/











































































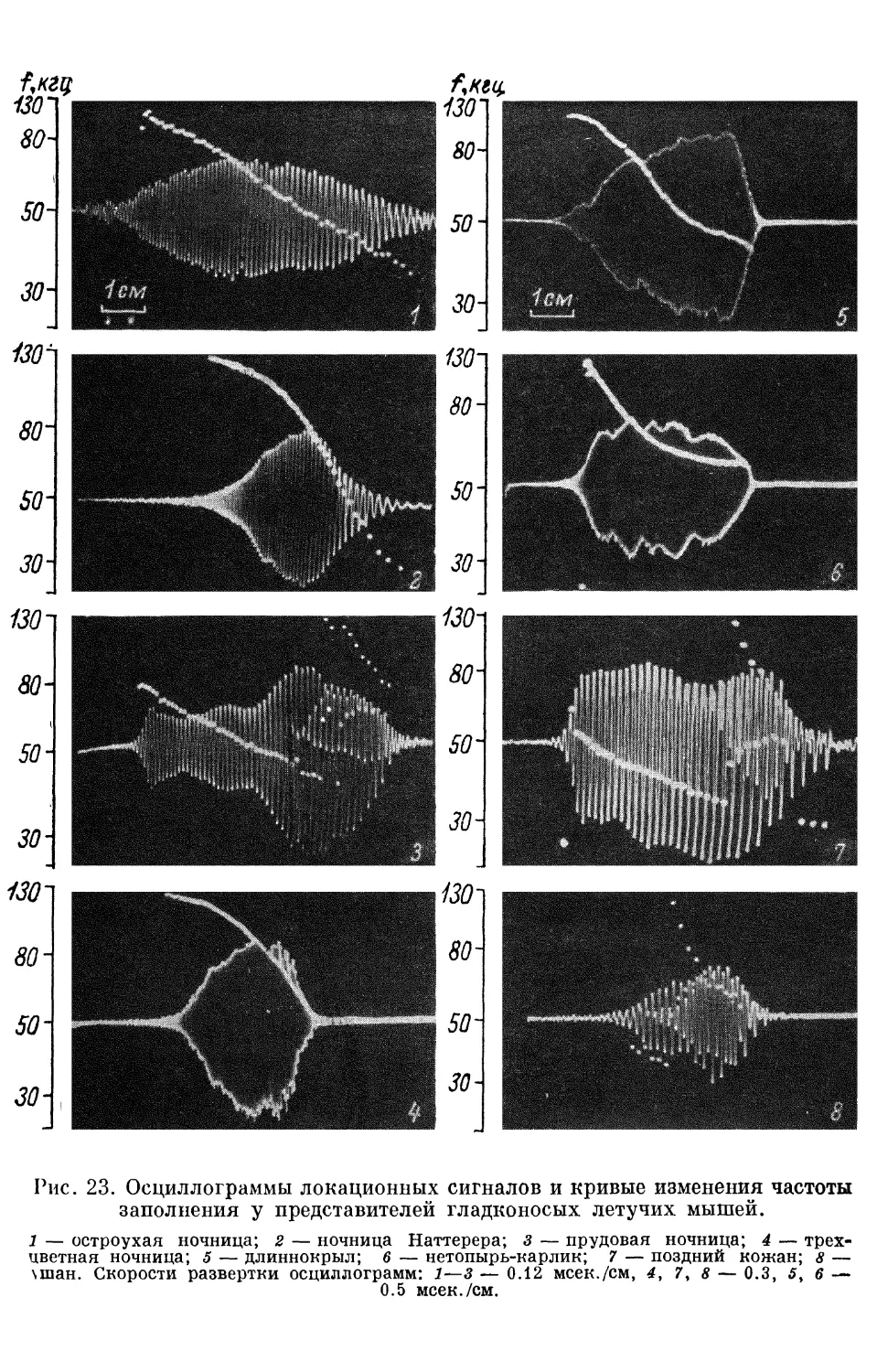




















































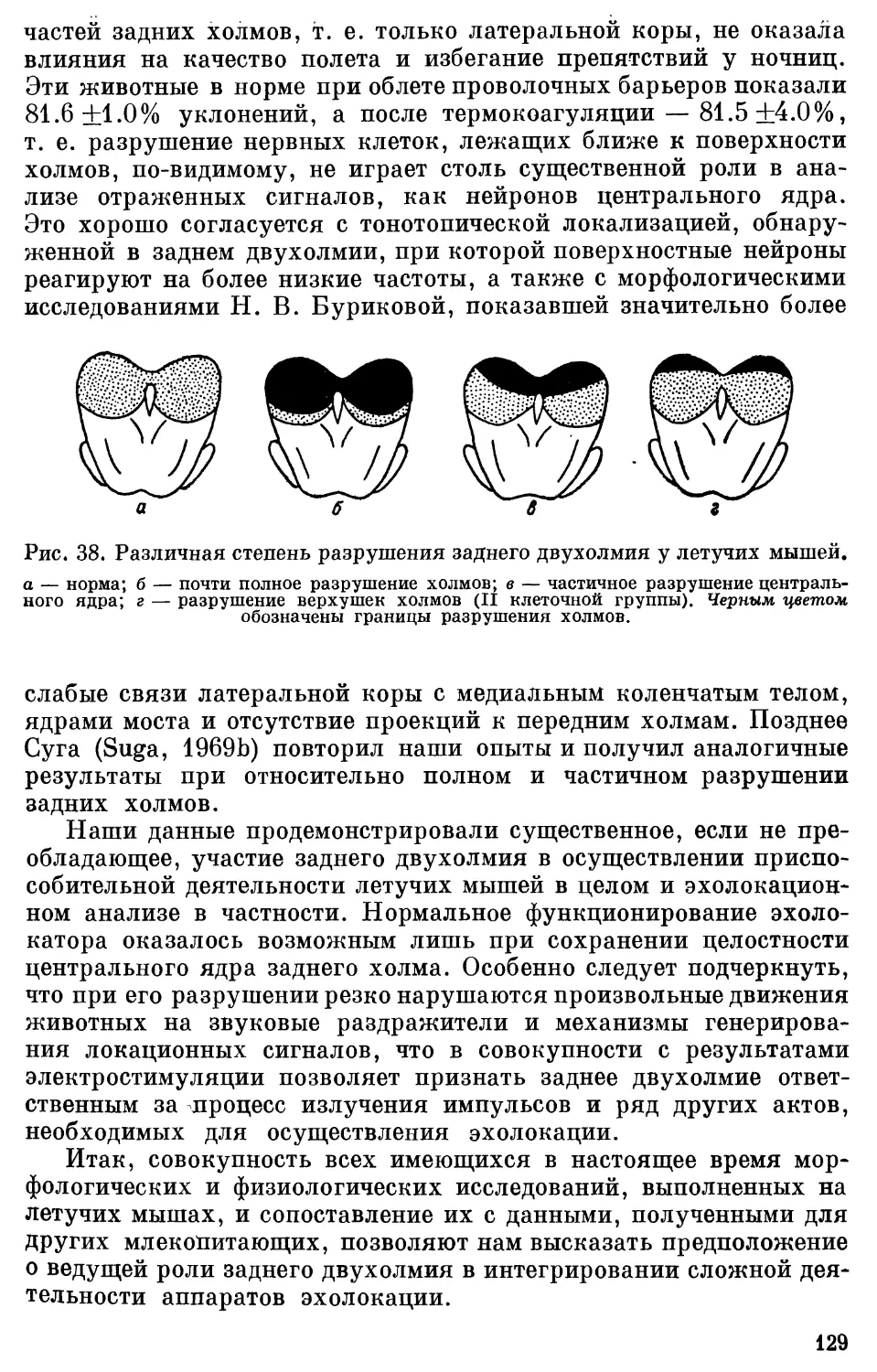




















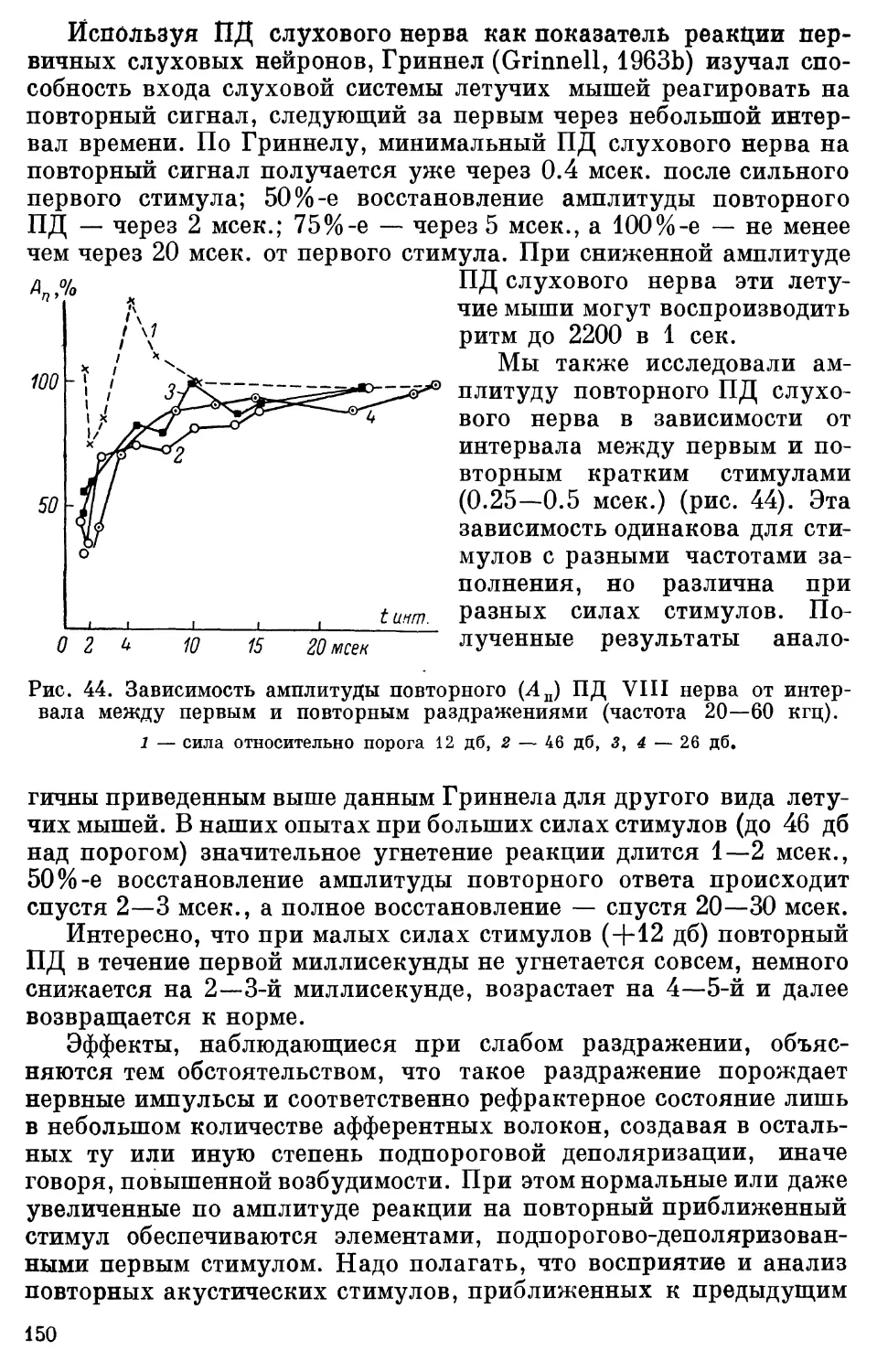




















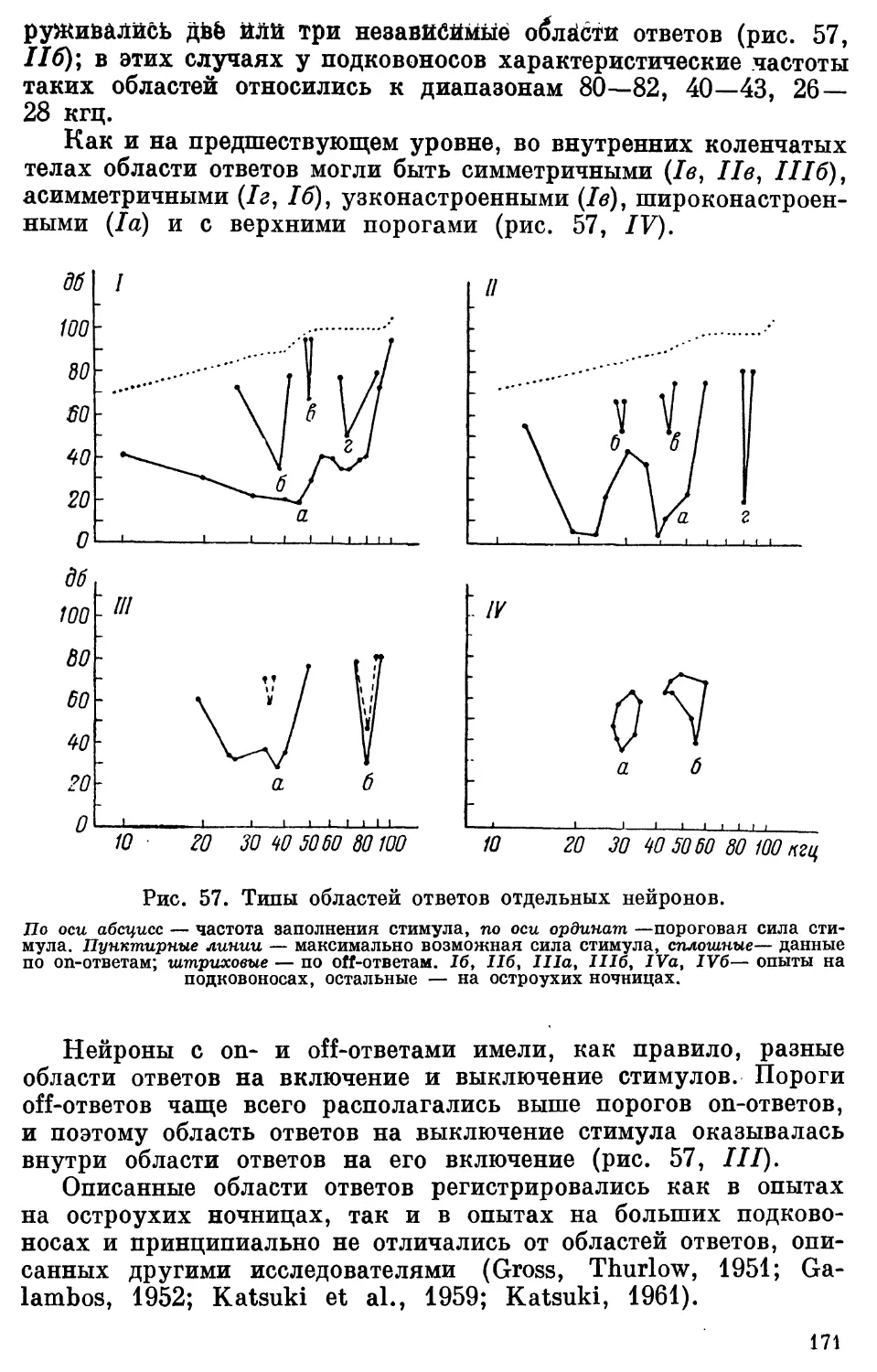
































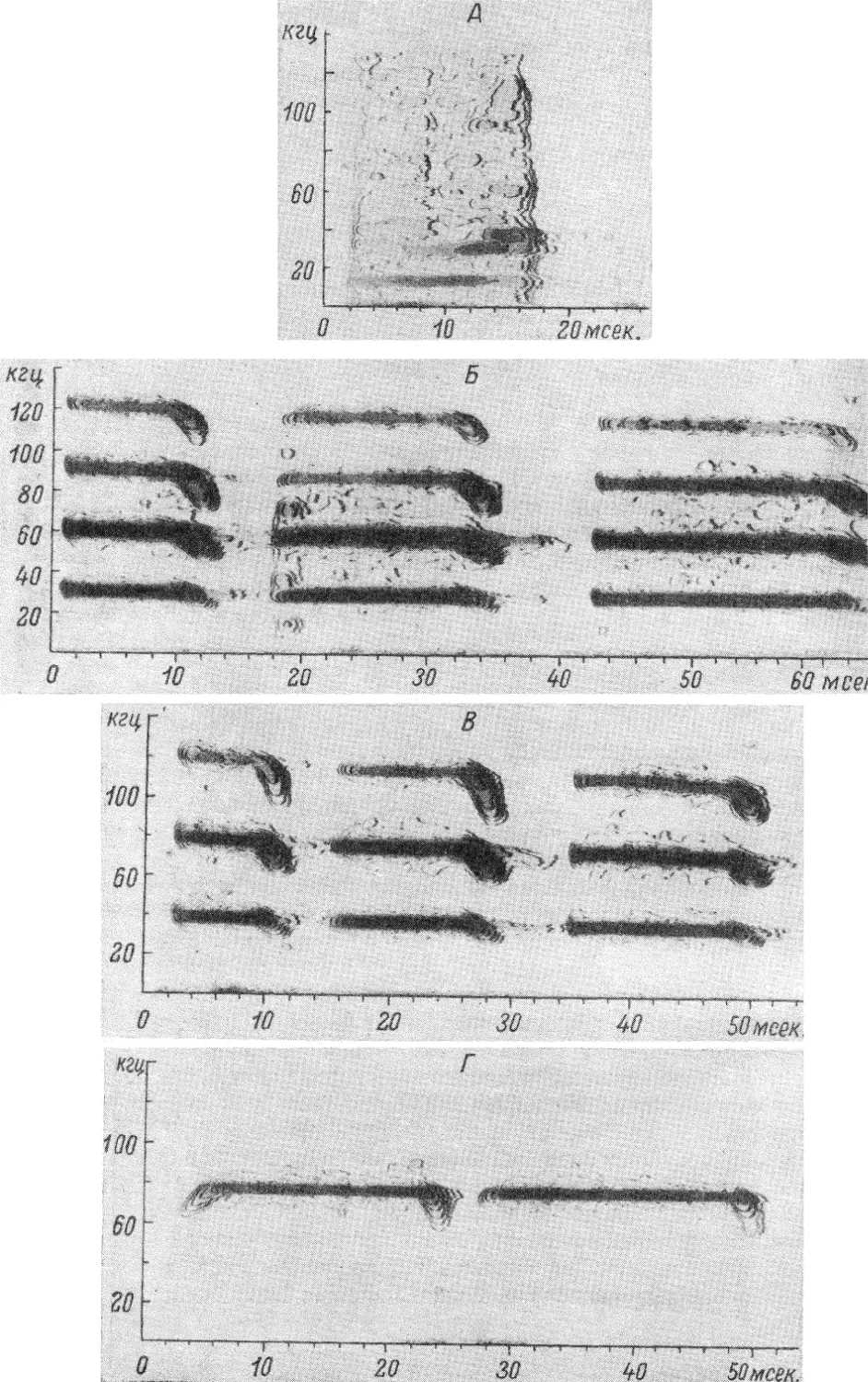



























































































































































































































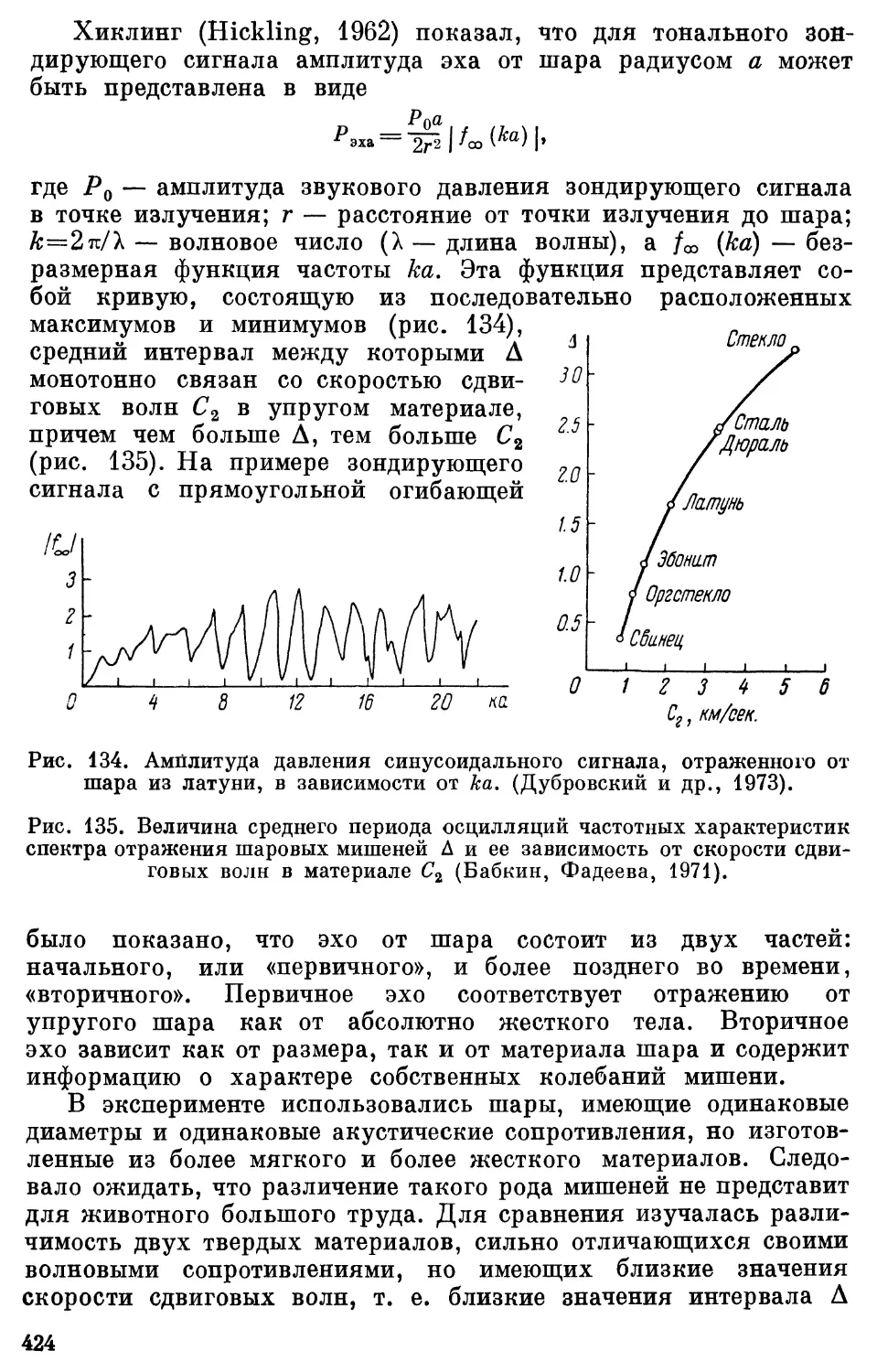




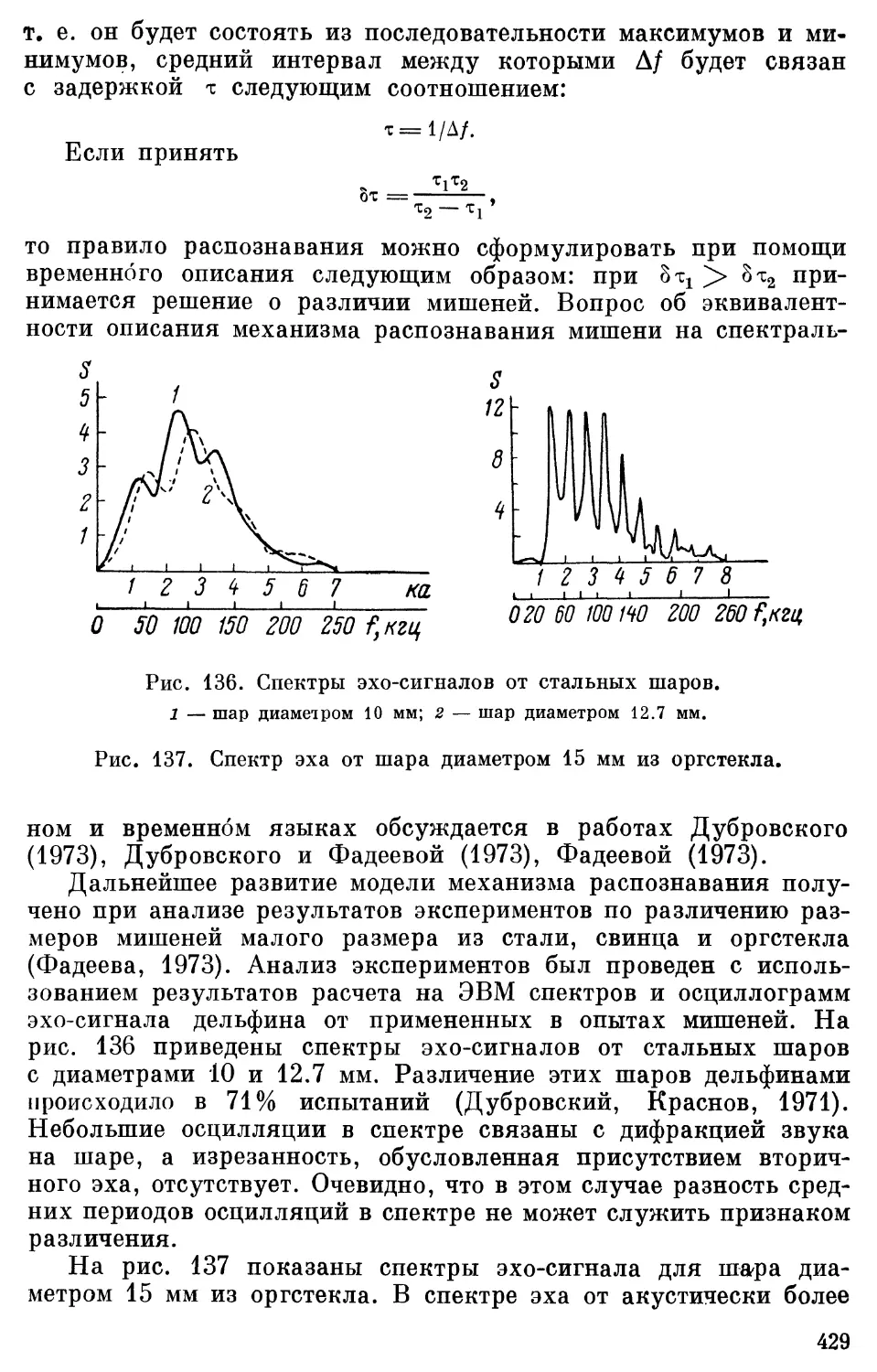
























































































Text
АКАДЕМИЯ НАУК СССР
ОТДЕЛЕНИЕ ФИЗИОЛОГИИ
Э.Ш.АЙРАПЕТЬЯНЦ
А.И.КОНСТАНТИНОВ
ЭХОЛОКАЦИЯ В ПРИРОДЕ
Издание второе, переработанное и дополненное
ИЗДАТЕЛЬСТВО «НАУКА» ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ ЛЕНИНГРАД -19 7 4
USSR ACADEMY OF SCIENCES
E. Sb. Airapetiantz and A. I. Konstantinov
ECHOLOCATION in NATURE
The monograph describes the vital activity of the echolocating animals, the history of the discovery of a specific means of spatial orientation, as well as of the mechanisms of radiation, perception and analysis of ultra-sound reflected by objects (echo). The principle underlying the functioning of echolocational system is discussed as well as the possibility of its utilization for engineering purposes. Special emphasis is laid on the latest investigations on bats and dolphins.
139 illustrations, 40 tables, 945 titles of references.
УДК 591.185.1:534.88
Эхолокация в природе. Изд. второе, переработ. и доп. Айра-петьянц Э. Ш., Константинов А. И. 1974. Изд-во «Наука», Ленингр. отд., Л. 1—512.
В книге описываются жизнедеятельность эхолоцирующих животных, история открытия особого средства пространственной ориентации, механизмов излучения, восприятия и анализа ультразвука, отраженного от предметов (эхо). Рассматривается принцип функционирования эхолокационной системы, возможность его применения для инженерных целей. Наиболее полно излагаются новейшие исследования на летучих мышах и дельфинах. Илл. 139, табл. — 40, библ. 945 назв.
A q^2 (01 )-74 711-73 0 Издательство «Наука» 1974
ПРЕДИСЛОВИЕ КО ВТОРОМУ ИЗДАНИЮ
Потребность в повторном издании монографии «Эхолокация в природе» обнаружилась спустя короткое время после выхода ее в свет. Тираж книги быстро разошелся, в чем нельзя не увидеть повышенного читательского спроса и возросшего интереса к сущности животной эхолокации, к результатам изучения специализированной функции головного мозга и особых средств пространственного анализа, в особенности у рукокрылых (летучих мышей) и китообразных (дельфинов).
В 1-м издании содержались все основные знания по эхолокации животных, добытые исследователями к моменту сдачи рукописи в издательство — к началу 1969 г. За истекшие годы всесторонне развивались работы в этом направлении как в ряде учреждений Советского Союза, так и за рубежом, преимущественно в США и ФРГ. Накопились существенного значения новые сведения, изложенные в многочисленных статьях и докладах на различных конференциях. С удовлетворением следует подчеркнуть, что теснее сближаются аспекты зоологических, физиологических (а среди последних на нейрональном уровне), биоакустических, инженерных исследований. Их результаты позволяют по совокупности обобщить современные представления о сложнейших механизмах и аппаратах мозга, выполняющих высокого уровня интегративную функцию: в микроинтервалах времени осуществить запуск, прием, обработку ультразвукового разнотипного самоизлучения. Мы близки к расшифровке содержащихся в акте эхолокации его подвижных компонентов, обеспечивающих в целом необходимую и достаточную информацию о пространственных атрибутах среды. Новые данные также приближают возможность рассмотреть в эволюционном аспекте морфофизиологическое переключение областей мозга эхолоцирующих животных в связи с длительно воздействующими экологическими факторами. Наконец, исходная бионическая задача изучения эхолокации за последние годы приобрела конкретность поисков, обогатилась рядом прикладных цррб с реальными подступами к инженерным моделям.
3
Научный коллектив сотрудников кафедры физиологии высшей нервной деятельности Ленинградского университета в прошедшие четыре года, продолжая разработку физиологии пространственного анализа, внес свой вклад в расширение знаний об эхолокации летучих мышей и дельфинов. Экспериментальный материал и его трактовка послужили основанием для защиты в 1972—1973 гг. докторской и трех кандидатских диссертаций и подготовки еще пяти кандидатских для их ближайшей защиты, а также к выпуску в свет сборника работ по эхолокации летучих мышей (Сравнительная физиология анализаторов, вып. 3, Изд. ЛГУ, 1973). За истекший, период новые материалы и теоретические обобщения доложены в Мюнхене на XXV международном конгрессе физиологов (1971 г.), на всесоюзных конференциях по бионике (1973 г.) и акустике (1973 г.).
При подготовке второго издания мы сочли полезным использовать некоторое увеличение объема для существенного дополнения материала (полученного в нашей и других лабораториях, в том числе и зарубежных) в виде новых глав и частичной переработки глав предыдущего издания.
В итоге глава 22 сохранилась в прежнем изложении, главы 1—3, 6, 9, 10, 12, 13, 15—21 подверглись изменению, добавлению текста, частью по структуре изложения, кроме того, включены семь новых глав: глава 4 «Общая функциональная схема регуляции эхолокатора», глава 5 «Особенности слухового восприятия акустических сигналов по параметрам частоты и длительности», глава 7 «Пространственный слух и точность локализации ультразвука», глава 8 «Развитие эхолокации у летучих мышей в онтогенезе», глава 11 «Точность измерения дальности и разрешающая способность эхолокатора», глава 14 «Сравнение возможностей эхолокационных систем и принципов их действия у гладконосых и подковоносых летучих мышей», глава 21 «Диапазон действия, помехоустойчивость и пределы распознавания» (у дельфинов). Последняя из перечисленных глав написана известным исследователем биоакустики дельфинов кандидатом физико-математических наук Н. А. Дубровским.
Текст «Заключения» не подвергся существенным изменениям. В нем лишь исключен абзац (стр. 348, 1-е изд.), где в лаконичной форме ставилась задача исследовать «становление эхолокации у летучих мышей в ранних фазах онтогенеза, точнее, в тот период, когда детеныш еще не летает и кормится молоком матери». Нам удалось эти исследования осуществить — результаты приведены в новой, 8-й, главе. Кроме этого, коротко изложены результаты наших начальных исследований в аспекте космической биологии — влияние длительной гравитационной перегрузки (гипервесомости) на пространственную ориентацию летучих мышей.
Библиографический аппарат расширен: внесено большое число источников — отечественных и зарубежных — в соответствии с их
изложением в тексте. Помимо этого, приложен список работ по проблеме эхолокации животных, опубликованных сотрудниками кафедры физиологии высшей нервной деятельности Ленинградского университета за период 1964—1973 гг.
В процессе оформления рукописи для второго издания нам оказали помощь сотрудники кафедры физиологии высшей нервной деятельности, в особенности по главе 5 — А. Г. Васильев, по главам 18, 19, 20, 21 — Б. Ф. Сергеев, по некоторым другим главам — Ю. И. Иваненко, В. Д. Жарская, И. А. Горлинский, А. К. Макаров, В. Н. Мовчан, Н. Н. Соколова, И. М. Стосман, В. И. Чи-лингирис. Технической подготовке книги содействовали В. П. Гаврилов, С. С. Головина, Н. Н. Саноцкая. Всем указанным товарищам по лаборатории выражаем свою признательность.
Настоящее издание могло быть осуществлено благодаря высокой оценке нашей монографии и горячим заботам о ее повторном издании покойного академика А. Н. Белозерского. С сердечной благодарностью и со светлой памятью о выдающемся ученом-биологе, талантливом организаторе советской науки произносим имя, олицетворяющее добро и идейность.
Нам вновь повезло — Александр Александрович Стрелков любезно согласился редакторски прочитать рукопись. Его дружеские конструктивные рекомендации, строгие требования профессора зоологии, научно-литературную критику мы приняли с искренней благодарностью.
Мы отдаем себе отчет в том, что весьма относительны пределы в совершенствовании нового издания научной книги-монографии, в особенности отображающей состояние знаний в молодом, активно развивающемся направлении. И все же мы старались как можно лучше представить нашему читателю оптимальный вариант существующих сведений и воззрений по эхолокации животных. Не нам судить об эффективности этих стараний.
Май 1973 г.
ПРЕДИСЛОВИЕ К ПЕРВОМУ ИЗДАНИЮ
За последние два десятилетия во многих областях естествознания, техники, философии возвысилась роль биологии. Старейшая наука о жизни с ее многочисленными разделами предстала в новом освещении перед взорами физиков, математиков, химиков, инженеров. Сведения о материальных основах носителей наследственности, различных способах получения информации живыми организмами, наконец, об устройстве мозга и принципах организации и согласованности действий миллиардов его нейронов стали предметом пристального внимания. Ряд закономерностей чисто биологического порядка был воспринят как ценнейший капитал для инженерной мысли.
Известно, что за указанный период в биологии не было ни открытий типа эволюционного учения (Ч. Дарвина) или учения о высшей нервной деятельности (И. П. Павлова) с их великим значением для всего естествознания, ни такого рода обобщений, которые непосредственно индуцировали бы новое движение в области техники. Независимо от биологии были найдены пути технического прогресса благодаря бурному развитию электроники, автоматики, телемеханики и созданию современных быстродействующих машин разнообразнейшего назначения. Более того, развитие этих наук предоставило естествознанию новые методические средства тончайшего анализа явлений, происходящих в органической и неорганической природе. Стало неотделимым от «большой теории» в естествознании эффективное инженерное творчество,
Однако оказалось, что ключ к решению ряда инженерных задач все же можно найти в биологии. То, что еще стремится изобрести человек, уже найдено животными в итоге тысячелетий, в процессе жестокой борьбы за существование, неумолимого естественного отбора. Инженеры увидели то, что искали, например, в ультрачувствителышх воспринимающих физиологических приборах (рецепторах), в сверхминиатюрных нейронных аппаратах, молниеносно осуществляющих переработку информации и bhhq-
сящих удивительно целесообразные решения. Подобные биологические прототипы инженерного искусства обнаружились теперь во множестве. Естественно открылась перспектива, оригинальный путь содружества между инженерами и биологами, — научный симбиоз в разработке общих проблем.
Так родилась бионика. Формальная дата ее рождения — сентябрь 1960 г.1 Она отпочковалась от кибернетики, вначале как ее спутник, затем очень быстро вступила на самостоятельный путь развития. Бионика за короткий срок привлекла к себе множество исследователей, но в то же время стала излишне популярным источником для рассказов о таинственных способностях животных или слишком легких технических проектов. В этом литературном тумане скрывалась опасность искажения целевого назначения бионики. Существуют различные толкования конкретных задач ее. Ограничимся высказыванием акад. А. И. Берга на I конференции по бионике (Москва, 1963 г.). Определяя задачи этой науки, он призывал исследователей к творческому отбору «живых прототипов». У природы следует заимствовать самые совершенные конструктивные схемы и механизмы биологических систем, удовлетворяющие запросам современной техники. Аксель Иванович Берг подчеркнул, что для бионики в особенности важны те системы, которые показывают «исключительно высокую гибкость и живучесть в сложных условиях существования» (1965, с. 3—10). Действительно, здесь открывается множество аспектов, неограниченный потенциал возможностей, и перед нами еще слишком молодая отрасль науки, чтобы быть вполне определенной.
У физиологов, изучающих головной мозг, вызывает глубокое удовлетворение выход знаний о мозге за пределы собственных задач биологии, психологии, медицины. Вместе с этим возникают на первых порах и некоторые трудности. Во-первых, обнаружилось, что некоторые сведения по физиологии мозга неконкретны в тех деталях, без которых машина не может быть имитирована; во-вторых, в этом аспекте не изучались физиологические механизмы и устройство нейронной организации. Преодоление этих трудностей несомненно принесет пользу и самой физиологии. В этом направлении проводятся исследования во многих научно-исследовательских институтах и на кафедрах вузов нашей страны (Москва, Киев, Ростов-на-Дону, Тбилиси, Горький, Ереван, Казань, Ленинград и т. д.).
В наших лабораториях (в Ленинградском университете, Институте физиологии им. И. П. Павлова — Колтуши, Мурманском морском биологическом институте — Дальние Зеленцы) по согласованной стратегической программе осуществляются в сравнительно-физиологическом плане исследования закономерностей про- 1
1 Первый симпозиум по бионике. Дайтон, США, 13 сентября 1960 t.
странственного анализа (или, по иной терминологии, пространственного восприятия, пространственной ориентации).
В этот сложный комплекс информации о среде входят в одном ряду показания формы, величины, местоположения, расстояния, перемещения предметов, их отношения между собой; в другом ряду — анализ положения собственного тела в среде. По компонентам указанных двух рядов формируются всевозможные биотические взаимосигнализации. Конечно, определенная роль сохраняется за врожденными, хотя и ограниченными пространственными атрибутами. Но следует особо подчеркнуть, что в анализе и синтезе (распознавании) множества дискретных параметров пространства и их соотношений решающая роль принадлежит приобретенным реакциям — временным связям. Они обеспечивают адаптивное пространственное поведение во всех фазах индивидуальной жизни. Наши исследования показали, что мозг не располагает специализированным анализатором пространства, подобно, например, слуховому, зрительному. Пространственный анализ требует участия ансамбля анализаторов — их интегративной деятельности. Особую роль выполняет нейронная организация двигательной области коры, где осуществляется конвергенция внешних и внутренних анализаторных систем. В определенных критических условиях деятельности мозга (разрушения внешних рецепторов — органов чувств, удаления участков мозга или одной его половины) вступает в действие викарирование — замещение (Айрапетьянц, 1960, 1967а, 19676, 1970).
Бионика несомненно заинтересуется и этим свойством конструкции мозга, в которой отражены высокая структурная надежность с многоступенчатым резервированием, автоматическая перестройка иерархических систем регулирования. В условиях аварии множества каналов возбуждения анализаторов или при необратимой потере миллиардов нейронов сохраняются достаточные ресурсы в хозяйстве мозга, чтобы запустить физиологически согласованные системы памяти, обеспечить кворум-элементы для реализации пространственного анализа. Следовательно, в мозговой организации заложен некий модус нейронной избыточности (не излишков) с огромной потенцией виртуальных механизмов — они и служат основой процесса замещения (Айрапетьянц, 1960, 1967а, 19676, 1970). Принцип викарирования универсален. Он действует также и в нормальной деятельности животных, с полной силой проявляясь в особых экологически обусловленных режимах и в экстремальных ситуациях.
Исследования механизмов пространственного поведения животных проводятся и с учетом их экологических особенностей. С этих позиций нас издавна интересовали ночные животные, и в первую очередь летучие мыши. Исследования по эхолокации летучих мышей, начатые в 1959 г. на кафедре физиологии высшей нервной деятельности Ленинградского университета, оказались первыми в Со
8
ветском Союзе. Нам пришлось не только упорно заниматься методическими условиями постановки опытов в поисках собственных подходов при разработке нервных механизмов эхолокации, но преодолевать и многие организационные трудности. При поддержке руководства Ленинградского университета и Физиологического института им. А. А. Ухтомского и особенно некоторых заинтересованных в результатах наших работ инженерных ведомств Ленинграда и Москвы мы смогли в относительно короткий срок стать на путь планомерного, систематического экспериментирования. В стартовой фазе экспериментов нам оказала существенное содействие и инструментальную помощь кафедра радиофизики Ленинградского университета. Пользуемся поводом выразить указанным организациям и их руководителям свою горячую признательность.
Вначале работа осуществлялась лишь авторами этой книги. Они же выступили с предварительными итогами исследований на I конференции по бионике (1963) с сообщением: «О роли эхолокации в анализе пространства летучими мышами». Постепенно вовлекались в дело новые сотрудники, образовалась рабочая группа, развернулись многоаспектные исследования с применением современной электронной аппаратуры. Параллельно проводился анатомо-гистологический анализ. Летом 1966 г. была осуществлена экспедиция в Нагорный Карабах, где в естественных условиях невдалеке от пещер с колониями нескольких тысяч летучих мышей различных семейств и видов удалось получить большой экспериментальный материал. Основные результаты сообщались на бионических и физиологических конференциях, совещаниях в Ленинграде, Москве, а осенью 1968 г. — в Вашингтоне на XXIV Международном конгрессе физиологов.
Что же такое эхолокация? Принцип, заложенный в основу этого необычного способа ориентации, предельно прост. Животные излучают серию акустических импульсов (звуковых или ультразвуковых) и, принимая отражения — эхо этих импульсов от встречающихся на их пути предметов, своевременно обнаруживают препятствия, опознают добычу и преследуют ее. Так отличается один предмет от другого по многим его свойствам и определяются направление движущегося предмета, скорость передвижения. Этот способ контактирования с внешней средой был назван эхолокацией по аналогии с работой радио- и гидролокационных систем, радаров, сонаров, созданных умом человека.
Исследования эхолокации в нашей лаборатории определились в следующих взаимодополняющих направлениях. В сравнительном аспекте у разных семейств и видов летучих мышей проводится биоакустическое изучение локационных сигналов, излучаемых животными в полете, при обнаружении препятствий, ловле добычи, распознавании геометрических фигур. Здесь выявляются также соотношения приобретаемых и врожденных сигнализаций. Далее приемами электрофизиологического анализа с использованием
микроэлектродной методики изучаются приемная (слуховая) система эхолокатора, нервные центры, участвующие в анализе ультразвуковых эхо-импульсов, свойства отдельных нейронов, их специализация по различным параметрам, а также средства помехозащищенности. И наконец, проводится детальное морфологическое исследование мозга в целом и различных его отделов для определения субстрата эхолокационного механизма. В итоге предстоит составить основы теории эхолокации, специфику интегративной деятельности анализаторов летучих мышей в анализе пространства.
В 1967 г. наше внимание привлекли дельфины, получившие в настоящее время такую большую популярность. Однако надо сказать, что описание поведения дельфинов оказалось запутанным в сетях новейших легенд о чудесах некоего «сверхживотного». Эти китообразные были принесены в очередную жертву антропоморфизма, как в свое время так же «пострадали» антропоиды, попав в чуждый им ранг интеллектуальных существ. Это обстоятельство само по себе извращает представления о весьма оригинальных, исключительно интересных по режиму поведения представителях водных млекопитающих. Можно уже сейчас на основании общих наблюдений и по результатам пока лишь разведочных экспериментов увидеть иную, чем у других млекопитающих, особенность динамики мозговой деятельности. Однако не следует ожидать каких-либо сенсационных открытий. В целом мозг дельфина выполняет свои функции в рамках первой сигнальной системы, т. е. не выходит за пределы предметного, конкретного, мышления, свойственного мозгу всех животных, не исключая даже столь высокоразвитых, как антропоиды.
Наша лаборатория поставила вполне определенную, хотя и нелегкую задачу — попытаться описать систему эхолокационного пространственного анализа у дельфинов и сопоставить ее с подобной системой летучих мышей, конечно, с учетом комплекса специфических черт ориентации в пространстве животных воздушной и водной сред. Результаты наших экспериментов на афалинах и белобочках излагаются в этой книге.
Таким образом, почти за десятилетний период раздумья, ме-тодических поисков, экспериментальных разведок и в процессе поэтапных обобщений фактического материала наша лаборатория успела накопить некоторый опыт в подходах к знанию об эхолокации. Как выше было сказано, проблема эта привлекает к себе почти всеобщий интерес. Однако вместе с этим в отечественной и в новейшей зарубежной литературе нет сводки о системе ультразвуковой сигнализации, о результатах опытов и наблюдений различных сторон эхолокации, а также более или менее полного описания форм поведения эхолоцирующих животных. Мы решились взяться за такой труд и в монографическом изложении представить основные сведения^
40
Проблема эхолокаций у животных связана с именем талантливого американского естествоиспытателя — зоолога, физиолога — Дональда Гриффина, известного советским читателям своими замечательными книгами «Эхо в жизни людей и животных» (1959 г.) и «Перелеты птиц» (1964 г.). На русский язык первая книга переведена в 1961 г., вторая — в 1966 г. Своими экспериментальными работами, выполненными в разное время совместно с Г. Пирсом, Р. Галамбосом, А. Новиком, А. Гриннелом и др., Д. Гриффин заложил основы дальнейших биофизических и собственно физиологических разработок данной проблемы. Результаты многолетнего изучения эхолокации летучих мышей и птиц Д. Гриффин обобщил в монографии «Listening in the dark» (1958) — «Слушая в темноте» (на русский язык не переведена). Естественно, что в нашей книге ссылки на его труд наиболее часты. С момента выхода в свет упомянутой монографии Д. Гриффина появилось множество оригинальных исследований. Они углубили и расширили первоначальные представления о физиологических механизмах, диапазоне действия и физической природе ультразвуковой импуль-сации и роли эхолокации в жизни различных животных. Особенно интенсивно начали развиваться исследования по гидроакустике китообразных. В 1961 г. вышла книга Уинтропа Келлога «Дельфины и сонар» («Porpoises and sonar»). Автор рассказывает о своих экспериментах и экспериментах других исследователей, установивших наличие эхолокации у дельфинов.
Были проведены симпозиумы и конференции, посвященные вопросам эхолокации. Отметим лишь некоторые из них. В апреле 1963 г. научно-исследовательским управлением ВМС США и Американским музеем естественной истории организуется на о. Бимини (Багамские острова) симпозиум по морской биоакустике. Из 23 докладов, представленных на этом симпозиуме, 5 относились к гидроакустике китообразных. В них дается подробный обзор имевшейся в то время информации о подводных звуках китообразных и рассматриваются также обобщающие методические вопросы, возникающие при изучении поведения этих животных. Труды симпозиума под редакцией видного биолога У. Н. Таволги вышли в свет в 1964 г. под названием «Морская биоакустика». На русский язык книга переведена в 1969 г.
Следующий симпозиум с тематикой исключительно по китообразным (их экологии, морфологическим особенностям, поведению, эхолокации, а также с описанием аппаратуры, используемой для гидрракустических исследований китообразных) был проведен в 1965 г. Материалы симпозиума «Киты, дельфины и морские свиньи» («Whales, dolphins and porpoises») опубликованы в 1966 г. под редакцией К. Норриса.
И наконец, последний симпозиум, непосредственно посвященный проблеме эхолокации животных, проходил в сентябре 1966 г. в итальянском городе Фраскати. Труды симпозиума «Сонарные
И
системы животных» («Animal sonar systems») опубликованы в двух томах под редакцией Р. Бюснеля (1967). Доклады включали исследования эхолокации летучих мышей, дельфинов, ластоногих и некоторые вопросы методического и технического порядка. В нашей книге излагаются наиболее важные материалы указанных симпозиумов.
Глава «Электрофизиологическая характеристика^нервных аппаратов слуховой системы летучих мышей» написана нами совместно с доктором медицинских наук Д. П. Матюшкиным, в течение ряда лет сотрудничавшим с нами на кафедре физиологии высшей нервной деятельности по эхолокационной проблеме.
Главы «Характеристика отряда китообразных» и «Механизмы генерирования звуков у китообразных» написаны кандидатом биологических наук В. М. Бельковичем, известным исследователем китообразных. Ему же мы выражаем признательность за ценные советы по второму разделу рукописи.
Профессорам П. И. Гуляеву и А. С. Мальчевскому, кандидату биологических наук П. П. Стрелкову выражаем благодарность за полезные советы при чтении рукописи.
Профессору Александру Александровичу Стрелкову — наша особая благодарность. Он был добрым оком рукописи — не один печатный лист в итоге оказался во благо экстирпирован, и для многих узкоспециализированных терминов по его совету мы нашли эквивалентное общенаучное выражение.
Академик В. А. Парин словом и делом постоянно поощрял наши новые исследования, способствовал их опубликованию — пользуемся поводом сказать ему большое спасибо.
В разработке исследований вопросов эхолокации в нашей лаборатории вместе с нами участвовали следующие сотрудники: Н. И. Ахмарова, Н. В. Бурикова, А. Г. Васильев, С. С. Головина, А. Г. Голубков, И. А. Ершова, В. Н. Зворыкин, В. И. Королев, Л. М. Котеленко, Г. А. Куликов, А. К. Макаров, Д. П. Матюшкин, О. А. Полунина, Б. В. Соколов, Н. Н. Соколова, И. М. Стос-ман, Л. И. Суховская.
По всей вероятности, книга привлечет внимание читателей широкого профессионального диапазона, но, надо полагать, не во всем они будут одинаково заинтересованы. Мы тщим себя надеждой, что эти противоречия и другие недостатки нашего труда окажутся терпимыми, и готовы с признательностью принять благотворную критику.
Май 1969 г.
ВВЕДЕНИЕ
Сущность эхолокации
Под словом «локация» понимается определение местоположения предметов, измерение их координат и параметров движения. В живой природе используются разнообразные формы и способы локации. У человека и большинства животных определение местоположения окружающих предметов осуществляется благодаря анализаторным системам дистантного действия, в основном зрительной и слуховой, причем эти системы в функциональном отношении у некоторых животных доведены до высочайшего совершенства. Достаточно вспомнить о необычайной остроте зрения у дневных хищных птиц или точности звуковой пеленгации добычи совами.
Для обнаружения объектов окружающей среды некоторые животные используют и другие виды информации. Глубоководные кальмары, например, помимо обычных органов зрения наделены особыми рецепторами, способными улавливать инфракрасные лучи, а своеобразные органы — «термолокаторы» — гремучих змей служат для поисков добычи, воспринимая тепловое излучение живых существ и реагируя на разность температур в тысячную долю градуса.
Приведенные примеры, несмотря на их разнообразие, представляют собой различные варианты так называемой пассивной локации, когда обнаружение объектов осуществляется только путем приема той энергии, которую непосредственно излучают или переизлучают сами исследуемые объекты.
Сравнительно недавно казалось, что более или менее чувствительными органами дистантного обнаружения как средствами пассивной локации ограничиваются возможности живой природы.
В самом начале XX в. человечество было вправе гордиться тем, что оно создало принципиально новый, активный способ локации, при котором невидимая прежде цель облучается потоком электромагнитной или ультразвуковой энергии и обнаруживается с по-
13
мощью той же энергии, ио уже отраженной от цели. Радио- и гидролокационные станции — эти приборы активной локации — пришли на смену различного рода «слухачам» — приборам пассивного обнаружения — ив настоящее время получили огромное развитие в решении народнохозяйственных, военных и космических проблем. В то же время несомненно, что принципы радиолокации подсказали биологам путь к решению вопроса о формах пространственной ориентации у некоторых животных, которые невозможно было объяснить функционированием хорошо известных анализаторов дистантного действия.
В результате кропотливых исследований с помощью новой электронной аппаратуры удалось установить, что ряд животных использует методы активной локации с применением двух видов энергии — акустической и электрической. Электрической локацией пользуются некоторые тропические рыбы, например мор-мирус, или водяной слоник, тогда как активная акустическая локация открыта у нескольких представителей наземных и водных позвоночных, стоящих на разных уровнях эволюционного развития.
Акустическая локация служит средством обнаружения объектов благодаря звуковым волнам, распространяющимся в данной среде.
По аналогии с радиолокацией различают две формы акустической локации: пассивную, когда обнаружение осуществляется только путем приема той энергии, которую непосредственно излучают или переизлучают сами исследуемые объекты, и актив н у ю, при которой анализ объекта основан на предварительном облучении его звуковыми сигналами с последующим восприятием этой же энергии, но уже отраженной от него. Первая форма акустической локации издавна обозначается как слух или слуховое восприятие, и звуковые колебания принимаются слуховым анализатором.
Вторую форму, т. е. активную акустическую локацию, американский ученый Д. Гриффин, впервые открывший ее у летучих мышей, назвал эхолокацией. Со временем термины «эхолокация», «акустическая локация» и «акустическая ориентация» стали в какой-то степени синонимами и широко используются в биологической литературе при описании активной формы локации у животных. Правда, в последние годы делаются попытки использовать термины «акустическая локация», «пассивная локация» для обозначения функций слуховой системы у сов, которые с высокой точностью производят локализацию местоположения своей добычи на слух во время ночной охоты (Ильичев, 1970; Payne, 1971). Этим хотят подчеркнуть ту огромную роль, которую играет слух в пищевом поведении сов, и сопоставить способы ориентации этих птиц с таковыми у летучих мышей, хотя это сопоставление неправомочно, ибо последние поднялись на следующую, качественно но-14
вую ступень акустической локации, применив активное зондирование пространства собственными акустическими сигналами.
Прежде чем перейти к характеристикам эхолокации, коротко остановимся на основных понятиях и определениях из области акустики, необходимых для понимания физических раздражителей слухового рецепторного аппарата.
Природа звуковых волн
Звуковые колебания. Когда мы говорим, что слышим звук, это означает, что где-то произошло колебание воздуха, которое достигло нашего слуха. Таким образом, звук есть колебательное движение частиц упругой среды, распространяющееся в виде волн в газах, жидкостях и твердых телах. Но так как органы слуха человека воспринимают только те колебания, частоты которых лежат в интервале примерно от 20 до 20 000 колебаний в 1 сек., то колебания этого диапазона и выделяются в особую группу звуковых, или акустических, колебаний, называемых также сокращенно звуком. Принято в качестве единицы частоты’[брать частоту такого колебательного процесса, при котором в 1 сек. совершается одно колебание. Эта единица частоты по имени немецкого физика Герца называется герцем, а тысяча колебаний в 1 сек. — килогерцем.
В акустике все колебания подразделяют на четыре категории: 1) дозвуковые колебания с частотами меньше 20 гц, или инфразвуки; 2) звуковые колебания — собственно звуки; 3) сверхзвуковые колебания с частотами выше 20 кгц, или ультразвуки; 4) гиперзвуковые колебания с частотами выше 109 гц (1000 мгц), или гиперзвуки.
Распространение звуковых волн. Явление распространения колебания в среде называется волной. Примером образования волн может служить действие камня, брошенного в воду. Область водяной поверхности, которая непосредственно возмущена падающим камнем, начинает колебаться, причем колебание распространяется от этой области к следующей, и мы получаем на поверхности воды волну. Сразу заметим, что при распространении колебаний колеблющиеся частицы не перемещаются с распространяющимся колебательным процессом, а колеблются около своих положений равновесия. Звук — такая же форма волнового движения, как и волны на воде, разбегающиеся от падающего камня. Но звуковые волны не сразу можно так реально представить себе, как волны на воде, потому что эти волновые движения происходят не на двухмерной поверхности пруда, а в трехмерном пространстве. Когда звук распространяется в среде, например в воздухе, давление в любой точке данной среды ритмически изменяется, то возрастая, то убывая. Если бы можно было сфотографировать распределение молекул воздуха вблизи источника звука, то оказалось бы, что в некоторых местах молекулы группируются тесно, а в других — свободно. В местах сгущения давление повышено, в местах разрежения — понижено. Графическое изображение изменения давления представляет собой синусоиду (рис. 1).
Основные свойства звуковых волн. Любые звуковые волны характеризуются скоростью распространения, длиной, частотой и интенсивностью. Легко заметить, что волны, вызванные падением камня на поверхность воды, движутся с постоянной скоростью. Это также относится и к звуковым волнам. Скорость звука (С) зависит от упругих свойств среды, в которой распространяется звуковая волна, и в меньшей степени — от температуры и некоторых других факторов. Например, скорость звука в воздухе при температуре 18° равна примерно 340 м/сек., а в морской воде при 0° — 1550 м/сек. Эти величины кажутся значительными, но во много раз уступают световым или радиоволнам, имеющим скорость 300 000 км/сек. Длина звуковой волны (X) — это расстояние между двумя последователь-
15
ними зонами максимума или минимума давления, возникающими при распространении звуковой волны. В нашем примере с камнем длиной волны будет расстояние между соседними гребнями или впадинами, где уровень воды соответственно выше среднего или ниже его.
Число волн, проходящих в 1 сек. через данный участок среды, называется частотой звука. Если волны короткие, то на заданном расстоянии их уложится большее число и за данный промежуток времени большее их число достигнет определенной точки, иначе говоря, частота их будет выше. Соотношение между скоростью звука С, его частотой / (или периодом Т) и длиной волны К можно выразить следующим простым уравнением:
С \ = — = СТ.
Так как скорость звука есть постоянная величина для определенной среды, то^длина волны изменяется обратно пропорционально частоте. Следовательно, чем выше частота звука, тем меньше длина звуковой волны. Например, длина волны звука с частотой 34 гц примерно равна 10 м, а частоте 50 гц соответствует длина волны
С
к = у = 340 000 мм/сек. :
: 50000 гц = 6.8 мм.
И наоборот, длине волны в 3.4 мм соответствует частота звука
С
f = -т- = 340 000 мм/сек. : к
: 3.4 мм = 100 000 гц, или 100 кгц.
Рис. 1. Изменение давления воздуха вблизи источника звука. (Griffin, 1961). X — длина волны.
Все приведенные выше примеры относились к звуковым волнам, распространяющимся в воздухе.
Другие соотношения частоты и длины волны получаются при распространении звука, например, в морской воде, в связи с тем что скорость звука в воде в 4.5 раза больше, чем в воздухе, и составляет 1550 м/сек. Поэтому частоте 50 кгц в морской воде будет соответствовать длина волны
Х = 1 550 000 мм/сек. : 50 000 гц = 3.1 см.
Понятие длины волны особенно важно при рассмотрении основных явлений, характерных для всех волновых процессов: дифракции и интерференции, отражения и преломления волн. Например, дифракционная картина становится заметной, когда длина волны делается сравнимой с размерами объекта, с которым встречается звуковая волна. При более коротких длинах волн происходят зеркальное отражение и образование резко очерченных звуковых теней, тогда как более длинные волны распространяются так, как будто препятствие отсутствует, другими словами, будут огибать его, не давая при этом заметных отражений.
16
Звук мы слышим за счет энергии, которую волна переносит от источника звуковых колебаний. Энергия, переносимая звуковой волной через единицу площади в единицу времени, представляет собой интенсивность, или силу звука. Однако измерение интенсивности связано с определенными трудностями и при решении практических задач в качестве измеряемого параметра звукового поля удобнее пользоваться давлением. При этом часто применяют термин «звуковое давление», понимая под этим изменения давления воздуха в данной точке поля, вызываемые звуковыми колебаниями. Предпочтение, отдаваемое давлению, объясняется тем, что из всех параметров звукового поля колебания давления можно измерить наиболее просто и точно, а также тем, что мы воспринимаем звук именно через колебания давления.
Звуковое давление, как и всякая другая переменная величина, характеризуется амплитудой (максимальным значением) и частотой звука. Звуковое давление измеряется в микробарах,
1 мкбар = 1 дин/см2 = 0.1 н/м2.
Минимальное давление звука, которое способно воспринять человеческое ухо, 0.0002 дин/см2. Эта величина является порогом слышимости. Интенсивность такого звука 10-16 вт/см2. Абсолютные значения звукового давления, при которых барабанная перепонка человека разрушается, составляют 2 -104 Дин/см2.
Вместо абсолютных значений звукового давления часто пользуются понятием относительного уровня звукового давления (L) или просто уровня силы звука. Оно выражается
L=201g-^-, дб, г о
где Р — среднее квадратичное значение звукового давления; Ро — порог слышимости человека 0.0002 дин/см2 (относительный 0).
Децибелы — логарифмические единицы, и их применение делает возможным сравнение Двух уровней звукового давления без знания абсолютного значения каждого из них. Два звука, отличающихся по силе в 10 раз, разнятся на 1 бел; если они отличаются в 100, 1000, 10 000 . . . раз, то имеют разницу в 2, 3, 4 . . . бел. Широко используется в десять раз меньшая едини-ница — децибел. Изменение интенсивностей в 10, 100, 1000, 10 000 раз составляет соответственно 10, 20, 30, 40 дб. Для измерения в децибелах отношений звуковых давлений следует учитывать, что интенсивность изменяется как квадрат звукового давления и логарифм отношения давлений должен быть умножен на 2.
Приведенные примеры круглых чисел, отличающихся порядками, позволяют производить логарифмирование в уме. Прирост на 3 дб равен увеличению звукового давления в ~1.5 раза; прирост на 6 дб — в 2 раза, а прирост на 10 дб — увеличению давления примерно в 3 раза. Число децибелов будет равно нулю при соотношении измеряемых величин, равных единице.
В качестве примера приведем средние уровни громкости (дб) для различных источников звука.
Тихая комната............................... 30
Спокойный разговор трех человек в комнате средних размеров................................. 48— 50
Шум оживленной улицы........................ 55— 60
Шум поезда метро при движении.............. 85— 90
Духовой оркестр.............................до 80—100
Шум авиационного мотора на расстоянии 1м... 110—120
Болевой порог .............................. 130
17
Сложные звуки. Чистые тона или звуки, которые графически можно представить в виде синусоидальных или гармонических колебаний, в природе встречаются довольно редко. Подавляющее большинство звуков, которые мы слышим (речь, музыка, шумы окружающей среды), являются сложными, т. е. состоят из нескольких или многих частот, придающих звуку его характерную окраску, или тембр.
Звуковые колебания, создаваемые роялем или кларнетом (рис. 2), носят значительно более сложный характер по сравнению с простыми колебаниями
Частотами,
и отличаются друг от друга по своей форме. Различие формы обусловлено их разным гармоническим составом, т. е. распределением амплитуд и фаз одних и тех же простых колебаний. Такое представление вытекает из общей теоремы Фурье, по которой любое сложное периодическое колебание периода Т представляет собой набор простых гармонических колебаний с кратными часто-1 1 тами /, равными у, 2 у ,
1 1
Зу, 4 у и т. д. Наиболее низкая частота называется основной частотой. Колебания с основной частотой (Д) есть первая гармоника, или основной тон, а колебания с частотами 2/1?3 Д, 4 /х — высшие гармоники (2-я, 3-я, 4-я), или обертоны.
Представление сложного звукового колебания в виде суммы простых гармонических колебаний, с кратными частотами, имеющими соответствующие значения амплитуд и фаз, называется разложением в спектр.
Рис. 2. Форма колебаний и звуковые спектры рояля (а; частота 128 гц) и кларнета (б; частота 275 гц).
Амплитуды отдельных гармоник в каждом спектре сильно различаются. В звуке рояля, например, заметны гармоники до 18-й, причем 15-я и 16-я практически отсутствуют, в то время как у кларнета налицо гармоники лишь до 12-й и отсутствуют 2-я и 4-я (рис. 2).
Сдвиги отдельных тонов по времени, т. е. изменение фаз этих тонов, не воспринимаются на слух, хотя и могут сильно изменять форму результирующего колебания.
Рассмотренные выше спектры линейчатые, и возникают они у периодических колебаний. Для колебаний, не имеющих периода, характерны сплошные спектры. Такие спектры имеют одиночные сигналы и разного рода шумы. Во всех случаях шумовые колебания состоят из огромного количества гармо
нических колебаний с разными частотами, равномерно распределенными по всему спектру.
Таким образом, у гармонического колебания спектр состоит из одной-единственной частоты. У периодического сложного колебания спектр представляет собой набор частот — основной и кратных ей. И наконец, непериодические колебания имеют спектры, в которых число различных частот неограниченно велико.
Ультразвук и его особенности
До недавнего времени считалось, что ультразвук редко встречается в природе. Однако исследования последних 20—30 лет привели к противоположному заключению. Сейчас можно определенно сказать, что человек живет в мире ультразвуков. Их излучают многие животные (позвоночные и беспозвоночные), моря и океаны, леса и джунгли. Многие явления природы — молния и гром, сильный ветер, песчаные бури, снежные бураны, горные обвалы — также служат источником ультразвуковых колебаний.
Интенсивность природных источников ультразвуковых колебаний варьирует в широких пределах. Так, самцы прямокрылых, сверчков и цикад излучают ультразвук, уровень силы которого на расстоянии 30 см от источника составляет около 90 дб. Джунгли, моря и океаны создают ультразвуки интенсивностью порядка 55 дб при частоте 20 кгц. Все природные источники образуют так называемый естественный фон ультразвуковых колебаний.
Помимо естественного фона, человек сам создает мощные ультразвуковые источники для различных целей. С начала XX в. и по настоящее время было проведено большое число исследований в области ультразвука. Энергия ультразвуковых колебаний быстро нашла практическое применение. Несомненно, что толчком к исследованиям в области ультразвука оказалась гибель «Титаника», который при первом же трансатлантическом рейсе в 1912 г. столкнулся с айсбергом и затонул. Вторым, еще более сильным толчком послужила проблема обнаружения немецких подводных лодок во время первой мировой войны. Именно в эти годы выдающийся французский физик Поль Ланжевен предложил использовать ультразвук для обнаружения, вражеских подводных лодок. Посылая в воду ультразвуковые импульсы, он измерял время между посылкой зондирующего импульса и получением отраженного эхо-сигнала от цели. Это изобретение вызвало к жизни новую область техники — гидроакустику, ставшую одним из эффективнейших средств обнаружения в военных операциях на море.
В 1928 г. советский инженер С. Я. Соколов предложил использовать тот же принцип отражения ультразвуковых волн от препятствий для обнаружения дефектов в твердых телах. В настоящее время с помощью ультразвука сверлят, сваривают металлы, очищают поверхность различных изделий, производят измерения толщины, определяют качество литья и сварных конструкций, разрушают горные породы, в медицине проводят ультразвуко-скопию тканей, глубокое их прогревание, массаж и т. д. (Бергман, 1957; Розенберг, 1957; Борисов, Марков, 1960; Горшков и др., 1965). Какие же особенности отличают ультразвук от других звуковых колебаний?
Как слышимые звуки, так и ультразвуки подчиняются одним и тем же акустическим законам, так как физическая природа их, как было указано выше, одинакова. Но условия излучения, распространения и восприятия звуков разных частот в соответствии с общими законами акустики оказываются различными. В силу именно этих различий излучение и восприятие звуков человеком ограничены слышимым диапазоном частот. Различна также и дальность распространения для звуков разной частоты.
Таким образом, особенности ультразвука обусловлены не тем, что он неслышим, а тем, что частота его выше и, слёдовдтельно, длина волны короче, чем у слышимого звука.
19
Как известно, звуковые волны распространяются во все стороны от источника звука. Например, звук заводского гудка одинаково радиирует во все стороны от завода. Шум улицы, проникающий через открытую форточку, практически одинаково слышен во всех точках комнаты, а не только против форточки. Это объясняется относительно большой длиной волны слышимых звуков, которая составляет от нескольких сантиметров до 15—20 м. В связи с этим окружающие нас предметы не являются препятствием для звуковых волн — они обтекают их, не давая звуковой тени.
Длина волны ультразвуковых колебаний, особенно высокочастотного диапазона, очень мала. В силу этого ультразвук, подобно свету, дает звуковые тени, или эхо, от сравнительно небольших предметов, сравнимых с длиной волны. Уточним значение некоторых слов, полезных для описания способности звуковых волн переносить информацию.
Эхо обычно означает четкое изолированное отражение звука от поверхности, расположенной на некотором расстоянии. Реверберация образуется многократными отражениями звука от близко расположенных поверхностей, когда отраженные волны перекрываются и смешиваются с исходными звуковыми волнами. В более общем смысле эхо означает звуковую волну, направление которой существенно изменилось в результате падения на некоторый предмет.
Следующее отличительное свойство ультразвука состоит в том, что его можно излучать в виде почти параллельного, узкого пучка. Реальные излучатели звука в слышимом диапазоне частот всегда излучают звук во всех направлениях. Так, во время разговора человек совсем не обязательно должен быть обращен лицом к своему собеседнику, чтобы тот его услышал. Как указывает Е. Я. Пумпер (1946), эти свойства ультразвука полностью согласуются с общей теорией дифракции волн, на основе которой сравнительно просто решается вопрос об излучении звука. Если линейные размеры плоского излучателя велики по сравнению с длиной волны звука, то излучается плоская волна, распространяющаяся в направлении, перпендикулярном к плоскости излучателя, в виде параллельного пучка. Если же размеры излучателя малы по сравнению с длиной волны звука, в этом случае излучатель по своим свойствам близок к точечному. Он излучает сферическую звуковую волну, радиирующую от него во все стороны. При всех других соотношениях между размерами излучателя и длиной волны звука имеют место промежуточные случаи: источники излучают расходящийся пучок волн, который будет приближаться к параллельному, если размеры излучателя возрастают или длина волны уменьшается; при уменьшении же размеров излучателя или увеличения длины волны пучок, наоборот, будет все больше расходиться.
Для звуковых колебаний в слышимом диапазоне частот практически всегда справедлив второй случай. Излучатели звука редко имеют размеры большие, чем несколько сантиметров, и низкие частоты обычно излучаются в виде сферических волн, только самые высокие среди них частоты могут иметь некоторую направленность.
Как раз обратный случай имеет место при излучении ультразвука. Так, при частоте 34 кгц длина волны ультразвука будет 1 см. Поэтому здесь сравнительно легко сделать излучатель большим по сравнению с длиной волны и получить направленное излучение в виде слабо расходящегося пучка.
Возможность образования ультразвуковых пучков позволяет не только концентрировать всю энергию ультразвука в нужном направлении, но и фокусировать ее в определенное место. Интенсивность ультразвука возрастает пропорционально квадрату амплитуды и частоты колебаний, в связи с чем при повышении частоты относительно легко могут быть получены ультразвуки огромной силы. Физические расчеты показывают, что при частоте 500—1000 кгц без труда удается получить силу звука порядка 10 вт/см2, что превосходит силу звука при пушечном выстреле в 1000 раз, а силу звука громкоговорителя — в 1-109 раз (Горшков и др., 1965).
20
И наконец, третье отличительное свойство ультразвука — его быстрое затухание. Даже если ультразвук излучается параллельным пучком, его амплитуда очень быстро уменьшается с расстоянием из-за больших потерь энергии в среде.
Величина затухания ультразвука характеризуется коэффициентом поглощения, который показывает степень уменьшения амплитуды звука в параллельном пучке с расстоянием. Обратная величина коэффициента поглощения звука дает то расстояние, на протяжении которого амплитуда звука убывает примерно в 3 раза. Чем больше коэффициент поглощения, тем меньше это расстояние, тем быстрее, следовательно, затухает звук. Но коэффициент поглощения звука пропорционален квадрату частоты. Следовательно, при увеличении частоты звука в 10 раз коэффициент поглощения возрастает в 100 раз. В 100 раз сократится и то расстояние, на котором амплитуда звука уменьшается в 3 раза. Таким образом, ультразвук, у которого частота выше, чем у слышимого звука, должен затухать на значительно меньших расстояниях от источника, чем последний.
Следует, однако, учесть, что при излучении слышимого звука трудно получить параллельный пучок. Поэтому здесь всегда будет иметь место уменьшение амплитуды еще и вследствие рассеивания. При низких частотах звука эта причина будет преобладать, но при распространении ультразвука на большие расстояния рассеивание мало по сравнению с затуханием (поглощением).
Именно благодаря большому затуханию ультразвуки не получили широкого применения для использования их в целях поиска в воздушной среде, где их полностью заменили электромагнитные волны. В то же время в воде, где затухание ультразвука в 1000 раз меньше и радиоволны не распространяются, в целях гидролокации применяются приборы, работающие исключительно на ультразвуковых и звуковых сигналах.
Итак, можно следующим образом обобщить особенности ультразвука, которыми он отличается от слышимого звука: 1) высокая частота и малая длина волны ультразвука создают благоприятные условия для получения четких отражений (эхо) от небольших предметов, которые звуковые волны слышимого диапазона огибают; 2) ультразвук можно излучать сравнительно узким, почти параллельным пучком, что очень трудно получить для слышимого звука; 3) ультразвук в силу большого поглощения распространяется на более короткие расстояния, чем слышимый звук.
История открытия эхолокации
Акустическая локация в живой природе была открыта сравнительно недавно, всего около 35 лет назад, однако этому событию предшествовала полная драматизма история многочисленных экспериментов, мучительных раздумий и горьких разочарований. История эхолокации ведет свое начало с 70-х годов XVIII столетия и связана с именем итальянского ученого Лазаро Спалланцани (Spallanzani, 1729—1799 гг.), уже тогда хорошо известного своими трудами в различных областях естествознания и философии. Это был ученый, поразительный по своему энциклопедическому таланту. Профессор в Родже, Модене, затем в Павии, он одновременно возглавлял кафедры логики, математики, естественных наук, греческого языка. Отметим лишь, что он был увлечен механизмами регенерации у животных и первый установил факт воспроизведения ящерицами отрезанных хвостов, лап тритонами. Он же впервые осуществил опыт искусственного
21
осеменения у млекопитающих. Спалланцани едва ли не первый физиолог, пытавшийся описать закономерности пищеварения у животных не методом вивисекции, а в хроническом эксперименте оригинальными приемами. В возрасте 64 лет он заинтересовался способами ориентации ночных животных.
В 1793 г. Спалланцани установил факт исключительной важности — летучие мыши свободно летают в темной комнате, где
Лазаро Спалланцани (1729—1799).
совы становятся совершенно беспомощными. Это послужило началом длительных и остроумных серий экспериментов, в результате которых Спалланцани узнал об ориентации летучих мышей столько же, сколько другие исследователи за 140 лет после его смерти. К сожалению, большая часть работ Спалланцани до сих пор остается неопубликованной, и мы должны быть благодарны голландскому ученому Дийкграафу (Dijkgraaf, 1949а, 1949b) и американским ученым Галамбосу и Гриффину (Galambos, 1942b; Griffin, 1958), которые подняли архивные материалы (в основном письма Спалланцани к его коллегам) и дали возможность позна
22
комиться широкому кругу биологов с замечательными опытами Спалланцани.
Первоначально Спалланцани полагал, что необыкновенная острота зрения позволяет летучим мышам избегать препятствий в темноте. Тогда на головы зверьков были надеты светонепроницаемые колпаки. Полет летучих мышей стал сразу неуверенным, и животные, натыкаясь на препятствие или стенку, падали на пол. Казалось, разгадка была близка, но когда ученый провел контрольные опыты с прозрачными колпаками, это не улучшило способность летучих мышей своевременно обнаруживать предметы на своем пути. Чтобы окончательно решить вопрос со зрением, Спалланцани ослепил несколько летучих мышей. К его удивлению, животные, оправившись после операции, летали так же хорошо, как и нормальные животные, и не испытывали никаких затруднений после лишения их зрения. Спалланцани немедленно разослал письма своим ученым коллегам некоторых европейских стран, в которых рассказывал о своих исследованиях, просил повторить эти эксперименты и сообщить ему о результатах. Большинство полученных ответов подтвердило его опыты, но в одном из них содержались новые, еще более удивительные сведения. Швейцарский энтомолог, орнитолог и ботаник Шарль Жюрин после ознакомления с результатами опытов Спалланцани провел ряд экспериментов с летучими мышами и в феврале 1794 г. сделал новое важное открытие. Если уши животных были плотно закупорены воском или другим материалом, то летучие мыши становились беспомощными и наталкивались на любые препятствия. На основании этого Жюрин пришел к заключению, что, по-видимому, органы слуха летучих мышей принимают на себя функцию зрения, давая возможность животным своевременно обнаруживать препятствия и избегать их. Краткий отчет об этих экспериментах Жю-рина был опубликован позднее, в 1798 г. (Jurine, 1798).
Узнав об опытах Жюрина, Спалланцани вначале отнесся к ним скептически, но как только смог получить новую партию летучих мышей весной 1794 г., сам повторил опыты своего швейцарского коллеги и убедился в его правоте. Спалланцани был дотошным исследователем. Чтобы избежать возможности механического воздействия ушных пробок на ориентацию летучих мышей, он выточил маленькие латунные трубочки и точно подогнал их по диаметру слухового прохода летучей мыши. Стоило заткнуть внешний конец такой трубочки, и летучая мышь оказывалась полностью дезориентированной; если же концы трубочек оставались открытыми, животные, даже ослепленные, летали свободно и легко избегали всевозможные препятствия. Незадолго до смерти, в 1799 г., Спалланцани сделал основной вывод: летучие мыши могут прекрасно обходиться без зрения, но всякое серьезное повреждение слуха для них равносильно гибели. Тем не менее ни Спалланцани, ни -Жюрин не смогли дать достоверного объяснения полученным
23
фактам, тем более что полет летучих мышей считался совершенно беззвучным. Поэтому их выводы о преобладающей роли слуха в ориентации летучих мышей, казавшиеся абсурдными, не нашли сторонников, были отвергнуты, высмеяны и впоследствии совершено забыты. Вот, например, высказывание по этому поводу одного из ярых противников «слуховой теории»: «Чтобы согласиться с заключениями, которые Жюрин черпает из своих опытов, что уши летучих мышей для них более существенны, чем глаза, при обнаружении предметов, требуется больше веры и меньше философского смысла, чем можно было бы ожидать от зоолога-философа (zootomical philosopher), которого можно было бы спросить, если летучие мыши видят своими ушами, то слышат они своими глазами?» (Montagu, 1809, цит. по: Griffin, 1958, с. 63).
Забвению выводов Спалланцани и Жюрина немало способствовала новая теория, выдвинутая блестящим французским зоологом Жоржем Кювье (Cuvier, 1795, 1800, 1805, 1836), который с легкостью отбросил выводы Жюрина и Спалланцани о роли слуха в беспрепятственном полете летучих мышей, сославшись на то, что методика опытов была крайне жестока и, вероятно, затыкание ушей оказывало гораздо большее влияние на животных, чем просто ограничение слуховой рецепции. В то же время он выдвинул свою, так называемую тактильную теорию, из которой следовало, что способность летучих мышей в темноте избегать препятствий основана на хорошо развитом у них чувстве осязания. И хотя Кювье знал о подобных экспериментах Спалланцани, в которых была показана несостоятельность этой точки зрения, он не придал этому никакого значения.
Благодаря авторитету Кювье тактильная теория была принята большинством естествоиспытателей без всяких экспериментальных доказательств. Более того, Кювье объявил, что им найден материальный субстрат предполагаемых исключительных тактильных свойств летучих мышей: наличие специфических нервных окончаний в их летательных перепонках. Это в значительной степени способствовало признанию тактильной теории Кювье.
F В дальнейшем Шобл (Shobl, 1871) подтвердил их наличие, однако в то же время другие авторы показали, что описанные Кювье структуры не являются нервными окончаниями, а представляют собой эластические балки, которыми очень богаты перепонки летучих мышей (Leidig, 1859; Великий, 1872; Stieda, 1872). Наш соотечественник, сторонник тактильной теории, Н. П. Забусов (1910) тщательно исследовал иннервацию летательных перепонок летучих мышей и объяснил механизм «шестого чувства» следующим образом. Каждый из многочисленных волосков, разбросанных по голой поверхности перепонок, представляет собой рычаг, длинное плечо^которого, выставляясь наружу, уже на значительном рас-стояншГможет воспринимать отраженную от препятствия воздушную волну. Колебание рычага передается прикорневому нервному
24
аппарату, а через него (в форме сигнала) достигает центральной нервной системы и вызывает соответствующую реакцию животного. Более сотни лет просуществовала в науке^тактильная теория Кювье, и за этот срок не было прибавлено ни одного нового факта к экспериментальному материалу Спалланцани и Жюрина. Поэтому, в то время как одни авторы продолжали пытаться обосновать физиологически и морфологически тактильную теорию (Jobert, 1872; Redtel, 1873; Забусов, 1910), другие начали вновь склоняться к старым, почти забытым представлениям Жюрина и Спалланцани о преобладающем значении слуха при пространственной ориентации летучих мышей (Rollinat, Trouessart, 1900; .Whitaker, 1906; Hahn, 1908; Sgonina 1935, и др.). Эти работы подтверждали экспериментальные данные Спалланцани и Жюрина, но не прибавили ничего нового для объяснения полученных фактов.
Впервые предположение о наличии звуковой локации у летучих мышей высказал X. Максим в 1912 г. (Maxim, 1912) — изобретатель скорострельного станкового пулемета. К мысли о звуковой локации Максима привел трагический случай гибели «Титаника», столкнувшегося с айсбергом. Максим предложил предупреждающее устройство, которое сигнализировало бы о приближении к айсбергу. Обосновывая предлагаемый принцип действия сигнального устройства, Максим высказал мысль, что и летучие мыши используют при полете в темноте звуковую локацию. Это была совершенно новая, прогрессивная постановка вопроса, очень близкая к истине. Однако Максим ошибочно считал, что при обнаружении предметов летучие мыши используют отражения очень низкочастотных звуков, порядка 15 гц и длиной волны 24 м, создаваемые взмахами крыльев и лежащие ниже слухового порога человека.
С другого конца к этому вопросу подошел английский нейрофизиолог X. Хартридж. В 1920 г. Хартридж, работая ночью в своем кабинете в Кембридже, заметил, что в открытое окно влетело несколько летучих мышей, преследовавших насекомых. Притворив окно, он стал наблюдать за летучими мышами, любуясь их быстротой и маневренностью. Его удивило, что они продолжали летать из комнаты в комнату даже тогда, когда свет был выключен, а дверь частично затворена. Будучи экспериментатором, он и тут остался верен себе. Оставляя щель в двери то шире, то уже, Хартридж заметил, что летучие мыши точно определяли свои возможности и ни разу не пытались пролететь в узкую щель, поднимаясь вдоль нее до самого верха. Животные совершенно явно располагали средствами, позволяющими им определить, достаточно ли широка щель, чтобы пролететь сквозь нее, не видя при этом двери и не прикасаясь к ней.
Хартридж в то время работал над проблемой зрения, и в его лаборатории имелась светонепроницаемая камера, в которой он проводил свои эксперименты. Таким образом, ему нетрудно было
25
продолжить исследования летучих мышей в темноте. Он снова подтвердил, что зрение летучих мышей не играет роли в преодолении препятствий, и, идя по пути исключения, в конце концов предположил, что парадокс летучих мышей «видеть ушами» может быть объяснен механизмом эхолокации с использованием звуков высокой частоты, лежащих выше порога слышимости человека и поэтому не воспринимаемых им. Хартридж выдвинул эту гипотезу (Hartridge, 1920) как возможное объяснение данного явления, но, будучи занят другими исследованиями, не подтвердил ее экспериментально.
Гипотеза Хартриджа первоначально не привлекла к себе внимания, и «спалланцаниевая проблема летучих мышей», как ее стали называть впоследствии, продолжала оставаться загадкой на протяжении еще 20 лет.
Объяснить тайну летучих мышей помогло появление новой электронной аппаратуры. В одной из лабораторий физического факультета Гарвардского университета в США Г. Пирс начал проводить исследования по изучению свойств ультразвуков, т. е. звуков, лежащих выше слухового порога человека. Под его руководством в 1937 г. был создан прибор — звуковой детектор, позволяющий улавливать звуки широкого диапазона частот. Он состоял из пьезокварцевого микрофона, расположенного в фокусе параболического рупора, узкополосного усилителя и самописца, при помощи которого осуществлялась запись зарегистрированного звука на ленте. Усилитель был осуществлен по схеме супергетеродина с промежуточной частотой, равной 175 кгц. Трансформация частоты осуществлялась после одного апериодического каскада предварительного усиления. После усиления на промежуточной частоте и фильтрации с помощью узкополосного магнитострикционного фильтра колебания вторично гетеродинировались в звуковую частоту, равную 1 кгц и слышимую человеческим ухом. На выходе усилителя можно было подключить ламповый вольтметр, громкоговоритель или самописец. Диапазон частот, воспринимаемый усилителем, лежал в пределах от 10 до 80 кгц. Исследование частотного состава ультразвука производилось с помощью быстрых поворотов рукоятки, меняющих резонансную частоту усилителя. В то же время отмечалось показание лампового вольтметра на выходе усилителя. Имея частотную градуировку усилителя, было нетрудно зарегистрировать распределение интенсивностей звука по частотам.
Именно этот прибор впервые зарегистрировал неслышимые звуки летучих мышей, когда в 1938 г. студент-биолог упомянутого выше университета Д. Гриффин принес в лабораторию Пирса полную клетку летучих мышей {Myotis lucifugus и Eptesicus fuscus). Вспоминая об этом, Гриффин писал: «Как только я поднес летучих мышей к аппарату Пирса, сразу же обнаружилось, что они издают множество звуков, но почти все эти звуки попадают в диа-26
пазон частот, лежащих выше порога слышимости человека» (Griffin, 1961, с. 17).
Пирс и Гриффин (Pierce, Griffin, 1938) провели частотный анализ звуков, излучаемых летучими мышами в полете, и установили,, что частоты этих звуков лежат в диапазоне 30—70 кгц при наибольшей интенсивности в области 45—50 кгц. Далее они обнаружили, что животные издают звук не непрерывно, а в виде дискретных импульсов, длительность которых составляет 1/1000—1/500 сек., или 1—2 мсек.
Уже эти первые сведения блестяще подтвердили гипотезу Картриджа о существовании неслышимых звуков, излучаемых летучими мышами при полете в темноте, и позволили по-новому взглянуть на результаты «абсурдных» опытов Спалланцани и Жюрина.
Однако установление только одного факта излучения ультразвука летучими мышами, несмотря на всю его важность, еще не объясняло способность животных беспрепятственно летать в полной темноте. Требовалось в условиях точного эксперимента доказать, что летучие мыши действительно используют ультразвук в целях ориентировки в пространстве и что они способны воспринимать эхо от этих звуков, отраженных от встречаемых на пути препятствий.
Используя барьеры вертикально натянутых проволок, Гриффин и Галамбос получили количественную оценку способностей летучих мышей преодолевать препятствия при частичном или полном выключении зрения, слуха и при закрывании рта. Результаты этих экспериментов представлены в табл. 1.
Эксперименты Гриффина и Галамбоса вновь подтвердили, что летучие мыши отлично ориентируются и без участия зрительной рецепции, но полное (двустороннее) или частичное (одностороннее) выключение слухового аппарата влечет за собой резкое ухудшение их способностей своевременно обнаруживать и избегать препятствия. Однако в этих опытах авторы пошли дальше своих предшественников. Они показали, что закрывание рта летучей мыши, лишающее ее возможности издавать эти высокочастотные звуки, оказывается столь же эффективным, как и плотное затыкание ее ушей. И в том и в другом случае мыши становились совершенно беспомощными и натыкались на стены комнаты и любые другие препятствия на их пути.
Используя аппарат Пирса, исследователи произвели запись ультразвука летучих мышей во время полета на ленте самописца. Выяснилось, что частота следования импульсов в разных ситуациях не остается постоянной, а меняется по мере приближения летучей мыши к препятствию (Galambos, Griffin, 1940, 1942).
При подготовке к взлету мышь излучает от 5 до 10 импульсов в секунду. В полете частота следования импульсов возрастает, достигая 30 импульсов. Наконец, когда мышь почти вплотную
27
Таблица 1
Способность летучих мышей (Myotis lucifugus) преодолевать барьеры проволок (диаметром 1.2 мм) в норме и при выключении зрения, слуха и источника ультразвука (Griffin, Galambos, 1941)
Число животных Вариант опыта Опыт Контроль
число пролетов % уклонений число пролетов % уклонений
28 Глаза закрыты 2016 76 3201 70
12 Оба уха закрыты 1047 35 1297 70
9 Глаза и уши закрыты (в контроле только глаза закрыты) 654 31 832 75
8 В уши вставлены стеклянные трубочки, концы которых закрыты (в контроле такие же трубочки, но открыты на конце) 580 36 636 66
12 Оба уха закрыты (в контроле одно ухо закрыто) .... 853 29 560 38
6 Глаза и одно ухо закрыты (в контроле только глаза закрыты) 390 41 590 70
7 Закрыт рот (в контроле с закрытыми глазами или интактные) 549 35 442 62
подлетает к препятствию или пытается сесть на стену, число импульсов в секунду может достигнуть 60. Факт регулярного изменения частоты следования импульсов ультразвука в зависимости от расстояния до препятствия при полете имел принципиальное значение, так как объективно показывал, что летучая мышь определяет расстояние до препятствия по отраженному ультразвуку. И хотя в общих чертах уже было ясно, что принцип использования ультразвука у летучих мышей близок к тем техническим принципам, которые лежат в основе радиолокационных устройств, или радаров, как их принято называть в Англии и США, тем не менее Галамбос и Гриффин не были удовлетворены этими результатами. Они считали, что их опыты достоверно показывают лишь то, что летучие мыши излучают ультразвук в виде импульсов, но еще не вполне было доказано, что мышь слышит ультразвук, отраженный от препятствия. Для решения этого вопроса необходимо было доказать, что летучие мыши действительно могут слышать ультразвук в том диапазоне, в котором они его излучают.
Методом отведения потенциалов улитки было показано, что животные способны воспринимать широкий диапазон частот — от 30 гц до 98 кгц (предел используемой аппаратуры) с максимумом ответов в области ультразвуковых частот порядка 30—50 кгц (Galaip-
28
bos, 1941b, 1942a). После этих опытов отпали последние сомнения в правильности взглядов о роли ультразвука в ориентировке летучих мышей при полете.1 Как уже отмечалось выше, по аналогии с радиолокационными техническими системами, широко используемыми в годы второй мировой войны, Гриффин (Griffin, 1944) предложил назвать акустический способ ориентации, основанный на посылке животным дискретных ультразвуковых импульсов с последующим восприятием эха, отраженных от предметов, эхолокацией.
Первоначально летучих мышей считали единственными представителями животного мира, использующими эхолокацию в целях ориентировки в пространстве. Но уже 50-е годы принесли новые неожиданные открытия. В 1953 г. звуковая локация была обнаружена у ночных птиц — гуахаро, гнездящихся в глубоких пещерах Венесуэлы (Griffin, 1953а), а несколько позднее — у стри-жей-саланган (Medway, 1959, 1962; Novick, 1959) и у рукокрылых из подотряда крыланов — Rousettus (Mohres, Kulzer, 1956; Novick, 1958b). Но наибольшую сенсацию вызвали первые сообщения о наличии эхолокации у обитателей водной среды — китообразных. Честь этого открытия принадлежит пионеру в изучении поведения и биологии дельфинов в неволе, куратору океанариума в Сан-Августино А. Мак-Брайду (McBride, 1956). В записях, датированных 1947 г., он отмечал, что дельфин афалина легко избегает лабиринты мелкоячеистых сетей, но запутывается в сетях с более крупной ячеей. Эти и ряд других наблюдений позволили ему высказать предположение, что дельфин так же, как и летучая мышь, использует эхолокационный аппарат для обнаружения препятствий. Об этом прозрении Мак-Брайда научный мир узнал лишь через девять лет, уже после его смерти, благодаря А. Шевиллу, который нашел и подготовил к печати его заметки. Экспериментальные доказательства эхолокационных способностей дельфинов были получены несколько позднее, благодаря усилиям большого числа исследователей, среди которых прежде всего следует отметить Келлога, Шевилла, Лоуренс и Норриса. Суть этих доказательств сводится к следующему.
1. В условиях ограниченной видимости (мутность воды экспериментального бассейна, проведение опытов в ночное время) или при полностью выключенной зрительной рецепции дельфины
1 Почти одновременно и независимо от американских исследователей к выводу об акустическом характере ориентировки летучих мышей пришел в 1932 г. голландский зоолог С. Дийкграаф (Dijkgraaf, 1943, 1946). Дийк-грааф полагал, что летучие мыши избегают препятствий, воспринимая эхо очень тихих, напоминающих тиканье часов, но постоянно слышимых во время полета звуков. Последующие исследования (Griffin, 1951) показали, что слышимые Дийкграафом звуки представляют собой низкочастотный слабый Юн, который механически сопровождает излучение каждого интенсивного ультразвукового импульса и не играет существенной роли для ориентации Животных в пространстве,
29
способны обнаруживать и избегать различные препятствия, отыскивать рыбу, бесшумно погруженную в воду, и даже дифференцировать предметы.
2. При решении любой из вышеуказанных задач животные излучают под водой серии коротких широкополосных сигналов-импульсов, воспринимаемых на слух как резкие щелчки. Частота следования щелчков нарастает по мере приближения животного к цели. Частотные составляющие спектра таких сигналов достигают почти 200 кгц. Максимум интенсивности наблюдается на частотах 20—25 кгц.
3. Диапазон звуков, воспринимаемых дельфинами, очень широк. Он целиком охватывает область звуковых частот, слышимых человеком, и простирается в область ультразвука до 150 кгц и выше.
Следовательно, у дельфинов имеются все предпосылки осуществлять пространственный анализ с использованием эхолокации как одного из основных компонентов в ансамбле различных анализаторных систем.
В 60-е годы список эхолоцирующих животных расширился еще на три представителя: два из них — землеройки (Gould et al., 1964) и мадагаскарский тенрек (Gould, 1965) из отряда насекомоядных и третий — калифорнийский морской лев из отряда ластоногих (Poulter, 1963а, 1963b, 1966).
Указанными видами ныне завершается перечень животных, которые при ориентации в оптически неблагоприятных условиях пользуются этим особым специфическим способом акустической сигнализации. Несмотря на то что эхолокация служит совершенно конкретным целям, ее роль в жизнедеятельности указанных представителей и степень совершенства оказываются весьма различными.
Только у летучих мышей и зубатых китообразных эхолокация есть основной, а у некоторых из них почти единственный способ дистантной ориентации. У других эхолоцирующих животных она служит лишь вспомогательным средством, уступая по функциональной значимости зрительной рецепции. Рассмотрим особенности акустической локации у различных представителей позвоночных, основываясь на знании сравнительного биоакустического анализа их локационных импульсов, особенностей восприятия и обработки отраженных сигналов и основных показателей их эхолокационных или сонарных систем.
ЭХОЛОКАЦИЯ В ВОЗДУХЕ
ТИПЫ ЛОКАЦИОННЫХ СИСТЕМ У ЛЕТУЧИХ МЫШЕЙ
С давних пор совершенно необоснованно летучие мыши пользуются дурной славой. «Вопрос о летучих мышах есть вопрос другого света, вопрос, который пахнет ересью. . . Все покрыто тайной, обманом и мраком в этих двусмысленных существах, представляющих собой высшую ступень противоестественности, мерзости и фантастичности. . . Летучая мышь — это химера, чудовищное невозможное существо, символ грез, кошмаров, призраков, больного воображения. . . Всеобщая неправильность и чудовищность, замеченная в организме летучей мыши, безобразные аномалии в устройстве чувств, допускающие гадкому животному слышать носом и видеть ушами, все это как будто нарочно приноровлено к тому, чтобы летучая мышь была символом душевного расстройства и безумия» (Туссенель, 1874).
Так описывал летучих мышей французский натуралист А. Туссенель в 70-х годах прошлого столетия, выражая тем самым широко распространенное мнение об этих безобидных животных.
Даже в наше”время, когда летучая мышь полностью «реабилитирована» многочисленными исследователями и раскрыта ее тайна беспрепятственно ориентироваться в темноте, все же для многих встреча с ней оставляет неприятное ощущение и летучие мыши продолжают подвергаться бессмысленному гонению и даже уничтожению.
Объясняется это прежде всего тем, что летучие мыши ведут сумеречно-ночной образ жизни, имеют необычный для млекопитающих внешний вид, а кроме того, о наших отечественных летучих мышах, не говоря уже о тропических и субтропических формах, слишком мало известно не только широким слоям населения, но даже специалистам, занятым в других областях биологии.
33
Основные систематические группы рукокрылых
Отряд рукокрылых (Chiroptera) один из наиболее многочисленных среди млекопитающих по количеству видов и многообразию жизненных форм. Судя по немногочисленным ископаемым остаткам, корни происхождения его уходят в дотретичное время, так как формы из раннего эоцена мало чем отличаются от современных представителей (Jepsen, 1970). Общепринято выводить рукокрылых от примитивных древесных насекомоядных (Insectivora) (Weber, 1928; Allen, 1939; Кузякин, 1950), однако промежуточные ископаемые формы, которые связывали бы эти два отряда, до сих пор не обнаружены.
Уже в самом названии отряда заключен основной признак, благодаря которому этих животных легко отличить от всех других представителей класса млекопитающих. Их передние конечности превратились в крылья за счет удлинения предплечья, метакарпальных костей и особенно фаланг пальцев (кроме первого), служащих каркасом для тонкой, эластичной, кожистой летательной перепонки, натянутой между ними, боками тела и задними конечностями.
В составе отряда насчитывается около 1000 видов, которые образуют две хорошо обособленные группы: подотряды крыланы (Megachiroptera) — 200 видов, обитающих в тропиках и субтропиках Восточного полушария, и летучие мыши (Microchirop-tera) — 800 видов, широко распространенных по всему земному шару. Оба подотряда, по-видимому, произошли от одних предков, но уже давно в результате разделения путей развития между ними появились значительные морфологические и биологические различия. Важно также подчеркнуть, что крыланы, несмотря на активность в ночное время, пользуются в основном визуальными средствами ориентации, тогда как летучие мыши при полете в темноте полагаются больше на ультразвуковую локацию, чем на зрение. Именно в силу этого летучие мыши представляют больший интерес для рассматриваемой темы, чем крыланы, о которых пойдет речь в одной из последующих глав, хотя последние считаются более древними в пределах отряда.
Летучие мыши — зверьки мелких размеров. Длина тела от 28 до 140 мм. По числу видов и разнообразию экологических форм они намного превосходят крыланов (Кузякин, 1950; Соколов, 1973). Подавляющее большинство летучих мышей насекомоядно и добывает свою пищу в стремительном полете, но встречаются формы, уклонившиеся от генеральной линии эволюции в отношении пищевой специализации. Так, имеются виды, которые, подобно крыланам, перешли к растительной пище — фруктам, нектару и пыльце цветов. Другие разнообразят свою диету рыбой, мелкими наземными позвоночными, а некоторые стали питаться кровью крупных животных.
34
Летучие мыши широко распространены в тропических и умеренных областях обоих полушарий, на север и на юг до границ древесной растительности.
В составе подотряда насчитывают 17 семейств.
Число родов
Ланцетоносы, или мышехвосты, — Rhinopomatidae................. 1
Мешкокрылы — Emballonuridae...................................... 8
Щелеморды — Nycteriidae.......................................... 1
Ложные вампиры — Megadermatidae ................................. 3
Щелегубы, или ложные подковоносы, — Hipposideridae............ 8
Подковоносы Rhinolophidae ....................................... 2
Мормопиды — Mormoopidae ......................................... 3
Американские листоносы — Phyllo stomatidae....................... 4
Десмодусы, или вампиры, — Desmodontidae.......................... 3
Рыбоядные — Noctilionidae ....................................... 1
Воронкоухи — Nataltdae .......................................... 1
Дымчатые летучие мыши — Furipteridae............................. 2
Американские присосконоги — Thyroptertdae ....................... 1
Мадагаскарские присосконоги — Myzopodidae........................ 1
Новозеландские летучие мыши — Myctacinidae....................... 1
Бульдоговые — Molossidae ........................................ 7
Гладконосые, или обыкновенные летучие мыши, — Vesperttlionidae. 23
Построение четких филогенетических взаимоотношений между отдельными, часто весьма специализированными семействами летучих мышей крайне затруднено. По ряду морфологических признаков, и в первую очередь по степени совершенства летательного аппарата, все семейства летучих мышей несколько условно подразделяют на 4 крупных таксона — надсемейства (Miller, 1907; Weber, 1928; Simpson, 1945; Grasse, 1955) или на 3 группы (Griffin, 1958). Последнее разделение в основном обусловлено особенностями строения плечевого пояса, в котором на месте прикрепления плечевой кости к лопатке у более быстролетных форм развиваются специальные приспособления в виде дополнительного сустава, что обеспечивает более жесткую фиксацию крыла.
Морфоэкологические сопоставления позволяют признать наиболее примитивными среди современных летучих мышей представителей 4 семейств: ланцетоносов, мешкокрылов, щелемордов и ложных вампиров. За исключением мешкокрылов, которые представлены в обоих полушариях, все остальные семейства встречаются исключительно в тропиках Старого Света. По мнению многих авторов, именно в пределах Восточного полушария и, возможно, даже более локально, в Юго-Восточной Азии (Andersen, 1905, 1918; Кузякин, 1950, 1958; Симкин, 1971а), возникли исходные формы отряда рукокрылых.
Более сложный, чем у рассмотренной группы, плечевой пояс обнаруживается у щелегубов и подковоносов — оба семейства Старого Света, а также у американских листоносов с несколькими близкородственными семействами фауны американских континентов. Представители всех этих семейств более специализированы и в экологическом отношении, а многие летучие мыши
35
американской фауны, помимо ловли насекомых, к тому же приспособились к питанию фруктами, цветами и мелкими позвоночными.
И наконец, наибольшего развития летательный аппарат получил у двух крупных семейств — круготропических бульдоговых и повсеместно распространенных гладконосых летучих мышей. Первые, сохранив ряд черт примитивной организации, достигли наивысшего среди рукокрылых совершенства летательного аппарата, что позволило им широко расселиться в тропиках и субтропиках Старого и Нового Света. Гладконосые также обладают достаточно совершенным строением плечевого пояса, а кроме того, универсальной морфологической организацией и физиологической пластичностью. Эти признаки, взятые в совокупности, обусловили широкое распространение их не только в тропиках обоих полушарий, но также в широтах умеренного и холодного климата.
После краткого ознакомления с основными систематическими группами отряда попытаемся в той же последовательности рассмотреть особенности эхолокационных систем у различных семейств летучих мышей, и в первую очередь параметры локационных сигналов, составляющих одну из основных характеристик любого локатора. Животные в этом отношении не должны представлять исключения. Данные параметры в дальнейшем могут оказаться дополнительным признаком для установления родственных отношений между видами, родами и семействами летучих мышей.
Методы регистрации и измерения основных параметров ультразвуковых сигналов
Исследование эхолокационных сигналов летучих мышей невозможно проводить без создания комплекса специальной измерительной аппаратуры, предназначенной для работы в ультразвуковом диапазоне частот. Такой комплекс включает в себя ультразвуковые микрофоны, микрофонные усилители, различные фильтры, необходимые для выделения определенных частот исследуемых сигналов, осциллографы и магнитофоны для записи и регистрации сигналов, а также специальные устройства для определения основных параметров импульсов: длительности, частоты повторения, частоты заполнения внутри импульса, величины ее девиации (изменения) и интенсивности. Создание такого комплекса приемной, измерительной и регистрирующей аппаратуры, отвечающей требованиям обеспечения высокой точности измерения, с минимальными искажениями сигналов, а также простоты и надежности в работе, связано с целым рядом трудностей. Во-первых, диапазон частот, который присутствует в импульсах летучих мышей, почти не применяется для технических задач, в связи с чем необходимая серийная аппаратура отсутствует. Во-вторых, как уже отмечалось во введении, ультразвук затухает в воздухе гораздо быстрее слышимого звука и, чтобы уловить сигналы летучих мышей на расстоянии нескольких метров, приемная аппаратура должна обладать очень высокой чувствительностью в широком диапазоне частот. И наконец, в-третьих, возникает большая сложность в проведении спектрального анализа сигналов летучих мышей, так как последовательность импульсов непериодична, а их
36
параметры изменяются в широких пределах за очень короткий промежуток времени. Следовательно, чтобы изучать эхолокацию у летучих мышей, а также и у других животных, необходимо прежде всего разработать и создать такую аппаратуру, которая удовлетворяла бы отмеченным выше требованиям.
Для того чтобы читатель имел общее представление о методах исследований основных параметров локационных сигналов летучих мышей, мы решили дать описание той регистрационной установки, которая была разрабо-
тана советскими инженерами и в настоящее время используется в наших
экспериментах (Макаров, 19736). Блок-схема указанного комплекса пред-
ставлена на рис. 3.
Прием ультразвуковых сигналов летучих мышей осуществляется конденсаторным микрофоном (М) с твердым диэлектриком. Диаметр капсюля микрофона составлял 10 мм. В качестве диэлектрика использовалась металлизированная с одной стороны мила-ровая пленка толщиной 15 мкм. Конденсаторный микрофон имел равномерную частотную характеристику в диапазоне 20—200 кгц с чувствительностью 1.7 мв/н«м2 + 1 дб. Нижний уровень его динамического диапазона определяется собственными шумами усили-
тельного тракта, а верхний
Рис. 3. Блок-схема установки, предназначенной для регистрации и анализа частотно-временных параметров ультразвуковых локационных сигналов.
А — блок-схема аппаратуры для записи сигналов на магнитофон (МУ-25) и кинорегистрации с экрана осциллографа О К-17. Внизу — осциллограммы 2 импульсов, пересеченные линией частотомера, по которой производился подсчет длительности и частоты заполнения. Метки через 10 мсек, служили для отсчета интервалов между импульсами. Б — блок-схема аппаратуры, используемой для обработки сигналов при их трансформировании в низкочастотный диапазон.
уровень составлял 167 дб. В ряде случаев для приема сигналов подковоносых летучих мышей использовался микрофон, имеющий резонансную частотную характеристику с максимальной чувствительностью 4.3 мв/н«м2 на частоте 80 кгц. Кроме того, иногда применялся микрофон, имеющий наибольшую чувствительность (7.8 мв/н-м2) в низкочастотной части ультразвукового Диапазона.
м С микрофона электрические сигналы по кабелю поступают на микрофонный усилитель (МУ). Для согласования капсюля конденсаторного микрофона, имеющего большое внутреннее сопротивление, с микрофонным кабелем использован предусилитель (КП), построенный по схеме катодного повторителя. Напряжение поляризации конденсаторного микрофона и питание предусилителя подавались от блока поляризации (БП), построенного по схеме стабилизированного выпрямителя.
4-КаскаДный широкополосный МУ имел линейную частотную характеристику с неравномерностью 3 дб в диапазоне 20—250 кгц. Коэффициент усиления 74 дб (5 -103 раз). Уровень собственных шумов усилителя равнялся 5 мкв. Коэффициент нелинейных искажений < 1 %.
Для определения частоты заполнения исследуемые сигналы через сш> тему фильтров (ФВЧ) подавались на измеритель девиации частоты (ИДЧ). ИДЧ измерял частоту заполнения в пределах импульса в каждый момент
37
времени, а показания его регистрировались на экране осциллографа одновременно с исследуемым сигналом. Принцип действия прибора основан на преобразовании частотной модуляции в амплитудную, т. е. на преобразовании высокочастотных колебаний частотно-модулированного (ЧМ) сигнала в последовательность импульсов, модулированных по амплитуде в соответствии с периодом высокочастотного заполнения. После детектирования такой последовательности импульсов на выходе детектора получается видеоимпульс, амплитуда которого в каждый момент времени пропорциональна периоду или обратно пропорциональна частоте заполнения в пределах исследуемого сигнала.
Измерение параметров локационных сигналов осуществлялось методом кинорегистрации сигналов с экрана двухлучевого осциллографа (ОК-17 или С1-12). Для этого на один луч подавался сигнал с выхода ИДЧ, а на другом луче воспроизводился сам измеряемый импульс. Появляющееся на экране осциллографа в каждый момент приема импульса совмещенное изображение самого сигнала и кривой его частотного заполнения регистрировалось на киноленту 35 мм кинокамерой (АКС-2).
Для определения длительности импульсов кривая частотного заполнения на экране осциллографа была промодулирована по яркости калиброванными метками с периодом 0.2 мсек. (рис. 3). Частота повторения импульсов определялась по яркостным меткам, воспроизводимым с периодом 10 мсек, по краю пленки. Определение частоты и величины девиации сигналов производилось по калибровочной шкале, нанесенной на экран осциллографа, где величина вертикального отклонения луча с сигналами ИДЧ отградуирована в соответствующих частотах.
Фоторегистрация сигналов с экрана осциллографа производилась киноаппаратом, работающим со скоростью 1 м/сек. в режиме непрерывной протяжки пленки. Съемка сигналов гладконосых летучих мышей, которые по длительности не превышают 10 мсек., производилась при развертке луча навстречу движению пленки. Это позволяло получить хорошее разрешение огибающей регистрируемых сигналов. Развертка осциллографа при этом работала в ждущем режиме и запускалась с приходом каждого исследуемого импульса. Для сигналов подковоносов съемка производилась без развертки луча осциллографа. Развертка сигнала на кинопленке осуществлялась только за счет ее движения, что при скорости съемки 2 м/сек. также давало хорошее разрешение сигнала.
В большинстве случаев в ходе опытов вместо киносъемки локационных сигналов летучих мышей с экрана осциллографа производилась только запись их на магнитную ленту. Для этого использовался серийный ультразвуковой магнитофон типа МУ-25 с полосой пропускания от 10 до 100 кгц, имеющий скорость протяжки пленки 152 см/сек.
Как показали наши измерения, магнитофон МУ-25 позволял записывать сигналы в диапазоне частот от 10 до 150 кгц с ослаблением верхних частот порядка 0.6 дб на каждые 10 кгц. При дальнейшей обработке записей отбирались наиболее характерные последовательности локационных сигналов, которые анализировались путем съемки их с экрана осциллографа вышеизложенным методом.
В последнее время для пересъемки коротких сигналов мы чаще стали пользоваться другим методом. Записи, полученные на МУ-25, воспроизводились со скоростью 15.2 см/сек. на магнитофоне МАГ-59. При этом частотный спектр импульсов трансформировался в 10 раз и укладывался в диапазон частот МАГ-59 — от 60 до 16 000 гц. С выхода магнитофона сигналы подавались на вход двухлучевого осциллографа С1-12 и ИДЧ. Измерение параметров сигналов производилось по изображению импульсов и кривой изменения частоты заполнения, снятому с экрана осциллографа на кинопленку фотоприставкой РФК-5.
Следует отметить, что при развертывании импульса на экране осциллографа неизбежно возникает потеря его переднего фронта, в результате чего часто возникают ошибки при измерении частоты к начале сигнала и его длп-38
ТелЬности. Чтобы устранить это явление, необходимо Задержать исслеДуё-мый сигнал по отношению к началу развертки. С этой целью на магнитофоне МАГ-59 были установлены две воспроизводящие головки с расстоянием 15 мм между ними, подключенные к 2 усилителям воспроизведения. Первая головка служила для запуска ждущей развертки осциллографа, вторая — для воспроизведения исследуемого сигнала.
При движении магнитной ленты возникала задержка сигнала на 80 мсек, относительно запускающего импульса. Запуск развертки осциллографа
также осуществлялся с задержкой после прихода запускающего импульса. Запускающие импульсы, кроме того, управляли сменой кадров в фотоприставке РФК-5 перед приходом каждого исследуемого сигнала. Таким образом, после пуска магнитофона дальнейшая фотосъемка каждого импульса с экрана осциллографа производилась автоматически. Необходимо также коротко остановиться на особенностях работы ИДЧ в
Рис. 4. Измерение частоты сигнала искаженной формы.
То — период основного тона; Ди/, — частоты, соответствующие Tt и Т2.
случае измерения частоты в сложных сигналах. Так как принцип работы ИДЧ основан на измерении периода синусоидального колебания, то при наличии гармонических составляющих, когда форма синусоиды будет искажена, показания ИДЧ могут не соответствовать частоте основного тона.
На рис. 4 показано, как происходит измерение частоты сигнала, когда в результате сильного искажения формы синусоиды волна дважды переходит через нулевую линию. ИДЧ в этом случае измеряет отдельно частоты fi и /2, соответствующие периодам и Т2. При изменении амплитуды и фазы гармонических составляющих на выходе ИДЧ появляются две линии частотного заполнения, так как соотношение соответствующих им периодов Т\ и Т2 меняется. Период основного тона То в этом случае равен сумме и Т2, а его частота /0 рассчитывается по формуле
/1 А + h
и в случае равенства Тг и Т2
/о —
А.
2 ’
Именно так определялись основные частоты сложных импульсов летучих мышей.
Эхолокация у примитивных летучих мышеи
Самым древним семейством среди летучих мышей считаются ланцетоносы, или мышехвосты (Rhinopomatidae). В определенном смысле они представляют как бы связующее звено с крыланами. Эти маленькие (длина тела 6—8 см) светло
39
серые зверьки отличаются от других летучих мышей длинным и тонким, свободным от межбедренной перепонки хвостом (рис. 5). Межбедренная перепонка недоразвита и лишь узкой каймой охватывает задние конечности животного. Крупные уши соединены между собой кожистой складкой по верху головы. На конце морды имеется небольшой округлый валик, который рассматривают как зачаток носового листка (Grasse, 1955).
Рис. 5. Ланцетоносы, или мышехвосты (род Rhinopoma). (Брем, 1893 г.).
Мышехвосты обитают в пустынных и полупустынных областях и распространены в Северной Африке и Южной Азии. Днем держатся в пещерах, расщелинах скал и зданиях, собираясь часто большими колониями, ночью добывают крупных летающих и нелетающих насекомых. В составе единственного рода выделяют 4 вида. Сведения об эхолокации этого интереснейшего с эволюционной точки зрения семейства ограничиваются лишь фрагментарным описанием характеристик локационных сигналов у одного вида — Rhinopoma microphyllum. Отмечено, что животные излучают через рот импульсы длительностью в среднем 4 мсек. Импульсы становятся более интенсивными при взлете и во время полета, причем частота их следования очень высокая — от 130 до 300 имп./сек. Сигналы не имеют частотной модуляции и спектр их простирается до 60 кгц (Mohres, Kulzer, 1955b).
Семейство щелемордов (Nycteridae) распространено в Африке и Южной Азии. Размеры мелкие и средние, длина тела 45—75 мм.
40
Название семейства обусловлено тем, что морда этих животных разделена продольной бороздой, от ноздрей назад по верху головы. Носовые листки в виде запятых изолированы друг от друга (рис. 6, а). Зверьки этого семейства обитают в сухих степях с одиночными деревьями, но встречаются и в дождливых тропических лесах. На день забираются в пещеры, здания, развалины, норы дикообразов и трубкозубов.
По относительно медленному полету и вылету на охоту за насекомыми в дневное время их легко отличить от других видов. Охотятся они, обычно летая вдоль стен домов, вокруг стволов деревьев, вблизи скал и ловят сидящих или медленно летающих насекомых.
Рис. 6. Nycteris macrotus (а) и Megaderma lyra (б). (Dobson, 1876).
Излучение локационных сигналов осуществляется не через рот, а через ноздри. Новик (Novick, 1958а) исследовал локационные сигналы у 3 видов: Nycteris grandis, N. arge, N. bispida — и показал, что они имеют очень близкие характеристики. Длительность сигналов короткая и варьирует в небольших пределах — от 0.4 до 1.4 мсек. Диаграмма частот состоит из основной частоты и сопровождающей ее 2—5-й гармоник. Амплитуды гармонических составляющих по ходу импульса изменяются. Общий спектр импульса 16—110 кгц. Основная частота соответствует примерно 20 кгц и изменяется у разных представителей от 18 до 24 кгц. Соответственно 2-я гармоника составляет 36— 48 кгц, третья — 54—72, 4-я — 72—96 кгц. В типичных импульсах основная энергия приходится на частоты около 40, 60 и 80 кгц, тогда как 1-я и 5-я гармоники очень слабые. Сигналы Nycteris имеют небольшую интенсивность — максимум 50 дин/см2 на расстоянии 5 см от плоскости носовых листков. Отмечены выраженная направленность излучения сигналов и полное отсутствие частотной модуляции.
41
Семейство Megadermatidae — ложные вампиры — тесно связано родственными отношениями с представителями предыдущего семейства, но отличается формой ушных раковин, сросшихся посредине, копьеобразным носовым листком и отсутствием хвоста. У большинства форм хорошо развиты глаза, что может свидетельствовать о важной роли зрительного анализатора при пространственной ориентации этих животных. Подобно Nycteridae, они медленно летают вдоль стен построек и скал, охотясь в основном за неподвижными или малоподвижными насекомыми и мелкими позвоночными, такими как лягушки, ящерицы, мелкие птицы и даже летучие мыши. Megadermatidae широко распространены в Африке и Юго-Восточной Азии. Типичный представитель этого семейства — Megaderma lyra (рис. 6,6) —-наиболее хорошо изучен с точки зрения эхолокационных возможностей (Mohres, Kulzer, 1957; Novick, 1958а; Mohres, 1967а).
Длительность излучаемых сигналов очень короткая —- 0.4— 1.8 мсек. Импульсы шумоподобного типа с низкой основной частотой — 17.0—19.6 кгц, которая остается постоянной на протяжении всего импульса, и наличием гармоник 2—4-го порядков. Интенсивность 3-й и 4-й гармонических составляющих, т. е. на частотах примерно 58.5 и 78.5 кгц, максимальна, тогда как 1-я гармоника очень слаба и часто вообще не обнаруживается.
Отмечено, что более длинные импульсы, как правило, имеют более низкий частотный диапазон (до 3-й гармоники включительно), чем короткие сигналы.
Все авторы единодушно отмечают, что общая интенсивность излучаемых импульсов у Megaderma очень слабая и их трудно записывать в полете. Новик оценивает общую интенсивность сигналов этого вида максимальными значениями 24—25 дин/см2 в 7 см от морды животного, тогда как Мёресу не удалось получить интенсивность более 1.0—1.6 дин/см2.
Излучение локационных сигналов у Megaderma осуществляется через ноздри направленным, сфокусированным пучком, и величина диаграммы направленности порядка 70° на уровне 0.5.
Частота повторения сигналов изменяется в зависимости от состояния животных. Если сидящая летучая мышь концентрирует внимание на каком-либо объекте, частота следования импульсов в среднем составляет 38—52 имп./сек. и в течение короткого периода иногда достигает 300 и более. Когда животное не проявляет большого интереса к окружению, частота следования импульсов не более 16—18 в секунду. Слабая интенсивность сигналов и относительно высокая частота их следования дают основания предполагать, что дальность действия локационной системы Megaderma, по-видимому, небольшая и не превышает 1—2 и.
А2
При охоте за неподвижными или малоподвижными крупными насекомыми или мелкими позвоночными такой диапазон действия эхолокации кажется вполне пригодным. В поисках добычи Megaderma обычно летает вдоль крупных объектов и хватает насекомых или позвоночных со стен домов и стволов деревьев. В таких условиях постоянная близость отражающей поверхности должна была бы вызывать перекрытие отраженного сигнала излучаемым импульсом, если длительность последнего не укоротить до минимума, что и наблюдается у этих летучих мышей.. Действительно, при длительности импульса 0.8 мсек, он распространяется на расстояние примерно 27 см, что позволяет животному летать в 13.5 см от объекта и не иметь перекрытия между посылкой и эхом. На таких расстояниях нецелесообразно использовать сигналы большой интенсивности, так как животное будет постоянно оглушать себя как собственным, так и отраженным сигналами; напротив, при использовании низкоинтенсивных импульсов легче отличить тонкие нюансы полезного эха. Кстати, как мы увидим дальше, все летучие мыши, подлетая к насекомому или препятствию на близкое расстояние, резко снижают интенсивность излучаемого сигнала.
У другого вида — Megaderma spasma, обитающего на Филиппинских островах, основные параметры сигналов оказались очень близки к описанным выше.
Мы видим, что локационные системы рассмотренных двух семейств этой группы очень близки по параметрам сигналов к представителям ланцетоносов. Те же шумоподобные импульсы с низкими основными частотами без признаков частотной модуляции, осложненные несколькими стабильными гармониками, используются этими животными для эхолокации, но в то же время здесь проявляются определенные черты специализации — импульсы становятся ультракороткими и резко снижаются по интенсивности.
Представители семейства мешкокрылов (Emballonurtdae) имеют мелкие и средние размеры (длина тела 3.7—10 см). В отличие от ланцетоносов они не имеют носового листка и большая часть хвоста у них включена в межбедренную перепонку, однако кончик его свободно располагается над этой перепонкой. Большинство видов в области плеча на летательной перепонке имеет специфическую кожную железу в виде своеобразного железистого мешка, который открывается на верхней поверхности крыльев. Отсюда и произошло название семейства. Мешок хорошо развит у самцов и выделяет красноватый секрет с сильным запахом.
Мешкокрылы обитают в различных ландшафтах тропических и субтропических областей Старого и Нового Света. Некоторые авторы считают, что мешкокрылов в экологическом и функциональном отношении следует рассматривать как своеобразный субстрат, на котором происходило формирование более молодых
43
групп летучих мышей обоих полушарий (Симкин, 1971). Мешкокрылы питаются в основном насекомыми, при этом наряду с эхолокацией важную роль в процессе ловли играет зрение.
Характеристики локационных сигналов исследованы у 6 видов следующих родов: у могильных мешкокрылов (Thaphozous и Emballonura) из Старого Света; балантиоптериксов (Balantiopte-гух), обыкновенных (Saccopteryx) и хоботковых (Rhinchonycteris) мешкокрылов из Нового Света.
Мёрес и Кульцер (Mohres, Kulzer, 1955b) дали следующие характеристики сигналам Thaphozous: длительность — 4.2 мсек., частота повторения — 60—160 в секунду, частотный спектр —
Рис. 7. Часть локационного сигнала Thaphozous, показывающая сложную рнутриимпульсную его структуру (Novick, 1958а).
25—80 кгц. Более детально исследование импульсов 2 видов этого рода провел Новик (Novick, 1958а). Импульсы Th, melano-pogan имели основную частоту в пределах 11—16 кгц, причем к концу сигнала наблюдалось небольшое падение частоты на 2— 3 кгц. Обычно уже в самом начале импульса появлялись гармонические составляющие 2-го и 3-го порядков, а в конце присутствовала дополнительно 4-я гармоника (рис. 7). Таким образом, частотный спектр сигналов был широким — от 11 до 68 кгц.
Амплитуды гармонических составляющих были всегда ниже основной частоты. Абсолютная интенсивность основной частоты в 7—15 см от рта животного была определена как 1650—5600 дин/см2, а 2-й гармоники — 440—930 дин/см2.
Параметры локационных сигналов у другого вида — Th, philtp-pinensis оказались близкими к вышеописанным. Длительность в среднем равнялась 4.6 мсек. (6.9—2.9 мсек.), основная частота изменялась от начала к концу импульса от 16—14 до 13—12 кгц и сопровождалась гармоническими составляющими 2—4-го порядка. Интенсивность сигналов была также высока — 1500—
44
3000 дин/см2. Следовательно, эти летучие мыши могут быть отнесены к разряду наиболее «громких» рукокрылых, как мы увидим дальше при сравнении с другими летучими мышами. Животные излучали локационные сигналы через рот.
У одного из представителей американских Emballonuridae характеристики сигналов оказались очень близкими к таковым у Thaphozous. Импульсы Balantiopteryx pltcata в полете варьировали по длительности от 2.6 до 4.2 мсек. Основная частота в начале импульсов была 17—21 кгц и понижалась к концу до 14— 17 кгц. Всегда присутствовали три гармонические составляющие, так что спектр сигналов достигал 60 кгц. Однако основная энергия была сосредоточена на низких частотах — 1-я и 2-я гармоники (Novick, 1962).
Локационные сигналы Saccopteryx bilineata исследовались несколькими авторами (Griffin, Novick, 1955; Novick, 1962; Pye, 1967, 1972, 1973; Grinnell, 1970). Несмотря на некоторую противоречивость полученных данных, можно сказать, что эти летучие мыши излучают 2 типа импульсов: один с постоянной частотой заполнения 20—33 кгц и длительностью 3.5—15 мсек.; второй — частотно-модулированный, со слабым изменением основной частоты от 20 до 18 кгц от начала к концу. В том и другом образцах сигналов всегда присутствует еще несколько гармонических составляющих, из которых обычно вторая доминирует. По-видимому, такого же характера сигналы присущи другому американскому виду мешкокрылов — Rhtnchonycteris naso.
Параметры сигналов у вида Emballonura nigrescens из Новой Гвинеи (Grinnell, Hagiwara, 1972а) несколько отличаются от характеристик сигналов видов, описанных выше. Длительность была очень короткой — 0.6—1.3 мсек., основная частота в течение первых 0.25 мсек, оставалась постоянной — 30.5 кгц, а затем в оставшиеся 0.5—0.6 мсек, падала до 18 кгц. В импульсах хорошо выражены 2-я и 3-я гармоники, причем 2-я (61—37 кгц) была более интенсивной. Частота следования импульсов составляла 150 имп./сек.
Итак, локационные сигналы мешкокрылов характеризуются в основном короткой длительностью, низкими основными частотами с присутствием стабильных гармонических составляющих 2-го и 3-го порядков на протяжении всего импульса. Иногда в конце появляется 4-я гармоника. Важно отметить, что в отличие от предыдущих семейств у мешкокрылов частота в пределах импульса редко остается постоянной. Как правило, она изменяется от начала к концу на 3—4 кгц, т. е. здесь впервые появляется слабо выраженная частотная модуляция. Исключительно высокая интенсивность, чрезвычайно быстрый темп и излучение сигналов через рот дополняют общую характеристику эхолокации этих животных.
45
Особенности эхолокации у щелегубов и подковоносов
Специализация акустической локации у представителей этих двух семейств шла по пути удлинения локационных сигналов, расширения участка с постоянной частотой и очищения спектра от дополнительных составляющих с преобладанием 2-й гармоники. В связи с этим обстоятельством мы решили рассмотреть данные семейства отдельно, хотя в систематическом от-
Рис. 8. Представители семейства ложных подковоносов (Hipposideridae). (Grasse, 1955).
а — Triaenops persicus, б — Rhinonicteris aurantia, в — Hipposideros tridens, г — Coelops frithii.
ношении их часто объединяют со щелемордами и ложными вампирами в надсемейство Rhinolophoidea.
Оба семейства — щелегубы, или ложные подковоносы (Hip-posideridae), и подковоносы (Rhinolophidae) — распространены в тропиках и субтропиках Старого Света. Наличие причудливых кожистых выростов на морде, лишенных шерсти (рис. 8,10), широкие листовидные, очень подвижные ушные раковины без козелка, своеобразное строение задних конечностей, непригодных для передвижения по твердому субстрату и служащих лишь
46
для подвешивания животного вниз головой, — вот основные признаки, свойственные представителям этих семейств. Отдельные виды значительно "отличаются друг от друга по размерам (длина тела 2.8—11 см), окраске, форме и строению носовых листков. Все летучие мыши указанных семейств питаются летаю* щими насекомыми.
У 14 видов ложных подковоносов исследованы параметры локационных сигналов. Полученные результаты представлены в табл. 2.
В большинстве случаев сигналы были записаны у животных, которых экспериментатор держал в руках, поэтому данные о дли-
Рис. 9. Спектрограммы локационных сигналов у разных видов ложных подковоносов. (Grinnell, Hagiwara, 1972а).
тельности сигналов следует относить к данной конкретной ситуации. Разброс этих данных значителен — 2.7—24 мсек.
Частота заполнения от начала сигнала была постоянна и варьировала у разных особей одного и того же вида в довольно широких пределах — от 56 до 150 кгц (рис. 9). Значения частот, приведенные в табл. 2, представляют собой 2-е высокоамплитудные гармоники импульсов, тогда как основные частоты (1-е гармоники) представлены в виде волн очень низкой амплитуды и не всегда обнаруживаются. В конце каждого импульса наблюдается понижение частоты, т. е. появляется частотно-модулированный участок. Величина модуляции у одних видов не превышает 2—6 кгц, у других достигает 40—50 кгц. Частота следования импульсов обычно составляла 50—75 в секунду. Во время полета отмечено снижение частоты постоянной составляющей на 1—2 кгц по сравнению с импульсами, излучаемыми животными в неподвижном состоянии. Сигналы излучаются животными через ноздри.
Сведения об акустической ориентации подковоносов появились в 50-х годах текущего столетия благодаря работам немецкого исследователя Мереса (Mohres, 1951, 1952, 1953). По Мёресу генерируемые "подковоносами сигналы превосходили по длитель-
47
Род и вид
Hipposideros
. diadema...............
. lankadiva ............
. commersoni . . . .
. brachyotus............
. obscurus..............
. speoris...............
. beatus ...............
. cyclops ..............
. cupidus...............
. calcaratus . . . .
. galeritus ............
. caff er...............
Asellia
A. tridens................
Aselliscus
A. tricuspidatus . . . .
fccjfcqtq taq taq taq tq
Таблица 2
Характеристики локационных сигналов ложных подковоносов
Длительность сигнала, мсек. Постоянная частота, кгц Заключительная частота, кгц Длительность ЧМ-участка Литературный источник
5.8-10.1 58—58.5 47 1.0 Grinnell, Hagiwara, 1972а
8-14 57-74 55-67 — Novick, 1958а
12—23.5 56-68 50—66 3—3.5 Pye, Roberts, 1970; Pye, 1972; Roberts, 1972
5-8 85—93 — — Novick, 1958a
8-10 95-109 — — Там же
6-10 100-120 — — » »
7-8 100-131 — — » »
17—24 101—109 — — » »
3-3.6 123—124 97 0.8 Grinnell, Hagiwara, 1972a; Roberts, 1972
4.4—5.6 128-129 88 1.0 Там же
2.9—4.6 144—145 120 1.0 » »
6—23 140-150 100 — Pye, Roberts, 1970; Roberts, 1972
9-10 120 50-70 Mohres, Kulzer, 1955a
7.8 115—120 95 1.0 Pye, Roberts, 1970
2.7—3.5 112 94 1.1 Grinnell, Hagiwara, 1972a
ности импульсы известных в то время гладконосых летучих мышей в 20—30 раз. Средняя продолжительность импульса большого подковоноса составляла 65.3 мсек., а во время полета животного регистрировались импульсы длительностью 90—110 мсек. В импульсах подковоносов отсутствует столь характерная для
Рис. 10. Большой подковонос (Rhinolophus ferrum-equinum).
(Фот. Г. Ю. Андреевой).
гладконосых частотная модуляция. Частота в пределах импульса постоянна и равняется у большого подковоноса 80 кгц, у малого подковоноса — 100 кгц. Частота следования импульсов состав ляет у сидящего животного 4 имп./сек., в полете — 5—6 имп./сек. Темп излучения сигналов тесно скоординирован с дыханием, и каждый выдох дает только один импульс. Мёресу не удалось измерить звуковое давление излучаемых подковоносами сигналов,
49
и он ограничился лишь общим выводом о том, что интенсивность излучения у подковоносов значительно выше, чем у исследованных в то время гладконосых летучих мышей.
Однако исследования Мереса из-за несовершенства регистрирующей аппаратуры и некоторых других причин оказались неполными и характеризовали только часть эхолокационного процесса подковоносов. Новик (Novick, 1958а) при анализе импульсов четырех тропических видов подковоносов отметил значительную вариабельность таких параметров, как длительность и интервал между импульсами. Им же было показано, что частота заполнения импульсов, оставаясь постоянной в средней части, меняется в терминальных участках каждого сигнала на несколько килогерц.
Гриффин (Griffin, 1962) обратил внимание на то, что одиночные сигналы большой длительности, описанные Мересом как единственные для эхолокации подковоносов, наблюдаются лишь в тех случаях, когда подковонос совершает спокойный курсирующий полет. Если же во время полета животное обнаруживает препятствие, частота следования импульсов увеличивается, а их длительность и интервалы между ними сокращаются. В этом отношении режим работы эхолокатора подковоноса приближается к тому, который наблюдается у гладконосых летучих мышей при подлете к препятствию или ловле насекомых.
Дальнейшие тщательные исследования эхолокационного процесса у нескольких видов подковоносов выявили два различных режима функционирования их акустических систем (Schnitzler, 1967а, 1967b, 1968; Айрапетьянц, Константинов, Матюшкин, 1968, 1969; Константинов, Соколов, 1969; Соколов, 1972а, 1973). Из шести видов подковоносов, отмеченных для фауны СССР, в нашей лаборатории были записаны и проанализированы локационные сигналы у пяти: малого (Rhinolophus hipposideros), Блаузиуса (Rh. blasii), Мегели (Rh. mehelyi), бухарского (Rh. bocharicus) и большого (Rh. ferrum-equinum).
При обзорной локации во время полета в просторном, уже знакомом помещении с места подковоносы обычно излучают длинные одиночные сигналы. Как показывают результаты измерений, длительность одиночных сигналов у подковоносов разных видов по средним показателям укладывается в диапазон 34— 50 мсек., но может варьировать в широких пределах — от 15 до 95 мсек. При этом у большого и бухарского подковоносов сигналы, как правило, длиннее, чем у других видов.
Интервалы между импульсами показывают еще большие изменения, чем длительности сигналов, и значения их зависят от состояния животного и окружающей обстановки. Спокойно висящий на одном месте подковонос излучает сигналы очень редко, через 200—300 мсек., но если его беспокоит какой-то посторонний шум или движение какого-либо предмета, одиночные сигналы следуют с интервалом 50—80 мсек. Такой же темп из
50
лучения сохраняется и в полете. Типичные осциллограммы одиночных сигналов разных видов представлены на рис. И.
Второй режим работы эхолокаторов подковоносов наблюдался при сближении животных с препятствием или при исследовании незнакомых предметов. В этих случаях вместо одиночных продолжительных сигналов все изученные виды подковоносов излучали серии, или пачки, более коротких импульсов (2—20 импульсов в каждой; рис. 12). Интервалы между пачками, состоящими из 2—3 импульсов, сохранялись примерно такими же, как между одиночными сигналами, но с увеличением количества импульсов в пачке они уменьшались.
Излучение каждой пачки независимо от ее длительности соответствовало выдоху животного, интервал между ними — вдоху. Импульсы в пределах каждой пачки отделялись друг от друга короткими паузами. G увеличением количества импульсов в пачке длительность их, равно как и пауз между ними, закономерно уменьшается, тогда как общая продолжительность самих пачек неуклонно возрастает.
Таблица 3
Характеристики локационных сигналов подковоносых летучих мышей (род Rhinolophus)
Вид Длительность импульсов, мсек. Частота заполнения средней части импульса, кгц Частота в конце импульса, кгц Интенсивность, Дб Литературный источник
Rh. arcuatus . . 17-34 55-69 51 Novick, 1958а
Rh. rouxi . . . 21-53 60—76 60 130 Там же
Rh. landeri . . . 14-72 115-122 80 — Pye, Roberts, 1970
Rh. subrufus . . 15—60 62—82 51 134 Novick, 1958а
Rh. hipposideros . . 16-62 116 80 94 Константинов, Соколов, 1969
Rh. mehelyi . . 20-90 НО 80 НО Там же
Rh. ferrum-equinum 18-95 81-83 67 120 » »
Rh. blasli . . . 15—65 91-93 —. 118 Соколов, 1972а
Rh. bocharicus . . 20—65 101-103 — 111 Там же
Rh. euryale . . 10-45 103-104 — 100 Schnitzler, 1968
Rh. fumigatus . . — 45-50 33 15-65 Roberts, 1972; Pye, Roberts, 1970
Rh. luctus . . . 30-42 42 —. 8-11.5 Roberts, 1972
Rh. alcyone . . 11-60 85—90 69 — Там же
В табл. 3 представлены параметры локационных сигналов у 13 видов подковоносов, полученные разными исследователями. Анализируя эти данные, можно заключить, что в целом у подковоносов наблюдается тенденция к дальнейшему увеличению
51
6
Рис. 11. Осциллограммы локационных сигналов подковоносов.
J — Rhinolophus hipposideros; 2 — Rh. mehelyi; 3 — Rh. bocharicus; 4, 5 — Rh. blasii; 6 — Rh. f er rum-equinum. Слева в каждом ряду — осциллограмма полного импульса и липин частотомера. По вертикали — частота, кгц. Период между калибровочными метками над каждой осциллограммой 20 мсек. Справа в каждом ряду — часть локационного сигнала при замедлении скорости воспроизведения в 128 раз.
Рис. 12. Локационные сигналы большого подковоноса, излучаемые в режиме одиночных имт гльсов и пачек.
Верхний ряд — последовательность одиночных импульсов; средний — последовательность пачек, состоящих из 2 импульсов; нижний ряд — учащение импульсов при облете препятствия. Линия над каждым импульсом показывает характер изменения несущей частоты. Период между калибровочными метками 10 мсек.
длительности сигналов, которая в одиночных импульсах при обзорной локации часто составляет более 50 мсек,, достигая величины 70—90 мсек., и даже при приближении животного к препятствию или насекомому не падает ниже 10—15 мсек.
Локационные сигналы подковоносов, как правило, заполнены одной высокой частотой, которая на протяжении большей части импульса сохраняется постоянной и лишь в конце снижается на 5—30 кгц у разных видов. Несомненно, что именно эта частота, значения которой проставлены в табл. 3, наиболее информативна для животного. Она представляет собой вторую гармоническую составляющую от слабой по амплитуде основной частоты. При этом в импульсах большого подковоноса последнюю вообще не удается обнаружить, в импульсах подковоноса Блазиуса она проявляется всегда, а у других видов появление ее нерегулярно.
Частота заполнения средней части импульсов подковоносов чрезвычайно стабильна и служит надежным диагностическим признаком вида. Например, у 8 взрослых особей большого подковоноса ее величина варьировала в пределах 80.6—81.5 кгц, а у 5 особей подковоноса Блазиуса —91.3—91.8 кгц. Интенсивность сигналов подковоносов у большинства видов относительно высокая, но встречаются и слабо интенсивные формы. Как и у щеле-губов, излучение локационных сигналов у подковоносов осуществляется через две ноздри, при этом носовой листок обеспечивает высокую направленность излучаемого ультразвука.
Локационные системы летучих мышей Нового Света
Фауна летучих мышей американских континентов весьма разнообразна. Помимо семейств, распространенных в обоих полушариях, здесь имеются семейства, эндемичные только для Нового Света. Основу фаунистического комплекса летучих мышей тропических районов Южной и Центральной Америки составляет обширная группа американских листоносов (сем. Phyllo-stomatidae). К ней тесно примыкают и, по-видимому, связаны родственными отношениями три семейства — Mormoopidae, No-ctilionidae и Desmodontidae, Систематическое положение первого из них до последнего времени оставалось неясным. Одни систематики (Miller, 1907; Simpson, 1945) относили его в ранге подсемейства Chilonycterinae к семейству американских листоносов, другие (Кузякин, 1950; Dalquest, Werner, 1954) рассматривали ►его отдельным семейством Chilonycteridae. Недавно была проведена тщательная ревизия представителей этой группы и их латинских названий (Smith, 1972). В результате все виды рассматриваемой группы были объединены в семейство Mormoopidae, по на
54
званию наиболее типичного рода Mormoops. Кроме указанного, в составе семейства был выделен еще один род — Pteronotus — с тремя подродами.
Не менее сложно обстояло дело и с семейством Noctilionidae. При одной из первых классификаций (Miller, 1907; Weber, 1928) его вместе с мышехвостами и мешкокрылами выделили в наиболее примитивное надсемейство Emballonuroidea, затем Симпсон (Simpson, 1945) перебросил его к гладконосым и буль-договым в надсемейство Vespertilionoidea. По-видимому, наиболее правильно размещение этого семейства в группе Phyllostomato-idea, как было сделано позднее другими систематиками (Bour-liere, 1955; Smith, 1972). Смит (Smith, 1972) полагает, что семейство Noctilionidae ближе всего стоит к Mormoopidae, но более специа л изиров ано.
Третье семейство — Desmodontidae — тесно связано по морфологическим признакам с американскими листоносами и нередко включается в него как подсемейство. Близки к американским листоносам и три небольших семейства американской фауны — Natalidae, Furipteridae и Thyropteridae.
Коротко остановимся на способах добывания пищи и особенностях эхолокации у исследованных представителей семейств рассматриваемой группы. Основные параметры локационных сигналов у 28 видов сведены в табл. 4.
Наиболее примитивным семейством в группе считается Мог-moopidae. Эти летучие мыши, как и подавляющее большинство рассмотренных выше семейств Старого Света, питаются летающими насекомыми. Локационные сигналы ими издаются через рот, причем губы складываются в своеобразный рупор. Как видно из табл. 4, у всех представителей рода Pteronotus условное обозначение характера изменения частоты в пределах каждого импульса (сигнальная формула) представляет собой ПЧ/ЧМ, т. е. частота в начале сигнала остается постоянной (ПЧ), а затем с середины или в конце его обнаруживается падение частоты на 2-—15 кгц (частотно-модулированный, или ЧМ-участок). По этому признаку сигналы трех рассматриваемых видов напоминают импульсы Saccopterix (из мешкокрылов). Длительность импульсов при обзорной локации относительно короткая — 2—8.6 мсек., частота вначале (менее х/3 длительности) сохраняется постоянной, а затем падает на 7—15 кгц. При сближении с препятствием или ловле насекомых импульсы укорачиваются и становятся исключительно частотно-модулированными за счет редукции передней части (рис. 13).
Только у одного представителя — Pteronotus parnellii rubiginosus сигналы значительно длиннее, до 30 мсек., и по частотным характеристикам поразительно напоминают импульсы подковоносов. При сближении с добычей сигналы становятся короче, но участок с постоянной частотой всегда сохраняется (рис. 14).
55
Таблица 4
Характеристики локационных сигналов 5 семейств летучих мышей Нового Света
Семейство и вид Тип питания Сигнальная формула Длительность, мсек. Частотный диапазон (/н~/к), кгц Литературный источник
Mormoopidae
Pteronotus parnellii rubiginosus (Chilonycteris rubiginosa) Насекомоядный пч/чм 13-30 32—28 (1-я) 64—56 (2-я) Griffin, Novick, 1955; Novick, 1971; Grinnell, 1970; Schnitzler, 1970a, 1970b
P. personatuspersonatus (Ch. per-sonata) » » 2.8—8.6 53—38 (1-я) 106—76 (2-я) Griffin, Novick, 1955; Novick, 1963a
P. personatus psilotis (Ch. psi-lotis) » » 3—5 21—17 (1-я) 42-34 (2-я) Novick, 1965, 1971
P. davyi » » 1—5 39-31.5 (1-я) 78-63 (2-я) Novick, 1963a, 1965, 1971; Grinnell, 1970
P. suaprensis » » 1.5—2.2 52—42 (2-я) Grinnell, 1970
Mormoops megalophylla » чм 4—5 48—37 Pye, 1967
Nоctil ionidae
Noctilio leporinus Рыбоядный пч/чм 3—13.8 58-30 (в конце 2-я) Suthers, 1965
N. labialis minor Насекомоядный чм? 4.1—13 62-41 (в конце 2-я) Griffin, Novick, 1955
Семейство и вид Тип питания Сигнальная формула
Phyllostomatidae
Macrotus mexicanus Насекомоядный ЧМ
Phyllo&tomus hastatus Плодо- и плото- »
ядный
Ph. discolor То же »
Phylloderma stenops » » »
Carollia perspicillata Плодоядный »
Centurio senex » »
Artibeus cinereus » »
A. jamaicensis » »
A. lituratus » »
Chiroderma villosum » »
Таблица 4 (продолжение)
Длительность, мсек. Частотный диапазон (/н —/к), кгц Литературный источник
2.0—3.7 38—28 (1-я) 76—56 (2-я) Novick, 1963а
0.5-4 65—25 Grinnell, 1970; Simmons, 1971а, 1971b; Peff, Simmons, 1972
0.5-4 50-25 Pye, 1967
0.8—2 70-35 Там же
0.5-1 40—22.5 Griffin, Novick, 1955;
(1-я) 80—55 (2-я) Pye, 1967; Grinnell, 1970
2 115-70 (2-я) Pye, 1967
— 90—60 (2-я) » »
2.7—3.2 53-32 (1-я) 106—56 (2-я) Novick,*!963a
1-3 65—42 Pye*1967
— 68—40 Там же
1-1.5 102-60 » »
Таблица 4 (продолжение)»
Семейство и вид Тип питания Сигнальная формула Длительность, мсек. Частотный диапазон (/н —/к), КГЦ Литературный источник
Anoura geoffroyi Нектароядный ЧМ 0.5-2 100-50 Руе, 1967
Glossophaga soricina » » 0.7—1 93—60 Novick, 1963а
G. longrostris » » 0.5-2 112-56 Руе, 1967
Leptonycteris nivalis Нектаро- и плодоядный » 2-8 50—25 (1-я) 100-50 (2-я) Novick, 1963а
Sturnira lilium Плодоядный » 1-1.3 96—50 Руе, 1967
S. tildae » » 1-1.3 96-50 Там же
Vampyrops helleri » » — 108—72 » »
Vampirum spectrum Плодо- и плотоядный » 0.5-1.8 110-65 Bradbury, 1970
Desmodontidae
Desmodus rotundus Питается кровью » 0.8-1.6 38—24 (1-я) 75-48 (2-я) Novick, 1963а
N at abidae
Natalus mexicanus Насекомоядный » 2.3—2.5 85-50 (1-я) 170—100 (2-я) Novick, 1962
Примечание. В графе «Частотный диапазон» в скобках указана соответствующая гармоника.
У всех видов в импульсах от начала до конца присутствуют две гармонические составляющие, причем несущей, доминирующей частотой является 2-я гармоника. В конце сигналов часто появляется
гармоническая составляющая.
третья
10 мсек. •------•
локационных сигналов Pteronotus personatus
13.
Рис.
Последовательность j во время сближения с насекомыми. (Novick, 1965).
У вида из другого рода — Mormoops — импульсы, по данным Пая (Руе, 1967), представляют собой ЧМ-сигналы. Однако описание их слишком фрагментарно.
Следующее семейство — N octilionidae — получило широкую известность благодаря тому, что один из двух его представителей — мексиканская рыбоядная летучая мышь (Noctilio lepori-nus) — в основном питается рыбой,
59
Noctilio leportnus распространены в южной Мексике, Центральной Америке, на Эквадоре, Тринидаде, Ямайке, в Бразилии, Панаме и на Кубе. Это довольно крупные летучие мыши, окрашенные в светло-палевые тона, размах их крыльев достигает 60—70 см (рис. 15). Характерная особенность Noctilio, связанная с пищевой специализацией, — своеобразное видоизменение в размере задних конечностей. Каждый палец с когтем на конце удлинен и сплюснут с боков. Все 10 пальцев задних конечностей направлены таким образом, что когда они погружены в воду, то представляют собой пару миниатюрных, хорошо отточенных «острог». С наступлением сумерек Noctilio вылетают из своих дневных убежищ и медленно летают на расстоянии нескольких сантиметров над поверхностью рек, озер или над морем. Когти задних конечностей оттянуты назад так, что «остроги» могут быть пущены в ход в доли секунды быстрым сокращением мышц пальцев задних лап. Пойманная добыча либо съедается на лету, либо откладывается в своеобразные защечные мешки, наличие которых позволяет летучей мыши сразу же продолжать охоту.
Уже первые наблюдения за рыбоядными летучими мышами в естественных условиях показали, что во время ночных охот они непрерывно излучают ультразвуковые сигналы высокой интенсивности (Griffin, Novick, 1955; Griffin, 1958). Сразу возникло предположение, что Noctilio использует эхолокацию для обнаружения рыбы под водой. И хотя с точки зрения физики все противоречило этой гипотезе, так как при переходе из воздуха в воду ультразвук на 99.9% отражается обратно, а 0.1% его энергии, пройдя в глубь воды, при отражении от подводной цели претерпевает точно такие же изменения, в результате чего лишь одна миллионная часть посланной энергии вернется обратно, Гриффин пытался обосновать возможность такого варианта для Noctilio. Ход его рассуждений подробно излагается в 1-м издании нашей книги, но так как последующие эксперименты с животными не подтвердили правильность данной гипотезы, не имеет смысла приводить их здесь.
Эксперименты, проведенные Сатерисом (Suthers, 1965) и позднее подтвержденные нашими исследованиями (Константинов, 1970), убедительно доказали, что рыбоядные летучие мыши не в состоянии обнаруживать свою добычу — мелкую рыбешку, скрытую под поверхностью воды, но четко реагируют на малейшее волнение воды, производимое жертвой, и на выступающие из воды части тела (например, спинной плавник) рыбы. Эхолокаторы этих животных отличаются высокой степенью разрешения малоразмерных целей, расположенных на поверхности воды.
С точки зрения эхолокационного процесса виды этого семейства проявляют большое сходство с представителями предыду-
60
Рис. 15. Рыбоядная летучая мышь (Noctilio leporinus}.
а — внешний вид (фот. А. И. Константинова); б — в полете над поверхностью воды (Slithers, 1965).
щего семейства. Как те, так и другие излучают локационные сигналы через рот, при этом губы вытягиваются, образуя своеобразный рупор, что повышает направленность излучения. Каждый импульс во время курсирующего полета состоит из 2 частей — от начала до середины частота (58—60 кгц) остается постоянной (ПЧ), а затем от середины к концу понижается по линейному закону на октаву до 30 кгц (ЧМ-часть). Наряду с указанными импульсами встречаются сигналы, где частотная модуляция в конце выражена слабее и не превышает 10—15 кгц. При за-
।______।______।______।______1______।
0 10 20 30 40 50 мсек.
Рис. 16. Спектрограммы импульсов Noctilio во время приближения к цели, выступающей из воды. (Suthers, 1965).
хвате рыбы длительность импульсов сокращается за счет редукции моночастотной части и сигналы становятся исключительно частотно-модулированными (рис. 16). В конце импульса всегда присутствует 2-я гармоника, которая так же, как и первая, изменяется по линейному закону.
По-видимому, близкие по частотным характеристикам сигналы использует и другой вид этого семейства — Noctilio la-bialis, который питается исключительно насекомыми.
Отличия локационных сигналов представителей этого семейства от предыдущего заключаются в повышении основной частоты, исчезновении второй гармонической составляющей на протяжении всего импульса, которая появляется лишь в конце, и в значительно большем размахе (до одной октавы) девиации частоты.
Семейство американских листоносов Phyllostomatidae (рис. 17) обширное и очень разнообразное по специализации жизненных форм. Оно включает 6 подсемейств и 49 родов. Американские листоносы, подобно крыланам Старого Света7 в основном при-
62
a
б
Рис. 17. Представители американских листоносов.
а — плодоядные листоносы: слева — Trachy ops, справа — Phyllostomus (Dobson, 1876);
б ~ нектароядные летучие мыши (Cockrum, Hayward, 1962).
способились к растительной пище (сочные плоды, крупные цветы и т. д.). Обилие последней в тропиках Южной и Центральной Америки обусловило пышный расцвет этой группы, несмотря на ряд черт морфологической примитивности. Однако несомненно предками их были насекомоядные формы, о чем свидетельствуют вторичное упрощение зубной системы, строение че-
та
.1
I
Рис. 18. Осциллограммы локационных сигналов ( некоторых представителей семейства Phyllostomatidae, (Novick, 1963а). *
а — локационный импульс плодоядной летучей мыши Artibeus; развертка 1.7 мсек.; основная частота в начале сигнала 56, в конце — 35 кгц. б — импульс нектароядной летучей мыши Glossophaga\ длительность развертки 1.7 мсек.; основная частота в начале сигнала 93, в конце — 57 кгц; 2-я гармоника 157 кгц появляется, когда основная частота падает до 79 кгц. в — импульс нектароядной летучей мыши Leptonycteris; длительность импульса 7.4 мсек.; видна резкая амплитудная модуляция.
репа и пояса передних конечностей. Копьеобразный носовой листок — характерная особенность большинства американских листоносов, но устроен он гораздо проще, чем носовая «насадка» у подковоносов Старого Света. Среди этой группы встречаются формы, которые питаются исключительно плодами фруктовых деревьев, другие — нектаром и пыльцой цветов, раскрывающихся только ночью, третьи фруктовую диету разнообразят мелкими позвоночными. Переход на новый тип питания несомненно отразился и на функционировании эхолокационных систем, которые приобрели черты адаптивных преобразований.
Первые сведения о параметрах сигналов нескольких видов американских листоносов появились давно (Griffin, Novick,
64
1955), однако последующее изучение их с использованием более совершенной аппаратуры позволило значительно уточнить частот* ные параметры этих импульсов (табл. 4).
Все изученные виды рассматриваемого семейства американ* ских листоносов в целях локации используют сигналы корот* кой длительности, в которых основная частота и ее 2-я гармоника плавно изменяются от начала и до конца каждого импульса. В самом конце или начиная с середины сигнала часто появляется
Рис. 19. Вампир обыкновенный {Desmodus rotundus). (Novick, been, 1970).-
3-я гармоника (рис. 18). Характерная особенность сигналов плодо- и нектароядных форм — их очень низкая интенсивность, порядка всего несколько дин/см2. У нектароядных форм пока трудно выявить полный спектр их сигналов, так как технические возможности записывающей аппаратуры ограничиваются в лучшем случае 200 кгц, а кроме того, остается неясной точная величина основных частот.
Многие виды излучают локационные сигналы через ноздри, и развитый носовой листок, вероятно, играет определенную роль в улучшении диаграммы направленности излучения или экранировки ушей от прямого воздействия зондирующего сигнала.
Как уже отмечалось выше, с американскими листоносами тесно связано семейство Desmodontidae — кровососущих летучих мышей, или вампиров, которые распространены в тропиках Центральной и Южной Америки. По образу жизни эти уникальные летучие мыши мало чем отличается от других рукокрылых. Размеры их мелкие, длина тела не превышает 5—8 см. У типич-
65
кого представителя вампиров Desmodus rotundus голова короткая и округленная, уши небольшие, нос широкий, украшенный толстокожим выростом, а нижняя губа рассечена (рис. 19). Поселяются они в темных пещерах, дуплах деревьев или на чердаках домов, часто образуя большие колонии, и с наступлением темноты вылетают на охоту. Однако по способу питания они настолько уклонились в сторону, что не имеют конкурентов не только среди рукокрылых, но даже во всем классе млекопитающих и напоминают скорее некоторых паразитических насекомых.
Как отмечает Ейзентраут (Eisenttfaut, 1957), название «кровососущий» не соответствует способу питания ни одного из видов вампиров, или десмодусов (от латинского названия семейства). Животное не высасывает кровь, так как для этого у него нет никаких специальных приспособлений, а с помощью длинных, острых зубов делает на коже жертвы глубокую рану и вытекающую кровь слизывает языком, при этом, возможно, слегка отсасывает губами. Своеобразно устроенный прикус вампира подобен острым ножницам. С молниеносной быстротой вампир вырезает небольшой участок кожи, в результате чего образуется ранка глубиной около 4 мм, из которой обильно сочится кровь. Питание кровью, естественно, не требует от животного пережевывания и размельчения пищи, поэтому количество зубов редуцируется до 20 и наибольшее развитие получают резцы и клыки.
В основном нападению вампиров подвергаются крупный и мелкий рогатый скот, лошади и мулы, которые в тропиках остаются всю ночь под открытым небом. Но и собаки, мелкие домашние животные и даже птицы также не гарантированы от их нападения. Наконец, в многочисленных сообщениях указывается, что десмодусы «цедят» кровь и у спящих людей. Излюбленным местом являются пальцы ног, по-видимому, в силу того что утомленный человек часто высовывает ноги из-под одеяла. При этом вампиры действуют настолько осторожно, что человек даже не чувствует укуса и не просыпается. Правда, для неслышного приземления на жертву у вампиров имеется специальное приспособление: маленькие припухлости в виде мягких подушечек на концах предплечий и на ступнях задних конечностей.
Сведения об эхолокации этих животных ограничиваются описанием локационных сигналов у одного вида — Desmodus rotundus (Novick, 1963а). Длительность их составляет 1—2.3 мсек. Основная частота в начале импульса соответствует 38—42 кгц и затем постепенно понижается, доходя в конце до 24—29 кгц. Уже в самом начале появляется 2-я гармоника с частотой 76—87 кгц, которая значительно превышает основную частоту по амплитуде. Когда основная частота падает примерно до 31 кгц, возникает 66
3-я гармоника. Таким образом, в начале импульса присутствуют две частоты, примерно 40 и 80 кгц; для середины импульса характерны три частоты — 30, 60 и 90 кгц, и для конца тоже три — 27, 54 и 81 кгц (рис. 20). 3-я гармоника в конце примерно такой же частоты, что и 2-я в начале импульса. Интенсивность излучаемых сигналов была очень слаба. Звуковое давление в нескольких сантиметрах от микрофона составляло 2 дин/см2.
Данное описание указывает, что нет никаких особенностей в импульсах десмодусов, которые отличали бы их от импульсов плодо- и нектароядных листоносов. Это обстоятельство лишний раз подтверждает филогенетическое родство этих двух групп.
Рис. 20. Осциллограммы Ориентационных импульсов Desmodus rotundus, (Novick, 1963а).
Длительность развертки в обойх случаях 1.7 мсек.; основная частота в начале сигнала 42, в конце — 25—28 кгц; в начале сигнала основная частота сопровождается 2-й гармоникой, в середине появляется 3-я гармоника.
В настоящее время ничего не известно о биологическом значении ультразвуковых импульсов десмодусов. Ясно одно, что служат они для эхолокации. Но используются ли они животным для того, чтобы своевременно обнаружить препятствие и уклониться в сторону, или найти нужную жертву, или, наконец, отыскать на жертве то место, где легче всего прокусить кожу и получить достаточное количество крови, — все эти вопросы остаются для будущих исследователей.
Итак, в противоположность ранее распространенному мнению о сложном шумоподобном характере сигналов американских листоносов и десмодусов теперь стало ясно, что локационные сигналы этих животных характеризуются короткой длительностью, присутствием на всем протяжении импульсов двух частотных составляющих, тогда как более высокие гармоники (3-го, иногда 4-го порядка) ограничены заключительными частями импульса. Обращает на себя внимание наличие частотной модуляции в сигналах, величина которой, как правило,
67
не достигает октавы, а также низкая интенсивность излучаемых импульсов.
Семейство Natalidae — небольшое семейство насекомоядных летучих мышей Нового Света. Параметры локационных сигналов исследованы у одного вида — Natalus mexicanus (Novick, 1962). Импульсы, излучаемые этими летучими мышами, в полете имели длительность в среднем 2.3 мсек. (2.2—2.4 мсек.), а в руках экспериментатора — 2.5 мсек. (1.5—2.8 мсек.). Основная частота в начале импульса варьировала от 72 до 91 кгц, но обычно равнялась 85 кгц. Наблюдалась четко выраженная частотная модуляция. В середине импульса частота падала до 53— 56 кгц, а к концу — до 35—42 кгц. В каждом импульсе присутствовала вторая гармоническая составляющая, которая начиналась почти с самого начала импульса и наблюдалась на всем его протяжении. Следовательно, в импульсе всегда присутствуют 2 частоты: в начале 85 и 170, а в конце 55 и НО кгц, причем вторая составляющая имеет более высокую амплитуду. Иногда в заключительных частях импульса появляется 3-я гармоника. По частотному параметру, на наш взгляд, сигналы Natalus близки к импульсам «высокочастотных» Phyllostomatidae. Импульсы излучаются через рот, сведения об их интенсивности отсутствуют.
Локационные системы бульдоговых и гладконосых летучих мышей
Первоначально возникнув в Восточном полушарии, представители этих семейств в настоящее время широко распространены в тропиках Старого и Нового Света, а гладконосые к тому же заняли экологически свободные ниши в областях с умеренным и холодным климатом. Исключительно насекомоядный тип питания, охота за быстролетающей добычей привели к совершенству летательного аппарата, что поставило этих летучих мышей в преимущественное положение перед другими рукокрылыми. Следовательно, мы вправе полагать, что их локационные системы также достигли наибольшего развития.
Семейство Molossidae — бульдоговые летучие мыши, — сохранив ряд черт примитивной организации (как зубная система, сильно развитая малая берцовая кость и др.), достигли наивысшего среди рукокрылых совершенства полета (Кузякин, 1950). По-видимому, последнее обстоятельство обусловило широкое распространение бульдоговых в тропиках Нового и Старого Света. Этим же можно объяснить наивысшую степень развития колониальности у бульдоговых. Их колонии насчитывают сотни и даже миллионы особей. Семейство объединяет 10 родов, из которых наиболее распространенным является род Tadarida, или складчатогубы (рис. 21). У 5 представителей этого рода были исследованы ярка
68
ционные сигналы. Приведенные в табл. 5 данные свидетельствуют о том, что складчатогубы для локации используют в основном короткие сигналы, длительность которых не превышает 10 мсек. Исключение составляла лишь одна летучая мышь, относящаяся к виду Tadarida luzonus, которая в руках экспериментатора излучала импульсы увеличенной длительности — 15—31 мсек. В тех же экспериментах у двух других особей этого вида во время полета были зарегистрированы импульсы не более 10 мсек. (Novick, 1958а). Думается, что сигналы увеличенной длительности
Рис. 21. Складчатогуб (Tadarida), (Novick, been, 1970).
стали результатом неадекватных для записи условий эксперимента или индивидуальными особенностями этой особи.
Частота заполнения сигналов складчатогубов представлена в низком и среднем ультразвуковом диапазоне, со слабо выраженными гармоническими компонентами в конце, которые не поддаются расшифровке. Импульсы имеют частотную модуляцию с падением частоты от начала к концу, величина которой около одной октавы.
Закон изменения частоты в сигнале, исходя из данных Новика (Novick, 1958а), по-видимому, нелинейный. Значительно быстрее частота меняется в первой половине импульса, а затем следует более плавное изменение. Отмечена амплитудная модуляция для многих импульсов складчатогубов. Животные излучают локационные сигналы через рот, и интенсивность их составляет порядка 100 дин/см2 на расстоянии 15 см от рта.
Семейство Vespertilionidae — гладконосые летучие мыши —
69
Таблица 5
Характеристики локационных сигналов летучих мышей семейств Molossidae и Vespertilionidae
Семейство и вид Длительность, мсек. Частота заполнения, кгц Интенсивность, дб Литературный источник
начало сигнала конец сигнала
Molossidae
Tadarida mexicana 3.8 51 35 — Griffin, 1958
3.1 45 25 — Vernon, Peterson, 1965
Т. yucatanica (1.4-4.2)
5.3 32 18 —. Griffin, 1958
Т. tragota 5.8 28 12.5 110—115 Novick, 1958a
Т. midas (3.1—8.1) (22-32) (11—14)
9 22.5 11.5 — Там же
Т, luzonus (6.9—10) 8 (21-25) 43 (10-12.8) 25.5 » »
Vespertilionidae (6.3-10) (35.5-47.5) (25—26)
Myotis lucifugus 2.3 78 39 109 Griffin, 1958
M. kenii 0.9 95 46 — Там же
M. nigricans 1.4 62 48 — » »
M. macrotarsus 2.3—3.7 43-59 23.5-33 130 Novick, 1958a
M. dasycneme 1.1 63 27 115 Константинов, Макаров, 1973
(0.6-1.4) (55-75) (25-30)
M. daubentonii 0.2—2.6 51-42 35-21 — Симкин, Патлякевич, 1969а
1.3 80 33 109 Константинов, Макаров, 1973
(1-1.5) (70-80) (30-35)
0.2—2.6 32—25 22-19 — Симкин, Патлякевич, 1969а
M. bechsteinii 2.5
M. mystacinus (1.0—5.0) 98-55 43—23 — Добрачев и др., 1968; Симкин, Патля-
1.1 87 30 117 кевич, 1969а Константинов, Макаров, 1973
(0.7-1.4) (70-110) (30-40)
0.3-4 120—55 51-27 — Добрачев и др., 1968; Симкин, Патля-
кевич, 1969а
Семейство и вид Длитель- Частота заполнения, кгц
ность, мсек. начало сигнала конец сигнала
М. emerginatus 1.1 (0.8-1.4) 0.35-2.4 130 (125-140) 120-100 60 (55-60) 52-41
М. myotis 1.7 103 33
(1-2.5) (70-130) (25—35)
0.7-3.3 107 43
М. oxygnathus 1.8 120 35
(1-3) (90—140) (25-40)
М. nattereri 1.2 130 30
(0.7-1.7) (110-140) (25-35)
М. capaccinii 1.2-5.6 50-70 30-40
Мiniopterus schreibersii 2.2 115 45
(1.5-3) (110—120) (45—50)
0.8-3.6 130—28 59-16
М. schreibersii escholtzii 1.5-3.7 67—104 52-35
М. australus 1.1—2.8 93—33 17-66
М. tristis 2.9—6.5 58—37.5 30-31
М. fuliginosus 1.9—3.3 — —
М. natalensis 2.7—3.6 103—69 45—50
Pizonyx vivesi 2.5 36 20
Pipistrellus subflavus 2.3 73 45
P. pipistrellus 2.5 116 56
(2-3) (110—130) (55-60)
1.0—3.9 110-80 42—54
P. nathusii 0.89-2.9 90-78 43-30
P. ceylonicus 1.7—3.3 42-31 24—26
P. coramandra 0.8-3.6 76—67 26—29
P, m. mimus 2.2—4.3 73—53 40—30
P, kuhlii 1.5 80 35-45
(0.7-6) (98-65)
P. papuanus 1-2 75 35
Таблица 5 (продолжение)
Интенсивность, дб Литературный источник
— Константинов, Макаров. 1973
— Добрачев и др., 1968; Симкин, Патля-кевич, 1969а
111 Константинов, Макаров, 1973
— Добрачев и др., 1968; Симкин, Патля-кевич, 1969а
111 Константинов, Макаров, 1973; Симкин, Патлякевич, 1969а
112 Там же
— Симкин, Патлякевич, 1969а
— Константинов, Макаров, 1973
138 135 115-120 Добрачев и др., 1968 Novick, 1958а Там же » » » » » »
— Griffin, 1958
НО Там же Константинов, Макаров, 1973
— Симкин, Патлякевич, 1970
95-105 110-115 Там же Novick, 1958а Там же » »
— Симкин, Патлякевич, 1970
— Grinnell, Hagiwara, 1972а
Семейство и вид Длительность, мсек. Частота заполнения, кгц
начало сигнала конец сигнала
Р. savii 1.5 (0.8—2.3) 0.7—2 96 (90-100) 80-62 40 (40-45) 45—30
Eptesicus ognevi 1.5 (1-2) 1.5 (0.7-2.5) 70 (60-80) 75-50 35 (30—35) 33-25
Е. serotinus 1.7 (1.5—2.5) 1.4-2.2 60 (55-70) 65—34 25 (25-30) 23—31
Е. superans 1.3 (0.4—3.5) 50-40 19-33
Е. tenuipinnis 0.9—1.9 62 37
Е, juscus 1—2.7 50 25
Vespertilio nilsonii 2—3 50 (65—45) 27 (22-35)
V. murinus 2 (1.5-3) 0.6-1.6 55 (50-60) 30—53 25 (25-30) 19—26
Nyctalus noctula 2.2 (1-4) 0.4-9 70 (60-90) 46—56 30 (25—35) 25—31
Tylonycteris pachypus 1.5-5.2 80 56—51
Lasiurus borealis 1.9 87 44
Antrowus pallidus 3.2 60 34
Plecotus rafinesquii 1.7 39 27
P, auritus 0.35-1.6 0.5 (0.4-0.6) 45 50 (40—60) 38 25 (24—26)
Barbastella barbastellus 0.38—3.2 53—41 33—28
Murina hilgendorfi 0.35—3.7 107—97 48—47
Таблица 5 (продолжение)
Интенсивность, дб Литературный источник
— Константинов, Макаров, 1973
103 Симкин, Патлякевич, 1970 Константинов, Макаров, 1973
— Симкин, Патлякевич, 1970
— Константинов, Макаров, 1973•
— Симкин, Патлякевич, 1970
— Там же
но Novick, 1958а Griffin, 1958
— Симкин, Патлякевич, 1970
— Константинов, Макаров, 1973* Симкин, Патлякевич, 1970
— Константинов, Макаров, 1973
130—145 Добрачев и др., 1968 Novick, 1958а Griffin, 1958
— Там же
— » »
92 Добрачев и др., 1968 Константинов, Макаров, 1973
— Добрачев и др., 1968
— Там же
обширная и повсеместно распространенная группа. В связи с адаптацией к суровым климатическим условиям в жизни Vespertilionidae в отличие от многих других семейств летучих мышей, обитателей тропиков, наблюдаются два периода: летней активной жизни и зимний, когда животные впадают в продолжительную спячку.
Все гладконосые — насекомоядные. Правда, некоторые из них, подобно Noctilio, разнообразят свою диету рыбой. Представители семейства вылавливают насекомых в различных условиях. Быстролетные виды (роды Miniopterus и Nyctalus) охотятся высоко над землей и в стремительном полете настигают добычу; нетопыри (род Pipistrellus) виртуозно выделывают «фигуры высшего пилотажа», мелькая среди деревьев и кустов в погоне за двукрылыми; мелкие ночницы (род Myotis) неторопливо кружатся над поверхностью водоемов, а ушаны (род Plecotus) предпочитают собирать нелетающих насекомых и гусениц на листьях и ветвях деревьев и кустарников. Состав поедаемых насекомых различными видами сильно варьирует — от жуков среднего размера и ночных бабочек до комаров и мошек. Разные способы и условия охоты, а также видовой состав поедаемых насекомых, т. е. различная пищевая специализация, несомненно наложили отпечаток как на внешний вид отдельных представителей (рис. 22), так и на функционирование локационных систем этих животных.
С точки зрения эхолокации семейство изучено наиболее полно. В настоящее время параметры локационных сигналов исследованы у 44 видов. Обстоятельные сведения о параметрах сигналов гладконосых летучих мышей фауны СССР можно найти в работах Симкина и Патлякевич (1969а, 1970) и Константинова и Макарова (1973).
Как видно из табл. 5, локационные сигналы гладконосых летучих мышей широко варьируют у различных представителей, но в общем характеризуются короткой длительностью, высокой или средней интенсивностью, относительно простым частотным заполнением с присутствием основной частоты и ее 2-й гармоники, которая почти всегда является подавленной, и, наконец, ярко выраженной частотной модуляцией в пределах каждого сигнала, величина которой часто превышает одну октаву.
Форма импульсов гладконосых летучих мышей может быть самой разнообразной не только у разных видов, но даже у одной и той же особи. Как правило, каждый из них начинается с незначительных амплитуд, которые либо постепенно увеличиваются, либо сразу же происходит подъем к одной или многим вершинам, после чего следует крутой спад амплитуды в конце. Более пологие фронты у импульсов отмечаются во время спокойного курсирующего полета животных, тогда как при приближении препятствию или насекомому сигналы по форме приближаются
73
к прямоугольному импульсу с чуть заваленными передним и задним фронтами.
Максимальная длительность излучаемого сигнала не превышает 10 мсек., но обычно, когда животное зондирует пространство, не встречая на пути препятствий, длительность импульсов у большинства видов составляет 1—3 мсек. При сближении с препятствием или насекомым импульсы все более и более укорачиваются. Минимальная зарегистрированная длительность равнялась всего 0.2—0.3 мсек.
Характерно быстрое изменение основной частоты заполнения каждого сигнала. Частота в начале сигнала достигает у некоторых видов 130—150 кгц и за 2—3 мсек, его звучания падает до 30— 40 кгц (рис. 23). Величина девиации, т. е. разница между максимальной и минимальной частотами, таким образом, может составлять более 100 кгц. Это особенно характерно для нескольких видов ночниц.
Наряду с видами, обладающими высокочастотными сигналами, примерно половина среди исследованных гладконосых летучих мышей имеет средний ультразвуковой диапазон частотного заполнения сигналов. У этих последних, как правило, в импульсах присутствует стабильная вторая гармоническая составляющая, которая, появляясь в середине, сопровождает основную частоту до конца сигнала (рис. 23).
Изменения основной частоты в импульсах большинства гладконосых летучих мышей происходят по линейному закону, но у некоторых видов нами отмечена линейность изменения частоты лишь в первой половине сигнала, а затем очень плавный ее спад.
Для всех видов характерно значительное изменение параметров локационных сигналов в зависимости от ситуации и тех задач, которые приходится решать животным. Частота следования импульсов во время спокойного курсирующего полета животного не превышает 10—20 имп./сек., при приближении к препятствию она возрастает до 50—60 имп./сек., тогда как во время перехвата насекомого темп излучения импульсов может достигать 150—200 в секунду. Соответственно наблюдается уменьшение длительности сигнала до 0.4—0.2 мсек, у некоторых мелких видов, при этом частота заполнения резко снижается и частотная модуляция становится нечетко выраженной. Именно поэтому мы имеем такой большой разброс значений длительности и частоты в начале импульсов.
Звуковое давление генерируемых импульсов, или их интенсивность, у многих представителей этого семейства составляет
Рис. 22. Представители гладконосых летучих мышей.
а — двухцветный кожан (фот. В. П. Дмитриевой); б — ушан (фот. А. И. Константинова); в — остроухая ночница (фот. А. И. Константинова).
75
?,кго f.Kiu.
Рис. 23. Осциллограммы локационных сигналов и кривые изменения частоты заполнения у представителей гладконосых летучих мышей.
1 — остроухая ночница; 2 — ночница Наттерера; з — прудовая ночница; 4 — трех-цветная ночница; 5 — длиннокрыл; 6 — нетопырь-карлик; 7 — поздний кожан; 8 — \шан. Скорости развертки осциллограмм: 1—3 — 0.12 мсек./см, 4, 7, 8 — 0.3, 5, 6 — 0.5 мсек./см.
60—100 дин/см* на расстоянии 10—15 см от рта животного. Но имеются виды, например из рода Plecotus, у которых интенсивность локационных сигналов исключительно низкая, как и у многих американских листоносов, тогда как у других, например у тропических длиннокрылов, интенсивность сигналов, по данным Новика (Novick, 1958а), достигает почти 2000 дин/см2, если, конечно, автор не ошибся в расчетах.
Гладконосые летучие мыши излучают локационные сигналы через рот, единственное исключение составляет род Ptecotus — ушаны, которые способны к производству импульсов как через рот, так и через ноздри.
Таким образом, для целей эхолокации представители этой последней группы используют относительно короткие ЧМ-сиг-налы, с большой величиной девиации основной частоты. В отличие от летучих мышей других подсемейств удельный вес дополнительных гармонических составляющих в сигналах животных этой группы резко снижен, а у многих видов в спектре представлена лишь основная частота.
Пути развития эхолокационных систем летучих мышей
Мы рассмотрели все известные на сегодняшний день типы локационных систем летучих мышей. При всем их многообразии можно выделить ряд общих черт и определенные линии специализации, а также проследить основные пути развития локационных сигналов у современных семейств.
Летучие мыши, у которых добывание пищи происходит в сложной акустической обстановке (наличие многочисленных эхо от близко расположенных предметов: стены домов, стволы, ветки, листва деревьев и кустарников) и к тому же объект добычи стационарен или малоподвижен, как правило, используют ультразвуковые, слабо интенсивные и обычно «зашумленные», т. е. с несколькими гармониками, сигналы. К ним относятся виды из различных надсемейств и семейств независимо от филогенетического развития, такие как Nycteridae, Megadermatidae, Desmodontidae и многие Phyllostomatidae, род Plecotus, а возможно, и ВагЪа-stella из Vespertilionidae. Из-за слабой интенсивности сигналов, по образному выражению Гриффина, всех их можно назвать «шепчущими» летучими мышами. Причины такой специализации у этих животных с акустической точки зрения мы уже рассматривали выше при описании семейства Megadermatidae.
В противоположность «шепчущим» летучие мыши, добывающие насекомых в стремительном полете, обладают локационными сигналами средней и высокой интенсивности. При больших скоростях полета основным требованием во время ночных охот должно быть своевременное обнаружение стоящих впереди пре-
пятствий или летящего насекомого. Поэтому использование более громких сигналов необходимо летучим мышам для получения эха с больших расстояний. Скорость полета этой группы летучих мышей значительно выше, а сам полет маневреннее в отличие от порхающего с частыми зависаниями на одном месте полета «шепчущих».
По параметру длительности сигналов, имеющей решающее значение при толковании принципов определения расстояния до цели по отраженному сигналу, у этих животных мы сталкиваемся с двумя функциональными типами эхолокационных систем.
Один из них представлен короткими сигналами длительностью в основном не более 5 мсек, и свойствен подавляющему большинству видов различных семейств.
По мере сближения животных с препятствием длительность сигналов прогрессивно уменьшается, что предотвращает наложение эха на зондирующий сигнал.
Второй функциональный тип основан на использовании длинных сигналов, обычно свыше 10 мсек., с обязательным перекрытием во времени излучаемого и отраженного импульсов. Несмотря на то что длительность сигналов также уменьшается по мере приближения животного к цели, участки перекрытия импульса и его эха присутствуют всегда, становясь непременным условием работы такого эхолокатора.
В Восточном полушарии этим типом локации несомненно пользуются ложные и настоящие подковоносы, а в Западном — некоторые представители Mormoopidae и, возможно, Nocttltonidae. Последние по параметрам сигналов представляют как бы переходную форму между рассматриваемыми двумя типами.
Сравнивая сигналы этих двух типов, мы замечаем еще одну закономерность. Как правило, короткие сигналы имеют более сложный спектр, обусловленный либо присутствием нескольких частотно-модулированных гармонических составляющих, либо большой величиной модуляции основной частоты. В то же время у летучих мышей, использующих длинные сигналы, спектральный состав гораздо проще и определяется одной или двумя частотами, которые на протяжении почти всего импульса остаются неизменными и только в конце наблюдается изменение частоты на величину, не превышающую одну октаву, а у большинства видов всего на несколько килогерц.
Длинные, относительно простые сигналы, сфокусированные в нужном направлении, имеют ряд преимуществ перед короткими для целей обзорной локации, но уступают последним при анализе деталей исследуемого объекта. Поэтому не случайно уменьшение длительности сигналов у летучих мышей, обладающих вторым типом локации, в сложных условиях ловли добычи идет за счет сокращения частей импульса с постоянной частотой, 78
тогда как частотно-модулированная часть сохраняется всегда неизменной.
Пока трудно сказать, почему для достижения одной и той же цели у насекомоядных летучих мышей развились два функционально различных типа эхолокации. Какие экологические и физиологические особенности видов способствовали такому разделению? До конца неясно, каковы преимущества и недостатки в использовании тех и других сигналов. Интересен тот факт, что второй тип локации, с применением длинных сигналов и постоянством частоты на протяжении большей части импульса, широко используется многочисленными представителями двух крупных семейств Старого Света, тогда как на американских континентах близкие по параметрам сигналы обнаружены всего у нескольких видов.
Возможно, такое неравномерное географическое распространение второго типа локации обусловлено более жестокой конкуренцией среди насекомоядных летучих мышей Восточного полушария. Бульдоговые и гладконосые летучие мыши с наиболее совершенным летательным аппаратом, чем у ложных и настоящих подковоносов, здесь представлены значительно шире в видовом отношении по сравнению с Америкой. Кузякин (1950) приводит следующие цифры: из 60 видов складчатогубов (из бульдоговых) в Америке обитает 14, из 80—100 форм ночниц — 20, из 100 с лишним видов кожанов и нетопырей — только 5. Развитие особого типа локации у ложных и настоящих подковоносов могло быть одним из тех факторов, которые обеспечили этим семействам не только выживание, но и расцвет в тропиках Восточного полушария. На американских континентах этот процесс ограничился лишь несколькими насекомоядными формами, так как многие другие виды перешли на другой тип питания и таким путем оказались вне конкуренции от проникших туда бульдоговых и гладконосых. Справедливо ли высказанное предположение, покажут дальнейшие исследования.
Попытаемся теперь на основе имеющихся в настоящее время данных представить в целом пути развития основных функциональных типов локационных систем у ныне существующих семейств летучих мышей (Константинов, 1973а).
Новик (Novick, 1958а), анализируя локационные сигналы летучих мышей 11 семейств, впервые обнаружил, что, несмотря на их разнообразие, представляется возможным связать их с образцами импульсов примитивных Emballonuridae. Более определенно на этот счет высказывается Симкин (1971а), рассматривая функциональный тип Emballonuridae как своеобразный субстрат, на котором происходило формирование более молодых групп летучих мышей Старого и Нового Света. Симкин справедливо замечает, что на первых этапах локационный сигнал, видимо, должен быть малоспециализированным, иметь постоянную частоту
79
и дополнительные гармоники. И далее, он приходит к выводу, что именно такой «первичный» тип сигналов обнаружен у одного из наиболее примитивных семейств — Emballonuridae. В своих рассуждениях он опирался в основном на первые, очень приблизительные данные Гриффина и Новика (Griffin, Novick, 1955) о неизменных частотных характеристиках импульсов этих животных. Последующие же работы (Novick, 1958а, 1962) наглядно продемонстрировали присутствие небольшой частотной модуляции в сигналах Emballonuridae либо от начала и до конца каждого импульса у большинства видов, либо в самом конце его. Несомненно, данный признак является прогрессивным по сравнению, например, с сигналами Nycteridae или Megadermatidae.
С другой стороны, никто не обращает внимания на характер локационных сигналов у истинно древнейшего из ныне живущих семейств летучих мышей — Rhinopomatidae. Несмотря на фрагментарное их описание, бросаются в глаза отсутствие частотной модуляции и не слишком короткая длительность импульса — 4 мсек. Верхняя граница спектра составляет 60 кгц, и по аналогии с другими примитивными семействами можно предполагать, что эта величина соответствует 4-й гармонике, а основная частота находится около 15 кгц. Если при последующем изучении описываемые параметры сигналов подтвердятся, то их можно рассматривать как тип, наиболее близко стоящий к гипотетическому «первичному» локационному сигналу, по крайней мере по частотным характеристикам. К сигналам Rhinopomatidae ближе всего подходят сигналы семейств Nycteridae и Megadermatidae, в импульсах которых также не обнаруживается частотной модуляции, основные частоты низкие и присутствует несколько стабильных гармоник. Отличия выражаются лишь в том, что в импульсах Nycteridae содержится 5 гармоник с преобладанием по интенсивности 2—4-й, а у Megadermatidae присутствуют лишь 2— 4-я гармоники, совершенно не выражена 1-я и во время полета часто отсутствует 4-я.
Следующий этап развития локационных систем представляют виды примитивного семейства мешкокрылов {Emballonuridae). Низкие основные частоты (11—30 кгц) и наличие стабильных гармонических составляющих 2-го и 3-го порядков свидетельствуют о том, что сигналы этих животных не слишком далеко продвинулись по сравнению с импульсами Rhinopomatidae. Однако прогрессивной чертой несомненно следует признать появление в сигналах мешкокрылов нисходящей частотной модуляции и разнообразие типов сигналов.
Большинство исследованных представителей мешкокрылов использует локационные сигналы короткой длительности с изменением основной частоты от начала к концу каждого из них на 2— 4 кгц. Именно такие сигналы могут быть поставлены у истоков направления, которое привело к организации широкополосного 80
ЧМ-сигнала с большим размахом девиации основной частоты у гладконосых летучих мышей (рис. 24, правая часть схемы). Промежуточным звеном на пути развития данного типа сигналов среди современных летучих мышей, по нашему мнению, следует считать локационные системы американских листоносов (сем. Phyllostomatidae). У насекомоядных и ряда плодоядных листоносов Нового Света основная частота в начале импульса
80-120
д Rhlnolophidae
Htpposidertdae Noctilionidae 58.
>100
Vespertilionidae
--------- X 30 en 577
Molosstdae 4^ 25
Embdllomiridae
= Megadermatldae /
"P /
—s Nycteritdae /
W /
NataliAae^
30
Desmodontldae 40 ------------x27
to
Phyllostomatidae^^
го^17
= Rhinopomatidae
Рис. 24. Направления развития основных типов локационных систем у летучих мышей.
Рядом с названием семейств поставлены типичные образцы частотного заполнения локационных сигналов (основная частота и гармонические составляющие). Жирными линиями выделены наиболее значимые для эхолокации частотные компоненты. Цифрами обозначена частота основного тона в начале и в конце сигнала, кроме сигналов подковоносов, для которых проставлены значения вторых гармоник.
повышается по сравнению с импульсами мешкокрылов до 40— 60 кгц и значительно расширяется ее девиация от начала к концу сигнала, хотя, как правило, еще не достигает октавы. Переход от насекомоядного типа к питанию фруктами, цветами, мелкими позвоночными и кровью животных и птиц, т. е. стационарными или малоподвижными объектами в сложной с точки зрения локации обстановке, приводит, как уже отмечалось выше, к адаптационным особенностям сигналов этих животных — сокращению их длительности и снижению интенсивности.
У Natalidae частота в начале сигнала увеличивается до 85 кгц, изменение ее к концу импульса достигает одной октавы, и в этом
81
отношении они приближаются к Vespertiltontdae. Однако стабильная высокоамплитудная 2-я гармоника, идущая от начала и до конца сигнала, а также появление в конце 3-й гармоники — типичные признаки сигналов Phyllostomatidae.
И наконец, у многих представителей гладконосых летучих мышей (Vespertilionidae) локационные сигналы заполнены только основной частотой, которая достигает максимального значения среди рукокрылых — 100 кгц и более в начале импульса. Вторые гармоники появляются у ряда специализированных форм, но достигают максимума всегда в заключительных частях сигналов. Величина частотной модуляции от начала и до конца каждого импульса часто превышает октаву, а у некоторых видов составляет две и даже три октавы.
Хотя бульдоговые летучие мыши (сем. Molossidae) достигли наибольшего совершенства полета среди рукокрылых, их локационные сигналы значительно уступают сигналам гладконосых. Величина частотной модуляции в импульсах достигает октавы, но основные частоты относительно низкие (50 кгц). 2-я гармоника приурочена к средней и заключительной частям импульса.
Итак, в процессе филогенетического развития формирование широкополосного ЧМ-сигнала прошло ряд последовательных этапов, что привело к созданию сигналов с огромной частотной девиацией. Напомним, что девиация частоты является одним из важнейших показателей в определении величины разрешающей способности локационной системы и чем; она выше, тем лучше разрешение по дальности и помехоустойчивость системы. Следует также отметить, что, как показывают результаты сравнительного анализа, расширение частотной модуляции идет одновременно с повышением основной частоты, уменьшением количества и функциональной значимости гармонических компонентов. Представители этого направления излучают локационные сигналы в основном через рот, с широкой диаграммой направленности, и только американские листоносы перешли на излучение сигналов через ноздри, чему, по-видимому, способствовал переход к новому типу питания.
Другой путь развития локационных систем привел к созданию локационного сигнала большой длительности с неизменяющимися частотными компонентами на протяжении большей части импульса, за исключением терминальных ЧМ-участков. Такие сигналы, свойственные в основном двум крупным семействам Старого Света — ложным и настоящим подковоносам (Hipposideridae и Rhinolo-phidae\ рис. 24, левая часть схемы), а также одному представителю американского семейства Mormoopidae, в эволюции могли возникнуть на базе относительно длительных сигналов с постоянной частотой заполнения, которые мы встречаем у 2 современных видов мешкокрылов (Saccopteryx и Rhinchonycteris). Сигналы этих видов характеризуются низкой основной частотой (20—30 кгц),
82
наличием нескольких гармонических составляющих, из которых 2-я гармоника становится несущей.
Как и при рассмотрении пути развития эхолокации первого типа, здесь также четко прослеживается процесс повышения частоты и очищения спектра от дополнительных составляющих, а кроме того, увеличение длительности сигнала. Импульсы Pteronotus parnellii rubiginosus из семейства Mormoopidae длительностью 20 мсек, имеют две гармонические составляющие на протяжении всего импульса с доминирующей 2-й гармоникой (64 кгц). В конце каждого импульса наблюдается падение частоты на 4 кгц. У многих ложных подковоносов в локационных сигналах, кроме слабой 1-й и мощной 2-й (~ 70 кгц) гармоник, других компонентов не встречается, а длительность импульсов возрастает до 30 мсек. Величина частотной модуляции в конце импульса увеличивается. У наиболее специализированных летучих мышей этого ряда настоящих подковоносов импульсы у многих видов заполнены лишь одной частотой, представляющей собой 2-ю гармонику (82— 100 кгц), тогда как 1-я и 3-я проявляются нерегулярно. Изменение частоты в конце составляет 15—30 кгц. Длительность сигналов достигает 60—90 мсек. Все члены данного ряда излучают локационные сигналы либо через рот, причем губы, вытягиваясь, образуют рупор, либо через ноздри.
Между рассмотренными двумя направлениями развития локационных сигналов следует выделить еще одно, как бы промежуточное. Свое начало оно берет, по-видимому, тоже от мешкокрылов. Сигналы одного из видов этого семейства — Emballonura nigrescens — характеризуются тем, что основная частота и две сопровождающие ее гармоники остаются неизменными от начала до середины импульса, а затем следует ЧМ-часть. Аналогичное изменение частотных компонентов в локационных сигналах происходит у ряда видов из семейств Mormoopidae и Noctilionidae. причем у представителей первого на протяжении всего импульса присутствуют две гармоники (1-я и 2-я), а у второго сигналы заполнены только основной частотой и 2-я гармоника появляется лишь в конце. В этом ряду также прослеживается указанная выше закономерность — повышение основной частоты, расширение величины частотной модуляции и сокращение числа гармонических компонентов.
Постоянство частоты в первой части импульса сближает сигналы этих видов летучих мышей с типом сигналов, которыми пользуются для эхолокации подковоносы. Кроме того, на близком расстоянии до цели происходит частичное перекрытие отраженного сигнала излученным (Novick, 1963b, 1965; Suthers, 1967). G другой стороны, такие признаки, как короткая длительность сигналов (1 — 13 мсек.), а также полная редукция моночастотной части импульса при сближении животного с добычей, в результате чего сигналы становятся исключительно частотно-модулированными, говорят
83
о сходстве этих локационных систем с ЧМ-системами первого направления.
Все сказанное позволяет прийти к заключению, что представители современных семейств летучих мышей в целях локации используют 4 типа локационных сигналов: 1) короткие шумоподобные сигналы без частотной модуляции; 2) короткие ЧМ-сигналы с разной степенью выраженности частотной девиации и разным количеством гармонических составляющих; 3) сигналы короткой или средней длительности с постоянным частотным составом в первой половине и частотной модуляцией во второй; 4) сигналы большой длительности с постоянными частотными характеристиками и заключительной ЧМ-частью.
Применяя филогенетический принцип при сопоставлении различных эхолокационных систем летучих мышей, удается наметить основные пути их развития в пределах подотряда Microchi-roptera и показать, как на базе сигналов первого типа примитивных семейств постепенно происходило формирование специализированных сигналов, свойственных более высокоорганизованным семействам.
Предлагаемая схема, естественно, не может считаться окончательной. Новые исследования, по-видимому, внесут в нее дополнительные коррективы и помогут с большей последовательностью, чем сейчас, представить пути функционального развития, которые прошли эхолокационные системы летучих мышей в процессе эволюции.
Последующее изложение будет посвящено описанию структурнофункциональных особенностей систем эхолокаторов, их возможностей, рабочих характеристик и принципов действия в основном у двух семейств: гладконосых и подковоносых летучих мышей, эхолокация которых достигла наибольшего совершенства по сравнению с другими семействами рукокрылых.
2
ЗВУКОИЗЛУЧАЮЩАЯ СИСТЕМА И ХАРАКТЕРИСТИКИ НАПРАВЛЕННОСТИ ИЗЛУЧЕНИЯ
Ультразвуковые локационные сигналы, как и звуки коммуникативного характера, излучаемые летучими мышами, своим происхождением обязаны функциям гортанных механизмов (Griffin, 1946, 1952, 1958; Mohres, 1953; Novick, 1955; Motta, 1956; Fischer, Vomel, 1961; Novick, Griffin, 1961). Для понимания механизмов генерирования ультразвуков необходимо обратиться к данным о строении гортани у летучих мышей.
Особенности строения гортани
Еще до открытия эхолокации при анатомическом исследовании гортани рукокрылых отмечались существенные отличия ее по сравнению с гортанью других млекопитающих: непропорционально большие размеры, частичное слияние и окостенение отдельных элементов, мощное развитие мускулатуры (Robin, 1881; Elias, 1907; Negus, 1929). Незначительные размеры суставов обеспечивают почти полную неподвижность гортанных элементов относительно друг друга, благодаря чему гортань в целом представляет собой как бы монолитное образование. Особенно характерны увеличение размеров, дорзальное срастание и частичное окостенение черпаловидных хрящей (с. arytenoidea), которые в виде двух тонких пластинок вытянуты в дорзо-вентральном направлении. Между ними располагается межчерпаловидный хрящ (с. intera-rytenoideus). Черпаловидные хрящи у летучих мышей полностью лишены вращательных движений в отличие от таковых у человека, у которого большая подвижность их играет важную роль в момент произнесения слов. Далее обращают на себя внимание увеличенный по высоте перстневидный хрящ (с. cricoidea), образующий основание всей гортани, и разрастающийся над ним в дорзо-вентральном направлении щитовидный хрящ (с. thyreoidea). Строение сустава
85
между ними обеспечивает некоторую подвижность щитовидному хрящу. Оба эти элемента у летучих мышей также частично окостеневают. Над входом в гортань располагается надгортанный хрящ (epiglottis), который кверху расширяется, а книзу сужается в стебелек, или ножку, сочленяясь с поверхностью щитовидного хряща. Гроссе (Grosser, 1900) отметил, что надгортанник у летучих мышей очень богат эластическими волокнами и имеет большую подвижность.
У гладконосых летучих мышей во время излучения локационных сигналов, по наблюдениям Фишера и Вёмеля (Fischer, Vomel, 1961), гортань несколько опускается, а надгортанник плоско прилегает к основанию языка. При таком положении гортани вход ее находится непосредственно в фокусе звуковой воронки (ротовой полости). У подковоносов в момент генерирования ультразвука надгортанник входит в небное отверстие носоглотки и таким образом происходит полное отделение трахеоназальных дыхательных путей. Небно-глоточная мышца сильно разветвляется, образуя мышечный сфинктер, который плотно зажимает надгортанник, входящий в небо. Благодаря этому создается прямой путь для воздушного потока и ультразвука в носовую полость и далее через ноздри наружу. У подковоносых летучих мышей в отличие от всех других млекопитающих имеется еще одна морфологическая особенность воздухоносного пути. На дорзальной стороне трахеи располагаются три хрящевые, частично окостеневшие колбообразные капсулы, открывающиеся в нее. Симметрично справа и слева от гортани лежат две парные капсулы, а ниже, примерно посредине, одна непарная. Этим капсулам приписывается роль резонаторов (Mohres, 1953), ответственных за исключительную чистоту тона, отмеченную для импульсов подковоноса, но, возможно, они выполняют и другие, пока еще неизвестные функции, выяснить которые позволят лишь измерения на модели.
Наряду с увеличенными размерами элементов скелета большое развитие у летучих мышей получает мышечный аппарат гортани. Все мышцы гортани поперечнополосатые, топографически разделяются на две группы — наружные и внутренние. К первой группе относится одна парная мышца — перстневидно-щитовидная (m. cricothyreoideus), достигающая у летучих мышей наибольших размеров. Она берет свое начало от наружной поверхности дуги перстневидного хряща, идет веерообразно вверх и назад и прикрепляется к нижнему краю щитовидного хряща, охватывая его изнутри и снаружи. Под воздействием этой мышцы щитовидный хрящ сближается с дугой перстневидного хряща и несколько наклоняется вперед. В результате расстояние между черпаловидными хрящами и углом щитовидного увеличивается, что приводит к натяжению голосовых связок.
Внутренние мышцы гортани в свою очередь подразделяются на две группы, одни расширяют голосовую щель, 86
а другие сужают ее. К первым относится задняя перстневидночерпаловидная мышца (ш. cricoarytenoideus posterior), ко второй — боковая перстневидно-черпаловидная (m. cricoarytenoideus lateralis), щитовидно-черпаловидная (m. thyreoarytenoi-deus) и поперечно-черпаловидная (m. arytenoideus transversus) мышцы.
Внутреннее строение гортани летучих мышей при фронтальном сечении напоминает форму песочных часов: в среднем отделе полость гортани сужена, кверху и книзу расширена. Средний отдел устроен наиболее сложно и отграничен от верхнего и нижнего двумя парами складок, идущими по боковым стенкам гортани. Верхняя складка, называемая преддверной, или ложной голосовой (plica ventricularis), представляет собой дубликатуру слизистой оболочки. В толще ее, ближе к внутреннему краю, лежит ложная голосовая связка. Нижняя складка — истинная голосовая (plica vocalis) — выступает сильнее верхней, т. е. ближе подходит к средней линии полости гортани. Щель между голосовыми складками — голосовая щель (rima glottadis) — самая узкая часть полости гортани и, в частности, у подковоносов составляет всего 2/5 внутреннего диаметра ее. Голосовая складка резко отличается от преддверной и внешним видом, и строением: край ее более острый, цвет белесоватый. Ближе к свободному краю ее лежит голосовая связка, а более латерально — голосовая мышца (m. vocalis).
У гладконосых летучих мышей хорошо развиты как истинные голосовые связки, так и ложные, имеющие диаметр 6—8 мкм, тогда как у подковоносов последние полностью отсутствуют. Как указывал еще Элиас (Elias, 1907), голосовые связки подковоносов исключительно короткие и напоминают сухожильные тяжи, почему и получили специальное название chorda vo-calia.
Итак, представленные морфологические данные говорят о том, что гортань летучих мышей как нельзя лучше приспособлена для производства звуков высокой частоты. В пользу этого свидетельствуют ограниченная подвижность гортанных элементов, наличие коротких голосовых связок и мощное развитие мышечного аппарата, обеспечивающего их быстрое и сильное натяжение.
Нервная регуляция голосового аппарата
Иннервация мышц гортани осуществляется в основном двумя веточками блуждающего нерва (n. vagus): верхним (n. laryngeus superior) и нижним (n. laryngeus reccurens) гортанными нервами. Верхний гортанный нерв подходит к гортани с латеральной поверхности и подразделяется на 2 веточки, одна из которых, сенсорная (внутренняя), обеспечивает иннервацию гортани, другая, моторная (внешняя) ветвь, иннервирует перстне
87
видно-щитовидную мышцу. Нижний, или возвратный, гортанный нерв проходит вдоль трахеи и иннервирует все другие мышцы гортани, кроме перстневидно-щитовидной (рис. 25).
У всех исследованных млекопитающих нижние гортанные нервы играют наиболее существенную роль в производстве различных звуков. Повреждение нижних гортанных нервов, напри
мер, у человека приводит к частичному параличу голосового аппарата, и нормальное качество голоса теряется. При повреждении же верхних гортанных нервов отмечается лишь незначительное изменение голоса.
У летучих мышей наблюдается обратная картина. Экспериментами американских исследователей (Griffin, 1958; Novick, Griffin, 1961) было показано, что перерезка обоих нижних гортанных нервов не вызывает у гладконосых летучих мышей изменений в излучении как локационных, так и коммуникационных сигналов. В то же время перерезка верхних гортанных нервов существенно
Рис. 25. Иннервация гортани летучей мыши. (Griffin, 1958).
1 — трахея; 2 — щитовидная железа; 3 — перстневидно-щитовидная мембрана; 4 — внутренние мышцы гортани; 5 — перерезанная грудинно-подъязычная мышца; 6 — верхний гортанный нерв; 7 — веточка подъязычного нерва; 8 — подъязычный нерв; & — веточка языкоглоточного нерва; 10 — языкоглоточный нерв; 11 —нижний гортанный нерв; 12 — блуждающий нерв.
влияет на частотные характеристики сигналов. Частота заполнения импульса снижается примерно до 10 кгц, и полностью исчезает внутриимпульсная девиация. В обоих случаях длительность сигналов и частота их следования не нарушались.
Мы повторили опыты с перерезкой нижних и верхних гортанных нервов на двух видах гладконосых летучих мышей — позднем кожане (6 животных) и остроухой ночнице (2 животных). Кроме того, впервые была предпринята попытка оценить влияние аналогичных перерезок на излучение локационных сигналов у больших подковоносов (8 животных). Результаты, полученные на гладконосых летучих мышах, совпали с данными американских исследователей. Двусторонняя перерезка нижних гортанных нер^
88
bob не изменила параметры локационных сигналов у исследованных видов, тогда как двусторонняя перерезка верхних гортанных нервов вызвала значительные изменения в частотном'заполнении импульсов, которые животные излучали во время старта и полета по экспериментальному манежу. Прежде всего частоты понизились до звукового диапазона и во время полета восприни-
Рис. 26. Осциллограммы импульсов летучих мышей в норме и после перерезки верхних гортанных нервов.
А — остроухая ночница; Б — большой подковонос. Период между калибровочными метками 0.3 мсек. 1 — норма, 2 — после операции.
мались экспериментатором на слух как «щебетание». Четко можно было уловить нарастание скорости повторения импульсов, когда животные совершали посадку на стены манежа.
Исследование спектральных характеристик показало, что ос* новная энергия в сигналах, например, остроухой ночницы прихо* дится на частоты 10—14 кгц, но в сигналах присутствуют многочисленные гармонические составляющие 2—4-го порядков. Полностью исчезает внутриимпульсная девиация, столь характерная для этих видов летучих мышей (рис. 26, А). Последующие наблюдения показали, что постепенно частотные характеристики сигналов восстанавливаются и спустя месяц импульсы почти не отли-
89
чаются от нормы. Вскрытие животных убедило нас в том, что регенерации нервного волокна не произошло. Следовательно, восстановление первоначальных параметров сигналов происходило за счет компенсаторной деятельности других нервов, осуществляющих иннервацию гортанной мускулатуры. В чем конкретно это выражалось — остается вопросом для будущего исследования.
У подковоносов билатеральная перерезка нижних гортанных нервов приводила к немедленной гибели животных. Смерть наступала через 10—15 сек., и всегда на полном вдохе. Этот факт свидетельствует о том, что функция нижних гортанных нервов у подковоносов в отличие от гладконосых летучих мышей тесным образом связана с регулированием дыхательных циклов и осуществление этого процесса невозможно без участия указанных нервов. При одностороннем удалении участка нервного волокна в поведении животных не было обнаружено каких-либо изменений, не нарушались и функции дыхания. Подковоносы во время полета продолжали излучать локационные сигналы обычной длительности, несущая частота заполнения (2-я гармоника) сохранялась на прежнем уровне (81—82 кгц). Отличия заключались лишь в том, что после операции в импульсах отчетливо проявилась 1-я гармоника, которая в сигналах интактных животных обычно подавлена.
Двусторонняя перерезка верхних гортанных нервов привела к значительному изменению частотных и амплитудных характеристик сигналов сразу после операции. Во время полета оперированных животных мы отчетливо слышали характерные «трески», издаваемые ими. Анализ осциллограмм (рис. 26, Б) показал резкие различия в форме излучаемых импульсов. Амплитудная модуляция оказалась настолько выраженной, что одиночный сигнал превратился в последовательность очень коротких импульсов (т=0.5 мсек.), связанных между собой участками с малой амплитудой. Усложнилась внутренняя структура сигналов, и вместо синусоидального импульс стал шумоподобным. Спектр такого сигнала (рис. 27) охватывал диапазон частот от 8 до 60 кгц с равномерно распределенной энергией по всему диапазону. Однако уже на 5-й день после операции форма многих записанных сигналов напоминала импульсы интактного животного, но их спектр имел четко выраженный полосовой характер с наличием четырех гармонических составляющих. Основная энергия приходилась на частоту 60 кгц. Наряду с такими импульсами отмечены сигналы более сложной структуры, в спектре которых можно было выделить шесть гармонических составляющих и максимум энергии приходился на частоты 20 и 40 кгц. Таким образом,
Рис. 27. Спектрограммы локационных сигналов большого подковоноса в норме и после перерезки гортанных нервов.
А — норма; Б — односторонняя перерезка нижнего гортанного нерва; В, Г — билатеральная перерезка верхнего гортанного нерва: В — на 2-й день, Г — на 10-й день.
91
и у подковоносых летучих мышей проявляется тенденция к восстановлению первоначальных параметров локационных сигналов, несмотря на выключение функций верхних гортанных нервов.
В результате проведенных экспериментов и сопоставления их с данными других исследователей становится ясно, что нервные механизмы, обеспечивающие производство звуков у летучих мышей, отличаются от таковых у человека и других млекопитающих. Основная роль в этом процессе у рукокрылых принадлежит верхним гортанным нервам, которые иннервируют перстневидно-щитовидные мышцы.
Вопрос о том, какие структуры мозга регулируют излучение ультразвуковых сигналов у летучих мышей, совершенно не освещен в литературе, поэтому предстояло провести специальные исследования. В поисках центрального аппарата, координирующего деятельность гортанных механизмов, мы исходили из того, что эхолокационная система летучих мышей, представляющая собой сложно согласованную систему излучения ультразвука и приема отраженных сигналов, должна иметь единый регулирующий центр. Из всех отделов слухового анализатора, о чем подробно будет сказано в следующей главе, наибольшего морфологического развития у летучих мышей достигают задние холмы четверохолмия среднего мозга (Зворыкин, 1959; Liitgemeier, 1962; Никитенко, 1969, 1971).
Уместно заметить, что еще В. М. Бехтерев (1895, 1906) на основании собственных экспериментов на ряде животных и данных других авторов пришел к заключению, что заднее двухолмие наряду с другими функциями выполняет роль высшего рефлекторного центра, приводящего к появлению голосовых реакций под влиянием слуховых раздражений.
Прус (Prus, 1899) считал, что в заднем двухолмии имеются центр для ассоциированных движений глаз и зрачка и центр голоса. Позднее А. Е. Афанасьевым (1946) было показано, что при подрезке задних бугров четверохолмия происходят изменение тонуса мышц позвоночника, исчезновение голоса и установка ушей в сторожевое положение.
На основании приведенных выше данных мы остановили свой выбор на заднем двухолмии и провели серию опытов с непосредственным воздействием на него электрической стимуляции.
В экспериментах были использованы 30 гладконосых летучих мышей (4 прудовых и 16 водяных ночниц). Летучая мышь под слабым гексеналовым наркозом закреплялась в специально сконструированном станке, жестко фиксировавшем голову животного. В черепе под задними холмами делалось отверстие для установки электрода. Раздражение производилось стальными изолированными электродами с диаметром кончика 40 мкм. В качестве раздражающих стимулов использовались прямоугольные электрические импульсы от стимулятора ЭС-4М длительностью 0.5—1 мсек., подаваемые либо одиночно с частотой 1 имп./сек., либо сериями по 10—40 имп./сек. Интервалы между раздражениями составляли 3—15 мин. Голосовые реакции регистрировались на экране осциллографа С1-4,
92
В результате проведенных экспериментов было показано, что электрическая стимуляция центрального ядра задних холмов вызывает у животных излучение ультразвуковых сигналов, внешне похожих на локационные сигналы данного вида летучих мышей (Айрапетьянц, Константинов, 19676, 1970а). В ряде опытов пороги вызванных ультразвуковых реакций составляли 5 в при раздражении одиночными импульсами, но иногда приходилось увеличивать напряжение до 20—30 в, чтобы получить ответ. В среднем латентный период от момента раздражения до момента излучения импульса равнялся 20—30 мсек. Обычно каждый стимул вызывал излучение 1—3 импульсов, каждый длительностью 2—3 мсек. При тетаническом раздражении (10—40 имп./сек.) получали серию ультразвуковых импульсов. При аппликации на холмы кристаллического стрихнина, вызывающего повышение возбудимости, последующее одиночное электрическое раздражение также сопровождалось излучением серии из 6—8 импульсов. Эта фаза повышенной возбудимости продолжалась в течение 1— 1.5 мин., после чего ответы надолго исчезали. В то же время аппликация на холмы хлористого калия, оказывающего депрессивное влияние, мгновенно прекращала излучение ультразвуковых и звуковых сигналов в ответ на электрические стимулы.
Для контроля производилось раздражение трех областей коры: височной, лобной и затылочной. При тех же параметрах раздражающих стимулов нам не удалось получить вызванных голосовых реакций у животных.
Отметим также, что при разрушении заднего двухолмия в наших опытах излучение локационных сигналов у ночниц либо полностью прекращалось, либо, наоборот, наблюдалось непрерывное их издавание в быстром темпе, не связанное с задачами ориентации в пространстве. Экстирпация височных областей коры у летучих мышей не вызывала изменений в характере следования импульсов при решении конкретной локационной задачи (Константинов, Соколова, 1970).
Таким образом, результаты наших исследований свидетельствуют о том, что заднее двухолмие у летучих мышей может регулировать рефлекторную деятельность гортанных механизмов, создающих ультразвуковую импульсацию. Недавно это было подтверждено опытами Суги и Шлегеля (Suga, Schlegel, 1972b) на аме* риканских видах гладконосых летучих мышей.
Механизмы излучения ультразвука
По поводу механизмов образования ультразвуковых сигналов у летучих мышей в литературе существуют три гипотезы. Одна из них, наиболее распространенная и хорошо аргументированная морфологическими и физиологическими исследованиями, сводится к тому, что ультразвук возникает в гортани
93
за счет вибрации голосовых связок (Galambos, Griffin, 1942; Griffin, 1952, 1958; Mohres, 1953). Было показано, что резонансная частота колебаний голосовых связок у Myotis lucifugus порядка 45 кгц, а непосредственная перерезка их препятствует излучению ультразвуковых сигналов. Перстневидно-щитовидные мышцы выполняют роль приводящих ремней, регулирующих степень натяжения голосовых связок. Вибрация последних по типу струны под влиянием выдыхаемого воздуха создает тон высокой частоты. Электрическая активность перстневидно-щитовидных мышц у гладконосых летучих мышей резко увеличивается на несколько миллисекунд перед излучением сигнала, а в момент излучения постепенно уменьшается (Novick, 1955). Следовательно, голосовые связки вначале получают максимальное натяжение, а затем происходит их быстрое расслабление. Этим объясняется наличие нисходящей частотной модуляции в сигналах гладконосых. У подковоносов голосовые связки в силу своей морфологической специализации, отмеченной выше, менее эластичны, чем у гладконосых, и, по-видимому, не способны к столь быстрому расслаблению. За 1—2 мсек, звучания локационного сигнала происходит натяжение голосовых связок до определенного уровня, и мощная мускулатура гортани удерживает их в этом состоянии довольно длительное время (от 10 до 100 мсек.).
Соответственно частота заполнения сигнала вначале быстро нарастает (восходящая частотная модуляция) и затем сохраняется на постоянном уровне. Перед окончанием импульса натяжение голосовых связок ослабевает, чем объясняется падение частоты в конце каждого сигнала (нисходящая модуляция).
Как показали морфологические исследования (Revel, 1962), внутренняя структура перстневидно-щитовидных мышц такова, что их сократительный аппарат обладает исключительно малой инерцией. Следовательно, даже при высоком темпе излучения локационных сигналов характер частотного заполнения их либо остается без изменений (подковоносы), либо несколько снижается (у гладконосых), т. е. функция мышечного аппарата обеспечивает полный цикл натяжения и расслабления голосовых связок для каждого нового импульса.
По второй гипотезе (Motta, 1956), вибрирующими элементами гортани при создании ультразвуков служат не голосовые связки, а черпало-надгортанные складки, натяжение которых определяет образование надгортанной щели и закрывание входа в гортань. Натяжение складок осуществляется в основном двумя группами мышц: подъязычно-надгортанной и задними перстневидно-черпаловидными мышцами. Вследствие натяжения черпало-надгортанные складки начинают вибрировать и, по мнению Мотты, способны создавать звуки и ультразвуки. Мотта прижигал каустиком черпало-надгортанные складки, небно-глоточные дужки, корень языка и наблюдал, как сказывается операция на облете летучими
94
мышами препятствий (аппаратуры для регистрации ультразвуковых импульсов у Мотты не было). В результате им было показано, что нарушение беспрепятственного полета наступает только при атрофии или перерезке черпало-надгортанных складок, тогда как другие хирургические вмешательства не нарушают ориентационных способностей у летучих мышей. В своей работе Мотта высказывает критические замечания по поводу выводов, сделанных Гриффином о главной роли голосовых связок в формировании ультразвука.
Наблюдения Гриффина над явлениями, связанными с надрезом верхних нервов гортани, по мнению Мотты, подтвердили правильность сделанных им выводов и о важной роли черпало-надгортанных складок, подвижность которых зависит от перстневиднощитовидных мышц, иннервированных верхними нервами гортани, в формировании ультразвуков.
Нам кажется, что, прежде чем отвергать или принимать гипотезу Мотты, необходимо проанализировать излучаемые ориентационные сигналы летучих мышей после перерезки черпалонадгортанных складок.
Третья гипотеза выдвинута немецкими учеными (Fischer, Vo-mel, 1961; Fischer, 1962). На основании гистологического исследования гортани ночницы и построения ее модели авторы приходят к выводу, что гортань летучих мышей очень примитивна. Многие мышцы сохранили первобытную функцию, подобную сфинктеру скелета жаберной дуги. Сама голосовая связка имеет очень простое строение и мало приспособлена к собственным колебаниям по типу струны, а следовательно, не способна формировать ультразвуки. Авторы считают, что образование ультразвуков у летучих мышей основано на совершенно других физических законах, чем у большинства млекопитающих, и происходит по типу «свистковых» звуков. При этом воздушная струя под большим давлением проходит через узкую щель с достаточно высокой скоростью. В воздушной струе образуются вихри, которые сменяются в быстрой последовательности и частотой смен определяют высоту тона. Авторы высказывают предположение, что краткость ультразвуковых импульсов, вероятно, следует объяснять быстрым открыванием и закрыванием голосовой щели. Для принятия этой гипотезы, на наш взгляд, требуются более основательные физиологические подтверждения.
Каким бы путем ультразвуковые колебания ни возникали в гортани летучей мыши, по форме волны они не являются синусоидальными, а носят комплексный характер из-за присутствия многочисленных гармонических компонентов (обертонов), хотя спектр их, по-видимому, различается у разных видов. Окончательное формирование частоты заполнения локационного сигнала и ее «очищение» от дополнительных гармоник, по мнению Пая (Руе, 1967), происходят в резонаторных полостях, заполненных возду-
95
ком, через которые проходит ультразвук непосредственно перед выходом наружу. У гладконосых таким широко настроенным резонатором служит ротовая полость; у подковоносов же — носовая полость, резонансные свойства которой определены более жестко. Степень совершенства организации звуковых каналов, по-видимому, и определяет большое разнообразие частотных характеристик сигналов у различных семейств и видов в подотряде летучих мышей.
Общим для всех летучих мышей является то, что локационные сигналы излучаются только в момент выдоха животного. На одном выдохе могут быть излучены один или серия сигналов, и частота дыхания тесно скоррелирована с частотой взмахов крыльев (Mohres, 1953; Schnitzler, 1971; Suthers et al., 1972).
Характеристики направленности излучения локационных сигналов
Как уже отмечалось выше, у летучих мышей имеются два канала для излучения локационных сигналов — ротовая полость и ноздри. При этом представители одних семейств используют при излучении только рот, другие — только ноздри, а третьи, как например ушаны, широкоушки и некоторые виды из семейства Phyllostomatidae, в зависимости от ситуации способны переключать излучение с одного канала на другой.
Линейные размеры рта, по нашим измерениям, у крупных отечественных видов гладконосых летучих мышей (остроухая ночница, рыжая вечерница) составляют примерно 15—20 мм, а у мелких (нетопыри, усатые ночницы) — около 10 мм. Длины волн ультразвуковых локационных сигналов лежат в диапазоне 2.4— 11 мм. Таким образом, имеются благоприятные соотношения между размером «излучателя» и длиной волны. Это дает возможность ожидать определенную направленность излучаемого ультразвука, что подтверждается экспериментальными и теоретическими исследованиями, выполненными для нескольких видов летучих мышей (Griffin, 1958; Mohres, 1967а; Schnitzler, 1968; Simmons, 1969; Strother, Mogus, 1970; Griffin, Hollander, 1973).
Направленность излучения эхолокационной системы у летучих мышей (как и любой радиолокационной станции) необходимо учитывать при определении ее чувствительности, разрешающей способности и помехозащищенности. Поэтому одной из задач наших исследований было установление диаграммы направленности излучающего аппарата у двух видов — остроухой ночницы и большого подковоноса.
Снятие точной диаграммы направленности излучения у гладконосых летучих мышей сопряжено с различными трудностями, если учитывать амплитуду и частотную модуляцию в импульсах, а также различную степень раскрытия ротовой полости. Кроме того, для ЧМ-импульса получается 96
пульсирующая диаграмма, ширина которой будет меняться в сйлу разйогб затухания той или иной частотной компоненты. Поэтому установление какой-то усредненной диаграммы в этом случае, по-видимому, лишено смысла и можно говорить лишь о диаграммах для определенных частот. У остроухой ночницы мы снимали диаграмму направленности излучения на частотах 80, 45 и 35 кгц. Животное закреплялось в специальный станок так, чтобы его голова была неподвижна. Станок с мышью устанавливался на поворотном столике, имеющем градуированную шкалу. На расстоянии 1 м от рта животного устанавливался конденсаторный микрофон, который принимал ультразвуковые сигналы летучей мыши. Воспринятый сигнал усиливался и подавался на вход двухлучевого осциллографа, на экране которого регистрировались огибающая сигнала и линия девиометра, показывающая его частотное заполнение.
Первоначально микрофон устанавливался на одной прямой с головой животного, и производилась киносъемка излучаемых импульсов. Затем положение мыши относительно микрофона смещали вначале в одну сторону, потом — в другую и через каждые 10° производили замеры амплитуды импульсов или фотографировали их с экрана осциллографа. Все измерения производились в горизонтальной плоскости.
Получение ультразвуковых сигналов от летучей мыши осуществлялось путем стимуляции ее постоянным электрическим током небольших напряжений. При этом многие животные издавали исключительно низкочастотные звуки, но две остроухие ночницы в ответ на электрические раздражения излучали типичные ультразвуковые сигналы, длительность которых была порядка 4—5 мсек., с изменением частоты заполнения от 90 кгц в начале сигнала до 30 кгц в конце. Именно этих животных мы и использовали для снятия диаграммы направленности излучения.
Выяснилось, что диаграмма направленности излучения у остроухой ночницы на различных частотах различна и с увеличением частоты сужается (рис. 28). Ширина луча наибольшая при частоте 30 кгц (конечные частоты заполнения импульса) и составляет 90° на уровне 0.5. При средней частоте заполнения 45 кгц ширина угла излучения равняется 70° и на высоких частотах — 80 кгц — составляет 60° при том же уровне. Некоторая асимметрия диаграмм и появление лепестков у второй ночницы, возможно, объясняются тем, что интенсивность излучения не всегда сохранялась постоянной, а кроме того, иногда животные незначительно меняли положение головы.
В противоположность гладконосым летучим мышам, как уже отмечалось, подковоносы излучают локационные сигналы через ноздри, которые окружены сложно устроенными носовыми выростами. В составе кожистого носового листка выделяют три основные части (рис. 29). Верхняя его часть — ланцет — представляет собой узкий кожный вырост, заостренный на конце и отогнутый в виде козырька. Его функция полностью н8 исследована. Предполагается, что он играет значительную роль при определении направления воздушных потоков (Mohres, 1953). Средняя часть — седло — состоит из небольшого, расположенного между ланцетом и подковой гребня, круто выступающего вперед. Его образуют 2 пластины, сросшиеся между собой, с овальной впадиной посредине. Подкова — широкая пластинка спереди и с боков, охватывающая
97
ноздри, которые имеют форму «запятых». Посредине подковы имеется прорезь, которая делит ее на 2 симметричные части. Постоянство формы всего носового выроста в центральных его частях обеспечивается хрящами, а в остальных частях — эластичными жировыми тканями.
С помощью скоростной киносъемки Мёрес наблюдал сильные и слабые углубления средней части подковы в момент излучения
а
Рис. 28. Диаграммы излучения ультразвуковых сигналов у ночницы, снятые в горизонтальной плоскости для разных частот.
а — 35, 45 и 80 кгц; б — 35 кгц.
импульсов, а также одновременное сужение и расширение дугообразных отверстий ноздрей. Свободные края подковы начинают вибрировать в момейгг интенсивного излучения сигналов, и темп вибрации совпадает с темпом излучения импульсов.
О функции сложного носового листка у подковоносов строились различные предположения. Одни рассматривали его как высокочувствительный орган осязания, другие считали, что он служит для приема отраженных сигналов. После установления факта излучения ориентационных сигналов у подковоносов через ноздри
98
было выдвинуто предположение, что подкова функционирует как
направленный излучатель, тогда как остальные части носового листка не имеют существенного значения для эхолокации (Mohres, 1950, 1953). Однако исследования, проведенные в нашей лаборатории (Макаров, Соколов, 1970; Соколов, Макаров, 1971), показали,
что все они в разной степени участвуют в формировании диаграммы
направленности излучения у подковоносов в горизонтальной и
вертикальной плоскостях.
Расположение ноздрей у подковоносов разных видов коррелируется с длиной волны излучаемого ими ультразвука. Так, у большого подковоноса частота средней части локационного сигнала составляет 81—82 кгц, что соответствует длине волны 4.25 мм, а расстояние между центрами обеих ноздрей равняется половине этой величины, т. е. 2.1 мм. Следовательно, излучающую систему подковоноса можно рассматривать как синфазный излучатель, состоящий из двух когерентных источников, расположенных на расстоянии половины длины волны рабочей частоты (рис. 30). Такая
большого подковоноса.
Рис. 29. Схема носового листка
Л — ланцет, С — седло, П — подкова. Стрелками отмечены щелевидные ноздри.
система имеет максимум излучения в направлении, перпендикулярном к ее оси (АВ), так как волны, излученные от обоих источников в этом направлении, не имеют разности хода и, следовательно, складываются друг с другом в одинаковой фазе, т. е. амплитуда излучения удваивается. В других направлениях появляется некоторая разность хода, возрастающая по мере увеличения угла <р между принятым и нулевым направлениями. Наибольшего значения, равного Х/2, она достигает под углом <р=90°. В этом направлении волны взаимно гасятся.
Нетрудно рассчитать теоретическую кривую характеристики направленности такой системы. Если расстояние от излучателя до произвольно выбранной точки значительно больше половины длины волны, то лучи, идущие в данном направлении от каждого источника, можно считать параллельными, т. е. значение разности хода будет равно величине отрезка АС (рис. 30). Величина интенсивности излучения под углом ср по отношению к норме в этом слу-нде определяется из соотношения
99
2
где I — расстояние между источниками; С — скорость звука; о) — частота (рад./сек.). Рассчитанная таким образом характеристика излучения для случая большого подковоноса представлена на рис. 31. Ширина ее на уровне 0.5 равна 60°.
Рис. 30. Схема интерференции волн ультразвука при излучении через два когерентных источника.
Сплошные линии — гребни волн; штриховые — впадины.
В случае, когда расстояние между двумя когерентными источниками меньше Х/2, диаграмма направленности излучения будет расширяться. Если расстояние между источниками больше Х/2, то, помимо основного лепестка, появятся боковые, соответствующие числу зон интерференции, а основной лепесток сузится.
Правильность теоретических расчетов была проверена экспериментальным путем. Методика оставалась той же, что и для остроухой ночницы, но использовались 2 микрофона, один из которых все время находился в 70 см от ноздрей животного и перемещался вместе с ним, другой оставался неподвижным. Характеристики излучения снимались путем одновременного измерения величины интенсивности сигнала на акустической оси и под некоторым углом к ней. Наблюдая на экране осциллографа изображения сигналов, принятых 2 микрофонами, с помощью аттенюатора выравнивали их по амплитуде. Величина затухания, вводимая аттенюатором, определяла величину ослабления сигналов для каждого ракурса. Применение 2 приемных трактов давало возможность нормировать интенсивность под любым углом относительно нулевого направления (направления максимума излучения) для каждого сигнала и позволяла производить измерение характеристик с точностью до 1° независимо от интенсивности излучения.
Ширина диаграммы направленности в горизонтальной плоскости при излучении локационных сигналов у большого подковоноса составила 55+2°. Диаграмма симметрична относительно аку-
ЮО
стической оси (рис. 31). Таким образом, экспериментальная диаграмма направленности близка к теоретической, что подтверждает правоту наших рассуждений по поводу механизмов ее формирования. Наблюдаемое различие в 5° можно объяснить фокусирующим влиянием подковы. Так, при закрывании одной ноздри характеристика направленности заметно расширяется, но в то же время остается значительно ^же (81 + 5°), чем от одного точечного источника, расположенного на плоскости. Излучение от одной
0°
180°
Рис. 31. Диаграммы направленности излучения большого подковоноса в горизонтальной плоскости.
1 — при излучении через одну ноздрю; 2 — расчетная диаграмма; 3 — у интактного животного.
ноздри равно по интенсивности излучению от другой, в 2 раза слабее суммарного излучения и направлено параллельно акустической оси в случае двух источников. Опыты показали, что удаление и верхней части седла не оказывает существенного влияния на ширину диаграммы направленности в горизонтальной плоскости.
Дополнительно у подковоносов снимались характеристики направленности излучения и в вертикальной плоскости. Диаграмма в этом случае оказалась резко асимметричной (рис. 32). Ширина основного лепестка составляла 44° на уровне 0.5, т. е. несколько $же, чем в горизонтальной плоскости. Вниз от него под углом 30° отходит лепесток, звуковое давление которого составляет 0.6— 0.7 звукового давления основного лепестка. Общая ширина диаграммы на уровне 0.5 равна 73+5°,
101
Отклонение направленности излучения вниз обеспечивается носовыми выростами — седлом и ланцетом, которые, кроме того, гасят верхние лепестки. Удаление этих выростов приводит к уменьшению амплитуды второго лепестка и появлению верхних лепестков, в результате диаграмма Становится симметричной (рис. 32). В то же время ширина основного лепестка остается без изменений. Как же можно представить себе механизм формирования диаграммы излучения в вертикальной плоскости? В этом случае она
1до°
Рис. 32. Диаграммы направленности излучения большого подковоноса в вертикальной плоскости.
1 — у интактного животного; 2 —у животного с залитыми верхними частями носовых* щелей; з — у животного после удаления седла и ланцета.
не может быть объяснена интерференцией от двух ноздрей, как в случае горизонтальной плоскости, так как в любой точке вертикальной плоскости волны от двух источников будут складываться в одинаковой фазе. Опыты подтвердили это — диаграмма излучения от одной ноздри совпадает с диаграммой интактного животного. Однако каждая ноздря представляет собой не точечный источник, а щель, верхний конец которой несколько расширен и открывается в углублении на границе между подковой и ланцетом. Между верхним и нижним концами щель сужена за счет складок основания седла. Длина ее от центра нижнего выхода до центра углубления на вершине составляет в среднем 2.85 мм. Таким образом, можно допустить, что каждая ноздря состоит как бы из двух когерентных источников, расположенных на расстоянии 2.85 мм.
102
Теоретический расчет показывает, что в этом случае ширина основного лепестка диаграммы за счет интерференции волн от двух точек составит 46° (на уровне 0.5). Полученная величина хорошо согласуется о экспериментальным значением. Заливка воском верхних частей носовых щелей привела к значительному расширению диаграммы, и разделение ее на лепестки исчезло (рис. 32).
Следовательно, формирование направленности излучения и в горизонтальной, и в вертикальной плоскостях происходит за счет интерференции от двух источников: в первом случае — от двух ноздрей, во втором — от двух точек каждой ноздри.
В упрощенном виде всю излучающую систему подковоносов можно рассматривать как четыре синфазных когерентных источника, расположенных по углам трапеции, на расстояниях, близких к половине длины волны локационного сигнала. Этим обеспечиваются хорошая направленность и высокая интенсивность излучения.
Носовые выросты окончательно формируют направленность излучения. Подкова несколько фокусирует излучение, является экраном, препятствующим распространению волн в каудальном направлении, и принимает участие наряду с нижней частью седла в оформлении щелей, в которые открываются ноздри. Верхняя часть седла частично отражает излучение и вместе с ланцетом участвует в формировании нижних лепестков в вертикальной плоскости. Ланцет, кроме того, гасит верхние лепестки.
Асимметрия направленности излучения в вертикальной плоскости, выявленная для подковоносов, имеет большое биологическое значение, позволяя животным постоянно контролировать высоту полета относительно данной поверхности.
Сравнение полученных данных для гладконосых и подковоносых летучих мышей показывает, что при одних и тех же частотах (80 кгц) измерения ширины диаграммы направленности излучения в горизонтальной плоскости оказываются близки. Однако у подковоносов за счет более совершенных механизмов ее формирования интенсивность излучения примерно на два порядка превосходит таковую у гладконосых летучих мышей. Ничего не известно о диаграмме направленности в вертикальной плоскости у гладконосых. Мы предполагаем, что контроль за высотой полета над землей у них может осуществляться благодаря расширению диаграммы на низких частотах в конце импульса. Кроме того, пульсирующая от импульса к импульсу диаграмма ЧМ-сигнала, возможно, обеспечивает летучим мышам лучшее разрешение объектов по расстоянию и их расположению относительно направления полета.
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ОСОБЕННОСТИ СЛУХОВОГО АНАЛИЗАТОРА ЛЕТУЧИХ МЫШЕИ
Функцию приемной антенны эхолокатора у летучих мышей выполняет слуховой анализатор. В его же центрах осуществляется полный анализ той информации, которую получает животное в виде отраженных ультразвуковых сигналов. Следовательно, его можно рассматривать в качестве приемного и анализирующего отдела эхолокационной системы.
Вопрос/) том, с какими морфологическими особенностями слуховой системы коррелирует способность летучих мышей воспринимать ультразвук, долгое время оставался неясным. Как отмечает Зворыкин (1959), «в умах исследователей укоренилось представление о существовании в данном вопросе парадоксального разрыва между функцией и формой. . . Казалось непонятным, каким образом примитивная в морфологическом отношении кора летучих мышей и значительно менее сложная, чем у человека, улитка могут определять восприятие уникальных по своей частоте ультразвуков, и наоборот, наличие у человека максимальной по своей сложности и совершенству слуховой коры и кортиева органа сочетается с отсутствием способности воспринимать ультразвуки» (с. 22).
За последнее десятилетие различные отделы слухового анализатора летучих мышей подверглись тщательному морфологическому изучению, и на основании обобщения результатов этих исследований удается выявить те специфические особенности, которые развились у этих животных в связи с переходом к ультразвуковой локации.
Наружное ухо
Ушная раковина у летучих мышей отличается от таковой других млекопитающих не только большими размерами относительно размеров головы, но и более сложным строением (Web
104
ster, 1966). Характерной особенностью летучих мышей, которой не обладают другие животные, является наличие хорошо развитого козелка (tragus) — узкого кожистого выроста копьеобразной или булавовидной формы, поднимающегося от основания ушной раковины и находящегося на пути звуковых и ультразвуковых волн. Исключение составляют лишь некоторые семейства рукокрылых (например, подковоносы), у которых козелок отсутствует. Это обстоятельство может иметь значение для акустических способностей животных, однако функция козелка остается пока неясной.
Размеры ушной раковины, ее форма и строение варьируют у разных представителей рукокрылых (Dobson, 1878; Grasse, 1955; Walker, 1964; Rosevear, 1965). По-видимому, величина ушной раковины обратно пропорциональна интенсивности излучаемых животными акустических сигналов. Так, например, длиннокрылы используют для эхолокации сигналы, при излучении которых, по данным Новика (Novick, 1958а), звуковое давление достигает 1000—2000 дин/см2, и их ушные раковины настолько малы, что почти не выступают над головой. Высота уха у длиннокрыла составляет всего 12 мм. У ночниц звуковое давление при излучении импульсов составляет 60—100 дин/см2. Ушные раковины этих летучих мышей средней величины и четко выделяются над головой животных. Так, у остроухой ночницы высота ушной раковины составляет в среднем 23.5 мм и выступает над головой животного на 18 мм. И наконец, у ушана величина ушной раковины равна или даже превышает длину тела животного (высота ушной раковины 35—42 мм при общей длине черепа 18 мм). Это животное излучает еще менее интенсивные сигналы — 30—35 дб относительно нулевого уровня.
Непосредственно под козелком на расстоянии 1.85 мм от его основания (промеры относятся к остроухой ночнице) лежит наружное отверстие слухового прохода. Длина слухового прохода от начала барабанной перепонки составляет 1.85 мм. Наружный слуховой проход имеет в сечении форму эллипса диаметром в начале 2.25x1 мм, в середине 1.8X 1.5 мм, а в конце, у барабанной перепонки, 1x1 мм. Таким образом, наружный слуховой проход имеет форму воронки.
Функция ушной раковины у летучих мышей, вероятно, подобна функции акустической линзы и заключается прежде всего в фокусировании акустических «лучей» в наружный слуховой проход, и этот эффект оказывается большим при высоких частотах, при малой длине звуковой волны. В связи с тем что ушные раковины разведены несколько в стороны, можно предположить, что направление максимальной акустической чувствительности у летучих мышей не строго прямолинейно.
Вероятно, существенное значение имеет взаимное расположение ушных раковин, К этому пункту мы еще вернемся и обсудим
105
его в связи с нашими электрофизиологическими данными, касающимися механизмов пространственного слуха.
Ушная раковина выполняет у летучих мышей и другую функцию — предохранительную. У всех млекопитающих ухо снабжено механизмами, предохраняющими его от сверхсильных звуков. Такое предохранение необходимо для чувствительных структур улитки, для ее волосковых клеток, так как эти клетки повреждаются сверхсильными раздражителями (Винников, Титова, 1961; Davis, 1964; Wever, 1966).
Вивер и Вернон (Wever, Vernon, 1961а) обнаружили в наружном ухе у летучих мышей два особых защитных механизма, которые вместе с сокращениями мышц среднего уха обеспечивают надежное предохранение улитки от сверхсильных раздражителей.
Первое из них — складывание ушной раковины. Этот эффект можно наблюдать при подаче очень сильного звука. Животное реагирует мгновенным наклоном вперед верхней половины ушной раковины. Существуют и другие движения ушной раковины, например резкое отклонение назад, однако, по мнению Вивера и Вернона, сгибание ушной раковины вперед обеспечивает наибольшее предохранение уха.
Второе предохранительное устройство в наружном ухе летучей мыши — закрытие наружного слухового прохода. Это происходит за счет плотного вхождения в слуховой проход полулунного бугорка, расположенного в 1.5 мм от барабанной перепонки на боковой стенке прохода.
При воздействии на ухо громкими звуками под действием соответствующих мышц полулунный бугорок закрывает вход в слуховой проход подобно пробке. При одновременном сгибании ушной раковины и закрытии наружного слухового прохода ослабляющий эффект на сильные раздражения очень значителен и колеблется в основном между 40—60 дб для звуков частотой выше 1000 гц и даже достигает 70 дб при 40 кгц.
Таким образом, защитные механизмы наружного уха, а также среднего уха, которые будут рассмотрены ниже, надежно предохраняют слуховой аппарат летучих мышей от нежелательных действий сильных звуковых и ультразвуковых раздражителей. Описание приводящей мускулатуры ушных раковин, обеспечивающих им большую подвижность, что имеет особенно важное значение при эхолокации у подковоносов, а также подробные сведения об иннервации наружного уха летучих мышей можно найти в работах ряда авторов (Macalister, 1872; Stanek, 1933; Schneider, Mohres, 1960; Schneider, 1961). -
Среднее ухо
Основная функция среднего уха — обеспечение акустической связи элементов внутреннего уха с воздушной средой. Это выполняют слуховые косточки среднего уха: молоточек,
106
наковальня и стремечко, связанные с одной стороны с барабанной перепонкой, а с другой — с овальным окном улитки. «Косточковый аппарат» производит усиление акустического давления сигналов, передаваемых из воздуха через барабанную перепонку. Это явление подробно исследовано в слуховой системе человека, но есть основания полагать, что аналогичные отношения свойственны и слуховой системе рукокрылых.
Морфология среднего уха у летучих мышей подробно описана в ряде работ (Hyrtle, 1845; Bondy, 1907;Stan6k, 1933; Henson, 1961), Дополнительное описание слуховых косточек представлено еще в четырех работах (Doran, 1878; Wassif, 1946, 1948; Tepaske, 1964), причем в последней из них слуховые косточки описаны у представителей всех 17 семейств летучих мышей, включая 77 родов.
При рассмотрении строения среднего уха у летучих мышей выявляется ряд существенных особенностей различных элементов, входящих в его состав. Небольшие размеры барабанной перепонки и особенно pars tympanicus, более вертикальное ее расположение по сравнению с другими млекопитающими, уменьшение размеров и утончение слуховых косточек, наличие глубоких выемок в них, соединение молоточка и наковальни под острым углом с наименьшими потерями при передаче энергии — вероятно, все это благоприятствует проведению высокочастотных звуков во внутреннее ухо (Wassif, 1946, 1948, 1950; Henson, 1961, 1970b).
Кроме функции обеспечения акустической связи и увеличения акустического давления аппараты среднего уха выполняют еще одну функцию — уменьшение величины энергии поступающего во внутреннее ухо при слишком большом уровне сигнала. Достигается это изменением сокращения мышц среднего уха, которые у летучих мышей получают большое развитие.
Тимпанальная мышца (m. tensor tympani) натягивает барабанную перепонку и у летучих мышей имеет веретенообразную форму, а сухожильный материал ее составляет лишь 16% объема всей мышцы, тогда как у кошки она состоит из центрального тяжа сухожильных волокон, которые занимают почти весь ее объем. Мышца стремечка (m. stapedius) удерживает косточковый аппарат среднего уха на месте и у летучей мыши по форме конусообразная, тогда как у кошки имеет форму веера.
Мышца, натягивающая барабанную перепонку у летучей мыши, имеет мышечную массу 0.142 мг, а мышца стремечка — 0.188 мг. Три слуховые косточки вместе с барабанной перепонкой весят 0.170 мг. Соответственно отношение масс барабанной мышцы к косточковому аппарату равно 0.82. Это же отношение у стре-мечковой мышцы составляет 1.1. В то же время, например, у кошки собственный вес мышцы, натягивающей барабанную перепонку, 5.81 мг, а стремечковой мышцы — 1.67 мг. Косточки и барабанная перепонка весят 12.4 мг. Отсюда отношение массы мышцы,
107
йатягивающей барабанную перепонку, к Массе косточкового аппарата у кошки составляет 0.47, т. е. почти в 2 раза меньше, чем у летучей мыши, а это же отношение для стремечковой мышцы равно 0.14.
Необычно большие размеры мышц среднего уха обусловлены сложными функциональными отношениями с барабанной перепонкой и косточковым аппаратом. Восприятие и проведение ультразвуков требуют от мышц создания высокого напряжения барабанной перепонки и системы слуховых косточек, а предохранение внутреннего уха от надпороговых сверхинтенсивных звуков, наоборот, их расслабления. Эти операции происходят в сотые доли секунды, а кроме того, высказывается предположение, что внутри-ушные мышцы обеспечивают настройку системы на определенные частоты (Blair, 1964).
Из всего вышесказанного ряд авторов (Griffin, 1958; Henson, 1961) заключает, что восприятие высокочастотных звуков у летучих мышей определяется в большей степени механизмами передачи вибрационной энергии через среднее ухо, нежели свойствами их внутреннего уха.
Уже в 1945 г. Хартридж высказал мысль о том, что ухо летучих мышей нуждается в предохранении от ею же издаваемых громких криков, для того чтобы затем услышать слабое эхо. Хартридж был убежден, что правильно рассчитанное сокращение мышц среднего уха обеспечит такое предохранение. В то время не было представлено никаких доказательств в пользу ;этой гипотезы, кроме аналогии с радарными и сонарными установками, в которых принимающее устройство включается в момент посылки сигнала.
Примерно в то же время Галамбос (Galambos, 1942а) при исследовании кохлеарных потенциалов у летучих мышей выявил интересные закономерности. Когда подавался интенсивный тон перед ухом анестезированного животного (летучей мыши), кохлеарные потенциалы были максимально высоки во время первых нескольких секунд, после чего их амплитуда обычно падала на 20—50%. Этот эффект исчезал совсем, когда летучая мышь получала дозу кураре, парализующего связи между моторными нервами и внутренними мышцами. Когда же анестезированная летучая мышь умертвлялась инъекцией гистологического фиксажа в кровяное русло (при этом парализовались соответствующие центры мозга, и таким образом устранялся рефлекс ушных мышц), получалось временное увеличение амплитуды кохлеарных потенциалов. Этот эффект наблюдался при частотах от 15 до 55 кгц. Поэтому был сделан вывод, что рефлекс мышц среднего уха у летучих мышей действительно обнаруживается на всем диапазоне частот, имеющихся в их ориентационных импульсах.
Однако долгое время гипотеза Хартриджа как таковая все же оспаривалась на том основании, что рефлекс внутриушных мышц 108
из-за его «центральной» инерции (инерции сократитёльных аппаратов самих мышц) не может эффективно перекрыть вход системы при локационном крике и затем сразу же полностью открыть этот вход для приема слабого эха.
Эффективное участие внутриушных мышц у летучей мыши в защите ее улитки от собственных локационных криков было экспериментально показано Хенсоном (Henson, 1964,1965). В хронических опытах на летучих мышах вида Tadarida brasiliensis mexicana регистрировались микрофонные потенциалы улитки и одновременно отводились потенциалы действия стремечковой мышцы среднего уха. При этом обнаружилось, что названная мышца активизируется при излучении каждого импульса. Начало этой мышечной реакции предшествует ему на 4—10 мсек., ее максимум совпадает с моментом излучения, а затем мышца очень быстро расслабляется (за 10 мсек.).
Ясно, что описанное сокращение мышцы не представляет собой рефлекса на излучаемый импульс. Его можно рассматривать как процесс, идущий параллельно с центральной активацией гортанного аппарата.
Описанные сокращения стремечковой мышцы (без участия барабанной мышцы), судя по микрофонным эффектам улитки, сильно (на 12—22 дб) ослабляют влияние излучаемого сигнала на вход слуховой системы. Что касается приходящего вслед за криком эха от окружающих предметов, то оно в этих условиях воспринимается улиткой практически без ослабления. Такое положение имеет место при относительно редком ритме следования локационных импульсов (до 50 в 1 сек.). При излучении сигналов с большей частотой следования защитная функция слабее. А при частоте сигналов 140 в 1 сек. сокращение мышцы становится непрерывным от начала до конца серии излучаемых сигналов. При этом прием эха от окружающих предметов ослабляется в равной мере, как и локационных импульсов (исключение составляет только последнее эхо, воспринимающееся незащищенным входом слуховой системы).
Внутреннее ухо. Микрофонные потенциалы улитки
Внутреннее ухо у летучих мышей обладает существенными структурными особенностями по сравнению с таковыми у остальных млекопитающих. Часть кохлеарного аппарата, расположенная ближе к среднему уху и имеющая отношение к наиболее высокочастотным сигналам (ультразвукам), у летучих мышей необычайно велика. Круглое окно здесь открывается в канал внутреннего уха иначе, чем у других млекопитающих, не в высокочастотном конце кохлеарного аппарата, а почти на миллиметр дальше вдоль его первого поворота.
Сведения о строении улитки рукокрылых содержатся в много-
109
численных работах (Hyrtle, 1845; Beauregard, 1894а, 1894b; Gray, 1907; Iwate, 1924; Zimmermann, 1926; Stanek,1933; Turkewitsch, 1933a, 1933b, 1935, 1937; Ikeda, Yokote, 1939; Stefanelli, 1944a, 1944b; Inuma, 1958; Pye A., 1964, 1966a, 1966b, 1967, 1970, 1973; Henson, 1970b; Firbas, Welleschik, 1973, и др.). Ее размеры и формы широко варьируют у летучих мышей, но в целом размер улитки относительно размера тела у представителей Microchiroptera значительно больший, чем у Мegachiroptera и других млекопитающих. Улитка поистине огромна у настоящих и ложных подковоносов и некоторых видов Pteronotus. У подковоносов, например, длина улиткового канала составляет примерно 20 мм, т. е. почти 2/3 его длины у человека. Число витков улиткового канала у большинства видов летучих мышей варьирует от 2.5 до 3.5. Наибольшего развития достигает базальный виток, расположенный ближе к среднему уху и имеющий прямое отношение к восприятию высокочастотных звуков. Хенсон (Henson, 1970b) отмечает, что базальный виток достигает максимального размера у тех летучих мышей, которые используют для эхолокации продолжительные сигналы с постоянной частотой заполнения в средней части.
У этих же видов обнаружены характерные изменения ширины базилярной мембраны, формирующей основу улиткового канала. Если у большинства летучих мышей, подобно всем другим млекопитающим, она прогрессивно расширяется от базального витка до апикального, то у подковоносов ширина базилярной мембраны около овального окна составляет 100 мкм, затем сужается до 80 мкм во втором витке и вновь расширяется, достигая вблизи геликотермы 120 мкм. У многих летучих мышей, но не у всех, на базилярной мембране в базальном витке имеются два дополнительных утолщения. У Rhinolophidae и Hipposideridae они достигают относительно крупных размеров — 20—50 мкм, тогда как у других видов выражены слабее. У летучих мышей и некоторых других млекопитающих эти утолщения встречаются в той части улитки, которая связана с восприятием тонов, имеющих наибольшее биологическое значение для животных. Так, например, у одного вида крыс наилучшая чувствительность слуха выявлена на низких частотах — 1—3 кгц и утолщения на базилярной мембране у них развиваются в апикальном (низкочастотном) витке улитки (Pye, 1966а, 1966b; Wever, 1966). В то же время настоящие и ложные подковоносы используют высокочастотные локационные сигналы с небольшой частотной модуляцией и имеют утолщения базилярной мембраны в основном витке улитки. Функциональная значимость их пока остается неясной.
В кортиевом органе рукокрылых существенных изменений не обнаружено. Правда, А. Пай (Руе, 1966а, 1967) было отмечено, что клетки Клаудиса у эхолоцирующих летучих мышей достигают значительно большей величины по сравнению с таковыми у неэхолоцирующих крыланов (100 мкм против 30—40). По другим 110
показателям внутреннее ухо летучих мышей не проявляет специализации к ультразвуковому диапазону воспринимаемых частот.
О деятельности этого отдела слухового анализатора обычно судят по так называемым улиточным потенциалам, в частности микрофонным потенциалам, точно повторяющим по форме действующий на ухо акустический сигнал (Wever, 1966). Последние возникают в волосковых (рецепторных) клетках кортиева органа и, по-видимому, имеют ближайшее отношение к раздражению чувствительных окончаний слухового нерва (Радионова, 1966; Wever, 1966).
Особенность микрофонных потенциалов у летучих мышей состоит в том, что они регистрируются не только в ответ на звуки, но главным образом в ответ на ультразвуки.
В 1942 г. микрофонные потенциалы улитки (МПУ) разных видов гладконосых летучих мышей изучал Галамбос (Galambos, 1942а). Он обнаружил МПУ в широком диапазоне — от 30 гц до 98 кгц (предел используемой аппаратуры), с максимумом амплитуды в районе 10—30 кгц и дополнительным ее повышением в районе 30—50 кгц.
Недавно эти исследования были повторены (Dalland et al., 1967) на 7 летучих мышах вида Eptesicus fuscus. Авторы, применяя длительные чистые тона, обнаружили МПУ в диапазоне от 0.3 до 100 кгц. Приняв условно за пороговую величину ответа МПУ в 1 мкв, они вычертили соответствующие частотно-пороговые кривые (ЧПК). На этих кривых минимум порога (максимум чувствительности) получился в районе 20—40 кгц, с дополнительным понижением около 60 кгц.
Сравнивая МПУ у летучих мышей и у других млекопитающих, эти же авторы показали, что пороги получения МПУ у летучих мышей выше (хуже), чем у кошек, на 20 дб. Однако пороги МПУ летучих мышей (для амплитуды 1 мкв) оказались несоответствующими порогам восприятия ультразвуков этими животными в естественных условиях (измеренным по условнорефлекторным методикам). В последней случае пороги гораздо ниже, особенно в высокочастотном (ультразвуковом) районе. Таким образом, МПУ или во всяком случае обычная методика регистрации МПУ (под наркозом), видимо, не могут дать точной характеристики натуральной чувствительности слухового анализатора летучих мышей. Впрочем, данные, полученные в методике регистрации МПУ у других животных, необходимо воспринимать также критически.
В той же работе анализировалась зависимость амплитуды МПУ от интенсивности и частоты акустических сигналов. Было найдено, что МПУ плавно возрастает при усилении сигналов в диапазоне до 70 дб. Максимальные амплитуды МПУ обнаруживались в районе "от 50 до 90 кгц.
111
У летучей мыши, пороги акустического восприятия которой были заранее измерены, авторы сравнили амплитуду МПУ на порогах для разных частот. При этом оказалось, что на разных частотах такие амплитуды МПУ различны. Эти данные авторы склонны объяснять тем, что связь между величиной МПУ и «пороговым» уровнем импульсации слухового нерва, вообще говоря, может быть неодинаковой для разных районов улитки. Возможно, что это действительно так. Но все же, как нам кажется, пока нельзя исключить и другого возможного объяснения описанных несоответствий: разными условиями исследования параметров МПУ и порогов слухового восприятия. И здесь надо иметь в виду не только то, что отведение МПУ осуществляется в условиях наркоза и иммобилизации животного, но и то, что отведение МПУ от круглого окна вряд ли обеспечивает равноценную регистрацию колебаний потенциалов, порождаемых в разных частях улитки. При этом не исключено, что особое положение круглого окна в улитке летучей мыши не улучшает, а ухудшает отведение потенциалов «высокочастотной» части ее кортиева органа.
Определяя пороги МПУ, Вивер и Вернон (Wever, Vernon, 1961b) исследовали угловую направленность входа слуховой системы Myotis lucifugus. При этом естественно обнаружилась наилучшая чувствительность к сигналам, направленным прямо в исследуемое ухо. При 60° смещения от этого направления потеря чувствительности составляла 5—7 дб. При подаче сигналов сзади от летучей мыши потеря чувствительности равнялась 23 дб. Таким образом, выявилось, что направленность приема на входах слуховой системы невелика, но вполне определенна. Экранирующее действие ушных раковин в связи с приведенными данными, видимо, можно оценивать как весьма слабое; это позволяет полагать, что основное значение в создании некоторой направленности приема акустических сигналов для каждого из ушей имеет экранирующее действие головы летучей мыши.
Известно, что, помимо микрофонных потенциалов, в улитке млекопитающих при достаточной интенсивности высокочастотных сигналов регистрируются еще и так называемые суммирующие потенциалы, как бы вычерчивающие огибающую стимула. Эти явления обнаруживаются и в улитке летучих мышей. Нам, в частности, их удавалось записывать от круглого окна у Myotis oxygnathus в ответ на ультразвуки с частотой 60—80 кгц.
В заключение надо заметить, что, несмотря на отмеченные особенности, характер биоэлектрических ответов улитки летучей мыши на адекватные стимулы вполне подобен наблюдаемому у других млекопитающих (Wever, 1966). Это позволяет думать, что механизм рецепции ультразвука у данных животных тождествен механизму звуковой рецепции у всех млекопитающих.
Что касается стационарных биоэлектрических явлений в улитке рукокрылых, таких как эндокохлеарный потенциал, а также мем
112
бранные потенциалы рецепторных клеточных элементов, то о них пока ничего не известно, и вопрос об их знаке и величине ждет экспериментального разрешения.
Проводниковый отдел
В литературе уже давно появлялись отдельные данные, правда довольно разрозненные и весьма неполные, касающиеся некоторых звеньев слуховых подкорковых центров летучих мышей (Hoffmann, 1908; Valeton, 1908; Polyak, 1926а; Hornet, 1933). Но исследователи того времени прочно стояли на позициях тактильной теории и ориентировку летучих мышей в пространстве относили к исключительному развитию поверхностного осязания. Попытка дать более полное описание слуховых подкорковых образований по количеству звеньев была предпринята немецким исследователем Шнейдером (Schneider, 1957). Тем не менее, как отмечает Зворыкин (1959), описания данного автора излишне кратки и содержат серьезные ошибки в идентификации звеньев слуховой системы подкорковых центров. В последнее время появились более полные и точные морфологические работы, позволяющие представить проводящие пути слухового анализатора рукокрылых.
Афферентные проводящие пути слухового анализатора у летучих мышей соответствуют общему плану их строения у млекопитающих. Первый уровень в восходящем слуховом пути составляют биполярные клетки спирального ганглия (ganglion spirale), являющиеся нейронами первого порядка. Их периферические отростки входят в кортиев орган и иннервируют волосковые клетки. Центральные отростки этих клеток выходят во внутренний слуховой проход и формируют слуховой нерв, представляющий слуховую ветвь VIII пары черепно-мозговых нервов. Слуховой нерв в продолговатом мозгу оканчивается на клетках кохлеарных ядер. Кохлеарные ядра представлены дорзальным ядром (nucleus со-chlearis dorsalis) или слуховым бугорком (tuberculum acusticum) и двумя вентральными — передним (nucleus cochlearis ventralis anterior) и задним (nucleus cochlearis ventralis posterior). Поляк (Polyak, 1926a, 1926b) считает, что у летучих мышей кохлеарные ядра занимают необычное экстрамедулярное положение и у некоторых видов даже выдаются во внутренний слуховой проход, тогда как у крыланов и других млекопитающих кохлеарные ядра располагаются на дорзальной поверхности медулярной пластинки. Отмечено, что наибольшее развитие у летучих мышей получают вентральные кохлеарные ядра, а дорзальное ядро сравнительно небольшое, тогда как у крыланов наблюдается обратная картина (Henson, 1970а).
От клеток кохлеарных ядер начинается как контрлатеральный, так и ипсилатеральный восходящий слуховой путь, но основная
113
масса волокон переходит на противоположную сторону. Аксоны клеток вентральных кохлеарных ядер образуют трапециевидное тело (corpus trapezoideus) и оканчиваются как в верхней оливе, так и в ядрах трапециевидного тела ипси- и контрлатеральной сторон. Аксоны клеток слухового бугорка в виде дорзальных слуховых полосок (stria cochlearis dorsalis) проходят под дном IV желудочка, и часть их оканчивается в оливах своей и противоположной сторон, другая входит в состав боковой петли.
Комплекс верхней оливы у млекопитающих состоит из нескольких ядер, куда входят латеральная олива (oliva superior lateralis), медиальная олива, или акцессорное ядро (oliva superior medialis, или oliva superior accessoria) и группа преоливарных ядер. У летучих мышей в противоположность высшим млекопитающим и человеку медиальная олива выражена слабо или вообще отсутствует, тогда как латеральная олива получает наибольшее развитие и вместе с крупным медиальным ядром трапециевидного тела по существу формирует верхнеоливарный комплекс (Зворыкин, 1959; Harrison, War, 1962; Harrison, Irving, 1966; Irving, Harrison, 1967). Интересно отметить, что подобное соотношение в строении олив этими же авторами обнаружено и у эхолоцирующих китообразных.
Аксоны клеток слухового бугорка, верхней оливы и трапециевидного тела образуют мощный пучок — латеральную петлю, которая направляется вперед и вверх от оливы и подходит к задним буграм четверохолмия, где в основном и заканчивается (рис. 33). Лишь небольшая часть волокон направляется во внутреннее коленчатое тело. По данным Зворыкина (1959), у летучих мышей, помимо вентрального и дорзального ядер латеральной петли, больших размеров достигает и интермедиальное ядро, дифференцируемое на две самостоятельные клеточные группы.
Заднее двухолмие (corpora quadrigemina posteriora) является высшим подкорковым слуховым центром и у летучих мышей располагается поверхностно между корой и мозжечком. По данным Никитенко (1971), относительная величина заднего двухолмия у летучих мышей почти в 100 раз превышает таковую у грызунов. Большие размеры сочетаются с увеличением количества в нем клеточных элементов и густотой их расположения по сравнению с аналогичными образованиями у других млекопитающих. Поляк (Polyak, 1926b) считал, что латеральные части задних холмов построены по слоистому принципу и напоминают строение коры. Однако результаты, полученные Зворыкиным (1959, 19636), полностью отрицают это. Автор отмечает ядерный принцип их построения и выделяет в составе каждого холма четыре клеточные группы. В каудальной части холмов, примыкающей непосредственно к мозжечку, располагается группа 1. Клеточная группа 2 занимает дорзо-латеральную часть заднего холма; группа 3 — всю центральную часть холма и вентрально граничит
114
с покрышкой среднего мозга. В месте соединения холмов располагается клеточная группа 4. По данным Суховской (1966), нейронный состав заднего двухолмия у 7 видов гладконосых летучих мышей остается тем же самым, что и у других животных, но количество нейронов в каждой клеточной группе значительно увеличено.
Обращает на себя внимание тот факт, что в различных клеточ ных группах заднего двухолмия летучей мыши, а особенно в кле-
Рис. 33. Срез через заднее двухолмие среднего мозга ночницы. Микрофот.
1 — задний холм; 2 — ядра латеральной петли; 3 — верхняя олива; 4 — трапециевидное тело; 5 — вентральное кохлеарное ядро.
точных группах 2 и 3, содержится большое количество звездчатых клеток, относящихся к типу ассоциативных нейронов. Увеличенное количество ассоциативных нейронов в заднем двухолмии летучей мыши указывает на сложную деятельность этого отдела мозга. Аксоны клеток заднего холма выходят на боковую поверхность и в составе ручки направляются во внутреннее коленчатое тело (corpus geniculatum mediale), где и заканчиваются. Волокна от внутреннего коленчатого тела оканчиваются в слуховой области коры (Fuse, 1920; Polyak, 1926а, 1926b).
Во многих из приведенных работ отмечается, что подкорковые слуховые центры у летучих мышей обнаруживают значительную гипертрофию (рис. 33) по сравнению с аналогичными центрами у других животных (Tamura, 1950; Schneider, 1957; Зворыкин, 1959; Schober, 1959—1960; Taniguchi, 1959; Mann, 1960; Lutge-meier, 1962; Никитенко, 1969).
Зворыкин (1959) впервые дал количественную оценку всей системы подкорковых слуховых образований летучих мышей
115
Й сопоставлений с таковой у человека. Он звено за звеном исследовал развитие этих структур на непрерывных сериях срезов, окрашенных по Нисслю. Было показано, что относительные размеры слуховых подкорковых центров у летучих мышей в 6— 100 раз больше, чем у человека. Только относительные размеры внутреннего коленчатого тела у летучей мыши и у человека оказались почти одинаковыми. Однако в целом различия между летучей мышью и человеком настолько велики, что делать выводы из такого сравнения часто бывает затруднительно. Поэтому наибольший интерес представляет сравнение развития структур слухового анализатора летучих мышей с аналогичными у близкородственной группы отряда рукокрылых — крыланов, не пользующихся локацией (Liitgemeier, 1962). При сравнении процентного отношения размеров слуховых центров к стволу мозга у нескольких представи телей гладконосых летучих мышей и крыланов (табл. 6) наиболее существенные различия наблюдаются в размерах кохлеарного ядра, верхней оливы и задних холмов, хотя и другие отделы (кроме дорзального кохлеарного ядра) увеличены у летучих мышей по сравнению с таковыми у крыланов. Лютгемейер отмечает, что в целом слуховая система у Microchiroptera составляет 14—23% ствола мозга, тогда как у Megachtroptera — всего только 5.3—8.5%, причем значение 8.5% относится к представителю рода Rousettus, который единственный среди крыланов пользуется примитивной акустической локацией.
Таблица 6
Сравнительное соотношение слуховых центров у. рукокрылых (Liitgemeier, 1962)
Подкорковые слуховые центры
Род и вид DCN VCN so TN IC mGB
Microchiroptera
Pipistrellus pipistrellus 0.80 3.05 3.98 0.60 11.6 3.18
Myotis daubentoni 0.65 2.32 3.48 0.39 10.9 3.01
Eptesicus serotinus 1.07 2.39 2.95 0.63 10.65 2.31
Myotis myotis 1.16 1.76 2.15 0.39 10.23 2.20
Megachtroptera
Rousettus leschenaulti 1.35 0.90 1.01 0.30 3.69 1.31
Pteropus giganteus 0.73 0.42 0.58 0.22 2.46 0.90
Примечание. ДСМ — дорзальное кохлеарное ядро, VCN — вентральное кохлеарное ядро, SO — верхняя олива, TN — ядро трапециевидного тела, IC — задний холм, mGB — внутреннее коленчатое тело.
116
Слуховая область корь!
По степени неокортикализации мозга рукокрылыё занимают одно из наиболее низких положений в сравнительном ряду плацентарных млекопитающих (Wirz, 1954; Liitgemeier, 1962; Mangold-Wirz, 1966; Никитенко, 1969, 1971; Henson, 1970а). Неокортекс у летучих мышей составляет 31—36% общей коры те-ленцефалона и характеризуется отсутствием борозд и извилин на наружной поверхности больших полушарий (лиссэнцефалия). Несмотря на то что величина височных областей неокортекса
Рис. 34. Цитоархитектонические формации коры полушарий летучей мыши. (Никитенко, 1969).
ВО — обонятельные луковицы; F — лобная область; FP — задняя лобная область; Р — теменная область; I — инсулярная кора; Та, Тг, Тр — поля височной области; О — затылочная область; РСх — древняя кора; С А — переднее двухолмие; Ср — заднее двухолмие.
у летучих мышей развита лучше по сравнению с другими областями (прецентральной, постцентральной, теменной и затылочной) (Никитенко, 1971; Курепина, 1967; рис. 34), представительство слухового анализатора в ней выражено слабо (Brodmann, 1909; Zunino, 1911; Rose, 1912; Suga, 1965a). В отличие от высших млекопитающих в слуховой коре летучих мышёй отсутствует четкая цитоар-хитектоническая дифференциация на поля. Отмечаются трудности и в расчленении слуховой коры на отдельные слои, что находит свое отражение при сравнении результатов различных исследователей. Одни авторы (Зворыкин, 1959) выделяют в ней 3 комплекса клеток, другие — 4 слоя (Никитенко, 1969), третьи — 5 (Zunino, 1911) и, наконец, четвертые — 6 слоев (Brodmann, 1909; Rose, 1912). Исследования нашей лаборатории (Добрякова, 1968; Айра-петьянц, Константинов, 1970а, 19706; Бурикова, 1971; Айра-петьянц, Бурикова и др., 1972, 1973) показали, что эта область имеет классический шестислойный тип строения. Среди других областей коры она легко отличается большой шириной (около 1 мм), разреженностью IV слоя, наличием крупных пирамид в слое
117
V, горизонтальной ориентацией клеток VI слоя (рис. 35)< Помимо этих признаков, позволяющих гомологизировать слуховую область летучих мышей с таковой у вышестоящих отрядов млекопитающих, в ней проявляется ряд признаков, связанных с низким положением этой группы в филогенетическом ряду. Это прежде всего примитивное строение верхнего этажа, сходное с уровнем развития слуховой коры на ранних стадиях онтогенеза вышестоящих отрядов: большая ширина слоя I, плотность и
Рис. 35. Поперечный срез слуховой области коры головного мозга летучей мыши. Ниссль. (Микрофот. Н. В. Буриковой).
сосочковидность слоя II, необособленность слоя IV и большое количество клеток неопределенной формы. Разделение же на слои нижнего этажа коры не вызывает затруднений, и клеточные элементы V и VI слоев имеют более правильную, типичную форму. Подобные примитивные признаки отмечены и для центральных областей коры летучих мышей (Хачатурян, 1969; Курепина, Хачатурян, 1970).
Таким образом, в отличие от слуховых подкорковых центров в строении корковой зоны у эхолоцирующих летучих мышей не удается выявить каких-либо особых черт специализации, отражающих ведущее значение слуха в ориентации этих животных.
Вопрос о наличии и функциональной значимости эфферентных связей слуховой коры рукокрылых до недавнего времени не подвергался экспериментальному изучению, хотя такие знания дают
118
возможность лучше оценить роль коркового звена в регулировании процесса слухового восприятия. Лишь совсем недавно в нашей лаборатории Бурикова (1971) с использованием методики Наута— Гигакс проследила дегенерацию аксонов и претерминальных разветвлений в различных структурах мозга после разрушения клеток слуховой коры. Полученные данные показали, что у летучих
Рис. 36. Схема распределения дегенерирующих волокон в образованиях ствола мозга после удаления участков слуховой коры у летучей мыши.
as — сильвиев водопровод; ret — сетчатая формация среднего мозга; пг — красное ядро; РР — основание ножки мозга; gi — межножковый узел; cgm — медиальное коленчатое тело; cba — переднее двухолмие; cbp — заднее двухолмие; прр — собственные ядра моста; рст — средние ножки мозжечка; nllv — вентральное ядро латеральной петли; п11г-т — промежуточное ядро латеральной петли; nlld — дорзальное ядро латеральной петли; ntr — ядро трапециевидного тела; osl — латеральное ядро верхней оливы; osm — медиальное ядро верхней оливы; pol — латеральное преоливарное ядро; рог — медиальное преоли-варное ядро; nvc — вентральное слуховое ядро; prf — парафлокулюс мозжечка; v. IV — четвертый желудочек мозга. Заштрихованный участок показывает локализацию места повреждения. Точками отмечены места, где были обнаружены дегенерировавшие волокна.
мышей от слуховой коры идет ряд проекционных путей к медиальному коленчатому телу, передним и задним буграм четверохолмия, латеральной петле, комплексу ядер верхней оливы, вентральному кохлеарному ядру и мозжечку (рис. 36). Локализация окончаний нисходящих слуховых волокон коры у рукокрылых в основном совпадает с таковой у кролика, кошки, собаки и обезьяны. Однако в отношении как количества, так и локализации дегене-рированных претерминалей у летучих мышей выявлен ряд особенностей,.
119
Прежде всего не удалось установить наличия комиссуральных связей между слуховыми областями коры обоих полушарий. В отличие от других животных, у которых аксоны клеток слуховой коры обильно представлены в медиальном коленчатом теле и заднем двухолмии, у летучих мышей в этих образованиях дегенерация выражена очень незначительно. Зато основная масса кортико-фугальных путей адресуется к структурам, расположенным достаточно низко по ходу афферентных слуховых волокон, таким как верхняя олива и вентральное кохлеарное ядро. Это свидетельствует о наличии путей от коры до волосковых клеток, состоящих из двух нейронов.
На основании сделанного обзора становится совершенно очевидным, что слуховой анализатор у летучих мышей в ходе эволюции подвергся значительным перестройкам и усложнению на всех уровнях, за исключением корковой слуховой проекции. Сопоставление различных его отделов у эхолоцирующих летучих мышей с подобными отделами у представителей крыланов, пользующихся в основном зрением, и других неэхолоцирующих млекопитающих дает возможность сделать более обоснованный вывод, что данные перестройки анализаторной системы связаны с рецепцией и анализом звуков ультразвукового диапазона частот.
4
ОБЩАЯ ФУНКЦИОНАЛЬНАЯ СХЕМА НЕРВНОЙ РЕГУЛЯЦИИ ЭХОЛОКАТОРА
Мы сосредоточили свое внимание на изучении функ* циональной значимости слуховой области коры и задних холмов, так как первая представляет собой верхний этаж слуховой системы и играет важную роль в анализе сложных акустических сигналов у высокоорганизованных млекопитающих и человека (Гершуни, 1963, 1967; Бару, 1966, 1967, 1972; Бару, Карасева, 1971), тогда как заднее двухолмие является основным подкорковым центром слуховых рефлексов (Бехтерев, 1895; Ades, 1959; Беленков, 1965; Меринг, 1967). Для решения этого вопроса обычно используется сочетание метода экстирпации определенных центров слуховой системы с методом условных рефлексов. Считается, что после выключения того или иного отдела его функции обнаруживаются в специфических дефектах поведения. Несмотря на ряд недостатков, экспериментально-хирургический метод оказался наиболее плодотворным в сопоставлении с электрофизиологическими и морфологическими методами исследования. Правильное применение этого метода предполагает обязательное проведение измерений состояния слуховой функции до и после повреждения отделов системы не по одному, а с помощью нескольких адекватных тестов.
О функциональной роли слуховой области коры
Многочисленные экспериментальные исследования, выполненные в основном на человеке и хищных млекопитающих (кошки, собаки), позволяют считать, что слуховая кора не имеет решающего значения в анализе частоты и интенсивности стационарных сигналов, но играет важную роль в оценке более сложных признаков звуковых сигналов: длительности, формы огибающей, временной последовательности и ритмического узора, а также ча
121
стично в осуществлении реакций, основанных на бинауральном взаимодействии (Альтман, 1972; Альтман, Дубровский, 1972; Бару, 1972). Особенно следует подчеркнуть, что разрушение слуховой коры приводит к избирательному повышению порогов обнаружения и различения коротких звуков (равных или меньших 10 мсек.). Это может свидетельствовать о том, что слуховая кора играет существенную роль в обработке информации, связанной с переходным процессом, фиксацией начального момента действия звука и существенных изменений его амплитуды, что чрезвычайно важно для всех операций, требующих отсчета времени и анализа временных характеристик сигнала.
Однако сведения о том, какую функцию выполняет слуховая кора у животных, стоящих на более низких ступенях филогенетического развития, в литературе практически отсутствуют. Поэтому неизвестно, применимы ли к ним те выводы о роли слуховой коры, о которых мы только что говорили.
По отношению к летучим мышам этот вопрос особенно актуален, учитывая короткую длительность локационных сигналов (а следовательно, и приходящего эха), а также быструю последовательность их излучения.
Двустороннее выключение слуховой коры в наших опытах проводилось на нескольких представителях гладконосых летучих мышей и на больших подковоносах с использованием различных приемов выключения и различных проверочных тестов.
В первой серии опытов на 6 гладконосых летучих мышах (3 ушана и 3 усатые ночницы) исследовалась их способность преодолевать проволочные барьеры (диаметр проволок 0.6 мм) в норме и после функционального выключения слуховой коры и путем аппликации на нее гамма-аминомасляной кислоты (ГАМК) концентрацией 10—20% по методу Буреша (Константинов, 1965а, 1965в). В соответствии с многочисленными электрофизиологическими исследованиями (см., например: Батуев, Сытинский, 1962; Буреш и др., 1962) ГАМК оказывает временное депрессивное влияние на корковые нейроны в месте ее приложения.
В норме у всех подопытных животных уклонение от препятствий при пролете сквозь проволочные барьеры составляло 75.6 + ±2.6%. Аппликация ГАМК не вызвала никаких изменений в поведении летучих мышей. Характер полета на протяжении всего опыта оставался таким же, как и в норме при той же быстроте и маневренности. Уклонение от препятствий составляло 80+2.6%. Статистический анализ полученных данных показал, что разница в обнаружении и избегании проволочных барьеров интактными и оперированными животными статистически недостоверна.
Во второй серии опытов, проведенной нами на 8 остроухих ночницах, изучалось влияние обширной двусторонней экстирпации слуховой области коры на преодоление животными барьеров из проволок уменьшающегося от опыта к опыту диаметра (1.12,
122
0.55, 0.20 мм). Дополнительно на 4 остроухих ночницах и 2 больших подковоносах проверялось сохранение ранее выработанного рефлекса и дифференцировки при распознавании плоских мишеней после аналогичной операции (Константинов, Соколова, 1970). Границы удаления слуховой коры определялись в соответствии с электрофизиологическими данными (Suga, 1965а; Котеленко, Матюшкин, 1969). После окончания экспериментов производился морфологический контроль путем исследования срезов толщиной 45—60 мкм, полученных на замораживающем микротоме. Макроскопический анализ показал, что удаление слуховой и части при-
лежащих областей коры было произведено полностью, иногда с повреждением подлежащего белого вещества мозга. Границы удаления слуховой зоны коры показаны на рис. 37. Все эксперименты проводились на предварительно энуклеи-рованных’животных.
В табл. 7 представлены результаты пролетов интактных и оперированных остроухих ночниц сквозь
Рис. 37. Границы обширного удаления слуховой области коры больших полушарий у остроухой ночницы.
проволочные барьеры. Как видно из сравнения этих данных, точность реакции у летучих мышей, а следовательно, и пороговая чувствительность слуховой системы при обнаружении не изменялись, разница статистически не достоверна, что полностью сов-
Таблица 7
Влияние двустороннего удаления слуховых зон коры у остроухих ночниц на пролет барьеров из проволок
Диаметр проволоки, мм Интактные животные После экстирпации слуховой коры
п °/0 уклонения п % уклонения
1.12 1063 80.29+1.9 517 78.21+2.3
0.55 1003 77.36 + 1.7 789 69.96+3.9
0.20 517 59.17+3.6 717 53.05+2.2
Примечание, п — цбщее число пролетов.
123
падает с результатами первой серии, а также с данными Суги (Suga, 1969а), использовавшего в качестве теста для оперированных летучих мышей тот же критерий.
Более сложной, чем обнаружение и преодоление препятствий, является для летучих мышей задача распознавания объектов одного класса (например, квадратов), которые отличаются либо по размерам, либо по материалу, либо по степени изрезанности их кромок. Процесс эхолокационного распознавания, с одной стороны, включает механизмы условнорефлекторной деятельности, а с другой — тонкое сравнение отраженных сигналов от каждой мишени и по минимальным различиям их спектральных характеристик — выбор положительной (подкрепляемой кормом) мишени. Таким образом, использование данного теста позволяло выявить влияние слуховой области коры на: 1) сохранение положительного условного рефлекса; 2) осуществление дифференцировки как формы условного внутреннего торможения; 3) способность улавливать небольшие различия в спектральных характеристиках приходящих сигналов от каждой мишени (см. главу 12); 4) осуществление сложного двигательного акта — посадки на фигуру с точной корректировкой расстояния до нее по отраженному сигналу.
Опыты показали, что распознавание предъявляемых мишеней и формирование целостной поведенческой реакции как у ночниц, так и у подковоносов сохраняются после обширного двустороннего удаления височных областей коры и проявляются уже на следующий день после операции. В первые дни отмечены некоторые двигательные расстройства, особенно при посадке на выбранную мишень, которые полностью исчезали спустя 3—5 дней после операции. Не было замечено изменений в частоте следования и в структуре ультразвуковых локационных сигналов, издаваемых летучими мышами при распознавании мишеней в норме и после операции. Результаты этих опытов представлены в табл. 8. В качестве положительной мишени во всех случаях был использован дюралюминиевый квадрат площадью 196 см2. Отрицательные, или дифференцировочные, мишени, кроме последней, по площади были равны положительному квадрату. Все они, за исключением одной (квадрат из плексигласа), были изготовлены из дюралюминия, и их поверхности подвергались одинаковой обработке пескоструйным аппаратом.
В специальной серии экспериментов у 2 ночниц вырабатывались двигательный условный рефлекс на чистый тон 750 гц и дифференцирование положения источника звука (справа—слева), вслед за этим проводилось удаление слуховой коры. У другой группы животных выработка условного рефлекса производилась уже после аналогичной операции. Было показано, что ранее выработанный рефлекс на звук сохраняется у оперированных животных на до-операционном уровне и может быть выработан вновь при отсутствии слуховых зон коры,
124
Таблица 8
Распознавание мишеней летучими мышами до и после удаления височных областей коры
Дифференцировочная мишень До операции После операции
ночницы подковоносы ночницы подковоносы
п % распознавания п % распознавания п % распознавания п °/0 распознавания
Квадраты: изрезанный со всех сторон (7г=15 мм) 212 92.9+1.9 136 88.3 + 0.7 65 100 31 85.0 + 0.8
изрезанный сверху (А=15 мм) ИЗ 92.6 + 3.5 — — 33 — — —
то же (7г=3 мм) 208 100 77 89.3 + 2.9 41 95.9 + 4.2 29 83.9+0
с двумя отверстиями по диагонали 150 92.1 + 3.8 — — 37 88.9+4.8 — —
из плексигласа 42 92.2 + 1.2 53 91 + 1.4 34 96.7+3.4 28 96.4+3.6
площадью 400 см2 .... 36 91 + 1.2 — — 25 91.5 + 1.6 — —
Примечание, п — общее число посадок на мишени; h — высота зубца на изрезанном квадрате.
Представляло интерес выяснить, не изменится ли электрическая активность на звуковое раздражение в оставшихся областях коры мозга у оперированных летучих мышей, что было бы проявлением компенсаторных процессов после удаления слуховых зон. Такие данные имеются для других животных (кошки), когда вызванные потенциалы на звук появлялись к моменту восстановления условных рефлексов после экстирпации слуховой коры в вентральной части задней сигмовидной извилины (Thompson, Sindberg, 1960; Maruyama, Kanno, 1961).
После полного восстановления положительного условного рефлекса на тон 750 гц и выработки его заново при отсутствии слуховых зон под гексеналовым наркозом исследовалась электрическая активность оставшихся областей путем регистрации суммарных ответов с поверхности коры. Ни в одном из электрофизиологических опытов не было получено вызванных потенциалов в лобной, теменной и затылочной областях коры в ответ на звуковые и ультразвуковые стимулы.
На основании вышеизложенных данных мы приходим к заключению, что у летучих мышей замыкание звуковых условных рефлексов независимо от того, является ли условным сигналом посторонний звук или эхо собственных импульсов, может осуществляться без участия слуховой коры на уровне подкорковых образований.
О функциональной роли заднего двухолмия
Заднее двухолмие — важнейший слуховой центр, которому издавна приписывались рефлекторные функции. У летучих мышей, как было отмечено выше, этот отдел мозга отличается резко увеличенными размерами и густотой клеточных элементов по сравнению с другими млекопитающими и даже представителями неэхолоцирующих рукокрылых — крыланов. В центральном ядре заднего холма у летучих мышей, по данным Суховской (1965, 1966), обнаружено большое количество звездчатых (ассоциативных) нейронов, и здесь же наблюдается наибольшее количество синапсов, расположенных как на теле, так ина дендритах клеток, т. е. имеются аксо-соматические и аксо-дендритические связи. Все это указывает на богатство ассоциативных связей в задних холмах и дает основание предполагать сложную функциональную деятельность этого подкоркового центра.
В соответствии с электрофизиологическими исследованиями в задних холмах обнаруживается вертикальная тонотопическая локализация, т. е. нейроны, лежащие в глубине холма, реагируют на более высокие ультразвуки, чем поверхностные нейроны (Grinnell, 1963а; Васильев, 1968а, 19686). По данным нашей лаборатории (Айрапетьянц, Константинов, Матюшкин, 1968, 1969), в нейронной массе задних холмов могут быть выделены своеобразные «нейронные шкалы» для оценок, во-первых, частоты и силы ультра-126
зйуковых сигналов, во-вторых, интервалов между сигналами, например между излученным сигналом и его эхом, и, в-третьих, направления прихода сигналов. Следовательно, заднее двухолмие выполняет центральную афферентную функцию, т. е. участвует в окончательном анализе афферентной импульсации, который частично осуществляется в нижележащих подкорковых центрах проводникового отдела слухового анализатора. С другой стороны, работами многих авторов, выполненных на различных млекопитающих, показано, что заднее двухолмие является важным рефлекторным центром, посредством которого слуховые раздражители, поступающие по афферентным волокнам, вызывают различные двигательные реакции без участия коры (Бехтерев, 1895, 1906; Калинина, 1962,1971; Меринг, 1963,1964; Беленков, 1965,1969;Белен-ков и др., 1969, и др.). Этому способствуют многочисленные эфферентные связи холмов с другими мозговыми структурами (Ramon у Cajal, 1902; Gehuchten, 1902; Клосовский, 1946; Johnson, 1954; Altman, Carpenter, 1961; Huberg-Hansen, 1964, и др.). Так, выявлены волокна, идущие к ретикулярной формации, сетчатому ядру покрышки, ядрам моста, некоторым черепно-мозговым ядрам (IV и VI), красному ядру и ядрам латеральной петли. Найдено небольшое количество волокон текто-спинального пути и показана тесная морфологическая и функциональная связь между задним и передним двухолмиями. От последнего начинается главная часть текто-спинального, текто-бульбарного и текто-тегментарного путей, а также пути к ретикулярной формации. Однако полученные данные относятся к животным, стоящим на разных филогенетических уровнях, и вряд ли можно ожидать, что у них эфферентные связи заднего двухолмия будут идентичными. Тем более экстраполировать полученные результаты на летучих мышей — относительно низкоорганизованных млекопитающих, как это делалось до недавнего времени, следует с большой осторожностью. Поэтому в лаборатории Э. Ш. Айрапетьянца в плане комплексного изучения структурно-функциональной организации эхолокационных систем проведены специальные исследования, посвященные данному вопросу.
Н. В. Бурикова (1973) на непрерывных сериях срезов мозга остроухой ночницы, импрегнированных по методу Наута—Гигакс, изучала эфферентные связи заднего двухолмия путем прослеживания дегенерированных волокон после односторонней коагуляции холмов. Ею показано, что полное повреждение холма вызывало дегенерацию волокон двух основных путей. Первый из них направлялся к медиальному коленчатому телу, где оканчивался преимущественно в центральных и латеральных отделах. Второй путь связывал холм с латеральным и частично вентральными ядрами варолиева моста. Кроме того, выявлены прямые пути к средним и глубоким слоям переднего двухолмия, а также к оральным отделам ретикулярной формации среднего мозга. Во всех выше-
127
указанных образованиях ствола мозга дёгёнераЦия НоЛокон Наблюдалась только на оперированной стороне. После коагуляции холма прослеживались пучки фрагментированных волокон в комиссуре между холмами, оканчивающихся в центральном ядре и в вентральных отделах латеральной коры противоположного холма. Обращает на себя внимание отсутствие прямых путей к коре, и в частности к слуховой и двигательным ее областям, а также эфферентных связей заднего двухолмия с нижерасположен-ными слуховыми центрами, которые выявлены у других исследованных млекопитающих.
Однако отсутствие убедительных данных об участии задних холмов в осуществлении эхолокационного процесса, а также прямых экспериментов по исследованию способности летучих мышей после повреждения заднего двухолмия к пространственной ориентации на основе акустических сигналов побудило нас к проведению соответствующих исследований.
В опытах на 5 ушанах и 5 остроухих ночницах было показано, что экстирпация большей части заднего двухолмия вызывает целый ряд глубоких, необратимых двигательных расстройств и приводит к полной потере способности животных ориентироваться в пространстве с помощью эхолокации (Константинов, 19656, 1965в; Айрапетьянц, Константинов, 19676,1970а). Как и в опытах с удалением слуховой коры, в этих исследованиях производился морфологический контроль за степенью повреждения холмов.
Важно отметить исчезновение полета и нарушение координации движения при ходьбе: заваливание животного на бок то в одну, то в другую сторону, круговые движения головой и вращение на одном месте. Полностью исчезала способность осуществлять эхолокационный анализ пространства. Летучие мыши не могли обнаруживать препятствия при побежке и избегать столкновения с ними. Излучение локационных сигналов либо полностью прекращалось, либо, наоборот, наблюдалось непрерывное их издавание в быстром темпе, не связанное с ориентацией. Отмечалось исчезновение ориентировочных реакций на звуковой раздражитель, на что обращалось внимание и в опытах на более высокоорганизованных животных (Меринг, 1964; Беленков и др., 1969).
При частичном разрушении холмов, т. е. при разрушении II клеточной группы или латеральной коры с небольшим захватом клеток центрального ядра (рис. 38), полет у животных (ушанов) сохранялся, но скорость его и маневренность значительно снижались. Крупные препятствия, такие как стены, мебель, летучие мыши облетали успешно, однако пролет сквозь проволочные барьеры оказывался невыполнимым. Если в норме процент уклонений составлял в среднем 75.5 ±0.9, то после операции он не превышал 24.0 ±4.2. В то же время термокоагуляция поверхностных 128
частей задних холмов, т. е. только латеральной коры, не оказала влияния на качество полета и избегание препятствий у ночниц. Эти животные в норме при облете проволочных барьеров показали 81.6+1.0% уклонений, а после термокоагуляции — 81.5+4.0%, т. е. разрушение нервных клеток, лежащих ближе к поверхности холмов, по-видимому, не играет столь существенной роли в анализе отраженных сигналов, как нейронов центрального ядра. Это хорошо согласуется с тонотопической локализацией, обнаруженной в заднем двухолмии, при которой поверхностные нейроны реагируют на более низкие частоты, а также с морфологическими исследованиями Н. В. Буриковой, показавшей значительно более
Рис. 38. Различная степень разрушения заднего двухолмия у летучих мышей. а — норма; б — почти полное разрушение холмов; в — частичное разрушение центрального ядра; г — разрушение верхушек холмов (II клеточной группы). Черным цветом обозначены границы разрушения холмов.
слабые связи латеральной коры с медиальным коленчатым телом, ядрами моста и отсутствие проекций к передним холмам. Позднее Суга (Suga, 1969b) повторил наши опыты и получил аналогичные результаты при относительно полном и частичном разрушении задних холмов.
Наши данные продемонстрировали существенное, если не преобладающее, участие заднего двухолмия в осуществлении приспособительной деятельности летучих мышей в целом и эхолокационном анализе в частности. Нормальное функционирование эхолокатора оказалось возможным лишь при сохранении целостности центрального ядра заднего холма. Особенно следует подчеркнуть, что при его разрушении резко нарушаются произвольные движения животных на звуковые раздражители и механизмы генерирования локационных сигналов, что в совокупности с результатами электростимуляции позволяет признать заднее двухолмие ответственным за процесс излучения импульсов и ряд других актов, необходимых для осуществления эхолокации.
Итак, совокупность всех имеющихся в настоящее время морфологических и физиологических исследований, выполненных на летучих мышах, и сопоставление их с данными, полученными для других млекопитающих, позволяют нам высказать предположение о ведущей роли заднего двухолмия в интегрировании сложной деятельности аппаратов эхолокации.
129
Схема регуляций эхолокатора как целостной системы
Попытаемся представить общую функциональную схему эхолокационного процесса у летучих мышей на основании имеющихся морфологических и физиологических данных (рис. 39). Под влиянием определенных факторов внешней среды и внутренних потребностей организма в нервных центрах возникает очаг возбуждения, что приводит к усилению двигательной активности животного. У летучих мышей это обычно проявляется в вылете на вечернюю охоту за насекомыми, причем уже в предстартовой фазе начинают действовать средства эхолокации, обеспечивая успешную ориентацию животных в пространстве. Импульсы возбуждения достигают координационного центра эхолокации, в нашем представлении заднего двухолмия, и отсюда адресуются на исполнительные органы, т. е. на излучающий и приемный аппараты эхолокатора. Однако результаты морфологических исследований Буриковой (1973} не выявили прямых нисходящих путей от задних холмов к соответствующим ядрам черепно-мозговых нервов. К последним возбуждение может поступать двояким путем, либо через переднее двухолмие, либо через ретикулярную формацию, с которыми заднее двухолмие рукокрылых имеет прямые нисходящие связи (Henson et al., 1968; Бурикова, 1973).
По Меринг (1967), переднее двухолмие наделено не только функцией аккомодации и конвергенции зрачковых реакций глаза, но многими исследователями рассматривается как структура, обеспечивающая выход слуховых и тактильных раздражений на моторные образования ствола, что выражается в появлении различных двигательных актов в ответ на раздражения разной модальности. Среди рукокрылых переднее двухолмие значительно лучше развито у летучих мышей, чем у крыланов (Lutgemeier, 1962), хотя последние пользуются в основном визуальными средствами ориентации. Все это позволяет предполагать, что, возможно, четверохолмие среднего мозга в целом участвует в осуществлении эхолокационного процесса у летучих мышей и задние холмы в основном разряжаются через передние, от которых идут многочисленные пути. Однако никаких экспериментальных доказательств на этот счет в литературе пока не имеется.
Наличие прямой связи задних холмов с ретикулярной формацией открывает второй путь для эфферентного потока импульсов. Ретикулярная формация играет существенную роль в выполнении ряда функций, являющихся составными частями целого рефлекторного акта. Ей придается значение дополнительного промежуточного звена между афферентными системами и двигательными центрами. Строение нейронов ретикулярной формации указывает на возможность избирательного восприятия или объединения и дальнейшей передачи разного рода возбуждений на другие
130
Рис. 39. Схема афферентных и эфферентных путей эхо локационной системы летучих мышей.
ta — слуховой бугорок; п. v. coch. — вентральное кохлеарное ядро; nt —ядро трапециевидного тела; оз — верхняя олива; nil — ядра латеральной петли; ci— задний холм; сз — передний холм; cgm —медиальное коленчатое тело; ret — ретикулярная формация; пр — ядра моста; сег — мозжечок; ти. з. —стремечковая мышца среднего уха;
t, t. — мышца барабанной перепонки. Цифры —ядра черепно-мозговых нервов.
группы нейронов. Аксоны клеток ретикулярной формации, направляясь рострально и каудально, отдают коллатерали, которые оканчиваются пуговками либо на телах ее же клеток, либо на ядрах черепно-мозговых нервов (Scheibel et al., 1955).
Для того чтобы произошло излучение локационного сигнала, нервные импульсы должны быть переданы на ядра блуждающего нерва (X пара) и по его волокнам — на ларингеальные нервы, которые иннервируют гортань, а также на ядра языкоглоточного нерва (IX пара), осуществляющего иннервацию глоточного сплетения. Далее, по описанному в главе 2 механизму, происходит излучение локационного сигнала.
Однако, прежде чем будет излучен локационный сигнал, должны сработать защитные механизмы приемного устройства. Иннервация мышц среднего уха осуществляется ветвью тройничного нерва (V пара), одной из веточек языкоглоточного нерва (IX пара) и лицевого нерва (VII пара) (Quay, 1970). Кроме того, последний иннервирует мышцы ушных раковин, сложные движения которых имеют важное значение для эхолокации, особенно у подковоносов (Schneider, Mohres, 1960). Кстати заметим, что для подковоносов пока не установлен синхронизированный контроль между излучением сигнала и сокращением мышц среднего уха.
Излученный локационный сигнал в ослабленном виде достигает улитки и по проводящим путям слухового анализатора поступает в высшие слуховые центры, где, по-видимому, сохраняется в качестве эталона, для сравнения с приходящим эхом. Возможно, благодаря этому импульсу осуществляется настройка всей системы на определенный частотный диапазон.
Вслед за окончанием излучения зондирующего импульса наступает следующая фаза эхолокационного процесса — восприятие отраженного сигнала. Воспринятый акустический сигнал во внутреннем ухе кодируется в афферентный поток нервных импульсов, который по проводящим путям слухового анализатора поступает в нейронную организацию задних холмов, где совершается обработка информации (декодирование и анализ).
В составе ручки заднего холма идет пучок волокон во внутреннее коленчатое тело, которое рассматривается в качестве релейного центра на пути афферентных импульсов, следующих к слуховой области коры. По Лютгемейеру (Lutgemeier, 1962), внутреннее коленчатое тело лучше развито у летучих мышей, чем у крыланов, но функциональное значение его для эхолокации не исследовано. А. Г. Васильев и Н. Г. Андреева (1971) выявили в нейронной организации внутреннего коленчатого тела большое количество единиц, обладающих узкими частотными настройками, особенно у подковоносов. Это может свидетельствовать об участии данного центра в обработке ультразвуковой информации. В то же время неясна причина, почему при относительно хорошо развитом внутреннем коленчатом теле у летучих мышей отмечается слабое
132
развитие слуховой коры. Такое несоответствие послужило основанием некоторым авторам выдвинуть предположение о существовании слуховых проекций от коленчатого тела к амигдалоидному комплексу, который ими рассматривается как важный подкорковый слуховой ассоциативный центр (Freeman, Williams, 1952). Однако морфологические связи между этими двумя отделами мозга у летучих мышей пока не выявлены.
Что касается роли слуховой коры в конкретном эхолокационном процессе, то этот вопрос нельзя считать окончательно решенным в настоящее время (Айрапетьянц, Бурикова и др., 1972а, 19726). Результаты экстирпации этого отдела, а также исследование электрических реакций слуховой коры и свойств ее отдельных нейронов свидетельствуют о том, что она не способна обрабатывать поток быстро следующих друг за другом отраженных сигналов и производить анализ каждого из них, что необходимо для эхолокации. В частности, обращает на себя внимание тот факт, что корковые первичные ответы оставляют за собой длительные периоды «невозбудимости» и восстановление амплитуды вызванного потенциала на повторный стимул в коре обычно начинается лишь спустя 200 мсек., а полное восстановление имеет место через 500—700 мсек, после первого стимула. Для большинства корковых нейронов характерны уменьшение количества импульсов в ответ на последующее раздражение и даже полное выпадение ответов. Первоначальная реакция нейрона может восстановиться лишь после значительного перерыва между стимулами или при смене их параметров.
В ходе ритмического раздражения с частотой, большей 1 — 2 стимула в секунду, наблюдается падение амплитуды первичных ответов в слуховой коре. Все это говорит о чрезвычайно низкой лабильности коркового слухового центра как системы, что, по-видимому, связано не с длительной рефрактерностью составляющих ее элементов, а с поступлением в кору залпа тормозящих импульсов, которые сопровождают возбуждающие (Котеленко, Матюшкин, 1969). Кроме того, в слуховой коре летучих мышей не обнаруживается четкой локализации частот, которая отмечается для более высокоорганизованных млекопитающих (Котеленко, Васильев, 1971). При морфологическом изучении слуховой коры рукокрылых также не выявлено каких-либо признаков совершенства в отличие от подкорковых образований, и ее строение в целом соответствует низкому уровню филогенетического развития этих животных. Все указанные выше признаки могут служить основанием для отрицания участия слуховой коры в осуществлении нового принципа деятельности слухового анализатора — эхолокации. Это тем более кажется вероятным, что слуховые области коры эхоло-Цирующих и неэхолоцирующих рукокрылых не различаются по своему строению.
Л далее, если принять во внимание, что задние холмы лету
133
чих мышей могут выполнять все высшие афферентные и эфферентные функции, связанные с процессом эхолокации, возникает мысль, что в эволюции становления и совершенствования особого акустического способа ориентации корковая субстанция осталась не вовлеченной в функциональную реконструкцию слухового анализатора.
Наряду с приведенной группой фактов имеются и другие, свидетельствующие о влиянии слуховой коры на функционирование слуховой системы в целом. Сюда относятся: 1) наличие прямых кортико-фугальных путей к первичным ядрам проводникового отдела слухового анализатора (Бурикова, 1971); 2) уменьшение латентного периода и увеличение амплитуды ответов в задних холмах на ультразвуковые стимулы при функциональном выключении коры (Куликов, 1970а, 19706, 1973а, 19736); 3) специфичность частотно-пороговых кривых слуховой коры, отображающая настройку на определенный диапазон частот локационного сигнала.
Становится очевидным, что все уровни слуховой системы, включая и ее входные звенья, в норме контролируются корковыми аппаратами и, возможно, не только слуховой областью. Вопрос лишь в том, каково удельное значение этих влияний в общей системе нисходящей эфферентации и какие компенсаторные механизмы включаются после прямого удаления корковых слуховых зон для сохранения эффективности эхолокационного анализа.
Мы рассмотрели структуру элементарного акта эхолокационного процесса. Практически же эхо локационный анализ совершается благодаря потоку импульсов, следующих друг за другом с интервалом в несколько миллисекунд. При этом каждый предыдущий, по-видимому, должен быть срочно обработан, чтобы с каждым последующим можно было получить большую полезную информацию, при необходимости изменить режим работы эхолокатора и правильно скоординировать положение крыльев во время полета. Последнее может быть быстро реализовано через прямой эфферентный путь от заднего двухолмия к собственным ядрам варолиева моста, ножки которого связаны непосредственно с мозжечком.
В заключение следует подчеркнуть, что мы далеки от мысли абсолютизировать значение заднего двухолмия в осуществлении эхолокационного анализа, о чем уже неоднократно упоминалось по ходу изложения. Некоторые элементы этого анализа, по-видимому, происходят в нижележащих подкорковых центрах (кохлеарных ядрах, оливах, ядрах латеральной петли) при прохождении через них афферентной импульсации. Каждый из них имеет прямые пути к ретикулярной формации, которая в этом случае составляет связующее звено между чувствующими и моторными образованиями ствола мозга.
ОСОБЕННОСТИ СЛУХОВОГО ВОСПРИЯТИЯ АКУСТИЧЕСКИХ СИГНАЛОВ ПО ПАРАМЕТРАМ ЧАСТОТЫ И ДЛИТЕЛЬНОСТИ
Границы частотного диапазона и зоны оптимального слуха
Первым доказательством того, что слуховая система летучих мышей способна реагировать на ультразвуковые частоты, была работа Галамбоса (Galambos, 1942а). Методом отведения микрофонных потенциалов улитки автор у нескольких видов гладконосых летучих мышей зарегистрировал ответы во всем диапазоне предъявляемых стимулов от 30 гц до 98 кгц, с максимумом амплитуды в районе 10—30 кгц и дополнительным ее новы* шением в районе 30—50 кгц. Впоследствии эта методика была использована рядом авторов (Wever, Vernon, 1961b; Henson, 1964, 1967; Vernon, Peterson, 1965, 1966; Dalland et al., 1967) для определения границ слухового восприятия у гладконосых, бульдоговых и американских листоносых летучих мышей. Применяя длительные чистые тона, авторы обнаружили ответы в улитке в диапазоне от 0.3—1 до 60—100 кгц у разных видов с неодинаковым распределением максимумов. Однако во всех случаях пороговые значения ответов были во много раз выше, особенно в ультразвуковом диапазоне, чем их величины на тех же частотах при использовании других методик, поэтому метод отведения микрофонных потенциалов улитки, по-видимому, не может дать точной характеристики натуральной чувствительности слуховой системы животных.
Лучшие результаты, т. е. более низкие пороговые значения ответов, дает метод отведения вызванных потенциалов от подкорковых слуховых центров. Однако и в этих электрофизиологических исследованиях экспериментаторы имеют дело не с активным, бодрствующим животным, для которого подаваемый стимул имел бы определенное сигнальное значение, а с препаратом, на котором изучаются свойства электрических реакций определенного Центра или отдельного нейрона. Кроме того, характер ответов и величины порогов в этом случае у летучей мыши сильно зависят
135
от температуры окружающего воздуха й тела животного (Rosen-blith et al., 1950; Henson, 1964, 1967; Harrison, 1965), от глубины наркоза (Neuweiler, 1970) и функционального состояния животных (Стосман, Константинов, 1972). Поэтому с целью выяснения функциональных возможностей биологической системы в целом следует больше полагаться на ответные реакции всего организма, чем на реакцию отдельных узлов этой системы. Для расчетов предельной дальности действия эхолокатора летучих мышей и ряда других задач знание реальных порогов их слуховой системы на частотах локационного диапазона имеет большое значение.
Нами была предпринята попытка исследовать частотно-пороговые характеристики слуха у летучих мышей методом пищевых условных рефлексов в ультразвуковом и звуковом диапазоне частот (Константинов и др., 1971). Последнее было продиктовано тем, что, с одной стороны, уже самые простые наблюдения за животными в неволе и при ловле насекомых убедительно свидетельствовали о хорошем восприятии ими низкочастотных звуков, а с другой — в единственной работе, выполненной с применением поведенческой методики (Dalland, 1965а, 1965b), указывалось на отсутствие реакций у Myotis lucifugus на звуки ниже 10 кгц, а у Eptesicus fuscus (1 животное) — ниже 2.5 кгц. И наконец, еще одна задача заключалась в определении верхней границы слуха, так как, по данным большинства электрофизиологических работ, ответы в слуховых центрах не выявляются на стимулы частотой выше 100—120 кгц, а пороги на этих частотах достигают 90—100 дб. Однако Гриннелу (Grinnell, 1963а) удавалось иногда регистрировать ответы в заднем холме у М. lucifugus на частоты 150—170 кгц и редко — 200 кгц, а в поведенческих опытах Дийкграафа (Dijk-graaf, 1957) трехцветная ночница хорошо реагировала на частоту 175 кгц; но сигналы частотой 400 кгц у нее не вызывали реакции.
Наши опыты проводились на двух видах — остроухих ночницах (Константинов и др., 1971) и больших подковоносах (Соколов, 1972а; Соколов, Соколова, 1972).
У Животных вырабатывался условный рефлекс на звуковой раздражитель (чистый тон длительностью 5 сек., частота которого в течение опытов менялась от 500 гц до 250 кгц), предъявляемый то слева, то справа от места старта в случайной последовательности. При правильном выборе направления животное получало пищевое подкрепление — мучных червей — из кормушки около излучателя, а в случае ошибки с середины пути возвращалось экспериментатором на место старта. Регистрировались число правильных ответов, число ошибок и определялся уровень дифференцировки в процентах — отношение правильных реакций к общему числу предъявлений сигнала. За пороговое значение принималась такая интенсивность сигнала, при которой летучие мыши проявляли еще четкую реакцию на звук и уровень положительных ответов был выше 75%. Ниже этих значений результаты считались сомнительными и не учитывались.
В качестве источников звука использовались генераторы ГЗ-ЗЗ или ЗГ-11, генерирующие звуковые колебания от 20 гц до 200 кгц (по шкале). В ходе опытов шкала частот была расширена и отградуирована до 250 кгц.
136
Сигнал со звукового генератора поступал на вход электронного ключа, формирующего из непрерывного тона определенной частоты импульсы различной длительности — от 5 сек. до 25 мсек, с завалом фронтов, от 5 до 0.01 мсек., либо прямоугольные импульсы. Для излучения низкочастотных звуковых сигналов от 500 гц (предел используемой аппаратуры) до 10 кгц применялись динамические громкоговорители типа 1-ГД-28 или игольчатые излучатели, а для излучения ультразвуков — конденсаторные излучатели нестандартного производства. Излучатели размещались на стенах манежа так, чтобы основной лепесток каждого из них перекрывал стартовое положение животного. Частотно-силовые характеристики излучателей были выравнены. В ходе опытов они включались в случайном порядке, интервалы между сигналами произвольно изменялись.
Измерение уровня звукового давления, создаваемого излучателями в месте расположения головы животного, производилось калиброванным конденсаторным микрофоном, с выхода которого сигнал поступал на усилитель и затем — на вольтметр.
В начале экспериментов 9 остроухих ночниц были разделены на две группы. На одной группе исследовались пороги слуха в низкочастотной области и первоначальным сигналом служил тон 500 гц. Для другой группы эксперимент начинался с тона частотой 20 кгц и затем устанавливались пороговые характеристики слуха в ультразвуковом диапазоне через 10 или 20 кгц.
У животных той и другой группы условный рефлекс на звук (или ультразвук) проявился уже в первый опытный день, после 8—10 сочетаний сигнала с пищевым подкреплением. Однако дифференцирование местоположения источника звука (справа—слева) протекало медленно и достигало 80—100% правильных ответов лишь к 7—10-му опыту, по 30—40 предъявлений сигнала в опыт. Сказывалось влияние хорошо развитого у летучих мышей рефлекса на место, отмеченное нами ранее (Константинов, 1964а, 1965в, 1966а): получив пищевое подкрепление, например справа, животное в следующий раз быстро направлялось в ту же сторону, несмотря на звучание сигнала слева. Таким образом, дифференцирование местоположения источника условного звукового сигнала устанавливалось одновременно с угашением рефлекса на место.
После закрепления дифференцировки переходили к определению пороговых величин слухового восприятия на данной частоте, постепенно уменьшая интенсивность стимула, что занимало от 5 до И дней. Усредненная частотно-пороговая кривая (ЧПК), полученная в результате проведенных экспериментов, представлена на рис. 40, А.
В области звуковых частот от 750 гц до 10 кгц пороги лежат около 10 дб. В сторону более низких частот наблюдается повышение порога на 15 дб при частоте 500 гц. Из-за методических трудностей мы не устанавливали пороги на частотах ниже 500 гц, однако еще относительно хорошая чувствительность слуха на данной частоте, а также опыты по обнаружению насекомых на слух, изложенные выше, дают основания полагать, что летучие мыши способна воспринимать и более низкочастотные звуковые колебания.
137
Наилучшая чувствительность слуха была зарегистрирована в области ультразвуковых частот от 20 до 100 кгц. Именно эти частоты представлены в широкополосных зондирующих сигналах остроухих ночниц, которые посредством эхолокации несут животным наибольшую информацию об окружающей среде.
Рис. 40. Частотно-пороговые кривые (ЧПК) слуха остроухой^ночницы (Л) и большого подковоноса (Б).
По оси абсцисс—частота (кгц); по оси ординат—уровень (дб) относительно 0.0002 дин/см2. 1 — поданным поведенческих опытов; 2 — по показателям заднехолмового ВП (2V4).
Минимальный порог (—13 дб) обнаружен на частоте 40 кгц, причем мы дважды исследовали эту область, вернувшись к ней после установления порогов на частоте 100 кгц. В обоих случаях была получена одна и та же величина порога. На этой же частоте были зарегистрированы минимальные пороги у другого вида ночниц — Myotis lucifugus (Grinnell, 1963а; Dalland, 1965b).
После 100 кгц наблюдается резкое повышение порогов на 30 дб уже при частоте 120 кгц, и затем ЧПК продолжает увеличиваться,
138
достигая порогового значения 64 дб на частоте 200 кгц. Пороги на частотах выше 200 кгц специально не определялись, но, судя по уровню правильных ответов, интенсивность предъявляемых сигналов была близка к пороговому значению слуха животных. Поэтому полученные данные включены в общую ЧПК. Опыты в этом диапазоне частот были поставлены на 2 остроухих ночницах (табл. 9).
Таблица 9
Реакции остроухих ночниц на ультразвуки 200—250 кгц
Частота сигнала, кгц Интенсивность сигнала в месте старта, дб Общее число предъявлений сигнала (п) Число правильных ответов животных % правильных ответов
200 64 34 28 81.7
210 70 14 11 78.5
220 70 47 33 70.2
220 80 30 22 73.3
230 80 86 62 72.0
240 85 58 43 74.1
250 90 102 75 73.5
Вплоть до предела используемого генератора — частоты 250 кгц — на все предъявляемые ультразвуки летучие мыши продолжали реагировать, показывая уровень положительных ответов выше 70%.
Правда, чтобы сохранить этот уровень для более высоких частот, интенсивность сигнала приходилось значительно увеличивать.
Уровень звукового давления сигнала при частоте 250 кгц составлял примерно 90 дб в месте старта животного. Дальнейшее повышение интенсивности на более высоких частотах с использованием другого генератора уже не имело смысла, так как, по данным ряда авторов, более сильные раздражители могли вызвать повреждение барабанных перепонок у животных (Dalland et al., 1967). Поэтому мы вправе допустить, что верхний предел воспринимаемых остроухими ночницами ультразвуковых частот если и превышает 250 кгц, то незначительно.
Аналогичные опыты по снятию ЧПК слуха в широком диапазоне частот были проведены на 6 больших подковоносах. Разница заключалась лишь в том, что ночницы совершали побежки со старта к излучателям, отнесенным на расстояние 40 см, а подковоносы подлетали к ним с расстояния 150 см.
Подопытные животные четко реагировали на сигналы от 500 гц До 200 кгц. Пороги для звуковых и ультразвуковых частот до 60 кгц были соизмеримы и варьировали в пределах 10—15 дб (рис. 40, Б). После 90 кгц кривая слышимости поднималась вверх, т. е. пороги резко возрастали и для частоты 200 кгц составляли
139
90—95 дб. Более высокие частоты на вызывали ответной реакции подковоносов. Большие перепады порогов обнаружились в области от 60 до 90 кгц и особенно на частотах, зарегистрированных в сигналах этого вида. Уже на 60 кгц пороги ответов возросли до 20 дб, а на 70 кгц — до 30 дб, однако на частоте 75 кгц пороги составили всего 1 дб. Для частоты 80 кгц пороги ответов вновь увеличились до 17 дб, но уже на локационной частоте, т. е. частоте средней части сигнала, измеренной у каждой подопытной особи методом нулевых биений, порог снизился до 3 дб. Значение локационной частоты варьировало у подковоносов от 80.6 до 81.3 кгц, следовательно, различие всего на 0.6—1.3 кгц приводило к изменению порога на 14 дб. Дальнейшее уменьшение порогов до —5 дб проявилось на частоте 83.5 кгц, а затем при повышении частоты наблюдалось ухудшение слуховой чувствительности. На частоте 85 кгц порог составил +6 дб, на 90 кгц--h 15 дб.
Таким образом, у подковоносов выявилась высокая частотная избирательность слуховой системы в области частот, близких к локационному диапазону. Частота 83.5 кгц, по нашим данным, в противоположность утверждению Шнитцлера (Schnitzler, 1968) отсутствует в зондирующих сигналах большого подковоноса независимо от того, остается он на месте или летит в сторону цели. Однако она имеет большое биологическое значение для подковоноса, так как благодаря эффекту Допплера во время полета животного со средней скоростью 5 м/сек. или приближения объекта локации частота отраженного сигнала должна увеличиваться по сравнению с излученной примерно на 2 кгц.
Уменьшение порогов на частоте 75 кгц, на наш взгляд, можно объяснить тем, что эта частота составляет середину частотно-моду-лированного заключительного участка импульса. Повышение чувствительности в этой области может облегчить выделение начала и конца излученного и принимаемого сигналов и, следовательно, служить в качестве своеобразной метки при оценке расстояния до цели. О большой информативности этого диапазона частот в обнаружении предметов окружающего пространства свидетельствуют также наши опыты с использованием ЧМ-помехи (Константинов, Мовчан, Макаров, 1973). Из всей области широкополосного шума (2—250 кгц) на эффективность обнаружения неподвижного шара подковоносами оказывает влияние лишь полоса частот 72— 75 кгц, соответствующая амплитудному пику ЧМ-части локационного сигнала большого подковоноса.
Полученные в условнорефлекторных экспериментах особенности ЧПК слуха подковоносов в ультразвуковом диапазоне частот хорошо согласуются с результатами электрофизиологических исследований (Васильев, 1968а; Айрапетьянц, Васильев, 1970а, 19706; Neuweiler, 1970; Neuweiler et al., 1971). Высокая частотная избирательность слуховой системы зарегистрирована и у других видов летучих мышей, использующих для эхолокации сиг
140
налы с постоянной частотой заполнения (Grinnell, 1967, 1970; Schnitzler et al., 1971; Васильев, 1973).
Таким образом, сравнение данных, полученных на остроухих ночницах и больших подковоносах в поведенческих опытах, прежде всего показывает, что по диапазону воспринимаемых частот летучие мыши оставляют далеко позади всех исследованных в настоящее время наземных млекопитающих. В звуковой области частот у тех и других видов зарегистрированы относительно низкие пороги, что соответствует необходимости обнаруживать насекомых по их жужжанию, обмениваться коммуникационными сигналами и своевременно избегать приближающейся опасности. В ультразвуковом диапазоне выявлены значительные отличия: у остроухой ночницы — примерно одинаковая чувствительность в широкой области частот (20—100 кгц), у подковоносов — обострение чувствительности только на определенные частоты в узких пределах (75 и 81—85 кгц), что хорошо объясняется различием в частотном заполнении их локационных сигналов. Как показали исследования нашей лаборатории, указанные различия проявляются на всех уровнях слухового анализатора, начиная от кохлеарных ядер и кончая слуховой областью коры, как при отведении вызванных потенциалов от соответствующих центров, так и при изучении свойств их отдельных нейронов (Васильев, 1969а; Айра-петьянц, Васильев, 1970а, 19706; Васильев, Андреева, 1971; Котеленко, Васильев, 1971).
С другой стороны, несмотря на сходство частотных настроек различных уровней слуховой системы в эхолокационном диапазоне, нельзя не отметить существенных различий в результатах условно-рефлекторных и электрофизиологических исследований, касающихся величины порогов слухового восприятия. На рис. 40 представлены усредненные ЧПК, снятые по Лг4 (отведение вызванных потенциалов от заднего холма среднего мозга) у наркотизированных остроухих ночниц (наши данные) и больших подковоносов (Neuweiler et al., 1971). Сравнивая их с ЧПК, полученными в поведенческих экспериментах, можно заметить, что у остроухих ночниц в диапазоне частот от 10 до 100 кгц пороги различаются на 20—80 дб. Заметное ухудшение чувствительности слуха начинается уже с частоты 60 кгц, тогда как 100 кгц практически составляет предел восприятия у остроухих ночниц. У больших подковоносов пороги в эхолокационном диапазоне, измеренные с помощью различных методик, также существенно различаются. По результатам электрофизиологических исследований (Neuweiler, 1970; Neuweiler et al., 1971), подковоносы практически не воспринимают частоту зондирующего сигнала. В условнорефлекторных экспериментах пороги, измеренные в этой точке, оказались достаточно низкими (+2 дб). И этот результат кажется нам логически оправданным: животное должно воспринимать эхо не только в полете, но и во время отдыха в неподвижном окружении, когда эффект Доп-
141
плера отсутствует и, следовательно, воспринимаемый отраженный сигнал имеет ту же частоту, что и излученный. Следует к тому же отметить, что описанные выше результаты электрофизиологических экспериментов были получены путем регистрации оп-ответов. Минимальные же пороги на выключение стимула (off-ответы) регистрировались именно на частоте 80.0—82.6 кгц (Neuweiler et al., 1971). Вероятно, в условнорефлекторных экспериментах в условиях регистрации реакции целостного организма при большой длительности раздражения это способствует снижению порогов на локационной частоте. Однако, несмотря на различие в абсолютных величинах порогов, в электрофизиологических исследованиях удалось выявить четкую корреляцию между чувствительностью слуха и частотным диапазоном локационных сигналов различных видов летучих мышей (Grinnell, 1963а, 1967, 1970; Васильев, 1967а, 19676,1968а, 1973; Васильев, Матюшкин, 1967; Айра-петьянц и др., 1968, 1969; Айрапетьянц, Васильев, 1970а, 19706, 1971; Neuweiler, 1970; Schnitzler et al., 1971; Grinnell, Hagiwara, 1972a).
Второе существенное различие результатов исследований, проведенных с применением описанных методик, касается восприятия частот звукового диапазона (ниже 10 кгц). Так, Ньювейлером и др. (Neuweiler et al., 1971) было показано, что начиная с 10 кгц у больших подковоносов происходит резкое возрастание порогов с уменьшением частоты стимула (рис. 40, 5). На частоте 1 кгц пороги достигают 100 дб, тогда как в наших данных пороги на низких частотах не превышают 10—15 дб. Причины столь больших расхождений пока неясны, однако совершенно очевидно, что в естественных условиях летучие мыши должны обладать хорошей чувствительностью слуха в звуковом диапазоне, так как, помимо излучения эхо локационных криков, летучие мыши издают различные звуковые сигналы, используемые ими для общения друг с другом. Часть спектров этих сигналов, по нашим данным, лежит ниже 15 кгц, и человек отчетливо воспринимает их даже на расстоянии нескольких десятков метров. Трудно допустить, чтобы сами летучие мыши слышали их хуже. Это подтверждают и наши данные, когда энуклеированные ночницы с расстояния 2—3 м (Константинов, 1969), а подковоносы с 6 м (Соколов, 19726) обнаруживали на слух без участия эхолокации ночных бабочек, частота взмахов крыльев которых не превышала 100 гц.
Зависимость порогов слуха от длительности сигнала
Как отмечалось выше, локационные сигналы гладконосых и подковоносов различаются не только по частотным, но и по временным характеристикам.
Так, остроухие ночницы используют в процессе ориентации в пространстве короткие сигналы, обычно не превышающие 3— 142
5 мсек., а при приближении к объекту локации их длительность может сокращаться до 0.6 мсек. Для других млекопитающих — собаки (Бару, 1967; Гершуни, 1967, и др.) и человека (Карасева, 1967; Бару, Карасева, 1971) — известно, что при укорочении сигнала до 1 мсек, пороги обнаружения возрастают, в среднем на 30 дб. Кроме того, в случае применения длительностей короче 10 мсек, значительно нарушается частотное различение сигналов. Поэтому было непонятно, как же остроухие ночницы могут получать исчерпывающую информацию об окружающей обстановке, используя столц короткие и, казалось "бы, трудно поддающиеся обработке сигналы.
Рис. 41. Зависимость порогов обнаружения сигналов от их длительности у остроухих ночниц (7) и больших подковоносов (2).
В противоположность ночницам подковоносы в ходе осуществления локации обычно не издают сигналов короче 10 мсек.
Представляло интерес провести исследование зависимости величины порогов обнаружения одиночных ультразвуковых сигналов от их длительности у активных ночниц и подковоносов (Соколова, 19736).
Для проведения опытов были выбраны частоты, которые лучше всего воспринимались ночницами (40 кгц) и подковоносами(83.5 кгц).
Полученные результаты выявили резкие различия в ходе кривых изменения порогов в зависимости от длительности стимула у исследованных видов летучих мышей (рис. 41). У ночниц пороги восприятия начинают возрастать лишь с 25 мсек, и к 0.1 мсек, увеличиваются на 25 дб. При этом наиболее круто их величина возрастает лишь для сигналов короче 0.5 мсек. Это и понятно, если вспомнить, что с импульсами длительностью от 5 до 0.6 мсек, ночницам обычно приходится сталкиваться во время приближения к цели. Стимулы короче 0.1 мсек., в нашем случае 0.025 мсек., не воспринимались ночницами. Уровень положительных реакций не превышал 50%, и, кроме того, полностью отсутствовала ориентировочная реакция на звук.
143
Пороговая кривая у больших подковоносов плавно поднимается уже при длительности стимула 250 мсек., и к 50 мсек, пороги увеличиваются на 10—15 дб. Затем происходит резкое увеличение порогов, которое, на наш взгляд, следует в первую очередь связать с тем, что большие подковоносы обычно не издают одиночных сигналов короче 60 мсек. Одиночные сигналы длительностью 5 мсек, вообще не вызывали ответной реакции подковоносов даже при уровне звукового давления 60 дб (предел используемой аппаратуры). Если же эти стимулы подавались сериями с частотой следования 40 имп./сек., подковоносы четко реагировали на них, пороги восприятия при этом равнялись 24 дб. Более того, при подаче сигналов сериями подковоносы могли воспринимать и еще более короткие импульсы, длительностью 2.5 мсек. При частоте следования 40 имп./сек. порог составил 52 дб, а при 70 имп./сек. — 26 дб.
Эти данные хорошо согласуются с тем, что в обычных условиях подковоносы, уменьшая длительность сигналов, увеличивают частоту их следования, группируя импульсы в серии-«пачки», что, как стало понятно, значительно снижает порог восприятия.
Таким образом, в отношении как частотных, так и временных настроек слуховой системы летучих мышей выявлена четкая корреляция с параметрами излучаемых сигналов.
Электрофизиологические исследования зависимости реакции заднего двухолмия от длительности раздражающего стимула проводились в нашей лаборатории путем регистрации on- и off-ответов (Васильев, 1967а, 1968а). Результаты этих измерений показали, что величина оп-ответов как у остроухих ночниц, так и у больших подковоносов не изменялась при укорочении стимула до 2 мсек., т. е. в этом случае не было выявлено какой-либо специфичности настроек слуховой системы.
В противоположность оп-реакциям off-ответы изменяются в зависимости от длительности стимула по-разному у ночниц и подковоносов. Если у первых укорочение сигнала до 10 мсек, не влияет на величину порога, то у вторых пороги off-реакций возрастают начиная с 50 мсек.
Исследование реакций отдельных нейронов позволило разделить их на 2 группы. В первой пороги восприятия и у ночниц, и у подковоносов не изменяются при уменьшении длительности до 0.5 мсек. Реакции же второй группы сходны с характеристиками off-ответов: у ночниц пороги начинают увеличиваться с 10 мсек., у подковоносов — с 50 мсек. Кроме того, в некоторых случаях у подковоносов off-ответы пропадали при стимулах 4.6 мсек, и более кратких.
Таким образом, характеристики отдельных нейронов, а также настройки по off-реакциям показывают наличие функциональных особенностей в слуховом анализаторе летучих мышей, связанных с использованием ими различных по длительности эхолокационных сигналов. Эти данные полностью соответствуют результатам условнорефлекторных опытов,
6
ЭЛЕКТРОФИЗИОЛОГИЧЕСКИЕ ХАРАКТЕРИСТИКИ НЕРВНЫХ АППАРАТОВ СЛУХОВОЙ СИСТЕМЫ ЛЕТУЧИХ МЫЩЕЙ
Электрофизиологические исследования нервных аппаратов слуховой системы эхолоцирующих летучих мышей ставят своей целью выяснить, чем эта система отличается от слуховых систем других млекопитающих, а также попытаться проникнуть в механизм обработки эхолокационной информации в мозгу летучих мышей. Электрофизиологические исследования нервных проводников и нервных центров слуховой системы летучих мышей ведутся сравнительно недавно. Первая работа в этом направлении относится к 1958 г. (Griffin, Grinnell, 1958), но уже в 60-х годах эти исследования получили широкое развитие (Grinnell, 1963а, 1963b, 1963с, 1963d, 1967; Suga, 1964а, 1964b, 1965а, 1965b, 1965с, 1967, 1968, 1969b; Suga, Griffin, 1964; Grinnell, Grinnell, 1965, и др.). В основном они посвящены электрофизиологическому анализу свойств слухового нерва, слуховых ядер продолговатого мозга, задних холмов и слуховой коры у некоторых американских видов гладконосых летучих мышей. В нашей стране подобные исследования проводятся в Ленинградском университете (Матюшкин, Васильев, 1966, 1968; Айрапетьянц, 1967а; Васильев, 1967а, 19676, 1968а, 19686, 1969а, 19696,1973а, 19736; Васильев, Матюшкин, 1967; Айрапетьянц, Константинов, Матюшкин, 1968, 1969; Матюшкин, Стосман, 1967, 1968; Котеленко, Матюшкин, 1969; Матюшкин, 1969, 1973; Айрапетьянц, Васильев, 1970а, 19706, 1971; Куликов, 1973а, 19736; Котеленко, 1973).
Уже сейчас накоплен большой экспериментальный материал по данному вопросу, который обобщен в настоящей главе.
Потенциалы действия слухового нерва, свойства первичных слуховых нейронов
Животное наркотизировалось внутрибрюшинным введением гексенала (0.08 мг/г веса мыши). Температура мыши в большинстве опытов поддерживалась на уровне 35—37° С. Мышь помещалась в звукозаглушенной
145
камере (размером 2X2X2 м). Ультразвуки получались с помощью конденсаторного излучателя (диаметром 6 см), располагавшегося на расстоянии 34 см спереди от головы животного. На излучатель подавались импульсы от генератора ГЗ-7А с помощью электронного ключа. Частота заполнения,
ZI
Рис. 42. Блок-схема раздражающей и регистрирующей установки и ее характеристика.
А — схема раздражающей и регистрирующей установки; Б: I — частотная характеристика излучателя; II — частотная характеристика усилителя; Bl 1 — осциллограмма электрического импульса, записанного с выхода усилителя; 2 — осциллограмма раздражающего сигнала, записанного после излучения конденсаторным микрофоном.
сила, длительность, завалы фронтов, частота повторения и угол подачи ультразвуковых сигналов могли изменяться соответственно задаче эксперимента в широких пределах (рис. 42).
Электрические потенциалы VIII нерва отводились от круглого окна униполярно манганиновым изолированным электродом и регистрировались с помощью усилителей УБШ-02 и катодного осциллографа ЭМОФ 2-01. Полоса пропускания частот усилителя была максимально расширена кверху (до 150 кгц).
146
В опытах выяснилось, что краткий ультразвук (0.25 мсек, при длительности фронтов по 0.1 мсек.) вызывает в слуховом нерве летучей мыши одиночный потенциал действия (ПД), который имеет двухфазную или трехфазную форму. При длительных и сильных ультразвуках вслед за основным ПД, возникающим при включении стимула, появляются дополнительные низкие колебания, сохраняющиеся на протяжении стимула, и иногда получается значительный ПД в ответ на выключение стимула.
Длительность ПД слухового нерва около 1 мсек., амплитуда (при данном способе отведения) не более 100 мкв, латентный период 0.5—1.4 мсек.
При отведении электрической активности от кохлеарных ядер ответы слухового нерва по форме являются функцией положения электрода. Как правило, on-ответы (реакции на включение стимула) состоят из быстрого колебания длительностью 0.5—1.5 мсек. Оно регистрируется трех- или двухфазным, начинаясь всегда с положительного отклонения (рис. 43, Л, 7, 2). В некоторых опытах подобные по форме ответы обнаруживаются и на выключение стимула (off-ответы).
Латентный период и амплитуда ПД слухового нерва зависят от силы ультразвука: чем сильнее ультразвук, тем больше ПД и тем короче его латентный период. При увеличении силы ультразвука от пороговой на 40—100 дб амплитуда ПД увеличивается от 0 до 100 мкв. Латентный период при этом укорачивается не более чем на 0.3 мсек. Рост амплитуды ПД слухового нерва при усилении ультразвука линейно зависит от силы ультразвука, выраженной в децибелах, и определяется увеличением количества возбуждаемых первичных слуховых нейронов. Поскольку волокна слухового нерва приблизительно однородны по диаметрам (данные на морских свинках и других млекопитающих — Davis, 1963), можно полагать, что между амплитудой ПД и количеством возбужденных волокон (первичных нейронов) имеется линейная зависимость.
Таким образом, увеличение количества возбужденных первичных слуховых нейронов при усилении краткого моночастотного ультразвукового сигнала происходит по логарифмическому закону.
По современной теории анализа частоты (высоты) тонов в улитке (Davis, 1963), пороговый по силе моночастотный ультразвук воздействует на небольшой район основной мембраны улитки и соответственно на относительно малую определенную группу волосковых клеток кортиева органа. При сверхпороговых силах ультразвука этот район расширяется, что приводит к увеличению числа возбужденных рецепторов. Не исключено также, что среди рецепторов одного и того же участка имеются разночувствительные (например, внешние и внутренние волосковые клетки), вовлекаемые последовательно при усилении деформации основной мембраны.
147
У части обследованных нами животных при усилении стимула за некоторый предел наблюдалось уменьшение ПД слухового нерва. Этот эффект получался на фоне роста микрофонного потенциала улитки (МПУ). Последнее обстоятельство, вероятно, свидетельствует о том, что в описываемых случаях имело место угнетение реакции в части первичных слуховых нейронов, связанное с явлениями какого-то тормозного взаимодействия в сфере первичных нейронов (Матюшкин, 1969).
Отмеченное выше укорочение латентного периода ПД при усилении раздражения, вероятно, определяется, во-первых, ускорением реакции рецепторных клеток в главном фокусе активности, во-вторых, тем, что при усилении стимула в реакцию могут вовлекаться более высокочастотные участки кортиева органа, возбуждающиеся быстрее, чем область главного фокуса колебаний.
В районе частот от 20 до 80 кгц, где пороги ответов были соизмеримыми, амплитуды ПД, вызываемых «припороговыми» (например, 10 дб над порогом) стимулами разной частоты, получались практически одинаковыми. Это значит, что количество первичных слуховых нейронов, отзывающихся на каждую из этих частот, приблизительно одинаково. Вместе с тем при больших силах стимулов (+40 дб над порогом) амплитуды ПД для «низкочастотных» ультразвуковых стимулов (20 кгц) были несколько больше (на 28%), чем для «высокочастотных» (80 кгц). Это, видимо, объясняется включением в область, раздражаемую сильным «низкочастотным» импульсом, районов «высокочастотной» настройки, затрагиваемых бегущей по основной мембране волной (см.: Bekesy, 1953).
Использовав ПД слухового нерва в качестве показателя реакции входа слуховой системы летучей мыши на ультразвуки, мы определили соответствующие частотно-пороговые кривые (ЧПК).
Выяснилось, что максимальная чувствительность к ультразвуку у обоих видов летучих мышей обнаруживается в диапазоне частот 10—30 кгц. В то же время на высоких частотах — 70—80 кгц для остроухих ночниц и 82—86 кгц для подковоносов — наблюдается дополнительное повышение чувствительности (рис. 43).
Наиболее низкие пороги для оп-ответов у остроухих ночниц отмечаются на частотах около 60 кгц, у подковоносов — на частотах 40+2 и 78—80 кгц (рис. 43, Б, /, II).
ЧПК отдельных первичных слуховых нейронов летучей мыши были изучены у ночницы (Frishkopf, 1964). Фришкопф изучал ЧПК 80 первичных слуховых нейронов. Области ответов этих нейронов оказались довольно узкими и асимметричными.
Частота акустического сигнала, к которой обнаруживалась наибольшая чувствительность у обследованных единиц (лучшая частота), была различной, но в общем все они располагались между 10 и 80 кгц, а у 2/3 из них — между 40 и 80 кгц.
148
У 80% нейронов обнаружилось торможение реакции, вызываемой стимулом лучшей частоты, при добавлении второго ультразвука с частотой, немного превышающей возбуждающие частоты.
Латентные периоды возбуждения и торможения были практически одинаковыми. В связи с последним обстоятельством Фриш-конф заключил, что торможение осуществлялось прямым воздействием «соседних» первичных слуховых нейронов на обследуемые.
Рис. 43. Осциллограммы электрических ответов слухового нерва (А) и их частотно-пороговые характеристики (Б).
А: 1—3 — записи с помощью макроэлектрода, расположенного в кохлеарных ядрах; 4 — около круглого окна; монополярное отведение. Сила стимула 70—75 дб, длительность 4 мсек. I, 2 — записи на ночнице, з, 4 — на подковоносе. Частота заполнения стимула для 1—4 — 35, 40, 30, 30 кгц; длительность фронта нарастания для всех стимулов 0.13 мсек.; акустическая задержка для 1—3 — 1 мсек., для 4 — 0 мсек. Позитивное колебание книзу. Б — частотно-пороговые кривые. По оси абсцисс — частота заполнения стимула; по оси ординат — сила стимула. Сплошные линии — измерения по оп-ответам, штриховые — по off-ответам. I — измерения, произведенные на остроухой ночнице;
II — на подковоносе.
Мы добавим, что, может быть, для объяснения скорости такого торможения необходимо допустить еще и его электротоническую природу.
Описанное периферическое латеральное торможение обеспечивает обострение частотных (тональных) настроек первичных слуховых нейронов летучей мыши, необходимое для тонкого тонального анализа ультразвуков у этих животных.
Можно думать, что полученные нами эффекты частичного «торможения» ПД слухового нерва летучей мыши при действии относительно сильных ультразвуков, а также off-эффекты в слуховом нерве связаны с рассмотренным латеральным торможением первичных нейронов.
149
Используя ПД слухового нерва как показатель реакции первичных слуховых нейронов, Гриннел (Grinnell, 1963b) изучал способность входа слуховой системы летучих мышей реагировать на
повторный сигнал, следующий за первым через небольшой интервал времени. По Гриннелу, минимальный ПД слухового нерва на повторный сигнал получается уже через 0.4 мсек, после сильного
первого стимула; 50 %-е восстановление амплитуды повторного ПД — через 2 мсек.; 75%-е — через 5 мсек., а 100%-е — не менее чем через 20 мсек, от первого стимула. При сниженной амплитуде
ПД слухового нерва эти летучие мыши могут воспроизводить ритм до 2200 в 1 сек.
Мы также исследовали амплитуду повторного ПД слухового нерва в зависимости от интервала между первым и повторным кратким стимулами (0.25—0.5 мсек.) (рис. 44). Эта зависимость одинакова для стимулов с разными частотами заполнения, но различна при разных силах стимулов. Полученные результаты анало-
Рис. 44. Зависимость амплитуды повторного (4Л) ПД VIII нерва от интервала между первым и повторным раздражениями (частота 20—60 кгц).
1 — сила относительно порога 12 дб, 2 — 46 дб, 3, 4 — 26 дб.
гичны приведенным выше данным Гриннела для другого вида летучих мышей. В наших опытах при больших силах стимулов (до 46 дб над порогом) значительное угнетение реакции длится 1—2 мсек., 50%-е восстановление амплитуды повторного ответа происходит спустя 2—3 мсек., а полное восстановление — спустя 20—30 мсек.
Интересно, что при малых силах стимулов (+12 дб) повторный ПД в течение первой миллисекунды не угнетается совсем, немного снижается на 2—3-й миллисекунде, возрастает на 4—5-й и далее возвращается к норме.
Эффекты, наблюдающиеся при слабом раздражении, объясняются тем обстоятельством, что такое раздражение порождает нервные импульсы и соответственно рефрактерное состояние лишь в небольшом количестве афферентных волокон, создавая в остальных ту или иную степень подпороговой деполяризации, иначе говоря, повышенной возбудимости. При этом нормальные или даже увеличенные по амплитуде реакции на повторный приближенный стимул обеспечиваются элементами, подпорогово-деполяризован-ными первым стимулом. Надо полагать, что восприятие и анализ повторных акустических стимулов, приближенных к предыдущим
150
на 1 мсек, и ближе, всегда опираются на вовлечение подпорогововозбужденных первичных чувствительных нейронов слуховой системы.
Суммируя изложенные данные о скорости срабатывания и скорости восстановления возбудимости, о предельных ритмах работы элементов слухового нерва летучих мышей, можно сказать, что этим последним, как и соответствующим рецепторным элементам, присуща весьма высокая лабильность. Несомненно, что такая лабильность необходима для осуществления эхолокации, при которой посылаемый сигнал и его эхо чаще всего чрезвычайно сближены во времени.
Электрические реакции
и свойства элементов кохлеарных ядер
Электрофизиологические исследования кохлеарных ядер летучих мышей немногочисленны (Suga, 1964а, 1964b, 1965b; Васильев, Стосман, 1970; Васильев, 1971). Большинство
Рис. 45. Типы областей ответов отдельных нейронов кохлеарных ядер.
По оси абсцисс — частота заполнения стимула; по оси ординат — сила стимула. Пунктирные линии — максимально возможная сила стимула. I — данные, полученные в опытах на остроухих ночницах; II, III — на подковоносах.
изученных нейронов на предъявление стимула отвечало тоническими разрядами (рис. 45), часть нейронов показала фазные ответы и ответы торможения. 13% нейронов обнаружило последействие, следующее за реакцией на сильные стимулы, достигающее иногда 0.6 сек. (Suga, 1964а). Латентные периоды ответов оказались равными 0.6—5.6 мсек.
Области ответов нейронов кохлеарных ядер можно разделить на широконастроенные и узконастроенные, с одной или двумя ха
151
рактеристическими частотами. Большинство нейронов имеет область ответов, значительная часть которой располагается ниже характеристической частоты (рис. 45, 7, а, г). У ночниц обнаружено 74% таких нейронов, у подковоносов — 62%. Другие нейроны имеют асимметричные области ответов, большая часть которых, напротив, располагается выше характеристической частоты (10% у ночници 15% у подковоносов; рис. 45,7, в). У остальных нейронов симметричные области ответов (7, б). Отметим, что у ночниц области ответов различных типов довольно равномерно распределены
Рис. 46. Зависимость порогов (А) и числа разрядов (Б) отдельного нейрона от частоты заполнения стимула.
А — область ответов отдельного нейрона. По оси абсцисс — частота заполнения стимула, по оси ординат — сила стимула. Сплошные линии — общая область ответов, штриховые — области off-ответов, пунктирные — области ответов нейрона, в которых регистрировались тонические ответы. Б — осциллограммы электрических ответов нейрона, область ответов которого представлена на А. Сила стимула 73 дб. Цифры слева — частота заполнения стимула.
по всему диапазону слышимых ими частот, тогда как у подковоносов области ответов второй и третьей групп (в порядке описания) расположены преимущественно в диапазоне 80—90 кгц.
Наиболее сложные области ответов обнаруживаются у подковоносов в диапазоне частот 70—90 кгц. Как правило, все нейроны отличаются высокой избирательностью к частоте заполнения стимула (рис. 45, 77). Большинство из них при большой силе стимула реагирует до частоты 78 или после 80 кгц, при этом чем ближе характеристическая частота к диапазону 78—80 кгц, тем уже область ответов. Некоторые нейроны (их было три) отвечали на частоты по обе стороны этой полосы (рис. 45, 777, а). Если же область отвс-
152
тов перекрывала диапазон 78—80 кгц, то характер реакций ней* рона был сложным (9 нейронов). Так, один из них (рис. 46) в полосе частот 40—67 и 80.5—90 кгц на воздействие стимула реагировал оп-ответами, в полосе частот 67—76 кгц предъявление стимула сопровождалось тоническими ответами, в диапазоне 78— 80 кгц ответы регистрировались только на включение и выключение стимула. В полосе частот 80—80.5 кгц снова наблюдались тонические ответы, плотность разряда которых выше, чем в диапазоне 67—78 кгц. Очевидно, что такие нейроны оказываются хорошими дискриминаторами частот (Q больше 800, коэффициент Q — отношение характеристической частоты к полосе частот на уровне 10 дб над порогом). Отметим, что если у таких нейронов не было on- и off-ответов в диапазоне 78—80 кгц, то у них оказывались две области ответов (рис. 45, III, б). Два нейрона, имевшие область ответов в полосе частот 50—90 кгц, отвечали только тоническими разрядами (рис. 45, III, а).
Используя ЧМ-стимулы, Суга (Suga, 1964а) нашел, что нейроны кохлеарного ядра летучей мыши реагируют на изменение модуляции любого направления. Но пороги их реакций на ЧМ-стимулы выше порогов для тональных сигналов лучшей частоты и равной длительности. Последнее обстоятельство автор объясняет тем, что кохлеарные нейроны воспринимают в ЧМ-сигналах только тот «небольшой» частотный компонент, на который они настроены.
Некоторые различия в характеристике зарегистрированных нейронов у обоих видов летучих мышей обнаруживаются при оценке настроенности нейронов по коэффициенту Q. Общие пределы границ изменения Q у остроухих ночниц 1—28, тогда как у подковоносов — 1—90. На этом уровне анализатора распределения областей ответов по Q у обоих видов летучих мышей подобны (рис. 47).
Рассмотренные электрофизиологические характеристики кохлеарных ядер летучих мышей во многом аналогичны известным характеристикам этих ядер у других млекопитающих. Вместе с тем у подковоносов обнаруживаются нейроны, настроенные на соответствующие частоты, используемые ими для эхолокации.
Для изучения особенностей реагирования на центрифугальную активацию при адекватных воздействиях исследовались реакции нейронов вентрального кохлеарного ядра на ультразвуковые стимулы после разрушения ипсилатеральной улитки (Куликов, 19726, 19736).
При этом были выявлены две группы нейронов, различающихся по значениям латентных периодов (ЛП), порогов их реакций и областям ответов. Распределение ЛП реакций нейронов первой и второй групп, а также различия реакций по ряду свойств свидетельствуют о том, что возбуждение элементов этих групп отражает активацию разных путей, входящих в вентральное кохлеарное ядро. Средний ЛП реакций нейронов первой группы приблизительно
153
на 1 мсек, больше ЛП реакций нейронов кохлеарного ядра летучих мышей на ультразвуковые раздражения при интактных входах. Такие малые значения ЛП реакций на контрлатеральную стимуляцию, по-видимому, подтверждают, что слуховые пути перекрещиваются ниже верхнеоливарного комплекса. Существование такого перекреста может иметь большое значение для быстрого взаимодействия входов при бинауральном слухе.
На основании сравнения полученных и литературных данных следует думать, что возбуждение нейронов второй группы с боль-
Рис. 47. Гистограммы частот коэффициента Q.
По оси абсцисс — значения коэффициента Q, по оси ординат — частота р* = —
(п— число нейронов в разряде; AQ — ширина разряда; N — общее число обследованных нейронов). I — данные, полученные в опытах на ночницах, II — на подковоносах.
шим ЛП отражает активацию оливокохлеарного пучка. Обнаружено, что этот канал центрифугальной активации характеризуется большим ЛП, высоким порогом, низкой частотной избирательностью, большим интервалом восстановления и значительным временем суммации.
При этом ярко выражено соответствие ряда характеристик реакций нейронов видовым особенностям локационных сигналов летучих мышей, что позволяет думать об их участии в функционировании слуховой системы при эхолокации.
Высокие значения порогов и интервала восстановления, а также резкое увеличение порогов при уменьшении длительности стимулов указывают на то, что в естественных условиях эти нейроны могут возбуждаться излучаемыми сигналами значительной длительности и с малой частотой следования. Большие значения ЛП реакций нейронов свидетельствуют о приуроченности их возбуждения к моменту прихода отраженных сигналов. Вышеуказанные условия излучения характерны для поисковой фазы эхолокации, в которой происходит обнаружение препятствий или объектов охоты.
154
Электрические явления в задних холмах четверохолмия и свойства нейронов задних холмов
Задние холмы четверохолмия как наиболее развитый и массивный отдел слуховой системы летучей мыши подверглись наиболее подробному электрофизиологическому исследованию.
Вызванные потенциалы задних холмов регистрировались нами с помощью униполярной методики. При этом активный электрод помещался на поверхности холма или углублялся в холм, а индифферентный электрод располагался на височной мышце с той же стороны. В таких условиях холмовой ВП на одиночный краткий (0.25—0.5 мсек.) тональный (или частотно-модулированный) ультразвук представляет собой положительное или положительноотрицательное колебание, перед которым (иногда заходя на его восходящую фазу) располагаются четыре быстрых ПД, обозначаемые Л\, N2, -ZV3, TV4 (рис. 48). Первый из этих ПД — N± — представляет собой уже описанный ПД VIII нерва, о чем свидетельствует, в частности, его ЛП. TV2, 7V3, TV4, вероятно, порождаются вторыми, третьими и четвертыми нейронами слухового пути соответственно. Наибольший ПД — TV4 — есть электрический ответ нейронов, отростки которых проходят в толще холма, поскольку он обнаруживается в холме и имеет максимум амплитуды в глубоких частях холма. Возможно, что это нейроны ядра латеральной петли, сильно развитого у летучих мышей. ЛП названных ПД (за вычетом акустической задержки) равны:
— 0.7 мсек., N2 — 1.6, N3 — 2.4, TV4 — 3.2 мсек, в среднем. Длительность каждого из этих ПД составляет около 1 мсек. Амплитуда наибольшего ПД — TV4 — достигает 100 мкв, прочие значительно меньше. Медленное колебание, возникающее в заднем холме (его знак извращается при погружении активного электрода в холм на глубине 1—2 мм), представляет собой алгебраическую сумму возбуждающих постсинаптических потенциалов (ВПСП), а также, видимо, и тормозящих (ТПСП), возникающих в холмовых нейронах при поступлении к ним залпа афферентных импульсов.
Длительность медленного колебания около 10 мсек., амплитуда иногда приближается к таковой у V4, но чаще меньше. Мы исследовали также реакции на более длительные тональные стимулы. При этом обнаружилось, что если ультразвуковой стимул имеет значительную длительность (например, 5—10 мсек, или более), то реакция на него оказывается более сложной. Начало действия такого «длинного» ультразвукового стимула всегда отмечается описанным комплексом, состоящим из JVj, N2, N3t N4 и медленной волны. Это реакция на «включение» раздражителя (on-эффект). На протяжении действия ультразвука обна-
155
Рис. 48. Осциллограммы электрических ответов заднего двухолмия и отдельных заднехолмовых нейронов. 1—2 __ отведение макроэлектродами; 3—9 — микроэлектродами. Второй луч (внизу) — запись раздражающего стимула с выхода усилителя раздражающего комплекса. Вертикальная черточка на верхнем луче — отметка раздражения. Акустическая задержка для 1—2_______1.7 мсек., для 3—9 — 1 мсек. Позитивные колебания книзу. 2 — запись биполярным электродом.
руживаются слабые суммарные электрические реакции в форме медленного положительного отклонения нулевой линии с беспорядочными низкоамплитудными ПД на его фоне. В момент прекращения действия (выключения) ультразвукового раздражителя выявляется off-эффект, особенно отчетливый в латеральных частях холмов. По форме и ЛП (отсчитываемому для off-эффектов от конца стимула) off-эффект сходен с вышеописанным on-эффекг м (рис. 48). Но так как для off-эффектов зависимость
Рис. 49. Зависимость амплитуды вызванных потенциалов заднего холма от силы раздражителя.
По оси абсцисс — сила стимула над порогом обнаружения ответа; по оси ординат — амплитуда ответа. 1 — кривые по медленному положительному колебанию; 2 — по JV4. I—III — данные по оп-ответам; IV—VI — по off-ответам. Ошибки средних значений каждой точки кривых не превышали 10 мкв.
амплитуды от силы ультразвука отличается от таковой для оп-эффектов и имеются различия в ЧПК для on- и off-ответов, можно утверждать, что on- и off-эффекты обеспечиваются не одними и теми же нервными структурами (Васильев, 1967а, 19676).
Амплитуда всех описанных компонентов суммарного ВП холма в некоторых пределах была тем больше, чем сильнее был стимул (тональный или частотно-модулированный ультразвук; рис. 49). Однако эта зависимость для А4 в on- и off-ответе и для медленной волны ВП была не вполне идентична.
Пороги оп-ответов в наших опытах не зависели или очень мало зависели от длительности стимулов, если последняя не уменьшалась за 0.5 мсек.? и от длительности фронта нарастания сти-
157
мула (которая в этих опытах не превышала' 0.5 мсек.). Амплитуда off-ответов также мало изменялась при указанных изменениях длительности стимулов, но значительно снижалась при уменьшении крутизны стимула (удлинении фронта его нарастания). off-ответы нарастали при удлинении стимулов и значительно снижались при удлинении времени нарастания стимулов (Васильев, 19686). Изменения медленного компонента ответа при вариации длительности сигнала и его фронта не были прослежены столь тщательно. Однако создалось впечатление, что этот компонент в общем изменяется в том же направлении, что и TV4.
При использовании ЧМ-сигналов, сходных с естественными локационными криками животных (с длительностью, равной 4 мсек., и частотой, линейно падающей от 80 до 37 кгц), off-ответы (TV4) были выражены слабо. Это, видимо, было связано с краткостью тональных компонентов сигнала, благоприятных для оп-ответов (80 или 60 кгц), и с невыгодной для off-ответа конечной частотой сигнала.
Если в качестве индикатора возбуждения слуховой системы использовался медленный компонент холмового ВП, отводимый с поверхности большим электродом, то ЧПК получались подобными вышеописанным. Однако при погружении точечного активного отводящего электрода в холм характер и сила реакции зависят от глубины погружения. Мы определяли при разных уг лублениях электрода частоту ультразвука, имеющую наибольший ответ при оптимальной силе стимуляции. При этом выявилось, что такие частоты для поверхностных слоев холма относительно низки; чем глубже исследуемый слой, тем они выше.
Такая же тонотопия в заднем холме летучей мыши обнаруживается и при исследовании ЧПК отдельных нейронов, о чем будет рассказано далее.
Рассмотрим способность слуховых центров, порождающих пик к приему повторного тонального сигнала (рис. 50). Она оказывается приблизительно такой же, как и у слухового нерва. Правда, в некоторых случаях здесь обнаруживаются те или иные отклонения (от описанного для ПД VIII нерва) хода восстановления амплитуды повторного ответа. Это имеет место на 4-й миллисекунде и далее. Иногда здесь наблюдается превышение повторного ТУ4 над первым, о чем писал ранее Гриннел (Grinnell, 1963b), называя это явление «аномальным восстановлением».1 Но может наблюдаться и обратное; в частности, при частоте ультразвука 50 кгц на 3—4-й миллисекунде после первого стимула иногда обнаруживается «вторичное» падение амплитуды повторного ответа (рис. 51).
1 По мнению Гриннела, «аномальное восстановление» получается лишь на больших частотах ультразвука. Мы получали эти явления на всех частотам локационного диапазона,
153
Повышение амйлитудьт повторного ответа здесь может быть объяснено облегчающими влияниями импульсов, порожденных первым стимулом и направляющихся в холм по длинным нейронным цепочкам, импульсов, «запаздывающих», таким образом, на 4 мсек.
Вторичное падение амплитуды повторного ответа У4 (на частоте 50 кгц) может быть объяснено только тормозящими импульсами, порождаемыми первым сигналом и приходящими к холму (точнее, к источнику А\) с запаздыванием на 2.5— 3.4 мсек. Заметим, что пороги для холмовых он-ответов на ультразвуки с с частотой^50 кгц обычно повышены по сравнению
с ^соседними частотами. В^районе этой же частоты преимущественно обнаруживаются off-ответы. Эти же факты тоже косвенно свидетельствуют о’наличии тормозящих импульсов в залпе,порождаемом ультразвуком с частотой, соответствующей середине
Рис. 50. Холмовые ВП'на одиночные и парные стимулы (60 кгц, 0.5 мсек., длительность нарастания фронта 0.1 мсек).
j(ceepxy вниз) — ВП на одиночный стимул и на пары стимулов с интервалами 1.75, 2.9, 3 мсек.; II (сверху вниз) — ВП на одиночный стимул и пары стимулов с интервалами 1.0, 5.0, 10 мсек. Над кривыми ВП — отметки раздражения и времени 2000 в секунду. III — зависимость амплитуды (А) ВП на повторный стимул от интервала. Частота заполнения стимулов 50 и 40 кгц, сила 106 и 40 102 дб.
ультразвукового сигнала летучей мыши. Если повторный стимул имеет малую силу, то, как и на входе слуховой системы, в задних холмах может не выявиться рефрактерного состояния. Объяснение этому факту уже давалось: он отражает собой замену канала, ответившего на первый стимул и вошедшего в рефрактерное состояние, каналом, подпорогово-возбужденным первым стимулом, облегченно срабатывающим на повторный.
У медленного компонента холмового ВП цикл восстановления возбудимости оказался более длительным, чем у быстрого (рис. 51).
159
Лабильность заднехолмового слухового центра летучей мыши исследовалась нами также в методике ритмической стимуляции. Здесь использовались в качестве раздражителей как тональные (моночастотные), так и ЧМ-сигналы.
Применяя длинный ряд тональных или ЧМ-сигналов, мы исследовали амплитуду реакции на повторные стимулы — 2-й, 5-й й 10-й стимулы — и сопоставляли их с реакциями на 1-й в ряду стимул.
При этом выявилось, что такие элементы реакции, как TV4 и положительное медленное колебание в холме, изменяются
при изменении ритма раздражения по-разному. Лучше всего воспроизводят ритм раздражения пики;У4. Их амплитуды в пределах ритмов от 1 до 80 в 1 сек. не уменьшаются. Более того, при ритмах около 80 в 1 сек. {их амплитуда даже несколько увеличена (до 20%) по сравнению с амплитудой Лг4 в ответе на одиночный стимул.
Рис. 51. Зависимость амплитуды вызванного потенциала на второй стимул от длительности стимулов и интервала между ними.
По оси абсцисс — интервал между стимулами; по оси ординат — отношение амплитуды ответов на второй стимул к амплитуде первого. 1,2 — данные по измерениям быстрого, •3, 4 —медленного позитивного колебания; 1,3 — длительность стимулов 0.5 мсек., 2, 4— 4.6 мсек. Частота заполнения стимула 50 кгц.
Медленный холмовой ВП при ритмах 2—5—10 в 1 сек. несколько увеличен против «нормы» для одиночной реакции (на 45— 20 и даже до 90%), но при ритмах, превышающих 10—20 в 1 сек., он уже явно снижен и иногда даже исчезает совсем. Таким образом, лабильность центров задних холмов, генерирующих медленный компонент ВП, ниже, чем у цепочек, порождающих пик TV4.
Необходимо отметить, что амплитуды вторых в ряду ВП падают меньше, чем амплитуды пятых и тем более десятых при соответствующих учащениях ритма. Иначе говоря, в ходе каждой серии ритмических стимулов и реакций на них свойства реагирующей системы центров задних холмов меняются, их лабильность падает.
Можно было бы думать, что в данном случае речь идет о характеристике средней лабильности подкорковых нейронов разных уровней и изменениях этой средней лабильности по ходу ритмического реагирования. Однако представляется более вероятным, что здесь перед нами сложные суммарные характеристики систем нейронов, так называемых нервных сетей. Лабильность таких
160
систем определяется не только лабильностью их элементов, но и характером взаимодействия нейронов (в особенности их тормозными взаимодействиями). Можно предположить, что резкое падение ВП при учащении стимуляции связано с выявлением и усилением тормозного компонента в афферентном потоке, следующего за возбуждающим. Вероятно, что чем выше расположен исследуемый центр (чем выше «порядок» его нейронов в сети), тем длительнее на него возбуждающее и тормозящее действие каждого афферентного залпа (из-за дисперсии импульсов во времени), тем ниже лабильность нервной сети, «заканчивающейся» на уровне данного центра. Можно думать, что если в ходе ритмической стимуляции имеет место усиление функции тормозящих нейронов сети, то лабильность сети в целом должна падать. К этим вопросам мы еще вернемся при обсуждении свойств отдельных нейронов заднего холма и при рассмотрении материалов, относящихся к слуховой коре летучей мыши.
В наших опытах не выявилось различий в показателях ла-бильностй центров при их стимуляции тональными и ЧМ-сиг-налами. Это, видимо, было связано с применением относительно низких ритмов раздражения.
Но еще раньше американские ученые (Grinnell, McCue, 1963) исследовали восстановление возбудимости подкорковых центров у Myotis lucifugus (по jV4) к тональному сигналу 40 кгц после применения кратчайшего (1 мсек.) сигнала (40 18 кгц). При этом
они нашли, что восстановление возбудимости в таких условиях идет быстрее, чем в случае применения двух однотонных сигналов. Полное восстановление амплитуды при этом получилось через 0.3 мсек, от конца 1-го стимула. Объяснение этого факта, по-видимому, должно быть сходным с приведенным выше для случая быстрого восстановления после слабого стимула.
С помощью внеклеточного отведения ПД во многих экспериментах исследовались реакции на ультразвуки нейронов задних холмов летучих мышей.
Реакции нейронов на кратчайшие ультразвуки (0.2—0.5 мсек.) чаще всего представляли собой одиночные ПД, реже — короткие серии ПД (по 2—3). На более длительные ультразвуковые стимулы нейроны холма чаще всего отвечали фазически, т. е. лишь короткими сериями ПД, связанными только с началом стимула (on-эффект) или иногда с выключением стимула (off-эффект). Тонические реакции на длительные стимулы (ритмический разряд на протяжении всего действия ультразвука) наблюдались редко. У некоторых нейронов, имеющих значительную фоновую активность, можно было наблюдать торможение активности при действии ультразвуков определенных параметров (рис. 52). ЛП реакций нейронов холма на круто нарастающие стимулы при оптимальной их силе варьировали в пределах 4.2— 17.2 мсек.
161
, 11мб Юмсел. ЮмсеКь
Рис. 52. Зависимость характера ответов отдельного нейрона от силы стимула.
Сила стимула записана на кадрах, частота заполнения 18 кгц. Акустическая задержка 1 мсек. Позитивные колебания книзу.
Таким образом, среди обследованных нейронов имелись единицы разных «порядков». Средний ЛП ответа холмовых нейронов составлял 8—10 мсек.
У других видов гладконосых летучих мышей Гриннел (Grinnell, 1963а) нашел еще больший разброс ЛП реакций нейронов задних холмов на ультразвуки: от 4 до 180 мсек. При этом он указывает, что нейроны с малыми ЛП чаще обнаруживаются в глубоких частях холма, а нейроны с большими — в поверхностных частях холма. В работе Суги (Suga, 1964b) этот разброс составил 3.9—26.2 мсек., при этом чаще всего величины ЛП реакции близки к 8—9 мсек. Мы в своих опытах не могли подтвердить закономерность, отмеченную Гриннелом. И в глубоких слоях холма, и вблизи его поверхности обнаруживались нейроны с весьма различными ЛП реакций на оптимальные для них стимулы. Последнее, вообще говоря, гармонирует с фактом вертикальной тонотопии в холме, описанной выше и обнаруживаемой также при исследовании тональных настроек холмовых нейронов. По-видимому, все тональные группы нейронов холма достаточно сложны по составу и включают нейроны разных порядков.
Надо заметить, что, по имеющимся данным, характер ответа нейрона на стимул (ЛП ответа, число ПД, их связь с началом, серединой или концом стимула, их стабильность), будучи различным при оптимальных условиях у разных нейронов, у каждого нейрона находится в некоторой зависимости от параметров стимула.
Например, при усилении ультразвукового стимула от порога до оптимальной величины у многих нейронов наблюдается некоторое укорочение ЛП реакций, иногда к тому же увеличение числа пд в ответе, всегда повышение стабильности реакции — вероятности ответа. То же имеет место при «оптимизации» других параметров стимула: частоты ультразвука, угла подачи сигнала и т. д.
При изменении параметров стимула в «неблагоприятном» направлении (слишком малая или слишком большая сила, неадекватная частота или угол стимуляции) наблюдаются противоположные вышеописанным изменения в ответах нейронов или даже замена этих ответов торможением спонтанной активности (при сверхсильной или неоптимальной по частоте ультразвука стимуляции) (рис. 52, 53).
Каждый из исследуемых нейронов заднего холма, вообще говоря, помимо средних или крайних параметров ^го ответа, характеризует условия его эффективного раздражения, например: а) ЧПК, иначе говоря, область его ответов в корреляционном поле параметров; частота раздражающего ультразвука, его сила; б) отношение к сложным, в частности к ЧМ-сигналам; в) отношение к крутизне нарастания и длительности стимулов;
163
г) лабильность — возможная частота повторения реакций или время восстановления нормальной реакционной способности после возбуждения; д) пространственная диаграмма порогов. Этот список характеристик можно было бы продолжить. Но получение необходимых данных в опыте,
даже только перечисленных, для одного и того же нейрона оказывается чрезвычайно затруднительным. Что касается выяснения отношения нейронов к сложным ультразвуковым сигналам, то оно ввиду бесконечного разнообразия вариантов сложных сигналов может быть лишь весьма приблизительным и касающимся лишь какого-то узкого класса таких сигналов.
Практически в опубликованных к настоящему времени работах у нейронов заднего холма летучих мышей исследовались лишь некоторые из названных характеристик, причем чаще всего у одних нейронов — одни, у других — иные.
Рассмотрим данные о ЧПК (областях ответа) у нейронов задних холмов гладконосых летучих мышей. Здесь обнаружилось большое разнообразие характеристик у разных холмовых нейронов. Области ответов одних нейронов лежали в узком частотном диапазоне, расположенном у разных представителей этой группы в разных участках шкалы частот ультразвуков (рис. 54, Л); об-
Рис. 53. Осциллограммы потенциалов действия двух (Z, II) нейронов при различных силах стимула.
Цифры по вертикали — сила стимула. Вертикальные черточки в начале развертки луча — отметка раздражения. Акустическая задержка 1 мсек. I — длительность стимула 4.6 мсек., частота заполнения стимула 48 кгц; II — длительность стимула 4.6 мсек., частота заполнения 22 кгц.
ласти ответов других занимали весь диапазон эхолокационных частот (рис. 54, Б). При этом у большинства нейронов можно было найти лучшую частоту раздражающего акустического сигнала, имеющую наименьший порог раздражения,
164
Характер распределения нейронов по остроте оказался у обоих видов летучих мышей подобным распределению на уровне кохлеарных ядер. При этом число узконастроенных нейронов у подковоносов оказалось больше, чем у остроухих ночниц, и обнару-
Рис. 54. Нейронные «шкалы» для оценки частоты и силы ультразвука.
А —частотно-пороговые кривые 10 холмовых нейронов, «настроенных» на разные частоты; Б -частотно-пороговые кривые нейронов, отзывающихся практически на все частоты исследуемого диапазона, но имеющих различные силовые пороги; В— частотнопороговые кривые холмовых и корковых нейронов с ограниченным диапазоном ответов на ультразвук; Г —электрограмма реакций холмового нейрона с ограниченным силовым диапазоном ответов. Длительность стимула 0.5 мсек., частота заполнения 65 кгц.
живались они преимущественно в эхолокационной полосе частот 70—90 кгц.
Узконастроенные нейроны часто обнаруживали рядом с областью ответа зону (или зоны) торможения спонтанной активности. Вероятно, эти тормозные зоны и обеспечивают «обострение» частотной настройки соответствующих нейронов. «Широкие» и «узкие» частотные настройки встречались приблизительно одинаково часто у нейронов с короткими и длинными ЛП с глубоким и поверхностным расположением в холме.
Мн уже указывали, что нейроны, принимающие ультразвуки
165
разной частоты, распределены в холме вертикально, и чем выше” частота, тем глубже ойй лежат (Grinnell, 1963а; Васильев, 1968а).
Среди нейронов с узким йли с широким частотным тональным диапазоном в задних холмах гладконосых летучих мышей обнаруживаются нейроны, имеющие и низкие, и высокие пороги возбуждения, а также имеющие более или менее высокий верхний порог (предел) области реагирования, за которым обнаруживается торможение. Это торможение формально можно назвать «запредельным».1 Его механизм, вероятно, связан с тормозящим действием на исследуемый нейрон другого нейрона или нейронов, активизируемых при усилении раздражения (рис. 53).
Количество нейронов с закрытой сверху запредельным торможением областью реагирования в задних холмах Myotis luci-fugus, по Гриннелу (Grinnell, 1963а), составляет около х/3 обследованных им. Суга (Suga, 1964b) на том же объекте не наблюдал таких нейронов. По данным нашей лаборатории, у остроухой ночницы в задних холмах обнаруживается 44% таких нейронов (по опытам на 98 нейронах — Матюшкин, Стосман, 1969). У некоторых нейронов холмов области ответов оказались весьма сложными, в особенности у нейронов с большой спонтанной активностью. Обнаруживались нейроны с двумя раздельными областями ответов, с отдаленными областями торможения, с раздельными областями для on- и off-реакций и т. п, (Васильев, 1968а).
Значительный интерес представляет вопрос о стабильности форм ЧПК (областей ответа) нейронов. Исследуя ЧПК нейронов холма многократно (до 25 раз подряд) и при подаче стимулов цод разными углами, мы обнаружили, что в общем эти характеристики довольно стабильны. Во всяком случае при многократном определении - области ответа обследованных заднехолмовых нейронов сохраняли свою ширину и местоположение на шкале частот.
Большой интерес представляют отношения отдельных нейронов задних холмов летучих мышей (гладконосых) к ЧМ-ультра-звукам, подобным крикам, издаваемым этими животными при эхолокации.
Реакции нейронов данной области на ЧМ-сигналы изучали многие исследователи (Grinnell, McCue,' 1963; Suga, 1964b, 1965b, 1968; Матюшкин, Котеленко, 1973) на различных представителях гладконосых летучих мышей. При этом было обнаружено, что большинство нейронов холма реагирует на ЧМ-сигналы, если последние затрагивают «область ответа» этих нейронов. Правда,
1 Термин «запредельное торможение» предложен и применялся И. П. Павловым главным образом для обозначения подобного торможения системной реакции, но иногда и для предполагаемого им клеточного процесса, аналогичного вышеописанному.
166
встречались нейроны, реагирующие на тональные и не реагирующие на ЧМ-сигналы. Нейроны с широкой частотной настройкой, отвечающие на ЧМ-сигналы, обычно были безразличными к направлению модуляции. Наоборот, узконастроенные нейроны обнаруживали избирательную чувствительность, к сигналам с определенным направлением частотной модуляции. Это, по Gyre (Suga, 1964а), было связано с положением тормозной области ответа. Если тормозная область располагалась с низкочастотной стороны области ответа, то нейрон был более чувствителен к сигналам, «пересекающим» область ответа от высоких частот к низким, и наоборот.
Особый интерес представляют нейроны, реагировавшие только на ЧМ-сигналы определенного вида и не реагировавшие на тональные сигналы (Grinnell, McCue, 1963; Suga, 1968).
Зависимость порогов и форм реакции нейронов заднего холма гладконосых летучих мышей от длительности стимулов исследовалась Гриннелом (Grinnell, 1963b) и Васильевым (1968а). В опытах Васильева у одних заднехолмовых нейронов пороги и формы реакции мало зависели от длительности тональных стимулов, если последняя превосходила 0.2 мсек, (для более коротких стимулов пороги были всегда повышены). Но у других нейронов при удлинении стимулов наблюдались падение порогов и иногда увеличение числа ПД в ответе.
Гриннел (Grinnell, 1963b) нашел ряд нейронов с резким (на десятки децибел) падением порогов при удлинении стимула и, что особенно интересно, один нейрон, реагировавший только в узком диапазоне длительностей стимула (0.5—4 мсек.). Можно предполагать, что реакция этого нейрона возникла с большим латентным периодом. Видимо, она могла тормозиться какими-то нейронными цепочками, включавшимися при стимулах, превышавших 4 мсек, по длительности.
Значение крутизны нарастания акустического стимула для формирования реакций нейронов заднего холма изучалось Васильевым (1968) на 16 нейронах. Было найдено, что у некоторых нейронов удлинение времени нарастания стимула не изменяет, а у других несколько увеличивает порог возбуждения. Порог «запредельного» торможения при этом часто снижался. Число ПД в ответе при удлинении фронта стимула чаще всего уменьшалось. Весьма интересно то, что обнаружилось небольшое число нейронов, отвечавших (on- и off-потенциалами действия) лишь при определенной крутизне стимула (например, от 0.1 до 0.65, как на рис. 55). В этом случае ограничение диапазона реакции нейрона несомненно связано с тормозными механизмами. Здесь можно предположить, что торможение данного нейрона (или какого-то низшего звена возбуждающей его нейронной цепочки) имеет два источника. Можно думать, что оно исходит, с одной стороны, от элементов, реагирующих только на очень е 167
крутой стимул (отсюда исключение ответа данного нейрона на «прямоугольный» стимул). С другой стороны, оно, видимо, исходит от каких-то нервных структур, реагирующих лишь на по-
20кгц 79дб ^6мсек.
Рис. 55. Осциллограммы электрических ответов отдельных нейронов заднего холма при различной длительности фронтов стимула.
Цифры по вертикали — длительность фронтов стимула, по горизонтали— частота заполнения, сила и длительность стимула. Вертикальные черточки в начале развертки луча —отметки раздражения. Акустическая задержка 1 мсек. Позитивные колебания книзу. На втором луче записаны раздражающие стимулы.
лого нарастающий стимул (отсюда подавление ответа нейрона при пологих стимулах).
Специальный интерес (в плане оценки механизмов эхолокации) представляют данные о способности нейронов заднего холма 168
летучей мыши реагировать на сигналы, следующие друг за дру-гом с короткими интервалами (как крик мыши и его эхо).
Оказалось, что время восстановления возбудимости у разных холмовых нейронов разное и варьирует в пределах от 0.5 мсек, до нескольких секунд. Правда, крайние варианты очень редки и основная масса единиц имеет время восстановления около 10—20 мсек. Отмечалось, что относительно короткие интервалы восстановления наблюдались при слабых, а длинные — при сильных сигналах, что характерно и для суммарной реакции.
При коротких интервалах между стимулами нейроны отвечали либо только на первый стимул, либо на оба, но иногда и только на второй. Последнее косвенно свидетельствует о том, что разные холмовые нейроны получают афферентные импульсы по весьма различным нервным цепочкам.
Несомненно, что в данном случае речь идет отнюдь не о вариациях рефрактерности нейронов как таковой. Длительная невоз-будимость заднехолмовых нейронов после их ответа на адекватный стимул является результатом воздействия на нейрон «тормозного» залпа, следующего тотчас за возбуждающим (Suga, 1964b). Заметим, что этот же тормозной залп укорачивает реакции нейронов на длинные акустические стимулы, делая эти реакции оп-ответами.
Оценивая в целом рассмотренные экспериментальные данные, касающиеся задних холмов летучих мышей, следует сказать, что они в основном качественно совпадают с полученными на других млекопитающих (Erulkar, 1959; Katsuki, 1961; Rose et al., 1963; Вартанян, 1966; Альтман, 1967; Марусева, 1967). Однако количественные характеристики этого отдела слуховой системы летучей мыши существенно отличаются от приводимых для таких млекопитающих, как кошка, морская свинка и крыса. Это касается не только ультразвуковых настроек нейронов у летучих мышей (определяемых входом системы), но и исключительных скоростей их реакций и восстановления возбудимости, остроты угловых характеристик. Все эти характеристики несомненно находятся в прямой связи с использованием летучими мышами ультразвуковой эхолокации, требующей от слуховых центров этих животных предельно высоких «разрешающих способностей».
Электрические явления во внутренних коленчатых телах
Исследования внутренних коленчатых тел летучих мышей единичны (Васильев, Андреева, 1971). Электрические ответы на включение стимула (оп-ответы), регистрируемые при макроэлектродном отведении от внутренних коленчатых тел, представляют собой негативно-позитивные комплексы (рис. 56,
169
А). Длительность негативного отклонения составляет 10—15, положительного — 20—25 мсек. ЛП первого колебания равен 5—8 мсек. В некоторых записях перед негативным колебанием регистрируется положительное отклонение малой амплитуды, имеющее ЛП 3—4 мсек., с накладывающимися на него быстрыми (1—1.5 мсек.) колебаниями того же знака и ЛП 4—5 мсек. Подобная форма ответов обнаруживается и у других млекопитающих (Ades, Brookhart, 1950; Гершуни, 1964).
Рис. 56. Электрический ответ внутреннего коленчатого тела (А) и частотнопороговые кривые (Б) по негативному колебанию.
А: длительность стимула 10 мсек., сила стимула 65 дб, частота заполнения 45 кгц, длительность фронта нарастания 0.1 мсек. Акустическая задержка 1 мсек. Вертикальная черточка в начале записи — отметка раздражения. Позитивное колебание книзу. В: по оси абсцисс — частота заполнения стимула; по оси ординат — пороговая сила стимула. Сплошные линии — данные по оп-ответам, штриховые — по off-ответам, пунктирные — максимально возможная сила стимула. I — опыт на остроухой ночнице, II — на подковоносе.
Регистрация порогов суммарных on- и off-ответов (негативного колебания) показывает, что они примерно такие же по форме, что и на предшествующих уровнях (рис. 56, I, II) у обоих видов летучих мышей. Это свидетельствует о том, что потоки импульсов об ультразвуковых раздражителях, в том числе и эхо-сигналах, передаются в вышележащие центры.
Измерение порогов разрядов отдельных нейронов — определение областей ответов отдельных нейронов — показывает, что и здесь все нейроны имеют характеристическую частоту, т. е. частоту, порог на которой оказывается минимальным. Некоторые нейроны имели две такие частоты (рис. 57, Па). При наличии нескольких характеристических частот у одного нейрона обна-170
руживалйсЬ двё ййй три независимые области ответов (рис. 57, 11б)\ в этих случаях у подковоносов характеристические частоты таких областей относились к диапазонам 80—82, 40—43, 26 — 28 кгц.
Как и на предшествующем уровне, во внутренних коленчатых телах области ответов могли быть симметричными (/в, Ив, Шб), асимметричными (1г, 16), узконастроенными (1в), широконастроенными (1а) и с верхними порогами (рис. 57, IV).
Рис. 57. Типы областей ответов отдельных нейронов.
По оси абсцисс — частота заполнения стимула, по оси ординат —пороговая сила стимула. Пунктирные линии — максимально возможная сила стимула, сплошные— данные по оп-ответам; штриховые — по off-ответам. 16, 116, Ша, Шб, IVa, IV6— опыты на подковоносах, остальные — на остроухих ночницах.
Нейроны с on- и off-ответами имели, как правило, разные области ответов на включение и выключение стимулов. Пороги off-ответов чаще всего располагались выше порогов оп-ответов, и поэтому область ответов на выключение стимула оказывалась внутри области ответов на его включение (рис. 57, III).
Описанные области ответов регистрировались как в опытах на остроухих ночницах, так и в опытах на больших подковоносах и принципиально не отличались от областей ответов, описанных другими исследователями (Gross, Thurlow, 1951; Ga-lambos, 1952; Katsuki et al., 1959; Katsuki, 1961).
171
Существенные различия в характеристике зарегистрированных нейронов у обоих видов летучих мышей обнаружились при оценке настроенности нейронов по коэффициенту Q. Общие границы
1 4 7 10 13 1619 22 25 26 31 40507090Q.
Рис. 58. Гистограммы частот коэффициента Q.
По оси абсцисс — значения коэффициента Q; по оси ординат — частоты. I — данные, полученные в опытах на ночницах (n=254), II — на подковоносах (п=251). п — число зарегистрированных нейронов у данного вида летучих мышей.
изменения Q у остроухих ночниц измерены от 1 до 50, у подковоносов — от 1 до 87 (рис. 58). Следовательно, число остронастроенных нейронов у подковоносов продолжает увеличиваться по сравнению с нижележащими центрами, тогда как у остроухих ночниц этот процесс не выражен.
Электрические явления в слуховой корковой области
Слуховая зона летучих мышей, точнее, симметричные слуховые корковые области, расположена на боковой поверхности правого и левого полушарий, как это показано на на рис. 59.
В этих областях у слабо наркотизированных летучих мышей могут быть зарегистрированы достаточно отчетливые вызванные потенциалы — первичные ответы (ПО) на ультразвуковые стимулы.
Форма ПО слуховой коры летучей мыши на краткий ультразвук (длящийся 1—4 мсек.) обычная: при униполярном поверхностном отведении это позитивно-негативное колебание (рис. 59), при углублении отводящего электрода в кору направление фаз ПО извращается, как и у других млекопитающих (Ройтбак, 1954). У остроухих ночниц, исследованных в наших опытах, латентный период ПО за вычетом акустической задержки составляет 7—13 мсек., амплитуда достигает 200—550 мкв. Длительность позитивной фазы 12—18 мсек., негативной фазы —
172
19—32 мсек. Это on-эффект. ЛП и амплитуда коркового ПО зависят от силы стимула. Эта зависимость, изученная нами для оп-эффектов, в большинстве случаев оказывается двухфазной: при увеличении силы стимула (от порога до некоторого предела) амплитуда ПО растет, а его ЛП укорачивается; при дальнейшем
Рис. 59. Электрофизиологические характеристики слуховой коры летучих мышей.
А — схема мозга летучей мыши (заштрихована область отведения ВП и реакции нейронов); Б — вызванный потенциал слуховой коры летучей мыши на ультразвуковой стимул (калибровка 100 мкв, 20 мсек.); В —частотно-пороговые кривые слуховой коры ночниц (1) и подковоносов (2). По оси абсцисс —частота заполнения стимула, по оси ординат — сила стимула. Ill, IV: сплошные линии —данные по оп-ответам, штриховые — по off-ответам.
увеличении силы стимулов амплитуда ПО падает, а его ЛП удлиняется. Наблюдаемое в данном случае (как и в рассмотренных случаях с холмовыми ответами) угнетение реакций сверхсильными стимулами аналогично запредельному торможению И. П. Павлова (1951). Вероятно, при стимуляции слуховых рецепторов в высшие отделы слуховой системы поступают не только воз-
173
бу ж Дающие, но и тормозящие ймйульсы. За это говорят данные, полученные на отдельных нейронах, которые будут обсуждены ниже. Можно полагать, что при малых силах раздражения количество тормозящих импульсов, поступающих в кору, невелико, а при больших силах велико, причем настолько велико, что оказывается достаточным для снижения эффекта возбуждающих импульсов (снижение суммированных возбуждающих постсинаптических потенциалов и соответственно импульсной активности в корковом этаже слуховой системы).
Отводимые большим поверхностным электродом корковые ПО при раздражении ультразвуками разной частоты (30, 60 и 80 кгц) сходны по форме и имеют соизмеримые амплитуды при равенстве сил раздражения относительно порогов. Это указывает на приблизительную равновеликость корковых проекций разных частот ультразвуков исследованного нами диапазона (соответствующего диапазону частот ультразвуковых импульсов, испускаемых мышами при эхолокации).
ЧПК, определенные по ПО, в общем подобны зарегистрированным на нижележащих уровнях, за исключением опытов на подковоносах. У них обнаружены ЧПК, содержащие три минимальных порога в диапазонах частот 27—28, 40—42 и 82—84 кгц. Если принять, что лучшей будет частота в диапазоне 82—84 кгц, то две другие равняются х/2 и х/3 ее. В этих же частотных полосах, как правило, регистрировались ответы на выключение стимула с минимальными порогами в диапазоне 78—82 кгц (рис. 59).
Корковые ПО в отличие от холмовых ответов тУ4 оставляют за собой длительный период «невозбудимости». Восстановление амплитуды ПО на повторное раздражение обычно начинается лишь спустя 200 мсек. (0.2 сек.), а полное восстановление имеет место лишь через 0.5 сек. после первого стимула. В ходе ритмического раздражения с частотой более чем 1—2 в 1 сек. обычно имеет место падение амплитуд ПО, так что амплитуда 5-го и тем более 10-го ответов меньше амплитуды 2-го ответа в серии.
Таким образом, лабильность коркового слухового центра как системы мала. Однако не следует думать, что здесь речь идет о низкой лабильности (длительной рефрактерности) элементов коркового центра. Длительный период невозбудимости вслед за ПО здесь несомненно связан с залпом тормозящих импульсов, сопровождающим возбуждающие. Об этом говорят многие факты, и в частности наблюдавшиеся нами «срывы» такого торможения — внезапные и резкие повышения амплитуд повторных ПО (вплоть до превышения ими ПО на первое раздражение) при сравнительно высоких ритмах стимуляции (10—12 в 1 сек.), обычно приводящих к сильному снижению корковых ПО (Котеленко, 1968). Суга (Suga, 1965а) исследовал реакции и свойства 140 корковых нейронов. При этом обнаружилось, что ЛП нейронов варьировали
174
от 8 до 22 мсек, (главным образом от 11 до 17 мсек.). Число ПД в ответе нейронов на ультразвук при постепенном усилении последнего увеличивалось, но чаще всего лишь до определенного предела, за которым начиналось уменьшение числа ПД (признаки запредельного торможения, по нашей терминологии).
Оказалось, что ЧПК, или области ответов, у разных корковых нейронов сильно различаются. Он выделил по этому признаку 4 типа нейронов: 1) нейроны с двумя областями ответа (высоко-и низкочастотной), разделенными зоной чувствительности; 2) нейроны с широкой областью ответа; 3) нейроны с узкими областями ответа (узконастроенные по частоте за счет латерального торможения); 4) нейроны с закрытыми сверху областями ответа (т. е. с запредельным торможением). Последних было очень мало (4 из 140). Суга считает, что корковые нейроны слуховой системы летучей мыши имеют области ответа не более широкие, чем у холмовых нейронов.
Аналогичные данные были получены Котеленко и Васильевым (1971) (рис. 60). Ими отмечаются некоторые особенности у подковоносых летучих мышей. В коре, как и во внутреннем коленчатом теле, получены нейроны с тремя областями ответов и расположением характеристических частот в диапазонах 26— 27, 40—42 и 80—83 кгц (рис. 60, IV, V). Далее, если у остроухих ночниц соотношение широко- и узконастроенных нейронов оказалось подобным нижележащим центрам слухового анализатора, то у подковоносов продолжается увеличение остронастроенных нейронов (рис. 61).
Весьма интересны данные Суги (Suga, 1965а) о реакциях разных корковых нейронов на частотно-модулированные ультразвуковые сигналы с модуляцией «разных направлений». Здесь автор выделил следующие категории нейронов: 1) «симметричные» нейроны (47%) — одинаково реагирующие на ЧМ-сигналы обоих направлений (это вместе с тем нейроны с двойными или с широкими областями ответа); 2) «асимметричные» нейроны (27%) — они реагируют преимущественно на сигналы с модуляцией частоты в одном каком-то направлении; 3) нейроны с верхними порогами (запредельным торможением) (11%) — они отвечают лишь на слабые ЧМ-сигналы; 4) нейроны, нечувствительные к ЧМ-сигналам (15%), хотя и чувствительные к монохроматическим, нейроны 2—4-й категорий имеют узкие области ответов; 5) нейроны, более чувствительные к ЧМ-сигналам, чем к монохроматическим (13%); 9 из 14 таких нейронов вообще не отвечали на монотонические сигналы; 6) нейроны, у которых ЧМ-сигналы вызывали ответы с резко увеличенным числом ПД, но пороги для этих и лучших монохроматических сигналов не различались (4%). Суга отмечает, что процент нейронов, особо реагирующих на ЧМ-сигналы (53%), в коре летучей мыши больше, чем в ее заднем холме.
175
Рис. 60. Типы областей ответов отдельных нейронов слуховой коры.
По оси абсцисс — частота заполнения стимула; по оси ординат— сила стимула. Пунктирные линии —максимально возможные силы стимулов, сплошные — измерения по оп-ответам, штриховые - по off-ответам. 1а, 16, На —данные опытов на остроухих ночницах, остальные — на подковоносах.
Рис. 61. Гистограммы частот коэффициента Q.
По оси абсцисс — значения коэффициента Q; по оси ординат— частоты. I—данные, полученные в опытах на ночницах (n=92), II — на подковоносах (п=112). п —число зарегистрированных нейронов у данного вида летучих мышей.
Для объяснения самых интересных из описанных категорий реакций (а именно категории 2, 4, 5) предложены схемы нейронных цепочек, приносящих к данным корковым нейронам возбуждающие и тормозящие импульсы. Некоторые из этих схем мы воспроизводим на рис. 62, 63. По Суге, сложные характеристики высших нейронов объясняются различными соотношениями областей ответов нижележащих нейронов, возбуждающих и «ла-терально» тормозящих данный нейрон. Если области латеральных торможений двусторонни и имеют низкие пороги, то «вышележащий» нейрон может быть нечувствителен к ЧМ-сигналу. Если с одной из «сторон» область торможения отсутствует или имеет высокий порог, то нейрон будет асимметричен (ответит лишь на сигнал, частота которого изменяется в направлении, показанном на рис. 63 сплошной стрелкой). Если два таких асимметричных нейрона с одинаковой тональной характеристикой имеют противоположную асимметрию и включены параллельно на некоторый нейрон высшего порядка, причем один из них возбуждает, а другой тормозит, то этот нейрон высшего порядка будет реагировать только на ЧМ-сигналы определенного направления (рис. 63), не отвечая на все прочие, даже монотона л ьные. Нас специально интересовал вопрос об «устойчивости» тональных настроек (форм областей ответа) у этих нейронов. Заметим, что в аналогичных исследованиях на нейронах задних холмов остроухой ночницы выявилась значительная устойчивость этих характеристик. Повторные промеры ЧПК (областей ответа) широконастроенных корковых нейронов в наших опытах не показали существенных изменений этих кривых. В то же время узконастроенные нейроны при повторных исследованиях, как правило, показывали измененные, иногда смещенные по частотной шкале, но всегда более широкие и даже предельно широкие области ответа.
Мы полагаем, что это расширение областей ответа нейронов в ходе опыта могло быть связано с постепенным ослаблением эффекта предшествующей однотонной (на лучшей частоте) сильной стимуляции, применяющейся при «выделении нейронного ответа из суммарной реакции» (при погружении микроэлектрода).
Такая стимуляция, видимо, обостряет настройку нейронов, реагирующих на данный тон за счет латерального торможения, тогда как разнотональная пороговая стимуляция в ходе измерений ЧПК этого эффекта не поддерживает.
Как бы то ни было, полученные факты (Котеленко, Матюшкин, 1969) свидетельствуют о потенциально широкой тональной настройке у многих корковых нейронов летучих мышей данного вида. Представляется вероятным, что это относится и к другим видам рукокрылых.
Потенциально широкая и изменчивая частотная настройка корковых нейронов свидетельствует об их связи со множеством
177
Стимул Канал В
а ъ
f, / е 1
/ Е 0
^3 i Е Е
гм /- ^Е 0.1
гм ^-Е Е
Рис. 62. Нейронная сеть, дающая узкую полоску ответа, немоно-тоническую spike-count-функцию и фазный оп-ответ. (Suga, 1965а).
На более низком уровне (например, улитковое ядро) нейроны а, Ь, с показывают тонические ответы. Области их ответов изображены кривыми в верхнем левом углу. Каждый из этих нейронов возбуждает соответственно нейроны с более высоким уровнем (А, В или С), но тормозит соседние нейроны через тормозные промежуточные нейроны. Таблица показывает эффект импульсов из нейронов а и на нейрон В и ответы нейрона В на каждый из чистых тонических импульсов Л,/2,/з- Кие — сильное и слабое возбуждение, I и i — сильное и слабое торможение, 0 — отсутствие эффекта. В результате этих взаимодействий нейрон В имеет узкую область ответа и области торможения (нижнее левое изображение), spike-count-функция становится немоното-нической при fa. Ответ на ЧМ-импульсы изменяется в зависимости от частотного изменения. Каналы торможения делают характеристику ответа фазным /з-ответом. Если есть потоки возбуждения и из соседних каналов, как показано точечными линиями (справа сверху), то область ответа нейрона В будет шире.
Рис. 63. Нейронная сеть, объясняющая нейроны, чувствительные к ЧМ-сигналам. (Suga, 1965а).
Когда нейроны а и Ъ, находящиеся на более низком уровне, имеют свойства, изображенные на графиках а и Ь, связаны с нейроном с, как показано верхней правой диаграммой (промежуточный тормозный нейрон), то нейрон с отвечает на ЧМ-сигналы, но не на чистые тонические импульсы. Остальные обозначения те же, что и на рис. 62.
разнонастроенных подкорковых нейронов (возбуждающие и тормозящие влияния которых на корковые нейроны могут приобретать в разных условиях разную силу). При таких неопределенных тональных настройках (иначе говоря, при таком «включении» в слуховую систему) соответствующие корковые нейроны, вероятно, не могут быть «опорными пунктами» в частотном анализе ультразвуков. Но эти нейроны коркового слухового центра у летучих мышей (как и у других млекопитающих), видимо, могут выступать в роли элементов, суммирующих разнотональные сигналы и порождающих обобщенные импульсы акустической модальности.
Рассмотренные нами данные об изменчивости тональных настроек корковых нейронов, конечно, нельзя расценивать как бесспорное опровержение данных Суги и его заключений о «свойствах» корковых нейронов у летучих мышей. Не исключено, что какая-то часть корковых нейронов рукокрылых стабильно демонстрирует свойства, описанные Сугой. Но в последнем можно сомневаться, и окончательное решение вопроса требует дальнейшей работы.
Говоря о свойствах слуховой коры летучих мышей, необходимо остановиться на исследовании характеристик кортико-фугальных влияний (Куликов, 1968, 1970а, 19706, 1972а, 19726, 1973а, 19736). С помощью физиологических тестов было выявлено наличие двух систем кортико-колликулярных связей.
Существование кортико-фугальных волокон поставило вопрос об их функциональной оценке. Для ее выяснения изучались изменения параметров N* на звуковые раздражения, поданные через разные интервалы времени после электрического раздражения слуховой коры. Было обнаружено, что электрическое раздражение вызывает уменьшение амплитуды и ЛП этого колебания.
Полученные данные совпадают для обоих видов летучих мышей. Таким образом, результаты исследования указывают на возможность кортикальной регуляции афферентного потока при звуковых раздражениях в слуховом анализаторе летучих мышей.
При дальнейшем исследовании роли кортикального уровня в деятельности эхолокационной системы изучалось влияние функционального выключения коры на пороговые, амплитудные и временные характеристики электрических реакций заднего двухолмия и улитки (ПД слухового нерва и микрофонного потенциала).
Функциональное выключение коры больших полушарий осуществлялось с помощью создания в височных областях очагов распространяющейся депрессии при двусторонней аппликации 20 %-го КС1. У остроухих ночниц при этом пороги на включение стимула (on-ответа) достоверно понижались в полосе частот
179
25—50 кгц. У больших подковоносов обнаружено увеличение порогов (on-ответа) в полосе частот 25—70 кгц. Контрольные опыты на обездвиженных животных дали аналогичные результаты.
Таким образом, одинаковые воздействия на кору приводят у этих видов летучих мышей к диаметрально противоположным изменениям порогов электрических реакций заднего двухолмия. При этом у остроухих ночниц понижение порогов располагается в полосе частот, соответствующих эхолокационным, а у больших подковоносов повышение обнаруживается в полосе частот, расположенной ниже эхолокационных. ЧПК заднего двухолмия, построенные по на выключение стимула (по off-ответам), не изменялись.
Очевидно, что полученные изменения порогов обусловлены результатом взаимодействия нисходящих влияний на более низких уровнях, чем заднее двухолмие. Поэтому в следующей серии изучалось влияние функционального выключения коры на пороги jN\ и микрофонного потенциала.
Как и у остроухих ночниц, так и у подковоносов было обнаружено понижение порогов в частотных полосах соответственно 25—70 и 38—50 кгц. ЧПК, зарегистрированные по микрофонному потенциалу, достоверно не изменялись. Данные опытов об изменении порогов Л;4 и N± подтвердились и при изучении зависимости амплитудных характеристик этих реакций от силы стимула до и после двусторонней аппликации КС1.
Проведение экспериментов под барбитуратовым наркозом, постановка контрольных опытов на обездвиженных животных, отсутствие изменений порогов off-ответов и микрофонного потенциала — все это позволяет считать, что обнаруженные влияния были реализованы через специфические системы нисходящих волокон.
Направленность изменений порогов, выявляемая на уровне заднего двухолмия, вероятно, улучшает избирательность слуховой системы к эхолокационным частотам, которые используются данными видами летучих мышей.
Эти изменения могут являться отражением реального функционирования эхолокационной системы у этих видов летучих мышей в режиме излучения сигналов с большой частотой следования, в связи с тем что нейроны слуховой коры не способны реагировать на быстроследующие раздражения. Такой режим излучения (до 200 имп./сек.) характерен при сближении летучих мышей с обнаруженными и распознанными объектами локации.
Некоторые соображения о принципах анализа ультразвуковых сигналов в слуховой системе
Попытаемся, исходя из рассмотренных материалов, нарисовать примерную схему работы данного анализатора по различению (определению): а) интенсивности ультразвукового
180
сигнала, б) его тональной характеристики (простой или сложной), в) его временных характеристик: длительности, крутизны фронтов, момента прихода (относительно какого-то предыдущего стимула).
Из этих элементов складывается как слуховая функция в обычном смысле этого слова, так и специфическая работа слухового анализатора (при сличении параметров излученного крика и его эха) в процессе ультразвуковой эхолокации. Нам представляется правдоподобным предположение, что названные элементы, или элементарные линии анализа (а—Ь), ультразвуков осуществляются одновременно в специальных сетях, в какой-то мере независимых друг от друга.
Вместе с тем имеющиеся фактические материалы позволяют предполагать, что принципы анализа на всех этих «элементарных линиях» весьма близки. Эти принципы были глубоко поняты еще И. П. Павловым, говорившим об обязательной «приуроченности динамики к структуре», отражении (кодировании) простых и сложных раздражителей возбуждением определенных клеток в мозговом конце анализатора, исключительной роли торможения в функции анализаторов. Эти принципы применительно к слуховому анализатору конкретизированы в ряде работ, выполненных на обычных лабораторных млекопитающих, а также в рассмотренных выше исследованиях на летучих мышах.
Предлагаемые схемы не претендуют на точное отражение действительности, они лишь демонстрируют, причем в упрощенной форме, один из возможных вариантов функционирования анализаторов летучей мыши при осуществлении той или иной «элементарной линии анализа». Нам представляется, что эти схемы могут быть легко согласованы с данными из области физиологической акустики (Гершуни, 1964; Гольдбурт, 1964; Katsuki, 1964).
Анализ интенсивностей коротких ультразвуков. Различие сил коротких ультразвуков любой частоты в слуховой системе летучих мышей, вероятно, опирается на различение количества рецепторов, возбуждаемых при разной силе ультразвука. Это следует из факта закономерного роста амплитуды ПД VIII нерва (А) при усилении раздражения, если последнее не чрезмерно.
О различительной способности системы в отношении силы ультразвуков можно грубо судить по усредненной кривой зависимости А от J (дб) с учетом вероятности этого или иного случайного отклонения А от каждого ее среднего значения. Если принять, что во всех точках этой кривой случайные отклонения А одинаковы (что близко к истине), то с вероятностью 0.95 можно утверждать, что изменение силы очередного краткого ультразвукового стимула (против предыдущей величины) на 2 дб будет отмечено в слуховой системе летучей мыши изменением количества возбуждающихся первичных нейронов. Можно думать, что дифференциальная чувствительность летучих мышей значительно
181
выше указанной величины. По данным Грййнёла (Grinnell, 1963aj/ она может составлять 0.2—0.5 дб.
Возможно, что при относительно длительных ультразвуках оценка силы последних отчасти опирается и на различение количества ПД в разряде нейронов (оно больше при более сильных стимулах). Но кодирование силы стимулов для^верхних этажей слуховой системы, вероятно, имеет другой механизм. Как уже говорилось, амплитуда холмовых и корковых ВП не столь строго следует силе стимулов. Таким образом, количество возбужденных нейронов оказывается непропорциональным силе стимула. Можно предполагать, что кодирование силы стимула и анализ силовой характеристики здесь опираются на часть нейронов, «представляющих» в высших центрах различные массы рецепторов улитки. Известно, что в задних холмах летучих мышей обнаруживаются разнопороговые нейроны, имеющие более или менее равное отношение ко всем частотам ультразвуков «эхолокационного диапазона». Такие нейроны могут быть элементами шкалы, на которой ведется отсчет силы раздражителя.
Особое значение здесь могут иметь нейроны с закрытой сверху областью ответа. Их возбуждение однозначно, хотя и глубоко кодирует соответствующую силу стимула. Можно предположить, что такие нейроны (или некоторые из таких нейронов) составляют элементы нейронной «шкалы», в которой соседние по порогам группы единиц имеют тормозное взаимодействие (как в тональной шкале). Эта гипотеза нам представляется правдоподобной. Ее’про-верка — дело будущего. |
Основные принципы частотного (тонального) анализа ультразвуков. Анализ (различение) частот ультразвука в улитке летучей мыши опирается на так называемое правило места, т. е. на распределение вдоль канала улитки и соответственно основной мембраны (несущей на себе кортиев орган с рецепторами) областей, отвечающих в порядке резонанса на разновысокие ультразвуки. «Правило залпов» (ритмов) здесь практически не имеет значения.
Известно, что в направлении от основания улитки к ее вершине последовательно расположены области, реагирующие на ультразвуки понижающейся частоты и далее на звуки понижающейся частоты, и так до вершины улитки. Таким образом, каждой частоте ультразвука соответствует особое место, «фокус» в кортие-вом органе, особые рецепторы.
Однако анализ частот (высот) тонов в улитке осложнен по меньшей мере тремя обстоятельствами: 1) поступлением ультразвука в канал улитки с его «высокочастотного» конца; это обстоятельство приводит к тому, что низкие ультразвуки и звуки, адресующиеся к верхним низкочастотным частям улитки, затрагивают «бегущей волной» высокочастотные части основной мембраны и высокочастотные рецепторы; 2) сильные ультразвуки приводят
182
к колебанию основной мембраны не только в фокусе данной частоты, но и в соседних районах (Bekesy, 1951); 3) рецептивные поля первичных слуховых нейронов относительно широки (Fri-sKko]Xf, 1964). Тут же заметим, что анализ высоты коротких моно-частотных ультразвуков, использованных в наших опытах (и тем более криков, издаваемых самими летучими мышами), затруднен относительно широким частотным спектром этих сигналов.
Отсюда ясна необходимость дальнейшего «обострения» частотного (тонального) анализа в слуховой системе летучих мышей, в ее центральных отделах.
По литературным данным (Katsuki, 1964), это обострение частотного анализа связано с «латеральным торможением». Сейчас известно, что тормозящее взаимодействие «соседних» каналов начинается на уровне нейронов VIII нерва и продолжается на высших уровнях, обеспечивая чрезвычайную «остроту» настроек отдельных нейронов в высших центрах слуховой системы летучих мышей (прежде всего задних холмов).
Возбуждение этих узконастроенных нейронов просто и надежно (однозначно) кодирует тональность стимула. Масса этих нейронов составляет своеобразную шкалу для оценки частот ультразвуков (частотный анализатор).
Схема анализа сложных, именно ЧМ-сигналов разного вида в конечном счете~также сводится к выделению и кодированию этих сигналов возбуждением специальных нейронов высших центров. Эта схема, предложенная и блестяще обоснованная Сугой (Suga, 1965а), базируется на элементах, которые только что рассмотрены. Настройки нейронов, принимающих сложные сигналы, складываются как сочетания настроек возбуждающих и тормозящих нейронов, воздействующих на данный «высший» нейрон. По-видимому, следуя этому принципу построения нервных сетей, можно нарисовать сеть, кодирующую (возбуждением выходного нейрона) любой сложный акустический сигнал.
О временном анализе в слуховой системе летучих мышей. На основании полученных и литературных данных можно предполагать, что отсчет малых интервалов между стимулами (например, криком и эхом) в слуховой системе летучих мышей осуществляется на «шкале» нейронов высших порядков, имеющих разные периоды «восстановления» возбудимости (исчезновение торможения), и в особенности нейронов, имеющих избирательное отношение к повторным стимулам, следующим с определенным интервалом после предыдущего стимула. Последняя категория нейронов может надежно кодировать величину соответствующих временных промежутков. Измерение длительности ультразвукового сигнала, вероятно, может осуществиться в высших слуховых центрах по тому же принципу с использованием on- и off-реакций низших отделов слуховой системы как пары сближенных стимулов.
183
Что касается оценки длительности фронтов стимула (его нарастания и спада), то и эта величина может оцениваться на «шкале» нейронов высших слуховых центров, избирательно реагирующих на сигналы с определенными длительностями фронтов.
Таким образом, все формы временного анализа в слуховой системе летучей мыши, по-видимому, могут быть связаны с деятельностью специальных «нейронных шкал» в высших центрах системы.
Представляет, конечно, большой интерес вопрос о том, каким образом начинается такой временной анализ (анализ интервалов между стимулами, длительностей и фронтов стимулов) в улитке и низших слуховых центрах, иначе говоря, как формируются названные нейронные шкалы.
Можно думать, что и здесь, как и в предыдущих случаях, основную роль играет взаимодействие возбуждающих и тормозящих сигналов, поступающих по различным нервным цепочкам к высшим нейронам, причем в данном случае особое значение имеют временные соотношения последних.
Например, можно себе представить нейрон, получающий вслед за раздражением и возбуждением два мощных и длительных тормозных залпа, идущих последовательно и разделенных коротким «светлым» промежутком, отстоящим от момента раздражения на время т. Такой нейрон при раздражении парами стимулов ответит на них двумя разрядами при интервале т (или при значительно больших интервалах). Здесь еще анализ временных интервалов ненадежен. Но можно представить себе более «высоко расположенный нейрон», возбуждающийся от первого лишь при суммации быстро следующих друг за другом импульсов. Этот нейрон возбудится уже только при интервале между стимулами т, т. е. он надежно закодирует своим импульсом (или серией импульсов) данный временной интервал между ультразвуковыми сигналами.
По такой же схеме можно построить гипотетические нервные сети, формирующие шкалы для анализа длительности сигнала и его фронтов.
Нельзя не отметить, что нарисованная нами картина работы слухового анализатора летучей мыши выглядит пока несколько статичной. Но мы, вероятно, исправим этот недостаток, если отметим, что в деятельности всех частей слухового анализатора летучей мыши, всех специализированных его каналов значительную роль играют уточнения настроек элементов в ходе их деятельности, борьба конкурирующих каналов и явления «доминантного» характера — все те моменты нейродинамики, которые так настойчиво подчеркивал А. А. Ухтомский (1954).
ПРОСТРАНСТВЕННЫЙ СЛУХ и точность ЛОКАЛИЗАЦИИ УЛЬТРАЗВУКА
Определение направления прихода отраженных сигналов в процессе эхолокации имеет важное значение для пространственной ориентации летучих мышей. Полученные к настоящему времени данные дают возможность оценить диаграмму приема слуховой системы летучих мышей и представить общую схему формирования направленности на различных мозговых уровнях. Кроме того, сведения о характеристиках пространственного слуха в ультразвуковом диапазоне позволяют высказаться о бинауральных механизмах, обеспечивающих локализацию эха этим животным.
Формирование направленности приема на разных уровнях слуховой системы
Угловую направленность входа слуховой системы изучали методом отведения микрофонных потенциалов улитки (МПУ) (Wever, Vernon, 1961b; Henson, 1964, 1967) и по суммарным реакциям слухового нерва (Кравцов, Звонов, 1969; Кравцов и др., 1969; Васильев, Стосман, 1970; Стосман, Этманова, 1971). При определении порога МПУ у Tadarida brasiliensis в зависимости от угла прихода стимула в горизонтальной плоскости было показано, что максимальная чувствительность обнаруживается при 30—40° ипсилатеральной стороны (Henson, 1964). Выявлена также четкая зависимость ширины диаграммы приема от частоты подаваемого стимула. Так, если для частоты сигнала 10 кгц, подаваемого на левое ухо и приходящего с направления 40°, порог ослаблялся по сравнению с правым ухом всего на 10 дб, то при частоте 30 кгц интегральная разница пороговых значений составляла уже 25 дб, а при 60 кгц увеличивалась до 40 дб.
Обострение диаграммы направленности приема при повышении частоты стимула показано и при изучении суммарных реакций
185
слухового нерва (Л\) у остроухой ночницы (Кравцов, Звонов, 1969) в горизонтальной, сагиттальной и фронтальной плоскостях. Например, для горизонтальной плоскости наибольшие значения амплитуды ответов на частоте 20—25 кгц отмечены в секторе 140° перед головой животного. Пороги ответов в этой области были соизмеримы и варьировали в пределах 5—10 дб. При повышении частоты стимула до 75—80 кгц обнаруживался ярко выраженный максимум дирекционной чувствительности в диапазоне углов 10—40° ипсилатеральной стороны.
Для большого подковоноса наибольшие пороги ответов зарегистрированы для направлений прихода сигнала частотой 30 кгц в секторе 0—60° ипсилатеральной стороны (разница в порогах не превышала 5 дб), который заметно сужался на частотах локационного диапазона (75—83 кгц) (Стосман, Этманова, 1971).
При исследовании формирования диаграммы направленности на уровне кохлеарных ядер проявляется влияние механизмов бинаурального взаимодействия. Тампонада контрлатерального входа приводила к снижению амплитуды вызванных потенциалов (ВП) этого центра на стимулы, подаваемые с ипсилатеральной стороны. 12% нейронов (из 100 обследованных) кохлеарных ядер проявляло активность при контрлатеральном раздражении (Васильев, Стосман, 1970; Стосман, Этманова, 1971; Стосман, 1973а).
В следующем звене слуховой системы — верхнеоливарном комплексе — максимум диаграммы направленности по сравнению с кохлеарными ядрами (рис. 64) смещался на противоположную сторону. Зона наилучшей чувствительности оставалась в пределах углов 0—40°, но теперь уже контрлатерально относительно регистрируемого входа. Это объясняется наличием перекреста афферентных путей слухового анализатора на уровне верхних олив.
Зависимости амплитуды вызванных потенциалов (TV4) и реакций нейронов задних холмов при изменении угла подачи ультразвукового раздражителя изучались рядом авторов (Grinnell, 1963с; Suga, 1964а; Grinnell, Grinnell, 1965; Матюшкин, Стосман, 1967, 1968, 1969; Васильев, 1968а, 19696; Матюшкин, Васильев, 1968; Grinnell, Hagiwara, 1972а; Стосман, 19736). Было показано, что при смещении источника раздражения в горизонтальной плоскости зоны минимальных порогов (наивысшей чувствительности) для всех частот захватывают центр полусферы и район, расположенный от него книзу и контрлатерально (относительно исследуемого холма). Последнее демонстрирует преимущественную связь заднего холма с контрлатеральным входом. Зона наименьшей чувствительности располагается на периферии, главным образом ипсилатерально и внизу (рис. 65). По сравнению с ней пороги вызванных потенциалов при раздражении с направления 90° с ипсилатеральной стороны оказались выше на 20—30 дб, а крутизна изменения порога в районе 15—45° составила 0.5 дб на 1° 186
30 кги, 60 кгц 80 кгц
Рис. 64. Диаграмма направленности приема на различных уровнях слуховой системы у остроухих ночниц (Л) и больших подковоносов (Б).
Слева — ипсилатеральная сторона по отношению к излучателю, справа — контрлатеральг ная сторона. I — на уровне кохлеарных ядер, II — верхних олив, 1Ц — на уровне заднего холма,
I
Рис. 65. Зависимость амплитуды вызванных потенциалов (ВП) от угла предъявления стимула постоянной силы у интактных и оперированных летучих мышей.
I — остроухие ночницы; II — большие подковоносы. Слева — осциллограммы ВП кохлеарного ядра при перемещении излучения. Вертикальные черточки в начале развертки луча — отметки раздражения. Акустическая задержка 1 мсек. Каждый кадр — суперпозиция 10 пробегов луча. Позитивное отклонение книзу. Справа: по оси абсцисс — угол предъявления стимула; по оси ординат — амплитуда ответа. Частота заполнения стимула: I — 60 кгц; II — 30 кгц; сила и длительность 30 дб и 4.6 мсек, соответственно. 1 — данные по измерениям оп-ответов; 2 — по off-ответам; з — амплитуда оп-ответов после ампутации ушной раковины и козелка; 4 — амплитуда оп-ответов после тампонады наружного слухового прохода. Сила стимула 30 дб над порогом обнаружения ВП прц предъявлении стимула под углом 0°.
и даже 1 дб на 1°. Таким образом, на уровне холмов диаграмма направленности сужается по сравнению со входом при одной и той же частоте раздражающего стимула. По данным Гриннела, перепад порогов TV4 достигает 40 и даже 68 дб. Это отчасти объясняется преимущественной связью соответствующего центра с контрлатеральным входом (улиткой). Кроме того, имеет место тормо-
жение импульсов ипсилатерального входа на пути к каждому холму за счет доминирующих влияний контрлатерального входа. Такой механизм, обостряющий диаграмму приема, выявляется уже в опытах с выключением контрлатерального уха (рис. 66). При этом кривая угловой настройки исследуемого холма «извращается», в частности происходит падение порогов (повышение чувствительности) к ипсилатеральному сигналу, что демонстрирует снятие «контрлатеральных» тормозных влияний. При выключении ипсилатерального уха и работе только одного контрлатерального кривая угловой настройки холма у ночниц почти не меняется.
пор.
--100
-50
90 75 60 45 30 15 0 15 30 45 60 75 90
Рис. 66. Диаграмма направленности приема для левого заднего холма (по TV4).
1 — при бинауральном слухе, 2 — при восприятии только ипсилатеральным левым ухом, 3 — только контрлатеральным. Заштрихованная область — зона торможения левого уха правым.
Исследование угловых характеристик отдельных нейронов заднего холма показало, что по сравнению с нейронами кохлеарных ядер в них наблюдается сужение пространственных диаграмм, что вызывает чрезвычайное обострение их дирекционной чувствительности. Так, по данным Суги (Suga, 1964а), скорость изменения порогов кохлеарных нейронов составляла 0.31 дб/град. в диапазоне 0—60°, тогда как холмовые нейроны показывали изменения порогов 0.41 дб/град. Увеличение дирекционной чувствительности по крайней мере на 0.1 дб/град., по мнению автора, объясняется бинауральными связями между этими слуховыми центрами. По результатам наших исследований, из 100 обследованных нейронов кохлеарных ядер основная масса имела широкую угловую настройку, в то же время из такого же количества нейронов задних холмов узконастроенных, или «направленных», оказалось более 50%.
Большое количество колликулярных нейронов, обладающих различной, весьма тонкой угловой чувствительностью, которая сформировалась на базе более грубых угловых настроек низших центров, может обеспечить системе приема своеобразную шкалу для оценки направлений прихода акустических сигналов
189
(Айрапетьянц, Константинов, 1970а; Константинов, Стосман, Гор-линский, 1973).
Таким образом, становится очевидным, что нейрональные механизмы ототопики летучих мышей хорошо приспособлены для обработки информации о положении цели в пространстве. Однако приведенные электрофизиологические данные не дают возможности оценить точность локализации источника ультразвука слуховой системой летучих мышей в целом. Поэтому предстояло выяснить этот вопрос индикаторами пространственной ориентации животного с использованием условнорефлекторной методики.
Точность локализации источника ультразвука
Серия этих опытов была проведена нами на 3 больших ночницах и 3 больших подковоносах (Константинов, Стосман, Горлинский, 1973).
На одном конце звукозаглушенной камеры размещались два конденсаторных ультразвуковых излучателя (диаметр корпуса 6 см), с которых в случайной последовательности подавался акустический сигнал, являющийся условным раздражителем для животных. Частота заполнения сигнала составляла 40 кгц (для ночниц) и 83.5 кгц (для подковоносов), т. е. были выбраны лучшие для каждого вида частоты; кроме того, ночницам также предъявлялся тон 83.5 кгц для сравнения с подковоносами. Длительность стимула была 5 сек., интенсивность в месте старта животного 60 дб.
В процессе опыта экспериментатор имел возможность последовательно уменьшать расстояние между излучателями от 100 до 9.5 см, что соответственно приводило к изменению угла между ними относительно места старта животного от 21 до 2°. Стартовая площадка с летучей мышью находилась на расстоянии 270 см от излучателей и располагалась так, что голова животного совпадала с акустической осью каждого из них. В момент слета животного со стартовой площадки сигнал автоматически выключался.
В ходе опытов у каждого животного вырабатывался условный рефлекс на подаваемый ультразвук и пространственное дифференцирование источника сигнала (справа—слева). Постепенно сближая излучатели, устанавливали предел слуховой системы летучих мышей в выборе направления прихода сигнала (рис. 67).
По мере уменьшения угла между излучателями относительно старта уровень правильных реакций у летучих мышей закономерно уменьшается. Этот факт указывает на то, что источники ультразвука дифференцировались подопытными животными именно по их пространственному положению. Полученные данные также свидетельствуют о том, что подковоносы несколько лучше справлялись с поставленной задачей по локализации ультразвука 83.5 кгц (процент положительных реакций был выше), чем ночницы при локализации тона 40 и 83.5 кгц. Это нашло свое логическое завершение при пороговых величинах углов. Предельная точность локализации источника ультразвука у подковоносов при взятом уровне положительных реакций 75% составила 4°, у ночниц — 5° независимо от частоты стимула.
190
Определение направления приходящих акустических Сигналов (направление на их источник) у летучих мышей, как и у прочих млекопитающих, обеспечивается бинауральным восприятием. Однако степень выраженности бинаурального взаимодействия у разных видов летучих мышей, по-видимому, различна. Как показали наши исследования (Константинов, 1965в; Константинов и др., 1967а, 19676), одностороннее повреждение слухового рецептора путем удаления большей части барабанной перепонки, приводящее к резкому снижению слуховой чувствительности животного (Bekesy, 1951), в разной степени отразилось на ориентацион-
Рис. 67. Изменение уровня положительных реакций у подковоносов (/) и ночниц (II) при уменьшении угла между излучателями.
I: 1 — при использовании длительного тонального сигнала (83.5 кгц), 2 — последовательность коротких импульсов; II: 1 — для сигналов частотой 40 кгц, 2 — частотой 83.5 кгц.
пых способностях ушанов, ночниц и подковоносов. Используя в качестве теста пролеты летучих мышей сквозь проволочные барьеры в норме и после односторонней перфорации барабанной перепонки, мы получили следующие результаты. У ушанов процент уклонений [при преодолении барьеров из проволок диаметром 0.6 мм в норме составлял 75.5+0.87, а после операции его значение уменьшилось до 29.0 + 1.9, что свидетельствует о полной потере способности этих животных обнаруживать препятствия. В то же время аналогичные нарушения слуховой системы у усатых ночниц в тех же условиях опытов не оказывали влияния (в норме процент уклонений 81.0 + 1.0, после перфорации — 80.0 + 1.7). Лишь при двусторонней операции процент уклонений у усатых ночниц понизился до 35.0+6.7.
Отсутствие различий при пролетах сквозь проволоки разного диаметра до и после операции показано и для остроухих ночниц (Константинов и др., 1967а, 19676). У подковоносов же одностороннее нарушение слуха почти не отразилось на проценте уклонений
191
Таблица 10
Ловля подвижных насекомых остроухой ночницей и подковоносом Меге ли в норме и после одностороннего нарушения слуховой рецепции
Вид Условия Число предъявляемых насекомых Число бросков на насекомое Число пойманных насекомых
Остроухая ночница Норма 10 14 10
Односторонняя перфорация 7 8 7
Подковонос Меге л и Норма 30 35 30
Односторонняя перфорация 6 60 1
при пролете проволок 0.55 и 0.2 мм, но чувствительность эхолокации значительно понизилась при пролетах через барьеры из проволок 0.12 мм.
Еще более существенные различия проявились при ловле насекомых (табл. 10).
По методике, описанной в главе 9, летучим мышам предъявлялись подвижные ночные бабочки. В норме, т. е. у предварительно энуклеированных животных с интактной слуховой системой, точность реализации добычи была одинаковой. Летучие мыши обычно с первого броска, иногда со второго захода, срывали насекомое с проволочки и съедали. После односторонней перфорации барабанной перепонки точность эхолокации и результативность ловли добычи у ночниц не изменились, но подковоносы практически оказались не способны поймать насекомое, хотя обнаруживали его и многократно атаковали.
Как показывают наши опыты, уже частичное нарушение бинаурального слухового восприятия полностью препятствует эхолокационному обнаружению мелких препятствий у ушанов, уменьшает чувствительность и точность эхолокации подковоносов и не оказывает влияния на восприятие отраженных сигналов у ночниц.
По данным Гриффина и Галамбоса (Griffin, Galambos, 1941), полное выключение одного из ушей путем тампонады слухового прохода у ночниц приводило к резкому ухудшению их ориентировочных способностей. В то же время немецкие исследователи (Moh-res, 1953; Schneider, Mohres, 1960) показали, что подковоносы нормально летали и правильно производили посадку после аналогичного ограничения слуховой рецепции. На основании этих экспериментов в литературе укоренилось представление о том, что подковоносы в отличие от гладконосых летучих мышей способны успешно ориентироваться в пространстве в условиях монаураль-ного приема отраженных сигналов.
В наших опытах по установлению точности локализации источ
192
ника ультразвука у двух подковоносов была произведена односторонняя тампонада слухового прохода. Животные продолжали летать по камере, садиться на стены, избегали крупные препятствия, но пространственная локализация звука оказалась на^ рушенной. Дифференцирование источников, разнесенных на 15° относительно старта животного, отсутствовало (50% положительных ответов). Летучие мыши хорошо реагировали на условный сигнал, но определить, с какого направления он приходит, не могли и садились либо между излучателями, либо несколько раз подряд под одним из них независимо от места подачи стимула. Если слуховой проход освобождался от тампона, то сразу же восстанавливался высокий уровень правильных ответов (90%). Следовательно, нарушение бинаурального восприятия у подковоносов приводит к резкому ухудшению их локализационных способностей. Об этом же свидетельствуют данные немецких исследователей (Flieger, Schnitzler, 1973).
В анализе направления прихода отраженного сигнала у подковоносов существенную роль могут играть быстрые движения ушных раковин, которые хорошо коррелируют с темпом излучения локационных сигналов (Schneider, Mohres, 1960; Griffin et al., 1962; Pye et al., 1962).Кроме того, в электрофизиологических исследованиях было показано, что при ампутации ушных раковин у подковоносов происходит выравнивание диаграммы направленности приема, снятой по (Стосман, Этманова, 1971), а изменение положения ушных раковин вызывает нарушение дирекционной чувствительности в задних холмах (Neuweiler, 1970).
Роль ушных раковин в восприятии отраженных сигналов в наибольшей степени проявляется у ушанов, что выделяет их среди других гладконосых летучих мышей. В наших опытах ушаны после ампутации одной ушной раковины вместе с козелком значительно хуже, чем в норме, избегали проволочные барьеры. Процент уклонений составил 42.5+3.7, тогда как в норме ушаны уклонялись от столкновения с проволокой на 75.5+0.8%. Для сравнения аналогичные опыты были проведены на усатых ночницах. Если в норме эти животные преодолевали проволочные барьеры с уклонением на 81.0 + 1.0%, то после ампутации одной, а затем и двух ушных раковин заметных ухудшений в ориентации не произошло. Процент уклонения составил соответственно 86.0 + 1.0 и 79.6+2.7. Эти результаты хорошо совпадают с электрофизиологическими исследованиями (Grinnell, Grinnell, 1965; Vernon et a]., 1966), в которых было показано, что загибание ушных раковин за голову ночницы не сказывается на пороговой чувствительности их слуховой системы при ультразвуковом раздражении; чувствительность понижалась не более чем на 5—10 дб, тогда как у ушанов изменение положения ушных раковин приводило к значительному изменению диаграммы направленности.
Коротко остановимся на возможных принципах определения
193
направления прихода отраженных сигналов у летучих мышей, реализуемых по механизму бинаурального взаимодействия. Как известно, в основе его могут лежать изменение фазового угла между синфазными тональными сигналами, приходящими к двум ушам (Д<р), разность по времени между моментом раздражения обеих улиток (АТ) и разность по интенсивности при раздражении каждой из них (А/) (Альтман, 1972; Альтман, Дубровский, 1972). Рассмотрим возможности применения каждого из этих способов для летучих мышей.
Допустим, что для данных животных минимально разрешаемое АТ не превышает аналогичный параметр слуховой системы человека и равно ~0.01 мсек. Тогда по имеющейся формуле (Альтман, Дубровский, 1972) можно построить график зависимости Д Т от угла прихода звука. Из-за малой величины интерауральной базы у летучих мышей (13—15 мм) точность локализации источника акустического сигнала составила бы в лучшем случае 20°, что находится в явном противоречии с нашими экспериментальными данными (4—5°). Другой фактор ототопики — бинауральное сравнение фаз приходящего сигнала дает однозначное определение направления на источник звука лишь в том случае, если длина волны в 2 раза превышает величину интерауральной базы. Такое условие для летучих мышей оказывается невыполнимым, так как длины волн излучаемого ультразвука составляют 10—2.5 мм. Следовательно, временные и фазовые бинауральные отличия в сигнале не могут нести летучим мышам информацию о положении цели в пространстве.
С другой стороны, быстрое затухание ультразвука в воздухе, экранирующие свойства головы и ушных раковин животных обеспечивают благоприятные условия для создания контрастных отличий в интенсивности полей на обоих ушах летучей мыши. Кроме того, обостренная дирекционная чувствительность подкорковых слуховых центров, соотнесенная с дифференциальными порогами по интенсивности, по данным нейрофизиологических исследований, может дать точность определения направления прихода ультразвука в 1°. Таким образом, из трех рассмотренных выше общепринятых факторов локализации звука лишь бинауральное сравнение интенсивности приходящего сигнала может объяснить полученный экспериментальный материал.
Для летучих мышей, использующих в качестве локационных сигналов ЧМ-импульсы с большим размахом девиации, теоретически возможен еще один принцип определения направления эха, основанный на анализе тембральных отличий сигнала, поступающего на оба уха. Однако этот вопрос требует специального исследования.
8
РАЗВИТИЕ ЭХОЛОКАЦИИ У ЛЕТУЧИХ МЫШЕЙ В ОНТОГЕНЕЗЕ
Вопрос о путях формирования эхолокации в процессе индивидуального развития у летучих мышей до последнего времени не подвергался систематическому изучению, несмотря на то что решение его позволяет глубже понять происхождение и эволюцию этого особого способа ориентации по крайней мере в пределах подотряда Microchiroptera. Сведения о ранних вокализациях у детенышей летучих мышей очень фрагментарны и носят чисто описательный характер. В ряде работ, посвященных изучению онтогенеза рукокрылых, отмечено, что до 10-дневного возраста детеныши непрерывно издают низкочастотные крики, которые, вероятно, служат им для связи с самками (Огг, 1954, 1970; Kul-zer, 1962; Jones, 1967; Davis et al., 1968; Kleiman, 1969). В одной из работ (Griffin, 1951) имеются указания на то, что звуки, издаваемые молодыми летучими мышами, лишь слегка увеличены по длительности и имеют гармоники высокой амплитуды в отличие от сигналов взрослых. Более подробно развитие вокализаций у молодых Myotis и Eptesicus исследовал Гульд (Gould, 1971). Им выделено несколько типов сигналов у детенышей этих видов и рассмотрен характер акустической связи между матерью и детенышем. Автор приводит некоторые физические параметры локационных или сонарных сигналов у детенышей летучих мышей, но специально не останавливается на динамике их становления по мере роста животных. Данные о становлении ультразвуковой рецепции в процессе индивидуального развития у животных, и в частности у летучих мышей, в литературе вообще отсутствуют. Указанные обстоятельства уже давно привели нас к мысли о необходимости проведения соответствующих исследований, и летом 1970 г. была организована экспедиция в Крымскую область, где в условиях, близких к естественным, мы смогли осуществить намеченную программу.
Основными задачами мы ставили выяснение этапов формирования частотно-временных характеристик локационных сигналов,
195
а также развитие функций слуховой системы у летучих мышей с разными типами локации от момента рождения до начала самостоятельной жизни. Вся работа была выполнена на 38 остроухих ночницах и 36 больших подковоносах в возрасте от 1 до 50 дней (Константинов, 1972а, 19726, 19736, 1973в; Константинов, Стосман, 1972, 1973; Стосман, Константинов, 1972).
Необходимым условием для проведения данной работы является знание точного возраста детенышей исследованных видов. Так как в литературе сведения о ходе развития остроухой ночницы и большого подковоноса в постнатальном онтогенезе носят слишком общий характер, требовалось провести дополнительные наблюдения за сроками прозревания, изменением поведения, состоянием молочных зубов, увеличением веса, размеров тела, предплечья и пальцев передних конечностей у детенышей разных возрастных групп. Совокупность всех этих данных и их сопоставление с таковыми у детенышей, которые воспитывались в неволе, позволили нам устанавливать возраст молодых зверьков, взятых из колонии, с точностью в 2—3 дня от момента рождения до 30 дней и с точностью до 5 дней в старших возрастных группах. Не вдаваясь в детали, ограничимся лишь описанием общего хода развития ночниц и подковоносов, так как более полное изложение постэмбрионального периода у изученных видов увело бы нас в сторону от основной темы.
Развитие детенышей у ночниц и подковоносов
Сроки появления на свет детенышей в колонии остроухой ночницы значительно варьировали. Начало родов происходило 10—15 мая, массовое размножение охватывало период с конца мая до середины июня, а некоторые самки приносили детенышей в конце июня. В колонии большого подковоноса роды начинались значительно позднее — в последней декаде июня, но протекали более дружно. Самки обоих видов приносили, как правило, по одному детенышу. Новорожденный детеныш ночницы голый, красного цвета, слепой, ушные раковины сморщены, слуховые проходы закрыты. В условиях клеточного содержания детеныши постоянно находились под крылом у матери в течение первых 7—10 дней. Детеныш подковоноса рождается значительно крупнее. Голова, плечи и часть спины до крестца покрыты густой серой шерстью, остальные части тела голые. Ушные раковины очень большие и закинуты назад, веки четко намечены, но глаза закрыты. Детеныш располагается на животе самки, держась ртом за паховые соски и своими длинными задними конечностями крепко обхватывает ее тело. К соску молочной железы детеныш прикладывается лишь время от времени на 15—20 мин. и затем снова принимает свою обычную позу.
В возрасте 10—12 дней ночницы прозревают, все тело со спины
196
покрывается короткой шерсткой, за исключением участка тела между лопатками, который еще долго не зарастает. Ушные раковины начинают вставать. При ходьбе детеныш опирается на предплечья и задние конечности, движения становятся направленными. Спустя неделю шерсть на голове и спине становится гуще, белым мехом покрывается живот. Под крыло матери детеныш забирается лишь во время кормления, остальную часть суток проводит либо на ней сверху, либо около. Начинается выпадение молочных зубов. В 25 дней детеныш быстро бегает, при этом расправляет крылья и подпрыгивает, как бы собираясь взлететь. Но в этом возрасте он еще не способен даже планировать и, подброшенный вверх, тут же падает на землю. Способность к полету проявляется к 30 дням, когда зверек срывается с поднятой руки и, часто махая крыльями, летит по наклонной траектории 3—4 м. В возрасте 40—45 дней полет становится более уверенным и продолжительным, но отличается от полета взрослых меньшей скоростью и маневренностью. В этом возрасте молодые ночницы, выросшие в неволе, проявляют живой интерес к ночным бабочкам, бьющим крыльями в руках экспериментатора, а вечером спускаются к кормушке вслед за взрослыми самками и пробуют мучных червей. В возрасте 55—60 дней они питаются почти исключительно насекомыми, но время от времени подбираются к матери и в течение 15—20 мин. сосут ее. Можно считать, что к концу второго месяца жизни остроухие ночницы становятся полностью самостоятельными.
Развитие детенышей большого подковоноса идет более быстрыми темпами и завершается раньше. В 7—10 дней детеныши прозревают, а в 15—17 дней уже начинают перелетать с одного предмета на другой. В 30-дневном возрасте молодые зверьки уже хорошо летают и предпринимают первые попытки ловить насекомых. К этому времени постоянные зубы прорывают кожу десны и выходят наружу (в отличие от большинства других видов летучих мышей генерация молочных зубов у подковоносов проходит в эмбриональный период). В возрасте 35—40 дней детеныши не уступают взрослым как при ловле насекомых, так и при преодолении проволочных барьеров.
Характер призывных криков и становление локационного сигнала
Запись звуков, издаваемых детенышами, производилась в день их отлова вместе с самками из больших колоний, расположенных на чердаках домов. Кроме того, регулярные наблюдения за развитием вокализаций, и в частности сигналов ориентации, проводились над детенышами, которые родились и воспитывались в неволе. Во время записи молодые зверьки были изолированы от самок и излучали сигналы либо во время передвижения по поверхности стола, либо во время их быстрого опускания вниз на руке, а старшие возрастные группы — перед слетом и во время полета. Звукозаписывающая аппаратура состояла из конденсаторного микрофона с полосой пропускания от 2 до 200 кгц, широкополосного усилителя и ультразву-
197
нового магнитофона МУ-25. Для анализа характеристик сигналов использовалась осциллографическая запись при замедлении скорости воспроизведения в 16 или 32 раза, а также спектрограммы, полученные на спектрографе «Кау Electric С°», тип 7029 А, с барабаном типа 669 и широким фильтром.1 Длительность сигналов и интервалы между ними определялись по специальным меткам, подаваемым на киноаппарат, работающий в непрерывном режиме при съемке сигналов с экрана осциллографа.
Ночницы в возрасте от 1 до 10 дней, взятые от самки, почти непрерывно в течение нескольких минут издают громкие крики, хорошо слышимые человеком. Интервалы между отдельными криками-импульсами обычно составляли 200 мсек., но иногда они следовали через 100—120 мсек, или более редко, через 300—400 мсек., в зависимости от ситуации. Биологическое значение этих криков мы определяем как призыв матери или врожденную сигнализацию о своем местоположении и называем их призывными криками, которые соответствуют isolation calls, по терминологии других авторов (например: Gould, 1970, 1971). Никаких других акустических сигналов у ночниц этого возраста мы не обнаружили.
Как видно на спектрограмме (рис. 68), спектр призывного крика новорожденной ночницы носит полосовой характер и охватывает широкий диапазон звуковых и ультразвуковых частот, но основная энергия сосредоточена в районе 16—20 кгц (более темные участки на спектрограмме). Отчетливо заметно понижение частоты каждой составляющей от начала к концу крика, особенно на более высоких частотах. На приведенной тут же осциллограмме (рис. 68) хорошо видны крутой передний фронт призывного крика и присутствие уже с самого начала двух кратных составляющих, одну из которых — 23.5 кгц — можно рассматривать в качестве 1-й гармоники или основной частоты, а другую — 47 кгц — ее 2-й гармоникой. В середине крика обычно появляется 3-я гармоника, а в конце картина становится настолько сложной, что оценить количество гармоник не представляется возможным. Удается проследить постепенное уменьшение частоты 1-й и 2-й гармоник к концу импульса. В середине величина основной частоты составляет 19 кгц, затем 17 кгц, а ближе к концу — 14.5 кгц. Таким образом, уже в первых акустических сигналах, издаваемых детенышами, основная энергия сосредоточена в области низких и средних ультразвуковых частот и наблюдается нисходящая частотная модуляция с падением основной частоты от начала к концу на 8—10 кгц. Интересно заметить, что, несмотря на общее сходство, призывные крики детенышей ночниц имели достаточно четкие индивидуальные отличия по длительности и спектральным характерис
1 Записи на спектрографе проводились в лаборатории биоакустики Института эволюционной морфологии и экологии животных АН СССР при содействии В. И. Маркова и А. Кузнецова, которым авторы выражают глубокую благодарность.
198
тикам. Это, по-видймому, помогает самкам отыскивать своих детенышей при возвращении с ночной охоты.
Характер вокализаций претерпевает значительные изменения к двухнедельному возрасту детенышей. На смену громким призывным крикам приходят более слабые (для слуха человека)
199
сигналы, которые излучаются отдельными сериями, по 5—10 импульсов через 100—120 мсек., с интервалами между сериями 200—300 мсек., во время передвижения зверьков. Меняется и поза животного при издавании сигналов. Если молодые зверьки, взятые от матери, при излучении призывных криков запрокидывают голову вверх, то в 14—15 дней они издают импульсы прямо перед собой. Длительность этих импульсов сокращается примерно наполовину по сравнению с призывными криками детенышей младшего возраста (табл. 11), частота основного тона повышается. Начиная с этого возраста акустические сигналы детенышей могут уже играть двоякую роль: сохраняют свое первоначальное значение для связи с самкой, а также приобретают свойства ориентационных сигналов.
Таблица И
Параметры сигналов ночниц разных возрастных групп
Возраст, дни Длительность, мсек. начало сигнала Частота, кгц конец сигнала девиация Число исследованных импульсов
1 37 (25—50) 23.5 (22—24) 14.5 (14-16) 9 130
10 30 (10—40) 25.5 (24—27) 17 (15-18) 8.5 25
15 14.4 (7—20) 31.6 (30-34) 17 (15-18) 14.6 28
17—18 8 (5.5-15) 35.5 (32-40) 19 (15-24) 16.5 33
20 4.2 (2.5-6) 42 (38-46) 25 (22-26) 17 35
25 3-0.8 48 (45-60) 33 (30-35) 15 40
30—33 3-0.8 60 (50-70) 34 (30-35) 26 52
35-37 3-0.8 72 (60-80) 35 (30-40) 37 43
40-43 3-0.8 96 (70—120) 38 (35-40) 58 97
45-50 3-0.8 120 (100—140) 36 (30-40) 84 40
Взрослые животные 1.8 3-0.8 120 (90-140) 35 (25-40) 85 620
По мере роста детенышей функция излучаемых сигналов становится все более однозначной — получение информации об окружающей среде с помощью эха. Происходит дальнейшее уменьшение длительности сигналов, которая к возрасту 25 дней лежит в пределах диапазона длительностей взрослых животных. К этому же времени стабилизируются заключительные частоты основного тона, тогда как повышение начальных частот в импуль
200
сах до уровня, отмеченного для сигналов взрослых ночниц, идет более медленно и завершается лишь к 45—50-дневному возрасту (табл. 11), т. е. к моменту, когда животные начинают охотиться за насекомыми.
Наряду с повышением основной частоты продолжается процесс постепенного исчезновения гармонических составляющих в спектре сигналов. Как видно из представленных осциллограмм сигналов ночниц разных возрастных групп (рис. 69), уже к 17— 20-му дню 2-я гармоника представлена лишь во второй части импульса, к 25-му дню она сдвигается в последнюю треть сигнала, а к
0 1 2 Змсе>
1__J__।__।_I----------1----------1
Рис. 69. Осциллограммы локационных сигналов у ночниц разных возрастных групп.
Возрастные группы (в днях): 1 — 15, 2—18—20, 5 — 25, 4 — 30, 5 — 45—50.
возрасту 30 дней либо совершенно исчезает, либо слабо прояв-
ляется в самом конце импульса.
Подобно ночницам, детеныши подковоноса уже с момента
рождения издают через рот резкие низкочастотные крики, содержащие комплексные волны с ярко выраженной амплитудной модуляцией. С точки зрения функциональной значимости их также можно классифицировать как призывные крики. Длительность их у новорожденного составляет в среднем 12 мсек. (5—2С мсек.), и при излучении импульсы сгруппированы в серии, или пачки, по 3—5 импульсов, с интервалами между ними в среднем 18 мсек, и между пачками — 120 мсек.
Среди нескольких сотен записанных криков нами обнаружено три одиночных сигнала, имеющих совершенно другую структуру. Длительность их составляла 30, 50 и 60 мсек., а спектр
201
Рис. 70. Осциллограммы трех типов призывных криков большого подковоноса в возрасте 10 дней.
1 — призывный крик, издаваемый через рот; 2 — смешанный сигнал — вначале излучение идет через рот, а затем, с середины — через ноздри; з — часть импульса, излучаемая через ноздри.
Л
я ф ЯС
я
я о я
Я
Я
я я
5
я о я о я
Я
34 та а
я о я
я Е
Я та я и
Е я и
я
Я
34
Я
й 1=1 ф я я Е Я
я я
а я
Я
я я
я
Я
Е и
й я Я Я я о 1=1 Я о я о я
я
?
та
Я
Я
Я
Я
ники. Образцы этих трех типов сигналов представлены на рис. 70. В пределах одной пачки можно было выделить один сигнал, из-лучаемый через рот, затем следующий, смешанный, и наконец последний — излучаемый через ноздри. Пачка могла начинаться в обратной последовательности. Постепенно именно «носовые», коммуникационные сигналы приобретают у подковоносов значение локационных, поэтому именно на них в дальнейшем мы и сосредоточили свое внимание.
До 20-дневного возраста в сигналах, излучаемых через ноздри, присутствуют 3 или 4 частотные составляющие и основная энергия сосредоточена в первой из них или в первой и во второй на частоте порядка 30 и 60 кгц (рис. 71). Подковоносы старшего возраста, перелетая с одного предмета на другой, пользуются исключительно «носовыми» сигналами, и крики, издаваемые через рот, обнаруживаются лишь в редких случаях, когда детеныши долгое время остаются одни. В спектре сигналов еще продолжают присутствовать 3 гармоники, но частоты 1-й и 2-й гармоник повышаются соответственно до 37—40 и 74—80 кгц. К 25—30-дневному возрасту в сигналах исчезают первая и третья составляющие, а доминирующей частотой становится 2-я гармоника, 81 — 82 кгц. Этот процесс сопровождается увеличением длительности одиночных сигналов и количества импульсов, входящих в пачку. Интересно отметить, что способность к издаванию призывных криков через рот у подковоносов сохраняется до одного года, но лишь в тех случаях, когда животное висит на месте, и никогда во время полета.
Итак, прослеживая характер изменения вокализаций в ходе индивидуального развития летучих мышей, можно прийти к заключению, что локационные сигналы, предназначенные для ориентации животных, формируются на базе призывных криков, обеспечивающих акустическую связь детеныша с матерью. Процесс функционального замещения «связных» криков на ориентационные у ночниц сопровождается изменением их параметров: сокращением длительности, увеличением частоты следования, повышением частоты основного тона, расширением девиации, «очищением» спектра от дополнительных гармонических составляющих. Изменение частотных характеристик сигналов в постнатальном онтогенезе у ночниц хорошо согласуется с направлением развития их у ныне существующих семейств летучих мышей, начиная от примитивных Emballonuridae и через ряд промежуточных стадий до наиболее развитых Vespertilionidae (Novick, 1958а; Симкин, 1971а; Константинов, 1973а), т. е. в определенной степени отображает филогенетические преобразования локационных сигналов. У подковоносов уже с первых дней жизни излучение призывных криков осуществляется через два канала — рот и ноздри. Возможность использовать носовую полость в качестве звукового канала, которую можно рассматривать как систему
203
A
сложноустроенных фильтров с более высокой добротностью, чем рот (Руе, 1967), имела решающее значение для того, чтобы именно «носовые» сигналы стали выполнять функцию локационных, а высокая 2-я гармоника оказалась несущей частотой сигнала. При этом длительность сигналов подковоносов в отличие от ночниц проявляет тенденцию в сторону увеличения. Указанные факты позволяют, на наш взгляд, рассматривать особенности эхолокации подковоносов как вторичное преобразование и предполагать, что в процессе эволюции развитие этого функционального типа локации взяло начало от гипотетического предка, использующего короткие сигналы, издаваемые через рот.
Развитие функций слуховой системы
Исследование электрических реакций, развивающихся при действии звуковых сигналов в слуховой системе незрело-рождающихся млекопитающих, интересно с нескольких точек зрения. Результаты количественной оценки отдельных параметров таких реакций должны не только дать представление об особенностях слуховой системы новорожденных животных, но и выявить основное направление изменений свойств исследуемого анализатора в процесс становления его функций. Для летучих мышей этот вопросе является особенно актуальным, если учесть, что слуховой анализатор этих животных выполняет специальные функции, связанные с анализом отраженных сигналов в процессе эхолокации.
При исследовании становления функций слуховой системы в качестве показателя реакции на ультразвуковые тональные стимулы были взяты быстрые колебания (/V4) суммарного ответа заднего холма. Детеныши летучих мышей под гексеналовым наркозом закреплялись на треугольной пластинке и помещались в звукозаглушенную камеру с постоянной температурой 38°. Акустические колебания, подаваемые от генератора ГЗ-ЗЗ, формировались электронным ключом в тональные посылки длительностью 0.2—10 мсек., с нарастанием длительности фронта 0.05 мсек, и воспроизводились через конденсаторный излучатель, расположенный в 34 см прямо перед головой животного. Регистрация вызванных потенциалов осуществлялась с помощью усилителя биопотенциалов УБП-1 и двухлучевого осциллографа С1-16, снабженного автоматической фотокамерой РФК-5.
Сопоставление данных, полученных при снятии ЧПК у летучих мышей различного возраста, позволило представить динамику становления слуховой чувствительности к ультразвукам в онтогенезе и установить наличие существенных возрастных изменений порогов и диапазона воспринимаемых частот.
Рис. 71. Спектрограммы акустических сигналов детенышей большого подковоноса разных возрастных групп.
А — один призывный крин, издаваемый через рот в возрасте 10 дней; Б —серия из трех импульсов, излучаемых в возрасте 10 дней через ноздри; В — серия из/грех импульсов, излучаемых в возрасте 20 дней через ноздри; Г — серия из трех импульсов, излучаемых в возрасте 30 дней через ноздри.
205
У ночниц, не достигших возраста 10 дней, и у подковоносов до 7 дней в заднем холме не удается зарегистрировать вызванных потенциалов на акустические стимулы в исследуемом диапазоне частот 8—150 кгц даже при максимальной интенсивности (100 дб) (Константинов, Стосман, 1973).
В возрасте 10—12 дней у ночниц отмечено появление нестабильных, низкоамплитудных позитивных потенциалов на стимулы 2—4 мсек, в узкой полосе частот 17—25 кгц. Иногда ответы возникали только при частоте стимула 18—20 кгц. Латентные периоды их составляли 5.5—6 мсек., а пороги — 76—85 дб. В 14—15 дней диапазон частот несколько расширяется — от 15 до 30 кгц, амплитуда ответов увеличивается до 10—30 мкв, но пороги продолжают оставаться высокими. Для возраста 17— 18 дней полоса ответов расширялась от 8 до 50 кгц и в некоторых случаях наблюдалось снижение порогов до 40 дб на частоте 30 кгц, но в основном пороги были высокими. В возрасте 20—25 дней ЧПК приобретают характер таковых у взрослых животных и по форме, и по диапазону частот. Вызванный потенциал представлял типичный позитивно-негативный ответ. Наиболее типичные индивидуальные ЧПК, снятые по TV4 у ночниц разных возрастных групп, приведены на рис. 72, А.
У подковоносов первые ответы в заднем холме были зарегистрированы в недельном возрасте на тональные стимулы частотой 20—30 кгц, а спустя 2—3 дня диапазон воспринимаемых частот расширялся от 10 до 50 кгц, хотя пороги были высокие. Амплитуда ответов колебалась от 20 до 40 мкв. В возрасте 12— 1бУдней наблюдалось расширение полосы воспринимаемых частот вверх по диапазону до 60—70 кгц, с понижением порогов до 30 дб на лучшей частоте (30 кгц). Амплитуда ответов увеличивалась до 100 мкв. К 20-дневному возрасту пороги снижались до 20 дб на частотах 10—30 кгц, но затем резко возрастали с увеличением частоты стимула. Детеныши этого возраста реагировали на ультразвуки вплоть до 100 кгц. Начиная с 25-дневного возраста и у более старших возрастных групп наблюдалось вторичное снижение порогов ответов в районе частот 70—75 кгц, которое всегда предшествует последующему повышению чувствительности у взрослых подковоносов на частоте 83.5 кгц (Neuweiler, 1970; Соколов, Соколова, 1972). Лучшие ЧПК слуха для детенышей подковоносов разного возраста представлены на рис. 72, Б.
Таким образом, развитие функций слуховой системы для восприятия ультразвука рабочего диапазона частот для каждого вида летучих мышей завершается к возрасту 25—30 дней. К этому же времени ответ на второй из двух предъявляемых стимулов проявляется при интервале между ними 1.5—2.5 мсек, и усваивается ритм раздражения 150—170 имп./сек., т. е. и по отсчету временных интервалов обнаруживается сходство со слуховой системой взрослых животных.
206
Представляет несомненно интерес сравнение данных, полученных на летучих мышах, с теми, которые имеются в литературе для других млекопитающих.
По результатам наших исследований, первые ответы в заднем двухолмии у летучих мышей могут быть получены лишь в возрасте 7—10 дней на звуковые раздражители интенсивностью 80—90 дб. Это хорошо согласуется, например, с данными на котятах, у которых ответы улитки и нейронов заднего двухолмия также удается зарегистрировать на 9—10-й день после рождения (Клявина, Марусева, 1963; Марусева, Малышева, 1970).
Рис. 72. Индивидуальные частотно-пороговые кривые, снятые по TV4 у ночниц (Л) и подковоносов (Б) разных возрастных групп.
Возрастные группы (в днях): А: 1 — 10, 2 — 15,3 — 17, 4 — 20, 5 — 25, 6 — взрослые; Б: 1 — 1,2 — 10, 3 — 15, 4 — 20, 5 — 25, 6 — взрослые.
Следует заметить, что в литературе для разных видов не-зрелорождающихся млекопитающих указываются как более ранние (Anggard, 1964; Pujol, Marty, 1968; Chaloupka et al., 1968), так и более поздние (Alford, Ruben, 1963; Myslivecek, Hasman-nova, 1963; Crowley, Reymond, 1966) сроки появления реакций на звук в слуховой системе. Причинами такого расхождения могут, очевидно, явиться неодинаковая интенсивность и различный характер звуковых раздражителей, применяемых исследователями. Так, например, зарегистрировать ответы улитки кролика на 5-й день после рождения удалось при использовании сигналов, интенсивность которых достигала 125 дб (Anggard, 1964). С другой стороны, нельзя также исключить возможность неодновременного созревания тех или иных структур слуховой системы у различных видов млекопитающих. В наших опытах более ранние сроки появления ответов в заднем холме были отмечены у подковоносов, которые в связи с затянутым эмбриональным периодом рождаются более развитыми по сравнению с ночницами.
207
Специального рассмотрения требует вопрос о возрастных изменениях ЧПК слуховой системы. Как и у других исследованных млекопитающих, у летучих мышей первые ответы ограничены узким частотным диапазоном звуковых раздражителей, который существенно расширяется к двухнедельному возрасту. Расширение диапазона происходит главным образом за счет постепенного увеличения способности животных реагировать на сигналы все более высокой частоты.
Более позднее появление реакций нервных центров на звуки высокой частоты определяется, по всей вероятности, структурными особенностями внутреннего уха незрелорождающихся животных. Слабая дифференциация опорных элементов кортиева органа, малые промежутки между волосковыми и поддерживающими клетками, значительная толщина базилярной мембраны, а также способ фиксации тенториальной мембраны, обнаруженные при гистологических исследованиях улитки новорожденных кроликов и крыс (Anggard, 1964; Crowley, Reymond, 1966), несомненно создают неблагоприятные условия для передачи высокочастотных колебаний. Однако у летучих мышей здесь проявляется ряд специфических особенностей. Уже первые ответы, зарегистрированные нами в холме, возникали только при действии сигналов, лежащих в диапазоне частот 20—30 кгц, тогда как, например, у котят первые реакции нейронов холма отмечались на стимулы 3—6 кгц (Марусева, Малышева, 1970). Именно эти й близкие к ним частоты у взрослых животных становятся лучшими, т. е. пороги ответов для них оказываются минимальными. Следовательно, у каждого вида или у определенной группы животных в раннем онтогенезе в первую очередь формируются совершенно определенные участки кортиева органа, которые предназначены для восприятия специфичного диапазона акустических раздражителей.
Исследование формирования диапазона восприятия ультразвука показывает, что этот процесс у обоих видов летучих мышей идет по одному и тому же пути — возникновение первых ответов на частотах 20—30 кгц с дальнейшим расширением полосы частот кверху и одновременным снижением порогов в диапазоне 10—50 кгц. Впоследствии у подковоносов происходит дополнительное изменение частотно-пороговых характеристик в районе 75—85 кгц, которое следует рассматривать как развившееся в процессе эволюции вторичное приспособление, обусловленное специфическими параметрами их локационных сигналов.
9
ОСОБЕННОСТИ ФУНКЦИОНИРОВАНИЯ ЭХОЛОКАЦИОННЫХ СИСТЕМ ЛЕТУЧИХ МЫШЕЙ ПРИ ПРЕСЛЕДОВАНИИ И ЛОВЛЕ ДОБЫЧИ
Как уже отмечалось выше, большинство видов летучих мышей является насекомоядными формами, причем многие из них настигают свою добычу во время полета. Наблюдения за процессом охоты, проведенные в естественной обстановке, показали, что темп ловли насекомых летучими мышами очень высок. За один час охоты летучая мышь совершает 1160—1280 бросков за пролетающими насекомыми (Gould, 1955, 1959). Даже если предположить, что в половине случаев животные промахивались, результативность охоты составит 580—640 насекомых в час, или примерно 10 в минуту. Эти цифры свидетельствуют о большом мастерстве летучих мышей, основанном на эффективной работе анализаторных систем дистантного действия в сочетании с высокими скоростями двигательных реакций.
Долгое время вопрос об участии того или иного анализатора этих животных в процессе ловли насекомых не подвергался экспериментальному анализу. Ряд наблюдений показывал, что обнаружение добычи летучими мышами может осуществляться путем восприятия тех звуков, которые производят сами насекомые (Кузякин, 1950; Mohres, 1950; Treat, 1955; Kolb, 1959, 1961).
Однако в 1953 г. было установлено, что в естественных условиях, подлетая к гасекомому, летучие мыши увеличивают частоту излучения локационных сигналов и одновременно изменяют их частотно-временные параметры (Griffin, 1953b, 1958). В последующих работах функциональная деятельность эхолокационной системы одного из видов ночниц (сем. Vespertilionidae) при ловле мелких, бесшумно летающих насекомых (дрозофил) или катапультируемых в воздух мучных червей подверглась тщательному изучению (Griffinet al., 1960; Cahlanderet al., 1964; Cahlan-der, 1967; Webster, 1967). Одновременно эхолокационный про
209
цесс при преследовании добычи был описан у представителей еще двух семейств: Mormoopidae (Novick, Vaisnys, 1964; Novick, 1965) и Noctilionidae (Griffin, 1962; Suthers, 1965). Результаты всех этих исследований убедительно доказывают, что реализация задачи наведения и захвата добычи при необходимости корректировки взаимных скоростей полета и маневров, а также высокой точности локализации малых объектов наиболее полно раскрывает возможности и характер деятельности эхолокационных систем летучих мышей.
Кроме того, было показано, что эхолокация может быть единственным способом дистантного обнаружения подвижной добычи в условиях, где деятельность других видов рецепции полностью исключается. В указанных работах исследователи делали основной упор на выявление закономерностей эхолокации при поимке насекомых летучими мышами. Однако в естественной обстановке, учитывая большую разреженность летающих в воздухе насекомых, быстрое затухание ультразвука и сложность обстановки, когда охота происходит в лесу или вблизи крупных объектов (скалы, стены домов и т. д.), трудно допустить, чтобы во вфемя поиска летучие мыши полагались только на эхолокацию и лишь отраженный от насекомого сигнал информировал животное о местоположении добычи, тем более что многие насекомые во время полета издают отчетливое жужжание, которое даже человек воспринимает на расстоянии нескольких метров. Поэтому естественно предположить, что гораздо чаще летучая мышь первоначально выслушивает звуки, издаваемые летящим насекомым, по ним определяет его присутствие и лишь затем с помощью эхолокационной системы осуществляет более точное наведение на добычу. Такой ход рассуждений требовал проведения специальных экспериментов в условиях, максимально приближающихся к естественным.
В 1966 г. нами была организована экспедиция в Нагорно-Карабахскую автономную область Азербайджанской ССР, где исследовался процесс ловли насекомых у нескольких отечественных видов — гладконосых и подковоносых летучих мышей (Константинов, Соколов, 1967; Константинов, 1969). Некоторые вопросы эхолокационного поведения подковоносов были уточнены и дополнены позднее в лабораторных условиях (Соколов, 19726)»
Конкретными задачами исследований было проверить возможность пассивного слухового обнаружения добычи летучими мышами и изучить динамику функционирования эхолокационных систем этих животных на разных этапах ловли насекомых. Опыты проводились на 6 видах: ночницах — остроухих и Нат-терера, нетопырях-карликах, подковоносах — больших, малых и Мегели.
210
Возможность пассивного (слухового) обнаружения насекомых
Процесс охоты летучих мышей включает в себя несколько последовательных этапов: поиск—обнаружение—преследование—захват добычи. Нет необходимости объяснять, что почти невозможно исследовать каждый отдельный цикл ловли насекомых охотящимися летучими мышами с тэм, чтобы увязать его поведение с деятельностью анализаторных систем. Поэтому требовалось ограничить пространство для охотящихся животных.
Под открытым небом был сооружен павильон в виде длинного коридора (15X3X3 м), который заканчивался шестиугольным расширением — «цирком» (5X6 м). Стены и потолок павильона были выполнены из тонкой капроновой сетки, которая ограничивала передвижение летучих мышей пределами павильона, но не препятствовала проникновению в него ночных насекомых, привлеченных светом лампы 500 вт. Ночью подопытные летучие мыши, которые отлавливались в ближайших пещерах, гротах или на чердаках домов и находились в отличном состоянии, летали по всему павильону, а мелкие виды, такие как нетопыри-карлики, ночницы Наттерера и малые подковоносы, великолепно охотились в основном в «цирке» за насекомыми, которые в большом количестве роились под лампой. Более крупные виды — остроухие ночницы, подковоносы большие и Мегели — пытались ловить ночных бабочек (бражников, совок), но последние в большинстве случаев быстро вылетали за пределы павильона и становились недоступными для летучих мышей.
Уже первые наблюдения за способами охоты летучих мышей показали, что обнаружение добычи происходит в основном на слух. Отдыхающие на сетке павильона животные внезапно взлетали и устремлялись вслед пролетающему мимо них насекомому, хотя до этого зондирующих сигналов они не излучали и эхолокационным способом не могли обнаружить насекомое. Летучие мыши, летающие по павильону, всякий раз резко изменяли траекторию своего полета и вплотную подлетали к экспериментатору, держащему в руках жужжащую стрекозу или бражника. При этом остроухие ночницы даже садились на него и выхватывали добычу прямо из рук, без всякого предварительного их обучения. Точно так же они реагировали и на "'спрятанных в картонную коробку насекомых, которые пытались "выбраться из нее.
Было отмечено, что положительная пищевая реакция животного на жужжащее насекомое проявляется на небольших расстояниях — от 1.5 до 3 м, редко 4 м — в зависимости от интенсивности жужжания разных видов.
Мелкие виды летучих мышей при охоте в «цирке» резко изменяли свой обычный круговой полет и устремлялись за добычей, которая до этого явно находилась вне радиуса действия их эхолокатора. Дальнейшее преследование и захват жертвы сопровождались излучением зондирующих импульсов, темп которых непрерывно увеличивался по мере сближения.
211
Тогда мы решили проследить реакцию летучих мышей на жужжащих подвижных насекомых и таких же, но обездвиженных. Летучим мышам одновременно предъявлялись два только что убитых насекомых и одно живое. Насекомые подвешивались на тонких проволочках перед входом в «цирк» на расстоянии 60—70 см друг от друга, а летучая мышь выпускалась с противоположного конца коридора. После каждого выпуска положение насекомых относительно друг друга менялось. В 74 случаях остроухие ночницы и нетопыри 70 бросков (94.5%) совершали на жужжащее насекомое и только 4 раза — на неподвижное. Предъявление только неподвижных насекомых, развешенных в разных местах павильона, не вызывало положительной пищевой реакции у животных. Летучие мыши иногда пролетали на расстоянии 20—30 см от насекомого, но всякий раз избегали контакта с ним, реагируя как на препятствие. Отрицательная реакция была и на раскачиваемых ветром мертвых насекомых. Забегая вперед, отметим, что после некоторой тренировки и чередования подвижных и неподвижных насекомых, предъявляемых в одном и том же месте, остроухие ночницы начинали хватать как тех, так и других, но первоначально у диких животных этого не наблюдалось.
Как и гладконосые летучие мыши, подковоносы обнаруживали насекомых в основном на слух и явно предпочитали ловить жужжащих насекомых, чем неподвижных. В 62 случаях одновременного предъявления одного живого и двух мертвых насекомых подковоносы показали 100%-ю реакцию на жужжащее насекомое. Но в отличие от гладконосых летучих мышей все исследованные нами подковоносы и в дальнейших опытах никогда не пытались схватить неподвижное насекомое, даже если последнее предъявлялось в одном и том же месте по нескольку раз. Более того, стоило бабочке сложить крылья и «замереть», как подковонос, которому оставалось лишь схватить добычу, прекращал преследование и отлетал в сторону. Трепещущее крыльями насекомое почти всегда привлекало внимание подковоносов.
В следующей серии экспериментов насекомое крепилось в стеклянном цилиндре с открытыми основаниями, который подвешивался на тонкой проволоке. Цилиндр лишь несколько ослаблял низкочастотные шумы, которые возникали при движении крыльев насекомого, но полностью исключал возможность отыскать его с помощью эхолокации. Тем не менее подковоносы Мегели обнаруживали добычу с расстояния около 3 м, подлетали к цилиндру и пытались сбить насекомое ударом крыла по стеклу. Цилиндр без насекомого или с обездвиженным насекомым не привлекал внимания подковоносов.
Позднее в лабораторных условиях удалось установить максимальное расстояние, на котором большой подковонос реагировал на мелкую бабочку — восковую моль (Соколов, 19726). Как только закрепленная на проволочке бабочка начинала ра-212
ботать крыльями, подковонос резко поворачивался всем корпусом в сторону добычи, лоцировал в направлении насекомого, а затем срывался в места, подлетал к добыче, схватывал ее и, подвесившись к потолку манежа, съедал. Когда дистанция между подковоносом и восковой молью не превышала 4.5 м, латентный период реакции был равен 1—2 мсек.
При возрастании расстояния от 4.5 до 6.5 м отмечалось увеличение латентного периода реакции до 5—10 сек. Летучие мыши при этом реагировали только на более крупные и активные экземпляры. На расстояниях, больших 6.5 м, заметная поведенческая реакция животных отсутствовала.
Установка войлочного экрана между подковоносом и насекомым, который полностью скрывал бабочку, был непроницаем для локационных сигналов животного, но в то же время не препятствовал распространению низкочастотных звуков, возникающих при движении крыльев насекомого, не повлияла на способность животного обнаруживать «жужжащую» добычу. Подковонос либо облетал экран и схватывал насекомое, либо подлетал ближе и с более удобной позиции атаковал бабочку. Если же бабочка за экраном не взмахивала крыльями, подковонос оставался на прежнем стартовом положении.
Подковоносы, так же как и гладконосые летучие мыши, сбивали настигнутых насекомых концом крыла, те падали в предварительно подставленную крыловую перепонку (у гладконосых в межбедренную перепонку) и уже оттуда доставались ртом. С пойманным насекомым животные обычно садились на сетку манежа и съедали у него в основном брюшко, тогда как крылья и головогрудь, особенно крупной добычи, оставляли нетронутыми.
Проведенные наблюдения свидетельствуют о преобладающей роли пассивного слухового восприятия на первом этапе охоты летучих мышей за насекомыми, т. е. в период поиска. Как было отмечено выше, диапазон слуховой чувствительности летучих мышей простирался в низкочастотную область вплоть до 30 гц. Следовательно, у животных имеются все предпосылки для восприятия низкочастотных звуков (ультразвукового излучения у используемых в наших опытах насекомых не было обнаружено), создаваемых вибрацией крыльев ночных насекомых во время полета.
Деятельность эхолокационных систем у гладконосых летучих мышей на разных этапах ловли насекомых
Динамика изменения локационных сигналов, излучаемых летучими мышами при сближении с добычей, изучалась в различных ситуациях. Сигналы нетопырей-карликов записыва
213
лись во время их ночной охоты за мелкими двукрылами, которые в большом количестве роились под лампой в «цирке» нашего павильона. Здесь же охотились малые подковоносы. Охота в «цирке» ничем не отличалась от таковой в естественной обстановке, и в этом отношении условия для изучения функций эхолокационных систем летучих мышей представлялись идеальными. Однако мелкие размеры насекомого и круговые полеты исследуемых животных не позволяли точно оценить расстояние сближения животного с целью и сделать привязку излучаемых сигналов с траекторией полета. Поэтому для других видов летучих мышей была разработана специальная методика, которая включала в себя некоторые элементы моделирования условий охоты.
На расстоянии 1 м перед входом в «цирк» на высоте 1.7—1.8 м от земли я а тонкой проволочке закреплялось живое насекомое. В основном это были ночные бабочки (совки, хохлатки, бражники), но также стрекозы, жуки-навозники и сверчки). Проволочка не препятствовала «полету» насекомого, яо удерживала его в определенном месте и лишала возможности садиться
Рис. 73. Общий вид экспериментального павильона.
Б, Ф — киноаппараты, М — микрофон, установленный позади насекомого, Л — осветительные лампы.
на потолок и стены павильона или вылететь за его пределы. В некоторых опытах вместо живых насекомых летучим мышам предъявлялись мертвые насекомые или имитаторы. Летучая мышь вылетала с противоположного ют насекомого конца коридора. Во время полета она обнаруживала добычу, приближалась к ней, срывала с проволоки и возвращалась обратно, так как вход в «цирк» был затянут тонкой капроновой сеткой. Пока летучая мышь поедала насекомое в дальнем конце павильона, экспериментатор успевал закрепить новое и подготовиться к регистрации очередного полета.
Ультразвуковые сигналы, излучаемые летучей мышью при подлете к цели, улавливались конденсаторным микрофоном, расположенным на расстоянии 1 м за насекомым. Воспринятый сигнал усиливался и, пройдя измеритель девиации, поступал на вход двухлучевого осциллографа (0К-17М и С1-4) и параллельно на стандартный ультразвуковой магнитофон (МУ-25). С экрана осциллографа импульс и его частотное заполнение фотографировались 35 мм кинокамерой с непрерывной протяжкой пленки со скоростью 1.5 м/сек. Одновременно с записью импульсов производилась киносъемка летящего животного двумя скоростными киноаппаратами (рис. 73, Ф и Б). Летучая мышь фотографировалась на фоне 2 белых экранов, на которых
214
были нанесены градуированные шкалы. По ним при дальнейшей обработке 2 кинопленок определялись траектория полета животного и расстояние до насекомого в каждый момент времени.
Так как скорости киноаппаратов, снимающих полет мыши и изображение ее сигналов на экране осциллографа, отличались друг от друга, необходимо было при обработке пленок совместить изображение каждого импульса с кадром, на который летучая мышь попадала в момент его излучения. Для этого использовалась система единого времени (СЕВ) — устройство, подающее на оба киноаппарата, фиксирующих полет летучей мыши, электрические метки в момент излучения мышью каждого импульса. Управляющие сигналы с указанной системы единого времени использовались также для запуска ждущей развертки осциллографа и одновременного запуска и остановки всех киноаппаратов и магнитофона.
Среди исследованных видов гладконосых летучих мышей наиболее полные сведения об эхолокационной активности удалось получить в опытах с остроухими ночницами. Было зафиксировано более 100 целенаправленных полетов, завершившихся захватом предъявляемого насекомого. Из них для анализа параметров локационных сигналов нами отобрано 30, в которых летучие мыши приближались к объекту по прямолинейной траектории.
Для выявления характерных закономерностей в изменении режима эхолокации, обусловленных присутствием насекомого, прежде всего исследовалась импульсная активность животного-при пролетах в пустом павильоне (рис. 74).
Каждая точка на графиках рис. 74 соответствует одному импульсу, излученному животным на определенном расстоянии при прямолинейной траектории полета от одного конца павильона до другого (т. е. до того места, где в последующих опытах помещалось насекомое). Как видно на графике (Л), темп излучения и параметры сигналов от начала и до конца пути почти не меняются. Частота следования импульсов обычно составляла 8—12 имп./сек., длительность сигналов 3—3.5 мсек., частота заполнения в начале сигналов около 120 кгц, в конце — 30—40 кгц. Такой режим работы эхолокатора летучей мыши свойствен обзорной локации при спокойном курсирующем полете животного.
Введение в эксперимент жужжащего насекомого, закрепленного на проволочке, вызывало значительные изменения в деятельности эхолокационной системы ночницы. Вылетая с исходного места, летучая мышь первоначально излучала зондирующие сигналы в том ритме, что и при полете в пустом павильоне. Но на расстоянии в среднем 2.3 м (1.8—3 м) от насекомого частота следования импульсов внезапно возрастала до 16—25 имп./сек., т. е. примерно в 2 раза по сравнению с исходным уровнем. Период между импульсами сокращался в среднем до 40 мсек. (30—70 мсек.), но основные параметры сигналов (длительность, частотное заполнение) оставались пока без изменений. Столь значительное учащение сигналов при отсутствии влияния посторонних факторов несомненно обусловлено тем, что летучая мышь обнаружила присутствие насекомого. Если исходный уровень излучения локационных сигналов до обнаружения насеко-
215
^кгц т^мсек. Т,мсек. Р^ги, %,мсек. Г, мсек.
Рис. 74. Изменение параметров локационных сигналов остроухой ночницы при пролете пустого манежа (А), при прямолинейном подлете к жужжащему (Б) и неподвижному (В) насекомому.
По оси абсцисс — расстояние; по оси ординат — период между импульсами (Г), длительность импульсов (т), частота заполнения импульсов (F). Б: первой стрелкой (справа налево) отмечено начало второй фазы эхолокационной активности, второй стрелкой — начало третьей фазы. Fu — частота в начале импульса; FK — частота в конце импульса.
мого обозначить первой фазой эхолокационной активности животного, то с момента учащения импульсов наступает вторая фаза. По времени она занимает менее 0.5 сек., и животное успевает излучить 3—5 импульсов, после чего эхолокация вступает в следующую, третью, фазу (рис. 74,-Б). Характеризуется она тем, что, во-первых, происходит еще более значительное учащение сигна-
лов и частота следования импульсов увеличивается сразу до 40 имп./сек., а затем доходит до 100 ймп./сек. перед самым захватом насекомого. Во-вторых, длительность сигналов сокращается одномоментно на 1/3 и далее постепенно уменьшается до 1—0.8 мсек. В-третьих, началу третьей фазы часто предшествует излучение импульса, амплитуда которого в несколько раз превышала амплитуду как предыдущего, так и последующего импульса (рис. 75). Появление такого сигнала может обозначать, что летучая мышь запеленговала объект атаки своим эхолокатором и с этого момента средства эхолокации становятся основными для животного в достижении цели. Важно отметить, что в дальнейшем наряду с сокращением длительности сигналов уменьшается и интенсивность их излучения. Это хорошо видно на представленных осциллограммах импульсов, следующих один «за другим по мере сближения животного с насекомым. Несмотря на то что расстояние до микрофона сокращается, амплитуда зондирующих сигналов уменьшается, но частотное заполнение сигналов не испытывает существенного изменения. Лишь в самых последних импульсах перед захватом насекомого происходит некоторое уменьшение величины девиации за счет снижения частоты в начале импульса до 100—80 кгц и незначительного ее повышения в конце
сб Я
8 О
о к
Я
217
(рис. 74). Переход второй фазы в первую происходил на расстоянии от насекомого в среднем 1.2 м (0.8—1.4 м).
Путем попеременного предъявления в определенном месте то подвижного (живого) насекомого, то неподвижного мы в течение одного опыта выработали у остроухих ночниц положительную реакцию (бросок) на подвешенное обездвиженное насекомое или такого же размера имитатор-камень. При этом в поведении животных не было замечено особых изменений. Как и при ловле насекомого, летучая мышь по прямой приближалась к неподвижному объекту и на расстоянии 8—10 см сбивала его крылом в межбедренную перепонку, после чего либо поедала насекомое, либо отбрасывала камень.
Анализируя деятельность эхолокационного аппарата летучей мыши в этом случае, мы установили полное отсутствие второй фазы, отмеченной при предъявлении живого насекомого. Приближаясь к неподвижному объекту, летучая мышь резко изменяла темп излучения сигналов лишь на расстоянии в среднем 1.2 м, и вслед за этим сразу начиналось характерное для третьей фазы уменьшение длительности сигналов и интервалов между ними (рис. 74). Следовательно, обнаружение неподвижного насекомого или его имитатора происходит на более близких расстояниях, чем обнаружение подвижного насекомого.
Выявленные отличия в эхолокации остроухой ночницы при ловле подвижного и неподвижного насекомого, и в частности отсутствие второй фазы в последнем случае, позволяют предполагать, что возникновение этой фазы обусловлено обнаружением добычи на слух, т. е. по жужжанию самого насекомого. Об этом свидетельствует расстояние в 2—3 м, на котором, по нашим данным, животные реагируют на насекомое без участия эхолокации, а также тот факт, что, кроме увеличения частоты повторения импульсов, их параметры остаются без изменений. Летучая мышь, услышав насекомое, начинает более активно лоцировать пространство, пока не получит отраженный сигнал от интересующего объекта. Именно тогда наблюдается переход эхолокации в третью фазу, т. е. на более высокий функциональный уровень с постепенным изменением всех параметров излучаемых сигналов.
Низкий уровень эхолокационной активности в поисковой фазе хорошо согласуется с тем, что вероятность обнаружения насекомого в естественной обстановке по отраженному сигналу крайне незначительна. Кроме того, если учесть, что некоторые виды ночных бабочек улавливают сигналы летучих мышей на значительно больших расстояниях — порядка 30 м (Roeder, 1965), то последним до определенного времени легче остаться незамеченными при небольшом темпе излучения сигналов. На близких расстояниях наведение на обнаруженный объект принимают на себя аппараты эхолокации.
218
Аналогичные фазные изменения в деятельности эхолокационной системы получены нами и для другого вида — ночницы Нат-терера.
Динамика изменения локационных сигналов у охотящихся нетопырей-карликов при сближении с объектом в общем подчиняется тем же закономерностям, что и для ночниц. Совершая круговые полеты в «цирке» манежа, нетопыри излучали импульсы, которые характеризовались следующими параметрами: длительность — 3—3.5 мсек., частота в начале сигнала — 110—130 кгщ частота в конце сигнала — 55—60 кгц. Частота следования их
Рис. 76. Осциллограммы импульсов нетопыря-карлика при ловле насекомых.
1 — импульс курсирующего полета; 11 — два импульса фазы приближения; III — три импульса заключительной фазы перед захватом насекомого.
была выше, чем у остроухих ночниц, и составляла 20—25 имп./сек. Обнаружение насекомых (обычно мелких двукрылых) происходило на небольших расстояниях, не превышающих 1 м. При сближении с насекомым темп излучения сигналов постепенно увеличивался, а длительность импульсов уменьшалась. Перед самым захватом добычи нетопырь излучал импульсы с частотой следования 220—230 за секунду. При этом интервалы между импульсами сокращались до 4.3 мсек., а длительность сигналов — до 0.3— 0.2 мсек., т. е. примерно в 10 раз по сравнению с импульсами, излучаемыми этим животным на стадии поиска. Частота заполнения в последних импульсах одного цикла охоты у нетопыря-карлика значительно снижается (рис. 76). Частота в начале импульса падает до 50—60 кгц, в конце — до 30—35 кгц, т. е. частотное заполнение уменьшается примерно в 2 раза по сравнению с нормой.
219
Более значительный диапазон изменений локационных сигналов у нетопырей-карликов по сравнению с остроухой ночницей, по-видимому, следует объяснять различной пищевой специализацией этих видов. В наших экспериментах нетопыри-карлики предпочитали ловить мелких двукрылых и небольших совок из семейства Noctuidae. Более крупных насекомых они не трогали, хотя и подлетали, привлеченные их жужжанием. Остроухие же ночницы мелких бабочек ловили с трудом, часто промахивались и срывали их с проволочки лишь с 5—6-го захода. Зато они великолепно справлялись с крупными бражниками, например с сиреневым (Sphinx ligustri) и большим ночным павлиньим глазом (Saturnia pyri) из семейства Saturniidae, а также с различными стрекозами, жуками, сверчками, кузнечиками и крупными совками. Не делали они попыток хватать только живых богомолов и одного из наиболее крупных бражников — мертвую голову (Acherontia atropos). Охота за мелкой добычей, к тому же летающей почти бесшумно, требует от эхолокационной системы животных более напряженной работы, что и проявилось в наших опытах у нетопырей-карликов. Примерно такой же высокий темп излучения локационных сигналов и резкое сокращение их длительности в заключительной фазе ловли дрозофил и москитов были зарегистрированы у одного мелкого вида американских ночниц — Myotis lucifugus (Griffin et al., 1960). Можно полагать, что мелкие виды летучих мышей во время охоты за бесшумно летающей добычей при обнаружении последней больше полагаются на эхолокацию, чем на пассивное восприятие звуков. Хотя не исключено, что гудение роя таких насекомых скорее привлечет внимание летучих мышей, чем одиночно летающие насекомые, и тогда на сравнительно ограниченном пространстве деятельность эхолокационной системы в сочетании с высокой маневренностью полета обеспечит им достаточное количество пищи. Следовательно, тактические приемы у разных видов гладконосых летучих мышей при ловле летающих насекомых могут быть различны. В случае остроухой ночницы речь идет об обнаружении и преследовании одиночной, достаточно крупной цели, тогда как для нетопырей и мелких ночниц задача, по-видимому, сводится к обнаружению группы насекомых. Отсюда ясно, что взаимодействие слухового восприятия и эхолокации будет выступать в том и другом случае в различных комбинациях, так же как различно будет и расстояние эхолокационного обнаружения добычи для разных видов.
Особенности эхолокации подковоносов при ловле насекомых
Сведения о деятельности эхолокационных систем подковоносых летучих мышей при охоте за насекомыми ограничивались лишь кратким замечанием о том, что большие подковоносы 220
при сближении с подброшенным вверх мучным червем увеличивают темп излучения сигналов, подобно гладконосым летучим мышам (Webster, 1968). В нашей лаборатории Б. В. Соколовым (19726) были проведены исследования функции эхолокации этого же вида при обнаружении, преследовании и захвате небольших бабочек (озимой совки и восковой моли), укрепленных на тонкой проволочке, как и в описанных выше экспериментах с остроухой ночницей. Первоначально устанавливался режим работы эхолокатора
Рис. 77. Динамика изменения периода следования (Г) и длительности (т) сигналов большого подковоноса в контрольных полетах (штрихпунктирная линия) и при подлете к насекомому (сплошная линия).
По оси абсцисс — расстояние до насекомого. Стрелкой обозначен момент обнаружения насекомого.
большого подковоноса при пролете участка манежа от места старта до микрофона без предъявления насекомого, т. е. устанавливалась норма. Во время таких пролетов подковоносы излучали одиночные сигналы длительностью 55—90 мсек. Период следования их варьировал от 70 до 120 мсек., т. е. за секунду излучалось 8—14 импульсов (рис. 77). Уровень звукового давления на расстоянии 10 см от головы животного равнялся в среднем 121 дб относительно 0.0002 дин/см2. Когда же на пути летучей мыши предъявлялось бьющееся насекомое, картина резко изменялась. Если на первых метрах полета подковоноса сигналы по своим параметрам не отличались от контрольных, то на определенном расстоянии от бабочки подковонос начинал излучать пачки из 2—3 сигналов, а затем следовала длительная серия из 8—10 импульсов, предпоследний или последний из которых совпадал с моментом захвата
221
насекомого. Соответственно изменялись и параметры сигналов. Их длительность на этом участке почти линейно падала с 55—90 до 8—12 мсек., а период следования с 70—120 до 10—15 мсек. На этом же участке наблюдалось изменение интенсивности сигналов. Уровень звукового давления сигналов уменьшался от 121 дб вначале до 91 дб к моменту, когда подковонос находился на расстоянии 30 см от насекомого. Таким образом, перепад уровней звукового давления может достигать 30 дб. После поимки добычи длительность и амплитуда сигналов резко возрастают, возвращаясь к норме. Такое изменение параметров сигналов подковоносов было обусловлено эхолокационным обнаружением ими добычи. Последний сигнал, не отличающийся от нормы, после которого наступало резкое уменьшение длительности, периода следования и амплитуды последующих сигналов, принимался за импульс обнаружения, а расстояние до насекомого, на котором подковонос начинал излучать этот сигнал, — за расстояние эхолокационного обнаружения. Эта величина для большого подковоноса изменялась от 1.2 до 2.3 м, когда предъявлялась восковая моль, и от 1.6 до 2.3 м — в случае более крупной озимой совки.
После захвата насекомого периоды и длительность импульсов вновь увеличивались. Таким образом, у большого подковоноса деятельность эхолокационной системы при ловле насекомых проявляет черты сходства с таковой у гладконосых летучих мышей.
Кроме большого подковоноса, как уже отмечалось выше, экспериментам были подвергнуты мелкие виды подковоносов — Мегели и малый. Проводя с ними опыты в полевых условиях,, мы пришли к заключению, что ловля насекомых этими видами основана главным образом на пассивном восприятии звуков, издаваемых ночными бабочками во время полета. Основой для такого заключения послужили два факта: отсутствие импульсов при приближении животных к подвижной бабочке и отсутствие пищевой реакции на насекомое, которое складывало крылья, даже в том случае, когда до него оставалось всего 15—20 см. Начиная полет в манеже, подковоносы излучали несколько зондирующих сигналов, но после обнаружения добычи замолкали, и импульсы возникали вновь лишь после того, как жертва была схвачена.
Сделанный вывод подтвердила и небольшая серия опытов, в которых была предпринята попытка заглушить звуки насекомого с помощью низкочастотного широкополосного шума. До включения шума подковонос Мегели совершил 14 бросков на подвижную бабочку из 16 предъявлений, тогда как после включения из 18 предъявлений он только 5 раз пытался схватить насекомое. В то же время остроухие ночницы в условиях шума продолжали совершать броски как на подвижных, так и неподвиж
222
ных насекомых с тем же успехом, что и в тишине, вероятно,
полагаясь в основном на эхолокацию.
Спустя несколько лет опыты с подковоносами Мегели были повторены в лабораторных условиях с использованием более совершенной звукоулавливающей аппаратуры (Соколов, 1972). В них ставилась задача проследить более детально режим работы эхолокационной системы этого вида при подлете к подвижной
бабочке и выяснить, на каком расстоянии подковонос прекращает излучение.
Обычно при пролете манежа подковонос Мегели излучал пачки, состоящие из 2 импульсов. Интервалы между пачками не превышали 25—30 мсек. Паузы между импульсами внутри пачки равнялись 5—7 мсек. Из-за разной величины интервалов между импульсами в пачке и между пачками импульсов график изменения периода следования сигналов имеет
Рис. 78. Динамика изменения периода следования (Т) и длительности (т) сигналов подковоноса Мегели в контрольных полетах (штрихпунктирная линия) и при подлете к насекомому (сплошная линия).
Остальные обозначения те же, что и на рис. 77.
пилообразный вид (рис. 78). Средняя длительность сигналов составляла 25 мсек., частота заполнения средней части импульса — 110 + + 2 кгц, уровень звукового давления — 110 дб. В выпусках, когда подковоносу Мегели предъявлялось насекомое, в начале полета параметры сигналов совпадали с контрольными. На некотором расстоянии от бабочки подковонос излучал один или несколько длительных (55—75 мсек.) одиночных сигналов постепенно уменьшающейся амплитуды. На последних 90—100 см от насекомого локационные сигналы исчезали (рис. 78). Изменение стандартного, в условиях опыта, способа излучения, а именно переход от пачек из 2 сигналов к одиночным импульсам, по-видимому, связано с эхолокационным обнаружением насекомого. Расстояние обнаружения бабочки, определяемое по этому признаку, варьировало в пределах 1.3—2.3 м. Импульсы вновь появлялись после захвата насекомого.
Таким образом, в отличие от всех ранее изученных видов
223
подковоносы Мегели после обнаружения насекомого не пытаются усилить работу своего эхолокатора, а наоборот, прекращают излучение импульсов. Появление длительных одиночных сигналов, возможно, помогает животному более точно определить координаты своей добычи. Как показали модельные опыты (Roeder, 1963; Соколов, 19726), интенсивность отраженного от бабочки ультразвукового сигнала меняется на 20—30 дб в зависимости от положения его крыльев в момент облучения. Частота взмахов крыльев у озимой совки и восковой моли составляет порядка 40 гц, и один взмах длится в среднем 25 мсек.
Можно ожидать, что длительный сигнал (более 50 мсек.) по крайней мере дважды отразится от бабочки с максимальной амплитудой, тогда как короткий сигнал (менее 10 мсек.) может попасть на насекомое в тот момент, когда оно дает минимальное отражение.
В таком случае биологическая целесообразность использования 1—3 длительных сигналов подковоносами Мегели после обнаружения добычи становится понятной. С их помощью животные дополнительно уточняют пространственные координаты летящей бабочки и в то же время остаются незамеченными, так как адаптация тимпанального органа бабочки к продолжительному чистому тону наступает очень быстро (Roeder, 1965). Последнее обстоятельство для мелких видов подковоносов становится решающим, учитывая, что пищей им служат в основном ночные бабочки, обладающие тимпанальными органами. Прекращая излучение локационных сигналов на расстоянии примерно 1 м до насекомого, подковоносы имеют больше шансов незаметно приблизиться к своей жертве и поймать ее.
Для многих гладконосых летучих мышей и больших подковоносов этот вопрос не стоит так остро. Большинство видов насекомых, которыми они питаются, не обладают тимпанальными органами и, следовательно, не способны воспринимать их ультразвуковые локационные сигналы. Б этом случае связь между хищником (летучей мышью) и его жертвой проявляется лишь односторонне, т. е. летучая мышь обнаруживает насекомое по его жужжанию, усиливает эхолокацию и, руководствуясь отраженными сигналами, настигает его. Выделить насекомое по отраженному сигналу среди других предметов окружающей обстановки помогают не только флюктуация амплитуды эха в зависимости от положения крыльев насекомого, но также и уменьшение интенсивности зондирующих сигналов по мере сближения. Последнее обстоятельство позволяет, во-первых, поддерживать величину воспринимаемого отраженного сигнала в оптимальных пределах независимо от расстояния, а во-вторых, лучше выделять эхо от насекомого по сравнению с эхом от более удаленных предметов, отражение от которых может оказаться ниже порога слухового восприятия.
224
В заключение коротко остановимся на функциональной значимости в пищевом поведении летучих мышей их других анализаторных систем — зрительной и обонятельной. В наших экспериментах после проведения описанных выше серий опытов ночницам и подковоносам была произведена двусторонняя энуклеация. Указанная операция не оказала влияния на способность животных обнаруживать и ловить фиксированных насекомых. Как и в норме, добыча схватывалась летучими мышами либо при первом же полете, либо со второго захода. Единственное отличие у ночниц обнаруживалось в том, что по сравнению с интактными животными они в поисковой фазе излучали импульсы в более быстром ритме и интервалы между сигналами составляли 50—80 мсек, при норме 100—130 мсек.
Вполне вероятно, что зрение может принимать некоторое участие на различных этапах ловли насекомых у таких насекомоядных видов летучих мышей, как например у рыжих вечерниц, которые вылетают на охоту еще засветло, или у ушанов, обладающих довольно крупными глазами и схватывающих добычу на листьях и ветвях деревьев, т. е. в непосредственной близости от себя, особенно в светлые лунные ночи. Тем не менее зрение в пищевом поведении насекомоядных летучих мышей следует рассматривать лишь как вспомогательный источник информации, без которого при осуществлении конкретного пищевого акта летучие мыши могут и обойтись.
Обоняние также не может иметь решающего значения при ловле насекомых. Так, гладконосые летучие мыши с его помощью могли обнаруживать на земле насекомых и выделять среди них съедобных лишь на расстояниях, не превышающих 12 см (Kolb, 1961). В полете выявляется ряд моментов, делающих такое обнаружение ненадежным. Это прежде всего сила и направление ветра, большие скорости полета, быстрота двигательных реакций животных и т. д. Подтверждением этому могут служить и наши эксперименты, в которых летучие мыши принимали за пищевой объект имитатор (камень) и не могли на расстоянии отличить съедобное насекомое от несъедобного.
Не исключено, однако, что зрение и обоняние участвуют в добывании пищи у тропических видов летучих мышей, особенно у нектароядных, фруктоядных, а также десмодусов, питающихся кровью позвоночных животных (Neuweiler, 1967; Chase, 1969; Chase, Suthers. 1969; Suthers et al., 1969; Suthers, 1970a).
10
ПРЕДЕЛЫ ОБНАРУЖЕНИЯ, ДАЛЬНОСТЬ ДЕЙСТВИЯ И ЧУВСТВИТЕЛЬНОСТЬ
ЭХОЛОКАЦИОННЫХ СИСТЕМ
Не подлежит сомнению, что развитие функций акустической локации в эволюции было обусловлено прежде всего необходимостью обнаружения препятствий в оптически неблагоприятных условиях и определения расстояния до них. Естественно, возникает вопрос, какими предельными возможностями обладает эхолокационная система летучих мышей при обнаружении, т. е. предметы какого размера еще замечаются животными и на каком расстоянии. Чтобы ответить на этот вопрос, необходимо выбрать какой-то критерий, по которому можно было бы судить о способности животного обнаруживать данный предмет или группу предметов и избегать столкновения с ними. В качестве такого критерия Хан (Hahn, 1908) предложил использовать барьеры из вертикально натянутых проволок, установленные на пути полета летучей мыши. Данная методика нашла широкое применение в ранних экспериментах по доказательству акустического способа ориентации рукокрылых (Griffin, Galambos, 1941; Galambos, Griffin, 1942) и продолжает оставаться на вооружении исследователей в настоящее время.
Пределы обнаружения
Использование металлических проволок обусловлено тем, что, во-первых, можно легко подобрать самые различные их диаметры, во-вторых, малейшее прикосновение к ним вызывает колебание и дребезжание соответствующей проволоки и поэтому облегчает подсчет ошибок, т. е. контактов с ними при пролете животного, и, в-третьих, можно подсчитать величину сигнала, отраженного от проволоки соответствующего диаметра. На пути животного устанавливают один или несколько таких барьеров и в условиях свободного полета животных подсчиты-226
вают общее число пролетов сквозь барьеры и число уклонений от проволок, т. е. тех пролетов, когда животные не касались ни одной из них. После этого определяется процент уклонений и проволоки заменяются меньшим диаметром. Расстояние между проволоками в ряду устанавливают по полному размаху крыльев летучей мыши изучаемого вида.
В настоящее время с использованием этой методики проведены опыты на 10 видах летучих мышей разных семейств, результаты которых сведены нами в табл. 12.
Полученные данные показывают, что с уменьшением диаметра проволок процент уклонений у летучих мышей постепенно снижается, однако пределы обнаружения у разных видов оказываются различными. Если проволоки диаметром от 2 до 0.32 мм обычно не вызывали трудностей при облете их летучими мышами, то при проволоках 0.2—0.15 мм у многих видов наступало заметное ухудшение ориентационных способностей. Куртис (Curtis, 1952) путем расчетов показал, что для Myotis lucifugus случайное уклонение при расположении проволок на расстоянии 30 см составляет приблизительно 35%. Это значение близко к нашим экспериментальным данным, полученным на ушанах и усатых ночницах при преодолении проволочных барьеров диаметром 0.6 мм в норме и после «оглушения» животных путем перфорации барабанных перепонок. У ушанов процент успешных пролетов после операции составил 30 при норме 76%, у усатых ночниц — 35 при норме 81 %. Таким образом, если животные в опыте показывают 35—30% и ниже успешных пролетов, они уже не способны обнаруживать препятствия и их преодоление происходит случайно.
При пролете через барьеры из более толстых проволок контакт животного с последними ограничивался лишь легким касанием крыла одной из них, тогда как при пролете проволок меньших 0.1 мм летучие мыши часто не успевали уклониться от препятствия, «врезались» на полном ходу в проволоку и после удара иногда падали на пол. Данные табл. 12 показывают, что Noctilio и Pizonyx уже не могли обнаруживать проволоки диаметром 0.1 мм, для Mtytis lucifugus это значение являлось пределом, тогда как Macorous и остроухие ночницы еще обнаруживали проволоки 0.08 мм. Особенно высокие результаты при проволоках 0.08 мм были зарегистрированы у подковоноса Мегели (77%). В дальнейшем Б. В. Соколовым (1972а) было показано, что подковонос Мегели еще обнаруживает проволоки 0.05 мм. Аналогичные предельные значения отмечены для южного подковоноса (Schnitzler, 1968). По данным Шнейдера и Мёреса (Schneider, Mohres, 1960), большие подковоносы надежно обнаруживали одиночные проволоки от 0.55 до 0.09 мм, расположенные вертикально в узкой щели (28 см), через которую животные пролетали, чтобы получить корм. Они хуже реагировали на проволоки 0.065 мм и не замечали тонкие перлоновые нити диаметром 0,03 мм. Авторы считают, что преде-
227
Таблица 12
Пролеты летучих мышей сквозь барьеры из проволок разного диаметра, % уклонений
Диаметр проволоки, мм
Вид CM 1.21—1.05 0.7-0.6 0.55-0.45 0.37—0.32 0.27-0.26 0.23-0.19 0.17-0.15 0.12-0.10 0.08-0.07 Литературный источник
Carollia perspicilata . . — — — — — 65 — 56 — — Griffin, Novick, 1955
Glossophaga coricina . . — 80 68 — — 75 — 89 — — Там же
Artibeus jamaicensis . . — 76 90 — — 85 — 72 — — » »
Macrotus mexicanus . . — — 79 77 79 76 70 53 52 46 Grummon, Novick, 1963
Noctilio leporinus . . . 91 — — 76 — — 60 — 20 — Suthers, 1967
Pizonyx vivesi .... 94 — — 79 — — 51 — 37 — Там же
Myotis lucifugus . . . — 85 77 — 72 52 — — 39 36 Curtis, 1952
-M. oxygnathus .... — 85 — 80 71 — 67 — 54 49 Константинов и др., 1967а
Plecotus auritus .... — 92 76 86 73 — 72 — 61 — Там же
.Rhinolophus mehelyi . . — 92 — 91 — — 82 — 82 77 » »
лом обнаружения для больших подковоносов являются проволоки 0.065 — 0.07 мм. Позднее было показано, что и большие подковоносы надежно обнаруживают проволоки 0.08 мм и не реагируют на проволоки 0.05 мм (Schnitzler, 1968; Соколов, 1972а), что хорошо согласуется с выводами Шнейдера и Мереса.
Для других видов летучих мышей не были установлены пределы обнаружения минимальных размеров препятствий, но высокий процент уклонений при пролете проволок 0.17 мм у Glossc-phaga и Artibeus позволяет допустить, что возможности их эхолокационных систем, по-видимому, не хуже, если даже не лучше, чем у подковоносов Мегели и южного.
Чтобы лучше представить, каким высоким разрешением обладают локационные системы летучих мышей, мы сопоставили диаметры предельно обнаруживаемых проволок с длиной волны излучаемого ультразвука у видов, для которых относительно точно установлены пределы обнаружения (табл. 13).
Таблица 13
Соотношение длины волны ультразвука и предельного диаметра проволок, обнаруживаемых летучими мышами
Вид Частота, кгц Длина волны X, мм Диаметр проволоки (d), мм X/d
Myotis lucifugus .... 45 7.6 0.12 63
М. oxygnathus Rhinolophus ferrum-equi- 85 4.0 0.08 57
num 82 4.2 0.07 60
Rh. mehelyi 110 3.1 0.05 62
Rh. euryale 102 3.3 0.05 66
Macrotus mexicanus . . . 80 4.2 0.07 60
Примечание. Для Myotis lucifugus и М. oxygnathus взяты средние частоты их ЧМ-сигналов, для Macrotus — величина сильной, 2-й, гармоники.
Близкое совпадение полученных данных у различных видов доказывает существование корреляции между длиной волны ультразвука и пределом обнаружения. Кроме того, эти данные показывают, что летучие мыши еще способны обнаруживать предметы, размер которых составляет всего 1/60 длины излучаемой волны. При таких соотношениях размера проволоки и длины волны наблюдается не прямое отражение от нее, а вторичные диафра-тированные волны, распространяющиеся во всех направлениях от малого объекта.
На основании проведенных расчетов можно примерно оценить разрешение эхолокаторов у всех изученных в настоящее время летучих мышей, В качестве иллюстрации подсчитаем, какими
229
предельными возможностями обладает эхолокатор Noctilio lepo-rinus. Средняя частота ее импульсов составляет 30 кгц (60—30 кгц), длина волны равна 11.3 мм, 11.3 : 61—0.18 мм, т. е. проволоки диаметром — 0.Л8 мм составят предел обнаружения для Noctilio. Полученная величина как раз укладывается между экспериментальными значениями, когда проволоки диаметром 0.21 мм обнаруживались животными при уровне уклонений 60%, а проволоки 0.10 мм не обнаруживались совсем (20%).
Методика преодоления проволочных барьеров очень проста и удобна для предварительной оценки эхолокационных способностей животных. Однако она имеет ряд недостатков, так как не учитывает скорость полета летучей мыши и ее маневренность, а кроме того, при малых сечениях проволок обнаружение их может происходить на столь близких расстояниях, когда животное уже не может уклониться от столкновения с ними.
Дальность действия
Приведенные выше данные позволили оценить предельные возможности эхолокации при обнаружении препятствий. Вторая часть задачи заключается в выяснении вопроса, на каком расстоянии летучие мыши обнаруживают проволоки разного диаметра.
Как показали американские исследователи (Grinnell, Griffin, 1958), подлет к препятствию у летучих мышей сопровождается возрастанием частоты повторения локационных сигналов. Начало стабильного изменения в периоде следования сигналов принято использовать в качестве критерия для оценки расстояния, на котором летучая мышь впервые обнаруживает препятствия. Следовательно, чтобы решить поставленную задачу, необходимо знать положение летучей мыши относительно препятствия в момент излучения каждого импульса. Для этой цели одновременно с записью локационных сигналов производится киносъемка животного при подлете к препятствию. Мы использовали в наших экспериментах с 9 остроухими ночницами (Голубков, Константинов, Макаров, 1969) ту же аппаратуру, что и в опытах с насекомыми.
Летучие мыщи стартовали в одном конце прямоугольного летного павильона (12.5 X 2.5 X 2,5м) и летели в сторону кормушки, расположенной в противоположном его конце (рис. 79). После нескольких тренировочных полетов у всех животных вырабатывалась прямолинейная траектория полета от места старта до места кормления. Вслед за этим подготовительным этапом начиналась серия основных экспериментов. На расстоянии 10 м от места выпуска летучих мышей в павильоне устанавливался металлический каркас, на который вертикально натягивались проволоки, используемые в качестве тестов для определения дальности эхолокации. Промежутки между проволоками составляли 40 см, что соответствовало размаху крыльев исследуемых животных. Каркдс с проволоками можно было передвигать 230
поперек павильона на 20 см в ту и другую сторону. В каждом опыте летучая мышь должна была обнаружить наличие препятствий и преодолеть их, прежде чем достичь места кормления. В опытах использовались проволоки нескольких диаметров: 4, 2, 1.5, 0.47, 0.21, 0.12 и 0.08 мм, предъявляемые последовательно. После каждого предъявления препятствия убирались, и животные в течение 1—2 дней летали в свободном павильоне, а затем эксперименты продолжались с проволоками следующего диаметра.
Первоначально исследовался режим работы эхолокатора летучих мышей при полете в свободном павильоне от места старта до места кормления. После выработки прямолинейной траектории полета все летучие мыши излучали импульсы примерно с одинаковым периодом следования на всей дистанции. Интервалы между импульсами в большинстве случаев составляли от 100 до 140 мсек., но иногда наблюдалось их падение до 80—70 мсек. Средняя величина периода следования импульсов в свободном павильоне равнялась 105 мсек. Длительность импульсов обычно колебалась у разных особей от 2 до 4 мсек., и каждый сигнал имел типичную для данного вида летучих мышей частотную модуляцию. Установка в павильоне пустого каркаса для проволок не повлияла на эхолокационную активность животных.
Введение заградительного барьера из проволок диаметром 4 мм вызывало заметные изменения как в поведении летучих мышей, так и в характере излучения локационных сигналов. В первых же выпусках все летучие мыши, на значительном расстоянии не долетая до препятствия, поворачивали обратно, чего не наблюдалось при полете в свободном павильоне. В последующих выпусках они все ближе подлетали к барьеру и наконец преодолевали его. Период следования сигналов при подлете животного к проволокам диаметром 4 мм менялся дважды.
Первое падение периода со 120—140 до 45—70 мсек, наблюдалось на расстоянии в среднем около 4 м, т. е. там, где первоначально летучие мыши совершали поворот. Затем на расстоянии в среднем 160 см происходило второе резкое сокращение периода следования на 70—80 мсек., с последующим его уменьшением до 10—20 мсек, в момент преодоления препятствия. Аналогичные двойные изменения периодов были отмечены нами при предъявлении летучим мышам проволок диаметром 2 мм. Разница состояла лишь в том, что менялось расстояние первого падения периода с уменьшением диаметра проволок, тогда как основное учащение импульсов в обоих случаях начиналось примерно на одной и той же дистанции.
На основании этих данных, сопоставленных с характером поведения животных, первое непродолжительное падение периода принималось за начало эхолокационного обнаружения препятствий летучими мышами.
На присутствие проволок меньшего диаметра летучие мыши реагировали одним резким сокращением периода между импуль-
231
Расстояние до препятствии
Периоды следодания импульсод, мсек.
сами, величина которого постепенно уменьшалась по мере приближения животного к препятствию. В экспериментах выявилась зависимость между диаметром проволок и расстоянием, на котором летучие мыши их обнаруживали. Средние значения расстояний, на которых животные впервые реагируют учащением импульсов на проволоки различного диаметра, приведены в табл. 14.
Таблица 14
Расстояние обнаружения проволок различного диаметра у двух видов ночниц
Myotis oxygnathus Myotis lucifugus
диаметр проволоки, мм дальность обнаружения, см диаметр проволоки, мм дальность обнаружения, см
4.0 400 (380—420) 3.0 215 (66-294)
2.0 330 (270—370) 1.07 185 (49—197)
1.5 250 (220—280) 0.65 150 (66-245)
0.47 173 (170—180) 0.46 120 (69—180)
0.21 110 (110—110) 0.28 105 (30—138)
0.12 85 (80—100) 0.18 90 (30-117)
0.08 49 (20—70)
Одновременно с увеличением частоты повторения импульсов во многих случаях наблюдалось сокращение их длительности до 1.5—1 мсек. Проволоки самого малого диаметра (0.08 мм) летучие мыши обнаруживали на расстоянии около 50 см перед барьером. Период между импульсами падал до 60—40 мсек. Излучив 1—2 импульса, животные преодолевали барьер, при этом часто задевая или обрывая проволоку.
Аналогичные эксперименты с другим видом ночниц {Myotis lucifugus) были проведены американскими исследователями (Grinnell, Griffin, 1958). При сравнении этих данных с нашими (табл. 14) наибольшие различия выявляются при пролетах животными проволок более крупного диаметра, тогда как тонкие проволоки обнаруживаются обоими видами летучих мышей примерно на одних и тех же расстояниях. Помимо видовой специфики животных и различных параметров излучаемых сигналов каждым из них, расхождения в результатах могут быть объяснены еще и тем,
Рис. 79. Летний павильон и графики зависимости периода следования импульсов при пролете летучими мышами барьеров из проволок разного диаметра (мм).
Стрелками на графиках обозначены принятые моменты учащения импульсов. Внизу для каждого графика построена теоретическая зависимость периода следования импульсов локатора с однозначным отсчетом.
233
что расстояние обнаружения измерялось несколько различными путями. Так, при проволоках диаметром 4 и 2 мм за расстояние обнаружения мы принимаем первое уменьшение периода следования импульсов, тогда как Гриннел и Гриффин не принимали его во внимание, хотя и отмечали его наличие во многих случаях. Кроме того, американские исследователи считали, что летучая мышь обнаружила препятствие тогда, когда она излучила импульс с более коротким промежутком по сравнению с последующими, и именно от этого импульса начинали отсчет. Мы же за исходную точку расстояния обнаружения брали последний импульс, излученный с большим промежутком, после которого начиналось учащение сигналов, считая, что отражение этого импульса дало возможность животному обнаружить препятствие и послужило началом учащения импульсов.
Проведенные эксперименты с проволочными барьерами позволили выяснить еще одну особенность эхолокационной системы гладконосых летучих мышей. В 1946 г. советский ученый Е. Я. Пум-пер высказал предположение, что летучая мышь издает каждый последующий ультразвуковой импульс сразу после того, как воспримет эхо предыдущего. Принятый после отражения сигнал является раздражителем, вызывающим посылку следующего зондирующего сигнала. Требовалось экспериментально проверить эту гипотезу, т. е. ответить на вопрос, в однозначном ли режиме действует эхолокационная система летучей мыши при определении дальности до препятствия. Период следования импульсного локатора с однозначным отсчетом определяется равенством
т ____ЗДщах
1 min — С ’
где /?тах — максимальная, однозначно определяемая дальность локатора; С — групповая скорость (340 м/сек. для звука в воздухе).
На рис. 79 для каждого экспериментального значения изменения периода следования импульсов ночниц построены теоретические кривые зависимости минимального периода следования (7\in) сигнала от минимальной дальности, полученные по приведенной формуле.
Анализ этих совмещенных графиков показывает, что для всех случаев расчетный минимальный период следования импульсов меньше экспериментальных значений, так Как ни в одном случае, даже при максимальной частоте следования импульсов, эти кривые не перекрещиваются. Следовательно, режим работы остроухой ночницы однозначен, т. е. каждый следующий локационный импульс излучается после того, как полуЧейа • информация -ёт предыдущего сигнала.
234
В последнее время в нашей лаборатории Б. В. Соколовым (1972а) были проведены аналогичные опыты с двумя видами подковоносов — большим и Мегели. Было показано, что при отсутствии препятствий большие подковоносы во время полета по манежу излучали одиночные сигналы длительностью 50—80 мсек, с периодом следования 80—130 мсек. При предъявлении препятствий на определенном расстоянии в зависимости от сечения проволок режим работы эхолокатора животных изменялся и вместо одиночных импульсов подковонос переходил на излучение пачек, состоящих первоначально из двух импульсов, затем 3—4 и перед самым пролетом проволок — из 10—18 импульсов. Именно переход на излучение пачек с сокращением длительности импульсов и периода их следования принимался автором за момент обнаружения препятствий подковоносами. Динамика изменения параметров локационных сигналов у подковоносов Мегели при обнаружении и преодолении барьеров в общих чертах была сходна с таковой у большого подковоноса. Однако в норме, т. е. при пролете манежа без препятствий, подковонос Мегели излучал пачки из 2 импульсов, а появление пачек из 3 сигналов принималось за момент обнаружения препятствий. Результаты опытов приведены в табл. 15.
Таблица 15
Расстояние (м) обнаружения проволочных барьеров подковоносами
Диаметр проволоки, мм Большой подковонос Подковонос Мегели
1.10 1.65 (1.4—1.9) 1.60 (1.4—1.9)
0.55 1.25 (1.1—1.6) 1.30 (1.1—1.4)
0.20 0.75 (0.6—1.1) 1.05 (0.9—1.2)
0.10 0.50 (0.3-0.65) 0.70 (0.6—0.9)
0.05 0.25 (0.2—0.3)
Расстояние обнаружения проволок различных диаметров оказалось примерно одинаковым у обоих видов, несмотря на то что уровень звукового давления сигналов подковоноса Мегели в 3 раза меньше, чем у большого подковоноса.
Соответствуют ли полученные значения реальным расстояниям обнаружения препятствий подковоносами — сказать трудно. Вполне возможно, что летучие мыши обнаруживают препятствие на больших расстояниях, но изменяют параметры зондирующих сигналов позднее, когда интенсивность эха от проволок достигнет какого-то определенного значения. Косвенным подтверждением этому могут служить результаты опытов Шнитцлера (Schnitzler, 1968), показавшего, что при проволоках диаметром 1.5, 2 и 3 мм
235
параметры сигналов у большого подкойойоса меняются на одних и тех же расстояниях — примерно 1.5 м. Правда, коэффициенты отражения от проволок данных сечений при частоте 81 кгц очень близки и составляют для 1 мм 0.7, для 2 мм — 0.7, для 3 мм — 2.0 (Морз, 1949; Griffin, 1958), однако многое зависит и от того, что брать за критерий обнаружения.
Итак, эксперименты с проволочными барьерами показали, что расстояние обнаружения зависит от размера предъявляемых препятствий (сечения проволоки) и, следовательно, от его отражательных свойств. Проволоки самого крупного сечения — 4 мм — остроухие ночницы обнаруживали на расстоянии 4 м. Этот результат наибольший для всех гладконосых летучих мышей, с которыми проводили исследования. Изучая дальность действия эхолокаторов подковоносов, Мёрес (Mohres, 1953) отметил, что предельное расстояние, на котором большой подковонос еще обнаруживал в стене отверстие размером 20x20 см, составляло 6.4 м. Однако трудно оценивать предельную дальность действия эхолокационных систем в экспериментальных комнатах, где отраженные сигналы от исследуемых предметов всегда идут на фоне более мощного отражения от стен, пола и потолка. Поэтому мы считаем, что полученные в опытных условиях расстояния обнаружения сильно занижены.
Путем расчета можно примерно оценить предельную дальность эхолокационных систем летучих мышей. Она определяется в основном следующими факторами: мощностью излучения, чувствительностью приемника, т. е. слуховой системы на локационных частотах, длиной волны излучаемого ультразвука и степенью его затухания в воздухе и, наконец, отражательными свойствами предметов.
Интенсивность излучения на расстоянии 10 см от головы животного у остроухой ночницы составляет 111 дб, у большого подковоноса — 122 дб; наилучшая чувствительность слуха на локационных частотах у ночницы варьирует от 0 (на 60 кгц) до —13 дб (40 кгц), у подковоноса равна —5 дб (83 кгц). Кривые ослабления ультразвука в воздушной среде для частот 81, 60 и 40 кгц (рис. 80) в зависимости от расстояния были рассчитаны нами следующим образом.
На расстоянии 10 см (гх) от головы животного интенсивность излучаемого сигнала принималась за единицу, тогда величина ослабления в децибелах (/) на любом большем расстоянии г2 в направлении максимума излучения определяется соотношением
'2
где а — коэффициент затухания в воздухе: для f=82 кгц составляет 0.7 1/м, для /=60 кгц — 0.58 1/м, для /=40 кгц — 0.35 1/м при температуре воздуха 20° и относительной влажности
236
30% (табличное значение — Griffin, 1958); е — основание натурального логарифма.
Например, на расстоянии 10 м для ультразвука с частотой 40 кгц величина ослабления составит
= дб,
т. е. на 55 дб уменьшится первоначальная интенсивность излученного сигнала. Зная параметры сигналов, пороги слуховой системы на лучших частотах и величину ослабления ультра-дб 140 120 100 00 60 40 20
0
Рис. 80. Кривые ослабления ультразвука при распространении его в воздухе. По оси абсцисс — расстояние, по оси ординат — ослабление.
звука при распространении в воздухе на разных расстояниях, можно подсчитать предельную дальность действия локационных систем исследуемых видов при условии, что преграда достаточно велика и дает зеркальное отражение падающей на нее волны.
По кривым ослабления находим, что сигнал с частотой 82 кгц при интенсивности излучения 122 дб распространится на 16.2 м, а с частотой 60 и 40 кгц при интенсивности 111 дб соответственно на 25 и 39.5 м. Половина этих расстояний может составить дальность действия эхолокаторов летучих мышей, так как сигнал, воспринимаемый животным, претерпевает двойное ослабление при прохождении до объекта и обратно. Однако порог слуховой системы на локационной частоте 82 кгц у большого подковоноса находится ниже нулевого уровня на 5 дб, за счет этого предельная дальность действия его эхолокатора составит 13.1 м. У ночниц сигнал частотно-модулированный и, как правило, большая часть его энергии сосредоточена в пределах 80—40 кгц. На 3 частотах в отдельности (80,60 и 40 кгц) с учетом порогов слуха от 0 до —13 дб дальность действия эхолокатора будет равна 6.7, 12.5, 25 м.
237
Йо так как сигнал Целиком дает информацию животному о расстоянии до предмета, то, по-видимому, среднее арифметическое этих трех величин будет больше соответствовать искомому значению предельной дальности действия эхолокатора остроухой ночницы. Ее величина равна 17 м.
Таким образом, проведенные расчеты приводят нас к неожиданному выводу, что теоретически предельная дальность действия у ночницы оказывается несколько выше, чем у подковоноса, несмотря на более мощное излучение у последнего, хотя обе эти величины получаются одного порядка и, по-видимому, совсем уравняются, если учесть диаграмму направленности излучения и приема у этих видов.
Максимальную дальность действия эхолокатора, например, остроухой ночницы можно рассчитать и другим способом, исходя из периода следования локационных сигналов при полете в свободном пространстве. Именно этот способ применяется в радиолокаторах с однозначным режимом отсчета, где период следования импульсов определяет радиус действия дальномера станции. У остроухой ночницы средняя величина периода следования (Г) сигналов при полете в большом манеже при отсутствии препятствий составляла 105 мсек. Тогда максимальная дальность ее однозначной локации определяется как
ТС 105 мсек. • 340 м/сек.
R max — 2 “ 2 =: 1^'8 м •
Дальше этого расстояния цели могут определяться по дальности уже неоднозначно, т. е. последующие зондирующие сигналы могут опережать прием эха от предыдущих, что не соответствует режиму эхолокационных систем гладконосых летучих мышей. Интересно отметить совпадение полученных данных, рассчитанных тем и другим методом для остроухой ночницы. Это лишний раз подтверждает, что период следования импульсов при обзорной локации не случайная величина, а соответствует предельным возможностям эхолокатора в целом для данного вида животного.
Однако в реальных условиях радиус действия эхолокационных систем летучих мышей будет, естественно, меньшим, так как всегда присутствует определенный шумовой фон и пороги слуха на локационной частоте повысятся, а кроме того, объекты локации отражают ультразвук не идеально.
Пороговая чувствительность эхолокатора
Опыты по обнаружению проволочных барьеров позволяют определить величину отраженного сигнала, которая вызывала реакцию животного. Иными словами, представляется возможным подсчитать пороговую чувствительность эхолока-238
Ционной системы летучей мыши в реальных условиях ее функционирования.
При условии, что цилиндр намного длиннее, чем длина волны звука (это условие соблюдается в наших экспериментах), интенсивность отраженного сигнала (/) на данном расстоянии от проволок до летучей мыши может быть вычислена по следующей упрощенной схеме (Голубков, Константинов, Макаров, 1969), выведенной из более сложного и общего уравнения (Морз, 1949):
X /0.1 \з
1=1°-*гг-2°\~г)е'
где /0 — интенсивность сигнала, излученного летучей мышью, измеренного на расстоянии 0.1 м от рта животного (/0=11.8-•10“2вт/м2); X — средняя длина волны ультразвука (Х=5.5 мм, что соответствует частоте 62 кгц); г — расстояние обнаружения, определяемое по первому резкому изменению периода следования импульсов; а — коэффициент затухания звука в воздухе, при X —5.5 мм а — 0.58 1/м (табличное значение); а —-у—относительная величина, характеризующая эффективность отражения провода диаметра (d) для данной длины волны (X); 2 а взято потому, что учитывается среднее арифметическое значение дальности обнаружения одновременно от двух проволок. Это объясняется тем, что наиболее вероятное направление полета летучей мыши будет приходиться на промежуток между двумя проволоками, а не точно на одну проволоку. Отражением от других проволок при пороговом сигнале можно пренебречь из-за селекции по дальности.
Важно отметить, что ультразвук, излучаемый ночницей на расстоянии более 10 см от рта животного, будет распространяться в виде сферических волн. Интенсивность сферической волны изменяется обратно пропорционально квадрату расстояния от источника. С другой стороны, эхо от цилиндрической проволоки может рассматриваться как цилиндрическая волна, интенсивность которой изменяется обратно пропорционально расстоянию от проволоки.
Следовательно, в целом интенсивность отраженного сигнала в нашем случае будет изменяться обратно пропорционально кубу расстояния от животного до проволоки. Поэтому в формулу введен коэффициент (-““У.
По формуле были рассчитаны интенсивность и уровень звукового давления относительно 2-10“5 н/м2 отраженных сигналов от проволок разного диаметра в той точке, где летучие мыши впервые резко изменяли период следования сигналов, принятой за момент обнаружения. Результаты расчетов представлены в табл. 16.
239
Таблица 16
Интенсивность и уровень звукового давления сигналов, отраженных от проволок разного диаметра в точке обнаружения
Диаметр проволоки, мм Дальность обнаружения, см Интенсивность отраженного сигнала, X 10~12 вт/м2 Уровень давления отраженного сигнала, Дб
4.0 400 8.3 9.2
2.0 330 16 18
1.5 250 145 21.6
0.47 173 58 17.6
0.21 НО 24 13.8
0.12 85 28 14.5
0.08 49 113 20.5
Отметим, что в предыдущих наших публикациях (Голубков, Константинов, Макаров, 1969; Айрапетьянц, Константинов, 1970а) при расчетах интенсивности отраженных сигналов была допущена ошибка, которая сейчас исправлена. Этим и объясняется расхождение данных в предыдущих работах с теми, которые представлены в табл. 16.
Таким образом, в реальных условиях эксперимента при шуме, создаваемом работой киноаппаратов и звукозаписывающей аппаратуры, а также многочисленных побочных отражениях от стен, пола, потолка, сетки павильона порог чувствительности приемной системы эхолокатора остроухой ночницы составил в среднем 15.5 дб (9.2—21.6 дб). Этот результат близок к соответствующим пороговым уровням отраженных сигналов, полученным путем аналогичных расчетов для других видов летучих мышей. Так, для Myotis lucifugus реакция на проволочные барьеры отмечена при уровне отраженного сигнала 23 дб, для Eptesicus fuscus — 17 дб (Griffin, 1958), а для большого подковоноса — 25—34 дб (Соколов, 1972а).
Все эти значения в несколько раз превышают экспериментальные пороги слуховой системы этих животных, полученные по условнорефлекторной методике, что вполне естественно, так как последние снимались в звукозаглушенной камере, где уровень шумов практически приближается к нулю.
Полученные величины пороговой чувствительности эхолокаторов позволяют еще раз скорректировать их предельные возможности по дальности в реальных условиях при идеальной отражающей поверхности препятствий. Пользуясь расчетами, аналогичными предыдущим, получаем, что для остроухой ночницы в этом случае максимальная дальность при частоте 40 кгц составит 15 м, 60 кгц — 9.7,80 кгц — 6 и в среднем 10.2 м, для подковоноса — 8,4 м,
ТОЧНОСТЬ ИЗМЕРЕНИЯ ДАЛЬНОСТИ И РАЗРЕШАЮЩАЯ СПОСОБНОСТЬ ЭХОЛОКАТОРА
Точность определения расстояния
Точность измерения дальности до предмета — следующая основная характеристика любой локационной системы, и требования к этому параметру, естественно, зависят от назначения данной системы. Так, например, для радиолокационной станции дальнего обнаружения объектов относительная точность по дальности считается удовлетворительной, если лежит в пределах 1—2% шкалы, что для дистанции 200 км дает ошибку в измерении 2—4 км. Ошибка для артиллерийских радиолокационных станций не превышает 20—25 м на дистанции до 40 км (Сиверс и др., 1959). Как было показано выше, эхолокаторы летучих мышей являются системами ближнего действия, радиус которых в идеальных условиях не превышает 10—17 м. Кроме того, преследование и захват летающих насекомых с помощью эхолокации требуют от животных высокой точности в определении координат подвижной цели во время сближения. То же самое относится к ситуации, когда летучая мышь совершает посадку на какую-либо поверхность. Уже на основании этого можно полагать, что точность дальнометрии эхолокационных систем у летучих мышей должна составлять очень малую величину. Экспериментальные данные по этому вопросу появились лишь в самое последнее время. Симмонс (Simmons, 1968а, 1968b, 1971а, 1971b), применив методику одновременного предъявления двух совершенно идентичных фигур (треугольников), разнесенных на разное расстояние относительно места старта животного, показал, что выбор близлежащей фигуры на уровне 75% правильных ответов летучие мыши (Eptesicus fuscus, Phyllostomus hastatus) могли осуществлять при разнице расстояния между ними в среднем 1.3 см. Отрицательная фигура, т. е. та, посадка на которую не подкреплялась пищей, всегда находилась на расстоянии 30 или 60 см от животного. Положительная фигура в начале опытов располагалась на 10 см ближе к животному (т. е. на расстоянии 20
241
или 50 см ), а затем постепенно, через 1 см, удалялась, т. е. уменьшалась разница по расстоянию между дифференцировочными фигурами. Так как опыты Симмонса остаются единственными, опубликованными в литературе, представлялось необходимым проверить их и сопоставить с результатами, полученными на других видах летучих мышей, чьи локационные сигналы характеризуются другими параметрами.
Опыты были проведены нами на 4 больших подковоносах и 10 остроухих ночницах, которые перед началом экспериментов были энуклеированы. Животные находились на стартовой площадке. На расстоянии 1 м от нее
Рис. 81. Схема опыта (Z) и результаты распознавания летучими мышами двух мишеней, расположенных на различном расстоянии относительно стартового положения животного.
I: знак плюс — положительная мишень, расположенная на расстоянии 1 м от старта; минус — отрицательная мишень, отставленная на расстояние 1 м-4-AZ (AZ — переменная величина) от старта. II: по оси абсцисс — разница в расстоянии между мишенями (Аг) и соответствующая временная задержка отраженных сигналов от ближней и дальней мишеней (AZ); по оси ординат — положительные ответы, а — изменение процента положительных ответов у летучих мышей при сближении цилиндров; б — то же при уголковых отражателях. 1 — результаты для остроухих ночниц; 2 — для подковоносов.
помещалась положительная мишень, посадка на которую подкреплялась пищей. Вторая, точно такая же мишень первоначально была удалена на 15 см дальше по сравнению с первой от места старта, а затем постепенно разница между ними уменьшалась. В каждом опыте взаимное расположение мишеней относительно стартовой площадки менялось в случайном порядке. Угол между фигурами и животным составлял 13°. Подковоносы совершали посадку непосредственно на фигуру, остроухие ночницы — на площадку, установленную за каждой из них. Выбор мишени животным осуществлялся
242
с места, после чего следовала прямолинейная траектория полета к одной из мишеней.
В первой серии опытов в качестве дифференцировочных мишеней использовались два одинаковых цилиндра (4=10 см, d=l см), во второй серии по причинам, изложенным ниже, они были заменены на два уголковых отражателя, представляющих собой трехгранную призму с открытым основанием в виде равнобедренного треугольника, каждая из сторон которого равнялась 7.3 см (рис. 81). Критерием дифференцировки мишеней служила разница в расстоянии, так как различия по интенсивности или угловым координатам составляли ничтожно малые величины, заведомо лежащие ниже порогов чувствительности приемной системы эхолокатора животных. В одном опыте каждое животное совершало от 20 до 50 подлетов и посадок на мишени.
Первая серия опытов с использованием цилиндров была начата на 8 остроухих ночницах, однако до конца опытов дошло лишь одно животное, остальные погибли раньше. Из усредненных графиков на рис. 81 видно что животные не испытывали трудностей в дифференцировке раздражителей, пока расстояние между ними составляло от 15 до 6 см. Затем процент правильных ответов постепенно снижался по мере уменьшения различий по расстоянию между мишенями. Пределом для ночниц оказалась разница по расстоянию в 2.3 см (при уровне положительных реакций 75%). Для подковоносов предельный уровень дифференцировки был получен при различии расстояния между мишенями в 4—4.5 см, т. е. примерно в два раза больше (Айрапетьянц, Константинов, 1973).
Сравнивая результаты опытов Симмонса на Eptesicus fuscus и наших на ночницах, мы получаем расхождение в 1 см, хотя внутриимпульсная модуляция у ночниц значительно больше, чем у американского вида кожанов (85 против 25 кгц), и следовало ожидать у ночниц лучших результатов. Наша рабочая гипотеза сводилась к тому, что расхождение в результатах обусловлено различными отражательными свойствами мишеней, используемых в тех и других экспериментах. Поэтому были разработаны и изготовлены новые мишени в виде уголковых отражателей. С применением этих мишеней опыты были повторены на 2 остроухих ночницах и 3 больших подковоносах (рис. 81, б).
У обеих ночниц в этой серии процент правильных ответов даже при отставлении мишеней на 1 см продолжал оставаться высоким — 85. Лишь при разнице в расстояниях 0.5 см животные не могли достоверно установить, какая из мишеней к ним находится ближе, и уровень дифференцировки не превышал 60%, а при одинаковом их расположении от места старта падал до 50%. По графику находим, что 75% распознавания приходится на 0.8 см между мишенями. Для подковоносов замена мишеней не вызвала изменений их дискриминационных способностей в точности измерения дальности. Как в первой, так и во второй серии опытов предельная разница в расстоянии между мишенями для них составила 4.3 см при уровне 75% положительных ответов.
243
Чтобы лучше понять, почему произошло улучшение порогов дифференцирования мишеней во второй серии у ночниц и различия отсутствовали у подковоносов, обратимся к анализу отражательных свойств цилиндров и уголковых отражателей. На нашей установке ультразвукового моделирования показано, что интенсивность отраженного сигнала на расстоянии 60 см от уголкового отражателя уменьшается на 20 дб, от цилиндра — на 27 дб относительно интенсивности падающей на мишень ультразвуковой волны. Отсюда, зная интенсивность излученного летучей мышью сигнала и рассчитав по кривой ослабления ультразвука (рис. 80) интенсивность падающей на мишень волны на расстоянии 1 м, мы подсчитали величину отраженного от мишени сигнала в точке старта животного.
Для большого подковоноса при частоте излучаемого сигнала 81 кгц отраженный сигнал от уголкового отражателя составил величину 71 дб, для цилиндра — 64 дб. Для остроухой ночницы при средней частоте сигнала с максимальной амплитудой 60 кгц величина отраженного сигнала от уголкового отражателя в месте старта животного составила 25 дб, от цилиндра — 18 дб.
Следовательно, при использовании цилиндров для ночниц интенсивность отраженного сигнала находилась около величины пороговой чувствительности ее эхолокатора (15.5 дб, по данным наших расчетов, приведенных выше), и увеличение отраженного сигнала на 7 дб при введении уголкового отражателя значительно облегчило задачу селекции мишеней по дальности. С другой стороны, подковоносы и в том и в другом случае имели дело с отраженными сигналами, интенсивность которых намного превышала пороговую чувствительность их эхолокатора. Последняя, по данным Б. В. Соколова (1972), составляла для большого подковоноса 28 дб. Кроме того, при использовании уголкового отражателя отпадала необходимость точной юстировки мишеней относительно места старта животного, так как величина отраженного сигнала от нее мало зависела от ракурса облучения (6 дб на 30°) по сравнению с его значением для цилиндра (6 дб на 3°). Таким образом, предельную величину точности измерения дальности для эхолокационной системы ночниц мы должны оценивать по результатам второй серии опытов, тогда как для подковоносов это не имеет существенного значения. Одновременно с нами аналогичные опыты с большими подковоносами были выполнены Симмонсом (Simmons, 19716). При расстоянии положительной мишени 30 см от старта животного этот автор получил предел измерения дальности для подковоносов 3 см. Путем теоретических расчетов им было показано, что в данном случае подковонос для анализа расстояния между мишенями использует заключительную ЧМ-часть.
В наших опытах положительная мишень всегда располагалась относительно старта животного на расстоянии 1 м. Следо-244
йаТёЛьно, отраженный от нее сигнал поступал к животному через время
после начала излучения зондирующего сигнала. Отраженный сигнал от второй мишени, отстоящей при первой дифференцировке на 15 см дальше, соответственно приходил через 6.8 мсек. Разница в приеме отраженных сигналов в этом случае составляла 0.9 мсек., или 900 мксек. При сближении мишеней относительно места старта животного уменьшалась и разница во времени прихода отраженных сигналов от мишеней примерно на 60 мксек, через каждый сантиметр. Величины временных различий (Л£) в отраженных сигналах для каждого экспериментального значения разницы в расстоянии между мишенями (Аг) указаны на графиках (рис. 81).
Таким образом, предельное разрешение приемной системы эхолокаторов при оценке двух сигналов, разнесенных по времени, составило для остроухих ночниц 50 мксек., для Eptesicus fuscus — 70, для большого подковоноса — 240 мксек.
Экспериментальные доказательства способности слуховой системы животных воспринимать эхо-сигналы, следующие через столь короткие временные интервалы, были представлены Симмонсом в следующей серии модельных опытов (Simmons, 1968а, 1968b, 1971а, 1971b).
Перед животными, предварительно участвовавшими в опытах по дифференцировке треугольников, устанавливался микрофон, которым воспринимались зондирующие сигналы, излучаемые летучей мышью.
Через систему двух параллельных фильтров сигнал поступал на два электростатических излучателя и, преобразованный в эхо-сигнал, переиз-лучался обратно к животному. Излучатели находились на одном уровне на расстоянии 30 см от места старта, перед каждым из них размещалась посадочная площадка. «Эхо-сигнал» с одного излучателя приходил к летучей мыши через 1.1 мсек, после излучения ею зондирующего сигнала, а «эхо-сигнал» от второго излучателя через линию задержки прибывал с отставанием на 600—30 мксек., что соответствовало разнице во времени прихода отраженных сигналов от мишеней, разнесенных по расстоянию от 10 до 0.5 см. Ближнее «эхо» переключалось в случайном порядке слева направо и наоборот электронным переключателем. Летучая мышь выбирала тот излучатель, от которого «эхо-сигнал» приходил раньше. Постепенно временная задержка между «эхо-сигналами» уменьшалась и устанавливался предел, когда летучая мышь уже не могла определить разницу во времени прихода сигналов от излучателей.
Результаты этих модельных опытов с искусственным эхом полностью совпали с результатами, полученными в экспериментах с мишенями. Предел обнаружения временной задержки для Eptesicus составил 70 мксек., что точно соответствовало предельной разнице в расстоянии между мишенями — 1.3 см. Следовательно, приемная система эхолокаторов летучих мышей действительно
245
способна разрешать временные интервалы между двумя эхо-сигналами менее 100 мксек.
Электрофизиологические исследования, выполненные на различных уровнях слуховой системы летучих мышей, показали, что ответ на второй из двух предъявляемых моночастотных стимулов проявляется при минимальном интервале между ними в 200—400 мксек. (Grinnell, 1963b; Suga, 1964а, 1964b), но в большинстве случаев при интервале 1—2 мсек. Однако эти значения явно не соответствуют возможностям эхолокационной системы в целом, так как даже в лучшем случае разрешение по дальности должно составлять 4—6.5 см, но никак не 0.7—1.5 см, полученные в поведенческих опытах. Поэтому вопрос о нервных механизмах, обеспечивающих высокую точность и соответственно
Рис. 82. Экспериментальные и теоретические кривые распознавания двух мишеней, расположенных на различном расстоянии, летучими мышами Eptesicus f ascus (Л) и Rhinolophus ferrum-equinum (Б), (llo: Simmons, 1971b).
По оси абсцисс — разница .в расстоянии между мишенями; по оси ординат — положительные реакции. J — усредненные данные опытов; 2 — расчетные значения для случая идеального приемника с испшььзованием ЧМ-сигналов, аналогичных сигналам летучих мышей; 3 — расчетные значения для случая, когда в качестве информативного признака используется наложение моночастотной и ЧМ-частей импульса подковоносов.
^вьадкую разрешающую способность по дальности эхолокационных .систем летучих мышей, пока остается открытым. Однако результаты поведенческих опытов и частотно-модулированный характер зондирующего сигнала (или по крайней мере заключительной части его у подковоносов) позволяют высказать предположение об использовании летучими мышами при обработке эхо-сигналов дополнительных способов, близких к оптимальным, благодаря которым можно значительно повысить эффективность эхолокационной системы.
Симмонсом была рассчитана автокорреляционная функция зондирующего сигнала Eptesicus, Phyllostomus и большого подковоноса, Опй.ающая этой автокорреляционной функции использовалась далее .для предсказания предела дальности идеального детектора с параметрами сигналов этих видов летучих мышей. Как видно
246
из приведенных графиков (рис. 82), построенные теоретические кривые порога дальности хорошо совпадают с экспериментальными кривыми, что подтверждает высказываемую гипотезу.
О разрешающей способности эхолокатора по дальности
В радиолокации разрешающей способностью станции по дальности 3D называют минимальное расстояние по дальности между двумя целями с одинаковыми угловыми координатами, при котором еще возможно их раздельное наблюдение. Исходя из данных, полученных нами при установлении точности измерения расстояния, попытаемся оценить потенциальную разрешающую способность по дальности эхолокационных систем летучих мышей. При этом следует иметь в виду, что последняя не может быть хуже величины точности измерения дальности.
В обычных импульсных радиолокационных станциях (РЛС) потенциальная разрешающая способность (3Dn) определяется прежде всего длительностью зондирующего импульса (ти);
где С — скорость распространения излучающей энергии.
Если гладконосые летучие мыши, как полагали Хартридж (Hartridge, 1946) и Гриффин (Griffin, 1958), оценивают расстояние до предметов исключительно импульсным методом, т. е. по временной задержке между излучаемым и приходящим сигналами, то величина разрешающей способности, например, для остроухой ночницы при длительности локационных сигналов 0.6— 4 мсек, составит 10.2—68 см. Однако полученные данные значительно превышают экспериментальное значение — 8 мм, предельное для данного вида при измерении расстояния между мишенями (уголковых отражателей). Следовательно, этот путь не соответствует реальным возможностям эхолокаторов гладконосых летучих мышей.
Однако наличие в локационных сигналах этих животных внутриимпульсной частотной модуляции, изменяющейся во многих случаях по линейному закону, наводит на мысль о возможности использования ими методов оптимальной обработки ЧМ-сигналов путем сжатия их в приемной (слуховой) системе (Strother, 1961).
В радиолокационных системах, работающих на этом принципе, зондирующие импульсы представляют собой ЧМ-сигналы, а приемное устройство включает в себя схему задержки, эффект которой зависит от частоты заполнения сигнала. Передний высокочастотный фронт импульса, поступающего на оптимальный фильтр
^247
в приемнике, задерживается больше, чем его последующие части с более низкой частотой. В результате все частотные составляющие импульса смещаются, т. е. импульс сжимается во времени, а его амплитуда резко возрастает по сравнению с входным сигналом. Радиолокационная система, использующая сжатие импульса, позволяет увеличить дальность действия сигналов без увеличения мощности излучения, а кроме того, в значительной степени повысить разрешающую способность локатора и при этом выделять перекрывающие друг друга отраженные сигналы.
Степень сжатия импульса полностью определяется пределами изменения частоты в импульсе (девиацией частоты Л/), и длитель-ность сигнала на выходе фильтра равна
Коэффициент сжатия импульса характеризуется отношением длительности зондирующего импульса (ти) к длительности сжатого импульса (тис)
ЬРС
Потенциальная разрешающая способность локатора с внутриим-пульсной частотной модуляцией зондирующего сигнала и сжатием импульса при обработке в этом случае определяется уже длительностью сжатого импульса
2 — 2ЯС/
Существует ли в действительности в слуховом анализаторе летучих мышей подобный фильтр с дисперсией частоты, или используются другие корреляционные методы (например, взаимо-корреляционный метод), сказать пока трудно, так как нет прямых экспериментальных доказательств по этому вопросу.
Расчеты потенциальной разрешающей способности по дальности по приведенной формуле у исследованных нами гладконосых летучих мышей, а также у Eptesicus., для которого имеются данные о точности измерения расстояния, представлены в табл. 17.
Как видно из этих данных, разрешающая способность у разных видов неодинакова и изменяется от 1.7 до 6.8 мм в зависимости от величины девиации частоты. Лучшая разрешающая способность (меньше 3 мм) обнаруживается в эхолокационных системах многих ночниц, длиннокрыла и нетопырей. Наименьшее разрешение получено среди гладконосых для кожанов и ушана, но при расчете не учитывалось присутствие сильной 2-й гармоники, которая у некоторых видов играет, по-видимому, значительную роль в получении информации.
248
Таблица 17
Девиация частоты и расчетные значения коэффициента сжатия и потенциальной разрешающей способности по дальности эхо локационных систем у исследованных видов летучих мышей
вид Д/, КГЦ Kc 8D , n MM
Myotis nattereri 100 120 1.7
М. oxygnathus 85 153 2.0
М. myotis 70 119 2.4
М. emarginatus 70 77 2.4
Miniopterus schreibersii 70 154 2.4
Pipistrellus pipistrellus 60 150 2.8
Myotis mystacinus 57 67 3.0
Pipistrellus savii 56 84 3.0
Nyctalus noctula 50 110 3.4
Myotis daubentonii 47 61 3.6
M. dasycneme 36 40 4.7
Eptesicus ognevi 35 53 4.9
E. serotinus 35 60 4.9
Vespertilio murinus 30 60 5.7
Plecotus auritus 25 13 6.8
Eptesicus fuse us 25 38 6.8
Rhinolophus ferrum-equinum .... 15 30 11.3
Примечание. При расчете KQ для подковоноса была взята длительность только заключительной ЧМ-части зондирующего сигнала.
Потенциальная разрешающая способность эхолокатора остроухой ночницы составила 2 мм, т. е. величину одного порядка с экспериментальным значением измерения дальности (Аг—8 мм) и пределом разрешения регулярных решеток (Лй=2.9 4- 4.1 мм). Близкие соотношения указанных величин обнаруживаются и для Eptesicus (&Z>n=6.8 мм, Аг13 мм). То, что потенциальные возможности любой системы всегда превышают реальные, — не требует специального объяснения.
О важном значении частотной модуляции в сигналах гладконосых может свидетельствовать и серия наших экспериментов, описанная в главе 2. После перерезки верхних гортанных нервов частота в зондирующем сигнале ночницы снизилась до 14 кгц, исчезла частотная модуляция, но длительность сохранилась в прежних пределах. Тем не менее ночницы не могли осуществить точную посадку на мишень и полностью нарушилась дифференцировка фигур (уголковых отражателей) по расстоянию. Последнюю удалось восстановить лишь при отставлении отрицательной мишени на 40 см относительно положительной (в норме предел различения фигур осуществлялся животными при Аг=1 см). В этом случае точность измерения дальности соответствовала
249
значению потенциальной разрешающей способности локатора, как если бы летучая мышь применяла только импульсный метод локации. После восстановления частотных параметров в локационных сигналах у оперированных животных предел разрешения по дальности снова составил 0.8—1 см.
Возможно, и у подковоносов разрешающая способность по дальности определяется модуляцией частоты в заключительных участках локационных сигналов. Полученное значение одного порядка с пределом разрешения мишеней по расстоянию для большого подковоноса (Аг равно 4 см в наших опытах и 3 см в опытах Симмонса). Тот факт, что заключительный ЧМ-участок импульса подковоносов играет исключительно важную роль в эхолокационном анализе, подтверждается и другими экспериментами.
Разрешающая способность эхолокатора по угловым координатам
Важной характеристикой любого радара или сонара, в том числе и у животного, считается величина его разрешающей способности по углу. Под последней понимается разность азимутов (угловое разрешение в горизонтальной плоскости) и углов места (угловое разрешение в вертикальной плоскости) предельно близких друг к другу целей, находящихся на одинаковой дальности, при которых еще возможно раздельное измерение азимута и угла места каждой цели.
В наших опытах была предпринята попытка определить разрешающую способность эхолокатора большого подковоноса в горизонтальной плоскости с использованием условнорефлекторной методики с пищевым подкреплением (Айрапетьянц, Константинов, 1973; Горлинский, 1972).
Летучим мышам предъявлялись четыре одинаковых металлических цилиндра диаметром 10 мм и высотой 180 мм, расположенных попарно на концах планки длиной 70 см, удаленной от места старта животного на 150 см (рис. 83). За положительную фигуру были выбраны два цилиндра с постоянным расстоянием между осями 10 мм, что составило относительно стартового положения подковоноса заведомо не разрешаемый им угол, равный 24'. Дифференцировочной фигурой служила вторая пара цилиндров, расстояние между осями которых последовательно менялось в процессе эксперимента от 210 до 80 мм, что соответствовало уменьшению угла между ними и стартовой площадкой, от 7°50' до 2°58'. Планка устанавливалась на такой высоте, чтобы подкова летучих мышей и середина цилиндров находились в одной горизонтальной плоскости. Три подковоноса обучались выбору с места старта и прямолинейному подлету к положительной фигуре. Последняя путем поворота планки менялась местами с отрицательной мишенью в случайном порядке. Перед началом опытов животные были энуклеированы.
Эксперименты показали, что выработка условного рефлекса на положительную фигуру у летучих мышей происходила за 1 — 250
3 дня, а первая дифференцировка (при максимальном угле между цилиндрами отрицательной мишени и стартом) закреплялась лишь через 7—13 опытов. Усредненные данные всего последовательно проведенного ряда дифференцировок приведены на графике (рис. 83). Процент правильных реакций подковоносов зако-
А
Рис. 83. Схема опыта по определению разрешающей способности сонара по углу (Л) и экспериментальная кривая углового разрешения эхолокатора большого подковоноса (Б).
По оси абсцисс — угол между животным и цилиндрами отрицательной фигуры; по оси ординат — положительные ответы.
номерно снижается по мере уменьшения угла между цилиндрами отрицательной фигуры и стартовой площадкой. Это свидетельствует о решающей роли его величины в распознавании подопытными жи-
вотными предъявленных им мишеней. Поэтому тот угол между
«отрицательными» цилиндрами и местом старта подковоноса, при котором он * перестает отличать две вышеупомянутые мишени, можно считать за разрешающую способность его эхолокатора по азимуту.
Разрешающая угловая способность большого подковоноса при уровне положительных реакций 75% в горизонтальной плоскости оказалась 4° 30'.
Недавно американскими исследователями Пэффом и Симмонсом (Peff, Simmons, 1972) была предпринята попытка оценить локационные возможности эхолокатора других семейств летучих мышей (гладконосых и американских листоносов). Объектами их исследований служили два вида, использующие короткие ЧМ-сигналы, — Eptesicus fuscus и Phyllostomus hastatus.
На расстоянии 30 см от стартового положения энуклеированного животного в горизонтальной плоскости по отношению к его голове располагались три одинаковые нейлоновые сферы диаметром 19 мм, поддерживаемые длинными и тонкими металлическими стержнями (рис. 84), Одна из этих
251
сфер постоянно находилась прямо перед летучей мышью, а две другие были разнесены от нее относительно стартовой площадки на углы д и б. Если величина последнего в процессе эксперимента поддерживалась постоянной и была равна 19°, то значение угла а варьировало от 37 до 19°.
По оси абсцисс — угловое разрешение; по оси ординат — положительные ответы.
Летучие мыши обучались переползать либо на левую, либо на правую посадочную площадки, расположенные несколько ниже горизонтальной плоскости, в сторону предъявления большего угла (А), который служил в данных опытах положительным стимулом и появлялся то справа, то слева от животного в соответствии с таблицей случайных чисел.
Было показано, что при 75% правильных реакций предел дифференцировки приходится между 4 и 6° разницы углов а и б для Phyllostomus и между 6 и 8° — для Eptesicus (рис. 84). Таким образом, полученные величины количественно характеризуют способность эхолокатора данных животных оценивать разницу в горизонтальных углах между смежно расположенными сферическими мишенями. Необходимо отметить, что несколько лучшие результаты у Phyllostomus по сравнению с Eptesicus связываются авторами с более широким частотным диапазоном сигналов первых и более узкой диаграммой направленности излучения в сравнении с Eptesicus.
На наш взгляд, решающую роль в процессе углового разрешения целей, основанного на определении угловых координат каждой из них, играют механизмы и характеристики приемной системы эхолокатора летучих мышей. В связи с этим остановимся на гипотетических возможностях определения направле
252
ния на цель эхолокатора большого подковоноса. Большая длительность его зондирующего импульса и взаимосвязанные с ним движения ушных раковин (Griffin et al., 1962; Руе et al., 1962) позволяют предположить использование этими летучими мышами для определения угловых координат целей известного в радиолокации принципа «равносигнальной зоны». Оценочные расчеты с привлечением данных о направленности приема большого подковоноса на уровне VIII нерва (Стосман, Этманова, 1971) и дифференциальной чувствительности по интенсивности слуха летучих мышей (Grinnell, 1963а) показывают, что в этом случае данные животные могли бы различать углы прихода эха с точностью 2—3°. Это значение хорошо совпадает с экспериментально полученными результатами.
Что касается гладконосых летучих мышей, то у них в связи с использованием коротких ЧМ-импульсов главную роль в угловом разрешении, вероятно, играет бинауральное сравнение интенсивностей и спектральных характеристик отраженного сигнала.
12
ДИАПАЗОН И ПРИНЦИПЫ
ЭХОЛОКАЦИОННОГО РАСПОЗНАВАНИЯ ПРЕДМЕТОВ
Теперь уже ни у кого не вызывает сомнения тот факт, что эхолокация у летучих мышей выполняет те же функции, что и зрение у большинства млекопитающих. С ее помощью многие виды рукокрылых обнаруживают препятствия, оценивают расстояние до них и преследуют свою добычу — летающих насекомых. Однако зрение позволяет животным не только обнаружить какой-либо предмет или группу предметов, но и различать их между собой, выделять нужный, а другие оставлять без внимания. Дает ли такие возможности летучим мышам эхолокационная система? Могут ли они только по различию отраженных сигналов судить о качестве предметов, оценивать их величину и форму? Этот вопрос, помимо чисто научного интереса, имеет принципиальное значение для техники связи и особенно для радиолокационного опознавания предметов.
Первые шаги в решении данного вопроса были предприняты еще Дийкграфом (Dijkgraaf, 1957), который в экспериментах на ушанах показал, что интактные животные этого вида с близкого расстояния (10—15 см) дифференцировали на уровне 76% диск от креста, имеющих одинаковую площадь. В 1964 г. мы провели небольшую серию опытов с тем же видом, используя в качестве дифференцировочных фигур куб, цилиндр и пирамиду одинакового объема (Айрапетьянц, Константинов, 1965; Константинов, 1965в). Животные безошибочно производили правильный отбор подкрепляемой фигуры, но после перфорации барабанной перепонки даже одного уха дифференцировочные способности полностью исчезали.
Американские исследователи (Griffin et al., 1965) провели эксперименты по распознаванию на другом виде летучих мышей — Myotis lusifugus. Авторы в темноте катапультировали в зоне полета летучих мышей мучных червей и металлические или пластиковые диски соответствующих размеров и установили, что щц-
254
ЬоТныё после длительной тренировки Вылавливали 98% мучных червей и избегали контакта с дисками на 80%. Последующие специальные исследования отраженных сигналов от всех трех видов целей не смогли выявить существенных различий, благодаря которым летучие мыши производили столь четкую дифференцировку (Griffin, 1967).
Таким образом, в этих первых работах были получены начальные сведения о возможности летучих мышей дифференцировать предметы, но требовались дополнительные доказательства, что распознавание может осуществляться исключительно с помощью эхолокации, без участия других дистантных анализаторов. В случае положительного ответа необходимо было выяснить, какова эффективность эхолокационного распознавания (пределы и диапазон), одинакова ли она у представителей разных семейств летучих мышей, чьи локационные сигналы резко отличаются друг от друга, как работает эхолокационная система при распознавании и, наконец, какие признаки отраженного сигнала наиболее существенны для животного, т. е. на чем основаны признаки распознавания объектов. На некоторых из перечисленных вопросов мы и сосредоточим свое внимание при дальнейшем изучении этой большой проблемы.
Одновременно с нами возможности и диапазон эхолокационного распознавания, а также особенности излучения локационных сигналов в этом процессе изучаются и другими исследователями на представителях семейств гладконосых, американских листоносов и рыбоядных (Suthers, 1965; Bradbury, 1968, 1970; Simmons, 1968а, 1968b; Симкин, Патлякевич, 19696; Симкин, 1970а, 19706, 19716; Simmons, Vernon, 1971; Симкин, Макаревич, 1972).
Наши исследования проводились в сравнительном плане в основном на отечественных видах летучих мышей: остроухой ночнице и большом подковоносе. Кроме того, одна из серий опытов была поставлена на рыбоядных летучих мышах Noctilio leporinus.
Пределы эхолокационного распознавания мишеней, отличающихся по размерам
В опытах, описанных ниже, при изучении способности животных распознавать предъявляемые мишени по параметрам отраженного сигнала нами использовались методика пищевых условных рефлексов и специально дифференцировочное торможение.
Опыты начинались с выработки пищевого двигательного рефлекса на определенную фигуру, которая в дальнейшем служила положительным или тестовым раздражителем, и посадка животного именно на эту мишень сопровождалась пищевым подкреплением (мучными червями).
Процедура закрепления положительного рефлекса для каждого животного независимо от типа применяемых мишеней занимала от 10 до 15 дней.
255
После этого начиналась серия основных экспериментов по распознаванию с введением новой мишени, которая была отрицательным или дифферен-цировочным раздражителем и отличалась наиболее контрастно (по сравнению с последующими предъявляемыми фигурами) от первой, т. е. положительной. Посадка животного на дифференцировочную мишень считалась ошибкой и не подкреплялась пищей. В большинстве опытов мишени (положительная и одна или редко две отрицательные) предъявлялись летучим мышам одновременно, и лишь в редких случаях использовалось последовательное предъявление.
У ночниц дифференцировка обычно вырабатывалась на 7—8-й опытный день, в редких случаях — на 13—15-й день, тогда как у подковоносов эта задача требовала не менее 15—17 опытов. Дифференцировка считалась закрепленной, а распознавание достоверным при уровне правильных ответов животного не менее 70—75%. Чтобы исключить возможность зрительного анализа, все животные перед опытом или в ходе опытов были энуклеированы. У гашение рефлекса на место или пищевого двигательного стереотипа обеспечивалось многократным перемещением мишеней относительно друг друга в одной плоскости или предъявлением их в разных местах манежа. Участие обоняния в распознавании исключалось самой методикой эксперимента. Таким образом, анализ предъявляемых раздражителей мог осуществляться лишь средствами эхолокации, путем сравнения отраженных сигналов от каждой мишени.
Дифференцирование цилиндрических мишеней. Опыты этой серии были проведены на 4 остроухих ночницах, 8 больших подковоносах и 2 рыбоядных летучих мышах.1
Положительной мишенью для ночниц служил плексигласовый цилиндр высотой 100 мм и диаметром 10 мм. Дифференцировочными раздражителями к нему использовались такой же высоты цилиндры, но большего диаметра — от 25 до 12 мм, т. е. постепенно приближающиеся к размеру положительного цилиндра.
Подковоносам первоначально предъявлялись пары аналогичных цилиндров, т. е. 10—20 и 10—25 мм, но в дальнейшем по причинам, изложенным ниже, положительной мишенью был взят цилиндр с диаметром 25 мм, а дифференцировочными к нему — 4, 8, 10 и 12 мм. Ночницам представлялась возможность производить выбор мишеней при полете в большом павильоне, подковоносам — первоначально в таких же условиях, а затем с места старта, отнесенного от предъявляемых раздражителей на расстояние 2 м. Опыты с рыбоядными летучими мышами проводились в большом экспериментальном помещении, посредине которого размещался бассейн с водой (4.3 X 2X0.3 м). Над поверхностью воды на 10 мм выступали плексигласовые цилиндры (положительный — диаметром 10 мм и отрицательные — от 20 до 11 мм), которые служили объектами для распознавания. При правильном выборе животное опускало задние конечности около цилиндра и выхватывало кусочки мяса или рыбы; скрытые от него под водой. Следует отметить, что, по результатам опытов Сатериса (Suthers, 1965) и нашим собственным наблюдениям (Константинов, 1970), пищевые объекты, расположенные под поверхностью воды, никогда не вызывают реакций рыбоядных летучих мышей. Поэтому в данных опытах условным сигналом служил именно цилиндр, выступающий из воды.
1 Эксперименты с рыбоядными летучими мышами проводились на базе департамента анатомии и физиологии Индианского университета (штат Индиана, США) в 1969 г. Животные и необходимое оборудование были предоставлены доктором Р. Сатерисом (R. Suthers), которому авторы, пользуясь случаем, выражают свою благодарность.
256
Анализируя полученные данные (табл. 18), мы видим, что у ночниц достоверный уровень распознавания сохраняется при разнице в диаметрах цилиндров 5 мм, тогда как у рыбоядных летучих мышей — при разнице 2 мм. Более высокий результат у последних хорошо согласуется с теми сложными задачами, которые приходится решать этим животным в естественной обстановке при распознавании, например, плавника рыбы, выставленного над водой, от различных других малоразмерных надводных целей.
Таблица 18
Распознавание цилиндров, отличающихся по диаметру, ночницами и рыбоядными летучими мышами
Разница в диаметрах цилиндров, мм Ночницы Рыбоядные мыши Разница в интенсивности отраженных сигналов, дб
число посадок на мишень уровень распознавания, % число бросков на мишень уровень распознавания, 0/о
15 112 85.7 3.8±0.6
10 269 85.6 214 85.0 3.0+0.4
7 277 84.8 100 85.0 2.4+0.3
5 236 77.0 48 85.4 1.9+0.3
4 133 62.8 — — 1.6+0.3
3 153 61.4 67 80.6 1.4+0.3
2 70 52.5 40 75.0 1.1+0.3
1 — — 60 51.6 0.6+0.3
Параллельно с экспериментами на остроухих ночницах по той же методике были начаты опыты с подковоносами. Первую дифференцировку цилиндров, отличающихся на 15 мм в диаметре, вырабатывали у 5 животных. На протяжении 23—25 опытов, в каждом из которых отдельное животное совершало от 15 до 40 подлетов и посадок на мишени, уровень положительных ответов в среднем не превышал 50%. Лишь у одного подковоноса после 18-го опыта уровень распознавания превысил 70%; из 133 посадок на мишени, полученных в 5 последующих опытах, правильное решение было в 99 случаях (74.4%).
Тогда мы видоизменили методику. За положительную мишень был взят цилиндр большего диаметра (25 мм). Дифференцировка вырабатывалась у трех новых подковоносов, но результат сохранился прежним. Уровень положительных ответов не превышал 50—55%. Становилось очевидным, что различия предъявляемых мишеней недостаточны для успешного распознавания их подковоносами. Поэтому положительной мишенью был оставлен тот же цилиндр (диаметром 25 мм), в качестве отрицательной взят цилиндр диаметром 4 мм. Опыты были продолжены на тех же трех подковоносах.
257
Уже во втором опыте, после предъявления этой пары цилиндров, уровень положительных ответов у всех животных превысил 80% и не снижался в последующих опытах. Его величина сохранялась примерно такой же и после замены отрицательной мишени цилиндром диаметром 8 мм. Но, как только мы снова взяли отрицательный цилиндр диаметром 10 мм, уровень дифференцировки резко понизился, у двух подковоносов в среднем до 52.6%, у одного — до 71% в первом опыте. На следующий опытный день этот подковонос показал 75% дифференцировки данной пары цилиндров из 49 предъявлений. При разнице диаметров цилиндров 13 мм их распознавание данным животным было уже недостоверно — 60% из 60 предъявлений (табл. 19). Данные таблицы свидетельствуют о том, что для большинства экспериментальных животных дифференцировка цилиндров, отличающихся по диаметру на 15 мм, — неразрешимая задача, но для двух подковоносов эта разница была все-таки пределом их дифференцировоч-ных способностей.
Таблица 19
Распознавание цилиндров, различающихся по диаметру, большими подковоносами
Разница в диаметрах цилиндров, мм Разница в интенсивности отраженных сигналов в каждой паре мишеней, дб Число посадок на мишень Число животных Уровень распознавания, %
21 9.2±0.4 260 3 88.7
17 5.4+0.4 . 238 3 84
15 3.8+0.6 173 2 74.7
939 5 50.7
13 3.2+0.4 60 1 60
При распознавании мишеней данного вида эффективность эхолокационной системы подковоносов оказалась намного ниже по сравнению с таковой у представителей других семейств.
На установке ультразвукового моделирования мы провели облучение каждого из использованных в опытах цилиндров с целью сравнения интенсивностей отраженных сигналов от положительной и отрицательной мишеней (по максимальной амплитуде). Длительность зондирующих импульсов составляла 1 мсек., частотный диапазон — 30—120 кгц.
На основании полученных экспериментальных данных мы смогли оценить различия в интенсивности отраженных сигналов для каждой пары предъявляемых мишецей (табл. 18, 19). Сопоставляя их с результатами распознавания для каждого вида животных, отмечаем, что предел дифференцировочных способностей у рыбоядных летучих мышей проявился при разнице
258
в интенсивности отраженных сигналов положительной и отри-цательной мишеней примерно 1 дб, у ночниц — 2 дб, а у подковоносов — 4—5 дб. В данном случае цилиндры были мишенями со слабыми отражательными свойствами по сравнению, например, с металлическими шарами, поэтому вряд ли следует рассматривать полученные данные как абсолютную величину дифференциального порога по интенсивности слуховой системы для каждого вида летучих мышей.
Дифференцирование шаровых мишеней подковоносами. В качестве тестовых мишеней были взяты стальные шары разного диаметра, отраженные сигналы от которых в воздушной среде отличаются только по интенсивности и идентичны по другим показателям при условии их небольших размеров (Freedman, 1963). Зная интенсивность излучения локационных сигналов большого подковоноса, расстояние, на котором осуществлялось распознавание, а также диаметр шара, нетрудно рассчитать величину отраженного сигнала для каждого диаметра шара.
Для случая плоской волны и абсолютно твердой сферы величина звукового давления отраженного сигнала (Рогр) рассчитывается по формуле, выведенной из более общего выражения (Hickling, 1962):
Рнчяаа р изл / г отр 2г •'°0’
где Ризл — величина звукового давления излучаемого сигнала на шаре; а — радиус сферы; г — расстояние от точки излучения до центра шара; а — коэффициент затухания звука в воздухе; для частоты 81—82 кгц его величина составляет 0.7 1/м (табличное значение); — функция, зависящая от волнового числа (К) и радиуса шара, рассчитана на ЭВМ.
Опыты проводились по следующей схеме. Животным, находящимся на стартовой площадке, предъявлялась пара шаров, на один из которых предварительно был выработан дифференцировочный условный рефлекс. Расстояние между шарами составляло 55.5 см. Они устанавливались в 2 м от стартовой позиции. Положительной мишенью служил шар диаметром 51 мм. В качестве дифференцировочных мишеней использовались стальные шары меньшего диаметра, постепенно приближающиеся к положительному: 20, 25, 30, 35, 38, 40, 45 мм. Расположение положительного шара (справа или слева от старта) менялось по таблице случайных чисел. Критерием дифференцировки считалась прямолинейная траектория полета подковоноса от места старта к выбранному шару. За уровень распознавания принималось 75% положительных реакций. Опыты были поставлены на 4 взрослых больших подковоносах.
Пределом распознавания для двух подковоносов (№№ 2 и 3) были шары с разницей в диаметре 13 мм (51 и 38 мм). По-видимому, такой же предел мог быть установлен для подковоноса № 1, но при проведении с ним опытов мы не располагали шаром диамет
259
ром 38 мм. Ни один из них не смог надежно, т. е. на уровне 75%, дифференцировать шары с разницей в диаметрах 11 мм и меньше. Только для одного подковоноса, № 4, предел дискриминационных способностей его эхолокатора составила пара шаров 51 и 30 мм, с увеличением диаметра отрицательного шара дифференцировка пропадала (табл. 20).
Таблица 20
Распознавание стальных шаров разного диаметра большими подковоносами
Диаметр отрицательных шаров, мм —!— , Дб Ротр Подковонос № 1 Подковонос № 2 Подковонос № 3 Подковонос № 4
п % п °/о п % п %
20 -8.3 34 94.1 59 93.2 107 80.3 57 78
25 -6.1 30 93.3 — — — — — —
30 -4.4 31 90.3 32 87.5 28 78.6 52 78.8
35 -3.3 65 78.5 37 89.2 40 82.5 62 50
38 -2.4 20 80 37 75.7 — —
40 -1.9 70 64.3 145 66.2 68 55.8 — —
45 -1 55 52.7 — — — — — —
Примечание. р£тр — величина отраженного сигнала от отрицательного шара; Р0тр — величина отраженного сигнала от положительного шара (диаметром 51мм); D* 4*
1 отр J<x>ax —------------соотношение интенсивностей отраженных сигналов предъявляемой
Ротр /ооаг пары шаров; п — число подлетов; % — уровень положительных реакций.
Проведенный расчет интенсивностей отраженного сигнала от каждого предъявляемого шара показал, что даже от самого малого (диаметром 20 мм) шара величина эха в месте старта животного составляла 17 дб, что значительно превышает порог слуха подковоноса на локационной частоте, полученный в поведенческой методике. Возможность возражения, что животные не дифференцировали шары каждой пары, а реагировали на эхо большей интенсивности от шара 51 мм, снималась дополнительной серией экспериментов, когда положительной мишенью служил шар 30 мм, а отрицательной — шар 51 мм. В этом варианте опытов с 2 подковоносами уровень положительных реакций у каждого из них составил 90%.
Таким образом, дифференциальный порог по интенсивности слуховой системы больших подковоносов в локационном диапазоне частот на основании полученных данных варьирует у разных особей от 4.4 до 2.4 дб. Эти величины хорошо совпадают с результатами опытов по распознаванию цилиндров, в которых предельная разница по интенсивности в отраженных сигналах для больших подковоносов составила 3.8 ±0.6 дб.
260
Дифференцирование квадратных мишеней. В первых опытах этой серии, проведенных на ночницах (Константинов, 1968; Константинов, Ахмарова, 1968), в качестве положительной мишени использовался квадрат размером 12.5 X X 12.5 см. В дальнейшем мы повторили их, сравнивая возможности ночниц и подковоносов, но с использованием дюралюминиевой квадратной пластины размером 15X15 см. При сопоставлении квадратов каждой пары в том и другом случае оказалось, что они не перекрываются, а хорошо дополняют ранее полученные результаты (табл. 21).
Таблица 21
Распознавание летучими мышами квадратов, различающихся по размерам
Разница между положительной и отрицательной мишенями Ночницы Подковоносы
по длине стороны, мм по площади, % число посадок на мишень уровень распознавания, 0 /о число посадок на мишень уровень распознавания, %
125 75.1 216 93.0
92 66.8 93 90.0 — —
51 50.0 81 88.3 — —
38 36.1 182 85.4 95 90.5
23 25.0 68 92.9 168 80.4
17 23.1 89 80.9 — —
15 17.1 161 83.1 205 81.8
9 11.2 75 75.6 108 77.8
6 9.3 46 80.4 — —
5 6.4 83 54.2 67 45.8
Ночницы и подковоносы распознавали предъявляемые мишени примерно одинаково. Уровень положительных ответов постепенно уменьшался по мере сближения квадратных пластин по линейным размерам. Пределом распознавания для подковоносов и большинства остроухих ночниц были квадраты, отличающиеся по стороне на 9 мм, а по площади — на 11.2%, но одна остроухая ночница распознавала мишени, отличающиеся на 6 мм по стороне (по площади — на 9.3%). При разнице в 5 мм между сторонами мишеней или 6.2% по площади летучие мыши не могли сделать правильный выбор и уровень распознавания составлял около 50%.
Результаты наших опытов близки к данным Симмонса (Simmons, 1968а), который изучал пределы распознавания нескольких пар треугольников приближающихся размеров на Eptesicus fuscus. Летучие мыши этого вида на уровне 75% положительных ответов дифференцировали треугольники, отличающиеся по площади на 12-16%,
261
Как показали исследования на установке ультразвукового моделирования, отраженные сигналы от квадратов, различных по площади, проявляют ярко выраженные различия в форме огибающей при облучении этих фигур ЧМ-сигналами (Л/=60 кгц) под любыми ракурсами, отличными от нулевого. А поскольку летучие мыши при распознавании всегда подлетали к фигурам под углом, то они вполне могли осуществлять выбор, основываясь на различии в структуре отраженных сигналов. В то же время различия в интенсивности отраженных сигналов от любой пары квадратов составляли слишком малую величину (меньше 1 дб), и, следовательно, данный признак не мог служить летучим мышам критерием для эхолокационного анализа, как в случае с цилиндром или шарами.
Распознавание мишеней, отличающихся по форме и материалу
В первой серии экспериментов у трех ночниц был выработан условный рефлекс на металлический квадрат площадью 156 см2, а затем на одном уровне с ним, на расстоянии 60 см, поместили металлический треугольник такой же площади. К 9-му опыту все летучие мыши производили дифференцировку фигур в среднем на 92%. Отдельные животные показывали еще более высокие результаты. Так, остроухая ночница № 38 в 5 опытах совершила 70 посадок, из них только 1 раз на отрицательную фигуру, показав уровень дифференцировки 98.6%.
Замена треугольника другим отрицательным сигналом (кругом) не вызвала никаких нарушений у тех же летучих мышей, и дифференцировка квадрата и круга сохранилась на том же высоком уровне. Летучие мыши четко дифференцировали все три фигуры, предъявляемые одновременно. Так, остроухая ночница № 38 из 45 посадок только 6 раз ошиблась и показала уровень дифференцировки 86.6%, а ночница № 39 из 52 посадок ошиблась лишь 4 раза, и уровень дифференцировки составил 92.3%.
Эту серию экспериментов мы закончили последовательным предъявлением фигур. При предъявлении только одной положительной мишени животные с первого подлета садились на нее. В то же время, если предъявлялась одна из отрицательных, летучие мыши неоднократно подлетали к ней, но всякий раз отлетали в сторону. Следовательно, условное эхо от положительной мишени прочно удерживается в памяти животных.
В последующих экспериментах выяснение диапазона и возможностей эхолокационного распознавания у летучих мышей было продолжено (Константинов, 1968; Айрапетьянц, Константинов, 1970а).
262
Двум остроухим ночницам и трем большим подковоносам наряду с положительным квадратом № 1 площадью 196 см2 последовательно предъяви лялись отрицательные квадраты (№№ 2—7), стороны которых были изрезаны в разной степени (рис. 85). Кроме того, в качестве отрицательных ми-
Рис. 85. Дифференцировочные фигуры, используемые в экспериментах по распознаванию, и уровни дифференцировки их ночницами и подковоносами.
По оси абсцисс — номера отрицательных фигур; по оси ординат — уровень распознавания. Все фигуры одинаковой площади. Квадрат № 1 — положительная фигура, остальные — отрицательные. № 12 сделан из плексигласа, 13 — из фанеры. Сплошная линия — результаты для ночниц, штриховая — для подковоносов.
шеней использовались вогнутый квадрат (№ 8), выпуклый (№ 9) и квадрат с 2 отверстиями диаметром 2 см по диагонали (№ 10). Все эти фигуры имели одинаковую площадь и были сделаны из алюминия. Высота зубцов по сторонам отрицательных квадратов была следующая: у квадратов №№ 2 и 5 — 15 мм, у квадратов №№ 3 и 6 — 7, у квадратов №№ 4 и 7 — 3 мм.
Опыты по дифференцировке начинались с введения отрицательной фигуры № 2, которая, на наш взгляд, легче всего отличалась от положительного квадрата. После того как уровень распознавания этой пары превысил 80% положительных ответов, у всех животных была произведена двусторонняя энуклеация, которая не отразилась на результатах опытов. Затем отрицательная мишень заменялась следующей в той последовательности, как они изображены на рис. 85. Как у ночниц, так и у подковоносов на все предъявляемые дифференцировки удалось получить достоверное различение. Однако уровень положительных ответов почти во всех случаях у ночниц оказался выше (в среднем по всем фигурам 94.5%) по сравнению с подковоносами, у которых в среднем по всем фигурам он составил 79.9%. Интересно отметить, что уменьшение величины изрезанности кромок квадратов не повлияло на способность распознавания их летучими мышами. Это лишний раз свидетельствует о том, что края фигуры играют существенное значение при формировании огибающей отраженного сигнала и
263
их изменения воспринимаются приемной системой эхолокатора, вызывая адекватную реакцию животного. Таким образом, экспериментально было доказано, что эхо локационные системы летучих мышей обоих семейств оказываются способны распознавать предметы одинакового размера, но различной конфигурации.
Способность летучих мышей распознавать одинаковые по размеру и форме, но отличающиеся по материалам мишени определялась с использованием квадратных пластин (15 X15 см) толщиной 4 мм. В качестве положительной мишени был взят дюралюминиевый квадрат, модуль упругости Юнга (Е) которого составляет 65-4 4-75-104 кг/см2 (табличное значение). Отрицательные квадраты были изготовлены из материалов с различным значением модуля упругости Юнга (табл. 22).
Таблица 22
Распознавание мишеней, отличающихся по материалу
Отрицательная мишень Е, ХЮ< кг/см2 Ночницы Подковоносы
число посадок на мишень уровень распознавания, % число посадок на мишень уровень распознавания, %
Плексиглас 2.9-4.1 152 98.6 373 84.9
Фанера 6.5 45 86.6 — —
Текстолит 10 90 81.5 121 82.8
Латунь 100 40 50 53 47.8
Железо 160 66 50 — —
Ночницы и подковоносы, участвующие в этих опытах, предварительно использовались в предыдущей серии. Закрепление новой дифференцировки (алюминий—плексиглас) не потребовало специального обучения животных, и уже во втором опыте уровень распознавания составил 80—90%. Мишени предъявлялись по степени приближения величины модуля Юнга.
Как показывают полученные данные, летучие мыши с высокой вероятностью распознавали дюралюминиевый квадрат от плексигласового, фанерного и текстолитового, отличающихся по модулю Юнга в 10—30 раз. Не могли они различать только материалы, близкие по упругим свойствам (металлы), и уровень распознавания в этих случаях не превышал 50%. Нам не удалось выяснить, по каким признакам производилось распознавание мишеней, выполненных из разных материалов, так как на нашей установке отраженные сигналы от всех мишеней были идентичны как по максимальной интенсивности, так и по форме огибающей. Возможно, при ударе ультразвуковой волны о пластины последние резонировали в разной степени, создавая различную окраску отраженных
264
сигналов. Во всяком случае распознавание пластин разного материала (кроме металлических) не вызывало затруднений у животных.
В другой серии опытов подковоносам предъявлялись одинаковые по размеру шары (диаметром 51 мм), изготовленные из различных материалов. При первой дифференцировке положительной мишенью являлся стальной шар, а отрицательной — из органического стекла. Эксперименты проводились на животных, предварительно участвовавших в распознавании шаров разного диаметра.
Из 150 подлетов к мишеням уже тренированные подковоносы показывали уровень распознавания не более 60%. Аналогичные результаты были получены при использовании плексигласового (положительного) шара и алюминиевого (отрицательного). Из 427 подлетов в 13 опытах подковоносы лишь 247 раз выбрали плексигласовый шар и в 180 случаях — алюминиевый. Уровень распознавания составил 57.5%. Таким образом, шаровые мишени, изготовленные из разных материалов, летучие мыши не способны различить по отраженному сигналу, В чем кроется секрет распознавания пластин и отсутствие такового при предъявлении шаров, отличающихся по материалу, предстоит решить в будущих исследованиях.
Пределы распознавания регулярных решеток
Опыты с данным видом мишеней были проведены на ночницах и подковоносах (Константинов, Малашин и др., 1973).
Регулярные решетки представляли собой вертикально расположенные в одной плоскости металлические стержни высотой 150 и диаметром 3 мм. Стержни крепились в отверстиях горизонтальной планки длиной также 150 мм, так что вся фигура была выполнена в форме квадрата (рис. 86). Расстояние между стержнями в каждой отдельной мишени было одинаковым, однако число их (п) менялось от 2 до И, и, следовательно, промежутки между ними у разных решеток оказывались различными. Регулярные решетки явились наиболее удобным видом мишеней для расчета спектра отраженного сигнала и его изменений по мере уменьшения или увеличения составляющих ее элементов.
Для обоих видов опыты начинались с предъявления животным решетки, состоящей из 7 стержней, которая в дальнейшем стала положительной мишенью (п+). При первой дифференцировке использовалась пара решеток — п+=7 и п~=2. Затем число элементов отрицательной решетки (п~) постепенно увеличивалось и серия заканчивалась предъявлением животным пары решеток — п+ = 7 и п~=11.
Как видно из графиков (рис. 87), по мере уменьшения разности числа элементов дифференцируемой пары решеток уровень положительных реакций животных постепенно снижается. Для ночниц пределом распознавания оказались решетки, отличающиеся всего на один элемент (п+—7 от п~=6 и п~~8), тогда как у подковоносов процесс дифференцировки двух решеток даже при наибольшей разнице их составляющих проходил сложно. У живот
265
ных часто наблюдались срывы, уровень правильных ответов в течение многих опытов колебался около 70% и только к 14—15-му опыту впервые превысил 75%. В дальнейшем подковоносы с вероятностью более 80% распознавали положительную решетку с п+, равным 7 элементам, от отрицательных решеток с п~, равным 2, 3 и 11 элементам. Пределы их эхолокационных возможностей составили решетки с п~ = 5 и п”=10 элементам.
Анализ траекторий полета животных показал, что до расстояния 1 —1.5 мм от места расположения мишеней летучие мыши в основном совершали прямолинейный полет. Начиная с указанных
Рис. 86. Остроухая ночница при посадке на регулярную решетку.
расстояний животное делало разворот и летело к фигуре, принятой за положительную. Если считать, что к моменту разворота летучая мышь принимает окончательное решение, то распознавание решеток наиболее часто происходит в диапазоне углов подлета 30—80°, причем распределение углов в этом диапазоне близко к нормальному.
Определим разницу в длине шага (Afe) той пары решеток, которая была пределом распознавания для каждого вида летучих мышей,
ДЛ = | V~ V| sin 0СР’
где hn+ и V — расстояния между элементами решеток (шаг решетки) положительной (п+=7) и отрицательных (п~); 6ср— среднее значение ракурса подлета при распознавании (0ср = 55°);
1 1 7
j , * — размер мишени; п — количество элементов,
266
Для остроухих ночниц:
ДА == | hrj — hQ | sin 55° = 4.1 И
ДА = | h7 — h8 | sin 55° = 2.9
Путем аналогичных расчетов находим, что для подковоносов Д/&=10.3 мм при п+ 2> п и \h = = 6.8 мм при п+ < п~. Следовательно, эхолокационное разрешение у ночниц оказывается более чем в 2 раза лучшим, чем у подковоносов.
Учитывая различия в частотном заполнении зондирующих сигналов тех и других летучих мышей, можно отметить, что полученные величины в общем удовлетворяют известному в теории разрешения сигналов соотношению
С
Sf'
где Д/ — девиация частоты в сигнале, С — скорость звука.
мм в случае п+ > п~ мм в случае п+ < п~.
-2 О J Ч 6
Рис. 87. Результаты распознавания регулярных решеток ночницами (7) и подковоносами (2).
По оси ординат — положительные ответы.
Исследование параметров локационных сигналов летучих мышей при распознавании
В различных сериях опытов мы производили регистрацию локационных сигналов у подопытных животных с целью выяснения динамики их изменений на различных участках пути и определения расстояния, на котором летучие мыши производили распознавание мишеней. Основные характеристики режима работы локационных систем ночниц и подковоносов были получены при различении квадратной дюралюминиевой мишени от других фигур.
До выработки условного рефлекса на мишень, пролетая по манежу, летучие мыши обычно излучали одиночные импульсы. Период их следования у остроухих ночниц составлял 100—120 мсек., а длительность — 2—3 мсек.; у больших подковоносов — 70— 120 мсек, при длительности 40—60 мсек. В процессе закрепления рефлекса, посадки на положительную мишень и становления диф
267
ференцировки наблюдались изменения в режиме работы эхолокатора.
У остроухих ночниц после выработки положительного рефлекса на квадрат частота следования импульсов при приближении животного к мишени не менялась до тех пор, пока расстояние не сокращалось до 1—1.5 м. Здесь наблюдалось резкое падение межимпульсного интервала до 40—50 мсек., что характерно для животных во время совершения предпосадочного маневра. Одновременно
Рис. 88. Измерение интервалов между импульсами при дифференцировке летучими мышами различных мишеней.
Каждая точка соответствует излученному импульсу. Штриховой линией показано место размещения мишеней, стрелками — первое изменение частоты следования импульсов.
уменьшалась длительность зондирующих сигналов. Именно на этом участке происходило изменение траектории полета, когда животное разворачивалось к мишени.
При распознавании двух мишеней изменение частоты следования импульсов отмечалось значительно раньше, на расстоянии 2.9—2.5 м, причем происходило не только уменьшение межимпульсного интервала, но и группировка импульсов в серии, по 2—3 импульса в каждой. Переход на излучение серии импульсов при распознавании мишеней был также отмечен и другими авторами (Симкин, Патлякевич, 19696; Симкин, 19706). При приближе
268
нии животного к фигурам в среднем на расстояние 1.2 м (0.6— 1.7 м) наблюдалось второе учащение сигналов, частота следования импульсов составляла 40—50 в секунду и затем постепенно уменьшалась вплоть до посадки его на выбранную мишень. Наиболее типичные случаи изменения частоты следования импульсов остроухих ночниц при курсирующем полете в манеже, посадке на одну мишень и при распознавании мишеней различной формы и размеров представлены на рис. 88.
Рис. 89. Изменение основных параметров импульсов ночницы при распознавании квадратных пластин.
По оси абсцисс — расстояние до мишени. Остальные обозначения те же, что и на рис. 74.
Следует отметить, что одновременно с началом первого учащения сигналов наблюдалось постепенное увеличение длительности импульсов, которая к расстоянию в среднем 1.5 м до мишеней достигала максимальной величины — 4.5 мсек.
Кроме того, происходило расширение девиации частоты внутри импульса за счет повышения начальных частот до 120—130 кгц против 100—110 кгц в начале полета (рис. 89).
Сравнивая изменения в локационных сигналах с поведением животных, мы пришли к заключению, что первое учащение импульсов связано с обнаружением мишеней и началом их распознавания. Сразу же после него начиналось одновременное увеличение длительности импульсов и девиации частоты. После излучения ночницей нескольких импульсов эти величины достигали своих
269
максимальных значений и с 1.5 м до мишеней начинали уменьшаться. Этот мрмент можно принять за окончание распознавания.
Таким образом, весь этот процесс происходит на отрезке пути от 2.9 до 1.5 м до мишеней. После его завершения уменьшались величины всех параметров сигналов и начиналось учащение, связанное с посадкой животного на выбранную мишень.
У больших подковоносов при распознавании происходило резкое уменьшение периода следования сигналов — от 80 до 40— 20 мсек, и их длительности — от 40—50 до 10 мсек, на расстоянии примерно 3 м до мишеней (рис. 90). Моменты начала уменьшения
Рис. 90. Изменение периода (7) и длительности (2) импульсов у подковоносов при распознавании фигур.
По оси абсцисс — расстояние; по оси ординат — период между сигналами и длительность. Стрелка — момент выбора положительной мишени.
периода и длительности, как правило, совпадали. На смену одиночным сигналам приходило излучение импульсов, сгруппированных в пачки. Сразу после начала учащения импульсов пачки состояли из 2—3 импульсов, далее — из 4—8 и к концу подлета — из 16—24 в каждой. Интервалы между этими сериями сигналов составляли в среднем 40 мсек. Иначе говоря, происходило такое же учащение сигналов, как и у остроухих ночниц при распознавании мишеней. Но сокращение периода в большей степени зависело от уменьшения длительности импульсов, чем интервалов между ними, как в случае с остроухими ночницами. Частота заполнения в большей части импульсов оставалась равной 82 кгц и не менялась на различных этапах полета.
У рыбоядных летучих мышей были изучены локационные сигналы при полете над бассейном, без погружения задних конечностей (8 выпусков), и во время распознавания цилиндров с захватом пищевого подкрепления около положительного цилиндра (20 выпусков). Пролетая над бассейном, рыбоядная летучая мышь до выработки условного рефлекса на цилиндр обычно излучала сигналы, которые группировались в серии, по 2—4 импульса в каждой из них. Интервалы между импульсами в серии составляли 30—40 мсек., а между сериями — 80—100 мсек. Длительность импульсов варьировала в диапазоне 6—10 мсек. Все импульсы
270
были смешанного типа, т. е. в первой части сигнала частота заполнения быстро повышалась и сохранялась постоянной на уровне 58—60 кгц, а затем во второй его части падала на октаву до 30 кгц. Иногда встречались импульсы, у которых моночастотная часть занимала до 80% его длительности, а в конце частота падала до 40—45 кгц, т. е. величина девиации не достигала октавы. По-видимому, эти импульсы были выделены Р. Сатерисом (Suthers, 1965) в тип сигналов с постоянной частотой.
После закрепления рефлекса и выработки дифференцировки в эхолокационной активности животных наблюдались характерные изменения. После старта со стены экспериментальной комнаты рыбоядная летучая мышь резко пикировала вниз к поверхности воды бассейна. На этом участке (2.4 м) импульсы излучались сериями с интервалами 25—35 мсек., а интервал между сериями составлял 70—80 мсек. Обычно за время подлета к бассейну летучая мышь излучала 3 серии по 3—8 импульсов в каждой. Длительность их не превышала 6—7 мсек. Затем следовал участок полета над водой на расстоянии 15—20 см над ее поверхностью. Уже в самом начале длительность импульсов возрастала до 8—10 мсек. Иногда отмечались сигналы увеличенной длительности — до 14— 17 мсек., которые в других ситуациях никогда не были зарегистрированы. Как правило, летучая мышь излучала одну серию в 4— 5 импульсов с интервалами 30—40 мсек., затем следовал промежуток 50—60 мсек., и с расстояния в среднем 2.2—3 м от расположения мишеней начиналась характерная фаза приближения к выбранной цели. Графики, представленные на рис. 91, наглядно демонстрируют закономерное уменьшение частоты следования и длительности сигналов с этого момента и вплоть до захвата рыбы, помещенной под водой около положительного цилиндра. Интервалы между импульсами сокращались с 30 до 5 мсек., длительность — с 6— 7 до 1—0.8 мсек. Постепенно сигнал из смешанного превращался в частотно-модулированный за счет редукции передней моночастот-ной части. После захвата рыбы период между импульсами сразу увеличивался до 40—50 мсек., длительность — до 6 мсек., сигналы становились смешанного типа и излучение их осуществлялось сериями. Сравнивая характер локационных сигналов при атаке положительной и отрицательной (в случае ошибки) мишеней с поведенческими реакциями животных, мы приходим к заключению, что процесс распознавания происходит на участке 3—2 м до мишеней, по-видимому, с помощью 4—5 импульсов.
Последнее учащение сигналов и сокращение длительности их в точности соответствуют стадии приближения рыбоядной летучей мыши к уже обнаруженной добыче (Suthers, 1965), в нашем случае — к выбранному цилиндру.
Таким образом, в динамике изменения локационных сигналов у 3 исследованных представителей летучих мышей в ходе распознавания мишеней обнаруживается много общих черт, несмотря на
271
различие сигналов по частотно-временным характеристикам. Все летучие мыши во время обнаружения и опознавания мишеней группируют сигналы в серии, или пачки. При приближении к выбранной фигуре во всех случаях увеличивается частота следова-
6
।-----1-------1------1------1------1-----j_______J О
мб б 4 3 2 1 О
Рис. 91. Графики зависимости частоты следования импульсов (вверху) и их длительностей (внизу) у Noctilio от расстояния до мишеней при курсирующем полете над бассейном (4) и при распознавании цилиндров (Б).
По оси абсцисс — расстояние.
ния и сокращается длительность сигналов. Расстояние, на котором происходило распознавание мишеней, при учете размеров последних оказалось наибольшим для рыбоядной летучей мыши, так как не требуется даже проводить специальных измерений, чтобы показать, что цилиндр, выступающий на 10 мм из воды, дает отраженный сигнал во много раз слабее, чем, например, квадратная пластина 15x15 см. Это и неудивительно, если вспомнить, что макси
272
мальная интенсивность зондирующих сигналов, по нашим измерениям, у рыбоядной летучей мыши составила 1240 дин/см2, тогда как у остроухой ночницы — не более 100, а у большого подковоноса — 270 дин/см2.
В заключение хочется отметить, что каких-либо особых характерных изменений в сигналах изученных видов, присущих именно процессу распознавания мишеней, по сравнению, например, с обнаружением препятствий или ловлей добычи нами не обнаружено. Единственной особенностью можно признать увеличение длительности сигналов у ночниц и рыбоядных летучих мышей в момент опознавания мишеней.
Механизмы формирования отраженного сигнала
Теоретические представления о механизмах формирования отраженных сигналов с внутриимпульсной частотной модуляцией от различных предметов можно найти в специальных работах, посвященных этому вопросу (Уфимцев, 1962; Фридман, 1963). На их основе сделаны попытки представить возможности распознавания пластин и решеток гладконосыми летучими мышами (Малашин, 1970а, 19706; Константинов, Малашин и др., 1973).
Рассмотрим вид отраженного, например, от решетки ЧМ-сиг-нала, длительность которого значительно превышает протяженность решетки. Будцм считать, что отраженные сигналы от отдельных элементов ее различаются только временем задержки. При этом огибающую результирующего отраженного сигнала можно представить в виде
П, . А АГ
где sm и; А/—внутриимпульсная частотная модуляция
ти С
зондирующего сигнала; ти— длительность зондирующего импульса; t — время; п — число элементов решетки; h — шаг решетки.
Из выражения (1) следует, что огибающая отраженного от решетки сигнала имеет сложный многолепестковый характер, причем с увеличением внутриимпульсной модуляции и числа элементов решетки роль боковых лепестков становится незначительной и основная часть энергии отраженного сигнала сосредоточивается в «главных» лепестках. При достаточно большом числе элементов п отраженные сигналы от регулярных решеток с числом элементов, различающимся на единицу, будут сколь угодно мало различаться между собой. То же самое можно сказать о частоте модуляции огибающей отраженного сигнала. Разность частот модуля-
273
ции огибающих сигналов, отраженных от решеток, определяется в этом случае выражением вида
Д/ 2
Воо = — • -7г • | hY — h2 |. (2)
Ти
Можно показать, что при облучении квадратной пластины или решетки с п=2 под произвольным углом 0 плоским фронтом падающей волны результирующий отраженный сигнал состоит из
Решетка п-2,
Решетка п=5
Решетка п = 5
t ра$&.~ 0.1 мсек./см
Рис. 92. Механизм формирования отраженного сигнала от решетки с п=2 (а) и осциллограммы отраженных сигналов (б, в).
2 раздельных импульсов, равных по величине и по форме совпадающих с зондирующим сигналом. Отдельные отраженные от пластины импульсы формируются на краях пластины, находящихся соответственно вблизи и вдали от излучателя, т. е. на расстояниях 1\ и г2 от излучателя (мыши). При таком представлении каждый из этих объектов эквивалентен двум идентичным источникам отражения .
Если длительность зондирующего сигнала ти больше радиальной < 2Z
протяженности объекта распознавания , что и имеет место в на-
21
шем случае, то через время происходит наложение (интерференция) отдельных отраженных импульсов, в результате чего формируется сложный суммарный сигнал, который имеет три характерных участка (рис. 92, а, б). В начале и конце сигнала имеются 274
OLL
до и от ти ДО
2Z 2Z
^и + тг соответственно- На участке 0 — происходит постепенное О и
увеличение числа суммируемых импульсов вследствие постепенного увеличения облучаемой поверхности объекта по мере продвижения по ней зондирующего импульса.
О . 21
На участке ти — ти + число суммируемых импульсов к моменту , 21 времени ти-|-уменьшается до нуля.
Средний участок, соответствующий временному интервалу
21 к -
— ти, представляет собой сумму всех отдельных импульсов, действующих (интерферирующих) одновременно, и может быть назван квазистационарным (рис. 92, а).
При облучении объекта ЧМ-сигналом отдельные отраженные импульсы, формируемые на различных дальностях от точки приема (мыши), имеют при наложении различные частоты в соответствии с величиной девиации частоты, приходящейся на данную протяженность (расстояние между источниками отражения). Нетрудно показать, что в этом случае огибающая сигнала, отраженного, например, от пластины или решетки из тг=2, будет, как это следует из выражения (1), изменяться на квазистационарном участке по гармоническому закону:
Д/ I
cos — • тг t = cos wZ. (3)
ти °
При предъявлении квадратных пластин, отличающихся линейными размерами, огибающая отраженного сигнала на квазистационарном участке от каждой из них будет характеризоваться различным числом периодов модуляции. В этом случае число периодов модуляции (N) огибающей при данном угле облучения (9), как вытекает из формулы (3), определяется выражением:
2Z sin 0 / 21 sin 0 \
(4)
которое связывает N с параметрами зондирующего импульса: девиацией частоты (Д/), длительностью сигнала (ти) и линейным размером (Z) пластины. Например, при ти=1.6 мсек., Л/=65 кгц, Z=15 см и 0=60° число периодов модуляции составит т¥=26. При тех же параметрах импульса, но при длине стороны мишени 16 см N=21. В эксперименте пределы распознавания пластин для ночниц и подковоносов оказались близки. Объяснить это, на наш взгляд, можно тем, что надежность распознавания по данному признаку невелика и мало зависит от параметров зондирую-ших сигналов.
275
Для регулярной решетки, состоящей из п=3 элементов, огибающая отраженного сигнала на квазистационарном участке будет определяться тремя слагаемыми вида (3), так как в этом случае 3 отдельных импульса будут находиться друг с другом в состоянии интерференции, образуя 3 пары комбинаций. При этом одна пара импульсов интерферирует между собой (сигналы от крайних наиболее удаленных друг от друга стержней) с частотой <1^ =
Д/ I
— —- и две пары — с частотой, в два раза меньшей, т. е. (о2 = ти С
1 Д/ I — О)з-2 С ’
Таким образом, огибающая результирующего отраженного сигнала на квазистационарном участке будет дополнительно про-модулирована по амплитуде частотой, в два раза меньшей основной частоты в случае решетки п — 2.
Для регулярной решетки, состоящей из п=5 элементам, огибающая отраженного сигнала на квазистационарном участке будет определяться уже 10 слагаемыми вида (3) (в этом случае 5 импульсов интерферируют между собой), из них: одна пара —
Д/ I 3 Д/ I
с частотой 0^ = —• —? две пары — с частотой , три
С - 4 ти С
2 Д/ I
пары — с частотой а)3 —— . —. —, четыре пары — с частотой а)4— 4 ти С
_j_ 1
V С •
Рассуждая аналогично предыдущему, можно показать, что сигналы от предельно распознаваемых остроухими ночницами решеток (п+ 7 и п"=6) в основном различаются тем, что в сигнале, отраженном от решетки п+=7, дополнительно присутствует шесть пар, образовавшихся в результате интерференции импульсов 1 Д/ I с частотой
Ь 6 Ти С
Для проверки изложенных теоретических рассуждений были проведены специальные эксперименты на установке ультразвукового моделирования. На ней воспроизводился сигнал, подобный излучаемому ночницей, но гораздо большей мощности, и соответственно принимался более интенсивный отраженный сигнал. В качестве объектов облучения использовались квадратная пластина (15 X15 см) и решетки, предъявляемые летучим мышам для распознавания. Отраженные сигналы регистрировались на фотопленку при ракурсах облучения, под которыми обычно летучая мышь производила распознавание. В качестве примера на рисунке представлены сигналы, отраженные от квадратной пластины и решеток (п равно 2, 3 и 5 элементам) при ракурсе облучения 55°. Результаты моделирования полностью подтвердили теоретические расчеты формирования отраженных ЧМ-сигналов и наглядно продемонстрировали отличие амплитудной модуляции огибающей от каждой отдельной мишени. Из приведенных фотографий видно, что огибающие отра
276
женных сигналов от квадратной пластины и решетки из п=2 элементам почти идентичны, но значительно отличаются от решеток с большим количеством составляющих. Последние в свою очередь хорошо различаются между собой. Кроме того, форма огибающей на квазистационарном участке отраженного сигнала от мишени сохраняется стабильной независимо от ракурса (40—90°) облучения ЧМ-сигналом с девиацией порядка 60—80 кгц (рис. 92, в), что облегчает животному распознавание положительной мишени в широком диапазоне углов подлета.
Модельные эксперименты мы проводили только с короткими ЧМ-сигналами типа сигналов ночниц. Можно предполагать, что механизм формирования отраженных сигналов в случае подковоносов будет во многом сходен с уже описанным. Отличия в основном будут обусловлены частотно-временными характеристиками их локационных сигналов. В случае решеток с п > 2 сформированные отраженные сигналы сложной формы удобно характеризовать различием в их спектрах. Как показывают расчеты, различия в спектрах отраженных сигналов предельно распознаваемых решеток для ночниц и подковоносов оказываются одинаковыми (порядка 2 кгц). Разницу, полученную в экспериментальном предельном разрешении у ночниц (ДД=3^-4 мм) и подковоносов (АД ^7~Ю мм), в этом случае можно объяснить различием их локационных сигналов.
Однако маловероятно, чтобы в процессе распознавания летучие мыши производили количественную оценку спектров отраженных сигналов. Скорее всего в качестве критерия они используют наиболее простой способ — качественный анализ формы огибающей отраженного сигнала. При этом в процессе обучения животное запоминает характерный рисунок эха от положительной мишени и в дальнейшем сравнивает его с рисунком эха от других предъявляемых мишеней.
В тех случаях, когда форма огибающей отраженных сигналов от двух мишеней (например, квадратной пластины и решетки из п =2) одинакова, летучие мыши могут распознавать такие мишени, основываясь на разнице в интенсивности отраженных сигналов.
Это было нами показано в опытах на двух энуклеированных ночницах, используемых ранее при распознавании квадратных пластин из разного материала. Положительной мишенью оставался алюминиевый квадрат 15x15 см, а в качестве отрицательной — 2 металлических стержня диаметром 3 мм, высотой 15 см, которые были расположены на аналогичной подставке, что и квадрат, на расстоянии 15 см друг от друга.
Уже в первом опыте уровень распознавания составил у одной ночницы 80 %, а у другой — 89 % , а в последующих двух опытах процент положительных ответов не падал ниже 92. Вместе обе ночницы совершили 197 посадок на предъявляемые мишени, из них 183 — на положительную. Уровень распознавания составил 92.8%.
277
После того как мы убедились в способности летучих мышей распознавать квадрат от двух металлических стержней, представляющих собой как бы боковые грани квадрата, было решено еще больше уподобить отрицательную мишень положительной. Для этого боковые стержни вверху были соединены третьим, таким образом получился контур квадрата. Летучие мыши в первом же опыте отреагировали снижением уровня распознавания на предъявление новой отрицательной мишени. У одного животного распознавание составило 77.4% (31 предъявление), у другого — 66.6% (69 предъявлений). Следовательно, верхний соединительный стержень принимает участие в формировании отраженного сигнала.
Однако уже в следующем опыте летучие мыши показывали высокие результаты — 80—90%, которые оставались стабильными на протяжении последующих 5 опытов. Одна остроухая ночница совершила 144 посадки на фигуры, из них 125 — на положительную (86.7 %), другая из 206 посадок 198 — на положительную мишень (96.1 %), при этом в последних двух опытах она показала 100% распознавания из 86 посадок. Траектория полета мышей при подлете к мишеням была самой различной, но чаще, особенно в конце данной серии, летучие мыши летели перпендикулярно плоскости мишени либо чуть выше ее. Постепенно у обеих мышей выработалась совершенно определенная траектория на фигуру, стоящую справа от нее. Если в данный момент там стояла положительная фигура, животные садились на нее, если отрицательная — пролетали дальше, делали поворот и садились на положительную. Таким образом, животные специально приспосабливали свой полет так, чтобы разница в суммарной интенсивности отраженных сигналов от той и другой мишени была наибольшей.
Итак, результаты наших экспериментов показали, что эхолокационные системы летучих мышей отличаются высокой эффективностью при распознавании объектов различных классов. Комплексное изучение предельных возможностей распознавания у животных в опыте и анализ отраженных сигналов позволили дать количественную оценку дискриминационных способностей эхолокаторов летучих мышей и выдвинуть ряд предположений о возможных принципах их функционирования. Какие реальные принципы заложены в эхолокационных системах этих животных — остается вопросом для будущих исследований. Однако совершенно очевидно, что эхолокатор летучих мышей оставил далеко позади перспективные широкополосные радиолокационные станции, которые все еще остаются на стадии разработки.
13
ПОМЕХОЗАЩИЩЕННОСТЬ ЭХОЛОКАТОРОВ
Летучие мыши в своей повседневной жизни непрерывно сталкиваются с необходимостью выделения полезного сигнала на фоне различных акустических помех естественного и биологического происхождения, а также помех, причинно связанных с излученными сигналами (собственных зондирующих сигналов, отражений от препятствий, не являющихся в данный момент объектами локации). Нет сомнений в том, что слуховой анализатор этих животных приспособлен к извлечению нужной информации из многообразного комплекса различных звуков.
Механизм, используемый летучими мышами во время эхолокации—выделения полезного сигнала при наличии помех, — представляет непосредственный интерес не только для биологов, но и для инженеров, работающих над этой основной проблемой в теории обнаружения.
В первых предварительных исследованиях, проведенных на отдельных представителях гладконосых летучих мышей (ушанах, мелких видах ночниц), было показано, что искусственно создаваемый низкочастотный шум (до 20 кгц) не влияет на способность животных избегать препятствий и ловить мелких насекомых, тогда как шум ультразвукового диапазона частот оказывает отрицательное воздействие на ориентацию летучих мышей (Griffin, Grinnell, 1958; Griffin et al., 1960).
Специальные опыты с ушанами (род Plecotus) показали, что летучие мыши этого вида способны избегать несколько рядов вертикально натянутых проволок с высоким процентом уклонений, даже когда диаметр проволок не превышал 0.3 мм.Интенсивность эха, получаемого летучими мышами, измеренная на расстоянии 100 мм от проволок, колебалась от 35 до 9 дб соответственно для проволок диаметром 1.07 и 0.28 мм. Когда ввели шумовой фон в полосе частот от 15 до 55 кгц и интенсивностью 70 дб, процент
279
уклонений от препятствия понизился, но весьма незначительно. Одновременно с введением шума наблюдалось повышение интенсивности ориентационных сигналов летучих мышей. Отношение интенсивности сигнала к шуму при этом составляло величину порядка 35 дб, однако это не мешало летучим мышам обнаруживать препятствия.
Таким образом, в этом эксперименте не удалось заглушить существенным образом сигналы летучих мышей, что послужило основанием для детального и углубленного изучения механизмов помехозащищенности у этих животных с использованием электрофизиологических и специальных биоакустических методов.
Особенности приема ультразвука в присутствии шумов
Хорошо известно, что когда чистый тон или частот-но-модулированные импульсы поступают на ухо животного в присутствии широкополосного шума, то ответы нейронов слуховых центров уменьшаются или просто исчезают. Громкий шум вызывает более значительное уменьшение амплитуды вызванных потенциалов. Этот феномен получил название маскировки. Такой же в общем эффект наблюдается, когда шум присутствует в интервале между двумя сигналами. Несомненно маскировка связана с торможением, длительностью периодов восстановления й в некоторой степени с адаптацией.
Для человека установлено, что изменения реакций нейронов на подаваемый сигнал в соответствии с изменениями в интенсивности широкополосного шума определяются соотношением сигнал/шум как 1:1. Другими словами, если фоновый шум увеличивается до определенной интенсивности, то и сила сигнала должна быть увеличена до такой же величины для достижения порога. Однако у летучих мышей отношение изменения в интенсивности сигнала к изменению интенсивности шума, необходимое для достижения порога, оказалось ближе к 3 : 2, чем 1 : 1 (Grinnell, 1963b, 1967). Нервные механизмы, обеспечивающие лучший эффект маскировки летучим мышам по сравнению с таковыми у человека, исследовались в нашей лаборатории с применением электрофизиологических методов (Васильев, 1968а; Матюшкин, Васильев, 1968; Котеленко, Матюшкин, 1969).
Значения силы сигналов (7С) и силы шума (7Ш) оценивались в децибелах относительно 0.0002 дин/см2. В качестве показателя эффективности маскирующего действия шума использовалось отношение силы сигнала к порогу маскирующего действия шума. При оценке сигналов и шума эта величина преобразуется в разность /с — 7Ш (дб). Чем больше эта разность, тем эффективнее маскирующее действие шума.
280
В результате экспериментов выяснилось, что маскирующее действие белого шума существенно различается в зависимости от способа сочетания сигналов и шума. Оно относительно слабо выражено в тех случаях, когда сначала включаются сигналы, а затем подбирается порогово-маскирующий шум. Наоборот, в тех слу-
Рис. 94. Зависимость показателя /с—7Ш от силы сигналов (Л), усредненная частотно-пороговая кривая (Б) и зависимость показателя /с~/ш (дб) от частоты заполнения сигнала при 10—20 дб над порогами (В).
чаях, когда сначала включается шум, а затем подбираются пороговые сигналы («преодолевающие» шум), маскирующее действие шума выражено сильно.
Этот существенный факт демонстрируется на рис. 93, где показаны величины 1С—1Ш при разных силах шума для случая «предшествования сигнала» (Л) и случая «предшествования шума» (JS).
В литературе по физиологической акустике принято рассматривать маскирующее действие шума в отношении сигнала как результат возбуждения шумами некоторых элементов слухового пути
281
(и вследствие этого возникновение в них рефрактерности) (Бибиков, Сухорученко, 1966). Такой подход, по-видимому, не может обеспечить объяснение факторов, показанных на рис. 93, а также аналогичного факта, описанного Гриннелом (Grinnell, 1963с) и состоящего в том, что на фоне шума сигнал пороговой величины (характерный для этих условий) вызывает резко увеличенный ответ так, как будто бы действие шума «преодолевается» сигналом полностью. Все эти факты можно объяснить, если предположить, что ультразвуковой сигнал и шум выступают в качестве раздражителей, вызывающих различные настройки слуховой системы, причем обнаруживающие «конкурентные взаимоотношения».
В дополнительной серии опытов мы специально исследовали взаимные влияния коротких сигналов и шумовых посылок, следующих друг за другом в том или ином порядке и не перекрывающихся во времени. При этом также обнаружилось, что не только предшествующая шумовая посылка ухудшает прием (повышает порог) для последующего сигнала, но и предшествующий тональный сигнал ухудшает прием шумовой посылки как акустического раздражителя.
Вопрос о механизме избирательной «настройки» слуховой системы тем или иным действующим стимулом уже затрагивался нами и будет обсуждаться в дальнейшем. Заметим, что, каков бы ни был механизм этой настройки, значение ее вообще и для эхолокации летучих мышей в частности должно быть существенным.
В естественных условиях эхолокационные ультразвуковые сигналы — крики летучих мышей — очень сильны. Они практически всегда преобладают над «шумами» и должны настраивать слуховую систему мышей на прием соответствующих эхо, одновременно понижая ее чувствительность к шумам.
Стремясь приблизиться к этим условиям, мы в своих последующих опытах при исследовании различных подробностей маскировки ультразвуковых сигналов длительным «фоновым» шумом начинали с подачи сигналов.
На рис. 94, А видно, что величина 1С—7Ш в случае предшествования сигналов тем больше, чем сильнее шум (в случае предшествования шума эта зависимость обратная, а величины показателя 7С—относительно больше). При достижении силы шума около 90 дб его маскирующее действие резко нарастает. Те же результаты могут быть представлены в виде графика зависимости величины 1с—1т от силы сигналов.
При этом обнаруживается, что по мере усиления маскируемых сигналов показатель 1С—увеличивается, т. е. маскирующий шум оказывается относительно более слабым, помехозащищенность падает.
Эту важную зависимость можно определить так: выделение сильного сигнала на фоне сильного шума происходит хуже выделения слабого сигнала на фоне пропорционально ослабленного 282
шума. Причина этой зависимости, вероятно, в том, что при сильных ультразвуковых сигналах возбуждаются не только рецепторы (и связанные с ними нейронные цепочки), настроенные на данный тон, но и соседние с ними, что может вызывать явления латерального «торможения» (Katsuki, 1964; Suga, 1964а, 1965а). Вообще известно, что усиление ультразвукового сигнала за некоторый предел (даже без шума) может приводить к снижению реакций слуховой системы летучей мыши (Grinnell, 1963с; Васильев, 1968а; Котеленко, Матюшкин, 1969).
Величина /с — при одной и той же силе шума и относительной (к порогу) величине ультразвукового сигнала зависит и от высоты (частоты заполнения) этого сигнала (/). На рис. 94, В показана эта зависимость и приведена усредненная ЧПК подопытных летучих мышей. Минимальное значение величина Zc — /ш имеет при 70 кгц, дополнительный минимум — при 30 кгц; эти точки соответствуют минимуму на ЧПК, но только на последней основной минимум находится при 30 кгц, а дополнительный — при 70 кгц.
Таким образом, эти данные демонстрируют особенно высокую помехозащищенность «высокочастотной» части ультразвукового диапазона слуховой системы, соответствующего характеристикам эхолокационных сигналов летучих мышей данного вида.
Далее оказалось, что величина маскирующего действия шума существенно зависит от направления прихода сигнала и шума.
В наших опытах исследовались: а) зависимость величины /с—Zra от угла подачи сигнала в горизонтальной и вертикальной плоскостях (при фронтальной подаче шума — из положения 0°); б) зависимость величины /с—/ш от угла подачи шума (в горизонтальной плоскости) при фронтальной подаче сигнала. Использовались короткие (2—4 мсек.) тональные сигналы в диапазоне 20— 60 кгц при силах, превышающих порог этих сигналов на 10—30 дб. Посылки белого шума, как уже указывалось, были длительными, составляли «фон» для сигналов, но включались с постепенным увеличением их силы от 0 до «порогово-маскирующей» величины.
Из рис. 95 видно, что наименьшие (наилучшие) значения разности 1С—1т обнаруживаются под малыми углами с контрлатеральной для данного (правого) холма стороны и снизу, а наихудшие значения — под углами 30—60° с ипсилатеральной стороны. Это значит, что для наиболее успешной конкуренции с фронтально подаваемым шумом тональные сигналы должны приходить с наиболее «выгодных» направлений — под углом 30° контрлатерально или снизу.
«Ипсилатеральное» направление с углами, равными 30—60°, как следует из полученных данных, наименее выгодно, что, вероятно, связано с мощным тормозящим компонентом в действии ипсилатерального сигнала на исследуемый заднехолмовой центр.
283
В опытах второй серии, где сигнал имел фиксированное фронтальное (или контрлатеральное — под углом 30°) направление, а направления фонового шумового раздражителя изменялись, были получены данные, подобные первой серии.
Результаты двух из таких опытов показаны на рис. 95, где видно, что при подаче сигнала с наивыгоднейшего направления
Рис. 95. Чувствительность слуховых центров правого заднего холма летучей мыши к ультразвуковым сигналам и показатели /с—/ш при разных углах подачи сигналов и шума.
А — чувствительность к сигналам при действии обоих ушей (К4-И), только ипсилатерального (И) и только контр латерального уха (К). Заштрихованы участки, демонстрирующие взаимнотормозящие действия правого и левого входов системы, /=30 кгц; Б — схема опыта с двумя излучателями для сигналов и для шума; В и Г — показатели 1С— при разных углах подачи шума (сигналы — под углом 30°, 0°): В — /=30 кгц; Г— 7о=30 дб.
(30° контрлатерально) наименьшее (наилучшее) значение 1С—/ш получается в тех случаях, когда шум подается под углом 60—75— 90° с ипсилатеральной стороны, а также под углом 75—90° с контрлатеральной стороны. При этом величина 1С—1т может составлять около 50 дб против 10—20 в случае подачи сигнала и шума с одного направления.
284
В других опытах этой серии при подаче сигнала под углом, равным 0°, и вариациях направления шума обнаружилась в общем та же закономерность.
Полученные данные нашей лаборатории позволяют определить основные условия, облегчающие выделение ультразвуковых сигналов из белого шума для слуховой системы летучих мышей: а) предварительная «настройка» слуховой системы на прием соответствующего ультразвукового сигнала; б) оптимальная частота заполнения этого сигнала; в) достаточная сила сигнала; г) поворот головы животного под определенными углами к направлению прихода сигнала и шума.
Настройка системы на прием первоначально предъявленных сигналов, сходная в некоторых отношениях с сенсорной доминантой, по-видимому, объясняется следовыми процессами в слуховых центрах — следовым возбуждением в нейронных цепях, избирательно возбужденных сигналом, и следовым торможением в «конкурирующих» нейронных группах, в частности в избирательно реагирующих на белый шум.
Напомним, что у летучих мышей данного вида в задних холмах имеются как нейроны, реагирующие лишь на ультразвуки определенной высоты, так и «широкополосные» нейроны, способные «суммировать» компоненты белого шума (Матюшкин, Васильев, 1968; Васильев, Матюшкин, 1967).
В связи со сказанным выше можно полагать, что сила эхо локационных сигналов, регулируемая при эхолокации летучей мышью, выступает как фактор преодоления шума лишь при относительно слабых шумах. Если шум достигает большой силы (в наших условиях 90 дб), то усиление сигнала неэффективно (рис. 95, «вертикальный» ход кривых).
Это косвенно свидетельствует либо о достижении системой «потолка» реакции на сигнал, либо о присоединении к возбуждающему действию сигнала его тормозящего влияния (о чем уже упоминалось выше).
В этих условиях, вероятно, можно повысить помехозащищенность системы лишь одновременным усилением сигнала и соответствующим понижением чувствительности. Последнее может быть обеспечено усилением защитной функции сокращений внут-риушных мышц.
Значение высоты (частоты) сигналов для помехозащищенности системы отчасти объясняется частотной характеристикой ее входа, но отчасти, видимо, какими-то особенностями ее нейронных сетей, обеспечивающими «предпочтение» частотам локационного диапазона.
Что касается значения для помехозащищенности углов прихода сигнала и шума, то оно объясняется определенной направленностью приема акустически^ стимулов (конструкцией и располо-
285
жением слуховых проходов и ушей), а также бинауральным взаимодействием, имеющим особое значение в условиях предварительной настройки сигналами, приходящими более или менее «односторонне».
Оценка помехозащищенности эхолокатора
В радиолокации под помехозащищенностью станции понимается ее способность сохранять тактико-технические характеристики при воздействии различных помех. Воздействие помех проявляется в понижении отношения энергии сигнал/шум на входе приемного устройства. Вследствие этого цели обнаруживаются с заданной вероятностью на меньших расстояниях или вовсе не обнаруживаются.
Первая попытка оценить помехозащищенность эхолокационной системы летучей мыши (ушана) была предпринята Гриффином с соавторами (Griffin et al., 1963) в экспериментах по обнаружению барьеров из вертикальных проволок в присутствии широкополосного шума. По результатам опытов было подсчитано, что в момент обнаружения проволоки отношение энергии отраженного сигнала к мощности шума на один период составляло 5 дб. Учитывая характер подлета животных к препятствию, а также способность их слуховой системы дискриминировать сигнал и шум по направлению прихода и эффекты бинаурального взаимодействия на уровне слуховых центров, авторы приходят к выводу, что эффективность эхолокатора летучих мышей близка к «идеальному» приемнику в соответствии с теорией обнаружения.
Нами были проведены две серии опытов с остроухими ночницами и большими подковоносами по изучению влияния широкополосного шума на эффективность распознавания и обнаружения мишеней.
В первой серии опытов 3 ночницам, предварительно энуклеированным, в качестве раздражителей предъявлялись объемные фигуры — куб, конус и цилиндр (высотой 15 см, с диаметром основания у 2 последних 15 см). У всех животных были выработаны от положительной мишени — куба — 100%-е дифференцировки на конус и цилиндр при одновременном их предъявлении. После этого опыты были повторены в условиях акустического широкополосного шума (0—150 кгц), интенсивность которого в месте расположения мишеней менялась от 40 до 75 дб.
Уже при первом включении шума интенсивностью 40 дб в поведении животных наблюдались резкие изменения. Они стали летать только до середины манежа, а затем поворачивали обратно, как будто перед ними находилось невидимое препятствие. Постепенно из выпуска в выпуск животные все ближе подлетали к фигурам и наконец начинали садиться на них, при этом четко выделяя положительную мишень. Увеличение интенсивности шума до 75 дб не оказало влияния на эффективность распознавания,
286
Положительные реакции животных сохранялись на 100%-м уровне, т. е. в этих экспериментах нам не удалось «заглушить» летучих мышей. Однако выявились интересные закономерности в изменении параметров локационных сигналов, особенно их интенсивности и длительности. В условиях тишины на расстоянии 2 м от микрофона интенсивность излучения у ночниц составляла всего 80 дб. При введении шума (40 дб) интенсивность сигналов значительно увеличилась и продолжала расти по мере увеличения интенсивности шумового поля. Таким образом, летучая мышь пыталась как бы «перекричать» шум. Длительность сигналов также увеличилась с 1.5—2 мсек, при полете в тишине до 5—6 мсек, в условиях подаваемого шума. Кроме того, наблюдалась тенденция сдвига частоты заполнения в область более высоких ультразвуков.
Полученные в этой серии данные продемонстрировали высокую лабильность и широкие адаптивные возможности эхолокатора ночниц к акустическим помехам. 100 %-я дифференцировка предметов в условиях шумового поля обеспечивалась за счет увеличения энергии локационных сигналов. Исследования выявили также, что эхолокационный аппарат летучей мыши не обладает инерционностью, так как выключение шума немедленно приводило к возвращению параметров сигналов к исходному уровню.
Вторая серия опытов была проведена на 3 больших подковоносах и 2 остроухих ночницах. Выяснялось влияние шума различной полосы частот и спектральной плотности на эффективность эхолокационного обнаружения шаровой мишени, и на основании полученных результатов дана оценка помехозащищенности системы в целом (Константинов, 1973а; Константинов, Мовчан, Макаров, 1973; Макаров, 1973а).
Методика экспериментов была следующая. Животное с места старта должно было обнаружить стальной шар диаметром 21 мм, подвешенный на тонкой нити на расстоянии 106 см, и по прямой траектории подлететь к нему. После посадки на шар животное получало пищевое подкрепление (мучного червя) и возвращалось на исходное место, а мишень перемещалась в другое положение. Были две фиксированные точки для предъявления шара — справа и слева от старта под углом 45°, и появление мишени в каждой из них в ходе опытов было равновероятным. Обнаружение считалось надежным при уровне положительных ответов не ниже 75%. Использование сферы явилось единственной удачной возможностью обеспечить стабильное эхо независимо от ракурса облучения и позволяло произвести точный расчет его интенсивности.
После выработки и закрепления у животных рефлекса на шар включался шум, амплитудные и частотные параметры которого последовательно менялись. Использовались следующие виды акустических помех: 1) белый шум, ограниченный полосой частот 10—250, 10—50, 40—70, 35—85 кгц; 2) ЧМ-помеха с различной шириной спектра.
Комплекс аппаратуры для создания помех состоял из генераторов шума, полосовых фильтров, усилителей мощности и излучающей системы, состоящей из 24 конденсаторных излучателей. Излучатели были направлены на стартовую площадку, располагались на двух вертикальных рамах, по
287
ставленных под углом 60° (рис. 96), и были завешены айустическй полупрозрачной тканью. Такое расположение излучателей позволило создать шумовое поле с неравномерностью +1.5 дб (учитывая характеристики направленности приема животного) в объеме, ограниченном углами 80° по горизонтали и 60° по вертикали.
В случае применения помех первого вида использовались два генератора типа Г2-1, подключенные к усилителям мощности, нагруженным через аттенюатор на конденсаторные излучатели. Полоса частот шума регулировалась подключением полосовых фильтров. В случае создания помех второго вида использовался ЧМ-генератор, модуляция частоты которого осуществлялась синусоидой с периодом 1 мсек., а величина девиации и несущая оставались заданными. Использование ЧМ-помехи позволило провести исследование помехозащищенности летучих мышей в узкой полосе частот, перекрывающих отдельные части локационного сигнала.
Первоначальное включение шума с полосой частот 10—250 кгц и интенсивностью в месте старта 12 дб (относительно нулевого уровня 0.0002 дин/см2) не повлияло на эффективность обнаружения цели животными. При дальнейшем увеличении интенсив-
Рис. 96. Схема опыта и блок-схема используемой аппаратуры при исследовании помехозащищенности эхолокации.
Шумовой комплекс: 1,2 — генераторы шума, 3 — двухканальный усилитель мощности, 4 — набор фильтров, 5 — аттенюатор, L — излучатели. Комплекс регистрации: М — микрофон, 6 — микрофонный усилитель, 7 — ультразвуковой магнитофон, 8 — измеритель девиации частоты, 9 — двух лучевой катодный осциллограф, 10 — кинорегистратор.
S — старт, t — мишень.
ности шума обнаружение оставалось таким же надежным, что и в тишине (рис. 97). Лишь при шуме 48 дб были отмечены изменения в поведении летучих мышей, которые выражались в увеличении времени, необходимого для обнаружения, а также в уменьшении надежности обнаружения. У подковоносов при шуме 51 дб процент положительных ответов стал меньше — 75, а при интенсивности 60 дб мыши вообще отказались летать в сторону шара, стараясь вылететь из зоны действия шума. Для ночниц пороговая величина шума составила 66 дб.
После определения величины широкополосного шума, при ко
288
тором животные переставали обнаруживать шар, были использованы шумы в полосах 10—50, 40—70, 35—85, 90—180 кгц. Полосовые шумы 10—50 и 40—70 кгц при интенсивности 55 дб не оказали заметного влияния ни на результативность обнаружения шара, ни на поведение животных во время опыта. При этом спектральная плотность их на И дб превышала плотность предельного широкополосного шума.
С другой стороны, шум в полосе 35—85 кгц, так же как и широкополосный шум (10—250 кгц), полностью исключал возможность обнаружения животными шума 49 дб и спектральной плотности 2 дб. После проведения дополнительной серии опытов с полосой 10—250 кгц оказалось, что порог восприятия эхо-сигналов на фоне широкополосного шума понизился на 3 дб и принял то же значение спектральной плотности, что и для полосы шума 35—85 кгц (для уровня 75% —hl дб).
Рис. 97. Влияние широкополосного шума на обнаружение шара и изменение интенсивности локационного сигнала у подковоносов.
По оси ординат (слева) — уровень обнаружения. Рш — интенсивность шума, — спектральная плотность шума, Рс—интенсивность излучаемого сигнала (на расстоянии 10 см от головы животного). 1 — уровень обнаружения шара, 2 — изменение интенсивности моночастотной части сигнала, 3 — изменение интенсивности ЧМ-части сигнала. Стрелкой отмечен момент включения шума. Штриховая линия — уровень обнаружения спустя 15 дней.
Для остроухих ночниц использование шумов с полосами 10— 50, 40—70, 35—85 и 90—180 кгц не оказало влияния на эффективность обнаружения даже при превышении плотности шума на 11 дб над пороговой и лишь в случае шума полосой 20— 100 кгц со спектральной плотностью, равной пороговому значению плотности широкополосного шума (10—250 кгц), удалось исключить возможность обнаружения.
Уточняя частотные границы шума, влияющие на эффективность обнаружения шара летучими мышами, мы использовали узкополосные помехи. Первоначально исследовалось влияние помехи с полосой 70—85 кгц, захватывающей не только частотный диапазон локационного сигнала подковоноса, но и частоты, возникающие в отраженном сигнале в результате эффекта Допплера (83.5 кгц). Оказалось, что летучие мыши не могли надежно обнаружить тестовую фигуру (уровень обнаружения становился меньше 75%)
289
при той же интенсивности и спектральной плотности, что и при шуме полосой 35—85 кгц. Тогда мы попытались выявить наиболее информативную часть локационного сигнала для осуществления животными поставленной задачи.
В первой серии проверялась значимость моночастотной части и ее допплеровского приращения. В опытах использовалась помеха в полосе 80—85 кгц, увеличение интенсивности которой вызвало падение уровня обнаружения до 75% лишь при спектральной плотности помехи 5 дб.
Во второй серии исследовалась информативность ЧМ-части с использованием помехи в полосе частот 70—80 кгц, которая
4 Л
6-
4
2
0
~2к
Т ,1—L...I—I—I—।__|__L_j_I__I_till
70 75 80 85
Рис. 98. Влияние узких полос помехи на маскировку отраженного сигнала.
F — частотный диапазон помехи; — спектральная плотность шума. Отрезками обозначена ширина полосы шума при той величине спектральной плотности, которая вызывала падение уровня обнаружения до 75%.
Рис. 99. Осциллограмма одиночного импульса при максимальном значении шума.
Импульс пересекает линия, показывающая его частотное заполнение.
оказала на эффективность обнаружения шара те же действия и имела те же значения спектральной плотности, что и шум широкой полосы. Аналогичный результат получился и при помехе 70— 75 кгц. В то же время использование помехи 75—82 кгц^при спектральной плотности 0 дб не повлияло на эффективность обнаружения, а снижение уровня обнаружения до 75% произошло при спектральной плотности Ц-6 дб.
В связи с тем что мы не располагали возможностью генерировать помехи в полосе уже чем 5 кгц, для уточнения границ наиболее информативной части сигнала использовалась помеха 72— 82 кгц. Выяснилось, что эта полоса частот, так же как и полоса частот 70—75 кгц, исключает возможность надежного обнаруже
290
ния цели при спектральной плотности помехи больше 1 дб (рис. 98).
В ходе опытов регулярно производились запись излучаемых локационных сигналов на ультразвуковой магнитофон и киносъемка их с экрана осциллографа. Как показал анализ проведенных записей, у подковоносов поиск и обнаружение тестовой фигуры со старта осуществлялись посредством одиночных сигналов длительностью порядка 50 мсек. Частота заполнения большей части импульса сохранялась постоянной и равнялась 81—82 кгц. В начале и в конце импульсов на протяжении 2—3 мсек, отмечалось снижение частоты в среднем на 10 и 17 кгц соответственно. Амплитуда моночастотной части сигнала колебалась незначительно, однако в конце импульса, имеющем частотную девиацию, как правило, наблюдался ее всплеск (рис. 99).
Интенсивность моночастотной части излучаемых сигналов для расстояния 10 см от головы животного в опытах без шума составляла 121 дб, а при интенсивности шума, полностью маскирующего эхо от шара, увеличивалась, достигая 127 дб. Еще более заметные изменения интенсивности наблюдались на ЧМ-части, которая увеличивалась от 107 (в тишине) до 122 дб (в шуме), с максимумом амплитуды на частоте порядка 74 кгц.
В условиях нашего эксперимента ЧМ-часть сигнала, соответствующая частотам 72—75 кгц, оказалась наиболее информативной. О значимости этой части сигнала говорит также и более быстрое, чем в моночастотной части, увеличение интенсивности ее амплитудного пика при возрастании уровня шума. Кроме того, это подтверждают и результаты электрофизиологических исследований (Schuller et al., 1971; Schuller, 1972).
У остроухих ночниц интенсивность излучаемых сигналов также зависит от интенсивности шума и изменяется от 90 (в тишине) до 122 дб при пороговом значении шума 58 дб. Эта величина интенсивности сигнала при дальнейшем увеличении шума остается постоянной, т. е. становится пределом для ночниц. Все части ЧМ-сигнала ночницы оказались значимыми для обнаружения цели. В случае применения узкополосных шумов «заглушить» летучую мышь не удается даже при использовании максимальной интенсивности, которая на 10—15 дб превышала пороговое значение в случае широкополосного шума.
Полученный в результате наших экспериментов уровень шума, полностью маскирующий эхо-сигналы от шара, и знание информативной части сигнала дают возможность количественно оценить помехозащищенность локационного аппарата летучих мышей. Она рассчитывалась нами как отношение интенсивности наиболее информативной части отраженного сигнала 72—75 кгц для подковоносов и 30—120 кгц для ночниц к интенсивности шума в данной полосе частот (при уровне обнаружения 75%). Величина отра
291
женного от шара сигнала, пришедшего к летучей мыши, была получена путем расчета (Hickling, 1962) и составила Ротр= =0.005 н/м2=48 дб. Этот результат был подтвержден экспериментально на установке ультразвукового моделирования. Интенсивность шума в указанных полосах определялась из соотношения
= 7Ш + 101g(A/),
где 7ш — спектральная плотность шума; Л/ — ширина эффективной полосы шума, и составила 34 дб.
Таким образом, в случае диффузного белого шума помехозащищенность эхолокационной системы большого подковоноса определяется: 48—34=+14 дб, тогда как у ночниц она оказалась значительно выше, составляя —17 дб. Лучшее значение помехозащищенности локационной системы ночницы, на наш взгляд, обусловлено большим размахом частотной модуляции (Л/=90 кгц) при длительности сигналов 4—5 мсек, и использованием всего частотного спектра для эффективной локации.
14
СРАВНЕНИЕ ВОЗМОЖНОСТЕЙ ЭХОЛОКАЦИОННЫХ СИСТЕМ И ПРИНЦИПОВ ИХ ДЕЙСТВИЯ У ГЛАДКОНОСЫХ
И ПОДКОВОНОСЫХ ЛЕТУЧИХ МЫШЕЙ
Результаты экспериментальных исследований и проведенные расчеты, изложенные в предшествующих главах, позволяют произвести оценку эхолокаторов животных в бионическом аспекте в сопоставлении с техническими системами, выявить сходство и различие между крайними типами эхолокации у летучих мышей на примере остроухой ночницы и большого подковоноса, а кроме того, в свете новых данных рассмотреть выдвинутые в разные годы гипотезы о принципах действия каждого из из них.
Прежде всего следует отметить, что у летучих мышей эхолокация есть средство дистантного анализа ближнего действия. Предельный радиус обзора эхолокаторов составляет 13—17 м, а в реальных условиях, если учесть присутствие различных акустических помех и далеко не идеальные отражательные свойства объектов локации, дальность их дейстия не превышает всего нескольких метров. Максимальное расстояние, на котором ночницы впервые замечали барьер из проволок диаметром 4 мм, равнялось в наших опытах 4 м. По данным Мереса (Mohres, 1953), большие подковоносы обнаруживали открытую дверцу клетки размером 20x20 см с расстояния 6.4 м. Эти цифры являются теми предельными значениями дальности действия эхолокаторов летучих мышей, которые удалось установить экспериментальным путем.
Относительно дальности обнаружения насекомого вопрос оказался сложнее, чем считалось ранее. Выяснилось, что в данном, конкретном случае летучие мыши чаще запеленговывают свою добычу по ее жужжанию, т. е. средствами так называемой пассивной локации. Пеленгация может осуществляться на расстоянии 3—4 м, а затем на более близкой дистанции средствами эхолокации производятся сопровождение и перехват добычи. Максимальная дальность обнаружения насекомого по отраженному сигналу ока
293
залась для подковоноса 2.3 м, тогда как для ночниц она составила 1.4 м. Полученные различия можно^объяснить тем, что при использовании зондирующего сигнала большой длительности подковоносу легче получить отраженный от насекомого сигнал максимальной амплитуды, чем ночнице, излучающей короткие импульсы. В зависимости от положения крыльев насекомого короткий сигнал может отразиться от него в тот момент, когда объект дает минимальное отражение.
^Однако на близких расстояниях широкополосные сигналы ночницы обеспечивают ей лучшее разрешение насекомого, чем подковоносу, что и демонстрируется в наших опытах, когда ночницы ловили как подвижных, так и неподвижных насекомых, а подковоносы отлетали от последних, не проявляя пищевой реакции. Особый интерес представляют тактические приемы мелких видов подковоносов при ловле ночных бабочек, обладающих тимпанальными органами. В отличие от всех других исследованных видов эти летучие мыши, приближаясь к насекомому, вначале переходят на импульсы увеличенной длительности, а затем, определив координаты своей жертвы, прекращают излучение и скрытно, т. е. молча, не обнаруживая себя, сближаются с ней и успешно атакуют. Это один из ярких примеров взаимодействия между хищником и жертвой, выработанного в процессе эволюции функций.
Ограниченные пределы рабочего пространства эхолокационных систем вполне удовлетворяют тем задачам, которые приходится решать летучим мышам в их повседневной жизни, но вместе с этим их реализация требует автоматизма срабатывания, быстрой скорости реакций на каждый приходящий сигнал в короткие интервалы времени и оптимальной обработки получаемой информации. Поэтому неудивительно, что эхолокаторы всех летучих мышей в своей деятельности являются полифункциональными системами. Анализ элементов окружающего пространства на основе отраженного сигнала одновременно обрабатывается по нескольким критериям. Обнаружение объекта связано с измерением дальности до него и определением угловых координат. Непрерывно, вплоть до полного сближения (перехват насекомого или посадка на предмет), производится эхолокационное слежение за объектом. В случае появления нескольких целей в поле зрения эхолокатора обеспечиваются их селекция по дальности и качественное распознавание по различию в параметрах отраженного сигнала. При этом анализ может осуществляться по линиям сравнения спектральных характеристик отраженных сигналов, формы огибающей или суммарной интенсивности. Следует подчеркнуть, что в своей способности к тонкому дифференцированию предметов по форме, размеру и качеству материала эхолокаторы летучих мышей оставили далеко позади самые современные РЛС.
Изменяющийся характер излучения зондирующих сигналов и способность к адаптации в зависимости от условий окружающей
294
обстановки являются важной особенностью эхолокационных систем и коренным образом отличают их от систем технических. В последних период следования и параметры локационных сигналов рассчитываются заранее, определяя назначения РЛС, и остаются неизменными при любых ситуациях. Природные эхолокаторы в этом отношении характеризуются широкой пластичностью. По мере сближения с объектом у всех летучих мышей наблюдаются закономерное увеличение частоты следования импульсов, уменьшение их длительности и интенсивности. При этом подковоносы переходят на режим излучения пачек относительно коротких сигналов, разделенных между собой небольшими паузами. По-видимому, увеличение количества сигналов в единицу времени обеспечивает больший приток информации и становится необходимым условием для более точного определения пространственных координат цели и расстояния до нее. При очень высоком темпе излучения у многих видов сокращение длительности сигналов сопровождается снижением частоты заполнения, которое идет за счет постепенной редукции передней высокочастотной части импульса. При тонких дифференцировках мишеней у ночниц проявляются особые адаптационные свойства их эхолокационной системы. На определенном этапе полета животные, наоборот, увеличивают длительность сигналов и несколько расширяют внутриимпульсную девиацию частоты, вследствие чего появляется возможность получать большую информацию о характере распознаваемых мишеней.
Однако высокая лабильность и широкие адаптивные возможности эхолокационных систем летучих мышей выступают в опытах с применением акустических шумов, полностью или частично перекрывающих частотный диапазон локационных сигналов. В столь сложной акустической обстановке как ночницы, так и подковоносы не остаются пассивными, а моментально переходят на более высокий уровень функционирования аппаратов эхолокации. Резко возрастает интенсивность зондирующих импульсов, достигая при максимальном шуме своего предельного значения — 127 дб для подковоноса и 122 дб для ночниц. У последних, кроме того, наблюдаются значительное увеличение длительности сигналов и некоторая тенденция сдвига частоты заполнения в область более высоких ультразвуков. Выявленные механизмы увеличения энергии сигнала в совокупности с рядом приспособлений слуховой системы отстраиваться от влияний акустических помех создают необходимые предпосылки для функционирования эхолокаторов животных даже в экстремальных условиях.
Наряду с общими свойствами между эхолокаторами ночниц и подковоносов выявляются и существенные различия. По результатам опытов представляется возможным произвести некоторые количественные сопоставления пределов их функционирования.
Уже в экспериментах по распознаванию мишеней разного раз-, мера и формы ночницы превосходили подковоносов как по уровню
295и
точности дифференцирования, так и по пределам распознавания. Например, предельная разница в диаметрах цилиндров для ночниц составила 5 мм, для подковоносов — 15 мм; пределы распознавания регулярных решеток при разнице в шаге решетки (ДА) у ночниц оказались 3—4 мм, у подковоносов — 7—10 мм. Однако более наглядно преимущества эхолокационных систем ночниц перед подковоносами проявляются при сравнении их основных характеристик, которые приведены ниже.
Остроухая ночница Большой подковонос
Потенциальная дальность действия м Максимальная дальность обнаружения насекомого (Z>max), м . . . 17 13
1.4 2.3
Точность измерения дальности (Аг), мм 8 40
Потенциальная разрешающая способность по дальности (о/)п), мм 2 17
Величина девиации частоты в сигнале (А/), кгц 85 15
Порог восприятия отраженного сигнала (/отр_), дб 15.5 30
Пороговая чувствительность по мощ-ности (Рпор_), ВТ 1.5-10-13 2.4-10-10
Величина помехозащищенности, дб —17 +14
Пороговая чувствительность эхолокатора по мощности была вычислена по формуле Люр=Лтр/25 (S — площадь уха летучей мыши; коэффициент 2 учитывает одновременное действие двух ушей). Такие важные показатели, как точность измерения дальности и разрешающая способность, пороговая чувствительность и помехозащищенность у ночниц оказываются во много раз лучше, чем у подковоносов.
В чем же секрет феноменальных возможностей эхолокатора ночниц? Ответ на этот вопрос, по-видимому, заключен в использовании животными локационных сигналов сложной структуры. Уже из общих положений теории информации вытекает, что чем сложнее сигнал, тем большую информацию из него можно извлечь. У ночниц, как, впрочем, и у многих других гладконосых летучих мышей, импульсы характеризуются большим размахом частотной модуляции, величина которой находится в прямой связи с высокими показателями эффективности работы эхолокатора. Наличие ЧМ-сигналов в свою очередь наводит на мысль о возможности применения методов оптимальной обработки путем сжатия отраженных импульсов в мозговых центрах приемного аппарата эхолокатора. В качестве одного из возможных механизмов сжатия сигналов, реализуемого в эхолокаторе летучей мыши, первоначально предполагалась ультразвуковая линия задержки (жесткий фильтр) (Strother, 1961, 1967). Однако такой фильтр работает только для
296
стандартного сигнала и в случае его использования затруднял бы животному изменять параметры зондирующих импульсов. Поэтому к настоящему времени большинство исследователей, разрабатывающих проблему эхолокации летучих мышей, склонны рассматривать их слуховую систему в качестве оптимального согласованного фильтра, или коррелометра, предназначенного для восприятия именно своего излучаемого сигнала и его отражения (Cahlander, 1967; McCue, 1966, 1969; Aites, Titlebaum, 1970; Simmons, 1971a, 19716; Константинов, 1973a; Лившиц, 1973). Точность сравнения излученного и принятого сигналов обеспечивается тем, что в момент излучения слуховая система в ослабленном виде все-таки воспринимает излучаемый сигнал, который может сохраняться в мозговых центрах до прихода соответствующего отраженного сигнала. Таким образом, вся система становится настроенной на прием определенного эха. При прохождении широкополосного отраженного сигнала через такой согласованный фильтр, или, что то же самое, при взаимокорреляционной обработке его, происходит сжатие импульса, т. е. импульс укорачивается до такой длительности, которая соответствует ширине его спектра.
Метод взаимной корреляции более эффективен по сравнению с автокорреляционным методом, каковым является диспергирующая линия задержки, рассматриваемая Стротером (Strother, 1961). Благодаря взаимной корреляции оптимальное сжатие сигнала присходит автоматически, а кроме того, легко преодолеваются взаимные помехи, связанные, например, с одновременным излучением локационных импульсов многих летучих мышей в большой колонии.
В свете изложенных представлений следует обсудить вопрос о методах дальнометрии, используемых в эхолокации гладконосых летучих мышей. По этому поводу в литературе было высказано несколько гипотез, многие из которых представляют теперь лишь исторический интерес и не подтверждаются экспериментальными данными. Тем не менее будет целесообразно коротко рассмотреть каждую из них.
По представлениям Гриффина (Griffin, 1944, 1958) и Харт-риджа (Hartridge, 1946), измерение дальности у этих животных основано на истинном эхо-принципе, т. е. на оценке расстояния по времени между моментами подачи звука и возвращения эха. В терминах радиолокации этот метод называется импульсным. Интервал времени между моментом излучения зондирующего импульса и моментом приема отраженного сигнала называют временем запаздывания (£3), или временной задержкой. Так как при эхолокации звуковые колебания дважды проходят расстояние (г) между животным и объектом, то временная задержка определяется
__2г
— С 9
297
где С — скорость распространения звука в воздухе (340 м/сек.).
Короткая длительность зондирующих сигналов (1.5—2 мсек.) гладконосых летучих мышей и изменение частоты следования импульсов по мере сближения с объектом подтверждали справедливость выдвигаемой гипотезы. Однако сразу возникло и затруднение при объяснении способности животных локализовать близко расположенные предметы. В то время еще не было получено экспериментальных доказательств сокращения длительности локационных сигналов одновременно с их учащением при подлете к препятствию или насекомому. Считалось, что при обычной длительности импульса 2 мсек., при скорости звука 340 м/сек. (34 см/мсек.) уже на расстоянии 34 см возникнет «мертвая зона», т. е. эхо вернется к животному раньше, чем закончится излучение зондирующего сигнала. Пытаясь найти выход из этого затруднения, Гриффин предположил, что наложение излучаемого сигнала и эха на близких расстояниях от объекта может не препятствовать их разграничению ввиду разницы по частоте в ЧМ-сигналах. Однако, если предмет находится рядом, эти различия должны быть очень не-значите л ьными.
Одновременно возникли и новые гипотезы, авторы которых исходили из противоположных предпосылок (Руе, 1960, 1961а, 1961b; Kay, 1961а, 1961b, 1962). По их представлению, перекрытие во времени излучаемого и отраженного ЧМ-сигналов является непременным условием режима работы эхолокатора на близкой дистанции. Суть этой гипотезы сводилась к тому, что отраженный ЧМ-сигнал с начальной частотой /х принимается в тот момент, когда излучаемый импульс имеет уже другую частоту — /2. В результате наложения возникает нота или тон биения Д/= =fr—f2, который есть функция времени и, следовательно, дает непосредственную меру расстояния.
Несмотря на то что аналогичный принцип используется в радиолокации и на основе его разрабатывается ультразвуковой прибор для ориентации людей, потерявших зрение (Кау, 1967), по отношению к эхолокации гладконосых летучих мышей он вряд ли может быть применим.
Прежде всего при длительности ЧМ-сигналов не более 2— 4 мсек, перекрытие отраженного сигнала излучаемым произойдет лишь на расстояниях 34—68 см. Каким же тогда методом даль-нометрии должны пользоваться животные на бблыпих расстояниях, лежащих в пределах радиуса действия их эхолокаторов? Пай выбрал компромиссное решение, считая, что в этом случае у летучих мышей реализуется эхо-принцип, тогда как Кей рассматривал возможным осуществление перекрытия не столько в воздухе, сколько в мозговых центрах слухового анализатора, хотя такое объяснение представляется чрезвычайно искусственным.
Последующие эксперименты показали, что при сближении с объектом (например, с насекомым) летучие мыши не только уве
298
личивают темп излучения импульсов, но и сокращают их длительность. Таким образом, даже на близких расстояниях эхо воспринимается в чистом виде, без перекрытия его зондирующим сигналом. Возможности нервных центров слуховой системы воспринимать отраженные сигналы, следующие друг за другом даже в предельном ритме (150—200 имп./сек.), а также короткие длительности импульсов (1—0.2 мсек.) были доказаны в нейрофизиологических исследованиях (Grinnell, 1963b),
Еще одну гипотезу при попытке объяснить способы восприятия предметов окружающей среды летучими мышами в те же годы выдвигал Нордмак (Nordmark, 1960, 1961). Его гипотеза основана на результатах исследования слухового восприятия у человека, проведенного в 1955 г. Терлоу и Смоллом (Thurlow, Small, 1955). Эти авторы обнаружили, что при раздражении серией коротких импульсов с частотой 100—150 имп./сек. человек воспринимает тон 100—150 гц и не улавливает несущей частоты. При подаче таких сигналов парами воспринимается тон, высота которого зависит от интервала. Чем короче интервал в паре, тем высота тона выше. Рассматривая зондирующий импульс и отраженный сигнал как пару, разделенную определенным временным интервалом, Нордмак полагал, что у летучих мышей расстояние до объекта определяется высотой тона, соответствующего межимпульсному интервалу. При сокращении расстояния от 4 м до 17 см высота тона повысится соответственно от 42.4 до 1000 гц. Таким образом, Нордмак сводит механизм эхолокации у гладконосых летучих мышей к восприятию низкочастотных тонов, возникающих в результате изменения временного интервала между посылаемыми импульсами и возвращенными эхо. Восприятие высокочастотных отраженных сигналов им отвергается, равно как и частотная модуляция в пределах импульса, которую он считает случайным следствием при генерации звуков. По данным этого автора, высота тона сохраняется даже в том случае, если сигналы достаточно длинные и имеет место перекрытие во времени зондирующего и отраженного импульсов. Это обстоятельство использовано им для объяснения по тому же принципу механизма эхолокации у подковоносов.
Гипотеза Нордмака не получила ни одного экспериментального подтверждения и вряд ли может быть принята для объяснения принципов эхолокации летучих мышей. Все имеющиеся в настоящее время морфофизиологические исследования говорят за то, что слуховой анализатор животных специально приспособлен к восприятию ультразвуков, а на низких частотах пороги восприятия резко увеличиваются.
Приведенные гипотезы были выдвинуты в то время, когда знания об эхолокации летучих мышей ограничивались сравнительно небольшим объемом экспериментального материала. Сейчас становится очевидным, что животные во время эхолокации не только производят измерение временной задержки между излучен
299
ным и отраженным сигналами, но повышают эффективность даль-нометрии за счет внутриимпульсной обработки сигналов. Об этом свидетельствуют следующие данные: 1) значительное снижение порогов ответов в слуховых центрах и сокращение времени восстановительных циклов при использовании ЧМ-сигналов по сравнению с применением тональных импульсов; 2) более высокая разрешающая способность и точность измерения дальности, чем можно ожидать при анализе только временной задержки; 3) способность к тонкому эхолокационному распознаванию целей и, наконец, 4) исключительная помехозащищенность эхолокатора ночниц, полученная в наших опытах.
Все эти факты в совокупности позволяют оценить принцип дальнометрии гладконосых летучих мышей как импульсно-частотный, включающий в себя методы оптимальной обработки отраженных сигналов.
О принципах эхолокации подковоносов в литературе также высказывались различные мнения. Большая длительность зондирующего сигнала, постоянство частоты заполнения и обязательное перекрытие отраженного сигнала излучаемым прежде всего навели на мысль о возможности использования подковоносами изменения частоты отраженного сигнала по сравнению с излучаемым, возникающего при движении в результате эффекта Допплера (Holl-mann, 1952; Kleesattel, 1952; Kay, 1961а; Pye, 1961b; Schnitzler, 1968).
Если обозначить через v скорость животного относительно неподвижного объекта, то частота принятого отраженного сигнала изменится по сравнению с частотой излучаемого импульса на величину
где / — скорость посылаемого звука; С — скорость звука в воздухе; Д/ — допплеровский сдвиг частоты. Изменение частоты может быть со знаком плюс, если объекты сближаются, и со знаком минус, если расстояние между ними увеличивается.
При средней скорости полета большого подковоноса 5 м/сек. и частоте зондирующего сигнала 81 кгц Д/ составит 2.38 кгц. Следовательно, при сближении с неподвижным предметом подковонос будет получать отраженный сигнал частотой 83.38 кгц.
Подтверждением тому, что эхолокационный аппарат подковоносов действительно учитывает допплеровское приращение частоты, свидетельствуют результаты экспериментов. Во-первых, при исследовании частотно-пороговых характеристик слуховой системы обнаружено значительное понижение порогов (на 20— 30 дб) на частоте 83—83.5 кгц по сравнению с близлежащими частотами (Neuweiler, 1970). Это облегчает животному дискриминировать отраженный сигнал от излучаемого по частотному признаку.
300
Во-вторых, по данным Шнитцлера (Schnitzler, 1968, 1971, 1973а, 1973b), подтвержденным и в наших опытах (Константинов, Макаров, Соколов, Соколова, 1973), подковонос меняет частоту зондирующего сигнала в зависимости от того, находится он на месте или летит (рис. 100). У сидящего подковоноса частота средней части сигнала оказывается на 2—3 кгц выше, чем у летящего животного. Объяснить такое саморегулирование частотного заполнения сигналов можно только тем, что животное стремится в том и другом случае получать отраженный сигнал одной и той же частоты. При локации с места эффект Допплера равен нулю, и, чтобы привести в соответствие частоту отраженного сигнала к настройке слуховой системы, следует повысить частоту зондирующего сигнала, что подковонос и делает. В полете частота отраженного сигнала повышается за счет допплеровских приращений, и для сохранения ее величины в определенных пределах подковонос снижает частоту зондирующего сигнала пропорционально скорости своего полета (рис. 100).
Различия наших данных с результатами немецких исследователей за-
Рис. 100. Изменение частоты зондирующего сигнала у большого подковоноса в зависимости от скорости полета.
По оси абсцисс — скорость полета животного, по оси ординат —частота. Гизд: сплошная линия — величина ПЧ-части сигналов, полученных в опыте, штриховая линия — ожидаемая величина ПЧ-части в случае компенсации допплеровских сдвигов в принимаемых сигналах; Глр: сплошная линия — принимаемая частота, рассчитанная по данным опыта, штриховая линия — ожидаемая величина ПЧ-части принимаемых сигналов.
ключаются лишь в количественной оценке частоты импульсов подковоносов. Шнитцлер (Schnitzler, 1968) для сигналов сидящего подковоноса приводит среднюю величину 83.5 кгц, тогда как в наших опытах при тех же условиях частота составляла 81 — 82 кгц. Так как точность измерений в тех и других опытах не вызывает сомнений, получаемую разницу, по-видимому, следует отнести к популяционной изменчивости частоты локационных сигналов у подковоносов.
Учет подковоносами допплеровских приращений частоты отраженного сигнала во время полета может быть использован ими для более точного определения скорости сближения с лоцируемым объектом и получения сведений об угловых координатах цели» Преимущество допплеровского локатора над импульсным проявляется в том, что он обеспечивает лучшую селекцию по скорости,
301
например селекцию движущихся целей на фоне неподвижных. Данное свойство эхолокатора подковоносов наглядно демонстрируется в наших опытах с предъявлением насекомых. Животные четко реагировали на подвижных бабочек и не обращали внимания или теряли из виду насекомых, которые прекращали движение крыльями.
Положительным качеством допплеровского локатора является также отсутствие характерной для обычных импульсных локаторов ближней непросматриваемой зоны, определяемой длительностью зондирующего импульса.
При наличии стабильности излучения и весьма узкополосных фильтров в приемнике на допплеровскую частоту, по теории радиолокации, возможно обеспечить оптимальную обработку без какого-либо преобразования частоты или получения биений в приемной системе. Оба эти условия, как было сказано, соблюдаются в эхолокаторах подковоноса; следовательно, можно ожидать, что обработка информации в слуховом анализаторе происходит оптимальным образом без дополнительных преобразований частотного спектра.
Однако допплеровские локаторы наряду с преимуществами имеют определенные недостатки, среди которых наиболее существенный состоит в том, что такой локатор не позволяет определить дальность цели (Теоретические основы радиолокации, 1970). Для подковоносов же это не менее важно, чем и для летучих мышей, обладающих другим типом локации.
Большая длительность сигналов и обусловленное этим наложение (перекрытие) отраженного сигнала излучаемым послужили основанием для отрицания истинного эхо-принципа в эхолокации подковоносов. Мёрес (Mohres, 1953), который первый установил особый тип локации подковоносов, выдвинул предположение о ведущей роли интенсивности отраженного сигнала в определении направления и расстояния этими животными.
Как указывалось выше, подковоносы имеют довольно узкую диаграмму направленности излучения (54° в горизонтальной плоскости). Максимум интенсивности находится прямо перед головой животного, по перпендикуляру к плоскости подковы. Таким образом, объект, расположенный точно по направлению полета подковоноса, даст максимальное отражение, а незначительное изменение направления сразу приведет к снижению интенсивности эха при условии сокращения расстояния до объекта.
Иначе говоря, по гипотезе Мереса, акцент ставится на степени точности оценки направления. Путем фиксирования угловых координат объекта из двух разнесенных точек удается довольно точно определять дальность до него. Однако этот метод прямой угловой засечки, как его обозначают в технической литературе, применим при условии большой базы (расстояния) между приемниками. Например, чтобы отличить расстояние в 6 м от 3 м.
302
подковоносу, имеющему интерауральную базу ~2 см, требуется учесть изменение угла настройки ушей в 5' (89°49' и 89°54' соответственно). Такое угловое разрешение явно не соответствует возможностям подковоносов, так как, по данным наших опытов, точность локализации местоположения источника звука для них составляет 4° (Горлинский, 1972; Константинов, Стосман, Гор-линский, 1973). Еще труднее приложить эту гипотезу к реальным условиям ловли насекомых, когда интенсивность отраженного сигнала независимо от расстояния будет значительно меняться от положения крыльев бабочки (Roeder, 1963; Соколов, 1972) и, следовательно, определение дальности указанным методом становится невозможным.
Новые экспериментальные данные, полученные за последние 5 лет, заставляют существенно пересмотреть ранние представления о принципе дальнометрии эхолокатора подковоносов. К ним прежде всего относятся установление второго режима с переходом на излучение пачек более коротких импульсов по сравнению с одиночными сигналами; закономерное уменьшение периода следования и длительности сигналов при сближении с объектом; наличие восходящей частотной модуляции в начале одиночного и первого в пачке импульсов и обязательное присутствие нисходящей частотной модуляции в конце каждого импульса; выделение в слуховых центрах ЧМ-сигнала амплитудным пиком. Все эти особенности зондирующих сигналов и режима их излучения позволили предположить возможность применения подковоносами того же принципа определения дальности, что и гладконосыми летучими мышами (Schnitzler, 1968; Константинов, Соколов, 1969; Айрапетьянц, Константинов, 1970а). Заключительные ЧМ-участки импульсов подковоносов, к тому же выделенные повышением амплитуды, напоминают ЧМ-сигналы гладконосых и могут служить хорошей меткой для отсчета времени между излучением и приемом, что явится мерой расстояния до объекта. К тому же эхо заключительной части как раз укладывается в тот небольшой интервал между окончанием одного импульса и началом следующего, т. е. в 2-миллисекундную паузу, разделяющую импульсы в пачке, и воспринимается в чистом виде. Тогда становится понятным, почему подковонос, приближаясь к препятствию или насекомому, увеличивает частоту следования импульсов и сокращает их длительность. Этим обеспечивается возможность получить большее количество ЧМ-меток в единицу времени и точнее определить расстояние до предмета.
В пользу выдвигаемой гипотезы говорят также следующие факты. ЧПК слуха подковоноса, снятая по суммарным электрическим ответам от заднего холма, показывает заметное снижение порогов ответов не только в области допплеровской частоты (83—83.5 кгц), но и в диапазоне частот 70—75 кгц (Neuweiler, 1970; Стосман? Константинов? 1972). Именно эти частоты входят
303
в состав заключительной ЧМ-части сигнала подковоноса. Одновременно в опытах с использованием узкополосных акустических шумов нами было установлено, что рассматриваемая часть сигнала является наиболее информативной для обнаружения неподвижной мишени сидящим подковоносом. И наконец, сравнение заднехолмовых off-ответов на стимулы длительностью 30 мсек, при моночастотном заполнении и с понижением частоты в конце на 16 кгц показало, что в последнем случае амплитуда off-ответов значительно возрастает (Schuller et al., 1971; Schuller, 1972). Это указывает на то, что ЧМ-часть сигнала хорошо выделяется слуховой системой подковоносов и играет важную роль в получении информации об окружающих предметах в процессе эхолокации. Возможно, благодаря оптимальной обработке ЧМ-части сигнала эхолокаторы подковоносов производят распознавание неподвижных предметов и обладают достаточно высокой точностью измерения дальности и разрешающей способностью. Однако, в силу того что величина девиации частоты в ней невелика по сравнению с импульсами гладконосых, локационные системы подковоносов по основным показателям значительно уступают эхолокаторам гладконосых. В то же время по отношению к техническим системам эхолокатор подковоноса представляет интересную комбинацию допплеровского и импульсного локатора, не имеющего себе равных среди современных РЛС.
Итак, рассмотренные принципы функционирования эхолокационных систем гладконосых и подковоносых летучих мышей позволяют прийти к заключению, что задача определения расстояния — дальнометрия по методам реализации отраженных сигналов — оказывается у них весьма сходной, несмотря на резкие различия в структуре зондирующих импульсов. Несомненно, что только сравнительный подход к изучению эхолокаторов этих животных явится наиболее перспективным для технического воплощения заложенных в них принципов в плане задач, выдвигаемых бионикой.
15
АКУСТИЧЕСКАЯ ЛОКАЦИЯ
У ПЕЩЕРНЫХ ПТИЦ И КРЫЛАНОВ
Открытие эхолокации у летучих мышей естественно поставило вопрос о возможном использовании такого же способа ориентации другими животными.
Первоначально наиболее реальными кандидатами в список эхолоцирующих животных считались многочисленные представители ночных птиц из отряда сов (Strigiformes). До сих пор, главным образом в научно-популярной литературе, проскальзывают сообщения о том, что совы якобы обладают эхолокаторами. Однако это явная ошибка. Полет сов совершенно бесшумный, и они не излучают звуков, которые хотя бы отдаленно напоминали локационные сигналы. Доказано, что совы во время охоты за грызунами используют так называемую пассивную локацию, т. е. обнаруживают свою жертву на слух. Точность пассивной локации сов высока и приближается к точности эхолокатора летучих мышей (Payne, 1962, 1965, 1971; Ильичев, 1970, 1972). Это обстоятельство было важной причиной того, что пространственный слух совы привлек внимание не только биологов, но также инженеров, акустиков, физиологов и отчасти математиков. Точный пространственный слух для сов крайне важен в условиях охоты за бегающими зверьками в сложной акустической обстановке природных биотопов. Полевые наблюдения показывают, что во время поискового полета ушастая сова летит медленно и бесшумно, слабо взмахивая крыльями, невысоко над землей (1—2 м). Ее лицевой диск при этом обращен вниз и вперед, голова совершает круговые движения. Замечено также, что в темные ночи охотящаяся сова бросается на шорох любого происхождения — звук сломанной веточки, шуршание листвы и т. д.
Используя дрессированных сипух, приученных отыскивать спрятанный в большом помещении источник звука, Пэйн (Payne, 1965) с помощью киносъемки в инфракрасных лучах установил,
305
что точность пассивного локатора этой совы достигает 1°. Примерно такая же точность пассивной локации обнаружена для других сов (ушастой совы, серой неясыти).
Частотный диапазон слухового восприятия сов определялся с помощью методики снятия микрофонных потенциалов улитки и условнорефлекторных реакций (Schwartzkopff, 1955). У ушастой совы (условнорефлекторные опыты) нижний порог слуха составил менее 0.1 кгц, верхний порог — 18, область оптимального слуха — около 6 кгц. У серой неясыти верхний порог достигал 21 кгц (микрофонные потенциалы). Область оптимального слуха составила 3—6 кгц.
Сопоставляя эти данные с характером звуков мышевидных грызунов, чаще всего наблюдаемых у них в естественной обстановке, обнаруживаем, что основные частоты некоторых сигналов колеблются от 3.7 до 5 кгц: у Microtus — от 1.3 до 1.5, у Рего-myscus — от 5.2 до 10.2 кгц (Houseknecht, 1968), совпадая с зоной максимальной слуховой чувствительности сов.
Ильичев (1970) отмечает, что особенности и степень развития локации сов четко скоррелированы с их образом жизни. Экология хорошо лоцирующих видов характеризуется, во-первых, мышеядным типом питания, во-вторых, ночным образом жизни, в-третьих, обитанием в труднопросматриваемых биотопах. Таким образом, общим фактором интенсификации слуха у сов является ограничение возможностей зрения, которое у многих дневных птиц играет ведущую роль в ориентации. Для добывания пищи в акустически неблагоприятных условиях (среди травы, кустов и т. д.) и для сохранения маскировки, по-видимому, было наиболее целесообразно вообще не издавать никаких сигналов, а ограничиться средствами пассивной локации.
Внимание исследователей привлекли и другие птицы, гнездящиеся в глубине пещер, где функции их зрительного анализатора бывают также ограничены. Уже первые наблюдения показали, что при полете в темноте они используют активную акустическую локацию. Автором этого открытия был Гриффин, который в 1953 г. провел серию наблюдений и экспериментов с птицами из отряда козодоеобразных — Caprimulgiformes — гуа-харо (Steatornis caripensis) в естественных условиях их обитания и в лабораторной обстановке (Griffin, 1953а).
«Жирная птица из Карипе»
В 1799 г. известный немецкий натуралист Александр фон Гумбольдт во время своего путешествия по Южной Америке посетил в северо-восточной Венесуэле близ города Карипе большую пещеру, которую местные жители прозвали «жировым рудником». В ней индейцы с незапамятных времен собирали «жировой урожай», иначе говоря, ловили невероятно 306
жирных птенцов птиц, во множестве обитающих в этой пещере. Птиц этих Гумбольдт впоследствии описал под латинским названием Steatornis caripensis («жирные птицы из Карипе»). Испанцы называли этих птиц «гуахаро», что означает «плачущий» или «стонущий», из-за пронзительных криков, которые они издают.
Рис. 101. Гуахаро, вылетающая из пещеры.
Гуахаро — крупные, размером с большого цыпленка, оранжево-коричневые птицы с размахом крыльев около 100 см (рис. 101). Рот гуахаро напоминает рот козодоя, но большой клюв имеет сходство с клювом ястреба.
В отчете Гумбольдта имелось два момента, которые впоследствии привлекли внимание Гриффина: во-первых, птицы гнездились и летали в темных отсеках пещеры, и, во-вторых, резкие, пронзительные звуки, издаваемые гуахаро, отражаясь от каменных стен горы, рождали отзвуки далеко за пределами пещеры.
В 1953 г. Гриффин вместе с орнитологом Фелпсом отправился в Карипе, чтобы на месте проверить возникшую у него гипотезу о возможности использования «жирными птицами» эхолокации при полете в темноте.
При наблюдениях в пещерах ставились две основные задачи: выяснить участие зрительного анализатора в ориентации этих
307
птиц и попытаться установить характер звуков, издаваемых ими при полете в темноте. Для проверки первой из них в самом отдаленном гроте пещеры (650 м от входа), где гнездилось большое количество птиц, исследователи потушили фонари и в течение 25 мин. оставались в темноте. Даже после наступления полной адаптации зрения в темноте ни один из них не обнаружил ни проблеска света. Не удовлетворившись этим наблюдением, они установили фотоаппарат с открытой диафрагмой, направленной в сторону выхода из грота, и экспонировали кадры фотопленки от 5 до 9 мин. Когда пленка была проявлена, стало ясно, что в глубь пещеры свет не проникал. Несмотря на это, гуахаро проявляли там оживленную деятельность. Раздраженные присутствием посторонних, они в полной темноте летали по залу пещеры, непрестанно издавая резкие, неприятные крики. Эти наблюдения показали, что зрение не может играть существенной роли в ориентации этих птиц при полете в темноте.
Во-вторых, необходимо было выяснить, в какой степени крики птиц могут служить им для ориентации. Для этих целей в пещере установили конденсаторный микрофон, усилители, портативный магнитофон с полосой пропускания от 50 гц до 15 кгц и катодный осциллограф с киноприставкой. Записанные на магнитофон крики гуахаро были нескольких типов. Одни из них являлись несомненно позывными криками сидящих на выступах скал птиц, другие — резкие, пронзительные — издавали возбужденные птицы, летая над исследователями. Но иногда выделялись совершенно другие звуки, которые напоминали быструю барабанную дробь и состояли из серии отдельных щелчков.
В сумерках гуахаро стали вылетать наружу. Картина звуков резко изменилась. Вместо неприятных пронзительных криков, которые издавали птицы внутри пещеры при вылете из сгущающейся темноты, шел ровный поток резких, быстро повторяющихся щелчков. Лишь изредка одна из сотен пролетающих птиц издавала продолжительный сигнал, похожий на те звуки, которые раздавались в глубине пещеры днем.
Тйпичные осциллографические записи этих щелчков показаны на рис. 102. Оказалось, что в то время как форма щелчков-импульсов в некоторой степени изменялась от записи к записи, длительность, частота импульсов и интервалы между ними сохранялись примерно постоянными.
Длительность этих импульсов составляла около 1 мсек, (крайние значения 0.3—1.5 мсек). Отдельный щелчок состоял из нескольких звуковых волн, имеющих частоту порядка 7 кгц. Ультразвуковых компонентов в импульсах не наблюдалось.
Гуахаро излучали импульсы короткими вспышками-сериями. Число импульсов в серии варьировало от 2 до 6 и более. Интервалы между импульсами в серии составляли в среднем 2.6 мсек. (1.7— 308
4.4 мсек.). Интенсивность импульсов-щелчков не была измерена, но автор отмечает, что они были слышны на расстоянии 100 шагов.
Таким образом, было установлено, что гуахаро во время полета в темноте производят громкие щелчки с низкой частотой заполнения, лежащей в пределах слуха человека. Но оставалось
еще доказать, что эти звуки используются птицами в целях эхолокации. Для окончательного решения этой проблемы были отловлены 3 птицы и каждой из них по очереди давали возможность пролететь несколько раз по темной комнате. Птицы велико
лепно ориентировались в темноте и избегали столкновения со стенами комнаты. После этого их слуховые проходы были плотно тампонированы. В каж-дом случае птица с закрытыми слуховыми проходами летела до противоположной стены и с шумом ударялась о нее. Однако ориентировка вновь восстанавливалась после удаления ушных пробок. В обоих случаях птицы издавали громкие
Рис. 102. Осциллограммы ориентационны сигналов гуахаро. (Griffin, 1958).
щелчки почти непрерывно.
Ситуация изменялась, когда в комнате включался свет. Издавание щелч-
ков почти совершенно прекращалось, но еще более поразительным было то, что птицы с тампонированными ушами на свету не теряли способности обнаруживать препятствия и избегать их. Очевидно, на свету гуахаро ориентировались с помощью зрения.
Гуахаро — плодоядные птицы и питаются в основном ко
сточковыми плодами, как правило, различных сортов пальм. Добывание плодов вряд ли требует участия эхолокации, поэтому можно предположить, что звуковая локация гуахаро используется лишь для того, чтобы вовремя обнаружить препятствие при ночных полетах за пищей и успешно ориентироваться в темноте глубоких пещер, которые служат им местом дневного убежища.
Эхолокация у стрижей-саланган
Гуахаро не единственный представитель пернатых, обитающих в глубине пещер и использующих акустический способ ориентации при полете в темноте. В Юго-Восточной Азии широко распространены стрижи-саланганы (род Collocalia), несколько видов которых гнездится в темных уголках глубоких
309
пещер, часто по соседству с летучими мышами. Птицы эти, подобно гуахаро, с давних пор преследовались туземным населением, так как их гнезда, построенные из смеси слюны, грязи, веточек и травинок, используются в пищу. Сбор съедобных гнезд на некоторых островах является основной профессией населения. Ежегодное истребление потомства заставило стрижей-саланган устраивать свои гнездовые колонии все дальше от входа в глубину пещер, на отвесных скалах и карнизах высоких подземных гротов.
В отличие от гуахаро саланганы — дневные насекомоядные птицы. Во время охоты за насекомыми они в основном руководствуются зрением, но когда направляются к своим гнездам в полумрак пещер, то начинают издавать низкочастотные, хорошо воспринимаемые на слух щелчки. Эти щелчки по своим параметрам были почти одинаковыми, но частота их повторения менялась, увеличиваясь по мере падения освещенности в пещере и достигая максимума при посадке на гнездо. Новик (Novick, 1959) приводит следующие параметры щелчков у цейлонской саланганы (Col-ilocalia brevirostris unicolor). Длительность каждого импульса-щелчка составляла 2—6 мсек. Основная частота была 4—5 кгц и сопровождалась множеством обертонов. Полный спектр сигнала получить не удалось. Частота следования импульсов была определена по нескольким 2—3-секундным записям серий импульсов, воспринимаемых на слух как «треск». Она варьировала от 5.5 до 10 имп./сек. Связь этих щелчков и «тресков» с эхолокацией была установлена экспериментальным путем, когда несколько экземпляров этих птиц исследовали в лабораторной комнате. С закрытыми глазами или в полной темноте они свободно летали по комнате, избегая различные большие препятствия, но непрерывно издавали щелчки, сливающиеся в «трески» при приближении к стене или другим предметам. При одновременном выключении зрения и слуха птицы становились беспомощными и сталкивались с любым препятствием. При хорошем освещении выключение слуха не влияло на ориентацию саланган, которая осуществлялась исключительно за счет зрения.
Медвей (Medway, 1959) исследовал щелчки у другого вида саланган — Collocalia maxima bowi.) Во время «треска» они издавали 5—6 импульсов в секунду. Длительность каждого из них не превышала 2.5 мсек., а частота заполнения колебалась от 1.5 до 5.5 кгц, с 2 максимумами амплитуды на частотах 2 и 4.5 кгц. Осциллограммы одиночных щелчков различных индивидуумов показывают распределение частотных пиков внутри импульсов. Эти индивидуальные вариации импульсов, возможно, облегчают каждой птице выделять свое собственное эхо среди криков и эха множества других птиц, летающих одновременно по пещере.
310
Специальная проверка показала отсутствие ультразвуковых частот в криках саланган Collocalia maxima, С. salangana, С. es-culenta (Cranbrook, Medway, 1965).
Эхолокационные щелчки издаются птицами только во время полета, при взмахивании крыльями. Сидящие птицы со сложенными крыльями щелчков и тресков не производят. Первые эхо-локационные крики у молодых саланган появляются лишь тогда, когда их крылья достигают размеров взрослых птиц (Harrison, 1966).
В 1967 г. Медвей записал и проанализировал эхолокационные сигналы у яванской саланганы (Collocalia fuciphaga) и обнаружил, что частоты заполнения их располагались примерно в том же диапазоне, что и у предыдущего вида, между 1.5 и 4.5 кгц (Medway, 1967). Кроме того, было показано, что птицы данного вида не могли в темноте избегать столкновений с деревянными, вертикально поставленными шестами, имеющими 1 см2 в поперечнике, но свободно маневрировали между ними при хорошем освещении. Расстояние между шестами составляло 15 см, т. е. почти вдвое меньше размаха крыльев саланган (27 см).
Более высокие результаты при экспериментах с преодолением препятствий были получены Гриффином и Сатерисом на другом виде саланган — Collocalia vanikorensis granti (Griffin, Sutters, 1970). В небольшой темной камере птицам последовательно были предъявлены 3 барьера: из пластиковых трубок (диаметром 8 мм), металлических стержней (диаметром 6.3 мм) и проволок в изоляции (диаметром 2 мм). При преодолении первых двух типов преград птицы показали соответственно 68 и 80% бесконтактных пролетов, но проволоки диаметром 2 мм вызывали трудности их обнаружения, и количество успешных пролетов составило лишь 43%. Различия с результатами опытов Медвея авторы объясняют тем, что С. vanikorensis в эхолокационных сигналах использовали более высокие частоты, что могло увеличить разрешающую способность их локационных систем. Наибольшее количество энергии в их сигналах (щелчках) было сосредоточено» между 4.5 и 7.5 кгц. Спектр нескольких щелчков распространялся даже до 16 кгц (рис. 103). Продолжительность высокоамплитудных частей сигналов обычно была 4—8 мсек., а интервалы между ними в среднем составляли 116 мсек., с минимумом 48 мсек, и максимумом 358 мсек., соответственно частота следования — 20—3 щелчка в секунду. Исследователям не удалось обнаружить какой-либо закономерности в изменении параметров локационных щелчков у данного вида саланган при приближении их к препятствию.
Итак, из имеющихся данных об акустической локации у птиц пока возможно сделать лишь следующие выводы. Во-первых, в оптически неблагоприятных условиях некоторые виды двух отрядов — Caprimulgiformes и Apodiformes — могут использовать
311
эхолокацию в целях беспрепятственного полета. Во-вторых, основная энергия в локационных сигналах (щелчках), издаваемых птицами, сосредоточена в слышимом для человека диапазоне частот и варьирует у разных представителей от 2 до 7.5 кгц. В-третьих, низкочастотное заполнение сигналов обусловливает
кгц
1------1------1_______I------1_______L
О 0.1 0.2 0.3 ОЛ 0.5
сек.
Рис. 103. Спектрограммы 8 ориентационных сигналов Collocalia vanikoren-sts при полете в темноте. (Griffin, Suthers, 1970).
Крики проанализированы с использованием различных полос: а — 300 гц на 3 дб, б —300 гц на 6 дб; в — 45 гц на 3 дб.
низкую разрешающую способность эхолокационных систем этих птиц и в функциональном отношении эхолокация у них, по-видимому, играет лишь соподчиненную роль при хорошо развитой зрительной рецепции.
Совершенно не исследованными остаются вопросы, связанные с механизмами излучения эхолокационных сигналов и спосо
312
бами восприятия и обработки отраженных импульсов. Не подвергались тщательному экспериментальному исследованию возможности и диапазон эхолокации птиц по таким показателям, как чувствительность, разрешающая способность, распознавание предметов и т. д.
Гуахаро и 4 вида стрижей-саланган — первые объекты в открытии эхолокации у птиц. Можно ли ожидать расширения этого списка в ближайшие годы? По этому поводу высказывается ряд предположений. Почти несомненно, что эхолокация будет обнаружена еще у нескольких видов пещерных стрижей-саланган.
Заслуживает внимания предположение Паултера (Poulter, 1969) о том, что Гумбольдтов пингвин использует эхолокацию при ловле рыбы, воспринимая звуки, возникающие за счет кавитационных процессов в водных струях, обтекающих тело птицы.
Многие перелетные птицы мигрируют ночью, и не исключено, что по крайней мере некоторые из них акустическим путем оценивают высоту своего полета или обнаруживают высокие преграды на пути. Это кажется тем более вероятным, что ночные мигранты при перелетах в условиях ограниченной видимости постоянно издают различные звуковые сигналы (Thorpe, 1961; Schwartzkopff, 1962). В частности, есть сообщения, что кроншнепы с помощью эхолокации могут ориентироваться в тумане, используя для этих целей специальные сигналы (Freeman, 1950).
Способы ориентации у крыланов
Крыланы, или, как их еще называют, летучие собаки, летучие лисицы, за внешнее сходство их голов с головами собак и лисиц (рис. 104) в отряде рукокрылых выделяются в подотряд Megachtroptera, о чем уже говорилось в главе 1. Подотряд включает единственное семейство Pteropodidae, которое разделяется на 3 или 4 подсемейства. Область распространения крыланов ограничивается тропиками и субтропиками Старого Света. Родина их — Африка, Юго-Восточная Азия, Австралия и некоторые острова южной части Тихого океана. Животные эти, мелких, средних и крупных размеров, ведут ночной образ жизни. Длина тела от 5 до 40 см. Наиболее крупные представители подотряда, например летучие лисицы (род Pteropus), в размахе крыльев достигают 170 см. На этих животных уже с давних времен смотрели как на истинных чудовищ. Их необоснованно называли отвратительными гарпиями и страшными вампирами: к числу этих животных относили и то воображаемое ужасное существо, которое якобы опускается на спящих людей и высасывает у них из сердца кровь.
На самом же деле все крыланы вегетарианцы. Пищей им служат плоды фруктовых деревьев, а некоторые мелкие формы кормятся на цветах, извлекая пыльцу и нектар.
313
Примитивное строение пояса передних конечностей, отсутствие у многих хвоста и хвостовой перепонки свидетельствуют о слабом развитии летных качеств крыланов. Полет их относительно медленный и лишен лой замечательной маневренности, которая отмечается для летучих мышей.
Рис. 104. Австралийская летучая лисица. (Breeden, 1964).
Дневку большинство видов крыланов проводит на высоких деревьях, прямо под открытым небом, либо в неглубоких нишах у входа в пещеры, на чердаках домов и в дуплах деревьев.
Как показали исследования Мёреса и Кульцера (Mohres, Kulzer, 1956; Kulzer, 1956), а позднее Новика (Novick, 1958b), большинство представителей крыланов при полете в темноте целиком руководствуется зрением. Когда глаза этих животных были закрыты, они испытывали серьезные затруднения. Если же крыланам затыкали уши в условиях хорошей освещенности, они
314
продолжали летать с той же легкостью, избегая препятствий! на своем пути.
В естественных условиях крыланы, прежде чем добраться до кормовых угодий, иногда пролетают большие расстояния — в 150 км и более (рис. 105). Предполагают, что основными визуальными ориентирами им служат реки. Но как же они находят фрукты на деревьях в темноте? Каким образом не сталкиваются с ветвями’и не повреждают свои крыловые перепонки о различного рода колючки и шипы? Сведений на этот счет очень мало. Совершенно неизвестно об условиях освещенности кормовых участков
Рис. 105. Вечерний вылет крыланоз. (been, 1970).
и о том, прилетают ли летучие собаки на кормежку безлунными ночами. Новик (Novick, 1958b) склонен полагать, что крыланы кормятся на деревьях, отстоящих друг от друга на некотором расстоянии, и не прилетают кормиться в самые темные ночи. Крыланы хорошо переносят отсутствие пищи в течение нескольких дней. Кроме того, в неволе они отказываются от еды в полной темноте и даже не пытаются покидать своего убежища. Видимо, в естественной обстановке благоприятные кормовые условия с требуемым освещением бывают достаточно часто, а кроме того, зрение летучих собак специально приспособлено к ночной ориентации.
Ньювейлер (Neuweiler, 1962) специально исследовал морфофункциональные особенности зрительного аппарата у индийской летучей лисицы. Он показал, что глаза у Рteropus giganteus по своей структуре характеризуются рядом специальных приспособлений для ночцого видения, часть из которых развита лучше,
315
чем у всех других животных, ведущих ночной образ жизни. Например, на 1 мм2 сетчатки насчитывается 672 400 палочек, что составляет наибольшую плотность зрительных клеток среди всех исследованных млекопитающих. На одну нервную клетку приходится 275—400 палочек, тогда как у совы Tyto alba это соотношение составляет всего 15 : 1. Острота зрения у летучей лисицы при полном освещении составила 18', т. е. примерно такую же величину, как и у других ночных животных, но много ниже, чем у человека и приматов (26'). Однако при слабой освещенности (от 10~4 до 10-6 млб) острота зрения крыланов значительно превосходила таковую человека. При 4.5 -10~5 млб минимальное различение у летучей лисицы составляло 48', тогда как у человека при тех же условиях только 1°20'. Абсолютный порог зрения по интенсивности освещенности у крыланов оказался таким же, как и у сов. Следовательно, у летучих собак имеются необходимые условия для того, чтобы хорошо ориентироваться в полете при слабом освещении в ночное время. В поисках пищи наряду со зрением большое значение имеет также и обоняние.
Крыланы издают различного рода писки и пронзительные крики, но эти звуки носят исключительно эмоциональный, предостерегающий или коммуникативный характер. С точки зрения возможности использования акустической локации во время полета Новиком (Novick, 1958b) были исследованы 12 видов крыланов, относящихся к 8 родам, и только у рода Rousettus удалось обнаружить издавание регулярных щелчков, которые носили характер эхолокационных сигналов. У представителей семи других исследованных родов — Eonycteris, Macroglossus, Ptenochirus, Pteropus, Eidolon, Cynopterus и Lissonycteris — не было зарегистрировано периодически повторяющихся звуковых или ультразвуковых сигналов во время полета, которые могли бы служить целям эхолокации.
Мёрес и Кульцер (Mohres, Kulzer, 1956) первыми заметили, что у Rousettus aegypticus каждый старт, полет и посадка сопровождаются отчетливо слышимыми металлическими звуками, напоминающими «тикание». При приближении животных к препятствию или при встрече их в полете интенсивность и частота этих звуков повышаются. Исследователи также установили зависимость между интенсивностью звуков и степенью освещенности помещения: чем темнее, тем интенсивнее издаются звуки. Уже эти наблюдения давали основание предполагать, что «тикающие» звуки используются животными для ориентации.
Дальнейшие опыты показали, что в условиях хорошего освещения помещения роузетусы великолепно ориентировались визуально при почти беззвучном полете. Но стоило выключить свет, раздавались четко слышимые щелчки, которые усиливались, если животное огибало препятствие. Роузетусов можно было заставить издавать звуки при каждом выключении света, но 316
крыланы сразу замолкали, как только свет снова зажигался. Если животному закрывали уши, то в полной темноте оно становилось совершенно беспомощным, а при свете ориентировалось с помощью зрения.
Несколько позднее Новик (Novick, 1958b) подтвердил акустическую локацию у данного вида роузетусов и показал, что по крайней мере еще два вида этого рода — R. amplexicaudatus и R. seminudus — используют эхолокацию во время полета. Он отмечает, что, когда роузетусы летают ночью от дерева к дереву во время кормежки в естественных условиях, часто можно слышать регулярно повторяющиеся щелчки, издаваемые ими. При этом темп повторения и интенсивность этих щелчков увеличивались, когда животное приближалось к ветке. Автору ни разу не удалось регистрировать такие звуки, когда животные отдыхали или находились в руках экспериментатора. По-видимому, так же как и пещерные птицы, роузетусы издают свои эхолокационные щелчки только в полете.
При описании параметров эхолокационных звуков роузетусов Мёрес и Кульцер сообщают, что длительность щелчков в среднем составляет 5.5 мсек, и они заполнены сложной смесью частот в диапазоне 6.5—100 кгц, с максимумом интенсивности 15—20 кгц. Более подробный анализ сигналов роузетусов приводит Новик (Novick, 1958b).
У трех исследованных видов длительность эхолокационных щелчков составляла 3—4 мсек. Начальная часть сигнала на протяжении 0.5—1 мсек, имела наибольшую амплитуду и сравнительно чистые тона заполнения. Частота этого сегмента сигнала составляла у R. aegyptlcus 11—17 кгц, у R. seminudus — 14— 18, у R. amplexicaudatus — 12 кгц. Следующие две трети сигнала представляли собой комплексные, не синусоидальные волны очень низкой амплитуды. Используют ли роузетусы эту часть своего сигнала при получении информации о предметах — остается неизвестным.
Первоначально предполагали, что эхолокационые сигналы роузетусы излучают через ноздри и образование их происходит в гортани животного. Однако дальнейшие исследования (Kulzer, 1956, 1960; Novick, 1958b) опровергли эту точку зрения. Наблюдения и киносъемка летающих животных показали, что рот у роузетусов закрыт во время полета, но губы растянуты и слегка приоткрыты по бокам. Именно через эти щели производится излучение локационных сигналов. Перерезка гортанных нервов не вызывала изменений в излучении сигналов или ухудшения ориентационных способностей животных. В то же время двусторонняя перерезка всех боковых веточек, идущих от проксимальной части подъязычного нерва (n. hypoglossus) и иннервирующих мышцы языка, исключила возможность издавания щелчков у роузетусов. Эти данные позволили предполагать, что основную роль в про-
317
изводстве ориентационных звуков у роузетусов играет язык, а позднее Кульцер (Kulzer, 1960) привел убедительные доказательства этому. Щелчок, производимый быстрым движением языка, как и всякий другой, создаваемый механическим путем, будет иметь широкий спектр, но основные составляющие располагаются в звуковом диапазоне, что и отмечено для локационных сигналов роузетусов.
Данные о диапазоне и чувствительности эхолокации роузетусов ограничиваются всего одной работой (Griffin et al., 1958). Один самец R. aegypticus (размах крыльев 75 см) испытывался на пролет ряда вертикальных препятствий, расположенных на расстоянии 53 см друг от друга, диаметр которых варьировал от 25 (для картонных трубок) до 0.28 мм (для металлических проволок).
Как показали результаты опытов, роузетус хорошо обнаруживал препятствия в темноте, пока их диаметр не становился меньше 1 мм, после чего затруднительно было говорить об успешной ориентации животных.
Сравнивая эти данные с аналогичными опытами, проведенными на стрижах-саланганах, о которых говорилось выше, можно видеть, что чувствительность эхолокации роузетусов значительно выше, чем у саланган.
Так, уклонение от препятствий на уровне 45%~у роузетусов обнаружилось при диаметре проволок 0.46 мм, тогда как у саланган 43% уклонений было зарегистрировано при проволоках диаметром 2 мм. Объяснить это можно только более высокой частотой заполнения эхолокационных щелчков у роузетусов, что приводит к увеличению разрешающей способности эхолокатора.
В этих же опытах было исследовано влияние шумового поля на преодоление препятствий роузетусом. Использовались два типа шумов: до 15 кгц с ослаблением нижних частот 24 дб на октаву и 25 кгц с таким же ослаблением верхних частот. Оказалось, что при шуме 15 кгц животное преодолевало проволоки диаметром 3 мм лишь на уровне 26.6% (60 пролетов), тогда как при шуме 25 кгц из 36 пролетов ни один не был выполнен успешно. Эти эксперименты лишний раз подтвердили, что частоты 15— 25 кгц являются наиболее эффективными для эхолокации роузетусов. При хорошем освещении шум не сказывался на ориентационных способностях животного и преодоление препятствий осуществлялось им на уровне 90%. Следовательно, в этом случае животное полностью полагалось на зрительное восприятие.
До сих пор остается непонятным, почему эхолокация возникла только у одного рода в пределах Megachiroptera. Зрительный аппарат роузетусов развит так же хорошо, как и у Pteropus (Neuweiler, 1967), и проявляет все черты адаптации к ночному образу жизни. Возможно, эхолокация у роузетусов развилась в связи
318
с тем, что эти животные в противоположность многим другим крыланам обитают в пещерах при слишком слабом освещении, чтобы воспользоваться зрением. Новик (Novick, 1958b) высказывает мысль о том, что и другие крыланы могли бы пользоваться эхолокацией, но для этого им недостает только способности издавать специализированные звуки. В этом отношении роузе-тусы сделали качественный скачок по сравнению с другими представителями крыланов, но их локационные способности ни в коей мере не могут быть сравнимы с ультразвуковой локацией летучих мышей. Эхолокация, по-видимому, возникла изолированно у данного рода крыланов и в общем для ограниченных целей — обеспечить беспрепятственный полет в темноте.
Нельзя не удивляться конвергентному сходству акустической локации у птиц и роузетусов. Как те, так и другие в качестве локационных сигналов используют примитивные широкополосные щелчки, основная энергия которых сосредоточена в слышимом диапазоне частот. Эти щелчки издаются наиболее простым механическим способом, который не имеет ничего общего с механизмами производства сложных сигналов у летучих мышей. Как птицы, так и роузетусы способны производить щелчки только в полете и, следовательно, в неподвижном состоянии не могут пользоваться локацией. И наконец, у них проявляется тесное взаимодействие и координированное переключение зрительной и эхолокационной рецепции, при этом последняя выполняет явно подчиненное значение.
Взаимодействие зрительного и эхолокационного анализа существует, вероятно, и у примитивных видов летучих мышей из семейств Nycterildae и Megadermatidae. У них хорошо развито зрение, и, по наблюдениям Новика (Novick, 1958а), эти летучие мыши вылетают из своих убежищ и охотятся за крупной добычей еще до захода солнца.
По мере совершенствования летательного аппарата увеличивается скорость передвижения животных и соответственно сокращается время реакции для . принятия решения. Вместе с этим усложняются ориентационные задачи, когда требуется по возможности на больших расстояниях обнаружить объекты и отдифференцировать их по степени значимости для конкретной ситуации. Тут уже эхолокационная система принимает на себя основную роль в осуществлении пространственного анализа у летучих мышей. Даже при хорошей освещенности молодые, а тем более взрослые животные продолжают пользоваться эхолокацией, не ограничиваясь возможностями зрительного анализатора. Если одну из систем эхолокационного аппарата выключить, ориентация животных резко нарушается, несмотря на сохранение условий для зрительного анализа. С другой стороны, выключение зрения, по нашим данным, а также данным других авторов, не влияет на способность летучих мышей обнаруживать
319
препятствия, ловить насекомых, распознавать различные мишени. Тем не менее тесная функциональная связь эхолокации со зрительной рецепцией проявляется и у них. Об этом свидетельствуют результаты наших опытов, полученные при изучении формирования направленного поведения у взрослых гладконосых летучих мышей (рыжих вечерниц) (Константинов, 1964а, 19646, 1965в, 1966а).
В условиях хорошо освещенного манежа у животных вырабатывался двигательный стереотип, который выражался в становлении прямолинейной траектории движения от места старта к месту кормления (кормушке). В каждом выпуске на всей дистанции следования животного регистрировалось общее количество излучаемых импульсов как показатель активности эхолокационной системы. Если в первых опытах после выработки прямолинейной траектории летучие мыши по пути к цели излучали до 140 импульсов (в среднем 90 импульсов), то уже спустя 2—3 опыта число сигналов уменьшалось в среднем до 27 импульсов, причем некоторые особи достигали кормушки, излучив всего 3—5 импульсов. Однако стоило изменить освещенность или хирургическим способом выключить зрительный аппарат, как тут же наблюдалось резкое усиление функций эхолокационной системы. Животные начинали излучать в 3—4, а в некоторых случаях в 10 раз больше импульсов, чем интактные или чем в условиях хорошего освещения на той же самой дистанции. Особенно наглядно процесс усиления эхолокации проявился у рыжей вечерницы № 6. В последнем опыте на свету эта летучая мышь излучала в каждом выпуске при достижении кормушки в среднем 8.8 импульсов, тогда как в темноте их количество достигало в среднем 93.5, т. е. увеличилось в 10.6 раза. Следовательно, при выпадении зрительной рецепции у летучих мышей происходит быстрое переключение аппаратов эхолокации на более высокий уровень в соответствии с общим принципом викариро-вания (замещения) утраченных функций за счет специфической активации деятельности оставшихся анализаторов (Айрапетьянц, 1960, 1961а, 19616).
16
ЭХОЛОКАЦИЯ У ГРЫЗУНОВ И НАСЕКОМОЯДНЫХ
Эхолокация у грызунов
Среди мелких млекопитающих, ведущих наземный образ жизни, наиболее простая форма акустической локации была обнаружена у белых лабораторных крыс. Розенцвейг, Рилей и Крич (Rosenzweig, Riley, Krich, 1955; Riley, Rosen-zweig, 1957) тренировали 10 слепых крыс в лабиринте, состоящем из двух коридоров. В каждом опыте один из них был закрыт в 20 см от его начала металлическим барьером и животным требовалось выбрать свободный коридор, чтобы достичь кормушки. Результаты опытов считались отрицательными, если животное заходило в прегражденный коридор на 4 шага и его вибриссы касались барьера. Условия опыта были подобраны так, чтобы полностью исключить у животных возможность пользоваться иными органами чувств, кроме слуха. Все крысы научились отличать незагороженный коридор от коридора, закрытого барьером, не дотрагиваясь до него. Семь из них в 20 поставленных опытах давали 18 верных решений. Некоторые видоизменения условий эксперимента, по мнению авторов, еще более убедительно доказали, что при выборе правильного решения задачи крысы пользовались эхолокацией.
Угол между барьером и стенкой коридора, обычно равный 90°, изменили на 45°, вследствие чего действия шести животных, использованных в этом опыте, изменились и стали незакономерными. Их поведение вернулось к норме, когда барьер вновь установили под углом 90°. Авторы полагают, что поведение животных менялось в связи с тем, что эхо их пеленгующих звуков отражалось не прямо к крысе, а в бок, в стенку коридора, что делало невозможным успешную акустическую ориентацию. Кроме того, три животных, у которых были перфорированы барабанные перепонки, вели себя так же незакономерно, как и в случае изменения угла барьера.
321
Первоначально авторы пришли к заключению, что крысы ориентировались в лабиринте, воспринимая эхо собственных ультразвуковых сигналов, не слышимых экспериментатором. Однако использование высокочастотного микрофона с высокой чувствительностью показало, что ультразвуки издаются крысами крайне редко и никак не могут определять их ориентационное поведение. Зато наблюдатели постоянно отмечали не ультразвуковые, а низкочастотные звуки, производимые крысами в лабиринте: фырканье, чихание, щелканье и скрежетание зубов, царапанье когтей. Поэтому окончательный вывод исследователей сводился к тому, что слепые крысы ориентируются в лабиринте, улавливая эхо перечисленных случайных звуков, дающих им представление о расположенных впереди препятствиях. Правда, в ряде опытов, давших положительные результаты, крысы не производили, казалось, вообще никакого шума. Однако авторы объясняют это лишь недостаточной остротой человеческого слуха. Результаты этих опытов хорошо согласуются с данными, полученными на слепых людях. Различные звуковые шумы, сопровождающие движение человека, — шум шагов, стук палки и т. д., отраженные от предметов, подсознательно воспринимаются обостренным слухом слепых. Эти отражения, или эхо, в известной степени способствуют своевременному обнаружению препятствий на пути слепого человека. При закрытых ушах или при сильном постороннем шуме эта способность у слепых пропадает.
Ограничиваются ли возможности грызунов лишь такого рода грубой акустической ориентацией, или по крайней мере некоторые из них обладают более совершенными методами акустической локации, в настоящее время остается неизвестным. Однако предпосылки для второго предположения имеются. Прежде всего многие виды грызунов активны в сумеречно-ночное время суток или обитают в оптически неблагоприятных условиях (Огнев, 1947; Наумов, 1948; Громов и др., 1963; Новиков и др., 1970). Следовательно, зрительная рецепция оказывается часто ограниченной или в некоторых случаях полностью выключенной для осуществления дистантной ориентации. Так, например, при посещении Саблинских пещер авторы постоянно отмечали следы мышевидных грызунов в глубине пещер, на расстоянии 1—1.5 км от входа, куда дневной свет никогда не проникает. Другой предпосылкой можно считать способность грызунов к издаванию ультразвуков.
В 1948 г. Шлейдт показал, что рыжая полевка испускает два типа звуков частотой заполнения 15.5 и свыше 21 кгц (Schleidt, 1948, 1951). Затем Каман и Остерман (Kahmann, Ostermann, 1951) обнаружили способность испускать высокочастотные звуки у многих мелких млекопитающих: землероек, сонь и особенно мышевидных грызунов. Эти авторы допускают возможность того, что, помимо выражения эмоций и взаимной связи отдель
322
ных особей, высокочастотные звуки этих мелких млекопитающих могут иметь значение ориентационных сигналов для целей акустической локации.
Впоследствии Андерсону (Anderson, 1954) удалось зарегистрировать ультразвуковые компоненты в визге крысы частотой 19 и 29 кгц, тогда как в спокойном состоянии крысы издавали высокочастотные звуки от 21.5 до 26.5 кгц. Севеллом (Sewell, 1970) было показано, что при агрессии животные издают звуки частотой порядка 50 кгц, в то время как частота заполнения звуков «подчинения» ниже и лежит в пределах 22—30 кгц. Наиболее высокочастотные акустические сигналы грызуны издают в первые дни после рождения. Так, новорожденные детеныши домовой, желтогорлой мыши и рыжей полевки издают звуки частотой 60—80 кгц, на которые быстро реагируют самки (Zip-pelius, Schleidt, 1965; Noirot, 1965). По существу этими работами и ограничиваются сведения об ультразвуковом диапазоне звуков, издаваемых грызунами. До сих пор не было предпринято попыток выявить наличие специальных акустических сигналов, которые могли бы быть использованы свободноживу-щими видами грызунов для целей эхолокации.
Третьей предпосылкой для предположения о наличии ультразвуковой эхолокации у грызунов следует считать способность слуховой системы многих из них воспринимать высокочастотные звуковые сигналы. Используя в качестве показателя реакции животного на звук рефлекс Прейера (движение ушных раковин) (Gould, Morgan, 1941; Dice, Barto, 1952; Schleidt, 1952), было показано, что домовая мышь воспринимает ультразвуки частотой почти до 100 кгц; серая крыса, орешниковая соня и мышь-малютка — до 60; садовая соня, морская свинка и рыжая полевка — до 50, а золотистый хомячок — до 23 кгц. Наряду с этим у других грызунов, даже близкородственных перечисленным видам, верхняя частотная граница лежит значительно ниже и не распространяется в ультразвуковую область: у черной крысы — до 20, обыкновенного хомяка — до 15—16 кгц. Это была лишь первая попытка оценить верхнюю границу слуха у диких грызунов. Дальнейшие исследования показали, что прейеровский рефлекс не совсем точный показатель чувствительности слуха у животных (Herman, Clark, 1963; Schleidt, 1965) и лучше для этой цели использовать методы условных рефлексов или электрические ответы улитки и слуховых центров.
При отведении микрофонных потенциалов улитки у морских свинок (Wever et al., 1963) и белых крыс (Iskii et al., 1965) ответы регистрировались вплоть до 100 кгц. Наилучшая чувствительность оказалась в диапазоне 10—15 кгц и оставалась хорошей до 55 кгц, после чего ЧПК поднималась круто вверх. При использовании аналогичной методики исследовался слух лесных И белых лабораторных мышей, а также одного из видов полевок
323
(Brown, 1970). Оказалось, что у всех исследованных животных имеются две области максимальной чувствительности слуха: одна — в звуковом, другая — в ультразвуковом диапазоне частот. У лабораторных мышей такими оптимальными частотами были 4, 10 и 40—45 кгц, у лесных мышей — 17 и 55, у полевок — 13 и 50 кгц. Ролз (Ralls, 1967), используя в качестве показателя реакции на звук вызванные потенциалы заднего двухолмия, установила, что дикие домовые мыши отвечали на ультразвуки до 80 кгц, пороги были наименьшими на частотах 15—30 кгц. Это хорошо совпадает с результатами поведенческих опытов, выполненных на том же виде (Birch et al., 1968). У двух видов из рода Peromyscus (Р. leucopus и Р. boylii), по данным Ролз, ЧПК оказались почти идентичными с ЧПК летучих мышей {Myotis lucifugus), снятыми по той же методике. Верхняя граница слуха у них составляла 90—100 кгц, с областью максимальной чувствительности на 40 кгц. В нашей лаборатории изучался диапазон воспринимаемых частот и зон наилучшей чувствительности слуха у трех видов сонь: орешниковых, лесных и сонь-полчков (Мовчан, 1973). В качестве показателя реакции на акустический стимул также использовались потенциалы задних холмов. Установлено, что диапазон слуха у исследуемых животных охватывает широкую область звуковых и ультразвуковых частот, однако верхний предел восприятия у разных видов различался от 70 до 110 кгц. С другой стороны, у всех исследованных видов наиболее низкие пороги отмечались в диапазоне частот 6—25 кгц. В этой области имеется зона небольшого повышения порогов в районе 10—14 кгц, которая делит ее на два наиболее чувствительных участка с оптимальными частотами 8 и 16 кгц. Кроме того, у лесных сонь и полчков было выявлено снижение порогов на частотах ниже 1 кгц.
Исходя из всей совокупности рассмотренных данных по частотной настройке слухового анализатора многих мелких грызунов, становится очевидным большое значение ультразвука в жизни этих животных.
И наконец, четвертой предпосылкой могут служить опыты, в которых животным предлагалось обнаруживать предметы только методом акустической локации, с исключением возможности использования других дистантных анализаторов. Каман и Остерман (Kahmann, Ostermann, 1951) описали опыты, проведенные с соней-полчком и золотистым хомячком. Соню-полчка обучали прыгать с площадки (место старта) на тонкую жердочку, отстоящую от животного на 40 см. Прыгнув на нее, зверек добирался до кормушки. Опыты проводились в темноте, и положение жердочки постоянно меняли. Тем не менее соня успешно обнаруживала ее в новом месте и без промаха совершала прыжок.
Второй опыт проводился с одним предварительно ослепленным золотистым хомячком. Зверек был посажен на высокий круглый столик, спуститься с которого можно было? ТОЛИНО 324
воспользовавшись стеклянной пластинкой, которая, наподобие ступеньки лестницы, располагалась ниже его поверхности. Положение стекла постоянно менялось по окружности. Слепой хомячок всегда опускался только в том месте, где высота прыжка сокращалась поставленным стеклом. По мнению авторов, зверек не мог обнаруживать площадку с помощью обоняния, так как стекло после каждого опыта менялось на новое. Результаты этих опытов давали основания предполагать использование
Рис. 106. Соня-полчок в опыте перед прыжком на скворешник. (Фот. В. Н. Мовчана).
животным звуковой локации во время обнаружения жердочки и площадки.
Мы попытались проверить предположение вышеуказанных авторов (Мовчан, 1973), Интактных сонь (полчков, лесных и орешниковых) обучали прыгать со стартовой площадки на домик (18x15x10 см), положение которого менялось по расстоянию (от 5 до 70 см) и по углу в случайном порядке (рис. 106). После выработки прочного пищевого двигательного рефлекса у животных проводилась двусторонняя энуклеация. Ни в одном из многочисленных опытов ослепленные животные не могли обнаружить домик даже на самом близком расстоянии (5—10 см). Лишь при касании вибриссами, т. е. контактным способом, они обнаруживали домик и перебегали в него. В процессе экспериментов нам не удалось отметить каких-либо акустических сигналов, издаваемых животными. Таким образом, наши эксперименты не подтвердили наличия эхолокации у соль. По всей ве-
325
роятности, в экспериментах немецких исследователей не удалось создать абсолютной темноты и сони-полчки обнаруживали жердочку с помощью зрения.
По усовершенствованной методике «круглого столика» проверялась также способность золотистых хомячков обнаруживать подставку, расположенную ниже поверхности столика (Кон.-стантинов и др., 1966).
Опыты проводились па 6 животных. Схема опытов была следующей. Животное помещалось на плоский плексигласовый диск (диаметром 15 см), укрепленный на вертикальном штативе. На этом же штативе под диском устанавливалась плексигласовая площадка шириной 6 см, выступающая за край диска на 7 см. Положение площадки легко менялось как в вертикальной, так и в горизонтальной плоскости вокруг диска. Животное должно было отыскать площадку, спрыгнуть на нее, после чего по приставной лесенке опуститься на пол установки, окруженной фанерным барьером, найти отверстие, ведущее к кормушке и, получив пищевое подкрепление, по узкому коридору вновь подняться к диску и выйти на него. Если животные прыгали мимо площадки на пол, они не получали пищевого подкрепления и вновь возвращались на диск. В каждом опыте одно животное совершало 10—17 прыжков на площадку или мимо нее.
Интактные золотистые хомячки в первых опытах показывали лишь 30—40% обнаружения площадки из всех совершаемых ими прыжков с диска. Но постепенно, к 5—6-му опыту, число прыжков на площадку увеличилось до 70%. На этом уровне была проведена двусторонняя одномоментная энуклеация. После операции время поисков площадки животными увеличилось в 2— 3 раза.. Хомячки бегали по краю диска, опускали голову вниз или свешивались целиком, удерживаясь на одной задней лапе, снова поднимались на диск и продолжали поиск. По нескольку раз они пробегали мимо площадки, опущенной на 6—8 см от поверхности диска. Количество правильных прыжков в первом опыте после энуклеации снижалось до 50%, но затем, уже во 2— 3-м опыте, возвращалось к исходным 70%. Два животных были энуклеированы до начала эксперимента. Тем не менее процесс обучения в нахождении площадки у них не отличался от интактных животных. Предполагая использование акустической локации хомячками при поисках площадки, мы пытались уловить какие-либо звуки, издаваемые ими. На слух мы воспринимали лишь слабые «пофыркивания» животных, когда они свешивались над самой площадкой. Попытка использовать в качестве приемника высокочастотный титанат-бариевый микрофон с резонансной частотой около 70 кгц не дала каких-либо результатов.
В этих же опытах мы обратили внимание на энергичные движения вибрисс у хомячков, когда они спускались на площадку. Первоначально полагая, что хомячок, низко свесившись с диска над площадкой, касается ее своими вибриссами, мы покрыли площадку листом закопченной бумаги, но никаких следов от вибрисс на ней не обнаружили. Следовательно, непосредствен-326
ного контакта с площадкой у животного не было. Тогда мы обрезали вибриссы на мордочке хомячка. Вслед за этим животные полностью утратили способность находить площадку и прыгать на нее. По-видимому, не акустическая локация, а хорошо развитая тактильная чувствительность обусловливала поведение ослепленных хомячков в наших и описанных выше опытах. