/

































































































































































































































































































































































































































Author: Шмидт-Ниельсен К.
Tags: физиология животных биофизика, биохимия и физиология животных и человека зоология
Year: 1982




Text
к. Ш.миФ'-Ниольсен ФИЗИОЛОГИЯ животных
К. Шмидт-Ниельсен
ФИЗИОЛОГИЯ
животных
ПРИСПОСОБЛЕНИЕ И СРЕДА
Second edition
Knut Schmidt-Nielsen James B. Duke Professor of Physiology, Department of Zoology, Duke University
ANIMAL PHYSIOLOGY
ADAPTATION AND ENVIRONMENT
Cambridge London New York Melbourne
К. Шмидт-Ниельсен
ФИЗИОЛОГИЯ животных
ПРИСПОСОБЛЕНИЕ И СРЕДА
В 2-х книгах
Книга первая
Перевод с английского
канд. биол. наук М. Д. ГРОЗДОВОЙ и
канд. физ.-мат. наук Г. И. РОЖКОВОЙ
лод редакцией
акад. Е. М. КРЕПСА
Москва «Мир» 1982
ББК 28.903
Ш 73 УДК 591.1
Ш73 Шмидт-Ниельсен К.
Физиология животных. Приспособление и среда, Книга 1: Пер. с англ./ Перевод Гроздовой М. Д., Рожковой Г. И.; Под ред. и с предисл. Е. М. Крепса. — М.: Мир, 1982. — 416 с., ил.
Книга известного американского физиолога посвящена вопросам сравнительной физиологии, рассматриваемым в экологическом аспекте. Русское издание выходит в двух книгах. В первую книгу входят разделы: кислород; пища и энергия; температура.
Предназначена для физиологов, зоологов, экологов, гидробиологов, специалистов по охране окружающей среды, для преподавателей и студентов университетов, педагогических, сельскохозяйственных и рыботехнических институтов.
21008—111
Ш041(01)—82 1,,—82, ч> *’
2007020000
ББК 28.903
591.4
Редакция литературы по биологии
© Cambridge University Press, 1975, 1979 © Перевод на русский язык, «Мир», 1982
ПРЕДИСЛОВИЕ РЕДАКТОРА ПЕРЕВОДА
Предлагаемая читателям книга профессора К. Шмидт-Ниель-сена «Физиология животных», хотя она и была написана как пособие для студентов, не похожа на все другие курсы физиологии. Это книга совсем особого рода. Чтобы написать такую книгу, надо быть не только крупным физиологом, мастером своего дела и опытным педагогом, но также компетентным зоологом и экологом, знатоком животных, позвоночных и беспозвоночных, и условий их существования.
Как известно, физиология — это наука о функциях живых организмов: о питании и пищеварении, о дыхании, переносе газов кровью, кровообращении и работе сердца, о выделении продуктов обмена веществ и функции почек, о работе мышц и движении, о деятельности нервной системы и функции рецепторов и т. д. В книге можно найти краткое, но очень четкое и ясное изложение всех этих вопросов. Однако есть немало хороших руководств по физиологии, и главная ценность книги не в этом. Проф. Шмидт-Ниель-сен ставит своей основной целью показать, как работают .у животных различные системы организма в тех реальных условиях, где эти животные обитают, — условиях подчас необычайно трудных и, казалось бы, мало приспособленных для жизни. Речь идет прежде всего о тех физиологических проблемах, которые возникают у представителей животного мира в специфической среде их обитания, и о том, как животные эти проблемы разрешают. Проблем этих неисчислимое множество.
Как животные адаптируются к недостатку воды, например в жаркой, безводной пустыне, или, наоборот, к избытку влаги? Как избегают они гибели от замерзания тканей где-нибудь на литорали арктического моря во время отлива или, наоборот, как избегают перегрева и необратимой денатурации тканевых белков под лучами палящего солнца Сахары? Каким образом различные формы получают достаточное количество кислорода, когда потребность в нем резко повышается (как, например, у птицы во время дальнего перелета) или когда кислорода в окружающей среде мало? Как приспосабливаются водные животные, дышащие жабрами, когда водоем пересыхает и они оказываются в воздушной среде?
А бесчисленные осмотические проблемы! Морские рыбы и рептилии живут в гипертоничной среде, пресноводные рыбы и амфибии— в гипотоничной, а надо поддерживать нормальную осмоти-
л6 Предисловие редактора перевода
ческую концентрацию крови и нужную концентрацию всех ионов. При этом покровы или жабры (Проницаемы для воды. Человеку тоже подчас надо выделять гипертоничную мочу, когда воды не хватает, а пустынным животным и подавно. Как справляется «корабль пустыни» — верблюд, совершая длинные переходы по жаре без воды, с возникающими физиологическими затруднениями? Подобных проблем множество, и во всех случаях Шмидт-Ниельсен ясно и убедительно разъясняет, как они решаются.
В книге собран огромный и по большей части новый, современный ^материал. Но было бы большой ошибкой думать, что книга проф. Шмидт-Ниельсена — просто компиляция. Нет лучшего учителя, чем сама природа, и автор сам прежде всего в природе изучал адаптацию животных к условиям существования. Автор — друпный исследователь, путешественник и экспериментатор, изучавший физиологические отправления очень различных организмов в самых разнообразных, нередко экстремальных экологических условиях. Им открыто немало важных и интересных физиологических адаптаций. Он исследовал терморегуляцию, водный и азотистый обмен у верблюдов в пустыне и у страусов в африканских степях, моллюсков, змей и ящериц, обитающих в пустыне. Это он открыл солевые железы у морских птиц и у морских рептилий и изучил их физиологию, исследовал почечную функцию у крабоядной лягушки и внес много нового в проблему осморегуляции у позвоночных. Он описал принцип противоточного обмена тепла, газов, солей, метаболитов и в почках, и в дыхательных путях птиц, и в плавательном пузыре рыб. Он изучал регуляцию температуры тела у однопроходных. Трудно переоценить вклад в сравнительную физиологию, сделанный лично Шмидт-Ниельсе-ном и его сотрудниками. Может быть, потому книга и читается с таким живым интересом, что в ней чувствуется энтузиазм и азарт исследователя и открывателя.
Автор приводит огромное количество фактов, открытых и изученных также и другими исследователями. 'Надо отдать ему справедливость, он очень строг в отборе материала, рассказывая только о точно установленных и хорошо проверенных фактах, которым всегда дает ясное толкование. Если мы сравним издание 1979 г. с первым изданием 1975 г., мы поразимся, как много нового сделано за какие-нибудь четыре года и как неутомим наш автор в замене несколько устаревших данных и толкований новыми, более точными или более надежными. В последнем издании мы находим новый материал по дыханию рыб и безлегочных саламандр, по кислородному обеспечению развивающихся яип птиц. В главе о пище и питании дается новый материал о морских восках и значении их в пищевых цепях в море, о фиксации азота у термитов и у кораллов. Мы находим пересмотренные сведения о влиянии на физиологические процессы высокого гидростатического давления в океане и пониженного барометрического давления на
Предисловие редактора перевода
7
больших высотах, о смерти от перегрева и выносливости к -высокой температуре, о концентрировании мочи у млекопитающих. В главах о движении и о сенсорных функциях пересмотрены старые представления и приведены новые факты — например, о рецепции электрических и магнитных стимулов и функции боковой линий у рыб. Даны новые трактовки ряду прежде известных, а также новых фактов, в частности связи нервной регуляции с регуляцией эндокринной — связи, которую Шмидт-Ниельсен всегда подчеркивал; добавлены новые интересные данные о нейросекреции.
Не только студенты, но и многие специалисты-физиологи удивятся, узнав немало нового и неожиданного для себя, как это произошло и со мной. Книга заставляет задуматься о путях развития животного -мира. Она показывает, с одной стороны, как необычайно разнообразна природа в решении своих проблем, как велика изменчивость в эволюции животного мира и, с другой стороны, как велик консерватизм в эволюции, как часто, найдя удачное решение какой-нибудь биологической проблемы, природа придерживается его в совершенно, казалось бы, несходных ситуациях, на самых разных уровнях развития животного мира. Ярким примером может быть принцип противоточной циркуляции. Он использован для разных целей и в плавательном пузыре рыб, и в легких птиц, и в почечных канальцах млекопитающих, и в ряде других органов и тканей.
Автор далек от мысли, что физиологам уже все ясно. Многое, конечно, стало ясным, но большое число вопросов еще ждет своего разрешения, требует дальнейших исследований; автор везде отмечает это и указывает, где можно ожидать волнующих новых открытий. Нам приятно отметить, что, когда автор располагает информацией о новых интересных советских работах, он не обходит их молчанием.
Автор снабдил книгу несколькими очень полезными приложениями. На Западе, как <и у нас, введено в обязательном порядке пользование новыми единицами измерения физических величин — системой СИ. В тексте книги все физические величины даны как в старой, привычной символике, так и в единицах системы СИ. В Приложении А приведены таблицы единиц измерения физических величин, как первичных, так и производных, в системе СИ и таблицы перевода между старыми и новыми системами. Это очень полезная информация. В Приложении Б — «О диффузии» — автор дает также полезную информацию о понятии диффузии и применении его к разным вопросам дыхания, в частности у водных организмов. Этот вопрос нередко бывает камнем преткновения для молодых физиологов. В следующих приложениях рассмотрены проблемы, касающиеся растворов, осмоса и осмотического давления, а также термодинамическое выражение температурных эффектов. Заканчивается книга таблицей современной классификации животного мира, несомненно очень нужной для
<3 Предисловие редактора перевода
всех интересующихся сравнительным и эволюционным аспектами ^биологической науки.
Мы признательны профессору К. Шмидт-Ниельсену за любезное разрешение перевести и издать его книгу «Физиология животных» на русском языке. Несомненно, книга эта будет с большим интересом встречена широким кругом читателей — студентами биологических факультетов и биологами разных профилей.
Академик Е. М. Крепе
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
Второе издание книги всегда должно быть лучше первого. Можно улучшить изложение того же материала, а можно и дополнить его новыми данными. Я постарался сделать и то и другое. Во втором издании заново отредактирована каждая глава, добавлен ряд новых существенных сведений; материал некоторых глав рассмотрен более глубоко.
Как и прежде, я придерживаюсь мнения, что понимание принципов гораздо важнее, чем простое ознакомление с множеством фактов, которые своей массой способны подавить любознательность молодого читателя. Я учитывал это при подборе нового материала.
Для читателей, занимающихся преподаванием физиологии, вероятно, представит интерес список внесенных изменений. Они состоят в следующем. В главы, посвященные дыханию, добавлен новый материал, касающийся дыхания рыб и безлегочных саламандр, а также снабжения кислородом птичьих яиц. Расширены и яснее написаны главы о крови и кровообращении. Глава о пище и питании содержит новые данные по морским воскам и их значению в цепи питания, по фиксации азота у термитов и коралловых полипов; значительно расширено также обсуждение вопроса о растительных ядах и химических средствах защиты.
В главу о метаболизме введено обсуждение вопросов о слое воды в океане с минимальным содержанием кислорода, а также о влиянии высокого давления и подъема на большие высоты. Полностью пересмотрен раздел о соотношениях между размерами тела и другими показателями.
Главы, касающиеся проблем терморегуляции и влияния температуры, содержат новые данные о толерантности к теплу и холоду и о гибели от перегрева. Обсуждаются новые представления о биологической роли лихорадочного состояния, а также результаты новейших исследований по терморегуляции у птиц и пчел. Сделано более ясным изложение материала в главах по осморегуляции и экскреции, четко представлены современные концепции о механизме концентрирования мочи в почках млекопитающих.
Три последние главы, посвященные мышцам, нервам и гормонам, полностью переработаны и значительно дополнены. Представлен обширный новый материал по ряду вопросов, начиная от амебоидного движения и кончая молекулярным механизмом мышечного сокращения. Значительное внимание уделено локомо*
10 Предисловие ко второму изданию
ции животных и биомеханике, так как способность к передвижению — это одна из важнейших особенностей животных.
В разделе о сенсорном восприятии приведены более современные данные, особенно в отношении рецепции электрических и магнитных раздражителей, а также функции боковой линии у рыб. Полностью переработан, расширен и приведен в соответствие с современными требованиями раздел, касающийся природы нервных импульсов и потенциалов действия.
Последняя глава — об интеграции физиологических процессов— тоже подверглась основательной переработке. Уже в первом издании мы подчеркивали тесную взаимосвязь между центральной нервной системой и функцией эндокринных желез; теперь эта тема дополнительно раскрывается в свете последних удивительных данных о нейроэндокринной функции. Вместо перечисления почти бесконечного числа гормонов, запомнить которые могли только студенты с хорошей механической памятью, мы приводим четкие таблицы, в которых резюмированы основные принципы современной эндокринологии.
Как и в первом издании, ряд основополагающих сведений, которые должны быть уже известны студентам, приведен в приложении; это обусловлено отнюдь не тем, что автор счел такую информацию второстепенной, — напротив, она настолько важна, что должна быть под рукой у тех, кому нужно будет вспомнить четкую формулировку фундаментальных положений.
Международная система единиц (система СИ), как и в первом издании, четко и точно представлена в приложении А. Кроме того, в тексте единицы СИ приводятся вместе с аналогичными традиционными единицами, чтобы облегчить перевод одних в другие.
Два важных раздела физиологии, касающиеся витаминов и размножения, изложены в книге очень поверхностно, а некоторые аспекты вообще не затронуты. Я полагаю, что частично указанные разделы достаточно известны студентам. Едва ли есть надобность приводить перечень витаминов или описание симптомов их недостаточности: с этим материалом студенты должны были уже ознакомиться в курсах общей биологии, домоводства и гигиены. Углубленный же разбор роли витаминов в обмене веществ относится к области биохимии и, таким образом, выходит за рамки данного учебника. Что касается проблемы размножения, то в отношении человека основные вопросы должны быть уже известны студентам, а размножение животных — настолько обширная тема, к тому же включающая массу данных по морфологии и биологии развития, что ее лучше рассматривать в отдельной книге.
Я надеюсь, что в результате внесенных изменений читатель получит от этого учебника больше пользы; буду рад, если студенты или коллеги сообщат мне, что еще следовало бы в нем улучшить.
Кнут Шмидт-Ниелъсен
МОИМ МОЛОДЫМ ЧИТАТЕЛЯМ
Этот учебник я написал со злости и от отчаяния; отчаивался я потому, что не мог порекомендовать своим студентам книги, где простыми словами рассказывалось бы то, что мне кажется самым увлекательным и важным в физиологии животных, где бы рассматривались проблемы и способы их решения, описывалось, как работает живой организм.
Около 20 лет я думал о создании такого учебника, прекрасно понимая, что физиология — это слишком большая область для того, чтобы один человек ею мог полностью овладеть и адекватно изложить. Я искал и не находил соавтора, на которого мог бы переложить часть ответственности. Однажды я страшно рассердился на самого себя и решил выполнить всю работу один. Может быть, недостатки учебника, связанные с чрезмерной сложностью задачи для автора, будут частично компенсированы определенным единством стиля и точки зрения. Для написания учебника я использовал свой годичный отпуск. Вернувшись к чтению лекций, я обнаружил, что лучше знаю физиологию по сравнению с тем, как я ее знал, когда принимался за мучительно трудную, но благодарную работу по созданию этой книги, в которую я вложил свою душу.
ОБ ЭТОЙ КНИГЕ
Эта книга — о животных, о жизненных проблемах, с которыми они встречаются, и о способах решения этих проблем. В ней освещены также те аспекты физиологии, которые мне лично кажутся особенно интересными. Книга рассчитана на студентов, которые стремятся понять, как работает живой организм и на что он способен.
В книге освещены обычные разделы физиологии: дыхание, кровообращение, пищеварение и т. д. Эти темы рассматриваются в 13 главах; основу для группировки материала составляют важнейшие факторы окружающей среды: кислород, пища и источники энергии, температура, вода. Я считаю такой план изложения очень важным, ибо нельзя стать хорошим физиологом и вообще хорошим биологом, не поняв зависимости функций живого организма от внешней среды.
Содержание книги элементарно, и она рассчитана на читателей с минимальной подготовкой. Я исходил из того, что студент знаком с теми основами знаний, которые можно получить в хорошей средней школе или во вводных курсах по биологии в колледжах. В других отношениях особой предварительной подготовки Практически не требуется. В книгу включено достаточно общих Сведений для того, чтобы можно было объяснить физиологические принципы в понятиях элементарной физики и химии. В ряде случаев более строгое описание дано в виде приложения (например, при рассмотрении растворов и осмотических явлений). Этими приложениями могут воспользоваться студенты, желающие лучше разобраться в вопросе, и преподаватели, считающие нужным ввести соответствующий материал в программу обучения.
Объем и сложность научной информации непрерывно возрастают, так что студенты уже сейчас перегружены материалом, который нужно запомнить. Кроме того, простое ознакомление с фактами не способствует пониманию общих принципов. В связи с этим я старался представить только те данные, из которых с достаточной убедительностью вытекали бы теоретические положения и выводы. Студенты увидят, что усвоение нескольких фундаментальных законов позволяет понять многие проблемы. Мне думается, что ясность общих представлений важнее, чем запоминание терминов, но поскольку ни одного представления нельзя выразить без слов, термины тоже необходимы. Однако для того, чтобы они помогали уяснить суть дела, они сами должны быть четко и логично определены.
Об этой книге 13
Чтобы не перегрузить студента информацией, для учебника приходится отбирать материал, поэтому многое будет сознательно спущено. Например, большинство студентов имеет достаточное представление о витаминах и о физиологии размножения, а соответствующая терминология вошла в повседневную речь, так что нет необходимости повторять эти разделы еще раз. Разумеется, знакомство с употребительными терминами еще не означает понимания существа процессов, протекающих в живых организмах. Очень важно создать у студентов целостное представление, согласующееся с современными данными, с законами физики и химии и с концепциями относительно потребностей организма, живущего в определенных условиях среды.
В этой книге неоднократно рассматривается вопрос о путях приспособления (адаптации) животных к условиям, казалось бы, совершенно непригодным для обитания. Сделана попытка сравнить различные подходы к решению жизненных проблем, использованные в разных группах животных. Пониманию общих принципов адаптации весьма способствует изучение форм с чертами анатомической или физиологической специализации. Но если мы не будем активно искать эти общие принципы, сравнительная физиология легко может превратиться в описание функций, свойственных необычным животным, — необычным не потому, что они редки, а потому, что они мало знакомы нам, так как отличаются от нас самих и от наших домашних и лабораторных животных — собак, кошек, крыс, лягушек. Напротив, объединение частных сведений, подводящее к формулировке общих принципов, позволит лучше понять, как функционирует организм любого животного.
В книге кое-где даны ссылки на источник сведений, и в конце каждой главы приводится список литературы. Это сделано не только для того, чтобы указать, откуда взято описание тех или иных фактов, но и для того, чтобы помочь студентам быстро найти дополнительную информацию по заинтересовавшему их вопросу. Обилие источников научной информации заставило прибегнуть к строгому отбору, и мнения о том, насколько этот отбор удачен, могут расходиться.
Чтобы открыть читателю путь к более специальным источникам сведений, после основной библиографии дается небольшой список работ для дополнительного чтения. Эти работы могут служить отправными точками для дальнейшего углубленного изучения того или иного вопроса. Стремять спасти студентов от чувства беспомощности, я сделал такие списки очень небольшими. За несколькими исключениями, здесь указаны достаточно свежие, современного уровня публикации.
Как и большинство авторов, я надеюсь, что благожелательные, а может быть, и не вполне благожелательные читатели укажут мне на допущенные ошибки и предложат те или иные улучшения, которые я мог бы сделать.
ЧТО ТАКОЕ ФИЗИОЛОГИЯ?
Физиология — это наука о функциях живых организмов: как они едят, дышат, передвигаются и вообще поддерживают свою жизнь. Выражаясь в более специальных терминах, физиология — это наука о пище и питании, пищеварении, дыхании, переносе газов кровью, кровообращении и функции сердца, выделении и функции почек, о мышцах и движении и т. д. У мертвого животного имеются структуры, способные при жизни выполнять эти функции; у живого — эти структуры работают. Физиология описывает также, как живой организм приспосабливается к условиям внешней среды: получает достаточно воды, чтобы выжить, или предохраняет себя от избытка воды, спасается от замерзания или от перегрева, передвигается в поисках более подходящих мест для жизни, питания или спаривания, а также каким образом животное получает информацию о внешней среде с помощью органов чувств. Наконец, физиология описывает регулирование всех этих функций, а именно — как происходит координация и интеграция их в живом, слаженно работающем организме.
Физиология не только описывает различные функции; она также задается вопросами «почему?» и «как?». Чтобы понять, как функционирует организм животного, нужно знать его строение и иметь элементарные сведения по физике и химии. Нельзя, например, разобраться в сущности дыхания, если ничего не знать о кислороде. Еще с древнейших времен жизнь от смерти отличали по наличию дыхательных движений, однако выяснение истинного смысла дыхания стало возможным только тогда, когда химики открыли кислород.
В изучении процессов жизнедеятельности огромную роль играет сравнительный подход. Сравнивая различных животных и исследуя, каким образом каждое из них выживает, приспосабливаясь к факторам внешней среды, мы получаем представление об общих закономерностях, которые без такого сравнения трудно было бы выявить. Ни одно животное не существует и не может существовать независимо от окружающей среды: оно использует ресурсы среды и в то же время должно преодолевать воздействие ее неблагоприятных факторов. Таким образом, как сравнительный, подход, так и изучение связи организма и среды позволяют проникнуть в сущность физиологических процессов.
При рассмотрении адаптации животного к среде нередко выявляется, что именно «хорошо» для животного. Это может пока
Что такое физиология? 15
заться слишком близким к телеологическому объяснению — к представлению о цели, что многие биологи считают некорректным с научной точки зрения. И все же всем нам свойственно задавать вопросы «почему?» и «для чего?» Как бы то ни было, животному необходимо выжить, и нет ничего предосудительного или антинаучного в том, что мы хотим узнать, как и почему это ему удается. Если бы животное не могло разрешить эту задачу, нам не пришлось бы его изучать.
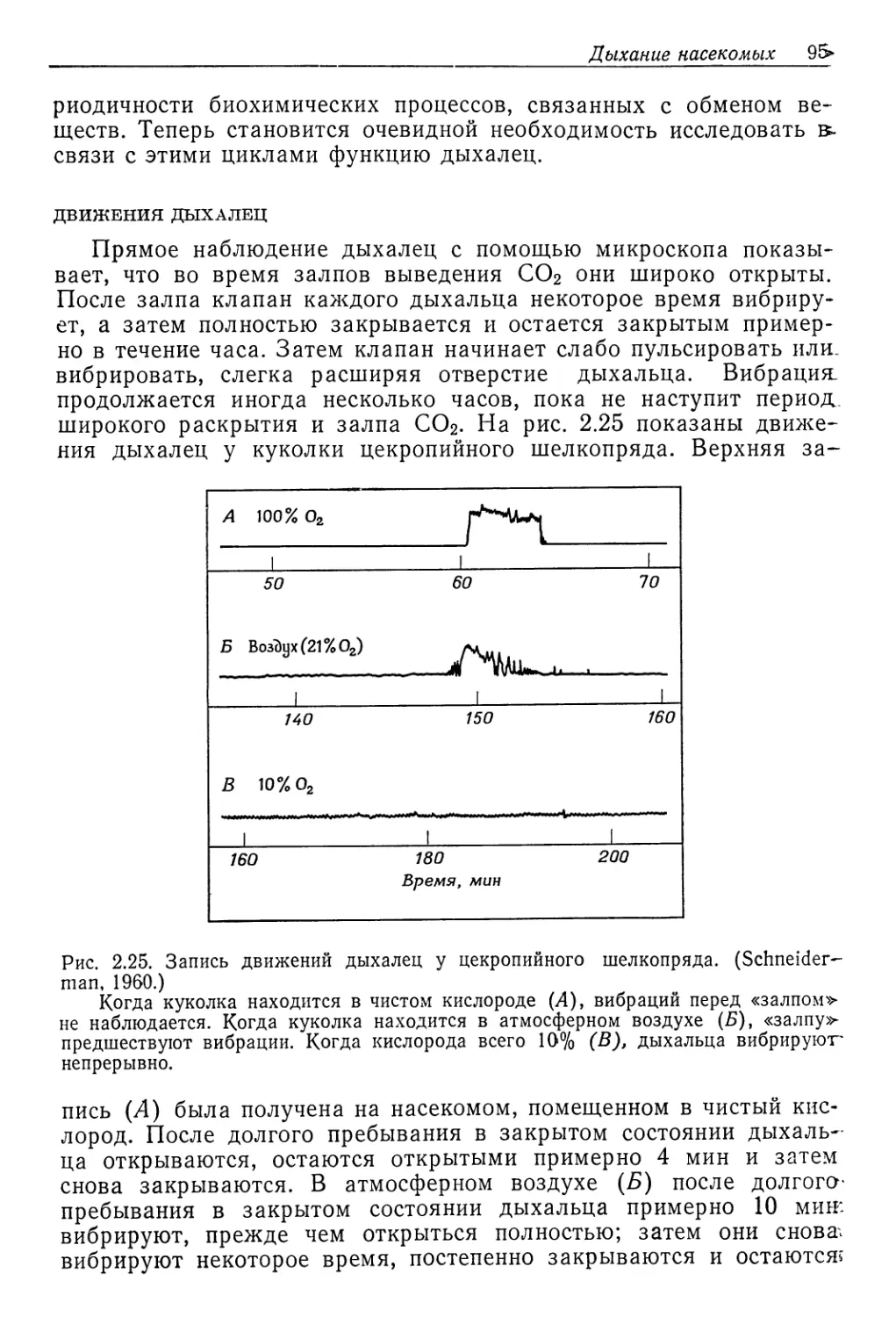
В этой книге сравнительная физиология излагается с экологических позиций. Описание начинается с того, как из внешней среды, водной или воздушной, животные получают кислород. Далее рассматривается роль крови в доставке кислорода тканям и описывается, как происходит перекачивание крови в организме. О снабжении энергией (пищей) речь идет в главе, посвященной питанию и пищеварению, а затем обсуждаются общие вопросы энергетического обмена. Воздействие такого важного внешнего фактора, как температура, рассматривается в двух главах. Затем списывается другой, не менее важный внешний фактор — вода. Еще одна глава посвящена движениям и локомоции и еще одна— способам получения информации об окружающей среде (органам чувств). В последней главе обсуждается вопрос, каким образом с помощью нервной и эндокринной систем осуществляется регуляция всех рассмотренных функций, координация их и интеграция всех систем в слаженно функционирующий целостный организм.
ЧАСТЬ ПЕРВАЯ
КИСЛОРОД
ГЛАВА 1
ДЫХАНИЕ В ВОДЕ
Все живые организмы используют энергию, которую они должны добыть из внешних источников. Большинство растений улавливает энергию солнечного света и использует атмосферную углекислоту (двуокись углерода) для синтеза сахаров и в конечном счете всех других сложных веществ, из которых состоит растение. Животные в свою очередь используют энергию химических соединений, которые они получают из растений либо непосредственно—> поедая растения, либо косвенным путем — поедая животных, питающихся растениями. Таким образом, используемая животными химическая энергия имеет своим первоисточником солнечное излучение.
Большинство животных удовлетворяет свои энергетические потребности путем окисления компонентов пищи. Некоторые животные способны использовать энергию органических веществ в отсутствие кислорода, однако полное окисление этих веществ доставляет примерно в 10—20 раз больше энергии. Пища животных состоит в основном из трех групп соединений: углеводов, жиров и белков. При полном окислении углеводов и жиров получаются только два конечных продукта — двуокись углерода (СО2) и вода; при окислении белков, кроме СОг и воды, образуются небольшие количества других конечных продуктов.
Процессы поглощения кислорода и выделения СО2 называют дыханием, причем это слово может относиться как к газообмену всего организма, так и к процессам, происходящим в клетках. Животные извлекают кислород из среды, в которой они живут, и выделяют в эту же среду СО2. Водные животные используют те небольшие количества кислорода, которые растворены в воде; наземные животные дышат кислородом, в изобилии содержащимся в атмосферном воздухе. Многие мелкие животные могут получать достаточное количество кислорода прямо через поверхность тела,
Газы в воздухе и воде 17
но большинство животных имеет специальные органы дыхания. Когда клетки используют кислород для окисления пищевых веществ, образуется двуокись углерода, которая перемещается в-противоположном направлении и выходит наружу через поверхность тела или дыхательные органы. Другой конечный продукт окисления — вода — просто включается в общий водный запас организма и не создает особых проблем.
Самый важный, а иногда и единственный физический процесс,, который обеспечивает перемещение кислорода из внешней среды к клетке, — это диффузия (т. е. движение газа в виде растворенного вещества в направлении от высоких концентраций к низким). Перемещение СО2 в обратном направлении также идет «вниз» по-градиенту концентрации. Диффузии часто помогает макродвижение (например, циркуляция крови), но именно градиенты концентраций создают основную движущую силу для перемещения: газов, участвующих в дыхании. Таким образом, чтобы понять механизм дыхания, надо получить представление о дыхательных газах, их растворимости и диффузионных процессах.
Жизнь, по-видимому, зародилась в море, так что удобнее сначала рассмотреть водное дыхание, а затем уже перейти к дыханию-в воздухе. Дышащие воздухом животные (в первую очередь позвоночные и насекомые) принадлежат к наиболее сложным организмам, однако большинство животных — водные, особенно много их среди не столь высокоорганизованных беспозвоночных. После краткого обзора дыхательных газов и небольшой порции основ физики мы в этой главе будем иметь дело с проблемами водного дыхания.
ГАЗЫ В ВОЗДУХЕ И ВОДЕ
СОСТАВ СУХОГО АТМОСФЕРНОГО ВОЗДУХА
Физиологически наиболее важные газы — это кислород, двуокись углерода и азот. Они присутствуют в атмосферном воздухе-в пропорциях, указанных в табл. 1.1. Кроме того, атмосфера содержит водяные пары в сильно варьирующих количествах.
То, что физиологи называют азотом, это на самом деле смесь азота примерно с 1% инертных газов, и ради точности их тоже надо было бы перечислить. Однако в физиологии принято объединять эти газы с азотом, главным образом потому, что в большинстве физиологических процессов азот и инертные газы одинаково безразличны для организма. Другая причина — то, что при анализе дыхательных газов обычно определяют количества О2 и СО2> а все остальное называют «азотом». Таким образом, для физиолога количество «азота» в воздухе равно 78,09 + 0,93%, т. е. 79,02%. Примерно 1% аргона представляет физиологический интерес только в некоторых весьма специфических случаях, напри-иер в связи с секрецией газов в плавательный пузырь у рыб. Пол-
48 Г лава 1. Дыхание в воде
Таблица 1.1
Состав сухого атмосферного воздуха (Otis, 1964). Нормальный атмосферный воздух содержит также водяные пары в весьма изменчивом количестве. На долю более редких инертных газов (гелия, неона, криптона и ксенона), взятых вместе, приходится всего лишь 0,002%.
Компонент Содержание, %
Кислород Двуокись углерода Азот Аргон 20,95 0,03 78,09 0,93
Всего 100,00
&1ый анализ состава газов в пробе воздуха можно произвести с помощью масс-спектрометра — дорогого и очень сложного прибора, который недоступен большинству физиологов.
Состав атмосферы чрезвычайно постоянен. Конвекционные токи обеспечивают интенсивное перемешивание до высот по меньшей мере 100 км, и никаких заметных изменений в процентном составе воздуха обнаружено не было, хотя давление на больших высотах сильно понижено. Утверждение, что верхние слои атмосферы обогащены легкими газами, особенно водородом и гелием, относится лишь к самым наружным слоям, которые не представляют никакого интереса для физиолога. Для наших целей газовый состав свободной атмосферы можно считать постоянным, если исключить водяные пары (Spitzer, 1949).
Состав воздуха поддерживается равновесием между потреблением кислорода в окислительных процессах (главным образом в окислении органических веществ до СО2) и освобождением кислорода в процессе ассимиляции СО2 растениями.
Опасение, что использование нами ископаемых топлив — нефти, угля и природных газов — может истощить запас кислорода в атмосфере и добавит в нее большие количества СО2, по-видимому, лишено оснований. В 1910 году чрезвычайно точный анализ дал для содержания кислорода цифру 20,948%, а в период с 1967 по 1970 г. повторные измерения давали величины 20,946±0,006%. Ученые, производившие эти очень точные анализы, вычислили, что если все известные запасы ископаемого топлива будут сожжены, то и тогда в асмосфере еще останется 20,8% кислорода (Ма-chia, Hughes, 1970). В физиологическом плане такое изменение не будет иметь никаких последствий.
Небольшое увеличение количества СО2 в результате сжигания всего топлива тоже оказало бы лишь ничтожный физиологический эффект, но это не значит, что оно было бы вообще безвредным.
Газы в воздухе и воде 19'
Даже незначительное изменение в концентрации двуокиси углерода повлияет на поглощение солнечной радиации в атмосфере, и может возникнуть непредсказуемый парниковый эффект, который со временем мог бы коренным образом изменить климатические условия на земной поверхности. Атмосфера более проницаема для. поступающего извне коротковолнового излучения, чем для длинноволнового, испускаемого Землей. Выходящее длинноволновое излучение поглощается в атмосфере главным образом двуокисью углерода и водяными парами. Подсчитано, что удвоение содержания СО2 в атмосфере приведет к повышению мировой температуры примерно на 1,3°, если содержание воды в атмосфере останется прежним. Однако при более высокой температуре атмосфера способна удерживать больше водяных паров, и это может усилить, экранирующий эффект и вызвать дальнейшее повышение температуры. С другой стороны, увеличение влажности атмосферы может усилить образование облаков, что в свою очередь приведет к отражению большей доли входящей солнечной радиации и тем самым вызовет обратный эффект. Сложность этих взаимоотношений-делает предсказания относительно парникового эффекта от повышенной концентрации СО2 весьма недостоверными (Sawyer, 1972; Baes et al., 1977).
Подчеркивая постоянство состава воздуха, мы должны добавить несколько слов о некоторых особых случаях. Например, в. микрообластях типа нор, занимаемых животными, состав воздуха сильно варьирует, причем содержание кислорода может опускаться там до 15% и ниже (Darden, 1972). Содержание СО2 там повышено, но не обязательно в той же степени; и все же оно может подниматься до 5%, что уже приводит к значительным физиологическим последствиям.
Воздух, находящийся в почве, в промежутках между ее частицами, часто содержит мало кислорода. Объясняется это тем, что> почва может содержать окисляющиеся вещества, которые иногда сильно истощают запас кислорода. Не только органические соединения, но и такие вещества, как сульфид железа, могут поглощать. О2 до тех пор, пока практически весь свободный кислород не будет израсходован. Такие окислительные процессы зависят от температуры, влажности и других факторов, а также от степени обмена почвенного воздуха с атмосферой. Например, дождь может блокировать почвенные поры в поверхностной зоне и одновременно вызвать повышение влажности и тем усилить процессы окисления; В результате микроатмосфера может коренным образом измениться.
ВОДЯНЫЕ ПАРЫ В ВОЗДУХЕ
Приведенные выше сведения о процентном составе атмосферы, относились к сухому воздуху, и теперь мы должны обратиться к. содержанию в воздухе воды. Давление водяных паров над свобод
20 Глава 1. Дыхание в воде
ной поверхностью воды изменяется с температурой (табл. 1.2). В точке замерзания давление пара равно 4,6 мм рт. ст. (0,61 кПа)1. Оно повышается с повышением температуры и при 100°C достигает 760 мм рт. ст. (101,3 кПа). Поэтому вода при 100°C закипает, если атмосферное давление равно 760 мм рт. ст. Если давление ниже, вода закипает при более низкой температуре— например, при 20 °C, если давление снижено до 17,5 мм рт. *ст. (2,34 кПа).
Любая смесь газсв типа атмосферного воздуха, находящаяся в равновесии со свободной поверхностью воды, содержит водяные пары, давление которых соответствует данной температуре, и поэтому часть объема воздуха, занятая водяными парами, с повы--шением температуры увеличивается (колонка 4 в табл. 1.2). При 37 °C — обычной температуре тела млекопитающих — давление водяных паров близко к 47 мм рт. ст. (6,28 кПа), и в этом случае пар занимает 6,2% объема воздуха (см. также рис. 9.9).
Воздух в легких человека и других наземных позвоночных всегда насыщен водяными парами при температуре тела, а атмосферный воздух обычно не насыщен. В случае его насыщения мы говорим, что относительная влажность (о. в.) равна 100%. Если в воздухе меньше водяных паров, влажность можно выразить в процентах от количества, необходимого для насыщения при данной температуре; например, относительная влажность 50% означает, что воздух содержит половину того количества воды, которое он
Таблица 1.2
Давление водяного пара
над свободной поверхностью воды при разных температурах
Температура, °C Давление водяного пара
мм рт. ст. кПа % от 1 атм мг Н2О на на 1 л воздуха
0 4,6 0,61 0,6 4,8
10 9,2 1,23 1,2 9,4
20 17,5 2,34 2,3 17,3
30 31,7 4,24 4,2 30,3
40 55,1 7,38 7,3 51,1
50 92,3 12,33 12,2 83,2
100 760,0 101,33 100,0 598,0
37 46,9 6,28 6,2 43,9
1 В физиологии по традиции используется единица «миллиметр ртутного столба» (мм рт. ст.). Она возникла в связи с применением ртутных манометров, и 1 мм рт. ст. при 0 °C называют также 1 торр. В Международной системе единиц (системе СИ) за единицу давления принят паскаль (Па), равный 1 ньютону на 1 квадратный метр (н-м“2). Таким образом, 1 мм рт. ст. =133,3 Па, или 0,133 кПа; 1 атм = 760 мм рт. ст.= 101,3 кПа. Другие подробности относительно системы СИ даны в Приложении А.
Газы в воздухе и воде 21
содержал бы, если бы был насыщен водяным паром при данной температуре.
Для некоторых целей относительная влажность — удобная мера, но если мы хотим знать общее количество водяных паров в воздухе, его можно выразить в миллиграммах воды на литр воздуха. Поскольку в холодном воздухе водяного пара очень мало даже при о. в. 100%, абсолютное количество воды в холодном воздухе невелико. Таким образом, если насыщенный наружный воздух зимой проникает в наши дома и нагревается, относительная влажность в помещении становится очень низкой (хотя абсолютная влажность воздуха не изменилась), и мы говорим, что воздух «очень сухой». При этом влажные поверхности, включая наши слизистые оболочки, быстро сохнут, что часто вызывает неприятные ощущения у чувствительных людей.
ВЫСОТА И АТМОСФЕРНОЕ ДАВЛЕНИЕ
Восхождение на высокие горы или подъем на большую высоту в негерметичном самолете вызывают серьезные физиологические сдвиги. На уровне около 3000 м влияние высоты на людей начинает проявляться в снижении физической работоспособности, а на высоте 6000 м (около 20 000 футов) большинство людей едва может выжить. Это происходит от недостатка кислорода, хотя содержание его в воздухе по-прежнему составляет 20,95%.
На уровне моря, где атмосферное давление равно 760 мм рт. ст. и 20,95% этой величины создает кислород, парциальное давление О2 в сухом воздухе равно 159 мм рт. ст. (21,2 кПа)1. На высоте 6000 м атмосферное давление вдвое меньше, чем на уровне моря, т. е. около 380 мм рт. ст. (50,7 кПа). Парциальное давление Ог здесь также вдвое меньше, т. е. около 80 мм рт. ст., или 10,6 кПа (20,95% от 380 мм рт. ст., т. е. от 50,7 кПа). Именно это уменьшение парциального давления кислорода и вызывает такие серьезные последствия.
Зависимость атмосферного давления от высоты представлена на рис. 1.1. На нижней шкале оси абсцисс указано парциальное
1 В газовой смеси общее давление является суммой тех давлений, которые каждый газ создавал бы, если бы он присутствовал один. В сухом атмосферном воздухе при стандартнОхМ барометрическом давлении (760 мм рт. ст.) парциальное давление кислорода (Ро2) равно 159,2 мм рт. ст. (20,95% от 760 мм рт. ст.), азота (^n2) — 600,6 мм рт. ст. (79,02% от 760 мм рт. ст.) и двуокиси углерода (Рсог) —0,2 мм рт. ст. (0,03% от 760 мм рт. ст.).
Атмосферный воздух никогда не бывает совсем сухим, и его водяные пары создают парциальное давление, соответствующее их содержанию в воздухе. Парциальные давления других газов при этом уменьшаются в соответствующей точной пропорции. Если при давлении 760 мм рт. ст. воздух содержит 5% водяных паров (Рн2о = 38 мм рт. ст.), общее давление остальных газов должно быть 722 мм рт. ст., а их индивидуальные парциальные давления должны быть пропорциональны их относительным концентрациям и в сумме должны составлять 722 мм рт. ст.
22 Глава 1, Дыхание в воде
давление кислорода во вдыхаемом воздухе. Нуль этой шкалы не совпадает с нулем атмосферного давления, а соответствует точке, где атмосферное давление чуть ниже 50 мм рт. ст. (6,67 кПа). Причина этого проста. При температуре нашего тела (37 °C) дав-
fl 50 100 150
Парциальное давление кислорода, ммрт.стп.
Рис. 1.1. Зависимость атмосферного давления от высоты. (Dejours, 1966).
1. Высота, на которой большинство неакклиматизированных людей теряет сознание от недостатка кислорода. 2. Наибольшая высота постоянных человеческих поселений. 3. Наибольшая высота, на которой акклиматизированные люди могут выживать несколько часов, вдыхая воздух. 4. Наибольшая высота, доступная для людей при дыхании чистым кислородом.
ление водяного пара составляет 47 мм рт. ст. (6,28 кПа). Значит» если человека поместить в атмосферу с давлением 47 мм рт. ст. (на высоту 19 000 м, или 63 000 футов), его легкие будут заполнены водяным паром и потому воздух или кислород не сможет войти в них.
Позднее мы вернемся к некоторым эффектам, вызываемым низким парциальным давлением кислорода у животных на больших высотах.
РАСТВОРИМОСТЬ ГАЗОВ В ВОДЕ
Газы растворяются в воде. Если свободную поверхность воды привести в контакт с газом, часть молекул газа будет проникать в воду и переходить в раствор. Это будет продолжаться до тех
Газы в воздухе и воде 23
пор, пока не установится равновесие, при котором в единицу времени равные количества молекул газа будут входить в воду и выходить из нее. Количество газа, которое будет при этом растворено в воде, зависит 1) от природы газа, 2) от давления газа в газовой фазе, 3) от температуры и 4) от присутствия других растворенных веществ. Для понимания физиологии необходимо знать основы этих зависимостей.
Растворимость различных газов в воде очень различна; данные для наиболее важных в физиологическом отношении газов приведены в табл. 1.3. Поскольку растворимость зависит как от температуры, так и от давления, значения этих двух переменных должны быть указаны.
Таблица 1.3
Растворимость газов в воде при 15 °C и давлении газа 1 атм. Под коэффициентом растворимости а подразумевается объем газа (в миллилитрах при СТДС), растворяющегося в 1 л воды в условиях, когда давление данного газа без водяного пара равно 1 атм. Количество газа выражают объемом, который данный газ без примеси Н2О будет занимать при О °C и давлении 1 атм; эти условия сокращенно обозначают СТДС: стандартные температура и давление, в сухом состоянии.
Кислород 34,1 мл О2 1
Азот 16,9 мл N2 1 на 1 л воды
Двуокись углерода 1019,0 мл СО2 J
Мы сразу видим, что азот растворяется вдвое хуже, чем кислород, а растворимость двуокиси углерода примерно в 30 раз больше, чем у кислорода, и в 60 раз больше, чем у азота. Такая высокая растворимость СО2 в воде позволяет приготовлять содовую воду или шампанское. Закупоренная бутылка содовой воды, давление в которой обычно в 2—3 раза больше атмосферного, содержит небольшое количество газа при этом давлении вверху горлышка, а остальная часть СО2 растворена в жидкости. Как мы .знаем, это должно быть немалое количество, так как множество пузырьков довольно долго продолжает подниматься к поверхности. Если бы при давлении втрое выше атмосферного был растворен азот, образовалось бы всего лишь несколько пузырьков, так как азота растворилось бы примерно в 60 раз меньше, чем СО2.
ВЛИЯНИЕ ДАВЛЕНИЯ И ТЕМПЕРАТУРЫ
Количество газа, растворенного в данном объеме воды, зависит от давления газа. Если это давление увеличить вдвое, вдвое больше газа будет растворено. Пропорциональность между дав
24 Глава 1. Дыхание в воде
лением и количеством растворенного газа известна под названием закона Генри и записывается следующим образом:
760 ’^н2о>
где a — коэффициент растворимости (см. табл. 1.3); уравнение дает нам число миллилитров газа (Vg, при СТДС), растворенных в воде при данном давлении (Pg), выраженном в мм рт. ст.
Растворимость газовых смесей зависит от парциального давления каждого газа, находящегося в газовой фазе. Каждый газ растворяется в соответствии со своим собственным парциальным давлением в газовой фазе независимо от присутствия других газов.
Растворимость газов уменьшается с повышением температуры. Большинство знает это по собственному опыту. Когда мы открываем бутылку теплой содовой воды или пива (боже упаси!), жидкость проявляет гораздо большее стремление пениться и выливаться, чем в холодном состоянии. Точно также, когда мы смотрим на котелок с водой, подогреваемый на плите, видно, что задолго до того, как начнется кипение, на стенках начинают образовываться мелкие пузырьки; эти пузырьки содержат газ, выталкиваемый из раствора по мере повышения температуры. Таким образом, растворимость газов в воде ведет себя прямо противоположно растворимости твердых тел, которые в большинстве своем в горячей воде растворяются лучше, чем в холодной (например, сахар).
Теперь мы можем рассмотреть растворимость газов более подробно. В качестве примера в табл. 1.4 представлены данные о растворимости кислорода. Обратите внимание, что эти цифры относятся к количеству О2, растворенного в воде, которая находится в равновесии с атмосферным воздухом (а не с чистым кислородом при давлении 1 атм, как в табл. 1.3). Можно видеть, что при повышении температуры от точки замерзания до 30 °C растворимость уменьшается примерно до половины исходного значения. Та-
Таблица 1.4
Количество кислорода, растворенного в пресной и морской воде в условиях равновесия с атмосферным воздухом. (Krogh, 1941)
Температура, °C В пресной воде, мл/л В морской воде, мл/л
0 10,29 7,97
10 8,02 6,35
15 7,22 5,79
20 6,57 5,31
30 5,57 4,46
Газы в воздухе и воде 25
кая пониженная растворимость О2 весьма существенна для многих водных животных.
В табл. 1.4 приведены также данные о растворимости атмосферного кислорода в морской воде, которая в среднем на 20% ниже, чем в пресной. Это связано с тем, что соли снижают растворимость газов. Такой эффект характерен для растворенных твердых тел, тогда как растворенные газы не влияют на растворимость других газов (в условиях, с которыми мы имеем дело в физиологии).
ПАРЦИАЛЬНОЕ ДАВЛЕНИЕ И НАПРЯЖЕНИЕ
Мы рассматривали растворимость газов, измеряемую количеством газа, которое переходит в раствор при определенном давлении этого газа. Теперь подойдем к этому с другой стороны. Возьмем пробу воды, в которой растворено определенное количество газа; это количество должно соответствовать определенному давлению газа в газовой фазе. Такое давление называют напряжением данного газа в воде1. Если в воде растворено несколько газов, напряжение каждого из них соответствует его парциальному давлению в атмосфере, с которой вода находится в равновесии. Таким образом, напряжение газа в растворе определяется как парциальное давление этого газа в атмосфере, находящейся в равновесии с данным раствором.
Когда давление газа над пробой воды снижается, газ стремится выйти из раствора. Если мы снизим давление газа примерно до половины первоначального, газ будет выходить из раствора, пока не наступит новое равновесие; это произойдет, когда количество растворенного газа уменьшится 'вдвое. Если давление газа снизить до нуля (что равносильно помещению воды в вакуум), то весь газ выйдет, или будет экстрагирован, из воды. Такая вакуумная экстракция служит одним из способов удаления из жидкости всего растворенного газа; на практике этот метод обычно используется, например, для анализа газов, содержащихся в пробах крови.
Поскольку растворенный в жидкости газ находится в равновесии со своей газовой фазой при данном парциальном давлении, мы можем сказать, что газ в жидкости находится под тем же самым парциальным давлением. Если мы возьмем сосуд с водой, которая пришла в равновесие с атмосферным воздухом, и введем
1 Физиологи часто говорят о напряжении газа в жидкости, а не о его парциальном давлении; они определяют напряжение газа в растворе как давление этого газа в атмосфере, с которой данная жидкость находится в равновесии. Главная причина такой замены терминов состоит в том, что растворенный газ как таковой не оказывает измеримого давления, и поэтому термин «парциальное давление» применяется здесь в несколько условном смысле. Однако часто оба термина используются как взаимозаменяемые.
26 Глава 1. Дыхание в воде
в воду небольшой пузырек газа, например азота, то кислород (так же как и СО2) будет диффундировать из воды в этот пузырек И’ равновесие наступит тогда, когда пузырек будет содержать. 20,95% кислорода. Часть азота вначале растворится в воде, так. как исходное давление азота в пузырьке равно 1 атм, а его напряжение в воде всего лишь 0,79 атм. Однако по мере проникновения в пузырек кислорода убыль азота будет снижаться и конечные концентрации внутри пузырька будут такими же, как в исходном уравновешивающем атмосферном воздухе. (Эти рассуждения верны только в том случае, если объем пузырька очень мал по сравнению с объемом воды, так что пузырек заметным образом не влияет на концентрации газов в воде.)
РАСТВОРИМОСТЬ ДВУОКИСИ УГЛЕРОДА И СКОРОСТЬ ЕЕ ДИФФУЗИИ
Растворимость СО2 в воде в 30 раз больше, чем растворимость О2 (см. табл. 1.3). Однако в связи с очень низким содержанием двуокиси углерода в атмосфере (0,03%) общее ее количество, растворенное в воде, очень мало. Зная коэффициент растворимости и концентрацию СО2 в атмосфере, это количество можно вычислить следующим образом:
Объем растворенной СО2=---foo2—— 0>3 мл СО2 на 1 л воды.
Наряду с растворенной двуокисью углерода природные воды содержат различные количества СО2 в составе бикарбонатов и карбонатов. Поэтому общее количество находящейся в воде СО2 может быть довольно большим и может варьировать в зависимости от присутствия катионов. Жесткая вода, например, содержит много растворенного бикарбоната кальция, Са(НСОз)2, что повышает общее количество двуокиси углерода. В морской воде, которая имеет слегка щелочную реакцию (pH ~8,2), общее содержание СО2 может варьировать в пределах от 34 до 56 мл на 1 л воды (Nicol, 1960), тогда как количество СО2 в виде растворенного газа в ней остается таким, какое необходимо для уравновешивания ю атмосферой, т. е. около 0,3 мл на 1 л; остальная СО2 существует главным образом в виде бикарбонат-ионов. Таким образом, несмотря на высокое содержание двуокиси углерода, напряжение ее в морской воде (и многих других природных водах) близко к ее давлению в атмосфере, т. е. к 0,23 мм рт. ст., или 30,7 Па (небольшие отклонения могут быть результатом процессов фотосинтеза и дыхания).
В дыхательных органах животных (например, жабрах и легких) газы диффундируют между средой и организмом — кислород входит в организм, а двуокись углерода выходит из него. Поэтому важно знать, насколько быстро газы диффундируют, т. е. знать скорость диффузии. Этот предмет заслуживает особого внимания,
Газы в воздухе и воде 27
так как многие биологи убеждены, что СО2 диффундирует намного быстрее, чем О2. На самом деле это не так.
Скорость диффузии газа обратно пропорциональна квадратному корню из его молекулярного веса; двуокись углерода тяжелее кислорода и поэтому диффундирует медленнее. Молекулярный вес двуокиси углерода 44, а кислорода — 32; квадратные корни из
Рис. 1.2. Диффузия кислорода и двуокиси углерода между воздухом и водой. При одинаковом парциальном давлении обоих газов в газовой фазе количество СО2, растворившейся у поверхности воды (при 15°C), в 29,8 раза больше, чем количество О2. Молекулы СО2 диффундируют медленнее (со скоростью, составляющей 0,86 от скорости кислорода), но благодаря их большей концентрации в поверхностном слое при одинаковых давлениях в газовой фазе в воду диффундирует в 25,6 раза больше СО2, чем О2 (29,8-0,86 = 25,6).
этих чисел — 6,6 и 5,7 соответственно. Значит, ввиду обратной пропорциональности скорости диффузии СО2 и О2 относятся как 5,7/6,6, что равно 0,86. Иными словами, СО2 диффундирует со скоростью, составляющей 0,86 скорости диффузии О2.
Когда двуокись углерода диффундирует между воздухом и водой, ее высокая растворимость в воде приводит к кажущемуся ускорению диффузии. Это можно пояснить с помощью рис. 1.2. Пусть мы имеем атмосферу, содержащую кислород с давлением 100 мм рт. ст. и двуокись углерода с давлением 100 мм рт. ст. (т. е. парциальные давления или концентрации этих двух газов равны). Каждый газ растворяется в поверхностном слое воды независимо от другого пропорционально своей концентрации в газовой фазе и коэффициенту растворимости. Количества О2 и СО2, растворившихся у поверхности, будут соответственно равны 4,5 и 134 мл на 1 л (при 15°C). Другими словами, у поверхности концентрация СО2 в растворе в 30 раз (или, точнее, в 29,8 раза) больше концентрации О2, хотя в газовой фазе их концентрации равны. Теперь двуокись углерода и кислород диффундируют с по--верхности в толщу воды. Количество СО2, растворенной у поверхности, намного больше, поэтому и большее ее количество продиф-
28 Глава 1, Дыхание в воде
фундирует внутрь. Сами по себе молекулы СО2 диффундируют со скоростью, составляющей 0,86 скорости молекул О2, так что количество продиффундировавшей СО2 по отношению к О2 равно 29,8-0,86=25,6.
Теперь мы поймем, что при диффузии двуокиси углерода и кислорода от равных концентраций в воздухе в толщу воды общее количество продиффундировавшей СО2 будет примерно в 25 раз больше. Аналогичным образом, если напряжения СО2 и О2 в воде равны (в нашем случае 100 мм рт. ст. у поверхности воды), двуокись углерода будет диффундировать в атмосферу быстрее. Важно отметить, что это кажущееся ускорение диффузии двуокиси углерода проявляется только тогда, когда диффузия происходит в воде или между газом и водой и мы исходим из напряжений или парциальных давлений газов, а не из их молярных концентраций. Эта быстрая диффузия связана с более высокой растворимостью двуокиси углерода; каждая же молекула СО2 диффундирует в соответствии с законами физики (т. е. несколько медленнее кислорода)1.
Зная физические основы диффузии газов, мы теперь можем перейти к вопросу о том, каким образом организм животного существует и функционирует в пределах этих ограничений.
ВОДНОЕ ДЫХАНИЕ
Ряд самых простых устройств для дыхательного газообмена мы находим у водных животных. Многие мелкие организмы получают кислород в результате его диффузии через поверхность тела и не имеют каких-либо специальных органов дыхания и кровообращения. Более крупные и более сложно организованные животные часто имеют специализированные поверхности для газообмена и кровеносную систему, которая транспортирует кислород быстрее, чем это могла бы обеспечить одна лишь диффузия.
ЖИВОТНЫЕ, НЕ ИМЕЮЩИЕ СПЕЦИАЛЬНЫХ ОРГАНОВ ДЫХАНИЯ
Простейшая геометрическая форма организма — сферическая. Для нашего рассмотрения важно, что сфера имеет при данном объеме наименьшую возможную поверхность; любое отклонение от сферической формы приводит к относительному увеличению поверхности. Если мы предположим, что сферический организм должен получать кислород путем диффузии через поверхность тела, то самый длинный путь для диффузии будет соответствовать рас* стоянию от поверхности до центра.
Для того чтобы кислород достиг центра, нужно, чтобы концентрация его у поверхности превосходила определенную величи
1 В случае газов молярные концентрации прямо пропорциональны давле* Ниям, и никакой путаницы со скоростью диффузии возникнуть не может.
Водное дыхание 29>
ну, поскольку по мере диффузии в глубь тела кислород потребляется в процессе обмена веществ. То напряжение О2 у поверхности,, которого достаточно для снабжения всего организма кислородом' путем диффузии, можно вычислить с помощью уравнения, выведенного Э. Н. Харви (Harvey, 1928), который хорошо известен? своими исследованиями люминесценции организмов в море:
F -2^2_.
°2— 6К
В этом уравнении Го2 — концентрация кислорода у поверхности, выраженная по отношению к атмосферной, Уо2 — потребление кислорода в миллилитрах на 1 г в 1 мин, г — радиус сферы в сан* тиметрах и К — константа диффузии (К указывает число миллилитров О2, которое продиффундирует за 1 мин через площадь 1 см2, если градиент равен 1 атм/см. Дальнейшее рассмотрение диффузии см. в Приложении Б.)
Взяв в качестве гипотетического примера сферический организм с радиусом 1 см, потреблением кислорода 0,001 мл «г-1 мин-1 и константой диффузии для О2 11 • 10~6 см2 «атм-1 мин-1 (такой,, как у соединительной ткани и многих других животных тканей),, мы найдем, что концентрация кислорода у поверхности, необходимая для снабжения им всего организма вплоть до центра путем диффузии, равна 15 атм. Очевидно, что данный организм не может быть обеспечен кислородом путем одной лишь диффузии при выбранной нами интенсивности обмена, которая, кстати, довольно низка даже для беспозвоночных. Отсюда вывод: чтобы хорошо снабжаться кислородом, организм либо должен быть намного меньше, либо должен иметь гораздо более низкую интенсивность обмена. Теперь рассмотрим меньший организм, взяв радиус 1 мм. Мы найдем, что необходимое давление кислорода у поверхности? составляет 0,15 атм. Поскольку хорошо аэрируемая вода находится в равновесии с атмосферой с 0,21 атм О2, такой организм может получить достаточное количество кислорода путем одной, только диффузии и будет вполне жизнеспособным.
Если мы рассмотрим реальных животных, мы увидим, что эти? вычисления дают достаточно разумные порядки величин. Организмы, которые снабжаются кислородом только путем диффузии (например, простейшие и плоские черви), в большинстве своем или очень малы — менее 1 мм, — или имеют очень низкий уровень метаболизма, как, например, медузы. Хотя медузы могут быть очень крупными, они иногда содержат менее 1% органического вещества: остальное — вода и соли. Среднее потребление О2 организмом медузы очень невелико, а метаболически активные клетки находятся у них около поверхности, где пути диффузии относительно коротки.
Организм, форма которого отличается от сферической, имеет большую относительную поверхность и более короткие пути диф
30 Глава 1. Дыхание в воде
фузии. Это обстоятельство использовано в строении многих сравнительно простых организмов. Они бывают уплощенными или нитевидными, образуют псевдоподии или имеют очень большую и сложную поверхность (таковы, например, коралловые полипы и губки) и благодаря этому могут получать достаточно кислорода путем диффузии, хотя размеры некоторых из них намного больше, чем те, для которых мы выше приводили расчеты.
ЖИВОТНЫЕ, ОБЛАДАЮЩИЕ ОРГАНАМИ ДЫХАНИЯ
Хотя мелкие организмы могут получать достаточно кислорода путем диффузии через поверхность, этого обычно нельзя сказать о более крупных животных. Разумеется, любая форма, отличная от сферической, имеет большую поверхность, и пути диффузии при этом укорочены. Однако в большинстве случаев этого недостаточно, и мы находим у таких животных специализированные органы дыхания с сильно увеличенными поверхностями. Часто эти органы имеют также более тонкие покровы, чем другие части тела, и это облегчает газообмен.
Если дыхательная поверхность вывернута наружу, образуя выпячивание, получившийся орган обычно называют жаброй. Вторично жабра может быть заключена в полость, как, например, у рыб, но это не меняет того факта, что жабры в основе своей — выпячивания.
Если поверхность тела образовала углубление или впячивание, получившуюся полость называют легким. Хорошим примером служат наши собственные легкие, хотя вторично они приобрели чрезвычайно сложную, разветвленную структуру. Существуют и более простые легкие; например, у легочной наземной улитки легкие мало чем отличаются от простого мешковидного углубления, в котором осуществляется газообмен. Термин «легкие» используется независимо от того, что является дыхательной средой — вода или воздух.
У насекомых дыхательная система устроена особым образом, Небольшие отверстия в поверхности тела насекомого соединяются с системой трубочек (трахеи), которые ветвятся и подходят ко всем частям тела. В этом случае дыхательный орган представляет собой комбинацию распределительной системы (трубочки) с газообменной системой, так как большая часть газа проходит че* рез стенки тончайших веточек этой системы и диффундирует непосредственно к клеткам.
Вообще говоря, жабры чаще служат для дыхания в воде, а легкие — для дыхания в воздухе. Но есть и исключения: голотурии имеют водные легкие, в которых, по-видимому, осуществляется большая часть газообмена. Жабры тоже могут быть модифицированы для использования в воздухе, но в целом они не слишком приспособлены для атмосферного дыхания. Например, боль
Водное дыхание 31
шинство рыб, если их вынуть из воды, скоро начинают страдать от удушья, хотя в воздухе намного больше кислорода, чем в воде. Дело в том, что в воде вес жабр хорошо скомпенсирован, а в воздухе они не выдерживают собственной тяжести, так как им не хватает механической прочности и жесткости.
Эффективный орган дыхания должен иметь, во-первых, большую поверхность и, во-вторых, тонкие стенки. Оба этих требования затрудняют создание механической жесткости, необходимой' для поддерживания жабры в воздухе. Кроме того, в воздухе поверхности рыбьих жабр легко слипаются. В связи с этим поверхность, открытая для воздуха, оказывается во много раз меньше той, которая доступна воздействию воды, и это сильно препятствует поглощению кислорода.
ВЕНТИЛЯЦИЯ ЖАБР
Если жабра будет извлекать кислород из совершенно неподвижной воды, то в пограничном, смежном с жаброй слое воды кислорода вскоре не останется. Поэтому для снабжения кислородом важно, чтобы вода обновлялась, и существуют различные* механические устройства для создания тока воды над поверхностью жабр. Ток воды можно усилить двумя способами: движением жабры в воде и перемещением самой воды.
Движение жабры в воде «практично» только для небольших организмов. Таким способом вентилируют жабры некоторые водные личинки насекомых, например личинки поденок (Ephemeri-dae). Трудность тут вот какая: если жабра перемещается в воде' таким образом, что ее основание служит точкой вращения, то сила, которая нужна, чтобы преодолеть сопротивление воды, будет слишком велика. Сопротивление возрастает пропорционально квадрату линейной скорости, и поэтому энергия, необходимая для перемещения жабры, возрастает в той же пропорции. Механическую прочность жабры тоже приходится увеличить, опять-таки пропорционально квадрату линейной скорости, как и силу, приложенную к основанию жабры. Крупная водная амфибия Nectu-rus все же перемещает свои жабры, но эти движения очень медленные.
Движение воды над дыхательной поверхностью — намного легче реализуемое решение. Такое движение может создаваться работой ресничек, как у простейших или в жабрах мидий и других, двустворчатых моллюсков. Губки прогоняют воду через свои отверстия при помощи жгутиков.
Более распространены механические приспособления, подобные насосу. Рыбы и крабы, например, прогоняют воду над своими жабрами именно таким способом. В принципе «дешевле» перемещать воду медленно над большой поверхностью, чем быстро над малой.
32 Глава 1. Дыхание в воде
У некоторых животных движению воды около жабр способствует перемещение всего тела. Это относится ко многим пелагическим рыбам; у крупных тунцов жаберные крышки почти неподвижны, и необходимая высокая скорость потока воды над жабрами достигается путем быстрого плавания в воде. Тунцы, вероятно, не 'могут выжить, если их лишить возможности плыть вперед, и когда этих рыб содержат в аквариумах, их обычно помещают в большие круглые бассейны, чтобы они могли беспрерывно двигаться, не встречая препятствий.
У кальмаров и осьминогов тоже имеется тесная корреляция между локомоцией и током воды над жабрами, но у этих животных взаимоотношения в некотором смысле обратные. Кальмар или осьминог вентилирует свои жабры, набирая воду в мантийную полость, а когда он выбрасывает воду через сифон, он продвигается в воде реактивным способом. В этом случае вентиляционная система была видоизменена для локомоции, но, как и у рыб, по мере того как с увеличением скорости плавания возрастает потребность в кислороде, последний автоматически поступает в большем количестве.
ДРУГИЕ ФУНКЦИИ ЖАБР
Жабры могут также выполнять функции, отличные от дыхания, и иногда трудно решить, какова основная или единственная функция данной жабры. Некоторые из так называемых анальных жабр у личинок комаров осуществляют осмотическую регуляцию: они всасывают из окружающей воды ионы, и сомнительно, чтобы они играли сколько-нибудь заметную роль в дыхании. У рыб и крабов жабры тоже участвуют в осморегуляции, но здесь совершенно ясно, что их главная функция — дыхание.
Чтобы оценить роль в газообмене какой-либо жабры или другого предполагаемого дыхательного органа, мы должны иметь сведения о количестве поглощаемого через этот орган кислорода, а желательно также и о количестве выделяемой СО2. Сравнивая эти величины с потреблением кислорода всем организмом, мы увидим, ответствен ли данный орган практически за все снабжение кислородом, за большую его часть, за малую часть или совсем незначительную долю.
Если организм имеет обособленные и хорошо развитые органы дыхания, обычно возникает необходимость в циркуляторной системе для доставки кислорода к различным частям тела. (Это не относится к насекомым, так как их трахейная система в принципе независима от циркуляторной.) Мы уже говорили о том, что жабры двустворчатых моллюсков создают движение воды с помощью ресничек. У этих животных, отцеживающих пищу из воды, жабры работают как сито, задерживая взвешенные в воде частицы, которые потом отправляются в рот. Таким образом, жабры здесь в
Водное дыхание 33
первую очередь играют роль в питании; имеют ли они существенное значение для газообмена — менее ясно. У двустворчатых моллюсков интенсивность обмена, как правило, очень низка, и возможно, что для необходимого газообмена им достаточно поверхности мантии.
ГАЗООБМЕН И ТОК ВОДЫ
Тот факт, что поверхность жабр должна быть достаточно большой, чтобы обеспечить необходимый газообмен, хорошо иллюстрируется данными о рыбах. Очень активные рыбы имеют наибольшие относительные площади жабр (рис. 1.3). У быстро плавающей макрели жаберная поверхность, отнесенная к единице веса тела, примерно в 50 раз больше, чем у такой медлительной придонной рыбы, как морской черт.
Для того чтобы газообмен отвечал потребностям, необходимы высокая скорость потока воды и тесный контакт между водой и жаброй. Этому способствует анатомическая структура жаберного аппарата. Жабры заключены в жаберную полость, которая обеспечивает защиту этих относительно хрупких органов и, что не менее важно, позволяет воде обтекать жаберную поверхность наиболее эффективным образом. Особые преимущества дает такое строение, при котором поток воды около жабры и поток крови внутри этого органа направлены противоположно друг другу. Это называется противотоком. Чтобы оценить важность такого устройства, нужно знать структуру жабры (рис. 1.4; фото 1.1).
У рыб жабры состоят из нескольких главных жаберных дуг с каждой стороны. От каждой жаберной дуги отходят два ряда жаберных нитей. Кончики нитей, принадлежащих смежным дугам, соприкасаются, заставляя воду течь между нитями. На каждой нити располагаются ряды плотно уложенных плоских жаберных пластинок. Газообмен осуществляется в этих пластинках по мере того, как вода протекает между ними в одном направлении, а кровь внутри них — в другом.
Этот принцип противотока имеет важное значение. Когда кровь уже готова покинуть жаберную пластинку, она встречает воду, из которой кислород еще не извлекался. Таким образом, эта кровь поглощает кислород из воды, в которой содержание Ог еще не понизилось, и в результате насыщение крови кислородом может достигнуть максимального уровня. По мере дальнейшего прохождения между пластинками вода встречает кровь со все более и более низким содержанием кислорода, и поэтому она будет все время отдавать и отдавать кислород. Таким образом, пластинки извлекают кислород из воды по всей своей длине, и вода, пройдя жабру, может потерять до 80—90% исходного количества О2 (Hazelhoff, Evenhuis, 1952). Это считается очень высокой степенью экстракции кислорода; у млекопитающих, например, кровь 3—1873
Рис. 1.3. У высокоактивных и быстро плавающих рыб площадь жабр больше, чем у медлительных придонных видов. Цифрами после каждого названия указана общая площадь жаберной поверхности, выраженная в произвольных единицах на 1 г веса тела рыбы. (I. Е. Gray, по данным Gray, 1954.)
Водное дыхание 35
извлекает лишь около четверти кислорода, поступающего в легкие.
Газообмен противоточного типа можно представить с помощью графика (рис. 1.5). Мы видим, как кровь по мере прохождения через жаберные пластинки все больше и больше насыщается кис-
Рис. 1.4. Жабры рыб состоят из нескольких жаберных дуг. Каждая дуга несет на себе по два ряда жаберных нитей. На каждой нити имеются тонкие параллельные дискообразные пластинки. Внутри этих пластинок кровь течет в направлении, обратном тому, в котором вода течет между пластинками. (Randall, 1968.)
лородом, и в конце концов напряжение Ог в ней приближается к напряжению О2 во входящей воде. Из выходящей воды большая часть кислорода извлечена, и его напряжение здесь намного меньше, чем в крови, покидающей жабру. Если бы вода и кровь текли в одном направлении, это было бы невозможно, так как напряжение О2 в крови тогда могло бы в лучшем случае достичь того же уровня, что и в вытекающей воде. Поскольку прокачивание воды через жабры требует энергии, благодаря увеличению экстракции
36 Глава 1. Дыхание в воде
кислорода противоток снижает также энергетические затраты на прокачивание.
Раньше думали, что в жабрах акул нет противотока, хотя акулы великолепные пловцы и должны были бы иметь превосходно работающие органы дыхания. Однако на самом деле противоток в жабрах акул есть (Grigg, 1970). Противоток обнаружен также
Фото 1.1. Жабра рыбы. Кончик одной жаберной нити форели. Вода течет между параллельно расположенными пластинками в направлении, перпендикулярном плоскости бумаги. (Фото предоставлено G. М. Hughes, Бристольский университет.)
в жабрах некоторых крабов, но у них эффективность извлечения кислорода из воды намного меньше, чем у рыб. Частично это может объясняться тем, что ток воды у крабов менее эффективен, но, возможно, это связано главным образом с тем, что у них выше жаберно-кровяной барьер для диффузии. Последнее объяснение, видимо, справедливо для европейского берегового краба (Carcinus), у которого экстракция кислорода, несмотря на наличие противотока, не превышает 7—23% (Hughes et al., 1969).
Так как в жабрах рыб пластинки располагаются очень тесно, возникла мысль, что это создает большое сопротивление и тока воды между пластинками практически нет, поскольку промежутки между ними могут не превышать 0,02 мм. Давление, которое гонит воду через жабры, часто бывает меньше 10 мм Н2О, и такого низкого давления кажется недостаточно для прогона воды через узкие промежутки. Хьюз (Hughes, 1966) тщательно изучил этот вопрос, основываясь на данных о ширине, высоте и длине проходов между пластинками. Используя модифицированную форму уравнения Пуазейля (см. гл. 4), он вычислил, что поток Перез жабры 150-граммового линя (Tinea) при давлении 5 мм Н2О должен быть равен 10,1 мл/с. Нормальный объем воды, прогоняемой через жабры этой рыбы, составляет 1—2 мл/с; из этого очевидно, что жаберные пластинки не создают большого сопротивления, так как они могли бы пропустить и больше воды, чем реально проходит у живой рыбы. Аналогичные расчеты, проведенные более чем для дюжины других видов рыб, показали, что во всех случаях венти
Водное дыхание 37
ляционные объемы, измеренные на живой рыбе, не превышают теоретически возможных.
Интересно, что наибольшие величины для потока воды найдены у некоторых антарктических рыб, очень необычных в том отношении, что в их крови нет гемоглобина. Позже мы вернемся к. этим интересным, рыбам и к тому, как они могут жить без «нор* мального» механизма транспорта Ог в крови.
Рис. 1.5. Противоток в жабрах рыб позволяет крови иметь на выходе из жабр почти такое же высокое напряжение кислорода, как у входящей воды. Вода отдает кислород на всем пути вдоль жаберной пластинки и, пройдя жабру, может потерять почти весь свой кислород.
Для того чтобы прогонять воду через жабры, костистые рыбы используют комбинированное действие ротовой полости и жаберных крышек при участии соответствующих клапанов для контроля потока. Модель такого насоса показана на рис. 1.6. На самом деле эта система состоит из двух насосов. Объем первого — ротовой полости — может быть увеличен опусканием челюсти и особенно дна рта, а объем второго — оперкулярной полости — увеличивается с помощью движения жаберных крышек при одновременном блокировании обратного тока воды вблизи краев кожными щитками, играющими роль пассивных клапанов. На схеме показан только один оперкулярный насос; в действительности имеются две оперкулярные камеры — по одной с каждой стороны.
Действие этих двух насосов таково, что ток воды через жабры поддерживается на протяжении почти всего дыхательного цикла. Вода продолжает течь несмотря на то, что во время какой-то части цикла давление в ротовой полости может сделаться меньше, чем во внешней среде; дело просто в том, что давление в оперку
38 Глава 1. Дыхание в воде
лярной полости поддерживается на еще более низком уровне, чем во рту.
Это видно по записям давления, произведенным в ротовой и оперкулярных полостях на протяжении дыхательного цикла (рис. 1.7). График показывает, что изменения давления синхронизированы с движениями рта и жаберных крышек. Кроме того, вид-
Рис. 1.6. Воду через жабры рыб прогоняет двойная система насосов. При помощи соответствующих клапанов эти насосы обеспечивают однонаправленный ток воды около жаберной поверхности. (Hughes, 1960.)
но, что разность между давлениями во рту и оперкулярных полостях (нижняя кривая) остается положительной в течение почти всего цикла и обеспечивает силу, проталкивающую воду через жабры. Только на короткое время создается небольшая инверсия давления. Детали кривых давления у разных рыб различны, но в основном эти кривые у всех до сих пор изученных видов сходны: на ^^яжении почти всего дыхательного цикла давление в ротовой полости больше, чем в оперкулярных полостях, и это обеспечивает практически непрерывный ток воды через жабры.
Некоторые рыбы не способны дышать таким способом. Ихтиологи давно знают, что крупные тунцы не выживают в неволе, если у них нет возможности беспрерывно плавать; их приходится помещать в большие кольцеобразные бассейны, где они могут плавать по кругу без остановки. Такие рыбы плавают, приоткрыв рот; видимые дыхательные движения у них отсутствуют, а вода непрерывно течет через жабры; это называют таранной вентиляцией.
Водное дыхание 34
Таранная вентиляция встречается не только у крупных, быстро* плавающих пелагических рыб. Многие рыбы при малых скоростях, используют для дыхания прокачивание, а при больших переходят к таранной вентиляции. Переход наблюдается в области скоростей от 0,5 до 1 м/с: при больших скоростях активные дыхательные движения прекращаются (рис. 1.8).
Рис. 1.7. Записи изменений давления в дыхательном насосе плотвы (Rutilus). Нижняя кривая — разность давлений в ротовой и оперкулярной полостях. Давление в 1 см водн. ст. соответствует примерно 0,1 кПа. (Hughes, Shelton, 1958.)
Переход к таранной вентиляции вовсе не означает, что дыхание «пускается на самотек»; это значит только, что вентиляцию обеспечивают теперь не мышцы оперкулярного насоса, а плавательные мышцы тела и -хвоста. Открытый рот увеличивает лобовое сопротивление, и это должно быть компенсировано усиленной работой мускулатуры. Однако возможно, что при высокой интенсивности дыхания, необходимой во время быстрого плавания, непрерывный ток воды при таранной вентиляции энергетически более выгоден, чем оперкулярное прокачивание.
Степень открытия рта, по-видимому, регулируется таким образом, чтобы обеспечивать достаточный ток воды через жабры, но не увеличивать лобовое сопротивление больше, чем это требуется для адекватной вентиляции. Этот вывод основан на результатах экспериментов, в которых макрелей заставляли двигаться с одной и той же скоростью, но постепенно снижали содержание кислорода в воде. Это приводило к постепенному раскрыванию рта, так
40 Глава 1. Дыхание в воде
что снижение подачи кислорода компенсировалось увеличением тока воды (Brown, Muir, 1970).
Когда вода протекает через жабры, взвешенные в ней твердые частицы нередко застревают там. Такие частицы можно удалить, изменив направление тока воды; это осуществляется путем расширения ротовой полости при сомкнутых губах, приводящего к рез-
Рис. 1.8. Когда скорость движения макрели в воде возрастает до 0,5—0,8 м/с, оперкулярный насос перестает действовать и рыба полностью переходит на дыхание с помощью таранной вентиляции жабр. Приведенные на графике данные были получены на 5 особях весом около 70 г. (Roberts, 1975.)
кому снижению давления во рту. Такой прием аналогичен кашлю, который служит для удаления мешающего материала из дыхательных путей млекопитающих.
Крабы прочищают свои жабры таким же способом. Ток воды через жабры направлен почти всегда в одну сторону, но время от времени прокачивание прекращается и происходит внезапная реверсия потока на несколько секунд. Частота таких реверсий сильно варьирует: они могут повторяться каждую минуту и раз в 10 мин или даже реже (Hughes et al., 1969). По-видимому, этк-внезапные реверсии потока служат для удаления частиц, застрявших в жабрах.
Физиологическая основа дыхательного газообмена, обрисованная в этой главе, связана с очень простыми физическими принципами. Мы видели, как используются эти принципы и каким образом физиологические функции животных приспособлены к требованиям физического окружения. В следующей главе мы увидим, как столь же простые физические принципы используются в процессе дыхательного газообмена у животных, дышащих воздухом.
Водное дыхание 41
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Baes С. F., Jr., Goeller Н. Е., Olson J. S., Rotty R. М. (1977). Carbon dioxide and climate: The uncontrolled experiment, Am. Sci., 65, 310—320
Brown С. E., Muir B. S. (1970). Analysis of ram ventilation of fish gills with application to skipjack tuna (Katsuwonus pelamis), J. Fish. Res. Bd. Can., 27, 1637—1652.
Darden T. R. (1972). Respiratory adaptations of a fossorial mammal, the pocket gopher (Thomomys bottae), J. Comp. Physiol., 78, 121—137.
De jours P. (1966). Respiration, New York, Oxford University Press, 244 pp.
Gray I. E. (1954). Comparative study of the gill area of marine fishes, Biol. Bull., 107, 219—225. >
Grigg G. C. (1970). Use of the first gill slits for water intake in a shark, J. Exp. Biol., 52, 569—574.
Harvey E. N. (1928). The oxygen consumption of luminous bacteria, J. Gen. Physiol., 11, 469—475.
Hazelhoff E. H., Evenhuis H. H. (1952). Importance of the «counter-current principle» for the oxygen uptake in fishes, Nature, bond., 169, 77.
Hughes G. M. (1960). A comparative study of gill ventilation in marine teleosts, J. Exp. Biol., 37, 28—45.
Hughes G. M. (1966). The dimensions of fish gills in relation to their function, J. Exp. Biol., 45, 177—195.
Hughes G. M., Knights B., Scammell C. A. (1969). The distribution of Po2 and hydrostatic pressure changes within the branchial chambers in relation to gill ventilation of the shore crab Carcinus means L., J. Exp. Biol., 51, 203—220.
Hughes G. M., Shelton G. (1958). The mechanism of gill ventilation in three freshwater teleosts, J. Exp. Biol., 35, 807—823.
Krogh A. (1941). The Comparative Physiology of Respiratory Mechanisms, Philadelphia, University of Pennsylvania Press, 172 pp.
Machia L., Hughes E. (1970). Atmospheric oxygen in 1967 to 1970, Science, 168, 1582—1584.
Nicol J. A. C. (1960). The Biology of Marine Animals, New York, Interscience, 707 pp.
Otis A. B. (1964). Quantitative relationships in steady-state gas exchange. In: Handbook of Physiology, sect. 3, Respiration, vol. I (W. O. Fenn and H. Rahn, eds.), pp. 681—698, Washington, D. C., American Physiological Society.
Randall D. J. (1968). Fish physiology, Am. Zool., 8, 179—189.
Roberts J. L. (1975). Active branchial and ram gill ventilation in fishes, Biol. Bull., 148, 85—105.
Sawyer J. S. (1972). Man-made carbon dioxide and the «greenhouse» effect, Nature, bond., 239, 23—26.
Spitzer L. (1949). The terrestrial atmosphere above 300 km. In: The Atmospheres of the Earth and Planets (G. P. Kuiper, ed.), pp. 211—247, Chicago, University of Chicago Press.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Altman P. L., Dittmer D. S. (eds.) (1971). Biological Handbooks: Respiration and Circulation, Bethesda, Federation of American Societies for Experimental Biology, 930 pp.
Bartels H., De jours P„ Kellogg R. H., Mead J. (1973). Glossary on respiration and gas exchange, J. Appl. Physiol., 34, 549—558.
Dejours P. (1975). Principles of Comparative Respiratory Physiology, Amsterdam, North-Holland, 253 pp.
Hoar W. S., Randall D. J. (eds.) (1969—1971). Fish Physiology, 6 vols, New York, Academic Press.
Hughes G. M. (1963). Comparative Physiology of Vertebrate Respiration, Cambridge, Mass., Harvard University Press, 145 pp.
42 Глава 1. Дыхание вводе
Hughes G. М., Morgan М. (1973). The structure of fish gills in relation to their respiratory function, Biol. Rev., 48, 419—475.
Johansen K. (1971). Comparative physiology: Gas exchange and circulation in fishes, Annu. Rev. Physiol., 33, 569—612.
Jones J. D. (1972). Comparative Physiology of Respiration, London, Edward Arnold, 202 pp.
Krogh A. (1939) The Comparative Physiology of Respiratory Mechanisms, Philadelphia: University of Pennsylvania Press, 172 pp. Reprinted by Dover Publications, New York, 1968.
Библиографические ссылки, относящиеся ко всем вопросам физиологии, читатель найдет в следующей книге:
Prosser С. L. (ed.) (1973). Comparative Animal Physiology, Philadelphia, Saunders, 1011 pp. [Имеется перевод: Сравнительная физиология животных. В 3-х томах./Под ред. Л. Проссера, — М.: Мир, 1977, 1978.]
ГЛАВА 2
ДЫХАНИЕ В ВОЗДУХЕ
Успешная эволюционная адаптация к воздушному дыханию и наземной жизни осуществилась в широком масштабе только у членистоногих и позвоночных. Хорошо приспособлены к жизни на суше также некоторые улитки (они встречаются даже в пустынях); кроме того, в различных наземных микробиотопах обитает небольшое число других беспозвоночных.
Атмосфера обеспечивает высокую и постоянную концентрацию кислорода, доступного для использования почти везде. Наибольшей помехой для дыхания в воздухе служит испарение воды.
Легкая доступность кислорода делает возможными высокую интенсивность обмена веществ и высокий уровень общей организации, как структурной, так и физиологической. Другое следствие жизни на суше — то, что малая теплопроводность и теплоемкость воздуха по сравнению с водой позволяет животным поддерживать значительную разницу между температурой собственного тела и температурой окружающей среды. Это почти невозможно для водных животных. Наземная жизнь определила появление теплокровных животных, таких, как птицы и млекопитающие. Однако существуют способы, с помощью которых отдельные водные животные, в особенности крупные и очень активные рыбы, также поддерживают температуру своего тела на уровне, значительно превышающем температуру воды (см. гл. 8).
Давайте сравним количества кислорода, доступного для использования в воде и в воздухе. Вода в равновесии с атмосферным воздухом при температуре 15°C содержит 7 мл О2 на 1 л (1000 мл). Эти 7 мл О2 весят 0,01 г, и такое количество находится в воде, вес которой в 100 000 раз больше. Значит, для того, чтобы получить заданное количество кислорода, мы должны пропустить через дыхательные органы массу воды, вес которой в 100 000 раз больше, чем вес необходимого кислорода1.
Между тем один литр воздуха содержит 210 мл О2, который весит 280 мг. Оставшаяся часть воздуха — 790 мл N2 — весит 910 мг. Чтобы получить заданные количества кислорода, мы должны пропустить только в 3,5 раза большую массу инертного газа. Это различие наглядно представлено на рис. 2.1.
1 Более корректно употреблять термин «масса», а не «вес». Поскольку в повседневной речи мы говорим, что для определения массы объекта мы его взвешиваем, стало привычным говорить «вес», когда следовало бы сказать «масса». (См. также примечание на стр. 54.)
44 Глава 2. Дыхание в воздухе
Такая огромная разница в массе инертной среды имеет одно важное следствие: в водных дыхательных системах движение воды почти всегда однонаправленное. Если бы поток воды входил и выходил в одном месте или двигался то туда, то сюда, то боль-
Воздух (1л)
(280 мг) (910 мг)
Вода (1л)
\ (1000000мг)
О2 (7мл-> Юмг)
Рис. 2.1. В воздухе имеется много кислорода (21^%), а биологически инертный газ азот содержится в количестве, которое по весу всего лишь в 3,5 раза больше количества кислорода. В отличие от этого в воде растворенный кислород составляет всего 0,7% по объему (при 15°C и Pq2 159 мм рт. ст.). Вес инертной среды в этом случае в 100 000 раз больше веса кислорода.
шую массу воды нужно было бы ускорять, затем останавливать, затем снова ускорять — уже в обратном направлении. Для беспрерывных изменений кинетической энергии воды требовались бы большие затраты энергии. В дыхательных органах, использующих воздух, например в легких, перемена направления потока обходится не слишком дорого, так как масса инертного газа всего в несколько раз больше массы используемого кислорода. Но, как мы увидим позже, когда будем рассматривать дыхание птиц и насекомых, у дышащих воздухом животных встречается и однонаправленное движение воздуха.
Кроме массы дыхательной среды, на количество работы, необходимое для ее перемещения, влияет еще один фактор. Вода имеет большую вязкость, чем воздух, и это увеличивает работу по перекачиванию, так как движущее давление должно возрастать пропорционально вязкости. Поскольку энергия, необходимая для перемещения жидкости, прямо пропорциональна давлению, она возрастает также и пропорционально вязкости.
Вязкость воды при 20°C равна 1 сПз (сантипаузу), а вязкость воздуха — 0,02 сПз (т. е. у воды она примерно в 50 раз больше,
Дыхание в воздухе 45
чем у воздуха). Соответственно возрастает и работа, требуемая для перекачивания.
Другое преимущество воздушного дыхания — высокая скорость диффузии кислорода в воздухе, которая при том же парциальном давлении (напряжении) здесь примерно в 10 000 раз больше, чем в воде (см. Приложение Б). При такой быстрой диффузии размеры элементов дыхательного органа могут быть совсем иными. Расстояния, на которые газ диффундирует, например, в легких, могут составлять несколько миллиметров, а диффузионные расстояния в жабрах рыб — это ничтожные доли миллиметра.
Чтобы не происходило чрезмерного испарения воды с дыхательной поверхности, наружный воздух не должен иметь к ней свободного доступа. Газообменные поверхности обычно располагаются в специализированных дыхательных полостях (легких), и это сильно ограничивает доступ воздуха. Обновление воздуха в такой полости часто очень точно регулируется и не бывает более интенсивным, чем это диктуется потребностью в кислороде.
Газообмен через всю поверхность тела обычно возможен только во влажных местообитаниях. Например, дождевые черви, у которых весь дыхательный газообмен осуществляется через поверхность тела, очень чувствительны к недостатку воды. Они живут во влажных местах, и если остаются неприкрытыми на поверхности земли, то быстро высыхают и гибнут.
Что касается потери воды, сопутствующей газообмену, то растения находятся, по существу, в гораздо худшем положении, чем животные. Для фотосинтеза растения нуждаются в двуокиси углерода и должны получать ее из воздуха, который, как мы уже видели, содержит всего лишь 0,03% СО2 (0,23 мм рт. ст.). Таким образом, диффузионный градиент, заставляющий СО2 переходить внутрь растения, чрезвычайно мал. Даже если бы растение могло поддерживать нулевую концентрацию двуокиси углерода в своих тканях, движущая сила для диффузии СО2 не превышала бы 0,23 мм рт. ст. Давление водяных паров в тканях зависит от температуры; при 25 °C оно равно 24 мм рт. ст. Таким образом, движущая сила для диффузии воды в сухой воздух равна 24 мм рт. ст. В случае влажного воздуха она меньше: например, при о. в. (относительной влажности) 50% она меньше вдвое. В любом случае давление пара, вызывающее диффузию воды наружу, примерно в 100 раз больше движущей силы для диффузии СО2 внутрь. Из-за такого неблагоприятного соотношения растения испытывают очень большую потребность в воде.
Ананас, например, облегчает свое положение следующим способом. Это растение открывает устьица ночью, когда влажность воздуха высока; при этом двуокись углерода диффундирует внутрь и запасается в растении (конечно, в темноте она не может быть использована для фотосинтеза). В дневное время устьица закрыты; это снижает потери воды, а имеющийся теперь сол
46 Глава 2. Дыхание в воздухе
нечный свет используется для фотосинтеза органических веществ из запасенной двуокиси углерода (Joshi et al., 1965; Szarek et al., 1973).
Ситуации, в которых находятся растения и животные, сопоставлены на рис. 2.2. У животных направленная внутрь движущая сила для кислорода соответствует 21% О2 в воздухе, т. е.
Растение
Животное
Воздух Лист
Воздух
Легкое
Двуокись углерода 0,23 мм рт. ст.
Кислород
159 мм рт. ст.
Рис. 2.2. Растениям трудно получать необходимое количество СО2 из атмосферы путем диффузии, не подвергая себя опасности огромной потери воды. Для животных получение кислорода без большой потери воды представляет меньшую проблему благодаря высокому парциальному давлению О2 в воздухе (оно в 700 раз больше, чем парциальное давление СО2).
составляет 159 мм рт. ст. При 25 °C направленная наружу движущая сила для отдачи воды в сухой воздух составляет 25 мм рт. ст., т. е. она в несколько раз меньше. Даже при высоких температурах, скажем при 37 °C, давление водяных паров мало по сравнению с давлением, заставляющим кислород входить внутрь. Таким образом, мы имеем большой контраст между неблагоприятным водным балансом, который приходится преодолевать растениям, и благоприятной ситуацией с кислородом, которая выпала на долю животных (по крайней мере, надеемся, что это так).
ОРГАНЫ ДЫХАНИЯ
У животных, дышащих воздухом, встречаются три главных типа дыхательных органов: жабры, легкие и трахеи.
Органы дыхания 47
Жабры. В целом жабры довольно плохо приспособлены для дыхания в воздушной среде и используются лишь немногими животными— главным образом теми, которые сравнительно недавно проникли в наземные местообитания и принесли с собой пережитки своего прежнего способа водного дыхания. Хорошим примером служат сухопутные крабы. Краб пальмовый вор (Birgus latro), почти полностью перешедший к наземной жизни (включая лазание по кокосовым пальмам), имеет достаточно жесткие жабры, пригодные и для дыхания в воздухе. Другой сухопутный краб, Cardiosoma, особенно интересен тем, что может неопределенно долго жить и на суше, и в воде, тогда как пальмовый вор через некоторое время погибает, если его держать погруженным в воду (Cameron, Mecklenburg, 1973). Еще одна группа воздуходышащих ракообразных с жабрами — это наземные равноногие (Isopoda), обычно называемые мокрицами; эти животные, как правило, предпочитают жить во влажных местах, и у видов, наиболее успешно приспособившихся к наземным местообитаниям, жабры заключены в полости, так что в функциональном плане их можно рассматривать как легкие.
У некоторых рыб, способных дышать воздухом (но не у всех), функционирующие жабры сохранились. Обыкновенный угорь (Anguilla vulgaris) хорошо выживает на воздухе, если ему обеспечить достаточное охлаждение и влажность. Тогда большая часть кислорода поступает через кожу и меньшая — через жабры, так .как жаберные нити имеют тенденцию слипаться и в жаберной полости лишь небольшая часть их поверхности остается в контакте с воздухом. В результате угорь не получает своего обычного количества кислорода, и потребление О2 уменьшается примерно вдвое (Berg, Steen, 1965).
Легкие. Различают два типа легких: диффузионные и вентиляционные. Диффузионные легкие характеризуются тем, что обмен воздухом с окружающей атмосферой осуществляется только путем диффузии. Легкие этого типа встречаются у относительно небольших животных, таких, как легочные улитки, скорпионы и некоторые равноногие раки. Вентиляционные легкие имеются только у позвоночных. Значительное и регулярное обновление воздуха в легких необходимо при больших размерах тела, сочетающихся с высокой интенсивностью метаболизма. Дыхательные системы позвоночных вентилируются входящими и выходящими (приливно-отливными) потоками воздуха. Однако у птиц эти системы намного сложнее, чем у млекопитающих, и устроены таким образом, что и при вдохе, и при выдохе воздух может течь через легкие в одном направлении (см. ниже раздел о дыхании птиц).
Трахеи. Этот тип органов дыхания характерен для насекомых. Он представляет собой систему трубочек, которые подводят кислород непосредственно к тканям, так что циркуляция крови для транспорта газов оказывается ненужной. Газообмен в трахейной
48 Глава 2, Дыхание в воздухе
системе может происходить только путем диффузии, но у многих крупных и особенно высокоактивных насекомых имеет место однонаправленное прокачивание воздуха через части трахейной си-
Рис. 2.3. Дыхательный цикл у лягушки (Rana). (Gans, 1969.)
А. Воздух набирается в ротовую полость путем опускания ее дна. Б. Воздух выпускается из легких через верхнюю часть ротовой полости. В. Ноздри закрываются, и воздух проталкивается в легкие. Г. Пока воздух удерживается в легких благодаря смыканию голосовой щели, цикл может быть повторен, начиная с нового набирания воздуха в рот.
стемы. Преимущество однонаправленного потока состоит в том, что он обеспечивает намного лучший газообмен, чем при поочередном всасывании и выталкивании воздуха.
ДЫХАТЕЛЬНЫЕ ДВИЖЕНИЯ
Легкие позвоночных вентилируются путем активного прокачивания воздуха. (Вентиляцию трахейной системы мы рассмотрим позже, так как насекомые отличаются от других животных в столь многих отношениях, что о них лучше говорить отдельно.) Вентиляция легких у позвоночных может осуществляться двумя способами: для наполнения легких используется либо нагнетательный насос, как у амфибий, либо всасывающий насос, как у большинства рептилий, птиц и млекопитающих.
Роль кожи в дыхании 49
Лягушка, чтобы наполнить легкие, набирает воздух в ротовую полость, а затем закрывает рот и ноздри и вытесняет воздух в легкие путем подъема дна ротовой полости. В деталях эта последовательность движений несколько сложнее (рис. 2.3). Благодаря такому механизму наполнения легких лягушка может несколько раз подряд набирать новые порции воздуха, не позволяя ему выходить, в результате чего она способна раздуться до значительных размеров.
В противовес общепринятому мнению Бентли и Шилд (Bentley, Shield, 1973) показали, что по крайней мере некоторые амфибии могут также использовать при дыхании механизм всасывания. Свойственно ли это всем амфибиям, неизвестно.
Нагнетание под действием положительного давления, аналогичное механизму амфибий, обнаружено у некоторых рептилий. Ящерица Sauromalus, обитающая в пустынях юго-западной части Северной Америки, часто прячется в трещинах скал, где, раздув свои легкие, она заклинивается так прочно, что ее невозможно вытащить. Единственный способ достать это животное, используемый индейцами, которые употребляют его в пищу,—проколоть его заостренной палочкой.
Обычный механизм наполнения легких у рептилий таков же, как у птиц и млекопитающих: легкие наполняются путем всасывания воздуха. Выдох может быть пассивным (упругая отдача после вдоха), или ему может активно содействовать мышечное сокращение. Использование принципа всасывающего насоса требует замкнутой грудной полости, в которой давление во время вдоха должно быть меньше, чем в окружающей атмосфере. У млекопитающих вдоху способствует сокращение мышечной диафрагмы. Птицы имеют перепончатую диафрагму, прикрепленную к стенкам тела с помощью мышц, но по своей функции она отличается от диафрагмы млекопитающих. Распространенное утверждение, что у птиц нет диафрагмы, неверно.
РОЛЬ КОЖИ В ДЫХАНИИ
Газообмен через кожу является нормой и играет важную роль у амфибий, кожа которых влажная и хорошо снабжена сосудами. Некоторые мелкие саламандры (Plethodontidae) совсем не имеют легких, и весь газообмен осуществляется у них через кожную поверхность, если не считать небольшого участия слизистой оболочки рта.
У лягушек относительное значение кожи и легких меняется на протяжении года (рис. 2.4). Зимой, когда потребление кислорода очень невелико, кожа поглощает больше кислорода, чем легкие. Летом, когда требуется много кислорода, поглощение его через легкие возрастает в несколько раз и намного превышает поглощение через кожу. Тот факт, что поглощение О2 через кожу пример
4—1870
50 Глава 2. Дыхание в воздухе
но одинаково в течение всего года, связан с постоянством концентрации этого газа в атмосфере, которое обеспечивает постоянную скорость диффузии. Если бы концентрация кислорода в крови оставалась одинаково низкой на протяжении всего года, диффузия через кожу не должна была бы сильно изменяться, так как скорости диффузии очень мало зависят от температуры. А по-
Рис. 2.4. У лягушек поглощение кислорода через кожу почти постоянно на протяжении всего года. Возрастающее потребление О2 в летнее время покрывается за счет сильно увеличивающегося поглощения его через легкие. (Dolk, Postma, 1927.)
скольку летом потребность в кислороде сильно возрастает, кожное дыхание становится недостаточным и дефицит должен быть покрыт за счет дополнительного поглощения О2 легкими, что и происходит на самом деле.
.Определяются ли сезонные изменения температурой? Этот вопрос изучался на жабах Bufo americanus, которых содержали при трех различных температурах: 5, 15 и 25 °C (рис. 2.5). При двух более высоких температурах через легкие поступало больше кислорода, чем через кожу, а при самой низкой температуре, наоборот, преобладало поглощение через кожу. Так же обстоит дело у лягушки. Для обмена СО2 кожа более важна при всех температурах. Таким образом, при низких температурах кожа играет более важную роль, чем легкие, как в отдаче СО2, так и в поглощении О2.
Саламандры семейства Plethodontidae необычны тем, что у них нет ни легких, ни жабр. Эти саламандры отнюдь не редкость— они составляют около 70% всех существующих видов саламандр. Это очень мелкие формы, встречающиеся как в наземных, так и в водных местообитаниях. Газообмен и роль крови в транспорте газов были изучены у Desmognathus fuscus — животного, которое во взрослом состоянии весит от 5 до 7 г.
Это животное дышит в основном кожей, хотя около 15% всего газообмена осуществляется через слизистую оболочку рта и
Роль кожи в дыхании 51
глотки. Кровь не отличается какими-либо особыми свойствами: и содержание гемоглобина, и его сродство к кислороду находятся в том же диапазоне, что и у других саламандр, как водных, так и наземных. Здесь нет специальных сосудов, которые несли бы насыщенную кислородом кровь от кожи к сердцу, и оксигенированная кровь, покидая кожу, смешивается с общей венозной кровью.
Рис. 2.5. Легочный и кожный газообмен у жабы Bufo americanus при различных температурах. (Hutchison et al., 1968.)
У других амфибий в сердце имеется частичное разделение оксигенированной и венозной крови, но у безлегочных саламандр такого разделения, по-видимому, нет, и поэтому кровь у них никогда не бывает полностью насыщена кислородом. Тем не менее широкое распространение безлегочных саламандр указывает на то, что они отлично адаптированы, несмотря на дыхательный аппарат, который с нашей точки зрения кажется весьма у есовершенным (Gatz et al., 1974).
Кожу рептилий в отличие от кожи амфибий чао о считают почти непроницаемой. Рептилии дышат легкими, и большинство их — наземные формы. Однако настоящие морские змея — действительно морские животные, и некоторые из них даже рождают в море живых детенышей. Они прекрасные пловцы и способны погружаться на глубину по меньшей мере до 20 м. Когда морская змея 4*
52 Глава 2. Дыхание в воздухе
Pelamis platurus находится под водой, она может поглощать кислород через кожу с интенсивностью, доходящей до 33% от стандартного уровня, и выделять до 9% обычного количества СО2. Хотя у этой змеи главным органом газообмена служат легкие, кожа, по-видимому, также оказывает существенную помощь, когда змея преследует мальков рыб, на которых она охотится (Graham, 1974).
У млекопитающих газообмен через кожу ничтожен. Существует часто повторяемая легенда об итальянских детях, которых для участия в религиозной процессии покрасили золотой краской; история дальше повествует, что все они умерли от удушья, потому что «кожа не могла дышать». Смерть от удушья здесь совершенно исключена, так как поглощение кислорода через кожу едва измеримо, а выделение двуокиси углерода составляет менее 1% от ее выведения через легкие (Alkalay et al., 1971). Позолоченные дети могли умереть от других причин. Правдоподобно объяснение, что золотая краска была приготовлена из амальгамы золота с ртутью, суспендированной в масле — обычной основе красок. Ртуть легко образует эмульсию с маслом и в таком виде быстро всасывается через кожу, так что дети вполне могли умереть от острого отравления ртутью.
У летучих мышей относительная поверхность кожи намного больше, чем у других млекопитающих; широкие, тонкие, лишенные волосяного покрова перепонки крыльев у них сильно васкуляризо-ваны и могли бы участвовать в газообмене. Действительно, через эти перепонки выделяется некоторое количество СО2. При 18 °C у летучей мыши Eptesicus fuscus через кожу крыльев выделяется 0,4% всей образующейся двуокиси углерода. Эта доля возрастает с температурой воздуха, и при 27,5 °C уже 11,5% СО2 выводится этим путем (Herreid el al., 1968). Однако поглощение кислорода через перепонки крыльев не настолько велико, чтобы оно имело какое-то значение; как уже говорилось, диффузия кислорода между водой и воздухом идет примерно в 25 раз медленнее, чем диффузия двуокиси углерода.
ЛЕГКИЕ МЛЕКОПИТАЮЩИХ
Переходя от одного класса позвоночных к другому по восходящей линии, мы увидим, что легкие становятся все более и более сложными. У амфибий легкое — это одиночный мешок, подразделенный несколькими гребнями, которые увеличивают поверхность. Легкие млекопитающих разделены на множество гораздо более мелких мешочков — альвеол, которые во много раз увеличивают поверхность, пригодную для газообмена. Измерения поверхности легких у лягушки показывают, что 1 см3 легочной ткани имеет общую газообменную поверхность 20 см2; соответствующая цифра для легких человека — 300 см2 на 1 см3 легочной ткани. Большая
Легкие млекопитающих 53
поверхность необходима для быстрого поглощения кислорода, которого требуется очень много при высокой интенсивности метаболизма, характерной для теплокровных животных.
ОБЪЕМ ЛЕГКИХ
У млекопитающих легкие занимают около 6% объема тела независимо от его веса (рис. 2.6). Если бы объем легких всегда составлял в точности одну и ту же часть всего объема тела, наклон линии регрессии на рис. 2.6 был бы точно равен 1. Линия регрессии, дающая наилучшее приближение, имеет наклон 1,02 (т. е. отклонение от строгой пропорциональности несущественно). Как и можно было ожидать, отдельные точки не попадают точно на линию, однако^ больших характерных отклонений нет. Это означа-
ла
ю
ч to х §
в ю
° 0,01
0,001
0,0001
0,01 oj 1,0 ю loo юоо юооа
Вес тела, кг
Рис. 2.6. График зависимости между весом тела и объемом легких у различных млекопитающих. Видно, что величина легких является определенной функцией размеров тела. (Tenney, Remmers, 1963.)
•ет, что мелкие млекопитающие, у которых интенсивность обмена высока, получают достаточно кислорода через легкие той же относительной величины, что и у крупных животных.
Следует отметить, что ныряющие животные, такие, как дель-.фин, ламантин и кит, укладываются в общую для млекопитающих схему в отношении размеров легких. Можно было бы думать, что .для более продолжительного пребывания под водой эти животные .должны иметь очень большие легкие, которые они могли бы на
54 Глава 2, Дыхание в воздухе
полнять воздухом перед погружением. Однако это не так; как мы увидим позже, время пребывания под водой у ныряющих животных не определяется запасами кислорода в легких.
Линия регрессии на рис. 2.6 может быть описана уравнением ^=0,0567.^’02,
где Vi — объем легких в литрах, а Л4ь — масса тела в килограммах L У млекопитающего с массой тела 1 кг ожидаемый объем легких будет 0,0567 л, или 56,7 мл. Если принять, что объем тела в литрах равен массе тела в килограммах (для практических целей это верно, ибо мы знаем, что человек близок к нейтральному равновесию в воде и, следовательно, имеет среднюю плотность около 1), объем легких получится равным 5,67% объема тела. (Тот, кто незнаком с арифметическими манипуляциями над степенными уравнениями, найдет краткое объяснение в Приложении В.)
ВДОХ и выдох
Газообмен в легких осуществляется в альвеолах; трахеи, бронхи и их разветвления — только соединительные трубки. В конце выдоха эти трубки наполнены «использованным» воздухом из альвеол, а когда начинается вдох, этот воздух втягивается обратно в альвеолы, прежде чем туда войдет свежий наружный воздух. Наличие воздуха в дыхательных путях уменьшает количество свежего воздуха, входящего в альвеолы, и поэтому объем дыхательных путей называют мертвым пространством. Объем воздуха, вдыхаемого за один вдох, называют дыхательным объемом. У здорового человека в покое дыхательный объем составляет около 500 см3. Так как мертвое пространство занимает около 150 см3, в альвеолы поступает только 350 см3 свежего воздуха. Таким образом, мертвое пространство составляет около 7з дыхательного объема в покое. При физической нагрузке относительная роль мертвого пространства меньше. Например, если человек, который глубоко дышит, вдыхает за один раз 3000 см3 воздуха, то мертвое пространство в 150 см3 составит лишь V20 дыхательного объема. Таким образом, в покое мертвое пространство составляет значи
1 Масса — фундаментальное свойство материи, а вес — это сила, с которой на нее действует определенное гравитационное поле. В гравитационном поле Земли масса и вес, если их выражать в килограммах, численно совпадают. В различных гравитационных полях масса остается постоянной, а вес меняется. Например, на Луне человек, весящий на Земле 70 кг, будет иметь ту же массу— 70 кг, а вес — в шесть раз меньший, чем на Земле, т. е. около 12 кг (более правильно писать кгс, что означает килограмм-сила). Из-за совпадения массы и веса на Земле между ними обычно не делают различия. Однако в нашем примере корректным будет такое утверждение: масса человека равна 70 кг независимо от того, где он находится — на Земле, на Луне или в космосе, а вес его на Земле будет 686 Н (ньютонов), на Луне—110 Нив космосе — О Н.
Легкие млекопитающих 55
тельную часть дыхательного объема, а при нагрузке — относительно несущественную.
Важной особенностью дыхания является то, что легкие никогда полностью не освобождаются от воздуха. Даже если человек выдохнет все, что может, в легких еще останется около 1000 см3 воздуха. Поэтому человек не может наполнить легкие только «свежим» воздухом: вдыхаемый воздух всегда смешивается с воздухом, оставшимся в альвеолах и мертвом пространстве.
При дыхании в состоянии покоя в легких человека перед началом вдоха может содержаться около 1650 см3 воздуха. Во время вдоха в легкие входят 350 см3 свежего воздуха и смешиваются в них с 1650 см3, которые уже находятся там. Таким образом, обновление воздуха происходит лишь на одну пятую часть. В результате этого состав газа в альвеолах остается достаточно постоянным, где-то на уровне 15% кислорода и 5% СОг. Такой состав альвеолярного воздуха сохраняется и при физической нагрузке; иными словами, усиленная вентиляция при нагрузке точно подстраивается к повышенному потреблению кислорода.
ПОВЕРХНОСТНОЕ НАТЯЖЕНИЕ
Каждый, кто выдувал мыльные пузыри, знает, что при открытом выходе для воздуха пузырь стремится сокращаться и выдавливать из себя воздух, пока не исчезнет совсем. В легких у позвоночных мы имеем нечто похожее: пузыреобразная форма и большая кривизна альвеол ведут к тому, что поверхностное натяжение во влажных внутренних поверхностях стремится заставить «пузыри» сжаться и исчезнуть. В результате могло бы произойти спадение альвеол, но эта тенденция сводится к минимуму благодаря присутствию на стенках альвеол веществ, сильно уменьшающих поверхностное натяжение.
Эти вещества — фосфолипиды, и за свое влияние на поверхностное натяжение они получили название сурфактантов. Сурфактанты обнаружены в легких всех позвоночных — млекопитающих, птиц, рептилий и амфибий. Количество сурфактантов в легких позвоночных, по-видимому, всегда превышает тот минимум, который требуется для покрытия всей легочной поверхности мономолеку-лярным слоем (рис. 2.7).
МЕХАНИЧЕСКАЯ РАБОТА ДЫХАНИЯ
Перемещение воздуха в легкие и обратно требует затрат энергии, и было бы интересно сравнить эти затраты с тем, что дает поглощаемый кислород. Проще всего было бы сравнить количество кислорода, расходуемое на работу насоса, с тем его количеством, которое организм получает за тот же период времени.
Определить работу, связанную с дыханием, довольно трудно,
56 Глава 2, Дыхание в воздухе
и тут возможны существенные ошибки. Большинство данных указывает на то, что затраты на дыхание у человека в покое составляют 1,2% от общего потребления кислорода. В покое легкие вентилируются примерно пятью литрами в минуту, а на дыхательные движения расходуется около 0,5 см3 О2 на 1 л вентилирующего воздуха.
Рис. 2.7. Зависимость между площадью поверхности легких и количеством сурфактанта, которое можно экстрагировать из легких различных позвоночных. Во всех случаях это количество больше теоретического минимума, необходимого для покрытия легочной поверхности мономолекулярным слоем. (Clements et al., 1970.)
Однако при усилении вентиляции затраты на дыхание возрастают. Если вентиляция увеличивается до 10 л/мин, эти затраты достигают 1 см3 О2 на 1 л воздуха, а при очень интенсивной вентиляции (50 л/мин) они возрастают до 2 см3 на 1 л воздуха (Otis, 1954). По другим оценкам, максимальные затраты на дыхание при нагрузке не превышают 3% от всего поглощенного кислорода, т. е. несколько меньше только что приведенной цифры (Margaria et al., 1960). Затраты на дыхание определяли также у собак; были найдены величины того же порядка, что и для человека.
А каковы затраты на дыхание у рыб, которым приходится перемещать гораздо более тяжелую и вязкую среду? Точные оценки затруднительны, и вопрос остается не вполне ясным. Высказывалось -предположение, что затраты на дыхание у рыб могут доходить до 30 и даже 50% от количества получаемого кислорода
Регуляция дыхания 57
(Schumann, Piiper, 1966). Эти цифры кажутся нереальными, и недавние оценки показали, что затраты намного ниже — несколько процентов от общего потребления кислорода. Если механическую работу дыхания вычислить из перепада давлений в жабрах и объема протекающей воды, то результат составит менее 1% —это гораздо более правдоподобная цифра. Без однонаправленного тока воды затраты на вентиляцию, вероятно, не могли бы быть столь малыми.
РЕГУЛЯЦИЯ ДЫХАНИЯ
Если потребность в кислороде возрастает, вентиляция дыхательных органов должна быть соответственно увеличена. Аналогичным образом, если концентрация кислорода в среде падает, это должно быть скомпенсировано усилением вентиляции, увеличением количества кислорода, извлекаемого из воздуха, или обоими этими способами.
У теплокровных позвоночных — млекопитающих и птиц — вентиляция легких очень точно регулируется в соответствии с потребностью в кислороде; при этом интересно, что главным фактором, ответственным за регуляцию, служит концентрация двуокиси углерода в легочном воздухе. Это легко продемонстрировать, добавляя СО2 к вдыхаемому воздуху: вентиляция легких быстро усиливается (рис. 2.8). В норме атмосферный воздух практически не содержит двуокиси углерода (0,03%), и если к вдыхаемому воздуху добавить 2,5% СО2, вентилирующий объем примерно удваивается. У млекопитающих и птиц эти эффекты сходны.
В действительности 2,5% СО2 — это не так уж много, поскольку легочный воздух у млекопитающих уже содержит около 5% СО2. Если концентрацию двуокиси углерода во вдыхаемом воздухе повысить до той, которая в норме имеется в легких, вентиляция увеличится в 7 раз. При существенно больших концентрациях СО2 действует как наркотик и поэтому вызывает аномальные реакции.
Кислород гораздо меньше влияет на вентиляцию. Если мы снизим концентрацию кислорода во вдыхаемом воздухе на 2,5% (с 21 до 18,5%), практически никаких изменений дыхания не последует.
Чувствительность дыхательной системы к концентрации СО2 часто используют пловцы, которые хотят подольше продержаться под водой. Дыша глубоко в течение некоторого времени, можно увеличить выход двуокиси углерода из легких и крови. Это устранит обычный стимул для вдоха, и в результате человек после такой гипервентиляции сможет дольше оставаться под водой, прежде чем он будет вынужден подняться на поверхность из-за потребности вдохнуть. Такая практика чрезвычайно опасна. Когда человек плавает под водой, запас кислорода в его крови постепенно истощается, но из-за отсутствия обычной концентрации СО2 субъ
58 Глава 2. Дыхание в воздухе
ективная потребность в дыхании не очень сильна. Поэтому он остается под водой; а так как содержание кислорода в крови будет падать, он может потерять сознание, даже не заметив опасности. Если в этом состоянии его тотчас не обнаружат и не извлекут из воды, он утонет. Такой ход событий действительно был причиной многих несчастных случаев, особенно в плавательных
Рис. 2.8. Влияние содержания СО2 во вдыхаемом воздухе на вентиляцию легких у кур (Johnston, Jukes, 1966.)
бассейнах, где хорошие пловцы, соревнуясь, предпринимали длительные подводные заплывы (Craig, 1961а, Ь).
Утверждали, что тюлени и киты менее чувствительны к двуокиси углерода, чем другие животные, и поэтому могут дольше оставаться под водой. Однако продолжительность ныряния, по-видимому, все же лимитируется запасом кислорода в организме, и вряд ли ее можно было бы увеличить простым снижением чувствительности к СО2.
Если мы хотим изучить реакцию на вдыхаемую окись углерода у разных животных, недостаточно измерять только частоту дыхания. У некоторых животных частота дыхания при избытке СО2 существенно возрастает, у других же она может почти или совсем не изменяться, но вместо этого значительно возрастает дыхательный объем. Такая реакция обнаружена, например, у ехидны (Tachyglossus) (Bentley et al., 1967). Между тем нам нужно знать минутную вентиляцию (т. е. произведение частоты дыхания на дыхательный объем).
Если мы будем определять увеличение минутной вентиляции, вызываемое избытком СО2, то окажется, что при концентрациях
Регуляция дыхания 59
Рис. 2.9. Увеличение содержания СО2 во вдыхаемом воздухе приводит к увеличению вентиляции легких в несколько раз. Обратите внимание на то, что тюлени более чувствительны к СО2, чем собаки. (Bentley et al., 1970.)
СО2 до 6% тюлень даже более чувствителен к ней, чем неныряющие животные, такие, как человек и собака (рис. 2.9). Это противоречит утверждениям, что ныряющие тюлени нечувствительны к двуокиси углерода.
СРАВНЕНИЕ ВОЗДУШНОГО И ВОДНОГО ДЫХАНИЯ
У многих водных беспозвоночных регуляция дыхания плохо развита или даже совсем отсутствует. Это особенно характерно для морских видов, которые обычно живут в хорошо аэрированной воде с относительно постоянным содержанием кислорода. Не
69 Глава 2. Дыхание в воздухе
которые низшие животные хорошо переносят недостаток кислорода; двустворчатые моллюски могут долгое время держать свои раковины закрытыми и в отсутствие вентиляции используют анаэробные метаболические процессы (см. гл. 6).
Для большинства водных животных главный стимул для дыхания— нехватка кислорода. Это типично для ракообразных, осьминогов, рыб и др. Влияние двуокиси углерода на водных беспозвоночных никогда не выражено сильно и может совсем отсутствовать. Напряжение СО2 (Рсо2) в природных водах почти всегда низко, и, как мы увидим в гл. 3, из-за большой растворимости СО2 в воде у водных животных не могут создаваться высокие величины Рсо2- Если бы стимуляция дыхания зависела у них от повышения Рсо2, то достаточное снабжение кислородом не было бы обеспечено.
Если мы сравним две родственные формы — морского рака омара (Homarus) и речного рака (Astacus), мы заметим характерное различие. У омара, морского животного, не происходит больших изменений вентиляции при уменьшении количества кислорода в воде (Thomas, 1954). В то же время пресноводный рак отвечает на снижение концентрации О2 усилением вентиляции. Эту разницу легко понять. Омар обитает в холодных водах, где кислорода всегда много, так что сложный механизм регулирования вентиляции был бы излишним; речной же рак легко может попасть в местообитания, бедные кислородом, где необходима усиленная вентиляция.
Рыбы, как правило, реагируют на уменьшение количества кислорода, а их реакция на изменения концентрации СО2 незначительна. В этом отношении они сходны с другими водными животными, а не с дышащими воздухом позвоночными. Насекомые в большинстве случаев весьма чувствительны к двуокиси углерода.
Можно ли делать обобщения относительно этих различий? Как мы уже упоминали, водным животным трудно основывать регуляцию дыхания на чувствительности к двуокиси углерода не только потому, что напряжение СО2 в природных водах низко, но и потому, что это ненадежная мера содержания кислорода. Морская вода сильно забуферена, так что напряжение СО2 в ней никогда не поднимается до сколько-нибудь существенных значений. Напротив, в застойной пресной воде концентрация СО2 может быть высокой, что обычно ассоциируется с низкой концентрацией О2_ По-видимому, водные животные не могут опираться на такую ненадежную вещь, как концентрация двуокиси углерода; в тех случаях, когда их дыхание регулируется, основой служит концентрация кислорода.
Теперь возникает вопрос: почему наземные воздуходышащие животные отказались от регуляции по кислороду и перешли на регуляцию по СО2 как главному показателю? Дело, вероятно, в том,
Рыбы, способные дышать воздухом 61
что при более легком извлечении кислорода из атмосферы концентрация СО2 в органах дыхания имеет тенденцию нарастать. Например, если какое-то млекопитающее снижает концентрацию кислорода во вдыхаемом воздухе с 21 до 16%, это ведет к одновременному повышению концентрации СО2 примерно до 5%, что сильно влияет на кислотно-щелочной баланс организма. Снижение концентрации О2 до 16% не влечет за собой существенных физиологических последствий, тогда как изменение концентрации СО2 всего лишь на 1% —скажем, с 4 до 5% —ведет к увеличению количества угольной кйслоты на 25%. Кроме того, с физиологической точки зрения, возможно, легче создать точную систему контроля, основанную на чувствительности к СО2 и на выявлении малых сдвигов концентрации ионов Н+, чем выработать систему,, чувствительную к небольшим изменениям концентрации О2. Так или иначе, мы всегда находим, что животные, дышащие воздухом^ намного чувствительнее к концентрации двуокиси углерода, чем к концентрации кислорода.
РЫБЫ, СПОСОБНЫЕ ДЫШАТЬ ВОЗДУХОМ
Каждый слышал о двоякодышащих рыбах, однако и многие другие рыбы могут дышать воздухом. Многие из них прибегают к этому способу дыхания только тогда, когда концентрация кислорода в воде низка; относительно немногие зависят от воздуха в такой степени, что погибают, если их удерживать под водой. Есть две главные экологические причины для использования воздушного дыхания в качестве дополнительного или единственного: 1) нехватка кислорода в воде и 2) периодические засухи. Двоякодышащая рыба, например, в засушливые периоды зарывается глубоко в ил, окружает себя капсулой и остается неактивной до» следующего влажного периода.
Большая часть воздуходышащих рыб — обитатели тропических пресных водоемов и эстуариев; истинно морские виды если и встречаются, то их очень мало. Пресные воды с дефицитом О2 гораздо чаще встречается в тропиках, чем в умеренных зонах. Это> объясняется тем, что в тропических водах много разлагающегося органического материала, температура высока и ускоряет деятельность бактерий; небольшие водоемы часто сильно затенены нависающими джунглями (что снижает фотосинтез и образование-кислорода в воде); температура мало изменяется на протяжении суток, в связи с чем тепловая конвекция, которая могла бы доставить богатую кислородом поверхностную воду к глубинным слоям, очень слаба. Однако не все дышащие воздухом рыбы обитают в тропиках. Хорошо известная ильная рыба (Amia calva) встречается в северной части США даже в тех местах, где озера зимой замерзают. В такие периоды эта рыба легко обходится без воздуха, так как низкая температура снижает потребление кислорода.
Таблица 2.1
Рыбы, которые могут использовать дыхание воздухом как дополнительное или зависят от него полностью. Большинство их — современные костистые рыбы; только три последние в этом списке — настоящие двоякодышащие рыбы
Орган, используемый для дыха- Рыба Место _ обитания Примечания
ния в воздухе
Жабра Symbranchus Южная Америка, Угреобразная рыба, не
пресные воды имеющая английского
названия
Кожа Anguilla Северная Америка, Обыкновенный угорь;
Европа размножается в море;
личинки мигрируют в пресную воду
Кожа Periophthalmus Побережья тропи- Довольно обычная рыба,
чеоких эстуариев которую часто называ-
ют илистым прыгуном
Рот и оперку- Electrophorus Южная Америка, Электрический угорь
лярные полости пресные воды
Рот и оперку- Anabas Юго-восточная Эту рыбу называют оку-
лярные поло- Азия, пресные нем-ползуном, но это
сти воды в действительности не
окунь, а родственник петушку (Betta) — сиамской бойцовой рыбке
Рот и оперку- Clarias Юго-восточная Сом, известный также
лярные поло- Азия (а также под названием шагаю-
сти Флорида, в ре- щего сома
зультате интродукции), пресные
воды
Рот и оперку- Gillichthys Тихоокеанское по-
лярные поло- бережье Север-
сти ной Америки
Желудок Plecostomus Южная Америка, Небольшой сомик, кото-
пресные воды рого часто содержат в
аквариумах
Желудок Anicistrus Южная Америка, Сом, защищенный круп-
пресные воды ными шипами и кост-
ными пластинками
Кишечник Hoplosternum Южная Америка, Тоже из «броненосных»
пресные воды сомов (сомик-гопло- стерн)
Плавательный Arapaima Южная Америка, Самая большая в мире
пузырь реки пресноводная рыба
Плавательный Amia Северная Америка, Ильная рыба; ареал про-
пузырь пресные воды стирается на север в
области, где озера всю зиму покрыты льдом; принадлежит к прими-
тивной группе костных рыб
Рыбы, способные дышать воздухом 63
Продолжение
Орган, используемый для дыхания в воздухе Рыба Место обитания Примечания
Плавательный пузырь Lepisosteus Северная Америка, пресные воды Панцирная щука, относится к примитивной группе костных рыб
Легкие Polypterus Африка, пресные воды Бишир; имеет легкие, но не является настоящей двоякодышащей рыбой (см. текст)
Легкие Lepidosiren Южная Америка,' пресные воды ।
Легкие Легкие Protopterus Neoceratodus Африка, пресные воды Австралия, пресные водоемы, реки Настоящие двоякодышащие рыбы.
В гл. 1 мы объясняли, почему жабры рыб мало приспособлены для дыхания в воздухе: у них нет необходимой жесткости, и они имеют тенденцию слипаться. Однако жабры все же могут извлекать из воздуха некоторое количество кислорода. Любая другая влажная поверхность тоже будет вносить некоторый вклад в газообмен, если эта поверхность контактирует с воздухом и снабжается кровью. Какой-то газообмен в'сегда может осуществляться через кожу и поверхность ротовой полости; кроме того, могут существовать другие, анатомически более специализированные органы, помогающие газообмену. Органы, которые обычно используются для дыхания воздухом, — это жабры, кожа, ротовая и оперкулярные полости, желудок, кишечник, плавательный пузырь и легкие.
Некоторые воздуходышащие рыбы перечислены в табл. 2.1 с кое-какими комментариями, помогающими составить о них представление. Нужно отметить, что все они, за исключением пяти последних, — вы'сшие костные рыбы (лучеперые — Actinopterygii). Ильная рыба (Amia) — представитель примитивной группы Но-lostei, а многопер (Polypterus) занимает особое место, так как он имеет легкое, но обе эти рыбы рассматриваются как примитивные представители Actinopterygii. Только последние три из перечисленных в табл. 2.1 рыб — настоящие двоякодышащие рыбы (Dipnoi), предположительно близкие к кистеперым (Crossopterygii; к этому подклассу относится знаменитая латимерия). На каждом из трех континентов — в Австралии, Африке и Южной Америке — имеется по одному роду настоящих двоякодышащих рыб.
64 Глава 2, Дыхание в воздухе
Таблица 2.2
Рыбы, для которых дыхание воздухом обязательно
Рыба Орган дыхания Место обитания
Protopterus Eepido siren Легкие Легкие Африка
Arapaima Hoplosternum Плавательный пузырь Кишечник Южная Америка
Ophiocephalus Глоточные полости Южная Азия и Африка
Electrophorus Рот Южная Америка
Потребность в дыхании воздухом зависит от количества кислорода в воде и температуры, так как потребность в кислороде с температурой возрастает. Поэтому не всегда можно сказать, нуждается или не нуждается в дыхании воздухом данная рыба. Однако некоторым рыбам воздух настолько необходим, что они не могут существовать даже в хорошо аэрируемой воде. Рыбы, перечисленные в табл. 2.2, — именно такие облигатные воздуходышащие формы; интересно, что австралийская двоякодышащая рыба Ateo-ceratodus сюда не относится.
На примере некоторых рыб, изученных физиологами, мы рассмотрим теперь, как функционируют различные механизмы и насколько они эффективны.
ОБЫКНОВЕННЫЙ УГОРЬ
Считается, что обыкновенный европейский и североамериканский угорь (Anguilla vulgaris) способен проползать значительные расстояния по суше и переселяться из одного водоема в другой, особенно ночью и по мокрой траве. Рыбаки часто держат живых угрей в ящиках по нескольку дней, просто прикрыв их влажным мешком, чтобы предохранить от высыхания.
Когда угорь находится вне воды, он держит свои жаберные полости наполненными воздухом. Этот воздух он обновляет примерно раз в минуту; между тем в воде при 20 °C угорь совершает около 20 дыхательных циклов в минуту.
Потребление кислорода в воздухе у угря примерно вдвое меньше, чем в воде при той же температуре. Когда угря вынимают из воды, к кислороду, извлекаемому из воздуха, вначале добавляется кислород, переходящий в кровь из плавательного пузыря; в течение первого часа его количество может быть примерно равным то
Рыбы, способные дышать воздухом 65
му, которое угорь получает из воздуха. Но если угря держать на воздухе дольше, у него постепенно накапливается значительная кислородная «задолженность» и в крови возрастает концентрация молочной кислоты. После возвращения в воду происходит быстрое восстановление, и через 2 ч концентрация молочной кислоты в крови уже снова нормальна.
Рис. 2.10. Когда угря вынимают из воды (при 7°C), потребление кислорода у него постепенно уменьшается и стабилизируется на каком-то сниженном уровне. Вначале кислород берется из плавательного пузыря, а когда весь этот запас израсходован, суммарного поступления кислорода через кожу и жабры хватает для поддержания некоторого сниженного уровня потребления Ог. (Berg, Steen, 1965.)
При более низкой температуре — 7 °C — потребление кислорода намного ниже, концентрация молочной кислоты в крови не повышается и кислородной задолженности, очевидно, нет. Проницаемость кожи для кислорода практически не должна зависеть от температуры, и при низких температурах общего поглощения Ог через кожу и жабры, по-видимому, достаточно для покрытия метаболических нужд (рис. 2.10).
Для угря относительное значение жабр и кожи в воде и на воздухе различно. В воде (при 20°C) около 90% всего кислорода поступает через жабры и около 10%—через кожу. На воздухе только треть кислорода поглощается через жабры, остальные две трети — через кожу. Хотя жабры и приносят угрю некоторую пользу в воздухе, их функция далеко не соответствует потребностям, и даже при содействии кожи общее количество получаемого кислорода не позволяет избежать кислородной задолженности, если температура не очень низка.
5—1873
66 Глава 2. Дыхание в воздухе
ИЛИСТЫЙ ПРЫГУН
Другой рыбой, получающей, подобно угрю, кислород из воздуха, главным образом через кожу, является хорошо известный илистый прыгун (Periophthalmus), который обычен у тропических побережий, особенно в илистых устьях рек. Эти рыбы достигают в длину около 20 см; они любят подолгу оставаться на воздухе и передвигаются, опираясь на грудные плавники. Если илистого прыгуна потревожить, он быстро скрывается в ямку, вырываемую в иле, на дне которой обычно имеется вода. Однако, если эту рыбу держать погруженной в воду, у нее возникают такие реакции кровеносной системы, которые указывают на частичную асфиксию.
SYMBRANCHUS
Эта южноамериканская болотная рыба не имеет общепринятого английского названия. Она примечательна тем, что, по-видимому, способна одинаково хорошо дышать и в воде, и на воздухе. Для дыхания она использует в первую очередь жабры, но стенки жаберной полости у нее сильно васкуляризованы и участвуют в газообмене. Когда рыба находится на воздухе, жаберная полость периодически наполняется и воздух задерживается там на 10—15 мин, пока примерно половина ^кислорода не будет извлечена.
Когда эта рыба находится в воде, артериальная кровь у нее всегда довольно слабо насыщена кислородом — не более чем на 50 или 60%. Наполнение жаберной полости воздухом неизменно приводит к существенному повышению количества артериального кислорода, которое затем достигает уровня насыщения (Johansen, 1968).
Когда Symbranchus дышит воздухом, артериальная концентрация СОг возрастает, что характерно для всех воздуходышащих, а после возвращения рыбы в водную среду снова быстро падает. Если концентрацию СО2 в воде увеличить, интенсивность дыхания уменьшается; таким образом, двуокись углерода здесь скорее тормозит, чем стимулирует дыхание, в противоположность тому, что наблюдается у других организмов, дышащих воздухом. Ослабление дыхания, вызванное высоким уровнем СО2 в воде, ведет к гипоксии, а это стимулирует дыхание воздухом. Таким образом, главным стимулом в регуляции дыхания у Symbrartchus служит недостаток кислорода, который заставляет рыбу переходить к воздушному дыханию.
ЭЛЕКТРИЧЕСКИЙ УГОРЬ
Южноамериканский электрический угорь (Electrophorus elect-ricus) известен благодаря своим сильным электрическим разрядам; эта рыба способна создавать напряжение до 550 В, достаточ
Рыбы, способные дышать воздухом 67
ное, чтобы оглушить или убить другую рыбу, а возможно, даже и человека. Электрический угорь дышит ртом и гибнет, если его лишить доступа к воздуху. В его ротовой полости, обильно васкуляризованной, имеется множество складок и сосочков, сильно увеличивающих поверхность.
Электрический угорь набирает воздух в рот с интервалами от нескольких секунд до нескольких минут. Количество кислорода в
§
Рис. 2.11. Воздух, удерживаемый в ротовой полости электрического угря, постепенно теряет кислород по мере использования его рыбой. Содержание СО2 в этом воздухе постепенно увеличивается, но не в такой степени, как убывает О2. После извлечения примерно трети кислорода рыба поднимается к поверхности для обновления воздуха. (Johansen et al., 1968.)
этом воздухе постепенно убывает, а содержание СО2 возрастает, хотя и не в той же пропорции (рис. 2.11). Когда кислорода становится меньше примерно на одну треть, рыба медленно поднимается к поверхности воды, чтобы обновить воздух.
Если у животного, дышащего воздухом, весь газообмен осуществляется между кровью и определенной массой воздуха, увеличение количества СО2 должно примерно соответствовать убыли О2. Поскольку у электрического угря дело обстоит не так, можно заключить, что двуокись углерода выходит еще где-то — возможно, через довольно рудиментарные жабры или, что более вероятно, через кожу.
Регуляцию дыхания у электрического угря можно исследовать, изменяя концентрации СО2 и О2 в той атмосфере, которой она дышит. Такой эксперимент показывает, что значительное повышение содержания СО2 в воздухе приводит лишь к умеренному ускорению дыхания, тогда как при снижении количества О2 темп дыхания ускоряется в несколько раз. И наоборот, подъем давления О2
5*
68 Глава 2, Дыхание в воздухе
выше нормального атмосферного уровня несколько подавляет дыхание (Johansen, 1968). Интересно, что у данной рыбы, перешедшей к облигатному воздушному дыханию, сохранился водный тип регуляции дыхания, зависимый от кислорода. Очевидно, это связано с тем, что СО2 выходит из организма через относительнопроницаемую кожу и не накапливается в концентрациях, достаточных для того, чтобы иметь физиологическое значение.
ПАНЦИРНАЯ ЩУКА
Панцирная щука (Lepisosteus osseus) — реликтовый представитель древней группы Holostei, который в настоящее время встречается только в Северной Америке. Это рыба с удлиненным рылом и жесткой чешуей, не популярная среди рыболовов-любителей. Часто можно видеть, как она высовывает голову над поверхностью воды, чтобы подышать (фото 2.1). Это большая рыба,.
Фото 2.1. Панцирная щука (Lepisosteus osseus}, вдыхающая воздух у поверхности воды. Около задней части головы виден пузырь воздуха; он только что вытолкнут из жаберной щели, и рыба готова набрать воздух путем опускания дна ротовой полости. Весь цикл смены воздуха занимает лишь немногим более 0,5 с, после чего рыба вновь уходит под воду. (Фото предоставлено Katharine Rahn, Буффало, штат Нью-Йорк.)
вес которой может превышать 10 кг; она широко распространена, причем обитает в северной части США даже там, где озера зимой замерзают на несколько месяцев.
Панцирная щука имеет жабры и, кроме того, использует для дыхания свой видоизмененный плавательный пузырь. Частота, с которой она поднимается к поверхности, чтобы набрать в пузырь свежего воздуха, зависит от температуры воды. При 22 °C
Рыбы, способные дышать воздухом 69х
интервал между «вдохами» у непотревоженной рыбы составляет в среднем 8 мин, но зимой, когда вода покрыта льдом, рыба, очевидно, не может дышать воздухом и полностью полагается на жабры. При температурах от 20 до 25°C воздушное дыхание покрывает примерно 70—80% всей потребности в кислороде; оставшуюся часть обеспечивают жабры. Зимой, когда рыба использует только жабры, выведение двуокиси углерода через плавательный пузырь, конечно, равно нулю. Летом выход СОг через пузырь достигает 10%; основная же масса СО2 выводится через жабры и кожу (Rahn et al., 1971). Это характерно для многих воздуходышащих рыб: хотя вдыхание воздуха важно для получения кислорода, двуокись углерода благодаря своей высокой растворимости выделяется главным образом в воду.
ДВОЯКОДЫШАЩИЕ РЫБЫ
Африканские и южноамериканские двоякодышащие рыбы (Protopterus и Lepidosiren) фигурируют в табл. 2.2 как формы, для которых дыхание воздухом обязательно. Они живут в застойных водоемах и в озерах, там, где длительные засухи могут приводить
Neoceratodus Protopterus Lepidosiren (Австралия) (Африка) (Южная Америка)
Рис. 2.12. Относительная роль жабр и легких в дыхательном газообмене трех видов двоякодышащих рыб при содержании их в воде со свободным доступом к воздуху. (Lenfant et al., 1970.)
к полному пересыханию их местообитаний. Рыбы проводят время в спячке до следующего влажного периода; тогда они выходят из своих капсул в иле и возобновляют нормальную жизнь.
70 Глава 2. Дыхание в воздухе
Австралийская легочная рыба Neoceratodus живет в реках и медленных ручьях. Она тоже впадает в спячку во время засушливых периодов, однако эта рыба значительно меньше использует легкое и дышит главным образом жабрами. Если держать ее в резервуа-.ре, из которого вода медленно вытекает, она начинает вести себя очень беспокойно и пытается погрузиться в остатки воды. Однако она все-таки наполняет свое легкое воздухом, что существенно помогает газообмену.
Рис. 2.13. Когда арфриканских и южноамериканских двоякодышащих рыб вынимают из воды, потребление кислорода у них почти не изменяется, а у австралийской двоякодышащей рыбы оно при этом резко снижается; насыщение ар • териальной крови кислородом у австралийской рыбы соответственно падает. (Lenfant et al., 1970.)
Если двоякодышащих рыб этих трех видов содержать в воде в условиях свободного доступа к воздушной среде, относительная роль жабр и легких будет у них такой, как показано на рис. 2.12. У австралийской рыбы главным органом газообмена служат жабры. У африканской рыбы большая часть кислорода поступает через легкое и лишь малая доля — через жабры. У южноамерикан
Дыхание птиц 71
ской двоякодышащей рыбы практически весь кислород поступает через легкие, а через жабры — только ничтожное количество. С двуокисью углерода дело обстоит иначе: как африканская, так и южноамериканская рыбы примерно половину ее выделяют через-легкие, остальное — через жабры. Таким образом, для обмена СО2, жабры существенны, хотя в поглощении кислорода главную роль играют легкие.
Сходные соотношения между тремя видами проявляются и в изменениях, которые происходят, когда этих рыб помещают в воздушную среду (рис. 2.13). Потребление кислорода африканской и южноамериканской рыбами практически не изменяется. В отличие от этого у австралийской двоякодышащей рыбы потребление О2 резко падает, так как в норме эта рыба дышит в воде и не может получать достаточно кислорода из воздуха.
Кривые насыщения артериальной крови кислородом (рис. 2.13, нижний график) показывают, что у тех двух рыб, для которых дыхание воздухом обязательно, уровень насыщения сохраняется или слегка увеличивается, если их вынуть из воды; у австралийской же рыбы содержание кислорода в артериальной крови падает почти до нуля — явное указание на то, что рыба начинает страдать от удушья.
Двоякодышащие принадлежат к наиболее изученным из тех рыб, которые способны дышать воздухом. Сведения относительно многих других недостаточны или практически отсутствуют. Исследование их могло бы оказаться очень плодотворным, хотя часто было бы сопряжено с техническими трудностями из-за малой величины объектов.
ДЫХАНИЕ ПТИЦ
СТРОЕНИЕ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
Органы дыхания у птиц сильно отличаются от соответствующих органов млекопитающих. Небольшие компактные легкие птиц сообщаются с объемистыми тонкостенными воздушными мешками — воздушными пространствами, которые расположены между внутренними органами и даже разветвляются внутрь костей черепа и конечностей. Эту обширную и сложную дыхательную систему рассматривают как приспособление, облегчающее полет. Однако мы можем сразу же сказать, что для полета такая система не необходима, поскольку летучие мыши — животные с легкими, типичными для млекопитающих, — тоже хорошие летуны и иногда даже мигрируют на большие расстояния.
Можно было бы думать, что летающие птицы должны потреблять очень много кислорода и что дыхательную систему птиц нужно рассматривать именно в этом свете. Однако величины потреб
72 Глава 2. Дыхание в воздухе
ления О2 птицами и млекопитающими в покое очень близки, и хотя нормальный полет требует 8—10-кратного увеличения расхода кислорода, многие млекопитающие тоже способны обеспечить такое увеличение. Наконец, во время полета летучие мыши потребляют примерно столько же кислорода, сколько и птицы с такой же массой тела. С другой стороны, хотя свойственная птицам ды-
Рис. 2.14. Самыми мелкими элементами легких у млекопитающих являются мешковидные альвеолы. В легких птиц тончайшие разветвления представляют собой открытые с обеих сторон трубки со сквозным проходом для воздуха. (Schmidt-Nielsen, 1972.)
хательная система и не служит необходимой предпосылкой для полета, она все же может давать существенные преимущества.
Можно сказать, что присутствие в теле воздушных пространств делает птицу легче, но только в весьма ограниченном смысле. Птице необходимы пищеварительная система, печень, почки и
Таблица 2.3
Объемы дыхательной системы у типичной птицы и типичного млекопитающего весом 1 кг. (По данным Lasiewski, Calder, 1971)
Птица Млекопитающее
Объем легких, мл 29, б1^ 53,5
Объем трахеи, мл 3,7 0,9
Объем воздушных мешков, мл 127,5 —
Общий объем дыхательной системы, мл Дыхательный объем, мл 160,8 54,4
13,2 7,7
Частота дыхания, мин-1 17,2 53.5
’) Это общий объем легкого птицы, которое содержит
только 9,9 мл воздуха.
Дыхание птиц 73
т. д.» и простое добавление к брюшной полости больших воздушных мешков не делает птицу ни на йоту легче. Если мы удалим этот воздух или удвоим его объем, птице придется нести во время полета в точности тот же вес. С другой стороны, если костный
Фото 2.2. Поперечный срез легкого курицы. Видны цилиндрические трубки (парабронхи), обеспечивающие однонаправленный сквозной ток воздуха, характерный для птиц. Диаметр каждой трубки в данном случае немногим менее 0,5 мм. (Фото предоставлено проф. H.-R. Duncker, Гиссен, ФРГ.)
мозг заменить равным объемом воздуха, кости будут весить меньше. Значит, наполненные воздухом кости действительно делают птицу легче, чего нельзя сказать о других больших воздушных пространствах.
Сравнивая объем отдельных частей дыхательной системы у птиц и млекопитающих, мы находим ряд резких различий (табл. 2.3). Объем легких типичной птицы составляет чуть больше половины объема легких млекопитающего той же величины. Напро
74 Глава 2, Дыхание в воздухе
тив, объем трахеи у птиц намного больше, чем у млекопитающих. Это легко понять, учитывая большую длину шеи у птиц, однако в дальнейшем мы увидим, что этому есть и другие объяснения. Воздушные мешки птиц велики, по объему они в несколько раз превышают легкие, а у млекопитающих воздушных мешков нет сов-
Рис. 2.15. В теле птицы имеется несколько больших тонкостенных воздушных мешков. Парные легкие невелики и располагаются вдоль позвоночника. Главный бронх, который проходит сквозь легкое, сообщается как с легким, так и с воздушными мешками. Вверху приведена схема такой системы, на которой для упрощения все передние мешки объединены в одно воздушное пространство, а все задние — в другое. (Schmidt-Nielsen, 1972.)
сем. Поэтому общий объем дыхательной системы у птиц примерно в три раза больше, чем у млекопитающих.
Различие между птицами и млекопитающими не ограничивается только воздушными мешками; сами легкие птиц по своему строению тоже радикально отличаются от легких млекопитающих. У последних самые тонкие веточки бронхов заканчиваются мешковидными альвеолами; у птиц же тончайшие разветвления бронхиальной системы (называемые парабронхами) допускают сквозной проход воздуха (рис. 2.14; фото 2.2). Таким образом, у птиц воздух может течь через легкие непрерывным потоком мимо
Дыхание птиц 75
газообменной поверхности; у млекопитающих же воздух должен входить в альвеолы и выходить из них. Это наиболее важное различие между дыхательными системами птиц и млекопитающих, и из него вытекают глубокие физиологические следствия.
Чтобы понять, как работает дыхательная система птицы, нам нужны некоторые дополнительные сведения по ее сложной анатомии; воспользуемся схемой легких и основных воздушных мешков (рис. 2.15). Нет надобности заучивать названия всех мешков хотя бы потому, что разные исследователи часто употребляют различные термины, и это просто запутает неспециалиста. Однако важно знать, что анатомически и функционально воздушные мешки образуют две группы: заднюю, включающую обширные брюшные мешки, и переднюю, которая состоит из нескольких мешков меньшего размера.
Трахея делится на два бронха; каждый бронх подходит к одному из легких и затем в буквальном смысле проходит сквозь него и заканчивается в брюшном мешке. Передние мешки подсоединяются к этому главному бронху в передней части легких; задние мешки присоединены к заднему отделу главного бронха. Главный бронх сообщается также с легкими, и, кроме того, некоторые из воздушных мешков непосредственно связаны с легочной тканью.
РАБОТА ДЫХАТЕЛЬНОЙ СИСТЕМЫ
Какова функция дыхательных мешков? Участвуют ли они в газообмене или, подобно кузнечным мехам, служат лишь для втягивания и выталкивания воздуха?
В морфологии воздушных мешков нет признаков того, что они играют сколько-нибудь важную роль в газообмене между воздухом и кровью. Их стенки тонки, непрочны, слабо васкуляризированы и не имеют ни складок, ни гребней, которые увеличивали бы поверхность. Простой эксперимент, который исключил их прямую роль в газообмене, был проведен в прошлом веке одним французским исследователем (Soum, 1896). Он закрыл проходы из больших абдоминальных воздушных мешков в остальную часть дыхательной системы и затем ввел в эти мешки окись углерода. Вообще птицы очень чувствительны к окиси углерода, но подопытные птицы не обнаружили никаких признаков отравления ею. Значит, окись углерода не переходила из воздушных мешков в кровь. Этот вывод можно обобщить; он справедлив и для других газов со сходными диффузионными свойствами, например кислорода.
В отношении функции воздушных мешков более правдоподобна гипотеза, что они, подобно мехам, служат для втягивания и выталкивания воздуха. Во время вдоха одновременно понижается давление и в передних, и в задних мешках, по мере того как они расширяются (см. схему на рис. 2.15). Значит, при вдохе воздух
76 Глава 2. Дыхание в воздухе
входит во все мешки (однако, как мы увидим далее, не все мешки наполняются наружным воздухом). Во время выдоха давление в мешках увеличивается, и воздух из них снова выходит.
Чтобы более детально проследить перемещение газа, можно использовать газовую смесь, отличную по составу от обычного воздуха. Для этой цели подходит чистый кислород, поскольку он
Обычный состав газа
Брюшной > Задние мешки
Задний грудной
(%О2)(%СО2)
17,5 А.З
17, А 3,6
Легкое (?) 3,9 (?)
[Передний грудной ~| 14,1 6,8
Передние ч мешки
| Межключичный ]
| Латеральный ключичный] 13,2
_________I_________I_________
Рис. 2.16. Когда страусу дают один раз вдохнуть чистый кислород, а затем он дышит обычным воздухом, кислород играет роль метки. Раньше всего он обнаруживается в задних воздушных мешках. Только во время второго или третьего цикла дыхания этот газ появляется в передних мешках. Частота дыхания— примерно 6 циклов в минуту. Обычный состав газа в воздушных мешках указан справа. (Schmidt-Nielsen et al., 1969.)
безвреден и концентрацию его легко регистрировать с помощью чувствительного к О2 электрода. Такие эксперименты удобно проводить на страусе, так как он дышит очень медленно (один цикл длится около 10 с) и поэтому легко прослеживать изменения в составе газа.
Когда страусу дают один раз вдохнуть вместо воздуха чистый кислород, к концу вдоха этот газ появляется в задних воздушных мешках (рис. 2.16). Это должно означать, что вдыхаемый кислород проходит туда прямо через главный бронх. Между тем в передних мешках концентрация кислорода во время первого вдоха никогда не возрастает. Эти мешки, однако, при вдохе тоже раздуваются — значит, они получают воздух откуда-то еще. К концу второго вдоха, во время которого птица вдыхает уже обычный воздух, количество кислорода в передних мешках начинает нарастать. Это может означать только то, что кислород, который теперь здесь появляется, раньше находился в какой-то другой части дыхательной системы (по-видимому, в задних мешках) и прошел в передние мешки через легкие.
Дыхание птиц 77
Представляют интерес концентрации газов, измеренные в воздушных мешках. Задние мешки содержат около 4% СО2 (см. рис. 2.16), а количество О2 по сравнению с атмосферным воздухом уменьшено здесь на ту же величину — с 21 до 17%. Однако в передних мешках концентрация СО2 выше — между 6 и 7% — и количество кислорода соответственно снижено до 13 или 14%. Эти различия в газовом составе могли бы навести на мысль, что задние мешки лучше вентилируются, а передние содержат более застойный воздух, в котором поэтому накапливаются более высокие концентрации двуокиси углерода. Однако такой вывод был бы неверным.
Вводя маркирующий газ, можно выяснить, насколько быстро обновляется воздух в мешках. После введения маркера непосредственно в мешок концентрация его понижается ступеньками, которые совпадают с последовательными дыхательными циклами, и скорость этого понижения характеризует быстроту обновления воздуха. Время, необходимое для того, чтобы количество газа-маркера уменьшилось до половины исходного уровня, можно назвать периодом полуобновления. У страуса этот период составлял от двух до пяти дыхательных циклов как для передних, так и для задних мешков; это означает, что обе группы мешков вентилируются примерно одинаково. В аналогичных опытах на утках тоже были найдены примерно равные периоды полуобновления для передних и задних мешков (Bretz, Schmidt-Nielsen, 1972). Поскольку эксперименты с прослеживанием потоков газа показали, что воздух проходит в передние мешки из легких (концентрация СО2 в них совместима с таким предположением), и поскольку обновление воздуха в них велико, можно заключить, что эти мешки служат резервуарами для приема из легких того воздуха, который должен быть выведен наружу при следующем выдохе.
Путь воздуха можно также определить, помещая в различных проходах миниатюрные датчики, чувствительные к движению воздуха. На рис. 2.17 схематически представлены результаты таких исследований, проведенных на утках. Здесь показано, как могла бы перемещаться некоторая порция воздуха. Во время вдоха (а) большая часть воздуха идет прямо в задние мешки. Хотя передние мешки при вдохе расширяются, они совсем не получают вдыхаемого наружного воздуха; вместо этого в них поступает воздух из легких. При выдохе (б) воздух из задних мешков не выходит через главный бронх, а направляется в легкие. При следующем вдохе (в) воздух из легких переходит в передние мешки, и наконец при втором выдохе (г) воздух из передних мешков выходит прямо наружу. Для того чтобы провести данную порцию газа через всю дыхательную систему, требуются два полных дыхательных цикла. Сказанное выше не означает, что эти два цикла чем-то отличаются друг от друга: они совершенно идентичны, и за
78 Глава 2. Дыхание в воздухе
каждой порцией газа следует другая порция, повторяющая тог же путь на один цикл позже.
Наиболее примечательная особенность такой системы — то,, что воздух всегда течет через легкие по направлению от задней их области к передней и что воздух проходит через легкие как при вдохе, так и при выдохе.
Рис. 2.17. Движение газа, поступившего за один вдох, через дыхательную систему птицы. Прохождение газа по всему пути занимает два полных цикла дыхания. (Bretz, Schmidt-Nielsen, 1972.)
Такой механизм весьма выгоден для газообмена между воздухом и кровью, так как он в принципе сходен с противоточным механизмом в жабрах рыб (см. рис. 1.4). Благодаря ему в оксиге-нированнной крови, покидающей легкие, парциальное давление кислорода может быть больше, чем в выдыхаемом воздухе. Это поясняет сильно упрощенная схема на рис. 2.18. Кровь, которая должна вот-вот покинуть легкие (на правой стороне схемы) обменивается с воздухом, который только что вошел в легкие ив задних мешков и в котором парциальное давление О2 еще не снизилось. По мере того как воздух проходит через легкие (на схеме— справа налево), он теряет кислород и насыщается двуокисью углерода. На всем своем пути этот воздух встречает кровь со все более низким напряжением О2 и поэтому отдает ей все больше и больше кислорода. Благодаря этому кровь может хорошо насыщаться кислородом и способна извлекать из легочного воздуха больше кислорода и отдавать в легких больше двуокиси углерода, чем у млекопитающих.
Дыхание птиц 79
На самом деле в легких птиц существует не идеальная противоточная система, а скорее система с поперечными потоками (рис. 2.19). Что касается напряжения О2 и СО2 в артериальной крови, то эта система дает результат, аналогичный описанному
Рис. 2.18. Газообмен в легком птицы на большой высоте. (Schmidt-Nielsen, 1972.)
На этой сильно упрощенной схеме показаны потоки крови и воздуха, проходящие через легкое в противоположных направлениях. Такой характер потоков позволяет оксигенированной крови иметь на выходе из легкого максимально возможное напряжение О2.
выше, но она не так эффективна, как настоящий противоточный газообменник.
Эффективность однонаправленного потока воздуха в легких птиц особенно существенна на больших высотах. Когда мышей и воробьев помещали в камеру с давлением воздуха 350 мм рт. ст. (чуть меньше 0,5 атм), соответствующим высоте 6100 м, мыши ле-
Рис. 2.19. В легком птицы кровь течет не по параллельным капиллярам, а скорее по сложной, мало упорядоченной сети. На этой упрощенной схеме показано, что выходящая из легкого кровь представляет собой смесь потоков, прошедших через разные части легкого и насыщенных кислородом в неодинаковой степени. Такую организацию можно назвать схемой с перекрестными потоками. (Scheid, Piiper, 1972.)
80 Глава 2. Дыхание в воздухе
жали на животе и едва могли ползать, в то время как воробьи все еще были способны летать (Tucker, 1968). У мышей и воробьев одинаковый вес тела, кровь имеет одно и то же сродство к кислороду и интенсивность обмена различается незначительно, так что наблюдаемую разницу нельзя связать с уровнями потребления О2 или с химическими свойствами крови. Наиболее правдоподобное объяснение — иная схема потоков воздуха в птичьих легких: она дает возможность крови извлекать кислород из воздуха, в котором его концентрация выше, чем в дыхательной системе млекопитающих, и вдобавок кровь извлекает его более эффективно благодаря однонаправленности и непрерывности тока воздуха через легкие. Эти эксперименты позволяют понять, почему в Гималаях можно видеть парящих над головой птиц в таких местах, где альпинисты без кислородных приборов едва способны ходить.
ДЫХАНИЕ ПТИЧЬИХ ЯИЦ
Проблема снабжения кислородом растущих эмбрионов и птенцов внутри твердой скорлупы птичьих яиц составляет интересную главу физиологии дыхания. Большая часть работ в этой области проведена на куриных яйцах, но основные принципы, вероятно, приложимы к любым птичьим яйцам, если не считать особенностей, связанных с колоссальными различиями в их размерах. Самое маленькое яйцо — яйцо колибри — весит менее 0,3 г, а яйцо страуса весит больше 1 кг. Самые крупные из известных яиц, принадлежащие ископаемой птице Aepyomis, весят в среднем около 10 кг. Таким образом, соотношение весов может превышать 40 000: 1.
Скорлупа яйца имеет твердый наружный слой из карбоната кальция, к которому с внутренней стороны примыкают две мягкие мембраны, называемые внешней и внутренней подскорлупо-выми оболочками. Твердый слой намного хуже пропускает газы, чем подскорлуповые оболочки, которые тоньше и гораздо более проницаемы. На тупом конце яйца имеется воздушная камера, которая располагается между внутренней и наружной подскорлупо-выми оболочками. Это воздушное пространство постепенно увеличивается, так как яйцо теряет воду в результате испарения.
В твердой скорлупе среднего куриного яйца имеется около 10 000 пор. Так как площадь поверхности яйца около 70 см2, это значит, что на 1 мм2 приходится в среднем по 1,5 поры. Диаметр каждой поры около 0,017 мм, так что общая площадь пор составляет 2,3 мм2 (Wangensteen et al., 1971). Весь газообмен между эмбрионом и окружающей атмосферой должен осуществляться через эти поры.
Потребление кислорода и выделение СО2 на протяжении всего периода инкубации (около 21 дня) возрастают. Поскольку диф
Дыхание птиц
фузионные характеристики яичной скорлупы остаются постоянными или почти постоянными, должны постепенно увеличиваться газовые градиенты между атмосферным воздухом и эмбрионом: парциальное давление О2 внутри яйца должно понижаться, а дав* ление СО2 — возрастать (рис. 2.20).
Рис. 2.20. В период инкубации куриного яйца в его воздушной камере концен-трация кислорода постепенно снижается, а концентрация СО2 соответственно* нарастает. (Romijn, Roos, 1938.)
В последний день инкубации положение резко меняется. Примерно за 28 ч до вылупления птенец прокалывает мембрану воздушной камеры, включается функция легких, и птенец начинает дышать воздухом из воздушного пространства (которое благодаря испарению воды увеличилось в объеме более чем до 10 см3). Спустя примерно 12 ч птенец начинает разбивать скорлупу (эту стадию называют проклевыванием), и главный барьер для газообмена теперь отпадает. В последующие часы — в период, когда птенец старается полностью освободиться, — газообмен уже всецело осуществляется через легкие.
На больших высотах, где давление О2 снижено, снабжение эмбрионов кислородом становится серьезной проблемой. Этот вопрос изучали на колонии белых леггорнов, которую содержали многие годы на исследовательской станции Уайт-Маунтэн в Калифорнии на высоте 3800 м (барометрическое давление 480 мм рт. ст., или 64 кПа). Когда колонию начинали создавать, цыплята выводились -только из 16% оплодотворенных яиц, тогда как на уровне моря они обычно выводятся из 90%. В дальнейшем выход цыплят постепенно увеличивался и после восьми поколений достиг максимума где-то около 60%.
6—1873
82 Глава 2. Дыхание в воздухе
Одно из следствий пониженного атмосферного давления — увеличение коэффициента диффузии для газов. На высоте 3800 м коэффициент повышен несколько больше, чем в полтора раза, и это способствует снабжению эмбриона кислородом. Однако при низком давлении повышается и коэффициент диффузии для водя-
Рис. 2.21. Трахейная система насекомого (блохи) состоит из тонких наполненных воздухом трубочек (трахей), по которым дыхательные газы транспортируются ко всем частям тела и обратно наружу. Трахеи сообщаются с наружным воздухом с помощью дыхалец. (Wigglesworth, 1972.)
ного пара, и это увеличивает опасность высыхания.
Яйца, откладываемые на большой высоте, мельче, а время инкубации больше, чем на уровне моря. Площадь пор на единицу поверхности скорлупы уменьшена, хотя толщина скорлупы не изменена. Уменьшение площади пор в сочетании с увеличением коэффициента диффузии газов обеспечивает примерно такую же проницаемость для газов, как на уровне моря. Таким об
разом, мы можем видеть, что уменьшение общей площади пор у яиц, откладываемых на большой высоте, служит необходимой адаптацией, поскольку иначе потери воды оказались бы
чрезмерными (Wangensteen et al., 1974).
ДЫХАНИЕ НАСЕКОМЫХ
Условия наземной жизни беспрерывно создают конфликт между потребностью в кислороде и потребностью в воде. То, что благоприятствует проникновению кислорода, способствует и потере воды. Насекомые — самая процветающая группа обитателей суши— имеют твердую кутикулу, которая очень мало проницаема для газов и покрыта восковым слоем, делающим ее практически непроницаемой также и для воды. Газообмен осуществляется через систему внутренних воздухоносных трубочек — трахей, которые сообщаются с наружной средой при помощи отверстий, называемых дыхальцами (рис. 2.21). Дыхальца обычно могут закрываться, что позволяет строго контролировать газообмен между воздухом в трахейной системе и наружной атмосферой. Трубочки ветвятся и заходят во все части тела. Самые ,тонкие веточки — трахеолы — имеют в диаметре около 1 мкм и менее и могут даже входить в отдельные клетки, например в мышечные волокна (фото 2.3).
Дыхание насекомых 83*
Трахейная система доставляет кислород прямо к тканям, а. двуокись углерода выводит в обратном направлении. Благодаря этому дыхание насекомых не зависит от системы кровообращения. Кровь у насекомых в отличие от крови позвоночных не играет роли в транспорте кислорода. Сравнение с позвоночными вполне уместно, так как, например, величины потребления кислорода у
Фото 2.3. Трахеолы насекомого. Трахеолы в тимпанальной мышце цикады Tibi-сеп sp. (Фото предоставлено David S. Smith, университет Майами.)
Почти все трахеолы (Тр) видны в поперечном разрезе; только одна срезана тангенциально, и здесь можно видеть спиральные утолщения ее стенки. Диаметр-трахеол около 0,5 мкм. Расстояние между двумя трахеолами, обозначенными Тр,. около 6,5 мкм (т. е. примерно соответствует диаметру эритроцитов человека). Светлые участки (М) — мышечные волокна в поперечном сечении; темные участки — митохондрии.
крупных бабочек и колибри во время полета близки, хотя механизмы снабжения кислородом различаются радикально.
Из этого не следует заключать, что циркуляторная система для насекомых не важна. Она выполняет много других функций. Одна ее роль очевидна: во время полета мышцы развивают большую мощность, и кислород для этого подводится через трахейную систему, а топливо должно доставляться кровью.
6
84 Глава 2, Дыхание в воздухе
ТРАХЕЙНАЯ СИСТЕМА
Трахеи приспособлены главным образом для дыхания в воздухе и должны были развиться в воздушной среде. Насекомые, вторично перешедшие к водному образу жизни, в принципе сохраняют воздушный тип дыхания, но их трахейные системы имеют много интересных особенностей, которые делают их пригодными для газообмена в воде.
Трахейная система может существенно отличаться от основного типа, для которого характерны 12 пар дыхалец — 3 пары на груди и 9 пар на брюшке. Часто дыхалец бывает меньше, и они могут даже совсем отсутствовать. Некоторые из возможных вариаций показаны на рис. 2.22.
Рис. 2.22. Основная схема строения трахейной системы насекомого (А) может подвергаться разнообразным видоизменениям (Б — Е). (Wigglesworth, 1972.) Объяснения в тексте.
В типичной системе крупные трахеи соединяются таким образом, что вся система оказывается состоящей из взаимосвязанных трубочек (рис. 2.22, Д). Часто к этому добавляются расширенные участки трахей, или воздушные мешки, которые могут сжиматься (Б). Объем этих мешков может изменяться при движениях тела, что ведет к насасыванию воздуха в трахейную систему или к его изгнанию. Это существенно, так как у крупных и очень активных насекомых одна лишь диффузия не обеспечила бы достаточного газообмена. Сами трахеи имеют внутри спиральные ребра и почти несжимаемы; таким образом, большие мешки необходимы для вентиляции. Если дыхальца будут открываться и закрываться синхронно с дыхательными движениями, но в противофазе по отношению друг к другу, можно получить однонаправленный поток воздуха через более крупные трахейные стволы.
Дыхание насекомых 85
На рис. 2.22, В показана модификация, характерная для многих водных насекомых. Большая часть дыхалец не функционирует, и только два самых задних открыты наружу. Они расположены таким образом, что насекомое, проколов кончиком брюшка поверхность воды, может осуществить контакт с атмосферой и обеспечить газообмен либо путем диффузии, либо более эффективным способом — с помощью дыхательных движений.
Трахейная система может быть совсем замкнутой, без каких-либо отверстий на поверхности тела, хотя она и заполнена воздухом (рис. 2.22, Г). В этом случае газообмен должен полностью осуществляться с помощью диффузии через кутикулу. Для многих мелких водных насекомых такого способа вполне достаточно: их кутикула относительно тонка, и они могут оставаться под водой, не испытывая потребности в контакте с атмосферой. И все же трахейная система здесь необходима для транспорта газов внутри самого тела, так как диффузия через ткани шла бы слишком медленно. Как мы уже знаем, кислород диффундирует в воздухе примерно в 300 000 раз быстрее, чем в воде; это ясно указывает на преимущества трахейной системы с точки зрения транспорта газов, когда циркуляция крови не приспособлена для этой цели.
Следующее усовершенствование того же плана снова демонстрируют водные насекомые (рис. 2.22, Д). Ветви замкнутой трахейной системы заходят в абдоминальные придатки, или «жабры», которые, имея относительно большую поверхность и тонкую кутикулу, позволяют осуществлять эффективный газообмен между водой и воздухом трахейной системы. Такие трахейные жабры есть, например, у личинок поденок (Ephemeroptera).
Еще одна разновидность показана на рис. 2.22, Е. Трахейные жабры расположены в полости прямой кишки, и насекомое вентилирует их, набирая воду внутрь кишки и выталкивая ее обратно. Такая система имеется у личинок некоторых стрекоз. Эти личинки могут использовать такой механизм и для локомоции: быстро выталкивая воду из кишки, они передвигаются реактивным способом, как кальмары и осьминоги.
ДИФФУЗИЯ И ВЕНТИЛЯЦИЯ
Многим мелким насекомым и сравнительно неактивным крупным насекомым для газообмена в трахейных системах достаточно одной лишь диффузии. Примером может служить крупная гусеница древоточца Cossus, изученная Крогом (Krogh, 1920). Эта гусеница весит около 3,4 г, а длина ее 60 мм. На теле у нее имеется 9 пар дыхалец, и тщательное измерение трахей, идущих от этих дыхалец, показало, что суммарная площадь поперечного сечения всех таких трахей близка к 6,7 мм2. Средняя длина трахей 6 мм. Любопытно, что по мере того, как трахеи делятся и разветв'
86 Глава 2. Дыхание в воздухе
ляются, суммарная площадь поперечного сечения системы остается постоянной, не изменяясь с удалением от дыхалец. Таким образом, для анализа диффузии через всю трахейную систему ее можно представить одной цилиндрической трубкой с поперечным сечением 6,7 мм2 и длиной 6 мм. Потребляемый животным кислород (0,3 мкл О2 в 1 с) сможет продиффудировать через трубку таких размеров, если разность парциальных давлений О2 между двумя концами будет равна И мм рт. ст. (1,5 кПа). Это означает^ что при давлении О2 в атмосфере 155 мм рт. ст. (20,7 кПа) напряжение О2 в тканях сможет достигать 144 мм рт. ст. (19,2 кПа). Очевидно, что для гусеницы Cossus достаточное снабжение кислородом гарантируется одной диффузией, даже если интенсивность обмена во время активности нужно будет увеличить в несколько раз.
Интуитивно может показаться, что диффузия через 6 мм воздуха должна быть медленной; однако кислород диффундирует в. воздухе в 300 000 раз быстрее, чем в воде, так что диффузия через 6 мм воздуха идет так же быстро, как диффузия через слой воды толщиной 0,02 мкм. Таким образом, наибольший барьер для доставки кислорода тканям скорее всего находится между тончайшими веточками трахеол и клетками. В очень активных тканях, таких, как летательные мышцы, на электронных микрофотографиях видно, что трахеолы подходят к митохондриям на расстояние до 0,07 мкм.
Количественное изучение разветвленной трахейной системы дает сведения еще об одном аспекте газообмена. По мере того как трахеи делятся и ветвятся, суммарная площадь стенок системы увеличивается. У гусеницы Cossus наибольшая трахея имеет диаметр около 0,6 мм, а самая тонкая трахеола — 0,001 мм. Поскольку общая площадь поперечного сечения трубок остается постоянной, площадь стенок трахеол будет в 600 раз больше площади стенок самой крупной трахеи той же длины. Таким образом, благодаря одному только уменьшению диаметра поверхность стенок увеличивается в 600 раз. Из этого следует, что практически вся поверхность, участвующая в газообмене, сосредоточена в тончайших веточках. Если мы учтем еще, что толщина стенок трахеол составляет менее одной десятой толщины стенок трахей, нам станет очевидно, что диффузия через стенки более крупных ветвей должна быть очень незначительной.
ДЫХАЛЬЦА
Выходы трахейной системы наружу — дыхальца — это очень сложные устройства, которые могут открываться и закрываться, позволяя тем самым изменять скорость газообмена. Их точное регулирование помогает избежать потери воды.
Дыхание насекомых &Т
При высокой температуре и усиленной активности насекомого дыхальца открываются чаще и шире — в соответствии с повышенной потребностью в кислороде. Дыхальца не обязательно открываются все одновременно; они находятся под контролем центральной нервной системы, и их поочередное открывание и закрывание позволяет регулировать поток воздуха через трахейную систему.
На вентиляцию трахейной системы и особенно на работу ды-
Время, недели
Рис. 2.23. Потеря воды организмом мучного червя в сухом воздухе. В те периоды, когда дыхальца были открыты (в результате добавления СО2 к воздуху), шотери воды возрастали в несколько раз. (Mellanby, 1934.)
Главным стимулом для открывания дыхалец, по-видимому, служит СО2. Если тонкую струйку этого газа направить на одно дыхальце, только оно и откроется; это показывает, что отдельное дыхальце может реагировать независимо от других. Необходимая концентрация двуокиси углерода довольно мала; например, у таракана заметный эффект вызывает уже 1% СО2 в воздухе, 2% держат дыхальца открытыми, а 3% заставляют их широко открываться.
Открывание дыхалец оказывает существенное влияние на потери воды. У мучного червя, который всю свою жизнь проводит среди сухой муки, управление дыхальцами играет очень важную роль. Если удерживать его дыхальца в открытом состоянии, добавив к воздуху СО2, потери воды немедленно возрастают в несколько раз (рис. 2.23).
Двуокись углерода не единственный фактор, влияющий на дыхальца; их можно также заставить раскрываться с помощью чистого азота (стимулом здесь служит нехватка кислорода). Это тоже можно было бы интерпретировать как эффект СО2, поскольку в отсутствие кислорода часто происходит накопление кислых продуктов обмена (например, молочной кислоты), которые повышают напряжение СО2. Таким образом, действие аноксии может быть связано с косвенным влиянием двуокиси углерода. Однако дыхание стимулируется даже умеренным дефицитом кислорода, поэтому весьма вероятно, что кислород оказывает и прямое влияние.
88 Глава 2. Дыхание в воздухе
ВЕНТИЛЯЦИЯ
Трахеи относительно жестки и поэтому обычно не сжимаются, однако спиральные складки стенок позволяют им немного укорачиваться наподобие гармошки. Это может служить для уменьшения их объема, что способствует активной вентиляции системы. У трахей с овальным или удлиненным сечением сжимаемость больше. Однако эффективная вентиляция обеспечивается тонкостенными расширениями — воздушными мешками, соединенными с более крупными трахеями, поскольку при сжатии этих мешков может выталкиваться большой объем воздуха. Выдох обычно бывает активной фазой, а вдох — пассивной.
У многих насекомых основные дыхательные движения совершает брюшко. Во время полета вентиляцию могут усиливать движения груди и синхронные с ними изменения давления, так что благодаря передаче кровяного давления даже голова может до некоторой степени активно вентилироваться. Объем вентиляции может быть довольно большим: при одном выдохе может опорожняться до одной трети и даже до половины всей емкости воздушной системы, что приводит в последнем случае к обновлению примерно половины воздуха. Это намного больше, чем у млекопитающих в покое, где за один цикл дыхания обновляется около одной десятой всего воздуха, содержащегося в системе, хотя максимальное возможное обновление близко к двум третям общего объема этой системы.
Дыхательные движения синхронны с открыванием и закрыванием дыхалец и тоже контролируются недостатком кислорода или избытком СО2. Избыток кислорода может полностью остановить дыхательные движения; поскольку высокая концентрация кислорода не должна влиять на образование СО2, кислород, по-види-мому, действует непосредственно на нервную систему.
Дыхательные движения управляются центрами в сегментированной брюшной нервной цепочке, и их могут выполнять даже изолированные сегменты. Координация между сегментами осуществляется высшими центрами, находящимися в протораксе. Голова, по-видимому, в этом не участвует.
У насекомых вообще меньше высших центров в голове, и декапитация почти не влияет на дыхание и многие другие функции.
ДЫХАНИЕ У ВОДНЫХ НАСЕКОМЫХ
Насекомые, живущие в воде, произошли от наземных форм и сохранили многие черты своих сухопутных предков. Почти все они пресноводные; очень немногие живут в солоноватой воде, а настоящих морских насекомых практически нет.
Некоторые водные насекомые сохранили воздушный тип дыхания через дыхальца (фото 2.4). Число дыхалец в этих случаях обычно меньше, и они располагаются на заднем конце тела, кото
Дыхание насекомых 89
рый приводится в контакт с атмосферным воздухом. Края дыха-лец гидрофобны и часто окружены гидрофобными волосками. Благодаря этому водная поверхность легко прорывается и дыхальце устанавливает контакт с воздухом.
Мелкие насекомые (например, личинки комаров) обходятся диффузионным газообменом между атмосферой и трахейной си-
Фото 2.4. Многие водные насекомые дышат воздухом. Личинка комара (слева) прокалывает поверхностную пленку воды с помощью дыхальцевой трубочки, которая открывается на кончике брюшка. Листовидные структуры справа — анальные папиллы, главная функция которых состоит в осморегуляции. Пока путем диффузии осуществляется газообмен, такая личинка неподвижно висит у поверхности. Жук-плавунец (справа) устанавливает контакт с поверхностью лишь на короткое время, чтобы только втянуть воздух в пространство под крыльями. В это воздушное пространство открываются дыхальца, и газообмен в трахеях продолжается под водой. (Фото предоставлены Thomas Eisner, Кор-нельский университет.)
стемой, но у более крупных личинок обновлению трахейного воздуха содействуют дыхательные движения. Трахеи часто весьма объемисты, так что находящийся в них воздух содержит достаточный запас кислорода для длительного пребывания под водой. Однако такое решение проблемы имеет жесткие пределы, так как
9Э Глава 2. Дыхание в воздухе
увеличение запасов воздуха приведет в конце концов к слишком большой плавучести и насекомое не сможет погружаться в воду..
НАРУЖНЫЕ ЗАПАСЫ КИСЛОРОДА
Многие водные насекомые несут запас воздуха на поверхности своего тела, и их дыхальца открываются в эту воздушную прослойку. Воздух удерживается с помощью несмачиваемых поверхностей, часто снабженных гидрофобными волосками. Взрослые жуки-плавунцы (Dytiscus) имеют большое воздушное пространство под крыльями; когда жук устанавливает контакт с водной поверхностью, это пространство вентилируется, а при погружении оно служит газовым резервуаром, в который открываются дыхальца. Аналогичный способ использует клоп-гладыш (Notanec-ta), который имеет запас воздуха на вентральной поверхности брюшка. Довольно большой воздушный пузырек придает клопу положительную плавучесть, так что ему приходится энергично плыть, чтобы удалиться от поверхности, и прицепляться к растениям или другим твердым объектам, чтобы оставаться под водой.
С помощью простого эксперимента можно показать, что воздушный пузырек нужен гладышу для дыхания и не имеет отношения к проблеме плавучести. Если клопа держать погруженным в воду, которая находится в равновесии не с воздухом, а с чистым азотом, он проживет всего лишь 5 минут. Если его держать в воде, насыщенной воздухом, он будет жить 6 часов. Если же его поместить в воду, находящуюся в равновесии с 100% О2 вместо воздуха, и перед погружением позволить ему наполнить свое воздушное пространство кислородом, он проживет всего лишь 35 минут (Egey 1915). Этот кажущийся парадокс проистекает из того, что пузырек служит двум целям: он содержит запас кислорода, который постепенно используется, и, кроме того, он играет роль диффузионной жабры, в которую кислород должен диффундировать из окружающей среды.
Посмотрим, что происходит в каждом случае. В первом опыте насекомому позволяют погрузиться с пузырьком атмосферного воздуха в воду, лишенную кислорода. Кислород из пузырька исчезает, потому что часть его используется организмом и еще большая часть быстро диффундирует в окружающую воду, не содержащую О2, так что через 5 минут кислорода больше не остается.
Во втором опыте насекомое также уносит с собой пузырек воздуха. Через некоторое время в результате потребления насекомым кислорода его содержание в пузырьке снизится, скажем, до 5%, тогда как содержание СО2 увеличится до 1%. Остальной газ в пузырьке (94%) —азот. Поскольку парциальное давление О2 в пузырьке теперь меньше, чем в воде, кислород будет диффундировать из воды в пузырек. С другой стороны, азота в пузырьке 94%,
Дыхание насекомых 91
а вода уравновешена нормальной атмосферой с 79% азота. Значит, парциальное давление азота в пузырьке выше, чем в воде; поэтому азот будет диффундировать наружу, объем пузырька постепенно будет уменьшаться, и в конце концов пузырек исчезнет (в описанном опыте он просуществовал 6 часов).
В третьем опыте и пузырек, и вода содержат только кислород. Пузырек уменьшается в соответствии с тем, как насекомое расходует кислород. Хотя вода насыщена кислородом, это не помогает, так как парциальные давления О2 в пузырьке и в воде одинаковы и добавочный кислород из воды не диффундирует. Поэтому пузырек исчезает намного раньше, чем при наличии также и азота, как во втором опыте.
Пузырек воздуха обладает одним невыгодным свойством: время его существования зависит от глубины, на которую опустилось насекомое. С увеличением глубины общее давление внутри пузырька возрастает, тогда как парциальные давления газов, растворенных в воде, остаются неизменными; поэтому пузырек исчезает быстрее. С каждым метром глубины парциальное давление азота в пузырьке увеличивается, и это ускоряет его диффузию в толщу воды.
На глубине 10 м общее давление погруженной массы воздуха равно 2 атм. При таком давлении газ быстро переходит в воду, поэтому на столь большой глубине воздушная масса может функционировать лишь очень короткое время. Поэтому насекомые, уносящие с собой пузырьки воздуха, могут погружаться лишь на ограниченную глубину. Они должны очень часто возвращаться на поверхность для обновления воздуха, и расстояние, которое им приходится преодолевать, требует дополнительной работы и расхода кислорода; общее время, проводимое на дне, должно быть коротким.
Для насекомого, которое использует пузырек воздуха как диффузионную жабру, общее количество доступного кислорода намного превышает исходный запас, содержавшийся в пузырьке при погружении. Азот выходит из пузырька медленнее, чем входит внутрь кислород, — главным образом потому, что из-за своей меньшей растворимости азот диффундирует между воздухом и водой вдвое медленнее, чем кислород. Благодаря начальному запасу азота воздушная масса существует настолько долго, что насекомое получает путем диффузии из окружающей воды в восемь раз больше кислорода, чем его первоначально содержалось в пузырьке. Это отношение 8 : 1 имеет место всегда — независимо от интенсивности метаболизма у насекомого, величины газообменной поверхности, начального объема воздушной массы и толщины пограничного слоя воды (в котором на количество доступного кислорода не будет влиять турбулентность или активная вентиляция) (Rahn, Paganelli, 1968).
92 Глава 2, Дыхание в воздухе
ПЛАСТРОН
Другой тип дыхательного устройства — это пластрон, У дышащих с помощью пластрона насекомых часть поверхности тела густо покрыта гидрофобными волосками, которые создают несма-чиваемую поверхность, удерживающую воздух. Это приспособление позволяет насекомому оставаться под водой неограниченное время. У клопа Aphelocheirus на каждом квадратном миллиметре поверхности пластрона имеется по 2 миллиона волосков длиной 5 мкм (Thorpe, Crisp, 1947). Насекомые, дышащие с помощью пластрона, имеют огромное преимущество перед теми, которые уносят с собой в качестве диффузионной жабры одиночный пузырек, так как вода не может проникать между гидрофобными волосками. В результате пластрон работает как несжимаемая жабра, в которую из воды диффундирует кислород. Поскольку объем остается постоянным, результирующая диффузия азота в такой жабре должна быть равна нулю и общее давление газа в воздушном слое должно быть отрицательным по отношению к напряжениям газов в окружающей воде. Это возможно благодаря тому, что поверхностная пленка воды подпирается гидрофобными волосками, и чтобы загнать воду в воздушные пространства между ними, требуется давление в 3,5—5 атм.
ВНУТРЕННИЙ ЗАПАС КИСЛОРОДА
Полужесткокрылые насекомые Виепоа и Anisops — родственники гладыша (Notonecta) — примечательны тем, что могут почти без усилий оставаться взвешенными в воде в течение нескольких минут. Это крайне необычно для насекомых и позволяет этим хищным клопам использовать среднюю зону воды в водоемах, бедных кислородом. В небольших временных водоемах, где нет рыб, средняя зона относительно свободна от хищников. Придонный слой опасен из-за прожорливых личинок стрекоз и жуков, другие хищники охотятся у поверхности, а в средних слоях и безопасно, и почти нет конкуренции из-за пищи.
И у Виепоа, и у Anisops в брюшке имеются крупные клетки, заполненные гемоглобином. Они служат для запасания кислорода, который расходуется во время пребывания под водой. Насекомые захватывают с собой лишь один небольшой пузырек воздуха, но пока используется кислород из гемоглобина, этот наружный запас расходуется очень медленно. Когда же кислород гемоглобина иссякает, внешний кислород быстро потребляется и насекомое теряет плавучесть, становясь тяжелее воды.
Долгосрочный запас кислорода дает этим насекомым некоторое преимущество. После возобновления запаса Ог у поверхности воды они активно уплывают вниз и затем долго остаются взвешенными в средних слоях воды в состоянии почти нейтральной плавучести, пока не израсходуется кислород. Насекомое снова
Дыхание насекомых 93
возвращается на поверхность только тогда, когда оно частично» использует наружный запас кислорода и начнет тонуть (Miller, 1964).
Если клопа Artisops поместить в воду, лишенную кислорода, время пребывания под водой сократится незначительно. Это показывает, что сравнительно небольшой пузырек воздуха здесь не-играет роли жабры, как у Notonecta. Но если с помощью окиси углерода вывести из строя гемоглобин, время погружения сильно* уменьшится, так как гемоглобин теперь не будет нести кислород. Даже если насекомое захватит с собой нормальный пузырек воздуха, это ничего не изменит: обычнный запас кислорода отсутствует, а наружная масса воздуха заметным образом не помогает.
ПРЕРЫВИСТОЕ, ИЛИ ЦИКЛИЧЕСКОЕ, ДЫХАНИЕ
Многим насекомым свойствен особый феномен, характеризующийся периодическим (циклическим) выделением больших количеств СОг. Кислород поглощается с более или менее постоянной скоростью, но двуокись углерода выводится лишь в течение коротких периодов, как бы залпами. Залпы выделения СО2 могут' длиться несколько минут и регулярно повторяться через длительные интервалы (до нескольких часов и даже дней). Запись такого залпа у куколки одного шелкопряда показана на рис. 2.24. По-
Рис. 2.24. Запись газообмена у куколки цекропийного шелкопряда (при 20 °C)’4 демонстрирует непрерывное поглощение кислорода. Сопутствующее выделение СО2 ничтожно, и только раз в несколько часов оно осуществляется большим* «залпом». (Punt et al., 1957.)
добные залпы могут возникать с частотой от одного раза в неделю до многих раз в течение часа, в зависимости от температуры* и интенсивности обмена веществ.
Циклическое дыхание наблюдали у тараканов, кузнечиков,, пчел, гусениц и куколок дневных и ночных бабочек, а также у*
94 Глава 2. Дыхание в воздухе
взрослых бабочек в период диапаузы. Явление это довольно обычное, но, по-видимому, оно характерно для насекомых с малой интенсивностью обмена. У некоторых насекомых его можно вызвать снижением температуры, приводящим к замедлению метаболических процессов (Schneiderman, 1960).
Количество двуокиси углерода, выделяемое в промежутках между залпами, ничтожно; кислорода за это время поглощается во много раз больше — в некоторых случаях в 20, а иногда и в 100 раз. Очевидно, что газообмен осуществляется через дыхальца, а не через кутикулу, так как, если все дыхальца залепить воском, не наблюдается ни поглощения кислорода, ни выделения СОг (т. е. кутикула почти непроницаема для газов). Это означает, что в течение межзалповых интервалов дыхальца позволяют кислороду проникать внутрь, практически не выпуская двуокись углерода наружу. Как это может быть? Этот и многие другие вопросы еще остаются без ответа.
КОЛИЧЕСТВО ВЫДЕЛЯЕМОЙ ДВУОКИСИ УГЛЕРОДА
У куколки цекропийного шелкопряда, которая весит 6 г, в течение одного залпа выделяется более 0,5 см3 СО2. Это больше всего объема трахейной системы, так что даже если бы все трахеи были заполнены только двуокисью углерода, то и тогда пришлось бы допустить, что во время короткого залпа этот газ выходит также из крови и тканей.
Точные измерения, произведенные на куколке другой бабочки, Agapema, показали, что объем трахей у нее составляет 60 мм3. Непосредственно перед залпом воздух трахей содержал 5,9% СО2. Это означает, что к началу залпа в трахейной системе находилось 3,5 мм3 СО2. Однако во время залпа выделилось 30 мм3 СО2; другими словами, только -10% двуокиси углерода могло выйти из воздуха трахей, остальная часть должна была выделиться еще откуда-то.
НЕТ ЛИ ПЕРИОДИЧНОСТИ В САМОМ МЕТАБОЛИЗМЕ?
Запись на рис. 2.24 показывает, что хотя поглощение кислорода идет непрерывно и в промежутках между залпами, во время залпа оно все же усиливается. В связи с такой периодичностью выделения СО2 и поглощения О2 возникает вопрос, не протекает ли обмен веществ у куколки циклически, с короткими «взрывами» образования СО2 и долгими периодами почти полной остановки метаболических процессов.
На этот вопрос можно ответить с помощью простого эксперимента. Если одно из дыхалец держать постоянно открытым, вставив в отверстие тонкую стеклянную трубочку, феномен цикличности исчезает. Следовательно, мы можем отвергнуть гипотезу о пе
Дыхание насекомых 9f>
риодичности биохимических процессов, связанных с обменом веществ. Теперь становится очевидной необходимость исследовать связи с этими циклами функцию дыхалец.
ДВИЖЕНИЯ ДЫХАЛЕЦ
Прямое наблюдение дыхалец с помощью микроскопа показывает, что во время залпов выведения СО2 они широко открыты. После залпа клапан каждого дыхальца некоторое время вибрирует, а затем полностью закрывается и остается закрытым примерно в течение часа. Затем клапан начинает слабо пульсировать или. вибрировать, слегка расширяя отверстие дыхальца. Вибрация продолжается иногда несколько часов, пока не наступит период, широкого раскрытия и залпа СО2. На рис. 2.25 показаны движения дыхалец у куколки цекропийного шелкопряда. Верхняя за-
Рис. 2.25. Запись движений дыхалец у цекропийного шелкопряда. (Schneiderman, 1960.)
Когда куколка находится в чистом кислороде (Л), вибраций перед «залпом» не наблюдается. Когда куколка находится в атмосферном воздухе (5), «залпу» предшествуют вибрации. Когда кислорода всего 10% (В), дыхальца вибрируют' непрерывно.
пись (Л) была получена на насекомом, помещенном в чистый кислород. После долгого пребывания в закрытом состоянии дыхальца открываются, остаются открытыми примерно 4 мин и затем снова закрываются. В атмосферном воздухе (5) после долгого* пребывания в закрытом состоянии дыхальца примерно 10 мин: вибрируют, прежде чем открыться полностью; затем они сновав вибрируют некоторое время, постепенно закрываются и остаются*
<96 Глава 2. Дыхание в воздухе
закрытыми до начала следующего периода вибраций. Третий график (В) показывает, что 10%-ная концентрация Ог заставляет дыхальца вибрировать непрерывно.
Во время долгого периода вибраций, когда дыхальца слегка приоткрыты, выведение двуокиси углерода ничтожно, но кислород поступает почти непрерывно. Таким образом, мы встречаемся с непонятным фактом: когда дыхальца вибрируют, скорость поступления О2 может в 100 раз превышать скорость выхода СО2. Каким образом дыхальца — единственные пункты газообмена — позволяют входить кислороду, но эффективно препятствуют выходу двуокиси углерода? Чтобы ответить на этот вопрос, нам нужны •сведения о концентрации газов внутри трахейной системы.
Насекомые, о которых шла речь, обычно не производят видимых дыхательных движений, и это позволяет предположить, что газообмен между трахейной системой и наружным воздухом осуществляется у них путем диффузии. Однако реальные концентрации газов, обнаруженные в трахейной системе, нельзя объяснить •одними лишь процессами диффузии.
Шнейдерману и его сотрудникам удалось осуществить отбор крохотных проб газа для анализа (рис. 2.26). Как и ожидалось,
'Рис. 2.26. Состав воздуха, взятого из трахейной системы в разные фазы циклического дыхания. Резкие подъемы концентрации О2 соответствуют моментам, когда дыхальца полностью открываются во время «залпа». Вес куколки 8,4 г; температура 25 °C. (Levy, Schneiderman, 1958.)
наименьшую концентрацию СО2 (около 3%) они обнаружили сразу после залпа, и в последующие 6 ч эта концентрация очень медленно возрастала до 6,5%. Концентрация кислорода во время залпа увеличивалась примерно до атмосферного уровня (18—20%), но после залпа быстро снижалась и оставалась постоянной на уровне 3,5% на протяжении всего межзалпового периода. Эти •удивительные цифры нельзя объяснить, если единственной движу
Дыхание насекомых 97
щей силой для движения газов через дыхальца является диффузия. Однако газы могли бы также перемещаться под действием давления, и Бак (Buck, 1958) предположил, что к интенсивному входу воздуха через дыхальца могло бы приводить небольшое отрицательное давление в трахейной системе.
Давление внутри трахейной системы можно измерить, если вставить в одно из дыхалец тонкий стеклянный капилляр и соединить его с манометром, оставив все остальные дыхальца интактными. Во время периодов вибраций давление в трахейной системе очень -близко к атмосферному, но слегка сдвинуто в отрицательную сторону. Этот небольшой отрицательный сдвиг вызывает интенсивный ток воздуха внутрь. Когда дыхальца закрываются полностью, давление в трахейной системе резко падает. Пока дыхальца остаются закрытыми, развивается значительное отрицательное давление, соответствующее быстрому спаду концентрации кислорода внутри трахейной системы сразу после закрытия дыхалец (рис. 2.26). Когда вибрации начинаются снова, давление постепенно нарастает, а когда они становятся регулярными, давление опять возвращается к уровню, близкому к атмосферному (Levy, Schneiderman, 1966b).
Быстрый спад давления при закрытых дыхальцах вызывается быстрым поглощением кислорода из замкнутой трахейной системы. Большая часть образующейся двуокиси углерода остается растворенной или забуференной в крови и тканях, и поэтому должно создаваться отрицательное давление. Если перед закрытием дыхалец поместить куколку в чистый кислород, чтобы трахейная система заполнилась одним только кислородом, развивается еще большее отрицательное давление. Именно этого и следует ожидать, когда кислород потребляется, а азот отсутствует.
Если мы объединим все имеющиеся данные, мы увидим, что они совместимы со следующей интерпретацией (рис. 2.27). На верхнем графике (рис. 2.27, А) показаны вибрации, за которыми следует период открытых дыхалец, а затем — период полностью закрытых дыхалец, после которого вибрации возобновляются. При закрытых дыхальцах и во время вибраций выход двуокиси углерода (Б) мал; большая часть СО2 выводится коротким залпом в период открытых дыхалец. Поглощение кислорода на записи выглядит как непрерывное; оно соответствует действительному потреблению О2 тканями и может быть измерено с помощью подходящих манометрических методов (Schneiderman, Williams, 1955).
Данные о составе трахейного воздуха (рис. 2.27, В) показывают, что концентрация двуокиси углерода падает во время залпа с 6,5 до 3% и снова медленно повышается в течение межзалпового периода. Изменения концентрации кислорода более резки. Когда дыхальца открываются, количество О2 быстро нарастает до 18%, что мало отличается от атмосферной концентрации. В тече
7—1 873
98 Глава 2. Дыхание в воздухе
ние последующего периода закрытых дыхалец концентрация Ог в трахеях неуклонно снижается, поскольку ткани потребляют кислород с постоянной скоростью. К моменту, когда начинаются вибрации, внутри трахейной системы уже создается отрицательное давление, и воздух теперь устремляется внутрь. Это ведет к час-
Рис. 2.27. Сводный график, отражающий события, происходящие синхронно с: движениями дыхалец при циклическом дыхании. (Levy, Schneiderman, 1966а.)
тичному возмещению израсходованного кислорода, однако азот тоже входит и постепенно накапливается. Если, скажем, в трахейной системе имеется 4% СО2 и 4% О2, то остальное должен составлять азот в концентрации 92%.
ЗНАЧЕНИЕ ЦИКЛИЧЕСКОГО ДЫХАНИЯ
Наземные животные уязвимы в отношении потери влаги, а насекомые часто живут в условиях очень большой сухости. Находящаяся в состоянии диапаузы куколка, которая может оставаться в своей оболочке почти целый год от одного лета до другого, не имеет возможности восполнить потерю воды. Она должна'быть очень экономной в расходовании воды, и в то же время ей нужен значительный газообмен для метаболических процессов. Поэтому
Дыхание насекомых 99
управление дыхальцами у нее должно определяться компромиссом между потребностью в кислороде и необходимостью избегать потери влаги.
Теперь мы можем понять смысл периодического открывания дыхалец. Потери воды зависят от давления пара внутри трахей (которое зависит от температуры), от влажности наружного воздуха и от состояния дыхалец. Если дыхальца закрыты, потери воды из трахейной'системы равны нулю; когда они полностью открыты, эти потери максимальны. Как мы видели, частичное прикрывание (вибрация) позволяет получить достаточное количество кислорода не только благодаря входящему потоку воздуха, но и благодаря тому, что высокая концентрация кислорода в атмосфере создает крутой градиент, вызывающий диффузию, направленную внутрь. Однако, когда концентрация О2 в трахейной системе нарастает и достигает примерно 6%, дыхальца широко раскрываются и позволяют этому газу выйти (залп двуокиси углерода). Если бы вместо этого двуокиси углерода позволялось выходить непрерывно через открытые дыхальца по мере ее образования, то непрерывно терялась бы и вода. Циклический характер газообмена, который на первый взгляд кажется непонятной ►аномалией, можно рассматривать как хитроумное приспособление для сохранения влаги.
Мы рассмотрели, каким образом в органах дыхания происходит газообмен и какие специальные приспособления обеспечивают необходимое поглощение кислорода и сопутствующее выведение СО2. В следующей главе мы ознакомимся с тем, каким образом выполняет свою роль кровь, транспортируя эти два важных газа между органами дыхания и тканями, в которых происходит обмен веществ.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Alkalay I., Suetsugu S., Constantine H., Stein M. (1971). Carbon dioxide elimination across human skin, Am. J. Physiol., 220, 1434—1436.
Bentley P. L, Herreid C. F., Schmidt-Hielsen K. (1967). Respiration of a monotre-me, the echidna, Tachyglossus aculeatus, Am. J. Physiol., 212, 957—961.
Bentley P, J., Shield J. W. (1973). Ventilation of toad lungs in the absence of the buccopharyngeal pump, Nature, bond., 243, 538—539.
Berg T., Steen J. B. (1965). Physiological mechanisms for aerial respiration in the eel, Comp. Biochem. Physiol., 15, 469—484.
Bretz W. L., Schmidt-Nielsen K. (1972). Movement of gas in the respiratory system of the duck, J. Exp. Biol., 56, 57—65.
Buck J. (1958). Cyclic CO2 release in insects. 4. A theory of mechanism, Biol. Bull., 114, 118—140.
Cameron J. N., Mecklenburg T. A. (1973). Aerial gas exchange in the coconut crab, Birgus latro, with some notes on Ge car coidea lalandii, Respir. Physiol., 19, 245-261.
Clements J, A., Nellenbogen J., Trahan H, J. (1970). Pulmonary surfactant and evolution of the lungs, Science, 169, 603—604.
Craig A. B., Jr. (1961a). Causes of loss of consciousness during underwater swimming, J. Appl. Physiol., 16, 583—586.
7’
100 Глава 2. Дыхание в воздухе
Craig А. В., Jr. (1961b). Underwater swimming and loss of consciousness, J.A.M.A., 176, 255—258.
Dolk H. E., Postma N. (1927). Ueber die Haut- und die Lungenatmung von Rana temporaria, T. Vergl. Physiol., 5, 417—444.
Ege R. (1915). On the respiratory function of the air stores carried by some aquatic insects (Corixidae, Dytiscidae and Notonecta), Z. Allg. Physiol., 17, 81—124.
Gans C., De Jongh H. J., Farber J. (1969). Bullfrog (Rana catesbeiana) ventilation: How does the frog breathe? Science, 163, 1223—1225.
Gatz R. N., Crawford E. C., Jr., Piiper J. (1974). Respiratory properties of the blood : of a lungless and gillless salamander, Desmognathus fuscus, Respir. Physiol., 20, 33—41.
Graham J. B. (1974). Aquatic respiration in the sea snake Pelamis platurus, Respir. Physiol., 21, 1—7.
Herreid C. F., Bretz W. L., Schmidt-Nielsen K. (1968). Cutaneous gas exchange in bats, Am. J. Physiol., 215, 506—508.
Hutchison V. H., Whitford W. G., Kohl M. (1968). Relation of body size and surface area to gas exchange in anurans, Physiol. Zool., 41, 65—85.
Johansen K. (1968). Air breathing in the teleost Symbranchus marmoratus, Comp.
Biochem. Physiol., 18, 383—395.
Johansen K. (1968). Air-breathing fishes, Sci. Am., 219, 102—111.
Joshi M. C., Boyer J. S., Kramer P. J. (1965). Growth, carbon dioxide exchange, and control of breathing in the electric eel, Electrophorus electricus, 7L. Vergl. Physiol., 61, 137—163.
Johnston A. M., Jukes M. G. M. (1966). The respiratory response of the decerebrate domestic hen to inhaled carbon dioxide-air mixture, J. Physiol., 184, 38—39P.
Joshi M. C., Boyer J. S., Kramer P. J. (1965). Growth, carbon dioxide exchange, transpiration, and transpiration ratio of pineapple, Botan. Gaz., 126, 174—179.
Krogh A. (1920). Studien uber Tracheenrespiration. 2. Ueber Gasdiffusion in den Tracheen, Pflugers Arch., 179, 95—112.
Lasiewski R. C., Calder W. A., Jr. (1971). A preliminary allometric analysis of respiratory variables in resting birds, Respir. Physiol., 11, 152—166.
Lenfant C., Johansen K., Hanson D. (1970). Bimodal gas exchange and ventilationperfusion relationship in lower vertebrates, Fed. Proc., 29, 1124—1129.
Levy R. I., Schneiderman H. A. (1958). An experimental solution to the paradox of discontinuous respiration in insects, Nature, Lond., 182, 491—493.
Levy R. I., Schneiderman H. A. (1966a). Discontinuous respiration in insects. 2. The direct measurement and significance of changes in tracheal gas composition during the respiratory cycle of silkworm pupae, J. Insect Physiol., 12, 83—104.
Levy R. I., Schneiderman H. A. (1966b). Discontinuous respiration in insects.
4. Changes in intratracheal pressure during the respiratory cycle of silkworm pupae, J. Insect. Physiol., 12, 465—492.
Margaria R., Milic-Emili G., Petit J. M., Cavagna G. (1960). Mechanical work of breathing during muscular exercise, J. Appl. Physiol., 15, 354—358.
Mellanby K. (1934). The site of loss of water from insects. Proc. R. Soc. Lond. B, 116, 139—149.
Miller P. L. (1964). The possible role of haemoglobin in Anisops and Buenoa (He-miptera: Notonectidae), Proc. R. Entomol. Soc. Lond. A, 39, 166—175.
Otis A. B. (1954). The work of breathing, Physiol. Rev., 34, 449—458.
Punt A., Parser W. J., Kuchlein J. (1957). Oxygen uptake in insects with cyclic CO2 release, Biol. Bull., 112, 108—119.
Rahn H., Paganelli С. V. (1968). Gas exchange in gas gills of diving insects, Res-, pir. Physiol., 5, 145—164.
Rahn H„ Rahn К- B., Howell B. J., Gans C., Tenney S. M. (1971). Air breathing of the garfish (Lepisosteus osseus), Respir. Physiol., 11, 285—307.
Romijn C., Roos J. (1938). The air space of the hen’s egg and its changes during the period of incubation, J. Physiol., 94, 365—379.
Scheid P., Piiper J. (1972). Cross-current gas exchange in avian lungs: Effects of reversed parabronchial air flow in ducks, Respir. Physiol., 16, 304—312.
Дыхание в воздухе 101
Schmidt-Nielsen К. (1972). How Animals Work, London, Cambridge University* Press, 114 pp.
Schmidt-Nielsen K., Kanwisher J., Lasiewski R. C., Cohn L E., Bretz W. L. (1969). Temperature regulation and respiration in the ostrich, Condor, 71, 341—352.
Schneiderman H. A. (1960). Discontinuous respiration in insects: Role of the spiracles, Biol. Bull., 119, 494—528.
Schneiderman H. A., Williams С. M. (1955). An experimental analysis of the discontinuous respiration of the cecropia silkworm, Biol. Bull., 109, 123—143.
Schumann D., Piiper J. (1966). Der Sauerstoffbedarf der Atmung bei Fischen nach Messungen an der narkotisierten Schleie [Tinea tinea), Pfliigers Arch., 288, 15-26.
Soum J, M. (1896). Recherches physiologiques sur 1’appareil respiratoire des oi-seaux, Ann. Univ. Lyon, 28, 1—126.
Szarek S. R., Johnson H. B., Ting I. P. (1973). Drought adaptation in Opuntia ba-silaris: Significance of recycling carbon through crassulacean acid metabolism, Plant Physiol., 52, 539—541.
Tenney S. M., Remmers J. E. (1963). Comparative quantitative morphology of the mammalian lung: Diffusing area, Nature, Lond., 197, 54—56.
Thomas H. J. (1954). The oxygen uptake of the lobster (Homarus vulgaris Edw.), J. Exp. Biol., 31, 228—251.
Thorpe W. H., Crisp D. J. (1947). Studies on plastron respiration. 1. The biology of Aphelocheirus [Hemiptera, Aphelocheiridae (Naucoridae)] and the mechanism of plastron retention, J. Exp. Biol., 24, 227—269.
Tucker V. A. (1968). Respiratory physiology of house sparrows in relation to high-altitude flight, J. Exp. Biol., 48, 55—66.
Wangensteen 0. D.t Wilson D., Rahn H. (1971). Diffusion of gases across the shell of the hen’s egg, Respir. Physiol., 11, 16—30.
Wangensteen 0. D„ Wilson D., Rahn H. (1974). Respiratory gas exchange of high altitude adapted chick embryos, Respir. Physiol., 21, 61—70.
Wigglesworth V. B. (1972). The Principles of Insect Physiology, 7th ed., London, Chapman and Hall, 827 pp.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Altman P..L., Dittmer D. S. (eds.) (1971). Biological Handbooks: Respiration and Circulation, Bethesda, Federation of American Societies for Experimental Biology, 930 pp.
Buck J. (1962). Some physical aspects of insect respiration, Annu. Rev. Entomol., 7, 27—56.
Comroe J. H., Jr. (1965). Physiology of Respiration: An Introductory Text, Chicago, Year Book, 245 pp.
Duncker H.-R. (1972). Structure of avian lungs, Respir. Physiol., 14, 44—63.
Fenn W. O.t Rahn H. (eds.) (1964, 1965). Handbook of Physiology, sect. 3, Respiration, vol. 1, pp. 1—926 (1964); vol. 2, pp. 927—1696 (1965).
Heath A. G., Mangum C. (eds.) (1973). Influence of the environment on respiratory function, Am. Zool., 13, 446—563.
Hughes G. A. (1963). Comparative Physiology of Vertebrate Respiration, Cambridge, Mass., Harvard University Press, 145 pp.
Johansen K., et al. (1970). Symposium on cardiorespiratory adaptations in the transition from water breathing to air breathing, Fed. Proc., 29 (3), 1116—1153. <
Jones J. D. (1972). Comparative Physiology of Respiration, London, Edward Arnold, 202 pp.
Miller P. L. (1964). Respiration: Aerial gas transport. In: The Physiology of Insec-ta, vol. Ill (M. Rockstein, ed.), pp. 557—‘615, New York, Academic Press.
Piiper J., Scheid P. (1977). Comparative physiology of respiration: Functional analysis of gas exchange organs in vertebrates, Int. Rev. Physiol., 14, 219—253.
Weibel E. R. (1973). Morphological basis of alveolar-capillary gas exchange, Physiol. Rev., 53, 419-495.
ГЛАВА 3
КРОВЬ
В двух предыдущих главах мы имели дело с дыхательными газами и газообменом между организмом и средой. Наше внимание было сосредоточено на двух самых важных газах — кислороде и двуокиси углерода.
Мы узнали, что для всех организмов, кроме самых мелких, одной диффузии недостаточно для перемещения газов внутри организма и что нужна какая-то механическая транспортная система. Почти все крупные животные, кроме тех, у которых потребность в кислороде очень мала, имеют распределительную систему, основанную на механическом круговороте внутри организма некоторой жидкости — крови. Насекомые не используют кровь для транспорта газов; у них для этой цели служат наполненные га-
Таблица 3.1
Наиболее важные функции крови
1. Транспорт питательных веществ от пищеварительного тракта к тканям, к местам накопления (жировой ткани, печени) и от них
2. Транспорт метаболитов (например, молочной кислоты от мышц к печени), который делает возможной метаболическую специализацию тканей
3. Транспорт веществ, подлежащих экскреции, из тканей к выделительным органам, из органов, где они образуются (например, мочевина — в печени), к почкам
4. Транспорт газов (кислорода и двуокиси углерода) между дыхательными органами и тканями; запасание кислорода
5. Транспорт гормонов (например, адреналина, вызывающего быструю реакцию, или гормона роста, вызывающего медленную реакцию)
6. Транспорт клеток, не выполняющих дыхательной функции (например, лейкоцитов у позвоночных); кровь насекомых не имеет отношения к дыханию, но она переносит кровяные клетки многих типов
7. Перенос тепла из глубоколежащих органов к поверхности для его рассеяния (что существенно для крупных животных с высокой интенсивностью обмена веществ)
8. Передача силы (например, для локомоции у дождевых червей; для разрыва кутикулы при линьке у ракообразных; для движения таких органов, как пенис, сифон двустворчатых моллюсков и т. п.; для разгибания ног у пауков; для ультрафильтрации в капиллярах почек)
9. Свертывание. Способность к свертыванию, присущая многим жидкостям, играющим роль крови или гемолимфы, служит для предотвращения потери крови
10. Поддержание «внутренней среды», подходящей для клеток в отношении pH, ионного состава, питательных веществ и т. д.
Кровообращение IQff
зом трубочки. И все же насекомые имеют кровь, которая перегоняется по телу, так как многие другие вещества необходимо транспортировать с большей скоростью, чем это может обеспечить одна лишь диффузия.
Действительно, кровь несет много функций, которые не сразу бросаются в глаза. Достаточно просмотреть перечень главных функций, выполняемых этой важной жидкостью (табл. 3.1).
Мы склонны думать, что основная функция крови — это транспорт газов, но в списке упомянуто целых шесть важных категорий веществ (в добавление к теплу), которые переносятся кровью. Очень существенной функцией является передача механической силы, хотя эту сторону часто упускают из виду. Из двух последних пунктов списка один касается высокоспецифического внутреннего свойства крови — ее способности свертываться, второй — общего химического состава крови как внутренней среды организма.
Большую часть этих функций могла бы выполнять почти любая водная среда. Например, никакие особые свойства не нужны для переноса метаболитов или продуктов, подлежащих экскреции. Исключение составляют газообмен и свертывание, которые связаны с чрезвычайно сложными химическими особенностями крови.
В этой главе нас будут интересовать в первую очередь роль крови в транспорте газов и те ее свойства, которые служат этой цели.
ПЕРЕНОС КИСЛОРОДА КРОВЬЮ
ДЫХАТЕЛЬНЫЕ ПИГМЕНТЫ
У многих беспозвоночных кислород переносится в крови или гемолимфе в виде простого физического раствора. Это помогает доставлять кислород с поверхности к различным частям тела, поскольку для всех организмов, кроме самых мелких, диффузия сама по себе — процесс слишком медленный.
Однако количество кислорода, которое может быть перенесено в виде простого раствора, невелико, и многие высокоорганизованные животные (а позвоночные — почти все без исключения) имеют кровь, способную обратимо связывать большие количества кислорода. В крови млекопитающих концентрация кислорода в растворенном состоянии составляет около 0,2 мл на 100 мл, а содержание О2, обратимо связанного с гемоглобином, может быть в 100 раз больше — около 20 мл на 100 мл крови. Таким образом, растворенный кислород имеет ничтожное значение по сравнению с кислородом, присоединенным к гемоглобину.
Вещества, которые служат переносчиками кислорода в крови,— это белки, содержащие металл (обычно железо или медь).
Таблица 3.2
Распространенные дыхательные пигменты и примеры животных, у которых они имеются
Пигмент Характеристика Молекулярный вес У каких животных встречается
Гемоцианин Медьсодержащий белок, находится в растворе 300 000— 9 000 000 Моллюски: хитоны, головоногие, переднежаберные и легочные брюхоногие, но не пластинчатожаберные Членистоногие: крабы, омары Паукообразные: Limulus, Euscorpius
Гемоэритрин Железосодержащий белок непорфириновой структуры, всегда находится внутри клеток 108 000 Сипункулиды: все исследованные виды Полихеты: Magelona Приапулиды: Halicryptus, Priapulus Плеченогие: Lingula
Хлорокруорин Железосодержащий белок, находящийся в растворе 2 750 000 Четыре семейства полихет: Sabellidae, Serpulidae, Chlorhaemidae, Amphateri-dae Одна лишь простетическая группа имеется у морских звезд (Luidia, Astro pecten)
Гемоглобин Железосодержащий белок, находится в растворе или внутри клеток; наиболее широко распространенный пигмент 17 000— 3 000 000 Позвоночные: почти все, за исключением лептоцефа-лид (личинки угрей) и некоторых антарктических рыб (Chaenichthys) Иглокожие: голотурии Моллюски: Planorbis, Ti-vela Членистоногие: насекомые (Chironomus, Gastr ophi- lus)', ракообразные: Daphnia, Artemia Кольчатые черви: Lumbricus, Tubifex, Arenicola, Spiror-bis (у некоторых видов имеется гемоглобин, у некоторых — хлорокруорин, другие не имеют в крови пигментов), Serpula (и гемоглобин, и хлорокруо-рин) Нематоды: Ascaris Плоские черви: паразитические сосальщики Простейшие: Paramecium, Tetrahymena Растения: дрожжи, Neuro-spora, корневые клубеньки бобовых растений (клевера, люцерны)
Перенос кислорода кровью 105
Они, как правило, окрашены, и поэтому их часто называют дыха-тельными пигментами. Самые распространенные из них перечислены в табл. 3.2.
ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ
У некоторых животных дыхательные пигменты растворены в крови; у других (например, у позвоночных) они заключены в клетки, а плазма крови не содержит растворенного пигмента. Данные о локализации наиболее обычных дыхательных пигментов в клетках или плазме крови приведены в табл. 3.3; кроме то-
Таблица 3.3
Молекулярные веса и локализация (в клетках или в плазме крови) дыхательных пигментов у различных животных
В клетках В плазме
Животйые Молекулярный вес Животные Молекуляр- i ный вес
Гемоглобин Млекопитающие ~68000l> Олигохеты
-Птицы ~68 ООО1) Lumbricus 2 946 000
Рыбы ~68 ООО1) Полихеты
Круглоротые Arenicola 3 000 000
Lampetra 19100 Serpula 3 000 000
My'xine 23100 Моллюски
Полихеты Planorbis 1 539 000
Notomastus 36000 Насекомые
Иглокожие Chironomus 31400
Thy one 23 600
Моллюски Area Насекомые 33 600
Хлорокруорин Gastrophilus 34 000 Полихеты
Spirographis 3400000
Гемоэритрин Sipunculus 66000
Phascolosoma 120 000
Гемоцианин Моллюски Helix (Головоногие) Rossia (кальмар) Octopus Eledone Членистоногие Limulus (Ракообразные) 6 680 000 3 316 000 2 785 000 2 791 000 1 300 000 397 000
Pandalus
Palinurus 447 000
Nephrops 812 000
Homarus 803 000
9 Мол. вес мышечного гемоглобина (миоглобина) равен 17 000.
106 Глава 3. Кровь
го, там указаны молекулярные веса пигментов для типичных представителей соответствующих групп животных.
Из этой таблицы сразу видно, что в тех случаях, когда пигменты заключены в клетки, их молекулярные веса относительно невелики — от 20 000 до 120 000; если же пигменты растворены в плазме, их молекулярные веса намного больше — от 400 000 до нескольких миллионов. Единственное исключение составляет гемоглобин комара Chironomus.
Крупные молекулы дыхательных пигментов, растворенных в плазме, — это на самом деле агрегаты молекул меньших разме-
Содержание гемоглобина г г на 100 мл
Рис. 3.1. Влияние содержания гемоглобина на вязкость крови. (Schmidt-Nielsen, Taylor, 1968.)
На графике представлены отношения вязкости крови к вязкости чистой воды (принятой за 1). I — вязкость крови козы с интактными эритроцитами. II — относительная вязкость гемолизированной крови. III — вязкость плазмы тех же проб крови. Сравнение точек А, В и С показывает, что у раствора гемоглобина вязкость примерно вдвое меньше, чем у интактной крови с тем же общим содержанием гемоглобина.
увеличивая числа белковых частиц, находящихся в растворе в плазме крови. При очень большом числе растворенных частиц возросло бы коллоидно-осмотическое давление плазмы, что в свою очередь повлияло бы на многие другие физиологические процессы, такие, как прохождение жидкости через стенки капилляров и ультрафильтрация (первичный процесс при образовании мочи в почках).
Высказывалось мнение, что при упаковке больших количеств гемоглобина внутри клеток вязкость крови оказывается ниже, чем
Перенос кислорода кровью 107
в случае раствора гемоглобина, обладающего той же способностью переносить кислород. Плазма млекопитающих содержит около 7% белков, а красные кровяные тельца, или эритроциты, содержат около 35% гемоглобина. Если бы весь гемоглобин эритроцитов был растворен в крови, а не находился внутри клеток, общая концентрация белков была бы около 20%, т. е. в три раза выше типичной концентрации белков в плазме млекопитающих.
Приводились аргументы в пользу того, что раствор с 20% белков будет очень вязким, сиропообразным — намного более вязким, чем кровь. Однако если мы сначала измерим вязкость какой-то пробы крови, а затем разрушим эритроциты с помощью ультразвука, чтобы освободить гемоглобин, то получившийся раствор гемоглобина будет иметь вдвое меньшую вязкость, чем кровь, в которой гемоглобин был заключен внутри клеток (рис. 3.1). Тем не менее это не полный ответ, поскольку вязкие свойства крови очень сложны; они будут обсуждаться позднее в гл. 4.
Если не говорить о вязкости, упаковка гемоглобина в клетки дает и другие преимущества. Пожалуй, самое важное из них то, что химическая среда внутри эритроцитов может отличаться от состава плазмы крови. На реакцию кислорода с гемоглобином наряду с неорганическими ионами сильно влияют и некоторые органические соединения, особенно органические фосфаты, и гемоглобин, локализованный внутри клеток, может быть обеспечен специфической, хорошо подобранной средой, отличной от плазмы.
РАЗМЕРЫ И ФОРМА ЭРИТРОЦИТОВ
Эритроциты млекопитающих — круглые, слегка вогнутые с обеих сторон диски. Имеется одно исключение: у всех представителей семейства верблюжьих эти клетки имеют овальную форму. Диаметр эритроцитов специфичен для каждого вида; у млекопитающих он варьирует в пределах от 5 до 10 мкм. Определенной связи между величиной эритроцитов и размерами тела животного нет: из млекопитающих самые мелкие эритроциты обнаружены у одного некрупного азиатского оленя, а у мыши и слона они примерно одинаковых размеров.
Эритроциты млекопитающих лишены ядер, хотя эти клетки живут около 100 дней и выполняют сложные метаболические функции. Эритроциты всех других позвоночных — птиц, рептилий, амфибий и рыб — содержат ядро и почти все имеют овальную форму. Часто они бывают очень крупными: например, у саламандр объем эритроцита в 100—200 раз больше, чем у млекопитающих. Имеет ли какое-нибудь значение наличие или отсутствие ядер в эритроцитах, не установлено. Не ясно также, играет ли какую-то функциональную роль величина эритроцитов.
Единственные позвоночные, у которых нет гемоглобина и эритроцитов,— это личинки угрей (лептоцефалиды) и некоторые антарктические рыбы семейства Chaenichthyidae.
108 Глава 3. Кровь
КРИВЫЕ КИСЛОРОДНОЙ ДИССОЦИАЦИИ
Свойство крови, наиболее важное для переноса кислорода — это способность обратимо связывать его с молекулой гемоглобина. Такое связывание можно записать в виде обычной обратимой химической реакции:
НЬ+О2 ньо2.
При больших концентрациях кислорода гемоглобин (НЬ) соединяется с кислородом, образуя оксигемоглобин (НЬО2)Г и реакция идет в правую сторону. При низких концентрациях кислород отделяется, и реакция идет в левую сторону. Если концентрация О2 снижается до нуля, гемоглобин отдает весь связанный им кислород.
Каждый атом железа в молекуле гемоглобина связывает одну молекулу кислорода, и когда при высокой концентрации (или высоком давлении) О2 все имеющиеся места связывания заняты, гемоглобин становится полностью насыщенным и не может больше принимать кислород. При каждой данной концентрации кислорода существует определенное количественное соотношение между ге-
Рис. 3.2. Кривые кислородной диссоциации для крови голубя. (Lutz et al., 1973.)
/ — кривая, полученная при нормальных для организма птицы условиях: температура 41 °C, Рсо235 мм рт. ст., pH 7,5; II— кривая, полученная после сдвига pH с 7,5 до 7,2 без изменения РСо2.
моглобином и оксигемоглобином. Если мы построим график зависимости количества оксигемоглобина от концентрации кислорода, мы получим кривую кислородной диссоциации, которая описывает, каким образом эта реакция зависит от концентрации или парциального давления О2. Одна такая кривая представлена на
Перенос кислорода кровью 1С9
рис. 3.2. Эта кривая показывает, что кровь голубя полностью насыщается кислородом (100% НЬО2) при парциальных давлениях О2 80—100 мм рт. ст. (10—13 кПа) и выше. При дальнейшем повышении давления кислорода гемоглобин больше не будет его принимать, и мы говорим, что гемоглобин полностью насыщен. При меньшем давлении О2 гемоглобин будет его отдавать, и при Роа =30 мм рт. ст. (4 кПа) половина пигмента будет находиться в форме оксигемоглобина, а другая половина — в форме гемоглобина. Такое характеристическое давление кислорода называют давлением полунасыщения (Р50). Если давление О2 снижать дальше, все больше кислорода будет отделяться, пока при нулевом давлении О2 весь гемоглобин не будет полностью деоксигенирован.
У других животных кривые кислородной диссоциации гемоглобина по форме сходны с кривой на рис. 3.2, но не идентичны ей. Кровь некоторых животных имеет более высокое сродство к кислороду- (т. е. не так легко его отдает), и кривые диссоциации у них сдвинуты влево. Кровь других животных отдает кислород легче (ее сродство к кислороду меньше), и кривые диссоциации сдвинуты вправо. Кроме этого, на связывание кислорода и тем самым на кривую диссоциации влияют и другие параметры (pH, ионная среда, органические фосфаты, температура). Эти различия имеют большое функциональное значение.
ВЛИЯНИЕ ТЕМПЕРАТУРЫ
Повышение температуры ослабляет связь между гемоглобином и кислородом, и эта связь легче разрывается. Поэтому при высоких температурах гемоглобин легче отдает кислород, и кривая диссоциации сдвигается вправо. С точки зрения физиологии это существенно, так как повышение температуры обычно сопровождается увеличением интенсивности обмена (а значит, и потребности в кислороде). Поэтому более легкая отдача О2 гемоглобином при повышенной температуре выгодна для организма. Это может иметь некоторое значение даже для теплокровных животных — например, в случаях лихорадки или при физической нагрузке, когда потребление кислорода возрастает.
ВЛИЯНИЕ ДВУОКИСИ УГЛЕРОДА И pH
Существенное влияние на кривую диссоциации оказывает pH плазмы крови. Увеличение количества СО2 или других кислот снижает pH плазмы, и это ведет к сдвигу кривой диссоциации вправо (рис. 3.2). Такой сдвиг означает, что повышенная концентрация W2 приводит к повышенной отдаче кислорода при любом заданном его давлении. Это явление известно как эффект Бора. В капиллярах тканей, где двуокись углерода поступает в кровь, гемоглобин отдает больше кислорода, чем это было бы при отсутствии влияния СО2 на его связывание. Таким образом, можно сказать,
110 Глава 3. Кровь
что эффект Бора облегчает доставку тканям дополнительного количества кислорода.
Эффект Бора не одинаково выражен у разных млекопитающих. За меру эффекта Бора можно принять сдвиг Ры при изменении pH на единицу. Тогда мы обнаружим, что эффект Бора зависит' от размеров тела и что сдвиг Р50 У мыши больше, чем у слона (рис. 3.3). Иными словами, гемоглобин мыши более чувствителен! к кислоте, чем гемоглобин слона. Позже мы увидим, что это важна для доставки кислорода тканям мелких животных, у которых удельная интенсивность обмена выше, чем у более крупных животных (Schmidt-Nielsen, 1972).
Влияние кислоты на кривую диссоциации у некоторых рыб выражено намного сильнее, чем у млекопитающих; этот более сильный эффект называют эффектом Рута (рис. 3.4). Его самое важное следствие касается функции крови при секреции кислорода в плавательный пузырь (гл. 11).
Для эффекта Рута характерно то, что при низком pH кровь не насыщается кислородом полностью ни при каком давлении О2-Это объясняется тем, что кровь у рыб содержит гемоглобин двух
Рис. 3.3. Связь между величиной эффекта Бора для гемоглобина и размерами тела. (Riggs, 1960.)
Гемоглобин мелких животных обнаруживает больший сдвиг Бора (т. е. более чувствителен к кислоте), чем гемоглобин крупных животных, и, следовательно, при данном Ро2 отдает больше кислорода.
разных типов: высокочувствительный к кислоте и нечувствительный к кислоте. В присутствии высокой концентрации СО2 первый совсем не связывает кислород, тогда как второй дает обычную кривую диссоциации, характерную для этого вида гемоглобина (Gillen, Riggs, 1973).
Перенос кислорода кровью 111
Зачем рыбам два различных гемоглобина? Легко понять значение молекул НЬ, чувствительных к кислоте, но для чего нужна -другая разновидность, которая к кислоте нечувствительна? Какая в этом необходимость? Дело, очевидно, в том, что у рыбы, когда она в критической ситуации развивает при плавании максимальные усилия, образуются большие количества молочной кислоты.
Рис. 3.4. Кривые кислородной диссоциации для крови рыбы Opsanus при 15 °C, (Root et al., 1939.)
I — кривая для цельной крови в отсутствие СО2; II— кривая для цельной'кро-ви при давлении СО2 25 мм рт. ст. (3,3 кПа).
Если бы в присутствии этой кислоты весь гемоглобин терял способность связывать кислород, рыба погибла бы от асфиксии. Однако наличие другого, нечувствительного к кислоте гомоглобина предотвращает эту беду.
ВЛИЯНИЕ ОРГАНИЧЕСКИХ ФОСФАТОВ
Перенос газов эритроцитами тесно связан с метаболизмом самих клеток. Благодаря выяснению роли органических фосфатов мы теперь понимаем многие особенности кривой кислородной диссоциации, которые раньше казались очень странными.
Эритроциты млекопитающих не имеют ядра, и в течение многих лет их рассматривали как нечто мало отличающееся от инертного мешка, набитого гемоглобином. Однако в эритроците имеет место активный углеводный обмен, который не только существен для жизнеспособности клетки, но и оказывает глубокое влияние на ее функцию при транспорте кислорода. Эритроцитам свойственно также высокое содержание аденозинтрифосфата (АТФ) и еще более высокий уровень 2,3-дифосфоглицерата (ДФГ). Присутствие этих органических фосфатов помогает объяснить, почему раствор чистого гемоглобина проявляет гораздо большее сродство к кислороду, чем цельная кровь (кривая диссоциации для НЬ
112 Глава 3, Кровь
в растворе сдвинута далеко вправо по отношению к кривой для крови). Если к раствору гемоглобина.добавить органические фосфаты, в особенности ДФГ, его сродство к кислороду сильно снижается и становится близким к уровню интактных клеток. Это снижение можно объяснить тем, что к гемоглобину присоединяется ДФГ, который изменяет его сродство к О2 (Chanutin, Curnish, 1967; Benesch et al., 1968a, b).
Открытие влияния ДФГ объясняет многие наблюдения, которые раньше казались совсем непонятными. Оно также объясняет, почему при исследовании цельной крови можно наблюдать ряд особенностей кривой диссоциации, которые не выявляются при изучении гемоглобина в растворе. Так, например, у высокогорных жителей уровень ДФГ в эритроцитах выше, чем у людей, живущих на уровне моря. Вследствие этого на больших высотах кривая диссоциации О2 сдвинута вправо. Этот сдвиг был хорошо известен, но механизм его оставался неясным до тех пор, пока не было установлено, что уровень ДФГ возрастает в ответ на гипоксию (Aste-Salazar, Hurtado, 1944; Lenfant et al., 1968).
Влияние ДФГ и других фосфатов позволило объяснить давно известный факт, что при хранении консервированной крови сродство ее к кислороду со* временем увеличивается. Оно также помогает понять, почему очищенные, освобожденные от ДФГ гемоглобины плода и взрослого человека обнаруживают одинаковое сродство к кислороду, хотя, у крови плода сродство к кислороду выше. Кроме того, теперь можно объяснить сдвиг вправо кривых диссоциации в период беременности, хотя беременная самка (в отличие от плода) все время имеет один и тот же гемоглобин.
КРИВЫЕ ДИССОЦИАЦИИ ГЕМОГЛОБИНА ЩЕОДА (ФЕТАЛЬНОГО НЬ)
У многих млекопитающих, включая человека, кривые диссо1-циации для крови плода сдвинуты влево от кривых для материнской крови (рис. 3.5). Это связано со способом, при помощи которого плод получает кислород из крови матери. Поскольку кровь плода имеет более высокое сродство к кислороду, чем материнская кровь, она может лучше захватывать О2. При одном и том же напряжении кислорода кровь плода будет содержать больше этого газа (иметь более высокий процент насыщения), чем материнская кровь; это облегчает извлечение кислорода кровью плода в плаценте. Различие кривых диссоциации для крови плода и матери отчасти обусловлено тем, что гемоглобин плода несколько отличается от материнского гемоглобина, а отчасти связано также с различием в органическом фосфате внутри эритроцитов. После рождения фетальный гемоглобин постепенно исчезает и замещается гемоглобином взрослого типа.
Смещение кривых диссоциации влево, означающее, что кровь активнее поглощает кислород, обнаружено также у куриных эмбрионов и у головастиков лягушки. У гемоглобина головастиков
Перенос кислорода кровью ИЗ»
сродство к кислороду выше, чем у взрослых лягушек, и, что особенно интересно, он нечувствителен к кислоте, тогда как гемоглобин взрослых особей обнаруживает выраженный эффект Бора и очень чувствителен к кислоте (рис. 3.6). Если мы вспомним о местах обитания головастиков, мы легко увидим преимущества
Рп , кПа
Рис. 3.5. Кривые кислородной диссоциации для крови беременной козы и плода. (Barcroft, 1935.)
Более высокое сродство к кислороду у крови плода способствует передаче ей О2 в плаценте. Пунктиром показаны пределы, в которых лежат кривые для небеременных взрослых коз.
этого: головастики нередко живут в застойных водоемах с низким содержанием кислорода, а иногда и с высоким содержанием СО2 в воде. Таким образом, высокое сродство к кислороду здесь выгодно. Поскольку низкая концентрация О2 и высокая концентрация СО2 очень часто сопутствуют друг другу, сильно выраженный эффект Бора был бы нежелателен, так как он уменьшал бы сродство крови к кислороду как раз тогда, когда высокое сродство крайне необходимо.
влияние высоты
Низкое атмосферное давление на большой высоте означает, что и парциальное давление кислорода здесь меньше, чем на уровне моря. У животных, которые постоянно живут в этих условиях относительного дефицита О2, в кривых диссоциации оксигемогло-
8—1873
fl 14 Глава 3. Кровь
бина проявляются соответствующие адаптации. Например, у ламы кривая располагается влево от обычной для млекопитающих области (рис. 3.7). Лама живет в высокогорной зоне южноамериканских Анд, часто выше 5000 м, и большое сродство ее гемоглобина к кислороду позволяет ее крови поглощать этот газ при низких давлениях. Такая адаптация к высоте — врожденное свой-
Рис. 3.6. Кривая кислородной диссоциации для крови головастика лежит левее, чем у взрослой лягушки (т. е. кровь головастика имеет большее сродство к О2). Кроме того, кровь головастика почти нечувствительна к кислоте, тогда как у крови взрослой лягушки сдвиг Бора хорошо выражен. (Riggs, 1951.)
ство: у лам, перевезенных в зоопарк, находящийся на уровне моря, кривые кислородной диссоциации указывают на такое же высокое сродство гемоглобина к Ог.
Когда человек поднимается на большую высоту, он постепенно все больше приспосабливается к низкому давлению кислорода; •он акклиматизируется к высоте, и его работоспособность, как физическая, так и умственная, улучшается. Однако эта адаптация не связана с повышением сродства крови к кислороду. Как это ни парадоксально, но кривые диссоциации по мере привыкания к высоте даже слегка сдвигаются вправо. Это результат других, более «сложных физиологических адаптаций (Lenfant et al., 1969).
Перенос кислорода кровью 115»
Смещение кривой диссоциации вправо (из-за увеличения уровня ДФГ), происходящее у человека на большой высоте,— это сдвиг, прямо противоположный изменению в крови ламы и других естественных обитателей больших высот. Возможно, это» трудно согласовать, но следует учитывать, что в организме срод-
ро2, кЛа
Рис. 3.7. Кривые кислородной диссоциации для крови ламы и викуньи располагаются слева от кривых для других млекопитающих (заштрихованная область). Большее сродство к кислороду у крови этих животных способствует поглощению О2 при низких давлениях, характерных для больших высот. (Hall et al.r 1936.)
ство гемоглобина к кислороду имеет большое значение в двух местах: 1) в легких, где гемоглобин поглощает кислород, и 2) в тканях, где гемоглобин его отдает. Что касается легких, то здесь высокое сродство к кислороду (сдвиг влево) усиливает поглощение О2; с другой стороны, отдача кислорода тканям облегчается при низком сродстве к нему (сдвиге вправо). По-видимому, сдвиг кривой диссоциации влево у ламы — это адаптация, предназначенная главным образом для лучшего поглощения кислорода при низких давлениях на большой высоте, а сдвиг вправо у человека в аналогичных условиях служит для лучшей отдачи кислорода тканям. Таким образом, любую кривую диссоциации можно рассматривать как результат компромисса между желательным высоким сродством к кислороду в легких и столь же желательным низким сродством в тканях.
В эритроцитах разных млекопитающих содержание ДФГ различно, и гемоглобин не одинаково чувствителен к уровню ДФГ. У человека, лошади, собаки, кролика, морской свинки и крысы уровень ДФГ в эритроцитах высок, а гемоглобин, будучи очищен 8*
a 16 Глава 3, Кровь
ют ДФГ, имеет большое сродство к кислороду. Напротив, у овцы, козы, коровы и кошки .концентрация ДФГ низка, а гемоглобин лишь слабо взаимодействует с ДФГ (Bunn, 1971). Эти различия пока трудно объяснить. Они не связаны явным образом ни с филогенетическими отношениями, ни с какими-либо характерными различиями в биологии тех и других видов.
КРИВЫЕ ДИССОЦИАЦИИ У НЫРЯЮЩИХ животных
Может показаться, что таким млекопитающим, как тюлени, которым свойственно нырять и для которых поэтому весьма обычно пребывание в условиях относительной нехватки кислорода, было бы выгодно иметь кровь с высоким сродством к О2. Однако это не так, и мы, подумав немного, поймем, почему они не получили бы ют этого особых преимуществ. Поскольку тюлени дышат обычным атмосферным воздухом, насыщение крови кислородом в легких происходит у них примерно в таких же условиях, как и у других млекопитающих, и для этого процесса не требуется какого-то особенно высокого сродства к кислороду. Во время ныряния такое повышенное сродство не способствовало бы переходу О2 из крови в ткани (прежде всего в центральной нервной системе и в сердечной мышце); напротив, оно, вероятно, даже затрудняло бы отдачу кислорода жизненно важным органам. В связи с этим кривые диссоциации О2 у ныряющих животных обычно мало отличаются от соответствующих кривых для близких к ним неныряющих видов.
КРИВАЯ ДИССОЦИАЦИИ И РАЗМЕРЫ ТЕЛА
Если кривые диссоциации различных млекопитающих свести на. одном графике, мы увидим, что у крупных животных эти кривые сдвинуты влево, а у мелких — вправо (рис. 3.8).
Как мы уже говорили, реакции связывания и отдачи кислорода гемоглобином играют важную роль в двух местах: в легких, где кислород поглощается, и в тканях, где кровь отдает его. Как видно из рис. 3.8, при нормальном для млекопитающих давлении О2 в легких, которое близко к 100 мм рт. ст. (13 кПа), кровь всех представителей этого класса насыщена на 100%. Таким образом, в легких кровь полностью нагружается кислородом, и в этом отношении большой разницы между видами нет. Поэтому рассмотрим теперь процесс отдачи кислорода в тканях.
У мелких млекопитающих кровь легче отдает кислород, так как она имеет меньшее сродство к нему. Обычно кровь отдает в тканях не весь свой кислород; приближенно можно сказать, что в среднем отдается около половины кислорода. Это соответствует состоянию полунасыщения гемоглобина (50% оксигемоглобина и 50% гемоглобина) —точке, с помощью которой удобно характеризовать положение кривой диссоциации. Соответствующее давле
Перенос кислорода кровью 117
ние Ог, которое мы можем рассматривать как среднее давление при отдаче, равно примерно 22,5 мм рт. ст. (3,0 кПа) для крови слона, 28 мм рт. ст. (3,7 кПа) для крови человека и около 45 мм рт. ст. (6 кПа) для крови мыши.
Такие различия в давлении при отдаче кислорода кровью можно объяснить различными метаболическими потребностями в
Рис. 3.8. Кривые кислородной диссоциации для крови млекопитающих разных размеров. (Schmidt-Nielsen, 1972.)
У мелких млекопитающих сродство к кислороду ниже. Это помогает доставке Ог тканям и поддержанию высокой интенсивности метаболизма, характерной для мелких животных.
кислороде. У мелких животных потребление Ог на 1 г веса тела больше, чем у крупных: например, у лошади весом 700 кг оно составляет 1,7 мкл-г-1-мин-1, а у 20-граммовой мыши — 28 мкл-•г-1-мин-1, т. е. среднее потребление кислорода одним граммом ткани у мыши примерно в 16 раз больше, чем у лошади.
Отсюда следует, что у мыши ткани должны снабжаться кислородом в 16 раз интенсивнее, чем у лошади, а это может быть достигнуто только при 16-кратном различии в диффузионных градиентах от капилляров к клеткам. Это схематически показано на рис. 3.9. Каждый капилляр снабжает кислородом окружающую ткань, и расстояние, на которое кислород должен диффундировать, определяется расстоянием между соседними капиллярами: середина этого расстояния — самая далекая точка, до которой кислород должен дойти. У лошади и у мыши величина эритроцитов примерно одинакова, а потому и капилляры у них сходны по своим
118 Глава 3, Кровь
размерам, и это упрощает анализ. Можно создать у мыши в 16 раз больший диффузионный градиент, варьируя две переменные: во-первых, можно сократить пути диффузии, уменьшив расстояния между капиллярами (т. е. увеличив плотность капилляров в тканях), и, во-вторых, можно увеличить давление кислорода в месте его отдачи (сдвинуть кривую диссоциации вправо).
Действительно, у мелких животных плотность капилляров больше. Например, у мыши на 1 мм2 поперечного сечения мышцы может приходиться около 2000 капилляров, а у лошади — менее 1000 (Schmidt-Nielsen, Pennycuik, 1961). Из рис. 3.8 видно, что в крови мыши давление Ог при его отдаче (Р50) примерно в 2,5 раза выше, чем у лошади. Из этого можно заключить, что более высокая интенсивность метаболизма и соответственно увеличение градиента для диффузии кислорода от капилляров к тканевым клеткам достигаются путем сокращения диффузионного пути в сочетании с повышенным давлением кислорода при его отдаче.
Рис. 3.9. Вокруг каждого капилляра можно выделить приблизительно цилиндрическую область, в которой ткань получает кислород путем диффузии из этого капилляра. Крутизна диффузионных градиентов зависит от радиуса тканевого цилиндра (R), радиуса капилляра (г) и разности парциальных давлений Ог в крови и в ткани.
ДЫХАТЕЛЬНАЯ ФУНКЦИЯ КРОВИ У РЫБ
Если мы захотим сравнить сродство к кислороду у крови различных рыб, мы сразу же натолкнемся на ряд проблем, которые трудно разрешить. Например, при какой температуре нужно производить сравнение? Если мы будем сравнивать кровь двух видов рыб, должны ли мы исследовать ее свойства при одной и той же
Перенос кислорода кровью 119
температуре или же при температурах, соответствующих обычным условиям жизни той и другой рыбы? Когда мы сравниваем тропических и арктических рыб, на этот вопрос нелегко ответить, но для рыб со сходными местообитаниями мы можем брать одну и ту же температуру.
Кривые диссоциации для гемоглобина различных рыб говорят о широком диапазоне величин сродства к кислороду и о характерных особенностях, связанных со степенью активности данного вида (Hall, McCutcheon, 1938). У макрели — очень активной рыбы, которая обычно плавает в насыщенной кислородом воде, — кривая диссоциации сдвинута вправо по сравнению с другими рыбами. Это согласуется с тем, о чем мы только что узнали: низкое сродство к кислороду облегчает отдачу кислорода тканям, а в ^хорошо аэрированной воде артериальная кровь и в этом случае будет полностью насыщена. Иглобрюх, у которого кривая очень сильно смещена влево, — медленно плавающая и относительно вялая донная рыба (Randall, 1970). Ее часто находят в не столь .хорошо аэрированной воде, и она очень устойчива к недостатку кислорода. Гемоглобин иглобрюха обладает чрезвычайно высоким сродством к кислороду, и это соответствует как среде, в которой рыба обычно живет, так и относительно низкой интенсивности ее метаболизма, причем здесь, по-видимому, более важны требования, предъявляемые к механизму насыщения крови кислородом, чем к механизму его отдачи в тканях.
Поскольку рыбы, особенно пресноводные, подвергаются воздействию значительных сезонных изменений температуры, может показаться, что зависимость сродства крови к кислороду от температуры для них невыгодна. В связи с этим интересно рассмотреть изменение кривых диссоциации у рыб по мере их адаптации к разным температурам, У американского сомика Ictalurus nebu-losus эта кривая зависит от температуры акклимации (рис. 3.10). Кровь рыбы, акклимированной к теплу, обнаруживает большее сродство к кислороду, чем кровь рыбы, акклимированной к холоду, если их исследовать при одной и той же температуре. А теперь сравним кривые, полученные при тех температурах, к которым рыбы акклимировались (две средние кривые из четырех). У рыбы, адаптировавшейся к 9 °C, кривая (снятая при 9 °C) сдвинута влево от кривой для рыбы, приспособившейся к 24 °C (снятой при 24°C). Акклимация приводит к сближению кривых, но у тепловодной рыбы, у которой интенсивность метаболизма выше, кривая все же остается сдвинутой вправо по отношению к кривой для рыбы, акклимированной к холоду.
Различия в свойствах крови и их сезонные изменения зависят от особенностей эритроцитов, а не гемоглобина. Если взять рыб, адаптировавшихся к теплу и к холоду, и исследовать их гемоглобины в растворе, измеримого различия в сродстве к кислороду не обнаружится.
120 Глава 3. Кровь
Рис. 3.10. Кривые кислородной диссоциации для сомиков Ictalurus, акклимиро-ванных к холоду и к теплу, снятые при температурах 9 и 24 °C; pH 7,8 (Crigg, 1969.)
КРИВЫЕ ДИССОЦИАЦИИ ДЫХАТЕЛЬНЫХ ПИГМЕНТОВ БЕСПОЗВОНОЧНЫХ
Ранее мы видели (табл. 3.2), что у беспозвоночных имеется ряд различных дыхательных пигментов. У многих из них сродство к кислороду очень велико, и в обычной системе координат кривые диссоциации будут сдвинуты далеко влево. Например, у гемоглобина двустворчатого моллюска Phacoid.es Pso для кислорода составляет около 0,2 мм рт. ст. (27 Па) (Read, 1962).
Гемоглобин личинки Chironomus тоже обладает чрезвычайно' высоким сродством к кислороду. На его кривую диссоциации влияют как температура, так и pH, и в области температур от 5 до 24 °C Ао изменяется у него от 0,1 до 0,6 мм рт. ст,- (от 13 да 80 Па). Для млекопитающих это были бы исключительно низкие величины. Личинки хирономид обычно живут в среде, где временами бывают очень низкие концентрации кислорода, и их гемоглобины, по-видимому, играют важную роль в первую очередь, тогда, когда кислорода недостаточно.
Это предположение подтверждается и тем фактом, что у бес-позвоночных, у которых имеется гемоглобин, его концентрация часто сильно возрастает при содержании животных в бедной кислородом воде. Так обстоит дело у дафний, артемии, личинок Chironomus и многих других.
Перенос кислорода кровью 121
Гемоэритрин химически отличается от гемоглобина тем, что не содержит порфириновой группы. Он найден только у нескольких морских форм (см. табл. 3.2), и у него обнаружена интересная корреляция между сродством к кислороду и естественными экологическими условиями. У двух представителей сипункулид — Dendrostomum и Siphonosoma — гемоэритрин имеется как в крови, так и в целомической жидкости. У Dendrostomum щупальца вокруг рта обильно снабжаются кровью; животное выставляет их над поверхностью песка, в котором оно живет, и они служат ему органом дыхания. Характерно, что у Dendrostomum целомическая жидкость имеет большее сродство к кислороду, чем кровь (рис. 3.11). Это означает, что кислород, поглощенный кровью в дыхательном органе, легко передается пигменту целомической жидкости. У Siphonosoma кровь и целомическая жидкость сильно не различаются, причем сродство к кислороду даже чуть выше у крови. Это соответствует образу жизни: ротовые щупальца Siphono-
Рис. 3.11. Кривые кислородной диссоциации для гемоэритрина из крови и целомической жидкости сипункулиды Dendrostomum zostericolum. (Manwell, 1960.)
зота использует не для дыхания, а для питания; животное нахо-дится в норе глубоко под поверхностью ила и поглощает кислород всей поверхностью тела. В связи с этим его кровь и целомическая жидкость имеют почти одинаковое высокое сродство к кислороду.
Сходное соотношение между сродством двух различных пигментов к кислороду обнаружено у боконервного моллюска Сгур-iochiton. Кровь этого животного содержит гемоцианин, а мышцы языка — миоглобин. Кровь имеет относительно низкое сродство к кислороду, а миоглобин — высокое (рис. 3.12). Это совершенно аналогично тому, что мы имеем у позвоночных. У Cryptochiton кислород передается от гемоцианина крови гемоглобину (миогло
122 Глава 3. Кровь
бину) мышц, у которого сродство к кислороду намного больше и кривая диссоциации смещена далеко влево — Р5о около 0,5 мм рт. ст. (0,07 кПа).
Рис. 3.12. Кривые кислородной диссоциации для гемоцианина (из крови) и миоглобина (из мышц радулы) боконервного моллюска Cryptochiton. (ManwelU 1958.)
ОБЛЕГЧЕННАЯ ДИФФУЗИЯ В РАСТВОРАХ ГЕМОГЛОБИНА
Очевидно, что гемоглобин, переносимый кровью, неизмеримо облегчает доставку кислорода тканям. Однако гемоглобин встречается не только в крови, но и в различных тканях, у высших животных— главным образом в мышцах. Может ли такой гемоглобин, находящийся внутри клеток и выключенный из циркуляции^ выполнять какую-либо функцию?
В тех случаях, когда потребность в кислороде может резка возрастать, увеличение кислородной емкости может способствовать сглаживанию колебаний. Например, при сокращении мышцы приток крови к ней может уменьшиться или на какой-то момент прерваться из-за механического сдавления сосудов сокращающейся мышцей. Тогда оксигенированный миоглобин будет отдавать свой кислород и, таким образом, будет функционировать как кратковременный резерв кислорода (Millikan, 1937). Однако сокращение мышц вряд ли может регулярно полностью прерывать движение крови; напротив, кровоток в напряженно работающих мышцах обычно сильно возрастает.
Когда выяснилось, что гемоглобин сильно ускоряет диффузию кислорода, это позволило по-иному интерпретировать присутствие гемоглобина в мышцах, а также его частое присутствие у животных, обитающих в местах, где хотя бы временами резко снижается напряжение кислорода. Некоторые из таких животных упомянуты
Перенос кислорода кровью 123
в табл. 3.2, и возможно, что они тоже пользуются преимуществами ускоренной, или облегченной, диффузии кислорода.
Шоландер (Scholander, 1960) обнаружил, что в отличие от азота, который диффундирует в растворе гемоглобина несколько медленнее, чем в воде, кислород проходит через раствор гемоглобина намного быстрее, чем через воду.
Шоландер поставил следующий эксперимент. Испытуемую жидкость (раствор гемоглобина, плазму крови или воду) абсорбировали сильно пористой мембраной. Затем этой мембраной отгораживали обычный воздух от вакуума. И кислород, и азот должны были теперь диффундировать из воздуха через мембрану в вакуум. Когда в мембране содержалась вода или плазма крови, отношение количеств кислорода и азота, продиффундировавших через жидкий слой, составляло около 0,5. (Именно такое отношение можно было предсказать, исходя из того, что воздух содержит примерно в четыре раза больше азота, чем кислорода, но азот растворяется вдвое хуже, чем кислород.)
Если давление воздуха над мембраной снизить до 0,5 атм, то в вакуум будет проходить вдвое меньше воздуха в единицу времени, но отношение между двумя газами останется прежним: объем кислорода будет составлять 0,5 от объема азота (рис. 3.13). Когда плазму заменяли раствором гемоглобина (полученным путем гемолиза), содержащим столько же гемоглобина, сколько его имеется в нормальной крови, количество проходящего через мембрану кислорода оказывалось примерно равным количеству проходящего азота. Это означает, что по отношению к азоту диффузия кислорода в присутствии гемоглобина ускоряется. При дальнейшем снижении давления в воздушной камере отношение скоростей все больше сдвигалось в пользу кислорода, и в конце концов диффузия кислорода ускорилась примерно в восемь раз по сравнению с его диффузией в отсутствие гемоглобина.
Облегчение диффузии связано со способностью гемоглобина обратимо связывать кислород. Гемоглобин, превращенный в метгемоглобин (путем окисления двухвалентного железа в трехвалентное), уже не может связывать кислород, и поэтому раствор метгемоглобина не ускоряет диффузию этого газа. Однако и миоглобин, и различные гемоглобины как позвоночных, так и беспозвоночных облегчают диффузию Ог.
Один из важных аспектов облегченной диффузии — то, что диффузия ускоряется только в том случае, если давление кислорода на конечном участке диффузионного пути близко к нулю. Действительно, максимальное ускорение наблюдается тогда, когда степень насыщения «верхнего» и «нижнего» слоев гемоглобина соответствует самому крутому участку кривой диссоциации.
Не облегчают ли диффузию также и другие дыхательные пигменты, например гемоцианин? Нет, не облегчают. Это, по-видимому, связано с большей величиной молекул гемоцианина (несколь-
Рис. 3.13. Диффузия кислорода и азота из воздуха в вакуум через слой плазмы крови всегда осуществляется в соотношении 0,5 : 1. Относительная скорость диффузии О2 через раствор гемоглобина больше и возрастает с уменьшением давления. Заштрихованной полоской справа указано давление водяных паров, которое было одинаковым по обе стороны мембраны. Подробности см. в тексте. (Scholander, 1960.)
Перенос кислорода кровью 12S
ко миллионов дальтон), которая сильно уменьшает их подвижность в растворе. Хотя точный механизм облегчения диффузии О2 еще не вполне понят, можно думать, что некоторую роль здесь играет обычное тепловое движение молекул гемоглобина в растворе. Если это движение замедлить (например, добавив желатину), облегчение уменьшается. Миоглобин, молекулярный вес которого (17 000) меньше, чем у гемоглобина (68000), еще больше-облегчает диффузию (грубо говоря, эффекты этих двух пигментов обратно пропорциональны квадратным корням из их молекулярных весов). Это указывает на то, что различие связано с тепловым движением молекул в растворе (Hemmingsen, 1965).
Главные моменты, характеризующие облегченную диффузию-кислорода в растворах гемоглобина, можно резюмировать следующим образом: 1) результирующая диффузия кислорода ускоряется, когда давление на выходе низко; 2) общий перенос кислорода складывается из эффекта простой диффузии и добавочного эффекта, обусловленного присутствием пигмента; 3) облегчение возрастает с увеличением концентрации гемоглобина; 4) скорость переноса обратно пропорциональна квадратному корню из молекулярного веса пигмента, так что сравнительно небольшие молекулы гемоглобина создают большее облегчение.
ФИЗИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ОБЛЕГЧЕННОЙ ДИФФУЗИИ
У позвоночных гемоглобин содержится в высокой концентрации главным образом в эритроцитах и в меньшем количестве — в красных мышцах. Сомнительно, чтобы облегченная диффузия: играла существенную роль в эритроцитах, так как для этого эффекта нужно, чтобы давление кислорода на конце диффузионного-пути приближалось к нулю. Эритроциты, в которых пути диффузии очень коротки, разве только в редких случаях оказываются' в среде со столь низким давлением О2 (если не говорить о крови в капиллярах напряженно работающих мышц).
Иначе обстоит дело с миоглобином. На некотором расстоянии: от капилляра напряжение кислорода в работающей мышце легко» может приблизиться к нулю. Это создает идеальные условия для облегченной диффузии, так как напряжение О2 в капилляре лежит где-то между артериальным и венозным уровнями, а в ткани (на конце диффузионного пути) почти равно нулю. Ускорение-диффузии в несколько раз, наблюдавшееся в лабораторных экспериментах, действительно в существенной мере улучшало бы! снабжение мышц кислородом. Эксперименты на грудной мышце-голубя подтвердили, что миоглобин может транспортировать значительную долю кислорода, потребляемого мышцей (Wittenberg; et al., 1975).
Кажется особенно примечательным то, что беспозвоночные, обладающие гемоглобином, часто живут в местах, где хотя бы вре
126 Глава 3. Кровь
менами напряжение кислорода бывает низким. Когда кислорода мало, напряжение его в тканях сильно снижается, и облегчение диффузии может играть очень важную роль. В таких случаях гемоглобин имеет особое значение, и многие животные приспосабливаются к длительным периодам нехватки кислорода путем увеличения количества гемоглобина в организме. Можно думать, что главная функция гемоглобина у этих животных — облегчение диффузии кислорода.
Действительно, многие нематоды, паразитирующие в кишечнике у овец, дышат более интенсивно, чем это, согласно расчетам, могло бы быть при максимальной скорости входной диффузии кислорода без эффекта облегчения (Rogers, 1949). Однако в клетках этих животных очень много гемоглобина, и он мог бы ускорять диффузию кислорода при тех очень низких парциальных давлениях О2, которые имеют место в кишечнике хозяина.
Относительно роли гемоглобина у растений выдвигалось немало гипотез, но теперь она достаточно ясна. Накапливающие азот растения, такие, как клевер и соя, имеют на своих корнях небольшие клубеньки, в которых содержатся бактерии, использующие атмосферный азот для синтеза азотсодержащих соединений (фиксация азота). Эти клубеньки содержат гемоглобин, называемый леггемоглобином, который необходим для связывания азота. Он имеет чрезвычайно высокое сродство к Ог (Рбо = О,О4 мм рт. ст., или 0,005 кПа), и поток связанного леггемоглобином кислорода превышает поток растворенного кислорода в 10 000—100 000 раз. Доставляемый кислород поддерживает образование аденозинтрифосфата (АТФ), который расходуется в сложном восстановительном процессе, приводящем к фиксации азота (Wittenberg, 1977).
ЭВОЛЮЦИОННЫЕ АСПЕКТЫ
Весьма любопытно, что имеющий столь сложное строение гемоглобин распространен так широко и при этом «разбросан» по разным группам организмов. В добавление к тому, что он имеется почти у всех позвоночных, гемоглобин тут и там встречается и в ряде других типов, включая простейших, а также у растений и бактерий. Часто его находят только у одного или нескольких представителей какой-либо обширной группы; иногда он есть у паразита, но отсутствует у родственных свободноживущих форм, и т. д. Но когда была установлена роль гемоглобина в ускорении диффузии, особенности его распространения сделались более понятными. Структурную основу гема — небелковой части молекулы гемоглобина — составляет порфириновое ядро, которое имеется у всех растений и животных в молекулах клеточных ферментов, принадлежащих к системе цитохромов. Относительно небольшие биохимические перестройки могут превратить эту исходную структуру в пигмент типа гемоглобина, обратимо связывающий кис
Перенос двуокиси углерода кровью 12Т
лород; чтобы такие мутации прочно закрепились, они должны давать какое-то эволюционное преимущество, которое способствовало бы их сохранению.
Самое очевидное и ближайшее преимущество — то, что гемоглобин улучшает доставку кислорода, облегчая его диффузию. После того как система синтеза гемоглобина сформировалась, дальнейшее улучшение транспорта кислорода может быть достигнуто путем добавления конвекционного транспорта (т. е. крови и циркуляторной системы),
Формирование полной циркуляторной системы — это сложное-изменение, которое потребовало бы не только относительно небольших химических модификаций порфирина, но и сложных морфологических и физиологических изменений во всем организме.-Такие радикальные изменения, конечно, возможны, но вряд ли они могут осуществиться в один или несколько этапов. Напротив,, появление гемоглобина в любой ткани сразу же обеспечивает организму выгоду, связанную с облегченной диффузией, и обусловливает селективное преимущество для закрепления нового признака. Следующим шагом могло быть создание условий для' включения гемоглобина в уже существующую кровеносную систему и тем самым — для дальнейшего улучшения доставки кислорода путем конвекции.
Из этой гипотетической последовательности событий вытекает,, что разрозненные случаи появления гемоглобина объясняются повсеместным присутствием железопорфириновых соединений, что* облегченная диффузия дает первый толчок для сохранения биохимического механизма синтеза гемоглобина и что добавление гемоглобина к уже существовавшей конвекционной системе логически становится следующим шагом в эволюционной истории.
ПЕРЕНОС ДВУОКИСИ УГЛЕРОДА КРОВЬЮ
По мере того как кровь проходит через ткани и отдает кислород, она попутно вбирает в себя СОг; когда она проходит через^ легкие, идет обратный процесс. Давайте проанализируем, что происходит в легких при отдаче двуокиси углерода.
Подходящая к легким венозная кровь млекопитающего содержит примерно 550 см3 связанной и свободной СОг на 1 л при напряжении около 46 мм рт. ст. (6,1 кПа). Когда кровь покидает легкие, она содержит около 500 см3 СО2 на 1 л при напряжении! около 40 мм рт. ст. Таким образом, кровь отдает лишь малую долю содержащейся в ней двуокиси углерода: различие между концентрациями СОг в артериальной и в венозной крови составляет всего около 50 мл на 1 л. Чтобы понять, как осуществляется транспорт двуокиси углерода, нам нужно лучше ознакомиться с* ее свойствами и поведением в растворе.
128 Глава 3. Кровь
Общее количество двуокиси углерода, переносимое кровью, намного больше того, которое было бы растворено в воде при том напряжении СО2, какое имеется в крови. Это связано с тем, что •большая часть двуокиси углерода переносится в форме других химических соединений, а не в виде свободного растворенного газа— СО2. Из таких соединений наиболее важны два: 1) ионы бикарбоната и 2) соединение СО2 с гемоглобином. (Отметим, что двуокись углерода присоединяется к концевым аминогруппам белковой молекулы, а не к месту связывания кислорода.)
Когда двуокись углерода растворяется в воде, она соединяется с водой и образует угольную кислоту (Н2СО3). Это можно записать следующим образом:
СО2 + Н2О ^=> Н2СО3 н+ + нсо3- (Н+ + СО32-).
Угольная кислота Н2СО3 диссоциирует на ион водорода и бикарбонат-ион (НСО3~). Она ведет себя как очень слабая кислота с кажущейся константой диссоциации порядка 8,0-10~7 при 37°С (р/С=6,1)1. Для полноты в приведенное уравнение включена дальнейшая диссоциация бикарбонат-иона еще на один ион водорода и карбонат-ион (СО32~), но при величинах pH, характерных для живых организмов, количество карбонат-ионов ничтожно, и обычно его можно не принимать во внимание.
При нормальном для крови млекопитающих pH большая часть всей двуокиси углерода находится в форме бикарбонат-ионов. Это можно легко вычислить из уравнения Хендерсона — Хассельбаха следующим образом:
1Н+] [А-] „
[НА] л’
рН-р/С— log^j-;
1 В действительности угольная кислота является более сильной кислотой с истинным значением рК около 3,8. Причина такого расхождения в значениях рК— то, что только 0,5% растворенной двуокиси углерода соединяется с водой и образует кислоту. Основная масса СО2 остается в виде растворенного газа, и только 0,005 гидратируется и образует Н2СО3. В физиологии принято считать, что вся двуокись углерода существует в виде угольной кислоты; в связи с этим угольная кислота выглядит более слабой, чем она есть в действительности. Если к истинному значению рХ для угольной кислоты (3,8) мы прибавим отрицательный логарифм гидратационной константы (log 0,005=—2,3), сумма будет 6,1, что соответствует кажущемуся значению рК для СО2, используемому в физиологии. В этой книге мы будем следовать трактовке, принятой в физиологии, и будем считать, что вся растворенная двуокись углерода образует Н2СО3.
Перенос двуокиси углерода кровью 129
для р/С=6,1 и при pH = 7,4
-log 1^-=рН-р/<=7,4-6,1=1,3;
log=-1,3=0,7-2;
[НА] _ [Н2СО3] __плк_ 1 [А“] [НСО3-] ’ 20 *
Это означает, что при pH 7,4 количественное отношение угольной кислоты к бикарбонат-ионам равно 1 :20, т. е. на каждую часть СО2, присутствующей в виде кислоты, приходится в 20 раз больше СО2, присутствующей в виде бикарбонат-ионов.
Ион водорода, появляющийся в результате ионной диссоциации угольной кислоты, взаимодействует с различными буферными веществами, содержащимися в крови, в результате чего СО2 оказывает лишь умеренное влияние на pH крови. Например, артериальная кровь может иметь в норме pH 7,45, а венозная — 7,42; таким образом, различие в pH, создающееся за счет поглощения кровью двуокиси углерода в тканях, не превышает 0,03.
Наиболее важными буферными веществами крови являются компоненты системы угольная кислота — бикарбонат, фосфаты и белки. Белки превосходно выполняют роль буферов, так как они содержат группы, которые могут диссоциировать и как кислоты, и как основания, в результате чего белки могут и отдавать, и принимать ионы водорода. Гемоглобин — это белок, содержащийся в крови в наибольшем количестве, и он играет главную роль в буферном действии; белкам плазмы принадлежит вторая по значению роль в этом отношении.
КРИВАЯ ДИССОЦИАЦИИ ДЛЯ ДВУОКИСИ УГЛЕРОДА
Количество двуокиси углерода, поглощаемое кровью, зависит от ее парциального давления. Таким образом, для крови можно построить кривую диссоциации СО2, аналогичную кривой кислородной диссоциации. Для этого откладывают по оси ординат общее количество двуокиси углерода в крови, а по оси абсцисс — парциальное давление СО2. В результате получаются кривые вроде представленных на рис. 3.14. На этом графике показаны слегка различные кривые диссоциации для оксигенированной крови и для крови, лишенной кислорода. Различие объясняется тем, что оксигемоглобин— несколько более сильная кислота, чем гемоглобин; поэтому насыщенная кислородом кровь несколько хуже связывает двуокись углерода. Это явление тесно связано с эффектом Бора, в котором просто выявляется другая сторона медали. Когда к крови добавлена двуокись углерода (кислота), она сдвигает равнове-
1Я73
130 Глава 3. Кровь
сие между оксигемоглобином и гемоглобином в сторону более слабой кислоты (гемоглобина). Это способствует освобождению большего количества кислорода, в чем, собственно, и состоит эффект Бора.
Точка, отмеченная на рис. 3.14 буквой Л, соответствует нормальной артериальной крови, а точка, отмеченная буквой V,— смешанной венозной крови (венозная кровь никогда полностью
Рсо2, *Па
Рис. 3.14. Кривые диссоциации для СО2, полученные в опытах с кровью млекопитающих. (Winton, Bayliss, 1955.)
Для оксигенированной и дезоксигенированной крови кривые несколько различаются. Линия А — V представляет собой физиологическую кривую диссоциации in vivo, отражающую различие между артериальной и венозной кровью, Прерывистая линия — количество СО2, удерживаемое в воде в случае физического раствора.
не освобождается от кислорода). Реальная кривая диссоциации СО2 для крови в живом организме — это А—V; ее называют функциональной кривой диссоциации.
На рис. 3.14 показана также кривая диссоциации для раствора бикарбоната натрия с тем же щелочным резервом, что и у крови. Наклон верхней части этой кривой представляет растворимость двуокиси углерода в водной среде. По мере того как давление СО2 падает, растворенная СО2 выделяется. Когда давление двуокиси углерода снизится примерно до 10 мм рт. ст., бикарбо-натный раствор начнет отдавать СО2 из бикарбонат-ионов, оставляя взамен карбонаты.
Перенос двуокиси углерода кровью 131
Если поместить раствор бикарбоната натрия в вакуум, то у этого бикарбоната может быть отнята половина всей двуокиси углерода, а в растворе вместо этого останется карбонат натрия (Na2CO3). Однако карбонат натрия не отдает двуокись углерода в вакуум, и получившийся в конце концов раствор содержит половину того количества СО2, которое первоначально присутствовало в форме бикарбоната. В отличие от этого помещенная в вакуум кровь отдает всю имевшуюся в ней двуокись углерода. Это происходит благодаря тому, что необходимые анионы образуются здесь из других веществ, главным образом белков. Если в вакуум поместить плазму крови, она по своему поведению будет больше напоминать раствор бикарбоната натрия: чтобы получилась нормальная кривая диссоциации, показанная на рис. 3.14, необходимо присутствие эритроцитов.
ДВУОКИСЬ УГЛЕРОДА ПРИ ВОДНОМ ДЫХАНИИ
Количество двуокиси углерода, образующейся в процессе метаболизма, тесно коррелирует с количеством потребляемого кислорода. Отношение между выделением СО2 и потреблением О2, называемое дыхательным коэффициентом, обычно находится в пределах между 0,7 и 1,0 (см. гл. 6). В последующем обсуждении мы будем рассматривать животное, у которого дыхательный коэффициент равен 1 (т. е. на каждую молекулу поглощенного О2 выделяется в точности одна молекула СО2).
Для животного, дышащего воздухом, например млекопитающего, у которого дыхательный коэффициент равен 1, концентрацию двуокиси углерода в выдыхаемом воздухе легко предсказать. Если концентрация О2 уменьшается с 21% во вдыхаемом воздухе до 16% в выдыхаемом, то выдыхаемый воздух неизбежно должен будет содержать 5% СО2. Если это же выразить на языке парциальных давлений данных двух газов, то можно сказать, что при снижении давления О2 в используемом для дыхания воздухе на 50 мм рт. ст. давление СО2 возрастет на ту же величину — на 50 мм рт. ст. (6,7 кПа). Каждый миллиметр изменения Ро2 приведет к равному, но противоположному изменению РСо2.
В случае газообмена в воде при дыхательном коэффициенте, равном единице, количества кислорода и двуокиси углерода, конечно, тоже будут равны, но изменения парциальных давлений этих двух газов будут очень различны, так как сильно различаются их растворимости. Если, например, рыба извлечет две трети содержащегося в воде кислорода, уменьшив Ро2 со 150 до 50 мм рт. ст. (с 20 до 6,7 кПа), то увеличение Рсо2 составит лишь около 3 мм рт. ст. (при условии, что вода не забуферена; в забуфе-ренной среде увеличение может быть еще меньшим). В зависимости от температуры снижение Ро2 на 100 мм рт. ст. может 9*
132 Глава 3. Кровь
означать, что, скажем, из 1 л воды было извлечено 5 см3 О2. Введение взамен этого пяти миллилитров СО2 повысит Рсо2 всего лишь примерно на 3 мм рт. ст., так как растворимость двуокиси углерода очень велика — приблизительно в 30 раз больше растворимости кислорода.
Это соотношение между напряжениями СО2, создающимися при дыхании в воздухе и в воде,
кПо
о 5 ю 15 20
представлено на рис. 3.15. На графике показаны Рсо2> которые получаются при данном уменьшении ко-ю личества кислорода в дыхательной среде. Да-же если бы рыба смогла 5 « извлечь из воды весь qS кислород, давление СО2 в воде поднялось бы только до 5 мм рт. ст. (0,67 кПа). Поскольку двуокись углерода диффундирует через жаберный эпителий почти так
Рис. 3.15. Соотношение между Ро2 и Рсо2 в воздухе и воде в случае, когда дыхательный коэффициент (ДК) равен единице. (Rahn, 1966.). Объяснение в тексте.
же легко, как и кислород, Рсо2 в крови рыб никогда не может превысить 5 мм рт. ст. (у на
земных позвоночных оно
примерно в 10 раз больше). Поэтому в отношении влияния на кислотно-щелочной баланс водное дыхание сильно отличается от воз-
душного. Это особенно важно для животных, которые используют то воздушное, то водное дыхание, таких, как амфибии и дышащие воздухом рыбы. У этих животных благополучный переход из одной среды в другую требует существенной перестройки (регулирования кислотно-щелочного баланса.
СОДЕРЖАНИЕ ДВУОКИСИ УГЛЕРОДА В КРОВИ И pH
Мы видели, что у водных и наземных животных напряжение (или парциальное давление) СО2 в артериальной крови очень различно. Общее содержание СО2 в крови тоже сильно варьирует: оно обычно невелико у рыб (часто менее 100 см3 на 1 л) и может быть очень высоким у черепах (более 1000 см3 на 1 л). Таким образом, у позвоночных возможны большие различия и по напряжению, и по общему содержанию двуокиси углерода в крови. Напротив, в отношении pH крови мы находим у разных позвоночных гораздо большее единообразие, чем можно было бы ожидать, учитывая значительные различия в содержании СО2. Это обусловлено соответствующей адаптацией кислотно-щелочного равнове
Перенос двуокиси углерода кровью 133
сия, достигаемой в первую очередь путем регуляции количества натрия (часто называемого свободным бикарбонатом или щелочным резервом) в крови.
У различных позвоночных, как теплокровных, так и холоднокровных, pH крови обычно варьирует где-то в пределах от 7,4 до 8,2. Но если мы построим график зависимости pH от температуры тела, мы получим более закономерное распределение (рис. 3.16). При температурах около О °C pH часто достигает значений выше 8,0 а при 40 °C оно ближе к 7,5. Однако при любой данной температуре pH крови различных животных укладывается в узкий диапазон порядка 0,2 единицы pH. Важно помнить, что диссоциация воды и, следовательно, pH нейтральной среды изменяются с температурой. Нейтральная точка соответствует 7,0 только при комнатной температуре (25°C); ее смещение в зависимости от температуры показано на рис. 3.16 линией с пометкой «вода».
Рис. 3.16. Величины pH крови позвоночных (представителей всех классов), измеренные при нормальной для каждого животного температуре тела. Штриховкой указан диапазон наблюдаемых значений; сплошной линией внизу представлена зависимость нейтрального pH воды от температуры. (Rahn, 1966.)
У всех позвоночных pH крови отклоняется от нейтральной точки примерно на 0,6 единицы pH, всегда в щелочную сторону. Таким образом, хотя позвоночные и различаются значительно в отношении pH крови, они удивительно единообразны в отношении отклонения от нейтральности, если измерять pH при обычной
134 Глава 3. Кровь
температуре тела. Такое же единообразие мы находим и в отношении внутриклеточного pH, что очень важно для активности метаболических ферментов (Alalan et al., 1976).
Большое различие условий для достижения надлежащего кислотно-щелочного равновесия при жизни в воде и на суше создает большие физиологические трудности для животных, которые в течение жизни постоянно переходят из одной среды в другую. У рыбы, обычно живущей в воде, кислотно-щелочной баланс приспособлен к напряжению СО2 в 2—3 мм рт. ст. Если эта рыба переходит на воздушное дыхание, напряжение СО2 в крови резко возрастает; это требует адаптаций всей буферной системы крови (в первую очередь изменения концентрации ионов натрия) или каких-либо вспомогательных механизмов для удаления двуокиси углерода. Амфибоидные животные выделяют значительные количества СО2 через кожу, и это помогает как двоякодышащим рыбам, так и водным амфибиям. Если животное выходит на сушу и становится полностью наземным, оно не может сохранить высокопроницаемую кожу из-за опасности обезвоживания. Если бы ему приходилось поддерживать низкую величину Рсо2 путем вентиляции легких, вентиляция должна была бы чрезмерно возрасти (по сравнению с тем, что необходимо для снабжения кислородом) и испарение стало бы серьезной проблемой. Таким образом, главной предпосылкой для перехода к постоянной жизни на суше является радикальная перестройка регуляции кислотно-щелочного равновесия.
КАРБОАНГИДРАЗА И СКОРОСТЬ
ВЗАИМОДЕЙСТВИЯ ДВУОКИСИ УГЛЕРОДА С ВОДОЙ
Реакции, происходящие при растворении двуокиси углерода в воде (см. уравнения, приведенные ранее в этой главе), совершаются не мгновенно. Когда СО2 попадает в воду и растворяется, начальная гидратация ее молекулы с образованием Н2СО3 протекает относительно медленно. Обратная реакция — высвобождение двуокиси углерода из Н2СО3— тоже идет не быстро: для него требуется от нескольких секунд до долей минуты. (Диссоциация Н2СО3 на Н+ и НСО3~ и обратный процесс совершаются, в физиологическом масштабе времени, мгновенно). Если мы учтем, что время пребывания крови в капилляре обычно составляет доли секунды, то встанет вопрос: как она успевает извлекать СО2 из тканей с той быстротой, с которой она это делает, и затем выделять эту СО2 в легочных капиллярах за то короткое время, что она там находится?
Эта проблема, по-видимому, разрешилась с окрытием фермента, который настолько ускоряет образование двуокиси углерода из Н2СО3, что этот процесс становится очень быстрым. Фермент
Перенос двуокиси углерода кровью 135
получил название карбоангидразы, которое не вполне корректно, так как ускоряется и соединение СО2 с водой, и ее освобождение из угольной кислоты (т. е. процесс катализируется в обоих направлениях). В плазме крови фермент отсутствует, но он содержится в высокой концентрации внутри эритроцитов. Он встречается и в других органах, особенно в железистых, таких, как почки, секреторный эпителий желудка, поджелудочная железа и слюнные железы, но сейчас это не будет нас интересовать. Следует, однако, заметить, что действие карбоангидразы высокоспецифично: кроме гидратации СО2 никаких других ее функций в организме не известно.
Через несколько лет после открытия карбоангидразы было синтезировано несколько мощных ингибиторов этого фермента, которые стали применяться как эффективные лекарственные препараты, особенно при определенных нарушениях функции почек. К большому удивлению физиологов, оказалось, что такие ингибиторы очень мало токсичны и что даже полное торможение карбоангидразы крови оказывает лишь незначительное влияние на транспорт СО2. Иными словами, фермент не играет особо важной роли в переносе двуокиси углерода, хотя подавление его активности и приводит к некоторым изменениям кислотно-щелочного равновесия.
Для выяснения возможной роли карбоангидразы могли бы оказаться полезными данные о ее количестве у различных животных. Мы знаем, что у мелких животных интенсивность метаболизма больше, чем у крупных. Мелкие животные потребляют на единицу веса тела больше кислорода и выделяют больше СО2; поэтому двуокись углерода должна высвобождаться в их легких за более короткое время. Анализ зависимости между активностью карбоангидразы в крови и весом тела показывает, что в крови мелких животных этот фермент действительно находится в значительно более высоких концентрациях (рис. 3.17). Это как будто согласуется с потребностями в переносе СО2, однако фермент присутствует в гораздо больших количествах, чем было бы нужно для этой цели. Между тем есть еще один процесс, для которого важна быстрая гидратация двуокиси углерода. Когда СО2 поступает в кровь тканевых капилляров, присутствие карбоангидразы способствует мгновенному образованию угольной кислоты, которая в свою очередь сразу же воздействует на диссоциацию чувствительного к кислоте оксигемоглобина. Эта чувствительность к кислоте (эффект Бора), очевидно, не успевала бы проявиться в период пребывания крови в капилляре, если бы гидратация молекулы СО2 не ускорялась ферментом. Значит, в отсутствие карбоангидразы эффект Бора не мог бы иметь большого значения.
Таким образом, можно думать, что карбоангидраза—фермент, специфически катализирующий гидратацию молекулы СО2, — возможно, не играет большой роли в транспорте двуокиси углерода,
(36 Глава 3. Кровь
но способствует эффективной доставке кислорода тканям. Поскольку ингибирование фермента у животных в норме не причиняет им большого вреда, этот фермент, по-видимому, важен главным образом в условиях, предъявляющих особые требования к механизму транспорта газов.
Рис. 3.17. Активность карбоангидразы в крови млекопитающих связана с размерами тела: она тем выше, чем меньше животное. (Magid, 1967.)
В других органах, где имеется карбоангидраза, она, по-видимому, всегда связана с функциями, для которых важны процессы переноса ионов, например в железистых тканях (в почках, поджелудочной железе, слюнных железах) и в цилиарном теле глаза. Ингибирование карбоангидразы оказывает глубокое влияние на эти процессы, что служит основой для широкого использования ее ингибиторов как в клинической медицине, так и в физиологических исследованиях.
Рассмотрев физиологические свойства крови и механизмы переноса ею газов, мы можем перейти к следующей главе, где речь будет идти о движении крови внутри организма.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Aste-Salazar Н., Hurtado А. (1944). The affinity of hemoglobin for oxygen at sea. level and at high altitudes, Am. J. Physiol., 142, 733—743.
Barcroft J. (1935). Foetal respiration, Proc. R. Soc. Lond., B, 118, 242—263.
Benesch R., Benesch R. E., Enoki У. (1968a). The interaction of hemoglobin and its subunits with 2,3-diphosphoglycerate, Proc. Natl. Acad. Sci. USA, 61, 1102—1106.
Кровь 137
Benesch R., Benesch R. E., Yu С. I. (1968b). Reciprocal binding of oxygen and diphosphoglycerate by human hemoglobin, Proc. Natl. Acad. Sci. USA, 59, 526— 532.
Bunn H. F. (1971). Differences in the interaction of 2,3-diphosphoglycerate with certain mammalian hemoglobins, Science, 172, 1049—1050.
Chanutin A., Curnish R. R. (1967). Effect of organic and inorganic phosphates on the oxygen equilibrium of human erythrocytes, Arch. Biochem. Biophys., 121, 96—102.
Gillen R. G., Riggs A. (1973). Structure and function of the isolated hemoglobins of American eel, Anguilla rostrata, X Biol. Chem., 248, 1961—1969.
Grigg G. C. (1969). Temperature-induced changes in the oxygen equilibrium curve of the blood of the brown bullhead, Ictalurus nebulosus, Comp. Biochem. Physiol., 28, 1203—1223.
Hall F. G., Dill D. B., Barron E. S. G. (1936). Comparative Physiology in high altitudes, J. Cell. Comp. Physiol., 8, 301—313.
Hall F. G., McCutcheon F. H. (1938). The affinity of hemoglobin for oxygen in marine fishes, J. Cell. Comp. Physiol., 11, 205—212.
Hemmingsen E. A. (1965). Accelerated transfer of oxygen through solutions of heme pigments, Acta Physiol. Scand. Suppl., 246, 1—53.
Lenfant C., Torrance J., English E., Finch C. A., Reynafarje C., Ramos J., Faura L (1968). Effect of altitude on oxygen binding by hemoglobin and on organic phosphate levels, J. Clin. Invest., 47, 2652—2656.
Lenfant C., Ways P., Aucutt C., Cruz J. (1969). Effect of chronic hypoxic hypoxia on the O2-Hb dissociation curve and respiratory gas transport in man, Respir. Physiol., 7, 7—29.
Lutz P. L.} Longmuir I. S., Tuttle J. V., Schmidt-Nielsen К (1973). Dissociation curve of bird blood and effect of red cell oxygen consumption, Respir. Physiol1., 17, 269—275.
Magid E, (1967). Activity of carbonic anhydrase in mammalian blood in relation to body size, Comp. Biochem. Physiol., 21, 357—360.
Malan A.f Wilson T. L., Reeves R. B, (1976). Intracellular pH in cold-blooded vertebrates as a function of body temperature, Respir. Physiol., 28, 29—47.
Manwell C. (1958). The oxygen-respiratory pigment equilibrium' of the hemocyanin and myoglobin of the amphineuran mollusc Cryptochiton stelleri, J. Cell. Comp. Physiol, 52, 341—352.
Manwell C. (1960). Histological specificity of respiratory pigments. 2. Oxygen transfer systems involving hemerythrins in sipunculid worms of different ecologies, Comp. Biochem. Physiol, 1, 277—285.
Millikan G. A. (1937). Experiments on muscle haemoglobin in vivo: The instantaneous measurement of muscle metabolism, Proc. R. Soc. Lond, B, 123, 218— 241.
Rahn H. (1966). Gas transport from the external environment to the cell. In: Development of the Lung (A. V. S. de Reuck and R. Porter, eds.), pp. 3—23 London, CIBA.
Randall D. J. (1970). Gas exchange in fish. In: Fish Physiology, vol. 4 (W. S. Hoar and D. J. Randall, eds.), pp. 253—292, New York, Academic Press.
Read К R. H. (1962). The hemoglobin of the bivalved mollusc, Phacoides pectina-tus Gmelin, Biol. Bull, 123, 605—617.
Riggs A. (1951). The metamorphosis of hemoglobin in the bullfrog, J. Gen. Physiol, 35, 23—44.
Riggs A. (1960). The nature and significance of the Bohr effect in mammalian hemoglobins, J. Gen. Physiol, 43, 737—752.
Rogers W. P, (1949). On the relative importance of aerobic metabolism in small nematode parasites of the alimentary tract. 2. The utilization of oxygen at low partial pressures by small nematode parasites of the alimentary tract, Aust. X Sci. Res, 2, 166—174.
138 Глава 3. Кровь
Root R. W., Irving L., Black E. C. (1939). The effect of hemolysis upon the combination of oxygen with the blood of some marine fishes, J. Cell. Comp. Physiol., 13, 303—313.
Schmidt-Nielsen K. (1972). How Animals Work, London, Cambridge University Press, 114 pp.
Schmidt-Nielsen K-, Pennycuik P. (1961). Capillary density in mammals in relation to body size and oxygen consumption, Am. J. Physiol., 200, 746—750.
Schmidt-Nielsen K-> Taylor C. R. (1968). Red blood cells: Why or why not? Science, 162, 274—275.
Scholander P. F. (1960). Oxygen transport through hemoglobin solutions, Science, 131, 585—590.
Winton F. R., Bayliss L. E. (1955). Blood and the transport of oxygen and carbon dioxide. In: Human Physiology (F. R. Winton and L. E. Bayliss, eds.), pp. 78— 118, Boston, Little, Brown
Wittenberg B. A., Wittenberg J. B., Caldwell P. R. B. (1975). Role of myoglobin in the oxygen supply to red skeletal muscle, J. Biol. Chem., 250, 9038—9043.
Wittenberg J. B. (1977). Facilitation of oxygen diffusion by intracellular leghemo-globin and myoglobin. In: Oxygen and Physiological Function (F. F. Jobsis, ed.), pp. 228—246, Dallas, Professional Information Library.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Altman P. L., Dittmer D. S. (eds.) (1971). Biological Handbooks: Blood and Other Body Fluids (3rd printing, with minor corrections), Bethesda, Federation of American Societies for Experimental Biology, 540 pp.
Bonaventura J., Bonaventura C.> Sullivan B. (1977). Non-heme oxygen transport proteins. In: Oxygen and Physiological Function (F. F. Jobsis, ed.), pp. 177— 220, Dallas Professional Information Library.
Howell B. J., Rahn H., Goodfellow D., Herreid C. (1973). Acid-base regulation and temperature in selected invertebrates as a function of temperature, Am. Zool., 13, 357—563.
MacFarlane R. G. (1970). The Haemostatic Mechanism in Man and Other Animals. Symposium of the Zoological Society of London, Academic Press, 248 pp.
Reeves R. B. (1977). The interaction of body temperature and acid-base balance in ectothermic vertebrates, Annu. Rev. Physiol., 39, 559-—586.
Riggs A. (1965). Functional properties of hemoglobins, Physiol. Rev., 45, 619— 673.
Rorth M., Astrup P. (eds.) (1972). Oxygen Affinity of Hemoglobin and Red Cell Acid Base Status, Proceedings, Alfred Benzon Symposium IV, Copenhagen, Munksgaard, 832 pp.
ГЛАВА 4
КРОВООБРАЩЕНИЕ
Главная роль циркулирующих в организме жидкостей состоит в том, что ойи быстро переносят различные вещества на такие расстояния, при которых диффузия была бы неэффективным или слишком медленным процессом. Поэтому циркуляция важна практически для всех животных величиной более нескольких миллиметров и совершенно необходима для всех крупных животных с интенсивным обменом веществ. Транспорт газов— кислорода и двуокиси углерода — существенная, но далеко не единственная функция циркуляторной системы.
Главные функции крови (перечисленные в табл. 3.1) распадаются на три основные категории: 1) массовый транспорт растворенных веществ и клеток; 2) перенос тепла; 3) передача силы. Функции, связанные с передачей силы, имеют отношение главным образом к движению отдельных органов, локомоции (передвижению животного в целом) и созданию давления для ультрафильтрации в почечных капиллярах; эти функции будут рассмотрены в другом месТе. В настоящей главе мы ознакомимся с переносом тепла и растворенных веществ (включая газы). Система, пригодная для конвективного транспорта кислорода, как правило, годится и для транспорта других растворенных веществ. Поэтому нас будет в первую очередь интересовать значение, циркуляции для переноса газов, в особенности кислорода.
Циркуляторные системы позвоночных хорошо изучены, поэтому с них мы и начнем. Функция большинства циркуляторных си-: стем беспозвоночных известна хуже; во многих случаях морфолог гия исследована весьма детально, а данные о функции часто недостаточны. В результате мы имеем мало возможностей для обобщений, и потому главное внимание в этой главе будет уделено высокоразвитым и более изученным циркуляторным системам позвоночных.
ОБЩИЕ ПРИНЦИПЫ
Работоспособная циркуляторная система состоит из одного или нескольких насосов и разного рода каналов, по которым, может течь кровь. Действие насоса, или сердца, основано на способности мышц сокращаться. Слой мышечной ткани, окружающей трубку или камеру, сокращаясь, может уменьшать ее объем. Та: ким путем могут создаваться насосы двух различных типов: tie*
140 Глава 4, Кровообращение
ристальтические насосы и камерные насосы с клапанами (рис. 4.1). В камерных насосах уменьшение объема может достигаться либо с помощью сократимых стенок (рис. 4.1,5), либо с помощью давления снаружи, со стороны других частей тела (рис. 4.1,5).
Перистальтические сердца встречаются главным образом у беспозвоночных; почти все без исключения позвоночные имеют серд-
Рис. 4.1. Три типа насосов, обеспечивающих движение крови в системе кровообращения. В случае перистальтического насоса (А) область сокращения перемещается вдоль трубки и толкает кровь перед собой. В случае обычного камерного насоса (Б) ритмические сокращения стенок выталкивают кровь из камеры. Клапаны препятствуют обратному току, и в результате кровь всегда движется в одном направлении. В камерном насосе другого типа (В) кровь выжимается из трубки под давлением окружающих тканей. В изображенном здесь случае давление создают мышцы, а клапаны определяют направление тока. К этому типу принадлежат венозные насосы в ногах человека.
це камерного типа с сократимыми стенками. Типичными насосами, работающими за счет внешнего давления, служат крупные вены в ногах человека. Стенки этих вен относительно тонки, и, кроме того, эти вены снабжены клапанами, препятствующими обратному току крови; мышцы ног, сокращаясь, сжимают вены, а клапаны заставляют кровь двигаться по направлению к сердцу. Такая перекачка сильно помогает перемещению крови из венозной системы ног против силы тяжести, которая способствует задержке крови в ногах.
Общие принципы 141
Поскольку мышцы могут создавать силу только путем укорочения, сердце не сможет засасывать в себя кровь, если для этого нет специального приспособления, какое имеется, например, у акул. Сердце акулы заключено в жесткую капсулу, и когда одна его камера сокращается, другая камера наполняется кровью путем всасывания, причем клапаны препятствуют обратному притоку крови с артериальной стороны. Подробнее это будет описано позже в этой главе.
Циркулирующая жидкость, или кровь, разносится от сердца по проводящим каналам и в конце концов возвращается обратно к сердцу. У позвоночных кровь заключена в систему эластичных трубок (артерий, капилляров, вен). К сердцу кровь возвращается, не выходя из этой системы, и поскольку кровь всегда остается внутри замкнутого объема, систему кровообращения позвоночных называют замкнутой. У многих беспозвоночных (например, насекомых, большинства ракообразных и многих моллюсков) кровь переходит из сердца в кровеносные сосуды, которые обрываются, так что кровь, прежде чем в конце концов вернуться к сердцу, более или менее свободно растекается между тканями. Такую систему называют незамкнутой. Некоторые особенности незамкнутой и замкнутой систем перечислены в табл. 4.1.
Таблица 4.1
Основные различия между замкнутыми и незамкнутыми системами кровообращения
| Замкнутые системы Незамкнутые системы
Давление в системе обычно велико Давление обычно небольшое
Создание высокого давления требует замкнутого пространства и сопротивления Создание высокого давления невозможно
Для поддержания высокого давления в промежутках между сокращениями сердца требуются эластичные стенки Поддержание давления невозможно
Кровь подводится прямо к органам Так же, как в замкнутых системах
Распределение потоков к различным органам может регулироваться Распределение крови регулировать трудно
Кровь возвращается к сердцу быстро Кровь возвращается медленно
Замкнутые системы кровообращения имеются у позвоночных, головоногих моллюсков (осьминога) и иглокожих, а незамкнутые— у большинства членистоногих, большей части моллюсков (кроме головоногих) и оболочников. Циркуляторная система оболочников интересна в нескольких отношениях. Это незамкнутая"
142 Глава 4. Кровообращение
система, в которой трубкообразное сердце проталкивает кровь с помощью перистальтической волны, проходящей от одного конца к другому. Такое сердце не имеет клапанов. Произведя серию сокращений— скажем, несколько дюжин или сотню, — сердце постепенно замедляет свою работу и останавливается. После паузы волна меняет направление, и кровь теперь перекачивается в другую сторону (Krijgsman, 1956). Это довольно необычное устройство: в большинстве циркуляторных систем сердце перекачивает кровь всегда в одну сторону и она течет только в одном направлении.
КРОВООБРАЩЕНИЕ У ПОЗВОНОЧНЫХ
РАСПРЕДЕЛЕНИЕ ВОДЫ В ОРГАНИЗМЕ II ОБЪЕМ КРОВИ
Тело позвоночного примерно на две трети состоит из воды. Точные цифры сильно варьируют, главным образом в связи с различиями в количестве жира. Жировая ткань содержит только около 10% воды, и у очень тучных особей, у которых жир может составлять больше половины веса тела, общее содержание воды сильно понижено. Если количество воды выражать в процентах от веса тела за вычетом жира, цифры будут варьировать намного' меньше. Большая часть воды заключена внутри клеток (внутриклеточная вода); меньшая доля находится снаружи от клеточных мембран (внеклеточная вода). Внеклеточная вода содержится частично в тканевых пространствах (интерстициальная жидкость) г а частично — в крови (вода плазмы). Таким образом, мы можем: считать, что вода в организме распределена между тремя ком-партментами и подразделяется соответственно на внутриклеточную воду, интерстициальную воду и воду плазмы (рис. 4.2).
Объем этих трех компартментов можно измерять различными способами, часто без всякого вреда для организма. Один из самых удобных способов — ввести подходящее вещество, которое равномерно распределялось бы в измеряемом компартменте. Тогда по данным о разведении этого вещества можно вычислить тот объем, по которому оно распределилось. Таким способом легче всего оценивать объем плазмы, объем межклеточной жидкости и общий объем воды в организме.
Общий объем крови нельзя измерить точно, выпуская всю» кровь, поскольку значительное ее количество все равно останется в сосудистой системе. Между тем метод разбавления дает надежные и воспроизводимые результаты. Одно из обычных веществ,, используемых для этой цели, — краситель, называемый синим: Эванса (известный также под названием Т-1824). Небольшая доза этого красителя вводится в виде водного раствора в кровяное* русло; краситель связывается белками плазмы и остается внутри сосудистой системы. Когда произойдет полное перемешивание (че-.
Кровообращение у позвоночных 143
рез несколько минут), берут пробу крови и измеряют концентрацию красителя в плазме. После этого по его разведению легко вычислить объем плазмы. Общий объем крови можно оценить, определив содержание в ней эритроцитов (гематокрит) путем центрифугирования пробы крови.
Рис. 4.2. Примерное содержание воды и твердых веществ в теле позвоночных, выраженное в процентах от веса тела без жира. Более подробные данные приведены в табл. 4.2.
Объем внеклеточной жидкости определяют путем инъекции вещества, которое проходит через стенки капилляров и распределяется по крови и межклеточному пространству, но не проникает внутрь клеток. Одно из таких веществ — полисахарид инулин (используемый также при исследовании функции почек; см. гл. 10). Инулин, имеющий мол. вес около 5000, равномерно распределяется по всему межклеточному пространству, но не включается в обмен веществ и не проникает в клетки. Поэтому степень его разбавления служит показателем величины всего внеклеточного объема. Объем интерстициальной жидкости можно вычислить как разность между общим внеклеточным объемом и объемом плазмы.
Конечно, общий объем воды в организме можно определить, забив животное и измерив содержание в нем воды путем высушивания. Однако метод разведения представляется менее варварским и более удобным. В настоящее время чаще всего для этой цели используют воду, меченную тритием — радиоактивным изотопом водорода. Делают инъекцию небольшого количества меченой воды; эта вода свободно диффундирует через стенки капилляров и клеточные мембраны и вскоре равномерно распределяется во всей жидкости тела. Чтобы вычислить объем, в котором рас
144 Глава 4. Кровообращение
пределилась введенная меченая вода, достаточно определить радиоактивность пробы любой жидкости тела. Не обязательно даже брать пробу крови: если перед экспериментом мочевой пузырь был опорожнен, проба свежей мочи даст ту же цифру, что и проба любой другой жидкости организма.
Таблица 4.2
Распределение воды в организме у представителей разных групп позвоночных. Все величины указаны в процентах от веса тела. Небольшие различия недостоверны и могут объясняться различием в методах исследования, в количестве жира у разных животных и т. п. Однако объем крови у костистых рыб действительно намного меньше, чем у других позвоночных.
Животное Общее количество воды, % Внутриклеточная вода, % Внеклеточная вода, % Объем крови, %
Минога1) 76 52 24 8,5
Амия2) Карп (пресновод- 71 58 13 6,8
ная рыба)3) Луциан (морская 71 56 15 3,0
рыба)3) 71 57 14 2,2
Лягушка-бык4) 79 57 22 5,3
Аллигатор5) 73 58 15 5,1
Гоферная змея5) 70 52 17 6,0
Голубь6) Виргинский фи- — — — 9,2
лин6) —. — — 6,4
Собака7) ’) Thorson, 1959. 2) Thorson, 1958. 3) Thorson, 1961. 4) Thorson, 1964. 5) Thorson, 1968. 6) Bond, Gilbert, 1958. 7) Hopper et al., 1944. * 63 37 19 9,1
В табл. 4.2 приведены данные об общем объеме воды и объе: ме внутриклеточной и внеклеточной воды для некоторых позвоночных. Общий объем воды в теле водных и наземных позвоночных одного порядка, около 70%, однако имеются отклонения в обе стороны. Распределение между внутриклеточным и внеклеточным компартментами также весьма однотипно, однако и тут есть, исключения.
У позвоночных количество крови обычно варьирует в пределах ox Jrдо. 10% рт веса тела; если не считать костистых рыб,, у которых оноДкак'Правило, составляет 2ггЗ% от. в.еса тела; У пластю
Кровообращение у позвоночных 145
ножаберных и круглоротых соответствующие величины лежат в-интервале от 6 до 10%, а у позвоночных, дышащих воздухом (амфибий, рептилий, птиц и млекопитающих), — в интервале от 5 до 10%. Таким образом, можно сказать, что в целом у позвоночных относительный объем крови составляет 5—10%, за исключением костистых рыб, у которых он существенно меньше. Пока еще не ясно, какое функциональное значение может иметь это различие.
СХЕМЫ КРОВООБРАЩЕНИЯ
Позвоночные, принадлежащие к одному классу, имеют однотипные схемы кровообращения, но различия между классами весьма значительны. По мере перехода позвоночных от водной жизни к наземной происходило усложнение циркуляции.
Рис. 4.3. В системе кровообращения рыб кровь из сердца проталкивается через* жабры и далее течет к тканям. У млекопитающих кровь проходит через легкие-и возвращается в сердце, прежде чем направиться в ткани под действием второго насоса. Благодаря такому устройству у млекопитающих в тканях может создаваться более высокое кровяное давление, чем в легких.
Устройство системы кровообращения у млекопитающих определяет два важных ее свойства, которые сразу видны из рис. 4.3. Во-первых, количество крови, протекающей через легкие, должн’о* быть равно ее количеству, протекающему через остальные части-тела (если не учитывать незначительных колебаний, которые могут вызываться небольшими изменениями ударного объема сердца в течение одного или нескольких его сокращений). Во-вторых, поскольку обе половины сердца сокращаются одновременно, весь, выталкиваемый .объем лкрови доджей размещаться за счет изменения-объема эластичных кровеносных сосудов.
10—1873
146 Глава 4. Кровообращение
Миксины
Пласгпиножаберные и костные рыбы
Двоякодышащие рыбы
Лягушки
Рис. 4.4. Основные схемы кровообращения у разных классов позвоночных. Темными участками
Кровообращение у позвоночных 14Т
Рептилии (кроме крокодилов)
Крокодилы
Млекопитающие
Птицы
представлены потоки венозной крови, белыми — потоки артериальной крови, богатой кислородом.
148 Глава 4. Кровообращение
У рыб и млекопитающих мы находим два крайних типа кровообращения. На рис. 4.4 показано постепенное разделение сердца на два отдельных насоса по мере перехода позвоночных от водной жизни к полностью наземному типу дыхания. Каждый класс позвоночных имеет свои характерные черты, и для объяснения различий между ними нужны некоторые комментарии.
вспомогательным сердцам относятся
Хрящ
Рис. 4.5. Каудальное сердце миксины отличается от других сердец позвоночных. Центральный хрящ изгибается под действием попеременных сокращений мышц, расположенных по обе стороны и служащих к тому же стенками двух камер. Когда мышцы одной стороны сокращаются, камера этой стороны наполняется кровью, а из другой камеры кровь выталкивается. Клапаны помогают обеспечить однонаправленный ток крови. (Gordon, 1972.)
Особенно интересны каудальные
КРУГЛОРОТЫЕ
Система кровообращения миксин отличается от циркуляторной системы всех других позвоночных. Это не замкнутая система, как у других позвоночных: в ней имеются большие кровяные синусы. Примечательная особенность ее состоит в том, что кроме основного (жаберного) сердца миксины имеют несколько добавочных сердец, главным образом в венозной системе (рис. 4.4,А). К этим воротное (портальное) сердце, получающее венозную кровь из кардинальной вены и от кишечника и накачивающее эту кровь в печень, кардинальные сердца, которые расположены по ходу кардинальных вен и помогают проталкивать кровь, и каудальные сердца — парные расширения каудальных вен. Все эти вспомогательные сердца находятся в венозной системе, а в продвижении вперед артериальной крови активное участие принимают жабры. Это достигается сокращением поперечнополосатых мышечных элементов в жабрах и жаберных протоках, которое помогает проталкивать кровь в артериальной системе (Johansen, 1960).
сердца миксин, так как по
своему устройству они отличаются от всех других сердец (рис. 4.5). Две камеры разделяет продольный стержень из хряща, а попеременные сокращения мышц с обеих сторон заставляют этот стержень изгибаться. Когда мышцы одной из сторон сокращаются, мышцы другой стороны обеспечивают давление для изгнания крови из своей камеры. Одновременно на сокращающейся стороне объем камеры увеличивается, и она наполняется кровью. Благода
Кровообращение у позвоночных 149
ря поочередным сокращениям обе камеры наполняются и опоражниваются в противофазе, а надлежащая работа клапанов обеспечивает однонаправленный ток крови.
Рис. 4.6. Перед двухкамерным сердцем рыбы расположен объемистый венозный синус, из которого кровь поступает в предсердие. Сердце выталкивает кровь в утолщенный участок артерии — артериальную луковицу (у костистых рыб) или артериальный конус (у пластиножаберных). (Randall, 1970.)
РЫБЫ
Схема кровообращения у рыб, одинаковая для костистых и пластиножаберных, показана на рис. 4.4, Б. Сердце состоит из двух последовательно соединенных камер — предсердия и желудочка. С венозной стороны перед сердцем находится расширенная камера — венозный синус (sinus veno-sus), который помогает поддерживать непрерывный приток крови к сердцу. У костистых рыб с артериальной стороны к сердцу непосредственно примыкает утолщенная мускульная часть брюшной аорты — артериальная луковица (bul-bus arteriosus; рис. 4.6). У пластиножаберных рыб имеется сходным образом расположенный утолщенный участок — артериальный конус (conus arteriosus), развившийся из сердечной мышцы. Он состоит из фиброзной ткани и снабжен клапанами, которые препятствуют обратному току крови в желудочек. Это особенно важно, так как сердце, расположенное в жесткой камере, может создавать отрицательные давления. Отрицательное давление способствует наполнению предсердия в результате «всасывания» крови из объемистого венозного синуса.
Последовательность со-
бытий, происходящих в сердце акулы на протяжении одного цикла сокращения, можно проследить по записям давления, приведенным на рис. 4.7. Во время сокращения желудочка давление
150 Глава 4, Кровообращение
крови в нем нарастает, и это давление передается в артериальный конус. Когда желудочек расслабляется, обратному току крови из конуса препятствуют клапаны, и поэтому высокое давление в конусе сохраняется и после того, как желудочек начал расслабляться.
Во время сокращения желудочка сердце уменьшается в объеме, и поскольку оно заключено в жесткую камеру, в этой камере
Рис. 4.7. Изменения давления в сердце и потока крови в вентральной аорте у’ Пластиножаберных на протяжении одного цикла сокращения. (Randall, 1970.) / — поток крови в аорте; II — давление в артериальном конусе; III — давление' 3 желудочке.
создается отрицательное давление. У акулы Порт-Джексона давление в желудочке во время его сокращения поднимается выше' 30 мм рт. ст. (4,0 кПа), а развивающееся при этом отрицательное давление в предсердии и венозном синусе может доходить до—4 мм' рт. ст. (—0,6 кПа) (Satchell, 1970). Если бы обширный синус и смежные вены не могли обеспечить равного по объему притока крови в предсердие при сокращении желудочка, то отрицательное* давление только препятствовало бы сокращению. Однако при наличии притока крови отрицательное давление не становится чрезмерным и просто служит для наполнения предсердия. Когда после; этого предсердие сокращается, уже расслабленный теперь желудочек начинает наполняться кровью из предсердия, так как обратному току крови в синус препятствуют клапаны. Таким образом,, в этой части цикла кровь просто перемещается из предсердия в; желудочек, причем объем содержимого перикарда остается неизменным (Randall, 1970).
Кровообращение у позвоночных 151
У позвоночных, дышащих воздухом, венозный синус и артериальная луковица имеют меньшее значение, а в сердце млекопитающих они и вовсе отсутствуют как отдельные структуры.
ДВОЯКОДЫШАЩИЕ РЫБЫ
Главное эволюционное приобретение двоякодышащих рыб — то, что в добавление к жабрам они имеют в качестве дыхательных органов еще и легкие (рис. 4.4, В). Жабры частично получают кровь, которая уже прошла через легкие. Если бы жабры были такими же, как у обычных рыб, это могло бы быть невыгодным моментом, так как двоякодышащая рыба, плавающая в бедной кислородом воде, тогда отдавала бы кислород из крови в воду, омывающую жабры. Однако у двоякодышащих рыб жабры подверглись дегенерации, и некоторые из жаберных дуг пропускают прямой сквозной поток крови _(Johansen et al., 1970).
Предсердие разделено перегородкой на две камеры, а желудочек тоже частично разделен. В этом отношении сердце двоякодышащих рыб несколько напоминает полностью разделенное сердце млекопитающих, птиц и крокодилов. В сердце двоякодышащих рыб даже больше структурных подразделений, чем у амфибий (Foxon, 1955). Из легких кровь возвращается в левое предсердие, а правое предсердие получает кровь из главного круга кровообращения. Частичное разделение желудочка не позволяет двум потокам крови сильно перемешиваться, так что оксигенированная кровь направляется главным образом в две первые жаберные дуги и голова относительно хорошо снабжается кислородом. Менее насыщенная кислородом кровь из правой половины сердца течет через задние жаберные дуги и попадает в спинную аорту и частично в легкие. Двоякодышащие рыбы сделали первый шаг к полному разделению легочного круга циркуляции крови и круга циркуляции через остальные части тела (Johansen et al., 1968).
Из анатомии двоякодышащих рыб не ясно, каким же образом в действительности течет кровь. Исследования на живой африканской рыбе Protopterus показали, что легочная циркуляция и общий круг кровообращения функционально в большой мере разделены, причем бедная кислородом кровь преимущественно направляется в легкие, а богатая кислородом — в сосуды всего тела. Особенно важно то, что это функциональное разделение достигает наивысшей степени непосредственно после вдыхания воздуха, когда содержание кислорода в легких максимально, тогда как позднее, в промежутке между вдохами, степень разделения уменьшается. Для эффективности газообмена в легких, которые наполняются свежим воздухом лишь через определенные интервалы времени, значение этого обстоятельства очевидно (Johansen et al., 1968).
152 Глава 4. Кровообращение
АМФИБИИ
Сердце современных амфибий (лягушек, жаб, саламандр) имеет два полностью разделенных предсердия и только один неразделенный желудочек (рис. 4.4,Г). Левое предсердие получает насыщенную кислородом кровь из легких, а правое — венозную кровь из большого круга кровообращения. Хотя желудочек не разделен, эти два потока крови почти не смешиваются, так что богатая кислородом кровь направляется в большой круг кровообращения, а кровь, бедная кислородом, течет отдельно в легочный круг.
Легочные артерии посылают ответвления также и в кожу; это имеет большое значение, так как влажная кожа амфибий играет значительную роль в поглощении кислорода. В артериальной луковице имеется продольный гребень (известный под названием, спирального клапана), который, по-видимому, существенно помогает разделению потоков крови.
РЕПТИЛИИ
У всех рептилий, кроме крокодилов, предсердия разделены полностью, а желудочек — только частично (рис. 4.4, Д). Даже и при этих условиях потоки насыщенной и бедной кислородом крови хорошо отделяются друг от друга и перемешивания крови поч^ ти нет; по существу,, мы имеем здесь хорошо развитую двойную циркуляцию. Таким образом, смысл неполного разделения желудочка у амфибий и рептилий (за исключением крокодилов) нельзя понять на основе одного лишь анатомического строения, которое само по себе позволяло бы предполагать смешение крови к желудочке.
Как правило, мелкие животные потребляют больше кислорода на единицу веса тела, чем крупные, и потоки крови у них разделены в значительно большей степени, чем можно было бы думать,, исходя из анатомических данных, и это справедливо как для.ам-; фибий, так и для рептилий. Чтобы оценить степень перемешивания, определяли содержание кислорода в крови, входящей в сердце, и в крови, выходящей из него по разным сосудам. Такие ис-> следования продемонстрировали почти полное разделение потоков крови (White, 1959; Johansen, 1962).
У крокодилов обе камеры сердца полностью разделены (рис. 4.4, Е). С этим полным разделением как будто не вяжется тот факт, что левая дуга аорты начинается от правого желудочка щ стало быть, должна получать венозную кровь. Однако существует отверстие, соединяющее две дуги аорты. Эти анатомические осо? бенности дали повод утверждать, что кровообращение у крокодил лов допускает смешение насыщенной и бедной кислородом крови; между тем более тщательное исследование показало; что потоки крови остаются разделенными и что обе дуги аорты несут несме?
Кровообращение у позвоночных 153
шанную оксигенированную кровь (White, 1956). Однако при нырянии кровообращение меняется: поток крови к легким уменьшается и значительная часть содержимого правого желудочка выталкивается в левую дугу аорты. Такой переброс позволяет изменять путь крови при погружении под воду и полностью или частично обходить легкие.
ПТИЦЫ И МЛЕКОПИТАЮЩИЕ
У птиц и млекопитающих сердце полностью разделено, а легочный и большой круги кровообращения полностью обособлены друг от друга (рис. 4.4, Ж, 3). Это приводит к одному важному следствию: в легочном и большом кругах кровообращения давление может быть различным. Сопротивление кровотоку в легочном круге намного меньше, чем в большом, и кровяное давление в легочном круге составляет лишь малую часть от давления в большом. Такого различия, конечно, не могло бы быть, если бы разделение сердца не было полным. Тем не менее, поскольку неполное разделение все-таки сохранилось и у амфибий, и у рептилий, оно может иметь другие преимущества, которые пока еще не очень ясны.
Между кровообращением птиц и млекопитающих есть некоторые различия, имеющие большое значение для сравнительной анатомии. Например, у млекопитающих сохранилась левая дуга аорты, а у птиц — правая. Физиологически важным различием является то, что почки всех позвоночных, кроме млекопитающих, получают венозную кровь от задней части тела {воротная система почек). Птицы сохранили эту воротную систему, а у млекопитающих она отсутствует. Это различие важно для понимания функции почек (гл. 10) (Johansen, Martin, 1965).
СЕРДЦЕ И ЕГО РАБОТА
Как правило, мелкие животные потребляют больше О2 на единицу веса, чем крупные, и поэтому сердце у них должно лучше снабжаться кислородом. Мы уже видели, что кислородная емкость крови у мелких и крупных животных примерно одинакова; значит, сердце у мелких животных должно перегонять кровь с большей скоростью. Чем это достигается — увеличением перепада давлений, увеличением ударного объема или учащением ритма сердца?
ВЕЛИЧИНА СЕРДЦА У ПОЗВОНОЧНЫХ
Размеры сердца некоторых млекопитающих представлены на рис. 4.8. Как и можно было ожидать, абсолютная величина сердца растет с увеличением размеров тела, но удивительно то, что
154 Глава 4. Кровообращение
у мелких и крупных животных относительная величина сердца* примерно одинакова. Если бы масса сердца была точно пропорциональна массе тела, наклон линии регрессии на рис. 4.8 должен был бы равняться 1; действительный наклон равен 0,98, что статистически неотличимо от точной пропорциональности. Эта линия описывается уравнением Л4С = 0,0059 УИТ°>98, которое означает, что
Рис. 4.8. Соотношение между величиной сердца и размерами тела у млекопитающих. (Stahl, 1965.)
Величина сердца почти пропорциональна величине тела и составляет примерно 0,6% от его общей массы как у мелких, так и у крупных животных. У самых различных приматов относительная величина сердца попадает в диапазон, характерный для других млекопитающих.
средняя масса сердца (Л4С) млекопитающего составляет 0,59% массы тела (Л4Т) независимо от размеров тела.
У птиц величина сердца может быть описана сходным уравнением: тИс = 0,0082 Л4т0,91 (Lasiewski, Calder, 1971). Из этого уравнения следует, что величина сердца не строго пропорциональна размерам тела: показатель степени при массе тела существенно меньше единицы; это означает, что у более крупных птиц относительная величина сердца в среднем несколько меньше, чем у мелких. Ожидаемый вес сердца у птицы весом 1 кг равен 8,2 г, а у млекопитающего такой же величины — 5,9 г.
Размеры сердца у амфибий и рептилий менее изучены, но имеющиеся данные указывают на то, что у рептилий вес сердца составляет 0,51% веса тела, а у амфибий — 0,46%. Эти цифры лишь немногим ниже, чем для млекопитающих, хотя интенсивность об
Кровообращение у позвоночных 155
мена у рептилий и амфибий составляет лишь около одной десятой интенсивности обмена у млекопитающих. У рыб размеры сердца .еще меньше — около 0,2% от веса тела.
Резюмируя, можно сказать следующее: сравниваем ли мы различные классы или животных разной величины внутри одного класса, мы видим, что большие различия в интенсивности метаболизма у позвоночных заметным образом не отражаются на величине сердца. Следовательно, различная потребность в кислороде должна отражаться в первую очередь на частоте сокращений сердца, так как ударный объем зависит от величины сердца, а количество кислорода, содержащееся в данном объеме крови, от размеров сердца не зависит.
ЧАСТОТА СОКРАЩЕНИЙ СЕРДЦА
Ритм сердца или частоту пульса обычно определяют как число сокращений в 1 мин. Частота пульса у взрослого человека в покое составляет около 70 ударов в 1 мин; при нагрузке она увеличивается в несколько раз.
Частота сокращений сердца определенно находится в обратной зависимости от размеров тела. У слона, который весит 3000 кг,
Рис. 4.9. Соотношение между частотой сокращений сердца и размерами тела у млекопитающих. (Stahl, 1967.)
Частота пульса больше у мелких животных. Если имеющиеся данные нанести на график в логарифмических координатах, они сгруппируются около прямой линии, которую можно представить логарифмическим уравнением (см. текст).
частота пульса в покое 25 ударов в минуту, а у трехграммовой землеройки — самого маленького из живущих ныне млекопитающих— более 600 (Morrison et al., 1959). Это означает, что у землеройки сердце совершает 10 полных циклов в секунду — скорость почти невероятная. Во время активности частота пульса становится еще больше: у колибри (Lasiewski et al., 1967) и у мелких ле
156 Глава 4. Кровообращение
тучих мышей во время полета (Studier, Howell, 1969) было зарегистрировано 1200 ударов в минуту.
Если построить график зависимости частоты пульса от массы тела в логарифмической шкале, точки расположатся близко к прямой линии (рис. 4.9). Уравнение этой линии fc = 241 УИТ-0’25. Наклон линии регрессии отрицателен, т. е. чем больше масса тела (Л4Т), тем ниже частота сокращений сердца (fc).
Кроме отрицательного знака наклона наиболее важная информация, заключенная в этом уравнении,—это численная характеристика наклона, т. е. показатель 0,25. Точно такую же цифру мы имеем для наклона линии регрессии на графике зависимости между массой тела и потреблением кислорода на единицу массы в покое, т. е. удельным потреблением кислорода (см. гл. 6).
Теперь мы знаем о двух важных закономерностях: 1) величина сердца составляет постоянный процент от массы тела; 2) частота сокращений сердца при переходе от крупных животных к мелким возрастает в точном соответствии с потребностью в кислороде.
Обратимся теперь к объему крови, перекачиваемой сердцем.
МИНУТНЫЙ ОБЪЕМ СЕРДЦА
Объем крови, перекачиваемой сердцем за 1 мин, обычно называют минутным объемом сердца, (В случае сердец с полностью разделенными желудочками это выражение относится только к одной половине сердца.) Минутный объем можно определить несколькими различными способами. Один из часто применяемых методов основан на принципе Фика — на том простом факте, что весь потребляемый животным кислород (Vog)1 переносится кровью, выталкиваемой из сердца. Следовательно, если мы знаем разницу между содержанием кислорода в артериальной и венозной крови, мы сможем вычислить минутный объем (Qc) из уравнения
^Оз^Фс (СаО2 — £вО2)-
Чтобы найти минутный объем, мы должны определить 1) потребление О2, 2) содержание О2 в артериальной крови и 3) содержание О2 в венозной крови. Пробу артериальной крови можно
1 В уравнениях, относящихся к газообмену, V обычно означает объем, а V (V с точкой)—объем в единицу времени. Соответствующий газ (в данном случае это О2) указывается для ясности в подстрочном индексе. Q означает количество крови, Q — кровоток в единицу времени, Qc — количество крови,, выбрасываемое сердцем (т. е. минутный объем сердца). С — концентрация газа в крови (Са — в артериальной, Св — в венозной).
Кровообращение у позвоночных 157
взять из любой артерии, но венозную кровь нужно брать только смешанную (т. е. из правой половины сердца или из легочной артерии). Венозная кровь из каких-либо других мест не подойдет, так как разные органы извлекают из проходящей через них крови неодинаковые количества кислорода. Пробу смешанной венозной крови легче всего брать из правой половины сердца, вводя в вену катетер и проталкивая его, пока он не дойдет до сердца. Это широко применяемая и относительно безвредная процедура.
Во время активности, когда организм потребляет больше кислорода, минутный объем сердца возрастает, но не пропорционально потреблению О2. Возросшую потребность в кислороде частично покрывает увеличение артерио-венозной разницы в содержании О2 (т. е. во время активности количество извлекаемого и^ крови кислорода увеличивается).
Определение минутного объема сердца на основе принципа Фика возможно только у тех животных, у которых артериальная кровь в сердце полностью отделена от венозной, как у млекопитающих и птиц. У амфибий и рептилий возможно смешение в сердце артериальной и венозной крови, и к ним этот принцип неприменим. Однако он применим к рыбам: пробу венозной крови можно брать из сердца или около него (но перед жабрами), а артериальной— из любой артерии после жабр.
Другие методы определения минутного объема основаны на иных принципах. Один из методов состоит в том, что в вену, ведущую к сердцу, в определенный момент инъецируют краситель,, концентрацию которого легко измерять. Вскоре краситель появляется на артериальной стороне, и путем интегрирования кривой, отражающей подъем и спад его концентрации в артерии, можно* вычислить минутный объем сердца.
Сходен с этим по принципу и метод терморазбавления. В вену вводят известный объем физиологического раствора; если этот раствор холоднее крови, температура артериальной крови на короткое время понизится. И в этом случае интегрирование соответствующей кривой даст сведения о потоке крови (т. е. о минутном объеме сердца).
РАСПРЕДЕЛЕНИЕ КРОВОТОКА
Кровь не распределяется равномерно по всем частям тела; некоторые органы получают ее в гораздо большем количестве, чем: другие, как в относительных, так и в абсолютных единицах. Распределение крови между важнейшими органами у человека в покое представлено в табл. 4.3.
Четыре лучше всего снабжаемых кровью органа — почки, печень, сердце и мозг—составляют только 5% массы тела, но эти7 органы получают более половины всей перекачиваемой крови^ Относительный приток крови к почкам больше, чем к любому дру-
158 Глава 4. Кровообращение
Таблица 4.3
Кровоток в важнейших органах у человека весом 70 кг в покое. (По данным Folkow, Neil, 1971)
_ Вес органа, Кровоток, Удельный
Орган кг л/мин кровоток,
л/(кг«мин)
Почки 0,3 1,2 4,0
Печень 1,5 ’ 3,5 1,4 3,6 0,9
Сердце 0,3 0,25 0,8
Головной мозг 1,4 J 0,75, 0,5
Кожа 2,5] 0,2 ] 0,08
Мышцы 29 66,5 0,9 2,0 0,03
Остальное 35 J 0,9 J 0,03
Всего 70 5,6
тому из главных органов (хотя для некоторых эндокринных органов, особенно щитовидной железы и надпочечников, эта величина может быть еще выше). На единицу веса почки получают более чем в сто раз больше крови, чем мышцы в покое.
При физической нагрузке минутный объем сердца возрастает и распределение кровотока изменяется. Это происходит в первую очередь потому, что сильно возрастает потребность мышц в кислороде, и потому, что нагрузка на сердце увеличивается из-за добавочной работы по перекачиванию крови.
Приток крови к коже также очень изменчив; он варьирует в соответствии с нуждами терморегуляции. На холоде кожные сосуды сужаются и кровоток через кожу минимален, а когда нужно увеличить отдачу тепла из-за высокой окружающей температуры или при повышенной теплопродукции в связи с мышечной работой, поток крови через кожу сильно возрастает. Этот вопрос будет рассмотрен подробнее в гл. 8.
РЕГУЛЯЦИЯ СЕРДЕЧНОГО РИТМА
Сердцу внутренне присуща способность сокращаться ритмично без какого бы то ни было внешнего стимула. Даже полностью отделенное от тела, сердце может продолжать биться довольно долгое время: изолированное сердце лягушки или черепахи ритмически сокращается в течение нескольких часов. Сердца млекопитающих продолжают биться не так долго, потому что они более чувствительны к температуре и снабжению кислородом. Поскольку изолированное сердце бьется само по себе, способность к ритмическим сокращениям должна быть его внутренним свойством. Превосходным доказательством независимой сократимости сердца служат данные о том, что сердце развивающегося куриного эм
Кровообращение у позвоночных 159»
бриона начинает биться, когда до него еще не доросли нервы, а что клетки сердечной мышцы в культуре ткани также сокращаются ритмически без всякого внешнего стимула.
У млекопитающих сокращение сердца начинается в небольшом, участке мышечной ткани эмбрионального типа, там, где полая вена впадает в правое предсердие. Это остаток венозного синуса, и: его называют синусным узлом. Сокращение быстро распространяется по мышечной стенке обоих предсердий и после небольшой, задержки переходит на мускулатуру желудочков. Когда волнам сокращения достигает границы между предсердиями и желудочками, участок ткани, известный под названием атриовентрикулярного пучка (пучка Гиса), проводит импульс к желудочкам, которые после небольшой задержки, связанной с проведением, сокращаются одновременно.
Синусный узел, где мышечная ткань начинает сокращаться^ называют также пейсмейкером сердца. Он запускает сокращения* и определяет их частоту присущей ему собственной ритмичностью. Как мы знаем, эта частота может изменяться в довольно широких, пределах: в ее регуляции участвуют два различных фактора: нервные импульсы, идущие к пейсмейкеру, и гормональные влияния. Ветвь блуждающего нерва, если ее стимулировать, замедляет’ ритм сердца. Это парасимпатический нерв, который воздействует* путем выделения ацетилхолина. Другой нерв — симпатический — ускоряет сокращения сердца. Когда этот нерв стимулируют, сердечный ритм учащается, так как из нервных окончаний в пейсмейкере выделяется норадреналин.
Норадреналин известен также как гормон. Когда он переходит* в кровь из мозгового вещества надпочечников, ритм сердца учащается. Такое влияние на сердце — один из главных эффектов^ адреналина, хорошо известный как часть синдрома «борьбы или: бегства». Аналогичной секреции замедляющего вещества — ацетилхолина, кроме как из окончаний блуждающего нерва, не обнаружено. Если бы ацетилхолин выделялся где-нибудь в другом1, участке тела, он не действовал бы на сердце по той простой причине, что в крови имеется фермент, быстро расщепляющий (гидролизующий) ацетилхолин.
На количество крови, перекачиваемой сердцем, может влиять-не только частота его сокращений, но и объем крови, выбрасываемый при одном сокращении — ударный объем. Сокращаясь, сердце выталкивает не всю кровь, находящуюся в желудочках. К концу сокращения в желудочках остается значительное количество-крови, которое может быть различным. На ударный объем влияют два основных фактора: один из них — это гормон адреналин, который увеличивает силу сокращения сердечной мышцы, так что* из сердца при каждом ударе выходит больше крови, а второй — количество крови в желудочках в начале сокращения. Действие^ последнего фактора определяется особенностью, присущей сердеч-
160 Глава 4. Кровообращение
ной мышце: она будет сокращаться с тем большей силой, чем больше она растянута перед сокращением.
Если венозный приток крови к сердцу по какой-то причине возрастает, то в желудочки входит больше крови, растяжение мышцы увеличивается и последующее сокращение выталкивает больший объем. Эта зависимость ударного объема сердца от венозного притока была открыта знаменитым английским физиологом Старлингом и известна под названием закона Старлинга. При мышечной нагрузке венозный приток к сердцу возрастает отчасти в результате «насосного» действия сокращающихся мышц на вены, а отчасти — из-за сжатия вен брюшной области; все это приводит к увеличению ударного и минутного объема сердца.
Итак, сердечные сокращения инициируются в синусном узле, или пейсмейкере. Минутный объем находится под влиянием трех регуляторных систем: нервной системы, гормонов и системы саморегуляции, основанной на том, что венозный приток оказывает воздействие на сердечную мышцу.
КРОВЕНОСНЫЕ СОСУДЫ
Кровеносные сосуды — это не просто трубки разных размеров. Они имеют эластичные стенки, и слой гладкой мускулатуры внутри этих стенок позволяет изменять диаметр сосуда.
Между артериями, капиллярами и венами имеются характерные различия. У артерий относительно толстые стенки, которые состоят из мощных, прочных слоев эластических волокон и гладкой мускулатуры. По мере того как артерии разветвляются и становятся все тоньше, отношение количества мышечной ткани к количеству эластической ткани стенок постепенно возрастает. Капилляры — самые узкие протоки — состоят из одного слоя клеток. Практически весь обмен веществами между кровью и тканями осуществляется через стенки капилляров. У вен стенки тоньше, чем у артерий, но и здесь повсюду имеются как эластические волокна, так и гладкая мускулатура.
Результаты некоторых измерений, произведенных на сосудистой системе собаки, представлены в табл. 4.4 (Burton, 1972). Всякий раз, когда более крупный сосуд ветвится, число протоков увеличивается, а диаметр их уменьшается. По мере того как ветвей становится больше, их суммарное поперечное сечение растет; в одной только брыжейке на уровне капилляров оно примерно в 800 раз больше, чем у аорты. Начиная с этого места сосуды объединяются во все более и более крупные вены, а их число и площадь поперечного сечения уменьшаются; поперечное сечение полой вены лишь ненамного больше сечения аорты. Из табл. 4.4 можно узнать еще один интересный факт: количество крови, находящееся в венозной системе, в несколько раз превышает количество крови в артериальной системе.
Кровообращение у позвоночных
161
Число и геометрические параметры кровеносных сосудов в брыжейке собаки
Таблица 4.4
Тип сосудов Диаметр, мм Число Общая площадь поперечного сечения, см2 Примерная длина, см Общий объем!), см3
Аорта 10 1 0,8 40
Крупные артерии Артериальные ветви Артериолы 3 1 0,02 40 2 400 40 000 000 3 5 125 20 . 5 0,2 190
Капилляры Венулы 0,008 0,03 1 200 000 000 80 000 000 600 570 0,1 0,2) 60
Вены Крупные вены Полая вена 2 6 12,5 2 400 40 1 30 И 1,2 5 20 40 680
9 Оценки общего объема основаны приведенные в предыдущей колонке. па более точном анализе, чем : оценки длины*
Кроме соотношения объемов в сосудистой системе нас должны интересовать еще давления и скорости тока крови в разных ее частях. Соответствующие данные для кровеносной системы человека приведены на рис. 4.10. Из первой колонки видно, что большая часть крови в каждый данный момент находится на венозной стороне.
Данные последней колонки показывают, что быстрее всего кровь течет в аорте. По мере увеличения суммарной площади поперечного сечения скорость резко уменьшается, и в капиллярах она доходит до уровня где-то между 7юо и Viooo от скорости в аорте. На венозной стороне движение крови снова ускоряется, но все же остается более медленным, чем в артериальной системе.
В средней колонке на рис. 4.10 представлены давления во всех частях системы. Конечно, на всем пути от аорты до крупных вен давление должно постепенно уменьшаться. Наибольший спад давления имеет место в самых мелких артериях, называемых артериолами, стенки которых содержат относительно много гладкой мышечной ткани; изменение их диаметра — наиболее важный фактор в изменении сопротивления потоку и тем самым — в регуляции распределения крови между различными органами.
Легочный круг кровообращения отличается от большого круга тем, что его общее сопротивление меньше; поэтому давление крови здесь может быть меньше и артерии могут иметь более тонкие стенки. В поперечном сечении легочная артерия не круглая, а овальная. Во время сокращения сердца, когда кровь выталкивается в легочную артерию, объем уплощенного сосуда значительно возрастает, по мере того как его сечение становится круглым;
11—1873
162 Глава 4, Кровообращение
только после этого стенки начинают растягиваться. Благодаря этому часть вытолкнутой крови заполняет расширившуюся артерию; таким образом, артерия служит как бы амортизатором и кровь не проходит через легочные капилляры толчками при каждом сокращении сердца (Melbin, Noordergraaf, 1971).
Обзем, мл Давление, мм рт ст Скорость, см/с II-
Аорта Артерии Артериолы Капилляры Венулы Вены Полая вена STOO К^ЗОО $50 Ш250 Ш300 ШШШШШ2 шзоо j 100 100-40 LWWI40-30 30-12 □ 12-10 200 □ Ю-5 Q2 с J 40 Ш 10-0,1 I Менее 0,1 8 Менее 0,3 ^0,5-5 L
Рис. 4.10. Объем, давление и скорость движения крови в различных частях со судистой системы человека (1 мм рт. ст.=0,13 кПа).
ФИЗИКА ТЕЧЕНИЯ ЖИДКОСТЕЙ ПО ТРУБАМ
Течение жидкости (или газа) может быть столь гладким и правильным, что в прямой трубе каждая частица вещества будет двигаться по прямой линии. Такое течение называют ламинарным, 'Если в искривленной трубе направление потока в каждой данной точке остается постоянным, поток тоже считается ламинарным. Путь, проходимый частицей вещества в ламинарном потоке, называют линией тока. Если частицы вещества движутся нерегулярно и образуют завихрения, поток называют турбулентным. В любой данной трубе поток из ламинарного переходит в турбулентный, если скорость превышает некоторую критическую величину. Последующие рассуждения будут относиться только к ламинарному потоку; динамика турбулентных потоков намного сложнее и мало» касается течения кропи в сосудах.
ТЕОРЕМА БЕРНУЛЛИ
Стационарное течение идеальной жидкости вдоль линии тока описывается теоремой Бернулли, названной так по имени швейцарского физика (1700—1782), ставшего профессором математики в возрасте 25 лет. Прежде всего мы должны заметить, что поток в трубе называется стационарным, если скорость в каждой точке трубы остается неизменной, даже если труба где-то сужается или расширяется и соответственно жидкость течет в одних участках быстрее, а в других медленнее.
Кровообращение у позвоночных 163
Теорема Бернулли гласит, что общая энергия жидкости (В) является суммой 1) потенциальной энергии, связанной с внутренним давлением, 2) потенциальной энергии, связанной с силой тяжести, и 3) кинетической энергии движущихся частиц:
£=(ру)+(^)4-(4"/72“2)- (О
В правой части этого уравнения первый член представляет энергию давления жидкости, равную произведению давления (р) на объем (и). Второй член — это гравитационная потенциальная энергия, равная произведению массы (т) на гравитационную постоянную (g) и на высоту (/i). Третий член — кинетическая энергия, зависящая от скорости (и) жидкости. Сумма этих трех величин и есть общая энергия жидкости. Содержание энергии в единице объема (В7) можно найти, разделив обе части уравнения на объем:
£'=р+Р£й+р^, (2)
где р —плотность жидкости.
В случае течения жидкости по горизонтальной трубе уравнение можно существенно упростить. Так как гравитационная потенциальная энергия от одного конца трубы к другому не меняется, соответствующий член остается постоянным, и его можно опустить. Кроме того, поскольку плотность жидкости тоже постоянна, эту константу можно исключить, так что в результате мы получим
E’^p-Y—. (3)
Если жидкость течет в трубе без трения, удельная энергия жидкости (В7) должна оставаться постоянной, так что при изменениях скорости (и), связанных с изменением диаметра, давление должно изменяться в противоположном направлении, т. е.
д4=-Др. (4)
Теперь рассмотрим трубу постоянного диаметра, в которой есть трение. При постоянном диаметре скорость (и) остается постоянной. Из-за сопротивления течению на продвижение воды через трубу должна затрачиваться энергия, и ее потери должны выражаться в падении давления (р), как это показано на рис. 4.11. Энергия, теряющаяся из-за трения, рассеивается в виде тепла, что проявляется в повышении температуры жидкости, однако в случае потока крови это несущественно.
Если диаметр трубы изменяется, то должна изменяться и скорость течения (и). Если труба горизонтальна, гравитационный 11*
164 Глава 4. Кровообращение
член в уравнении (2) опять-таки можно опустить. Теперь представим себе на минуту, что энергия не рассеивается в результате трения и ее удельное содержание остается постоянным. Тогда мы увидим, что скорость и давление будут изменяться в противопо-
Рис. 4.11. Когда жидкость течет по горизонтальной трубе, сопротивление приводит к постоянному уменьшению давления по длине трубы.
ложных направлениях, согласно выражению (4), которое означает, что при возрастании скорости давление в жидкости должно па-
Рис. 4.12. Когда жидкость протекает через узкий участок трубы, скорость ее движения возрастает. Увеличение скорости сопровождается снижением давления.
Часто утверждают, что жидкость всегда течет по направлению от большего давления к меньшему. Рис. 4.12 показывает, что это-не всегда так, поскольку в более узкой части трубы давление меньше, чем далее вниз по течению. На самом деле жидкость всегда течет от точки, где ее общая энергия больше, к точке, где ее общая энергия меньше.
В предыдущем обсуждении мы игнорировали гравитационный член в уравнении (2). Если мы имеем U-образную трубку вроде изображенной на рис. 4.13, то в покое жидкость в ней будет иметь одну и ту же удельную энергию во всех участках. Хотя давление
Кровообращение у позвоночных 165
в нижней (горизонтальной) части трубки больше, чем в верти-кальных частях, жидкость не течет. Обратившись к уравнению (2), мы увидим, что с уменьшением высоты (/г) гравитационная потенциальная энергия на единицу объема (pgh) уменьшается и со-
Рис. 4.13. Жидкость в U-образной трубке остается в покое. В результате воздействия силы тяжести на жидкость давление в нижней части трубки выше, чем в вертикальных участках.
ответственно должна возрастать энергия давления (р). Мы связываем это увеличение энергии давления с гидростатическим дав* лением жидкости, понимая под этим выражением увеличение давления с глубиной по отношению к давлению на поверхности.
КРОВЯНОЕ ДАВЛЕНИЕ
Посмотрим, как можно применить описанные выше принципы. В ступнях ног у человека венозное давление повышено за счет высоты столба крови (рис. 4.14). Артериальное давление здесь тоже превышает давление в сердце — также в соответствии с весом столба крови. Высокое давление в венозной системе ног растягивает вены, поэтому сокращение мышц и действие венозных клапанов становится существенной подмогой для возвращения крови к сердцу.
Другое следствие наличия гравитационного члена — это то, что для снабжения кровью головы необходимо определенное артериальное давление. Из-за влияния гравитации артериальное кровяное давление в голове снижено примерно до 50 мм рт. ст. (7 кПа). У жирафа, который держит голову на высоте около 2 м над уровнем сердца, давление, необходимое для кровоснабжения мозга, намного выше, чем у человека.
166 Глава 4. Кровообращение
Измерения кровяного давления у жирафа показали, что систолическое давление доходит у него до 260 мм рт. ст. (35 кПа) (Van Citters et al., 1968). (У человека нормальное систолическое давление около 100 мм рт. ст., или 13 кПа.) Чтобы выдерживать столь высокие давления, артерии у жирафа имеют необычайно
Рис. 4.14. Артериальное и венозное давление у человека при различном положении тела. Цифрами указано давление в разных точках по отношению к давлению в правом предсердии (1 мм рт. ст.=0,13 кПа). (Burton, 1972.)
толстые стенки, а венозная система везде оснащена клапанами, которые облегчают возвращение крови из конечностей. Когда жираф высотой 4,5 м опускает голову, чтобы напиться, клапаны в шейных венах препятствуют обратному притоку крови к голове и возрастанию гидростатического давления в мозгу, которое иначе произошло бы при опускании головы.
ДИАМЕТР ТРУБОК И ТОЛЩИНА ИХ СТЕНОК
У полого цилиндра натяжение стенок (71) равно произведению давления, действующего на стенки (р), на радиус (г):
Т=р.г.
Это соотношение было выведено Бернулли, но обычно его называют законом Лапласа. Оно означает, что при данном давлении
Кровообращение у позвоночных 167
натяжение стенок возрастает прямо пропорционально радиусу. Для того чтобы стенки могли выдерживать это натяжение, их толщина должна соответственно увеличиваться. Поэтому крупные артерии должны иметь более толстые стенки, чем мелкие.
У вен, в которых давление крови намного ниже, стенки тоньше. Опять-таки мелкие вены могут выдерживать венозное давление, имея более тонкие стенки, чем у крупных вен. В капиллярах давление, конечно, выше, чем в венах, но благодаря очень малому радиусу стенки капилляров, состоящие из одного слоя клеток, все же достаточно прочны. Таким образом, малый радиус капилляров позволяет им иметь достаточно тонкие стенки, допускающие быстрый обмен веществами между кровью и тканями.
вязкость
Сопротивление потоку в трубе обусловлено внутренним трением в жидкости, т. е* ее вязкостью. С этим свойством приходится иметь дело каждому: все знают, что вода и сироп вытекают из бутылки не одинаково быстро. Мы говорим, что вода имеет маг лую вязкость, а сироп — большую.
Величина потока через трубу пропорциональна разности давлений на ее концах и обратно пропорциональна сопротивлению:
С=-к-
Член, отражающий сопротивление (7?), зависит от размеров трубы и природы жидкости (от ее вязкости). Эту зависимость исследовал французский физик Пуазейль (1799—1869), который заинтересовался течением крови в капиллярах. Он работал с трубками различных диаметров и нашел, что поток через трубку прямо пропорционален давлению и четвертой степени радиуса трубки и обратно пропорционален длине трубки и вязкости жидкости. Формально это выражается уравнением Пуазейля, которое можно записать следующим образом:
\ д г4 л
где Q — поток в единицу времени, Др — перепад давлений, г — радиус трубки, I — ее длина и т] — вязкость1.
Самое поразительное — это колоссальное влияние, которое оказывает на поток радиус трубки. Однако здесь мы будем больше интересоваться вязкостью. Из уравнения видно, что с увеличением вязкости ток жидкости пропорционально замедляется.
Для многих целей вязкость любой жидкости удобно выражать по отношению к вязкости воды, приняв последнюю за единицу. Плазма обладает большей вязкостью, чем вода, главным образом
1 Вязкость жидкости изменяется с температурой. При понижении температуры с 37 до О °C вязкость воды увеличивается примерно в 2,6 раза.
168 Глава 4. Кровообращение, из-за 7% растворенных белков: относительная вязкость плазмы равна примерно 1,8. Предположим, что мы измеряем ток воды и плазмы через некоторую трубку: если мы примем поток воды за 1, то поток плазмы будет равен 71,8- Уменьшим теперь радиус трубки наполовину, оставив ее длину и давление прежними. Согласно уравнению Пуазейля, поток воды снизится до (’А)2, т. е. до Vie своей прежней величины. То же самое произойдет и с потоком плазмы, так что отношение потоков по-прежнему останется равным 71.8- Это значит, что вязкость плазмы по отношению к воде (относительная вязкость) не зависит от размеров трубки.
Если вместо плазмы мы возьмем цельную кровь, мы обнаружим, что вязкость ее выше, чем у плазмы, и возрастает с увеличением количества эритроцитов (рис. 3.1). Однако кровь ведет себя не совсем так, как можно было ожидать. Ее относительная вязкость меняется с изменением радиуса трубки, по которой она течет. С уменьшением радиуса относительная вязкость крови уменьшается (т. е. кровь становится больше похожей на воду и проходит легче).
Такую аномальную жидкость называют неньютоновой жидкостью-, из-за сложности ее поведения поток крови в сосудах трудно описать, особенно учитывая тот факт, что кровеносные сосуды эластичны и радиус их отнюдь не остается неизменным. Конечный результат, однако, очень важен: через капилляры кровь протекает легче и ее эффективная вязкость ниже, чем это можно было бы ожидать на основании размеров кровеносных сосудов.
Теперь мы можем вернуться к вопросу, который был затронут в главе 3: как влияет на вязкость крови то, что гемоглобин заключен внутри клеток? Из-за своего неньютонова поведения цельная кровь проходит через артериолы и капилляры (где сосредоточено основное сопротивление потоку) легче, чем проходили бы эквивалентные растворы гемоглобина (Snyder, 1973).
Другая интересная особенность течения крови через капилляры связана с тем, что диаметр капилляров часто значительно меньше, чем размеры эритроцитов. Вопреки ожиданиям оказалось, что току крови это не мешает: эритроциты легко деформируются и без труда проходят через капилляр (Skalak, Branemark, 1969). Это создает очень своеобразный тип течения (корпускулярное течение), при котором каждый эритроцит действует как поршень: он обеспечивает быстрое обновление жидкости в пограничном слое вдоль стенок капилляра, тем самым способствуя обновлению диффундирующих веществ в этом слое.
КАПИЛЛЯРНАЯ СИСТЕМА
Общее число капилляров в теле огромно. В одной только брыжейке собаки число капилляров превышает миллиард (см. табл. 4.4). Для точного подсчета числа капилляров особенно под-
Кровообращение у позвоночных 169
ходят мышцы, так как капилляры в них идут между мышечными волокнами, параллельно им. Поэтому на поперечном срезе мышцы сравнительно легко подсчитать число капилляров на единицу площади. Это число очень велико, но обычно не все капилляры открыты и наполнены кровью. В покоящейся мышце морской свинки на поперечном сечении в 1 мм2 насчитывается около 100 открытых капилляров, через которые течет кровь. Когда мышца работает, суженные артериолы расширяются и кровь идет по большему числу капилляров: при максимальном напряжении на 1 мм2 поперечного сечения может приходиться более 3000 открытых капилляров. У обычного карандаша поперечное сечение стержня составляет около 3 мм2. Вообразите себе примерно 10000 тонких трубочек, идущих параллельно друг другу внутри этого стержня!
У мелких млекопитающих плотность расположения капилляров больше, чем у крупных. Это согласуется с более высокой интенсивностью метаболизма и большей потребностью в кислороде у мелких животных (см. гл. 3).
Обмен веществами между жидкостью внутри капилляров и содержимым тканевых межклеточных пространств определяется двумя главными процессами. Стенки капилляров очень тонки — 1 мкм толщиной— и состоят из одного слоя клеток. Вода и растворенные вещества с малым молекулярным весом (газы, соли, сахара, аминокислоты и т. д.) могут сравнительно легко диффундировать через стенки капилляров, и, кроме того, существующее внутри капилляров давление заставляет большие количества жидкости просачиваться через стенки наружу в результате фильтрации. Однако молекулы таких размеров, как у большинств/а белков (молекулярный вес около 70 000 или более), не проходят через стенки капилляров. Белки остаются внутри капилляра, а через стенки выдавливается относительно свободный от белков фильтрат. Концентрации в этом фильтрате растворенных низкомолекулярных веществ близки к соответствующим концентрациям в плазме, но не равны им, так как белки оказывают некоторое влияние на распределение ионов (эффект Гиббса—Доннана).
Если мы будем рассматривать стенку капилляра как полупроницаемую мембрану, мы увидим, что небелковые вещества, свободно проходящие через нее, не влияют на осмотическое давление. Однако белки удерживаются внутри капилляра и создают некоторый осмотический эффект — так называемое коллоидное осмотическое давление, В плазме млекопитающих это давление составляет около 25 мм рт. ст.; оно стремится присасывать воду из окружающей тканевой жидкости обратно в капилляр. Когда гидростатическое (кровяное) давление внутри капилляра больше коллоидного осмотического давления, жидкость выталкивается через стенки капилляра наружу; когда же внутреннее гидростатическое давление опускается ниже коллоидного осмотического, жидкость засасывается внутрь. Кровяное давление в капилляре быва
170 Глава 4. Кровообращение
ет различным, но на артериальном конце оно обычно выше, а на венозном ниже коллоидного осмотического давления. Вследствие этого на артериальном конце капилляра жидкость профильтровывается наружу, а на венозном конце входит обратно (рис. 4.15). Такое представление о фильтрации и обратном возвращении жидкости через стенки капилляров впервые было выдвинуто Старлингом (Starling, 1896).
Коллоидное осмотическое давление (25 мм рт ст.)
Гидростатическое давление Гидростатическое давление/
(АО мм рт. ст ) (15 мм рт. cm.) I
Плазма |
\
Коллоидное осмотическое давление (25 мм рт. ст)
Рис. 4.15. Стенка капилляра полупроницаема, и кровяное давление заставляет жидкость выходить наружу путем ультрафильтрации (А). Белки плазмы остаются внутри капилляра и противодействуют процессу ультрафильтрации. Кровяное давление по ходу капилляра снижается, и когда оно опускается ниже уровня коллоидного осмотического давления, создаваемого белками, возникает обратный ток жидкости внутрь капилляра, вызываемый осмотическими силами (Б). Стенка капилляра свободно пропускает соли и другие растворенные вещества с небольшими молекулами, поэтому движение жидкости происходит так, как если бы этих веществ не было.
Количество жидкости, выходящее через стенки капилляров, и Количество, входящее обратно благодаря коллоидному осмотическому давлению, сильно варьирует. Обычно выходной поток больше входного, и избыточная жидкость остается в интерстициальных пространствах между тканевыми клетками. Эта жидкость — лимфа— медленно переходит в тонкие лимфатические сосуды, или лимфатические капилляры. В настоящее время считается, что этот поток не есть результат чисто пассивного дренажа. Мелкие лим
Кровообращение у позвоночных 171
фатические сосуды подвержены постоянным ритмическим сокращениям, благодаря насосному действию которых создается небольшое отрицательное давление, и пока количество жидкости, фильтрующейся через стенки капилляров, невелико, лимфатическая система способна удалить всю избыточную жидкость (Guyton, 1976).
Мелкие лимфатические сосуды постепенно сливаются друг с другом и в конце концов объединяются в более крупные лимфатические протоки. Эти протоки в свою очередь впадают в относительно крупные вены, так что жидкость возвращается в кровеносную систему. Лимфатическую систему можно рассматривать как часть сосудистой системы, но циркуляции лимфы как таковой нет; скорее можно сказать, что это дренажная система, которая возвращает в кровь избыток жидкости, просочившейся из капиллярной системы.
Если бы в плазме не было белков, любое гидростатическое давление, превышающее давление в тканевых пространствах, заставляло бы жидкость профильтровываться наружу и не было бы силы, заставляющей жидкость возвращаться обратно. Таким образом, белки плазмы необходимы для удержания жидкой части крови в сосудистой системе.
У млекопитающих коллоидное осмотическое давление плазмы выше, чем у других позвоночных (табл. 4.5). Удивительно, что
Таблица 4.5
Коллоидное осмотическое давление плазмы крови различных позвоночных (1 мм рт. ст.=0,13 кПа). (По данным Altman, Dittmer, 1971)
Животное Давление, мм рт. ст. Животное Давление, мм рт. ст.
Корова 21 Аллигатор 9,9
Овца 22 Черепаха 6,4
Собака 20 Лягушка 5,1
Кошка 24 Жаба 9,8
Курица 11 Треска 8,3
Голубь 8,1 Камбала 8,5
у птиц оно очень невелико, хотя артериальное кровяное давление у них обычно намного выше, чем у большинства млекопитающих. У некоторых птиц кровяное давление превышает 200 и даже 250 мм рт. ст. (27—33 кПа), тогда как у млекопитающих оно чаще всего около 100 мм рт. ст. Эти различия не очень понятны. Можно предположить, что либо у птиц должно быть более высокое сопротивление току крови на уровне артериол, в связи с чем капиллярное давление у них не очень велико, либо стенки капилляров у них обладают не такими свойствами, как у млекопитаю
172 Глава 4. Кровообращение
щих. Более высокое сопротивление в артериолах кажется бессмысленным, ибо зачем тогда птицам иметь высокое начальное давление в артериях?
У других позвоночных — рептилий, амфибий и рыб — коллоидное осмотическое давление тоже сравнительно невелико. Это согласуется с тем фактом, что и артериальное кровяное давление у них в общем ниже, чем у млекопитающих.
КРОВООБРАЩЕНИЕ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ
Во время мышечной активности потребность в кислороде повышена, а значит, и количество кислорода, которое кровь должна доставить тканям, тоже должно быть больше. Для удовлетворения этой возросшей потребности есть два пути: увеличение объема крови, перекачиваемой сердцем (минутного объема сердца), и увеличение количества кислорода, доставляемого данным объемом крови. Артериальная кровь уже полностью насыщена и не может больше поглощать кислород, но содержание кислорода в венозной крови в норме составляет больше половины его содержания в артериальной крови. Увеличение отдачи кислорода кровью — очевидный путь получения большего количества Ог из каждого ее объема.
Рассмотрим сначала процесс увеличения экстракции кислорода из крови. Вся масса мышц худощавого человека, составляющая почти половину его веса, потребляет около 50 мл О2 в 1 мин. Такое количество кислорода доставляется потоком крови объемом примерно в 1 л (т. е. при превращении артериальной крови в венозную содержание в ней кислорода снижается с 200 мл на 1 л до 150 мл на 1 л). Поскольку из артериальной крови извлекается четвертая часть кислорода, мы говорим, что экстракция составляет 25%. При большой физической нагрузке кровоток в мышцах у здорового человека может составлять 20 л в 1 мин (у хорошо тренированных спортсменов — даже больше), а экстракция кислорода в мышцах увеличивается до 80 или 90%; иными словами, в венозной крови, идущей от напряженно работающих мышц, остается очень мало кислорода (Folkow, Neil, 1971).
Второй путь увеличения доставки кислорода — это увеличение минутного объема сердца. Оно может быть достигнуто учащением ударов сердца и увеличением ударного объема. Благодаря интересу, связанному с медициной и спортом, относительно человека имеется намного больше сведений, чем относительно других млекопитающих. В покое сердце человека бьется с частотой около 70 ударов в минуту, а ударный объем составляет около 70 мл (для каждой стороны), так что минутный объем равен примерно 5 л. При большой физической нагрузке работа сердца может легко возрасти в пять раз и более (если притом экстракция кислорода утраивается, то это будет соответствовать 15-кратному увеличе
Кровообращение у позвоночных 173
нию доставки кислорода). Большая часть прироста минутного объема сердца связана с увеличением частоты пульса, которая может возрасти до 200 ударов в минуту, однако увеличивается и удапный объем, который может превысить 100 мл.
'Рис. 4.16. Распределение всего потока крови (минутного объема) (А) и потребления кислорода (Б) между мышцами (заштрихованные участки столбиков) и другими частями тела (светлые участки). Приведены данные для человека в покое (I), для среднего человека при тяжелой мышечной нагрузке (II) и для спортсмена высокого класса при тяжелой нагрузке (III). (Folkow, Neil, 1971.)
На рис. 4.16 показано распределение кровотока у человека в покое и при нагрузке. У спортсмена приток крови к мышцам в экстремальных условиях может увеличиваться в 25—30 раз; приток крови к остальным частям тела при этом слегка уменьшается. Потребление кислорода мышцами у спортсмена может возрастать в 100 раз; это возможно только благодаря примерно трехкратному увеличению экстракции кислорода.
174 Глава 4. Кровообращение
ЦИРКУЛЯЦИЯ ЖИДКОСТЕЙ У БЕСПОЗВОНОЧНЫХ
Многие беспозвоночные имеют хорошо развитые циркуляторные системы; примером могут служить кольчатые черви, иглокожие, членистоногие и моллюски. Большинство их имеет незамкнутые системы. У головоногих (осьминогов и кальмаров) системы кровообращения замкнутые, а у представителей других классов моллюсков— незамкнутые. Следует заметить, что у насекомых — высокоорганизованных, сложно устроенных животных, способных поддерживать очень высокий уровень метаболической активности, — системы циркуляции тоже незамкнутые.
КОЛЬЧАТЫЕ ЧЕРВИ (АННЕЛИДЫ)
Некоторые аннелиды имеют хорошо организованные системы циркуляции, а их кровь часто содержит дыхательные пигменты, растворенные в плазме. Наиболее обычный пигмент — гемоглобин, но у ряда кольчатых червей имеется хлорокруорин или гемоэритрин.
У дождевого червя Lumbricus органом дыхания служит вся поверхность тела, и она снабжена густой сетью капилляров (рис.
Рис. 4.17. Система кровообращения дождевого червя. (Meglitsch, 1972.) Спинной сосуд способен сокращаться и накачивать кровь в крупные сосуды. Газообмен осуществляется через наружные покровы, которые обильно васкуляризированы. Особые сосуды, идущие от кожи, снабжают оксигенированной кровью брюшную нервную цепочку.
4.17). Циркуляторная система имеет два продольных сосуда: спин’ ной, в котором кровь движется вперед, и брюшной, в котором она течет в противоположном направлении. Нервная цепочка у многих аннелид, включая дождевого червя, снабжается кровью через дополнительные продольные сосуды, и часть насыщенной кислоро
Циркуляция жидкостей у беспозвоночных 175
дом крови направляется от покровов прямо в этот важный орган. Таким образом, замкнутая система сосудов имеет явное преимущество в том отношении, что позволяет строго определять, куда именно должна течь кровь.
У аннелид нет одного четко обособленного сердца, но некоторые сосуды имеют расширения, способные сокращаться. Наиболее важный из кровеносных сосудов — спинной, в котором кровь проталкивается вперед с помощью перистальтических волн. Кроме того, сосуды, соединяющие с обеих сторон тела главный спинной еосуд с главным брюшным, тоже сократимы и служат вспомогательными сердцами.
ИГЛОКОЖИЕ
Иглокожие (например, морские звезды, морские ежи и голотурии) имеют три системы, наполненные жидкостью: целомическую систему, кровеносную, или гемальную, и водно-сосудистую, или амбулакральную. Последняя функционирует главным образом при локомоции: это гидравлическая система, которая приводит в движение трубчатые ножки и заполнена жидкостью, близкой по составу к морской воде.
Между стенками тела и пищеварительным трактом у иглокожих имеется большой целом, наполненный целомической жидкостью, которая, по-видимому, играет важную роль в переносе питательных веществ от пищеварительного тракта к другим частям тела (Farmanfarmaian, Phillips, 1962).
Гемальная, или кровеносная, сосудистая система иглокожих содержит отдельную жидкость, в составе которой у некоторых голотурий есть гемоглобин. У голотурии Cucumaria он обладает довольно высоким сродством к кислороду: давление полунасыщения варьирует в пределах от 5 до 10 мм рт. ст. (в зависимости от температуры); удивительная особенность этого гемоглобина — отсутствие эффекта Бора в интервале pH от 7,5 до 6,5 (Manwell, 1959). Эти свойства, по-видимому, весьма выгодны в бедной Ог илистой среде, в которой живут голотурии.
Взаимоотношения между гемальной и целомической системами не вполне понятны, и роль гемальной системы в дыхании и доставке тканям кислорода не ясна (Martin, Johansen, 1965). Амбулакральная система обслуживает в первую очередь локомоцию, и она лучше изучена.
моллюски
Все моллюски, за исключением осьминогов и кальмаров, имеют незамкнутые системы кровообращения. Кровь многих моллюсков содержит гемоцианин, а у некоторых имеется гемоглобин. Способность крови переносить кислород коррелирует с размерами животного и особенно с его активностью.
176 Глава 4. Кровообращение
Моллюски обычно обладают хорошо развитым сердцем, и частота его сокращений регулируется у них в соответствии с физиологическими потребностями в кислороде. Увеличение венозного притока крови к сердцу ведет к увеличению как амплитуды, так и частоты биений. Сердце моллюска продолжает сокращаться, если даже отделить его от нервной системы; оно имеет внутренне присущий ему ритм, задаваемый пейсмейкером. Однако оно находится также под влиянием нейросекретов, которые изменяют частоту его сокращений. Ацетилхолин тормозит сердце, а серотонин оказывает возбуждающее действие.
У двустворчатых моллюсков активное движение ноги осуществляется при помощи крови. Обширные синусы в кровеносной системе ноги управляются клапанами, которые помогают регулировать и величину ноги, и ее движения. Это особенно важно для роющих видов, которые используют ногу для продвижения через донный материал.
Головоногие моллюски (осьминоги и кальмары) обладают высокоразвитой замкнутой системой кровообращения с четко дифференцированными сетями артерий, вен и капилляров. Это связана с высокой организацией и активностью этих животных и с важной ролью крови в функционировании жабр (газообмен) и почек.
Тот факт, что головоногие имеют замкнутую циркуляторную систему, означает, что кровь всегда остается внутри сосудов, поэтому она четко отделена от тканевой жидкости. На долю крови приходится около 6% веса тела, а на долю тканевой жидкости— около 15%. Эти величины поразительно близки к соответствующим цифрам для позвоночных. В незамкнутых системах моллюсков, не относящихся к головоногим, кровь не отделена от тканевой жидкости и свободно растекается по внеклеточному пространству; в связи с этим количество крови велико и может составлять до 50% сырого веса тела (без раковины) (Martin et al., 1958)^
НАСЕКОМЫЕ
У насекомых обычно имеется крупный кровеносный сосуд, который тянется вдоль спинной стороны тела. Задний отдел этого сосуда играет роль сердца; он снабжен рядом отверстий с клапанами, через которые кровь может входить внутрь. Передняя часть сосуда, которую можно было бы назвать «аортой», способна сокращаться и может создавать перистальтические волны, проталкивающие кровь вперед. Этот главный кровеносный сосуд разветвляется и идет до головы и здесь оканчивается. Ответвления от аорты снабжают кровью большую часть тела, где кровь свободно растекается между тканями и медленно просачивается обратно к сердцу.
У многих насекомых имеются продольные перепонки, придающие току крови между тканями определенное направление. В ан
Циркуляция жидкостей у беспозвоночных МТ
тенны и конечности кровь может входить вдоль одной стороны, а выходить из них — вдоль другой. Таким образом, кровь направляется по определенным путям, хотя она и не течет внутри обособленных кровеносных сосудов (рис. 4.18).
Поскольку кровеносная система у насекомых незамкнутая,, кровяное давление у них может быть лишь ненамного выше дав^ ления в тканях. Кровь не играет здесь особой роли в дыхании;
Рис. 4.18. В хорошо развитой системе кровообращения насекомого имеется» главный спинной сосуд, несущий кровь от сердца. Хотя эта система незамкнутая, благодаря наличию продольных перепонок, особенно в ногах, кровь идет по определенным путям. Стрелками указаны направления потоков. (Wiggles* worth, 1972.)
С — спинная диафрагма с мышцами; И — нервная цепочка; /70— пульсирующие* органы; П — перепонки в конечностях.
ее главная функция — разносить по телу питательные вещества и? метаболиты и обеспечивать транспорт гормонов, которые у насекомых очень важны для физиологической координации (роста,, линьки и т. п.). У некоторых насекомых циркуляция крови существенна также для распределения тепла, особенно для регулирования теплоотдачи у высокоактивных летающих насекомых (Heinrich, 1971).
Хотя сердце и спинной сосуд — главные компоненты кровеносной системы насекомых, многие из них имеют еще дополнительные «насосы», или сердца. Они особенно важны для поддержания циркуляции в придатках (крыльях, ногах и антеннах). Току крови в крыльях способствуют пульсирующие органы внутри самих крыльев, расположенные в каналах, по которым кровь возвращается к телу (Thomsen, 1938). Течению крови в антеннах по1-могают сократимые органы, расположенные в основаниях антенн. Они работают, всасывая кровь из кровяных синусов, имеющихся в голове. У многих насекомых есть также ножные сердца, способствующие циркуляции крови в конечностях. Дополнительные сердца насекомого и их сокращения независимы от спинного сердца!
12—1873
•178 Глава 4. Кровообращение
они могут сокращаться с разной частотой, одно из них может ос-тановиться, тогда как другие будут продолжать биение.
Наполнение спинного сердца насекомого осуществляется путем всасывания. Имеется группа мышц (крыловидные мышцы), расхо-
дящихся во все стороны
Рис. 4.19. Сердце насекомого — это сократимый участок главного спинного сосуда. Наполнение сердца кровью осуществляется путем всасывания с помощью мышц, прикрепленных к сердцу снаружи и растягивающих его. На данном рисунке изображен препарат таракана Blaberus. (Nutting, 1951.)
от сердца; при их сокращении сердце расширяется и втягивает кровь в свою полость через снабженные клапанами отверстия (остия). Последующее сокращение сердца проталкивает кровь вперед в направлении к голове, а клапаны препятствуют вытеканию крови через остия (рис. 4.19).
ПАУКООБРАЗНЫЕ
Циркуляторная система пауков и скорпионов сходна с системой насекомых, но она может играть большую роль в дыхании. В крови некоторых скорпионов был найден гемоцианин, и как скорпионы, так и пауки имеют органы дыхания, через которые проходит кровь. Это не должно казаться удивительным, Tai как у паукообразных нет трахейной системы, характерной для насекомых.
Сердце у паукообразного расположено в дорсальной части брюшка. Оно наполняется через снабженные клапанами
отверстия (остия) и опорожняется в артерии. У пауков особые артерии идут к конечностям, где относительно высокое кровяное давление существенно для локомоции. В ногах пауков нет мышц-разгибателей, и для разгибания ног используется давление крови (см. гл. И).
Циркуляция жидкостей у беспозвоночных 179
РАКООБРАЗНЫЕ
Циркуляторные системы этих животных чрезвычайно разнообразны. У мелких представителей они плохо развиты; крупные ракообразные, в особенности десятиногие (омары, крабы, речные раки), имеют хорошо развитые циркуляторные системы и кровь с дыхательным пигментом (гемоцианином).
Рис. 4.20. Кровообращение у омара (Л). У крупных ракообразных богатая кислородом кровь из жабр направляется прямо в сердце, которое накачивает ее далее в ткани (Б). У рыб сердце расположено иначе — перед жабрами (см. рис. 4.3). (Meglitsch, 1972.)
Система кровообращения у ракообразных состоит из тех же основных морфологических структур, что и у других членистоногих, но она отличается от системы насекомых в одном важном физиологическом отношении: ракообразные имеют жабры, а в связи с этим — и хорошо выраженные пути к жабрам.
У крупных ракообразных спинное сердце лежит в перикардиальном синусе, из которого кровь поступает в сердце через остия. От сердца обычно отходят главные артерии, идущие в переднем и заднем направлениях. По мере ветвления артерий кровь покидает сосуды и начинает течь между тканями к системе вентральных синусов. Из них кровь направляется в жабры, а затем по особым сосудам-—назад к сердцу.
Благодаря такому устройству сердце ракообразного непосредственно получает оксигенированную кровь, которая затем нака-12*
180 Глава 4. Кровообращение
чивается в ткани (рис. 4.20). Это противоположно тому, что имеет место у рыб: у них в сердце поступает отдавшая кислород венозная кровь, которая идет отсюда в жабры и далее в ткани. Однако сама ткань сердца у рыб снабжается кровью, насыщенной О2, через одну из ветвей жаберной системы циркуляции, так что оксигенированная кровь из жабр поступает прямо в сердечную мышцу.
СВЕРТЫВАНИЕ КРОВИ И ГЕМОСТАЗ
Потерю крови при разрыве кровеносных сосудов помогают предотвращать несколько механизмов. Большая кровопотеря ведет к снижению кровяного давления и тем самым замедляет вытекание крови из поврежденного участка. Поврежденные сосуды сжимаются и таким образом уменьшают поток крови. Однако самый важный механизм — это закупорка кровеносных сосудов в месте повреждения пробкой, состоящей из свернувшегося белка и .клеточных элементов крови (фото 4.1). Такая пробка (кровяной сгусток, или тромб) полностью останавливает кровотечение при незначительных повреждениях, но если разорваны крупные сосуды, ее недостаточно.
Механизм свертывания, или коагуляции, крови хорошо изучен у млекопитающих, особенно у человека, так как этот процесс имеет большое значение в медицине. Для того чтобы механизм свертывания был эффективным, он должен действовать быстро, а в то же время кровь внутри сосудистой системы не должна загустевать. Поэтому крови должна быть внутреннее присуща способность свертываться, и соответствующий механизм должен быть готовым включиться, как только это будет нужно. С другой стороны, иметь такой механизм — все равно что сидеть на бомбе; необходимы все предосторожности против его случайного срабатывания.
У позвоночных кровяной сгусток состоит из белка фибрина — нерастворимого фибриллярного белка, образующегося из фибриногена— растворимого белка, который содержится в нормальной плазме в количестве около 0,3%. Для превращения фибриногена в фибрин необходим катализатор — фермент тромбин, и кровь не свертывается внутри сосудистой системы именно потому, что в циркулирующей крови этого фермента нет. Однако тромбин может быстро образовываться, так как его предшественник — протромбин— в плазме уже имеется. Для инициации свертывания необходимо, чтобы из протромбина образовался тромбин. Но это только последний шаг в сложной последовательности биохимических реакций, которую медленно расшифровывали, изучая больных с различными дефектами механизма свертывания (например, гемофилией). Всего идентифицировано 12 факторов свертывания крови, которым даны номера от I до XIII (термин «фактор VI»
Свертывание крови и гемостаз 181
теперь не используется). Несколько конечных этапов показано на следующей схеме:
Тромбопластин (III)
Са2+ (IV)
I
Протромбин (II) ---------> Тромбин
(в плазме) ।
Фибриноген (I) --> Фибрин
(в плазме) (тромб)
Многие этапы механизма свертывания могут показаться излишним усложнением, и они сильно затруднили выяснение действительного хода событий. Биологическое значение такой сложно-
Фото 4.1. Эритроцит человека, захваченный фибриновой сетью в кровяном сгустке. Клетка, диаметр которой около 6 мкм, имеет форму двояковогнутого диска, характерную для эритроцитов млекопитающих. (Фото предоставлено Е. Bernstein, Gillette Company Research Institute; рисунок на обложке Science, 1971, vol. 173, № 3993.)
сти, по-видимому, состоит в том, что механизм свертывания крови работает как биохимический усилитель (MacFarlane, 1964). Он обычно приводится в действие при контакте крови с инородной поверхностью или поврежденными тканями. Это инициирует цепь ферментативных реакций, в которой фермент, образовавшийся на
182 Глава 4. Кровообращение
первом этапе, служит катализатором или активатором для следующего этапа, и т. д. Таким образом создается «ферментативный каскад», завершающийся образованием кровяного сгустка, когда растворимый фибриноген переходит в нерастворимый фибрин.
Такая цепь ферментативного усиления позволяет сгустку образовываться быстро и в то же время обеспечивает значительный порог безопасности, предотвращающий спонтанное свертывание крови внутри сосудистой системы. Аналогия с обычной электронной системой усиления здесь очевидна. Если нам нужен низкий уровень шума, мы используем несколько ступеней с малым усилением на каждой ступени, а не одну ступень с большим усилением. Это минимизирует вероятность того, что случайный шум в системе сможет запустить конечную ступень, и гарантирует достаточную степень безопасности.
Для большинства беспозвоночных механизм гемостаза так же важен, как и для позвоночного животного. Тот факт, что многие из них имеют незамкнутые системы кровообращения, осложняет дело, поскольку в такой системе сжатие кровеносных сосудов ничем не может помочь. С другой стороны, в незамкнутой системе кровяное давление всегда ниже, и это уменьшает вероятность потери большого объема жидкости.
Два гемостатических механизма позвоночных — свертывание крови и местное сужение сосудов — имеют свои аналоги и у беспозвоночных. Самый простой механизм у беспозвоночных — это агглютинация (склеивание) клеток крови без участия белков плазмы (Gregoire, Tagnon, 1962). Вслед за агглютинацией начинается образование клеточных сетей, которые сжимаются и помогают затянуть рану. К этому часто добавляется сокращение мышц стенки тела, способствующее закрытию раны.
У многих членистоногих, особено у ракообразных, описано и настоящее свертывание, вызываемое ферментативными превращениями нестабильных белков крови. Механизм свертывания у беспозвоночных, там, где он есть, биохимически отличен от соответствующего механизма позвоночных. Например, у позвоночных свертывание тормозится гепарином — мукополисахаридом, который можно выделить из печени млекопитающих. Гепарин не оказывает никакого влияния на систему свертывания крови мечехвоста (Limulus) и почти не влияет на кровь ракообразных (Needham, 1970).
Сведения относительно механизмов свертывания крови у беспозвоночных очень неполны, но имеющиеся данные указывают на то, что такие механизмы, вероятно, возникали в ходе эволюции много раз независимо друг от друга.
В главах 1—4 мы рассмотрели дыхательные газы и их транспорт. Теперь мы обратимся к вопросам, касающимся доставки энергии, и другим связанным с этим проблемам.
Кровооб ращение
183
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Altman Р. L., Dittmer D. S. (eds.) (1971). Biological Handbooks: Respiration and Circulation, Bethesda, Federation of American Societies for Experimental Biology, 930 pp.
Bond C. F., Gilbert P. W. (1958). Comparative study of blood volume in representative aquatic and nonaquatic birds, Am. J. Physiol., 194, 519—521.
Burton A. C. (1972). Physiology and Biophysics of the Circulation: An Introductory Text. 2nd ed., Chicago, Year Book, 217 pp.
DeLong К. T. (1962). Quantitative analysis of blood circulation through the frog heart, Science, 138, 693—694.
Farmanfarmaian A., Phillips J. H. (1962). Digestion, storage and translocation of nutrients in the purple sea urchin (Strongylocentrotus purpuratus), Biol. Bull., 123, 105—120.
Folkow B., Neil E. (1971). Circulation, New York, Oxford University Press, 593 pp.
Foxon G. E. H. (1955). Problems of the double circulation in vertebrates, Biol. Rev., 30, 196—228.
Gordon M. S. (1972). Animal Physiology: Principles and Adaptations, 2nd ed., New York, Macmillan, 592 pp.
'Gregoire C., Tagnon H. J. (1962). Blood coagulation, Comp. Biochem. (B), 4, 435—482.
Guyton A. C. (1976). Symposium: Interstitial fluid pressure and dynamics of lymph formation, Fed. Proc., 35, 1861—1885.
'Heinrich B. (1971). Temperature regulation of the sphinx moth, Manduca sexta. 2. Regulation of heat loss by control of blood circulation, J. Exp. Biol., 84, 153—166.
Hopper J., Jr., Tabor H., Winkler A. W. (1944). Simultaneous measurements of the blood volume in man and dog by means of Evans blue dye, T1824, and by means of carbon monoxide. 1. Normal subjects, J. Clin. Invest., 23, 628—635.
Johansen K. (1960). Circulation in the hagfish, Myxine glutinosa L., Biol. Bull., 118, 289—295.
Johansen K. (1962). Double circulation in the amphibian Amphiuma tridactylum, Nature, bond., 194, 991—992.
Johansen K. (1968). Air-breathing fishes, Sci. Am., 219, 102—111.
Johansen K. (1970). Air breathing in fishes. In: Fish Physiology, vol. 4 (W. S. Hoar and D. J. Randall, eds.), pp. 361—411, New York, Academic Press.,
Johansen K„ Lenfant C., Hanson D. (1968). Cardiovascular dynamics in the lungfishes, Z. Vergl. Physiol., 59, 157—186.
Johansen K., Martin A. W. (1965). Comparative aspects of cardiovascular function in vertebrates. In: Handbook of Physiology, sect. 2, Circulation, vol. 3 (W. F. Hamilton and P. Dow, eds.), pp. 2583—2641, Washington, D. C., American Physiological Society.
Krijgsman B. J. (1956). Contractile and pacemaker mechanisms of the heart of tunicates, Biol. Rev., 31, 288—312.
Lasiewski R. C., Calder W. A., Jr. (1971). A preliminary allometric analysis of respiration variables in resting birds, Respir. Physiol., 11, 152—166.
Lasiewski R. C., Weathers W. W., Bernstein M. H. (1967). Physiological responses of the giant hummingbird, Patagona gigas, Comp. Biochem. Physiol., 23, 797—813.
MacFarlane R. G. (1964). An enzyme cascade in the blood clotting mechanism and its function as a biochemical amplifier, Nature, bond., 202, 498—499.
Manwell C. (1959). Oxygen equilibrium of Cucumaria miniata hemoglobin and the absence of the Bohr effect, J. Cell. Comp. Physiol., 53, 75—84.
Martin A. W., Harrison F. M., Huston M. J., Stewart D. M. (1958). The blood volumes of some representative molluscs, J. Exp. Biol., 35, 260—279.
Martin A. W., Johansen K. (1965) Adaptations of the circulation in invertebrate animals. In: Handbook of Physiology, sect. 2, Circulation, vol. 3 (W. F. Ha
184 Глава 4, Кровообращение
milton and Р. Dow, eds.), pp. 2545—2581, Washington, D. C., American Physiological Society.
Meglitsch, P. A. (1972). Invertebrate Zoology, 2nd ed., New York, Oxford University Press, 834 pp.
Melbin J., Noordergraaf A. (1971). Elastic deformation in orthotropic vessels: Theoretical and experimental results, Circulation Res., 29, 680—692.
Morrison P„ Ryser F, A., Dawe A. A, (1959). Studies on the physiology of the masked shrew Sorex cinereus, Physiol. Zool., 32, 256—271.
Needham A. E. (1970). Haemostatic mechanisms in the invertebrata. Symposia of the Zoological Society of London, No. 27 (R. G. MacFarlane, ed.), pp. 19—44, London, Academic Press.
Nutting W. L. (1951). A comparative anatomical study of the heart and accessory structures of the orthopteroid insects, J. Morphol., 89, 501—597.
Randall D. J. (1970). The circulatory system. In: Fish Physiology, vol. 4 (W. S. Hoar and D. J. Randall, eds.), pp. 133—172, New York, Academic Press.
Satchell G. H. (1970). A functional appraisal of the fish heart, Fed. Proc., 29, 1120—1123.
Skalak R., Branemark P. 1. (1969). Deformation of red blood cells in capillaries, Science, 164, 717—719.
Snyder G, K. (1973). Erythrocyte evolution: The significance of the Fahraeus-Lind-qvist phenomenon, Respir. Physiol., 19, 271—278.
Stahl W. R, (1965). Organ weights in primates and other mammals, Science, 150, 1039—1042.
Stahl W, R, (1967). Scaling of respiratory variables in mammals, J. Appl. Physiol., 22, 453—460.
Starling E. H, (1896). On the absorption of fluids from the connective tissue spaces, J. Physiol., Lond., 19, 312—326.
Studier E, H., Howell D. J. (1969). Heart rate of female big brown bats in flight, J. Mammal., 50, 842—845.
Thomsen E. (1938). Ueber den Kreislauf im Flugel der Musciden, mit besonderer Berucksichtigung der akzessorischen pulsierenden Organe, Z. Morphol. OekoL Tiere, 34, 416—438.
Thorson T. B. (1958). Measurement of the fluid compartments of four species of marine Chondrichthyes, Physiol. Zool., 31, 16—23.
Thorson T. B, (1959). Partitioning of body water in sea lamprey, Science, 130, 99—100.
Thorson T. B. (1961). The partitioning of body water in Osteichthyes: Phylogenetic and ecological implications in acquatic vertebrates, Biol. Bull., 120, 238— 254.
Thorson T. B. (1964). The partitioning of body water in Amphibia, Physiol. Zool., 37, 395—399.
Thorson T. B, (1968). Body fluid partitioning in Reptilia, Copeia, 1968 (3), 592— 601.
Van Citters R, L., Kemper W, S.t Franklin D. L. (1968). Blood flow and pressure in the giraffe carotid artery, Comp. Biochem. Physiol., 24, 1035—1042.
White F. N, (1956). Circulation in the reptilian heart (Caiman sclerops), Anat. Rec., 125, 417—431.
White F. N. (1959). Circulation in the reptilian heart (Squamata), Anat. Rec., 135, 129—134.
Wigglesworth V, B. (1972). The Principles of Insect Physiology, 7th ed., London, Chapman and Hall, 827 pp.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Altman P. L„ Dittmer D. S. (eds,) (1971). Biological Handbooks: Respiration and Circulation, Bethesda, Federation of American Societies for Experimental Biology, 930 pp.
Кровообращение 185
Burton А. С. (1972). Physiology and Biophysics of the Circulation: An Introductory Text, 2nd ed., Chacago, Year Book, 217 pp.
Caro C, G., Pedley T. J., Schroter R. C., Seed W. A. (1978). The Mechanics of Circulation, Oxford, Oxford University Press, 540 pp. [Имеется перевод: Каро К., Педли Т.э Шротер Р., Сид У. Механика кровообращения. — М.: Мир, 1981.]
Chien S., Usami S.t Dellenback R. J,, Bryant C. A. (1971). Comparative hemorheology: Hematological implications of species differences in blood viscosity, Biorheology, 8, 35—57.
Folkow B., Neil E. (1971). Circulation, New York, Oxford University Press, 593 pp. Hamilton W. F., Dow P. (eds.) (1962, 1963, 1965). Handbook of Physiology, sect. 2, Circulation, vol. 1, pp. 1—758 (1962); vol. 2, pp. 759—1786 (1963); vol. 3, pp. 1787—2765 (1965).
MacFarlane R. G. (ed.) (1970). The Haemostatic Mechanism in Man and Other Animals. Symposium of the Zoological Society of London, No. 27, London, Academic Press, 248 pp.
Martin A. W. (1974). Circulation in invertebrates, Annu. Rev. Physiol., 36, 171— 186
Merrill E. W. (1969). Rheology of blood, Physiol. Rev., 49, 863—888.
Taylor M. G. (1973). Hemodynamics, Annu. Rev. Physiol., 35, 87—116.
ЧАСТЬ ВТОРАЯ
ПИЩА И ЭНЕРГИЯ
ГЛАВА 5
ПИЩА, ТОПЛИВО И ЭНЕРГИЯ
Животные нуждаются в пище 1) как в источнике энергии для поддержания жизни и осуществления своих функций и 2) как в материале для построения и обновления своих клеточных структур и метаболического аппарата.
Большинство растений использует энергию солнечного света и двуокись углерода из атмосферы для синтеза сахаров и —непрямым путем — всех сложных соединений, входящих в состав растения. Все животные используют в качестве источников энергии и строительных материалов химические соединения; животные должны получить эти соединения извне — либо непосредственно из растений, поедая их, либо поедая другие органические материалы. В конечном счете та химическая энергия и те органические вещества, которые нужны животным, имеют своим первоисточником растения и тем самым — косвенно — энергию солнечного света.
Добывание и поглощение пищи называют питанием. Практически вся пища, как растительного, так и животного происхождения, состоит из очень сложных соединений, которые не могут включаться в состав организма или использоваться как топливо, не будучи расщеплены на более простые соединения. Эти сложные вещества обычно даже не могут всасываться в организм без такого расщепления, называемого перевариванием пищи (пищеварением).
Разнообразные химические вещества могут использоваться для получения энергии, но, кроме того, у животных есть специфические потребности в веществах, которые они не могут сами синтезировать, — в аминокислотах, витаминах, некоторых минеральных солях и т. д. Как потребность в пище для обеспечения энергией, так и потребности в специфических пищевых компонентах составляют предмет науки о питании.
Способы питания 187
В этой главе мы рассмотрим пищу и ее потребление, начав с механизмов питания, предшествующих пищеварению, которое делает сложные вещества доступными для организма, а затем кратко охарактеризуем некоторые требования в отношении качества пищи и специфические пищевые потребности. И наконец, мы обсудим, каким образом растения могут защищать себя, вырабатывая вещества, которые делают их несъедобными для многих животных.
СПОСОБЫ ПИТАНИЯ
Для добывания пищи существуют различные механические способы, и они определяют природу пищи, которую данное животное может получать и использовать. В табл. 5.1 перечислены основные механизмы питания и приведены примеры животных, использующих тот или иной способ добывания пищи.
МЕЛКИЕ ЧАСТИЦЫ
Микроскопические водоросли и бактерии могут поступать непосредственно внутрь клеток (например, в пищеварительную вакуоль амебы). Многие животные, даже достаточно крупные, улавливают микроорганизмы при помощи ресничек. Ряд животных, в особенности оболочники и некоторые брюхоногие моллюски, задерживают мелкие взвешенные частицы с помощью слоя слизи; затем эта слизь заглатывается, и организм переваривает содержащуюся в ней пищу.
Голотурии добывают пищу при помощи щупалец. Они зарываются в ил и выставляют свои щупальца над его поверхностью. Щупальца улавливают мелкие частицы, и животное время от времени втягивает их в рот, где с них снимается материал, который можно переварить.
Для мелких планктонных ракообразных, например веслоногих, а также для бокоплавов, губок и двустворчатых моллюсков характерны разнообразные фильтрующие устройства. Очень немногие позвоночные (от рыб до млекопитающих) также используют для получения пищи фильтрацию, однако их фильтры обычно отцеживают пищу более крупную, чем те микроскопические частицы, которые обыкновенно используются беспозвоночными.
Многие пелагические рыбы питаются планктоном. Сельди и скумбрия имеют жаберные гребни, устроенные таким образом, что они работают как сито, улавливающее планктон, главным образом мелких ракообразных. Некоторые из самых больших акул— гигантская акула и китовая акула — питаются исключительно планктоном, который отцеживается из воды, поступающей в рот и протекающей через жабры. По произведенным оценкам, гигант-
188 Глава 5. Пища, топливо и энергия
Таблица 5.1
Способы, используемые различными животными для получения пищи, сгруппированные в соответствии с характером пищи
_ Животные, использующие Тип пищи Способ питания данный способ
Мелкие частицы Образование пшцеваритель- Амебы, радиолярии ных вакуолей Использование ресничек Инфузории, губки, дву- створчатые моллюски, головастики Образование слизистых ло- Брюхоногие моллюски, обо- вушек лочники Использование щупалец Голотурии Использование щетинок и Мелкие ракообразные (Ha-т. п., фильтрация пример, дафнии), сельде- вые, усатые киты, фламинго, буревестники Крупные частицы Заглатывание пассивных Детритоядные, дождевые или массы масс черви Соскабливание, жевание, Морские ежи, улитки, насе-сверление комые, позвоночные Ловля и заглатывание Кишечнополостные, рыбы, жертвы змеи, птицы, летучие мыщи Жидкости или Высасывание сока растений, Тли, пчелы, колибри мягкие ткани нектара Высасывание крови Пиявки, клещи, насекомые, летучие мыши (вампиры) Сосание молока и сходных Детеныши млекопитающих, с ним выделений птенцы Наружное пищеварение Пауки Поглощение через поверх- Паразиты, ленточные черви ность тела Растворенный ор- Поглощение из разбавлен- Водные беспозвоночные (ве-ганический мате- ных растворов роятно, это не основной риал способ питания) Питательные веще- Использование внутрнкле- Парамеции, губки, коралло-ства, образуемые точных симбиотических вые полипы, гидры, пло- симбионтами водорослей ские черви, двустворчатые моллюски
ская акула за 1 ч отцеживает планктон из 2000 тонн воды (Matthews, Parker, 1950).
Некоторые морские птицы тоже кормятся планктоном. Один из буревестников — прионус — имеет ряд пластинок, лежащих вдоль края верхней половины клюва. Он питается ракообразны* ми, отцеживая их из поверхностной воды примерно тем же способом, который более широко используют беззубые киты. Фламинго также питаются планктоном, отцеживая из воды мелкие организмы с помощью клюва.
Способы питания 189>
Беззубые киты полностью специализировались для питания планктоном. Их фильтрующий аппарат состоит из множества роговых пластинок, прикрепленных к верхней челюсти и свисающих вниз с обеих сторон. Когда кит плывет, вода обтекает пластинки и течет между ними, а планктон застревает в волосовидной «бахроме» на их краях. К китам, питающимся планктоном, относится самое крупное из современных животных — синий кит, вес которого может достигать более 100 тонн. Интересно; что и самые крупные киты, и самые крупные акулы кормятся планктоном, а не являются плотоядными; тем самым они избегают лишних звеньев-в пищевой цепи.
МАССИВНАЯ ПИЩА
При питании крупными частицами или массами пищи используются чрезвычайно разнообразные механизмы и структуры. Некоторые животные заглатывают среду, в которой они живут, и переваривают содержащийся в ней органический материал — мелкие живые организмы или мертвое органическое вещество. Такие животные часто пропускают через свой пищеварительный тракт большие количества относительно инертной массы и утилизируют любой органический материал, который она содержит.
Многие животные используют для добывания пищи (нередко растительного происхождения) такие механические приемы, как жевание и соскабливание. Круг этих животных столь широк, что перечислять их было бы бессмысленно. Стоит только вспомнить об огромном числе насекомых и других беспозвоночных, а также о многих позвоночных, питающихся растительной пищей. В эту группу входят и некоторые плотоядные. Они ловят добычу и, прежде чем проглотить ее, разрывают, кромсают или разжевывают.
Другие плотоядные, поймав добычу, заглатывают ее целиком. Среди беспозвоночных таких сравнительно^ мало, среди позвоночных же они многочисленны и включают представителей всех классов — рыб, амфибий, рептилий, птиц и млекопитающих.
Рот и ротовая полость могут быть снабжены челюстями и зубами различного типа, служащими для механического дробления пищи на более мелкие частицы (жевания); это делает пищу более доступной для воздействия пищеварительных ферментов.
У некоторых животных механическое измельчение пищи происходит в особых органах с толстыми мышечными стенками — мускульных желудках, где пища перетирается. У дождевого червя, например, есть мускульный мешок для такого перетирания. Птицы, у которых нет зубов, имеют аналогичный орган — зоб — и часто заглатывают небольшие камешки, помогающие им в измельчении пищи. Это особенно важно для птиц, питающихся зернами, так как эта пища твердая и грубая, и без перетирания требова
t90 Глава 5. Пища, топливо и энергия
лось бы много времени для ее размягчения в пищеварительном тракте. Перед мускульной камерой может находиться простая накопительная камера, в которой начинается смачивание или размягчение пищи, но не происходит сколько-нибудь существенного перетирания.
В начале пищеварительного тракта обычно имеется большой специальный отдел — желудок, который служит накопительным органом и в котором осуществляется также переваривание пищи. У всех позвоночных, за немногими исключениями, желудок играет главную роль в переваривании белков, и наиболее важным ферментом желудка является расщепляющий белки фермент пепсин, который лучше всего работает в растворе с высокой кислотностью. Желудок позвоночных имеет также специальные клетки, выделяющие соляную кислоту.
За желудком идет полая трубка — кишечник, в котором происходит дальнейшее переваривание и который играет особенно важную ;роль во всасывании продуктов пищеварения. Эти продукты— в основном простые сахара, глицерин, жирные кислоты и аминокислоты. Длина кишечника сильно варьирует, и обычно его можно подразделить на несколько отделов. Задний отдел практически не участвует в переваривании пищи и служит главным образом для ее всасывания. Оставшийся материал периодически выбрасывается через анальное отверстие в виде фекалий.
“ЖИДКОСТИ
Многие жидкости могут служить пищей. Животные, питающиеся жидкостями, часто высокоспециализированы и приспособлены к (своим пищевым источникам, и в ряде случаев имеется взаимная выгода и для потребителя, и для поставщика пищи. Например, насекомые получают от цветков нектар и в свою очередь, перелетая с цветка на цветок, производят опыление. Во многих случаях питающиеся жидкостями животные — паразиты; таковы тли, которые пьют сок растений, а также пиявки, комары и клещи, сосущие кровь других животных.
Все млекопитающие в начале своей жизни кормятся жидкостью, так как в течение некоторого периода они получают исключительно молоко, производимое матерью. Не так хорошо известно, что и некоторые птицы вскармливают своих птенцов выделениями, сходными с молоком. У голубя эти выделения образуются в зобе и известны под названием зобного молочка; они отрыгиваются для кормления еще не вылетевших из гнезда птенцов. Интересно, что образование зобного молочка стимулируется пролактином — тем же самым гормоном, который стимулирует выработку молока молочными железами млекопитающих. Биологические преимущества такого способа вскармливания молоди состоят в том, •что он позволяет самим родителям питаться любой подходящей
Способы питания 191
для них пищей и освобождает их от необходимости искать специальный корм (например, насекомых) для молоди. Этот способ» уменьшает также зависимость питания молоди от случайных обстоятельств.
Это преимущество выступает особенно наглядно у императорских пингвинов, которые могут кормить своих птенцов «молоком», выделяемым пищеводом (Prevost, Vilter, 1962). Императорский’ пингвин размножается в середине зимы. Когда в марте приходит антарктическая зима, птицы покидают море и идут по льду к местам размножения, где самка откладывает единственное яйцо, которое в течение двух месяцев будет согревать ее супруг, стоя на-льду в холоде и мраке зимней ночи. Тем временем самка идет обратно к морю для питания и позже возвращается к голодающему самцу, чтобы кормить птенца. Однако если самка задерживается, то все еще голодающий самец начинает, кормить птенца «молоком», и птенец при этом не только выживает, но может* даже прибавлять в весе. По содержанию белка и жира «молоко»' голубя и пингвина (табл. 5.2) близко к молоку млекопитающих, (хотя в молоке многих млекопитающих намного больше углеводов).
Таблица 5.2
Состав «молока» голубя и императорского'пингвина’ в процентах от общего количества сухого остатка. Для сравнения приведен состав кроличьего молока;
(Prevost, Vilter, 1962)
Компонент Голубь Пингвин Кролик
Белки 57,4 59,3 50,6
Липиды 34,2 28,3 34,3
Углеводы 0 7,8 6,4
Минеральные
вещества 6,5 4,6 8,4
Всего 98,1 100,0 99,7
Особый случай питания жидкостями мы наблюдаем у пауков. Добыча паука часто бывает размером с его собственное тело и: даже больше, и обычно она покрыта жестким хитином, который нелегко содрать. Чтобы получить пищу из пойманного насекомого, паук прокалывает его своими пустотелыми челюстями и накачивает в его тело пищеварительные соки. Эти соки растворяют ткани, и паук затем высасывает внутренность жертвы.
Некоторые паразиты (например, ленточные черви) живут в-очень питательной среде, и пищеварительный тракт у них полностью редуцировался. Поскольку их хозяева сами обеспечивают
192 Глава 5. Пища, топливо и энергия
переваривание пищи, у этих паразитов нет ни пищеварительной системы, ни соответствующих ферментов, которые иначе были бы необходимы, и все питательные вещества эти паразиты поглощают прямо через поверхность тела.
Фото 5.1. Насекомое, питающееся жидкостью. Жуки-хищники (Staphylinidae) питаются различными беспозвоночными. Жук пронзает жертву своими острыми мандибулами и вводит в нее ферменты, которые переваривают ее изнутри. Затем жук высасывает разжиженную пищу через волосковое сито, расположенное у входа в ротовую полость. (Фото предоставлено J. F. Lawrence, Музей сравнительной зоологии, Харвардский университет.)
РАСТВОРЕННЫЙ ОРГАНИЧЕСКИЙ МАТЕРИАЛ
Могут ли водные животные использовать органический материал, растворенный в воде, — этот вопрос остается дискуссионным. В начале нашего века немецкий биолог Пюттер высказал мысль, что растворенные органические вещества, хотя они и содержатся в воде в очень низкой концентрации, все же могут непосредственно использоваться морскими животными. Пюттер предположил даже, что такой растворенный материал имеет большее значение, чем планктон, поскольку .планктонные организмы так далеко разбросаны друг от друга .в воде, что их улавливание часто бывает невозможным.
Играет ли существенную роль поглощение растворенного органического материала, трудно было выяснить экспериментальным путем, так как нужно было бы анализировать чревычайно раз
Способы питания 193
бавленные растворы. Когда появилась возможность метить органические вещества радиоизотопами, удалось ясно показать, что многие водные беспозвоночные действительно могут поглощать и глюкозу, и аминокислоты из очень сильно разведенных растворов (Stephens, 1962, 1964).
В таких экспериментах несложно установить, что какое-то количество вещества поглощается; это не вызывает сомнений. Трудно показать, что данного вещества поступает в организм больше, чем за то же время выходит из организма наружу. Чтобы для животного была какая-то польза, поступление должно превышать потери. Одновременное поглощение и выделение свободных аминокислот организмами в морской воде сравнили Иоганнес и др. (Johannes et al., 1969). В экспериментах на одном виде турбел-лярий они показали, что выход свободных аминокислот в воду примерно в четыре раза превышает одновременное поглощение аминокислот, помеченных 14С. Другой вопрос, который остается открытым, — это вопрос об участии в обсуждаемом поглощении бактерий. Бактерии могли бы поглощать растворенные органические вещества, а затем заглатываться животным. Они способны эффективно извлекать глюкозу или ацетат из растворов, содержащих эти вещества в концентрации всего лишь 1—10 мг/л; для обычных пресноводных водорослей это невозможно (Wright, Hob-bie, 1966).
Хотя имеющиеся данные неопределенны, нет явных указаний на то, чтобы какое-либо водное беспозвоночное существовало главным образом за счет поглощения органического материала из сильно разведенного раствора, в котором оно живет. Сложные приспособления для питания тоже говорят против сколько-нибудь важной роли растворенных органических веществ.
СИМБИОНТЫ КАК ИСТОЧНИК ПИТАТЕЛЬНЫХ ВЕЩЕСТВ
У некоторых беспозвоночных внутри клеток имеются симбиотические водоросли. Эти водоросли существенны для снабжения хозяина питательными веществами, так как обеспечивают наиболее короткий из возможных путей между фотосинтезом и животным. В свою очередь водоросли используют аммиак, выделяемый животным-хозяином, для синтеза белка. Такие симбиотические отношения между животным и растением встречаются у простейших, губок, коралловых полипов, гидр, плоских червей и двустворчатых моллюсков.
При взаимоотношениях такого рода животное получает органические вещества (в первую очередь углеводы) из живых клеток растения, а не путем разрушения и переваривания этих клеток. В этом отношении симбиотическое снабжение питательными веществами в принципе отличается от всех механизмов питания, об-13—1873
194 Глава 5. Пища, топливо и энергия
суждавшихся выше, так как все они включают разрушение части или всего организма, служащего источником пищи.
Важное значение симбиоза было продемонстрировано на гидрах путем сравнения животных, содержащих водоросли (зеленые гидры) и не содержащих их. Присутствие симбиотических водорослей сильно удлиняет время выживания гидр в условиях голодания, а если дается ограниченное количество пищи, зеленые гидры растут быстрее. Зеленые гидры поглощают на свету меньше кислорода, чем в темноте, и меньше, чем гидры без симбиотических водорослей (Muscatine, Lenhoff, 1965; Pardy, Dieckmann, 1975).
Внутри животных-хозяев встречаются в основном водоросли двух типов: зеленые (зоохлореллы) и бурые (зооксантеллы) (табл. 5.3). Если под углеводами мы будем понимать не только* сахара, но и их производные, такие, как сахароспирты и глицерин, то можно будет сказать, что основная масса передаваемого» материала имеет углеводную природу. Растворимые углеводы, выделяемые в организм животного-хозяина, всегда отличны от основных внутриклеточных углеводов водоросли. Например, зоохлореллы синтезируют и накапливают в своих клетках сахарозу, а в организм хозяина экстретируют мальтозу или глюкозу.
Количество органического материала, получаемого от симбионта, довольно значительно. Водоросли в зеленой гидре отдают
Таблица 5.3
Выделение углеводных соединений из симбиотических водорослей в организм животного-хозяина. (Smith, et al., 1969)
Водоросли Тип животных-хозяев Вид животного-хозяина Выделяемый углевод или родственное соединение
Зоохлореллы Простейшие Paramecium bursaria Мальтоза
Губки Spongilla lacustris Глюкоза
Кишечнополостные Плоские черви Chlorohydra viridissi-ma (дикий тип) Ch. viridissima (мутант) Convoluta roscoffen- 1 Мальтоза Глюкоза
Моллюски sis ’ Placobranchus iantho- 1 bapsus J Tridacna crispata Не известно
Зооксантеллы Кишечнополостные Моллюски Pocillopora datnicor-1 nis Anthopleura elegantis-sima Zoanthus confertus Fungia scutaria Tridacna crocea . Глицерин
Пищеварение 195
организму хозяина 45—50% углерода, связываемого в процессе «фотосинтеза. Изолированные водоросли выделяют углеводы, главным образом мальтозу, в окружающую среду в количестве, которое зависит от pH среды. При pH 4,5 эти водоросли отдают до 85% связанного при фотосинтезе углерода, а в щелочной среде выделение снижается до нескольких процентов. Таким образом, не исключено, что организм хозяина может контролировать количество углеводов, выделяемых водорослью, регулируя свое внутриклеточное pH.
Если зооксантеллу выделить из ее хозяина и инкубировать в морской воде, она будет выделять некоторое количество углеводов, но это количество возрастет, если к воде добавить гомогенизированную ткань хозяина. В случае зооксантеллы, взятой из Tridacna (гигантский двустворчатый моллюск), было получено 16-кратное увеличение отдачи углеводов в присутствии гомогената клеток хозяина. Однако это не универсальное явление: на зоохлореллу, взятую из гидры, гомогенаты ее хозяина не влияют (Smith et al., 1969).
ПИЩЕВАРЕНИЕ
Пища животных состоит из органических веществ, большая часть которых относится к трем главным группам соединений — к белкам, жирам и углеводам. Эти три типа органических соединений безраздельно доминируют в составе практически всех растений и животных. Большинство их имеет довольно большие молекулы. Даже у самых мелких (сахаров) молекулярные веса составляют несколько сотен; молекулы жиров крупнее; многие белки имеют молекулярные веса от 100 000 до нескольких миллионов, а молекулы крахмала и целлюлозы не имеют определенных размеров, так как это полимеры, составленные из малых углеводных единиц. И в тех случаях, когда пища используется как топливо, и тогда, когда она служит для построения и поддержания структур тела, большие молекулы пищи вначале дробятся на более простые компоненты; последние затем всасываются и либо включаются в состав тела, либо расщепляются для получения энергии.
Главная задача пищеварения — расщепить большие и сложные молекулы, имеющиеся в пище, таким образом, чтобы их фрагменты могли всосаться и были пригодны для использования в организме. Такое дробление достигается в пищеварительном тракте с помощью ферментов.
ВНУТРИКЛЕТОЧНОЕ И ВНЕКЛЕТОЧНОЕ ПИЩЕВАРЕНИЕ
Пищеварение может осуществляться или внутри, или вне клеток. У одноклеточных животных пищеварение обычно по необходимости внутриклеточное. Простейшие набирают пищу в пищева-13*
196 Глава 5, Пища, топливо и энергия
рительную вакуоль и секретируют в эту вакуоль ферменты, расщепляющие углеводы, жиры и белки. Аналогичное внутриклеточное переваривание происходит у губок и в какой-то мере у кишечнополостных, гребневиков и турбеллярий. В сочетании с внеклеточным пищеварением оно встречается также у ряда более* сложно устроенных животных. Например, у двустворчатых моллюсков мелкие пищевые частицы нередко захватываются и перевариваются клетками пищезарительпой железы.
Некоторые животные, питающиеся более крупными кусками пищи, например кишечнополостные, имеют частично внутриклеточное, а частично внеклеточное пищеварение. Переваривание начинается в пищеварительной (гастроваскулярной) полости; затем фрагменты частично переваренной пищи захватываются внутрь клеток, образующих стенки этой полости, где перевариваются окончательно.
Внеклеточное пищеварение имеет одно очевидное преимущество: оно позволяет заглатывать большие куски пищи, тогда как внутриклеточное пищеварение ограничено частичками, достаточно-малыми для поглощения их отдельными клетками организма.
Внеклеточное пищеварение обычно сочетается с хорошо развитым пищеварительным трактом, в котором выделяемые ферменты могут воздействовать на пищевой материал. Пищеварительный тракт может иметь одно отверстие, как у кишечнополостных, офиур и плоских червей. У этих животных все непереваренные материалы выбрасываются через то же отверстие, которое служит ртом. У более сложных животных пищеварительный тракт имеет два отверстия: рот и анус. Это позволяет осуществить «конвейерный» процесс переваривания. Пища, поступающая внутрь через рот, передвигается дальше и подвергается действию ряда пищеварительных ферментов; растворимые продукты переваривания всасываются, а непереваренный материал в конце концов выбрасывается через анус, не мешая приему пищи. При таком способе прием пищи может продолжаться во время пищеварения, и прохождение пищи по пищеварительному тракту может осуществляться без перерывов.
Все кишечнополостные — плотоядные. Они имеют хватательные приспособления — щупальца, которые помогают им ловить и парализовать добычу. Щупальца снабжены специализированными клетками — нематоцистами, которые, соприкоснувшись с подходящей добычей, выбрасывают тонкую полую нить, пронзающую жертву. Через эту нить из клетки выходит яд, .который парализует добычу. Затем щупальца заталкивают жертву в гастральную полость для переваривания.
У плоского червя (планарии) рот ведет в гастроваскулярную полость, разветвления которой расходятся по всему телу. Благодаря своей разветвленности эта полость не только служит для пищеварения, но и доставляет пищу ко всем частям организма.
Пищеварение 197
Система ветвей увеличивает также общую поверхность гастровас-кулярной полости, что способствует всасыванию переваренной пищи. У планарий внеклеточное пищеварение помогает измельчению пищи, но большая часть пищевых частиц захватывается клетками, выстилающими полость, и переваривается внутриклеточно.
ФЕРМЕНТАТИВНОЕ ПЕРЕВАРИВАНИЕ ПИЩИ
Молекулы большинства пищевых веществ либо очень крупны (белки), либо нерастворимы в воде (жиры), либо и крупны, и нерастворимы (крахмал, целлюлоза). Прежде чем элементы пищи смогут всосаться и использоваться организмом, они должны быть переведены в растворимое состояние и расщеплены на более мелкие составные части.
Белки, крахмал и целлюлоза — полимеры, образованные из более простых строительных блоков путем синтеза с отщеплением воды; жиры — это эфиры жирных кислот и глицерина, опять-таки синтезируемые путем отщепления воды. Разделение этих пищевых веществ на более простые компоненты связано с поглощением воды и поэтому называется гидролизом. Реакция гидролиза протекает самопроизвольно с выделением энергии в форме тепла. Такой спонтанный гидролиз пищевых веществ происходит с неизмеримо малой скоростью, но его можно ускорить катализаторами. Вырабатываемые живыми организмами катализаторы называются ферментами. Ферменты необходимы для всех пищеварительных процессов. Практически все метаболические процессы внутри клеток также катализируются ферментами.
Все ферменты — белки; многие из них были выделены в чистом виде, и их действие изучено очень подробно. Многие ферменты получили свои названия по субстратам, на которые они действуют, с добавлением окончания -аза. Пищеварительные ферменты часто действуют на обширную группу субстратов, и некоторые из них больше известны под старыми традиционными названиями, хотя они имеют более точные названия согласно принятой международной номенклатуре. Поскольку детально изучены пищеварительные ферменты лишь небольшого числа организмов, мы будем в большинстве случаев употреблять традиционные названия, а не следовать биохимической номенклатуре.
После субстрата, на который действует фермент, или реакции, которую он катализирует, наиболее важным параметром фермента является его активность, т. е. скорость, с которой протекает катализируемая им реакция. На эту активность влияет много факторов; при изменении структуры ферментного белка он обычно становится неактивным. Высокая температура, сильнодействующие химикаты, другие ферменты, расщепляющие белки, и тяжелые металлы, связываемые белками, — все эти факторы могут инактивировать фермент.
198 Глава 5. Пища, топливо и энергия
На активность всех ферментов сильно влияет pH раствора. Большинство ферментов проявляет наибольшую активность в узком диапазоне pH; это так называемый оптимум pH для данного фермента. Два хорошо известных фермента, расщепляющих белки,— это пепсин, образующийся в желудке позвоночных, и трипсин, вырабатываемый поджелудочной железой позвоночных.
Для пепсина оптимум pH соответствует растворам с очень высокой кислотностью — он близок к pH 2, а для трипсина опти-
Рис. 5.1. Влияние температуры на расщепляющий белки фермент (протеазу) из асцидии Halocynthia. Как показывают кривые, для действия фермента существует температурный оптимум, но этот оптимум тем ниже, чем длительнее эксперимент. Объяснения в тексте. (ВеггШ, 1929.)
мальны слабощелочные растворы с pH около 8. Оптимум pH у данного фермента не обязательно одинаков для всех субстратов, на которые он действует. Например, для пепсина оптимум pH равен 1,5 в случае переваривания яичного альбумина, 1,8 — в случае казеина и 2,2 — в случае гемоглобина (White et al., 1964). По обе стороны от оптимального pH активность снижается и при большом удалении от него падает до нуля. Пепсин еще оказывает некоторое действие при pH 4 и полностью восстанавливает свою активность, когда pH возвращается к оптимуму. Однако при pH 8 пепсин разрушается и инактивируется необратимо.
Скорость ферментативных реакций сильно зависит от температуры; с повышением температуры она увеличивается. Но так как нагревание обычно ведет к свертыванию белков, оно также инактивирует и ферменты; при температурах выше 50 °C или около того большинство ферментов полностью утрачивает активность. Та
Пищеварение 199
ким образом, умеренное повышение температуры ускоряет реакцию, но если температуру повышать дальше, увеличению скорости реакций начнет противодействовать термическое разрушение ферментов. При какой-то температуре скорость будет максимальной, но, поскольку термическое разрушение с течением времени нарастает, мы обнаружим, что температурный оптимум зависит от длительности эксперимента. В длительном опыте повышенная температура будет воздействовать на фермент дольше и наблюдаемый температурный оптимум будет поэтому ниже; в кратковременном опыте с тем же самым ферментом оптимум будет выше. Таким образом, температурный оптимум не является точной характеристикой фермента; он зависит от длительности опыта (рис. 5.1).
Большинство ферментов быстро инактивируется при температурах выше 45—50 °C. Это та область температур, где лежит верхний предел жизни для большинства сложных организмов, за исключением некоторых покоящихся стадий и определенных термофильных бактерий. Однако небольшое число ферментов термоустойчиво; примером могут служить расщепляющий белки фермент папаин из плодов дынного дерева и сходный с ним фермент из ананаса. Эти ферменты применяют для размягчения мяса, так как их протеолитическое действие продолжается во время варки до температур 70—80 °C, хотя при температуре кипения воды они быстро инактивируются.
ПЕРЕВАРИВАНИЕ БЕЛКОВ
Белки — это полимеры, состоящие из аминокислот, соединенных между собой пептидными связями. Аминокислота — относительно простая органическая кислота, у которой атом углерода, смежный с кислотной группой (карбоксильной группой —СООН), несет на себе аминогруппу (—NH2). Пептидные связи образуются в результате отщепления воды между карбоксильной группой одной аминокислоты и аминогруппой другой. Имеется около 20 «обычных» аминокислот, которые в различных пропорциях содержатся практически во всех живых организмах. Это означает, что аминокислоты, получающиеся при гидролизе белков, служат строительными блоками, пригодными для синтеза почти любого специфического белка.
Ферменты, расщепляющие белки, делятся на две группы в соответствии с тем, на какую часть молекулы белка они действуют. Экзопептидазы гидролизуют концевые пептидные связи в длинной пептидной цепи; эндопептидазы воздействуют на внутренние участки пептидной цепи. Две наиболее известные пищеварительные эндопептидазы позвоночных — это пепсин и трипсин.
Пепсин секретируется желудком в виде неактивного предшественника— пепсиногена. Желудок выделяет также соляную кис-
200 Глава 5. Пища, топливо и энергия
Рис. 5.2. Переваривающий белки фермент пепсин разрывает пептидные связи между дикарбоновой и циклической аминокислотами. Трипсин действует на пептидные связи, смежные с аминокислотами, имеющими две аминогруппы. Подробности см. в тексте.
лоту, которая обеспечивает его содержимому низкое pH. В кислой среде с pH ниже 6 пепсиноген аутокаталитически активируется, превращаясь в активный фермент—пепсин. Пепсин специфически гидролизует пептидные связи между аминокислотой, содержащей фенильную группу (тирозином или фенилаланином), и ка-какой-нибудь дикарбоновой аминокислотой (глутаминовой или аспарагиновой). Таким образом, пепсин действует только на некоторые из пептидных связей в длинной белковой цепи, а не на все, и в результате получается набор более коротких цепей — фрагментов, которые дальше пепсином не расщепляются.
Трипсин выделяется поджелудочной железой в виде неактивного трипсиногена. Он активируется в кишечнике с помощью фермента энтерокиназы, секретируемой железами в стенках кишки. Трипсиноген активируется также активным трипсином (поэтому активация его идет все быстрее по мере того, как образуется все больше трипсина). Это называют аутокаталитической активацией.
Трипсин лучше всего действует при слабощелочных pH между 7 и 9. Он гидролизует пептидные связи, смежные с основными аминокислотами (т. е. с теми, в которых две аминогруппы, — лизином или аргинином) (рис. 5.2).
Белковые фрагменты и пептидные цепочки, образовавшиеся под действием пепсина и трипсина, перевариваются далее с помощью экзопептидаз, которые действуют только на концевые пептидные связи. Карбоксипептидаза, выделяемая поджелудочной железой, гидролизует концевые связи, смежные со свободной -карбоксильной группой, а аминопептидаза, выделяемая кишечни
Пищеварение 201
ком, гидролизует связи, смежные со свободной аминогруппой. И наконец, дипептидаза гидролизует пептидные связи тех фрагментов, которые состоят только из двух аминокислот.
У беспозвоночных переваривание белков, как внеклеточное, так и внутриклеточное, в принципе протекает так же, как у позвоночных, за исключением того, что беспозвоночные обычно не имеют пепсиноподобных ферментов, действующих в кислых растворах. Во всех других отношениях действие ферментов сходно, хотя они и не идентичны по структуре ферментам позвоночных.
Однако нельзя сказать, что пепсиноподобные ферменты свойственны исключительно позвоночным. Личинка обыкновенной комнатной мухи (Musca domestica) выделяет протеазу, активную в диапазоне pH от 1,5 до 3,5 с оптимумом при 2,4. Имеет ли этот фермент значение для пищеварения, зависит от того, может ли для него создаваться подходящая кислая среда, а это пока не выяснено (Greenberg, Paretsky, 1955; Lambremont et al., 1959).
ПЕРЕВАРИВАНИЕ ЖИРОВ
Переваривание жиров сходно у позвоночных и беспозвоночных. Обычные жиры как растительного, так и животного происхождения состоят из эфиров, образованных одной молекулой глицерина (трехатомного спирта) и тремя молекулами жирных кислот с длинной цепью. Они практически нерастворимы в воде и поэтому нелегко гидролизуются ферментами. Поджелудочная железа позвоночных выделяет гидролизующий жиры фермент липазу, но для приведения его в контакт с жирами необходима помощь детергентов. Эту функцию выполняют желчные кислоты, выделяемые печенью. Под действием желчных кислот и механических движений кишечника жиры эмульгируются, а образующиеся при их гидролизе жирные кислоты (тоже относительно нерастворимые) с помощью желчных солей удерживаются в растворе и благодаря этому могут всасываться. Глицерин (это нормальный промежуточный продукт метаболизма практически во всех клетках) растворим в воде и поэтому легко всасывается и включается в обмен веществ. Однако жиры могут частично всасываться кишечным эпителием и без предварительного гидролиза: мелкие капельки, образующиеся при эмульгировании с помощью желчи, непосредственно захватываются эпителиальными клетками.
Некоторые жироподобные вещества, в том числе воска, не гидролизуются обычными липазами. Воска — это эфиры одной молекулы алифатического высокомолекулярного спирта с одной молекулой жирной кислоты; если бы они могли гидролизоваться, их компоненты усваивались бы и давали большое количество энер-. гии. Из восков наиболее известен пчелиный воск; за одним исключением, он не переваривается позвоночными и потому не имеет питательной ценности.
2С2 Глава 5. Пища, топливо и энергия
Хотя позвоночные не вырабатывают ферментов, расщепляющих пчелиный воск, он играет важную роль в питании небольшой группы птиц — южноафриканских медоуказчиков. Эти птицы, родственники дятлов, известны своим умением приводить капского медоеда или человека к гнездам диких пчел. Птица привлекает к себе внимание, поднимая треск и шум, и если за ней последовать, она приведет к пчелиному гнезду. Там она подождет, пока гнездо грабят, а затем будет есть оставшийся воск. Медоуказ-чики предпочитают воск меду и едят чистый воск так же охотно, как и соты. Эти птицы могут переваривать воск благодаря тому, что в их кишечном тракте живут симбиотические бактерии, которые вырабатывают ферменты, необходимые для расщепления воска. Практически все другие животные, как позвоночные, так и беспозвоночные, не способны переваривать воск. Только гусеница большой восковой огневки (Galleria)—обычный паразит пчелиных ульев — может существовать за счет пчелиного воска (Friedmann, Kern, 1956а, b).
Предположение о том, что за переваривание воска медоуказ-чиком ответственны симбиотические бактерии, было подтверждено путем введения домашним курам чистых культур бактерий, взятых из пищеварительного тракта медоуказчиков. В норме куры совершенно не способны переваривать воск, но если скармливать им воск вместе с такими культурами, они могут его переваривать и усваивать (Friedman et al., 1957).
Переваривание воска птицами с помощью симбиотических бактерий является редкой диковиной. Напротив, как мы увидим далее в этой главе, симбиотическое переваривание целлюлозы — одного из самых обычных растительных материалов — представляет огромную важность для травоядных животных.
Хотя воска не играют существенной роли в питании обитателей суши, они занимают чрезвычайно важное место в пищевых цепях морских животных наряду с обычными жирами и маслами.
Воска имеются у большого числа разнообразных морских организмов: головоногих и других моллюсков, креветок, актиний, коралловых полипов и многих рыб. Первичными производителями восков, по-видимому, являются мелкие планктонные ракообразные, особенно веслоногие. У некоторых из них воска могут составлять до 70% сухого веса тела. Веслоногие питаются фитопланктоном, который не содержит воска. Диатомеи и панцирные жгутиковые аккумулируют масляные шарики, состоящие в основном из триглицеридов. Однако жирные кислоты воска веслоногих очень напоминают характерные жирные кислоты, свойственные фитопланктону, и есть основание заключить, что эти последние непосредственно используются веслоногими для выработки восков.
Роль, восков в морских цепях питания неизмеримо более важна, чем это представлялось несколько лет назад, так как планктонные ракообразные — главное связующее звено между микро
Пищеварение 203
скопическими фотосинтезирующими водорослями и крупными обитателями моря. По произведенным оценкам, благодаря такому звену примерно половина всего органического вещества, образующегося на Земле в процессе фотосинтеза, на какое-то время превращается в воск (Benson et al., 1972).
Рыбы, питающиеся веслоногими (например, сельди, анчоусы, сардины), имеют в своем пищеварительном тракте липазы для расщепления восков (Sargent, Gatten, 1976). Спирты этих восков окисляются до жирных кислот, которые затем входят в состав обычных нейтральных жиров — триглицеридов. У ряда других рыб количество восковых липаз намного меньше, и вопрос о том, насколько хорошо они могут переваривать воска, остается открытым. Поскольку воска содержатся в'жирах и маслах очень многих морских животных, даже у китов, трудно сказать, могут ли они использоваться в процессах метаболизма и служить энергетическим резервом, или они просто накапливаются в результате пассивного поглощения и затруднений с их использованием. Этот важный вопрос сейчас интенсивно изучается.
ПЕРЕВАРИВАНИЕ УГЛЕВОДОВ
В отношении переваривания углеводов между позвоночными и беспозвоночными большой разницы нет. Простые сахара, такие, как глюкоза и фруктоза, всасываются без каких-либо изменений и непосредственно используются в обычных метаболических процессах. Дисахариды, например сахароза (сахар растительного происхождения) или лактоза (содержащаяся в молоке), расщепляются до моносахаридов, и только после этого они могут всасываться и использоваться. Фермент сахараза выделяется в кишечнике, но ее нет в клеточном аппарате животных. Поэтому, если сахарозу ввести в организм позвоночного путем инъекции, она будет полностью выведена с мочой в неизмененном виде.
Множество растений накапливает в качестве основного энергетического резерва крахмал. Это полимер, состоящий из остатков глюкозы. Он относительно нерастворим, но гидролизуется ферментом амилазой (от латинского amylum — крахмал), выделяемым слюнными железами человека (и некоторых других, но не всех, млекопитающих) и в большем количестве — поджелудочной железой.
Хотя крахмал плохо растворим, его легко переваривают многие животные. Однако амилаза действует на него эффективнее, если он подвергался нагреванию, и это основная причина того, что люди готовят крахмалистую пищу (картофель, хлеб и т. д.‘) горячим способом. Вполне возможно, что у многих животных, легко переваривающих сырой крахмал, начальному расщеплению крахмала помогают бактерии, после чего он лучше поддается действию амилазы.
204 Глава 5. Пища, топливо и энергия
СИМБИОТИЧЕСКОЕ ПЕРЕВАРИВАНИЕ ЦЕЛЛЮЛОЗЫ
Наиболее важным структурным материалом растений является целлюлоза (клетчатка) —полимер глюкозы, который нерастворим и чрезвычайно устойчив к химическим воздействиям. Ферменты, расщепляющие целлюлозу — целлюлазы — в пищеварительных соках позвоночных отсутствуют, и все же многие позвоночные переваривают целлюлозу, и она служит для них главным источником энергии. Настоящие целлюлазы были найдены в кишечном тракте некоторых беспозвоночных, питающихся древесиной и сходными растительными продуктами, но во многих случаях переваривание целлюлозы осуществляют симбиотические микроорганизмы, живущие в пищеварительном тракте своих хозяев.
ПЕРЕВАРИВАНИЕ ЦЕЛЛЮЛОЗЫ У БЕСПОЗВОНОЧНЫХ
Вопрос о том, кто вырабатывает целлюлазу — само животное или симбиотический микроорганизм, — часто трудно решить с полной определенностью. Проиллюстрируем эти трудности несколькими примерами.
Улитки. С давних пор считалось, что виноградная улитка, Helix pomatia, переваривает целлюлозу и действительно выделяет целлюлазу. Однако тщательный анализ экстрактов пищеварительных желез и кишечной стенки не выявил никакой целлюлазной активности, хотя присутствие целлюлазы в содержимом кишечника продемонстрировать нетрудно (Florkin, Lozet, 1949). Это, по-видимому, указывает на важную роль кишечных микроорганизмов, хотя сам факт отсутствия целлюлазы в экстрактах пищеварительных органов не обязательно исключает возможность того, что животное может вырабатывать этот фермент.
Корабельный червь. Другой организм, который считают способным переваривать целлюлозу, — это корабельный червь (Teredo); на самом деле это сверлящий дерево двустворчатый моллюск. В его кишке была обнаружена настоящая целлюлаза, и кишечные экстракты высвобождали сахар из целлюлозы, хотя никаких переваривающих целлюлозу бактерий или простейших из пищеварительного тракта выделить не удалось (Greenfield, Lane, 1953). В данном случае можно не согласиться с тем, что видимое отсутствие таких микроорганизмов является доказательством их отсутствия в живом животном. Экстракты кишечника, проявлявшие целлюлазную активность, были приготовлены из кусочков кишки, содержимое которой не удалялось, и поэтому источник активности остался неуточненным.
Чешуйница. Есть, однако, сообщения и о настоящих целлюлазах, полученных из ряда животных в условиях, которые, по-видш мому, исключают какое бы то ни было участие симбиотических микроорганизмов. Чешуйница Ctenolepisma lineata переваривает целлюлозу и может выживать на рационе из одной лишь целлю
Пищеварение 205
лозы, хотя ее и недостаточно для длительного питания. Кишка этого насекомого содержит много микроорганизмов, но никаких переваривающих целлюлозу видов из нее выделено не было. Еще важнее то, что путем отмывания яиц в растворе хлористой ртути и этанола и выкармливания нимф на стерилизованных овсяных хлопьях и витаминах в асептических условиях были выращены особи, свободные от бактерий. Такие особи переваривали целлюлозу, меченную 14С, и выдыхали ИСС>2. Наконец, целлюлаза была обнаружена в экстрактах средней кишки. Ввиду этого можно, видимо, считать установленным, что чешуйница действительно вырабатывает целлюлазу (Lasker, Giese, 1956).
Термиты. В случае термитов, питающихся почти исключительно древесиной, роль симбиотических бактерий несомненна. Кишечный тракт переваривающего древесину термита буквально набит различными жгутиковыми и бактериями. Некоторые виды жгутиковых были выделены и выращивались в чистой культуре, и это позволило тщательно изучить их роль в переваривании целлюлозы.
Симбиотические жгутиковые, выделенные из термитов, являются строгими анаэробами (т. е. они могут выживать только при отсутствии свободного кислорода). Эту чувствительность к кислороду можно использовать для удаления жгутиковых из организма животного-хозяина и получения термитов, свободных от простейших. Если термитов подвергать воздействию кислорода под давлением 3,5 атм, жгутиковые избирательно гибнут примерно за полчаса, а термиты выживают без всякого вреда для себя (Cleveland, 1925). Но такие «дефаунированные» термиты не могут выжить, если кормить их, как обычно, древесиной. При этом бактерии в их кишечниках все еще имеются, из чего можно заключить, что за переваривание целлюлозы ответственны не они, а симбионты-простейшие. Если термитам ввести соответствующих жгутиковых, они снова начинают переваривать целлюлозу (Hungate, 1955).
Гипотеза о том, что целлюлозу переваривают жгутиковые, получила дальнейшее подтверждение при исследовании культур этих организмов. Одно из таких жгутиковых (Trichomonas ter-mopsidis), которое выращивали в культуре в течение трех лет, послужило источником экстрактов, содержащих целлюлазу (Trager, 1932). Более того, это простейшее вообще не может использовать никаких углеродных соединений, кроме целлюлозы, так что нет никаких сомнений относительно его роли в переваривании целлюлозы термитами (Trager, 1934).
Хотя древесная целлюлоза и может дать значительное количество энергии, древесина содержит очень мало азота. Поскольку термиты на этой пище процветают, было высказано предположение, что в их кишечной флоре имеются азотфиксирующие бактерии, которые служат добавочным источником белка. Эта гипотеза
206 Глава 5. Пища, топливо и энергия
казалась правдоподобной. Многие ранние попытки продемонстрировать фиксацию азота в общем азотном балансе термитов не увенчались успехом (Hungate, 1941), но теперь ясно, что фиксация азота при участии микроорганизмов действительно имеет место (Benemann, 1973; Breznak et al., 1973). Возможно, что фиксация азота даже более широко распространена у сверлящих дерево животных, чем предполагалось раньше. У некоторых видов корабельных червей (Teredo и других) фиксация азота связана с кишечными бактериями; эти бактерии были выделены и было показано, что они способны фиксировать азот (Carpenter, Culliney» 1975).
ПЕРЕВАРИВАНИЕ ЦЕЛЛЮЛОЗЫ ПОЗВОНОЧНЫМИ
Многие млекопитающие растительноядны, и пищевой рацион большинства таких форм делает необходимым переваривание целлюлозы. Жвачные животные, к которым относятся наши важнейшее поставщики мяса и молока (крупный рогатый скот, овцы, ко> зы), имеют специализированный пищеварительный тракт, в высокой степени приспособленный для симбиотического переваривания целлюлозы. Однако и многие нежвачные животные тоже нуждаются для переваривания целлюлозы в микроорганизмах-симбионтах, хотя их анатомические адаптации не таковы, как у настоящих жвачных.
Жвачные, Желудок жвачного состоит из нескольких отделов: (рис. 5.3), или, если выражаться точнее, настоящему пищеварительному желудку предшествует несколько крупных камер, из которых первая, самая большая, называется рубцом. Рубец служит бродильным чаном, в котором пища, смешанная со слюной, подвергается интенсивной ферментации. В рубце содержатся в больших количествах и бактерии, и простейшие. Эти микроорганизмы расщепляют целлюлозу и делают ее доступной для дальнейшего» переваривания. Продукты брожения (главным образом, уксусная» пропионовая и масляная кислоты) всасываются и используются организмом; двуокись углерода и метан (СН4) выводятся наружу путем отрыжки (через рот).
Жевание жвачки состоит в отрыгивании и повторном измельчении непереваренного волокнистого материала, который потом заглатывается снова. Этот процесс и дал группе жвачных их название. При повторном поступлении в рубец пища подвергается дальнейшему брожению. Раздробленные частицы пищи постепенно переходят в следующие отделы желудка, и в четвертом отделе (который соответствует пищеварительному желудку других млекопитающих) они подвергаются действию обычных пищеварительных, соков.
Продукты брожения из рубца — главный источник энергии.. Было подсчитано, что органические кислоты, образующиеся в руб
Пищеварение 207
це, доставляют у крупного рогатого скота до 70% всей необходимой организму энергии. С другой стороны, метан безвозвратно теряется для жвачных, и эта потеря существенна. Корова, получавшая ежедневно 5 кг сена, выделяла каждый день 191л метана; этот метан заключал в себе более 10% всей энергии, содержавшейся в суточном рационе (Kleiber, 1961).
Продукты брожения — это в основном органические кислоты с короткой цепью, и обильная слюна, выделяемая жвачными, ней
трализует эти продукты в рубце. Слюна жвачного мало чем отличается от простого разбавленного раствора бикарбоната натрия и служит одновременно буфером и подходящей ферментационной средой для микроорганизмов.
Количество слюны, выделяемой жвачными, внушительно. По произведенным оценкам, суммарное выделение слюны за день у •овец и коз составляет 6— 16 л, а у крупного рогатого скота 100—190 л (Bailey, 1961; Blair-West et al., 1967). Поскольку овца или коза весит около 40 кг, а корова — около 500 кг, ежедневная выработка слюны
Рис. 5.3. У жвачных животных перед настоящим желудком (сычугом) имеется ряд других отделов. Первый и самый большой из них — рубец — служит гигантским бродильным чаном, способствующим перевариванию целлюлозы.
может составлять примерно треть веса тела. А поскольку тело на две трети состоит из воды, можно считать, что около половины всей воды организма каждый день
проходит через слюнные
железы (и рубец).
Среди простейших, находящихся в рубце, имеются инфузории, внешне сходные со свободноживущими формами, такими, как па-
рамеции. Количество их достигает нескольких сотен тысяч на кубический миллилитр содержимого рубца. Многие микроорганизмы из рубца культивировали в лаборатории, и экстракты из чистых культур проявляли целлюлазную активность (Hungate, 1942). Эти инфузории — строгие анаэробы; они должны получать энергию за -счет процессов брожения. Такие процессы доставляют относитель
но мало энергии самим микроорганизмам, но это означает, что
208 Глава 5. Пища, топливо и энергия
продукты брожения сохраняют ценность для животного-хозяина, которое в свою очередь использует их в процессе окислительного метаболизма. Поскольку симбиотическое переваривание целлюлозы— это единственный способ, который делает ее доступной для млекопитающих, относительно малые потери в пользу симбиотических микроорганизмов количественно несущественны; важно то, что эти организмы и в некоторых других отношениях способствуют питанию хозяина.
Микроорганизмы рубца могут синтезировать белок из неорганических азотных соединений, таких, как соли аммония (McDonald, 1952). Особенно важно то, что к пище жвачных можно добавлять мочевину (которая обычно является экскретом, выводимым с мочой) и таким образом увеличивать синтез белка. Это используют в молочной промышленности: так как синтетическая мочевина недорога, добавлять ее в рацион молочных коров выгоднее, чем использовать более дорогой корм, богатый белком.
Содержимое рубца коровы может весить до 100 кг, и из них около 2 кг составляет общий вес простейших, в которых в свою очередь содержится 150 г белка. Исходя из скорости размножения этих простейших, можно вычислить, что каждый день в книжку (третий отдел желудка жвачных) переходит 69% их популяции и что это дает корове более 100 г белка в день (Hungate; 1942).
Синтез белка микроорганизмами в рубце имеет особое значение, когда животное получает низкокачественный корм. Было обнаружено, что верблюд, получающий почти лишенную белка пищу (плохое сено и финики), практичёски не выделяет с мочой мочевину. Образование мочевины в процессе обмена веществ продолжается, но вместо того, чтобы экскретироваться, этот «ненужный» продукт возвращается в рубец — отчасти через стенки рубца, а отчасти со слюной. В рубце мочевина гидролизуется до СО? и аммиака, а последний используется для ресинтеза белка. Таким способом верблюд, получающий плохой корм, может повторно-использовать значительную часть того малого количества белкового азота, которое имеется в его распоряжении (Schmidt-Nielsen et al., 1957).
Аналогичное повторное использование азота мочевины животными, в рационе которых мало белка, наблюдалось у овец (Houpt^ 1959); в определенных условиях и кролик (который не относится к жвачным) может в значительной мере использовать мочевину в своем азотном обмене (Houpt, 1963).
Если к рациону жвачного добавлять неорганический сульфат,, микробный синтез белка улучшается и, что особенно важно, сера сульфата включается в весьма важные аминокислоты — цистеин-и метионин (Block et al., 1951). Таким образом, микроорганизмы рубца вносят вклад как в синтез белка, так и в качество белка. Поскольку микрофлора рубца может синтезировать все незамени-
Пищеварение 209
Рис. 5.4. Желудок ленивца (Bradypus tri-dactylus) имеет сложное строение и очень напоминает желудок жвачных. (Grasse, 1955.)
мне аминокислоты, жвачные в своем питании не зависят от их наличия в корме и качество белка, который они получают извне, не имеет существенного значения.
Еще одно преимущество пищеварения жвачных с точки зрения питания — это то, что микроорганизмы синтезируют ряд важных витаминов, в том числе некоторые витамины группы В; в частности, витамин В12 жвачные в естественных условиях получают только от микроорганизмов.
Неоквачные животные. У многих нежвачных травоядных млекопитающих перевариванию целлюлозы тоже содействуют микроорганизмы. Целлюлозосодержащая пища обычно объемиста, а брожение идет относительно медленно и занимает много времени. Поэтому для брожения требуется большое пространство, и тот отдел пищеварительного тракта, где оно происходит, имеет большие размеры. У некоторых животных желудок велик и состоит из нескольких отделений, а пищеварение обнаруживает явные черты сходства с пи
щеварением жвачных. У других основное сбраживание целлюлозы осуществляется в крупном ответвлении от нижней части кишечника — слепой кишке.
Многокамерные желудки имеются не только у некоторых нежвачных копытных, но и у очень далеких от них животных, таких, как ленивец (Denis et al., 1967) и обезьяна лангур (Bauchop, Martucci, 1968) (рис. 5.4). Даже среди сумчатых есть животные, у которых желудок сходен с рубцом. Одно из них — кенгуру куок-ка, величиной с зайца и весом от 2 до 5 кг. Его большой желудок, который дает приют микроорганизмам, участвующим в переваривании целлюлозы (Moir et al., 1956), может содержать почти 0,5 кг влажного материала. Для животного, весящего около 3 кг, это составляет 15% от веса тела — соотношение, сходное с тем, которое часто находят у жвачных.
Микробное брожение в слепой кишке в значительной мере сходно с брожением в рубце, но у рубца есть три преимущества перед слепой кишкой. Первое — это то, что рубец находится в пе-14—1873
.210 Глава 5. Пища, топливо и энергия
редней части желудочно-кишечного тракта, так что продукты пищеварения могут проходить отсюда через длинный кишечник для дальнейшего переваривания и всасывания.
Второе преимущество — то, что механическое измельчение пищи может доводиться до высокой степени, так как грубые непереваренные частицы могут отрыгиваться обратно снова и снова. Результат хорошо виден при сравнении фекалий крупного рогатого скота (жвачных) и лошадей (нежвачных). Фекалии лошадей содержат интактные грубые фрагменты пищи; фекалии коров — хорошо перетертая, однородная масса, содержащая мало крупных фрагментов.
Третье и, возможно, самое большое преимущество пищеварения жвачных с микробным брожением в передней части желудочно-кишечного тракта — это возможность повторного использования азота мочевины, который иначе был бы выброшен и потерян для организма. Как мы уже видели ранее, это важно для жвачных, получающих низкосортный корм. Подобные жвачным крупные сумчатые кенгуру и валлаби, у которых микробное брожение тоже происходит до поступления пищи в пищеварительный желудок, используют такой же механизм. Когда в начале засушливого периода содержание азота в растениях снижается, валлаби начинает использовать мочевину повторно и продолжает делать это на протяжении всего долгого сухого периода, приобретая тем самым относительную независимость от качества имеющейся пищи (Kin-near, Main, 1975).
Улучшение переваривания растительного материала с помощью брожения встречается не только у млекопитающих. Например, белая куропатка на полуострове Аляска в течение нескольких зимних месяцев питается одними лишь ивовыми почками и побегами. У большинства птиц семейства куриных имеются две слепые кишки для сбраживания целлюлозы. У белой куропатки продуктами брожения являются этанол и уксусная, пропионовая, масляная и молочная кислоты в различных соотношениях. Брожение в слепой кишке дает до 30% энергии основного обмена— величина весьма внушительная (McBee, West, 1969).
'Копрофагия. Невыгода локализации сбраживания целлюлозы в заднем отделе кишечного тракта может быть скомпенсирована следующим интересным способом. У многих грызунов, а также у кроликов и зайцев из содержимого слепой кишки формируются фекалии особого рода, которые животное снова заглатывает, так что пища проходит через весь пищеварительный тракт второй раз. У них имеется два вида фекалий: хорошо известный обычный твердый помет, характерный для грызунов, и более мягкие и крупные светлые экскременты; последние животное не роняет, а поедает прямо из анального отверстия. В прямой кишке они лежат отдельно от обычного помета, и их повторное заглатывание позволяет более полно переваривать и утилизировать пищу.
Питание 21L
Копрофагия (от греч. copros—кал и phagein — есть) обычна для грызунов и имеет огромное значение для их питания. Например, крысам при исключении копрофагии требуется добавочный пищевой источник витамина К и биотина. Кроме того, быстрее развивается и недостаточность других витаминов. Еще интереснее то, что даже при разнообразных добавках к рациону скорость роста крыс все же снижается примерно на 15—25%. Даже если
предоставить крысам возможность поедать обычные крысиные фекалии, без естественной копрофагии рост животных остается замедленным (Barnes et al., 1963).
У кроликов исключение копрофагии ведет к снижению способности переваривать пищу, а также утилизации белков и накопления азота. После возобновления копрофагии способность переваривать целлюлозу вновь возрастает (Thacker, Brandt, 1955).
Специальные мягкие фекалии, которые кролик повторно заглатывает, образуются в слепой кишке. Эти фекалии не разжевываются и не смешиваются в желудке с другой пищей; они имеют тенденцию располагаться
Рис. 5.5. В желудке кролика проглоченная пища заполняет пилорический отдел (слева), где находятся пищеварительные железы. Вторично заглатываемые комочки фекалий локализуются у основания (справа), где они остаются обособленными от пищевого материала в течение всего времени ферментации. (Harder, 1'949; Grasse, 1955.) отдельно в области дна желудка
(рис. 5.5). Такие фекалии покрыты оболочкой и подвергаются в
желудке брожению в течение многих часов; один из продуктов брожения — молочная кислота (Griffiths, Davies, 1963). Таким образом, область около дна желудка служит бродильной камерой,, аналогичной рубцу овец и коров, и это создает животному значи-
тельные преимущества с точки зрения питания.
ПИТАНИЕ
Механические свойства пищи должны соответствовать способу питания животного, а ее химическая природа — механизмам пищеварения. Если эти требования соблюдены, можно поставить следующий вопрос: подходит ли данная пища для удовлетворения потребностей организма? Именно этим вопросом занимается наука о питании.
14'
212 Г лава 5, Пища, топливо и энергия
Эту проблему удобно подразделить на две части: вопрос о количестве пищи и вопрос о качестве пищи. Любой организм нуждается 1) в энергии для внешней активности и самоподдержания и 2) в поступлении определенных веществ, необходимых для роста и обновления структур тела. К таким специфическим веществам относятся аминокислоты, витамины и некоторые другие органические соединения, а также различные минеральные вещества и другие элементы, некоторые — в столь ничтожных количествах, что трудно установить, действительно ли они нужны.
В последующем обсуждении мы сначала займемся энергетическими потребностями организма и теми компонентами пищи, которые доставляют энергию. Затем мы перейдем к специфическим веществам, необходимым организму. Наконец, мы упомянем ряд менее желательных компонентов, которые могут встречаться в пище, и отметим их биологическое значение.
СНАБЖЕНИЕ ЭНЕРГИЕЙ; ТОПЛИВО
Основная масса органического материала, используемого в пищу, состоит из белков, углеводов и жиров. Главные продукты переваривания этих соединений — аминокислоты (из белков), различные простые сахара (содержащиеся в пище или получаемые при переваривании крахмала), жирные кислоты с короткой цепью (в основном продукты сбраживания целлюлозы) и с длинной цепью (от расщепления жиров).
