/




























































































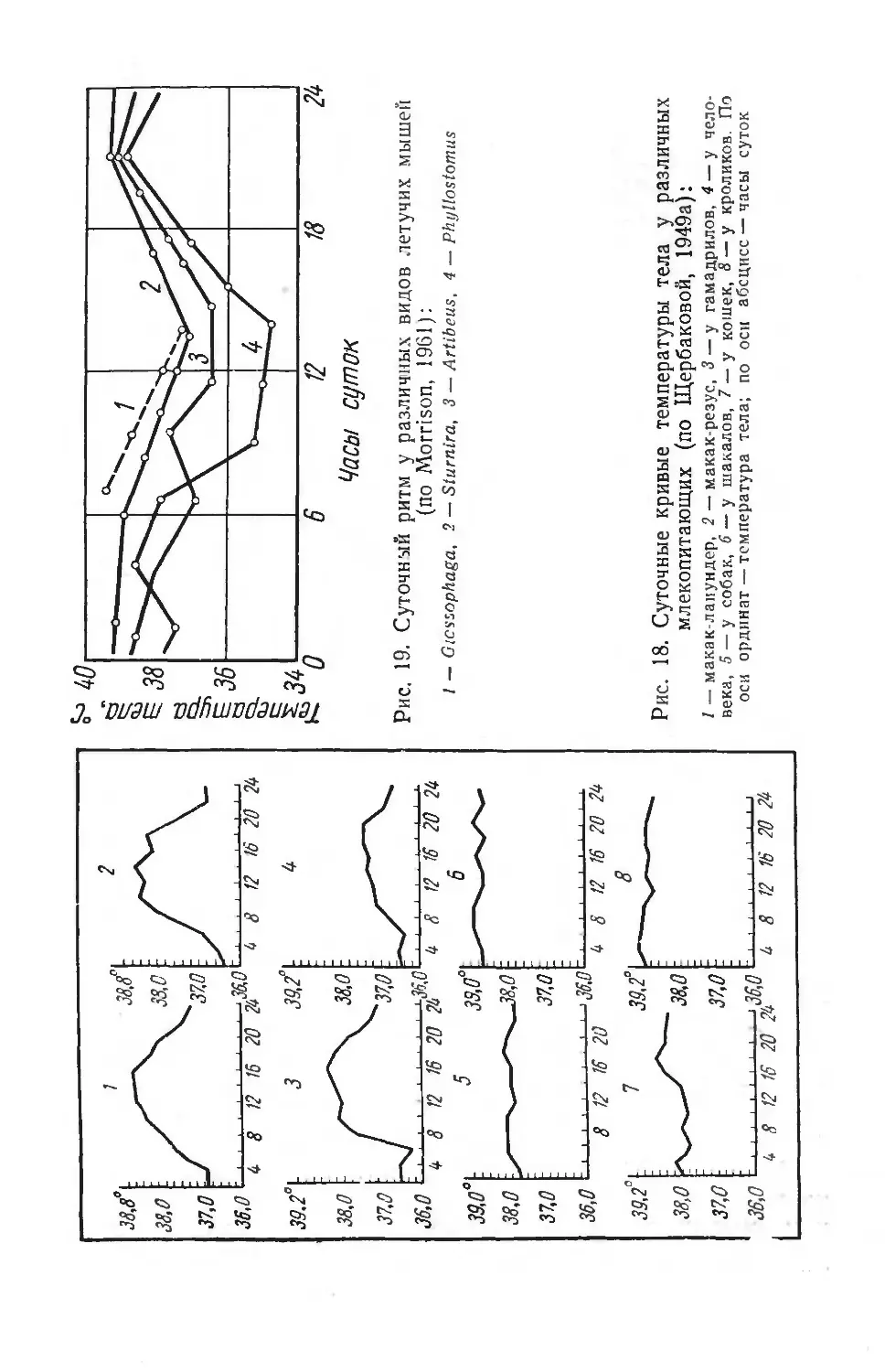
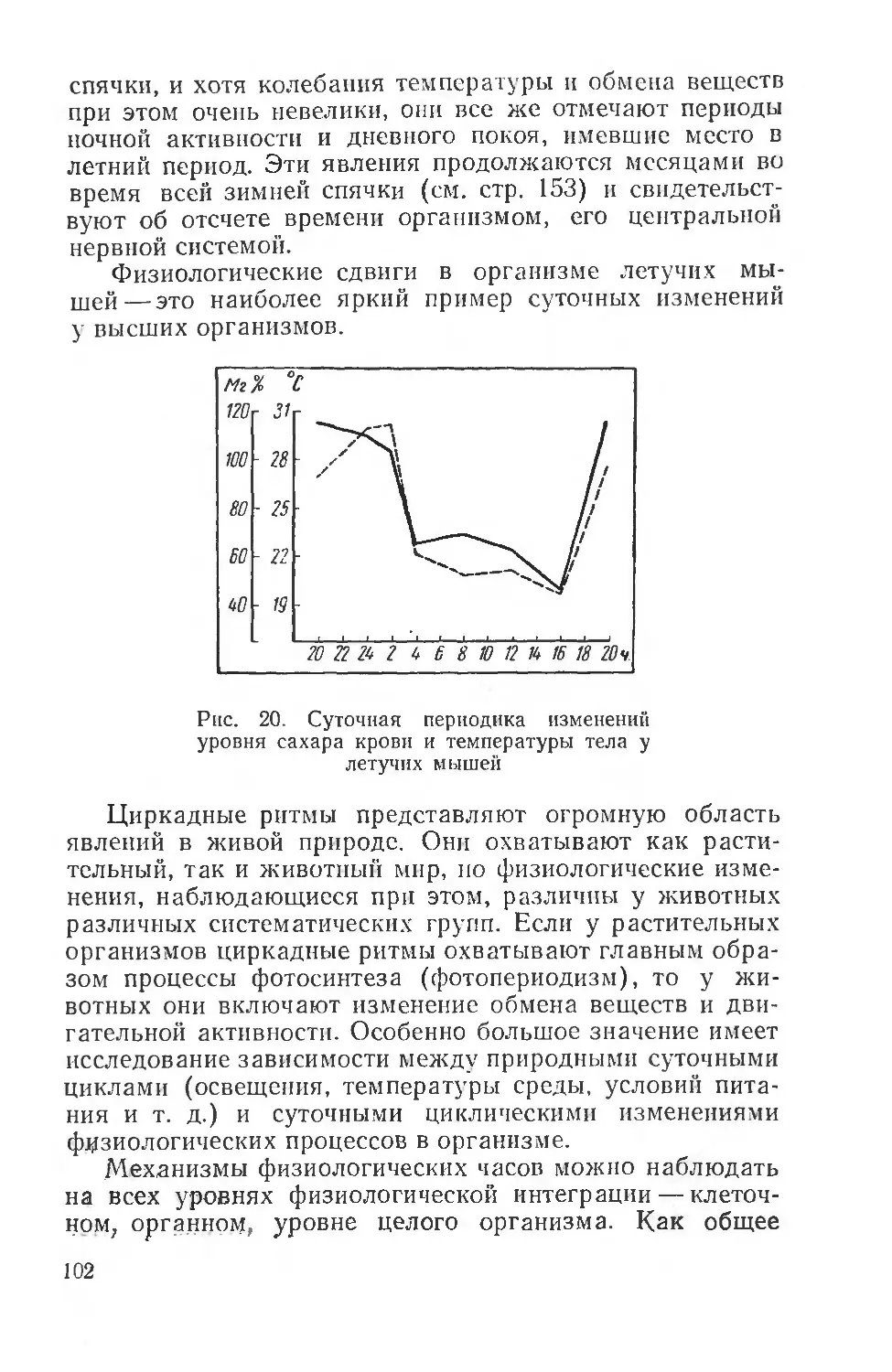









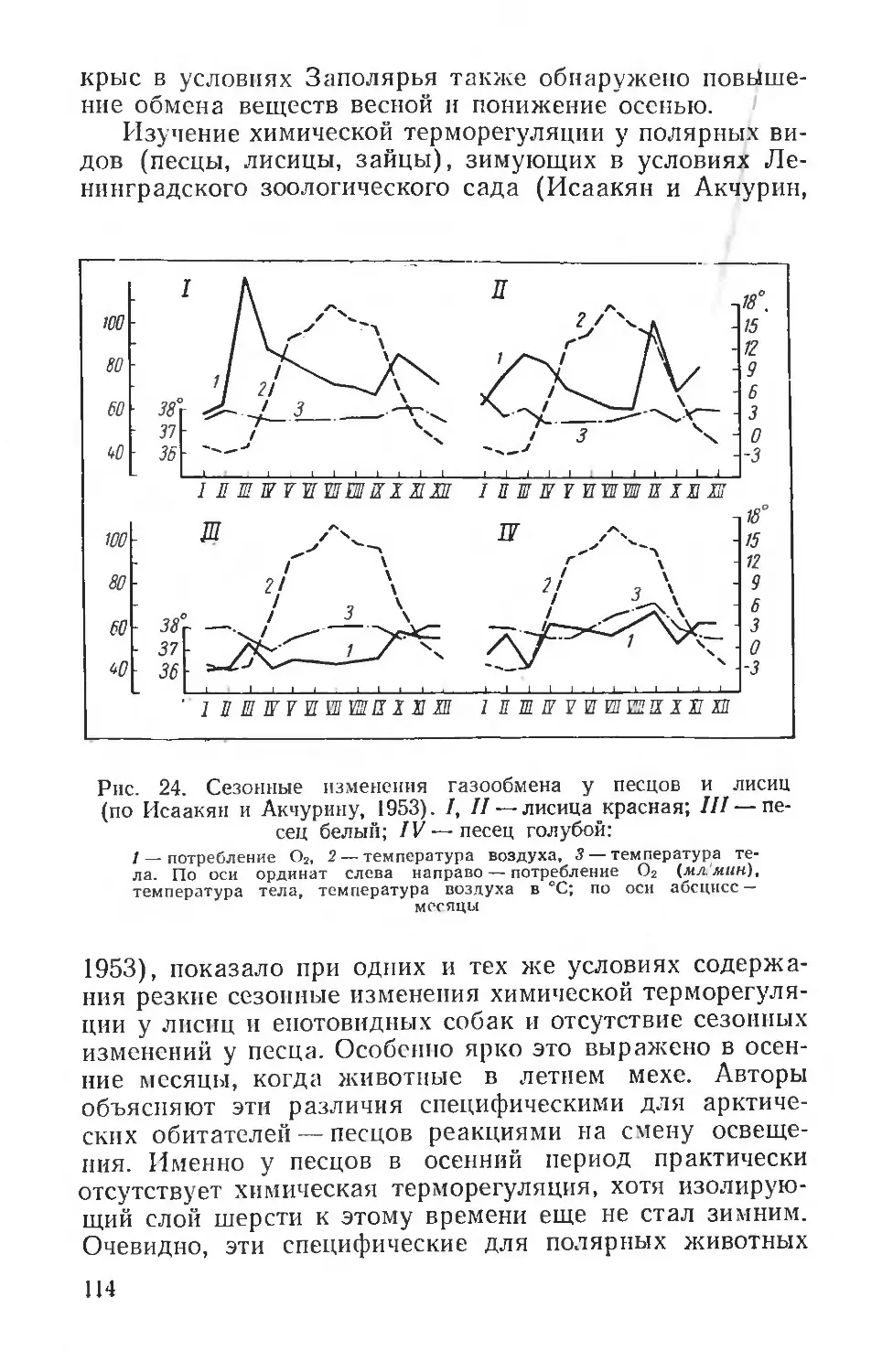





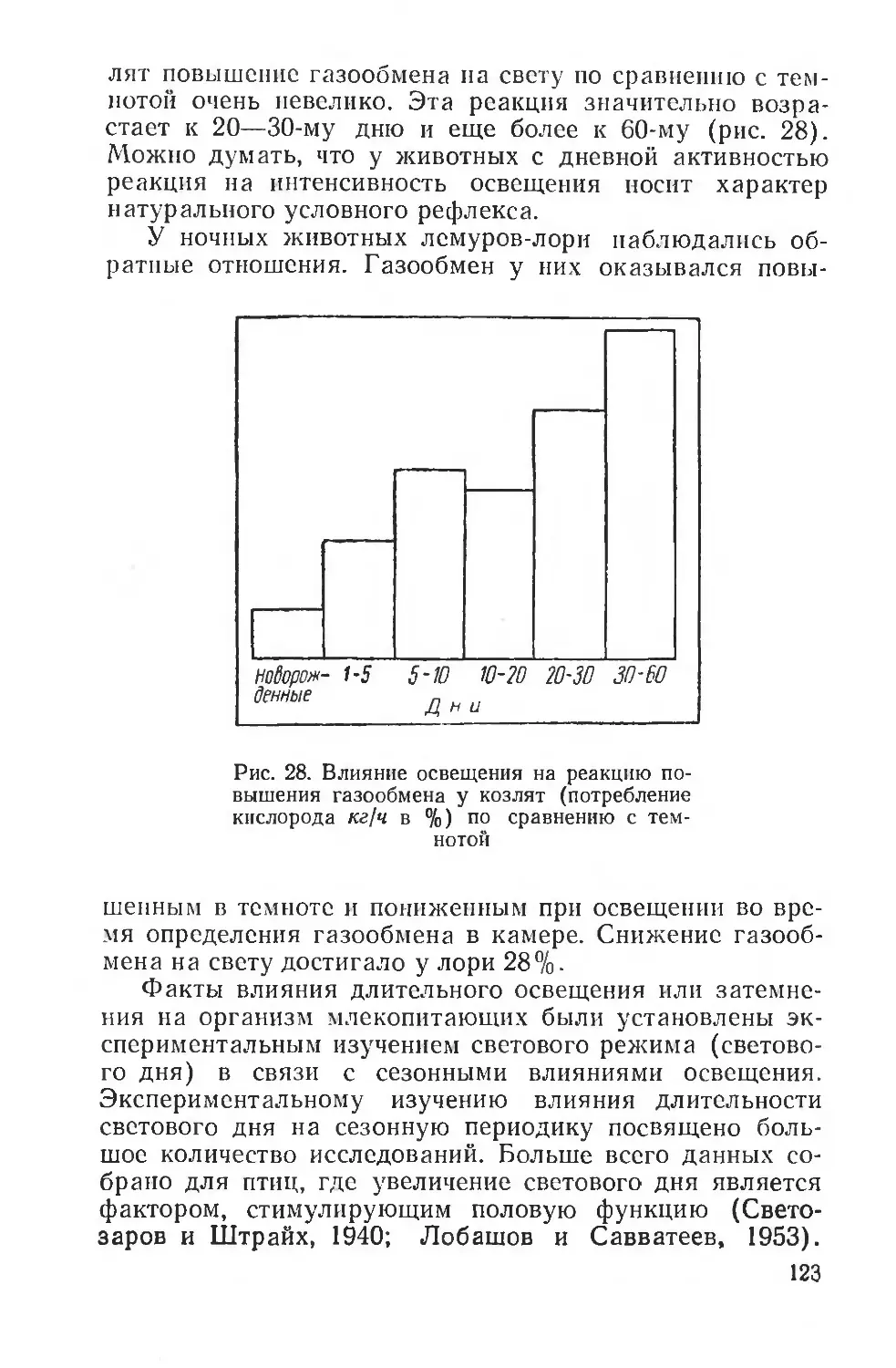














































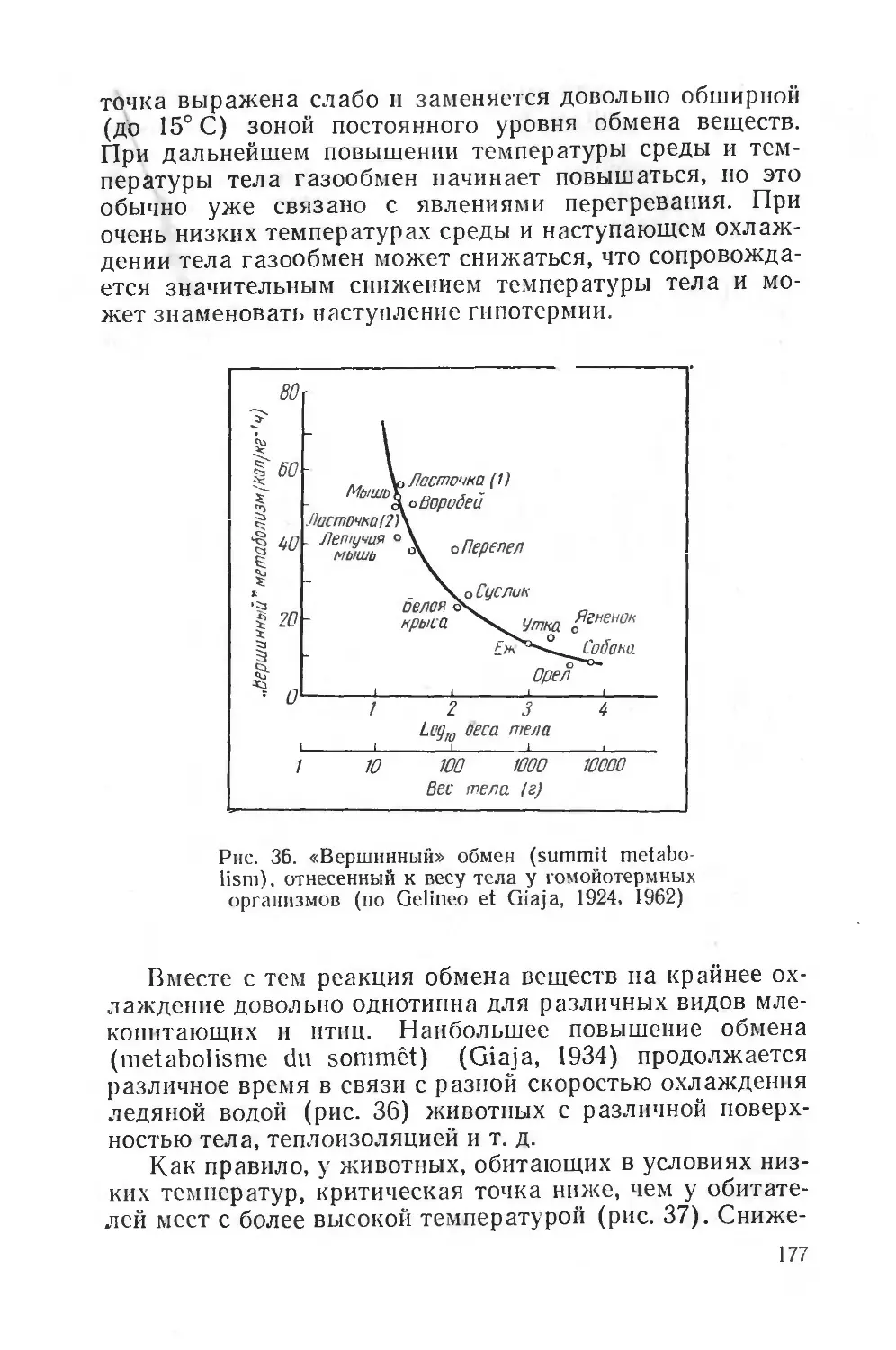
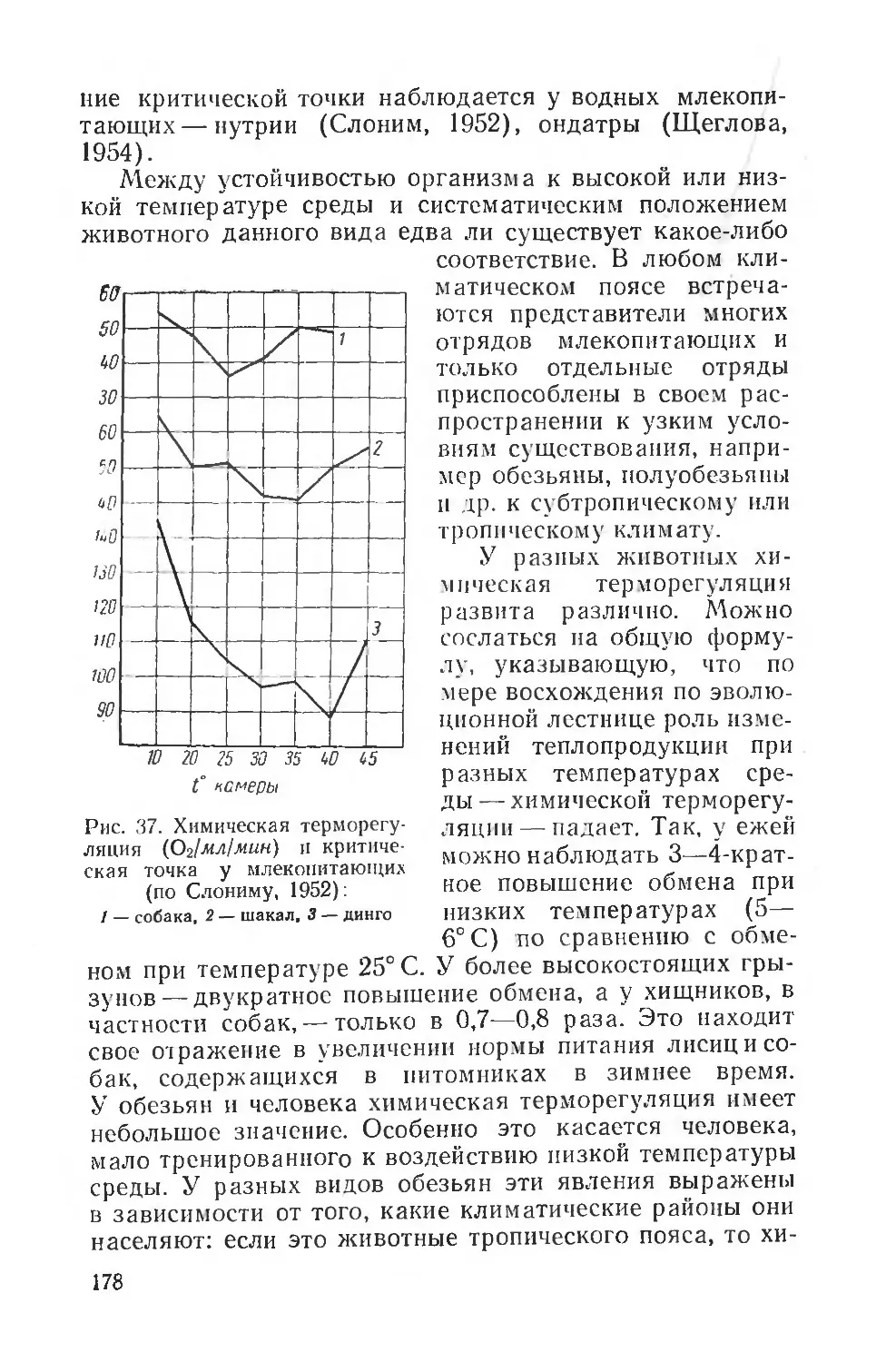



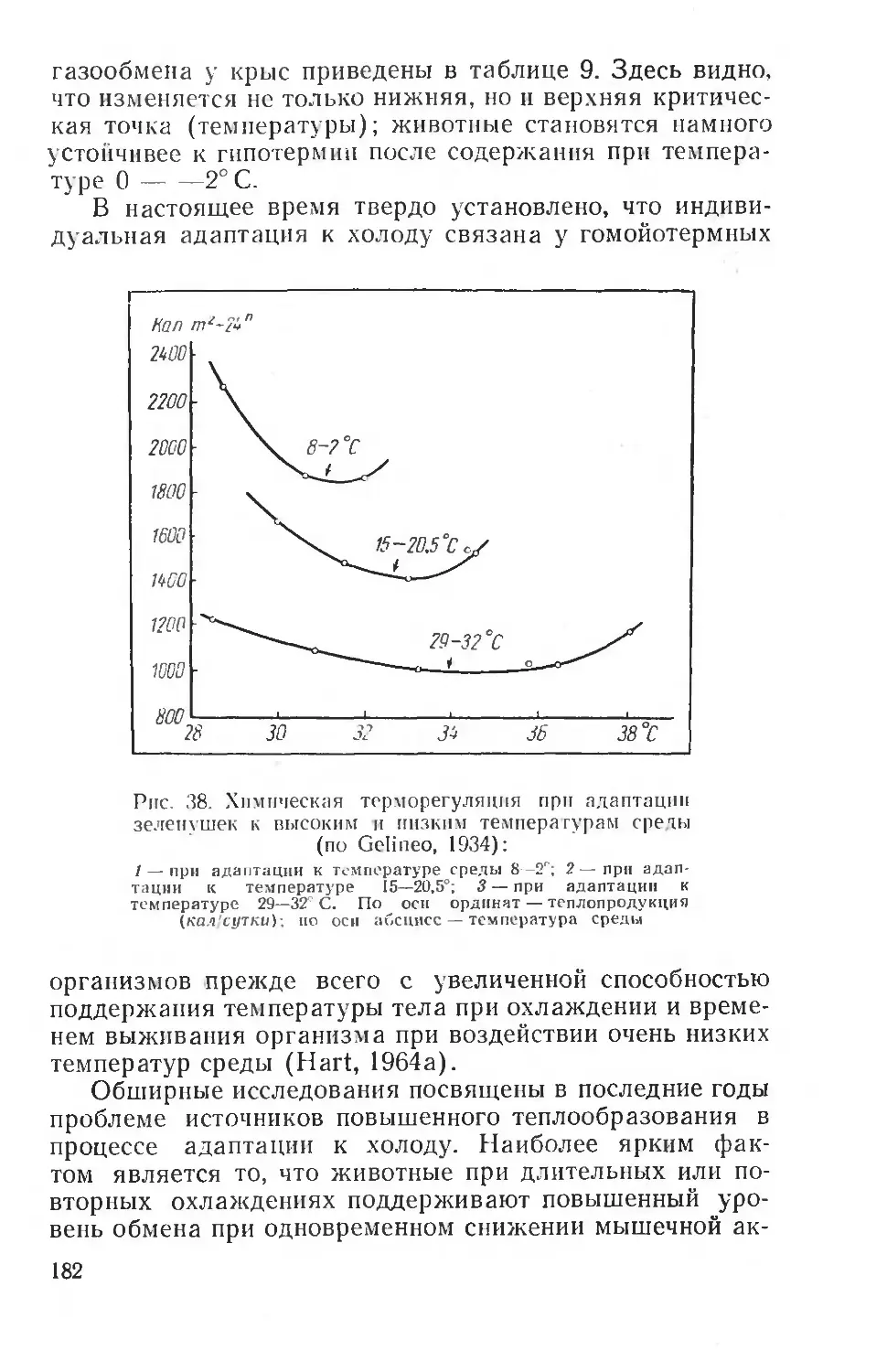



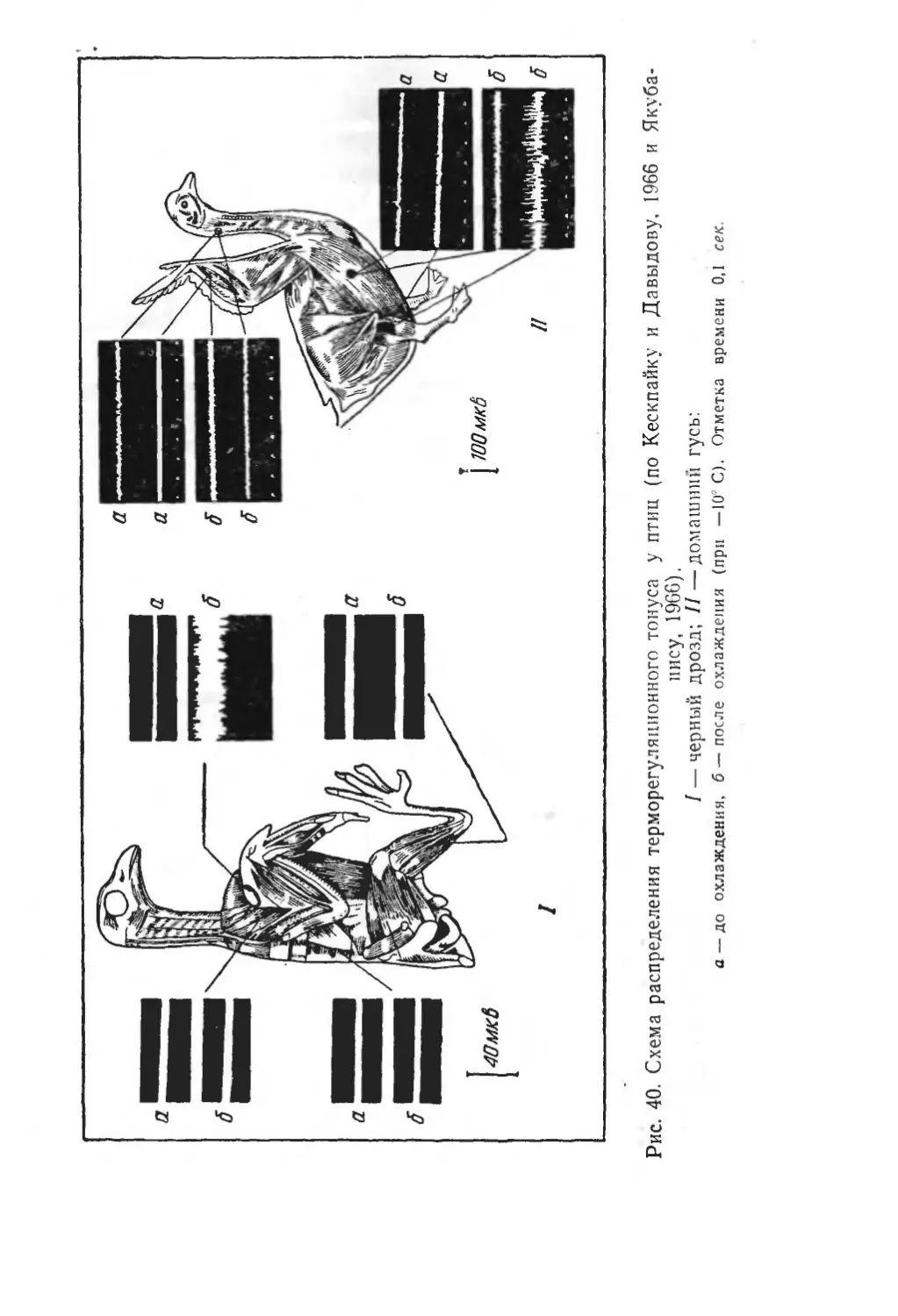










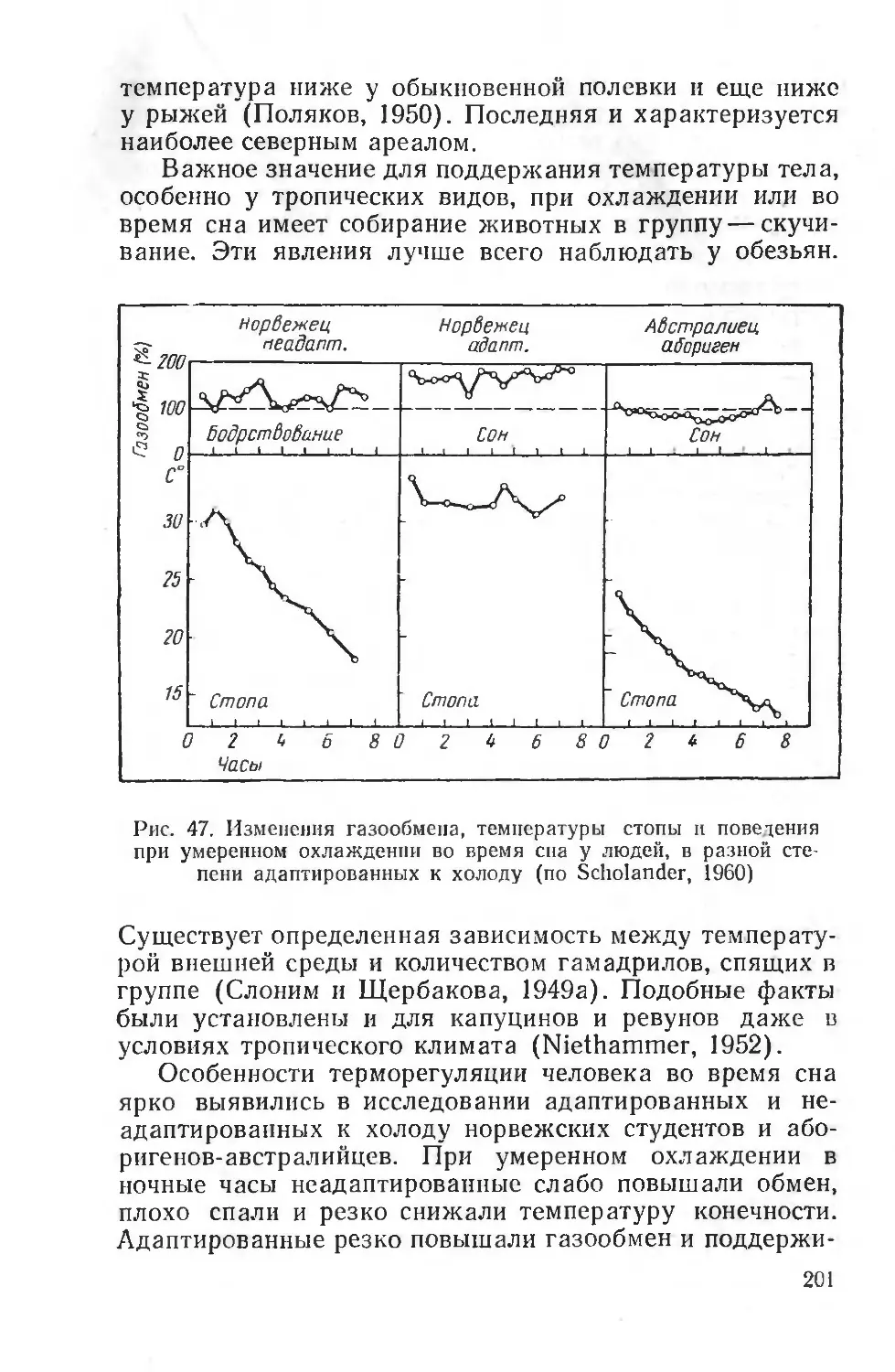



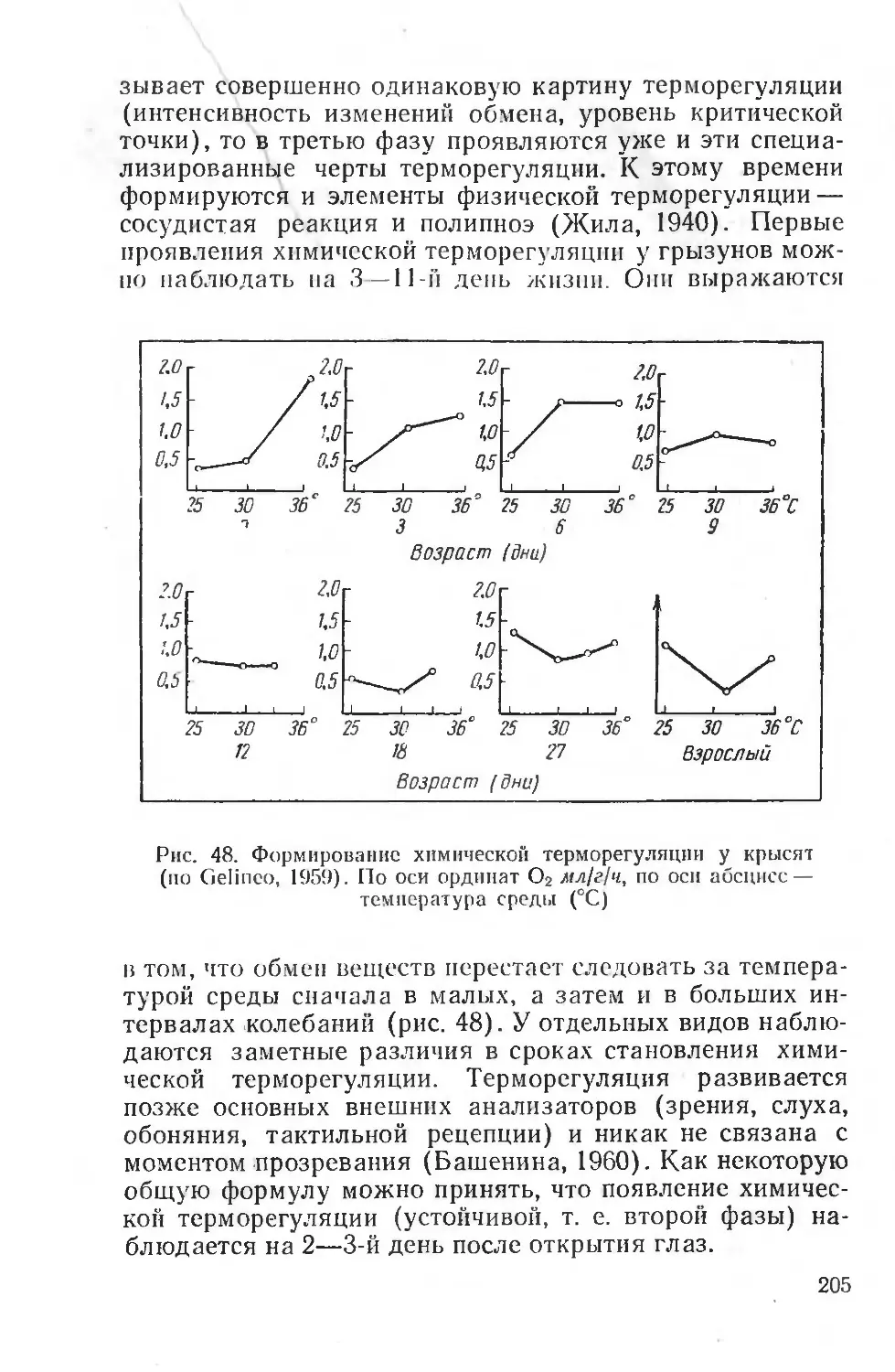






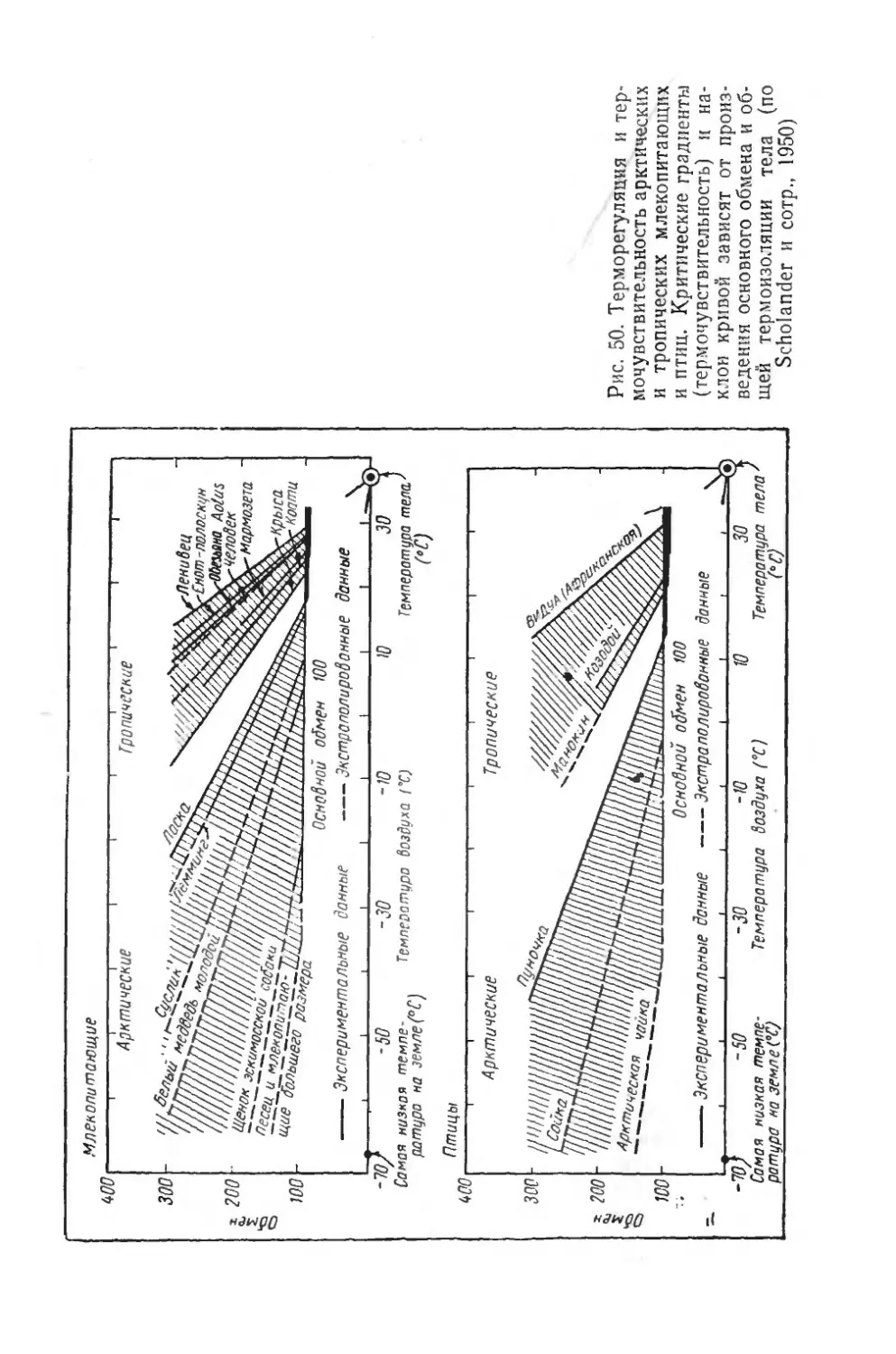

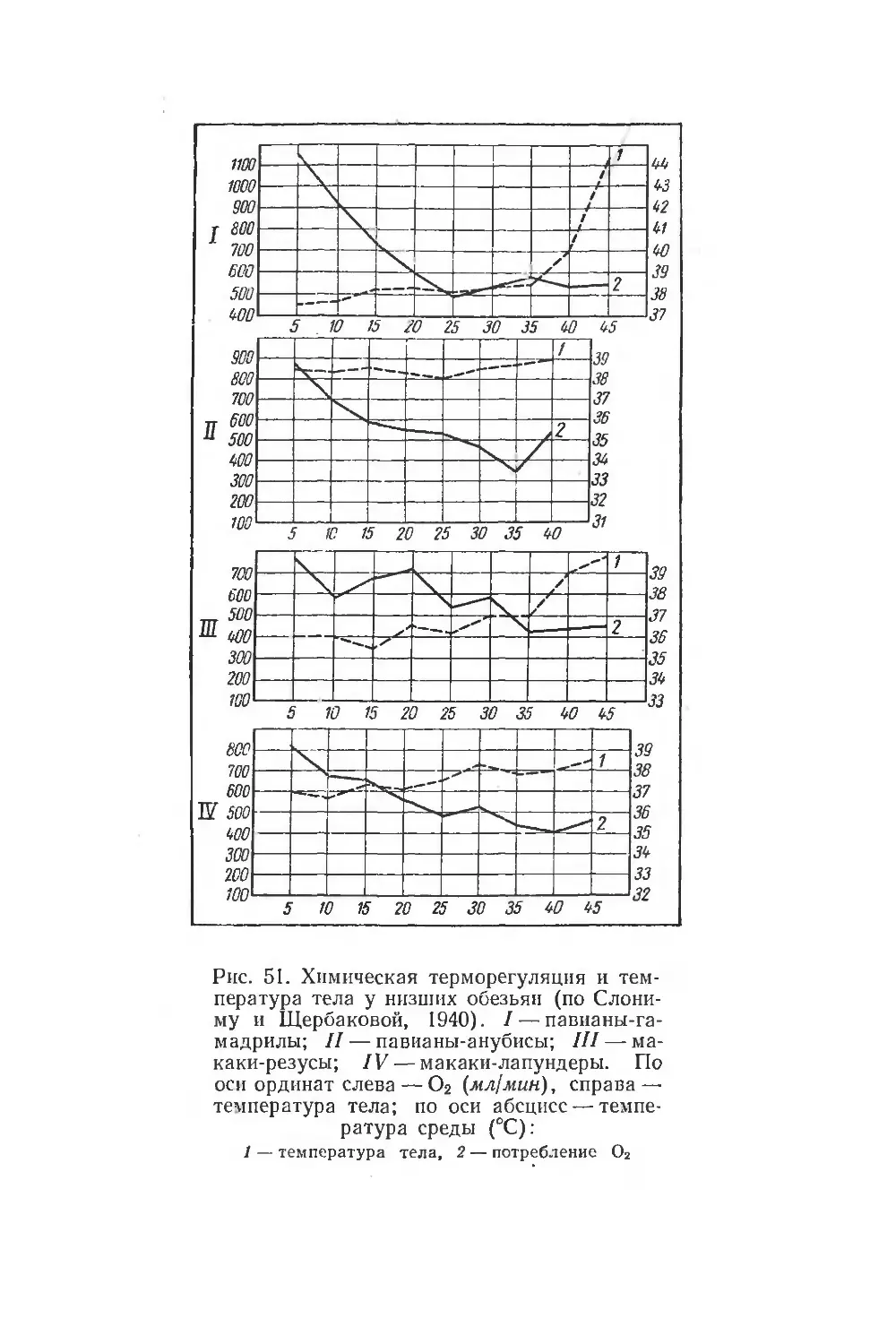










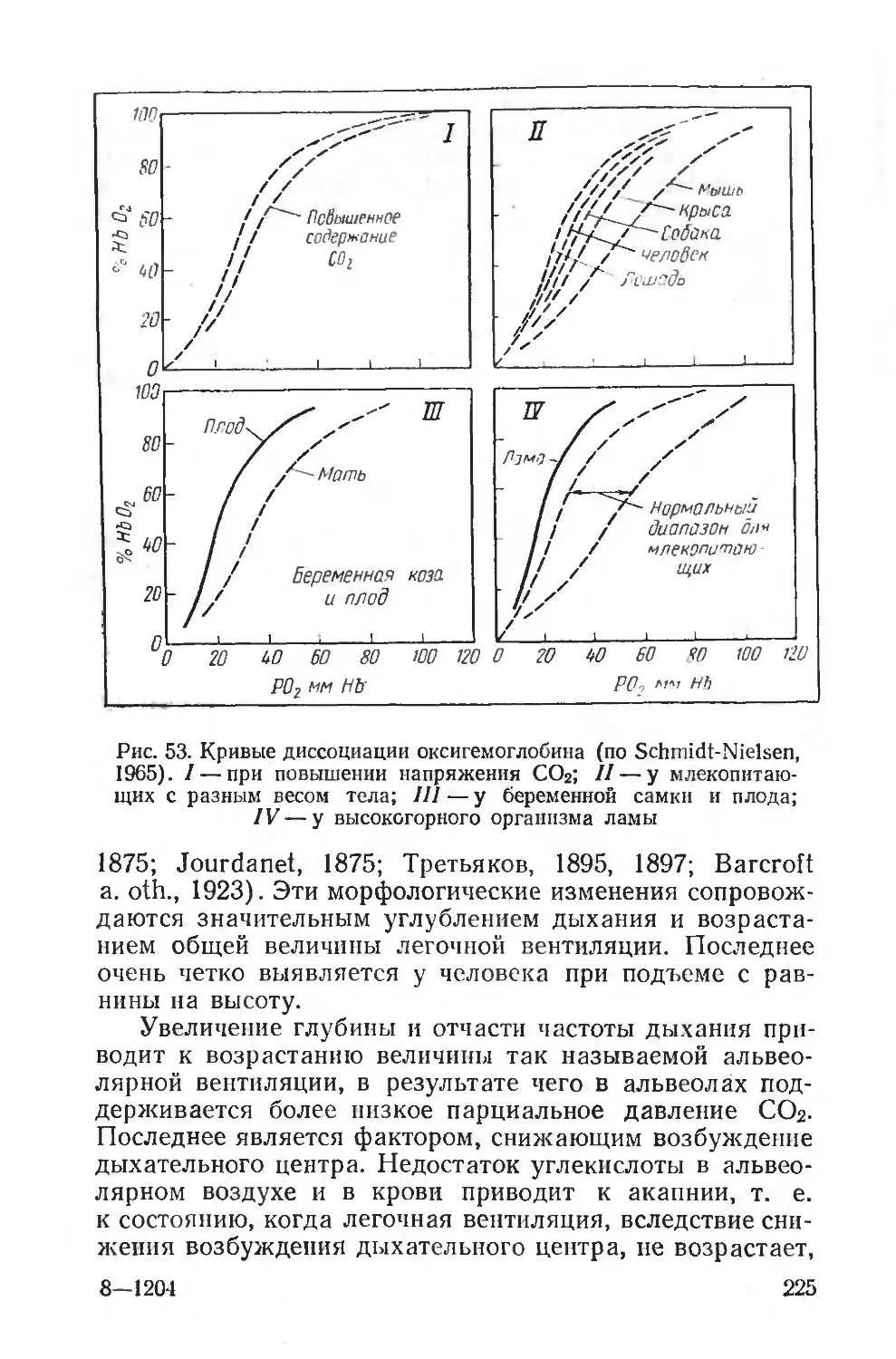


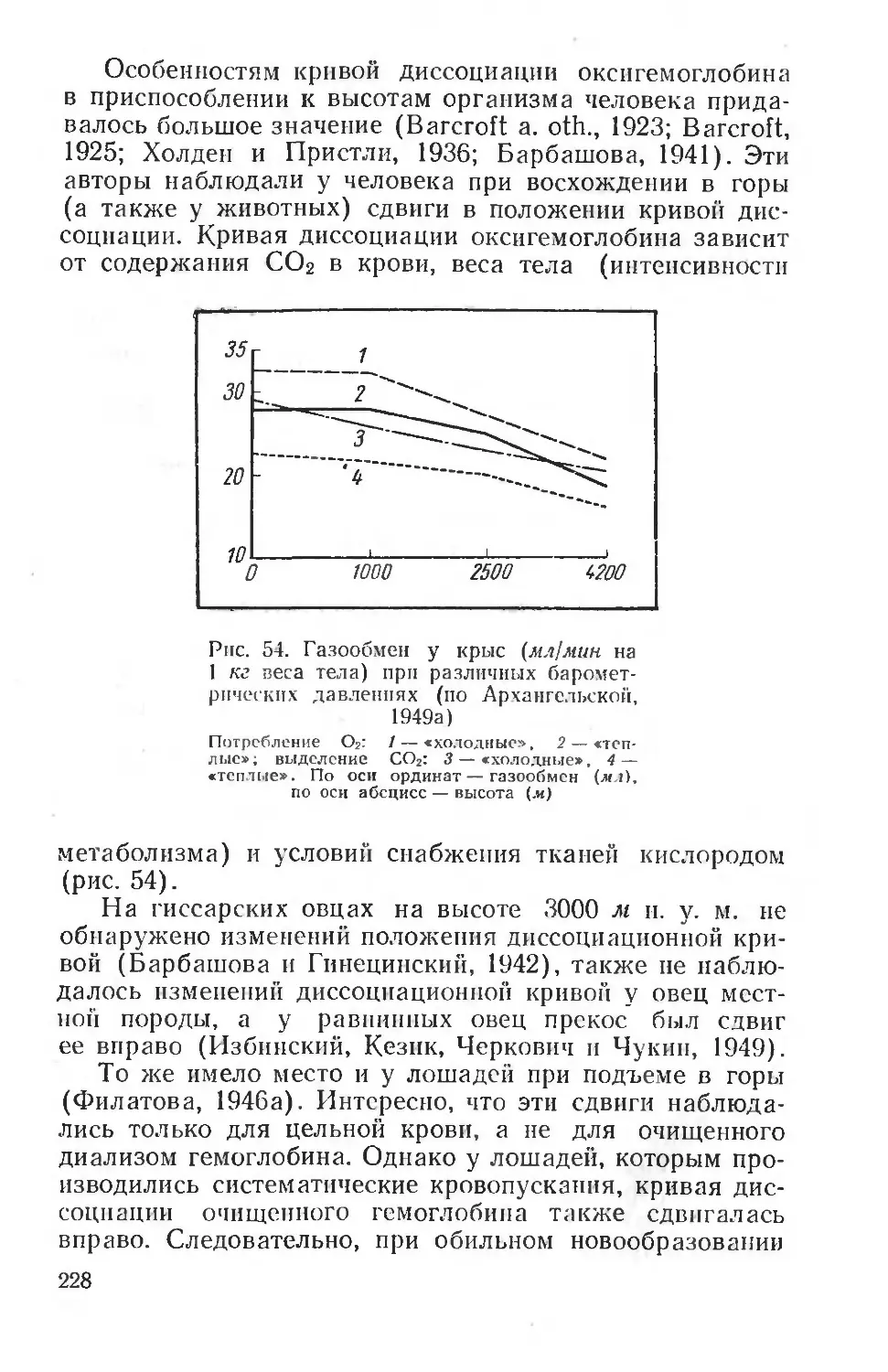












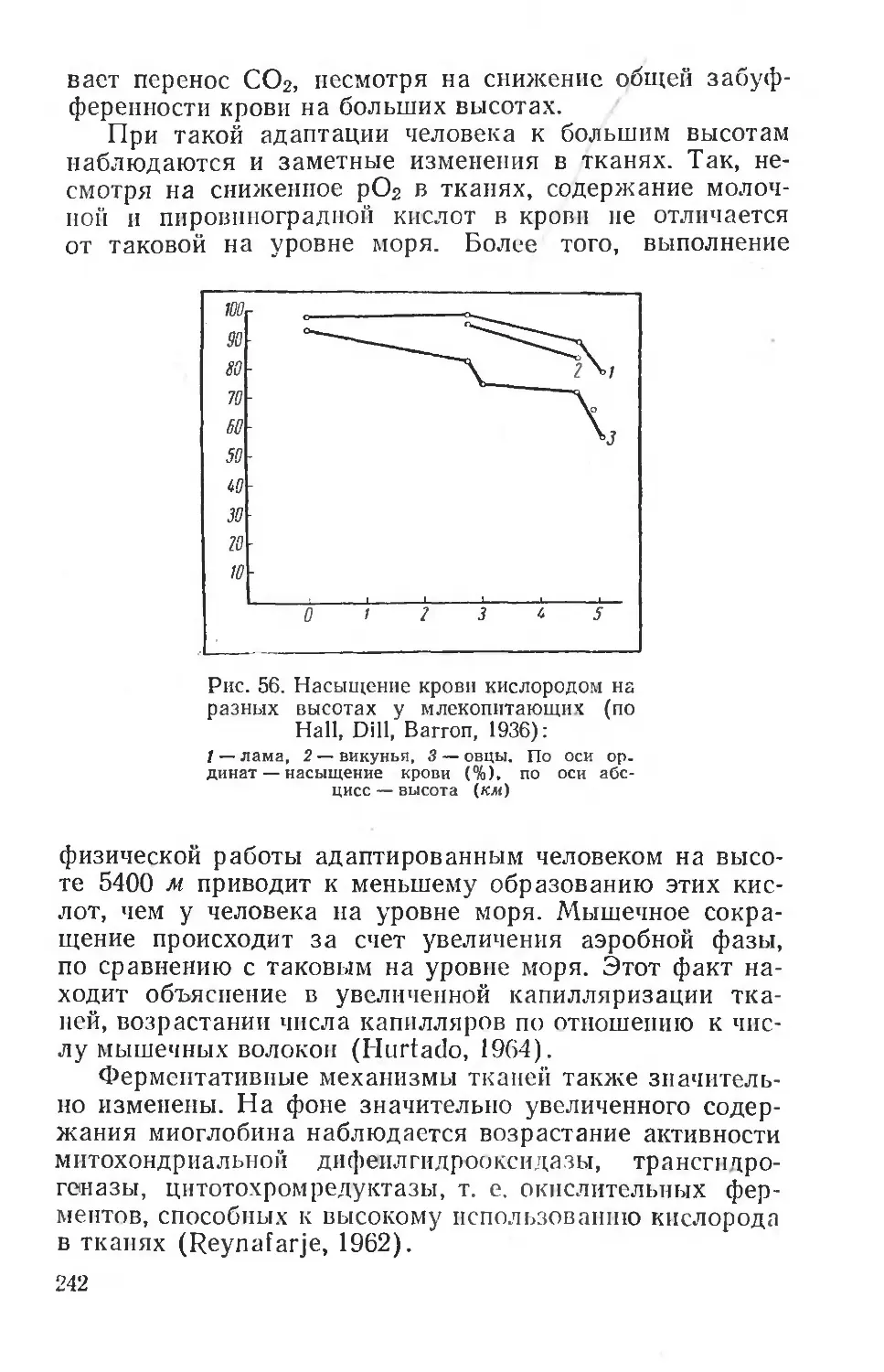






















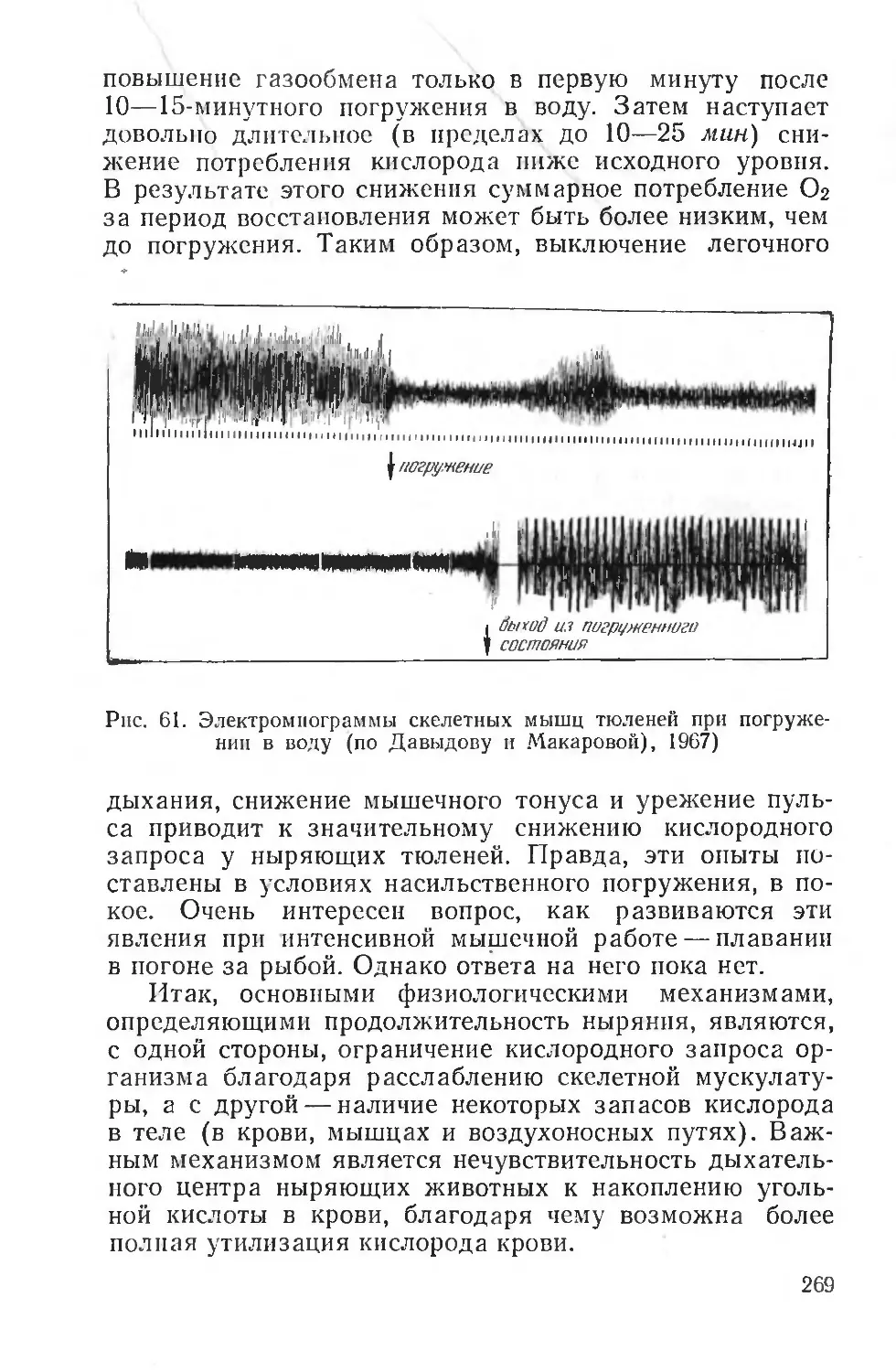







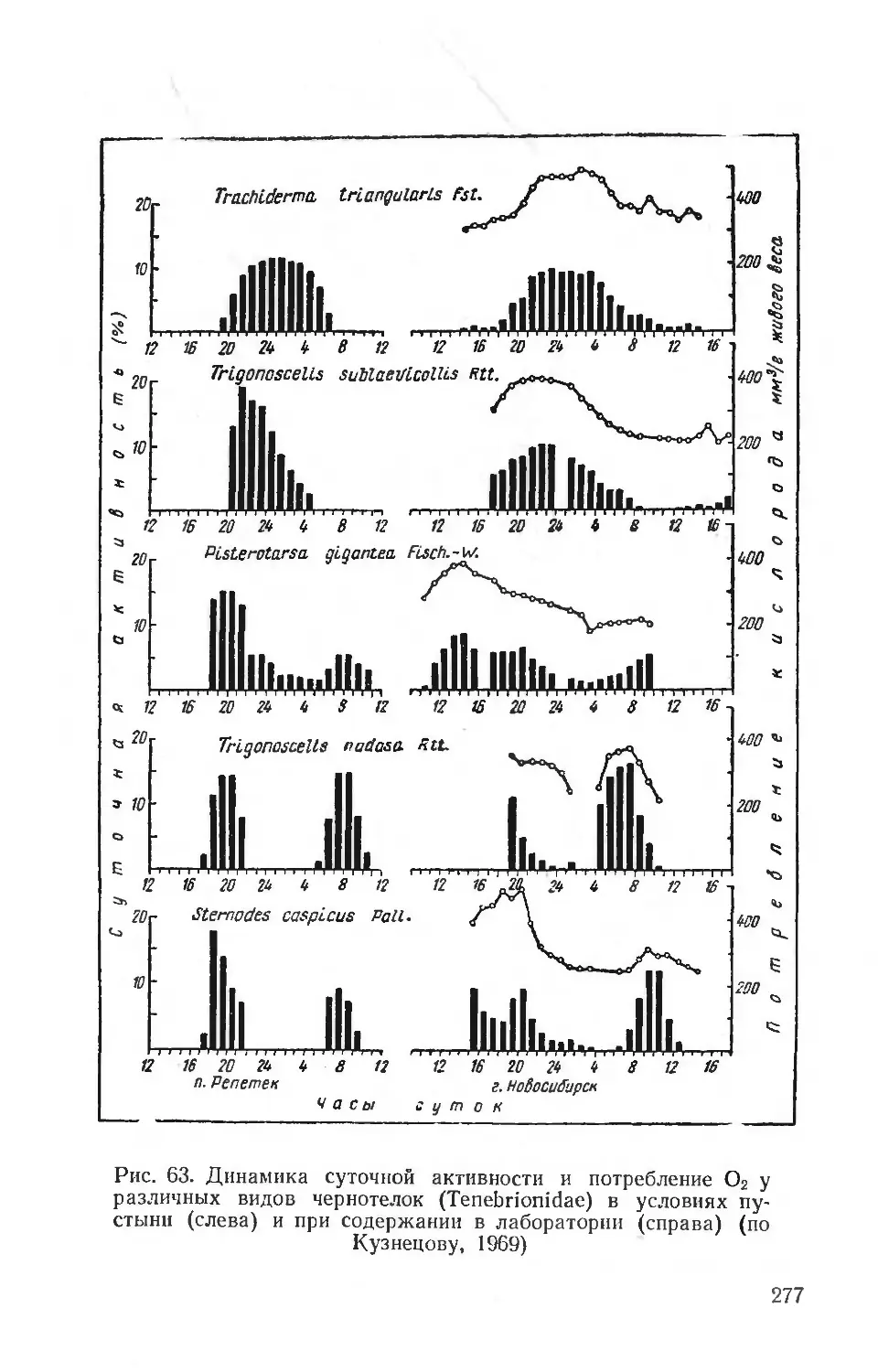





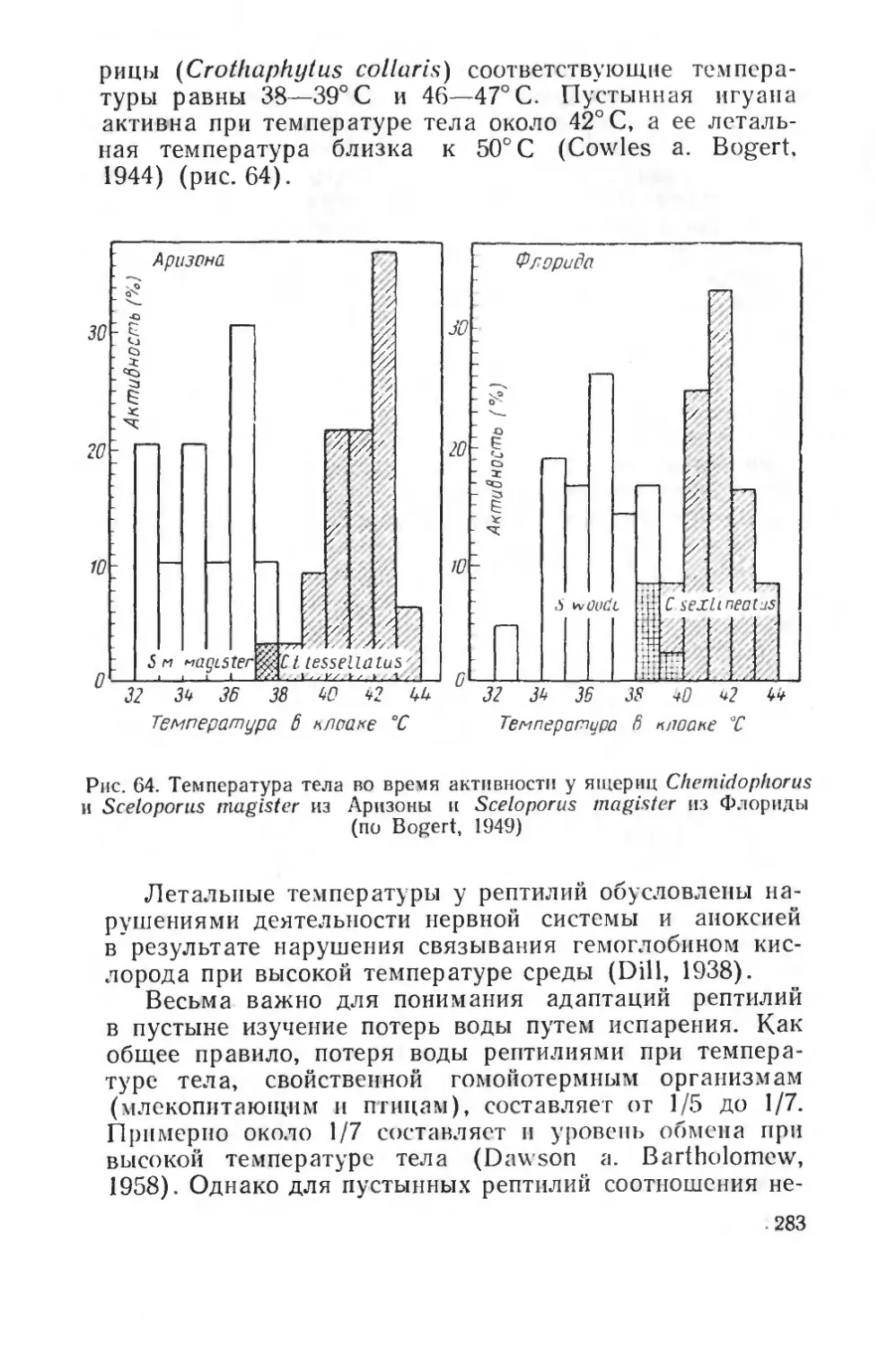


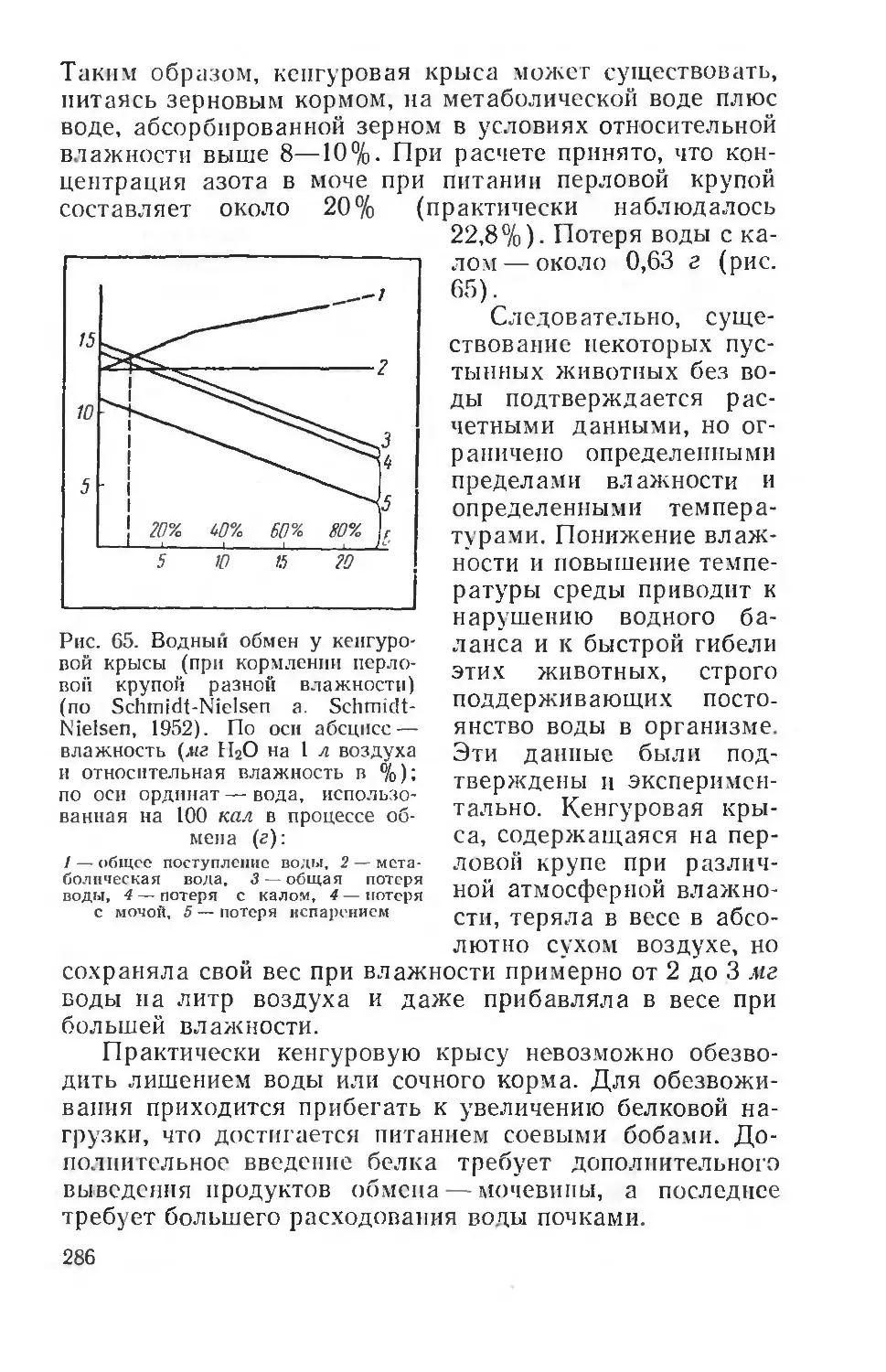























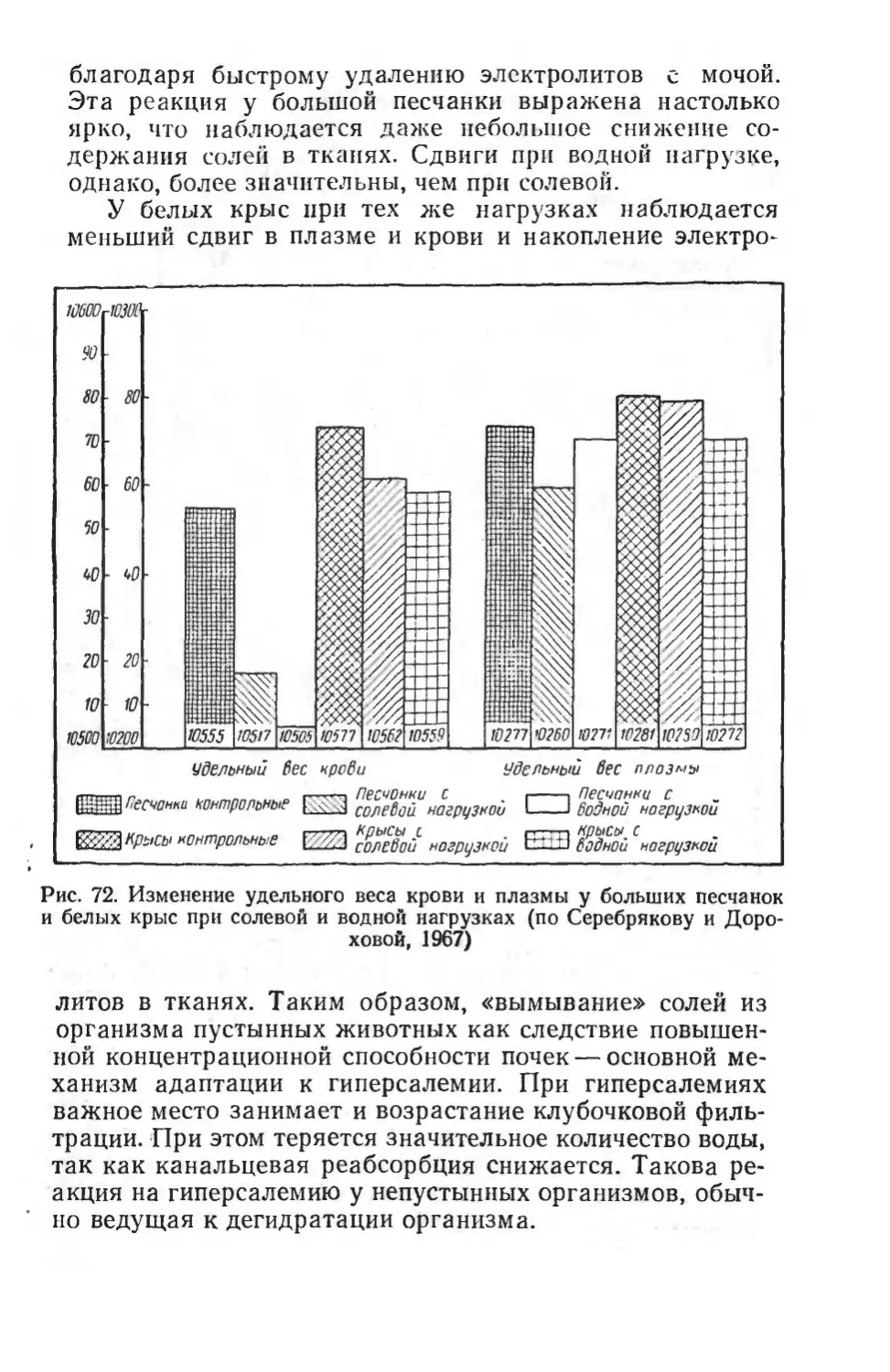





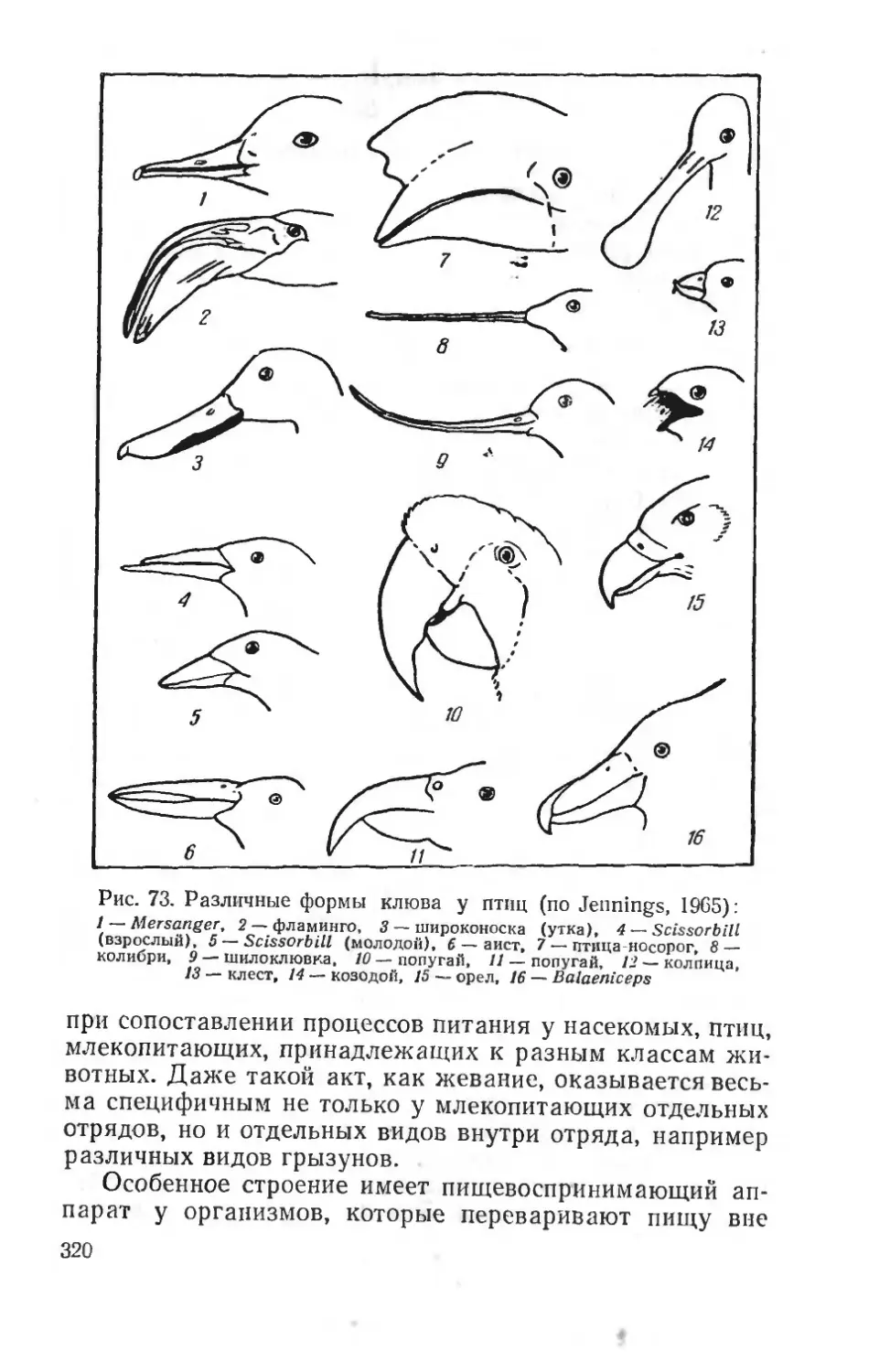





















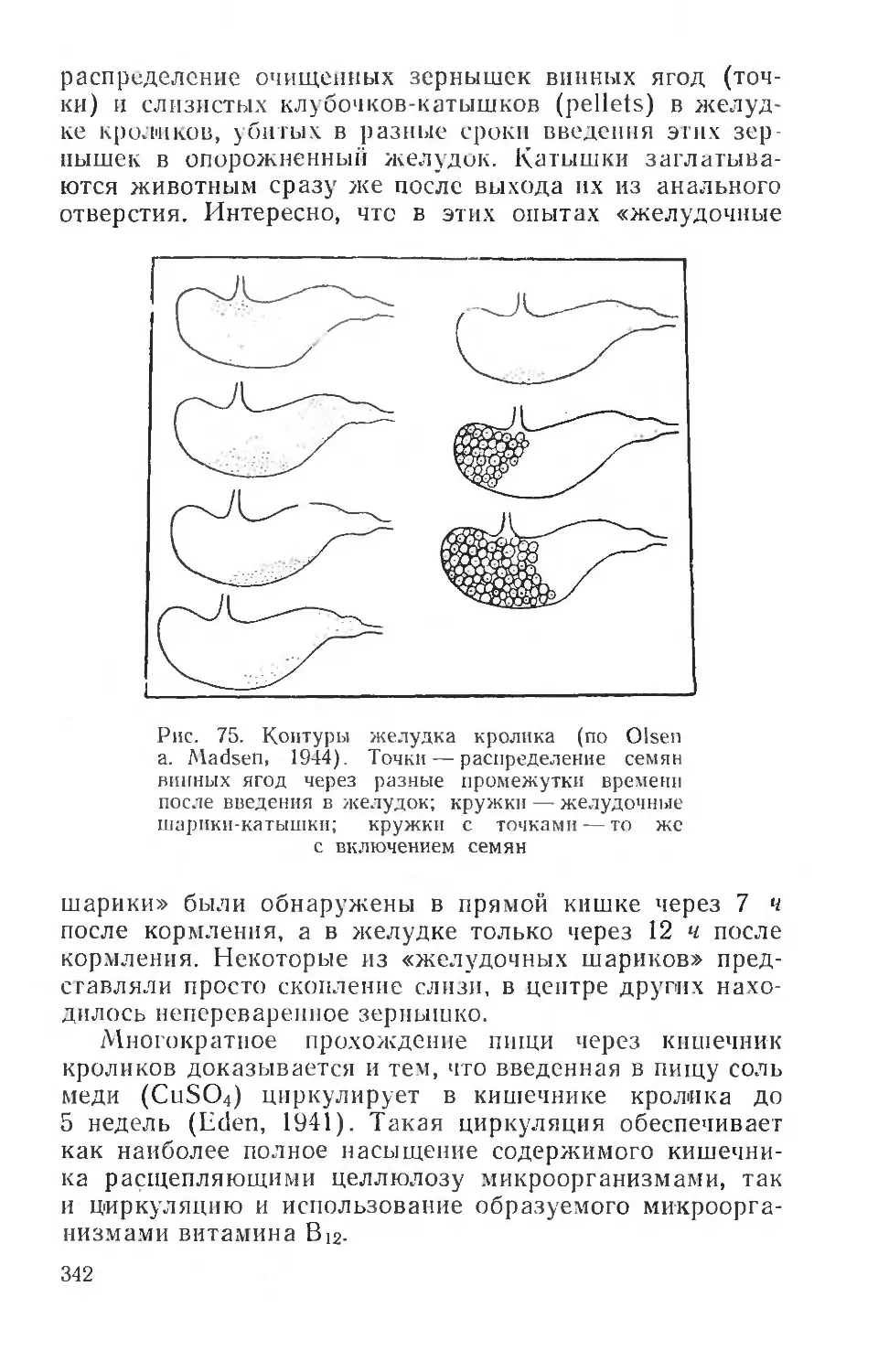




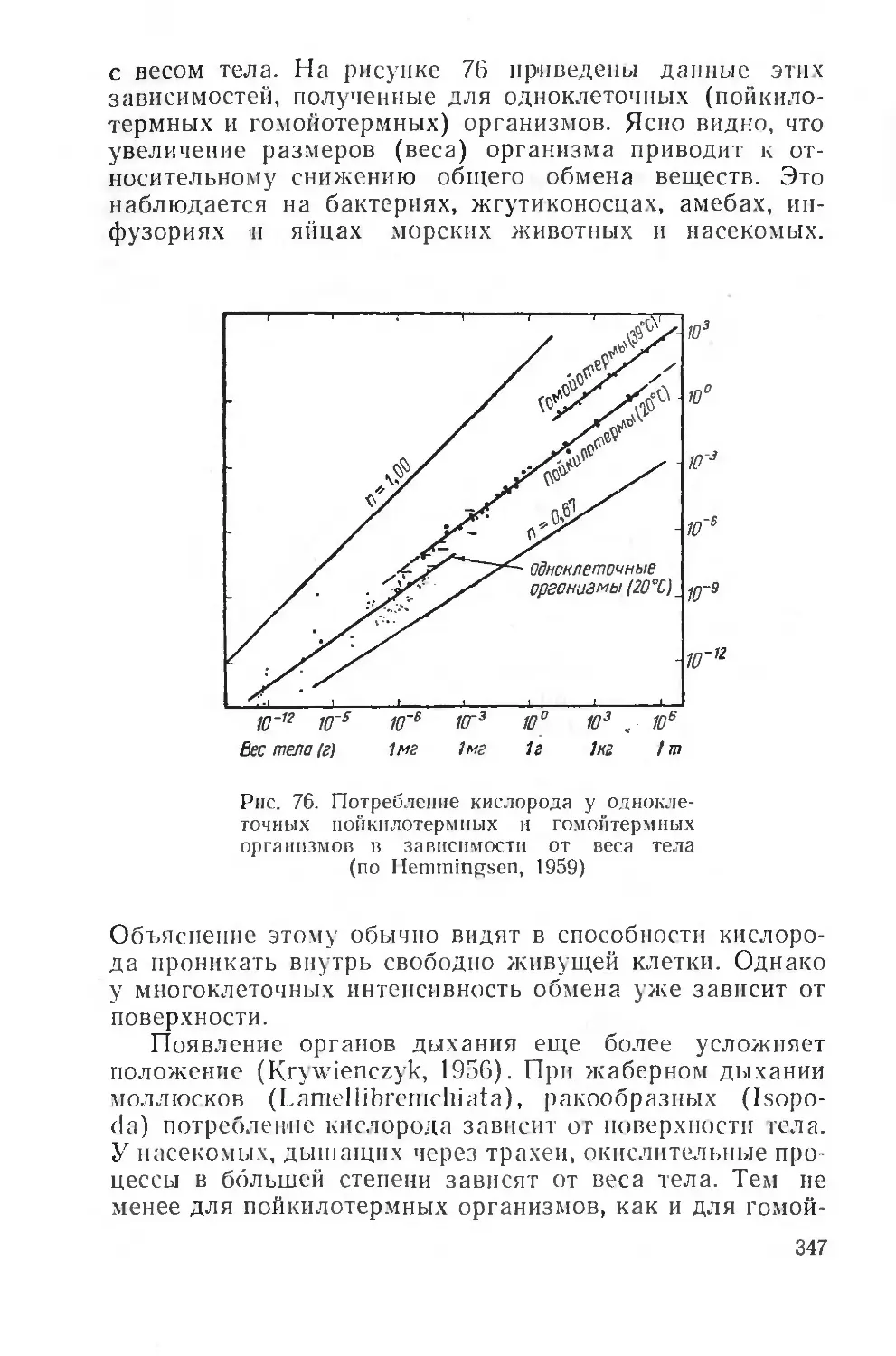

































































































Text
А. Д. СЛОНИМ ЭКОЛОГИЧЕСКАЯ
ФИЗИОЛОГИЯ животных
Допущено Министерством высшего и среднего специального образования СССР в качестве учебного пособия для студентов биологических специальностей
университетов
Издательство «Высшая школ
АННОТАЦИЯ
В книге дается краткий обзор современного состояния физиологии адаптации к природным факторам среды и микроэволюции физиологических функций в связи с этими факторами. Эти проблемы приобретают все большее значение при народнохозяйственном освоении новых территорий, в связи с задачами повышения продуктивности биоценозов, с проблемами завоевания космоса.
Современное состояние проблемы в целом не позволяет в одинаковой мере подробно осветить явления физиологической адаптации на всех этапах эволюции животного мира; наиболее полно в этом отношении изучены высшие животные и человек. Поэтому в книге основное внимание уделено высшим организмам.
Книга состоит из двух частей. Первая охватывает общие явления адаптации организма, изученные иа различных уровнях физиологической интеграции. Сюда же следует отнести и исследования поведения организма как неизменной составляющей физиологических адаптаций.
Вторая часть охватывает специальные разделы экологической физиологии, посвященные реакции организма на сложные природные факторы среды.
Изложение предмета в книге отличается от традшц/онной формы, так как рассматриваются не отдельные функции, а их взаимоотношения, явления замещений функций в процессе адаптаций как обусловленной генетически, так и возникающей в процессе индивидуальной жизни. Особое внимание обращено на сопоставление физиологических особенностей и реакций организмов близких видов как основного пути изучения микроэволюции.
2-10-6
111-112-70
От автора
Стремление к познанию физиологических основ существования животных и человека в сложных, а иногда и крайних (экстремальных) условиях быстро возрастает.
Непрерывный рост населения земного шара, задачи повышения жизненного уровня миллионов людей требуют хозяйственного освоения новых территорий, ранее почти не использованных человеком—Арктики, Антарктиды, жарких пустынь—аридной зоны, а также повышения биологической продуктивности уже давно населенных человеком территорий. Это требует прежде всего знаний о возможностях организма животных и человека к существованию в новых условиях, о путях и предельных возможностях приспособления (адаптации) к ним. Особое значение эти проблемы имеют для СССР, где расширение народнохозяйственного использования природных ресурсов происходит наиболее интенсивно, а задачи биологических исследований в природе разнообразны, как и сама природа страны.
Автор сделал попытку объединить в одной доступной студенту-биологу, физиологу и врачу книге состояние основных проблем физиологии в приложении к существованию животных, а человека — к его деятельности. Столь широкий охват самой проблемы адаптации животного организма потребовал специального подбора материала, отражающего, с одной стороны, фактическую сторону дела, а с другой — само положение учения об адаптации в общей проблеме эволюции. К этому автора побуждало твердое убеждение, что поскольку функциональные особенности организма в процессе эволюции являются более подвижными по сравнению с морфологическими, их изучение наиболее плодотворно на уровне микроэволюции вообще и адаптивной микроэволюции в частности.
3
Книга ориентирует читателя иа изучение экологической физиологии высших организмов: птиц, млекопитающих и человека, но автор старался использовать и важные для понимания этих вопросов данные физиологии низших позвоночных и беспозвоночных.
В отличие от обычного курса физиологии, где излагается в определенном порядке функционирование отдельных органов и систем, в книге сначала даются общие представления о физиологических адаптациях и их механизмах, а затем о специальных формах адаптаций, соответствующих отдельным факторам среды.
В основу плана построения книги положены курсы лекций, читанные автором в разные годы студентам-физиологам Л\осковского, Ленинградского, Казахского и Новосибирского университетов.
Автор не считал нужным избегать спорных вопросов, а их возникает более чем достаточно при попытке применить данные лабораторного эксперимента к явлениям, протекающим в живой природе. Если эти необъяснснные сегодня факты привлекут внимание будущих исследователей и прежде всего нашей вступившей в науку молодежи, задачу книги автор будет считать выполненной.
ВВЕДЕНИЕ
ПРЕДМЕТ И ЗАДАЧИ ЭКОЛОГИЧЕСКОЙ ФИЗИОЛОГИИ
Под экологической физиологией понимают совокупность знаний о физиологических основах приспособлений (адаптаций) к природным факторам среды и к сложному сочетанию их в различных физико-географических условиях. При этом особое значение приобретают сведения о влиянии упомянутых выше факторов как на видовые особенности физиологических функций и поведения животных, так и на особенности отдельных популяций видов в связи с особенностями их существования в разных ареалах, в разные сезоны года, в зависимости от тех или других изменений условий среды.
Как физиологическая дисциплина экологическая физиология представляет ветвь эволюционной физиологии — предмета, ставящего своей задачей изучение происхождения физиологических функций, их эволюцию в связи с общей эволюцией органического мира. Экологическая физиология пользуется физиологическими методиками исследования и физиологическим экспериментом. Задачей экологической физиологии (в отличие от проблем эволюционной физиологии) является исследование адаптаций — совокупности физиологических явлений в их взаимной связи, суммирующей, замещающей (вика-рирующей) или еще более сложно интегрирующей взаимоотношения отдельных органов и систем, отдельных более или менее сложных элементов поведения и регуляций физиологических функций.
Экологическая физиология тесно связана с проблемами экологии как самостоятельной науки о приспособлении видов и популяций в широком биологическом плане,— науки, опирающейся на данные генетики, зоогеографии, климатологии, не говоря уже о морфологии и систематике животных.
Проблемы эволюционной физиологии, так же как и проблемы экологической физиологии, были четко декла
5
рированы в нашей стране на основе материалистического учения II. М. Сеченова, II. П. Павлова, являющегося дальнейшим развитием эволюционного учения Ч. Дарвина. Принципиальные положения этого учения служат основой как при собирании материала, так и его обобщении в плане решения многих теоретических и практических задач. Исследование влияния природных факторов среды на протекание физиологических функций получило широкое распространение и за рубежом, особенно в плане изучения отдельных физико-географических и ландшафтных зон. Это направление получило право гражданства как «физиология внешней среды» (environmental physiology),— термин, прямо не переводимый на русский язык, но часто употребляемый в английской и американской литературе.
В связи с задачами изучения природы отдельных физико-географических зон в зарубежной литературе появились такие главы физиологии, как «пустынная физиология» (desert physiology), «горная физиология» (mountain physiology), «арктическая физиология» (arctic physiology) и т. п. Все эти разделы науки являются частными проблемами экологической физиологии, использующими для исследований ее приемы и методы.
Первые попытки специальных эколого-физиологических сопоставлений в отечественной литературе принадлежат Е. М. Крепсу (1936). Эколого-физиологические исследования терморегуляции были выполнены А. Д. Слонимом и О. П. Щербаковой (1935, а, б, в; 1940). Затем физиологические методы в экологических исследованиях были применены А. С. Шаталиной (1935), Д. Н. Кашкаровым и Е. П. Коровиным (1936), Н. И. Калабуховым (1933, а, б), А. И. Израэлем (1936). А. И. Израэль (1936) опубликовал первый обзор как собственных и отечественных, так и иностранных исследований. Эти работы в основном преследовали цели изучения изменений физиологических функций под влиянием определенных факторов среды.
Экологическая физиология животных получила особое развитие при изучении водных организмов. Достижения ее в этой области широко используются гидробиологами. Экологическая физиология занимается изучением и млекопитающих, в том числе сельскохозяйственных животных. Эколого-физиологические исследования про
6
водятся на любых уровнях филогенетического развития животных.
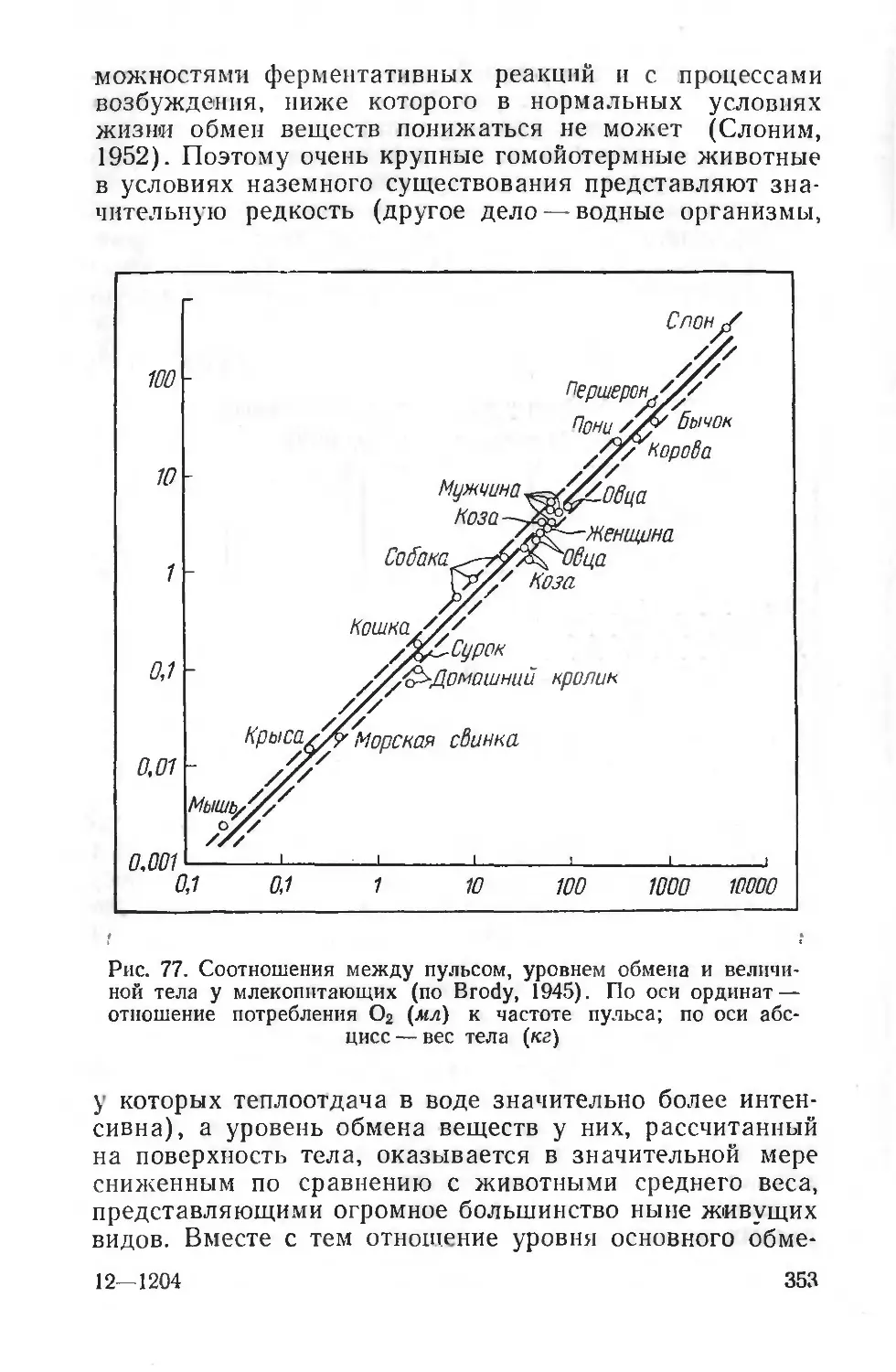
Если физиолог может использовать разных животных, например крысу, кошку, черепаху, с целью получения представлений о развитии тех или иных функций па разных этапах эволюции, то в экологической физиологии для сопоставления необходимо иметь, по возможности, близкие между собой виды, но живущие в разных условиях, историю отпочкования которых от эволюционного древа можно было бы проследить. Сопоставление физиологических особенностей у одних и тех же животных, которые закрепились на разных территориях и существуют в разных условиях, позволяет глубоко проникнуть в закономерности формирования рефлекторной деятельности, понять особенности биохимических процессов, т. е. дать анализ глубокого воздействия определенного комплекса факторов среды на организм в процессе эволюции.
В СССР развитие как экологической, так и эволюционной физиологии неразрывно связано с исследованиями школы И. П. Павлова.
Л. А. Орбели создал предпосылки для изучения эволюции физиологических процессов не только на основе сопоставления их у видов, находящихся на разных ступенях эволюционной лестницы, но и используя физиологический эксперимент и данные патологии человека. Им обоснованы приемы выявления более древних и более новых элементов в деятельности центральной нервной системы (например, мозжечка, лимбической коры) в регуляции функций организма современных животных.
Учение о кортикальных регуляциях физиологических функций, разработанное К. М. Быковым и его сотрудниками, открыло широкие перспективы анализа взаимоотношений сложного поведения и регуляций физиологических функций животных и человека. Само понятие о среде обитания значительно усложнилось благодаря выделению как физических, так и сигнальных ее факторов, действующих на организм.
Возможность образования условных рефлексов, изменяющих как функцию отдельных органов и систем (железы пищеварительного тракта, гладкая мускулатура, почка и др., а также система крови, сердечно-сосудистая система, дыхание), так и общую жизнедеятельность организма и ее проявление — общий обмен веществ, азоти
7
стый и углеводный обмен, открыли совершенно новые пути в изучении взаимоотношений между организмом и внешней средой.
Этот путь исследования позволил понять и многие стороны происхождения патологических явлений, вызванных нарушениями деятельности головного мозга.
Наконец, Д. А. Бирюковым (1948, 1960) разработано представление о значении адекватности раздражителя (в том числе и природного) как для выявления врожденной — безусловнорефлекторной деятельности, так и образования условных рефлексов. По представлениям Д. А. Бирюкова и его сотрудников, нервная система животного отражает не только уровень развития на филогенетической лестнице животного мира, но и весьма тонко экологические условия существования данного вида. Это выражается как в особенностях сложнорефлектор-пой деятельности, так и в свойствах нервной системы, ее возбудимости, силе и подвижности нервных процессов.
Все эти исследования намечают основные пути познания реакции животного на внешнюю среду в связи с уровнем его организации и условиями существования в природе. Открылись возможности выделения генетически обусловленных (врожденных) адаптаций организмов и адаптаций, приобретаемых в процессе индивидуального развития в различных условиях.
Эколого-физиологическое исследование очень редко может ограничиться изучением только одного какого-либо органа или системы. Как правило, приспособление (адаптация) какой-либо системы к условиям существования сопровождается изменениями и сопряженных функций. Так, например, адаптации организма к недостатку кислорода обязательно охватывают систему легочного, или внешнего дыхания, сердечно-сосудистую систему и систему крови. Очень часто системные адаптации дублируются или замещают друг друга. Только сопоставление функций отдельных систем и их реакций на изменение во внешней среде может пролить свет на действительные механизмы, которые обеспечивают животному существование в необычных, измененных, а иногда и крайних условиях среды.
В работе по экологической физиологии практика выявила два основных приема. Первый из них — использование массового полевого (обычно экспедиционного)
8
материала — животных, отлавливаемых непосредственно перед постановкой опытов. Физиолог в этом случае получает животных непосредственно из природы и одновременно использует большое количество сведений об их жизни в природе, обрабатывая их сообразно поставленной задаче, и ставит опыт в соответствии с этими природными условиями.
Приближение условий экспериментирования к естественным (например, создание нор, укрытий, соответствующего освещения, изоляции и т. д.) устраняет многие трудности и в сочетании с полевыми наблюдениями обещает много существенного в понимании физиологических реакций животного. Эти приемы особенно часто применяются при изучении поведения.
Особое значение в полевой работе физиологов приобретают дистантные методы исследования. Сейчас в экологии п физиологии применяется метод меченых атомов, позволяющий точно исследовать расселение организмов на данной территории. Большое значение приобретают исследования с помощью регистрации по радио физиологических функций (пульса, дыхания, электрических явлений в тканях и органах, движений и т. п.).
Другой важнейший прием работы — содержание животных в неволе, в лаборатории. При этом открываются возможности использовать животных, история которых известна по возможности полнее (сроки и место вылова, условия содержания и т. д.).
В последнее время все чаще и чаще стараются пользоваться животными, родившимися и выросшими в лаборатории. Это очень важно для исследования происхождения физиологических особенностей, так как здесь физиолог знает всю историю данного организма и может создать те или иные условия воздействия на него.
Специальным и очень важным разделом экологической физиологии является экологическая физиология человека. Необходимость хозяйственного освоения новых территорий часто заставляет человека заселять ранее безлюдные территории. Плотность населения зависит от социально-экономических условий, но заболеваемость и физиологическое состояние человека определяется часто природными неблагоприятными факторами среды. Полностью изолировать от них человека, в том числе детей, нет никакой возможности, а частичная и посильная
9
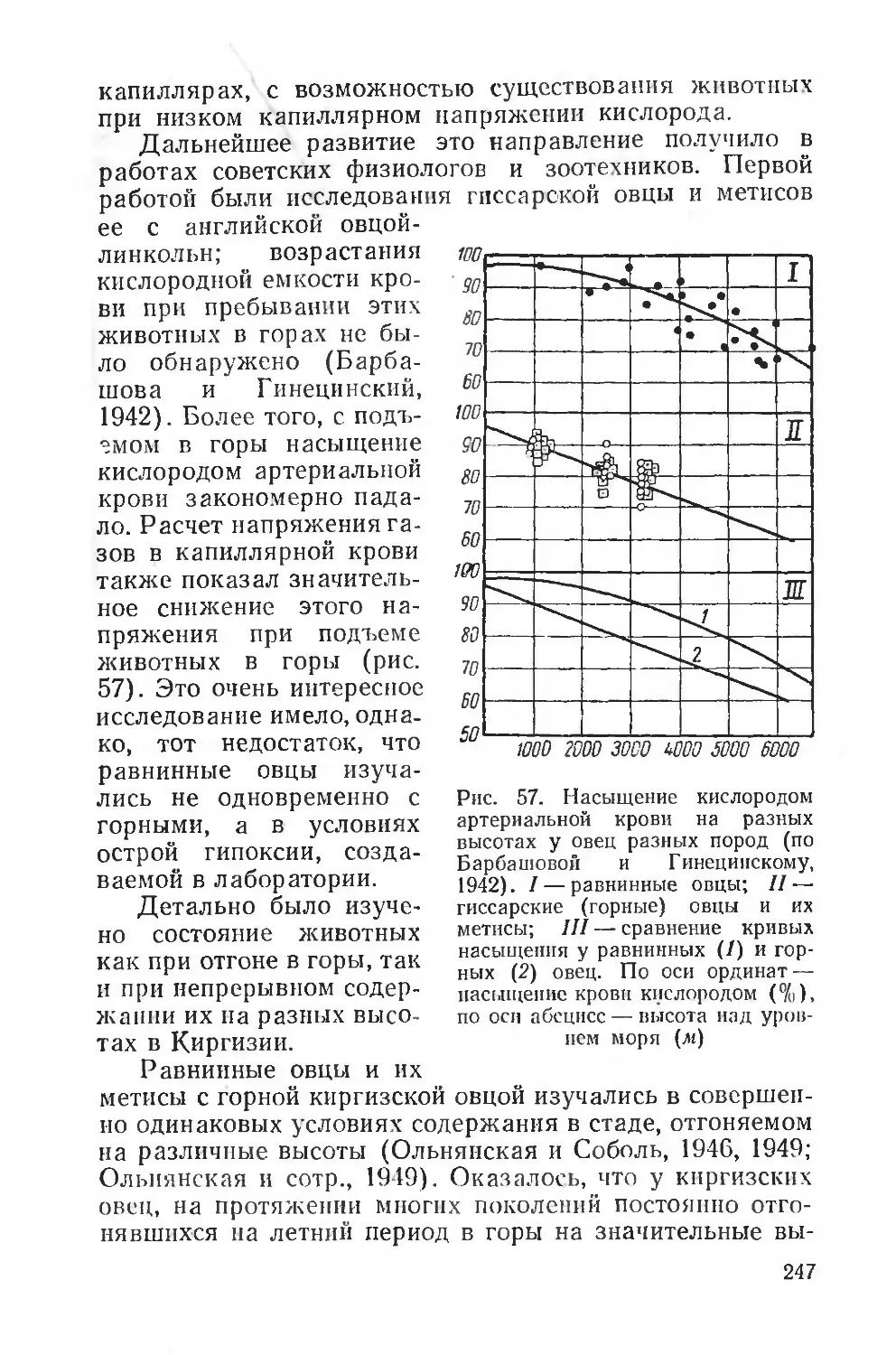
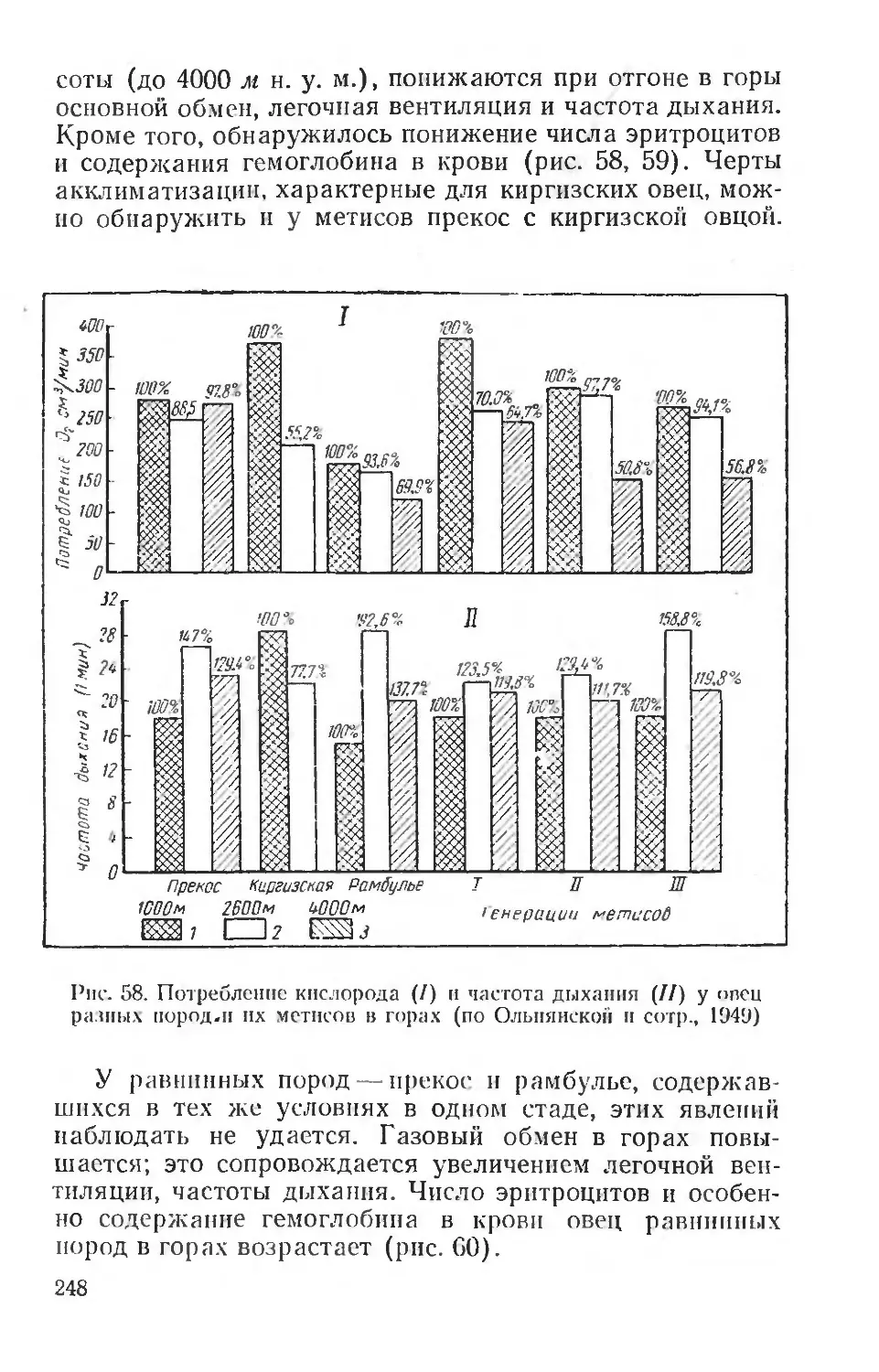
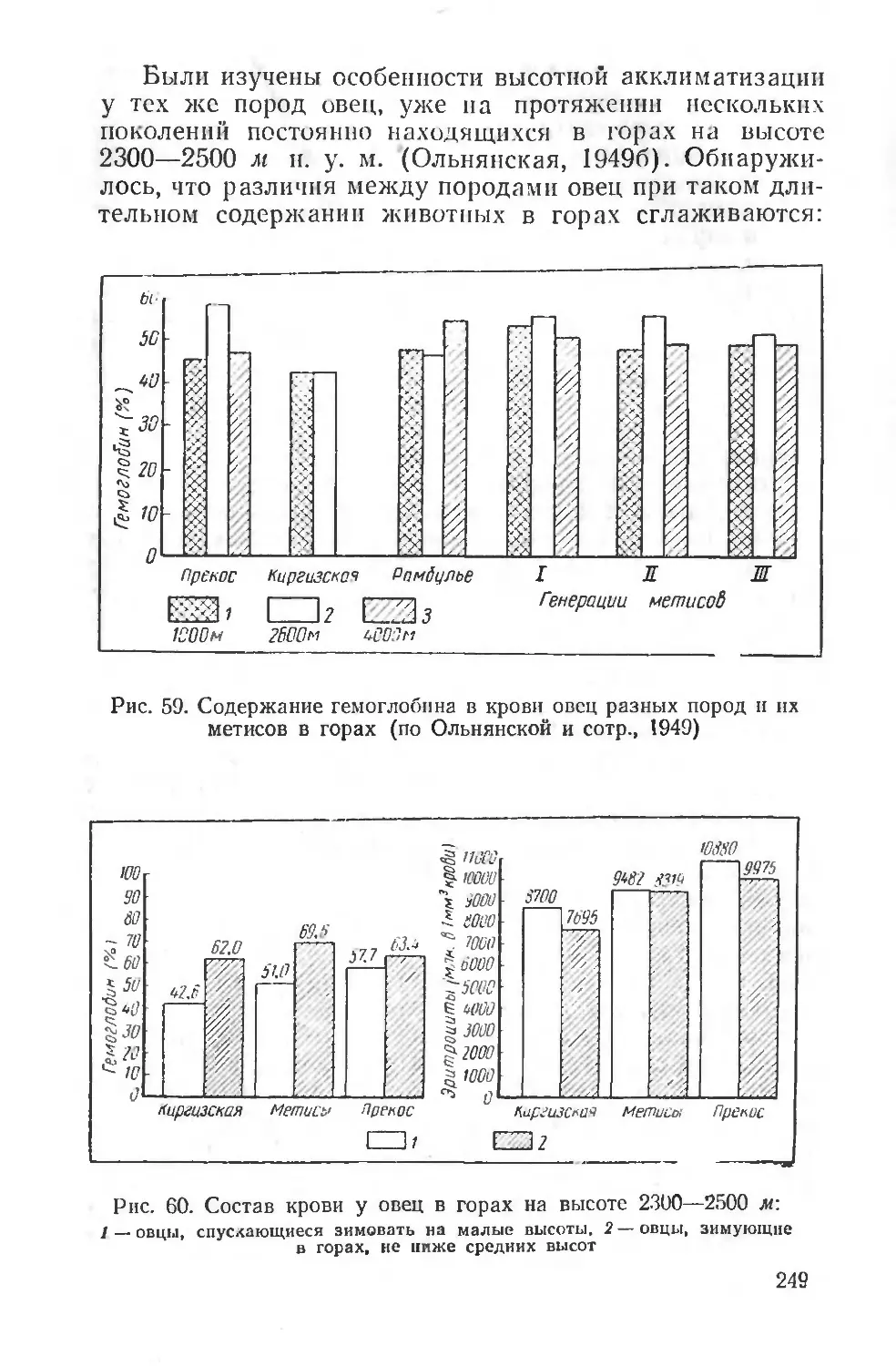
борьба с влиянием этих неблагоприятных факторов может вестись только на основе знания механизмов их сложного и длительного воздействия. Следовательно, экологическая физиология человека является основой для разработки гигиенических мероприятий, методов физического воспитания человека, широкого планирования оздоровительных мероприятий, строительства новых городов и их планировки и т. д.
Такие проблемы, как физиолого-гигиеническое обеспечение жизни человека в условиях Крайнего Севера, жаркого и пустынного климата, в условиях высокогорья и т. д., имеют в своей основе глубокое изучение общебиологических закономерностей влияния среды на организмы. Неудивительно поэтому, что многие вопросы экологической физиологии, например проблема суточного ритма, некоторых сезонных изменений, лучше всего разрабатываются на человеке, хотя, разумеется, физиологические особенности этих реакций у человека имеют свою очень сложную специфику. Особые условия среды, создаваемые человеком в культурных условиях жизни, сильно осложняют многие вопросы приспособления к природным факторам, но вместе с тем позволяют путем выбора соответствующих объектов исследования решать многие общие для человека и животных вопросы. Например, такие исследования, как влияние климата на людей, работающих на открытом воздухе или в закрытых помещениях, влияние переездов из одних климатических зон в другие в различные сезоны — все это вопросы, близкие гигиенисту, врачу-курортологу и вместе с тем дающие большой материал для эколого-физиологических сопоставлений.
В последние годы экологическая физиология человека получила широкое развитие во всем мире благодаря деятельности международной организации — Интернациональной Биологической программы (International Biological Programme), поставившей задачей всестороннее изучение здоровья и деятельности человека в различных природно-климатических районах земного шара. Программа предусматривает проведение исследований по единой методике, с тем чтобы результаты, полученные в разных странах различными исследователями, могли быть сопоставлены и позволили обосновать как научные, так и практические выводы. Специальный симпозиум в Киото (Япония) в 1965 г. подробно обсуждал эти во
10
просы. Следует отметить весьма важные способы регистрации двигательного поведения, способы разложения суточной деятельности человека на отдельные элементы, определения энергетических затрат, кожной температуры, температуры разных областей тела, определения потоотделения и водного баланса, разработку систем специальных климатических камер, позволяющих унифицировать метеорологические условия исследования человека, адаптированного к разным климатическим зонам.
С другой стороны, эколого-физиологическая характеристика многих видов животных, например наиболее близких к человеку обезьян, позволяет решить ряд вопросов терморегуляции человека, особенностей приспособления к интенсивной мышечной деятельности и т. д. Можно поэтому говорить и об эколого-физиологических аспектах исследования и в области физиологии человека, в особенности когда речь идет о воздействии природных физических агентов или более сложных факторов среды.
Практическое значение эколого-физиологических исследований очень велико. Кроме фактических данных, важных для гигиенических мероприятий, касающихся жизни и деятельности человека, огромное значение они приобретают для сельскохозяйственной физиологии, физиологии полезных и вредных для сельского хозяйства животных. Вопросы породного районирования и увеличения продуктивности животных, научное обоснование норм кормления в разных условиях и условий содержания, специальные приемы по выращиванию молодняка и т. д.— вот далеко неполный список вопросов, непосредственно опирающихся на данные эколого-физиологических исследований. Не менее важна и борьба с вредными для сельского хозяйства и здоровья человека животными. Практической проблемой является, например, научное обоснование приманочного метода борьбы с вредными грызунами, прогнозирование их размножения и планирование мероприятий по борьбе с вредными животными, защита посевов от повреждений и т. п.
Важное значение имеют эколого-физиологические исследования и для разведения полезных представителей дикой фауны, для восстановления и увеличения численности охотничье-промысловых животных. В то же время эколого-физиологическое исследование требует анализа наблюдаемых в природной обстановке явлений на всех
11
уровнях физиологической интеграции, на уровне целостного организма, на органном и системном, на тканевом и клеточном, и наконец, молекулярном.
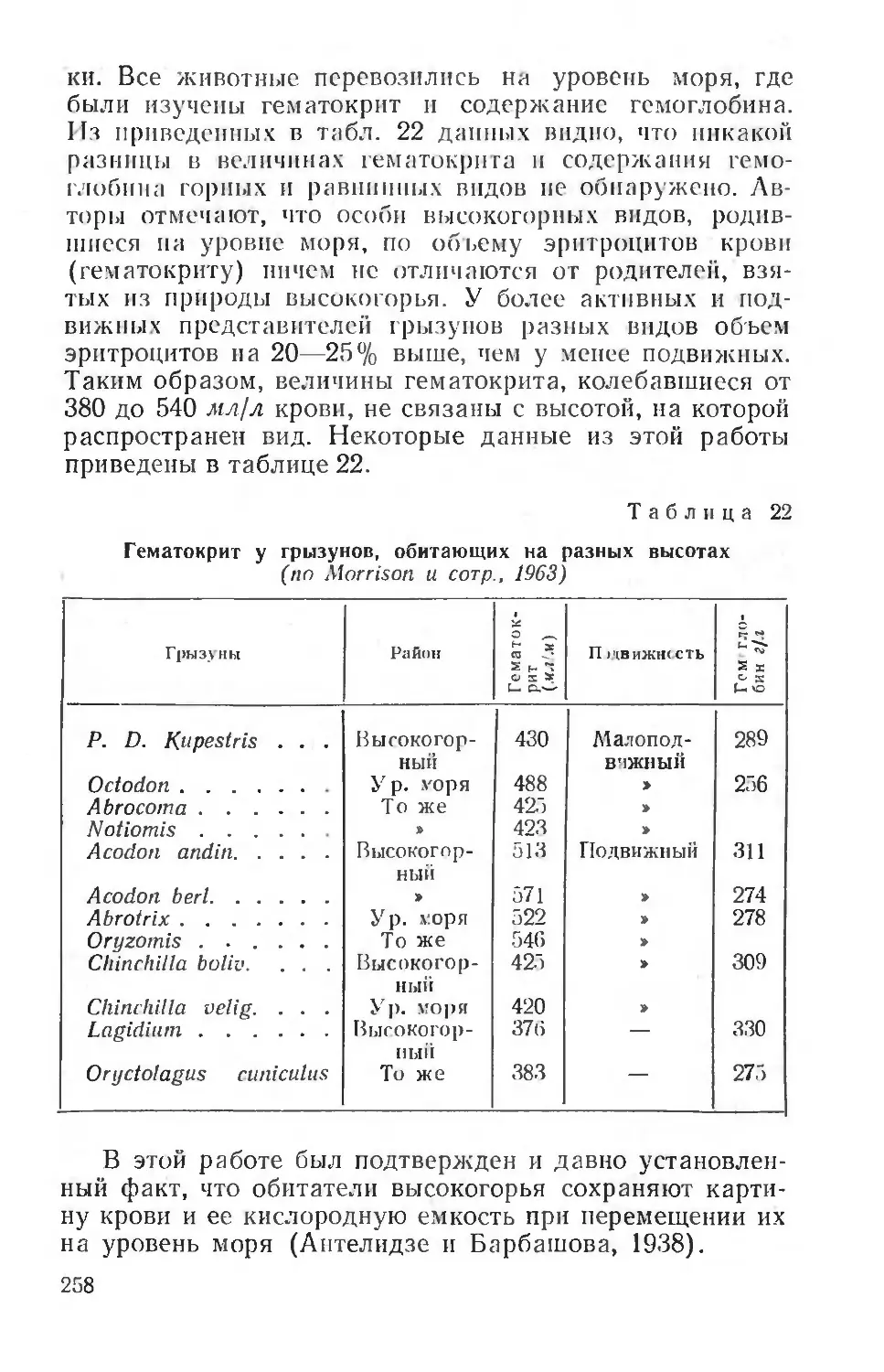
При изучении любого животного объекта исследователь сталкивается, во-первых, с элементами, генетически обусловленными филогенезом вида, во-вторых, с элементами, приобретенными в разные периоды индивидуального развития. Последние охватывают натуральные условные рефлексы различной сложности и прочности (элементы запечатлевания — импринтинга), более глубокие изменения гормональных отношений и тканевых процессов и, наконец, некоторые результаты естественного (а у домашних животных и искусственного) отбора.
Экологи под популяцией понимают совокупность животных, принадлежащих к определенному виду и населяющих территорию с однородными условиями существования. Морфологические и физиологические особенности популяции зависят от целого ряда внешних воздействий на организм и от того, в какой мере данный вид обладает большей или меньшей вариабильностыо своих признаков, определяющих существование животных на данной территории. Физиологический анализ особенностей популяции чрезвычайно важен для решения ряда практических вопросов, так как популяция характеризует и представляет данный вид в его естественных отношениях к другим видам, к тем или другим факторам среды. Поэтому интересен анализ тех факторов, которые определяют те или другие особенности популяции. Изучение этих особенностей заставляет обратиться к формированию рефлекторной деятельности в разные периоды онтогенеза. Безусловные и образующиеся на их основе условные рефлексы возникают в разные периоды постнатального развития. Поэтому чрезвычайно важно установить, в какой мере формирование тех и других зависит от факторов среды и насколько прочными будут вновь образованные условные рефлексы или сформулированные безусловные, насколько определяющими — факторы среды, действующие на разных этапах постнатального развития, для всего последующего развития отдельных особей, популяции в целом.
Глава I
ПРИЕМЫ И МЕТОДЫ
ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИХ ИССЛЕДОВАНИИ
ИССЛЕДОВАНИЕ ФИЗИОЛОГИЧЕСКИХ ФУНКЦИЙ
В ПРИРОДНЫХ УСЛОВИЯХ И В ЭКСПЕРИМЕНТЕ
Эколого-физиологическое исследование, направленное на изучение физиологии животных и человека в естественных условиях для установления физиологического состояния организма или деятельности отдельных его систем, требует не только специальных приемов экспериментирования, но и разработки специальной аппаратуры. Последняя должна отвечать требованиям современной физиологической науки и обеспечивать сохранение или воспроизведение специфических для данного вида организмов реакций, действующих на организм факторов и форм поведения.
Если рассмотреть с этой стороны обстановку обычной физиологической лаборатории и приемы физиологического экспериментирования па целом организме, то нельзя не признать почти полную их непригодность для работы на огромном большинстве диких животных. Всевозможные типы камер, станков, клеток и других фиксирующих или ограничивающих движения животных приспособлений, изолирующих его от окружающей среды и стремящихся выделить только некоторые из факторов этой среды, неизбежно нарушают природные связи животного и требуют создания новых, искусственных, что и выражается в более или менее длительном приучении животных к условиям эксперимента. Обычно это удается, и только ценой нарушения естественных связей животного с внешней средой физиологи получают возможность работать на диких организмах.
Для домашних или полностью одомашненных видов обстановка жилья человека, включая и лабораторию,
13
является более естественной средой существования, и здесь задача физиолога значительно упрощается.
Практически эколого-физиологическое исследование идет по двум направлениям. В первом случае работа ведется в полевых условиях, когда исследователь использует природную обстановку в качестве экспериментального фона. Главной методической задачей здесь является регистрация как элементов двигательного поведения, так и состояния физиологических функций, по возможности без изменений условий среды. Идеальными методами исследования являются в этом случае дистантные методы, начиная от точного и хорошо поставленного визуального наблюдения и кончая дистантной регистрирующей радиоаппаратурой. Примером таких работ могут служить исследования по образованию условных рефлексов у суслика в естественной обстановке (Реймов, 1958). 'Здесь визуальным методом изучались особенности двигательных реакций на различные ориентировочные, пищевые и оборонительные раздражители, подаваемые экспериментатором вблизи естественного места обитания — норы. Трудность постановки таких опытов с дикими животными очевидна, но опа целиком компенсируется ценностью полученных фактов. Дистантные методы исследования и регистрации, основанные на использовании радиоэлектроники, нашли свое применение в спортивной физиологии, космической физиологии и начинают играть важную роль в эколого-физиологических исследованиях как на сельскохозяйственных, так и па диких животных (в кн. Биотелеметрия, 1965) (рис. 1, 2).
Весьма важной в экспериментировании на животном организме является проблема фиксирования животного. Наиболее простые методы фиксирования, как-то: привязывание за лапки, повал (для копытных), станок для собак и т. д., широко применяются в физиологической практике и останавливаться на их описании нет никакой необходимости. Важно заметить, что применение всех этих методов к диким животным сильно изменяет их физиологическое состояние, часто вызывает неугасающую оборонительную реакцию, затрудняющую получение необходимых данных даже и в состоянии относительного покоя животного. Поэтому разработаны такие методы фиксации, которые являются наиболее адекватными для отдельных видов животных и, не вызывая длительной
14
rr
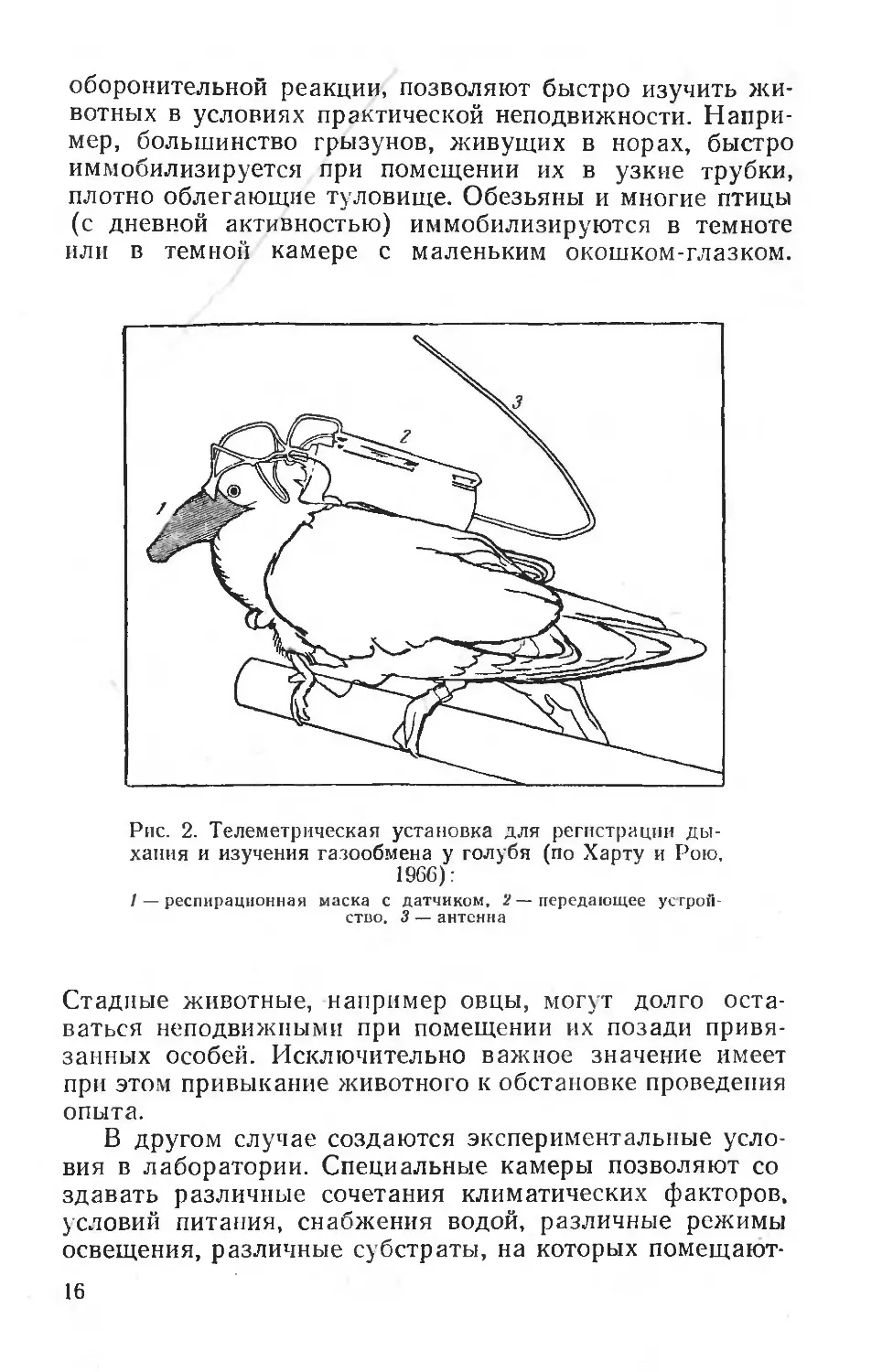
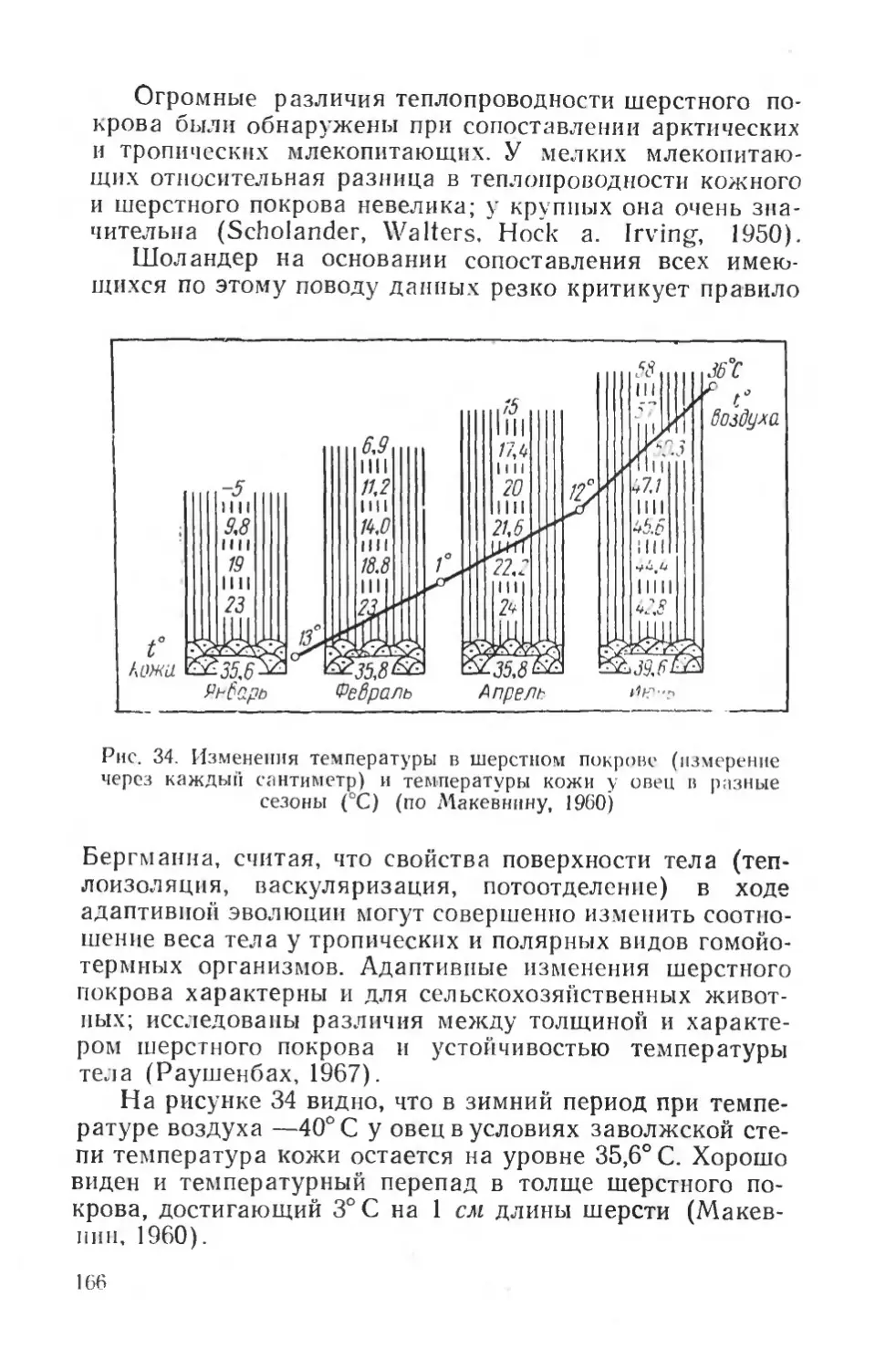
Рис. 1. Телеметрическая установка для регистрации электромиограмм и температуры тела (по Бакалову, 1968). I — правая часть — передающее, левая — регистрирующее устройства; // — процесс регистрации
15
оборонительной реакции, позволяют быстро изучить животных в условиях практической неподвижности. Например, большинство грызунов, живущих в норах, быстро иммобилизируется при помещении их в узкие трубки, плотно облегающие туловище. Обезьяны и многие птицы (с дневной активностью) иммобилизируются в темноте или в темной камере с маленьким окошком-глазком.
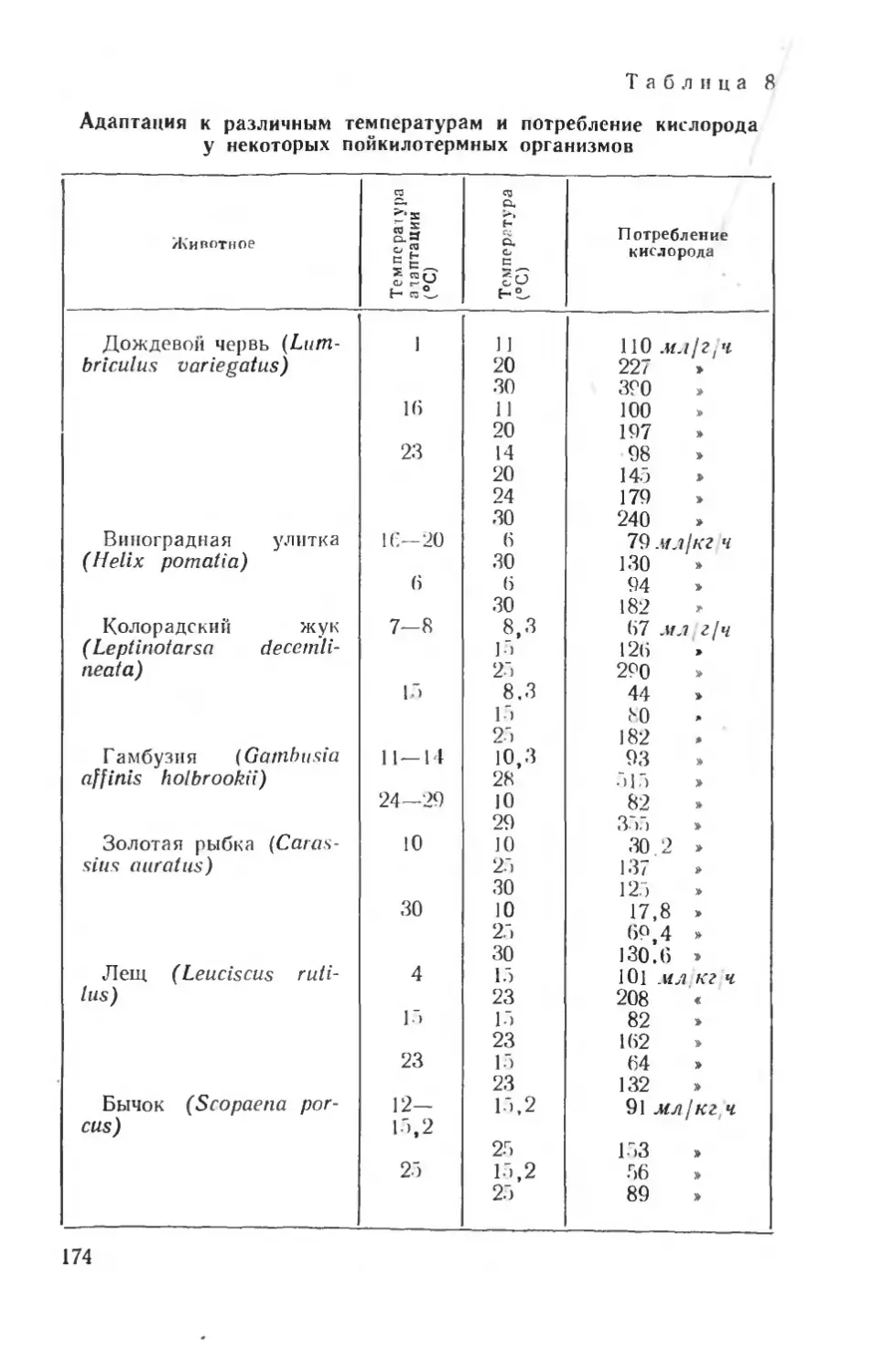
Рис. 2. Телеметрическая установка для регистрации дыхания и изучения газообмена у голубя (по Харту и Рою, 1966):
I — респирационная маска с датчиком, 2—передающее устройство. 3 — антенна
Стадные животные, например овцы, могут долго оставаться неподвижными при помещении их позади привязанных особей. Исключительно важное значение имеет при этом привыкание животного к обстановке проведения опыта.
В другом случае создаются экспериментальные условия в лаборатории. Специальные камеры позволяют со здавать различные сочетания климатических факторов, условий питания, снабжения водой, различные режимы освещения, различные субстраты, на которых помещают
16
ся животные. Таким образом имитируются условия подземного существования, комплекс, засухи, влияние высоких и низких температур, специальные режимы ограничения доставки пищи и воды, снижения содержания кислорода или барометрического давления в окружающей среде. Наконец, исключительное значение при изучении адаптации приобретает исследование двигательного поведения. Среди функций, которые наиболее часто используются для эколого-физиологических характеристик, наибольшее значение имеет изучение общего обмена веществ как функции, отражающей общую жизнедеятельность организма. Эта функция в равной мере имеет значение как для высших организмов, так и для различных видов беспозвоночных. Важное место занимает исследование питания, которое включает пищедобыва-тельную деятельность и ферментативную и двигательную активность пищеварительного аппарата. Эти проблемы
включают как вопросы изучения поведения, так и вопросы, непосредственно связанные с эколого-биохимическими исследованиями. Третья группа вопросов включает
изучение поддержания различных сторон гомеостазиса, особенно важных для характеристики высших организ
мов— позвоночных, но имеющих и немаловажное значение для видов, населяющих территории, физические параметры которых подчас представляются совершенно не
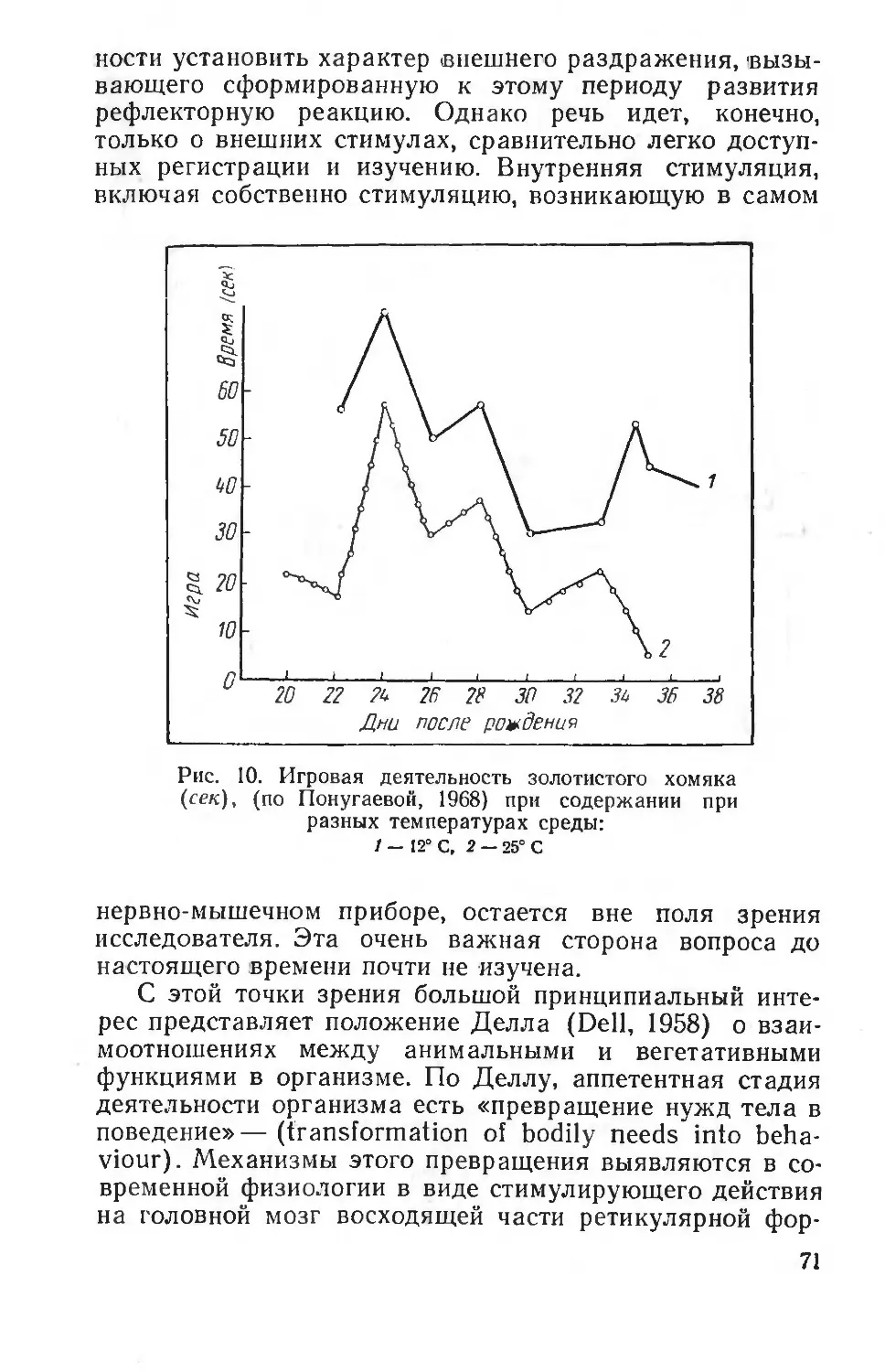
совместимыми с жизнью.
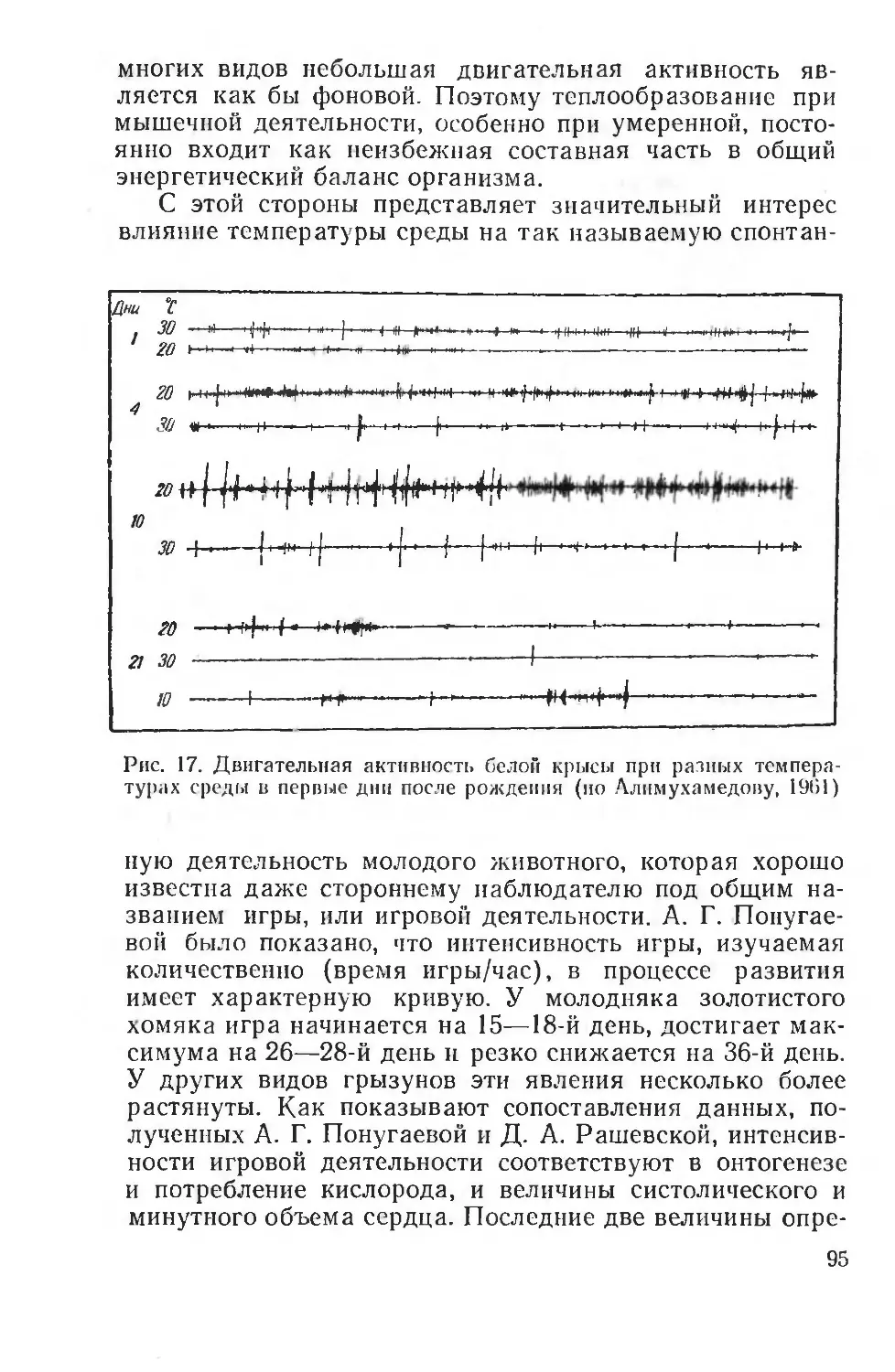
Методические приемы эколого-физиологических исследований приобретают за последние годы все большее разнообразие, а самые исследования большую глубину. Физиологи все больше начинают регистрировать не только феноменологическую сторон ' явлений адаптации, но
и стремятся понять лежащие в их основе механизмы. В этом отношении любопытно замечание американского физиолога Шмидт-Нильсена (Schmidt-Nielsen, 1967), указывающего, что использование в работах физиолога и в лабораторной практике необычных животных, т. е. животных — обитателей различных экологических зон, на
разных этапах эволюции привело к значительным открытиям в области физиологии и патологии. Вместе с тем
эта тенденция требует замены работы в кратковремен-IX экспедиционных условиях, создания стационарных ’^сЬбораторш'Г'В' различных природных условиях. Сейчас уж\создана целая сеть специальны^ лабораторий как
17
в СССР, так и за рубежом, ориентированных на изучение человека и животных в условиях Арктики, тропиков, пустынь, а также ряд морских биологических станций.
ИЗУЧЕНИЕ ПОВЕДЕНИЯ В ПРИРОДНЫХ И ЛАБОРАТОРНЫХ УСЛОВИЯХ
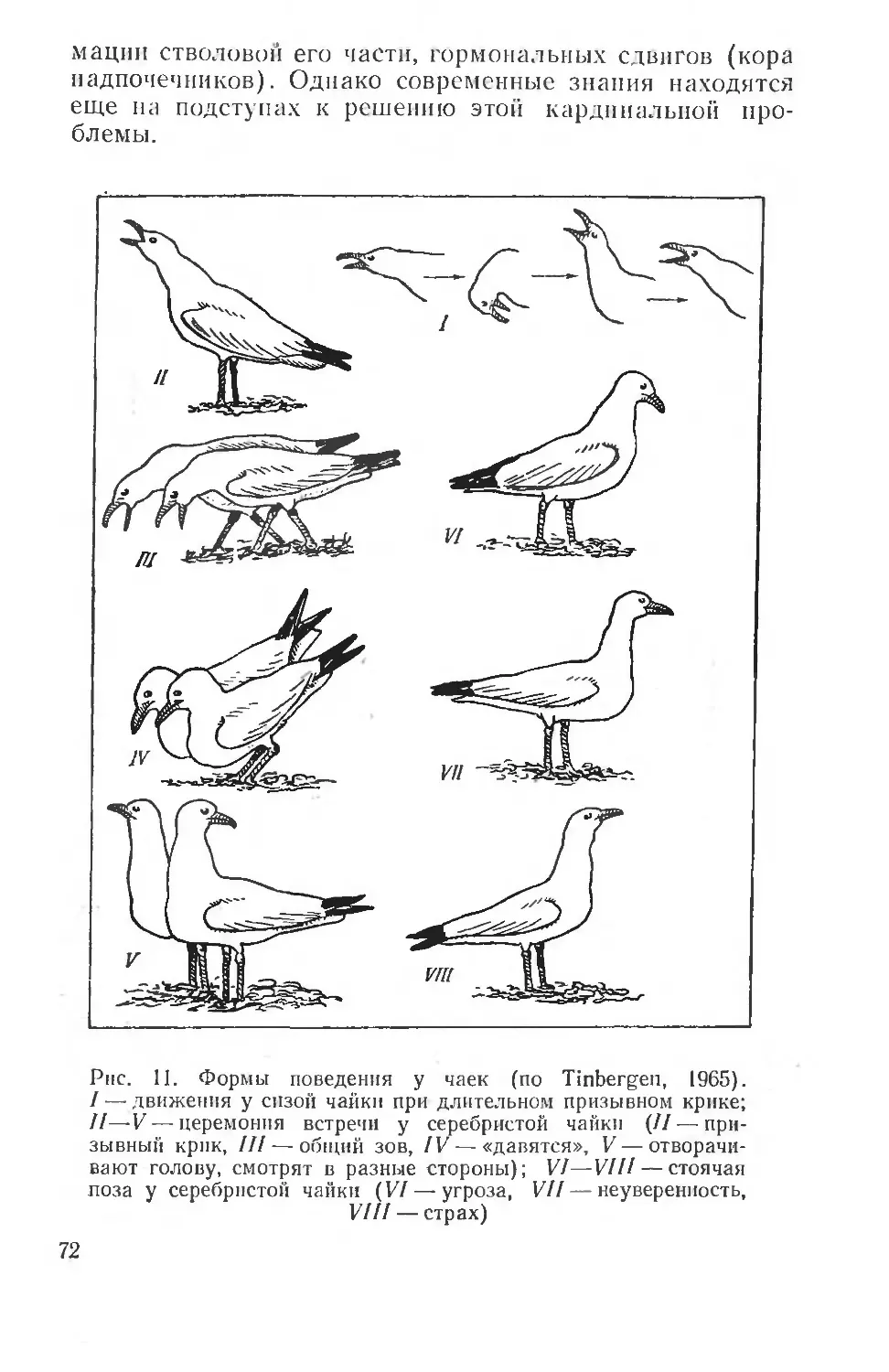
Наряду с исследованием физиологического состояния организма и отдельных его систем важное значение для понимания особенностей распределения и существования животных в природе приобретает изучение двигательного поведения, которое в последнее время выделяется в самостоятельную науку — этологию, по включает элементы как физиологического, так и психологического анализа, детальную характеристику сложных форм поведения в природе, количественный анализ этого поведения и его воспроизведение при изолированном воздействии сложных природных факторов (как биотических, так и абиотических). Следует заметить, что развитие этологических исследований в настоящее время находится главным образом на стадии собирания и систематизации материала. Теоретические основы этологических исследований еще очень неопределенны (см. стр. 61), поэтому здесь изложены те методы изучения поведения, которые лежат в основе понимания адаптивной эволюции организмов и неразрывно связаны с регуляцией вегетативных функций.
В изучении поведения отдельных видов и их популяций важное место приобретает задача выявления специфичности действия раздражителей, происхождения отдельных форм поведения в онто- и филогенезе, внутренней (эндогенной) стимуляции поведения, и наконец, процесса замещения поведенческих реакций изменениями физиологических процессов или наоборот.
Наиболее общей характеристикой поведения животного организма является распределение двигательной активности на протяжении суток — так называемая суточная периодика двигательной активности. Обычно исследование состоит в том, что все движения животного, помещенного в клетку, регистрируются на протяжении суток. Иногда регистрация усложняется путем дополнительной записи на кимографе движений, связанных с
18
\ кормушкой, с выходом из гнездовой камеры и т. д. Распределение периодов активности и покоя служит важной характеристикой состояния возбуждения и торможения в центральной нервной системе, отражением воздействия внешних условий на эти отношения, выражением уже выработанного на протяжении прошлого индивидуального опыта или врожденного жизненного цикла.
Получаемые таким путем графики активности (акто-граммы) характеризуют распределение активности во времени, но совершенно не отражают интенсивность мышечных движений. Кроме того, до настоящего времени не разрешен окончательно вопрос о регистрации сложных форм мышечной деятельности животных, например роющей деятельности, полета, специальных двигательных актов.
Анализ актограмм не представляет трудности. Полученные отношения наносятся на линейную или круговую (24-часовую) шкалу. При этом в целях суммирования получаемых в отдельные дни данных движения суммируются в пределах от 15 мин до 1 ч.
Значительно труднее осуществляется регистрация суточных особенностей активности животных в природе. Исследователями в полевой обстановке применен ряд остроумных методов исследования, сочетающих наблюдение за поведением животных с некоторыми аппаратурными приемами регистрации. Наиболее простым методом является суточный учет попаданий животных в ловушки. Этот метод, применяемый обычно для грызунов, дает хорошие результаты только при высокой численности этих животных. Используется также методика изучения прикопки нор в разные часы суток. Имеющиеся в нашем распоряжении лабораторные данные свидетельствуют о большом соответствии и совпадении во времени, например роющей деятельности, с общей суточной активностью животного. Поэтому эти простые методы можно считать вполне приемлемыми для регистрации суточного распределения активности.
Применяются, конечно, и визуальные наблюдения за активностью животных, например при изучении колоний сусликов, песчанок, стад копытных и т. д.
Совершенно особое положение при изучении двигательного поведения (пищевого, оборонительного, полового и т. д.) занимает визуальная регистрация во времени
19
отдельных элементов деятельности животного — методика хронометража поведения. По происхождению методика хронометража относится к области изучения протекания трудовых процессов у человека, но ее точность и легкая применимость к изучению любых процессов, протекающих во времени, сделали ее универсальным ме тодом исследования.
При постановке хронометражных наблюдений можно точно выявить так называемые фиксажные точки, т. е. такие элементы движения или сложного поведения, которые периодически повторяются и могут быть выделены с большой точностью из общего сложного комплекса движений животного. Такими фиксажными точками при изучении пищевых реакций в условиях природы могу г быть время начала и конца приема пищи, начала и конца ходьбы, бега, периодических переходов с места на место, выхода из норы или ухода в нору, начала и конца элементов игровой деятельности молодняка и т. д. Если такое хронометрирование производить на протяжении суток (или на протяжении периодов активности животного), то при нанесении на суточную шкалу времени получается «фотография активности животного». Обработка такой фотографии ведется так же, как и суточной актограммы, по часам суток или суммарно в процентах к общему изученному времени. Важно отметить, что суммирование фотографий дня возможно только после обработки в процентах или по часам суток.
Другой способ регистрации текущей деятельности основан на фиксировании состояния активности изучаемого объекта через определенные промежутки времени (Ъмин, 10 мин, 1 ч, 2 ч и т. д.). Этот способ позволяет единовременно изучать поведение на большом числе животных, например при пастьбе, при отдыхе в стаде. Так, единовременно на стаде в 20 голов отмечается, сколько животных пасется, сколько переходит с места на место и т. д. Этот способ позволяет судить о поведении целой группы и при достаточном материале оказывается очень ценным для выявления зависимостей поведения от внешних факторов. Таким путем было установлено влияние различных факторов среды (температуры, ветра, снежного покрова и т. д.) на характер и особенности пастьбы северного оленя. Эти данные использованы для изучения пастьбы других животных.
20
Хронометражные наблюдения приходится применять не только в полевых условиях, но иногда и в лабораториях. Таким путем удалось зарегистрировать время акта еды у различных животных с различными типами и особенностями питания в условиях зоопарка и в лаборатории.
Несмотря на свою трудоемкость, хронометраж является незаменимым методом изучения поведения животных в разных физиологических состояниях и его значение для эколого-физиологических исследований очень велико.
Но далеко не всегда двигательная деятельность животных в природе доступна для наблюдения и особенно для ее количественной характеристики. Между тем только последняя позволяет сопоставить ее с другими формами деятельности или с изменением физиологического состояния. Поэтому исследователь часто пытается воспроизвести различные формы поведения в экспериментальной обстановке (в лабораторной или полевой), чтобы облегчить точную регистрацию поведения в целом или отдельных его элементов.
Наиболее просто воспроизводится в условиях эксперимента пищевое поведение. Так, например, изучение скорости поедания определенных видов пищи может производиться в строго постоянных условиях голодания и характеризует как пищевую возбудимость животного, так и сложнорефлекторные отношения питания и пнщедобы-вательной деятельности.
Скорость приема пищи можно изучать в связи с ее добыванием, например с выкапыванием ее грызунами из-под определенного слоя земли. В этом случае скорость пищедобывательной реакции характеризует пищевую возбудимость животного, а также наличие образованных ранее условных рефлексов с обонятельного анализатора Такой метод работы может быть применен в садках или небольших вольерах. Разумеется, всякая фиксация животного здесь исключается, а время регистрируется визуально с отсчетом по секундомеру или методом контактной регистрации.
Более сложно воспроизведение в условиях лаборатории таких форм деятельности, как грызение, роющая деятельность, запасание корма, гнездостроение. Поскольку эти формы деятельности врожденные и стимулируются определенным сочетанием внешних раздражителей —
21
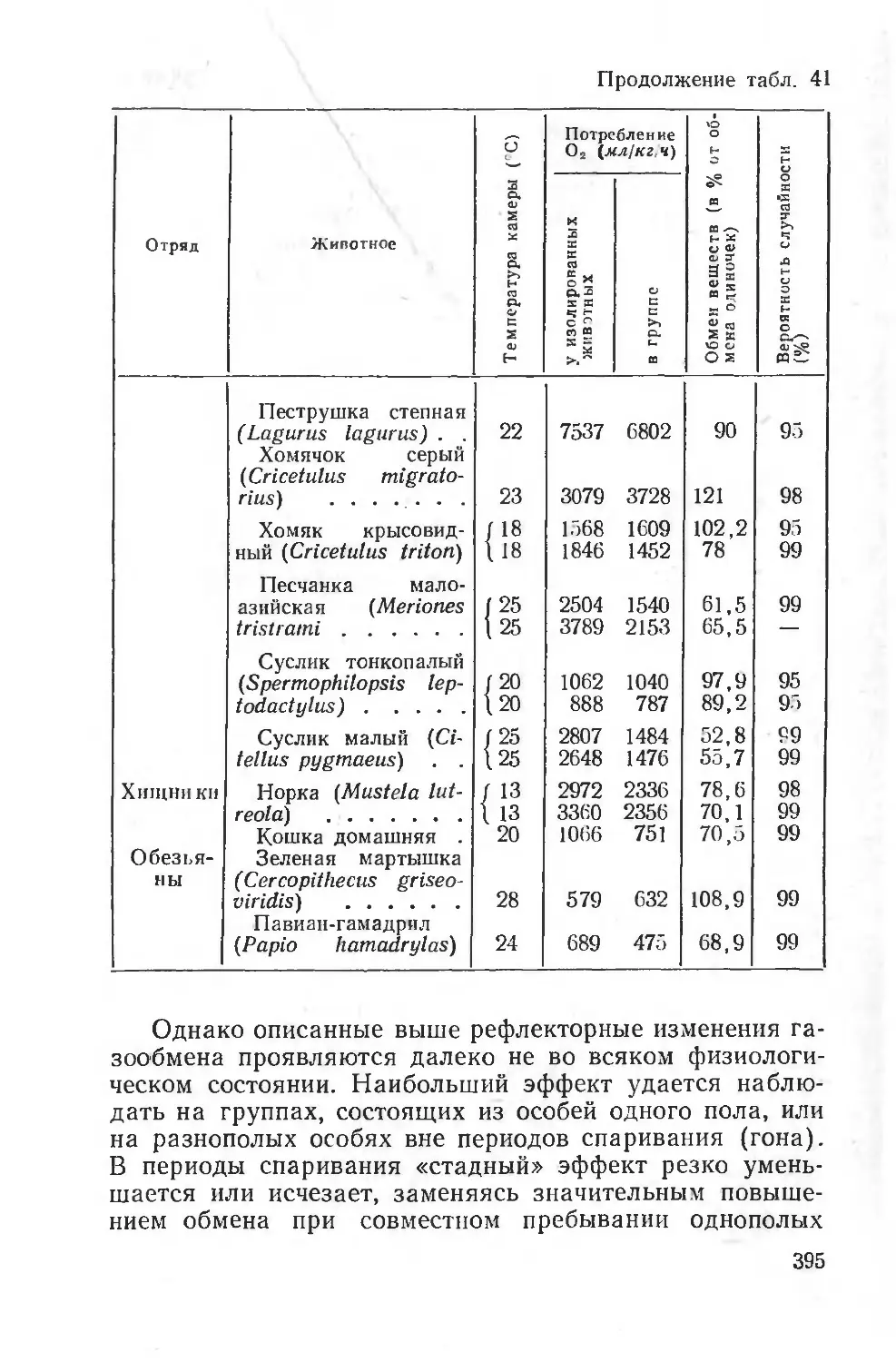
объектов запасания, грызения, материалов для постройки гнезд, наличием сыпучего субстрата (земли, песка и т. д.) и эндогенных факторов (эндокринные железы, условия питания, суточный цикл и т. д.); исследователь имеет возможность имитировать эти природные раздражители и длительное время изучать двигательное поведение животных. При этом можно изучать влияние отдельных факторов среды как абиотических (температура, барометрическое давление и т. д.), так и биотических (питание, наличие особей того же или другого вида и т. д.) (Попугаева, 1960; Слоним, 1961). Опыт показывает, что такие формы деятельности, как запасание корма или грызение, производятся животными на протяжении многих часов подряд в определенные отрезки суточного и сезонного цикла.
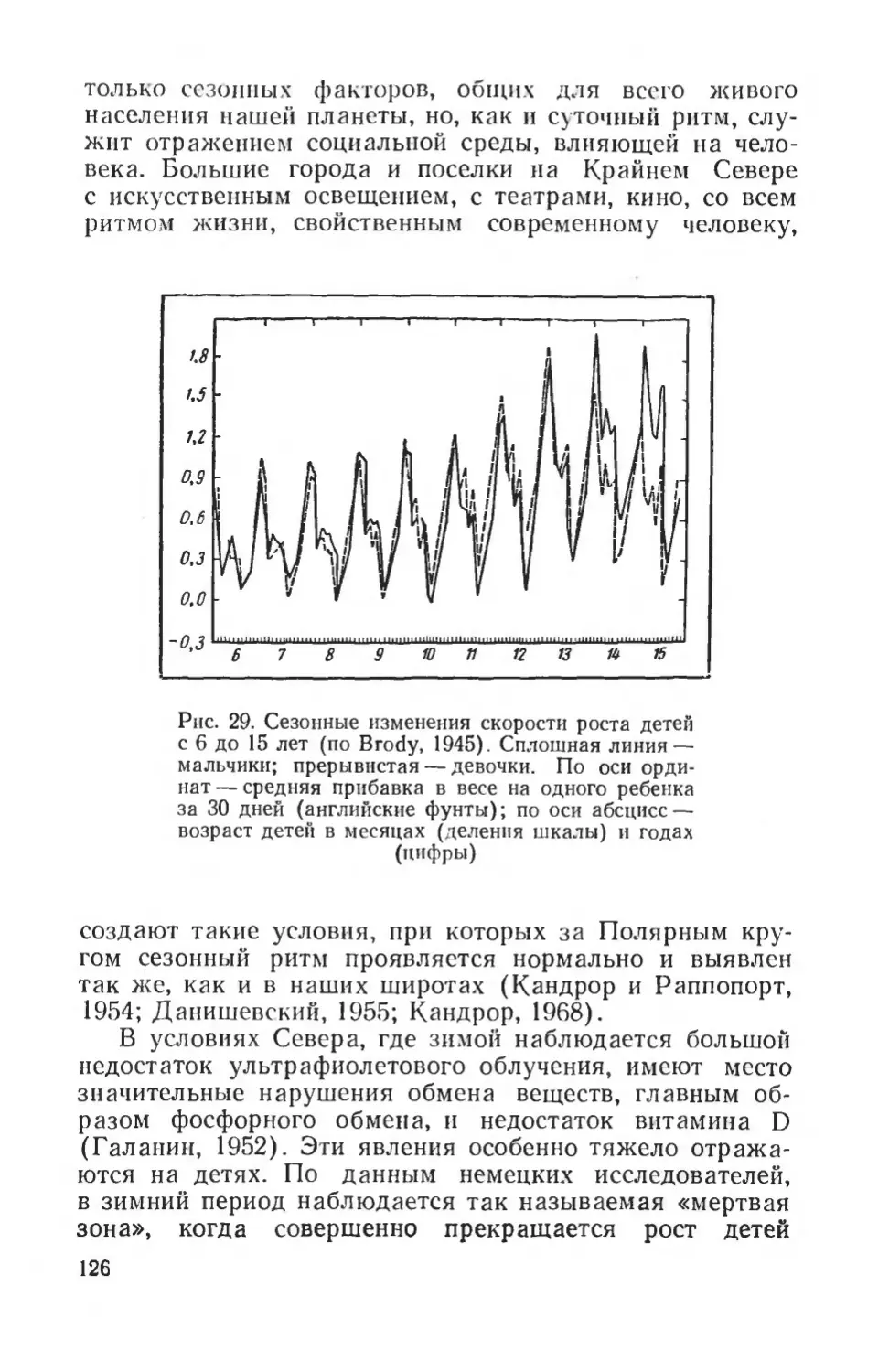
Легко воспроизводится игра молодняка при подборе определенных пар из помета грызунов. Во время игры могут регистрироваться определенные ее элементы (движения, поза), их распределение во времени, частота повторений, общая продолжительность и т. д. Таким образом дается путем визуального наблюдения (хронометража) количественная характеристика этой формы деятельности животных.
Воспроизведение отдельных форм поведения в лаборатории сильно облегчается благодаря методу муляжей. Под муляжем в данном случае понимают создание искусственных моделей пищевых объектов, особей противо положного пола, яиц, пор и убежищ, на которые экспери ментальное животное реагирует специфическим образом. Такие модели могут быть гипертрофированы в определенном направлении, например темно пли ярко окрашены, увеличены в размерах целиком или отдельные их части. Применение муляжей не только облегчает постановку экспериментов с воспроизведением различных форм деятельности, но и позволяет выделить наиболее важные элементы в сложном комплексе раздражителей, которые вызывают ту или иную деятельность. Например, при показе самцу трехиглой колюшки нормальной самки с раздутым икрой брюшком возникает значительно более слабая специфическая реакция, нежели при показе грубого макета с силыю раздутым брюшком. Подобные реакции наблюдаются и у многих птиц. Из пищевых моделей хорошо известна меловая болтушка, имитиру
22
ющая молоко; она вызывала в классических исследованиях И. С. Цитовича положительную пищевую двига тельную реакцию и слюноотделение у щенков собаки.
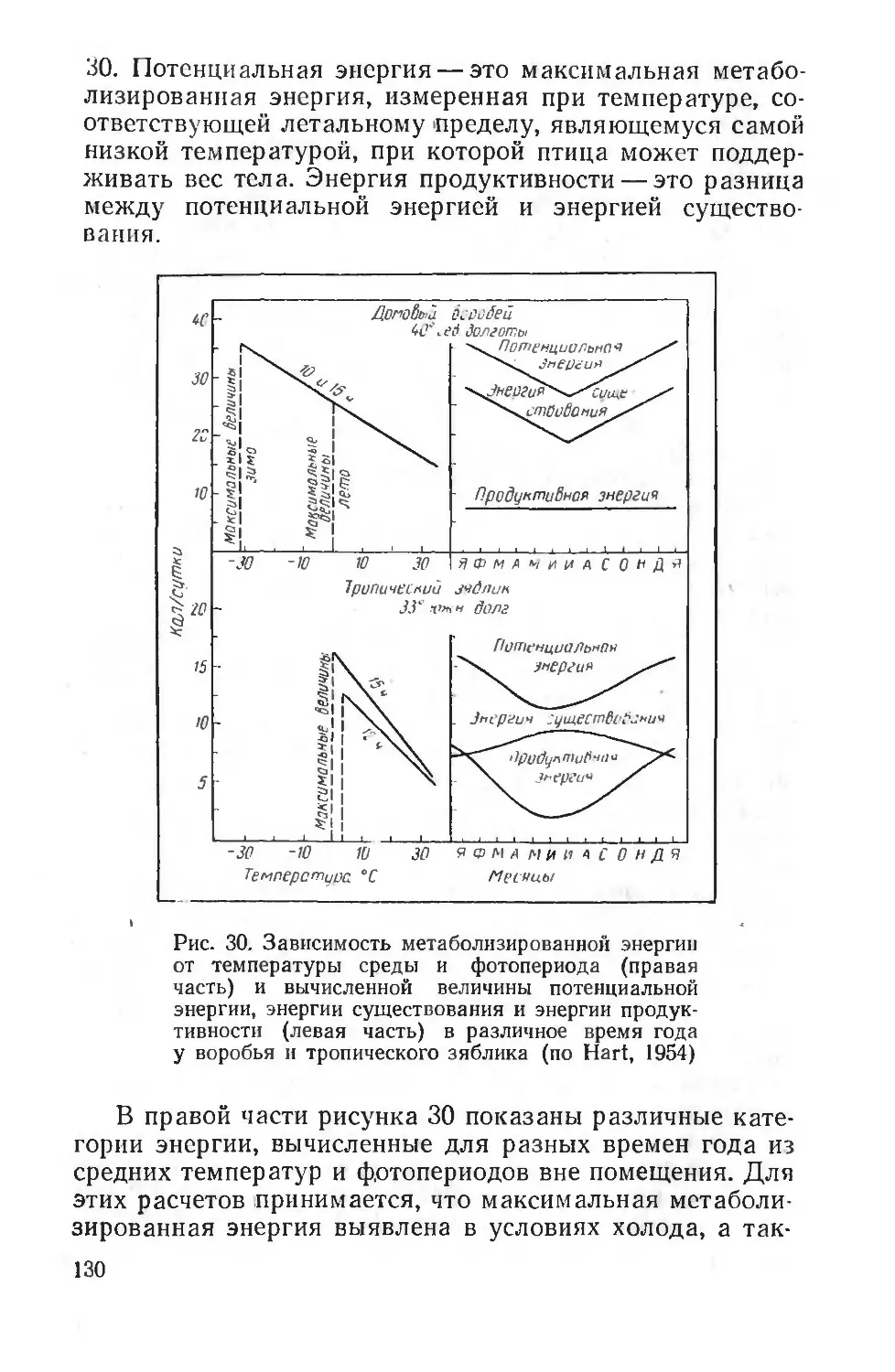
Использование моделей применялось для изучения стадных реакций грызунов, моллюсков, птиц. Примером могут служить деревянные модели уток, применяемые охотниками.
Весьма важной задачей в изучении поведения является установление генетического происхождения отдельных его форм или элементов. Для решения этого вопроса применяется метод изоляции (изолированного выращивания) или как его часто называют в иностранной литературе метод Каспар — Хаузеров. Немедленная после рождения или вылупления из яйца изоляция животного от тех или иных факторов внешней среды (особей того же вида, определенных сортов пищи, температурных условий, смены освещения) приводит к выпадению возможности образования соответствующих условных рефлексов на эти факторы (см. также стр. 70). Тем не менее в условиях такой изоляции в определенные сроки развиваются врожденные элементы поведения и регуляции физиологических функций. Так, например, изолированные от мясной пищи щенки домашней собаки начинают на 18—21-й день жизни тянуться за куском мяса и выделяют при этом слюну. Изолированные от мясной пищи котята проявляют на 30—32-й день охотничью реакцию на мышей и птиц (Слоним, 1961). Таким образом удается установить, что данная форма деятельности врожденная. С другой стороны, ягнята и козлята, изолированные от грубого корма и питавшиеся только молоком, будучи выпущены на луг, не паслись (Рахимов, 1958).
Такое разделение на врожденные и приобретенные формы поведения весьма важно для понимания происхождения адаптивных его форм. Метод изоляции весьма широко распространен в физиологии и очень хорошо позволяет понять формирование сложных адаптивных реакций и сложных форм деятельности в онтогенезе (см. гл. II).
МЕТОДЫ СОСТАВЛЕНИЯ ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИХ ХАРАКТЕРИСТИК
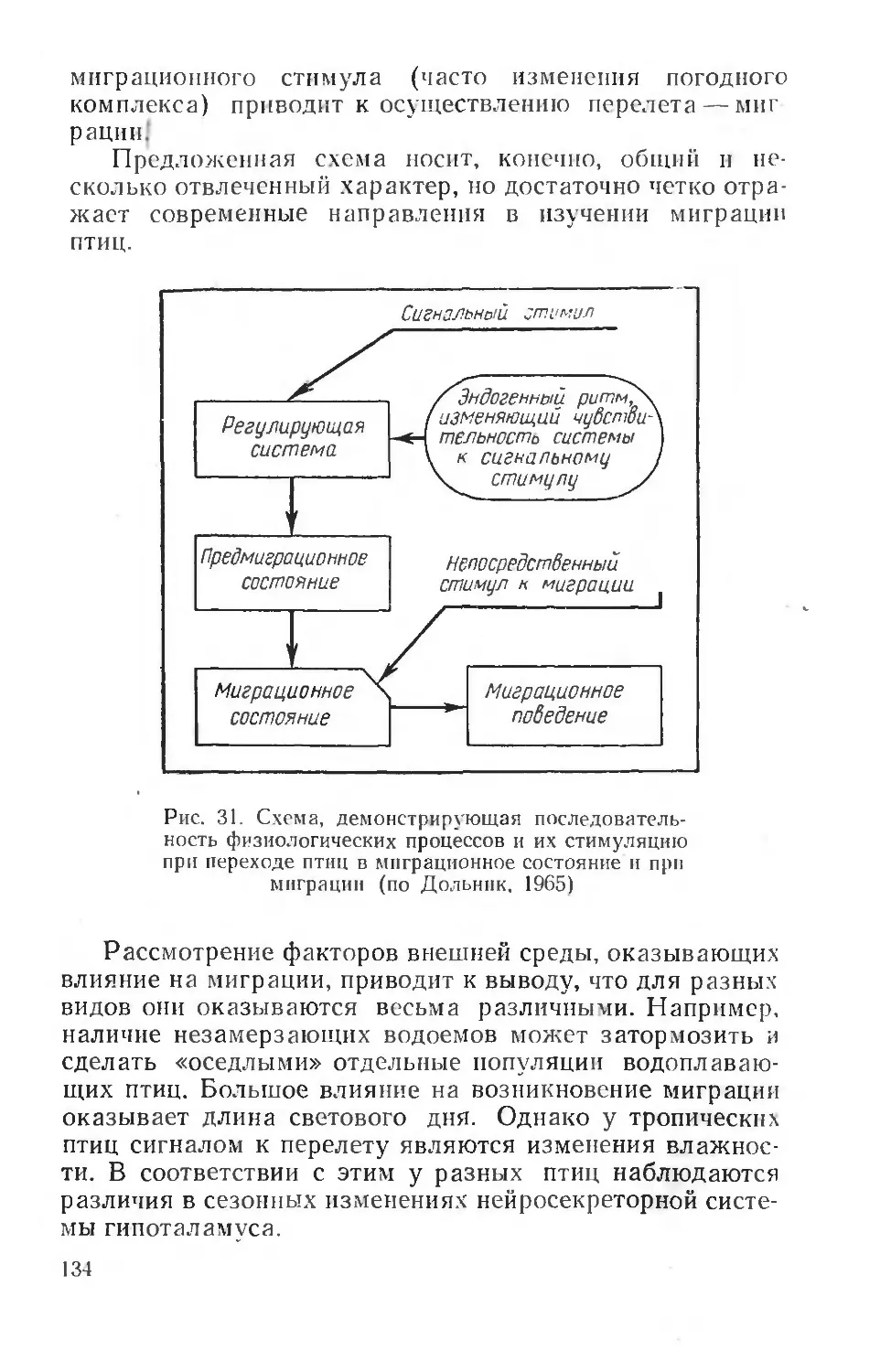
При выявлении физиологических основ приспособления животных к факторам среды вообще и к экстремаль
23
ным в частности физиолог неизбежно использует ряд методических приемов эксперимента и оценки наблюдаемых при этом физиологических данных. Наиболее часто применяется для эколого-физиологической характеристики животных ряд методов. Как правило, для экологофизиологической характеристики организма не могут быть использованы параметры только одной его функции или системы. Например, при изучении организма в условиях недостатка кислорода необходимо исследование не только легочного или внешнего дыхания, но и дыхательной функции крови, окислительных ферментов тканей, не говоря уже о кровообращении. При изучении приспособления организма к условиям недостатка воды (пустыня) необходимо изучение всех функций органов, участвующих в поддержании водного баланса — легких, почек, пищеварительного тракта и кожи.
Физиолог, собравший все необходимые данные, как бы рисует схему, характеризующую деятельность отдельных органов и систем, их взаимозамещение, дублирование, что только и обеспечивает надежность регулирования жизнедеятельности организма как физиологической системы. Однако оценка каждого органа и системы, да и организма в целом, происходит на основании определенных приемов, к изложению которых мы и переходим.
Наиболее часто физиологическая характеристика организма производится путем изучения: а) уровня физиологической активности животного или отдельных его систем; б) физиологического (а иногда и патологического) сдвига под влиянием изолированного или сложного воздействия на организм; в) времени и картины восстановления после сдвига, вызванного в эксперименте; г) времени выживания при воздействии экстремальных факторов среды.
Уровень физиологического состояния и морфо-физиологическая характеристика организмов являются предметом изучения сравнительной физиологии, но представляют и большой эколого-физиологический интерес. Так, например, частота сердечных сокращений у кролика и зайца относится, как 3: I. Это обеспечивает значительно большую резервную мощность сердца у зайца по сравнению с кроликом, реализуемую при выполнении мышечной работы. Это явление обеспечивается и морфологическими различиями сердца обоих видов (Аршав
24
ский, 1958; Hesse, 1921). Подобные явления можно обнаружить и при сопоставлении дыхательной функции крови животных, в разной мере адаптированных к интенсивной мышечной работе (Крепе, 1937), гипоксии и т. д. При изучении животных в покое необходимо учитывать периодические изменения, постоянно имеющие место в организме (см. гл. IV).
Для регистрации физиологических сдвигов, наблюдаемых при изменении условий среды, питания и т. д. отклонение функций отдельных систем или реакций целостного организма служит часто применяемым приемом эколого-физиологического исследования. Для изучения этих сдвигов необходимо знание нормальных («должных») величин, характерных для данного организма,что касается не только таких величин, как уровень общего обмена (газообмена), но и показателей общего и местного кровообращения, водно-солевых констант, температуры тела и т. д. Установление этих констант представляет значительные трудности, так как у многих организмов эти величины очень лабильны. Поэтому в большинстве случаев исследователи проводят ряд контрольных исследований, совпадающих во времени (по часам суток, сезонам года и т. д.) с постановкой основного эксперимента. Сдвиги различных функций позволяют вскрыть устойчивость данного организма к воздействующему фактору и выявить физико-химические и физиологические механизмы, лежащие в основе этой устойчивости. Таковы механизмы устойчивости беспозвоночных к переохлаждению, устойчивости высших животных к кислородному голоданию, недостатку воды. Подробно эти вопросы освещены в соответствующих главах.
Регистрация времени картины восстановления после острого воздействия на организм широко применяется при изучении не только физиологических, но и патологических сдвигов. Сюда относятся исследования восстановления температуры тела и других функций после гипотермии, гипертермии, нарушения водного баланса, недостатка кислорода после интенсивной мышечной работы и т. д. Все эти сдвиги могут рассматриваться как образование известного «долга» в организме при таком нарушении гомеостазиса. Время и характер кривой погашения такого долга учитываются как критерий устой
25
чивости организма к воздействию данного фактора или к выполнению данного вида деятельности в определенных условиях среды.
В экологии, а отчасти и в экологической физиологии критерием устойчивости организма считается время выживания в экстремальных условиях среды. Так, время выживания при недостатке кислорода служит одним из важнейших приемов изучения «высотного потолка», т. е. установление устойчивости к пониженному барометрическому давлению (см. гл. VI). Время выживания, конечно, не позволяет дать физиологический анализ наблюдаемых здесь проявлений устойчивости, но является итоговым критерием оценки всех физиологических процессов организма и отчасти отражает его морфологические особенности.
Большое место в изучении устойчивости организмов, а главное их способности избегать неблагоприятных факторов среды занимает изучение отталкивающих (ге-pellants) и привлекающих (attractants) агентов, которые изменяют двигательное поведение животных. К этим факторам следует отнести не только различные химические агенты, искусственно синтезируемые человеком, но и ряд биологически активных веществ, раздражающих отдельно анализаторы и направляющих поведение животных (Киршенблат, 1968).
ЭКОЛОГО-ФИЗИОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
человека
Несмотря на то, что жизнь современного человека в значительной мере ограждена от воздействия крайних и неблагоприятных природных факторов, проблема его адаптации в разных климатических условиях стоит очень остро. Если для дикой фауны распространение и численность животных определяется экологическими факторами, с одной стороны, и адекватностью реакций организма на эти факторы — с другой, то человек благодаря своей деятельности населяет территории, не только совместимые с его жизнью, но и несовместимые. Завоевание Арктики и Антарктиды, космические полеты — все это многочисленные примеры нормального существования человека в экстремальных условиях среды, никак не совместимой с жизнью.
26
Однако повседневная жизнь и трудовая деятельность человека не могут быть изолированы от влияния природных факторов среды как климатических, так и условий питания, обеспечения микроэлементами, витаминами и т. д.
Поэтому за последние десятилетия вслед за интенсивным хозяйственным освоением ранее безлюдных районов земного шара наблюдается повышенный интерес к особенностям физиологии человека в различных экологических условиях. Кроме того, природные факторы среды широко используются для профилактики и лечения различных заболеваний, и это смыкает интересы врачей-курортологов и климатологов с интересами гигиенистов при проектировании новых городов и поселений, курортного строительства и т. д.
Эколого-физиологические исследования па человеке обычно проводятся в следующих направлениях.
Наиболее распространенным приемом исследования является физиологическая характеристика аборигенного и приезжего населения данного климатического района. В этом случае целью исследования является установление «должных» показателей физиологического состояния человека в данном районе. Подобные исследования в СССР широко проводятся в условиях жаркого климата (Авазбакиева, 1958; Юнусов, 1950; Кафарова, 1967 и др.), малых, средних высот и высокогорья (Миррахи-мов, 1965, 1966; Сиротинин, 1950; Колчинская, 1965 и мн. др.), в условиях муссонов Дальнего Востока (Матюхин, 1965) и в других районах. Такое изучение идет обычно в условиях определения основного обмена и включает исследование сердца и сосудистой системы, легочного дыхания, картины красной и белой крови, температуры тела, температуры кожи и реакций терморегуляции, основного обмена и других функций.
Специальным аспектом таких исследований является изучение возрастных особенностей физиологии человека в данных условиях среды и жизни. Это особенно важно в связи с проблемой развития детей в условиях Субарктики, жаркого климата и т. д. (Юнусов, 1947; Багдаса-рова, 1965; Кафарова, 1966 и мн. др.).
Большое место в исследованиях на человеке занимает изучение суточной и сезонной динамики физиологических функций. По существу, без этих знаний трудно
27
говорить об установлении каких-либо физиологических констант. Ряд исследований посвящен проблеме переездов в широтном и долготном направлениях и применяется для экспериментального изучения сезонного ритма и вопросов акклиматизации (см. гл. IV, V, VI и VII).
Наконец, исследование физиологических изменений при переездах в новые природно-климатические условия представляет большой интерес для климатологии и курортологии.
В настоящее время разработана международная программа исследований (International Biological Programme), которая позволит сопоставить результаты физиологических исследований па человеке в различных районах земного шара.
Важной стороной исследований на человеке является и изучение патологических явлений. Данные этого раздела пауки — географической патологии, также служат для установления пределов устойчивости человека к крайним природным факторам среды.
Совершенно еще мало изученной областью является физиология труда человека в специальных природных условиях. Несмотря на большие возможности создания искусственной среды, оптимальной для работающего человека, полное выключение природных факторов, особенно таких, как температура, барометрическое давление и отчасти световой режим, в горах и в высоких и низких широтах практически невозможно. Кроме того, в этих условиях возникают и специальные формы труда, например дальние зимние переезды, плавание на судах в тропиках, строительные работы, экспедиционные работы и зимовки и т. д. Здесь обычные формы труда человека комбинируются с подчас экстремальными факторами среды, что накладывает глубокий отпечаток на протекание физиологических процессов. Отдельные формы труда всегда отражаются в картине адаптации человека к природным факторам среды.
Глава II
О ФИЗИОЛОГИЧЕСКИХ МЕХАНИЗМАХ ПРИРОДНЫХ АДАПТАЦИИ
КЛАССИФИКАЦИЯ АДАПТАЦИИ
И ИХ ЗНАЧЕНИЕ ДЛЯ ФУНКЦИИ
РЕГУЛИРУЕМЫХ СИСТЕМ
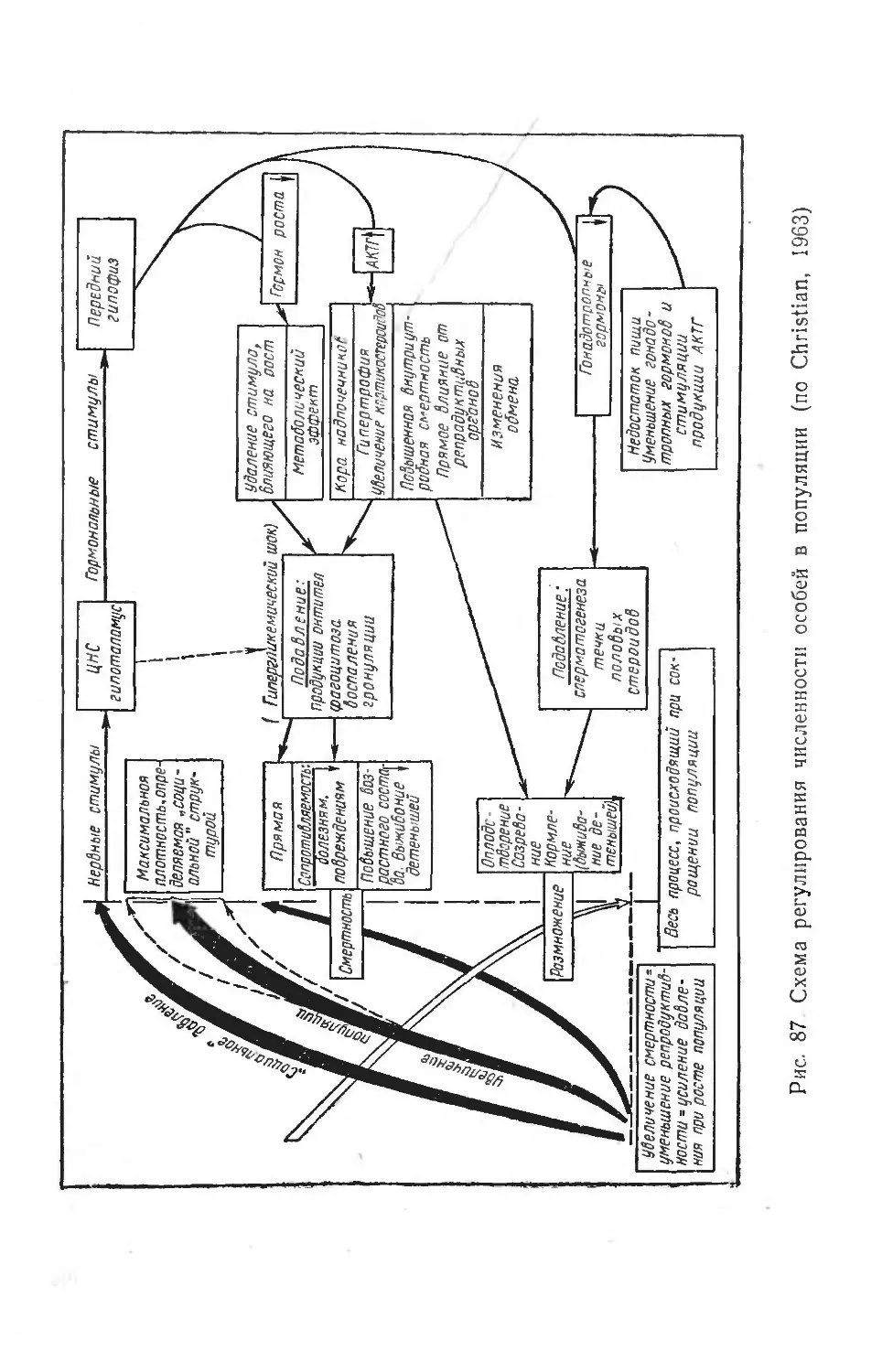
Одной из важнейших проблем современной физиологии является выявление физиологических механизмов, лежащих в основе приспособления — адаптации организма к воздействующим на него агентам внешней среды или к изменениям физиологического состояния.
Под физиологической адаптацией следует понимать совокупность физиологических особенностей, обусловливающих уравновешивание организма с постоянными или изменяющимися условиями среды. В зависимости от длительности и повторяемости этих изменений адаптации могут носить циклический характер и быть более или менее стойкими. Самый термин «адаптация» характеризует только феноменологию явления и не предполагает какого-либо объяснения механизмов, лежащих в его основе.
За последние годы предложено несколько классификаций физиологических адаптаций. Эти классификации обычно учитывают стадии развития процесса и в зависимости от этого включают вопрос о его обратимости.
Хензель и Хильдебрандт (Hensel a. Hildebrandt, 1964) предлагают классификацию адаптаций, основанную на времени воздействия на организм. Они выделяют три типа адаптаций:
1. Острые изменения регуляций функций, возникающие в ответ на внешние или внутренние сдвиги, продолжительностью от нескольких секунд до нескольких минут, а иногда и часов.
29
2. Слабые адаптивные ответы организма на изменения во внешней среде; они включают понятия акклима-ции и акклиматизации. Продолжительность этих сдвигов— от часов или месяцев до нескольких лет.
3. Адаптации в эволюционном аспекте — превращение и селекция генетически адаптированных типов — чрезвычайно медленный процесс, вовлекающий ряд поколений и во времени растянутый на миллионы лет.
Этой классификацией авторы пытались заменить классификацию канадского физиолога Харта (Hart, 1955), также подразделяющую (по отношению к воздействию холода) все явления на акклимацию— острый и обратимый процесс; акклиматизацию — процесс, протекающий в течение всей индивидуальной жизни; и адаптацию — процесс, длящийся на протяжении многих поколений.
Однако эти классификации не дают анализа происхождения адаптаций в онто- и филогенезе, а главное — не предлагают отделить врожденные их элементы от приобретенных в течение индивидуальной жизни.
С целью изучения природных адаптаций (Слоним, 1962) была предложена классификация адаптаций в зависимости от их происхождения в онто- и филогенезе и от их значимости для жизни отдельной особи, популяции или вида в целом. Исходя из наличия в адаптации врожденных и наследственно закрепленных элементов, а также элементов, приобретенных в процессе индивидуального развития, все адаптационные явления было предложено разделить на три группы.
Первая группа явлений включает индивидуальные адаптации, возникающие на протяжении постнатального развития. Сюда относится формирование условных рефлексов и более сложных стереотипов, возникающих при воздействии факторов среды на взрослый организм. Эти явления могут носить несколько иной характер при воздействии в определенные, главным образом ранние этапы постэмбрионального развития (см. стр. 74). К группе индивидуальных адаптаций нужно также отнести изменения гормональных отношений (типа стресс, неспецифических явлений адаптации) и тканевых процессов. Все эти изменения в организме (особенно при относительно коротких воздействиях) практически обра
30
тимы и сравнительно легко обнаруживаются в эксперименте.
Вторая группа явлений включает видовые, наследственно закрепленные адаптации. Они обусловлены наследственно закрепленными особенностями нервной системы и гормональными и тканевыми регуляциями и в значительной мере всей динамикой морфологических изменений, возникших в процессе онтогенеза особи данного вида. Эти адаптации охватывают отдельные системы органов с характерным для каждого типа адаптаций замещением одного органа и системы другой.
Адаптивные особенности врожденных актов поведения обеспечивают у высших организмов контакт с кормящей самкой в гнездовой период развития, закономерности расселения молодняка (распад гнезда и гнездовых отношений) и т. д. Эти наследственно программированные рефлекторные акты и сложные гормональные отношения весьма специализированы и очень различаются даже у близких в таксономическом отношении видов. Адаптивное значение таких физиологических реакций при сопоставлении их с факторами среды обитания обычно не вызывает сомнений. Они составляют основной фонд знаний в области экологической физиологии.
Третья группа — популяционные адаптации возникают в процессе формирования популяции в данных конкретных условиях ее существования. Исследование этих адаптаций и динамики их образования представляет наибольший интерес для экологии в целом, так как характеризует поведение вида в разных условиях существования. Популяционные адаптации по своей генетической структуре очень сложны. Они отражают наследственные формы адаптации и накладывающиеся на них влияния среды на всех этапах как пренатального, так и постнатального развития, включая явления импринтинга (см. гл. III). Кроме того, они включают, разумеется, и все строго генетические отношения, связанные с естественным (а иногда и искусственным) отбором.
Приспособительные изменения физиологических реакций, возникающие в ответ на воздействие различных факторов среды, могут зависеть от особенностей структуры и функции клеток организма, целых систем органов и, наконец, регуляций, связанных с поддержанием общего уровня физиологических реакций животного.
31
Одной из основных особенностей адаптации как процесса, позволяющего организму продолжать существование в измененной среде, является поддержание жизнедеятельности и некоторых сторон гомеостазиса, свойственных организмам данного вида, данного уровня развития нервных и гормональных его механизмов. В соответствии с эволюционным уровнем развития животного можно говорить о разных типах адаптации, охватывающих различные уровни регулируемых систем — клеточный, тканевой, органный и уровень целостного организма. В последнем случае в процесс адаптации вовлекаются, кроме изменений собственно вегетативных функций, и изменения двигательного поведения.
Важнейшими адаптациями организмов к условиям среды являются термическая, осмотическая, окислительно-восстановительная и пищевая (ферментативная). Они свойственны, по существу, всем без исключения живым существам, включая и растительные организмы.
Однако по своим механизмам адаптивные изменения физиологических функций могут быть достаточно четко разграничены в зависимости от наличия у них тех или других гомеостатических механизмов. Это позволяет разделить особенности адаптации гомойо- и пойкило-термных организмов, гомойо- и пойкилоосмотических организмов, водных и наземных организмов и т. д.
Процесс эволюции живых существ на протяжении нескольких миллионов лет включал в себя «химическую эволюцию» (Prosser, 1964). За этот период организмы приобрели способность использовать высокую потенциальную энергию фосфатов в процессе обмена веществ, генетическое кодирование при помощи нуклеиновых кислот, специфические протеины в качестве катализаторов (ферментные системы), избирательную проницаемость клеточных мембран, избирательную способность к удержанию отдельных ионов (калий). Именно эти тканевые механизмы поддержания жизнедеятельности на клеточном уровне и легли в основу адаптивной эволюции организмов.
Однако уровни, на которых используются эти элементарные химические механизмы, могут быть совершенно различными. Кроме организменных уровней регуляции существуют и «надорганизменные».
32
В соответствии с зависимостью жизнедеятельности организма от данного фактора внешней среды можно различать организмы «зависимые» (conforming organisms) и «регулирующие» (regulating organisms). Лучше всего различие между «зависимыми» и «регулирующими» организмами может быть обнаружено при сопоставлении зависимости интенсивности общего обмена веществ от окружающей организм температуры. Чем выше температура среды (до известного критического предела), тем интенсивнее протекает обмен веществ у пойки-лотермного организма. Вместе с температурой среды нарастает и температура тела. Вместе с тем, при длительном воздействии высокой температуры наступает адаптация. Обмен веществ нарастает уже несколько меньше. У гомойотермных организмов па фоне постоянного, по сниженного обмена веществ наблюдается и постоянная температура тела.
Адаптация имеет место в обоих случаях, но у гомойо-термов она проявляется па уровне целостного организма (терморегуляция), а у пойкилотермпых организмов — на уровне клеточных систем.
Тканевые адаптации у млекопитающих и птиц обнаруживаются по отношению к колебаниям температуры тканей, снабжению их кислородом, содержанию воды и ионного состава, содержанию углекислоты. Кроме того, несомненно, клеточную природу имеет устойчивость определенных организмов к ядам (например, устойчивость насекомоядных к змеиному яду и др.).
Наиболее ярко выражены тканевые адаптации к понижению температуры. До настоящего времени остается загадкой, каким образом лишенные какой-либо теплоизоляции конечности морских птиц (чаек, бакланов, пингвинов и др.) не замерзают при очень низких температурах воздуха. Каким образом осуществляется тканевой обмен веществ, отдача кислорода кровью в тканевых капиллярах при температуре, близкой к 0°С, а иногда и ниже 0°С, когда все тканевые ферментативные системы оказываются инактивными, а оксигемоглобин гомойотермных организмов не способен к отщеплению кислорода даже при высоких напряжениях СО2. На многие из этих вопросов нельзя в настоящее время дать достаточно убедительный ответ, однако само исследование адаптационных изменений клеточных систем открывает широ-
2—1204
33
кис перспективы для понимания физиологического механизма экологических адаптаций животных.
КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ
У ГЮЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ
В отличие от гомойотермпых организмов, где адаптация связана с вовлечением механизмов, поддерживающих температурный гомеостазис, пойкилотермные организмы не относятся к «регулирующим» свое физиологическое состояние, а «настраиваются» на определенный уровень жизнедеятельности условиями внешней среды (Hensel a. Hildebrandt, 1964).
Тем не менее уже на тканевом уровне у многих беспозвоночных можно наблюдать клеточные сдвиги в процессе индивидуальной адаптации. Так, при воздействии низких температур среды наблюдалось снижение содержания воды в мышцах краба (Pachygrapsus crassips) одновременно с повышением потребления кислорода (Roberts, 1957). Однако в противоположность этим данным в мышцах бедра лягушки (Rana esculenta) «свободная» и «связанная» вода возрастает при адаптации к низким температурам (Stangenberg, 1955).
При адаптации к холоду наблюдается «перемещение» веществ в органах. У арктической рыбы скульпины (Myoxcephalus quadricorius) при переходе от температур -;-2° С к +20° С в мышцах снижается содержание фосфолипидов, а содержание липидов в печени возрастает. Содержание К и Na у рыб при миграции из океана в реки может изменяться в направлении, обратном изменению содержания этих элементов в среде.
Проссер и сотрудники (Prosser a. oth., 1957) указывают, что при адаптации золотой рыбки (Carassius аи-ratus) к низкому напряжению кислорода в воде наблюдается понижение окислительных процессов в мышечной ткани (от 0,157 мл О2 на 1 г сухой ткани до 0,78 мл О2).
Чрезвычайно ярко выражены клеточные и тканевые адаптации у некоторых насекомых при воздействии холода, что неразрывно связано с явлениями переохлаждения клеток и тканей.
Эти явления хорошо изучены для зимоспящих организмов, впадающих в состояние оцепенения.
34
Поскольку все членистоногие (Arthropoda) иойкило-термны и полностью подчиняются воздействию окружающей среды, выживание при температурах ниже нуля может протекать двумя путями:
1) организм выдерживает замерзание,
2) организм избегает его. Первое предполагает наличие специальных защитных свойств, которыми обладают немногие артроподы; последнее достигается переохлаждением.
Как известно, вода обычно приходит в состояние переохлаждения, степень которого зависит от ее физического состояния в данный момент и от поверхности тел, находящихся в ней или содержащих ее. Ткани членистоногих обычно охлаждаются значительно сильнее, чем аналогичная масса воды. Это зависит от ряда обстоятельств:
а) точка замерзания воды, 0°С, не имеет особого значения, поскольку жидкости тела содержат растворы, понижающие точку замерзания их до уровня ниже точки замерзания чистой воды;
б) точка замерзания организма, являющаяся наивысшей температурой, при которой любая из жидкостей тела может замерзнуть (без какого бы то ни было переохлаждения), находится несколько ниже 0°С. Разницу можно определить по концентрации и химической природе растворов. Эта температура является также точкой плавления, т. е. наивысшей температурой, при которой лед может сохранять свое состояние в системе;
в) при охлаждении ниже точки замерзания организм переохлаждается до температуры, при которой внезапно наступает замерзание. Это — точка переохлаждения;
г) кратковременное обогревание или «отдача» наблюдается, когда начинается замерзание, и вызывается освобождением тепла кристаллизации. Этот первый этап замерзания пропорционален степени переохлаждения и идеально подогревает систему почти вплоть до точки замерзания (концентрация растворов, вызываемая образованием льда, несколько понижает первоначальную точку замерзания). Достигаемая при этом наивысшая температура называется точкой «отдачи», но этот термин не имеет биологического значения. Однако он отражает некоторые физические характеристики системы. В хорошо изолированном большом количестве разбавленного
2*
35
раствора точка «отдачи» может приблизиться к точке замерзания.
Таким образом, величина переохлаждения равна разнице между точкой переохлаждения и точкой замерзания. Поскольку последняя у членистоногих обычно приближается к —1°С, величина переохлаждения и точка переохлаждения практически совпадают. Однако некоторые виды, особенно те, которые перезимовывают на открытых местах в условиях холодного климата, имеют значительную концентрацию защитных растворов, вроде глицерола, могущих понизить точку замерзания примерно на 20°С. Точка переохлаждения понижается на такую же величину.
Большинство членистоногих при замерзании погибает. Точка переохлаждения является для них минимальной летальной температурой и поэтому факторы, устанавливающие ее положение, имеют большое значение. С чего начинается замерзание и как? С понижением температуры до пределов ниже точки замерзания молекулярное строение воды становится все более близким к строению льда. Движение молекул замедляется, они сближаются и вероятность их ориентации в кристаллическую решетку льда становится все более явной. Предполагается, что вначале образуются скопления молекул, которые затем расширяются или съеживаются в ответ на различные воздействия и, в частности, на температуру. Когда такое скопление достигает критических размеров, оно превращается в ядро кристалла. Затем уже замерзание системы обусловливается ростом ядра путем наращения. Если ядра образуются только из молекул воды, ядерные образования считаются гомогенными. Однако в биологических тканях всегда имеются инородные, т. е. неводные поверхности, которые и образуют более или менее приемлемые участки для образования ядер. Ядерные образования на таких поверхностях называются гетерогенными. В общем, чем больше инородная поверхность напоминает кристаллическую решетку льда, тем более эффективной является она как ядрообразующий агент.
Неизвестно, какие вещества определяют замерзание у членистоногих, по некоторые факты известны. Все зимоспящие особи вида обычно переохлаждаются примерно одинаково, что дает основание предполагать на-
36
лпчие единого типа ядрообразующего агента. Когда появляется возможность вести наблюдение за приблизительным началом замерзания, выясняется, что оно случайно распределяется по телу, свидетельствуя о гуморальном или, иначе говоря, общем распределении. Воздействие на кутикулы различных веществ показало, что ядерные образования скорее внутреннего происхождения, а не кожного.
Большинство наземных видов членистоногих способно к значительному переохлаждению во время зимней спячки; в среднем температура понижается до —20—25° С, нередко до —30—35° С. Точка переохлаждения ниже —35° С предполагает более низкую, чем обычно, точку замерзания, так как предел переохлаждения для чистой воды находится примерно около —40° С. Зимующие личинки Bracon cephi могут охлаждаться до —47° С, но охлаждаются лишь до —30° С, поскольку у них достаточно глицерола для понижения точки замерзания на — 17° С (Salt, 1959).
КЛЕТОЧНЫЙ И ТКАНЕВОЙ УРОВНИ АДАПТАЦИЙ
У ГОМОЙОТЕРМНЫХ ОРГАНИЗМОВ
Поскольку для гомойотермпых организмов характерно поддержание постоянства внутренней среды, явления клеточных и тканевых адаптаций, казалось бы, должны занимать незначительное место в общей картине приспособления к различным факторам среды и условиям существования. Однако (как это будет показано в последующем изложении) представление о гомеостазисе как о строгом постоянстве физических и химических условий во внутренней среде организма (крови, тканях, отдельных системах органов) может служить лишь схемой, практически никогда не выявляющейся в условиях природных отношений организма. Это положение в равной мере относится к температуре отдельных частей тела, pH тканей, содержанию воды в клеточных элементах и в межклеточных пространствах, напряжению кислорода в тканевых капиллярах и т. д., это касается также и поддержания постоянства химического состава организма — процесса питания, который далеко неодинаково обеспечивает расход вещества в отдельных органах и системах.
37
Так, голодание неодинаково сказывается на весе отдельных органов и систем (Пашутин, 1902).
В последнее время изучалась возможность изменения тканевого дыхания у разных животных при голодании. Оказалось, что полное голодание вызывает не только значительное падение веса отдельных тканей, но и ограничение жизнедеятельности тканей, что находит свое
Рис. 3. Изменения тканевого дыхания при голодании (по Слони-му, 1968). I — краснощекий суслик; II— белая крыса
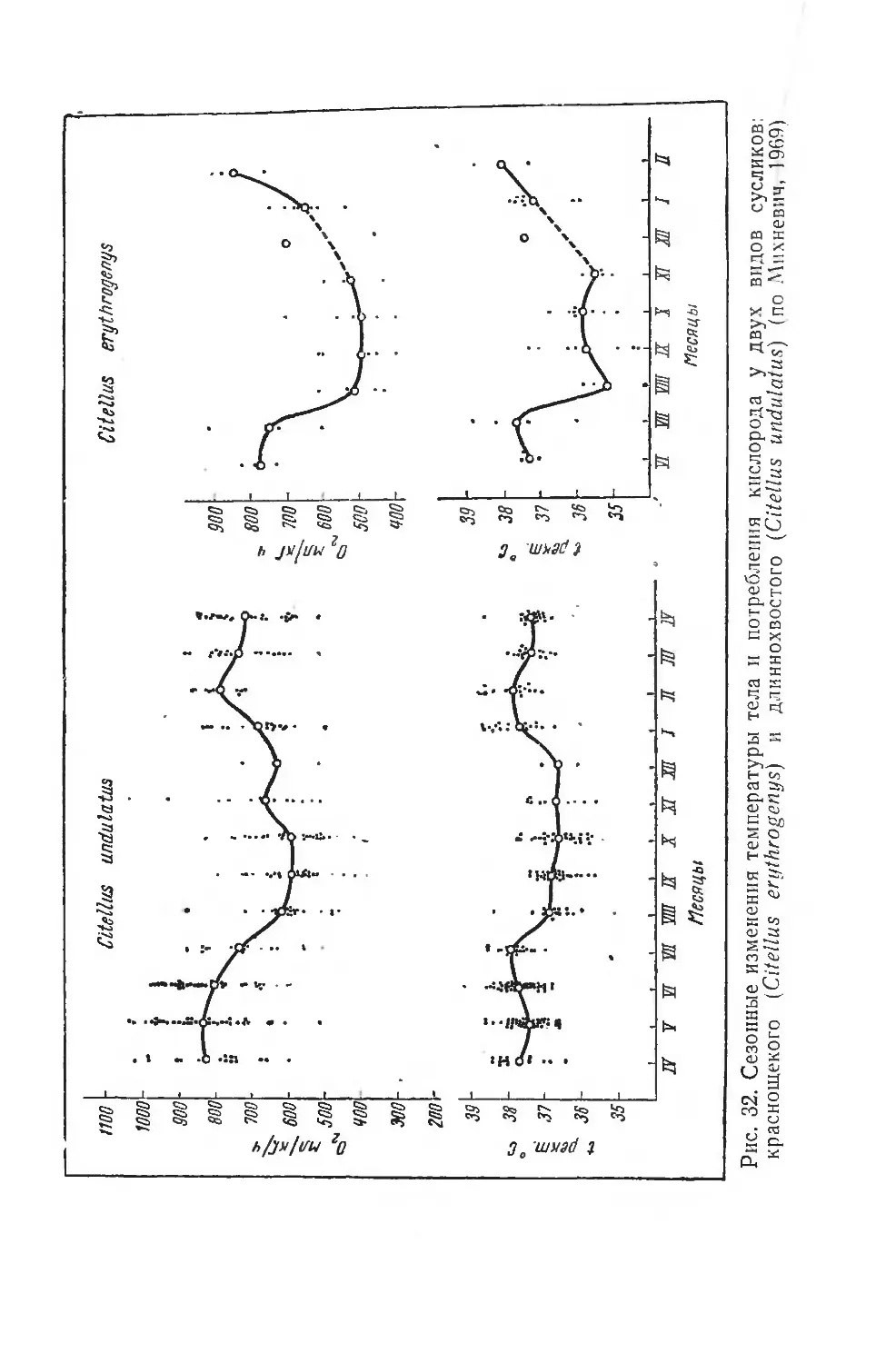
отражение в снижении их энергетического расхода. На рисунке 3 приведены некоторые данные, полученные на голодающих крысах и краснощеких сусликах.
Неоднородная реакция дыхания тканей при голодании указывает па особую тканевую специфичность и на существование специальной регуляции обмена веществ при голодании и при питании. В пользу этого говорит и тот факт, что наличие жирового депо в значительной мере сглаживает изменения тканевого дыхания при голодании.
В истории изучения клеточных адаптаций значительную роль сыграл постулат Пфлюгера (Pfluger, 1878), установившего, что количество энергии, вырабатываемой клетками тканей тела организма, не зависит от доставки к ним кислорода или пищевых веществ. Это положение Пфлюгера оказало большое влияние на последующие работы и развирало мысль о том, что потребление кисло* 38
рода клеточными изолированными элементами, а также изолированными тканями не зависит от общего уровня обмена животного. Ткани пойкилотермных и гомойотерм-ных животных (лягушка, кролик, мышь, собака) при исследовании in vitro потребляют при одинаковых температурах равное количество кислорода на 1 г веса ткани (Grate, 1928). Отсюда был сделан весьма важный вывод, что уровень общего обмена веществ определяется не свойствами самих клеток и тканей, а сложным нервно-гуморальным аппаратом регуляции их деятельности. Последующие исследования ввели значительные поправки в эти широко известные данные Графе. Изолированная мышечная ткань крыс и мышей обладает при одинаковых температурных условиях более высоким уровнем обмена веществ, нежели ткань более крупных грызунов — кроликов (Krebs, 1950; Bertalanffy a. Estvick, 1953). В этих же исследованиях обнаружилось, что в пределах одного вида изолированная мышечная ткань животных разного веса потребляет одинаковое количество кислорода. Следовательно, положения Графе оказываются справедливыми для животных разного веса в пределах одного вида, но не подтверждаются для животных разных видов с разной относительной поверхностью тела. Берталанфи и Пироцинскнй (Bertalanffy a. Pyrozynski, 1953) указывают, что внутривидовые различия в уровне основного обмена, связанные с относительной поверхностью тела, обусловливаются влиянием нервной и гуморальной систем. У разных же видов можно обнаружить заметные и притом стойкие различия тканевого дыхания, зависящие от внутриклеточных особенностей тканей.
Ряд исследований установил неодинаковую реакцию тканей тела на охлаждение и нагревание. У белых мышей наблюдалось значительное повышение потребления кислорода тканью печени после кратковременного (30-минутного) пребывания животных при низкой температуре (4-5°С), у шестинедельных лабораторных крыс наблюдается после 10 и 26-дневного пребывания при температуре 4-5° С повышение потребления кислорода всеми тканями тела, за исключением ткани головного мозга (Weiss, 1954). Когда таких же крыс содержали 5 дней при температуре 4-12° С, повышения потребления кислорода тканями не наблюдалось. Таким образом, несмотря на последующую изоляцию тканей от организма,
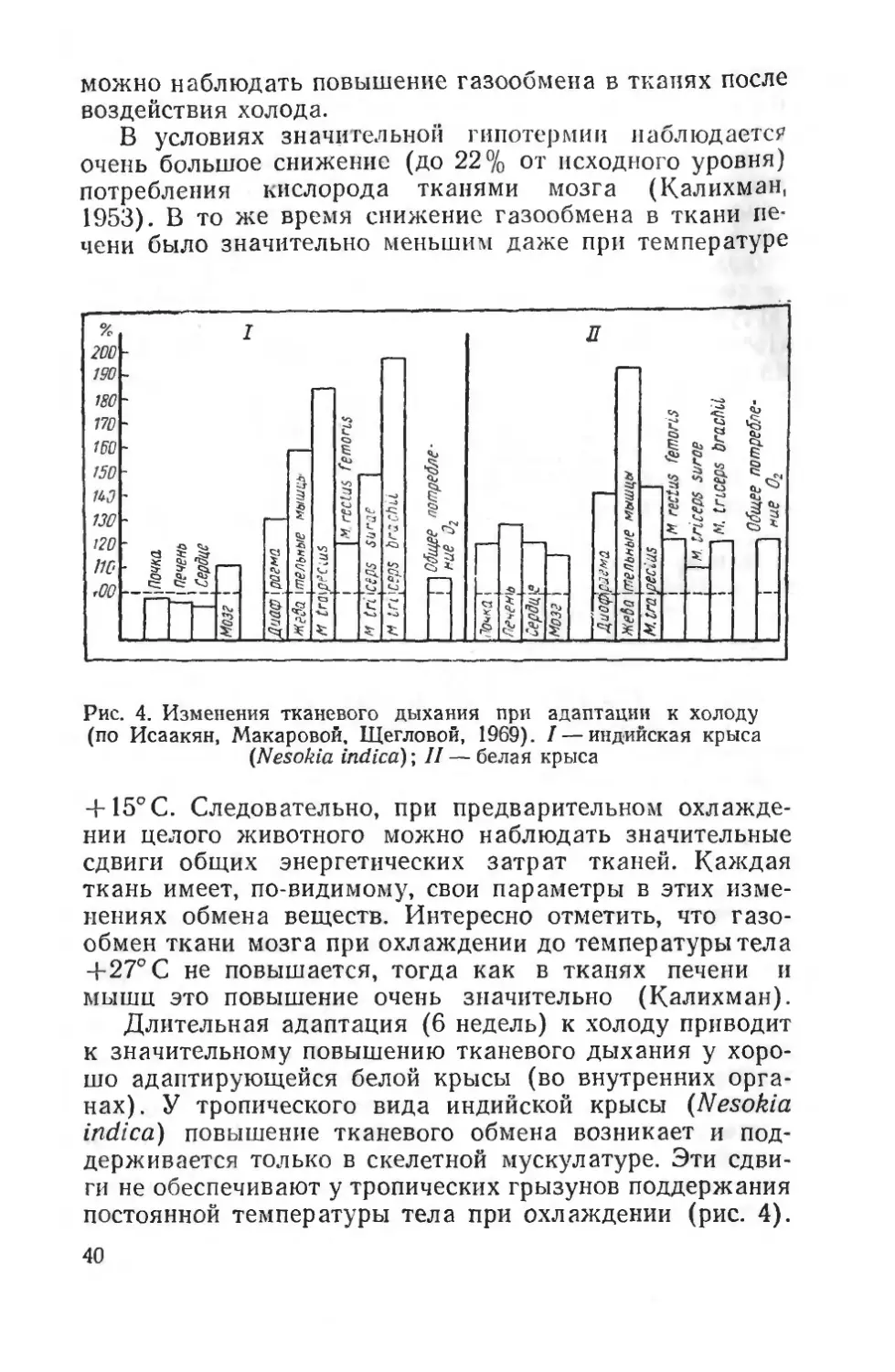
39
можно наблюдать повышение газообмена в тканях после воздействия холода.
В условиях значительной гипотермии наблюдается очень большое снижение (до 22% от исходного уровня) потребления кислорода тканями мозга (Калихман, 1953). В то же время снижение газообмена в ткани печени было значительно меньшим даже при температуре
Рис. 4. Изменения тканевого дыхания при адаптации к холоду (по Исаакян, Макаровой, Щегловой, 1969). I — индийская крыса (Nesokia indica); II — белая крыса
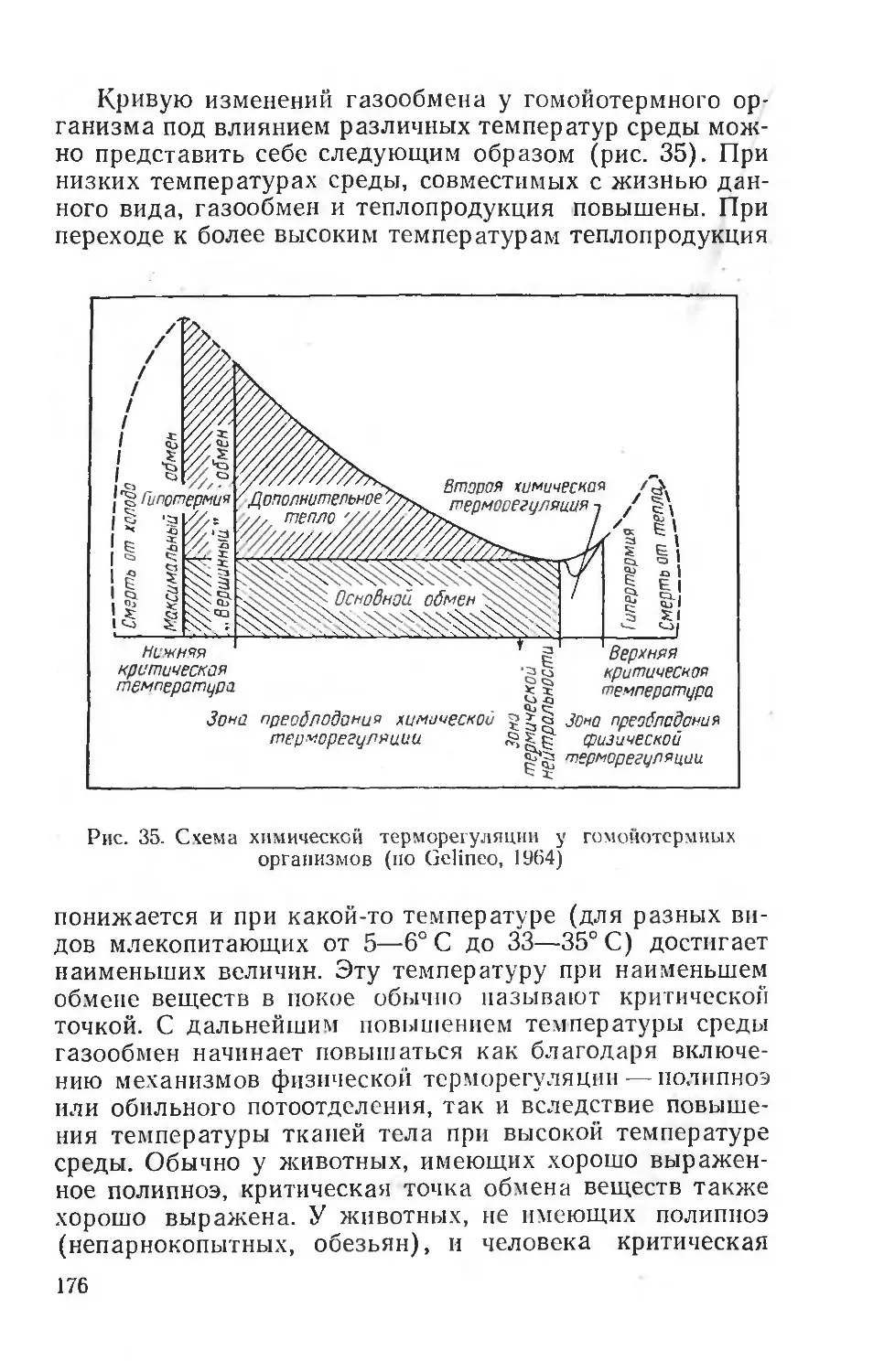
+ 15°С. Следовательно, при предварительном охлаждении целого животного можно наблюдать значительные сдвиги общих энергетических затрат тканей. Каждая ткань имеет, по-видимому, свои параметры в этих изменениях обмена веществ. Интересно отметить, что газообмен ткани мозга при охлаждении до температуры тел а 4-27° С не повышается, тогда как в тканях печени и мышц это повышение очень значительно (Калихман).
Длительная адаптация (6 недель) к холоду приводит к значительному повышению тканевого дыхания у хорошо адаптирующейся белой крысы (во внутренних органах). У тропического вида индийской крысы (Nesokia indica) повышение тканевого обмена возникает и поддерживается только в скелетной мускулатуре. Эти сдвиги не обеспечивают у тропических грызунов поддержания постоянной температуры тела при охлаждении (рис. 4).
40
Изучение реакций организма на тканевом и клеточном уровнях открывает широкие перспективы для понимания стойких изменений в организме, возникающих под влиянием факторов внешней среды. Это хорошо иллюстрируется изменениями клеточных элементов скелетной мускулатуры при воздействии холода.
Общеизвестно, что скелетная мышца по своей микроструктуре не однородное образование; отдельные ее волокна обнаруживают различную способность к длительным тоническим сокращениям, чувствительность к ацетилхолину, отличаются по содержанию миоглобина и т. д.
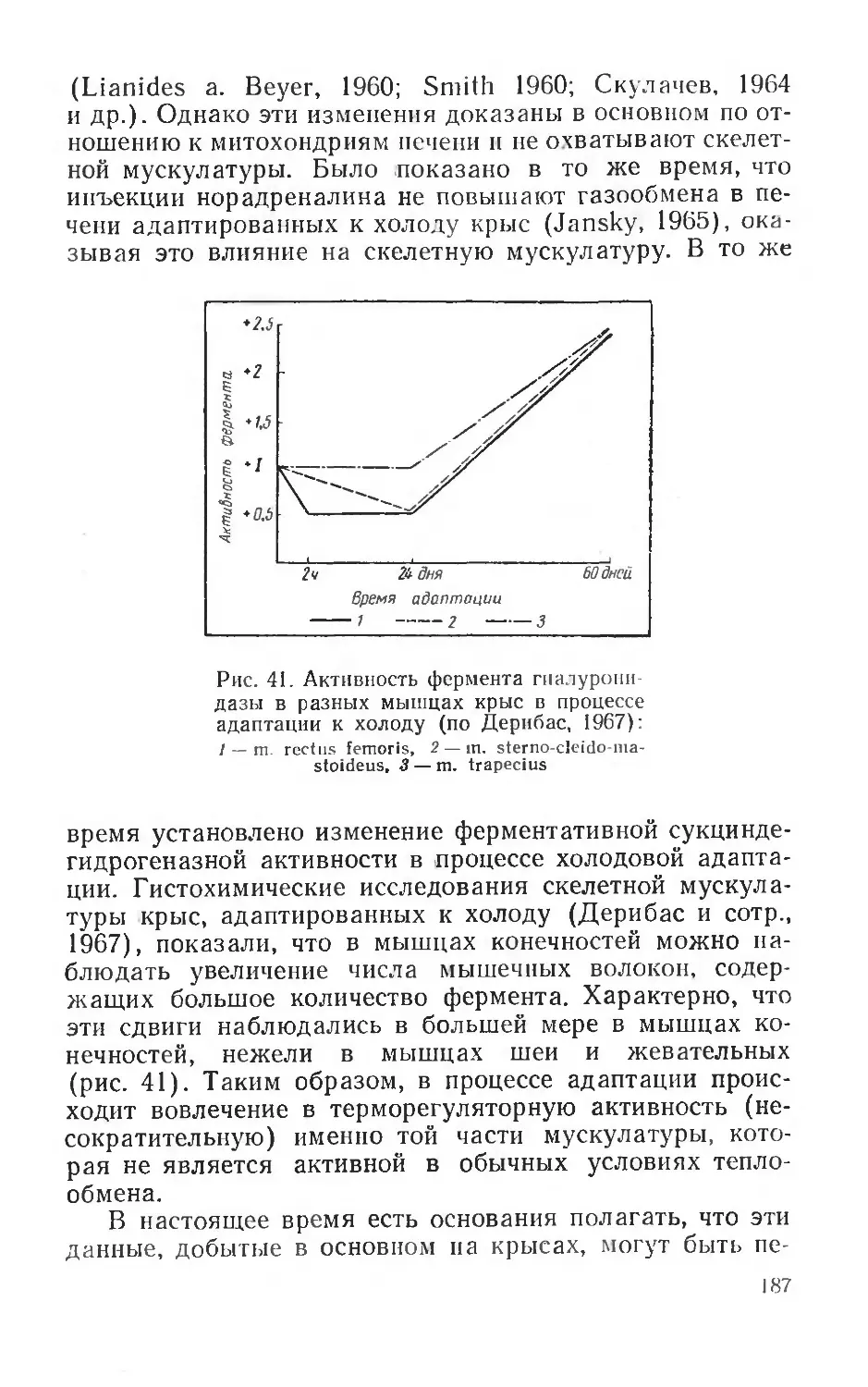
Оказалось, что сильно реагирующие при охлаждении организма мышцы — трапецевидная и мышца бедра у крыс (см. стр. 184) —не являются однородными по толщине мышечных волокон, снабжению этих волокон капиллярами и по содержанию окислительных ферментов (сукциндегидрогеназа). По содержанию окислительных ферментов мышцы состоят как бы из трех клеточных популяций. Степень васкуляризации волокон, содержащих большое количество фермента, в полтора раза выше, чем волокон, в которых фермента мало. Было выявлено и распределение волокон с различным содержанием окислительного фермента. В мышцах крысы обнаружено своеобразное, расположенное в центре мышцы ядро, интенсивно окрашенное и содержащее много фермента, и периферическая часть с малым содержанием фермента. Такое распределение волокон с разным содержанием фермента не является, однако, постоянным. Экспозиция крыс в течение 60—100 дней при температуре + 5° С привела к некоторому изменению состава клеточных популяций, количество интенсивно окрашенных волокон возросло (Дерибас, Филипченко, Шошен-ко, 1966).
Механизм тканевых адаптаций физиологических процессов можно проследить не только по изменениям общей жизнедеятельности клеточных элементов, общим выражением которой является потребление кислорода. Немаловажное значение приобретают в этом случае и отдельные стороны обмена веществ, и связанные с ним накопления отдельных потенциалов клеточных энергетических превращений. Так, например, под влиянием холода как в эксперименте на крысах (Page et Babineau,
41
1954; Pavlovic, 1954), так и у животных — обитателей полярных областей (Шварц, 1959; Ливчак, 1960) возрастает содержание гликогена в печени. Под влиянием охлаждения возрастает количество аскорбиновой кислоты у крыс и обезьян (Dugal et Therien, 1947; Dugal et Fortier, 1952). У обского лемминга и большой узкочерепной полевки (полярные виды) по сравнению со степной пеструшкой (степной вид) наблюдалось повышенное содержание аскорбиновой кислоты в тканях почки, печени (Ливчак, 1960). В этой же работе установлены прямые взаимоотношения между потреблением кислорода на 1 г ткани целого животного и содержанием аскорбиновой кислоты в ткани почки. Эти данные хорошо иллюстрируются следующей таблицей.
Таблица 1
Интенсивность потребления кислорода и содержание аскорбиновой кислоты в почках у грызунов
(по Ливчак, 1960)
Вш ЖИВ ЛНОГО П )тр?бл?ние при 163 С (мг'ч) К нцен-;ация А К в почках (мг%)
Обский лемминг (Lemmas obensis) 3,6 П,2
Узкочерепная полевка (Microtus gregalis) . 4,9 13,4
Степная пеструшка (Lagurus lagu-rus) 7,0 20,1
В условиях гомойотермии значительные колебания температуры тканей отсутствуют, однако в ряде случаев приходится считаться с непосредственным влиянием холода на тканевые процессы.
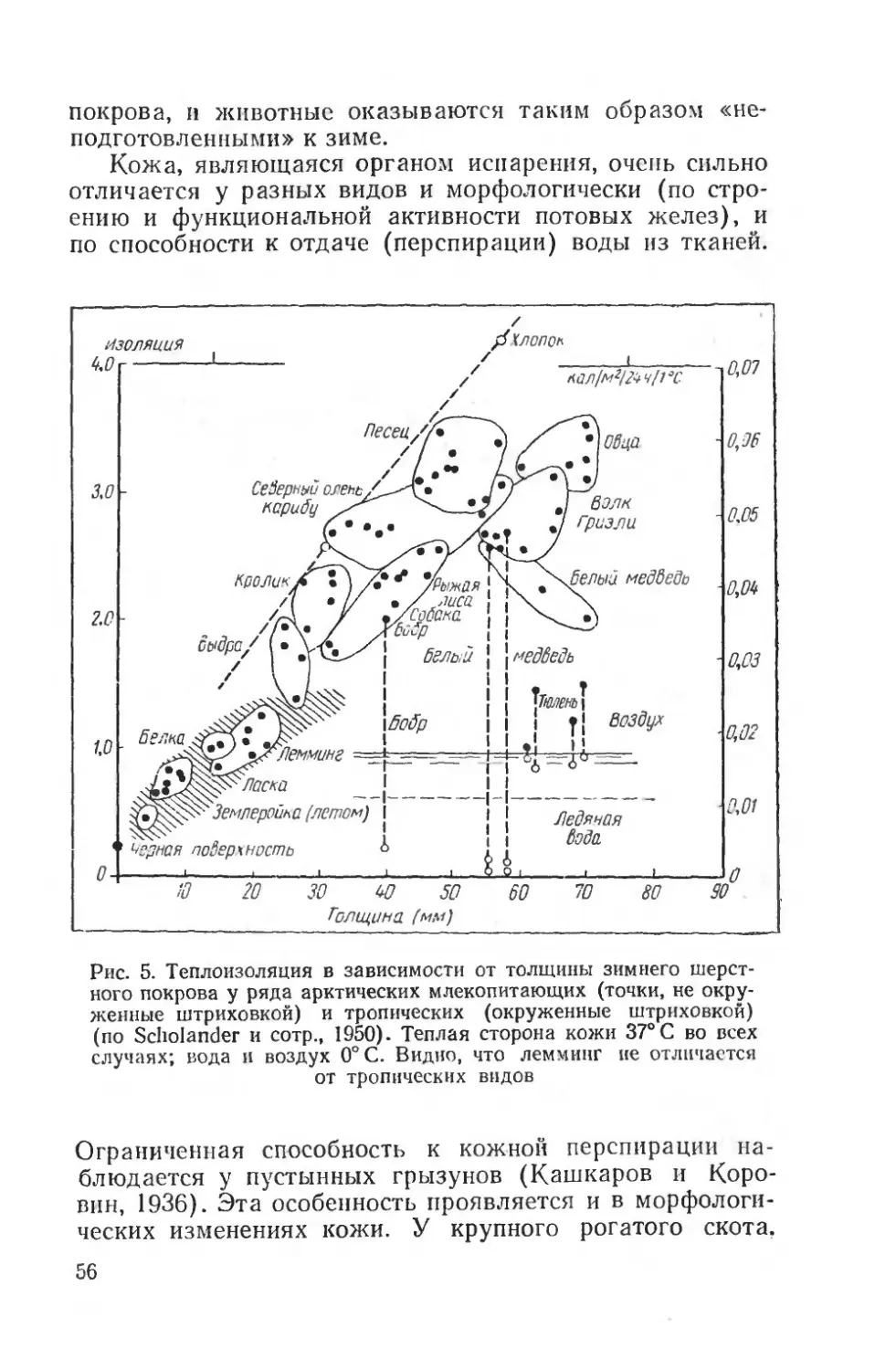
Понижение температуры конечностей имеет место у большинства полярных млекопитающих и птиц. Это находит свое отражение в интересных данных об особенностях точки плавления жира в разных частях тела полярных животных, (Irving, Schmidt-Nielsen a. Abraham-sen, 1957). Оказалось, что жир тканей внутренних органов имеет точку плавления +38—50° С, в суставных сумках стопы и пальцев — ниже +10° С (у лисицы, со
42
баки и северного оленя около 0—5°С), что совпадает с данными для зимоспящих животных. У тропических животных точка плавления жира конечностей много выше.
Имеются довольно многочисленные данные, характеризующие термоустойчивость живых тканей у пойкило-термных организмов (Ушаков, 1955, 1956). Обнаружены химические особенности белков мышечного волокна. Это доказывается различной устойчивостью мышц к нагреванию при действии аденозинтрифосфатазы. К сожалению, данных для млекопитающих и особенно для мышц в целом организме, функционирующих при различных температурах (например, для мышц конечностей или мышц туловища), пока пет.
Для свойств пищеварительных ферментов 1 были установлены как факты изменения их видовых особенностей (например, у плотоядных и растительноядных организмов), так и при длительном питании всеядных животных растительной или животной пищей (Уголев, 1958а). Это оказалось ярко выражено в особенностях ферментов, расщепляющих растительный и животные углеводы (ферментативной активности по крахмалу и по гликогену).
Весьма значительные изменения в химических процессах в тканях наблюдаются в связи с особенностями снабжения их кислородом. С этой стороны особый интерес представляют данные о содержании миоглобина, указывающие на то, что в мышцах млекопитающих, устойчивых к функциональной гипоксии (горные животные, ныряющие животные, животные, выполняющие интенсивную мышечную работу) количество миоглобина повышено (Hurtado, Rotta, Alerino, Pons, 1937; Irving, 1939; Robinson, 1939; Clark, Criscuolo, Coulson, 1952; Верболо-вич и сотр., 1955, 1956).
Миоглобин во всех случаях является веществом, способным связать дополнительное количество кислорода в самой мышечной клетке, активировать окислительные процессы с использованием кислорода в момент ограничения доставки его извне (например, при нырянии) или
1 Различия в термоустойчивости различных пищеварительных и тканевых ферментов установлены в работах П. А. Коржуева и X. С. Коштоянца (1934), П. А. Коржуева (1936), Т. И. Привольне-ва (1953), А. И. Комковой и Б. П. Ушакова (1955).
43
при интенсивной мышечной работе, когда снабжение кислородом через дыхательно-циркуляторную систему оказывается меньшим, чем кислородный запрос работающих мышц.
Однако тканевые клеточные процессы в условиях ограничения доставки кислорода изменяются не только в связи с увеличением содержания миоглобина как вещества, создающего известное депонирование кислорода. Как уже указывалось выше, постулат Пфлюгера предусматривает независимость интенсивности тканевого дыхания от содержания кислорода в окружающей! среде. Следовательно, процесс приспособления к недостатку кислорода должен приводить с этой точки зрения к более полному поддержанию уровня окислительного метаболизма независимо от условий снабжения кислородом.
Потребление кислорода кусочками тканей сердца, печени, головного мозга, диафрагмы, взятыми от акклиматизированных мышей (поднимавшихся систематически в барокамере), немного повышается в среде атмосферного воздуха и резко возрастает при снижении давления кислорода до 1,5—1,7% (Барбашова, 1960). В последнем случае потребление кислорода тканью сердца составляет 202% для мышей и 218% для крыс, почечной тканью соответственно 190 и 191%, тканью головного мозга — 212 и 219% и диафрагмой у крыс—185%- Следовательно, у акклиматизированных лабораторных крыс и мышей возрастает способность тканей поддерживать высокий уровень потребления кислорода в бедной кислородом среде. Такой высокий уровень потребления кислорода имеет место только при крайнем ограничении его доставки в ткани. Близкие данные были получены в работе Сэндстрема и Михаэлса (Sundstroem a. Michaels, 1942) и некоторых других авторов, хотя, по-видимому, для различных тканей условия, в которых поддерживается относительно повышенный при недостатке кислорода окислительный метаболизм, могут быть различны.
Имеются данные, свидетельствующие об изменениях в состоянии тканевых окислительно-восстановительных систем (цитохрома, цитохромоксндазы). Содержание цитохрома, по данным той же работы 3. Барбашовой, в мышцах далеко не всегда возрастает в процессе акклиматизации к высоте. Содержание сукциноксидазы у акклиматизированных кошек, крыс и мышей, однако, воз
44
растает по сравнению с контрольными животными. Особенно резко возрастает активность этого фермента в условиях пониженного содержания кислорода в среде, окружающей мышечную ткань. При достаточном снабжении ткани кислородом активность сукциноксидазы у акклиматизированных и неакклиматпзированных животных не отличается (Крепе, Вержбинская, Ченыкаева, Чирковская, Гавурина, 1956). Таким образом, между характером потребления кислорода в бедной кислородом среде и активностью сукциноксидазы существует прямой параллелизм, характеризующий состояние тканей животных, акклиматизированных и неакклиматизированных к пониженному барометрическому давлению.
Кроме изменений цитохромной системы под влиянием высотной акклиматизации, в тканях обнаружены сдвиги ряда других ферментных систем. Так, обнаружено увеличение желтого фермента и дегидразы (Delachaux et Tissieres, 1946), повышение активности сукциндегидроге-на п трифосфатазы (Tappan, Reynafarje, van Potter a. Hurtado, 1957). Резко возрастает содержание в тканях карбоангидразы (Вержбинская, 1946; Стрельцов и Хазен, 1946; Пигарева, 1947 и др.). Фермент карбоангидраза находится в тесной связи с интенсивностью обмена веществ. Значение его заключается в ускорении удаления СОг из тканей. Это особенно ярко выражено в хрусталике глаза, где кровеносные сосуды отсутствуют, и в сетчатке глаза, где васкуляризация выражена слабо, несмотря на интенсивный обмен веществ (Четвериков, 1948; Kauth a. Sommer, 1953).
Под влиянием повторной и длительной гипоксии может изменяться также соотношение между анаэробной и аэробной фазами метаболического процесса в тканях, особенно в мышечной ткани. Повышение анаэробного гликолиза в эмбриональных тканях развивающегося в условиях гипоксии куриного яйца наблюдала Т. Ф. Комарова (1952). Такое же явление было обнаружено в мышцах акклиматизированных к высоте животных (Бар-башова, 1960), а также после тренировки к скоростным, сопровождающимся острой кислородной недостаточностью мышечным напряжениям (Яковлев, 1950, 1955).
Повторное воздействие гипоксии, например при ане-мизации конечности, приводит к изменениям местной тканевой возбудимости (хронаксии), указывающим на
45
наличие тканевых процессов адаптации к гипоксии. Резкое падение возбудимости становится значительно менее выражении после 15-мииутной анемизации конечности (руки человека), производившейся ежедневно в течение месяца. В этом случае имеет место очень сложная картина адаптации как рецепторов тканей, так и, возможно, самих мышечных элементов.
Однако едва ли приведенные выше еще отрывочные данные в настоящее время могут исчерпать всю сложную картину тканевых адаптаций млекопитающих, возникающую под влиянием факторов внешней среды. Устойчивость целого организма к гипоксии, наблюдающаяся в результате высотной акклиматизации, устойчивость тканей животного к низким температурам, свойственная многим арктическим животным и птицам, далеко выходят за пределы значения описанных выше частных физико-химических механизмов. Следовательно, различные стороны явлений такой устойчивости, хотя и однородные по их внешним проявлениям, не могут быть объяснены известными физиологическими механизмами.
Адаптации к изменению содержания воды и осмотического давления складываются из процессов удержания внутриклеточной и межклеточной воды. Содержание воды в тканях большой песчанки представляется в следующем виде (см. табл. 2).
Таблица 2
Содержание воды в органах большой песчанки при питании растительным кормом разной влажности
(по Арешевой и Щегловой, 1961)
Влажность естественных керм о в Вес тела (г) Сердце Печень Почка Мышцы Кожа
73—76% 43-47% 90,3 93,5 81,4 79,2 74,2 73,2 79,9 79,0 78,1 77,7 64,5 67,1
При уменьшении влажности корма с 73—76% ДО 43— 47% содержание воды в тканях ряда органов — печени, почке, сердце, скелетной мускулатуре — уменьшается. В то же время в коже содержание воды несколько возрастает. Это указывает на несомненную роль последней в качестве водного депо организма.
46
Содержание воды в мышцах овец и верблюдов в условиях пустыни Каракумы претерпевает значительные сезонные изменения (Кузнецов, 1947). Так, содержание воды в феврале равнялось в среднем 76,65%, в мае — 77,65% (до 89,05%), в июне — 77,82%, в ноябре — 76,28%- Увеличение влажности корма в весенние месяцы сопровождается и возрастанием содержания воды в скелетных мышцах. Следовательно, скелетная мускулатура, составляющая значительную часть тела (по весу), является у этих животных водным депо. Его значение у разных млекопитающих в процессе приспособления к условиям недостатка воды еще неясно. Очень важен вопрос об образовании в процессе обмена веществ так называемой метаболической воды.
У сусликов обнаружено повышенное образование метаболической воды в процессе окислительного метаболизма (Ковальский, Шульгина и Ржешевский, 1951). По их данным, этот процесс может быть искусственно усилен в условиях ограниченной доставки воды в организм.
Итак, тканевые процессы гомойотермных организмов подвержены глубокой адаптации, направленной подчас на изменение ряда физико-химических клеточных механизмов, но обеспечивающей существование животного в специальных, а иногда трудных для жизни условиях среды.
Тканевые (клеточные) адаптации связаны не только с прямым воздействием факторов внешней среды, но и отражают особенности регуляции функций отдельных систем целого организма. Так, например, ткани зимоспящих животных нечувствительны к глубокому охлаждению только при выключении терморегуляции, т. е. в состоянии зимней спячки. Однако у бодрствующего животного сильное охлаждение вызывает такие же картины нарушения функций, как и у других млекопитающих.
ОРГАННЫЕ II СИСТЕМНЫЕ АДАПТАЦИИ
У ПОЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ
Органные адаптации и формирование систем органов на разных этапах эволюции настолько связаны между собой, что выявление собственно физиологических адаптаций возможно только на сравнительно небольших стадиях филогенетического развития.
47
Уровни адаптации у беспозвоночных практически те же, что и у позвоночных, но изучение их также необходимо проводить на близких в систематическом отношении видах, четко разделенных различными условиями существования.
В этом отношении хорошо изучена функция дыхания, которая у водных, наземных и переходных форм ярко отражает процессы адаптивной эволюции. Адаптивная эволюция, однако, протекает на фоне филогенетической и неразрывно с ней связана. Так, например, жаберный аппарат, обеспечивающий дыхание в водной среде, встречается у полихет, личинок насекомых, личинок хвостатых амфибий (Urodela), не говоря уже о рыбах. Сама классификация адаптивных изменений дыхания, возникших в процессе эволюции, необычайно сложна, так как дыхание осуществляется и в водной среде, и в воздушной. При этом имеет место как процесс диффузии (градиент концентрации), так и сочетание диффузии с вентиляцией, обеспечивающее интенсивный процесс диффузии при низком градиенте. Эти особенности газообмена дополняются многочисленными изменениями циркуляции крови и лимфы. Само дыхание может происходить при большем или меньшем кислородном долге.
По существу, все классы беспозвоночных животных, за исключением Coelenterata имеют своих представителей на суше. При переходе животных на сушу возникают воздушные жабры, функционирующие во влажной среде. Такое строение жабр имеют крабы, а лесные вши (Iso-poda) включают жаберные виды, адаптированные к водной, влажной и сухой среде. Классификация дыхательных образований на жабры и легкие затруднялась вследствие многих адаптивных модификаций, не имеющих специальных названий. У водных личинок насекомых Chironomus, например, существуют жаберные выросты, называемые «кровяными жабрами», так как первоначально предполагали, исходя из их строения, что они имеют дыхательную функцию. Часто они являются не адаптацией дыхания, а ионообменными (например, хлоридными) структурами и, возможно, они не должны называться «жабрами». «Книжки» получили свое название благодаря их листообразному строению и потому, что они предназначены для дыхания в воздушной среде (пауки, скорпионы). Однако среда, в которой осу
48
ществляется дыхание, не может быть использована для классификации.
Эволюция функции дыхания, охватывающая все без исключения живые системы, приобретает особое значение в связи с существованием в водной или воздушной среде. Кроме того, дыхательной функции принадлежит основная роль в поддержании гомеостазиса как в смысле обеспечения цепи биохимических превращений в организме, так и в плане поддержания деятельности специальных органов, например выделительной системы. Эволюция дыхания направлена от существования организмов в воде к существованию их в воздушной среде. На это основное направление эволюции накладываются в разных случаях определенные величины кислородного запроса и условия его удовлетворения в более богатой среде.
В водной среде дыхание осуществляется как процесс диффузии в воде, интенсивность которого обеспечивается разностью напряжения газов во внутриклеточных участках окисления и в окружающей среде. Экологически вся дыхательная среда ограничивается общими характеристиками растворения газов в толще воды. При максимальном внутриклеточном окислении интенсивность дыхания ограничивается общей массой диффузии устойчивой протоплазмы, проникающей между поверхностью клетки и окисляющими участками, и уровнем, на котором па поверхности клетки сохраняются обычные концентрации дыхательных газов, свойственных окружающей среде.
Эти концентрации лежат в основе всех дыхательных адаптаций животных — свободно существующих одноклеточных, зиготы многоклеточных или самих многоклеточных организмов.
Основная тенденция дыхательной адаптации заключается в возможно большем уменьшении устойчивой массы протоплазмы и в возможно большем увеличении восстановления поверхностной концентрации. На эволюционной шкале возрастания сложности дыхания физические адаптации идут перед химическими. На физическом уровне захват поверхности, наряду со смещением протоплазмы или смещением жидкостей тела, сталкивается с внутренними проблемами; выбор среды или смещение среды встречаются с внешними проблемами.
49
Передаточные пигменты внутриклеточного Ог и транспортирующие пигменты О2 жидкостей тела эволюционируют на химическом уровне. Все дыхание как внутреннее, так и внешнее остается в основном водным в том смысле, что О2 переходит в водный раствор до того, как пройти поверхностные мембраны любого животного.
Типичным органом, служащим для увеличения дыхательной поверхности водных организмов, являются выросты на теле — образования, функционирующие как жабры. У всех, кроме самых малых организмов, жабры требуют некоторых транспортных процессов, и первыми организмами такого рода с явно выраженными жабрами и специальным транспортом являются кольчатые черви Annelidae. На этом уровне развития дыхательный пигмент уже вполне сформирован как функциональный компонент транспортируемой жидкости. Водное диффузионное дыхание у Annelidae осуществляется наружными жабрами, что вообще характерно для животных; но у всех филогенетических образований, стоящих выше Annelidae, водное диффузионное дыхание дополняется вентиляционным дыханием, и жабры заключаются в бронхиальную камеру. Среди филогенетических образований, из которых возникли виды с воздушным дыханием, водные дыхательные системы сохранились у тех, которые обитают во влажной среде. Поверхность жабр уменьшается у видов, живущих как на суше, так и в воде, а у наземных видов строение жабр становится более жестким; водные вентиляционные механизмы уменьшаются или исчезают, а транспортные механизмы усиливают свою активность, и кожный покров становится менее проницаемым для воды и газов.
Органом увеличения дыхательной поверхности у наземных животных служит инвагинированная полость, функционирующая как легкое. У всех, кроме беспозвоночных очень малого размера, функция легкого сопровождается и транспортирующим процессом. Первым классом с явно выраженными легкими являются амфибии. Очень малое количество классов имеют виды с диффузионным дыханием в сухой среде. Это прежде всего паукообразные (Arachnida) и насекомые (Insecta). У первых сохраняется дыхательный орган водного типа в виде «книжки» или жабр. Такое диффузионное дыхание требует внутренней обменной поверхности с регули-50
руемым наружным отверстием и кожным покровом, непроницаемым для воды. У насекомых и хордовых, которые перешли от влажного типа дыхания к сухому (ксерическому), поверхность легких становится все более сложной и диффузия дополняется вентиляционным типом дыхания. Вентиляция предшествует диффузии через поры только у тех немногих видов насекомых, которые имеют наивысший обмен; именно в этом случае атмосферное диффузионное дыхание достигает наивысшего уровня специализации при отсутствии специального транспорта. Из числа хордовых у всех наземных амфибий вентиляционный тип дыхания поддерживает диффузионное дыхание и предшествует диффузии при переходе дыхания от влажного воздуха к сухому.
У личинок рыб и амфибий и у некоторых взрослых амфибий существует дыхание типа диффузии в воде через кожный покров или в сочетании с наружными жабрами. Для рыб типична непрерывная эпноическая циркуляционная вентиляция внутренних жабр; однако у видов с факультативным воздушным дыханием эпноическая вентиляция жабр иногда замещается воздушным диффузионным дыханием жабрами наряду с периодической буккофарингеальной вентиляцией pulmonate плавательного пузыря. У амфибий наблюдается последовательная замена наружных жабр, вентилируемых внутренних жабр легкими и дыханием типа атмосферной легочной вентиляции, дополняющей диффузию через кожный покров.
Сравнительное использование легких и кожного покрова для дыхания у многих видов амфибий позволяет предположить, что легкие имеют столь же важное значение для плавучести в качестве «гидростатического легкого», как и для дыхания. Вентиляционный тип дыхания использует буккофарингеальный механизм рыб и амфибий, но легочная вентиляция у амфибий носит периодический и возрастающе-убывающий характер, а эпноическая вентиляция имеет место только в назобуккофарин-геальной полости. Во время апноэ легкое отключается от атмосферы. Такой контроль за диффузионным обменом с атмосферой является существенной особенностью полностью развитого атмосферного дыхания. Преобразование легочной поверхности в альвеолы с увеличивающейся легочной васкуляризацией начинается у амфибий и ох
51
ватывает всех, включая рептилий, но васкуляризация кожного покрова соответственно уменьшается и у рептилий диффузия через кожный покров исчезает.
Явный переход от водного типа дыхания к воздушному осуществлен у амфибий. Переход от влажного дыхания ко ксерическому имеет место у рептилий. Эпноиче-ская буккофарингеальная вентиляция сохраняется у рептилий, как и у амфибий, но сама легочная вентиляция становится тораколюмбальной и у амфибий сохраняет лишь периодический характер вентиляции. Сохраняется также замкнутая легочная система, но во время легочной вентиляции буккофарингеальная вентиляция приостанавливается во время апноэ; за пределами легких какого-либо значительного дыхательного обмена не наблюдается. Только у крокодилов (Crocodilia) полное отделение системной циркуляции от легочной развивается в направлении транспорта жидкости под высоким давлением к тканям. У рептилий также происходит некоторая дифференциация при отделении участков легочной диффузии от вентиляционной емкости через проницаемые воздушные мешки.
Дальнейшее развитие альвеолярной структуры продолжается у птиц и млекопитающих. Поверхность дыхательного обмена значительно увеличивается, и легочная васкуляризация возрастает с целью вместить весь объем циркулирующей венозной крови при уровнях давления, характерных для преобладающего системного давления рептилий. Легочная вентиляция становится эпноической. Легочная система становится открытой для воздействия атмосферы, так как глотка закрывается только при глотании или при фонации, кашле и дефекации.
Альвеолярные образования вмещают объем газа, выталкиваемый из вентиляционного объема. Этот объем сообщается с объемом вентиляционного обменного газа, проходящего через поры. Его способность к разбавлению остается постоянной на таком узком диапазоне, что устойчивая дыхательная среда у позвоночных может быть обнаружена только при водном типе дыхания. Дифференциация вентиляционной способности наиболее явно заметна — структурно — в дискретных воздушных мешках у птиц. У них вентиляция самих легких является, по-видимому, циркуляционной. Такая вентиляция в отли
52
чие от вентиляции у млекопитающих обеспечивает почти атмосферную концентрацию Ог у альвеолярных отверстий. Физиологические характеристики этой системы вполне аналогичны характеристикам трахеальной вентиляции у летающих насекомых. Поэтому атмосферная циркуляция, эпноическая вентиляция системы, открывающейся в трубчатое диффузионное легкое с регулируемыми порами, является высшей дыхательной специализацией у животных.
Морфологические и механические адаптации дыхательных систем в процессе переноса сопровождаются гистологическими и химическими адаптациями. У позвоночных тенденция эволюции явно направлена в сторону специализации гемопоэза и молекулярной характеристики гемоглобина. Возрастающая специализация дыхательного обмена сопровождается увеличивающейся лабильностью и устойчивостью объемов плазмы и форменных элементов. Жизнь эритроцитов у гомойотермных организмов значительно короче, чем у пойкилотермных. У гомойотермных организмов регуляция эритроцитов более чувствительна к дыхательным переменным, что выражается в преобладающем уровне О2 в тканях. Эритроциты становятся меньше, количество их возрастает и концентрация гемоглобина в них возрастает в соответствии с увеличением обмена в дыхательных системах. Наиболее ярко адаптация к дыхательной специализации обнаруживается в химической характеристике гемоглобина. Молекулярные свойства гемоглобина наряду со свойствами внутримолекулярной среды, которая обусловливает ее деятельность, несомненно являются адаптацией для большинства организмов. Они в равной мере относятся к развитию дыхательных условий всех живых существ и отдельного животного.
ОРГАННЫЕ И СИСТЕМНЫЕ АДАПТАЦИИ
У МЛЕКОПИТАЮЩИХ И ПТИЦ
Приспособления органов и систем к разным условиям существования млекопитающих и птиц часто хорошо выражены морфологически. Густота и теплопроводимость шерстного и перьевого покровов, распределение жира, длина ушей, хвоста, конечностей, вес сердца, лег-
53
кпх, почек п т. д. могут служить наиболее яркими примерами. Морфологические и физиологические адаптации отдельных органов оказываются настолько мощными, что другие системы организма могут иногда совершенно ие изменяться в связи с приспособлением к крайним условиям среды. Таковы, например, изменения структуры почек у пустынных видов млекопитающих. Приспособительные реакции органов могут охватывать все системы, включая систему крови.
Морфологические адаптации органов пищеварения хорошо выражены, проявляются в особом строении ротовой полости, желудка, длине кишечника. В качестве физиологических адаптаций можно привести многочисленные примеры изменении активности ферментов в связи с характером питания (Уголев, 1958) и двигательной функции кишечника у близких видов грызунов, особенностей водного обмена в кишечнике у верблюда в связи с существованием в аридной зоне (жаркие пустыни). Эти системные физиологические адаптации обеспечивают существование организма в естественных для вида условиях питания, снабжения водой и т. д.
Система крови также находится в тесной связи с экологической специализацией животного. Это касается прежде всего ее дыхательной функции (содержания эритроцитов, гемоглобина, кривой диссоциации оксигемоглобина, активности угольной ангидразы). Увеличение способности крови связывать кислород и переносить его тканям наблюдается главным образом в процессе приспособления к гипоксии (к условиям пониженного барометрического давления), интенсивной мышечной работе, нырянию (Барбашова, 1941, 1960; Крепе, 1936). При частичном понижении температуры тканей (например, при очень низкой температуре конечностей) перенос кислорода кровью обеспечивается соответствующим изменением содержания гемоглобина в крови. У северного оленя в зимний период наблюдается повышенное (с 77 до 90% по Сали) содержание гемоглобина в крови. Одновременно температура тела конечностей оленя падает до 4-4— 10°С. Очевидно, высокая концентрация гемоглобина в крови обеспечивает снабжение кислородом тканей при таких исключительно низких температурах среды.
Органы выделения изменяются у млекопитающих различной экологической специализации главным образом в
54
связи с характером обмена веществ и особенностями водного обмена в частности. В этом отношении большое значение имеют морфологические особенности — соотношения между толщиной коркового и мозгового слоев почки и соответственно длиной канальцевого аппарата почки. У пустынных форм млекопитающих (грызунов) обнаруживается крайне увеличенный корковой слой с длинными канальцами, покрытыми утолщенным цилиндрическим эпителием. Этим морфологическим отличиям соответствует и большая способность к концентрации мочи, достигающая соотношений первичной мочи ко вторичной, как 1000/1 вместо 100/1 для животных — обитателей орошаемых районов (Schmidt-Nielsen а. Schmidt-Nielsen, 1952). У некоторых хищных (например, львов) наблюдаются аналогичные отношения, хотя по характеру водного обмена они не отличаются от непустынных млекопитающих (Sperber, 1944). К сожалению, с этой стороны изучены лишь немногие важные в эколого-физиологическом отношении виды млекопитающих.
У птиц органы выделения в значительной мере адаптированы к выведению повышенной концентрации солей. Особенно ярко выражены эти явления у птиц — обитателей пустынной зоны (Dawson a. Schmidt-Nielsen, 1964).
Очень большие морфологические изменения в связи с условиями обитания претерпевают у млекопитающих кожа и ее придатки. Следует отметить общую закономерность, проявляющуюся в величине шерстного покрова и его теплоизоляционных свойствах в связи с сезонами года, а главное в связи с ареалом данного вида (Калабухов, 1950; Scholander, Hock, Walters a. Irving, 1950) (рис. 5). Увеличенные теплоизолирующие свойства шерстного покрова характерны для полярных млекопитающих, а также для зимнего периода. Сезонная смена мехового покрова происходит под непосредственным влиянием режима освещения и, в частности, ритмической смены его на протяжении суток. Поэтому раннее наступление зимы или перевод (в целях акклиматизации) животных из северных районов в более южные может вызывать нарушения терморегуляции. Относительно длинный день в южных условиях не вызывает соответствующей температуре среды смены мехового
55
покрова, и животные оказываются таким образом «неподготовленными» к зиме.
Кожа, являющаяся органом испарения, очень сильно отличается у разных видов и морфологически (по строению и функциональной активности потовых желез), и по способности к отдаче (перспирации) воды из тканей.
Рис. 5. Теплоизоляция в зависимости от толщины зимнего шерстного покрова у ряда арктических млекопитающих (точки, не окруженные штриховкой) и тропических (окруженные штриховкой) (по Scholander и сотр., 1950). Теплая сторона кожи 37°С во всех случаях; вода и воздух 0°С. Видно, что лемминг не отличается от тропических видов
Ограниченная способность к кожной перспирации наблюдается у пустынных грызунов (Кашкаров и Коровин, 1936). Эта особенность проявляется и в морфологических изменениях кожи. У крупного рогатого скота.
56
акклиматизируемого в условиях жаркого сухого климата, обнаружено возрастание числа потовых желез и увеличение их функциональной активности (Раушенбах, 1952, 1958). Последняя после акклиматизации приближалась к активности, свойственной аборигенному крупному рогатому скоту Средней Азии.
Кожа и ее сосудистый аппарат как орган терморегуляции имеет у различных видов крайне специализированный характер. Так, например, поверхность кожи уха кролика и зайца чутко реагирует на изменение температуры (а следовательно, и интенсивность теплоотдачи) на мышечную работу, производимую животным. Нарушение симпатической иннервации уха не только устраняет эту тонкую регуляцию при мышечной работе, но и ограничивает продолжительность работы вследствие наступления перегревания. Если, например, интактный кролик способен пробежать в третбаие около трех часов, то после односторонней перерезки симпатического ствола в шейной области кролик прекращает работу при той же температуре тела (около +42°С) после 1,5-часового бега той же интенсивности (Склярчпк, 1963). У крыс таким органом является хвост. К сожалению, дикие виды млекопитающих с этой стороны совершенно не изучены.
У птиц перьевой покров и его теплоизоляционные свойства также сильно варьируют в зависимости от условий обитания и сезонов года (King a. Earner, 1961; Шилов, 1968).
Разнообразие в характере и интенсивности мышечной деятельности обеспечивается столь же разнообразными изменениями мышечной системы, носящими адаптивный характер. Они ярко выражены в виде большей или меньшей степени развития соответствующей мускулатуры у роющих, бегающих, прыгающих форм (Огнев, 1951; Гамбарян, 1953). Однако физиологические особенности отдельных высокоспециализированных мышечных групп почти совершенно не изучены.
Поскольку в мышечной ткани осуществляются главным образом окислительные процессы, между развитием скелетной мускулатуры, интенсивностью потребления кислорода и интенсивностью мышечной деятельности имеют место самые тесные зависимости. Особенно ярко они могут быть прослежены при рассмотрении строения и
57
функции сердечно-сосудистой системы и главным образом сердца.
Лучше других отделов кровеносной системы изучено сердце, особенно его морфология и отношение веса к общему весу тела, обычно выражаемое в % о- Строение и размеры сердца отражают не только видовые особенности мышечной деятельности, но и общий уровень обмена веществ, связанный с химической терморегуляцией (Шв арц, 1959), а также увеличенный кровоток в связи с недостатком кислорода в горах (Машковцев, 1935). Значение видовых особенностей строения сердца для характеристики различных видов млекопитающих в связи с их мышечной деятельностью установлено многими исследователями (Strohl, 1910а, 19106; Hesse, 1921; Машковцев, 1935; Шварц, 1959 и мн. др.).
Как правило, относительные размеры и вес сердца уменьшаются с увеличением размеров тела (внутри одного вида). Многочисленные примеры этого явления приведены в цитированной выше книге С. С. Шварца. Что касается изменений веса сердца в связи с природными климатическими условиями, то на основании имеющегося сейчас уже большого материала, приводимого в книге Шварца, и сопоставления его с данными по химической терморегуляции у палеарктических, субарктических и арктических видов (Слоним, 1952; Быков и Слоним, 1960) имеется полная зависимость между интенсивностью химической терморегуляции и относительным весом сердца. У палеарктических и субарктических видов лучше всего развита химическая терморегуляция, а размеры сердца увеличены. У арктических видов размеры сердца могут не отличаться от южных форм.
Легочное дыхание, как и кровообращение, находится в тесной зависимости от интенсивности окислительных процессов, от особенностей содержания кислорода в окружающей среде. Уже давно установлено увеличение размеров грудной клетки у человека в горах (Forbes, 1870; Barcroft, 1925). Увеличенные размеры легких, больший диаметр бронхов и сильно развитая бронхиальная мускулатура характеризуют породы овец и коз, разводимых в условиях гор Заилийского Алатау (Лебедева, 1954). У земноводных, обитающих в горах, наблюдается большее развитие легочного дыхания, нежели у видов, обитающих на равнине (Шварц, 1959).
58
НЕРВНЫЕ II ГОРМОНАЛЬНЫЕ МЕХАНИЗМЫ АДАПТАЦИИ
Огромное значение в формировании реакций животных на внешние природные факторы среды имеют нервные и гормональные механизмы. Но если нервные механизмы сравнительно хорошо изучены как в аспекте исследования видовых особенностей поведения, так и в аспекте исследования регуляций физиологических функций, то гормональным механизмам уделялось до сего времени очень небольшое внимание. Тем не менее имеется некоторый материал, характеризующий специально гормональные адаптации, разумеется, тесно связанные с особенностями нервной системы.
По существу нервные и гормональные особенности организмов следует рассматривать как высший уровень физиологических интеграций, суммирующих клеточные и тканевые приспособления с органными и создающих ту характерную для вида совокупность физиологических регуляций, которая и составляет предмет эколого-физиологических исследований.
Величина и структура многих эндокринных желез находится в прямой связи с величиной тела. Это касается таких органов, как щитовидная железа, надпочечники, инсулярный аппарат поджелудочной железы, гипофиз (Oboussier, 1948; Padour, 1950; Erlant, 1951). Под влиянием низких температур у крольчат увеличивается вес сердца и резко возрастает вес надпочечников. При этом гипертрофии подвергается как корковый, так и мозговой их слои.
У полевок в зимний период наблюдается значительное возрастание размеров щитовидной железы и надпочечников (Шварц, 1959). Однако это явление неодинаково выражено у Microtus gregalis и М. Middendorffi. У первого вида щитовидная железа значительно отстает в размерах при резком увеличении надпочечников.
По-видимому, различие в гормональном статусе отдельных видов обусловливается активностью адреналовой системы и содержанием в крови кортикостероидов.
Исследования Лишака, Эндреци (Lissak, Endreszi, 1960) установили определенные различия в содержании кортикостероидов в крови серой дикой и белой лабораторной крыс. Однако авторы указывают и на большие
59
индивидуальные колебания в содержании кортикальных гормонов в крови вены надпочечников.
Агрессивная реакция лактирующей самки крысы может быть устранена введением эстрогенных гормонов — эстрона (Endreszi, Lissak, Telegdi, 1957). Гидрокортизон в свою очередь снимает тормозящее действие эстрона. Авторы полагают наличие антагонистического влияния на центральную нервную систему этих эндокринных систем. По данным Лишака и его сотрудников, повреждение определенных подкорковых образований (архикор-текса) устраняет эту агрессивную реакцию лактирующей крысы. Следовательно, имеется связь между гормональным статусом и возбуждением определенных областей нервной системы. По данным той же лаборатории, после удаления аммониева рога у кошки пропадал на длительное время рефлекс охоты на мышей. Разрушение nuclei amigdalarum (Karli, 1956) устраняет у серой крысы агрессивную реакцию на белых мышей, которых интактные серые крысы убивают совершенно стереотипным образом, прокусывая брюшную стенку. Таким образом, многие характерные для вида акты поведения, иногда и достаточно сложные, обусловливаются функцией определенных центральных нервных образований и определенным гормональным статусом животного.
Имеются, хотя и немногочисленные, данные о видовой специфичности центральных аппаратов регуляции вегетативных функций. Так, например, обнаружены различия в возбудимости дыхательного центра к повышенной концентрации углекислоты во вдыхаемом воздухе у овец гибридов архаро-каракуль по сравнению с романовскими и каракульскими овцами (К- П. Иванов, А. Р. Макарова и Д. А. Рашевская). В то же время реакция на недостаток кислорода во вдыхаемом воздухе у всех сравниваемых животных была однотипной. Следовательно, имеется видовая специфика возбудимости дыхательного центра, вероятно, связанная с особенностями мышечной деятельности животных.
С другой стороны, нельзя не отметить фактов, когда введение определенных гормональных веществ в организм не оказывает влияния на характерные черты его поведения. Так, введение адреналина не изменяет замедленной, характерной для ленивца двигательной реакции (см. гл. IX).
Глава III
ВРОЖДЕННОЕ И ПРИОБРЕТЕННОЕ ПОВЕДЕНИЕ (ОСНОВЫ ЭТОЛОГИИ)
АДАПТИВНОЕ ПОВЕДЕНИЕ
Самостоятельная жизнь организма и его развитие в постэмбриональный период совершенно невозможны без готовых к моменту рождения или вылупления элементов поведения различной сложности. Эти врожденные формы поведения направлены на осуществление первого контакта с кормящей самкой или кормящими родителями, сохранение контакта с молодняком себе подобных, поиски адекватных объектов питания. Кроме того, врожденное поведение включает и ряд защитных реакций, свойственных данному виду организмов и направленных на сохранение особи.
Подобно другим адаптивным физиологическим реакциям, врожденное поведение подвержено закономерной эволюции на протяжении индивидуальной жизни. Это связано с развитием мозга, гормональных систем, а также с образованием на протяжении всей жизни или определенных ее этапов многочисленных условных рефлексов, многие из которых также характерны для всех особей данного вида и отличаются различной степенью прочности. Очень часто врожденное поведение называют инстинктом, подразумевая под этим ту его часть, которая обусловлена генетически и не определяется условиями жизни организма, т. е. образованием условных рефлексов или обучением. Однако выделение собственно врожденного поведения представляет большие трудности и потому сам термин инстинкт, или инстинктивное поведение, не всегда точно отражает происхождение тех или иных актов поведения животных или даже человека. Вместе с тем сложные акты поведения проявляются не только вместе с регуляциями отдельных функций и тканевых процессов (см. гл. II), но в известной мере могут
61
замещать их. Так, например, пустынный грызун — большая песчанка (Rhombotnis opimus) при воздействии высокой температуры повышает общую двигательную деятельность и роющую в частности. Роющая деятельность замещает в данном случае недостаточную в этих условиях терморегуляцию (Щеглова и Смирнов, 1964). Подобные же явления известны для птиц с различной дальностью миграций — перелетов. У дальних мигрантов наблюдается меньшая реакция на охлаждение, чем у ближних. У оседлых птиц реакции на холод выражены резче, чем у перелетных (Benedict, 1936; King a. Farner, 1961) .^Следовательно, целый ряд поведенческих актов является в полном смысле слова адаптивным; они не только предохраняют организм от вредных влияний внешней среды, но и дополняют недостаточные для этого реакции поддержания постоянства внутренней среды — гомеостазиса.
ПОНЯТИЕ ОБ ИНСТИНКТЕ
Наибольшее эколого-физиологическое значение имеют различные формы адаптивного поведения как сформированного в процессе эволюции и носящего характер наследственных видовых признаков, так и поведения, приобретенного в процессе индивидуального опыта. Анализ этих отношений иногда представляет значительные трудности и не случайно до настоящего времени не существует признанного всеми исследователями определения термина инстинкт как характеристики видового поведения организма.
Сложность и трудность исследования этой проблемы привели к тому, что врожденные формы деятельности стали изучаться особой сформировавшейся на Западе (ФРГ, ГДР, Англии, Голландии) наукой — этологией. Этологию определяют как науку о целостном поведении, включающем как врожденные, так и приобретенные в индивидуальной жизни элементы поведения. В действительности до настоящего времени изучались главным образом врожденные акты поведения, тогда как приобретенные являлись предметом изучения высшей нервной деятельности.
И. П. Павлов считал понятие инстинкта и безусловного рефлекса синонимами. С этим можно, однако, со
62
гласиться только отчасти, так как существует ряд форм инстинктивной деятельности, проявление которых возникает как бы спонтанно (самопроизвольно), без всякого регистрируемого раздражителя, действующего из внешней или внутренней среды организма. В настоящее время есть все основания считать, что проявление отдельных врожденных форм деятельности в большей или меньшей мере может зависеть от действия внешних или внутренних стимулов. Такие формы деятельности, как игры молодняка, в значительной мере независимы от внешних раздражителей.
Этой стороне вопроса различные исследователи-этологи придают разное значение, а отсюда и довольно многочисленные попытки дать современное определение инстинкта. Приведем некоторые из этих определений: инстинктом называют не зависящие от воспитания действия животного; инстинкт есть врожденное поведение; инстинктивное поведение построено на безусловных рефлексах; инстинктивное поведение состоит из цепей рефлексов; инстинктивное поведение основано на специфических для данного вида нормах движения (эндогенные автоматизмы), а также па специфичных для вида нормах реагирования; инстинкт есть нервная интеграция внутренних элементов возбуждения и внешних элементов возбуждения под влиянием обучения. Большинство же этологов определяет инстинкт приблизительно так: инстинктивное поведение есть специфическое видовое поведение, которое должно рассматриваться с тех же позиций, что и специфическое видовое строение тела и его функции.
Это определение очень близко к определению понятия инстинкта, данного Ч. Дарвиным о том, что инстинкт есть результат определенных наследственных свойств мозга (...some inherited modification of the brain).
Наибольшие трудности представляет, как об этом упоминалось выше, возможность установить, какие раздражители и состояния организма способны вызывать проявления врожденной деятельности — инстинкта. Французский исследователь Пьерон (Pieron, 1956) предложил рассматривать следующие раздражения как способные вызывать инстинктивные ответы: условные раздражения, не оказывающие внешне никакого действия, но вызывающие у животного состояние возбуждения;
63
раздражения, непосредственно вызывающие рефлексы и частичные реакции; раздражения, непосредственно вызывающие направленное поведение; раздражители, вызывающие устранение или ускорение действия; предостерегающие или «патические» раздражители, вызывающие устранение или бегство; «разрешитепи», освобождающие специфические для данного вида инстинктивные движения; заключительные раздражения, вызывающие прекращение инстинктивного акта; комбинации раздражителей, действующих как «сигналы» или «символы».
Для определения инстинктивной формы поведения Торп (Thorpe, 1956) предлагает три критерия: эта инстинктивная деятельность должна возникать спонтанно (эндогенно), быть унаследованной и организованной внутри. Эта крайняя, противостоящая И. П. Павлову точка зрения рассматривает инстинкт не как рефлекторную деятельность, а как спонтанную в самом прямом смысле слова.
Крэг (Craig, 1918) первый ввел в понятие инстинкт представление о накоплении в организме специфической энергии, обеспечивающей протекание реакции. Поэтому разрешающая ситуация приводит к освобождению этой энергии, а само поведение приобретает направленность — аппетенцию и оканчивается завершающим актом (соп-summatory action).
Дальнейшее направление исследования акцентировало внимание на двух существенных сторонах проблемы.
Лоренц (Lorenz, 1937) ввел понятие о наследственных видовых координациях как основе инстинктивных движений. Эти движения вызываются только специфическими стимулами, соответствующими условиям жизни данного вида и «подходящими, как ключ к замку». Однако это, по мнению многих этологов, только одна сторона вопроса.
Другая связана непосредственно с накоплением в организме специфической для проявления данной формы деятельности энергии (Lorenz, 1937). Накопление этой гипотетической энергии приводит к появлению реакции даже при отсутствии внешних или внутренних стимулов или к так называемой «реакции в пустоте» (Leerlaufreak-tion). Сама структура инстинктивного поведения складывается из отдельных фаз. Первая из пих—аппетент-ное поведение. Под последним понимают неспецифиче-
64
ское состояние возбуждения организма. Затем наступает собственно инстинктивное действие, заканчивающееся «завершающим актом» (consumrnatory action). Этот акт благодаря афферентной сигнализации изменяет состояние определенных областей мозга, и инстинктивная деятельность прекращается. Она может возобновиться лишь
борьба
Преследование Кусание Угроза и тд.
потомстве
Рытье испытание материалов Сверление
Склеивание и т. д.
Зигзагообразный танец Ведет самку к гнезду Показывает вход
Дрожанцр
Оплодотворение икринок и т д
Промывка икринок
Зхрана икринок и т д.
Рис. G. Схема иерархической организации полового инстинкта трехпглой колюшки (самца) (по Tinbergen, 1952)
после накопления новой порции гипотетической специфической энергии в нервных центрах (overflow activities— переливание через край) или при воздействии специфического раздражителя из внешней или внутренней среды. Можно представить, что в процессе жизнедеятельности происходит снижение порога возбудимости отдельных центров активности и, таким образом, инстинктивная деятельность возникает в ответ на подпороговые раздражения.
Важное место в этологии занимает вопрос о центральной локализации определенных форм врожденного поведения — инстинкта.
Тинберген (Tinbergen, 1952) разработал схему взаимного торможения центров в определенной соподчинен-ности— иерархии. При этом высшие центры инстинктов тормозят низшие. На рисунке 6 показана схема иерар
3—1204
65
хической зависимости инстинкта спаривания у трехиглой колюшки (Gasterosteus aculeatus), проанализированная Тинбергеном.
СЛОЖНЫЕ ФОРМЫ ПОВЕДЕНИЯ
И ИХ ПРОИСХОЖДЕНИЕ
Этологическая терминология, основанная на представлениях о неделимости поведения и о невозможности его нейрофизиологического, нейрохимического и структурного анализа, резко отличается от физиологической, используемой в учении о высшей нервной деятельности и нейрофизиологии.
Однако при изучении сложного поведения организма, складывающегося из врожденных и приобретенных элементов, как физиологи, так и этологи обращают внимание на одни и те же явления, обозначая их разными терминами.
В таблице 3 сделано некоторое сопоставление терминов, применяемых в физиологии высшей нервной деятельности, и терминов, применяемых этологами. Разумеется, некоторые из них приводятся весьма условно, но они все же позволяют ориентироваться хотя бы в тех аспектах, в которых проводится объективное изу юнце поведения животных.
Таблица 3
Некоторые терминологические сопоставления из области высшей нервной деятельности и современной этологии
1 Термины, приня'ые в физиол гии 1 высшей нервш й деятельности Термины этологические
Доминанта Адекватный раздражитель «Натуральный рефлекс» Безусловные рефлексы Натуральные условные рефлексы Искусственные условные рефлексы Уровень возбуждения, тонус нервной системы Обратная афферентация Аппетенция Врожденный реализующий механизм Запечатлевание (импринтинг) Врожденное поведение, инстинкт Облигатное обучение Факультативное обучение Зарядка центров Реафференция
66
Многие расхождения между отдельными исследователями носят часто терминологический характер. Это касается, например, вопросов обучения и образования условных рефлексов, образования прочных условных рефлексов и явлений так называемого запечатлевания, проявлений доминанты и аппетенции, не говоря уже о терминах условнорефлекторного — «разумного» и безусловнорефлекторного — «инстинктивного» поведения. Все эти различия в терминах не имеют в своей основе глубокого расхождения в понимании явлений, и это сильно затрудняет сопоставление фактов, полученных даже в близких методических условиях. Другое дело — глубокие методологические расхождения, касающиеся проблемы так называемой «реакции в пустоте» (Leerlaufreaktion), т. е. реакции, не вызванной факторами, действующими внутри или вне организма. Эта позиция многих этологов (Lorenz, 1937, 1957, 1960; Tinbergen, 1955; Eibl-Eiblsfeldt, 1958) не может быть не признана фина-листской и здесь какие бы то пи было терминологические сопоставления, конечно, совершенно бесполезны.
Современное представление о рефлекторной деятельности, изучаемой на уровне целого организма, включает детальное изучение как форм двигательной активности и ее чередования (поведение), так и ряда вегетативных процессов, которые вместе с отдельными формами поведения составляют целостные рефлекторные акты.
Уже сразу после рождения или вылупления из яйца организм способен реагировать на определенный комплекс раздражителей. Так, зрелые искусственно вынутые из яйцевых оболочек личинки жука тополевого листогры-за (Metasoma populi) задерживаются на листьях кормовых растений — тополя, ольхи и сразу покидают листья некормовых — например, березы или жасмина. Однако и на этих некормовых растениях они задерживаются дольше, чем на искусственных предметах, например бумаге, кинопленке и т. п. (Уждавини. 1957) (рис. 7). Следовательно, существует врожденная избирательная способность (врожденное узнавание и различение) дифференцировать непищевые и пищевые объекты. Влажная поверхность бумаги привлекает этих личинок больше, чем сухая. Значит, существует и определенная градация элементов этой дифференцировки от
3*
67
более общих к более частным свойствам анализируемого субстрата.
Эти врожденные элементы «узнавания» дополняются образованием многочисленных условных рефлексов (временных связей). Значительный материал в этом направлении собран в лаборатории М. Е. Лобашова на пчелах и шелковичных червях.
Рис. 7. Схема установки для изучения передвижения личинок ио субстрату (по Уждавини, 1958): 1 — стержень для помещения личинки и субстрата, 2 — углубление стержня, 3 —- миллиметровая сетка на поверхности столика
Обширный материал, свидетельствующий о значении слияния врожденного и приобретенного поведения у птиц, собран Н. М. Промптовым (1956). Он показал, что поведение птиц и многие черты их нения в период размножения зависят не только от врожденных особенностей организма (видовые характеристики), но и от условий выкармливания своими или другими видами птиц. Особенно ярко выражены в этом отношении голосовые реакции птиц, выращенных родителями другого вида. Эти реакции являются довольно прочными, но могут изменяться на протяжении индивидуальной жизни.
У млекопитающих весьма сложные формы поведения можно наблюдать уже сразу после рождения до осуществления какого-либо контакта с факторами внешней среды. Наиболее общей реакцией, существующей практически у всех видов млекопитающих и вызывающей не только приближение новорожденного, но и пищевую реакцию, является положительная реакция на теплую поверхность.
Особенно важно отметить специфическое значение определенных факторов среды для появления отдельных 68
и притом специализированных форм поведения при экспозиции животных па разных стадиях онтогенеза. В этом случае — вне всякого образования условного рефлекса — возможно проявление чисто безусловного рефлекса, и такой адекватный раздражитель является в собственном смысле слова реализующим механизмом этологов (ange-borene auslosende Mechanismus). Так, например, у щенков, содержавшихся от рождения и до месячного возраста на молочном рационе, свойственная хищным положительная реакция на мясо появляется только на 20—21-й день после рождения, хотя мяса эти животные никогда до этого не если (рис. 8).
Другим более сложным примером может служить игровая деятельность у золотистого хомяка. Появление этой формы врожденной деятельности на определенном этапе онтогенеза не зависит от факторов внешней среды, но в то же время может усиливаться при образовании соответствующих условных рефлексов и усиливаться и тормозиться внешними температурными влияниями (рис. 9, 10). Таким образом, хотя собственно «реализатором» этой формы врожденной деятельности является только партнер по игре, предшествующие раздражения (температурные), условные рефлексы на обстановку (систематическое отсаживание в клетку для игры), время опыта несомненно имеют в этом случае немаловажное значение. Однако здесь имеет место и определенная «зарядка» центров, которая как бы отражает накопленный нервными центрами потенциал. Если представление об аккумуляции энергии центрами является в данном случае только гипотетическим и ничего не объясняющим, то само состояние повышенной возбудимости и работоспособности выполняющих эту форму деятельности центральных и периферических аппаратов не вызывает никакого сомнения.
Если характеризовать с этой стороны врожденные формы деятельности, то на основании уже многих исследований (Попугаева, 1959, 1960; Рахимов, 1958; Ужда-вини, 1955, 1958; Щеглова, 1958, 1958а; Шепелева, 1960, 1963), можно утверждать, что существуют определенные сроки постнатального развития, когда эти реакции вызываются ничтожными по силе (подпороговыми) внешними раздражениями из внешней среды нли даже возникают «спонтанно». В последнем случае нет никакой возмож-
69
1
Рис. 8. Двигательная реакция на мясо у щенков домашней собаки (по Уждавини, 1958); вверху—1-й день после рождения, в середине— 2-й день, внизу — 21-й день
Рис. 9. Изменение некоторых функций в онтогенезе у золотистых хомяков (по Понугаевой, 1968):
1 — газообмен, 2 — пульс, 3 — кислородный пульс, 4 — игровая деятельность. По оси абсцисс — возраст (дни); по левой оси ординат слева — частота пульса, справа — газообмен (О2 мл'кг ч); по правой оси ординат слева — длительность игровой деятельности (сек), справа — кислородный пульс (О2 мл'кг веса на 1 удар пульса)
70
ности установить характер «внешнего раздражения, 'вызывающего сформированную к этому периоду развития рефлекторную реакцию. Однако речь идет, конечно, только о внешних стимулах, сравнительно легко доступных регистрации и изучению. Внутренняя стимуляция, включая собственно стимуляцию, возникающую в самом
Рис. 10. Игровая деятельность золотистого хомяка (сек), (по Понугаевой, 1968) при содержании при разных температурах среды:
1 — 12° С, 2 — 25° С
нервно-мышечном приборе, остается вне поля зрения исследователя. Эта очень важная сторона вопроса до настоящего времени почти не изучена.
С этой точки зрения большой принципиальный интерес представляет положение Делла (Dell, 1958) о взаимоотношениях между анимальными и вегетативными функциями в организме. По Деллу, аппетентная стадия деятельности организма есть «превращение нужд тела в поведение»—(transformation of bodily needs into behaviour). Механизмы этого превращения выявляются в современной физиологии в виде стимулирующего действия на головной мозг восходящей части ретикулярной фор-
71
нации стволовой его части, гормональных сдвигов (кора надпочечников). Однако современные знания находятся еще на подступах к решению этой кардинальной проблемы.
Рис. II. Формы поведения у чаек (по Tinbergen, 1965). / — движения у сизой чайки при длительном призывном крике; II—V — церемония встречи у серебристой чайки (// — призывный крик, III — общий зов, IV — «давятся», V — отворачивают голову, смотрят в разные стороны); VI—VIII — стоячая лоза у серебристой чайки (VI— угроза, VII— неуверенность, VIII — страх)
72
Несмотря на то, что индивидуальные элементы поведения (благодаря образованию многочисленных условных рефлексов) могут различаться у разных особей одного и того же вида, существует четко выраженная
Рис. 12. Типичные формы поведения у красной лисицы (по Tembrock, 1964). I— нападение; II— пассивно-оборонительная реакция (тенденция к бегству); III — наложение состояний 1 и II (полное торможение двигательной активности); IV— оборонительное поведение против укуса сзади; V— наложение состояний I и IV (нападение с согнутой спиной); VI — ритуализированное поведение подчинения
специфика в формах поведения, реакциях па определенные раздражители у разных, даже близких в систематическом отношении видов. Эта называемая этологами «морфология поведения» служит важным таксономическим признаком и представляет одну из наиболее характерных черт сложных форм поведения. На рисунках 11, 12 приведены примеры такого поведения у представителей птиц и млекопитающих. Изучение этих особенностей
73
поведения у отдельных популяций одного и того же вида представляет огромный интерес для решения экологофизиологических проблем.
ЗАПЕЧАТЛЕВАНИЕ (ИМПРИНТИНГ)
Под запечатлеванием, или импринтингом, подразумевают реакции, образующиеся у млекопитающих и птиц в ранний постнатальный (чувствительный) период развития.
Эти явления были впервые описаны Лоренцом (1937) для птиц в связи с реакцией следования. Если сразу после вылупления (для зреловылупляющихся — выводковых птиц) перед молодняком продемонстрировать какой-либо одушевленный или неодушевленный предмет (птицу или ее модель, шар, горящую лампочку, контуры фигуры и т. п.), то у птенцов наступает стойкая реакция следования, продолжающаяся в течение всего периода постнатального развития (рис. 13). При этом сочетаются раздражения зрительного, слухового, температурного, химического и других анализаторов с двигательными центрами центральной нервной системы, т. е. с врожденными элементами поведения — ходьбой, следованием за удаляющимся предметом и т. д. Важной особенностью образования таких прочных рефлексов является быстрота формирования связи (однократное раздражение), а также невозможность переделки в довольно значительное время после рождения. Так, например, ягненок, двигающийся за человеком, остается совершенно неподвижным при виде удаляющейся от него овцы. Такие особенности импринтинга позволили некоторым исследователям характеризовать образующиеся в чувствительный период связи как своеобразное завершение эмбрионального развития. Действительно, следование за движущимся предметом есть врожденная реакция организма; следование за данной особью (овцой, птицей и т. д.) формируется сразу после рождения или вылупления.
Типичные формы импринтинга характерны для зре-лорождающихся животных, у которых общение с кормящей или оберегающей (для птиц) самкой не подкрепляется кормлением. Импринтинг может возникать не только на зрительные, но и на слуховые раздражения и,
74
вероятно, при раздражении обонятельного и вкусового анализаторов (Sluckin, 1961).
Чувствительный период, в течение которого возможно запечатлевапие, колеблется от 6—8 ч до 4—5 дней. Наибольшее количество исследований посвящено импринтингу у домашней птицы — цыплят, утят, гусят.
Рис. 13. Аппарат, применяемый для изучения импринтинга, состоит из беговой дорожки, по окружности которой движется манок утки (по Gess, 1959). Утенок следует за манком. Контролирующая аппаратура на переднем плане
Импринтированная реакция следования не сохраняется пожизненно, а исчезает на 70—72-й день после вылупления. К этому моменту молодняк перестает следовать за взрослой самкой.
Очень мало изучен импринтинг у млекопитающих. У них вслед за периодом молочного питания можно выделить период, характеризующийся чрезвычайно развитой пищевой ориентировочной реакцией, сроки развития и проявления которой различны для разных видов. Зрелорождающиеся животные (ягнята, козлята) уже
75
появляются с резко выраженной ориентировочной реакцией. У щенков домашней собаки эта обобщенная, генерализованная ориентировочная реакция появляется к концу первого месяца жизни. Генерализация ориентировочной реакции характеризуется тем, что на любое воздействие внешней среды животное отвечает пищевой реакцией с ярко выраженными двигательными и секреторными компонентами. В этот период, благодаря широкому общению с факторами внешней среды, формируется чрезвычайно большое количество новых натуральных условных рефлексов. Так, отползающие от гнезда молодые животные как бы знакомятся с окружающим миром путем образования многочисленных положительных и тормозных оборонительных условных рефлексов.
Однако дело не сводится только к образованию определенного комплекса — стереотипа положительных и тормозных условных рефлексов. В этот ранний период рефлексы отличаются исключительной прочностью, что может рассматриваться как явление запечатлевания. Это касается многих сторон рефлекторной деятельности: рефлексов на терморегуляцию, рефлексов, регулирующих протекание водного обмена, функций воспроизведения, оборонительных рефлексов и т. д. Если взять помет обыкновенной полевки (Microtus arvalis) и содержать молодняк при высокой температуре и ограниченном количестве влаги, то они вырастают и становятся половозрелыми животными, отличаясь, однако, малым весом тела, сниженным обменом веществ, пониженной способностью к размножению. Если затем этих животных поставить в условия благоприятного питания и снабжения водой, то восстановить вес и все те функции, которые свойственны животным, выращенным в нормальных условиях, уже нс удается. Если воздействие таких же неблагоприятных факторов производится в более поздние сроки развития, можно наблюдать такое же падение веса и обмена веществ, но эти изменения оказываются обратимыми, т. е. при помещении этих животных в благоприятные условия среды можно наблюдать полное восстановление нарушенной функции.
Таким образом, существует период, когда влияние факторов среды оказывается чрезвычайно сильным и прочным. Это сказывается на всей последующей физиологической характеристике отдельных особей и всей
76
популяции. Такая особенность развития используется в разных областях биологии, как метод так называемого направленного воспитания. Таким приемом воздействуют на развивающийся организм (например, холодный метод выращивания молодняка сельскохозяйственных животных, главным образом крупного рогатого скота). Помещение телят сразу после рождения в условия низких температур и последующее их содержание при таких температурах среды приводит к формированию стойких изменений обмена веществ, терморецепции, скорости роста и т. д. Так животноводы, исходя из этих физиологических данных, получают очень мощные средства воздействия па развивающийся организм, на физиологические его особенности в последующий период. С другой стороны, экологи, исходя из этих представлений, получают возможность оценивать динамику численности животных в природе.
Учитывая факторы среды (температурные, кормовые и другие условия на данной территории) и развивающиеся под их влиянием популяции животных, можно довольно точно предсказать состояние последующих поколений, возможности их дальнейшего размножения и теоретически обоснованно прогнозировать размножение животных, что имеет огромное практическое значение, особенно когда речь идет о таких организмах, как вредители сельского хозяйства (насекомые, грызуны и пр.).
Несмотря на столь важное значение запечатлевания для млекопитающих, оно еще мало изучено. Имеется описание запечатлевания у ягнят; первые кормления их молоком создают прочные связи с кормящей самкой или дающим корм человеком. Это описано и дтя дикого барана-архара (Слоним, 1961). У морских свинок сразу после рождения наблюдается хорошо выраженное запе-чатлевание на окраску стен ящика, где происходили роды.
Нельзя, по-видимому, ограничивать явления запечатлевания только реакцией следования и только начальным «чувствительным» (sensitive) периодом. Есть все основания считать, что в миграционной активности животных (перелетах птиц, миграциях рыб) имеют место и элементы запечатлевания. Однако с такой точки зрения эти явления еще совершенно не изучены.
Какова роль запечатлевания в развитии человека?
77
Общеизвестно, что воспитание, т. е. вся сумма педагогических и других воздействий в раннем возрасте откладывает отпечаток на всю последующую деятельность человека. Л1ногне приобретенные в детстве особенности не поддаются переделке впоследствии. По-видимому, здесь имеется общая с животными физиологическая основа — образование чрезвычайно прочных связей или запечатлевание, которые практически не удается затем изменить.
Следовательно, физиология отдельных особей при учете условий их развития в онтогенезе характеризует не только особенности данного вида, по и особенности данной популяции. Эта характеристика популяции во многом определяется историей развития отдельных особей в раннем постнатальном периоде.
РОЛЬ АНАЛИЗАТОРОВ В ФОРМИРОВАНИИ СЛОЖНОРЕФЛЕКТОРНОЙ ДЕЯТЕЛЬНОСТИ
Факторы внешней среды, воздействующие на организм изолированно или в сложном их сочетании (что всегда имеет место в природных условиях), улавливаются и оцениваются анализаторами — чрезвычайно сложной системой, трансформирующей энергию внешнего воздействия в рефлекторный ответ животного. В эколого-физиологическом аспекте интересны роль и значение анализаторов и пути их изучения не в строго ограниченных условиях эксперимента, а в сложных условиях, когда целый ряд факторов среды воздействует на организм одновременно. Обычно в эксперименте воздействуют на животное, выполняющее присущий ему род деятельности. Так, при изучении роли различных анализаторов в питании и в пищедобывательной деятельности летучей мыши бесполезно вести исследование животного, находящегося в покое. Ведь в природе летучая мышь питается на лету, и только на фоне этой деятельности можно изучать роль того или другого анализатора. Крот питается при одновременной интенсивной роющей деятельности, и т. д.
Изучение этих вопросов осложняется явлением так называемого «взаимодействия анализаторов». Например, в пищедобывательной деятельности огромную роль играет сложный комплекс химического, термического, механического и других анализаторов. Они сформирова
78
лись отчасти уже в процессе филогенеза, отчасти — в процессе онтогенеза.
Прежде всего необходимо изучить отдельные, наиболее важные постоянно действующие на организм факторы внешней среды. При анализе специфических реакции животных приходится иметь дело с неспе-пифической реакцией, которая зависит от силы раздражения и от общих свойств нервной системы (силы уравновешенности и подвижности), и с очень тонкими, сложными специализированными реакциями, свойственными лишь животным данного вида,— реакциями, которые, как было показано выше, образуются в его онто- и филогенезе. Зрительный анализатор, реагирующий на лучистую энергию видимой части спектра, имеет для организма исключительное значение. В зависимости от того, дневное, сумеречное или ночное животное, интенсивность освещения оказывает в большей или меньшей степени возбуждающее или тормозящее влияние на его деятельность, прежде всего на двигательную активность.
Хорошо известно, что если у дневных животных (а их очень много среди млекопитающих и птиц) свет повышает двигательную активность, лежащую в основе пищевой и всякой другой деятельности, то для ночных животных таким же возбуждающим фактором является затемнение, создание сумерек. Эти рефлекторные ответы, возникающие при раздражении зрительного анализатора, имеют свои особенности для каждого вида организмов и тесно связаны с его экологией.
Как развиваются эти рефлексы, как они проявляются в процессе онтогенеза и какие из них являются врожденными и приобретенными? У зрелорождающегося млекопитающего, например козленка или новорожденной обезьяны, освещение уже с самых первых часов после рождения вызывает заметное повышение общего газообмена у совершенно неподвижного животного. Это повышение невелико. Если животное после рождения оставить в условиях непрерывного круглосуточного освещения, т. е. не менять режим освещения в зависимости от состояния бодрствования и покоя, то через несколько дней можно убедиться, что смена дня и ночи не оказывает никакого влияния на газообмен. Если же родившаяся обезьянка находится в условиях нормальной смены дня и ночи, то наблюдавшийся в самые первые
79
часы жизни врожденный рефлекс на освещение увеличивается и через несколько дней затемнение вызывает резкое понижение газообмена, а освещение — резкое его повышение. По-видимому, врожденный рефлекс па освещение очень быстро дополняется условными рефлексами, связанными с активностью матери, с активностью самого детеныша в дневные часы. Так формируется хорошо выраженный у обезьяны рефлекс на дневное освещение, который охватывает функции дыхания, двигательную активность, обмен веществ, процессы выделения. Как правило, сразу же после рождения у животного наблюдаются элементы врожденного рефлекса, которые затем при наслаивании условных рефлексов создают сложнорефлекторный ответ, хорошо выраженный у более взрослых животных. Подобное явление можно наблюдать и у козлят, где повышение обмена под влиянием освещения сразу после рождения незначительно, а затем постепенно увеличивается.
Образующийся условный рефлекс может изменить значение силы воздействующего па организм раздражителя. Например, для обезьян наиболее сильным фактором, снижающим обмен в состоянии покоя, является не абсолютная темнота, а сумерки, наиболее возбуждающим фактором для птиц—-не яркий свет, который включается внезапно, а постепенный переход от темноты к свету — рассвет. Это влияние сигнальных факторов среды. Ведь именно динамика постепенного затемнения в условиях приближающейся ночи или постепенно нарастающего освещения в процессе рассвета и служит сигнальным раздражителем для предстоящего покоя или деятельности животных; силовые отношения раздражителей здесь несколько нарушаются. В этом случае никак нельзя прямо связывать абсолютную силу воздействующего раздражителя с получаемым физиологическим эффектом. К сожалению, относительно других врожденных и постепенно развивающихся реакций со зрительного анализатора сведений очень мало. Известна еще оборонительная реакция «на глубину» у новорожденной обезьяны. Обезьяна, которую поднимают над пропастью, цепляется за ручку, за барьер, за брюхо матери. Эта оборонительная реакция, никогда не подкреплявшаяся, есть врожденная реакция, возникающая при раздражении только одного зрительного анализатора.
80
С пуховой анализатор и звуковая среда представляют исключительный интерес при анализе эколого-физиологических отношений. II в этом случае имеют место как неспецифические ответы, так и высокоспециализировап-ные реакции. Если, например, воздействовать на собаку звуками различной интенсивности, то можно наблюдать характерные сдвиги обмена веществ, которые являются критерием общей жизнедеятельности организма. Характерно, что здесь ярко выражены силовые отношения, о которых речь шла выше У животного с более сильным типом нервной системы наблюдается повышение обмена веществ в ответ на воздействие раздражителей средней интенсивное!и. При увеличении силы раздражителя это повышение может смениться понижением обмена. Последнее возникает быстрее у животного с более слабой работоспособностью нервной системы. Если взять очень сильный раздражитель (трещотку), то можно получить резкое падение обмена веществ.
Чем слабее действующий раздражитель и сильнее нервная система, тем в порядке ориентировочной реакции можно получить больший эффект повышения общего обмена веществ. Эта песпецпфическая реакция характерна для всех видов животных, по, по-видимому, каждый вид и отчасти каждая особь имеют свои параметры. Поскольку речь идет об ориентировочном рефлексе, о воздействии раздражителя, с которым животное не встречалось, эта реакция, как и всякий ориентировочный рефлекс, угасает.
Кроме этого неспецифического воздействия со слухового анализатора (которое может иметься и у человека), можно наблюдать и резкую специфическую реакцию на совершенно определенный звуковой раздражитель. К таким определенным раздражителям прежде всего относятся звуки, связывающие организмы между собой. Это, выражаясь языком этологов, звуки биотического происхождения. Например, стадные звуки, которые характерны для стада копытных или обезьян. Рефлексы на такие раздражители — врожденные, безусловные. Только что родившийся ягненок немедленно отвечает на крик матери или на имитацию крика. Эта реакция не обладает свойством угасания, следовательно, ее нельзя назвать ориентировочной. Реакция собаки на лай другой собаки, как показывают очень тонкие исследования с примене
81
нием плетизмографической методики, практически неугасима.
Есть большое количество факторов звуковой среды не биотического характера, но тоже высокоспецифических для данного вида животных. Например, плеск воды оказывает влияние на сердечную деятельность и дыхание животных, которые ведут водный образ жизни. Треск сучьев вызывает неугасимую реакцию учащения сердечной деятельности у зайцев и кроликов. Очень легко образуются при этом и условные рефлексы, отличающиеся исключительной прочностью, и если они образовались, то угасить их практически невозможно (Бирюков, 1960).
Очень сложны реакции, возникающие при раздражении температурного анализатора. Во-первых, всегда существует какой-то температурный фон, на котором животное осуществляет свою деятельность, и этот фон оказывает определенное влияние на организм, создает тот уровень, на котором протекают жизненные процессы. Во-вторых, температура может воздействовать как раздражитель в случае, когда происходит суточная или сезонная смена температуры. Важно то обстоятельство, что, как и в других анализаторах, здесь имеются необычайно широкие возможности' к адаптации, а в температурном анализаторе это особенно ярко выражено. Так, длительное воздействие температуры среды приводит к тому, что животное перестает реагировать на нее, а у представителей одного и того же вида можно встретиться с совершенно одинаковым уровнем физиологических процессов и обмена веществ, дыхания, двигательной активности и других функций при самых разнообразных термических условиях. Это особенно хорошо выражено у гомойотермпых организмов и, по существу, такие механизмы позволяют животным вести действительно активное существование, в значительной мере независимое от сезонных изменений температуры среды.
Воздействие температур тесно связано с безусловной рефлекторной деятельностью, и для многих гомойотерм-ных организмов прослежено формирование ее в онтогенезе. Если дети рождаются недоношенными, то они неспособны поддерживать температуру тела, но постепенно эта реакция формируется, и при достижении нормальной физиологической зрелости наблюдается хорошо выраженная терморегуляция. Для многих птиц, млеко
82
питающих (хищников, грызунов) такое состояние при рождении нормально. У них терморегуляция и реакция на температурные раздражители развиваются уже в процессе постнатального развития. Здесь большое значение приобретает формирование многочисленных натуральных рефлексов. Эги натуральные рефлексы образуются уже с первого дня рождения, а такие факторы, как воздействие ветра, освещения, безразличные в самом раннем постнатальном периоде, становятся сигнальными факторами, дополняющими температурные воздействия на организм и вызывающими реакцию терморегуляции.
Таким образом, температурный анализатор лишь с трудом можно выделить в обычных условиях жизни взрослого организма. Здесь- термический раздражитель всегда дополняется раздражителем кожного анализатора, раздражителем зрительного анализатора, так как реакция на сложные климатические воздействия никогда не сводится к воздействию температур, а обязательно включает и элементы воздействия светового режима, ветра, влажности и других факторов.
Очень большое значение в оценке отношений животных к внешней среде имеют химические анализаторы. Обонятельный анализатор у многих видов животных является ведущим при осуществлении разнообразных форм деятельности. Из общей физиологии хорошо известно, что обоняние складывается из химического воздействия па обонятельные элементы (ольфактивпая чувствительность) и из очень сложного воздействия (менее специализированного) на окончания nervi trigemini (тригеминальная чувствительность). В последней большое значение имеет движение воздуха в носовых ходах (принюхивание) и отчасти специфическое влияние химических веществ, раздражающих рецепторы. Изучение обонятельного анализатора чрезвычайно важно для решения целого ряда вопросов. Например, только за последнее время стало понятным, каким образом сухопутные животные, например антилопа-сайгак, способны находить воду иногда за несколько десятков километров. Исследования последнего времени указывают на возможность своеобразной гидрорецепции, рецепции влажности воздуха. Очень велика роль обонятельного анализатора при пищедобывательной деятельности, особенно когда животное добывает корм, зарытый в землю. Работа обоня-
83
телыюго анализатора осуществляется здесь па фоне интенсивной мышечной деятельности, что заставляет применять ряд специальных приемов исследования.
Для изучения обонятельной рецепции существует метод ольфактомстрпи, но практика его применения сводится к изучению обоняния у человека. Эта форма исследования оказывается непригодной для животных, так как отдельные анализа горы постоянно взаимодействуют между собой. Если взять раствор сахарозы, настопько слабый, что он не создает ощущения сладкого во рту у человека, то его питье не вызывает повышения содержания сахара в крови. Но если дать человеку сосать стеклянную бусинку и запивать этим же раствором, то этот раствор вызывает гипергликемическую реакцию и воспринимается как сладкий. Здесь очень ярко видно, что при отсутствия раздражения всего ротового анализатора, в частности механорецепции, связанной с сосанием, нет ощущения сладкого и нет соответствующего рефлекторного повышения содержания сахара в крови. Онтогенетически и филогенетически эти явления имеют глубокие корни. Именно сладкое вызывает акт сосания у детей и у обезьян. Чувствительность ротового анализатора к сладкому в процессе сосания резко возрастает.
На примере обонятельного анализатора можно хорошо продемонстрировать значение условиорефлектор-ной деятельности в его формировании. В отечественной зоологической литературе проходила оживленная дискуссия по вопросу о том, какое значение имеет обонятельный анализатор для добывания пищи, выкапывания семян, орехов из земли и т. п. у грызунов. Этот вопрос очень важен с практической точки зрения. Результатом таких исследований является целый ряд мероприятий чисто агротехнического порядка. Отдельные исследователи установили важную роль обонятельного анализатора в пищедобывательной деятельности, в выкапывании грызунами семян. Другие исследователи пришли к выводу, что обонятельный анализатор не имеет никакого значения для их пищедобывательной деятельности. Оказалось, что все зависит от того, какие условные рефлексы образованы у животного. Например, если животное получало пищу на поверхности, то оно выкапывать семян и орехов не станет, даже если голодно^ а орехи зарыты
84
в землю. Переделка этого 5 же выработанного пищевого стереотипа оказывается очень трудной (рис. 14).
Велика роль обонятельного анализатора и в протекании ориентировочного рефлекса. В ответ па воздействие обонятельного раздражителя большинство животных отвечает специфической реакцией — принюхиванием. Кстати, у разных видов животных принюхивание различно.
Рис. 14. Схема установки для изучения скорости выкапывания грызунами пищевых приманок (по Радько, 1957): 1—схема расположения приманок, 2 — регистрирующие приборы, 3— экспериментальный ящик
Так, у собак наблюдается фиксированное принюхивание к отдельным предметам. Для копытных характерно поднятие головы кверху и нюхание воздуха. Это создает условия для восприятия запахов издалека, что очень важно в условиях жизни дикого стада. Оказалось, что с этими особенностями связаны различия в реакциях обмена веществ. В то время как у собак раздражение обонятельного анализатора не вызывает изменений или понижает газообмен, у копытных (например, коз) обмен повышается. Это связано с характером ориентировочной двигательной реакции, возникающей в ответ на раздражение обонятельного анализатора. У собак при этом имеет место фиксированная поза (обнюхивание), у копытных — описанное выше нюхание воздуха, что связано с мышечной деятельностью и вызывает передвижение стада с места на место.
85
Следующий комплекс анализаторов связан с важным фактором среды, вне которого вообще нельзя представить какую-либо деятельность и существование животных,—.это фактор пространственных отношений. Известно, что в оценке пространства, окружающего организм, принимает участие целый ряд анализаторов. Ведущим для большинства организмов является зрительный анализатор. Велика роль и обонятельного, слухового, а также мышечного анализаторов; с последним связано перемещение животного в пространстве. О влиянии расстояния па организм и его физиологических механизмах высказывался еще И. М. Сеченов.
Описанный выше опыт с новорожденной обезьяной, поднимаемой над пропастью, указывает, что существует уже врожденный рефлекс на пространство, который никогда не преодолевался в индивидуальном опыте. Однако этот врожденный рефлекс дополняется огромным количеством условных рефлексов. Поэтому у взрослых животных реакция на расстояние всегда высоко специфична для данного вида и даже для данной особи. Хорошей иллюстрацией этих возникающих как в фило-, так и в онтогенезе отношений может служить характеристика пищевых натуральных условных рефлексов, вызываемых раздражением на расстоянии видом пищи. Если вызывать натуральный условный рефлекс слюноотделения на раздражение видом и запахом большого и маленького куска мяса, то всегда можно обнаружить строго определенные взаимоотношения: маленький кусок мяса вызывает обильное слюноотделение, а большой—• почти никакого. Казалось бы, это противоречит основному положению о значении силы действующего на организм раздражителя. Ведь и изображение па сетчатке, если взять зрительный анализатор, и интенсивность обонятельного раздражения при большом куске мяса больше, чем при малом. Однако если обратиться к индивидуальному опыту животного, то подкрепление большими кусками мяса, как правило, не имело место, в то время как подкрепление маленькими кусками мяса для собак обычно. Отсюда понятна такая якобы парадоксальная реакция, находящаяся в обратном отношении с физическим параметром воздействия раздражителя.
На каком же расстоянии оказывает влияние пищевой раздражитель? С какого расстояния у собаки можно
86
наблюдать натуральное условнорефлекторное слюноотделение? Оказывается, если отнести мясо на 3—4 м, то никакого эффекта на слюнные железы у собаки этот раздражитель не оказывает. Начиная с 3 м и ближе, можно уже наблюдать слюноотделение, причем оно закономерно нарастает по мере приближения пищевого объекта. Если мясо положить за стеклянную перегородку, как это делал И. С. Цитович, эффект не меняется. Оказывается, существует то же отношение расстояния. Чем ближе подносится пища, тем сильнее она действует как пищевой раздражитель. Следовательно, здесь имеет место раздражение не обонятельного анализатора, а анализатора пространства. Не случайно условные рефлексы сначала назывались рефлексами на расстояние. В какой мере этот рефлекс на расстояние различен у разных животных? Решение этого вопроса позволяет понять рефлекторный механизм пространственных отношений в жизни животных. Для каждого вида можно определить то максимальное расстояние, на котором осуществляется натуральное условнорсфлекторное слюноотделение. Это относится к раздражению мясом или другой пищей:
для собак — 3 м для лисиц — 2,5—3 м
для обезьян —около 4 м для ежа — не больше 10 см
Несколько иные реакции наблюдаются у копытных. Например, у романовской овцы натуральный условный рефлекс при раздражении видом или запахом пищи (зеленой травы) как бы отсутствует. Но зато у этих овец можно наблюдать резкое слюноотделение при виде другой овцы, поедающей корм даже на расстоянии около 300 м. У овец не каракульской породы, разводимых в пустынях и полупустынях Средней Азии, наблюдается значительное натуральное условнорефлекторное слюноотделение при раздражении видом и запахом травы. Это зависит от того, что романовская овца пасется на лугах с густым травостоем, а каракульская — с крайне разреженным; отсюда разная условнорефлекторная реакция па зеленый корм как на пищевой раздражитель. Таким образом, реакция на расстояние как на раздражитель содержит и безусловные и условнорефлекторные компоненты, и эти условнорефлекторные компоненты различны у представителей разных видов, как отличаются у них и натуральные условные рефлексы.
87
Можно привести еще пример: когда собаке показывают мясо на корнцанге, ш слюны выделяется меньше, чем если мясо держат в руке. Это происходит потому, что рука человека является тем природным раздражителем, который в течение жизни собаки много раз подкреплялся приемом пищи. Известно, что щенки домашней собаки отползают очень недалеко от матери и от места, где они постоянно находятся, а лисята производят более длинные экскурсии. У щенков домашней собаки хорошо выраженная ориентировочная реакция примерно через полтора месяца после рождения сменяется резко выраженной пассивно-оборонительной реакцией. Щенок хотя и отходит от матери, от логова, по всякие внешние раздражители вызывают у пего не положительную ориентировочную реакцию, не контакт с воздействующими на него агентами, а наоборот, пассивно-оборонительную реакцию. Животное либо тут же куда-то забивается, прижимается к полу, к степе, либо быстро бросается иа свое привычное место.
Очевидно, что у животных, развивающихся в стаде, возникают совершенно иные реакции на пространство. Сразу после рождения, когда животное следует за матерью, преодолеваются большие расстояния. В этом случае стадный рефлекс является ведущим, а новорожденное животное следует за себе подобными в стаде.
Деятельность животного во внешней среде определяется не только воздействием таких раздражителей, сформированных на их основе условных рефлексов и реакций запечатлевания. Большую роль играет и образ жизни животного, отражающий как врожденные особенности, так и приобретенный стереотип поведения. Здесь большое значение приходится придавать и энергетическим взаимоотношениям, пищевым связям животного с окружающим миром.
Отдельные виды животных обладают совершенно различными уровнями интенсивности мышечной деятельности и, следовательно, разнообразными энергетическими затратами. Этим объясняется различная степень «прожорливости» того или иного животного. Как формируется разная по интенсивности мышечная деятельность? Почему отдельные формы мышечной деятельности у животных характерны для данного вида и почему проявляются в течение многих часов подряд? Кто не
88
видел в зоологическом саду, как животное (особенно хищники) ходят из угла в угол? Это повторяется на протяжении многих часов и длится годы. Поведение такого животного трудно объяснить. Если открыть клетку, животное не попытается уйти в открытую дверь. По-видимому, здесь имеют место очень сложные отношения между проявлением этой деятельности и раздражениями, которые идут из внутренней среды организма. Загадкой остается и такая форма мышечной деятельности, как игра молодых животных, которая является неизбежным этапом в жизни каждого животного.
Можно привести следующие опыты, проведенные на большой песчанке. Животное многие часы подряд в течение дня грызет деревянные бруски, причем грызение не является пищевой реакцией, потому что животное дерева не поедает. Сумма взвешенных стружек и исходных брусков совпадает, а животное совершает огромную деятельность с повышением энергетических затрат на 200% и более по сравнению с состоянием покоя. Оказалось, что такая деятельность поддерживается только в условиях определенного жизненного стереотипа—животное в течение многих часов грызет эти бруски дерева только в TOxM случае, если в кормушке есть корм: оно грызет и питается. Образуется прочный условный рефлекс на кормление с включением в стереотип грызущей деятельности, присущей грызунам. По-видимому, при воздействии экстеро- и интероцептивных раздражителей образуется такой условнорефлекторный стереотип, который поддерживает эти формы мышечной деятельности (рис. 15).
Однако разнообразие форм поведения не может быть объяснено у различных видов животных только образованием условнорефлекторного стереотипа. Поведение животных различных видов отличается довольно четкими количественно видовыми особенностями. Это особенно ярко проявляется при изучении количественной характеристики поведения в связи с процессами питания и обмена веществ.
К фактора.м внешней среды, воздействующим на животных, относится и ландшафт — совокупность физико-географических условий в широком смысле слова, который является и совокупностью воздействующих на организм физических и химических раздражителей. Таким
89
образом раскрывается физиологическая сторона понятия о ландшафте как факторе, определяющем животное население данной территории. Так, в лесу на организм воздействуют зрительные раздражители, особая влажность, своеобразие содержания кислорода и углекислоты в дневные и ночные часы, здесь целый ряд обонятельных раздражителей, связанных с растительностью и др. Весь этот комплекс раздражителей и представляет собой воздействие ландшафта на организм.
Рис. 15. Опыт изучения грызущей деятельности у большой песчанки (по Щегловой, 1958)
Лучше всего изучено влияние ландшафта на человека. Это имеет огромное практическое значение, и врачи давно научились воздействовать на здоровье человека, особенно больного с повышенной чувствительностью, тем или иным ландшафтом: степным, горным, морским и т. д.
Все эти явления представляют большой интерес при изучении поведения животных, причем для них большее значение имеет влияние не макроландшафта, а прежде всего микроландшафта. Так, для полевки микроландшафтом является копна соломы. Каждому виду животных соответствует сугубо специфическая форма микроландшафта.
90
КОЛИЧЕСТВЕННАЯ ХАРАКТЕРИСТИКА
И ЭНЕРГЕТИКА ПОВЕДЕНИЯ
Внимание к этой стороне вопроса было привлечено в связи с изучением суточного ритма двигательной активности и влияния на него различных факторов. При этом оказалось возможным изучение суммарной двигательной активности животного за определенный отрезок времени и распределение этой активности на протяжении суток (Szymanski, 1918, 1920; Щербакова, 1937, 1938, 1949; Калабухов, 1938, 1940, 1950; Черкович, 1950, 1958 и мн. др.).
Большой фактический материал позволяет утверждать, что общая суммарная двигательная активность (ходьба, бег, прыжки) и их энергетический эквивалент остаются при одинаковых условиях у отдельных особей изо дня в день постоянными. Это впервые было показано О. П. Щербаковой (19496) на обезьянах, причем удалось отметить определенные взаимоотношения между горизонтальными прыжками и вертикальными. Общая величина выполненной работы в кг]м и м/кг при одинаковых условиях исследования (температура среды, освещение) оставалась постоянной. Суммарная активность животного и ее энергетический эквивалент являются довольно постоянным признаком, характеризующим данный вид и даже данную особь. На высших обезьянах — шимпанзе и на лемурах подобные отношения в течение суток были установлены Р. С. Персон и Н. Е. Мярков-ской (1953).
Тот факт, что суммарный энергетический эквивалент мышечной работы, выполняемой животным в течение суток, остается постоянным, заставил обратиться к изучению влияния навязанных мышечных нагрузок на спонтанную деятельность животного. Первым исследованием на эту тему явилась работа М. Р. Майзелис (1953), проведенная на больших песчанках. Оказалось, что непроизвольная, даже кратковременная, мышечная работа значительно сокращает спонтанную активность животного. Это уменьшение активности может продолжаться несколько суток; наблюдается снижение газообмена и снижение веса в дни сокращенной двигательной активности.
91
На фоне общего снижения двигательной активности выявились некоторые ее особенности. В течение суток можно было наблюдать определенные циклы, характеризующие такие движения, как бег и ходьба, питание и чистка меха; чередование этих элементов оставалось неизменным. Сокращалось только их время и увеличивались промежуточные периоды покоя. Некоторые эле-
Рис. 16. Длительность игровой активности золотистых хомяков (по Склярчик, 1965). / — контроль; // — бег в течение 600 мин\ III—-бег в течение 3600 мин. Белые столбики— до бега в третбапе; заштрихованные—-после бега в третбане. Линии — размахп колебаний (минимум и максимум), столбики— средние величины:
1 — дни с большой длительностью игровой актив ности в первом исследовании. 2 — дни с меньшей длительностью игровой активности в первом исследовании
менты активности, связанные с питанием, мочеиспусканием, дефекацией, существенно не изменились. Впоследствии А. Г. Понугаева и Е. Л. Склярчик установили такие же отношения в изменении поведения после мышечных нагрузок и по отношению к циклам игры у молодых животных (золотистые хомяки) (рис. 16).
Следовательно, в поведении животного можно различать как облигатные, так и факультативные для каждого вида элементы. Облигатные элементы не могут быть изменены даже количественно (конечно, у нормальных особей данного вида); они характеризуются своеобразием не только качественным, но даже и количественным — удельным весом в общей двигательной активности
92
животного. По-видимому, пищевые рефлексы и пищедо-бываиие не могут всегда считаться облигатными элементами поведения. Это видно хотя бы из опытов В. Г. Кассиля, А. М. Уголева и В. Н. Черниговского (Черниговский, 1962), установивших, что в зависимости от концентрации раствора и его состава (глюкоза 8% и 40%-ные растворы), а также в зависимости от состояния пищевой возбудимости количество выпитого раствора меняется. При этом 40%-иый раствор глюкозы использовался как пищевое вещество. Значит, акт питья может изменяться в зависимости от состояния пищевой возбудимости и наоборот.
Рассмотрение полученного материала убеждает в том, что между временем еды и калорийностью съеденного корма существуют обратные соотношения. Чем ниже калорийность пищи, тем больше время еды. При расчете на 1 кг веса тела \ растительноядных травоядных форм поступает 0,04—0,14 кал!кг/мин, у плотоядных 1,03— 9,66 кал/кг/мин, у всеядных 1,4—5,5 кал/кг/мин. Такова энергетическая основа пищевого поведения с количественной стороны. Разумеется, эти отношения, полученные в условиях неволи, совершенно элиминируют природную пищедобывательную деятельность. Однако наблюдения в условиях пастбища свидетельствуют также об определенных отношениях времени пищедобывания у разных видов в разных условиях (Трайб, 1959; Стопим, 1962).
Таким образом, количественная сторона поведения определяется и энергетическими отношениями, отношениями поверхности тела и, по-видимому, именно эти условия определяют видовое постоянство.
Количественная характеристика поведения как общего (двигательного), так и специального (пищевого, пи-щедобывательного) в значительной мере зависит от температуры внешней среды. Это хорошо выявлено во многих исследованиях пастьбы сельскохозяйственных животных (Трайб, 1959). В лабораторных условиях изменения таких врожденных форм поведения, как роющая деятельность, грызение под влиянием температуры внешней среды, выявились в работах А. Г. Попугаевой (1960) и А. И. Щегловой (1958). Для грызущей деятельности большой песчанки, например, установлена оптимальная зона температур +20—22° С, при которых животные сгрызают наибольшее количество древесины. Высокие
93
температуры тормозят роющую деятельность у грызунов различных видов. Следовательно, как для общей двигательной активности, так и для специализированных ее форм существует определенный температурный оптимум.
Каков же физиологический механизм, лежащий в основе этого оптимума? Многие исследователи наблюдали сокращение времени пастьбы и сокращение времени всей пищедобывательной деятельности сельскохозяйственных животных. Это обычно объясняют необходимостью снизить теплопродукцию в условиях нагревания, что и достигается уменьшением приема пищи. Возможно также, что повышенная теплопродукция во время двигательной активности является фактором, повышающим температуру тела, и в порядке регуляции теплового баланса наступает разлитое торможение в центральной нервной системе, ограничивающее двигательную активность. Пъ-видимому, между температурой внешней среды и величиной двигательной активности гомойотермпого организма существуют более сложные отношения.
А. А. Алимухамедовым (1960) и А. А. Алимухамедо-вым и К. П. Ивановым (1963) было установлено, что температурная реакция проявляется ранее, нежели развивается собственно терморегуляция. Новорожденные грызуны, взятые из гнезда, где они находились с матерью, реагируют на небольшое охлаждение сначала повышением произвольной двигательной активности, а затем ее понижением. В последующие после рождения дни картина резко меняется (рис. 17). Начальная фаза повышения обмена при охлаждении все более растягивается во времени, а затем тепло, которое в первые дни и часы после рождения вызывало возбуждение животного, начинает его угнетать. Появляется реакция, свойственная взрослому гомойотермному животному,— реакция, которая входит как обязательный компонент в поддержание температуры тела (терморегуляцию) в естественных условиях жизни. Очень трудно согласиться с высказываниями некоторых исследователей (Hart, 1960; Scholander, 1960), указывающими на то, что способность к поддержанию температуры тела при охлаждении следует рассматривать как явление адаптации только в состоянии полного покоя животного. Многие гомойотерм-ные организмы практически не остаются в покое длительное время (мышевидные грызуны, землеройки). Для
94
многих видов небольшая двигательная активность является как бы фоновой. Поэтому теплообразование при мышечной деятельности, особенно при умеренной, постоянно входит как неизбежная составная часть в общий энергетический баланс организма.
С этой стороны представляет значительный интерес влияние температуры среды на так называемую спонтан-
. 30 •—*>---ц‘4* — < in .......... j w—«f-ii-i.i.нr»।•>.......
20 !•!' '»i ... -.в---------и—м---------------------i
20 м<-'|'|> "»>м||'«Ц|,4 .. «. и»..। .Im
30 —h-h—i—।------------> <------*----1 • > * H———
го i>
10
30 -ь----U-H-fj-—»--------bU—I----p-1—ft-<>)>....I--------H-++
20 ----------------------------------------'— -----------'-------—
21 30 ---------------------------------—I-----------------------------
Ю —------1------rr—-----------H*—1.....------------—-------------— * У
Рис. 17. Двигательная активность белой крысы при разных температурах среды в первые дин после рождения (ио Ллпмухамедову, 19(51)
ную деятельность молодого животного, которая хорошо известна даже стороннему наблюдателю под общим названием игры, или игровой деятельности. А. Г. Попугаевой было показано, что интенсивность игры, изучаемая количественно (время игры/час), в процессе развития имеет характерную кривую. У молодняка золотистого хомяка игра начинается на 15—18-й день, достигает максимума на 26—28-й день и резко снижается на 36-й день.
У других видов грызунов эти явления несколько более растянуты. Как показывают сопоставления данных, полученных А. Г. Понугаевой и Д. А. Рашевской, интенсивности игровой деятельности соответствуют в онтогенезе и потребление кислорода, и величины систолического и минутного объема сердца. Последние две величины опре-
95
делились как прямым путем по Фику, так и расчетом так называемого кислородного пульса (см. рис. 10).
Исследования А. Г. Попугаевой показали, что при содержании помета хомяка при температуре среды + 10— 15° С на игровую деятельность приходилось около 85% от общего времени опыта; у помета хомяков, содержавшихся при температуре +23—25° С, эта величина составляла только 65%.
Весьма важные данные получены в термоградиент-ириборе при изучении игры. «Холодные» хомячки, содержавшиеся при температуре +10—15°С, играли в температурной зоне +12—16° С. «Теплые» хомячки, содержавшиеся при +23—25° С, играли в зоне +17—34° С. Интересно, что после мышечной работы — игры как теплые, так и холодные животные располагались и засыпали в термоградиентприборе при одинаковой температуре +22—23° С. Следовательно, предварительное содержание молодняка в различных температурах не только оказывает влияние па уровень спонтанной активности, по и позволяет выявить существенные различия в термопреферен-думе у этих животных. Выполненная мышечная работа резко снижает термическую чувствительность организма (Слоним, 1952, 1962, 1962). Низкая температура среды (известный термический градиент теплоотдачи) является одним из важных факторов, стимулирующих мышечную активность. II хотя некоторые исследователи (Hardy, 1961; Hart, 1962) считают, что мышечная дрожь по своему калоригенпому эффекту более эффективна, нежели произвольные мышечные движения, она только в случаях крайне интенсивного охлаждения используется животными (едва ли кто-нибудь сознательно доведет себя до дрожи и откажется сделать несколько энергичных движений, если ему придется некоторое время пробыть на холоде). В этих условиях и человек, и животные начинают двигаться, и снижение температурной чувствительности играет, по-видимому, немаловажную роль.
Изложенные выше данные, а также специальные исследования соотношений между температурой тела и явлениями утомления при мышечной работе животных (Склярчик, 1963) позволяют высказать гипотезу о термостатической регуляции мышечной работоспособности и интенсивности двигательной активности животных. Во всяком случае, для общей количественной оценки двига-
96
тельного поведения животных температура внешней среды имеет преимущественное значение как в момент самого исследования, так и в предшествующий исследованию период (температурная адаптация).
Однако как стимуляцию, так и угнетение общей двигательной активности можно наблюдать не только при воздействии разных температур среды. А. И. Щегловой было установлено, что ограничение дачи воды и снижение содержания воды в корме резко повышает двигательную активность у пустынного грызуна большой песчанки. При этом распределение двигательной активности ее в течение суток не изменяется (песчанка — животное с дневной активностью). У ночного вида — полуденной песчанки (Pallasiomys meridianus) содержание на такой скудной ио содержанию воды пище вызывает значительное снижение активности. Такие же данные были получены А. И. Щегловой и у лабораторных крыс. Ограничение двигательной активности при сухоядении у желтого суслика наблюдала Л. Г. Филатова (1944).
Таким образом, такой казалось бы неспецифический раздражитель, как изменение осмотической концентрации крови и тканей, вызывает совершенно различные изменения поведения у животных с разной экологической специализацией. Если ограничение движений в случае с желтым сусликом является пассивной адаптацией, вызывающей экономию потери воды, то увеличение активности у дневного вида большой песчанки, наоборот, связано, вероятно, с миграцией или увеличением роющей деятельности, что приводит к добыче частей растений, содержащих воду.
Количественное изучение поведения животного, проведенное в различных условиях, может дать очень много для понимания биологической значимости отдельных его форм, а иногда и вскрыть отдельные физиологические механизмы, лежащие в его основе. Однако только одна количественная характеристика не дает возможности вскрыть ряд зависимостей поведения различных видов от их экологической специализации. В этом плане значительные услуги оказывает исследование структуры отдельных элементов поведения — рефлекторных актов, включающих как двигательные, так и вегетативные функции.
4—1204
Глава IV
ПЕРИОДИЧЕСКИЕ ИЗМЕНЕНИЯ
ФИЗИОЛОГИЧЕСКИХ ПРОЦЕССОВ В ОРГАНИЗМЕ
ОТСЧЕТ ВРЕМЕНИ В ОРГАНИЗМЕ (ФИЗИОЛОГИЧЕСКИЕ ЧАСЫ)
Одним из самых существенных факторов, оказывающих влияние на организм, является время; физиологический механизм отсчета времени чрезвычайно сложен. Этот отсчет осуществляется деятельностью каждого отдельного органа, его обменом веществ, элементами движения, секреторными циклами и т. д. По существу, каждая отдельная физиологическая система, каждый орган имеют свой отсчет времени, который может быть охарактеризован периодами покоя и деятельности.
Однако при изучении жизни целого ряда животных исследователь встречается с отсчетом времени целым организмом, причем этот отсчет может регистрировать микро- и макроинтервалы. Можно различать отсчеты времени в организме длительные, протяженные и короткие, в течение которых часто периоды покоя и деятельности чередуются.
Отсчет длительных периодов времени связан прежде всего с суточными и сезонными циклами, которые накладываются на явления индивидуального развития животных, протекающие, как было показано выше, по собственным закономерностям, также связанным подчас с еще более значительными периодами времени. Сложные или более простые комбинации этих факторов, их сочетание и определяют протекание повторяющихся физиологических реакций животных во времени и ритмику физиологического состояния организма — физиологические «часы».
Постоянно повторяющиеся физиологические изменения, протекающие во времени, можно наблюдать на любых отрезках его, начиная от микроинтервалов, связап-
98
пых с элементарными процессами возбуждения живых клеток и тканей, и кончая такими большими интервалами, как суточная и сезонная периодика. Для физиологии целого организма в экологическом аспекте наибольшее значение имеют макроинтервалы времени. Хорошо известно, что многие физиологические функции и поведение организма претерпевают строго циклические изменения, приуроченные к таким же циклическим изменениям во внешней среде. Наиболее ярко проявляется в этом смысле суточный ритм, происхождение которого несомненно связано с суточным космическим циклом, с суточными циклическими изменениями освещения, температуры, отчасти влажности и с суточными циклическими изменениями поведения других организмов. Суточный ритм широко распространен не только у высших млекопитающих, но и у беспозвоночных. Широко известно, что отсчет времени на протяжении суток имеет место у насекомых (часы лета и активности у комаров), актиний, где эта активность выражается в раскрывании щупалец во время приливов и свертывании при отливах. Последнее наблюдается даже у актиний, помещенных в аквариум с постоянным уровнем воды.
Отсчет времени представляет собой широко распространенное общее физиологическое явление. У млекопитающих и птиц оно получает яркое выражение не только в двигательном поведении, но и в глубоких изменениях физиологических функций, в частичных изменениях гомеостазиса. Так, у дневных птиц в течение суток можно наблюдать характерные фазы перехода от покоя к деятельности на рассвете и от деятельности к покою с наступлением сумерек (большая синица). Одновременно резко изменяются температура тела (на 4-3—4°С), дыхание, частота сердечных сокращений. Изменения вегетативных функций наблюдаются даже тогда, когда животное остается в полное покое. Эти факты представляют наибольший интерес для исследования, так как позволяют выявить циклические и закономерные изменения центральной регуляции физиологических функций животных в состоянии покоя.
Суточные циклы у млекопитающих протекают очень различно и в значительной мере соответствуют поведению животных, образу их жизни (дневные, ночные, сумеречные животные). Однако соотношение между дви-
4*
99
гателыюй активностью — поведением и циклическими изменениями физиологических функций у разных видов резко отличается (рис. 18).
Наиболее ярко выражена зависимость между двигательной активностью и состоянием вегетативных функций у представителей отряда летучих мышей (Chirop-tera). Многие виды палеарктических видов летучих мышей в период дневного покоя (летом в сезон активности) являются пойкилотермными организмами с температурой тела, почти равной температуре окружающей среды, с редким пульсом, редким дыханием и с очень низкой возбудимостью отдельных анализаторов (рис. 19 и 20). Такое животное не в состоянии быстро реагировать. Например, в ответ на раздражение пучком света, направленным на нее, летучая мышь улетает не сразу, а после длительного периода разогревания, во время которого резко увеличивается обмен веществ — теплопродукция, повышается температура тела, учащается дыхание, возрастает частота пульса и повышается возбудимость нервного и мышечного аппаратов. Только по окончании периода разогревания летучая мышь может взлететь.
Очень глубокие различия в физиологическом состоянии наблюдаются и в разные часы суток; у летучих мышей они связаны с периодом их суточной активности. Многие виды летучих мышей летают только в течение нескольких вечерних или ночных часов, когда вертикальные токи воздуха поднимают с земли огромное количество насекомых, которыми эти животные питаются.
Для летучих мышей, обитающих в наших широтах, характерна вечерняя и ночная гомойотермия и утренняя и дневная пойкилотермия. В основе этих изменений физиологического состояния лежит так называемый циркадный ритм, т. е. врожденные периодические колебания центральных и периферических аппаратов, обеспечивающих гомеостазис.
Циркадные ритмы могут возникать и поддерживаться непосредственно без периодического влияния на организм факторов внешней среды. Это можно показать на следующем примере. Так, если отделить летучую мышь от стада и поместить в изолированную камеру, то в часы активности всего стада в пещере у нее можно наблюдать описанные выше явления разогревания — калорифика-ции. Последние наблюдаются даже во время зимней
100
Часы суток
Рис. 19. Суточный ритм у различных видов летучих мышеи (по Aiorrison, 1961):
/ — G icszophaga, 2 — Sturnira, 3 — Artibeus. 4 — Phyllostomus
Рис. 18. Суточные кривые температуры тела у различных млекопитающих (по Щербаковой, 1949а):
/ — макак-лапундер, 2 — макак-резус, 3 — у гамадрилов, 4 — у человека, 5 — у собак, 6 — у шакалов, 7 — у кошек, 8 — у кроликов. По оси ординат — температура тела; по оси абсцисс — часы суток
спячки, и хотя колебания температуры и обмена веществ при этом очень невелики, они все же отмечают периоды ночной активности и дневного покоя, имевшие место в летний период. Эти явления продолжаются месяцами во время всей зимней спячки (см. стр. 153) и свидетельствуют об отсчете времени организмом, его центральной нервной системой.
Физиологические сдвиги в организме летучих мышей—это наиболее яркий пример суточных изменений у высших организмов.
Рис. 20. Суточная периодика изменений уровня сахара крови и температуры тела у летучих мышей
Циркадные ритмы представляют огромную область явлений в живой природе. Они охватывают как растительный, так и животный мир, но физиологические изменения, наблюдающиеся при этом, различны у животных различных систематических групп. Если у растительных организмов циркадные ритмы охватывают главным образом процессы фотосинтеза (фотопериодизм), то у животных они включают изменение обмена веществ и двигательной активности. Особенно большое значение имеет исследование зависимости между природными суточными циклами (освещения, температуры среды, условий питания и т. д.) и суточными циклическими изменениями физиологических процессов в организме.
Механизмы физиологических часов можно наблюдать на всех уровнях физиологической интеграции — клеточном; органном, уровне целого организма. Как общее
102
правило, клеточные ритмы в значительной мере находятся в большей зависимости от циклических факторов внешней среды, чем ритмы целого организма, особенно высшего. У человека образование множественных суточных ритмов практически невозможно.
Клеточные процессы во многом протекают ритмично. Такие явления, как синтез нуклеиновых кислот, объединение молекул РНК и белка в рибосомы, синтез белка на рибосомах повторяются в клетках ритмически. Существует и циркадный суточный ритм в образовании нуклеиновых кислот в печени мышей (Halberg, Halberg, Barnum, Bittner, 1959). Имеются данные о суточном ритме синтеза некоторых ферментов в крови. Можно предполагать, что регулирование во времени происходит с участием обратной связи. Накопление продуктов обмена ведет к торможению процесса. Время от начала до конца цикла является своеобразным маятником, регулирующим весь ход процесса. Наконец, хорошо выражен суточный ритм митотической активности клеток.
Большое значение в формировании физиологических часов имеют гормоны. У насекомых здесь большую роль играет нейросекреторная система, в частности подглоточного ганглия. Инерция суточного ритма черного таракана связана с инерцией выделения нейросекрета (Harker, 1960). Здесь выделение гормона под влиянием темноты поддерживает ночную активность и связано с активностью многих внутрисекреторных образований (прилежащих тел, околосердечной железы).
У высших животных и у человека механизм отсчета времени связывается с гипоталамусом. Это влияние гипоталамуса гуморально распространяется на кору надпочечников, что приводит к суточному циклу образования кортикостероидов. Значительную роль в суточном цикле играет кора головного мозга, или высшие отделы центральной нервной системы, связанные с образованием условных рефлексов на время. Последние могут быть образованы на разные интервалы времени у всех высших организмов, но только у обезьян и человека (кроме летучих мышей, о которых речь была выше) включают кроме элементов двигательной активности сосудистых реакций и значительные сдвиги температуры тела, обмена веществ и т. д. Однако эти изменения сложнорефлекторные, т. е. они включают как условные рефлексы
103
на время, так и непосредственное влияние режима освещения на возбудимость гипоталамической области.
ЭКСПЕРИМЕНТАЛЬНОЕ ИЗУЧЕНИЕ СУТОЧНЫХ
(ЦИРКАДНЫХ) РИТМОВ
Исследованиями последних лет (обзоры: Бюннипг, 1961; Эмме, 1967) установлено, что суточный ритм физиологических процессов, в особенности двигательной активности, является одной из основных особенностей жизнедеятельности всех живых существ. Лучше всего дневная суточная активность выражена у большинства видов воробьиных птиц, а из млекопитающих — у обезьян и человека. Столь же ярко выражена суточная ночная активность у летучих мышей и ночных птиц.
Если исключить летучих мышей, о которых подробно речь шла выше, то наиболее яркие и столь же хорошо выраженные сдвиги имеют место у обезьян, суточные колебания температуры тела которых составляют 4— 5° С. У человека суточные изменения температуры тела достигают 1°С.
В течение суток претерпевают закономерные колебания также частота дыхания, состав альвеолярного воздуха, состав крови, мочи, потоотделение, работа сердца (резко меняется характер .электрокардиограммы), мышечная и умственная работоспособность, отделение желудочного сока и т. д. Сейчас уже собрано большое количество фактов, касающихся этой стороны динамики физиологических функций человека.
У ряда животных почти не удается наблюдать суточные изменения физиологических функций (хищные, собаки), у грызунов резко изменяется периодика двигательной активности, но это не сопровождается значительными изменениями физиологических функций, как у обезьяны, человека и особенно у летучих мышей.
Каков механизм образования и поддержания суточной периодики? Как формируются эти суточные циклы физиологического состояния, представляющие собой фон, на который воздействуют все раздражители и который определяет в значительной мерс ответы организма на эти раздражители? Самые старые и достаточно научно аргументированные теории считали суточный ритм
104
врожденной реакцией; суточную активность, изменения физиологических функций — отражением космического цикла на животный организм. Опыты на человеке и отчасти на животных подтверждали это положение. У человека в условиях эксперимента практически невозможно изменить суточный ритм. Даже переход на работу в ночные часы не изменяет обычного суточного ритма. Данные, полученные при изучении таких постоянных представителей ночного труда, как сторожа, показали, что иногда наблюдаются какие-то элементы нарушения суточного ритма, но далеко не у всех и не всегда. Было, например, показано, что напряженный труд ответственных работников на железнодорожном транспорте в течение всей Великой Отечественной войны, т. е. в условиях совершенно нерегулярного режима сна и бодрствования, не изменил суточных колебаний температуры тела; они остались в пределах нормы.
Между тем имеются уже довольно старые данные (Osborne, 1909) и более новые (Кандрор, 1954) о том, что при переезде в экваториальном направлении (из Лондона в Мельбурн и из Владивостока в Москву) суточный ритм человека постепенно перестраивается в соответствии с географической долготой места.
При изменении режима сна и бодрствования в условиях полярной ночи датский физиолог Линдхардт (Lind-hardt, 1907) наблюдал полное извращение суточной кривой температуры тела и ритма дыхания. Однако это происходит лишь при полном изменении всей окружающей человека обстановки.
Опыты с изоляцией человека от окружающей среды п изменением продолжительности суток путем изменения скорости движения стрелок часов были поставлены американским исследователем Клейтманом (Kleitman, 1939). Оказалось, что 21-часовые сутки для человека являются пределом укорочения естественных суток, создающих часть функций (температуры тела и др.). Предел удлинения суток — 28-часовой ритм. У разных людей наблюдаются большие различия в пределах суточного ритма. Это выявилось в исследованиях Лоббана (Lobban, 1957), когда члены семьи, выехавшей в условия полярного дня в Гренландию, совершенно неодинаково реагировали на искусственно созданное извращение суточного режима сна и бодрствования. По-видимому, для человека боль
105
шое значение имеют типологические особенности нервной системы, подобно тому, как это выявлено в эксперименте у обезьян (см. также Кандрор, 1968).
Наиболее обстоятельные исследования суточного ритма и прежде всего поддерживающих его факторов были проведены в эксперименте на обезьянах (Simpson a. Galbraith, 1905а; 1905; Щербакова, 1949) и на птицах (Hilden a. Stenback, 1917). При этом выяснилось, что
Рис. 21. Влияние режима питания и освещения на суточные кривые температуры тела у обезьян (по Щербаковой, 1938, 19496): 1 — нормальный режим питания и освещения, 2 — извращенный режим питания и освещения, 3 — двухфазовый суточным режим, 4 — режим освещения днем, питания — ночью. По оси ординат — температура тела; по оси абсцисс — часы дня
извращение суточного режима питания и освещения приводит к полному изменению суточной кривой температуры тела, дыхания, pH мочи, не говоря уже о двигательной активности животных. На рисунке 21 представлены примеры извращения суточного режима у низших обезьян — павианов-гамадриллов и макаков-резусов. Специальными опытами было выяснено, что ведущим рецептором, при раздражении которого происходит извращение суточного ритма, является зрительный, а ведущим фактором — освещение. В тех же исследованиях О. П. Щербаковой (1949) удалось с применением двухфазового суточного режима смены освещения и питания получить двухфазовую суточную кривую температуры тела. Удалось получить и измененные соответствующим образом кривые при 36-часовых «сутках» с периодами
106
освещения и затемнения по 18 ч. Изменение только режима питания (кормушки в этом случае подсвечивались, а помещения для обезьян затемнялись) не приводило к заметным изменениям суточной температурной кривой.
В то же время американские ученые Гриффин и Уэлш (Griffin a. Welsh, 1937) нашли возможность изменить суточную кривую активности у летучих мышей Myotis lacifugus при искусственном освещении и подкармливании в неволе в дневные часы.
Для формирования суточного ритма имеют значение и некоторые специфические для данного вида животных раздражители. Так, например, у обезьян-гамадрилов суточный ритм температуры тела можно изменить в зависимости от распределения стадных звуков во время суток. Если обезьяну поместить в изолированном домике внутри большой вольеры, то можно получить изменение суточного ритма, не связанное с освещением в домике, а в зависимости от стадных звуков животных, находящихся в вольере. В этом случае суточный ритм поддерживается, благодаря контакту между изолированным животным и стадом, раздражением слухового анализатора (Черкович, 1953а).
У большинства сельскохозяйственных животных (коров, лошадей, овец, коз) суточный ритм обеспечивает периодическое изменение двигательной активности. Суточные изменения температуры тела, дыхания, кровообращения этих животных крайне невелики, хотя, как показывают специальные исследования, вполне достаточны для того, чтобы оказать влияние на продуктивность животных, в частности на молочную продуктивность в связи с режимом содержания животных.
Следовательно, суточный ритм образуется у разных животных при раздражении различных анализаторов.
Как же формируется суточный ритм в онтогенезе? Об этом сравнительно мало данных. У детенышей обезьян двигательная активность приурочивается ко времени кормления. Врожденным признаком здесь является способность к концентрации периода возбуждения и торможения, характерная для обезьян. Молодая обезьянка после каждого кормления обязательно длительное время активна и длительное время остается в покое. В процессе развития после рождения это проявляется в период, когда начинает формироваться нормальный суточный
107
ритм. На определенном этапе онтогенеза способность к концентрации возбудительного и тормозного процесса во времени формируется как безусловная реакция, как врожденное свойство нервной системы, а факторы внешней среды являются теми условиями, которые и определяют образование суточного ритма как условного рефлекса на время.
Все факты, которыми в настоящее время мы располагаем для доказательства того положения, что суточный ритм есть очень сложный динамический стереотип, концентрируются вокруг вопроса о возможности изменения суточного ритма в зависимости от условий среды. Установлено, что у обезьян с сильным, уравновешенным типом нервной системы полное извращение суточного ритма в связи с изменением условий кормления и освещения происходит на 3—4-й день, т. е. чрезвычайно быстро. Для обезьян со слабым типом нервной системы, с большей инерцией нервных процессов характерно затяжное извращение суточного ритма. Встречаются обезьяны, у которых невозможно извратить суточный ритм даже после 25—30 дней. Типологические особенности играют ведущую, очень важную роль при изменениях суточного ритма — суточного динамического стереотипа (Слонпм и Черкович, 1959).
Можно образовать суточный ритм и при различных формах деятельности животного. Так, если обезьяну кормить в определенные часы три раза в день, то можно наблюдать повышение обмена веществ, приуроченное к этим часам кормления, даже при отсутствии каких-либо пищевых раздражителей (Черкович, 1958).
Между прочностью суточного (циркадного) ритма и расселением организмов можно наблюдать интересные взаимоотношения. Так, например, существуют виды крыс, населяющие определенные области (туркестанская крыса, черная крыса)—они не способны так легко менять вновь образованный стереотип. По-видимому, в законах расселения организмов эта пластичность и подвижность нервной системы играют исключительную роль. Переделка суточного стереотипа практически не удается ни у туркестанских, ни у черных крыс (видов с ограниченным распространением); у серых крыс она происходит легко. Здесь четко выступают глубоко закрепленные видовые особенности нервной системы. Они определяли и опре
108
деляют законы расселения, ареал этих видов, с ними связан и целый ряд других физиологических особенностей каждого вида.
Знания об особенностях происхождения и влияния на организм суточного ритма получили довольно широкое практическое применение в зоотехнии, прикладной физиологии и в клинике. Создание путем изменения режима освещения двухфазового суточного ритма повышает яйценоскость у кур, увеличивает отложение жира при откорме свиней. Есть основания думать, что и молочная продуктивность крупного рогатого скота повышается при создании двухфазовых суток с дробными периодами отдыха.
Многое объясняет ритм и при изучении режима рабочего дня человека, при изучении явления утомления. Оказывается, что режим труда, в частности режим труда студентов с огромной перегрузкой во время экзаменов, сопровождается значительным нарушением суточного ритма. В поздние ночные часы, когда должно было наступить разлитое сонное торможение, сопровождающееся снижением температуры тела, температура держится еще на очень высоком уровне (до 12—1 ч ночи), т. е. имеется длительный застойный процесс возбуждения, не сменяющийся постепенно тормозным процессом, как это имеет место в норме. Соответствующий отдых и отчасти медикаментозная терапия позволяют устранить эти нарушения суточного ритма и вернуть его к норме.
Это доказано и для процесса сна. Между сном и суточным ритмом существуют известные соотношения. Сон наступает в определенные фазы суточного ритма при известной сниженной возбудимости центральной нервной системы. По этому поводу можно привести некоторые любопытные факты. Для людей, работающих с постоянным многофазным рабочим днем, например на железнодорожном транспорте, где работа протекает в любые часы дня и ночи, где отсутствует нормальный суточный режим, характерен нормальный физиологический ритм (кривая температуры тела, частота пульса, дыхания и т. д.), но в то же время образуется удивительная способность засыпать в любое время суток. Такой рабочий, приходя на работу из дома, где он провел перед этим 36 ч, получает наряд вести паровоз точно через 3 ч. У него образован условный рефлекс засыпать в любое
109
время, как только он ляжет в постель. Рабочий ложится спать в специальной комнате отдыха, через 3 ч поднимается, едет, потом опять идет в комнату отдыха и опять засыпает до следующей поездки. Физиологические исследования показывают, что такой сон не сопровождается значительным понижением физиологических функций; это не тот сон, который имеет место ночью, однако он наступает в любое время. Сопоставляя сои и явления суточного ритма, можно отмстить, что сон представляет торможение в первую очередь всей двигательной активности, тогда как суточный ритм охватывает все без исключения физиологические функции: изменение дыхания, температуры тела, кровяного давления и т. д.
СЕЗОННЫЕ ИЗМЕНЕНИЯ ФИЗИОЛОГИЧЕСКИХ
ФУНКЦИЙ
Сезонные изменения включают глубокие сдвиги в организме под влиянием изменений питания, температуры окружающей среды, лучистого солнечного режима и под влиянием периодических изменений эндокринных желез, связанных главным образом с размножением животных. Сам вопрос о факторах внешней среды, определяющих сезонную периодику, чрезвычайно сложен и до настоящего времени полного разрешения не получил; в формировании сезонных циклов приобретают большое значение сдвиги в функциях половых желез, щитовидной железы и др., имеющие очень устойчивый характер. Изменения эти, хорошо устанавливаемые морфологически, очень устойчивы в своем последовательном развитии для разных видов и сильно усложняют анализ влияния физических факторов, вызывающих сезонную периодику.
Сезонные изменения в организме включают и реакции поведения. Они заключаются либо в явлениях миграций и кочевок (см. ниже), либо в явлениях зимней и летней спячки, либо, наконец, в разнообразной деятельности по постройке нор и убежищ. Между глубиной нор некоторых грызунов и зимним понижением температуры существует прямая зависимость.
Огромное значение для суммарной суточной активности животного имеет режим освещения. Поэтому сезонную периодику невозможно рассматривать вне широтного распространения организмов. На рисунке 22 при-
110
вёдены сезоны размножения у птиц в разных шпротах северного и южного полушарий. Видны четко сдвинутые на более ранние месяцы сроки размножения при передвижении с севера на юг в Северном полушарии и почти зеркальное отображение этих отношений в Южном. Подобные же зависимости известны и для млекопитающих, например для овец. Здесь рассматриваются в основном
I И Ш IV Г W Е7 ИГ К X XL ZU I X Ш IV V VT
50° - 70°C-5(f-6O°C-«f-SCTC 30°-tf°C ?0°-30°c-K)°-?D°O o-nfc*
Illi
0° 10°K) II — —
Ю°-20°Ю ! -
2O~30°K)^ ——----------------------------
зо° l/jx——-------------------------------------------
----------
I К Ш V V ET H3VXIX X XI XX 1 К Ш IV V VL
Рис. 22. Влияние широты па сезоны размножения у птиц (по Hart, 1964). Площадь гистограмм на всех широтах одинакова. По оси абсцисс — месяцы года
физиологические изменения в организме, происходящие в умеренном климате средних широт Северного полушария. Наибольшие изменения в организме в течение сезонов года касаются системы крови, общего обмена веществ, терморегуляции и отчасти пищеварения. Исключительное значение для бореальных организмов имеет накопление жира как энергетического потенциала, расходуемого на поддержание температуры тела и мышечную деятельность.
П1
Наиболее заметные изменения двигательной активности в разные сезоны можно наблюдать у дневных животных, что несомненно связано с режимом освещения. Лучше всего эти отношения изучены у обезьян (Щербакова, 1949). При содержании обезьян в течение круглого года при постоянной температуре среды суммарная суточная активность зависела от продолжительности светового дня: возрастание активности имело место в мае
Рис. 23. Сезонные изменения двигательной активности у обезьян (по Щербаковой, 1949в). По оси ординат слева направо — температура воздуха в °C (7), количество движений (столбики), продолжительность дня в часах (2)
и июне. Повышение суммарной суточной активности наблюдалось еще в декабре и январе. Последнее никак нельзя отнести за счет влияния светового дня и связано, вероятно, с весенними проявлениями в природе в условиях Сухуми (рис. 23).
В этих исследованиях обнаружилась также значительная сезонная периодика температуры тела у обезьян. Наиболее высокая температура в прямой кишке наблюдалась в июне, наиболее низкая — в январе. Эти сдвиги не могут объясняться изменениями температуры во внешней среде, так как температура помещения оставалась постоянной. Очень вероятно, что здесь имело место влияние радиационного охлаждения, благодаря сниженной температуре стен помещения.
В естественных условиях (Хрусцелевский и Копылова, 1957) яркую сезонную динамику двигательной актив-
112
пости проявляют полевки Брандта в Юго-восточном Забайкалье. У них наблюдается резкое сокращение активности— выхода из нор в январе, марте, ноябре и декабре. Причины такой картины поведения довольно сложны. Они связаны с характером протекания беременности обычно весьма активных самок, со сроками восхода и захода солнца, высокими температурами летом и низкими зимой. Суточная активность, изученная в условиях природы, много сложнее и не всегда отражает картину, получаемую исследователем с помощью актографической методики.
Столь же сложные отношения обнаружены (Леонтьев, 1957) для полевки Брандта и монгольской песчанки в Амурской области.
У норок (Терновский, 1958) наблюдаются значительные изменения двигательной активности в зависимости от сезонов года. Наибольшая активность имеет место весной и летом, что, по-видимому, связано с длительностью светового дня. Однако при понижении температуры активность снижается, равно как и при осадках. У всех без исключения стадных копытных наблюдаются сезонные изменения стадности, что ярко выражено у лосей. У северного оленя стадные отношения (группирование, следование друг за другом) более заметны осенью, нежели летом пли весной (Салганский, 1952).
Лучше всего изучены сезонные изменения обмена веществ (основного обмена). Еще в 1930 г. японский исследователь Ишида (Ishida, 1930) обнаружил значительное повышение основного обмена у крыс в весенний период. Эти факты были затем подтверждены многочисленными исследованиями (Kayser, 1939; Sellers, Scott a. Thomas, 1954; Kocarcv, 1957; Gelineo a. Heroux, 1962). Установлено также, что зимой основной обмен у крыс гораздо ниже, чем летом.
Очень яркие сезонные изменения основного обмена обнаруживаются у пушных зверей. Так, основной обмен у песцов летом по сравнению с зимой повышен па 34%, а у серебристо-черных лисиц — на 50% (Фирстов, 1952). Эти явления несомненно связаны не только с сезонным циклом, но и с имеющим место в летнее время перегреванием (см. гл. V) и отмечены разными исследователями у песцов, енотовидных собак (Слоним, 1961). У серых
113
крыс в условиях Заполярья также обнаружено повышение обмена веществ весной и понижение осенью.
Изучение химической терморегуляции у полярных видов (песцы, лисицы, зайцы), зимующих в условиях Ленинградского зоологического сада (Исаакян и Акчурин,
Рис. 24. Сезонные изменения газообмена у песцов и лисиц (по Исаакян и Акчурину, 1953). /, // — лисица красная; III — песец белый; IV — песец голубой:
1 _ потребление О2, 2 — температура воздуха, 3 — температура тела. По оси ординат слева направо — потребление О2 (ял мин), температура тела, температура воздуха в °C; по оси абсцисс— месяцы
1953), показало при одних и тех же условиях содержания резкие сезонные изменения химической терморегуляции у лисиц и енотовидных собак и отсутствие сезонных изменений у песца. Особенно ярко это выражено в осенние месяцы, когда животные в летнем мехе. Авторы объясняют эти различия специфическими для арктических обитателей — песцов реакциями на смену освещения. Именно у песцов в осенний период практически отсутствует химическая терморегуляция, хотя изолирующий слой шерсти к этому времени еще не стал зимним. Очевидно, эти специфические для полярных животных
114
реакции не могут объясняться только физическими свойствами кожных покровов: они результат сложных видовых особенностей нервного и гормонального механизмов терморегуляции. Эти реакции у полярных форм сочетаются с теплоизоляцией (Шоландер и сотрудники, см. стр. 208).
Большой материал по сезонным изменениям газообмена у различных видов грызунов (Калабухов, Ладыгина, Майзелис и Шилова, 1951; Калабухов, 1956, 1957; Михайлов, 1956; Скворцов, 1956; Чугунов, Кудряшов и Чугунова, 1956 и др.) показал, что у незимоспящих грызунов можно наблюдать повышение обмена веществ осенью и понижение зимой. Весенние месяцы характеризуются повышением обмена веществ, а летние относительным понижением. Такие же данные на очень большом материале были получены для обыкновенной полевки и рыжей полевки в Московской области.
Схематически можно представить сезонную кривую изменений обмена веществ у не впадающих в зимнюю спячку млекопитающих следующим образом. Самый высокий уровень обмена веществ наблюдается в весеннее время в период половой активности, когда животные после зимнего ограничения питания приступают к активной пищедобывательной деятельности. Летом уровень обмена вновь несколько снижается в связи с высокой температурой, а осенью несколько повышается или держится на летнем уровне, постепенно понижаясь к зиме. Зимой наступает некоторое понижение основного обмена, а к весне он опять резко повышается. Эта общая схема изменений уровня газообмена на протяжении всего года для отдельных видов и в отдельных условиях может значительно варьировать. Это особенно относится к сельскохозяйственным животным. Так, основной обмен у не-лактирующих коров (Ritzman a. Benedict, 1938) в летние месяцы даже на 4—5-й день голодания оказался выше, чем зимой и осенью. Кроме того, очень важно отметить, что весеннее повышение обмена веществ у коров не связано с беременностью и лактацией, с условиями содержания в стойле или на пастбище. При стойловом содержании газообмен весной оказывается выше, чем при пастбищном осенью, хотя пастбищное содержание само по себе повышает газообмен в покое в течение всего пастбищного сезона (Калитаев, 1941).
115
В летний период газообмен у лошадей (в покос) возрастает почти на 40% по сравнению с зимним. Одновременно возрастает и содержание эритроцитов в крови (Магидов, 1959).
Очень большие различия (30—50%) в энергетическом обмене зимой и летом отмечаются у северного оленя (Сегаль, 1959). У каракульской овцы, несмотря на протекание беременности в зимний период, имеет место значительное снижение газообмена. Случаи снижения обмена веществ зимой у северного оленя и у каракульской овцы несомненно связаны с ограничением питания в зимний период.
Изменению основного обмена сопутствуют и сдвиги в химической и физической терморегуляции. Последнее связано с повышением теплоизоляции (insulation) шерстного и перьевого покровов в зимнее время. Снижение теплоизоляции летом сказывается как на уровне критической точки (см. гл. V), так и на интенсивности химической терморегуляции. Так, например, величины теплоотдачи в летнее и зимнее время у разных животных относятся: у белки, как 1 : 1; у собаки 1 : 1,5; у зайца 1 : 1,7. В зависимости от сезонов года теплоотдача с поверхности тела значительно изменяется в связи с процессами линьки и обрастания зимней шерстью. У птиц электрическая активность скелетной мускулатуры (благодаря отсутствию несократительного термогенеза) зимой и летом не изменяется; у млекопитающих, например серой крысы, эти различия очень значительны (рис. 25).
Сезонные изменения критической точки обмена веществ обнаружены в последнее время у полярных животных в условиях Аляски (Irving, Krogh a. Monson, 1955)—у красной лисицы они составляют летом +8°, зимой —13°; у белки — летом и зимой -;-20°С; у дикобраза (Erethizoon, dorsatum) летом +7°, а зимой —12°С. Эти изменения авторы также связывают с сезонными изменениями теплоизоляции меха.
Обмен веществ у полярных животных в зимний период даже при температуре —40° С повышается сравнительно незначительно: у лисицы и полярного дикобраза— не более 200% от уровня обмена при критической точке, у белки — около 450—500%. Подобные же данные были получены в условиях Ленинградского зоосада па песцах и лисицах (Ольнянская и Слоним, 1947). Сме
116
щение критической точки обмена веществ от температуры -!-30‘ С до +20° С наблюдалось у серой крысы зимой (Синичкина, 1959).
Изучение сезонных изменений газообмена у степных пеструшек (Lagurus lagurus) показало (Башенииа, 1957), что зимой критическая точка у них, в отличие от других видов полевок, необычайно низка — около 23° С. Критическая точка обмена веществ у полуденных песчанок смещается в разные сезоны, а у гребенщиковой остается постоянной (Мокриевич, 1957).
Рис. 25. Электрическая активность мышц во время холодового воздействия па серых крыс (справа) и вечернего дубоноса (слева) зимой и летом (по Hart, 1964). Переход от комнатной температуры к холодному помещению осуществляется в точке 0
Наиболее высокие величины потребления кислорода при температурах среды от 0 до 20° С наблюдались у желтогорлых мышей, отловленных летом, а наиболее низкие— в зимний период (Калабухов, 1953). Данные для мышей, отловленных осенью, занимали среднее положение. Эта же работа позволила обнаружить очень интересные изменения теплопроводности шерсти (снятых с животных и высушенных шкурок), сильно возрастающей летом и понижающейся зимой. Некоторые исследователи склонны приписывать этому обстоятельству ведущую роль в изменениях обмена веществ и химической терморегуляции на протяжении разных сезонов года. Разумеется, такие зависимости отрицать нельзя, но и у лабораторных животных (белых крыс) имеется ярко выраженная сезонная динамика даже при постоянных температурах среды (Исаакян и Избинский, 1951).
В опытах па обезьянах и диких хищных обнаружено (Слоним и Безуевская, 1940), что химическая терморе
117
гуляния весной (апрель) более интенсивна, нежели осенью (октябрь), несмотря на то, что температура окружающей среды была в обоих случаях одинаковой (рис. 26). Очевидно, это результат предшествующего влияния зимы и лета и соответствующих изменений
Рис. 26. Сезонные изменения химической терморегуляции у обезьян и хищных (по Слониму и Безуевской, 1940). /, II — павиан-гамадрил;
III, IV — макак-лапундер; V — динго; VI,
VII — шакал; VIII — енотовидная собака:
1 — при температуре среды 20° С, 2 — при температуре среды 35° С. По оси ординат — потребление О2 (мл/мин)
в эндокринных системах организма. В летний период наблюдается снижение интенсивности химической терморегуляции, в зимний — увеличение.
Своеобразные сезонные изменения химической терморегуляции обнаружены у впадающего в зимнюю и летнюю спячку желтого суслика и незимоспящего тонкопалого суслика (Калабухов, Нургельдыев и Скворцов, 1958). У тонкопалого суслика сезонные изменения терморегуляции выражены более отчетливо, чем у желтого (разумеется, в состоянии бодрствования). Зимой обмен тонкопалого суслика резко повышен. У желтого суслика
118
летом химическая терморегуляция нарушается уже при + 15—5° С. Сезонные изменения терморегуляции у него почти отсутствуют и заменяются длительной зимней и летней спячкой (см. ниже). Столь же мало выражены сезонные изменения терморегуляции у впадающего в летнюю и зимнюю спячку тарбаганчика.
Сопоставление сезонных изменений химической терморегуляции и биологического цикла животных (Н. И. Калабухов и сотр.) показало, что сезонные изменения выражены слабо как у зимоспящих видов, так и у видов, проводящих зиму в глубоких норах и мало подвергающихся воздействию низкой температуры наружного воздуха (например, большая песчанка).
Таким образом, сезонные изменения терморегуляции сводятся в основном к возрастанию теплоизоляции зимой, снижению интенсивности реакции обмена веществ (химической терморегуляции) и к смещению критической точки в зону более низких температур среды.
Несколько изменяется и термочувствительность организма, что связано, по-видимому, со сменой шерстного покрова. Такие данные установлены Н. И. Калабуховым для песцов (1950) и желтогорлых мышей (1953).
У серых крыс, обитающих в средней полосе, предпочитаемая температура зимой составляет от 21 до 24°, летом—25,9—28,5°, осенью — 23,1—26,2° и весной — 24,2°С (Синичкина, 1956).
В естественных условиях у диких животных сезонные изменения потребления кислорода и теплопродукции могут в значительной степени зависеть от условий питания. Однако экспериментальное подтверждение пока отсутствует.
Значительно изменяется по сезонам года кроветворная функция. Наиболее яркие сдвиги в этом отношении наблюдаются у человека в Арктике. Весной здесь можно наблюдать большое нарастание числа эритроцитов и гемоглобина (НЬ) крови, что связывают с переходом от полярной ночи к полярному дню, т. е. с изменениями инсоляции. Однако даже в условиях достаточной инсоляции в горах Тянь-Шаня у человека зимой наблюдается несколько сниженное количество гемоглобина в крови. Резкое повышение НЬ наблюдается весной. Количество эритроцитов уменьшается весной и возрастает летом (Авазбакиева, 1959). У многих грызунов, например у
119
песчанок, в летний период содержание эритроцитов снижается, а в весенний и осенний периоды — повышается (Калабухов и сотр., 1958). Механизм этих явлений до настоящего времени неясен. Здесь имеют место и изменения питания, витаминного обмена, ультрафиолетовая радиация и т. д. Не исключены также и влияния эндокринных факторов, причем особенно важная роль принадлежит щитовидной железе, стимулирующей эритропоэз.
Наибольшее значение в поддержании сезонного ритма имеют гормональные сдвиги, представляющие как самостоятельные циклы эндогенного происхождения, так и связанные с воздействием важнейшего фактора среды— режима освещения. При этом в настоящее время уже намечается схема взаимоотношений гипоталамус — гипофиз — кора надпочечников.
Сезонные изменения гормональных отношений выявлены у диких животных в природных условиях на примере изменения веса надпочечников (играющих, как известно, большую роль в приспособлениях организма к специфическим и неспецифическим условиям «напряжения»— стресса).
Сезонная динамика веса и активности надпочечников имеет очень сложное происхождение и зависит как собственно от «напряжения» в связи с условиями жизни (питание, температура среды), так и от размножения (Шварц и сотр., 1968). В этом плане интересны данные об изменениях относительного веса надпочечников у не-размпожавшихся полевых мышей (рис. 27). В период усиленного питания и оптимальных температурных условий вес надпочечников резко возрастает. Осенью с похолоданием этот вес начинает снижаться, по с установлением снежного покрова стабилизируется. Веспой (апрель) начинается увеличение веса надпочечников в связи с ростом организма и половым созреванием (Шварц, Смирнов, Добрипский, 1968).
Морфологическая картина щитовидной железы у многих видов млекопитающих и птиц подвержена значительным сезонным изменениям. В летнее время отмечается исчезновение коллоида фолликул, уменьшение эпителия, уменьшение веса щитовидной железы. В зимнее время имеют место обратные отношения (Riddle, Smith a. Benedict, 1934; Watzka, 1934; Miller, 1939; Hoehn, 1949).
120
Столь же четко выражена сезонная изменчивость в функции щитовидной железы у северного оленя. В мае и июне наблюдается ее гиперфункция с усилением секреторной деятельности эпителиальных клеток. Зимой, особенно в марте, секреторная деятельность этих клеток прекращается. Гиперфункция сопровождается уменьшением объема железы. Сходные данные получены и у овец, но закономерность выражена гораздо менее резко.
Рис. 27. Сезонные изменения относительного веса надпочечника (л«г/г) у неразмножавшихся полевых мышей (из Шварца, Смирнова и Добринского, 1968)
В настоящее время имеются многочисленные данные, указывающие на наличие устойчивых сезонных колебаний содержания тироксина в крови. Наиболее высокий уровень тироксина (определяемого по содержанию йода в крови) наблюдается в мае и июне, наиболее низкий — в ноябре, декабре и январе. Как показали исследования (Sturm a. Buchholz, 1928; Curtis, Davis a. Philips, 1933; Stern, 1933) между интенсивностью образования тироксина и уровнем газообмена у человека на протяжении сезонов года есть прямой параллелизм.
Имеются указания на тесную связь между охлаждением тела и выработкой гормона щитовидной железы и тиреотропного гормона гипофиза (Uotila, 1939; Войтке-вич, 1951). Эти отношения также имеют весьма важное значение в формировании сезонной периодики.
По-видимому, значительная роль в сезонной периодике принадлежит и такому неспецифическому гормону, как адреналин. Большое количество данных свиде-
121
тсльствует о том, что адреналин способствует лучшей акклиматизации как к теплу, так и к холоду. Особенно сильно действуют комбинации препаратов тироксина и кортизона (Hermanson a. Hartmann, 1945). Хорошо акклиматизированные к холоду животные обладают большим содержанием аскорбиновой кислоты в корковом слое надпочечников (Dugal a. Fortier, 1952; Dugal, 1953).
Адаптация к низкой температуре среды сопровождается повышением содержания аскорбиновой кислоты в тканях, повышением содержания гемоглобина в крови (Gelineo et Raiewskaya, 1953; Raiewskaya, 1953).
В последнее время накоплен большой материал, характеризующий сезонные колебания содержания кортикостероидов в крови и интенсивности их выделения при инкубации коры надпочечника in vitro.
Роль режима освещения в формировании сезонного ритма признается подавляющим большинством исследователей. Освещение, как было установлено еще в середине прошлого столетия (Moleschott, 1855), оказывает значительное влияние на интенсивность окислительных процессов в организме. Газообмен у человека и животных под влиянием освещения повышается (Moleschott u. Fubini, 1881; Арнаутов и Веллер, 1931).
Однако до последнего времени совершенно не изученным оставался вопрос о влиянии освещения и затемнения на газообмен у животных с разным образом жизни. И лишь при изучении влияния интенсивности освещения на газообмен у обезьян (Иванов, Макарова и Фуфачева, 1953) выяснилось, что он всегда выше на свету, чем в темноте. Однако не у всех видов эти изменения оказались одинаковыми. У гамадрилов они были выражены наиболее резко, далее шли макаки-резусы и меньше всего влияние освещенности сказалось на зеленых мартышках. Различия можно было понять только в связи с экологическими особенностями существования перечисленных видов обезьян в природе. Так, обезьяны-гамадрилы— обитатели безлесных нагорий Эфиопии; макаки-резусы — обитатели леса и сельскохозяйственных культурных участков, а зеленые мартышки — густого тропического леса.
Реакция на освещение появляется в онтогенезе относительно поздно. Так, например, у новорожденных коз
122
лят повышение газообмена па свету по сравнению с темнотой очень невелико. Эта реакция значительно возрастает к 20—30-му дню и еще более к 60-му (рис. 28). Можно думать, что у животных с дневной активностью реакция на интенсивность освещения носит характер натурального условного рефлекса.
У ночных животных лемуров-лори наблюдались обратные отношения. Газообмен у них оказывался повы-
Рис. 28. Влияние освещения на реакцию повышения газообмена у козлят (потребление кислорода кг)ч в %) по сравнению с темнотой
шенным в темноте и пониженным при освещении во время определения газообмена в камере. Снижение газообмена на свету достигало у лори 28%.
Факты влияния длительного освещения или затемнения на организм млекопитающих были установлены экспериментальным изучением светового режима (светового дня) в связи с сезонными влияниями освещения. Экспериментальному изучению влияния длительности светового дня на сезонную периодику посвящено большое количество исследований. Больше всего данных собрано для птиц, где увеличение светового дня является фактором, стимулирующим половую функцию (Свето-заров и Штрайх, 1940; «Лобашов и Савватеев, 1953).
123
Полученные факты указывают как на значение общей длины светового дня, так и на значение смены фаз освещения и затемнения.
Хорошим критерием влияния режима освещения и длительности светового дня для млекопитающих служит протекание овуляции. Однако именно у млекопитающих такого прямого влияния света на овуляцию у всех без исключения видов установить не удается. Многочисленные данные, полученные на кроликах (Smelser, Walton a. Whethem, 1934), морских свинках (Dempsey, Ateyers, Young a. Jennison, 1934), мышах (Kirchhof, 1937) и на сусликах (Welsh, 1938), показывают, что содержание животных в полной темноте не оказывает никакого влияния на процессы овуляции.
В специальных исследованиях имитировались «зимние условия» охлаждением (от —5 до 4-7° С) и содержанием в полной темноте. Эти условия не оказали влияния на интенсивность размножения у обыкновенной полевки (Microtus arvalis) и скорость развития молодняка. Следовательно, сочетанием этих основных факторов внешней среды, определяющих физическую сторону сезонных влияний, нельзя объяснить зимнее угнетение интенсивности размножения, хотя бы для грызунов этого вида.
На хищных было обнаружено значительное влияние света на функцию размножения (Беляев, 1950). Уменьшение светового дня приводит к более раннему созреванию меха у норок. Изменение же температурного режима не оказывает никакого влияния па этот процесс. У куниц дополнительное освещение вызывает наступление периода спаривания и рождения детенышей на 4 месяца раньше обычного срока. Изменение режима освещения не оказывает влияния на основной обмен (Беляев, 1958).
Однако сезонную периодику нельзя представлять себе только как результат влияния факторов внешней среды, на что указывает большое количество экспериментов. В связи с этим возникает вопрос, существует ли сезонная периодика у животных, изолированных от воздействия природных факторов. У собак, которые в течение круглого года содержались в отапливаемом помещении при искусственном освещении, можно было наблюдать характерную для собак сезонную периодику (Maignon
124
el Guilhon, 1931). Сходные факты были обнаружены в опытах на лабораторных белых крысах (Избинский и Исаакян, 1954).
Другой пример чрезвычайной прочности сезонной периодики касается животных, привезенных из южного полушария. Так, например, австралийский страус в заповеднике Аскания Нова откладывает яйца в условиях нашей зимы, несмотря на сильный мороз, прямо в снег в сезон, соответствующий лету в Австралии (М. М. За-вадовский, 1930). Австралийская собака динго щенится в конце декабря. Хотя эти животные, как и страусы, в течение многих десятков лет разводятся в северном полушарии, никаких изменений в их природном сезонном ритме не наблюдается.
У человека изменение обмена веществ протекает по той же схеме, что и у незимоспящих животных. Имеются наблюдения, полученные в естественной обстановке с попыткой извращения естественного сезонного цикла. Наиболее простой способ такого извращения и наиболее достоверные факты получены при изучении переездов из одной местности в другую. Так, например, переезд в декабре— январе из средней полосы СССР в южную (Сочи, Сухуми) вызывает эффект повышения сниженного «зимнего» обмена в течение первого месяца пребывания там в связи с новыми условиями юга. По возвращении весной на север наступает вторичное весеннее повышение обмена. Таким образом, при зимней поездке на юг можно в течение года наблюдать два весенних подъема уровня обмена у одного и того же человека. Следовательно, извращение сезонного ритма имеет место и у человека, но только в условиях изменения всего комплекса природных факторов внешней среды (Иванова, 1954).
Особый интерес представляет формирование сезонных ритмов у человека на Крайнем Севере. В этих условиях, в особенности при жизни на небольших станциях, резко нарушается сезонная периодика. Недостаточная мышечная деятельность вследствие ограничения прогулок, часто невозможных в условиях Заполярья, создает практически полное выпадение сезонного ритма (Слоним, Ольнянская, Руттенбург, 1949). Опыт показывает, что создание благоустроенных поселков и городов в Заполярье восстанавливает его. Сезонный ритм у человека является до некоторой степени отражением не
125
только сезонных факторов, общих для всего живого населения нашей планеты, но, как и суточный ритм, служит отражением социальной среды, влияющей на человека. Большие города и поселки на Крайнем Севере с искусственным освещением, с театрами, кино, со всем ритмом жизни, свойственным современному человеку,
Рис. 29. Сезонные изменения скорости роста детей с 6 до 15 лет (по Brody, 1945). Сплошная линия — мальчики; прерывистая — девочки. По оси ординат — средняя прибавка в весе на одного ребенка за 30 дней (английские фунты); по оси абсцисс — возраст детей в месяцах (деления шкалы) и годах (цифры)
создают такие условия, при которых за Полярным кругом сезонный ритм проявляется нормально и выявлен так же, как и в наших широтах (Кандрор и Раппопорт, 1954; Данишевский, 1955; Кандрор, 1968).
В условиях Севера, где зимой наблюдается большой недостаток ультрафиолетового облучения, имеют место значительные нарушения обмена веществ, главным образом фосфорного обмена, и недостаток витамина D (Галанин, 1952). Эти явления особенно тяжело отражаются на детях. По данным немецких исследователей, в зимний период наблюдается так называемая «мертвая зона», когда совершенно прекращается рост детей
126
(рис. 29). Интересно, что в Южном полушарии (в Австралии) это явление наблюдается в месяцы, соответствующие лету в Северном полушарии. Сейчас дополнительное ультрафиолетовое облучение рассматривается как один из важнейших приемов коррекции нормального сезонного ритма в условиях северных широт. В этих условиях приходится говорить не столько о сезонном ритме, сколько о специфическом недостатке этого природного необходимого фактора.
Сезонная периодика представляет также огромный интерес для животноводства. Ученые склоняются сейчас к тому, что значительная часть сезонных периодов должна изменяться сознательным воздействием человека. Речь идет прежде всего о сезонном режиме питания. Если для диких животных недостаток питания приводит иногда к гибели значительного количества особей, к уменьшению численности представителей их в данной местности, то по отношению к культурным сельскохозяйственным животным это совершенно недопустимо. Питание сельскохозяйственных животных не может базироваться на сезонных ресурсах, а должно дополняться на основе хозяйственной деятельности человека.
Сезонные изменения в организме птиц тесно связаны с характерным для них инстинктом перелета и в основе их лежат изменения энергетического баланса. Однако, несмотря на перелеты, у птиц наблюдаются как сезонные изменения химической терморегуляции, так и изменения теплоизоляционных свойств перьевого покрова (инсу-ляция).
Хорошо выражены изменения обмена веществ у домового воробья (Passer domesticus), энергетический баланс которого при низких температурах поддерживается большей теплопродукцией зимой, чем летом. Результаты, полученные при измерении потребления пищи и обмена, показывают уплощенный тип кривой химической терморегуляции, обычно встречающийся в тех случаях, когда теплопродукция оценивается на основе потребления пищи на протяжении нескольких дней, а не исходя из потребления кислорода в кратковременном эксперименте.
В последнее время было установлено, что максимальная теплопродукция у воробьиных птиц зимой выше, чем летом. У дубоносов, голубей Columba livia и скворцов Sturnus vulgaris время выживания в холодные периоды
127
зимой было большим главным образом в результате возросшей способности поддерживать более высокую теплопродукцию. На длительность периода до наступления смерти влияют также состояние оперения—линька и продолжительность нахождения в неволе, но сезонный эффект при этом всегда ярко выражен. У находящихся в клетке птиц потребление пищи зимой возрастало на 20—50%. Но потребление пищи зимой у содержащихся в клетке вьюрков (Fringilla montefringilla) и у домашних воробьев, находящихся на воле, не увеличивалось (Rautenberg, 1957).
Значительная ночная гипотермия, наблюдаемая зимой у только что пойманных птиц, отсутствует у дубоносов и черноголовой гаички. Ирвинг (Irving, I960) пришел к выводу, что холодными ночами северные птицы охлаждаются ниже уровня своей дневной температуры тела примерно настолько же, насколько и птицы умеренных областей.
Увеличение веса оперения, наблюдаемое у некоторых птиц в зимнее время, позволяет предположить наличие теплоизоляционной адаптации, которая могла бы частично покрыть изменения метаболизма на холоду. Однако исследования Ирвинга на нескольких видах диких птиц в зимнее и летнее время, а также Дэвиса (Davis, 1955) и Харта (Hart, 1962) дают мало оснований для предположения о том, что увеличение метаболизма при падении температуры на Г было в эти сезоны различным. Установлено, что у голубей теплопродукция, измеренная при 15° С, зимой была ниже, чем летом. Однако величина этих сезонных изменений была небольшой и в диапазоне критических температур не наблюдалось никаких сдвигов. Данные о сдвигах уровня критической температуры были получены для кардинала (Richmondena cardi-nalis) (Lawson, 1958).
Вальгрен (Wallgren, 1954) изучал энергетический обмен у желтой овсянки (Emberiza citrinelid) при 32,5° С и при —11° С в разное время года. Обмен в состоянии покоя не обнаружил сезонных изменений; при —11° С в июне и июле обмен был значительно выше, чем в феврале и марте. Эта инсулятивная адаптация частично объясняется большей толщиной и «распушиванием» оперения и большим сужением сосудов в зимнее время (так как оперение было самым густым в сентябре — пос-
128
ле линьки, а максимальные изменения обмена — в феврале).
Теоретически изменениями оперения можно объяснить понижение летальной температуры примерно на 40° С.
Исследования, проведенные на черноголовой гаичке (Parus atricapillus), также указывают на наличие низкой теплопродукции в результате теплоизоляционной адаптации в зимнее время. Частота пульса и частота дыхания имели сезонные сдвиги, причем зимой при 6° С понижение было большим, чем летом. Критическая температура, при которой дыхание резко возрастало, зимой также сдвигалась к более низкому уровню.
Увеличение основного обмена при термонейтральных температурах, резко выраженное у млекопитающих и птиц, подвергнутых воздействию холода в течение нескольких недель, не играет заметной роли во время адаптации к зиме. Единственное свидетельство значительного сезонного изменения основного обмена было получено на домовых воробьях, но нет оснований предполагать, что оно играет сколько-нибудь значительную роль у птиц, живущих на воле. У большинства исследованных видов вообще не наблюдается никаких изменений. Кинг и Фар-нер (King a. Earner, 1961) указывают, что большая ин теиспвпость основного обмена в зимнее время была бы невыгодной, так как птице потребовалось бы ночью увеличить расход своих энергетических резервов.
Наиболее характерными сезонными сдвигами у птиц являются их способность изменять свою теплоизоляцию и поразительное свойство поддерживать более высокий уровень теплопродукции в условиях холода. На основе измерения потребления пищи и -выделений при различных температурах и фотопериодах были сделаны оценки энергетических потребностей для существования и продуктивных процессов в различное время года. Для этой цели птицы были размещены в отдельных клетках, где у них измерялась метаболизированная энергия (максимальный приток энергии минус энергия выделений при различных температурах и фотопериодах). Наименьшая метаболизированная энергия, потребная для существования при определенных температурах и фотопериодах испытания, называется «энергией существования». Ее корреляция с температурой показана в левой части рисунка
5—1204
129
30. Потенциальная энергия — это максимальная метаболизированная энергия, измеренная при температуре, соответствующей летальному 'пределу, являющемуся самой низкой температурой, при которой птица может поддерживать вес тела. Энергия продуктивности — это разница между потенциальной энергией и энергией существования.
Рис. 30. Зависимость метаболизированной энергии от температуры среды и фотопериода (правая часть) и вычисленной величины потенциальной энергии, энергии существования и энергии продуктивности (левая часть) в различное время года у воробья и тропического зяблика (по Hart, 1954)
В правой части рисунка 30 показаны различные категории энергии, вычисленные для разных времен года из средних температур и фотопериодов вне помещения. Для этих расчетов принимается, что максимальная метаболизированная энергия выявлена в условиях холода, а так
130
же для продуктивных процессов при более высоких температурах. У домового воробья потенциальная энергия подвергается сезонным изменениям вследствие сезонных изменений пределов выживаемости. Энергия существования также изменяется в соответствии со средней температурой вне помещения. Благодаря сезонным изменениям потенциальной энергии и энергии существования, энергия продуктивности остается постоянной в течение всего года. Некоторые авторы указывают, что возможность домового воробья жить в дальних северных широтах обуслов лена его способностью растягивать свой максимальный энергетический баланс на всю зиму и метаболизировать столько же энергии во время короткого дневного фотопериода зимой, сколько и в течение длительных фотопериодов летом.
У белогорлого воробья (Z. albicallis) и у юнко (/. hue-malis) при 10-часовом фотопериоде количество метаболизированной энергии меньшее, чем при 15-часовом, что является серьезным недостатком зимнего времени (Seibert, 1949). Эти наблюдения были сопоставлены с фактом миграции обоих видов на юг в зимнее время.
В отличие от домового воробья, тропический сине-черный зяблик {Volatinia jacarina) мог сохранять энергетический баланс примерно до 0° С при 15-часовом фотопериоде и до 4° С при 10-часовом. Фотопериод ограничивал энергетику в большей степени при понижении температуры, в чем состоит отличие этих птиц от домашнего воробья. Благодаря воздействию фотопериода потенциальная энергия была наиболее низкой в зимнее время, когда энергия существования наивысшая. Следовательно, энергия продуктивности была также наиболее низкой в то время года. Эти физиологические характеристики не позволяют данному виду существовать в зимнее время в северных широтах.
Хотя для терморегуляции энергетические потребности в холодное время года оказываются максимальными, различные виды активности птиц распределяются, по-ви-димому, в течение всего года равномерно и поэтому кумулятивные эффекты ничтожны. Распределение установленных энергетических запросов для различной активности в течение всего года лучше всего описано для трех воробьев S. arborea (West, 1960). У этого вида наибольшее количество энергии продуктивности потенциально прихо-,5*
131
дилось на летнее время. Поэтому деятельность, требующая энергетических затрат, как, например, миграция, гнездование и линька, равномерно распределяется между апрелем и октябрем Дополнительные затраты на свободное существование являются тем неизвестным, которое может или не может повысить теоретический потенциал. Однако вполне возможно, что потенциальная энергия может быть использована в любое время года, по крайней мере на кратковременные периоды — на время полета.
СЕЗОННЫЕ ИЗМЕНЕНИЯ ПОВЕДЕНИЯ (МИГРАЦИИ И КОЧЕВКИ)
Сезонная периодика, характерная для животных и прежде всего для дикой фауны, определяется в первую очередь изменением условий существования. Например, изменение условий питания вызывает миграции животных на большие или меныппе расстояния. Так, на несколько десятков километров млекопитающие спускаются с гор на равнину, где они легче находят пищу, не покрытую глубоким слоем снега (Формозов, 1946; Касимович, 1955). В условиях тундры миграции па сотни километров замечены у северных оленей, лосей. На более короткие расстояния мигрируют некоторые грызуны — серый хомячок, серая крыса. У птиц миграция может быть на тысячи километров (перелет).
Миграционная деятельность животных приурочена, как правило, к определенным сезонам года, и такие явления, как перелет птиц, миграция крупных млекопитаю щих (лосей, бизонов, северного оленя) уже издавна связывались с сезонными изменениями условий питания, температуры, длины светового дня и т. д. Действительно, можно указать на вертикальные миграции животных в горах, которые связаны с изменением снежного покрова, изменениями температуры. Вместе с тем миграционное поведение в ряде случаев является врожденным видовым признаком. Так, например, скрещивание европейского немигрирующего зубра с американским мигрирующим бизоном приводит к тому, что гибриды — зубробизоны, проявляют яркую тенденцию к сезонным миграциям.
Большое разнообразие форм миграций и их продолжительности не позволяет, конечно, даже в общей форме определить физиологическую картину миграционного со
132
стояния, общего для всех организмов птиц, млекопитающих, рыб. Однако некоторые общие черты здесь все же могут быть отмечены. К ним следует отнести накопление жира в предмиграционный период, изменения в функции нейросекреторной системы, изменения реакции терморегуляторного аппарата, стадных отношений и т. д.
Миграции птиц. В настоящее время принято считать, что перелеты — мт рации птиц вызываются как экзогенными, так и эндогенными ф.акторами. Внешними факторами издавна считались понижение температуры среды, замерзание водоемов, уменьшение количества корма. Однако только этими факторами совершенно невозможно объяснить закономерные сдвиги в организме птиц и возникающие на их основе миграционные явления.
Само явление миграции у птиц по существу довольно четко разделяется на 2 ряда проявлений активности. Первое — это стимуляция и поддержание собственно миграционной активности, второе — ориентация птиц при перелете. Как те, так и другие явления имеют строго выраженный видовой характер, по в известной степени постоянно коррегируются факторами внешней среды (погодой, картиной местности, наличием корма и т. д.).
Миграционные явления у птиц, их возникновение и зависимость от влияний внешней и внутренней среды организма довольно четко представлены на следующей схеме (рис. 31). Из этой схемы видно, что факторы внешней среды (сигнальные стимулы ') оказывают непосредствен ное влияние на регулирующую систему. Последняя в равной мере зависит как от внешних стимулов, так и ог эндогенного (главным образом сезонного) ритма. Изменения этой системы вызывают появление предмиграцион-ного состояния, которое ведет к образованию стай, усиленному питанию и жироотложению, изменению в поведении и т. д. Предмигранионное состояние на каком-то этапе его развития переходит в миграционное, характеризующееся своеобразным поведением птицы (перелетное беспокойство — Zugunruhe, немецк.), изменением суточной периодики, состоянием центральной нервной системы. Миграционное состояние при одновременном воздействии
1 Слово «сигнальный» здесь нс следует понимать в смысле услов-порефлекторный, а лишь как сигнал из внешней пли внутренней среды.
133
миграционного стимула (часто изменения погодного комплекса) приводит к осуществлению перелета — миг рации.
Предложенная схема носит, конечно, общий и несколько отвлеченный характер, но достаточно четко отражает современные направления в изучении миграции птиц.
Сигнальной стимул
Рис. 31. Схема, демонстрирующая последовательность физиологических процессов и их стимуляцию при переходе птиц в миграционное состояние и при миграции (по Дольник. 1965)
Рассмотрение факторов внешней среды, оказывающих влияние на миграции, приводит к выводу, что для разны:< видов они оказываются весьма различными. Например, наличие незамерзающих водоемов может затормозить и сделать «оседлыми» отдельные популяции водоплавающих птиц. Большое влияние на возникновение миграции оказывает длина светового дня. Однако у тропических птиц сигналом к перелету являются изменения влажности. В соответствии с этим у разных птиц наблюдаются различия в сезонных изменениях нейросекреторной системы гипоталамуса.
134
Изучение состояния нейросекреторного аппарата гипоталамуса, оказывающего огромное влияние на синтез гормонов, представляет значительный интерес для понимания механизмов возникновения миграционных состояний вообще и миграции птиц в частности. Как известно, нейросекреторный аппарат гипоталамуса имеет хорошо выраженный цикл сезонных изменений, которые в значительной мере зависят от длины светового дня. Освобождение секрета соответствует наступлению миграционной активности.
Чрезвычайно противоречивы данные о роли различных желез внутренней секреции в миграционных явлениях. Наиболее старые данные имеются в отношении половых желез. Они могут стимулироваться длинным световым днем, но увеличения их активности не наблюдалось в период миграции. В настоящее время эта так называемая гонадная гипотеза уже не является ведущей. Также не получила подтверждения довольно старая гипотеза о роли щитовидной железы. Поддержанная в свое время рядом физиологов эта теория предполагала, что осеннее угнетение активности щитовидной железы приводит к ослаблению интенсивности теплообразования, нарушениям терморегуляции и вызывает перелетное беспокойство у птиц. Годовой цикл активности щитовидной железы с минимумом зимой и максимумом летом, однако, не может объяснить изменений физиологического состояния в весенний и осенний периоды, т. е. во время перелета.
Предложена гипотеза о значении периодических нарушений соотношений гормонов — синергистов и антагонистов. Имеется в виду годичный цикл половых гормонов с одной стороны, и годичный цикл гормона инсулярного аппарата поджелудочной железы — с другой. Эта гипотеза по существу всеобъемлющая, конечно, ничего не объясняет, так как не позволяет воспроизвести миграционное поведение в эксперименте. Таким образом, гормональные механизмы явлений миграции представляются еще в достаточной мере неопределенными, хотя они несомненно играют роль в природе.
Большое место в изучении миграции птиц заняло в по следние годы исследование обмена веществ. Наиболее яркими в период миграции являются изменения жирового обмена. Они сводятся к тому, что в результате возросшего питания (гиперфагии) резко увеличивается отложение
135
жира в организме птиц. Изучение дыхательного газообмена показывает в этот период высокие величины ды нательного коэффициента, свидетельствующие о переходе углеводов в жир (Дольник, 1966). Нужно заметить, что накопление жира предшествует перелету не только птиц, но и летучих мышей (впрочем, как зимней спячке) и является итогом питания в летний период. При перелетах птиц жир расходуется и накапливается на остановках, когда они кормятся.
Отмечаются и сдвиги углеводного обмена. Последний возрастает во время линьки и угнетается при подготовке к перелету.
Весьма примечательны и следующие факты. Отощавшие во время перелета птицы имеют значительное количество гликогена в печени. При голодании он расходуется. Собственно миграционное поведение птиц представляет весьма сложное явление, включающее как врожденные, так и приобретенные элементы. Наиболее удобным объектом для изучения механизмов стимуляции перелета является перелетное беспокойство. Последнее может рассматриваться как своеобразный инстинкт, возбуждаемый как внутренними, так и наружными сигналами. Многими исследователями такие сигналы рассматриваются как «реализующий механизм» этой формы инстинктивного поведения.
Ориентация при перелетах птиц. Эта проблема изучалась специалистами орнитологами на протяжении последнего столетия, но физиологические механизмы ориентации еще очень мало выявлены. Существует два основных метода изучения ориентации птиц. Первый — давно использованный метод завозов. При этом птицы завозились в отдаленные места в темных ящиках без возможности воспринимать какие-либо ориентиры на местности и затем изучались пути и скорость их возвращения. Другой метод — ориентация в клетках во время перелетного беспокойства. При этом птица в своих движениях ориентируется по направлению к зимовке.
Сейчас уже широко применяются методы радиотелеметрии, когда птица, снабженная миниатюрным радиопередатчиком, «сообщает» о пути, проделываемом ею при перелете. Довольно широко попользуется и радарная техника, позволяющая регистрировать движение стаи в целом.
136
Представления о механизмах ориентации птиц при перелетах еще далеко не сформировались, несмотря на большие работы в этом направлении. Наиболее старые взгляды опираются на роль наземных ориентиров при перелетах— очертаниях материков, гор и т. д. Эти представления, однако, совершенно не объясняют механизмов перелетов над морем.
В настоящее время подверглась значительной критике и почти оставлена магнитная теория ориентации птиц. Создавая различные условия распределения магнитных сил вокруг птицы в эксперименте, в условиях природной магнитной аномалии, исследователи не могли, однако, обнаружить отклонения птиц от нормального направления к зимовке. Также не подтвердились предположения о значении сочетания магнитных сил с силами вращения земли (Кариолисовыми силами).
Большое место в современных представлениях об ориентации птиц занимают вопросы ориентировки по положению Солнца и по времени суток, а также и положению звезд (так как многие виды птиц летят преимущественно ночью). Имеются даже данные, что существует врожденная ориентация птпц по звездам, так как птицы-ночные мигранты, выращенные в неволе при полной изоляции (никогда не видевшие неба), точно ориентируются в направлении перелета.
Все это позволяет предполагать и наличие у птиц способности к навигации, т. е. к сопоставлению координат местности и координат направления (цели) перелета. Исследования в этом направлении не могут считаться сколько-нибудь законченными.
Миграции рыб. Миграционное поведение рыб изучено значительно меньше, чем миграции птиц, но представляет огромный не только теоретический, но и практический интерес, особенно в связи со строительством гидротехнических сооружений на реках и нарушением путей миграций.
Различают пассивные миграции молодняка рыб из рек в море и анадромные (активные миграции) из моря в реки, связанные с нерестом.
Физиологические проблемы, возникающие при рассмотрении этих явлений, включают вопросы происхождения миграционного импульса и формирования миграционного поведения, вопросы изменений осморегулирующих
137
систем организма в связи с изменениями солености среды. Пассивное перемещение молоди рыб из рек в море не сопровождается специальными физиологическими изменениями. Здесь, однако, имеет место формирование стаи — процесс, включающий как врожденные поведенческие акты подражания, так и условнорефлекторные механизмы (Герасимов, 1965).
Образование стаи связано и с «групповым эффектом», т. е. с закономерным снижением интенсивности общего обмена веществ, подобно тому, что имеет место у наземных позвоночных (Штефан, 1958; Алексеева, 1959; Коштоянц, Мамокина, Александрюк, 1960 и др.).
Очень характерно то, что одиночно живущие виды рыб в период миграционной активности образуют стаи. При формировании стаи большое значение имеет не возраст, а размеры рыб. Имитационное поведение и прежде всего реакция следования и подражания проявляется па особь одинаковой величины. Роль различных анализаторов (зрительного, слухового) в формировании стаи рыб еще недостаточно изучена.
Физиологический механизм анадромных миграций, по мнению ряда исследователей (Гербильский, 1957, 1965; Баранникова, 1965, 1968), основан на глубоких изменениях нейросекреторной активности ядра гипоталамуса, а также на функции эндокринных желез, изменениях почечной функции и клеток жабр, экскретирующих натрий. Функциональные морфологические изменения нейросекреторной системы гипоталамуса, аденогипофиза связаны с увеличенной выработкой гонадотропного гормона. В результате возникают изменения в образовании половых продуктов и двигательного поведения.
Изложенная схема, разумеется, еще далека от возможности объяснить все особенности миграционного поведения и связанных с ним у анадромных рыб изменений регуляции водно-солевого гомеостаза. Однако именно по этой схеме идет в настоящее время накопление фактов в этой важной области физиологии адаптаций.
ФИЗИОЛОГИЧЕСКИЕ ИЗМЕНЕНИЯ ПРИ ЗИМНЕЙ
СПЯЧКЕ
Зимнее угнетение жизнедеятельности организма, носящее сезонный циклический характер, особенно ярко вы
138
ражено у млекопитающих, впадающих в зимнюю и летнюю спячку. Под спячкой понимают состояние резко пониженной жизнедеятельности животного, сопровождающееся изменением поведения: оно перестает двигаться и принимает ооычно характерную позу, соответствующую позе сна. Наиболее ярким проявлением спячки является снижение температуры тела. Многие исследователи считают, что в основе физиологического механизма спячки лежит временное нарушение или выключение терморегуляции, так называемое состояние гетеротермии. Тщательное сопоставление уровня температуры тела у зимоспящих и незимоспящих млекопитающих по возможности близких в систематическом отношении видов обнаруживает у зимоспящих более низкий уровень температуры тела (см. табл. 4).
Таблица 4
Температура тела у зимоспящих и незимоспящих млекопитающих (в состоянии бодрствования)
3им< спяшн? t c Незимоспяшпе t c
Еж европейский (Eri-naceus europaeus) . . . около 35,0 Крот европейский (Talpa euro раса) . . . 39,4
Сурок альпийский (Marmota baibacina) . . 36,0 Серая крыса-пасюк (Rattus norvegicus) . . 37,6
Суслик крапчатый (&-iellus suslicus) 36,5 Заяц-русак (Lepus europeus) 38,0
Хомяк обыкновенный (Cricetus cricetus) . . . 34,7 Полевая мышь (Apode-inus agrarius) 38,5
Соня-полчок (Glis Elis) 35,5 Морская свинка (Pro-cauia cobaya) 38,6
Соня орешниковая (Muscardinus avellanari-us) 35,7 Собака (Curtis fainilia-ris) Шакал (Cauls aureus) 38,5 38,5
Барсук (Meles tneles) Медведь закавказский (Ursus arctos caucasicus) Енотовидная собака (Nyctereutes procyonoi-des) 37,8 37,0 37,8 Лисица (Vulpes vul-pes) 39,0
Многие исследователи указывают, что температура тела млекопитающих, впадающих в спячку, отличается большим непостоянством даже в период бодрствования,
139
Рис. 32. Сезонные изменения температуры тела и потребления кислорода у двух видов сусликов' краснощекого (Citellus erythrogenys) и длиннохвостого (Citellus undulatus) (по .Михневич, 1969)
например у ежей, сонь, сусликов. Гак, суточные колебания температуры тела ежа могут достигать около 2,5° С (Herter, 1933) , хомяка — от 2,1 до 7,0° С (Horwath, 1878; Kayser, 1939а, 19396, 1940а, 19406). Таким образом, у зимоспящих форм и в период бодрствования наблюдается более низкая температура тела, подверженная более значительным колебаниям под влиянием изменяющихся температур среды. У бодрствующих сусликов при круглогодичном содержании при комнатной температуре можно было наблюдать закономерные снижения уровня обмена веществ и температуры тела в месяцы, соответствующие периоду спячки в природе (рис. 32). У наземных хищных (см. табл. 4)—барехка, медведя, енотовидной собаки, для которых характерен лишь продолжительный зимний сон, температура тела также несколько снижена по сравнению с другими хищными млекопитающими (Слоним, 1952).
В период зимней спячки температура тела снижается, как правило, до уровня температуры окружающей среды, лишь на доли градуса превышая ее. Другим не менее характерным проявлением спячки является резкое снижение обмена веществ (потребления кислорода, выделения углекислоты) в 10 и более раз по сравнению с состоянием бодрствования. Уровень обмена зависит от температуры среды, глубины спячки, но вообще он остается довольно постоянным па протяжении многих месяцев.
О величинах снижения теплопродукции во время зимней спячки у разных млекопитающих можно судить по таблице 5.
По-видимому, понижение обмена наступает в одинаковой мере у всех видов вне зависимости от величины поверхности тела. Следовательно, в состоянии зимней спячки закон поверхности тела теряет свое значение.
Наблюдается резкое замедление частоты дыхания, а иногда и прекращение его. В этом случае обмен газов осуществляется благодаря так называемым кардпопнев-матпческпм движениям, когда сердечные сокращения, также редкие, вызывают толчок, который способствует перемешиванию воздуха в воздухоносных путях, достаточный для того, чтобы удовлетворить резко сниженный кислородный запрос. Так, например, у ежа частота дыхания в бодрствующем состоянии достигает 40—50 в минуту, а при впадении в спячку падает до 6—8 и даже до
141
Таблица 5
Теплопродукция у некоторых зимоспящих в летний (при бодрствовании) и зимний период (в спячке)
(по Kayser, из Eisentraut, 1956)
Зим осп я nine Летом Зимой
Вес тела G) Калории (кг, ч) Вес тела (г) Калории (кг.ч)
Сурок обыкновенный (Martnota bobac) . . . 1868 2.1 2146 0,086
Еж европейский (Eri-naceus europaeus) . . . 684 3.5 600 0,075
Суслик малый (Citel-lus pygmaeus) 227 4.5 275 0,085
Соня-полчок (Gils glis) 127 5.0 130 0,069
Соня орешниковая (Muscardinus avellanarius) 19 13,0 23 0,170
1 в минуту (Suomalaincn, 1944). У сурка частота дыхания понижается с 25 до 4—5 в 1 мин (Benedict а. Lee, 1938). Подобные же данные имеются для зимоспящих сусликов, летучих мышей и т. д. Во время зимнего сна у хищных (барсука, медведя) частота дыхания заметно не снижается.
Тщательно изучены изменения дыхания орешниковой сопи (Muscardinus avellanarius) при зимней спячке и пробуждении (Томилин, 1958). Во время глубокой зимней спячки обнаружены дыхательные паузы продолжительностью до 11 мин. Такие паузы чередуются с сериями дыхательных движений. При пробуждении (повышении температуры тела) длительность дыхательных пауз возрастает, а число дыханий между паузами уменьшается; при понижении температуры имеют место обратные соотношения. При температуре среды около —5,5° С наступает пробуждение, и температура тела достигает 34,5— 36,0° С, т. е. температуры тела бодрствующего животного.
Очень сильно изменяется во время зимней спячки кровообращение. Частота сердечных сокращений падает у ежа с 230 до 24 в минуту (Suomalainen a. Sarajas, 1951). У суслика частота пульса урежается в 8—10 раз
142
(Johnson, 1929). Минутный объем сердца может у суслика снижаться в 65 раз (Popovic, 1964). Резко замедляется во время спячки кровоток. Если время оборота крови у бодрствующего животного — кролика составляет 7— 8 сек, то у спящего сурка — 3—4 мин. Большое количество крови депонируется в селезенке. Кровяное давление падает с 70—72 мм Hg до 16 мм Hg в сонной артерии (Dubois, 1896).
Большой интерес представляют морфо-физиологические особенности сердца зимоспящих. Сердце зимоспящих
Рис. 33. Электрокардиограмма европейского ежа (по Suonialainen и Sarajas, 1951) I, II, III — отведения:
I — в состоянии бодрствования, 2 — в зимней спячке
млекопитающих способно к ритмическим сокращениям даже при 0° С, тогда как у незимоспящих его остановка происходит уже при температурах около 15° С (Lyman а. Chatfield, 1956; Sarajas, 1960; Johansson, 1967, 1968). При этом у зимоспящих никогда не наблюдается явлений фибрилляции желудочков при выходе из состояния гипотермии, что является обычным осложнением при попытках вывести незимоспящее животное из гипотермии, вызванной охлаждением. Эта очень интересная особенность сердца зимоспящих получила свое объяснение недавно. Оказалось, что клетки миокарда зимоспящих совершенно не иннервируются адренергическими волокнами. Эта иннервация ограничивается лишь сосудами миокарда и повышение содержания катехоламинов, имеющее место при пробуждении, не вызывает явлений фибрилляции желудочков (Nielsen a. Owman, 1968). Во время спячки резко уменьшается количество мочи, а концентрация ее 4значительно возрастает.
143
Терморегуляция и температурная чувствительность не полностью выключена. Резкое охлаждение зимоспяще/о животного обязательно вызывает пробуждение. Можно думать, что в условиях зимней спячки и значительного выключения терморегуляционной активности сохраняется «сторожевой пункт» в центральной нервной системе, раздражение которого вызывает пробуждение.
Глубоким изменениям подвергается также качественная сторона обмена веществ. Так как удовлетворение пониженных энергетических потребностей организма во время спячки осуществляется за счет накопленных жиров, то в это время наиболее интенсивные изменения происходят в жировом обмене. В теле летучей мыши до зимней спячки было найдено 21,2 г жирных кислот на 100 г веса тела, а после 58 дней спячки — лишь 12,6 г (Reach, 1910).
Различные жиры, отложенные к концу лета, расходуются на протяжении зимней спячки неодинаково. Зимняя спячка сравнительно мало отражается на количестве внутреннего жира у тарбаганов; в этот период сильно расходуется подкожный жир. К концу спячки у летучей мыши (Myotis myotis) точка плавления жира оказалась повышенной (Mann, 1936). Изменяется также иодное число, кислотность жира и другие его химические и физические свойства.
Особую роль в протекании зимней спячки и в пробуждении играет скопление бурого жира, или бурой жировой ткани. До недавнего времени этой ткани приписывали роль своеобразной эндокринной железы. Однако в последние годы благодаря химическим и гистохимическим исследованиям роль бурого жира становится более ясной (Joel, 1964). Анатомически бурый жир располагается в области сердца, диафрагмы и в межлопаточной области вдоль позвоночника. Цвет бурой жировой ткани обусловлен скоплением сосудов, богатством митохондрий и цитохрома. В межлопаточной бурой жировой ткани суслика содержание цитохромов в 4 раза больше, чем в ткани мозга или мышц.
Бурый жир встречается не только у зимоспящих организмов, но у них он подвержен значительным сезонным колебаниям. Особенно значительно убывание бурого жира в процессе пробуждения от зимней спячки. Так, из общего содержания липидов в бурой жировой ткани сусли
144
ка в процессе пробуждения исчезает около 45%, а по весу эта ткань теряет около 22%.
Потребление кислорода бурой жировой тканью очень значительно. У суслика (Citellus tridecimlineatus) оно здесь гораздо выше, чем в ткани почек того же животного (Hook a. Barron, 1941). Особенно ярко этот факт проявляется при инкубировании срезов бурой жировой ткани при разных температурах и при добавлении гормонов и ферментов (см. табл. 6).
Таблица G
Потребление кислорода срезами бурой жировой ткани и органов суслика при разных температурах и добавлении гормонов и ферментов
(по Joel, 1965)
Тк.ШИ II орг; цы Ии куб ’НИИ при 37° С (.ц.Ю., 100 мг свежей ткани час) Инкубация при 8° С (% от скорости при 37° С) Количество нар инкубаций
Бурая жировая ткань без добавления гормонов 539 ±15 31 ±3 12
+ норадреналин . . . 719 ±32 38+3 12
+ норадреналин + сукцинат 1260 ±60 23 ±5 Г)
Белая жировая ткань без добавления гормонов 10 ± 1 9±1 7
+ норадреналин . . . 14+2 >±3 7
Печень 228 ±18 20 ±1 4
Диафрагма 214 ± 18 20±2 7
Почка 418 ±24 8.2±0,4 .)
Наблюдается резкое уменьшение содержания гликогена в бурой жировой ткани после пробуждения. Таким образом, бурая жировая ткань представляется в настоящее время субстратом повышенных окислительных процессов при пробуждении и источником повышенного теплообразования (подробно см. гл. V). Высказывается предположение, что в процессе разогревания используется бурый жир, непосредственно прилегающий к жизненно важным органам — сердцу, спинному мозгу. На ранних стадиях
145
разогревания (пробуждения) кровообращение оказывается особенно интенсивным в этих отделах бурой жировой ткани, что сопровождается и начальным повышением температуры. В последующие стадии разогревания бурая жировая ткань является, вероятно, источником значительного повышения содержания жировых кислот в крови.
Во время спячки дыхательный коэффициент падает до очень низких величин, что, однако, связано, по имеющимся данным, с удержанием некоторого количества углекислоты кальцием и магнием костей.
Существуют многочисленные доказательства значительных изменений углеводного обмена во время зимней спячки. Уже давно (Valentin, 1857; Bernard, 1859) установлено, что содержание гликогена в печени в период спячки увеличивается, а в период пробуждения падает. Содержание сахара в крови во время спячки резко снижается, доходя до 33 лгг%, и повышается в период пробуждения до 180 мг% и более (Dubois, 1896); количество гликогена на протяжении спячки закономерно падает в печени, сердце, поперечнополосатой мускулатуре (Leduc et Lyman, 1952). Между уровнем сахара крови и температурой тела во время спячки наблюдаются прямые соотношения: чем ниже температура тела, тем больше выражена гипогликемия (Endrews, 1930; Фердманн и Файн-шмидт, 1932; Suomalainen, 1944). При пробуждении уровень молочной кислоты в крови нарастает (Фердманн и Файншмидт). Таким образом, для зимней спячки характерно своего рода блокирование углеводных резервов в организме. С пробуждением начинается их интенсивное расходование. Этому способствует энергичная мышечная деятельность — дрожание, связанное с пробуждением. Что касается обмена азота в организме зимоспящего, то в период спячки он резко снижается.
Своеобразным глубоким изменениям во время зимней спячки подвергаются процессы регуляции постоянства внутренней среды (гомеостазиса).
Еще в конце XIX столетия было установлено (Dubois, 1898), что в крови спящих животных (сурков) обнаруживается повышенное содержание углекислоты. Это дало повод этому исследователю рассматривать спячку (да и сон вообще), как проявление наркоза, вызванного углекислотой (карбонаркоза). Резкое нарастание содержания
146
углекислоты связано с понижением возбудимости дыхательного центра. В последние годы установлен ф.акт значительного повышения щелочных резервов крови у спящего ежа (Филатова, 1946). В поддержании желочно-кислотного равновесия в период зимней спячки играет важную роль и выделение значительного количества кислого желудочного сока. Благодаря этому освобождается значительное количество щелочных оснований крови, связывается дополнительное количество угольной кислоты и уменьшается возбудимость дыхательного центра. По-видимому, в основе резкого сокращения частоты дыхания и уменьшения легочной вентиляции во время спячки лежат оба механизма — понижение возбудимости дыхательного центра и увеличение щелочных резервов крови (Персон, 1950). Изменяются и физико-химические свойства крови— удлиняется время свертывания. Механизм происхождения этих явлений пока не выяснен.
Изменения в нервной системе и соответственно рефлекторной деятельности имеют в основе выключение высших отделов центральной нервной системы: коры, подкорковых узлов, области промежуточного мозга. Все же сложные координации физиологических процессов, связанные с деятельностью продолговатого мозга, сохраняются. Больше того, во время спячки проявляются ярко выраженные рефлексы, которые отсутствуют в бодрствующем состоянии животного. Все эти рефлексы относятся к поддержанию позы в состоянии спячки. Обычно спящее животное принимает характерную позу: ежи сворачиваются в клубок благодаря сокращению специальной мускулатуры; для летучих мышей характерно подтягивание на задних конечностях (во время спячки они висят вниз головой). Установлена точная локализация этих сложных рефлексов в области продолговатого мозга. Таким образом, можно считать, что глубокая спячка характеризуется отнюдь не выключением, а усилением деятельности отдельных участков продолговатого мозга (Mersbacher, 1903).
У многих млекопитающих во время спячки сохраняется способность производить некоторые движения. Особенно ярко это выражено у летучих мышей. Если висящую головой вниз в состоянии глубокой спячки летучую мышь поместить в клеточку, а затем перевернуть ее таким образом, что животное окажется головой вверх, то можно на-
147
олюдать медленное переползание оез одновременного повышения температуры тела и обмена, т. е. без пробуждения животного. Многие исследователи обращают внимание на перелеты летучих мышей с места на место в пещерах во время зимней спячки. По-видимому, здесь имеют место самые различные степени пробуждения животных. Однако, несмотря на значительные передвижения внутри зимних убежищ, летучие мыши во время спячки нищи не принимают. Вопрос этот требует дополнительных исследований и представляет большой физиологический интерес для изучения уровней двигательных координаций.
Относительно высшей нервной деятельности во время зимней спячки сказать что-либо трудно. Опыты с попытками выработать условные рефлексы во время зимней спячки не проводились. Ранее же образованные условные рефлексы во время спячки выпадают.
Имеются данные о том, что результаты дрессировки животных исчезают даже после кратковременной зимней спячки. Так, хомяки, приученные брать пищу из рук человека, теряли эту способность даже после короткой спячки и к ним возвращалась их природная дикость (Portig, 1950).
У ежа после кратковременной спячки наблюдалось исчезновение искусственного двигательного условного рефлекса на время кормления (в утренние часы) (Филатова, 19496). У летучей мыши (ночница, Nyctalus noctula) хорошо выработанный двигательный условный рефлекс на место кормления исчез после зимней спячки, продолжавшейся только одни сутки со снижением температуры тела животного до 3,5° С (Эйзептраут, 1956). Исчезновение всех явлений приручения после кратковременной зимней спячки наблюдал этот же автор и у ушастого ежа. В этом отношении зимняя спячка сильно отличается от влияний гипотермии, после которой (Калихман, 1949; Malmejac et al., 1956) глубоких изменений условнорефлекторной деятельности не обнаруживается. Механизм такого рода явлений заслуживает самого тщательного изучения.
Однако во время зимней спячки у летучих мышей, по крайней мере, сохраняется суточный (циркадный) ритм обмена веществ. Он проявляется только в определенных температурных условиях, но отражает режим по
148
коя и бодрствования, имевший место в летний период (Попугаева и Слонам, 1953).
Следовательно, можно думать, что образованные условные рефлексы на время частично сохраняются и во время самой зимней спячки, ио исчезают при пробуждении. Но, если предположить, что кора головного мозга во время спячки выключена, то замыкание этой связи, очевидно, осуществляется за счет нижележащих центров го ловного мозга.
Вопрос о возможности сохранения памяти во время зимней спячки в последние годы вновь привлек внимание исследователей в связи с электрофизиологическими данными о выпадении во время зимней спячки электрической активности мозга, лежащей в основе циклического возбуждения— реверберации. Реверберация, по мнению многих исследователей, является механизмом удержания кратковременной, образной памяти. В опытах над европейскими сусликами (Citellus citellus), у которых осенью были образованы навыки прохождения через лабиринт, было показано, что эти навыки хорошо сохраняются после длительной зимней спячки на 32 й день после пробуждения. Это приводит к общему выводу, что долговременная память связана со структурными изменениями в нервной системе и сопровождается возрастанием содержания РН1\ и белковых субстанций в мозг) (Mihailovic, Petrovic-MTiic, Protic a. Divac, 1968). Значительное повышение содержания РНК в гиппокампе было обнаружено при пробуждении из спячки у краснощекого суслика (Citellus erythrogenys)-, после полного пробуждения содержание РНК не отличалось от содержания во время спячки (Семешина, 1969). Таким образом, различные формы памяти в разной мере сохраняются как при искусственной гипотермии, так и при естественной зимней спячке.
ФАКТОРЫ, ВЫЗЫВАЮЩИЕ СПЯЧКУ, ЗАСЫПАНИЕ
И ПРОБУЖДЕНИЕ
Понижение температуры внешней среды (если исключить летнюю спячку)—основной ф,актор, вызывающий проявление спячки на фоне остальных необходимых условий. Смена температуры среды — ее повышение — относится к важнейшим факторам, вызывающим пробуж
149
дение. У субтропических видов зимняя спячка наступает при относительно очень высокой температуре среды. Так, у африканского и палестинского видов ежей впадение в зимнюю спячку наблюдалось при температуре среды от 19 до 23° С (Herter, 1957).
Обычно зимняя спячка протекает при 1,5—16° С (Herter, 1934; Eisentraut, 1956). Если температура становится ниже этих пределов, наступает пробуждение животных. Указанный температурный режим поддерживается в течение зимы в норах, пещерах и других убежищах и играет важную роль в протекании зимней спячки у различных видов. В природных условиях наступлению зимней и летней спячки предшествуют миграции и переселения животных, постройка специальных зимовочных нор, закрываемых земляной пробкой, сосредоточение животных в группы и т. д. Все это имеет значение для создания постоянной сравнительно низкой — с небольшими колебаниями, температуры среды, т. е. условий, в которых может осуществляться спячка. Натуралистами-зоологами описаны различные формы убежищ и нор, в которых проходит зимняя спячка (Калабухов, 1936; Eisentraut, 1956 и др.). Иногда, например у хомяка, постройка зимовочной норы сопровождается и накоплением обильных запасов корма (Свириденко, 1957).
Такие факторы среды, как освещение, не оказывают непосредственного влияния на спячку. Возможно, здесь имеет некоторое значение влияние его на образование витамина D. Ограничение питания ускоряет впадение в зимнюю спячку, особенно в сочетании с влиянием низкой температуры среды. В этих условиях у ежей при искусственном охлаждении удается наблюдать спячку даже в летние месяцы (Mimachi u. Weinland, 1910; Herter, 1957 и мн. др.).
Важным биохимическим аспектом изучения зимней спячки является изучение роли накопления аскорбиновой кислоты в организме в различные сезоны и соотношение ее содержания с витамином Е — токоферолом. У грызунов, впадающих в зимнюю спячку, происходит закономерное повышение содержания токоферола в печени и аскорбиновой кислоты в надпочечниках (Калабухов, 1962; Бондаренко, Калабухов, Козакевич, Скворцов, 1968). Так как токоферол (Т) тормозит уровень обмена веществ в тканях и тем самым подавляет выведение и
150
расход аскорбиновой кислоты (АК), то их соотношение АК/Т служит важным регулятором физиологического' состояния зимоспящих животных. Самый процесс впадения в спячку и выход из нее рассматривается как результат влияния соотношения АК/Т. Этот индекс закономерно изменяется у зимоспящих животных, снижаясь в период 'впадения в спячку и при выходе из .нее. В летний период содержание АК резко возрастает, и индекс значительно повышается.
Зимняя спячка сопровождается характерными изменениями водно-электролитного обмена. В начале спячки и конце ее резко возрастает содержание в легких и в почках гиалуронидазы — фермента, обеспечивающего функцию удержания воды в организме (Григоренко, 1967). Установлено и уменьшение содержания воды в легких в тот же период (Высочина, 1967). Большие запасы жира обеспечивают у сусликов поддержание водного баланса при лишении их воды.
Огромное значение для возникновения и протекания зимней спячки имеет состояние гормональных систем организма. Основную роль здесь играет щитовидная железа, гормон которой — тироксин — повышает обмен веществ и усиливает процессы химической терморегуляции. Падение содержания тироксина осенью и одновременная дегенерация щитовидной железы определяют фон, на который оказывают влияние внешние воздействия.
В период подготовки к спячке резко падает содержание адреналина в крови — с 112 до 70 лгг% (Suomalainen, 1944). Однако, по мнению некоторых авторов, эти колебания находятся в пределах физиологических изменений, имеющих место на протяжении года. Тем не менее снижение уровня сахара в крови трудно связать с чем-либо иным, кроме понижения содержания адреналина в крови.
Введение инсулина вызывает состояние, близкое к спячке, а гипертрофия островков Лангерганса подтвер ждает значение этого гормона для зимней спячки. Несомненное влияние на протекание зимней спячки оказыва ют гормоны гипофиза и половые.
Однако гормональные сдвиги сами по себе не могут привести к состоянию зимней спячки. Они приводят организм зимоспящего в состояние готовности, т. е. такое состояние, когда внешний стимул — изменения температу
151
ры, питания и т. д. могут вызвать появление «дремоты» (см. ниже) и впадение в зимнюю спячку.
У зимоспящих организмов наблюдаются ярко выраженные сезонные изменения тонуса вегетативной нервной системы. В период спячки повышается чувствительность парасимпатической вегетативной нервной системы (Кратиков, 1947). Одновременно понижается чувствительность симпатической нервной системы (к адреналину). Удаление верхних шейных симпатических узлов приводит к более быстрому впадению в спячку и к большей ее продолжительности (Арбузов, 1952). Пробуждение сопровождается появлением симпатико-миметических веществ в крови.
В период глубокой зимней спячки у сусликов красно-щекого (Citellus erythrogenys) и длиннохвостого (С. ип-dulatus) наблюдается низкая секреторная активность ги-поталамо-гипофизариой нейросекреторной системы, повышающаяся к весне и уже резко повышающаяся к моменту пробуждения (Юрисова и Поленов, 1967). При этом повышается как образование нейросекрета, так и его выведение в кровь. Это приводит к повышению выведения антпдпуретического гормона (АДГ), возрастанию гиалуронидазной активности тканей.
Сроки наступления этих изменений у упомянутых двух видов сусликов различны. У краснощекого суслика снижение нейросекреции супраоптического ядра гипоталамуса наступает раньше, чем у длиннохвостого, что соответствует срокам впадения в зимнюю спячку. Таким образом, возможен центральный механизм регуляции гормональных отношений при зимней спячке.
Переходная стадия от бодрствования к спячке начинается с расстройства высшей нервной деятельности, выражающегося в том, что животное хорошо реагирует на сигнальные раздражители, но соотношение между условными и безусловными рефлексами нарушается. Стадия перехода от бодрствования к спячке была названа сонным опьянением (Schlaftrunken; Merzbacher, 1903 и др.) и состоянием дремоты (Пашутин, 1902). Еж перед впадением в зимнюю спячку реагирует на пищевые сигналы, по пищу почти не принимает. Появляется резкая оборонительная реакция, растормаживаются дифференцировки. Изменяется и двигательное поведение животного, характеризующееся качаниями из стороны в сторону. Наконец,
152
животное окончательно перестает принимать пищу и засыпает. Температура тела при этом снижается на 5—8° С, по еще поддерживается на высоком уровне.
Явления сонного опьянения удается вызвать экспериментально путем образования условного рефлекса. Если ежа, подготовленного к зимней спячке, в поздний осенний период систематически помещать в охлажденное помещение, можно выработать условный рефлекс на охлаждение (состояние дремоты). После ряда таких сочетаний (около 15—17), поместив ежа в урочное время и в то же помещение, но нс охлаждая его при этом, можно наблюдать дремоту— сонное опьянение, возникшее как условный рефлекс. При неподкреплспип этот рефлекс через несколько дней угасает (Исаакяи, 1946).
Пробуждение — переход от спячки к бодрствованию, сопровождается быстрым повышением температуры тела, очень большим повышением обмена веществ и соответствующими изменениями дыхания и кровообращения. Особенно быстро и последовательно протекает пробуждение у летучих мышей. Процесс сопровождается сначала сокращением мышц дистальных отделов конечностей, а затем судорожными сокращениями охватывается вся поперечнополосатая мускулатура. Резко повышается содержание сахара в крови (Архангельская, Канфор, Слоним, Черкович, 1949).
Зависимость продолжительности зимней спячки от температуры тела равна логарифму величин температуры тела в пределах от 2 до 25°С (Twente a. Twente, 1965). Однако эти отношения не остаются постоянными на протяжении всей зимней спячки.
Изучение двигательного поведения во время спячки у американского суслика (Citellus lateralis) с вживленными термопарами показало, что температура тела осенью во время спячки постепенно падает, а весной постепенно повышается. Таким образом, наблюдается закономерный сезонный цикл изменений температуры тела во время спячки (Pengelley a. Fisher, 1961; Twente a. Twente, 1967). Обнаружена и сезонная периодика изменений чувствительности при введении солевых растворов норадреналина. При введении норадреналина период пробуждения сокращается как при учащении пробуждений, так и в связи с потерей животными веса в конце зимней спячки (Даудова и Ипатьева, 1968).
153
Большой интерес представляет химическая динамика, лежащая в основе повышенного теплообразования при пробуждении. Концепция разобщения между тканевым дыханием и окислительным фосфорилированием (отношение P/О) привлекла внимание многих исследователей в связи с проблемой адаптации к холоду и природой несократительного термогенеза в скелетной мускулатуре (Скулачев, 1962; Гусева, 1964; Маслов, 1964; см. также гл. V). Разобщение между дыханием и окислительным фосфорилированием можно наблюдать у летучих мышей Myotis a. austroriparius при инкубации митохондрий печени только при температурах 13 и 21° С (Horwitz, Nelson a. Popovic, 1967). Понижение отношения P/О связывается авторами с дифференцированной температурой чувствительностью ферментов по отношению к различным субстратам. Некоторые (Даудова и Ипатьева, 1968) высказывают сомнение в значительной роли явлений разобщения в разогревании при пробуждении. Механизм, вызывающий изменения в этих процессах, до сих пор остается неясным.
Пробуждение наступает обычно в различные сроки: от 15—20 мин до 2 ч в зависимости от глубины спячки и условий среды. Во всяком случае, оно происходит много быстрее, чем переход от бодрствования к спячке, продолжающийся 10—14 ч, а иногда (у ежей, сусликов, сурков) и больше (Suomalainen a. Suvanto, 1953).
В природных условиях пробуждение вызывается повышением температуры среды. Это доказывается специальными экспериментами над сусликом (Citellus latera-lus), когда период спячки в интервале температур от +2 до +25° был тем короче, чем выше температуры среды (Twente a. Twente, 1965). Само пробуждение сопровождается повышением содержания норадреналина в крови и может быть вызвано введением гормона интраперитонеально. При этом резко возрастает частота пульса, но не наступает первичного повышения теплообразования. Вероятно, норадреналин оказывает влияние на возбудимость гипталамической области мозга (Twente a. Twente, 1968).
ФОРМЫ ЗИМНЕЙ СПЯЧКИ. ЛЕТНЯЯ СПЯЧКА
Имеющийся материал позволяет выделить 3 группы зимоспящих млекопитающих и соответственно три типич-154
ные формы спячки как по факторам, ее вызывающим, так и по характеру ее протекания.
Первая форма зимней спячки тесно связана с понижением температуры среды. По этому типу протекает спячка всех видов сонь — грызунов, распространенных в СССР на юге. Они засыпают сразу при понижении температуры среды. В горных местностях при понижении температуры можно наблюдать, как они засыпают каждую ночь, что позволяет брать их из гнезда неподвижными.
На сонях Muscardinus avellanarius был проделан интересный опыт с продлением зимней спячки. Они были завезены на высокогорную метеорологическую станцию и содержались в ящике па снегу; спали 345 дней, после чего проснулись совершенно нормальными. Это — факультативный тип спячки, вызванной исключительно низкой температурой внешней среды.
Вторая форма спячки связана с накоплением жира. В эту группу входит большинство зимоспящих животных: суслики, сурки, ежи. Эти животные впадают в спячку только при особых условиях: накоплении жира в большом количестве и — для некоторых видов—аскорбиновой кислоты.
Третья форма спячки — зимний сон. Зимний сон имеет место у некоторых хищных (у медведей, барсуков) и представляет переходную форму между состоянием зимоспящего и незпмоспящего организма. Температура тела во время зимнего сна понижается очень незначительно— только на 1.5—2° С. Обмен веществ понижается примерно на 50—70%. Резко уменьшается двигательная активность, что ведет к большой экономии энергии. Наблюдается некоторое урежение частоты дыхания и сердечных сокращений. По существу, такой сон является как бы удлинением ежесуточного сна. Барсук, например, к началу зимы все меньше и меньше двигается и наступают дни, когда он совсем перестает выходить из норы и питаться.
Вопрос о том, каким же образом осуществляется обмен веществ, поддержание жизни во время зимнего сна, очень давно интересовал исследователей. У барсуков зимой в период бодрствования был обнаружен высокий обмен веществ (Пашутин, 1902). В условиях субтропической зимы (Сухуми) у барсуков удалось наблюдать очень
155
характерную особенность: животное, как правило, отказывается от пищи, ио исследования газообмена показывают, что в те периоды, когда оно просыпается и удастся его покормить, полностью выпадает специфическое динамическое действие пищи. Следовательно, во время зимнего сна нарушаются не только питание, но и .процессы ассимиляции (Слоним и Щербакова, 1949).
Наряду с зимней спячкой у многих животных встречается летняя — эстнвация. Летняя спячка характеризуется теми же явлениями, что и зимняя, только начинается и протекает она при высокой температуре среды. Наблюдаются те же фазы — дремота, падение температуры тела, ограничение физиологических функций. Летняя спячка, как правило, связана с высыханием эфемерной растительности в пустынях и полупустынях, с наступлением неблагоприятных условий для питания животных. Летняя спячка встречается у насекомоядных, грызунов и даже у полуобезьян лемуров (Мадагаскар) в засушливое время года.
Механизм летней спячки совершенно не изучен. Известно только, что она может быть вызвана у животного, уже подготовленного к спячке, накопившего достаточное количество жира (а для некоторых видов грызунов—и достаточное количество аскорбиновой кислоты, которую они плохо синтезируют). Определенную роль здесь играют и внешние раздражители. Если взять взрослого ожиревшего желтого суслика (Citellus fulviis) и иммобилизовать его, растянув за лапки, то можно наблюдать резкое падение температуры. Животное тут же засыпает, не делая попыток к бегству. Сон продолжается в течение нескольких часов. Если дать такому животному поголодать, пробудив его, то у него снижается содержание аскорбиновой кислоты в крови и вновь ввести его в спячку не удается (Исаакяи и Фельбербаум, 1949; Филатова, 19496).
В условиях эксперимента малый суслик впадает в летнюю спячку при полном водном голодании (содержании на сухом корме) (Калабухов, 1936). Наблюдения на желтом суслике этих фактов не подтвердили: при содержании на сухой диете желтые суслики действительно резко ограничивают двигательную активность, но не снижают температуры тела и реагируют на внешние раздражители характерным свистом. Таким образом, вопрос о том,
156
какие факторы непосредственно вызывают состояние летней спячки, остается открытым.
Спячка встречается у представителей многих отрядов млекопитающих. Низшие млекопитающие — утконос, ехидна не впадают в зимнюю спячку, хотя в холодное время года у них наблюдается ограничение активности (Martin, 1901; Eisentraut, 1956). В условиях охлаждения и недостаточного питания имеет место некоторое подобие холодовой летаргии (Krieg, 1924). Эти явления можно наблюдать и у сумчатой крысы (Dydelphis Virginia па).
У насекомоядных явления зимней спячки представлены очень широко. Она хорошо выражена у среднеазиатского ушастого ежа, не говоря уже о еже европейском (Eiseittraut, 1956). Виды ежей, обитающих в Сахаре, в зимнее время при небольшом понижении температуры впадают в спячку (Herter, 1952).
Зимняя спячка рукокрылых (летучих мышей) охватывает представителей всех палеарктических и нсоарктиче-ских вптов. У тропических форм, например летучих собак (Megachiroptera), се нет.
Спячка наблюдается у большого количества видов грызунов-—сусликов (большинство видов), сурков, сонь, хомяков и т. д. У многих из них, например сусликов, летняя спячка непосредственно переходит в зимнюю (Кала-бухов, 1936; Кашкаров и Коровин, 1936).
Существуют немногочисленные указания о летней спячке у некоторых полуобезьян (лемуров на Мадагаскаре). В неволе у лемура-потто эти полевые наблюдения не подтвердились (Эйзентраут).
У представителей американских неполнозубых (Хе-nar’thra) настоящей зимней спячки не наблюдается, хотя температура тела у многих представителей низка и отличается значительными колебаниями.
Что касается хищных, то, как об этом уже говорилось, они в зимний период погружаются на некоторое время в зимний сон. не сопровождающийся снижением температуры тела.
В условиях зимней стужи и затрудненной добычи пищи многие грызуны, хищные, особенно мелкие, остаются многие дни в покое. Однако эти явления нельзя аналоги -зировать не только с зимней спячкой, но даже с явлениями зимнего сна.
157
В онтогенезе способность к впадению в зимнюю и летнюю спячку появляется сравнительно поздно. На примере многих видов сусликов можно наблюдать, что первыми в зимнюю и летнюю спячку укладываются взрослые зверьки прошлых лет рождения. Молодняк текущего года рождения длительное время остается активным и впадает в спячку только по мере накопления достаточного количества жира. Так, молодняк горной популяции малого суслика отличается более поздним сроком залегания в спячку и более поздним накоплением жира. У молодых животных с еще несформироваипой терморегуляцией легко наступает состояние гипотермии, но они не впадают в спячку в полном смысле слова.
У летучих мышей переход от гомойотермип к пойки-лотермии нарушается, если удалить большие полушария мозга (Слоним, 1938). Летучая мышь остается после такой операции или активной (теплой) пли в состоянии покоя (холодном) многие часы. Сходные данные нарушения суточной активности и поведения (полета) были описаны Мерцбахером (Merzbacher, 1903). Ряд обстоятельств говорит о том, что зимняя спячка не есть приспособление специально к переживанию зимы, а лишь к более или менее неблагоприятным условиям существования. В условиях Заполярья с его чрезвычайно коротким летом и очень продолжительной зимой зимняя спячка была бы, конечно, неосуществима, поскольку за короткий летний период животное не могло бы накопить нужного энергетического потенциала — необходимого количества жира. На средние широты падает наибольшее количество впадающих в спячку видов. Особенно ярко это выражено в тех климатических условиях, где есть колебания температуры и влажности, например в условиях полупустынь и сухих степей. Здесь воздействуют два фактора — уменьшение доставки воды в организм — выгорание растительности, ведущее к летней спячке, и неблагоприятный зимний период (недостаток корма), ведущий к зимней спячке. В этих условиях спячка некоторых видов продолжается около 8 месяцев.
Теории зимней с п я ч к и. Теории зимней спячки довольно разнообразны и отражают отдельные стороны этого явления, установленные в разное время различными авторами.
158
Наиболее простая теория объясняет явления зимней спячки недостаточностью терморегуляции животного, недостаточным поддержанием других сторон гомеостазиса. Эта теория разработана иа летучих мышах (Irving а. Krog, 1954) и получила довольно широкое развитие в работах по искусственной зимней спячке (Лабори и Гюге-иар, 1956). В отечественной литературе эта теория защищалась В. С. Галкиным (1947). Совершенно очевидно, что эта теоретическая концепция никак не может объяснить всей сложной цепи явлений, связанных с зимней спячкой. Зимняя спячка является очень сложным и тонко регулируемым состоянием животного, а отнюдь не простым подчинением его условиям внешней среды. Летучие мыши, как было показано выше, не составляют в этом отношении никакого исключения (Слоним, 1945; Пону-гаева и Слоним, 1953).
Другие теории основывались на изменении содержания определенных веществ в организме: сахара в крови (Dworkin a. Finney, 1927); аскорбиновой кислоты (Suo-malainen, 1944), содержания СО2 в крови (Dubois, 1896). Эта точка зрения также не может объяснить явлений впадения в зимнюю спячку и в равной мере механизмов пробуждения.
Важное место в теоретических представлениях о механизме зимней спячки занимают изменения эндокринных систем. Эти точки зрения были высказаны в разное время Адлером (Adler, 1926) —на основании изменений щитовидной железы, Кайзером (Kayser, 1953) —на основании изменений ряда эндокринных желез, наблюдаемых при зимней спячке. Однако ни недостаток тироксина, гормонов гипофиза, ни удаление надпочечников и т. д. не ведут к развитию картины, типичной для зимней спячки. Эндокринные железы определяют готовность животного к впадению в спячку и к пробуждению, но не определяют от начала до конца всей цепи этих явлений.
В последнее время все большее признание получает нервная теория зимней спячки. Эта точка зрения была высказана впервые еще Мерцбахером (Merzbacher, 1903), Проссером (Prosser, 1950), а в нашей литературе К- М. Быковым (1942), Н. И. Калабуховым (1950), А. Д. Слонимом (1952) и др. В пользу этой точки зрения можно привести ряд ф,актов, указывающих на то, что разрушения центральной нервной системы как в области
159
больших полушарий мозга, так и в области промежуточного мозга ведут к изменению протекания зимней спячки. Пробуждение сурка из спячки возможно только в случае целости промежуточного мозга (Dubois, 1896). Повреждение его нарушает процесс пробуждения; это связано с нарушением центров терморегуляции. Однако в самое последнее время (Maiап, 1968) доказана возможность пробуждения и спинномозговых животных.
Наиболее простая точка зрения (Prosser, 1962) исходит из изменений чувствительности центральных, регулирующих тепло аппаратов. Именно изменения состояния центров терморегуляции, по мнению Проссера, и являются причиной снижения температуры тела и приводят к изменениям физиологического состояния, характерного для зимней спячки.
ХОЛОДОВОЕ ОЦЕПЕНЕНИЕ У ПТИЦ
(ТОРПИДНОЕ СОСТОЯНИЕ)
Некоторые птицы — колибри, стрижи, козодои характеризуются удивительной способностью к впадению в сноподобное — торпидное, состояние с понижением температуры тела и резким ограничением жизненных процессов, напоминающих зимнюю спячку млекопитающих. Это состояние было впервые описано Егером (Yaeger, 1949) под названием зимней спячки стрижей и в последние годы подверглось многочисленным исследованиям (Cowles a. Dawson, 1951; Howell a. Bartholomew, 1959, Lasiewski a. Dawson, 1964).
Эта группа птиц обладает низким основным обменом по сравнению с другими видами того же веса, большой устойчивостью к высоким температурам среды. Такая своеобразная гетеротермия лучше выражена у колибри— самых мелких птиц, у которых состояние гомойотермии и пойкилотермии носит характер закономерных суточных колебаний. При помещении в темноту у колибри шести видов наблюдалась полная зависимость интенсивности газообмена от температуры среды в пределах от 0 до 20° С и значительное повышение обмена при более высоких температурах, когда птицы переходили в гомойотерм-ное состояние. В торпидном состоянии Q,o у колибри приближается к 4,1, а уровень обмена — к таковому у спящих летучих мышей или рептилий. Частота сокращений
160
сердца в активном состоянии колеблется между 600 и 1000 в минуту в покое. В торпидном состоянии эти величины не превышают 100 в минуту. Частота дыхания в активном состоянии колеблется между 400 и 180 и зависит от температуры среды. Дыхание имеет большое значение для терморегуляции: с повышением температуры среды отдача воды испарением возрастает. Даже при температуре среды 26° С суточные колебания температуры тела у колибри (Amazilia) достигают 8° С (от 32 до 40° С). Переход от покоя к активности и полету сопровождается повышением температуры тела на 5—6° С со скоростью повышения температуры 0,5° С в минуту (Morrison, 1962).
Разогревание колибри происходит за счет больших грудных мышц, в которых обнаружено множество крупных митохондрий, тесно связанных с узкими миофибриллами. Величина миофибрилл возрастает с уменьшением веса тела птицы (диаметр до 18 мк). Мышечные волокна содержат огромное количество сукциндегидрогеназы; содержание фермента выше, чем у всех изученных крупньп птиц. Все эти особенности связаны с крайне малым размером тела и особенностями разогревания (Lasiewski, Galey, Vasquez, 1965).
Значительно устойчивее температура тела у более крупных птиц — козодоев и стрижей. Однако обнаружена значительная гетеротермия у птенцов стрижей и у молодых летающих птиц (Кескпайк и Давыдов, 1967). Еще более резко выражена такая гетеротермия у козодоев. У малого козодоя (Chordeiles minor) в активном состоянии наблюдается слабо выраженная химическая терморегуляция, однако в темноте даже при температуре среды 15—20° С уровень газообмена в 3 раза меньше, чем в состоянии гомойотермип; частота сердечных сокращений снижается с 250 до 50—60 в минуту (Lasiewski а. Dawson, 1964).
Важнейшее условие самостоятельного выхода птиц из торпидного состояния — степень их упитанности. При длительном голодании птицы из состояния холодового оцепенения не выходят. Все имеющиеся до сих пор факты позволяют думать, что холодовое оцепенение (тор-пидное состояние) у птиц имеет биологическое значение, предохраняя их от излишних потерь тепла (у колибри). Однако детально роль этих физиологических особенностей некоторых видов птиц представляется еще неясной.
6—1204
Глава V
ТЕМПЕРАТУРА СРЕДЫ ОБИТАНИЯ
ОБЩЕЕ ВЛИЯНИЕ ТЕПЛА И ХОЛОДА
НА ЖИВЫЕ СИСТЕМЫ
Температурный диапазон, на фоне которого проходит жизнь в естественных условиях существования, довольно широк. Не будет преувеличением, если определить его примерно в 100° С, при диапазоне температур от —50° С до +50° С. Низкие температуры довольно обычны в условиях Арктики. Высокие температуры свойственны жарким пустыням тропических областей.
Кроме того, для многих континентальных областей (Сибирь, Центральная Азия) характерны огромные колебания температур среды в течение суток — от минусовых до высоких. Здесь адаптивные изменения в организме чрезвычайно сложны, и их изучение представляет особенный интерес для понимания законов термической адаптации. Однако все эти условия не только совместимы с жизнью низших и высших организмов, но, что особенно важно, температура тела млекопитающих и птиц поддерживается на почти постоянном уровне, несмотря на большие колебания температуры внешней среды.
Это оказывается возможным благодаря сочетанию физиологических и морфологических адаптаций: тканевых, адаптаций поверхности тела (относительной к весу величины) организмов, изменения теплопродукции, теплоотдачи (сосудистой терморегуляции, оперения, шерстного покрова, теплоотдачи дыханием), поведения (изменения позы, суточной (циркадной) активности, постройки нор и убежищ, скучивания организмов, миграций и кочевки).
У пойкилотермных животных огромное значение имеют адаптации ферментных систем, обеспечивающие протекание жизненных процессов в условиях экстремальных температур.
162
В основе реакции живых систем на изменения температуры внешней среды лежит изменение скорости химических реакций под влиянием охлаждения и нагревания. Охлаждение приводит к уменьшению энергии превращения веществ, к снижению обмена веществ и понижению скорости многих ферментных реакций. Для каждого фермента в организме имеется свой температурный оптимум— температура, выше и ниже которой скорость синтеза веществ или их расщепления снижается. Высокие температуры также ограничивают ферментные реакции и тем самым приводят организм в состояние теплового окоченения и далее к гибели. Собственно говоря, ограничение жизнедеятельности организма при охлаждении сводится не к угнетению отдельных его систем, а включает угнетение двигательной активности животного, что приводит в известных пределах к состоянию, известному как холодовое оцепенение. Такие отношения существуют у пойкилотермных организмов или у гомойотермных с выключенным аппаратом терморегуляции (например, в состоянии зимней или летней спячки, гипотермии или фармакологического выключения терморегуляции).
Регулирование тепла в организме (отдельные элементы которого имеют место и в пойкилотермном организме) не только сильно расширяет температурный диапазон высокого уровня жизнедеятельности организма, но и обеспечивает далеко идущее постоянство этого уровня. Поэтому температурные адаптации гомойотермных организмов, сохраняя некоторые элементы, свойственные пой-килотермным (например адаптации активности ферментов, использование радиационных потерь тепла, испарения с поверхности кожи и слизистых и др.)> коренным образом различаются. Здесь в основе поддержания теплового баланса лежат соотношения продукции тепла и потери — отдачи его в окружающую среду. Соотношение скорости продукции и отдачи тепла создает условия для поддержания термического гомеостазиса — поддержания постоянной температуры тела. Это же постоянство является непременным условием нормальной деятельности большинства систем организма высших позвоночных — млекопитающих и птиц, особенно деятельности их центральной нервно?! системы, достигающей высокого уровня развития. Следовательно, и холод и тепло действуют на организм гомойотермного животного не непосредствен
6*
163
но, угнетая или возбуждая клеточные и тканевые элементы, а опосредованно, через нервную систему, гормональный аппарат, т. е. через аппарат терморегуляции.
МОРФОЛОГИЧЕСКИЕ АДАПТАЦИИ
К ТЕПЛУ И ХОЛОДУ
Существование в различных температурных диапазонах в связи с климатическими условиями, образом жизни и типом питания накладывает значительный отпечаток на многие стороны морфологии животных.
Наиболее яркая зависимость между климатическими (температурными) условиями и морфологией организмов была отмечена Бергманном (Bergmann, 1848) еще в середине прошлого столетия. Правило Бергманна заключается в том, что нойкилотермпые организмы, близкие в систематическом отношении, по мере удаления от полюсов к экватору становятся крупнее, а гомойотермпые— мельче. Так как с увеличением веса животного уменьшается поверхность тела (относительно единицы веса), то такая закономерность позволяет крупным гомойотерм-ным животным Севера уменьшать потери тепла, а мелким южным формам увеличивать теплоотдачу поведением.
Увеличение размеров пойкилотермных организмов в связи с более высокой температурой обитания не находит такого простого объяснения и обычно связывается с более интенсивным ростом этих организмов в условиях высоких температур среды, с лучшими условиями питания. Впрочем, и по отношению к гомойотермным организмам правило Бергманна встречает ряд возражений и поправок. Лучше всего это правило было доказано при сопоставлении веса тела таких животных, как медведи, волки, лисицы и многие копытные. В тропических странах представители тех же родов имеют более мелкие размеры тела, чем в умеренном климате и в условиях Севера (Арктики и Антарктики). Правило Бергманна находит свое подтверждение и для птиц. Оно явилось предпосылкой для обоснования физиологического закона поверхности тела Рише— Рубнера.
Увеличение поверхности тела обеспечивает более интенсивную отдачу тепла в окружающее организм пространство даже в условиях небольшой разницы между температурой кожи и температурой окружающей среды.
164
Увеличение теплоотдачи, наряду с уменьшением веса тела, обеспечивается и изменением его формы. Как правило, поверхность тела у южных видов млекопитающих увеличена за счет более длинных ушей, длинного хвоста, длинных конечностей. Это правило Аллена полностью подтверждается при сопоставлении формы тела близких видов лисиц, некоторых грызунов.
По существу, правило Аллена является весьма важным дополнением к правилу Бергманна. Это в последнее 'время показано па примере изучения терморегуляции у африканского слона — весьма крупного животного, уши которого содержат большое количество кровеносных сосудов, а количество протекающей крови составляет до 30% от всей циркулирующей крови (Luck a. Wright, 1965). Увеличенная теплопродукция в связи с интенсивной мышечной работой приводит также к выявлению правила Аллена, например у зайцев, хотя одновременно со храняется и значение правила Бергманна.
Экспериментальные данные свидетельствуют о том, что животные одного вида, выращиваемые в условиях высоких температур, изменяют на протяжении многих поколений свой рост, т. е. увеличивают поверхность тела. Так было при переселении лисиц из Англии в Австралию. Самнер обнаружил (цит. по Hesse, 1924), что белые мыши, выращенные при температуре ±6,1° С, были крупнее выращенных при 4-26,3° С. Эти изменения стойко отли чади пх от основного штамма. Серые крысы, обитающие в холодильниках при постоянной температуре —15° С, оказались более крупными, чем обитающие на элеваторах (Шепелева, 1950). Вес тела серых крыс на элеваторе на 70 г меньше, чем в холодильнике (на 21,5%). У тропических животных наблюдается менее густой шерстный покров и отсутствует подшерсток (правило Ренша).
Однако этими общими морфологическими закономерностями не исчерпывается вся сложность приспособления животных к тропическому и жаркому климату. Немаловажную роль играет цвет шерстного покрова. Светлая окраска многих тропических животных, особенно обитающих в условиях открытых пространств, предохраняет их от прямого нагревания солнечными лучами. У лесных форм — обитателей тропического леса, эта защитная окраска отсутствует и наблюдается увеличение содержания пигмента.
165
Огромные различия теплопроводности шерстного покрова были обнаружены при сопоставлении арктических и тропических млекопитающих. У мелких млекопитающих относительная разница в теплопроводности кожного и шерстного покрова невелика; у крупных она очень значительна (Scholander, Walters, Hock a. Irving, 1950).
Шоландер на основании сопоставления всех имеющихся по этому поводу данных резко критикует правило
Рис. 34. Изменения температуры в шерстном покрове (измерение через каждый сантиметр) и температуры кожи у овец в разные сезоны (°C) (по Макевнину, I960)
Бергманна, считая, что свойства поверхности тела (теплоизоляция, васкуляризация, потоотделение) в ходе адаптивной эволюции могут совершенно изменить соотношение веса тела у тропических и полярных видов гомойотермных организмов. Адаптивные изменения шерстного покрова характерны и для сельскохозяйственных животных; исследованы различия между толщиной и характером шерстного покрова и устойчивостью температуры тела (Раушенбах, 1967).
На рисунке 34 видно, что в зимний период при температуре воздуха —40° С у овец в условиях заволжской степи температура кожи остается на уровне 35,6° С. Хорошо виден и температурный перепад в толще шерстного покрова, достигающий 3°С на 1 см длины шерсти (Макев-пип, 1960).
166
Не меньшее значение, чем шерстный покров, имеет и распределение жира в организме. У ряда обитателей юга жировые отложения не занимают всей поверхности тела, как это имеет место у обитателей холодного климата, а расположены локально (например, курдюк овец, разводимых на юге, горб верблюда и индийского рогатого скота зебу). С другой стороны, жировые отложения у тюленей расположены по поверхности тела и могут составлять до 25% от веса тела. Толщина этого жирового слоя подвержена значительным сезонным колебаниям, причем вес шкуры с жировым слоем может падать весной па 60—70%- В весенний период наблюдается физиологическое голодание этих животных, направленное на уменьшение теплоизоляции в летний период (Шепелева, 1967).
ЛУорфологические адаптации выражены также у гомойотермных организмов и в связи с различной интенсивностью обмена веществ на севере и на юге. С изменением уровня обмена веществ связана и величина сердца у крупных и мелких видов, населяющих разные климатические зоны. У мелких видов гомойотермов в условиях жаркого климата обнаружен меньший вес сердца, чем у обитателей умеренного и холодного климата (правило Гессе). Вес сердца имеет непосредственную связь с уровнем обмена веществ, более высоким у тех обитателей холодной зоны, которые имеют меньшую теплоизоляцию поверхности тела, а следовательно, большую теплоотдачу. С этим же связана и большая величина и длина кишечника в условиях Севера (правило Ренша), а у птиц и большая поверхность крыльев, обусловленная дальностью перелетов.
Имеются многочисленные указания на то, что распределение жира, в том числе и полостного, также имеет адаптивный характер. Так, у сибирского и монгольского крупного рогатого скота наблюдается мощное отложение околопочечного жира в сальнике и других частях брюшной полости. Последнее связывают с поеданием животными на зимних пастбищах мерзлого корма и значительным охлаждением при этом брюшных внутренностей. Даже у человека, например у профессиональных ныряльщиц за губками и жемчугом (ама), обнаружено увеличение отложений подкожного жира (Remmie, Covino, Howel, Song, Kang, Hong, 1962).
167
ТЕРМИЧЕСКИЕ АДАПТАЦИИ
У ПОЙКИЛОТЕРМНЫХ ОРГАНИЗМОВ
Вопросы адаптации пойкилотермных организмов к различным температурам довольно широко изучены в большом количестве исследований, направленных на решение нескольких основных проблем. Наиболее общей из них является состояние холодового оцепенения и порогов появления активности у наземных и водных форм. Поскольку у всех без исключения пойкилотермных организмов имеет место подчиненность жизненных процессов температурным условиям среды, исследование холодового оцепенения имеет не только большое теоретическое, но и практическое значение и позволяет установить влияние температурного фактора среды на основные процессы — питание, размножение и расселение этих организмов. Это же явление представляет очень удобный объект для рассмотрения как индивидуальных, так и видовых популяционных адаптаций.
Для огромного большинства пойкилотермных организмов период похолодания и понижения температуры среды до 0°С и ниже ведет пе только к холодовому оцепенению, прекращающему двигательную активность, по и резко снижает энергетический обмен. Вместе с тем это холодовое оцепенение (chill-coma) в значительной мере зависит от предшествующих температурных условий. Уже давно (Mellanby, 1939) установлено, что предшествующее содержание насекомых (имаго и в личиночных стадиях) в течение одного дня приводит к значительному снижению температурного порога, при котором наступает холодовое оцепенение (см. табл. 7).
Большой материал, собранный различными исследователями, указывает, что в процессе температурной адаптации (тепловой и холодовой) изменяется не только температура холодового оцепенения, но и летальная температура (Salt. 1964).
Вместе с тем адаптация к теплу и к холоду у пойкилотермных организмов оказывает очень различное влияние у разных видов на потребление кислорода и температурный коэффициент Qio. Этот коэффициент на протяжении уже более 50 лет привлекает внимание исследователей, так как его приложение к процессам, происходящим в целостном организме (общему обмену веществ),
168
Таблица 7
Температура холодового оцепенения у пяти видов после адаптации к трем различным температурам среды
(по Mellanby, 1939)
Вины Температуоы адаптации
14—’.7° С 30° с 36° с
Blatta orientalis . . . Cimex lectularius . . Rhodnius prolixus . . Lucilla sericata, личинка Lucilla sericata, имаго ползающ. . Calliphora erythrocep-hala, личинка Calliphora erythrocep-hala, имаго ползающ. 2.0 4,5 8,6 2,0 3,5 1.0 1.0 7,5 7,0 10,5 4,5 6.0 1,0 5, 1 9,5 7,5 12.0
или отдельным его системам (например, частоте дыхания или сердечных сокращений) позволяет сопоставлять закон физико-химических моделей с отдельными физиологическими процессами. Так, показано (Scholander, Flagg, Walters a. Irving, 1953), что арктические морские беспозвоночные и рыбы при сопоставлении с близкими в систематическом отношении тропическими видами имеют одинаковый газообмен при температурах 15—20° С. Все же наземные беспозвоночные (8 ракообразных, 4 насекомых, 1 паук) не обнаружили холодовой адаптации, кроме узкого диапазона температур от 10 до 20° С. Противоречивые данные между изменениями температуры холодового оцепенения и Qio общего обмена веществ до сих пор не нашли своего объяснения.
Вместе с тем эти данные свидетельствуют о том, что индивидуальные адаптации, и генетически обусловленные, у пойкилотермных организмов резко отличаются. Есть все основания предполагать, что ограничение возможности индивидуальной адаптации пойкилотермных животных являются важным фактором, ограничивающим географическое распространение видов.
Исследование изменений газообмена в процессе индивидуальной адаптации приобрело за последние годы
169
большое значение, так как позволяет понять механизмы, лежащие в основе описанных выше сдвигов порога холо дового оцепенения и летальных температур. Рядом исследований показано, что пойкилотермные организмы более устойчивы к высоким температурам, если они были предварительно адаптированы к ним (Gelineo, 1964).
Различные пойкилотермные организмы по-разному реагируют на экспозицию при температуре, при которой они живут в течение нескольких дней или недель. У ярко выраженных стенотермных организмов тепловую адапта цию обнаружить не удается. У некоторых эвритермных организмов, живущих при почти постоянной температуре, реакция термогенетической системы вначале либо увеличивается, либо падает. Если животное, адаптированное к высокой температуре, поместить в условия более низкой температуры, то в новой среде потребление кислорода возрастет. С другой стороны, при перенесении животного из холода в тепло потребление кислорода после определенного периода времени уменьшается.
Однако у некоторых видов не наблюдается изменений окислительных процессов при температурной адаптации. Рыба Scorpaena porcus и насекомое тополевый листогрыз (Metasoma populi) не изменяют уровня обмена после температурной адаптации.
Однако чаще наблюдается другой тип адаптации: повышение или понижение потребления кислорода при новой температуре. Во всяком случае, уровень потребления кислорода — как повышенный, так и пониженный — сохраняется после окончания процесса адаптации. У краба Pachygrapsus crassipes, адаптированного к трем различным температурам среды, при более высокой температуре адаптации потребление кислорода оказывается выше, чем при более низкой. Таким же образом адаптируются черви, моллюски Helix fluviatilis, Ancylus fluviatilis, ракообразные Emerita talpoida и Astacus fluviatilis, насекомые— черный таракан (Periplaneta americana) и другие пойкилотермные организмы.
Из рыб подобным же типом адаптации обладает Lampetra fluviatilis. При одной и той же температуре морская рыба Tautogolabrus adspersus потребляет зимой больше кислорода, чем летом. Классические работы (Wells, 1935) показали, что у Fundulus parvipinnis, адаптированных к 20—24° С, интенсивность общего обмена
170
при 12—20° С понижается на 30—40% по сравнению с организмами, адаптированными к 10—12° С, и что у Gillichthys mirabilis наблюдается та же реакция. Это было подтверждено другими исследователями на многих видах рыб.
По мнению некоторых исследователей, рептилии не обладают видовыми особенностями адаптации к холоду и потому их распространение на север ограничивается ог 52° до 66° широты в северном полушарии и до 44° в южном. Также у лягушки Rana sylvatica не обнаружено различий в активности, сезонах размножения или скорости эмбрионального развития между особями, выловленными около Нью-Йорка и на Аляске (Hock, 1964). Поэтому можно считать, что именно температурный фактор ограничивает распространение отдельных видов рептилий и амфибий на Север и что индивидуальной и популяционной адаптации у них обнаружить не удается. Температура около 15° С является нижним пределом активности для видов, обитающих на Аляске.
Совершенно иные температурные пределы активности зарегистрированы для южных видов рептилий. Так, у ночной змеи Aspis cerastes из Сахары температура снижения активности наблюдается уже при 32—34° С (Saint-Girons et Saint-Girons, 1956). Такие же данные имеются для субтропических и тропических видов ящериц. Однако у близких видов ящериц США и Мексики (разница в средней температуре на 19° С, в высоте над уровнем моря 2200 м) температура среды, при которой наблюдается двигательная активность, колеблется всего лишь от 32,9 до 36,9° С. Температура тела в период активности у близких ящериц из Аризоны и Флориды вовсе не отличалась (Bogert, 1943) (рис. 64).
Механизмы, лежащие в основе видовых адаптаций пойкилотермных организмов к температуре внешней среды, изучены еще совершенно недостаточно.
Обширные исследования показали, что теплоустойчивость клеток и тканей пойкилотермных организмов тесно связана с ареалом видов и отчасти с физиологическим состоянием организма (Ушаков и сотр., 1963, 1964, 1967). Было установлено, что в зимний и отчасти весенний периоды предварительное содержание при низких или высоких температурах таких животных, как лягушки, черепахи, моллюски, гидры, актинии, не приводит к сущест
171
венным изменениям теплоустойчивости, т. е. к изменению температуры, при которой происходят изменения (необратимые) белковых структур протоплазмы. Значительно большие сдвиги удается обнаружить в летний период, а также под воздействием гормонов. В то же время наблюдается значительная разница в теплоустойчивости клеток и тканей у организмов близких видов, живущих в разных экологических условиях.
Особо следует отметить разнообразие теплоустойчивости клеток пойкилотермных организмов на ранних стадиях эмбрионального развития. Установлены некоторые факты соответствия между теплоустойчивостью отдельных тканей и клеток (возбудимость соматической мускулатуры, мерцательного эпителия, подвижности сперматозоидов, а также отдельных тканей по витальной окраске) и температурой тела в природе температурному префе-рендуму двух видов лягушек — Rana temporaria и Rana ridibunda (Ушаков, 1964).
Все эти фактические данные указывают на значение температуры среды для функционирования протоплазмы у пойкилотермных организмов, что в свою очередь связано с целостью составляющих ее белковых комплексов. Однако в целом организме индивидуальная адаптация к различным температурам среды зависит нс только от непосредственного термического воздействия па клетки и ткани, а от влияния гормональных систем и воздействия на нервную систему (см. Precht, Hensel, Christofersen, 1955). В этом плане выявляются и изменения общего газообмена у различных пойкилотермных организмов.
Так, адаптивные изменения обмена у зеленой лягушки (Rana esculenta) заключаются в типичных различиях в потреблении кислорода при трех различных температурах. Адаптации накладываются на повышенный его уровень во время размножения. То же установлено и для травяной лягушки (Rana temporaria). У ящерицы Lacerta melisetlensis после адаптации наблюдается тот же уровень обмена. Однако у ящерицы Urosaurus ornatus, адаптированной к 35° С, наблюдается значительно более низкое потребление кислорода при 15° С, чем у адаптированной к 8° С; у ящериц Uta stansburiana, Sceloporsus occidentalis наблюдается та же реакция.
После адаптации к холоду пойкилотермные организмы при одной и той же температуре потребляют больше
172
кислорода, чем адаптированные к теплу. Золотая рыбка, адаптированная к 10° С, потребляет больше кислорода при 25° С, чем адаптированная к 30° С (Kanungo a. Prosser, 1959). Важно также и то, что у пойкилотермных организмов, способных к адаптации к различным температурам среды, потребление кислорода зависит не только от температуры тела, но и от физиологических состоянии, возникающих в процессе адаптации. Ящерица Lacerta tn. galvagnii потребляет одинаковое количество кислорода (196 мл/кг/ч) при 13, 19 или 25° С в том случае, если температура тела равна температуре адаптации (Gelineo а. Gelineo, 1955). Это свойство пойкилотермных организмов, характеризуемое тем, что интенсивность процессов окисления регулируется температурой живого существа, основано на сдвигах в энзиматических системах в период адаптации и напоминает дыхание изолированных тканей, интенсивность которого возрастает, если гомойотермные организмы жили в холодных условиях (табл. 8).
Период адаптации у пойкилотермных организмов короче, чем в гомойотермных. Обычно он достигает двух недель.
Данные, характеризующие популяционные изменения у пойкилотермных организмов, обитающих на разных широтах, очень ограниченные. Потребление кислорода морской звездой (Astarte montagui) из Гренландского моря (2° С) при температуре от 0 до 14° С было большим, чем у особей того же вида из более теплого Датского пролива (10°С). Гренландские особи при температуре окружающей среды (2° С) потребляют то же количество кислорода, что и датские формы при 10° С. Также моллюски Mytilus edtilis, обитающие вблизи Копенгагена (5° С), и особи того же вида из Средиземного моря вблизи Неаполя (15° С) потребляют одинаковое количество кислорода. Эти формы обладают наивысшей возможной адаптацией к температуре, так же как ящерицы и некоторые рыбы (Gelineo, 1959). Термогепетическая система этих форм допускает одинаково интенсивное протекание жизненных процессов как в холодной, так и в теплой среде, независимо от различных уровней температуры тела. Сопоставление температурных коэффициентов активности ферментных систем у этих видов представляет особый интерес.
173
Таблица 8
Адаптация к различным температурам и потребление кислорода у некоторых пойкилотермных организмов
Животное Температура а таптаций (°C) Температура (°C) Потребление кислорода
Дождевой червь (Lum-briculus variegatus) Виноградная улитка (Helix pomatia) Колорадский жук (Leptinotarsa decemli-neata) Гамбузия (Gambusia affinis holbrookii) Золотая рыбка (Caras-sius auratus) Лещ (Leuciscus ruti-lus) Бычок (Scopaena porous) 1 16 23 16—20 6 7—8 15 11—14 24—29 10 30 4 I-'. 23 12- 13,2 25 11 20 30 11 20 14 20 24 30 6 30 6 30 8,3 15 25 8.3 15 25 10,3 28 10 29 10 25 30 10 25 30 15 23 15 23 15 23 15,2 25 15,2 25 110 227 » 390 100 197 98 145 » 179 240 79 мл/кг ч 130 94 » 182 67 мл г/ч 126 » 290 > 44 » 80 182 93 515 » 82 » 35.5 » 30.2 » 137 » 125 17,8 » 69,4 » 130.6 > 101 .ил кг ч 208 '« 82 162 64 > 132 91 мл/кг-'Ч 153 » 56 » 89
174
Продолжение табл. 8
Животное Температура алеп’сщий (°C) Температура (°C) Потребление кислорода
Ящерица (Lacerta meli-sellensis melisellensis) 11—13 20-24 13,3 24,8 30,2 13,3 25 30 284 мг]кг[Ч 525 » 607 > 158 256 369
ТЕРМИЧЕСКИЕ АДАПТАЦИИ
У ГОМОЙОТЕРМНЫХ ОРГАНИЗМОВ
Физиологические механизмы терморегуляции гомойотермных организмов включают как изменения теплообразования, так и изменения теплоотдачи. В эти процессы вовлекаются почти все органы и системы и только в итоге направленного изменения функции каждой системы температура тела млекопитающих и птиц остается на постоянном уровне или в рамках закономерных физиологических колебаний.
Кроме того, большую роль в поддержании термического гомеостазиса играют реакции поведения как более простые — приближение или удаление от источников тепла или холода, смена температурной зоны (уход в тень, в нору, выход на солнце, скучивание особей (одного или разных видов), так и более 'сложные — постройка гнезд, нор, убежищ и т. д.
На примере изучения реакций животных при различной температуре среды в природных и экспериментальных условиях можно хорошо проследить взаимоотношения индивидуальных и видовых адаптаций и сопоставить лежащие в их основе физиологические механизмы.
Наиболее изучены адаптивные изменения реакций обмена веществ в организме млекопитающих и птиц на изменение температуры внешней среды. Эти реакции в физиологии уже давно получили наименование химической терморегуляции.
175
Кривую изменений газообмена у гомойотермного организма под влиянием различных температур среды можно представить себе следующим образом (рис. 35). При низких температурах среды, совместимых с жизнью данного вида, газообмен и теплопродукция повышены. При переходе к более высоким температурам теплопродукция
Вторая химическая терморегуляция
si
^1
Си \
V!
Нинхняя критическая температура
Зона преобладания химической терморегуляции s Ц
s Гипотермия
Дополнительное
Верхняя критическая температура
Зона преобладания
физической терморегуляции
Рис. 35. Схема химической терморегуляции у гомойотермных организмов (по Gelineo, 1964)
понижается и при какой-то температуре (для разных видов млекопитающих от 5—6° С до 33—35° С) достигает наименьших величин. Эту температуру при наименьшем обмене веществ в покое обычно называют критической точкой. С дальнейшим повышением температуры среды газообмен начинает повышаться как благодаря включению механизмов физической терморегуляции — полипноэ пли обильного потоотделения, так и вследствие повышения температуры тканей тела при высокой температуре среды. Обычно у животных, имеющих хорошо выраженное полипноэ, критическая точка обмена веществ также хорошо выражена. У животных, не имеющих полипноэ (непарнокопытных, обезьян), и человека критическая
176
точка выражена слабо п заменяется довольно обширной (до 15° С) зоной постоянного уровня обмена веществ. При дальнейшем повышении температуры среды и температуры тела газообмен начинает повышаться, но это обычно уже связано с явлениями перегревания. При очень низких температурах среды и наступающем охлаждении тела газообмен может снижаться, что сопровождается значительным снижением температуры тела и может знаменовать наступление гипотермии.
Рис. 36. «Вершинный» обмен (summit metabolism), отнесенный к весу тела у гомойотермных организмов (по Gelineo et Giaja, 1924, 1962)
Вместе с тем реакция обмена веществ на крайнее охлаждение довольно однотипна для различных видов млекопитающих и птиц. Наибольшее повышение обмена (metabolisme du sommet) (Giaja, 1934) продолжается различное время в связи с разной скоростью охлаждения ледяной водой (рис. 36) животных с различной поверхностью тела, теплоизоляцией и т. д.
Как правило, у животных, обитающих в условиях низких температур, критическая точка ниже, чем у обитателей мест с более высокой температурой (рис. 37). Сниже-
177
Рис. 37. Химическая терморегуляция (СЬЛил/лшн) и критическая точка у млекопитающих (по Слониму, 1952):
1 — собака, 2 — шакал, 3 — динго
ние критической точки наблюдается у водных млекопитающих— нутрии (Слоним, 1952), ондатры (Щеглова, 1954).
Между устойчивостью организма к высокой или низкой температуре среды и систематическим положением животного данного вида едва ли существует какое-либо соответствие. В любом климатическом поясе встречаются представители многих отрядов млекопитающих и только отдельные отряды приспособлены в своем распространении к узким условиям существования, например обезьяны, полуобезьяны и др. к субтропическому или тропическому климату.
У разных животных химическая терморегуляция развита различно. Можно сослаться на общую формулу, указывающую, что по мере восхождения по эволюционной лестнице роль изменений теплопродукции при разных температурах среды— химической терморегуляции— падает. Так, у ежей можно наблюдать 3—4-кратное повышение обмена при низких температурах (5— 6° С) по сравнению с обме
ном при температуре 25° С. У более высокостоящих грызунов— двукратное повышение обмена, а у хищников, в частности собак, — только в 0,7—0,8 раза. Это находит свое отражение в увеличении нормы питания лисиц и собак, содержащихся в питомниках в зимнее время. У обезьян и человека химическая терморегуляция имеет небольшое значение. Особенно это касается человека, мало тренированного к воздействию низкой температуры среды. У разных видов обезьян эти явления выражены в зависимости от того, какие климатические районы они населяют: если это животные тропического пояса, то хи
178
мическая терморегуляция приближается по интенсивности к таковой у человека.
Первое исследование, характеризовавшее особенности популяций, взятых из разных широт (Руттенбург, 1950), показало, что в зоне температур от 0 до 35° С популяции серой крысы (Rattus norvegicus) из Мурманска, Ленинграда, Севастополя и Сухими резко отличаются и по интенсивности химической терморегуляции и по критической точке обмена веществ.
С другой стороны, популяции черной крысы из Мурманска и Севастополя, а также александрийской крысы из Ленинграда и Севастополя совершенно не отличались. Этот весьма важный факт указывает на малую вариа-бильность популяционных адаптаций у подвидов с более узким географическим распространением — черной и александрийской крысы — по сравнению с широко распространенной под всеми широтами серой крысой.
Южные формы (черная и александрийская), заносимые на север морскими судами, образуют популяции в отапливаемых зданиях, кухнях и т. д. и, таким образом, холод и отсутствие способности к адаптации ограничивают их широкое распространение в условиях Севера.
Уровень обмена и химическая терморегуляция значительно отличались у двух популяций желтогорлых мышей (Apodemus flavicollis), выловленных под Харьковом и в Крыму (Калабухов и Ладыгина, 1953). У крымской популяции наблюдалось более низкое потребление кислорода при температуре от 0 до 25° С.
Географические отличия в теплообмене сказываются очень ярко даже у популяций полевок Microtus arvalis, разделенных с севера на юг расстоянием около 100 км. Северная популяция полевок обладала более высоким обменом, чем южная.
Между интенсивностью химической терморегуляции и устойчивостью организма к холоду нет прямой зависимости. Например,- интенсивность терморегуляции у крысы выше, чем у кролика (Gelineo, 1948); вместе с тем общая устойчивость организма кролика к холоду выше, чем у крысы. Так, кролик способен поддерживать химическую терморегуляцию до —45° С, тогда как крыса только до — 20° С. Это различие зависит от величины теплоотдачи с поверхности тела кролика и крысы, которая у крысы,
179
конечно, оказывается значительно большей на сдиницх массы тела.
Повышение интенсивности обмена и химической терморегуляции при переходе с юга на север не распространяется на организмы, живущие за Полярным кругом. Полярные животные, с трудом добывающие корм в условиях зимы, не могли бы существовать, если бы так резко увеличивался их обмен веществ при низких температурах. В зимних условиях на побережье Ледовитого океана песцы, например, не добывают двойного количества пищи, а довольствуются питанием, уменьшенным по сравнению с летним. Отсюда исключительно теплый мех — приспособление, которое ограничивает повышение обмена веществ при действии холода. Лучше всего выражена химическая терморегуляция у животных, обитающих в средних широтах в континентальном климате, а в условиях тропиков или в условиях Крайнего Севера она развита слабо.
Большую роль в интенсивности реакций обмена веществ на охлаждение играет теплоизоляция кожных покровов (мех и подкожный жировой слой). Эти особенности могут резко уменьшать реакцию на охлаждение. Это имеет место у животных в зимнем мехе или, например, у полярных видов (Scholander, Hock, Walters а. Irving, 1950). Однако только этот фактор (см. ниже) не может объяснить своеобразия химической терморегуляции ни в зимний период, ни в условиях Арктики. Но, несмотря на то, что отдельные виды обладают характерными чертами приспособления к определенным климатическим областям, некоторые из них имеют очень обширное географическое распространение, не ограниченное определенной климатической зоной. Подобная особенность — способность к существованию в разных температурных условиях—получила в экологии название эвритермии. Эвритермпыми организмами среди млекопитающих и птиц являются многие грызуны (серая крыса, серый хомячок), красная лисица, заяц-русак, волк, домовый воробей, серая ворона и др. (Hesse, 1924). В основе эвритермии лежит способность животного приспосабливаться к измененным температурным условиям — способность к быстрой и медленной стойкой акклиматизации.
Особенно ярко выражены эти особенности терморегуляции у животных, приспособленных к континентально-ISO
му климату, где имеются огромные суточные перепады температуры (суточная амплитуда колебаний температуры в условиях Казахстана 30—32° С). Эти значительные перепады приводят к формированию соответствующих реакций терморегуляции. Животные, которые здесь обитают (лисица-караганка, степной хорек и др.), показывают очень хорошую картину терморегуляции и приспособлены к любым температурам внешней среды благодаря чрезвычайно подвижной реакции обмена веществ — химической терморегуляции и полипноэ. Эти реакции обмена и их подвижность осуществляются за счет чрезвычайно быстрого образования условных рефлексов, их быстрого дифференцирования и угасания (Ольнянская и Слоним, 1947; Исаакян, 1949; Слоним, 1952).
Географические различия в химической терморегуляции у птиц изучены очень мало. Наблюдаются значительные различия критической точки у различных видов гусей, гнездящихся в Северном полушарии (Якубанис, 1967).
Влиянию индивидуальной адаптации на химическую терморегуляцию посвящено большое количество исследований. Классическими многолетними исследованиями (Gelineo, 1934а, б, 1936, 1948) было показано, что как интенсивность химической терморегуляции, так и температура критической точки изменяются под влиянием длительного (3—4-недельного) содержания при разных температурах внешней среды. Джелппео назвал эти сдвиги теплопродукции, изученные им у птиц, белых крыс и сусликов, адаптационными изменениями обмена веществ и терморегуляции. По Джелинео, величина теплопродукции складывается из двух компонентов: а) уровня обмена веществ, который при всех температурах среды после длительного пребывания в условиях низких температур повышается, а после пребывания при высоких, наоборот, падает; б) химической терморегуляции, которая после пребывания в условиях низких температур возрастает, а после пребывания при высоких — падает. Эти отношения хорошо видны на рисунке 38, где приведены данные для зеленушки (Chrysomitris spinus) после содержания ее при температурах среды —2°—8° С; 15°—20,5° С и 29°— ( + 32°) С. На этом рисунке видно, что критическая точка сдвигается после адаптации к более высоким температурам с 30° С к 34° С. Данные адаптационных изменений
181
газообмена у крыс приведены в таблице 9. Здесь видно, что изменяется но только нижняя, но и верхняя критическая точка (температуры); животные становятся намного устойчивее к гипотермии после содержания при температуре О ----2° С.
В настоящее время твердо установлено, что индивидуальная адаптация к холоду связана у гомойотермных
Рис. 38 Химическая терморегуляция при адаптации зеленушек к высоким и низким температурам срезы (по Gelineo, 1934):
1 — при адаптации к температуре среды 8 2Р; 2 — при адаптации к температуре 15—20,5°; 3 — при адаптации к температуре 29—.32 С. По оси ординат — теплопродукция (кал сутки)-. ио оси абсцисс — температура среды
организмов прежде всего с увеличенной способностью поддержания температуры тела при охлаждении и временем выживания организма при воздействии очень низких температур среды (Hart, 1964а).
Обширные исследования посвящены в последние годы проблеме источников повышенного теплообразования в процессе адаптации к холоду. Наиболее ярким фактом является то, что животные при длительных или повторных охлаждениях поддерживают повышенный уровень обмена при одновременном снижении мышечной ак-
182
Таблица 9
Теплопродукция и ее особенности у крыс, адаптированных к трем различным температурам среды
(по Celineo, 1934)
Температура адаптации 30—32° с 16-20° С 0—2» С
Термическая нейтральность 32—34,5 30—32 29—30
Объем в покое (кг/кал/л12/24 ч) .... 364 640 911
Избыточное теплообразование на ГС (кг/кал/24 ч) 29 40 55
«Вершинный обмен» (кг/«ал/л12/24 ч) . . . . 746 1817 3100
Низшая критическая точка (°C) 24 2 -10
Температура, при которой обмен максимален (°C) 2 -10 -10
Максимальный уровень обмена (кг/кал/л2/24 ч) 1514 2313 3100
тивности, т. е. видимой дрожи или определяемой с помощью специальной аппаратуры высокой электрической активности скелетной мускулатуры (Heroux, 1962; Hart, 19646; Бисерова и Гембчински, 1964; Иванов, 1965). Так возникло представление о несократителыюм термогенезе, т. е. таком повышении обмена веществ, которое не связано с усиленной функцией скелетной мышцы как сокращающегося органа. Прямым доказательством существования несократительного термогенеза явилось влияние норадреналина на общий газообмен у адаптированных к холоду крыс и на газообмен мышц тех же крыс, изученный in situ (Hsieh, Carlson, Gray, 1957; Depocas, 1960; Heroux, 1962; Ткаченко, 1968). Норадреналин в данном случае не является основным веществом, поддерживающим высокий метаболизм в целом организме. Приведенные выше данные (Исаакян, Макарова, Щеглова, 1968; Jansky a. Hart, 1968) свидетельствуют о значительном повышении газообмена во внутренних органах, что также приводит к возрастанию роли несократительного термо
183
генеза. Возрастает и кровоток во внутренних органах и жировой ткани (Jansky a. Hart, 1968).
Несократительный термогенез ярко выражен в раннем постнатальном периоде (исследования на морских свинках) и связывается в этом случае с развитием бурой жировой ткани в межлопаточной области (см. также гл. IV), затем он в нормальных условиях развития животного заменяется сократительным термогенезом, а при холодовой адаптации вновь проявляется (Briick, u. Wiinnenberg, 1965, 1966; Moore a. Simmonds, 1966).
Сократительный термогенез и его распределение в организме лучше всего может быть изучено при отведении игольчатыми электродами биотоков с разных групп мышц у животного, подвергающегося охлаждению (Слоним, 1964, 1966; Иванов, 1965; Баженов, 1968; Давыдов и Кузьмо, 1968 и др.). Как общую схему здесь можно принять, что наибольшую электрическую активность при охлаждении обнаруживают мышцы шеи, жевательные мышцы и отчасти мышцы спины и передних конечностей. Мышцы задних конечностей вовлекаются в терморегуля-циопную активность в последнюю очередь при очень интенсивном охлаждении. В эту схему, характерную для всех млекопитающих, необходимо, однако, ввести поправки для водных организмов (нутрия, ондатра), у которых наибольшая реакция наблюдается в мышцах спины и конечностей, т. е. в мышцах, непосредственно подвергающихся охлаждению в воде (Давыдов и Кузьме, 1968). У грызунов терморегуляционный тонус выражен вообще относительно слабо; он заменяется в жевательных мышцах активными мышечными сокращениями при охлаждении (рис. 39).
Такая же картина распределения мышечного терморегуляционного тонуса наблюдается у птиц. На рисунке 40 приведены данные, характеризующие электрическую активность мышц груди и мышц бедра у представителей воробьиных и водоплавающих. У последних мышцы бедра являются наиболее активными в отношении сократительного теплообразования.
В процессе адаптации наблюдаются некоторые изменения в химической динамике скечетной мускулатуры. Они касаются взаимоотношений между процессами дыхания и окислительного фосфорилирования в процессе теплообразования (так называемый коэффициент Р/О)
184
Рис. 39. Схема распределения терморегуляционного тонуса у грызунов(по
Иванову, 1964; Слони-му, 1964). /—белая мышь; //— крыса; /// — кролик: а—до охлаждения, б— после охлаждения
Рис. 40. Схема распределения терморегуляционного тонуса у птиц (по Кескпайку и Давыдову, 1966 и Якуба-пису, 1966),
I— черный дрозд; II—домашний гусь:
а — до охлаждения, б — после охлаждения (при —10° С). Отметка времени 0,1 сек.
(Lianides a. Beyer, 1960; Smith 1960; Скулачев, 1964 и др.). Однако эти изменения доказаны в основном по отношению к митохондриям печени и не охватывают скелетной мускулатуры. Было показано в то же время, что инъекции норадреналина не повышают газообмена в печени адаптированных к холоду крыс (Jansky, 1965), оказывая это влияние на скелетную мускулатуру. В то же
Рис. 41. Активность фермента гиалуронидазы в разных мышцах крыс в процессе адаптации к холоду (по Дерибас, 1967): / — гл. rectus femoris, 2 — in. sterno-cleido-nia-stoideus, 3 — m. trapecius
время установлено изменение ферментативной сукцинде-гидрогеназной активности в процессе холодовой адаптации. Гистохимические исследования скелетной мускулатуры крыс, адаптированных к холоду (Дерибас и сотр., 1967), показали, что в мышцах конечностей можно наблюдать увеличение числа мышечных волокон, содержащих большое количество фермента. Характерно, что эти сдвиги наблюдались в большей мере в мышцах конечностей, нежели в мышцах шеи и жевательных (рис. 41). Таким образом, в процессе адаптации происходит вовлечение в терморегуляторную активность (несократительную) именно той части мускулатуры, которая не является активной в обычных условиях теплообмена.
В настоящее время есть основания полагать, что эти данные, добытые в основном па крысах, могут быть пе-
187
ренессны на большинство млекопитающих, но не на птиц, где явления несократительного термогенеза до настоящего времени не обнаружены. В то же время у обезьян Saimiri sciurea при адаптации к холоду обнаружены изменения общего газообмена, окислительных ферментов в тканях, скорости окисления а-глицероф.осфата в бурой жировой ткани, митохондриях печени, совершенно аналогичные тем, которые описаны для лабораторных крыс (Chaffee, Allen, Brewer, Horvath, Mason a. Smith, 1966).
Изменения основного обмена (измеренного при определенной температуре) при адаптации к холоду представляются в следующем виде. В течение первых пяти дней у крыс наблюдается значительное повышение газообмена, затем газообмен продолжает медленно повышаться до 15—20-го дня, адаптация несколько снижается после 44-го дня к 60-му (рис. 42) (Cottle a. Carlson, 1954; Щеглова, 1967). Однако совершенно иные картины адаптационных изменений газообмена были получены при длительном содержании с температурой около 5° С представителей субтропических видов грызунов — индийской крысы незокин (Nesokia indica) и большой песчанки (Rhombomis opitnus) (рис. 43). Оказалось, что грызуны— обитатели субтропиков и тропиков при температуре около 5° С не обнаруживают никаких адаптационных изменений обмена веществ. Температура тела остается пониженной и не поднимается в процессе адаптации, как это имеет место у крыс. Следовательно, изменения газообмена являются весьма важными звеньями в процессе термической адаптации и их отсутствие указывает на невозможность образования адаптированных к изменившимся условиям популяций.
В противоположность адаптации к холоду содержание этих животных в тепле приводит к стойкому снижению обмена, более выраженному, чем у крыс. Отсюда понятна возможность расселения этих организмов далеко на юг и возможность образования стойких популяций с изменением обмена веществ и химической терморегуляции (рис. 43, 44).
Следовательно, адаптационные изменения газообмена к холоду образуются только в том случае, если температурные условия среды не ведут к значительным нарушениям терморегуляции, например к значительному понижению температуры тела. При склонности многих мыше-
1К«
Рис. 42. Теплопродукция у белой крысы в процессе адаптации к холоду (по Cottle a. Carlson, 1954)
Рис. 43. Изменение веса тела, потребления О2 (мл!г!ч) и ректальной температуры у лабораторной крысы и тропической крысы незокии (по Исаакян, Макаровой, Щегловой, 1969)
189
Рис. 44. Динамика физиологических функций у остриженных кроликов во время двухнедельной адаптации к 28° С и последующей 3-недельной адаптации к 6° С. I—вес тела (г); II — потребление пиши (г/день); III — образование кала (г!день); IV — электрическая активность мышц (mkV/ЗО мин); V — потребление кислорода при 28°С (мл1мин13$ мин)’, VI — температура тела (°C) (по Heroux, 1966)
190
видных грызунов (полевок) снижать уровень температуры тела и обмена при действии низкой температуры адаптационные изменения обмена полностью отсутствуют. Так, у содержавшихся в холоде пеструшек обмен под влиянием охлаждения оказывался ниже, чем у тех же пеструшек, содержавшихся в тепле (Гладкина, Мейер, Мокеева, 1958). Вероятно, только благодаря адаптационным изменениям теплообмена некоторые исследователи обнаруживают крайнюю нестойкость эколого-физиологических изменений, связанных с термическими условиями среды обитания.
АДАПТИВНЫЕ ОСОБЕННОСТИ ТЕПЛООТДАЧИ И СЛОЖНЫЕ ФОРМЫ ТЕРМОРЕГУЛЯЦИИ
Отдача тепла во внешнюю среду происходит тремя путями. Во-первых, конвекцией — нагреванием окружающего воздуха, когда нагретый воздух расширяется и поднимается вверх, а новые порции холодного воздуха поступают на его место и омывают тело животного. Таким образом, чем больше разница температур кожи и окружающего воздуха, тем интенсивнее происходит теплоотдача. Во-вторых, радиацией, которая заключается в том, что между излучением более холодных окружающих предметов и более теплой поверхностью тела животного устанавливается известное равновесие, при котором животное отдает больше тепла с длинноволновой радиацией и получает меньше тепловой энергии от окружающих предметов. Этот второй путь также очень важен. Некоторые исследователи считают, что около 50% всего тепла в обычных условиях среды отдается путем тепловой радиации. В-третьих, испарением воды с поверхности кожи, слизистых, легких. Это основной путь отдачи тепла при высокой температуре среды, когда отдача тепла конвекцией и радиацией невозможна. Значение таких физических путей отдачи тепла в природных условиях может резко изменяться как для животных, так и для человека.
Теплоотдача с поверхности тела осуществляется по правилу Ньютона:
где Тъ — температура тела животного; Та — температура окружающего воздуха; К—константа; / — устойчивость
191
к потере тепла (теплоизоляция), И — теплопродукция. Зная величину критической точки и температуру тела, можно вычислить величину теплоизоляции, т. е. температурный перепад, который не оказывает влияния на теплопродукцию животного и человека.
По американским данным (Irving, 1964), для человека эта температура оказывается
Ть — Та=37—27=10 С;
для арктического грызуна лемминга
7'fc-—7'я = 37—17 = 20° С;
для арктического хищника песца
7'6 —7Я = 39 —( —41 ) = 80° С.
По данным таких расчетов, арктические животные могут практически не изменять обмена веществ при очень низких температурах за счет весьма ограниченной теплоотдачи. Но схема построена на основе весьма грубых расчетов и может быть только иллюстрацией значения теплоизоляции.
Прежде всего в условиях основного обмена, т. е. у голодных н неподвижно сидящих животных никогда не поддерживается ректальная температура (температура тела) на постоянном уровне. Это касается не только мелких животных — мышевидных грызунов, воробьиных птиц и т. д., но даже таких крупных, как верблюды, лошади, овцы. Более постоянная температура у представителей семейства собак (Canidae), например домашних собак, лисиц, песцов. В табл. 10 приведены колебания температуры тела в зоне температур от 15 до 30° С при 2—3-часовой экспозиции животных в условиях основного обмена.
Таким образом, расчеты американских исследователей (Scholander a. oth, 1950; Irving, 1964; Hart, 1964 и др.), несмотря на их привлекательную простоту, не отражают всей сложности процессов, происходящих в организме при короткой экспозиции животного организма в разных температурных условиях среды.
Вторым возражением против этой схемы является вопрос о длительности воздействия холода. В многочисленных исследованиях (Слоним, 1952, 1962) было показано, что первые применения холодовых воздействий (если раздражитель не очень силен) вызывают обычно лишь слабо
192
Таблица 10
Колебания температуры тела у некоторых видов млекопитающих при экспозиции в зоне температур от 15 до 30° С (по Слониму, 1952)
Животное Колебания ректальной температуры ( С)’ Ав гор
Утконос {Ornithorhync-
has anatinus) Ехидна {Echidna acu- 3,5 Мартин, 1902
leata) Летучая мышь {Myo- З.о »
tis myotis) Еж европейский {Eri- Более 20 Слонпм, 1952
naceus europaeus) . . . Полевка общественная 8,0 »
{Microtus socialis) . . Ондатра {Ondatra zi- 2,2 »
bethica) Индийская крыса {Ne- (5,2 »
sokia indica) 8,5 »
Крыса лабораторная 2.8 Слепим, 1952
Большая песчанка •
{Rhombomys opimus) 4,2 »
Собака домашняя . . 1.2 »
Шакал {Canis aureus) Песец {Alopex Iago- 1.0 »
pus) 0,2
Барсук {Meles moles) Гамадрил {Papio ha- 0.5
madrylis) Макак-резус {Macaca 2.2 »
mulatto.) 2,3 »
выраженную реакцию обмена веществ. Примером может быть собака — животное, у которого вообще химическая терморегуляция достаточно хорошо выражена. Еще более это заметно у человека при сравнительно коротком воздействии слабых холодовых раздражителей. Лучше выявляется реакция химической терморегуляции у грызунов— лабораторных крыс, мышей. У многих представителей диких грызунов, особенно субтропических, например незокии (Nesokia indica), александрийской крысы (Rattus rattus alexandrinus), туркестанской крысы (Rattus turkestanicus), большой песчанки (Rhombomis opi-
7—1204
193
mas), песчанок малоазиатской (Meriones tristrami) и краснохвостой (Meriones erythrourus), первые применения холодового раздражителя вызывают слабо выраженное повышение газообмена, даже несмотря на значительное снижение температуры тела. Слабо выражена химическая терморегуляция и у тропических видов обезьян, например макак-резусов и макак-лапундер.
Отсутствие значительной реакции терморегуляции при кратковременных воздействиях характерно для всех гомойотермных организмов.
Значительное изменение реакции терморегуляции можно наблюдать при ориентировочной реакции животных (резкое охлаждение может вызывать такую реакцию), эмоциональном возбуждении и т. д. В этой связи следует отметить значительные колебания температуры тела, главным образом функциональную гипертермию, часто наблюдаемую при измерении температуры у диких и мало приученных к этой процедуре животных, например обезьян, лемуров и даже рептилий (Britton a. Klein, 1939; Слоним, 1952). При повторных измерениях (приучении животных) эта гипертермия может исчезать, а при воздействиях холода возрастает реакция на это воздействие, т. е. повышение газообмена.
Весьма важное место в регуляции теплоотдачи занимает изменение легочного дыхания — его резкое учащение при воздействии па организм тепла и замедление при воздействии холода. Реакция дыхания — полипноэ, изучалась неоднократно и служит часто простым и надежным критерием «напряжения терморегуляции», так как обычно появляется тогда, когда возможности регулирования температуры тела снижением обмена оказываются исчерпанными (Слоним, 1952).
При понижении температуры среды у животных, хорошо поддерживающих температуру тела и приспособленных к охлаждению, наблюдается углубление и уреже-ние дыхания (Калабухов, 1944; Раушенбах, 1958). Эта реакция, как бы обратная термическому полипноэ, еще мало изучена. Она сводит до минимума потери тепла испарением с поверхности легких и ведет к повышению коэффициента утилизации кислорода в легких. Таким образом уменьшаются потери тепла во внешнюю среду. Отдача тепла легкими — изменением дыхания — при высоких температурах особенно выражена у хищников, хо
194
тя полипноэ наблюдается в какой-то мере и у всех остальных млекопитающих. Нет полипноэ только у обезьян, непарнокопытных и у человека. Между реакцией полипноэ и потоотделением у животных существует обратная зависимость: если функционируют потовые железы и они хорошо развиты, то полипноэ отсутствует и наоборот. Между особенностями протекания полипноэ у млекопитающих различных отрядов установлены существенные различия. Так, полипноэ хищных выражено резче; частота дыхания достигает огромных величин (до 600 дыханий в 1 мин), дыхание очень поверхностное. Однако у хищных полипноэ не может продолжаться очень долго, так как требует большого расхода энергии и ведет в конечном счете к явлениям перегревания. Особенность полипноэ хищных (особенно у Canidae) заключается в том, что оно возникает при почти не изменившейся под влиянием нагревания температуре тела (Richet, 1898; Апгер а. Нат-niouda, 1933; Нестеровский и Слоним, 1935: Веселкин, 1945). Полипноэ сопровождается отделением жидкой слюны (Парфенов, 1906; Александров, 1954; Уждавини, 1960).
Для полипноэ характерны значительная акапния и сдвиги щелочно-кислотного равновесия. Обнаружено, что у собак в молодом возрасте при полипноэ наблюдается большее падение содержания СОг в крови, чем у старых. Оно сопровождается значительным учащением пульса. Нарушенный водный баланс организма оказывает влияние на полипноэ. В опытах на собаках установлено, что газообмен, легочная вентиляция и частота дыхания при недостатке воды в организме снижаются. У грызунов полипноэ всегда возникает на основе значительного повышения температуры тела, хотя дыхание и становится более поверхностным. По-видимому, у них полипноэ также не может долго поддерживать температуру тела при нагревании, и животные гибнут от гипертермии (Stigler, 1930).
Полипноэ у насекомоядных (ежей) изучено Веселкиным (1945) и Филатовой (1949в). Частота дыхания у ежей при нагревании увеличивается до 240 в 1 мин. Температура тела при этом возрастает с 34,5 до 40,0° С. У жвачных полипноэ отличается сравнительно небольшой частотой дыхания — обычно до 200 в 1 мин — и очень большой продолжительностью этой реакции. У крупного
7*
195
рогатого скота, овец, коз полипноэ может продолжаться всю жаркую часть дня и особенно усиливается при инсоляции. Для жвачных при полипноэ характерно усиление теплообразования в межреберных мышцах, компенсирующееся теплоотдачей; таким образом, кровь, направляющаяся к коже, проходит в большом количестве через межреберные артерии в межреберные мышцы (Goodall a. Young, 1954) с последующим разветвлением в коже на спине и на боках (тип «чудесной» сети) (рис. 45).
При резких термических воздействиях обычно наблюдаются очень яркие проявления сосудистой регуляции. Особенно это выражено у человека, но отчасти наблюдается и у животных. В последнее время установлены и специальные механизмы — «теплообменники» в конечностях. При этом артерия, окруженная венами, отдает тепло в венозную кровь и таким образом в зависимости от просвета вен в большей или меныпей степени нагревает самую конечность. Подобные «теплообменники» существуют, по-вндпмому, в ряде органов животных: хвосте у грызунов, конечностях у морских млекопитающих, ушах у жвачных и т. д. Схема такого органа представлена на рисунке 46.
На основании большого экспериментального материала показано, что метаболическая реакция при охлаждении начинается лишь после того, как исчерпаны возможности физической терморегуляции, в том числе и теплоизоляции, сосудистой реакции и т. д. (Hart, 19646). Однако эта схема едва ли может быть принята в связи с наличием кортикального механизма терморегуляции, сильно изменяющего эти взаимоотношения на первых этапах адаптации к теплу и холоду (Быков и Слоним, 1960).
ДВИГАТЕЛЬНОЕ ПОВЕДЕНИЕ
И ТЕМПЕРАТУРА СРЕДЫ
Двигательные реакции — поведение — имеют огромное значение в приспособлении животных к температурным условиям окружающей среды. Животное определенным образом изменяет свое поведение ib зависимости и от метеорологических условий среды. Например обезьяны в жаркие часы дня проделывают большее количество горизонтальных прыжков, чем вертикальных. При низкой
196
Рис. 15. Кровоснабжение кожи теленка (по Goodall а Young, 1954):
а — поверхностные сосудистые анастомозы, б — глубокие сосу-анстые анастомозы; 1 — сальная железа, 2 — волосяной фолликул. { — волосяная луковица, 4 — вена, 5 — артерия, 6 — потовые железы
Рис. 46. Схема артериовенозного «теплообменника» (по Scholander, 1960). Изменения температуры крови в артериях и венах в лапе собаки. Поперечный разрез через артерию (4) и оплетающие ее венозные пазухи (В). I — в тепле; II — на холоде
197
температуре количество вертикальных прыжков увеличивается. Этот, казалось бы, странный факт быт установлен точным наблюдением, и очевидно, что прод> кция тепла уменьшается или увеличивается в зависимости от характера прыжков у животных (Щербакова, 1949в). В зависимости от температуры среды животное принимает определенное положение тела. Например, скручивание собаки кольцом на холоде связано с уменьшением отдачи тепла поверхностью тела в этой позе.
Роль поведения животных в осуществлении поддержания температуры тела настолько велика, что некоторые исследователи (Стрельников, 1958) предлагают термин «экологическая терморегуляция» для характеристики всей совокупности изменений в организме под влиянием термических условий внешней среды. Такие явления, как гнездостроение, собирание в группы (групповая терморегуляция), а также сложный стереотип поведения в зависимости от погодных комплексов являются примерами сложной реакции организмов на изменение температуры среды.
Совокупность физической и химической терморегуляции в широком смысле слова позволяет животным поддерживать температуру тела в условиях различных температур внешней среды. Терморегуляция является совокупностью сложнейших физиологических процессов, включающих как изменения двигательного поведения, так и изменения уровня так называемых вегетативных функций. Изменения теплопродукции и теплоотдачи, центрального и периферического кровообращения, потоотделения и слюноотделения (при полипноэ), возбудимости нервной системы сопровождаются совершенно закономерными изменениями поведения, позы, активности и покоя, специальными формами деятельности. Схематически эти сложнейшие физиологические отношения могут быть представлены как изменения физиологических функций и сложного поведения животного организма, а для человека— также и его сознательной деятельности (см. табл. 11).
Совокупность изменений поведения и регуляций физиологических функций, характерная для каждого вида в определенных условиях существования, позволяет выделить определенные жизненные формы, характеризующиеся приспособленностью к условиям данного биотопа
198
Таблица 11
Элементы терморегуляции у гомойотермных организмов (по Hensel, 1955)
Показатели Регуляция физиологических функций Изменения поведения и деятельности (человек)
Температура среды Теплообразование Теплоотта'а Изменение повеления Испарение воды Геометрический фактор (поверхность) Тонус мышц и холодовая дрожь. Химический тонус мышц. Обмен веществ во внутренних органах Кровенаполнение сосудов кожи; положение перьев и шерсти (пиломоторная реакция), линька Секреция пота; полипноэ; секреция носовых и ротовых желез Изменение позы Выбор макро- И микроклиматических зон. Выбор солнца, тени и т. д. (искусственное отопление и охлаждение) Активные движения. Специальное динамическое действие (прием холодной и горячей пищи) Постройка гнезд, нор; поиски пещер, щелей и других укрытий; изоляция убежищ сухими листьями, вентиляция пчелиного улья (вентиляция постройки, жилища) Увлажнение поверхности кожи (увлажнение одежды) Групповые скопления животных (групповая терморегуляция)
(например, жизненные формы грызунов, которые отличаются различной чувствительностью к температурам внешней среды). Устойчивость к охлаждению и перегреву различных животных из разных микроареалов различна. Н. И. Калабухов (1954) выделяет 4 жизненные формы грызунов лесостепей и степей левобережья Украины, различно реагирующие на понижение и повышение температуры среды (см. табл. 12).
199
Таблица 12
Чувствительность к влиянию температуры (по Калабухову, 1954)
Темпераiypa Жизненные формы
• степей песков ПГШ 41 леса
Высокая - + +
Низкая -— + + —
Примечание: + обозначает наличие влияния, — отсутствие влияния.
Для обитателей песков и степи высокие температуры являются адекватными раздражителями, а на жизнедеятельность обитателей пашен они оказывают угнетающее влияние. Лесные формы мало чувствительны к низким температурам, но очень чувствительны к высоким. Низкая температура оказывает значительное влияние на обитателей песков и пашен, но мало влияет на обитателей степи. Эта схема, конечно, нуждается в дальнейшем уточнении, так как многое в этих влияниях зависит от образа жизни животных, устройства нор, особенностей их подвижности. Тем не менее такие поиски эколого-физиологических зависимостей необходимы для уточнения физиологических особенностей организмов, живущих в природе в разных условиях.
Повышение и понижение температуры обусловливает и глубину нор грызунов одного и того же вида. Так, глубина нор обыкновенной полевки в различное время года разная и это, по-видимому, зависит от температурных условий. В нежаркое время года норы располагаются на глубине 5—25 см\ в жаркий и бесснежный холодный периоды года глубина их доходит до 60—80 см. При значительном и устойчивом снеговом покрове норы устраиваются на поверхности почвы непосредственно под снегом. Температура в норах разных видов, по-видимому, определяет и возможности распространения данного вида. Так, температура в норах на протяжении с мая по декабрь самая высокая у общественной полевки, причем как в предгорьях Карабаха, так и в степном Крыму эта 200
температура ниже у обыкновенной полевки и еще ниже у рыжей (Поляков, 1950). Последняя и характеризуется наиболее северным ареалом.
Важное значение для поддержания температуры тела, особенно у тропических видов, при охлаждении или во время сна имеет собирание животных в группу — скучи-вание. Эти явления лучше всего наблюдать у обезьян.
Рис. 47. Изменения газообмена, температуры стопы и поведения при умеренном охлаждении во время сна у людей, в разной степени адаптированных к холоду (по Scholander, 1960)
Существует определенная зависимость между температурой внешней среды и количеством гамадрилов, спящих в группе (Слоним и Щербакова, 1949а). Подобные факты были установлены и для капуцинов и ревунов даже в условиях тропического климата (Niethammer, 1952).
Особенности терморегуляции человека во время сна ярко выявились в исследовании адаптированных и неадаптированных к холоду норвежских студентов и аборигенов-австралийцев. При умеренном охлаждении в ночные часы неадаптированные слабо повышали обмен, плохо спали и резко снижали температуру конечности. Адаптированные резко повышали газообмен и поддержи
201
вали температуру конечности на постоянном уровне во время сна. Австралийские аборигены хорошо спали, не повышали обмена веществ и еще более резко снижали температуру конечности. Таким образом, последний тип адаптации связан со значительными нарушениями температурного гомеостазиса (Scholander, 1960) (рис. 47).
СЛОЖНОРЕФЛЕКТОРНЫЙ МЕХАНИЗМ
ТЕРМОРЕГУЛЯЦИИ И ЕГО ЗНАЧЕНИЕ ДЛЯ АДАПТАЦИИ К ТЕПЛУ И ХОЛОДУ
Представления об адаптации организма к теплу и холоду складывались на основе исследования длительного и реже повторного воздействия этих факторов на организм. Как правило, при этом описываются итоговые влияния тепла или холода, позволяющие животным существовать в крайних условиях среды и даже поддерживать в широких пределах температуру тела.
Однако, как уже отмечалось, в ряде случаев па применение холодового или теплового раздражителя впервые организм отвечает атипичной реакцией, обычно более слабой, чем на последующие воздействия. Затем наступает стадия возрастания реакции и, наконец, стадия постепенного ее снижения, т. е. возникновение устойчивого состояния. По поводу этих явлений до сих пор нет единообразного толкования и описываются они главным образом феноменологически.
В последнее время рядом зарубежных исследователей обращено внимание на «помехи», которые возникают в процессе реагирования, главным образом человека, на термические раздражители (Glasser, 1962; Glasser а. Griffin, 1962; Yoshimura a. Ida, 1963; Leblanc, 1966). Все эти фактические данные вновь привлекают внимание к вопросам высшей нервной деятельности и ее влияния на сложное сочетание вегетативных и поведенческих актов, составляющих терморегуляционный ответ организма.
В то время как образование условного рефлекса на вегетативную реакцию организма (метаболизм, сосудистые ответы, сердечная деятельность, потоотделение и т. д.), как правило, повышает рефлекторные ответы организма (Быков, 1942; Ольнянская, 1950, 1965; Рогов, 1951; Пшоник, 1952; Слоним, 1952; Быков и Слоним, 1960 и мн. др.), длительное воздействие факторов среды или
202
большое количество повторных сочетаний приводит к их уменьшению или даже полному исчезновению (Слоним, 1962; Slonim, 1964). Таким образом, кортикальные влияния на терморегуляцию занимают определенное место в общих явлениях индивидуальной адаптации на определенных стадиях ее развития. Сейчас есть все основания предполагать, что кортикальный (условнорефлекторный) механизм терморегуляции имеет наибольшее значение на первых этапах индивидуальной адаптации гомойотерм-иого организма. Действительно, наиболее яркие проявления кортикальной регуляции обмена веществ, сосудистой деятельности, потоотделения могут исчезать при длительных или повторных термических воздействиях. Это наблюдали для газообмена (Слоним, 1952, 1962), реакции сосудов (Орлов, 1961), электрической активности скелетной мускулатуры (Ольнянская и сотр., 1966).
Все описанные выше явления очень характерны для индивидуальной адаптации, возникающей в результате эксперимента. Гораздо сложнее влияют естественные сочетания отдельных факторов среды, определяющие в конечном счете теплоотдачу и тепловое состояние организма.
Имеющийся в настоящее время экспериментальный материал позволяет говорить, что сочетания термического влияния среды и элементов производственной работы у человека, сочетание воздействия природных метеорологических факторов (освещения, движения воздуха, конвекционной и радиационных потерь тепла) образуют в центральной нервной системе прочный динамический (жизненный) стереотип, соответственно которому направляются терморегуляторные реакции и нарушение которого вызывает их торможение — полное или частичное. Так, описаны случаи выпадения терморегуляционных реакций в темноте (Тгошр, 1956; Kamyama, 1963) и усиление их при солнечном освещении. В условиях увеличенных радиационных потерь тепла (свыше 50% общих теплопотерь) наступает охлаждение организма при малой реакции сосудов и метаболизма (Марголина и Попугаева, 1949; Малышева, 1959). Движение воздуха может вызвать реакцию на охлаждение без сколько-нибудь значительных потерь тепла с поверхности кожи (Давыдов, 1953). Эти прочные и весьма сложные для анализа
203
физиологические реакции, хотя и являются сигнальными, практически сохраняются в течение всей жизни.
Физиологический механизм таких прочных реакций животных и человека еще совершенно не изучен. Некоторые исследователи (Бирюков, 1960) рассматривают их как своеобразные натуральные рефлексы. Можно думать, что в формировании их имеют большое значение реакции, образованные на определенных (в том числе ранних) этапах онтогенеза по типу импринтинга, или запечатле-вания. Все это требует еще дополнительных исследований.
О ФОРМИРОВАНИИ ТЕРМИЧЕСКИХ АДАПТАЦИИ
В ОНТОГЕНЕЗЕ
Формирование терморегуляции в онтогенезе проходит через три характерные фазы. Первая из них — фаза пой-килотермии. У многих незрелорождающихся гомойотер-мов млекопитающих и птиц эта фаза ярко выражена сразу после рождения. Животное не только не в состоянии поддерживать температуру тела на постоянном уровне, но и почти не реагирует на охлаждение или нагревание характерными для гомойотермов изменениями обмена. Температура тела полностью отражает температуру внешней среды. К таким формам относятся многие грызуны (мышевидные), все зимоспящие млекопитающие (грызуны, насекомоядные, летучие мыши), птенцовые птицы. В этих условиях огромное значение имеет нагревание теплом матери в гнездовых условиях развития. Вне присутствия матери молодняк охлаждается, но часто не погибает, а сначала (первая фаза) лишь резко ограничиваются уровень обмена веществ и подвижность (Антошкина, 1939а и б; Жила, 1940; Слоним, 1945; Gelineo, 1954; Adolph, 1957 и ми. др.). Во второй фазе появляется интенсивная химическая терморегуляция, однако температура тела все еще подвержена большим колебаниям в результате слабого функционирования сосудистой терморегуляции. Эта фаза продолжается до конца гнездового периода развития, до перехода молодняка к самостоятельному существованию (Понугаева, 1953). Затем (третья фаза) молодняк приобретает черты терморегуляции, свойственные взрослой форме. Если во вторую фазу развития молодняк тропических и полярных видов пока
204
зывает совершенно одинаковую картину терморегуляции (интенсивность изменений обмена, уровень критической точки), то в третью фазу проявляются уже и эти специализированные черты терморегуляции. К этому времени формируются и элементы физической терморегуляции — сосудистая реакция и полипноэ (Жила, 1940). Первые проявления химической терморегуляции у грызунов можно наблюдать па 3 —11-п день жизни. Они выражаются
Рис. 48. Формирование химической терморегуляции у крысят (по Gelinco, 1959). По оси ординат О2 мл1г)ч, по оси абсцисс — температура среды (°C)
в том, что обмен веществ перестает следовать за температурой среды сначала в малых, а затем и в больших интервалах колебаний (рис. 48). У отдельных видов наблюдаются заметные различия в сроках становления химической терморегуляции. Терморегуляция развивается позже основных внешних анализаторов (зрения, слуха, обоняния, тактильной рецепции) и никак не связана с моментом прозревания (Батенина, 1960). Как некоторую общую формулу можно принять, что появление химической терморегуляции (устойчивой, т. е. второй фазы) наблюдается на 2—3-й день после открытия глаз.
205
У животных, рождающихся зрелыми (например зайцы, копытные, обезьяны), а также у человека первая фаза развития терморегуляции приходится на внутриутробный (а у выводковых птиц — внутрияйцевой) период развития. У некоторых видов, например копытных, температура тела сразу становится уже довольно постоянной. Однако специфические для вида реакции терморегуляции развиваются в более поздний период (например у обезьян-гамадрилов и макак). В более поздний период формируются и такие специфические реакции, как, например, реакции на движение воздуха, являющиеся у взрослых организмов специфическим стимулом для терморегуляции. Есть основания думать, что специфические черты терморегуляции каждый вид приобретает в сроки появления самостоятельности: после распада гнезда, появления самостоятельной игровой деятельности (в стаде копытных) и т. д.
На скорость развития терморегуляции у млекопитающих и птиц оказывает влияние ряд факторов; важнейший из них — питание. Чем меньше детенышей в помете у грызунов, тем интенсивнее кормление и тем скорее наступает поддержание постоянной температуры тела. Другим весьма важным фактором является температура среды. При содержании крысят в помещении с температурой 34° С формирование терморегуляции замедляется. У тундровой красной полевки (Clethrionotnis rutilus) формирование терморегуляции происходит чрезвычайно быстро: в период с 12-го на 14-й день наблюдается появление чрезвычайно устойчивой температуры тела (Morrison, Ryser a. Strecker, 1954). У большой песчанки химическая терморегуляция проявляется в лаборатории при температуре среды 18—20° Сна 5—6-й день, а в условиях пустыни в Средней Азии при температуре 25—30° С — только на 13—14-й день (Алимухаммедов, 1960). Таким образом, влияние температуры среды и ее колебаний на скорость развития терморегуляции несомненно как у зрелорожда-ющихся животных (копытных), так и у незрелорождаю-щихся (грызунов).
Среда, в которой происходит развитие молодняка, оказывает большое влияние на все последующие особенности терморегуляции. Эти явления, протекающие по типу импринтинга (см. гл. III), впервые установлены животноводами в практике так называемого холодного выра-
206
щивания молодняка. Формирующиеся при этом особенности терморегуляции оказываются очень прочными и характеризуются более высоким обменом веществ, интенсивным кровообращением, большой чувствительностью таких животных к высоким температурам среды (Пшеничный, 1955; Рогаль, 1955).
Содержание при низких температурах ягнят, козлят и морских свинок сразу после рождения стимулирует главным образом формирование сосудистой терморегуляции (Калашников, 1958, 1962). Включение навязанной мышечной работы (бег в трстбане) оказывает противоположное влияние па обмен веществ, который при такой холодовой экспозиции повышен. Таким образом, прочное влияние условий развития на формирование терморегуляции бесспорно и представляет большой научный и практический интерес.
Практически зрелорождающиеся животные, например копытные, в молодом возрасте значительно отличаются от взрослых. Так, у ягнят полипноэ сопровождается значительным учащением пульса, тогда как у взрослых овец полипноэ сопровождается увеличением минутного объема сердца за счет увеличенного систолического объема (Юнусов, 1960). Близкие явления обнаружены при сравнении молодых и взрослых домашних лам (Давыдов, Лобанов, ^Макарова, Рашевская, 1967). Даже учащение дыхания, отсутствующее у взрослых непарнокопытных, наблюдается у молодняка пони в первый месяц после рождения, когда еще не сформирована функция потоотделения (Давыдов, Лобанов, Абакарова, Рашевская, Юнусов, 1967). Все это указывает на позднее в онтогенезе формирование специфических видовых особенностей терморегуляции и значение ранних стадий постнатального развития для формирования адаптаций.
Таким образом, в ранние периоды развития на организм можно оказать значительное влияние, приводящее к необратимым изменениям. Есть основания думать, что эти явления лежат в основе природных расселений животных, занимающих подчас все новые и новые области. При акклиматизации животных человеком в условиях Севера, например, процесс идет успешно только тогда, когда формирование терморегуляции в ранний период постнатального онтогенеза происходит в условиях низкой температуры.
207
КРАЙНИЕ ТИПЫ АДАПТАЦИИ К ТРОПИЧЕСКОМУ И ПОЛЯРНОМУ КЛИМАТУ
Климатические особенности Арктики и Антарктики с чрезвычайно низкой среднегодовой температурой, крайне продолжительной зимой и совсем коротким летом создают совершенно особые условия существования гомойотермных организмов. Еще более ста лет назад были измерены температуры тела у полярных животных (песца, полярного зайца-беляка) при температурах окружающей среды около —50°С (Раггу et Lyon, 1825). Эти исследования обнаружили, что постоянство температуры тела может поддерживаться при разнице внутренней температуры (тела) и среды, достигающей 80° С (см. табл. 13).
Таблица 13
Температура тела полярных млекопитающих и температура среды
(по Hesse, 1924)
Температура (°C)
Bui тела с репы Разиина
Песец (Alopex lago-pus) 39,6 —35,0 74,6
Волк (Canis lupus) Заяц-беляк (Lepus ti- 40,5 —32,8 73,3
tnidus) 38,3 —29,4 67,7
Снежная куропатка (Lagopus mutus) . . . 43,3 -37,3 0,6
Постоянство температуры тела у некоторых полярных видов даже в условиях сильных холодов (до —25° С) было обнаружено в работе с песцами и лисицами (Ольнян-ская и Слоним, 1947); при этом оказалось, что теплопродукция (газообмен) в условиях значительного охлаждения не повышается. Слабое повышение газообмена при охлаждении на фоне несколько снижающейся температуры тела было обнаружено у серой крысы (пасюка) в условиях Заполярья — Кольский полуостров (Руттенбург, 1950, 1953). На рисунке 49 приведены данные изменений газообмена и температуры тела у некоторых обитателей
208
Л Б
39-90-
38-70-
20 15 Ю 5 0 5 10 15 20
Ы) 39 38 37 36
А Б
А Б
II
-110--90-
-70--50--3Qt-
20 15 10 5 0 5 10 15 20
39-90
38-70
37 -50
36 *~30*——1—1—1—।—।—।—।—
20 15 10 5 0 5 10 15 20
+
Рис. 49. Изменения газообмена и температуры тела при разных температурах среды у арктических млекопитающих (по Ольнянской и Слониму, 1947). / — песец белый; II — песец голуб( й; III — заяц-беляк; IV — заяц-русак:
а — температура тела (”С), б — потребление кислорода (мл)
Заполярья. Пределы выживаемости некоторых млекопитающих и птиц при низких температурах видны из табл. 14.
Таблица 14
Выживаемость некоторых животных при низких температурах (по Hart a. oth., 1957)
Живо гные Температура (°C) Врэмя выживания
Крупные млекопитающие Арктики —40 Неопре теленно
Кролик —35 3,5—6,5 ч
Крупный рогатый скот — 13 2 нетели
Крыса . —35 4->—120 мин
Утка — 10 384 ч
Г олубь — 10 16—30 ч
Воробей -30 3—5 ч
Продолжительность выживания при интенсивном охлаждении зависит главным образом от теплоизоляции, создаваемой шерстным и перьевым покровом животных. От этой теплоизоляции в значительной мере зависит и интенсивность повышения обмена веществ при охлаждении. У арктических животных повышение обмена наблюдается только при очень низких температурах среды: у ласки, суслика и лемминга при —15—18° С и ниже; эскимосской собаки —24° С; песца —40° С; белого медведя —50° С (Scholander, Hock, Walters a. Irving, 1950). Исключительно постоянная температура тела наблюдалась как в состоянии сна, так и в периоды бодрствования у таких видов, как эскимосская собака, песец. При температуре среды —80° С температура тела оставалась постоянной на уровне 38° С. По этим же данным критическая точка у этих животных лежит ниже —50° С. Температура кожи у эскимосской собаки под мехом составляет около 30° С при температуре среды около —50° С. Однако температура носа, конечностей (например, у северного оленя) может не превышать в этих условиях 10° С (Irving, 1951). Вместе с тем при одинаковой теплоизоляции шерстного покрова устойчивость к охлаждению может быть различной. Так, при сравнении трех мелких млекопитающих Аляски — тундровой рыжей полевки (Clethrionotnys rutilus dawsoni), тундровой полевки (Microtus oeconomus
210
macjarlani) и длиннохвостой землеройки (Sorex cincreus hollisteri), несмотря на одинаковую величину теплоизоляции, наиболее устойчивой к холоду оказалась рыжая полевка. Это особенно ярко было выражено при охлаждении в условиях движения воздуха. При этом характерными являются п позы животных при охлаждении, что также определяет устойчивость температуры тела (Morrison a. Tietz, 1957).
Очень низкая температура критической точки обнаружена у горного козла (Creamnos americanus) на Аляске (Krogh a. Monson, 1954). При температурах среды от 20 до —20° С газообмен оставался постоянным, при — 30° С наблюдалось повышение его на 23%. а при — 50° С потребление кислорода повысилось на 30%. Температура — 20° С является средней для обитания этого животного.
На основании ряда исследований Шоландером предложена схема (рис. 50) изменений теплопродукции у тропических и арктических млекопитающих и птиц. Эта схема показывает, что у устойчивых к холоду гомойотермных организмов теплопродукция с понижением температуры среды нарастает очень медленно. Это связывается Шоландером с высокой теплоизоляцией (см. выше). Тропические формы обладают большей реакцией на охлаждение. Поэтому они не могут длительное время существовать в условиях, когда температура снижается ниже — 10° С. Интенсивная химическая терморегуляция (по существу приближающаяся к величинам вершинного обмена — metabolisme de sommet) быстро приводит к истощению энергетических ресурсов и к гибели (Scholander a. oth, 1950).
У тропических видов — обезьян и хищных наблюдается ряд особенностей химической терморегуляции и ее взаимоотношений с физической, а также ряд особенностей теплоотдачи (Слоним и Щербакова, 1935; Слоним, 1937, 1941, 1952). Так, например, у представителей тропических и субтропических хищных —шакалов, динго — была обнаружена более высокая температура критической точки, свидетельствующая о том, что полное выключение химической терморегуляции наблюдается при более высокой температуре внешней среды. На рис. 37 приведены эти данные, полученные для домашней собаки, шакала и динго, содержавшихся на протяжении нескольких
211
ЧОО
Млекопитающие
। ।
Арктические
Гропические
300
7/&ъ
>&?
200
Ленивец
Снот -полоскун
КлОбпьвна Adas
Yfr- человек
- Мармозеза
100
Крыса г коати
Основной одмен ЮО -----Экспериментальные ванные ----------Экстраполированные данные
-10/ -50
Самая низкая темпера тура на земле (°С)
-30 -10
Температура воздуха t’C)
10 30
Температура тела. (°C)
200
300
100
Птицы
ЧОО
10) -50
Самая низкая температура на земле СС)
-30 -ю
Температура воздуха (°C)
10 30
Температура тела (°C)
Рис. 50. Терморегуляция и термочувствительность арктических и тропических млекопитающих и птиц. Критические градиенты (термочувствительность) и наклон кривой зависят от произведения основного обмена и общей термоизоляции тела (по Scholander и сотр., 1950)
месяцев в одинаковых температурных условиях. На этих же объектах обнаружено, что интенсивность химической терморегуляции у тропических форм ниже, чем у обитателей умеренного климата. Например у шакала — обитателя субтропиков процент изменений обмена веществ при переходе от 10 к 25°С составляет около 4%, тогда как у собаки (с одинаковой примерно длиной шерсти) достигает 7%. То же имеет место и у тропической собаки динго.
Подобные же данные получены при сравнении обезьян— павианов и гамадрилов — обитателей плоскогорий Эфиопии (200 м н. у м.) и обезьян-макак (резус и ла-пундер). У первых обнаружена очень интенсивная химическая терморегуляция (до 5% изменений обмена на 1°С), а у макак значительно более слабо выраженная (до 2% на 1°С). Критическая точка была резче выражена у макак при более высокой температуре внешней среды.
Следует отметить, что критическая точка лучше выражена у животных, обладающих механизмом полипноэ. В этом случае пониженная, по сравнению с низкими температурами, теплопродукция при дальнейшем повышении температуры среды или при увеличении длительности экспозиции сразу возрастает (Слоним, 1952). Так обстоит дело у всех хищных, жвачных (Brody, 1945), грызунов и насекомоядных (Веселкин, 1945; Слоним, 1952). У животных, обладающих потоотделением, например обезьян, лошади и у человека, критическая точка заменяется зоной постоянного уровня обмена, отражающего включение сначала сосудистой, а затем и потоотделительной реакции. На рисунке 51 приведены данные, полученные для разных видов обезьян. Проявление критической точки при разных температурах среды обусловлено тем, что непосредственно перед включением полипноэ обмен веществ более или менее длительное время оказывается пониженным. Это явление так называемой второй химической терморегуляции было описано (Kestner, 1924) и подробно изучено на разных видах (Слоним и Щербакова, 1938; Слоним, 1952). У тропических форм это понижение обмена при высоких температурах выражено лучше, чем у нетропических, что можно наблюдать у шакалов, динго, обезьян-макак. Вторая химическая терморегуляция, как правило, особенно хорошо выявляется при длительном умеренном нагревании, и это ограничение про-
213
ЯЙ7 W00 900 800 ЮО 600 500 400 - 1
/' LA
L?
i / 4/
I / 4/7 39
2
- — JO 17
5 0 '5 20 2 5 30 35 Ы) 45
опп /
800 700 600 500 400 300 zoo 100 Ч——.. JJ
38 37
п 2 35 34 33 3?
31
L 10 15 20 5 30 Л г
1 39 38 J7
/ии 600 500 400 300 200 1ПП /
/
ш 2 Of
3d 35 ?4
33
5 10 15 2 0 25 30 3i 4/7 45
рпп 39
1
tuu £ПГ 3 QI
IV 500
2 35 3b
чии ъпг
dUU ОПП
LUU 1ПП QQ
/ии 5 10 15 20 25 30 3 5 4/7 45
Рис. 51. Химическая терморегуляция и температура тела у низших обезьян (по Слони-му и Щербаковой, 1940). I— павианы-гамадрилы; II — павианы-анубисы; III — макаки-резусы; IV — макаки-лапундеры. По оси ординат слева — О2 (мл/мин), справа — температура тела; по оси абсцисс — температура среды (°C):
1 — температура тела, 2 — потребление О2
дукции тепла, несомненно, имеет большое значение и в процессе осуществления животным мышечной деятельности.
АДАПТАЦИИ К ПРИРОДНЫМ ТЕМПЕРАТУРНЫМ УСЛОВИЯМ У ЧЕЛОВЕКА
Не только по своему филогенезу от обезьяноподобных предков, но и по физиологическим особенностям терморегуляции человек как гомойотермный организм должен быть отнесен к тропическим видам. Относительно слабое развитие химической терморегуляции, яркая и охватывающая большие области тела сосудистая реакция и хорошо развитое потоотделение с большим количеством эккринных потовых желез, характеризуют терморегуляцию у человека. Температура тела человека подвержена суточным колебаниям в пределах 1°С и не является стабильной в разных частях тела.
Физиологические изменения в организме человека под длительным влиянием холода близки к тем, которые наступают в организме экспериментальных животных. Наблюдается общее повышение газообмена, уменьшение электрической активности скелетной мускулатуры при охлаждении, возрастание реакции газообмена в мускулатуре при введении норадреналина, возрастание устойчивости температуры тела при охлаждении (Davis a. oth., 1965; Leblanc, 1966; Кандрор, 1968). Однако особое место занимают изменения чувствительности конечностей к холоду и изменения кожной вазоконстрикции. Как показали детальные исследования у рыбаков, руки которых подвергаются длительному систем этическому охлаждению, наблюдается общая пониженная реакция к холоду в результате изменения общей чувствительности к нему (Leblanc, 1960, 1962). В этих же исследованиях обнаружено, что явления адаптации у рыбаков могут сохраняться в течение 15 лет после прекращения работы. Следует заметить, что экспериментальные адаптации у крыс, о которых речь была выше, быстро исчезают после прекращения холодового воздействия. Все это позволяет заключить, что у человека холодовая адаптация, связанная с характером его деятельности, представляет своеобразную «память», отраженную в центральной нервной системе; тут играют важную роль кортикальные механизмы терморегуляции и их своеобразная динамика.
215
В то же время адаптация к условиям Субарктики и Арктики не аналогична адаптации человека к холоду даже в условиях его повседневного воздействия на организм, например при холодовом закаливании или производственной работе, хотя контакты с природой имеют большое значение. Например, в условиях Арктики многие исследователи наблюдали у местных жителей повышение уровня основного обмена. Однако это повышение скорее было связано с ношением тяжелой одежды, чем с непосредственным воздействием холода на терморецепторы. Тем не менее у человека основной обмен может повышаться под влиянием длительного охлаждения (Кандрор, 1968) в условиях Арктики у лиц, постоянно работающих на открытом воздухе. У людей, не работавших па открытом воздухе, основной обмен в таких суровых климатических условиях не изменялся.
В зимнее время основной обмен эскимосов повышается на 25%, объем плазмы крови — па 25—45% и объем эритроцитов — па 15—20%. Летом все эти сдвиги исчезают, что происходит, как полагают авторы, в результате дезакклиматизации (Brown, Bird, Bong, Delahayc, Green, Hatcher a. Page, 1954). С другой стороны, не обнаружено никаких отличий критической точки обмена веществ у лопарей (27°С), по сравнению с обитателями умеренного пояса (Scholander, 1957). Автор считает, что все явления приспособления лопарей к низким температурам среды происходят за счет использования теплой одежды. Вопрос о собственно холодовой акклиматизации у народов Севера остается, таким образом, открытым.
По-видимому, в условиях Арктики большое значение имеет и своеобразная диета, включающая значительное количество белков и жиров. Исключительное значение, кроме того, имеет, по-видимому, для человека и режим мышечной деятельности. При ограничении движений и пребывания на открытом воздухе основной обмен в условиях Заполярья у человека по сравнению с условиями средних широт понижен (Слоним, Ольнянская и Руттеп-бург, 1949). Однако процесс адаптации у человека усиливается при сочетании собственно климатических воздействий с умеренной мышечной деятельностью. Так, в санаториях под влиянием таежного климата основной обмен повышается при одновременном снижении частоты дыхания и пульса. Увеличивается и скорость восста-
216
повления температуры кожи после местного охлаждения.
При акклиматизации человека к условиям Севера различают три фазы, последовательно наступающие одна за другой (Данишевский, 1955): а) начальная фаза акклиматизации, когда ярче всего проявляются реакции организма на новые для него климатические условия; б) фаза уравновешивания и перестройки механизмов уравновешивания организма с внешней средой. При этой фазе наблюдаются случаи «полома» механизмов уравновешивания и явления дезадаптации и в) фаза устойчивой акклиматизации.
В первый период акклиматизации в условиях Арктики у человека наблюдается тенденция к понижению кровяного давления. Причины этого явления неясны.
Одним из критериев акклиматизации к условиям Севера можно считать скорость восстановления кожной температуры после стандартного охлаждения. Эта скорость особенно велика у коренного населения Севера — чукчей, эскимосов, якутов (Капдрор, Солтысский, 1959). У приезжих из умеренного климата — при условии выполнения работы па открытом воздухе — картина восстановления кожной температуры после охлаждения приближается к картине аборигенов только после трех лет такой акклиматизации. В зимнее время сосудистая реакция более выражена, чем в летнее.
Однако акклиматизация к условиям полярных областей не ограничивается непосредственными изменениями терморегуляции под влиянием только пониженных температур среды. Не меньшее значение имеют и особенности светового и ультрафиолетового режима в условиях как полярного дня, так и полярной ночи. Полярная ночь оказывает значительное и притом отрицательное воздействие на организм человека. Световое голодание приводит к увеличению случаев заболевания детей рахитом. Наблюдается снижение содержания в крови лейкоцитов и гемоглобина. Изменяются иммунобиологические реакции как детей, так и взрослых, что выражается в увеличении заболеваниями скарлатиной, корью в зимние месяцы. Наблюдается снижение неспецифического иммунитета, особенно у тех, кто недавно прибыл в Заполярье.
Вопрос об акклиматизации человека в условиях Заполярья решается, по-видимому, исключительно в плане
217
современных гигиенических мероприятий, позволяющих не только создать для человека достаточный тепловой комфорт, по и компенсировать световое и ультрафиолетовое голодание. Такие же вопросы, как физиология размножения и развития, требуют еще значительных физиолого-гигиенических исследований для создания нормальных физиологических отношений в организме человека при этих своеобразных условиях внешней среды.
Большое место в рассматриваемой проблеме занимают исследования терморегуляции у человека в условиях жаркого климата. По вопросу приспособления человека к существованию в условиях тропиков имеется значительная литература. Большинство исследователей приходит к выводу, что в процессах приспособления к тропикам у людей различных рас нет существенных различий (Stigler, 1920; Morrison, 1956; Ladell, 1964 и др.). Обычно принимают, что тропический климат с его строго постоянной температурой среды (с колебаниями в году до 1°С и отсутствием суточных колебаний) может обеспечить нормальный теплообмен человека без всякой одежды в тени и в полном покое. Всякая деятельность в этих условиях связана с дополнительной продукцией тепла и требует увеличения теплоотдачи путем потоотделения. Имеется довольно большое количество фактов, свидетельствующих о том, что потоотделение в условиях жаркого климата увеличено, и способность к потоотделению в процессе акклиматизации возрастает. Этим и объясняется то, что ходьба в условиях тропиков по горизонтали с 20 кг нагрузки у хорошо потеющего человека перегревания не вызывает.
Значительные изменения в условиях тропиков претерпевает у человека кровообращение. Большинство исследователей обнаруживает стойкое снижение кровяного давления и возрастание минутного и ударного объема сердца. Однако и у человека дыхательному аппарату принадлежит в теплоотдаче значительная роль. Исследования температуры выдыхаемого воздуха показали, что последняя зависит не только от температуры внешней среды, но и от одежды исследуемого, т. е. от величины общей теплоотдачи организма.
Таким образом, несмотря на отсутствие у человека собственно механизма полипноэ, теплоотдача дыханием 218
занимает даже в условиях жаркого климата (особенно сухого) значительное место.
Температура тела в условиях тропиков часто бывает повышенной, причем наблюдается обратная зависимость между интенсивностью потоотделения и температурой тела (Ladell, 1964).
Собственно процессы адаптации человека к жаркому климату сводятся в основном к снижению температуры тела, возрастанию периферического кровообращения. Увеличенное кровоснабжение кожи обеспечивает не только большую теплоотдачу с поверхности тела, но и усиленную работу потовых желез (Lewis, 1942; Юнусов, 1950). Наиболее ярко воздействие тропического климата проявляется в увеличении минутного объема сердца, сопровождающегося учащением сердечной деятельности. Часто увеличение кровообращения коррелируется с повышением температуры тела.
Важное место при адаптации в тропиках занимают изменения в крови. Большинство исследователей отмечает увеличение содержания воды в плазме, особенно ярко выраженное в первый период воздействия высокой температуры (Юнусов, 1961). Активная реакция крови не изменяется, хотя есть некоторая тенденция к се сдвигу в щелочную сторону.
Наиболее неясным представляется изменение общего обмена веществ. Как правило, большинство исследователей находит в условиях тропиков лишь незначительное снижение основого обмена, что отчасти связывается с особенностями питания в условиях высокой температуры. Тем не менее ряд исследователей в строгих условиях изучения основного обмена наблюдали его снижение в тропиках как у местного населения, так и у хорошо адаптированных приезжих (Ozorio de Almeida, 1919; Knipping, 1923). Имеются указания на то, что интенсивность химической терморегуляции у человека, адаптированного к высокой температуре, снижена. Значительно возрастает в условиях тропиков энергетический расход на мышечную работу. Это связывается, однако, с включением в активность большого количества систем (кровообращение, дыхание, потоотделение), обеспечивающих поддержание температуры тела.
Таким образом, несмотря на то, что человек, по мнению ряда исследователей (Слоним, 1952; Scholander,
219
1958 и др.), является тропическим организмом, интенсивная работа его в тропических условиях крайне затруднена и требует специальных искусственных мер охлаждения. Можно сделать и более общий вывод, что существование человека в условиях различных климатических поясов от Арктики и Антарктики до экватора обеспечивается отнюдь не физиологическими особенностями его терморегуляции, а создаваемым человеком микроклиматом — одеждой и жилищем (Бартон и Эд-холм, 1957). Тем не менее факт адаптации человека к различным температурным условиям несомненен и обеспечивается физиологическими механизмами, близкими к таковым у высших млекопитающих.
Глава VI
НЕДОСТАТОК КИСЛОРОДА И ЕГО ВЛИЯНИЕ НА ОРГАНИЗМ
ТИПЫ ГИПОКСИИ И МЕХАНИЗМЫ
ЕЕ ВОЗНИКНОВЕНИЯ
Протекание окислительных процессов связано с непрерывным поступлением кислорода в организм—'Процессом, который обеспечивается легочным, жаберным, кожным, трахейным и другими видами внешнего дыхания. Внешнее дыхание на разных этапах эволюции в разной мере непосредственно обеспечивает тканевое при разном участии циркуляторного аппарата (кровообращения пли лимфообращения). Все эти процессы обеспечивают кислородный запрос организма, т. е. такое поступление кислорода в организм, которое отвечает затрачиваемой организмом энергии. Всякое усиление жизнедеятельности связано с увеличением кислородного запроса и наоборот. Поэтому в природных условиях состояние гипоксии, т. е. несоответствие между доставкой кислорода и кислородным запросом, часто наступает не только при изменении содержания кислорода в окружающей среде, но и при изменении деятельности организма. Наиболее простым примером может здесь служить мышечная деятельность, резко повышающая интенсивность обмена веществ, а отсюда и кислородный запрос организма.
Известная классификация гипоксий, предложенная английским физиологом Баркрофтом, включала: гипоксическую гипоксию, связанную с нарушением поступления кислорода в организм, циркуляторную — с нарушением кровообращения, гистотоксическую — с нарушением тканевого дыхания при выключении дыхательных ферментов, анемическую — при недостатке крови, застойную — при нарушениях кровообращения.
221
В плане эколого-физиологического сопоставления можно выделить следующие типы гипоксий, связанные с особенностями среды обитания организма или с его деятельностью. Эти формы гипоксий в различной мере компенсируются животным организмом и поэтому адаптивные сдвиги оказываются у разных видов выражены в различной степени: при недостатке кислорода в горах и на высотах; при мышечной деятельности; при нырянии; при оборонительных реакциях (скручивание тела).
Следует заметить, что различные организмы в разной степени компенсируют во времени недостаток кислорода и образующийся при неполностью удовлетворенном кислородном запросе так называемый кислородный долг. Адаптивные реакции в организме в связи с этим могут занимать большее или меньшее место в регуляции жизнедеятельности и поведения животного и быть в различной степени выражены.
ПОСТУПЛЕНИЕ КИСЛОРОДА В ОРГАНИЗМ
И ЕГО ПЕРЕНОС К ТКАНЯМ
Разность в напряжении кислорода в воде и жидкостях организма как основной путь снабжения кислородом при клеточном дыхании дополняется в процессе эволюции рядом механизмов, имеющих адаптивный характер. Наиболее важный этап в эволюции здесь — появление дыхательных пигментов, постепенно приобретающих исключительное значение в снабжении кислородом тканей.
Огромное разнообразие морфологических и физиологических адаптаций, обеспечивающих обмен газов между организмом и средой, связано с различным содержанием (напряжением) газов во внешней среде, с одной стороны, и кислородным запросом организма, с другой. Дополнительные движения тела или отдельных его частей (дыхательные движения) способствуют поддержанию высокой разницы в содержании газов вне и внутри организма. Для экологической физиологии особенно важно соответствие всех этих дополняющих морфологических и функциональных механизмов с уровнем кислородного запроса, с его изменениями в организме и отдельных его частях.
До настоящего времени известно очень немного случаев настоящей секреции газов, т. е. продвижения их 222
через мембраны против высокой концентрации. Эти случаи известны для плавательного пузыря некоторых видов рыб (закрытопузырных) и некоторых водных организмов. По существу таким образом создаются химически свободные запасы кислорода в организме. Однако в подавляющем большинстве случаев в процессе эволюции происходит образование более или менее сложных химических систем связывания как кислорода, так и СО2. Эти химические механизмы связывания газов могут замещать как физико-химические механизмы внешнего дыхания, так и собственные механизмы дыхания (вентиляцию, смешивание или проталкивание воды и т. д.). Химические механизмы могут носить адаптивный характер и па уровне клетки.
Рис. 52. Физиологические изменения, протекающие на разных уровнях в процессе адаптации к гипоксии (по Барбашовой, 1964)
3. И. Барбашова (1960, 1964) предлагает схему «борьбы за кислород», которая заключается в том, что ткани, поставленные в условия ограниченного снабжения кислородом, «акклиматизируются» и оказываются способными дышать более интенсивно в бедной кислородом среде. В основе этого явления лежит, по-видимому, обогащение тканей окислительными ферментами. Однако эта «борьба за кислород» может происходить не только
223
на тканевом уровне, но и на уровне транспортной системы— крови, что выражается в увеличении ее дыхательной функции, т. е. способности переносить О2 и СО2. Кроме того, «борьба за кислород» вовлекает и функции дыхания и кровообращения. Эта схема в настоящее время может быть положена в основу изучения физиологических изменений, наступающих в организме при гипоксии (рис. 52).
РЕАКЦИИ ОРГАНИЗМА НА ГИПОКСИЮ
Поскольку в основе явлений гипоксии лежит несоответствие между величинами кислородного запроса организма и способностью транспортных систем в его удовлетворении, общие реакции организма на гипоксию могут иметь двоякую направленность: а) включение физиологических механизмов, увеличивающих доставку кислорода тканям и б) приспособление самих тканей к существованию в бедной кислородом среде или ограничение кислородного запроса путем снижения жизнедеятельности организма или отдельных его систем. Такая схема физиологических изменений позволяет разделить все случаи приспособлений к недостатку кислорода на реактивный тип приспособления, связанный с увеличением транспорта кислорода тканям, и тканевой тип, связанный с приспособлением к недостатку кислорода тканей тела (Campbell, 1926—1927, 1927а, 1927; Барбашова, 1941, 1960; Гинецинский и Барбашова, 1942, 1945; Ольнянская, 1950; Слоним, 1952; Филатова, 1957, и др.).
Физиологическое влияние высоты на организм складывается из уменьшения парциального давления кислорода (вследствие снижения общего барометрического давления с высотой) и понижения температуры окружающей среды. Кроме того, на больших высотах значительную роль играет и ультрафиолетовая солнечная радиация, на равнине в значительной мере задерживаемая атмосферой и изменяющая чувствительность организма к гипоксии.
Длительное пребывание в условиях недостатка кислорода вызывает значительные изменения дыхания. Из морфологических изменений следует указать на увеличение размеров грудной клетки. Последнее установлено для человека, постоянно живущего на высотах (Forbes, 1870,
224
Рис. 53. Кривые диссоциации оксигемоглобина (по Schmidt-Nielsen, 1965). I — при повышении напряжения СО2; II— у млекопитающих с разным весом тела; III — у беременной самки и плода;
IV — у высокогорного организма ламы
1875; Jourdanet, 1875; Третьяков, 1895, 1897; Barcroft a. oth., 1923). Эти морфологические изменения сопровождаются значительным углублением дыхания и возрастанием общей величины легочной вентиляции. Последнее очень четко выявляется у человека при подъеме с равнины на высоту.
Увеличение глубины и отчасти частоты дыхания приводит к возрастанию величины так называемой альвеолярной вентиляции, в результате чего в альвеолах поддерживается более низкое парциальное давление СО2. Последнее является фактором, снижающим возбуждение дыхательного центра. Недостаток углекислоты в альвеолярном воздухе и в крови приводит к акапнии, т. е. к состоянию, когда легочная вентиляция, вследствие снижения возбуждения дыхательного центра, не возрастает,
8—1204
225
несмотря на развивающиеся явления кислородного голодания. При этом вследствие понижения содержания СОг в крови ухудшаются условия снабжения тканей кислородом из-за уменьшения степени диссоциации оксигемоглобина. Чувствительность к акапнии у разных организмов весьма сильно различается. Так, у человека акапния быстро приводит к появлению кислородного голодания п горной болезни. У млекопитающих, обладающих механизмом полипноэ, всегда сопровождающимся значительным вымыванием углекислоты из альвеолярного воздуха и из крови, этого почти ие наблюдается.
Изменение кровообращения при пребывании на высотах сводится к значительному увеличению объема выбрасываемой сердцем крови (минутного объема сердца), при этом учащается и пульс.
Некоторые считают отсутствие реакции со стороны сердечно-сосудистой системы в процессе приспособления к высотам благоприятным критерием акклиматизации.
Кровяное давление, как правило, в горах возрастает; то же наблюдается и в опытах в барокамерах. Однако во многих горных местностях (например, система Тянь-Шань) эти явления не наблюдаются, в Альпах и на Кавказе, напротив, они выражены очень ярко.
Яркие морфологические изменения наблюдаются в сердце. Впервые они были обнаружены у альпийской горной куропатки (Lagopusy Strohl, 1910) и выражаются в увеличении веса сердца и гипертрофии миокарда. Это подтверждено рядом дальнейших исследований у млекопитающих животных и птиц (Hesse, 1921; Машковцев, 1935; Шварц, 1959), а также у человека (Миррахимов, 1964).
Особое место в изменениях кровообращения занимают изменения кровообращения мозга. В процессе акклиматизации к высотам у крыс наблюдается стойкое (на 30—40%) увеличение содержания крови в мозге (Крепе и сотр., 1956). Кроме того, увеличивается кровоснабжение также печени и мышц, но эти изменения выражены менее ярко, чем увеличение кровообращения мозга.
Огромным количеством исследователей установлено, что в процессе акклиматизации к пребыванию в горах наблюдается увеличение содержания гемоглобина и эритроцитов в крови. Эти изменения в картине красной крови— следствие увеличения кроветворения, а при корот
226
ких воздействиях и перераспределения крови, благодаря чему наблюдается возрастание кислородной емкости всей циркулирующей крови. Усиленное кроветворение наступает вследствие раздражения костного мозга; перераспределение крови — благодаря опорожнению кровяных депо: селезенки, печени, сосудов кожи. В табл. 15 приведены некоторые полученные в работах разных авторов данные об изменении содержания эритроцитов в крови у разных животных в связи с пребыванием на высотах.
Содержание эритроцитов и гемоглобина в крови определяет ее кислородную емкость. Кислородная емкость крови имеет огромное значение для экономного переноса кислорода, но ее величина не всегда определяет способность животного к существованию в условиях гипоксии.
Таблица 15
Количество эритроцитов в крови животных на различных высотах (млн. в 1 мл)
(по Барбашовой, 1941)
Животное Высота (.и н. у. м.) Количество эритроцитт в (млн. в 1 мл) Авторы
Овца Кролик .... Крыса Мышь 10,53 4,55 8,97 11,39 11,72 — — — 6,58 — — 14,37 - — 15,72 Holl и соТр. То же Lintzel и Roder То же
Другой стороной дыхательной функции крови является характер диссоциации оксигемоглобина, определяющий особенности переноса кислорода из легких в ткани. Кривая диссоциации зависит, с одной стороны, от природных особенностей гемоглобина животного данного вида, а с другой, от среды, в которой происходит реакция окисления гемоглобина и восстановления его (pH плазмы, содержание СОг, содержание С1 в плазме). Кроме того, в последнее время к числу факторов, оказывающих влияние на кривую диссоциации оксигемоглобина, относят также интенсивность гликолиза в эритроцитах и расщепление в них же бикарбонатов карбоангидразой. Следовательно, эритроцит как физико-химическая система оказывает также значительное влияние на дыхательную функцию крови (Ван-Лир и Стикней, 1967).
В* 227
Особенностям кривой диссоциации оксигемоглобина в приспособлении к высотам организма человека придавалось большое значение (Barcroft a. oth., 1923; Barcroft, 1925; Холден и Пристли, 1936; Барбашова, 1941). Эти авторы наблюдали у человека при восхождении в горы (а также у животных) сдвиги в положении кривой диссоциации. Кривая диссоциации оксигемоглобина зависит от содержания СОг в крови, веса тела (интенсивности
Рис. 54. Газообмен у крыс (мл/мин на 1 кг веса тела) при различных барометрических давлениях (по Архангельской, 1949а)
Потребление О2: / — «холодные». 2 — «теплые»; выделение СО2: 3 — «холодные», 4 — «теплые». По осн ординат—газообмен (лгл), по оси абсцисс — высота (.и)
метаболизма) и условий снабжения тканей кислородом (рис. 54).
На гиссарских овцах на высоте 3000 м н. у. м. не обнаружено изменений положения диссоциационной кривой (Барбашова и Гинецинский, 1942), также не наблюдалось изменений диссоциационной кривой у овец местной породы, а у равнинных овец прекос был сдвиг ее вправо (Избпнский, Кезик, Черкович и Чукин, 1949).
То же имело место и у лошадей при подъеме в горы (Филатова, 1946а). Интересно, что эти сдвиги наблюдались только для цельной крови, а не для очищенного диализом гемоглобина. Однако у лошадей, которым производились систематические кровопускания, кривая диссоциации очищенного гемоглобина также сдвигалась вправо. Следовательно, при обильном новообразовании
228
гемоглобина могут изменяться его природные свойства, что, несомненно, имеет место для гемоглобина животных различных видов, для гемоглобина плода и новорожденного. Сдвиг диссоциационной кривой, как и абсолютная величина (%) насыщения кислородом гемоглобина артериальной крови, не могут служить критериями устойчивости его к условиям высокогорья. Это показывают приводимые далее наблюдения на горных породах овец и многие наблюдения на человеке.
Сдвиг кривой диссоциации влево приводит к более полному насыщению кислородом крови, по в то же время затрудняет отдачу его тканям. Поэтому имеет значение не только абсолютное положение диссоциационной кривой НЬОг, но и форма последней и ее конфигурация, определяющая положение точек зарядного и разрядного напряжения. Уровень потребления кислорода при гипоксии закономерно не изменяется. Большой материал, собранный исследователями на человеке, главным образом при восхождении в горы, а также экспериментальные исследования гипоксии на лабораторных животных позволяют утверждать, что постоянство кислородного запроса в условиях гипоксии сохраняется. Снижение потребления кислорода при высоких степенях гипоксии обычно вызывает накопление продуктов неполного окисления в организме и последующее возрастание потребления кислорода после возвращения к нормальным условиям дыхания (Giaja, 1938; Шик, 1939).
Способность удерживать потребление кислорода во время гипоксии на одном уровне рассматривается многими авторами (Крепе, Вержбинская, Ченыкаева, Чир-ковская, Гавурина, 1956) как приспособление к условиям кислородного голодания. Некоторые отмечают явление так называемого гипоксического парадокса, когда при недостатке кислорода кислородный запрос возрастает (Opitz u. Schneider, 1950). В основе лежит проявление действия гипоксии еще до того, как в клетках произойдут гипоксические изменения обмена. Потребление кислорода тканями остается на прежнем уровне или даже слегка повышается, в то время как организм дает отчетливые симптомы кислородного голодания вследствие понижающегося напряжения кислорода в крови и тканях.
Это явление объясняется двояко. Во-первых, рефлексы, вызывающие соответствующие компенсаторные ре
229
акции (увеличенное дыхание и кровообращение), возникают и развиваются при таком напряжении кислорода, которое еще не настолько низко, чтобы вызвать непосредственное нарушение аэробных процессов в клетках. Поэтому при наличии выраженного увеличения дыхания, кровообращения и других функций, характеризующих
Рис. 55. Средние градиенты давления рО2 от трахеального воздуха до смешения венозной крови у уроженцев Лимы (уровень моря) и Марокоча (4540 м н. у. м.). Альвеолярный воздух, артериальная кровь и смешанная венозная кровь (из легочной артерии) получены одновременно в покое лежа. рО2 капиллярной вычислено (по Hurtado, 1964)
определенную фазу гипоксии, потребление кислорода может несколько повышаться. Во-вторых, имеется и биохимический аспект этих сдвигов, так как сама по себе величина напряжения кислорода является важным условием деятельности различных ферментных систем. Известно, что при определенных условиях пониженное напряжение кислорода может вызвать усиление аэробного гликолиза без изменений в величине потребления кислорода. Реакцией, очень чувствительной к понижению напряжения кислорода, является, например, синтез ацетилхолина и некоторые другие биохимические процессы.
230
Низкая температура среды повышает требования организма к минимальному давлению кислорода, при котором могут в полной мере осуществляться окислительные процессы. Это зависит не от возрастания кислородного запроса, а от своеобразных изменений регуляции обмена гипоксии. Факт повышения устойчивости организма к гипоксии при высоких температурах среды и понижения при низких был установлен Е. А. Карташевским (1908) и детально изучен Н. А. Архангельской (1949а, 19496).
В исследованиях Н. А. Архангельской было установлено, что при понижении барометрического давления снижение газообмена у крыс зависит от того, при какой температуре они содержались до опыта. У животных, длительно содержавшихся при температурах среды 6 — 8° С, снижение обмена при гипоксии выражено при более высоком давлении О2, чем у содержавшихся при 26— 28° С.
На рисунке 54 видно, что уровень потребления кислорода у «холодных» крыс при всех изученных уровнях барометрического давления выше, чем у «теплых». Снижение барометрического давления вызывает у «холодных» крыс понижение потребления кислорода, начиная с давления, соответствующего 1000 м н. у. м., а у «теплых» крыс снижение обмена наступает на высоте 2500 м н. у. м. Следовательно, влияние оказывает не только температура среды, но и ее последействие.
Таблица 16
Зависимость между температурой среды, потреблением О2 и «начальным» давлением О2 (по Giaja et Marcovic, 1950)
Температура среды (°C) Потребление Оа (мл,'кг,ч) Начальнг е давление (лглг) Бгрокоэффиииент /О, (мл/кг. Ч\ ХрО2 мм Hg/
— 1 3970 138 28
3 3306 149 22
11,5 2716 125 22
27,5 1648 104 17
30,7 1405 89 13,5
Исследованиями югославского физиолога Джайя (Giaja, 1938, 1953, 1957) и его школы выявлены опреде-
231
ленные зависимости между уровнем потребления кислорода и интенсивностью охлаждения тела животного в условиях недостатка кислорода. Установлено понятие начального давления, т. е. того давления кислорода, ниже которого начинает снижаться газообмен (при разных температурах среды) и понятие барокоэффициента (Bq), представляющего отношение между потреблением О2 в мл/кг/ч, и напряжением О2 в мм Hg (табл. 16).
Под влиянием гипоксии (вдыхание смеси с 12,5 %-ным содержанием кислорода) можно наблюдать уменьшение электрических ответов в мышце кролика (Иванов, 1957). Такое снижение электрической активности сопровождается понижением общего газообмена. Переход от гипоксического состояния к нормальному повышению энергетических затрат сопровождается повышением электрических явлений в покоящейся мышце. Таким образом, поскольку электрические явления в мышце определяются импульсацией из центральной нервной системы, нет оснований считать снижение обмена веществ при гипоксии лишь проявлением подчинения тканевых процессов недостатку кислорода и заменой части окислительных превращений анаэробными. Последнее несомненно имеет место, но сопровождается и глубокими изменениями трофической функции центральной нервной системы.
Температура тела даже при умеренной гипоксии почти всегда снижается. Многие факты понижения кислородного запроса при гипоксии могут, по-видимому, быть поставлены в прямую зависимость от этого снижения температуры тела и тканей и, по-видимому, от общего снижения возбудимости нервной системы. Последнее выражается в снижении величин рефлекторных ответов при глубокой гипоксии. Нарушается также и условно-рефлекторная деятельность. Особенно страдает при этом тормозной процесс (растормаживание дифференцировок). При гипоксии резко нарушается желудочное пищеварение— отделение желудочного сока (Разенков, 1945). Систематическая тренировка собаки в барокамере ведет к нормализации кривой желудочного сокоотделения на высоте (Малкиман, 1948). Лучше всего такой эффект наблюдается у собак с желудочком по Павлову, хуже—• с желудочком по Гейденгайну. Повторные гипоксические состояния (подъем в барокамере) приводят к образованию весьма стойких условных дыхательных рефлексов (Архангельская и Сегаль, 1954), а также к условнореф-
232
лекторному изменению содержания эритроцитов и НЬ и к другим реакциям, связанным с острой гипоксией.
Специфическая роль эндокринных регуляций и, в частности, щитовидной железы в приспособлении организма к гипоксии остается еще не ясной. В настоящее время можно считать установленным, что тиреоидэктомия приводит к повышению устойчивости организма к недостатку кислорода (Streuly, 1918; Веселкин, 1942 и др.). С другой стороны, имеются указания на то, что гипоксические состояния, например отравление окисью углерода, могут быть причиной гипертиреоидизма (Raab, 1934; Schultze, 1936). По некоторым данным (цит. по: Grandjean, 1948), на высотах понижается интенсивность действия тиреотропного гормона гипофиза. Ряд клиницистов наблюдали в горах снижение симптомов гипертиреоидизма (Grandjean, 1948). В последнее время появились определенные указания на зависимость устойчивости организма к острой гипоксии от функции щитовидной железы и коры надпочечников. Угнетение этих функций или тиреоидэктомия и адреналэктомия приводят к увеличению времени выживания животных при пониженном барометрическом давлении (Debias, 1966).
Адаптация к высоте угнетает активность щитовидной железы, а искусственное введение тироксина замедляет адаптацию. У адаптированных крыс тироксин оказывает большее влияние на потребление корма (увеличивает его), чем у животных на уровне моря (Weihe 1966).
Экспериментальный подъем животных в барокамере приводит к заметным изменениям поведения. Они выражаются в прекращении активности на высоте. Адаптация мышей к высоте постепенно уменьшает этот эффект и активность становится нормальной. Вместе с тем не наблюдается никакой зависимости между реакцией крови и изменениями активности. Так, если животным вводить эритропоэтины и вызвать без подъема на высоту увеличение гематокрита с 45 до 70% эритроцитов (что имеет место и при высотной адаптации), то такое возрастание гематокрита не снимает снижения активности мышей при подъеме па высоту. Следовательно, одна дыхательная функция крови не обеспечивает адаптации к высоте, а является лишь одним из ее компонентов. Двигательная активность на высоте при адаптации восстанавливается раньше, чем наступают изменения в крови (Klighley, Lowy a. McCune, 1968).
233
УСЛОВИЯ СУЩЕСТВОВАНИЯ ЖИВОТНОГО ОРГАНИЗМА В ГОРАХ И ТИПЫ АДАПТАЦИИ К ГОРНЫМ УСЛОВИЯМ
Физические факторы, действующие на животный организм в горах, не ограничиваются пониженным парциальным давлением кислорода, пониженным общим атмосферным давлением, а включают также пониженную температуру воздуха и почвы, повышенную солнечную радиацию, значительные суточные колебания относительной влажности воздуха и, наконец, особенности минерального питания, связанного с малым содержанием солей в воде и почве. Все эти факторы и их сочетание отличаются значительным разнообразием, зависящим от рельефа местности и ориентировки отдельных склонов и долин в отношении стран света. К этому разнообразию собственно физических и химических параметров среды следует добавить еще и значение собственно рельефа местности, передвижение по которой представляет для одних и тех же видов (например, для пасущихся животных) разную по интенсивности мышечную работу.
Животное население горных районов, а отчасти и человек не остаются постоянно в течение разных сезонов года. Изменение температуры, глубина снежного покрова в горных условиях столь значительны, что многие организмы лишь периодически в весенне-летние месяцы мигрируют в горы, спускаясь в предгорья и долины в зимнее время. Все животное население гор можно разделить (Насонов, 1923) на несколько групп в зависимости от степени их вертикальной «оседлости». Животные, постоянно обитающие в горах на одних и тех же высотах и не мигрирующие в долины, были названы стеногипсох-тонными. В противоположность им мигрирующие организмы— эвригипсохтонными. Эта классификация, забытая экологами, однако, совершенно необходима, когда делается попытка эколого-физиологических сопоставлений отдельных видов и изучения формирования адаптаций животных к горам в процессе эволюции отдельных видов. В известной мере эти отношения касаются и физиологических изменений у человека в горах, так как вертикальные миграции человека известны с незапамятных времен и связаны с отгоном скота па различные высоты в связи с высыханием растительности в предгорьях и долинах.
234
По мнению II. В. Насонова, собственно стеногипсох-тонными животными являются некоторые виды моллюсков (главным образом голые слизни), населяющих отдельные высокогорные долины и образующих большое количество эндемичных видов. Более подвижными в этом отношении являются насекомые, расселение которых (например, жужелиц) охватывает значительно большие области по вертикали. Это касается также и птиц. Значительно меньшие передвижения по вертикали проделывают довольно многочисленные в горах виды грызунов, из которых некоторые (пищухи, сурки) представляют типичные для данной горной системы высокогорные или среднегорные виды. Наконец, наибольшие миграции (да и лучше всего изученные) можно наблюдать у копытных. В этом отношении дикие копытные в известной мере предвосхитили режим выпаса домашних, отгоняемых человеком в горы на летнее время (так называемая летов-ка овец и крупного рогатого скота).
Исходя из этих особенностей распространения животных и человека в горах можно выделить три случая физиологических адаптаций к горным условиям: адаптация равнинных организмов, в своей эволюции не мигрировавших в горы; адаптации мигрирующих организмов и, наконец, адаптации организмов, постоянно обитающих на высотах. К сожалению, не все эти группы изучены в одинаковой степени физиологами, но и имеющийся материал уже позволяет наметить ряд важных сопоставлений.
АДАПТАЦИИ РАВНИННЫХ ОРГАНИЗМОВ В ГОРАХ
И В ЭКСПЕРИМЕНТЕ
Исследования реакции на недостаток кислорода как важнейший фактор воздействия на организм высокогорья широко представлены на равнинных организмах, к числу которых должен быть отнесен и человек. При этом используется как подъем в барокамерах в эксперименте, так и перемещение в горы на разные высоты.
Практически на основании анализа довольно большого материала реакций равнинных организмов обычно делят горные условия на 3 группы в зависимости от высоты над уровнем моря. Малые высоты включают подъем в горы от 200 до 1000 м, средние высоты (среднегорье)
235
от 1000 до 2500—3000 .л и большие высоты от 3000 м над уровнем моря и выше. Практически жизнь в естественных условиях высокогорья простирается до 5000—5500 м. Выше проникает только человек, если иметь в виду высокогорные экспедиции, и часто с применением дополнительной подачи кислорода для дыхания.
Наиболее простым и весьма важным критерием устойчивости организма к недостатку кислорода является определение высотного потолка. Подьем в барокамере с определенной скоростью пли время выживания на определенной высоте (барометрическом давлении) позволяют сравнить различных животных по их устойчивости к острому или хроническому кислородному голоданию.
Наиболее систематические исследования высотного потолка были проведены на разных животных Н. Н. Си-ротининым (1950). Большая устойчивость в гипоксии имеет место у коз. Автор высказывает предположение, что это обусловлено горным происхождением этих животных. Свиньи, по тем же данным, очень чувствительны к недостатку кислорода — их высотный потолок не превышает 6000—7000 м н. у. м. Низкий высотный потолок наблюдается у домашней кошки; он не превышает 10 000 м. Очень низкий потолок — 8000 м и у медведя.
Как правило, пойкилотермные организмы обладают гораздо большей устойчивостью к гипоксии, нежели гомойотермные. Это также ярко выявляется в опытах в барокамере. В этой связи значительный интерес представляет очень высокий высотный потолок у летучих мышей — животных, дневной сон которых сопровождается резким понижением температуры тела (см. выше).
Более высоким, чем другие животные равнины, высотным потолком обладают организмы, в течение жизни подвергающиеся гипоксическим состояниям,— ныряющие и животные, способные сворачиваться клубком (скручиваться).
Очень высоким потолком обладают некоторые копытные и в частности овцы и козы.
Подъем в барокамере вызывает изменения в ряде физиологических функций, направленных на борьбу с развивающейся гипоксией. Наиболее ярко эти явления были продемонстрированы в исследованиях Л. Г. Филатовой (1961) (см. табл. 17).
236
Таблица 17
Физиологические изменения в организме под влиянием подъема в барокамере на 6000 м (по Филатовой, 1958)
Животное Эритроциты* Гемоглобин (%) Частота дыхания (в 1 мин} Частота пульса (в 1 мин}
л> после % ЛО пгсле % Л"’ п' слг % ло I п еле 1 %
Морская свинка . . . 5,66 6,08 107 80,8 81,3 100,6 106 139 131 289 292 101
Белая крыса .... 7,68 7,75 101 80,1 84,9 106,1 88 127 144 460 466 100
Кролик 4,40 5,39 122 61,0 71,0 120,0 162 133 82 240 243 101
Ондатра (Ondatra zi-bethica) 5,97 6,28 105 90,1 91,5 101,5 77 98 121 303 319 105
Суслик желтый (Citel-lus fulvus) 5,21 5,59 107 73,0 73,8 101,1 47 93 198 274 321 117
Еж ушастый (лето) (Erinaceus auritus) . . 9,18 9,99 109 74,7 81,0 108,4 35 89 254 201 270 134
Еж ушастый (зима) 9,01 11,01 122 71,0 84,0 118,3 45 30 68 280 239 85
Серый хомячок2 (Cri-cetulus migratorius) . . 8,29 6,68 80 102,0 86,0 84,2 116 202 172 600 600 100
1 Эритроциты изучались сразу после подъема, частота дыхания и пульса во время подъема (на 30-й минуте). 2 Этот вид встречается на разных высотах и широко распространен в южных районах Евразии.
Из таблицы видно, что отдельные физиологические функции изменяются при острой гипоксии у различных животных неодинаково. Типично равнинные формы, например ушастый еж, желтый суслик или лабораторные крысы и кролики, обнаруживают некоторое повышение содержания эритроцитов. У ныряющей формы — ондатры наблюдается небольшая реакция дыхания и изменение содержания эритроцитов.
Существует обратная зависимость между изменениями отдельных функций при гипоксии. Так, например, у крыс почти не изменяется количество эритроцитов, процент гемоглобина и частота пульса, но резко увеличивается частота дыхания. У желтого суслика почти не изменяется состав крови, но резко возрастает частота дыхания и пульса. У морских свинок картина очень близка к тому, что и у крыс. У серого хомячка повышается только частота дыхания, кислородная емкость крови даже несколько снижается.
Большая разница наблюдается в реакции ежей в летний и зимний периоды (в бодрствующем состоянии). Интересно резкое увеличение кислородной емкости крови (реакция в зимний период) при совершенно неизменных дыхании и пульсе. Таким образом, можно видеть не только глубокие различия в отдельных реакциях, но и их взаимное замещение при коротком влиянии кислородного голодания.
На лабораторных животных, поднятых в горы, и на животных, выращенных на разных высотах, проведено сравнительно мало исследований. Среди них особое место занимает работа южноамериканских исследователей, проведенная на собаках. При исследовании собак, выросших на высоте 4530 м н. у. м. и на уровне моря, было обнаружено у первых сравнительно небольшое повышение числа эритроцитов, более значительное увеличение гемоглобина крови и очень большое увеличение миогемоглобина (Hurtado, Rotta, Merino a. Pons, 1937). Последнее было выражено слабее в сердечной мышце и сильнее в скелетной мускулатуре (табл. 18).
Данные Хуртадо и сотрудников не были подтверждены Полем (Poel, 1949), работавшим на акклиматизированных в барокамере крысах. Только в сердечной мышце наблюдалось после 152 ежедневных подъемов в барокамере (высота 7620 м и. у. м.), увеличение содер-
238
Таблица 18
Эритроциты, гемоглобин крови и мышечный гемоглобин у собак, выросших на разных высотах
жания миоглобина. Такие же данные были получены для скелетной мускулатуры собак (Bowen a. Eads, 1949). В дальнейшем у крыс, акклиматизируемых в атмосфере газовых смесей на протяжении четырех поколений, не было обнаружено увеличения миоглобина мышц, равно как и увеличения активности цитохромоксидазы (Крепе и сотр., 1956). Только при интенсивной мышечной тренировке нескольких поколений крыс в высокогорье на уровне около 4000 м н. у. м. установлено возрастание содержания миоглобина в скелетной мускулатуре на 50—70% (Clark, Criscuolo a. Coulson, 1952).
Следовательно, возрастание гемоглобина связано в основном с воздействием высоты (при длительной акклиматизации) на равнинные организмы, а возрастание миоглобина — главным образом с интенсивной мышечной деятельностью, протекающей в условиях пониженного парциального давления кислорода. Различия между этими формами гипоксических состояний по их влиянию па химическую динамику снабжения тканей кислородом еще требуют дополнительных исследований. В этой связи интересно отметить, что у диких видов млекопитающих и птиц наблюдается небольшое увеличение НЬ крови но сравнению с домашними животными и значительное увеличение мышечного гемоглобина (Верболович и Остаток, 1956). Так, гемоглобина у зайца на 20—22% больше, чем у кролика, а у дикой степной кошки па 10—27% больше, чем у домашней. Соотношение содержания гс-
239
моглобпна и миогемоглобина и определяет, по-видимому, приспособление к условиям мышечной работы животных.
Реакция организма человека на подъем в барокамере была описана ранее. При подъеме в горы на малые и даже средние высоты многими исследователями не обнаруживалось заметных и достоверных возрастаний числа эритроцитов и содержания гемоглобина в крови. Только начиная с 2000 м н. у. м. в условиях Швейцарских Альп можно наблюдать некоторую реакцию крови на
пребывание па высоте.
Таблица 19
Гематологическая картина жителей разных высот
(по Hurtado, 1964)
Показатели Лима (на уровне м<>ря) Мар- коч (4540 м н. у. м.)
Число испытуемых Число испытуемых
Общий объем крови (л) Кровь смЧкг веса тела Общий объем плазмы (л) Плазма с мН кг веса тела Общий гемоглобин (г) Гемоглобин (г/кг веса тела) Эритроциты (млн/мм3) Гематокрит (% эритроцитов) Гемоглобин (а/100 с.н3 крови) 20 20 20 250 250 250 4,77 79,6 2,52 42,0 756 12,6 5,11 46,6 15,64 20 20 20 83 83 83 5,70 100,5 2,23 39,2 1166 20,7 6,44 59,5 20,13
Наибольший материал по длительной адаптации человека к горам собран в Андах (Monge, 1943; Hurtado, 1964), а также в горах Тянь-Шаня (Миррахимов, 1964а, 19646). В таблице 19 приведены данные, характеризующие состав крови у жителей Лимы (Перу) па уровне моря и жителей Марокоча па высоте 4540 м н. у. м.
240
Из таблицы видно, что у постоянных жителей, обитающих на высоте свыше 4000 л/, увеличены все основные показатели красной крови: общий объем крови и плазмы, гемоглобин, количество эритроцитов и гематокрит. Следовательно, при достаточно сильном гипоксическом воздействии резко проявляется реакция крови, направленная на дополнительное снабжение организма кислородом. Эти явления, отражающие раздражение костного мозга, отсутствуют у новорожденных (Reynafarjc, 1959) и появляются лишь спустя несколько дней после рождения ребенка.
У родившихся и постоянно проживавших на высоте 4540 м н. у. м. наблюдается устойчивое повышение легочной вентиляции на 20—30% по сравнению с величинами, обычно наблюдающимися на уровне моря. Однако, несмотря на такую гипервентиляцию, насыщение артериальной крови кислородом у жителей Марокоча не превышает 80%• Дыхательный центр обладает повышенной возбудимостью, так как гипервентиляция сохраняется и после нормального насыщения крови кислородом.
Особенно интересны данные об изменении в этих условиях проницаемости для газов легочных мембран. Так, градиент между парциальным давлением кислорода в альвеолярном воздухе и напряжением кислорода в крови при такой акклиматизации снижается. Восстановление оксигенации крови при такой акклиматизации снижается, а после мышечной деятельности — ускорено. Состав альвеолярного воздуха изменен на 30—50%• Форма диссоциационной кривой оксигемоглобина не отличается от таковой на уровне моря. Количество эритроцитов увеличено, что обеспечивает повышенную кислородную емкость крови. Однако, несмотря па это, напряжение кислорода в тканевых капиллярах снижено с 57 мм на уровне моря до 37 мм па высоте 4540 м и. у. м., в смешанной крови соответствующие отношения 42 и 37 мм (рис. 55).
Кривая диссоциации оксигемоглобина несколько сдвинута вправо, что обеспечивает более легкую отдачу кислорода тканям. Напряжение СО2 в крови несколько снижено, но одновременно снижено и содержание бикарбонатов, что обеспечивает нормальные величины pH крови. Повышенное содержание эритроцитов обсспечп-
241
вает перенос СО2, несмотря на снижение общей забуф-ференности крови на больших высотах.
При такой адаптации человека к большим высотам наблюдаются и заметные изменения в тканях. Так, несмотря на сниженное рО2 в тканях, содержание молочной и пировиноградной кислот в крови не отличается от таковой на уровне моря. Более того, выполнение
Рис. 56. Насыщение крови кислородом на разных высотах у млекопитающих (по Hall, Dill, Barron, 1936):
/ — лама, 2 — викунья, 3 — овцы. По оси ординат — насыщение крови (%), по оси абсцисс — высота (кл<)
физической работы адаптированным человеком на высоте 5400 м приводит к меньшему образованию этих кислот, чем у человека на уровне моря. Мышечное сокращение происходит за счет увеличения аэробной фазы, по сравнению с таковым на уровне моря. Этот факт находит объяснение в увеличенной капилляризации тканей, возрастании числа капилляров по отношению к числу мышечных волокон (Hurtado, 1964).
Ферментативные механизмы тканей также значительно изменены. На фоне значительно увеличенного содержания миоглобина наблюдается возрастание активности митохондриальной днфеилгидрооксидазы, трансгндро-гоназы, цптотохромредуктазы, т. е. окислительных ферментов, способных к высокому использованию кислорода в тканях (Reynafarje, 1962).
242
В противоположность этим фактам, свидетельствующим о картине глубокой адаптации человека на больших высотах, был получен ряд данных, свидетельствующих о малой реакции со стороны дыхательно-циркуляторной системы и системы крови при адаптации к средним высотам. Следует помнить, что постоянное пребывание человека на больших высотах возможно только в южных широтах, на Тянь-Шане и Памиро-Алае, в МНР и в Тибете, а также в горах Южной Америки. В более северных широтах жизнь человека и его круглогодичное пребывание возможно лишь на высотах, не превышающих 1500—2000 м. На этих высотах производилось исследование влияния постоянного пребывания человека в горах Кавказа, Альпах и других горных местностях. Здесь постоянно наблюдались заметные сдвиги в составе крови, а также изменения дыхания и кровообращения (Grandjean, 1948). В горах Южной Америки — Андах, впервые было обращено внимание (Monge, 1943) на то, что значительное повышение содержания эритроцитов и гемоглобина в крови не является признаком адаптации человека к данной высоте, а является «хронической горной болезнью». Монге была выдвинута теория адаптации к высотам, привлекшая внимание к тканевым процессам. Он считал, что адаптация включает прежде всего повышенное использование кислорода тканями и возрастание транспортных средств доставки кислорода лишь дополняет такие изменения. Эти явления были особенно детально изучены па средних и даже на больших (до 3500 м н. у. м.) высотах рядом исследователей (Слоним и сотр., 1949; Аваз-бакиева, 1958; Филатова, 1960; Миррахимов, 1964а, 19646).
У постоянных жителей Киргизии и Казахстана на высотах 1650—1800 м н. у. м. частота и глубина дыхания, частота пульса, содержание эритроцитов и НЬ в крови не превышает верхней границы нормы (Авазбакиева, 1958). На высоте 1700 и 2600 м н. у. м. (озеро Иссык-Куль) у человека в состоянии покоя не наблюдается увеличения числа эритроцитов, содержания гемоглобина, а также увеличения частоты дыхания, пульса и повышения кровяного давления (Филатова, 1954а). Однако при выполнении мышечной работы кровяное давление повышается значительнее, чем на высоте около 900 м.
243
Значительное снижение основного обмена в горах Тянь-Шаня было отмечено на высотах 760—2400 м и. у. м. на 315 здоровых людях (Фанталис, I960).
Небольшое повышение содержания эритроцитов и НЬ крови обнаружено у жителей высокогорья Тянь-Шаня на высотах от 2100 до 2200 м (Миррахимов, 1964): увеличенным оказался и объем эритроцитов, и содержание гемоглобина. У постоянных жителей Тянь-Шаня обнаружена брадикардия. Систолическое кровяное давление не было изменено, а диастолическое имело тенденцию к повышению. Особенно тщательно были исследованы изменения сердечно-сосудистой системы человека в горах Киргизии. Изучены изменения кровяного давления, скорости кровотока, особенности капиллярного кровообращения у постоянных жителей малых и средних высот и у приезжих — мало акклиматизированных. Максимальное кровяное давление во многих случаях характеризуется низкими величинами, минимальное несколько повышено. Пульс в покое замедлен, а граница сердца несколько расширена. Функциональные нагрузки характеризуются быстрым возвращением состояния сердечнососудистой системы к исходному уровню.
Совершенно особое место при этом заняло изучение кислородного запроса организма как в покое в условиях основного обмена, так и во время выполнения мышечной работы. В упомянутых выше исследованиях, проведенных на малых и средних высотах Тянь-Шаня, постоянно наблюдалось снижение основного обмена, особенно четко выраженное, если исследование производилось с учетом погодных условий — температуры внешней среды (Миррахимов, 1965). Эти факты были созвучны отсутствию реакции со стороны кроветворной, дыхательной и сосудистой систем и затем детально проанализированы при изучении адаптации к высоте у людей, прибывших из различных районов СССР. Оказалось, что такой аре-активный (тканевой) тип адаптации свойствен людям, прибывающим из местностей, расположенных на малых высотах (Авазбакиева, 1968). Другими словами, снижение реакции возникает, по-видимому, в результате предварительно длительно воздействовавшей на организм умеренной гипоксии.
В этом отношении значительный интерес представляют и исследования энергетического расхода при выпол-244
нении мышечной работы в горах. Большинство прежних исследований, проведенных главным образом на альпинистах при кратковременных подьемах в горы, обнаруживало повышение газообмена и энергетических затрат на выполнение дозированной мышечной работы в горах. В последние годы, однако, показано, что у постоянных жителей гор кислородный запрос на выполнение мышечной работы значительно ниже, чем у постоянных жителей равнины. При этом оказалось, что у горцев в Тянь-Шане при мышечной работе насыщение крови на 5 об.% меньше, чем у жителей равнины. Артериальное давление при выполнении одинаковой мышечной работы у горцев ниже. Таким образом, сниженный энергетический обмен и кислородный запрос являются одним из критериев высотной адаптации у человека при переезде из одной горной местности в другую (Дубинина, Ковалева, Мирра-химов, 1967).
При подъеме в горы с уровня моря исследователи наблюдали другую картину. При переезде из Ленинграда на Кавказ (Пятигорск — 550 м н. у. м.) газообмен повышался; дальнейший подъем в горы до высоты 2100м вызывал еще большее повышение газообмена, причем не только в состоянии покоя, но и во время мышечной работы. При подъеме и пребывании на высоте 2100 м количество эритроцитов в крови нарастает (Быков, Мартинсон, 1933). По данным С. М. Бедаловой (1959а, 19596), у человека в условиях средних высот Малого Кавказа (1200—1600 м н. у. м.) наблюдается учащение пульса, понижение кровяного давления. При кратковременном пребывании учащение пульса выражено более резко (возрастание с 80 до 90 ударов в 1 мин). Увеличивается также венозное давление и скорость кругооборота крови.
При одпомесячной адаптации детей в горах Малого Кавказа кислородная емкость крови увеличивалась, повышался газообмен и легочная вентиляция. Переезд и в этом случае происходил с уровня моря (Салахов, 1960; Багдасарова, 1968).
В условиях горы Арагац были обследованы группы людей, проживающих постоянно на разных высотах: 960, 2000 и 3250 м н. у. м. В этом случае в системе гор Малого Кавказа также не обнаружено отклонений основного обмена от нормы (Захарян, 1965).
245
Таким образом, до настоящего времени вопрос о кратковременной картине адаптации человека в различных горных системах остается открытым. Решение его потребует еще выявления последовательных и повторных влияний факторов, воздействующих на человека в горах на разных высотах. Высказываются предположения о значении функции щитовидной железы. В горах Тянь-Шаня некоторые исследователи находили ее функцию пониженной.
Нет никаких данных о значении содержания микроэлементов для высотной адаптации человека и животных.
У человека скорость обмена железа (л/г/Ре/сутки/кг), равная на уровне моря 0,37, возрастает уже через 2 ч после прибытия на высоту (4500 м н. у. м.) до 0,54; через 1—3 недели — до 0,91, а через 6 месяцев адаптации к высоте держится на уровне 0,73. У аборигенов высокогорья эта величина равна только 0,49, а после спуска на уровень моря падает до 0,16 (Reynafarje, 1966). Возрастает всасывание железа в пищеварительном тракте; эта величина изменяется параллельно содержанию гемоглобина в крови. При спуске на уровень моря эти показатели изменяются в обратную сторону. У человека, как поднявшегося в горы, так и родившегося там и проживающего постоянно, спуск на уровень моря приводит к быстрой дезадаптации, к появлению отношений, характерных для жителей равнины.
АДАПТАЦИИ ОРГАНИЗМОВ, МИГРИРУЮЩИХ В ГОРЫ
Переезды человека с одних высот на другие в настоящее время уже не всегда носит сезонный характер. Но пастухи периодически перегоняют сельскохозяйственных животных в горы, особенно овец.
Установлено, что насыщение артериальной крови у овец падает с высотой (Hall, Dill a. Barron, 1936). В то же время постоянные обитатели высокогорья — лама и грызун вискаша поддерживают напряжение О2 в артериальной крови па высоком уровне (рис. 56). При подъеме в горы содержание эритроцитов у овец сначала увеличивается, а затем, на средних высотах, снижается. Авторы уже тогда связали это явление с возможностями глубокого снижения напряжения кислорода в тканевых
246
капиллярах, с возможностью существования животных при низком капиллярном напряжении кислорода.
Дальнейшее развитие это направление получило в работах советских физиологов и зоотехников. Первой работой были исследования гпссарскон овцы и метисов ее с английской овцой-
линкольн; возрастания кислородной емкости крови при пребывании этих животных в горах не было обнаружено (Барбашова и Гинецннский, 1942). Более того, с подъемом в горы насыщение кислородом артериальной крови закономерно падало. Расчет напряжения газов в капиллярной крови также показал значительное снижение этого напряжения при подъеме животных в горы (рис. 57). Это очень интересное исследование имело, однако, тот недостаток, что равнинные овцы изучались не одновременно с горными, а в условиях острой гипоксии, создаваемой в лаборатории.
Детально было изучено состояние животных как при отгоне в горы, так и при непрерывном содержании их па разных высотах в Киргизии.
Равнинные овцы и их
Рис. 57. Насыщение кислородом артериальной крови на разных высотах у овец разных пород (по Барбанювой и Гинециискому, 1942). / — равнинные овцы; II— гиссарские (горные) овцы и их метисы; III — сравнение кривых насыщения у равнинных (7) и горных (2) овец. По оси ординат — насыщение крови кислородом (%), по оси абсцисс — высота над уровнем моря (xi)
метисы с горной киргизской овцой изучались в совершен-
но одинаковых условиях содержания в стаде, отгоняемом на различные высоты (Ольнянская и Соболь, 1946, 1949; Ольиянская и сотр., 1949). Оказалось, что у киргизских овец, на протяжении многих поколений постоянно отгонявшихся на летннй период в горы на значительные вы
247
соты (до 4000 м н. у. м.), понижаются при отгоне в горы основной обмен, легочная вентиляция и частота дыхания. Кроме того, обнаружилось понижение числа эритроцитов и содержания гемоглобина в крови (рис. 58, 59). Черты акклиматизации, характерные для киргизских овец, можно обнаружить п у метисов прекос с киргизской овцой.
Рис. 58. Потребление кислорода (/) и частота дыхания (//) у овец разных пород,и их метисов в горах (по Ольпянской и corp., 1949)
У равнинных пород — прекос и рамбулье, содержавшихся в тех же условиях в одном стаде, этих явлений наблюдать не удается. Газовый обмен в горах повышается; это сопровождается увеличением легочной вентиляции, частоты дыхания. Число эритроцитов и особенно содержание гемоглобина в крови овец равнинных пород в горах возрастает (рис. G0).
248
Были изучены особенности высотной акклиматизации у тех же пород овец, уже па протяжении нескольких поколений постоянно находящихся в горах на высоте 2300—2500 м и. у. м. (Ольнянская, 19496). Обнаружилось, что различия между породами овец при таком длительном содержании животных в горах сглаживаются:
Рис. 59. Содержание гемоглобина в крови овец разных пород и их метисов в горах (по Ольнянской и сотр., 1949)
Рис. 60. Состав крови у овец в горах на высоте 2300—2500 л:
1 — овцы, спускающиеся зимовать на малые высоты, 2 — овцы, зимующие в горах, не ниже средних высот
249
снижение газообмена и легочной вентиляции наблюдается у овец всех пород как горных, так и равнинных и их метисов. Число эритроцитов у овец, постоянно находящихся в горах, было ниже, чем у спускавшихся ежегодно в долину.
Таким образом, при постоянном содержании животных в горах преобладают явления тканевой акклиматизации, а транспортные ее механизмы выражены крайне слабо. Такая форма акклиматизации выявляется не сразу после рождения. У ягнят, родившихся на высоте 3000 м и. у. м., в первые 10 дней после рождения можно наблюдать увеличенное количество эритроцитов и гемоглобина в крови, сравнительно высокий уровень газообмена и учащенное дыхание (Ольнянская, 1949в). В дальнейшем эти особенности исчезают, и картина крови ягнят киргизской горной породы начинает к 20-му дню жизни приближаться к картине крови их родителей. Следовательно, высотная акклиматизация без реакции со стороны транспортных систем организма не следствие какого-то недоразвития последних, а, наоборот, форма акклиматизации, приобретаемой животными в более поздние периоды онтогенеза. Это позволяет сделать вывод о том, что овцы в своем филогенетическом прошлом являются животными равнинными, лишь впоследствии населившими горные районы (Насонов, 1923; Цалкин, 1950).
Выводы Р. П. Ольнянской и сотрудников были подтверждены большим количеством исследований в горах Тянь-Шаня и Памиро-Алая (Арав и Смоличев, 1947; Жуков и Кожевников, 1947; Арав и Мартынова, 1949; Росляков, Евдаков, Басенко, 1953; Басенко, 1959). В этих исследованиях еще раз было показано, что типично равнинные— овцы каракульские при отгоне в горы обнаруживают значительные сдвиги в дыхании и кровообращении. Основной обмен повышается при этом на 29% (Арав, 1960). По этим же данным, наблюдаемые изменения связаны с пастьбой на склонах гор, которая сопровождается повышением обмена на 100% при подъеме и на 52% при спуске. По мере отгона в горы от средних к большим высотам у каракульских овец наблюдается повышение недоокисленных продуктов обмена в моче (vacat — О).
Влияние пребывания в горах на физиологические функции крупного рогатого скота в последнее время
250
изучено в ряде исследований. При этом обнаружились различия в реакциях со стороны животных, разводимых на равнине и в горах. Так, в условиях Киргизии уменьшение числа эритроцитов при одновременном увеличении содержания в них гемоглобина наблюдали у коров при отгоне на горные пастбища (с 1550 до 2800 м н. у. м.) (Одынец, Докукин и Фаиталис, 1956). Кривые диссоциации оксигемоглобина при этом практически не изменялись. Однако на большой высоте наблюдалось снижение насыщения артериальной крови кислородом. Последнее сопровождалось небольшим повышением частоты дыхания и частоты пульса, быстро исчезавшим при возвращении на исходную высоту над уровнем моря.
Проводились исследования высотной акклиматизации у крупного рогатого скота в условиях Северного Кавказа (Костин и Гастева, 1957; Костин, 1958а, 19586). При сравнении двух пород: красного степного равнинного скота и кубано-черноморского, издавна разводимых в горах, оказалось, что как молодняк, так и взрослые животные кубано-черноморской породы реагируют на отгон с равнины на высоту 1800 м и. у. м. снижением газового обмена и легочной вентиляции. У молодняка красностепной породы также снижаются легочная вентиляция и газообмен, а у взрослых животных эти функции повышаются.
Картина высотной акклиматизации лошадей подвергалась за последнее десятилетие тщательному изучению, главным образом в условиях Киргизии и Казахстана. Лошади содержались в стойле, а на высоте 2300 м и. у. м. — на пастбище (Попугаева и Слоним, 1949). Содержание эритроцитов в крови колебалось как на высоте, так и при стойловых условиях содержания от 5,6 до 9,03 млн., содержание гемоглобина — от 60 до 78% по Сали. Очень интересные данные получены при сравнительном изучении ряда пород лошадей в условиях гор и равнин Киргизии (Чашкин, 1959). Наиболее высокое содержание гемоглобина и эритроцитов оказалось у тяжеловозов, хуже всего акклиматизирующихся в горах. Хорошо акклиматизированные в горах киргизская, новокиргизская н донская породы отличались низким содержанием эритроцитов и гемоглобина. В процессе акклиматизации наблюдалось снижение содержания
251
эритроцитов у лошадей равнинных пород; если этого не происходило, животные погибали.
Общее количество циркулирующей крови, однако, оказывалось у хорошо акклиматизируемых пород довольно высоким (киргизская — 8,4%, новокиргизская 8,1 %, донская — 7,8% веса тела). Общая суммарная поверхность эритроцитов па 100 кг живого веса оказалась наименьшей у киргизских лошадей — 4970 л/2, а у равнинных чистокровных — 6170 л/2. При подъеме в горы на высоты 1600—2500 м н. у. м. наибольшее учащение дыхания и пульса обнаружили киргизские лошади, а наименьшее— донские и чистокровные верховые. Таким образом, имеет место как бы замещение дыхательной функции крови увеличением дыхания и кровообращения. Подобные факты в большой мере характеризуют адаптацию картины крови у лошадей на высотах 400, 700, 2000 и 3000 м н. у. м. В опытах сравнивались три группы лошадей казахской породы: а) равнинная, никогда в горы не поднимавшаяся; б) предгорная, ежегодно отгонявшаяся в горы до 2000 м н. у. м. и в) горная, содержавшаяся круглый год на высоте от 2000 до 3000 л* н. у. м. (Раушенбах, 1958а) (см. табл. 20).
Таблица 20
Физиологические изменения у лошадей при отгоне их в горы
(по Раушенбаху, 1958а)
Группа Количество ЖИВОТНЫХ Район исследования Высота (ж н. у. м.) Чаете а дыхания (за 1 мин) Частота пульса (за 1 мин) Число эритроцитов (млн./жлг) Гемоглобин по Сали (%)
Равнинная 16 Низовья реки Чу 400 14 43 7,08 63
Предгорная 22 Северный склон 700 12 43 6,50 62
Киргизского Алатоо 2000 12 47 7,38 70
Горная 12 Кунгей-Ллатоо 2000 14 42 6,79 65
3000 13 40 6,53 65
Полученные данные свидетельствуют о том, что у равнинной группы лошадей при подъеме в горы наблю-
252
дастся возрастание числа эритроцитов и повышение содержания НЬ в крови (до 7,08 млн.) и 63% НЬ по Сали. У предгорной группы также наблюдалось повышение этих данных, а у горной при подъеме с 2000 до 3000 м н. у. м. никаких сдвигов не обнаружено.
Различные влияния горных систем на реакции животных на одинаковых высотах над уровнем моря были изучены в горах Северного Кавказа при отгоне овец на средние высоты в летнее время. В условиях Северного Кавказа при отгоне тонкорунных овец на пастбища на высоту 2000 м н. у. м. увеличивается легочная вентиляция на фоне неизменного потребления кислорода. Состав красной крови за период акклиматизации не изменился. Контрольная группы, оставленная на равнине, также не обнаружила изменений состава крови. У полугрубошер-стных овец, кроме возрастания легочной вентиляции к концу периода акклиматизации (через 30 дней после подъема), наблюдалось и увеличение потребления кислорода. Таким образом, в условиях Северного Кавказа у исследованной группы овец было обнаружено увеличение легочного дыхания (Ольнянская, Давыдов, Романовская, 1969).
На основании сопоставления большого материала, добытого на отгоняемых в горы, по по своему происхождению равнинных животных (Ю. О. Раушенбах, 1958, 1967) считает возможным разделить весь процесс адаптации в генетическом аспекте на стадии.
Первой стадией адаптации к высоте является усиление легочного дыхания и кровообращения. Вторая (по данным Ю. О. Раушенбаха) характеризуется увеличением кислородной емкости крови при уменьшении реакции со стороны дыхания и кровообращения. Третья характеризуется увеличением кислородной емкости крови при уменьшении реакции со стороны дыхания и кровообращения и снижением кислородного запроса тканей. Различные животные и разные возрастные группы по-разному проходят эти стадии. Молодые животные легче достигают третьей стадии и поэтому легче акклиматизируются в горах.
Что же касается вопроса о различном влиянии отдельных горных систем на картину высотной адаптации, то этот вопрос как для человека, так и для животных остается открытым.
253
АДАПТАЦИИ ГОРНЫХ ОРГАНИЗМОВ
Расселение организмов в горах по вертикали зависит от основных физических факторов, характеризующих горный климат, а также от условий питания.
На основании имеющихся данных можно утверждать, что расселение пойкилотермных организмов не ограничивается самим фактором понижения парциального давления кислорода (Нооск, 1964). Многие низшие позвоночные выдерживают в барокамере понижение давления до 20 000—25000 м н. у. м. Это касается ящериц, лягушек, жаб, ужей.
В настоящее время известно 5 видов ящериц и 4 вида моллюсков, существующих на высоте 3600 м н. у.' м. Лишь один вид ящериц известен как обитающий до высоты 5500 м н. у. м. Фактором, ограничивающим расселение на больших и даже средних широтах, является для пойкилотермных организмов температура. По существу, только время солнечного сияния в летние месяцы позволяет этим животным сохранять активность, необходимую для питания и размножения. Это особенно важно для животных с хищным типом питания, так как во многих случаях и объекты питания отсутствуют на больших и средних высотах.
В известной мере все сказанное выше относится и к ряду высших животных — млекопитающих и птиц. Это особенно касается животных с ночным образом жизни и ночным питанием, когда низкая температура в горах и ущельях, плохо прогреваемых солнцем, создает неблагоприятную ситуацию для питания мелкими беспозвоночными и насекомыми.
Высокогорная фауна представлена по преимуществу растительноядными организмами или организмами, питающимися этими последними. Можно полагать, что верхней границей распространения по вертикали в этих случаях является сниженное парциальное давление кислорода.
Старое представление о том, что кровь горных животных, а также и человека, всегда содержит увеличенное количество эритроцитов и гемоглобина, сначала получило как будто подтверждение на видах и популяциях, постоянно живущих па разных высотах. Например, на двух популяциях горных лесных мышей (Apodemus sil-
254
vaticus ciscaucasicus Ogn.) и равнинных (Л. silvaticus mosquensis Ogn.) было установлено, что подвид на высотах до 1800 м н. у. м. имеет более высокое содержание эритроцитов и гемоглобина — эритроцитов на 12%, НЬ па 6,3% (Калабухов, 1935). Однако изучение двух популяций малого суслика, взятых на равнине (Ростовская обл.) и в горах (горы Кавказа), не подтвердило этого факта как явлений, общего для всех видов. Как процент содержания гемоглобина, так и количество эритроцитов оказались у этих двух популяций приблизительно одинаковыми (Калабухов, 1950).
У нескольких видов грызунов была также изучена реакция на «подъем» в барокамере до 3000 м н. у. м. в течение 72 ч. Изучались грызуны, взятые из природы в разных районах и на разных высотах.
Таблица 21
Изменение числа эритроцитов у грызунов после подъема в барокамере
(по Калабухову, 1937, 1950)
Ви i животного Измерения после подъема (%) Местообитание
Лесная мышь (Apode-mus sylvaticus) .... 109 Московская обл.
Лесная мышь (Apode-mus sylvaticus) .... 121 Северный Кавказ (1500 м н. у. м)
Полевая мышь (Apo-
detnus agrarius) .... 84 Северный Кавказ (равнина)
Рыжая полевка (Cle-thrionomys glareolus) 95,1 Московская обл.
Серый хомячок (Crice-tulus migratorius) . . . 90,4 Северный Кавказ (равнина)
Желтогорлая мышь (Apodemus flavicollis) 127,2 Северный Кавказ (равнина)
Малый суслик1 (Citel-lus pygtnaeus) ..... . 98,3 Северный Кавказ (горный)
1 Изучался при разрежении в барокамере, соответствующем 500 Л1 н. у. м.
255
Из всех изученных видов грызунов только два — лесная мышь и желтогорлая мышь — реагировали па понижение барометрического давления увеличением содержания эритроцитов. Примерно такие же результаты получены и при изучении изменении в крови гемоглобина. Интересно, что у горной популяции малого суслика снизилось содержание эритроцитов и гемоглобина «mi высоте». У рыжей полевки и серого хомячка в ответ на понижение барометрического давления обнаружилось снижение числа эритроцитов. Интересно, что одинаковый эффект повышения числа эритроцитов наблюдался у лесных мышей, взятых из природы в Московской обл. и на Северном Кавказе. Следовательно, реакция на понижение барометрического давления является очень стойким видовым признаком, мало меняющимся в зависимости от места обитания. Этот факт представляет большой интерес в смысле выявления врожденных реакций видов животных, не зависящих от условий обитания.
Однако указанные особенности, по-видимому, все же не связаны с распространением этих видов грызунов. Так, серый хомячок поднимается на Кавказе в горы до высоты 1600 м, а в Средней Азии (Семиречье) —до высоты 2500 м н. у. м.
Что касается желтогорлой мыши, то предел ее распространения в горы, по-видимому, ограничивается 800— 1000 м н. у. м. Между способностью вида существовать в горных условиях и способностью увеличивать кислород-тую емкость крови при кратковременной 72-часовой гипоксии никакой зависимости не наблюдается.
Несколько иную трактовку дала в последнее время этим данным 3. И. Барбашова (1960), которая отмечает, что в результате подъемов в барокамере содержание эритроцитов у разных видов как бы уравнивается, приближаясь к уровню около 9,2—9,5 млн. При этом у полевой мыши количество эритроцитов даже снижается. 3. И. Барбашова делает вывод, что повторная гипоксия вызывает уравнивание физиологических констант применительно к особенностям действия раздражителя. Это замечание хотя и снимает при характеристике исследованных видов всякую эколого-физиологическую специфику, позволяет думать, что малые и даже средние высоты не оказывают стимулирующего влияния на кроветворение у постоянно здесь обитающих видов.
256
Последние годы принесли в этом направлении ряд новых фактов. В Баркрофтской лаборатории на высоте 3800 л/ и. у. м. (White Mountain Research Station) были поставлены опыты с сопоставлением реакции на острую гипоксию у белой крысы, рожденной на этой высоте, и золотистого суслика (Citellus lateralis), живущего в диком состоянии на этих же высотах. У сусликов оказалась большая устойчивость к острой гипоксии, нежели у хорошо адаптированных крыс. Это, однако, необходимо связать и со способностью сусликов впадать в зимнюю спячку и с норным образом жизни (Bullard a. Kollias, 1966), который также повышает устойчивость животных к гипоксии (Hall, 1964).
Исследования физиологического состояния животных — постоянных обитателей высокогорья, пока еще довольно немногочисленны. Они проведены исключительно на американских видах, высоко поднимающихся в горы Северной и особенно Южной Америки.
Основной обмен был изучен (Cook a. Hannon, 1954) у трех популяций оленьей мыши (Peromiscus sp.), обитающей на трех высотах Калифорнии: на уровне моря, на высоте 1220 м и 3800 м н. у. м. Мыши отлавливались на месте и исследовались на уровне моря. Разумеется, температура среды па высотах, откуда были выловлены животные, ниже, чем на уровне моря. Исследования обнаружили более низкий уровень обмена веществ у животных, взятых с большой высоты, нежели у животных со средней высоты и с уровня моря. К сожалению, данные о составе крови в этих исследованиях отсутствовали.
Специально этому вопросу посвящены исследования (Morrison, Kerst, Rosemann, 1963), выполненные на грызунах высокогорья Чилийских Анд. Были изучены — высокогорная полевка (Acodon andinus), кроликообразная вискаша (Lagidiutn viscacia), взятые из природы на высотах от 4200 до 4700 м н. у. м., и два вида шиншилл — боливийская (Chinchilla boliviana) с высоты 2800 м и Chinchilla veligera, обитающая в Чили на малых высотах.
Кроме этих видов, изучен ряд грызунов, не поднимающихся в горы: длиннохвостая рисовая крыса (Orysomys longicaudatus), шиншилловая крыса (Abro-сота Bennetti), слепыш (Notiomys megalonix), полевая мышь (Acodon abrothrix), а также лабораторные кроли
9—1204
257
ки. Все животные перевозились на уровень моря, где были изучены гематокрит и содержание гемоглобина. Из приведенных в табл. 22 данных видно, что никакой разницы в величинах гематокрита и содержания гемоглобина горных и равнинных видов не обнаружено. Авторы отмечают, что особи высокогорных видов, родившиеся па уровне моря, по объему эритроцитов крови (гематокриту) ничем нс отличаются от родителей, взятых из природы высокогорья. У более активных и подвижных представителей грызунов разных видов объем эритроцитов па 20—25% выше, чем у менее подвижных. Таким образом, величины гематокрита, колебавшиеся от 380 до 540 мл]л крови, не связаны с высотой, на которой распространен вид. Некоторые данные из этой работы приведены в таблице 22.
Таблица 22
Гематокрит у грызунов, обитающих на разных высотах (по Morrison и сотр., 1963)
Грызуны Район Гематокрит (мл’м) П >чвижи< сть Гем глобин ij.i
Р. D. Kupestris . . . Высокогорный 430 Малоподвижный 289
Octodon Ур. .горя 488 » 256
Abrocoma То же 423
Notiomis 423
Acodon andin Высокогорный 513 Подвижный 311
A codon berl » 571 » 274
Abrotrix Ур. у.оря 522 278
Oryzomis ...... То же 546
Chinchilla boliv. . . . Высокогорный! 425 309
Chinchilla velig. . . . Ур. моря 420
Lagiditun Высокогорный 376 — 330
Oryctolagus cuniculus То же 383 — 275
В этой работе был подтвержден и давно установленный факт, что обитатели высокогорья сохраняют картину крови и ее кислородную емкость при перемещении их на уровень моря (Антелидзе и Барбашова, 1938).
258
В специальном исследовании П. А. Коржуева и Н. Н. Булатовой (1953), охватившем ряд равнинных и горных копытных в условиях Московского зоопарка, у горных животных отмечено увеличение количества и размеров эритроцитов при одинаковом общем содержании НЬ. Наибольшее количество эритроцитов обнаружено у винторогого козла (млн.):
Козел домашний................... 13,5
Козел випторогий................. 25,8
Сибирский козерог................ 15,2
Козел горный сибирский........... 19,8
У яков, спустившихся с высоты 2000 м н. у. м., наблюдается повышение содержания гемоглобина в крови на 17% по сравнению с таковым у яков, содержавшихся на равнине 3000 м н. у. м. Колебания в содержании эритроцитов и гемоглобина у яков различных возрастов очень велики. Так, содержание эритроцитов колеблется от 6,07 до 10,18 млн., а процент гемоглобина по Сали от 69,10 до 81,7 (Арав, 1950а). Яки, спущенные с Восточного Памира (около 4000 м и. у. м.) на высоту около 700 м, сохраняли эту картину крови в течение двух месяцев. При этом у животных разного возраста нарастало число эритроцитов от рождения к трехмесячному возрасту и снижалось к годовалому и старше. Как домашний як (Poephagus grunniens), так и дикий (Р. ти-tus) чрезвычайно приспособлены к существованию в условиях высокогорья, обладают относительно большими по сравнению с горным скотом размерами сердца и легких.
Три поколения лам, прожившие па уровне моря, сохранили основные отличия в морфологии крови (большое количество мелких эритроцитов) (Антелидзе и Барбашова, 1938). Кривая диссоциации ламы сдвинута влево (см. рис. 38) с зарядным напряжением около 55 мм НЬ и разрядным около 24 мм НЬ. Столь низкое зарядное напряжение обеспечивает насыщение артериальной крови при снижении парциального давления кислорода в горах. Однако вискаша {Viscacia viscacia)—грызун, высоко поднимающийся в горы Южной Америки, обладает кривой диссоциации, весьма близкой к кривой человека. !
Реакция высокогорных южноамериканских грызунов на подъем в барокамере со стороны частоты дыхания
9*
259
и пульса отсутствует. Только у равнинных мышей и грызуна Akodon дыхание участилось при подъеме на высоту 4500 м н. у. м. на 22%. Учащение пульса на 25% наблюдалось у мышей с равнины и равнинного вида Phyllolis (Morrison a. Elsner, 1962). Однако высокогорные виды имеют при нормальном давлении более частые пульс и дыхание.
Далеко неоднородный материал, полученный на диких животных — обитателях высокогорья, однако, неопровержимо свидетельствует о том, что обычные нормы адаптации равнинных организмов и организма человека совершенно неприложимы для характеристики этих видов. Картина красной крови, содержание гемоглобина и реакции дыхания и кровообращения не могут объяснить удивительной устойчивости к гипоксии таких животных, как крупные представители семейства Cameli-dae ламы, викуньи, альпака, а также многочисленных видов высокогорных грызунов. Исследования над несколькими видами перуанских высокогорных грызунов показали, что содержание гемоглобина в крови у них также не превышает нормальный уровень у животных, обитающих на равнине (Tappan a. Reynafarje, 1957). Это привело к выводу о необходимости изучения тканевых окислительных ферментов и содержания миоглобина в тканях.
Оказалось, что у высокогорных видов перикот {Phyllolis}, травяной полевки {Akodon}, шиншиллы {Chinchilla sahamai}, а также у камелид (ламы и викуньи) обнаружены значительные особенности в содержании миоглобина. У перикот, взятых с высот 3900— 4500 м н. у. м., обнаружена удвоенная концентрация миоглобина в диафрагме и мышцах конечности. Более активные высокогорные полевки Akodon не отличались от перикот.
Миоглобин мышцы сердца у домовых мышей {Mas tnusculus} на высоте 4500 м был выше, чем на уровне моря, но миоглобин диафрагмы и мышц ноги — одинаковый. У викуньи и альпака миоглобин сердца был более высокий, а миоглобин диафрагмы и ноги значительно более высокий, чем у исследованных грызунов.
Следовательно, тканевые изменения более выражены у животных — аборигенов высокогорья, чем изменения крови.
260
Большой интерес в этом плане представляют исследования обмена железа как количественного критерия скорости новообразования эритроцитов.
Хотя высокогорные животные — ламы, викуньи, алпа-ка и не обнаруживают в высокогорье значительных сдвигов содержания гемоглобина и эритроцитов, самый характер обмена железа и срок жизни эритроцита у них резко отличаются от таковых у хорошо акклиматизированного человека — уроженца больших высот. Срок жизни эритроцита у этих животных почти вдвое короче, чем у человека (60 дней). Общий объем эритроцитов у них составляет 25—31 см? [кг веса тела, что гораздо меньше, чем в тех же условиях у человека. Следовательно, кругооборот эритроцитов и их новообразование у этих видов животных резко повышены, костный мозг функционирует очень интенсивно и компенсирует быстрое разрушение эритроцитов. Можно полагать, что малая реакция высокогорных камелид — ламы и др. на гипоксию связана с большей поверхностью эритроцита вследствие малой величины последних. Это обеспечивает даже при одинаковой капилляризации больший контакт с кислородом в легких. Сдвинутые влево кривые диссоциации также указывают на эту зависимость. Следовательно, самые высокие степени адаптации организма к недостатку кислорода следует видеть в особенностях структуры и жизни эритроцитов, сродства гемоглобина к кислороду и свойств ткани утилизировать этот кислород. Несомненное значение в эволюции на определенных ее этапах приобретают адаптации кислородного запроса, особенно хорошо выраженные при повторных воздействиях гор на мигрирующие на разные высоты организмы.
ФИЗИОЛОГИЧЕСКИЕ ОСОБЕННОСТИ НЫРЯЮЩИХ ОРГАНИЗМОВ
Физиологические исследования ныряющих организмов, в частности млекопитающих, касаются главным образом особенностей их легочного дыхания, естественно прекращающегося на продолжительное время во время ныряния. Способность пырять довольно характерна для различных видов млекопитающих. Продолжительность пребывания млекопитающих под водой колеблется от 50 сек до 2 ч (Irving, 1939) (табл. 23).
261
Таблица 2с
Продолжительность ныряния млекопитающих (по Irving, 1939; Harrison, 1960)
Объект Время Объект Время
Утконос (Ornithor-hynchus anatinus) . . 10 мин Кашалот (Physeter catodon) 1 Ч — 1 Ч
Морской слон (На-lichocrus grypus) . . 6 мин 48 сек Бутылконос (Hyper oodon ampullatus) 15 мин 2 ч
Нерпа (Pysa hispid а) 15 мин Гренландский кит (Balaena mysticetus) 1 ч
Тюлень-тевяк (На-lichoerus grypus . . 15 мин Блювал (Balaenop-tera musculus) . . . 49 мин
Водяная крыса (Аг-vicola terrestris) . . 2 мин Японский кит (Eu-balacna glacialis) . . 50 мин
Ондатра (Ondatra zibethica) 17 сек 12 мин Большой полосатик (Balaenoptera niuscu-lus) 30 мин
Бобр (Castor fiber) Кролик (Oryctola- 15 мин Финвал (Balaenoptera nodosa) .... 20 мин
gus cuniculus) . . . Собака домашняя 3 ми н 4 мин Горбатый кит (Me-gaptcra nodosa) . . 30 мин
Бегемот [Hippopo- 25 сек Человек (искатели жемчуга и губок) 2 мин
tatnus amphibius) . . 50 сек 30 сек
Морж (Odobcnus rosmarus) . ... , 16 мин 20 сек
Сравнение приведенных величии показывает, что наибольшей устойчивостью к прекращению дыхания обладают китообразные: некоторые из них способны находиться под водой от одного до двух часов. Насекомоядные (выхухоль), грызуны (ондатра), а также ластоногие (тюлени) способны оставаться под водой очень ограниченное время.
Среди водных млекопитающих, способных к нырянию, т. е. обладающих повышенной устойчивостью к гипоксии, можно встретить представителей многих отрядов. Так, из низших млекопитающих ныряет утконос, из насекомоядных— выхухоль, из грызунов — водяная крыса, ондатра, нутрия, бобр, из наземных хищных — выдра, из нежвачных парнокопытных — бегемот. Ныряют, разумеет-
.262
ся, все ластоногие и китообразные и ныне почти вымершие сиреиовые.
Физиологические особенности ныряющих животных касаются изменении дыхательного аппарата, изменении дыхательной функции крови, содержания миогемоглобина, а также некоторых особенностей регуляции дыхания и кровообращения в условиях гипоксии.
/Морфологические отличия легких заключаются в том, что объем их у пыряющих организмов значительно превосходит объем легких у наземных форм. Например, у дельфинов легкие составляют по весу 27% веса тела, тогда как у наземных млекопитающих только около 12% (Клейненберг, 1956). Количество легочных альвеол почти в три раза превосходит таковое у человека (150 млн. у человека и 437 у морской свиньи). Сильно развита у дельфинов хрящевая ткань, окружающая не только трахею, но и бронхи и бропхиоли и не позволяющая легким спадаться. Такая структура легких в условиях ныряния и повышения даь тения предохраняет животное от кессонной болезни (Scholander, 1940). Вес сердца дельфина несколько меньше, чем у волка того же веса (около 30 кг) (Клейненберг, 1956). Скорость кровотока у ныряющих организмов понижена. Дыхательная функция крови некоторых ныряющих животных отличается несколько повышенной кислородной емкостью.
В табл. 24 приведены некоторые данные сравнения дыхательной функции крови человека, собаки и ныряющих млекопитающих.
Таблица 24
Дыхательная функция крови (по Irving, 1939)
Объск г Кр >вь (м.1 о2;юо.«.?) Эритр ни ы (.«л Oj/100 лог)
Человек 20,7 45
Собака 21,8 —
Тюлень 29,3 61,3
Морской лев 19,8 68
Бобр 17,7 40
Ондатра 25,0 50
Дельфин 20,5 57,7
Кит 14,1 —
263
Таблица 25
Запасы кислорода в теле тюленя к моменту ныряния (вес животного 29 кг)
(по Scholander, 1940)
Место связывания О2 Общее количество Оа (лгл)
Альвеолярный воздух (350 мл с 16 об. % О2) 50
Кровь (4500 мл с 25 об. % О2) 1100
Мышцы (6000 г с 4,5 об. % О2) 270
Ткани (кроме мышц) 100
Весь запас О2 . . . 1520
С уверенностью можно говорить о большой кислородной емкости крови только у некоторых ныряющих форм. У многих она нисколько не выше, чем у наземных организмов. Одно увеличение дыхательной функции крови не может обеспечить возможности ныряния. Содержание миогемоглобина в мышцах типичных ныряющих животных, особенно китов, резко повышено. У тюленя количество его в 7 раз выше, чем у быка (Robinson, 1939). Это же обнаружено и в сердечной мышце (Theorell, 1934). Эти факты были затем подтверждены для китов и тюленей (Scholander, 1940; Коржуев, 1949). Таким образом, огромные массы мышц крупных морских млекопитающих во время ныряния оказываются снабженными кислородом за счег восстановления мышечного гемоглобина. Есть предположение, что у китообразных мышечный аппарат снабжается кислородом во время ныряния именно за счет миогемоглобина.
Регуляция дыхания и кровообращения у ныряющих форм отличается исключительно малой чувствительностью дыхательного центра к накоплению СОг. Это было установлено еще в прошлом столетии (Bert, 1878; Richet, 1898) и подтверждено многочисленными современными исследователями. Отсутствие реакции на накопление СОг характерно и для таких ныряющих форм, как тюлени, бобры, ондатры, т. е. животных, далеко отстоящих в систематическом ряду друг от друга (Irving, 1939). Напряжение СОг в альвеолярном возду-
264
хе во время ныряния резко возрастает, но это не служит раздражителем для дыхательного центра. Следовательно, кислород, имеющийся в легких, может быть использован в большей мере, чем у не ныряющих организмов. Однако в последнее время прямая нечувствительность дыхательного центра к накоплению углекислоты подвергается сомнению. У уток при децеребрации и трахеотомии (т. е. при выключении влияний с высших отделов центральной нервной системы и назо-фарингеальных рецепторов) наблюдается ярко выраженная реакция при вдыхании воздуха с 6—10% СОг. Следовательно, имеет место постоянное торможение дыхательного центра, возрастающее при раздражении интраторакальных рецепторов (Jukes, Jukes a. Banihfa, 1968).
У ныряющих животных во время ныряния постоянно наблюдается уменьшение частоты пульса. По-видимому, это связано с уменьшением кровоснабжения мускулатуры, которая у всех ныряющих животных (кроме китообразных) во время ныряния расслабляется.
Вопрос об изменениях сердечной деятельности при нырянии на разных глубинах был изучен детально (Harrison, 1960). При небольших погружениях длительностью не более 15 мин у тюленей наблюдалась лишь скоропроходящая брадикардия, сменявшаяся тахикардией. Если период брадикардии продолжался 3 мин, то «восстановительная» тахикардия—до 5—10 мин. Обнаруживается и связь брадикардии с возрастом животного. У старых тюленей эти явления развиваются медленнее, чем у молодых. Более глубокие погружения (от 24 до 91 лг) осуществлялись в специальной камере, наполненной водой, где повышалось давление. При этом у всех животных (как старых, так и молодых) наступает сразу брадикардия (пульс от 4 до 15 в минуту), сменяющаяся через 2—3 мин тахикардией, доходящей до 30 пульсовых ударов в минуту. Интересно, что в воздушной среде аналогичные повышения давления вызывают лишь очень слабый эффект со стороны сердца. Только при очень больших повышениях давления в воздушной среде (соответствующих глубине погружения 60 м) наблюдались брадикардия и одновременное урежение дыхания.
Минутный и систолический объем сердца у тюленей сразу после 10 минутного пребывания под водой оказывается резко повышенным (минутный с 5,5 л до 14,7 л
265
и систолический с 38 мл до 100 мл) (Давыдов и Макарова, 1964). Столь значительное повышение систолического объема сердца свидетельствует об особенностях сердечной мышцы. Расширение сердечной мышцы у человека и у наземных животных не превышает двукратного объема систолы во время работы по сравнению с покоем.
Регуляция щелочно-кислотного равновесия у ныряющих животных отличается, кроме особенностей невозбу-димости дыхательного центра к накоплению СОг, повышенными щелочными резервами крови. Суммарный запас кислорода к моменту ныряния складывается из содержания его в легких, крови и мускулатуре. Кроме того, имеется некоторое количество его в подкожном жире, где может раствориться кислорода в 5 раз больше, нежели в воде (Irving, 1939). Полное использование кислорода из легких благодаря отсутствию реакции на накопление СОг, использование запасов кислорода из миогемоглобина и, наконец, значительные изменения кровообращения с преимущественным снабжением мозга обусловливают способность животного к существованию в условиях дчительного выключения акта дыхания. По-видимому, раздражителем дыхательного центра этих животных служит недостаток кислорода, оказывающий влияние на рецепторы каротидного синуса.
В конечном счете, продолжительность ныряния определяется запасами кислорода в организме и способностью тканей, в том числе и нервной системы, функционировать в условиях резкой гипоксемии. У ныряющих животных содержание кислорода в тканях и крови значительно.
Совершенно особое значение в физиологии ныряющих животных имеют сетевидные сосудистые сплетения, так называемая чудесная сеть. Эти образования встречаются у китообразных в грудной клетке вдоль сонной и позвоночной артерий. По мнению некоторых исследователей, чудесная сеть обеспечивает кровоснабжение мозга во время ныряния. При этом резко сокращается кровообращение в мускулатуре и возрастает в центральной нервной системе. Эти явления, возникающие при нырянии, в последнее время стали характеризоваться как своеобразный «рефлекс ныряния». Последний предполагает трансформацию кровообращения в «сердечно-мозговую
2G6
цепь», направляющую все резервы Ог к сердцу и мозгу, почти полностью приостанавливая кровообращение даже в интенсивно работающих мышцах.
Рефлекс начинается с раздражения рецепторных окончании носа и через бульбарные механизмы резко усиливает реакцию, возбуждаемую через хеморецепторы сосудов. Кожные артериовенозные анастомозы обеспечивают быстрое возвращение крови к сердцу без потерь Ог в тканях. Такой круюоборот дает возможность полностью использовать резервы О2 мозгом и сердцем в период ныряния. При всплытии наблюдается быстрое увеличение минутного объема сердца (до 50 раз против уровня покоя) (Folkow, 1968).
Развитие пыряния в онтогенезе совершенно не изучено. Известно, что молодняк тюленей — бельки, остается длительное время (до 1 месяца) после рождения на льду. Однако наблюдалось нормальное плавание белька гренландского тюленя, непосредственно перед этим извлеченного оперативным путем из матки (В. К- Шепелева). Неизвестно, в какой мере это относится также и к грызунам— бобрам, ондатрам.
Известно также непродолжительное произвольное ныряние у детеныша тюленя, извлеченного из матки (Уждавипи, Шепелева, 1966). Время ныряния не превышало 1 мин. Детеныши гренландского тюленя сразу после рождения способны оставаться под водой (при насильственном погружении) лишь самое минимальное время (2—5 мин). В более старших возрастах (стадии «хохлуша» и «серка») возможность погружения возрастает до 10 мин-, отдельные животные оставались неподвижно под водой до 7,5 мин (Давыдов и Макарова, 1964).
Нервные регуляции ныряния следует рассматривать как систему рефлекторных актов, связанных с раздражением определенных рецепторных образований на поверхности кожи. /Можно полагать, что большое значение в этих актах имеет терморсцепция, так как благодаря значительной теплопроводности воды раздражения возникают при каждом погружении. У лягушек, как было показано выше, имеет преимущественное значение специальная хеморсцепция.
Особенности центральной нервной регуляции у ныряющих животных проявляются в описанном виде рас-267
слабления мускулатуры. Это установлено для утки (Bert, 1878), ондатры, бобра и тюленя (Irving, 1938). Утки при однократном погружении в воду способны выживать в течение 5—15 мин (Richet, 1898). После многократных погружений способность выживать под водой возрастает до 12—17 мин. Децеребрация утки (Huxly, 1913) не устраняет типичной задержки дыхания при погружении головы и клюва в воду, как не устраняет общего для ныряющих форм замедления пульса. Экстирпация лабиринтов снижает этот эффект, но не устраняет его. По-видимому, он осуществляется в этом случае с рецепторов шейных мышц.
Есть основания полагать, что у ныряющих и наземных млекопитающих и у птиц нет принципиальной разницы в реакциях при нырянии. Эти различия носят количественный характер и зависят от особенностей как рецепторов, так и центров, регулирующих дыхание. Ваго-томированный утенок становится таким же чувствительным к асфиксии, как цыпленок (Richet, 1899). Ритм дыхания у ныряющих организмов отличается прерывистостью. Тип периодического дыхания (Чейн-Стокса) характерен в норме для таких организмов, как бегемот, но не встречается у тюленей.
Изменения мышечного тонуса во время ныряния были установлены электромиографическими исследованиями, проведенными над молодняком гренландского тюленя (А. Ф. Давыдов и А. Р. Макарова). Погружение в воду (без погружения головы) вызывает очень небольшое снижение электрической активности скелетной мускулатуры. Погружение тела с головой вызывает резкое ограничение электрической активности, сопровождающееся, как уже указывалось выше, урежением пульса. Так как понижение тонуса возникает сразу же после погружения, нет оснований думать, что это следствие накопления СОг, как это имело место в исследованиях И. С. Репина (1959а), и недостатка кислорода, как в опытах К- П. Иванова (19596). Можно думать о существовании специального механизма, снижающего мышечный тонус у ныряющих животных (тюленей), обусловливающий сниженный кислородный запрос в период ныряния (рис. 61).
Исследование потребления кислорода во время ныряния (А. Ф. Давыдов и А. Р. Макарова) обнаружило
268
повышение газообмена только в первую минуту после 10—15-минутного погружения в воду. Затем наступает довольно длительное (в пределах до 10—25 мин) снижение потребления кислорода ниже исходного уровня. В результате этого снижения суммарное потребление Ог за период восстановления может быть более низким, чем до погружения. Таким образом, выключение легочного
Рис. 61. Эчектромиограммы скелетных мышц тюленей при погружении в воду (по Давыдову и Макаровой), 1967)
дыхания, снижение мышечного тонуса и урежение пульса приводит к значительному снижению кислородного запроса у ныряющих тюленей. Правда, эти опыты поставлены в условиях насильственного погружения, в покое. Очень интересен вопрос, как развиваются эти явления при интенсивной мышечной работе — плавании в погоне за рыбой. Однако ответа на него пока нет.
Итак, основными физиологическими механизмами, определяющими продолжительность ныряния, являются, с одной стороны, ограничение кислородного запроса организма благодаря расслаблению скелетной мускулатуры, а с другой — наличие некоторых запасов кислорода в теле (в крови, мышцах и воздухоносных путях). Важным механизмом является нечувствительность дыхательного центра ныряющих животных к накоплению угольной кислоты в крови, благодаря чему возможна более полная утилизация кислорода крови.
269
ФИЗИОЛОГИЧЕСКИЕ ГИПОКСИИ У НАЗЕМНЫХ ОРГАНИЗМОВ
Ограничение доставки кислорода вследствие уменьшения возможностей дыхания или повышения кислородного запроса ткапей занимает большое место при разных формах деятельности животных. К первой группе явлений относятся все случаи замирания животных в связи с реакциями нападения (например, у хищных) пли оборонительными (у грызунов). Ограничение дыхания связано здесь с торможением дыхательного центра и обычно не ведет к значительной гипоксии в организме. Так, уменьшение частоты и глубины дыхания можно наблюдать у кошки, выслеживающей добычу (мышь или птицу), лисицы и т. д. Есть основания думать, что эти состояния сопровождаются значительным уменьшением мышечного тонуса и в результате — понижением кислородного запроса. Замирание грызунов при действии оборонительных раздражителей относится к числу часто наблюдаемых в природе явлений. Это своеобразное проявление пассивно-оборонительной реакции может продолжаться очень длительное время у таких видов, как домовая мышь, черная крыса, садовая соня и т. д. Специальное исследование газообмена (Шарипова, 1953) на садовой соне обнаружило резкое его понижение при экспозиции животных в камере на ветке под водой.
Наиболее яркое выражение среди сухопутных млекопитающих эти реакции приобретают у ежей как видов, отличающихся своеобразной оборонительной реакцией скручивания. Скручивание у ежа сопровождается не только изменением позы, но и резким увеличением вредного пространства дыхательных путей вследствие погружения отверстий рта и носа в полость, ограниченную сокращающейся кольцевой кожной мышцей. В таком состоянии скручивания еж находится во время сна, зимней спячки и во время оборонительной реакции па сильные раздражители. Специальному исследованию функциональной гипоксемии, развивающейся во время скручивания, были посвящены работы Л. Г. Филатовой (19466, 1958). Физиологическая гипоксемия, вызываемая скручиванием, наблюдается у ежей как при осуществлении короткой оборонительной реакции, так и при длительном состоянии скручивания во время зимней спяч-
270
кп. В ранний весенним период, непосредственно вслед за пробуждением после зимней спячки, у таких ежей имеет место очень низкое насыщение артериальной крови кислородом (Филатова, 19466). Одновременно в ранний ве
сенний период у ушастого ежа основного обмена.
Повышению насыщения артериальной крови с 43,7 до 68% соответствует понижение теплопродукции со 128,0 до 87,9 кал на 1 кг веса в сутки. Специальные расчеты и определения кривых диссоциации оксигемоглобина в крови ежей показали, что содержание СО2 в альвеолярном воздухе у ежей после спячки должно достигать около 12%,что намного превышает эти величины у млекопитающих. При напряжении углекислоты 40 мм разрядное напряжение еще очень высоко и условия отдачи кислорода тканям сильно затруднены. При напряжении СО2, равном 100 мм Hg, разрядное напряжение падает и, таким образом, на фоне общей гипоксемии может осуществляться удовлетворительное снабжение тканей кислородом. Так как после зимней спячки ежи оказываются активными, охотятся и много времени пребывают в рас
очень высокий уровень
Рис. 62. Потребление кислорода (мл/мин) у ежей (по Филатовой, 1944, 1958). / — в раскрученном состоянии; // — в скрученном состоянии
крученном состоянии, то такое
напряжение кислородг! в крови является следствием длительного снижения возбудимости дыхательного центра при зимней спячке. Только низкая возбудимость дыха
тельного центра в это время может позволить животному существовать без явлений тяжелой одышки в этих резко измененных условиях.
С пробуждением от спячки эта низкая возбудимость сохраняется еще некоторое время, обусловливая дли
271
тельную и стойкую гипоксемию у бодрствующих животных. Для полного обоснования такого толкования фактов необходимо было изучить рефлекторные изменения дыхания и обмена веществ, которые возникают у ежей при скручивании. При изучении газообмена и дыхательной функции крови у ежей в скрученном и раскрученном состоянии было установлено следующее (Филатова, 1958). Уже кратковременное 15-минутное скручивание вызывает у ушастого ежа резкое снижение потребления кислорода (в среднем на 43,5%) по сравнению с раскрученным состоянием (рис. 62). При этом наблюдается прогрессирующая гипоксемия. Процент насыщения кислородом артериальной крови тем ниже, чем дольше продолжается состояние скручивания. В то время, как в раскрученном состоянии насыщение артериальной крови у ежей равно почти 100%, в скрученном состоянии насыщение кислородом артериальной крови падает при увеличении продолжительности скручивания. Резкое снижение потребления кислорода при скручивании — результат расслабления скелетной мускулатуры, деятельность которой находится в состоянии функционального антагонизма с кожно-мышечным аппаратом ежа.
Глава VII
ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ
К АРИДНОЙ ЗОНЕ (ЖАРКИЕ ПУСТЫНИ)
УСЛОВИЯ СУЩЕСТВОВАНИЯ В ПУСТЫНЕ
Существование животных в аридной зоне — в условиях жарких пустынь, связано с весьма своеобразными приспособлениями к резко колеблющимся в течение годового и суточного цикла физическим условиям среды, резким колебаниям условий питания, снабжения водой, минерального состава воды и пищи. На эти общие для всего животного населения пустынь факторы, однако, по-разному реагируют организмы, находящиеся на разных степенях эволюции, с разной экологической специализацией.
Факторы, действующие на организм в пустыне, не только многообразны, но подчас вызывают противоположные реакции организма (например, па воздействие как крайне высоких, так и крайне низких температур среды), поэтому именно для пустынных организмов характерен огромный диапазон физиологических адаптаций и параметров устойчивости.
Наиболее ярко выражены в условиях аридной зоны следующие факторы:
1. Резкие колебания температуры воздуха. В условиях пустынь северного полушария они могут достигать 29° С для суточных и 50° С для сезонных колебаний. Эти колебания температуры вызывают не только значительные колебания жизнедеятельности организмов (очень многие млекопитающие пустыни впадают в зимнюю или летнюю спячки), но и резкие, циклически повторяющиеся изменения поведения и регуляции физиологических функций.
2. Резкие колебания температуры почвы. Характерны перепады от 12 до 20° С в зимний и от 20 до 70° С в летний периоды. Последние приводят к значительным суточным колебаниям жизнедеятельности (ночной активности)
273
пойкилотермпых организмов — беспозвоночных и низших позвоночных, главным образом рептилий.
3. Резкие колебания инсоляции. Вызывают возникновение ряда защитных морфологических изменений животных: окраски, динамики хроматофоров.
4. Сезонные колебания в растительном покрове. Выражаются в появлении эфемерной растительности в весенний (и отчасти осенний) период и замены ее солянками в летник. Эти изменения резко меняют кормовую базу, отсюда пищевое поведение животных.
5. Сезонные колебания содержания воды в почве и солей в естественных и искусственных водоемах. Это весьма важный фактор для крупных млекопитающих и многих птиц пустыни.
Основными факторами, воздействующими в аридной зоне в жаркий период года, являются высокая температура, недостаток (а иногда и полное отсутствие) воды и повышенное содержание солей в воде и почве.
Экстремальные условия существования в пустыне приводят к появлению ярко выраженной миграционной активности у птиц и некоторых млекопитающих, способности к зимней и летней спячке. Большинство пойкило-термных организмов также приобретает в пустыне специфические физиологические особенности.
АДАПТАЦИИ ПУСТЫННЫХ БЕСПОЗВОНОЧНЫХ
Удивительная способность многих беспозвоночных — насекомых, пауков, скорпионов, существовать в пустыне при полном отсутствии воды издавна привлекала внимание биологов, и физиологические исследования над этими организмами были начаты гораздо раньше, чем на млекопитающих и птицах.
Механизмы адаптации позволяют разделить всех пустынных членистоногих на две большие группы. Первая группа энергично испаряет воду через кожные покровы — интегумент. Сюда относятся пустынные виды многоножек (Myriapoda), древесные мокрицы (Hemilepis-tus). Это типично ночные виды с ярко выраженной положительной реакцией на темноту. Ночью повышается относительная влажность воздуха, снижается его температура, что ограничивает испарение с поверхности те-274
ла. Многие из этих видов населяют норы грызунов, где условия испарения более благоприятные. Вторая группа включает многие виды насекомых, пауков, скорпионов — они в значительной мере независимы от испаряющей силы воздуха благодаря непроницаемому панцирю — интегументу. Эта группа не обязательно ведет ночной образ жизни, и последний, если он наблюдается, связан с условиями питания.
Итак, адаптации членистоногих к пустыне складываются из ограничения потерь воды через хитиновый покров и путем ограничения испарения воды при трахейном дыхании, т. е. ограничении ее потерь через спираку-лы. Главной особенностью поведения пустынных видов пауков и насекомых является хорошо выраженная роющая деятельность. Например, мокрица (Hemilepistus reaumuri) роет вертикальные норы диаметром 5 мм и глубиной до 10 см при 35° С на сухой почве и при 45° С, если почва увлажнена (Verrier, 1932). Многие пустынные насекомые хорошо морфологически адаптированы к рытью, например, жуки-чернотелки (Tenebrionidae). Хорошо роют песок также низшие насекомые (Thysanu-га). Для многих характерно рытье в горизонтальном направлении, что способствует быстрому засыпанию насекомого песком. Многие скорпионы и сольпуги очень глубоко зарываются в почву, проделывая длинные глубокие ходы.
Хорошо используется насекомыми в пустыне ориентация по странам света. Термиты в условиях тропиков создают свои постройки в направлении с севера на юг, благодаря чему подвергаются меньшему облучению. Личинки американского муравьинового льва (Myrmeleon immaculatus) изменяют расположение своих ловчих воронок на протяжении дня.
Замечено, что муравьи мало приспособлены к условиям пустыни и встречаются там только во влажных местах (Cloudsley-Thompson, 1961).
В лабораторных экспериментах было установлено, что некоторым насекомым (Triboleum confusum, Т. costane-ит) свойствен своеобразно обратимый преферендум на влажность. Положительная реакция на пониженную влажность сменяется отрицательной под действием ветра, а главное в состоянии «высушивания» и голодания (Roth a. Willis, 1951).
275
Очень существенной стороной поведения пустынных членистоногих является реакция на освещение; отсюда их своеобразный суточный режим. Очень многие пустынные артроподы выходят из нор вечером и ночью при снижениях температуры. Крупные размеры тела жуков-чернотелок и их длинные конечности позволяют им существовать на большей территории, нежели, например, муравьи. Но нагревание нервно-мышечного аппарата у насекомых мало зависит от их размеров и, таким образом, все дело сводится к микроклиматическим различиям (Cloudsley-Thompson, 1961). Перемещение с голого песка на растительность в зависимости от температуры описано многими исследователями (Chapman, 1955; Кашкаров и Коровин, 1936 и др.).
Активность разных пустынных видов на протяжении суток, зарегистрированная в актографе, различна. Так, сумеречной активностью обладают сколопендры (Sco-lopendra), некоторые скорпионы (Buthus и Scorpio). Европейские виды (Enscorpius italicus и Е. carpaticus) обладают ночной активностью.
Большое разнообразие суточной активности от дневной до ночной выявлено В. А. Кузнецовым (1969) на Tenebrionidae в Каракумах. Отдельные виды чернотелок стойко сохраняют особенности суточного ритма активности при длительном содержании в лаборатории (рис. 63).
Суточный ритм и связанные с ним изменения температуры, в свою очередь, зависят от сезонного ритма. Как общее правило, насекомые в пустыне активны в зимние месяцы в дневные часы, а летом в вечерние и ночные (Bodenheimer, 1954). У многих насекомых изменяется на протяжении года также и цикл размножения.
Характерные особенности приобретает в пустыне и протекание спячки и диапаузы. Диапауза представляет состояние, при котором прекращаются питание, рост, резко снижается обмен веществ, процессы размножения. Это состояние характеризуется повышенной устойчивостью насекомого к воздействию факторов внешней среды и особенно термических. Диапауза у насекомых наблюдается на любых стадиях развития, но особенно на ранних (личиночных), хотя может охватывать и стадии нимфы и имаго. Большинство исследователей считают диапаузу связанной с низкими зимними температурами.
276
мм3/г живого веса
часы суток
Рис. 63. Динамика суточной активности и потребление О2 у различных видов чернотелок (Tenebrionidae) в условиях пустыни (слева) и при содержании в лаборатории (справа) (по Кузнецову, 1969)
277
В аридной зоне встречаются случаи облигатной диапаузы, когда развитие приостанавливается на 6 месяцев даже при высокой влажности среды. В других случаях диапауза может быть факультативной, т. с. зависит в значительной мере от температуры среды. Впрочем, факторы, вызывающие диапаузу, еще далеко не ясны и здесь положение близко к тому, что известно для зимней спячки у млекопитающих.
Особое место занимает у пустынных членистоногих механизм удержания воды. У пустынной мокрицы (Не-milepistus reaumuri) потери воды на 5 мг[см2/ч ниже, чем у мокрицы умеренного климата (Porcellio seaber) (Cloudsley-Thompson, 1939). Испарение воды прямо пропорционально недонасыщению водяными парами окружающего воздуха. Такие же данные известны для стоножек (сколопендр).
Большое значение в поддержании водного баланса некоторых насекомых пустыни имеет реабсорбция воды в кишечнике. Так называемая криптонефрия заключается в том, что вода, выводимая мальпигиевыми сосудами, вместе с экскретами поступает в задний отдел кишечника, где подвергается обратному всасыванию. Это хорошо выражено у пустынных чернотелок (Tenebrionidae) (Гиляров, 1964).
Особенно хорошо способность удерживать воду в зависимости от экологии вида установлена для видов скорпионов — обитателей пустынь Европы и Северной и Тропической Африки. Пустынные виды отличаются еще и очень большой устойчивостью к высокой температуре.
Некоторые насекомые способны адсорбировать влагу из недонасыщенного воздуха. Впервые эти факты были получены для колорадскою жука (Leptinotarsa decimli-neata), а затем для мучных червей — личинок жука Те-nebrio molitor. Классические исследования (Buxton, 1930, 1932; Mellanby, 1932) четко наметили наблюдаемые здесь зависимости. Так, например, если мучных червей содержать при сравнительной влажности 80% или ниже, они теряют в весе; но они набирают его при 90% как при 23°, так и при 30° С. Среднее увеличение веса идет за счет воды, содержание которой возрастает от 57 до 65% общего веса насекомых в течение 23 дней содержания их при 30°С и относительной влажности 90%. Равновесие достигается при относительной влажности 88% и 278
определенных температурах, но при более низких величинах относительной влажности личинки теряли в весе. Следовательно, этот процесс зависит от относительной влажности, а не от дефицита насыщения. Анестезия насекомых 35% СОг прекращает поступление воды из воздуха. Предполагают, что при этом выключается особая активность кутикулы, а не открытие спиракул.
Своеобразное равновесие между влажностью окружающей среды наблюдается у других отрядов насекомых— прямокрылых, двукрылых, а также у некоторых родов клещей (Ixodes, Ornithodorus). Важную роль в поддержании водного баланса у насекомых играет метаболическая вода, образующаяся в процессе окисления углеводов, особенно жиров.
Рядом исследователей установлено влияние влажности окружающей среды на окислительные процессы. Впервые Бергер (Berger, 1907) показал, что мучные черви при голодании сохраняют процент воды и сухого вещества постоянными. Данные были подтверждены Бэкстоном (Buxton, 1930), установившим это постоянство в среде от 0 до 64% влажности. Механизм такого поддержания постоянства внутренней среды насекомого был детально изучен Мелланби. На основании того, что у мучного червя при 23О,С повышается обмен веществ при недостатке влаги и понижается во влажной среде, Мелланби предложил гипотезу о дополнительном окислении, направленном на поддержание водного обмена. В дальнейшем, однако, было высказано предположение, что повышение окислительных процессов связано с усилением вентиляции через спиракулы. Здесь, однако, важно вспомнить и старое предположение Бэкстона (Buxton, 1930) о том, что насекомые способны к удержанию водяных паров в воздухоносных путях, прежде чем этот воздух покидает их. Эти явления в настоящее время хорошо изучены на пустынных млекопитающих. Многие аридные беспозвоночные способны выдерживать значительные потери воды (высушивание). Это относится к некоторым видам муравьев, личинкам комаров (Chiro-nomidae) и даже ракообразным (Artemia). У личинок хирономид во влажной среде содержание воды в тканях может возрасти с 3 до 32% за 6 ч при 25° С.
Непосредственная адаптация тканей к высокой температуре также весьма значительна. Многие западно
279
африканские виды активны в зоне температур от 18 до 30° С, тогда как близкие к ним североамериканские — от 16 до 28°С (Wallwork, 1960). Летальные температуры соответствуют в этих случаях 37 и 40° С, а термический преферендум от 24 до 31° С и от 21 до 26° С.
Ряд величин летальных температур известен для пустынных насекомых различных отрядов — от 38 до 50° С (Cloudsley-Thompson, 1964). Наиболее высокие величины наблюдались у скорпионов (Leiurus quiquestriatus) и у «верблюжьего паука» — camel spider (Galeodes grati-ti). Очень высокая устойчивость к высокой температуре и у яиц. Так, яйца ракообразного Artemia salina могут выдерживать в сухом состоянии температуру 83°С в течение 4 ч.
Механизмы, лежащие в основе устойчивости беспозвоночных к очень высоким температурам, совершенно не изучены. Аридные и адаптированные к существованию в бедной влагой среде беспозвоночные отличаются исключительной устойчивостью к голоданию. Подчас голодание может продолжаться несколько месяцев.
АДАПТАЦИИ РЕПТИЛИЙ К ПУСТЫНЕ
Широко распространенные в аридной зоне рептилии представляют весьма интересный пример адаптаций организма, включающих как элементы подчинения факторам внешней среды (conforming organisms), так и элементы регуляции (regulating organisms), предохраняющие животное от чрезмерных сдвигов во внутренней среде. В пустыне встречаются представители всех отрядов — змей, ящериц и черепах, питающихся как растительной, так и в большинстве животной пищей.
У многих представителей рептилий, главным образом ящериц, обнаружены явления видовых температурных адаптаций, выражающихся в различных температурах, при которых наступает холодовое окоченение. Для ящериц характерно то, что активность наступает лишь при определенной температуре тела, которая поддерживается на постоянном уровне в течение всего времени, пока животное активно (колебания 0,56 — до 0,005°С). Такое своеобразное поддержание температуры тела достигается реакциями поведения, четко выраженной предпочитаемой температурой. Температура тела пустынных 280
рептилий в состоянии активности в значительной мере поддерживается и солнечным облучением. При этом некоторые виды способны изменять свою окраску и тем самым регулировать поглощение и отражение лучей различной части спектра.
Поскольку поверхность почвы в пустыне может в дневные часы нагреваться свыше 70° С, рептилии в это время суток укрываются в норах или на ветках растений (Кашкаров и Коровин, 1936). Многие виды пустынных ящериц—агамы, кругоголовки зарываются в песок, глубокие слои которого имеют более низкую температуру, нежели поверхность, и таким образом изолируются от прямого воздействия солнечных лучей.
Однако реакции поведения дополняются у пустынных рептилий мощным механизмом поддержания температуры путем испарения воды с поверхности верхних дыхательных путей — полипноэ. Испарение с поверхности тела, как и кожные железы у рептилий отсутствуют.
Термическое полипноэ лучше всего изучено у ящериц. При этом обнаружено, что между интенсивностью полипноэ, средней температурой активности и летальной температурой существуют определенные прямые и обратные зависимости (табл. 26).
Таблица 26
Реакция на нагревание у трех видов ящериц (по Dawson a. Templeton, 1964)
Вичы Температура
Средняя активности Летальная Наступления полипноэ Возрастания частоты пульса на 1%
Сцинк (Eumeces obso-letus) Ящерица (Crotaphytus collaris) Игуана (Dipsosaurus dorsalis) 33 38 39-42 43 46,5 48—50 Отсутствует 42,5-43,0 43 2 4,5 7
281
Очень важное значение в поддержании постоянной температуры тела имеет так называемый париетальный глаз. При удалении его температура тела ряда видов ящериц повышалась при нагревании как в природной обстановке, так и в лаборатории (Steblin a. Eakin, 1958). Такие данные были получены В. Н. Долининым (1968) при заклеивании париетального глаза пластинкой станиоля. Эти факты указывают на рефлекторный характер поддержания температуры тела у пустынных ящериц, но изучены совершенно недостаточно.
Изменение окраски ящериц под влиянием солнечного облучения является одним из важных механизмов поддержания температуры тела. Отражаются от поверхности тела преимущественно инфракрасные (тепловые) лучи. У европейских видов отражение составляет около 20% магниевого (MgO) стандарта. Однако исследование ряда видов с различным характером обитания указывает на исключительную роль отражения лучей у пустынных видов. Так, отражающая способность для лучей от 320 до 1100ммк. составляет для тропической ящерицы-игуаны {Iguana iguana) 6,2%, а для пустынной {Phery-nosotna plathyshinos)—35%. Резкое уменьшение поглощения солнечного тепла наступает у ящериц благодаря побелению поверхности кожи спины, наступающему одновременно с повышением температуры тела. У некоторых видов наступает и побеление кожи живота, что, вероятно, предохраняет животное от отраженного излучения поверхности субстрата.
Тканевые адаптации у рептилий в пустыне выражаются в большей термоустойчивости мышечных белков, активности адезинтрифосфатазы (Ушаков, 1963; Schmidt-Nielsen a. Dawson, 1964).
Повышение температуры тела приводит у рептилий к возрастанию обмена веществ, учащению ритма сердечных сокращений. Реакция на температуру среды зависит от дневного и ночного образа жизни животного. Так, летальная температура у дневных видов приходится на 49—50° С, а у ночных — 43—44° С (Saint-Girons et Saint-Girons, 1956). Как правило, температурный предел устойчивости к нагреванию коррелирует с уровнем температуры тела во время активности животных. У пустынных видов ящериц рода Sceleropus температура активности 35° С, а летальная 43° С. У воротниковой яще-
282
рицы (Crothaphytus collaris) соответствующие температуры равны 38—39° С и 46—47° С. Пустынная игуана активна при температуре тела около 42° С, а ее летальная температура близка к 50° С (Cowles a. Bogert. 1944) (рис. 64).
Рис. 64. Температура тела во время активности у ящериц Chemidophorus и Sceloporits magister из Аризоны и Sceloporus magister из Флориды (по Bogert, 1949)
Летальные температуры у рептилий обусловлены нарушениями деятельности нервной системы и аноксией в результате нарушения связывания гемоглобином кислорода при высокой температуре среды (Dill, 1938).
Весьма важно для понимания адаптаций рептилий в пустыне изучение потерь воды путем испарения. Как общее правило, потеря воды рептилиями при температуре тела, свойственной гомойотермным организмам (млекопитающим и птицам), составляет от 1/5 до 1/7. Примерно около 1/7 составляет и уровень обмена при высокой температуре тела (Dawson a. Bartholomew, 1958). Однако для пустынных рептилий соотношения не
283
сколько иные. Воротниковая ящерица испаряет в 5 раз меньше воды по сравнению с белой мышью, но только в 2 раза меньше по сравнению с кенгуровой крысой {Dipodomis merriami) (Schmidt-Nielsen, 1950).
Пребывание в норах — особенность экономии воды рептилиями. Более низкая температура среды, а следовательно, и более низкий обмен, более высокая относительная влажность, а следовательно, и более низкая испаряющая сила воздуха создают здесь для рептилий наиболее благоприятные микроклиматические условия.
Однако низкий уровень обмена (теплопродукции) создает благоприятные условия для существования рептилий и вне норы, так как в состоянии активности при одинаковой с пустынными млекопитающими температуре тела животному угрожает поступление тепла извне и наполовину меньшее образование тепла в теле. Этим можно объяснить богатую и разнообразную фауну рептилий в пустынях Старого и Нового Света.
Очень низки потери воды испарением через кожные покровы и при дыхании у черепах пустыни. У Gopherus agassizii потерн воды оказались намного меньшими, чем у Terrapene Carolina, обитающего во влажной среде (Schmidt-Nielsen a. Bentley, 1966).
Особенно ярко эти зависимости проявляются при сравнении небольшого каймана {Caiman sclerops) с пустынными ящерицами {Iguana iguana и Sauromalus obe-sus). У ящериц испарение воды кожей составляло лишь 5% от этой величины у каймана (Bentley a. Schmidt-Nielsen, 1966). Данные такого рода совершенно изменяют укоренившееся представление о малой проницаемости кожи рептилий. Сравнительно высокие потери воды испарением, однако, хорошо компенсируются малыми потерями воды с мочой и калом. Моча рептилий содержит мочевую кислоту как основной конечный продукт азотистого обмена. Для пустынных рептилий этот тип азотистого обмена является весьма благоприятным, так как при этом теряется минимальное количество воды. Это достигается секрецией гиперосмотической — по сравнению с плазмой — мочи.
Многие рептилии пустыни к тому же обладают удивительной .способностью хорошо переносить увеличение концентрации хлоридов крови. Высокая концентрация
284
летом наблюдается и в полевых условиях. Регуляция водного обмена у пустынных рептилий сходна с той, что имеет место у млекопитающих.
ВОДНЫЙ ОБМЕН У ПУСТЫННЫХ МЛЕКОПИТАЮЩИХ
Приспособительные механизмы к недостатку воды у грызунов пустыни хорошо развиты. Так, мышей Ре-rognathus flavescens и Р. fasciatus можно держать на сухой диете много недель (Bailey, 1923). Мышь Ре-rognathus fallox можно держать на сухом ячмене около 3 лет (Stefens, 1906). Следовательно, некоторые млекопитающие действительно способны существовать только на одной метаболической воде, т. е. на воде, которая освобождается в тканях в результате окислительных процессов в ходе обмена веществ. Необходимо отметить, что определенное содержание воды в теле стойко удерживается животными, несмотря на разные условия снабжения водой.
Произведен точный расчет баланса воды кенгуровой крысы при кормлении ее перловой крупой (сухой) из расчета 25 г в день (Schmidt-Nielsen a. Schmidt-Nielsen, 1951). Большим удобством этого расчета оказалось то обстоятельство, что 25 г сухой перловой крупы точно содержит 100 кал. Принято, что 25 г перловой крупы соответствует 13,4 г метаболической воды. При различной влажности и условно принятой температуре 25° С содержание воды ib корме увеличивается с возрастанием влажности воздуха. Влажность воздуха изменяет потери воды испарением и количеством воды, абсорбированной кормом.
Метаболическая вода сухого корма остается, разумеется, постоянной при всех степенях влажности среды и корма. Количество суммарной воды, принимаемой организмом, возрастает с увеличением влажности среды. Количество испаряемой воды с повышением влажности снижается. Снижаются также величины (относительные) потерь воды с мочой (на 1 г сухого зерна) и с калом.
Сопоставление кривых приводит к выводу, что при температуре 25°С относительная влажность ниже 8% приводит к дефициту воды у кенгуровой крысы. При влажности 20% имеется положительный баланс воды, а при более высокой влажности количество ее еще выше.
285
Таким образом, кенгуровая крыса может существовать, питаясь зерновым кормом, на метаболической воде плюс воде, абсорбированной зерном в условиях относительной влажности выше 8—10%. При расчете принято, что концентрация азота в моче при питании перловой крупой составляет около 20% (практически наблюдалось
Рис. 65. Водный обмен у кенгуровой крысы (при кормлении перловой крупой разной влажности) (по Schmidt-Nielsen a. Schmidt-Nielsen, 1952). По оси абсцисс — влажность (мг Н2О на I л воздуха и относительная влажность в %); по оси ординат — вода, использованная на 100 кал в процессе обмена (г):
1 — общее поступление воды, 2 — метаболическая вода, 3 — общая потеря воды, 4 — потеря с калом, 4 — потеря с мочой, 5 — потеря испарением
22,8%). Потеря воды с калом— около 0,63 г (рис. 65).
Следовательно, существование некоторых пустынных животных без воды подтверждается расчетными данными, но ограничено определенными пределами влажности и определенными температурами. Понижение влажности и повышение температуры среды приводит к нарушению водного баланса и к быстрой гибели этих животных, строго поддерживающих постоянство воды в организме. Эти данные были подтверждены ц экспериментально. Кенгуровая крыса, содержащаяся на перловой крупе при различной атмосферной влажности, теряла в весе в абсо-
лютно сухом воздухе, но сохраняла свой вес при влажности примерно от 2 до 3 мг воды на литр воздуха и даже прибавляла в весе при большей влажности.
Практически кенгуровую крысу невозможно обезводить лишением воды или сочного корма. Для обезвоживания приходится прибегать к увеличению белковой нагрузки, что достигается питанием соевыми бобами. Дополнительное введение белка требует дополнительного выведения продуктов обмена — мочевины, а последнее требует большего расходования воды почками.
286
Опыты показали, что в этом случае кепгу >овая крыса может терять в весе до 34%. Однако процент содержания воды в теле при этом не падает и составляет около 67,2 (Schmidt-Nielsen, Schmidt-Nielsen a. Brokaw, 1948).
Среди биологов значительную дискуссию постоянно вызывал вопрос о запасах воды в теле у пустынных животных. В результате тщательных исследований (Schmidt-Nielsen a. Schmidt-Nielsen, 1951) кенгуровой крысы было установлено, что содержание ее в течение 52 дней на сухом корме не изменило веса тела. Содержание воды в теле при этом было равно 66,1%, т. с. совершенно не изменилось в результате длительного лишения воды. Следовательно, думать о каких-то запасах воды в теле таких животных не приходится.
При исследовании большой песчанки в условиях различного снабжения водой (различная влажность корма— саксаула) выяснилось, что понижение влажности естественного корма приводит к небольшому снижению содержания воды в различных тканях тела (2—2,5%) (Арешева и Щеглова, 1961). Следует учесть, что прямое сопоставление в эксперименте пустынного грызуна — большой песчанки и влаголюбивого — белой крысы, крайне затруднено. При ограничении водного режима белые крысы резко уменьшают количество поедаемого корма и наступает не только водное, но и общее голодание. 5 суток содержания на сухом корме приводит у большой песчанки к снижению веса на 11,3%, а у белых крыс на 26,3%. Песчанки в этих условиях резко сокращают расход воды. По данным тех же авторов, удельный вес мочи больших песчанок на влажном корме колеблется от 1,012 до 1,017, т. е. он весьма невысок. При содержании на сухом корме удельный вес возрастает до 1,042—1,047, т. е. приближается к удельному весу мочи верблюда, столь хорошо использующего воду. Очень длительное содержание песчанок на сухом корме, однако, оказалось невозможным, так как у них из-за резкого снижения влажности кала наступала закупорка кишечника и гибель.
Сопоставление водного обмена с энергетическим у разных видов млекопитающих пустыни было сделано Л. Г. Филатовой (1944, 1949а). Величины водного обмена, отнесенные к весу тела и энергетическому обмену, колеблются у исследованных видов по весу от 0,014 до
287
Таблица 27
Максимальная концентрация мочи у некоторых млекопитающих (Schmidt-Nielsen, 1949)
Объект Электролиты (%) Мочевина (%)
Человек 2,2 6
Крыса лабораторная 3,5 15
Кенгуровая крыса 7,0 23
0,061, а по обмену (О2) от 0,56 до 1,60. Наиболее низкий уровень водного обмена наблюдается у тушканчика Северцева (Allactaga severtzovi), серого хомячка (Crice-tulus migratorius), желтого суслика и ушастого ежа (Hemiechinus sp.); наиболее высокий — у морской свинки, белой крысы и степного хорька (Putorius ewersman-ni). Ограничению подвергаются как потери воды мочой (ренальные), так и потери воды дыханием и испарением с поверхности кожи — экстраренальные. Так, самый низкий процент экстр арен ал ьных потерь воды, обеспечивающих охлаждение животного путем испарения, у тушканчика Северцева. Вместе со слабо выраженной химической термогуляцисй это обстоятельство и определяет образ его жизни как типичного пустынного ночного животного. Самое большое количество воды на 1 г веса тела потребляет гребенщиковая песчанка, обитание которой приурочено к берегам арыков или других водоемов.
Следовательно, пустынные организмы в различной степени ограничивают расход воды. Такие формы, как гребенщиковая песчанка, распространенная вдоль ирригационных сооружений, не отличаются по характеру водного обмена от непустыпных форм. Такие же отношения были обнаружены у пластипчатозубой крысы — незокии, обитающей на поливных землях (Щеглова, 1952а). У ряда пустынных форм образуется очень небольшое количество мочи, обладающей высокой концентрацией. Так, у кенгуровой крысы концентрация мочевины в моче (взятой прямо из мочевого пузыря) равна 19,7—22,8% сухого остатка с концентрацией электролитов, соответствующей 8,7% раствора NaCL У белой крысы эта концентрация почти в 2 раза меньше, а у человека — почти
288
' в 5 раз меньше. У тушканчиков Аравийского п-ова эта концентрация оказалась еще более высокой. Таким образом, расход воды на выделение продуктов обмена у пустынных животных минимальный (Schmidt-Nielsen, 1949); см. табл. 27. Самая высокая концентрационная способность почек установлена для австралийской пустынной прыгающей мыши (Nomis alexis). Осмотическая концентрация мочи у этих животных при содержании их па сухом зерне достигала 9370 мм/Осм/л, т. е. почти в три раза превышала эти величины для кенгуровой крысы, а в среднем вдвое превышала их (MacMillen а. Lee, 1969). По-видимому, эти величины являются рекордными для млекопитающих.
Детальное изучение механизмов выделительной функции у кенгуровой крысы было сделано Хауэллом и Тершем (Howell a. Gersch, 1935) и Шмидт-Нильсеном с сотр. (Schmidt-Nielsen, Schmidt-Nielsen, Brokaw a. Schneider-mann, 1948). В канальцах сосочков при содержании животного на сухой пище обнаружена более высокая концентрация ферроцианида натрия, а также значительная реадсорбция воды из мочевого пузыря при кормлении сухой пищей. Однако в последнее .время факты реадсорбции в мочевом пузыре подвергаются сомнению.
Между систематическим положением млекопитающих и способностью к концентрации мочи нет прямой зависимости. У самого примитивного из ныне живущих млекопитающих— ехидны в состоянии дегидратации можно наблюдать концентрацию мочи выше 2300 мм/Осм!л (Bentley a. Schmidt-Nielsen, 1966). В то же время «возвращение» к примитивной почке имеет место у горного грызуна (Aplodontia rufa), обитающего в условиях изобилия воды. Слабую концентрирующую способность почек у водяной полевки наблюдали Л. И. Иванова и А. П. Филюшина (1969). Это подтверждено п морфологическими исследованиями почек (Pfeiffer et al., 1960; Dicker a. Eggleton, 1964; Nungesser a. Pfeiffer, 1965). В противоположность этому, реабсорбционная способность у кенгуровой крысы в 10 раз превышает эту способность у непустынных видов. Обнаружено выделение канальцами мочевины, чего никогда не наблюдали у других млекопитающих.
Очень яркие особенности гормональной регуляции почечной функции были обнаружены Эймсом и Вап-Дей-
10—1204
289
ком (Ames a. Van Dyke, 1950). Оказалось, что концентрация антидиуретического гормона в моче во много раз выше у кенгуровой крысы, нежели у белой крысы (50 МЕ/лм мочи у кенгуровой крысы и 6 МЕ/л/л — у белой). Гипофиз кенгуровой крысы содержит больше антидиуретического гормона (0,9 МЕДиг), чем гипофиз лабораторной белой крысы (ОЗМЕ/л/г).
Как известно, при ограничении воды содержание антидиуретического гормона у животных нарастает. Содержание антидиуретического гормона в гипофизе кенгуровой крысы при нормальном для нее водном режиме такое же, как у белой крысы после 72-часового водного голодания.
Таким образом, у кенгуровой крысы удерживание воды телом увеличено.
Эти физиологические данные могут быть дополнены и морфологическими. Так, у большинства представителей млекопитающих обнаруживается замечательная зависимость между длиной почечных канальцев и способностью концентрировать мочу. Это положение было впервые установлено Петером (Peter, 1909) и затем Спербером (Sperber, 1944), изучившими большое количество видов млекопитающих. Спербером же было установлено, что относительная толщина мозгового слоя почек связана с условиями обитания животного; обитатели пустыни (грызуны, антилопы, жирафы, верблюды, львы и т. д.) имеют значительно большую толщину этого слоя, нежели обитатели орошаемых местностей. Так, у льва толщина мозгового слоя почек около 34 мм, а у свиньи только 6,5 мм. Сравнивая травоядных, плотоядных и насекомоядных, можно заметить, что плотоядные имеют очень длинную петлю Генле, а травоядные — короткую. Исключение из этого представляют пустынные грызуны, также обладающие очень длинной петлей. В этом отношении пустынные грызуны больше походят на плотоядных, чем на травоядных (рис. 66).
Травоядные и всеядные организмы имеют специальный тип нефрона, называемый кортикальным нефроном. Эти нефроны характерны гем, что проксимальный отдел канальца не опускается в мозговое вещество и участок этот очень короток. Такая форма нефронов свойственна свиньям, мышам, слонам. Корковое вещество занимает у них 9/10 всей почки.
290
Между развитием петли Гейле и способностью концентрировать мочу в онтогенезе существует прямая зависимость. У новорожденных эта способность очень мала (Schmidt-Nielsen, 1958). Крупные животные пустыни— верблюд, осел, каракульские овцы, джейран и сайгак изучены значительно меньше, нежели пустынные
Среда
обитания
Водная
Отряд
Однопроходные и сумчатые
Насекомоядные
Besm ап а
Psammomys
___ Тип питания
(с
Плодоядный
В Vespertilio Летучие мыши
Насекомоядный
Зерноядный
Приматы
Рис. 66. Строение почки (соотношение толщины коркового и мозгового вещества у млекопитающих) (по Sperber, 1944)
грызуны. Тем не менее есть основания для верблюда наметить некоторую схему его адаптации к условиям аридной зоны.
Удельный вес мочи верблюда не отличается от удельного веса у других копытных (в среднем он равен 1,030— 1,060) (см. табл. 21). Верблюд обладает большой жаж-доустойчивостыо благодаря способности ограничивать и без того малое количество мочи; колебания принятой per os воды достигают от 1 до 64 л, однако его почки не способны к большой концентрации мочи (Бахтозина,
10'
291
1960а, 19606). Температура тела верблюда в Сахаре нередко повышается с 33 до 40° С. При высокой темпе ратуре начинается интенсивное потоотделение, предохраняющее животное от дальнейшего нагревания (Schmidt-Nielsen, 1954). Высокая температура тела уменьшает количество тепла, принимаемого излучением, и позволяет лучше сохранить постоянную температуру тела на этом высоком уровне, сохраняя при этом воду. По данным этого же автора, в плазме поддерживается строго
Рис. 67. Суточные колебания температуры тела при разном питьевом режиме у верблюда, осла и человека в пустыне (по Schmidt-Nielsen, 1957):
в — дача воды ad libitum, д — содержание без воды. По оси ординат: верхние три кривые — температура тела, нижняя — температура среды (°C); по оси абсцисс — даты опытов (июнь 1954 г.)
постоянная концентрация солей и белка за счет поступления плазмы из межмышечных пространств. Особое значение имеет густая шерсть, предохраняющая от нагревающего действия солнечных лучей и от чрезмерного испарения. Специальными опытами показано, что остриженный верблюд теряет больше влаги, чем покрытый шерстью (Schmidt-Nielsen, 1954; Schmidt-Nielsen, Schmidt-Nielsen, Houpt a. Jarnum, 1956; Schmidt-Nielsen, Schmidt-Nielsen, Jarnum, Houpt, 1957) (рис. 67).
При питании сочной растительностью верблюд совершенно не пьет воды (это часто бывает в Сахаре зимой, а в Средней Азии весной). Летом на сухом подножном корму верблюд подходит к колодцу не чаще чем 1 раз в четыре дня. В самое жаркое время года верблюд может находиться на сухой диете (в работе) около 7 дней, теряя почти четверть веса тела без всяких серьезных нарушений физиологического состояния и работоспособности. В течение летнего дня верблюд теряет около 2,2%
292
(на сухой диете), в то время как осел в тех же условиях— около 7,7% 'веса тела.
Обмен веществ возрастает с повышением температуры тела при Qi0 = 2.06. При дегидратации обмен веществ снижается у верблюда до 77% первоначального уровня. Частота дыхания возрастает при гипертермии (Q;0 = = 10,8), хотя типичного полипноэ не наблюдается (Бах-тиозина, 1960; Schmidt-Nielsen, Crawford, Hewsome, Rawson a. Hammel, 1967).
Лишение лактирующей верблюдицы воды (при наличии зеленого корма, содержащего 65—75% воды) на протяжении 23 дней не оказало влияния на количество и состав молока (Туленбеков, 1958). Значительная устойчивость верблюда к потерям воды подтверждается еще и тем, что при полном водном голодании удельный вес крови, колеблющийся в норме от 1,045 до 1,057 возрастает только на 12-й день до 1,069. Следовательно, несмотря на лишение воды, содержание ее в крови остается постоянным (Бахтиозина, 1959, 1961а, 19616).
Обнаружены небольшие различия содержания воды в крови каракульских овец в разные сезоны (Ковальский и Падучева, 1951). Наименьшее количество (307 .иг%) наблюдалось в апреле, наибольшее — в феврале (401 л/г%). При различном режиме водопоя — ежедневном и через день — авторы не наблюдали определенно направленных изменений хлоридов в крови. Содержание хлоридов в крови верблюдов составляет около 400—520 л/г % (Бахтиозина, 1954). При водном голодании содержание хлоридов в крови несколько возрастает (до 510—600 л/г%). Это сопровождается резким сокращением мочеотделения и уменьшением выделения хлоридов мочой, которое падает, по данным Б. X. Бахтио-зиной, с 3000—3600 до 900 мг в сутки.
Однако в пустыне на солевое равновесие в организме одновременно воздействуют два фактора: значительные потери воды, связанные с испарением (терморегуляцией), и значительное содержание солей в воде и в распространенных в пустыне растениях. Эти влияния отражаются на водно-солевом обмене сельскохозяйственных животных.
В плазме крови пустынных млекопитающих поддерживается постоянная осмотическая концентрация NaCl (Schmidt-Nielsen a. Schmidt-Nielsen, 1952). Это положе
293
ние в общем подтверждается и при изучении животных в условиях пустыни и в опытах с водным голоданием. Наибольшие сдвиги содержания хлористого натрия в крови были найдены у каракульских овец (Кузнецов, 19516). Содержание хлористого натрия повышалось в летние месяцы по сравнению с зимними (с 459 до 683 лг%). Автор связывает эти изменения с поеданием большого количества соли с растительной пищей. Уровень NaCl в крови верблюдов стойко удерживается как при различном режиме снабжения водой на пастбище, так и при экспериментальном водном голодании (Бах-тиозина, 1961а, 19616). Несколько большие изменения (в пределах 15%) наблюдаются при переводе верблюдов с пастбищного на стойловое содержание (Бестужев, 1951). У лошади отмечены такого же рода изменения в крови (Протасеня, 1939; Коропов, 1946). Количество же хлоридов в мышцах подвержено очень большим колебаниям в зависимости от водного режима и содержания солей в корме. Так, содержание воды в мышцах изменяется по сезонам года (Кузнецов, 1951а). Влажность мяса в марте — 76,65%, в мае — 77,65 (до 79,0), в июне — 77,82, в ноябре — 76,28%. Следовательно, по содержанию хлоридов и влаги скелетная мускулатура овец в пустыне обнаруживает закономерные колебания. Мясо туркменских животных содержит воды от 76,6 до 81,7%, тогда как у животных умеренного климата оно содержит не более 75—76% (Кузнецов, 1947, 1949, 1951а).
Обильное содержание NaCl и воды позволяет рассматривать мускулатуру животных как своеобразное водное депо. Интересными в этой связи могут быть сопоставления с содержанием воды в мясе архаров (Бута-рин, 1949), у которых оно равно 71,04—73,52% (а у гибридных архаромериносов 55,97%). У горных животных, в отличие от пустынных, ткани значительно обеднены водой.
Исследование водного обмена (мл!кг/сутки) у разных пород овец в Кении (Африка) показало, что самым высоким водным обменом обладают каракульские овцы (114 мг), затем следует черноголовая сомалийская (107 мг), далее меринос (95 мг); самый низкий обмен наблюдался у овец-катумани (Macfarlane, 1965). Среди подобранных близнецов крупного рогатого скота у евро-294
пейского (Bos taurus) водный обмен был выше, нежели у зебувидного (Bos indicus) (Macfarlane a. Howard, 1966). Это наблюдалось как при стойловом, так и при пастбищном кормлении.
Распределение воды в организме крупных млекопитающих в условиях аридной зоны хорошо показано на
Рис. 68. Распределение воды в теле мериносовых овец в сухих тропиках (21° ю. ш.) и умеренном климате (35° ю. ш.). Ярко выражено возрастание количества воды клеточной и межклеточной
(по Macfarlane, 1965)
рис. 68. Из него видно, что содержание воды относительно возрастает во всех тканях: в плазме, в интерстициальной соединительной ткани, в клетках и тканях паренхиматозных органов. Таким образом, в организме создаются запасы воды, обеспечивающие терморегуляцию при высоких температурах внешней среды. Однако это влияние высокой температуры — фактор весьма важный в аридной зоне, но не единственный. Другим фактором является одновременный недостаток воды.
ТЕРМОРЕГУЛЯЦИЯ ПРИ НЕДОСТАТКЕ ВОДЫ
И ТИПЫ АДАПТАЦИЙ К УСЛОВИЯМ ПУСТЫНИ
Трудные условия для теплоотдачи у млекопитающих в пустыне приводят к уменьшению теплопродукции. Впервые понижение основного обмена было установлено .у большой песчанки (Rhombomis opitnus) по сравнению с гребенщиковой песчанкой (Meriones tamaricscnus).обитателем орошаемых земель и белыми и серыми крысами
295
одинакового веса (Щеглова, 1949). Этот факт был подтвержден детальными исследованиями на молодняке песчанок и белых и серых крыс. Оказалось, что у молодняка песчанок весом до 50—60 г основной обмен более чем в два раза ниже, чем у крысят того же веса. Соответственно этим величинам у песчанок также крайне низок и минутный объем крови сердца (Рашевская, 19b8). Крайне низкий уровень основного обмена был установлен и для обитателя высокогорной пустыни Невада оленьей мыши (Peromyscus maniculatus sonorien-sis); он связывался с большим содержанием жира в теле (52%). Эта точка зрения получила развитие в работах ряда американских исследователей (McNab a. Morrison, 1963; Lee, 1963; Hudson, 1964; Hayward, 1965). Снижение основного обмена по сравнению с контрольными животными (крысами) достигало у так ix видов, как Peromis-cus crinitus и Heterocephalus glaber, от 25 до 60%. Для очень суровых аридных условий снижение окислительных процессов в летний период позволяет организму довольствоваться метаболической водой, окисляя при этом минимальные количества жира (McNab, 1968) при условии, что вода не используется для терморегуляции.
В противоположность грызунам у верблюда установлен довольно колеблющийся уровень обмена веществ в зависимости от уровня температуры тела и величины дегидратации (Бахтиозина, 1958). Снижение обмена коррелирует с потерей воды, которая у верблюда может достигать 25% без каких-либо патологических нарушений (у непустынных млекопитающих не более 12—15%) (Schmidt-Nielsen, Crawford, Newsome a. Rawson, 1963; Schmidt-Nielsen, Crawford, Newsome, Rawson a. Hammel, 1967). Таким образом, снижение обмена веществ в условиях пустыни является твердо установленным фактом, но механизм его остается неясным.
Высокая температура среды, наблюдаемая в пустыне в летние месяцы, создает условия для перегревания организма, так как отдача тепла проведением и излучением становится невозможной. Единственный путь охлаждения организма — испарение воды с поверхности кожи, слизистых. Однако недостаток воды в пустыне вызывает значительные нарушения водного баланса, поэтому такой путь отдачи тепла сильно затруднен. В результате возникает ситуация, казалось бы, несовместимая с
296
жизнью. Выживание в этих условиях обеспечивается физиологическими адаптациями, лежащими в основе устойчивости как пойкилотермных, так и гомойотермных организмов в пустыне. Эти адаптации могут быть разбиты на 3 основные группы.
Первая группа характеризуется изменением поведения— главным образом суточного жизненного цикла. Большинство обитателей пустыни ведет ночной или сумеречный образ жизни (Кашкаров и Коровин, 1936). Так, типичными ночными видами являются многие насекомые, особенно жуки-чернотелки (Tenebrionidae), многие жужелицы, прямокрылые, бабочки, ряд рептилий (геккон). Ночной образ жизни характерен и дтя многих грызунов. На рисунке 69 приведены актограммы некоторых пустынных грызунов; на них видно, что для большей части исследованных животных характерен покой в наиболее жаркие часы дня. В это время грызуны укрываются в норах, где температура не превышает 18—25° С.
Вторая группа адаптаций, свойственная крупным обитателям пустыни, характеризуется значительной устойчивостью организма к нагреванию и недостатку воды. К этой группе следует отнести прежде всего вербтюда.
В условиях пустыни верблюд полностью лишен какого-либо убежища: он никогда не уходит в тень, несмотря на самые высокие температуры воздуха и почвы. По своему пастбищному режиму он отличается от всех других встречающихся в пустыне сельскохозяйственных животных. Пастбищный режим верблюда характеризуется небольшими затратами времени на поедание корма (по сравнению с коровами и лошадьми) и еще меньшей затратой времени на водопой (Бестужев, 1951). Малые затраты времени на эти процессы позволяют верблюду сохранить значительное количество энергии, не расходуя ее на пастьбу и связанные с ней процессы терморегуляции. Курдючные овцы, разводимые в пустьинях Казахстана с незапамятных времен, приближаются по характеру пастбищного режима к верблюду. Режим пастьбы в жаркое время года в пустынях Средней Азии характерен и для каракульской овцы. Эти животные заканчивают утреннюю пастьбу, когда солнце еще не успело нагреть остывшую за ночь поверхность земли, и ложатся па еще холодные участки. В таком положении они остаются всю жаркую часть дня, отдавая тепло в ненагретую
297
Рис. 69. Суточная двигательная активность некоторых видов пустынных грызунов (по Bodenhei-mer, 1957). I—Microtus guntheri; II — Meriones tristrami; III—Aco-mys dimidiatus-, IV — Cricetulus migratorius, V — Spermophilus sp; VI — Dipodillus dasyurus; VII—Mus musculus praetextus;
VIII — Spalax typhlus ehrenbergi. По оси абсцисс — часы суток; по оси ординат — двигательная активность (в условных единицах)
солнцем землю. Характерно для каракульских овец также упрятывание головы в тень другой овцы, благодаря чему образуются своеобразные цепочки или круговые скопления этих животных. По-видимому, инсоляция создает гораздо большие затруднения в регулировании теплового баланса овец в пустыне, нежели такое ограничение теплоотдачи при собирании в группы.
Своеобразна регуляция теплоотдачи конвекцией и испарением у крупного американского пустынного грызуна Lepus alleni. Оказалось, что около 25% всей поверхности тела приходится на уши, не покрытые шерстью. При очень высокой температуре происходит сокращение сосудов уха и испарение с их поверхности прекращается. Такая реакция имеет место при роющей активности этого грызуна. При температуре среды ниже температуры тела теплоотдача может возрастать в 2— 3 раза. При более высоких температурах животное нагревается. Таким образом, организм весом в 2—2,5 кг способен значительно экономить воду для терморегуляции (Dawson a. Schmidt-Nielsen, 1966).
У каракульских овец питье холодной воды приводит к довольно значительному охлаждению тела. Питье холодной колодезной воды приводит к снижению ректальной температуры овцы на 1,5° С (Миропольский, 1939; Макарова, Попугаева и Щеглова, 1955; Сафонов, 1958).
Таким образом, значительная группа организмов существует в пустыне в условиях периодической дегидратации, периодического повышения температуры тела. Последнее особенно ярко выражено у верблюда и осла. Суточные колебания температуры тела у осла и верблюда при различных режимах снабжения водой резко отличаются. При ограничении дачи воды повышение температуры тела оказывается более значительным. У верблюда величина суточных колебаний температуры тела может достигать 6° С.
Третий тип адаптаций к пустыне связан с миграциями. К этому типу следует отнести многие виды птиц. Большинство птиц, встречающихся в пустыне, залетает в орошаемые районы (Miller, 1951); например, значителен прилет птиц к пресному водоему в Каракумах (Репетек, Туркмения) (Аманова, 1968). К этому же типу следует отнести многих диких копытных — джейрана, сайгака, кулана, совершающих огромные переходы
299
и таким образом добывающих воду для поддержания температуры тела.
АДАПТАЦИИ ПТИЦ К ПУСТЫНЕ
Отсутствие потовых желез у птиц, а потому очень ограниченное испарение воды с поверхности тела определяет преимущественное охлаждение испарением при дыхании. Термическое полипноэ у птиц наступает, как правило, при повышении температуры тела до 42,5— 43,5° С. Количество тепла, отдаваемое при полипноэ (при температурах среды около 40°С), у большинства птиц составляет от 30 до 50% от общей теплопродукции. С повышением температуры среды теплоотдача возрастает, хотя температура тела птицы при этом поднимается на 2—4° С (Dawson a. Schmidt-Nielsen, 1964). Эта способность организма птиц имеет в условиях пустыни, однако, ограниченное значение, так как не обеспечивает собственно экономии воды. Кроме того, самое учащение дыхания при полипноэ является дополнительным источником расхода энергии и источником теплообразования. У многих птиц полипноэ дополняется особыми колебаниями глотки — gular fluttering, наблюдаемое у водоплавающих птиц и у птиц пустыни (козодоев Phalenoptillis nutallii).
Механизм gular flattering, требующий очень малых затрат энергии, обеспечивает значительную отдачу тепла, достигающую при 44°С 170% общей теплопродукции организма у козодоя Ph. nutallii (Bartholomew, Hudson, Howell, 1962). Нет, однако, никакой уверенности в том, что механизм этот в сочетании с вообще низкой температурой тела у козодоев (см. также гл. IV) специфичен для птиц пустыни. Можно только думать о некоторой повышенной устойчивости у них к колебаниям температуры тела. По существу, значительное повышение испарения у птиц пустыни, например у кардинала (Richmon-dena cardinalis), начинается с температуры 38° С и выше. Здесь начинают функционировать механизмы повышенной теплоотдачи, о которых речь была выше.
Высокую теплоотдачу во время полета обеспечивает и фу [кция воздушных мешков (Salt a. Zenteen, 1960).
Потери воды пустынными птицами не столь ограничены, как у пустынных млекопитающих и рептилий. Все
300
же они несколько меньше, чем у домашней птицы. По сравнению с пустынными грызунами потери воды птицами вообще и пустынными в частности очень велики и превосходят потери грызунов от 5 до 12 раз. В противоположность большим потерям воды у птиц с дыханием, потери при мочеотделении очень ограничены, что не связано с высокой концентрирующей способностью почки птиц, выделяющей продукты азотистого обмена в виде мочевой кислоты. Концентрирующая способность в отношении мочевины у птиц немного меньше этой величины у пустынных млекопитающих.
Способность пустынных видов птиц переносить потери воды не отличается от непустынных видов. Потеря в весе около 15% позволяет восстановить водный баланс, Весьма интересные особенности обнаружила М. Аманова (1968) при изучении распределения воды в тканях пустынной птицы скотоцерки (Scotocerca inquieta) в Каракумах. Оказалось, что относительный вес сердца у этих птиц много меньше, чем у пеночек (теньковки и веснички). Наблюдались закономерные уменьшения сердечного индекса в летний период. В то же время в летний период ткани скотоцерки содержали больше воды, нежели птиц, менее связанных с пребыванием в пустыне. Особенно эти различия были заметны в летний период.
Малый вес многих птиц пустыни (менее 40 г) и сопутствующий этому весу высокий обмен вызывают значительное образование метаболической воды в организме. У некоторых мелких птиц при температуре среды около 25° С потребность в воде за счет метаболических процессов может быть покрыта па 75% (Dawson, 1958).
Большие колебания температуры тела наблюдаются у страуса — до 4°С. Испарение воды возрастает до 0,4 г/мин при 5° С и 4,5 г/мин при 45° С. При высоких температурах наблюдается взъерошивание перьев, что предохраняет от нагревания кожи. Таким образом, по характеру терморегуляции страус — обитатель аридных степей (саванн) близок к верблюду (Crawford a. Schmidt-Nielsen, 1967).
Питание птиц в пустыне связано с поглощением дополнительного количества солей. Птицы резко отличаются от млекопитающих наличием солевых назальных желез (Schmidt-Nielsen, 1965). Можно предположить, что секреция солевых желез связана с реабсорбцией во
301
ды в клоаке, необходимой для образования мочи. Пустынный воробей саванн (Passerculus sandwichensis beldingi) отличается большой устойчивостью к высоким концентрациям соли. Концентрация солей в моче у этого вида пустынного воробья равна 527 м)Экв1л, а у непустынного—175 м!Экв!л (Poulson a. Bartholomew, 1962). Несмотря на это, в эксперименте пустынные виды птиц не могут существовать без воды. В природе поступление воды происходит за счет питания суккулентами, откуда и избыток поступающей в организм соли.
Пустынная авиафауна крайне бедна. Так, из 274 видов птиц, размножающихся в Калифорнии, только 17 размножаются в пустыне и 23 вида частично встречаются в пустыне. Около 60% всей авиафауны пустыни не являются собственно пустынными видами. Следует отметить, что физиологические особенности птиц, населяющих пустыни СССР (да и вообще пустыни Старого Света), совершенно не изучены.
РОЛЬ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА В АДАПТАЦИИ
К АРИДНОЙ ЗОНЕ
Особенности пищеварения и водного обмена пустынных млекопитающих
В связи с тем, что существование в пустыне связано, с одной стороны, с большой экономией воды и что, с другой стороны, поедаемый многими животными корм содержит воду в крайне ограниченном количестве, у пустынных млекопитающих имеется целый ряд особенностей пищеварения. Они сводятся в общем к тому, что обильно выделяющиеся благодаря сухой пище пищеварительные соки подвергаются полному обратному всасыванию в кишечнике. Интересна в этом отношении высокоспециализированная реакция слюнных желез у каракульских овец.
У каракульской овцы, хорошо приспособленной к существованию в пустыне, при поедании сухого корма (по сравнению с влажным) не наблюдается общего для всех млекопитающих увеличения слюноотделения из подчелюстной и подъязычной желез (Файзиев, 1957, 1958). Таким образом, у каракульских овец уменьшены потери
302
воды при испарении слюны с поверхности слизистой рта и языка. Интересно, что приспособления к условиям пустыни идут в этом направлении и дальше. У каракульских овец не происходит выделения слюны во время введения в полость рта сухого песка (отмывающая слюна). Это, вероятно, связано с большим количеством песка, заглатываемого животными с пищей во время пастьбы в пустыне. У новорожденных же каракульских овец введение сухого корма и песка в рот вызывает увеличение слюноотделения. Очевидно, эта адаптивная особенность взрослых каракульских овец появляется в более поздний период развития. У большой песчанки сухость корма вызывает возрастание слюноотделения, как и у других млекопитающих (Арешева и Щеглова, 1961). Выявилось также и обильное слюноотделение при всыпании сухого песка в рот (отмывающая слюна). Как показали специальные наблюдения А. И. Щегловой, песчанки в жаркое время дня поедают большую часть корма, предварительно затаскивая его в норы, где влажность воздуха значительно выше. Прежде чем есть, они тщательно отряхивают песок, прилипший к растениям. Следовательно, наличие отмывающей слюны в этом случае не может вести к значительным дополнительным потерям воды.
Уменьшенное слюноотделение у каракульской овцы компенсируется обильным выделением других пищеварительных соков. Этот процесс имеет значение как для осуществления переваривания сухой пищи, так и для выделения продуктов обмена веществ в связи с крайне ограниченным выделением мочи. Так, объем всасываемой воды в кишечнике каракульской овцы достигает весной 7,5 л, а летом 10,7 л (весной в два, а летом в три раза превышает количество воды, принятой per os) (Исхаков, 1954, 1960).
Таким образом пустынные животные увеличивают выделение пищеварительных соков, не подвергающихся испарению. Увеличенная тем самым циркуляция воды в организме при уменьшенном диурезе обеспечивает выделение продуктов обмена веществ. У каракульской овцы наблюдается увеличение количества химуса при поедании сухих кормов и уменьшение его при поедании влажных. При содержании па влажном и сухом кормах процент воды в верхних отделах пищеварительного тракта
303
остается практически неизменным. У больших песчанок уменьшенное количество воды, поступающее с пищей, компенсируется дополнительным поступлением воды с пищеварительными соками. Однако при этом наблюдается значительное различие содержания воды в кале. При влажном корме процентное содержание воды в кале большой песчанки равно 66,9%, а при сухом корме — 49,1%. Так как в слепой кишке количество воды при разных режимах не изменяется, то, очевидно, ограничение поступления воды вызывает увеличение обратного всасывания в толстых кишках (Арешева и Щеглова, 1961).
У кенгуровой крысы при содержании на сухом корме (перловая крупа) содержание воды в кале составляет около 45%, тогда как у белой крысы — около 68%.
Интенсивный обмен воды между кровью и желудочно-кишечным трактом был установлен у верблюда и каракульских овец. При полном двухнедельном водном голодании объем проходящего химуса снизился у верблюда с 86 до 60 л. Содержание же сухого вещества было до водного голодания 2,8—4,3%, а после голодания 1,3— 3,6%. Обнаружено выделение краски нейтральрот в рубце и сетке. Следовательно, в кишечнике верблюда даже в условиях водного голодания содержится ее большое количество. Повышение температуры среды приводит к значительному выделению кальция в просвет кишечника. Также возрастает и выделение азотистых веществ в кишечник при повышении температуры среды (Лутфулли-на, 1956а, 19566; Базанова и Степапкина, 1957; Степан-кина, 1958).
Значительное увеличение (с 10 до 23%) золы в сухом веществе кала обнаружено у большой песчанки, а также значительное увеличение мочевины в желудке при водном голодании у крыс и особенно больших песчанок (Бельская и Войтенко, 1968). Между содержанием мочевины в крови и выведением ее в желудок при усиленной его секреции, вызванной гистамином, существуют обратные взаимоотношения. Большая песчанка очень мало чувствительна к накоплению мочевины в крови при дегидратации. Даже повышение ее содержания до 200 мг1°1о не вызывает у этих животных каких-либо патологических явлений (Арешева и Щеглова, 1968).
304
Все это позволяет говорить о частичном замещении функции почек деятельностью пищеварительного аппарата как механизма, связанного с адаптацией к аридной
РОЛЬ КОЖИ И ЛЕГКИХ В АДАПТАЦИИ К ПУСТЫНЕ
Весьма важное место в ограничении потерь воды у пустынных организмов занимают адаптивные особенности отдачи воды кожей и легкими при дыхании. Эти потери составляют значительную часть так называемых незаметных потерь веса (perspiratio insensibilis), которые издавна являются одним из путей изучения общего обмена веществ. Кожно-легочные потери складываются из потерь воды при дыхании путем почти полного насыщения водяными парами вдыхаемого воздуха, испарения воды с поверхности (потоотделение и кожная перспирация) и, наконец, из разности веса поглощаемого кислорода и выделяемой при кожном и легочном дыхании углекислоты. Эта разность учитывается при определении кожно-легочных потерь воды путем одновременного определения газообмена и незаметных потерь веса.
Кожно-легочный механизм экономии воды у пустынных организмов был впервые обнаружен А. Й. Щегловой (1949). Кожно-легочные потери воды у большой песчанки значительно ограничены по сравнению с лабораторной белой крысой. Важно и то, что при повышении температуры среды у белой крысы кожно-легочные потери воды резко возрастают, а у пустынного вида — большой песчанки это возрастание незначительно. В итоге при высоких температурах среды (30°С) кожно-легочные потери воды у белой крысы почти в 2 раза превышают таковые у большой песчанки (рис. 70). Песчанка лишь в минимальных количествах расходует воду на терморегуляцию. А так как химическая терморегуляция у большой песчанки также выражена слабо, то, очевидно, существование этого вида в пустыне обусловлено своеобразной формой поведения — роющей деятельностью, обеспечивающей в глубоких норах такой термический режим, который не нарушает постоянства температуры тела (Щеглова, 1949).
Ограничение водного обмена было затем подтверждено на желтом суслике, тушканчике Северцева (Филатова, 1944). У этих видов оказатся более низкий водный
305
обмен и более низкие экстраренальные потери воды, нежели у гребенщиковой песчанки — обитателя орошаемых земель и белой крысы. В дальнейшем установлено (Щеглова, 1949, 1952а, 19526, 1959), что ограничение водного обмена можно наблюдать у многих пустынных
Рис. 70. Кожно-легочные потери воды у большой песчанки и белой крысы при температурах среды 10°, 20°, 30° С. Черные столбики — большая песчанка
грызунов — тонкопалого суслика, малоазиатской песчанки, малого суслика. При сравнении трех видов сусликов — тонкопалого, желтого и малого, самый низкий уровень водного обмена оказался у тонкопалого суслика, несколько больший у желтого и самый высокий — у малого (Щеглова, 19526). Различия могут быть объяснены не только особенностями расселения этих видов, но и
сроками залегания в спячку желтого и малого сусликов. Первый залегает в спячку очень
рано, когда содержание воды в растениях пустыни снижается. Малый суслик залегает в спячку поздно и долго питается содержащей воду степной растительностью.
Подобные же отношения у пустынных видов грызунов Нового Света (Schmidt-Nielsen a. Schmidt-Nielsen, 1950, 1952). Потери воды кожей и легкими у пустынных видов составляют около 2/3 потерь воды у лабораторных животных. Обращают на себя внимание низкие величины потерь воды у диких видов грызунов, особенно домовой мыши, которая может находиться без воды неопределенно долгое время (Bing a. Mendel, 1931). Этот факт интересен как пример приспособления к недостатку воды
в условиях, весьма далеких от пустынных.
Ограничение кожно-легочных (экстрарепальных) потерь воды у обитателей пустыни определяется особенно
306
стями кожного покрова и испарения воды легкими. Так как грызуны не обладают потовыми железами, то роль кожи в кожно-легочных потерях воды невелика. Выдыхаемый воздух у непустынных млекопитающих насыщен водяными парами практически на 100%- Небольшое недона'сыщение может быть лишь при очень поверхностном дыхании.
Дефицит насыщения выдыхаемого воздуха был впервые обнаружен у верблюда в Южном Казахстане (Бах-тиозина, 1948), а затем тщательно изучен как в условиях нормального снабжения водой, так и при водном голодании (Бахтиозина, 1960а, 19606). Обнаружилось, что верблюды при достаточном снабжении водой в жаркие летние месяцы способны выдыхать воздух с относительной влажностью, не превышающей 69—86% при исходной величине 88—95%. После прекращения водного голодания относительная влажность выдыхаемого воздуха возвращалась к исходной на 5—6-й день. Относительная влажность вдыхаемого воздуха была в этих опытах от 26,8 до 55% (табл. 28). Недонасыщение выдыхаемого воздуха водяными парами при водном голодании было ярко показано на курдючных овцах (в Казахстане в летний период). При низкой относительной влажности, не превышающей 45—47%, и температуре около 31—33°С относительная влажность выдыхаемого воздуха до водного голодания превышала 90 % • При 4-дневном водном голодании относительная влажность выдыхаемого воздуха снизилась с 78 до 62 %.
При редком водопое (1 раз в три дня) у каракульских овец наблюдается снижение потерь воды легкими (Падучева, Ковальский и Шраер, 1957). По сравнению с ежедневным водопоем снижение потерь воды через легкие может достигать в апреле 1,4%, а в июле 23,2%. Авторы объясняют это уменьшением легочной вентиляции и падением окислительных процессов.
Было высказано предположение, что длинные носовые ходы некоторых пустынных животных, смачиваемые выделяемой слизистой жидкостью, могут благодаря ее испарению охлаждать область морды и таким образом конденсировать водяные пары. Однако это соображение лишено какой-либо энергетической основы, так как количество испаренной с поверхности жидкости должно быть равно количеству удержанной, т. е. не вызывать
307
Таблица 28
Относительная влажность вдыхаемого и выдыхаемого верблюдом воздуха при водном голодании (Бахтиазина, 1960а)
День опыта Температурi окружаю ш го воздуха (°C) Относительная влажность воздуха
вдыхаемого (%) выхыхаемаг» (%)
Нормальные условия
1 25,7 55,0 95,0
2 25,6 38,0 90,0
3 24,2 54,2 93,0
4 22,4 52,4 91,8
5 24,8 42,6 88,8
Период водного голодания
1 28,3 31,2 82,2
2 29,9 32,4 79,8
3 27,6 36,8 86,2
4 26,6 39,5 74,6
5 24,7 38,1 79,8
6 26,0 40,8 72,7
7 25,4 34,0 78,3
8 25,7 35,0 73,0
9 25,7 36,5 69,5
10 23,3 41,6 70,4
11 25,6 26,8 69,3
Восстановительный период
1 26,0 37,7 73,0
2 26,2 35,0 83,8
3 24,1 37,7 79,8
4 23,8 40,1 82,4
5 24,9 29,5 71,3
6 25,7 32,7 91,6
7 25,0 32,0 91,0
какого-либо энергетического эффекта. Было высказано и предположение (Слоним, 1962), что здесь может иметь место активное удержание водяных паров слизью, скапливающейся по ходу верхних дыхательных путей. Пос
308
леднее требовало для подтверждения изучения ферментативных механизмов, изменяющих физико-химические свойства муцинов. Были необходимы и специальные патофизиологические исследования легочной ткани и верхних дыхательных путей, в том числе и межклеточных образований, играющих (Гинецинский, 1962) важную роль в перемещениях воды в организме.
Специальные исследования активности тканевой гиалуронидазы в легочной ткани и ткани почек (Григоренко, 1967, 1968), показали, что у впадающего в зимнюю спячку краснощекого суслика (Citellus erythrogenys) и у большой песчанки наблюдается взаимно противоположно направленная динамика активности этого фермента. У песчанки наибольшая активность в засушливый период года, у суслика — в период впадения в зимнюю спячку и сразу после пробуждения. В почках наблюдаются те же взаимоотношения. Эти фактические данные позволяют думать, что проницаемость межклеточных образований для воды и состояние кислых мукополисахаридов могут играть существенную роль в динамике удержания воды в легких. Введение антидиуретического гормона вызывает однозначное изменение активности фермента в легких и почках.
С другой стороны, гистохимические, ультрамикроско-пические и гистологические методы исследования (Соболева, 1967, 1968) осветили и морфологическую картину в легких пустынных животных. Оказалось, что у верблюда и большой песчанки строение легких отличается значительной длиной воздухоносных путей. Эпителий, выстилающий бронхиоли, цилиндрический, лишь в самых конечных отделах переходит в плоский — альвеолярный. Такое строение легочной ткани обеспечивает меньшие потери воды при поддержании высокого напряжения водяных паров в альвеолах.
Особенную роль приходится отвести большим клеткам альвеолярного эпителия у всех млекопитающих, принимающим на себя функцию фагоцитов-макрофагов. Превращение этих клеток в макрофаги сопровождается изменениями, регистрируемыми гистохимически: исчезает активность неспецифической эстеразы и проявляется накопление липазы. Такое явление сопровождает дедифференцировку клеток и превращение их в макрофаги.
309
Рис. 71. Микрофотография легкого большой песчанки после дегидратации. 1 — клетки на разных стадиях дедифференцировки, 2 — в просвете альвеолы пино-цит (по Соболевой, 1964)
После содержания больших песчанок на безводной диете большие клетки альвеолярного эпителия берут на себя новую функцию — пнноцитов, т. е. клеток, задерживающих воду. При этом клетки оказываются бедными липидами, белком и мукополисахаридами. Цитоплазма содержит большое количество неокрашивающихся вакуолей. Клетки эти часто наблюдаются не только в стенке, но и в просвете альвеол. У белых крыс при содержании на безводной диете на 6—7-й день также наступают начальные стадии дедифференцировки альвеолярного эпителия, но до образования пиноцитов дело, по-видимому, не доходит (рис. 71). У верблюда после 20 дней лишения воды также появляются большие комплексы ли-пофагов, в которых можно предполагать начальные стадии развития пиноцитов.
Кроме того, в этих же исследованиях были обнаружены значи
тельные изменения в жировой ткани, расположенной подплевраль-но, заключающиеся в нарушении структуры жировой ткани и на
коплении кислых мукополисахаридов. При дальнейшем
водном голодании жир почти исчезает и заменяется новой тканью, состоящей из неправильных многоугольных и округлых клеток с уплощенными ядрами, передвигающимися к периферии. В цитоплазме этих клеток мало белка, нет липидов и мукополисахаридов и содержится большое количество вакуолей. Это позволяет думать о
наличии системы пиноцитов, расположенных в подплевральной области и удерживающих воду.
Изложенные данные позволяют выявить пути удержания воды легкими при дыхании и понять интимные механизмы этого явления. Окончательное решение этих вопросов требует еще дальнейших исследований.
310
АДАПТАЦИИ К ЗАСОЛЕНИЮ (ВОДА, ПОЧВА, РАСТИТЕЛЬНОСТЬ)
Наряду с ограниченным снабжением водой, организмы в условиях пустыни встречаются с значительным увеличением концентрации солей в питьевой воде, почве и растительности, что оказывает наибольшее влияние на растительноядные организмы, но играет роль и для плотоядных. Практически это касается не только экологофизиологических сопоставлений, но имеет значение и для организации содержания сельскохозяйственных животных в пустыне, обоснования водного режима сельскохозяйственных животных, использования пастбищ, различно удаленных от источников питьевой воды с различной концентрацией солей (табл. 29). Имеются литературные данные, указывающие на предельно допустимые концентрации солей в воде, пригодной для использования человеком и животными (Бокина, 1968).
Таблица 29
Характеристика питьевой воды, используемой для водопоя сельскохозяйственных животных в пустыне
(по Минервину, 1955)
Характеристика воды Плотный г ста гск (г/л)
Пресные воды, пригодные в течение круглого года Слабосоленые (солоноватые), пригодные к круглогодовому использованию Соленые, годные только осенью или зимой . . Соленые, годные только зимой Чрезмерно засоленные, непригодные До 2 » 6 > 13 . 13—16 Свыше 16
Содержание овец на протяжении тысячелетий в пустыне привело к значительному привыканию этих животных к высоким концентрациям солей. Этому способствовало и то обстоятельство, что содержание солей в колодцах обычно повышается от весеннего периода к зимнему. Режим изменения содержания солей в питьевой воде сочетается и с изменением растительного корма, с переходом от питания в весенний период эфемерами, содержащими мало солей, к питанию летом и осенью солянками
311
с большим количеством солей. Поэтому в пустыне в весенние месяцы можно наблюдать значительное уменьшение поступления солей в организм животных, а в летние и осенние месяцы — большой их избыток (Минервин, 1955). Все это приводит к сезонным колебаниям содержания солей в тканях животных в пустыне.
Однако природа физиологических адаптаций к гипер-салемии стала только в последние годы предметом специальных исследований. Впервые Бартолемью и Хадсон (Bartholomew a. Hudson, 1959) обнаружили, что некоторые грызуны способны извлекать воду из концентрированных растворов хлористого натрия. Так, суслики Citellus leucurus способны поддерживать постоянство веса при употреблении воды с концентрацией солей в 1,4 раза больше, чем морская вода. При этом они обладают способностью к большой концентрации мочи и осмотическое давление ее возрастает в 6,4 раза.
Солевые нагрузки вызывают не только местные реакции со стороны выделительных органов, по и значительные сдвиги в нейросекреторных и гормональных реакциях организма. Особенно интересна в этом отношении стимуляция гипоталамической нейросекреции, в свою очередь стимулирующая образование антидиуретическо-го гормона, который повышает концентрационную способность почек (Поленов, 1962).
Установлено, что в период ограниченного поступления воды в организм (конец лета) у краснощекого суслика наблюдается усиленное выведение нейросекрета из супраоптического ядра гипоталамуса, который идет на синтез антидиуретического гормона — вазопрессина. Эти изменения соответствуют периоду питания растительностью с повышенным содержанием солей. Особенно выявляется солевая нагрузка при питании солянками,характерными для пустыни второй половины лета. К концу лета в Сахаре концентрация солеи в растениях, являющихся основной пищей типичного грызуна пустынь Северной Африки и Ближнего Востока — песчаной крысы (Psammotnis obesus), очень велика (табл. 30).
Сопоставление концентрации солей в таких растениях с концентрацией в морской воде указывает на ту нагрузку, которую в течение длительного времени несет регулирующий осмотическое равновесие физиологический аппарат этих животных. По тем же данным концен-
312
Таблица 30
Концентрация ионов в трех видах солянок (по Sclimidt-Nieslen, 1964)
Объект- Na K Ca Щавелевая кислоia
Arthrocnemum sp. ... 723 38 872 0
Traganutn nudatum . . 706 153 61 421
Sueda mollis 796 51 344 555
Морская вода 470 10 548 0
трация электролитов в моче песчаной крысы в естественных условиях обитания может достигать 1920 м1Экв1л. Это в 4 раза превышает концентрацию солей в морской воде. Растворы поваренной соли 5, 8 и 10%-ные не вызывают потери веса животного в течение 4—10 дней при питании сухим кормом и питье исключительно этой воды.
До настоящего времени остается совершенно не ясным механизм адаптации пустынных животных к высокому содержанию щавелевой кислоты в корме. Щавелевая кислота ядовита для огромного большинства организмов, так как вызывает выпадение ионов кальция из растворов и тем самым вызывает симптомы гипокальци-немии. Возможно, что в кишечнике песчаной крысы происходит разложение щавелевой кислоты бактериями. Известна подобная же устойчивость овец к введению в желудочно-кишечный тракт щавелевой кислоты. Непосредственное влияние введения растворов поваренной соли в организм пустынных грызунов — больших песчанок было детально изучено Б. Р. Дороховой (1967,1968). Животным в течение 10 дней через желудочный зонд вводился ежедневно 7,7%-ный раствор СаС1 и КС1 из расчета 10 мл раствора на 250 г веса тела в сутки. Контрольным животным вводилась дистиллированная вода, т. е. вызывалась гипергидратация (рис. 72). Оказалось, что как водная, так и солевая нагрузка вызывает снижение удельного веса плазмы крови, общего белка плазмы, гемоглобина и показателя гематокрита, что свидетельствует о поступлении воды в кровь. Следовательно, высокая концентрационная способность почки позволяет животным пустыни реагировать на введение солей в организм не сгущением крови, а, наоборот, разжижением,
313
благодаря быстрому удалению электролитов с мочой. Эта реакция у большой песчанки выражена настолько ярко, что наблюдается даже небольшое снижение содержания солей в тканях. Сдвиги при водной нагрузке, однако, более значительны, чем при солевой.
У белых крыс при тех же нагрузках наблюдается меньший сдвиг в плазме и крови и накопление электро-
Крысы контрольные солевой нагрузкой EZ3 водной нагрузкой
Рис. 72. Изменение удельного веса крови и плазмы у больших песчанок и белых крыс при солевой и водной нагрузках (по Серебрякову и Дороховой, 1967)
литов в тканях. Таким образом, «вымывание» солей из организма пустынных животных как следствие повышенной концентрационной способности почек — основной механизм адаптации к гиперсалемии. При гиперсалемиях важное место занимает и возрастание клубочковой фильтрации. При этом теряется значительное количество воды, так как канальцевая реабсорбция снижается. Такова реакция на гиперсалемию у непустынных организмов, обычно ведущая к дегидратации организма.
Глава VIII
АДАПТАЦИИ К ПИТАНИЮ, ПИЩЕВАЯ СПЕЦИАЛИЗАЦИЯ И ОБМЕН ВЕЩЕСТВ
ТИПЫ ПИТАНИЯ животных ОРГАНИЗМОВ
Вещества, превращаемые животными в энергию роста, размножения, двигательной активности и т. д., а также используемые ими в процессе жизнедеятельности и для поддержания структур собственного тела, могут быть разделены на небольшое количество групп. Это биополимеры — белки, углеводы, жиры, а также витамины, вода, неорганические соли. Однако процессы питания, переваривания, всасывания и усвоения пищевых веществ у различных животных отличаются исключительным разнообразием. Таким же разнообразием отличаются и пищевые связи животных с внешним миром. Эти связи и их классификация основаны на изучении характера принимаемой животными данного вида пищи.
При изучении пищевых связей в экологии принято различать эврифагию, т. е. питание разнообразной пищей (часто и растительной, и животной) и стенофагию — питание однородной пищей. Иногда различают монофа-гию, т. е. питание одним видом пищи, олигофагию — немногими видами пищи и полифагию — питание разнообразной пищей (всеядность).
, Однако, несмотря на все эти многообразные формы пищевых (трофических) связей животных организмов с внешним миром, конечный их итог — обеспечение полного энергетического баланса с физиологическим состоянием и особенностями деятельности животного. Оно достигается целым рядом физиологических особенностей пищеварения и всасывания, ассимиляции и диссимиляции, а также специальными формами пищедобывательной деятельности и акта еды. Вся эта совокупность двигательных и вегетативных процессов и составляет
315
сущность процесса питания — исходного этапа осуществления обмена веществ, этапа, в значительной мере определяющего характер обмена и его особенности.
За основу формирования пищевых связей с внешним миром в физиологии давно уже принята следующая схема. Обмен веществ вызывает соответствующее обеднение энергетических потенциалов организма. Это приводит к повышению пищевой возбудимости, к возбуждению тех нервных образований, которые И. П. Павлов называл пищевым, или химическим, центром. Интеграция всех раздражений и, следовательно, регуляция пищевого поведения связана с концентрацией возбуждения в областях холинэргического уровня, т. е. в сетевидной формации таламуса и обонятельных долях мозга (Soulairac, 1963). В результате возбуждения химического центра возникает пищедобывательная деятельность животного, а если есть пища, то и самый пищевой акт. Животное добывает пищу или поедает ее до тех пор, пока возбудимость пищевого центра не снижается — тогда животное перестает добывать пищу и перестает есть. Наступает период пищеварения, всасывания и ассимиляции. Эта элементарная схема лежит в основе изучения всякого пищевого рефлекса и рефлекторной пищедобывательной деятельности. Если попытаться приложить схему к условиям существования различных видов животных, то она оказывается совершенно недостаточной для пояснения наблюдаемых явлений.
Количество потребляемой пищи прямо связано с энергетическими потребностями организма. Однако в природе постоянно наблюдаются случаи, когда животные принимают значительно больше пищи, нежели требуют их энергетические затраты, т. е. они повышают свой энергетический потенциал главным образом за счет накопления жира. С другой стороны, эти жировые запасы расходуются в неблагоприятные для добывания пищи времена года или в условиях выкармливания детенышей. Это наблюдается, например, у медведя, самки которого рожают и выкармливают детенышей зимой, не выходя из берлоги. Лактирующие самки тюленей некоторое время после родов не принимают пищи совсем. Таким образом, нельзя считать, что питание обеспечивает сохранение энергетического и вещественного баланса в организме в каждый данный момент. Явления избы
316
точного питания и голодания постоянно чередуются в естественных условиях и сопровождаются анатомическими изменениями. Так, у крыс после искусственно вызванной гиперфагии, длившейся 6—12 месяцев, значительно возрастала длина кишечника, особенно тонкий отдел (Lesby a. Levin, 1966). Можно считать, что только на протяжении очень больших отрезков времени (порядка годового цикла) пищевые связи с внешним миром уравновешивают все жизненные потребности животного организма. По существу, все эти явления лежат в основе энергетики зимней спячки и зимнего сна.
От состояний накопления и расходования энергетического потенциала следует отличать такое состояние баланса вещества, когда в организме задерживаются главным образом азотсодержащие вещества (ретенция азота), что связано с ростом и развитием организма, формированием половых продуктов и внутриутробным развитием плода. К этой же группе явлений относятся лактация и рост шерстного покрова после линьки или стрижки.
К обычным источникам питания животных относятся зеленые или сухие растения, другие животные, а также микроорганизмы (бактерии и грибки). Эти основные группы содержат почти бесчисленное множество различных видов пищевых веществ и столь же различны типы механизмов питания.
При рассмотрении аппаратов питания необходимо наметить какую-то классификацию, но она не может основываться на обычной таксономической классификации животных, поскольку способ питания любого вида зависит от характера пищи (растительного или животного происхождения, размера частиц относительно веса данного животного), особенностей среды обитания, а также уровня организации животного. Животные разных видов, обитающие вместе в любой среде, могут добывать и поедать одни и те же пищевые вещества с помощью способов питания и органов, которые, хотя и отличаются в деталях, однако действуют на основе одних и тех же основных принципов. Также возможно обратное положение, когда животные близких видов потребляют совершенно различную пищу и разными способами как за счет изменений исходных механизмов питания, так и при помощи совсем других систем.
317
Эти положения могут быть проиллюстрированы на примере животных любого местообитания, рассмотрения различных механизмов питания, при помощи которых они получают пищу. На морском побережье, например, животные с организацией очень различных уровней и сложности — губки, кольчатые черви, различные моллюски и ракообразные, а также организмы, имеющие оболочку, питаются примерно одинаково, отфильтровывая из морской воды мелкие растения и животных, личинок и органический детрит. Структуры, используемые этими разными видами животных при поедании пищи, неоднородны, однако все они действуют на основе одного и того же принципа: пропускают морскую воду через какой-то аппарат фильтрационного типа, который задерживает находящиеся в ней пищевые вещества и передает их органам пищеварения.
Контрастом этим неродственным видам животных, питающихся одинаковым образом, могут служить также прибрежные представители одного типа или класса, сильно отличающиеся по способу питания и поедаемой пищи.
Особенно яркие примеры дают моллюски гастроподы, у которых некоторые члены класса малоподвижные травоядные животные, пасущиеся на морских водорослях, другие — хищные и охотятся за другими моллюсками, а третьи отфильтровывают пищевые вещества из морской воды или ловят свободноплавающие частицы пищи с помощью приспособлений. Эти приспособления полностью отличаются от аппаратов, которые используются травоядными и хищными представителями.
Все эти факты не позволяют создать классификацию механизмов питания на основе сопоставления таксономических признаков животных. Были предложены классификации, основанные на других критериях, и наиболее пригодной и исчерпывающей оказалась составленная С. М. Йонджем (1954). Она охватывает все типы беспозвоночных и Chordata (подтипы Hemichorda, Urochorda, Craniota) и основана в основном на учете размера частиц поедаемой пищи. Несмотря на то, что предлагаемая классификация разработана прежде всего для беспозвоночных, она образует удобную основу для краткого обзора аппаратов и способов питания большинства животных.
318
Классификация способов и аппаратов питания животных
(по Ионджу, 1954)
Аппараты поедания малых, частиц пищи
1. Псевдоподиальный.
2. Жгутиконосцевый.
3. Цилиарный.
4. Фильтрующий (тентакулярный).
5. Мукоидный.
Способы поедания больших частиц пищи
1. Проглатывание неподвижной пищи.
2. Соскребывание и рытье.
3. Схватывание добычи.
а) Схватывание и проглатывание.
б) Схватывание и жевание перед проглатыванием.
в) Схватывание и наружное переваривание до проглатывания.
Способы поедания жидкостей или мягких тканей
1. Прокалывание и высасывание.
2. Высасывание.
3. Абсорбирование всей поверхностью тела.
В соответствии с этими особенностями питания и пи-щедобывания необычайно разнообразной оказывается и морфология аппарата пищедобывания и пищеварения. В качестве примера приведем различия в строении клюва у различных видов птиц: хищных, насекомоядных, зерноядных, всеядных (рис. 73). Формы клюва соответствуют не только этим большим группам питания, но и отражают особенности питания рыбой, насекомыми (в земле, под корой деревьев и т. п.), мелкими плавающими организмами и т. д. У представителей одного класса сильно отличаются как аппарат пищедобывания, так и акт еды. Как общее правило, в отношении пищевых адаптаций наибольшим адаптивным изменениям в процессе эволюции подвергаются пищедобывательная деятельность и акт еды, далее следуют особенности строения желудка и желудочного пищеварения. Пищеварение в кишечнике подвержено значительно меньшим адаптивным видовым различиям. Это можно наглядно видеть
319
Рис. 73. Различные формы клюва у птиц (по Jennings, 1965): 1 — Mersanger, 2 — фламинго, 3— широконоска (утка), 4 — Scissorbill (взрослый), 5—Scissorbill (молодой), б—аист, 7— птица-носорог, 8 — колибри, 9 — шилоклювка, 10 — попугай, 11 — попугай, 12 — колпица, 13 — клест, 14 — козодой, 15 — орел, 16 — Balaeniceps
при сопоставлении процессов питания у насекомых, птиц, млекопитающих, принадлежащих к разным классам животных. Даже такой акт, как жевание, оказывается весьма специфичным не только у млекопитающих отдельных отрядов, но и отдельных видов внутри отряда, например различных видов грызунов.
Особенное строение имеет пищевоспринимающий аппарат у организмов, которые переваривают пищу вне 320
желудочно-кишечного тракта; выделение ферментов (с соками) может растворять твердые пищевые вещества, которые потом всасываются. Такой гни питания встречается у многих хищных насекомых (например, жужелп цы), а также у крабов.
К этому типу питания относится и адсорбция нищевых веществ всей поверхностью тела. Помимо простейших, такой тип питания свойствен многим экто- и эндопаразитам: червям, насекомым и их личинкам.
ТИПЫ ПИЩЕВАРЕНИЯ
Под пищеварением понимается совокупность процессов, обеспечивающих ферментативное расщепление (чаще всего гидролитическое) полимерных пищевых веществ па простые составные части, в основном мономеры (Уголев, 19G7).
До настоящего времени в зависимости от условий действия ферментов на пищевой субстрат установлено три типа пищеварения: дистантное, внеклеточное пищеварение, внутриклеточное и пристеночное (контактное, или мембранное).
У высших животных протекают процессы пищеварения всех трех типов и есть все основания думать, что они могут дополнять друг друга в зависимости от характера субстрата и условий выделения ферментов, а также от концентрации отдельных ферментов.
Все эти формы пищеварения изучены у разных видов с разной экологической специализацией неодинаково.
Наиболее изученное дистантное внеклеточное пищеварение характеризуется тем, что самый процесс расщепления происходит главным образом в специальных полостях пищеварительного аппарата. Реже ферменты выделяются во внешнюю среду, воздействуя на пищевые вещества, которые затем в расщепленном виде так или иначе всасываются животным. К последним случаям следует отнести внеклеточное пищеварение у некоторых насекомых — жужелиц, муравьиного льва и др. Общим признаком внеклеточного пищеварения служит значительное расстояние этих полостей (желудка, кишечника, зоба) от секретирующих клеток желез. Этот тип пищеварения является объектом исследования давно, начиная с классических исследований И. П. Павлова и его предшественников.
11—1204
321
Внутриклеточное пищеварение имеет наибольшее значение у одноклеточных животных, однако часто встречается и у высших форм как в виде фагоцитоза, так и в виде пииоцитоза — поглощения отдельными клетками (например, эпителием) жидких веществ.
Характерная черта внутриклеточного пищеварения — образование вакуоли, т. е. отграниченного от протоплазмы клетки пузырька (пространства), куда поступают ферменты, расщепляющие твердые или жидкие субстраты пищи. Эта способность была установлена сравнительно недавно путем электронно-микроскопических исследований. Таким образом, внутриклеточное пищеварение имеет большое сходство с внеклеточным — полостным.
Пристеночное (контактное, мембранное) пищеварение было открыто всего около 10 лет назад А. М. Уголе-вым. Оно осуществляется ферментами, фиксированными на клеточной мембране. Клеточные мембраны осуществляют, таким образом, не только перенос веществ из окружающей среды в клетку, но и участвуют в процессе пищеварения (расщепления), ориентируя определенным образом активные центры ферментов, подобно тому, как при гетерогенном катализе. Пристеночное пищеварение осуществляется на специальных поверхностных структурах, например на щеточной кайме тонкой кишки. Оно может происходить и на всей поверхности клеток у одноклеточных организмов.
В качестве общей схемы можно указать, что дистантное пищеварение приобретает наибольшее значение, когда собственно пищевое вещество отграничено от среды воздействующего на него фермента различными трудно проницаемыми барьерами, например оболочками растительных клеток, соединительной тканью и т. д. Внутриклеточное пищеварение обеспечивает пищевыми веществами непосредственно данную клетку. Пристеночное пищеварение осуществляет одновременное направленное действие фермента на пищевой объект и создает условия для всасывания, т. е. переноса через оболочку пищеварительного тракта или для одноклеточных организмов оболочку клетки.
Эколого-физиологическое значение описанных выше типов пищеварения изучено еще совершенно недостаточно. Наиболее важный в этом отношении факт касается зависимостей в развитии полостного и пристеночного
322
пищеварения у млекопитающих в постиатальиом онтогенезе. В этот период жизни млекопитающих как пиноци-тозная активность клеток кишечника, так и полостное пищеварение (содержание ферментов) очень ограничены, а между тем осуществляется интенсивное переваривание молока и всех его составных частей, в частности гидролиз крахмала. В этот период пищеварение осуществляется за счет действия ферментов на внешней поверхности клеточных мембран (Уголев, 1967). Эту функцию кишечник животных начинает осуществлять уже к концу эмбрионального периода развития, предупреждая, таким образом, наступление фазы молочного питания. Важно и то, что у морских свинок (зрелорождающихся) амилолитическая активность поджелудочной железы оказывается уже значительной к моменту рождения. Развитие отдельных сторон (типов) пищеварения является врожденным признаком, индуцируемым генетическим аппаратом клеток пищеварительных желез и гуморальными стимулами. Пока еще очень мало данных о возможностях смещения взаимоотношений между полостным и пристеночным пищеварением под влиянием диеты в более поздние фазы онтогенеза.
Приспособительный характер пристеночного пищеварения можно иллюстрировать еще и следующим фактом. У круглых червей — кишечных паразитов аскарид структура щеточной каймы кишечника такая же, как и у высших животных. Ферменты тоже абсорбированы на этих поверхностях. Важно и то, что у спороцист некоторых кишечных паразитов щеточная кайма покрывает эктодермальную поверхность тела, т. е. эктодерма является аналогом кишечной поверхности высших животных.
АД ХПТАЦИЯ ПИЩЕВАРИТЕЛЬНЫХ ФЕРМЕНТОВ
Разнообразие объектов питания приводит к образованию в организме большого количества ферментов, активность которых направлена на расщепление сложных пищевых веществ. Эти ферменты группируются следующим образом: амилолитические, расщепляющие углеводы различной сложности; протеолитические, расщепляющие белки, и липолитические — жиры.
В лаборатории И. П. Павлова давно уже был установлен кардинальный факт, что выделение пищеварительных
11*
323
соков по количеству и качеству, т. е. по содержанию в них ферментов, зависит от химических особенностей поедаемого пищевого продукта. Зависимость была установлена в опытах на собаках, но сохраняет свое значение в процессах питания животных на всех этапах эволюции. Однако в связи с различными условиями среды, продолжительностью пребывания пищи в разных отделах пищеварительной системы и т. п. адаптации довольно сложны и разнообразны. Кроме того, для огромного количества пойкплотермных организмов ферментативные системы характеризуются оптимумом активности, соответствующим температуре тела животного, которая зависит от температуры среды обитания в период активности. Для гомойотермных организмов температурный оптимум активности ферментов находится в сравнительно узком диапазоне температур от 37 до 40° С.
При рассмотрении адаптаций пищеварительных ферментов следует различать их по происхождению — видовые генетические адаптации, свойственные видам с определенным типом питания, и индивидуальные, возникшие в процессе питания каким-либо одним видом пищи.
Всеядные организмы, как общее правило, обладают значительно большим набором ферментов, нежели плотоядные или растительноядные. У первых преобладают протеазы, у вторых — карбогидразы (амилазы).
Однако наблюдается и более тонкая специализация ферментов, что выявляется особенно ярко в процессе индивидуального развития организма. Например, многие насекомые в личиночной стадии имеют более богатый набор ферментов в кишечнике, чем в стадии имаго. Это особенно заметно для гусениц бабочек, пчел, у которых остаются только амилолитические ферменты. В желудочном соке новорожденных млекопитающих постоянно есть химозин — фермент, створоживающий молоко; он отсутствует у взрослых. В соке желудочных и поджелудочных желез многих рептилий, амфибий и млекопитающих содержится хитиназа, расщепляющая хитиновые оболочки насекомых (Frankignol et Jeuniaux, 1965). Таким образом, имеется значительное соответствие между изменениями в объектах питания и пищеварительными ферментами в онтогенезе. Большую роль в процессе расщепления пищевых веществ играют и ферменты гканей, служащих объектом питания. Это особенно касается
324
растительных тканей и имеет большое значение для растительноядных насекомых, особенно для полифагов.
У млекопитающих ферментный состав желудочного сока имеет определенную направленность. Легче всего у каждого животного перевариваются белки, которыми в норме питается данный вид. Это было четко установлено еще в лаборатории И. П. Павлова для желудочного сока собаки по отношению к белкам животного и растительного происхождения. Последние переваривались желудочным соком собаки с большим трудом, т. е. требовали большего количества фермента. Ферменты пищеварительных соков различных животных не идентичны по своей активности в отношении одинаковых пищевых веществ у видов с различной экологической специализацией.
Имеются указания на то, что даже кристаллические препараты пепсина быка, свиньи и цыпленка неодинаковы по свое?! активности в отношении различных белков. Протеиназы гидролизируют нативные белки животного другого вида или денатурированные белки того же вида.
Пищеварительная роль и содержание, а также активность ферментов слюны у разных животных сравнительно невелики. В слюне некоторых животных, например лошади, вовсе не удается обнаружить присутствующего у других животных амилолитического фермента. Почти нет этих ферментов также и в слюне хищных млекопитающих. Небольшая амилолитическая активность обнаружена в слюне кошек, лисиц и очень незначительная —в слюне морских свинок, белых крыс и низших обезьян (Уголев, 1958). В таблице 31 приведены некоторые данные сравнения амилолитической активности у млекопитающих.
Для слюны обезьян характерно наличие в ней амилазы и очень большая чувствительность состава слюны к химическим и механическим раздражителям полости рта (Малиновский, 1953). Высокая амилолитическая способность слюны связана с растителыюядностью обезьян и многообразием поедаемых ими растительных продуктов.
Значительная ясность в эти вопросы с эколого-физиологической точки зрения внесена работами А. М. Уго-лева (1957, 1958а, 19586, 1958в). Была изучена сравни-
325
Т а б л н ц а 31
Амилолитическая активность слюны у некоторых млекопитающих (Уголев, 1958а)
Животные Амилолитическая активность слюны (опретеление п > Вольге-муту)
Кошка домашняя Лисица Морская свинка Крыса белая Макак-резус 20-160 До 10 320—1 280 5 000—20 000 10 000—100 000
тельная активность ферментов пищеварительных соков (слюны, желудочного, поджелудочного и кишечного) у многих видов млекопитающих по отношению к крахмалу и гликогену как основным продуктам, близким по химической структуре, но различным по происхождению— животному или растительному. Оказалось, что ферменты пищеварительных соков (например, слюны) различны по своей активности в отношении крахмала (фптолптическая активность) и гликогена (зоолитиче-ская активность). В отношении желудочного сока и его фермента пепсина выявилось, что постоянно наблюдаемое более медленное переваривание растительных белков у собаки может быть изменено, если, например, накануне накормить ее хлебом. Такой удивительный факт не может быть объяснен концентрацией фермента, так как п-ри этом замедляется переваривание белков мяса и ускоряется переваривание растительных. Следовательно, здесь имеет место определенное приспособление ферментного состава пищеварительных соков к роду пищи, не количественное — в отношении содержания фермента, а качественное — в отношении направленности действия этих ферментов. Пепсин желудочного сока и его свойства очень тонко регулируются в зависимости от режима питания животного. Очевидно, пищевой режим животного определяет не только количество желудочного сока, выделяемого на различные пищевые вещества, но и спектр его ферментативной активности.
Индивидуальные адаптации пищеварительных ферментов обнаружены у многих организмов. Так, у тара-
326
кана при содержании его на пшеничных отрубях вырабатывается больше протеиназы, чем при кормлении белым хлебом. При содержании на картофеле у насекомых резко повышается активность амилазы. Подобные же данные известны для .моллюсков.
Особенно хорошо явления адаптации ферментов желудочных желез изучены на собаке. Впервые факты этого рода были установлены X. С. Коштоянцем (1928) и В. А. Музыкантовым (1929). Оказалось, что длительное содержание собаки на мясном или углеводном пищевом режиме приводит к значительному изменению количества и состава желудочного сока, отделяемого па различные пищевые вещества. Составу сока и особенностям его отделения, изменяющимся при длительном кормлении определенными веществами, соответствуют и изменения морфологии желез желудка (Лозовский, 1948). Значительное изменение морфофизиологических особенностей желудка в связи с пищевым режимом можно наблюдать и у человека. Характерно, что длительные приспособительные сдвиги в составе и характере желудочного сокоотделения повторяют приспособительные сдвиги, наблюдаемые при однократном кормлении этими же веществами.
Сравнение состава сока у разных видов млекопитающих показывает значительные особенности, связанные как с отличиями возрастного питания, так и с характером питания данного вида. Однако эти различия намного меньше соответствующих различий в ферментном составе слюны. Протеолиз в кислой среде, расщепление белков мяса и молока и только частичное расщепление жира— вот общая практически для всех млекопитающих функция ферментов желудочного сока. У жвачных она заменяется частичным перевариванием растительных белков и белков, составляющих тела симбионтных микроорганизмов (бактерий и инфузорий). У крыс в без-железистом отделе желудка происходят процессы, аналогичные рубцовым, и образующиеся летучие жирные кислоты (триацетат, трипропионат и трибутират) могут служить источниками энергии, что выяснилось при добавлении этих веществ в корм (Little a. Mitchell, 1965).
Механизм регуляции отделения желудочного сока и его пищеварительных свойств осуществляется нервной системой и многочисленными гуморальными связями,
327
хорошо изученными в физиологии животных и человека. Основной опыт с изучением нервной регуляции деятельности желез желудка — опыт мнимого кормления — оказался эффективным для всех исследованных до сих пор млекопитающих. Так, по данным павловской лаборатории (Тихомиров и Савич, 1911), секреция желудочного сока из сычуга у козы происходит непрерывно. Можно наблюдать усиление ее в часы обычного кормления. Блуждающий нерв является основным секреторным первом желудка млекопитающих. Мнимое кормление мясом вызывает выделение желудочного сока, активного по белкам животного происхождения, а мнимое кормление хлебом — сока, активного по отношению к растительным белкам (Уголев, 1958). Отсюда можно сделать вывод, что не только истинное, но и мнимое кормление собаки хлебом 'или мясом оказывает влияние на специфическую зоо- или фитолитическую активность желудочного сока на следующий день. Тончайшее приспособление желез желудка к роду пищи происходит рефлекторно при раздражении рецепторов полости рта жевательного аппарата.
Секреция поджелудочной железы мало изучена как в сравнительно-физиологическом, так и в эколого-физиологическом отношениях. Есть указания, что поджелудочная железа различных животных выделяет сок, по химическому составу одинаковый. Три основных фермента— трипсин, липаза и амилаза находятся в поджелудочном соке в одинаковых соотношениях, изменяется лишь концентрация их в связи с родом пищи (Разен-ков, 1948).
Как известно, поджелудочная железа является источником не только ферментов поджелудочного сока, но и ферментов крови — трипсина, амилазы и липазы. Количество и соотношение последних могут изменяться в зависимости от пищевого режима организма. Была исследована активность амилазы крови у лабораторных мышей, крыс, морских свинок, кроликов, хомяков, кошек, собак, африканских белых хорьков и ежей (Уголев, 1958). Оказалось, что независимо от принадлежности к тому или другому отряду у растительноядных видов амилолитическая активность крови преобладала над гликогенолитической, у всеядных обе были равны, а у плотоядных превалировала гликогенолитическая актив
328
ность. Аналогичные данные были получены и при исследовании водных экстрактов поджелудочной железы у разных животных. Что касается регуляции деятельности поджелудочной железы, то как нервные пути (блуждающие нервы), так и гуморальный механизм (секретин) не отличаются специфичностью даже в пределах разных классов животных.
Из всего изложенного можно сделать вывод, что изменение ферментативной активности как в пределах отряда млекопитающих, так и в пределах одного вида при разных пищевых режимах является основным механизмом приспособления (в широком смысле этого слова) к роду питания. Детально эти механизмы еще не изучены.
АДАПТАЦИИ ДВИГАТЕЛЬНОЙ ФУНКЦИИ ПИЩЕВАРИТЕЛЬНОГО ТРАКТА
Различные типы питания, особенно поедание пищи твердой (костей), инкапсулированной (семян), а также разные типы пищедобывательной деятельности связаны с особенностями строения и двигательными функциями полости рта, желудка и отчасти кишечника. Наибольшую специализацию приобретают при этом функции полости рта и желудка; движения тонкого и толстого кишечника довольно однотипны у разных видов млекопитающих.
Но время общего прохождения пищи по желудочно-кишечному тракту у разных видов животных представляет характерные особенности. Как правило, оно больше у растительноядных форм (обладающих и большей длиной кишечной трубки) и меньше у плотоядных.
Акты ж е в а пи я и г р ы з е н и я. Жевание у разных видов чрезвычайно разнообразно. Лучше всего изучена механическая сторона жевания у человека и отчасти собаки (Рубинов, 1958). Применяемая обычно графическая методика регистрации жевательных движений (так называемая мастикациография) позволяет установить время и интенсивность смыкания и размыкания зубных рядов, растирающих и перемалывающих движений челюстей. Для каждого вида пищи можно наблюдать своеобразное сочетание этих компонентов жевательного акта; такая структура акта жевания наблю
329
дается у человека. Акт жевания собаки резко отличается; отсутствуют длительные смыкания челюстей, характеризующие растирающие и размалывающие движения.
У жвачных характер жевания и жевательного рефлекса изменяется в зависимости от количества захваченного корма, его физических и химических свойств. Введение в рот сладкого, соленого, кислого и горького изменяет протекание жевательного рефлекса (Плетнев, 1953). Жевательные рефлексы у сельскохозяйственных животных (главным образом, овец) тщательно изучены (Плетнев и Щербакова, 1952). Число жевательных движений зависит от пищевой возбудимости и «грубости» корма. Чем грубее корм и голоднее животное, тем больше число жевательных движений во время еды. Определенной степени возбудимости пищевого центра соответствует и определенное число жевательных движений. Это устанавчивается наблюдениями в разные сроки после последнего кормления животного. У непарнокопытных жевание осуществляется очень долго с чередованием работы двух противоположных сторон.
У насекомоядных (ежа) жевание носит характер разламывающих движений. Оно характеризуется таким же чередованием работы челюстей.
У грызунов разных видов процесс жевания очень сложен и приобретает характер грызения. Оно представляет самостоятельную форму деятельности, обеспечивающую не только функцию питания, но и играющую большую роль в жизнедеятельности отряда. Грызение у кроликов вызывает длительное повышение газообмена, хорошо выраженное и после прекращения такой мышечной работы (Исаакян, 1953). Эти данные были подтверждены на большой песчанке (Щеглова, 1959); оказалось, что грызение у нее может занимать до 30% суточного времени. Грызение же древесины не связано с приемом пищи, а входит в общий жизненный стереотип этих животных.
Движения губ и щечной мускулатуры 1принимают непосредственное участие в акте еды. У представителей разных видов млекопитающих роль движений губ различна. Большое значение они имеют у обезьян — низших и особенно у антропоидных. Губы принимают непосредственное участие в акте питья у копытных, обезьян, от*
330
части грызунов. Питье хищных, насекомоядных осуществляется лакающими движениями языка. Роль щечной мускулатуры главным образом заключается в осуществлении акта питья путем создания отрицательного давления в полости рта. Какие-либо объективные исследования этих мышечных аппаратов неизвестны.
Функция защечных мешков. Парные защечные мешки есть у некоторых представителей отряда грызунов и у низших обезьян. Они располагаются в передней полости рта (cavum oris anterior) и несут функции запасания пищи. У грызунов защечные мешки по существу являются органом запасания корма. У новорожденных обезьян при первом кормлении из рожка можно наблюдать заполнение защечных мешков. Есть основания думать, что их функция начинает осуществляться с (Самого начала питания через пищеварительный канал (Архангельская, 1958). Иннервация защечных мешков грызунов и обезьян совершенно не изучена.
Важное значение для млекопитающих приобретает обратная перистальтика верхней части желудочно-кишечного тракта, связанная с актом рвоты, отрыгиванием принятой нищи и отрыгиванием жвачки. Акт рвоты — важнейший оборонительный рефлекс, свойственный многим низшим позвоночным (рыбам, амфибиям, рептилиям), млекопитающим (хищным, приматам, жвачным; у непарнокопытных отсутствует), птицам. Он возникает при сильных раздражениях желудка, всего кишечника и его тонкого отдела. Этот оборонительный рефлекс в норме легче всего возникает в ответ на чрезмерное растягивание желудка и является своего рода охранительным при избыточном поедании пищи. Однако самый акт рвоты биологически используется не только как охранительный рефлекс. Так, у хищных (например, волка) принятая пища отрыгивается при запасании ее или при кормлении молодняка в логове. Многими хищными (собаками, кошками) отрыгиваются непереваренные остатки пищи (к примеру кости значительной величины). У грызунов акт рвоты отсутствует и заменяется своеобразным задерживанием ядовитых веществ, попавших в желудок с пищей.
Двигательная функция пищеварительного тракта. Движения желудка и кишечника в эколого-физиологическом отношении изучены крайне недо-
331
статочпо; имеются данные для собаки, сельскохозяйственных животных и для человека.
Наиболее простые отношения в моторике желудка наблюдаются у организмов с плотоядным и всеядным типом питания. В качестве общей схемы здесь можно принять следующее. Пища, поступающая в желудок, в течение длительного времени остается неподвижной в его фундальной части, не смешиваясь с выделяющимся при этом желудочным соком. Сокращения продвигают ее по направлению к pylorus. Однако здесь благодаря работе так называемого препилорического сфинктера происходит отбрасывание части пищевых масс назад и перемешивание их с желудочным соком. Препилорический сфинктер есть у собаки, низших обезьян, а также человека. У зерноядных грызунов пища также долго задерживается в желудке, где она, однако, остается совершенно неизмененной, подвергаясь перевариванию только в тонком кишечнике.
Двигательная функция рубца жвачных состоит из периодических сокращений, которые стимулируются как условиями наполнения рубца, так и самим актом приема корма. Своеобразная моторика рубца связана с его функцией и потому зависит от характера питания. Двигательная функция рубца возникает независимо от перехода животного от питания молоком к грубым кормам (Тул-баева, 1959). Возникновение жвачки может быть связано с раздражением грубыми кормами стенок преджелудков жвачных (опыты на овцах и козах). Так, у каракульских овец движения рубца зависят не только от наличия корма, но и от состояния пищевой возбудимости. При наполненном рубце и при длительном голодании движения прекращаются (Хайрутдинов и Воробьева, 1954).
Двигательная функция кишечника в эколого-физиологическом аспекте почти не изучена.
ПРИЕМ ПИЩИ И ТИПЫ ПИЩЕДОБЫВАТЕЛЬНОЙ ДЕЯТЕЛЬНОСТИ
Разнообразие типов питания сопровождается и значительным разнообразием поведенческих актов, приводящих организм в контакт с объектами питания. Во многих случаях эта деятельность вызывается рефлектор
332
но при раздражении видом, запахом пищи, звуками, издаваемыми живой добычей, и т. п.; во многих случаях здесь имеют значение химические изменения в крови («голодная» кровь) или раздражение отдельных органов, например желудка.
В настоящее время наиболее приемлемой можно считать схему, согласно которой обеднение крови отдельными химическими веществами приводит к раздражению специальных областей мозга. Изменения состояния этих регулирующих центров служат стимулом для возникновения общего возбуждения животного и проявлений специальных форм пищедобывательной деятельности.
Гипоталамическая область является объектом многочисленных исследований в этом направлении. В гипоталамической области расположены специфические нервные центры регуляции пищевого поведения, его стимуляции и торможения (Anand, 1968). Различают два центра: латеральный — «пищевой центр», вызывает пищевое поведение, стремление (drive) к объекту питания и поддержание состояние голода, медиальный — центр насыщения, тормозит это поведение, подавляя активность пищевого центра. Активность «центра насыщения» определяется главным образом изменениями внутренней среды при поступлении пищи. К факторам внутренней среды, влияющим на центр насыщения, относятся уровень содержания (использования) глюкозы в крови, состояние специфического динамического действия, уровень содержания жира в депо. Растяжение желудочно-кишечного тракта служит дополнительным источником сигналов, возбуждающих центр насыщения и прекращающих прием пищи. Таким образом, состояния голода и насыщения, обусловливающие стремление животного к пище, зависят от взаимодействия гипоталамических центров.
Стремление животного к определенному виду пищи осуществляется благодаря сочетанному возбуждению гипоталамических центров и интеграции этого возбуждения в лимбической системе. Гипоталамическая область является и центром осморецепции, регулирующей прием воды (Andersson, 1967). Это доказывается введением гипертонического раствора поваренной соли в переднюю медиальную часть гипоталамуса или в полость III желудочка (Andersson, 1953). При этом животное (коза) выпивает огромное количество воды.
333
Эти факты свидетельствуют о центральной гипоталамической регуляции приема пищи и воды, которая, однако, непрерывно коррелируется сигналами из внутренней среды организма, а также сигналами из высших отделов центральной нервной системы — коры головного мозга. Последнее может быть доказано образованием многочисленных условных рефлексов., не только направляющих и изменяющих пищевое поведение, но и всю деятельность пищеварительного аппарата.
Важное значение имеют и натуральные условные рефлексы, образованные на разных этапах индивидуального развитая. Так, был описан факт, что щенки, выращенные на молоке, до 6-месячного возраста крайне медленно и «неуверенно» поедали мясо (Цитович, 1911). Эти факты были подтверждены позднее (Бреслав, 1952). Также у ягнят и козлят, выращенных на молочном рационе, до 5-месячного возраста отсутствует акт пастьбы; вид и консистенция обычного корма и у таких искусственно выращенных щенят, и у ягнят не вызывают возбуждения пищевого центра, имеющего условнорефлекторное происхождение.
Насыщение животного или вливание голодному животному «сытой» крови, т. е. крови накормленного животного, вызывает отказ от еды и торможение выделения слюны и желудочного сока (Разенков, 1927).
Большую роль играют интероцептивные раздражения, исходящие из желудка. Так, скорость поедания хлеба у собак по мере насыщения падает, представляя довольно характерную кривую. Промывание желудка непосредственно перед едой новокаином не изменяет скорости акта еды. Раздувание желудка собаки резиновым баллоном (до 500 см3) резко затормаживает акт еды, меньшее раздувание не оказывает на него влияния (Бельская, 1949). Раздувание желудка вызывает раздражение иптероцеиторов его мышечной оболочки.
Роль интероцепторов в регуляции питания и питья ярко показана в исследованиях А. М. Уголева и В. Н. Черниговского (1959). Было установлено, что ритм потребления крысами пищи (в опытах — 40% раствора глюкозы) изменяется после перерезки блуждающих нервов под диафрагмой. Если ставить опыт на сытых животных, эта закономерность не наблюдается, так как в этом случае раствор глюкозы используется в качестве питья,
334
а пе пищи. Суточное количество глюкозы, принятой после перерезки нервов под диафрагмой, оставалось практически неизменным. После двусторонней денервации синокаротидных клубочков потребление глюкозы уменьшалось, а потребление воды (8%-ного раствора глюкозы) резко возрастало. Авторы делают вывод, что рецепторы пищеварительного канала играют преимущественную роль в регуляции режима питания (покрытия энергетических трат организма), а синокаротидная зона — в регуляции поступления воды. Однако этот пример не единственный в области регуляции двигательного поведения в связи с изменениями во внутренней среде организма.
Введение гипертонического раствора хлористого натрия в желудок собаки через фистулу вызывает отказ собаки от питья концентрированных (2,5%, 3,5% NaCl) растворов поваренной соли в молоке (Кассиль, Уголев и Черниговский, 1959). Двусторонняя перерезка блуждающего нерва под диафрагмой снимала этот эффект более чем в половине случаев.
Таким образом, афферентная импульсация с желудка, обнаруживаемая и электрофизиологически, имеет, по-видимому, прямое отношение не только к механорецепции желудка, но и к хеморецепции, в свою очередь изменяющей характер пищевого поведения животного.
Что касается стимуляции пищедобывательной деятельности в природных условиях — стимуляции акта еды, то здесь зависимости очень сложные, физиологический анализ их — дело будущего. Так, совместное пребывание крыс стимулирует количество потребляемого корма (Soulairac et Soulairac, 1954).
Существует ли контроль (регуляция) за калорической стоимостью пищевого материала, поступающего в организм? Бробек (Brobeck, I960) предполагал, что тепло, образующееся в процессе переваривания и ассимиляции принятой пищи, регистрируется термочувствительными структурами промежуточного мозга и таким образом специфическое динамическое действие является важным звеном в регуляции пищевого поведения. Этим взглядам противостояли теории глюкостатическая, липостатическая и протеостатическая, пытавшиеся объяснить насыщение организма и угнетение пищевого поведения накоплением в организме белков, углеводов пли жиров (Мауег, 1955).
335
Ма ньон и Та л лои (Magnen et Та I Ion, 1968) убедительно показали, что кормление крыс ad libitum жирами или углеводами приводит к потреблению их соответственно калорическому эквиваленту. Насколько эти факты могут восстановить значение термогенной теории Бробека, должны показать будущие исследования.
Высказанные выше в значительной мере противоречивые точки зрения на механизм поддержания аппетита и регулирования приемов пищи все же позволяют сделать некоторые обобщения.
Рис. 74. Схема, показывающая общий ход регуляции для четырех основных систем питания. Каждый квадратик перекрывает соответствующий квадратик с другого края, предшествующий и последующий эффекты образуют систему замкнутого контура. Влияния могут оказываться в любую сторону. Прочное соединение всех контуров указывает на центральную роль питания во всех компонентах регуляции (по Gamilton, 1965)
Изменения в крови имеют значение для самого акта приема пищи. Наступающие при этом растяжение и механическое раздражение желудочно-кишечного тракта и изменения обмена веществ (включая и специфическое динамическое действие) определяют новый уровень состояния крови. Это состояние опять-таки вызывает после-дующее состояние пищевого возбуждения и характерный для данного вида цикл пищедобывательной деятельности и приема пищи (рис. 74).
При изучении времени, необходимого для осуществления пищедобывательной деятельности, было устаиов-
336
лево, что животные с хищным типом питания затрачивают на поедание пищи намного меньше времени, чем растительноядные (Слоним, 1962). Особенно продолжительна пастьба копытных в разных условиях и при различной питательности пастбищного корма, его разреженности и т. д. Особенный интерес в этом отношении представляет зимняя пастьба (тебеневка) оленей, связанная с выкапыванием корма из-под снега и требующая больших затрат энергии на пищедобывание (Макарова и Сегаль, 1958).
В эксперименте регламентирование времени поедания пищи приводит к увеличению усвояемости пищи, к ожирению (опыты на крысах) (Leveill а. О’Неа, 1967). Описано также образование стереотипа пищевого поведения у обезьян (Быков и Слоним, 1960). Значение режима питания для огромного большинства организмов еще мало изучено.
Широко распространенное представление о том, что объект питания раздражает своими физическими или химическими свойствами один или несколько анализаторов, в результате чего последовательно наступают акт захватывания пищи, акт еды и процессы пищеварения, очень схематично и практически не имеет реального воплощения в природе. Наиболее близка такая схема к всеядному характеру питания домашней собаки. Всеядность, возникшая в процессе одомашнивания, привела собаку к потере специализированных реакций хищника, к крайнему упрощению пищедобывательного рефлекса (Слоним,1962).
СИМБИОНТНОЕ ПИТАНИЕ И ПИЩЕВАРЕНИЕ
Под симбионтиым питанием и пищеварением понимают участие животных или растительных организмов в обеспечении цитация (или расщепления отдельнЫ/Х пищевых веществ) животного-хозяина. Хотя эти симбионт-ные отношения лучше всего выражены в организме некоторых насекомых (термитов) и млекопитающих (жвачных), их можно наблюдать и у животных, стоящих на более низких ступенях эволюции.
Довольно распространено у низших организмов — кораллов, турбсллярпй, некоторых моллюсков симбионт-
337
нос питание при помощи зооксантелы и зоохлореллы. Одноклеточные зоохлореллы образуют благодаря фотосинтезу крахмал, который затем поглощается подвижными клетками — амебоцитами. Зоохлореллы поглощаются полипами.
Особенно большое значение приобретает симбионтное питание при усвоении целлюлозы. У большинства высших организмов нет фермента, расщепляющего целлюлозу— целлюлазы. Только некоторые организмы, в том числе дождевые черви, способны самостоятельно усваивать целлюлозу. Большинство же улиток (Helix) переваривает целлюлозу при помощи бактерий. Из насекомых жуки-усачи (Cerambycidae), златки (Buprestidae) и жуки-точильщики (Anobiidae) имеют в кишечнике целлюлазу и могут питаться древесиной. Другая группа жуков— рогачи (Lamellicornia), личинки которых тоже питаются древесиной, содержат в кишечнике бактерии, расщепляющие целлюлозу.
Однако симбионтное питание не ограничивается использованием целлюлазы. У жвачных огромная масса инфузорий и бактерий, населяющая рубец, переваривается в сычуге и нижележащих отделах кишечного канала; так удовлетворяется около 1/3 общей потребности животного в белке.
Микроорганизмы рубца способны синтезировать белок из таких простых азотистых соединений, как мочевина, аммонийные соли и даже аммиак. Это служит основанием для использования синтетической мочевины при кормлении сельскохозяйственных животных. Есть основания полагать, что в слепой кишке грызунов происходят аналогичные процессы, и поедание цекотрофов кала служит источником белка, так как в основном це-котрофы состоят из тел микробов и инфузорий. При содержании животных в стерильных условиях для них требуется большее количество белка. То же наблюдается и при исключении цекотрофии.
Пребывание пищи в рубце жвачных характеризуется интереснейшим среди высших организмов примером симбиоза бродильных бактерий, инфузорий с макроорганизмом жвачного. Рубцовое «пищеварение» связано с деятельностью микроорганизмов, расщепляющих клетчатку, и с образованием глюкозы, молочной кислоты и летучих жирных кислот (уксусной, пропионовой, масляной, мура-
338
вьииой и др.). При этом выделяется и значительное количество тепла. Если бактериальная флора рубца обеспечивает химическое расщепление клетчатки путем брожения, то инфузории механически расщепляют клетчатку, содействуя тем самым более полному ее расщеплению. Брожение происходит с образованием большого количества нелетучих жирных кислот и СО2. И те, и другие всасываются в кровь, а углекислота частично отрыгивается. Непрерывная секреция паротндных слюнных желез обеспечивает поддержание в рубце слабощелочной реакции (pH около 8,0), что является оптимальным для протекания бродильных процессов. Фактически размножение микробов и инфузорий приводит к тому, что перевариванию в сычуге и кишечнике подвергается огромное количество микробных тел и простейших, составляющих основной источник усвояемости жвачными азота (Lously, 1949; Renaux, 1955; Ennison, a. Lewis, 1962). Кроме того, некоторые микробы синтезируют аминокислоты из мочевины, а также витамины.
Около 40% энергетического обмена восполняется в организме жвачных за счет микрофлоры и простейших рубца. Кормление корнеплодами, содержащими много углеводов, усиливает процесс перехода растительных белков в животные белки тел микроорганизмов в рубце. Переход на грубые корма у новорожденных жвачных сопровождается появлением инфузорий в рубце. Так, по старым данным, появление инфузорий в рубце ягнят п телят можно наблюдать только с началом кормления сеном. Закономерная динамика нарастания количества инфузорий обнаруживается к 6-месячному возрасту животного с последующим падением числа простейших к 7—8 месяцам, когда их количество достигает величин, характерных для взрослого. Корм задерживается в рубце в течение 5—6 ч (овцы). Реакция содержимого рубца при скармливании углеводов становится кислой, при скармливании грубого корма — щелочной. У овец бродильные процессы приобретают наибольшее развитие через 4 ч после начала пастьбы. Кормовая свекла увеличивает степень брожения и вызывает увеличение количества инфузорий (Ташенов, 1956). По данным той же работы, наименьшее брожение вызывает скармливание животным силоса. Это может объясняться тем, что в силосе уже есть большое количество перебродивших про
339
дуктов расщепления углеводов. Процессы нитрификации в рубце протекают крайне энергично. При этом усваивается как неорганический азот (например, азот аммиака), так и азот мочевины (Виноградова-Федорова, Ми-хина, Павлов, Солдатенков и Трофимова, 1935; Имше-нецкий, 1953; Ташенов, 1956).
Таким образом, в преджелудках жвачных, поскольку у млекопитающих нет ферментов, расщепляющих клетчатку, осуществляется расщепление клетчатки вместе с населяющими преджелудки бактериями и отчасти инфузориями. Здесь обеспечивается переваривание до 45% всей клетчатки, поступившей с пищей. Сетка и книжка служат, по существу, фильтрами, обеспечивающими более полное разложение грубой пищи в рубце и препятствующими переходу ее в сычуг.
Как было сказано выше, переваривание клетчатки обеспечивается не только процессами, происходящими в рубце у жвачных (расщепление клетчатки микроорганизмами), но и расщеплением клетчатки в слепой кишке непарнокопытных и специальным механизмом цекотро-фии у некоторых видов грызунов. Было установлено расщепление целлюлозы в слепой кишке лошади и доказана роль бактерий в этом процессе (Scheunert, 1906).
Переваривание клетчатки у непарнокопытных (лошади) обеспечивается интенсивно развитой слепой кишкой, вмещающей до 37 л содержимого (вдвое больше желудка). По характеру процессов слепая кишка может быть уподоблена рубцу с той разницей, что интенсивность процессов брожения и нейтрализации образующихся при этом жирных кислот происходят за счет выделения кишечной слизи. Частично процессы брожения протекают в толстой (ободочной) кишке. Пища задерживается в слепой кишке лошади до 72 ч, причем переваривается около 40—50% всей клетчатки, содержащейся в химусе. Здесь происходит переваривание химуса ферментами частично растительного, частично животного происхождения, поступающими из вышележащих отделов кишечного канала (Никулин, 1954). Однако и у жвачных слепая кишка является местом интенсивного брожения. Мигрирующие из лимфатических желез в просвет кишечника лимфоциты регулируют развитие флоры кишечника и тем самым процессы брожения (Синельников и Ивкина, 1954).
340
По-видимому, сходные явления длительного задерживания пищевой массы (химуса) в слепой кишке происходят и у некоторых видов грызунов, питающихся зелеными частями растений (полевок, песчанок). Толстая и прямая кишка составляет у домовой мыши 18,6% общей длины кишечника, а у обыкновенной полевки — 38,3% (Рубин, 1953); в заднем отделе кишечника и слепой кишке идут процессы брожения и усвоения клетчатки.
Первые исследования значения слепой кишки для пищеварения у кролика принадлежат Цунтцу (Zuntz, 1905). Морфологические особенности слепой кишки указывают на чрезвычайное развитие этого органа у ряда видов грызунов (травоядные). Наибольшего развития слепая кишка достигает у травоядных форм — полевок. Удаление слепой кишки в асептических условиях приводило к быстрой гибели полевок, снижению веса тела у черной крысы, но не оказывало практически никакого влияния на состояние крыс-пасюков (В. К. Шепелева). Как известно, пасюк в значительной мере питается животной пищей и усвоение клетчатки имеет для этого вида наименьшее значение в ряду грызунов. Отсутствует слепая кишка у зимоспящих видов — сонь, однако для этих животных характерно исключительно зерноядное питание, главным образом орехами и семенами хвойных. У домашних кроликов многие исследователи (Morot, 1882) уже давно обнаружили поедание собственного кала (копрофагию) и скопление кала в области дна желудка. При лишении возможности поедания части собственного кала кролики погибали в течение нескольких месяцев. Копрофагия играет важную роль в пищеварении многих грызунов: крыс, морских свинок и хомяков (Krzywanek, 1927). Кал кроликов состоит из двух частей: более плотной части — мелких шариков (pellets), которые поедаются животными, и более крупных, составляющих обычный кал (Briiggeman, 1937). Было высказано предположение (Morot, 1882), что два вида кала связаны в своем происхождении с однократным или двукратным прохождением его по кишечнику. Исследованиями Мадсена (Madsen, 1939) было установлено, что в кишечнике кролика формируются особые слизистые комки, которые заглатываются животными и откладываются в области дна желудка. На рисунке 75 показано
341
распределение очищенных зернышек винных ягод (точки) и слизистых клубочков-катышков (pellets) в желудке кроликов, убитых в разные сроки введения этих зернышек в опорожненный желудок. Катышки заглатываются животным сразу же после выхода их из анального отверстия. Интересно, что в этих опытах «желудочные
Рис. 75. Контуры желудка кролика (по Olsen a. Madsen, 1944). Точки — распределение семян винных ягод через разные промежутки времени после введения в желудок; кружки — желудочные шарики-катышки; кружки с точками — то же с включением семян
шарики» были обнаружены в прямой кишке через 7 ч после кормления, а в желудке только через 12 ч после кормления. Некоторые из «желудочных шариков» представляли просто скопление слизи, в центре других находилось непереваренное зернышко.
Многократное прохождение пищи через кишечник кроликов доказывается и тем, что введенная в пищу соль меди (CuSO4) циркулирует в кишечнике кролика до 5 недель (Eden, 1941). Такая циркуляция обеспечивает как наиболее полное насыщение содержимого кишечника расщепляющими целлюлозу микроорганизмами, так и циркуляцию и использование образуемого микроорганизмами витамина Bi2-
342
Вторичное пережевывание выделяемых шариков-катышков кроликами и зайцами рассматривалось некоторыми исследователями, как так называемая псевдожвачка, а роль всех этих явлений в усвоении целлюлозы начисто отрицалась. Так как шарики-катышки поедаются животными прямо из анального отверстия, в нормальных условиях видеть эти образования невозможно. Специальные формы эксперимента — деревянные воротники, позволили некоторым исследователям (Olsen a. Aladsen, 1944; Harder, 1950 и др.) не только отделить эти образования от кала, но и установить значение поедания и двукратного прохождения части пищи через кишечник. Олсен и Мадсен обнаружили, что одевание воротника на кроликов ведет к падению веса и привес в процессе развития уменьшается. Они рассматривали эти явления как результат уменьшенного снабжения организма витаминами, которые образуются в результате деятельности бактерий и поедаются животными. Однако более детальные исследования (Harder, 1950; Frank, Hadeler a. Harder, 1951; В. К- Шепелева) установили несомненную роль слепой кишки в происхождении упомянутых выше шариков-катышков.
Хардер приводит довольно противоречивые данные о влиянии выключения цекогрофпи на рост и поддержание веса у грызунов. У лабораторных мышей надевание воротника, исключающего поедание шариков-катышков, быстро приводит к гибели; у кролика в зимний период можно наблюдать значительное падение веса, в летний период при поедании зеленого растительного корма падения в весе почти не происходит. По-видимому, здесь имеют значение и растительные ферменты расщепления клетчатки, содержащиеся в зеленом корме.
Некоторые исследователи считают копрофагию у кроликов (домашних и диких), а также зайцев нормальным физиологическим явлением и критерием полноценности пищи в отношении содержания витаминов группы В (Anghi, 1957; Rodel, 1957).
У бобров поедание кала наблюдается главным образом при питании травой, в меньшей мере — картофелем и еще меньше при введении кефирной подкормки (Шпд-ловская, I960). Автор считает, что копрофагия (цекотро-фпя) обеспечивает витаминный обмен в организме, главным образом витаминов группы В (В12). Поедание так
343
называемого мягкого кала (цекотрофа) бобрами происходит непосредственно из клоаки.
На основании детального исследования поедания катышков, их химического состава выдвинута теория цеко-трофии, т. е. значения образующихся в слепой кишке шариков-цекотрофов в пищеварении грызунов (Harder, 1950). Цекотрофы содержат значительное количество белка, большое количество бактериапьных тел (кал — 2700 млн. бактериальных тел, цекотрофы — 95G0 млн.). Цекотрофы, формирующиеся в слепой кишке, поедаются затем животными сразу после выхода из анального отверстия и откладываются в слепом конце желудка. Оттуда отдельными порциями они поступают в кишечный канал и поддерживают на протяжении всего пищеварительного канала расщепление целлюлозы. Наиболее развит (да и лучше всего изучен) механизм цекотрофии у кролика, хотя есть все основания придавать ему большое значение у всех без исключения травоядных грызунов.
ВЫПАДЕНИЕ ПИЩЕВЫХ РЕФЛЕКСОВ
И ФИЗИОЛОГИЧЕСКОЕ ГОЛОДАНИЕ
Питание животных, как сказано выше, отнюдь не всегда ведет к полному уравновешиванию прихода — расхода вещества и на коротких отрезках времени (иногда в пределах целых сезонов) можно наблюдать или накопление или расходование пищевых резервов организма. С этой стороны особенный интерес представляет физиологическое состояние, когда организм на длительное время полностью (или частично) прекращает прием пищи и текущие энергетические затраты осуществляются исключительно за счет ранее накопленного вещества.
Впервые на состояние физиологического голодания обратил внимание В. В. Пашутин (1902), рассматривавший явления зимней спячки и зимнего сна. Как уже указывалось (Слоним, 19616), одно из первых проявлений зимней спячки заключается в полном выпадении пищевых рефлексов, прекращении питания и пищедобыва-телыюп деятельности. По существу, нельзя проводить полную аналогию между состоянием зимней спячки и
344
состоянием физиологического голодания, так как в спячке одновременно с прекращением приемов пищи наблюдается и резкое ограничение общего обмена веществ, т.е. сокращение расходования пищевых ресурсов. Гораздо ближе к состоянию подлинного голодания зимний сон некоторых хищных млекопитающих (медведя, барсука, енотовидной собаки), не сопровождающийся значительным снижением температуры тела и обмена веществ. Отсюда очень большая потеря в весе этих животных, резкие сдвиги во всей регуляции обмена веществ и пищеварения. Некоторое снижение обмена при зимнем сне возникает, ио-видимому, за счет почти полного выпадения специфического динамического действия пищи (обнаруживаемого в том случае, если животное удается покормить; Слоним и Щербакова, 19496).
Состояние органов пищеварения во время физиологического голодания почти совершенно не изучено. Известно, что при глубокой зимней спячке (температуре тела около 17—18°С) у сусликов выделяется кислый желудочный сок — pH 3,5—3,8. Это явление связывается с необходимостью поддержать щелочно-кислотное равновесие в организме при резком ограничении легочного дыхания и мочеотделения (Персон, 1950; Valentin, 1857).
Удалось изучить некоторые особенности функции желудка у нескольких видов тюленей во время лактации, когда эти животные в течение длительного времени (до 6 недель) не принимают пищи (Шепелева, 1968). На только что убитых на льдинах Гренландского моря тюленях изучались pH содержимого желудка, характер содержимого и титруемая кислотность. У непитающихся л актирующих самок тюленя-хохлача (Cystophora crista-ta) pH желудочного содержимого колеблется от 5,0 до 6,0. В отдельных случаях можно наблюдать и очень высокие величины pH, связанные с забрасыванием дуоденального содержимого. Следовательно, у непитающихся животных в процессе голодания устанавливается стойкая щелочная реакция в желудке. Поддержание щелочно-кислотного равновесия, несмотря на обильное выделение молока и связанную с этим потерю щелочей, происходит за счет изменений дыхания и мочеотделения. У морского зайца (Erignatus barbatus), по данным той же работы В. К. Шепелевой, при питании обнаружено
345
уже значительное содержание кислот в желудке и pH желудочного содержимого колебался от 1,0 до 4,0. Следовательно, состояние физиологического голодания приводит не только к выпадению пищевых и ппщедобыва-тельных рефлексов (отказу от пищи), ио и к глубокому изменению реакции пищеварительных желез. Эго голодание особого рода, не сопровождающееся повышением возбудимости пищевого центра. Центральный физиологический механизм такого голодания можно рассматривать как проявление отрицательной индукции с других центров и формирование на такой основе соответствующей «отрицательной пищевой доминанты». Однако эти общие физиологические соображения требуют для своего подтверждения специальных исследований.
ЭНЕРГЕТИЧЕСКИЙ РАСХОД ОРГАНИЗМА
Пойкилотермные животные. Потребность в пище и ее удовлетворение тесно связаны с энергетическими затратами, в свою очередь зависящими от ряда факторов внешней среды, из которых главное место занимает температура. Кроме того, энергетические затраты животного зависят от размеров тела, подвижности, ряда сложных биологических циклов (суточного, сезонного, лунного и т. д.), стадии развития в онтогенезе, условий питания, а также от наличия других организмов того же вида. Эти очень сложные связи не могут быть сведены к какой-либо элементарной схеме, тем более, что безусловно не существует никакой зависимости между уровнем энергетических затрат (потреблением кислорода) и уровнем эволюционного развития (Hemmingsen, 1950).
Наиболее общая зависимость, которая здесь наблюдается,— это правило поверхности тела, установленное впервые для гомойотермных организмов и затем широко исследованное для пойкилотермных (Hemmingsen, 1950). В основе этого правила (закона) лежит факт, что между весом тела и уровнем энергетического обмена есть логарифмическая зависимость. Так как поверхность тела представляет собой экспоненциальную функцию веса тела, то при изображении такой кривой в логарифмическом масштабе можно наблюдать прямую, наклон которой зависит от возрастания уровня обмена в связи
346
с весом тела. На рисунке 76 приведены данные этих зависимостей, полученные для одноклеточных (пойкилотермных и гомойотермных) организмов. Ясно видно, что увеличение размеров (веса) организма приводит к относительному снижению общего обмена веществ. Это наблюдается на бактериях, жгутиконосцах, амебах, инфузориях hi яйцах морских животных и насекомых.
Рис. 76. Потребление кислорода у одноклеточных пойкилотермных и гомойтермных организмов в зависимости от веса тела (по I lemmingsen, 1959)
Объяснение этому обычно видят в способности кислорода проникать внутрь свободно живущей клетки. Однако у многоклеточных интенсивность обмена уже зависит от поверхности.
Появление органов дыхания еще более усложняет положение (Krywienczyk, 1956). При жаберном дыхании моллюсков (Lamellibrcmchiata), ракообразных (Isopo-da) потребление кислорода зависит or поверхности гела. У насекомых, дышащих через трахеи, окислительные процессы в большей степени зависят от веса тела. Тем не менее для пойкилотермных организмов, как и для гомой-
347
отермпых, обмен веществ в покое может быть рассчитан по формуле
M = KWb,
где М— общий обмен веществ (кал), V7— вес тела, b — константа, отличающая наклон кривой, полученной в эксперименте, от кривой соотношений веса (по оси абсцисс) и теоретического потребления кислорода (по оси ординат) в логарифмических выражениях. Константа К показывает уровень окислительных процессов, на котором ведется отсчет. Для млекопитающих она равна примерно 10. Величина b обычно меньше 1,0 и у млекопитающих равна 0,70. У пойкилотермных величина b также меньше 1,0. Так, для амфибий она равна 0,738, для моллюсков и равноногих раков (Isopoda) около 0,66, для насекомых около 1, что означает, что закон поверхности тела для них практически отсутствует (Bertalanffy, 1949). Впрочем, у насекомых разного веса эти отношения оказываются различными. Так, у жуков-чернотелок (Те-nebrionidae) метаболизм возрастает на единицу веса до 5 мг, затем быстро снижается до 45 мг и остается постоянным до 140 мг (Chauvin, 1956). У насекомых с полным превращением b приближается к 1,0, а у насекомых с неполным превращением этот коэффициент ниже. Так, у тараканов и палочников он равен примерно 0,8 (Enger a. Savalov, 1958). Для очень мелких беспозвоночных весом менее 40 мг коэффициент b равен 0,95 (Zeuthen, 1953). У более крупных ракообразных значение b снижается, т. е. выявляется правило поверхности тела.
В последнее время выявлены зависимости между интенсивностью дыхания и содержанием азота в теле животных (Zeuthen, 1953). С увеличением содержания азота интенсивность газообмена снижается. Это хорошо выявлено у рыб, ракообразных и насекомых.
Механизмы, лежащие в основе 'изложенных выше зависимостей, неясны. В исследованиях дыхания тканей установлена обратная корреляция между интенсивностью Q О2 птиц и размерами тела (Bertalanffy a. Pirozynski, 1951). Некоторые ткани (печень, мозг, мышцы) дышат с одинаковой интенсивностью у рыб разного веса (Vern-berg, 1954).
Вместе с тем наблюдается четкая зависимость между количеством митохондрий на вес ткани и общим обменом
348
веществ. Это же относится и к содержанию некоторых окислительных ферментов (сукциноксидазы, дегидрогеназы) (Prosser a. Brown, 1962).
Таким образом, у пойкилотермных организмов такая же тенденция к зависимости уровня обмена от поверхности тела, как и у гомойотермных. Однако очень большое значение приобретает механизм дыхания — снабжение кислородом отдельных тканей, что приобретает особое значение и в связи с тем, что большинство пойкилотермных организмов подчиняет уровень окислительных процессов содержанию кислорода в окружающей среде (конформация дыхания), т. е. потребление кислорода зависит от его доставки (Tang, 1931). Особенно ярко это выражено у беспозвоночных, не имеющих дыхательных пигментов. Впрочем, у многих форм пигмент насыщается в зависимости от содержания кислорода в среде и потому тип дыхания становится зависимым (конформируемым).
Г о м о й о т е р м н ы е животные. Общая величина энергетических затрат организма складывается из затрат энергии в полном покое, дополнительного расхода энергии на пищеварение, всасывание, ассимиляцию, дополнительного расхода на терморегуляцию, расхода на поддержание положения тела, сопротивление току воды и т. п. затраты энергии в полном покое — основной обмен.
Понятие об основном обмене, разработанное главным образом для человека, лишь с большим трудом может быть применено к животным и особенно к тем формам, у которых процессы пищеварения и всасывания идут непрерывно. Именно для этих животных состояние голодания (состояние «натощак») не является физиологическим и достигается уже в результате настоящего голодания с распадом собственных тканей тела. Так обстоит дело, например, у всех копытных. Некоторые грызуны (кролики, зайцы) при лишении их пищи также очень долго сохраняют в желудке пищевые массы. По-видимому, это касается и многих других травоядных млекопитающих. Изучение основного обмена v всех этих животных возможно только на какой-то стадии пищеварения и всасывания и, следовательно, сравнивать их с хищными, обезьянами и человеком как с организмами с фазовым типом пищеварения и всасывания очень трудно.
349
Другой стороной рассматриваемого вопроса является мышечная деятельность. Для многих мелких видов млекопитающих и птиц исключить двигательную активность практически невозможно. Многие мышевидные грызуны, некоторые мелкие хищные (хорьки, ласки, виверровые), воробьиные птицы очень редко остаются в состоянии покоя, а при исключении мышечной деятельности у них значительно снижается температура тела.
Учение об основном обмене — следствие работ дерпт-ских физиологов середины XIX столетия Биддера и Шмидта, установивших у собаки удивительно постоянный уровень метаболизма изо дня в день. Эти факты были подтверждены большим числом исследователей.
Под основным обменом всегда понимают величину энергетических затрат (в единицу времени), отнесенную к весу тела животного. Таким образом, величины основного обмена всегда отражают интенсивность тканевых окислительных превращений. Общеизвестно, что уровень обмена веществ, рассчитанный на единицу веса тела, резко падает с увеличением веса тела животного, с уменьшением его относительной поверхности. Поскольку последняя является важным фактором, определяющим величину теплоотдачи животных, возник важнейший биологический принцип, получивший название закона поверхности тела. Впервые положение о значении поверхности тела для уровня обмена веществ было сформулировано более 100 лет назад (Sarrus et Rameaux, 1837, 1838; Rameaux et Sarrus, 1838, 1839). Эти исследователи впервые сделали вывод, что уровень теплоотдачи зависит от величины «свободной поверхности животного», а потребление кислорода пропорционально количеству вдыхаемого воздуха. Отсюда был сделан вывод, что потребление кислорода пропорционально потере тепла и, следовательно, зависит от соотношений веса и поверхности тела. Интересно, что эти авторы включили в число факторов, определяющих поверхность тела, еще и общую поверхность легких и количество дыханий. Саррю и Рамо представили взаимоотношения возрастания веса и снижения при этом числа пульсовых ударов и частоты дыханий, продукции и потери тепла по формуле:
У = а\\Г7 2/3,
350
где У— величины перечисленных физиологических функций, а — константа для каждого животного, U’’—вес тела (2). Таким образом, первые положения о законе поверхности связывались с уровнем основных физиологических функции животных. Дальнейшие исследования ряда физиологов подтвердили эти положения о связи величин обмена веществ и величин, характеризующих легочное дыхание и кровообращение.
Важным этапом в развитии представлений о роли поверхности тела организмов в регуляции их жизнедеятельности явились положения Бергманна (Bergmann, 1847—1848). Бергманн, впервые употребивший термины «гомойотермный» и «пойкилотермпый», обратил внимание на то, что первые от полярных стран к тропическим (у близких в систематическом отношении видов) имеют все меныпие размеры (вес) тела; у пойкилотермных организмов, наоборот, тропические формы обладают большими размерами тела, чем обитатели северных широт. Эти закономерности важны не только для эколого- и зоогеографических сопоставлений, но и служат основанием для физиологического истолкования поверхности тела как важнейшего фактора, определяющего обмен веществ в организме.
Правило Бергманна было использовано для физиологического исследования обмена веществ на собаках (Rubner, 1883) и кроликах (Richet, 1889). На сельскохозяйственных животных и птицах было выявлено значение поверхности тела для интенсивности охлаждения и повышения обмена веществ при низких температурах среды (Kleiber, 1932).
Формулировка закона поверхности тела принадлежит Рубнеру (Rubner, 1883). Он обосновывал закон поверхности тела тем, что при расчете на единицу поверхности тела обмен веществ собаки остается почти постоянным, тогда как при расчете на единицу веса он резко изменяется. Положения основывались на исследовании теплопродукции в полном покое и в голодном состоянии у 7 собак весом от 31 до 3 кг (табл. 32). Такие же явления наблюдаются и при сопоставлении теплопродукции у животных разных видов (табл. 33).
На рисунке 77 приведены данные величин основного обмена (Benedict, 1938), нанесенные на полулогарифмическую шкалу. Из схемы видно, что обмен веществ, рас-
351
Таблица 32
Теплопродукция у собак различною веса (по Rubner, 1883)
Собаки (№) Вес тела (кг) Тсплопротукция (кал за 24 ч)
на 1 кг на 1 м~ поверхности тела
1 31,20 35,68 1036
2 24,00 40,91 1112
3 19,80 15,87 1207
4 18,20 46,20 1097
5 9,61 65,16 1183
6 6,.'.О 66,07 1153
7 3,19 88,07 1212
Таблица 33
Теплопродукция у животных разных видов
Объект В.с тела (кг) Теплопротукция (кал за 24 ч)
на 1 кг веса тела на 1 м* п верхно-сти тела
Лошадь 441 11,3 948
Свинья 128 19,1 1078
Человек 64,3 32,1 1042
Собака 15,2 51,5 1039
Кролик 2,3 75,1 776
Г усь 3,5 66,7 969
Курица 2,0 71,0 943
считанный на единицу поверхности тела, возрастает очень медленно от самых маленьких животных (мышь, воробей и т. д.) к средним (до 100 кг), но затем у крупных животных (копытных — пони, першерона, быка, слона) — очень резко. Следовательно, относительно очень малая поверхность тела не является фактором, снижающим уровень окислительных процессов. Для протекания жизненных процессов гомойотермного организма существует какой-то минимум, связанный, по-видимому, с воз-
352
мощностями ферментативных реакций и с процессами возбуждения, ниже которого в нормальных условиях жизни обмен веществ понижаться не может (Слоним, 1952). Поэтому очень крупные гомойотермные животные в условиях наземного существования представляют значительную редкость (другое дело — водные организмы,
Рис. 77. Соотношения между пульсом, уровнем обмена и величиной тела у млекопитающих (по Brody, 1945). По оси ординат — отношение потребления О2 (мл) к частоте пульса; по оси абсцисс — вес тела (кг)
у которых теплоотдача в воде значительно более интенсивна), а уровень обмена веществ у них, рассчитанный на поверхность тела, оказывается в значительной мере сниженным по сравнению с животными среднего веса, представляющими огромное большинство ныне живущих видов. Вместе с тем отношение уровня основного обме-
12—1204
353
на и частоты пульса у гомойотермпых организмов удивительно постоянны (см. рис. 76) и нанесенные на логарифмическую шкалу (по весу тела) представляют такую же прямую зависимость, как и величина тела и потребление кислорода у пойкилотермных.
Были изучены газообмен и некоторые физиологические функции у самых маленьких животных — землероек весом около 3,5 г. В таблице 34 дано сравнение самых маленьких млекопитающих с животными среднего и крупного веса.
Таблица 34
Основной обмен у различных млекопитающих (по Morrison a. Pearson, 1947)
Объект Вес (кг) Основной <бмен (кал) на:
кг/сутки м*/сутки
Бык 500 12 1090
Человек 63 2 920
Макак 4,2 49 670
Морская свинка .... 0,41 86 710
Мышь белая 0,021 170 330
Землеройка (Blarina) 0,020 370 1020
Землеройка (Sorex) . . 0,0031 830 1570
Согласно этим данным, землеройки двух исследованных родов Blaritia и Sorex сильно отличаются по своим физиологическим особенностям от близких им по весу тела грызунов. Авторы объясняют это чрезвычайной прожорливостью землероек, не могущих оставаться без пищи дольше 10—11 ч.
У крупных животных ритм сердечных сокращений уменьшается в связи с весом животного (Brody, 1945; Аршавский и Пяткин, 1958). Изменение сердечной деятельности крупного рогатого скота в онтогенезе связано с развитием и возбудимостью антигравитационной мускулатуры. Зависимости между весом и поверхностью тела и частотой пульса и дыхания не менее сложны, чем эти же зависимости при основном обмене веществ.
Закон поверхности тела может быть прослежен на разных этапах развития млекопитающих; его нельзя 354
установить, вероятно, только для летучих мышей (Слоним, 1952). У Chiroptera летательные перепонки, периодически наполняющиеся кровью и периодически — во время состояния гомойотермип и полета — отдающие значительное количество тепла, в состоянии пойкилотер-мии и дневного сна не принимают сколько-нибудь значительного участия в теплоотдаче. В таблице 36 приведена теплопродукция у летучих мышей (2 вида) в состоянии гомойотермип в сопоставлении с теплопродукцией желтогорлой мыши (Apodemus jlavicollis), близкой ей по весу.
Таблица 35
Газообмен и теплопродукция у летучих мышей и желтогорлой мыши
(по С лини му, 1952)
Вит Вес тела (г) Поверхность тела (с.и5) Вы пихаемый СО2 (слР.'.иии) Гсплоот ia<ia (кал)г)
на 1 кг па I .w2
Подковонос (Rhynolop-lius terrium equinum) . . 22 451,4 0,95 12,90 27,5
Большая ночница (Myotis myotis) .... 26 504,6 1,02 11,99 23,8
Желтогорлая мышь (Apodemus flavicollis) 20 66,3 2,3 345,0 520,0
Несмотря на большую поверхность тела летучих мышей, их основной обмен при температуре среды около 30° С в гомойотермном состоянии оказывается более низким, чем у мыши. Это объясняется тем, что температура большей части поверхности тела летучей мыши очень низка — почти совпадает с температурой внешней среды. Условия кровообращения в летательных перепонках летучей мыши таковы, что в покое (при определении основного обмена) отдача тепла происходит почти исключительно с поверхности туловища.
Правило Бергманна, хорошо подтвержденное современными исследователями (Allen, 1906; Botticher, 1915; Rensch, 1932, 1939; Лукин, 1940), в известной мере отражает значение климатического фактора для формирова
12*
355
ния высших организмов. Вместе с тем вся организация животного — его относительный вес и длина кишечника, поверхность легких, вес сердца, печени и т. д. тесно связана с основными жизненными процессами в организме, с их интенсивностью. Эти особенности развиваются в процессе эволюции или параллельно, или в тесной зависимости от основных энергетических потребностей животного организма. Способность организма отдавать и терять тепло в окружающую среду определяет в значительной мере тепловой баланс организма, а потому величина поверхности тела в установлении уровня общей жизнедеятельности гомойотермных организмов несомненно играет выдающуюся роль.
Рассмотрение большого материала внутри отдельных отрядов (близких в систематическом отношении видов) позволяет выявить значение закона поверхности тела как общей закономерности, свойственной всем без исключения организмам. Однако на эту зависимость накладываются влияния экологического порядка, связанные с особенностями питания, пищедобывательной деятельностью и с длительным воздействием температуры среды обитания.
Глава IX
АДАПТАЦИЯ К ПЕРЕДВИЖЕНИЯМ
И МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ
ОКРУЖАЮЩЕЕ ОРГАНИЗМ ПРОСТРАНСТВО
КАК ФАКТОР СРЕДЫ
Существование организмов в окружающей среде невозможно без мышечной активности, т. е. без преодоления этого пространства или без двигательных актов, приводящих организм в контакт с пищей, изменяющих содержание газов в теле, без оборонительных реакций и т. п. Поэтому, строго говоря, далеко не все мышечные двигательные реакции животных связаны с перемещением тела в окружающем пространстве, но все они направлены на преодоление этого пространства. Последнее можно наблюдать даже у сидячих организмов — губок, коралловых полипов, актиний, асцидий.
Двигательные акты, обусловливающие перемещение животных в окружающей среде, чрезвычайно многообразны. Передвижение в воздушной среде — полет, в водной— плавание, в плотной (земля)—роющая деятельность, передвижение по поверхности — скольжение, бег, ходьба, ползание — вот далеко неполный перечень типов передвижения многоклеточных организмов. Учитывая и сложные акты передвижения у высших форм, и двигательные акты простейших (жгутиковых, реснитчатых, амебовидных), едва ли можно создать какую-либо рационально построенную классификацию.
Однако, как правило, физиологические особенности мышечной деятельности различных по уровню организации и систематическому положению организмов тесно связаны с их образом жизни, особенностями пищедобывательной, оборонительной, миграционной и воспроизводительной деятельности. Именно эти основные биологические стимулы легли в основу формирования не только структуры двигательных актов, но и обеспечения их
357
энергетическими ресурсами, циркуляцией, дыханием и т. д. Поэтому адаптивные изменения мышечной деятельности и двигательной активности охватывают большинство видов и в какой-то мере создают морфологически однотипные жизненные формы. Это можно видеть прежде всего при изучении структуры конечностей, типы которых (бегающий, роющий, прыгающий, плавающий, летающий) в известной мере сходны у видов, принадлежащих не только к различным отрядам, по и к разным классам животных.
На морфологических особенностях строения конечностей отражается, однако, не только субстрат (среда), по и самый параметр перемещения, характерная для вида скорость, попутная деятельность, характер стимулов, определяющих эту форму деятельности (ориентировочная, оборонительная, пищевая и т. д.).
ОСОБЕННОСТИ ПЕРЕДВИЖЕНИЯ
(ПРЕОДОЛЕНИЯ ПРОСТРАНСТВА) В ВОДНОЙ СРЕДЕ
Передвижение в водной среде (плавание) лучше всего изучено у водных позвоночных (рыб и млекопитающих), так как их движения, связанные с деформациями тела, лучше всего могут быть подвергнуты биодинамическому и математическому анализу (Cray, 1933; Bainbridge, 1961, Шулейкин, 1966 и др.).
При изучении этих явлений можно наметить три аспекта: 1) изучение морфологии рыбы —формы тела, обеспечивающей наименьшее сопротивление в различных условиях обитания; 2) изучение механики движения — математический анализ двигательных актов, построенный на данных киноциклосъемки; 3) изучение энергетических затрат при плавании с различной скоростью, явлений, связанных с производимой животным мышечной работой. Сочетание всех этих исследований позволяет вычислить для плавающих организмов коэффициент полезного действия и найти оптимальное значение скоростей в связи с величиной организма и особенностями биодинамики.
Эколого-морфологические особенности водных организмов выражены очень ярко. Они касаются в равной мере как формы тела рыб, живущих в различных условиях, разного типа питания, так и различных по этим
358
Рис. 78. Различные типы формы тела рыб (по Никольскому, 1963): 1—стреловидный (с; рган), 2—торпедовидный (скумбрия), 3—сплющенный с боков, ле-щевиднын (обыкновенный лещ), 4— тип рыбы луны (луна-рыба), 5—тип камбалы (речная камба-
ла), 6— змеевидный (угорь), 7—лентовидный (сельдяной король), 3—шаро-ви 1НЫЙ (кузовок), 9—плоский (скат)
же признакам морских млекопитающих. На рисунке 78 приведена различная форма тела рыб; передвижение их в воде значительно отличается и связано с условиями среды и скоростью плавания. Так, торпедовадная форма обеспечивает наиболее быстрое передвижение в открытой воде, сплющенная с боков — возможность плавания в водоемах с густой растительностью; впрочем, этот тип формы тела встречается и среди живущих в толще воды, и среди придонных и донных. Змеевидный тип тела характерен для рыб зарослей и зарывающихся в ил. Медленно плавающие рыбы имеют лентовидную форму тела, шаровидную и плоскую.
Особое положение занимают в этом своеобразном ряду летучие рыбы, с громадной скоростью движущиеся в воде и завершающие этот двигательный акт своеобразным полетом в воздухе. Здесь большое значение приобретают не только змеевидные движения туловища, но и большие боковые плавники, обеспечивающие своеобразное парение в воздухе. Характерно, что до момента полного отрыва летучей рыбы от воды скорость благодаря движениям хвоста продолжает нарастать. В воздухе летучая рыба держится около 10 сек, пролетая за это время около 100 м (Никольский, 1963).
Своеобразной адаптацией рыб к существованию в водной среде и к перемещению на различные глубины является способность к изменению удельного веса тела. Это осуществляется при помощи плавательного пузыря, сокращение которого позволяет изменять количество газа в пузыре или давление в нем. Наполнение плавательного пузыря происходит либо за счет заглатывания воздуха (у открытопузырных и у всех рыб на ранних стадиях постэмбрионального развития) или за счет выделения кислорода и СОг по типу секреции специальным образованием — красным телом. У рыб, живущих в толще воды и быстро передвигающихся по вертикали (акулы, скумбриевые), плавательный пузырь отсутствует вовсе. Плавательный пузырь связан у рыб со слуховым лабиринтом и давление в пузыре регистрируется в центральной нервной системе.
Приведенными примерами, конечно, не исчерпывается ряд морфологических адаптаций рыб, обитающих в морях, реках и других водоемах (Никольский, 1963; Строганов, 1964).
360
Скорость перемещения рыб в воде очень различна. Наибольшей скоростью обладают некоторые акулы, тунцы, меч-рыбы, лососевые. Так, тунец перемещается со скоростью 20 м/сек, голубая акула— 10 м/сек, лосось — 5 м/сек (Никольский, 1963). Поскольку скорость движения зависит от длины тела, для сравнения рыб различного размера принят так называемый коэффициент скорости, рассчитываемый по формуле—где V — абсо-
Г-
лютная скорость, J L — корень квадратный от длины рыбы. Получаемые коэффициенты колеблются от 70 (акулы, тунцы) до 5 (луна-рыба). В таблице 36 приведены некоторые данные скоростей плавания рыб разных видов.
Таблица 36
Скорость передвижения (плавания) водных организмов
Животное MjCCK
Барракуда (Sphyraena picuda) Голубая акула (Carcharias glaucus) Тупец (Thunnus thunnus) Лосось (Salmo salar) Форель (Salmo trutta morpha fario) Щука обыкновенная (Esox Indus) Треска (Gadus morhua) Плотва (Rutilus rutilus) Лещ (Abramis brama) Налим (Lota lota) Морской окунь (Sebastes marinas) 12,2 10,1 6,2 6,0 3,6 3,5 2,5 1,2 0,9 0,6 0,4
Стандартная скорость плавания рыб принимается обычно как 4 А. У мелких рыб при бросках эта скорость может намного превышать отношение к длине тела. Так как сопротивление движению пропорционально квадрату линейных размеров, а мощность (масса) мышц — кубу линейных размеров, крупные рыбы движутся при прочих равных условиях быстрее мелких.
Некоторые морские млекопитающие отличаются большой скоростью плавания. Так, для кита-сервала описана скорость 6 м/сек, для дельфинов до 10 м/сек и более.
361
Скорость передвижения в воде зависит от формы тела, характера движений туловища, хвоста и плавников, а также от гидродинамических особенностей тела рыбы, т. е. от сопротивления, оказываемого водой при продвижении вперед. Между скоростью плавания рыбы и ее гидродинамическими свойствами существует значительный параллелизм (Шулейкин, 1966). Так, наименьшее сопротивление при движении оказывает вода телу дельфина, наибольшее медленно плавающей рыбе — морскому языку.
Гидродинамические свойства плавающих организмов определяются, однако, не только формой тела, но особенностями его поверхности. Помимо гладкой поверхности, почти не создающей трения «I вызывающей минимальное образование пограничного слоя движущейся вместе с плывущим телом воды, здесь большое значение имеет выделение слизи. Она выделяется специальными бокаловидными железами и имеет большое значение для увеличения коэффициента полезного действия плавающего животного. Мягкий демпферный слой подкожной жировой клетчатки у дельфинов также выполняет функцию гашения турбулентности.
Плавание осуществляется благодаря движениям тела, при которых поперечные колебания головы оказываются меньшими по сравнению с колебаниями хвоста. Большой движущей силой обладает хвост; он перемещается из стороны в сторону, проделывая синусоидный путь. Само тело рыбы в зависимости от вида в большей или меньшей степени проделывает синусоидные движения. Силы, действующие при сокращении туловища и движениях хвоста, изгибают хвост, но тем не менее хвост образует угол с движением тела по воде. Поэтому на хвост действуют и сила сокращения, и упор о воду. Равнодействующая этих сил направлена вперед. Когда хвост перемещается влево, равнодействующая направлена вправо и наоборот, но средняя сила, измеренная на всем протяжении сокращения цикла, направлена точно вперед.
Мощность общей работы, выполняемой телом плывущей рыбы, является произведением скорости и силы сопротивления движению. Эта сила в свою очередь складывается из силы сопротивления воды движению сложно изгибающегося тела рыбы и силы внутреннего сопротивления изгибу тела. Кроме того, затрачивается энергия 362
на сопротивление распространяющейся вдоль тела волны.
Специальные исследования показали, что каждый сегмент тела рыбы обладает различной мощностью, и это соответствует и анатомической толщине расположенных на отдельном сегменте мышц (Шулейкин, 1968). Мышечный тяж, идущий вдоль тела рыбы, обладает в различных его участках (сегментах) разными биохимическими особенностями. Следовательно, плавание рыбы представляет исключительно интересный объект для изучения эволюции адаптаций мышечной системы к выполнению различных нагрузок.
Энергетический обмен у рыб при плавании был объектом многих работ отечественных и иностранных исследований. Поскольку многие рыбы существуют в проточной воде, плавание как способность противостоять течению является у них функцией непрерывной. Отсюда возникло понятие о так называемом активном обмене — обмене в толще движущейся воды (Ивлев, 1960). Эти величины активного обмена составляют около 200% от уровня обмена веществ в неподвижной воде.
Исследование мышечной деятельности (плавания) рыб производится в специальных гидродинамических камерах, где можно создавать различную скорость течения воды, поддерживать постоянную температуру и измерять содержание кислорода в воде (Fry a. Hart, 1948; Blazka a. oth., 1960; Brett, 1964), содержание углекислоты. Подобно исследованиям, проведенным на наземных организмах, главным образом на человеке, энергетическая оценка движения — плавания производится путем вычитания из величин газообмена, наблюдаемых во время плавания, величин так называемого стандартного обмена— более низких величин газообмена в состоянии полного покоя рыбы. Подобно фактам, хорошо изученным на человеке, потребление кислорода при плавании с достаточно большой скоростью не сопровождается полным удовлетворением кислородного долга, что установлено как при сопоставлении теоретически вычисленных величин потребления кислорода с наблюдаемыми фактически, так и при изучении величин кислородного долга (Brett, 1964; Smith, 1965). С возрастанием скорости газообмен повышается, однако имеется определенный энергетический оптимум скорости, когда газообмен, рассчи-
363
Плавание
Покой
<з
* Oj
1200
Запрос и обеспечение
Накопление
ь Кислородный''
О, /«
1000
Обмен во время работы
800
600
№0
200
too-
100-\
80-
Стандартный обмен
20
У1" Утомление ^Нормальная—" б нагрузка
Погашение долга
Коллапс
Возобновление работы
Обмен во время работы
Время юмле> /мин)
2
Кислородный
Стандартный обмен
—L
1
Время восстановления
। Спонтанная । активность
Скорость плавания (1/сек)
5
В
%
1
?
4
5
Рис. 79. Взаимоотношения между потреблением кислорода, скоростью движения (плавания), временем работы до утомления, кислородным долгом и восстановлением у красной нерки при температуре воды 15° С (по Brett, 1964)
танный на пройденный путь, оказывается наименьшим. Этим величинам ряд исследователей придают большое значение, предполагая, что именно эти наиболее «экономные» скорости используются рыбами в процессе дальних миграций.
Одной из характерных черт мышечной энергетики рыб является очень длительная (после работ, проведенных до утомления) ликвидация кислородного долга, продолжающаяся более 3 ч. Нагрузка, при которой рыба может продолжать работу в устойчивом состоянии (steady state), не превышает величины 4 линейных ее размеров (4 Л) в секунду (Brett, 1964). На рисунке 79 приведены отношения потребления кислорода, скорости утомления и восстановления у нерки (Oncorhynchus nerka) весом 50 г.
Таблица 37
Потребление кислорода и скорость плавания при различных температурах среды у нерки
(по Brett, 1964)
Температура сре чы (°C) Вес тела Потребление О5 (мг!кг ч) при работе Потреблен не О2 (мг кг, ч) стандартный обмен Критическая скорость плавайия
о!>т сек 1'сек
5 37,6 514 41 1,76 3.26
10 32,9 627 60 1Л0 3,65
13 55,2 895 71 2,52 4,12
20 62,6 852 120 2,43 3,90
24 32,2 848 196 2,23 3,75
В последнее время для изучения энергетических затрат плавающих рыб стали применять косвенный метод расчета по содержанию жира в теле. Эти пути исследования имеют большое значение при изучении миграций.
Весьма интересные данные по плаванию рыб получены при различной температуре воды. Максимальная скорость плавания с повышением температуры воды возрастает. При этом стандартный метаболизм увеличивается с 41 до 196 мг1кг1ч\ расход же на плавание — с 514 до 948 мг!кг!ч, т. е. менее чем вдвое. Эти результаты полу
365
чены после двухнедельной адаптации к температурам от 5 до 24° С и указывают па значительную независимость коэффициента полезного действия скелетной мускулатуры рыбы от температуры среды и от предшествующих температурных воздействии, да и скорость плавания не возрастает с температурой выше 15°С.
Несмотря на большой интерес, проявляемый в последнее время к изучению морских млекопитающих, особенно дельфинов, какие-либо данные о физиологии плавания отсутствуют. Имеются только предположительные расчеты, исходя из наблюдений за скоростью и весом тела, свойствами поверхности тела.
ПЕРЕДВИЖЕНИЕ НАЗЕМНЫХ ОРГАНИЗМОВ
Мышечная активность наземных млекопитающих отличается исключительным разнообразием и включает, кроме собственно перемещения тела, многие формы «деятельности».
Низшие млекопитающие — однопроходные, сумчатые и неполнозубые — представляют в этом отношении крайне пеструю картину. Среди однопроходных утконос сравнительно малоподвижен на суше и более подвижен в воде; он хорошо плавает, ныряет и роет землю. Мышечная сила утконоса (особенно спинных мышц) очень значительна, что позволяет ему быстро передвигаться во время плавания и при зарывании в землю. Ехидна — ночное наземное животное, более подвижна. Для нее характерны ходьба, бег и особенно рытье земли.
Сумчатые млекопитающие включают очень разнообразные формы, начиная от малоподвижных сумчатых кротов и кончая сумчатыми копытными. К последней группе относятся кенгуру. Наряду с малыми скоростями у сумчатых наблюдаются и большие, которые развивает кенгуру, способный делать прыжки 6—10 м длиной. Бег и прыжки у большого кенгуру настолько интенсивны, что по скорости передвижения (до 90 км/ч) с ним не может сравниться ни одно животное.
Чрезвычайно большой интерес представляет мышечная деятельность представителя американских неполнозубых (Edentata) —ленивцев. Их отличительная особенность— исключительная медленность движений, причем каждый шаг может продолжаться до минуты и более.
366
Как древесные животные, ленивцы висят вн'из головой и в такой же позе совершают переходы по ветвям. Для ленивцев характерна низкая температура тела —31,5— 36,0° С. У беременной самки ленивца значительно более постоянная температура тела, не понижающаяся даже при охлаждении (наружная температура 10°С); разница между температурой тела беременной и небеременной самки достигает 12° С. При введении американскому ленивцу (Bradypus tridactilis) экстрактов щитовидной железы и гипофиза не обнаружилось повышения двигательной активности и повышения температуры тела. Таким образом, медлительность, с которой совершаются мышечные сокращения у ленивца, не зависит от гормональных влияний, а связана, по-видимому, с особенностями его нервной системы.
Насекомоядные (Insectivora)—большей частью ночные животные — обладают очень небольшой скоростью движения. Тем не менее мышечная деятельность насекомоядных довольно разнообразна. Так, для ежа и землероек характерны ходьба, бег и роющая деятельность; для крота — исключительно развитая роющая деятельность, для выхухоли — плавание.
Чрезвычайно большую мышечную работу выполняет крот, питание которого осуществляется только под землей путем выкапывания дождевых червей и личинок насекомых. Эта исключительно интенсивная мышечная деятельность крота находит свое отражение в высоком содержании НЬ в теле — до 11,8 а на 1 кг веса тела, тогда как у полевок всего около 7,0 г.
У ежа сильно развита кожная мускулатура, имеющая первостепенное значение при осуществлении оборонительного рефлекса — скручивания в шар. Это поперечнополосатая мускулатура большой мощности. Еж имеет три мышечные системы: поддерживающую позу, стягивающую кожный покров и тело ежа в шар, и стягивающую кожу над головой. Исследование роющей деятельности ежей разных видов — европейского и ушастого — показало, что ушастый еж вырывает в два раза меньше земли, чем обыкновенный, но роет вдвое дольше и при этом затрачивает в 1,5 раза меньше энергии на 1 кг вырытой земли. По-видимому, значительная способность к длительному рытью, свойственная степным формам, накладывает отпечаток на физиологические реакции этих близ-
367
ких видов животных. В таблице 38 приведены некоторые из данных, полученных при сравнении двух видов ежей.
Длительное рытье создает и особый тип выполнения этой работы — с небольшой интенсивностью потребления кислорода в единицу времени и с более экономным расходованием энергии.
Таблица 38
Роющая деятельность обыкновенного и ушастого ежей (по Понугаевой, 1960)
Потребление О2 (ли8)
Еж обыкновенный (Erinaceus europaeus) 9
Еж ушастый (Erina-ceus auritus) .... 9
292 1650 20
261 1253 35
1835 3107
1629 2143
Рукокрылые, или летучие мыши (Chiroptera),— отряд млекопитающих, приспособленных к полету и к покою при висении головой вниз в состоянии повышенного тонуса задних конечностей. Медленные переползания по горизонтали сравнительно редко наблюдаются в природе. У летучих мышей можно наблюдать ползание вверх по вертикальной стене задним концом тела вперед. Полет осуществляется благодаря исключительно большой летательной перепонке, представляющей складку кожи, натянутую между туловищем и задними и передними конечностями, а также пальцами передних конечностей и хвостом. Очень развита у них грудная мускулатура при сравнительно слабой тазовой. В зависимости от строения летательных перепонок различают острокрылые, длиннокрылые виды и ширококрылые, относительно тупокрылые (Огнев, 1951). Первые являются весьма быстрыми летунами, вторые — медленными. А. П. Кузякин (1950) различает три типа полета у летучих мышей: гребной, порхающий и планирующий. Схемы движений при этих трех 368
формах полета приведены на рисунке 80 (кадры киносъемки). Гребной полет характерен для большой ночницы, малого подковоноса. При порхающем полете животное удерживается на месте и поднимается вверх. Такой полет позволяет летучей мыши-ушану подхватывать спящих насекомых. Эта форма полета свойственна не только медленно летающим летучим мышам (ширококрылым), но и быстро летающим (например, длиннокрыл
Рис. 80. Типы полета у летучих мышей (по Кузякину, 1950). / — гребной; // — порхающий; III — планирующий
южноевропейск'пй — Miniopleriis schreibersii). Скорость полета летучих мышей, по-видимому, не превышает 8— 10 м[сек.
Грызуны (Rodentia) —отряд млекопитающих, насчитывающий наибольшее количество видов, населяют все климатические зоны земного шара. По количеству не только видов, но и особей грызуны занимают, несомненно, доминирующее место среди млекопитающих.
369
У грызунов встречаются всевозможные проявления мышечной деятельности. Так, у летяг наблюдаются элементы полета по типу затяжных прыжков, близких к прыжкам белки. Среди грызунов немало плавающих и ныряющих, многие развивают большую скорость бега, например тушканчики, зайцы. У египетского тушканчика (длина тела от конца морды до основания хвоста 130 мм) задние конечности имеют длину 161 мм, что
Роет землю — Выбрасывает
Лесная мышь । । । । ।
землю J I г
Разравнивает землю |
Отдыхает (сидит) -|—।
Очищает шкурку -|—I
Слепец ।
Роет землю Выбрасывает землю
Отдыхает (сидит) Очищает шкурку
Рис. 81. Распределение роющей деятельности грызунов во времени (по Понугаевон, 1960)
позволяет ему делать прыжки в 2,5 м. При этом развивается скорость до 40 км/ч. Многие грызуны хорошо лазают (белки, черные крысы и др.). Большинству грызунов свойственна интенсивная роющая деятельность и интенсивное грызение как пищевых, так и непищевых объектов. Если прибавить к этому многочисленные позы, принимаемые грызунами в связи с ориентировочными оборонительными реакциями (например, поза столбика у суслика), то совершенно очевидно большое многообразие мышечной деятельности, связанное с разнообразными условиями жизни этого отряда.
На весьма распространенной у этого отряда роющей деятельности можно наблюдать смену форм движений, что позволяет грызунам выполнять сравнительно тяжелую работу на протяжении многих часов подряд. На рисунке 81 приведены данные, сопоставляющие режим роющей деятельности лесной мыши и слепца. Ясно видно, что интенсивная роющая деятельность слепца в зна-370
чительной мере более непрерывна, чем у лесной мыши. Здесь имеет значение и прямой отдых, и смена интенсивной мышечной работы на более легкую (очистка шкурки), что ускоряет процесс восстановления (эффект Сеченова) .
Многообразие форм мышечной деятельности связано с многообразием морфологических приспособлений: длины ног, хвоста, ушей, особенностей строения кисти и т. д. Эти данные прекрасно изложены в книге С. II. Огнева «Экология млекопитающих». Для грызунов характерна большая пластичность их мышечной деятельности.
А. Н. Северцов (1922) считал, что изменения в поведении организмов имеют огромное приспособительное значение для сохранения вида в целом в новых условиях существования. Действительно, многие грызуны, например крысы, способны приспосабливаться в своих движениях к окружающей обстановке; например, александрийская крыса (Rattus rattus alexandrinus}—лазать по канатам, проводам и т. п. Кролики, живущие в Западной Европе на земле в норах, при переселении в леса Австралии легко переходят к древесному образу жизни. Необычайная пластичность двигательных функций отличает нс только отдельных индивидуумов, по и отряд в целом.
Таблица 39
Максимальная скорость движения (бега) у грызунов
В п 1 Скорость (км ч) Автор
Заяц-беляк (Lepus Н-midus) Суслик малый (Citel-lus pygmaeus) .... Бурундук (Eutamias sibiricus) ...... . Дикобраз (Hystrix leucura) . 55—со 12 11,5 3 Огнев, 1940 Cottam, 1942 >
Как показано в таблице 39, многие грызуны способны развивать при движении — беге очень большие скорости.
Хищные млекопитающие (Carnivora) представляют со стороны их мышечной деятельности большой интерес,
371
поскольку некоторые их представители (собаки и кошки) служат наиболее распространенными лабораторными животными.
Наземные хищные отличаются необычайным приспособлением к выполнению различной по интенсивности мышечной работы. У них прекрасно развиты механизмы распределения крови, регуляции дыхания, изменения дыхательной функции крови, обеспечивающие очень интенсивный и дифференцированный перенос кислорода. У хищных встречаются очень разнообразные формы мышечной деятельности (рытье, грызение, прыжки, бег, ползание, плавание). По скорости .выполнения движений хищные неоднородны. Так, барсук, медведь обладают медленными движениями, а волк, собака, лисица развивают при беге очень большую скорость. Максимальная скорость бега у волка достигает 45 км/ч, причем такая скорость может поддерживаться 15 км подряд. У лиси-сы скорость бега не превышает (по тем же данным) 35 км/ч (Зверев, 1948).
Среди копытных встречаются представители с большой скоростью движения: кабарга, сайга, передвигаясь прыжками, могут развивать скорость 70-80 км/ч (табл. 40).
Таблица 40
Максимальная скорость движения (бега) у копытных
Вп 1 животного Скорость движения (Км/ч) Автор
Джейран (Casella sub-gutturosa) 56—64 Зверев, 1948
Сайга (Saiga tatarica) Архар (Ovis ammon 72
polii) 50—60 »
Олень благородный (Cervas elaphus) . . . 56 Cottam a. Williams,
1943
Большерог (Ovis canadensis) . 48 >
С другой стороны, здесь встречаются такие животные, как верблюд, с очень характерными медленными движениями, но своеобразным режимом работы и отды
372
ха. Любопытные данные в этом отношении приводит Н. М. Пржевальский (1888), который указывает, что для верблюда характерна способность очень интенсивно работать в течение недель и месяцев, но отдых, требующийся после этого, продолжается от 2 до 3 месяцев, а иногда и до года. Опыт монголов по содержанию этих животных показывает, что такой режим необходим для восстановления рабочего потенциала животного. Подобные же факты известны из практики спорта и у человека, когда для устранения явлений перетренировки требуется длительный отдых.
Очень большой интерес представляют наиболее близкие к человеку млекопитающие — обезьяны. Переходные формы между приматами и прочими млекопитающими— полуобезьяны (тупайи, лемуры) Prosimiae — отличаются крайне однообразным типом движений, мало подвижны. Эти по преимуществу ночные лазающие животные с очень узким ареалом, по существу, связаны с очень ограниченными условиями обитания. Физиология почти не изучена. Низшие обезьяны по сравнению с человеком и хищниками отличаются чрезвычайным непостоянством условий внутренней среды. Такне константы, как температура тела, уровень сахара в крови, дыхание подвержены у низших обезьян закономерным изменениям в течение суток (что связано с суточными изменениями возбудимости центральной нервной системы), а также с большими колебаниями при выполнении даже кратковременной мышечной деятельности.
Средний путь, проделываемый обезьяной в большой клетке (объемом 7,7 я3), составляет около 10—12 км в сутки, т. е. около 1 км!ч активности (Щербакова, 1949). Обезьяны-гамадрилы способны за 6 ч пробежать в третбане около 38 км (Г. М. Черкович). При этом, однако, наблюдалось значительное утомление, выразившееся в повышении температуры в ночные часы. Моторная функция обезьян чрезвычайно разнообразна и складывается из сложного сочетания врожденных и приобретенных в течение индивидуальной жизни движений. Среди обезьян не все виды отличаются богатством движений. Особенно разнообразны движения у древесных видов (макак). Наземные виды (павианы) отличаются более однообразными движениями и несколько большим постоянством физиологических функций.
373
Наиболее важный момент в эволюции моторики обезьян — переход их к передвижению на задних конечностях. При этом передние конечности освобождаются для более сложной, но у низших обезьян еще достаточно примитивной деятельности. Передвижение обезьян происходит в основном на четырех ногах (особенно убегание от опасности); движения кисти, а также ряд других, связанных с жизнью стада и воспитанием детенышей, происходят при стоянии на двух ногах. Важно отметить, чго скорость передвижения па двух ногах в 4 раза меньше скорости передвижения па четырех. В эволюции такое падение скорости компенсировалось развитием передних конечностей и образованием более сложных жизненно важных форм движения (поиски и лучшее выбирание пищи, сложные прыжки и т. п.).
Полное отсутствие физиологических данных о мышечной деятельности высших человекообразных обезьян не позволяет продолжать обзор и связать его непосредственно с особенностями физиологии мышечной деятельности человека. Можно отметить малую интенсивность и скорость мышечной деятельности у высших обезьян, являющихся животными также с узким ареалом, с трудом приспосабливающихся к изменениям условий среды, питания и существования.
Мышечная деятельность человека отличается значительным разнообразием, причем характерны большие возможности тренировки скорости и выносливости. Вся мышечная работа человека протекает при относительно неглубоких изменениях постоянства внутренней среды, а чрезмерная чувствительность высокоорганизованной нервной системы человека ко всяким колебаниям последней не мешает проведению работы в крайних пределах.
По своим возможностям к осуществлению мышечной деятельности человек, разумеется, уступает многим из рассмотренных животных форм, но по многообразию форм движений он их значительно превосходит.
ФИЗИОЛОГИЧЕСКИЕ АДАПТАЦИИ
ПРИ МЫШЕЧНОЙ ДЕЯТЕЛЬНОСТИ
Поскольку всякая мышечная деятельность животного сопровождается увеличенным энергетическим расходом, наиболее общей чертой является здесь адаптация
374
физиологических систем, направленных на обеспечение организма кислородом. Поэтому наибольшие сдвиги в процессе адаптации происходят в функции дыхания, кровообращения и в дыхательной функции крови.
Между величиной выполняемой животным работы как в короткие промежутки времени, так и на протяжении, например суток, и общим энергетическим расходом существуют прямые соотношения. Однако только для очень немногих форм деятельности известны энергетические характеристики; большинство сложных актов поведения и длительной работы животных с этой стороны еще совершенно не изучены.
Мышечная деятельность по своей интенсивности и общей величине выполненной работы распределяется в течение жизни животного совершенно неравномерно. Особенно интенсивна мышечная деятельность в определенные периоды жизни и в определенные сезоны. Так, большая мышечная работа производится животными при кочевках, миграциях и перелетах. С другой стороны, например, период беременности у самок многих грызунов, сопровождается резким ограничением мышечной активности. Большую мышечную работу производят животные при постройке нор и укрытий, а также в процессе запасания корма. Наконец, резкие возрастные отличия, связанные с повышенной игровой деятельностью молодняка, характеризуют большинство млекопитающих.
Мышечная деятельность по своей интенсивности и суммарному эффекту (в кг/м работы) величина постоянная для отдельных видов. Особенно это характерно для многих хищных, которые даже в условиях клеточного содержания в зоопарках выполняют большую мышечную работу, ритмически передвигаясь по клетке в течение многих часов в сутки (многие куньи, рыси, гиены, львы и т. д.). Физиологический механизм, лежащий в основе этой «произвольной» деятельности, совершенно не ясен. Во всяком случае, он никак не связан с пищедобывательной, оборонительной или какой-либо другой формой деятельности. Происхождение этой деятельности также неясно, как и происхождение игровой деятельности молодого животного.
Физиологические изменения, происходящие при мышечной работе, а равно и приспособление к выполнению
375
мышечной работы различной интенсивности, скорости и продолжительности изучены главным образом на человеке в связи с проблемами физиологии труда и спорта. При трактовке многих фактов, касающихся мышечной деятельности животных, как домашних, так и диких, приходится опираться на этот большой и хорошо систематизированный материал (Atzler, 1927; Бейнбридж, 1927; Кекчеев, 1931; Конради, Слоним, Фарфель, 1934; Виноградов, 1941; Крестовников, 1951; Зпмкин, 1956; Виноградов, 1958).
При мышечной работе наблюдается увеличение содержания эритроцитов и НЬ в крови. Это обычно связывается с сокращением селезенки и выбрасыванием крови из кровяных депо в периферическое кровяное русло. Объем такой депонированной крови может содержать до 40% всей крови. По-видимому, у разных млекопитающих роль отдельных депо различна. У человека это в основном кожа, у собаки — селезенка и сосуды печени. Сдвиг в содержании эритроцитов объясняется тем, что депонированная кровь содержит больше эритроцитов, чем циркулирующая. Кроме того, усиливается и новообразование крови, что выражается в возрастании числа ретикулоцитов.
Наиболее ярки и доступны для наблюдения изменения частоты пульса. При мышечной работе частота пульса возрастает параллельно ее интенсивности, т. е. параллельно потреблению кислорода. При очень интенсивной мышечной работе последнее происходит не только во время выполнения работы, но и после ее окончания. Соответственно этому и пульс, учащенный во время работы, приходит к исходным величинам спустя несколько минут (иногда 50—60) после окончания работы. Установлено, что у тренированных людей и животных пульс учащается меньше, чем у нетренированных. У тренированного человека при мышечной работе резко увеличивается объем каждой систолы—ударный объем сердца, а частота пульса сохраняется на более низком уровне.
Факт уреженного пульса у тренированного человека, объясняемый обычно повышением тонуса блуждающего нерва, можно наблюдать и у животных, выполняющих очень интенсивную мышечную работу и хорошо приспособленных к ней. Так, частота пульса у зайца составляет около 70 в 1 мин, у кролика, не выполняющего 376
столь интенсивной работы, как заяц, частота пульса в покое — около 180 в 1 мин (Аршавский, 1958). Такие же соотношения были обнаружены при сравнении частоты пульса каракульских и романовских овец и гибридов 1 генерации архара и каракульской овцы. Частота пульса у каракульских овец была около 100, а у гибрида в покое — около 70. Это находит свое отражение и при сравнении лошадей различных пород. Так, частота пульса у шотландского пони весом 284 кг около 42,9 в 1 мин, а у першерона весом 646 кг — также около 42,8 (Brody, 1949). Таким образом, несмотря на разницу в весе, приспособленный к интенсивной работе пони имеет относительно замедленный пульс. Частота пульса у очень крупных животных невелика — у слона она равна 31 в 1 мин (Brody, 1945).
Кровяное давление во время мышечной работы возрастает. Это касается прежде всего систолического давления, возрастание которого находится в прямой зависимости от интенсивности мышечной работы. Диастолическое кровяное давление падает или поддерживается на постоянном уровне. Разность между величинами систолического и диастолического давления — пульсовое кровяное давление, как правило, возрастает. Существует определенная зависимость между величиной пульсового кровяного давления и минутным объемом сердца.
Изменения дыхания при мышечной деятельности сопровождаются увеличением частоты и глубины дыхания, что позволяет увеличить минутный объем дыхания или легочной вентиляции. Как правило, учащение дыхания при мышечной работе сопровождается углублением его; однако есть основания считать, что при тренировке и в дыхании наблюдаются такие же изменения, как и в минутном и ударном объеме крови. Частота и глубина дыхания зависят от интенсивности выполняемой работы. У тренированных людей увеличиваются и частота и глубина, у нетренированных — главным образом частота дыхания.
Частота дыхания у животных, приспособленных к интенсивной мышечной деятельности, также несколько ниже, чем у домашних, мало приспособленных. Так, частота дыхания у гибридов архарокаракуль оказывается более редкой, а объем выдоха большим, чем у овец каракульской или романовской породы.
377
Замечательным свойством дыхательной деятельности и сердечнососудистой системы тренированного организма является способность к поддержанию содержания кислорода в артериальной крови на постоянном уровне даже при выполнении довольно интенсивной (но, конечно, не предельной, истощающей) мышечной работы. Измеренное с помощью осигемометра насыщение артериальной крови у тренированного человека показывает почти постоянный уровень как при интенсивной, так и при умеренной работе. У нетренированного человека в первые минуты после начала работы насыщение кислородом артериальной крови значительно падает (Войтке-вич, 1952). К сожалению, проверка этих данных в приложении к диким животным, способным к выполнению интенсивной мышечной деятельности, не производилась.
Поддержание постоянства внутренней среды и прежде всего постоянства состава крови и реакции осуществляется за счет выведения продуктов мышечного обмена (молочной кислоты, СОг) из организма почками и потом, а СО2 — легкими. Однако огромное значение для поддержания pH крови имеют буферные системы крови, которые лучше всего могут быть изучены суммарно как щелочные резервы крови. Щелочные резервы крови (способность крови связывать угольную кислоту) у человека и животных разной тренированности к мышечной работе также различны. У тренированных они повышены и сохраняют этот высокий уровень и при выполнении мышечной работы (Конради, Слоним, Фарфель, 1935). Повышенная резервная щелочность встречается у животных, выполняющих интенсивную мышечную работу. Северный олень обладает большой кислородной емкостью и более высокими щелочными резервами. Кривая диссоциации оксигемоглобина сдвинута вправо, что обеспечивает интенсивный перенос кислорода и отдачу его тканям (Крепе, 1936; Антелидзе и Барбашова, 1938). Количество крови у северного оленя превышает наблюдающееся у домашних животных (от 11 до 14% к весу тела), тогда как у других сельскохозяйственных животных эти отношения не превышают 10% (Городецкий, 1959). Высокие щелочные резервы крови наблюдаются также у таких животных, как лошадь — 72%, осел — 66%, собака — 56% и, наконец, у человека — около 60%.
378
Низкие щелочные резервы характерны для животных с малой активностью: кролик — 48%, кошка — 46%, мышь белая — 45%, морская свинка — 36% (Меньшикова и Писарева, 1945). Любопытно, что низким щелочным резервом обладает романовская овца (по данным тех же авторов, около 45%). Это можно объяснить малой мышечной активностью овец этой породы, разводимых почти исключительно в стойловых условиях.
Содержание угольной ангидразы — фермента, расщепляющего водный раствор угольной кислоты, находится в обратной зависимости от величин щелочных резервов крови.
Существуют многочисленные исследования содержания сахара в крови при мышечной деятельности (Конради, Слоним, Фарфель, 1935; Виноградов, 1941 —1958). В качестве общей схемы, одинаково приемлемой для человека и животных, можно принять следующие положения. При коротких, хотя и интенсивных мышечных нагрузках, наблюдается возрастание содержания сахара в крови, особенно ярко выраженное в начале работы. Эю явление отражает картину мобилизации гликогена, расходуемого в первую очередь как энергетический материал для мышечной деятельности. При более продолжительной (хотя и не предельной по напряжению) мышечной деятельности наблюдается снижение содержания сахара в крови. Это явление отражает, вероятно, процессы начинающегося истощения запасов гликогена в организме. Оно связано также и с ресинтезом углеводов из молочной кислоты, происходящим на протяжении продолжительной и не слишком тяжелой мышечной работы. По-видимому, при более или менее продолжительной работе можно наблюдать как начальную фазу повышения содержания сахара в крови, так *и фазу понижения, связанную с нарастающим истощением организма. Эти факты, полученные на человеке, нашли свое подтверждение и при исследованиях, проведенных на лошадях как в процессе коротких беговых испытаний, так и при длительных переходах.
Мышечная деятельность — полет, сопровождается у летучих мышей разогреванием, повышением температуры тела и возрастанием содержания сахара в крови. Во время длительного полета содержание сахара в крови падает.
379
По интенсивности работы и по возможностям обеспечения ее выполнения доставкой кислорода все мышечные работы могут быть разделены на 2 группы: истощающая работа, протекающая с нарастанием кислородного долга и требующая его устранения после окончания работы (или перехода на более легкую работу, например с бега на шаг), и работа, когда количество продуктов мышечного метаболизма (молочной кислоты), образующееся б единицу времени, не превышает того количества, которое может быть устранено окислением или восстановлением в 1исходные вещества (гликоген). Состояние такого равновесия получило название устойчивого состояния (steady state). Кроме величины выполняемой работы, оно зависит еще от мощности дыхательной и циркуляторной систем организма.
Однако такое состояние наблюдается лишь при сравнительно нетяжелых, а главное не очень интенсивных мышечных работах. При очень тяжелых, истощающих работах потребность работающих мышц в кислороде превышает возможности доставки его с помощью аппаратов дыхания и кровообращения. В этих условиях в результате несоответствия между кислородным запросом тканей и его доставкой образуется кислородный долг. Нарастание этого долга на протяжении самой работы ограничивает ее продолжительность, а кислородный долг ликвидируется после работы в результате усиленного дыхания и восстановительных процессов в мышцах.
Выше уже указывалось, что по особенностям выполнения мышечной работы организм можно характеризовать по силе, быстроте (скорости) и выносливости. Каждое из этих качеств в различной степени представлено в отдельных мышечных спортивных упражнениях, каждое из них в различной степени представлено у животных с разной экологической специализацией.
Картины кривых диссоциации оксигемоглобина у лошади и верблюда различны (Крепе, 1936). Кривая диссоциации лошади характеризовалась значительной крутизной падения, что в ее разрядной части (при рО2 40) обеспечивает быструю отдачу кислорода оксигемоглобином в тканевых капиллярах. Такой тип кривой диссоциации обеспечивает обильное снабжение тканей кислородом при резко возросшем кислородном запросе. Тип кривой диссоциации у верблюда оказался более
380
пологим, с меньшей возможностью отдачи кислорода тканями и с большей способностью к фиксации его кровью. Первый тип кривой обеспечивает быстроту и большую интенсивность мышечной деятельности, второй— медленную устойчивую работоспособность на длительное время (рис. 82).
Характер мышечной деятельности и работоспособность животного в значительной мере определяются также и морфологическими особенностями скелетной мускулатуры. В организме животного имеются мышцы двух
Рис. 82. Диссоциация оксигемоглобина у животных с разными типами мышечной деятельности (по Крепсу, 1936). По оси ординат — содержание оксигемоглобина (%); по оси абсцисс — напряжение кислорода
(мм II):
/—лама. 2— верблюд, 3 — лошадь, 4 — северный олень
родов: мышцы белые, главным образом быстро сокращающиеся, и мышцы красные — тонические, обладающие свойствами более медленного сокращения и более развитыми свойствами поддержания мышечного напряжения— тонуса. Последнее особенно важно при поддержании позы и характеризуется малым энергетическим расходом, малой утомляемостью и малой интенсивностью центральной импульсации (Виноградов, 1958).
У лошадей количество желудочного сока, отделяемого во время мышечной работы, уменьшается, но после окончания работы быстро восстанавливается (Курилов, 1955). Практически это находит свое отражение в том, что при мышечной работе интенсивностью около
381
2 .млн. кг/м в сутки наблюдается снижение перевариваемости сено-овсяных рационов. Последнее по клетчатке составляло 9%, а по жиру 7% (Чалюк, 1950). Однако это положение может быть правильным только для тех организмов, у которых процесс еды происходит или по окончании пищедобывательной деятельности (например, у хищника после охоты) или у животных, питающихся на одном месте после поисков корма (например, питающихся падалью, всеядных организмов).
Движения обезьян, направленные на добывание пищи (движения передних конечностей), всегда сопровождаются отделением слюны (Уголев, Волкова, Корневищ 1953). У лисицы при раскапывании норы грызунов наблюдалось увеличенное слюноотделение (Уждавини).
Мышечная деятельность оказывает очень большое влияние на терморегуляцию организма. Поскольку мышечные сокращения связаны со значительным теплообразованием и при выполнении работы в условиях средних температур (не говоря уже о высоких), всегда имеется положительный баланс тепла. Это находит свое отражение в том, что температура тела, как правило, при интенсивной мышечной деятельности повышается. У человека такое повышение температуры тела может наблюдаться при работе значительной интенсивности, у животных — в зависимости от интенсивности работы и видовых особенностей животного. Наибольших сдвигов температура тела во время работы достигает у летучих мышей, являющихся практически пойкилотермными организмами в периоды отсутствия работы (полета).
Очень сильно повышается температура тела при мышечной работе у низших обезьян. Повышение температуры тела после бега в третбане может достигать 2,5— 3°С. Особенно ярко это выражено у молодых животных— подростков, отличающихся большой подвижностью.
МЫШЕЧНАЯ ДЕЯТЕЛЬНОСТЬ У ПТИЦ
Мышечная деятельность у птиц изучается с разных сторон: такие явления, как сезонные миграции, самый механизм полета, ориентация при полете представляет огромный интерес для биологии и механики. Однако до настоящего времени наши знания о физиологических
382
механизмах, действующих .при выполнении мышечной работы в полете, чрезвычайно ограничены.
Самый полет птиц с биодинамической стороны изучался неоднократно, начиная с работ французского физиолога Марея (Магеу, 1890) и кончая последними исследованиями (Гладков, 1960).
Значительное количество исследований посвящено изучению энергетических резервов птиц, связанных с перелетом, в том числе и резервов жира, являющегося в этом случае источником образования механической энергии (Дольник, 1965). Установлена определенная зависимость между резервами жира и дальностью перелета у отдельных видов. Однако эти исследования позволяют лишь косвенно судить об энергетике полета птиц, а добытый материал по своему объему и достоверности не идет ни в какое сравнение с данными, имеющимися для млекопитающих.
Только в самые последние годы метод радиотелеметрии позволил получить первые прямые данные о физиологических изменениях, происходящих у птиц во время полета. В некоторых более ранних исследованиях было установлено, что подъем крыла у голубя сопровождается вдохом, тогда как у чайки, кряковой утки число взмахов крыла на одно дыхание может колебаться от 3 до 9. У небольших птиц число дыханий много меньшее, чем движений крыла (Tomlinson, 1957; Loz, Berios a. Coch-zan, 1962).
Однако главным приемом изучения физиологических изменений во время полета птиц оставались расчеты, исходившие из сопоставлений мышечной работы птиц с работой млекопитающих. Так, было рассчитано, что полет голубя со скоростью 50 км/ч должен сопровождаться увеличением теплопродукции в 10 раз, а легочной вентиляции в 20 раз, чтобы обеспечить отдачу тепла и сохранение температуры тела птицы. Однако исследования испарения, проведенные с помощью двойной изотопной метки (Litson, Gordon a. Me Clintock, 1955), не подтвердили этого расчета.
Непосредственное определение легочной вентиляции было проведено с применением радиотелеметрии и регистрации изменений давления и сопротивления в маске, одетой на головку птицы (Hart a. Roy, 1966). Оказалось, что легочная вентиляция возрастает при полете более
383
чем в 20 раз, при неизменном объеме дыхания. После полета в восстановительном периоде глубина возрастает одновременно с понижением частоты дыхания. Минимальный объем дыхания при -максимальной частоте был найден во время термического полипноэ. Следовательно, легочная -вентиляция во время полета почти в 2,5 раза превышает метаболические потребности. Легочная вентиляция в покое составляла 7,2 л{ч, в полете—147, после полета — 20,4. Соответственно продукция тепла в покое — 2,69 кал{ч, в полете — 22,0 и после полета 8,2 кал/ч. Частота дыханий в покос составляла 26 в 1 мин, в полете — 487, при ходьбе 77, а после полета . 156. Частота дыхания при полипноэ достигала 437 дыханий в минуту. Частота пульса в покое в среднем составляла около 166 в мин (птицы весом около 400 г), а во время полета возрастала незначительно — не более чем в 2 раза против величин покоя. Приведенный материал еще недостаточен для обобщений, касающихся птиц вообще, но он совершенно ясно свидетельствует об исключительной роли легочной вентиляции в регуляции тепла у птиц во время полета. При этом огромное значение приобретают и воздушные мешки, служащие главным путем отдачи тепла (Zeuthen, 1942; Hart a. Roy, 1967).
Глава X
СТАДНЫЕ И ПОПУЛЯЦИОННЫЕ ОТНОШЕНИЯ И ИХ ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ
ГРУППОВЫЕ РЕАКЦИИ ЖИВОТНЫХ
И подражательная деятельность
Из факторов внешней среды, оказывающих значительное влияние на физиологическое состояние организма, его поведение, а также реакции на отдельные действующие агенты, следует особенно выделить организмы того же вида или реже других видов. Изучение этих влияний составляет большой раздел экологии — влияние биотических факторов среды.
Под биотическими факторами в экологии понимают совокупность влияний на организм других организмов, входящих в состав данного биоценоза. Последний включает всю совокупность организмов (животных, растительных, микроорганизмов), существование которых взаимосвязано при осуществлении питания, защиты от врагов, размножения и т. д. (Кашкаров, 1944; Наумов, 1963).
В экологии установлены многочисленные и вместе с тем очень важные для существования видов и их популяций биотические факторы. Это отношения, существующие между хищником и жертвой, между объектом питания и питающимся. В понятие биотического фактора входят сложные взаимоотношения между двумя видами, населяющими один и тот же узкий ареал, сложные отношения между возрастными группировками внутри отдельных популяций, внутри всего животного населения данной территории. В группу биотических факторов входят также отношения животных одного и того же вида между собой — колониальные, стадные и групповые отношения.
В принятых в настоящее время в физиологии представлениях об «уровнях» изучения физиологических
13—1204
385
функций, их сложного взаимного влияния выделяют и так называемый надорганизменный уровень исследования, а возникающие при этом связи характеризуются трудно переводимым английским термином — mass physiology (Christian, 1950). Исследование взаимного влияния организмов включает теперь уже хорошо очерченное учение об агрегациях (скоплениях) животных, подражательных реакциях (имитационных) и подражательном поведении, учение о доминировании особей, специализации стада и нервных и гормональных регуляциях численности популяций.
Рассмотрение всех этих направлений в физиологических исследованиях, проводимых на надорганизменном уровне, уместно начать с группы подражательных реакций, которые, как уже указывалось выше, относятся к важнейшим компонентам сложных форм поведения.
Отношения -между животными — себе подобными или даже животными разных видов — складываются из очень широко распространенной подражательной деятельности. Например, если взять группу взрослых крольчат и группу щенков и поставить перед ними миску с морковью, то голодные кролики бросятся к ней. Сюда же бросятся и щенки, но есть морковь, конечно, не будут, а начнут нюхать ее, облизывать и повторять движения, близкие к тем, которые проделывают пожирающие морковь крольчата. В основе таких явлений лежат подражательные рефлексы — рефлексы условные, где на основе рецепции звуков и вида акта еды воспроизводится акт еды животного, даже другого, далеко отстоящего вида.
Впервые детальное изучение подражательных реакций, лежащих в основе стадных взаимоотношений, было осуществлено В. Я- Кряжевым (1935, 1955). Им был установлен основной факт, заключающийся в том, что если собаку кормить в присутствии другой собаки, то у последней выделяется слюна. Эти взаимоотношения между собакой-актером и собакой-зрителем основаны на условных рефлексах, где сигнальными раздражителями являются звук и вид самого акта еды. В зависимости от силы действующего раздражителя (пищевой возбудимости «зрителя») и типологических особенностей его нервной системы реакция может принимать различный характер.
386
А. Г. Понугаева (1953 б), изучая в близкой к опытам В. Я. Кряжева методической постановке реакцию ушастого ежа на поедание пищи другим ежом, обнаружила усиленное слюноотделение у «зрителя» наряду с весьма агрессивной реакцией по отношению к «актеру». Таким образом, такого рода взаимоотношения у животных разных видов неоднородны и отражают черты безусловнорефлекторного видового поведения.
Значительное ускорение образования условных рефлексов в условиях общения В. Я. Кряжев объясняет повышением возбудимости нервной системы животных в группе. Из его опытов несомненно явствует, что в условиях группы (по крайней мере собак) процессы возбуждения преобладают над процессами торможения. Это выражается в невозможности добиться четко выраженных дифференцировок, полного угасания условных рефлексов.
Если голодную обезьяну держать в камере перед группой поедающих пищу обезьян, то у первой можно наблюдать резкое снижение газообмена (Макарова, 1953).
В тех же условиях у обезьян резко снижается содержание сахара в крови, т. е. проявляется реакция, противоположная той, которая имеет место при акте еды. По-видимому, для столь возбудимых животных, как обезьяны, такой раздражитель сверхсильный, он вызывает запредельное торможение и вместо повышения обмена, связанного с актом еды и последующим специфическим динамическим действием пищи (рефлекторной его фазой), можно видеть его понижение.
Самый простой случай реакции подражания обычно связывался с образованием натурального условного рефлекса на звук, запах, вид поедающего пищу животного. Однако в исследованиях В. Я- Кряжева (1955) показано, что изоляция животных от себе подобных не выключает таких рефлекторных актов. Эти явления наблюдаются у животных, воспитанных в самой строгой изоляции, что позволяет считать подражательную деятельность в значительной степени врожденной и только дополняемой целым рядом условных рефлексов. На звук и вид акта еды и поедаемой пищи может образоваться условный рефлекс и у изолированного от других особей животного.
13*
387
В процессе подражания (имитации) также может быть образовал целый ряд условных рефлексов. Выше уже было подчеркнуто значение имитационного условного рефлекса при пастьбе копытных.
Л. Г. Воронин (1947) выработал двигательные пищевые условные рефлексы у детеныша обезьяны, хотя детеныш, находящийся с матерью, пищевого подкрепления не получал (питаясь исключительно материнским молоком), а лишь подражал матери. Следовательно, в стаде эти формы поведения приобретают исключительный вес и значение.
Однако стадные отношения охватывают не только различные формы двигательного поведения животных. Они сильно изменяют и протекание основных физиологических (вегетативных) функций. Особенно ярко это проявляется у отдельных особей, например при удалении или приближении стада овец или коз к одиночному животному или группе (Понугаева и Трубицына, 1955). При удалении стада от одиночно стоящей овцы пли козы у последней наблюдается резкое повышение газообмена (реакция следования). При приближении стада к стоящему одиночно животному изменения газообмена отсутствуют. В основе происходящих здесь физиологических изменений лежат мышечные реакции, связанные с поддержанием определенного расстояния между членами агрегации. По-видимому, от видовых особенностей двигательных реакций и зависит протекание физиологических процессов у животных разных видов. Добытые в этом направлении факты полностью оправдывают такую постановку вопроса.
Следование за движущимся предметом особенно ярко проявляется у многих копытных в определенные периоды постнатального развития. Если в первые дни жизни новорожденный не следует за овцой-маткой или за кормящим его человеком (при выращивании в искусственных условиях), то через некоторое время он обязательно начнет следовать за ними. Давно описаны случаи, когда теленок бизона приходил в лагерь вслед за лошадью охотника или за самим охотником. То же отмечено и у жеребенка зебры. У молодняка гну наблюдается следование за всяким удаляющимся предметом («способность догонять все, что удаляется»; Hediger, 1952). При воспитании молодняка овец разных пород и муфлонов
388
(в неволе) также наблюдалось неотступное следование молодняка за кормящим их человеком (Goethe, 1939).
В соответствии с этими врожденными физиологическими особенностями копытных разных видов можно предполагать, что и газообмен, и такие физиологические реакции, как пульс или дыхание, изменяются при дистантном раздражении новорожденного копытного видом удаляющейся матери или движущегося предмета.
Повышение обмена у новорожденных ягнят сразу после рождения наблюдалось при удалении кормящей овцы-матки. Это явление очень хорошо выражено у ягнят каракульской породы. Приближение овцы-матки к ягненку обычно не вызывает изменения газообмена, а также пульса и дыхания. Только на фоне очень повышенной пищевой возбудимости у каракульских ягнят можно наблюдать повышение газообмена при приближении к ним овцы-матки. Иная картина наблюдается у ягнят гибридов (архарокаракуль). Здесь как приближение, так и удаление матери вызывает у новорожденного значительное повышение газообмена и частоты пульса. Реакции эти на протяжении индивидуального развития не остаются постоянными. Различия в реакциях на удаляющуюся и приближающуюся овцу у новорожденных ягнят каракульской овцы и ягнят гибридов архарокаракуль интересно сопоставить с особенностями врожденного поведения ягненка овцы и ягненка архара.
Как известно, домашние овцы-матки никогда не оставляют своих ягнят с самого рождения и до взрослого состояния. При этом как ягненок следует за матерью, так и овца следует за ягненком. Совершенно иную картину можно наблюдать у многих видов диких копытных. У отнятого от самки сразу после родов ягненка архара наблюдалось полное отсутствие следования за движущимся предметом и человеком в первые 2—3 дня после рождения. Лишь на 5—6-й день ягненок архара неотступно следовал за человеком, который его постоянно кормил молоком.
СТАДНЫЕ II АТРЕГАЦИОИНЫЕ РЕАКЦИИ
Подражательное поведение и возникающие на его основе явления, однако, не исчерпывают всех реакций животных на себе подобных и являются скорее лишь
389
частной и весьма специализированной формой врожденной физиологической реакции. И тем не менее есть все основания думать, что именно на основе этого подражательного поведения, фиксирующего расстояние между особями, и создается объединение животных в группы — агрегации (Слоним, 1965). Само же существование животных в группе приводит к ряду физиологических изменений в организме каждого члена группы. Лучше других при этом выявляются реакции общего обмена веществ и эндокринные, главным образом со стороны коры надпочечников.
Изменения газообмена в группе лучше всего выявляются в состоянии полного мышечного покоя. Если взять группу лабораторных белых крыс и поместить их в условия низкой температуры, то они образуют скопление: садятся друг на друга, постоянно меняют положение (крайние особи протискиваются внутрь группы) (Hill, 1913). Отмечено, что при групповом скоплении газообмен у них резко снижен. Это явление тогда же было истолковано Хиллом как проявление терморегуляции, как следствие ограничения теплоотдачи в связи с тем, что суммарная поверхность тела животных, собранных вместе, значительно меньше отдающей тепло поверхности животных, посаженных врозь. Это, если можно так выразиться,— коллективная терморегуляция. Однако уже расчеты Хилла обнаружили недостаточность такого объяснения, так как уменьшение поверхности тела было меньше, нежели соответственное падение теплообразования. Хилл считал, что здесь имеет место снижение двигательной активности животных.
Хорошо выраженный двигательный аффект в группе был недавно обнаружен Ю. Кескпайком и А. Ф. Давыдовым (1966). Оказалось, что с развитием терморегуляции у птенцов появляется хорошо выраженное «скучива-ние» при умеренном охлаждении, когда родители покидают гнездо. На рисунке 83 показаны позы птенцов и их реакция через 20 мин на изменение температуры камеры. Птенцы в возрасте четырех суток «скучиваются» при температуре 20° С и отодвигаются друг от друга при температуре 33° С. В первый день после вылупления скучи-вание наблюдается даже при температурах 34 и 35° С. Двигательная реакция на охлаждение появляется ранее, чем формируется терморегуляция. Последняя как бы за
390
меняет при определенных температурах реакцию окучивания. На рисунке 84 показаны те же отношения у птенцов разного возраста по наблюдениям, произведенным через 10 мин после вылета самки из гнезда.
Явление скучивания (агрегации) широко распространено и среди пойкилотермных организмов: массовые скопления лягушек и жаб, насекомых во время зимовки, массовые скопления моллюсков (Allee, 1931; Понугаева,
Рис. 83. Положение птенцов дрозда в гнезде при различной температуре среды (°C) через 15—20 мин после экспозиции в камере.
1, II — 1-е сутки; /// — 4-е сутки
1953). Для таких скоплений характерно резкое суммарное понижение обмена веществ в покое, достигающее у представителей отдельных видов 40% и более (Понугаева, 1949, 1952, 1953, 1960). Факт этот был обнаружен у моллюсков, рыб и лягушек, т. е. у животных, не обладающих терморегуляцией. Интересно, что у моллюсков понижение обмена веществ можно обнаружить не только при помещении их в группу живых моллюсков, но и среди вываренных раковин. Очевидно, это связано не только с химическими раздражителями, особенно интенсивными в водной среде.
М. Штефан (1957), а затем К. А. Алексеева (1959) на рыбах'подтвердили данные Хилла, Элли и А. Г. Пону-
391
гаевой. Было установлено, что удаление переднего мозга рыб влечет за собой выпадение стадного влияния на газообмен. Последний оказался неодинаково выраженным у разных видов рыб. У водных организмов исследователи неоднократно обнаруживали химические вещества, выделяющиеся в окружающую среду и могущие действовать на обмен веществ (телергоны) (Киршен-блат, 1968). (Имеются многочисленные указания па
Рис. 84. Положение птенцов различного возраста в гнезде через 10 мин после вылета самки: / — односуточные; II— двухсуточные; III — трехсуточные; IV — четырехсуточные; V — пятисуточные;
VI — шестисуточные
вещества, стимулирующие и задерживающие рост растений, действующие через корневую систему.)
Особенно ярко изменение обмена веществ в стаде или группе проявляется у млекопитающих со стадным или колониальным образом жизни. Для получения «стадного» эффекта непосредственный термический контакт животных друг с другом не обязателен (Понугаева, 1960). Например, обезьяны собираются на ночь (особенно при низких температурах среды) в большие группы (Слоним и Щербакова, 1949). Снижение газообмена в группе обезьян можно наблюдать, если посадить животных в различные камеры. Контакт животных обеспечивается с по-392
мощью стадных звуков, и обмен веществ в такой разделенной группе снижается.
У сусликов, живущих колониями, можно наблюдать яркий «стадный» эффект при отсаживании их в общую газообменную камеру. Физиологический механизм рефлекторного влияния на обмен веществ в группе подобных организмов подвергается тщательному изучению с целью определения роли различных анализаторов и сроков возникновения стадных рефлексов. В онтогенезе стадные рефлексы (изменения газообмена) возникают у грызунов сравнительно поздно (на 30-й день). Можно считать установленным, что этих рефлексов нет ни в момент рождения животного, ни в последующий период гнездового развития (Попугаева, 1960). Понижения газообмена при групповом скоплении не наблюдается у представителей ряда млекопитающих, ведущих во взрослом состоянии одиночный образ жизни. Помещение таких животных, как серый хомячок (Mesocricetus griseus), незокия (Nesokia indica), ушастый еж {Hemiechinus attritus'), в среду себе подобных вызывает у них повышение газообмена (Попугаева, 1960).
В таблице 41 приведены основные величины изменений газообмена.
Изменения обмена веществ в группе и во время спячки не исчезают. Исследования, проведенные в пещере «Адзаба» возле Сухуми, в Саблинской пещере под Ленинградом на различных видах летучих мышей, обнаружили снижение обмена веществ в группе и в состоянии дневного сна у летучих мышей летом и во время зимней спячки (Понугаева, 1949; Понугаева и Слоним, 1953). Последний факт представляет большой интерес, так как поясняет значение скоплений животных в их зимних убежищах во время спячки. В этих условиях скучивание животных обеспечивает сниженный энергетический расход, а следовательно, лучшие условия выживания во время прекращения питания в зимний период. Большие зимние скопления грызунов в копнах сена и соломы (полевая мышь — Apodemus agrarius, курганчиковая мышь — A4us musculus hortulanus и др.) также способствуют лучшему выживанию в этих резервациях. Ограничение газообмена в группе мелких видов может достигать 50% по сравнению с одиночными особями.
393
Таблица 41
Изменение газообмена у изолированных животных и в группе (по Понугаевой, 1960)
Отряд Жив >тное Температура камеры CC Потребление O2 (мл,кг/ч) Обмен веществ (в °А от сб-мена одиночек) Вероятность случайности । (%)
у изолированных животных я группе
Р у КО- Подковонос малый
крылые (в покое) (Rhino tophus hipposidcros) . . 23 1169 838 72 —
Ночница большая
(в покое) (Myotis myotis) 14 4366 2364 54 —
Длиннокрыл обык-
новенный (в покое) (Miniopterus schrei-bersi) 23 1846 1084 59
Насеко- Еж ушастый (Не- 16 134
моя тные miechinus auritus) . . 1698 2280 95
Грызуны Полевка обыкновенная (Microtus aril al is) 20 5689 4196 73,8 99
Полевка общест-
венная (Microtus so-cialis) 25 6218 3756 60,4 99
Полевка рыжая (Clethrionomys gla-reolus) 22 2366 2771 117 99
Крыса пасюк (серая) (Rattus norvegi-cus) 20 4615 2529 54,8 99
Крыса пасюк альбинос (Rattus norvegi-cus var. albino . . . 20 3413 2704 79,2 95
Незокия (Nesokia indica) ..... . 16 1390 1375 98,5 95
Мышь домовая /20 4412 4061 92 96
(Mus musculis) . . (20 4173 4127 98,8 95
Мышь белая . . . Соня лесная (Dyro- 22 8168 6309 77,2 99
mys nitedula) .... 25 3767 3166 84 99
394
Продолжение табл. 41
Отряд Жипотное Температура камеры (rC) Потребление O2 (мл/кг,ч) Обмен веществ (в % от обмена одиночек) Вероятность случайности (%)
у изолированных ЖИВОТНЫХ в группе
Пеструшка степная (Lagurus lagurus) . . 22 7537 6802 90 95
Хомячок серый (Cricetulus migrato-rius) ....... 23 3079 3728 121 98
Хомяк крысовид- f 18 1568 1609 102,2 95
ный (Cricetulus triton) 118 1846 1452 78 99
Песчанка мало- азийская (Meriones 125 2504 1540 61,5 99
tristrami t 25 3789 2153 65,5 —
Суслик тонкопалый (Spermophilopsis lep- 120 1062 1040 97,9 95
todactylus) 120 888 787 89,2 95
Суслик малый (Ci- 1 25 2807 1484 52,8 99
tellus pygmaeus) . . 125 2648 1476 55,7 99
Хищники Норка (Mustela lut- / 13 2972 2336 78,6 98
reola) I 13 3360 2356 70,1 99
Кошка домашняя . 20 1066 751 70,5 99
Обезья- Зеленая мартышка
II ы (Cercopithecus griseo-viridis) 28 579 632 108,9 99
Павиан-гамадрил (Papio hamadrylas) 24 689 475 68,9 99
Однако описанные выше рефлекторные изменения газообмена проявляются далеко не во всяком физиологическом состоянии. Наибольший эффект удается наблюдать на группах, состоящих из особей одного пола, или на разнополых особях вне периодов спаривания (гона). В периоды спаривания «стадный» эффект резко уменьшается или исчезает, заменяясь значительным повышением обмена при совместном пребывании однополых
395
Рис. 85. Схема постановки опытов с изучением стадных рефлексов на газообмен. I—группа; II—разделенная группа; III — одиночка
особей, что особенно ярко проявляется у малого суслика в весеннее время.
Специальной формой эксперимента, когда животные помещались в отдельные клеточки в общей камере, удалось установить наличие стадного эффекта. Однако когда часть животных была заменена источниками теплового излучения равной интенсивности, то стадный эффект оставшегося в камере животного исчез. Следовательно, изменения газообмена не могут быть сведены к процессам ограничения теплоотдачи, т. е. к процессам терморегуляции (рис. 85).
Физиологические сдвиги, происходящие в группе себе подобных организмов, не ограничиваются только изменением газообмена. Например, в группе овец можно наблюдать учащение пульса и дыхания при удалении других овец. У взрослых овец этот эффект выражен ярче, чем у ягнят (Понугаева и Трубицына, 1955). Так, частота сердечных сокращений у новорожденного ягненка, оставшегося без овцы-матки на 30—40 увеличивалась на 5 ударов у серой каракульской породы, на 12 у черных и на 14 у
гибридов. В момент удаления матки у серых пульс учащался на 17, у черных на 13 и у гибридов на 2 удара; при приближении матки у серых пульс повышался на 3 удара, у черных на 9, у гибридов архарокаракуль на 12. Изменения дыхания были незначительны. На 5-й день после рождения характер изменения пульса и дыхания остался тем же (изучалась реакция только у гибридов). Так, у находящегося с овцой-маткон ягненка частота пульса была 218 ударов, во время ухода матки частота
396
пульса увеличилась до 256 ударов. Когда ягненок остался один, пульс понизился до 200 ударов, при приближении овцы-матки увеличился до 238.
Анализаторы, раздражение которых вызывает описанные эффекты, по-видимому, достаточно разнообразны и до сих пор у грызунов изучены мало; вероятно,
температуре внешней среды (по Понугаевой, I960)./ — песчанка; II— общественная полевка; III — обыкновенная полевка:
1 — одиночные животные, 2 — животные в группе. По оси ординат — потребление Оз (мл'мин/г)' по оси абсцисс — температура среды
все анализаторы принимают участие при изменении реакции газообмена в группе. Установлена роль зрительного, обонятельного и, по-видимому, звукового анализаторов. Опыт показывает, что если этих животных рассадить по отдельным камерам, имеющим прозрачные стенки, так, чтобы общение осуществлялось только за счет звукового и зрительного анализаторов, а обонятельный будет выключен, то в первое время стадная реакция исчезает, но через некоторое время вновь обнаруживается. Очевидно, нарушенный динамический стереотип через некоторое время восстанавливается на основе других анализаторов. Однако здесь имеют место большие видовые различия.
Чрезвычайно важен вопрос о роли температурного анализатора в изменениях обмена веществ в стаде. Из рисунка 86 видно, что эти изменения наиболее ярко вы
397
ражаются при низких температурах среды, при средних они уменьшаются, а при высоких сравнительно невелики, по не исчезают полностью. Следовательно, стадный эффект имеет много общего с «групповой» терморегуляцией, но не аналогичен ей.
Стадные изменения обмена веществ обнаруживаются не только у бодрствующего (гомойотермного) животного, и во время зимней спячки. Так, у спящей группы летучих мышей газообмен оказывался пониженным по сравнению с одиночной особью в том же состоянии, при той же температуре среды и температуре тела.
Имеются немногочисленные данные о физиологических механизмах, лежащих в основе наблюдаемых изменений газообмена. Роль щитовидной железы в проявлении стадного эффекта у белых крыс изучалась путем подавления ее функции при введении в организм р-ме-тилтиоурацила — 25,35 и 50 мг ежедневно. Во всех случаях наблюдалось понижение газообмена в зависимости от дозы. Однако на фоне этого сниженного уровня наблюдался отчетливо выраженный стадный эффект (около 15% снижения обмена в группе). Кроме того, были поставлены исследования с полной одномоментной экстирпацией щитовидной железы. В ряде случаев наблюдалось увеличение стадного эффекта на фоне выключения (полного и частичного) функции щитовидной железы. Особенно яркий эффект получен при очень низкой температуре среды (7°С в летний период). Здесь стадный эффект почти отсутствовал у нормальных крыс и составлял около 15% снижения обмена у крыс, получавших метилтиоурацил. Таким образом, в отличие от специфического динамического действия пищи, выпадающего при угнетении функции щитовидной железы, стадный эффект при этом сохраняется и даже становится более значительным.
Снижение газообмена в 70% случаев сопровождается снижением величины электрической активности скелетной мускулатуры. Есть все основания предполагать, что в основе наблюдаемых изменений газообмена лежат сдвиги в состоянии мышечного тонуса, изменения возбуждения мышечных волокон скелетной мускулатуры. Каковы при этом изменения в других органах и в частности во внутренних органах и в центральной нервной системе, должны показать будущие исследования.
398
Стадные и групповые реакции связаны с эндокринными системами. В группе животных постоянно действуют факторы, которые ^ерез нервную систему вызывают изменения в функции эндокринных желез, особенно гипофизарно-адренокортикальной и репродуктивной систем (Christian, 1963). В основе этих явлений лежит реакция стресса, т. е. неспецифических стадий адаптации, особенно ярко выраженных при экспозиции животных в группе на сравнительно короткое время. В опытах Кристиана подсаживание полевки Microtus agrestis в клетку, где уже жила пара полевок этого же вида, приводило к борьбе, покусам животных н к значительной гипертрофии надпочечников и селезенки, атрофии тимуса, увеличению межпозвоночных хрящей, т. е. к явлениям стресса, что было ярче выражено у подсаженных особей, нежели у тех, которые жили здесь ранее.
При сопоставлении числа рубцов от укусов (в процессе борьбы) с перечисленными выше морфофизиологическими изменениями в организме; никакой связи между этими явлениями установить не удалось (Clarke, 1953; Chitty a. oth., 1956; Christian, 1959). Таким образом, физиологические изменения в группе не связаны пи с мышечной деятельностью, пн с травмами, которые животные получают в процессе борьбы.
Физиологические механизмы, лежащие в основе явлений стресса, возникающего в этих условиях, были тщательно изучены и описаны Кристианом (Christian, 1963). Они заключаются в возбуждении гипофизо-адренокортикальной системы, усилении распада белков, подавлении роста костей, соединительной ткани, продукции коллагена, в подавлении митотической активности, что связано и со снижением выделения гормона роста. Повышение активности коры надпочечников (выделение кортикоидов) может оказывать угнетающее влияние и на щитовидную железу. Однако, по-видимому, снижение обмена веществ в группе связано не только с этим угнетением тиреоидальной функции, а и с изменением состояния скелетной мускулатуры и особенно с двигательным поведением животного.
Изучение ориентировочно-исследовательской реакции по «поведению на открытой площадке» (open field behaviour) показало, что изолированное содержание белых крыс ведет к снижению ориентировочно-исследова-
399
тельской активности по сравнению с белыми крысами, содержащимися в группе. Ориентировочно-исследовательская активность как у изолированных, так и у содержащихся в группе животных несколько выше в зимний период (январь—февраль), чем в весенне-летний.
Меняется в зависимости от сезона и активность гипофизарно-надпочечниковой системы. Однако направленность этих изменений различна у «групповых» и изолированных животных (Старыгин, 1969). В зимнее время уровень кортикостерона в крови у содержащихся в группе животных в 2—2,5 раза выше, чем у изолированных. С наступлением весны активность гипофизарно-надпочечниковой системы у изолированных животных повышается, а у групповых несколько снижается. В результате в мае уровень кортикостерона у групповых животных оказывается в 2 раза ниже, чем у изолированных.
СТАДНЫЕ РЕАКЦИИ И «СОЦИАЛЬНОЕ» ДОМИНИРОВАНИЕ
Стадные реакции и физиологическое состояние животных в стаде неразрывно связаны со своеобразной структурой стада или группы, которую в зарубежной литературе называют «общественной» или «социальной». Эти явления носят характер либо более простой иерархии «доминирования» или «подчинения», либо более сложной специализации, что имеет место особенно у «социальных» видов насекомых.
Лучше всего формы доминирования и подчинения описаны для низших обезьян (Hartmann, 1932; Войтонис, 1948; Тих, 1950). Более простые формы изучены в работах Дэвиса и Кристиана (Davis a. Christian, 1959) на домовых мышах и Эйкнесса (Eiknes, 1959) на собаках. В группе домовых -мышей, помещенных в общую клетку, возникает борьба, вскоре прекращающаяся. Вместо борьбы устанавливается своеобразная иерархия, когда одна мышь доминирует, а другая подчиняется. Такой иерархии подчинены все члены этой группы. Установление иерархии сопровождается гипертрофией коры надпочечника, наибольшей у животных, оказавшихся в самом низу иерархической лестницы. Гипертрофия уменьшается по мере подъема вверх по иерархической лестнице и у доминирующих животных полностью отсутствует или вы-400
ряжена очень слабо. Наблюдается подавление секреции гонадотропных гормонов в связи с увеличенной секрецией гипофиза и кортикостероидов. В этих условиях у доминирующих мышей увеличивались и органы размножения (Davis a. Christiarl, 1957).
Доминирующие по поведению собаки отличались меньшей секрецией кортикостероидов по сравнению с собаками, занявшими подчиненное положение. Такие же данные были получены для самцов серых крыс (Rattus norvegicus) методом изучения содержания эозинофилов в крови. Здесь особенно выявились изменения в крови животных, которых подсаживали в колонии с уже сформировавшимися отношениями между их членами (Barnett, 1958).
Кристиан считает, что повышение адренокортикальной активности и снижение функции размножения у грызунов происходит в результате массированного раздражения нервной системы у подчиненных животных, т. е. у тех из них, которые уклоняются от борьбы с доминирующими.
Факты и сопоставления Кристиана были в известной мере подтверждены на обезьянах-резусах. При содержании этих животных в- группах наблюдалось значительное увеличение гидрокортикоидов в моче (Mason, 1959).
Своеобразные групповые отношения хорошо изучены и на многих видах птиц, особенно домашних курах. Здесь доминируют, как правило, петухи. Их доминирование прерывается периодом насиживания и выращивания выводка цыплят. Затем куры возвращаются в свои группы.
Особенно интересно для рассматриваемой проблемы распределение птиц на определенной территории, также изменяющееся в связи с репродуктивным циклом. Опыт, поставленный на изолированном необитаемом коралловом острове (Mebride, 1967), показал четкое разделение территории у ряда образовавшихся групп птиц.
Физиологические механизмы, лежащие в основе «социального» доминирования или подчинения, не представляются ясными. Они включают свойства нервной системы, в свою очередь зависящие от особенностей гормонального аппарата. Свойства нервной системы грызунов (Федоров, 1962) значительно варьируют как по силе, так и по уравновешенности нервных процессов. Это касается как лабораторных популяций крыс, так и диких видов
401
(пасюк, большая песчанка). Отсюда, можно Полагать, возникают различия в стадном поведении и формировании своеобразной иерархии (доминирования и подчинения). Однако эта проблема ждет еще будущих исследователей.
НЕРВНЫЕ И ГОРМОНАЛЬНЫЕ МЕХАНИЗМЫ
РЕГУЛИРОВАНИЯ ЧИСЛЕННОСТИ В ПОПУЛЯЦИИ
Изменения физиологического состояния и появление симптомов адаптации по Селье оказалось возможным наблюдать и в естественных популяциях млекопитающих. Наиболее четкие данные были получены для серой крысы, ряда видов полевок (Microtus), мышей (Рего-niiscus) и оленей (Cervus nippon). Наиболее убедительным было сопоставление городских и сельских популяций серых крыс. При искусственном уменьшении численности популяции в отдельных городских кварталах (являвшихся как бы островами) наблюдалось немедленное и пропорциональное плотности уменьшение веса надпочечников (на 33%) (Christian a. Davis, 1955).
В сельской местности при отлове получены такие же данные, как и для домовых мышей и полевок в естественных популяциях. В естественных популяциях был и эффект внесения новых (чужих) особей в популяцию. При этом наблюдалось общее снижение численности популяции (Calhoun, 1948) и уменьшение веса надпочечников. Возможно, что здесь имело место значительное возрастание смертности средн вновь внесенных особей, а также среди «подчиненной» части аборигенов популяции.
Имеются данные о зависимости между естественными колебаниями численности и весом надпочечников у полевок (Christian, 1959). Сезонные изменения численности также могут быть сопоставлены с весом надпочечников. Так, резкое сокращение плотности популяции полевок от сентября к ноябрю сопровождается снижением веса надпочечников. С ноября до марта, несмотря на стрессирующие зимние условия, вес надпочечников, а также низкая плотность популяции поддерживались на одинаковом уровне. Существует обратная зависимость между плотностью естественной популяции и содержанием эозинофилов в крови полевок (Louch, 1958).
402
Зависимость веса надпочечников от изменения плотности естественных популяций была установлена также для мышей Peromiscus и для ондатр (Beer a. Meyer, 1951), а также для оленей Cervus nippon в результате их естественной гибелц (Christian a. oth., 1960).
Интересно, что ограничение питания на фоне постоянной плотности популяции не приводит к возрастанию веса надпочечников. В то же время недостаток питания приводит к уменьшению интенсивности размножения, что, возможно, является следствием уменьшения активности гипофиза.
Первыми исследованиями о регулировании численности в популяциях были работы Крю и Мирской (Creu a. Mirskaja, 1931) и Ретцлафа (Retzlaff, 1938), установивших, что численность животных на определенной территории (плотность популяции) приводит к торможению интенсивности размножения. Интенсивность размножения мышей была обратно пропорциональна численности, состоящей из 1, 4, 8 или 12 пар. Уже в этих работах было выявлено, что наиболее агрессивные самки оказывались и наиболее плодовитыми. Ретцлаф также заметил, что неагрессивные подчиненные самки чаще подвергались инфекциям и погибали. Интересно, что понижение температуры среды, при которой содержались мыши, с 20 до 4° С приводило к снижению числа детенышей в помете. Это также связывачось с тем, что при низких температурах мыши скучиваются и это оказывает влияние на их плодовитость. В последующих работах было показано, что эстральные циклы самок мышей в группах оказывались более короткими.
Скученность мышей в клетке в течение трех недель приводит к 30%-ному увеличению площади поперечного сечения коркового слоя надпочечников и к 80%-ному увеличению площади мозгового слоя (Bullough, 1952). Эти сдвиги сопровождались 60%-ным сокращением митотической активности эпидермиса, которая стимулируется секрецией кортикостероидов. Содержание полевок в группе снижает уровень гликогена в печени (Chitty, 1955).
Таким образом, эмпирически были установлены зависимости между плотностью популяции млекопитающих— грызунов, изменениями в надпочечниках и репродуктивной функцией. Последняя сопровождается уменьшением веса семенников как абсолютного, так и
403
Рис. 87 Схема регулирования численности особей в популяции (по Christian, 1963)
по отношению к тоже уменьшающемуся весу тела. Эти явления сопровождаются уменьшением веса тимуса.
Все изложенные выше физиологические изменения, возникающие в экспериментальных условиях в популяциях различной плотности, позволили Кристиану высказать положение о саморегуляции численности популяции млекопитающих. Схема такой саморегуляции представлена на рисунке 87. В левой части рисунка указаны факторы, оказывающие угнетающее влияние на численность животных. Это факторы «социального давления», возникающего внутри популяции на основе агрегационных реакций, возрастания численности смертности взрослых животных. В правой части рисунка приводится последовательность развивающихся при этом физиологических и патологических явлений. Все эти явления возникают после достижения популяцией максимальной плотности и поэтому действуют по принципу механизма обратной связи. Изложенная схема, построенная на экспериментальных популяциях, получила некоторое подтверждение на естественных популяциях.
За последние годы накоплен значительный материал, характеризующий соотношения между состоянием естественных популяций млекопитающих и эндокринного аппарата. Так, установлен значительный параллелизм между численностью популяции и весом надпочечника и числом взрослых (зрелых) самок у полевки Microius pennsylvanicus (Christian a. Davis, 1966). Установлена также и часть коркового вещества надпочечника (внутренняя юкстомедуллярная часть), связанная с секрецией эстрогенов, которая увеличена при созревании самок полевок. Все это приводит к установлению корреляций между весом надпочечников и величиной популяции.
В естественных популяциях приобретает, однако, значение не только факт воспроизведения — рождаемость, но и смертность в данной популяции. Последняя зависит от сопротивляемости организма, в частности сопротивляемости инфекциям. В частности на мышах в лаборатории и на диком американском грызуне Arctomys топах было изучено влияние введения АКТГ на протекание гломерулонефрита, очень частого заболевания среди лабораторных и диких популяций грызунов. Нагрузка этим гормоном увеличивала смертность животных. Эта смерт
405
ность стояла в прямой связи с «социальной» структурой популяции (Christian, Lloyd a. Davis, 1965).
Совершенно определенные данные о значении перенаселения для смертности оленей Cervas nippon были получены на популяции, интродуцированной в штате Мэриленд (США). Наблюдалась гибель до 60% стада при явлениях гломерулонефрита, а также расстройства водноэлектролитного обмена в связи с нарушениями симпатоадреналовой и адренокортикальной регуляций. Численность высших организмов в природе может зависеть не только от физических факторов среды, не от условий питания, а от плотности популяции.
Вопрос о влиянии факторов среды и условий существования на популяционные отношения очень мало изучен. По данным Кристиана, условия питания не оказывают влияния на иерархические отношения в группе крыс и не влияют на процессы размножения.
Интересные факты добыты при изучении солевого питания животных Гленном, Ауманном и Эмленом (Glenn, Aumann, Emlen, 1965); они показали, что между плотностью популяций грызунов в природе и содержанием натрия в почве существует тесная корреляция. При низком содержании натрия в почве численность полевок была 30 особей на акр, при среднем — до 400, а при высоком— до 1000. Эти положения добыты на основании анализа большого количества литературных источников. Поставленные в лаборатории эксперименты подтвердили эти положения. Животные с высокой плотностью популяции по сравнению с популяциями малой плотности выбирали большие концентрации солевых растворов. Натриевый дефицит в плотных популяциях авторы рассматривают как возникающие в них нарушения адренокортикальной регуляции.
Эти факты получили свое отражение и развитие в работе А. Ф. Баженовой и М. Г. Колпакова (1969), установивших в условиях высокогорья Памира расселение полевок Alticola в районах с высоким содержанием натрия в почве и растениях. При переводе полевок в условиях эксперимента на рацион с меньшим содержанием натрия выявлена возможность адаптации и сохранения концентраций натрия и калия в крови. Следовательно, при высокой плотности популяции механизм этой адаптации нарушается.
ЛИТЕРАТУРА
Авазбакиева М. Ф., 1958. Влияние климата Казахстана и Киргизии на организм человека. Алма-Ата.
Авазбакиева М. Ф., 1959. К вопросу о сезонных изменениях крови, кровообращения и основного обмена у человека. Совет, по экол. физиол., тез. докл.. Изд-во АН СССР, вып. 2., М.— Л.: 3.
Александров И. С., 1954. Влияние на слюнные железы веществ, возбуждающих центральную нервную систему. Автореф. дисс., Л.
Аманова ЛЕ, 1968. Интерьерные особенности скотоцерки в Каракумах. Всес. научн. коиф. по изуч. и освоению пустынных тер-рит. Ср. Азии и Казахст. Секц. 2. Подсекция «Животный мир». Тез. докл., Ашхабад: 37.
Анохин П. К., 1968. Биология и нейрофизиология условного рефлекса. Изд-во «Медицина», М.
Антелндзе Б. и Барбашова 3., 1938. Сравнительная характеристика дыхательной функции крови некоторых копытных животных. Физиол. жури. СССР, т. 25, вып. 4.
Антошкина Е. Д., 1939 а. Онтогенетическое развитие терморегуляции. Физиол. жури. СССР, т. 26, вып. 1.
Антошкина Е. Д., 1939 б. Онтогенетическое развитие терморегуляции. Сообщ. 2. Физиол. жури. СССР, т. 26, вып. 1.
Арав И. М., 1950 а. Картина красной крови у яка Памира. Сообщ. Таджик, фпл. АН СССР, вып. 26, Душанбе: 31.
Арав И. М., 1951. Сравнительная характеристика картины крови у равнинных и горных пород овец. Тр. Таджик, фил. АН СССР, вып. 28, Душанбе: 133.
Арав И. М., 1960. Обмен веществ у каракульских овец в горах. Тез. докл. 2-й конф, физиол., биохим. и фармакол. Ср. Азии, Фрунзе: 39.
Арав И. М. и Мартынова В. Н., 1959. Результаты исследований по физиологии гиссарских овец при отгонно-пастбищной системе содержания. Всес. совещ. по физиол. и биохим. с.-х. животных. Тез. докл. Изд-во АН СССР, М. — Л.: 128.
Арав И. М. и С м о л и ч е в Е. П., 1947. Акклиматизация сельскохозяйственных животных в горах. Сообщ. 2. Некоторые особенности крови в горах. Изв. Таджик, фил. АН СССР, № 14, Душанбе: 23.
Арбузов С. Я., 1952. Влияние химических веществ на зимнеспящих животных. Усп. совр. биол., т. 33, вып. 1.
А р е ш е в а 3. С., С к л я р ч и к Е. Л., Ш е г л о в а А. И., 1968. Возрастные изменения содержания воды в теле у большой песчанки. В кн.: «Сравнительная и возрастная физиология», 186. Изд-во «Наука», Л.
Арешева 3. С. и Щеглова А. И., 1961. Распределение воды в организме некоторых грызунов. Тез. докл. III конф, по эволюц. физиол., Л.: 8.
407
Арешева 3. С. и Щеглова А. И., 1968. Содержание мочевины в крови некоторых грызунов при разных водных режимах. В кн.: «Сравнительная и возрастная физиология», Изд-во «Наука», Л.: 7.
Арнаутов П. Д. и Веллер Е. Г., 1931. Влияние естественного и искусственного света на газообмен у человека. Русск. физиол., т. 14, вып. 2.
Архангельская Н. А., 1949а. Влияние низких и высоких температур среды на устойчивость к гипоксической гипоксии. Сб. «Опыт изуч. регуляц. физиол. функц.». Изд-во АН СССР, М. — Л.: 107.
Архангельская Н. А., 1949 б. Влияние функционального выключения и экстирпации коры головного мозга на устойчивость организма к гипоксической гипоксии. Сб. «Опыт изуч. регуляц. физиол. функц.» Изд-во АН СССР, М. — Л.: 123.
Архангельская Н. А., К а н ф о р И. С., Слоним А. Д. и Черкович Е. М., 1949. Материалы по изучению гомеостазиса у летучих мышей. Реф. науч.-исслед. работ АМН СССР, сер. медико-биол. наук, № 7, М.: 163.
Архангельская Н. А. и Сегаль Н. А., 1954. Регуляция дыхания в условиях недостатка кислорода. Бюлл. эксп. биол. и мед., № 23.
Аршавский И. А., 1958. К сравнительно-онтогенетической характеристике некоторых черт физиологии кроликов и зайцев в связи с особенностями их экологии. Совещ. по экол. физиол., тез. докл., вып. 1, изд-во АН СССР, М. — Л.: 3.
Аршавский И. А., и Пятки и Е. М., 1958. К механизму изменения естественного ритма сердца у крупного рогатого скота в процессе постнатального онтогенеза. Совещ. по физиол. и патол. нервн. сист. животных и человека ранних возрасти, этапов развития. М.: 7.
Багдасарова Т. А., 1965. Об адаптации детского организма в горах Малого Кавказа. Тр. IX Всес. съезда физиол., т. П. Изд-во АН СССР, М. — Л.
Баженов Ю. И., 1968. Мышечная деятельность и химическая терморегуляция. Автореф., Л.
Баженова А. Ф. и Колпаков М. Г., 1969. Состояние натриевого гомеостаза у травоядных животных, адаптированных к условиям высокогорья. В кн.: Сборн. реферат, к V конф, физиол. Ср. Азии и Казахст., стр. 6. Новосибирск.
Базанова Н. У. и С теп ан кин а И. М., 1957. Физиологические особенности деятельности пищеварительной системы овей и верблюдов в зависимости от породы, возраста, условий кормления и содержания. Вопр. физиол., с.-х. животн. Изд-во АН СССР, Л.: 216.
Баранникова И. А., 1965. Функциональные основы миграционного поведения анадромных рыб. В кн. «Биохимическое значение и функциональная детерминация миграционного поведения животных». Изд-во «Наука», М. — Л.: 32.
Барбашова 3. И., 1941. Материалы к проблеме акклиматизации к низким парциальным давлениям кислорода. Изд-во АН СССР, М, —Л.
Барбашова 3. И., 1960. Акклиматизация к гипоксии и ее физиологические механизмы. Изд-во АН СССР, М. — Л.: 12.
408
Б ар баш ова 3. И. и Гинецинский А. Г., 1942. Особенности приспособления к высоте у гиссарских овец. Изв. АН СССР, сер. биол. № 5: 295.
Бартон А. и Эд холм О., 1957. Человек в условиях холода. Изд-во ИЛ, М.
Бахтиозина Б. X., 1948. К изучению водного обмена у верблюдов. Вести. АН КазССР, № 10 (43), Алма-Ата.
Бахтиозина Б. X., 1954. О физиологии водного обмена у верблюда. Автореф. дисс., Л.
Бахтиозина Б. X., 1959. Регуляция водного обмена у верблюда. Уч. зап. Каз. гос. пед. ин-та., сер. естеств.-геогр., т. 17, Алма-Ата: 30.
Бахтиозина Б. X., 1960 а. Количественная характеристика водного обмена у верблюда. Изв. АН КазССР, сер. мед. и биол., вып. 1 (13): 90.
Бахтиозина Б. X., 1960 б. Изменение объема воды при водном голодании верблюда. Изв. АН КазССР, сер. мед. и биол., вып. 2(14): 101.
Бахтиозина Б. X., 1961 а. Содержание хлоридов в моче и крови у верблюда при водном голодании. Изв. АН КазССР, сер. мед. и биол., вып. 1 (15): 37.
Бахтиозина Б. X., 1961 б. Влияние водного голодания на состав крови верблюдов. Изв. АН КазССР, сер. мед. и биол., вып. 1 (15): 27.
Башенина Н. В., 1957. К вопросу о кормлении степных пеструшек при разведении их в неволе. Зоол. журн., т. 36, вып. 12.
Башенина Н. В., 1960. Материалы по онтогенезу терморегуляции обыкновенной полевки и степной пеструшки. Сб. «Фауна и экология грызунов», вып. 6, изд-во МГУ: 111.
Бедалова С. М., 1959а. Изменение венозного давления, скорости кровотока и капиллярного кровообращения у человека под влиянием горного климата Малого Кавказа. Совещ. по экол. физиол., тез. докл., вып. 2. Изд-во АН СССР, М. — Л.: 18.
Бедалова С. М., 1959 б. Изменение пульса и кровяного давления у человека под влиянием горного климата Малого Кавказа. Совещ. по экол. физиол., тез. докл., вып. 11. Изд-во АН СССР, М. — Л.: 17.
Бейнбридж Ф. А., 1927. Физиология мышечной деятельности. ГИЗ, М.
Бельская И. П., 1949. О влиянии интероцептивных раздражений на акт еды. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР, М. — Л.: 266.
Бельская И. П. и Войтенко Н. И., 1967. К вопросу об участии пищеварительного тракта грызунов в адаптации их к обезвоживанию сухоядением. Матер, к III Всес. совещ. по экол. физиол., биохим., морфол. Т. «Биохимич., фармакол. и токсикол. аспекты исслед. адаптаций», Новосибирск: 15.
Беляев Д. К-, 1950. Роль света в управлении биологическими ритмами млекопитающих. Журн. общ. биол., т. 11, № 1.
Бестужев А. Г., 1951. Кормление двугорбых верблюдов. Алма-Ата.
Бирюков Д. А., 1960. Экологическая физиология нервной деятельности. Медгиз, Л.
409
Бисерова А. Г. и Гембчински М., 1964. Об изменениях холодовой дрожи при повторных охлаждениях организма. В ки.: «Теплообразование в организме». Изд-во «Наукова думка»: 38.
Б оки на А. И., 1968. Физиологические основы гигиенического нормирования солевого состава питьевых вод. Автореф. дисс., М.
Бондаренко А. И., Калабухов Н. И., Козаке-вич В. П. и Скворцов Г. Н., 1968. Сезонные изменения содержания аскорбиновой кислоты и токоферола у грызунов. Жури, эволюц. биохим. и физиол. Изд-во «Наука», т. IV, № 2: 137.
Бреслав И. С., 1952. Газообмен и условные пищеварительные рефлексы у собак. Тр. ин-та физиол. им И. П. Павлова АН СССР, т. I: 103.
Булатова Н. А., 1953. Некоторые особенности дыхательной функции крови высокогорных животных. Автореф. дисс., М.
Бухарин Н. С., 1949. Краткие итоги работы по созданию новой породы овец архаромеринос. Изв. АН КазССР, т. 1, вып. 5, Алма-Ата.
Быков К. М., 1942. Кора головного мозга и внутренние органы. Киров; 1952, изд. 2-е. Медгиз, М. — Л.
Быков К. М. и Мартинсон Э. Э., 1933. Материалы по физиологии горного климата. Арх. биол. наук., т. 32, вып. 1—2.
Быков К. М. и Слон и м А. Д., 1960. Исследование слож-ио-рефлекторной деятельности животных и человека в естественных условиях существования. Изд-во АН СССР, М. — Л.
Бюннинг Э., 1961. Ритмы физиологических процессов («физиологические часы»), Медгиз, М.
Ван Лир Э. и Стик ней К., 1967. Гипоксия. Изд-во «Медицина».
Басенко Е. П., 1959. Некоторые вопросы экологии сельскохозяйственных животных. Объед. научи, сессия по пробл. повыщ. продукт, животн. в Казахст., тез. докл., Алма-Ата: 168.
Верболович П. А., Валитова М. С., Гроссман Ю. Г., Куровская Н. И. и Пухова Н. Н., 1955. Влияние условий существования на химический состав мышц животных. Тез. докл. VIII Всес. съезда физиол., биохим. и фармакол. М„: 113.
Верболович П. А. и Остапюк 3. А., 1956. О содержании гемоглобина в крови диких и домашних животных. I конф, физиол., биохим. и фармакол. Средн. Азии и Казахстана, тез. докл., Ташкент: 56.
Вержбинская Н. А., 1946. Угольная ангидраза в мозгу млекопитающих. Изд. АН СССР, сер. биол., № 1: 135.
Веселкин П. Н., 1942. О влиянии пониженного барометрического давления на животных с экспериментальным гипотиреозом. Билл. эксп. биол. и мед., т. 14, вып. 4.
Веселкин П. Н., 1945. Бюлл. эксп. биол. и мед., т. 14, вып. 4.
Веселкин П. Н., 1945. Тепловая одышка. Л.
Виноградов М. И., 1941. Очерки по энергетике мышечной деятельности. Изд-во ЛГУ, Л.
Виноградов М. И., 1958. Физиология трудовых процессов. Изд-во ЛГУ.
Виноградова-Федорова Т. В., Михина Т. Н., Солдатенков П. Ф., Павлов Г. Н., Трофимова А. И., 1935. 410
К вопросу о роли инфузорий жвачных в усвоении азота мочевины. Физиол. жури. СССР, т. 19, вып. 3.
Войткевич А. А., 1951. Зависимость функции щитовидной железы от центров переднего мозга. Журн. общ. биол., т. 12, № 5.
Войткевич В. II., 1952. Оксигемометрические исследования насыщения артериальной крови в здоровом и больном организме. Автореф. днсс., Л.
Войтоннс Н. Ю., 1948. Предыстория интеллекта. Изд-во АН СССР, М.
Воронин Л. Г., 1947. К вопросу об имитационных способностях у низших обезьян. Физиол. журн. СССР, т. 33, № 3.
Высочи на Т. К., 1967. Ионные сдвиги в организме мелких грызунов при экспериментально вызванной дегидратации. Сб. «Гормональные механизмы адаптации. Сезонная периодика в организме. Адаптация водно-солевого обмена». Матер. III Всес. совещ. по экол., физиол., биохнм. и морфол. Изд-во СО АН СССР. Новосибирск: 154.
Галанин II. Ф., 1952. Лучистая энергия и ее гигиеническое значение. Л.
Галкин В. С., 1947. Об изучении нервных механизмов физиологических и патологических реакций. Сб. «Механизм патол. реакций». вып. 9—10., Л.: 5.
Гамбарян О. И., 1953. Адаптивные особенности передних конечностей слепца. Зоол. сб., вып. 8. Изд. АН АрмССР, Ереван: 103.
Гербильский Н. Л., 1965. Биологическое значение и функциональная детерминация миграционного поведения рыб. В кн.: «Биологическое значение и функциональная детерминация миграционного поведения животных». Изд-во «Наука», М. — Л.: 23.
Гиляров М. С., 1964. Основные направления приспособлений насекомых к жизни в пустыне. Зоологич. журн., т. 43, вып. 3, стр. 443.
Г и н е ц и н с к и й А. Г., 1963. Физиология водно-солевого равновесия. Изд-во АН СССР, М. — Л.
Г л а д к и н а Т. С„ Мейер М. Н. и М о к е е в а Т. М„ 1958. Некоторые морфо-физпологические отличия подвидов степной пеструшки и их реакции на температурные условия. Совещ. по экол., физиол., тез. докл. вып. 1, изд-во АН СССР, М. — Л.: 15.
Гладков Н. А., 1960. Полет птиц. Изд. МОИН, М.
Григоренко В. Е., 1967. Гиалуронидазная активность тканей у животных с разной экологической специализацией. Матер, к III Всес. конф, по экол. физиол., биохим. и морфол., вып. «Биохимия», Новосибирск: 12.
Гуменер П. И., 1962. Изучение терморегуляции в гигиене и физиологии труда. Медгиз, М.
Гусева Л. А., 1964. Изменение некоторых окислительных процессов при адаптации к холоду. Сб. «Теплообразование в организме», Киев: 58.
Давыдов А. Ф., 1958. Наблюдения по терморегуляции у северного оленя. Сб. «Вопросы физиологии и патологии нервной системы», Л.: 47.
Давыдов А. Ф., Лобанов Н. В., Макарова А. Р., Рашевская Д. А. и Юнусов С., 1967. Особенности терморегуляции у представителей семейства Equidae в онтогенезе в условиях высокой температуры. В кн.: «Физиол.-генетич. исслед. адаптаций у животных». Изд-во «Наука»: 5.
Давыдов А. Ф., Лобанов Н. В., Макарова А. Р.,
411
Рашевская Д. Л. и Юнусов С., 1967. Возрастные изменения легочного дыхания и кровообращения в условиях тепловой нагрузки у муфлонов, тонкорунных овец и их гибридов. В кн.: «Физиол,-генетич. исслед. адаптаций у животн.», изд-во «Наука»: 21.
Давыдов А. Ф., Макарова А. Р., 1964. Изменения терморегуляции и кровообращения у новорожденных тюленей прп переходе к водному образу жизни. Физиол. журн. СССР, т. 50, № 7: 894.
Данишевский Г. М., 1955. Акклиматизация человека на севере. Медгиз, М.
Даудова Г. М. и Ипатьева Н. В., 1968. Изменение температуры в различных участках тела сусликов Citellus major во время искусственного пробуждения от зимней спячки. Журн. эволюц. биохим. и физиол. Изд-во «Наука», т. 4, № 4: 348.
Дейвис Т. и Р. Джой. 1965. Естественное и искусственное приспособление человека к холоду. В кн.: «Биометеорология». Изд. Гидрометеорол., Л.
Дерибас В. И., Ливчак Г. Б., Филипченко Р. Е. и Шошенко К. А., 1966. К вопросу о функциях «красных» и «белых» волокон в скелетных мышцах белых крыс. Тез. докл. Всес. конф, по терморегуляции, Л.
Долинин В. А., 1967. Исследование основного обмена и водных потерь у пустынных видов рептилий. Реф. покл. к III Всес. совещ. по экол. физиол.. биохим., морфол., т. «Видовые и прпродно-климат. адаптации», Новосибирск: 33.
Дольник В. Р., 1965. Физиологические основы миграций птиц. Сб. «Биологическое значение и функциональная детерминация миграционного поведения животных». Изд-во «Наука», М. — Л.: 12.
Дорохова Б. Р., 1967. Исследование водно-солевого обмена у белых крыс и больших песчанок (Rhomb omis opimus) при водной и солевой нагрузках. Матер, к III Всесоюзн. совещ. по экол. физиол., биохим., морфол., т. «Гормон, механизмы адаптации», Новосибирск: 159.
Дорохова Б. Р. и Серебряков Е. П., 1967. Изменение водно-солевого обмена у белой крысы и большой песчанки (Rhombo-mis opimus') в условиях дегидратации. Матер, к III Всес. совещ. по экол. физиол., биохим., морфол., т. «Гормональные механизмы адаптации...», Новосибирск.
Дубинина Ж. С., Ковалева Р. И. и М и р р а х и-м о в М. М., 1967. К вопросу о регуляции дыхания на высоте. Сб. «Физиол. и патол. организма в усл. высокогорья». Фрунзе: 7.
Дэн Су-и и Иванов К. П., 1958. О влиянии режима освещения на периодику физиологических функций у «ночных» животных (лемуров). Совещ. по экол. физиол., тез. докл., вып. 1. Изд-во АН СССР, М—Л.: 17.
Жила Е. С., 1940. Материалы к сравнительной физиологии терморегуляции. Терморегуляция у новорожденных животных (грызуны. хищники, приматы). Физиол. журн. СССР, т. 28, вып. 4.
Жуков А. П. и Кожевников В. А., 1947. Сравнительная характеристика красной крови в горах. Изд. Таджик, фпл. АН СССР, № 14, Душанбе: 731.
Журавлев И. II., 1948. О питьевой возбудимости. XIII совещ. по физиол. пробл., тез. докл.. Л.: 41.
Журавлев И. Н„ 1952. Анализ двигательных пищевых реакций в свете учения И. П. Павлова о пищевом центре и анализаторах. 412
Сб. «50 лет учения акад. И. П. Павлова». Изд-во АН СССР, М. — Л: 231.
Завадовский И. М., 1930. Исследования по экспериментальной зоологии летом 1923 г. Сб. «Аскания Нова», ГИЗ, М.: 343.
3 а х а р я и А. Б., 1965. Основной обмен у лиц, проживающих в условиях высокогорья. Арагац. В кн.: «Матер, симпозиума «Газообмен в условиях высокогорья», Фрунзе: 21.
Зверев М. Д., 1948. К вопросу о быстроте бега некоторых животных. Тр. Алма-Ат. гос. запов., вып. 7: 145.
3 и м к и н Н. В., 1956. Физиологическая характеристика силы, быстроты и выносливости. Изд-во «Физкультура и спорт», М.
Иванов К. П., 1954. Регистрация дыхательных движений у грызунов. Физиол. журн. СССР, т. 40, вып. 3.
Иванов К. П., 1959. Потребление кислорода и изменение тонуса скелетных мышц при гипоксии. Физиол. журн. СССР, т. 46, вып. 8.
Иванов К- П., 1965. Мышечная система и химическая терморегуляция. Изд-во «Наука», М. — Л.
Иванов К- П. и АлимухаммедовА. А.» 1963. О физиологических механизмах химической терморегуляции в онтогенезе. Физиол. журн. СССР, т. 49, № 4.
Иванов К. П., Макарова А. Р. и Ф уф а ч ев а А. А., 1953. Влияние интенсивности освещения на газообмен у обезьян. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2, Изд-во АН СССР, М. —Л.,: 94.
Израэль А. И., 1936а. Значение факторов высокогорья в физиологии человека и животных. Труды САГУ, серия VIII, вып. 24.
Израэль А. И., 1936 б. Физиология лошади на горных перевалах Тянь-Шаня. Тр. Среднеаз. гос. ун-та, сер. Villa, вып. 28, Ташкент.
Имшепецкпй А. В., 1953. Белок микроорганизмов в пищеварительном тракте жвачных животных. Микробиология целлюлозы. Госиздат., М.
И с а а к я п Л. А., 1946. Филогенез кортикального механизма терморегуляции у млекопитающих. Дисс. АМН СССР, Л.
И саакян Л. А., 1949. О кортикальном механизме терморегуляции у некоторых отрядов млекопитающих. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР, М. — Л.: 21.
И саакян Л. А., 1953. Об условнорефлекторном механизме терморегуляции у ежей. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2. Изц-во АН СССР, М. — Л.: 46.
Исаакян Л. А. и Акчурин Р. И., 1953. Сезонные изменения газообмена у хищных млекопитающих. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2. Изд. АН СССР, М.— Л.: 217.
Исаакян Л. А. и Избинекий А. Л., 1951. Материалы к вопросу о сезонных изменениях газообмена и терморегуляции. Бюлл. эксп. биол. и мед., т. 32. вып. 5, № 11.
Исаакян Л. А. и Ф е л ь б е р б а у м Р. А., 1949. Физиологические исследования желтого суслика (Citellus fulvus) при впадении в летнюю спячку. Сб. «Опыт изуч. период изменений физиол. функций в организме». Изд-во АМН СССР, М. — Л.: 194.
Исхаков 3. А., 1954. Изучение пищеварительной деятельности каракульских овец в условиях пустыни. Автореф. дисс.
Исхаков 3. А., 1960. Пищеварительная деятельность кара
413
кульских овец в условиях пустыни. Сб. «Биология каракульской ов-цы». Изд-во АН Уз. ССР, Ташкент: 63.
Калабухов Н. И., 1933 а. Анабиоз у животных при температурах ниже 0°. 1. Действие низких температур на летучих мышей. Бюлл. МОИП, т. 47, вып. 3.
Калабухов Н. И., 1933 б. Физиологическая закономерность в изменении эритроцитов у млекопитающих. Зоол. журн., т. 12, вып. 1.
Калабухов Н. И., 1935. Физиологические особенности горных и равнинных подвидов лесной мыши (Apodemus silvaticus). Докл. АН СССР, нов. сер., т. 2. вып. 1.
Калабухов Н. И., 1936. Спячка животных. Бпомедгиз, ДА.
Калабухов Н. II., 1937. Особенности реакции некоторых видон равнинных грызунов на понижения атмосферного давления. Зоол. журн., т. 16,: 48.
Калабухов II. И., 1938. Некоторые экологические особенности близких видов грызунов. 1. Особенности реакции лесных мышей (Apodemus silvaticus п A. flavicollis) и сусликов (Cytellus pygtnaeus и С. suslica) на интенсивность освещения. Зоол. жури., т. 17, вып. 3.
Калабухов Н. И., 1940. Суточный цикл активности животных. Усп. совр. биол., т. 12, вып. 1.
Калабухов Н. И., 1944. О зависимости темпа дыхания млекопитающих от температуры. Докл. АН СССР, т. 53, № 9.
Калабухов И. II., 1950. Эколого-физиологические особенности животных и условия среды. 1. Дивергенция некоторых эколого-физиологических признаков близких форм млекопитающих. Харьков.
Калабухов Н. И., 1953. Сезонные изменения реакции желто-горлых мышей на воздействие условий среды. Бюлл. МОИП, отдел, биол. т. 58, вып. 3.
Калабухов Н. II., 1956. Сезонные изменения реакции краснохвостой Meriones erythrourus и большой Rhombomis opimus песчанок на влияние температуры среды. Тр. Ин-та биол. АН ТуркмССР, вып. 4: 3.
Калабухов Н. И., 1957. Предпочитаемая температура млекопитающих и ее связь с другими особенностями их терморегуляции. «Грызуны и борьба с ними», вып. 5, Саратов: 3.
Калабухов Н. И. и Ладыгина К. М., 1953. Возникновение эколого-физиологических особенностей у млекопитающих под воздействием внешней среды. Зоол. журн., т. 32, вып. 2.
Калабухов Н. И., Ладыгина Е. М., М а й з е л и с М. Р. и Шилова Т. И., 1957. Сезонные изменения в организме некоторых видов мышей и хомячков. Тез. докл. II экол. конф., ч. 3—4, Киев: 92.
Калабухов Н. И., Нургельдыев О. Н. и С к в о р-ц о в Г. Н., 1958. «Жизненные формы» грызунов песчаных и глинпс-стых пустынь Туркмении. Зоол. жури., т. 37, вып. 3.
Калашников В. П„ 1958. О влиянии мышечной тренировки на терморегуляцию у новорожденных козлят при низких температурах среды. Сб. «Опыт изуч. регуляции физиол. функций», т. 4. Изд-во АН СССР, М. —Л.: 158.
Калита ев М. В., 1941. О легочном газообмене и теплопродукции у дойных коров в условиях стойлового и пастбищного режимов. Докл. ВАСХНИЛ, вып. 4, М.: 37.
К а л и х м а н А. А., 1953. Изменения общего газообмена и окислительных процессов в тканях при общем охлаждении теплокровного 414
организма. Сб. «Опыт изуч. действия низких температур на теплокр. организм». Изд. АМН СССР, М.: 61.
Кандрор И. С., 1954. Изменения суточной температурной периодики у человека при переездах на дальние расстояния в широтном направлении. Сб. «Опыт изуч. регуляции физиол. функций», вып. 3. Изд-во АН СССР, М. — Л.: 185.
Кандрор II. С., 1958. О нормировании пищевого рациона с учетом сезонов и климатических поясов. Вопр. питания, вып. 3: 9.
Кандрор И. С., 1968. Очерки по физиологии и гигиене человека на Крайнем Севере. Изд. «Медицина», М.
Кандрор И. С. и Раппопорт К. А., 1954. Основной обмен у человека в холодном климате. Сб. «Опыт изуч. регуляции физиол. функций, вып. 3. Изд-во АН СССР, М. — Л.: 153.
Кандрор И. С. и Солтысский Е. Н., 1959. Акклиматизация и заболеваемость в холодном климате. Сб. «Физические факторы внешней среды», М.: 365.
Канфор И. С., 1959. О сложнорефлекторной регуляции углеводного обмена. Автореф. дисс., М.
Карташев ск ий Е. А., 1908. О влиянии недостатка кислорода на обмен веществ и теплопроизводство в животном организме. Изв. ВМА, т. 16: 259.
К а ф а р о в а Р. 3., 1966. Возрастные особенности терморегуляции и их сезонные изменения в условиях жаркого климата. Автореф. дисс., Баку.
Каш кар ов Д. В., 1944. Основы экологии животных. Учпедгиз., Л.
Кашкаров Д. И. и Коровин Е. П., 1936. Жизнь пустыни. Биомедгпз, М.
Кекчеев К. X., 1931. Физиология труда. М.
Кескпайк Ю. и Давыдов А., 1966. Теплообмен и поведение птиц в гнездовой период развития. Изв. АН Эст. ССР, т. 15, сер. биол., № 1: 99.
Киршенблат Я. Д., 1968. Телергоны. Химические средства воздействия животных. Изд-во «Наука», М.
Клейненберг С. Е., 1956. Млекопитающие Черного и Азовского морей. Изд-во АН СССР, М.
Климов Н. М., 1949. К физиологии желудочного пищеварения у северных оленей. Сов. зоотехн., № 6.
Ковальский В. В. и Падучева А. Л., 1951. Вопросы водного обмена у каракульских овец. Каракулеводство и звероводство. № 1: 26.
Ковальский В. В., Шульгина Н. С. и Ржешев-ский И. К., 1951. Роль наследственности в приспособительных изменениях водного обмена у близких видов. Укр. биохимич. журн., т. 23, № 3.
Колчинская А. 3., 1965. Гипоксия в детском возрасте. Изд-во «Медицина», Киев.
Комарова Т. Ф., 1952. Процессы дыхания и гликолиза в мозгу куриных эмбрионов в зависимости от условий среды. Автореф. дисс., Л.
Комкова А. И. и Ушаков Б. П., 1955. Температурная инактивация аденозинтрифосфатазы мышц травяной и озерной лягушек. Докл. АН СССР, т. 102, № 6.
415
Конради Г. П.» Слоним А. Д. и Ф а р ф е л ь В. С., 1935. Общие основы физиологии труда. Биомедгиз., М.
Коржуев П. А., 1936. Влияние высокой температуры на трипсин теплокровных и холоднокровных позвоночных животных. Физиол. журн., СССР, т. 21, № 3.
Коржуев П. А. и Коштоянц X. С., 1934. Материалы к сравнительной физиологии пищеварительных ферментов. Зоол. жури., т. 13, вып. 3.
Коропов В. М., 1946. О водном голодании у лошадей. Ветеринария, № 8—9: 30.
Коштоянц X. С., 1923. Влияние различных пищеварительных режимов на секреторную деятельность желудочных желез. Жури, экспер. мед., т. 16(1): 109.
Коштоянц X. С., 1937. Основы сравнительной физиологии, т. 1. Изд-во АН СССР.
Коштоянц X. С., 1947. Основы сравнительной физиологии, т. 1.
К р а т и н о в А. Г., 1947. Спячка млекопитающих и сезонная динамика функционального состояния вегетативной нервной системы. VIII Всес. съезд физиол., биохим. и фармакол., докл., М.: 295.
Крепе Е. М., 1936. Дыхательная функция крови. Бюлл. ВИЭМ, № 3—4, М.
Крепе Е. М., Вержбинская Н. А., ЧеныкаеваЕ. Ю., Чирковская Е. В., Гавурина Ц. К-, 1956. О приспособлении животных к хронической гипоксии (влияние приспособления к хронической гипоксии на «потолок» и на высоту газообмена при пониженном содержании кислорода). Физиол. журн. СССР, т. 42, № 1.
Крестовников А. Н., 1951. Очерки по физиологии физических упражнений. Изд-во «Физкультура и спорт», М.
Кряжев В. Я., 1935. Пищевые рефлексы «стадного характера». Физиол. журн. СССР, т. 18, № 4.
Кряжев В. Я., 1955. Высшая нервная деятельность животных в условиях общения. Медгиз, М.
Кузнецов В. А., 1947. Сезонные колебания влажности
и хлоридов в мясе каракульских овец. Тр. Туркм. с.-х. ин-та, т. 5, Ашхабад.
Кузнецов В. А., 1949. О мясе туркменского одногорбого верблюда. Изв. Туркм. фил. АН СССР, № 3, Ашхабад: 29.
Кузнецов В. А., 1951а. Сезонные колебания влажности и хлоридов в мясе каракульских овец. Тр. Туркм. с.-х. ин-та, т. 5, Ашхабад: 41.
Кузнецов В. А., 1951 б. Мясная продуктивность туркменской каракульской овцы. Автореф. дисс., Л.
Кузякин А. П., 1950. Летучие мыши. Изд-во «Советская наука», М.
Курилов Н. В., 1955. Физиологические основы диетического кормления лошадей. Автореф. дисс., М.
Лабори А. и Гюгенар П., 1956. Гибернотерапия (искусственная зимняя спячка) в медицинской практике. Медгиз, М.
Лазовский Ю. М., 1948. Функциональная морфология в норме и патологии. Изд-во АМН СССР, М.
Лебедева Н. Г., 1954. Особенности в строении органов дыха-
416
ния некоторых новых пород овец и коз. Тр. ин-та эксп. биол. АН КазССР, т. 2, Алма-Ата: 78.
Леонтьев А. Н., 1957.. К суточной активности монгольской песчанки и полевки Брандта. Пзв. Иркутск, науч.-исслед. прогиво-чумн. ин-та Спб. и Д. Востока, вып. 16, Иркутск: 78
Ливчак Г. Б., 1960. Некоторые эколого-физиологические
и биохимические особенности полярных полевок. Автореф. дисс., Л.
Л ишак К. и Эндреци Э., 1967. Иейро-эндокринпая регуляция адаптационной деятельности. Изд. АП, Будапешт.
Лобашев М. Е. и Савватееи В. Б., 1953. Изменение безусловных рефлексов в онтогенезе у кур методом условных рефлексов. Сообщ. 1. Изменение суточного стереотипа и времени кладки яиц. Тр. Ин-та физиол. им. И. II. Павлова АН СССР. Изд-во АН СССР, М. — Л.: 503.
Лукин Е. И., 1940. Дарвинизм и географические изменения организмов. Изд-во АН СССР, М. — Л.
Лутфуллипа М. С., 1956а. Изучение суммарной деятельности главных пищеварительных желез верблюда. Тр. Ин-та биол. АН Каз. ССР, т. 3, Алма-Ата: 40.
Лутфуллина М. С., 19566. К вопросу водного обмена у верблюдов. Тр. Ин-та биол. АН Каз. ССР, т. 3, Алма-Ата: 104.
Ма гидов Г. А., 1959. Сезонные и возрастные изменения в обмене веществ у лошадей. Всес. совещ. по физиол. и биохим. с.-х. животп., тез. докл. Изд. АН СССР, М. — Л.: 151.
М а й з е л и с М. Р., 1953. Влияние мышечной деятельности на суточную периодику активности у грызунов. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2, Изд. АН СССР, М. — Л.: 162.
Макарова А. Р., 1953. О влиянии приемов пищи и режимов питания на газообмен. Автореф. дисс., Л.
Макарова А. Р., П о н у г а е в а А. Г. и Щеглова А. И., 1955. Исследование газообмена и дыхания каракульской овцы в условиях полупустыни. Тез. докл. II совещ. по физиол. с.-х. животн., АН СССР: 34.
Макарова А. Р. и Сегаль А. Н., 1958. Физиологическая характеристика зимней пастьбы северного оленя. Сб. «Опыт изуч. регуляции физиол. функций», т. 4. Изд. АН СССР: 29.
Малиновский О. В., 1953. Безусловная секреция околоушной железы обезьяны макака резус. Физиол. журн. СССР, т. 39, вып. 1.
М а л к и м а н И. В., 1948. Материалы к вопросу о влиянии пониженного барометрического давления на секрецию желудочных желез. Сб. «Влияние пониженного барометр, давл. на процессы пи-щевар.», М.: 19.
Маслов С. П., 1964. О самостоятельном значении специфической терморегуляторной функции мышц. Сб. «Теплообразование в организме», Киев: 130.
Малышева А. Е., 1959. О некоторых функциональных сдвигах при работе в условиях охлаждения. Конф, по физиол., теплообмена и гигиене промышл. микрол., тез. докл., М.,: 31.
Матюхин В. А., 1965. О физиологии и патологии человека в условиях дальневосточного муссонного климата. Автореф. дисс., Л.
Машковцев А. А., 1935. Влияние горного климата на конституцию млекопитающих. Тр. Лабор. эвол. морфол. АН СССР, т. 2, вып. 3, М.
14—1204
417
Меньшикова А. В. ч Пигарева 3. Д., 1945. К сравнительной физиологии угольной ангидразы. Тр. Ин-та им. И. П. Павлова АП СССР, т. 1. Изд. АН СССР, М. —Л.: 84.
Минер вин В. II., 1955. Пастбищно-водопойный режим овец в пустыне. Изд. АН ТуркмССР, Ашхабад.
Мпропольский С. В., 1939. Газообмен и терморегуляция у каракульской овцы в пустынях Средней Азии. Вопр. экол. и биохим., № 5—6.
Миррахимов М. М., 1964а. О картине периферической крови в условиях высокогорья Тянь-Шаня и Памира. Фрунзе.
Миррахимов М. М., 19645. Об акклиматизации человека к высокогорью Средней Азии. Автореф. дисс., Л.
Миррахимов М. М., 1965. Основной обмен в высокогорье. Матер, сими. «Газообм. в усл. высокогорья» и конф. «Высокогорн. климат и больной организм», Фрунзе: 26.
Миррахимов М. М., 1964. Очерки о влиянии горного климата Средней Азии на организм. Изд. «Кыргызстан», Фрунзе.
Михайлов В. М., 1956. Сезонные изменения некоторых эколого-физиологических признаков емуранчиков и тарбаганчиков. Тр. Ростовск. н/Д. противочумн. ин-та, т. 2: 161.
Мокриевич Н. А., 1957. Сезонные изменения некоторых эколого-физиологических особенностей полуденной (Meriones meridi-anus Pall.) и гребенщиковой песчанок (Meriones tamariscinus Pall.) в Волжско-Уральских песках. Сб. «Грызуны и борьба с ними», вып. 5, Саратов,; 29.
Музыкантов В. А., 1929. Об особенностях секреторной деятельности желудочных желез при молочной диете. Сб. «Оздоровление труда», вып. 9, М.: 42.
Нас им о вич А. А., 1955. Роль режима снежного покрова в жизни копытных животных на территории СССР. Изд-во АН СССР, М.
Насонов Н., 1923. Географическое распределение горных баранов. Изд-во АН СССР, Л.
Наумов Н. П., 1963. Экология животных. Изд-во «Высшая школа».
Нестеровой ий Г. В. и Слоним А. Д., 1935. Условно-рефлекторный механизм тепловой одышки. Бюлл. ВИЭМ, вып. 3—4, М.
Никольский Г. В., 1963. Экология рыб. Изд-во «Высшая школа», М.
Никулин В. Н., 1954. Закономерности анатомического строения желудочно-кишечного тракта некоторых млекопитающих. Рефер. докл. Рязаиск. с.-х. ин-та, вып. 1, Рязань: 28.
Огнев С. И., 1951. Экология млекопитающих. МОИП, М.
О д ы н е ц Р. Н., Докукин А. Ф. и Ф а н т а л и с И. А., 1956. Изменение дыхательной функции крови у лактпрующих коров при отгоне их на горные пастбища. Тр. Ин-та зоол. и паразитол. АН КиргССР, вып. 5, Фрунзе: 9.
Ольнянская Р. П., 1949. К физиологии высотной акклиматизации у новорожденных ягнят. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР, М. — Л.: 164.
Ольнянская Р. П., 1950. Кора головного мозга и газообмен. Изд-во АМН СССР, М.
418
Ольнянская Р. П., Давыдов Л. Ф., Романовская Г. Д., 1959. Об изменениях легочного дыхания, газообмена и состава красной крови у овец при акклиматизации их к горным условиям Северного Кавказа. Совещ. по экол. физиол., тез. докл., вып. 2, изд-во АН СССР, М. — Л.: 73.
Ольнянская Р. П., Из б и некий А. Л., Кез и к Р. С., Черковнч Г. М. и Чу кин К- А., 1949. Кровь и ее дыхательная функция в горах Тянь-Шаня на различных высотах. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР, М. — Л.: 135.
Ольнянская Р. П. и Слоним А. Д., 1947. О приспособлении организмов к очень низким температурам среды. Изв. АН СССР, сер. биол., № 2.
Ольнянская Р. П. и Соболь Е. М., 1946. Газообмен и легочное дыхание у овец на различных высотах. Тр. Кирг. фил. АН СССР, т. 4, Фрунзе: 141.
Ольнянская Р. П. и Соболь Е. М., 1949. Легочное дыхание и газообмен у овец в горах Тянь-Шаня на различных высотах. Сб. «Опыт изуч. регуляции физиол. функции», Изд-во АН СССР, М. — Л.: 146.
Опыт изучения периодических изменений физиологических функций в организме. 1949, изд-во АМН СССР, М.
Опыт изучения регуляций физиологических функций в естественных условиях существования организмов: т. 1 — 1949, т. 2 — 1952, т. 3—1954, т. 4 — 1958, т. 5—1961, т. 6 — 1963. изд-во АН СССР, М. — Л.
Орлов В. В., 1961. Плетизмография. Изд-во АН СССР, М. — Л.
Падучева А. Л., Ковальский В. В. и Ш р а й е р Б. С., 1957. Приспособительные изменения и нечувствительные потери воды у каракульских овец в различных температурных условиях и при разреженном водопое. Докл. АН СССР, т. 113, № 4.
Парфенов Н. Ф., 1906. Специальный случай работы слюнных желез у собаки. Тр. Общ-ва русских врачей в СПб, год 73, сент.— дек., стр. 30.
Пашутин В. В., 1902. Курс общей и экспериментальной патологии (Патологическая физиология), СПб.
Персон Р. С., 1950. Желудочная секреция во время спячки как фактор регулирования щелочно-кислотного равновесия крови. Докл. АН СССР, т. 72, № 5.
Персон Р. С. и Мярковская Н. Е., 1953. Материалы по суточной периодике некоторых представите чей приматов. Опыт изуч. регуляции физиол. функций, т. 2, Изд-во АН СССР, М. — Л.: 183.
Писарева 3. Д., 1947. Угольная ангидраза в крови и в мозгу в эмбриногенезе кроликов и морских свинок. Докл. АН СССР, т. 58, № 7: 1535.
Плетнев А. В., 1953. Жевательные и жвачные рефлексы. Совещ. по физиол. с.-х. животн., тез. докл. Изд. АН СССР, М. — Л.: 40.
Плетнев А. В. и Щербакова Г. Ф., 1952. Жевательные рефлексы у сельскохозяйственных животных. Докл. АН СССР, т. 84, № 3.
Поляков И. Я., 1950. Травопольная система земледелия
и вредные мышевидные грызуны. Журн. общ. биол., т. 9, № 1.
Понугаева А. Г., 1949. Стадная форма существования и су-
14* 419
точная периодика у летучих мышей. Опыт изуч. периодпч. изменений физиол. функции в организме. Изд-во АМН СССР: 104.
Пон у гаева А. Г., 1952. О взаимоотношениях стадных и оборонительных рефлексов на теплообмен. Тр. Ин-та физиол. им. И. П. Павлова АН СССР, т. 2. Изд-во АН СССР, М.— Л.: 490.
П о н у г а с в а А. Г., 1953. Рефлекторная регуляция газообмена при групповых скоплениях грызунов в гнездовой период развития. Тр. Ин-та физиол. им. И. П. Павлова АН СССР, т. II. Изд-во АН СССР, М. — Л.: 490.
Попугаева А. Г., 1959. Изучение реакции запасания корма у золотистых хомяков в онтогенезе. Совещ. по экол. физиол., тез. докл. вып. II, изд-во АН СССР, М.— Л.: 81.
Понугаева А. Г., 1960. Физиологические исследования инстинктов у млекопитающих. Изд-во АН СССР, 1960, М. — Л.
Попугаева А. Г. и Марголина О. И., 1949а. Химическая терморегуляция у человека при низких температурах среды. Сб. «Опыт изуч. регуляции физиол. функций» Изд-во АН СССР, М. — Л.: 40.
Понугаева А. Г. и Марголина О. И., 19496. Кортикальный механизм химической терморегуляции у человека. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР, М. — Л.: 40.
Понугаева А. Г. и Слоним А. Д., 1953. Суточный ритм теплопродукции у летучих мышей во время зимней спячки. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2. Изд. АН СССР, М. — Л.: 155.
Понугаева А. Г. и Трубицына Г. А., 1955. Стадные изменения газообмена у овец. Тр. Ин-та физиол. им. И. П. Павлова АН СССР, т. 4. Изд-во АН СССР, М. —Л.: 171.
Пржевальский Н. М., 1888. От Кяхты на истоки Желтой реки. Гл. 1. Как путешествовать по Центральной Азии. СПб.
Привольнев Т. И., 1953. Влияние температуры солнечного света на активность каталазы рыб. Изв. Всес. научи.-иссл. ин-та озерн. и речи. рыбн. хоз., т. 28: 127.
Промптов А. Н., 1956. Очерки по проблеме биологической адаптации поведения воробьиных птиц. Изд-во АН СССР, М. — Л.
Проссер Л. П. и Браун Ф.. 1967. Сравнительная физиология животных. Изд-во «Мир». М.
Протасеия Т. П„ 1939. Физиологические и биохимические сдвиги при разных степенях водного голодания лошадей. Сов. ветеринария, № 2: 63.
Пшеничный П. Д., 1955. Основы учения о воспитании сельскохозяйственных животных. Изд-во АН УССР, Киев.
П шони к А. Т., 1952. Кора головного мозга и рецептивная функция организма. Изд-во «Советская наука», М.
Ра зенков И. П., 1927. Исследование вазомоторных свойств крови. Изд-во Ком. акад. М.
Разенков И. П., 1945. Пищеварение на высотах. Медгиз, М. — Л.
Разенков II. И., 1948. Новые данные по физиологии и патологии пищеварения. М.
Рахимов К., 1959. О формировании пищевых рефлексов в постиатальпом онтогенезе у жвачных. Автореф. дисс., Л.
Раушенбах Ю. О., 1952. О природе устойчивости сельскохозяйственных животных к специфическим факторам среды. Тр. сове-420
щания по биол. основ, повыш. продукт, животновод., Изд. АН СССР, М. — Л.: 71.
Раушенбах Ю. О., 1958а. О физиологической природе устойчивости животных к гипоксическим условиям высокогорья. Сб. «Опыт изуч. регуляции физиол. функций», вып. 4. Изд-во АН СССР, М. — Л.: 71.
Раушенбах Ю. О., 19586. О природе устойчивости сельскохозяйственных животных к высокой температуре среды. Сб. «Опыт изуч. регуляции физиол. функций», вып. 4, Изд-во АИ СССР, М. — Л.: 84.
Раушенбах Ю. О., 1967. Видовые различия в структуре реакции на высокие и низкие температуры у домашних животных. Реф. докл. к III Всес. совет, по эколог., физиол., биохим., морфол., т. «Видовые и природно-климатич. адаптации», Новосибирск: 91.
Рашевская Д. А., 1968. Возрастные изменения кровообращения у некоторых видов грызунов. В кп.: «Сравнительная и возрастная физиология». Изд-во «Наука-», стр. 272.
Рейм о в Р., 1958. Полевое изучение высшей нервной деятельности малого суслика (Citellus pygmaeus) в целях усовершенствования борьбы с ним. Автореф. дисс., Л.
Репин И. С., 1959. Блокирующее действие СО2 на терморегулярный мышечный тонус и влияние ее на физическую теплорегуляцию. Ежегод. Ин-та эксп. мед. за 1958 г., Л., 119.
Рогаль И. Г., 1955. О холодном методе выращивания молодняка сельскохозяйственных животных и его физиологическом обосновании. Журн. общ. биол., т. 16, № 4.
Рогов А. А., 1951. О сосудистых условных и безусловных рефлексах человека. Изд. АН СССР, М. — Л.
Росляков А. К., Евдаков А. Н., Басенко Е. П., 1953. Газовый обмен у овец в зависимости от физиологического состояния организма и условий внешней среды. Тр. Ин-та эксп. биол. АН КазССР, т. 1, Алма-Ата: 108.
Рубин Ф. Е., 1953. Эколого-морфологический очерк некоторых видов грызунов Белоруссии. Автореф. дисс., Минск.
Рубинов И. С., 1958. Физиология и патофизиология жевания и глотания. Медгиз, Л.
Руттенбург С. О., 1950. Физиологические особенности некоторых видов крыс в связи с их географическим распространением. Автореф. дисс., Л.
Руттенбург С. О., 1953. Химическая терморегуляция у некоторых видов крыс в связи с их географическим распространением. Сб. «Опыт изуч. регуляции физиол. функций», т. 2. Изд-во АН СССР, М, —Л.: 35.
Салахов А. X., 1960. Изменение морфологического состава крови у практически здоровых людей при пребывании в Чухугорте. Сб. тр. Азерб. мед. ин-та, т. 7, Баку: 136.
Сал га некий А. А., 1952. Опыт выпасного содержания пятнистых оленей без изгороди. Каракулеводство и звероводство № 5: 52.
Сафонов А. Ф., 1958. Характер внешнего дыхания у при-аральскпх тонкорунных овец. Тр. Ин-та биол. АН КазССР, т. 4, Алма-Ата: 49.
Светозаров Е. и Штрайх Г., 1940. Свет и половая периодичность у животных. Усп. совр. биол., т. 12, вып. 1.
421
С в и р п д е п к о II. Л., 1957. Запасание корма животными. Пзд-во АН УССР, Киев.
Се вер нов А. Н., 1945. Эволюция и психика. Собр. соч., т. 3. Пзд-во АН СССР, М. —Л.: 289.
Сегаль А. II., 1959. Зависимость некоторых физиологических функции у северного оленя от температуры воздуха в условиях .лес-пых пастбищ Карелии. Совещ. по экол. физиол., вып. 2. Изд-во АН СССР, М.— Л.: 95.
Синельников Е. И. и Ивкина К. Л., 1954. Регуляция нормального состава бактериальной флоры кишечника лимфатическими образованиями. Тр. научн. совещ. по физиол. и патол. пищевар. Изд-во АН СССР. М. — Л.: 23.
С и н и ч к и п а А. А., 1956. Особенности реакции серых крыс на влияние сезонных изменении условий среды. Тр. Ин-та «Микроб», вып. 2, Саратов.
Синичкина А. А., 1959. Особенности реакции серых крыс на сезонные изменения среды. Совещ. по экол. физиол., тез. докл., вып. 2. Изд-во АН СССР, М. — Л.: 96.
Сироти нин Н. Н., 1941. Об аноксической аноксии. Кчин. мед., т. 19, вып. 3: 5.
С и р о т п и и н Н. Н., 1950. Об эволюции адаптаций к гипоксии. Мед. журн. АН УССР, т. 20, вып. 6, Киев.
Скворцов Г. II., 1956. Сезонные изменения некоторых эколого-физиологических особенностей тушканчиков мохноногого (Dipus sagitta) и тарбаганчика (Alactagulus acontion) в условиях Туркмении. Автореф. дисс., М.
Склярчик Е. Л., 1963. Минутный объем сердца и газообмен у кроликов и зайцев в покое и при мышечной деятельности. Сб. «Опыт изуч. регуляции физиол. функций», т. 6, Изд-во АН СССР, М. — Л.: 16.
Скула ч ев В. П., 1962. Соотношение окисления и фосфорилирования в дыхательной цепи. М.
Скулачев В. П., 1964. Разобщение окисления и фосфорилирования в дыхательной цепи. Изд-во АН СССР, М.
Слоним А. Д., 1937. К физиологии терморегуляции у некоторых тропических и горных животных. Физиол. журн. СССР, т. 22: 89.
С л о и и м А. Д., 1941. Температура среды обитания и регуляция тепла в организме млекопитающих. Усп. совр. биол., т. 14: 52.
Слоним А. Д., 1945. Суточная и сезонная периодика активности и терморе’«ляцпп у летучих мышей. Изв. АН СССР, сер. биол., № 3: 303.
Слоним А. Д., 1952. Животная теплота и ее регуляция в организме млекопитающих. Пзд-во АН СССР, М. — Л.
Слоним А. Д., 1958. О влиянии факторов внешней среды на физиологические функции. Материал I кон. физиол., биохим. и фар-макол. Ср. Азии и Казахст. Ташкент: 207.
Слоним А. Д., 1961. Основы общей экологической физиологии млекопитающих. Пзд-во АН СССР, М. — Л.
Слоним А. Д., 1962. Частная экологическая физиология мпе-копптающих. Пзд-во АП СССР, М. — Л.
Слоним А. Д., 1964. О физиологических механизмах природных адаптаций животных и человека. Изд-во «Наука», М. — Л.
Слоним А. Д., 1966. Физиология терморегуляции и термиче
422
ской адаптации у сельскохозяйственных животных. Изд-во «Наука», М. — Л.
Слоним А. Д., 1967. Инстинкт. Изд-во «Наука», Л.
Слоним А. Д. и Безус вс кая Р. Я., 19 И). Сезонные
изменения терморегуляции. Физиол. журн. СССР, т. 28, вын. 2—3.
С л о н н м А. Д„ О л ь и я н с к а я Р. И. в Р у т т е н-бург С. О., 1949. Опыт изучения динамики физиологических функций человека в условиях Заполярья. Сб. «Опыт изуч. период, изменений физиол. функций». Изд-во АН СССР, М.— Л.: 207.
Слоним А. Д., Понугаева А. Г., Марголина О. И., Руттепбург С. О., Лупинская 3. А. и Избив ский А. Л., 1949. Опыт изучения физиологии высотной акклиматизации человека в горах Тянь-Шаня. Сб. «Опыт изуч. регуляции фп-зиол. функций». Изд-во АН СССР, М. — Л.: 180.
Слоним А. Д. и Черкович Г. М., 1959. Роль типологических особенностей нервной системы в формировании суточного ритма физиологических функций у обезьян. Конф, по вопр. о роли типа нервн. сист. в обмен., компенсат. и восстан. реакц. организма. Pv р. докл., Киев: 84.
Слоним А. Д. и Щербакова О. П., 1935а. Материалы к сравнительной физиологии терморегуляции. Сообщ. 1. Терморегуляция у обезьян гамадрилов. Бюлл. ВИЭМ, № 11 —12.
Слоним А. Д. и Щербакова О. П., 19356. Материалы к сравнительной физиологии терморегуляции. Сообщ. 2. Терморегуляция у обезьян макак. Бюлл. ВИЭМ, № 11 —12.
Слоним А. Д. и Щербакова О. П. 1935в. Материалы к сравнительной физиологии терморегуляции. Сообщ. 3. Терморегуляция у хищников. Бюлл. ВИЭМ, № 11 — 12.
Слоним А. Д. и Щербакова О. П., 1938. О последующем влиянии высоких температур среды на газообмен и температуру тела. Физиол. журн. СССР, т. 25: 827.
Слоним А. Д. и Щербакова О. П., 1940. Физиология терморегуляции у низших обезьян. Архив биол. наук, т. 55: 3.
Слоним А. Д. и Щербакова О. И., 1949а. Наблюдения над ночным сном у обезьян. Сб. «Опыт изуч. период, изменений физиол. функций», Изд-во АМН СССР, М.: 156.
Слоним А. Д. и Щербакова О. П., 19496. Обмен веществ и физиологические особенности зимнего сна у барсука. Сб. «Опыт изуч. период, изменений физиол. функций в организме». Изд-во АМН СССР, М.: 167.
Смирнов В. С. и Шварц С. С., 1957. Сезонные изменения относительного веса надпочечников у млекопитающих в природных условиях. Докл. АН СССР, т. 115, № 6.
Соболева А. Д., 1967. Гистохимические механизмы адаптации легких к условиям пустынного существования (сухоядение). Материалы к III Всес. конф, по эколог., физиол., биохим. и морфолог. Вып. «Морфология», Новосибирск: 72.
Степа нк и на М. К., 1958. Об участии пищеварительного тракта некоторых пород овец и верблюда в межуточном обмене азота. Автореф. дисс., Алма-Ата.
Стрельников И. Д., 1958. Экологическая терморегуляция у некоторых наземных беспозвоночных (насекомые) и позвоночных (рептилии и млекопитающие). Совещ. по экол. физиол., тез. докл., вып. 1. Изд-во АН СССР, М. — Л.: 61.
423
Стрельцов В. В. и Хазен II. М., 1946. Активность карбоангидразы в крови и некоторых тканях организма при пониженном барометрическом давлении. Бюлл., Эксп. биол. и мед., т. 21, вып. 4.
Ташенов К. Т., 1956. Активность фаупы рубца овец. Тр. Ин-та биол. АН Каз. ССР, т. 3, Алма-Ата: 7.
Терновский Д. В., 1958. Биология и акклиматизация американской норки на Алтае. Новосибирск.
Томилин А. Г., 1958. Некоторые данные о терморегуляции у орешниковой сони (Muscardltius avellanurus). Изменение характера дыхания и температуры тела во время спячки и бодрствования. Зоол. жури., т. 37, выи. 1.
Трайб Д. Е., 1959. Поведение пасущихся животных. Сб. «Новое в физиологии домашних животных», т. 2, Сельхозгиз, М. — Л.: 227.
Третьяков Н., 1895. К вопросу о влиянии разреженного воздуха Памирских высот. Военно-мед. журн.: 183.
Третьяков Н., 1897. К вопросу об акклиматизации. Дисс. СПб.
Ту л баев П., 1959. О рефлекторной деятельности многокамерного желудка коз в онтогенезе в связи с типом питания. Автореф. дисс., Л.
Уголев А. М., 1957. Адаптивные свойства желудочного сока. Докл. АН СССР, т. 113, № 1.
Уголев А. М., 1958а. К сравнительной физиологии ферментативных свойств пищеварительных соков. Совещ. по экол. физиол., тез. докл., вып. 1. Изд-во АН СССР, М. — Л.: 64.
Уголев А. М., 19586. Приспособление пищеварительных желез к качеству пищи. Автореф. дисс., М.
Уголев А. М., 1958в. Материалы о деятельности слюнных желез у грызунов. Сб. «Опыт изуч. регуляции физиол. функций», т. 4, Изд-во АН СССР, М. — Л.: 173.
Уголев А. М., 1961. Пищеварение и его приспособительная эволюция. Изд-во «Высшая школа», М.
Уголев А. М., 1963. Пристеночное (контактное) пищеварение. Изд-во АН СССР.
Уголев А. М., 1967. Физиология и патология пристеночного (контактного) пищеварения. Изд-во «Наука», Л.
Уголев А. М., Волкова Е. А. и Кор не виц Г. В., 1953. Натуральные пищевые условные рефлексы у обезьян. Сб. «Опыт изуч. регуляции физиол. функций», т. 2. Изд-во АН СССР, М, —Л.: 147.
Уголев А. М. и Черниговский В. Н., 1959. О роли интероцепторов в формировании поведения высших животных. Докл. АН СССР, т. 126, № 2.
Уждавини Э. Р., 1955. О формировании натуральных пищевых рефлексов в онтогенезе у собаки. Автореф. дисс., Л.
Уждавини Э. Р., 1958. Врожденные пищевые рефлексы у щенков. Сб. «Опыт изуч. регуляции физиол. функций». Изд-во АН СССР: 101.
Уждавини Э. Р., 1960. Материалы к сравнительной физиологии слюнных рефлексов. Тр. Ии-та физиол. им. И. П. Павлова АН СССР, т. 9, Изд-во АН СССР, М. — Л.: 139.
Уждавини Э. Р. и Шепелева В. К-, 1966. Очерки развития врожденного поведения. Изд-во «Наука», М. — Л.
424
Ушаков Б. П., 1955. Теплоустойчивость соматической мускулатуры земноводных в связи с условиями существования вида. Зоол. журн., т. 34, вып. 3.
Ушаков Б. П., 1956. Теплоустойчивость клеточных белков холоднокровных животных в связи с видовым приспособлением к температурным условиям существования. Журн. общ. биол., т. 17, № 2.
Ушаков Б. П., 1963. О классификации приспособлений животных и растений и о роли цитоэкологии в разработке проблемы адаптации. Сб. «Проблемы цитоэкологии животных». Изд. АН СССР, М. — Л.
Ушаков Б. П., 1964. Исследование теплоустойчивости клеток и протоплазматических белков пойкилотермных животных в связи с проблемой вида. «Клетка и температура среды», в khz Тр. межд. симпозиума по цитоэкологии. Л., 1963.
Ушаков Б. П., 1967. Заметка о корреляции между индивидуальной изменчивостью показателей теплоустойчивости клеток животных в онто- и филогенезе». Изд-во «Наука», Л.
Файзиев С., 1957. Безусловные и натуральные условные слюнные пищевые рефлексы у овец романовской и каракульской пород. Автореф. дисс., Л.
Файзиев С., 1958. Об особенности оборонительного слюноотделения у овец каракульской породы. Сб. работ аспир. АН УзССР. Ташкент: 219.
Ф а н т а л и с И. А., 1960. Изменение некоторых физиологических показателей у людей в горных районах Киргизии. 2-я конф, физиол. биохим. и фармакол. Ср. Азии, тез. докл., Фрунзе: 362.
Федоров В. К-, 1959. Современное состояние проблемы наследования изменений высшей нервной деятельности животных. Журн. высш, нервн. деят., т. 9, вып. 6.
Фер дм ан Д. Л. и Файн шмидт О. Н., 1932. До биохимии зымовой сплячкы. Наук. зап. Укр. биохимичн. институту, т. 5, вып. 2.
Физиология человека в пустыне, 1952. Изд-во ИЛ, М.
Филатова Л. Г., 1944. Эколого-физиологические исследования у млекопитающих — обитателей полупустынь Киргизии. Тез. дисс., Фрунзе. Ф.
Филатова Л. Г., 1946а. Дыхательная функция крови при повторной длительной гипоксемии. Журн. Биол. Ин-та Кирг. фил. АН СССР. 1, Фрунзе, 173.
Филатова Л. Г., 19466. Особенности легочного дыхания и физиологическая гипоксемия у ежа. Бюлл. эксп. биол. и мед., т. 22, вып. 5.
Филатова Л. Г., 1949а. Водный обмен у животных — обитателей полупустынь Киргизии. Сб. «Опыт изуч. регуляции физиол. функций». Изд. АН СССР, М. — Л.,: 220.
Филатова Л. Г., 19496. Суточный ритм у грызунов и насекомоядных и опыт его экспериментального изучения. Сб. «Опыт изуч. измен. период. функций организма», Изд-во АМН СССР, М. — Л„: 116.
Филатова Л. Г., 1948в. Терморегуляция млекопитающих — обита гелей полупустыни. Сб. «Опыт изуч. регуляц. физиол. функций». Изд-во АН СССР, М. — Л.: 83.
425
Филатова Л. Г., 1954. Опыт изучения влияния горного климата Киргизии па организм человека. Уч. зап. биол.-почв. фак-та Кирг. гос. ун-та, вып. 4, Фрунзе: 29.
Филатова Л. Г., 1957. Исследование высотной акклиматизации животных п человека в условиях Тянь-Шаня. Автореф. дисс., Л.
Филатова Л. Г., 1958. Реакция на гипоксемию у млекопитающих с различной экологической специализацией. Совещ. по экол. физиол., тез. докл., вып. 1. Изд. АН СССР, М. — Л.: 75.
Филатова Л. Г., 1960. Акклиматизация к гипоксии. Фрунзе.
Филатова Л. Г., 1961. Исследования по физиологии высотной акклиматизации животных и человека. Изд-во Кирг. гос. ун-та, Фрунзе.
Фирстов А. А., 1952. Биологические особенности в основном обмене у пушных зверей. Тр. Моск, пушно-мех. ин-та, т. 3, М.: 212.
Формозов А. Н., 1946. Снежный покров в жизни млекопитающих и птиц СССР. Изд. МОИП, М.
Хайрутдинов X. Ш., Воробьева А. В., 1954. Двигательная реакция рубца каракульских овец на различные внешние факторы. Докл. АН УзССР, № 10, Ташкент.
Холден Дж. и Пристли Д ж., 1936. Дыхание. Биомед-гиз., М.
X р у с ц е л е в с к и й В. П. и Копылова О. А., 1957. Материалы по экологии полевки Брандта. Изв. Ирк. гос. науч.-исслед. противочумного ин-та Спб. и Д. Востока, т. 16, Иркутск: 69.
Цалкии В. II., 1950. Горные бараны СССР. Изд. МОИП, М.
Ц и т о в п ч И. С., 1911. Происхождение и образование натуральных условных рефлексов. Дисс. СПб.
Ч а л ю к Е. А., 1950. Влияние работы на перевариваемость сено-овсяных рационов. Тр. Всес. иауч.-иссл. ни-та коневодства, вып. 18.
Чашкпн И. И., 1959. Современное состояние новокиргизской породы лошадей и дальнейшее ее совершенствование. Тр. Кирг. науч.-иссл. ип-та животновод., вып. 14, Фрунзе: 136.
Черкович Г. М„ 1950. Экспериментальное изучение суточной периодики некоторых физиологических функций у обезьян. Автореф. дисс., Л.
Черкович Г. М., 1953. Слуховой анализатор и роль стадных отношений в суточной периодике физиологических функций у обезьян. Сб. «Опыт изуч. регуляции физиол. функций», вып. 2. Изд-во АП СССР, М, —Л.: 187.
Черкович Г. М., 1958. Получение неврозов у обезьян при нарушении стереотипа натуральных раздражителей. Совещ. по экол. физиол., тез. докл., вып. 1. Изд-во АН СССР, М. — Л.: 76.
Черниговский В. II., 1962. Значение интероцептивной сигнализации в пищевом поведении животных. Изд-во АН СССР, М. — Л.
Черниговский В. Н., III е х т е р С. Ю. и Я роше в-с к п й А. Я., 1967. Регуляция эритропоэза. Изд-во «Наука», Л.
Четвериков Д. А., 1948. Карбоангидраза тканей глаза в онтогенезе. Пзв. АН СССР, сер. биол., № 4: 461.
Чубин скип С. М., 1965. Биоклиматология. Изд-во «Медицина», М.
426
Чугунов Ю. Д., Кудряшова II. И., Чугунова II. Д., 1956. Особенности терморегуляции степных п горных малых сусликов в условиях Северного Кавказа. Тр. Науч.-неся. противочумп. ин-та Кавказа и Закавказья, вып. 1. Ставрополь: 265.
Шарипова Р. Р., 1953. Исследование газообмена при тонических напряжениях. Сб. «Опыт изуч. регуляции физиол. функции-», т. 2. Изд-во АН СССР, М. — Л.: 109.
Шаталина А. С., 1935. Влияние климата на состав крови. Журн. «Соц. наука и техн.», Ташкент, № 3.
Шварц С. С., 1959. Некоторые вопросы проблемы впда у наземных позвоночных животных. Тр. Ип-та биол. Уральск, фил. АН СССР, вып. II, Свердловск.
Шварц С. С., 1960. Принципы и методы современной экологии животных. Изд-во АН СССР, Свердловск.
Шварц С. С., Смирнов В. С. и До бри некий Л. Н., 1968. Метод морфофизиологических индикаторов в экологии наземных позвоночных. Тр. ин-та экологии растений и животн., Свердловск.
Шепелева В. К., 1950. Некоторые особенности питания пасюка (Rattus norvegicus) и черной крысы (Rattus rattus). II экол. конф., тез. докл., ч. 2, Киев: 121.
Шик Л. Л., 1939. Цит. по: Кандрор и Раппопорт, 1957.
Штефан М., 1957. Сравнительно-физиологическое исследование группового эффекта у рыб. Автореф. дисс., МГУ, М.
Шулейкин В. В., 1966. Физика моря. Гл. IX. Изд-во «Наука», М.
Щеглова А. И., 1949. Терморегуляция п кожно-легочные потери воды у большой песчанки (Rhombomis ophnus Licht). Co. «Опыт изуч. регуляции физиол. функций» Изд-во АН СССР, М. — Л.: 71.
Щеглова А. И., 1952а. Физиологические особенности некоторых видов грызунов в связи с их экологией. Автореф. дисс., Л.
Щеглова А. И., 19526. Приспособление водного обмена у некоторых видов сусликов к условиям жизни. Докл. АН СССР, т. 83, № 5.
Щеглова А. II., 1958. Опыт физиологического анализа грызущей деятельности большой песчанки. Сб. «Опыт изуч. регуляции физиол. функций», т. 4, Изд-во АН СССР, М. — Л.: 184.
Щеглова А. И., 1959. Значение водного обмена в жизнедеятельности млекопитающих. Совещ. по экол. физиол., тез. докл. Изд-во АН СССР, М. — Л.: 125.
Щеглова А. И. и Смирнов П. К., 1963. Роющая деятельность некоторых грызунов в разных условиях температуры среды и солнечной радиации. Сб. «Опыт изуч. регуляции физиол. функций», вып. 6. Изд-во АН СССР, М. — Л.: 110.
Щербакова О. П., 1937. Материалы к изучению суточной периодики физиологических процессов у высших млекопитающих. Сообщ. 1. Нормальная суточная периодика физиологических процессов. Бюлл. эксп. биол. и мед., т. 4, вып. 4.
Щербакова О. П., 1949. Сезонные и суточные изменения двигательной активности у обезьян. Сб. «Опыт изуч. период, измен, физиол. функций организма». Изд-во АМН СССР, М. — Л.: 143.
Щербакова О. П., 1949а. Суточная периодика физиологических процессов у некоторых отрядов млекопитающих. Сб. «Опыт
427
изуч. периодич. измен, физиол. функций организма». Изд-во АМН СССР. М. —Л.: 15.
Щербакова О. П., 19496. Сезонные и суточные изменения двигательной активности у обезьян. Сб. «Опыт изуч. период, измен, физиол. функций», Изд-во АМН СССР. М.: 143.
Эмме А. М., 1967. Биологические часы. Изд-во «Наука», Новосибирск.
Юнусов А. Ю., 1950. Физиологическая характеристика потоотделения у детей. УзГИЗ, Ташкент.
Юнусов А. Ю., 1961. Физиология крови человека и животных в жарком климате. Медгиз УзССР, Ташкент.
Юнусов А. Ю. и Коротько Г. Ф., 1962. Функции органов пищеварения в жарком климате. Медгиз УзССР, Ташкент.
Юнусов С., 1960. Газообмен и минутный объем сердца у овец и муфлона. Сб. «Регуляция обмена тепла и др. физиол. функц. у с.-х. животн. в условиях высоких температур». Краснодар: 301.
Юнусов С., 1961. Физиологическое состояние у овец при polypnoe. Тр. Аидпжанск. гос. пед. ии-та, т. VIII, Андижан.
Юрисов а М. Н. и Поленов А. Л., 1967. Сравнительный эколого-гистофизиологический анализ состояния гипоталамо-гипофи-зарной нейросекреторной системы у двух видов сусликов. Матер. III Всес. совещ. по экол. физиол., биохи.м., морфол., вып. «Морфол. исслед.», Новосибирск: 176.
Яковлев Н. Н., 1950. Последовательность биохимических изменений в мышцах при тренировке и растренировке. Физиол. журн. СССР, т. 36, № 6.
Яковлев И. Н., 1955. Очерки по биохимии спорта. Изд-во «Физкультура и спорт», М.
Adler L., 1926. Der Winterschlaf. Boethe’s Handb. d. norm. u. pathol. Physiol., Bd. 17, Teil III, Correlationen.
Adolph E. F., 1957. Ontogeny of physiological regulations in the rat. Quart. Res. Biol., vol. 32 : 89.
Alexander R. Me. N., 1967. Functional Design in Fishes. Hutchinson Univers. Library, London.
Allee W. C., 1931. Animal aggregation. Chicago Press.
Allen J. A., 1906. The influence of physical conditions on the genesis of species. Ann. Rep. of Smith’s Inst.: 375.
Ames R. G. a. H. B. Van Dyke, 1950. Antidinretic hormone in the urine and pituitary of the kangaroo rat. Proc. Soc. Exp. Biol. a. Med., vol. 75:417.
Anand В. K-, 1968. Limbic system in innate animal behavior. Proc, of the XXIV Int. Congr. of Physiol. Sci., vol. VI, Washington, D. C.
Andersson B„ 1956. Hypothalamic control of the water intake. In: XX Int. Physiol. Congr., Bruxelles, vol. 1 : 126.
Andersson B., 1967. Studies on the thirst mechanism. Acta Biol. Exper. (Warsaw), vol. 27, No. 3 : 269.
Andersson В. a. M. McCann, 1956. The effect of hypothalamic lesions on the water intake of the dog. Acta Physiol. Scand., vol. 35 : 312.
Anghi C. G., 1957. Die Coecotrophie als Indicator der richtigen Kaninchenfiitterung. Kaninchenziichter, No. 13, Jg. 61: 121.
428
Animal drives. 1965. Ed. by G. A. Cicala D. Van Nostrand C°, Inc.
An n i s о n E. F. a. D. Lewis, 1962. Metabolism in the Rumen. London Methuen and C°.
Airep G. W. a. M. Ham mouda, 1933. Observation on panting. J. Physiol. London, vol. 67 : 87.
A tfc I e r E., 1927. Korper und Arbeit. Leipzig.
Bailey V., 1923. Sources of water supply for desert animals. Sci. Monthly, vol. 17 : 66.
Bainbridge R., 1961. Problems of fish locomotion. Symp. Zool. Soci No. 5.» Vertebrate locomotion, London : 13.
Bainbridge R., 1963. Caudal fin and body movement in the propulsion of some fish. J. Exptl. Biol., vol. 40 : 23.
Barbashova Z. I.» 1964. Cellular level of adaptation. Handb. of Physiol., Environment, U. S. A.: 37.
Barcroft J. S„ 1925. The respiratory function of the blood. I. Lessons from high altitude. Cambridge.
Barcroft J. S., С. A. В i n g e r, A. W. Bo ok, J. H. Boggart, H. S. Forbes, G. Harrop, J. C. Meakins a. A. C. Redfield, 1923. Observations upon the effect of high altitude on the physiological processes of the human body. Phil. Trans. Roy. Soc. B., vol. 211: 353.
Barnett С. H., 1955. Competition among wild rats. Nature, 175 : 126.
Barnett С. H., 1958. Physiological effects of «social stress» in wild rats. I. The adrenal cortex. J. Psychosomat. Res., 3: 1.
Bartholomew G. A. a. J. W. Hudson, 1959. Effects of sodium chloride on weight and drinking in the antelope ground squirrel. J. Mammology, vol. 40 : 354.
Bartholomew G. A., J. W. Hudson a. T. R. H о w e 11, 1962. Body temperature oxygen consumption, evaporative water loss, and heart rate in the poorwill. Condor 64 : 117.
Beer J. R. a. R. K- Meyer, 1951. Seasonal changes in the endocrine organs and behavior patterns of the musk rat. J. Mam-mol. 32 : 173.
Benedict F. G., 1938. Vital energetics, a study in comparative basal metabolism. Carneg. Inst., Washington, Publ. No. 503.
Benedict F. G. a. R. C. L e e, 1938. Hibernation and marmot physiology. Carneg. Inst., Washington, Publ. No. 497.
Bentley P. J. a. K. Schmidt-Nielsen, 1966. Cutaneous Water Loss in Reptiles. Science 151, No. 3717 : 1547.
Berger B., 1907. Uber die Widerstandsfahigkeit der Tenebrio-larven gegen Austrockung. Arch. Ges. Physiol. 118 : 607.
Bergmann H., 1847—1848. Heber die Verhaltnisse der Wiir-meokonomie der Tiere zu ihrer Grosse. Gottingen.
Bernard CL, 1859. De la matiere glicogene consideree comme condition de developement de certain tissues. C. R. Acad. Sci. t. 48 : 673.
Bertalanffy L., 1949. Relation between total and tissue metabolism and body size and growth type. Nature 116 : 156.
Bertalanffy L. a. R. Estwick, 1953. Tissue respiration of musculature in relation to body size. Amer. J. Physiol. 173, 1.
Bertalanffy L. a. W. Y. Pirozynski, 1953. Tissue respiration, growth and basal metabolism. Biol. Bull. 105, 1.
429
Bing F. С. а. С. В. Mendel, 1931. The relationship between food and water intakes in mice. Amer. J. Physiol. 98 : 169.
В 1 a z к a P„ M. Volf a. M. С e p e 1 a, 1960. A new type of respirometer for the determination of the metabolism of fish in an active state. Physiol. Bohemosl. 9 : 553.
В о d e n h e i m e r F. S., 1954. Problems of physiology and ecology of desert animals. In: Biology of Deserts. Ed. by J. L. Ckiudsley — Thompson. London, Inst. Biol. : 162.
Botticher II., van., 1915. Untersuchungen fiber den Zusam-rnenhang zwischen Klima und Korpergrosse bei homdothermen Thieren. Zool. Jarb., Bd. 40 : 1.
Bowen W. J. a. H. J. E a d s, 1949. Effect of 18 000 ft simulated altitude on myoglobin content of dogs. Amer. J. Physiol. 159:77.
Brett J. R., 1965. The respiratory metabolism and swimming performance of young sockeye salmon. J. Fish. Res. Bd. Canada, vol. 22 : 623.
Brett J. R., 1967. Swimming Performance of Sockeye Salmon (Oncorhynchus nerka) in relation to Fatigue Time and Temperature. J. Fish. Res. Bd. Canada, 24 : 1731.
Britton S. W. a. R. F. К I e i n, 1939. Emotional hyperglicemia and hyperthermia in tropical mammals and reptiles. Amer. J. Physiol. 125 : 730.
В г о b e с к J. R., 1960. Food and temperature. Recent Progr. Hormone Res. 16: 439.
Brody S„ 1945. Bioenergetics and Growth. New-York.
Brown С. M., G. S. Bird, L. M. В о u g, D e 1 a h a у e, G. E. Green, J. D. H a t c h e r a. Page, 1954. Metabolism clinical and experimental. Metabolism 333.
Brfick К. a. B. Wfinnenberg, 1966. Influence of ambient temperature in the process of replacement of nonshivering by shivering thermogenesis during postnatal development. Fed. Proc. 25, 4.
Bullard R. W. a. J. К о 1 1 i a s, 1966. Functional characteristics of two high — altitude mammals. Fed. Proc. 25, 4: 1288.
В u 11 о u g h W. S., 1952. Stress and epidermal mitotic activity. J. endocrinol. 8 : 265.
Buxton P. A., 1930. Evaporation from the meal — worm (Tene-brio: Coleoptera) and atmospheric humidity. Proc. Roy. Soc. London Ser. B. 116: 560.
Buxton P. A., 1932. Terrestrial insects and humidity of the environment. Biol. Rev. Cambridge Phil. Soc. 7 : 275.
Calhoun J. B., 1948. Mortality and movement of brown rats in artificially supersaturated populations. J. Wildlife Meng. 12 : 167.
Campbell J., 1926—1927. Prolonged alterations of oxygen pressure in the inspired air with special reference to tissue oxygen tension, tissue carbon dioxide tension and haemoglobin. J. Physiol. (London) 62: 211.
Campbell J., 1927. Note on some pathological changes in the tissues during attempted acclimatization to alterations of O2 pressure in the air. Brit. J. Exp. Pathol. 8 : 347.
Chaffee R. R., J. R. Allen, M. Brewer, S. M. Horvath, C. Mason a. R. E. S m i t h, 1966. Cellular physiology of cold and heat—exposed squirrel monkeys (Saimiri Sciurea). J. Appl. Physiol. 21 : 151.
430
Chapman R. F., 1955. Some temperature responses of numphs of Loiusta migratoria migratoria migratorioides (R. S. F.) with special reference to aggregation. J. Exptl. Biol. 32: 126.
Chauvin R., 1956. Physiologic de 1’insectes. Inst. Nat. de la Recherche Agronomique, Paris.
Chitty D., 1955. Adverse effects of population density upon the viabilitylof later generations. In: The Number of Men and Animals, ed. by Olivei a. Boyd, Edinbourgh : 57.
Chitty D. a. C. R. Austin, 1957. Environmental modification of oestrus.in the vole. Nature 179 : 592.
Christian J. J., 1950. The adrenalo-pituitary system and population cycles in mammals. J. Mammol. 31 : 247.
Christian J. J., 1959. Adrenocortical splenic and reproductive responses to inanition and to grouping. Endocrinology 65 : 189.
Christian J. J., 1963. Endocrine Adaptive Mechanisms and the Physiologic Regulation of Population Growth in Physiological Mam-mology. Ed. by Meyer a. Gelder, Acad. Press New York a. London, vol. 1 : 189. I
Clark R. T., J. D. Criscuolo a. С. K. Coulson, 1952. Effects of 20 000 ft simulated altitude on mioglobin content of animals with and without exercise. Feder. Proc. 11 : 25.
Clarke J. R., 1953. The effect of fighting on the adrenals, thymus, and spleen of the vole (Microtus agrestis). J. Edocrinol. 9 : 114.
Cl ou d s I ey Th om p so n J. L., 1959. Studies in diurnal rhythms. IX. The water relations of some nocturnal tropical arthropods. Entomol. Exptl. Appl. 2 : 249.
Cloudsley-Thompson J. L., 1961. Rhythmic activity in animal. Physiology and Behaviour, Acad. Press, London.
Cook S. F. a. J. P. H a n п о n, 1954. Metabolic differences between three strains of Peromyscus maniculatus. J. Mammal. 35, 4.
Cottle W. H. a. L. D. Carlson, 1954. Adaptive changes in rats exposed to cold. Caloric exchange. Amer. J. Physiol. 178 :305.
Cowles R. В. a. С. M. В о g e r t, 1944. A preliminary study of the thermal requirements of desert reptiles. Bull. Am. Museum Nat. Hist. 83 : 261.
Cowles R. B. a. W. R. D a w s о n, 1951. A cooling mechanism in the Texas nighthawk. Condor 53 : 19.
Craig W., 1918. Appetites and Aversions as Constituents of Instinct. Biol. Bull. 34,2 : 91.
C r a w f о r d E. C., Jr., a. K. Schmidt-Nielsen, 1967. Temperature regulation and evaporative cooling in the ostrich. Am. J. Physiol. 212. 2: 347.
Crew F. A. a. L. M i r s к a j a, 1931. The effect of density on an adult mouse population. Biologia gencralis 7 : 239.
Curtis G. M., С. B. D a v i s a. F. J. P h i 1 i p s. 1933. Significance of the iodine content of human blood. J. Amer. Medic. Ass. 101 :901.
Davis D. E. a. J. J. Christian, 1957. Relations of adrenal weight to social rank of mice. Proc. Soc. Exp. Biol. Med., 94:728.
Davis E. A., 1955. Seasonal changes in the energy balance of the English sparrow. Auk 72 : 385.
D a \v son W. R., 1958. Relation of oxygen consumption and evaporative water loss to temperature in the cardinal. Physiol. Zool. 31, 1 : 37.
431
Dawson W. R. a. G. A. Bartholomew, 1958. Metabol/c and Cardiac Responses to Temperature in the Lizard Dipsosaurus dersalis. Physiol. Zool. 31, 2 : 100. /
Dawson W. R. a. K- Schmidt-Nielsen, 1964. Terrestrial animals in dry heat: desert birds. In: Handb. of Physiol. Sect. 4. Adaptation to Environment. Washington D. C. (
Dawson W. R. a. K. S c h m i d t - N i e 1 s e n. 1966. Effect of Thermal Conductance on Water Economy in the Antelope JaZk Rabbit, Lepus alleni. J. of Celhil. Physiol. 67, 3 : 463.
D e b i a s D. A., 1966. Thyroid-adrenal relationship in altitude tolerance. Fed. Proc. 25, 4 : 1227.
D e 1 a c h a u x A. et A. T i s s i e r e s, 1946. L’adaptatipn tissulaire a 1’hypoxie dosee. Helvet. med. acta 13 : 333.
Dempsey Z. W., N. J. M e у e r s, W.C. Young a. I. Jen-n i s о n, 1934. Absence of light and the reproduction cycle in the guinea pig. Amer. J. Physiol. 109, 2.
D e p о c a s F., 1960. Calorigenesis from various organ systems in the whole animal. Feder. Proc. 19. Suppl. 5 : 25.
Dicker S. E. a. M. G. E g g 11 о n, 1964. Renal function in the primitive mammal Aplodontia rufa with some observations in squirrels. J. Physiol. 170 : 186.
Dill D. B„ 1938. Life. Heat and Altitude. Cambridge, Harvard Univ. Press : 211.
Dubois. R., 1894. Variations du glycogene du foie dans 1’etat du torpeur et pendant le passage de la torpeur a 1’etat de veille. C. R. Soc. Biol. 64: 219.
Dubois R., 1896. Physiologie comparee de la marmotte. Lion.
Du gal L. R., 1953. Les glandes endocrines et le froid. XIX Int. Congr. Physiol., Resumes et commun., Montreal: 121.
D u g a 1 L. R. a. G. F о r t i e r, 1952. Ascorbic acid and acclimatization to cold in monkeys. J. Appl. Physiol. 5 : 143.
D u g a 1 L. R. a. AL T h e r i e n, 1947. Ascorbic acid and acclimatization to cold environment. Canad. J. Res. Sect. E. 25: 1.
Dworkin S. a. W. H. F i n n e y, 1927. Artificial hibernation in the woodshuck (Aretomis monax). Amer. J. Physiol. 80:75.
Eden A., 1941. Studies on the excretion of copper in the rabbit. J Agric Sci 31 * 145
Eibl-Eiblsfeldt J. a. S. Kramer, 1958. Ethology, the comparative study of animal behaviour. Quart. Rev. of Biol. 33, 3 : 181.
Eisentraut M., 1956. Der Winterschlaf mit seinen okologischen und physiologischcn Begleiterscheinungen. VerL G. Fischer.
E n g e r a. S a v a 1 о v, 1958. Metabolism of tropical arthropods. J. Insect. Physiol. 2 : 232.
Folkow B. 1968. Circulatory adaptation to diving in ducks XXIV Intern. Congr. Physiolog. Sci. v. 6 p. 23 Washingion D. C.
Forbes D., 1870. On the Aymara Indians of Bolivia and Peru. From: Barcroft J., 1925.
Frank L, U. Hadeler a. W. Harder, 1951. Zur Ernah-rungsphysiologie der Nagetiere. Pfliigers Archiv, 253: 173.
Frankignoul AL a. Ch. J e u n i a u x, 1965. Effect of chitin on chitinase secretion by rodents. Life Sci. 4, 17 : 1669.
Fry F. E. J. a. J. S. Hart, 1948. Cruising speed of goldfish in relation to water temperature. U. Fisheries Res. Board Canada 7 : 169. 432
Gelineo S., 1934a. Influence du milieu thermique d’adaptation sur la thermogenese des homeothermes. Ann. Physiol. Physicochim. Biol. 10 \ 1083.
Gelineo S., 1934b. Influence du milieu thermique d’adaptation sur la coturbe de la thermoregulation. C. rend. Soc. Biol. 40: 117.
G elineo S., 1936. Der Sauerstoffverbrauch bei Behaglichkeits-temperatui\ nach exogener Hyperthermie. Biologia generalis 12 : 234.
Gelineo S., 1948. Contribution a la connaissance de la calori-facation di| lapin. Bull. Mathem. et Naturclles Science 24 XII.
Gelineo S„ 1959. Temperature d’adaptation et consummation d’oxygene cbez la scorpene truie — Scorpaena porcus L. Bull. Acad. Serbe Sci. 2j : 37.
Gelineo S., 1964. Organ systems in adaptation: the temperature regulating system. In: Handb. of Physiol., sect. 4. Adaptation to Environment, Amer. Physiol. Soc. Washington : 259.
Gelineo S. a. A. G e 1 i n e o, 1955. Les echanges respiratoires des lizards noirs de Dalmatie, adaptes a differentes temperatures. Compt. Rend. Soc. Biol. 149 : 387.
Gelineo S. et R. Kocarev, 1957. Sur ia depence d’oxygene chez le rat rassasie au cours de la lutte centre le froid. Bull, de 1’Acad. Serbe Sci., Sci. naturelles 18, 5.
Gelineo S. et T. Raiewskaya, 1953. Influence d’un regime alimentaire de mais sur la concentration de I’hemoglobine a des temperatures differentes. C. R., Soc. Biol. 147 : 136.
G i a j a J., 1938. Sur 1’analyse de la fonction de la aolorification de 1’homeotherme par la depression barometrique. C. R. Soc. Biol. 127 : 1355.
G i a j a J., 1953. Hypothermie, hybernation et poikilothermie experimentalle. Biol. Mcdicale 42, 6.
Giaja J. a. S. G e 1 i n e o, 1931. La thermogenese aux temperatures inferieures a celle de la resistance homeotherme. Ann. Physiol. Physicochim. Biol. 7: 152.
Giaja J. a. L. M a г к о v i c - G i a j a, 1950. Le spectre de la thermogenese de 1’organisme refroidi. Arch, des Sci. Physiol. 9:89.
Glasser E. M. a. J. P. G r i f f i n, 1962. Effect of the cerebral cortex upon habituation. J. Physiol. (London) 160 : 429.
Glenn D., A u m a n n J. a. T. E m 1 e n. 1965. Relation of population density to sodium availability and sodium selection by microti-nae rodents Nature v. 208 p. 198.
Goethe F., 1939. Auf Jugendleben des Muffelwildes. Zool. Garten, N. F. 11 : 1.
Goodall A. M. a. S. H. Y о n g, 1954. The vascular supply of the skin of ayrshire calves and embryos. J. Agric. Sci. 1 :44.
Grafe C., 1928. Die nervose Regulation des Stoffwcchsels. In: Oppeiih. Handb. d. Biochem. Bd. 8.
Grandjean E., 1948. Physiologic du climat de la montagne. J. de physiol. 49, 1 : 52.
Gray J., 1933. Studies in animal locomotion. J. Exp. Biol. 10: 88.
Griffin D. R. a. J. H. W e 1 s h, 1937. Activity rhythms in bats undes constant external conditions. J. Mammal. 18 : 337.
H a 1 b e r g F., E. H a 1 b c r g, С. P. Bar n u ni a. .1. .1. В i t-t n e r, 1959. Physiologic 24-hour periodicity in human beings and
433
mice, the lighting regimen and dayly routine. Photoperiodism and related phenomena in plants and animals. Ed. by Withrow, A. A. A. S. Washington.
Hall F., D. D i 11 a. G. E. В a г г о n, 1936. Comparative physiology in high altitude. J. Cell. Comp. Physiol. 8: 301.
Hamilton C. L., 1965. Control of food intake. In: Physiological Controls and Regulations, Ch. 13. Ed. by Saunders C°. Philadelphia, London. I
Handbook of Physiology, 1964. Sect. 4. Adaptation to the Environment. Am. Physiol. Soc. Washington D. C. '
Harder W., 1950. Zur A4orphologie und Physiologle des Blind-darmes der Nagetiere. Zool. Anzeiger. Suppl. 14 : 95.
Hardy J. P., 1961. Physiology of temperature regulation.
Physiol. Rev. 41 : 521.
Harker J., 1960. Endocrine and Nervous Factors in Insect Circadian Rhythms. In: Biological clocks, Cold Spring Harbor Symposia on Quantitative Biology, XXV, N. Y.
Harrison S. J., 1960. Experiments with diving seals. Nature 188: 1068.
Hart J. S., 1956. Seasonal changes in insulation of the fur. Canad. J. Zool. 34, 1.
Hart J. S„ 1959. The relation between thermal history and cold resistance in certain species of rodents. Canad. J. Zool. 31, 2.
Hart J. S., 1960. Energy metabolism during exposure to cold.
Feder. Proc. 19, 4, part II : 15.
Hart J. S., 1962. Seasonal acclimatization in four species of wild birds. Physiol. Zool. 35 : 35—224.
Hart J. S., 1962a. Heat production mechanisms. Proc, of the Int.
Union of Physiol. Sci. XXII Int. Congr., Leiden 1 :398.
Hart J. S., 1964. Insulative and metabolic adaptations to cold in vertebrates. Symp. Soc. Exp. Biol. 18:31. Feedback Homeostasis and Mechanisms. Cambr. Univ. Press.
Hart J. S. a. O. Z. R о y, 1966. Respiratory and cardiac responses to flight in pigeons. Physiol. Zool. 39 : 291.
Hart J. S. a. O. Z. Roy, 1967. Temperature regulation during flight in pigeons. Amer. J. Physiol. 213 : 1311.
Hartmann F. A. a. K. A. Brownell 1949. The adrenal glands. Ed. by Lea a. Febiger, Philadelphia.
Hayward J. S., 1965. Metabolic rate and its temperature-adaptive significance in six geographic races of Peromyscus. Can. J. Zool., v. 43, p. 309.
Hediger IL, 1952. Beitriigc zur Saugetier-Soziologie. Structure et Physiol, des Soc. Animalcs, Colloq. Internal. Paris.
Hem mingsen A., 1950. Energy metabolism as related to body size and respiration surfaces and its evolution. Rep. Steno Hosp. 4 : 7-58.
Hensel H., 1955. Temperatur und Leben. Von H. Precht Christophersen u. H. Hensel. Berlin.
Hermanson V. a. F. A. Hartmann, 1945. Protection of adrenalectomized rats against high temperature. Amer. J. Physiol. 144 : 108.
Heroux. O., 1962. Seasonal adjustments in captured wild rats. IL Survival time, pelt insulation, shivering and metabolic and pressor responses to noradrenaline. Canad. J. Biochem. Physiol. 40 : 537.
434
Heirter К., 1933. Dressurversuche mit Igeln. Z. Vergl. Physiol.
Heitor K., 1932. Der Tcinperalursinn flor Siingetiere. Leipzig.
Her ter K., 1957. Das Verhalten der Insectivoren. Handb. d.
Zool. 8, Lii. 9, Berlin.
Hess E. H., 1959. Two conditions limiting critical age for imprinting. J. Comp. Physiol. Psychol. 52 : 515.
Hess W. R., 1949. Das Zwischenhirn. Basel.
Hesse R., 1921. Das Herzgewicht der Wirbeltiere. Zool. Jahrb. Abt. Zool. u Physiol. 38, 3 : 243.
Hesse R., 1924. Tiergeographie und okologische Grundlage. Jena.
H i 1 d e п A. a. K. S. S t e n b a c k, 191G. Zur Kentniss der Tages-schwankungen der Korpertemperatur bei den Vogeln. Skand. Arch, f. Physiol. 34 : 382.
Hill A. V., 1913. Heat production in animals living together. J. Physiol. London 46, 2 : 411.
Hock R., 1964. Animals in high altitudes: reptiles and amphibians. Handb. of Physiol. Sect. 4. Adaptation to Environment, Washington, D. C. : 841.
Hoehn E. O., 1949. Seasonal changes in the thyroid gland and effects of thyroidectomy in the mallard in relation to moult. Amer. J. Physiol. 158 : 337.
Hoock R. J., 1951. Temperature and metabolism of bears. Proc. Second Alaska Science Confer.: 310.
Hoock R. J., 1964. Animals in high altitudes: reptiles and amphibians. Handb. of Physiol. Sect. 4. Adaptation to the Environment. Am. Physiol. Soc., Washington : 841.
Hook W. E. a. E. S. Barron, 1941. The respiration of brown adipose tissue and kidney of the hibernating and nonhibernating ground-squirrel. Amer. J. Physiol. 133 : 56.
Horwath A., 1878. Beitrag zur Lehre fiber den Winterschlaf. Verh. d. Phys. med. Gesellschaft zu Wiirzburg, Bd. 12, 1.
Horwitz B. A., L. Nelson a. V. Popovic, 1967. Effect of temperature on oxidative phosphorylation in hibernators and nonhiber-nators. J. Appl. Physiol. 22 : 639.
Howell A. B., 1965. Speed in animals. Hapn. Publ. Comp.
Howell A. B. a. J. Gersch, 1935. Conservation of water by the rodent Dipodomis. J. Mammol. 16 : 1.
H о w e 1 1 T. R. a. G. A. Bartholomew, 1959. Further experiments on torpidity in the poor-will. Condor 61 : 180.
Hsein A. C. L. a. L. D. Carlson, 1957. Role of the Thyroid in AJetabolic Response to Low Temperature. Am. J. Physiol. 188, 1 : 40.
Human Adaptability and its Methodology, 1966. Papers at the Symp. in Kyoto, Japan. Ed. for IUPS a. IBP by Jap. Soc. of Promot. for Sci., Tokyo.
Hudson J. W., 1964. Temperature regulation in the round-tailed ground squirrel, Citellus tereticaudus. Ann. Acad. Sci. Fenn. A. IV, v. 30, p. 219.
Hurtado A., 1964. Animals in high altitudes: resident man. Handb. Physiol. Sect. 4. Adaptation to the Environment, Washin gton D. C. : 843.
Hurtado A., A. R о 11 a, С. M e r i n о a. J. P о n s, 1937. Stu
435
dies of myohaemoglobin at high altitudes. Amer. J. Med. Sci. 194 : 708.
Huxley F. M., 1913. On the reflex nature of apnoea in the duck in diving. Quart. J. Physiol. 6 : 147.
Irving L., 1939. Respiration in diving mammals. Physiol. Rev. 19, 1.
Irving L., 1951. Physiological adaptation to cold in arctic and tropic mammals. Fed. Proc. 10 : 543.
Irving L„ 1960. Birds of Anaktuvak Pass, Kobuk and Old Crow. U. S. Nat. Mureum Bull. 217.
Irving L., 1964. Terrestrial animals in cold: birds and mammals. Handb. of Physiol. Sect. 4. Adaptation to Environment. Washington, D. C.
Irving L. a. J. Krog, 1954. Temperature of skin in the Arctic as a regulator of heat. J. Appl. Physiol. 7 : 355.
Irving L., H. Krogh a. M. Monson, 1955. The Metabolism of Some Alaskan Animals in Winter and Summer. Physiol. Zool. 28: 173.
Irving L., a. K. Schmidt-Nielsen a. N. S. Abraham-sen, On the melting points of animal fats in cold climates. Physiol. Zool. 30 : 93.
Ishida K., 1930. Uber den Einfluss der Jahreszeit auf den respi-ratorischen Gaswechsel weisser Ratten. Mifteil. Mediz. Acad. Kioto, 4: 117.
Jukes M. G. M., M. M. R. Jukes a. W. Banihfa, 1968. The respiratory response of the duck to inhaled carbon dioxide XXIV Intern. Congr. Physiolog. Sci. v. 7 p. 23 Washington D. C.
Jansky L., 1965. Adaptability of heat production mechanisms in homeotherms. Acta Univer. Carolinae Biologica vol. 1 : 1, Praha.
Jansky L. a. J. S. H a r t, 1968. Cardiac output and organ blood flow in warm- and cold-acclimated rats exposed to cold. Can. J. Physiol. Pharmacol. 46 : 653.
Jaeger E. S., 1949. Further observations of the hibernation of the poor-will. Condor 51 : 105.
Jennings J. B., 1965. Feeding Digestion and Assimilation in Animals. Pergamon Press.
Joel C. D., 1965. The physiological role of brown adipose tissue. Handb. of Physiol. Sect. 5. Adip. tissue. Am. Physiol. Soc. Washington.
Johansson B., 1960. Brown fat and its possible significance for hibernation. Bull. Museum Comp. Zool. Harvard Coll. 124 : 233.
Johansson B. W., 1967. Heart and circulation in hibernators.
Mammalian Hibernation III, New York, o. 200.
Johansson B. W., 1968. Electrocardiographic changes in depressed metabolism. Proc, from the Conference in Depressed Metabolism, Washington, D. C.
Johnson G. E., 1929. Hibernation of the thirteen-lined ground squirrel Citellus tridecemlineatus. III. The rise in respiration, heart beat and temperature in waking from hibernation. Biol. Bull. 57 : 107.
Jourdanet D., 1875. L’influence de la pression de Pair sur la vie de 1’homme. Paris.
Kami у am a K., 1956. On effects of environment condition on vasomotor reflex. Papers in Meteorol. a. Geophysics 7, 2.
Kanungo M. S. a. C. L. Prosser, 1959. Physiological and biochemical adaptation of goldfish to cold and warm temperatures.
436
I. Standard and active oxygen consumption of cold- and warm acclimated goldfish at various temperatures. J. Cellul. Comp. Physiol. 54 : 259.
Karli P„ 1956. The norway rats killing response to the white mouse: an experimental analysis. Behaviour, Leiden 10, 1—2:81.
Kauth H. u. H. Sommer, 1953. Das Ferment im Tierkorper, IV. Biol. ZbL 72, 3—4.
Kayser Ch., 1939. Evolution saisonniere de la thermoregulation chimique chez les hibernants reveilles. C. R. Soc. Biol. 131 : 893.
Kayser Ch., 1940a. Les echanges respiratoires des hibernants. 3-e partie: Echanges respiratoires des hibernants a 1’etat de sonuneil hibernal. Ann. Physiol. 16: 128.
Kayser Ch., 1940b. Les echanges respiratoires des hibernants. 4-e partie: Essai d’analyse du mechanisme du sommeil hibernal. Ann. Physiol. 16 : 314.
Kayser Ch., 1953. L’hibernation des mammiferes. Ann. Biol. 29 : 109.
Kayser Ch., 1961. The physiology of natural hibernation. Per-gamon Press.
King J. R. a. D. S. F a r n e r, 1960. Energy metabolism, thermoregulation and body temperature. In: Biology and Comp. Physiol, of Birds, ed. by Marshall, vol. 2. Acad. Press, N.—Y. : 215.
Kirchhoff H., 1937. Jahreszeiten und Belichtungen in ihrem Einfluss auf weibliche Genitalfunktionen. Arch. f. Gynakol. 163.
Kleiber M., 1932. Body size and metabolism. Hilgardia 6, 315.
К 1 e i t m a n N., 1939. Sleep and wakefulness. Univ, of Chicago Press. Chicago.
Klighley, G. P. Low у a. C. McCune, 1968 Polycythemia as ineffective component of altitude acclimation. XXIV Int. Congr. Physiol. Sci. Washington D. C.: 232.
Krebs H. A., 1950. Body size and tissue respiration. Biochem. et Biophys. Acta 4.
Krieg H., 1924. Biological Reisestudien on Siidamerika, HI.
Chilenische Beutelratten. Z. Morph. Okol. 3 : 169.
Krogh H. a. M. Л4 о n s о n, 1954. Notes on the Metabolism of a mountain goat. Amer. J. Physiol. 178 : 515.
Krueger H. M., J. B. Saddler, G. A. Chapman, I. J. Tinsley a. R. K- Lowry, 1968. Bioenergetics, Exercise and Fatty Acid of Fish Amer. Zool. 8, 1.
Krywienczyk J., 1952a Korpergrosse, Korperzeiten und Energiebilanz. IVte Mitteilung. Korpergrosse, O2-Konsum und Kriech-geschwindigkeit bei Prosobranchieren. Z. Vergl. Physiol. 34 : 6.
Krywienczyk J., 1952b. Korpergrosse, Korperzeiten und Energiebilanz. V Mitteilung. Korpergrosse, O2-Konsum und Kriechge-schwindigkeit bei Wasserpulmonaten. Z. Vergl. Physiol. 34: 14.
Krzywanek F. W., 1928. Uber die Temperatur im Panzer des Schafes. Pfliigers Arch. Physiol. 222, 89.
L a d e 1 1 W., 1964. Terrestrial animals in humid heat: man. Handb. Physiol. Sect. 4. Adaptation to Environment. Washington, D. C.:625.
Lasi ewski R. C., 1963. Oxygen consumption of torpid, resting, active and flying humming birds. Physiol. Zool. 36 : 122.
L a s i e w s к i R. C. a. W. R. Dawson, 1964. Physiological responses to temperature in the common lighthawk. Condor 66 : 477.
437
Lasiewski R. C., F. R. G a I e у a. C. Vasquez, 1965. Morphology and physiology of the pectoral muscles of humming birds. Nature''206 : -104.
Leblanc J., 1962. Local adaptation to cold of Gaspe fishermen. J. Appl. Physiol. 17 : 950.
Leblanc J., 196G. Adaptive mechanisms in humans. Ann. of N—Y Acad. Sci. 134, 2 : 721.
L e b 1 a n c J., J. A. H i 1 d e s a. O. Heroux, 1960. Tolerance of Gaspe fichermen to cold water. J. Appl. Physiol. 15 : 1031.
Leduc E. H. a. Ch. P. Lyman, 1952. Changes in blood sugar and tissue glicogen in the Hamster during arousal from hibernation. Anat. Rec. 113: 112.
Lee A. K-, The adaptation to arid environments in wood rats of the genus Neotoma. Univ. CaliL Pubis Zool., v. 64, p. 57.
Les by J. E. a. R. S. Levin, 1966. Anatomic adaptation of the alimentary tract of the rat to the hyperphagia of chronic alloxan diabet. Nature 210, 5034.
Leveill G. A. a. E. K. O'Hea, 1967. Influence of Periodicity of Eating on Energy Metabolism in the Rat. J. Nutr. 93, 4: 541.
L i a n i d e s S. P. a. R. E. Beyer, 1960. Oxidative phosphorylation in liver mitochondria from cold-exposed rats. Am. J. Physiol. 199 : 836.
Lindhardt J., 1917. Physiological investigation in the Dane-mark expedition. Meddelelseron Gronland, Kopenhavn.
Litson N. L, G. B. Gordon a. R. McClintock, 1955. Measurement of total carbon dioxide production by means of D2O1S. J. Appl. Physiol. 7 : 704.
Little L. O. a. G. E. M i t c h e 11, 1965. Rumen metabolites as metabolizable energy sources for rats. Life Sci. 5 : 905.
Lobban M. C., 1957. Studies on physiological diurnal rhythms in polar regions. Advancem. Sci. 13 : 413.
L о о s 1 i J. K-, H. H. Williams, W. E. Thomas, F. H. Ferris a. L. A. M a g n a r d, 1949. Synthesis of aminoacids in the rumen. Science 110, 5.
Lord R. D., F. С. В e 11 г о s e a. W. W. Cochran, 1962. Radiotelemetry of the respiration of a flying duck. Science 137 : 39.
Lorenz K., 1937. Uber die Bildung des Instinktbegriffes. Natur-wissenschaften 25 : 199.
Louch C. D., 1958. Adrenocortical activity in two meadow vole populations. J. Mammol. 39 : 109.
Liman С. P. a. P. O. Chatfield, 1956. Physiology of hibernation in mammals. The Physiology of Induced Hypothermia, Washington, p. 80.
Alacfarlane W. V., 1965. Water metabolism of desert ruminants. In: Studies in Physiol., Springer-Verlag N—Y : 191.
Macfarlane W. V. а. В. H о w a r d, 1966. Water content and turnover of identical twin Bos indicus and Bos taurus in Kenya. J. Agris. Sci. 66 : 297.
MacMillen R. E. a. A. K. Lee, 1967. Australian Desert Mice: Independence of Exogenous Water. Science, v. 158, N 3799, p. 383.
MacMillen R. E. a. A. K- Lee, 1969. Water metabolism of Australian hopping mice. Comp. Biochem. Physiol., v. 28, p. 493.
M c N a b. В. K., 1968. The influence of fat deposits on the basal
438
rate of metabolism in desert homoiotherms. Comp. Biochcm. Physiol., v. 216, p. 337.
McNab В. К. a. P. R. A'lorrison, 1963. Body temperature and metabolism in subspecies of Peromyscus from arid and mesic environments. Ecol. Monorg., v. 33, p. 63.
Madsen H., 1939. Does the hare chew the cud? Nature 143: 981.
de Magnen J. et S. T a 1 I о n, 1968. Equivalence des apports caloriques ghicidiques et lipidiques dans la regulation de la prise alimentaire chez 1c rat. Arch, des Sci. Physiol. 22, 1 : 127.
M a i g п о n F. et H. G u i 1 h о n, 1931. L’influence des saisons sur les changes respiratoires chez le chien. Cr. Acad. Sci. 192 : 1410.
Malmejac J., G. N e v e г r e, P. Plane, M. Montero et С. M a 1 m e j a c, 1956. Action de hypothcrmie «provoquee» sur les differentes parties du systeme nervoeux: cortex, centres bulbo-protube-rantiel, ganglions vegetatifs. XX Int. Congr. de Physiol., Resumes et commun. Bruxelles : 612.
Mammalian Hibernation. Bull. Museum Comp. Zool., Cambridge, Mass. U. S. A.
M a rey E. J., 1890. Le vol des oiseax. Ed. Masson, Paris.
Martin C. J., 1901. Thermal adjustment and respiratory exchange in monotremes and marsupials. Proc. Roy. Soc. (London) 69 : 352.
Mason J. W., 1959. Central nervous system factors in the regulation of endocrine secretion. Hormone Research 15 : 345.
Mayer J., 1955. Regulation of energy intake and the weight: the glucostatic theory and the lipostatic hypothesis. Ann. N. Y. Acad. Sci. 63: 15.
Mellanby K., 1932. Effects of temperature and humidity on the metabolism of the fasting ber-bug (Cimex lectularius). Hemiptera Parasitology 24: 419.
Merzbacher L„ 1903. Die Nervendegeneration wahrend des Winterschlafes. Die Beziehungen zwischen Temperatur und Wintersch-laf. Pfliig. Arch. 100 : 568.
Mihai lovic Lj., В. P e t г о v i c - M i n c, S. Protic, I. D i-v a c, 1968. Effects of Hibernation on Learning and Retention. Nature, v. 218, N 5136, p. 197.
Miller A. H., 1951. An analysis of the distribution of the birds of California. Univ. Calif., Berkley, Publ. Zool. 50:531.
MillerS., 1939. A study of the phlsiology of the sparrow thyroid. J. Exp. Zool. 80, 2.
Mimachi H. u. E. W e i n 1 a n d. 1910. Beobachtungen am Igel in der Periode dor Nahrungsaufnahme. Z. Biol. 55 : 1.
Mole schott J„ 1855. Uber den Einfluss des Lichtcs anf die Menge von Tierkorper ausgeschwidenen Kohlensaure. Wien. med. Wochcnschr. 43 : 1.
Moleschott J. ii. S. F u b i n i, 1881. Uber den Einfluss gemischten und farbigen Lichtes auf die Ausscheidung der Kohlensau-re bei Tieren. Moleschotts Untersuchen 12, 2 : 266.
Monge C., 1943. Chronic mountain sickness, Physiol. Rev.
Moore R. E. a. M. A. Simmonds, 1966. Decline with age in the thermogenic response of the voung rat to 1-noradrenaline. Fed. Proc. 25, 4.
Morot Ch., 1882. Des pelotes stomacales des leporides. Memoires Soc. Ctr. Med. Veter. 12, Paris : 1.
439
Morrison P. R. a. W. J. T i e z t, 1957. Cooling and thermal conductivity in three small alaskan mammals. J. Mammol. v. 38, N 1, p. 78.
Morrison P. R. 1962. Modification of body temperature by activity in brazilian humming birds. Condor 64 : 315.
Morrison P. a. R. Elsner, 1962. Influence of altitude on heart and breathing rates in some Peruvian rodents J. appl. Physiol, v. 17, p. 467.
Morrison P. R., К. К e r s t a. M. R о s e n m a n n, 1963. Hematocrit and Hemoglobin Levels in some Chilean Rodents from High and Low Altitude. Int. J. Biometeor. 7 : 45.
Morrison P. R. а. О. P. Pearson, 1947. The metabolism of very small mammals. Science 104 : 287.
Morrison P. R„ F. A. R у s e r a. F. S t г e с к e r, 1954. Growth and the development of temperature regulation in the tundra redback vole. J. Mammal. 35 : 3.
Nielsen К. С. а. С. О w m a n, 1968. Difference in cardiac adrenergic innervation between hibernators and non-hibernating mammals. Acta Physiol. Scand., Suppl. 316.
Niethammer G., 1952. Tiergeographie. Fortschr. Zool. 9 : 368.
Nungesser W. С. a. E. W. Pfieffer, 1965. Water balance and maximum concentrating capacity in the primitive rodent, Aplodo-ntia rufa. Comp. Biochem. Physiol. 14 : 289.
О b о u s s i e r B., 1948. Uber die Grossenbeziehungen der Hypo-physe und ihre Teile bei Saugetieren und Vogeln. Arch. f. Entw. Meehan, d. Organismen 143 : 172.
Olsen H. a. H. Madsen, 1944. Investigation on the pseudorumination in rabbits. Videnskabelige Meddelelser. 107, Kopenhavn : 37.
Opitz E. u. M. Schneider, 1950. Uber die Sauerstoffversor-gung des Gehirns und den Mechanismus von Mangelwirkungen. Erg. Physiol. 46 : 126.
Ozorio de Almeida B., 1919. Le metabolisme minimum et le metabolisme basal de 1’homme tropicale de race brauche. J. Phys, et Path. 18: 958.
Pa dour L., 1950. Differenzen endokrinen Driisen bei Saugetieren verschiedener Korpergrosse mit besondcrer Beriicksichtigung der Lan-gerchanschcn Inseln. Zool. Jahrb., Abt. Physiol. 62: 3.
Page E. a. L. M. В a b i n e a u, 1954. Tissue glicogen and glucose absorption in rats adapted to cold. Canad. J. Biochem. Physiol. 32, 4.
Parry et Lyon, 1825. Temperature des animaux aux grands froids. Ann. de Chim. et de Physique 28 : 223.
P a v 1 о v i с. V., 1954. Adaptation to environmental temperature and liver glycogen fractions in rats —Mus decumanus. Arch. Biol. Sci., Beograd 6, 3.
Pengel ley E. T. а. К- C. F i s h e r, 1963. Rhythmical arousal from hibernation in the golden — mantled ground squirrel Citellus lateralis tescorum. Can. J. Zool. 39 : 105.
Peter K-, 1909. Die Nierenkaniilchen des Menschen und andere Siiugetiere. Untersuchungen fiber Ban und Entwicklung der Niere. Jena.
P flii ger E., 1878. Uber Warme mid Oxydation der lebenden Materie. Pflfig. Arch. 188 : 4.
440
Physiological Mammoiogy, 1963. Vol. I. Mammalian populations. Ed. by V. V. Alayer a. R. G. van Gelder, Academic Press.
Pier on H., 1958. De 1’actinie a 1’homme. I. Paris.
Pool W. E., 1949. Effect of anoxic anoxia on myoglobin concentration in striated muscle. Amer. J. Physiol. 156 : 44.
Popovic V., 1964. Cardiac output in hibernating ground squirrels. Am. J. Physiol. 207 : 1345.
Portig F., 1950. Bemerkungen zur Uberwinterung des Hamsters (Cricetus cricetus). In: Nene Ergebn. u. Probl. d. Zoolog. : 756.
Poulson T. L. a. G. A. Bartholomew, 1962. Salt balance in the savannah sparrow. Physiol. Zool. 35: 109.
P r e c h t IL, C h r i s t о p h e r s e n J. a. H. H c n s e 1, 1955. Tem-peratnr und Leben. Springier — Verlag, Berlin.
Prosser. C. L., 1964. Perspectives in adaptation theoretical aspects. In: Handb. of Physiol. Sect. 4. Adaptation to Environment, p. II, Washington, D. C.
Prosser C. L. a. F. В г о w n, 1962. Comparative Animal Physiology. W. B. Saunders Company Philadelphia — London.
Raab W., 1934. Morbus Basedov nach Kohlenoxydvergiftung. Wien. Klin Wchschr., Jahrg. 47 : 1482.
Raiewskaya T., 1953. Influence de la temperature d’acclimatisation sur la concentration de hemoglobine. Comp. Rend. Soc. Biol. 147: 1981.
Rameaux et Sarrus, 1838—1839. Rapport sur un memoire adressee a 1’Academie Royale de Medicine. Bull, de 1’Acad. Roy. de Med. 3 : 1094.
Rautenberg W., 1957. Vergleichende Untersuchungen fiber den Energieaushalt des Bergfinken (Fringilla montefringilla) und des Haussperling (Passer domesticus). J. Ornithol. 98 : 36.
Reach F., 1910. Beitrag zur Physiologic des Winterschlafes. Bioch Z. 26 : 331.
Readings in ecology, 1965. Prentice—Hall, Inc. New Jersey.
Rennie D. W., B. G. С о v i n о, B. J. H о w e 11, S. H. S о n g, B. S. К a n g a. S. K- Hong, 1962. Physical insulation of Korean diving women. J. Appl. Physiol. 17 : 961.
Rensch B„ 1932. Uber die Abhangigkeit der Grosse des relati-ven und Oberflachenstruktur der Landeschneckschalen von den Um-weltfactoren. Z. f. Morphol. Okol. 25: 16.
Rensch B., 1939. Klimatische Auslese von Grossenvarianten. Arch. f. Naturg. N. F. 8 : 1.
Re t z 1 a f f E. G., 1938. Studies in population physiology with the albino mouse. Biologia Generalis 14 : 238.
Reynafarje C., 1966. Iron metabolism during and after altitude exposure in man and in adapted animals (Camelids). Fed. Proc. 25, 4.
Reynafarje В. a. P. Morrison, 1962. Myoglobin Levels in Some Tissues from Wild Peruvian Rodents Native to High Altitude. J. Biol. Chem. 237, 9 : 2861.
R i c h e t Ch., 1889. Chaleur animal. Paris.
Riche t Ch., 1898. De 1’influence de 1’education sur la resistance du canard a 1’asphyxie. C. R. Soc. Biol. (Paris) 50 :481.
R i c h e t Ch., 1899. De la resistance des canard a 1’asphixie. J. Physiol, et Pathol. Gencr. 1 : 641.
441
Riddle О., G. C. S m i t h a. F. G. В с n e d i c t, 1934. Seasonal and temperature factors and their determination in pigeon on percentage metabolism change per degree of temperature change. Amer. J. Physiol. 107 : 334.
Ritz man E G. a. F G. Benedict, 1938. Heat production in sheep. Carnegie Inst. Washington, Publ. 494.
Robinson D., 1939. The muscle hemoglobin of seals as an oxygen store in diving. Science 90, 2334.
R 6 d e 1 E., 1957. Zum Problem der Coecotropie als Anzeiger richtiger Kaninchcnfiittenmg. Kaninchenzuchter 19, 61 : 181.
Roth L. M. a. E. R. Willis, 1951. The effect of dessication and starvation on the humidity behaviour and water balance of Tribo-lium confusum a. T. castancuin. J. Exptl. Zool. 118:337.
Rubner M., 1883. Uber den Einfluss der Korpergrosse auf Stoff und Kraftwechsel. Z. f. Biol. 19: 535.
Saint-Girons H. et M. C. Saint-Girons 1956. Cycle d’activite et thermoregulation chez les reptiles (lizards et serpents). Vie et Milieu 7 : 133.
Salt G. W., 1964. Respiratory evaporation in birds. Biol. Rev. 39 : 113—136.
Salt G. W. a. E. Z e u t h e n, 1960. The respiratory system. In: Biology and Comparative Physiology of Birds, ed. by A. J. Marshall. New York, Acad. Press, vol. 1 : 363.
Sarajas H. S. S.. 1960. On the cardiac responce in hibernation and induced hypothermia. Functional, pathologic and metabolic aspects. Mammalian Hibernation, Bull. Museum Comp. Zool., v. 124, Cambridge, Mass., p. 337.
Sarrus et Rameaux, 1837—1838. Application des sciences accessoires et principalment des mathematiques a la physiologic generale. Bull, de I’Acad. de Med. 2 : 538.
Scheunert A., 1906. Zum Mechanismus der Magenverdauung. Pfliig. Arch. 114 : 64.
Schmidt-Nielsen B., 1949. Reduced evaporative water loss from the lungs of certatin desert mammals. Feder. Proc. 8 : 139.
Schmidt-Nielsen K-. 1949. Efficiency of excretory system in desert mammals. Feder. Proc. 8 : 140.
Schmidt-Nielsen K-, 1954. Heat regulation in small and large desert mammals. Biol, of Deserts, J. L. Cloudsley — Thompson, Ed., Inst, of Biol., London : 182.
Schmidt-Nielsen K-, 1956. Animals and Arid Conditions: Physiological Aspects of Productivity and Management. The Future of Arid Lands, Amer. Assoc, for Advancem. of Sci., Washington, D. C.: 368.
Schmidt-Nielsen K-> 1958. Animals and arid conditions. In: Climatology and microclimatology. Proceedings of the Canberra Symposium. Paris, Unesco : 217.
Schmidt-Nielsen K-, 1965. Animal Physiology. Prentice — Hall, Inc. New Jersey.
Schmidt-Nielsen К., E. C. Crawford Jr., A. E. New-some, K. S. R a w s о n a. H. T. H a m m e 1, 1967. Metabolic rate of camels: effect of body temperature and dehydration. Am. J. Physiol. 212, 2 :341.
S c h m i d t - N i e 1 s e n K. a. W. R. Dawson, 1964. Terrestrial
442
animals in dry heat: desert reptiles. Handb. of Physiol. Sect. 4. Adaptation to the Environment. Am. Physiol. Soc. Washington : 467.
Schmidt-Nielsen К. a. B. Schmidt-Nielsen, 1951. A complete account of the water metabolism in kangaroo rats and an experimental verification. J. Cell. Comp. Physiol. 38 : 165.
Schmidt-Nielsen К. a. B. Schmidt-Nielsen. 1952. Water metabolism in desert mammals. Physiol. Rev. 32, 2.
Schmidt-Nielsen К, B. Schmidt-Nielsen a. A. Brokaw, 1948. Urea excretion in desert rodents exposed to high protein diets. J. Cell. Comp. Physiol. 32 : 361.
Schmidt-Nielsen В., K. Schmidt-Nielsen, A. Brokaw a. A. S c h n e i d e r m a n n, 1948. Water conservation in desert rodens. J. Cell. Comp. Physiol. 32 : 331.
S c h m i d t - N i e 1 s e n K-. B. S c h m i d t -N i с 1 s e n, S. A. J a r-num a. T. R. H о u p t, 1957. Body temperature of the camel and its relation to water economy. Am. J. Physiol. 188 : 103.
S c h m i d t - N i e I s e n В., K. S c h m i d t -N i e 1 s c n, T. R. H o-ii p t a. S. A. J a r n u m, 1956. Water balance of camel. Amer. J. Physiol. 185 : 185.
Scholander P. F., 1940. Experimental investigation on the respiration in diving mammals and birds. Norse. Vid. Akad. 1 : 31.
Scholander P. F., 1955. Evolution of climatic adaptation in homeotherms. Evolution 9, 1 : 15.
Scholander P. F., 1958. Studies of man exposed to cold. Feder. Proc. 17 : 1054.
Scholander P. F„ 1961. Habituation processes. Ann. Acad. Reg. Sci. Upsalieu 5 : 1.
Scholander P. F., Flagg G. S., V. Walters a. L. I r-v i n g, 1953. Climate adaptation in arctic and tropical poikilotherms. Physiol. Zool. 26 : 67.
Scholander P. F., R. Hock, V. Walters a. L. Irving, 1950. Adaptation to cold in arctic and tropical mammals and birds in relation to body temperature insulation and basal metabolism rate. Biol. Bull. 99 : 259.
Schultze E., 1936. Kohlenoxyd und Schilddrusc. Arch. exp. Pathol, n. Pharm. 180 : 639.
Seibert H. C„ 1949. Differences between migrant and non-mig-rant birds and water intakes at various temperatures and pholoperiods. Auk 66 : 128.
Sellers E. Л., I. W. S с о 11 a. N. T h о m a s, 1954. Electrical activity of skeletal muscle of normal and acclimatized rats on exposure to cold. Amer. J. Physiol. 177 : 371.
Simpson S. a. J. J. Galbraith, 1905. An investigation into the diurnal variation of the body temperature of nocturnal and other birds and a few mammals. J. Physiol. (London), 33:225.
S luckin W., 1965. Imprinting and early learning. Aldine Publ. Comp. Chicago.
Smelser G. R., Walton a. E. O. Whet hem, 1934. The effect of light on ovarian activity in the rabbit. J. Exp. Biol. 11,4.
Smith R. E., 1960. Mitochondrial control of oxidative phosphorylation in cold-acclimated rats. Feder. Proc. 19, suppl. 5 : 64.
Soulairac A., 1963. Neurological factors in the control of the appetite. Intern. Rev. Neurobiol. 5, New York-London, Acad. Press.
443
S о u 1 a i г a с A. a. M. L. S о п 1 a i r a c, 1954. Effets du groupment sur le comportement alimentaire du rat. Conipt. rend. Soc. biol. 148, 3—4 : 304.
Sperber J., 1944. Studies on the mammalian kidney. Zool. Bidrag Iron Uppsala 22 : 249.
S t e b 1 i n s R. C. a. R. M. E a к i n, 1958. The role of the “third eye” in reptilian behavior. Am. Museum Novitates 1870 : 1.
Stern F., 1933. Der Blutjodspiegel bei Hautkrankheiten. Arch. Derm. u. Syph. 167 : 285.
Stigler R., 1920. Rassenphysiologische Studien. Wiener Med. Wochensch: 1263.
Stigler R., 1930. Vergleichende Untersuchungen der physiolo-gischen Warmeregulation. Die Warmeregulation der Nagetierc bei cxo-gcn gesteigerter Korpertemperatur. Ach. exper. Pathol, u. Pharmacol. 155: 257.
Streuly H., 1918. Das Verhalten von Schilddriisenlosen und milzlosen Tieren bei O2—Mangel. Zugleich ein Beitrag zur Theorie der Bergkrankheit. Bioch. Z. 87 : 359.
Strohl J., 1910. Die Massenverhaltnisse des Herzens in Hoch-gebirge. Zool. Jahrb. Abt. Zool. u. Physiol. 30, 5 : 1.
Strohl J., 1910a. Le poids relatif du coeur et 1’effet des grands altitudes. Etude comparative sur deux especes de lagopedes, habitant 1’une les Hautes-Alpes, 1’autre les plaines de la Laponie. C. R. Acad. Sci. 150 : 1257.
Sturm A. u. B. Buchholz, 1928. Beitrage zur Kenntnis de' Jodstoffwechsels. IV Mitteil. Jodverteilung im menschlichen und tieri-schen Organismus in ihrer Bezichung zur Schilddriise. Dtsch. Arch, klin. Med. 161 : 227.
Sundstroem E. S. a. G. Michaels, 1942. The adrenal cortex in adaptation to altitude, climate and cancer. Berkley.
S u о m a 1 a i ne n P., 1944. Uber die Physiologie des Winterschla-fes. Sitzber. Finn. Akad. d. Wis>ensch., Helsinki.
Suomalainen P. a. S. Sarajas, 1951. Heart-beat in the hibernating hedgehog. Nature 168 ; 211.
Szymanski J. S., 1918. Versuche uber Aktivitat und Ruhe bei Sauglingen. Pf lug. Arch. 172 : 424.
Szymanski J. S., 1919. Aktivitat und Ruhe bei Tieren und Menschen. Z. f. Allg. Physiol. 18: 105.
Tang. P. S., 1933. O2 consumption and O2 pressure. Quart. Rev. Biol. 8: 260.
Tappan D. V. a. B. Reynafarje, 1957. Tissue pigment manifestation of adaptation to high altitudes. Am. J. Physiol. 190 : 99.
T a p p a n D. V., B. R e у n a f a r j e, R. v a n P о 11 e r a. A. H u r-t a d o, 1957. Alteration in enzymes and metabolites resulting from adaptation to low oxygen tensions. Amer. J. Physiol. 190 : 93.
Tembrock G., 1964. Verhaltungsforschung. Eine Einfuhrung in die Tier — Ethologie. Veb. G. Fischer Verlag, Jena.
Thorpe W. H., 1956. Learning and instinct in animals. London.
Tinbergen N., 1955. The study of instinct. Oxford Univ. Press.
Tomlinson J. T., 1957. Pigeon wing beats synchronized with breathing. Condor 59 : 401.
Tromp S. W., 1963. The influence of weather and climate on urinary volume pH, 17-ketosteroids, hexosamines, Cl, K, Na and urea. Int. J. Biometeor. 7 : 59.
444
Twente J. W. a. J. A. Twente, 1965. Regulation of hibernating periods by temperature. Proc. Nat. Acad. Sci. 54 : 1058.
Twente J. W. a. J. A. T w e n t e, 1967. Seasonal variation in the hibernating behaviour of Citellus lateralis. Mammal. Hibern. III. Elsevier N. Y. : 47.
Twente J. W. a. J. A. Twente, 1968. Effects of epinephrine upon progressive irritability of hibernating Citellus lateralis. Comp. Biochem. Physiol., v. 25, p. 475.
U о t i 1 a U. U., 1939. The role of the pituitary stalk in the regulation of the anterior pituitary with special reference to the thyrcotropic hormone. Endocrinology 25 : 635.
Valentin G., 1857—1888. Beitrage zur Kenntniss des Wintcr-schlafes der Murmeltiere.
Vaughan В. E. a. N. Pace, 1956. Changes in myoglobin content of the high altitude acclimatized rat. Amer. J. Physiol. 183: 549.
Vernberg F. Y., 1954. O2 consumption by tissues of teleosts of different size. Biol. Bull. 106 : 360.
V e r r i e r M. L., 1932. Etude des’rapports de la forme de 1’habitat et du comportement de quelques crustaces isopodes. Bull. Biol. France — Belg. 66 : 200.
Wai Igren H., 1954. Energy metabolism of two species of the genus Emberiza as correlated with distribution and migration. Acta Zool. Fennica 84 : 1.
Wall work J., 1960. Observations on the behaviour of some oribatid mites in experimentally controlled temperature gradients. Proc. Zool. Soc. London 135 : 619.
Watzka M., 1934. Physiologische Veranderngen der Schilddrusc. Z. mikr.-anat. Forsch. 36 : 67.
W e i h e W. H., 1966. Influence of age, physical activity and ambient temperature on acclimatization of rats to high altitude. Fed. Proc. 25, 4 : 1342.
Weiss A. K., 1954. Adaptation of rats to cold air and effects on tissue oxygen consumption. Amer. J. Physiol. 177, 2:201.
Wells N. A., 1935. Variations in the respiratory metabolism of the Pacific killfish Fundulus parvipinnis due to size, season, and continued constant temperature. Physiol. Zool. 8 : 318.
Welsh J. H., 1938. Diurnal rhythms. Quart. Rev. Biol. 13: 123.
West G. C., 1960. Seasonal variation in the energy balance of the three sparrow in relation to migration. Auk 77 : 306.
W i n d h e m С. H. a. J. F. xM о r r i s о n, 1956. Heat regulation of Masarwa (bushman). XX Congr. Int. Physiol., Resum. de commun., Bruxelles : 981.
Y о s h i m u r a H. a. T. I d a, 1952. Studies on the reactivity of skin vessels to extreme cold. Jap. J. Physiol. 2 : 177.
Yonge С. M., 1954. Feeding mechanism in the Invertebrata. Tabulae Biologicae 21, 3, 22.
Zeuthen E., 1942. The ventilation of respiratory tract in birds. Det. Kgl. Danske Vidensk. Biol. Medd. 17 : 1.
Zeuthen E., 1953. O2 uptake as related to size of organisms. Quart. Rev. Biol. 28 : 1.
Zuntz N., 1905. Die Bedeutung des Blinddarms fiir die Ver-dauung bcim Kaninchen. Arch. f. Anat. u. Physiol., Physiol. AbteiL. Leipzig, 1 : 403.
445
ОГЛАВЛЕНИЕ
Стр.
От автора................................................ 3
Введение. Предмет и задачи экологической физиологии . . 5
Глава I. Приемы и методы эколого-физиологических исследований ........................................... 13
Исследование физиологических функций в природных условиях и в эксперименте..................... 13
Изучение поведения в природных и лабораторных условиях.......................................... 18
Методы составления эколого-физиологических характеристик ......................................... 23
Эколого-физиологические исследования человека . . 26
Глава II. О физиологических механизмах природных адаптаций ............................................. 29
Классификация адаптаций и их значение для функции регулируемых систем........................... 29
Клеточный и тканевой уровни адаптаций у пойкилотермных организмов................................ 34
Клеточный и тканевой уровни адаптаций у гомойотермных организмов............................... 37
Органные и системные адаптации у пойкилотермных организмов ....................................... 47
Органные и системные адаптации у млекопитающих и птиц............................................ 53
Нервные и гормональные механизмы адаптации ... 59
Глава III. Врожденное и приобретенное поведение (основы этологии).......................................... 61
Адаптивное поведение.............................. 61
Понятие об инстинкте.............................. 62
Сложные формы поведения и их происхождение . 66
Запечатлевание (импринтинг) ...................... 74
Роль анализаторов в формировании сложно-рефлекторной деятельности............................... 78
Количественная характеристика и энергетика поведения . .......................................... 91
Г лава IV. Периодические изменения физиологических процессов в организме................................. 98
Отсчет времени в организме (физиологические часы) 98
Экспериментальное изучение суточных (циркадных) ритмов .......................................... 104
Сезонные изменения физиологических функций ... 110
Сезонные изменения поведения (миграции и кочевки) 132
446
Стр.
Физиологические изменения при зимней спячке . . . 138
Факторы, вызывающие, спячку, засыпание и пробуждение ......................................... . 149
Формы зимней спячки. Летняя спячка............... 154
Холодовое оцепенение у птиц (торпидное состояние) 160
Г лава V. Температура среды обитания.................. 162
Общее влияние тепла и холода на живые системы . . 162
Морфологические адаптации к теплу и холоду . . . 164
Термические адаптации у пойкилотермных организмов ............................................. 168
Термические адаптации у гомойотермпых организмов 175 Адаптивные особенности теплоотдачи и сложные формы терморегуляции................................ 191
Двигательное поведение и температура среды .... 196
Сложнорсфлекторный механизм терморегуляции и его значение для адаптации к теплу и холоду.......... 202
О формировании термических адаптаций в онтогенезе 204 Крайние типы адаптации к тропическому и полярному климату . , .................................. 208
Адаптации к природным температурным условиям у человека......................................... 215
Глава VI. Недостаток кислорода и его влияние на организм 221
Типы гипоксии и механизмы се возникновения .... 221
Поступление кислорода в организм и его перенос к тканям........................................... 222
Реакции организма па гипоксию.................... 223
Условия существования животного организма в горах и типы адаптаций к горным условиям............... 234
Адаптации равнинных организмов в горах и в эксперименте . . .................................... 235
Адаптации организмов, мигрирующих в горы ... 246
Адаптации горных организмов...................... 254
Физиологические особенности ныряющих организмов 261
Физиологические гипоксии у наземных организмов . . 270
Г лава VII. Физиологические адаптации к аридной зоне (жаркие пустыни)................................. 273
Условия существования в пустыне.................. 273
Адаптации пустынных беспозвоночных............... 274
Адаптации рептилий к пустыне..................... 280
Водный обмен у пустынных млекопитающих .... 285
Терморегуляция при недостатке воды и типы адаптаций к условиям пустыни......................... 295
Адаптации птии к пустыне......................... 300
Роль пищевари.сльного тракта в адаптации к аридной зоне......................................... 302
Особенности пищеварения и водного обмена пустынных млекопитающих................................ 302
Роль кожи и легких в адаптации к пустыне......... 305
Адаптации к засолению (вода, почва, растительность) 311
447
Стр.
Г iaea VIII. Адаптации к питанию, пищевая специализация и обмен веществ................................... 315
Типы питания животных организмов................. 315
Типы пищеварения................................. 321
Адаптация пищеварительных ферментов.............. 323
Адаптации двигательной функции пищеварительного тракта........................................... 329
Прием пищи и типы пшпедобывательной деятельности ............................................. 332
Спмбиоитное питание и пищеварение.............. 337
Выпадение пищевых рефлексов и физиологическое голодание . . ................................... 344
Энергетический расход организма.................. 346
Глава IX. Адаптация к передвижениям и мышечной деятельности ........................................ 357
Окружающее организм пространство как фактор среды . ,........................................ 357
Особенности передвижения (преодоления пространства) в водной среде .............................. 358
Передвижение наземных организмов.............. 366
Физиологические адаптации при мышечной деятельности ........................................... 374
Мышечная деятельность у птиц.................. 382
Глава X. Стадные и популяционные отношения и их физиологические механизмы.............................. 385
Групповые реакции животных и подражательная деятельность .................................... 385
Стадные и агрегационные реакции.................. 389
Стадные реакции и «социальное» доминирование . . 400
Нервные и гормональные механизмы регулирования численности в популяции . ....................... 402
Литература............................................. 407
Абрам Данилович Слоним экологическая физиология животных
Редактор Л. П. Петровская. Художник Н. В. К р а с о в и т о в а.
Художественный редактор С. Г. Абелин. Технический редактор Л. Л. Муравьева. Корректор Р. К. Иванова
ьХ^дано в набор 1I/V 1970 г. Поди, к печати 19/XI 1970 г.
С/формат 84Х108‘/з2 Объем 14 печ. л. 23,52 усл. п. л. Уч.-изд. л. 25,18
Изд. № E-19I Тираж 10 000 экз. Цена 1 р. 07 к.
План выпуска литературы издательства «Высшая школа» (вузы и технику-
мы) на 1970 год. Позиция № 111—112.
Москва, К-51. Неглинная ул., д. 29/14, Издательство «Высшая школа»
Московская типография № 8 Главполиграфпрома Комитета по печати при Совете Министров СССР, Хохловский пер., 7. Зак. 1204.