/









































































































Author: Работнова Т.А.
Tags: география растений фитогеография флора географическое распространение растений биология биологическая флора
Year: 1975




Text
БИОЛОГИЧЕСКАЯ
ФЛОРА
Московской области
БИОЛОГИЧЕСКАЯ
ФЛОРА МОСКОВСКОЙ ОБЛАСТИ
Под редакцией профессора Т. А. РАБОТНОВА
Выпуск второй
ИЗДАТЕЛЬСТВО МОСКОВСКОГО УНИВЕРСИТЕТА
1975
УДК 581.9 (471. 311)
BIOLOGICAL FLORA OF THE MOSCOW REGION
Chief Editor prof. T. A. Rabotnov
Part 2
Edited by
I. A. Gubanov and M. G. Vakhrameeva
Книга содержит разносторонние сведения о 20 видах растений, широко распространенных в Подмосковье и других областях СССР. Многие из них широко известны как лекарственные и пищевые растения: лещина, горец, горицвет, рябина, черемуха, чабрец, бузина и др. Охарактеризованные в книге виды клевера — важнейшие кормовые растения. Включены материалы о некоторых подмосковных орхидеях и хохлатках, которые из-за своей декоративности сильно истребляются и поэтому нуждаются в охране.
Печатается по постановлению Редакционно-издательского совета Московского университета
Редакторы выпуска И. А. Губанов и М. Г. Вахрамеева
Рецензенты:
докт. биол. наук А. Г. Воронов, канд. биол. наук В. С. Новиков, канд. биол. наук К. В. Киселёва
© Издательство Московского университета, 1975 г.
г 21006—076 077(02)—75
149—75
ПРЕДИСЛОВИЕ
Второй выпуск справочника «Биологическая флора Московской области» написан по тому же общему плану, что и первый, выпущенный Издательством Московского университета в 1974 г. Во втором выпуске охарактеризовано 20 видов подмосковных растений. В его составлении приняли участие сотрудники Московского государственного университета им. М. В. Ломоносова, Московского государственного педагогического института им. В. И. Ленина, Главного ботанического сада АН СССР, Ботанического института им. В. Л. Комарова АН СССР, Всесоюзного научно-исследовательского института информации и технико-экономических исследований по сельскому хозяйству, Всесоюзного научно-исследовательского института лекарственных растений, Первого Московского медицинского института им. И. М. Сеченова, Тамбовского педагогического института, Центральной лаборатории охраны природы МСХ СССР и Лаборатории лесоведения АН СССР.
Как и в предыдущем выпуске, статьи-монографии настоящей книги не равноценны. В каждой из них отдельные разделы написаны значительно полнее и обстоятельнее, чем другие. Тем не менее коллектив авторов и редакторов надеется, что помещенные в выпуске монографии позволят составить довольно разностороннее представление об охарактеризованных таксонах. Две статьи второго выпуска посвящены целым родам: хохлатке и чабрецу. В них изложены подробные сведения о всех зарегистрированных на территории Московской области видах растений этих родов. Остальные статьи характеризуют отдельные виды. Большая часть их посвящена родам, имеющим в Подмосковье только по одному представителю (гудайера, лещина, горицвет, рябина, черемуха, пролесник, борщевик, бузина, молокан). Следовательно, такие статьи охватывают род полностью. Прочие монографии характеризуют наиболее распространенные и практически важные виды полиморфных родов (любка двулистная, горец змеиный, клевер луговой, клевер розовый).
Второй выпуск отредактировали и подготовили к печати И. А. Губанов и М. Г. Вахрамеева под общим руководством
3
Т. А. Работнова. Научное рецензирование и отчасти первичное ре дактирование некоторых разделов провели Т. А. Работнов, Т. И. Серебрякова и В. Н. Тихомиров. Номенклатурные цитаты для все? видов, охарактеризованных в выпуске, составили по единому плану (или дополнили и уточнили) И. А. Губанов и В. Н. Тихомиров. На звания грибов и других низших растений в первом и втором выпу сках выверила Г. Д. Успенская, а названия насекомых и други; членистоногих — А. Д. Никитина. Авторы и редакторы принося' благодарность всем лицам, принявшим участие в создании настоя щей книги или способствовавшим появлению ее в свет. Особук признательность выражаем официальным рецензентам первого второго выпусков В. Н. Вехову, А. Г. Воронову, К- В. Киселеве?
В. С. Новикову, А. К. Скворцову.
Поскольку «Биологическая флора Московской области» мыс лится как многотомное издание, редакция ее крайне заинтересован в оценке первых выпусков. Заранее благодарим коллег и коллект! вы, нашедших время прорецензировать наш труд и сообщить свс мнение как по содержанию выпусков и качеству публикуемых м; териалов, так и по оформлению. Наш адрес: Москва, 117234, Лени; ские горы, МГУ, биологический факультет, кафедра геоботаники.
М. Г. Вахрамеева и Л. В. Денисова
ГУДА Й ЕРА ПОЛЗУЧАЯ
Номенклатура
Goodyera repens (L.) R. Br. in Ait., Hort. Kew., ed. 2, 5 (1813) 198; Кауфм., Моск, фл., изд. 1 (1866) 479, изд. 2 (1889) 494; Цингер, СО. свед. (1885) 423; Шмальг., Фл. 2 (1897) 449; Сырейщ., Илл. фл. I (1906) 268; он же, Опр. раст. Моск. губ. (1927) 97; Невский во Фл. СССР 4 (1935) 639; Назаров в Маевск., Фл., изд. 7 (1940) 245; Голицын, там же, изд. 8 (1954) 693, изд. 9 (1964) 680; Скворцов в Опр. паст. Моск. обл. (1966) 124.— Satyriutn repens L., Sp. pl. (1753) 11-15.— Гудайера ползучая, гудьера ползучая, ползучка. (Сем. Orchi-diiccae.)
Латинское название рода дано в честь английского ботаника Джона Гудайра (J. Goodyer), работавшего в XVII в. (Watt, Renwick, 1915).
Goodyera R. Br.— широко распространенный, в основном горно-лесной азиатский род. Содержит 25 видов, встречающихся преимущественно в тропической и умеренной Азии и Америке, несколько видов отмечено в Австралии, Новой Каледонии и Самоа. В Европе — 1 вид, в СССР — 1 вид.
Морфологическое описание
Многолетнее травянистое растение со шнуровидным плагио-тропным корневищем, которое ветвится, образуя на растущих концах ортотропные побеги или розетки листьев. Корневище обычно не проникает глубже, чем на 2,5—5 см. Все корни придаточные, немно-,'очисленные, достигают 2,5 мм в диаметре и от 2,5 до 8 см в длину. Размещаются в основном в подстилке, не достигая минерального лоя почвы. Корни густо покрыты корневыми волосками, которые к'тречаются маленькими изолированными группами и на корневищах (Mollison, 1943). Более многочисленные крупные корни образуются на участках корневища, несущих-генеративные побеги.
Стебель выходит из середины листовой розетки, очень крепкий, <руглый, высотой от 6—7 до 20 см (редко 25—30), в верхней полотне, а иногда и по всей длине коротко-железистоопушенный, несет
5
мелкие линейно-ланцетные плотно прилегающие листья. Подземные побеги кончаются 3—8 листьями, образующими розетку или несколько отстоящими друг от друга.
Розеточные листья удлиненно-яйцевидные или эллиптические, заостренные, 1,8—3,5 см длиной и 1—2 см шириной, к основанию закрученные или клиновидно суженные в короткий широкий черешок. По жилкованию гудайера резко отличается от других орхидных — листья имеют ясно заметную сетку из 3—7 продольных и многочисленных поперечных белых или желтоватых жилок. Белый рисунок листа объясняется присутствием воздушных полостей (Fuller, 1962).
Соцветие — колос (2) 4—7 (15) см длины, цветки расположены однобоко или слегка спирально. Прицветники зеленоватые, заостренные, обычно длиннее завязи. В соцветии 10—20 (до 30) цветков. Цветки мелкие, белые или желтовато-белые, снаружи коротко-же-лезистоопушенные. Наружные листочки околоцветника овальные или продолговато-овальные, средний сливается вместе с двумя листочками внутреннего круга, образуя шлем. Лепестки 3—4 мм длины, 0,75—2,5 мм ширины, губа без шпорца, сильно вытянутая, вогнутая, на конце с треугольным носиком до 1 мм длины (Флора СССР, 1935). Гиностений короткий; клювик прямой, заостренный, короткодвураздельный; пыльник прямой; поллинии в числе 2, сидячие, обратнояйцевидные, связанные довольно большой округлой или ланцетной железкой. Завязь небольшая (значительно меньше, чем у других орхидей), сначала удлиненно-кругловатая, потом веретенообразная, почти сидячая, железисто-коротковолосистая, при основании слегка скрученная. Плод — одногнездная коробочка с продольным гребешком внутри. Семена очень мелкие, коричневые (Downie, 1943 а, Ь). Вес 1000 семян 0,002 г (Кожевников, 1950).
У гудайеры наблюдается полиэмбриония (Поддубная-Арноль ди, 1953). Диплоидный набор хромосом при неоднократных иссле дованиях оказался различным: во Франции — 28—30, в Скандина вии и Нидерландах — 30, в Голландии — 40, в Японии — 32 (Хромо сомные числа цветковых растений, 1969; Schotsman, 1970). Связт между морфологией, числом хромосом и географическим распро странением популяций гудайеры в Европе не выяснена (Schotsman 1970).
Географическое распространение
Гудайера ползучая — голарктический вид, ареал которого тесн< связан с ареалом хвойных пород и охватывает Скандинавию, Цент ральную и Атлантическую Европу, Балканский п-ов, Малую Азию Афганистан, Гималаи, Северную Америку. В СССР она встречаете; в лесах европейской части, Кавказа, Западной и Восточной Сибирц Казахстана. Она является самым северным представителем род; (Hulten, 1968). Карта ее распространения в СССР приведена в ра боте А. И. Толмачева (1954). В Московской обл. встречается изред
6
Кл и Северном, Клинско-Дмитровском, Западном, Восточном и Приокском флористических районах, а также собрана в Серебряном бору и Москве (Ворошилов и др., 1966).
Широкое и прерывистое распространение, а также малая вариабельность свидетельствуют о значительной древности гудайеры ползучей как вида (Толмачев, 1954). В ледниковый период гудайера сохранялась в реликтовых сосновых рощах, а на юге в горных ельниках. По мере отступания ледника она переселялась в смешанные и березовые леса, расширяя свой ареал. Развитие широколиственных лесов привело к ее исчезновению из ряда мест в Европе (Zlegcnspeck, 1936).
Онтогенез и сезонный ритм развития
Семя гудайеры состоит из семенной оболочки и недифференцированного эмбриона. Различные стадии развития эмбриона подробно изучены И. Вейратом (Veyrat, 1956). Эмбрион имеет сравнительно большой запас питательных веществ в виде масел, содержащихся почти во всех клетках, за исключением меристематических. Прорастание семени происходит только при наличии грибов из рода Rhizo-rlonia (Downie, 1943 а, с). Водная вытяжка из гифов гриба стимулирует развитие эмбриона, спиртовая вытяжка вызывает быстрое, по ненормальное развитие (Downie, 1950).
Первые два — четыре года проросток ведет подземный образ жизни и является микотрофом. Лишь на пятый год на корневище, которое к этому времени имеет 3 членика (междоузлия), появляют-первые зеленые листья и растение, по крайней мере частично, юрсходит к автотрофному способу питания. В условиях хорошего (снещения зеленые листья появляются значительно раньше, даже П1 второй год (Ziegenspeck, 1936; Fuller, 1962). Более старые участ-<п корневища с этого времени начинают отмирать. Цветение, видимо, может наступить не ранее пятилетнего возраста (Fuller, 1962).
При ветвлении корневища из пазушной почки в первый год шзвивается 3 маленьких членика корневища, на следующий год щс 3 членика. При этом ассимиляция отсутствует, поступление штательных веществ из старого корневища ограничено, а питание, io-видимому, в основном происходит за счет гриба-симбионта. На четвертый год образуется 2 членика, несущих чешуйки, и третий лсник с корнями и несколькими большими чешуйчатыми листьями. 1а пятый год междоузлия становятся толще, вырастают настоящие пстья. На шестой год корневище начинает частично отгнивать. На едьмой-восьмой год корневище становится положительно фототро-ичным и отрицательно геотропичным, появляются ортотропные по-еги, несущие соцветия. В подземной (плагиотропной) части корне-щца восьмого года жизни гифы грибов отсутствуют, происходит еиленное развитие придаточных корней.
После цветения побег отмирает, его замещают побеги, развива-щиеся из боковых почек. До цветения побеги гудайеры развива
7
ются моноподиально, после отмирания цветущего побега наблюдается симподиальное ветвление, затем побеги нарастают моноподиально до следующего цветения (Ziegenspeck, 1936). Иногда в неблагоприятных условиях, особенно при затенении, развитие побегов происходит более замедленно, они ведут подземный сапротроф-ный образ жизни. В некоторых случаях даже особи целиком могут вести сапротрофный образ жизни (возможно, они находятся в состоянии покоя), но при улучшении условий они быстро образуют надземные вегетативные и генеративные побеги.
Отмечено и обратное явление, когда побеги могут достичь поверхности почвы и образовать маленькие зеленые розетки листьев в первый же год. Розетки сохраняются в течение зимы, на следующий сезон растут, а на третий год могут дать цветочную стрелку (МоШ-son, 1943). По-видимому, скорость развития зависит от степени освещенности местонахождения. На корневищах имеются спящие почки, сохраняющие жизнедеятельность в течение ряда лет. По мере отмирания старых участков корневища они могут просыпаться и развиваться, если этому не препятствует отсутствие гриба.
Гудайера — единственная в СССР зимнезеленая орхидея. Листья у нее зимуют и отмирают лишь весной следующего года, а иногда и на третий год, поэтому стебель у основания имеет много вялых, но хорошо сохранивших форму листьев. Цветочную стрелку гудайера выбрасывает рано, в конце мая —начале июня, но цветение начинается не ранее середины июля и продолжается в условиях Подмосковья в течение трех недель. Плоды созревают через 1,5—2 месяца.
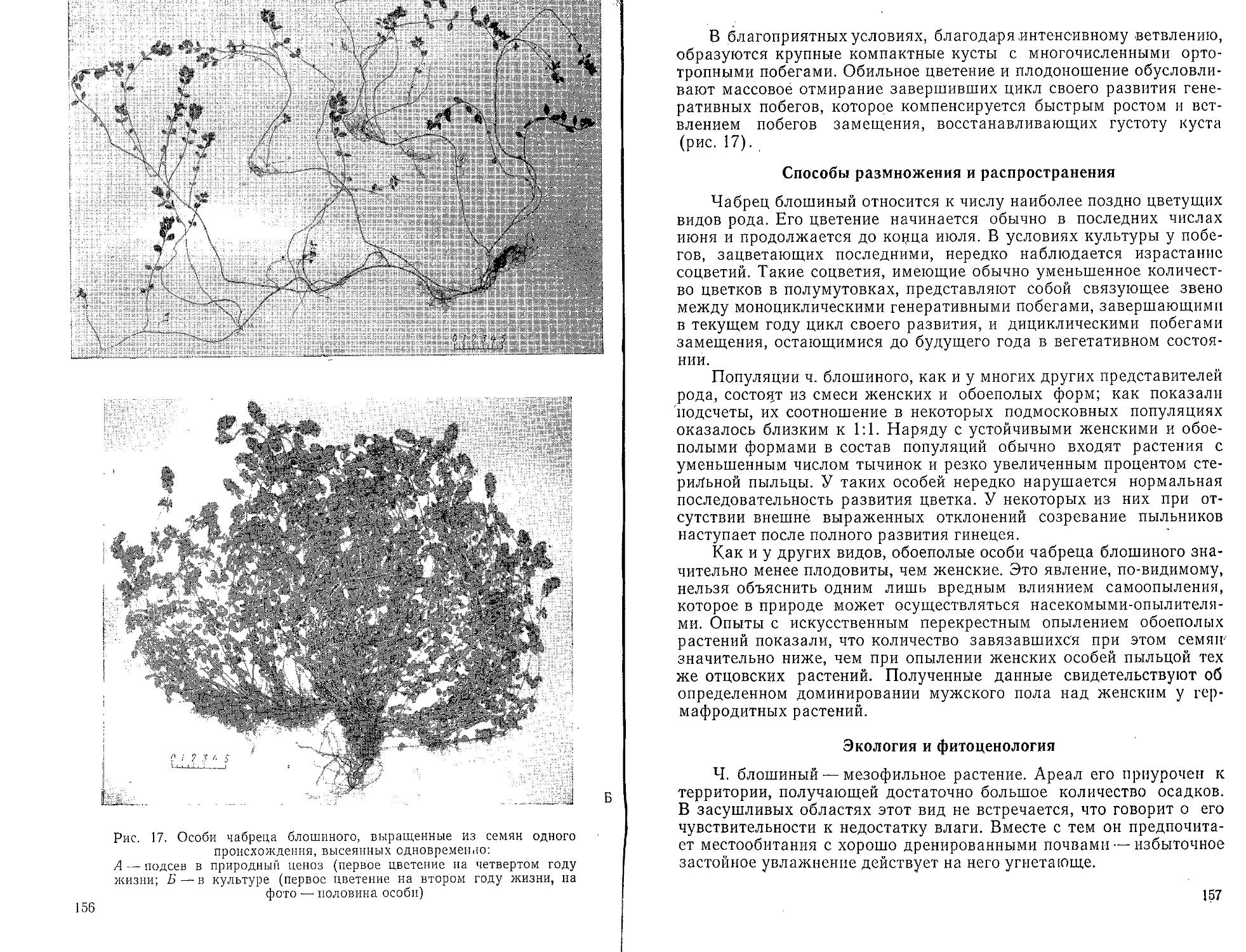
Способы размножения и распространения
Для гудайеры характерно только перекрестное опыление. Цветки имеют сладковатый запах,'привлекающий насекомых. Перекрестное опыление обеспечивается своеобразным расположением пестика и тычинок и строением околоцветника, листочки которого расположены так, что нектар можно достать только через узкую щель. Поллиний расположен около нижней лопасти, а нижняя губа несет кармашек с липким секретом. Насекомое, берущее нектар, касается кармашка с секретом, к нему прилипает поллиний. При посещении следующего цветка поллиний с насекомого прикрепляется к рыльцу.
Семена распространяются при помощи ветра. Семенное возобновление возможно лишь в присутствии гриба-эндофита, поэтому вегетативное размножение имеет не меньшее значение. Новые места гудайера заселяет при помощи семян, но размножение в освоенных местообитаниях происходит в основном вегетативным путем (Ziegenspeck, 1936).
Вегетативно гудайера размножается при помощи подземных побегов, развивающихся из почек на корневище. Через несколько лет старые участки корневища отмирают, а оставшиеся молодые участки корневища становятся самостоятельными растениями. В ре
8
зультате часто образуются куртины значительных размеров (Molli-son, 1943). Вегетативное размножение облегчается большой самостоятельностью отдельных частей корневища, которые сами могут переваривать грибы и переходить к автотрофному питанию.
Экология
Гудайера предпочитает умеренно влажные местообитания, избегая как избыточно влажные и заболоченные, так и сухие местообитания. Пониженные температуры, по-видимому, переносит хорошо, так как растет.в суровых условиях на высоте до 2000 м над уровнем моря. К тому же ее листья остаются зелеными даже под снегом.
Особенно чувствительна гудайера к свету: при сильном затенении она переходит на сапротрофный способ питания и может существовать несколько лет в виде подземных побегов, но затем отмирает, если не улучшатся условия. При осветлении местообитаний (вы-падение^деревьев, вырубка и т. п.) появляются зеленые цветущие особи. Этим объясняется неожиданное появление многочисленных генеративных побегов гудайеры на вырубках. В Подмосковье гудайера встречается при освещенности от 1,5 до 11 тыс. люкс, но наиболее многочисленна в местообитаниях с освещенностью 3—6 тыс. люкс.
Гудайера относительно мало требовательна к почвам — встречается на подзолистых, дерново-подзолистых и торфянистых почвах, на суглинках, песках, известняках, на кислых и близких к нейтральным почвах. В Подмосковье чаще встречается на сильно- и среднеподзолистых почвах (pH водное 4,4—4,9; pH солевое 3,4—3,6).
Биологическая продуктивность
В подмосковных ельниках-зеленомошниках в местах наибольшей численности надземная масса гудайеры достигает 125 кг/га сырого и 34,9 кг/га воздушно-сухого веса, а подземная масса — 76 кг/га сырого и 34,9 кг/га воздушно-сухого веса.
Фитоценология
Растет гудайера по тенистым хвойным (еловым, сосновым, пихтовым, кедровым, лиственничным) и смешанным лесам, предпочитая мшистые, богатые гумусом участки. Произрастает куртинами, часто вместе с грушанками и некоторыми орхидеями (из родов Lis-tera, Neottianthe, Corallorhiza и др.).
В Московской обл. встречается в ельниках и сосняках зеленомошных, борах на дюнах, ельниках-черничниках и брусничниках, смешанных сосново-еловых и елово-мелколиственных лесах, однако в чистых мелколиственных лесах, выросших на месте сведенных хвойных лесов, как правило, исчезает. Иногда растет большими
9
скоплениями, насчитывающими сотни розеток, но никогда не доминирует в травяном ярусе. У нас на открытых местах не встречается, но в Шотландии отмечена на пустошах (Downie, 1943 с).
Консортивные связи
Симбионтами гудайеры являются грибы из рода Rhizoctonia. Чаще встречается R. goodyera-repentis, который живет как сапро-троф на опавшей сосновой хвое, не проникая в глубокие слои почвы (Downie, 1943 а, Ь, с). Гриб стимулирует прорастание семян гудайеры, пронизывая базальные клетки гифами в виде завитков (Molli-son, 1943). В культуре семена заражаются грибом через 5 дней после посева, рост эмбриона начинается немедленно после проникновения гиф в его клетки. Через некоторое время гриб теряет свою жизненность и переваривается гудайерой, что подтверждается наличием участков корней и корневищ, свободных от него. Обычно нет гриба в корневой меристеме. Надземные части также грибом не заражены (Mollison, 1943).
Внутривидовая систематика
В Подмосковье можно выделить несколько форм гудайеры, отличающихся размером и окраской листьев: пестролистную, мелколистную и крупнолистную. Все три формы встречаются в одних и тех же тийах леса, выявить их приуроченность к каким-либо особым условиям не удалось. Известны попытки описания географических рас гудайеры ползучей в ранге разновидностей, но общего признания выделенные таксоны не получили (Fernaid, 1899; Schot-sman, 1970). Э. Хультен (Hulten, 1968) считает этот вид мало изменчивым.
Н. Г. Царевская
ЛЮБКА ДВУЛИСТНАЯ
Номенклатура
Platanthera bifolia (L.) L. C. Rich., Mem. Mus. Paris, 4 (1818) 47; Кауфм., Моск, фл., изд. 1 (1866) 472, изд. 2 (1889) 488; Цингер, Сб. свед. (1885) 419; Сырейщ., Опр. раст. Моск. губ. (1927) 96; Невский во Фл. СССР 4 (1935) 656; Назаров в Маевск., Фл., изд. 7 (1940) 246; Голицын, там же, изд. 8 (1954) 696, изд. 9 (1964) 683; Скворцов в Опр. раст. Моск. обл. (1966) 125.— Orchis bifolia L., Sp. pl. (1753) 939, p.p.— Platanthera solstitialis Boenningh. in Reich, f., Fl. Germ. Exc. (1830) 120; Сырейщ., Илл. фл. 1 (1906) 259, 4 (1914) 47.— Habenaria bifolia R. Br. in Ait., Hort. Kew, ed. 2, 5 (1813) 193; Шмальг., Фл. 2 (1897) 462.— Любка двулистная, ночная фиалка. (Сем. Orchidaceae.)
Морфологическое описание
Многолетнее травянистое растение, характеризующееся наличием особых запасающих органов—-корневых клубней, образованных из придаточных корней (Федоров и др., 1962). Ко времени цветения имеются два клубня, из которых один молодой, гладкий, наполненный запасными веществами; другой — старый (прошлогодний), сморщенный, истративший значительную часть своих запасных веществ на развитие стебля с листьями и цветками (Irmisch, 1850; Флора СССР, 1935). Клубни неразделенные, сравнительно большие (0,6—2,8 см длиной и 0,4—1,5 см шириной), несколько удлиненные, почти свеклообразные, со шнуровидным окончанием (Hegi, 1909; Рзазаде, 1952).
В зависимости от возраста имеет от 1 до 10 поглощающих корней. По нашим данным, удельный вес этих корней значительно меньше, чем у шнуровидного окончания клубня, соответственно 651 н 1013 мг/см3, что объясняется, видимо, различиями в их анатомическом строении, так как они выполняют разные функции. .
И. Г. Серебряков (1962) включает любку в класс наземных травянистых поликарпических растений с ассимилирующими побегами несуккулентного типа. Многодетность особи обеспечивается
почти полным обновлением всего растения за счет образования замещающих корневых клубней. Ю. А. Лукс (1970) считает, что наземные орхидеи с корневыми клубнями следует отнести к особой жизненной форме наземных травянистых поликарпических растений с корневыми клубнями, ежегодно полностью или почти полностью обновляемых.
Любка имеет два сближенных, почти супротивных, эллиптических, тупых, к основанию суженных в крылатый черешок зеленых листа. Листья длиной 4—17 см (с черешком) и 0,5—6 см шириной, голые, блестящие, цельнокрайние, однотонные. Лист имеет 2—12 жилок, дугообразно расположенных, сближенных при основании и у верхушки пластинки и соединенных тонкими перемычками, поперечно или косо направленными. Более крупные жилки выступают в виде ребер на нижней стороне листа.
Цветоносный стебель прямостоячий, высотой 43± 13 см, желто-зеленый, полый, при основании с буроватыми влагалищными листьями. Соцветие довольно рыхлое, почти цилиндрическое, 12±6 см длиной, многоцветковое, несет 14±6 цветков. Прицветники травянистые, ланцетные или узколанцетные, туповато-заостренные, длиной 1,4±0,3 см, шириной 0,4±0,1 см.
Цветки обоеполые, с приятным запахом, особенно сильным вечером, ночью и при пасмурной погоде. Околоцветник трехчленный из двух кругов, венчиковидный, зигоморфный, снабжен тонким нитевидным шпорцем, на дне которого скапливается нектар. Листочки околоцветника белые, на концах зеленоватые.
Из 6 тычинок развита только 1 в наружном круге. На месте 2 передних тычинок внутреннего круга — 2 маленьких стаминодия. Пыльник с узким связником состоит из двух гнезд, открывающихся продольной щелью. Пыльца в каждом гнезде склеена в поллинии.
Гинецей паракарпный из трех плодолистиков. Завязь нижняя, сидячая, скрученная, одногнездная, с тремя париентальными плацентами, на которых расположены многочисленные мельчайшие семяпочки. Рыльце имеет вид железистой вогнутой площадки, расположено ниже основания гнезд пыльника.
Плод — коробочка длиной 1,6±0,2 см, раскрывается шестью продольными щелями. Семена темно-коричневые, очень мелкие, без эндосперма, с широкой прозрачной сетчатой оболочкой и недифференцированным шаровидным зародышем.
Географическое распространение
Любка двулистная широко распространена в Европе и на значительной части Азии (Ziegenspeck, 1936). Самое северное ее местонахождение— в Норвегии, на 70°9' с. ш. В целом северная граница ареала в Европе совпадает с границей распространения деревьев. Она растет высоко в горах: в Баварии до высоты 1700 м над уровнем моря, в Центральных Альпах местами поднимается до 2300 м. В средиземноморских районах любка очень редка: отмечена в горах
12
Северной Африки, на севере Испании, в горах Италии и в Балкан-, ских странах севернее Греции. На восток доходит через Сибирь и Среднюю Азию до Китая и Японии, на юге — до Гималаев.
На территории СССР любка двулистная встречается в европейской части, на Кавказе, в Западной Сибири, на Алтае, в Саянах и на Дальнем Востоке. В Московской обл. обычна во всех районах.
Онтогенез и сезонный ритм развития
Благодаря ежегодной смене клубней большой жизненный цикл любки двулистной представляет непрерывную последовательность поколений особей, ежегодно сменяющих друг друга.
Всходы любки ведут подземный образ жизни. Уже в первую осень проросток имеет микоризу (Burgeff, 1936). Ось молодого проростка (протокорм) сильно заражена эндофитом, не проникающим только в область, близкую к точке роста, из которой развивается несколько поглощающих корней. Первый клубень образуется у основания второго листа возле точки роста. У клубня вырастает длинная ножка и он погружается в почву. В конце вегетационного сезона протокорм, его листья и корни отмирают, остается только клубень, из которого в следующем году формируются листья и придаточные корни. Инфекция никогда не проникает в клубень, корни заражаются из почвы. Образование первого более или менее выраженного запасающего клубенька и первого зеленого листа происходит на третий год жизни (Ziegenspeck, 1936).
У ювенильных растений только один лист и один (реже два) поглощающий корень. Лист узкий, длиной 4,95±0,37 см (без черешка) и шириной 0,96±0,11 см, с 2—6 жилками. Имматурные растения крупнее, с одним, реже двумя листьями и двумя-тремя поглощающими корнями. Листья длиной 7,12±0,58 см и шириной 1,7 ± 0,16 см, с 8—10 жилками.
Взрослые вегетативные растения с одним, чаще двумя листьями и четырьмя-пятью поглощающими корнями. Листья имеют более 10 жилок, длину 9,65±0,57 см и ширину 2,91 ±0,18 см. По размерам они близки, а иногда даже больше листьев генеративных особей. Вполне возможно, что в эту группу входят и особи постгенеративные.
Любка может зацветать на шестой-седьмой год (Ziegenspeck, 1936). Генеративные растения имеют листья длиной 10,08±0,49-см и шириной 3,18±0,18 см, с числом жилок более 10. Поглощающих корней у них 6. Судя по наличию у некоторых генеративных особей остатков цветоносных побегов прошлого года, .любка может цвести, по крайней мере, два года подряд.
Сенильные особи мы не наблюдали, однако было найдено несколько экземпляров взрослых вегетативных и генеративных особей, которые не заложили замещающего клубня на следующий год. По-видимому, эти особи можно считать сенильными.
13
Изменение биоморфологических признаков любки двулистной в зависимости от возрастного состояния
Возрастные состояния особей
Показатели ювенильные имматурные взрослые вегетативные генеративные
Длина листа, см 4,95±0,11 7,12±0,58 9,65 + 0,57 10,08±0,49
Ширина листа, см 0,96±0,11 1,70±0,16 2,91 ±0,18 3,18±0,18
Площадь листа, см2 3,86 + 0,63 9,7± 1,3 23,3 + 2,54 26,01+2,4
Число жилок 2—6 8-10 более 10 более 10
Число поглощающих корней 1,46±0,31 2,38±0,29 4,36±0,29 6,62 + 0,41
Средняя длина поглощающих корней, см 1,51 ±0,53 1,48 + 0,15 2,38±0,25 3,41 ±0,37
Длина шнуровидного окончания клубня, см 1,8±0,66 1,8±0,46 3,38 + 0,49 4,0± 1,13
Глубина укоренения, см 3,43±0,46 4,23± 1,0 6,99±0,49 8,38± 1,27
Диаметр распространения корней, см 2,1 ±0,48 2,03+0,32 3,76±0,41 5,5±0,69
Соотношение воздушно-сухого веса надземной и подземной частей 2,85 + 0,46 3,16 + 0,84 4,02 + 0,29 1,76+0,36
Число устьиц на 1 - мм2 листа 66,±3,4 69+2,8 79±4,1 86 + 4,8
Протяженность жилок на 1 см2 листа 26,5+1,3 26,5 + 1,4 28,2 + 1,6 28,1 + 1,8
Удельная плотность листьев, мг/см2 21,4 24,2 22,1 25,3
В таблице приведены данные об изменении различных биоморфологических показателей при переходе особей любки из одного возрастного состояния в другое.
Мы изучали численность и спектр возрастных состояний популяций любки двулистной в различных условиях местообитания в Одинцовском р-не Московской обл.: в ельнике и сосняке- зеленомошных, в злаково-разнотравных березняках, на щучково-белоусо-во-лапчатковом лугу и на просеке, поросшей душистым колоском, лапчаткой прямостоящей и подростом ели и березы. Установлено, что на всех участках любка двулистная образует популяции нормального типа. Однако на более освещенных участках —на лугу, просеке и в разреженном березняке — процент генеративных особей выше (29—41,7%), что свидетельствует о том, что эти условия для любки двулистной ближе к оптимальным.
14
Способы размножения и распространения
Для любки двулистной характерно перекрестное опыление, ооычно осуществляемое ночными бабочками (Дарвин, 1928). Вскоре после опыления околоцветник вянет, а колонка и завязь разрастаются. Увядший околоцветник сохраняется до созревания плода. Довольно много цветков остаются неопыленными.
Созревание семян происходит приблизительно через 1,5—2 месяца, а растрескивание плодов — приблизительно через 2—2,5 месяца после опыления. Мелкие необычайно легкие семена любки образуются в огромном числе и широко распространяются воздушными течениями. Легкость и летучесть обеспечиваются наличием нежной однослойной сетчатой семенной кожуры и воздушной полости.
Чрезвычайно мелкие размеры и недифференцированность зародыша, а также весьма незначительное количество запасных веществ, которыми могут располагать проростки, затрудняют прорастание и дальнейшее развитие проростков. Симбиоз орхидей с грибами, образующими эндомикоризу, возмещает недостаток запасных веществ в семенах и облегчает образование проростков.
Семенная продуктивность высокая: в среднем число цветков в соцветии 14, плодов — 11, семян в одной коробочке 5384± 1684, средняя семенная продуктивность особи — 59224+ 18418 семян. Средний вес семян в одной коробочке 5,14 + 0,41 мг.
Ежегодно особи любки образуют только один замещающий клубень с ростовой почкой будущего года, т. е. коэффициент вегетативного размножения равен 1:1. Т. Ирмиш (Irmisch, 1850) подробно показал на примере любки двулистной процесс заложения и развития замещающих корневых клубней и ростовых «почек». Он отмечает наличие только одного замещающего корневого клубня, что подтверждают и наши данные. Однако Ю. А. Лукс (1970) неоднократно наблюдал у этого растения образование одного-двух дочерних клубней помимо одного основного замещающего.
Способы питания
Любка двулистная — микотрофное растение, ее микориза относится к эндотрофному типу (Горбунова, 1956). Помимо сапротроф-ного питания, благодаря наличию зеленых листьев, она еще и фотосинтезирует как обычный автотроф (Терехин,1965).
После проникновения гриба в семя и внедрения его в клетки базальной части зародыша зародыш переходит к паразитному способу питания (Ziegenspeck, 1936; Горбунова, 1956; Терехин, Кфме-лина, 1969). У зародышей всех орхидей, в том числе и любки, местом, через которое внедряется гриб, является точка прикрепления подвеска.
Если агрессивность внедряющегося гриба невелика, то его мицелий хорошо проникает в клетки зародыша, где образует клубки гиф, но быстро подвергается в них внутриклеточному переварива
15
нию (Ziegenspeck, 1936; Магру, 1949), в результате чего растение получает питательные вещества. Любка двулистная осенью до зимы и ранней весной (пока растение не имеет листьев) питается за счет корней, переваривающих грибные гифы. Вполне вероятно, что переваривание в основном играет защитную роль, и обмен веществ происходит непрерывно через активные поверхности живого гриба в клетках хозяина (Лобанов, 1963).
С большинством орхидных микоризу образуют несовершенные грибы из рода Rhizoctonia (Burgeff, 1936; Bernard, 1899), ярко выраженные аэробы, страдающие от недостатка кислорода. В качестве источника углерода они могут использовать многие органические вещества. В исследованных нами подземных органах любки также обнаружены гифы микоризного гриба, который проникает только в поглощающие корни, а клубни от него свободны, что подтверждает точку зрения Н. Бернара (Bernard, 1911). Гифы гриба обнаружены и в корнях взрослых растений. Следовательно, любка является частичным сапротрофом в течение всей жизни. Н. Бургеф (Burgeff, 1909) утверждал, что взрослое растение любки двулистной связывают с почвой около 3200 гиф.
Экология
Любка двулистная отличается довольно большой экологической амплитудой. В отношении освещенности она довольно требовательна, явно .предпочитая хорошо освещенные места, однако может выдерживать и значительное затенение. В условиях Одинцовского р-на Московской обл. встречается при относительной освещенности от 11,2 до 87,5 и даже 100%. Содержание хлорофилла в листьях — от 0,7 до 1,3 мг/г — также говорит о светолюбии этого вида.
К влажности почвы любка довольно безразлична, избегает лишь совсем сухих и застойно-увлажненных местообитаний. К реакции среды также не очень требовательна, встречается как на кислых, так и на нейтральных почвах. В Московской обл. чаще растет на подзолистых и дерново-подзолистых суглинистых и супесчаных почвах с pH 4—6.
По способу перезимовки любку можно отнести к корнеклубневым геофитам.
Фитоценология
Любка двулистная не обнаруживает привязанности к определенным растительным сообществам. В Западной Европе она встречается в лиственных лесах, прежде всего в буковых (Ziegenspeck, 1936). На Карпатах также приурочена к буковым лесам, которые, как и в Альпах, включают в себя также явор, ель, граб. Растет она и в еловых, дубовых и смешанных елово-широколиственных лесах, нередко и на лугах, особенно суходольных. В Московской обл. любка двулистная встречается в разнообразных сообществах: в ельни
16
ках-зеленомошниках (преимущественно близ «окон»), в березняках, елово-березовых лесах, в сосняках, на лугах и вырубках.
Существенного воздействия на окружающие виды растений любка двулистная, по-видимому, не оказывает. Однако, поселившись на какой-то территории, стойко удерживает свои позиции, даже оказавшись в неблагоприятных условиях (Ziegenspeck, 1936).
Охрана вида
Любка двулистная — изящное растение с довольно крупным соцветием, обладающим приятным запахом. Эти декоративные свойства приводят, к сожалению, к уничтожению растения в огромных количествах. В окрестностях Москвы, на тех участках, где недавно она была обильна, теперь встречаются лишь единичные экземпляры. Популяции ее неуклонно «стареют», например, в Одинцовском р-не ювенильные особи, по нашим подсчетам, составляют от 2 до 16%, имматурные — 2—11, а взрослые вегетативные — 50—74%.
Причины сокращения численности любки, по-видимому, следующие: отрицательное воздействие антропогенного фактора (прежде всего постоянное уничтожение значительного числа генеративных особей), трудность, прорастания семян (обязательное присутствие определенных микоризных грибов), отсутствие вегетативного размножения. Достаточно высокие темпы сокращения численности особей указывают на необходимость организации охраны этого вида. Любка двулистная должна быть отнесена, по крайней мере, к растениям ограниченного, регулируемого сбора.
11, Л, Аксенова и М. Г. Вахрамеева
ЛЕЩИНА ОБЫКНОВЕННАЯ
Номенклатура
Corylus avellana L., Sp. pl. (1753) 998; Кауфм.,Моск. фл., изд. 1 (1866) 441, изд. 2 (1889) 456; Цингер, Сб. свед. (1885) 391; Шмальг., Фл. 2 (1897) 427; Сырейщ., Илл. фл. 2 (1907) 46; он же, Опр. раст. Моск. губ. (1927) 108; Бобров во Фл. СССР 5 (1936) 264; Назаров в Маевск., Фл., изд. 7 (1940) 267; Бобров, там же, изд. 8 (1954) 177, изд. 9 (1964) 180; Tutin in Fl. Eur. 1 (1964) 60; Ворошилов в Опр. раст. Моск. обл. (1966) 134.—Лещина обыкновенная, орешник обыкновенный, о. лесной. (Сем. Betulaceae.)
Латинское название рода происходит от «coris» — шлем, чепец (по форме плюски). Видовое название «avellana» лещина получила от имени города Avella (Авеллино) в Италии, где встречалась в больших количествах (Koch, 1872),
Морфологическое описание
Кустарник 4—5 м высотой с яйцевидной или плоскошаровидной кроной. Обладает мощной корневой системой, достигающей глубины 1—1,5 м, основная масса корней располагается в гумусовом горизонте (до 10—15 см). Оси лещины диаметром до 3—5 см (иногда 12—15 см) расходятся дугообразно и образуют опрокинутый конус. Многолетние побеги покрыты гладкой серо-коричневой корой, однолетние имеют тонкую серую опущенную кору, на двулетних побегах появляются пробковые бородавочки. Древесина рассеянно-сосудистая, белого цвета со светло-коричневым или слегка розовым оттенком, слабым блеском или тусклая. Годичные слои различаются плохо. Древесина умеренно тяжелая и умеренно прочная (Вихров, 1959).
Почки шарообразные (5X5 мм), слегка сдавленные, красновато-бурые, гладкие, сидят несколько в стороне от листового рубца, с 6—8 широкими кроющими чешуями, окруженными коричневой каймой, располагающимися на почке двурядно и косо. Листовой рубец полукруглый с 5 следами сосудисто-волокнистых пучков, размещенных в одну линию.
18
Листорасположение спиральное, может быть обозначено как 7з и */2 (Codaccioni, 1962). Листья сидят на коротких (0,5—1,2 см) железисто-щетинистых черешках. Прилистники продолговато-яйцевидные, покрыты волосками, рано опадают. Листовая пластинка обратнояйцевидная с неравно- или дважды зубчатым краем и не-равнобоко-сердцевидным основанием. Форма листовой пластинки меняется по мере роста листа и в зависимости от положения листа па побеге. М. Д. Киндякова (1969) предложила формулу для массового определения площади листьев лещины, закончивших свой рост: s = 0,8321.h. Средняя длина листовой пластинки 7—13 см, ширина 6—10 см. Молодые листья лещины рассеянно-опушенные, с возрастом верхняя поверхность становится голой, а на нижней опушение сохраняется главным образом по жилкам.
Лещина — однодомное растение с раздельнополыми цветками. Одиночные мужские цветки расположены на внутренней стороне кроющих чешуек и собраны в длинные густые цилиндрические повислые сережки около 5 см длиной. Каждый цветок состоит из кроющей чешуйки, двух прицветных чешуек и четырех тычинок, расщепленных до основания. Тычинки с короткими тычиночными нитями, несущими одногнездные пыльники с пучком волосков на верхушке, производят огромное количество пыльцы — в одной сережке до 4 млн. пыльцевых зерен (Нейштадт, 1953). Существует корреляция между размером пыльцевых зерен и составом растительного сообщества, размеры их варьируют от 22,5 до 36 мк (Dabrowsky, 1957). Теоретическая дальность полета пыльцы 121 км (Нейштадт, 1953).
Женские цветки сидят у основания кроющей чешуи двуцветных дихазиев, средний цветок, как правило, не развивается (Eichler, 1878; Willis, 1966). Ко времени распускания мужских сережек женские цветки выпускают красные рыльца, свешивающиеся в виде кисточек. Каждый женский цветок имеет по два прицветника и невзрачный приросший к верхушке завязи околоцветник из двух листочков с маленьким мелкозубчатым отгибом. Столбик с двумя нитевидными красными рыльцами, завязь нижняя, образована двумя плодолистиками (Имс, 1964). Некоторые авторы считают, что завязь лещины двугнездная, однако О. Хагеруп (Hagerup, 1942) доказал, что она имеет только одно гнездо с четырьмя семяпочками. Позднее при развитии плода плаценты и ножки семяпочек настолько сближаются, что образуется узкая полость между ними, которая разделяет гнездо завязи на две части. В это время завязи выглядят так, как будто они имеют перегородку и центральную плаценту. Изучение эмбрионального развития семяпочки показало, что на одном и том же растении могут встречаться завязи с 2 и 4 семяпочками, что зависит от времени исследования. На ранних стадиях в завязи находятся чаще 4 семяпочки, а к моменту оплодотворения их остается 2 (остальные недоразвиты и плохо заметны). Как правило, только 1 семяпочка из 4 развивается.и образует семя.
После оплодотворения завязь разрастается в плод-орех, окру
19
женный светло-зеленой листовидной оберткой-плюской, образованной сросшимися прицветниками (Сапанкевич, 1956). Орехи разнообразны по форме (Geitler, 1943): яйцевидные, продолговато-яйцевидные, плоско-округлые, широкоэллиптические, вальковатые, гладкие или с намечающимися продольными бороздками; по окраске от буровато-желтых до темно-бурых, матовые. В нижней части орех имеет беловатое пятно —место его прикрепления к плюске, которая у основания короткобархатистая или щетинисто-железистая, длиннее или короче ореха, сверху открытая. Она сильно варьирует по форме, длине, степени рассеченности. С. Н. Агаркова (1968) выявила корреляционную зависимость между формой ореха и содержанием в них ядра (г —+0,62). Содержание ядра в орехах удлиненной формы выше (до 50,9% по весу), чем в орехах округлой формы.
Географическое распространение
Ареал лещины обыкновенной занимает европейскую часть СССР (включая Крым), Кавказ, Центральную и Западную Европу, Малую Азию, северную часть Африки. Самая северная точка ареала— в Норвегии (67°56' с. ш.), южная — на севере Сирии (37° с. ш.). Западные пределы ареала — б°30' з. д., восточная граница проходит вдоль Уральского хребта, не переходя его.
На территории СССР ареал лещины имеет следующие очертания. Южная граница идет от Тирасполя на Кировоград, Днепропетровск, Ростов-на-Дону, устье Сев. Донца, станицу Усть-Медведин-скую, сёло Лапоть Саратовской обл. Далее по правобережью Волги граница поднимается до Куйбышева, по р. Самарке идет до ее среднего течения, поворачивает на восток в Оренбургскую обл., где проходит через поселки Малый Боклинский, Пономаревский, Шарлых-ский, затем переходит в Башкирскую АССР и южнее селения Ме-леуз поднимается до истоков р. Белой. Отсюда восточная граница идет по западным предгорьям Уральского хребта до устья р. Уфы, далее вверх по ее долине примерно до среднего течения. Затем граница идет на Красноуфимск (Свердловская обл.), Кунгур (Пермская обл.), Оханск, вниз по р. Каме до пристани Галево, на Воткинск (Удмуртская АССР), Советск (Кировская обл.), станцию Шарья, Буй, Грязновец, Черновец, Белозерск, Тихвин, Ленинград, Выборг.
В. В. Иванов (1951) обнаружил крупное местонахождение лещины в 200 км южнее сплошной границы распространения. Лещина росла в дубраве на левом берегу р. Урала близ селения Амангель-ды, что, возможно, свидетельствует о том, что в недалеком прошлом граница лещины проходила значительно южнее современной.
Палеогеографические сведения. Наиболее древние ископаемые остатки Corylus относятся к древнетретичному периоду. Л. А. Смолянинова (1929) указывает, что уже в олигоцене был найден плод, сходный С плодом нашей лещины. Остатки С. avellana были найдены у нас в позднетретичных отложениях на Алтае и в плиоценовых
20
отложениях верхнего Днепра и Верхней Волги. Основываясь на данных пыльцевого анализа, М. И. Нейштадт (1953) рассматривает историю расселения лещины, начиная с Днепровско-Валдайской межледниковой эпохи, когда лещина была широко распространена и образовывала густые заросли, граница которых проходила севернее Онежского озера. Затем в связи с начавшимся в конце межледниковой эпохи похолоданием лещина исчезает из состава леса. Е. М. Лавренко (1938) считает, что основным убежищем ее в этот момент был северо-балканский центр консервации теплолюбивой флоры, откуда лещина, первая из широколиственных пород, появилась в Центральной Европе после отступления ледника. Относительная суровость субарктического периода мешает ее распространению. В раннем голоцене, характеризующемся теплым климатом, лещина достигает пышного расцвета в Прибалтике и на 600 км продвигается на восток. К этому времени появляется второй центр распространения лещины в восточных Карпатах, откуда она начинает продвигаться на северо-восток и сливается с прибалтийским потоком в районе р. Припяти. Ранний голоцен считают временем, когда лещина была наиболее распространена. Она встречалась в это время на значительной территории Европы, в Карпатах поднималась на 300 м выше своей современной высотной границы. В Европе этот период называют фазой орешника. В конце раннего голоцена лещина продвигается на север, юг и восток. В среднем голоцене она достигает своей современной южнойтраницы, на востоке переходит Каму, на севере продвигается на 300 км дальше своей современной северной границы, доходя до среднего течения Сев. Двины. Одновременно уменьшается ее роль в западных районах. В позднем голоцене сильно развившиеся широколиственные породы снижают роль лещины в лесах. Если в раннем голоцене встречались и лещиновые леса, то в позднем голоцене лещина играет лишь роль подлеска (Нейштадт, 1953). В конце позднего голоцена начинается отступление лещины на юг и общее сокращение ареала, продолжающееся и поныне.
Онтогенез
Односеменные плоды лещины с твердым сухим деревянистым околоплодником сидят скученно по 2—5 на концах коротких листоносных побегов. Размеры орехов 18—20X12—15 мм. Вес 1000 орехов 0,64—1,5 кг (Джиаукштас, 1957). При сборе семена имеют всхожесть в среднем 75%, которая сохраняется, однако, недолго (Кощеев, 1950; Озол, Зуковска, 1955). На песчаных почвах грунтовая всхожесть осеннего посева 70%. Околоплодник построен склеренхимными волокнами с толстыми одревесневшими стенками, а снаружи и внутри — с двумя слоями каменистых клеток; с внутренней стороны покрыт тонкой пленкой, состоящей из нескольких рядов сдавленных тонкостенных клеток; толщина околоплодника 0,7—
21
2 мм (Заборовский, 1962). Скорлупа составляет от 47 до 52% веса ореха.
Семя состоит из тонкой светло-коричневой пластинчатой кожуры и белого зародыша с двумя толстыми семядолями, между которыми едва заметен корешок с зародышевой почкой. Благодаря густой сети проводящих пучков в наружнем слое кожура морщинистая. У набухшего семени она легко разделяется на два слоя: наружный— коричневый, с пучками спиральных сосудов и внутренний— светлый, в виде прозрачной пленки. В июле семя молочного цвета размером 2—3 мм, плод 12X10 мм, в августе в зрелом плоде почти вся полость занята семенем. Обычно даже на одном кусте можно обнаружить плоды разной степени спелости; бессемянные плоды попадаются редко — менее 5% (Заборовский, 1962). Скорлупа и семенная кожура водопроницаемы, наиболее интенсивно поглощают воду первые четверо суток. Зрелые семена без околоплодника быстро прорастают в вегетационных сосудах, в околоплоднике— лишь через 3 месяца. При весеннем посеве необходима стратификация при температуре 3—4° в течение 3—4 месяцев (Заборовский, 1962).
Вопросам прорастания семян лещины под воздействием различных факторов посвящен ряд работ. Показано, что покой семян может быть нарушен охлаждением до 5° в течение 30 дней (Bradbeer, Colman, 1967). Установлено, что в семенах, охлаждаемых при температуре 4° в течение 50 дней и затем содержащихся при 20° в течение 5 дней, происходит резкое увеличение интенсивно'сти метаболизма нуклеиновых кислот в семядолях (Wood, Bradbeer, 1967). Резко сокращают период покоя семян гиббереллин, кинетин и тиомочевина (Frankland, Wareing, 1966; Bradbeer, Pinfield, 1967; Pinfield, 1968).
При прорастании семядоли, заключенные в оболочку плода, остаются в почве и сохраняются до осени. Стебелек в нижней части несет мелкие недоразвитые низовые листья в виде двух очередных узколанцетных чешуек, утолщающихся к основанию. Первый лист почти округлый, по краю мелко-выемчато-зубчатый, на коротком черешке, с мелкими линейно-ланцетными прилистниками при основании. Второй лист более крупный, широкояйцевидный, при основании сердцевидный, неравномерно выемчато-зубчатый и реснитчатый по краю, на верхушке зубчиков железистые шипики. Третий и четвертый лист такие же, но более крупные. Прицветники мелкие, ланцетно-шиловидные. Стебли и черешки листьев усажены короткими, бело-сероватыми шелковистыми простыми волосками с примесью железистых. Корневая система стержневая, вверху веретеновидно утолщенная с обильными мелкими боковыми корнями. Развитие лещины в первый год жизни заканчивается формированием побега высотой 12—18 (до 40) см с 4 листьями (редко бывает недоразвившийся пятый).
Морфогенез лещины изучен подробно (Серебряков и др. 1954). В благоприятных условиях на 3—4 год начинается ветвление глав-
22
пой оси, однако главная ось еще отчетливо заметна. С 4—5 лет происходит ослабление роста главной оси, прирост постепенно сокращается: 20 см в 4 года, 15 см в 6 лет, 11 см в 8 лет, 5 см в 10 лет. На 12—14 год прирост главной оси составляет всего 1—2 см. Главная ось живет не более 13—16 лет. Боковые ветви с 4—5 лет мало уступают по росту главной оси, а к 9—10 годам обгоняют ее.
Ветвление главной оси происходит не только в надземной, но и в подземной части, где побеги образуются из спящих почек. У хорошо развитых кустов в условиях Подмосковья число скелетных осей может составлять 60—80 (до 150); диаметр таких кустов достигает 1,5 м. В первые годы жизни корневая система орешника имеет стержневой характер, но на 2—4 год появляются придаточные корни в надсемядольной сфере главной оси. С появлением новых скелетных осей появляются придаточные корни и в зоне перехода побегов от горизонтального к вертикальному положению. К 10 годам корневая система окончательно приобретает поверхностный характер. Рост орешника в различных условиях в первые 10 лет жизни исследовали М. С. Двораковский и Н. А. Захарова (1955). Они отмечают более быстрый рост лещины на супесях по сравнению с суглинками (разница в высоте около 10 см, а в диаметре — около 2 мм). Рост лещины неравномерен — на 4-м и особенно 6-м году жизни наступает резкий скачок прироста в высоту.
На хорошо освещенных участках лещина начинает плодоносить с 4—6 лет (Шиманюк, 1967). Наиболее высокие урожаи дает с 10— 15-летнего возраста (Круссер, 1940). Общая продолжительность жизни, по данным различных авторов, от 80—100 (Флора СССР, 1936; Biisgen, 1927; Харитонович, 1968) до 200 лет (Biisgen, 1913).
Сезонный ритм развития
Лещина — один из наиболее раноцветущих наших кустарников. В условиях Подмосковья она цветет в марте — апреле, до распускания листьев. В Белоруссии цветение наступает через 11—13 дней после перехода среднесуточной температуры через 0 (Шиманюк, 1967). Средняя продолжительность цветения 9—10 дней. В. А. Штамм (1953) отмечает, что нижние ветви лещины, бывшие под снегом, зацветают раньше верхних, что, видимо, связано с тем, что под снегом температура выше и ровнее зимой, чем над ним.
Раннее цветение возможно благодаря тому, что все основные органы заложены в почках с предыдущего года. На побегах три рода почек: из одних развиваются мужские соцветия, из других — побеги, несущие женские цветки, из третьих — вегетативные побеги. Заложение мужских сережек начинается обычно после прекращения активного роста побега — в июне. В начале или середине июля сформированные сережки уже выходят из почек. Сережки (их в группе бывает 1—6) очень быстро растут, достигают 1,3—4 см длины и в таком состоянии зимуют. Каждая сережка содержит до 200 цветков и более. Мужские цветки начинают формироваться в пери
23
од выхода сережек из почек. Заканчивается дифференциация к наступлению устойчивых заморозков. Располагаются они в пазухах прицветных чешуек, сросшихся с . кроющей чешуей (Сапанкевич, 1956).
В то время когда мужские соцветия уже вполне сформированы (в середине сентября), начинают закладываться женские соцветия. Разница во времени заложения мужских и женских соцветий 2—3 месяца. Дифференциация женских цветков лещины происходит ранней весной незадолго до начала цветения (Киндякова, 1958). Завязь и семяпочки начинают развиваться после попадания пыльцы на рыльце пестика. В целом развитие яйцевого аппарата в отличие от других покрытосеменных происходит очень поздно (Навашин, 1951 а, б, в). Завязь срастается с околоплодником, обвертка (плюска) начинает развиваться с началом листораспускания. Число цветков в одном соцветии от 8 до 25, на старых ветвях иногда до 50.
Лещина — анемофильное растение. Опыление происходит в апреле, а оплодотворение через 3—3,5 месяца, в конце июня — начале июля (Киндякова, 1958). Процесс оплодотворения лещины подробно изложен в работе С. Г. Навашина (1951 в). Созревание плодов обычно происходит через 140 дней после опыления.
Рост побегов, листьев и корней начинается весной при полном оттаивании почвы и среднесуточной температуре 10° (в конце апреля — начале мая) и продолжается 4 месяца. Рост побегов происходит двумя периодами: с максимумом в конце мая — начале июня и в середйне июля — середине августа. Активный рост листьев с приростами 1,0—3,2 см2 продолжается 16—20 дней. Рост корней происходит также двумя периодами (с минимумом во время летней почвенной засухи) и продолжается до замерзания почвы. Наибольший среднесуточный прирост осевых корней 5 мм, боковых— 1,8 мм (Киндякова, 1966).
По данным 3. Ф. Сапанкевич (1956), вегетативная почка состоит из почечных чешуй и зачаточных листьев, в пазухе которых лежат меристематические бугорки. Эти бугорки закладываются в конце июня и к осени образуют уже 2—5 крохотных почечных чешуек. В таком состоянии они зимуют. С мая следующего года меристематические бугорки полностью дифференцируются в развитые пазушные почки и в таком виде проводят вторую зиму. Весной они образуют вегетативные побеги. При изучении вегетативных почек замечена интересная особенность развития листа: в течение всего периода развития почек в пазухе кроющих чешуй, которые А. В. Кожевников (1950) называл будущими прилистниками, развиваются зачаточные листья, в конце лета и осенью они выглядят перистораздельными, густо покрытыми длинными одноклеточными волосками. Главные и боковые жилки их заканчиваются удлиненной несколько расширенной кверху красной железкой.
В процессе последующего развития доли лопастей зачаточного листа постепенно срастаются, образуя гофрированную поверхность
24
простого листа. Ко времени цветения срастание достигает до ’Д от общей поверхности пластинки листа. Только в период распускания почек (в конце апреля — начале мая) листовая пластинка становится цельной, но она остается еще гофрированной и часто сложенной вдвое. К этому времени красные железки исчезают. Кроющие чешуйки сохраняются некоторое время в виде прилистников, но скоро опадают. Такое развитие листа автор объясняет вторичным упрощением сережкоцветных, к которым относится лещина.
Способы размножения и распространения
Лещина хорошо размножается семенным и вегетативным путем. Женские цветки нередко находятся в верхних частях побега, а мужские— на нижних, поэтому пыльца, видимо, не попадает на расположенные выше рыльца, что предотвращает самоопыление в пределах одной особи (Смолянинова, 1929). Плодоносит лещина регулярно, но неравномерно. На каждые 10 лет приходится 3 неурожайных года (Флора СССР, 1936). С одного куста можно собрать 50—250 г плодов в неурожайные годы и не менее 2,5 кг в урожайные (Козлов, 1970). С 1 га зарослей лещины собирают 0,6—1 т плодов (Гроздов, 1960; Шиманюк, 1967). Внесение суперфосфата под кусты лещины повышает урожайность в 3 раза (Джиаукштас, 1957).
Периодичность плодоношения нередко связана с суровыми зимними условиями, вызывающими гибель мужских соцветий. Одной из причин неурожая может быть также массовое опадение плодов во время весенне-летней засухи (Киндякова, 1964). В благоприятных условиях в культуре хорошие урожаи бывают почти ежегодно.
По мере созревания плоды выпадают из плюски на землю. Плоды широко потребляются человеком и некоторыми животными, преимущественно грызунами, поэтому лишь небольшая часть их может превратиться во всходы. В условиях Подмосковья наилучшее естественное возобновление происходит на достаточно богатых почвах под разреженным древостоем.
Вегетативное размножение лещины встречается широко и происходит различными способами — пневой порослью, отводками, корневыми отпрысками (Озол, Зуковска, 1955; Гроздов, 1960; Willis, 1966; Шиманюк, 1967). В искусственных условиях лещина размножается семенами, реже отводками с надрезом, зелеными черенками, взятыми из нижней части растений. Размноженные отпрысками и отводками особи плодоносят на 3—4 год. Высокая способность к -порослевому размножению приводит к тому, что лещина на лесосеках размножается так энергично, что мешает естественному возобновлению основных древесных пород. Вполне возможным является ведение хозяйства на лещину со сроком рубки 10—15 лет (Флора СССР, 1936).
25
Экология
Экологическая пластичность лещины довольно значительна. Она предпочитает умеренно влажные почвы, однако встречается и в довольно засушливых условиях, но не переносит избыточное увлажнение и особенно заболачивание (Сукачев, 1938; Кощеев, 1950; Джиаукштас, 1957). За день лещина испаряет 3,33 г воды с 1 дм2 листовой поверхности при прямом солнечном свете, температуре 22—24° и относительной влажности воздуха 39—52% (Schim-рег, 1889). Дневная транспирация 1 г свежих листьев равна 4,2 г (Lyr и. а., 1967). Средняя интенсивность транспирации в Серебряноборском лесничестве Московской обл. составляет 37,5±4,9 мг на 1 г за час при 18,3°. Отмечен очень высокий коэффициент корреляции, (0,87±0,06) между транспирацией лещины и температурой воздуха (Иванов и др., 1951).
Лещина довольно теневынослива — может произрастать под пологом сомкнутых дубрав и хвойно-широколиственных лесов, но в таком случае плодоносит слабо. На вырубках, опушках, в окнах хорошо развивается и обильно плодоносит. В неблагоприятных почвенных условиях минимальная освещенность, которую переносит лещина, равна 15,3%, а на плодородных почвах — 2% (Lyr. и. а., 1967). Лещина — морозоустойчивая порода, однако в очень суровые зимы наблюдается частичное обмерзание побегов, например, в суровые зимы 1939/40 и 1941/42 гг. К почве лещина довольно требовательна., Предпочитает свежие и влажные плодородные суглинки и супеси с pH почвенного раствора, близкой к нейтральной (Харитонович, 1968): деградированные черноземы, темно-серые и серые лесостепные почвы, слабооподзоленные и дерново-подзолистые легкие суглинки и супеси, подстилаемые неглубоко залегающей мореной в лесной зоне. В степи лещина удовлетворительно растет в зоне выщелоченных, мощных и обыкновенных черноземов, преимущественно в более обеспеченных влагой местоположениях. На засоленных почвах не растет.
Мощная поверхностная корневая система позволяет лещине расти по крутым склонам разной экспозиции и довольно высоко подниматься в горы. В Вогезах лещина растет на 800 м, в Альпах — на 1250 м, в Боснии — на 1600 м, а на Кавказе — до 2100—2300 м над уровнем моря (Нейштадт, 1953).
Фитоценология
Лещина обыкновенная растет преимущественно в широколиственных, а также в хвойно-широколиственных лесах, образуя хорошо выраженный ярус подлеска. Встречается также в сосняках и ельниках высших бонитетов, в горных буково-каштановых лесах (в Закавказье). Часто поселяется на вырубках, вытесняя коренные породы. В Литовской ССР наибольшие скопления лещины в спелых и перестойных дубняках: елово-ясеневом, елово-снытевом и елово-кис
26
личном (Джиаукштас, 1957). В степных районах лещина растет по опушкам балочных лесов, по оврагам, обрывам, поймам рек.
Отличаясь прекрасными- почвоулучшающими свойствами за счет быстрого разложения опада, лещина обогащает почву минеральными солями, особенно кальцием.
Внутривидовая систематика
Отличаясь большой изменчивостью, в особенности плодов и листьев, лещина образует несколько разновидностей. А. Редер (Rehder, 1927) различал следующие: var. aurea Kirch.— листья желтые; var fusco-rubra Dipp.— листья пурпурные или коричнево-красные; var. heterophylla bond.— листья перистолопастные с острозубчатыми долями; var. pendula Dipp.—с поникающими ветвями; var. contorta Bean.— с изогнутыми и скрученными ветвями; var. grandis Lam.— с большими орехами, обычно полушаровидными; var. pontica Winke.— листья округлые, опушенные снизу, черешки короткие, орехи большие, но плюска длиннее орехов. К. Дальгрен (Dahlgren, 1941) отмечал разновидность лещины var. zimmermannii Hahne.— с необычной формой листьев (мелкозубчатые, более темные, чем у типичной формы), предполагая, что это связано с мутациями в почках. Л. А. Смолянинова (1931) Выделяла разновидности: var silvestris DC., var. ovata Willd., var. brachychlamus Spach. Б. И. Федорако (1940) отмечал наличие трех форм лещины в башкирской лесостепи: мелкоплодной скороспелой, крупноплодной скороспелой, крупноплодной поздней.
Известно очень много культурных сортов, разводимых преимущественно на юге европейской части СССР, на Кавказе и в Средней Азии с промышленной целью. Они отличаются тонкой скорлупой и более высокой урожайностью по сравнению с дикорастущей лещиной.
Консортивные связи
Большую роль в распространении лещины играют грызуны (Hagerup, 1942). Они же часто препятствуют семенному возобновлению, поедая значительную часть плодов.
Лещина довольно чувствительна к различным грибным заболеваниям. Наиболее распространенные болезни лещины (Ванин и др., 1931): пятнистость листьев (возбудители — Gloeosporium coryli Sacc., G. perexiguum Sacc., Cercospora coryli Montem., Septoria avellanae Berk, et Br., Leptothyrium corylinum Fuck., Phyllostricta corylaria Sacc., Ph. coryly West), мучнистая роса (возбудители — Microsphaera alni Wint., Phyllactinia suffulta Sacc.), ржавчина (возбудитель — Pucciniastrum corili Komar.), розовая плесень (возбудитель — Trichothecium roseum Link.), рак корней (возбудитель— Agrobacterium tumefaciens (Smith et Town.) Conn.). Кроме
27
того, повреждения вызывают Dtaporthe sulphurea Fuck., Nectria cinnabarina Fr., Sterium hirsutum Pers., Monilia fructigena Pers.
На лещине зарегистрировано 26 энтомовредителей (Гусев, Римский-Корсаков, 1951). Листья повреждаются долгоносиками, листоедами, растительными клещами, орешниковой галлицей, различными молями и тлями; листовые почки — орешниковым почковым клещиком; сережки — орешниковой сережковой галлицей; плоды — орешниковой плодожоркой и орешниковым долгоносиком. Многочисленные вредители встречаются на ветвях и стволах — ложнощитовки, златки, орешниковый усач, различные короеды.
Хозяйственное значение
Лещина обыкновенная находит широкое применение как орехоплодное, почвозащитное и почвоулучшающее растение. Плоды содержат до 54—71% ценных растительных масел, 12—18% белков, 7,3—15% углеводов (Круссер, 1940; Киндякова, 1970). Ореховое масло идет в пищу, употребляется в живописи (оно не меняет цвета красок и быстро высыхает), в лакокрасочной промышленности, парфюмерии. Во многих странах Европы промышленные посадки лещины занимают тысячи гектаров.
Молодые побеги с листьями идут на корм скоту. Из листьев приготовляют мочегонный чай, а кора обладает сосудосуживающим действием и может использоваться при расширениях вен (Йорданов и др., 1972). Ветви пригодны для топлива и изготовления рисовального угля, употребляются также на обручи, грабли, для плетения корзин.
Лещина широко применяется при облесении оврагов, склонов, откосов, так как благодаря мощной поверхностной корневой системе обладает высокими почвозакрепляющими свойствами. Довольно широко лещину используют в полезащитном лесоразведении, лесных посадках и в зеленом строительстве (Шиманюк, 1967).
В. Jl. Тихонова
ГОРЕЦ ЗМЕИНЫЙ
Номенклатура и систематическое положение
Polygonum bistorta L., Sp. pl. (1753) 360; Кауфман, Моск, фл., изд. 1 (1886) 429, изд. 2 (1889) 444; Цингер, Сб. свед. (1885) 379; Шмальг., Фл. 2 (1897) 389; Сырейгц., Илл. фл. 2 (1907) 72; он же, Опр. раст. Моск. губ. (1927) 115; Комаров во Фл. СССР 5 (1936) 681; Федченко в Маевск., Фл., изд. 7 (1940) 284; Шишкин, там же, изд. 8 (1964) 208; Ворошилов, там же, изд. 9 (1964) 207; Chater in Fl. Eur. 1 (1964) 80; Ворошилов в Опр. раст. Моск. обл. (1966) 143—. Горец змеиный, змеевик, раковые шейки.
Принадлежит к секции Bistorta Adans, р. Polygonum L. сем. Polygonaceae. Описан из Швейцарии, тип в Лондоне.
Латинское название рода происходит от древнегреческого polygonon: poly —много; gbny (род. падеж gonatos)—колено, узел, так как у многих видов резко выделяются узлы стеблей. Видовое название от bis — дважды, torta — скрученная, так как корневище дважды изогнутое. Русское название «змеиный, змеевик» обязано корневищу, внешне напоминающему свернутую змею.
Морфологическое описание
Многолетний травянистый поликарпик с коротким, толстым, несколько сплюснутым, деревянистым, змеевидно-изогнутым корневищем длиной 5—10 см, толщиной 1—1,5 (3) см, черным или темнобурым снаружи, розовым или буровато-розовым на изломе, в верхней части одетым остатками листьев и покрытым рубцами (следами прикрепления розеточных листьев). Главный корень рано Отмирает, корневая система генеративных особей состоит из многочисленных тонких придаточных корней, отходящих от нижней части корневища. На корнях обнаружена эндотрофная микориза (Ячевский, 1933).
Стебли генеративных побегов одиночные или их несколько, прямостоячие, цилиндрические, голые, высотой (10)30—80(150) см, с 5—6 узлами, слегка утолщенные под узлами. Прикорневые и ниж-
29
ние стеблевые листья длиной 4—20(30) см, шириной 1—7 см, низбе-гающие в длинный крылатый черешок, который равен или превышает по длине пластинку листа. Пластинки листьев продолговато-яй- 1 цевидные или широколанцетные, у основания закругленные или сердцевидные, с волнистыми краями, сверху зеленые, снизу сизые, голые или короткрволосистые. Верхние стеблевые листья мельче и уже, очередные, ланцетовидные, самые верхние — линейные, почти сидячие. Прилистники срастаются в трубки, образуя влагалища-раструбы, охватывающие нижнюю часть междоузлий.
Обычно генеративный побег не ветвится, но в культуре часто наблюдается появление небольших соцветий в пазухах верхних стеблевых листьев. Иногда после цветения на соцветии развиваются боковые побеги 5—10 см длиной с мелкими листьями. Соцветия на концах генеративных побегов овальные или цилиндрические, колосовидные, густые. Цветки мелкие (около 3,5 мм), сидят по 2 в пазухах пленчатых прицветников. Околоцветник неопадающий, простой, лепестковидный, пятичленный, почти до основания раздельный, розовый, реже белый или темно-красный. Тычинок 8. Пыльцевые зерна ; крупные, достигают 61,7X51,7 мк, от желтовато-серой до темно-желтой окраски, относятся к группе трехбороздно-трехпоровых зерен ; (Пыльцевощанализ, 1950; Ананова, 1958). Пестик с верхней трехгранной завязью, тремя столбиками и маленькими головчатыми ' рыльцами. 1
Плод — лизикарпный орешек с трехчленной оболочкой (Каден, '' 1965),'в очертании яйцевидный, в разрезе трехгранный, с тупыми | толстыми ребрами и равными между собой гранями, с носиком. Стенки плода твердые, гладкие, блестящие, красновато- или темно-коричневые. Длина орешка с носиком 3,15—4,4 (в среднем 3,8) мм, .j ширина 1,85—2,55 (в среднем 2,19) мм, длина носика 0,20—0,45 мм. Вес 1000 семян 2,4—4,6 г (Золотницкая, Авакян, 1950).
Число хромосом, по разным определениям, 24, 44, 46, 48 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Северная граница ареала горца змеиного в СССР идет от мыса 1 Святой Нос на Баренцевом море к р. Шуе на южном берегу Белого моря, захватывая весь п-ов Канин, острова Колгуев, Вайгач и южный о-в Новой Земли. В Западной Сибири граница пересекает п-ов Ямал, идет через Гыданский п-ов к Енисейскому заливу, через п-ов 1 Таймыр (до 75°) до Хатангского залива. Восточная граница этого вида, вероятно, здесь опускается вниз по р. Хатанге и ее притокам, по Нижней Тунгуске, пересекает верховья р. Лены и вдоль западного берега Байкала достигает государственной границы СССР. 1
Южная граница на территории Казахстана проходит севернее оз. Зайсана (змеевик встречается также в горах Тарбагатая). От р. Иртыша граница поднимается к р. Чулыму, пересекает р. Обь и । 30 1
выходит к р. Томи, минуя Барабинскую степь, выходит по р. Уй к Таре, далее вдоль Иртыша к р. Тоболу и его притокам и к Челябинску. (Отмечены отдельные нахождения в окрестностях Кустаная.) От Челябинска граница опускается, огибая Южный Урал, по левому берегу р. Урал к Оренбургу, далее по р. Белой, через Заволжье к Жигулям, вниз по Волге к Вольску, на Балашов и далее на запад по границе степи через Змиев к Киеву, Хмельницкому, Тернополю, на Карпаты (в Закарпатье отсутствует). Южнее имеются отдельные нахождения в горной части Крыма.
На западе ареал отступает от Балтийского моря и через Каунас, Вильнюс и Белоруссию идет к Зап. Двине, оз. Ильмень и Ладожскому оз., далее к востоку от Беломорско-Балтийского канала. В Прибалтике, Карелии, Псковской и Ленинградской областях змеевик встречается лишь спорадически (Гаммерман, Шасс, 1954).
За пределами СССР горец змеиный известен в Скандинавии и горах Европы. В ископаемом состоянии найден в постплиоцене (Флора СССР, 1936).
В условиях Московской обл. горец встречается довольно часто на влажных лугах, в кустарниках, на лесных полянах и просеках, на опушках. Особенно часто в Северном, Клинско-Дмитровском, Западном и Заокском ботанико-географических районах. В силу своей декоративности он усиленно уничтожается и стоит в списке растений, которые в недалеком будущем могут оказаться полностью уничтоженными (Ворошилов и др., 1966).
Онтогенез
В природных условиях горец змеиный размножается семенным и вегетативным путем.
В опытах М. М. Костроминой (1970) как свежесобранные, так и хранившиеся в течение нескольких лет семена горца' совершенно не прорастали. Анатомическое изучение плодовой оболочки показало, что она содержит толстостенные механические клетки, которые, по-видимому, затрудняют проникновение влаги. Обработка семян кипящей водой, гиббереллином, накалывание, скарификация не вызывали прорастания. Стратификация в течение 5 месяцев позволила получить всходы (2—18%).
Наши наблюдения в условиях Московской обл. (ВИДР) показали, что плоды горца легко осыпаются, не достигнув зрелости. Среди собранных семян много щуплых и пустых со светло-коричневой окраской. Проращивание семян в лабораторных условиях при постоянном температурном режиме (2—5°) позволяет наиболее полно выявить всхожесть семян. Прорастание семян начинается через 1—1,5 месяца после закладки, происходит растянуто, недружно, в большей степени зависит от погодных условий в период созревания. Всхожесть колеблется от 0 до 64%. При хранении семян в комнатных условиях всхожесть снижается.
31
Тип прорастания у горца — надземный. При прорастании гипокотиль, выносит над поверхностью почвы плодовую оболочку, которая вскоре сбрасывается. Семядоли обратнояйцевидные, эллиптические, с длинными черешками, сросшимися в трубчатое влагалище. Первый лист обычно ланцетный, выходит через верхнее отверстие влагалища, не разрушая его. Второй лист округлый, на верхушке островатый, у основания слегка сердцевидный, выходит через разрыв влагалища у его основания. Проростки горца — розеточного типа.
При выращивании горца в условиях теплицы двухмесячные растения уже имеют корневище диаметром 2—4 мм, шаровидное, коричневого цвета. В полевых условиях развитие растений из семян идет медленно и большая часть их погибает. К концу первого вегетационного периода растения горца имеют розетку из 2—5 мелких листьев (до 8 см в длину) и корневище длиной до 1,3 см, диаметром 0,8 см, с хорошо развитыми придаточными корнями. Корневища розового цвета и уже имеют характерную изогнутую форму.
Виргинильный период продолжается в культуре один — три года. На втором году жизни примерно у половины экземпляров в пазухе одного из первых розеточных листьев (Серебряков, 1952) образуется генеративный побег. Остальные цветут на третьем и даже на четвертом году жизни. Растения, полученные от посева семян в грунт, на втором году жизни обычно еще не образуют столонов и имеют одно корневище весом в среднем 9 г. При выращивании через рассаду у растений второго года жизни весной или в начале лета начинают образовываться столоны, на конце одного из них развивается дочернее корневище; вес корневищ одного экземпляра достигает 20 г (Киченко, Тихонова, 1968).
Переход в генеративное состояние и начало вегетативного размножения не связаны между собой.. Мы наблюдали еще не цветущие на третьем году жизни растения, имеющие по 3 дочерних корневища. В то же время некоторые цветущие растения не имели столонов и дочерних корневищ. Обычно растения третьего года жизни способны и к генеративному и к вегетативному размножению.
Рассадные особи в условиях питомника проявляют очень сильную индивидуальную изменчивость. Так, число розеточных листьев колеблется от 3 до 64 (в среднем 24), число генеративных побегов от 0 до 7. Материнское корневище достигает 9—11 см в длину, дочерние— от 0,5 до 4—6 см. Число дочерних корневищ варьирует от 0 до 13. Длина столонов обычно 1—3 см, реже до 6—10 см, но тогда они изогнуты и прилегают к корневищу, поэтому вся подземная часть культивируемого горца очень компактна. Вес сырых корневищ с корнями у трехлетнего семенного экземпляра 73 г (21—178 г), без корней — 60 г (18—162), надземных органов — 29 г (1—86).
В условиях питомника генеративные особи ежегодно цветут и образуют семена. Длительность генеративного периода неизвестна. В природе общая продолжительность жизни растений измеряется десятками лет (Ларин и др., 1951).
32
Сезонный ритм развития
Горец змеиный относится к моноподиальным растениям, у которых преобладающая часть ассимилирующих листьев находится в розетке (на главной оси). Генеративные побеги развиваются из пазушных почек первых розеточных листьев текущего года (Серебряков, 1952). Зимующих листьев нет. По строению корневища его относят к группе короткокорневищных, подгруппе полурозеточных растений (Голубев, 1956, 1958). Генеративные побеги горца моно-циклические. Внутрипочечный рост стебля протекает по типу верхушечного с акропетальной последовательностью, внепочечный рост только интеркалярный, так как верхушка стебля завершается соцветием. Побег в почке сформирован заранее (Серебряков, 1952).
В природных условиях в разнотравном луговом сообществе в пойме р. Вологды вегетация у горца в 1924 г. началась в середине апреля, цветение 25 мая, семена созрели во второй половине июля, в начале августа растения уже закончили вегетацию. Часть растений вообще не цвела (Шенников, 1927). В условиях питомника растения начинают вегетировать обычно в апреле, цветут в июне, плоды созревают в конце июня — июле или в июле — августе (в зависимости от зоны). Цветение длится от 2 до 6 недель (Киченко, Тихонова, 1968; Иванова и др., 1970; Костромина, 1970). В условиях культуры в Московской обл. и г. Минске отмечено вторичное цветение в августе— сентябре. Оно недружное, растянутое, не прекращается даже после первых заморозков, так как невысокие генеративные побеги, находящиеся в гуще розеточных листьев, не страдают от заморозков. Вегетация растений заканчивается обычно в октябре.
По нашим наблюдениям в питомнике одно соцветие цветет 12— 13 дней, один цветок — 6—7 дней. Цветение протекает в две волны, в первую зацветает около 56% бутонов. На 7—8-й день вытягиваются цветоножки оставшихся бутонов и начинается вторая волна цветения. На 1 см оси соцветия расположено 35—40 цветков. Опыление насекомыми. Из общего числа цветков только 35—40% дают еемена.
Горец змеиный — геофит. Корневища у него погружены в почву до 12—18 см. На корневище значительный запас спящих почек, обеспечивающих высокий коэффициент вегетативного размножения.
Способы размножения и распространения
Семенная продуктивность горца змеиного почти не изучена. Одно растение дает до 50 семян (Сорные растения СССР, 1934). Они поедаются птицами и медведями. Семена имеют плохую всхожесть. Прорастают сразу после осыпания или весной (Ларин и др., 1951). Всходы горца в природе часто гибнут (Красовская, 1949). В условиях питомника у вегетативно размноженных растений первого — третьего года жизни семенная продуктивность составляет 1—1,3 г, у более старых — до 5 г.
2 Зак. 255
33.
В природе у горца преобладает вегетативное размножение за счет специализированных органов — столонов. На одном растении их может образоваться до 50 (Сорные растения СССР, 1934). Длина столонов достигает 15—20 и даже 30 см.
В культуре была изучена возможность вегетативного размножения дочерними корневищами, отрезками корневищ разного размера, столонами с хорошо развитой верхушечной почкой, столонами со спящими почками, молодыми побегами (Киченко, Тихонова, 1968). Лучшие результаты дала осенняя посадка молодых корневищ и отрезков молодой части корневищ длиной 3—5 см с верхушечной почкой. Они дают высокий процент укоренения (до 100%) и наиболее интенсивный прирост. При семенном размножении лучшие результаты дал осенний посев в грунт (Московская обл.), в областях с достаточным летним увлажнением — летний (г. Новосибирск). Большая часть всходов погибает в первый вегетационный сезон и в первую зиму.
Экология
Наибольшего проективного покрытия горец достигает на почвах с влажно- и сухолуговым увлажнением (Раменский и др., 1956). Он растет на гипновых болотах, выносит близкое залегание грунтовых вод. К затоплению поверхностными водами менее приспособлен, отрицательно реагирует на заиление (Ларин и др., 1951). В поймах крупных рек горец встречается в притеррасных частях, в долинах мелких рек — на лугах прирусловой части. В условиях культуры повышенное количество осадков в летнее время увеличивает продуктивность корневищ горца.
Горец плохо переносит затенение, приурочен к ценозам с хорошей освещенностью. В условиях Московской области не страдает от заморозков. Растет на почвах разного богатства: небогатых (подзолистых, дерново-подзолистых, подзолисто-глеевых, торфяных и перегнойно-торфянистых) и довольно богатых (луговых и лесостепных суглинках, выщелоченных черноземах) (Раменский и др., 1956). Хорошо отзывается на внесение азотных удобрений. Встречается на почвах различной кислотности (pH 5,0—7,5), наиболее часто на слабокисль!х почвах (pH 6,0—6,5).
Горец хорошо растет только на слабозадерненных почвах, при увеличении задернения, а также при уплотнении почвы его способность к вегетативному размножению снижается, уменьшается численность, растения угнетаются. Плохо выносит выпас и частое скашивание (Ларин и др., 1951).
Биологическая продуктивность
По данным Н. П. Евсеенко (Калининская и Брянская области)-, на пойменном лугу при численности горца 28—36 особей на 1 м2 урожай сырых корневищ с корнями составил 326—472 г/м2, на опушке леса при 32—48 особях — 344—520 г/м2; на суходольном лугу при
34
JIG—56 особях на 1 м2 урожайность колебалась от 392 до 680 г/м2. Сырой вес корневищ с корнями одной особи составлял в изученных ассоциациях 10—13 г; на лугу с сырой глеевой почвой в Калининской обл.— от 38 до 54 г (Киченко, Тихонова, 1968).
В условиях питомника продуктивность резко возрастает. С трехлетних плантаций, заложенных корневищами, можно получить до 17 т/га сырых корневищ с корнями или 9,4—9,8 т/га корневищ без корней. Вес сырых подземных органов трехлетнего вегетативно размноженного растения составляет в среднем 320 г, двулетнего — от 8() до 200 г (на продуктивность в сильной степени влияют погодные условия вегетационного сезона).
Фитоценология
Горец змеиный встречается в- основном на лугах различного типа, иногда доминирует. Часто растет на болотах, лесных опушках, в изреженных сырых еловых лесах. В горах приурочен к субальпийским и альпийским лугам.
Консортивные связи
Опыляется в основном пчелами, хороший нектаронос (Глухов, 1955). Листья часто поражаются ржавчиной Puccinia bistorta DC.
Хозяйственное значение
Корневища горца издавна используют в медицине и ветеринарии. Они содержат до 25% дубильных веществ, щавелевую кислоту (1%), оксиметилантрахиноны, оксалат кальция, витамин С, каротин, парарабин и т. д. Корневища и препараты из них применяют внутрь как кровоостанавливающее, вяжущее и противовоспалительное средство при заболеваниях кишечника и дизентерии; наружно — при воспалении слизистой оболочки рта (Атлас лекарственных растений СССР, 1962; Турова, 1967).
Корневища горца используют для дубления кож (Варлих, 1912; Вульф, 1932; Шлыков, 1932; Чернышев, 1934; Первухин, 1961), для окраски шерсти (Телятьев, 1971), в ликеро-водочной промышленности (Пряно-ароматические растения СССР, 1963). Вымоченные корневища теряют горечь и могут добавляться к муке. Заготовку корневищ для лекарственных и технических нужд следует проводить в фазу бутонизации, когда в них содержится максимальное количество дубильных веществ (Полякова, 1967).
Надземные органы горца содержат флавониды, дубильные вещества (5—7%), витамин С, каротин, минеральные элементы. Листья и молодые побеги можно употреблять в пищу в сыром, варёном, сушеном и квашеном виде (Медведев, 1957). Горец — важное кормовое растение в тундре (Ларин и др., 1951).
2*
А. П. Пошкурлат и И. А. Губаноё
ГОРИЦВЕТ ВЕСЕННИЙ
Номенклатура и систематическое положение
Adonis vernalis L., Sp. pl. (1753) 547; Цингер, Сб. свед. (1885) 35; Шмальг., Фл. 1 (1895) 11; Бобров во Фл. СССР 7 (1937) 535; Невский в Маевск., Фл., изд. 7 (1940) 374; Голицын, там же, изд. 8 (1954) 87; Маценко, там же, изд. 9 (1964) 88; Tutin in Fl. Eur. 1 (1964) 222.— Горицвет весенний, адонис весенний, стародубка, черногорка.
Относится к ряду Vernales Bobr. секции Consiligo DC. р. Adonis L., входящего в подсем. Ranunculeae сем. Ranunculaceae (Адольф, 1930; Флора СССР, 1937). Описан из Европы, тип в Лондоне.
. Родовое название отражает ярко-красную окраску цветков некоторых видов этого рода. Латинское «adonis» связано с именем мифического Адониса, будто бы превращенного в кровяно-красный цветок, русское «горицвет» также указывает на «огненность» цветков. Видовое название «весенний» отражает раннее цветение данного вида.
Морфологическое описание
Многолетнее травянистое растение с хорошо развитой корневой системой, состоящей из корневища и придаточных корней. Начиная со второго года жизни особи, первоначальная стержневая корневая система постепенно заменяется мочковато-кистевой. Образующаяся густая сеть ветвящихся придаточных корней захватывает большой объем почвы. Основная масса их сосредоточена в верхних горизонтах, до глубины 80—100 см, но наиболее длинные корни достигают 220 см, имея общую длину до 300 см. От центра особи корни распространяются на 50—60 (до 110) см. Преобладающий диаметр корней' на глубине 10 см — 1—1,5 мм (от 0,5 до 2 мм). По мере старения меняется цвет корней: молодые — белые; двух-, трехлетние — светло-коричневые; более старые — темно-коричневые или почти черные.
Корневища короткие, темно-коричневые, грушевидной или цилиндрической формы, у крупных особей — многоглавые. Погружены в почву на 6—8 см.
36
Надземных побегов у каждой особи от 1 до 20 (иногда Польше). Они ветвятся до третьего, редко до четвертого порядка (рис. 1). Стебли прямые, высотой 20—70 см, слегка ребристые, молодые опушены, оолее старые — голые, у основания коричневатые, выше зеленые.
Листорасположение очередное. Нижние листья побега представляют коричневые чешуи, выше по стеблю расположены влагалищные листья с неразвитой пластинкой. Вся верхняя часть побега, включая разветвления, олиствена сидячими ассимилирующими листь-вми пятиугольной формы, паль-чаторассеченными на дважды-перистые узкие линейные доли
или слегка опушены, сизовато-зеленые.
Рис. 1. Схема максимального ветвления горицвета весеннего (цифрами обозначен порядок ветвления осей)
(до 1 мм шириной). Листья голые
Рис. 2. Цветение горицвета весеннего (фото В. С. Жмыховой)
Цветки одиночные, крупные, расположены на концах побегов (рис. 2), состоят из 5 чашелистиков, 15—22 лепестков, многочислен-
ных тычинок и пестиков. Чашелистики зеленые, иногда с фиолетовым или коричневым оттенком, слегка опушены. Лепестки ярко-желтого цвета. Тычинки желтые, свободные, до 6 мм длиной, расположены спирально, с пыльниками, вскрывающимися продольной трещиной по бокам широкого связника. Пестики светло-зеленые, спирально расположены на вытянутом цветоложе. Завязь верхняя, столбик вытянутый, рыльце малозаметное.
Плод — многоорешек, округлой или конусовидной формы, длиной 20—25. мм, шириной 10—15 мм. Отдельные плодики-орешки односеменные, овальные, размером 5X3 мм, с хорошо развитым спинным килем и носиком, с ячеисто-морщинистой поверхностью, густо опушенные.
Набор хромосом 2п = 16 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Ареал горицвета весеннего довольно обширен (рис. 3). В долготном направлении он простирается от Пиренейского п-ва । до бассейна верхнего течения Енисея (Stary, 1952; Meusel u. а., 1965).
На территории СССР распространение горицвета связано с ле- j сос'гепью, лишь на юге он незначительно заходит в степи. Приурочен вид в основном к возвышенностям: Волыно-Подольской, Приднепровской, Среднерусской, Приволжской, Белебеево-Бугульминской. В сибирской части ареала горицвет широко распространен в Южном Зауралье, далее через Барабинскую низменность, Обскую и Са-'] лаирскую дренированную равнину доходит до Салаирского кряжа, | который служит восточной границей сплошного распространения i этого вида. В низменностях горицвет представлен ограниченно или I выпадает совсем'(Гаммерман, Шасс, 1954). |
Помимо основного ареала имеются изолированные фрагменты! и отдельные местонахождения горицвета весеннего. В нашей стране I известны следующие крупные фрагменты (рис. 3): в Крыму — крым-1 ский; в Предкавказье — анапский, ставропольский и северо-пред-1 горный; севернее Приволжской возвышенности — сергачский; в I Предуралье — кунгурский и красноуфимско-месягутовский; в Сиби-1 ри — кузнецкий в Кузнецкой котловине и .минусинский в верхнем! течении Енисея. Наиболее восточные находки горицвета весенне-1 го — бассейн Лены возле Олекминска. Как фрагментарные, так и от-1 дельные местонахождения связаны с интразональными участками! лесостепи (Пошкурлат, 1970). I
В Московской обл. горицвет встречается только в самой юго-1 восточной части — в Серебряно-Прудском р-не. Здесь проходит се-1 верная граница ареала вида, поэтому растение должно строго охра-1 няться. I
38
Рис. 3. Ареал горицвета весеннего: массовое распространение, 2 — отдельные местонахождения
Онтогенез
Латентный период горицвет переживает в состоянии плодиков (семян). Период покоя у них длится 60—70 дней (Пошкурлат, 1969). По мнению В. Кульпа (Kulpa, 1960), прорастание семян тормозится ингибиторами в виде аммиачных соединений, находящихся в около-плоплоднике. Он экспериментально показал возможность устранения тормозящих веществ путем переменного намачивания. Возможно, на этом и основана естественная стратификация. И. А. Иванова (1966) считает, что задержка в прорастании вызвана недоразвитием зародыша, которому необходим определенный срок для дифференциации, а также низкой ферментативной активностью.
Оптимальная температура для прорастания семян 20—30° (Kulpa, 1960). Прорастанию способствует свет, семена прорастают при заделке в почву не глубже 1—2 см (Gawlowska, 1958; Kulpa, 1960; Пошкурлат, 1969). Средняя всхожесть в лабораторных условиях 50% (Kulpa, 1960; Аксельрод, 1961), полевая всхожесть 0—5% (Пошкурлат, 1969). Семена сохраняют всхожесть в течение одного года, на второй год всхожесть утрачивается почти полностью.
Рис. 4. Схема прорастания семени горицвета весеннего (по Kulpa, 1960)
Прорастание семян надземное (рис. 4). Проросток имеет типичное строение двудольного растения. Семядоли вначале узкие, потом приобретают овальную или почти округлую форму, что харак
40
тернО для всех лютиковых (Irmisch, 1856). Поверхность семядолей голая, размеры 7X5 или 7X7 мм. Первый ассимилирующий лист морфологически отличается от всех последующих наличием длинного черешка (2—2,5 см) с расширенным влагалищем. Пластинка его нальчато рассечена на 3 доли. Главная и боковые жилки заканчиваются широкими воронковидными гидатодами.
Ювенильные растения имеют неразветвленный побег высотой 4 -20 см, с одной чешуей внизу, одним черешковым листом и несколькими сидячими листьями. С возрастом увеличивается число чешуй (до 3-х генераций) и число сидячих листьев (Пошкурлат, 1965). Корневая система в этот период переходная от стержневой к кистевой. Корневище цилиндрическое, вертикальное, длиной от 0,5 до 20 мм, диаметром 1,5—7 мм. Общая продолжительность ювенильного периода в культуре 1 год, в природе —от 7 до 12 лет, в ксеро-фнтных условиях — до 30 лет.
Генеративный период можно разделить на три возрастных этапа: восходящего развития, максимального и нисходящего. Цветок у молодых растений не всегда бывает полноценным. На этапе максимального развития цветок достигает наиболее крупных размеров (6—7 см в диаметре) и продуцирует наибольшее число плодиков. На этапе нисходящего развития размеры цветков постепенно уменьшаются (до 1,5—2 см в диаметре), а у самых старых генеративных растений бутоны остаются нераскрытыми. На нисходящем этапе развития продуцируется значительное число неполноценных плодиков, что объясняется нарушением процесса оплодотворения и развития зародышевого мешка (Ефремова, 1968, 1971).
В начале генеративного состояния каждое растение горицвета весеннего имеет лишь один надземный побег. С течением времени число надземных побегов увеличивается за счет одновременного прорастания постепенно увеличивающегося числа почек возобновления. На максимальном этапе растение имеет несколько плотно расположенных побегов. Прорастание нескольких почек возобновления приводит к ветвлению корневища, оно становится многоглавым. Возникает большое число придаточных корней — на один побег приходится 40—50 корней. Постепенно формируется все большее число почек обогащения, и отдельные побеги достигают максимального ветвления — развивают до 6—7 осей второго порядка, большинство из которых имеет по 3—4 оси третьего порядка.
Признаки старения проявляются в сокращении ветвления в результате неполного развития почек обогащения. Почки трогаются в рост, но остаются с короткими осями и этиолированными листьями, затем перестают трогаться в рост, а у еще более старших побегов перестают закладываться совсем.
На этапе нисходящего развития самые слабые побеги становятся вегетативными, в то время как часть побегов еще долгое время продолжает оставаться генеративными. За счет разрастания периферических и отмирания центральных участков ранее плотные
41
особи превращаются в клоны, состоящие из отдельных неравноценных партикул. Клоны могут достигать размера 70 X100 см.
Перерывы в цветении наблюдаются у растений восходящего и нисходящего этапа. У первых продолжительность их не больше 1—2 лет. С возрастом перерывы прекращаются. Снова появляются перерывы только у более старых растений нисходящего этапа развития. Чем старше растение, тем продолжительнее перерывы, они могут длиться до 7—8 лет. Помимо возрастного состояния для ’энергии цветения имеют значение погодные условия предыдущих лет, влияющие на формирование почек возобновления. Кроме перерывов в цветении наблюдаются и перерывы в отрастании всего экземпляра, что бывает только у растений нисходящего этапа (вторичный покой). Продолжительность таких перерывов может быть от 1 года до'5 лет.
В сенильное состояние горицвет переходит постепенно, начиная еще с нисходящего этапа, когда часть более слабых партикул клона уже не производит репродуктивных побегов. После полного прекращения цветения наступает сенильное состояние всего клона. К этому времени полностью клон сохраняется редко. Большей частью встречаются только остатки клона, состоящие из бывших более крупных партикул. Побеги сенильных растений с очень ограниченным ветвлением: осей второго порядка мало и они короткие. Участки корневищ крупные, но сильно разрушенные.
Вследствие неодновременного отмирания партикул к концу жизни сохраняется одна из них, образующая уже только один побег. В бо’лее молодом состоянии одиночные сенильные растения развиваются еще на старых крупных участках корневищ, но по мере старения и ограниченной способности корневища разрастаться в толщину у сенильных растений образуются очень тонкие корневища. После разрушения старого крупного корневища в глубокой старости сенильные растения на тонких цилиндрических корневищах развивают слабые неветвящиеся надземные побеги высотой 4—6 см. Тонкие корневища могут получаться и в результате партикуляции материнского корневища.
Максимального развития горицвет весенний достигает примерно к 40—50 годам, а общая продолжительность жизни предположительно не меньше 150 лет.
Сезонный ритм развития
Время прорастания плодиков горицвета весеннего в природе связано с достаточным количеством влаги в осеннее и весеннее время. Осенью всходы появляются с конца августа до начала октября; весной — с конца апреля до первой декады мая (Winkler, 1893; Каменецкая, 1949; Пошкурлат, 1966; Сарычева, 1966). В Челябинской обл. отмечено только весеннее прорастание (Строкова, 1970). Отсутствие осенних всходов на Южном Урале можно объяснить боль-шой.сухостью осени.
Численность всходов зависит от погодных факторов, а также от
42
экологических условий места, куда попадают плодики. На лесных полянах, а также на участках, затененных, травостоем, всходы не появляются. Во вторую половину мая и в июне наблюдается большая гибель всходов либо из-за недостатка влаги в весенне-летний период, либо от грибковых заболеваний, поражающих всходы в дождливые сезоны (Пошкурлат, 1966, 1969).
При летне-осеннем прорастании горицвет находится в состоянии проростков в течение нескольких месяцев, уходя в зиму с семядолями, при весеннем прорастании — около двух недель. Переход в ювенильное состояние происходит всегда только в первой половине мая.
У горицвета весеннего формируется два типа почек — возобновления и обогащения. Из первых развиваются надземные побеги, а из вторых — оси второго и третьего порядков. Почки возобновления как меристематические бугорки закладываются с марта по май в пазухах кроющих чешуй, как дочерние образования на материнской почке возобновления. По сведениям Ю. А. Дударь (1969), в условиях Ставрополья заложение меристематических бугорков наблюдается только в июле.
Органогенез почки возобновления продолжается от заложения меристематического зачатка до развития надземного побега 24—27 месяцев, захватывая три вегетационных периода, с марта первого года развития почки до мая третьего года развития. Примерно 2—4 месяца почка возобновления находится в состоянии недифференцированного меристематического бугорка, 8 месяцев уходит на формирование многолетней части оси и трех генераций чешуй; 2 месяца (июнь'—июль второго года развития) —на органогенез однолетней части оси, цветка, четвертой генерации чешуй, листовых зачатков и ночек обогащения; 5 месяцев — на внутрипочечный рост и 6 месяцев (по 3 в каждый год формирования — с ноября по февраль) находится в состоянии внутриклеточной дифференциации. Наиболее активный органогенез происходит в июле второго года развития.
Вегетация надземного побега продолжается 5 месяцев (с мая по сентябрь): с конца апреля — бутонизация, в течение мая — цветение, до середины июня — созревание плодов, до конца июня — осыпание плодов и в середине сентября — конец вегетации (Борисова-Гул енкова, 1960).
У горицвета весеннего и генеративные, и вегетативные побеги однотипные — моноциклические. Их рост можно отнести, по И. Г. Серебрякову (1952), к третьему, линейному типу. Время развития побегов, особенно линейного роста, его активность и продолжительность у разновозрастных групп различно. Раньше всего начинают развиваться более сильные генеративные побеги растений максимального этапа (конец апреля — начало мая), позже всего — наиболее старые сенильные растения (конец мая — начало июня). Интенсивность роста генеративных побегов следует правилу одновершинной кривой. Молодые генеративные побеги интенсивнее растут в первой декаде мая (прирост до 2 см в сутки). Чем старше генератив
43
ный побег, тем больше удлиняется его ось, и период интенсивного роста передвигается на более позднее время — вторую, реже третью декаду мая.
У вегетативных побегов наблюдается два-периода интенсивного роста (Дохман, 1948). Длительность этих периодов определяется возрастом растений. У ювенильных и имматурных растений интенсивный рост происходит в первой половине мая. Чем старше становится растение, тем на более поздние сроки сдвигается период интенсивного роста, но не позже первой декады июня.
Между периодами интенсивного роста и формированием мата-меров у горицвета существует определенная корреляция. У генеративных побегов интенсивный рост оси первого порядка происходит в период цветения. Рост и цветение осей второго порядка происходит в период формирования плодов на оси первого порядка. Рост осей третьего порядка происходит в такой же последовательности — они растут после прекращения роста осей второго порядка. Интенсивный рост вегетативных побегов происходит после окончания цветения и развития генеративных. Рост побегов всех возрастов заканчивается к концу осыпания плодов и интенсивного роста окружающих растений, т. е. ко второй декаде июня.
Молодые корни появляются ранней весной (конец апреля) до активного роста надземных побегов, но самое большое число корней вырастает и интенсивный рост их происходит в августе и в начале сентября, после формирования почек возобновления. Годичный прирост'старых корней 2—5 см, молодых— 10, иногда 15 см. Корневище формируется из симподиально нарастающих многолетних базальных участков побегов. Годичное нарастание корневища в длину около 0, 5 см.
Время отрастания и продолжительность развития побегов в пределах одного экземпляра не одинаковы. Раньше всего завершают рост более сильные побеги, расположенные на конечных, более молодых участках корневищ, и позже всего — более слабые побеги средних частей корневищ. В первой фазе распускаются цветки на осях первого порядка более сильных побегов; во второй — происходит одновременно цветение средних осей второго порядка и цветков более слабых побегов на осях первого порядка; в третьей фазе цветения, которая приходится на третью декаду мая, доцветают цветки запоздавших в развитии нижних осей второго порядка. Соответственно разновременному цветению формирование, созревание и осыпание плодиков на разных побегах происходят неодновременно.
Горицвет весенний — типичный геофит.
Способы размножения и распространения
Размножается горицвет только семенным путем. Каждый цветок бывает раскрыт в течение 8—10 дней (крайние колебания — от 6 до 11), открываясь ежедневно в 5—9 часов утра и закрываясь в 2— 4 часа дня. Периодическое движение лепестков происходит за счет
44
неравномерного роста их стенок. За сутки длина лепестков увеличивается на 1—2 мм. На открывание и закрывание цветка влияют температура и освещение. Цветки открываются при температуре не ниже 11 —13° (Оловянникова, 1949). В пасмурную погоду цветки могут не открываться совсем. Хорошо выражена протерогиния — тычинки созревают только на второй — четвертый день после раскрытия цветка, а пестики готовы к восприятию пыльцы сразу после раскрытия. В пасмурную погоду возможно самоопыление (Оловян-пикова, 1949).
Межвидовые гибриды встречаются в местах перекрытия ареалов разных видов рода Adonis. Так, в Воронежской обл. в Л пекинском р-не часто встречаются гибриды A. vernalisXA. wolgensis Stev. В Кемеровской обл. в Беловском р-не, близ с. Бочаты, были обнаружены гибриды A. vernalisXA. sibiricus Patr.
Семенная продуктивность горицвета очень изменчива. Она зависит от климатических условий различных провинций ареала, от годичной изменчивости погодных условий каждой провинции, от экологических условий произрастания популяций, а также ее возрастного состава. Наибольшая урожайность отмечена в южной части Заволжья (в Куйбышевской обл.) — 16 312 семян на 100 м2. Наименьшее число семян оказалось на севере Среднерусской возвышенности (на некосимом участке заповедной Стрелецкой степи)— всего 1167. Плодики горицвета не переходят в почвенный запас. Специальные исследования показали полное отсутствие в почве цельных плодиков, обнаружены только остатки эндокарпия (Голубева, 1962; Пошкурлат, 1969).
Экология
Горицвет весенний распространен на хорошо дренированных элементах рельефа с абсолютными высотами от 150 до 480 м. В южных районах он произрастает на склонах северной и северо-западной экспозиции, в более северных районах предпочитает юго-восточную и юго-западную экспозиции, еще дальше к северу переходит на склоны южной экспозиции. Наиболее обычен в нижней трети склонов.
Для горицвета как ранневесеннего растения имеет большое значение насыщенность почвы влагой тающего снега. Этим, вероятно, объясняется произрастание этого растения в местах, где задерживается снег: на опушках леса, полянах, среди кустарника.
В европейской части СССР горицвет произрастает преимущественно на карбонатных черноземах, реже на выщелоченных черноземах и темно-серой лесной почве. В сибирской части ареала он встречается большей частью на выщелоченных, реже на обыкновенных черноземах, серой лесной и лугово-черноземовидной почвах. Амплитуда содержания гумуса в почвах с зарослями горицвета — от 2,8 до 15,1%. Кислотность же почвы всюду остается в пределах нейтральной или слабокислой (pH от 6 до 8,1).
45
Растение светолюбивое. В условиях недостаточного освещения развивается плохо или выпадает совсем. Растет в разных климатических условиях. От поздневесенних заморозков гибнут цветки. В малоснежные морозные зимы иногда подмерзают корневища.
Биологическая продуктивность
В зависимости от почвы и других экологических условий урожайность надземных частей колеблется в большом интервале — от 0,5 до 8,4 кг сырой массы на 100 м2.
На общее жизненное состояние и накопление биомассы большое влияние оказывает почвенная влага. Этим объясняется лучшее состояние горицвета в нижней трети склона в области смыкания подвешенных и поднимающихся капиллярных вод.
Фитоценология
A. vernalis распространен, как уже отмечено выше, преимущественно в лесостепи, отчасти заходит в северную часть настоящих степей. В более южных вариантах степей европейской части СССР он растет отдельными массивами в местообитаниях с соответствующими микроусловиями. Произрастает горицвет в основном на остеп-ненных лугах и в луговых степях с мезофильным разнотравьем, в меньшей степени — в типчаковых и ковыльных степях.
Для большинства популяций горицвета европейской части ареала характерна высокая численность, наличие подроста, среднее накопление биомассы. Высота растений колеблется от 20 до 40 см. Среднее число побегов одной особи от 3 до 10.
Для сибирской части ареала-характерна меньшая численность. Популяции западной части сибирского ареала, примерно до Ишима, имеют высокий процент генеративной фракции и чрезвычайно низкий— виргинильной. В восточной части ареала, начиная от Бара-бинской низменности, численность подроста значительно возрастает и в восточных провинциях (Верхнеобской и Кузнецкой) она достигает довольно высоких показателей. Наиболее крупные особи генеративной фракции (10—30 побегов высотой 40—60 см) развиваются на Зауральском плато и в восточных провинциях.
На жизненное состояние популяций сильно влияет хозяйственная деятельность человека, в частности, выпас скота, заготовка лекарственного сырья, сенокошение. При большой пастбищной нагрузке наблюдается сильное измельчение растений. Неправильные приемы заготовок сырья в течение продолжительного времени тоже приводят к угнетению популяции.
Отсутствие сенокошения способствует большей задержке и накоплению влаги, что содействует большему накоплению биомассы. Однако сомкнутый травостой и ветошь мешают развитию подроста. При сенокошении, наоборот, снижается запас влаги в почве, что резко сказывается на развитии растений горицвета. Более разре-
46
жснный травостой косимой степи создает нужные условия для подроста.
Несмотря на слабую возобновляемость массивов горицвета, они сохраняются довольно долго за счет большой продолжительности жизни особей (Цибанова, 1961). В связи с медленным развитием новые территории горицвет захватывает чрезвычайно редко и осваивает их долго (и то лишь при сочетании благоприятных климатических условий).
Хозяйственное значение
В надземных частях горицвета весеннего обнаружено более 20 сердечных гликозидов, из которых 10 карденолидов выделено в индивидуальном состоянии: цимарин, адонитоксин, строфантидин, К-строфантин-р, строфандогенин, 3-ацетилстрофандогенин, ацетил-идонитоксин, адонитоксол и др. (Зоз и др., 1967). Кроме того, из надземных и подземных частей выделен флавоновый гликозид, адо-нивернит, кумарин вернадин, спирт адонит (Атлас лекарственных растений, 1962; Зоз и др., 1967). Гликозиды горицвета обладают высокой биологической активностью и нашли широкое применение в медицине и ветеринарии. Их используют для. лечения хронической недостаточности сердечной деятельности и неврозов сердца; в сочетании с бромом назначают при повышенной нервной возбудимости, бсссонице, эпилепсии (Варлих, 1912; Землянский, 1958; Царев, 1964; Турова, 1967; Йорданов и др., 1972; Машковский, 1972).
Горицвет весенний известен как медонос (Глухов, 1955) и как декоративное растение (Clapham и. а., 1958). Его использовали для окраски шелковых тканей в желтые тона.
Наличие гликозидов обусловливает ядовитость всех органов-‘горицвета, в связи с чем он может вызывать отравления при поедании его животными. Обычно домашние животные это растение не трогают (Полякова, 1939), однако'ранней весной могут быть случаи отравления, особенно у ягнят (Захарьин, 1935; Вильнер, 1966). Кроме того, наблюдаются отравления лошадей при поедании горицвета в сене (Kummer, 1952).
Как важное лекарственное растение г. весенний вводят в культуру. Дикорастущие заросли его требуют бережного обращения, а в ряде районов должны строго охраняться.
О. В. Смирнова и В. А. Черемушкина!
РОД ХОХЛАТКА — Corydalis Medic.
Во флоре Московской обл. род Corydalis Medic, (из сем. Fuma-riaceae) представлен 4 ^видами, относящимися к подроду Capnites DC.
Два из них—С. bulbosa (L.) DC. и С. marschalliana (Pall.) Pers.—входят в секцию Radix-cava Irmisch, которая включает ложноодносемядольные виды (ложноодносемядольность предположительно возникла в результате недоразвития второй семядоли). Хох-латкр этой секции — эфемероиды, геофиты, подземные части которых представлены моноподиально нарастающим полым клубнем. Генеративные побеги развиваются в пазухах сближенных чешуевидных листьев и несут два очередных сближенных срединных листа. Прицветники цельные. Придаточные корни рассеяны по всему 'клубню (Irmish, 1862; Prantl, Kiindig, 1891; Velenovsky, 1910; Ryberg, 1959).
Два других вида — С. solida (L.) Swartz и C. intermedia (L.) Merat.—относятся в секции Pes-gallinaceus Irmisch. В эту секцию включены также ложноодносемядольные раннецветущие эфемерои- ды, геофиты, но с клубнем, не имеющим полости и сменяющимся ежегодно. Молодые клубни окружены разрушающимися остатками 'клубней прошлых лет. Нарастание симподиальное. Генеративные побеги развиваются из терминальной почки ив некоторых случаях дополнительно из почки в пазухе верхнего чешуевидного листа. Последний отделен от нижних сближенных чешуевидных листьев длинным междоузлием и обычно размещен над поверхностью почвы (или в подстилке). Столбик сохраняется при плодах (Irmish, 1862; Ryberg, 1950, 1955).
1 Разделы «Номенклатура» составлены В. Н. Тихомировым.
48
1. ХОХЛАТКА ЛУКОВИЧНАЯ
Номенклатура
Corydalis bulbosa (L.) DC. in Lam. et DC., Fl. Fr., ed. 3, 4 (1815) 637,-—C. bulbosa subsp. bulbosa: Mowat, Chater in Fl. Eur. 1 (1964) 253.— Fumaria bulbosa a. cava L., Sp. pl. (1753) 699 (pro var. typ.).— F. cava (L.) Miller, Gard. Diet., ed. 8 (1768) №7.— Corydalis cava (L.) Schweigg. et Koerte, Fl. Erlang. 2 (1811) 44; Кауфм., Моск, фл., изд. 1 (1866) 40, изд. 2 (1889) 28; Цингер, Сб. свед. (1885) 51; Шмальг., Фл. 1 (1895) 40; Петунн., Крит, обзор 1 (1896) 58; Сырейщ., Илл. фл. 2 (1907) 167; он же, Опр. раст. Моск. губ. (1927) 137; Попов во Фл. СССР 7 (1937) 659; Федченко в Маевск., Фл., изд. 7 (1940) 378; Васильченко, там же, изд. 8 (1954) 327, изд. 9 (1964) 320; Скворцов в Опр. раст. Моск. обл. (1966) 173.— Хохлатка луковичная, х. полая.
Описана из Европы. Тип в Лондоне.
Морфологическое описание
Многолетнее, травянистое, поликарпическое, клубневое растение. Корневая система состоит из эфемерных .придаточных корней длиной 2—10 мм и диаметром менее 1 мм. Боковых корней не образуется. Клубень взрослых растений залегает на глубине 5—12 см. Он 2—2,5 см высотой и 1—1,5 см шириной, продолговато- или округло-треугольный, с полостью внутри. Полость имеется у всех генеративных растений, но у более молодых она плохо заметна (Michalet, 1859; Irmish, 1862; Troll, 1957; Ryberg, 1959).
У растения на протяжении всей или почти всей жизни функционирует только верхушечная почка, которая формирует розеточный побег. На годичном участке этого побега развиваются 3—4 срединных ассимилирующих листа с черешками длиной 8—14 ем. Пластинка их трижды тройчато-рассеченная, длиной 2,8 см и шириной 3,5 см, доли ее на коротких черешках или сидячие, цельные или крупнозубчатые, заостренные, реже острые или тупые. Листья нежные, светло-зеленые, с сизоватым налетом. У цветущих растений листья на розеточном побеге не всегда развиваются, в таком случае функцию ассимиляции осуществляют два сближенных черешковых листа, расположенных в верхней части генеративных побегов. Число генеративных побегов у одной особи колеблется от 1 до 3- (редко 4), высота 20—30 см.
Соцветие — цилиндрическая рыхлая кисть акропетального типа (из 5—13 цветков), заканчивается стерильным рудиментом оси. Прицветники цельные, травянистые, зеленые с фиолетовым оттенком, в 2—3 раза длиннее цветоножки. 2 очень мелких чашелистика рано опадают. Венчик 18—28 мм длины, состоит из 4 лепестков, расположенных в двух кругах. Отгиб наружных лепестков широкий, с большой выемкой, нижний лепесток с широким малозаметным бу
49
горком, шпора толстая, тупая, с изогнутым концом, по величине равна лепестку. Существуют фиолетово-розовая и белоцветковая формы, которые встречаются по всему ареалу. Над шпорцем при основании тычинки имеется нектарник. Завязь синкарпная, из 2 плодолистиков, столбик 1 прямой, рыльце зубчатое, 2 тычинки, возникшие вследствие расщепления тычинок внутреннего круга и срастания их с тычинками наружного круга, имеют широкие нити, разделенные наверху на 3 доли. Средняя доля несет обычный четырехгнездный пыльник, боковые — неполные двугнездные пыльники! В отличие от близкородственного вида С. marschalliana у С. bulbosa найдена только трехбороздчатая пыльца (Fischer, 1890). Диаграмма цветка составлена Эйхлером (Eichler, 1878).
Плод — стручковидная, острая, поникающая коробочка, в зрелом состоянии буроватая от выступающих семян, с толстыми стенками, длиной 10—12 мм и шириной 3—4 мм, длина цветоножки 3—4 мм. Семена с эндоспермом, округлые, черные, блестящие, мелкоточечные, крупные, до 3 мм в диаметре, с придатком лентовидной формы (Hegi, 1919; Ryberg, 1955, 1959; Troll, 1964). Число хромосом (2 и) равно 16, в Чехословакии найдена тетраплоидная форма (Mi-chalkova, 1967; Хромосомные числа цветковых растений, 1969).
Географическое распространение
С. bulbosa — европейский атлантическо-субатлантический вид (Meilsel, 1969). В Западной Европе распространена от восточной границы Франции до Польши. На севере достигает побережья Балтийского моря, отдельные местообитания отмечены на юге Скандинавии. Южная граница ареала проходит по Италии и северной части Балканского п-ова. Небольшой изолированный участок описан на севере Пиренейского п-ова (Engler, 1879; Meusel, 1943, 1969). В пределах европейской части СССР произрастает по всей Украине: сплошной ареал доходит до Днепра, восточнее известны изолированные местонахождения по р. Суле, Северному Донцу и на Донецком кряже. После небольшого перерыва этот вид встречается в Могилевской, Смоленской, Московской, Тульской и Калужской областях. Кроме этого, он отмечен в Горьковской, Саратовской, Тамбовской и Ростовской областях (Клеопов, 1941; Флора СССР, 1937). Карта ареала составлена Мейзелем (Meusel, 1969).
В Московской обл. встречается очень редко в Клинско-Дмитров-ском, Западном и Восточном ботанико-географических районах (Ворошилов и др., 1966).
Онтогенез
Семена хохлатки луковичной прорастают в начале апреля в юго-западных районах СССР и в конце апреля — начале мая в центральных районах европейской части СССР. Тип прорастания надземный. Единственная семядоля имеет длинный черешок, в подзем-
50
Пой части покрытый эпидермальными выростами, выполняющими функцию корневых волосков, и продолговато-эллиптическую пластинку (1—1,2 см длины, и 0,8 см ширины). Главный корень небольшой (несколько миллиметров длины), не ветвится. Клубень у проростка формируется вследствие развития гипокотиля и к концу первого года вегетации достигает 3—4 мм в диаметре (Ryberg, 1959; Lukasiewicz, 1962).
На второй год жизни растение развивает первый срединный пссимилирующий лист ювенильного типа с небольшой 3—5-раздель-пой пластинкой. Листовая серия ювенильных растений состоит из 2—3 чешуевидных и 1 (редко 2) срединного ассимилирующего листа. Главный корень отмирает, возникают первые придаточные корни, расположение корней рассеянное. Число придаточных корней увеличивается с возрастом растения. В образовании клубня принимает участие не только гипокотиль, но и стеблевая часть. Форма клубня шаровидная. Ювенильное состояние длится 1—3 года.
Рис. 5. Схема онтогенеза хохлатки луковичной и хохлатки Маршалла: 1—проросток, 2—молодое вегетативное растение, 3— средневозрастное генеративное, 4— сенильные растения
В имматурном и молодом вегетативном состоянии происходит постепенное увеличение размеров растения (рис. 5). Пластинка листа изменяется от дважды до трижды тройчато-рассеченной. Клубень имеет овально-округлую форму, на поперечном разрезе в центре его появляются участки отмирающих паренхимных клеток сердцевины. Полость появляется у 3—4-летних экземпляров (Lukasiewicz, 1962).
Генеративные побеги развиваются из почек в пазухах чешуевидных листьев. Молодые генеративные растения обычно имеют 1 генеративный побег, розеточные ассимилирующие листья, как правило, не развиваются. Форма клубня овальная или продолговато-треугольная, полость небольшая. В средневозрастном генеративном состоянии увеличивается число генеративных побегов от 2 до 4 (8), клубни крупные, неправильно-треугольной формы с небольшими выростами, полость ясно заметна.
51
В старом генеративном состоянии наблюдается интенсивное развитие полости, что приводит к разделению мощных клубней на несколько самостоятельных жизнеспособных частей и образованию клона. Деление клубня может сопровождаться отмиранием верхушечной почки, в некоторых случаях она сохраняется на одной из партикул. На участках клубня, лишенных верхушечной почки, трогаются в рост спящие пазушные почки. Возможно развитие и придаточных почек (Ryberg, 1959). Старые генеративные растения обычно образуют 1—2 генеративных побега, клубень треугольно-конусовидной формы, почти полностью разрушен, диаметр его живой части составляет около 5 мм, общий диаметр 3—5 см (Ryberg, 1955).
В старом вегетативном состоянии от клубня остается только ’/3 часть. В некоторых случаях верхушечная почка отмирает и в рост трогаются спящие пазушные почки. У растения развиваются 2—3 чешуевидных и 1 срединный лист с 5—7-раздельной пластинкой. У сенильных растений верхушечная или заменившая ее пазушная почка формируют 1—2 чешуевидных и 1 срединный лист ювенильного типа.
У экземпляров с нормальной или пониженной жизненностью онтогенез заканчивается в фазе одноосного побега, у экземпляров с повышенной жизненностью формируется недолговечный клон, состоящий обычно из старых вегетативных и сенильных особей.
. Сезонный ритм развития
Заложение и полное развитие листовых зачатков у монопо-диально нарастающего розеточного побега осуществляется в течение двух лет. В начале июня на конусе нарастания дифференцируется 1—2 листовых примордия, число их к осени достигает 6. В течение осени и начала весны 3 наружных примордия дифференцируются в чешуевидные листья, полностью прикрывающие почку. Весной в пазухах чешуевидных листьев ясно заметны небольшие почки. У внутренних примордиев начинается дифференциация листовой пластинки, но обычно лишь 1 или 2 из них превращаются в срединные ассимилирующие листья, остальные остаются в полудифферен-цированном состоянии и выполняют роль почечных чешуй. Через , год после заложения первых листовых примордиев, в июне, из пазушных почек первых трех чешуевидных листьев наблюдается развитие генеративного побега. Обычно развиваются не все заложив-шиеся побеги. Преобладание получает побег в пазухе второго чешуевидного листа. Одновременно с заложением генеративных побегов происходит формирование на конусе нарастания новых ли- 1 стовых примордиев.
Генеративный побег полностью формируется к сентябрю. В это время можно ясно различить два ассимилирующих листа, полностью сформированные прицветники и зачатки цветков. Окончательное формирование цветка, микро- и мегаспорогенез наблю- , даются в конце осени и начале зимы. Таким образом, хохлатка ,
луковичная относится к первой группе видов, выделенных И. Г. Серебряковым (1947 а),
В октябре появляются новые придаточные корни. У хохлатки луковичной два типа придаточных корней: первые появляются осенью и отмирают летом следующего года после окончания вегетации, вторые возникают весной и отмирают летом того же года после окончания вегетации. Придаточные корни возникают в камбиальном слое и вследствие этого отмирающие корни выходят из более глубоких слоев клубня, чем новые.
В январе — феврале верхушечная почка заметно увеличивается в размерах. В марте начинается интенсивный надземный рост генеративного побега. В начале апреля побег появляется на поверхности почвы, увеличивается в размерах и зацветает. Продолжительность цветения 2—2,5 недели. Хохлатка луковичная — весеннезеленый вид (Walter, 1968). Цветки опыляются шмелями и некоторыми длиннохоботковыми бабочками (Velenovsky, 1910; Трофимов, 1939). Самоопыление возможно, но происходит редко, в связи с тем что пыльца созревает раньше, чем рыльце готово к опылению (Кернер, 1902).
Семена созревают в начале — середине мая, одновременно отмирают полностью надземные побеги. Продолжительность ассимиляционного периода в европейской части СССР 1—1,5, в Западной Европе 2—2,5 месяца (Hegi, 1919; Ryberg, 1955).
Способы размножения и распространения
Преобладающее значение имеет семенное размножение. Созревающие семена, снабженные особым придатком-элайосомом, распространяются муравьями. Семена осыпаются с недоразвитым зародышем, состоящим из нескольких недифференцированных клеток. До начала зимы зародыш не успевает полностью сформироваться. Семена очень чувствительны к недостатку влаги и не прорастают, если в течение зимы содержатся сухими (Ryberg, 1950, 1955). Жидкость, содержащаяся в элайосоме, частично предохраняет семя от высыхания. Прорастают семена весной через год после созревания.
Вегетативное размножение наблюдается в конце жизненного цикла. Оно представляет собой сенильную партикуляЦию.
Экология
Хохлатка луковичная — эумезофит, произрастает на богатых свежих почвах, суглинистых и супесчаных. Плохо переносит тяжелые почвы. Она обычна на равнинах, но местами поднимается до высоты 1800 м (например, в Альпах). Освещенность в местообитаниях этого вида колеблется от 25 до 80% от полной. Сосущая сила зеленых листьев равна 8,7 атм, пожелтевших-—6,4—10,0 атм (Hegi, 1919; Ryberg, 1955; Walter, 1968).
53
Фитоценология
С. bulbosa относится к временным доминантам, входит в весеннюю синузию эфемероидов, образуя вместе с С. solida аспект в широколиственных лесах на территории европейской части СССР (Алехин, 1951). Она встречается в зарослях кустарников, у изгородей, в садах, в лиственных (в частности буковых) лесах Европы, входит под полог прикарпатских дубовых и дубово-грабовых, приднепровских грабово-дубовых и среднерусских дубово-липовых насаждений. В дубравах Тульских и Калужских засек растет вместе с черемшой и зубянками. В Московской обл. встречается, в частности, в обедненных широколиственных лесах юго-восточных склонов Клинско-Дмитровской гряды (Алехин, 1951; Растительный покров СССР, 1956; Walter, 1968; Ярошенко, Грабарь, 1969).
Хохлатка луковичная относится к группе видов, популяции которых характеризуются ярко выраженным левосторонним спектром: большая часть особей относится к виргинильному периоду. Такой характер спектра связан с интенсивным семенным возобновлением, относительно небольшой протяженностью генеративного периода и почти полным отсутствием в популяциях особей сенильного периода. Основной отпад происходит в конце генеративного периода. Например, в популяции, описанной в Закарпатье в средневозрастных дубово-буковых насаждениях, при общей численности хохлатки около 100 особей на 1 м2 на долю виргинильных приходится 80%, генеративных—17%. Старые вегетативные и сенильные особи присутствуют в популяции в очень небольших количествах (2—3%).
Внутривидовая систематика
У С. bulbosa выделены следующие формы: f. angustifolia Beck — доли листа узкие, заостренные (эта форма наиболее распространена); f. incisa Р. Junge — нижние прицветники сильно рассечены, по форме напоминают конечные доли стеблевых листьев; f. paniculata Р. Junge — в пазухах чешуевидных листьев возникает по два и более генеративных побегов (Hegi, 1919).
Хозяйственное значение
В клубнях растения содержится ряд алкалоидов: бульбокап-нин, коритуберин, коридин, изокоридин, корикавин, корикавидин (Атлас лекарственных растений, 1961). Бульбокапнин обладает снотворными свойствами, но лечебного применения пока не нашел. В народной медицине клубни употребляли при лечении глистных заболеваний и как успокаивающее средство.
54
2. ХОХЛАТКА МАРШАЛЛА
Номенклатура
Corydalis marschalliana (Pall.) Pers., Syn. 2 (1807) 269; Цин-rep, Сб. свед. (1885) 50; Шмальг., Фл. 1 (1895) 40; Петунн., Крит, обзор 1 (1896) 59; Сырейщ., Илл. фл. 2 (1907) 168; он же, Опр. раст. Моск. губ. (1927) 137; Попов во Фл. СССР 7 (1937) 659; Федченко в Маевск., Фл., изд. 7 (1940) 378; Васильченко, там же, изд. 8 (1954) 327, изд. 9 (1964) 320; Скворцов в Опр. раст. Моск. обл. (1966) 172.— Fumaria marschalliana Pall, in Nov. Act. Petrop. 10 (1797) 315 — Corydalis bulbosa (L.) DC. subsp. marschalliana: Chater in ITeddes Repert. 69 (1964) 56; Monat, Chater in Fl. Eur. 1 (1964) 253.— Хохлатка Маршалла.
Описана из Крыма. Тип в Берлине.
Морфологическое описание
Морфологически этот вид очень близок к С. bulbosa. Хохлатка Маршалла — многолетнее травянистое поликарпическое клубневое растение. Корневая система представлена эфемерными придаточными корнями, размеры и размещение их существенно не отличаются у обоих видов. Клубень небольшой, 1—1,5 см высотой и 0,8—1,2 см шириной, конической или полусферической формы, с полостью внутри, залегает на глубине 8—15 см.
Многолетние части растения представлены укороченным розеточным побегом, нарастающим моноподиально, с пазушными генеративными побегами. На годичном приросте розеточного побега развивается 3—5 чешуевидных и 3—7 срединных ассимилирующих листьев. Из почек в пазухах чешуевидных листьев формируются генеративные побеги 20—35 см высотой с 2 сближенными очередными листьями в верхней части побега.
Ассимилирующие листья розеточного побега на длинных черешках (10—14 см), листья на генеративном побеге имеют короткие черешки (1—2 см). Листовая пластинка приблизительно в 1,5— 2 раза больше, чем у хохлатки луковичной, почти всегда дважды тройчато-рассеченная. Срединная доля крупная, цельная, продолговато-удлиненная, закругленная, реже заостренная. Боковые доли обычно делятся на большую внутреннюю и более мелкие наружные дольки.
Соцветие — довольно рыхлая, цилиндрическая, многоцветковая кисть акропетального типа (рис. 6). Прицветники зеленые, цельные, продолговатые или- яйцевидные, крупнее, чем у С. bulbosa. Цветки до 25 мм длины, отличаются от цветков С. bulbosa только окраской венчика и формой рыльца. Окраска венчика желтая, желтовато-белая, на Кавказе встречаются особи с розовыми наружными и желтыми внутренними лепестками. Рыльце дисковидное, по краю мел-кобородавчато-городчатое.
55
Рис. 6. Взрослое генеративное растение хохлатки Маршалла
Пыльцевые зерна 32—38 мк в диаметре, бугорки экзины крупные, редко расположенные, ясно выступающие, в плане имеют вид крупных угловатых пятен с рассеянными зернами и мелкими бугорками между ними. Встречаются отклоняющиеся пыльцевые зерна — трехбороздчатые, а также с фрагментированными, слившимися, неправильно расположенными бороздами (Сагдуллаева, 1959; Ryberg, 1959).
56
Плод — стручковидная коробка, более узкая, чем у хохлатки луковичной. Семена округлые, черные, мелкоточечные, блестящие, с мясистым придатком, диаметром около 3 мм.
Географическое распространение
С. marschalliana входит в комплекс кв.ерцетальных светолюбивых элементов, связанных в своем происхождении с субксерофиль-ными дубравами Средиземноморья (Клеопов, 1941). Европейскую часть СССР подобные виды заселили в течение послерисского периода (Гроссет, 1962).
Ареал С. marschalliana не сплошной. Один крупный участок занимает север Балканского п-ова от Албании до Черного моря, за-1’’ хватывая Молдавию. Второй участок расположен в Восточной Европе. Западная граница в европейской части СССР проходит по Калужской, Сумской и Днепропетровской областям. Данные о произрастании вида в Орловской и Брянской областях отсутствуют. На север этот вид не заходит дальше Владимирской и юга Горьковской областей, самая восточная точка ареала — северная часть Пензенской обл. Ареал хохлатки Маршалла доходит до Азовского моря, далее узким языком спускается к Кавказу, захватывая частично юго-восточную часть Ростовской обл. и Ставропольского края. На Кавказе она встречается повсеместно, довольно широко распространена в лесном поясе горного Крыма. Третий изолированный участок ареала находится в Малой Азии и Иране (Гроссгейм, 1950; Ryberg, 1955; Meusel, 1943, 1969). Карта ареала составлена Мейзе-лем (Meusel, 1969).
В Московской обл. встречается очень редко лишь в южных районах (Ворошилов и др., 1966; Губанов, Алексеев, 1975).
Онтогенез
Онтогенез С. marschalliana почти не отличается от онтогенеза С. bulbosa. Строение проростков обоих видов идентично (рис. 7). Все признаки возрастных состояний — число листьев, степень рас-членности пластинки листа, форма клубня и степень развития полости— у обоих видов полностью совпадают (см. рис. 5).
Как и хохлатка луковичная, рассматриваемый вид в течение всего онтогенеза существует в стадии моноподиально нарастающего одноосного побега и лишь у экземпляров с повышенной жизненностью наблюдается факультативное образование клона вследствие распадения клубня на ряд жизнеспособных участков.
Сезонный ритм развития
Сезонный ритм обоих видов существенно не различается. Заложение и развитие листовых зачатков у моноподиально нарастающего розеточного побега осуществляется в течение двух лет, заложе-
57
CM
Рис. 7. Проростки хохлатки Мар-' шалла
ние и развитие генеративного побега происходит в течение одного года. Все сезонные изменения у обоих видов осуществляются приблизительно в одни и те же сроки, изменяясь только в зависимости от широты местности.
Заложение генеративного побега хохлатки Маршалла более детально исследовано в условиях Ставропольского края (Скрипчинский, 1969). Недифференцированный конус нарастания в пазухе чешуевидного листа можно ясно различить уже в конце зимы, в начале апреля начинается заложение соцветия. Цветок в виде бугорка появляется уже в мае и заканчивает свое развитие к сентябрю. В это время генеративный побег сформирован уже полностью с листьями и вполне развитым соцветием. В то же время образуются молодые корни, рост их
продолжается всю осень и, возможно, часть зимы. Пыльца и пыльники созревают значительно позднее. Заложение пыльников происходит в конце октября, в ноябре наблюдается редукционное деление. С конца января по март происходит подземный рост растения. Сначала увеличивается в размерах почка, затем почечные чешуи раскрываются и начинает расти генеративный побег. В марте — начале апреля генеративный побег появ
ляется над поверхностью почвы, через некоторое время растение
зацветает.
Опыление производится шмелями и некоторыми длиннохоботковыми бабочками. Пчелы часто прокусывают шпорец цветка, добывая нектар. Семёна созревают в первой половине мая. Период вегетации длится у этого весеннезеленого вида 1—1,5 месяца.
Способы размножения и распространения
Хохлатка Маршалла размножается почти исключительно семенным путем. Как и х. луковичная, она относится к группе мирме-кохоров (Sernander, 1906). Зародыш в опадающих семенах не дифференцирован. В лабораторных условиях семена, как правило, не прорастают, так как не выдерживают Длительного пребывания в сухом состоянии. Высеянные сразу же после созревания (в мае) в условиях Ставрополья семена прорастают в январе — мае следующего года, всхожесть в среднем 16,4% (Скрипчинский, 1963).
58
Реальная семенная продуктивность х. Маршалла; определенная нами в Воронежской обл., составила следующую величину: в одном плоде в среднем 5—6 семян, на одном побеге — 40—48 семян, на одной средневозрастной генеративной особи —96 семян. В проле-сково-снытевой дубраве Теллермановского опытного лесничества запас жизнеспособных семян этого вида — 480—500 на 1 м2.
. Экология
Хохлатка Маршалла — эумезофит, произрастает в равнинных и предгорных районах на почвах разного механического состава: глинах, суглинках, песках и известняках. Тяготеет к старым широколиственным лесам на почвах нейтральной реакции, богатых гумусом. Освещенность в местообитаниях этого вида изменяется от 30 до 40% от полной (Алехин, 1951).
Биологическая продуктивность
На темно-серых лесных песчаных почвах в снытевых дубравах Воронежской обл. воздушно-сухой вес генеративных особей колеблется от 0,75 до 2,34 г. Вес ювенильных особей составляет 0,03 г, сенильных— 1,22 г. В снытевых дубравах Воронежского заповедника биомасса популяции, включая надземную и подземную ее части, составляет 13,32 г/м2.
Фитоценология
С. marschalliana является временным доминантом в травяном покрове дубрав, довольно часто господствуя в синузии эфемероидов (Алехин, 1947, 1951). В снытевых дубравах европейской части СССР она образует весенний аспект вслед за Scilla sibirlca Andrews и С. solida (L.) Swartz. В качестве содоминанта она встречается в осоково-снытевых и снытево-осоковых дубо-липняках, но при значительном обилии Carex pilosa Scop, исчезает полностью. Она встречается в гирканских смешанных широколиственных лесах в нижнем поясе Талышского хребта, в частности в лесах из шелковой акации, а также в дубово-грабовых, дубовых и буковых лесах горного Крыма (Гроссгейм, 1950; Растительный покров СССР, 1956).
Популяции этого вида исследованы в снытевых и осоково-снытевых дубравах Воронежской обл. на темно-серых лесных почвах в насаждениях разного возраста. Как и предыдущий вид, С. marschalliana характеризуется популяциями левостороннего типа: в возрастном спектре преобладают особи виргинильного периода (74—90% от общей численности), среди генеративных особей господствуют молодые, сенильные растения во многих популяциях не встречены.
Наиболее высокая жизненность особей (максимальный вес, высота побегов и семенная продуктивность) отмечена в 200—250-летних снытевых дубравах Теллермановского опытного лесничества.
59
Обйгая численность хохлатки в этих насаждениях невелика (13— 27 особей на 1 м2), что обусловлено конкурентными отношениями с другими эфемероидами. Численность генеративных особей при этом максимальная, что свидетельствует о высокой приживаемости вида в ценозе. Максимальной численности вид достигает в осоково-сны-тевом дубо-липняке в Воронежском заповеднике, где при общей численности 60 особей хохлатки на 1 м2 наибольшее число их находится в ювенильном и молодом вегетативном состояниях. Подобный молодой облик популяции определяется интенсивным семенным размножением, небольшой длительностью генеративного и особенно сенильного периодов, практически полным отсутствием вегетативного размножения.
Внутривидовая систематика
На Кавказе часто встречаются "формы С. marschalliana, выделяемые в особые разновидности: var. rosea-purpurea Rupr.— лепестки розовато-пурпурные, нижний пестик желтый; var. pseudocava Grossh. ( = С. cava auct. fl. Talysch.)—боковые сегменты листьев надрезаны на более узкие, чем в типе, дольки (Гроссгейм, 1950).
Хозяйственное значение
В клубнях хохлйтки Маршалла обнаружены те же алкалоиды, что и в клубнях х. луковичной (Атлас лекарственных растений, 1962). В народной медицине растение использовалось как лекарственное при сердечно-сосудистых заболеваниях.
В прошлом веке х. Маршалла широко культивировалась в садах как декоративное растение (Ryberg, 1955).
3. ХОХЛАТКА ПЛОТНАЯ
Номенклатура
Corydalis solida (L.) Swartz in Svensk Bot., 8 (1817) t. 531.— Кауфм., Моск, фл., изд. 1 (1866) 40, изд. 2 (1889) 28; Цингер, Сб. свед. (1885) 51; Шмальг., Фл. 1 (1895) 41; Петунн., Крит, обзор 1 (1896) 58; Сырейщ., Илл. фл. 2 (1907) 168, 4 (1914) 82; он же, Опр. раст. Моск. губ. (1927) 137; Mowat, Chater in Fl. Eur. 1 (1964) 256; Скворцов в Окр. раст. Моск. обл. (1966) 172.— Fumaria bulbosa у. solida L., Sp. pl. (1753) 699.— F. solida Ehrh., Beitr. 6 (1791) 146.— F. halleri Willd., Prodr. fl. Berol. (1787) 229,— Corydalis halleri (Willd.) Willd., Enum. hort. Berol. 2 (1809) 740; Попов во Фл. СССР 7 (1937) 671; Федченко в Маевск., Фл., изд. 7 (1940) 377; Васильченко, там же, изд. 8 (1954) 326, изд. 9 (1964) 319.— Хохлатка плотная, х. Галлера.
Описана из окрестностей Берлина. Тип в Берлине.
60
Морфологическое описание
Многолетнее травянистое поликарпическое растение. Главный корень отмирает в первый вегетационный период, со второго года жизни у растения развивается корневая система придаточного типа. Корни сосредоточены в базальной части клубня, эфемерны, сменяются ежегодно, длина их 2—5 см. Обычно развиваются боковые корни первого порядка. Клубень небольшой, овально-шаровидный, светло-коричневый, 0,8—1,5 см в диаметре, располагается на глубине 5—10 см, ежегодно возникает из камбия, расположенного под почкой возобновления.
В основании годичного побега находятся 2—3 сближенных чешуевидных листа, междоузлие верхнего чешуевидного листа удлиненное. Верхний чешуевидный лист крупный, до 1 см длины, линейно-ланцетный, вогнутый. В верхнёй части побега под соцветием располагаются на коротких черешках 2 сближенных, очередных, срединных, тонких, зеленовато-сизых, ассимилирующих листа с дважды или почти трижды тройчато-рассеченной пластинкой. Сегменты (доли) листа на длинных черешочках, почти до основания или до 'Д— ’/г надрезанные на линейнопродолговатые, слегка клиновидные, тупые, реже обратнояйцевидные дольки. Дольки листа цельные или на верхушке с 2—4 тупыми зубцами. Форма, размеры и степень рассеченно-сти как долей листа, так и прицветников очень сильно варьирует у этого вида, что приводило к выделению большого числа разновидностей и форм.
Соцветие — плотная, многоцветковая (15—25 цветков), верхушечная кисть акропетального типа (рис. 8). У более мощных экземпляров дополнительный генеративный побег развивается из почки в пазухе верхнего чешуевидного листа. Обычно он слабо развит, в некоторых случаях вместо него формируется укороченный веге- Рис. Взрослое генеративное расте-тативный побеге 1—2листьями. ' ние хохлатки плотной
61
Прицветники клиновидно-обратнояйцевидные, спереди надрезаны на линейные дольки или зубчатые. В некоторых случаях нижние прицветники цельные, верхние слабозубчатые или рассеченные. Цветоножки нитевидные, нижние почти равны прицветникам, до 10 мм длины,, верхние короче. Цветок устроен так же, как и у других видов рода. Чашелистики мелкие, быстро опадающие. Венчик розово-фиолетовый (окраска его может очень резко варьировать от почти белой до темно-фиолетовой и темно-розовой)', 15—20 см длины, отгиб наружных лепестков довольно широкий, на верхушке широковыемчатый, нижний лепесток с заметным бугорком при основании, шпора почти прямая или чаще слегка дуговидно согнутая вниз, тупая, равна лепесткам или чуть длиннее их, толстая. Изредка встречаются цветки с 2 шпорцами, с 1 шпорцем и 1 рудиментом и цветки с образованиями промежуточного типа (Goebel, 1913).
Пыльцевые зерна — 30—34 мк в диаметре, 3-бороздчатые или 6- и 12-морщинные, бугорки пыльцевых зерен относительно крупные, редко расположенные, высокие, в плане в виде крупных угловатых, иногда неправильных пятен. Сэкзина несколько толще нэк-зины, поверхность ее слабоволнистая. Завязь одногнездная, содержит от 5 до 7 кампилотропных семяпочек (Eichler, 1878; Fischer, 1890; Schnarf, 1929; Сагдуллаева, 1959; Коробова-Семенченко, 1960). Рыльце дисковидное, по краю мелкобороздчато-зубчатое. .
Плод — стручковидная коробочка, поникающая или отклоненная, продолговатая, 10—12 мм длины и 3—4 мм ширины, на верхушке заостренная в носик. Семена черные, блестящие, около 2 мм в диаметре, придаток узколанцетовидный, пленчатый, недлинный, отстоящий.
Диплоидное число хромосом, по разным определениям, равно 16, 24 или 32 (Ryberg, 1950; Хромосомные числа цветковых растений, 1969).
Географическое распространение
Хохлатка плотная распространена в Европе от западной границы Франции до Урала. Южная граница ареала проходит по Италии, Сицилии и Балканскому п-ову. Изолированные местонахождения зарегистрированы на Корсике, Сардинии, на севере Африки и в Малой Азии. Северная граница ареала проходит в Нормандии, на севере ФРГ, в ГДР, южной части Польши, а также по побережью Балтийского моря,-Ленинградской, Вологодской и Архангельской областям. Отдельные местонахождения известны на севере Польши и в Швеции (Ryberg, 1955; Meusel, 1969). Восточная граница сплошного ареала расположена в предгорьях Среднего и Южного Урала (Горчаковский, 1968).
Изолированный участок ареала находится в низовьях Енисея (Красноярский, Минусинский и Канский районы) и в Даурии (окрестности г. Нерчинска). Даурские и енисейские популяции С. soli-da относятся к особой разновидности var. subremota М. Pop. и пред-
62
ставляют промежуточную форму между типичной С. solida и С. ге-то1а Fisch. Последний вид отличается более узкими долями листа, узким цветком, у нижнего лепестка близ цветоножки нет бугра (Попов, 1957).
Карта ареала С. solida составлена Мейзелем (Meusel, 1969), а для северной части Западной Европы — Хультенем (Hulten, 1950).
В Московской обл. хохлатка плотная широко распространена практически во всех районах.
Онтогенез
Строение проростков у всех хохлаток подрода Capnites ничем принципиально не отличается. Проросток С. solida характеризуется лишь несколько меньшими размерами семядоли (рис. 9). '
На второй год жизни в ювенильном состоянии у растения формируется удлиненный побег за счет растяжения междоузлия верхнего чешуевидного листа. Верхушечная почка оказывается поднята
над поверхностью почвы на 2—3 см. В ювенильном состоянии у растения образуется 2—3 чешуевидных и 1 срединный трехраздельный лист. В пазухе одного из нижних чешуевидных листьев формируется крупная почка, которая является почкой возобновления. После окончания вегетации надземная и подземная части побега отмирают до почки возобновления. Растение переходит к симподиаль-ному нарастанию (Irmisch, 1862).
У С. solida происходит ежегодная смена клубней. Новые клубни возникают внутри старых вследствие деятельности нового камбиального кольца, формирующегося в основании развивающейся пазушной почки (Irmisch, 1862; Jost, 1890; Ryberg, 1959). При-
Рис. 9. Начальные этапы онтогенеза хохлатки плотной:
1 — проросток, 2 — ювенильное растение, 3— имматурное
рода клубней взрослых растений этого вида долго обсуждалась в литературе. В настоящее время М. Риберг (Ryberg, 1959) пришел к заключению, что клубни хохлатки плотной имеют гипокотильную
63
структуру особого рода, что сближает этот род с х. луковичной и х. Маршалла. Замещающий клубень образует слабовыраженный ложностержневой корень (заметен только у молодых растений) и несколько. придаточных корней, формирующихся у его основания.
Дальнейшее развитие растения приводит к появлению в имма-турном состоянии дважды тройчато-рассеченных срединных листьев. В молодом вегетативном состоянии развиваются 2 срединных листа: первый трижды, а второй дважды тройчато-рассеченный. Клубень округлый, до 1 см в диаметре. Междоузлие верхнего чешуевидного листа достигает 5—7 см длины.
Зацветает хохлатка плотная на 4—6 год (Ryberg, 1950; Никитина, 1955). Генеративные побеги развиваются из верхушечных почек. Молодые генеративные растения обычно имеют один генеративный побег. У более мощных экземпляров из пазушной почки верхнего чешуевидного листа развивается розеточный побег с 1—2 срединными листьями (рис. 10).
Рис. 10. Схема онтогенеза хохлатки плотной и хохлатки промежуточной: 1 — проросток, 2 — ывйшпъшж растение, 3 — молодое вегетативное, 4 — средневозрастное генеративное, 5 — старое генеративное
В средневозрастном генеративном состоянии закладываются 2 почки возобновления в пазухах базальных чешуевидных листьев. Каждая из них формирует свой клубень, но некоторое время оба дочерних клубня остаются скрытыми под оберткой из разрушающихся клубней прошлых лет (Goebel, 1913; Трофимов, 1952; Ryberg, 1959). У такого растения имеется 2 генеративных побега. Средневозрастные особи с повышенной жизненностью развивают по 2 генеративных побега из каждого дочернего клубня: один из верхушечной почкй, другой из почки в пазухе верхнего чешуевидного листа. У экземпляров с пониженной жизненностью деление клубня в средневозрастном генеративном состоянии не наблюдается. Они отличаются от молодых особей лишь несколько большими размерами, числом цветков, придаточных корней и большим количеством J разрушающихся остатков клубней прошлых лет.
К моменту полного обособления дочерних клубней от материнского число их может достигать 4, а в очень редких случаях — 6 и 8,
64
образуется клон. Размеры дочерних клубней по мере их возникновения уменьшаются по сравнению с материнским, соцветия содержат меньше цветков, а в некоторых случаях цветки недоразвиваются. Дочерние особи переходят в более старое по сравнению с материнской возрастное состояние (рис. 11). В течение жизненного цикла появляется 2—3 вегетативно возникших поколения. Продолжительность существования клона составляет в среднем 4—6 лет.
Рис. 11. Старое генеративное растение хохлатки плотной
Особи редко доживают до сенильного состояния, обычно лишь самые мощные растения образуют сенильные клоны, которые состоят из большого числа мелких, часто уродливых клубней, окруженных разрушающими остатками материнских клубней. Наземные по
3 Зак. 255 65
беги образуют листья ювенильного типа. Массовое образование клонов отмечено на северном пределе ареала этого вида, в Швеции (Ryberg, 1959). Семенйое возобновление при этом подавлено.
Сезонный ритм развития
Монокарпический побег развивается по моноциклическому типу. Полный цикл развития осуществляется в течение 22 месяцев. Зачаток почки будущего монокарпического побега появляется в августе в пазухе нижнего чешуевидного листа уже сформированной почки возобновления. С августа по май следующего года этот зачаток почки остается недифференцированным. В июне в почке возобновления начинается интенсивная деятельность камбия, почка заметно увеличивается в размерах, достигая 2,25 мм в высоту, в это же время наблюдается заложение чешуевидных листьев, срединных листьевщ стебля. К концу июля почка имеет 4 мм в высоту и 3,2 мм в ширину, в ней заметно уже зачаточное соцветие (0,7 мм высотой) с частично дифференцированными прицветниками и цветками.
Дочерний клубень в это время достигает базальной части материнского клубня. Доли рассеченного листа в виде бугорков. К середине августа на молодом клубне возникают придаточные корни 2—6 мм длиной. В середине августа — сентябре в пазухах нижних чешуевидных листьев закладываются зачатки будущих почек.
Оптимальная температура для нормального подземного развития Хохлатки плотной в течение летних месяцев 15—18°, осенью 3—5°. При отсутствии или недостаточной длительности осеннего холодного периода растения образуют карликовые побеги. Оптимальная продолжительность осеннего охлаждения — 2 месяца (Горы-шина, 1961, 1965; Митина, 1969)'. Пониженная летняя температура задерживает развитие корней.
В. начале сентября цветки в соцветии уже полностью дифференцированы, но семяпочки представлены недифференцированными бугорками. В ноябре зародышевые мешки находятся на двуядерной стадии развития,- В это время в пыльниках имеются одноядерные пыльцевые зерна. Зародышевый мешок развивается по нормальному типу. Полностью сформированные семяпочки обнаружены в декабре— январе (Трофимов, 1952; Ryberg, 1959; Коробова-Семенченко, 1960). К февралю — марту полностью сформированное соцветие достигает 1,5—2 см высоты, кроющие листья бледно-желтые. В это время побег возобновления выходит на поверхность почвы. Для окончательного формирования соцветия и цветения хохлатке необходимо воздействие пониженных температур. Осенне-зимний покой у нее довольно глубокий, с сентября по декабрь. С декабря по март наступает вынужденный.покой. В эксперименте зацветали растения, взятые в природе не раньше декабря (Трофимов, 1952; Серебряков, 1959; Горышина, 1965).
В средней полосе европейской части СССР хохлатка плотная зацветает в середине апреля. Многолетние наблюдения в Воронеж
66
ском заповеднике позволяют более детально охарактеризовать фенологию цветения. Самое раннее начало цветения —5 апреля, самое позднее — 26 апреля, соответственно самые ранние и поздние сроки массового цветения — 9 и 28 апреля, конца цветения — 17 апреля и 5 мая (Гоббе, 1964). Продолжительность цветения — около двух недель.
Начало весенней вегетации может происходить при значительных колебаниях температуры почвы и воздуха, но начало цветения связано с температурой почвы не ниже 3—3,5°. Суточные колебания температуры воздуха во время цветения достигают 20—25°. Цветки обладают ароматом, содержат много нектара и характеризуются большим разнообразием окраски венчика, который бывает белым, розовым, голубоватым,- светло- и темно-сиреневым, малиновым и красным. Опыление цветков осуществляется длиннохоботковыми шмелями и некоторыми бабочками, короткохоботковые шмели прокусывают шпорец, добывая нектар. В дубравах Белгородской обл. (лес на Ворскле) прокусанные цветки составляют 50—70% от общего числа (Горышина, 1965).
Семена созревают в первой половине мая, цветоносы полегают в процессе созревания семян. В это же время наблюдается массовое пожелтение листьев. Продолжительность ассимилиционного периода—3—5 недель (Hegi, 1919; Ryberg, 1959; Горышина, 1969).
С. solida — весеннезеленый вид, почки возобновления которого полностью формируются к осени (Серебряков, 1947 a; Walter, 1968). С момента заложения почки возобновления до полного разрушения сформированного ею клубня проходит 4 года (Bersillon, 1958) .
Способы размножения и распространения
В популяциях С. solida, как и у двух предыдущих видов, господствует семенное возобновление. Хохлатка плотная — мирмекохор, семена опадают с йедоразвитым зародышем (Никитина, 1955, 1957; Ryberg, 1955, 1959). Реальная семенная продуктивность в дубравах Воронежской обл. в 1973 г. составила 3—5 семян на 1 побег. Семенная продуктивность этого вида в дубняке снытевом в Белгородской обл. составляет в среднем 749 семян на 1 м2.
Вегетативное размножение отмечено в конце жизненного цикла. Существенной роли в самоподдержании популяции оно не играет.
- Экология
Хохлатка плотная—эумезофит, произрастает на почвах разного механического состава: глинах, суглинках и супесях. Широко распространена на равнинах, в горах поднимается до 1900 м над уровнем моря (Hegi, 1919). Отличается высоким светолюбием. Для нормальной продуктивности фотосинтеза необходима освещенность более 50% от полной. (В эксперименте растение погибает при 5%
3*
67
йолной освещенности.) Устойчива к низким й высоким температурам.
В течение периода вегетации растение не испытывает недостатка влаги. Содержание воды в листьях составляет 90,9% от сырого веса листа. Дневной ход содержания воды в листьях отличается незначительными колебаниями. Активная деятельность корневой системы наблюдается даже при низкой температуре почвы, о чем свидетельствует обильная гуттация (Коореп, 1962; Горышина, 1965). В течение периода вегетации оводненность листьев держится на высоком уровне, затем стремительно падает в связи с увяданием. С появлением новых придаточных корней влажность клубней достигает 65%. Высокая влажность клубней сохраняется в течение всей зимы и связана с поступлением воды в корни в зимнее время.
Листья лишены всяких приспособлений, уменьшающих испарение: кутикула слабо развита, опушение отсутствует. Палисадная паренхима представлена одним слоем клеток небольшой толщины, губчатая паренхима рыхлая с крупными межклетниками. Устьица расположены на обеих сторонах листа, на 1 мм2 листа в среднем 170 устьиц. Механическая ткань слабо развита. Площадь листьев у одного растения в период максимального развития составляет в среднем 37 см2. Площадь листьев, приходящаяся на 1 га в период максимального развития, составляет 6432 м2, в среднем за вегетационный период — 3220 м2.
Образование хлорофилла начинается при 0°. Содержание хлорофилла в листьях в среднем за вегетационный период достигает 2,28 мг на 1 г сырого веса листа. Наибольшая величина потенциальной интенсивности фотосинтеза отмечена в период массового цветения и соответствует 134 мг СОг/г-час. Дневные кривые потенциального фотосинтеза одновершинны, имеют максимум в 9—11 часов. Наблюдаемый фотосинтез по характеру сезонных и дневных изменений повторяет ход потенциальной интенсивности фотосинтеза этого вида. Максимальная величина наблюдаемого фотосинтеза во время цветения хохлатки составляет 106 мг СОг/г-час. '
Хохлатка плотная отличается высокой интенсивностью дыхания. В дневные часы при 14—20° оно достигает 1,4—2,0 мг СОг на 1 г сырого веса в час. Наиболее интенсивное дыхание отмечено в начале вегетации, наиболее интенсивный фотосинтез — в конце вегетации. Сезонный пик дыхания приурочен к фазе бутонизации. Транспирация в дневные солнечные часы достигает 1200—1600 мг на 1 г сырого веса листа в час. Дневной ход транспирации представляет одновершинную кривую с максимумом в дневные часы. Низкая температура почвы не снижает интенсивности транспирации. Суммарный расход воды на транспирацию за 1 день равен 5,8—9,4 г на 1 г сырого веса листа, за вегетационный период — 220 г на 1 г сырого веса листа или 35 т/га. Водный запас в листьях полностью сменяется за 1 час (Горышина, 1965).
68
Биологическая продуктивность
На темно-серых лесных песчаных почвах в снытевых и снытево-осоковых дубравах Воронежской обл. воздушно-сухой вес генеративных особей колеблется от 0,30 до 0,95 г, вес ювенильных особей 0,01 г, сенильных 0,38 г. Максимальный сухой вес листьев — 0,22 ц/га, средний — 0,15 ц/га. Суммарная продуктивность фотосинтеза составляет 15,6 г на г сухого вещества листа или 242 кг/га, расход на дыхание-—41,1 г на г сухого вещества листа или 90 кг/га.
Участие С. solida в травостое снытевого липо-дубняка по сухому весу составляет 15%, по площади листьев — 13%. Число клубней в снытевой дубраве под Харьковом — 183—188 на 1 м2 (Карпенко и др., 1966). Биомасса в течение года изменяется следующим образом: сухой вес подснежных ростков составляет 5 кг/га; надземных частей весной — 25 кг/га, подземных — 57 кг/га; подземных частей летом — 92 кг/га (Горышина, 1969). Биомасса популяции хохлатки плотной в дубравах Воронежского заповедника, включая надземную и подземную части, составила 8, 60 г/м2.
Фитоценология
С. solida является временным доминантом в травяном покрове широколиственных и хвойно-широколиственных лесов (Алехин, 1951). Обладает более широким ценоареалом, чем два предыдущих вида. В Средней и Атлантической Европе вид встречается в холмистых листопадных лесах, кустарниковых зарослях на юрских известняках, изредка (в Арденах) на пестрых песчаниках. Он отмечен на песчаных почвах во многих группировках, принадлежащих союзу Alneto-Ulmion Br.— Bl. и Fraxineto-Carpinion Tuxen и обычно произрастает вместе с Anemone ranunculoides L. (Lefort, 1951).
Вместе с другими эфемероидами образует весенний аспект в приднепровских дубовых и дубово-грабовых лесах. В дубовых лесах Закарпатья встречается вместе с С. bulbosa. Весенний аспект С. solida образует во всех широколиственных лесах европейской части СССР от запада Украины до Предуралья (Петров, 1947, 1955). В черноолыпаниках Припятского Полесья, в ясенево-ольхо-пых и дубово-ясенево-ольховых сообществах встречается вместе с Anemone ranunculoides L.
Проникает в хвойно-широколиственные леса Московской и соседних областей, встречается в липово-хвойных лесах Среднего Урала (Игошина, 1943; Горчаковский, 1948). Вместе с другими эфемероидами образует ранневесеннюю синузию в травяном покрове лиственничников Средней и Восточной Сибири (Алтай, Саяны, отроги Кузнецкого Алатау).
Популяции этого вида исследованы в Закарпатье в дубовых лесах и в дубовых и дубово-липовых насаждениях Воронежской Обл. на серых и темно-серых лесных почвах. Как и два предыдущих Пида, С. solida характеризуется популяциями левостороннего спект
69
ра: виргинильные особи составляют от 60 до 90% от общей численности популяции. Численность особей этого вида достаточно велика во всех исследованных местообитаниях (50—80 особей/м2), наиболее высокая численность отмечена в дубовых лесах Закарпатья (87,2 особей/м2).
По данным Т. К. Горышиной (1969), численность С. solida в ли-по-дубняке снытевом в Белгородской обл. составляет 234 особи/м2, в дубняке осоковом — 47 особей/м2, причем большая часть особей (187 и 43 соответственно) находится в виргинильном периоде. Резкое снижение общей численности хохлатки плотной наблюдается в осоковых ассоциациях, при этом возрастание обилия Carex pilosa Scop, приводит к укорочению жизненного цикла хохлатки и усиленному отмиранию генеративных особей. По данным М. В. Митиной (1965), наблюдается обратная корреляционная зависимость между обилием С. solida и Carex pilosa.
Наиболее высокая жизненность особей хохлатки плотной (максимальный вес, высота побегов, наибольшая семенная продуктивность) отмечена в 250-летних снытевых дубравах Теллермановского опытного лесничества на темно-серых лесных почвах. Высокая жизненность у этого вида сочетается с большой численностью — 72,34 особи/м2.
Внутривидовая систематика
С. solida — очень изменчивый вид, у которого довольно трудно определить систематическую значимость наблюдаемых форм. В частности, степень рассеченности прицветников не является существенным признаком.
Хозяйственное значение
В клубнях найдены алкалоиды бульбокапнин и протопин.
4. ХОХЛАТКА ПРОМЕЖУТОЧНАЯ
Номенклатура
Corydalis intermedia (L.) Merat., Nouv. Fl. Env. Par. (1812) 272; Шмальг., Фл. 1 (1895) 41; Попов во Фл. СССР 7 (1937) 665; Федченко в Маевск., Фл., изд. 7 (1940) 377; Васильченко, там же, изд. 8 (1954) 327, изд. 9 (1964) 320; Mowat, Chater is Fl. Eur. 1 (1964) 253; Скворцов в Опр. раст. Моск. обл. (1966) 172.— Fumaria bulbosa [3. intermedia L., Sp. pl. (1753) 699.— Corydalis fabacea (Retz.) Pers., Syn. 2 (1807) 269; Сырейщ., Илл. фл. 4 (1914) 82; он же, Опр. раст. Моск губ. (1927) 137.— Fumaria fabacea Retz., Prodr; fl. Scand., ed. 2 (1975) 167.— Хохлатка промежуточная, x. средняя.
Описана из Западной Европы.
70
Морфологическое описание.
Многолетнее травянистое поликарпическое .растение. Клубень подземный, ежегодно сменяющийся, маленький, 8—15 мм в диаметре, шаровидный, светло-бурый, по строению и способу образования ничем существенно не отличается от клубня С. solida. Корневая система состоит из ежегодно сменяющихся, эфемерных придаточных корней.
Листовая серия взрослых растений состоит из 3—4 чешуевидных и 2 срединных листьев. Междоузлие верхнего чешуевидного листа удлиненное (4—5 см). Срединные ассимилирующие листья длинночерешковые, дважды тройчато-рассеченные, сегменты на длинных, доли на коротких черешочках. Доли листа обратнояйце-видно-клиновидные, надрезанные на продолговато-линейные, слегка клиновидные, тупые крупные дольки. Консистенция листьев нежная, цвет зеленовато-сизый.
Соцветие — короткая, малоцветковая, до цветения поникающая верхушечная кисть. У большинства экземпляров в отличие от С. solida из почки в пазухе верхнего чешуевидного листа развивается дополнительный генеративный побег. Прицветники цельные, крупные, обратнояйцевидные, туповатые или короткозаостренные. Цветоножки тонкие, даже при плодах в 2—3 раза короче прицветников. Чашелистики очень мелкие, пленчатые, зубчатые. Венчик мелкий, около 15 мм длины, тускло-пурпурный, отгиб наружных лепестков довольно широкий, на верхушке с большой выемкой. Шпора широкая, прямая, тупая, слегка длиннее лепестка. Нижний лепесток с едва заметным бугром. Рыльце дисковидное, по краю мелкоборо-давчато-зубчатое, без рожков при основании.
Плод — коробочка, прямостоячая или отклоненная, продолговатая, 10—15 мм длины и 4—5 мм ширины, заостренная в плоский носик, переходящий в столбик. Семена черные, блестящие, 2 мм в диаметре. Каринкула пленчатая, мешковидная, длинная, отстоящая (Hegi, 1919; Fedde, 1936; Ryberg, 1955).
Число хромосом (2н) равно 16 (Ryberg, 1950; Хромосомные числа цветковых растений, 1969).
Географическое распространение
Хохлатка промежуточная распространена в Центральной и частично в Восточной Европе. Северная граница ареала проходит в Швеции (изолированные местонахождения отмечены в Финляндии), западная-—по Бельгии, Голландии, центру Франции, южная — по северу Италии и Хорватии. В пределах европейской части СССР встречается в Белоруссии и центральных районах РСФСР, на востоке достигает Горьковской обл. (Hegi, 1919; Fedde, 1936; Флора СССР, 1937; Ryberg, 1955). Карта ареала составлена X. Мейзелем (Meusel, 1969).
В Моск. обл. отмечена в ряде районов как редкое растение (Ворошилов и др., 1966). По-видимому, распространена значительно
71
шире, чем представлена в гербарных коллекциях, так как легко просматривается в природных фитоценозах (Губанов, Алексеев, 1975).
Онтогенез и сезонный ритм развития
В онтогенезе хохлатки промежуточной не выявлено существенных отличий от онтогенеза х. плотной, за исключением более редкого образования клонов в сенильном периоде.
Сезонный ритм развития в основных чертах повторяет ритм развития х. плотной. Цветение, созревание плодов и отмирание надземных побегов у х. промежуточной происходит на 1—1,5 недели раньше, чем у х. плотной.
Как и многие другие виды рода, х. промежуточная — эфемероид, геофит.
Способы размножения и распространения
Хохлатка промежуточная размножается семенным путем, вегетативное размножение почти полностью отсутствует. Семенная продуктивность в широколиственных лесах Воронежской обл. выше, чем у х. плотной. Число семян на одном побеге в среднем 15,92.
Экология, биологическая продуктивность и фитоценология
Хохлатка промежуточная — эумезофит, произрастает на почвах разного механического состава: супесчаных, суглинистых и'глинистых. Охотно селится на известняках. Растет на равнинах и в горах, в Альпах поднимается до 2 000 м над уровнем моря.
Вес взрослых особей в широколиственных лесах Воронежской обл. составляет 0,5 г. Общая биомасса популяции 5,4 г/м2, только надземной части — 2,3 г/м2.
X. промежуточная выступает как временный доминант или со-доминант синузии дубравного широкотравья. Она образует так называемый «хохлатковый тип» в дубово-буковых лесах Центральной Европы (Querceto-carpinetum corydaletosum). Встречается на лугах и в лесных островах среди луговой растительности под липой или лещиной (Tuxen, 1937; Ellenberg, 1938). В европейской части СССР встречается вместе с х. плотной и х. Маршалла в дубовых и дубоволиповых лесах, но, как правило, не господствует в них.
Популяции этого вида, изученные в Воронежской обл., ничем практически не отличаются от популяций х. плотной. Численность особей на 1 м2 10 экземпляров.
Внутривидовая систематика
Хохлатка промежуточная отличается значительно большим постоянством признаков, чем другие виды секции. В естественных фитоценозах и в культуре легко образует гибриды с другими видами рода. Большая часть гибридов оказывается стерильной (Ryberg, 1950).
М. Г. Вахрамеева
РЯБИНА ОБЫКНОВЕННАЯ
Номенклатура
Sorbus aucuparia L., Sp. pl. (1753) 477; Кауфм., Моск, фл., изд. 1 (1866) 170, изд. 2 (1889) 171; Цингер, Сб. свед. (1885) 171; Сырейщ., Илл. фл. 2 (1907) 279; он же, Опр. раст. Моск. губ. (1927) 157; Комаров во Фл. СССР 9 (1939) 376; Пояркова в Маевск., Фл., изд. 7 (1940) 423, изд. 8 (1954) 103, изд 9 (1964) 104; Скворцов в Опр. раст. Моск. обл. (1966) 192; Warburg, Karpati in Fl. Eur. 2 (1968) 68.— Pyrus aucuparia (L.) Gaertn., Fruct. Sem. pl. 2 (1788) 45; Шмальг., Фл. 1 (1895) 353.-—Рябина обыкновенная.
Сем. Rosaceaе, подсем. Pomoideae.
Описана из Северной Европы, тип в Лондоне.
Морфологическое описание
Крупный кустарник или дерево 4—15 (реже 20) м высотой и диаметром до 30—40 см, с яйцевидной или шарообразной кроной; ствол покрыт серой корой. Молодые побеги красновато-бурые с серым налетом, нередко опушенные. Древесина с хорошо заметным серовато-бурым или красновато-бурым ядром и белой заболонью, с тонкими годичными слоями (Абраменко, Фишер, 1933).
Почки рябины крупные, конически заостренные, 0,8—2,0 см длины, с 4—6 кроющими чешуями; листовые рубцы с 5 листовыми следами. Генеративные почки окружены основаниями 3—4 черешков листьев, вегетативные — лишь одним основанием, выполняющим функцию кроющей чешуи. Генеративные почки (и побеги) всегда терминальные, а вегетативные — боковые (Серебряков, 1952).
Корневая система состоит из неглубоко идущего стержневого корня, который на глубине 20—25 см резко обрывается, образуя «лапу» с расходящимися в разные стороны ответвлениями, и большого количества боковых корней, расположенных в основном на глубине до 20 см (Исаченко, 1954)-. Скелетные корни в своей периферической части ветвятся довольно слабо. У взрослых деревьев диаметр корневой системы примерно в 2 раза превышает диаметр кроны. Общая глубина проникновения корней около 100 см.
73
Листья сложные, непарноперистые, в общем очертании продол-1 говато-эллиптические, до 30 см длины и 20 см ширины, с черешками длиной 2—4 см и опадающими прилистниками; состоят из 4—7 (реже 9) пар продолговатых или. продолговато-ланцетных, в нижней части цельнокрайних, выше пильчатых, сверху матово-зеленых, снизу сизых или сероватых листочков. Конечный листочек эллиптиче-ски-овальный, часто к основанию более суженный, иногда сросшийся с 1—2 верхними боковыми листочками.
Цветки с пятичленным околоцветником, 0,8—1,5 см в диаметре, снабжены опадающими прицветниками, отличаются резким запахом 3-метиламина, собраны в многоцветковые щитковидные соцветия до 10—11 см в диаметре. Чашелистики с железистыми ресничками по краям, опушенные, иногда голые; лепестки беловатые, округлые, при основании суженные в ноготок; тычинок 20, равных по длине лепесткам; завязь трехгнездная, с 3 (реже 2—5) столбиками, образована 2—5 плодолистиками; в цветке имеется голый бороздчатый диск.
Плод — олигомерное яблоко (Каден, 1965), круглый или немного сплюснутый, 8—42 мм в диаметре, при созревании от желтоватооранжевого до красного цвета, горьковато-кисло-сладкого терпкого вкуса, содержит 3—5 семян (по 1—2 в гнезде). Семена светло- или темно-коричневые, блестящие, узкие, слегка изогнутые, суженные у места прикрепления к плаценте, 4—4,5 мм длиной, 1,9—2,1 мм шириной и 1 мм толщиной.
Диплоидный набор хромосом равен 34 (Diill, 1961; Хромосомные числа цветковых растений, 1969).
Географическое распространение
Рябина обыкновенная широко распространена на всей территории Европы, включая Исландию, Скандинавию, приатлантические районы и Балканы, а также в Малой Азии (Корреп, 1888; Hegi, 1923; Коновалов, 1952; Петров, 1957; Rubner, 1960; Diill, 1961).
В пределах СССР северная граница ареала проходит по Кольскому п-ову, через Хибины, восточнее оз. Имандра (67°50' с. ш.), к бухте Индиги, по долине Печоры между устьями Усы и Цильмы, затем по Уральскому хребту. Южная граница от г. Хотина (Черновицкая обл.) идет через Волыно-Подольскую возвышенность к Днепру, между Каневым и Черкассами, далее через Дубны, Полтаву, Харьков, Балашов, Саратов, затем по северу Общего Сырта и р. Сакмаре к Южному Уралу.
Изолированно от общего ареала рябина встречается в Крыму и на Кавказе (Габриэлян, 1958 а, б; Попов, 1960). В Крыму она представляет собой реликт периода материкового оледенения, сохранившийся в настоящее время только в наиболее высокой части гор на их северных склонах (Попов, 1960).
74
Онтогенез
Созревшие семена сохраняют всхожесть в течение года (Кап-пер, 1952). Наивысшую всхожесть (47%) имеют семена, посеянные через 3—5 дней после сбора (Савченко, 1963). Для весеннего посева семена необходимо стратифицировать, лучше всего при 1—2° в течение 7 месяцев в торфяной крошке, увлажненной до полной влаго-емкости (Савченко, 1966).
Всходы рябины имеют сравнительно тонкую пурпурную подсемядольную часть. Семядоли овальные, 7—8 мм длиной и 4 мм шириной, на верхушке закругленные, на коротких черешках. Первый лист глубоко трехлопастной или трех-, пятираздельный, второй — обычно тройчатый, третий — непарноперистый с 5 листочками. Листочки яйцевидные, по краю пильчато-зубчатые, с перистыми жилками. Надсемядольное междоузлие небольшое (3 мм длиной), опушенное (Васильченко, 1960).
В первые годы жизни, особенно в культуре, рябина имеет большие приросты — до 50 см в год. После 20—25 лет прирост резко снижается (Хохлов, Ковалева, 1950; Эйзенрейх, 1959; Харитонович, 1968). На открытых местах высокий темп прироста сохраняется довольно долго.
В Московской обл. в березово-еловом лесу на дерново-подзолистой суглинистой почве рябина имеет в первые годы жизни следующие средние размеры: в 3 года — высота 13,5 см, диаметр стебля 2 мм, годичный прирост 3,6 см; в 5 лет — соответственно 31 см, 3,2 мм, 7,9 см; в 7 лет — 59 см, 4,2 мм, 12,8 см; в 10 лет — 106 см, 5,7 мм, 21 см. М. В. Пайбердин (1970) для условий Поволжья (Марийской АССР) приводит следующие данные таксационных показателей рябины в березняке липовом на свежей супесчаной почве:
Возраст, годы Средняя высота, м Средний диаметр ствола, см Число стволов на 1 га Запас древесины, м3/га Урожай плодов, кг/га
10 2,3 1,6 1300 1,30 30
12 2,9 1,9 1100 1,43 69
14 3,6 2,8 974 2,93 83
16 4,2 3,4 934 4,53 105
18 4,6 4,1 833 5,83 116
20 5,0 4,7 726 6,53 116
22 5,3 5,3 650 6,43 129
24 5,7 5,8 450 5,63 166
26 6,1 6,2 320 4,03 179
28 6,5 6,7 200 3,00 188
И. Г. Серебряков (1954) отмечает более раннее старение главной оси у рябины по сравнению с другими древесными породами. Быстрый рост рябины в первые 8—10 лет связан с недоразвитием ее
75
боковых ветвей. Мощное развитие ветвей происходит позже. Оно приводит к ослаблению роста главной оси (т. е. ее старению), что влечет за собой пробуждение спящих почек в основании ствола и рост новых осей параллельно материнскому. В связи с этим возникает совершенно необычное многоствольное или кустовидное дерево, для которого характерно одновременное развитие стволов нескольких или, по крайней мере, двух поколений.
Изменение характера корневой системы с возрастом изучал во Владимирской области на среднесуглинистых среднеподзолистых почвах Е. М. Петров (1957). У однолетних сеянцев корни идут вглубь до 63 см, боковые корни расходятся в стороны на 17—26 см. У 2—7-летних особей основная масса корней расположена на 3—5 см ниже подстилки. Диаметр корневой системы превышает диаметр крон не менее чем в 3—4 раза. У взрослых деревьев диаметр корневой системы 10—12 м, а диаметр кроны 6—6,4; корни проникают в почву на глубину до 1 м, но основная масса корней находится на глубине 40 см.
Плодоносить рябина начинает в зависимости от условий в различном возрасте: в посадках в благоприятных условиях — уже в 5 лет, в менее благоприятных условиях — в 10—15 лет (Петров, 1957; Пайбердин, 1970). Продолжительность жизни, по данным различных авторов, от 50—80 лет (Hegi, 1923; Петров, 1957) до 140— 150 лет (Абраменко, Фишер, 1933)
Сезонный ритм развития
В условиях Подмосковья рябина цветет во второй половине мая. Распускаются почки при среднесуточной температуре 6—7°. Зацветает рябина при среднесуточной температуре 15—16° (Петров, 1957). Из цветочных (точнее, смешанных) почек появляется сначала 3—4 листа, а затем соцветие. По данным В. И. Долгошева (1947), в Подмосковье первые развернувшиеся листья рябины появляются 7. V (25. IV—21. V) , начало цветения 24. V (7. V—8. VI), первые зрелые плоды 11. III (17. VII—29. VIII), полная осенняя раскраска листьев большинства особей 28. IX (15. IX—19. X), конец листопада большинства особей 18. X (25. IX—24. X).
Рост побегов заканчивается довольно рано — к концу цветения длина прироста составляет 93% от общей длины годичного прироста (Петров, 1957). Рост междоузлий по всей их длине происходит равномерно в течение всего периода растяжения. Длина междоузлий на годичном побеге изменяется по типу одновершинной кривой (Серебряков, 1952).
Рост корней начинается весной при температуре 3—4°. Максимальный их рост происходит во второй половине мая — первой половине июля. Оптимальная температура для роста корней 8—16°.
Способы размножения и распространения
Рябина хорошо размножается семенным и вегетативным путем (Петров, 1957; Эйзенрейх, 1959; Пятницкий и др., 1963). Характерно
76
для нее перекрестное опыление (цветки прототипичны), однако очень редко происходит и самоопыление. Возможна партенокарпия, при этом образуются бессемянные плоды (Петров, 1957). Известны межвидовые (с Sorbus alpina и A. melanocarpa) и межродовые (с Mespilus germanica и Crataegus sanguinea) гибриды, введенные в культуру сорта «Бурка», «Ликерная», «Десертная», «Гранатная».
В соцветии рябины до 100—200 цветков (Габриэлян, 1958 6; Diill, 1961), иногда до 550 (Лозовский, 1936). Среднее число цветков на одном дереве — 3806. Продолжительность цветения 10—12 дней. Иногда наблюдается вторичное цветение. Средний процент завязывания плодов — 23,8. Через 15 дней после цветения остается 39,5% завязей, через 35 дней — 34,4% (Лозовский, 1936).
В 1 кг 2200 плодов и в них 220—265 тыс. семян, вес 1000 семян 3,5—4,5 г (Хохлов, Ковалева, 1950; Kantor, 1956; Петров, 1957). Объем 1000 семян — 8,1—8,5 см3 (Савченко, 1966). Существует тесная зависимость между весом плодов и диаметром ствола — коэффициент корреляции 0, 62—0,97 (Федоров, 1971).
Урожай плодов в лесу в значительной мере зависит от состава насаждения: в сосняке-брусничнике — 3—20 кг/га, в сосняке липовом— 15—40, в березняке и осиннике липовом— 18—160, в липняке— 12—50, ельнике липовом — 20—50, на вырубке — до 400 кг/га (Федоров, 1964). В насаждениях со значительной примесью рябины урожай еще выше — 600—1000 кг/га (Ковалев, 1934; Пайбердин, 1970). Плодоносит рябина довольно регулярно, хорошие урожаи бывают через 1—2 года. Семенному размножению и расселению способствуют птицы, благодаря которым рябина появляется не только в лиственных и смешанных, но и в хвойных лесах (Харитонович, 1968).
Вегетативное размножение рябины (корневыми отпрысками, пневой порослью, укоренением побегов) встречается обычно чаще, чем семенное. Е. М. Петров (1957) указывает для Татарской АССР 84% порослевых и 16% семенных рябин, для Тамбовской обл.— соответственно 72% и 28%. Сходное соотношение наблюдается и в Подмосковье. Очень распространенная у рябин пневая поросль образуется обычно на крупных корнях первого порядка, расположенных близ поверхности почвы. Придаточные почки на корнях часто располагаются группами. Под пологом древостоя у молодых рябин часто возникают стелющиеся побеги, самостоятельно укореняющиеся и образующие отводки. После рубки рябина успешно возобновляется пневой порослью.
Экология
Рябина обыкновенная отличается большой экологической амплитудой, встречаясь в самых разнообразных местообитаниях. По отношению к влаге она мало требовательна и произрастает как в засушливых условиях, так и при достаточном увлажнении (Коновалов, 1952;Харитонович, 1968). Предпочитает, однако, хорошоувлаж-
77
ненные места, но избегает избыточно увлажненные почвы (Rubner, 1960). Интенсивность транспирации рябины в условиях Подмосковья колеблется в зависимости от температуры следующим образом: от 133 (при 7°) до 731 (при 26°) мг/г за час; транспирационное число 477 (Иванов и др., 1951).
В молодом возрасте переносит значительное затенение, встречаясь под пологом темнохвойных лесов, позже становится более светолюбивой. Обильно цветет и плодоносит лишь на хорошо освещенных местах — вырубках, гарях, опушках. По зимостойкости занимает одно из первых мест среди лиственных древесных пород (Лапин, Сиднева, 1971). В экспериментальных условиях корневая система рябины переносит успешно без повреждений охлаждение до —14,5°.
По отношению к почвам мало требовательна. Растет как на подзолистых, дерново-подзолистых суглинистых и супесчаных почвах, так и на серых и темно-серых, а также на каменистых горных почвах, избегая засоленные и торфяно-болотные почвы (Харитонович, 1968). Лучше всего растет на влажных гумусированных почвах (Rubner, 1960). В лесной зоне хорошо растет на дерново-подзолистых и серых лесных почвах, лучше на легких и средних суглинках, на песках растет плохо (Петров, 1957; Rubner, 1960). В Московской обл. чаще встречается на дерново-подзолистой почве с pH водным 4—6 и pH солевым 3—5.
Рябина обыкновенная поднимается в горы до высоты 2300— 2400 м (Эйзенрейх, 1959; Габриэлян, 19586). Хорошо переносит длительное воздействие дыма в городах (Эйзенрейх, 1959)'.
Фитоценология
Рябина обычно растет одиночно или небольшими группами, очень редко доминируя в растительных сообществах. Б. А. Быков (1965) относит ее к субдоминантам 5 яруса, указывая, что рябина иногда доминирует в хвойных лесах, например в сообществах: Picea abies — Sorbus aucuparia — Dryopteris spinulosa; Abies sibirica — Sorbus aucuparia — Equisetum silvaticum. Б. В. Гроздов (1950).описал в Смоленской обл. на свежих относительно плодородных почвах ельник рябиновый. В Подмосковье также иногда можно встретить небольшие участки леса с рябиной в качестве доминанта (обычно на месте небольшой вырубки или крупного «окна»).
В Московской обл. распространена широко в подлеске и нередко в нижнем древесном ярусе в лесах самого разнообразного состава, чаще- в хвойно-мелколиственных, но форму дерева имеет лишь на опушках, близ дорог, на прогалинах.
Рябина относится к почвоулучшающим породам благодаря обильному и легко минерализующемуся опаду (Шиманюк, 1967). В культуре считается ценной породой, сопутствующей дубу, ели, сосне.
78
Консортивные связи
Большое значение в жизни рябины имеют насекомые-опылители и птицы (особенно дрозды-рябинники), последние способствуют естественному распространению рябины. Плоды рябины — важнейший корм боровой'дичи. Нередко отмечают повреждения рябины грызунами (мышами, белками, зайцами), а также лосями (Muller-Stoll, 1947; Эйзенрейх, 1959; Динесман, 1968). Выпас рябина переносит плохо.
Одной из распространенных болезней рябины является кольцевая пятнистость листьев, вызываемая вирусом (Kegler, 1959; Krali-kova, 1961). Наиболее распространенные болезни, вызываемые грибами (свыше 25 видов): пятнистость, ржавчина и мучнистая роса листьев, засыхание ветвей, пятнистость коры, гниль ствола и корней, пятнистость плодов (Ванин, 1931; Muller-Stoll, 1947; Ванин и др., 1950; Эйзенрейх, 1959; Muller ш а., 1956).
На рябине отмечено около 30 видов энтомовредйтелей: на листьях — пилильщики, гусеницы совок, пядениц, белянок, паутинной моли, растительные клещи; на цветках — галлицы; на плодах— рябиновый большой и малый семееды, яблоневый семеед, рябиновая моль, яблоневый плодовый пилильщик; на ветвях и стволах— щитовки и короеды (Muller-Stoll, 1947; Гусев, Римский-Корсаков, 1951; Muller и. а., 1956; Заборовский, 1962).
Внутривидовая систематика
Внутривидовое разнообразие рябины обыкновенной очень велй-ко. Выделяют много форм (от 8 до 12) различного таксономического ранга в зависимости от размеров, формы и строения кроны, строения и опушения листьев и особенно от формы, окраски, размеров и вкуса плодов (Корреп, 1888; Hedlund, 1901; Вольф, 1923; Hegi, 1923; Rehder, 1927; Bailey, 1930; Ковалев, 1934; Hulten, 1949; Коновалов, 1952; Raatikainen, 1960; Dull, 1961).
Некоторые формы рябины вошли в культуру. Особенно большую популярность приобрела у нас невежинская рябина (var. rossi-са Spath.), происходящая из с. Невежина Владимирской обл. Многие ее сорта — кубовую, желтую, сахарную, красноплодную — широко применяют в плодоводстве (Петров, 1957).
Хозяйственное значение
Рябина обыкновенная использовалась с древности как плодовая культура (Ковалев, 1934), особенно две ее разновидности: var. moravica и var. rossica, дающие урожай до 600 кг/га. Плоды рябины богаты сахарами, органическими кислотами, пектиновыми веществами, витаминами А, С, Р и т. д. (Петров, 1957; Федоров, 1964; Сильянова, 1957), благодаря чему они широко применяются в пищевой промышленности и имеют лекарственное значение (Эйзен-
79
рейх, 1959). Плоды рябины рекомендуют также вводить в рацион питания детей (Donath, 1954).
Богаты биологически активными веществами и листья, которые могут служить хорошим витаминным кормом для сельскохозяйственных животных, особенно птиц (Курьянов, 1967; Федоров, 1971). Кора рябины содержит значительное количество таннинов и может быть использована в дубильном производстве.
Твердая и хорошо полирующаяся древесина применяется в мебельной промышленности. Будучи устойчивой к дыму и благодаря высокой декоративности (особенно осенью) рябина широко используется в городских посадках. При создании защитных лесных полос (полезащитных, почвозакрепляющих, противопожарных) также используется рябина (Каппер, 1952).
М. Г. Вахрамеева
ЧЕРЕМУХА ОБЫКНОВЕННАЯ
Номенклатура
Prunus padus L., Sp. pl. (1753) 473; Кауфм., Моск, фл., изд. 1 (1866) 148, изд. 2 (1889) 146; Цингер, Сб. свед. (1886) 153; Шмальг., Фл. 1 (1895) 314; Сырейщ., Илл. фл. 2 (1907) 279; он же, Опр. раст. Моск. губ. (1927) 167; Скворцов в Опр. раст. Моск. обл. (1966) 207; Webb in Fl. Eur. 2 (1968) 80.— P. racemosa Lam., Fl. Fr. 3 (1778) 107.— Padus'racemosa (Lam.) Gilib., Fl. Luthuan. 5 (1785) 231; Юзепчук и Пояркова в Маевск., Фл., изд. 7 (1940) 453; Комаров во Фл. СССР 10 (1941) 576; Пояркова в Маевск., Фл., изд. 8 (1954) 135, изд. 9 (1964) 134.—Слива-черемуха, черемуха обыкновенная. (Сем. Rosaceae.)
Морфологическое описание
Кустарник или дерево до 10 (реже 17) м высоты, с диаметром ствола до 10 (реже 40) см, с узкой довольно густой кроной. Корневая система сначала стержневая, позже приобретает поверхностный характер (Барыкина, 1960).
Ствол покрыт матовой черно-серой корой с хорошо заметными чечевичками. Молодые побеги оливковые или вишнево-красные. У молодых деревьев (до 6—8 лет) ветви отходят вверх под острым углом (Селиванова-Городкова, 1957а), с возрастом они пригибаются под действием собственной тяжести книзу.
Древесина ствола мягкая, легко поддается обработке, хорошо полируется, с желтовато-белым ядром, иногда красновато-беловатым, нередко отграниченным от желтовато-беловатой заболони (Абраменко, Фишер, 1933; Никитин, Суворов, 1949). Ранняя древесина окрашена светлее, чем поздняя. Годичные слои ясно заметны. Удельный вес 0,55—0,82 (Отенов, 1970). Объемный вес древесины при 15% влажности 0, 52 г/см3, временное сопротивление сжатию вдоль волокон 333 кг/см3 и статистическому изгибу 539 кг/см3, твердость (торцовая) 250 кг/см3 (Никитин, Суворов, 1949).
Почки ростовые и смешанные (последние, возможно, правильнее считать цветочными) (Байкина, 1946; Артюшенко, Соколов,
81
1955). Верхушечные почки всегда ростовые, цветочные обычно закладываются на побегах прошлого года и всегда боковые. Почки конические, заостренные, голые, спирально расположенные, длина их 0,5—1,3 см, причем размеры их убывают к основанию побега. Кроющих чешуи у верхушечных почек 8—12, у боковых (пазушных) 6—8 (Ефимова, 1955). Наружные чешуи с пленчатым краем, мельче внутренних, состоят из толстостенных мертвых клеток, многие из которых содержат друзы, под наружным эпидермисом чешуй расположена пробковая ткань (Ефимова, 1955). Внутренние чешуи не имеют пробки, но в них есть мощные межклетники, заполненные воздухом, мощная кутикула и толстостенные клетки. Нередко мож- . но наблюдать постепенный переход от чешуй к листьям. '
Листья очередные, тонкие, слегка морщинистые, яйцевидно-ланцетные или продолговато-эллиптические, заостренные, 3—10, реже -15 см длиной, с пильчатыми зубцами по краю, на которых находятся легко опадающие красно-бурые железки. Черешки листьев короткие (1—1,5 см длиной), с 1—3 железками. Прилистники шиловидные, очень рано опадающие. Осенью листья приобретают красную или желтую окраску.
Соцветие — кисть, 8—12 см длиной, довольно густое, поникающее, из 15—50 цветков. Цветки с характерным запахом, на цветоножках, в несколько раз превышающих чашечку, их длина 0,5—1,5 см. Гипантий чашевидный, снаружи голый, внутри мохнатый (Флора СССР, 1941). Доли чашечки трехгранные, по краям с железками. Лепестки венчика в числе 5, обычно белые, реже розовые (Бойченко, 1968; Вашкулат, 1959; Харитонов, 1958; Сюзев, 1906), быстро опадающие. Тычинок около 20, с желтыми пыльниками. Завязь образована одним плодолистиком с зеленым голым столбиком. Плод — сочная голая однокостянка (Каден, 1965), черная, очень редко белая. Косточка округло-яйцевидная, извилисто-выемчатая. Вес 1000 семян 42—68 г. В 1 кг 4 тыс. плодов и 17 тыс. косточек (Соколов, 1954). Иногда плоды имеют удлиненную форму (Корчагин, 1951).
Диплоидный набор хромосом равен 32 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Черемуха обыкновенная относится к евразиатским элементам. Ее ареал занимает территорию почти всей Европы и значительной части Азии (Корреп, 1888; Hegi, 1923; Hulten, 1950; Соколов, 1954; Селиванова-Городкова, 1957 б; Meusel и. а., 1965).
В пределах СССР черемуха распространена широко, отсутствует лишь в тундре и на юге европейской части СССР. Северная граница ареала почти совпадает с границей леса, проходит по Кольскому п-ову (69° с. ш.), берегам оз. Имандра, затем несколько отклоняется к югу до 68° 40' с. ш. Далее граница вновь поднимается несколько севернее, приближаясь к Полярному кругу, проходит че-82
рез реки Цильму, Пешу, пересекает Печору близ устья Цильмы, идет далее через Уральский хребет к Енисею, где у Усть-Курейки достигает 66° 40' с. ш. Южная граница проходит по линии Каменец-Подольск, юг Киевской обл., Полтава, Харьков, пересекает Дон у устья р. Медведицы, далее через Саратов, Общий Сырт к Оренбургу, через р. Урал на Орск, к южной части Мугоджар. В Западной Сибири черемуха встречается в березовых колках Барабинской степи, на северо-западе Алтая, по Иртышу и до г. Зайсан. Восточная граница проходит немного восточнее Енисея. Островное местонахождение отмечено в западной части Кавказа, преимущественно в субальпийском поясе (Соколов, 1954).
За пределами СССР черемуха встречается в Португалии, южной части Испании, в Пиренеях, на Аппенинском п-ове, в Северной и Центральной Европе, захватывает северную часть Балканского п-ова. Довольно широко распространена в Малой Азии, встречается в Северной Турции, Гималаях, Афганистане. Самая северная точка, где черемуха плодоносит, лежит в восточной части Финляндии — 70° 20' с. ш. Есть сведения о натурализации черемухи обыкновенной в США, в штате Пенсильвания (Long, 1927).
Онтогенез
Плоды черемухи созревают в августе — сентябре. К этому моменту семена еще не дозрели, хотя зародыши уже дифференцированы на две округлые семядоли с маленькой почечкой между ними, зачаточный стебелек и первичный корешок (Байкина, 1946). Для прорастания семян необходима стратификация в течение 80—90 дней при 1—5°.
Семена черемухи прорастают в условиях Московской обл. в конце мая — начале июня. Проросток имеет овальные семядоли 6—7 мм длиной, 3,5—4 мм шириной, сверху матовые темно-зеленые, снизу блестящие, сидячие, мясистые, около 1—1,5 мм толщиной (Васильченко, 1960). Надсемядольная часть красноватая, покрыта мельчайшими густыми волосками, подсемядольная часть утолщенно цилиндрическая, книзу несколько расширенная и более светлая, кверху красноватая. Первые листья супротивные яйцевидные, снизу светлые, сверху матово-зеленые, по краю мелко пильчато-зубчатые, на верхушке острые, на коротких черешках. Жилкование листьев выражено в виде утолщенной снизу сильно выдающейся средней жилки и тонких боковых жилок, углубленных сверху и выдающихся снизу, которые у края пластинки заворачиваются кверху и соединяются друг с другом с образованием крупных удлиненных петель. Ответвления боковых жилок входят в верхушки краевых зубчиков пластинки. Ближайшие последующие листья сходны с первыми, часто на черешках с двумя железками. Прилистники щетиновидно-линейные мелкозубчатые.
В первые годы жизни черемуха растет не очень быстро. Так, в ельнике кислично-зеленчуковом на дерново-сильноподзолистой поч
83
ве (Одинцовский р-н Московской обл.) 4-летний подрост черемухи • достигает высоты 20—23 см и 2,3 мм в диаметре, средний годичный J прирост его 5 см, несет 5 листьев; в 8 лет подрост достигает высоты ( 60 см, диаметра 5,3 см, годичный прирост 7—10 см, листьев— 15— 20; к 10 годам высота — 1 м, диаметр 6 — 8,5 мм. %. 12—15 летам черемуха в условиях достаточной освещенности начинает плодоносить. В более благоприятных условиях черемуха, может зацветать значительно раньше — в 4—6 лет (Байкина, 1946).
Данные о продолжительности жизни черемухи довольно разно- ‘ речивы: от 100 лет (Кбрреп, 1888) до 150—200 лет (Отенов, 1971).
Сезонный ритм развития
Для черемухи характерна высокая степень одновременности зацветания (высокий коэффициент эфемерности). Колебание в сроках зацветания отдельных особей составляет 0,7—1,3 дня (Батманов, 1962). Этот разрыв увеличивается лишь при наступлении похолодания. По годам амплитуда начала цветения составляет 42 дня (Тюрин, 1959, 1969). Для Подмосковья самый ранний срок зацветания-25 апреля, самый поздний — 5 июня, средний—15—16 мая± 7 дней. И. Г. Серебряков (1952) указывает срок зацветания черемухи в Подмосковье 17 мая— 11 июня. Определяющим фактором зацветания черемухи является температура; скорость продвижения J фазы цветения составляет 2,4 суток на 100 км (Малышева, 1968). 1 Созревание плодов происходит примерно через два месяца после на- ! чала цветения: в Подмосковье в среднем через 57 дней, в Архангельске через 52 дня (Шиманюк, 1964).
Побеги будущего года закладываются полностью еще летом. Формирование почек начинается до весеннего развертывания листьев (Серебряков, 1947а, б). Сначала закладывается 12—13 почечных i чешуй и 2 предлиста. Этот процесс протекает около двух месяцев и j почти совпадает с периодом роста побегов. Заложение листьев про- 1, исходит с первой декады июня и заканчивается в октябре. Формиро- . вание соцветий начинается в июне, этому предшествует формирова- $ ние 2—3 зачатков листьев, которые впоследствии недоразвиваются (Серебряков, 1947 а).
И. Г. Серебряков (1947 6) подробно изучил процесс заложения и развития листьев черемухи. Величина пластохрона в одних и тех же почках прогрессивно возрастает по мере затухания органообразующей деятельности конуса нарастания. В период с 17 мая по 11 июня величина пластохрона составила 2,8 дня. В период весеннего набухания почек и первых этапов роста листа (апрель — май) происходит настолько интенсивное деление клеток эпидермиса, что их !• площадь уменьшается почти вдвое. У теневых листьев интенсивность • деления клеток ниже, поэтому у них наблюдается гораздо большее увеличение площади каждой клетки — в 4,5 раза, а у световых — в 2 раза; соответственно число клеток эпидермиса у световых листьев возрастает в 525 раз, у теневых — в 168 раз. В процессе внутрнпо-
84
чечного развития листа черемухи увеличение пластинки идет за счет краевого меристематического роста, протекающего по симультаннодивергентному типу. С начала набухания почек и выхода листьев из почки увеличение пластинки идет за счет ее равномерного поверхностного роста, обязанного своим происхождением интенсивному делению клеток эпидермиса и мезофилла и в значительной степени увеличению размеров самих клеток. В целом развитие листа протекает по так называемому параллельному типу — элементы закладываются одновременно на всем протяжении срединной жилки, все его части являются одновозрастными. Для черемухи характерен так называемый поверхностный рост листа — по выходе из почки рост поверхности пластинки в длину и ширину протекает строго равномерно по всей площади.
Цветение одной кисти длится 6—7 дней (Байкина, 1946). Бутоны обычно распускаются утром. Образование зародышевого мешка проходит по обычному типу. Вскрывание пыльников происходит одновременно с распусканием цветков.
Способы размножения и распространения
Черемухе свойственно семенное и вегетативное размножение. Соотношение между особями семенного и порослевого происхождения в лесах Московской обл. примерно 1:2. Цветет ежегодно, но интенсивность цветения и плодоношения значительно колеблется. Вероятность хорошего урожая после обильного составляет 34% (Дол-гошев, 1959). Средний урожай плодов в условиях Марийской АССР составляет около 40 кг/га (Данилов, 1964).
Способы вегетативного размножения разнообразны: корневыми отпрысками, пневой порослью, отводками (Hegi, 1923; Селиванова-Городкова, 1957 а; Барыкина, 1960; Пятницкий и др., 1963; Отенов, 1970). Широко распространено размножение с помощью горизонтально направленных подземных корневищ, которые обычно образуются из придаточных почек на гипокотиле или реже на главном корне. Очень часто побеги от таких корневищ принимают за корнеотпрысковые. Р. П. Барыкина (I960) считает, что образование подземных корневищ, по-видимому, связано с редукцией системы главного побега у кустовидных форм черемухи. У прямоствольных форм подземные корневища заменяются отпрысками, которые образуются на поверхностных горизонтальных корнях, выходящих за пределы кроны дерева.
С. С. Пятницкий (1963) отмечает у черемухи наличие «шагающих» отводков, особенно на влажных местообитаниях, где образуются длинные побеги, дугообразно свисающие до поверхности почвы, на концах таких побегов образуются придаточные корни. Из боковых почек на месте укоренения образуются мощные побеги, которые часто по толщине превосходят материнский побег. Эти дочерние побеги также могут пригибаться к земле и укореняться. В итоге образуется система отводков, как бы шагающих в стороны от мате
85
ринского дерева. Укореняются ветви, которые расположены на высоте до 4 м.
Процесс укоренения ветвей подробно изучила Е. А. Селиванова-Городкова (1957а). С возрастом эти ветви утолщаются, при затенении теряют ассимилирующие и репродуктивные органы и получают совершенно новую функцию, являясь опорой стволу. Наиболее характерным местообитанием черемухи являются периодически подмываемые почвы речных долин, где ствол черемухи нередко сильно наклоняется в сторону максимального развития кроны и нуждается в опоре, чем и являются эти укоренившиеся опорные ветви.
Экология
Черемуха обыкновенная характеризуется довольно большой экологической амплитудой. В молодом возрасте она теневынослива, но с возрастом становится светлолюбивой. Цветет и плодоносит в «окнах», по опушкам и на полянах, где может достигать максимальных размеров — 15 м высоты и 30—40 см в диаметре.
Отличается исключительной зимостойкостью (Шиманюк, 1967; Артюшенко, Соколов, 1955; Селиванова-Городкова, 1957б). Из лиственных деревьев в наших лесах раньше других одевается листвой, не страдает при этом от весенних заморозков (Артюшенко, Соколов, 1955).
Предпочитает хорошо увлажненные дренированные места, встречаясь .часто по берегам рек й озер. Сухость воздуха переносит плохо. К почвам довольно требовательна, предпочитает плодородные почвы, хорошо растет на свежих суглинистых почвах. Встречается часто на молодом аллювии в уремах (Селиванова-Городкова, 1957 6), а также на почвах, бедных известью (Hegi, 1923). В условиях Московской обл. обычна на почвах с pH 4—5,5. Опад черемухи способствует улучшению подстилки, так как ее листья богаты кальцием, калием и натрием (Гроздов, I960).
В горы поднимается довольно высоко — до 1265 м в Скандинавии, до 2000 м на Кавказе и до 2200 м в Альпах (Hegi, 1923; Селиванова-Городкова, 19576).
Фитоценология
Черемуха обыкновенная довольно широко распространена в в хвойных, смешанных и лиственных лесах на хорошо увлажненных участках, особенно по берегам рек, в поймах (Hegi, 1923; Селиванова-Городкова, 1957 6; Смирнов, 1961). Как правило, она входит в состав второго яруса или подлеска. Встречается черемуха и в березовых колках Барабинской степи (Флора СССР, 1941). Е. А. Селиванова-Городкова (1957 6) считает, что черемуха нередко встречается в местонахождениях, которые являются очагами реликтовой третичной флоры в широколиственных лесах Правобережья Украины и Предуралья, в приволжских дубравах, в черневой тайге Алтая, в
86
сложных лесных насаждениях Западных Саян, в кустарниково-степном поясе Тянь-Шаня.
Б. А. Быков (1965) называет черемуху в качестве доминанта 5 яруса преимущественно в уремных лесах с кустарниковым подлеском из черной смородины, спиреи иволистной, малины и черной бу-зины.'Кроме того, она доминирует в сообществах Prunus padus — Calamagrostis langsdorfii, P. padus — Filipendula ulmaria и др.
Черемуха обладает довольно значительной средообразующей способностью (Сукачев, 1938). Атмосферные осадки, промывшие опавшие листья черемухи, имеют слабощелочную реакцию, нейтрализуют почву. Листья быстро минерализуются, обогащая почву К, Са и N (Шиманюк, 1967), поэтому черемуха считается желательной в большинстве лесных насаждений (с господством сосны, дуба и других пород), так как создает высококачественную подстилку.
Консортивные связи
Черемуха относится к строго перекрестноопыляемым растениям, опыление происходит с помощью мух, муравьев и некоторых других мелких насекомых (Байкина, 1946). Плодами черемухи питаются и способствуют ее распространению черный дрозд, дубонос и некоторые другие птицы (Оленев, 1955).
Черемуха подвержена заболеваниям, вызываемым грибами: деформацию плодов вызывает Exoascus pruni Fuck. var. padi Jacz. (Ванин, 1931); неукоренившиеся и наиболее затененные ветви поражаются грибами Nectria cinnabarina Fries и Stereum purpureum Fries. (Селянинова-Городкова, 1957). Черемуха является промежуточным хозяином ржавчинного гриба Pucciniastrum padi, повреждающего шишки ели (по этой причине не рекомендуется вводить черемуху в посадки ели).
Зарегистрировано около 30 видов насекомых и клещей, повреждающих черемуху обыкновенную. Листья объедают гусеницы бабочек-пядениц, личинки пилильщиков, минируют листья моли и минирующая златка, черемуховые клещики образуют галлы; повреждаются листья также листовертками и тлями, паутинные гнезда образуются личинками пилильщиков-ткачей, черемуховой моли и гусеницами златогузок и боярышниц. Плоды повреждаются личинками пилильщиков и долгоносиков; цветочные^ почки повреждает гусеница серой кармашковой пяденицы; цветки повреждаются черемуховым цветоедом. Вредителями стволов и веток являются черемуховый побеговый долгоносик, личинки различных короедов, персиковая ложнощитовка (Гусев, Римский-Корсаков, 1951; Оленев, 1955).
Внутривидовая систематика
В ряде флористических сводок (Hegi, 1923; Флора СССР, 1941; Hulten, 1948) самостоятельным таксоном выделяют низкорослую разновидность var. petraea Tausch. (-Padus borealis Schiib.), pacnpo-
87
страненную в горах и на севере ареала. Многие авторы (Сюзев, 1906; Харитонов, 1958; Вашкулат, 1959; Александров, 1967; Бойченко, 1968) выделяют розовоцветную разновидность— var. roseiflora Sinz., чаще указываемую для более северных местонахождений (Кировская, Архангельская, Пермская области). Морфологически разновидности сходны с белоцветной, но розовую окраску сохраняют постоянно, лишь несколько варьируя по годам (Вашкулат, 1959). Кроме того, А. А. Корчагин (1951) выделил разновидность с удлиненными плодами — var. dolichocarpa A. Korz. А. П. Шиманюк (1967) указывает позднецветковую форму — var. tardiflora Alech., выделенную В. В. Алехиным в прибрежной полосе Ветлуги и зацветающую на месяц позднее обычной формы. Ф. А. Александров (1967) отмечает присутствие в лесах Кировской обл. махровой, по-лумахровой и крупноцветной форм черемухи, перспективных для использования в озеленении.
Выделено много культурных форм черемухи (Hegi, 1923; Rehder, 1927; Bailey, 1937; Соколов, 1954).
Хозяйственное значение
Черемуха обыкновенная имеет довольно разнообразное применение в народном хозяйстве. Она широко используется в садах й парках в качестве прекрасного декоративного дерева, особенно розовоцветковая форма. Благодаря почвоулучшающим особенностям (Степанов, 1938; Шиманюк, 1967) черемуху желательно вводить в культуры сосны, дуба, лиственницы (но не ели). Способность укореняться может быть широко использована при облесении склонов, оврагов (Селиванова-Городкова, 1957 6).
Черемуха — хороший весенний медонос и перганос (Соколов, 1954). Плоды черемухи содержат сахар, яблочную и лимонную кислоты, витамин С (Знаменский, 1932; Флора СССР, 1941; Шиманюк, 1967). Их широко употребляют в пищу, особенно на Урале и в Сибири. Кора обладает красильными свойствами (Ефимова, 1954; Смирнов, 1961). Древесина пригодна для столярных изделий (Соколов, 1954).
Широко известна черемуха как лекарственное растение. В медицине ее плоды применяют при желудочно-кишечных заболеваниях (Атлас лекарственных растений, 1962). Различные органы черемухи обладают токсическим действием на болезнетворных микроорганизмов и простейших (Сацыперова, 1951; Граменецкая, 1952; Стуккей, 1955), а также на червей и насекомых (Драбкин, 1954; Оленев, 1955). Благодаря этому препараты из черемухи могут использоваться при лечении долго не заживающих ран (Разумович, Горфункель, 1956).
Н. П. Крылова и Т. А. Работное 1
КЛЕВЕР ЛУГОВОЙ
Номенклатура
Trifolium pratense L., Sp. pl. (1753) 768; Кауфм., Моск, фл., изд. 1 (1866) 128, изд 2 (1889) 124; Цингер, Сб. свед. (1885) 132; Шмальг., Фл. 1 (1895) 238, р.р.; Петунн., Крит, обзор 1 (1896) 122; Сырейщ., Илл. фл. 2 (1907) 294, 4 (1914) 112; он же, Опр. раст. Моск. губ. (1927) 172; Федченко в Маевск., Фл., изд 7 (1940) 465; Бобров во Фл. СССР 11 (1945) 248; он же в Тр. БИН АН СССР сер. I, 6 (1947) 300; Борисова в Маевск., Фл., изд. 8 (1954) 151, изд. 9 (1964) 153; Тихомиров в Опр. раст. Моск. обл. (1966) 213; Coombe in Fl. Eur. 2 (1968) 168.— Клевер луговой, к. красный.
Сем. Бобовые — Leguminosae (Papailionaceae).
Морфологическое описание
Многолетнее травянистое растение. Корневая система стержневая, со многими вторичными корнями (Бобров, 1947; Wilsie, 1962). На заливных лугах в пойме Оки в Московской обл. корневая система достигает глубины 30—40 см (Покровская, 1960 а; Крылова, 1964). В условиях европейской лесостепи глубина проникновения корней — около 120 см (Голубев, 1962). Культурные формы клевера в благоприятных условиях на хорошо дренированных и аэрированных почвах развивают корневую систему до 1—2 м (Сергеев, 1951, 1958; Fergus, 1960) и даже до 3 м (Weaver, 1926). На влажных материковых лугах корневая система поверхностная, углубляется на (>— 12 см (Linkola, Tiirikka, 1936).
Клевер луговой имеет прямостоячие или слегка изогнутые надземные побеги. Число побегов в кусте значительно варьирует и зависимости от условий местообитания и составляет от 1 до 10 и даже 18 на растение (Покровская, 1960а; Крылова, 1964). Центральный побег укорочен и в течение почти всего жизненного цикла клевера сохраняется в виде прикорневой розетки листьев, вокруг которой расположены боковые генеративные и вегетативные побеги (мопо-
1 Раздел «Консортивные связи» написала Г. Д. Успенская.
89
подвальный тип возобновления). На генеративных побегах насчитывается 6—7 ясно различимых междоузлий и 2 сближенных (Крылова, 1964). Число узлов у клевера — признак непостоянный (Лисицын, 1951). Встречаются особи с числом узлов от 9 до 18 (Покровская, 1960 а). Листья тройчатые, цельнокрайние, верхние листья на коротких, нижние — на более длинных черешках. Стебли и листья опушены. Прилистники срослись с листовыми черешками.
Соцветие — головка, у основания которой имеются два небольших сидячих листа. Длина трубки венчика у клевера лугового от 3,83 до 9,7 мм (Абрамова, I960; Крылова, 1964). Плод — односемянный, реже двусемянный боб. Семена желтоватого, буроватого или фиолетового цвета, яйцевидной формы (Сергеев и др., 1963). Средний вес 1000 семян— 1,71 г, колебания от 0,96 до 2,03 г (Филимонов, 1953; Лебедев,'1956). Наибольший абсолютный вес семян клевера лугового в пойме Оки — 1,37 г (Крылова, 1964).
Как культурные сорта, так и дикорастущие формы клевера лугового разного географического происхождения являются диплои-дами и имеют 2й—14 (Арутюнова, 1940; Шутова, 1965).
Географическое распространение
Клевер луговой в диком виде широко распространен во многих районах Евразии, а как культурное растение — во всем мире. Это важная кормовая культура в Европе, западной части Центральной Азии, США, Канаде, Новой Зеландии и Австралии (Ларин и др., 1951).
Дикорастущий клевер луговой делится на две расы: атлантическую Т. pratense proles atlanticum Bobr. и континентальную Тр. proles continentale Bobr. (Флора СССР, 1945). В обширном ареале культурного клевера на территории СССР принято выделять три части (Лисицын, 1951).
1. Юго-западная часть занята двуукосным клевером. Сюда относится вся Украина, Курская и Брянская области, южная половина Белоруссии.
2. Дальше к северо-востоку расположен пояс смеси двуукосных клеверов с одноукосным. Сюда относится центральная и западная части Ленинградской обл., а также северная половина Белоруссии, западная и юго-западная части Орловской и Брянской областей.
3. Дальше на северо-восток и на север распространен в основном одноукосный клевер.
Дикий клевер луговой, как и культурный, является сложной популяцией, но с преобладанием скороспелых рас (Лисицын, 1961). В Московской обл. он распространен повсеместно. На окских лугах по времени заложения соцветий у дикорастущего клевера лугового выделяют две формы: одни особи являются типичными аборигенами средней полосы, другие представляют собой естественные гибриды дикорастущих форм с культурными и даже одичалые культурные клевера (Покровская, 1960 а).
90
Онтогенез и сезонный ритм развития
Основными морфологическими показателями при выделении возрастных этапов у клевера принято считать состояние главного побега и главного корня, а также наличие или отсутствие боковых генеративных побегов и придаточных корней разных порядков. В развитии клевера лугового выделяют следующие возрастные этапы (Покровская, 1960 а):
I — ювенильный и отчасти предгенеративный (кущения нет, хотя в пазухах листьев имеются почки возобновления);
II — предгенеративный (крупные розетки, в пазухах листьев которйх находятся укороченные побеги или почки);
III — начально-генеративный (ясно выражены боковые генеративные побеги, но они немногочисленны);
IV — генеративный (имеется большое число генеративных побегов при одновременном существовании верхушечной почки);
V — этап вегетативного размножения (полное отмирание верхушечной почки и начало разрушения главного корня);
VI — сенильный (постепенное отмирание и разрушение главного корня).
Наблюдения за фиксированными особями в пойме Оки позволили выделить в жизненном цикле клевера лугового следующие возрастные группы особей: всходы, ювенильные, имматурные и взрослые виргинильные, генеративные, сенильные (Крылова, 1964).
О длительности сохранения жизнеспособности у семян клевера лугового, погребенных в почве, в литературе мало сведений. В опыте Дювеля она составила 39 лет (Toole, Brown, 1946). По данным Госса (Goss, 1933), семена клевера лугового сохраняют всхожесть после 20 лет хранения в почве. По данным А. М. Овеснова (1963), в почве Сылвенского залива Пермского водохранилища после 5 лет затопления содержались единичные всхожие семена клевера лугового. При хранении в лаборатории семена могут сохранять всхожесть длительный срок — от 20—28 лет (Мещеряков, 1957) до 81 года (Turner, 1933). По данным Е. И. Якушевой (1947), после 10-летнего срока хранения всхожесть семян клевера понижается на 60%, а после 30-летнего могут быть получены лишь единичные всходы.
У проростков клевера лугового гипокотиль низкий, шириной около 0,8 мм, зеленоватый. Семядоли эллиптические, темно-зеленые, слегка мясистые, длиной 5 мм, шириной 3 мм, на черешке длиной 5—8 мм. Первый лист на длинном (30—40 мм) черешке, второй лист тройчатый с эллиптическими, на верхушке выемчатыми, волосистыми листочками. Прилистники листьев узколанцетныс, в некоторых случаях всходы имеют 3 семядоли (Васильченко, 1937). Па лугах в пойме Оки у всходов в течение одного-двух месяцев после появления сохраняется первый простой листочек, который является точным признаком возникновения особи в данном вегетационном сезоне. Всходы наряду с простым округло-сердцевидным листом пс-
91
Сут 1—4 тройчатых листа, уже на первом листе легко просматривается опушение (Крылова, 1964).
Ювенильные особи клевера лугового представляют собой маленькие розетки листьев с хорошо выраженным главным корнем. Кущение отсутствует, но в пазухах листьев отчетливо просматриваются почки возобновления. Переход от состояния всходов к ювенильной фазе выражается в утолщении узлов кущения и гипокотиля. Высота ювенильных особей на окских лугах от 3 до 11 см. Обычно в этот период развиваются третий — пятый настоящие листья. В чистых посевах эта фаза наступает на 60—65 день после посева (Исаин, 1959). Нарастание стебля и листьев на данном этапе идет за счет бокового ветвления верхушечного конуса роста, которое протекает у клевера по типу моноподиального ветвления (Лебедев, 1958). В связи с бактериотрофией — получением от клубеньковых бактерий связанного азота — молодые особи развиваются на лугах быстро, если нет каких-либо других ограничивающих условий.
По Т. М. Покровской (1959, 1960 а), ювенильные особи клевера лугового относятся к растениям, находящимся на I возрастном этапе. В Московской обл. на лугах поймы Оки при двуукосном использовании и при поздних сроках скашивания к ювенильным относится большинство особей клевера второго года жизни. Продолжительность пребывания в ювенильном состоянии и устойчивость ювенильных особей зависят от природных и антропогенных факторов: метеорологических условий, состава и степени развития окружающего травостоя, режима использования. Ювенильная фаза заканчивается с началом кущения.
Благоприятные условия увлажнения, богатство почв доступным для растений фосфором и калием и раннее скашивание сокращают продолжительность пребывания особей в ювенильном состоянии. Наблюдения за фиксированными особями на окских лугах показали, что они способны пребывать в ювенильном состоянии не больше одного вегетационного периода, превращаясь в дальнейшем в имма-турные и взрослые виргинильные особи. Такие особи, как правило, характеризуются морфологическими признаками растений, находящихся, по Т. М. Покровской (1960 а), на втором этапе формирования куста.
Наибольшее число ювенильных особей клевера лугового ежегодно отмечалось при раннем скашивании основного укоса, что объясняется равномерным обсеменением во втором укосе. В опыте на окских лугах внесение фосфорно-калийных удобрений при двуукосном использовании не устраняет колебаний в численности ювенильных особей: в среднем на 1 м2 в 1959 г. их было 10, в 1960 — 0, в 1961 —4, в 1962 — 0. Отмирание ювенильных особей, как и всходов, может происходить в результате резкой смены условий произрастания после скашивания основного укоса и более выражено в сухие, чем во влажные, годы.
Кущение у к. лугового наступает после образования третьего — пятого настоящего листа (Семенов, 1947; Лебедев, 1958). По нашим
92
Наблюдениям, взрослая виргинйльная особь клевера лугового имеет кроме прикорневой розетки 1—2 (при внесении удобрений до 6) боковых вегетативных побега в виде крупных розеток листьев. В на зухах каждого листа отчетливо видны укороченные побеги или почки (II этап по Т. М. Покровской, 1960 а). Каждая розетка чаще всего состоит из 5 листьев с длинными черешками. Главный корень выражен отчетливо, но проникает на небольшую глубину.
Продолжительность пребывания во взрослом виргинилыюм состоянии изменяется в зависимости от внешних условий. Положительное влияние фосфорно-калийных удобрений на мощность вегетативных побегов отчетливо проявляется даже в засушливые годы. Среди взрослых виргинильных особей клевера лугового при двуукосном использовании преобладают растения третьего года жизни. Нередко во влажные годы у трехлетних особей в основном укосе происходит усиленное кущение, а к моменту второго укоса большинство их переходит в генеративную фазу.
При раннем скашивании в течение одного вегетационного периода у клевера лугового происходит совмещение II (предгенера-тивного) и III (начально-генеративного) возрастно-морфологических этапов, выделенных Г. М. Покровской. Иногда при раннем скашивании с внесением РК можно наблюдать прохождение в течение одного вегетационного периода II, III и даже IV (генеративного) периодов жизни особи. Однако жизненное состояние и продуктивность клевера лугового при ранних сроках скашивания, как правило, ниже, чем при позднем скашивании.
Клевер луговой на природных лугах в благоприятных условиях зацветает на 3—4 год жизни растения. В менее благоприятных условиях зацветает не ранее чем в возрасте 6—8 лет (Linkola, Tiirikka, 1936). В благоприятных условиях это моно- или дикарпичное растение. В малоблагоприятных условиях клевер — поликарпик с вероятной длительностью жизни до 20 лет, с выраженными перерывами в цветении, но его генеративные особи развиты слабо. Внесение фосфорно-калийных удобрений ускоряет переход клевера в генеративное состояние, но сокращает длительность генеративного периода.
Генеративная особь в благоприятных условиях имеет большей частью 3—6 (иногда до 18) генеративных побегов при одновременном сохранении вегетативной центральной розетки. Т. М. Покровская (1959) пишет, что в этот период (IV возрастно-морфологический этап) на конусе нарастания отмечается три зачатка листьев и значительное погружение его («втягивание до 0,5 см»), что может быть связано с прекращением роста, оси первого порядка. Главный корень морфологически выражен хорошо, хотя наблюдается отмирание первичной ксилемы. Образование побегов во втором укосе у генеративных особей клевера происходит в результате роста центральной розетки и развития спящих боковых почек, а также из пазушных почек в нижних узлах скошенных генеративных побегов.
93
Аналогичные данные по наблюдениям на окских лугах приводятся ! Т. И. Серебряковой (1953, 1956) и А. Н. Лихачевым (1959). '
Характеризуя ритм побегообразования генеративной особи клевера лугового, Т. И. Серебрякова (1956) отмечает большое число весенне-летних листьев у главной розетки осенью и наличие в верхушечной почке главной розетки 5—6 зимующих зачатков листьев розетки будущего года. Перезимовавшие прошлогодние листья весной отмирают, сменяясь к июлю листьями, сформировавшимися из прошлогодних зачатков. В течение июля весенние листья розетки засыхают, а вновь заложившиеся развертываются с последующим формированием зачатков листьев розетки будущего года. По данным Т. И. Серебряковой (1956), боковые розетки — будущие генеративные побеги — начинают развертываться в пазухах засыхающих листьев в конце июля — августе, последовательно снизу вверх. Вторичное цветение клевера лугового после скашивания идет за счет । побегов третьего порядка, пазушных в нижних оставшихся узлах срезанного генеративного побега. При низком срезании генеративные побеги удаляются почти нацело и вторичного цветения не происходит.
По данным Г. М. Денисовой (1953), у клевера лугового, как и у других стержнекорневых бобовых, общей закономерностью в побегообразовании является формирование боковых побегов из спящих почек, расположенных на старых участках материнского побега. Боковые побеги в своем развитии аналогичны материнскому побегу.
По 'наблюдениям в пойме Оки, чем значительнее в основном укосе выражен максимум репродуктивной мощности, тем слабее происходит образование новых генеративных побегов во втором укосе. При раннем скашивании или весеннем подкашивании, осо-бенйо при внесении фосфорно-калийных удобрений, значительная часть особей клевера лугового переходит в генеративное состояние на третий год жизни, образуя по 4—6 генеративных побегов. При повторном цветении на четвертый год жизни у таких особей образуется небольшое число генеративных побегов — в среднем 1—2. Возможны случаи, когда трехлетние особи при раннем скашивании вступают в генеративную фазу осенью. На следующий (четвертый) год у таких растений в основном укосе образуется 7—8 новых генеративных побегов, т. е. максимум репродуктивной мощности выражен довольно хорошо.
Раннее скашивание травостоя способствует также пробуждению почек возобновления на пеньке генеративного побега. Возобновление побегов из почек на пеньке скошенного генеративного побега — довольно распространенное явление во втором укосе, но сохранение почки возобновления в течение зимы и образование из нее на следующий год нормально развитого генеративного побега происходит очень редко.
Период максимума репродуктивной мощности у особей клевера лугового сменяется к весне следующего года переходом особей в фазу постгенеративной вегетации или полным отмиранием (Крыло-
94
ва, 1964; Работнов, 1955, 1960). Отмирание особей после прохождения генеративной фазы наступает в позднеосенний и зимний периоды. Переход в фазу постгенеративной вегетации наступает обычно на пятый или четвертый годы жизни.
У сенильных особей клевера лугового сохраняется только центральная розетка листьев, жизненное состояние угнетенное. У отдельных особей можно различить лишь основание центральной розетки, состоящее из слабых мелких листочков. У других особей видны лишь два боковых слабых листа центральной розетки. Почки возобновления в пазухах многочисленных отмерших листьев центральной розетки встречаются очень редко. Главный корень почти полностью разрушен. Продолжительная постгенеративная вегетация у клевера встречается значительно реже, чем кратковременная. Из зафиксированных в 1961 г. 60 генеративных 3-—4-летних особей в 1962 г. (после перезимовки растений) обнаружено лишь 6 особей в фазе постгенеративной вегетации. Из 49 генеративных особей, зафиксированных в 1962 г., в 1963 г. в фазе постгенеративной вегетации обнаружено лишь 4 особи. При этом все особи в фазе постгенеративной вегетации встречены только при раннем скашивании основного укоса. По нашим наблюдениям, на окских лугах в состоянии постгенеративной вегетации особи клевера лугового пребывают не более одного вегетационного периода.
Способы размножения и распространения
Данные о семенной продуктивности клевера лугового на природных лугах приводятся в работах Перттула (Perttula, 1941), Т. А. Работнова (1950), И. В. Каменецкой (1952). Различные условия местообитания обусловливают значительную разницу семенной продуктивности, приводимой этими авторами. Число семян на одну генеративную особь по данным Перттула (Южная Финляндия) составляет 15,2; по данным Т. А. Работнова (Северный Кавказ) — от 13 до 50; по данным И. В. Каменецкой (Стрелецкая степь) — 106. По нашим наблюдениям, на природных лугах Московской обл. наибольшая семенная продуктивность клевера лугового во втором укосе 56—60 семян на особь (Крылова, 1964).
Максимальное число живых семян на особь клевера при позднем скашивании не превышает 40 даже в годы, благоприятные для обсеменения. Внесение фосфорно-калийных удобрений оказывает положительное влияние на жизненное состояние и семенную продуктивность, но не обеспечивает устойчивости в семенной продуктивности. Урожай семян клевера лугового даже при позднем скашивании основного укоса подвержен колебаниям по годам, но отсутствие обсеменения отмечается лишь в редкие годы, например, в пойме Оки в 1962 г.
Несмотря на колебания в урожае семян по годам, условия для семенного размножения клевера при раннем скашивании основного укоса являются достаточно благоприятными. На окских лугах мак
95
симальный урожай семян во втором укосе получен в 1962—1963 гг., когда в среднем на 1 м2 насчитывалось до 3825 семян. В другие годы (1959—1961). число семян колебалось от 18 до 338 на 1 м2 на участках, где не вносили удобрения, и до 509 — при внесении фосфорио-калийного удобрения. Посевы культурного двуукосного клевера во втором укосе дают больший урожай семян при скашивании первого укоса в фазу бутонизации по сравнению с более поздним скашиванием (Мирвис, 1959; Морозов, 1960).
Несомненно, что среди факторов, определяющих высокую семенную продуктивность клевера, во втором укосе большое значение имеет формирование мелких цветков, которые лучше опыляются насекомыми, что отметил еще Ч. Дарвин. По наблюдениям на Витебской опытной станции (Морозов, 1960), на цветущем клевере во втором укосе бывает больше насекомых-опылителей, в частности в 2—3 раза больше шмелей.
Несомненное значение имеет и снижение повреждений семян клевера насекомыми во втором укосе. По наблюдениям на Ганусов-ской опытной станции (Мирвис, 1959), при скашивании первого укоса на семена получено здоровых головок 2%, частично поврежденных— 38%, уничтожено насекомыми 57%. При скашивании первого укоса в фазе бутонизации и второго укоса на семена получено здоровых семян 47%, частично поврежденных — 43%, уничтожено 10%. По данным Витебской опытной станции (Морозов, 1960), только от фитономуса, клубенькового долгоносика и особенно клеверного долгоносика потери семян в первом укосе составляют 17—27%, во втором укосе — 2—6, редко — 10%.
При различных режимах использования положительное влияние фосфорно-калийных удобрений на урожай семян клевера прослеживается не ежегодно, так как определяется главным образом численностью генеративных особей, а колебания их численности по годам внесением удобрений не устраняется.
Как урожай, так и осыпаемость семян клевера в условиях естественного луга изменяются по годам. Количество осыпавшихся семян при двуукосном использовании лугов и даже при позднем скашивании первого укоса невелико. Наибольшее число осыпавшихся семян клевера лугового в пойме Оки отмечено при самообсеме-нении во втором укосе — до 150—160 на 1 м2 при раннем скашивании первого укоса.
По наблюдениям О. И. Соколовой (1934), в условиях Московской обл. среди семян дикорастущего клевера лугового хорошую всхожесть имели 70,6%, среднюю — 22%, плохую — 7,4%. В южной части Финляндии всхожесть семян клевера в природных условиях составляет 21—40% (Perttula, 1941). По нашим наблюдениям (пойма Оки), содержание жизнеспособных семян в урожае у клевера при различных режимах использования колебалось по годам от 55 до 90%. Отчетливого влияния внесения удобрений на изменение содержания жизнеспособных семян у клевера не прослеживается.
Для клевера лугового характерно высокое содержание в уро
96
жае твердых семян, особенно во втором укосе, где их доля составляет от 55 до 67%. Твердые семена сохраняют жизнеспособность в течение длительного времени. Так, в опытеХмелара (Chmelar, 1941), в благоприятных для прорастания семян условиях 77% твердых семян проросло в течение первого года, за 10 лет проросло 90%, за 20 лет — 91%, за 26 лет — 92%.
При двуукосном использовании клевера лугового запас его семян в почве на окских лугах, по нашим наблюдениям, не превышает 25 семян на 1 м2. Наибольший запас отмечен при позднем скашивании луга, где вносились фосфорно-калийные удобрения, и при раннем скашивании первого укоса с обсеменением во втором укосе — соответственно 236 и ПО семян на 1 м2.
По наблюдениям на заливных лугах поймы Оки, число появляющихся всходов клевера зависит от урожая и осыпаемости семян в предыдущий год, поэтому колеблется от года к году. Так, после частичного самообсеменения клевера при обычном двуукосном использовании его в 1959 г. число всходов в 1960 г. было в 4—8 раз больше, чем в 1961, 1962 гг. Наиболее устойчиво семенное размножение клевера лугового при самообсеменении его во втором укосе после раннего скашивания основного укоса. При поздних сроках скашивания на приживаемости всходов клевера отрицательно сказываются длительное затенение и конкуренция со взрослыми растениями за питательные вещества и влагу. Большое значение в появлении всходов и устойчивости клевера в травостое имеет содержание в урожае твердых семян (Шайн, 1949).
Как показали наши наблюдения на окских лугах, а также исследования Г. М. Денисовой (1953), вегетативное размножение клевера лугового—чрезвычайно редкое явление. В менее благоприятных условиях, например в северных районах, оно, по-видимому, более распространено.
Экология
Клевер луговой широко распространен на лугах в пределах лесной и лесостепной зон, а также в лесном и субальпийском поясах гор. В Московской обл. он наиболее распространен на хорошо дренированных краткопоемных лугах в долинах рек и на суходольных лугах.
Судя по шкалам Л. Г. Раменского, наибольшее участие в травостоях принимает при увлажнении в пределах 58—72 ступеней (свежевлажное луговое увлажнение), 9—11 ступеней переменности увлажнения (умеренная переменность) и 9—18 ступеней богатства почвы (от небогатых до богатых почв). Наиболее хорошо растет на суглинистых слабокислых или нейтральных почвах (pH в водной вытяжке не ниже 5,5).
Отрицательно реагирует на засоление и на ухудшение аэрации почвы, связанное с заболачиванием, задернением, уплотнением почвы, образованием сплошного мохового покрова. Не выносит дли
4 Зак. 255
97
тельного (свыше 10—15 дней) затопление полыми водами ив особенности застоя воды, а также перекрытия мощным наилком.
Дикорастущий клевер достаточно устойчив к низким температурам, а также к колебаниям температуры ранней весной (сравнительно «весностоек»). Клевер луговой довольно требователен к условиям освещения. Умеренно устойчив к выпасу.
Биологическая продуктивность
На природных лугах для клевера лугового характерно резкое колебание его продуктивности по годам. Так, на краткопоемном, неежегодно заливаемом лугу в пойме Оки продуктивность клевера в первом укосе в течение II лет (1954—1964 гг.) изменялась от 0,6 до 313,2 г с 8 м2, т. е. примерно в пределах от 1 до 400 кг/га. Резкое колебание продуктивности обусловлено различием' в самообсемене-нии клевера в отдельные годы/ в появлении и приживаемости его всходов, а также своеобразным жизненным циклом клевера. Это своеобразие выражается в том, что в благоприятные годы большая часть особей клевера переходит в генеративное состояние и пышно развивается — клевер может стать доминантой («клеверный» год), но его пышно развитые особи на следующий год или через год отмирают, что сопровождается резким снижением продуктивности.
Важным фактором, обеспечивающим увеличение продуктивности клевера на лугах, является внесение удобрений. В зависимости от содержания в почве легкорастворимых Р2О5 и КгО продуктивность резко возрастает при внесении калийных, фосфорных или ка-лийно-фосфорных удобрений. Так, на краткопойменном лугу в долине Оки наибольший эффект дали калийные, а на субальпийском лугу Северного Кавказа — фосфорные удобрения (Работнов, 1948 6, 1963, 1973). При ежегодном (в течение 10 лет) внесении калийных удобрений суммарный урожай клевера лугового на окских лугах за период с 1955 по 1964 г. возрос в 2 раза. Особенно значительное увеличение по сравнению с контролем имело место в «клеверные» года (Работнов, 1973). Внесение калийных и фосфорно-калийных удобрений не устраняет цикличности в биологической продуктивности клевера лугового. На внесение азотного и полного (NPK) минерального удобрения этот вид реагировал отрицательно.
Устойчивость и продуктивность клевера возрастают при подсеве его семян. По наблюдениям в пойме Оки продолжительность жизни подсеянного клевера 4 года. Урожай его сена (при норме подсева семян 6 кг/га) в сумме за 2 года составил около 40 ц/га (Крылова, 1963).
Фитоценология
. Участие клевера лугового в травостоях лугов резко изменяется от года к году (Работнов, 1955, 1972). Массовое участие вплоть до доминирования в отдельные годы и последующее исчезновение кле
98
вера обычно связано с тем, что в годы, благоприятные для развития клеверов, большая часть особей переходит в генеративное состояние (см. выше в разделе «Биологическая продуктивность»). Такое высокое участие клевера в травостое может длиться один или два года, так как его пышно развитые особи, закончив свой жизненный цикл, отмирают. Так, на окском краткопойменном лугу в 1948 г. на площадке в 5 м2 из 308 особей клевера 288 (93,5%) были в генеративном состоянии. К 1949 г. 75% из них отмерли, 22% были в вегетативном состоянии и только 3% цвели повторно. К 1950 г. все особи отмерли (Rabotnov, 1960). В годы массового развития .клевера успевают обсемениться в первом или во втором укосе, из семян появляются всходы и через несколько лет вновь повторяется «клеверный» год. Интервалы между клев'ерными годами в пойме Оки близ с. Дедино-ва варьируют в зависимости от почвенных и метеорологических условий от 1 до 3 и более лет. Таким образом, клевер луговой — типичный ценоциклофлюктуент.
В литературе описаны многочисленные полидоминантные ассоциации, в состав доминантов которых входит клевер луговой. Все они являются флюктуционными состояниями каких-то других ассоциаций, в которых клевер периодически входит в состав доминантов. Наибольшее значение в формировании луговых фитоценозов это растение имеет на краткопоемных (часто неежегодно заливаемых) лугах, где он периодически доминирует совместно со злаками, некоторыми бобовыми (мышиный горошек, чина луговая, клевер розовый, а при пастбищном использовании клевер белый) и видами разнотравья. Из злаков совместно с клевером доминируют виды умеренно влажных достаточно богатых почв (пырей ползучий, овсяница луговая, тимофеевка луговая, полевица белая, лисохвост луговой, мятлик луговой и др.), а из разнотравья — герань луговая, тысячелистник обыкновенный, щавель пирамидальный, колокольчик скученный, порезник горный и др.
Консортивные связи
В процессе эволюции между Г. pratense и шмелями (р. Bom-bus) выработались тесные консортивные отношения. Шмели являются чуть ли не единственными опылителями клевера. Значительно меньшая роль принадлежит пчелам (р. Apis). Это обусловливается особенностями ротового аппарата этих насекомых и строением цветка клевера. За один рабочий час шмель опыляет более 1500, а пчела— до 800 цветков (Алпатов и др., 1933).
Симбиотические отношения сложились у клевера с почвенной микрофлорой, что проявилось в образовании клубеньков на корнях бактериями р. Rbizobium. По специфичности в отношении растения-хозяина и некоторым физиолого-биохимическим признакам различают ряд видов, из которых Rh. leguminosarum и Rh. trifolii известны на корнях к. лугового, фиксация молекулярного азота осущест
4* 99
вляется этими бактериями только в симбиозе с корнями бобового растения. Подобные консортивные взаимоотношения проявляются во всех фитоценозах, по число клубеньков зависит от условий произрастания.
Комплекс микроорганизмов ризосферы корня клевера специфичен для определенных почвенно-климатических зон и состоит из разных видов микроскопических грибов и бактерий.
Паразитические консортивные взаимоотношения дикорастущего клевера лугового с микромицетами отрицательно влияют на растения. Поражая листья и стебель, они ограничивают ассимиляционную способность растения, вызывая преждевременное опадение листьев, ломкость стебля, что ведет к снижению урожая и ухудшению качества сена. На Т. pratense известно свыше 50 возбудителей заболеваний (Купревич, 1954),ноне все они встречаются в Московской обл. и неодинаково их значение. Наиболее распространена Peronospora trifolii-pratensis A. Gust.— возбудитель ложной мучнистой росы, часто развивающийся в низинах, достигая обильного развития при влажной и теплой погоде. Возбудители бурой пятнистости Pseudo-pesiza trifolii (Biv.— Bern.) Fuck., ржавчины Uromyces trifolii (Hedw. f.) Lev., концентрической пятнистости Stemphylium sarei-naeforme (Kav.) Wilt., аскохитоза Ascochyta trifolii Bond, et Trus., черной пятнистости Polythrincium trifolii Kunz, церкоспороза Cer-cospora zebrina Pass., стагоноспороза Stagonospora meliloti Petrak., мучнистой росы Erysiphe communis f. trifolii Rab. и др. встречаются повсеместно и наибольшего развития достигают к середине лета. Некоторые из них продолжают жить на отаве (Успенская, 1959). Очаговое распространение имеет возбудитель антракноза Kabatiella caulivora (Kirchn) Karak, Это зависит от условий произрастания клевера (Миняева, 1952). Сходные паразитические связи имеются между Т. pratense и возбудителем бактериальной пятнистости Pseudomonas trifoliorum (Jones et al.) Stapp. Особенно часто они возникают на клеверах, растущих в поймах рек (Успенская, Духо-вич, 1958).
Отмечено, что на растениях Т. pratense одновременно могут развиваться U. trifolii и St. sarcinaeformis, illtrifolii и E. communis f. trifolii. Комбинации возбудителей на одних и тех же экземплярах растений могут быть различными. Основными факторами в возникновении таких консорций являются .физиолого-биохимическое состояние высшего растения, отсутствие антагонистических взаимоотношений грибов при их совместном развитии в тканях растений, соответствующие экологические условия.
Подобные отрицательные консортивные связи существуют и у возделываемого красного клевера. Паразитические консортивные связи дикорастущих клеверов способствуют развитию заболеваний культурного клевера, так как больные невозделываемые клевера являются источниками инфекции.
100
Хозяйственное значение
Хозяйственное значение клевера лугового исключительно велико. Он является ценнейшим сенокосно-пастбищным видом и одним из основных компонентов травосмесей, высеваемых в полевых севооборотах. Роли клевера как кормового растения посвящена обширная отечественная и зарубежная литература (см., например, Ларин и др., 1951; Сергеев и др., 1963; Мухина, 1971; Залыгин, 1971).
Н. П. Крылова1
КЛЕВЕР РОЗОВЫЙ
Номенклатура
Trifolium hybridum L., Sp. pl. (1753) 766; Кауфм., Моск. фл. изд. 1 (1866) 129, изд. 2 (1889) 126; Цингер, Сб. свед. (1885) 133; Шмальг., Фл. 1 (1895) 244,4 (1914) 114; он же, Опр. раст. Моск. губ. (1927) 172; Федченко в Маевск., Фл., изд. 7 (1940) 464; Бобров во Фл. СССР, 11 (1945) 212; он же в Тр. БИН АН СССР, сер. 1, 6 (1947) 228; Борисова в Маевск., Фл. изд. 8 (1954) 149, изд 9 (1964) 151; Тихомиров в Опр. раст. Моск, обл, (1966) 212; Coombe in Fl. Eur. 2 (1968) 162.— Клевер розовый, к. гибридный, к. шведский.
Сем. Бобовые — Leguminosae (Papilionaceae).
Морфологическое описание
Многолетнее травянистое моно- или дикарпичное растение с снмподпальным типом возобновления побегов (Серебряков, 1952) и неглубокой корневой системой (Дмитриев, 1948; Понятовская, 1956). Главный корень стержневой, длиной 30—40 см, хорошо развиты боковые корни (Дмитриев, 1948; Ларин и др., 1951; Лапин, I960; Крылова, 1964). Рост главного, корня при повышении уровня грунтовых вод у клевера розового ослабляется, но отмечается интенсивное образование придаточных корней (Голубев, 1960). i В культуре система главного корня развита значительно лучше, но придаточные корни формируются в меньшем числе (Покровская, 1960б). У тетраплоидов корневая система развита лучше, чем у. диплоидов (Goral, Г964).
Надземные побеги прямостоячие или слегка изогнутые, высотой от 30 до 100 см, часто полые внутри, имеют склонность к полеганию (Gilbert, 1956; Rauschert, 1965). Число побегов в кусте колеблется от 1—2 до 6—8, число междоузлий на стебле 7—11, чаще 8—9 (Кры-} лова, 1964). Культурные формы имеют большее число генеративных | побегов (Покровская, 1960 б). Листья с прилистниками, в верхней ;
1 Разделы «Экология» и «Фитоценология» написал Т. А. Работнов, раздел «Консортивные связи» — Г. Д. Успенская.
102
части заостренными. Листья гладкие, тройчатые, с эллиптическими листочками. Края листочков вырезанно-зубчатые. Прикорневые листья собраны в розетку, приподнятые, длина черешков у прикорневых листьев от 1,5 до 6 см, а у стеблевых — от 0,5 до 12,5 см.
Соцветие — шаровидная головка до 20—25 мм в диаметре, расположено на длинном цветоносе. Среднее число соцветий на одном побеге 4—5. При внесении удобрений количество соцветий возрастает (Крылова, 1964). Венчик 8—10 мм длиной, в начале цветения белый, позднее бледно-розовый или розово-красный. Длина трубки венчика 2,6—2,9 мм (Абрамова, 1960; Крылова, 1965).
Плод — нерастрескивающийся боб. Односемянных бобов у клевера розового развивается в среднем 20,9%, двусемянных — 43,4, трехсемянных — 28,5, четырехсемянных — 7,0, пятисемянных — 0,2% (Абрамова, 1960). Семена зеленовато-желтого или зеленовато-коричневого цвета, гладкие, блестящие, округло-сердцевидной формы, длиной около 1 мм. Зародышевый корешок занимает около половины длины семени. Вес 1000 семян в условиях Московской обл. от 0,43 до 0,64 г.
Диплоидное число хромосом 16 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
В СССР дикорастущий клевер розовый является обычным растением в лесной и лесостепной зонах европейской части. За пределами СССР он распространен на естественных лугах в области умеренного климата в Европе и Азии, встречается в Алжире. Возделывается как кормовое растение в США и Канаде (Ларин и др., 1951; Hughes, 1953; Gilbert, 1956; Покровская, 1960 6).
Онтогенез и сезонный ритм развития
На основании анализа состояния главного побега и главного корня, развития боковых генеративных побегов и придаточных корней разных порядков у клевера розового выделяют те же возрастные этапы формирования куста (Покровская, 1960а, б), что и у к. лугового (см. выше).
Наблюдения, проведенные за фиксированными особями, дали основание для выделения у к. розового ювенильных, имматурных, генеративных и сенильных особей (Крылова, 1964). Благодаря бак-териотрофии (получению связанного азота от клубеньковых бактерий) развитие молодых растений в благоприятных условиях идет быстро.
Всходы имеют высоту от 1,5 до 5 см. Наряду с простым округлосердцевидным листом у них есть от 1 до 3 тройчатых листьев, стебель очень тонкий. В условиях заливных лугов Московской обл. появлению всходов благоприятствует отсутствие мощного наилка, летних заморозков, регулярных продолжительных разливов, умерен
103
ное увлажнение. По наблюдениям на лугах поймы Оки, максимум появления всходов приходится на весну. Осенний максимум выражен довольно слабо и только при поздних сроках скашивания основного укоса. Появление и отмирание всходов летом происходит в значительных размерах. Распределение всходов единичное и групповое (от 2—3 до 13—22).
Ювенильные особи — растения, имеющие маленькие розетки листьев и хорошо выраженный главный корень. Высота их на окских лугах от 5 до 15 см. Обычно в этот период происходит образование третьего — пятого листов. Кущение отсутствует, но в пазухах листь-ещотчетливо просматриваются почки возобновления. Переход от состояния всходов к ювенильной фазе у клеверов выражается прежде всего в утолщении узлов кущения и гипокотиля. По Т. М. Покровской (1906 б), ювенильные особи относятся к растениям, находящимся на первом возрастно-морфологическом этапе развития. Ювенильными являются главным образом особи второго года жизни. Продолжительность пребывания в ювенильном состоянии определяется сроками перехода особей в фазу формирования центральной розетки листьев с удлиненными нижними междоузлиями и обычно не превышает одного вегетационного периода.
Во влажные годы даже при двуукосном использовании или позднем скашивании у большинства особей второго года жизни к моменту основного укоса намечается переход к формированию центральной розетки с удлиненными нижними междоузлиями, что характерно для имматурных особей. При внесении фосфорно-калийных удобрений продолжительность ювенильной фазы заметно сокращается. В отдельных случаях (при раннем скашивании или в негустом травостое) особи второго года жизни к моменту основного укоса формируют генеративный побег.
Отмирание ювенильных особей, так же как и всходов, происходит главным образом летом, после основного укоса, усиливаясь в засушливые годы. Большое число их гибнет в позднеосеннее и зимнее время. В условиях достаточного увлажнения почвы, например на лугах центральной поймы, значительных изменений численности ювенильных особей в зависимости от погодных условий сезона не наблюдается. Существенное влияние на изменение их численности оказывают сроки скашивания. Наибольшее число всходов отмечено при весеннем подкашивании травостоя и внесении фосфорно-калий-ного удобрения. Этот режим использования луга способствует наименьшему выпадению ювенильных растений клевера.
Имматурная особь представляет собой розетку листьев с удлиненными нижними междоузлиями, высотой от 10 до 17 см, у основания которой находятся укороченные боковые вегетативные побеги. При двуукосном использовании и позднем скашивании среди имматурных встречаются особи как третьего, так и второго года жизни. При весеннем подкашивании и ранних сроках скашивания к имма-турным относится большинство особей второго года жизни. Внесение удобрений способствует более быстрому переходу имматурных
104
особей во взрослое состояние. По Т. М. Покровской (1960 6), II этап в жизни к. розового, характеризующий предгенеративный период развития, на лугах наблюдается не всегда, так как происходит быст рый переход оси первого периода к генерации.
К- розовый обычно зацветает на третий год жизни. При позднем скашивании трехлетние растения цветут только во влажные годы или при внесении фосфорно-калийных удобрений. Продолжительность генеративного периода 2 (реже 3) года. В благоприятных экологических условиях возможно завершение генеративного периода в течение одного года.
У отдельных особей клевера при внесении удобрений и.раннем скашивании максимум репродуктивной мощности наступает на третий год жизни, когда образуется до 7 генеративных побегов. Во влажные прохладные годы максимум репродуктивной мощности выражен сильно и у особей четвертого года жизни, у которых насчитывается 4—6 генеративных побегов, у отдельных особей — до 13— 15 (Крылова, 1964). В сухие годы у особей четвертого года жизни обычно бывает по 2 генеративных побега.
На заливных лугах Московской обл. к. розовый, как правило, дикарпичное растение. При систематическом двуукосном использовании, когда травостой в основном укосе скашивается в фазу цветения клевера, среди генеративных особей встречается примерно равное количество особей третьего и четвертого года жизни. При весеннем подкашивании и раннем скашивании основного укоса, особенно на удобренных участках, большая часть генеративных особей представлена особями третьего года жизни.
Цветение к. розового на заливных лугах Московской обл. в пойме Оки наступает в сухие годы 10—15 июня, в умеренно сухие — около 25 июня и в годы с влажным прохладным лётом — в начале июля. Период цветения — созревания семян растянут. При двуукосном использовании к моменту основного укоса на 100 генеративных побегов насчитывается 130 бурых соцветий, 192 цветущих и 166 бутонизирующих (среднее за 4 года) (Крылова, 1964).
В сухие годы к. розовый отличается слабой отавностью. Побегообразование в отаве усиливается во влажные годы, особенно при внесении удобрений.
Те особи клевера, у которых генеративная фаза не сопровождалась быстрым разрушением главного корня, вступают в период затухания репродуктивной мощности. Большинство особей проходит период затухания репродуктивной мощности в возрасте пяти лет, образуя в основном укосе один боковой генеративный побег. Пятилет-ние особи у к. розового встречаются чаще, чем у к. лугового. При раннем скашивании основного укоса период затухания репродуктивной мощности может наступить у четырехлетних растений во втором укосе. Образования генеративных побегов во втором укосе у особей пятого года жизни не наблюдалось (Крылова, 1964).
Образование большого числа побегов в период максимума ре-продуктивности мощности приводит к быстрому отмиранию особи в
105
течение последующего осенне-зимнего периода независимо от ее возраста. Внесение удобрений ускоряет завершение жизненного цикла.
О периоде постгенеративной вегетации в жизненном цикле к. розового данные противоречивы. Т. М. Покровская (1959) сенильный период у этого вида не выделяет. Наблюдения, проведенные за фиксированными особями на лугу в пойме Оки, показали, что сенильный период в жизненном цикле этого растения выражен отчетливо (Крылова, 1965). Переход в фазу постгенеративной вегетации происходит на пятый год жизни в середине вегетационного периода (после скашивания основного укоса) или в конце его.
У к. розового нами выявлена кратковременная и продолжительная поСтгенеративная вегетация. Кратковременная наступает после прохождения генеративной фазы в основном укосе и заканчивается в конце данного вегетационного сезона. У многих особей резкого ослабления жизненного состояния в фазе кратковременной постгенеративной вегетации не обнаружено. Продолжительную постгенеративную вегетацию можно выделить для особей, вступивших в эту фазу после перезимовки, а также для особей, находящихся в постгенеративной вегетации более одного вегетационного периода. В этой фазе особи к. розового встречаются гораздо реже, чем в фазе кратковременной постгенеративной вегетации. Наблюдения на заливных лугах в пойме Оки показали, что продолжительность периода постгенеративной вегетации у этого растения обычно 3—5 месяцев. Фазу постгенеративной вегетации проходят, главным образом, те особи, которые в генеративном состоянии характеризовались не самым высоким жизненным состоянием.
Продолжительность жизни к. розового в условиях Московской обл. 4—5, очень редко 6 лет. Как показали наши опыты, скашивание в различные сроки не оказывает влияния на продуктивное долголетие клеверов, но изменяет сроки наступления отдельных возрастных фаз.
Способы размножения и распространения
- Клевер розовый размножается почти исключительно семенами. Вегетативное размножение не имеет существенного значения.
Семенная продуктивность у этого растения значительно колеблется по годам в зависимости от метеорологических условий и режима скашивания. По нашим пятилетним наблюдениям в пойме Оки, при скашивании основного укоса в фазу массового цветения клевера одна особь образовала от 0 до 44 семян, при весеннем подкашивании травостоя в сочетании с основным укосом в фазу созревания семян клевера — от 24 до 230 семян, при скашивании основного укоса в фазу бутонизации и обсеменении клевера во втором укосе — от 0 до 224 семян; при скашивании основного укоса в фазу созревания семян — от 93 до 240 семян. Внесение фосфорпо-калий-. ных удобрений способствовало росту семенной продуктивности (она
106
соответственно составила: 0—90, 66—396, 1—360, 84—414), по не устраняло ее колебаний по годам.
Высокая семенная продуктивность к. розового во втором укосе обусловлена благоприятными условиями для образования семян в связи с ослаблением конкуренции со стороны злаков и разнотравья, хорошим световым режимом и формированием во втором укосе более мелких цветков. Средняя длина венчика без флага во втором укосе 1,99 мм, максимальная — 2,27 мм, минимальная —1,2 мм (в первом укосе — соответственно 2,88 7,0, 1,0 мм) (Крылова, 1964, 1965). Извёстно, что во втором укосе более мелкие цветки клевера лучше опыляются насекомыми, а семена в меньшей степени повреждаются вредителями (Морозов, 1960; Мирвис, 1959).
В отдельные годы к. розовый частично обсеменяется даже при двуукосном использовании: урожай составляет около 200—250 семян на 1 м2 (на некоторых участках до 1500 семян на 1 м2). При позднем скашивании травостоя и небольшом участии клевера в сложении травостоя урожай его семян составляет 450—500 семян на 1 м2, а при значительном участии — 4000—5000 семян. При систематическом весеннем подкашивании травостоя или при обсеменении в отаве после раннего скашивания первого укоса минимальный урожай составляет 1500—3000, максимальный-—7500—9000 семян.
Среди жизнеспособных семян к. розового обычно преобладают твердые. При обсеменении в отаве доля твердых семян подвержена значительным колебаниям по годам — от 23 до 70%. Обилие твердых семян обеспечивает ежегодное равномерное семенное размножение клевера, даже если урожай семян в предыдущем году был незначительным, что отмечено и для к. лугового (Шайн, 1947, 1949).
По наблюдениям на окских лугах, число осыпавшихся семян к. розового в сухие годы колеблется от 70—90 на 1 м2 при двуукосном использовании до 700 при позднем скашивании основного укоса. При обсеменении клевера в основном укосе после весеннего подкашивания число осыпавшихся семян может достигать 1200 и даже 2300 на 1 м2. Во влажные годы число осыпавшихся семян колеблется от 100—140 при позднем скашивании основного укоса до 460 при обсеменении клевера во втором укосе после раннего скашивания основного укоса.
Большое .число всходов клевера появляется при весеннем подкашивании с внесением фосфорно-калийных удобрений, при позднем скашивании основного укоса и при обсеменении.во втором укосе. В отдельные годы число всходов при обсеменении в основном укосе после весеннего подкашивания (фон — РбоКбо) было в 6 раз, и при обсеменении клевера во втором укосещочти в 12 раз больше, чем при обычном двуукосном скашивании (Крылова, 1964, 1965), Влияние весеннего подкашивания на семенное размножение клевера может быть настолько значительным, что после трех лет ежегодного подкашивания число всходов может достигать 800—1000 па I м2.
Запас семян в почве на природных лугах невелик. В Московской обл. на лугах в пойме Оки в среднем отмечено 10—60 семян па
107
1 м2 (Работнов, 1956 а). В отдельные годы запас семян существенно возрастает. При различных режимах использования луга формируется неодинаковый запас в почве семян к. розового: при обычном двуукосном использовании луга, когда первый укос скашивается в фазу массового цветения,— 60—160 семян на 1 м2; при обсеменении в основном укосе после весеннего подкашивания — до 4950 семян; при обсеменении во втором укосе — 70—160; при скашивании основного укоса в фазу обсеменения клевера — .1650—2160. Наибольшее накопление семян в почве происходит при обсеменении клевера в основном укосе после весеннего подкашивания (Крылова, 1964, 1965).
Экология
По шкалам Л. Г. Раменского оптимальными условиями для клевера розового на лугах являются: увлажнение в лесной зоне свежелуговое— влажнолуговое (62—67 ступени), а в степной зоне — сыро-свежелуговое (83—89 ступени); умеренная переменность увлажнения (9—11 ступени); довольно богатые почвы (11—14 ступени); отсутствие наилка или очень слабое его отложение. Хорошо растет на лугах с уровнем грунтовых вод 40—90 см и даже выше (Раменская, 1951). На пойменных лугах устойчив к затоплению полыми водами продолжительностью 1—2 недели (не дольше -1 месяца), имеются сведения, что к. розовый выдерживал затопление до 46 дней (Янсон, 1968).
Удовлетворительно переносит как зимние морозы, так и весенне-осенние заморозки. Оптимальная реакция почвы pH 6—7, однако может произрастать и на кислых почвах (pH 4—5). Предпочитает богатые перегноем среднеувлажненные супесчаные почвы или суглинки, хотя при достаточной влажности может расти на других почвах (Раменская, 1951; Янсон, 1968). Выносит лишь слабо интенсивный выпас.
Биологическая продуктивность
Продуктивность клевера розового на лугах резко колеблется по годам. Это определяется особенностями его жизненного цикла (массовым отмиранием пышно развитых особей в связи с завершением ими жизненного цикла), а также различиями в обсеменении и в приживаемости всходов в отдельные годы. Так, на краткопойменном почти ежегодно заливаемом лугу в пойме Оки в условиях, благоприятных для произрастания к. розового, по наблюдениям за 10 лет, его продуктивность в «клеверные» годы достигала 344—520 г/10м2 (т. е. 3—5ц/га),ав «межклеверные» годы падала до 0. Внесение фосфорно-калийных удобрений в «клеверные» годы повышало продуктивность клевера в 3 раза (10,7—15,7 ц/га) (Работнов, 1973).
Эффективным приемом повышения устойчивости и урожайности к. розового на кратко- и среднезаливаемых лугах с устойчивым
108
режимом увлажнения является весеннее подкашивание с внесением фосфорно-калийных удобрений, которое способствует увеличению урожая сена клевера примерно. вдвое по сравнению с внесен нем удобрений при двуукосном использовании и не снижает общей урожайности луга (Крылова, 1963, 1964, 1965).
Фитоценология
В Московской обл. клевер розовый широко распространен на заливных лугах с хорошим режимом увлажнения, на луговых дерновых почвах, при отсутствии перекрытия сколько-нибудь значительным наилком, в различных фитоценозах. На суходольных лугах участие его в травостое обычно невелико.
К. розовый, как и к. луговой,— типичный ценоциклофлюктуент, способный доминировать в благоприятных для него условиях лишь периодически, в отдельные («клеверные») годы, когда большая часть его особей переходит в генеративное состояние, а затем на следующий год или через год отмирает. В связи с этим период доминирования длится 1—2 года. После массового отмирания особей участие клевера в луговых фитоценозах резко снижается, формируется новая популяция из семян, осыпавшихся в «клеверные» годы или из имевшихся в почве, пока вновь не возникнут условия, благоприятные для перехода особей в пышно развитое генеративное состояние.
В годы доминирования к. розовый обычно входит как доминант в состав полидоминантных фитоценозов. На пойменных лугах он часто доминирует совместно с более требовательными к воде злаками: лисохвостом луговым, полевицей белой, щучкой.
Консортивные связи
Для Т. hybridum характерны связи с насекомыми-опылителями, симбиотические и паразитические консортивные отношения, подобные тем, что описаны для Т. pratense. В. Ф. Купревич (1954) отмечал на клевере розовом 22 вида грибов-паразитов. Из них наиболее распространены Peronospora trifolii-hybridi Gauman., Erysiphe communis f. trifolii, Urotnyces trifolii-repentis (Cast.) Liro, Polythrinciuin trifolii и др. (Успенская 1974). Последние три вида грибов иногда развиваются на одном и том же растении. По М. С. Дунину (1946), их можно отнести к разряду возбудителей сопряженных болезней, когда виновник одного заболевания способен развиваться на тех же участках растения, где уже паразитирует другой. В разбираемом случае пустулы .ржавчины и спороношения возбудителя черной пятнистости всегда развиваются на нижней поверхности листьев, пораженных мучнистой росой. Эти отрицательные связи приводят к преждевременному засыханию и опадению листьев.
109
Хозяйственное значение
Клевер розовый относится к числу ценных сенокосных растений (Ларин и др., 1951; Лапин, 1960; Янсон, 1970; Власова, 1971; Илющенко, 1971). Этот вид — один из основных компонентов травосмесей на кормовых угодьях с повышенной влажностью почв, в частности на освоенных низинных болотах. Оптимальный срок уборки на сено — фаза начала цветения (Раменская, 1951). По содержанию протеина превосходит к. луговой, хотя по поедаемости уступает последнему.
Для улучшения долголетних сеяных или природных лугов рекомендуется подсев к. розового (6—8 кг/га семян) в дернину. Такой подсев дает положительные результаты и на пастбищах, особенно сеяных (Лапин, 1960). По данным Г. К- Абрамовой (1965), долговечность к. розового определяется условиями его выращивания. При интенсивной культуре он удерживается в травостое до 2—3 лет, тогда как в естественных ценозах он произрастает 5 и более лет.
К. розовый — весьма ценный медонос (Глухов, 1955).
О. В. Смирнова и Н. А. Торопова1
ПРОЛЕСНИК МНОГОЛЕТНИЙ
Номенклатура и систематическое положение
Mercurialis perennis L., Sp. pl. (1753) 1035; Кауфм., Моск, фл., изд. 1 (1866) 106, изд. 2 (1889) 104; Цингер, Сб. свед. (1885) 390; Шмальг., Фл. 2 (1897) 418; Сырейщ., Илл. фл. 2 (1907) 337; он же, Опр. раст. Моск. губ. (1927) 180; Федченко в Маевск., Фл., изд. 7 (1940) 496; Пояркова во Фл. СССР, 14 (1949) 296; Проханов в Маевск., Фл., изд. 8 (1954) 297, изд. 9 (1964) 290; Ворошилов в Опр. раст. Моск. обл. (1966) 266; Tutin in Fl. Eur. 2 (1968) 212.— Пролесник многолетний (перелесник, собачья капуста).
Род Mercurialis L., насчитывающий около 10 видов, относится к сем. Euphorbiaceae, подсем. Crotonoideae Pax, трибе Crotoneae Pax, подтрибе Mercurialinae Pax.
В Московской обл. встречается только М. perennis L., описанный из Европы. Тип в Лондоне.
Морфологическое описание
Многолетнее травянистое поликарпическое двудомное растение. Многолетняя система побегов взрослого растения состоит из участков длинных гипогеогенных корневищ, несущих чешуевидные листья, и сближенных базальных частей ортотропных побегов, образующих в основании парциального куста своеобразное симпо-диальное корневище (Серебряков, Серебрякова, 1965).
Корневая система взрослых особей состоит из придаточных корней, образующихся на симподиальных корневищах парциальных кустов и на некоторых узлах гипогеогенных корневищ. Придаточные корни довольно толстые (диаметр 2 мм), извилистые, густо покрыты длинными корневыми волосками, сохраняющимися и па опробковевших участках корня. При проникновении корпя в глубь
1 При составлении раздела «Онтогенез» использованы данные Г. П. Рысп-ной (начальные этапы онтогенеза); раздела «Способы'размножения и распространения» — Г. П. Рысиной и отчасти К. В. Зворыкиной.
111
почвы между старыми корневыми’ волосками могут появляться новые, старые в это время засыхают и опадают (Mukerji, 1936). Растущие придаточные корни белые, опробковевшие — светло-бурого цвета. Длина их в среднем 12—15 см (от 6 до 53 см). В пределах парциального куста образуется от 10 до 25 придаточных корней. На них развиваются боковые корни первого — третьего порядков, длиной 1—7 см и диаметром 0,5—0,9 мм. На 1 см придаточного корня приходится 0,7—1 боковых корней первого порядка.
В европейской части СССР максимальная глубина проникновения корней пролесника (25—30 см) отмечена на темно-серых лесных почвах и выщелоченных черноземах на известняке. Рабочая глубина корневой системы составляет на дерново-подзолистых почвах под хвойно-широколиственными лесами 5—10 см, на серых и темно-серых лесных почвах под широколиственными лесами — 10— 15 см и определяется главным образом глубиной гумусового горизонта.
Плагиотропные гипогеогенные корневища пролесника расположены в приповерхностном слое почвы, обычно на глубине 3—5 см. Длина годичного прироста их 15—30 см, число узлов 2—4, диаметр 2—4 мм. Корневища в течение всей жизни покрыты многочисленными простыми волосками, морфологически напоминающими корневые волоски. Молодые корневища белые, хрупкие, сочные, многолетние— опробковевшие, бурые. Чешуевидные листья на гипогеоген-ном корневище супротивные, широкотреугольной формы, 3—4 мм длины. Большая часть почек, развивающихся в их пазухах, переходит в разряд спящих (Кернер, 1902; Юрцев, 1951; Шик, 1953; Голубев, 1965; Любарский, 1967). Нормально развитые почки возобновления развиваются в пазухах последней пары чешуевидных листьев гипогеогенного корневища, в месте перехода плагиотропной части побега в ортотропную.
Надземные побеги зеленые, прямостоячие, 15—45 см высоты, не-, ветвистые; побеги женских растений обычно выше, чем мужских. Стебли голые или почти голые (в верхней части слабо опушенные), ребристые. Общее число узлов на надземной части побега 6—10. Наиболее длинными обычно являются третье и четвертое междоузлия, верхние междоузлия короткие.
Листья супротивные, на первом — третьем узлах наземного побега развиваются чешуевидные листья, на остальных — 4—6 пар срединных ассимилирующих. Чешуевидные листья широкотреугольные, 2—3 мм длины. Срединные листья черешковые, пластинки их овально-ланцетные или ланцетно-эллиптические, заостренные или острые, с клиновидным (верхние — иногда с более или менее округлым) основанием, пильчато-городчатые, темно-зеленые (иногда с голубоватым отливом), негусто шершаво-волосистые. Длина их 3—10 см, ширина 1,5—4,5 см. Черешки голые или волосистые, желобчатые, с завернутыми вверх краями, длиной 4—4,5 см. Часть срединных ассимилирующих листьев занимает промежуточное положение между чешуевидными и вполне развитыми срединными листьями. Верхние
112
срединные листья сближенные, более узкие, мелкие, почти сидячие, по форме и размерам приближаются к катафиллам.
Прилистники светло-зеленые, перепончатые, удлиненно-овально-ланцетные, 1—2 мм длины, сохраняющиеся до конца вегетации. У мужских особей листья длиннее и уже, у женских — более округлые. На концах зубчиков пластинки листа имеются гидатоды.
Цветки пролесника однополые. Имеются сообщения о находках обоеполых цветков, а также мужских цветков на женских экземплярах (Saunders, 1883; Williams, 1925, 1926), особенность эта сохраняется и при-перенесении обоеполых экземпляров в культуру.
Тычиночные цветки сидячие или на коротких ножках (1—2 мм), собраны в малоцветковые (4—7) клубочки (ложные колосья), расположенные в пазухах перепончатых кроющих листьев. Клубочки образуют прерывистый колос, обычно по длине превышающий листья. В начале цветения колосья вверхстоячие, после выбрасывания пыльцы поникают и отмирают. Обычно на одном побеге развивается 4—8 мужских соцветий в пазухах верхних пар листьев. Околоцветник трехраздельный, доли его яйцевидно-овальные, заостренные, голые, вогнутые, гладкие, 2 мм длины, тычинок от 8 до 20, обычно соединенных в 2 или 3 пучка. Пыльники двугнездные, тычиночные нити тонкие, длинные. Пыльцевые зерна серно-желтые, слабобугорчатые, эллипсовидные, 39x20 мк. После выбрасывания пыльцы доли пыльника синеют.
Пестичные цветки собраны в короткие (в 3—4 раза короче листьев) кистевидные соцветия в пазухах прицветников, длиной 2 мм. Ось соцветия тонкая, 1,5—3 мм (до 5,5 мм) длины, при плодах несколько удлиняется. Женские соцветия обычно 3—4-цветковые, иногда развивается лишь один терминальный цветок. Обычно на одном побеге имеется 3—4 (1—7) женских соцветия, развивающихся в в пазухах четвертой — седьмой-пары листьев. Околоцветник трехраздельный, доли его несколько сросшиеся, овальные, заостренные, голые, 2 мм длины. Завязь дву-,. редко трехгнездная, щетинистоволосистая и негусто покрытая сосочками. Столбика 2, нитевидные, у основания сросшиеся, с внутренней стороны усаженные сосочками. Со стороны швов завязи расположены 2 нитевидных расширенных к основанию нектарника, которые представляют собой редуцированные тычинки. Семяпочка анатропная с брюшным швом. Пролесипку свойственна порогамия (Сырейщиков, 1907; Веттштейн, 1912; Williams, 1925, 1926; Durand, 1957; Каден, 1964, 1965).
Плод — двучленная синкарпная дробная коробочка, сплюснуто-шаровидная, 6—8 мм ширины и 4—5 мм длины, морщинистая, шершаво-пушистая. Околоплодник тонкий, покрыт бородавками и щетинками. Семена округлые или овальные, на верхушке острые, 3 мм в диаметре. Вес 1000 семян 8 г. Семенная кожура толстая, жесткая, семя имеет беловатый присемянник, развивающийся около рубчика. Зародыш мелкий, эндосперм занимает большую часть семени.
113
Число хромосом (2п), по данным разных авторов, составляет 42—64—84. Отмечены разные числа хромосом у мужских (66) и женских (64) экземпляров (Mukerji, 1936; Хромосомные числа цветковых растений, 1969).
Географическое распространение
Пролесник многолетний входит в западную (европейскую) группу неморальных видов с центром распространения в средней и восточной частях Европы, отчасти в Малой Азии и на Кавказе. Ареал вида охватывает большую часть Европы, Северную Африку, Малую Азию и Иран. Северная граница ареала проходит по югу Норвегии, середине Швеции, северу ФРГ и ГДР, западная — по Шотландии и Ирландии, южная — по Северной Африке и Малой Азии, восточная — по Ирану и предгорьям Урала (Hegi, 1931; Mukerji, 1936; Lippmaa, 1938; Boatman, 1956). Северная граница ареала близка к полярному кругу, так как самое северное местообитание отмечено на западном побережье Норвегии на 65° 26' с. ш.
В СССР пролесник встречается почти по всей территории европейской части, за исключением самых северных районов, и на Кавказе (Колхидская и Гирканская провинции). Самые восточные местообитания этого вида отмечены в Предуралье: в окрестностях пос. Мензелинска, близ г. Бирска и с. Благовещенского в Башкирской АССР (Горчаковский, 1968). В Московской обл. этот вид встречается не часто и приурочен к широколиственным и хвойно-широколиственным лесам. По данным Ю. Д. Клеопова (1941), пролесник многолетний является потомком тургайской флоры, связанным в своем происхождении с Восточной Азией.
Онтогенез
Семена пролесника созревают в июне. Жизнеспособные зрелые семена содержат нормально развитые зародыши до 1 мм длины и массивный эндосперм. В природных местообитаниях семена, посеянные в июне, прорастали в сентябре — октябре. Однако прорастают они не одновременно, большая часть семян зимует, а прорастает весной следующего года. Итак, период покоя семян, прорастающих весной, составляет около 9 месяцев. Период прорастания семян растянут по крайней мере на 15 месяцев (Boatman, 1956).
До начала прорастания заметных изменений в зародыше не происходит, он имеет тот же вид и размеры, что и в свежесобранных семенах. При прорастании эндосперм разбухает и кожура, разрывается. Через несколько дней появляется корешок. Плоские семядоли приобретают зеленовато-желтую окраску. Положение семядолей относительно поверхности почвы у пролесника может быть различно, наиболее часто встречается подземный тип прорастания (Рысина, 1968).
Проросток пролесника имеет две овальные семядоли, утолщенный гипокотиль, длиной до 1,5 см, и главный корень 5—6 см. Об-
114
втягиваются в почву при-
Рис. 12. Первичный куст про-лесника многолетнего и пмми-турном возрастном состоянии
ласть .корневой шейки и главный корень покрыты корневыми волосками, буреющими к концу первого вегетационного периода. Корневые волоски сохраняются несколько лет и по ним хорошо .чпмсгпп граница между гипокотилем и корнем. Главный корень слабо ветвится, первые два придаточных корня появляются на гипокотиле между основаниями семядолей, они обычно достигают большей длины, чем главный, и ветвятся. К моменту образования второй пары срединных ассимилирующих листьев в пазухах семядолей закладываются почки возобновления. Осенью 'семядоли обычно опадают, двулетние растения с хорошо сохранившимися семядолями являются редкостью. У однолетних растений к зиме побег первого порядка отмирает выше семядольного узла, а из пазушных почек начинают развиваться побеги второго порядка. С этого момента начинается симподиальное возобновление пролесника.
После полного отмирания семядолей растение переходит в ювенильное состояние. Группу ювенильных составляют однолетние и двух-, трехлетние растения. Побеги второго и следующих порядков кроме срединных имеют 1—2 пары чешуевидных листьев, в в пазухах нижней пары закладываются почки возобновления. Сближенные основания ортотропных побего! даточными корнями и формируют своеобразное симподиальное корневище, по числу порядков побегов которого можно определить абсолютный возраст растения. В ювенильном состоянии у пролесника увеличивается число боковых и гипокотильных корней, начинают появляться придаточные корни стеблевого происхождения. Гипокотиль и главный корень сохраняются долго, но впоследствии их трудно обнаружить среди множества более крупных придаточных корней.
В имматурном состоянии в рост одновременно трогается несколько почек, образуя побеги, значительно отличающиеся от побегов ювенильных растений по размерам. Соответственно резко увеличивается и запас почек возобновления. Формируется первичный куст (рис. 12). Имматурное состояние длится два-три года.
Следующим этапом развития особи является молодое вегетативное
состояние, когда кроме ортотропных побегов, составляющих первичный материнский куст, появляются подземные нлагиотршшые побеги (отбеги) — гипогеогенные корневища. У молодых особей длина их невелика — около 10 см. Из верхушечной ночки этих кор
115
невищ развиваются надземные ортотропные (парциальные) побеги, такие же, как и ортотропные побеги в составе первичного куста. Парциальные побеги дают начало парциальным кустам, которые развиваются подобно первичному кусту. Корневая система первичного куста существует 4—8 лет. Первичный куст в совокупности с парциальными кустами и побегами образует куртину (рис. 13). Мо-
Рис. 13. Куртина пролесника многолетнего лодое вегетативное состояние у пролесника очень непродолжительно, 1—2 года, иногда пролесник зацветает одновременно с образованием первого парциального побега, практически минуя молодое вегетативное состояние.
Пролесник зацветает на 5—7 год жизни. Генеративный период самый продолжительный в его жизненном цикле. В это время начинается вегетативное размножение путем отделения от материнской особи систем парциальных кустов вследствие разрушения участков корневищ. К генеративным растениям относятся куртины и системы парциальных кустов вегетативного происхождения.
116
Основным структурным элементом взрослой особи пролссиика является парциальный куст. Развитие его начинается с появлением вегетативного или генеративного парциального побега из терминальной почки плагиотропного корневища. Побеги следующих порядков возникают в пазухах нижних чешуевидных листьев ортотропного побега, в месте перехода плагиотропной части побега в ортотропную -(Шик, 1953). Придаточные корни развиваются в непосредственной близости к почкам возобновления. Парциальные кусты 2—5-летнего возраста имеют вегетативные и генеративные побеги и образуют плагиотропные корневища — отбеги, являясь центрами формирования новых парциальных кустов. У 2—б-летних кустов наблюдается образование Силлептических побегов. Парциальные кусты 6—7-летнего возраста развивают только ортотропные побеги, отбеги же не образуются. В дальнейшем, на 8—'9-й год, парциальный куст начинает отмирать: прекращается образование ортотропных ассимилирующих побегов и придаточных корней, старые корпи постепенно разрушаются. В некоторых случаях у отмирающих парциальных кустов из спящих пазушных почек развиваются небольшие ортотропные побеги ювенильного типа.
Отмирающий парциальный куст представляет собой систему сближенных оснований побегов прошлых лет (подиумов) и служит для закрепления особи и связи с соседними более молодыми парциальными кустами. Следующим этапом является разрушение подиумов. Онтогенез парциального куста длится в среднем 10 лет. В возрасте 2—7 лет парциальные кусты способны переходить в состояние вторичного покоя. Наземные побеги не развиваются, но в кусте сохраняются жизнеспособные почки, обеспечивающие развитие куста в последующиетоды.
Подразделение генеративного периода пролесника основано на соотношении активно растущих плагиотропных корневищ-отбегов и коммуникационных корневищ, молодых растущих и отмирающих парциальных кустов.
К молодым генеративным растениям относятся куртины и системы парциальных кустов, у которых преобладают активно растущие части. У молодых генеративных особей, в частности у молодых куртин, часто наблюдается недоразвитие соцветий.
Средневозрастные генеративные растения представлены мощно развитыми системами парциальных кустов. Признаки старения у этих особей проявляются в уменьшении числа растущих корневищ и понижении способности к захвату новой площади. В дальнейшем е увеличением возраста у пролесника не только уменьшается число растущих корневищ, но, и сокращается число ассимилирующих ортотропных побегов — растение переходит в старое генератпппое состояние.
В это время все интенсивнее происходит разрушение систем парциальных кустов. Старые вегетативные особи обычно состоят in двух или нескольких парциальных кустов с небольшим числом ор то тропных побегов. У последних резко уменьшаются размеры и число
117
срединных листьев. Растущих корневищ эти особи, как правило, не образуют, запас почек возобновления у них невелик, образования придаточных корней не происходит.
К сенильным относятся особи пролесника, представленные одним-двумя парциальными кустами, у которых развивается один ортотропный побег ювенильного типа, корневая система почти полностью разрушена. Это последний этап в жизненном цикле растения (Смирнова, Торопова, 1973).
Сезонный ритм развития
Заложение и формирование почек возобновления, которые дают начало как ортотропным побегам, так и побегам с плагиотропной частью, начинается еще в период роста и формирования материнского побега (июнь — июль). Внутрипочечная фаза развития длится около года. Дальнейшая дифференциация и рост побегов продолжаются после окончания цветения и плодоношения материнских побегов в июне следующего года. В течение июля формируется побег, достигающий к концу июля 1—5 см длины. В составе побега имеется 6—7 узлов, из которых 2—4 несут чешуевидные листья, а верхние— зачатки зеленых листьев. Удлинение побегов происходит за счет вытягивания нижних междоузлий, в одних случаях незначительного (1—1,5 см)—это будущие ортотропные побеги, в других-— очень большого (20—30 см)—это побеги с плагиотропной частью. Верхние узлы остаются сближенными и образуют подобие почки, хотя1 и незакрытой сверху чешуевидными низовыми листьями. Почка загнута назад, и побег раздвигает частицы почвы оказавшимся впереди участком стебля.
Почки плагиотропных и ортотропных побегов закладывают соцветия одновременно. Хорошо заметные соцветия впервые обнаруживаются в конце июля. В начале сентября большинство ортотропных побегов поднимается над уровнем почвы на 10—20 мм, листья зеленые, свернуты в трубочку, сближены. Цветки слабо дифференцированы, 0,8—1,3 мм. В конце сентября в мужских цветках хорошо заметен развитый археоспорий, в женских — зачаток пестика. В начале ноября в мужских цветках наблюдаются материнские клетки пыльцы, в женских — зачатки семяпочек. В середине февраля заметны вполне сформированные пыльцевые зерна. Цветки достигают 1,3—1,8 мм (Шик, 1953).
Таким образом, пролесник многолетний относится к первой группе растений, выделенной И. Г. Серебряковым (1947 а): к осени побег возобновления сформирован полностью, включая соцветие. По положению почек возобновления в течение зимы он относится к' хамефитам, хотя часть временно покоящихся почек находится под землей. В теплые зимы наблюдается рост побегов под снегом (Кожевников, 1931).
В середине апреля происходит дальнейшее развитие побегов за счет растягивания междоузлий и роста листьев. Растение зацветает прежде, чем полностью разовьются листовые пластинки и прекра-118
тится рост побега. Рыльца становятся восприимчивыми за 2 дня до вскрывания пыльников. В некоторых случаях мужские цветки начинают пылить значительно раньше, чем созревают рыльца (Mukerji, 1936). Пролесник по преимуществу анемофил, роль насекомых в опылении этого вида относительно невелика. Цветет в конце апреля—в мае; семена созревают и рассеиваются в середине июня.
Активная вегетация и рост надземных побегов происходят в светлую фазу, но вегетация в целом продолжается до конца августа— октября. После смыкания древесного полога начинаются ак-тивные ростовые процессы в подземной сфере (рост корней п корневищ). Первые придаточные корни появляются в начале, июня. Процесс образования корней длится около двух месяцев, рост ах продолжается до конца вегетационного периода. Первые боковые корни образуются в середине — конце июня.
Содержание углеводов в корневищах пролесника увеличивается от середины июля к середине октября в 1,5—2 раза, главным образом за счет высокомолекулярных углеводов, содержание же легкорастворимых сахаров уменьшается (Любарский, 1967). В конце вегетационного периода (конец августа'—сентябрь) надземные побеги отмирают до базальной части. Основание побега с почками возобновления помещается под землей. Во время летних засух надземные побеги отмирают в начале августа. Таким образом, побеги пролесника, не учитывая внутрипочечной фазы, являются моноцик-лическими (озимыми).
Продолжительность ассимиляционной деятельности — около 5 месяцев. Пролесник относится к апериодическим видам, по данным И. Г. Серебрякова (1959), обладает кратким периодом вынужденного покоя и близок к выделенному Дильсом (Diels, 1918) Leuco-jum-типу. Продолжительность периода, в течение которого у пролесника отсутствует видимый рост, составляет в средней полосе европейской части СССР около 6 месяцев.
Способы размножения и распространения
В популяциях пролесника вегетативное размножение преобладает над семенным. При созревании коробочка вскрывается, разбрасывая заключенные в ней семена на 2—4 м. Кроме саморазбра-сывания семена распространяются дождевыми потоками, сильным ветром, сонями и муравьями. Пролесник является мирмекохором (Sernander, 1906; Mukerji, 1936). Распространяя пролесник, муравьи используют в пищу придаток (воздушная элайосома), развивающийся у основания семени.
Урожайность семян пролесника сильно варьирует от года к году и от местообитания к местообитанию. На одном побеге может развиваться от 3 до 30 (42) семян, в среднем от 3,7 до 9,2 (Mukerji, 1936; Рысийа, 1968).
От 30 до 60% семян оказываются нежизнеспособными (недоразвитые или уродливые зародыши), часть семян уничтожается
II!)
птицами и другими животными (до 70—90%), повреждается грибными заболеваниями или прорастает в неблагоприятных условиях. Поэтому в сплошных зарослях пролесника проростки появляются нечасто даже в годы, благоприятные для прорастания семян, большая часть проростков гибнет вследствие конкуренции со взрослыми растениями. По данным А. В. Кожевникова (1929), в Погонно-Лосином острове из 600 исследованных побегов пролесника только у 11 % были обнаружены вполне развитые соцветия.
Вегетативное размножение наблюдается, главным образом, в генеративном периоде. Оно происходит раз в 4—8 лет, когда гипо-геогенные корневища, соединяющие самые старые парциальные кусты, полностью разрушаются (перегнивают). Обычно из одной материнской особи образуются 2—3 дочерних. При перегрызании гипогеогенных корневищ роющими животными происходит травматическое вегетативное размножение, но в связи с некоторой ядовитостью растения это отмечается редко.
' Скорость разрастания особей пролесника — 15—30 см/год (Кернер, 1902; Boatman, 1956; Смирнова, Торопова, 1974). В Ирландии при смешанном (вегетативном и семенном) распространении пятно пролесника перемещалось со скоростью 60 см/год (Boatman, 1956).
Экология
Пролесник произрастает на разных высотах: от 300—400 м над уровнем моря до 1200—4000 м в горах Центральной Европы. Он встречается на песчаных, глинистых, суглинистых почвах, на известняках, почвах вулканического происхождения, гранитах и сланцах и даже на гравии. Исключение составляют торфяники. Этот вид характеризуется как кальцефил и нитрофил (Кац, 1943; Ellenberg, 1963). Однако повышенное содержание нитратов при прочих благоприятных условиях оказывает незначительное воздействие на рост его побегов (Martin, 1968).
Увлажнение почв в местообитаниях пролесника варьирует от влажно- до сыролугового (Раменский и др., 1956). Встречается на почвах с pH от 4,0 до 8,5, предпочитает богатые почвы с pH от 5,8 до 7,2 (Olsen, 1923; Никишин, 1939; Martin, 1968). Корни проникают на самую большую глубину, достигая при этом наибольшей длины и веса, на нейтральных или слабощелочных почвах.
Пролесник — чуткий показатель условий аэрации почв. При поверхностном заболачивании его корни не проникают в почву глубже 10 см, выживают лишь поверхностные корни (Salisbury, 1920, 1925; Mukerji, 1936). В хорошо аэрируемых глинистых почвах на известняках корни сильно ветвятся и проникают на глубину до 20 см, в плохо аэрируемых — до 6 см. Наилучший рост корней отмечен на почвах с влажностью 25—35%.
Пролесник встречается на почвах с содержанием воды от 12 до 70%, однако, если весной влажность почвы в его местообитаниях
120
превышает 35—55%, он отмирает. Наибольший сухой нес и макси мальное содержание золы наблюдаются при влажности 40 Г>0%, На влажных почвах рост лимитируется плохой аэрацией. При улучшении аэрации почв высота побегов увеличивается с 8 до 33 ем, площадь листьев на побеге — с 18 до 150 см2. Распространение пролесника ограничено площадями с высокой скоростью диффузии к недорода. Наименьшее содержание кислорода в почве, при котором выживают его корни, 0,2%. Периодическое затопление ограничивает развитие пролесника (Wardle, 1959).
Повышенная концентрация ионов М.п++ токсична для иролее-ника, при этом, появляются маленькие черные пятна на листьях. Повышенное содержание ионов железа Fe++ вызывает отмирание кончиков корней.
Пролесник — представитель теневой флоры. Освещенность в его местообитаниях под пологом бука составляет в светлую фазу 25—45% от полной, в теневую — 4—6%, под пологом дуба — соответственно 60—90 и 10%. При освещенности около 90% от полной совершенно подавляется рост пролесника. Амплитуда колебаний освещенности в местообитаниях женских растений меньше, чем мужских; освещенность менее 3,6% служит порогом для развития женских особей. Наиболее благоприятна для растений обоих полов освещенность в 40—50% от полной в светлую фазу (Martin, 1968).
Содержание золы в побегах значительно выше в начале вегетационного периода (апрель) и заметно уменьшается к осени, что коррелятивно связано с образованием подземных корневищ и корней (Mukerji, 1936). В более затененных местообитаниях содержание золы значительно больше в женских растениях, чем в мужских, в более освещенных — наоборот. Содержание золы в листьях в разные сезоны колеблется от 7 до 18%, в среднем составляет 12—15%.
Биологическая продуктивность
Сухой вес одного надземного побега пролесника колеблется от 0,15 до 0,95 г в зависимости от условий местообитания. Сухой вес побегов на 1 м2 в сложных сосняках Подмосковья составляет 13 г (Рысин, Золотова, 1968). По данным Е. Л. Любарского (1967), в широколиственных лесах окрестностей г. Казани воздушно-сухой вес листьев у одной особи составляет в среднем 0,85 г (от 0,24 до 1,46 г), вес корневищ — 2,09 г, вес 1 см корневища — 0,02 г.
Ассимиляционная поверхность особи пролесника составляет 286 (107—514) см2; площадь листьев, приходящихся на 1 см корневища,— 3,2 (2,0—5,1) см2; площадь листьев, приходящихся на 1 г корневища,— 149 (99—202) см2 (Любарский, 1967).
В снытево-пролесковой дубраве в Воронежском заповеднике биомасса популяции пролесника составляет 196 г/м2 (воздушно-сухой вес), надземной части — 64 г/м2, подземной— 132 г/м2. В иро-лесково-снытевой дубраве Теллермановского лесничества Воронежской обл. эти показатели соответственно равны: 120 г/м2, 44 г/м2, 76 г/м2.
121
Фитоценология
Пролесник многолетний — довольно распространенный доми-нат травяного покрова широколиственных и хвойно-широколиственных лесов Европы на почвах, достаточно увлажненных и наиболее богатых гумусом. Он является характерным растением свежих, влажных и сырых дубрав (Д2—Д4). Б. А. Быков (1965) определяет его как субдоминант-коннектор 2 яруса.
Пролесник доминирует в буковых, дубовых, ясеневых лесах Западной Европы, а также в лесах с господством вяза, березы, липы, граба, тисса, сосны (Mukerji, 1936; Martin, 1968). Он часто господствует во взрослых дубово-грабовых, дубовых и буковых лесах Карпат, Кодр, приднестровской части Молдавии, Крыма и Кавказа (Поплавская, 1948; Kaida, 1962; Ярошенко, Грабарь, 1969). Он обычен в темнохвойных лесах Карпат (Попов, 1949), в дубовых и липовых лесах на равнинной территории европейской части СССР, на темно-серых и серых суглинистых и супесчаных почвах. В южных дубравах пролесник приурочен к понижениям рельефа при отсутствии заболачивания (Высоцкий, 1913; Иваненко, 1923; Коновалов, 1929; Гроссет, 1930; Воробьев, 1953; Елагин, 1963; Курнаев, 1968).
Пролесниковая дубрава является крайним членом экологического ряда по увеличению богатства почв (Алехин, 1951). На южном пределе хвойно-широколиственных лесов в европейской части СССР пролесник иногда выступает как доминант, преимущественно в елово-липовых лесах (Кац, 1943; Алехин, 1947; Порфирьев, 1959; Рысин, Рысина, 1965), чаще же как сопутствующий вид. В некоторых типах елово-широколиственных лесов пролесник вместе с липой и елью образует особые коренные парцеллы (Дылис, 1969). Пролесник господствует в травяном ярусе некоторых типов сложных сосняков в Московской обл. и в низинных сероолыпаниках (Работнов, 1939).
В широколиственных и хвойно-широколиственных лесах пролесник образует либо чистые заросли, либо встречается вместе с другими видами, чаще всего вместе со снытью. Семенное возобновление у него подавлено. В сплошных зарослях проростки встречаются редко, ювенильные особи практически отсутствуют. Во всех исследованных популяциях этого вида господствуют генеративные особи, сенильные встречаются в небольшом числе; виргинильные, кроме проростков, полностью отсутствуют. Возобновление популяций неопределенно долгое время осуществляется за счет вегетативного размножения. Семенное возобновление эпизодическое, обычно на вырубках или при иных нарушениях травяного покрова.
Увеличение числа побегов и веса пролесника неблагоприятно сказывается на росте и количестве подроста ясеня. Отрицательное влияние пролесника проявляется в раннем затенении подроста ясеня и раннем достижении последним точки компенсации (Wardle, 1959).
122
I
I внутривидовая систематика.
д
У Mercurialis perennis L. выделены три разновидности, стойко : удерживающие свои признаки в культуре (Mukerji, 1927);
var. genuine, Muttel.— Aarg.— листья широкоэллиптические, длина их в 2-—3 раза больше ширины, верхняя сторона листа почти голая, на нижней стороне опушение редкое по жилкам;
var. salisburfana Mukerji — глубоко надрезанный (зубчатый) край листовой пластинки, очень короткий черешок, более короткие и вверх направленные рыльца; растения темно-зелёные и более сильно опушенные;
var. leiocarpa Mukerji (syn. M. leiocarpa Sieb. et zucc.) — отличается голой завязью и коробочкой.
Кроме трех разновидностей Мукерджи (Mukerji, 1936) указывает еще 6 форм, поведение которых в культуре неизвестно: .
f. silvatica Hoppe — листья удлиненные, эллиптически-ланцет-ные; центральная и боковые жилки листьев значительно толще и крупнее, чем у типичных форм;
f. ovalifolia Hausskn.— листья овальные или овально-ланцетные, верхняя часть стебля поникающая; черешки значительно короче, чем у типичных форм; эта форма обитает на солнечных склонах среди кустарников или других сухих местообитаниях;
f. angustifolia Murr.— растет, как и предыдущая, на солнечных местах, но листья более узкие, ланцетные, более толстые и сильнее опушены;
f. robusta Grossh.— листья необычно большие, 10—15 см длины и более 5 см ширины; растения очень высокие и более грубые, чем обычно, иногда достигают 60—65 см в высоту, они обычно встречаются под старыми бука!ми или дубами по влажных тенистых местах;
f. saxicola Beck — листья удлиненно-эллиптические, длина их более чем в 4—5 раз превышает ширину, постепенно суживающиеся к верхушке; встречается обычно на меловых холмах;
f. variegata Mukerjie-—листья покрыты желтыми и белыми пятнами.
По нашим наблюденимя, f. silvatica, f. angustifolia, f. robusta встречаются на территории центра европейской части СССР.
Хозяйственное значение
Пролесник — ядовитое растение, содержит алкалоид меркурпа-лин. Попадая в корм скоту, он вызывает отравление, молоку придает синий цвет. В народной медицине пролесник используется как слабительное. В Курской обл. использовался для приготовления голубой краски (Rendle, 1925; Маевский, 1954; Ларин и др., 195(>; Землинский, 1958).
И. Ф. Сацыперова ’
БОРЩЕВИК СИБИРСКИЙ
Номенклатура
Heracleum sibiricum L.,Sp. pl. (1753) 249; Кауфм., Моск, фл., изд. 1 (1866) 211, изд. 2 (1889) 213; Цингер, Сб.свед. (1885) 203; Пе-тунн., Крит, обзор 1 (1896) 196; Сырейщ., Илл. фл. 2 (1907) 418; он же, Опр. раст. Моск. губ. (1927) 199; Федченко в Маевск., Фл., изд. 7 (1940) 557; Манденова во Фл. СССР 17 (1951) 231; Шишкин в Маевск., Фл., изд. 8 (1954) 406, изд. 9 (1964) 393; Тихомиров в Опр. раст. Моск. обл. (1966) 254.— Н. sphondylium L. 6. sibiricum (L.) Schmalh., Фл. 1 (1895) 413.— H. sphondylium L. svb&p' sibiricum (L.) Simonkai, Enum. Fl. Transs. (1887) 266; Brummitt in Fl. Eur. 2 (1968) 366,—Борщевик сибирский, пупырь, свиные дудки (Umbelli-ferae — Apioideae — Pastinaceae).
Латинское название дано Плинием растению в честь древнегреческого героя Геракла. К- Линней присвоил это название роду из семейства зонтичных, очевидно, обратив внимание на крупные размеры растений (Манденова, 1950). Русское название «борщевик», широко распространенное во многих районах нашей страны, возникло из-за использования местным населением многих видов этого рода в качестве приправы при приготовлении борща. Видовое название было дано К- Линнеем, описавшим вид по образцу, собранному в Сибири.
Тип в Лондоне.
Морфологическое описание
Двулетнее или многолетнее полурозеточное длительно вегетирующее травянистое растение со стержнекорневой системой (по классификации В. Н. Голубева, 1965), моно- или поликарпик. Двулетние растения имеют простую стержнекорневую систему, проникающую
1 Разделы «Морфологическое описание» и «Внутривидовая систематика» написаны совместно с В. Н. Тихомировым.
124
вглубь до 1 м. Диаметр стержневого корня у основания—10--15 мм, на глубине 50 см — 2—4 мм. Многолетние растения имеют короткое вертикальное корневище и систему главного корня. Корневище од ноглавое (у монокарпиков) или многоглавое (у поликарпиков), образуется из остатков ортотропных ди- или полициклических побегов, соединенных с сильно разросшимся одревесневшим гипокотилем. От него отходят 2—3 крупных и несколько мелких придаточных корней, которые располагаются в поверхностном слое почвы. Главный корень разветвляется на 3—5 крупных и несколько мелких боковых ветвей. Все боковые корни первоначально отходят косо вниз, а затем принимают вертикальное положение. Диаметр стержневого корня у основания от 7,9 до 21,0 мм, на глубине 50 см— 1,5 до 5,0 ем. Диаметр наиболее крупных боковых корней в начале их ответвления 1—3 см. Окраска корней коричнево-бурая. Максимальная глубина проникновения— 110—230 см, ширина простирания — 30—100 см (Голубев, 1962).
Стебель ортотропный, 50—190 см высоты, 0,6—1,5 см в диаметре, ребристый (число ребер 20—22), внутри полый, в верхней части слабо ветвистый, опушен по ребрам вниз направленными или оттопыренными, длинными, жесткими, белыми волосками. Боковые ветви возвышаются над главной осью, реже они ниже главной оси. Нижнее (первое) и верхнее (четвертое или пятое) междоузлия укороченные (2—10 см длиной), средние (второе — четвертое) —длинные (20—52 см).
Листья очередные, жесткие, сверху более темные, с нижней стороны по жилкам и по краю листа покрыты более короткими, чем на стебле, с волосками, в общем очертании овально-яйцевидные, тройча-то- или перисторассеченные (с двумя парами сегментов), прикорневые (их 5—6) на длинных черещках, стеблевые (3—4) — короткочерешковые. Черешки тонкие, отстоящие, желобчатые, с узкими длинными влагалищами. Прикорневые листья, собранные в розетку, самые крупные: длина пластинки 10—40 см, ширина 18—26 см, длина черешка 20—35 см, длина влагалища 10—14 см. Размеры стеблевых листьев и их черешков в акропетальной последовательности уменьшаются, размеры же влагалищ (по отношению к длине черешка) увеличиваются (у самых верхних листьев пластинка листа отходит от влагалища). Сегменты перистолопастные, раздельные или рассеченные, в очертании широкояйцевидные, конечный — округло-яйцевидный или округлый; все они по краю неравномерно пильчатозуб-чатые, с перисто-петлевидным жилкованием. Дольки (сегменты) последнего порядка сильно варьируют по форме — от яйцевидных до линейно-ланцетных и линейных.
Соцветие — сложный зонтик. Центральный зонтик чашевидный, в поперечнике 7—18 см, с 15—30 неравными по длине лучами, опушенными редкими жесткими белыми волосками. Лучи, несущие зонтички, расположены в 4 круга. Число лучей в круге уменьшается в центростремительном направлении (от 5—10 в наружном до 2 5 во внутреннем). Зонтички первого круга несут 20—25 цветков, вто
125
рого— 15—20, третьего — 12—15, четвертого — 8—12. Диаметр зон-тичков 1—2 см. Боковые зонтики (их обычно 2—3, редко до 5) отличаются от центрального менее крупными размерами (6-—12 см) и меньшим числом лучей (12—20). Листочки обертки (в числе 2—5) линейные, неравные, длиной 0,4—0,6 см, опушенные, опадающие. Листочки оберточки (их 5—8) линейно-шиловидные или линейные, равные, длиной 0,2—0,3 см, опушенные, неопадающие.
Цветки на длинных неравных цветоножках, обоеполые, актиноморфные (только краевые в зонтичках первого круга слегка зигоморфные), до 0,4 см в диаметре. Чашечка редуцирована — зубцы ее незаметны. Венчик из 5 свободных зеленовато-желтых обратнояйцевидных лепестков 0,1—0,2 см длиной; лепестки с цельной верхушкой, загнутой внутрь примерно до х/2 длины лепестка, оттянутые в короткий ноготок. Тычинок 5, они свободные, в бутонах загнутые внутрь, при раскрытии венчика распростертые, чередующиеся с лепестками, зеленовато-желтые; тычиночные нити по длине почти равны лепесткам, пыльники двугнездные, прикрепленные к нити серединой, раскрываются продольно. Пыльцевые зерна продолговатой формы, с тремя закрытыми экваториальными порами; тапетум — клеточный с двуядерными клетками, тип образования тетрад пыльцы— симультанный (Кордюм, 1967). Гинецей синкарпный, завязь голая, полунижняя, двугнездная; плацентация центрально-краевая; в каждом гнезде закладывается по две, но развивается по одной висячей анатропной семяпочке; стилодии равны подстолбиям или вдвое длиннее их, с головчатыми рыльцами.
Плод — сухой вислоплодник из двух мерикарпиев с двураздельной от основания колонкой. Мерикарпий 4,9—9,8 мм длины, 3,6— 7,2 мм ширины, 0,4—0,8 мм толщины, сильно сжатый со спинки, в очертании овальный, широкоовальный, обратнояйцевидный или широко-обратнояйцевидный, полый, соломенно-желтый или желтовато-светло-коричневый; ребра только первичные, спинное и промежуточные— тонко-нитевидные, краевые — ширококрылатые; секреторные канальцы в ложбинках одиночные, коричневые на просвет, к основанию утолщенные, неравные, достигающие V2—4/s всей длины плода; на комиссуральной стороне два более коротких и широких канальца. Экзокарпий однослойный, паренхиматический. Наружная зона мезокарпия паренхиматическая, внутренняя — склеренхимная, образующая гипэндокарпий. Эндокарпий в зрелом плоде неразличим. Реберные проводящие пучки мелкие, в краевых ребрах разделенные на два тяжами склеренхимы. Канальцы глубоко расположенные, прилегающие к гипэндокарпию. Фуникулус с расширенным в тангентальном направлении проводящим пучком. Эндосперм нуклеарного типа, обильный, выполняющий всю полость семени, плотный, на комиссуральной стороне почти плоский. Зародыш мелкий, расположен в микропилярной части семени (в верхней трети его), погружен в эндосперм. Он дифференцирован на две свободные продолговатые семядоли (Васильченко, 1941), гипокотиль, зачаток зародышевого корешка и мельчайшую почечку (апекс стебля).
126
Семена боковых зонтиков в большинстве случаев беззародышоные или с очень мелким, почти не дифференцированным зпро дышем.
Диплоидное число хромосом равно 22 (Кордюм, 1967; Хромо сомные числа цветковых растений, 1969).
Географическое распространение
Ареал охватывает Скандинавию, всю Восточную Европу, вклю чая европейскую часть СССР, Предкавказье, Западную Сибирь, Монголию. Борщевик сибирский следует отнести к группе растений с евразиатским типом ареала. Он встречается от лесотундры до Средиземноморья, в горах поднимается до лесного пояса. Произрастает на материковых лугах лесной зоны, в прирусловой и центральной частях поймы, среди кустарников по берегам рек, на опушках и на лужайках влажных лесов, в светлых березово-осиновых и березово-еловых лесах, вдоль обочин дорог, на межах, мусорных местах около селений.
В палеоботанической литературе имеются сведения о нахождении б. сибирского в постплиоценовых отложениях Ленинградской обл. (Данилевский, 1928).
Онтогенез
По литературным данным, семена б. сибирского не имеют периода покоя (Ларин и др., 1956). Наши исследования показали: период покоя у семян этого растения вполне выражен, длится 2—3 месяца. Он обусловлен недоразвитием зародыша. Дозревание зародыша происходит при низких температурах (+2—5°). При выдерживании семян в течение трех месяцев при +2—5° или под снегом лабораторная всхожесть колеблется от 35 до 78%, всходы начинают появляться на 5—12 день. При подзимнем посеве в грунт (в октябре) происходит естественная стратификация семян, всходы появляются в течение месяца после стаивания снега. Полевая всхожесть колеблется от 20 до 70%. Срок появления всходов растянут до 30 дней. Часть семян (до 25%) прорастает только через год. Срок сухого хранения семян при комнатной температуре не более двух лет, после трех лет всхожесть составляет менее 10%.
Для б. сибирского характерен надземный тип прорастания. Семядоли продолговатые (10—15 мм длины, 3—5 мм ширины), па верхушке острые, книзу постепенно суженные в черешок длиной до 20 мм. Подсемядольная часть слабо развита (Васильченко, 1941). Кроме семядолей и заключенной между ними почечки проростки имеют короткий гипокотиль и главный корень длиной до 12 см. Из верхушечной почки в последующем развивается укороченный веге-
127
Рис.'14. Возрастные этапы борщевика сибирского: проростки, 2 — ювенильное растение, 3 — имматурное, 4 — взрослое вегетативное, 5 — генеративное
тативный побег, несущий одновременно от 3 до 6 листьев, собранных в розетку. Первые 3 листа простые, округлые, неясно-пятилопастные, при основании сердцевидные, по краю городчато-зубчатые на длинных черешках. Рассеченность пластинок последующих листьев постепенно усложняется.
В пределах виргинильного периода у б. сибирского можно выделить 4 возрастных этапа (рис. 14): проростки, ювенильный, имма-турный и взрослый вегетативный (Работнов, 1956 6). Одновозрастные растения в разных экологических условиях могут находиться на разных возрастных этапах. Все же можно считать, что ювенильное и имматурное состояния длятся менее одного сезона, хотя в отдельных случаях имматурное состояние может затянуться до 3 лет. Во взрослом вегетативном состоянии растения могут находиться от 4 до 10 лет. Первое цветение в естественных ценозах наступает на третий год жизни, в культуре — на второй. В пойме Оки в возрасте четырех лет зацвела Уз растений от общего их числа (Работнов, 1956 6). Половозрелые особи цветут не ежегодно: около 17% из них цветет повторно, примерно 50% после цветения переходит в вегетативное состояние на несколько лет, а затем вновь зацветают. Отдельные особи цветут 2—3 года подряд, потом переходят в вегетативное состояние на несколько лет и вновь зацветают. У б. сибирского встречаются моно-, ди- и поликарпичные (с 3—4-кратным цветением) особи; последние преобладают. Таким образом, он может быть отнесен к группе растений, переходных от моно- к пойикарпич-ности со слабо выраженной кратностью цветения. Средняя продолжительность генеративного периода 3—5 лет (Работнов, 19566). Первые признаки старения проявляются в образовании дупла в корневище, которое постепенно увеличивается. В результате происходит расчленение материнской особи на 2 (редко 3) самостоятельные особи, расположенные в непосредственной близости друг от друга. Длительность сенильного периода составляет в среднем 1,5 года.
Переход от одного возрастного этапа к другому сопровождается морфологическими изменениями надземных и подземных частей растения. Так, для проростков характерно наличие семядольных листьев. Ювенильные растения имеют простые округлые трехлопастные листья, имматурные — трехраздельные, а взрослые вегетативные особи-—тройчатые или перистые листья. Генеративные растения отличаются от взрослых вегетативных особей наличием удлиненного цветоносного побега или следов от него и иногда двух-трех розеток листьев. Сенильные растения имеют несколько розеток листьев, следы бывших цветоносных побегов и отличаются от генеративных растений наличием дупла в корневище. Корневая система б. сибирского с изменением возраста растений постепенно из простой стержневой превращается в многоосевую стержнекорневую или стержнекисте-корневую с коротким одноглавым или многоглавым вертикальным корневищем. Общая продолжительность жизни б. сибирского не первышает 25 лет (Работнов, 1956 6).
5 Зак. 255
129
Сезонный ритм развития
Всходы появляются с середины апреля до первой половины мая. При благоприятных условиях около 50% из них сохраняется в течение первого года жизни; основная масса особей гибнет в ювенильном состоянии. К осени большинство особей успевает достигнуть имматурного состояния. В течение первого года жизни -у растений развивается розеточный вегетативный побег с шестью очередными листьями, из которых в розетке одновременно находится не более трех. Первый лист появляется на 10—12-й день после развертывания семядолей. Он развивается медленно,-лишь через месяц достигает максимальных размеров и отмирает в среднем через 2,5 месяца после появления. Размеры листьев и темпы их роста увеличиваются по мере увеличения порядкового номера. Только первые 3—4 листа успевают фактически пройти полный жизненный цикл, остальные уходят под снег в зеленом состоянии.
Взрослые особи трогаются в рост рано, в апреле. Из терминальной почки, которая начинает формироваться в августе предыдущего. года, развиваются ди- или полициклические вегетативные побеги, находящиеся в фазе розетки от двух до пяти лети являющиеся лишь фазой в развитии монокарпического генеративного побега. В течение сезона развивается две генерации листьев: весенне-летняя и летне-осенняя. Листья первой генерации (1—3 низовой и 4—7 срединной формаций) появляются с апреля по июнь и успевают пройти полный'жизненный цикл. Листья второй генерации (8—И срединной формации) появляются с июля по сентябрь и уходят в зеленом состоянии под снег. Длительность жизни листьев низовой формации не превышает одного месяца, листьев срединной формации — от 58 до 90 дней. Одновременно в розетке в течение всего сезона находится от двух до пяти листьев.
Корневая система взрослых вегетативных растений второго года жизни такая же, как у однолетних растений к концу вегетации, т. е. стержнекорневая простая. В начале вегетации происходит развитие всасывающей части корневой системы. Затем наблюдается рост стержневого корня в глубину, утолщение и разрастание боковых корней. Одновременно в течение вегетационного сезона происходит утолщение и сокращение базальной части корня, обусловленное процессом контрактации, и сокращение укороченного стебля предыдущего года развития, что приводит к сближению листовых рубцов и узлов.
У растений, вступивших в генеративный период, весной из терминальной почки развиваются 2—3 розеточных (срединной формации) листа, а затем удлиненный ортотропный генеративный побег, несущий 4—5 стеблевых листьев.
По ритму цветения б. сибирский следует отнести к растениям со среднелетним сроком цветения: цветение начинается в июне и продолжается в среднем 26 дней. Первыми зацветают краевые цветки в зонтичках, расположенных в центральном зонтике. Распускание
130
цвётков во всех зонтичках происходит в центростремительном, а увядание— в центробежном порядке. Длительность цветения одного цветка колеблется от 2 до 5 дней, зонтичка — от 3 до 5 дней, центрального зонтика — от 10 до 12 дней. Распускание цветков в боковых зонтиках происходит после увядания цветков в центральном зонтике.
Цветки б. сибирского следует отнести к цветкам с утренним типом распускания: большинство (62,4%) распускается с 6 до 8 часов утра. Раскрытие цветка начинается с отгибания лепестков и тычинок (мужская фаза цветка), пыльники растрескиваются на сутки раньше, чем рыльце становится готовым к восприятию пыльцы. Женская ф.аза цветка характеризуется удлинением столбиков, отклонением их в противоположные стороны и появлением нектара на стилоплодиях. Во время прохождения женской фазы тычинки усыхают и отваливаются.
Плоды в центральном зонтике созревают с июля по первую половину августа, в боковых — в августе — сентябре. Созревшие плоды плотно сидят на плодоносцах, часто сохраняются на растении до первого снега.
Б. сибирский после скашивания дает хорошую отаву. Скашивание способствует дополнительному развитию двух-трех листьев, образующих розетку, а иногда появлению новых генеративных побегов из имеющихся розеточных побегов. Число таких вновь зацветших растений не превышает 5% от числа особей, находившихся в цветущем состоянии во время скашивания (Ларин и др., 1956). Цветение вновь развившихся побегов запаздывает на 1,5—2 месяца, семена на этих побегах развиваются, но созреть не успевают. Выпас скота способствует повреждению терминальных почек, а также пробуждению и быстрому развитию спящих почек, из которых к осени развиваются новые вегетативные розеточные побегиг
Б. сибирский является гемикриптофитом: он уходит под зиму с розеткой зеленых листьев, которые после выпадения снега отмирают. Терминальная почка укрыта остатками влагалищ старых листьев, она находится на уровне поверхности почвы. Боковые (зимующие) почки располагаются у корневой шейки в пазухах отмерших листьев. Они погружены в поверхностный слой почвы. В течение зимнего периода внутри терминальной почки происходит формирование вегетативного побега или вегетативной части смешанного генеративного побега. К началу весны в терминальной почке бывают полностью сформированы первые 3 листа низовой формации и 2—3 листа срединной формации, а также зачатки будущего цветоносного стебля. Пазушные почки в виде крохотных бугорков могут находиться в спящем состоянии в течение нескольких лет. В год, предшествующий цветению, 1 (реже 2—3) почка начинает увеличиваться в размерах и в год цветения или на следующий после цветения она развивается в вегетативный побег, который является лишь фазой монокарпического генеративного побега.
S*
131
Способы размножения и распространения
Семенное размножение. Для б. сибирского характерно наличие обоеполых и функционально мужских цветков. Первые расположены в центральном и боковых зонтиках, вторые — только в боковых. Обоеполые составляют до 90% от общего числа цветков в зонтиках. Это перекрестноопыляемое энтомофильное растение со следующими приспособлениями для такого типа опыления: цветки собраны в крупные сложные зонтики, протерандричны и обильно выделяют нектар, привлекающий многих насекомых. Данные о специфических особенностях опыления и оплодотворения б. сибирского отсутствуют. Очевидно, эти процессы сходны с описанными для других видов зонтичных (Borthwick, 1931; Кордюм, 1967).
Нам удалось получить межвидовые гибриды при искусственном опылении Н. sosnowskyi Manden. пыльцой Н. sibiricum.
Для б. сибирского характерно наличие одного, редко двух генеративных побегов. Каждый генеративный побег несет центральный и 2 или 3 боковых зонтика. В центральном зонтике число цветков колеблется от 212 до 575, в каждом боковом — от 177 до 394. В центральном зонтике завязывается от 80 до 100% цветков, в боковых'— от 1 до 10%. Семенная продуктивность одной плодущей особи колеблется, по нашим наблюдениям, от 1000 до 1800 выполненных мерикарпиев, по данным Т. А. Работнова (1949), от 5000 до 8000 мерикарпиев, по данным И. В. Ларина и др. (1956), составляет 400 мерикарпиев.
Б. сибирский цветет и плодоносит нерегулярно. По наблюдениям Т. А. Работнова (1956 6), только 1,5% особей от общего числа, находившихся в генеративном состоянии, цвело и плодоносило 5 лет подряд, около 3% —в течение трех лет и 13,5% —два года подряд.
По способу распространения семян б. сибирский относится к группе баллистов-анемохоров (Левина, 1967). Рассеивание мерикарпиев происходит только при раскачивании цветоносного стебля, который к периоду массового созревания плодов становится сухим и упругим, как и лучи зонтика и плодоножки. Резкие порывы ветра, а также сотрясения от соприкосновения растений с человеком или животными способствуют рассеиванию мерикарпиев. Так как мерикарпии плоские, легкие, с небольшими крыльями, они замедленно падают и рассеиваются обычно в радиусе 60 см от растения, отдельные плодики встречаются на расстоянии до 2 м от плодоносящей особи (Левина, 1957). Мерикарпии, попавшие на участки с разреженным травяным покровом, после перезимовки под укрытием листьев и остатков растений весной, если достаточно влаги, дружно прорастают (всхожесть до 70%). Мерикарпии, пролежавшие в поверхностном слое почвы в течение года, могут прорасти на второй год (всхожесть их составляет не более 10%). При более длительных сроках пребывания мерикарпиев в почве наблюдаются только единичные случаи их прорастания.
Периодическое скашивание лугов, в состав фитоценозов кото
132
рых входит б. сибирский, способствует затягиванию сроков перехода от одного возрастного состояния к другому и сокращает естественное возобновление. Частое скашивание лугов в течение одного сезона вызывает несв'бевременное цветение растений и прекращение обсеменения.
Вегетативное размножение. Б. сибирский размножается вегетативно только путем партикуляции в сенильный период материнской особи на 2—3 дочерние. Вегетативное размножение малоэффективно. Возникшие вегетативным способом особи находятся в непосредственной близости к материнским растениям. Как правило, они редко зацветают, находятся в вегетативном состоянии 3—4 года, а затем отмирают.
Экология
Б. сибирский — луго-лесной эумезофит (Раменский и др., 1956). Он использует влагу атмосферных осадков и в меньшей степени ка-пиллярно-поднимающуюся почвенную воду. По классификации И. Н. Бейдеман (1954) его можно отнести к омброфитам и условно к трихогидрофитам. Он предпочитает участки с глубоким уровнем залегания грунтовых вод, но мирится и с грунтовыми водами на уровне от 18 до 85 см.. Хорошо развивается при умеренно переменном увлажнении, но встречается и на почвах со среднеобеспеченным водным питанием.
Легко переносит непродолжительное (10—15 дней) затопление полыми водами. Устойчив к заилению: хорошо развивается в условиях умеренного отложения (около 2 м) и при сильном отложении (2—4 см) наилка. Даже при катастрофической мощности наилка в 10—15 см можно встретить единичные экземпляры борщевика (Раменский и др., 1956). В то же время он неустойчив к замерзанию почвы, насыщенной водой. Притертая ледяная корка, образующаяся на почве после осеннего паводка, способствует выпадению борщевика из луговых ценозов (Работнов, 1971).
Б. сибирский — светолюбивое растение, но мирится с частичным затенением. В условиях недостаточного освещения увеличивается длительность пребывания особей в виргинильном состоянии. Произрастает в довольно широких пределах тепловых условий. Начинает вегетировать сразу же после стаивания снега, прекращает вегетацию лишь после выпадения снега или наступления устойчивых заморозков. Вполне холодостоек и морозоустойчив. Первые весенние и осенние заморозки задерживают рост листьев, но не повреждают их.
Б. сибирский — эутроф. Он встречается массово на лугах с богатыми и довольно богатыми почвами с нейтральной или слабокислой средой (pH 6,0—7,5). В то же время заходит на слабосолончаковые почвы со щелочной реакцией (pH 7,5—8,3) и на мезотрофные слабокислые (pH 5,5—6,5) почвы. Единично его можно встретить на бедных кислых почвах (pH 5,0—5,5) и даже на средне- и сильносо
133
лончаковых почвах со щелочной реакцией (pH 7,5—8,4) (Раменский и др., 1956).
Внесение удобрений на луга, где встречается б. сибирский, способствует особенно пышному его развитию и даже переходу в доминанты (Работнов, 1965). При этом глубина проникновения корней в почву на удобренных участках уменьшается более чем в 3 раза, да-, же преобразуется морфологический тип корневой структуры — она становится кистекорневой (Демин, 1969).
Биологическая продуктивность
Урожай надземной массы б. сибирского в полидоминантных луговых ценозах в пойме Оки, где травостой отличается большим разнообразием разнотравья и бобовых, колеблется от 2 до 30 г на 10 м2 и составляет от 0,3 до 5,6% от общего урожая, полученного от одного укоса. При ежегодном внесении NPK. продуктивность увеличивается в 5—10 раз (Работнов, 1965).
В опытных посевах урожай зеленой массы составил в первый год 25—30 т/га, во второй — 60, в третий — 40, в четвертый — 24 (Ларин и др., 1956). Урожай мерикарпиев достигал 1,35 кг с 10 м2.
Фитоценология
Б. сибирский может быть ассектатором, доминантой, соэдифи-катором и эдификатором. Он входит в состав полидоминантных луговых сообществ, принимая участие в образовании первого яруса. Как ассектатор часто встречается на пойменных разнотравно-крупнозлаковых (с костром) лугах в южной подзоне лесотундры европейской части СССР и Западной Сибири (Шенников, 1938), а также на материковых суходольных лугах лесной зоны европейской части СССР: в клеверно-душисто-колосковых сообществах, разнотравных сильно замоховелых белоусниках (Понятовская, Сыроком-ская, 1970) и др. Кроме того, как ассектатор он встречается на остепненных лугах (Ярошенко, Юрова, 1970) и в луговых степях северного варианта, входя в состав злаково-разнотравных сообществ (Кучеров и др., 1971).
Борщевик становится доминантой в пойменных лугах лесной, лесостепной и степной областей европейской части СССР, где он входит в состав разнотравно-злаковых лугов среднего уровня центральной поймы, в частности разнотравных белополевичников (Шенников, 1938; Никитина, 1966). Он является субэдификатором или эдификатором в пойменных среднезональных (центральная пойма) крупноразнотравных лугах, где образует борщевиковые луга или крупнозонтичные луга с примесью порезника и бутеня (Шенников, 1938).
При благоприятных условиях (богатые органическими удобрениями и достаточно влажные почвы) он легко вытесняет ряд видов, становится преобладающим растением и существенно изменяет
134
состав травостоя. По данным Т. А. Работнова (1958), травостой злакового пойменного луга спустя 4 года после подсева б. сибирского превратился в разнотравно-злаковый с преобладанием борщевика.
Консортивные связи
Цветки б. сибирского обильно посещаются представителями отрядов перепончатокрылых, двукрылых, полужесткокрылых и жесткокрылых. Основными опылителями можно считать перепончатокрылых, так как они в основном посещают цветки в часы растрескивания пыльников.
Листья и стебли борщевика поражаются мучнистой росой. Плоды часто повреждаются личинками моли, которые выедают эндосперм семени; они иногда повреждают до 80% созревших мерикарпиев. В зимний период полевки выедают борщевик на лугах. Выпас скота приводит к выпадению б. сибирского из ценоза (Работнов, 1956 6).
Внутривидовая систематика
Растения борщевика сибирского сильно варьируют по форме листьев — по всему ареалу встречаются особи, у которых сегменты листьев разделены на широкояйцевидные, яйцевидные, ланцетные, линейные доли. На этом основании описан ряд разновидностей, в том числе var. longifolium Koch с узкими ланцетными или линейноланцетными долями листьев последнего порядка, но, по-видимому, эти формы не имеют таксономического значения.
Хозяйственное значение
Корни содержат следующие кумариновые соединения: бергап-тен, изобергаптен, пимпинеллин, сфондин и умбеллиферон (Колесников и др., 1961; Комиссаренко и др., 1961; Ognyanov et al., 1966); из которых бергаптен, пимпинеллин и умбеллиферон обладают противогрибковым действием (Лесников, 1969). Кроме того, в них находится эфирное масло, выход и состав которого не изучены (Горяев, 1952).
В листьях обнаружено от 105,4 до 538,5 мг% аскорбиновой кислоты (Гурцевич, Шиврина, 1942; Ефремов, 1943; Муравьева, Вань-ковский, 1947) и от 3,2 до 12,8 мг% каротина от веса сырых листьев (Савинов, 1948).
Плоды по составу кумариновых соединений отличаются от корней только наличием в них изопимпинеллина (Svendsen, Ottesrcd, 1957). Они также содержат от 0,3 до 3,0% эфирного масла, одним из компонентов которого является октиловый спирт, используемый в парфюмерии (Карпенко, 1931; Путохин, Смирнова, 1935). Летучие фракции плодов обладают фунгицидным действием (Дуброва,
135
1952). Содержание аскорбиновой кислоты в плодах составляет 42 мг% (Клобукова-Алисова, 1958).
Б. сибирский относится к луговым сорнякам, хотя он и обладает определенной кормовой ценностью. В вегетативном состоянии в растении находится (в пересчете на абсолютно сухой вес) до 27,7% протеина, 15,2% растворимых углеводов и 13,9 золы; содержание же клетчатки не превышает 16,4%. К периоду цветения содержание растворимых углеводов повышается до 23,2% и клетчатки — до 32,3%, количество же протеина снижается до 12,4%, а золы — до 8,7%. Наиболее богаты питательными веществами листья, которые при ворошении и сгребании сена легко превращаются в труху и теряются. В результате в сено попадают только грубые стебли, которые содержат мало протеина (5,2%) и золы (5,8%), но много клетчатки. Кроме того, свежие стебли плохо сохнут и легко плесневеют, что вызывает порчу сена. Наиболее целесообразно борщевик использовать для силосования. Он легко силосуется в чистом виде и даже с трудносилосуемыми травами, в засилосованном виде хорошо поедается крупным рогатым скотом. Свежие листья и молодые побеги хорошо поедаются крупным и мелким рогатым скотом, а также кроликами; свиньи едят не только листья, но и корни (Ларин и др., 1956).
Молодые побеги в разных районах нашей страны издавна употреблялись в пищу в сыром и соленом виде для салатов, из них варили супы и приготовляли начинку для пирогов. Черешки листьев после сцятия кожицы мариновали, а из корней гнали спирт (Роллов, 1908; Медведев, 1957; Клобукова-Алисова, 1958).
По данным М. М. Глухова (1955), борщевик — хороший медонос, его цветки выделяют много нектара.
Б. сибирский использовали в русской народной медицине. Его препаратами лечили сердечные заболевания, чесотку, опухоли (Роллов, 1908), плохое пищеварение, эпилепсию и др. заболевания (Зем-линский, 1958). В Болгарии надземную часть и корни употребляют внутрь при нарушении пищеварения, для возбуждения аппетита и в качестве спазмолитического средства, главным образом при катаре желудка и кишечника, дизентерии и поносах. Наружно отвары из этих же частей растения применяют при кожных заболеваниях (Иорданов и др., 1970).
Ранее использовали борщевик для окраски по алюминиевой протраве шерсти и шелка в желтый цвет (Клобукова-Алисова, 1958).
Е. Е. Гогина
РОД ЧАБРЕЦ (ТИМЬЯН) — Thymus L.
Обширный род сем. губоцветных (Labiatae), относящийся к трибе Satureieae. Имеет средиземноморское происхождение, распространен в Европе, Азии и Северной Африке. Отличается большим разнообразием трудно дифференцируемых форм, которым придается различное таксономическое значение. Следствием этого является неупорядоченность номенклатуры и критическое состояние систематики рода.
Во флоре Московской обл. насчитывается лишь три вида этого рода: Т. marschallianus Willd., Т. pulegioides L. и Т. serpyllum L. Указывавшийся раньше для области Т. loevyanus Opiz (Ворошилов и др., 1966), по нашим данным, представляет естественный гибрид между Т. pulegioides и Т. marschallianus (Гогина, 1974). Виды чабреца встречаются на территории области лишь в немногих разобщенных местообитаниях и относятся к числу редких растений. Это объясняется тем, что все три вида находятся здесь у пределов своего распространения, будучи при этом представителями разных флористических элементов во флоре области.
Т. marschallianus принадлежит к числу характерных растений степной и лугово-степной евразиатской зон, по Московской обл. проходит северная граница его естественного распространения. 7'. pulegioides входит в группу видов, распространение которых связано с листопадными лесами Западной и Центральной Европы. Во флоре области он представляет западный субатлантический элемент, тогда как Т. serpyllum отражает ее северные связи: основной ареал последнего вида лежит в северной части европейской бореальной области, где он предпочитает песчаные стации.
Все три вида представляют собой низкорослые ароматические полукустарнички с восходящими или стелющимися побегами и имеют лекарственное значение.
137
1. ЧАБРЕЦ (ТИМЬЯН) МАРШАЛЛА
Номенклатура и систематическое положение
Т. marschallianus Willd., Sp. pl. 3 (1800) 1141; Петунн,, Крит, обзор 2 (1900) 128; Сырейщ., Илл. фл. 3 (1910) 79, 4 (1914) 141; он же, Опр. раст. Моск. губ. (1927) 219; Иконников-Галицкий в Маевск., Фл., изд. 7 (1940) 625; Клоков во Фл. СССР 21 (1954) 511; Борисова в Маевск., Фл., изд. 8 (1954) 521, изд. 9 (1964) 503; Ворошилов в Опр. раст. Моск. обл. (1966) 283.— Т. serpyllum L. var. marschallianus Ledeb., Fl. Ross. 3 (1849) 346; Кауфм., Моск, фл., изд. 1 (1866) 374, изд. 2 (1889) 386; Цингер. Сб. свед. (1885) 342; Шмальг., Фл. 2 (1897) 311.— Т. pannonicus All., Auct. Syn. Meth. Stirp. Hort. Taurin. (1773) 6, p. p.; Jalas in Fl. Eur. 3 (1972) 182, p. p.— Ч. Маршалла.
Во «Флоре СССР» (1954) отнесен к секции Verticillati Klok.; во «Flora Europaea» — к секции Serpyllum (Miller) Bentham. Описан из Таврической губернии. Тип в Берлине, в гербарии Вильденова.
Зарубежные исследователи рода Thymus в последние годы приходят к выводу о неправомерности использования названия Т. marschallianus. Необходимость его замены обосновывается прежде всего тем, что аутентичные экземпляры из Крыма, находящиеся в гербарии Вильденова (Берлин — Далем) под этим названием, относятся к родству Т. callieri Borb. ex Vel. (Schmidt, 1973). Для переиме-нования'предлагается использовать название Т. pannonicus АП., которое было опубликовано без описания. В качестве его лектотипа избран приведенный в работе К. Клюзиуса (Clusius, 1583) рисунок венгерского растения, идентичного Т. marschallianus в привычном понимании (Jalas, 1971 b). Отказ от названия Т. marschallianus связывается также с изменением понимания объема этого вида, который предложено объединить с близкородственным Т. kosteleckya-nus Opiz (Schmidt, 1968; Jalas, 1971 а), отличающимся опушенно-стью всех частей растения.
Несмотря на серьезность высказанных соображений, следует отметить, что они не свободны от некоторых неясностей и допускают ряд возражений. Так, по данным Ю. Л. Меницкого (1973), один из аутентичных экземпляров в гербарии Вильденова (не имеющий, к сожалению, указания о происхождении) вполне' соответствует диагнозу Т. marschallianus. Не бесспорна и типификация названия Т. pannonicus, которое было первоначально применено к опушенной форме из Южных Альп, а затем употреблялось в весьма различных значениях (Hegnauer, 1948). Подробный критический разбор приведенных взглядов дан в работе Ю. Л. Меницкого (1973).
Имеются возражения и против объединения опушенной и гололистной рас. Б. Павловский (Pawlowski, 1966) считает его неправильным, подчеркивая различия в их географическом распространении. Как правило, опушение в роде Thymus, действительно, не является надежным признаком, поэтому вполне понятно возникновение
13S
сомнений в самостоятельности вида Т. kosteleckyanus. Однако на протяжении большей части своего обширного ареала Т. marschallianus представлен гололистной формой. Вычленение опушенных рас наблюдается в основном по периферии ареала. Заслуживают ли эти географические расы выделения в самостоятельные таксоны и в каком ранге, должны показать дальнейшие исследования. Вопрос о видовой самостоятельности Т. kosteleckyanus и соответственно об отказе от закрепившегося названия Т. marschallianus правильнее поэтому отложить до более подробного обоснования необходимости такого изменения.
Морфологическое описание
Чабрец Маршалла, как и большинство других видов рода, представляет собой типичный симподиально нарастающий полукустарничек. Главный корень, имеющий стержневую форму, сохраняется в течение всей жизни растения. Глубина его проникновения, по данным Г. М. Зозулина (1959), достигает в условиях Центральночерноземного заповедника 77—87 см, по данным Б. С. Кожухова (1931) — 120 см. Наряду с этим на прилегающих к поверхности почвы частях скелетных осей обычно образуются более или менее обильные придаточные корни.
Взрослые особи образуют сложную систему побегов, в которой хорошо различимы многолетняя одревесневшая базальная часть, прилегающая к поверхности субстрата, и прямостоячие или восходящие травянистые побеги текущего года. Внешний облик растений, соотношение числа цветоносных и вегетативных побегов и общее их число сильно изменяются в зависимости от условий местообитания. Цветоносные стебли достигают в среднем высоты 25 см, в поперечном сечении округло-четырехгранные, опушение их равномерное, к соцветию усиливающееся (от короткого ретрорзного до длинного отстоящего).
Листья простые, без прилистников, к основанию клиновидно суженные, сидячие или почти сидячие, значительно варьируют по форме (от линейно-ланцетных до продолговато-эллиптических), светло-зеленые, тонкие, с малозаметными боковыми жилками, у основания по краю реснитчатые. Длина листьев колеблется от 15 до 25 мм, ширина — от 2,5 до 5 (7) мм. (Как и у других видов рода, размер листьев постепенно увеличивается к соцветию, поэтому принято использовать для сравнения листья в средней части стебля.) Поверхность листьев голая или рассеянно-волосистая, с мелкими, малозаметными точечными железками и одноклеточными шипико-видными волосками.
Соцветие образовано накрест супротивными ложными полумутовками (из дихазиев с сильно укороченными цветоножками), колосовидное, нередко с расставленными нижними полумутовками, заканчивающееся недоразвитой почкой, во время плодоношения заметно удлиняющееся. Цветение распространяется по соцветию
139
снизу вверх, нередко, однако, одна или две нижние ложные мутовки зацветают с опозданием, что, по-видимому, свидетельствует об их происхождении из укороченных и придвинутых к основанию соцветия генеративных побегов обогащения. Зацветание в пределах каждой полумутовки происходит в строгом соответствии с порядком цветков в дихазии: первым зацветает центральный цветок, последними цветки на осях высших порядков.
Число цветков в одной полумутовке зависит от ее положения на оси соцветия: нижние полумутовки обычно наиболее многоцветковые, в условиях культуры в них может развиваться до 13—17 цветков, верхние обычно содержат по одному цветку. Прицветные листья сходны по форме со стеблевыми, в верхней части соцветия постепенно мельчают, нижняя пара обычно отогнута вниз. У основания цветоножек расположены мелкие, узкие, ресничатые по краю прицветники.
Чашечка двугубая, с десятью жилками, около 2—2,5 мм длины во время цветения, при плодах увеличивается до 3—3,5 мм, с отстоящей опушенной трубочкой и волосистым кольцом в зеве. Верхняя губа широкая и до одной трети рассечена на остротреугольные, почти одинаковые, по краю ресничатые зубчики; нижняя губа разделена до основания на два шиловидных длиннореснитчатых зубчика.
Венчик двугубый, с двунадрезанной верхней и трехлопастной нижней губой и короткой трубкой, от бледно-лилового до белого, с более темными крапинками в основании нижней губы. Размеры венчика и чашечки колеблются в зависимости от пола цветка и его положения в полумутовке. Венчик обоеполых цветков крупнее, чем женских. По данным Л. М. Злобиной (1967), средняя длина его у обоеполых цветков равна 5,45 мм, у женских — 4,48 мм. Центральные цветки в полумутовках заметно крупнее цветков, расположенных на осях последних порядков.
Тычинки выдаются из трубки венчика, двусильные. Пыльники интрорзные, с двумя гнездами, смещенными книзу вследствие неравномерного утолщения связника. Пыльца шестибороздная, к полюсам несколько вытянутая, 31,7X36,8 мк (размеры определены по гербарным образцам, собранным в Сумской обл.).
Гинецей обычного для губоцветных строения. В каждом гнезде завязи находится одна апотропная семяпочка, развитие которой вызывает выпячивание верхней части гнезда. Четыре образующихся вздутия перерастают основание столбика, который становится ги-нобазическим. От зрелого плода (ценобия) отделяются четыре сухие закрытые и невскрывающиеся односеменные части, которые ввиду их морфологического своеобразия нередко именуют не орешками, а эремами (Каден, Закалюкина, 1965).
Нижняя часть сросшихся плодолистиков представляет собой мясистый диск (гинобазис), функционирующий во время цветения в качестве нектарника. Развитие плодов вызывает формирование в ткани гинобазиса особых внутренних железок, раскрывающихся ко времени созревания эремов (Гогина, 1971а). Их порошковидный 140
секрет, представляющий собой, по-видимому, сильно трансформированные эфирные масла, покрывает вею поверхность зрелых эремов, которые довольно долго не осыпаются, так как этому препятствует волосистое кольцо, перегораживающее зев чашечки.
Зрелые эремы чабреца Маршалла имеют шаровидную форму и окраску от темно-коричневой до черной. Длина их в среднем 0,6 мм, однако она может заметно изменяться в зависимости от положения цветка в полумутовке и количества развивающихся зачатков. Наибольшей величины достигают одиночные эремы в центральных цветках полумутовки. Женские особи дают в среднем немного более крупные плоды. В условиях культуры вес 1000 эремов женской формы равнялся 191,5 мг, вес 1000 эремов обоеполой формы— 141,6 мг (Гогина, 1971 б).
По данным О. Ф. Билимович (1935), околоплодник эремов у чабрецов, в том числе у ч. Маршалла, состоит из наружного эпидермиса, паренхиматического слоя, механического слоя и внутреннего эпидермиса. Тонкостенные, спавшиеся в сухом состоянии клетки наружного эпидермиса имеют полигональную форму. При увлажнении они быстро ослизняются и пузыревидно вздуваются, приклеивая при подсыхании эрем к частицам'почвы. Паренхиматический слой образован несколькими рядами клеток, которые у зрелых плодов сильно смяты и трудно различимы. Механический слой представлен одним рядом клеток полигональной формы с сильно утолщенными стенками и маленькой полостью, вытянутой параллельно боковым стенкам. Внутренний эпидермис образован одним рядом тонкостенных сильно сжатых клеток. Клетки 1—2-рядного эндосперма имеют многоугольную форму и содержат алейроновые зерна и масла. Крахмал в клетках эндосперма и зародыша отсутствует. Зародыш прямой, с утолщенными округлыми семядолями и слабо дифференцированной осевой частью.
Диплоидное число хромосом по многочисленным определениям равно 28 (Хромосомные числа цветковых растений, 1969).
Как и другие виды рода, Т. marschallianus склонен к межвидовой гибридизации, давая при этом фертильное потомство. Он хорошо скрещивается с встречающимися в Подмосковье Т. pulegioid.es и Т. serpyllum. При скрещивании с первым из них процент завязавшихся гибридных семян в наших опытах был более высоким.
Географическое распространение
Т. marschallianus имеет весьма обширный ареал, который протягивается от Австрии и Чехии на западе до Алтая и Средней Азии на востоке. Для него характерна ясная широтная приуроченность к степной и лесостепной зонам (понтический тип ареала).
В Московской обл. северная граница естественного распространения ч.абреца Маршалла проходит по долине Оки, где он приурочен к островным участкам степной растительности и остепненным
141
лугам (Приокско-Террасный заповедник, выходы известняков в окрестностях Коломны). Эти местонахождения несколько оторваны от ареала сплошного распространения вида, охватывающего окское правобережье (рис. 15).
Рис. 15. Распространение видов чабреца в Московской обл.:
1 — Thymus serpyllum L. s. str., 2 — T. pulegioides L., 3 — T. marschallianus Willd., 4— T. loevyanus Opiz.
В более теплые и сухие периоды граница распространения вида, по-видимому, проходила севернее. На это, в частности, указывает произрастание чабреца Маршалла на остепненных лугах, развитых по высокому правому берегу долины Москвы и прилегающим водораздельным участкам, а также на вторичных местообитаниях в этом же районе, в частности на луговинах вдоль шоссе Москва — Рязань на протяжении между Боровским курганом и Коломной. Спорадические местонахождения Т. marschallianus вдоль насыпей железных дорог имеют, очевидно, заносный характер.
142
Онтогенез и сезонный ритм развития
Семена Т. marschallianus не нуждаются даже в кратковременном периоде покоя или дозревания. При комнатной температуре они начинают дружно прорастать на третий день после посева, отдельные— на второй день. Энергия прорастания весьма высока —в первые 7—10 дней прорастает основная масеа семян, лишь единичные эремы остаются в состоянии покоя в течение нескольких месяцев. По данным Л. М. Злобиной (1967), семена чабреца Маршалла, собранные с женских растений, имели несколько более высокую лабораторную всхожесть (95%), чем с обоеполых-форм (89%). В наших опытах семена обеих форм обладали одинаково высокой всхожестью (более 95%).
Полевая всхожесть семян значительно ниже лабораторной и существенно изменяется в зависимости от глубины их заделки. Максимальной (46%) оказалась всхожесть семян на поверхности почвы, хотя в этом варианте опыта прорастание в наибольшей степени зависело от колебания погодных условий. При заделке эремов на глубину 5 мм полевая всхожесть снижалась до 34%, семена, присыпанные слоем почвы толщиной в 10 мм, практически не прорастали (всхожесть 1,6%, лишь по трещинкам почвы). В естественных местообитаниях появление массовых всходов обусловлено погодными причинами. В годы с обильными осадками оно практически совпадает со временем обсеменения материнских растений, в сухие или холодные годы задерживается до осени или следующей весны.
По наблюдениям на экспериментальном участке Главного ботанического сада АН СССР, всходы чабреца Маршалла при весеннем посеве к осени успевают хорошо раскуститься. Ветвление начинается примерно через полтора месяца после появления всходов, когда они имеют около 4 пар листьев. Вскоре после этого семядоли, которые имеют почти округлую, при основании слабо сердцевидную форму, подсыхают. Первые ветви возникают'в пазухах семядолей и в нижних узлах всходов. К осени они достигают размера материнской оси и начинают в свою очередь ветвиться. Ветви третьего порядка нс успевают удлиниться; лучше всего они развиты в средней части осей второго порядка. Покоящимися остаются лишь мелкие недифференцированные почки в первых сближенных узлах боковых осей.
В природных фитоценозах ветвление нередко начинается лишь на второй год жизни растения и протекает весьма замедленно. Всходы из эремов разной величины вначале хорошо различаются но мощности развития, однако к концу вегетационного периода эти различия сглаживаются. Листья всходов мелкие, 2—3 мм длины, и имеют отчетливо выраженный черешок. Материнская ось всхода первоначально растет ортотропно, но примерно через два месяца поникает и продолжает свой рост в плагиотропной направлении. Нарастание материнской оси происходит моноподиально, по тормозящее влияние верхушечной почки выражено слабо, вследствие чего развитие дочерних осей отличается большой независимостью, и
143
главная ось вскоре теряется в массе хорошо развитых ветвей второго порядка.
Рост побегов продолжается до глубокой осени, но темп его значительно снижается, вследствие чего на осях возникает зона укороченных междоузлий, совпадающая с границей годичных приростов. Растения имеют открытые почки и уходят в зиму с зелеными листьями. В почках возобновления сформирована чрезвычайно малая часть побега будущего года — лишь зачатки двух-трех пар листьев, зачатки генеративных органов в них отсутствуют. Подснежного развития почек не наблюдается. Перезимовавшие листья возобновляют фотосинтез сразу же после стаивания снега. Развертывание новых листьев и весенний рост побегов начинаются примерно через две недели после этого. Листья осенней генерации и первые весенние листья, развившиеся из заложившихся осенью зачатков, отличаются меньшими размерами, лучше выраженным черешком и более грубой консистенцией.
В начале вегетационного периода в условиях культуры трогаются в рост почти все точки роста, за исключением почек в переходной зоне между годичными приростами и в основании ветвей. Возникающие из них многочисленные побеги растут ортотропно и сильно различаются по степени развития, которая зависит от размера их прошлогоднего прироста. Первые нижние ветви всхода дают в следующем году наиболее сильные оси, последние — наиболее слабые, нередко остаются вегетативными.
Большая часть побегов в начале июня переходит к цветению, успев дать пазушные побеги обогащения (как генеративные, так и вегетативные). Ко времени зацветания прирост прошлого года обычно одревесневает, а в узлах полегших осей, чаще у границ годичных приростов, при благоприятных условиях увлажнения образуются придаточные корни. К началу фазы плодоношения перезимовавшие листья и первые листья весенней генерации подсыхают, вследствие чего куст осветляется, и трогаются в рост покоившиеся пазушные почки на границе годичных приростов и в основании ветвей, из которых возникают плагиотропные побеги второй генерации — побеги возобновления. Одновременно усиливается рост отставших в развитии осей (рис. 16).
С окончанием фазы плодоношения закончившие развитие побеги отмирают, после чего моноподиальное нарастание скелетных осей сменяется симподиальным. Отмирает почти весь весенний прирост этого года. Таким образом, истинный прирост куста-—нарастание его многолетней части — происходит за счет плагиотропных побегов возобновления, возникающих из пазушных почек с выраженным периодом покоя. Обильно ветвясь, эти побеги образуют к осени густую зеленую поросль в основании засохших отплодоносивших ветвей. Дальнейшее развитие куста весной следующего года происходит в основном за счет их отрастания. Следовательно, большинство побегов чабреца Маршалла развивается по озимому типу, причем ежегодно отмирает их большая часть, образовавшаяся весной.
6 Зак. 255
145
Кроме этого основного типа побегов могут быть выделены весенние эфемерные побеги обогащения и побеги, не успевшие зацвести в начале вегетационного периода. Часть таких отставших побегов отмирает, часть заканчивает свое развитие на третий вегетационный период. Такие побеги с удлиненным циклом развития принимают существенное участие в создании скелетной системы куста. В конце лета, после плодоношения рост их заметно усиливается, они полегают на поверхность почвы и начинают усиленно ветвиться. Побеги этого типа обычно имеют хорошо развитую стелющуюся часть и могут достигать значительной длины, способствуя расползанию куста. Как правило, у них ясно различимы нижняя неветвящаяся часть, развившаяся весной в условиях повышенного затенения и угнетения, и образовавшаяся в конце лета интенсивно ветвящаяся часть. При благоприятных условиях некоторые из таких отставших побегов успевают зацвести в конце вегетационного периода, создавая слабо выраженную волну вторичного цветения.
Два периода активного роста побегов, различающиеся по его направленности, обусловливают сезонный диморфизм куста. Побеги обогащения, образующиеся из пазушных почек, не имеющих периода покоя, большей частью укорочены до розеток листьев в-, узлах весеннего прироста материнской оси. В благоприятных условиях, в особенности в культуре, они могут удлиняться и переходить к цветению, что более обычно для побегов в верхних, приближенных к соцветию узлах. Такие побеги отличаются удлиненными первыми междоузлиями, листья их заметно мельче и уже, чем на материнской оси.
Различные типы побегов не всегда строго разграничены. Вегетативные побеги обогащения обычно отмирают вместе с материнской осью, но иногда они трогаются в рост. По мере своего удлинения такие побеги достигают поверхности почвы и принимают на себя функции побегов возобновления. В этих случаях весенний прирост материнской оси отмирает неполностью, сохраняется ее отрезок, расположенный ниже такой изменившей тип развития боковой ветви.
Климатические и фитоценотические условия, в которых происходит развитие растения, существенно сказываются на его внешнем облике. Под их влиянием изменяются соотношения типов побегов и степень их развития. Благоприятная температура и хорошие условия увлажнения во второй половине лета стимулируют рост побегов второй генерации. Вследствие этого оси, продолжающие свой рост на следующий год, имеют отчетливо выраженную стелющуюся прошлогоднюю часть, которая нередко укореняется. В разреженных фитоценозах, при повышенной освещенности ветвление побегов второй генерации очень интенсивно, в результате чего усиливается конкуренция между ними, возрастает число побегов с удлиненным циклом развития и формируются крупные расползающиеся кусты.
В сомкнутых фитоценозах побеги второй генерации сильно
146
удлиняются, но мало ветвятся, что увеличивает рыхлость куста, развивающегося на следующий год.
При повышенной сухости или низких температурах осеннее отрастание подавлено, поэтому стелющаяся прошлогодняя часть осей укорочена и не укореняется; это ослабляет развитие побегов второй генерации на границе годичных приростов в конце следующего вегетационного периода. В этих условиях возобновление происходит главным образом за счет развития почек в основании осей, растения слабее ветвятся, что приводит к более равномерному развитию побегов (побеги с удлиненным циклом развития формируются в меньшем числе) и куст приобретает более компактную форму.
Эти особенности побегообразования обусловливают высокую лабильность структуры куста Т. maschallianus, в которой находят отражение погодные условия не только текущего, но и предыдущего года. Немалое значение в отношении степени выраженности побегов различного типа имеют и наследственные различия отдельных рас и особей. В сравнительно влажном климате Московской обл. пауза в развитии, наступающая в начале фазы плодоношения, несколько сглажена. В более засушливых, условиях пробуждение покоящихся почек и вторичная активизация роста побегов задерживается, вследствие чего период летнего покоя выражен более резко.
Будучи типичным представителем Жизненной формы полукустарничков, чабрец Маршалла характеризуется преобладанием ба-зисимподиального нарастания скелетных осей и отмиранием большей части годичных побегов (преимущественно весеннего прироста). Плагиотропная часть куста образована древеснеющими побегами второй генерации, ортотропная — травянистыми весенними побегами.
Ежегодная смена направления роста осей, совпадающая с сезонным ходом изменения климата, улучшает условия перезимовки для незащищенных точек роста и должна рассматриваться как приспособление растения к переживанию неблагоприятного времени года. Чабрец Маршалла, как и большинство других видов рода, относится к группе гемикриптофитов.
Закономерности развития структуры куста осложняются наличием переходных форм между отдельными типами побегов.
Незащищенные покровами, открытые почки тимьяна нередко отмирают под влиянием неблагоприятных условий (засуха, заморозки, механические повреждения), что особенно часто наблюдается у молодых, не успевших окрепнуть растений и всходов. Вследствие этого моноподиальное нарастание обычно сменяется симподпальпым до вступления растений в генеративную фазу. В условиях культуры, однако, при благоприятной перезимовке отмирают лишь завершившие цикл своего развития отплодоносившие оси.
Виргинильный период у культивируемых особей значительно укорочен. При весеннем посеве растения зацветают на второй период вегетации (иногда в конце первого). В естественных местообитаниях развитие протекает значительно медленнее, ветвление юве
6:
147
нильных растений сильно ослаблено, цветение начинается лишь на 4—5 году жизни.
Выраженность отдельных скелетных осей в общей структуре куста и длительность их жизни изменяются в зависимости от условий среды. Факторы, благоприятствующие генеративному развитию растений, ускоряют процесс смены скелетных осей, в особенности если создаются условия, неблагоприятные для осеннего отрастания и укоренения побегов. В условиях, способствующих укоренению скелетных осей и задержке генеративного развития, продолжительность жизни скелетных осей увеличивается до нескольких лет, и многолетняя плагиотропная часть куста хорошо выражена.
В культуре отдельные экземпляры чабреца достигали лишь 5— 8-летнего возраста. Гибель растений вызывалась в основном неблагоприятными условиями перезимовки ( вымоканием, вымерзанием и т. п.). Наиболее чувствительными к неблагоприятным условиям оказались особенно обильно плодоносившие, хорошо развитые растения. В природных местообитаниях продолжительность жизни растений, по-видимому, может быть значительно более продолжительной.
Способы размножения и распространения
Чабрецу Маршалла, как и другим представителям рода, свойственна гинодиэция, или женская двудомность. Отдельные популяции существенно различаются по процентному соотношению женских и обоеполых форм. Наряду с константными половыми формами в составе популяций встречаются особи с неустойчивым, изменяющимся половым состоянием. Первые цветки в соцветиях таких особей могут быть обоеполыми, а раскрывшиеся позднее — чисто женскими. Существенных различий во времени зацветания разных половых форм у этого вида не наблюдалось, по признакам вегетативной сферы они тождественны (Гогина, 1970).
Цветение начинается в первых числах июня и продолжается обычно около месяца. Цветки раскрываются, как правило, утром; при пасмурной погоде раскрывание их может запаздывать. Продолжительность цветения отдельного цветка зависит от его пола и времени опыления. Обоеполые цветки, характеризующиеся протеран-дричностью развития, цветут в среднем на день-два дольше женских. В течение одного-двух дней после расцветания они находятся в мужской фазе развития. Гнезда пыльников уже в бутонах имеют продольную щель, вдоль которой по мере развития пыльника формируется ряд чувствительных клеток. Прикосновение к ним вызывает активное выдавливание слипшихся пыльцевых зерен из пыльника. Эта особенность лучше выражена у крупноцветковых видов рода, а у мелкоцветковых, к которым относится и Т. marschallianus, она несколько ослаблена. На второй-третий день постепенно удлиняющийся столбик перерастает сначала задние, короткие, а затем и передние тычинки и выдвигается за край верхней губы венчика. После
148
этого раскрываются лопасти его рыльца и цветок вступает в женскую фазу развития. Растущий столбик прижат к верхней губе венчика и минует поэтому сближенные пыльники коротких тычинок сверху, со стороны их утолщенного связника, что также препятствует самоопылению. (У интерсексуальных форм эти закономерности развития цветка нередко оказываются нарушенными.) Продолжительность женской фазы зависит от времени опыления. В разгар цветения при хорошей погоде, когда посещения цветков насекомыми-опылителями учащаются, она короче и равна 'в среднем двум дням. Это относится в равной мере и к цветкам женских особей, у которых выпадает мужская фаза развития. В условиях изоляции продолжительность цветения значительно увеличивается: женские цветки сохраняются до 8—10 дней, обоеполые — до 10—12 дней. Опыты с изоляцией обоеполых соцветий чабреца показали, что про-терандрия, как правило, надежно предохраняет его цветки от самоопыления (завязывание семян наблюдалось лишь в единичных условиях).
В естественных условиях, благодаря продолжительному цветению отдельных экземпляров и соцветий, опыление собственной пыльцой вполне возможно, так как цветки посещаются многими видами насекомых. П. Кнут (Knuth, 1899) перечисляет 72 вида опылителей тимьянов. По нашим данным их наиболее часто посещают пчелы и муравьи, которые обследуют почти все цветки каждого цветущего растения. Лишь женские особи при этом гарантированы от самоопыления. Пониженная плодовитость обоеполых особей, возможно, является следствием частичного самоопыления, производимого насекомыми при свободном переопылении. Подсчеты показали, что семенная продуктивность женских особей более чем в два раза превышает продуктивность обоеполых (у женских особей она составляла 68,6%, у обоеполых — 31,1%). В среднем в условиях культуры число эремов в одном соцветии обоеполой особи равнялось — 164, в одном соцветии женской особи — 269.
Хотя созревание отдельного эрема происходит довольно быстро, приблизительно в течение четырех-пяти недель, окончание периода плодоношения растянуто примерно на месяц, оно заканчивается в конце июля: Созревшие эремы некоторое время задерживаются в чашечке благодаря волосистому кольцу, перегораживающему ее зев. Их высыпание происходит при резком сотрясении или раскачивании цветоносного стебля. Основная масса осыпавшихся эремов оказывается при этом в непосредственной близости от материнского растения. В литературе нет данных о запасе семян тимьянов в почве; возможно, тонкие покровы эремов не могут обеспечить долгое сохранение жизнеспособности семян.
Для сохранения численности особей в ценозах решающее значение имеет генеративное размножение.
В редких случаях возможно разделение куста на отдельные пар-тикулы, которое происходит при отделении укоренившихся скелетных осей в результате механических повреждений. В целом же вид
149
характеризуется ограниченной способностью к вегетативному размножению.
Мелкие эремы чабрецов, как уже упоминалось, не содержат значительных запасов питательных веществ и отличаются дружным прорастанием. Поэтому судьба всходов сильно зависит от колебания условий среды. Однако, несмотря на большой процент отпада, который может сильно изменяться по годам, семенное размножение осуществляется достаточно эффективно, вследствие чего общая численность популяций чабреца в местах естественного произрастания оказывается довольно стабильной.
Экология и фитоценология
Как и другие степные растения, чабрец Маршалла относится к группе ксерофитов. Однако приуроченность этого вида к сухим местообитаниям объясняется не столько его сухолюбием, сколько невысокой конкурентной способностью и засухоустойчивостью. Изолированные популяции чабреца, имеющие бесспорно заносное происхождение, могут существовать значительно севернее его основного ареала, например в Финляндйи (Jalas, Kaleva, 1970).
Особенности сезонного развития также свидетельствуют о его ограниченном сухолюбии. К наступлению засушливого периода в местах естественного произрастания тимьян обычно уже находится в фазе плодоношения, которая сопровождается приостановкой ростовых процессов. К этому времени растения сбрасывают большую часть листьев, что значительно уменьшает их транспирацию. Вторая активизация роста совпадает обычно с увеличением количества осадков в конце вегетационного периода. В условиях засухи наблюдается массовый отпад всходов и отмирание концов побегов у ювенильных особей. О чувствительности этого вида к недостатку влаги свидетельствует также значительное улучшение его роста в условиях достаточного водоснабжения.
Вместе с тем избыточное, застойное увлажнение губительно действует на чабрец Маршалла, даже если оно не слишком продолжительно. При таких условиях, создающихся преимущественно ранней весной или осенью, в коллекции тимьянов, собранной в Главном ботаническом саду АН СССР, резко возрастала гибель растений.
~ Будучи весьма чувствительным к условиям освещения, ч. Маршалла предпочитает разреженные сообщества травянистых ксерофитов, хотя изредка заходит на опушки и поляны немногочисленных лесных ценозов степной зоны. Наилучшего развития растения достигают при полном солнечном освещении. Экземпляры, выросшие в густых ценозах, отличаются ослабленным ветвлением, удлиненными междоузлиями и уменьшением количества цветоносных стеблей. В сомкнутых лесных насаждениях чабрец Маршалла не встречается.
Ареал вида отчетливо связан с областью распространения черноземных почв (с их многочисленными разностями), однако не избе
150
гает он и каменистых субстратов со слабо развитым почвенным покровом; в северной части ареала чабрец Маршалла встречается на луговых-почвах. В местах своего естественного произрастания на юге Московской обл. в пойме Оки он растет на хорошо дренированных почвах, заливаемых полыми водами лишь на короткий срок; в окрестностях Коломны встречается на неглубоких дерновых почвах, подстилаемых известняком. В местообитаниях с повышенной кислотностью почв отсутствует.
На территории Приокско-Террасного заповедника, по наблюдениям П. А. Смирнова (1958), встречается редко, отдельными группами в составе разнотравно-типчаковых сообществ. В окрестностях Коломны (близ с. Коробчеево) он сохранился лишь на небольшом участке естественной растительности по краю выработанного известнякового карьера. Вместе с ним здесь растут Aster amellus, Anemone sylvestris, Salvia verticillata, Coronilla varia, Achillea nobilis И др.
Хозяйственное значение
В ряде областей СССР чабрец Маршалла используют в лечебных целях в качестве антисептического и отхаркивающего средства. Главным действующим веществом является тимол, входящий в состав эфирного масла. Отдельные расы и особи далеко не однородны по составу и содержанию эфирного масла в надземных органах. Соотношение компонентов эфирного масла и его качественные показатели могут существенно изменяться в зависимости от фенологического состояния растений. Наибольший выход масла они дают в фазе цветения. Его содержание в отдельных органах растений также различно: наибольшее— в соцветиях, наименьшее — в стеблях. Общее содержание масла в абсолютно сухом сырье этого вида колеблется от 0,27 до 1, 68% (Казакевич, Соболевская, 1928; Васильев, Кисис, 1935; Киченко, 1852). Чабрец Маршалла находит применение в пищевой промышленности, где используется как пряность.
2. ЧАБРЕЦ (ТИМЬЯН) БЛОШИНЫЙ
Номенклатура и систематическое положение
Т. pulegioides L., Sp. pl. (1753) 592; Клоков во Фл. СССР 21 (1954) 487; Ворошилов в Опр. раст. Моск. обл. (1966) 283; Jalas in Fl. Eur. 3 (1972) 182,— T. chamaedrys Fries, Nov. Fl. Sues. 2 (1814) 35; Петунн., Крит, обзор 2 (1900) 128; Иконников-Галицкий в Маевск., Фл., изд. 7 (1940) 624.—Г. serpyl-lum L. var. chamaedrys auct: Кауфм., Моск. Фл., изд. 1 (1866) 374, изд. 2 (1889) 385; Цингер, Сб. свед. (1885) 342; Шмальг., Фл. 2 (1897) 311.— Т. glaber Mill., Gard. Diet., ed. 7 (1759) № 6; Сырейщ.,
151
Илл. Фл. 3 (1910) 79, 4 (1914) 141; он же, Опр. раст. Моск. губ. (1927) 218.— Т. pulegioides L. subsp. ucrainicus Klok. et Shost. в BicH. Кшвськ. бот. саду 16 (1932) 6.— T. ucrainicus (Klok. et Shost.) Klok. во Фл. СССР 21 (1954) 486; Борисова в Маевск., Фл., изд. 8 (1954) 521, изд. 9 (1964) 502.— Ч. блошиный, ч. пулегевидный.
Во «Флоре СССР» (1954) отнесен к секции Goniothymus Klok. (подсекция Goniotrichi Borb., ряд Pulegioides Klok et Shost.); во «Flora Europaea» (1972)—к секции Serpyllum (Miller) Bentham. Описан по материалу из Франции (Монпелье) К. Линнеем, который, однако, во втором издании «Species plantarum» отнес этот вид к роду Cunila под названием С. thymoides. Тип хранится в гербарии. Линнея в Лондоне.
Многими авторами включался в состав обширного сборного вида Т. seryllutri L. s. 1. Название Т. pulegioides L. было вновь введено в употребление и утвердилось в литературе благодаря работам Роннигера (Ronniger, 1923, 1924). Против использования этого названия возражал Клоос (Kloos, 1925), поскольку один из экземпляров, на которых основывался Линней при описании вида, имеет изросшее соцветие. Хиландер (Hylander, 1945), однако, справедливо считает это возражение неубедительным. Подобное явление нередко наблюдается у тимьянов и, по нашему мнению, не может служить достаточной причиной для отказа от линнеевского названия вида.
На протяжении своего ареала Т. pulegioides распадается на ряд мелких трудно различимых разновидностей и форм, систематическое значение которых оценивается весьма противоречиво. Так, М. В. Клоков (Флора СССР, 1954) выделяет восточную расу Т. ри-legioides в особый вид — Т. ucrainicus (Klok. et Schost.) Klok. Ее отличия от западноевропейских форм, однако, весьма незначительны, поэтому их объединение, произведенное Б. Павловским (Pawlowski, 1966) и П. Шмидтом (Schmidt, 1971), следует признать правильным.
Морфологическое описание
По типу побегообразования Т. pulegioides сходен с чабрецом Маршалла. Он также представляет собой симподиально нарастающий полукустарничек и имеет главный корень стержневого типа, сохраняющийся в течение всей жизни растения. У этого вида, однако, более сильно развиты плагиотропная часть осевой системы и придаточные корни. Глубина проникновения главного корня, по-видимому, не превышает 30—40 см (по наблюдениям за культивируемыми растениями). Высота цветоносных стеблей весьма изменчива, в среднем около 20 см. В поперечном сечении стебли имеют острочетырехгранную форм'у. Пары супротивных граней каждого междоузлия различаются по ширине — две из них более узкие и вогнутые. Короткое ретрорзное опушение развито только по ребрам. Листья короче междоузлий, ясночерешковые/яйцевидной или яйцевидно-эллиптической формы, по черешку до основания пластинки
152
ресничатые, с камптодромным жилкованием и мало заметными жилками. Длина листьев с черешком в среднем равна 10—15 мм, ширина 4—10 мм. Количество устьиц на верхнем эпидермисе листа значительно меньше, чем на нижнем, что отличает этот вид от других представителей рода (Hegnauer, 1948; Kaleva, 1967).
Соцветие в начале цветения головчатое, позже вытягивающееся и рыхлеющее, нередко прерванное. Нижние полумутовки содержат до 15—17 цветков, верхние полумутовки обедненные. Зацветание цветков в соцветии и в пределах полумутовок подчинено тем же закономерностям, что и у чабреца Маршалла. Нижние прицветные листья по форме сходны со стеблевыми, верхние уменьшенные.
Чашечка обычной для рода формы, колокольчатая, во время цветения от 2,5 до 3,5 мм длины, ко времени плодоношения увеличивающаяся до 3,5 — 4,5 мм. Зубчики верхней губы остротреугольные, по краю ресничатые. Трубка чашечки в нижней половине опушена отстоящими волосками, сверху в основании верхней губы оголенная. Венчик 4—6 мм длины (у женских цветков до 2—4 мм), розово-лиловый. Строение цветка и его частей аналогично описанному у чабреца Маршалла, но тычинки и столбик сильно пространственно разобщены и более отчетливо выражена протерандрия. Пыльца про-долговато-шаровидная, шестибороздная, ее диаметр по продольной оси 24,8, по экваториальной — 22,9 мк (Sorsa, 1966).
Зрелые эремы имеют округлую форму и окраску от светло-коричневой до коричневой. Как и у чабреца Маршалла, они заметно варьируют по размеру, в среднем их диаметр равен 0,7—0,8 мм.
Диплоидное число хромосом равно 28 (Хромосомные числа цветковых растений, 1969). У этого вида известен, однако, также редко встречающийся цитотип с 2n = 30 (Trela-Sawicka, 1968).
Географическое распространение
Ареал Т. pulegioides охватывает умеренную зону Западной и и Центральной Европы, обнаруживая ясную связь с областью распространения листопадных лесов. Этот вид встречается как на равнинной территории, так и в горах, поднимаясь на наибольшую высоту в южной части своего ареала. Он распространен от Пиренейского п-ова и южной части Англии на западе до центра европейской части СССР на востоке (Московская обл.). На Скандинавском п-ове заходит довольно далеко на север, более обычен в южной Швеции. На юг дальше всего проникает по горным системам Балканского и Пиренейского полуостровов.
Вследствие сильных антропогенных изменений растительного покрова Европы значительная часть местообитаний этого вида имеет вторичный характер. Его первичными местообитаниями являются, по-видимому, лесные опушки и поляны, откуда он распространился в сообщества, возникшие на месте сведенных лесов. Многие его местонахождения являются результатом заноса. Это, по-види
153
мому, единственный представитель рода, проникший в Северную Америку, куда он был занесен из Европы (Roussine, 1961).
В Московской обл. Т. pulegioid.es относится к числу редких растений. Наибольшее число его местонахождений известно из западной части области. На юге он заходит в пределы ареала тимьяна Маршалла (долина Оки в окрестностях Коломны, р. Лопасня). Ряд сообществ, в которых произрастает ч. блошиный на территории области, имеет очевидный вторичный характер. В прошлом он встречался и в пределах современной Москвы (Сокольники, берег р. Москвы у Симонова монастыря), но в настоящее время там: отсутствует (рис. 15).
Онтогенез и сезонный ритм развития
По строению эремов и особенностям их прорастания ч. блошиный весьма сходен с чабрецом Маршалла. Условия окружающей среды оказывают столь же сильное влияние на быстроту и мощность развития его всходов.
В условиях культуры всходы начинают интенсивно ветвиться на ранних фазах развития. Как и у предыдущего вида, первые ветви появляются в пазухах семядолей приблизительно через месяц — полтора после прорастания, когда главная ось всхода имеет лишь 3—4 пары листьев. Немногим позже направление роста материнской оси изменяется, она полегает на поверхность почвы. Дальнейший рост ее происходит плагиотропно и сопровождается непрерывным образованием ветвей, которые в свою очередь начинают ветвиться. Ветвление продолжается обычно до образования осей третьего-чет-вертого порядков. Наиболее мощные ветви образуются в нижних узлах главной оси всхода. Покоящимися остаются, как правило, лишь почки в первых узлах боковых осей.
Первые листья всходов отличаются мелкими размерами (2— 3 мм), последующие довольно быстро сравниваются с листьями взрослых растений. К концу вегетационного периода растения образуют густые дерновинки, достигающие 30 см в диаметре. В естественных местообитаниях развитие всходов сильно замедлено. Их ветвление обычно ограничивается образованием ветвей в пазухах семядолей, но может протекать и значительно интенсивнее, поскольку на темпе роста растений сказываются даже мелкие различия в условиях существования.
К осени скорость роста побегов снижается, вследствие чего на осях возникает зона сближенных междоузлий с покоящимися пазушными почками; ее выраженность зависит от климатических условий года. Ч. блошиный имеет зимнезеленые листья и открытые почки возобновления с небольшим числом зачатков.
Большим сходством отличается и сезонный ритм развития побегов у обоих видов. Он также характеризуется двумя периодами активного роста: весенним ортотропным и летне-осенним пдагио-
154
тропным. У ч. блошиного могут быть выделены те же основные типы побегов, но их роль в создании структуры куста оказывается иной. Тип развития побегов у этого вида может легко изменяться под влиянием внешних условий, что отражается на облике растений. Перезимовавшие точки роста весной образуют ортотропные цветоносные и вегетативные побеги, их соотношение в сильной степени зависит от условий внешней среды. Наиболее сильные скелетные оси и пазушные ветви, заложившиеся в прошлом году во второй половине постфлоральной активации роста, завершаются обычно терминальным соцветием. Вегетативные ветви образуются чаще всего из почек, тронувшихся в рост в предыдущем году в начале периода вторичного роста материнской оси, т. е. в средней части годичного прироста, и из почек вблизи границы годичных приростов. В последнем случае они слабее.
Во время цветения вегетативные побеги замедляют свой рост. Он вновь усиливается в конце фазы плодоношения, что вызывает полегание этих осей. Такие побеги с удлиненным циклом развития, которые могут быть названы побегами замещения, имеют большое значение в формировании структуры куста ч. блошиного, способствуя увеличению ее плагиотропной части. Обычно они развиваются по дициклическому типу и приурочены к средней части годичного прироста материнской оси, вследствие чего у этого вида преобладает мезосимподиальное нарастание скелетных осей. Более многочисленны у тимьяна блошиного и генеративные побеги обогащения. Побеги возобновления, развивающиеся из покоившихся почек и образующие вторую генерацию побегов, выражены слабее, чем у чабреца Маршалла. Они возникают обычно в зоне укоренения полегших осей у границ годичных приростов и в основании скелетных осей. Повреждение материнской оси или ее отмирание заметно стимулирует их образование.
В условиях затенения и угнетения характерны подавленность ветвления и удлинение цикла развития побегов (до 3, реже до 4 лет). В результате возникают длинные, мало разветвленные стелющиеся оси, которые, расползаясь, образуют на поверхности почвы рыхлую сеть с далеко расставленными олиственными ортотропными побегами на концах. В этих условиях иногда наблюдается нарушение обычного порядка зацветания осей — точка роста материнской оси может остаться вегетативной, тогда как пазушные ветви переходят к цветению. В подобных случаях дольше сохраняется мо-ноподиальный тип нарастания скелетных осей. Этот признак, имеющий в принципе весьма важное значение для систематики, не может, следовательно, считаться у ч. блошиного абсолютно стабильным (рис. 17).
У подобных ценотических форм одревесневшие части осей почти не утолщаются, что увеличивает возможность их отчленения. Если' такие обособившиеся части имеют достаточно развитые придаточные корни, они переходят к самостоятельному существованию, что делает возможным вегетативное размножение вида.
155
Рис. 17. Особи чабреца блошиного, выращенные из семян одного происхождения, высеянных одновременно:
А — подсев в природный ценоз (первое цветение на четвертом году жизни; Б—в культуре (первое цветение на втором году жизни, на фото — половина особи)
156
В благоприятных условиях, благодаря ,интенсивному ветвлению, образуются крупные компактные кусты с многочисленными ортотропными побегами. Обильное цветение и плодоношение обусловливают массовое отмирание завершивших цикл своего развития генеративных побегов, которое компенсируется быстрым ростом и ветвлением побегов замещения, восстанавливающих густоту куста (рис. 17).
Способы размножения и распространения
Чабрец блошиный относится к числу наиболее поздно цветущих видов рода. Его цветение начинается обычно в последних числах июня и продолжается до коцца июля. В условиях культуры у побегов, зацветающих последними, нередко наблюдается израстанис соцветий. Такие соцветия, имеющие обычно уменьшенное количество цветков в полумутовках, представляют собой связующее звено между моноциклическими генеративными побегами, завершающими в текущем году цикл своего развития, и дициклическими побегами замещения, остающимися до будущего года в вегетативном состоянии.
Популяции ч. блошиного, как и у многих других представителей рода, состоят из смеси женских и обоеполых форм; как показали подсчеты, их соотношение в некоторых подмосковных популяциях оказалось близким к 1:1. Наряду с устойчивыми женскими и обоеполыми формами в состав популяций обычно входят растения с уменьшенным числом тычинок и резко увеличенным процентом стерильной пыльцы. У таких особей нередко нарушается нормальная последовательность развития цветка. У некоторых из них при отсутствии внешне выраженных отклонений созревание пыльников наступает после полного развития гинецея.
Как и у других видов, обоеполые особи чабреца блошиного значительно менее плодовиты, чем женские. Это явление, по-видимому, нельзя объяснить одним лишь вредным влиянием самоопыления, которое в природе может осуществляться насекомыми-опылителями. Опыты с искусственным перекрестным опылением обоеполых растений показали, что количество завязавшихся при этом семян1 значительно ниже, чем при опылении женских особей пыльцой тех же отцовских растений. Полученные данные свидетельствуют об определенном доминировании мужского пола над женским у гермафродитных растений.
Экология и фитоценология
Ч. блошиный—мезофильное растение. Ареал его приурочен к территории, получающей достаточно большое количество осадков. В засушливых областях этот вид не встречается, что говорит о его чувствительности к недостатку влаги. Вместе с тем он предпочитает местообитания с хорошо дренированными почвами — избыточное застойное увлажнение действует на него угнетающе.
157
Большое значение для распространения этого вида имеют и условия освещения. Отличаясь повышенным светолюбием, он предпочитает открытые места, встречаясь преимущественно в травянистых сообществах на месте сведенных лесов, по лесным опушкам и обочинам дорог. Он произрастает на почвах разнообразного состава, имеющих нейтральную или умеренно кислую реакцию.
Будучи конкурентно слабым растением, ч. блошиный в сомкну-: тых сообществах оказывается оттесненным на участки с нарушенным растительным покровом. Он нередко селится на колониях поч-венных муравьев, что, возможно, объясняется не только особенностями его экологии, но и тяготением муравьев к чабрецу.
Внутривидовая систематика
Высокий наследственный полиморфизм Т. pulegioides проявляется даже в пределах относительно небольшой Московской обл. Здесь представлены хорошо различающиеся популяции с крупными и широкими и с более мелкими и узкими листьями., Эти различия, сохраняющиеся и в условиях культуры, возможно, объясняются отдаленными следами гибридизации данного вида с тимьяном ползучим, так как отдельные участки их ареалов, несомненно, могли соприкасаться в прошлом.
Способность к межвидовым скрещиваниям проявляется у тимьяна блошиного в весьма высокой степени, известны его гибриды с видами других секций, даже с Т. vulgaris L. (Schratz.Growm, 1968).
Хозяйственное значение
В литературе, посвященной лекарственным и эфирномасличным растениям, как правило, приводятся описания лишь двух признанных лекарственными видов — Т. vulgaris L. и Т. serpyllum L. Чабрец блошиный в ней незаслуженно не упоминается. Это объясняется сохранением устаревших систематических воззрений на объем вида Т. serpyllum, который понимается в самом широком смысле. Как показывает, однако, ряд исследований (Hegnauer, 1948; Messer-schmidt, 1965; Лякавичюс, Ясконис, 1968), именно Т. pulegioides представляет собой среди форм, входящих в комплекс Т. serpyllum L. s. 1., наиболее перспективный для лекарственного использования вид. По данным А. А. Лякавичюса и Ю. А. Яскониса (1968), он содержит примерно в десять раз больше эфирного масла, чем Т. serpyllum L. При этом внутривидовые формы чабреца блошиного заметно различаются по содержанию эфирного масла. Максимальный выход масла достигал в литовских популяциях 1,21% от абсолютно сухого веса сырья, минимальный — 0,11 %. Общее количество фенолов, входивших в состав масла, равнялось 38—42%. По данным В. Мессершмидта (Messerschmidt, 1965), содержание эфирного масла в различных по происхождению образцах этого вида колебалось
158
от 0,19 до 0,68%, причем обнаружена и значительная изменчивость •состава его компонентов (тимол—1—4%, карвакрол — 4,9—32,5%, борнеол — 0,1—14,9%, гераниол — 2,5—9,6%). Эфирное масло ч. блошиного находит применение в медицине как антисептик и средство от кашля (входит в состав «пертуссина»), в пищевой и парфюмерной промышленности.
3. ЧАБРЕЦ (ТИМЬЯН) ПОЛЗУЧИЙ
Номенклатура и систематическое положение
Т. serpyllum L., Sp. pl. (1753) 590; Кауфм., Моск, фл., изд. 1 (1866) 373, изд. 2 (1889) 385; Цингер, Сб. свед. (1885) 341, р. р., Шмальг., Фл. 2 (1897) 310; Петунн., Крит, обзор 2 (1900) 128; Сы-1рейщ., Илл. фл. 3 (1910) 78; он же, Опр. раст. Моск. губ. (1927) 218; Иконников-Галицкий в Маевск., Фл., изд. 7 (1940) 625; Клоков во Фл. СССР 21 (1'954) 537; Борисова в Маевск., Фл., изд. 8 (1954) 522, изд. 9 (1964) 502; Ворошилов в Опр. раст. Моск. обл. (1966) 283,— Т. serpyllum L. subsp. serpyllum: Jalas in Fl. Eur., 3 (1972) 182.— 4. ползучий.
Во «Флоре СССР» (1954) отнесен к секции Euserpyllum Klok., ряду Euserpylla Klok. et Shost.; во «Flora Europaea» (1972) — к секции Serpyllum (Miller) Bentham. Описан, по-видимому, по образцам из Швеции. Типовой экземпляр, к сожалению, утрачен (Schmidt, 1969).
Краткость данного Линнеем описания, допускающая различные толкования, послужила основанием для чрезмерного расширения объема вида. Значение названия было ограничено Миллером, который связал его с распространенным в Швеции растением. Эта точка зрения в настоящее время общепринята. В таком понимании Т. serpyllum L. emend Mill, представляет собой отчетливо очерченный вид с двумя хорошо различающимися подвидами: типовым subsp. serpyllum [Г. serpyllum L. subsp. angustifolius Celak., Prodr. Fl. Boh-men. 2 (1871) 351.— T. serpyllum L., Sp. pl. (1753) 590 p. p.; Клоков во Фл . СССР 21 (1954) 537] и северным subsp. tanaensis (Hyl.) Jalas in Acta Bot. Fenn. 39 (1947) 20. [T. subarcticus Klok. et Schost. в Журн. Инет. бот. АН УССР 9/17 (1936) 194; Клоков во Фл. СССР 21 (1954) 538].
Морфологическое описание
В отличие от двух предыдущих видов чабрец ползучий имеет моноподиально нарастающие многолетние скелетные оси, стелющиеся по поверхности почвы, которые обычно сильно ветвятся и укореняются. Ортотропные неветвящиеся генеративные побеги отходят от узлов этих осей. Как правило, они развиваются на прошлогодних участках осей из перезимовавших в укороченном состоянии
159
пазушных ветвей, реже — как побеги обогащения на приросте текущего года.
Длительно сохраняющийся главный корень имеет стержневую форму, глубина его проникновения в почву не превышает обычно 20—30 см, но на песках глубина проникновения может достигать, 50 см (Pigott, 1955). Весьма важную роль в жизни растения играют многочисленные придаточные корни, образующиеся на стелющихся,, участках осей. По форме роста ч. ползучий относится к группе низ-' ких шпалерных полукустарничков. Высота его генеративных органов не превышает обычно 7—10 см, нередко встречаются особи с более низкими цветоносными стеблями (до 2—3 см). Стебель в поперечном сечении округло-четырехгранный, равномерно опушенный отстоящими или книзу отклоненными волосками, более длинными под соцветием.
Листья эллиптические, удлиненно-эллиптические или узко-обратнояйцевидные, 5—7 (до 10) мм длины, 1,5—3 (до 4) мм ширины, суженные в короткий черешок, в основании цветоносных стеблей почти сидячие, на вегетативных побегах несколько более узкие, по черешку и в основании пластинки реснитчатые, с толстоватыми, выступающими снизу жилками и мало заметными железками. Прицветные листья несколько шире, по краю более высоко реснитчатые.
Соцветие головчатое, с короткими цветоножками и небольшим числом цветков (1—3, редко 5) в полумутовках, к концу цветения мало удлиняющееся. В среднем в соцветии насчитывается около 24 цветков.' Чашечка 3,5—4,25 мм длины, с треугольными верхними зубчиками, более или менее реснитчатыми по краю. Трубка чашечки опушена реснитчатыми волосками, под зубцами сверху оголенная. Венчик 6—8 мм длины, розовато-лиловый с крапчатым рисунком на нижней губе. Строение цветка в принципе аналогично описанному выше у других видов. Пыльца шестибороздная, вытянуто-шаровидная, с поперечником по экваториальной оси 24,5 мк, по продольной— 26,4 мк (Sorsa, 1966).
Зрелые эремы округлые, коричневые, около 0,7 мм в диаметре.
Диплоидное число хромосом по неоднократным определениям равно 24. Имеются, однако, сообщения о существовании форм с 2 п=20 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Ареал Т. serpyllum s. str. не выходит за пределы Европы. Имевшиеся в нашем распространении материалы из Сибири, хотя и относились к близкородственным видам, не были тождественны ему. Наши наблюдения подтверждают вывод П. Шмидта (Schmidt, 1969) о том, что Т. serpyllum представляет собой чисто европейский вид, не переходящий за Урал; по-видимому, он не встречается восточнее района Сыктывкара. Северный его подвид заходит за Полярный круг, достигая северной оконечности Скандинавского п-ова, граница сплошного распространения типового подвида проходит значитель-
но
но южнее, поднимаясь наиболее далеко к северу в районе устья Северной Двины. Южнее 50° с. ш. ч. ползучий встречается лишь в изолированных местонахождениях, крайние из которых расположены у границ Венгрии и Югославии. Наиболее далеко на запад Т. serpyllum проникает в юго-восточной части Англии. Центр распространения вида находится в северной части бореальной Европы. Разнообразие его форм заметно возрастает по окраине ареала.
В Московской обл. Т. serpyllum встречается редко (рис. 15). Большая часть известных его местонахождений находится на севере области (окрестности Дмитрова, долины Дубны, Сестры, Лутос-ни и т. д.). В конце прошлого века он встречался и на территории современной Москвы (в окрестностях с. Останкино). Наиболее характерными его местообитаниями являются осветленные сосновые леса и поляны на песчаной почве. Некоторые местонахождения, несомненно,— результат заноса,
Онтогенез и сезонный ритм развития
Строение эремов и особенности их прорастания аналогичны описанным у других видов. Сходны и начальные фазы развития растений, однако в дальнейшем у ч. ползучего обнаруживается принципиально отличный характер нарастания скелетных осей. В природных условиях темп развития всходов обычно замедлен, но вследствие большой их чувствительности к условиям среды может быть весьма различным в неоднородных по строению фитоценозах. У культивируемых растений ветвление начинается на очень ранних фазах развития, примерно через месяц после появления всходов, когда материнская ось имеет лишь 3—4 сформированных узла выше семядолей. Первыми начинают рост ветви в пазухах семядолей. Они быстро сравниваются в размерах с материнской осью, которая полегает примерно через два месяца после появления всходов. Эти ветви растут плагиотропно под большим углом к материнской оси и в свою очередь рано переходят к ветвлению, вследствие чего образуется радиально разрастающаяся по поверхности почвы дерновина. Длина первых междоузлий на осях второго порядка соответствует длине ближайших междоузлий на материнской оси. У нижних боковых осей первые узлы сближены, почки в их пазухах почти не дифференцированы и остаются в состоянии покоя или дают ослабленные короткие ветви. Первые мелкие листья всходов, не превышающие 2—3 мм длины, довольно рано сменяются листьями обычного размера. К концу вегетационного периода непрерывное ветвление приводит к образованию осей 4—5 порядков; скелетные оси успевают к этому времени хорошо укорениться в узлах.
В условиях природных ценозов всходы ветвятся значительно слабее, чаще всего в течение первого вегетационного периода жизни не успевают перейти к ветвлению. Появление всходов приурочено обычно к окончанию фазы плодоношения.
При весеннем посеве в культуре отдельные особи зацветают
161
уже на первом году жизни. Способность переходить к цветению в течение первого же вегетационного периода, без перезимовки, проявляющаяся даже у этого северного вида, связана, по-видимому, с
Рис. 18. Особи чабреца ползучего подмосковной популяции:
1—'однолетняя; 2 — двулетняя; 3 — трехлетняя; 4 — в начале генеративного периода; 5—двулетний участок моноподиально нарастающей скелетной оси (границы приростов обозначены стрелками) : а — ортотропные моноциклические генеративные побеги прошлого года, б — текущего года, в — моноподиально нарастающие вегетативные пазушные ветви прошлого года, г — текущего года; 6 — смежные пазушные ветви, развивающиеся по разному типу: д — закончившаяся на втором году жизни терминальным соцветием, е — моноподиально нарастающая
южным, средиземноморским происхождением рода. В отличие от предыдущих видов, у ч. ползучего на первом году жизни зацветают лишь пазушные ветви второго порядка, образовавшиеся в средней
162
части материнской оси; нижние, наиболее мощные ветви и материнская ось остаются вегетативными. Как и другие виды рода, зимует с зелеными листьями и имеет вследствие продолжительного роста побегов открытые почки с малым числом зачатков. Осенне-зимняя приостановка роста вызывает на всех осях формирование зоны сближенных междоузлий.
Массовое цветение в условиях культуры происходит на втором году жизни. В природе растения зацветают значительно позже — на пятом-шестом году (рис. 18).
Цветение ч. ползучего начинается в первых числах июня и длится около месяца. Цветки характеризуются хорошо выраженной протерандричностью. Созревание эремов заканчивается в течение одного — полутора месяцев.
С наступлением генеративной фазы отчетливо проявляются характерные особенности побегообразования, отличающие этот вид. К этому времени происходит основная дифференциация осей, выделяются главные скелетные оси, нарастающие моноподиально и стелющиеся по поверхности почвы. Функционально они равноценны, хотя представляют собой оси разных порядков ветвления. Тип развития постоянно образующихся пазушных ветвей зависит от времени их появления и положения на материнской оси. Все перезимовавшие оси примерно через две недели после выхода из-под снега начинают свой рост в ортотропном направлении. При этом скелетные оси и пазушные ветви, успевшие значительно удлиниться в предыдущем году, остаются в вегетативном состоянии и продолжают нарастать моноподиально. Ветвление их в начальный весенний период роста обычно подавлено.
Пазушные ветви на последнем, наиболее поздно сформировавшемся в предыдущем году участке материнской оси, перезимовавшие в укороченном, розеточном состоянии, вступают в фазу генеративного развития. Именно из них возникают отходящие рядами от материнской оси характерные цветоносные стебли, которые имеют розетку мелких сближенных прошлогодних листьев в основании и более крупные и расставленные листья на участке весеннего прироста.
Рост вегетативных осей приостанавливается в период цветения и вновь усиливается с окончанием плодоношения. В это время формируется вторая, слабее выраженная, зона укороченных междоузлий. Возобновление активного роста сопровождается изменением его направления. Верхушки побегов поникают, касаются поверхности почвы и продолжают свой рост плагиотропно. Это изменение направления роста находит отражение в слабой волнистости многолетних скелетных осей. Их годичный прирост складывается, таким образом, из двух участков — более короткого весеннего и более длинного летне-осеннего. Пазушные ветви, образовавшиеся вблизи зон приостановки роста и в особенности в начале второго периода роста, развиваются по полициклическому типу. Удлиняясь, они выполняют функцию дальнейшего развития скелетной системы расте
163
ния, его вегетативного разрастания. Образовавшийся весной отрезок оси несколько приподнят над поверхностью почвы и поэтому слабее укореняется, обычно он успевает одревеснеть до конца вегетационного периода. Пазушные ветви в средней части второго прироста развиваются, как правило, по моноциклическому, озимому типу и служат генеративному размножению вида.
Осенний прирост осей в большей своей части одревесневает и укореняется только весной. На границах приростов ветвление подавлено, пазушные почки крайне мелки и находятся в состоянии покоя. Эти закономерности лучше выражены у растений, развивающихся в условиях ценоза, где интенсивность ветвления уменьшена и вследствие этого ограничено разнообразие типов осей.
Таким образом, по особенностям побегообразования этот вид обнаруживает много общих черт с крымскими моноподиально нарастающими видами рода, структура которых была проанализирована В. Н. Голубевым (1968).
В благоприятных условиях трогаются в рост покоящиеся пазушные почки в базальной части скелетных осей и на границах приростов, из них развиваются побеги возобновления со сближенными узлами в основании. Этот процесс, особенно хорошо выраженный у молодых растений в условиях культуры, обычно совпадает по времени со вторым периодом активного роста побегов и может привести к образованию вторичной разветвленной системы осей,-накладывающейся на уже сформированную.
Несмотря на отсутствие прямых данных о максимальной продолжительности моноподиального роста скелетных осей, можно предположить, что он не беспределен и через несколько лет завершается образованием терминального соцветия или отмиранием точки роста. Об этом говорит отсутствие скелетных осей старше пятилетнего возраста в просмотренном нами материале. Продолжительность цикла развития осей при этом, очевидно, изменяется в зависимости от их места в побеговой системе растения. Наиболее долговечны первые скелетные оси низшего порядка.
Описанные закономерности осложняются существованием осей переходного типа, которые нередко развиваются в пределах одного годичного прироста материнской оси. Такие оси представляют собой связущее звено между многолетними моноподиально нарастающими осями и моноциклическими генеративными побегами. Продолжительность их вегетативного роста весьма непостоянна, поэтому стелющаяся часть их может быть разной длины. Цветение таких побегов иногда запаздывает и они Могут давать пазушные генеративные побеги обогащения. В подобных случаях в основании ортотропных цветоносных стеблей отсутствуют розетки мелких сближенных листьев. Терминальные цветоносные стебли обычно заметно крупнее, чем у пазушных моноциклических побегов. После плодоношения они отмирают на значительную часть своей длины вместе с побегами обогащения и представляют собой, таким образом, видоизмененную форму характерных для полукустарничков удлиненных
164
генеративных побегов. Гетероциклические побеги переходного типа обычны у хорошо развитых экземпляров тимьяна ползучего, хотя их число весьма непостоянно.
Определенное детерминирующее влияние на тип развития побегов оказывают, по-видимому, и придаточные корни. Многолетние скелетные оси обычно хорошо укореняются придаточными корнями, зацветающие рано моноциклические генеративные побеги лишены корней. Различиями в укоренении объясняются, возможно, и те случаи, когда в зоне образования переходных побегов в пазухах супротивных листьев развиваются побеги разного типа. Такие побеги-близнецы обычно развиваются из различно ориентированных пазушных почек. Вследствие накрест-супротивного листорасположения на стелющихся осях чередуются латерально и дорзовент-рально расположенные пары почек. В последнем случае у побегов, развивающихся из нижних прижатых к почве почек, создаются лучшие условия для укоренения.
Нормальный ход побегообразования у ч. ползучего нередко нарушается из-за отмирания не защищенных покровными чешуями точек роста скелетных осей или при отчленении отдельных осей от общей системы побегов, обусловленном механическими причинами. В первом случае происходит перевершинивание осей: отмершую верхушечную почку замещает обычно ближайший боковой побег более высокого порядка — побег продолжения. На следующий год он нередко переходит к цветению, что создает ложное впечатление зацветания скелетной оси.
Многолетние части стелющихся осей ч. ползучего весьма хрупки, вследствие чего часто обламываются. Случаи нарушения связи отдельных осей с системой главного корня способствуют вегетативному размножению вида. Оно вызывает пробуждение покоящихся почек в основании куста и усиливает рост ближайших к месту обрыва боковых ветвей. На обособившихся частях осей происходит усиленное образование и углубление имеющихся придаточных корней, приобретающих вторичностержневой характер. Более резко это выражено в тех случаях, когда обрыв оси происходит вблизи ее нарастающего конца. Такие отделившиеся части обнаруживают очень большое внешнее сходство с молодыми растениями семенного происхождения. Усиленное образование придаточных корней в свою очередь влияет на развитие и длительность жизни главного корня. Чрезвычайно большое количество интенсивно ветвящихся корней, появляющихся в условиях культуры, вызывает уменьшение относительной мощности развития главного корня. Следствием этого, видимо, является его раннее отмирание, наблюдавшееся у отдельных, наиболее крупных экземпляров на третьем году жизни. В условиях ценоза у растений, на стелющихся осях которых образуются лишь неглубокие тонкие придаточные корни, главный корень сохраняется неопределенно долго, и его относительная длина значительно больше.
Как и другие виды рода, Т. serpyllum обладает высокой чувст
165
вительностью к условиям окружающей среды. Под влиянием внеш-‘ них условий существенно изменяется величина годичного прироста скелетных осей, интенсивность их ветвления и разнообразие типов побегов. В естественных условиях годичный прирост осей у взрослых растений равен в среднем 10—12 см. В условиях культуры он возрастает до 20—25 см, в результате чего возникают крупные и густые куртины округлой формы. Наибольшие из них достигают на третьем году жизни 80—90 см в диаметре. На строении многочисленных скелетных осей, формирующих наземное тело растения, сказываются как различия в климатических условиях отдельных лет, так и неоднородность внутренней среды фитоценоза. Неблагоприятная погода в отдельные периоды роста побегов, встреча растущей оси с моховой дерниной или другим конкурентно мощным растением подавляют ветвление и корнеобразование на соответствующем участке оси, замедляют темп ее роста. Изменение окружающей обстановки в благоприятную сторону, напротив, стимулирует рост и ветвление побегов.
Различные сочетания условий освещения, увлажнения и температуры, так же как и колебания напряженности конкурентных отношений в ценозе, создают весьма пеструю картину разнонаправленных влияний, сказывающуюся на общем облике растения, для которого характерны многочисленные местные нарушения интенсивности роста и ветвления побегов. Такая гибкая система побегообразования, позволяющая растению полнее использовать окружающую среду и лучше приспособиться к ней, облегчает ему борьбу за существование, способствует освоению им новых территорий. Но именно эта высокая структурная подвижность создает большие трудности в работе ботаников, поставивших перед собой задачу разобраться во всем многообразии форм этого вида. Особые сложности при этом возникают у систематиков, которые до последнего времени базировались в основном на гербарных материалах, включающих лишь отдельные и не всегда типичные части побеговой системы растения. Следствием этого явилось выделение неоправданно большого числа таксонов внутри этого вида.
Основными направлениями эволюции побеговой системы Т. serpyllum являются: увеличение продолжительности жизни скелетных осей и переход к моноподиальному типу нарастания, специализация осей, приводящая к резкому сокращению размеров ежегодно отмирающей части годичного прироста, преобладание плагиотропного направления роста побегов, усиление роли адвентивного корнеоб-разования и вегетативного размножения. Особый интерес представляет сближение жизненной формы этого полукустарничка со шпалерными кустарничками типа Sibbaldia procumbens L. Об этом говорит отчетливая дифференциация его осей на два основных типа — многолетние вегетативные и моноциклические генеративные. Побеги последнего тип§ не ветвятся и имеют сокращенную вегетативную часть, вследствие чего могут быть функционально приравнены к олиственным соцветиям, отмирание которых не за
166
трагивает многолетней основы куста. О принадлежности чабреца ползучего к полукустарничкам свидетельствует, однако, существование осей переходного типа, которые представляют собой связующее звено между этими принципиально различными типами побегов.
Способы размножения и распространения
Тенденция к возрастанию роли вегетативного размножения, наметившаяся в эволюции чабреца ползучего, связана,' очевидно, с продвижением его предковых форм на север, где они встретились с климатическими условиями (меньшая интенсивность солнечного освещения, большая влажность), не благоприятствующими успеху семенного размножения. Этим объясняется, по-видимому, большая устойчивость распределения признаков пола у этого вида. В отличие от двух рассмотренных выше видов Т. serpyllum. не обладает женской двудомностью. При обследовании его подмосковных популяций было обнаружено лишь одно растение с неполностью редуцированным числом тычинок в цветках. Эту особенность оно сохранило и в культуре.
Семенная продуктивность ч. ползучего в условиях культуры оказалась довольно высокой. Искусственное самоопыление вызывало резкое уменьшение количества полноценных эремов, что говорит о его неполной самосовместимости. Это свойство должно ограничивать самооплодотворение, возможность которого, несмотря на небольшие размеры соцветий, создается дружным и обильным цветением растений. Ч. ползучий легко гибридизирует с другими видами рода, даже филогенетически удаленными.
Экология и фитоценология
Как и другие виды рода, ч. ползучий весьма чувствителен к условиям освещения и произрастает на открытых участках или в осветленных, преимущественно сосновых, лесах с негустым травяным покровом. В Московской обл. его фитоценотическая роль незначительна.
Почти на всем протяжении своего ареала тяготеет к местообитаниям с песчаной и супесчаной подзолистой почвой, лишь изредка, обычно по краю ареала, заходя на выходы скал. Вследствие своей псаммофильности, ч. ползучий не отличается требовательностью к плодородию почв и способен мириться скорее с сухостью, чем с застойным увлаженением субстрата. Он предпочитает местообитания с нейтральной или щелочной реакцией, но может расти и на умеренно кислых почвах,
167
Хозяйственное значение
Чабрец ползучий дает невысокий выход эфирного масла — от 0,12 до 0,27% от абсолютно-сухого веса сырья (Лякавичюс, Яско-нис, 1967). Качество его масла ниже, чем у чабреца блошиного (Hegnauer, 1948). Употребляемое в литературе по лекарственным растениям название Т. serpyllum охватыва'ет комплекс видов, среди которых тимьян ползучий в узком понимании является одним из наименее ценных и перспективных. Рекомендацию этого вида в.качестве основного источника лекарственного сырья надо признать неправильной.
В районах массового распространения дружно и обильно цветущий чабрец ползучий может использоваться в качестве медоносного растения. С одного гектара сообщества с его господством может быть получено около 380 кг нектара (Лякавичюс, Ясконис, 1967).
Н. А. Аксенова
БУЗИНА КРАСНАЯ
Номенклатура и систематическое положение
Sambucus racemosa L., Sp. pl. (1753) 270; Кауфм., Моск, фл., изд. Г (1866) 222, изд. 2 (1889) 224; Цингер, Сб. свед. (1885) 209; Шмальг., Фл. 2 (1897) 2, р. р.; Сырейщ., Илл. фл. 3 (1910) 201; он же, Опр. раст. Моск. губ. (1927) 242; Федченко в Маевск., Фл., изд. 7 (1940) 675; Пояркова, там же, изд. 8 (1954) 535; она же во Фл. СССР 23 (1958) 431; она же в Маевск., Фл., изд. 9 (1964) 518; Ворошилов в Опр. раст. Моск. обл. (1966) 304.— Бузина красная, б. обыкновенная, б. кистистая.
Относится к сем. Caprifoliaceae, колену Sambuceae. Описан из Европы. Тип в Лондоне.
Морфологическое описание
Бузина красная — сильно ветвистый кустарник или деревце до 5 м высоты, со сбежистым, толстым у основания стволом и поверхностной корневой системой. Кора на стволах и старых ветвях серовато-бурая, слабо опробковевшая, с продольными морщинами, у молодых побегов бурая или более менее окрашенная в фиолетовые или пурпурные тона. Молодые побеги округлые или слегка ребристые, блестящие, с ясно заметными крупными щелевидными чечевичками, между которыми видны мелкие чечевички. Сердцевина побегов мягкая, губчатая, светло-бурая, древесина зеленая, очень легкая.
Почки супротивные, конечные часто недоразвитые. Боковые почки на небольших ножках; цветочные — шарообразные, листовые— продолговато-яйцевидные, рыхлые, голые, зеленовато-бурые, красно-бурые или красноватые, 8—15 мм длиной и 5—8 мм шириной. У почек из-под чешуек на вершине нередко проглядывают недоразвившиеся листья. Чешуйки почек голые, реснитчатые, располагаются крестообразно, при основании почек по одной-две развитых чешуйки, верхушки которых заканчиваются коротким мягким -острием.
Листовой рубец полукруглый, большой, щитковидный, у основания несколько приподнятый, серовато-беловатый, с 3—5 следами.
169
Супротивно лежащие листовые рубцы соединены мозолистой перетяжкой. Листовая подушка несколько возвышается над поверхностью побега.
Прилистники чаще в виде крупных, иногда стебельчатых, мясистых железок, реже очень мелкие, ланцетовидные, зеленоватые, быстро опадающие. Листья супротивные, непарно-перистые, с 3—8 яйцевидными, продолговато-яйцевидными или эллиптическими листочками, длинно заостренными на верхушке и клиновидными или округлыми при основании, 5—8 см длиной и 3—4 см шириной. Листья голые или слегка волосистые, с пильчато-зубчатым краем, сидят на общем черешке 5—И см длиной с двумя железками при основании нижней пары. Листовая пластинка сверху зеленая, снизу сизоватая, с неприятным запахом.
Желтовато-белые актиноморфные цветки бузины собраны в плотные яйцевидные или конические соцветия 2,5—6 см длины и 2,5—5,5 см ширины на тонком и коротком (0,8)1—3(4,5) см длины цветоносе. Нижние веточки соцветия большей частью отогнуты, осевые части его голые или усажены (иногда густо) мелкими сосочками. Зубцы чашечки около 0,5 мм длиной, большей частью языковидные, тупые, „на верхушке нередко зазубренные, реже треугольные, острые. Венчик беловато- или зеленовато-желтый, 4—5 мм, с эллиптическими или продолговато-яйцевидными тупыми или острыми долями, более короткими, чем завязь, наверху иногда с небольшими зубчиками. Завязь около 2 мм длиной, столбик шаровидный или шаровидно-конический, с тремя красными рыльцами. Тычинки в два раза короче лепестков, с шаровидными пыльниками, превышающими нити или равными им по длине.
Плод бузины — синкарпная, сочная, блестящая, ярко-красная костянка, шаровидной формы, 3—5 мм в диаметре, с желтой сочной мякотью в плотных яйцевидных или конических гроздьях по 54— 217 штук. В плоде обычно 3 косточки 2—4 мм длиной, 0,9—2 мм шириной, 0,7—1,1 мм толщиной. Косточки трехгранно-яйцевидные, светло-желтые, матовые, с мелкоморщинистой поверхностью. В 1 кг 435 тысяч семян, 1000 семян весят 2—2,6 т (Артюшенко, 1962). Жизнеспособность семян 86% (Некрасов, 1973). Плоды бузины несъедобны, по характеру плодоношения ее относят к видам с устойчивым типом плодоношения.
Диплоидное число хромосом равно 36 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Родина бузины — Западная Европа, где она растет в диком виде в горах.
В Советском Союзе бузина красная в диком виде встречается только в Верхне-Днестровье, как одичалая распространена от Прибалтики до Кавказа, растет также в Сибири и на Дальнем Востоке.
170
В Московской обл. бузина встречается в старых парках, Лесах, вблизи жилья, распространена по всей области (Ворошилов и др., 1966).
Онтогенез
Семя бузины красной трехгранно-яйцевидное, в поперечнике трехгранно-овальное, 3X2X1 мм. Семенная кожура представлена слоем облитерированных клеток интегумента, плотно прилегающих к эндокарпу, но не срастающихся с ним. Эндокарп состоит из двух слоев одревесневших волокон и одного слоя каменистых клеток неодинаковой длины, благодаря чему поверхность семени мелко гофрированная. В семенах имеется хорошо развитый эндосперм, клетки которого богаты маслом и белком. Небольшой зародыш с семядолями неравной длины расположен в верхней части семени.
При весеннем посеве семян после стратификации всходы появляются через 43 дня (Бородина и др., 1970). Всходы бузины красной имеют сравнительно тонкую, бледно-зеленую, голую подсемядольную часть, овальные, слегка яйцевидные семядоли 7—8 мм длины и 5—6 мм ширины, закругленные на верхушке и слабо выемчатые, на коротких черешках. С верхней стороны семядоли темно-зеленые, с нижней светлее, жилки слабо заметны. Первые листья супротивные, коротко- и широкояйцевидные, при основании сердцевидные, с остро-мелкозубчатым краем, на длинных черешках. Третий и четвертый листы более вытянуты, овально-яйцевидные. Надсемядольное междоузлие около 2 мм длины (Васильченко, 1960).
Растет бузина красная быстро, уже однолетки ее можно высаживать на постоянное место; на третий год бузина начинает плодоносить.
Ветвление у бузины симподиальное, ежегодно ее побеги теряют верхушечную точку роста, в том числе и вегетативные, и постоянно возобновляются за счет боковых почек. Иногда у бузины отмирает верхушечный побег до нескольких десятков сантиметров длиной. Равномерный рост междоузлий на побегах бузины протекает по всей длине в течение периода растяжения, междоузлия на побегах бузины могут достигать 40 см длины. Характер листьев в пределах годичного побега изменяется: чем выше по стеблю, тем более расчленены низовые листья. На* границе со срединными нередко образуются своеобразные переходные формы листьев (Серебряков, 1952).
Сезонный ритм развития
Олиствение и цветение у бузины красной проходят одновременно во второй половине апреля — начале мая. Плодоношение в Московской обл. наблюдается с конца июля по сентябрь, причем
171
плоды долго сохраняются на кустах. Средний урожай с одного растения 300 г (Некрасов, 1973).
В сентябре у бузины начинают окрашиваться листья, в конце месяца проходит листопад.
Способы размножения и распространения
Бузина размножается семенами,, отводками и черенками, дает обильную поросль от пня.
Плодоносит бузина регулярно и обильно. При заготовке плоды ее освобождают от сочной мякоти растиранием с последующей отмывкой. Выход чистых семян из сырья 3—5% (Заборовский, 1962). При проращивании в лабораторных условиях семена набухают быстро, но не прорастают месяцами. При посеве весной без предварительной подготовки к концу первого года семена бузины дают незначительную грунтовую всхожесть, большее число всходов , появляется лишь следующей весной. Осенние посевы бузины также не всегда дают следующей весной хорошую грунтовую всхожесть. В Ленинграде семена бузины, собранные в разные сроки и сразу же высеянные в грунт, дали весной следующую грунтовую всхожесть: семена сбора 1 августа — 55%, сбора 1 сентября — 75%, сбора 15 сентября — всего 40% (Заборовский, 1962). В опытах П. Н. Ваш-кулата (1952), проведенных в районе Куйбышева, свежесобранные семена бузины красной имели лучшие показатели при посеве в'период 20 июля— 1 августа, а сентябрьские посевы давали низкую всхожесть или вообще утрачивали к осени способность быстро прорастать. Для весенних посевов лежалых семян Г. X. Абианц (1939) рекомендует стратификацию их во влажном песке при температуре 0—5° в течение 4 месяцев, в некоторых случаях 6—7 месяцев. Продолжительность сохранения всхожести у семян б. красной около двух лет. Хранят их в стеклянной или металлической таре.
Норма высева семян бузины 0,7—2 г на 1 погонный метр, глубина заделки 0,5—1 см (Хохлов, Ковалева, 1950). Средний выход сеянцев — 24 с 1 погонного метра (Мисник, 1949).
Плоды бузины охотно поедаются птицами, способствующими ее распространению.
Экология
Б. красная не очень требовательна к почве, хорошо растет на разнообразных почвогрунтах, является прекрасной почвоулучшающей породой: листья ее очень богаты зольными веществами. Предпочитает свежие плодородные суглинистые почвы, заметно реагирует на азотистые удобрения, особенно пышно разрастается на местах, где сбрасывают огранические остатки и мусор. На рыхлых плодородных почвах бузина отличается таким бурным ростом, что через 4—5 лет куст редеет снизу и теряет свою пышность. В таких
172
случаях его сажают на пень й через год-два снова получают пышный куст.
Бузина переносит самое сильное отенение. К температурным условиям она мало требовательна, отличается жаростойкостью и зимостойкостью, однако при росте в сырых местах побеги ее нередко подмерзают.
Фитоценология
На родине бузина растет в горных лесах. В одичалом состоянии— повсеместно в лесах и зарослях кустарников, предпочитая сосновые и еловые леса, по сорным местам. В Московской обл. бузину можно встретить в самых разнообразных местах.
Внутривидовая систематика
Бузина красная в природе варьирует в основном по форме и окраске листьев. А. И. (Колесников (1958) указывает для нее следующие формы: f. plumosa — б. перистая; f. laciniata (f. serratifo-lia) —б. разрезнолистная; f. tenuifolia— б. тонколистная; f. plumo-sa-aurea— б. перистая золотистая. А. И. Пояркова (во Флоре СССР, 1958) называет также f. ornata— с разрезанными листьями; f. таг-morata и f. aureo-variegata— с пестрыми листьями; f. semperflo-rens — почти непрерывно цветущая; f. spectabilis — с чисто-белыми цветками; f. purpurea — с розовым снаружи венчиком; f. flaves-cens — с золотисто-желтыми плодами. Кроме того, встречаются: f. violacea — с фиолетово-красной при распускании, Затем сизоватой листвой; f. viridis — со светлой ярко-зеленой листвой; f. fructi-fla-vis — с желто-оранжевыми плодами.
Консортивные связи
Консортивные связи бузины изучены мало. Известно, что она как хороший медонос активно посещается насекомыми. Плоды бузины охотно поедаются и распространяются птицами, преимущественно славками, дроздами, зарянками (Рычин, 1972).
Цветкам бузины наносят вред личинки бузинной галлицы — Arnoldia sambuci, черноногой галлицы — Placochaela nigripes и жимолостной галлицы — Syndiplosis lonicerarum (Заборовский, 1962).
Хозяйственное значение
Бузина красная имеет немалое хозяйственное значение. Почти все части растения используются в народной медицине: чай из соцветий и сок из ягод — как потогонное средство, плоды и кора — как слабительное, чай из сушеных цветков и плодов считается хорошим средством от катаров, ревматизма и сыпи (Строгий, 1934).
173
Ё Незрелых плодах и листьях бузины содержится синильная кислота (Гринер, 1960). В соцветиях имеется энзим эмульсин; в листьях и коре ветвей и корней до 2,5% сахарозы и эмульсина. Плоды содержат жирное, красновато-желтое масло, в состав которого входят пальмитиновая, олеиновая, арахиновая и некоторые другие кислоты, фитостерин, инвертин, 8,5% сахаров, 9,4% сырого протеина, 2,8% пектина. В семенах содержится жирное масло, обладающее послабляющим свойством с глицеридами кислот линолевой, линоленовой, масляной, пальмитиновой (Флора СССР, 1958).
В листьях и ветвях бузины содержатся дубильные вещества: в листьях они приурочены к кожице, в паренхиме коры и сердцевине также имеются крупные, 25—164 мк в длину и 18—120 мк в ширину, вместилища дубильных веществ (Раздорский, 1949).
Плодами бузины хорошо отмывать руки от зелени или соков, сок ягод ее хорошо растворяет смолы. Ягоды бузины можно использовать для приготовления мармелада (Машкин, Голицын, 1952). Л4асло из семян б. красной в некоторых странах Европы используется в технических целях, из плодов получают спирт.
Бузина — ядровая порода с желтовато-белой заболонью и буровато-желтым ядром. Древесина ее умеренно тяжелая, умеренно высыхающая, прочная, посредственно твердая, умеренно хрупкая, удельный вес ее 0,6—0,65. Она хорошо полируется, поэтому находит применение в токарном производстве. Большую ценность для поделок представляют корневые наплывы бузины. Сердцевина бузины красной очень широкая, занимает более половины поперечника побега, рыжая, рыхлая, находит применение в микроскопии и при изготовлении физических приборов. Молодые ветви идут на изготовление детских игрушек.
Быстрый рост и неприхотливость бузины заслуживают большого внимания для использования ее при лесоразведении, в мелиоративных целях, для закрепления оврагов и крутых склонов. Ее хорошо использовать в качестве подлеска, так как бузина привлекает на гнездовья массу птиц. Очень интересна бузина с декоративной точки зрения, так как она теневынослива, прекрасно переносит обрезку, почти не страдает от дыма и газов (Рычин, 1972) и относится к кустарникам, наиболее эффективно снижающим городской шум (Галактионов и др., 1967).
Благодаря продолжительному пребыванию плодов на ветках бузина очень декоративна как в естественном виде, так и в штамбовых посадках (Овсянников, 1930). Бузину рекомендуют использовать для создания живых изгородей (Керн, 1926), в качестве подлеска в полезащитных насаждениях (Шиманюк, 1967), в парках, групповых и одиночных посадках.
A. M. Былова
МОЛОКАН ТАТАРСКИЙ
Номенклатура и систематическое положение
Mulgedium tataricum (L.) DC., Prodr., 7,1 (1838) 248; Сырейщ., Опр. раст. Моск. губ. (1927) 271; Ильин и др. в Маевск., Фл., изд. 7 (1940) 768; Кирпичников, там же, изд. 8 (1954) 630.—,Son.chus tata-ricus L., Mant. 2 (1771) 572.— Lactuca tatarica (L.) C. A. Mey., Ver-zeichn. Pfl. Cauc. (1831) 56; Кирпичников во Фл. СССР 29 (1964) 282; он же в Маевск., Фл., изд. 9 (1964) 616; Тихомиров в Опр. раст. Моск. обл. (1966) 344.— Молокан татарский, молоканник, осот голубой.
Относится к сем. Compositae (Asteraceae), подсем. Cichorioideae Kitam., колену Cichoreae Spreng., подколену Lactucina Less. Тип в Лондоне.
Морфологическое описание
Молокан татарский — травянистое, многолетнее, поликарпиче-ское, облигатно-корнеотпрысковое растение (Серебряков, 1962). Корневая система у него широко распростертая (особенно на рыхлых почвах) и глубоко проникающая. Одна особь может занимать площадь до 10 м2 (Былова, 1960 а) и углубляться до 4—5, а иногда до 7 м (Мамонов, 1955; Смирнов, 1956).
От стержневого корня на разной глубине отходят боковые, в свою очередь неоднократно ветвящиеся. Одни из них тонкие, всасывающие, менее 1 мм толщиной, другие — более толстые, составляющие основной скелет подземной части растения. Поверхностные крупные боковые корни толщиной в 1,5—2 мм отходят от главного корня на глубине 10—50 см и располагаются в почве почти горизонтально, слабо ветвятся и постепенно приближаются к дневной поверхности. Достигнув длины нескольких десятков см (20—280), боковой корень делает крутой изгиб и растет вертикально вниз, углубляясь до 1—2 м и обильно ветвясь по всей длине. Изгиб корня оказывается в среднем на 6 см ближе к поверхности почвы, чем ме
175
сто его отхождения. От выпуклой стороны Изгибов, которые всегда утолщены примерно вдвое, отходят боковые корни следующего порядка, имеющие подобный же характер нарастания, ветвления и расположения в почве. У одного растения могут образоваться за год корни семи порядков. На утолщенных изгибах боковых корней закладываются придаточные почки, меньше их на горизонтальных и вертикальных участках корня. На вертикальных корнях почки могут быть на глубине 2—3 м (Мамонов, 1956 а). Из этих почек развиваются ортотропные побеги — корневые отпрыски, поэтому крупные боковые корни называют корнями размножения или корнями распространения. У растений молокана, растущих на необрабатываемой почве, корни размножения в небольшом числе или совсем отсутствуют, но зато лучше развиты крупные боковые корни, спускающиеся отвесно вниз и расположенные близко от главного корня, иногда сплетающиеся с ним в своеобразный жгут (Былова, 1958). У растений, растущих на пахотных землях, главный корень, как правило, отсутствует, и корневая система представлена корнями размножения нескольких порядков.
От подземных частей корневых отпрысков отходят придаточные корни, среди которых различают более крупные, подобные боковым корням размножения, несущие почки, и более мелкие, тонкие и короткие всасывающие корни. Последние эфемерны, развиваются и функционируют при повышенной влажности почвы в любое время вегетационного периода и засыхают при пересыхании верхних слоев почв,ы. Очень редко на них возникают почки.
Подземные отрезки корневых отпрысков являются вертикальными корневищами. По годичным приростам древесины установлено, что они могут существовать в течение 11 лет (Былова, 1962). Верхняя часть их осенью отмирает вместе с надземными побегами. Из почек вблизи отмершего конца на следующий год развиваются новые побеги. Так осуществляется симподиальное нарастание корневища. На плотных почвах, где молодые растущие отпрыски встречают механические препятствия, корневища изгибаются, иногда скручиваясь спиралью. Корневище имеет почти одинаковую с надземным стеблем толщину, в среднем 3—7 мм.
Молодые однолетние корневища белые или слегка желтоватые, несут белые сочные чешуевидные низовые листья, которые засыхают по мере роста надземной части побега, при этом корневище покрывается перидермой, становится коричневым. Появляющиеся ин-вагинально, реже экстравагинально придаточные корни сосредоточены главным образом вблизи узлов. Длина корневищ зависит от глубины залегания почек, из которых развивается побег, и может быть 55—60 см.
Из почек на корневищах и корнях вырастают надземные моно-циклические, монокарпические побеги полурозеточного типа. Специализированных вегетативных розеточных побегов у взрослых растений молокана нет, но так как побеги вырастают в любое время вегетационного периода, то возникшие осенью побеги не успевают
176
завершить малый жизненный цикл и к зиме отмирают в фазе розетки.
Высота генеративных побегов молокана обычно 15—80 см, иногда 150—190 см. Средняя толщина стеблей 5—6 (7) см, они почти округлые, слегка бороздчатые, внутри полые, зеленые, слегка сизоватые, иногда с фиолетовым оттенком от антоциана, в верхней части до цветения густо, позднее — слабо опушены простыми белыми спутанными волосками. В затенении побеги молокана ветвятся только в верхней части, в условиях хорошего освещения — по всей длине. Ветви направлены косо вверх, но верхние побеги обогащения нередко отклонены горизонтально. Стебель олиствен по всей длине. Размеры метамеров побега (длина междоузлий и листьев) подчиняются правилу одновершинной кривой изменения метамеров. Формула листорасположения 2/5.
Листья зеленые, сизоватые, довольно толстые, мясистые, жестковатые, простые, без прилистников; розеточные и нижние стеблевые сужены в черешок, средние и верхние — сидячие, полустебле-объемлющие. Размеры листьев и их рассеченность уменьшаются снизу вверх. Нижние и средние стеблевые листья перисто-выемча-тонадрезные или струговидные, перисто-лопастные с острыми лопастями, слегка повернутыми к основанию листа, голые или опушенные, верхние — ланцетные или продолговатые с редкими одиночными зубцами или цельнокрайние, имеют более или менее густое белое войлочное клочковатое опушение, как и молодые листья розеток. Край листа редкозубчатый, верхушка острая или притупленная, как и зубцы заканчивается хрящеватым шипиком, жилкование сетчатое, перистое. Выражена фотометричность листьев, но в гораздо меньшей степени, чем у Lactuca serriola Torner.
Соцветия — корзинки средней величины, диаметром в 20—28 мм, состоят из 20 (16—23) язычковых цветков. Ось соцветия голая, ям-чатая, обертка большей частью четырехрядная, у бутонов цилиндрической или усеченно-конической формы. Листочки обертки травянистые, зеленые или окрашены антоцианом в буровато-фиолетовый цвет, снаружи тонко опушенные, наружные острые, внутренние по краю с пленчатой, обычно волнистой каймой, на верхушке туповатые. Корзинки расположены на оси первого порядка и на побегах обогащения второго и третьего порядков, образуя как бы сложное широкометельчатое соцветие.
Цветки пахнут ванилью, имеют сиренево-голубой язычковый венчик из 5 лепестков. Длина трубки венчика 7—8 мм, отгиба — 9—10 мм. В трубке венчика скапливается нектар. Хохолок короче венчика, состоит из большого числа почти одинаковых белых, мягких, ломких, зазубренных волосков. Из трубки венчика более чем наполовину выходит голубая, с синими полосами по месту срастания пыльников тычиночная трубка длиной 3—3,5 мм, образованная пятью сросшимися вдоль пыльниками. Пыльники интрорзные. Свободные белые тычиночные нити прирастают к трубке венчика. Сквозь пыльниковую трубку выставляется столбик с синим двураз
7 Зак. 255
177
дельным рыльцем. Выметающие волоски короткие, равномерно покрывают верхнюю часть столбика. Пыльца белая. Завязь нижняя, белая, гинецей паракарпный из двух плодолистиков.
Плод — семянка 5 (4,5—6) мм длины и 0,75—1 мм ширины. Плоды продолговато-линейные или эллиптические, слегка сжатые, иногда немного изогнутые, черные, реже желтовато-коричневые или оливковые с 5—7 продольно выступающими невысокими ребрышками, довольно густо покрытые очень короткими жесткими сосочковидными волосками, на верхушке постепенно оттянутые в носиковидное сужение, обычно более светлое, чем семянка, и заканчивающееся воронковидным диском, несущим хохолок, в 1,2—2 раза превышающий семянку, легко отделяющийся. Абсолютный вес семянок 0,476 г (Бажанов, 1922, 1929).
Число хромосом по данным различных авторов определено как 2п= 18 (Хромосомные числа цветковых растений, 1969).
Географическое распространение
Ареал молокана татарского охватывает значительную часть центра Евразии. Он распространен в Центральной Европе, на Балканском п-ве (в Сербии и Болгарии), в Турции (около Босфора), в Малой Азии, Иране, Афганистане, в южных и юго-восточных районах СССР, в Монголии, северной части Китая, Западном Тибете, северных штатах Индии (Шмальгаузен, 1897; Флора СССР, 1964). Изображение ареала в пределах СССР дано в книге «Районы распространения важнейших сорных растений в СССР» (1935), всего ареала — в неопубликованной диссертации А. М. Быловой (автореферат Былова, 1962).
В пределах СССР молокан особенно широко распространен в Среднем и Нижнем Поволжье, Заволжье, на Южном Урале (в Башкирской АССР, в Оренбургской, Куйбышевской, Саратовской, Волгоградской областях) и в Северном Казахстане. В Московской обл.'это — редкое заносное растение. Он встречен по берегу Сенежского озера на песке (Сырейщиков, 1927) и на железнодорожной насыпи Ярославской железной дороги за Яузой (Александров, 1923). Нами найден в Москве на ул. Шаболовке и на ул. Бурденко (в палисаднике), на площади Гагарина (на газоне), на ул. Усачева (у забора около рынка), на Трубецкой ул. и в других местах.
Продвижение молокана на север по грунтовым дорогам, железнодорожным путям и рекам вполне очевидно (Дмитриев, 1937; Красовский, 1949; Ларюнов, Макодзеба, 1957). Только заносом можно объяснить редкие местонахождения этого растения значительно севернее границы сплошного распространения: под Калугой, Москвой, Островом, близ Кирова, Архангельска, Котласа, у с. Самаровского на Оби и в других местах.
Ареал молокана охватывает многие ботанико-географические зоны: смешанные и лиственные леса, лесостепь, северные и южные степи, полупустыни и пустыни. В лесной зоне он растет по обрыви
178
стым высоким берегам рек. В Средней и Центральной Азии он растет также по берегам водоемов (рек, озер, арыков), у колодцев, на мокрых солонцах, а также на барханных песках в пустынях Гоби и Алашань, в песках Тенгери, на бугристых песках Большие и Малые Барсуки и Муюн-Кум,ы (Пташицкий, 1910). В степной и полупустынных зонах встречается на водораздельных пространствах, занимая преимущественно пахотные земли. Молокан может произрастать и высоко в горах (П. П. Горбуновым собран на Памире на высоте 3400м).
Онтогенез
Семя молокана заполняет всю семянку, кроме носика, плотно прилегает к околоплоднику. Семенная кожура тонкая, белая. Зародыш продолговатый, прямой, с длинными вдоль сложенными семядолями, коротким гипокотилем, корешком и недифференцированной точкой роста. Все клетки зародыша содержат много масла.
Семена могут прорастать летом и осенью без периода покоя, либо весной после перезимовки. Они сохраняют всхожесть в течение двух лет (Бажанов, 1922, 1929). При достаточном увлажнении всходы молокана могут появляться летом, но так как в местах произрастания его лето, как правило, засушливое, то всходы появляются обычно весной и осенью. Весенние всходы в массе гибнут летом во время засухи и объедаются животными. Гибнут всходы и во время перезимовки.
Лабораторная всхожест’ семян 40—70%, всхожесть стратифицированных семян выше, чем хранящихся зимой в комнатных условиях, и прорастают они быстрее и дружнее (Былова, 1965). Полевая всхожесть сильно зависит от влажности почвы и глубины заделки. При орошении всхожесть возросла с 2,2 до 22,4% (Владимиров, 1947, 1948). Предельная глубина заделки семян 2—3 см, с глубины 5 см и более всходы не появляются. При благоприятных условиях прорастание начинается на третий день после посева. Прорастание надземное. На третий — пятый день появляется корешок, а через день-два начинает вытягиваться гипокотиль, вынося вверх семядоли, которые освобождаются от околоплодника и быстро зеленеют.
У проростка овальные или яйцевидные семядоли длиной в 6— 10 (12) мм и шириной 4—5 (6) мм тонким цилиндрическим гипокотилем выносятся на высоту 8—12 мм над уровнем почвы. Семядоли горизонтально отклоненные, сизоватые, голые, короткочерешковые, с хорошо заметной средней жилкой. Первый лист тоже голый, сизоватый, овальный или продолговато-обратнояйцевидный, сужен в черешок, с остроконечной верхушкой, по краю с редкими зубцами или без них. Второй и последующие листья с более заметными зубцами образуют розетку из 5—6 листьев 1,5—3 см длиной. У проростков хорошо развит главный корень, обильно ветвящийся в базальной части, при повреждении он замещается одним из боковых. В конце лета и осенью у проростков на главном корне на глубине
7*
179
5—20 мм обнаруживаются бугорки — скрытые внутри корня почки или несколько уже развитых почек возобновления 1—2 мм длиной, с белыми мясистыми чешуями.
Весной из одной почки на корне за-счет запасов инулина во всех паренхимных тканях корня развивается новый розеточный побег, а розеточный побег проростка, гипокотиль и основание главного корня за зиму отмирают. Растение переходит в ювенильное состояние. У проростков, появившихся весной, семядоли засыхают к середине или концу июня, развиваются более крупные листья, и растение уже в возрасте двух месяцев становится ювенильным.
У ювенильных растений розеточный побег имеет 3—7 листьев до 5 см длиной. Стержневой корень толщиной в 1,5—2 мм у основания и 0,5 мм у верхушки углубляется до 80—100 см и ветвится по всей длине, но поверхностные крупные корни размножения не развиты.' Они могут появляться у ювенильных растений на первом году жизни только при сильном увлажнении (в опыте). Почки возобновления закладываются на главном корне глубже, чем у проростков, а именно на глубине 3—5 см. Тонкие боковые корни летом в верхнем пересохшем горизонте почвы (до глубины примерно 20 см) засыхают, отмирают, но после увлажнения почвы дождем на этом же отрезке главного корня появляются новые придаточные тонкие корни, густо покрытые корневыми волосками.
Сходный с ювенильными облик имеют растения, возникшие вегетативным путем от мелких корневых черенков. От черенка отходит обычно один крупный стержневой корень и 1—2 розеточных побега. Если они появились во второй половине лета или осенью, то в текущем году не зацветают.
Длительность виргинильного периода у молокана не выяснена. На регулярно обрабатываемой почве, где не успевают развиться генеративные побеги, этот период может быть очень длительным.
Взрослые растения молокана семенного или вегетативного происхождения, имеющие сильно развитую корневую систему и большой запас питательных веществ в ней, растущие на нетронутых или редко обрабатываемых почвах, ежегодно развивают генеративные побеги. У старых растений молокана, обнаруженных на 17—20-летней залежи, совершенно отсутствовали боковые корни размножения, а сохранялся только стержневой корень, сильно разрушенный в базальной части, несущий пеньки отмерших побегов и отпрыски. Растения, находящиеся в состоянии покоя, развивали не достигающие дневной поверхности отпрыски или розеточные побеги один раз в 3—5 лет (Смирнов, 1956).
В течение всего вегетационного периода происходит нарастание корневой системы, заложение почек (Милащенко, 1961), развитие их в побеги, поэтому у одного растения одновременно можно видеть почки в виде бугорков, только что тронувшиеся в рост почки, подземные отпрыски, розеточные и генеративные побеги. В любое время вегетационного периода в корнях и корневищах молокана обнаруживается инулин (в гораздо меньшем количестве — крахмал).
180
Запасы его в подземных оргайах истощаются лишь при многократных систематических подрезках отрастающих побегов, начиная с весны.
Сезонный ритм развития
По степени сформированное™ побега в почках возобновления молокан относится, по И. Г. Серебрякову (1952), к 3-й группе. Соцветия закладываются и формируются весной по мере роста побега. Скученные бутоны корзинок на вершине растущих побг"ов появляются в последней декаде мая. Единичные корзинки раскрываются в середине июня, но пора массового цветения — конец июня и первая половина июля. Постепенно затухая, цветение длится до середины октября, что связано с разными сроками появления побегов, но в поздно цветущих корзинках семянки не вызревают.
Первой раскрывается верхушечная корзинка на побеге, а затем корзинки на осях второго и третьего порядков. Раскрывание корзинок происходит рано утром, спустя 15—20 минут после восхода солнца. Солнечным летним утром полностью раскрывается корзинка за •30—40 минут, в пасмурное утро — за 2 часа, прохладным осенним утром — более чем за час (пример термонастии). По мере отгибания листочков обертки и венчиков до горизонтального положения вырастают и тычиночные нити, выдвигая пыльниковые трубки, из которых высовываются столбики с двулопастным рыльцем со сложенными лопастями, которые позднее расходятся и скручиваются спиралью. Выталкиваемая столбиками пыльца попадает на поверхность рылец. Специальными опытами с заключением корзинок в марлевые изоляторы установлено, что хотя возможно самоопыление, но завязывания плодов при этом не происходит (Былова, 1960 а, 1965). Основными опылителями молокана являются мухи-журчалки, пчелы и осы, достающие нектар из узких трубочек венчика или собирающие пыльцу. Максимум их наблюдается при солнечной погоде в 9—10 часов утра. Раскрытыми остаются корзинки в течение одного дня: в жаркие летние дни — до 10—И часов, а в пасмурные и прохладные осенние — до 18—19 часов. Посещение корзинок насекомыми ускоряет их закрывание.
По отцветании листочки обертки поднимаются вверх и сближают венчики, которые бледнеют и засыхают. Под защитой обертки летом в течение 10—12 дней созревают семянки. В одной корзинке созревает в среднем 16 семянок и 6 остаются недозрелыми. При созревании семянок листочки обертки подсыхают, скручиваются и отгибаются вниз. На оси соцветия семянки с белыми хохолками расходятся и образуют в совокупности почти сферу. Единичные плодоносящие корзинки встречаются в конце июня, к концу первой декады июля уже наблюдается перевес числа плодоносящих корзинок над цветущими, а в первой половине августа большинство растений уже обсеменяется.
181
После плодоношения генеративные побеги молокана начинают засыхать и к зиме надземные их части отмирают полностью, как и розеточные побеги. Все почки возобновления оказываются на большей или меньшей глубине под поверхностью почвы (криптофит).
Способы размножения и распространения
Семенная продукция молокана очень велика. Один среднеразвитый генеративный побег может дать 2200 зрелых семянок, а отдельные лучше развитые побеги до 4000—5000 (Суднов, 1946; Пенин, 1959). На поле со средней засоренностью молоканом, где на 1 м2 насчитывается 80—90 побегов, образуется 191 400 семянок, а на участках с еще большей засоренностью — несколько сотен тысяч.
Семянки обладают большой парусностью, легко срываются ветром и разносятся по воздуху на расстояние до 5 км. На значительно меньшее расстояние распространяются они водяными потоками во время дождя, а также растаскиваются муравьями. Несмотря на огромную семенную продукцию, семенное размножение подавлено, так как мала полевая всхожесть семян и велика гибель проростков. Только во влажные годы и в условиях орошения семенное размножение может играть более существенную роль.
Основной способ размножения — вегетативный. Естественное вегетативное размножение наблюдается при перегнивании базальных частей корней размножения и отделении их от материнского растения. Этому процессу способствует человек. При обработке почвы легко дробятся хрупкие корни, лишенные механических тканей, и корневища молокана. Образующиеся черенки способны к регенерации. Они образуют придаточные корни, закладывают новые почки или развивают уже имеющиеся почки в побеги.
Молокан может регенерировать на очень ранних этапах своего развития. При наличии достаточного количества влаги (в опыте) окореняются отделенные от корня семядоли с небольшим кусочком гипокотиля, а также семядоли, отделенные от проростков в возрасте нескольких дней (Былова, 1960 6). Придаточные корни появляются у таких черенков на десятый — двенадцатый день. Они развиваются на месте среза гипокотиля, а у семядолей — вблизи главной жилки.
При регенерации молокана от черенков взрослых растений решающим фактором является влага. Весной при наличии в почве большого запаса зимне-весенней влаги при неглубокой заделке могут окореняться и развивать побеги даже очень маленькие отрезки корней в 1,5—2 см. Корневые черенки длиной в 7—9 см могут отрастать на песчаной почве с глубины 55—57 см. Черенки, вынесенные при обработке почвы на поверхность, очень быстро высыхают и не окореняются. Образующиеся летом черенки, оказавшись в пересохшем пахотном слое, также не приживаются. Хуже отрастают черенки и при глубокой заделке в почву, особенно на тяжелых почвах.
182
Регенерация молокана от оставшихся в поч; °, слабо поврежденных или стержневых корней может происходить в течение всего вегетационного периода, так как в данном случае растения всасывают воду из подпочвы и имеют достаточный запас питательных веществ в корнях. Корневые черенки окореняются легче и дают больше отпрысков, чем черенки из корневищ.
Экология
Молокан — ксерофит с глубоко проникающей корневой системой, способной доставать воду из подпочвы и запасающий воду в мощно развитой корневой системе, богатой паренхимными тканями и пронизанной в коровой части густой сетью млечников. Листья тоже ксерофильного типа; они покрыты толстой кутикулой и восковым налетом, средние и верхние стеблевые несколько фотометрич-ны. Транспирационный коэффициент в полтора раза выше, чем у пшеницы, и равен в среднем 603 (Бажанов, 1915), поэтому молокан перехватывает воду у культурных растений и сильно иссушает почву. Хорошо переносит атмосферную и почвенную засуху и быстро развивает придаточные корни при увлажнении почвы дождем.
Типичные местообитания молокана характеризуются чаще всего сухостепным и среднестепным увлажнением (Раменский и др., 1956). Растет на участках, где грунтовые воды залегают на глубине от 120 см до нескольких метров (Никитин, 1957). Переносит затопление водой (Левина, 1956).
Молокан светолюбив, о чем свидетельствует распространение его на открытых местах. Произрастая в затенении в колках или в зарослях и посадках кустарников, он сильно вытягивается, побеги слабо ветвятся и могут достигать 150—190 см.
Благодаря погруженности в почву зимующих органов и почек возобновления молокан переносит большие морозы и может расти в областях, где зимние минимумы температуры почвы достигают минус 40° при маломощном снеговом покрове. Вегетация его начинается в конце апреля и заканчивается с первым снегопадом и морозами, во время которых надземные побеги отмирают. Весенние заморозки переносит, при этом в листьях вырабатывается много антоциана и они окрашиваются в красновато-бурый цвет.
Отношение молокана к богатству и засоленности почвы сильно варьирует (Раменский и др., 1956). Произрастание молокана на сильно засоленных почвах отмечают многие авторы (Бажанов, 1922, 1929; Суднов, 1946; Красовский, 1949; Казакевич, 1951; Бахтизин, Рахимов, 1958; и др.). А. И.Мальцев (1933) относит его к солелюбивым растениям. Он растет на почвах разного механического состава— от песчаных до тяжелых суглинков, часто является пионером на крутых обнаженных склонах оврагов, прибрежных речных и морских песках.
183
Фитоценология
Молокан встречается как в естественных фитоценозах, так и в агроценозах. Он растет в оазисах Туркмении как сопутствующий вид в тростниковой ассоциации (Никитин, 1957). На Волго-Уральском междуречье встречен также в зарослях тростника, в составе лугово-болотной растительности по днищу пересохшей речки вместе с Typha angustifolia L., Bulboschoenus maritimus (L.) Palla, Alisma plantago-aquatica L., Gypsophila trichotoma Wend, и др., по берегу р. Малого Узеня в разнотравно-вейниковой ассоциации (Левина, 1956); в житняковой (из Agropyron pectiniforme Roem. et Schult.) и разнотравной ассоциации по берегам лиманов Прикаспийской низменности (Манохина и др., 1956); в ассоциации Agropyron ramosum Richt. (Фурсаев и др., 1956). П. Н. Красовский (1949), Л. И. Казакевич и Б. М. Смирнов (1950) считают прибрежную растительность для молокана его естественным фитоценозом, откуда он заносится в посевы и другие места. Встречается молокан в небольшом числе в осиновых колках Северного Казахстана и по их опушкам в густых зарослях Spiraea crenata L. и S. hy pericifolia L. E. K. Kox (1952) отмечает высокое обилие молокана в изреженных посадках сосны в ассоциациях Роа angustifolia L. и Artemisia austriaca Jack. М. С. Гиляров (1951) считает, что молокан, как и многие другие сорные растения, поселился сначала на выбросах грызунов, откуда потом распространился на прилегающие территории.
Молокан растет на разновозрастных залежах, на нарушенных и пахотных землях, засоряя абсолютно все культуры на полях, в садах, на бахчах, огородах, лесополосах, а также часто растет вдоль каналов, дорог, на железнодорожных насыпях. На молодых залежах растет часто большими куртинами или латками, занимающими площадь в несколько м2 (иногда десятки м2). Внутри куртин он образует чистые или почти чистые заросли с проективным покрытием до 90—95%, где на 1 м2 может приходиться до 250 побегов молокана. По мере старения залежи обилие снижается, но он может сохраняться в сильно угнетенном, покоящемся состоянии в течение 17—20 лет (Смирнов, 1956).
При большом обилии, когда молокан образует густую сеть корней, пронизывающих большие объемы почвы, он сильно иссушает почву, поглощает большое количество элементов минерального питания, его густые заросли затеняют соседние растения, поэтому он играет значительную средообразующую роль и обладает большой конкурентной способностью. Угнетающе действует молокан не только на другие сорняки и культурные травянистые растения, но и на рост молодых одно- и двулетних дубков, посеянных на лесных полосах на юго-востоке европейской части СССР, снижая годовой прирост и уменьшая олиственность их побегов (Былова, 1960 а, б, 1962). На пырейных залежах корни молокана, лишенные механических тканей, легко пронзаются острыми «шильцами» корневищ пырея.
В посевах молокан может доминировать как над другими сор
184
няками, так и над культурными растениями. Скорость разрастания молокана может быть 2,5—3 м (Владимиров, 1939) и даже 10 м за один год. Загущенные посевы ржи, пшеницы и кормовых трав угнетают молокан и сильно снижают его обилие (Мальцев, 1933; Судков, 1946; Владимиров, 1948, 1956).
Консортивные связи
В районах массового произрастания молокана, например в Волгоградской обл., главными опылителями его являются пчелы из разных родов и мухи-журчалки. На бутонах и распустившихся корзинках можно встретить жуков-нарывников. Они пожирают иногда целиком все цветки в корзинке и даже верхушки листочков обертки. Нередко посещают раскрытые корзинки молокана дневные бабочки, доставая длинными хоботками нектар из трубок венчиков. На листьях и стеблях молокана можно встретить галлы, иногда листья минированы; на побегах бывает многа тлей (в Москве), на бутонах корзинки найдены яички флерницы.
Молокан поражается ржавчиной Puccinia minussensis Thiim. Поражение местное (Бажанов, 1929). Обычно гриб сильно поражает корни и корневища при их поранении (при подрезках). Вблизи места поранения возникает клубневидное утолщение, на котором образуется масса почек, развивающихся затем в надземные хилые, зараженные побеги. Почки и побеги, зараженные ржавчиной, покрыты рыжевато-бурым налетом спор.
Стравливание молокана и вытаптывание копытными животными не угнетает его заметно, но при неумеренном выпасе, приводящем к сильному уплотнению почвы, он выпадает из травостоя, ибо сильного уплотнения почвы не выносит.
Внутривидовая систематика
Б. А. Федченко выделял форму м. татарского с коротким, почти отсутствующим носиком у семянки — f. pumilum В. Fedtsch., а С. И. Коржинский — особую разновидность с недоразвитыми семянками— var. anomala Korsh.
Хозяйственное значение
Молокан, обладая большой конкурентной способностью, благодаря огромной энергии вегетативного разрастания и способности к легкой регенерации является злостным, трудноискоренимым корнеотпрысковым сорняком, прочно удерживающим за собой занятые территории. Меры борьбы с ним должны быть направлены прежде всего на истощение запасов инулина в его подземных органах (Казакевич, Захарова, 1935; Казакевич, 1945; Мамонов, 1956 6). При систематических подрезках отрастающих побегов запасы инулина полностью можно истощить к середине июня. Известны удачные
185
опыты использования гербицидов для борьбы с этим растением (Остроумов, Смагина, 1935; Мамонов, 1956а; Милащенко, 1961).
Молокан — растение среднего кормового достоинства (Ларин и др., 1956; Раменский и др., 1956). Он хорошо поедается крупным рогатым скотом, давая хорошую нажир'овку при нагуле и способствуя увеличению удоя молока у коров. Молодые побеги его поедаются верблюдами, лошадьми, козами, овцами, кроликами (Бажанов, 1929; Конуров, 1940 и др.). Жители г. Камышина и окрестных селений собирают для скота молодые сочные побеги молокана во второй половине лета, когда пастбища оскудевают. Сено молокан дает плохое, так как при сушке часто плесневеет и чернеет, а высушенные листья крошатся.
Есть указания, что семена молокана обладают ядовитыми свойствами.
ЛИТЕРАТУРА
А б и а н ц Г,- X. 1939. Стратификация семян красной бузины. «Лесное хоз-во», № 5.
Абраменко С. Н., Фишер Б. Р. 1933. Определитель важнейших древесных пород СССР. М., Гослестехиздат.
Абрамова Г. К. I960. Некоторые биологические особенности клевера розового при возделывании его в полевых условиях Ленинградской области. В кн.: «Вопросы сенокосно-пастбищного хозяйства». М., Изд-во МСХ СССР.
Абрамова Г. К- 1965. Долговечность клевера розового в зависимости от условий произрастания. «Вести, с.-х. науки», № 2.
Агаркова С. Н. 1968. Изменчивость морфологических и хозяйственноценных признаков у дикорастущей лещины в Тамбовской области. «Бюлл. научн. информ. Центр, генетич. лаб.», вып. 14. Мичуринск.
Адольф Н. А. 1930. Многолетние виды рода Adonis Dill (подрод Consili-go DC.) в СССР. «Тр. по прикладной ботанике, генетике и селекции», т. 23, вып. 1.
Аксельрод Д. М. 1961. Биологические особенности и приемы возделывания горицвета весеннего. В кн.: «Дикорастущие и интродуцируемые полезные растения в Башкирии», вып. 1. Уфа.
Александров Л. П. 1923. Заметки о редких находках подмосковной флоры. «Бот. мат-лы Гербария Гл. бот. сада РСФСР», т. 4, вып. 19—20.
Александров'Ф. А. 1967. Формы черемухи обыкновенной в Кировской области и перспективы использования их в озеленении. «Тр. Ин-та экологии раст. и жив. Уральского ФАН СССР», вып. 54. Свердловск.
Алехин В. В. 1947. Растительность и геоботанические районы Московской и сопредельных областей. Изд-во МОИП.
Алехин В. В. 1951. Растительность СССР в основных зонах, изд. 2. М., «Сов. наука».
Алпатов В. В. и др. 1933. Опыление красного клевера и пути клеверного семеноводства. М.
Ананова Е. Н. 1958. Морфология пыльцы Polygonum bistorta. ДАН СССР, т. 118, № 1.
Артюшенко 3. Т. 1962. Род Бузина — Sambucus L. В кн.: «Деревья и кустарники СССР», т. 6. М.— Л., Изд-во АН СССР.
Артюшенко 3. Т., Соколов С. Я- 1955. Формирование почек и развитие годичных побегов у некоторых древесных пород. «Тр. Бот. ин-та АН СССР», сер. 6, вып. 4.
Арутюнова Л. Г. 1940. Морфология хромосом некоторых видов клевера. ДАН СССР, т. 27, № 18.
Атлас лекарственных растений СССР. 1962. М., Медгиз.
Бажанов С. С. 1915. Наблюдения над сорной растительностью в 1913 го
187
ду на Бузулукском опытном поле и в его окрестностях. «Тр. Бюро по прикладн. бот.», т. 8, № 3.
Б а ж а н о в С. С. 1922. Сорные травы Самарской губернии и борьба с ними. Самара.
Бажанов С. С. 1929. Mulgedium tataricum DC (молокан, молочай, синий молочай, осот татарский) как сорное растение на полях. «Тр. Средне-Волжского с.-х. ин-та», вып. 5. Самара.
Байкина А. Д. 1946. Материалы по биологии цветения и эмбриологии черемухи. «Сов. бот.», № 1.
Барыкина Р. П. 1960. О морфогенезе подземных органов размножения черемухи. «Биол. науки», № 4.
Батманов В. А. 1962. Фенологическая карта СССР «Начало цветения черемухи». В кн.: «Сибирский географ, сб.», вып. 1. М., Изд-во АН СССР.
Бахтизин Н. Р., Рахимов Э. М. 1958. Сорные растения Башкирской АССР и меры борьбы с ними. Уфа.
Бейдеман И. Н. 1954. Развитие растительности и почв в низменности восточного Закавказья. В кн.: «Вопросы улучшения кормовой базы в степной, полупустынной и пустынной зонах СССР». М,— Л., Изд-во АН СССР.
Билимович О. Ф. 1935. Значение околоплодника Labiatae для их систематики. «Тр. Воронежск. ун-та», вып. 7.
Биологическая флора Московской области. 1974, вып. 1. Изд-во МГУ.
Б о б р о в Е. Г. 1936. История и систематика рода Corylus. «Сов. бот.», № 1.
Бобров Е. Г. 1947. Виды клеверов СССР. «Тр. Бот. ин-та АН СССР», сер. 1, вып. 6.
Бойченко А. М. 1968. Розовоцветная черемуха в Зауралье. «Тр. Ин-та экологии раст. и жив. Уральского ФАН СССР», вып. 60. Свердловск.
Борисова-Гуленкова М. А. 1960. Ритм сезонного развития растений луговой степи. «Бюлл. МОИП», отд. биол., т. 65, вып. 6.
Бородина Н. А., Комаров И. А., Лапин П. И. и др. 1970. Семенное размножение интродуцированных древесных растений. М., «Наука».
Бы к,о в Б. А. 1965. Доминанты растительного покрова Советского Союза, т. 3. Алма-Ата, «Наука».
Былова А. М. 1958. О корневой системе Mulgedium tataricum DC. «Бот. журн.», т. 43, № 4.
Былова А. М. 1960а. Молокан татарский. В кн.: «Биология сорных растений». М., Учпедгиз,
Былова А. М. 19606. Регенерация Mulgedium tataricum DC. «Уч. зап. Моск. гос. пед. ин-та», № 142 (ф-т естествозн., вып. 4).
Былова А. М. 1962. Биология молокана татарского (Mulgedium tataricum (L.) DC.). Автореф. канд. дисс. М.
Былова А. М. 1965. Семейное и вегетативное размножение молокана татарского. «Уч. зап. Моск. гос. изд. ин-та», № 212 (химия, ботаника, зоология и гистология).
Ванин С. И. 1931. Болезни сеянцев и семян лесных пород. М.—Л., Сель-колхозгиз.
Ванин С. И., Журавлев И. И., Соколов Д. В. 1950. Определитель болезней древесных пород и кустарников, применяемых для полезащитных насаждений. М.—Л., Гослесбумиздат.
Вар лих В. К. 1912. Русские лекарственные растения. Изд. 2. СПб.
Васильев В. Ф., Киси с А. Д. 1935. Эфиромасличные растения Воронежской области. «Зап. Воронежск. с.-х. ин-та», вып. 1 (16).
Васильченко И. Т. 1937. Определитель всходов сорных растений СССР. М.—Л., Сельхозгиз.
Васильченко И. Т. 1941. О филогенетическом значении морфологии прорастания у зонтичных (Umbelliferae). «Сов. бот.», № 3.
Васильченко И. Т. 1960. Всходы деревьев и кустарников. М.— Л., Изд-во АН СССР.
Вашкулат П. Н. 1952. К биологии прорастания семян бузины. «Агробиология», № 5.
188
ВашкулатП. Н. 1959. Аномалии генеративных органов у Padus racemosa Gilib. «Бот. журн.», т. 44, № 6.
Веттштейн Р. 1912. Руководство по систематике растений, т. 2, ч. 2. М.
Вильнер А. М. 1966. Кормовые отравления сельскохозяйственных животных, изд. 4. Л., «Колос».
Вихров В. Е. 1959. Диагностические признаки древесины главнейших лесохозяйственных и лесопромышленных пород СССР. М., Изд-во АН СССР.
Владимиров И. Ф. 1939. Сорная растительность и борьба с нею. Куйбышев'.
Владимиров И. Ф. 1947. Сорные растения и борьба с ними. Куйбышев.
Владимиров И. Ф. 1948. Молокан татарский (Mulgedium. tataricum DC) и меры борьбы с ним. «Докл. ВАСХНИЛ», вып. 11.
Владимиров И. Ф. 1956. Влияние растительного покрова на всхожесть и развитие сорняков. В кн.: «Бот. сб. работ Куйбышевского отделения Всесоюзн. бот. об-ва». М.— Л., Изд-во АН СССР.
Властова К. С. 1971. Тетраплоидный розовый клевер. «Сб. научн. тр. Белорусск. НИИ земледелия», т. 15.
Вольф Э. Л. 1923. Sorbus aucuparia L. var aquilonaris Egb. Wolf. «Бот. мат-лы Гербария Главн. бот. сада РСФСР», т. 4, вып. 23—24.
Воробьев Д. В. 1953. Типы лесов европейской части СССР. Киев, Изд-во АН УССР.
Ворошилов В. Н., Скворцов А. К-, Тихомиров В. Н. 1966. Определитель растений Московской области. М., «Наука».
В у л ь ф Е. В. 1932. Дубильные растения. Л., Госхимтехиздат.
Высоцкий Г. Н. 1913. О дубравах в Европейской России и их областях. «Лесн. журн.», т. 43, № 1—2.
Габриэлян Э. Ц. 1958а. Кавказские представители рода Sorbus L. «Тр. Бот. ин-та АН Арм ССР», т. 11.
Габриэлян Э. Ц. 19586. К анатомии плодов и морфологии цветков кавказских представителей рода Sorbus L. «Изв. АН Арм ССР», сер. биол. науки, № 7.
Галактионов И. И., В у А. В., Осин В. А. 1967. Декоративная дендрология. М., «Высшая школа».
Г а м м е р м а н А. Ф., Ш а с с Е. Ю. 1954. Схематические карты распространения важнейших лекарственных растений СССР. М.— Л., Изд-во АН СССР.
Гиляров М. С. 1951. Роль степных грызунов в происхождении полевой почвенной энтомофауны и сорно-полевой растительности. ДАН СССР, т. 79, № 4.
Глухов М. М. 1955. Медоносные растения, изд. 6. М., Сельхозгиз.
Гоббе Л. А. 1964. Типы весен и сроки весеннего развития растений в лесу Воронежского заповедника. «Тр. Воронежск. заповеди.», вып. 14.
Гогииа Е. Е. 1970. О некоторых особенностях цветения тимьянов. «Бюлл. ГБС АН СССР», вып. 77.
Гогина Е. Е. 1971а. О гинобазисных железках у Thymus и некоторых других растений сем. Labiatae. «Бот. журн.», т. 56, № 12.
Гогина Е. Е. 19716. По поводу различий в семенной продуктивности обоеполых и женских особей у Thymus. «Бюлл. ГБС АН СССР», вып. 82.
Гогина Е. Е. 1974. О гибридной природе подмосковной расы Thymus laevyanus Opiz. «Бюлл. ГБС АН СССР», вып. 92.
Голубев В. Н. 1956. К биоморфологии природных растений Подмосковья с запасающими органами побегового происхождения. «Уч. зап. Моск. обл. пед. ин-та», т. 41 (тр. каф. ботаники, вып. 1).
Голубев В. Н. 1958. О короткокорневищных растениях. «Бюлл. МОИП», отд. биол., т. 63, вып. 3.
Голубев В. Н. 1960. К экологии й морфологии Trifolium hybridum L. «Бот. журн.», т. 45, № 3.
Голубев В. Н. 1962. Основы биоморфологии травянистых растений центральной лесостепи. «Тр. Центр.-Черноземн. заповеди.», вып. 7.
Голубев В. Н. 1965. Эколого-биологические особенности травянистых растений и растительных сообществ лесостепи. М., «Наука».
189
Голубев В. Н. 1968. О морфогенезе моноподиальных полукустарников Крымской Яйлы. «Бюлл. МОИП», отд. биол., т. 73, вып. 4.
Голубева И. В. 1962. Некоторые данные о запасе живых семян в почвах под луговостепной растительностью. «Бюлл. МОИП», отд. биол., т. 67, вып. 5.
Горбунова Н. П. 1956. О взаимоотношении гриба и высшего растения в микоризах. «Усп. совр. биол.», т. 42, вып. 2.
Горчаковский П. Л. 1948. Синузия весенних раннецветущих-растений в горных лиственных лесах Сибири. ДАН СССР, т. 60, № 3.
Горчаковский П. Л. 1968. ’Растения европейских шйроколиственных лесов на восточном пределе их ареала. «Тр. ин-та экологии раст. и жив. Уральск. ФАН СССР», вып. 59. Свердловск.
Горышина Т. К- 1961. О температурном режиме ранневесенних растений в дубовом лесу. «Бот. журн.», т. 46, № 9.
Горышина Т. К. 1965. О характере осенне-зимнего покоя у различных сезонных групп травянистых растений дубрав. «Бюлл. МОИП», отд. биол., т. 70, вып. 5.
Горышина Т. К. 1969. Ранневесенние эфемероиды лесостепных дубрав. Изд-во ЛГУ.
Горяев М. М. 1952. Эфирные масла флоры СССР. Алма-Ата, Изд-во АН КазССР.
Граменицкая В. Г. 1952. О фитонцидных свойствах некоторых растений.. В кн.: «Фитонциды, их роль в природе и значение для медицины». М., Изд-во АМН СССР.
Гроздов Б. В. 1950. Типы леса Брянской, Смоленской и Калужской областей. Брянск.
Гроздов Б. В. 1960. Дендрология, .изд. 2. М,—Л., Гослесбумиздат.
Гроссгейм А. А. 1950. Флора Кавказа, изд. 2, т. 4. М.— Л., Изд-во АН СССР.
Гроссет Г. Э. 1930. Лес и степь в их взаимоотношениях в пределах лесостепной полосы Восточной Европы. Воронеж.
Гроссет Г. Э. 1962. Возраст термофильной реликтовой флоры широколиственных лесов Русской равнины, Южного Урала и Сибири в связи с палеогеографией плейстоцена и голоцена. «Бюлл. МОИП», отд. биол., т. 64, вып. 3.
Губанов И. А., Алексеев Ю. Е. 1975. Новые местонахождения редких и интересных растении в Московской области. «Бюлл. МОИП», отд. биол., т. 80, вып. 3.
Губанов И. А., Б. А. Старостин, В. Н. Тихомиров. 1972. Флора и растительность Московской области (История изучения и аннотированная библиография). Изд-во МГУ.
Гусев В. И., М. Н. Римский-Корсаков. 1951. Определитель повреждений лесных и декоративных деревьев и кустарников европейской части СССР. Л., Гослесбумиздат.
Гуцев ич С. А., Ш и в р и н а А. Н. 1942. Съедобные дикорастущие травы. Л.
Данилов М. Д. 1964. Урожаи дикорастущих плодово-ягодных растений в Марийской АССР и их использование. В кн.: «География плодоношения лесных древесных пород, кустарников, ягодников, значение их урожаев в народном хозяйстве и жизни фауны». М„ изд. МОИП.
Даниловский И. В. 1928. Фауна и возраст известковых туфов на правом берегу р. Луги близ д. Вяз. «Изв. Геол, комит.», т. 47, № 6.
Дарвин Ч. 1928. Приспособления орхидных к оплодотворению насекомыми. Поли. собр. соч., т. 4, ки. 1.
Двораковский М. С., Захарова Н. А. 1955. Сравнительная характеристика подроста обыкновенной лещины в различных экологических условиях. «Бюлл. МОИП», отд. биол., т. 60, вып. 3.
Демин А. П. 1969. Влияние удобрений на морфологическую структуру подземных органов некоторых луговых растений. «Бюлл. МОИП», отд. биол., т. 74, вып, 6.
Денисова Г. М. 1953. Побегообразование и ритм сезонного развития луговых растений низовий Северной Двины. Автореф. канд. дисс. М.
190
Джиаукштас П. И. 1957. Использование лещины в широколиственных лесах Литовской ССР. М., изд. Ин-та леса АН СССР.
Динесмаи Л. П 1961. Влияние диких млекопитающих на формирование древостоев. М., Изд-во АН СССР.
Дмитриев А. М. 1948. Луговодство с основами луговедения, изд. 2. М., Сёльхозгиз.
Дмитриев Г. О. 1937. Главные сорняки Башкирии и борьба с ними. Уфа.
Долгошов В. И. 1947. Календарь природы Подмосковья. В кн.: «Очерки природы Подмосковья и Московской обл.». Изд-во МОИП.
Долгошов В. И. 1959. Некоторые особенности плодоношения лесных пород. «Природа», № 1.
До хм а и Г. И. 1948. К экологии видов рода Adonis L. Эколого-фитоцено-тическое обоснование ритма развития и некоторых морфологических особенностей строения видов. «Бюлл. МОИП», отд. биол., т. 53, вып. 2.
Драбкин Б. С. 1954. О механизме действия фитонцидов черемухи. «Биохимия», т. 19, вып. 4.
Дуброва Г. Б. 1952. Дейс'твие фитонцидов некоторых растений на плесневые грибы. В кн.: «Фитонциды, их роль в природе и значение для медицины». М., Изд-во АМН СССР.
Дударь Ю. А. 1969. Особенности развития монокарпического побега горицвета весеннего (Adonis vernalis L.). «Тр. Ставропольск. НИИ сельск. хозяйства», вып. 6.
Дунин М. С. 1946. Иммуногенез и его практическое использование. «Тр. ТСХА», вып. 40.
Д ы л и с Н. В. 1969. Структура лесного биогеоценоза. М., «Наука».
Ел аги и И. Н. 1963. Типы леса нагорной части Теллермаиовского опытного лесничества и их хозяйственное значение. В кн.: «Биогеоценотические исследования в дубравах лесостепной зоны». М., Изд-во АН СССР.
Ефимова М. А. 1954. Деревья и кустарники зимой. Л., Учпедгиз.
Ефимова М. А. 1955. Анатомическое строение кроющих чешуек почек липы, черемухи, березы и черной смородины. «Уч. зап. Ленингр. пед. ин-та», т. 109.
Ефремов В. В. (ред.). 1943. Витаминные растения и их использование в пищу. М.
Ефремова Л. Д. 1968. О ритме развития горицвета весеннего (Adonis vernalis L.) в Курском заповеднике. В кн.: «Рефераты докл. по морфологии раст.». Изд-во МГУ. '
Ефремова Л. Д. 1971. Состояние репродуктивных органов у Adonis vernalis L. на различных этапах генеративного развития. «Растит, ресурсы», т. 7, вып. 2.
Забровский Е. П. 1962. Плоды и семена древесных и кустарниковых пород. М., Гослесбумиздат.
3 а л ы г и н С. П. 1971. Красный клевер. М., «Сов. Россия».
Захарьин М. Н. 1935. Ядовитые и вредные растения. Саратов.
Земли некий С. Е. 1958. Лекарственные растения СССР, изд. 3. М., Медгиз.
Злобина Л. М. 1937. Цветение и плодоношение тимьяна (Thymus marschallianus Willd.). «Ботаника. Исследования» (сб. работ Белорусск. отд. ВБО), вып. 9.
Знаменский И. Е. 1932. Дикие съедобные растения. Л.
3 о з И. Г., К о м и с а р е н к о Н. Ф., Черных Н. А. 1967. Буфадиенолид-карденолидсодержащие растения флоры СССР. «Растит, ресурсы», т. 3, вып. 2.
Зозулин Г. М. 1959. Подземные части основных видов травянистых растений и ассоциаций плакоров Средне-Русской лесостепи в связи с вопросами формирования растительного покрова. «Тр. Центр.-черноземн. заповеди.», вып. 5. Курск.
Золотницкая С. Я-, Авакян А. А. 1950. Атлас и определитель семян лекарственных растений. Ереван.
Иваненко Б. И. 1923. Условия произрастания и типы насаждений Погон-но-Лоснного острова. М.
191
Иванов В. В. 1951. Лещина в долине реки Урала. «Бот. журн.», т. 36, № 3.
Иванов Л. А., С и л и н а А. А., Жм у р Д. Г., Це л ь н и к е р Ю. Л. 1951. Об определении транспирационного расхода древостоем леса. «Бот. журн.», т. 36, № 1.
Иванова А. И. 1966. О внутреннем строении семян лютиковых. «Билл. ГБС АН СССР», вып. 61.
Иванова Е. В., Пашина Г. В., Кухарева Л. В. 1970. Полезные травянистые растения. Минск, «Урожай».
Игошина К. Н. 1943. Остатки широколиственных ценозов среди пихтовоеловой тайги Среднего Урала. «Бот. журн.», т. 28, № 4.
Илющенко Э. И. 1971. Значение клевера розового в подборе травосмесей для пойменных земель. «Научн. тр. СибНИИ сельск. хозяйства», т. 1 (16).
И м с А. 1964. Морфология цветковых растений. М., «Мир».
Инспекторов М. М., Стенковская М. Л. 1951. Сорные растения и борьба с ними. Омск.
Исаин В. Н. 1959. К биологии клевера' (Trifolium pratense L.). «Докл. ТСХА», вып. 46.
Исаченко X. М. 1954. Лесоводственные свойства и хозяйственное значение деревьев и кустарников. М.— Л., Гослесбумиздат.
Иорданов Д., Николов П., Бойчинов А. 1972. Фитотерапия, изд. 3. София.
Каден Н. Н. 1964. К вопросу о дробных плодах. «Бот. журн.», т. 49, № 7.
Каден Н. Н. 1965. Типы плодов растений средней полосы европейской части СССР. «Бот. журн.», т. 50, № 6.
Каден Н. Н., Зака люк ин а Т. П. 1965. Морфология-гинецея и плода бурачниковых и губоцветных. «Вести. Моск, ун-та», сер. биол., почвовед., № 3.
Казакевич Л. И. 1945. Истощение подземных частей молокана при подрезках. «Бюлл.. Ин-та зернового хоз-ва Юго-Востока СССР», № 3. Саратов.
Казакевич Л. И. 1951. Сорные растения и способы их уничтожения. Саратов. >
Казакевич Л. И., Захарова Т. А. 1935. Борьба с сорняками при орошении. В кн.: «Итоги по опытной работе ВИЗХ за 1934 г.». Саратов.
Казакевич Л. И., Смирнов Б. М. 1950. Как очистить поля от сорняков. Саратов.
Казакевич Л. И., Соболевская О. Ю. 1928. Дикорастущие душистые растения Нижнего Поволжья и их эфирные масла. «Журн. опытн. агрономии Юго-Востока», т. 5, № 2.
Каменецкая И. В. 1949. Влияние метеорологических условий на семенное возобновление травянистых растений Стрелецкой степи. «Бюлл. МОИП», отд. биол., т. 54, вып. 4.
Каменецкая И. В. 1952. Семенное возобновление растительности северной степи (на примере Стрелецкой степи Курской обл.). Автореф. канд. дисс. Л.
Каппер О. Г. 1952. Рябина обыкновенная или горькая (для защитных лесных полос). «Лесн. хоз-во», № 8.
Карпенко Ю. О. 1931. Пахуч! ростлини Украши та добування з них ете-рово! оли. Харк1в — Кшв.
Кауфман Н. Н. 1866, 1889. Московская флора, или описание высших растений и ботанико-географический обзор Московской губернии, изд. 1—2. М.
Кац Н. Я- 1943. На пути к познанию структуры лесных фитоценозов. «Бот. журн.», т. 28, № 4.
Керн Э. Э. 1926. Иноземные древесные породы, их лесоводственные особенности и лесохозяйственное значение. Л.
Кернер фон Марилаун А. 1902. Жизнь растений, т. 2. СПб.
Киндякова М. Д. 1958. К биологии цветения и плодоношения лещины обыкновенной — Corylus avellana L. «Уч. зап. Мордовск. ун-та», вып. 7, сер. естеств. и с.-х. наук. Саранск.
Киндякова М. Д. 1964. О причинах нерегулярного плодоношения обыкновенной, лещины в Мордовской АССР. В кн,: «География плодоношения лесных
192
древесных пород, кустарников и ягодников, значение их урожаев в народном хозяйстве и жизни фауны». М., изд. МОИП.
Киндякова М. Д. 1966. Динамика роста побегов, листьев и корней лещины обыкновенной в годовом цикле. «Уч. зап. Горьков, пед. ин-та», вып. 57, сер. биол. наук.
Киндякова М. Д. 1969. Определение площади листа лещины обыкновенной. «Тр. Горьков, пед. ии-та», вып. 100, сер. биол. наук.
Киндякова М. Д. 1970. О сроках сбора урожая лещины обыкновенной в Ярославской области. «Растит, ресурсы», т. 6, вып. 2.
КиченкоВ. И. 1952. Дикорастущие чабрецы района Кавказских Минеральных вод. «Уч. зап. Пятигорск, фарм. ин-та», вып. 1.
Киченко В. И., Тихонова В. Л. 1968. Испытание горца змеиного в культуре. «Бюлл. ГБС АН СССР», вып, 71.
Клеопов Ю. Д’ 1941. Основные черты развития флоры широколиственных лесов европейской части СССР. «Мат-лы по истории флоры и растит. СССР», т. 1. М.— Л., Изд-во АН СССР.
К л о б у к о в а-А л и с о в а Е. Н. 1958. Дикорастущие полезные и вредные растения Башкирии, т. 1. М.— Л., Изд-во АН СССР.
Ковалев Н. В. 1934. Рябина как северное плодовое дерево. В кн.: «Проблемы северного растениеводства», вып. 4. Л.
Ковалев Н. В., Костина К. Ф. 1935. К изучению рода Primus Focke. (Вопросы систематики и селекции). «Тр. по прикладной бот., генетике и селекции», сер. 8, № 4.
Кожевников А. В. 1929. К фитосоциологической характеристике липовой части 38-го квартала Погонно-Лосиного острова. «Тр. по лесному опытному делу», вып. 6. М.
Кожевников А. В. 1931. О перезимовке и ритме развития весенних растений липового леса. «Бюлл. МОИП», отд. биол., т. 40, вып. 1—2.
Кожевников А. В. 1950. Весна и осень в жизни растений, изд. 2. Изд-во МОИП.
Кожухов Б. С, 1931. Некоторые типы корневых систем растений степных участков ЦЧО. В кн.: «Степи Цеитральио-Черноземной области». М.— Л., Сельхозгиз.
Козлов Г. В. 1970. Некоторые морфологические и биологические особенности лещины в Ленинградской области. «Сб. тр. аспирантов и молодых научн. сотр. ВИРа», вып. 17.
Колесников А. И. 195,8. Декоративные формы древесных пород. М., Изд-во Мин. коммун, хоз-ва РСФСР.
Колесников Д. Г., Комиссаренко Н. Ф.', .Чернобай В. Т. 1961. Кумарины борщевика сибирского (Heracleum sibiricum L.). «Мед. пром. СССР», № 6.
К о м и с с а р е н к о Н. Ф., 3 о з И. Г., Чернобай В. Т., Колесии-ков Д. Г. 1961. Кумарины плодов борщевиков и таксономия. «Биохимия», т. 26, № 6.
Коновалов И. Н. 1952. Род. Рябина — Sorbus L. В кн.: «Деревья и кустарники СССР», т. 3. М.— Л., Изд-во АН СССР.
Коновалов Н. А. 1929. Типы леса подмосковных опытных лесничеств. «Тр. по лесному опыта, делу Центр, лесной опыта, станции», вып. 5—6. М.— Л.
Конуров И. Г. 1940. Сорные растения и борьба с ними в АССР НП. Энгельс.
Кррдюм Е. Л. 1967. Цитоэмбриология семейства зонтичных. Киев, «Нау-кова думка».
К о р о б о в а-С е м е н ч е н к о Л. В. 1960. О подснежном развитии Corydalis halleri Willd. «Биол. науки», № 1.
Корчагин А. А. 1951. Новая (длинноплодиая) разновидность черемухи Padus racemose. (Lam.) Gilib. «Бот. журн.», т. 36,.№ 2.
Костромина М. М. 1970. К биологии горца змеиного (Polygonum bis-torta L.). В кн.: «Эколого-морфологические и биохимические особенности полезных растений дикорастущей флоры Сибири». Новосибирск, «Наука».
193
Котт С. А. 1961, 1969. Сорные растения и борьба с ними, изд. 3—4. М., Сельхозгиз — «Колос».
Кох Е. К. 1952. Типология лесных посадок в пределах трассы государственной лесной полосы Саратов — Камышин и их засоренность. «Уч. зап. Сара-товск. ун-та», т. 29.
Кощеев А. Л. 1950. Распространение и лесоводственные свойства древесных пород и кустарников для полезащитных насаждений. М.— Л., Гослесбум-издат.
Красовская С. А. 1949. Изучение жизненного цикла цветковых растений в питомнике и в природе в Башкирском заповеднике. «Научно-методические зап. Главк. Управл. по заповедникам», вып. 12. М.
Красовский. П. Н. 1949. О распространении молокана татарского (Mulgedium tataricum (L.) DC.) на Урале. Пенза.
Круссер О. В. 1940. Биохимия лещины. В кн.: «Биохимия культурных растений», т. 7. М.— Л..
Крылова Н. П. 1963. К вопросу о повышении устойчивости бобовых в травостоях природных заливных лугов. «Бюлл. МОИП», отд. биол., т. 68, вып. 4.
Крылова Н. П. 1964. Закономерности семенного размножения и устойчивости бобовых растений природных заливных лугов (на примере Дединовского расширения поймы р. Оки). Автореф. канд. дисс. М.
Крылова Н. П. 1965. Семенное размножение клеверов красного и розового на природных заливных лугах в пойме р. Оки. «Уч. зап. Ульяновского пед. ин-та», т. 20, вып. 6. Саратов.
Купревич В. Ф. 1954. Болезни клевера и люцерны. М.—Л., Изд-во АН СССР.
КурнаевС. Ф. 1968. Основные типы леса средней части Русской равнины. М., «Наука».
Курьянов М. А. 1967. Биологически активные вещества в плодах рябины и аронии. «Тр. Центр, генетической лаб.», т. 9. Тамбов.
Кучеров Е, В. и др. 1971. Характеристика естественных кормовых угодий южной части западного склона Южного Урала. В кн.: «Дикорастущие и интродуцируемые полезные растения в Башкирии», вып. 3. Уфа, изд. БашФАН СССР.
Лавренко Е. М. 1938. История флоры и растительности СССР по данным современного распространения растений. В кн.: «Растительность СССР», т. 1. М.— Л., Изд-во АН СССР.
Л а п и н Е. М. 1960. Дикорастущий розовый клевер. Вологда.
Лапин П. И., Сиднева С. В. 1971. Сезонный ритм развития у видов рода Sorbus при интродукции. «Бюлл. ГБС АН СССР», вып. 79.
Ларин И. В. н др. 1951, 1956. Кормовые растения сенокосов и пастбищ СССР, тт. 2, 3. М.— Л., Сельхозгиз.
JIapioHOB Д. К., Макодзеба!. А. 1957. Бур’яни i боротьба з ними. Кшв.
Лебедева В. И. 1958. Органогенез вегетативных и генеративных органов красного клевера. «Вести. Моск, ун-та», сер. биол., почв., № 3.
Левина Р. Е. 1957. Способы- распространения плодов и семян. Изд-во МГУ.
Левина Р. Е. 1967. Плоды. Морфология, экология, практическое значение. Саратов.
Левина Ф. Я- 1956. Растительность лиманов севера Волго-Уральского междуречья. В кн.: «Природа и кормовые особенности растительности лиманов Волго-Уральского междуречья». М.— Л., Изд-во АН СССР.
Лесников Е. П. 1969. Аитифунгальиые свойства высших растений. В кн.: «Антифунгальные свойства высш. раст.». Новосибирск, «Наука».
Лисицин П. И. 1951. Избранные сочинения, т. 1. М., Сельхозгиз.
Лихачев А. Н. 1959. Формирование отавы некоторых луговых растений при сенокосном и пастбищном использовании. «Уч. зап. Моск, городск, пед. ин-та», т. 100 (каф. ботаники, вып. 5).
Лобанов Н. В. (ред.). 1963. Микориза растений. М., Сельхозгиз.
194
Лозовский Т. А. 1936. Производственно-биологические особенности рябины как плодового дерева в связи с ее селекцией и сортоиспытанием. «Тр. по прикладн. бот., генетике и селекции», сер. 8, № 5.
Лукс Ю. А. 1970. К вопросу о естественном вегетативном размножении некоторых наземных орхидей с корневыми клубнями. «Тр. Никитского бот. сада», т. 43.
Любарский Е. Л. 1967. Экология вегетативного размножения высших растений. Казань.
Лякавичус А. А., Ясконис Ю. А. 1967, 1968. О роли тимьяна (Thymus L.) в Литве. «Тр. АН ЛитССР», сер. В (1-е сообщ.—1967, № 1; 2-е сообщ.— 1968, №2).
М а г р у Ж- 1949. Симбиоз у орхидей и картофеля. М., ИЛ.
Маевский П. Ф. 1940, 1954, 1964. Флора средней полосы европейской части СССР, изд. 7—9. М.—Л., Сельхозгиз — «Колос».
Мальцев А. И. 1933. Главные сорняки зерновых культур СССР и меры борьбы с ними, изд. 2. М.— Л., Сельхозгиз.
Малышева Г. С. 1968. Географические закономерности зацветания Padus racemosa Gilib. на северо-западе Русской равнины. «Бот. журн.», т. 53, № 12.
Мамонов Н. Н. 1956а. Борьба с молоканом и березкой в условиях орошения на каштановых почвах Заволжья. Автореф. канд. дисс. Саратов.
Мамонов Н. Н. 19566. Глубокая обработка почвы — средство борьбы с молоканом и березкой на орошаемых землях. «Земледелие», № 1.
Манохина Л. А., Ларин И. В., Акимцева 3. С. 1956. Влияние лиманного орошения на луговую, пустынную и степную растительность лимана Утиного. В кн.: «Природа и кормовые особенности растительности лиманов Волго-Уральского междуречья». М.— Л., Изд-во АН СССР.
Манденова И. П. 1950. Кавказские виды рода Heracleum. Тбилиси.
Машкин С. И., Голицын С. В. 1952. Дикорастущие и разводимые деревья и кустарники Воронежской области. Воронеж.
Машковский М. Д. 1972. Лекарственные средства, изд. 7, чч. 1, 2. М., «Медицина».
Медведев П. Ф. 1957. Пищевые растения СССР. В кн.: «Растительное сырье СССР», т. 2. М.— Л., Изд-во АН СССР.
Меницкий Ю. Л. 1973. Надвидовые таксоны рода Thymus L. (Labiatae). «Бот. журн.», т. 58, № 6, 7.
Мещеряков Д. П. 1957. О длительном сохранении всхожести семенами бобовых. «Изв. ТСХА», вып. 2.
Милащенко Н. 3. 1961. Борьба с осотами в связи с применением ржаных паров и гербицидов в условиях Омской области. Автореф. канд. дисс. Омск.
Миняева О. М. 1952. Антракноз клевера в условиях нечерноземной зоны. М., Сельхозгиз.
Мирвис Д. А. 1959. Какой укос у двуукосного клевера следует оставлять на семена. «Тр. Ганусовской с.-х. опыта, станции», вып. 1. Минск.
Мисник Г. Е. 1949. Производственная характеристика семян деревьев и кустарников городских насаждений. М., Изд-во Мин. коммун, хоз-ва РСФСР.
Митина М. Б. 1965. Влияние осоки волосистой на распределение и обилие эфемероидов в дубовом лесу. «Вести. Ленингр. ун-та», № 15, сер. биол., вып. 3.
Митина М. Б. 1969. Об ассимиляционной деятельности доминантов сезонных синузий в снытевой дубраве лесостепной зоны. В ки.: «Механизмы взаимодействия растений в биогеоценозах тайги». Л., «Наука».
Морозов С. Ф. 1960. О получении семян клевера со второго укоса. «Агробиология», № 6. .
Муравьева В. И., Банковский А. И. 1947. Исследование растений, применяемых в народной медицине, на содержание аскорбиновой кислоты. «Тр. ВИЛАР», вып. 9. М.
М у х и и а Н. А. 1971. Клевер красный. Л., «Колос».
Навашин С. Г. 1951а. К. истории развития халазогамных Corylus avellana L. «Избранные тр.», т. 1. М.— Л., Изд-во АН СССР. .
195
Навашин С. Г. 19516. Новые данные по эмбриологии орешника Coryllus avellana L. «Избранные тр.», т. 1. М.—Л., Изд-во АН СССР.
Навашин С. Г. 1951в. Об оплодотворении у вяза и орешника. «Избранные тр.», т. 1. М.— Л., Изд-во АН СССР.
Нейштадт М. И. 1953. Расселение лещины обыкновенной по территории европейской части СССР в послеледниковое время. «Бот. журн.», т. 38, № 3.
Некрасов В. И. 1973. Основы семеноведения древесных растений при интродукции. М., «Наука».
Нестерович Н. Д., Чекалине кая И. И., Сироткин Ю. Д. 1967. Плоды и семена древесных растений. Минск, «Наука и техника».
Никитин А. А., Суворов И. П. 1949. О свойствах древесин черемух СССР. «Тр. Бот. ин-та АН СССР», сер. 5, вып. 2.
Н и к и т и Н--В. В. 1957. Сорная растительность Туркмении. Ашхабад.
Никитина К. К. 1955. К вопросу о длительности ювенильного периода жизни некоторых растений. В кн.: «Тез. докл. на науч. конф. Саратовск. пед. ин-та, посвященной итогам научно-исслед. работы за 1954 г.». Саратов.
Никитина К. К. 1957. К биологии ранневесенних растений. «Уч. зап. Саратовск. пед. ин-та», вып. 28.
Никитина С. В. 1966. Пойменные луга реки Мезени в нижнем ее течении. «Докл. ТСХА», вып. 126.
Овеснов А. М. 1963. Всхожие семена в почве, затопленной водами Пермского водохранилища. В кн.: «Мат-лы по биол. и гидрологии волжских водохранилищ». М,— Л., Изд-во АН СССР.
Овсянников В. Ф. 1930. Лиственные породы. Хабаровск.
Оз о л А. М., Зуковска 3. Я- 1955. Распространение лещины в Латвийской ССР. «Тр. Ин-та биологии АН ЛатвССР», т. 3.
О лен ев Н. О. 1955. О фитонцидах черемухи. «Природа», № 4.
ОловянниковаИ Н. 1949. Экология и ритм цветения растений Стрелецкой степи. «Научно-методич. зап. Главн упр. по заповеди.», вып. 12. М.
Остроумов Б. В., С маги и а Е. К. 1935. Флорпикрин как средство борьбы с молоканом синим и как фактор повышения урожайности яровой пшеницы. «Химизация соц. земледелия», № 9—10.
О тенов Т. О. 1970. Особенности образования корневых отпрысков у черемухи. «Вести. Каракалпакск. ФАН УзССР», № 2.
О те нов Т. О. 1971. Интродукция видов рода Padus Mill, в Каракалпакию. Автореф. канд. дисс. Ташкент.
Пайбердин М. В. 1970. Об использовании рябины обыкновенной и ее таксационных показателей. «Леей, жури.», № 4.
Пенин М. М. 1959. Борьба с осеменением и распространением молокана. «Сельское хоз-во.Поволжья», № 8.
Первухин Ф. С. 1961. Введение в культуру травянистых дубильных растений в условиях Северо-Запада СССР. «Тр. Бот. ин-та АН СССР», сер. 5, вып. 7.-
Петров А. П. 1947. Сезонная динамика травяного покрова широколиственных лесов. «Уч. зап. Казанск. пед. ин-та», ф-т естеств., № 5.
Петров А. П. 1955. Кайбицкие дубравы. «Уч. зап. Казанск. ун-та», т. 115, кн. 8.
П е т р о в Е. М. 1957. Рябина. М., Сельхозгиз.
Петунников А. Н. 1896, 1900, 1901. Критический обзор московской флоры: ч. 1 — «Бот. зап., изд. при Бот. саде СПб. ун-та», т. 6, вып. 13; ч. 2 —«Тр. СПб. о-ва естествоисп.», т. 30, вып. 3, отд. бот.; ч. 3— «Тр. СПб. о-ва естёство-исп.», т. 31, вып. 3, отд. бот.
Поддубная-Арнольди В. А. 1953. Выращивание орхидей из семян. «Тр. ГБС АН СССР», т. 3.
Покровская Т. М. 1959. Этапы формирования куста лугового клевера в условиях Московской области. ДАН СССР, т. 124, № 5.
Покровская Т. М. 1960а. Возрастно-морфологические особенности клеверов Московской области. Автореф. канд. дисс. М.
196
Покровская Т. М. 19606. Возрастно-морфологические этапы формирования куста шведского клевера (Trifolium hybridum L.) в условиях Московской области. «Биол. науки», № 2. '
Полякова Л. В. 1957. Динамика содержание дубильных веществ в некоторых представителях рода Polygonum L. в зависимости от фаз вегетации. В кн.: «Полезные растения природной флоры Сибири». Новосибирс, «Наука».
Полякова О. М. 1939. К вопросу о поедаемости дикорастущих степных —• растений крупным рогатым скотом. «Тр. Новочеркасского зооветеринарного ин-та», вып. 5. Ростов-на-Дону.
Понятовская В. М. 1956. Корневые системы важнейших кормовых злаков и бобовых Калининградской области. «Тр. Бот. ин-та АН СССР», сер. 3, вып. 10.
Поплавская Г. И. 1948. Растительность горного Крыма. «Тр. Бот. ин-та АН СССР», сер. 3, вып. 5.
Попов К. П. 1960. Рябина Крыма и перспективы их использования в культуре. Автореф. канд. дисс. Симферополь.
Попов М. Г. 1949. Очерк растительности и флоры Карпат. Изд-во МОИП.
П о п о в М. Г. 1957. Флора Средней Сибири, т. 1. М.— Л., Изд-во АН СССР.
Порфирьев В. С. 1959. Хвойно-широколиственные леса Волжско-Камского края и Предуралья и вопросы их классификации. В кн.: «Мат-лы по классификации растительности Урала». Свердловск.
ПошкурлатА. П. 1965. Анатомо-морфологический анализ растений вир-гинильного периода Adonis vernalis L. «Тр. Центр.-Черноземн. заповеди.», вып. 9.
Пошкурлат А. П. 1966. Материалы стационарных наблюдений за развитием горицвета весеннего Adonis vernalis L. «Бюлл. МОИП», отд. биол., т. 71, вып. 2.
Пошкурлат А. П. 1969. Семенное размножение весеннего горицвета (Adonis vernalis L.). «Биол. науки», № 7.
Пошкурлат А. П. 1970. Распространение горицвета весеннего и задача охраны его ресурсов. «Растит, ресурсы», т. 4, вып. 1.
Пошкурлат А. П. 1973. Горицветы СССР. Автореф. докт. дисс. М.
Пряно-ароматические растения СССР и их использование в пищевой промышленности. 1963. М., Пищепромиздат.
Пташицкий М. И. 1910. Растительность бассейна среднего течения реки Или. СПб.
Путохин Н., Смирнова В. 1935. Материалы по обследованию эфирных масел дикорастущих растений Куйбышевской предгорной зоны. М.— Куйбышев.
Пыльцевой анализ. 1950. М., Госгеолиздат.
П я т н и ц к и й С. С., К о в а л е н к о М. П., Л о х м а т о в Н. А. и др. 1963. Вегетативный лес. М., Сельхозиздат.
Работнов Т. А. 1939. Типы сероольховых насаждений северо-западной части Московской области. «Бот. журн.», т. 24, № 1.
Работнов Т. А. 1949. Луговые сорняки и меры борьбы с ними. М., Сель-хозгиз.
Работнов Т. А. 1950. Жизненный цикл многолетних травянистых растений в луговых ценозах. «Тр. Бот. ин-та АН СССР», сер. 3, вып. 6.
Работнов Т. А. 1955. Разногодичная изменчивость лугов. «Бюлл. МОИП», отд. биол., т. 60, вып. 3.
Работнов Т. А. 1956а. Некоторые данные о содержании всхожих семян в почве луговых сообществ. В кн.: «Акад. В. Н. Сукачеву к 75-летию со дня рождения». АА— Л., Изд-во АН СССР.
Работнов Т. А. 19566. О жизненном цикле борщевика сибирского (Не-racleum sibiricum L.). «Бюлл. МОИП», отд. биол., т. 61, вып. 2.
Работнов Т.-А. 1958. Изменение состава травостоя пойменного луга при внедрении борщевика сибирского (Heracleum sibiricum L.). ДАН СССР. т. 121, № 4.
197
Работнов Т. А. 1960. О флористической и ценотической полночленностй ценозов. ДАН СССР, т. 130, № 3.
Работнов Т. А. 1963. Биологические и экологические основы рационального использования и улучшения сенокосов и пастбищ. В кн.: «Природные сенокосы и пастбища». М.— Л., Сельхозиздат.
Работнов Т. А. 1965. О динамичности структуры полидоминантных луговых ценозов. «Бот. журн.», т. 50, № 10.
Работнов Т. А. 1971. Влияние образования льда в почве и на ее поверхности на луговые растения. «Бюлл. МОИП», отд. биол., т. 76, вып. 1.
Работнов Т. А. 1972. О наименовании ассоциаций со сменяющимися в течение вегетационного сезона или от года к году доминантами. «Вести. Моск, унта», сер. биол., почв., № 2.
Работнов Т. А. 1973. Влияние минеральных удобрений на луговые растения и луговые фитоценозы. М., «Наука».
Раздорский В. Ф. 1949. Анатомия растений. М„ «Сов. наука».
Разумович М. Б., Горфункель Д. А. 1956. Лечебные и дезорирую-щие свойства фитонцидов черемухи при лечении гнойных ран. «Уч. зап. Брестск. пед. ин-та», вып. 1.
Районы распространения важнейших сорных растений в СССР. 1935. М.— Л., Сельхозгиз.
Раменская М. Л. 1951. Естественные луга Карело-Финской ССР. «Изв. Карело-Финского ФАН СССР», № 3.
Раменский' Л. Г., Цаценкин И. А., Чижиков О. Н., Антипин Н. А. 1956. Экологическая оценка кормовых угодий по растительному покрову. М., Сельхозгиз.
Растительный покров СССР. 1956, т. 1. М.— Л., Изд-во АН СССР.
Рзазаде Р. Я- 1952. Род Platanthera L. С. Rich. В кн.: «Флора Азербайджана», т. 2. Баку, Изд-во АН АзССР.
Роллов А. X. 1908. Дикорастущие растения Кавказа, их распространение, свойства и применение. Тифлис.
Рыси'н Л. П., Золотова Ф. П. 1968. К методике определения продуктивности надземной части травяного покрова. В кн.: «Сложные боры хвойно-широколиственных лесов и пути ведения лесного хозяйства в лесопарковых условиях Подмосковья». М., «Наука».
Рысин Л. П., Рыси на Г. П. 1965. Почвенный запас семян травянистых растений в лесу и факторы, влияющие на их прорастание. В кн.: «Леса Подмосковья». М., «Наука».
Рыси на Г. П. 1968. К характеристике семенной продуктивности некоторых лесных травянистых растений. В кн.: «Сложные боры хвойно-широколиственных лесов и пути ведения лесного хозяйства в лесопарковых условиях Подмосковья». М., «Наука».
Р ы ч и н Ю. В. 1972. Древесно-кустарниковая флора, изд. 2. М., «Просвещение».
Савинов Б. Г. 1948. Каротин (провитамин А) и получение его препаратов. Киев.
Савченко А. И. 1963. Выращивание сеянцев рябины обыкновенной. «Лесн. хоз-во», № 8.
Савченко А. И. 1966. Предпосевная подготовка семян и выращивание сеянцев рябины обыкновенной. В кн.: «Ботаника. Исследования» (сб. работ Белорусок. отд. ВБО), вып. 8. Минск.
Сагдуллаева А. Л. 1959. Морфология пыльцы семейства маковых (Ра-paveraceae). «Проблемы бот.», т. 4. М.— Л., Изд-во АН СССР.
Сапанкевич 3. Ф. 1956. Заложение и развитие почек лещины обыкновенной. «Тр. Брянского лесохоз. ин-та», т. 7.
Сарычева 3. А. 1966. Динамика растительного покрова луговых степей северо-восточной части лесостепи Украины по исследованиям в заповеднике «Михайловская целина».- Автореф. канд. дисс. Киев.
Сацыперова И. Ф. 1951. О динамике накопления фитонцидов в плодах черемухи обыкновенной. ДАН СССР, т. 79, № 3.
198
С ел ив анов а-Г о р о д ков а Е. А. 1957а. Об опорных ветвях черемухи обыкновенной. ДАНСССР, т. 115, № 5.
Селиванова-Городкова Е. А. 19576. Черемуха и ее биологические особенности. «Бот. журн.», т. 42, № 8.
Семенов А. Л. 1947. Красный клевер. Минск.
Сергеев П. А. 1951. Красный клевер. В кн.: «Многолетние травы в луго-пастбищных севооборотах». М., Сельхозгиз.
Сергеев П. А. 1958. Приемы повышения устойчивости урожаев клевера красного. «Земледелие», № 1.
Сергеев П. А., Шайн С. С., Константинова А. М. и др. 1963. Клевер, изд. 2. М., Сельхозиздат.
Серебряков И. Г. 1947а. О ритме сезонного развития подмосковных лесов. «Веста. Моск, ун-та», № 6.
Серебряков И. Г. 19476. Некоторые данные по истории развития листьев черемухи и липы. «Вести. Моск, ун-та», № 7.
Серебряков И. Г. 1952. Морфология вегетативных органов высших растений. М., «Сов. наука».
Серебряков И. Г. 1954. О морфогенезе жизненной формы дерева у лесных пород средней полосы европейской части СССР. «Бюлл. МОИП», отд. биол., т. 59, вып. 1.
Серебряков И. Г. 1959. Период покоя у некоторых травянистых и древесных растений Подмосковья. «Уч. зап. Моск, городск. пед. ин-та», т. 100 (каф. ботаники, вып. 5).
Серебряков И. Г. 1962. Экологическая морфология растений. М., «Высшая школа».
Серебряков И. Г., Доманская Н. П., Родман Л. С. 1954. О морфогенезе жизненной формы кустарника на примере орешника. «Бюлл. МОИП», отд. биол., т. 59, вып. 2.
Серебряков И. Г., Серебрякова Т. И. 1965. О двух типах формирования корневищ у травянистых многолетников. «Бюлл. МОИП», отд. биол., т. 70, вып. 1.
Серебрякова Т. И. 1953. Побегообразование и ритм сезонного развития растений заливных лугов Средней Оки. Автореф. канд. дисс. М.
Серебрякова Т. И. 1956. Побегообразование и ритм сезонного развития луговых растений. «Уч. зап. Моск. гос. пед. ии-та», т. 97 (каф. ботаники, вып. 3).
Сильянова ГО. И. 1969. О накоплении витамина С в рябине. «Тр. Ка-занск. с.-х. ин-та», т. 41, вып. 1.
Скрипчинский В. В. 1963. Прорастание семян некоторых дикорастущих декоративных растений в естественных условиях. «Бюлл. ГБС АН СССР», вып. 50.
Скрипчинский В. В. 1969. Особенности роста и развития весенних эфемероидов в условиях Ставрополя и Ленинграда. «Тр. Ставропольск. НИИ сельск. хоз-ва», вып. 6.
Смирнов Б. М. 1956. О периоде покоя у корнеотпрысковых сорняков. «Агробиология», № 3.
Смирнов Н. Н. 1961. Дикорастущие ягоды Сибири и Дальнего Востока. Омск.
Смирнов П. А. 1958. Флора Приокско-Террасного государственного заповедника. «Тр. Приокско-Терр. заповеди.», вып. 2. М.
Смирнова О. В., Торопова Н. А. 1974. О сходстве жизненных циклов и возрастного состава популяций некоторых длиннокорневищных растений дубрав. «Уч. зап. Моск. гос. пед. ин-та».
Смольянинова Л. А. 1929. Обзор литературы по роду Corylus. «Тр. по прикладной бот., генетике и селекции», т. 21, вып. 5.
Смольянинова Л. А. 1931. Сорта и культура лещинного орешника в Абхазии. «Тр. по прикладной бот., генетике и селекции», т. 24, вып. 4.
Соколов С. Я. 1954. Род черемуха — Padus Mill. В кн.: «Деревья и кустарники СССР», т. 3. М.— Л., Изд-во АН СССР.
Соколова О. И. 1934. О всхожести семян дикорастущих кормовых трав. «Соц. растениеводство», сер. А, № 11.
199
Сорные растения СССР, т. 2. 1934. Л., Изд-во АН СССР.
Степанов Н. Н. 1938. Следует ли вводить в лесные культуры крушину и черемуху. «В защиту леса», № 3.
Строгий А. А. 1934. Деревья и кустарники Дальнего Востока. М.— Хабаровск, Дальгиз. .
Строкова Н. П. 1970. Семенное возобновление горицвета весеннего в условиях Челябинской области. В кн.: «Вопросы биол. раст.», вып. 2. Челябинск.
Стуккей К. Л. 1955. О протистоцидных свойствах летучих фитонцидов листьев черемухи и лавровишни. «Бюлл. эксперим. биол. и медицины», т. 39, вып. 4.
С у д н о в П. Е. 1946. Осоты и меры борьбы с ними. Ставрополь.
Сукачев В. Н. 1938. Дендрология с основами лесной геоботаники, изд. 2. Л., Гослестехиздат.
Сырейщиков Д. П. 1906, 1907, 1910, 1914. Иллюстрированная флора Московской губернии, чч. 1—4. М.
Сырейщиков Д. П. 1927. Определитель растений Московской губернии. М.
Сырейщиков Д. П. 1931. Виды и формы, новые для Московской области, и критические заметки. «Бюлл. МОИП», отд. биол., т. 40, вып. 1—2.
Сырокомская И. В., Понятовская В. М. 1960. Эколого-биологическая характеристика некоторых луговых ассоциаций. «Тр. Бот. ,ин-та АН СССР», сер. 3, вып. 12.
С юз ев П. В. 1906. О розовоцветной черемухе. «Изв. СПб. бот. сада», т. 6, вып. 1.
Телятьев В. В. 1971. Лекарственные растения Восточной Сибири. Иркутск.
Терехин Э. С. 1965. О терминах «сапрофит», «полусапрофит» и «полупаразит». «Бот. журн.», т. 50, № 1.
Терехин Э. С., К а мел ин а О. П. 1969. Эндосперм в Orchidaceae (к вопросу о редукции). «Бот. журн.», т. 54, № 5.
Толмачев А. И. 1954. К истории возникновения и развития темнохвойной тайги. М.— Л., Изд-во АН СССР.
Трофимов Т. Т. 1939. К вопросу о ритме развития ранневесенних растений. «Научно-метод. зап. Комитета по заповеди. РСФСР», вып. 5. М.
Т р о ф и м о в Т. Т. 1952. Вегетативное размножение хохлатки Галлера (плотной) (Corydalis halleri Willd.). «Бюлл. МОИП», отд. биол., т. 57, вып. 3.
Турова А. Д. ,1967. Лекарственные растения СССР и их применение. М., «Медицина».
Тюрин А. В. 1959. О зацветании Padus racemosa (Lam.) Gilib., Caragana arborescens Lam. и Betula verrucosa Ehrh. в Подмосковье за 74 года. «Бот. журн.», т. 44, № 11.
Тюрин А. В. 1969. О зацветании черемухи обыкновенной Padus racemosa (Lam.) Gilib. в Свердловске за 93 года. «Бот. журн.», т. 54, № 4.
Успенская Г. Д. 1959. Пятнистости листьев клевера в Московской области. «Биол. науки», № 1.
Успенская Г. Д. 1974. Консортивные связи микромицетов и бобовых растений в окрестностях Звенигородской биостанции МГУ. «Тр. ЗБС МГУ», вып. 1.
Успенская Г. Д., Духович В. М. 1958. Бактериальные пятнистости клевера. «Вести. Моск, ун-та», сер. биол., почв., № 2.
Федорако Б. И. 1940. О формовом разнообразии лещины в лесостепной зоне западного Предуралья. «Природа», № 10.
Федоров А. А., Кирпичников М. Э., Артюшенко 3. Т. 1962. Атлас по описательной морфологии высших растений. Стебель и корень. М.— Л, Изд-во АН СССР.
Федоров П. Н. 1964. Содержание аскорбиновой кислоты, сорбита, каро-' тина и витамина Р в плодах некоторых форм обыкновенный рябины, урожайность плодов в различных условиях местопроизрастания. В кн.: «География плодоношения лесных древесных пород, кустарников и ягодников, значение их урожаев в народном хозяйстве и жизни фауны», изд. МОИП.
200 '
Федоров П. Н. 1971. Биохимическая характеристика форм рябины обыкновенной южной полосы лесной зоны Среднего Поволжья. Автореф. канд. дисс. Л.
Филимонов М. А. 1953. Повышение посевных качеств семян кормовых трав. М., Сельхозгиз.
Флора СССР. 1935, 1936, 1937, 1941, 1945, 1949, 1951, 1954, 1958, 1964. Тт. 3, 4, 5, 7, 10, 11, 14, 17, 21, 23, 29. М,— Л., Изд-во АН СССР.
Ф у р с а е в А. Д., Щ е г о л е в а А. Д., Миловидова И. Б. 1956. Искусственные лиманы Заволжья, их почвы и растительность. В кн.: «Природа и кормовые особенности растительности лиманов Волго-Уральского междуречья». М.— Л„ Изд-во АН СССР.
Харитонов А. М. 1958. Розовая черемуха. «Сад и огород», № 1.
Харитонович Ф. Н. 1968. Биология и экология древесных пород. М., «Лесн. пром.».
Хохлов С. С., Ковалева А. С. 1950. Деревья и кустарники Нижнего Поволжья. Саратов.
Хромосомные числа цветковых растений. 1969. Л., «Наука».
Царев С. Г. 1964. Лекарственные растения в ветеринарии. М., Россель-хозцздат.
Цибанова Н. А. 1960. К биологии горицвета (Adonis vernalis L.). «Тр, Центр.-Черноземн. заповеди.», вып. 6. Курск.
Цингер В. Я. 1885. Сборник сведений о флоре.Средней России («Уч. зап. Моск, ун-та», отд. ест.-истор., вып. 6 за 1886 г.). М.
. Чернышев П. Я. 1934. Дубильные растения нашей страны. М.-—Л.
Шайн С. С. 1949. Значение твердосемянности многолетних бобовых для повышения устойчивости их травостоя. В кн.: «Вопросы кормодобывания». М., Сельхозгиз.
Шенников А. П. 1927. Фенологические спектры растительных сообществ. «Тр. Вологодской обл. с.-х. опытн. станции», вып. 2.
Шенников А. П. 1938. Луговая растительность СССР. В кн.: «Растительность СССР», т. 1. М.— Л., Изд-во АН СССР.
Шик М. М. 1953. Сезонное развитие травяного покрова дубрав. «Уч. зап. Моск. гос. пед. ин-та», т. 73 (каф. ботаники, вып. 2).
Шиманюк А. П. 1964. Биология древесных и кустарниковых пород СССР, изд. 2. М., «Просвещение».
Шиманюк А. П. 1967. Дендрология. М., «Лесн. пром.».
Шлыков Г. Н. 1932. Дубильные растения СССР. Л.— М., Сельколхозгиз.
Шмальгаузен И. Ф. 1895, 1897. Флора Средней и Южной России, Крыма и Северного Кавказа, тт. 1, 2. Киев.
Штамм В. А. 1953. О причинах разновременного зацветания лещины. «Бюлл. ГБС АН СССР», вып. 16.
Шутова 3. П. 1965. Новые данные по цитологии красного клевера. «Бот. журн.», т. 50, № 3.
Эйзенрейх X. 1959. Быстрорастущие древесные породы. М., ИЛ.
Юрцев Б. А. 1951. Некоторые данные о корневых системах травянистых многолетников подмосковных широколиственных лесов. «Бюлл. МОИП», отд. биол., т. 56, вып. 4.
Якушева Е. И. 1947. Продолжительность жизнеспособности семян некоторых бобовых трав. «Селекция и семеноводство», № 1.
Я н с о н Ф. И. 1968. Клевер розовый. М., «Колос».
Янсон Ф. И. 1970. Возделывание клевера розового на корм и семена. Автореф. докт. дисс. Таллин.
Ярошенко П. Д., Грабарь В. А. 1969. Смена растительного покрова Закарпатья. Л., «Наука».
Ярошенко П. Д., Юрова Э. А. 1970. Об остепненных лугах Владимирского ополья. «Бюлл. МОИП», отд. биол., т. 74, вып. 4.
Ячевский А. А. 1933. Основы микологии. М.— Л., Сельхозгиз.
В a i 1 е у L. Н. 1957. Standard cyclopedia of horticulture. N. Y.
Bersillon G. 1958. Organisation et structure du tubercule de Corydalis solida Sw. «Compt. Rend. Acad. (Paris)», t. 246, № 18.
201
Bernard N. 1899. Sur la germination du Neottia nidus avis. «Compt. Rend.' Acad. (Paris)», t. 128.
Bernard N. 1911. Sur la function fungicide des bubles d’Ophrydees. «Ann. Sc. Nat. (Paris)», ser. 9, t. 14.
Boatman D. J. 1956. Mercurialis perennis L. in Ireland. «Journ. Ecol.», vol. 44, N 2.
Borthwick H. A. 1931. Development of the macrogametophyte and, embryo of Daucus carota. «Bot. Gaz.», vol. 92.
Bradbeer J. W., Colman B. 1967. Studies in seed dormancy. I. The metabolism of (2-I4C) acetate by chilled seeds of Corylus avellana L. «New Phytol.», vol. 66, N 1.
Bradbeer J. W., Pin field N. J. 1967. The effects of gibberellin on the dormant seeds of Corylus avellana L. «New Phytol.», vol. 66, N 4.
В u r g e f f H. 1909. Die Wurzelpilze der Orchideen, ihre Kultur und ihr Le-ben in der Pflanze. Jena.
Burgeff H. 1936. Samenkeimung der Orchideen und Entwicklung ihrer Keimpflanzen. Jena.
Biisgen M. 1911. Cupuliferae. In: «Lebensgeschichte der Bliitenpflanzen Mit-teleuropas». Bd. 2, Abt. 1. Stuttgart.
В ii s g e n M. 1927. Bau und Leben unserer Waldbaume. Jena.
С 1 a p h a m A. R., T u t i n T. G., Warburg E. Fl 1958. Flora of the British Isles. Cambridge.
Ch m el ar F. 1941. Kliceni tezce bobtnajicich (tvrdych) zrn jeteie cerveneho (Trifolium pratense) vlozenych do xlicidla pfed 26 lety. «Sborn. Ceskosl. Akad. Ze-medSlsk.», vol. 19, N 1—3.
Clusius C. 1583. Rariorum aliquot stirpium, per Pannonicam, Ausfram, et vicinas quasdem provincias observatarum Hisforia. Antwerpen.
Codaccioni M. 1962. Recherches morphologiques et onthogeniques sur quelques cupuliferes. «Rev. Cytol. Biol. Veget.», t. 25, f. 1—2.
Dabrowski M. 1957. Zmiennosc wielkesci ziarn pylkow leszczyny (Corylus avellana L,) na tie zespoiow lesnych Bialowieskiego Parku Narodowego. «Acta Soc. Bot. Polon.», vol. 26, N. 2.
Dahlgren K. 1941. Eine neue Corylus Varietat. mit Ascidienblattern und ilber Corylus avellana L. var. Zimmermannii Hahne. «Sv. Bot. Tidskr.», Bd. 35, H. 4.
Diels L. 1918. Das Verhaltnis von Rhythmik und Verbreitung bei den Peren-nen des europaischen Sommerwaldes. «Вег. Deutsch. Bot. Ges.», Bd. 36.
Donath T. 1954. Die Edelberesche und ihre Einfiihrung in die Sauglingser-nahrung. «Arch. Gartenbau», Bd. 2, H. 3.
Downie D. G. 1943a. On the germination and growth of Goodyera repens. «Trans. Proc. Bot Soc. Edinburg», vol. 33, p. 1.
Downie D. G. 1943b. Notes on the germination of some British Orchids. «Trans. Proc. Bot. Soc. Edinburg», vol. 33, p. 2.
Downie D. G. 1943c. Source of symbiont of Goodyera repens. «Trans. Proc. Bot. Soc. Edinburg», vol. 33, p. 4..
Downie D. G. 1950. The germination of Goodyera repens (L.) R. Br. in fungal extract. «Trans. Proc. Bot. Soc. Edinburg», vol. 35, p. 2.
Dull R. 1961. Die Sorbus-Arten und ihre Bastarde in Bayern und Thiiringen. «Вег. Bayer. Bot. Ges.», Bd. 34.
Durand B. 1957. L’organisation de I’androcee des Mercurialis. «Compt. Rend. Acad. (Paris)», t. 244, N 5.
E i c h 1 e r A. W. 1878. Bliitendiagramme, t. 2. Leipzig.
Ellenberg H. 1939. Uber Zusammensetzung, Standort und Stoffproduktion bodenfeuchter Eichen- und Buchen-Mischwaldgesellschaften Nordwestdeutsclands. «Mitt. Flor-Soz. Arb. Niedersachsen», H. 5.
Ellenberg H. 1963. Die Vegetation Mitteleuropas mit den Alpen. «Einfiih-rung in die Phytologie», Bd. 4, T. 2.
Engler A. 1879. Versuch einer Entwicklungsgeschichte der Pflanzenwelt, insbesondere der extratropischen Florengebiete seit Tertiarperiode. T. I, 2. Leipzig.
Fedde F. 1936. Papaveraceae. In A. Engler, K. Prantl «Die natiirlichen Pflanzenfamilien», 2 Aufl., Bd. 17.
202
Fernaid M. 1899. The Rattlesnake-plantains of New England. «Rhodora», vol. 1, N 1. .
Fergus E. N., Hollowell E. A. 1960. Red clover. «Advances in Agronomy», vol. 12.
Fischer H. 1890. Beitrage zur vergleichenden Morphologic der Pollen-Kor-ner. Breslau.
Flora Europaea. 1964, 1968, 1970, vol. 1, 2, 3. Cambridge.
Frankland B., WareingP. 1966. Hormonal regulation of seed dormancy in hazel (Corylus avellana L.) and beech (Fagus sylvatica L.). «Journ. Exp. Bot.», v. 17, N 52.
Fuller F. 1962. Goodyera und Spiranthes. In: «Die neue Brehm-Biicherei». Berlin.
Gawlowska J. 1958. Ochraria naturalnych zasobow a mozliwosci uprawy milka wiesennego (Adonis vernalis L.). «Ochrona przyrody», roc. 25.
Geitler L. 1943. Fruchtformen der Hasel in Wildpopulationen. «Wien. Bot. Zeitschr.», Bd. 92, H. 1—2.
G i 1 b e r t H. 1956. Forage crops. N. Y.
G о e b e 1 K. 1913. Organographie der Pflanzen. T. 1. Jena.
G 6 r a 1 S. 1964. Badania dynamiki wzrostu systemu korzeniowego ditetraplo-idalnych form Trifolium pratense L. i T. hybridum L. «Roczniki Nauk Rolniczych», t. 88, ser. A, zc. 3.
Goss W. L. 1933. Buried seed experiment. «Monthly Bull. Dep. Agric., California», vol. 22, N 6.
Hagerup O. 1942. The morphology and biology of the Corylus — fruit. «Det. Kgl. Danske VidensK. Selsk., Biol. Med.», Bd. 17, N 6.
Hedlund T. 1901. Monographic der Gattung Sorbus. «Kongl. Sv. Vet. Akad. Handl.», Bd. 35, N 1.
Hegi G. 1909, 1913, 1923, 1931, Illustrierte Flora von Mitteleuropa. Bd. 2, H. 1; Bd. 4, H. 2, 5; Bd. 5, H. 1. Mflnchen.
Hegnauer R. 1948. Beitrag zur chemischen und morphologischen Kenntnis der Schweizerischen Thymus-formen. «Вег. Schweiz. Bot. Ges.», Bd. 58.
Hui ten E. 1949. On the races in the Scandinavian Flora. «Sv. Bot. Tidskr.», vol. 43, N 2, 3.
Hui ten E. 1950. Atlas over vaxternas utbredning i Norden. Stockholm.
Hui ten E. 1968. Flora of Alaska and neighboring territories. Stanford.
Hylander N. 1945. Nomenklatorische und systematische Studies tiber nor-dische Gefasspflanzen. «Uppsala Univ. Arsskr.», Bd. 1, N 7.
I r m i s c h T. 1850. Zur Morphologie der monocotylischen Knollen- und Zwie-belgewachse. Berlin.
Irmisch T. 1856. Uber einige Ranunculaceen. «Bot. Zeit.», T. 14.
I r mis ch T. 1862. Uber einige Fumariaceen. «Abh. Naturf. Ges. Halle», Bd. 6.
J al as J. 1971a. Notes on Thymus L. (Labiatae) in Europa. I. Supraspecific classification and nomenclature. «Journ. Linn. Soc. London (Bot.)», vol. 64, N 2.
J a 1 a s J. 1971b. Notes on Thymus L. (Labiatae) in Europe. 2. Comments on species and subspecies. «Journ. Linn. Soc. London (Bot.)», vol. 64, N 3.
J a 1 a s J., Kaleva K. 1970. Supraspecifische Gliederung und Verbreitungs-typen in der Gattung Thymus L. (Labiatae). «Feddes Repent.», Bd. 81, H. 1—5.
Jost L. 1890. Die Erneuerungswiese von' Corydalis solida Sw. «Bot. Zeit.», vol. 48.
Kaida A. 1962. Laialehised metsad Eestis. In: «Salumatsade majandamise Kiisimusi.». Tartu.
Kaleva K. 1967. Notes on some epidermal characteristics in Thymus L. (Labiatae). «Ann. Bot. Fenn.», vol. 4, N 1.
Kantor I. 1956. Jerab obecny (Sorbus aucuparia L.) jeho vyznam a pecteni sadebniho materialu. «Sborn. Vysoke Skoly Zem. Lesn. Fak. Brne», cis. 3.
Kegler H. 1959. Das Ringflecken mosaik der Eberesche (Sorbus aucuparia L.). «Phytopathol. Zeitschr.», Bd. 37, H. 2.
К 1 о о s A. W. 1926. Het geschlacht Thymus in Nederland. «Nederland. Kruidk. Arch.» (Jg. 1925).
203
Knuth P. 1899. Handbuch der Bliitenbiologie, Bd. 2. Leipzig.
Koch K. 1872. Dendrologie. T. 2. Erlangen.
Koponen T. 1962. Some observations on the cold resistance of Corydalis solida (L.) Sw. «Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo», vol. 17, N 1.
Ko p pen F. 1888. Geographische Verbreitung der Holzgewachse des euro-paischen Russlands und des Kaukasus. T. I. St.-Pet.
К r a 1 i к о v a K. 1961. Krufkovitost jarabiny v CSSR. «Biologia», roc. 16, cis. 11.
Kulp a W. 1960. Biologia kielkowania miika wiosennego (Adonis vernalis L.). «Roczniki Nauk Rolniczych», ser. A, t. 81, N 2.
Kummer H. 1952. Vergiftungen bei Pferden durch Adonis in Luzernecheu «Tierarztl. Umschau», Jg. 7, N 21/22.
Lefort F. 1951. Les Corydalis de la flore luxembourgeoise. «Bull. Soc. Bot. Belgique», t. 83, f. 2.
Linkola K., Tiirikka A. 1936. Uber Wurzelsysteme und Wurzelausbrei-tung der Wiesenpflanzen auf verschiedenen Wiesenstandorten. «Ann. Bot. Soc. Zool.-Bot. Fenn. Vanamo», t. 6, N 6.
Lippmaa T. 1938. Areal und Alterbestimmung einer Union (Galeobdolon-Asperula-Asarum-Union) sowie das Problem der Charakterarten und des Konstan-ten. Tartu.
Long B. 1927. Naturalized occurence of Prunus padus in America. «Rhodora», vol. 25, N 298.
Lukasiewicz A. 1962. Morpholigiczno-rozwojowe typy bylin. Poznan.
Lyr H., Polster H., Fiedler H. 1967. Geholzphysiologie. Jena.
Martin M. H. 1968. Conditions affecting the distribution of Mercurialis pe-rennis L. in certain Cambridgeshire woodlands. «Journ. Ecol.», vol. 56. N 3.
Messerchmidt W. 1965. Gas- und dunschichtchromatographie Untersu-chungen der atherischen Ole einiger Thymusarten. 2. «Planta Medica», Bd. 13, N 1.
Meusel H. 1943. Vergleichende Arealkunde. 2. Berlin.
Meusel H. 1969. Chorologische Artengruppen der Mitteleuropaischen Eichen-Hainbuchwalder. «Feddes Repert.», Bd. 80, H. 2—3.
Meusel H., Jager E., Weinert E. 1965. Vergleichende Chorologie der zentraleuropaischen Flora. Jena.
M i c h a 1 e t E. 1859. Sur le development et la vegetation des Corydalis solida Sw. et C. cava Schweigg. et Koert. «Bull. Soc. Bot. Fr.», vol. 6.
Michal к ova V.. 1967. Tetraploide Population der Art Corydalis cava (L.) Schweigg. et Koerte (Hohler Lerchensporn) in Wald Dubnik bei Sered. «Acta Fac. Rer. Natur. Univ. Comenianae, Bot.», Bd. 15.
Molli son J. E. 1943. Goodyera repens and its endophyte. «Trans. Proc. Bot. Soc. Edinburg», vol. 33.
Mukerji S. K. 1927. New variety of Mecrurialis perennis L. «Journ. Bot.», vol. 65.
Mukerj.i S. K. 1936. Contributions to the autecology of Mercurialis perennis L. «Journ. Ecol.», vol. 24, N 1—2.
Muller FL, Stritzke S., Treutler I., Donatt E. 1956. Die Edelbere-sche. Ein neues Obstgeholz. Berlin.
Muller-Stoll W. R. 1947. Die stifle Eberesche, Sorbus aucuparia L. var. als Obstbaum und Vitamintrager. «Die Pharmazie», Jg. 2, H. 2.
Ognyanov L, Gentscheva G., Georgiev V., Panov P. 1966. Na-turcumarine. I. Cumarine in Heracleum verticillatum Pane, und H. sibiricum L. «Planta medica», vol. 14, N 1.
Olsen S. 1925. Studies on the hydrogen-ion concentration of the soil and its significance to the vegetation, especially to the natural distribution of plants. «Compt. Rend. Trav. Lab. Carlsberg», vol. 15, N 1.
Pawlowski B. 1966. Observationes ad Thymus polonicos et nonnullis ucrai-nicos pertinentes. «Fragm. Fl. Geobot.», vol. 12, N 4.
Pert tula U. 1941. Untersuchungen uber die generative und vegetative Ver-mehrung der Bliitenpflanzen in der Wald-Hain-Wiesen und Hainfelzen-Vegetation. «Ann. Acad. Sci. Fenn. Ser. A, Biol.», vol. 58.
204
P i g о 11 C. D. 1955. Thymus L. «Journ. Ecol.», vol. 43, N 1.
Pin field N. J. 1968. The effect of gibberellin on the metabolism of ethanol-soluble constituents in the cotyledons of hazel seeds (Corylis avellana L.). «Journ. Exp. Bot.», vol. 19, N 60.
Prantl K., Kiindig J. 1891. Papaveraceae. In: A. Engler, K. Prantl «Die naturlichen Pflanzenfamilien», T. 3. Abt. 2. Leipzig.
Raatikainen T. 1960. Studies on Finnish populations of Sorbus aucuparia L. «Arch. Soc. Zool. Bot. Fenn. Vanamo», vol. 15, N 1—2.
Rabotnov T. A. 1960. Some problems in increasing the proportion of leguminous species in permanent meadows. In: «Proceed. Eigth Intern. Grassland Congr.», 1960, Session 11 A.
Rauscherts. 1965. Wiesen-und Weidepflanzen. Leipzig.
R e h d e r A. 1927. Manual of cultivated trees and shrubs. N. Y.
Rendle A. 1925. Development of blue pigment in Mercurialis. «Journ. Bot.», vol. 63, N 1.
Ronniger K. 1924. Beitrage zur Kenntnis der Gattung. Thymus. I. Die Bri-tisch Arten und Formen. «Feddes Repert.», Bd. 20.
Ronniger K. 1924. Beitrage zur Kenntnis der Thymus—Flora Balkanhalb-insel. «Feddes Repert.», Bd. 20.
Roussine N. 1961. Note sur les especes du genre Thymus aux Etats Unis d’Amerique. «Nat. Monspeliensia», ser. Bot., f. 13.
Rubner K. 1960. Die Pflanzengeographischen Grundlagen des Waldbaues. Berlin.
Ryberg M. 1950. Studies in the taxonomy and fertility of some Scandinavian Corydalis species of the sect. Pes-gallinaceus Irmisch. «Acta Horti Berg.», vol. 15, N 10.
Ryberg M. 1955. A. taxonomical survey of the genus Corydalis Ventenat. «Acta Horti Berg.», vol. 17, N 5.
Ryberg M. 1959. A morphological study of Corydalis nobilis, C. cava. C. solida and some allied species with special reference to their underground organs. «Acta Horti Berg.», vol. 19, N 3.
Salisbury E. J. 1920. The significance of the calcicolous habit. «Journ. Ecol.», vol. 8, N 3.
Salisbury E. J. 1925. The incidence of species in relation to soil reaction. «Journ. Ecol.», vol. 13.
S a 1 m о n С. E. 1930. Notes on Sorbus. «Journ. Bot.», vol. 68.
Saunders I. 1883. Monoecious and hermaphrodite of Mercurialis perennis. «Journ. Bot.», N 21.
Schimper A. 1889. Pflanzengeographie auf physiologischer Grundlage. Jena.
Schmidt P. 1968. Beitrag zur Kenntnis der Gattung Thymus L. in Mittel-deutschland. «Hercynia», vol. 5, N 4.
Schmidt P. 1969. Zur taxonomischen Abgrenzung und Verbreitung von Thymus serpyllum L. emend. Mill. «Wiss. Zeitschr. Halle Univ.», Bd. 18, N 6.
Schmidt P. 1971. Die Thymus-Arten in Nordosten des europaischen Teiles der UdSSR. «Feddes Repert.», Bd. 81, H. 8—9.
Schmidt P. 1973. Ubersicht fiber die mitteleuropaischen Arten der Gattung Thymus L. «Feddes Repert.», Bd. 83, H. 9—10.
S c h n a r f K. 1929. Embryologie der Angiospermen. Berlin.
Schotsman H. D. 1970. Contribution a la caryologie des angiospermes de la Sologne et du Vai de Loire. «Bull. Cent. Etud. Rech. Biarritz», t. 8, N 1.
Schratz E., Grown G. 1968. Experimentalle Bastardierung in der Gattung Thymus. «Planta Medica», vol. 16, N 3.
Sernander R. 1906. Entwurf einer Monographie der europaischen Myrme-kochoren. «Kongl. Sv. Vet. Akad. Handl.», Bd. 41, N 7.
Sorsa P. 1966. Pollen morphological studies in the genus Thymus L. (Labia-tae). «Ann. Bot. Fenn.», vol. 13, N 2.
Stary F. 1952. Farmakobotanicka monografie Adonis vernalis L. «Preslia», t. 24, N 1. '
205
Swendseh A., Ottesred E. 1957. Cumarine der Friichte von Heracleum panaces L. und H. sibiricum L4 «Pharm. Acta Helv.», Bd. 32, N 12.
Toole E. H.. Brown E. 1946. Final results of the Duvel buried seed experiment. «Journ. Agricult. Res.», vol. 72, N 6.
Trela-Sawicka Z. 1968. Cytological investigations in the genus Thymus L. «Acta Biol. Cracov. (Bot.)», vol. 11, N 1.
Troll W. 1957. Praktische Einfiihrung in die Pflanzenmorphologie. T. 2. Jena.
T г о 11W. 1964. Die Infloreszenzen. Bd. 1. Jena.
Tiixen R. 1937. Die Pflanzengesellschaften Nordwestdeutschlands. «Mitt. Flor. Soz. Arbet. Niedersachsen», H. 3.
VelenovskyJ. 1907, 1910. Vergleichende Morphologie der Pflanzen. T. 2, 3. Prag.
Veyret I. 1956. Embrydgenie des Orchidacees. Les lois du developpement chez le Goodyera repens R. Br. «Bull. Soc. Bot, France», t. 103, N 9—10.
W a 11 e r H. 1968. Die Vegetation der Erde. Bd. 2. Jena.
W a r d 1 e P. 1959. The regeneration of Fraxinus excelsior in woods with fields layer of Mercurialis perennis. «Journ. Ecol.», vol. 47, N 2.
Watt L., Renwick J. 1915. Notes on the occurence of Goodyera repens R. Br. in Scotland. «Glasgow Nat.», vol. 7, N 1—4.
Williams LA. 1925. 1926. Monoecious form of Mercurialis perennis. «Journ. Bot.», vol. 63, 64.
Willis J. C. 1966. Dictionary of the flowering plants and ferns. Cambridge.
W i I s i e C. 1962. Red clover and alske clover forage. Iowa.
Winkler A. 1893. Einige BemerKungen fiber die Keimung von Adonis ver-nalis L. «Deutsche Bot. Monatschr.», Jg. 11, N 1.
Wood A., Brad beer J. 1967. Studies in seed dormancy. 2. The nucleic acid metabolism of the cotyledons of Corylus avellana L. seeds. «New Phytolog.», vol. 66, N 1.
Ziegenspeck H. 1936. Orchidaceae. In: «Lebensgeschichte der Blutenpflan-zen Mitteleuropas», Bd. 1, Lf. 4. Stuttgart.
СОДЕРЖАНИЕ
Предисловие..................................................... 3
М. Г. Вахрамеева и Л. В. Денисова. Гудайера ползучая .... 5
Н. Г. Царевская. Любка двулистная................................И
Н. А. Аксенова и М. Г. Вахрамеева. Лещина обыкновенная ... 18
В. Л. Тихонова. Горец змеиный....................................29
А. П. Пошкурлат и И. А. Губанов. Горицвет весенний .... 36
О. В. Смирнова и В. А. Черемушкина. Род Хохлатка — Corydalis
Medic......................................................48
М. Г. Вахрамеева. Рябина обыкновенная...........................73
М. Г. Вахрамеева. Черемуха обыкновенная........................81
И. П. Крылова и Т. А. Работнов. Клевер луговой..................89
Н. П. Крылова. Клевер розовый.................................102
О. В. Смирнова и Н. А. Торопова. Пролесник многолетний . . . 111
И. Ф. Сацыперова. Борщевик сибирский...........................124
Е. Е. Гогина. Род Чабрец (тимьян)—Thymus L.....................137
Н. А. Аксенова. Бузина красная '...............................169
А. М. Былова. Молокан татарский................................175
Литература.....................................................187
Биологическая флора Московской области, вып. 2.
Тематический план 1975 г. № 149
Редактор Е. Г. Зверева
Переплет художника М. М. Носовой Технический редактор В. И. Овчинникова
Корректоры
. Л. С. Клочкова, Н. В. Тютина
Сдано в набор 11/Ш 1975 г.
Подписано к печати 4/VIII 1975 г.
Л-21861. Формат бОХЭО'Дб
Бумага тип. № 2
Физ. печ. л. 13,0
Уч.-изд. л. 14,31. Изд. № 2405
Зак. 255. Тираж 2070 экз.
Цена 1 р. 54 к.
Издательство Московского университета.
Москва, K~S, ул. Герцена, 6)7.
Тульская типография «Союзполиграфпрома» при Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли, г. Тула, проспект им. В. И. Ленина, 109