Окисление этих продуктов пищеварения доставляет практически всю химическую энергию, необходимую организму животного. Все это весьма простые органические вещества, состоящие главным образом из углерода, водорода и кислорода в различных соотношениях, если не считать аминокислот, содержащих также азот (в составе аминогруппы —NH2). Если аминокислоты используются для получения энергии, то сначала отщепляется аминогруппа (дезаминирование), а затем оставшиеся относительно небольшие фрагменты (различные органические кислоты с короткой цепью) вступают на обычные пути промежуточного обмена.
Все эти разнообразные органические соединения в широких пределах взаимозаменяемы с точки зрения энергетического обмена, хотя здесь есть некоторые ограничения. Например, мозг человека нуждается в углеводном топливе (глюкозе), тогда как большинство других органов может использовать жирные кислоты, которые составляют главное топливо для мышечного метаболизма млекопитающих. В отличие от этого летательным мышцам плодовых мушек {Drosophila) нужны углеводы, и когда источники углеводов истощаются, мушки не могут летать, хотя запасенный жир может использоваться для других процессов, требующих энергии (Wigglesworth, 1949). Однако не всем насекомым для полета нужна глюкоза: саранча использует для своих длительных перелетов главным образом жиры (Weis-Fogh, 1952).
Питание 213
Чтобы животное могло находиться в стационарном состоянии, общие затраты энергии должны покрываться эквивалентным поступлением пищи. Если количество пищи недостаточно, часть энергетических потребностей покрывается за счет внутренних резервов организма, в первую очередь — запасенного жира. Если поступление пищи превышает расход энергии, большинство животных запасает излишки в виде жира, независимо от состава пищи. По этой причине домашний скот, например быки и свиньи, может жиреть при кормлении зерном или кукурузой, которые содержат в основном крахмал. Однако некоторые животные запасают главным образом углеводы в виде гликогена. Это существенно, так как гликоген может служить источником энергии в отсутствие кислорода (гл. 6).
РЕГУЛЯЦИЯ ПОТРЕБЛЕНИЯ ПИЩИ
У многих животных потребление пищи удивительно точно регулируется в соответствии с энергетическими затратами (Mayer, Thomas, 1967). Если в связи с физической активностью потребность в энергии возрастает, потребление пищи соответственно увеличивается. У млекопитающих регуляторные механизмы находятся в мозгу, в области гипоталамуса, но в контроле сиюминутного уровня голода и потребления пищи большую роль играют и определенные периферические влияния. Один из важных факторов— это уровень сахара в крови. Было показано, что приступы голода совпадают с сокращениями желудка и что последние особенно часты тогда, когда уровень глюкозы в крови падает. Кроме того, для возникновения чувства насыщения и для прекращения приема пищи очевидное значение имеет степень наполнения желудка.
Эти факторы могут играть лишь второстепенную роль в регуляции среднего потребления пищи за длительный период и в поддержании постоянства веса тела. Увеличение объема пищи путем добавления в нее непереваримых материалов существенно влияет на наполнение желудка и на его подвижность, но животные быстро приспосабливаются к этому и увеличивают потребление пищи. Крысы, переведенные на менее калорийную пищу, уже на следующий день обеспечивают себе нужное число калорий, поглощая большее количество корма. У крыс, которых переводят на более концентрированную в отношении калорий пищу, количество потребляемых калорий несколько дней остается на повышенном уровне, но потом снижается до обычной нормы (рис. 5.6).
Механизмы этой регуляции еще не вполне изучены, но известно, что важную роль играет гипоталамическая область мозга. Если определенные участки гипоталамуса разрушены, потребление пищи сильно изменяется и может как снижаться, так и возрастать. Тщательно локализованные повреждения гипоталамуса могут настолько повышать потребление пищи, что животное бу
214 Глава 5. Пища, топливо и энергия
дет есть непрерывно и разжиреет так, что будет не способно двигаться.
Регуляция потребления пищи существует не только у высших позвоночных. Золотые рыбки, которые продолжают расти в тече-
0 5 10 15 20 5 10
Время, Ъни
Рис. 5.6. Изменение количества пищи, съедаемой за сутки взрослыми крысами (сплошные кривые) в случаях, когда калорийность пищи (на единицу ее веса) уменьшена (А) или увеличена (Б). Прерывистые кривые — доставляемое съеденной пищей количество энергии, выраженное как эквивалентное количество обычной пищи (1 килокалория=4,184 килоджоуля). (Hervey, 1969.)
ние всей жизни (хотя и со снижающейся скоростью), использовались в сходных экспериментах с пищей, обедненной калориями. В таких опытах рыбки начинали потреблять больше корма. Понижение температуры воды с 25 до 15 °C приводило к уменьшению потребления пищи в два-три раза. Таким образом, золотые рыбки, по-видимому, стремятся получить нужное количество калорий, так как снижение температуры на 10° приводит к снижению метаболизма и энергетических потребностей как раз примерно в два-три раза (Rozin, Mayer, 1961).
ПОТРЕБНОСТЬ В СПЕЦИФИЧЕСКИХ ПИТАТЕЛЬНЫХ ВЕЩЕСТВАХ
Пищевой рацион, доставляющий необходимое количество энергии, может быть совершенно неподходящим как источник «сырых материалов» для роста, а также для построения и поддержания клеточных структур и метаболического аппарата.
Питание 215
У большинства животных энергетические нужды могут быть полностью удовлетворены одной только глюкозой, но глюкоза ничего не дает с точки зрения других потребностей. Последние можно разделить на три основные категории: потребность в белках и аминокислотах, в витаминах и родственных им веществах и в минеральных веществах и редких элементах.
БЕЛКИ И АМИНОКИСЛОТЫ
Когда животное растет, белок непрерывно синтезируется и включается в структуры тела. Однако у взрослого организма количество белка остается более или менее постоянным на протяжении всей жизни. В связи с этим может показаться, что после достижения организмом его окончательных размеров получение белка с пищей становится менее существенным. Это не так: недостаточное снабжение белками ведет к серьезным расстройствам.
Почему так получается? Казалось бы, будучи однажды включены в состав некоторого органа (например, мышцы), белки должны оставаться там как часть неизменной структуры. Поэтому, когда биохимик Рудольф Шёнхеймер обнаружил, что белки тела постоянно разрушаются и синтезируются вновь, это явилось большой неожиданностью. Шёнхеймер вводил в рацион взрослых крыс аминокислоты, меченные тяжелым изотопом азота 15N, и эти аминокислоты неизменно оказывались включенными в тканевые белки. Таким образом было показано, что клеточные компоненты, которые ранее рассматривались как стабильные структуры, имеют высокую скорость оборота белков (Schoenheimer, 1964).
Не у всех белков скорость оборота одинакова. У человека, например, период полуобновления белков сыворотки крови — около 10 дней, а для структурных белков соединительных тканей это время значительно больше. После того как эти структурные белки сформированы, они остаются относительно стабильными по сравнению с белками таких метаболически активных тканей, как кровь, печень, мышцы и т. д.
Новый белок синтезируется из отдельных аминокислот, а не из частично расщепленных фрагментов других белковых молекул. У взрослого человека синтезируется и разрушается около 400 г белка в день, что намного превышает общее суточное потребление белков, которое в среднем варьирует в пределах от 100 до 150 г. Таким образом, в процессе синтеза используются не только аминокислоты пищи, но частично и те, которые получаются при расщеплении белка в организме.
Каждый организм содержит большое число различных белков — структурных компонентов клетки, ферментов и т. п. При этом каждый вид животных или растений имеет белки, которые специфичны только для этого вида, и даже особи в пределах одного вида могут иметь белки с характерными отличиями. Напри
216 Глава 5, Пища, топливо и энергия
мер, группы крови человека связаны с различиями в структуре некоторых белков крови.
В белках обычно встречается всего лишь около 20 различных аминокислот, но поскольку молекула белка может содержать от 100 и менее до многих тысяч аминокислот, практически нет предела числу различных белков, которые могут быть построены из этих 20 основных строительных блоков.
Обычно организм получает нужные ему аминокислоты с белками пищи, но их можно заменить химически чистыми аминокислотами, взятыми в подходящих соотношениях. Однакд нет надобности получать извне все эти 20 различных аминокислот. Часть из них может образовываться в организме из других аминокислот, но есть такие, которые не могут синтезироваться организмом животного и должны поступать с пищей. Эти последние называются незаменимыми аминокислотами.
Термины «заменимые» и «незаменимые» указывают лишь на то» должны ли эти аминокислоты обязательно содержаться в пище, но ничего не говорят о том, насколько они важны для организма. Заменимые аминокислоты на самом деле так же важны, как и другие структурные компоненты белков. Они даже, по-видимо-. му, настолько важны, что рискованно полагаться на их содержание в пище, и организм не может обойтись без механизма для их синтеза.
Из 20 с небольшим обычных аминокислот для крыс незаменимыми являются 10 (табл. 5.4) (Rose, 1938). Исключение любой из них (кроме аргинина) вызывает глубокие расстройства. При
Таблица 5.4
Классификация аминокислот по отношению к пищевым потребностям крыс
Незаменимые Заменимые
Лизин Изолейцин Глицин Глутаминовая
Триптофан Треонин Аланин кислота
Гистидин Фенилаланин Метионин Валин Серин Норлейцин Пролин Цитруллин
Лейцин Аргинин1* Аспарагиновая кислота Тирозин Цистеин
*) Аргинин может синтезироваться, но скорость его синтеза недостаточна для того, чтобы обеспечить нормальный рост.
отсутствии аргинина рост крыс замедляется, но аргинин все-таки-может синтезироваться в организме, хотя и не так быстро, как это нужно для оптимального роста. У человека и других млекопитающих потребности такие же, за исключением того, что взрос
Питание 217
лым людям не нужен гистидин, хотя он, по-видимому, требуется всем другим изученным высшим животным (Rose, 1949). Особенно поражает то, что у рыб, многих насекомых и даже простейших потребности в незаменимых аминокислотах практически перекрываются (Tarr, 1958; Taylor, Medici, 1966).
Растения в отличие от животных синтезируют белки из неорганического азота (аммонийных солей или нитратов), а некоторые микробы могут использовать даже молекулярный азот. Азотфик-сирующие бактерии содержатся в корневых клубеньках клевера и других бобовых, которые в связи с этим особенно важны для повышения белкового выхода сельскохозяйственной продукции.
Мы уже упоминали, что термиты и другие сверлящие древесину насекомые используют азотфиксирующих бактерий кишечника, чтобы обеспечить себе добавку к тому чрезвычайно малому количеству белка, которое имеется в их пище. Возможно, что пользу из азотфиксирующих бактерий извлекает гораздо большее число животных, которые сами не способны утилизировать молекулярный азот. С этим связана, в частности, высокая биологическая продуктивность коралловых рифов, поражавшая биологов в течение многих лет. Для тропических водных масс характерно очень низкое содержание растворенных питательных веществ, и, что особенно важно, низкое содержание связанного азота часто бывает главным фактором, лимитирующим продуктивность фитопланктона. .Как выяснилось, при обтекании водой мелководных коралловых рифов в ней заметно возрастает содержание азота, и за. это ответственна главным образом азотфиксирующдя сине-зеленая водоросль Colothrix. Без ее помощи в снабжении азотом было бы трудно объяснить высокую продуктивность рифовых сообществ (Webb et al., 1975).
В конечном счете все животные белки имеют растительное происхождение. Это относится даже к белкам, синтезируемым из аммиака и мочевины в рубце жвачных, так как без микробов этот синтез не имел бы места. Микробный синтез белка в рубце имеет одно интересное следствие: он делает жвачных независимыми от содержания в пище незаменимых аминокислот, поскольку они синтезируются микроорганизмами. Жвачные не нуждаются в получении таких аминокислот с пищей, хотя для метаболизма они столь же необходимы (Loosli et al., 1949; White et al., 1964).
ДОБАВОЧНЫЕ ПИЩЕВЫЕ ВЕЩЕСТВА: ВИТАМИНЫ
В начале этого века было установлено, что диета из углеводов, жиров и белков не удовлетворительна, если в ней нет незначительных количеств других компонентов, которые получили название витаминов. Необходимые количества их очень малы — они измеряются миллиграммами или микрограммами. По этой причине такие соединения называют также добавочными пищевщми
218 Глава 5. Пища, топливо и энергия факторами. Открытие первого витамина было связано с изучением болезни бери-бери (полиневрит), которая вызывается отсутствием тиамина (витамина Bi). Вначале витамины обозначались буквами алфавита, но по мере того, как выяснялись их химическая структура и биохимическая роль, становилось все более принятым использовать вместо этого их химические названия. Подробный перечень витаминов с указанием их роли в обмене веществ можно найти в любом руководстве по вопросам питания,, и поэтому здесь мы его приводить не будем.
Потребности в витаминах не одинаковы у разных животных.. Например, аскорбиновая кислота (витамин С) нужна человеку,, но не нужна другим млекопитающим, кроме некоторых приматов,, летучих мышей и морских свинок. Остальные млекопитающие* синтезируют аскорбиновую кислоту сами и могут не получать ее с пищей, тогда как рыбам этот витамин нужен (Hegsted, 1971).
Удивительные различия существуют в отношении холестерола^ который позвоночные легко синтезируют. Для человека холестерол даже считают нежелательным компонентом пищи, так как он играет важную роль в развитии артериосклероза. Однако насекомые не могут синтезировать холестерол и поэтому должны получать его с пищей; для насекомых он является витамином (Horning, 1958). Все до сих пор изученные насекомые не способны синтезировать холестерол из нестероидных предшественников и нуждаются в пище, содержащей стеролы. Это относится и к ракообразным, хотя они занимают среди исследованных животных одно из первых мест по концентрации холестерола в тканях. Некоторым другим беспозвоночным тоже нужны источники стеролов; среди простейших одни нуждаются в стеролах, другие — нет.
Важными источниками витаминов часто служат симбиотические организмы. Например, витамин К, который необходим позвоночным для нормального свертывания крови, у млекопитающих синтезируется бактериями пищеварительного тракта. Его физиологическая роль была выяснена в опытах на птицах, которым давали пищу без холестерола с целью изучения биосинтеза этого вещества. У таких птиц возникали обильные кровотечения, и была установлена связь их с недостатком определенного витамина. Только после этого было выяснено, что тот же витамин нужен и млекопитающим. Но это явилось следствием случайного открытия его значения для птиц, которые должны получать его с пищей.
Симбиотический синтез витаминов особенно важен для жвачных, которые, по-видимому, не нуждаются в некоторых витаминах группы В. Именно микробный синтез обеспечивает независимость жвачных от пищевого источника этих веществ (McElroy, Goss, 1940).
Сведения о потребности в витаминах у низших позвоночных и у беспозвоночных пока недостаточны. Соответствующие исследования трудны, так как они требуют использования диет, состав-
Питание 219
ленных из очищенных компонентов, а многие животные отказываются от такой пищи в лаборатории. В связи с этим питание основательно изучается главным образом у простейших, насекомых и высших позвоночных. При этом, чтобы достоверно установить пищевые потребности, необходимо знать, не служат ли существенным источником витаминов симбиотические микроорганизмы. Это можно выяснить только при выращивании стерильных особей, а тогда процедура становится очень сложной и дорогой.
МИНЕРАЛЬНЫЕ ВЕЩЕСТВА И МИКРОЭЛЕМЕНТЫ
Какие химические элементы необходимы для жизни? Это выяснено лишь в отношении нескольких лабораторных животных. Из 90 с лишним встречающихся в природе элементов только 25 определенно необходимы для крыс и/или цыплят.
Четыре самых распространенных элемента — кислород, углерод, водород и азот. На их долю приходится до 96% веса тела млекопитающих (табл. 5.5). Почти все оставшиеся ^4% составляют семь следующих элементов (в порядке уменьшения количества): кальций, фосфор, калий, сера, натрий, хлор и магний. Требуются еще пятнадцать элементов, но их общее количество в теле млекопитающего составляет по весу менее 0,01%.
Таблица 5.5
Приблизительный элементарный состав тела человека весом 70 кг
Элемент Количество, г Процент от веса тела Элемент Количество, г Процент от веса тела
Кислород 45 500 65,0 Калий 245 0,35
Углерод 12 600 18,0 Сера 175 0,25
Водород 7 000 10,0 Натрий 105 0,15
Азот 2 100 3,0 Хлор 105 0,15
Кальций 1 350 1,93 Магний 35 0,05
Фосфор 785 1,12 —
Всего 70 000 100,00
Вода и органические компоненты тела — белки, нуклеиновые кислоты, жиры и углеводы — содержат большую часть кислорода, водорода, углерода и азота, а также часть серы и фосфора. Минеральные компоненты скелета вместе с растворенными ионами составляют оставшиеся несколько процентов. Количественно самые важные ионы в организме животного — это натрий, калий, хлор и бикарбонат. Как мы увидим в гл. 9, поддержание нормальных концентраций ионов натрия и хлора необходимо для осмотическо
220 Глава 5. Пища, топливо и энергия
го равновесия. В то время как натрий обычно является главным катионом крови и межклеточных жидкостей, калий — преобладающий катион внутри клеток, где натрий содержится в соответственно меньшей концентрации. Хлора обычно больше во внеклеточной и меньше — во внутриклеточной жидкости. Как было показано в предыдущих главах, бикарбонат-ион тесно связан с дыханием и транспортом СО2 и имеет большое значение в регуляции pH организма.
Кальций, магний, фосфат и сульфат — тоже очень важные ионы, необходимые для многих физиологических процессов. Значительные изменения их концентрации приводят к серьезным расстройствам или даже к смерти. Ионы кальция нужны для нормального функционирования клеток, проведения нервных импульсов, сокращения мышц и свертывания крови. Если концентрация кальция в крови млекопитающего снизится примерно до половины своей нормальной величины (около 5 мэкв/л), возникнут тяжелые или даже смертельные тетанические судороги.
Кроме того, кальций — важный компонент скелета и многих других жестких механических структур. Кости и зубы позвоночных состоят в основном из фосфата кальция (гидроксиапатита). Минерализованные структуры беспозвоночных, например двустворчатых моллюсков (раковины) и коралловых полипов, состоят в основном из карбоната кальция.
Не все опорные образования у животных минерализованы. Наружный скелет насекомых состоит из органического материала — хитина. У ракообразных наружный скелет тоже может быть полностью органическим, но у многих крупных форм (таких, как крабы и омары) он инкрустирован карбонатом кальция. Кроме карбоната кальция, скелеты беспозвоночных часто содержат и карбонат магния, а нередко также незначительные количества сульфата кальция. Особый случай представляет скелетный мате-, риал радиолярии Acantharina (Protozoa), который целиком состоит из сульфата стронция (Odum, 1951, 1957).
Опорные структуры многих растений часто содержат, кроме целлюлозы, аморфную окись кремния (кремнезем). Из животных некоторые губки имеют скелеты, составленные из кремневых спикул (SiO2).
МИКРОЭЛЕМЕНТЫ
Рассмотрим теперь те 15 элементов, которые составляют менее 0,01% тела млекопитающего. Эти элементы содержатся в столь малых количествах, что ранние исследователи могли выявить только их присутствие в организме, так как аналитические методы не были достаточно тонкими для точного анализа. Поэтому о таких элементах упоминалось как о встречающихся в «следовых» количествах, и они получили название следовых элементов, или
Латание 22i
микроэлементов. С усовершенствованием аналитических методов, особенно с введением радиоизотопов, удалось не только определить более точно содержание этих элементов, но и выяснить их физиологическую и биохимическую роль.
Микроэлементы можно разбить на 3 группы: 1) те, о которых известно, что они необходимы организму; 2) те, которые влияют на обмен веществ, но, возможно, не являются необходимыми; и 3) те, которые очень часто встречаются в живых организмах, но, по-видимому, попадают туда лишь случайно. Некоторые микроэлементы требуются в столь малых количествах, что трудно придумать пищевой рацион, полностью лишенный их, и вполне вероятно, что по мере дальнейшего исследования некоторые элементы второй, а возможно, даже и третьей группы будут переходить в первую группу.
Простое наличие микроэлемента ничего не говорит нам о его* функции; поэтому одно лишь выявление его присутствия мало информативно: практически любой элемент периодической таблицы может быть обнаружен в каком-то ничтожном количестве.
Роль микроэлементов выясняют в основном двумя способами: 1) применяя синтетические рационы, составленные из хорошо очищенных компонентов и не содержащие изучаемого элемента, и* 2) изучая болезни человека и животных, предположительно обусловленные недостатком данного микроэлемента. Такие болезни' часто ассоциируются с исключительно низкими концентрациями определенных элементов в почве того или иного района. Первый метод имеет тот недостаток, что трудно исключить из рациона все' следы какого-то элемента и что высокоочищенный рацион легко становится дефектным в других отношениях. Например, если в> пище будет нехватать не только изучаемого' микроэлемента, но и какого-то витамина, у животного не наступит ожидаемого улучшения, если добавить один лишь микроэлемент. В связи с этим исследование микроэлементов было затруднено, пока прогресс в изучении витаминов не позволил разработать подходящие экспериментальные рационы.
Железо [26] L Присутствие железа в живых организмах было известно так давно, .что его обычно не считают микроэлементом: Тем не менее оно необходимо организму. В периодической системе железо входит в группу «переходных» элементов (см; рис. 5.7), которая включает девять известных микроэлементов: Шесть других расположены в периодической- таблице в непосредственной близости к тем неметаллам, которые содержатся -в живых организмах в наибольших количествах.
Железо — составная часть гемоглобина и ряда внутриклеточных ферментных систем, прежде всего цитохромов. Общее коли-
1 В квадратных скобках после названия элемента указан его атомный номер
-222 Глава 5. Пища, топливо и энергия
чество железа в организме не очень велико: у взрослого человека его примерно 4 г. Из этого количества 70% содержится в гемоглобине, 3,2% в миоглобине, 0,1% в цитохромах и 0,1%—в каталазе, а остальное — в резервных соединениях, главным образом в печени (O’Dell, Campbell, 1971). У взрослых млекопитающих
Рис. 5.7. Периодическая система элементов, встречающихся в природе. Элементы в заштрихованных клетках известны как необходимые животным.
потребность в получении с .пищей железа очень мала, так как железо, освобождающееся :при разрушении гемоглобина, накапливается в печени и снова используется для синтеза гемоглобина. Растущим животным железа нужно больше, а взрослым самкам надо возмещать то, что 'Они теряют при процессах, связанных с размножением (при росте утробного плода, менструациях).
Кобальт [27]..Это единственный в своем роде микроэлемент, ионы которого, насколько известно, не нужны как таковые (в свободном виде) никаким животным. Однако всем высшим животным кобальт необходим в форме витамина Bi2) который нужен для образования крови. Недостаток его ведет к тяжелой анемии. У жвачных витамин Bi2 синтезируется в рубце бактериальной флорой, если в пище есть достаточное количество кобальта.
Первое убедительное свидетельство того, что кобальт является незаменимым микроэлементом, пришло из Австралии, где тяжелая болезнь крупного рогатого скота и овец причинила большой экономический ущерб. Добавка небольшого количества кобальта предотвращает эту болезнь. Вводить кобальт в удобрения дорого, так как для этого (нужны довольно большие его количества. Вместо этого, как выяснилось, можно давать каждой овце содержащий кобальт керамический шарик, который остается в рубце и медленно выделяет кобальт в течение нескольких лет.
Питание 223
Никель [28]. Никель в периодической таблице соседствует с кобальтом, и полагают, что он необходим организму. Известно» что никель влияет на рост перьев у цыплят, однако точно не установлено, существен ли этот элемент для жизни. Дело- в том» что небольшие количества никеля почти всегда сопутствуют железу, и поэтому чрезвычайно трудно полностью исключить его из диеты.
Медь [29]. Медь широко распространена в почве, но в определенных областях концентрации ее так низки, что и растения^ и животные страдают от ее недостатка. Один из наиболее заметных симптомов — анемия. Медь не входит в состав молекулы гемоглобина, но недостаток меди препятствует образованию этого пигмента. Конкретный механизм этого эффекта неизвестен.
Медь является также составной частью многих> ферментов, наиболее важный из которых — цитохромоксидаза, содержащая и-железо, и медь1.
Цинк [30]. Цинк входит в состав многих важных ферментов» включая карбоангидразу и несколько пептидаз. С помощью очищенного рациона без цинка можно вызвать характерные симптомы.
Ванадий [23]. Ванадий сильно ускоряет рост крыс, содержащихся в изоляции в пластмассовых клетках на высокоочищенном-аминокислотном рационе. Необходимое количество — около 1 мкг в день на крысу — лежит в диапазоне концентраций, находимых обычно в естественной пище и в тканях высших животных (Schwarz, Milne, 1971).
Интересно, что некоторые виды оболочников накапливают в крови очень высокие концентрации ванадия, хотя у других оболочников нет необычных концентраций этого элемента. Смысл этого” обогащения ванадием, если таковой имеется, остается неясным.
Хром [24]. Хром представляет собой сравнительно недавнее добавление к списку необходимых микроэлементов. Он нужен для того, чтобы инсулин мог максимально способствовать поглощению глюкозы тканями (Mertz, 1969). Пока нет данных о том, чтобы хром был компонентом какой-либо^ ферментной системы или действовал как активатор фермента.
Молибден [42]. В молибдене нуждаются все азотфиксирующие микроорганизмы, высшие растения и животные. Он входит в состав фермента ксантиноксидазы и, таким образом, играет роль в* окислении пуринов до мочевой кислоты.
Марганец [25]. Марганец широко распространен в почве и-содержится во всех растениях и животных. Сравнительно не таю давно было установлено, что он является необходимым компонентом диеты. Недостаток марганца приводит к нарушению развития костей, и, кроме того, этот элемент необходим, для функциони-
1 Кроме того, медь — основной металл кровяного» пигмента гемоцианина.— Прим, ред.
224 Глава 5. Пища, топливо и энергия
рования яичников и семенников. Его биохимическая роль стала известна только недавно. Он действует как активатор определенных ферментных систем, но связь этой функции с симптомами его недостаточности не вполне ясна.
Кремний [14]. Давно было известно, что кремний, который по распространенности в земной коре стоит вслед за кислородом, необходим растениям. Однако то, что он нужен также для роста и нормального развития животных, было установлено лишь в последние годы (Carlisle, 1972). У цыплят, не получающих кремния, развиваются аномалии костей, в том числе черепа, — очевидно, в результате какого-то влияния на коллаген костной ткани (Carlisle, 1977).
Олово [50]. Олово, относящееся к той же группе периодической системы, что и углерод с кремнием, составляет одно из самых поздних добавлений к списку необходимых микроэлементов. Неорганические соли олова нужны для нормального роста крыс, содержащихся на высокоочищенных рационах с другими известными микроэлементами (Schwarz et al., 1970).
Мышьяк [33]. Этот элемент, по-видимому, необходим крысам -и предположительно другим животным. При очень низком содержании мышьяка в пище у крыс вырастает грубая шерсть и они развиваются медленнее, чем контрольные животные. Наблюдаются и другие нарушения, в частности увеличение селезенки и низкие величины гематокрита. По каким-то неизвестным причинам у самцов явления недостаточности всегда проявляются сильнее, чем у самок (Nielsen et al., 1975).
Селен [34]ч. Из-за своей токсичности селен имеет практическое значение не только как необходимый компонент пищи животных. В некоторых областях селен содержится в почве в таких высоких концентрациях, что растения становятся ядовитыми для скота, и пасущиеся животные могут страдать от острого отравления зеленом. Однако в то же время селен — необходимый организму -микроэлемент; проявлениями его недостаточности могут быть некроз печени и мышечная дистрофия. Биохимическая роль селена, по-видимому, связана с функцией витамина Е, который необходим для роста и нормального размножения крыс.
Фтор [9]. До недавнего времени фтор не считался необходимым элементом, хотя некоторые его физиологические эффекты были хорошо известны; однако положение изменилось, когда было обнаружено, что он нужен для нормального роста крыс (Schwarz, Milne, 1972).
Давно было известно, что .фтор присутствует в небольших количествах в зубах и костях, где его ионы включены в кристаллическую решетку фосфата кальция (апатита)—главного минерального компонента. Так ли необходим фтор в этой роли, не ясна, .но он делает зубы более устойчивыми к кариесу. В связи с этим для снижения частоты кариеса было рекомендовано добавление
Ядовитые вещества и химическая защита 225
фторида к водопроводной воде. Эффективность этого средства несомненна, а добавляемые количества столь малы, что никаких токсических эффектов не наблюдается.
Однако в некоторых географических районах фтор содержится в природных источниках воды в таких высоких концентрациях, что он вызывает изменение зубной эмали, известное под названием «пятнистая эмаль». С косметической точки зрения этот эффект нежелателен, но «пятнистые» зубы особенно устойчивы к кариесу. Еще более высокие уровни поглощения фтора ведут к деформации костей и серьезному отравлению; такие случаи имели место вблизи плавильных заводов и других предприятий, выбрасывающих фтор в атмосферу.
Иод [53]. Физиологическое значение иода связано с тем, что он входит в состав гормонов щитовидной железы. Эти гормоны ускоряют клеточный метаболизм и необходимы как регуляторы интенсивности обмена веществ и нормального роста позвоночных. Давно известно, что случаи зоба (увеличения щитовидной железы) у человека очень часты в тех географических районах, где содержание иода в почве особенно низко. Увеличение щитовидной железы, по-видимому, является реакцией на недостаток иода, и введение небольших его количеств с пищей (обычно в виде малых добавок йодистого натрия к обычной пищевой соли) позволило практически ликвидировать эту довольно серьезную в прошлом проблему.
Районы с недостатком иода — это большей частью внутренние гористые местности, удаленные от океана. Морская вода содержит небольшое количество иода, и он попадает в атмосферу с брызгами от волн, осаждается с дождем и удовлетворяет нужды как животных, так и человека даже на расстояниях в несколько сотен километров от побережья.
Другие микроэлементы, В живых тканях встречается также много других следовых элементов (например, литий, рубидий, бериллий, стронций, алюминий, бор, германий, бром), и возможно, что они имеют функциональное значение, хотя определенных свидетельств этого нет. Некоторые из них весьма токсичны, но даже и в этом случае они могут быть необходимыми в следовых количествах. Кроме того, в настоящее время нет причин думать, что у всех животных потребности в микроэлементах в точности одинаковы.
ЯДОВИТЫЕ ВЕЩЕСТВА И ХИМИЧЕСКАЯ ЗАЩИТА
Один из способов не быть съеденным — это стать несъедобным. Этот простой принцип широко используется и растениями, и животными. Растения — первичные продуценты в пищевой цепи, и .поскольку они не могут бегать, они остаются на месте и их легко поедать.
15—1873
226 Глава 5. Пища, топливо и энергия
Ядовитые вещества — очень распространенное средство защиты от поедания, и поразительное множество растений несъедобно для животных. Но «хищники» в свою очередь выработали способы обхода этой защиты или противодействия ей, так что едва ли есть хоть одно растение, совершенно не имеющее врагов (Whittaker, Feeny, 1971). В качестве примера такой адаптации можно привести бабочку-капустницу, гусеницы которой питаются на определенных крестоцветных (капусте, горчице и т. п.). Эти растения содержат горчичные масла, которые ядовиты или смертельны для других бабочек, но хорошо переносятся гусеницами капустницы и даже служат аттрактантами, привлекающими взрослых бабочек для откладки яиц на этих растениях.
Животные тоже используют яды и токсины для защиты; у некоторых (например, змей, пауков, скорпионов) яд служит как для целей обороны, так и для поражения добычи.
Некоторые животные не только ухитряются «обойти» химическую защиту растений, но даже используют токсичные компоненты поедаемых растений, чтобы сделать себя невкусными или несъедобными для своих врагов.
РАСТИТЕЛЬНЫЕ ЯДЫ
Разнообразные химические компоненты растений влияют почти на все физиологические системы. Они могут действовать на кожу, на пищеварительный тракт, на кровь, на различные части нервной системы, влиять на обмен веществ, на питательную ценность пищи, а также на гормональную и репродуктивную системы животного. Разнообразие веществ и их эффектов делает их трудно-поддающимися классификации, но их можно объединить в несколько основных групп частично по химическому сходству, а частично по сходству действия. Приведенные ниже примеры проиллюстрируют огромное разнообразие таких соединений и их физиологических эффектов.
АЛКАЛОИДЫ
Алкалоиды принадлежат к группе органических соединений, которые имеют между собой мало общего, кроме того, что все они содержат азот. Среди них есть токсичные соединения, оказывающие сильное воздействие на центральную нервную систему позвоночных (например, опиум — сырой продукт из опийного мака — содержит полезные для медицины алкалоиды морфин и кодеин). Широко используемым алкалоидом является никотин; другие хорошо известные и высокотоксичные алкалоиды — это стрихнин и компоненты стрельного яда кураре.
Ядовитые вещества и химическая защита 227
гликозиды
Гликозидами называют соединения, которые при гидролизе дают одну или несколько молекул сахаров и ряд других соединений. Один из самых известных гликозидов — дигиталис, широко применяемый при лечении болезней сердца. Он содержится в наперстянке, и его передозировка опасна для жизни.
Ряд растений содержит гликозиды, из которых при гидролизе образуется синильная кислота (HCN). Из таких гликозидов хорошо известен, например, амигдалин, содержащийся в горьком миндале и в косточках абрикосов, слив и вишен. Семена яблони тоже содержат небольшие количества этого высокотоксичного соединения. Для большинства из нас это несущественно, однако сообщалось, что один человек, обожавший яблочные семечки как деликатес, накопил их полную чашку и после того, как съел их все за один прием, умер от отравления цианидом (Kingsbury, 1964).
Во многих тропических странах один из главных продуктов питания — кассава (маниок) —выделяет так много синильной кислоты, что даже приготовление клубней для еды связано с риском смертельного отравления. Собранные клубни измельчают в воде и, после того как крахмальные зерна осядут на дно, ядовитую жидкость сливают. У некоторых индейцев Центральной Америки процесс приготовления кассавы считается столь опасным, что эту работу поручают только старикам — видимо, потому, что считают их отжившими свой век.
Интересно, что для одних людей синильная кислота (цианид) сильно пахнет горьким миндалем, а для других—совершенно лишена запаха. Это различие не обусловлено неспособностью ощущать запах горького миндаля, так как бензальдегид, который для чувствующих цианид пахнет очень похоже на него, имеет характерный запах «горького миндаля» и для людей, не ощущающих запах цианида. Те кто не чувствует запаха цианида (таких людей довольно много), должны быть очень осторожны при использовании этого опасного вещества, поскольку обоняние не может предостеречь их.
МАСЛА И СМОЛЫ
Разного рода масла и смолы широко распространены в растениях и вызывают много различных физиологических эффектов. Одна из таких смол является действующим началом в марихуане— препарате конопли (Cannabis), который благодаря своему влиянию на центральную нервную систему изменяет состояние сознания и вызывает галлюцинации.
Другие масла и смолы раздражают кожу или действуют на желудочно-кишечный тракт, вызывая рвоту, понос и ряд других симптомов. Некоторые, вещества раздражают слизистую носа и 15*
228 Глава 5. Пища, топливо и энергия
рта и таким образом быстро «отучают» поедать соответствующие растения.
ЩАВЕЛЕВАЯ КИСЛОТА
Щавелевая кислота содержится во многих растениях из весьма различных семейств. Ее острое токсическое действие обусловлено тем, что она осаждает кальций в виде нерастворимой соли — оксалата. При остром отравлении концентрация кальция в крови снижается, что ведет к неврологическим симптомам, судорогам и сильному повреждению почек.
В связи с этим представляет интерес растение Halogeion, обычное для западных районов США. Свыше 30% сухого веса этого растения могут составлять растворимые соли щавелевой кислоты. Оно чрезвычайно токсично для пасущихся животных и явилось причиной гибели тысяч овец.
К тому же семейству Chenopodiaceae относятся два растения, часто употребляемые в пищу человеком, — шпинат и ревень. Ни одно из них обычно не едят в количествах, достаточных для острого отравления. Однако листья ревеня, которые иногда используют как овощную пищу, содержат больше щавелевой кислоты, чем стебли, что привело в нескольких случаях к смерти. Если каждый день есть много шпината, осаждение оксалата кальция в кишечнике приведет к уменьшению количества кальция, способного всасываться, и может со временем вызвать симптомы недостаточности этого элемента.
ИНГИБИТОРЫ ФЕРМЕНТОВ
Некоторые растения, включая ряд представителей семейства бобовых, содержат вещества, подавляющие действие трипсина — важного протеолитического фермента поджелудочной железы. Такие ингибиторы трипсина обнаружены в соевых бобах, лимских бобах, арахисе и ряде растений из других семейств.
Большинство ингибиторов трипсина чувствительно к нагреванию, и поскольку люди обычно варят пищу, эти ингибиторы не имеют большого значения для человека. Однако они снижают использование белка животными, которые едят сырые или необработанные соевые бобы и сходные продукты, так что эти вещества существенны для животноводства. Их вредное действие можно уменьшить или устранить путем тепловой обработки (Rackis, 1965).
ВЕЩЕСТВА, ВЛИЯЮЩИЕ НА ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Капуста, горчица, редис и ряд других растений семейства крестоцветных обязаны своим острым вкусом горчичным маслам, ко
Ядовитые вещества и химическая защита 229
торые действуют как раздражающие вещества и даже могут вызвать смертельное отравление. Кроме того, многие из этих растений содержат вещества, подавляющие функцию щитовидной железы и вызывающие ее увеличение (зоб). Когда в рационе лабораторных животных много листьев капусты или репы, у них развивается зоб.
Антитиреоидные вещества растений химически родственны лекарственным препаратам вроде тиомочевины, применяемым при* лечении гиперфункции щитовидной железы. Хорошо известно, что-некоторые люди находят фенилтиомочевину чрезвычайно горькой; другие находят ее безвкусной. Неспособность ощущать вкус тио-мочевины наследуется как рецессивный признак. Одно из антитиреоидных веществ репы и капусты также безвкусно для одних и горько для других, причем чувствующие горечь и не чувствующие ее соответствуют ощущающим и не ощущающим вкус фенилтиомочевины (Boyd, 1950). Это указывает на возможную физиологическую роль соответствующего гена.
ВЕЩЕСТВА, ВЛИЯЮЩИЕ НА РАЗМНОЖЕНИЕ ЖИВОТНЫХ
Одна из стратегий защиты, используемых растениями, состоит в отрицательном воздействии на размножение своих врагов с помощью веществ, «обманывающих» эндокринную систему животного. От этого могут страдать как насекомые, так и позвоночные.
Такие вещества могут, например, нарушать нормальное развитие насекомых, которые в личиночной стадии проходят определенное число линек, прежде чем превращаются во взрослую форму (имаго). Развитие личинок регулируется так называемым ювенильным гормоном, но для превращения в имаго, напротив, необходимо отсутствие этого гормона. Ряд американских вечнозеленых деревьев синтезирует вещества, которые действуют как ювенильный гормон (см. гл. 13); результат очень прост: насекомые не переходят во взрослое состояние, и тем самым их размножение предотвращается.
Было создано много синтетических веществ, сходных по своему действию с ювенильным гормоном; некоторые из них даже намного более эффективны, чем естественные гормоны. При определенных условиях их можно использовать как сильные инсектициды, оказывающие минимальное воздействие на других животных, в том числе домашних, которые нечувствительны к гормонам насекомых. Один из недостатков таких инсектицидов — то, что они эффективны только в течение короткого периода, когда личинки превращаются в имаго.
Более разностороннее влияние оказывают синтезируемые растением вещества, которые действуют как антагонисты ювенильно
230 Глава 5. Пища, топливо и энергия
го гормона (Bowers et al., 1976). Они эффективны на протяжении значительно более долгого периода и нарушают нормальное развитие, а также ряд других процессов, регулируемых ювениль-лым гормоном. Они полностью или частично подавляют выработку этого гормона; это ведет не только к преждевременному метаморфозу, но также к нарушению диапаузы, откладки яиц, образования половых аттрактантов и т. д. Эти вещества, вероятно, приобретут большее значение, когда будет получено больше сведений об их физиологическом действии.
Размножение позвоночных тоже подвержено влиянию растительных веществ, действующих на эндокринную систему. Некоторые растения, обычно поедаемые животными на пастбищах, особенно представители семейства бобовых, содержат биологически активные вещества с эстрогенным действием, имитирующим нормальную активность эстрогенов животных. Эти эстрогены нарушают цикл размножения у овец и крупного рогатого скота и могут играть роль в регулировании численности диких грызунов.
Вредные эффекты растительных эстрогенов широко изучались в Австралии, где они влияют на размножение овец. У животных, поедающих подземный клевер, плодовитость падает из-за нарушения процессов, с которыми связано зачатие, так что число суягных овец резко сокращается (Shutt, 1976).
Эстрогенные вещества растений имеют значение не только для животноводства; их роль в природе гораздо шире. В безводных районах Калифорнии однолетние растения в засушливые годы вырабатывают фитоэстрогены. Когда их листья поедаются перепелами, эти вещества тормозят размножение и предотвращают появление потомства, которому не хватило бы пищи. Во влажные годы растения энергично растут и концентрация эстрогенов в них низка, и в такие годы перепела размножаются в изобилии. Богатые урожаи растительной массы и зерна в такие годы гарантируют выживание увеличившейся популяции. Тесная связь между размножением животных и продуктивностью растений известна давно, и механизм ее, по крайней мере в приведенном примере,— это хорошо отрегулированный естественный биологический контроль численности популяции (Leopold et al., 1976).
ИСПОЛЬЗОВАНИЕ РАСТИТЕЛЬНЫХ ЯДОВ ЖИВОТНЫМИ
Обычный ваточник (Asclepias) и другие растения, принадлежащие к тому же семейству, синтезируют сердечные гликозиды. Кроме влияния на сердце животных, поедающих эти ядовитые растения, гликозиды действуют также на мозговые центры, контролирующие рвоту. Когда животное поедает растение, рвота обычно начинается раньше, чем успеет всосаться летальная доза яда, так что эффект сводится лишь к очень неприятному желудочному расстройству.
Ядовитые вещества и химическая защита 231
Несмотря на свою ядовитость, ваточники служат единственной пищей личинок целой группы насекомых, в которую входит бабочка-данаида Danaus plexippus. В природе различные насекомо-ядные позвоночные избегают этих бабочек и их гусениц по той простой причине, что в них накапливаются ядовитые вещества’ ваточников. Если птица съест всего лишь одну ярко окрашенную1 данаиду, она будет жестоко страдать и больше никогда не попробует это повторить. Таким образом, у этих бабочек выработалась-способность не только питаться ядовитыми растениями, но и использовать эти яды для защиты от врагов (Brower et al., 1967).
Такое использование токсичных веществ, получаемых из пищи, широко распространено среди насекомых, но оно не ограничено только этим классом. Ряд морских животных питается различными кишечнополостными, которые от других врагов хорошо защищены своими стрекательными капсулами, или нематоцистами. Одно из таких кишечнополостных, со свойствами которого человек познакомился на собственном мало приятном опыте, — это сифо-нофора Physalia, которая может даже вызвать смерть пловца, близко с нею соприкоснувшегося. Некоторые голожаберные моллюски, питающиеся этим животным, заглатывают кусочки его ткани и переваривают их. При этом нематоцисты не перевариваются и не разряжаются; они включаются в ткани моллюска и служат ему для защиты.
ИСПОЛЬЗОВАНИЕ ЖИВОТНЫМИ СОБСТВЕННЫХ ЯДОВ
Животные используют также разнообразные яды и токсины собственного производства. Хорошо известные примеры — змеи, скорпионы, пауки, пчелы и осы. К менее обычным средствам защиты относится синильная кислота, которую выделяют многоножки из группы двупарноногих, будучи потревожены (Eisner et al., 1963); еще более эффектный пример — аэрозоль с температурой 100°C, выбрасываемый жуками-бомбардирами (Aneshansley et al., 1969).
Жуки-бомбардиры (фото 5.2) выпускают свою защитную струю из пары желез, открывающихся на кончике брюшка. Струя видна как легкая дымка, и разряд сопровождается слышимым звуком «взрыва». Секретирующие железы — двухкамерные. Большая, внутренняя камера содержит раствор гидрохинонов и перекиси водорода; в наружной камере находится смесь ферментов — каталаз и пероксидаз. Когда жук производит разряд, он выжимает жидкость из внутреннего резервуара в наружную камеру. Происходит мгновенная взрывная реакция; перекись водорода разлагается ферментами, и гидрохиноны окисляются с выделением огромного количества тепла. Под давлением образующегося газа смесь выстреливается в виде тонкодисперсного аэрозоля, который оказы-
232 Глава 5. Пища, топливо и энергия
Фото 5.2. Жук-бомбардир. Это насекомое защищается, выбрасывая струю химического раствора с температурой 100 °C. Струя направляется прямо на врага (например, на муравья, вцепившегося в одну из ног). Для фотосъемки жука удерживали на месте с помощью металлической проволоки, приклеенной к его спине. Струя направлена на «врага» — пинцет, которым схвачена одна из ног. Жуки-бомбардиры встречаются во всех главных районах мира, кроме Океании. Показанный здесь вид, Stenaptinus insignis, привезен из Кении; длина его тела около 20 мм. (Фото предоставлено Т. Eisner и D. Aneshansley, Корнельский университет.)
вает сильное отталкивающее действие на врагов. Жук может быстро производить ряд залпов и направлять кончик своего брюшка практически в любую сторону, с которой его атакуют.
❖
❖
Резюмируя все сказанное в этом разделе, мы можем заключить, что огромное число животных и растений использует химические средства защиты, либо вырабатывая собственные яды, либо беря на вооружение вещества, производимые другими организ
Ядовитые вещества и химическая защита 233
мами. Вероятно, лишь небольшая доля организмов не защищается каким-нибудь таким способом по крайней мере от некоторых потенциальных врагов.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Aneshansley D. J., Eisner Т., Widom J, М., Widom В. (1969). Biochemistry at 100 °C: Explosive secretory discharge of bombardier beetles (Brachinus), Science, 165, 61—63.
Bailey С. B. (1961). Saliva secretion and its relation to feeding in cattle. 3. The rate of secretion of mixed saliva in the cow during eating with an estimate of the magnitude of the total daily secretion of mixed saliva, Br. J. Nutr., 15, 443—451.
Barnes R. H., Fiala G., Kwong E. (1963). Decreased growth rate resulting from prevention of coprophagy, Fed. Proc., 22, 125—133.
Bauchop T., Martucci R. W. (1968). Ruminant-like digestion of the langur monkey, Science, 161, 698—700.
Benemann J. R. (1973). Nitrogen fixation in termites, Science, 181, 164—165.
Benson A. A., Lee R. F., Nevenzel J. C. (1972). Wax esters: Major marine metabolic energy sources, Biochem. Soc. (Lond.) Symp., No. 15, pp 175—187.
Berrill N. J. (1929). Digestion in ascidians and the influence of temperature, J. Exp. Biol., 6, 275—292.
Blair-West J. R., Coghlan J. P., Denton D. A., Wright R. D. (1967). Effect of endo-crines on salivary glands. In: Handbook of Physiology, sect. 6, Alimentary Canal, vol. II, Secretion (C. F. Code, ed.), pp. 633—664, Washington, D. C., American Physiological Society.
Block R. J., Stekol J. A., Loosli J. K. (1951). Synthesis of sulfur amino acids from inorganic sulfate by ruminants. 2. Synthesis of cystine and methionine from sodium sulfate by the goat and by the microorganisms of the rumen of the ewe, Arch. Biochem. Biophys., 33, 353—363.
Bowers W, S., Ohta T,, Cleere J. S., Marsella P. A, (1976). Discovery of insect antijuvenile hormones in plants, Science, 193, 542—547.
Boyd W. C. (1950). Taste reactions to antithyroid substance, Science, 112, 153.
Breznak J. A., Brill W. J., Mertins J. W., Coppel H. C, (1973). Nitrogen fixation in termites, Nature, Lond, 244, 577—580.
Brower L. P., Brower J. V. Z., Corvino J. M. (1967). Plant poisons in a terrestrial food chain, Proc. Natl. Acad. Sci. USA, 57, 893—898.
Carlisle E. M. (1972). Silicon: An essential element for the chick, Science, 178, 619—621.
Carlisle E. M. (1977). A silicon requirement for normal skull formation, Fed. Proc., 36, 1123.
Carpenter E. J., Culliney I. L. (1975). Nitrogen fixation in marine shipworms, Science, 187, 551—552.
Cleveland L. R. (1925). Toxicity of oxygen for protozoa in vivo and in vitro: Animals defaunated without injury, Biol. Bull., 48, 455—468.
Denis G., Jenuiaux Ch., Gerebtzoff M. A., Goff art M. (1967). La digestion stomaca-le chez un paresseux: L’unau Choloepus hoffmanni Peters, Ann. Soc. R. Zool. Belg., 97, 9—29.
Eisner T., Eisner H. E., Hurst J. J., Kafatos F. C., Meinwald J. (1963). Cyanogenic glandular aparatus of a millipede, Science, 139, 1218—1220.
Florkin M., Lozet F. (1949). Origine bacterienne de la cellulase du contenu intestinal de 1’escargot, Arch. Int. Physiol., 57, 201—207.
Friedmann H., Kern J. (1956a). The problem of cerophagy or wax-esting in the honey-guides. Q. Rev. Biol., 31, 19—30.
Friedmann H., Kern J. (1956b). Micrococcus cerolyticus, nov. sp., an aerobic lipolytic organism isolated from the African honey-guide, Can. J. Microbiol., 2„ 515—517.
234 Глава 5. Пища, топливо и энергия
Friedmann Н., Kern Hurst J. Н. (1957). The domestic chick: A substitute for the honey-guide as a symbiont with cerolytic microorganisms, Am. Nat., 91, 321— 325.
Grasse P.-P. (1955). Traite de Zoologie: Anatomie, Systematique, Biologie, vol. 17, fasc. 2; Mammiferes, pp. 1173—2300, Paris, Masson.
Greenberg B., Paretsky D, (1955). Proteolytic enzymes in the house fly, Musca domestica (L.), Ann. Entomol. Soc. Am., 48, 46—50.
Greenfield L. J., Lane С. E. (1953). Cellulose digestion in Teredo, J. Biol. Chem., 204, 669—672.
Griffiths M., Davies D. (1963). The role of the soft pellets in the production of lactic acid in the rabbit stomach, J. Nutr., 80, 171—180.
Harder W. (1949). Zur Morphologie und Physiologie des Blinddarmes der Nagetie-re, Verh. Dtsch. Zool., 2, 95—109.
Hegsted D. M. (1971). Fish require dietary vitamin C, Nutr. Rev., 29, 207—210. Hervey G. R. (1969). Regulation of energy balance, Nature, bond., 222, 629—631. Horning M. G. (1958). The sterol requirements of insects and of protozoa. In: Cholesterol, Chemistry, Biochemistry and Pathology (R. P. Cook, ed.), pp. 445— 455, New York, Academic Press.
Houpt T. R. (1959). Utilization of blood urea in ruminants, Am. J. Physiol., 197, 115—120.
Houpt T. R. (1963). Urea utilization by rabbits fed a low-protein ration, Am. J. Physiol., 205, 1144—1150.
Hungate R. E. (1941). Experiments on the nitrogen economy of termites, Ann. Entomol. Soc. Am., 34, 467—489.
Hungate R. E. (1942). The culture of Eudiplodinium neglectum, with experiments on the digestion of cellulose, Biol. Bull., 83, 303—319.
Hungate R. E. (1955). Mutualistic intestinal protozoa. In: Biochemistry and Physiology of Protozoa, vol. II (S. H. Hutner and A. Lwoff, eds.), pp. 159—199, New York, Academic Press.
Johannes R. E., Coward S. J., Webb K. L. (1969). Are dissolved amino acids an energy source for marine invertebrates? Comp. Biochem. Physiol., 29, 283— 288.
Kingsbury J. M. (1964). Poisonous Plants of the United States and Canada. Englewood Cliffs, N. J., Prentice-Hall, 626 pp.
Kinnear J, E., Main A. R. (1975). The recycling of urea nitrogen by the wild tarn-mar wallaby (Macropus eugenii), a «ruminant-like» marsupial, Comp. Biochem. Physiol., 51A, 793—810.
Kleiber M. (1961). The Fire of Life. An Introduction to Animal Energetics, New York, Wiley, 454 pp.
Lambremont E. N., Fisk F. W., Ashrafi S. (1959). Pepsin-like enzyme in larvae of stable flies, Science, 129, 1484—1485.
Lasker R., Giese A. C. (1956). Cellulose digestion by the silverfish Ctenolepisma lineata, J. Exp. Biol., 33, 542—553.
Leopold A. S., Erwin M., Oh J., Browning B. (1976). Phytoestrogens: Adverse effects on reproduction in California quail, Science, 191, 98—100.
Loosli J. K., Williams H. H., Thomas W. E., Ferris F. H„ Maynard L. A. (1949). Synthesis of amino acids in the rumen, Science, 110, 144—145.
Matthews L. H., Parker H. W. (1950). Notes on the anatomy and biology of the basking shark (Cetorhinus maximus (Gunner)), Proc. Zool. Soc. Lond., 120, 535—576.
Mayer J., Thomas D. W. (1967). Regulation of food intake and obesity, Science, 156, 335—337.
McBee R. H., West G. C. (1969). Cecal fermentation in the willow ptarmigan, Condor, 71, 54—58.
McDonald I. W. (1952). The role of ammonia in ruminal digestion of protein, Biochem. J., 51, 86—90.
McElroy L. W., Goss H. (1940). A quantitative study of vitamins in the rumen contents of sheep and cows fed vitamin-low diets. I. Riboflavin and vitamin K, J. Nutr., 20, 527—540.
Пища, топливо и энергия 235
Mertz W. (1969). Chromium occurrence and function in biological systems, Physiol. Rev., 49, 163—239.
Moir R. J., Somers M., Waring H. (1956). Studies on marsupial nutrition. I. Ruminant-like digestion in a herbivorous marsupial (Setonix brachyurus Quoy and Gaimard), Aust. J. Biol., Sci., 9, 293—304.
Muscatine L., Lenhoff H. M. (1965). Symbiosis of hydra and algae. 2. Effects of limited food and starvation on growth of symbiotic and aposymbiotic hydra, Biol. Bull., 129, 316—328.
Nielsen F. H., Givand S. H., Myron D. R. (1975). Evidence of a possible requirement for arsenic by the rat, Fed. Proc., 34, 923.
O'Dell B. L., Campbell B. J. (1971). Trace elements: Metabolism and metabolic function, Comp. Biochem., 21, 179—266.
Odum H. T. (1951). Notes on the strontium content of sea water, celestite radio-laria and strontianite snail shells, Science, 114, 211—213.
Odum H. T. (1957). Biogeochemical deposition of strontium, Univ. Texas Publ. Inst. Mar. Sci., 4, 38—114.
Pardy R. L., Dieckmann C. (1975). Oxygen consumption in the symbiotic hydra Hydra viridis, J. Exp. Zool., 194, 373—378.
Prevost J., Vilter V. (1962). Histologie de la secretion oesophagienne du Manchot Empereur. In: Proceedings, 13th International Ornithological Congress, Vol. 2, pp. 1085—1094, Baton Rouge, La., American Ornithologists’Union.
Rackis J. J. (1965). Physiological properties of soybean trypsin inhibitors and their relationship to pancreatic hypertrophy and growth inhibition of rats, Fed. Proc., 24, 1488—1500.
Rose W. C. (1938). The nutritive significance of the amino acids, Physiol. Rev., 18, 109—136.
Rose W. C. (1949). Amino acid requirements of man, Fed. Proc., 8, 546—552.
Rozin P., Mayer J. (1961). Regulation of food intake in the goldfish, Am. J. Physiol., 201, 968—974.
Sargent J. R., Gatten R. R. (1976). The distribution and metabolism of wax esters in marine invertebrates, Biochem. Soc. (Lond.) Trans., 4, 431—433.
Schmidt-Hielsen B., Schmidt-Nielsen K., Houpt T. R., J ar num S. A. (1957). Urea excretion in the camel, Am. J. Physiol., 188, 477—484.
Schoenheimer R. (1964). The Dynamic State of Body Constituents, New York, Hafner, 78 pp.
Schwarz K„ Milne D. B. (1971). Growth effects of vanadium in the rat, Science, 174, 426—428.
Schwarz K., Milne D. B. (1972). Fluorine requirement for growth in the rat, Bioinorg. Chem., 1, 331—338.
Schwarz K., Milne D. B., Vinyard F. (1970). Growth effects of tin compounds in rats maintained in a trace-element-controlled environment, Biochem. Biophys. Res. Commun., 40, 22—29.
Shutt D. A. (1976). The effects of plant oestrogens on animal production, Endeavour, 35, 110—113.
Smith D, Muscatine L., Lewis D. (1969). Carbohydrate movement from autotrophs to heterotrophs in parasitic and mutualistic symbiosis, Biol. Rev., 44, 17— 90.
Stephens G. C. (1962). Uptake of organic material by aquatic invertebrates. 2. Uptake of glucose by the solitary coral, Fungia scutaria, Biol. Bull., 123, 648— 659.
Stephens G. C. (1964). Uptake of organic material by aquatic invertebrates. 3. Uptake of glycine by brackish-water annelids, Biol. Bull., 126, 150— 162.
Tarr H. L. A. (1958). Biochemistry of fishes, Annu. Rev. Biochem., 27, 223— 244.
Taylor M. W., Medici J. C. (1966). Amino acid requirements of grain beetles, J. Nutr., 88, 176—180.
Thacker E. J., Brandt C. S. (1955). Coprophagy in the rabbit, J. Nutr., 55, 375— 385.
236 Глава 5, Пища, топливо и энергия
Thompson Т. Е., Bennett L (1969). Physalia nematocysts: Utilized by mollusks for defense, Science, 166, 1532—1533.
Trager W. (1932). A cellulase from the symbiotic intestinal flagellates of termites and of the roach, Cryptocercus punctulatus, Biochem. J., 26, 1762—1771.
Trager W. (1934). The cultivation of a cellulose-digesting flagellate, Trichomonas termopsidis and of certain other termite protozoa, Biol. Bull., 66, 182— 190.
Webb K. L., DuPaul W. D., Wiebe W., Sottile W., Johannes R. E. (1975). Enewe-tak (Eniwetok) atoll: Aspects of the nitrogen cycle on a coral reef, Limnol. Oceanorg, 20, 198—210.
Weis-Fogh T. (19*52). Fat combustion and metabolic rate of flying locusts (Schi-stocerca gregaria Forskal), Philos, Trans, R. Soc. bond., B, 237, 1—36.
White A., Handler P., Smith E. L. (1964). Principles of Biochemistry, 3rd ed., New York, McGraw-Hill, 1106 pp.
Whittaker R. H., Feeny P. P. (1971). Allelochemics: Chemical interactions between species, Science, 171, 757—770.
Wigglesworth V. B. (1949). The utilization of reserve substances in Drosophila during flight, J. Exp. Biol., 26, 150—163.
Wright R. T., Hobbie J. E. (1966). Use of glucose and acetate by bacteria and algae in aquatic ecosystems, Ecology, 47, 447—464.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Biicherl W., Buckley E. E., Deulofeu V. (eds.) (1968, 1971). Venomous Animals and Their Venoms, vol. 1, 707 pp. (1968); vol. 2, 687 pp. (1971); vol. 3, 537 pp. (1971), New York, Academic Press.
Code C. F., Heidel W. (eds.) (1967, 1968). Handbook of Physiology, sect. 6, Alimentary Canal, vols. I—V, pp. 1—2896, Washington, D. C., American Physiological Society.
Crawford M. A. (ed.) (1968). Comparative Nutrition of Wild Animals, Symposium, Zoological Society of London, No. 21, London, Academic Press, 429 pp.
Eisner T. (1970). Chemical defense against predation in arthropods. In: Chemical Ecology (E. Sondheimer and J. Simeone, eds.), pp. 157—217, New York, Academic Press.
Fenchel T. M., Jargensen В. B. (1976). Detritus food chains of aquatic ecosystems: The role of bacteria, Adv. Microbl. Ecol., 1, 1—58.
Florkin M., Stotz E. H. (eds.) (1971). Comprehensive Biochemistry, vol. 21, Metabolism of Vitamins and Trace Elements, Amsterdam, Elsevier, 297 pp.
Goreau T. F., Goreau N. J., Yonge С. M. (1973). On the utilization of photosynthetic products from zooxanthellae and of a dissolved amino acid in Tridacna maxima f. elongata (iMollusca, Bivalvia), J. Zool. Lond., 169, 417—454.
Halver J. E. (ed.) (1972). Fish Nutrition, New York, Academic Press, 714 pp.
J ar gensen С. B. (1975). Comparative physiology of suspension feeding, Annu. Rev. Physiol., 37, 57—79.
Labov J. B. (1977). Phytoestrogens and mammalian reproduction, Comp. Biochem. Physiol., 57A, 3—9.
Liener I. E. (1969). Toxic Constituents of Plant Foodstuffs, New York, Academic Press, 500 pp.
McDonald P., Edwards R. A., Greenhalgh J. F. D. (1973). Animal Nutrition, 2nd ed., New York, Hafner, 487 pp.
Stephens G. C. (1975). Uptake of naturally occurring primary amines by marine annelids, Biol. Bull., 149, 397—407.
Underwood E. J. (1977). Trace Elements in Human and Animal Nutrition, 4th ed.,-New York, Academic Press, 545 pp.
ГЛАВА 6
ЭНЕРГЕТИЧЕСКИЙ ОБМЕН
В предыдущих главах речь шла о пище и питании, а в этой главе мы рассмотрим, каким образом пища доставляет организму энергию. Для осуществления разнообразных функций нужна химическая энергия, и процессы использования этой энергии у животных обычно называют энергетическим обменом1.
Животные получают энергию главным образом путем окисления питательных веществ, и поэтому мерой энергетического обмена может служить потребление кислорода. Значительная часть этой главы будет посвящена потреблению О2, которое мы часто будем считать показателем интенсивности энергетического обмена. Однако потребление кислорода не всегда может служить таким показателем, так как в организме возможны и другие пути получения химической энергии, не связанные с использованием О2. Например, некоторые животные способны жить при полном отсутствии свободного кислорода; они тоже используют химическую энергию для процессов жизнедеятельности, но получают ее иным способом. Мы говорим, что такие животные вырабатывают энергию путем анаэробного обмена. Анаэробный метаболизм свойствен очень небольшой группе животных, которые обитают в среде, бедной кислородом, и (или) вынуждены переносить на протяжении длительного периода полное отсутствие кислорода. Необходимую им энергию эти животные получают без использования окислительных процессов.
В живом организме для процессов и реакций, требующих затраты энергии, источником ее служит одно и то же вещество — аденозинтрифосфат (АТФ). Это вездесущее соединение, высвобождающее энергию при гидролизе своей «высокоэнергетической» фосфатной связи, является, по-видимому, непосредственным источником химической энергии для таких процессов, как сокращение мышц, движение ресничек, свечение светляков, разряды у электрических рыб, перенос веществ в клетках, всевозможные биосин
1 Термин «обмен» (обмен веществ, метаболизм) употребляют в разных значениях. Например, выражение «обмен (или метаболизм) железа» относится ко всем аспектам физиологической роли железа: к его потреблению, всасыванию и накоплению, к синтезу гемоглобина и цитохромов, выделению и т. д. Термин «промежуточный обмен» стал означать всю совокупность биохимических реакций и взаимопревращений, происходящих в организме. Таким образом, термин «обмен» довольно неопределенный. В этой главе под «обменом» будет подразумеваться энергетический обмен.
238 Глава 6. Энергетический обмен__________________
тетические реакции и т. д. АТФ образуется на различных экзергонических этапах окисления питательных веществ, а также хотя и в меньшем количестве — в ходе анаэробных процессов, идущих с выделением энергии. АТФ — это универсальный посредник, обеспечивающий поток химической энергии от питательных веществ к метаболическим процессам, требующим ее затраты, как у аэробных, так и у анаэробных организмов.
ИНТЕНСИВНОСТЬ МЕТАБОЛИЗМА
Под интенсивностью метаболизма (обмена) понимают величину энергетического обмена в единицу времени. Интенсивность обмена в принципе можно определять тремя различными способами.
Первый способ состоит в том, что вычисляют разность между энергетической ценностью всей потребленной пищи и энергетической ценностью всех экскретов (главным образом мочи и кала). Такой способ основан на предположении о постоянстве состава организма, поэтому он неприменим при работе с растущими животными или с животными, у которых изменяется (в ту или другую сторону) запас жира или иных веществ. Этот метод технически сложен, и для точности результатов здесь необходим достаточно длительный период наблюдений; нужно убедиться, что организм животного не претерпел существенных изменений в размерах и химическом составе.
Второй способ основан на определении общей теплопродукции организма. Здесь учитывается все использованное «топливо», и в принципе этот метод наиболее точен. Практически определения производят, помещая организм в калориметр. При этом могли бы быть получены очень точные результаты, но технически вся процедура очень сложна. При расчете общей теплопродукции необходимо вводить поправки на нагрев потребляемой пищи, испарение воды и т. п. Кроме того, если животное производит внешнюю работу, не переходящую в тепло, эта работа должна быть добавлена к найденной величине теплопродукции. Представьте себе, например, что человек, помещенный в большой калориметр, перекладывает тонну кирпичей, поднимая их с пола на стол. Потенциальная энергия этих кирпичей увеличилась за счет энергетического метаболизма человека, и ее прирост мог бы быть преобразован в тепло, если бы все кирпичи свалились обратно на пол.
Третий способ определения интенсивности обмена основан на измерении количества кислорода, используемого в процессах окисления, при условии, что известно, какие вещества подвергались окислению (и отсутствует анаэробный обмен).
^Измерять потребление кислорода технически несложно, и такой метод настолько широко применяется для оценки интенсивности обмена, что эти два понятия часто употребляются как взаимозаменяемые. Последнее, разумеется, неправильно; например.
Интенсивность метаболизма 239
полностью анаэробные организмы совсем не потребляют кислорода, но это вовсе не значит, что их метаболизм равен нулю!
Почему количество потребляемого кислорода может все-таки служить мерой интенсивности обмена? Дело в том, что количество тепла, выделяющееся при использовании каждого литра кислорода в метаболических процессах, почти не зависит от того, окисляются ли белки, жиры или углеводы (табл. 6.1, столбец в). /Максимальная величина (5,0 ккал1 на 1 л О2 при окислении углеводов) и
Таблица 6.1
Продукция тепла и потребление кислорода в процессах метаболизма основных питательных веществ. Величины для белков зависят от того, будет ли конечным продуктом распада мочевина или мочевая кислота. Отношение количества образовавшейся СО2 к количеству потребленного О2 называется дыхательным коэффициентом (ДК).
(По данным Lusk, 1931; King, 1957)
Питательное вещество (а) ккал/г (б) л О2/г (в) ккал/л О2 (а) СО2 ДК= -т— о2
Углевод 4,2 0,84 5,0 1,00
Жир 9,4 2,0 4,7 0,71
Белок (- ^мочевина) 4,3 0,96 4,5 0,81
Белок (->мочевая кислота) 4,25 0,97 4,4 0,74
минимальная (4,5 ккал на 1 л О2 при окислении белков) различаются всего лишь на 10%; поэтому при расчетах в качестве меры интенсивности обмена обычно используют среднюю величину — 4,8 ккал на 1 л О2. Максимальная ошибка, возможная при использовании этой усредненной цифры, составит 6%, но если окислению подвергается обычная смесь питательных веществ (углеводов, белков, жиров), то ошибка будет незначительной. К тому же определения интенсивности обмена в большинстве случаев вообще не отличаются большой точностью — не из-за недостатков имею
1 Традиционная единица измерения тепла — калория (кал)—не входит теперь в Международную систему единиц (СИ), но повсеместно употребляется в таблицах, книгах и различных публикациях по физиологии. Поэтому было бы непрактичным отказаться от нее в этом учебнике. Единица энергии в системе СИ —Джоуль (Дж), а калория определяется как 1 кал = 4,184 Дж (1 ккал = 4,184 кДж). Далее, 1 кал — это количество энергии, необходимое для того, чтобы нагреть 1 г воды с 14,5 до 15,5 °C, а 1 ккал =1000 кал. Иногда, особенно при изложении начальных сведений или в научно-популярной литературе, калории и килокалории путают между собой. Нередко килокалории называют «большими калориями» и обозначают «Кал». Поскольку калория — это точно определенная единица измерения, эпитеты «большая» и «малая» здесь лишены смысла. Дополнительные сведения об использовании единиц измерения и их обозначений! <см. в Приложении А.
240 Глава 6. Энергетический обмен
щейся аппаратуры, а из-за большой изменчивости таких факторов, как двигательная активность животного и другие физиологические функции.
Хотя использование 1 л кислорода приводит к примерно одинаковому высвобождению энергии для всех трех главных групп питательных веществ, при окислении 1 г этих веществ энергетический выход существенно различен. Например, окисление 1 г жира дает в 2 раза больше энергии, чем окисление 1 г углеводов (табл. 6.1, столбец а).
Хорошо известно, что жиры обладают высокой энергетической ценностью; биологически важное следствие этого состоит в том, что накопление энергии в виде жиров связано с меньшей прибавкой веса и объема по сравнению с использованием других классов соединений. Любопытно, что энергетическая ценность спирта тоже очень высока — около 7 ккал/г. Поэтому люди, стремящиеся похудеть, должны избегать спиртных напитков; кроме того, употребление спиртного быстро ослабляет решимость следить за своей диетой, и это дополнительно ухудшает дело.
Приводимая величина энергетической ценности белков — 4,3 ккал/г — отличается от величины 5,3 ккал/г, получаемой химиками при сжигании белка. Причина этого — то, что в организме белки не окисляются полностью. Млекопитающие выделяют азот в виде мочевины CO(NH2)2 — органического соединения, при сжигании которого выделяется энергия, эквивалентная упомянутому различию. Это различие следует иметь в виду, если энергетический баланс вычисляют, исходя из величин энергетической ценности, определяемых при сжигании белка в калориметрической бомбе.
Количество кислорода, необходимое для окисления 1 г жира, более чем вдвое превышает то, которое нужно для окисления углеводов или белков (табл. 6.1, столбец б). В сочетании с высокой калорийностью жиров это и приводит к тому, что энергия окисления в расчете на 1 л О2 у жиров, углеводов и белков примерно одинакова (см. выше).
В последнем столбце табл. 6.1 приведено отношение количества образующейся в процессе метаболизма СО2 к количеству потребляемого О2. Это отношение, называемое дыхательным коэффициентом (ДК) или показателем дыхательного газообмена, играет важную роль в физиологии обмена веществ. По величине ДК можно судить об основных субстратах, используемых в обмене. Обычно ДК колеблется в пределах от 0,7 до 1,0. Величина, близкая к 0,7, свидетельствует о преобладании жирового обмена, а близкая к 1,0 — о том, что используются главным образом углеводы. В случае промежуточных значений ДК судить о природе окисляемых веществ труднее: это могут быть белки, смесь жиров и углеводов или же смесь всех трех субстратов. Можно, однако, определить количество используемых белков по выделению азота
Запасание энергии: жир и гликоген
241
(мочевины у млекопитающих, мочевой кислоты у птиц и рептилий) и из полученных данных вычислить, какая доля потребляемого 02 и образующейся СО2 связана с белковым обменом. Остальная часть газов будет связана с использованием углеводов и жиров,, и далее можно будет вычислить вклад каждого из этих субстратов. Таким образом, определив потребление кислорода, выделение: СО2 и выведение азота, можно рассчитать, сколько по отдельности белков, жиров и углеводов было использовано в энергетическом обмене за данный промежуток времени.
Можно ли определять интенсивность метаболизма по выделению СО2, а не по потреблению кислорода? Теоретически — да, на практически это сложно, и результат будет менее точным по двум причинам: во-первых, в организме имеется большой пул СО2; во-вторых, энергетический эквивалент одного литра СО2 при окислении разных субстратов различен.
Количество двуокиси углерода, постоянно присутствующей в организме (пул СО2), легко подвержено изменению. Например, при гипервентиляции легких большое количество СО2 выходит с выдыхаемым воздухом. Разумеется, это выведение СО2 не отражает интенсивности обмена. После гипервентиляции наступает период пониженной вентиляции, во время которого пул СО2 возвращается к норме, и этот период пониженного выделения СО2 опять-таки ничего не говорит об уровне обмена. На содержание двуокиси углерода в организме влияют и другие обстоятельства. Например, при тяжелой физической работе в мышцах образуется много молочной кислоты. Переход ее в кровь дает такой же эффект, как добавление кислоты к раствору питьевой соды: в крови освобождается двуокись углерода, и тогда она выводится через легкие в избыточном количестве. В этом случае выделение СО2 тоже не отражает интенсивности метаболизма.
Вторая причина меньшей пригодности СО2 (по сравнению с О2) для оценки интенсивности обмена состоит в том, что различные питательные вещества дают разное количество энергии в рас-счете на 1 л образовавшейся СО2. Углеводы дают 5,0 ккал на 1 л СО2, а жиры — 6,7 ккал. Таким образом, для жиров этот показатель на !/з выше, чем для углеводов, и если мы не знаем, использовались ли в обмене жиры или углеводы, ошибка при расчетах может быть весьма значительной.
По этим двум причинам выделение двуокиси углерода не может быть надежным показателем интенсивности обмена.
ЗАПАСАНИЕ ЭНЕРГИИ: ЖИР И ГЛИКОГЕН
У большинства взрослых животных потребление пищи и расходование энергии примерно уравновешивают друг друга. Если же расход энергии больше притока ее с пищей, то источником дополнительной энергии служат компоненты тела, главным образом 16—1873
242 Глава 6. Энергетический обмен______________________________
жиры. Наоборот, если потребление пищи превышает энергетические потребности, то избыток запасается — независимо от состава пищи — в виде жира. Например, свиньи жиреют, питаясь в основном углеводами (зерном). В период накопления жира, когда большое количество углеводов превращается в жиры, ДК превышает 1,0. Это обусловлено тем, что жиры содержат относительно меньше кислорода, чем углеводы, и в окислительных процессах частично используется «избыточный» кислород из углеводов; поэтому потребление О2 при дыхании снижается, а отношение выделяющейся СО2 к поглощаемому О2 возрастает.
Поскольку калорийность жиров в два раза выше, чем углеводов, жиры служат более подходящей формой запасания энергии. .Из этого правила, однако, есть исключения. У малоподвижных животных, например у устриц и других двустворчатых моллюсков, вес тела не имеет существенного значения, и они запасают энергию в форме гликогена. Кроме того, гликоген накапливают многие кишечные паразиты, например аскариды. Гликоген, вероятно, служит для них более удобным запасным веществом, чем .жир, так как кишечные паразиты часто оказываются в условиях недостатка или даже полного отсутствия кислорода, а из гликогена можно получать энергию путем его анаэробного расщепления до молочной кислоты. Таким образом, поскольку двустворчатые моллюски часто надолго замыкают свои раковины, а кишечные пара-литы живут в среде, практически лишенной кислорода, этим животным выгодно накапливать гликоген, а не жир. Неподвижным .животным и паразитам нет особой надобности уменьшать вес тела, и с этой точки зрения запасание гликогена вместо жира не является для них невыгодным.
Однако для подвижных животных очень важно уменьшить вес тела. Это имеет особенно большое значение для перелетных птиц, которые иногда пролетают более 1000 км без остановки. Количество жира у них достигает 40—50% от веса тела; если бы их «топливо» было тяжелее, то тело оказалось бы слишком тяжелым и дальние перелеты были бы невозможны просто потому, что большая часть энергии расходовалась бы на поддержание тела в воздухе.
Накопление гликогена вместо жира приводит к значительно большему увеличению веса не только потому, что энергетическая ценность углеводов ниже, но и потому, что гликоген откладывается в клетках вместе со значительным количеством воды. По приближенной оценке, отложение гликогена в клетках печени и мышц сопровождается накоплением 3 г воды на каждый грамм гликогена. Судя по некоторым данным, количество воды может быть еще больше — до 4—5 г на 1 г гликогена (Olsson, Saltin, 1968).
Чтобы выразить вес «топлива» по отношению к его энергетической ценности, проще всего вычислить его изокалорический вес, т. е. вес, приходящийся на единицу энергии. Для интересующих
Запасание энергии: жир и гликоген
243
нас веществ изокалорический вес выражается следующими величинами (в граммах на килокалорию):
Жир 0,11
Белок 0,23
Крахмал 0,24
Гликоген+Н20 ~1,0
При таком способе выражения сразу же становится очевидным, что гликоген (если учесть связанную с ним воду) как источник энергии примерно в 10 раз тяжелее, чем жир. И все же отложение гликогена является важной формой запасания энергии. Оно дает два преимущества: во-первых, гликоген в отличие от жира очень быстро мобилизуется для использования и, во-вторых, — и это, вероятно, еще важнее — он может доставлять энергию в аноксических условиях. Такие условия часто создаются в мышцах при тяжелой физической работе, когда поступление кислорода с кровью не покрывает потребности в нем. Однако для запасания энергии в больших количествах и на долгий срок гликоген не подходит, и для этой цели используется жир.
Накопление вместе с гликогеном воды позволяет, возможно, объяснить загадочное явление, с которым сталкиваются многие лица, переходящие на диету для похудания. В первые несколько дней они быстро теряют в весе, часто намного больше, чем можно было бы ожидать, учитывая только потерю жира организмом. В этот начальный период происходит распад резервного гликогена и выделяется избыток воды. Таким образом, значительная часть потери веса обусловлена выведением воды. Спустя несколько дней, даже если потребление пищи ниже энергетических расходов организма, запасы гликогена восстанавливаются. Когда организм переходит на использование резервного жира, вес уже не снижается так быстро; он может даже опять увеличиться, хотя энергетический баланс остается отрицательным и жировые запасы уменьшаются. Из-за таких неожиданных изменений веса, которые редко получают правильное объяснение, многие люди, перешедшие на разгрузочную диету, в первые дни становятся сверхоптимистичными, а затем быстро разочаровываются: несмотря на все ограничения в питании, потеря веса значительно замедляется или даже происходит его прибавка.
ВЛИЯНИЕ КОНЦЕНТРАЦИИ КИСЛОРОДА
НА ИНТЕНСИВНОСТЬ МЕТАБОЛИЗМА
Считается, что потребление кислорода (или интенсивность обмена) не зависит от содержания кислорода в среде в широком диапазоне его концентраций. Если, например, заменить обычный воздух чистым кислородом, млекопитающее будет потреблять кислород в прежнем количестве, хотя внешняя концентрация О2 при 16*
244 Глава 6. Энергетический обмен
такой замене увеличится почти в 5 раз. То же самое наблюдается и в обратной ситуации: если уменьшить концентрацию кислорода в 2 раза, например снизив вдвое общее атмосферное давление (что будет эквивалентно подъему на высоту около 6000 м над уровнем моря), то потребление кислорода также практически не изменится.
^о2> кПа
Рис. 6.1. Потребление кислорода у рыб зависит от его парциального давления <₽О2 ) в воде. У иглобрюха и скала оно уменьшается по мере снижения Ро2 лишь в небольшой степени, и только при очень низких уровнях Ро2 потребление кислорода быстро падает. У третьей рыбы — Opsanus — потребление кислорода прямо пропорционально изменению Ро2 в воде на всем диапазоне измерений. (Hall, 1929.)
Однако такая независимость от внешней концентрации кислорода не является универсальной, и возможны прямо противоположные случаи. На рис. 6.1 показано, как влияет концентрация кислорода в воде на потребление его некоторыми рыбами. У двух рыб (скапа и иглобрюха) потребление кислорода снижается примерно на 7з, а при очень низкой концентрации кислорода (менее 15 мм рт. ст.) оба эти вида не выживают. Иначе ведет себя Ор-.sanus: у этой рыбы потребление кислорода снижается пропорционально его концентрации, так что эти два параметра связаны
Концентрация кислорода и интенсивность метаболизма 245
линейной зависимостью. Opsanus способен длительное время выживать в воде, лишенной кислорода; в этих условиях, когда потребление О2 падает до нуля, весь энергетический обмен становится анаэробным.
Сходным образом реагируют на изменение уровня кислорода в среде и некоторые беспозвоночные (например, обычный омар).
Рис. 6.2. Потребление кислорода у омара тем больше, чем выше температура, но при каждой данной температуре оно находится в линейной зависимости от концентрации О2 в воде. (Thomas, 1954.)
Как видно из рис. 6.2, чем выше температура, тем больше омар потребляет кислорода, однако при любой температуре потребление О2 находится в линейной зависимости от концентрации кислорода в среде1. Если же изучать потребление кислорода в широком диапазоне температур, то для такого животного, как золотая рыбка, мы получим кривые вроде представленных на рис. 6.3: при относительно высоком напряжении кислорода потребление О2 не зависит от его количества, тогда как при более низких величинах
1 В этой книге количества кислорода выражаются в единицах объема (объем сухого газа при О °C и давлении 760 мм рт. ст.). В системе СИ единица измерения количества любого вещества — моль, и количество О2 должно выражаться не в литрах или миллилитрах, а в молях и миллимолях (сокращения: моль, ммоль). Например, 1 л 02 при 0°С и 760 мм рт. ст. = 44,64 ммоль. В настоящее время происходит постепенный переход к употреблению единиц системы СИ, но многие традиционные единицы более привычны и будут какое-то время еще использоваться. Там, где это разумно, мы будем предпочитать единицы СИ (см. Приложение А).
246 Глава 6. Энергетический обмен
напряжения 02 имеет место линейная зависимость. Точка перегиба кривой, где потребление кислорода становится независимым от его концентрации, при более низких температурах сдвигается влево, в сторону меньших величин напряжения О2— у золотой рыбки, например с <20 мм рт. ст. при 5°C до ~40 мм рт. ст. при 35 °C.
PQ э мм ртп стп
Рис. 6.3. При низких концентрациях кислорода его потребление золотой рыбкой прямо пропорционально содержанию его в воде, но в области более высоких значений РОо этой зависимости нет. (Fry, Hart, 1948.)
Вернувшись к рис. 6.1 и 6.2, мы увидим, что прямолинейные графики, представленные здесь для рыбы Opsanus и омара, — это, возможно, всего лишь начальные участки кривых того же типа, что и кривые для золотой рыбки. Если это так, мы можем ожидать, что и кривые для омара и Opsanus при какой-то неизвестной более высокой концентрации кислорода достигнут плато.
Даже у млекопитающих, у которых интенсивность метаболизма почти не меняется при изменении количества кислорода в среде, встречается иногда более выраженная зависимость от О2, чем обычно полагают. Например, потребление кислорода изолированными мышцами крысы и кролика обратимо подавляется при низких концентрациях О2, причем другие клеточные функции не изменяются (Whalen, 1966). Это позволяет понять, почему ныряющие животные после ныряния не увеличивают потребления О2 настолько, чтобы погасить кислородную задолженность, которая должна была бы возникнуть у них во время пребывания под водой (см. ниже в этой главе). У плывущего животного больше всего кислорода потребляют мышпы, и зависимость уровня метаболизма этой ткани от напряжения кислорода должна приводить именно к тому,
Концентрация кислорода и интенсивность метаболизма 247
что наблюдается у многих ныряющих животных. Может быть, независимость обмена от кислорода — это (вопреки обычному представлению) частный случай, а зависимость — общее правило.
АККЛИМАЦИЯ К НИЗКОМУ УРОВНЮ 02
Многие быстро плавающие активные рыбы очень чувствительны к снижению содержания кислорода в воде. Это относится к большинству лососевых, например к гольцу (Salvelinus fontinalis). Однако для этой рыбы летальный уровень О2 смещается, если она предварительно находилась в условиях низкой, но еще совместимой с жизнью концентрации кислорода: пребывание в обедненной кислородом воде делает гольца более устойчивым к гипоксии (рис. 6.4). Такое повышение устойчивости можно объяснить уве-
Рис. 6.4. Если гольцов предварительно выдерживать в воде с пониженным содержанием кислорода, они становятся более устойчивыми к недостатку О-и минимальный уровень О2 для выживания у них снижается. (Shepard, 1955.)
личением способности извлекать кислород из воды. Если бы ткани действительно становились более выносливыми к аноксии, то рыбы, приспособившиеся к низкому уровню О2, в полностью анаэробных условиях выживали бы дольше, чем рыбы, не подвергнутые предварительной акклимации; между тем такого различия не наблюдается (Shepard, 1955). Повышенное извлечение кислорода из воды могло бы достигаться увеличением объема воды, протекающей через жабры, увеличением способности жабр извлекать кислород, повышением кислородной емкости крови, усилением работы сердца или какой-то комбинацией этих способов.
248 Глава 6. Энергетический обмен
АНАЭРОБНЫЙ ОБМЕН
Животные, переносящие длительное отсутствие кислорода, по необходимости должны получать энергию анаэробным путем. К таким животным относятся многие кишечные паразиты, а также обитатели бескислородного ила на дне озер и прудов, двустворчатые моллюски, надолго закрывающие свои створки, и другие организмы.
Обычный путь анаэробной выработки энергии — это расщепление углеводов до молочной кислоты. Например, 1 моль глюкозы может в анаэробных условиях расщепляться с образованием 2 молей молочной кислоты (C6Hi2O6—>2С3Н60з). Этот процесс, называемый гликолизом, часто происходит в мышцах млекопитающих, когда потребность в энергии велика, а снабжение ткани кислородом недостаточно. При гликолизе, свободная энергия (AF) которого составляет 50 ккал на 1 моль глюкозы, высвобождается только около 7% энергии, получаемой при полном окислении глюкозы (691 ккал/моль).
В большинстве случаев исходным субстратом гликолиза служит не глюкоза, а гликоген. Молярный энергетический выход при распаде гликогена на 7 ккал больше, чем при распаде глюкозы, так что образование молочной кислоты ведет к высвобождению 57 ккал. Поскольку универсальным источником энергии для процессов, требующих ее затраты, служит АТФ, можно расчитывать энергетический выход реакций аэробного и анаэробного обмена по количеству образующегося АТФ. При гликолизе, т. е. образовании 2 молей молочной кислоты из 1 моля глюкозы, синтезируются 2 моля АТФ (а если субстратом служит гликоген, то на 2 моля молочной кислоты образуется 3 моля АТФ). В отличие от этого при полном окислении глюкозы до СО2 и воды образуются 36 молей АТФ (6 — при образовании 2 молей пировиноградной кислоты и 30 — при полном окислении этой кислоты в цикле Кребса).
Таким образом, в полностью анаэробных условиях выработка АТФ за счет энергии гликолиза невелика, но кишечные паразиты, вероятно, не очень нуждаются в экономии. Однако те животные, которые лишь временами переходят на гликолиз, могут использовать молочную кислоту как субстрат для дальнейшего окисления (в цикле Кребса) и таким образом в конечном счете полностью реализуют энергетическую ценность исходного углевода.
Выносливость некоторых животных по отношению к аноксии поразительна. Сообщалось, что карась (Carassius carassius) может просуществовать в замерзшем озере 5V2 месяцев, хотя в воде подо льдом из-за гниения отмерших растений нет кислорода и присутствует сероводород (Blazka, 1958). (Сероводород ядовит, так как он присоединяется к цитохромоксидазе и инактивирует ее, однако без кислорода цитохромоксидаза не функционирует, и поэтому действие яда не проявляется.) При этом в тканях карася
Концентрация кислорода и интенсивность метаболизма 249 почти не накапливается молочной кислоты; значит, здесь происходят какие-то иные метаболические процессы. Было высказано предположение, что в этом случае в конечном результате образуются жирные кислоты, о чем свидетельствует накопление жира у рыб, длительно находящихся в анаэробных условиях. Существуют и другие реакции анаэробного обмена; уже изучен ряд путей выработки энергии у различных живых существ, способных переносить длительное или постоянное отсутствие свободного кислорода (Hochachka, Mustafa, 1972).
СЛОИ ВОДЫ С МИНИМАЛЬНЫМ СОДЕРЖАНИЕМ КИСЛОРОДА
На средних глубинах в океанах есть зоны с очень низким содержанием кислорода. Обширная зона такого рода имеется на северо-востоке тропической области Тихого океана. Около побережья центральной и южной Мексики подобный слой занимает глубины примерно от 100 до 900 м; концентрация растворенного кислорода здесь обычно ниже 0,1 см3/л, причем эта величина относительно мало подвержена сезонным изменениям (Douglas et •al., 1976). И в этой обедненной кислородом воде обитает поразительное количество живых существ!
Некоторые животные, особенно рыбы, совершают ежедневные миграции к поверхности, поднимаясь в сумерках и возвращаясь на глубину перед восходом солнца, причем их путь по вертикали составляет 300 м и больше. Удивительно, что рыбы, обитающие в этом бедном О2 слое, часто имеют плавательный пузырь, наполненный газом с высоким содержанием кислорода. Днем, находясь па глубине, эти рыбы, вероятно, получают энергию с помощью анаэробных процессов. Их кровь отличается малой кислородной емкостью, а гемоглобин обладает настолько низким сродством к кислороду, что на глубине его насыщение О2 должно составлять не более 1% (Douglas et al., 1976).
Мелкие ракообразные, обитающие в слое с минимальным содержанием О2, очень эффективно извлекают кислород из воды. Изучение 28 видов, обитающих в этом слое вблизи южной Кали-•форнии, показало, что все они обладают способностью поглощать кислород даже при самых низких величинах Ро2> с которыми они встречаются в природе, иногда вплоть до 4 мм рт. ст. Исключением оказались два паразитических вида и одно животное, ежедневно совершающее вертикальные миграции в слои, обогащенные кислородом (Childress, 1975).
Судя по имеющимся данным, наличие богатой фауны в слое € минимальным содержанием О2 обусловлено способностью обитателей этого слоя использовать кислород даже при его очень низкой концентрации, так что эти животные существуют в основном за счет аэробного обмена. Однако исследование интереснейших биологических проблем, связанных с такими областями моря,
250 Глава 6. Энергетический обмен
еще только начинается, и пока мы не можем сказать, что здесь играет главную роль — уникальная способность использовать кислород в очень низких концентрациях или же способность длительно существовать за счет анаэробного метаболизма.
Последняя возможность отнюдь не исключена, поскольку в принципе существует несколько биохимических механизмов анаэробного обмена. Анаэробный гликолиз с образованием молочной кислоты обычно представляется нам самой типичной формой анаэробного метаболизма, так как он свойствен высшим позвоночным. Однако гликолиз — малоэффективный процесс, высвобождающий лишь несколько процентов того количества энергии, которое можно получить при полном окислении субстрата. Для синтеза АТФ при длительной или постоянной гипоксии есть много других возможностей. Большой пул свободных аминокислот, имеющийся у многих беспозвоночных, может быть важным потенциальным источником энергии, несравненно более эффективным, чем образование молочной кислоты (Hochachka et al., 1973).
ПРОБЛЕМЫ, СВЯЗАННЫЕ
С НЫРЯНИЕМ МЛЕКОПИТАЮЩИХ И ПТИЦ
Все водные птицы и млекопитающие сохранили легкие и дышат воздухом, однако для успешного приспособления к водной среде им нужен ряд специальных механизмов. Наиболее очевидно то, что для продолжительного пребывания под водой необходимо эффективное использование ограниченного запаса кислорода, однако важную роль играют и другие функциональные особенности.
Таблица 6.2
Водные млекопитающие. (Можно видеть, что они есть в большинстве основных отрядов; некоторые отряды целиком состоят из водных животных)
Отряд Представители, ведущие водный образ жизни
Однопроходные Утконос
Сумчатые Нет
Насекомоядные Ку тор а
Летучие мыши Нет
Приматы Нет
Грызуны Бобр, ондатра
Зайцеобразные Нет
Китообразные Киты (все виды)
Хищные Калан
Ластоногие Тюлени (все виды)
Сирены Морские коровы, ламантин, дюгонь (все виды)
Копытные
непарнокопытные Тапир
парнокопытные Бегемот
Ныряние у млекопитающих и птиц 251
Таблица 6.3
Водные и сухопутные птицы. Многие семейства почти целиком перешли к водному образу жизни (если не считать размножения); в других семействах мало или совсем нет водных представителей
В основном водные В основном неводные
Пингвины Бескилевые (страусы,
Гагары нанду и др.)
Поганки Соколы
Буревестники и альба- Куры, фазаны и т. п.
тросы Голуби
Пеликаны Попугаи
Цапли Совы
Утки Козодои
Болотные птицы Колибри
Чайки Дятлы
Чистиковые Воробьиные (вьюрки,
скворцы, вороны и др.)
Приспособление к водному образу жизни встречается почти во всех основных отрядах млекопитающих, за исключением летучих мышей (Chiroptera), зайцеобразных (Lagomorpha — зайцы и кролики), приматов и сумчатых (табл. 6.2). Из списка водных и сухопутных птиц (табл. 6.3) видно, что в филогенетическом отношении эти группы крайне разнообразны. Однако физиологические пути адаптации к водной среде у них весьма сходны. Тот факт, что все ныряльщики используют одни и те же механизмы, в известной степени свойственные и неныряющим позвоночным, наводит на мысль, что ныряльщики воспользовались уже ранее имевшимися адаптациями и развили их.
Чтобы понять, какие трудности стоят перед ныряющими животными, представим себе человека, который ныряет в соответствующей экипировке или без нее. Некоторые из проблем, встающих перед человеком, не возникают у животных, другие же оказываются общими. Все дышащие воздухом животные при нырянии должны обеспечить себя кислородом, но для тех, которые опускаются достаточно глубоко, существуют дополнительные трудности, которые можно разделить на четыре группы:
1) кессонная болезнь;
2) токсичность кислорода;
3) наркотический эффект газов;
4) прямое воздействие высокого давления.
КЕССОННАЯ БОЛЕЗНЬ
Этот опасный синдром известен под несколькими названиями: воздушная эмболия, болезнь водолазов, кессонная болезнь. Он
252 Глава 6. Энергетический обмен _____________________________
возникает у людей, поднимающихся на поверхность после длительного пребывания на глубине более 20 м; чем больше глубина или время погружения, тем сильнее проявляется болезнь. Она обусловлена выделением в тканях и крови пузырьков газа, образующихся совершенно так же, как образуются пузырьки в бутылке содовой воды, когда ее открывают. В обоих случаях пузырьки возникают при понижении давления над жидкостью, насыщенной газом при большем давлении. Кессонная болезнь возможна также при быстром подъеме на высоту (на воздухоплавательных аппаратах или самолетах) в негерметизированной кабине; в этом случае опасность кессонной болезни возникает при снижении давления примерно на 0,5 атм (на высоте более 6000 м).
В газированной воде растворена двуокись углерода. Газ, вызывающий кессонную болезнь, — это всегда азот (если только водолаз не дышал искусственной газовой смесью, содержащей другие инертные газы, например гелий). Даже на большой глубине напряжение СО2 не намного превышает физиологическую норму (около 40 мм рт. ст.), а для развития кессонной болезни требовалось бы перенасыщение газом примерно при 2 атм. Кислород не вызывает кессонной болезни, так как он быстро используется в тканях; кроме того, как мы увидим позже, при давлении свыше 1 атм он настолько токсичен, что повышенные концентрации его недопустимы.
С увеличением глубины на каждые 10 м давление в воде возрастает примерно на 1 атм. Для того чтобы на глубине 10 м обычный водолазный костюм был наполнен воздухом, абсолютное давление в нем должно быть на 1 атм выше нормального атмосферного давления, т. е. составлять 2 атм. Когда водолазу, находящемуся на глубине 10 м, подают обычный воздух, давление кислорода в нем составляет 0,4 атм, а азота — 1,6 атм. В результате растворение азота в крови, а затем постепенно и в воде тканей происходит при повышенном давлении. Кроме того, в жире, который у здорового худощавого мужчины составляет около 15% веса тела, растворяется примерно в 5 раз больше азота на единицу веса, чем в воде. В организме такого мужчины весом 70 кг содержится около 50 л воды и 10 кг жира; значит, общее количество азота, растворенного в жире, примерно равно тому количеству, которое растворено в воде.
Для полного насыщения организма азотом на глубине требуется довольно много времени. Поэтому кратковременный спуск на глубину сравнительно неопасен, тогда как после длительного пребывания под водой, когда ткани почти полностью насыщены азотом под повышенным давлением, опасность возрастает. После возвращения водолаза на поверхность для удаления азота требуется несколько часов, так как азот лишь медленно выходит из тканей, растворяется в крови и выводится через легкие.
Ныряние у млекопитающих и птиц 25$
Чем выше давление, при котором происходило растворение^ газа, тем больше опасность образования пузырьков. Движение,, мышечное напряжение, усиленное кровообращение способствуют’ образованию пузырьков, действуя так же, как встряхивание бутылки с пивом или газированной водой перед открыванием. Пузырьки часто образуются в суставах, причиняя сильную боль.. В кровяном русле пузырьки попадают в мелкие сосуды и закупоривают их; если это происходит в центральной нервной системе^, то это особенно опасно и может привести к внезапной смерти. Единственный способ лечения кессонной болезни — это быстрое повышение давления с целью вызвать растворение образовавшихся пузырьков. Для этого водолаза вновь опускают на ту глубину, с которой он вернулся, или же помещают в рекомпресси-онную камеру, где можно создать давление воздуха заданной величины.
Для предупреждения кессонной болезни подъем водолаза на поверхность должен происходить медленно и постепенно. Например, вполне допустимо, если водолаз поднимется до половины той глубины, на которой он работал, и пробудет здесь некоторое время, пока не выделится значительная часть азота, — скажем, 20— 30 мин. Затем можно подняться выше и вновь остановиться на какое-то время. Широко применяются специальные таблицы, в которых точно указана допустимая скорость подъема с той или иной-глубины.
Если ныряльщик использует баллон со сжатым воздухом, он должен дышать воздухом под давлением, равным давлению воды на данной глубине, иначе его грудная клетка будет сдавлена водой. Поэтому аквалангисты подвергаются такой же угрозе развития кессонной болезни, как и водолазы в обычных водолазных костюмах.
Опасность развития кессонной болезни можно уменьшить, используя вместо азота другие газы. С этой целью часто применяют гелий, который имеет то преимущество перед азотом, что он менее растворим в воде и жире и скорость его диффузии в несколько, раз больше. Большая скорость диффузии позволяет сократить время подъема на поверхность; однако по той же причине насыщение тканей гелием происходит быстрее, и поэтому при кратковременных погружениях применение его менее выгодно.
До сих пор мы рассматривали только водолазов, снабжаемых воздухом под давлением. Ныряльщики, неоднократно спускающиеся без водолазных костюмов на значительную глубину, тоже подвергаются опасности развития кессонной болезни. Каждый раз,, когда ныряльщик наполняет легкие воздухом и опускается на. глубину, давление воды сжимает его грудную клетку и давление: воздуха в легких возрастает. Если он опускается на глубину 20 м, давление воздуха в легких достигает 3 атм. Один или два
254 Глава 6. Энергетический обмен. спуска на такую глубину не представляют опасности, но многократные спуски уже рискованны.
Описан достоверный случай кессонной болезни у ныряльщика, не пользовавшегося водолазным снаряжением. Речь идет о датском враче, тренировавшемся в бассейне глубиной 20 м. Он применял прием «падения на дно», состоящий в том, что ныряльщик, набрав воздуха, бросается вниз с края бассейна; на глубине 2—3 м его грудная клетка настолько сжимается, что далее он идет ко дну со все возрастающей скоростью. За пять часов врач 60 раз опускался на дно, находясь там по 2 мин, и после этого у него появились боли в суставах, затрудненное дыхание, расстройство зрения и боли в животе. Его поместили в рекомпрессионную камеру при давлении 6 атм, и все эти симптомы быстро исчезли. Постепенно давление в камере снизили до 1 атм, для чего потребовалось 19 ч 57 мин, согласно соответствующим таблицам морского флота США. В данном случае лечение оказалось вполне успешным, однако далеко не всегда дело кончается так благополучно (Paulev, 1965).
Если у людей, многократно ныряющих с задержкой дыхания, развивается кессонная болезнь, то как же избегают ее тюлени и киты, без конца погружающиеся на глубину? Нередко они всплывают на поверхность всего лишь на несколько секунд, а при нырянии достигают значительной глубины.
Лучший ныряльщик среди тюленей — антарктический тюлень Уэддела, который опускается на глубину 600 м и более и способен провести под водой до 43 мин (Kooyman, 1966). Среди китов глубже всех ныряет кашалот (Physeter catodon), для которого известен рекорд в 1134 м глубины. Этот любопытный рекорд документирован тем, что на такой глубине был порван трансатлантический кабель, а когда для ремонта кабель подняли на поверхность, оказалось, что в нем запутался нижней челюстью погибший кашалот. Судя по тому, как кабель обмотал челюсть, можно было представить себе, что кашалот, плавая у дна, зацепился за кабель и пытался освободиться.
Почему киты и тюлени могут глубоко нырять и быстро подниматься на поверхность и у них не возникает при этом кессонной болезни? Возможны три ответа на этот вопрос: 1) им не вредит образование пузырьков в крови и тканях; 2) у них есть какие-то механизмы, предотвращающие образование пузырьков даже при перенасыщении газом, или 3) у них не возникает перенасыщения газом. Судя по имеющимся данным, хотя и скудным, наиболее вероятен третий из вариантов.
Самое существенное отличие тюленей и китов от человека, ныряющего в водолазном снаряжении,’—то, что животные лишены непрерывного притока воздуха. Однако у человека многократные погружения с наполненными воздухом легкими тоже в конце кон
Ныряние у млекопитающих и птиц
255
цов вызывают кессонную болезнь. В связи с этим очень важно, что тюлень делает в начале каждого погружения выдох, а не остается под водой с легкими, наполненными воздухом, как человек. Выдыхают ли воздух в этой ситуации киты, остается неизвестным, но у них чрезвычайно большая и широкая трахея. Возможно, что это — приспособление, позволяющее киту быстро вентилировать-легкие при кратковременном подъеме на поверхность.
Не менее, а может быть, и еще более важен тот факт, что объем легких у китов невелик по сравнению с объемом верхних дыхательных путей (Scholander, 1940). Когда кит опускается на глубину, возросшее давление сжимает его легкие и выдавливает воздух в обширную трахею, которая защищена от сжатия костными кольцами. Если объем несжимаемой трахеи составляет Vio объема легких, то на глубине 100 м легкие будут полностью сжаты и весь воздух перейдет в трахею. Если в легких нет воздуха, то и азот не может переходить в кровь (в отличие от того, что происходит у человека). Азот не попадает в кровь также и потому, что кровоток в легких кита на глубине падает до минимума; таким образом, даже если в легких и было некоторое количество воздуха, то, прежде чем кит достигнет глубин, где легкие сжимаются,, очень мало азота успеет попасть в кровь.
ТОКСИЧНОСТЬ КИСЛОРОДА
Чистый кислород при давлении 1 атм вреден для большинства-теплокровных животных. Человек может благополучно дышать-чистым кислородом в течение 12 ч, но после 24 ч возникают неприятные ощущения за грудиной и признаки нарастающего раздражения легких. Крысы, если держать их в атмосфере чистого» кислорода, погибают с симптомами сильного раздражения легких через несколько дней. Если давление О2 выше 1 атм, то раздражению органов дыхания предшествуют неврологические симптомы. Продолжительное вдыхание кислорода под давлением 2 атм вызывает судороги, а мышечная работа повышает чувствительность к О2. Давление кислорода в 3 атм человек может выносить не дольше нескольких часов, а более длительное воздействие уже небезопасно.
Значение токсичности кислорода в связи с водолазным делом: очевидно. Если водолаз спускается на глубину 40 м и дышит сжатым воздухом, то общее давление этого воздуха будет 5 атм. Поскольку кислород составляет V5 часть воздуха, парциальное давление О2 составит 1 атм (что близко к границе токсичности). Понятно, что погружение на большие глубины тем опаснее, чем больше давление О2 превышает 1 атм.
„Единственный реальный путь предупреждения токсического воздействия кислорода на водолазов, работающих на глубинах 40 м и более, состоит в том, чтобы снизить содержание кислорода во
256 Глава 6. Энергетический обмен
вдыхаемом ими воздухе. На глубине 40 м (5 атм) водолаз мог бы использовать для дыхания газовую смесь, содержащую 10% О2 и 90% N2. В этом случае парциальное давление О2 будет 0,5 атм, что не представляет никакой опасности. При спуске на •большую глубину содержание кислорода в газовой смеси следовало бы еще больше снизить, а для уменьшения риска кессонной болезни при декомпрессии азот можно было бы частично или полностью заменить другим газом, например гелием.
НАРКОТИЧЕСКОЕ ДЕЙСТВИЕ
БИОЛОГИЧЕСКИ ИНЕРТНЫХ ГАЗОВ
Еще одно ограничение в отношении глубины, на которую может спуститься водолаз, связано с прямым физиологическим действием инертных газов. Азот под давлением в несколько атмосфер вызывает наркотический эффект, подобный действию закиси азота (веселящего газа); разница состоит в том, что эффект азота проявляется лишь при высоких давлениях.
Наркотический эффект развивается не сразу, но на глубине 100 м он настолько выражен, что применение азотно-кислородщях смесей практически невозможно. Поэтому необходимо использовать какие-то другие газы, лучше всего опять-таки гелий. Для этого годился бы и водород, однако он взрывоопасен, что сильно усложняет обращение с ним. В конце концов и гелий под очень высоким давлением начинает оказывать значительное побочное действие на организм, и это ограничивает глубину, на которую может спускаться водолаз даже в наилучшем снаряжении; речь не идет, конечно, о случаях, когда человек находится внутри прочной конструкции вроде батискафа или жесткого скафандра, где давление остается на низком уровне.
Для ныряющих животных проблемы наркотического эффекта инертных газов, так же как и токсического действия кислорода, не существует по той простой причине, что они не дышат во время пребывания под водой.
ОБЕСПЕЧЕНИЕ КИСЛОРОДОМ ВО ВРЕМЯ НЫРЯНИЯ
Человек, ныряющий без специального снаряжения, должен прежде всего обеспечить себя кислородом. Обычно он наполняет легкие воздухом, чтобы увеличить свой запас кислорода и иметь возможность дольше оставаться под водой. Однако теоретически имеется целый ряд других способов для достижения той же цели (табл. 6.4). Некоторые из них по физиологическим причинам -неосуществимы или же, насколько известно, не используются животными. У большинства ныряющих животных имеется сочетание нескольких адаптаций, и в результате способность к нырянию у них достигла — по человеческой мерке — поразительной степени.
Ныряние у млекопитающих и птиц 257
Таблицы 6.4
Возможные способы увеличения времени пребывания ныряющих животных под водой. В скобках указаны те варианты, которые мало распространены, маловероятны или по физиологическим причинам неосуществимы
Увеличение запаса кислорода
[Увеличение объема легких]
Увеличение объема циркулирующей крови и повышение содержания в ней гемоглобина
Увеличение количества мышечного гемоглобина
[Неизвестные способы запасания кислорода]
Использование анаэробных процессов
Образование молочной кислоты
[Использование неизвестных акцепторов водородных ионов]
Снижение потребления кислорода
Снижение интенсивности обмена
Водное дыхание
Кожное дыхание (лягушка, морские змеи)
Дыхание через пищевод или прямую кишку (некоторые черепахи)
[Вдыхание -воды в легкие (испробовано только в эксперименте) ]
ЗАПАСАНИЕ КИСЛОРОДА
Самый простой способ обеспечить себя кислородом при нырянии, казалось бы, состоит в том, чтобы набрать в легкие побольше воздуха. Однако объем легких у ныряющих животных в общем примерно такой же, как у других млекопитающих, а некоторые из лучших ныряльщиков имеют сравнительно небольшие легкие. Это нетрудно объяснить, если немного подумать. Во-первых, чем больше воздуха в легких, тем больше азота переходит в жидкости тела и тем больше опасность развития кессонной болезни. Во-вторых, большие легкие, наполненные воздухом, вначале затрудняют погружение под воду. Правда, этот аргумент имеет, возможно, второстепенное значение, так как после погружения на некоторую глубину давление воды вызовет сжатие легких и животное беспрепятственно сможет оставаться на глубине.
Посмотрим, какие источники кислорода использует при нырянии тюлень и на какое время хватает этого кислорода. Данные по этому вопросу получены норвежским физиологом Шоландером (Р. F. Scholander, 1940), сравнившим запасы кислорода у небольшого тюленя и у человека (табл. 6.5). Здесь больше всего поражает малое количество кислорода в легких тюленя. Вместо того чтобы вдохнуть воздух перед нырком, тюлень, по-видимому, выдыхает его и уходит под воду с минимальным запасом воздуха в легких.
17—1873
258 Глава 6. Энергетический обмен
Этим и обусловлена разница в содержании кислорода в легких у тюленя и человека, так как последний перед нырянием вдыхает воздух.
Совсем иначе обстоит дело с содержанием кислорода в крови, Хотя в приводимом примере тюлень весил вдвое меньше человека, общий объем крови у них был примерно одинаков. Кроме того, кровь тюленя обладала большей кислородной емкостью, так что общее количество кислорода в его крови было больше, чем у человека.
Таблица 6.5
Запасы кислорода у тюленя и человека. Приведены лишь приближенные величины (например, в крови на самом деле меньше кислорода, так как значительная часть крови, находящаяся
в венозной системе, не полностью насыщена О2). (Данные о тюлене из Scholander, 1940)
Локализация кислорода Количество кислорода мл
Тюлень (30 кг) Воздух в легких (350 мл, 16% О2) Кровь (4,5 л; 25 мл О2 на 100 мл) Мышцы (6 кг; 4,5 мл О2 на 100 г) Вода в тканях (20 л; 5 мл О2 на 1 л) Всего Мл О2 на 1 кг веса тела Человек (70 кг) Воздух в легких (4,5 л; 16% О2) Кровь (5 л; 20 мл О2 на 100 мл) Мышцы (16 кг; 1,5 мл О2 на 100 г) Вода в тканях (40 л; 5 мл О2 на 1 л) Всего Мл О2 на 1 кг веса тела 55 1125 270 100 1550 52 720 1000 240 200 2160 31
Вообще, высокая кислородная емкость крови характерна для хороших ныряльщиков. У человека этот показатель (20 мл О2 на 100 мл крови) выше, чем у многих других наземных млекопитающих, однако у тюленей и китов он варьирует в пределах от 30 до 40 мл на 100 мл крови. Но увеличение кислородной емкости крови имеет физиологические границы. Эритроциты и так практически до предела наполнены гемоглобином, а при увеличении содержания эритроцитов до уровня, превышающего примерно 60% от общего объема крови, чересчур возросла бы вязкость крови; в результате для проталкивания крови через сосуды потребовалась бы чрезмерно напряженная работа сердца.
Увеличить общее количество гемоглобина в организме, не повышая вязкости крови, можно лишь путем увеличения объема цир
Ныряние у млекопитающих и птиц 259
кулирующей крови. Такое решение проблемы мы находим у многих хорошо ныряющих животных, однако увеличение объема крови тоже имеет пределы.
Большое количество кислорода у тюленя запасается также в мышцах. Красный цвет мышц у млекопитающих обусловлен присутствием мышечного гемоглобина (миоглобина). Последний находится внутри клеток и потому сохраняется в мышечной ткани, даже если из нее вытекла вся кровь. Содержание гемоглобина в мышцах гораздо ниже, чем в крови, но поскольку масса мышечной ткани очень велика — около 20% веса тела1, миоглобин связывает значительное количество кислорода. У тюленей и китов мышцы содержат гораздо больше миоглобина (и мясо этих животных поэтому заметно темнее, чем у наземных млекопитающих), так что у 30-килограммового тюленя накапливается примерно столько же кислорода в мышцах, сколько у человека весом 70 кг»
Во всей воде организма растворено некоторое количество кислорода; оно зависит от общего содержания воды в теле (которое у человека и тюленя примерно одинаково), но в целом невелико. Жир тоже способен растворять кислород, но роль его как кислородного депо незначительна, так как кровеносная сеть в жировой ткани слабо развита и при нырянии циркуляция крови здесь, вероятно, вообще прекращается.
Ориентировочные подсчеты (в табл. 6.5) показывают, что запас кислорода на 1 кг веса тела у тюленя намного (хотя и не вдвое) больше, чем у человека.
При интенсивности метаболизма, обычной для состояний покоя, этого запаса тюленю должно хватить не больше чем на 5 мин. Однако тюлень может находиться под водой по крайней мере втрое дольше. У человека общий запас О2 был бы достаточен для поддержания метаболизма на уровне покоя, вероятно, в течение примерно 4 мин, но это больше того времени, которое человек способен провести под водой (обычно около 2 мин, если не брать в расчет редких рекордов). Дело в том, что человек не приспособлен к полному расходованию всего свободного кислорода, и поэтому максимальное время пребывания под водой у него меньше, чем можно было ожидать, исходя из запасов О2 в организме. Но почему тюлень может находиться под водой намного дольше?
Хотя a priori нельзя исключить, что у тюленя есть какое-то неизвестное депо кислорода, в свете имеющихся данных это кажется крайне маловероятным. То, что мы знаем о водных млекопитающих, позволяет полностью объяснить возможности даже лучших ныряльщиков, не постулируя наличия дополнительных, пока еще неизвестных механизмов.
1 Приведенная цифра невелика из-за большого количества жира в организме тюленя. В расчете на массу тела за вычетом жира доля мышечной ткани значительно больше.
17*
260 Глава 6. Энергетический обмен
ИЗМЕНЕНИЯ В СИСТЕМЕ КРОВООБРАЩЕНИЯ
Одна из самых характерных физиологических реакций, наблюдаемых у тюленя при нырянии, — это внезапное урежение сердечных сокращений в 10 и более раз по сравнению с нормой (рис. 6.5). Оно имеет рефлекторную природу, так как если бы оно было связано с постепенным расходованием кислорода, то развивалось бы
Время, мин
Рис. 6.5. Когда тюлень ныряет, частота сокращений сердца у него быстро падает от примерно 140 до менее чем 10 ударов в минуту. Начало и конец пребывания под водой показаны вертикальными пунктирными линиями. (Elsner, 1965.)
Несмотря на замедление ритма сердца, артериальное давление остается на прежнем уровне. Это возможно лишь в том случае, если возрастает сопротивление периферических сосудов вследствие их сжатия. При уменьшенном минутном объеме сердца кровь поступает только в жизненно важные органы, к которым относится центральная нервная система и само сердце. Происходит полное перераспределение кровотока в организме, причем мышцы и органы брюшной полости почти или совсем не получают крови. Тюлень как бы становится меньше: кислород сохраняется только для небольшой части тела, поступая в наиболее нуждающиеся в нем органы. В результате имеющийся в крови к началу погружения под воду кислород расходуется медленно, и время пребывания под. водой примерно соответствует времени полного использования этого кислорода.
Более точные данные о распределении кровотока при нырянии были получены путем введения радиоактивного изотопа рубидия. Рубидий быстро обменивается с калием внутри клеток, и это позволяет оценить количество крови, поступающее в тот или иной орган. На рис. 6.6 представлены результаты одного опыта на утке. Можно видеть, что во время ныряния приток крови увеличивается
Ныряние у млекопитающих и птиц 261
главным образом к голове, сердцу и глазам, и в наибольшей степени уменьшается кровоснабжение почек, которые в обычных условиях обильно снабжаются кровью (Johansen, 1964).
В мышцы — и активные, и бездействующие — кровь при нырянии почти не поступает. Работу мышц всецело обеспечивают анаэробные процессы, ведущие к образованию молочной кислоты. Из-за отсутствия кровотока в мышцах эта молочная кислота накапливается в самой мышечной ткани, а содержание ее в крови
тракт
Рис. 6.6. Кровоснабжение различных органов у ныряющей утки. (Johansen, Темные столбики — кровоснабжение при нормальном дыхании воздухом, светлые — во время пребывания под водой. Уровень кровоснабжения определяли с помощью радиоактивного рубидия; высота столбиков соответствует накоплению рубидия в ткани, выраженному в процентах от введенного количества изотопа в расчете на 1 г ткани.
. 262 Глава 6. Энергетический обмен_____________________________
остается на низком уровне. Сразу же после возвращения на поверхность кровообращение в мышцах возобновляется и содержание молочной кислоты в крови резко возрастает. (Способность мышц тюленя к работе в анаэробных условиях не представляет собой ничего уникального; молочная кислота образуется и в мышцах человека при напряженной физической работе или спортивных состязаниях.)
Миоглобин доставляет ныряющему тюленю некоторое количество кислорода, но запас этого кислорода очень быстро исчерпывается. Количество Ог, запасаемое миоглобином, слишком мало, чтобы объяснить продолжительность пребывания тюленя под водой; главную роль играют изменения в кровообращении и анаэробный обмен в мышцах.
Урежение ритма сердца при нырянии отмечено у большого числа животных (например, у тюленей, китов, бобров, ондатр, бегемотов, уток). У некоторых из них оно происходит более постепенно, чем это показано на рис. 6.5, но в целом это явление, видимо, характерно для всех ныряющих животных.
Тем не менее мы не можем считать брадикардию обязательной физиологической реакцией на погружение в воду, так как многие наблюдения, сделанные при насильственном погружении подопытных животных в воду, требуют пересмотра в свете новых данных.
Эксперименты на аллигаторах показали, что погружение под воду ведет к замедлению сердечного ритма с 40 и более до 3 ударов в минуту. Однако если сигнал от животного регистрируется с помощью радиотелеметрии, удается получить информацию о- свободно передвигающихся животных, ныряющих в отсутствие человека. В телеметрических исследованиях, проведенных на кайманах (Caiman crocodilus), было показано, что при работе экспериментатора с животными у них при погружении под воду развивалась типичная для ныряния брадикардия: частота сокращений сердца падала с 28 до 5 в минуту. Однако в отсутствие человека ритм сердца у животных, находящихся в покое, постепенно замедлялся на протяжении нескольких часов до 10—12 ударов в минуту. Когда далее эти кайманы погружались в воду по собственной воле, ритм сердца урежался у них всего на 1—2 удара в минуту или не изменялся вовсе. Но если в комнату входил экспериментатор, кайманы уходили под воду, а частота сердечных сокращений падала у них до 5 ударов в минуту (Gaunt, Gans, 1969). Эти результаты были подтверждены на аллигаторах (Alligator mississippiensis), свободно плававших в озере: кратковременное непринудительное погружение под воду совсем не вызывало у них урежения сердечного ритма (Smith et al., 1974). Поэтому важно проводить наблюдения в условиях, обеспечивающих минимум внешнего вмешательства.
Интересно, что у рыб отмечено нечто аналогичное подводной брадикардии, но в противоположной ситуации. Если рыбу вынуть
Ныряние I/ млекопитающих и птиц 263
из воды, ей не хватает кислорода и ритм сердца у нее тотчас же замедляется. Это наблюдалось как у костистых рыб, так и у пластиножаберных. В отличие от рыб, ведущих целиком водный образ жизни, у илистого прыгуна (Periophthalmus), который дышит воздухом (см. гл. 2), ритм сердца урежается, когда он прыгает в свою наполненную водой нору. В этом смысле илистый прыгун ведет себя как наземное животное, нырнувшее в воду (Gordon et al., 1969).
На рис. 6.7 приводятся записи дыхания и содержания газов в крови у тюленя во время ныряния. Верхний график (А) показывает, что количество кислорода в крови постепенно снижается,
Рис. 6.7. Дыхательный газообмен у тюленя весом 29 кг во время 12-минутного погружения в воду в условиях эксперимента. (Scholander, 1940.)
А — концентрация О2, СО2 и молочной кислоты в крови; Б — вентиляция легких;
В — потребление О2 и выделение СО2 при дыхании.
264 Глава 6. Энергетический обмен______________________________
а содержание СОг соответственно возрастает. Уровень молочной кислоты в период пребывания под водой изменяется мало, но после выныривания происходит его резкий подъем, как только возобновляется дыхание. Эта молочная кислота, которая свидетельствует о кислородной задолженности, постепенно выводится из крови. Большая часть ее используется для ресинтеза гликогена в печени и мышцах, а меньшая часть полностью окисляется до СОг и воды.
На среднем графике (рис. 6.7, Б) показаны изменения дыхательного объема легких: во время ныряния он равен нулю, а после выныривания сразу же резко возрастает.
На нижнем графике (рис. 6.7, В) представлено поглощение кислорода в легких. Этот показатель тоже резко возрастает тотчас после выныривания. Выведение двуокиси углерода почти все время соответствует потреблению Ог, но может быть чуть-чуть ниже. Однако сразу же после выныривания выведение СОг сильно возрастает, так как поступающая в кровь молочная кислота вытесняет двуокись углерода из бикарбонатов крови, подобно тому как уксус вытесняет ее из питьевой соды.
СНИЖЕНИЕ ИНТЕНСИВНОСТИ ОБМЕНА
Как видно из рис. 6.7,Б, при нырянии создается кислородная задолженность, а после этого наступает фаза повышенного потребления кислорода. Рассчитанная величина кислородного долга представлена в виде заштрихованной площади (соответствующей обычному уровню обмена у тюленя). Повышенное потребление Ог говорит о том, что возникшая задолженность погашается; при этом степень повышения должна соответствовать величине долга. Однако если сравнить добавочную площадь под повышенным участком кривой с заштрихованной областью, то компенсация окажется неполной. Это может быть связано с тем, что процессы, погашающие кислородную задолженность, сильно растянуты во времени и их трудно выявить на фоне нормальных небольших колебаний интенсивности метаболизма в покое. Возможно также, что уровень обмена при нырянии действительно снижается, так что истинная кислородная задолженность меньше, чем это представлено на рисунке.
Дальнейшие наблюдения показали, что последнее объяснение весьма правдоподобно. Так, у утки повышенное потребление кислорода после ныряния составляет около ’/4 или !/з от ожидаемой величины кислородной задолженности; это можно наблюдать на протяжении ряда последовательных ныряний, и при этом никаких признаков нарастания кислородной задолженности не отмечается.
Снижение обмена ниже обычного уровня покоя кажется неожиданным, так как привыкли считать этот уровень весьма постоянным. Однако это не всегда так. Вспомним, что при нырянии кро
Ныряние у млекопитающих и птиц 265
воснабжение многих органов резко уменьшается или прекращается вовсе. Почки, которые в норме обильно снабжаются кровью и потребляют много кислорода, во время ныряния практически лишены притока крови, и при этом в них сразу прекращается клубочковая фильтрация и образование мочи (Murdaugh et al., 1961). Другие органы при нырянии тоже фактически отключаются от
Рис. 6.8. У ныряющих черепах истинную интенсивность обмена можно определить по величине теплопродукции. Во время ныряния у черепахи, перед тем дышавшей атмосферным воздухом, уровень обмена постепенно снижается примерно до 0,06 ккал на 1 кг веса тела в час — до того же уровня, который устанавливается у черепахи, вдыхавшей перед нырянием чистый азот. (Jackson, Schmidt-Nielsen, 1966.)
кровоснабжения, и можно думать, что они почти перестают функционировать; поэтому и уменьшается кислородная задолженность, которую нужно компенсировать после выныривания.
Измерить истинную интенсивность метаболизма у птиц или млекопитающих, находящихся под водой, не так легко. Однако для таких исследований удобны водные черепахи, которые легко переносят длительное пребывание под водой и недостаток кислорода. Очевидно, что в анаэробных условиях потребление кислорода не может служить мерой интенсивности обмена, но последнюю точно отражает величина теплопродукции. Как показано на рис. 6.8 пунктирной линией, у черепахи уровень обмена, определяемый по теплопродукции, во время ныряния постепенно снижается.
После двух часов пребывания под водой выделение тепла достигает минимума, близкого к тому, который устанавливается при погружении после дыхания чистым азотом. Эта величина теплопродукции целиком определяется анаэробными процессами, продолжающимися при полном отсутствии кислорода.
Если же черепаха перед погружением под воду дышала чистым кислородом, то при нырянии теплопродукция вначале очень велика, но потом постепенно снижается по мере исчерпания запасов О2. Таким образом, у черепахи уровень обмена явно зависит
266 Глава 6. Энергетический обмен______________________________
от количества имеющегося кислорода. Эти результаты полностью согласуются с тем фактом, что потребление кислорода покоящейся мышцей не является постоянной величиной, а зависит от кровотока (т. е. от снабжения кислородом) (Whalen et al., 1973).
КОЖНОЕ И РЕКТАЛЬНОЕ ДЫХАНИЕ
Мы уже видели, насколько важную роль играет кожное дыхание у лягушек и как безлегочные саламандры поглощают весь необходимый им кислород через кожу (гл. 2). Что касается водных птиц и млекопитающих, то нет никаких оснований предполагать, что они могут получать сколько-нибудь существенное количество кислорода этим путем. Прежде всего, это достаточно крупные животные, и относительная величина их поверхности соответственно мала. Кроме того, это теплокровные организмы, и потребность в кислороде у них очень велика. Наконец, их кожа по своей структуре резко отличается от тонкой, влажной и обильно васкуляризованной кожи земноводных.
В случае ныряющих рептилий дело может обстоять иначе. Водные черепахи обладают толстым и, вероятно, непроницаемым данцирем, но при длительном пребывании под водой газообмен у них мог бы осуществляться через слизистую рта. Некоторые •черепахи набирают воду в прямую кишку, но какую роль это играет в газообмене, неясно; скорее всего они используют воду для регулирования своей плавучести, подобно тому как на подводных лодках набирают воду в балластные цистерны.
Хорошо известно, что морские черепахи — великолепные пловцы и ныряльщики. Тем не менее совершенно неожиданным оказалось сообщение, что зеленые черепахи (Chelonia rnydas) могут перезимовывать, впадая в спячку, под водой на дне Калифорнийского залива (Felger et al., 1976). При этом они находятся на глубине от 10 до 15 и более метров, где остаются неподвижными несколько месяцев. Поскольку рыбаки узнали о существовании этих черепах и о том, как легко их обнаруживать с помощью современных приспособлений, неизбежным результатом будет теперь их чрезмерный вылов, который быстро приведет к резкому уменьшению популяции этих удивительных животных. Существуют ли зеленые черепахи, находясь под водой, за счет анаэробных процессов или же как-то получают то небольшое количество кислорода, которое необходимо им в период спячки, — это еще предстоит выяснить.
Морские змеи Pelamis platurus, которые, охотясь на мелких рыб, ныряют на глубину около 20 м, обычно держатся на поверхности, но голову опускают в воду, где высматривают добычу. У этих животных до !/з необходимого кислорода поступает через кожу и этим же путем выводится свыше 90% СО2. Такой способ дыхания не только дает возможность змее дольше оставаться не
Ныряние у млекопитающих и птиц
267
подвижной с опущенной в воду головой, но и оказывается полез-ним, когда змея одолевает и заглатывает крупную добычу: это может продолжаться до 20 мин и не позволяет дышать воздухом (Graham, 1974).
ДЫХАНИЕ ЖИДКОСТЬЮ
Что будет, если наземное животное попробует дышать водой? Мы знаем, что человек на это не способен, он тонет. Известно также, что в 1 л воды содержится во много раз меньше кислорода, чем в 1 л воздуха. В связи с этим естественно возникает вопрос: а что если бы мы увеличили содержание кислорода в воде, например путем насыщения им воды под высоким давлением?
Такой опыт был сделан. При насыщении воды чистым кислородом под давлением 8 атм в 1 л воды растворяется около 200 мл О2 (при 37°C); иными словами, в 1 л воды будет примерно столько же кислорода, сколько в 1 л обычного воздуха. Необходимо учесть еще одно обстоятельство: для дыхания нужно использовать не чистую воду, а сбалансированный солевой раствор, сходный по составу с плазмой крови, иначе в легких вода будет быстро переходить в кровь из-за разности осмотического давления, что приведет к гемолизу эритроцитов и значительному снижению концентрации солей. Именно эти последствия поступления воды в легкие обычно служат причиной смерти при утоплении в пресной воде. Поэтому искусственное дыхание часто не дает результата при попытках спасти утонувшего в пресной воде и более эффективно при утоплении в морской воде.
Опыты с полным погружением в перенасыщенную кислородом воду с успехом проводились на мышах и собаках, причем животные выживали в такой воде в течение нескольких часов. Когда мышей помещают в барокамеру и затем погружают в воду, перенасыщенную О2, они сначала стремятся всплыть; но если этому воспрепятствовать, они начинают вдыхать жидкость, но не тонут, а продолжают дышать оксигенированным физиологическим раствором.
Основной недостаток такого способа дыхания — то, что при этом резко возрастает затрата энергии, так как вязкость воды примерно в 50 раз больше вязкости воздуха. Кроме того, в.воде происходит смывание слоя сурфактантов, покрывающих в.-норме внутреннюю поверхность легких; это ничем не мешает во время самого опыта, но при возобновлении воздушного дыхания легкие обнаруживают тенденцию спадаться.
Более серьезная проблема связана с необходимостью выведения СО2 с той же скоростью, с какой происходит поглощение кислорода. Для поддержания Рсо2 в крови на уровне нормы, т. е. 40 мм рт. ст., количество жидкости, входящей в легкие и выходящей из них, должно быть увеличено в несколько раз. Это четко
268 Глава 6. Энергетический обмен_____________________________
выявилось в опытах на животных, дышащих жидкостью: при полном насыщении артериальной крови кислородом содержание СОг в крови сильно возрастало. При использовании водных растворов эту проблему очень трудно разрешить (Kylstra, 1968).
Другой подход состоит в замене воды определенными синтетическими жидкими веществами, в частности фторуглеродными соединениями. Кислород обладает очень высокой растворимостью в этих органических жидкостях, что позволяет обойтись без процедуры насыщения кислородом под высоким давлением. Однако растворимость СОг не так велика, и это затрудняет практическое применение фторуглеродов. Все же они были испытаны в опытах на животных, и было показано, что животные могут дышать ими без особых осложнений в течение нескольких часов. Однако кажется маловероятным, что эти жидкости найдут практическое применение.
Дыхание жидкостью в принципе обладает одним преимуществом, которое может оказаться ценным в определенных, сугубо специфических условиях. Поскольку вдыхается жидкость, а не газ, нет необходимости в разбавлении кислорода инертными газами, такими, как азот или гелий. Теоретически это позволило бы дышать жидкостью при очень высоких давлениях, так как из-за отсутствия инертных газов опасность перенасыщения ими организма и последующего развития кессонной болезни в принципе исключена: И все же возможность практического использования этого преимущества представляется весьма ограниченной.
ВЛИЯНИЕ ВЫСОКОГО ДАВЛЕНИЯ
Используя для дыхания смесь гелия с кислородом, водолазы-профессионалы работают в настоящее время на глубинах примерно до 200 м (20 атм). Глубина погружения в экспериментальных условиях достигает около 600 м и может быть увеличена еще больше путем применения специальных газовых смесей, включающих водород. Как уже говорилось, газы, инертные при умеренном давлении, могут стать причиной кессонной болезни, для предотвращения которой необходимы продолжительные периоды декомпрессии; при высоком давлении эти газы обладают также наркотическим действием. Но, кроме того, высокое давление могло бы само по себе оказывать отрицательное действие, независимое от эффекта газов. При изучении животных, дышащих воздухом, не так легко отличить прямое действие высокого давления от действия газов, поскольку эти два фактора невозможно изменять независимо друг от друга. Таким образом, вопрос этот достаточно сложный.
Как влияет высокое давление на ныряющих тюленей и китов, способных, как мы уже видели, опускаться на глубину более 600 м (тюлень Уэддела) и 1100 м (кашалот)? Судя по имеющим
Ныряние у млекопитающих и птиц 269
ся данным, эти животные не страдают от кессонной болезни, так как у них не происходит большого перенасыщения тканей азотом. Однако химики и биохимики прекрасно знают, что давление сильно влияет на протекание химических реакций и структуру сложных молекул, и раньше думали даже, что огромное давление в глубинах океанов исключает там возможность всякой животной жизни. Однако при изучении океанского дна в конце концов выяснилось, что животный мир существует даже на самых больших глубинах, превышающих 10 000 м (1000 атм).
Если животных, обитающих на мелководье, поместить в условия высокого давления, не изменяя других факторов (температуры и количества кислорода), то в зависимости от вида эти животные будут реагировать по-разному. У многих видов давление от 50 до 100 атм вызывает увеличение потребления кислорода и нередко повышение двигательной активности. Однако более высокие давления, порядка нескольких сотен атмосфер, обычно действуют угнетающим образом и могут вызывать гибель (MacDonald, 1975). Вполне возможно, что на тюленей и китов гидростатическое давление само по себе не влияет; но, вероятно, самые глубокие из зарегистрированных погружений под воду — это особые, вероятно редкие, случаи. Когда водолазы опускаются на максимальную глубину (около 60 атм), прямое действие высокого давления на организм, возможно, становится весьма существенным.
Действие высокого давления на биологические системы можно в значительной части объяснить влиянием его на структуру белков и ионизацию слабых кислот. Увеличение давления способствует диссоциации слабых кислот и оснований; кроме того, давление влияет на константы скоростей химических реакций.
Вероятно, наиболее важным с биологической точки зрения воздействием на белки является изменение их третичной структуры, т. е. «укладки» полипептидных цепей, стабилизируемой слабыми связями и другими слабыми взаимодействиями. Давление оказывает значительное влияние на третичную структуру, от которой в свою очередь зависят многие биологически важные свойства белковых молекул — ферментов, сократимых белков мышц, структурных белков мембран и т. д. (Somero, Hochachka, 1976).
В интактном организме эти эффекты могут проявиться в нарушении нервной проводимости и функции нервной системы, мышечного сокращения, переноса кислорода в крови, функциях мембран и механизмов транспорта ионов. Чрезвычайно чувствительны к изменению давления сложные биохимические процессы экспрессии генов, связанные с синтезом информационной РНК, а значит, и процессы эмбрионального развития.
Животные, обитающие на больших глубинах, физиологически приспособлены к высокому давлению, при котором они обычно живут. В настоящее время функциональная адаптация их струк
270 Глава 6, Энергетический обмен
турных белков, ферментов и других систем интенсивно изучается. Первые результаты уже позволяют судить о том, какие типы биохимической адаптации необходимы для жизни в условиях высоких давлений (Hochachka, 1975).
ИНТЕНСИВНОСТЬ МЕТАБОЛИЗМА И РАЗМЕРЫ ТЕЛА
МЛЕКОПИТАЮЩИЕ
Среди водных млекопитающих, о которых говорилось выше, есть и огромные киты, способные оставаться под водой около двух часов, и такие маленькие ныряльщики, как кутора, которая редко остается под водой дольше чем полминуты. В целом, чем крупнее водное животное, тем больше времени оно может провести под водой. Чем это объяснить? Дело в том, что потребление кислорода на 1 г веса тела у мелких млекопитающих намного выше, чем у крупных.
Таблица 6.6
Потребление кислорода млекопитающими разной величины
Животное Вес (масса) тела (М^), кг Общее 1 потребление О2 <УО2>- */« Потребление'Од на 1 кг веса (VV’ Л/(КГ’Ч)
Землеройка1) 0,0048 0,0355 7,40
Мышь-малютка2) 0,0090 0,0225 2,50
Малый прыгун3) 0,0152 0,0273 1,80
t Мышь4) 0,025 0,041 1,65
' Суслик5) 0,096 0,09 1,03
Крыса4) 0,290 0,25 0,87
Кошка4) 2,5 1,70 0,68
Собака4) 11,7 3,87 0,33
Овца4) 42,7 9,59 0,22
Человек4) 70 14,76 0,21
Лошадь4) 650 71,10 0,11
Слон4) 3833 268,00 0,07
*) Hawkins et al., 1960.
2) Pearson, 1960.
3) Bartholomew, MacMillen, 1961.
*) Brody, 1945.
e) Hudson, 1962.
В табл. 6.6 приводятся данные о потреблении кислорода различными млекопитающими. Самое крупное из них — слон — в миллион раз больше, чем самое мелкое — землеройка (фото 6.1), и понятно, что слон потребляет намного больше кислорода. Однако сравнение величин общего потребления кислорода непоказательно.
Интенсивность метаболизма и размеры тела 271
Зато если мы сделаем пересчет на единицу массы тела, т. е. найдем удельное потребление кислорода1 (последний столбец таблицы), то получим поразительную картину зависимости между размерами тела и потреблением О2.
Фото 6.1. Эта моментальная фотография, изображающая землеройку и брошенного ей в качестве пищи таракана, хорошо иллюстрирует относительные размеры одного из самых мелких млекопитающих и сравнительно крупного насекомого.
Мы увидим, что потребление кислорода в расчете на 1 г неуклонно снижается по мере увеличения размеров тела. Это еще яснее видно при графическом представлении данных (рис. 6.9). Здесь для оси абсцисс выбрана логарифмическая шкала, иначе все величины для средних и мелких животных сосредоточились бы в левой части графика. Для оси ординат, напротив, использована Линейная шкала, и мы сразу можем видеть, что у землеройки один грамм ткани потребляет примерно в 100 раз больше кислорода, чем у слона. При такой огромной разнице в потреблении Ог необходимо, чтобы доставка его тканям, а значит, и кровоток у землеройки были в 100 раз больше (в расчете на 1 г ткани), чем у слона. Соответственно должны различаться и другие физиологические показатели — функция сердца, дыхание, потребление пищи и т. д.
Если данные табл. 6.6 представить на графике с логарифмической шкалой по обеим осям, все точки будут лежать примерно на одной прямой (рис. 6.10). Эта линия регрессии отражает правило, согласно которому у млекопитающих потребление кислорода на единицу веса тела равномерно уменьшается с увеличением размеров тела; наклон прямой служит количественным выражением степени этого уменьшения.
По этому графику можно найти «ожидаемое» потребление кислорода для млекопитающего любой величины. Например, у типич-
1 Слово «удельный», стоящее перед названием физической величины, означает «деленный на массу». Таким образом, удельное потребление кислорода означает потребление кислорода на единицу массы (веса).
272 Глава 6, Энергетический обмен
кого млекопитающего весом 1 кг потребление О2 должно составлять около 0,7 л/ч. Если же окажется, что американская норка весом 1 кг потребляет в два раза больше кислорода, чем предсказывает график, то можно будет сказать, что для норки данная величина вдвое больше, чем мы могли бы ожидать для типичного млекопитающего такой величины.
Рис. 6.9. Удельное потребление кислорода у различных млекопитающих. Потребление О2 на единицу веса тела быстро увеличивается по мере уменьшения размеров тела. Обратите внимание, что на оси абсцисс логарифмическая шкала, а на оси ординат — линейная.
Найденную зависимость можно представить не только графически, но и в виде уравнения
Vo2M = 0,676-Л1ь-»>2\
где Уоа/Л1ь — удельное потребление кислорода, выраженное в литрах О2 на 1 кг веса (массы) тела в 1 ч [л/(кг-ч)], а Мъ — вес (масса) тела в килограммах. Это общее уравнение основано на ре-
Рис. 6.10. Если зависимость между удельным потреблением кислорода различ-ными млекопитающими и весом их тела представить в логарифмических координатах. то получится прямая регрессии с наклоном —0,25 (данные из табл. 6.6).
Интенсивность метаболизма и размеры тела 27$
зультатах изучения гораздо большего числа животных, чем приведено в табл. 6.6, причем их веса варьировали от нескольких, граммов до нескольких тонн. Данные для отдельных видов могут больше или меньше отклоняться от найденной прямой, которая представляет собой просто вычисленную линию, в наибольшей степени соответствующую всем имеющимся данным. Некоторые авторы получали несколько иные величины констант (например, —0,27 для показателя степени, или наклона), но эти различия слишком малы, чтобы быть статистически достоверными (Kleiber, 1961).
Приведенное выше уравнение удобнее использовать, если записать его в логарифмической форме:
lg VO2/Mb- 1g 0,676-0,251g Mb.
Это уравнение имеет общую форму у=Ь + ах (уравнение прямой с наклоном а, причем в данном случае этот наклон отрицательный» а=—0,25) (см. также Приложение В).
Если мы вернемся к табл. 6.6 и рассмотрим общее потребление Оз разными животными (столбец 2), то, разумеется, найдем, что слон потребляет больше кислорода, чем мышь. Представив данные этого столбца в виде логарифмического графика, мы получим прямую линию с положительным наклоном 0,75. Уравнение для линии регрессии будет
VO2==0,676Alb0,7S.
Это уравнение можно получить из предыдущего, умножив обе его части на массу тела (Л4ь1,0). Опять-таки при арифметических расчетах (в случае, если нет электронного калькулятора) удобнее использовать это уравнение в логарифмической форме:
lgyO2==lg0,676 + 0,751gAlb.
Зависимость между потреблением кислорода и размерами тела изучалась на различных животных, как позвоночных, так и беспозвоночных. Как правило, холоднокровные позвоночные потребляют меньше кислорода, чем теплокровные, но опять-таки данные практически укладываются на прямой, имеющей наклон 0,75 (рис. 6.11)* 1. У многих беспозвоночных потребление кислорода со
1 Хотя интенсивность обмена у животных чаще всего определяют по потреблению кислорода, многие авторы пересчитывают свои данные на величину теплопродукции, исходя обычно из того, что потребление 1 л кислорода (при стандартных температуре и давлении) эквивалентно выделению 4,8 ккал. На рис. 6.11—6.13 интенсивность обмена выражена на шкале слева в килокалориях в час, а справа — в литрах кислорода в час.
В системе СИ единица энергии (в данном случае тепла) — джоуль (Дж);
1 кал=4,184 Дж. Теплопродукция 1 ккал/ч= 1,1622 Дж/с, или (поскольку
1 Дж/с=1 Вт) 1,1622 Вт. Интенсивность обмена, соответствующая потреблению 1 л О2 в 1 ч, составляет 5,5787 Вт, если принять, что 4,8 ккал эквивалентно 1 л Ог. Другие коэффициенты для пересчета приведены в Приложении А.
18—1873
274 Глава 6, Энергетический обмен
ответствует той же или сходной линии регрессии, но есть и исключения из этого общего правила. Например, у некоторых насекомых, у легочных улиток и у нескольких других групп беспозвоночных величины потребления Ог укладываются на линию регрессии, наклон которой близок к 1,0. Такой наклон означает, что потребление кислорода прямо пропорционально массе тела (т. е.
Рис. 6.11. Для самых разнообразных организмов зависимость потребления кислорода от массы тела (представленная в логарифмических координатах) соответствует линии регрессии с наклоном 0,75. Обратите внимание, что каждое деление на осях координат соответствует 1000-кратному изменению величин. (Hemmingsen, 1960.)
если одно животное вдвое больше другого, то оно потребляет в два раза больше О2, и т. д.).
Потребление кислорода микроорганизмами разной величины тоже практически соответствует линиям со сходным наклоном, и даже у некоторых деревьев имеется то же соотношение между потреблением О2 и размерами (Hemmingsen, 1960). Тот факт, что зависимость между потреблением кислорода и размерами тела одинакова у столь разнообразных организмов, позволяет рассматривать этот феномен как общебиологическую закономерность.
Интенсивность метаболизма и размеры тела 275
Многие физиологи, изучавшие эту проблему, пытались найти рациональное объяснение установленной зависимости. Почти 100 лет назад немецкий физиолог Макс Рубнер исследовал интенсивность метаболизма у собак разной величины. Он нашел, что у более мелких собак она в расчете на единицу веса тела выше, чем у крупных; это полностью соответствует всему сказанному ранее. Рубнер выдвинул разумную идею, согласно которой наблюдаемая зависимость может быть обусловлена тем, что у мелких животных отношение поверхности тела к объему больше, чем у крупных. Так как температура тела у собак различной величины одинакова, для поддержания ее выработка тепла в процессе метаболизма должна быть пропорциональна его потере. Поскольку у мелких собак относительная поверхность больше, чем у крупных,, они должны вырабатывать больше тепла на единицу веса. Рубнер-вычислил теплопродукцию на квадратный метр поверхности тела и нашел, что она составляет — как у крупных, так и у мелких собак — примерно 1000 ккал на 1 м2 поверхности в сутки. Таким образом, он, казалось бы, подтвердил свою гипотезу о том, что интенсивность обмена определяется размерами поверхности тела и необходимостью поддержания постоянной температуры; этот вывод вошел в науку как правило Рубнера.
К сожалению, однако, одинаковый наклон линий регрессии, приведенных на рис. 6.11, нельзя связать с необходимостью компенсации потерь тепла, поскольку для рыб, крабов или буковых деревьев не существует проблемы поддержания температуры тела, а между тем у них мы находим ту же зависимость между уровнем обмена и размерами, что и у млекопитающих. Кроме того, если бы интенсивность обмена была действительно пропорциональна поверхности тела, то наклон линий регрессии на рис. 6.11 составлял бы 0,67!. На самом же деле он близок к 0,75. Тот факт, что при изучении холоднокровных позвоночных, а также многих беспозвоночных (и даже растений, по крайней мере некоторых) получены линии регрессии с таким же наклоном, как и у теплокровных животных, показывает, что требования терморегуляции не могут лежать в основе сходства этих линий.
Закономерную зависимость между интенсивностью обмена и размерами тела и величину 0,75, характеризующую наклон линий регрессии, объяснить не так просто. Заметим, однако, что практически невозможно представить себе ряд млекопитающих разной величины, для которых эта линия регрессии имела бы наклон 1,0
1 У двух геометрически подобных тел разной величины площади поверхностей относятся между собой как квадраты соответствующих линейных размеров, а объемы — как кубы линейных размеров. Поэтому отношения площадей будут равны отношению объемов, взятых в степени 2/з, или 0,67. Это правило, которое относится к любым геометрически подобным телам, нетрудно понять, если посвятить несколько минут сравнению ребер, граней и объемов кубов различной величины.
18*
276 Глава 6. Энергетический обмен______________________________
{т. е. интенсивность метаболизма была бы прямо пропорциональной весу тела). Клейбер (Kleiber, 1961) рассчитал, что если бы у- быка удельная (на единицу веса) интенсивность обмена была такой же, как у мыши, то для отдачи тепла, уравновешивающей «го выработку, пришлось бы поднять температуру поверхности тела выше точки кипения. Наоборот, если бы удельная интенсивность обмена у мыши снизилась до уровня, характерного для быка, то для сохранения теплоты тела мыши понадобился бы меховой покров толщиной не менее 20 см.
’ Очень важно иметь в виду, что не только потеря тепла, но и -многие другие физиологические процессы зависят от площади поверхности. Невозможно было бы «сконструировать» жизнеспособный организм без тщательного учета роли его различных поверхностей. От величины и свойств поверхностей зависит множество физиологических процессов; например, поглощение кислорода & легких или жабрах определяется площадью поверхности этих •органов; диффузия кислорода из крови в ткани происходит сквозь -стенки капилляров, т. е. опять-таки зависит от их поверхности; всасывание пищи в кишечнике зависит от поверхности ее слизистой и т. д. В сущности, все клетки имеют поверхность, через которую должны поступать внутрь кислород и питательные вещества и выходить наружу продукты метаболизма. Внутриклеточная жидкость отличается по составу ионов от внеклеточной, и это различие опять-таки поддерживается за счет процессов, зависящих от свойств и величины поверхности клеток. Из всего этого видно, что нельзя рассматривать обмен веществ в целом вне связи с системой различных поверхностей. Гораздо труднее, однако, объяснить, почему эта связь проявляется настолько постоянным образом, что наклоны линий регрессии, как уже говорилось, очень часто равны 0,75 или очень близки к этой величине.
ПТИЦЫ
До сих пор мы рассматривали интенсивность обмена у млекопитающих. А как обстоит дело со второй основной группой теплокровных— птицами? У воробьиных (воробьев, вьюрков, ворон и др.) уровень метаболизма, как правило, выше, чем у других птиц, и поэтому мы соответственно разделим этот класс на две группы. Данные об интенсивности обмена известны в общей сложности для 58 видов, не относящихся к семейству воробьиных,— для различных птиц весом от 0,003 кг (колибри) до 100 кг (страусы). Полученная линия регрессии (рис. 6.12) описывается уравнением.
УОа=-_ 0,679 .Мь0’723.
Это уравнение очень сходно с тем, которое получено для млекопитающих: Го2 = 0,676-Afb0,75 (Vo2 выражается в литрах Ог в
Интенсивность метаболизма и размеры-тела 277
1 ч, а Мъ — в килограммах). Это означает, что если какое-то млекопитающее и какая-то птица, не относящаяся к воробьиным, имеют одинаковые размеры, то и уровни метаболизма у них скорее всего тоже будут примерно одинаковыми.
Что касается воробьиных, то здесь имеются данные для 36 видов птиц весом от 0,006 до 0,866 кг. Полученное для них уравнение имеет вид V’oa == 1,11 • ЛГь°>724. Таким образом, наклон линии
Рис. 6.12. Потребление кислорода (интенсивность обмена) у птиц в покое тем выше, чем больше размеры тела. Угол наклона прямой свидетельствует о том, что птица любой величины, не относящаяся к отряду воробьиных, потребляет примерно столько же кислорода, сколько млекопитающее такого же размера. Воробьиные в целом характеризуются более интенсивным метаболизмом. (La-siewski, Dawson, 1967.)
Потребление
регрессии как для воробьиных, так и для остальных птиц одинаков, но если сравнить сходных по величине представителей обеих групп, то окажется, что у воробьиных интенсивность обмена примерно на 65% выше, чем у птиц другой группы (65% соответствует различию между 1,11 и 0,679). Конечно, у отдельных видов возможны большие или меньшие отклонения от линии регрессии, так как последняя проведена таким образом, чтобы получилось наилучшее соответствие всем имеющимся данным (Lasiewski, Dawson, 1967).
278 Глава 6. Энергетический обмен
СУМЧАТЫЕ И ОДНОПРОХОДНЫЕ
У большинства сумчатых температура тела несколько ниже (около 35°C), чем у плацентарных млекопитающих (около 38°C). Об этом факте часто упоминают, нередко интерпретируя его как свидетельство того, что сумчатые по своей физиологии занимают промежуточное положение между «низкоорганизованными» однопроходными (у которых температура тела еще ниже — около 30 °C) и «высокоорганизованными» плацентарными млекопитаю-
Рис. 6.13. Интенсивность обмена в покое у сумчатых всегда несколько ниже, чем у плацентарных млекопитающих. Однако у обеих групп животных зависимость уровня обмена от размеров тела выражается линиями регрессии с одинаковым углом наклона. (Dawson, Hulbert, 1970.)
щими. Однако нет оснований a priori рассматривать более низкую температуру тела как признак более примитивной физиологической организации, так как сумчатые способны поддерживать постоянство своей температуры в широком диапазоне внешних условий. Если считать, что чем ниже температура, тем ниже уровень организации, птицы окажутся «выше» млекопитающих, так как температура тела у них выше (обычно 40—42°C).
Ввиду этих различий в температуре тела интересно будет сравнить уровни энергетического обмена у разных групп животных и выяснить, существует ли у сумчатых та зависимость между интенсивностью обмена и размерами тела, которая свойственна другим теплокровным позвоночным. Исследование австралийских сумчатых с весом тела от 0,009 до 54 кг показало, что эта зависимость у них сохраняется, но в целом на несколько более низком уровне (Dawson, Hulbert, 1970).
Уравнение линии регрессии для сумчатых имеет вид Voa= = 0,409-Л4ь0’75 (все величины в тех же единицах, что и в уравнении для плацентарных млекопитающих). Как видно из рис. 6.13, дан-
Размеры тела и соотношения различных параметров 279 ные укладываются на прямую с таким же наклоном, как у линии регрессии для высших млекопитающих (Йоа=0,676-Мь0’75). Между тем уровень обмена у сумчатых заметно ниже, и интересно, что это сочетается с пониженной на 3° температурой тела; однако мы не можем сказать, которое из этих двух отличий является причиной и которое — следствием.
Если вычислить, какой была бы интенсивность обмена у сумчатых, если бы температура тела у них поднялась до 38°C, то окажется, что этот «скорректированный» уровень был бы близок г уровню плацентарных млекопитающих. Такой расчет1 скорректированных интенсивностей обмена для трех групп млекопитающих (однопроходных, сумчатых и плацентарных) и для птиц, не относящихся к воробьиным, дает очень сходные величины. Воробьиные же птицы стоят особняком: интенсивность метаболизма у них в целом более чем на 50% выше по сравнению с остальными теплокровными, даже в пересчете на более низкую температуру тела, чем обычно им свойственная (Dawson, Hulbert, 1970).
РАЗМЕРЫ ТЕЛА И СООТНОШЕНИЯ
РАЗЛИЧНЫХ ПАРАМЕТРОВ
В предыдущем разделе мы рассмотрели зависимость между интенсивностью метаболизма и размерами тела и вывели общие уравнения, описывающие эту зависимость. Полученные уравнения показывают, как изменения величины, или «масштаба», животного отражаются на уровне энергетического обмена.
Вопрос о результатах изменения величины, или масштаба, очень важен для инженерной науки — настолько важен, что его изучение выделилось в особую дисциплину. Инженер повседневно сталкивается с проблемами изменения масштаба, строит ли он более высокие здания, более длинные мосты, более крупные суда или переходит от небольшой рабочей модели к объекту натуральной величины.
В физиологии также существует понятие масштабных соотношений (scaling), которое мы определили как структурные и функциональные последствия изменений величины, или масштаба, у сходно организованных животных. Анализируя имеющиеся данные, мы находим, что от размеров тела зависит множество функциональных показателей. Мы уже встречались с рядом ограничений, которые накладывает на интенсивность обмена величина тела млекопитающего; существует и много других ограничений, определяющих «конструкцию» животного организма тех или иных размеров. Всем известно, что у крупного животного, например слона, кости относительно массивнее, чем у мелкого, например мыши,
1 О том, как можно вычислить влияние изменения температуры на уровень обмена, см. в гл. 7.
280 Глава 6. Энергетический обмен___________________________
и это вполне понятно, так как кости должны поддерживать вес животного, а вес возрастает пропорционально кубу линейных размеров. В результате для поддержания возросшего веса диаметр костей у крупного животного должен быть не только абсолютно, но и относительно больше, чем у мелкого.
В определенном отношении к размерам тела находится и ряд других структурных и функциональных параметров. Например, сердце лошади во много раз больше сердца мыши, однако у обоих видов вес сердца составляет около 0,6% от веса всего тела (Мъ). В общей форме это можно представить так:
Вес (масса) сердца — 0,006 -Мь1’0.
Это уравнение, основанное на измерениях веса сердца у сотен различных животных, показывает, что вес сердца у млекопитающих пропорционален весу тела (показатель степени равен 1,0). Конечно, могут быть и некоторые отклонения, но в целом уравнение отражает общую тенденцию.
Объем легких у млекопитающих тоже прямо пропорционален размерам тела, что видно из рис. 2.6 (стр 53). Линия регрессии на графике имеет наклон 1,02; это означает, что у всех млекопитающих размеры легких и всего тела изменяются практически в одной пропорции. Общее правило состоит в том, что относительный объем легких у всех млекопитающих примерно одинаков и составляет 63 мл на 1 кг веса тела (опять-таки с некоторыми отклонениями). Если мы возьмем площадь внутренней поверхности легких, а не их объем, то мы найдем, что эта величина прямо пропорциональна количеству потребляемого животным кислорода, а не массе тела (рис. 6.14). Это вполне понятно, так как потребление кислорода, как мы видели, пропорционально весу тела в степени 0,75, поэтому и поверхность легких, т. е. площадь, через которую происходит диффузия Ог, должна определяться подобным же образом и зависеть от массы тела в степени 0,75.
В табл. 6.7 приведен ряд физиологических параметров, соотношение которых с размерами тела описывается аналогичными уравнениями. Как уже было сказано, объем легких прямо пропорционален массе тела. Дыхательный объем легких (т. е. объем вдыхаемого и выдыхаемого воздуха) в покое тоже прямо пропорционален массе тела (табл. 6.7, строка 5). Разделив уравнение для дыхательного объема на уравнение для общего объема легких, мы получим формулу для соотношения этих двух величин. Показатели степени, в которой в эти уравнения входит масса тела, практически одинаковы; поэтому полученным в результате деления показателем степени 0,01 можно пренебречь как величиной крайне малой по сравнению с ошибками измерений, на основании-которых выведены исходные уравнения. Таким образом, мы получим частное от деления 0,0062 на 0,063, т. е. почти 0,1. Эта безразмерная величина показывает, что дыхательный объем у мле-
Размеры тела и соотношения различных параметров
281
копитающих в покое будет, как правило, составлять 7ю объема легких независимо от размеров тела. Хотя возможны отклонения от этого общего правила, оно ценно тем, что позволяет делать определенные прогнозы. Например, при изучении какого-нибудь животного, например крысы или собаки, мы можем, исходя из общих
Потребление Oz , л/ч
Рис. 6.14. Общая площадь поверхности легких у млекопитающих пропорциональна потреблению кислорода, о чем свидетельствует наклон линии регрессии, связывающей эти два параметра, который равен 1,0. (Tenny, Remmers, 1963.)
сопоставив затем эти ожидаемые величины с данными прямых измерений, мы увидим, насколько данное животное соответствует общему правилу или отклоняется от него.
Изучая далее уравнения, приведенные в табл. 6.7, мы можем найти и другие интересные взаимосвязи, представляющие прогностическую ценность. Например, объем крови (л) у млекопитающих всегда составляет 5,5% от массы, или веса, тела (кг) независимо от его размеров (степень 0,99 можно считать несущественно отличающейся от 1,0). Как уже говорилось, вес сердца (т. е. величина насоса, обеспечивающего ток крови) также пропорционален весу тела. Но поскольку кровь снабжает ткани кислородом в соответствии с метаболическими нуждами, а интенсивность метаболизма зависит от веса тела в степени 0,75, скорость работы насоса (частота сердечных сокращений) также должна зависеть от размеров тела, и мы видим, что это действительно так. Частота сокращений сердца у мелких животных очень высока; она снижается с увеличением размеров животного, на что указывает отрицательный наклон (—0,25) линии регрессии (табл. 6.7, последняя стро-
282 Глава 6. Энергетический обмен
Таблица 6.7
Зависимость физиологических параметров от массы тела (ЛГь, в килограммах) у млекопитающих1).
(По данным Adolph, 1949; Drorbaugh, 1960; Stahl, 1967)
Потребление О2 (л/ч)
Потребление О2 на 1 кг [л/(ч-кг)]
Вентиляция легких (л/ч) Объем легких (л) Дыхательный объем (л) Объем крови (л) Вес сердца (кг)
Частота дыхания (мин-1)
Частота сокращений сердца (мин-1)
= 0,696 -Л1ь0’75
= 0,696 .‘Л/ь-°>25
= 20,0 .Л4Ь0’75
= 0,063 -Мь1’02
= 0,0062-Alb1’01
= 0,0055-Л1ь0’99
= 0,0058-Л1Ь°’99
= 53,5 -AV0’28
= 241 .Afb'0’25
’) Сопоставляя эти уравнения с аналогичными уравнениями, приводимыми в других книгах или статьях, нужно обращать внимание на используемые единицы измерения. В настоящей главе такими единицами везде служат литры О2, часы и килограммы. Пересчет литров в миллилитры и часов в минуты и секунды очень прост. Дело обстоит несколько сложнее, если масса (вес) тела выражается не в килограммах, а в граммах. Для примера рассмотрим уравнение
Vq2=3,8 Л4^о,75, где единицами
для интенсивности обмена
измерения служат миллилитры, часы и граммы. Если от граммов перейти к килограммам, то что произойдет с коэффициентом 3,8? Для перевода в килограммы мы должны разделить массу в граммах (А!^) на 1000, но, поскольку число 1000 будет далее возведено в степень 0,75, коэффициент 3,8 необходимо умножить на это же число (1000°,7о = 177,83).
В результате уравнение примет вид Vq =3,8-177,83• ./Иь°»75= =676Мьо,75 (мл, ч, кг), или 0,676 М^о,1$ (л, ч, кг). Эти последние единицы и используются во всех уравнениях, приводимых в настоящей главе.
ка). Таким образом, частота сердечных сокращений, а тем самым и минутный объем сердца определяются потребностью в таком объеме кровотока, который обеспечил бы доставку кислорода, соответствующую уровню обмена.
Уравнения, приведенные в табл. 6.7, относятся к параметрам, прямо пропорциональным весу тела (показатель степени 1,0) или зависящим от интенсивности обмена (показатель степени 0,75). Однако не все анатомические и физиологические параметры определяются таким образом. Например, размеры почек и печени выражаются следующими уравнениями:
Масса почек=0,021-Мь0’88;
Масса печени = 0,082 • Мь°>87.
Оба органа играют очень важную роль в обмене веществ, но их размеры не пропорциональны массе тела: они относительно тем меньше, чем крупнее животное. Но они не пропорциональны и интенсивности обмена, поскольку они зависят от массы тела в
Энергетическая цена локомоции
.283
степени, показатель которой больше 0,75. Таким образом, мы видим, что у животных размеры органов и физиологические параметры не всегда находятся в простой зависимости от массы тела. В целостном, хорошо функционирующем организме имеют место более сложные взаимоотношения.
ЭНЕРГЕТИЧЕСКАЯ ЦЕНА ЛОКОМОЦИИ
Очевидно, что при плавании, полете или беге расходуется больше энергии, чем в покое, но каковы энергетические соотношения между этими видами локомоции? Ходьба и бег нам ближе и понятнее, чем плавание или полет, и поэтому мы больше знаем о первых двух способах передвижения. К тому же человек — самый лучший экспериментальный объект, так как он может активно сотрудничать с исследователем, а иногда даже сильно заинтересован в успехе работы. Это имеет большое значение в опытах с максимальными нагрузками: когда животное отказывается бежать быстрее или дольше, как узнать — не хочет оно или не может? Много исследований проведено также на собаках, так как они обычно понятливы, готовы «сотрудничать» и с ними легко работать;. временами их поведение даже кажется сознательным. Иначе дело обстоит с изучением полета — ведь человек почти не имеет опыта летания за счет собственных мышечных усилий; по сравнению с рыбами и китами он оказывается также неуклюжим и неэффективным пловцом.
Сравнивая разные способы локомоции, следует учитывать, что главные различия между передвижением в воде, по земле или в воздухе обусловлены физическими особенностями этих сред. При этом основную1 роль играют два фактора: насколько среда поддерживает тело животного и каково ее сопротивление движению.
Большинство водных животных находится практически во взвешенном состоянии (нейтральная плавучесть): окружающая вода полностью поддерживает их вес, так что они не тратят усилий на преодоление силы тяжести. В совершенно ином положении находятся летающие и бегающие животные: они должны сами поддерживать весь свой вес. У бегающих животных есть прочная опора под ногами, тогда как летающие должны все время удерживать себя в воздухе — в среде с низкой плотностью и низкой вязкостью.
Сопротивление среды имеет важное значение для плавающих животных, так как вода характеризуется высокой плотностью и вязкостью. Бегающие и летающие формы пользуются тем преимуществом, что передвигаются в среде малой плотности и вязкости. Физические особенности среды оказывают глубокое влияние на структурные приспособления животных к тому или иному способу локомоции. Например, водные животные имеют обтекаемую форму тела и продвигаются вперед с помощью хвоста, плавников или их
284 Глава 6. Энергетический обмен.
производных. У птиц тело и крылья также обладают обтекаемой формой и действуют на основе тех же общих принципов гидро-и аэродинамики, что и хвост рыб. Однако чем меньше размеры животного, тем в меньшей степени обтекаемость формы снижает сопротивление воздуха, и потому мелкие насекомые уже не обладают этой особенностью в отчетливом виде.
Бегающие животные используют свои конечности как рычаги, перемещающие тело по твердому субстрату. Сопротивление воздуха не играет существенной роли при беге, и форма тела у таких животных не бывает в строгом смысле обтекаемой. Обтекаемая форма гораздо важнее для птиц, так как они обычно движутся намного быстрее наземных млекопитающих, а сопротивление воздуха возрастает примерно пропорционально квадрату скорости. Большинство животных — четвероногие, но некоторые крайне своеобразные виды, например человек и кенгуру, — двуногие.
БЕГ
Рассмотрим сначала потребление кислорода животным (например, крысой), когда оно бежит с той или иной скоростью (рис. 6.15). Потребление кислорода с увеличением скорости воз-
Скорость бега, км/ч
Рис. 6.15. Потребление кислорода у бегущих белых крыс возрастает линейно» с увеличением скорости бега. (Taylor et al., 1970.)
растает, причем эта зависимость линейна в широком диапазоне-скоростей. (При очень больших скоростях кривая может подниматься круто вверх; в этом случае «кислородная цена» бега увеличивается значительно больше, чем скорость, — это известно-каждому, кто участвовал в спринте.) Если продолжить прямую-
Энергетическая цена локомоции
285
линию назад до пересечения с осью ординат, соответствующего? нулевой скорости бега, то отсеченная на ординате величина окажется выше уровня потребления О2 в покое. Значит, если бы мы начертили кривую для всего диапазона скоростей от нуля до максимума, то прямой линии у нас бы не получилось. Однако в области средних скоростей эта зависимость линейна у множества разнообразных животных, включая человека. Человеку ходьба
Рис. 6.16. Затрата энергии у человека при ходьбе или беге по ровному месту* со спуском и с подъемом. (Margaria et al., 1963.)
с умеренной скоростью обходится «дешевле» в смысле кислородной цены движения, чем бег (рис. 6.16). По мере увеличения скорости кислородная цена ходьбы становится все выше, и в той точке», где кривая для ходьбы пересекается с кривой для бега, энергетически выгоднее перейти на бег. Рис. 6.16 отражает также тот хорошо известный факт, что движение в гору требует большей затраты энергии, чем по ровному месту или под гору.
Вернемся теперь к рис. 6.15 и рассмотрим линейную зависимость между скоростью движения и его кислородной ценой. То» что линия регрессии при экстраполяции не достигает уровня покоя, можно объяснить некоторой затратой энергии на поддержание определенной позы при беге. Действительно, интенсивность, обмена у спокойно стоящего человека близка к той величине, которую отсекает на оси ординат кривая, описывающая его бег. Это-проще всего понять таким образом, что разница между величиной* отсекаемой на оси ординат, и обменом в покое представляет собой
• 286 Глава 6. Энергетический обмен
цену поддержания позы, необходимой для движения, и эту разницу можно было бы назвать эффектом позы. Отсюда следует, что линейное увеличение потребления О2 выше отсекаемого на ординате уровня отражает кислородную цену возрастания скорости движения; это тот избыток кислорода, который расходуется на движение при данной скорости. Таким образом, метаболическая цена движения выражается наклоном кривой, а поскольку кривая прямолинейна, эта цена (наклон) остается постоянной и не меняется при изменении скорости бега.
Если мы посмотрим, как зависит потребление кислорода от скорости бега у различных млекопитающих (рис. 6.17), мы увидим, что чем меньше животное, тем круче наклон соответствующей .линии регрессии (т. е. тем быстрее растет потребление кислорода с увеличением скорости бега).
Существует простой способ сравнения цены бега у различных животных; нужно вычислить количества энергии, необходимые данным животным для переноса единицы массы тела на единицу расстояния [например, для перемещения 1 кг на расстояние 1 км;
Рис. 6.17. У самых разнообразных млекопитающих потребление кислорода во время бега возрастает линейно с увеличением скорости. Чем крупнее животное, тем меньше прирост потребления О2 в расчете на единицу веса тела. (Taylor et al., 1970.)
ответ выражается в л О2/(кг-км)]. Возвращаясь к рис. 6.15 и 6.17, мы видим, что наклоны линий регрессии соответствуют именно этой величине. Наклон кривой (определяемый как отношение приращения ординаты к приращению абсциссы) представляет собой отношение прироста потребления О2 [(л/(кг-ч)] к приросту скорости (км/ч). В результате сокращения общих множителей в чис
Энергетическая цена локомоции 287
лителе и знаменателе мы получим выражение величины наклона в л О2/(кг-км), т. е. именно в тех единицах, в которых, как указывалось в начале абзаца, выражается цена движения.
Теперь мы можем непосредственно сравнить цену бега на 1 км в расчете на 1 кг веса тела у ряда животных, представленных на рис. 6.17. Получаемые результаты иллюстрирует график на рис. 6.18. Сразу видно, что энергетическая цена бега законо-
Рис. 6.18. Энергетическая цена бега у млекопитающих в зависимости от размеров тела. (Schmidt-Nielsen, 1972а.)
«Чистая цена» означает цену перемещения 1 кг массы тела на расстояние 1 км; ее вычисляют из повышения интенсивности обмена при беге (исходя из наклона соответствующей линии регрессии). Общая цена отражает общую интенсивность, обмена при беге и поэтому несколько выше «чистой» цены.
мерно снижается с увеличением размеров тела. Таким образом, для бегающих млекопитающих энергетически относительно «дешевле», а потому и выгоднее, иметь большие размеры.
Вопрос об энергетической цене локомоции можно рассмотреть и в другом аспекте. До сих пор мы обсуждали вопрос о том, насколько больше кислорода потребляется при беге по сравнению с неподвижным состоянием. Однако, в сущности, при движении животного энергией должен быть обеспечен весь метаболизм. Например, пасущаяся овца должна съедать достаточно пищи, чтобы полностью покрыть все энергетические потребности, а не только те, которые добавились в связи с локомоцией как таковой. Если мы учтем общие метаболические затраты при движении, то мы получим более высокие величины, так как в них войдет и количество кислорода, потребляемое в неподвижном состоянии.
ПЛАВАНИЕ И ПОЛЕТ
Энергетическую цену плавания или полета можно выражать в тех же величинах, что и в случае бега, и при этом тоже оказывается, что цена движения зависит от размеров тела.
288 Глава 6. Энергетический обмен.
Если при беге потребление кислорода возрастает неуклонно и линейно с увеличением скорости (рис. 6.15), то у птиц при полете дело обстоит иначе. Опыты на волнистых попугайчиках (фото 6.2) показали, что существует оптимальная скорость полета, при которой птица потребляет наименьшее количество кислорода (рис. 6.19); при ускорении или замедлении полета потребление
Фото 6.2. Волнистый попугайчик (Melopsittacus undulatus) сфотографирован во время полета в аэродинамической трубе. Для того чтобы определить потребление кислорода при полете, выдрессированную птичку снабжают пластиковой маской с трубкой, через которую отводится весь выдыхаемый воздух. (Фото 'предоставлено V. A. Tucker, Duke University.)
кислорода будет возрастать. Поэтому кривая, отражающая зави--симость метаболической цены полета от его скорости, имеет U-образную форму.
Хотя у волнистого попугайчика потребление кислорода в единицу времени будет наименьшим при горизонтальном полете со скоростью 35 км/ч, однако при расчете затрат энергии на единицу расстояния наиболее экономичной будет несколько большая скорость полета, для волнистого попугайчика — 40 км/ч. Это связано с тем, что чем больше скорость, тем быстрее птица преодолевает данное расстояние. Наиболее экономичную скорость полета можно вычислить, но можно также найти ее и непосредственно из графика, проведя из начала координат касательную к кривой. Точка касания дает минимальное отношение потребления О2 к скорости полета, т. е., как ранее указывалось, цену движения [в л Ог/(кг-•км)]. (Цена движения, определяемая с помощью касательной к кривой на рис. 6.19, является общей ценой перемещения, а не ценой движения как такового, которую мы вычислили для млекопитающих. Однако для птиц это не имеет существенного значения,
Энергетическая цена локомоции 289
так как интенсивность их метаболизма во время полета примерно в 10 раз выше, чем в покое, и вычитание величины для покоя не скажется заметным образом на результате. У млекопитающих разность между общей и «чистой» ценой движения значительно больше.)
Рис. 6.19. Зависимость потребления Ог волнистым попугайчиком от скорости полета. (Schmidt-Nielsen, 1972b, по данным Tucker, 1968.)
Минимальное потребление кислорода наблюдается при скорости около 35 км/ч, но цена перелета на определенное расстояние будет минимальной при скорости около 40 км/ч (см. текст).
На рис. 6.20 сопоставлены данные о цене движения при плавании, полете и беге. Интересно, что при одном и том же весе тела полет оказывается гораздо более экономичным способом преодоления больших расстояний, чем бег. На первый взгляд это может показаться странным: интуитивно нам кажется, что при полете нужны будут значительные усилия уже для одного того, чтобы просто держаться в воздухе. Вспомним, однако, что перелетные птицы преодолевают без отдыха более 1000 км, тогда как трудно представить себе небольшое млекопитающее, например мышь, которое пробежало бы это расстояние без остановок для еды и питья.
Плывущая рыба тратит еще меньше энергии, чем летящая птица. Опять-таки это может показаться странным, если учесть большую вязкость воды и ее значительное сопротивление движущемуся телу. Однако следует принять во внимание два обстоятельства. Во-первых, обычно рыбы не плавают слишком быстро, а обтекаемая форма их тела хорошо приспособлена к передвижению в среде с относительно большой плотностью и вязкостью. Во-вторых, рыбы находятся практически во взвешенном состоянии: их тело целиком поддерживается водой и они не тратят усилий на преодоление силы тяжести.
19—1873
290 Глава 6. Энергетический обмен
Иначе обстоит дело, если животное менее приспособлено для плавания. Если вычислить цену передвижения плывущей утки, она окажется примерно равной цене бега млекопитающего тех же размеров. Для человека плавание еще менее экономично. По сравнению с рыбой он с трудом передвигается в воде; форма и особенности строения тела делают человека плохим пловцом, и цена его движения в воде в 5—10 раз больше цены передвижения по
°ис. 6.20. Сравнение энергии, необходимой для переноса единицы веса тела на расстояние 1 км при беге, полете и плавании. (Schmidt-Nielsen, 1972а.)
земле. Неприспособленность человека к плаванию ясно видна из того, что если представить цену его движения в воде на графике (рис. 6.20), то соответствующая линия окажется намного выше линии регрессии для бега.
ВЛИЯНИЕ БОЛЬШИХ ВЫСОТ
Основные сведения о реакции организма на условия больших высот, и в частности на понижение давления, были получены на человеке. Как уже говорилось в гл. 1, процентный состав атмосферы на больших высотах не меняется, но снижение барометрического давления оказывает резкое физиологическое воздействие.
Люди, не привыкшие к большим высотам, при подъеме в горы примерно на высоте 3000—4000 м начинают испытывать физическую слабость; но спустя несколько дней происходит акклиматизация1 и их самочувствие улучшается. Самое высокогорное из постоянных поселений человека — это поселок шахтеров в централь-
1 В настоящее время принято употреблять термин «акклиматизация», когда речь идет о приспособлении к определенному климату, например к сезонным изменениям от лета к зиме. Скажем, рыба, оставшаяся зимой в ледяной воде, акклиматизируется к этой воде. Слово же «акклимация» означает адаптацию организма к искусственно созданным условиям. Например, про золотую рыбку, помещенную на два месяца в воду с температурой 10 °C, говорят, что она акклимируется к 10 °C.
Влияние больших высот 291
них Андах в Южной Америке, расположенный на высоте почти 5000 м. Опыт покорителей Эвереста показал, что даже акклиматизированные люди лишь с большим трудом могут подняться без дополнительного обеспечения кислородом на высоту 8600 м (28 200 футов). По данным многих высокогорных экспедиций, процесс акклиматизации прогрессивно усиливается с увеличением высоты, однако при подъеме выше 6000 м над уровнем моря состояние организма начинает ухудшаться, причем быстрота и степень этого ухудшения тем значительнее, чем выше подъем. Это в конце концов заставляет альпинистов спускаться (Pugh, 1964).
Симптомы, возникающие при низком давлении кислорода, называют горной болезнью. Тяжесть этого состояния варьирует у разных людей, однако даже на умеренных высотах оно проявляется в виде ощущения слабости, нежелания двигаться и работать. Затем наступает головная боль, тошнота, иногда рвота, а также ухудшение умственной деятельности — точности суждений, способности выполнять простые арифметические расчеты и т. п. Сон и отдых тоже становятся затруднительными, что еще больше ухудшает дело.
За исключением кессонной болезни, которая может развиться при быстром подъеме на негерметизированном самолете или на воздушном шаре, все остальные известные эффекты высоты обусловлены низким парциальным давлением кислорода и недостаточностью кислородного снабжения тканей. Вдыхание чистого кислорода на высоте около 12 000 м, где барометрическое давление около 150 мм, снимает все симптомы горной болезни. Если бы человек на этой высоте дышал атмосферным воздухом, он выжил бы не дольше нескольких минут.
Снабжение тканей кислородом включает пять последовательных этапов:
1) вентиляцию легких;
2) диффузию кислорода из альвеолярного воздуха в кровь;
3) перенос кислорода кровью;
4) диффузию кислорода из крови в ткани;
5) использование кислорода в тканях.
Акклиматизация к высоте связана с изменением всех этих этапов.
Непосредственная реакция на нехватку кислорода состоит в усилении вентиляции легких путем увеличения как дыхательного объема, так и частоты дыхания. Усиленная вентиляция ведет к повышенному выведению СО2 из крови и соответственно к сдвигу pH крови в щелочную сторону, что ослабляет стимуляцию дыхательного центра. В результате далее наступает уменьшение вентиляции и снижение доставки кислорода; это запускает весь описанный процесс сначала, так что начинаются циклические переходы от повышенного дыхания к пониженному и обратно.
19*
292 Глава 6. Энергетический обмен
При акклиматизации к высокогорью реактивность дыхательного центра и регулирование концентрации бикарбоната в крови изменяются таким образом, что кислотно-щелочное равновесие и нормальная величина pH крови поддерживаются при более низком Рсо2. Чем больше развиваются эти приспособительные изменения, тем легче осуществляется повышенная вентиляция легких.
У уроженцев равнин диффузионная способность легких в условиях высокогорья не меняется. У горцев же объем легких больше,, чем у жителей равнин, и диффузионная способность легких также повышена, поскольку она зависит от размеров легочной поверхности. Это различие обусловлено усиленным ростом легких у горцев в детском возрасте. Аналогичное явление наблюдается у молодых крыс, выращенных в условиях пониженного давления. Таким образом, различие в диффузионной способности, по-видимому, связано с акклиматизацией в процессе индивидуальной жизни.
Перенос кислорода кровью тоже изменяется; это касается как кислородной емкости крови, так и сродства ее к О2. У человека на высотах около 4000 м над уровнем моря число эритроцитов увеличивается с 5 до 8 млн. на 1 мм3. Это явление наблюдается и у постоянных обитателей высокогорья, и у уроженцев низменностей, живущих на больших высотах.
Вопрос о сродстве крови к кислороду обсуждался в гл. 3. У ламы и викуньи — животных, обитающих высоко в горах,— сродство гемоглобина к О2 выше, чем у других млекопитающих, и это облегчает захват кислорода в легких. С другой стороны, у человека, оказавшегося в горах, иногда происходит небольшой сдвиг этого показателя в противоположную сторону; это не способствует захвату кислорода в легких, но несколько облегчает отдачу его в тканях.
Диффузия кислорода из крови в ткани может возрасти при укорочении диффузионного пути или при повышении концентрации миоглобина в тканях (облегченная диффузия). При акклиматизации были отмечены оба явления: может возрастать и число капилляров на единицу объема ткани (сокращение пути диффузии), и концентрация миоглобина (ускорение диффузии).
Последний этап — использование кислорода на тканевом уровне. Однако вопрос о том, каким адаптивным изменениям подвергаются ферментные системы и какую роль играют эти изменения в акклиматизации к высоте, еще плохо изучен.
На рис. 6.21 суммированы данные, показывающие значение перечисленных пяти этапов транспорта О2 у человека. Здесь сравниваются соответствующие показатели у постоянных жителей Лимы, расположенной на уровне моря, и у жителей Морокочи — шахтерского городка, находящегося на высоте 4540 м. Атмосферное давление в Морокоче составляет в среднем 446 мм рт. ст., а на уровне моря — 760 мм. В табл. 6.8 приведены данные о пар
Влияние больших высот 293
циальном давлении кислорода (Роа) У этих двух групп жителей. Разница в Ро2 между воздухом в трахее (т. е. увлажненным атмосферным воздухом) и венозной кровью примерно вдвое больше на уровне моря, чем на высоте. Несмотря на это, Ро2 смешанной венозной крови на высоте было только на 7,3 мм рт. ст. ниже; это показывает, что доставка кислорода тканям у акклиматизи-
Рис. 6.21. Средние величины парциального давления кислорода у жителей Лимы (уровень моря) и Морокочи (Перу, высота 4540 м). (Hurtado, 1964.)
рованных уроженцев высокогорья осуществляется почти так же эффективно, как и у тех, кто живет на уровне моря (Hurtado, 1964).
Птицы намного лучше переносят условия больших высот, чем млекопитающие, и альпинисты неоднократно сообщали, что на тех высотах, где им приходилось жестоко страдать от нехватки кислорода, они видели пролетавших над ними птиц. Эта особенность птиц обусловлена несколькими причинами. Прежде всего,
Таблица 6.8
Парциальное давление кислорода (в мм рт. ст.) в воздухе трахеи и в венозной крови у коренных жителей Лимы (на уровне моря) и Морокочи (4540 м).
Средние величины для 8 здоровых людей из той
и другой группы. Те же данные, что и на рис. 6.21.
(Hurtado, 1964)
Воздух трахеи Смешанная венозная кровь Разность
Лима 147,2 42,1 105,1
Морокоча 83,4 34,8 48,6
Разность 63,8 7,3 56,5
294 Глава 6. Энергетический обмен
дыхательная система птиц обладает большей эффективностью, так как воздух проходит через легкие не только при вдохе, но и при выдохе. Сродство крови к кислороду у птиц примерно такое же, как у млекопитающих, но благодаря принципу противотока достигается большее насыщение крови в легких (гл. 2). Вторая причина — исключительная устойчивость птиц к алкалозу; когда кровь теряет много СО2, связывание кислорода кровью увеличивается благодаря выраженному эффекту Бора. Наконец, последняя причина состоит в том, что у птиц (в отличие от млекопитающих) при низком Ро2 крови поддерживается нормальное кровоснабжение мозга; это частично предохраняет птиц от тяжелых нарушений функции центральной нервной системы, которые возникают на больших высотах у млекопитающих.
Подводя итоги, мы можем сказать, что энергетический обмен у животного тесно связан не только с потреблением пищи, но и с дыханием и обеспеченностью кислородом. Переходя далее к рассмотрению еще одного фактора внешней среды — температуры,— мы снова увидим, что принципы, установленные в предыдущих главах, сохраняют свою силу, ибо существует тесная взаимосвязь между функционированием всех физиологических систем.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Adolph Е. F. (1949). Quantitative relations in the physiological constitution of mammals, Science, 109, 579—585.
Bartholomew G. A., MacMillen R. E. (1961). Oxygen consumption, estivation, and hibernation in the kangaroo mouse, Microdipodops pallidus, Physiol. Zool., 34, 177-183.
Blazka P. (1958). The anaerobic metabolism of fish, Physiol. Zool., 21, 117— 128.
Brody S. (1945). Bioenergetics and Growth: With Special Reference to the Efficiency Complex in Domestic Animals, New York, Reinhold, 1023 pp., Reprinted (1964), Hafner, New York.
Childress J. J. (1975). The respiratory rates of midwater crustaceans as a function of depth of occurrence and relation to the oxygen minimum layer off southern California, Comp. Biochem. Physiol., 50A, 787—799.
Dawson T. L, Hulbert A. J. (1970). Standard metabolism, body temperature, and surface areas of Australian marsupials, Am. J. Physiol., 218, 1233—1238.
Douglas E. L., Friedl W. A., Pickwell G. V. (1976). Fishes in oxygen-minimum zones: Blood oxygenation characteristics, Science, 191, 957—959.
Drorbaugh J. E. (1960). Pulmonary function in different animals, J. Appl. Physiol., 15, 1069—1072.
Elsner R. (1965). Heart rate response in forced versus trained experimental dives in pinnipeds, Hvalradets Skrifter, No. 48, pp. 24—29.
Felger R. S., Cliff ton K., Regal P. J. (1976). Winter dormancy in sea turtles; Independent discovery and exploitation in the Gulf of California by two local cultures, Science, 191, 283—285.
Fry F. E. L, Hart J. S. (1948). The relation of temperature to oxygen consumption in the goldfish, Biol. Bull., 94, 66—77.
Gaunt A. S., Gans C. (1969). Diving bradycardia and withdrawal bradycardia in Caiman crocodilus. Nature, Lond., 223, 207—208.
Влияние больших высот 295
Gordon М. S., Boetius I., Evans D. Н., McCarthy R., Oglesby L. C. (1969). Aspects of the physiology of terrestrial life in amphibious fishes. 1. The mudskipper, Periophthalmus sobrinus, J. Exp. Biol., 50, 141—149.
Graham J. B. (1974). Aquatic respiration in the sea snake Pelamls platurus, Respir. Physiol., 21, 1—7.
Hall F. G. (1929). The influence of varying oxygen tensions upon the rate of oxygen consumption in marine fishes, Am. J. Physiol., 88, 212—218.
Hawkins A. E., Jewell P. A., Tomlinson G. (1960). The metabolism of some British shrews, Proc. Zool. Soc. Lond., 135, 99—103.
Hemmingsen A. M. (1960). Energy metabolism as related to body size and respiratory surface and its evolution, Rep. Steno Hosp., 9 (2), 1—110.
Hochachka P. W. (ed.) (1975). Pressure effects on biochemical systems of abyssal and midwater organisms: The 1973 Kona Expedition of the Alpha Helix, Comp. Biochem. Physiol., 52, (IB), 1—199.
Hochachka P. W., Fields J., Mustafa T. (1973). Animal life without oxygen: Basic biochemical mechanisms, Am. Zool., 13, 543—555.
Hochachka P. W., Mustafa T. (1972). Invertebrate facultative anaerobiosis, Science, 178, 1056—1060.
Hudson J. W. (1962). The role of water in the biology of the antelope ground squirrel Citellus leucurus, Univ. Calif. Publ. Zool., 64, 1—56.
Hurtado A. (1964). Animals in high altitudes: Resident man. In: Handbook of Physiology, sect. 4, Adaptation to the Environment (D. B. Dill, E. F. Adolph and C. G. Wilber, eds.), pp. 843—860, Washington, D. C., American Physiological Society.
Jackson D. C., Schmidt-Nielsen K. (1966). Heat production during diving in the fresh water turtle, Pseudemys scripta, J. Cell Physiol., 67, 225—232.
Johansen K. (1964). Regional distribution of circulating blood during submersion asphyxia in the duck, Acta Physiol. Scand., 62, 1—9.
King J. R. (1957). Comments on the theory of indirect calorimetry as applied to birds, Northwest Sci., 31, 155—169.
Kleiber M. (1961). The Fire of Life: An Introduction to Animal Energetics, New York, Wiley, 454 pp.
Kooyman G. L. (1966). Maximum diving capacities of the Weddell seal, Lepto-nychotes weddelli, Science, 151, 1553—1554.
Kylstra J. A. (1968). Experiments in water-breathing, Sci. Am., 219, 66—74.
Lasiewski R. C., Dawson W. R. (1967). A re-examination of the relation between standard metabolic rate and body weight in birds, Condor, 69, 13—23.
Lusk G. (1931). The Elements of the Science of Nutrition, 4th ed., Philadelphia, Saunders, 844 pp.
MacDonald A. G. (1975). Physiological Aspects of Deep Sea Biology, London, Cambridge University Press, 450 pp.
Margaria R., Cerretelli P., Aghetno P., Sassi G. (1963). Energy cost of running, J. Appl. Physiol., 18, 367—370.
Murdaugh H. V., Jr., Schmidt-Nielsen B., Wood J. W., Mitchell W. L. (1961). Cessation of renal function during diving in the trained seal (Phoca vitulina), J. Cell. Comp. Physiol., 58, 261—266.
Olsson K- F., Saltin B. (1968). Variation in total body water with muscle glycogen changes in man. In: Biochemistry of Exercise, Medicine and Sport Series, vol. Ill, pp. 159—162, Basel, Karger.
Paulev P. (1965). Decompression sickness following repeated breath-hold dives, J. Appl. Physiol., 20, 1028—1031.
Pearson О. P. (1960). The oxygen consumption and bioenergetics of harvest mice, Physiol. Zool., 33, 152—160.
Pugh L. G. С. E. (1964). Animals in hihg altitudes: Man above 5,000 meters — mountain exploration. In: Handbook of Physiology, sect. 4, Adaptation to the Environment (D. B. Dill, E. F. Adolph and C. G. Wilber, eds.), pp. 861—868, Washington, D. C., American Physiological Society.
Schmidt-Nielsen K. (1972a). Locomotion: Energy cost of swimming, flying, and running, Science, 177, 222—228.
296 Глава 6. Энергетический обмен
Schmidt-Nielsen К. (1972b). How Animals Work, Cambridge, Cambridge University Press, 114 pp.
Scholander P. F. (1940). Experimental investigations on the respiratory function in diving mammals and birds, Hvalradets Skrifter, No. 22, pp. 1—131.
Shepard M. P. (1955). Resistance and tolerance of young speckled trout (Salveli-nus fontinalis) to oxygen lack, with special reference to low oxygen acclimation, J. Fish. Res. Bd. Can., 12, 387—446.
Smith E. N., Allison R. D., Crowder W. E, (1974). Bradycardia in a free-ranging American alligator, Copeia, 1974, 770—772.
Somero G. N., Hochachka P. W. (1976). Biochemical adaptations to pressure. In: Adaptation to Environment: Essays on the Physiology of Marine Animals (R. C. Newell, ed.), pp. 480—510, London Butterworth.
Stahl W. R. (1967). Scaling of respiratory variables in mammals, J. Appl. Physiol., 22, 453—460.
Taylor C. R., Schmidt-Nielsen K., Raab J. L. (1970). Scaling of the energetic cost of running to body size in mammals, Am. J. Physiol., 219, 1104—1107.
Tenney S. M., Remmers J. E. (1963). Comparative quantitative morphology of the mammalian lung: Diffusing area, Nature, bond., 197, 54—56.
Thomas H. J. (1954). The oxygen uptake of the lobster (Homarus vulgaris Edw.), J. Exp. Biol., 31, 228—251.
Tucker V. A, (1968). Respiratory exchange and evaporative water loss in the flying budgerigar, J. Exp. Biol., 48, 67—87.
Whalen W. J. (1966). Intracellular Po2: A limiting factor in cell respiration, Am. J. Physiol., 211, 862—868.
Whalen W. J., Buerk D., Thuning C. A. (1973). Blood-flow-limited oxygen consumption in resting cat skeletal muscle, Am. J. Physiol., 224, 763—768.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Andersen H. T. (ed.) (1969). The Biology of Marine Mammals, New York, Academic Press, 511 pp.
Bennett P. B., Elliott D. H. (1975). The Physiology and Medicine of Diving and Compressed Air Work, London, Bailliere Tindall, 566 pp.
Brody S. (1964). Bioenergetics and Growth: With Special References to the Efficiency Complex in Domestic Animals, reprint ed., N. Y., Hafner, 1023 pp.
Frisancho A. R. (1975). Functional adaptation to high altitude hypoxia, Science, 187, 313—319.
Heath D., Williams D. R. (1977). Man at High Altitude, Edinburgh, Churchill Livingston, 292 pp.
Heatwole H„ Seymour R. (1975). Diving physiology. In: The Biology of Sea Snakes (W. Dunson, ed.), pp. 289—327, Baltimore, University Park Press.
Hemmingsen A. M. (1960). Energy metabolism as related to body size and respiratory surfaces, and its evolution, Rep. Steno Hosp., 9 (2), 1—110.
Kleiber M. (1961). The Fire of Life: An Introduction to Animal Energetics, New York, Wiley, 454 pp.
Kooyman G. L. (1972). Deep diving behaviour and effects of pressure in reptiles, birds, and mammals, Symp. Soc. Exp. Biol., No. 26, pp. 295—311.
Kylstra J. A. (1975). Liquid breathing and artificial gills. In: The Physiology and Medicine of Diving and Compressed Air Work, 2nd ed. (P. B. Bennett and D. H. Elliott, eds.), pp. 155—165, London, Bailliere Tindall.
Newell R. C. (ed.) (1976). Adaptation to Environment: Essays on the Physiology of Marine Animals, London, Butterworth, 539 pp.
Schmidt-Nielsen K. (1972). How Animals Work, Cambridge, Cambridge University Press, 114 pp.
Schmidt-Nielsen K. (1975). Scaling in biology: The consequences of size, J. Exp. Zool., 194, 287—308.
Sleigh M. A., MacDonald A. G. (eds.) (1972). The Effects of Pressure on Organisms, Symposia, Soc. for Exper. Biol., No. 26, Cambridge, 516 pp.
ЧАСТЬ ТРЕТЬЯ
ТЕМПЕРАТУРА
ГЛАВА 7
ВЛИЯНИЕ ТЕМПЕРАТУРЫ
В предыдущей главе мы рассматривали энергетический обмен у животных и влияние на него таких переменных, как концентрация кислорода, размеры тела и двигательная активность. Теперь мы познакомимся с тем глубоким влиянием, которое оказывает на живые организмы и их метаболические процессы температура.
Активные проявления животной жизни возможны только в узком диапазоне температур — от нескольких градусов ниже точки замерзания чистой воды (О °C) примерно до + 50 °C1. По сравнению с космическими температурами эти границы поистине крайне узки. Однако температурные условия, пригодные для обитания животных, имеются и повсеместно в океанах, и на большей части земной суши хотя бы в какое-то время года. Рыбы и множество беспозвоночных живут в арктических водах, где круглый год держится температура около —1,8°С. Противоположная крайность—• это немногочисленные виды животных, обитающие в горячих источниках при температуре около 50 °C. Некоторые примитивные растения способны жить при еще более высоких температурах, а термофильные бактерии успешно размножаются при температуре, близкой к точке кипения воды.
Оказавшись вне диапазона температур, совместимых с активной жизнью, многие животные способны выживать, переходя в состояние оцепенения. Более того, некоторые животные могут переносить температуру жидкого воздуха (около —190 °C) или даже жидкого гелия (—269°C). Устойчивость к высоким температурам не так велика, хотя некоторые животные в фазе покоя в
1 Речь идет о температуре самого организма, а не окружающей его среды. Когда человек выходит на мороз, температура его тела остается на обычном уровне около 37 °C.
298 Глава 7. Влияние температуры
этом отношении весьма выносливы, а споры бактерий могут выживать при температурах выше 120 °C.
У большинства животных, в том числе у всех водных беспозвоночных, температура тела очень близка к температуре окружающей среды. В отличие от этого у птиц и млекопитающих температура почти постоянна и мало изменяется даже при больших колебаниях внешней температуры. Есть также очень немногочисленные виды как низших позвоночных, так и беспозвоночных, которые временами способны поддерживать значительную разницу между температурой тела и окружающей среды.
Классифицировать животных по этим свойствам совсем не просто. Обычно птиц и млекопитающих по традиции называют теплокровными, а всех остальных — холоднокровными. Эти термины настолько укоренились, что удобнее продолжать ими пользоваться, хотя они неточны и могут даже вводить в заблуждение. Холоднокровные животные вовсе не обязательно холодны: температура тела у тропической рыбы, у ящерицы в пустыне или у сидящего на солнце насекомого может быть выше, чем у млекопитающего. С другой стороны, некоторые млекопитающие и птицы впадают в зимнюю спячку или в оцепенение, и тогда температура их тела может снижаться почти до точки замерзания воды без всякого вреда для организма. Было бы довольно странно называть их в этом состоянии теплокровными.
Соответствующие научные термины — пойкилотермные вместо «холоднокровные» и гомойотермные (или гомеотермные) вместо «теплокровные» — также неточны. Слово «пойкилотермный» (от гр.еч. пойкилос — изменчивый) указывает на то, что температура холоднокровного животного изменяется вместе с температурой среды. Рыба, например, имеет такую же температуру, как и вода, В которой она плавает, а дождевой червь — температуру той почвы, в которой он находится. Но как быть с глубоководными рыбами? Вся их жизнь проходит в тех слоях воды, где колебания температуры ничтожны, так что это поистине животные с практически постоянной температурой тела. Было бы логично называть таких рыб гомойотермными, однако это слово употребляют только в отношении птиц и млекопитающих, температура тела которых, в сущности, колеблется в пределах нескольких градусов, а во время зимней спячки может даже снижаться почти до 0°С. Тех животных, у которых температура тела только временами поддерживается на высоком и хорошо регулируемом уровне, а в другие периоды зависит от окружающей среды, часто называют гетеро-термными (от греч гетерос — различный).
Гомойотермные животные (птицы и млекопитающие), как правило, имеют высокую температуру тела и остаются активными как в условиях холода, так и в тепле. Большинство пойкилотерм-ных животных по мере снижения температуры становятся все менее и менее активными. Есть, однако, примечательные исклю
Влияние изменений температуры 299
чения. Например, ящерица, обогреваясь на солнце, может поддерживать свою температуру на уровне, который значительно выше температуры окружающего воздуха. Для того чтобы отличать такую ящерицу от птиц и млекопитающих, согреваемых теплом внутреннего, метаболического происхождения, пользуются терминами эктотермные и эндотермные организмы.
У эндотермных животных высокая температура тела поддерживается внутренним образованием тепла; эктотермные животные зависят от внешних источников тепла, прежде всего от солнечной радиации. Но и эти понятия следует употреблять с известными оговорками. Например, у тунца, как мы увидим- далее, температура мышц может на 10—15° превышать температуру окружающей воды. Хотя источником тепла здесь служит метаболизм мышц, нельзя все же считать тунца таким же эндотермным животным, как птицы и млекопитающие.
В отношении насекомых применение данной терминологии еще больше усложняется. Например, насекомое может греться на солнышке и в то же время вырабатывать дополнительно тепло за счет интенсивных мышечных сокращений. Очевидно, что этот случай не соответствует полностью какому-то одному из двух понятий (эндо- или эктотермия). То же самое можно сказать и относительно тех птиц и млекопитающих, которые в определенных случаях используют тепло солнечных лучей, чтобы снизить собственную теплопродукцию.
Выбор терминологии определяется прежде -всего удобством, и иногда действительно трудно найти термины, приложимые в любой ситуации. Вопрос состоит не в том, правильна или неправильна какая-то терминология, а в том, насколько она полезна для данной цели. Вместе с тем необходимо всегда четко определять используемые понятия, чтобы не было никаких сомнений в их точном смысле.
ВЛИЯНИЕ ИЗМЕНЕНИЙ ТЕМПЕРАТУРЫ
НА ФИЗИОЛОГИЧЕСКИЕ ПАРАМЕТРЫ
Изменение температуры очень сильно сказывается на многих физиологических процессах. С известными ограничениями можно сказать, что повышение температуры ускоряет большинство процессов. Рассмотрим, например, ее влияние на потребление кислорода, так как последнее служит удобным показателем общей метаболической активности организма.
В диапазоне температур, переносимых данным животным, потребление кислорода часто довольно регулярным образом возрастает с повышением температуры. Обычно при повышении ее на 10° потребление Ог увеличивается в 2—3 раза.
Увеличение скорости какого-либо процесса при повышении температуры на 10° обозначают как Qi0. Если скорость увеличивается
300 Глава 7, Влияние температуры
вдвое, то Qio=2, если втрое, — <2ю=3 и т. д. Это относится не только к потреблению кислорода, но и к скорости любого процесса, зависящего от температуры (см. Приложение Г).
Если животное способно выдерживать изменение температуры в широких пределах, то потребление им кислорода может возрастать при повышении температуры во много раз. Если, например, <2ю=2, то по сравнению с 0°С при 10°С потребление О2 возрастет
Рис. 7.1. Скорость многих процессов, зависимых от температуры, неуклонно возрастает по мере ее повышения. Подробности см. в тексте.
вдвое, при 20 °C — в 4 раза, при 30 °C — в 8 раз; если же <2ю=3, то при тех же изменениях температуры оно увеличится соответственно в 3, 9 и 27 раз.
Построив по этим цифрам график (рис. 7.1), мы получим круто поднимающиеся вверх линии, называемые экспоненциальными кривыми, так как математически они описываются экспоненциальными функциями1 общего вида
у=Ь-ах.
Если скорость какого-то процесса при температурах 72 и Tt обозначить соответственно Т?2 и и использовать принятый сим
1 См. также Приложение В.
Влияние изменений температуры 301
вол Qio, то уравнение будет выглядеть так:
-Сю 10 •
Экспоненциальными уравнениями описываются скорости очень многих процессов, например радиоактивного распада, роста и т. д. Для расчетов удобнее использовать эти уравнения в логарифмической форме:
1g lg b+x- 1g а. или
Теперь видно, что 1g R2 линейно возрастает с изменением температуры (Т2 — Л). Следовательно, если мы отложим логарифм скорости против температуры, мы получим прямую линию. Так
Рис. 7.2. Если скорость температурозависимого процесса вроде представленного на рис. 7.1 отложить по оси ординат в форме логарифма, то зависимость скорости от температуры изобразится прямой. Обратите внимание, что абсцисса имеет линейный масштаб и только ордината — логарифмический. Объяснения в тексте.
обычно и поступают, используя так называемую полулогарифмическую бумагу, где ордината имеет логарифмический масштаб, а абсцисса — линейный. График приведенного уравнения представлен на рис. 7.2.
302 Глава 7; Влияние температуры
Таблица 7.1
Потребление кислорода колорадским жуком (Leptinotarsa decemlineata) при различных температурах от 7 до 30 °C. Перед опытом жуков выдерживали при 8°C. (По данным Marzusch, 1952)
Температура, °C Потребление О2, мкл/(г-ч) Температурные интервалы, °C $10
7 61
10 81 7—10 2,57
15 126 10—15 2,41
20 200 15—20 2,52
25 290 20—25 2,10
30 362 25—30 1,56
Очевидно, что для нахождения Qw нет необходимости измерять скорости при температурах с интервалом ровно 10°. Можно
Рис. 7.3. Потребление кислорода колорадским жуком возрастает с увеличением температуры (сплошная линия). Пунктиром показано, какой вид имела бы кривая, если бы величина Qio была постоянна и равна 2,5. Данные взяты из табл. 7.1.
взять любые две температуры, лишь бы они достаточно разнились между собой для надежности измерения температурного эффекта. Чтобы вычислить Qio из скоростей при двух разных температурах,
Влияние изменений температуры 303
можно преобразовать уравнение и использовать одну из двух его форм:
lg <310 = (!g ^2 — lg ^1) т •
или
10
Q ___( #2 \ Т2 Т1
Далее вычисляют Qio для интервала между Т2 и Д Однако очень часто Qio не остается постоянным на всем диапазоне температур, совместимым с жизнью животного, и потому необходимо точно указывать, в каких условиях производились измерения.
Рассмотрим реальный пример. В табл. 7.1 приведены данные о потреблении кислорода колорадским жуком в интервале от 0 до 30 °C. На всем этом интервале потребление О2 с повышением температуры возрастает, и в среднем Qi0 для данного интервала
Рис. 7.4. Если потребление кислорода колорадским жуком отложить по оси ординат в виде логарифма, то зависимость этого процесса от температуры в диапазоне от 7 до 20 °C отобразится прямой. При температурах выше 20 °C фактический прирост меньше, чем следовало бы ожидать при QI0=2,5 (пунктирная линия). Данные взяты из табл. 7.1.
равно 2,17. Но если мы вычислим Qi0 для более узких .температурных интервалов, использованных в опыте, то мы увидим, что до 20°C Qio остается достаточно постоянным, составляя примерно 2,5, но при дальнейшем повышении температуры Qio падает (табл. 7.Г,
304 Глава 7, Влияние температуры, последний столбец). Если мы представим эти данные графически (рис. 7.3), то мы ясно увидим, что полученные результаты для температур выше 20°C отклоняются от правильной экспоненциальной функции.
На практике довольно хлопотно вычислять Qio для каждой серии определений; кроме того, неудобно строить экспоненциальную кривую в арифметических координатах для сопоставления с экспериментальными данными. Если вместо этого мы отложим наши цифры на логарифмической шкале, мы получим прямую для тех интервалов, в которых эффект температуры остается постоянным и Qio не меняется (рис. 7.4). Прямая соответствует величине Qio, равной 2,50, и можно видеть, что данные, полученные для температур до 20 °C, полностью укладываются на эту прямую. Однако при температурах выше 20 °C кривая потребления кислорода все больше и больше отклоняется от прямой линии (т. е. потребление О2 все еще возрастает с температурой, но величина температурного эффекта снижается). При дальнейшем повышении температуры это отклонение становится еще более резким, и в конце концов достигается верхний температурный предел для жизни.
ЭКСТРЕМАЛЬНЫЕ ТЕМПЕРАТУРЫ;
ТЕМПЕРАТУРНЫЕ ПРЕДЕЛЫ ДЛЯ ЖИЗНИ
Животные значительно различаются в отношении диапазона переносимых температур. Одни могут существовать только в очень узком диапазоне, для других этот диапазон намного шире. Кроме того, температурная толерантность иногда меняется с течением времени, и в этом отношении возможна некоторая адаптация: после длительного пребывания животного в условиях, близких к пределу его толерантности, этот предел может сдвинуться. Некоторые животные особенно чувствительны к крайним температурам в определенные периоды жизни, особенно на ранних стадиях развития.
Обсуждая вопрос о толерантности к крайним температурам, следует различать ту температуру, при которой данный организм может выжить, и ту, при которой он может пройти весь жизненный цикл. Следует также осознать, что нельзя точно определить, какая температура для данного организма детальна, так как большое значение имеет время температурного воздействия. Так, например, животное может выносить какую-то высокую температуру в течение нескольких минут, но погибнет, если ее воздействие продлится несколько часов.
Важно еще заметить, что речь идет о температуре тела самого животного, а не о температуре окружающей среды. Для подавляющего большинства водных организмов эти две величины почти равны между собой, но для наземных, как уже говорилось, они могут сильно различаться. У ящерицы, сидящей на солнце, тем
Экстремальные температуры; пределы для жизни 305
пература тела благодаря нагреву солнечными лучами может на 10—20° превышать температуру окружающего воздуха. Так называемые теплокровные животные — млекопитающие и птицы,— как правило, способны переносить лишь небольшие колебания температуры тела, но в то же время они способны существовать в очень широком диапазоне внешних температур — от Арктики до самых жарких пустынь.
ТОЛЕРАНТНОСТЬ К ВЫСОКОЙ ТЕМПЕРАТУРЕ
Ни одно животное не может пройти весь жизненный цикл при температуре выше 50 °C. Некоторые растения более устойчивы, чем животные; например, одноклеточная сине-зеленая водоросль Synechococcus встречается в горячих источниках с температурой, достигающей 73—75 °C. Это, по-видимому, верхний температурный предел для жизни фотосинтезирующих организмов. Термофильные бактерии проявляют еще большую термостойкость: было обнаружено, что они живут и растут в горячих источниках йеллоустонского национального парка при температуре около 92°C (на высоте Йеллоустона это точка кипения воды) (Bott, Brock, 1969; Brock, 1970).
Как уже говорилось в начале главы, покоящиеся стадии у некоторых животных исключительно устойчивы к высоким температурам. Личинка мухи Polypedilum (из Нигерии и Уганды) переносит обезвоживание и в высушенном состоянии выживает при 102°C в течение одной минуты, после чего способна расти и претерпевать нормальный метаморфоз (Hinton, 1960). Другим примером крайней толерантности могут служить яйца пресноводного рачка Triops (из Судана): в течение зимы и начала лета они находятся в сухом иле, где иногда прогреваются на солнце до температуры 80 °C. В лабораторных условиях они выживали при температуре лишь на 1° ниже точки кипения. Если температуру кипения воды повышали, увеличивая давление, то жизнеспособность яиц сохранялась при 103±1°С в течение 16 ч, но при нагреве до 106°C они погибали за 15 мин (Carlisle, 1968).
Эти примеры ясно показывают, что невозможно точно установить верхний температурный предел для жизни.
ОПРЕДЕЛЕНИЕ ЛЕТАЛЬНЫХ ТЕМПЕРАТУР
Если группу животных подвергнуть воздействию температуры, близкой к пределу их толерантности, то некоторые из них погибнут, а некоторые выживут. Как же точно определить летальную температуру для данного вида?
Обычно летальной называют такую температуру, при которой 50% животных погибает, а 50% выживает; ее часто обозначают как Z.T50. Если бы эту величину определяли методом проб и оши-20—1873
306 Глава 7. Влияние температуры
бок, то пришлось бы проводить очень много опытов, прежде чем один из них кончился бы гибелью ровно половины животных. Вместо этого сначала устанавливают летальную температуру приблизительно, а далее поступают так, как показано на рис. 7.5.
Рис. 7.5. Для определения А750, т. е. температуры, летальной для 50% организмов взятой группы, исследуют выживание в определенном диапазоне температур; по полученным данным строят график и из него находят температуру, соответствующую 50%-ному выживанию.
Несколько групп животных (например, четыре) подвергают воздействию ряда температур в течение одного и того же периода времени (например, 2 ч) и откладывают на графике, какой про-
Врёмя до гибели 50% осооей, ч
Рис. 7.6. Влияние времени воздействия на температурную толерантность становится очевидным, если LT^ отложить против длительности опыта, в котором определяли эту величину.
цент животных выжил в каждой группе. После этого путем интерполяции легко найти LT50.
Такой метод позволяет определить летальную температуру только для той длительности воздействия, которая испытывалась
Экстремальные температуры; пределы для жизни 307
в эксперименте; большая продолжительность воздействия снизит* выживаемость, а меньшая — повысит ее. Оценить эффект времени воздействия (экспозиции) можно с помощью тестов, подобных представленным на рис. 7.5, но с разными периодами экспозиции,, например 1, 2, 4 и 8 ч. Если полученные величины ЛТ50 отложить против времени воздействия, то при логарифмической шкале по оси абсцисс все точки часто будут лежать на одной прямой (рис. 7.6). Именно ради удобства использования логарифмической шкалы и были выбраны указанные в нашем примере периоды экспозиции. В результате такого эксперимента получается кривая, дающая полное представление о температурной толерантности изучаемого организма.
ЛЕТАЛЬНАЯ ТЕМПЕРАТУРА И ПРИЧИНА ГИБЕЛИ ПРИ ПЕРЕГРЕВЕ
Хотя верхний предел активной животной жизни близок к 50 °C, многие животные погибают при воздействии значительно меньших температур. В особенности это относится к водным животным и в первую очередь к морским, которые в природных условиях обычно не подвергаются воздействию слишком высоких температур. Ведь даже в тропических морях температура редко достигает 30 °C, хотя в мелких закрытых бухтах и лагунах она может быть и выше.
В ином положении находятся животные, обитающие в приливной (литоральной) зоне. Во время отлива они подвергаются воздействию теплого воздуха и солнечных лучей, так что их температура может заметно повыситься. Этому нагреву может до некоторой степени противодействовать испарение воды, но скудность ее запасов ограничивает возможности этого способа охлаждения.
Четкая корреляция между устойчивостью к нагреву и местом обитания наблюдается у брюхоногих моллюсков литоральной зоны (табл. 7.2). Моллюски, живущие в верхней полосе литорали и потому дольше других находящиеся в воздушной среде, значительно устойчивее тех, которые обитают в нижней полосе, вблизи минимальной отметки уровня воды.
Среди рыб наиболее устойчивы к нагреву, вероятно, карпозу-бики, обитающие в теплых источниках пустынь Калифорнии и Невады. Вид Cyprinodon diabolus живет в источнике, известном под названием Чертовой Дыры, где по меньшей мере за последние 30 000 лет температура (33,9°C), вероятно, почти не изменялась (Brown, Feldmeth, 1971). Верхний температурный предел для выживания этой крошечной рыбки, взрослые особи которой весят менее 200 мг, составляет около 43 °C — это наивысший предел, известный для рыб.
Напротив, некоторые арктические и антарктические животные обладают поразительно малой теплоустойчивостью. Антарктиче-20*
308 Глава 7, Влияние температуры
Таблица 7.2
Брюхоногие моллюски, обитающие у верхней границы литорали, способны выносить повышение температуры почти на 10° большее, чем те, которые живут около нижнего уровня воды. Приведенные в таблице величины соответствуют той максимальной температуре, которую животное выдерживает в течение часа. (Fraenkel, 1968)
Место обитания Вид Температура, °C
Полоса прибоя, Tectarius vilis 48,5
верхняя часть литораль- Planaxis sulcatus 48
ной зоны Nodilittorina granularis 47
Littorina brevicola 47
Мелкие временные водоемы в зоне прибоя Paesiella raepstorttiana 47
Средняя часть литорали N er it a japonica 46
Ner it a albicilla 44
Нижняя часть литорали — Lunella coronata 43
затененные участки и ме- Drupa granulatus 42
ста, прикрытые камнями Purpura clavigerus 42
Monodonta labis 42
Нижняя часть литорали, у границы воды Tegula lischkei 39
ские рыбы рода Trematomus особенно чувствительны к теплу: верхний предел для них около 6 °C. В природе эти рыбы живут при температуре воды —1,9 °C, причем сезонные колебания составляют всего лишь 0,1° (Somero, DeVries, 1967).
Исключительно низкая толерантность этих животных к теплу имеет большое значение для понимания механизма тепловой смерти. Высказывались предположения, что здесь играют роль следующие факторы:
1) денатурация белков, коагуляция их в результате нагрева;
2) термическая инактивация ферментов, идущая со скоростью, превышающей скорость их синтеза;
3) недостаток кислорода;
4) различие в температурном коэффициенте (Qio) для взаимосвязанных метаболических реакций;
5) изменение структуры мембран.
Рассмотрим тепловую денатурацию белков. Действительно, многие животные погибают при той температуре, при которой денатурируются многие белки, а именно при 45—55 °C. Трудно, однако, себе представить, чтобы термическая денатурация какого-либо белка происходила при +6°С — температуре, летальной для Trematomus.
Вторая предполагаемая причина — тепловая инактивация отдельных ферментных систем, особо чувствительных к повышенным
Экстремальные температуры; пределы для жизни 309'
температурам. Но опять-таки трудно представить себе существование столь термолабильных ферментов, что они инактивировались бы при +6 °C. К тому же исследование некоторых ферментных систем Trematomus показало, что их активность возрастает по мере повышения температуры приблизительно до 30 °C (Somero, DeVries, 1967).
Третью возможную причину тепловой смерти — недостаток кислорода в условиях, когда потребность в нем из-за повышенной температуры возрастает, — удалось во многих случаях исключить. Например, насекомые в атмосфере чистого кислорода не обнаруживали большей толерантности к перегреву. Точно так же форель—рыба, обитающая в холодной воде,—погибала в теплой воде при обычной для нее летальной температуре и тогда, когда содержание О2 в воде увеличивали в несколько раз путем аэрации чистым кислородом.
Четвертое предположение — о роли различий в Qi0— согласуется со многими экспериментальными данными. Если различные реакции промежуточного обмена изменяются при повышении температуры по-разному (т. е. имеют разные величины Qio), то это приведет к избыточному накоплению или нехватке отдельных промежуточных продуктов. Рассмотрим следующую схему:
А—>В—>С—>D
Если при повышении температуры реакция С—>D ускоряется в большей степени, чем реакция В—>С, то в этих условиях будет не хватать вещества С. Если же это вещество необходимо и для другого процесса (С—>-Е), то последний окажется заторможенным. Различие в температурной чувствительности сотен ферментов, участвующих в промежуточном обмене, может, таким образом, привести к полному нарушению нормального биохимического равновесия в организме.
Хотя в ряде случаев различия в температурной чувствительности ферментных систем могут быть правдоподобной причиной гибели (например, в случае антарктической рыбы, погибающей при + 6°C), это не обязательно единственный фактор. Определенную роль могла бы играть любая из упомянутых выше причин, и если речь идет о животных, погибающих только при очень высокой температуре, то здесь скорее всего действует несколько факторов одновременно.
Последняя из предполагаемых причин — изменение структуры мембран — включает, по существу, самые разнообразные случаи. В настоящее время общепризнано, что мембрана состоит из бимолекулярного слоя липидов, в который встроены или к которому
310 Глава 7. Влияние температуры
прикреплены функционально активные белки, и любое изменение взаимодействий между последними должно отражаться на функциональных свойствах мембраны. Изменение температуры (т. е. уровня кинетической энергии) оказывает глубокое влияние на такие взаимодействия; это может быть связано с изменением конформации белков, взаимодействий белков с липидами, липидов между собой и т. д. — короче говоря, с изменением молекулярных структур, которые зависят от слабых связей, легко изменяющихся при сдвиге температуры. Возникающие при этом нарушения в функционировании мембран играют, по-видимому, важнейшую роль в повреждении организма при перегреве.
УСТОЙЧИВОСТЬ К НИЗКИМ ТЕМПЕРАТУРАМ
Влияние низкой температуры не менее сложно, чем высокой. Некоторые организмы выдерживают глубокое промерзание, но для большинства животных оно гибельно. Известны виды, сохраняющие жизнеспособность после погружения в жидкий азот (—196 °C) и даже жидкий гелий (—269°C, или около 4 К); с другой же стороны, некоторые животные настолько чувствительны к охлаждению, что погибают при температуре намного выше 0°С. Это хорошо знают те, кто держит тропических рыбок в комнатных аквариумах: стоит отключить подогрев, и все рыбки могут погибнуть в первую же прохладную ночь. Например, гуппи (Lebistes reticulatus), которых содержали при 23 °C и выше, не переносят охлаждения примерно до 10 °C (Pitkow, 1960). По-видимому, их гибель наступает в результате холодового торможения дыхательного центра и последующей аноксии, так как увеличение концентрации кислорода в воде повышает их устойчивость к холоду, а уменьшение, наоборот, снижает ее.
УСТОЙЧИВОСТЬ К ОХЛАЖДЕНИЮ И ЗАМОРАЖИВАНИЮ
Животные, обитающие в умеренных и холодных зонах, выдерживают длительные периоды зимы, когда температура падает намного ниже точки замерзания воды. Они избегают холодового повреждения с помощью двух механизмов — переохлаждения и толерантности к замораживанию. Под переохлаждением понимают снижение температуры жидкости ниже точки ее замерзания без образования льда; толерантность к замораживанию означает способность переносить замерзание воды и образование льда в организме. Животных, не переносящих образования льда в теле, называют чувствительными к замораживанию, а тех, которые выживают в этих условиях,— толерантными к замораживанию.
Как происходит переохлаждение? Если воду или водный раствор охладить до температуры ниже точки замерзания, это не
Экстремальные температуры; пределы для жизни 311
обязательно ведет к образованию кристаллов льда1. Чистую воду можно охладить намного ниже О °C без какого-либо образования льда. Вероятность замерзания такой переохлажденной воды зависит от трех основных факторов: температуры, наличия центров образования льда и времени. При отсутствии чужеродных частиц, служащих центрами кристаллизации, чистую воду легко переохладить до —20 °C, а при особых предосторожностях — почти до —40 °C. Как только появился первый кристаллик льда, замерзание всей жидкости идет дальше очень быстро. Некоторые растворенные вещества не только снижают точку замерзания, но и влияют на ту степень переохлаждения, которую может выдержать раствор.
Рассмотрим теперь, каким образом животное, находящееся при температуре намного ниже точки замерзания жидкостей тела, может остаться в переохлажденном состоянии. Такая способность действительно имеет большое значение для выживания. Например, она может быть очень важна для животных, чувствительных к замораживанию, в случае внезапного наступления ночных холодов. Как показали опыты с пресмыкающимися и некоторыми другими позвоночными, у которых температура замерзания жидкостей тела —0,6 °C, они выдерживают переохлаждение до —8 °C без замерзания (Lowe et al., 1971).
Некоторые животные способны выживать даже при интенсивном образовании льда в их теле. Личинка комара Chironomus без вреда для себя выдерживает многократное замораживание при —25 °C и оттаивание. Казалось бы, трудно установить, переходят ли эти крошечные животные в состояние переохлаждения или же в их организме образуется лед. Однако оказалось возможным определить точное количество льда в теле животного, не причиняя ему вреда; для этого использовали остроумный метод, основанный на том, что превращение воды в лед сопровождается изменением объема. Определяя удельный вес замороженных личинок, нашли,
1 Термин «точка замерзания» может иметь несколько различных значений. В нашем случае мы определяем точку замерзания как температуру, при которой в жидкости образуется очень малое количество льда, находящееся в термодинамическом равновесии с жидкостью при данной температуре.
Точка замерзания чистой воды — 0 °C, однако водные растворы, содержащие соли и другие растворенные вещества, замерзают при более низкой температуре. Рассмотрим раствор, имеющий температуру несколько ниже 0 °C, в котором присутствуют отдельные кристаллы льда. Если этот раствор немного нагреть, часть льда растает; если же его несколько охладить, некоторое количество воды превратится в лед. В последнем случае концентрация растворенных веществ в оставшейся жидкости возрастет, и в результате температура, при которой сможет происходить дальнейшее образование льда, понизится. Этот процесс будет продолжаться до тех пор, пока не замерзнет весь раствор, однако точку полного замерзания трудно точно установить, и ее редко учитывают. Чтобы найти точку замерзания, как мы ее определили выше, обычно замораживают весь образец раствора, а затем его медленно нагревают и отмечают тот момент, когда вот-вот исчезнут последние мельчайшие кристаллики льда. Правильнее'было бы называть эту температуру точкой плавления, но этот термин редко используется в данном контексте.
312 Глава 7. Влияние температуры _________________________
что при —5 °C переходит в лед 70% их воды, а при —15 °C — 90%. Особей с установленным количеством льда в организме подвергали оттаиванию и проверяли на жизнеспособность (Scholander et al., 1953а).
Хотя многие беспозвоночные в высоких широтах обычно переносят в зимнее время очень низкие температуры, они крайне редко попадают в такие переменчивые условия, в каких находятся обитатели приливной зоны в субполярных областях. Зимой здесь регулярно происходят два полных цикла смены температур выше и ниже точки замерзания. Во время отлива животные на литорали промерзают так, что большая часть воды в их организме превращается в лед, а когда снова наступает прилив, они оттаивают. В течение шести и более часов кряду эти животные могут находиться в воздушной среде при температуре —30 °C, и при этом их собственная температура приближается к температуре окружающего воздуха. При —30 °C более 90% воды в организме замерзает» а в оставшейся жидкости концентрация растворенных веществ во много раз увеличивается. Это означает, что клетки таких организмов должны быть приспособлены не только к потере воды, превращающейся в кристаллы льда, но и к чрезвычайно большому повышению осмотического давления.
Изучение животных литоральной зоны показало, что им свойственно именно промерзание, а не переохлаждение, причем образование льда приводит к резкой деформации их мышц и внутренних органов. Однако кристаллы льда, как правило, находятся вне клеток, а клетки сморщиваются и, по-видимому, совсем не содержат льда. При оттаивании ткани вновь приобретают нормальный вид в течение нескольких секунд (Kanwisher, 1959).
Многие растения тоже обладают высокой толерантностью к замерзанию, и в холодные зимние дни вода в них превращается в лед, не вызывая необратимых повреждений. Толерантность к замораживанию зависит от сезона; растение в зимний период может выдержать охлаждение до —196 °C, тогда как летом погибает, если температура снижается на несколько градусов ниже точки замерзания (Weiser, 1970).
Замерзание воды в организме зависит не только от степени охлаждения, но и от длительности воздействия низкой температуры. На рис. 7.7 такая зависимость показана на примере зимующих личинок хлебного пилильщика Cephus cinctus. Если личинок содержали в условиях постоянной низкой температуры, то при температурах выше —15 °C образования льда в них не происходило. При более низких температурах личинки замерзали — тем быстрее, чем ниже была температура; так, при —30°C личинки промерзали за 1,2 с, тогда как при —17 °C для промерзания 50% личинок в большой выборке требовалось более года.
Давно уже известно, что глицерин защищает эритроциты н сперму млекопитающих от повреждения при замораживании. Гли
Экстремальные температуры; пределы для жизни 313
церин широко используют с этой целью; его добавляют в надлежащей концентрации к человеческой или бычьей сперме, которую затем замораживают, и спермин сохраняют в таком состоянии жизнеспособность в течение нескольких лет. Без глицерина замороженные спермин погибают.
Глицерин содержится в высокой концентрации у некоторых на-
Температура, °C
Рис. 7.7. Зависимость между температурой среды и средним временем, в течение которого промерзает 50% личинок хлебного пилильщика. (Salt, 1966.)
было высказано предположение, что именно этим объясняется способность насекомых выживать при низкой температуре. Возможны два механизма повышения устойчивости насекомых к холоду благодаря наличию глицерина: 1) глицерин мог бы защищать от повреждения льдом ткани тех насекомых, которые при низких температурах промерзают, и 2) понижая точку замерзания жидкостей тела и увеличивая тем самым возможную степень переохлаждения, он мог бы способствовать тому, чтобы в организме вообще не образовывался лед.
У многих насекомых концентрация глицерина возрастает перед наступлением зимы. Например, у паразитической осы Bracon cephi она к зиме достигает 5 молей на 1 кг воды (около 30%), а к весне вновь уменьшается. Это огромное количество растворенного вещества снижает температуру замерзания гемолимфы до —17,5 °C. Точка переохлаждения снижается в еще большей степени, и в результате личинка Bracon может быть переохлаждена (без обра
314 Глава 7. Влияние температуры
зования льда) до —47,2 °C. Зависимость между точками переохлаждения и замерзания (точнее, оттаивания) личинки показана на рис. 7.8. Сплошная линия представляет собой линию регрессии, вычисленную из всех полученных данных; прерывистой линией показано, как пошла бы кривая, если бы разность между точками замерзания и переохлаждения была постоянной и составляла 29°.
Рис. 7.8. Зависимость между наблюдаемым переохлаждением и точкой плавления для личинок паразитической осы Bracon cephi (Salt, 1959.)
Сплошная линия — линия регрессии, вычисленная из всех полученных данных; пунктиром показана постоянная степень переохлаждения на 29°.
Почти все экспериментальные точки лежат ниже прерывистой линии, т. е. почти у всех исследованных особей степень переохлаждения превышала 30°, причем она возрастала по мере снижения температуры оттаивания.
Однако глицерин не единственный фактор, защищающий насекомых от мороза и повреждающего действия замерзания. Из 11 видов насекомых, собранных в Альберте (Канада), три вида оказались толерантными к замерзанию, хотя у одного из них содержалось менее 3% глицерина, а у другого глицерина вообще не было. Остальные восемь видов, у большинства из которых было больше 15% глицерина, погибали при замораживании. Очевидно,, одного лишь глицерина недостаточно для защиты насекомых от
Экстремальные температуры; пределы для жизни 315
повреждения при замерзании. Имеет место также значительная степень адаптации: насекомые, подвергнутые воздействию умеренного холода, в последующем проявляют повышенную устойчивость к замерзанию, и у них наблюдается большее переохлаждение. Это относится как к накапливающим, так и к не накапливающим глицерин насекомым. Таким образом, толерантность к холоду нельзя полностью объяснить присутствием глицерина; имеют значение и какие-то другие компоненты гемолимфы (Somme, 1964, 1967).
АНТИФРИЗЫ РЫБ
Осмотическая концентрация жидкостей тела у костистых рыб составляет около 300—400 мосмоль; это соответствует температуре замерзания примерно от —0,6 до —0,8 °C. Температура морской воды в приполярных областях часто снижается до —1,8 °C, и тем не менее там обитает множество различных рыб1. Почему же эти рыбы не замерзают, когда плавают в воде при —1,8 °C? Не снижена ли у них по сравнению с обычными рыбами точка замерзания жидкостей? Или они на протяжении всей жизни находятся в переохлажденном состоянии? По-видимому, реализуются обе эти возможности.
Эта проблема изучалась в Хеброн-фиорде в северном Лабрадоре, где летом температура поверхностных слоев воды бывает на несколько градусов выше точки замерзания, а у дна остается равной —1,73 °C в течение всего года. В этом фиорде обитает несколько видов рыб, как в поверхностных, так и в придонных водах, и они служат превосходным материалом для изучения проблемы адаптации к холоду.
Летом точка замерзания крови у всех рыб в Хеброн-фиорде примерно такая же, как и у других костистых рыб, а именно около —0,8 °C (рис. 7.9). Для поверхностных рыб фиорда при этом не возникает никаких проблем, тогда как глубинные рыбы, очевидно, должны находиться в переохлажденном состоянии. Рыб, выловленных на глубине, в лаборатории можно переохладить (без промерзания) до той температуры, при которой они живут в море (—1,73 °C). Если же к такой рыбе прикоснуться куском льда, то в ее теле сразу начнется образование льда и она почти тотчас же погибнет. Это позволяет заключить, что придонные рыбы всю жизнь существуют в состоянии переохлаждения.
1 Содержание солей в морской воде соответствует концентрации, равной почти 1,0 осмолю на 1 л. Молярная депрессия точки замерзания воды равна 1,86 °C, поэтому морская вода не замерзает до тех пор, пока ее температура не снизится до —1,86 °C. Когда вода, морская или пресная, начинает замерзать, лед сначала плавает на поверхности. Пресная вода имеет наибольшую плотность при +4 °C, и когда на поверхности озера образуется лед, температура воды у дна может все еще оставаться на уровне +4 °C. Морская вода не обладает этой особенностью, и поэтому в полярных морях, поверхность которых покрыта льдом, температура воды повсюду равна примерно —1,8 °C.
316 Глава 7. Влияние температуры
С беспозвоночными дело обстоит проще. Морские беспозвоночные, как правило, находятся в осмотическом равновесии с окружающей их водой и благодаря этому не замерзают до тех
пор, пока остаются в воде.
Зимой придонные рыбы Хеброн-фиорда находятся в тех же условиях, что и летом, и температура замерзания остается у них примерно равной —0,8 °C. Температура воды тоже остается неизменной (—1,73°С), и
рыбы существуют в переохлажденном состоянии, так как промерзание тотчас же их убило бы.
Рыбы, обитающие в верхних слоях, зимой соприкасаются со льдом, и будь они просто В пере-
точка замерзания плазмы крови, °C
охлажденном состоянии, они бы неминуемо промерзли. Но этого не происходит, так как зимой точка замерзания у них снижается примерно до уровня температуры воды, в которой они плавают. При этом количество главных ионов, растворенных в крови,— натрия и хлора — у них воз-
Рис. 7.9. Поверхностным рыбам Хеброн-фиорда (Лабрадор) летом не грозит промерзание. Между тем рыбы, обитающие на больших глубинах, сталкиваются с этой проблемой, так как они находятся в воде с температурой —1,73 °C, а температура замерзания их крови и других жидкостей тела равна —1 °C; в результате эти рыбы всегда находятся в переохлажденном состоянии. (Scholander et al., 1957.)
растает лишь ненамного; основную роль в снижении точки замерзания играет накопление вещества-антифриза, химическая природа которого в период, когда проводились описанные исследования, была не известна
(Gordon et al., 1962).
Позднее удалось выделить и детально изучить антифриз, содержащийся в крови антарктической рыбы Trematomus (фото 7.1). По своей химической природе он представляет собой гликопротеид, который находится в крови в трех формах с молекулярными весами соответственно 10 500, 17 000 и 21 500. В низких концентрациях (6 г/л) этот гликопротеид более эффективно предотвращает образование льда, чем хлористый натрий (Komatsu et al., 1970). Это кажется удивительным, так как молекула гликопротеида в несколько сот раз больше, чем молекула NaCl. Иными ело-
Экстремальные температуры; пределы для жизни 317
вами, если растворить равные по весу количества того и другого вещества в воде, то молярная концентрация гликопротеида будет в несколько сот раз ниже молярной концентрации NaCl. Степень депрессии точки замерзания обычно зависит от числа растворенных частиц, так что в расчете на молярную концентрацию эффективность гликопротеида в предотвращении образования льда необычайно высока.
Фото 7.1. Антарктическая рыба Trematomus. В ее крови содержится гликопротеид, обладающий свойствами антифриза. Благодаря этому рыба может жить в морской воде с температурой —1,8 °C, хотя осмотическое давление ее крови недостаточно, чтобы в отсутствие антифриза предотвратить образование льда при этой температуре. (Фото предоставлено A. L. DeVries, Калифорнийский университет в Сан-Диего.)
На рис. 7.10 показано, как снижается точка замерзания в присутствии глюкозы, хлористого натрия и антифриза в зависимости от их концентрации. Глюкоза и хлористый натрий понижают точку замерзания прямо пропорционально концентрации, но поскольку хлористый натрий диссоциирует на два иона — Na+ и С1“, он вдвое эффективнее глюкозы. Что касается антифриза, то даже в ничтожной концентрации он предотвращает образование льда в воде в 200—500 раз более эффективно, чем хлористый натрий.
Гликопротеид рыбы Trematomus целиком построен из повторяющихся субъединиц, состоящих из двух аминокислот — аланина (23%) и треонина (16%) —и присоединенного к треонину дисахарида— производного галактозы. Это вещество, по-видимому, является первым природным гликопротеидом, структура которого полностью выяснена (DeVries et al., 1970; Shier et al., 1972).
316 Глава 7. Влияние температуры
Возможны по крайней мере два механизма, с помощью которых гликопротеиды могли бы понижать точку замерзания водных растворов. Один из них — это структурирование воды в результате образования водородных связей с гликопротеидом; второй механизм состоит в том, что гликопротеид тормозит рост образующих-
Моли но 1кг HZO
Рис. 7.10. Вещество со свойствами антифриза, выделенное из антарктической рыбы Trematomus, в водном растворе с необыкновенной эффективностью препятствует образованию льда. В расчете на молярную концентрацию это вещество в сотни раз эффективнее, чем можно было бы ожидать. (DeVries, 1970.)
По оси абсцисс — моляльная концентрация вещества, по оси ординат — кажущаяся осмоляльная концентрация, вычисленная по снижению точки замерзания раствора.
щая тем самым присоединение новых молекул воды к кристаллической решетке. На существование второго механизма указывает то, что если раствор, содержащий гликопротеид, все же заморозить, то температура плавления образовавшегося льда будет намного выше той, которая была нужна, чтобы раствор замерз. Таким образом, «точка плавления» не снижается, что противоречит предположению об образовании структурированной воды. Этот •факт легче объяснить тем, что гликопротеиды снижают температуру, допускающую рост кристаллов, по-видимому, за счет того, что •его многочисленные ОН-группы препятствуют ориентации молекул Н2О на поверхности кристалла (DeVries, 1974) .
Гипотеза о торможении роста кристаллов льда согласуется с данными, полученными при изучении многих арктических и антарктических рыб. У придонных и глубинных рыб, которые никогда не соприкасаются со льдом, точка замерзания жидкостей тела близка к —0,8°C, т. е. эти рыбы существуют в переохлажденном на Г состоянии. У рыб, обитающих в верхних слоях воды и со
Физиологическая адаптация к перемене температуры 319"
прикасающихся с морским льдом при температуре —1,8 °C, точка замерзания тоже близка к —0,8°C, но плазма крови у них гораздо устойчивее к замораживанию благодаря торможению роста ледяных кристаллов. Плазма у этих рыб содержит больше белка, чем у других позвоночных, а белки затормаживают рост кристаллов-льда (Hargens, 1972).
Высокомолекулярные антифризы были обнаружены у многих рыб, принадлежащих к 11 филогенетически отдаленным друг or друга семействам (Duman, DeVries, 1975). Обнаружены существенные различия в химической структуре антифризов у рыб разных семейств, так что способность к синтезу таких веществ, по-видимому, выработалась у них независимо.
ФИЗИОЛОГИЧЕСКАЯ АДАПТАЦИЯ К ПЕРЕМЕНЕ ТЕМПЕРАТУРЫ
Пределы температурной толерантности у животных не являются строго фиксированными. Воздействие сублетальной температуры часто ведет к некоторой адаптации, и в результате летальная ранее температура становится для животного переносимой.
Нередко пределы температурной устойчивости неодинаковы зимой и летом. В зимнее время животное часто не страдает и даже остается активным при таком холоде, который летом оказался бы для него смертельным; и, наоборот, устойчивость к высоким температурам зимой ниже, чем летом. Такое изменение температурной' толерантности при изменении внешних условий называется акклиматизацией. Аналогичное явление можно наблюдать и в лаборатории, если содержать животных некоторое время при заданной' температуре. Для того чтобы отличать адаптивные (приспособительные) изменения, наблюдаемые в лаборатории, от природной: акклиматизации, реакцию животных на экспериментальные условия часто называют аккламацией (Hart, 1952; Prosser, 1973).
Возможна акклиматизация или акклимация к различным факторам внешней среды — температуре, напряжению кислорода, характеру пищи, влажности. Здесь мы рассмотрим главным образом реакции на перемену температуры: они изучены лучше, чем реакции на изменение других внешних факторов.
ГЕОГРАФИЧЕСКИЕ РАЗЛИЧИЯ И СЕЗОННЫЕ АДАПТАЦИИ
Очевидно, что животные Арктики живут, дышат и передвигаются при температурах, близких к точке замерзания, в условиях, в которых сходные животные тропической зоны погибли бы или по крайней мере полностью утратили бы активность. Арктические-рыбы и ракообразные активны в ледяной воде и погибают при температурах выше 10—20 °C. Сравнимые тропические формы, напротив, погибают при температуре ниже 15—20 °C. Таким образом,.
320 Глава 7. Влияние температуры
существуют явные различия в температурной толерантности животных разных географических зон (Scholander et al., 1953b).
Географическое распространение близко родственных животных, например различных видов лягушек рода Rana, тесно связано с их температурной выносливостью. Данные о четырех видах североамериканских лягушек, представленные на рис. 7.11, показы-
ВиЪ "Время размножения
Rana clamiians
4- ..о -]- Июнь
Rana palustris 4------------о 4-
Rana pipiens 4“ о 4"
Rana sylvatica -о 4-^
I I I_________I___I____I—J_____I
СереЪина апреля "Начало апреля Конец марта
О 5 10 15 20 25 30 35
Температура, °C
Рис. 7.11. Зависимость между температурной толерантностью и временем размножения у четырех видов североамериканских лягушек. (Мооге, 1939.) Горизонтальными отрезками показан диапазон температур, в котором возможно нормальное развитие; кружочки — средняя температура воды в период икрометания; крестики — летальные температуры.
Бают, что виды, продвинувшиеся дальше всех на север, являются .и самыми холодоустойчивыми.
Корреляция между ареалом вида и температурной толерантностью распространяется и на температуру воды в период размножения, а также на верхнюю и нижнюю температурные границы развития икры. Для лягушек, обитающих в более холодном кли-.мате, характерны более низкие минимальные и максимальные температуры, при которых возможно размножение и эмбриональное развитие (табл. 7.3).
Из четырех приведенных в таблице видов наиболее широкий ареал у Rana pipiens. Северные популяции этого вида отличаются от южных популяций таким же образом, как северные виды от южных. Вид R. pipiens представлен расами, у которых физиологические реакции на температурные условия различны. Ясно, что это связано с необычайно широкой областью распространения вида (Мооге, 1949).
Рассмотрим теперь некоторые встречающиеся в природе, а также вызванные экспериментально изменения в температурной толерантности. На значительной части земной поверхности сезонные изменения температуры достаточно велики, и в одно время года животное легко переносит такую температуру, которая гибельна для него в другое время года. Например, летом верхняя летальная температура для сомика Icialurus (Ameiurus) nebulosus почти 36 °C, зимой же эта рыба погибнет, если температура воды
Физиологическая адаптация к перемене температуры. 321
Таблица 7.3
Сравнение условий размножения и развития у четырех видов лягушек (Rana). (Moore, 1949)
R. sylva-tica R. pipiens R. palustris R. clami-tans
Последовательность размножения 1 2 3 4
Температура воды в период размножения, °C 10 12 15 25
Северная граница распространения, °с. ш. 67 60 55 50
Нижний предел температуры для развития икры, °C 2,5 6 7 11
Верхний предел температуры для развития икры, °C 24 28 30 35
Длительность периода между стадиями 3 и 20 при 18,5 °C, ч 87 116 126 138
превысит 28 °C (рис. 7.12). Эти изменения в температурной толерантности возникают в ответ на естественную сезонную смену условий, и потому их следует рассматривать как пример естественной акклиматизации.
Нижняя летальная температура тоже изменяется в зависимости от сезона. Например, если жуков Pterosticus brevicornis из Аляски заморозить летом (для этого нужна температура —6,6°C),
Рис. 7.12. Верхняя летальная температура для сомика Ictalurus (Ameiurus) nebulosus меняется в зависимости от сезона. Точки на графике соответствуют температуре, при которой за 12 ч погибает 50% особей. (Fry, 1947.)
то они все погибают. Зимой же эти жуки выживают при температуре ниже —35 °C и полном промораживании (Miller, 1969). Особенно интересна способность этих жуков выдерживать промерзание, так как часто утверждают, что толерантность к промерзанию свойственна только личинкам и куколкам насекомых, но не взрослым формам. Ясно, что последнее не является общим правилом.
21—1873
322 Глава 7. Влияние температуры
Таким образом, у животных имеются генетические различия в температурной устойчивости, связанные с их географическим распространением, и сезонные изменения (акклиматизация); последние можно моделировать в лабораторных условиях, подвергая животных соответствующим термическим воздействиям (акклнма-ция), но при этом определяемые генетически границы устойчивости остаются без изменений.
МАКСИМАЛЬНЫЙ ДИАПАЗОН ТОЛЕРАНТНОСТИ
Если границы толерантности зависят от тех температурных условий, в которых находилось животное ранее, то возникает вопрос, насколько эти границы могут быть расширены путем мед-
Рис. 7.13. Сплошная линия очерчивает полный диапазон температурной толерантности золотой рыбки. Если золотую рыбку подвергают акклимации при температуре, соответствующей какой-то точке пунктирной диагонали, то верхний температурный предел выживания акклимированных особей можно найти на верхней сплошной линии, а нижний предел — на нижней сплошной линии. Например, у рыбки, содержавшейся при температуре 30 °C, верхний летальный предел составляет 38 °C, а нижний — 9 °C. (Fry et al., 1942.)
ленной и постепенной акклимации. Есть ли какой-то предел, который не может быть перейден? Для того чтобы выяснить это, несколько групп животных содержат при разной (но постоянной для данной группы) температуре в течение периода, достаточного* для полной акклимации. После этого можно определить верхние и нижние границы температурной устойчивости для каждой труп
Физиологическая адаптация к перемене температуры 323
пы и результаты, полученные после акклимации к ряду температур, представить в виде графика. На рис. 7.13 приведен такой график, дающий полную характеристику термической толерантности золотой рыбки.
Золотая рыбка способна переносить колебания температуры в исключительно широком диапазоне, и поэтому площадь, ограниченная кривой толерантности, очень велика. У рыб с более узкой
Рис. 7.14. Границы температурной толерантности кеты, обитающей в холодных водах, значительно уже, чем у сомика Ictalurus. Последний по широте своей толерантности сходен с золотой рыбкой (рис. 7.13). (Brett, 1956.)
толерантностью, например у кеты (Oncorhynchus keta), ограничиваемая кривой площадь гораздо меньше. Кета, подобно другим лососевым, — типичная холодноводная рыба, и ее верхний температурный предел не может быть поднят выше 24 °C. Это видно из рис. 7.14, где показаны границы температурной толерантности кеты и сомика Ictalurus nebulosus, который сходен по диапазону устойчивости с золотой рыбкой. Площадь, ограниченная кривой толерантности, у Ictalurus почти вдвое больше, чем у кеты. Для измерения этой площади мы можем взять в качестве единицы квадрат со стороной, равной 1 градусу на каждой из осей координат. Тогда для Ictalurus величина площади составит 1162 таких квадрата [или, точнее, градуса в квадрате (°C)2], а для кеты — 468 квадратов.
При исследовании температурной толерантности других рыб большая часть данных укладывалась между приведенными край
21
324 Глава 7. Влияние температуры
ними величинами. Как сомик Ictalurus, так и кета выживают при температуре замерзания воды, тогда как у многих тропических рыб кривые толерантности не опускаются ниже О °C, а замыкаются при более высоких температурах. Полная картина температурной толерантности выяснена только для относительно небольшого числа рыб, но создается впечатление, что в целом пресноводные рыбы обладают более широкой толерантностью, чем морские.
Толерантность пресноводных рыб варьирует в пределах от 140 (°C)* 2 для лосося до 1220 (°C)2 для золотой рыбки (Brett,. 1956). Из трех изученных видов морских рыб толерантность оказалась наибольшей у менидии (Menidia menidia)—715 (°C)2, а наименьшей у иглобрюха (Spheroides maculatus) —550 (°C)2; камбала Pseudopleuronectus americanus занимала промежуточное положение (Hoff, Westman, 1966).
Для удобства сравнения эти данные, а также результаты, полученные на других видах, сопоставлены в табл. 7.4. Большая
Таблица 7.4
Температурная толерантность различных пресноводных и морских рыб, выраженная в градусах в квадрате (объяснение см. в тексте). Для сравнения в таблицу включен омар
Вид
Толерантность, (°C) 2
Золотая рыбка (Carassius auratus) 12201^
Сомик Ictalurus nebulosus 11621)
Калифорнийская гирелла (Girella nigricans) 8002)
Менидия (Menidia menidia) 7153)
Камбала Pseudopleuronectes americanus 6853 4>
Американская палия (Salvelinus fontinalis) 6254)
Иглобрюх (Spheroides maculatus) 5503)
Кета (Oncorhynchus keta) 468х)
Омар (Homarus americanus) 8305^
’) Brett, 1956.
2) Doudoroff, цит. по McLeese, 1956.
3) Hoff, Westman, 1966.
4) Fry et al., 1946.
5) McLeese, 1956.
температурная толерантность сомиков и золотых рыбок, очевидно, обусловлена тем, что они живут в мелких пресных водоемах, которые гораздо легче прогреваются, чем быстрые холодные речки, где обитают лососи. В конце таблицы упоминается омар как важное промысловое животное; если бы он относился к рыбам, его* следовало бы поместить не в последней строке, а значительно выше.
Физиологическая адаптация к перемене температуры 325
Необходимо дальнейшее изучение температурной толерантно» сти рыб, так как вопрос о границах адаптивных возможностей в этом отношении становится все более актуальным в связи с проблемой термического загрязнения земного шара под влиянием производственной деятельности человека, особенно в результате работы электростанций и атомных реакторов.
СКОРОСТЬ АККЛИМАЦИИ
Повышение устойчивости к высокой температуре происходит у рыб сравнительно быстро. Сомик Ictalurus при подъеме температуры с 20 до 28 °C полностью приспосабливается к новым условиям менее чем за 24 ч (рис. 7.15). Для процесса температурной акклн-
Рис. 7.15. В воде с достаточным содержанием кислорода (I) температурная акклимация сомика Ictalurus происходит за 24 ч; при низкой концентрации О» (II) акклимация идет крайне медленно. (Fry, 1947.)
мации требуется достаточное снабжение кислородом. Хотя сомик легко переносит недостаток кислорода, дефицит последнего, по-видимому, полностью подавляет термическую акклимацию. Как правило, процесс акклимации идет тем быстрее, чем выше температура, так что он, очевидно, связан с метаболическими процессами; однако механизмы, ответственные за изменение температурного предела для выживания, остаются неясными, хотя известно, что важную роль здесь играют изменения ферментных систем (Hochachka, Somero, 1973).
Если сомику Ictalurus nebulosus предоставить выбор той или иной температуры воды, поместив его в условия температурного градиента, то этот выбор будет определяться предшествовавшими температурными условиями (табл. 7.5). Рыбки, акклимированные к 7°C, сначала выбирают 16°C, но вскоре начинают перебираться в более теплую воду. Спустя 10 ч они предпочитают 25 °C, а спустя 24 ч — уже 29—30°C, т. е. такую же температуру, какую через 24 ч выбирают и рыбки, акклимированные к 32 °C.
326 Глава 7, Влияние температуры
Таблица 7.5
Сомики Ictalurus nebulosus, акклимированные к различным температурам от 7 до 32 °C, вначале расходятся в выборе предпочтительных температурных условий, но спустя 24 ч все они одинаково отдают предпочтение температуре около 29 °C. (Crawshaw, 1975)
Температура акклимации, °C Предпочитаемая температура, °C
вначале спустя 10 ч спустя 24 ч
7 16 25 -/29
15 21 25 -/29
24 26 29 —/29
32 31 30 —/29
Если рыбу переместить из более теплой воды в более холодную, то потеря толерантности к высокой температуре и увеличение устойчивости к низкой будут происходить довольно медленно. Например, если для рыбы Pimephales promelas понизить температуру с 24° до 16 °C, то полная адаптация произойдет только через 10—20 дней (Brett, 1956).
Итак, мы видим, что толерантность у рыб зависит от предшествовавших температурных условий и на быстроту адаптации к новому температурному режиму влияет обеспеченность кислородом. Существенное значение имеют и другие факторы, в частности возраст и размеры рыб, качество воды. Особенно интересно то, что у золотых рыбок, даже если их содержат в лаборатории в условиях полного постоянства температуры и питания, сохраняются заметные сезонные изменения температурной толерантности — зимой повышается устойчивость к холоду. Этот феномен связывают с фотопериодом (Hoar, 1956). Как правило, длинный световой день, подобный летнему, увеличивает устойчивость к теплу, а короткий, подобный зимнему, — устойчивость к холоду (Roberts, 1964).
ТЕМПЕРАТУРНАЯ АККЛИМАЦИЯ И ИНТЕНСИВНОСТЬ МЕТАБОЛИЗМА
До сих пор мы рассматривали летальные температуры и адаптацию как результат воздействия предшествовавших температурных условий. Адаптация, однако, не ограничивается изменением температурных пределов для жизни; существуют и другие компенсаторные механизмы, которые противодействуют прямому влиянию перемены температуры. Каждый, кто ловил рыбу зимой, знает, что рыба в этот период не кажется заметно более медлительной или сонной, чем летом. Экспериментальные исследования подтверждают это впечатление; действительно, по истечении некоторого
Физиологическая адаптация к перемене температуры 327
времени эффект понижения температуры в значительной мере может компенсироваться соответствующими изменениями интенсивности обмена. Это касается не только рыб, но и многих других, пойкилотермных животных.
Предположим, что у рыбы, адаптированной к зимней температуре 5°C, температурная толерантность лежит в пределах от 0 до
Рис. 7.16. Предполагаемое изменение интенсивности обмена (потребления кислорода) у рыбы, акклимированной к температуре воды 5 °C (а). При последующей акклимации такой рыбы к 25 °C изменение интенсивности обмена в случае полной компенсации оисывается прямой б, а при полном отсутствии компенсации— пунктирной линией а'. Шкала по оси ординат логарифмическая.
25°C и величина Qio на всем этом интервале равна 2,0. Интенсивность обмена у этой рыбы описывается прямой а на рис. 7.16. Допустим далее, что мы акклимируем эту рыбу к летней температуре 25 °C и что при этом верхний летальный предел отодвигается у нее к 40 °C. Если не произойдет никаких компенсаторных явлений в метаболической регуляции, то интенсивность обмена должна будет возрасти в соответствии с пунктирной линией а'. Если же эффект перемены температуры будет полностью скомпенсирован, то уровень метаболизма при 25 °C останется таким же, каким он был при 5 °C. Если не изменится и Qi0 (2,0), то потребление кислорода этой рыбой будет описываться прямой б. В итоге изменение температуры будет полностью скомпенсировано и интенсивность метаболизма вернется к исходному уровню.
Помимо двух рассмотренных случаев (отсутствие компенсации и полная компенсация) возможны и другие типы адаптации
~328 Глава 7. Влияние температуры
к длительному изменению температуры. В общем виде они представлены на рис. 7.17. Если у животного не происходит компен-•сации и потребление кислорода продолжается со скоростью, соответствующей острому эффекту перемены температуры, то интенсивность обмена описывается прямой а (которая идентична прямой а' на рис. 7.16). С другой стороны, при полной компенсации потребление кислорода после адаптации животного к новым тем-
Рис. 7.17. При изменении температуры от 7\ до Т2 процесс компенсации может протекать по-разному в соответствии с изображенными на рисунке кривыми. (Precht et al., 1955.) Подробности в тексте.
пературным условиям возвращается к исходному уровню, и поэтому потребление кислорода описывается прямой б, которая соответствует полной компенсации. Любой ответ, укладывающийся на прямую, расположенную между линиями а и б, соответствует состоянию частичной компенсации эффекта перемены температуры. Возможен также случай «сверхкомпенсации», когда потребление кислорода изменяется в соответствии с прямой 7. Наконец, описано несколько случаев, когда сдвиг температуры (по крайней мере в какой-то части температурного диапазона) вызывал обратный "(противоположный компенсации) эффект, описываемый прямой 5.
На самом деле у животных редко встречаются эти идеальные типы ответа: обычно их реакция намного более сложна. Примером могут служить реакции двух водных животных — форели и европейского речного рака — на снижение температуры на 10° и последующее, после полной адаптации, повышение температуры на 10° (рис. 7.18).
Крайне мала или даже вовсе отсутствует компенсация у рачка-бокоплава Talorchestia megalophthalma из района Вудс-Хоул (штат Массачусетс). Если измерять количество потребляемого рачками кислорода при одной и той же температуре, оно окажется
Физиологическая адаптация к перемене температуры 329
одинаковым зимой и летом. Таким образом, здесь практически нет акклиматизации или компенсации сезонных изменений температуры. В результате зимой метаболизм у этих рачков настолько снижается, что они теряют подвижность и впадают в оцепенение, закопавшись в песок в маленькие норки. Если их потревожить зимой, то реакция окажется крайне вялой, но стоит поместить рачка
Рис. 7.18. Влияние изменения температуры на уровень метаболизма у речного рака и форели. (Bullock, 1955.)
Первоначально животные были акклимированы к 15 °C. Когда температура воды понизилась на 10°, интенсивность обмена у форели быстро достигла нового плато; у рака этот процесс шел медленнее. При повышении температуры вновь до 15 °C у форели наблюдалась сверхкомпенсация с последующим воз* вращением к исходному уровню обмена; рак реагировал на это изменение температуры очень медленно.
в тепло — и он вскоре оживится, как летом (Edwards, Irving, 1943 b).
В отличие от этого песчаный крабик (Emerita talpoida) —дру-гое ракообразное из того же местообитания — компенсирует зимнее понижение температуры усилением метаболизма. Зимой при 3°С он потребляет в 4 раза больше кислорода, чем летом при той же температуре. В холодную погоду он остается активным, но поскольку песчаные берега в районе Вудс-Хоула зимой крайне негостеприимны, он уходит из литоральной зоны на глубину (Edwards, Irving, 1943 а). Таким образом, песчаный крабик приспосабливается к сезонной смене температур и зимой остается активным, а не впадает в оцепенение. Верхняя предельная температура для песчаного крабика также зависит от времени года. Зимой для него детальны температуры выше 27 °C, а летом — выше 37 °C.
33 Э Глава 7. Влияние температуры
Мы рассмотрели ряд важных физиологических эффектов тепла и холода — не только летальное действие очень высоких или очень низких температур, но и приспособительные реакции животных в нормальном для них температурном диапазоне. Теперь мы перейдем к вопросу о том, каким образом животные приобретают независимость от изменений температуры окружающей их среды.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Bott Т. L., Brock Т. D. (1969). Bacterial growth rates above 90 °C in Yellowstone hot springs, Science, 164, 1411—1412.
Brett J. R. (1956). Some principles in the thermal requirements of fishes, Q. Rev.
Biol., 31, 75—87.
Brock T. D. (1970). High temperature systems, Annu. Rev. Ecol. Syst., 1, 191— 220.
Brown J. H., Feldmeth C. R. (1971). Evolution in constant and fluctuating environments: Thermal tolerances of desert pupfish (Cyprinodon), Evolution, 25, 390—398.
Bullock T. H. (1955). Compensation for temperature in the metabolism and activity of poikilotherms, Biol. Rev., 30, 311—342.
Carlisle D. B. (1968). Triops (Entomostraca) eggs killed only by boiling, Science, 161, 279—280.
Crawshaw L. I. (1975). Attainment of the final thermal preferendum in brown bullheads acclimated to different temperatures, Comp. Biochem. Physiol., 52A, 171—173.
DeVries A. L. (1970). Freezing resistance in antarctic fishes. In: Antarctic Ecology, vol. 1 (M. Holdgate, ed.), pp. 320—328, New York, Academic Press.
DeVries A, L. (1974). Survival at freezing temperatures. In: Biochemical and Biophysical Perspectives in Marine Biology, vol. 1 (D. C. Malins and J. R. Sargent, eds.), pp. 289—330, New York, Academic Press.
DeVries A. L., Komatsu S. K., Feeney R. E. (1970). Chemical and physical properties of freezing point-depressing glycoproteins from antarctic fishes, J. Biol. Chem., 245, 2901—2908.
Duman J. G., DeVries A. L. (1975). The role of macromolecular antifreezes in cold water fishes, Comp. Biochem. Physiol., 52A, 193—199.
Edwards G. A., Irving L. (1943a). The influence of temperature and season upon the oxygen consumption of the sand crab, Emerita talpoida Say, J. Cell. Comp. Physiol., 21, 169—182.
Edwards G, A., Irving L. (1943b). The influence of season and temperature upon the oxygen consumption of the beach flea, Talorchestia megalophthalma, J. Cell. Comp. Physiol., 21, 183—189.
Fraenkel G. (1968). The heat resistance of intertidal snails at Bimini, Bahamas, Ocean Springs, Mississippi and Woods Hole, Massachusetts, Physiol. Zool., 41, 1—13.
Fry F. E. I. (1947). Effects of the environment on animal activity, University of Toronto Studies, Biological Services, No. 55, Publ. Ontario Fish. Res. Lab., 68, 1—62.
Fry F, E. I., Brett I. R., Clawson G. H. (1942). Lethal limits of temperature for young goldfish, Rev. Can. Biol., 1, 50—56.
Fry F. E. I., Hart I. S., Walker K. F. (1946). Lethal temperature relations for a sample of young speckled trout (Salvelinus fontinalis), University of Toronto Studies, Biological Series, No. 54, Publ. Ontario Fish. Res. Lab., 66, 1—35.
Gordon M, S., Amdur В. H., Scholander P. F. (1962). Freezing resistance in some northern fishes, Biol. Bull., 122, 52—56.
Hargens A. R. (1972). Freezing resistance in polar fishes, Science, 176, 184— 136.
Физиологическая адаптация к перемене температуры 331
Hart J. S. (1952). Geographic variations of some physiological and morphological characters in certain freshwater fish, University of Toronto Studies, Biological Series, No. 60, Publ. Ontario Fish. Res. Lab., 72, 1—79.
Hinton H. E. (1960). A fly larva that tolerates dehydration and temperatures of —270 °C to +102 °C, Nature, Lond., 188, 336—337.
Hoar W. S. (1956). Photoperiodism and thermal resistance of goldfish, Nature, Lond., 178, 364—365.
Hochachka P. W., Somero G. N. (1973). Strategies of Biochemical Adaptation, Philadelphia, Saunders, 358 pp. [Имеется перевод: Хочачка П., Сомеро Дж. Стратегия биохимической адаптации. — М.: Мир, 1977.]
Hoff J. G., West man J. R. (1966). The temperature tolerances of three species of marine fishes, J. Mar. Res., 24, 131 —140.
Kanwisher J. (1959). Histology and metabolism of frozen intertidal animals, Biol. Bull., 116, 258—264.
Komatsu S. K., DeVries A. L, Feeney R. E. (1970). Studies of the structure of freezing point-depressing glycoproteins from an antarctic fish, J. Biol. Chem., 245, 2909—2913.
Lowe С. H., Lardner P. J., Halpern E. A. (1971). Supercooling in reptiles and other vertebrates, Comp. Biochem. Physiol., 39A, 125—135.
Marzusch K. (1952). Untersuchungen fiber die Temperaturabhangigkeit von Le-bensprozessen bei Insekten unter besonderer Berficksichtigung winterschlafen-der Kartoffelkafer, Z. Vergl. Physiol., 34, 75—92.
McLeese D. W. (1956). Effects of temperature, salinity and oxygen on the survival of the American Lobster, J. Fish. Res. Bd. Can., 13, 247—272.
Miller L. K. (1969). Freezing tolerance in an adult insect, Science, 166, 105— 106.
Moore J. A. (1939). Temperature tolerance and rates of development in the eggs of Amphibia, Ecology, 20, 459—478.
Moore J. A. (1949). Geographic variation of adaptive characters in Rana pipiens Schreber, Evolution, 3, 1—24.
Pitkow R. B. (1960). Cold death in the guppy, Biol. Bull., 119, 231—245.
Precht H., Christopher sen J., Hensel H. (1955). Temperatur und Leben, Berlin, Springer-Verlag, 514 pp.
Prosser C. L. (1973). Comparative Animal Physiology, 3rd ed., Philadelphia, Saunders, 966 pp.
Roberts J. L. (1964). Metabolic responses of fresh-water sunfish to seasonal photoperiods and temperatures, Helgol. Wiss, Meeresunter, 9, 459—473.
Salt R. W. (1959). Role of glycerol in the cold-hardening of Bracon cephi (Gahan), Can. J. Zool., 37, 59—69.
Salt R. W. (1966). Relation between time of freezing and temperature in supercooled larvae of Cephas cinctus Nort, Can. J. Zool., 44, 947—952.
Scholander P. F., Flagg W., Hock R. J., Irving L. (1953a). Studies on the physiology of frozen plants and animals in the arctic, J. Cell. Comp. Physiol., 42 (Suppl. 1), 1-56.
Scholander P. F., Flagg W., Walters V. , Irving L. (1953b). Climatic adaptation in arctic and tropical poikilotherms, Physiol. Zool., 26, 67—92.
Scholander P. F., Van Dam L.f Kanwisher J. W., Hammel H. T., Gordon M. S\ (1957). Supercooling and osmoregulation in arctic fish, J. Cell. Comp. Phy-' siol., 49, 5—24.
Shier W. T., Lin Y., DeVries A. L. (1972). Structure and mode of action of glycoproteins from an antarctic fish, Biochim. Biophys. Acta, 263, 406—413L
Somero G, N., DeVries A. L. (1967). Temperature tolerance of some antarctic fishes, Science, 156, 257—258.
Samme L. (1964). Effects of glycerol on cold-hardiness in insects, Can. J. Zool.» 42, 87—101.
Samme L. (1967). The effect of temperature and anoxia on haemolymph composition and supercooling in three overwintering insects, J. Insect. Physiol., 13» 805—814.
332 Глава 7. Влияние температуры
Weiser С. J. (1970). Cold resistance and injury in woody plants, Science, 169, 1269—1278.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Asahina E. (1969). Frost resistance in insects, Adv. Insect Physiol., 6, 1—49.
Hochachka P. W., Sotnero G. N. (1973). Strategies of Biochemical Adaptation, Philadelphia, Saunders, 358 pp. [Имеется перевод: Хочачка П., Сомнер Дж. Стратегия биохимической адаптации. — М.: Мир, 1977.]
Precht H.t Christophersen J., Hensel H., Larcher W. (1973). Temperature and Life, New York, Springer-Verlag, 779 pp.
Somero G, N., Hochachka P. W. (1976). Biochemical adaptations to temperature. In: Adaptation to Environment: Essays on the Physiology of Marine Animals (R. C. Newell, ed.), pp. 125—190, London, Butterworth.
ГЛАВА 8
ТЕРМОРЕГУЛЯЦИЯ
В главе 7 рассматривалось влияние температуры на физиологические процессы у тех многочисленных животных, у которых температура тела практически равна температуре окружающей среды. Независимость от этого внешнего фактора должна, видимо, создавать большие преимущества, и в этой главе мы рассмотрим животных, у которых температура тела поддерживается на более или менее постоянном уровне независимо от условий среды. Мы начнем с птиц и млекопитающих, так как в этом отношении они достигли наибольшего совершенства; далее мы перейдем к другим животным, способным в известной мере регулировать температуру своего тела независимо от внешней температуры.
Прежде всего необходимо будет разъяснить, что мы понимаем под таким отнюдь не простым термином, как температура тела. Далее придется вспомнить о том, что температура какого-либо тела остается постоянной лишь тогда, когда приток тепла и его потеря (отдача) равны между собой. Для понимания последних двух процессов необходимо знакомство с элементарной физикой переноса тепла.
В условиях холода тепловой баланс животного может быть стабилизирован путем уменьшения отдачи тепла и (или) путем увеличения его притока (выработки). Большинство птиц и млекопитающих весьма успешно справляются с этой задачей, однако некоторые млекопитающие и несколько видов птиц в определенных условиях «пасуют»: температура тела у них стремительно снижается и они впадают в оцепенение или зимнюю спячку. Но и в этом случае регуляция температуры тела не прекращается полностью; напротив, спячка представляет собой хорошо регулируемое и управляемое состояние.
В слишком теплой среде для поддержания постоянной температуры тела требуются противоположные меры: организм должен препятствовать повышению своей температуры и нередко бывает вынужден снижать ее путем испарения влаги.
Терморегуляция у птиц и млекопитающих настолько эффективна, что колебания температуры тела на протяжении почти всей жизни этих животных не превышают нескольких градусов. Однако •и некоторые представители других классов обладают удивительной способностью поддерживать температуру своего тела на уровне, отличном от температуры среды. Таковы наземные позвоночные (например, ящерицы), многие насекомые, особенно высоко-
334 Глава 8. Терморегуляция
активные, и даже, как это ни странно, некоторые рыбы, у которых температура отдельных областей тела может приближаться к температуре «теплокровных» животных.
ТЕМПЕРАТУРА ТЕЛА У ПТИЦ
И МЛЕКОПИТАЮЩИХ
ЧТО ТАКОЕ ТЕМПЕРАТУРА ТЕЛА?
Чтобы тепло, выработанное организмом животного, могло быть отдано внешней среде, оно должно быть сначала перенесено к поверхности тела. Поэтому температура у поверхности тела должна быть ниже, чем внутри, иначе переноса тепла не будет. Отсюда с необходимостью вытекает, что температура разных частей организма не может быть одинаковой.
Если мы посмотрим, где в организме млекопитающего происходит выработка тепла, то мы увидим, что в одних местах его образуется больше, чем в других. У человека на долю органов грудной и брюшной полостей, составляющих менее 6 % массы тела, приходится 56 % всей теплопродукции организма (табл. 8.1).
Таблица 8Д
Выработка тепла важнейшими органами человека в покое (вес тела 65 кг, общая теплопродукция 1872 ккал в сутки = 78 ккал/ч = 90,65 Вт).
Вес главных внутренних органов составляет около 5 кг, но они обеспечивают более 72% общей теплопродукции (Aschoff et а!., 1971)
Вес органа Продукция тепла
Орган % от веса кг тела % от общей ккал/ч теплопродук- ции
Почки 0,29] 0,45 6,0 7,7
Сердце 0,29 0,45 8,4 10,7
Легкие Мозг Органы брюшной об- 0,60 1,35 2,50. 5,03 0,9 2,1 3,8 7,7 3,4 12,5 4,4 16,0 - 72,4
ласти (кроме почек) 26,2 33,6 ,
Кожа 5,00 ] 7,8 1 1,5 1,9 ]
Мышцы 27,00 ^59,97 41,5 92,3 12,2 15,7 > 27,6
Другие органы 27,97 J 43,0 ) 7,8 10,0 J
Всего 65,00 100,0 78,0 100,0
Если сюда присоединить и головной мозг, который у человека велик по размерам и вырабатывает много тепла, то мы получим уже 72 %, т. е. более двух третей общей теплопродукции.
Таким образом, мы можем подразделить тело на внутреннюю .часть, где вырабатывается основное количество тепла, и гораздо
Температура тела у птиц и млекопитающих 335
большую по размеру «периферию», включающую кожу и мышцы и продуцирующую лишь небольшую долю общего тепла тела. При физической работе, когда интенсивность метаболизма возрастает в 10 и более раз, положение меняется, поскольку это усиление обмена происходит главным образом в мышцах (включая диафрагму и другие дыхательные мышцы). Для того чтобы при физической нагрузке внутренняя температура тела оставалась постоянной, перенос тепла к поверхности тела должен возрастать более чем в 10 раз по сравнению с состоянием покоя.
РАСПРЕДЕЛЕНИЕ ТЕМПЕРАТУРЫ В ТЕЛЕ
Температура внутренних органов поддерживается на довольно постоянном уровне, но это не значит, что она повсюду одинакова. Органы, вырабатывающие больше тепла, могут быть теплее других, хотя они охлаждаются кровью (т. е. оттекающая от них венозная кровь теплее, чем артериальная). Разница в температуре может достигать 0,5°. Поэтому нельзя говорить о какой-то одной температуре всех внутренних органов, хотя для практических целей в качестве таковой часто используют глубокую ректальную температуру.
У человека при сбалансированном теплообмене температура поверхности тела всегда ниже внутренней температуры. Это означает, что артериальная кровь, притекающая к поверхностным тканям, теряет часть тепла, * и оттекающая от них венозная кровь холоднее, чем артериальная. Именно таким образом в основном осуществляется перенос тепла к поверхности тела, или, другими словами, так охлаждаются внутренние органы. В зависимости от обстоятельств (температуры среды и потребности в теплоотдаче) температура кожи подвержена значительным колебаниям. Кроме того, подлежащие ткани, в том числе значительная часть мышечной массы, могут иметь температуру значительно более низкую, чем внутренние органы (рис. 8.1).
Что такое средняя температура тела? Ее можно вычислить из результатов многочисленных измерений температуры в разных участках тела при условии, что имеется какой-то метод «взвешивания» вкладов различных измерений. Однако такой расчет средней температуры не имеет большого смысла с физиологической точки зрения. Как мы уже видели, внутренняя температура не отражает теплового состояния организма в целом, периферическая температура колеблется в широких пределах, а та глубина, на которую она распространяется, может очень резко изменяться. Изменение периферической температуры тела указывает на сдвиг в общем количестве тепла в организме, хотя внутренняя температура может оставаться при этом постоянной. Когда человек переходит из помещения с температурой 35 °C в обычную комнату ч(20°С), снижение периферической температуры может соответст
336 Глава 8. Терморегуляция
вовать потере тепла в количестве 200 ккал (>800 кДж). Чтобы представить себе величину этой потери, достаточно сказать, что она примерно равна 3-часовой выработке тепла организмом в покое.
Рис. 8.1. Температура различных областей тела человека при температуре воздуха 20 °C (слева) и 35 °C (справа). (Aschoff, Wever, 1958.)
Как показывают изотермы (линии, соединяющие точки с одинаковой температурой), при 35°C внутренняя температура (заштрихованная область), равная 37 °C, распространяется на ноги и руки. При 20 °C область внутренней температуры ограничена только туловищем и головой, а по всей длине рук и ног отмечается постепенный спад температуры.
«НОРМАЛЬНАЯ» ТЕМПЕРАТУРА ТЕЛА
У ПТИЦ И МЛЕКОПИТАЮЩИХ
СУТОЧНЫЕ КОЛЕБАНИЯ ВНУТРЕННЕЙ ТЕМПЕРАТУРЫ
Внутренняя температура у человека и других млекопитающих, а также у птиц подвержена регулярным суточным колебаниям, обычно в пределах 1—2°. У дневных животных температурный максимум наблюдается днем, а минимум — ночью; у ночных животных— наоборот. Однако такой суточный ритм не связан непосредственно со сменой активности и покоя, поскольку он сохраняется и в том случае, если животное все время находится в полном покое.
Температура тела у птиц и млекопитающих 337
У многих млекопитающих и птиц суточные колебания температуры тела соответствуют периодам света и темноты. Например^, если тауи (Pipilo aberti, сем. овсянковых) содержать в условиях. 12 ч — темнота и 12 ч — свет, то колебания температуры птицьь будут синхронны с циклом освещенности. При комнатной температуре 23 °C внутренняя температура у тауи ночью составляет
Рис. 8.2. Когда птицу тауи (Pipilo aberti) содержали при постоянной температуре 23 °C, то температура ее тела изменялась в соответствии с циклом освещенности. Когда в 6.00 часов включали свет, температура тела поднималась, на 3°; при выключении света в 18.00 часов она возвращалась к исходному уровню. Если температуру помещения снижали до 5 °C, цикличность изменения' температуры тела сохранялась, но на несколько более высоком уровне. (Dawson, 1954.)
около 39 °C, а при включении света быстро возрастает почти до* 42 °C (рис. 8.2). При другой температуре помещения, например при 5 °C, внутренняя температура совершенно так же следует за сменой дня и ночи, хотя в целом она примерно на 0,5° выше, чем в предыдущем случае.
Аналогичные суточные циклы были отмечены у многих других птиц, принадлежащих к различным семействам. У дневных птиц пик температуры всегда наблюдается днем, а у ночных — ночью. Примером ночной птицы может служить нелетающий обитатель Новой Зеландии — киви (Apteryx). Внутренняя температура у киви ниже, чем у большинства других птиц, составляя в среднем 36,9 °C в дневное время. Ночью — в период активности этой птицы— температура возрастает в среднем до 38,4 °C (т. е. ночная температура примерно на 1,5° выше дневной) (Earner, 1956).
Как ритм колебаний температуры, так и соответствующий ритм интенсивности метаболизма могут быть обращены при обращении периодов темноты и света. Это показывает, что весь цикл определяется освещенностью (Dawson, 1954). Однако, если содержать животных круглосуточно в условиях постоянной и равномерной освещенности, суточные колебания температуры в основном сохраняются. А поскольку правильный ритм поддерживается без каких-либо внешних регулирующих факторов, значит, он присущ самому организму и представляет собой истинно эндогенный ритм. Он является самоподдерживающимся в том смысле, что продолжает-22—1873
ч338 Глава 8. Терморегуляция
ся дни, недели и даже месяцы, хотя обычно период его постепенно отклоняется от точного 24-часового. Поэтому для того, чтобы измерение температуры тела действительно имело смысл, желатель-1 но иметь информацию об активности животного, времени суток и нормальном для данного вида цикле температуры тела.
РАЗЛИЧИЯ в ТЕМПЕРАТУРЕ ТЕЛА У РАЗНЫХ ГРУПП ЖИВОТНЫХ
Пренебрегая разницей в 1—2°, можно считать, что каждой из основных групп теплокровных позвоночных свойственна постоянная, «нормальная» для данной группы внутренняя температура чела, однако между группами существуют характерные различия (табл. 8.2).
Таблица 8.2
Приближенные величины нормальной и летальной внутренней температуры у некоторых крупных групп млекопитающих и птиц. Цифры для летальных температур основаны на наблюдениях, сделанных в весьма разнообразных условиях. Интервал между нормальной и летальной температурами довольно постоянен и составляет примерно 6°
Группа животных
Нормальная внутренняя температура, тела, °C
Летальная внутренняя температура тела, °C
Однопроходные (ехидна) 30— 31
Сумчатые 35 —362 *)
Насекомоядные (ёж) 34—36
Человек 37
Высшие млекопитающие 36—383)
Птицы (киви) 384>
Птицы, кроме воробьиных 39—402>
Птицы, воробьиные 40—4121
371) 40—415 *>
416)
43 42—447 8>
468)
479, ю)
’) Schmidt-Nielsen et al., 1966.
2) Dawson, Hulbert, 1970.
’) Morrison, Ryser, 1952.
4) Farner, 1956.
6) Robinson, Morrison, 1957.
«) Shkolnik, Schmidt-Nielsen, 1976.
7) Adolph, 1947.
8) Robinson, Lee, 1946.
®) Calder, 1964.
,0) Dawson, 1954.
Часто трудно бывает установить, какая температура тела яв-•ляется для данного животного нормальной. Однако, пренебрегая изменениями температуры, обусловленными активностью животного и влиянием среды, можно все же сказать, что, как правило, температура тела у большинства птиц составляет 40±2°С, у плацентарных млекопитающих 38±2 °C, у сумчатых 36±2 °C и у однопроходных 31 ±2°C. У мелких птиц температура тела может быть
Температура тела у птиц и млекопитающих 339»
несколько выше, чем у крупных (McNab, 1966), но у млекопитающих нет ясной зависимости между размерами тела и температурой-(Morrison, Ryser, 1952); в отношении сумчатых этот последний вопрос исследован недостаточно.
В связи с тем, что так называемые «примитивные» группы — насекомоядные, сумчатые и особенно однопроходные — обладают* относительно низкой температурой тела, возникает ряд интересных эволюционных проблем. Эти группы считаются очень древними; значит, у них, вероятно, было больше времени, чем у теплокровных более позднего происхождения, для эволюции в направлении высокой температуры тела, если бы это качество было «желательным» и давало преимущества. Почему же они остались на* своем «примитивном» уровне? Может быть, они были не способны эволюционировать в этом направлении? Но, судя по тому, что эта группа уже так долго существует, она, без сомнения, достаточно' жизнеспособна.
Мы, собственно, не можем полностью оценить, в чем состоит* преимущество одной температуры тела перед другой. Во всяком: случае, было бы ошибкой считать, что относительно низкая температура служит признаком «примитивной» и потому несовершенной терморегуляции. Высказывалось мнение, что ехидна — яйцекладущее млекопитающее — занимает промежуточное положение между холоднокровными и теплокровными животными и* поэтому не способна полностью регулировать свою температуру. На самом же деле ехидна отлично регулирует температуру своего* тела, и ее внутренняя температура остается постоянной при снижении температуры среды по меньшей мере вплоть до О °C, хотя: высокие температуры она переносит плохо (Schmidt-Nielsen^ 1966).
В табл. 8.2 (последний столбец) приведены примерные значения летальной температуры для различных групп теплокровных, позвоночных. Создается впечатление, что летальная температура^, как правило, примерно на 6° выше нормальной внутренней температуры. Например, ехидна погибает при температуре тела 37°СГ тогда как для других млекопитающих эта температура является нормальной, а для птиц — намного ниже нормы. От группы к группе граница выживания неизменно отстоит от нормы на 6°, ш мы не знаем, чем определяется постоянство этой величины.
ТЕМПЕРАТУРА ТЕЛА ЖИВОТНЫХ, ОБИТАЮЩИХ В ХОЛОДНОМ КЛИМАТЕ
Какова температура тела у птиц и млекопитающих — обитателей Арктики? Отличаются ли они в этом отношении от других теплокровных?.
Чтобы выяснить этот вопрос, измеряли температуру тела у ряда птиц и млекопитающих Аляски, помещая их в условия весьма различной, но низкой температуры. Исследованные птицы принадлежали к 30 разным видам (вес тела от 0,01 до 2 кг). При; 22"
-349 Глава 8, Терморегуляция в
температуре воздуха от 4-20 до —30 °C средняя температура тела составляла у них 41,1 °C, т. е. лежала в том же диапазоне, что л у птиц умеренного и тропического климата.
Арктических млекопитающих (22 вида, вес тела от 0,1 до 1000 кг) помещали в условия холода вплоть до —50 °C, а один вид (песцов) даже до —80 °C. При этом температура тела у всех •исследованных животных оставалась в нормальных для млекопитающих пределах. Средняя температура тела для всех видов составила 38,6 °C, что примерно на 0,5° выше, чем средняя величина для млекопитающих умеренной зоны, ранее установленная по данным о большом числе животных; однако ввиду ограниченного числа обследованных арктических видов разницу в 0,5° нельзя считать достоверной (Irving, Krog, 1954).
Итак, мы можем заключить, что птицы и млекопитающие Арктики поддерживают температуру своего тела на том уровне, который свойствен каждому из этих классов в целом, несмотря на •существование в одном из самых холодных районов мира.
ЖИВОТНЫЕ В ЕСТЕСТВЕННЫХ УСЛОВИЯХ
В противоположность обилию данных о температуре тела у лабораторных животных и животных, содержащихся в неволе, систематических исследований на животных в природных условиях проведено очень мало. Разработка методов измерения температуры с помощью датчиков, передающих информацию по радио (телеметрия), дала возможность регистрировать состояние животных в естественных условиях.
Имплантация такого телеметрического датчика животному позволяет следить за температурой его тела в течение целого года. При исследовании этим методом пасущихся овец оказалось, что колебания внутренней температуры у них составляют менее 1° независимо от погоды — будет ли солнце, дождь или буря. На протяжении всего года, включая лето и морозную зиму, максимальный диапазон колебаний температуры был 1,9° (от 37,9 до 39,8 °C) (Bligh et al., 1965).
ЛИХОРАДКА
Лихорадка — это повышение температуры тела, обычно вызванное бактериальной или вирусной инфекцией. Температура может повышаться и при инъекции убитых бактерий: в бактериях -содержатся особые вещества — пирогены, — которые и вызывают лихорадочное состояние. Организм ведет себя подобно термостату, у которого так изменили регулировку, что он поддерживает температуру на несколько градусов выше прежней.
Лихорадка, несомненно, не обусловлена недостаточной эффективностью терморегуляции. Если человека в лихорадочном состоянии подвергнуть охлаждающим воздействиям, то для поддержа
Температура тела у птиц и млекопитающих 341
ния повышенной температуры у него увеличится выработка тепла; согревание вызовет обратную реакцию. Таким образом, терморегуляция сохраняется полностью, но температура поддерживается на более высоком уровне.
Представление о том, что лихорадка — это изменение настрой->ки хорошо работающего «термостата», подтверждается также следующим опытом. Если собаке ввести пироген, то у нее начинается лихорадка. Однако если приложить тепло локально к гипоталамусу, где находится центр терморегуляции, то повышение температуры тела будет предотвращено. Организм ведет себя в этом случае так, как если бы «заданный» повышенный уровень температуры был уже достигнут. Это показывает, что действие бактериальных пирогенов состоит не в повреждении регуляторного механизма в гипоталамусе, а в том, что возникает «потребность» в определенной гипоталамической температуре, и когда последняя устанавливается, уже не требуется дальнейшего согревания (Andersen «et al., 1961).
Как у млекопитающих, так и у птиц при бактериальной инфекции развивается лихорадочное состояние, но полезно это или вредно, остается неясным. Интересная попытка разрешить этот спорный вопрос была сделана в эксперименте на холоднокровном животном — ящерице Dipsosaurus dorsalis. Когда эту ящерицу помещали в среду, где имелся температурный градиент, она обычно выбирала такие условия, чтобы температура тела у нее была на уровне около 38,5 °C. Однако после введения взвеси подходящих бактерий ящерица предпочитала уже более теплые места, стремясь к тому, чтобы температура была примерно на 2° выше.
Сходные реакции на бактериальные пирогены наблюдаются у рыб и амфибий (Reynolds et al., 1976; Kluger, 1977): они выбирают более теплые места и таким образом поддерживают более высокую температуру тела. Следовательно, «лихорадочная» реакция свойственна всем позвоночным, а не только теплокровным — птицам и млекопитающим.
Вернемся теперь к вопросу о том, полезна лихорадка или вредна. Для его решения был поставлен следующий опыт на ящерице Dipsosaurus. Пяти группам ящериц ввели суспензию бактерий Aeromonas hydrophila, а затем поместили эти группы в нейтральные условия (38 °C) и в условия пониженных (34 и 36 °C) и повышенных (40 и 42 °C) температур. Обнаружилась поразительная корреляция между температурой и выживанием: при самой низкой температуре все животные погибли меньше чем за 4 дня; по мере повышения температуры выживаемость росла и была наибольшей при 42 °C. Примерно эту же температуру предпочитают и сами инфицированные животные, если имеют возможность выбора (Kluger et al., 1975). Таким образом, в описанном случае повышенная температура тела, несомненно, оказывала благоприятный эффект.
342 Глава 8. Терморегуляция
Свойственная птицам и млекопитающим способность к поддержанию постоянной температуры тела, а также их способность, к перестройке на новый уровень хорошо регулируемой температуры (как при лихорадочном состоянии) требует исключительна мощных физиологических механизмов теплообмена. Прежде чем перейти к их рассмотрению, необходимо ознакомиться с основами физики теплообмена.
ТЕМПЕРАТУРА, ТЕПЛО И ПЕРЕНОС ТЕПЛА
В предыдущем разделе речь шла о температуре, а понятие^ «тепло» упоминалось только вскользь. Необходимо ясно понимать разницу между этими двумя физическими величинами и отдавать себе отчет в том, что не всегда измерение температуры позволяет получить информацию о количестве тепла.
Температуру измеряют обычно в градусах Цельсия (°C), хотя в физической химии и в термодинамике используют абсолютную’ температуру в кельвинах (К)1.
В биологии тепло измеряют обычно в калориях. Калория — эта-количество тепла, необходимое для повышения температуры 1 г воды на 1°. (Калория не входит в систему СИ, но этот термин настолько распространен, что, несомненно, будет еще употребляться какое-то время. Для перевода калорий в джоули — единицы системы СИ — пользуются соотношением: 1 кал = 4,184 Дж.)
Чтобы нагреть 1 г воды, имеющей комнатную температуру (25°C), до кипения (100°C), требуется 75 кал тепла, а чтобы в такой же мере (на 75°) нагреть 100 г воды, требуется 7500 кал (7,5 ккал). В обоих случаях начальная и конечная температуры одинаковы; значит, изменение температуры само по себе не позволяет судить о количестве затраченного тепла. Но если известна-количество воды, то по данным об изменении температуры можно вычислить количество затраченного тепла, ибо, по определению, тепло, необходимое для повышения температуры 1 г воды на Г, составляет 1 кал. Количество тепла, необходимое для нагревания 1 г данного вещества на 1°, называется удельной теплоемкостью этого вещества2. Удельная теплоемкость воды равна 1,0 кал/ /(г-°С) и по сравнению с другими веществами очень высока. Удельная теплоемкость каучука равна 0,5, дерева — 0,4, большинства металлов — 0,1 и ниже. Удельная темплоемкость воздуха 0,24 кал/(г-°C), а поскольку плотность его 1,2 г/л (при 20°C),, теплоемкость 1 л воздуха составляет 0,3 кал.
1 Абсолютный нуль соответствует —273,15 °C. Температура по Цельсию (Тс> и абсолютная температура (Тк) связаны соотношением: ^=^+273,15.
2 Рекомендуется термин «удельный» перед названием физических величин применять только в значении «деленный на массу» (Council of the Royal Socie*
Температура, тепло и перенос тепла 343
Для того чтобы повысить температуру тела животного, требуется несколько меньше тепла, чем для нагрева такого же по весу количества воды. Средняя удельная темплоемкость тканей млекопитающего составляет примерно 0,8. Таким образом, для нагрева тела млекопитающего весом 1000 г на 1° требуется около 800 кал. Точное значение удельной теплоемкости животного организма не вполне постоянно. Например, удельная темплоемкость тела мыши варьирует в пределах 0,78—0,85, в среднем составляя 0,824 (Hart, 1951). Главный в количественном отношении компонент тела — вода с удельной теплоемкостью 1,0; другие компоненты — «белки, кости, жиры — несколько снижают эту величину. Жир играет особенно большую роль, так как его количество может в широких пределах изменяться, а удельная теплоемкость жира всего лишь около 0,5. Во многих случаях можно приближенно считать удельную теплоемкость организма животного равной 0,8, тем «более что для расчета изменений в количестве тепла необходимо знать среднюю температуру тела, а точное определение этой величины, как мы уже видели, крайне затруднительно.
•ФИЗИКА ПЕРЕНОСА ТЕПЛА
Для того чтобы температура тела оставалась постоянной, абсолютно необходимо, чтобы отдача тепла была равна его притоку. Для поддержания постоянства температуры тела животного скорость отдачи тепла должна равняться скорости его выработки в процессе метаболизма. Как мы уже видели, выработка метаболического тепла при физической работе легко может возрасти более чем в 10 раз, и если потеря тепла не увеличится в такой же пропорции, температура тела животного будет быстро повышаться. Кроме того, условия теплоотдачи в огромной степени зависят от внешних факторов, таких, как, например, температура воздуха и ветер. Чтобы разобраться в физиологических механизмах, регулирующих теплопродукцию и теплоотдачу, необходимо вспомнить элементарную физику теплообмена.
Если физические тела имеют разную температуру, то тепло передается от более нагретого тела к более холодному. Этот перенос тепла осуществляется путем теплопроводности или путем излучения. Тело может отдать часть тепла этими способами только в том случае, если температура окружающей среды или какой-то ее части ниже температуры поверхности тела. Есть, однако, и третий путь отдачи тепла, а именно испарение воды. Эти три способа переноса тепла — теплопроводность1, излучение и испаре
1 Передача тепла от твердой поверхности к соприкасающемуся с ней потомку жидкости (или газа) происходит путем теплопроводности. Движение масс -В жидкости (газе), называемое конвекцией, обеспечивает обновление жидкости (газа) в пограничном слое, вследствие чего описание передачи тепла путем теплопроводности усложняется (см. далее в этой главе).
344 Глава 8. Терморегуляция
ние — являются единственными средствами удаления избыточного» тепла, выработанного в процессе метаболизма.
ТЕПЛОПРОВОДНОСТЬ
Передача тепла путем теплопроводности осуществляется между физическими телами, находящимися в контакте друг с другом^ независимо от того, твердые они, жидкие или газообразные. Теплопроводность связана с прямой передачей кинетической энергии, молекул, а эта энергия всегда передается от более нагретого тела к менее нагретому.
Предположим, что у нас есть проводящий тепло стержень, у которого один конец холодной, а другой — теплый (рис. 8.3)-Тогда скорость переноса тепла (Q) можно выразить уравнением
Q=kA
где k — коэффициент теплопроводности материала; А — площадь,, через которую передается тепло (нормальная к направлению по-
Рис. 8.3. Проведение тепла однородным стержнем зависит от его поперечного сечения, материала, из которого он изготовлен, и градиента температуры.
тока тепла); Т2 и Т\ — температуры двух участков, разделенных расстоянием I. Дробь (Т2—1\)/1 характеризует градиент температуры, т. е. разность температур на единицу расстояния по длине стержня.
Температура, тепло и перенос тепла 345
Приведенное уравнение можно выразить простыми, интуитивно понятными словами. Поток тепла возрастает с увеличением 1) теплопроводности (k) материала, из которого сделан проводник тепла, 2) поперечного сечения (Л) проводника и 3) разности температур между двумя его сечениями (Т2 и Увеличение расстояния (Z) между двумя сечениями проводника с температурами Т2 и Т[ (если последние постоянны) снижает скорость передачи тепла.
Коэффициент теплопроводности (А) показывает, насколько хорошо данный материал передает тепло. В табл. 8.3 приведены величины k для ряда распространенных материалов. Металлы, как известно, обладают высокой теплопроводностью; стекло и дерево хуже проводят тепло; теплопроводность воды и тканей человека еще несколько ниже, но того же порядка величины. То, что вода и ткани нашего тела сходны по своей теплопроводности, вполне понятно, так как большинство тканей примерно на 2/з—% состоят из воды. Очень плохо проводят тепло воздух и мех животных; это означает, что они обладают высокими термоизоляционными свойствами (препятствуют теплообмену). Малая теплопроводность меха обусловлена в основном тем, что между волосками задерживается большое количество воздуха. Другие материалы, удерживающие много воздуха (например, войлок, шерстяные ткани, пух), тоже плохо проводят тепло, т. е. служат превосходными изоляторами.
Таблица 8.3
Коэффициенты теплопроводности (k) ряда распространенных материалов. (Hammel, 1955; Hensel, Bock, 1955; Weast, 1969)
Материал k, кал/(с-см-°С) Материал k, кал/(с*см«°С)
Серебро Медь Алюминий Сталь Стекло Почва (сухая) 0,97 0,92 0,50 0,11 0,0025 0,0008 Каучук Дерево Вода Ткани человека Воздух Мех животных 0,0004 0,0003 0,0014 0,0011 0,000057 0,000091
К сожалению, приведенное выше простое уравнение теплопроводности применимо только в тех случаях, когда перенос тепла происходит через плоский объект, например через стену. Но поверхность тела у большинства животных искривлена, и поэтому теплообмен через нее описывается значительно более сложным образом. Для практических целей достаточно принять, что поток тепла зависит от градиента температуры и площади поверхности, однако в случаях, когда требуется более точная количественная оценка теплообмена у животного, необходимо более строго учитывать физику переноса тепла.
346 Глава 8. Терморегуляция
Перенос тепла в жидкости или газе почти всегда значительно ускоряется благодаря конвекции, под которой понимают движение масс жидкости или газа. Представим себе холодную жидкость, соприкасающуюся с теплой твердой поверхностью. Тепло передается жидкости, и ее слои, прилегающие к твердой поверхности, нагреваются. Если жидкость находится в движении, то теплая жидкость около поверхности сменяется холодной и в результате отдача тепла поверхностью ускоряется. Таким образом, конвекция, т. е. перемещение масс жидкости (или газа), способствует потере тепла твердой поверхностью, но основой теплообмена между твердым телом и жидкостью остается теплопроводность.
Конвекция в жидкости (газе) может создаваться в результате разности температур или под действием внешней механической силы. Нагревание или охлаждение жидкости обычно изменяет ее плотность, а это в свою очередь вызывает перемещение ее масс. Например, если теплая твердая поверхность соприкасается с холодной жидкостью, то нагретые слои последней расширяются и поднимаются вверх, а их место занимают холодные слои. В этом случае причиной конвекции служит разность температур, и такую конвекцию называют свободной или естественной. Этот термин применим и тогда, когда жидкость теплее твердой поверхности, гак что прилегающие к последней слои жидкости охлаждаются, становятся более плотными и опускаются вниз1. Разумеется, свободная конвекция может иметь место как в воде, так и в воздухе; она в значительной мере способствует отдаче тепла живыми организмами.
Движение масс жидкости или газа может быть вызвано также внешними силами, например ветром, течениями или, наконец, вентилятором. Конвекцию, вызванную внешними силами, а не изменением плотности, называют принудительной.
Поскольку конвекция связана с перемещением масс жидкости (газа), она подчиняется довольно сложным законам гидродинамики, связывающим такие параметры, как вязкость и плотность,, а также теплопроводность. Потеря тепла, обусловленная конвекцией, зависит не только от площади поверхности твердого тела. Имеют значение кривизна поверхности и ее ориентация. Для учета этих переменных требуются довольно сложные математические выражения, что создает большие трудности при определении количества тепла, теряемого организмом животного. Однако, как мы увидим далее, для измерения теплообмена между животными и средой можно применять ряд практических способов, позволяющих обойтись без точного анализа физической стороны процесса.
1 Плотность воды при +4 °C выше, чем при температуре замерзания. Это» имеет большое значение в пресноводных водоемах, но в большинстве физиологических ситуаций не играет роли. Аномалия плотности вблизи точки замерзание не свойственна морской воде (см. также стр. 315).
Температура, тепло и перенос тепла 347
ИЗЛУЧЕНИЕ
Передача тепла путем излучения происходит при отсутствии непосредственного контакта между телами. Все физические тела при температуре выше абсолютного нуля излучают электромагнитные волны. Длина волн и интенсивность излучения зависят от температуры излучающей поверхности (и как мы увидим далее, ют ее излучающей способности). Кроме того, все тела воспринимают излучение от окружающих объектов. Электромагнитная радиация свободно проходит через вакуум, а для наших целей мы можем считать и атмосферный воздух полностью прозрачным для электромагнитных волн.
Интенсивность излучения, испускаемого телом, пропорциональна четвертой степени абсолютной температуры его поверхности. Для потока теплового излучения это выражается законом Стефана — Больцмана:
где Т — абсолютная температура излучающей поверхности (в кельвинах, К); о —константа Стефана — Больцмана '[1,376-10“12 кал/(с-см2-К4), или 5,67-10“8 Вт/(м2-К4)]. Поскольку количество излучаемой энергии или тепла возрастает пропорционально четвертой степени абсолютной температуры, с повышением температуры поверхности излучение увеличивается очень быстро.
Спектр длины волн испускаемого излучения зависит от температуры поверхности: чем горячее поверхность, тем короче излучаемые волны. Если тело нагревается, то по мере повышения его температуры излучение все более сдвигается в коротковолновую область спектра. Излучение раскаленного куска железа начинает становиться видимым (появляется красный свет), когда его температура достигает примерно 1000 К. При дальнейшем нагреве он начинает излучать и более короткие волны (т. е. еще больше лучей видимой области). Таким образом, с повышением температуры цвет излучения сдвигается от красного к желтому и белому. Солнечная радиация, имеющая пик в видимой части спектра, соответствует температуре поверхности около 6000 К; в ней содержится также значительная доля ближних ультрафиолетовых лучей. Тела с температурой, близкой к физиологической, испускают главным образом лучи средней инфракрасной области. Например, пик инфракрасного излучения кожи живого человека (Тс^ЗОО К) лежит в области 10 000 нм. Поскольку видимый свет соответствует .диапазону от 450 до 700 нм, излучение нашей кожи недоступно для глаза (тот свет от кожи, который мы видим, является отраженным, и в темноте его нет).
Зависимость интенсивности и спектра излучения от температуры излучающей поверхности иллюстрируют кривые на рис. 8.4.
348 Глава 8. Терморегуляция
Мы уже упоминали об излучающей способности. К этому понятию проще всего подойти, рассмотрев сначала поглощающую1' способность, т. е. способность поверхности поглощать падающее-на нее излучение. По определению, черное тело поглощает полностью все падающее излучение независимо от длины волн и ничего не отражает. Таким образом, поглощающая способность чер-
Рис. 8.4. Как спектр, так и интенсивность теплового излучения тела зависит-от температуры его поверхности. Чем выше температура поверхности, тем короче волны и тем выше интенсивность излучения. На рисунке показано спектральное распределение энергии теплового излучения Солнца (6000 К), раскаленной плиты (1000 К) и человеческого тела (300 К). (Hardy, 1949.)
ного тела равна 100%. (Хотя мы обычно мыслим себе «черное» применительно к видимому свету, физическое определение включает все длины волн).
В противоположность черному телу поверхность, которая отражает все падающее на нее излучение, имеет поглощающую способность, равную нулю, и является идеальным отражателем. К этому состоянию приближается отполированная до блеска металлическая поверхность (например, серебряное зеркало). Поскольку любое излучение, падающее на тело, либо отражается им, либо поглощается, отражающая и поглощающая способности тела должны в сумме составлять единицу (или 100%). Например, если тело отражает 30% падающего излучения, то оно поглощает 70% г и т. д.
Как правило, поглощающая (а значит, и отражающая) способность зависит от длины волны падающего излучения. В области видимого света мы воспринимаем это как цвет предметов; напри
Температура, тепло и перенос тепла 349»
мер, мы видим желтым тот предмет, который отражает главным образом желтые лучи и поглощает остальные компоненты видимого света. По отношению к средней части инфракрасного спектра, которая представляет для нас наибольший интерес в связи с тепловым излучением тел при физиологических температурах,, почти все поверхности являются черными. Кожа человека, например, поглощает практически 100 % падающего на нее инфракрасного излучения и, таким образом, является по отношению к нему-черным телом независимо от ее видимого цвета.
Теперь, когда мы познакомились с поглощающей способностью», рассмотрим, что такое излучающая способность. Эти две характеристики численно равны между собой, что можно проиллюстрировать следующим образом. Представим себе предмет, находящийся в вакууме внутри полого шара, температура стенок которого повсюду одинакова. Излучение стенок падает на этот предмет и частично отражается им, а частично поглощается. Аналогичным образом и сам предмет испускает излучение, которое:-частью отражается, а частью поглощается внутренней поверхностью шара (отраженное излучение в свою очередь либо снова> падает на предмет, либо попадает на другую точку стенки шара). Когда система находится в равновесии, температура предмета* равна температуре окружающей сферической поверхности и количество поглощаемого им излучения в точности равно количеству испускаемого. Если бы это было не так, температура предмета♦ отличалась бы от температуры поверхности шара. Последнее же-физически невозможно, ибо система не может генерировать энергию и работать как perpetuum mobile. В качестве примера из* практики возьмем блестящий металлический кофейник с отражающей способностью 95 % (а поглощающей — 5%). Его излучающая способность также равна 5% (т. е. кофейник с блестящей поверхностью отдает тепло путем излучения очень медленно).
Представим себе теперь, что предмет внутри полого шара почти черный и его поглощающая способность равна 99%. Очевидно, что в равновесном состоянии у этого предмета будет такая же-температура, как и у находящегося в тех же условиях отполированного серебряного предмета, хотя поглощающая способность-последнего может составлять всего лишь 1%. Причина в том, что в равновесной системе количество поглощаемого излучения в точности равно количеству испускаемого. Конечная температура у' обоих предметов будет одинаковой, но для ее достижения предмету с хорошо отражающей поверхностью потребуется больше времени.
Повторим еще раз, что тела с высокой поглощающей способностью обладают столь же высокой излучающей способностью. Тело, абсолютно «черное» по отношению к данной длине волн, будет также идеальным излучателем волн этой же длины.
350 Глава 8. Терморегуляция
Кожа человека, мех животных и всякого рода другие неметаллические поверхности обладают высокой поглощающей способностью в средней инфракрасной области — в диапазоне от 5000 до 10 000 нм. В этой области кожа человека независимо от степени ее пигментации является практически черной. Те различия в цвете кожи, которые мы замечаем, относятся к видимой области спектра, а поскольку в этой области кожа не излучает, особенности пигментации не влияют на ее излучение.
Вследствие высокой поглощающей способности в средней инфракрасной области спектра излучающая способность кожи в этой области тоже близка к 100%. При этом пигментированная и не-пигментированная кожа теряет путем излучения примерно одинаковое количество тепла; та и другая излучает как практически черное тело. Это относится и к меховому покрову животных, излучение которого не зависит от цвета меха в видимом свете.
Незнание этого простого физического факта породило ряд ложных представлений относительно значения той или иной окраски животных. Например, высказывалось предположение, что животные с темным мехом быстрее теряют тепло путем излучения, чем светлоокрашенные животные. Но так как излучение в инфракрасной области никак не связано с видимой окраской, различие в таком излучении можно было установить только путем прямых измерений в инфракрасной области. Данные таких измерений не выявили предполагаемых различий.
Однако цвет кожи или мехового покрова может существенно влиять на поглощение радиации, пик которой лежит в видимой области спектра. На эту область приходится примерно половина энергии солнечного излучения (рис. 8.4), и для теплового баланса организма важно, поглощается этот свет или отражается. Под прямыми солнечными лучами темноокрашенный кожный или меховой покров поглощает больше лучистой энергии, чем светлый.
Расчет суммарного переноса тепла путем излучения. Если между двумя поверхностями происходит теплообмен путем излучения, то каждая из них будет излучать согласно закону Стефа-/на— Больцмана и для суммарного переноса тепла (Qr) между .ними мы получим
QR=a8182 (Т24— Tfi А,
тде о — константа Стефана — Больцмана, 81 и 82 — излучающие способности двух поверхностей, Ti и Т2—абсолютные температуры этих поверхностей и А — эффективная излучающая площадь1. Если внешняя среда является однородной сферой, то А — это просто общая «видимая» в направлении лучей поверхность. Если же
1 Приведенное уравнение для суммарного переноса тепла основано на допущении, что излучающая способность одной или обеих поверхностей близка к единице или равна ей.
Температура, тепло и перенос тепла ЗЕ t
внешняя среда неоднородна, и особенно если в ней имеется точечный источник излучения (например, солнце), то интегральный учет поверхностей, участвующих в теплообмене, усложняется. С этой точки зрения природные ситуации чрезвычайно сложны, и здесь очень трудно дать точное математическое описание переноса тепла. Тем не менее, поняв в принципе физические основы теплообмена путем излучения, мы можем легко избежать ошибок вроде того вывода о роли пигментации, о котором упоминалось выше.
Применяемое на практике упрощение расчета. Хотя перенос тепла путем излучения зависит от четвертой степени абсолютной температуры, уравнение переноса все же можно упростить, если разница в температуре поверхностей не слишком велика. В диапазоне температур около 20° ошибка при упрощении уравнения Стефана — Больцмана мала, и ею часто можно пренебречь; суть упрощения состоит в том, что приближенно теплообмен путем излучения считают пропорциональным разности температур между двумя поверхностями. При малых разностях температур ошибка сравнительно невелика, но она быстро возрастает с увеличением разности.
Мы будем пользоваться этим упрощением, считая радиационный теплообмен пропорциональным разности температур. У теплокровного животного отдача тепла в холодной среде осуществляется как путем теплопроводности, так и путем излучения. Поскольку скорость обоих процессов пропорциональна разности температур (Г2—Л), суммарная скорость теплоотдачи также пропорциональна Т2—Л, или
Q=C(T2-Tt),
где все константы, входящие в уравнение теплообмена, объединены в один фактор пропорциональности С.
Мы еще вернемся к этому упрощенному уравнению при расчете потери тепла теплокровным животным в условиях холода.
ИСПАРЕНИЕ
Испарение воды связано с затратой большого количества тепла. Чтобы 1 г воды, имеющей комнатную температуру, превратить в пар с той же температурой, требуется 584 кал (2443 Дж). Это поразительно много тепла: вспомним, что для нагревания 1 г воды от точки замерзания до точки кипения расходуется только 100 кал (418 Дж); таким образом, для испарения воды нужно почти в 6 раз больше тепла, чем для нагрева ее от 0 до 100 °C.
Количество тепла, необходимое для фазового перехода воды из жидкости в пар, называют теплотой парообразования (ЯУД Эта величина несколько изменяется в зависимости от температуры, при которой происходит фазовый переход; так, при. CL°G Z/v.=
352 Г лава 8. Терморегуляция
= 595 кал на 1 г воды, при 22 °C Я¥ = 584 кал/г, а при температуре кипения (100 °C) Я¥ = 539 кал/г.
В физиологии обычно используют величину 580 кал на 1 г воды, что приближенно соответствует теплоте перехода воды в пар на поверхности кожи потеющего человека при температуре около 35 °C.
Измерять потерю тепла при испарении воды очень удобно: достаточно знать количество испарившейся воды. В условиях жары организм человека охлаждается путем испарения пота со всей поверхности кожи, тогда как у собаки испарение влаги идет главным образом в дыхательных путях и на языке. Разумеется, количество тепла, расходуемого на испарение 1 г воды, в обоих случаях одинаково (т. е. площадь испаряющей поверхности и ее локализация не имеют для нас значения).
Воздух, выдыхаемый млекопитающими и другими животными, насыщен водяным паром, и поэтому даже при отсутствии перегрева в дыхательных путях всегда испаряется значительное количество влаги. Это, конечно, необходимо учитывать при всех расчетах общего теплового баланса организма; к последнему вопросу мы сейчас и перейдем.
ТЕПЛОВОЙ БАЛАНС
Как мы уже не раз подчеркивали, для поддержания постоянной температуры тела необходимо, чтобы потеря тепла была уравновешена его притоком. Однако температура тела не всегда остается постоянной. Предположим, что потеря тепла не вполне равна его выработке в процессе метаболизма, а намного меньше. Тогда температура неизбежно будет повышаться. Это означает, что часть образующегося тепла остается в организме, и повышение температуры соответствует этому накоплению тепла. Если же средняя температура тела снижается, что имеет место при теплоотдаче, превышающей теплопродукцию, то эту избыточную потерю тепла мы можем рассматривать как расходование тепловых запасов. Количество накопленного тепла зависит от изменений средней температуры тела, от массы тела и от удельной теплоемкости тканей (последнюю у птиц и млекопитающих обычно принимают равной 0,83).
Теплообмен между организмом и средой осуществляется тремя описанными выше способами, а именно путем теплопроводности (включая конвекцию), излучения и испарения. Обычно все эти процессы ведут к отдаче тепла, но это не всегда так. Если температура воздуха выше температуры поверхности тела, поток тепла, передаваемого путем теплопроводности, будет направлен к телу, а не от него. При наличии внешнего источника сильного теплового излучения результирующий поток тепла тоже может быть направлен к телу. Испарение почти всегда ведет к потере тепла
Тепловой баланс 353
организмом, но в некоторых необычных условиях возможен и обратный процесс. Это, например, имеет место, когда охлажденное тело соприкасается с горячим влажным воздухом1.
Можно объединить описанные факторы, определяющие теплообмен, в простом уравнении2:
//общ ==: —//р “
где //общ — выработка тепла в процессе метаболизма (всегда положительный член), Нт — теплообмен путем теплопроводности и конвекции ( + в случае теплоотдачи), Яр— теплообмен путем излучения (радиации) ( + в случае теплоотдачи), НИ — теплообмен путем испарения (+ в случае теплоотдачи) и На — аккумуляция тепла в организме (плюс в случае накопления, минус в случае потери).
Три пути теплообмена — теплопроводность, излучение и испарение — зависят от ряда внешних факторов, из которых самым важным является температура. Очевидно, что при снижении температуры среды теплоотдача возрастает, а при ее повышении уменьшается; если же внешняя температура выше температуры поверхности тела, передача тепла как путем теплопроводности, так и путем излучения может идти в направлении из среды в организм. При этом общий приток тепла в организм будет складываться из метаболической теплопродукции и тепла, получаемого извне. В этом случае для поддержания постоянной температуры тела (//а = 0) нужно настолько увеличить испарение, чтобы оно отнимало все избыточное тепло.
1 Когда человек с температурой кожи около 35 °C входит в турецкую парную баню, где воздух почти полностью насыщен водяным паром при температуре выше 40°, на его коже сразу же конденсируется вода. В этом случае перенос тепла происходит в противоположном (по сравнению с обычным) направлении. В финской бане (сауне) воздух обычно сух, и влага, которая вскоре появляется на коже вошедшего в нее человека, представляет собой пот.
2 Важно отметить, что в это уравнение не входит работа. Для расчета теплового баланса организма уравнение в таком виде вполне корректно. Однако если для определения интенсивности метаболизма и вычисления теплопродукции используют величину потребления кислорода, то следует учитывать внешнюю работу, так как та доля потребления О2, которая обеспечивает выполнение этой работы, не проявится во внутренней теплопродукции организма. Например, летящая птица расходует около 20% метаболической энергии на придание ускорения воздуху, в котором она движется (в конечном итоге эта внешняя работа превращается в тепло в хвосте воздушных вихрей, образующемся позади птицы). Следовательно, у летящей птицы только 80% потребляемого кислорода используется для выработки тепла, и только эта часть входит в уравнение теплового баланса, как то метаболическое тепло (ЯОбщ), которое рассеивается в ходе процессов, представленных в правой части уравнения. Все это означает также, что нельзя использовать калорический эквивалент кислорода (4,9 ккал на 1 л потребленного О2) при расчете теплопродукции животного, выполняющего внешнюю работу.
23_1R73
354 Глава 8. Терморегуляция
Физиологические реакции на жару и холод разнятся во многих отношениях, и поэтому удобнее обсуждать их по отдельности. Мы рассмотрим сначала, как осуществляется терморегуляция на холоде, а затем — в тепле.
ТЕРМОРЕГУЛЯЦИЯ В УСЛОВИЯХ ХОЛОДА
Для поддержания постоянной температуры тела животное должно находиться в таком стационарном состоянии, когда выработка тепла в процессе метаболизма (//) уравновешивается теплоотдачей (Q). Допустим на минуту, что отдача тепла (в холодной среде) происходит только путем теплопроводности (включая конвекцию) и излучения, а потерей тепла в результате испарения влаги можно пренебречь1. Для описания отдачи тепла используем упрощенное уравнение, выведенное в предыдущем разделе и выражающее зависимость теплоотдачи (Q) от окружающей температуры:
H=Q=C(Tb-Ta).
Это уравнение просто означает, что скорость выработки тепла в организме равна скорости теплоотдачи, которая в свою очередь пропорциональна разности между температурами тела и окружающей среды (Тъ—Та). Множитель С служит показателем теплопроводности, и мы к нему вернем'ся несколько позже.
Каким образом может поддерживаться стационарное состояние? В приведенном уравнении величина Та (температура окружающей среды) не зависит от животного, если только оно не перемещается в иную среду. Для приспособления к неблагоприятной внешней температуре животное может изменять остальные величины: собственную теплопродукцию (Я), теплопроводность (С) и температуру тела (7ь). Но так как последняя из этих величин по условию должна быть постояйной, у животного остаются только две возможности: изменить собственную теплопродукцию (Н) или же теплопроводность (С). (Третья возможность — изменение температуры тела — реализуется при зимней спячке, которую мы рассмотрим позже.)
ПОВЫШЕНИЕ ТЕПЛОПРОДУКЦИИ
Если снижение теплопродукции (интенсивности метаболизма) возможно только до определенного минимального уровня, то повышение уровня метаболизма, напротив, может использоваться в
1 При средних и низких температурах испарение влаги происходит главным образом в дыхательных путях, и связанная с ним потеря соответствует лишь нескольким процентам метаболической теплопродукции.
Тепловой баланс 355
очень широких пределах. Основные средства для увеличения теплопродукции следующие: 1) мышечная активность, 2) непроизвольное сокращение мышц (дрожь) и 3) так называемый недро-жевой термогенез. Последний связан с повышением интенсивности обмена без заметного сокращения мышц.
Рис. 8.5. Температура в термоизолированной камере поддерживается на заданном уровне (Ть) с помощью нагревателя, обеспечивающего приток тепла со скоростью Н. При понижении внешней температуры (Та) внутренняя температура (Ть) останется постоянной, если приток тепла будет увеличиваться пропорционально разности Ть — Та. Эта схема представляет собой модель терморегуляции у млекопитающих.
Чтобы понять значение повышенной теплопродукции, рассмотрим простую физическую модель животного (рис. 8.5). Представим себе камеру с теплоизолирующими стенками, в которой имеется электрическая спираль, излучающая тепло со скоростью Н. В результате этого нагрева температура внутри камеры (Ть) будет выше, чем в окружающей среде. Если понизить внешнюю температуру (Та), то для поддержания внутренней температуры придется увеличить подачу тепла (Я); при этом возрастание Н должно быть прямо пропорционально увеличению разности между внутренней и внешней температурами, пока теплоизоляция (теплопроводность стенок) остается неизменной.
Точно так же обстоит дело с животными в условиях холода. Чем ниже окружающая температура, тем больше должна быть интенсивность обмена, чтобы температура тела оставалась постоянной. Как видно из рис. 8.6, при температурах ниже определенной точки, называемой нижней критической температурой (Тнк), интенсивность метаболизма возрастает линейно со снижением температуры. Выше этой критической точки выработка тепла остается постоянной, так как она не может быть ниже уровня покоя.
23*
356 Глава 8. Терморегуляция
Для того чтобы сравнить между собой ряд различных животных, удобно принять уровень их метаболизма в покое за 100%. Именно это сделано на рис. 8.7; из рисунка видно, что для большинства млекопитающих тропической зоны критические точки лежат в пределах от +20 до +30°C; при температурах ниже критической интенсивность обмена быстро возрастает по мере
Рис. 8.6. Потребление кислорода карликовым опоссумом (Cercaertus nanus) при различных температурах среды. (Bartholomew, Hudson, 1962.)
При температурах ниже определенного уровня, называемого нижней критической температурой (ТНк), потребление Ог линейно возрастает по мере снижения внешней температуры.
снижения температуры. Для млекопитающих Арктики характерны значительно более низкие критические температуры, а у такого хорошо защищенного от холода животного, как песец, уровень метаболизма заметно повышается только при температуре воздуха ниже —40 °C.
Наклон прямых на рис. 8.7 соответствует теплопроводности (С) в приведенном выше уравнении1. Как правило, тропические млекопитающие характеризуются высокой теплопроводностью, а арктические — низкой (т. е. хорошей теплоизоляцией). Таким образом, у тропических животных из-за высокой теплопроводности (малой теплоизоляции) обмен веществ должен резко возрастать даже при умеренном снижении температуры. Например, у обезьяны, имеющей температуру тела 38 °C и нижнюю критическую температуру 28 °C, интенсивность обмена должна увеличиваться вдвое при снижении температуры до 18 °C (на 10° по сравнению с критической). С другой стороны, арктическая земляная белка, у которой критическая температура близка к 0°С,
1 В случаях, когда данные измерений представлены в процентах от теплопродукции в состоянии покоя, принятой за 100%, наклон кривых соответствует теплопроводности не в абсолютных, а в относительных единицах.
Тепловой баланс 357
не нуждается в удвоении интенсивности метаболизма до тех пор, пока температура среды не упадет почти до —40 °C. Песец же с его критической температурой —40 °C даже при самых низких температурах Арктики (до —70 °C) повышает интенсивность обмена менее чем на 50% •
"Интенсивность обмена (нормализованная)
температура на Земле
тела
Рис. 8.7. Интенсивность обмена у различных млекопитающих в зависимости от температуры воздуха. (Scholander et al., 1950а.)
Нормальный уровень метаболизма в покое при отсутствии холодового стресса принят за 100%. Выражая повышение обмена при понижении температуры через эту нормализованную величину, можно сравнивать между собой весьма разнообразных животных.
Различие между животными тропиков и Арктики очень ясно выступает на рис. 8.7. У арктических млекопитающих ширина термонейтральной зоны.1 намного больше, а повышение уровня метаболизма при воздействии холода намного меньше, чем у тропических. Однако график на рис. 8.7 построен так, что в определенном смысле может ввести в заблуждение. Представим себе, что мы изучаем двух млекопитающих одинаковой величины и с одинаковой теплопроводностью, но у одного интенсивность обмена в термонейтральной зоне вдвое выше, чем у другого. Если в этом случае мы будем .откладывать на графике интенсивность обмена не в процентах, а в абсолютных величинах (рис. 8.8), то увидим, что нижняя критическая температура у этих животных различна, но при температуре ниже критической они будут тратить на согревание себя одинаковое число калорий. Из этого видно, что критическая температура (ширина термонейтральной
1 Термонейтральной зоной называют тот диапазон температур, в котором интенсивность метаболизма не зависит от температуры.
358 Глава 8. Терморегуляция
зоны) сама по себе еще не позволяет судить о том, насколько хорошо животное приспособлено к холоду. Для оценки взаимосвязи между энергетическим обменом и терморегуляцией нужны данные об интенсивности метаболизма в покое или, еще лучше, о величине теплопроводности.
Рис. 8.8. Если два животных с одинаковой температурой тела и одинаковой теплопроводностью отличаются друг от друга по интенсивности метаболизма в покое, то у них будет различна и нижняя критическая температура (верхний график). Если же метаболизм покоя принять за 100% (нижний график), то создастся ложное впечатление, будто у этих животных различная теплопроводность.
ТЕПЛОПРОВОДНОСТЬ, ТЕПЛОИЗОЛЯЦИЯ И ТОЛЩИНА МЕХА
Теплопроводность в том смысле, как мы употребляли до сих пор этот термин, является мерой потока тепла от животного в окружающую среду. Сюда включается перенос тепла от внутренних частей тела к поверхности кожи и от кожи через меховой покров во внешнюю среду. Низкая теплопроводность озна
Тепловой баланс 359
чает высокие теплоизоляционные свойства. По существу, теплоизоляция есть величина, обратная теплопроводности1.
Рассмотрим животных, обитающих в холодном климате, у которых мех служит основным препятствием для утечки тепла. Теплоизоляционные свойства меха разных животных весьма неодинаковы. Для некоторых из них соответствующие величины, отложенные против толщины меха, представлены на рис. 8.9. Как и следовало ожидать, теплоизоляционные свойства меха возрастают с увеличением его толщины, достигая максимума у более крупных животных с толстым слоем меха, таких, как песец. У мелких животных ясно видна корреляция между толщиной меха (и его теплоизоляционными свойствами) и размерами животного. Чтобы иметь возможность передвигаться, мелкие животные должны обладать тонким и легким меховым покровом. В особенности это относится к самым мелким млекопитающим — мышевидным грызунам и землеройкам. Из-за относительно плохой теплоизоляции этим животным приходится подыскивать себе микроклиматические условия (например, жить в норах) или впадать в спячку, чтобы можно было не поддерживать высокой температуры тела.
Интересно, что у белого медведя мех, будучи грубым и рыхлым, создает, несмотря на свою толщину, довольно плохую теплоизоляцию. Еще важнее то, что при погружении этого меха в ледяную воду он почти утрачивает свои теплоизоляционные свойства, и тепло проходит через него в воде в 20—25 раз быстрее, чем на воздухе. Если же слои воды перемешиваются (когда зверь плывет), потеря тепла возрастает еще больше — примерно в 50 раз. Это связано с тем, что вода приходит в контакт с поверхностью кожи, вытесняя из меха весь воздух. В этих условиях медведя спасает подкожный жир, который и обеспечивает теплоизоляцию (см. далее в этой главе).
У тюленя с его относительно тонким мехом теплоизоляцию тоже создает толстый слой подкожного жира. Поэтому разница в теплоизолирующих свойствах шкуры тюленя в воздухе и в воде невелика. В воздушной среде шкура тюленя с прилегающим слоем жира обеспечивает лишь ненамного большую теплоизоляцию, чем шкурка лемминга (слой толщиной 60—70 мм, состоящий в основном из жира, примерно так же защищает от охлаждения, как мех толщиной 20 мм). При погружении тюленя в воду теплоизоляция его снижается, но не так резко, как у белого медведя.
Хорошо известно, что густота меха у животного зависит от времени года: зимой мех гуще и, по-видимому, лучше сохраняет
1 Теплопроводность (поток тепла в единицу времени через единицу площади
в расчете на Г разности температур) выражается в единицах Вт/(м2-°С) или кал/(с-см2-°C). Теплоизоляция как величина, обратная теплопроводности, выражается в единицах м2-°С/Вт (или с-см2’°С/кал).
360 Глава 8. Терморегуляция
10
Ю
20
О
30 40 50 60
Толщина меха, мм
Рис. 8.9. Коэффициент теплоизоляции меха зависит от его толщины. Поскольку у мелких животных шерсть по необходимости короче, теплоизоляция у них хуже, чем у крупных животных. Коэффициент теплоизоляции меха на воздухе обозначен светлыми кружочками, в воде — треугольниками. Этот коэффициент измеряют в воде или в воздухе при О °C, поддерживая при этом с внутренней стороны меха температуру 37 °C. Наклонная прерывистая линия соответствует коэффициенту теплоизоляции для слоя ваты различной толщины. На рисунке показано, что теплоизоляционная ценность меха толщиной 4 см составляет примерно 1 м2*°С/Вт, что соответствует 41 840 с*см2*°С/кал. Мех толщиной 1 см обладает в 4 раза меньшей теплоизоляционной ценностью (10 460 с*см2*°С/кал). Следовательно, коэффициент теплоотдачи (величина, обратная коэффициенту теплоизоляции) составит 0,000096 кал/(с*см2-°C), что хорошо согласуется с данными, приведенными в табл. 8.3. (Scholander et al., 1950b.)
тепло, чем более легкий летний мех. Измерения показали, что сезонные изменения наиболее значительны у крупных животных, а у мелких грызунов они выражены слабее. У черного медведя, например, теплоизоляционные свойства меха летом на 52% ниже, чем зимой (т. е. зимняя шкура в 2 с лишним раза лучше со
Тепловой баланс 36Г
храняет тепло, чем летняя). В субарктической зоне.наименьшая, величина сезонных изменений — летнее снижение теплоизоляции онных свойств шкурки всего лишь на 12% — была найдена у ондатры.
теплоизоляция у птиц
Мы уже видели, что у млекопитающих теплопроводность покровов такова, что интенсивность обмена в условиях холода должна возрастать пропорционально холодовому стрессу. Интенсивности обмена при низких температурах укладываются на прямую, которая при экстраполяции достигает точки, соответствующей температуре тела; значит, модель, приведенная на рис. 8.5» адекватно описывает млекопитающее в условиях холода.
С птицами дело обстоит сложнее. Некоторые из них ведут себя так же, как и млекопитающие (т. е. мы находим здесь сходную зависимость уровня метаболизма от внешней температуры). Однако у других птиц наблюдается несколько иная картина. Например, у голубя и калифорнийской кукушки метаболическая теплопродукция при низкой температуре воздуха возрастает в меньшей степени, чем можно было ожидать (рис. 8.10). У этих птиц кривая интенсивности обмена при экстраполяции достигает не температуры тела (примерно 40°C), а значительно большей величины — выше 50 °C.
Рис. 8.10. Потребление кислорода у голубя и калифорнийской кукушки при низкой температуре воздуха увеличивается, однако при экстраполяции линии регрессии мы не получаем температуры тела животного в отличие от того, что имеет место у млекопитающих. (Calder, Schmidt-Nielsen, 1967.)
Этому может быть только одно объяснение: к птицам непри-, менимо простое уравнение Н=С(Тъ—Тя), С остается посто-
янным при низкой температуре. Характер линии регрессии может указывать на то, что теплопроводность (С) постепенно уменьша
362 Глава 8. Терморегуляция
ется по мере снижения внешней температуры (7\). Механизм этого уменьшения теплопроводности у птиц при температуре ниже критической остается не совсем ясным. Изменение наклона линии регрессии могло бы быть связано со снижением температуры тела, но это не согласуется с имеющимися данными о постоянстве внутренней температуры тела у птиц.
Каким же образом птицы уменьшают теплопроводность (увеличивают теплоизоляцию)? Один из возможных способов состоит в том, что птица взъерошивает перья, втягивает в них лапки и нахохливается, превращаясь в шар из перьев. Второй способ связан с заметным охлаждением периферических тканей при поддержании постоянства внутренней температуры. Падение периферической температуры тела и (или) увеличение толщины периферического слоя приводит к нескольким последствиям: в этом слое снижается кровоток; в нем уменьшается выработка тепла; в результате утолщения этого слоя соответственно уменьшается объем внутренней области. Каждый из этих сдвигов способствует как снижению теплоотдачи, так и уменьшению (по сравнению с теоретическим ожиданием) прироста интенсивности обмена, но Все же позволяет поддерживать внутреннюю температуру на постоянном уровне (хотя и в уменьшенном внутреннем объеме).
ОКУЧИВАНИЕ (ВЗАИМНЫЙ ОБОГРЕВ)
Среди пингвинов Антарктики самые удивительные — большие императорские! пингвины (Aptenodytes forsteri), которые не только живут в болёе холодных условиях, чем какие-либо другие птицы, но и обладаю^. уникальной особенностью: выводят птенцов среди зимё1.
С приближением Зимы императорские пингвины покидают открытую воду и направляются,, к своим постоянным местам размножения на ледяном шельфе,. проходя по морскому льду 50— 100 километров. Там !саь<кд откладывает одно яйцо, самец помещает его себе ,на лап1р1 и обогревает, оставаясь на месте до выведения птенцов,; а_самйа возвращается в море на кормежку. Самен стоит на льду-более двух месяцев на морозе, который может достигать —30 и даже —4(£°С, щ-ри сильнейшем ветре. Самка обычно возвращается с полным желудком к моменту вылупления птенцов. Она выкармливает птенца, в то время как самец отправляется к морю кормиться.
Императорские пингвины могут кормиться только в море; во время длительного голодания самцы существуют за счет больших запасов подкожного жира. Период голодания длится иногда более 100 дней, и к концу его самцы теряют до 40% первоначального веса.
Сколько расходуется энергии на длинный путь до места размножения и обратно и сколько —для поддержания температуры
7 епловой баланс 363-
тела в условиях антарктического холода? И сохраняют ли-пингвины в этих условиях постоянную температуру тела? , Эти вопросы сравнительно недавно были подвергнуты подробному изучению (Le Maho et al., 1976; Pinshow et al., 1976).
Оказалось, что пингвины сохраняют нормальную температуру тела 38 °C от начала до конца периода размножения. Сколько же топлива тратят птицы на обогрев и сколько на переход? Крупный самец, выходя из моря, может весить 35 кг, а за период голодания теряет около 15 кг. Чтобы выяснить, сколько энергии нужно птице для передвижения, в лабораторных условиях пингвинов приучали ходить на тредбане и во время ходьбы измеряли потребление ими кислорода. Результаты показали, что на 200-километровый переход к местам размножения и обратно к морю может расходоваться около 1,5 кг жира. Возникает вопрос: достаточно ли того, что остается, для поддержания постоянной температуры тела в течение нескольких месяцев голодания при инкубации яйца?
Можно было бы предположить, что пингвины обладают какими-то физиологическими особенностями, отличающими их от других птиц, — например, что термонейтральная зона у них распространяется на область крайне низких температур (т. е. у них очень малая теплопроводность). Это, однако, не так. По теплопроводности пингвины не отличаются от других птиц такой же величины, а их нижняя критическая температура равна —10 °C, т. е. намного выше температур, обычных для антарктической зимы. Если в лаборатории содержать пингвина при температурных условиях, близких к естественным, то интенсивность обмена у него будет такой, что для поддержания температуры тела во время зимнего голодания ему потребовалось бы около 25 кг жира; это намного превышает жировые запасы даже крупнейших самцов.
Это видимое противоречие связано с особенностями поведения пингвинов в местах размножения; они стоят здесь плотной толпой, насчитывающей иногда несколько тысяч особей. Интенсивность метаболизма у таких сгрудившихся пингвинов не измеряли, однако хорошим показателем ее может служить потеря веса, обусловленная, видимо, использованием жировых запасов; Одиночный императорский пингвин теряет на холоде до 0,2 кг веса в день; скученные пингвины теряют вдвое меньше, т. е. около 0,1 кг. Ясно, что скучивание помогает экономить энергию, и это нетрудно понять: если пингвин-одиночка охлаждается со всех сторон, то в толпе значительная часть поверхности тела соприкасается с другими пингвинами (фото 8.1). Когда два тела имеют одинаковую температуру, передачи тепла не происходит. Таким образом, скучивание пингвинов, без сомнения, необходимо для их выживания и успешного размножения. Почему императорские пингвины размножаются именно зимой — это особый вопрос, на который еще нет ответа.
364 Глава 8. Терморегуляция
Другие животные тоже иногда скучиваются для сохранения тепла, хотя это может проявляться не в такой впечатляющей форме, как у пингвинов. Скучивание уменьшает величину открытой поверхности, уменьшая тем самым воздействие холода и метаболические расходы на выработку тепла. Это имеет особое значение для птенцов и новорожденных млекопитающих, живу-
фото 8.1. Сгрудившиеся пингвины. (Фото предоставлено Ivon LeMaho, Лионский университет, Франция.)
Птенцы императорского пингвина сфотографированы на покрытом льдом гнездовье в Антарктике. Птенцы прижимаются друг к другу и таким образом уменьшают поверхность, открытую холодному воздуху. Такое поведение значительно уменьшает затраты энергии на поддержание высокой температуры тела в суровом климате. На фото запечатлена группа примерно из 50 птиц; несколько птенцов на мгновение подняли головы, чтобы посмотреть на фотографа.
щих вместе как выводок: во-первых, они дольше могут сохранять тепло в отсутствие родителей; во-вторых, сокращение расхода метаболической энергии на выработку тепла способствует ускорению роста.
ТЕПЛОИЗОЛЯЦИЯ У ВОДНЫХ МЛЕКОПИТАЮЩИХ
Многие тюлени и киты обитают в ледяных водах арктических и антарктических морей. При этом потери тепла в воде намного выше, чем в воздухе при той же температуре, вследствие высокой
Тепловой баланс 365
теплопроводности воды и ее большой теплоемкости. Коэффициент теплопроводности у воды примерно в 25 раз больше, чем у воздуха, а из-за конвекции, как свободной, так и принудительной, охлаждающее действие воды обычно еще сильнее — в некоторых случаях оно в 50 и даже в 100 раз больше, чем у воздуха.
Фото 8.2. На этом поперечном срезе замороженного тюленя виден толстый слой подкожного жира, занимающий в данном случае 58% всей площади среза; остальные 42% приходятся на долю мышц, костей и внутренних органов. (Фото предоставлено Р. F. Scholander, Калифорнийский университет в Сан-Диего.)
Как же поддерживают киты и тюлени свой тепловой баланс в этих условиях? Видимо, это им вполне удается, поскольку и тех и других гораздо больше в холодных водах, чем в тропиках. Так как они не в состоянии изменить температуру воды (не мигрируя в более теплые моря), выбор возможных путей приспособления у них ограничен. Они могли бы 1) существовать при пониженной температуре тела, 2) повысить интенсивность обмена или 3) улучшить теплоизоляцию организма, сократив таким образом потери тепла.
В отношении температуры тела киты и тюлени сходны с другими теплокровными животными: она обычно близка у них к 36— 38 °C (Irving, 1969). Поэтому следует рассмотреть другие пути адаптации к существованию в ледяной воде.
Интенсивность обмена была измерена у ряда тюленей и некоторых дельфинов (морских свиней; но не у крупных китов — это явно неудобное экспериментальное животное). У большинства исследованных животных уровень метаболизма в покое примерно вдвое превышал величину, которой можно было ожидать,
356 Глава 8. Терморегуляция
исходя из размеров тела (Irving, 1969). У гренландского тюленя ^Phoca grenlandica) — одного из арктических видов — интенсивность обмена не изменялась при охлаждении воды вплоть до
Рис. 8.11. Температура поверхности кожи живого тюленя, погруженного в ледяную воду, почти равна температуре воды. У тюленей теплоизоляция обеспечивается в основном толстым слоем подкожного жира. (Irving, Hart, 1957.)
точки замерзания; иными словами, даже самая холодная вода не отнимала столько тепла, чтобы возникла необходимость в повышении теплопродукции. Таким образом, нижняя критическая температура для этого вида тюленей лежит ниже точки замерзания воды, и ее не удалось определить (Irving, Hart, 1957).
Очевидно, задача приспособления решается главным образом, третьим способом — с помощью эффективной теплоизоляции. И у тюленей, и у китов имеется толстый слой подкожного жира, препятствующий потере тепла (фото 8.2). В пользу этого говорят и измерения температуры кожи; на ее поверхности температура
'Тепловой баланс 367
оказалась практически равна температуре воды (рис. 8.11). Но если это так, то тело может отдавать воде лишь очень немного тепла. Именно подкожный жир обеспечивает градиент температуры: последняя на глубине около 50 мм (соответствующей толщине жирового слоя) уже почти равна внутренней температуре тела.
Наземные арктические млекопитающие имеют иной градиент температуры: поверхность кожи под слоем меха у них всего лишь на несколько градусов холоднее внутренних областей тела (Irving, Krog, 1955); следовательно, их кожа защищена теплоизоляционным слоем снаружи. Что касается белого медведя, мех которого в воде теряет в основном свои теплоизолирующие свойства, то у него имеется толстая прокладка подкожного жира, защищающая от холода при плавании в ледяной воде. По-видимому, именно наличие этого жира позволяет белому медведю вести полуводный ббраз жизни (0ritsland, 1970).
Если тюлени и киты обеспечены такой прекрасной теплоизоляцией, то как им удается избегать перегрева при потеплении воды или ускорении метаболизма при быстром плавании? Как
Температура, °C
Рис. 8.12. Температура поверхности кожи у тюленей в воде и воздухе. Сплошной линией показано равенство температур кожи и окружающей среды. (Hart, Irving, 1959.)
можно видеть на рис. 8.12, у тюленя, перемещенного из водной среды в воздушную, температура кожи значительно .повышается. Такое повышение необходимо для увеличения теплоотдачи, так как охлаждающая способность у воздуха ниже, чем у воды. Оно обусловлено усиленным притоком крови через подкожный жир к поверхностному слою кожи, хорошо снабженному кровеносными сосудами. Эта система кожных сосудов позволяет точно регулировать количество тепла, поступающего к поверхности кожи и отводимого таким образом во внешнюю среду.
368 Глава 8. Терморегуляция
Итак, главное отличие водных млекопитающих от наземных в отношении способов теплоизоляции состоит в том, что у водных животных изолирующий слой (подкожный жир) расположен глубже той поверхности, через которую происходит отдача тепла. Поэтому кровь может притекать к этой поверхности в обход теплоизоляции, и в теплой воде или при большой физической нагрузке тепло может отводиться минуя теплоизоляционный слой. Наземные же млекопитающие, у которых этот слой лежит снаружи от кожной поверхности, не способны столь эффективно регулировать потерю тепла со всей поверхности кожи, так что в случае необходимости им приходится прибегать к другим способам теплоотдачи (рис. 8.13).
Рис. 8.13. Когда возрастает потребность в отдаче тепла, теплоизоляция, обеспечиваемая подкожным жиром, может быть «отключена» благодаря притоку крови к коже. Иначе дело обстоит с мехом, который расположен поверх кожи, так что «обойти» его невозможно.
РАСПРЕДЕЛЕНИЕ ТЕПЛОИЗОЛИРУЮЩЕГО МАТЕРИАЛА
Следует учесть, что покрытые мехом животные Арктики не могут в равной мере защищать от холода все части тела: должны оставаться какие-то открытые участки поверхности для отда
Тепловой баланс 369
чи тепла, особенно при физической нагрузке. Поскольку большая часть тела у них хорошо защищена от холода мехом, им нужны места со слабой теплоизоляцией (на ногах, морде и т. п.), где в случае необходимости могла бы происходить усиленная отдача тепла.
Таблица 8.4
Соотношение поверхностей, покрытых мехом различной толщины, у южноамериканского гуанако (Lama guanicoe). Примерно 40% поверхности тела покрыто толстым слоем меха, а около 20% почти не имеют мехового покрова, что позволяет в широких пределах регулировать теплоотдачу. (Morrison, 1966).
Толщина мехового покрова, мм
Доля всей поверхности кожи, %
Шерсти почти нет 1
Короткая шерсть 4
Шерсть средней длины 15
Длинная шерсть 30
19
20
20
41
Данные относительно участков тела, покрытых мехом различной толщины, указывают на то, что некоторые животные обладают широкими возможностями регулировать свою теплоотдачу
Рис. 8.14. Распределение меха различной толщины на теле гуанако. Площади различным образом заштрихованных участков на схеме пропорциональны поверхности соответствующих участков тела. (Morrison, 1966.)
путем изменения позы. Такие данные для самца гуанако представлены в табл. 8.4 и в виде схёмы на рис. 8.14 (Morrison, 1966).
Поскольку теплопроводность меха обратно пропорциональна длине шерсти, понятно, что почти голые участки кожи способны
24—1873
<370 Глава 8. Терморегуляция
рассеивать гораздо больше тепла, чем участки, покрытые густым мехом. Поэтому, если животное выпрямляет ноги и вся их поверхность оказывается открытой, то через эту «форточку» может выветриваться более половины отводимого во внешнюю среду тепла. Если же гуанако лежит или спокойно стоит так, что обращенные друг к другу поверхности ног прикрыты, то площадь, через которую происходит отдача тепла, уменьшается вдвое. Наконец, когда животное ложится и сворачивается так, чтобы по возможности укрыть все не защищенные мехом места, теплоотдача с охлаждающейся поверхности уменьшается до минимума, а именно в 5 раз по сравнению с теплоотдачей у распрямившегося животного. Короткошерстные участки кожи есть и у других животных; изменяя их положение и приток крови к ним, животное может регулировать потерю тепла через эти участки, чем достигается большая гибкость в регуляции теплоотдачи.
ТЕПЛООБМЕННИКИ
Ласты и хвостовые плавники тюленей и китов лишены подкожного слоя жира и плохо защищены от охлаждения. В то же время эти придатки имеют развитую сеть кровеносных сосудов и обильно снабжаются кровью. Таким образом, эти сравнительно тонкие структуры с большой поверхностью могут терять много тепла и способствовать охлаждению организма. Возникает вопрос: как можно избежать избыточной утечки тепла, когда тепло необходимо сохранять? Если бы температура крови, оттекающей от этих придатков, была близка к 0°С, то внутренние области тела быстро охлаждались бы.
Оказывается, чрезмерное охлаждение крови в ластах предотвращается- благодаря особенностям структуры их кровеносной -сети, устрбенной таким образом, что сосуды действуют как теплообменники. В плавниках китообразного каждая артерия полностью окружена венами (рис. 8.15), так что приходящая сюда теплая артериальная кровь охлаждается окружающей ее со всех сторон венозной кровью. В результате артериальная кровь поступает в периферические участки уже в охлажденном состоянии, и эти участки отдают воде очень мало тепла. В то же время венозная кровь оказывается нагретой еще до того, как она воз* вращается, во внутренние области тела. Если теплообмен высокоэффективен, то венозная кровь может согреваться почти до температуры артериальной крови и тогда практически не будет охлаждать тело. Теплообменник такого типа называют противоточным, так как кровь в нем движется двумя противоположно направленными потоками (Scholander, Schevill, 1955).
На рис. 8.16 показана схема противоточного теплообменника, которая помогает понять принцип его действия. Допустим, что вода с температурой 40 °C течет по медной трубке, попадает да
Тепловой баланс 37Е
лее в спираль, помещенную в баню с ледяной водой, и возвращается по второй трубке, находящейся в непосредственном контакте с первой, так что между трубками легко происходит теплообмен. После выхода из спирали вода имеет температуру, близкую к температуре бани, т. е. к О °C. Протекая дальше около
Рис. 8.15. В плавниках дельфина (морской свиньи) каждая артерия окружена сетью вен. Благодаря этому венозная кровь, прежде чем попасть во внутренние части тела, согревается в результате передачи тепла от артериальной крови. (Schmidt-Nielsen, 1970.)
трубки с теплой водой, холодная вода забирает тепло, охлаждая при этом теплую воду. Спустя некоторое время устанавливаются стационарные уровни температур (вроде указанных на схеме), которые зависят от условий теплообмена между трубками и от их длины.
Когда животное плывет в теплой воде, главная проблема для него состоит в отдаче, а не в сохранении тепла. Анатомическое устройство теплообменника таково, что при увеличении притока крови и артериального кровяного давления центральная артерия расширяется и сдавливает окружающие вены. Теперь венозной крови приходится оттекать по другим венам, расположенным ближе к поверхности ласта. В результате теплообменник не работает и артериальная кровь отдает свое тепло воде, тогда как венозная кровь возвращается во внутреннюю область тела без предварительного согревания и поэтому способствует охлаждению тела. Таким образом, кровеносная система ластов может
24*
372 Глава 8. Терморегуляция
функционировать по-разному и способствовать либо сохранению, либо отдаче тепла.
Интересно, что такие противоточные теплообменники есть и у некоторых других животных. Например, они имеются в придатках тела у ламантинов, обитающих в водах тропиков и субтропиков. Они, казалось бы, не нужны животным, постоянно находящимся в теплой воде; но дело в том, что ламантины очень медлительны
Рис. 8.16. Модель противоточного теплообменника.
В этом устройстве происходит передача тепла от притекающей воды к оттекающей; после достижения системой стационарного состояния температура воды на выходе и входе различается не более чем на 1°. Объяснение в тексте.
и для своих размеров имеют относительно низкую интенсивность обмена, поэтому уменьшение потери тепла имеет для них существенное значение.
Теплообменники в конечностях имеются не только у водных животных. Даже у человека в руках и ногах происходит некоторый теплообмен между главными артериями и прилегающими крупными венами, расположенными в глубине тканей. В условиях холода большая часть венозной крови возвращается по этим венам, тогда как в тепле венозная кровь идет по поверхностным венам, лежащим под кожей, тем самым избегая теплообмена с артериальной кровью (Aschoff, Wever, 1959).
Теплообмен в ногах играет очень важную роль у птиц, особенно у тех, которые стоят или плавают в воде. Если бы кровь, притекающая к покрытым тонкой кожей поверхностям ног, не проходила через теплообменник, потери тепла были бы очень велики, а благодаря теплообменнику они сводятся к минимуму. Так, например, у чайки, лапы которой находились два часа в ледяной воде, потеря тепла через кожу лап составила всего лишь 1,5% от общей теплопродукции организма (Scholander, 1955).
Тепловой баланс 373
Сосудистые теплообменники имеются даже в конечностях некоторых тропических животных. У ленивцев артерия, идущая к передним лапам, разделяется на несколько десятков тонких параллельно расположенных артерий, соприкасающихся с примерно таким же числом вен. Такой теплообменник может показаться излишней роскошью для тропического животного; однако в дождливую и ветреную ночь потери тепла у спящего в кроне де-
Рис. 8.17. При температуре воды ниже 24 °C потеря тепла за счет охлаждения лап составляет у утки лишь небольшую долю от общей теплопродукции организма. При температуре ниже О °C переход тепла в лапы (и соответственно отдача его воде) возрастает пропорционально снижению температуры воды. (Kilgore, Schmidt-Nielsen, 1975.)
рева ленивца могут быть очень большими. Значение теплообменника для ленивца было показано экспериментально. Лапу животного помещали в ледяную воду; при этом температура крови ниже теплообменника резко снижалась, а выше его венозная кровь была почти такой же теплой, как и внутренняя область тела (Scholander, Krog, 1957).
Высказывалось предположение, что сосудистые пучки в передних лапах ленивцев и лори играют совершенно другую роль, а именно связаны со способностью этих лазающих животных долго и крепко держаться за ветку дерева (Buettner-Janusch, 1966; Suckling et al., 1969). Однако сосудистые пучки расположены в проксимальной части передних конечностей, тогда как мышцы, обеспечивающие хватание — в дистальной. То, что кровь проте
374 Глава'8. Терморегуляция
кает по пучку артерий, соединяющихся далее в один общий сосуд, едва ли может как-то влиять на мышцы, расположенные более дистально. Содержание кислорода и питательных веществ в-крови при этом не изменяется, и только может несколько понизиться кровяное давление, что определенно не способствует увеличению силы мышц.
В условиях холода животное сталкивается с двумя противоположными проблемами: как сохранить тепло и как предохранить периферические части тела, в частности ноги, от замерзания. При не очень низких наружных температурах конфликта не возникает: уменьшение кровотока и охлаждение артериальной крови в теплообменнике позволяет свести потери тепла к минимуму.
Когда, однако, температура опускается намного ниже точки замерзания, требуется доставка дополнительного тепла в периферические ткани. Вспомним, например, голые лапы уток и других птиц. До тех пор, пока не возникает угрозы обморожения, потери тепла в лапах очень невелики, но на морозе эти потери возрастают пропорционально снижению внешней температуры (Kilgore, Schmidt-Nielsen, 1975). Повышенная теплоотдача заставляет организм усилить выработку тепла, что обнаруживается в виде отчетливого перегиба на кривой метаболизма (рис. 8.17).
ТЕРМОРЕГУЛЯЦИЯ ПРИ ИЗБЫТКЕ ВНЕШНЕГО ТЕПЛА
Как мы видели, птицы и млекопитающие вынуждены увеличивать выработку тепла, чтобы предохранить себя от охлаждения, если температура среды падает ниже определенного уровня, называемого нижней критической температурой. Посмотрим теперь, что происходит при внешних температурах выше этого критического уровня.
В этом случае метаболическая теплопродукция (в покое) остается постоянной в границах так называемой термонейтралъ-ной зоны' (см. рис. 8.6). Понятно, что для поддержания постоянной температуры тела выработка тепла должна быть равна его отдаче. Вернувшись к знакомому уже нам уравнению теплового баланса Н=С(Тъ—Та), мы заметим, что если Н и Ть остаются постоянными, а Та изменяется, то и теплопроводность С также должна изменяться.
В данном уравнении «теплопроводность» соответствует суммарной отдаче тепла организмом, и она может быть увеличена несколькими способами. Прежде всего может увеличиться приток крови к коже, что ускорит перенос тепла из внутренних областей тела к поверхности. Второй способ состоит в том, что животное вытягивается и открывает дополнительные участки поверхности, особенно те места, где нет или мало шерсти.
Тепловой баланс 275
Вернемся опять к основному уравнению теплового баланса (стр. 353):
Яобщ=±Ят±Яр±Яи±Яа.
Чем выше температура среды, тем менее благоприятны условия для отдачи тепла путем теплопроводности и конвекции (Ят) или путем излучения (Яр). Поскольку теплопродукция организма (Яобщ) не изменяется (или даже слегка возрастает), в уравнении теплового баланса должны увеличиваться два других члена — потеря тепла за счет испарения (Яи) и аккумуляция тепла (Яа).
Рассматривая терморегуляцию в условиях холода, мы не учитывали потерю тепла за счет испарения, так как она составляла лишь малую долю общей теплоотдачи. Однако при высокой температуре окружающей среды испарение играет ключевую роль в тепловом балансе. Представим себе, что мы повысили температуру среды до температуры тела. Теперь уже не будет отдачи тепла путем теплопроводности, и результирующий теплообмен путем излучения тоже приблизится к нулю. Для того чтобы температура тела оставалась в этих условиях постоянной (т. е. аккумуляция тепла Яа оставалась равной нулю), вся теплопродукция организма (ЯОбщ) должна компенсироваться путем испарения воды.
Этот вывод был проверен на ряде птиц и млекопитающих. Когда их содержали при температуре, равной температуре тела, те из них, которые оказались способны предотвратить повышение собственной температуры (а это могут не все виды), испаряли влагу в количестве, эквивалентном выработке тепла в- процессе метаболизма. На рис. 8.18 видно, что именно так обстоит дело, например, у обитающего в пустыне флоридского зайца.
Разумеется, температура среды может оказаться и выше температуры тела животных, что нередко бывает, например, в пустынях. Тогда в организм переходит тепло из окружающей среды: ют нагретого воздуха путем теплопроводности, от нагретой поверхности земли и в особенности от солнца — путем излучения. В этих условиях все тепло, как вырабатываемое в процессах метаболизма, так и получаемое от окружающей среды, должно отводиться с помощью испарения. Накопление тепла в организме не является удачным решением проблемы, поскольку, как мы уже знаем, животные обладают лишь ограниченной толерантностью к перегреву тела. И в'се же, как мы увидим дальше, даже умеренное повышение температуры тела может играть большую роль в адаптации обитателей пустынь к жаркому климату.
Для того чтобы оценить поступление тепла из окружающей среды, подсчитаем, сколько воды (в виде пота) испаряет человек на жаре в пустыне. Интенсивность обмена в покое составля-
376 Глава 8. Терморегуляция
ет у человека около 70 ккал в 1 ч, и для отдачи такого количества тепла путем испарения требуется 120 мл (0,12 л) воды. Известно также, что в жаркий день на солнце скорость потоотделения у человека колеблется от 1 до 1,5 л в 1 ч. Для упрощения расчета примем скорость потоотделения за 1,32 л/ч. Если темпе-
Рис. 8.18. Охлаждение путем испарения у флоридского зайца (в процентах от одновременной метаболической теплопродукции). (Dawson, Schmidt-Nielsen, 1966.)
Испарение возрастает по мере снижения разности между температурой тела (Ть) и температурой воздуха (Та). Когда эта разность приближается к нулю (т. е. при равенстве температур тела и воздуха), отдача всего образующегося в организме тепла происходит путем испарения.
расходуется на отведение тепла, поступающего из внешней среды. Таким образом, сумма тепла, получаемого путем теплопроводности от горячего воздуха и путем излучения — от горячего грунта и от солнца, компенсируется испарением 1,2 л воды; следовательно, в данном случае из внешней среды поступает ровно в 10 раз больше тепла, чем вырабатывается в организме человека в состоянии покоя. Повышение температуры тела на 1° соответствовало бы накоплению примерно 60 ккал, что эквивалентно теплоте испарения всего лишь 0,1 л воды. Отсюда видно, насколько велика может быть тепловая нагрузка на организм, когда температура среды выше температуры тела.
Ясно, что такие условия оказывают значительное влияние на организм. Если бы человек испарял влагу с указанной выше скоростью и при этом был лишен воды для питья, ему угрожала бы смерть от обезвоживания уже в конце первого жаркого дня в пустыне. Тем не менее во всех пустынях мира имеется богатый и разнообразный животный мир, хотя в большинстве пустынь нет
Тепловой баланс 377
открытых источников воды. Посмотрим, как животные адаптируются к таким условиям.
ЗНАЧЕНИЕ РАЗМЕРОВ ТЕЛА
Если мы положим на солнцепеке большую глыбу льда и маленькую льдинку, ясно, что льдинка растает гораздо быстрее. Если мы туда же положим огромный камень и мелкую гальку, то последняя нагреется намного раньше, чем камень. Дело в том, что мелкие предметы имеют большую по отношению к объему поверхность. Если же мы разобъем большой камень или льдину на мелкие кусочки, то появится множество новых поверхностей, открытых для солнечных лучей и теплого воздуха. В результате увеличения поверхности вся масса камня или льда соответственно нагреется или растает быстрее.
Для тел одинаковой формы зависимость между поверхностью и объемом очень проста. Если какой-то куб разделить на кубики с длиной ребра в 10 раз меньшей, то суммарная площадь поверхностей всех полученных кубиков будет в 10 раз больше, чем поверхность исходного куба; если линейные размеры маленьких кубиков будут в 100 раз меньше, чем у большого куба, общая поверхность увеличится в 100 раз, и т. д. Это правило применимо для всех геометрически подобных тел. Чем меньше линейные размеры любого предмета, тем больше его поверхность по отношению к объему. Форма тела у крупных и мелких млекопитающих достаточно сходна для того, чтобы их поверхность находилась в правильной зависимости от их объема, т. е. на них распространяется указанное выше правило.
Вернемся к животному, находящемуся в условиях высокой температуры. Его организм получает тепло из двух источников: из внешней среды и от собственного метаболизма. Приток внешнего тепла, осуществляющийся путем теплопроводности, конвекции и излучения от земли и солнца, идет только через поверхность тела, и поэтому внешняя тепловая нагрузка прямо пропорциональна этой поверхности. Как было показано в гл. 6, у млекопитающих выработка тепла хотя и не совсем пропорциональна поверхности тела, но приближенно можно считать ее таковой. Поэтому общая тепловая нагрузка на организм—сумма собственной теплопродукции и тепла, поступающего извне, — приближенно пропорциональна величине поверхности тела. В результате мелкие животные с их относительно большей поверхностью оказываются в смысле тепловой нагрузки в гораздо худшем положении, чем крупные.
Если человек в жаркой пустыне теряет с потом 1 л воды в час (т. е. примерно 0,6 л/ч на квадратный метр поверхности тела), то, исходя из соотношения поверхностей тела, мы можем приближенно предсказать, сколько влаги должны испарять в
378 Глава 8. Терморегуляция
аналогичных условиях другие животные, чтобы компенсировать, тепловую нагрузку. Мы можем построить теоретическую кривую, показывающую, какое количество испаряемой воды обеспечила бы достаточное охлаждение того или иного животного (рис. 8.19). Это количество является экспоненциальной функцией массы тела, и в логарифмических координатах кривая будет прямой линией. График на рис. 8.19 построен в линейном масштабе, чтобы показать, как круто вверх идет кривая и как много воды приходилось бы испарять мелким животным, если бы у них не была
Рис. 8.19. Для того чтобы млекопитающее могло сохранять постоянство температуры тела в условиях жаркого климата пустыни, оно должно испарять влагу в количестве, пропорциональном тепловой нагрузке. Поскольку мелкие животные обладают большой относительной поверхностью, величина тепловой нагрузки и соответственно теоретически рассчитанная интенсивность испарения (по отношению к весу тела) быстро возрастают с уменьшением размеров тела. Расчет приведенной кривой основан на предположении, что тепловая нагрузка пропорциональна площади поверхности тела. (Schmidt-Nielsen, 1964.)
иных способов избегать перегрева. Мелкие грызуны весом от 10 до» 100 г должны были бы за 1 ч терять в результате испарения влаги 15—30% своего веса. Поскольку для млекопитающих потеря воды в количестве 10—20% от веса тела смертельна, ясно, что такой способ охлаждения нереален. Поэтому живущие в пустыне мелкие грызуны должны избегать жары, что они и делают, прячась в течение дня в подземных норах.
Пример крупного животного: верблюд. Как видно из рис. 8.19, большие размеры тела дают животному существенное преимущество. Однако у достаточно крупных животных дальнейшее увеличение размеров тела уже не дает большой выгоды. Теоретически, если бы верблюд во всем, кроме размеров, был подобен че
Тепловой баланс 379
ловеку, то ему приходилось бы испарять только вдвое меньше воды (на единицу веса тела), чем человеку. Но верблюд не похож на человека, и он использует гораздо меньше воды для нужд терморегуляции: как мы увидим, у него есть ряд приспособлений, уменьшающих приток тепла из внешней среды.
Посмотрим сначала, что происходит у верблюда с такими двумя переменными в уравнении теплового баланса, как теплота испарения (Яи) и аккумуляция тепла (#а). Об аккумуляции тепла можно судить по повышению температуры тела. В обычных
Рис. 8.20. У верблюда, получающего достаточно воды, температура тела на протяжении суток изменяется в пределах примерно 2°. При отсутствии воды для питья дневные колебания температуры тела могут достигать 7°. Это оказывает большое влияние на использование воды для нужд терморегуляции (см. текст). (Schmidt-Nielsen, 1963.)
условиях у верблюда, ежедневно получающего воду в достаточном количестве, температура тела колеблется в пределах менее чем 2°, а именно от 36 до 38 °C. Однако при лишении питьевой воды суточные колебания значительно возрастают (рис. 8.20) (Schmidt-Nielsen et al., 1957). К утру температура тела может снижаться до 34 °C, а к концу дня достигать максимума — почти 41 °C. Столь значительное повышение дневной температуры означает аккумуляцию тепла в организме. Для верблюда весом 500 кг повышение температуры на 7 °C эквивалентно 2900 ккал, что соответствует экономии 5 л воды. В прохладную ночь накопленное тепло может быть рассеяно путем излучения и теплопроводности, без расходования воды.
380 Глава 8. Терморегуляция
Высокая температура тела в течение дня не только позволяет аккумулировать тепло, но дает еще одно преимущество: при повышении температуры тела уменьшается температурный градиент между организмом и средой (определяющий приток тепла из внешней среды). Это уменьшение притока тепла в организм, так же как и аккумуляция тепла (если не в большей степени), спо-
Рис. 8.21. Изменение температуры тела у верблюда, лишенного питьевой воды, намного снижает поступление тепла из внешней среды, а значит, и затрату воды для терморегуляции (путем испарения). (Schmidt-Nielsen et al., 1957.)
собствует сбережению воды. Это иллюстрирует рис. 8.21, где представлены данные о тепловом балансе некрупного взрослого верблюда в условиях пустыни Сахары. У животного, получающего достаточно воды, количество влаги, испарявшейся за 10 самых жарких часов дня, составляло 9,1 л, что соответствует отдаче 5300 ккал тепла. У того же животного, лишенного питья, испарение воды за 10 ч снизилось до 2,8 л, что соответствовало отдаче 1600 ккал. Таким образом, при обезвоживании организма использование воды уменьшилось более чем в 3 раза.
Основная причина того, что верблюд в пустыне расходует гораздо меньше воды, чем человек (верблюд — 0,28 л/ч, а человек, весящий примерно в 4 раза меньше, — около 1 л/ч), состоит в том, что у верблюда повышается температура тела и, следовательно, происходит аккумуляция тепла и снижается поступление тепла извне. У человека же температура тела остается на почти постоянном уровне — около 37 °C, т. е. тепло практически не накапливается. Кроме того, температурный градиент между орга
Тепловой баланс 381
низмом и средой у человека намного круче, чем у верблюда, так как средняя температура поверхности кожи у потеющего человека примерно 35 °C.
Существенно и то, что верблюд покрыт толстым слоем шерсти, обладающим высокими теплоизоляционными свойствами. Наличие такой изоляции снижает поступление тепла из внешней среды в организм. Значение меха легче всего проверить, если верблюда остричь. В одном из таких опытов расход воды после стрижки (при прочих равных условиях) увеличился почти на 50%.
Итак, главные причины меньшей потери воды у верблюда (по сравнению с величиной, вычисленной с учетом одних только размеров тела) состоят в следующем: 1) у верблюда происходит аккумуляция тепла, связанная с повышением температуры тела; 2) повышение температуры тела уменьшает приток тепла из внешней среды; 3) меховой покров существенно замедляет поступление тепла из внешней среды.
У верблюда есть еще одно преимущество перед человеком: он переносит гораздо большую степень обезвоживания организма. У человека потеря воды в количестве около 10—12% от веса тела уже близка к летальному пределу обезвоживания, тогда как верблюд может без видимого вреда перенести вдвое большую потерю воды. В результате верблюд может без питья идти по пустыне в течение 6—8 дней, а человек погибнет в этих условиях в течение одного дня.
В конце концов, однако, верблюду приходится пить, чтобы восполнить потерю воды; при этом он способен выпивать ее в количестве, превышающем */з веса его тела. Эта поразительная способность к питью породила легенду, будто перед длинным переходом через пустыню верблюд набирает воду в некие резервуары; на самом же деле он, подобно другим млекопитающим, пьет, чтобы восполнить недостаток воды и восстановить ее нормальное содержание в организме, и нет никаких данных в пользу того, что он пьет «в запас», предвидя последующую жажду.
Пример маленького животного: суслик. Как мы уже видели, мелкие грызуны из-за своих малых размеров и относительно большой поверхности должны быть не способны выносить жару и вести дневной образ жизни в пустыне. Однако мелкие суслики североамериканских пустынь опровергают это предположение. Этих мелких грызунов, вес которых варьирует от величины менее 100 г до нескольких сот граммов, часто можно видеть в дневное время. Они быстро перебегают с места на место, часто прячутся в норке, но очень скоро, нередко через несколько минут, появляются снова. В разгар лета в самые жаркие часы дня они менее активны, но утром и под вечер часто вылезают из нор.
Как правило, в местах обитания этих сусликов нет открытых источников воды. Каким же образом они могут оставаться актив-
382 Глава 8. Терморегуляция
яыми в дневное время? Видимо, существенную роль играет аккумуляция тепла, но она не идет с суточной цикличностью, как у верблюда. Находясь вне своей норы на дневной жаре, суслики •очень быстро начинают перегреваться. По чувствительности к повышению температуры тела они не отличаются от других млекопитающих и погибают при подъеме температуры до 43 °C. Однако 42,4 °C они переносят благополучно. Как только суслик начинает перегреваться на жаре, он тотчас возвращается в относительно прохладную нору, где вскоре охлаждается благодаря большой относительной величине поверхности тела. Часто уходя в прохладную нору и выходя обратно, суслик быстро аккумулирует тепло и быстро сбрасывает его избыток, обходясь таким образом без испарения воды. Это и позволяет ему оставаться активным в течение дня.
-ИСПАРЕНИЕ: ПОТООТДЕЛЕНИЕ И УЧАЩЕННОЕ ДЫХАНИЕ
Как мы знаем по собственному опыту, когда возникает потребность в охлаждении, человек потеет. В отличие от человека у собак очень мало потовых желез, и они охлаждаются главным •образом путем учащения дыхания: чтобы увеличить йспарение в гверхних дыхательных путях, они начинают дышать очень быстро и поверхностно (panting). Некоторые животные увеличивают испарение еще одним способом: они смачивают свой мех слюной .и облизывают лапы, охлаждаясь таким образом за счет испарения слюны.
Количество тепла, необходимое для испарения 1 кг воды (580 ккал, или 2426 кДж), не зависит, разумеется, от источника влаги или места, где она испаряется. Посмотрим, нет ли у различных способов испарения влаги каких-то особенностей, которые делали бы один способ предпочтительнее другого для того или иного животного. Ознакомимся для этого с характерными чертами каждого из способов.
У человека повышенное испарение влаги практически полностью осуществляется путем потоотделения. Испарение со всей поверхности тела является эффективным способом охлаждения, тем более что при отсутствии шерстного покрова вода испаряется очень быстро. Однако и у некоторых животных, покрытых шерстью, особенно у крупных (быки, крупные антилопы и верблюды), испарение происходит главным образом путем потоотделения. Хотя у верблюда мех очень густой, это не препятствует испарению, так как воздух в пустыне очень <сух; пот испаряется настолько быстро, что это вводило в заблуждение некоторых авторов, со-эбщавших, будто верблюды никогда не потеют. С другой стороны, у ряда менее крупных копытных — овец, коз и многих небольших газелей — испарение усиливается за счет частого поверхностного дыхания. Это относится и к исследованным хищ
Тепловой баланс 38$
никам. Третий способ — смачивание слюной и облизывание — свойствен многим австралийским сумчатым, в том числе крупным кенгуру, а также ряду грызунов, включая обычную лабораторную крысу. Этот способ не очень эффективен и, видимо, используется главным образом как срочная мера в случаях, когда температура тела угрожает приблизиться к летальному уровню.
Помимо того что смачивание слюной и облизывание менее эффективны, чем потоотделение и частое дыхание, этот способ охлаждения и значительно реже встречается, так что мы не будем' обсуждать его далее. Вместо этого сравним два других способа охлаждения и посмотрим, в чем могут заключаться их преимущества и недостатки.
Птицы в отличие от млекопитающих не имеют потовых желез.. Усиленное испарение влаги достигается у них либо путем учащения дыхания, либо посредством быстрых колебаний тонкого дна ротовой полости и верхней части глотки [так называемое-гулярное трепетание (gular flutter)]. По-видимому, оба этих приема служат эффективными способами охлаждения, и птицы используют либо один, либо другой, либо комбинацию их обоих. (Bartholomew et al., 1968).
Очевидная разница между учащением дыхания и потоотделением как средствами испарения влаги состоит в том, что при* учащенном дыхании животное само создает ток воздуха над, влажной поверхностью, ускоряя таким образом испарение. В этом, отношении потеющие животные находятся в худшем положении.. Второе различие — то, что пот (по крайней мере у человека) содержит значительное количество солей, и при обильном потоотделении потеря солей иногда так велика, что может возникнуть-их дефицит в организме. Именно поэтому рекомендуется добавлять в пищу больше поваренной соли в условиях сильной жары ш избыточного потоотделения. Что касается животных, прибегающих к учащенному дыханию, то они не теряют электролитов, выделяемых железами ротовой и носовой полости (если только» слюна не капает изо рта), так что и в этом отношении они находятся в лучшем положении.
Но учащенное дыхание имеет два очевидных недостатка. Во-первых, усиленная вентиляция легких может привести к избыточному выведению СОг и вызвать сильный алкалоз; во-вторых, увеличение вентиляции требует дополнительной мышечной работы, что связано с увеличением теплопродукции, а тем самым и: тепловой нагрузки на организм. .Развитие алкалоза можно частично предотвратить переходом на более поверхностное дыхание-(уменьшение дыхательного объема), с тем чтобы при учащенном дыхании усиленно вентилировалось главным образом мертвое пространство верхних дыхательных путей. Тем не менее при очень-сильном учащении дыхания у животных неизменно развивается выраженный алкалоз; значит, увеличение вентиляции не огра-
384 Глава 8. Терморегуляция
ничивается только мертвым пространством (Hales, Findlay, 1968; Hales, Bligh, 1969).
Усиление мышечной работы при учащенном дыхании было бы существенным недостатком этого способа, если бы не то любопытное обстоятельство, что мышечная работа (а значит, и теплопродукция) сильно уменьшается благодаря эластическим свойствам дыхательной системы. Когда собака, чтобы охладиться, начинает часто дышать, число дыхательных движений почти сразу возрастает у нее с 30—40 до относительно постоянного уровня 300—400 в минуту. При средней тепловой нагрузке частота дыхания у собаки не устанавливается на каком-то промежуточном уровне; вместо этого животное дышит с высокой частотой короткие интервалы времени, перемежая их с периодами обычного дыхания.
Смысл этого станет нам ясен, если мы вспомним, что вся дыхательная система эластична и, подобно другим эластичным телам, обладает собственной частотой колебаний. При вдохе значительная часть мышечной работы затрачивается на растяжение упругих элементов, которые при выдохе сами собой сжимаются, подобно тому как отскакивает в противоположном направлении теннисный мяч. Для поддержания колебаний дыхательной системы в ее собственном ритме (с резонансной частотой) требуются минимальные мышечные усилия. Соответственно и выработка тепла оказывается незначительной, так что тепловая нагрузка на организм почти не возрастает (Crawford, 1962).
Если бы при учащенном дыхании не использовался резонанс упругой системы, то, как показали расчеты, в связи с добавочной работой мышц выделялось бы больше тепла, чем вообще можно рассеять с помощью такого дыхания. По-видимому, и птицы с целью охлаждения могут переходить на резонансную частоту дыхания, что дает им такую же выгоду, как и млекопитающим (Crawford, Kampe, 1971).
Хотя нам известны некоторые функциональные различия между потоотделением и учащенным дыханием, мы все же не имеем ясного представления о том, почему одни животные охлаждаются так, а другие — иначе. Поскольку многие животные используют оба способа охлаждения, можно думать, что каждый способ обладает какими-то достоинствами, которые мы еще не вполне можем оценить.
Самое важное преимущество учащенного дыхания, возможно, состоит в следующем. Когда животное внезапно подвергается тепловому стрессу (как, например, африканская газель, спасающаяся бегством от хищника) и у него сильно повышается внутренняя температура тела, учащение дыхания могло бы предохранить от перегрева мозг — наиболее чувствительный к повышению температуры орган. На первый взгляд это кажется невероятным, поскольку мозг интенсивно снабжается артериальной кровью. Де-
Тепловой баланс 385
.ло, однако, обстоит 'следующим образом (см. рис. 8.22). У газелей и других копытных большая часть крови притекает к мозгу по внутренней сонной артерии, разделяющейся у основания черепа на сотни мелких артерий; последние, .прежде чем войти в мозг, вновь соединяются в общий ствол. Эти мелкие артерии проходят сквозь большой венозный синус, через который протекает
Рис. 8.22. Газель способна поддерживать температуру мозга на уровне ниже внутренней температуры тела. Артериальная кровь, прежде чем достигнуть мозга, проходит по мелким артериям через синус с охлажденной венозной кровью, оттекающей от носовой полости, где происходит испарение влаги. (Taylor, 1972.)
кровь, идущая от стенок носовой полости и потому охлажденная. Таким образом, кровь в мелких артериях тоже охлаждается и только после этого достигает мозга; в результате температура мозга может быть на 2—3° ниже температуры внутренних областей тела и крови сонных артерий.
Именно такая разница температур была обнаружена у газели Томсона — небольшой восточно-африканской газели, весящей от 15 до 20 кг. Когда эта газель бежала в течение 5 мин со скоростью 40 км/ч, температура ее артериальной крови повысилась из-за увеличения выработки тепла с 39 °C (нормальная для газели температура) до 44°C, но температура мозга не достигла и 41 °C, т. е. осталась на вполне безопасном уровне (Taylor, Lyman, 1972). Трудно представить себе систему охлаждения, которая предохраняла бы от перегрева все тело в условиях, когда животное бежит на жаре со скоростью, требующей 40-кратного увеличения интенсивности обмена, однако избирательное охлаждение крови, идущей к мозгу, позволяет избежать наиболее опасных проявлений перегрева.
25—1873
386 Глава 8. Терморсгу ыция
Такое же охлаждение крови, притекающей к мозгу, обнаружено и у ряда домашних копытных; возможно, что этот механизм имеется у всех животных, охлаждающихся с помощью учащенного дыхания. Но вряд ли подобный механизм существует у человека, поскольку у него охлаждается вся поверхность тела, а к учащенному дыханию как способу охлаждения он не прибегает.
Уже давно известно, что некоторые рептилии, оказавшись в условиях сильного перегрева, начинают дышать .чаще и с открытым ртом, что весьма напоминает учащенное дыхание (panting) у теплокровных животных. Довольно трудно было установить, какую роль в терморегуляции играет такое усиленное дыхание у рептилий, хотя оно, несомненно, увеличивает испарение влаги в дыхательных путях. Правда, количество испаряемой воды недостаточно для того, чтобы ‘существенно понизить температуру тела по сравнению -с внешней температурой; по-видимому, это лишь зачаточная форма того механизма, который используется у птиц и млекопитающих. Однако учитывая способность млекопитающих избирательно охлаждать мозг, можно иначе оцепить и значение учащенного дыхания у ящериц.
Рис. 8.23. Если ящерицу Sauromalus перенести из среды с температурой 15 °C в условия 45 °C, то у нее быстро повышается температура как мозга, так » клоаки. Однако после достижения 41 °C кривые температур расходятся: мозг остается на 2° холоднее клоаки и на 3° холоднее воздуха. (Crawford, 1972.)
Показано, что у пустынной ящерицы Sauromalus obesus учащение дыхания играет некоторую роль в общем тепловом балансе организма. При внешней температуре 45 °C тело ящерицы оказывается лишь ненамного холоднее (44,1 °C), но температура мозга остается на уровне 42,3 °C, т. е. почти на 3° ниже температуры окружающей среды. Если эту ящерицу перенести из помеще
Зимняя спячка и оцепенение 387
ния с температурой 15 °C в условия 45 °C, то 'сначала температура как тела (клоаки), так и мозга быстро повышается (рис. 8.23), но температура 'мозга, достигнув 41—42 °C, устанавливается на этом уровне, тогда как температура, измеряемая в клоаке, продолжает расти, приближаясь к уровню температуры среды.
У Sauromalus сонные артерии проходят очень близко к поверхности глотки — настолько близко, что они видны, когда животное открывает рот. Артериальная кровь протекает здесь под влажной поверхностью, где происходит испарение, и охлаждается, прежде чем попасть в мозг. Это еще один пример животного, способного поддерживать в разных частях тела разную температуру, причем на хорошо регулируемом уровне.
ЗИМНЯЯ СПЯЧКА И ОЦЕПЕНЕНИЕ
Поддержание высокой температуры тела в условиях холода ценой увеличения интенсивности обмена в несколько раз — это весьма дорогостоящий путь. Мелкие животные исходно обладают высоким уровнем метаболизма, и дальнейшее его повышение может быть для них слишком большой «растратой», особенно при отсутствии или недостатке пищи. Самый простой и единственный логичный выход из положения состоит в том, чтобы прекратить борьбу за поддержание высокой температуры тела — пусть опа снижается. При этом не только исчезает проблема повышения теплопродукции, но и сберегаются энергетические резервы, поскольку охлажденные ткани потребляют меньше топлива. В этом, •собственно, и заключается сущность явления, называемого зим-ней спячкой или гибернацией1.
Многие млекопитающие и несколько видов птиц регулярно впадают в спячку каждую зиму. Это означает, что температура нх тела падает почти до уровня окружающей среды; очень сильно снижаются метаболизм, дыхание, частота сокращений сердца и многие другие функции; животное находится в торпидном (оцепенелом) состоянии и слабо реагирует на внешние стимулы, например на шум или прикосновение. Прекратив активный образ жизни и резко снизив интенсивность обмена, животное переживает долгую зиму. Готовясь к зимней спячке, животные обычно накапливают много жира (т. е. создают большие запасы топлива). Эти запасы, которые можно уже не тратить на согревание тела, используются в течение длительных неблагоприятных периодов.
В спячку впадают главным образом мелкие животные; это понятно, так как при свойственной им интенсивности метаболизма они нуждаются в обильном питании. Например, впадают в спячку
1 Термин «гибернация» (от лат. hiberna — зима) означает также перезимовывание, т. е. неактивное состояние, пойкилотермных животных, например насекомых или улиток.
25*
.388 Глава 8. Терморегуляция
многие грызуны: хомяки, перогнаты, сони. Обитатели высоких широт, питающиеся насекомыми, едва ли могут зимой найти себе пищу; поэтому насекомоядные (например, ежи) и летучие мыши тоже впадают в зимнюю спячку. Спячка свойственна и некоторым сумчатым Австралии, в частности карликовому опоссуму (Bartholomew, Hudson, 1962). К числу впадающих в спячку птиц относятся колибри — самые мелкие из всех птиц, а также питающиеся насекомыми стрижи и некоторые птицы-мыши (африканский род Coitus).
Понятию зимняя спячка (или гибернация) нелегко дать вполне удовлетворительное определение. В физиологии так называют состояние оцепенения со значительным снижением интенсивности обмена. Таким образом, медведей, которые могут спать на протяжении большей части зимы, большинство физиологов все же не отнесет к истинно гибернирующим животным. Температура тела зимой у них падает только на несколько градусов, уровень метаболизма и другие физиологические показатели снижаются лишь в умеренной степени, а самки нередко рождают зимой детенышей. Иными словами, зимняя «спячка» медведей не соответствует тому, что физиологи обычно понимают под истинной гибернацией.
Другой термин — эстивация (от лат. aestas — лето)—означает неактивное состояние животных в течение лета; определение этого термина еще менее четко. Эстивацией можно назвать «сонное», неактивное состояние улиток при наступлении засухи или состояние сусликов, которые в самые жаркие месяцы исчезают в своих норах, оставаясь там вялыми и неподвижными. Колумбийские суслики начинают впадать в эстивацию в жарком августе, остаются неактивными на протяжении всей осени и зимы и вылезают из нор только к следующему маю. Что это—зимняя или летняя спячка? И когда одно состояние переходит в другое? По существу, между этими состояниями нет четкого физиологического различия. Кроме того, многие зимнеспящие животные (например, летучие мыши) могут впадать в торпидное состояние ежедневно, и при этом у них снижается температура тела и интенсивность обмена. Таким образом, это физиологическое состояние вполне сходно с гибернацией, хотя продолжается не недели и месяцы, а всего лишь несколько часов.
Ввиду отсутствия точных определений описанные состояния трудно разграничить, и поэтому мы будем рассматривать спячку и оцепенение как единый физиологический феномен.
Ранее полагали, что зимняя спячка и оцепенение возникают как результат недостаточности терморегуляторной системы в условиях холода и, таким образом, отражают определенную «примитивность» организации и слабость механизмов физиологического контроля. Сейчас, однако, стало совершенно ясно, что спячку нельзя объяснить недостаточностью терморегуляции; напро
Зимняя спячка и оцепенение 389
тив, это хорошо регулируемое физиологическое состояние, и кажущееся сходство между находящимся в спячке млекопитающим и холоднокровным животным (например, лягушкой или ящерицей) обманчиво. Как мы увидим, гибернацию отнюдь нельзя считать результатом неэффективности физиологических механизмов.
ТЕМПЕРАТУРА ТЕЛА И ПОТРЕБЛЕНИЕ КИСЛОРОДА
У некоторых животных, в частности у летучих мышей, могут быть и ежедневные периоды оцепенения, и более длительная спячка. На понижение температуры летучие мыши реагируют одним из двух характерных способов. Это видно из рис. 8.24, где
Рис. 8.24. Температура тела у двух видов североамериканских летучих мышей. (Herreid, Schmidt-Nielsen, 1966.)
При похолодании эти животные могут либо оставаться активными и сохранять нормальную (около 34 °C) температуру тела, либо впадать в оцепенение, и тогда температура снижается примерно до уровня температуры воздуха.
представлены данные о температуре тела у двух видов североамериканских рукокрылых. При температурах среды ниже примерно 30 °C эти животные либо впадают в оцепенение и их температура в пределах 1—2° совпадает с температурой воздуха, либо остаются активными и поддерживают нормальную температуру тела между 32 и 36 °C. Даже при температуре воздуха ниже 10°С эти
399 Глава 8. Терморегуляция
летучие мыши могут либо оставаться активными, либо впадать в оцепенение.
Оба вида имеют сравнимые размеры тела. Более крупный вид — бурый кожан (Eptesicus fuscus) весом около 16 г — почти повсеместно распространен в США и по крайней мере в северных частях своего ареала впадает в зимнюю спячку. Несколько мельче другой вид — мексиканский складчатогуб (Tadarida mexica-
Рис. 8.25. Потребление кислорода у летучих мышей, находящихся в активном состоянии, возрастает по мере снижения температуры воздуха; мыши, впавшие в оцепенение, потребляют во много раз меньше О2, чем активные (те же виды, что и на рис. 8.24). (Herreid, Schmidt-Nielsen, 1966.)
па)\ эти животные весом примерно 10 г обитают в сухом и жарком климате юго-запада США и в Мексике; зимой они мигрируют к югу и, насколько известно, спячка им не свойственна.
Хотя в естественных условиях первый вид впадает в спячку, а второй — нет, их реакция на понижение температуры воздуха поразительно сходна. У особей, сохраняющих в условиях похолодания нормальную температуру тела, наблюдаются сдвиги, типичные для гомойотермных животных. По мере снижения температуры воздуха увеличивается потребление кислорода, что отражает рост энергетических расходов на поддержание теплоты тела. Как видно из рис. 8.25, минимум потребления Ог соответствует температуре воздуха 35°C. Однако для особей, впадающих в оцепенение, характерна очень низкая интенсивность обмена. Например, при 15 °C оцепенелые животные потребляют в 40 раз меньше кислорода, чем активные. Иными словами, первые расходуют энергетические резервы в 40 раз медленнее, чем вторые, и соответственно запасов жира может хватить им на больший (в 40 раз) срок.
Зимняя спячка и оцепенение 391
Экономию энергии, обусловленную переходом в торпидное состояние, тщательно проанализировал в своих работах Таккер (Tucker, 1965а, Ь, 1966). Потребление кислорода в период оце: пенения определить нетрудно, но нужно также учитывать количество энергии, необходимое для согревания тела после выхода из этого состояния. Исследуя калифорнийского перогната (Perog-nathus californicus), Таккер нашел, что в случаях кратковремен-
Часы суток
Рис. 8.26. Потребление кислорода на протяжении суток у перогната при ограниченном пищевом рационе (1,5 г зерна в день). (Tucker, 1965а.)
В течение 9 ч животное находилось в состоянии оцепенения, потребляя очень мало кислорода, после чего наблюдался пик поглощения Ог, знаменующий возвращение в активное состояние. Температура воздуха была 15 °C.
ного оцепенения расход энергии на согревание составляет существенную часть общих энергетических затрат. Перогнат весит около 20 г; он легко впадает в оцепенение при любой температуре воздуха от 15 до 32°C. Если температура опускается ниже 15 °C, то нормальное состояние оцепенения оказывается нарушенным и животное уже не в силах выйти из него самостоятельно; другими словами, при охлаждении ниже 15 °C перогнат не способен выработать столько тепла в организме, чтобы начался процесс согревания.
Перогнат особенно легко впадает в торпидное состояние при ограниченном пищевом рационе. При постепенном снижении количества пищи периоды оцепенения у него становятся все более продолжительными. Благодаря этому вес перогната не снижается, даже если количество пищи уменьшается втрое по сравнению с тем, которое он обычно съедает, находясь в активном состоянии. На рис. 8.26 представлена кривая потребления кислорода этим
392 Глава 8. Терморегуляция
животным при ограниченном пищевом рационе. В данном случае перогнат получал 1,5 г зерна в день и при таком скудном питании ежедневно находился в состоянии оцепенения примерно 9 ч. При этом он не терял в весе, хотя получал меньше половины той пищи, которая была бы нужна ему в активном состоянии.
С чего начинается процесс оцепенения? Происходит ли дело таким образом, что сначала теряется тепло и соответственно падает температура тела, а в результате снижается уровень метаболизма, что в свою очередь ускоряет падение температуры тела? Или же сначала уменьшается интенсивность обмена, далее температура тела начинает падать, это приводит к дальнейшему снижению теплопродукции и т. д.?
Имеющиеся данные о выработке и .потере тепла на всех стадиях торпидного состояния позволяют ответить на этот вопрос. Оказалось, что переход в такое состояние может быть просто результатом того, что прекращается терморегуляторное повышение уровня обмена при падении температуры воздуха ниже критической точки (для перогната — около 32,5°C). При любой температуре ниже этой точки снижение обмена до уровня, характерного для состояния покоя в термонейтральной зоне, само по себе (без изменения теплопроводности) 'Приведет к падению температуры тела. Иными словами, особого механизма, повышающего потерю тепла, не требуется. Как только температура тела начинает падать, метаболизм замедляется и животное впадает в оцепенение, причем как теплопродукция, так и температура тела продолжают снижаться.
Пробуждение у перогната возможно при любой температуре выше 15 °C; для этого теплопродукция организма должна достигнуть максимума для данной температуры, что связано с увеличением потребления О2 в 10—*15 раз по сравнению с минимумом при той же температуре. Таким образом, пробуждение — активный процесс, который требует значительной траты энергии на протяжении довольно долгого промежутка времени, пока температура тела не достигнет нормального уровня.
Представим себе, что при 15 °C перогнат впадает в оцепенение, а затем тотчас же пробуждается. Дает ли это какую-либо экономию энергии или же согревание обходится слишком дорого: Снижение температуры тела занимает около двух часов, а общее потребление кислорода за это время составляет 0,7 мл на 1 г веса тела. Если животное после этого сразу же «просыпается», то согревание до нормальной температуры тела занимает 0,9 ч и требует потребления 5,8 мл О2 на 1 г. Таким образом, при переходе в оцепенение и немедленном выходе из него потребляется 6,5 мл О2 на 1 г на протяжении 2,9 ч. В то же время для поддержания нормальной температуры тела в течение 2,9 ч при температуре воздуха 15 °C перогнат использовал бы 11,9 мл О2 на 1 г. Значит, благодаря оцепенению он затрачивает лишь 55% той энергии, ко
Зимняя спячка и оцепенение 393
торая нужна для поддержания нормальной температуры в течение того же времени (Tucker, 1965b).
Более длительные периоды оцепенения, например 10 ч, дают еще большую экономию энергии. Если к ранее подсчитанным затратам мы прибавим количество кислорода, потребленного за 7,1 ч пребывания в оцепенении при 15 °C, а именно 1,2 мл/г, то для всего цикла длительностью в 10 ч, включающего переход в торпидное состояние и выход из него, мы получим 7,7 мл О2 на 1 г. Это меньше 20% «цены» поддержания нормальной температуры тела в течение 10 ч при 15 °C (составляющей более 40 мл О2 на 1 г.).
Можно видеть, что во всех рассмотренных случаях наибольшая затрата энергии связана с периодом пробуждения. При 10-часовом цикле на этот период приходится 75% всего расхода энергии.
Широкое распространение этого феномена — перехода в торпидное состояние при неблагоприятных условиях — позволяет предполагать, что он имеет фундаментальное значение для выживания. К числу животных, способных впадать в оцепенение, относится ехидна (Tachyglossus aculeatus)—очень отдаленный родственник современных млекопитающих. В обычных условиях ехидна превосходно поддерживает нормальную температуру даже при падении температуры воздуха ниже нуля (Schmidt-Nielsen et al., 1966). Однако если ее содержать при 5 °C без пищи, она быстро впадает в оцепенение. При этом ритм сердца урежается у нее с 70 до 7 ударов в минуту, температура тела падает примерно до +5,5°, а потребление кислорода составляет около 0,03 мл/(г-ч), т. е. оно теперь примерно в 10 раз меньше, чем в состоянии покоя в обычных условиях (Augee, Ealey, 1968).
МЕХАНИЗМЫ РЕГУЛЯЦИИ
Оцепенение и спячка — строго контролируемые физиологические состояния. Как мы только что видели, продолжительность ежедневного оцепенения у перогната определяется количеством пищи и потребностью в экономии энергии. Очевидно, что не только переход в оцепенение, но и длительность этого состояния, и выход из него должны строго регулироваться.
В естественных условиях начало зимней спячки у животных приурочено к определенному времени года, но оно вовсе не обязательно обусловлено холодом или недостатком пищи. Годичные циклы гибернации зависят от длины светового дня и гормональных изменений. Часто невозможно бывает, например, заставить впасть в спячку животное (которому она свойственна) в начале лета и особенно в период размножения.
Оцепенелое животное с его низкой температурой тела кажется совершенно инертным; оно не способно к координированным
394 Глава 8. Терморегуляция
движениям, едва отвечает на раздражение органов чувств. Внешне оно напоминает пойкилотермное животное в охлажденном состоянии. Всем нам известно, как «немеют» пальцы при охлаждении кистей рук; дело в том, что при температуре ниже 10—15 °C прекращается проведение нервных импульсов и исчезает чувствительность. (Если мы еще можем шевелить холодными пальцами, то лишь потому, что соответствующие мышцы расположены выше кисти, в предплечье.)
Если у находящегося в спячке животного нервы при пониженной температуре не действуют, то и вся нервная система не может координированно работать, скажем, при 5 °C. Тем не менее дыхание и многие другие физиологические функции остаются вполне координированными, хотя и замедленными. Если температура среды опускается до нуля и ниже, то некоторые животные, впавшие в спячку, погибают. Другие же реагируют одним из двух способов, причем эти реакции определенно свидетельствуют о сохранности ряда центральных нервных функций. Указанные реакции состоят либо в том, что животное выходит из состояния спячки, либо в том, что у него прекращается снижение температуры тела в результате регулируемого повышения теплопродукции, так что температура животного остается на каком-то низком, но постоянном уровне, например 5 °C.
Такая хорошо регулируемая теплопродукция, несомненно, требует эффективной координации центральных нервных функций. В качестве примера можно привести европейского ежа, температура которого во время морозов поддерживается на уровне от +5 до +6 °C. Это позволяет животному избежать обморожения и в то же время сберечь «топливо», которое, таким образом, не тратится ни на энергетически дорогостоящий процесс полного пробуждения, ни на поддержание высокой температуры тела, свойственной активному состоянию. Для животного, которому морозы могут угрожать несколько раз в течение зимы, повторные пробуждения могли бы оказаться энергетически слишком невыгодными; поддержание же температуры тела на самом низком уровне, еще предохраняющем животное от замерзания, — гораздо более экономная стратегия.
Аналогичная регуляция температуры тела во время оцепенения была обнаружена у вест-индского колибри Eulampis jugula-ris, который, подобно другим колибри, легко впадает в торпидное состояние. У оцепенелого Eulampis температура тела обычно близка к температуре воздуха, однако если последняя падает ниже 18 °C, то температура тела больше не снижается и остается на уровне 18—20 °C; значит, теплопродукция при внешних температурах ниже 18 °C должна возрастать (рис. 8.27), и это возрастание должно быть пропорционально снижению температуры.
Eulampis для нас особенно интересен еще и как пример того, что состояние оцепенения свойственно не только обитателям хо-
Зимняя спячка и оцепенение 395
лодного климата, но и тропическим теплокровным животным. Кроме того, четкая регуляция интенсивности обмена при двух разных уровнях температуры тела свидетельствует, что оцепене-
Рис. 8.27. Потребление кислорода у тропического колибри Eulampis. (Hainsworth, Wolf, 1970.)
У особи, находящейся в активном состоянии, потребление О2 возрастает линейно по мере понижения температуры воздуха. Особь, впавшая в оцепенение, потребляет мало кислорода, но если температура воздуха падает ниже 18 °C, то возрастает выработка тепла, так что температура тела остается на уровне 18—20 °C, хотя птичка не выходит из торпидного состояния.
механизма. Наконец, интересно отметить, что общая теплопроводность (наклон линии регрессии на рис. 8.27) у колибри в тор-пидном и в активном состоянии одинакова.
ВЫХОД ИЗ ТОРПИДНОГО СОСТОЯНИЯ («ПРОБУЖДЕНИЕ»)
Повышение температуры тела при пробуждении связано, как мы видели, с наибольшими затратами энергии по сравнению с другими этапами цикла. В этот период животное начинает очень сильно дрожать, его мышцы сокращаются и запасы топлива используются, по-видимому, с максимальной скоростью. В процессе разогревания важную роль играют, однако, не только мышцы, но и особый вид жировой ткани, называемый бурым жиром.
Помимо обычного, или светлого, жира у многих млекопитающих имеются большие или меньшие участки особого бурого жира; последний отличается от светлого жира не только цветом, но и характером обмена. Бурая жировая ткань содержит особенно
396 Глава 8. Терморегуляция
много цитохрома, благодаря чему он может очень интенсивно потреблять кислород; светлая жировая ткань, напротив, метаболически мало активна. Особенно много бурого жира у зимнеспящих животных, и уже давно высказывалась мысль, что между зимней спячкой и количеством бурого жира существует прямая связь. Природа этой связи долго оставалась неизвестной, пока биохимические исследования не показали, что бурая жировая
Рис. 8.28. У выходящего из зимней спячки хомяка температура передней части тела (защечных мешков) возрастает быстрее, чем температура задней части (ректальная). (Lyman, 1948.)
ткань обладает способностью к интенсивному потреблению кислорода и, следовательно, к быстрой выработке тепла. При выходе из торпидного состояния температура бурого жира, особенно в местах его скопления между лопатками, выше температуры других областей тела.
У выходящего из состояния спячки животного температура различных частей тела не одинакова, так как процесс согревания идет отнюдь не равномерно. На рис. 8.28 приведены данные о температуре в защечных мешках и в прямой кишке у «пробуждающегося» хомяка. Передняя часть тела, где расположены такие жизненно важные органы, как мозг и сердце, согревается гораздо быстрее, чем задняя. Повышение температуры сердца не просто существенно: оно должно быть начальным этапом пробуждения, так как сердце обеспечивает циркуляцию крови и доставку кислорода во все остальные органы. Основная масса бурого жира также находится в передней части тела. Измерения показывают, что согревание задней части тела начинается только после того, как температура передней части приблизится к норме. На этот-то момент, когда все тело животного быстро приобретает нормальную температуру, и приходится пик поглощения кислорода; после этого потребление О2 снижается и устанавливается на^обычном уровне.
Зимняя спячка и оцепенение 397
Тот факт, что температура разных частей тела повышается неодновременно, приводит к неизбежному выводу: на начальных стадиях пробуждения кровоток почти исключительно направлен к жизненно важным органам передней части тела и только после их разогрева он заметно усиливается и в задних частях.
Объем кровотока в различных органах можно определить с помощью радиоактивных индикаторов; такие исследования показали, что скелетные мышцы передней части тела у животного, пробуждающегося от спячки, получают в 16 раз больше крови, чем у бодрствующего животного в обычном состоянии. Это подтверждает, что мышечная ткань играет важную роль в повышении теплопродукции при выходе из спячки. В начале этого периода мышцы задней части тела получают примерно в 10 раз меньше крови, чем мышцы передней части. В то же время в бурой жировой ткани объем кровотока больше, чем даже в самых активных мышцах, что говорит об участии этой ткани в теплопродукции. Как и можно было ожидать, оказалось, что желудочно-кишечный тракт, в особенности тонкий кишечник, относится к органам, хуже всего снабжаемым кровью при пробуждении от спячки (Johansen, 1961).
Как ни странно, бурая жировая ткань, имеющая столь важное значение для зимнеспящих млекопитающих, по-видимому, отсутствует у ряда регулярно впадающих в спячку птиц, в частности у колибри, стрижей и козодоев (Johnston, 1971).
ТЕМПЕРАТУРА ТЕЛА
У «ХОЛОДНОКРОВНЫХ» ЖИВОТНЫХ
По традиции птиц и млекопитающих называют теплокровными, а всех остальных животных — холоднокровными. При всех недостатках этой терминологии она общепринята и удобна, поэтому мы ею воспользуемся за неимением других названий, достаточно полных и точных. Температура тела у большинства холоднокровных животных в основном зависит от температуры окружающей среды. Однако у некоторых так называемых «холоднокровных» она может поддерживаться и действительно поддерживается на более высоком уровне, чем температура воды или воздуха, где эти животные обитают. То, чего они могут в этом отношении достичь, определяется простыми физическими принципами, хотя отдельные адаптации поистине свидетельствуют об изобретательности природы.
Вернемся к уравнению теплового баланса (стр. 353) и переставим его члены так, чтобы в левой части оставалось только количество аккумулированного тепла (Яа). Тогда в правой части окажутся общая теплопродукция (//общ) и теплообмен путем излучения (Яр), теплопроводности и конвекции (Ят) и испарения (Яи). Если температура тела выше температуры окружающей
398 Глава 8. Терморегуляция
среды, то путем теплопроводности (Ят) и испарения (Яи) тепло выводится из организма, т. е. эти члены становятся отрицательными. В этом случае уравнение принимает вид
яа=яобщ±яр-ят-ни.
Ниже будут рассмотрены способы увеличения аккумуляции тепла (Яа), необходимой для повышения температуры тела.
ВОДНЫЕ ЖИВОТНЫЕ
В случае водных животных ситуация довольно проста, поскольку в воде не происходит испарения и нет значительных источников излучения (инфракрасные лучи водой быстро поглощаются). В результате уравнение принимает вид Яа = ЯОбщ—Ят. Отсюда видно, что аккумуляция тепла может быть увеличена путем изменения только двух параметров: должна либо возрасти общая теплопродукция, либо снизиться до минимума отдача тепла путем теплопроводности.
Из-за большой теплопроводности и теплоемкости воды мелкие животные быстро теряют тепло и не могут поднять температуру тела существенно выше температуры окружающей среды. Даже если такие животные интенсивно вырабатывают тепло (высокая интенсивность обмена) и могли бы увеличить теплопродукцию еще более, им для этого пришлось бы повысить потребление кислорода.
Здесь-то и заключена суть проблемы. Ведь для интенсивного’ потребления кислорода требуется большая поверхность жабр и нужна кровь для транспорта О2. Но кровь, протекая по жабрам, неизбежно будет охлаждаться до температуры воды: жаберная мембрана, которая должна быть достаточно тонкой, чтобы обеспечить диффузию кислорода, практически не создает препятствия для отдачи тепла. Поэтому в случае, если между жабрами и тканями нет теплообменника, животное не сможет существенно повысить температуру тела.
Такой теплообменник имеется у некоторых крупных, быстро плавающих рыб (например, тунцов и акул), благодаря чему они могут контролировать температуру в отдельных областях тела; в частности, они поддерживают высокую температуру, не зависящую от температуры окружающей воды, в своей плавательной мускулатуре.
Теплообменники, сохраняющие тепло в плавательных мышцах тунца, в принципе аналогичны противоточным теплообменникам в плавниках кита, но анатомически они устроены несколько иначе. У обычных рыб плавательная мускулатура снабжается кровью через периферические ветви спинной аорты, идущей вдоль позвоночника. У тунца дело обстоит иначе.
Сосуды, снабжающие кровью темные красные мышцы (кото
Температура тела у «холоднокровных» животных 399
рые тунец использует при равномерном быстром плавании), расположены по бокам тела непосредственно под кожей. От этих главных стволов отходит множество тонких параллельных сосу-
Рис. 8.29. У тунца кровь, идущая к плавательным мышцам, проходит через теплообменник, в результате чего мышцы сохраняют тепло, хотя температура .артериальной крови равна температуре воды. (Carey, Teal, 1966.)
дов; при этом образуется плотное сплетение, в котором артерии перемешаны с венами, несущими кровь в противоположном направлении (рис. 8.29 и фото 8.3). Холодный конец этого теплообменника расположен у поверхности тела, а теплый — глубоко в мышцах. Артериальная кровь, оттекающая от жабр, имеет тем
400 Глава 8. Терморегуляция
пературу воды. Протекая по тонким артериям, окруженным ве-нами, эта кровь поглощает тепло венозной крови, оттекающей or мышц. Ускорению теплообмена способствует малый диаметр со-судов — порядка 0,1 мм. Когда венозная кровь собирается в более крупные вены, расположенные под кожей, она уже охлажде-
Фото 8.3. Теплообменник тунца. (Фото предоставлены Е. D. Stevens, University of Guelph, Онтарио.)
Слева — поперечный срез 2-килограммового тунца Kalsuwonis pelamis. Видно,, что тело этого сильного пловца состоит в основном из мышц. Красные мышцы, имеющие высокую температуру, на фото выглядят почти черными. Основной теплообменник расположен непосредственно под позвоночным столбом, почти точно в центре снимка. Справа — поперечный разрез сосудистого теплообменника; видны артерии (небольшие, с толстыми стенками), перемежающиеся с примерно таким же числом вен (большего диаметра, тонкостенные). Диаметр артерий около 0,04 мм, диаметр вен 0,08 мм, длина тех и других сосудов около 10 мм.
на — ее тепло вернулось к мышцам через согретую артериальную кровь. В результате разность между температурой мышц тунца и температурой воды, в которой он плавает, может достигать 14 °C.
Повышение температуры мышц дает то преимущество, что увеличивает их мощность1. Благодаря большой мощности мышц тунец способен плавать с большой скоростью относительно независимо от температуры воды, и это позволяет ему успешно пре-
1 Сила, развиваемая сокращающейся мышцей, относительно мало зависит от температуры; поэтому почти не зависит от нее и работа, производимая при одиночном сокращении (силаХрасстояние). Однако при более высокой температуре мышца сокращается быстрее, и в результате увеличения числа сокращений в Единицу времени повышается мощность (работа'в единицу времени).
Температура тела у «холоднокровных» животных
401
следовать такую быстро плавающую добычу, как пелагические рыбы (например, макрель) или кальмары.
У некоторых (хотя далеко не у всех) крупных акул имеются аналогичные теплообменники, обеспечивающие поддержание относительно высокой температуры мышц (Carey, Teal, 1969; Carey et al., 1971).
Значительная разница в температуре отдельных частей тела' дает тунцу еще одно преимущество. Благодаря специальным теплообменникам поддерживается высокая температура печени и органов пищеварения. Ведь большая мощность мышц требует и надлежащего обеспечения топливом, а значит, и быстрого пищеварения; последнее лучше всего достигается повышением температуры желудочно-кишечного тракта.
Сравним теперь этих «теплых» рыб с морскими млекопитающими и посмотрим, в чем основные различия и черты сходства между ними. Как мы уже знаем, тюлени и киты имеют на поверхности толстый теплоизоляционный слой (подкожный жир); потеря тепла в конечностях у них снижена благодаря теплообменникам; но главное состоит в том, что они дышат воздухом, и поэтому кровь у них не охлаждается на дыхательных поверхностях до температуры окружающей воды. «Теплым» рыбам, напротив, не нужна теплоизоляция поверхности тела, поскольку у них вся артериальная кровь, оттекающая от жабр, уже имеет температуру воды. Но они способны регулировать температуру отдельных областей тела благодаря соответственно расположенным эффективным теплообменникам, которые сохраняют локально вырабатываемое тепло.
НАЗЕМНЫЕ ЖИВОТНЫЕ
В уравнение теплового баланса наземных животных входят все члены, приведенные на стр. 398. Для повышения температуры тела (т. е. увеличения аккумуляции тепла) этим животным важно уменьшить испарение и потерю тепла путем теплопроводности, а также довести до максимума приток тепла с излучением и собственную теплопродукцию. Обычно единственным источником излучения является солнце, и в отсутствие солнечного света только повышение собственной теплопродукции может обеспечить согревание тела. Разумеется, внешний и внутренний источники тепла (солнечная радиация и обмен веществ) могут использоваться одновременно, но животному выгоднее получать тепло извне, чем расходовать ресурсы организма.
Тепло солнечных лучей особенно интенсивно используют насекомые и пресмыкающиеся. Поглощать больше лучистой энергии этим животным помогают окраска тела и ориентация его по отношению к солнцу. Многие рептилии способны изменять свою окраску путем концентрирования или диспергирования пигмен-26—1873
402 Глава 8. Терморегуляция
тов в особых клетках кожи. Поскольку примерно половина энергии солнечного излучения приходится на область видимого света, темный цвет кожи существенно увеличивает поглощение солнечной энергии (уменьшая отражение). На поглощение ближнего инфракрасного света изменение окраски влияет лишь незначительно, так как для этой области спектра поверхность тела уже близка к «черной» (см. выше раздел «Излучение»).
Второй способ усилить поглощение солнечного тепла состоит в том, чтобы увеличить освещенную солнцем поверхность. Для этого животное располагает свое тело под прямым углом к солнечным лучам, распластывается и вытягивает конечности. Таким путем ящерица, например, может поднять свою температуру до уровня значительно выше температуры воздуха. Прогревшись в нужной степени, ящерица предотвращает дальнейшее согревание, изменяя окраску кожи на более светлую и меняя позу — располагаясь более или менее параллельно солнечным лучам; в конце концов животное может уйти в тень. Большое значение имеет также температура субстрата; например, ящерица может согреться, прижавшись к теплому камню и увеличив таким образом приток тепла.
Греясь на солнце, ящерицы могут очень сильно повышать температуру своего тела. Например, в горах Перу на высоте 4000 м, где прохладно даже летом, ящерица Liolaemus остается активной и выходит из укрытия даже при температурах ниже 0 °C. Одну из этих ящериц, весом 108 г, поместили утром на солнце, когда температура воздуха еще была —2 °C. Спустя час температура воздуха поднялась до +1,5°C, а измеряемая в клоаке температура тела ящерицы возросла с +2,5 до 33°C (рис. 8.30). Таким образом, тело было уже на 30° теплее окружающего воздуха, и это позволяло животному находиться в активном состоянии, несмотря на холод.
Когда ящерица согрелась, температура ее тела далее остается на более или менее постоянном уровне около -35 °C, хотя температура воздуха продолжает постепенно повышаться. Животное достигает этого тем, что снижает поглощение солнечной энергии, изменяя положения тела; поддерживаемую при этом температуру тела называют предпочитаемой или эккритической температурой.
Разумеется, для животного важно согреваться и переходить к полной активности как можно быстрее. С другой стороны, когда животное уходит в тень, ему выгодно было бы замедлить неизбежное охлаждение. Может ли ящерица каким-то образом ускорить свое согревание или замедлить охлаждение? Согревание внутренних областей тела может быть ускорено путем увеличения притока крови к нагретым участкам кожи; но как обстоит дело с замедлением охлаждения?
Этот вопрос был исследован на нескольких видах ящериц. Проблема охлаждения имеет особое значение для галапагосской
Температура тела у «холоднокровных» животных 40?
морской игуаны, которая питается в воде, имеющей температуру 22—27°C. Выйдя из воды, игуана устраивается на камнях вблизи полосы прибоя и греется на солнце. Когда же она погружается в море, где кормится водорослями, ее температура начинает падать, приближаясь к уровню температуры воды, который ниже эккритического. Этому животному было бы выгодно замедлить
Рис. 8 30. Перуанская горная ящерица Liolaemus появляется утром, когда температура воздуха ниже нуля; подставляя себя солнечньш лучам, она быстро согревается до температуры, обеспечивающей полную активность. (По данным Pearson, 1954.)
охлаждение в воде, так как это позволило бы удлинить период, в течение которого оно может активно есть и достаточно быстро двигаться, чтобы не стать добычей хищника. А после выхода на сушу, наоборот, выгодно было бы согреваться как можно быстрее.
Сравнение скоростей согревания и охлаждения морской игуаны показало, что как в воде, так и в воздухе она согревается примерно вдвое быстрее, чем охлаждается (рис. 8.31). Если холодное животное поместить в воду с температурой на 20° выше температуры тела, то скорость его нагрева окажется в 2 раза больше скорости охлаждения в условиях, когда теплое животное помещают в воду, которая на 20° холоднее тела. В воздушной среде скорость обоих процессов ниже, чем в воде, но опять-таки нагрев идет примерно вдвое быстрее, чем охлаждение при той же разности температур.
Различие в скоростях согревания и охлаждения в основном, по-видимому, обусловлено изменениями в циркуляции крови, особенно в коже. При согревании частота сокращений сердца высока и возрастает с повышением температуры; при охлаждении же сердце сокращается вдвое реже, чем при той же температуре тела в стабильных условиях. Такая способность регулировать скорость согревания и охлаждения путем изменений в цир-26*
•404 Глава 8. Терморегуляция
куляции крови, безусловно, полезна для животного, которое в поясках пищи периодически погружается в прохладную воду.
На солнце греются не только ящерицы, но и другие пресмыкающиеся. Черепахи, змеи и даже такие в наибольшей степени связанные с водной средой рептилии, как крокодилы, часто принимают солнечные ванны. Какую роль это играет в согревании тела и терморегуляции, в большинстве случаев не известно.
Рис. 8.31. Скорости согревания и охлаждения галапагосской морской игуаны (вес тела 652 г) в воде (треугольники) и в воздушной среде (кружочки). Начальная разность температур животного и внешней среды (ДТ) во всех случаях составляла 20 °C. (Bartholomew, Lasiewski, 1965.)
Животное может также повышать температуру своего тела, повышая собственную теплопродукцию (т. е. интенсивность метаболизма). Известно, что к этому способу прибегают самки некоторых змей в период, когда они согревают яйца. В частности, это наблюдали в нью-йоркском зоопарке, где самка питона длиной 2,7 м высиживала кладку из 23 яиц. Поскольку при этом она свернулась вокруг яиц, термопару для измерения температуры помещали между плотно прижатыми друг к другу кольцами змеиного тела. После снижения температуры воздуха с 33 до 25 °C у змеи возросло потребление кислорода, а температура тела оставалась на 4—5° выше температуры среды (рис. 8.32). Повышенное потребление О2 было, по-видимому, обусловлено сильными сокращениями мускулатуры, напоминающими дрожь у млекопитающих. При температуре воздуха 25,5 °C змея, согревавшая кладку яиц, поглощала в 9,3 раза больше кислорода, чем обычно. При дальнейшем снижении температуры до 21 °C животное уже не могло дополнительно повысить интенсивность обмена: темпе
Температура тела у «холоднокровных» животных
405
ратура тела падала, потребление кислорода снижалось. После 30-дневного согревания кладки вес змеи уменьшился с 14,3 до 10,3 кг, т. е. почти на 30%, — видимо, в результате расходования резервного «топлива» на выработку тепла (Hutchison et al., 1966).
Рис. 8.32. Когда самка питона, выводя потомство, согревает кладку яиц, ее температура поднимается выше температуры окружающей среды благодаря сильным мышечным сокращениям, напоминающим дрожь у млекопитающих. Проведенная на рисунке диагональ соответствует равенству температур тела змеи и воздуха. (Vinegar et al., 1970.)
ЛЕТАЮЩИЕ НАСЕКОМЫЕ
Большинство насекомых при низких температурах становятся вялыми и не могут летать. Однако некоторые из них способны разогревать свои летательные мышцы и сохранять активность в условиях очень холодного воздуха. Летательные мышцы насекомых, расположенные в .грудном отделе, могут вырабатывать большое количество тепла в результате процесса, сходного с дрожью у позвоночных. Такое разогревание мышц перед взлетом свойственно в основном крупным насекомым, таким, как саранча, крупные ночные бабочки и шмели, а также осам и пчелам, способным *к быстрому и длительному полету.
Шмель может полететь только в том случае, если температура его груди (торакса) достигает по меньшей мере 29—30 °C (Hein
406 Глава 8. Терморегуляция
rich, 1972). При меньшей температуре скорость сокращения мышц недостаточна для полета, при котором крылья должны совершать 130 взмахов в секунду.
Благодаря способности поддерживать высокую торакальную температуру шмель собирает нектар даже при 5°C. Однако он не может постоянно поддерживать высокую температуру, если не потребляет пищу по меньшей мере с такой же скоростью, с какой он расходует свои энергетические ресурсы. Шмель, весящий 0,5 г, может поглощать до 50 мл кислорода в час (100 л О2 на 1 кг веса тела в 1 ч), что соответствует использованию 60 мг сахара. Расход такого количества топлива, по-видимому, оправдывает себя, так как в холодную погоду (когда большинство насекомых теряет активность) конкуренция со стороны других потребителей нектара невелика.
Благодаря малым размерам тела насекомые по сравнению с млекопитающими имеют то преимущество, что могут не только очень быстро согреться, но и быстро подвергнуться охлаждению. Поэтому им легко привести температуру своего тела в соответствие с энергетическими ресурсами, и насекомое может поддерживать высокую температуру тела только тогда, когда приход энергии покрывает ее расход; иными словами, у насекомых есть-большая свобода выбирать, когда им быть «теплокровными».
Шмели повышают температуру своего тела также при обогреве расплода. В этом случае матка держит брюшко тесно прижатым к соту, и поскольку температура брюшка поддерживается на уровне 31—35 °C, расплод находится в тепле. Даже при температуре наружного воздуха 5 °C шмелиные матки сохраняют температуру в сотах на уровне около 20 °C (Heinrich, 1974). При этом потребление кислорода шмелиной маткой возрастает пропорционально снижению температуры воздуха, так что метаболическая кривая (рис. 8.33) сходна с соответствующей кривой для млекопитающих (см., например, рис. 8.7).
Процессы терморегуляции были тщательно изучены еще у одного насекомого — у бабочки-бражника Manduca sexta (его личинка— табачная гусеница, имеющая рог). Бражники довольна крупные: они весят 2—3 г; питаются нектаром, который сосут, повисая в воздухе перед цветком, подобно колибри. Ведя ночной образ жизни, бражники нередко летают при очень низких температурах воздуха; такой полет был бы невозможен, если бы они не могли разогревать себя. Для «зависания» в воздухе температура летательных мышц должна быть не ниже 35°C; бражники же летают и кормятся на цветках даже при понижении температуры воздуха до 10 °C. Дело в том, что перед полетом они разогревают свою грудь. Чем ниже начальная температура, тем медленнее происходит разогрев (рис. 8.34). Во время разогрева бражник, по-видимому, дрожит, так как у него слегка вибрируют крылья.
Температура тела у «холоднокровных» животных 407
Тепло образуется в результате сокращения летательных мышц, но бабочка при этом не машет крыльями, так как мышцы, поднимающие крылья, и мышцы, опускающие их, сокращаются одновременно, а не поочередно, как при полете. Можно было бы предположить, что скорость разогрева должна возрастать по ме-
Рис. 8 33. Потребление кислорода двумя шмелиными матками (Bombus vosne-senskii) в периоды непрерывного обогревания сота. Вес груди, где находятся мышцы, вырабатывающие практически все тепло, составляет около 0,2 г; общий вес матки (в среднем 0,5 г) варьирует в зависимости от наполнения медового зобика. (Heinrich, 1974.)
ре повышения температуры тела, поскольку выработка тепла при этом увеличивается. Но одновременно возрастают потери тепла, и поэтому кривые, приведенные на рис. 8.34, почти прямолинейны.
Грудь бражника покрыта длинными, мохнатыми чешуйками, и это помогает сохранять тепло при разогреве. Во время полета температура грудного отдела поддерживается на уровне 40—41 °C в широком диапазоне внешних температур. Ввиду интенсивной выработки тепла в летательных мышцах летящему бражнику, чтобы избежать перегрева груди, нужно иметь возможность увеличивать теплоотдачу. Это достигается путем усиленного обмена кровью между грудью и брюшком.
Отдаче тепла брюшком способствует большая поверхность этой части тела и ее слабая теплоизоляция. Если перевязать большой спинной кровеносный сосуд, идущий от сердца (расположенного в брюшке), и тем самым сделать невозможным охлаждение летательных мышц, то бражник быстро перегревается и перестает летать даже при температуре воздуха 23 °C. Конечно, причиной этого могло бы быть недостаточное снабжение летательных мышц кровью и соответственно «топливом». Однако простой опыт позволяет отвергнуть это предположение: если у бражника с перевязанным сосудом удалить чешуйки, покрывающие грудь, то теплоотдача значительно возрастает и насекомое начинает летать снова (Heinrich, 1970).
408 Глава 8. Терморегуляция
Медоносные пчелы регулируют не только собственную индивидуальную температуру, но и температуру во всей колонии. Летом температура в улье поддерживается на уровне около 35°C с очень небольшими отклонениями. Такой уровень оптимален для развития расплода. Быстрое размножение обеспечивает интенсивный рост популяции в наиболее благоприятном сезоне; по-
Рис. 8.34. Повышение температуры груди у бражника Manduca sexta происходит тем быстрее, чем выше начальная температура. (Heinrich, Bartholomew, 1971.)
этому поддержание температуры, оптимальной для развития яиц и личинок, очень важно с экологической точки зрения. Если в улье становится чересчур тепло, пчелы разбрызгивают над сотами воду; испарению ее способствуют токи воздуха, которые пчелы создают трепетанием своих крылышек. При температурах ниже 30 °C пчелы обычно не пьют воды, но при более высокой температуре потребление ими воды в огромной мере возрастает, так как она используется для охлаждения (Free, Spencer-Booth, 1958).
При низкой температуре воздуха пчелы скучиваются, и температура внутри их скопления поддерживается на уровне 20—30 °C. Теплее всего в центре скопления, и пчелы, находящиеся с краю, пробиваются в более теплую середину, оттесняя согревшихся особей к периферии. В большом диапазоне температур воздуха, вплоть до —17 °C, температура внутри скопления пчел поддерживается на уровне от 18 до 32 °C. Отмечается даже тенденция к повышению этого уровня при сильных морозах. Снижение температуры наружного воздуха на каждые 5° сопровождается повышением температуры в скоплении пчел примерно на 1° (Southwick, Mugaas, 1971). В этом отношении семья пчел ведет себя как единый крупный организм, поддерживающий свою температуру на высоком, хорошо регулируемом уровне при таких внешних условиях, в которых быстро погибла бы одиночная пчела.
Температура тела у «холоднокровных» животных 409
* * *
Итак, мы обсудили вопросы, связанные с температурой; в этой последней главе было показано, какую важную роль играет вода в поддержании нужной температуры тела. Теперь мы можем перейти к более детальному рассмотрению этого важного вещества и его физиологической роли.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Adolph Е. F. (1947). Tolerance to heat and dehydration in several species of mammals, Am. J. Physiol., 151, 564—575.
Andersen H. T., Hammel H. T., Hardy J. D. (1961). Modifications of the febrile response to pyrogen by hypothalamic heating and cooling in the unanesthetized dog, Acta Physiol. Scand., 53, 247—254.
Aschoff J., Gunther B., Kramer K. (1971). Energiehaushalt und Temperaturregu-lation, Munich, Urban and Schwarzenberg, 196 pp.
Aschoff J., Wever R. (1958). Kern und Schale im Warmehaushalt des Menschen, Naturwissenschaften. 45, 477—485.
Aschoff J., Wever R. (1959). Warmeaustausch mit Hilfe des Kreislaufes, Dtsch. Med. Wochenschr., 84, 1509—1517.
Augee M. L., Ealey E. H. M. (1968). Torpor in the echidna, Tachyglossus aculeatus, J. Mammal., 49, 446—454.
Bartholomew G. A., Hudson J. W. (1962). Hibernation, estivation, temperature regulation, evaporative water loss and heart rate of the pigmy possum, Cer-caertus nanus, Physiol. Zool., 35, 94—107.
Bartholomew G. A., Lasiewski R. C. (1965). Heating and cooling rates, heart rate and simulated diving in the Galapagos marine iguana, Comp. Biochem. Physiol., 16, 575—582.
Bartholomew G. A., Lasiewski R. C., Crawford E. C., Jr. (1968). Patterns of panting and gular flutter in cormorants, pelicans, owls and doves, Condor, 70, 31— 34.
Bligh J., Ingram D. L., Keynes R. D., Robinson S. G. (1965). The deep body temperature of an unrestrained Welsh mountain sheep recorded by a radiotele-metric technique during a 12-month period, J. Physiol., 176, 136—144.
Buettner-Janusch J. (1966). Origins of Man, New York, Wiley, 674 pp.
Calder W. A. (1964). Gaseous metabolism and water relations of the zebra finch, Taeniopygia castanotis, Physiol. Zool., 37, 400—413.
Calder W. A., Schmidt-Nielsen K. (1967). Temperature regulation and evaporation in the pigeon and the roadrunner, Am. J. Physiol., 213, 883—889.
Carey F. G., Teal J. M. (1966). Heat conservation in tuna fish muscle, Proc. Natl.
Acad. Sci. USA, 56, 1464—1469.
Carey F. G., Teal J. M. (1969). Mako and porbeagle: Warm-bodied sharks, Comp. Biochem. Physiol., 28, 199—204.
Carey F. G., Teal J. M., Kanwisher J. W., Lawson K. D. (1971). Warm-bodied fish, Am. Zool., 11, 137—145.
Council of the Royal Society (1975). Quantities, Units and Symbols, 2nd ed., London, The Royal Society, 55 pp.
Crawford E. C., Jr. (1962). Mechanical aspects of panting in dogs, J. Appl. Physiol., 17, 249—251.
Crawford E. C., Jr. (1972). Brain and body temperatures in a panting lizard, Science, 177, 431—433.
Crawford E. C., Jr., Kampe G. (1971). Resonant panting in pigeons, Comp. Biochem. Physiol., 40A, 549—552.
410 Глава 8. Терморегуляция
Dawson W. R. (1954). Temperature regulation and water requirements of the brown and abert towhees, Pipilo fuscus and Pipilo aberti, Univ. Calif. Publ. Zool., 59, 81—124.
Dawson T. J., Hulbert A. J. (1970). Standard metabolism, body temperature and surface areas of Australian marsupials, Am. J. Physiol., 218, 1233—1238.
Dawson T., Schmidt-Nielsen K. (1966). Effect of thermal conductance on water economy in the antelope jack rabbit, Lepus alleni, J. Cell. Physiol., 67, 463— 472.
Farner D. S. (1956). The bodv temperatures of the North Island kiwis, Emu, 56, 199—206.
Free J. B., Spencer-Booth У. (1958). Observations on the temperature regulation and food consumption of honeybees (Apis mellifera), J. Exp. Biol., 35, 930— 937.
Hainsworth F. R., Wolf L. L. (1970). Regulation of oxygen consumption and body temperature during torpor in a hummingbird, Eulampis jugularis, Science, 168, 368—369.
Hales J. R. S., Bligh J. (1969). Respiratory responses of the conscious dog to severe heat stress, Experientia, 25, 818—819.
Hales J. R. S., Findlay J. D. (1968). Respiration of the ox: Normal values and the effects of exposure to hot environments, Respir. Physiol., 4, 333—352.
Hammel H. T. (1955). Thermal properties of fur, Am. J. Physiol., 182, 369— 376.
Hardy J. D. (1949). Heat transfer. In: Physiology of Heat Regulation and the Science of Clothing (L. H. Newburgh, ed.), pp. 78—108, Philadelphia, Saunders.
Hart J. S. (1951). Calorimetric determination of average body temperature of small mammals and its variation with environmental conditions, Can. J. Zool., 29, 224—233.
Hart J. S. (1956). Seasonal changes in insulation of the fur, Can. J. Zool., 34, 53—57.
Hart J. S., Irving L. (1959). The energies of harbor seals in air and in water, with special consideration of seasonal changes, Can. J. Zool., 37, 447—457.
Heinrich B. (1970). Thoracic temperature stabilization by blood circulation in a free-flying moth, Science, 168, 580—582.
Heinrich B. (1972). Temperature regulation in the bumblebee Bombus vagans: A field study, Science, 175, 185—187.
Heinrich B. (1974). Thermoregulation in bumblebees. 1. Brood incubation by Bombus vosnesenskii queens, J. Comp. Physiol., 88, 129—140.
Heinrich B., Bartholomew G. A. (1971). An analysis of pre-flight warm-up in the sphinx moth, Manduca sexta, J. Exp. Biol., 55, 223—239.
Hensel H., Bock K. D. (1955). Durchblutung und Warmeleitfahigkeit des menschli-chen Muskels, Pfliigers Arch., 260, 361—367.
Herreid C. F., II, Schmidt-Nielsen K. (1966). Oxygen consumption, temperature and water loss in bats from different environments, Am. J. Physiol., 211, 1108— 1112.
Hutchison V. H., Dowling H. G., Vinegar A. (1966). Thermoregulation in a brooding female Indian python, Python molurus bivittatus, Science, 151, 694—696.
Irving L. (1969). Temperature regulation in marine mammals. In: The Biology of Marine Mammals (H. T. Andersen, ed.), pp. 147—174, New York, Academic Press.
Irving L., Hart J. S. (1957). The metabolism and insulation of seals as bare-skinned mammals in cold water, Can. J. Zool., 35, 497—511.
Irving L., Krog J. (1954). Body temperatures of arctic and subarctic birds and mammals, J. Appl. Physiol., 6, 667—680.
Irving L., Krog J. (1955). Temperature of skin in the arctic as a regulator of heat,. J. Appl. Physiol., 7, 355—364.
Johansen K. (1961). Distribution of blood in the arousing hibernator, Acta Physiol. Scand., 52, 379—386.
Т ерморегуляция 411
Johnston D. W. (1971). The absence of brown adipose tissue in birds, Comp. Biochem. Physiol., 40A, 1107—1108.
Kilgore D. L., Jr., Schmidt-Nielsen K. (1975). Heat loss from ducks' feet immersed in cold water, Condor, 77, 475—478.
Kluger M. J. (1977). Fever in the frog Hyla cinerea, J. Thermal Biol., 2, 79— 81.
Kluger M. J., Ringler D. H., Anver M. R. (1975). Fever and survival, Science, 188, 166—168.
Le Maho Y., Delclitte P., Chatonnet J. (1976). Thermoregulation in fasting emperor penguins under natural conditions, Am. J. Physiol., 231, 913—922.
Lyman С. P. (1948). The oxygen consumption and temperature regulation of hibernating hamsters, J. Exp. Zool., 109, 55—78.
McNab В. K. (1966). An analysis of the body temperatures of birds, Condor, 68, 47—55.
Morrison P. (1966). Insulative flexibility in the guanaco, J. Mammal., 47, 18— 23.
Morrison P. R., Ryser F. A. (1952). Weight and body temperature in mammals, Science, 116, 231—232.
Qritsland N. A. (1970). Temperature regulation of the polar bear (Thalarctos ma-ritimus), Comp. Biochem. Physiol., 37, 225—233.
Pearson О. P. (1954). Habits of the lizard Liolaemus multiformis multiformis at high altitudes in Southern Peru, Copeia, 1954 (2), 111 —116.
Pinshow B., Fedak M. A., Battles D. R., Schmidt-Nielsen K. (1976). Energy expenditure for thermoregulation and locomotion in emperor penguins, Am. J. Physiol., 231, 903—912.
Reynolds W. W., Gasterlin M. E., Covert J. B. (1976). Behavioural fever in teleost fishes, Nature, 259, 41—42.
Robinson K. W., Lee D. H. K. (1946). Animal behaviour and heat regulation in hot atmospheres, Univ. Queensland Papers, Dept. Physiol., 1, 1—8.
Robinson K. W., Morrison P. R. (1957). The reaction to hot atmospheres of various species of Australian marsupial and placental animals, J. Cell Comp. Physiol., 49, 455—478.
Schmidt-Nielsen K. (1963). Osmotic regulation in higher vertebrates, Harvey Leet., 58, 53—93.
Schmidt-Nielsen K. (1964). Desert Animals: Physiological Problems of Heat and Water, New York, Oxford University Press, 277 pp.
Schmidt-Nielsen K. (1970). Animal Physiology, 3rd ed., Englewood Cliffs, N. J., Prentice-Hall..
Schmidt-Nielsen K., Dawson T. J., Crawford E. C., Jr. (1966). Temperature regulation in the echidna (Tachyglossus aculeatus), J. Cell. Physiol., 67, 63— 72.
Schmidt-Nielsen K., Schmidt-Nielsen B., Jarnum S. A., Houpt T. R. (1957). Body temperature of the camel and its relation to water economy, Am. J. Physiol., 188, 103—112.
Scholander P. F. (1955). Evolution of climatic adaptation in homeotherms, Evolution, 9, 15—26.
Scholander P. F., Hock R., Walters V., Johnson F., Jrving L. (1950a). Heat regulation in some arctic and tropical mammals and birds, Biol. Bull., 99, 237— 258.
Scholander P. F., Krog J. (1957). Countercurrent heat exchange and vascular bundles in sloths, J. Appl. Physiol., 10, 405—411.
Scholander P. F., Schevill W. E. (1955). Countercurrent vascular heat exchange in the fins of whales, J. Appl. Physiol., 8, 279—282.
Scholander P. F., Walters V., Hock R., Irving L. (1950b). Body insulation of some arctic and tropical mammals and birds. Biol. Bull., 99, 225—236.
Shkolnik A., Schmidt-Nielsen K. (1976). Temperature regulation in hedgehogs from temperate and desert environments, Physiol. Zool., 49, 56—64.
Southwick E. E., Mugaas J. N. (1971). A hypothetical homeotherm: The honeybee hive, Comp. Biochem. Physiol., 40A, 935—944.
412 Глава 8. Терморегуляция
Suckling J, A., Suckling Е. Е., Walker А. (1969). Suggested function of the vascular bundles in the limbs of Perodicticus potto, Nature, Lond., 221, 379— 380.
Taylor C, R. (1972). The desert gazelle: A paradox resolved. In: Comparative Physiology of Desert Animals (G. M. O. Maloiy, ed.), Symposia, Zoology Society of London, No. 31, pp. 215—227, London, Academic Press.
Taylor C. R., Lyman С. P. (1972). Heat storage in running antelopes: Independence of brain and body temperatures, Am. J. Physiol., 222, 114—117.
Tucker V. A. (1965a). Oxygen consumption, thermal conductance, and torpor in the California pocket mouse Perognathus californicus, J. Cell. Comp. Physiol., 65, 393—403.
Tucker V. A. (1965b). The relation between the torpor cycle and heat exchange in the California pocket mouse Perognathus californicus, J. Cell. Comp. Physiol., 65, 405—414.
Tucker V. A. (1966). Diurnal torpor and its relation to food consumption and weight changes in the California pocket mouse Perognathus californicus, Ecology, 47, 245—252.
Vinegar A., Hutchison V. H„ Dowling H. G. (1970). Metabolism, energetics and thermoregulation during brooding of snakes of the genus Python (Reptilia, Boidae), Zoologica, 55, 19—50.
Weast R. C. (1969). Handbook of Chemistry and Physics, 50th ed., Cleveland, Chemical Rubber.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Fischer К. C., Dawe A. R., Lyman С. P., Schonbaum E., South F. E., Jr. (eds.) (1967). Mammalian Hibernation, vol. Ill, Edinburgh, Oliver and Boyd, 535 pp.
Gates D. M. (1962). Energy Exchange in the Biosphere, New York, Harper and Row, 151 pp.
Gates D. M., Schmerl R. B. (eds.) (1975). Perspectives of Biophysical Ecology, New York, Springer-Verlag, 609 pp.
Hardy J. D., Gagge A. P., Stolwijk J. A. J. (eds.) (1970). Physiological and Behavioral Temperature Regulation, Springfield, Ill., Thomas, 944 pp.
Heinrich B. (1974). Thermoregulation in endothermic insects, Science, 185, 747— 756.
Heller H. C., Glotzbach S. F. (1977). Thermoregulation during sleep and hibernation, Int. Rev. Physiol., 15, 147—188.
Maloiy G. M. O. (ed.) (1972). Comparative Physiology of Desert Animals. Symposia, Zoological Society of London, No. 31, London, Academic Press, 413 pp.
Monteith J. L. (1973). Principles of Environmental Physics, London, Edward Arnold, 241 pp.
Newburgh L. H. (ed.) (1968). Physiology of Heat Regulation and the Science of Clothing (reprinted), New York, Hafner, 457 pp.
Precht H., Christophersen J., Hensel H., Lurcher W. (1973). Temperature and Life, New York, Springer-Verlag, 779 pp.
Schmidt-Nielsen K. (1964). Desert Animals: Physiological Problems of Heat and Water, New York, Oxford University Press, 277 pp.
Whittow G. C. (ed.) (1970, 1971, 1973). Comparative Physiology of Thermoregulation: vol. 1. Invertebrates and Nonmammalian Vertebrates (1970), 333 pp.; vol. 2. Mammals (1971), 410 pp.; vol. 3. Special Aspects of Thermoregulation (1973), 278 pp., New York, Academic Press.
ОГЛАВЛЕНИЕ
Предисловие редактора перевода ..................................... 5
Предисловие ко второму изданию......................................... 9'
Моим молодым читателям.................................................11
Об этой книге...........................................................12
Что такое физиология?...................................................14
Часть первая. Кислород
Глава 1. Дыхание в воде....................
Газы в воздухе и воде..................
Водное дыхание.........................
Цитируемая литература......................
Дополнительная литература...................
Глава 2. Дыхание в воздухе...............
Органы дыхания......................
Дыхательные движения................
Роль кожи в дыхании.................
Легкие млекопитающих................
Регуляция дыхания...................
Рыбы, способные дышать воздухом .
Дыхание птиц........................
Дыхание насекомых...................
Цитируемая литература...................
Дополнительная литература ..............
Глава 3. Кровь..........................
Перенос кислорода кровью ....
Перенос двуокиси углерода кровью .
Цитируемая литература...................
Дополнительная литература ..............
16
17
28
41
41
43-
46-
47
49*
52
57
61
71
82
99»
101
102'
103» 127
136
138’
Глава 4. Кровообращение .
Общие принципы ....
Кровообращение у позвоночных Циркуляция жидкостей Свертывание крови и
Цитируемая литература Дополнительная литература
у беспозвоночных . гемостаз . . . .
139
139 142*
173
180
183-
184
вторая. Пища и энергия
Глава 5. Пища, топливо и энергия ....
Способы питания .........................
Пищеварение..............................
Часть
18&
187
195-
414 Оглавление
Питание ..........................................................211
Ядовитые вещества и химическая защита.............................225
Цитируемая литература.................................................233
Дополнительная литература.............................................236
Глава 6. Энергетический обмен.........................................237
Интенсивность метаболизма.........................................238
Запасание энергии: жир и гликоген.................................241
Влияние концентрации кислорода на интенсивность метаболизма . . 243
Проблемы, связанные с нырянием млекопитающих и птиц .... 250
Интенсивность метаболизма и размеры тела..........................270
Размеры тела и соотношения различных параметров...................279
Энергетическая цена локомоции.....................................283
Влияние больших высот.............................................290
Цитируемая литература.................................................294
Дополнительная литература.............................................296
Часть третья. Температура
Глава 7. Влияние температуры.........................................297
Влияние изменений температуры на физиологические параметры . . 299
Экстремальные температуры; температурные пределы для жизни . . 304
Физиологическая адаптация к перемене температуры.................319
Цитируемая литература................................................330
Дополнительная литература............................................332
Глава 8. Терморегуляция..............................................333
Температура тела у птиц и млекопитающих..........................334
Температура, тепло и перенос тепла ............................. 342
Тепловой баланс................................................. 352
Зимняя спячка и оцепенение.......................................387
Температура тела у «холоднокровных» животных.....................397
Цитируемая литература................................................409
Дополнительная литература .......................................... 412
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении, качестве перевода и другие просим присылать по адресу:
129820, Москва, И-110, ГСП 1-й Рижский пер., д. 2, издательство «Мир».
КНУТ ШМИДТ-НИЕЛЬСЕН физиология животных. ПРИСПОСОБЛЕНИЕ И СРЕДА
Книга 1
Ст. научн. редактор Ю. И. Лашкевич
Мл. редактор 3. Е. Кожанова
Художник В. Г. Ежков
Художественный редактор Л. М. Кузнецова
Технический редактор Н. И. Борисова
Корректор В. И. Постнова
ИБ № 2999
Сдано в набор 24.07.81.
Подписано к печати 15.12.81.
Формат 60Х90’/1б.
Бумага типографская № 1.
Гарнитура литературная. Печать высокая.
Объем 13 бум. л. Усл. печ. л. 26. Усл. кр.-отт. 26. Уч.-изд. л. 27,61. Изд. № 4/1263.
Тираж 10 000 экз. Зак. 1873. Цена 2 р. 30 к.
ИЗДАТЕЛЬСТВО «МИР» Москва, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома при Государственном комитете СССР по делам издательств, полиграфии и книжной торговли. Москва, 113105, Нагатинская ул., д. 1.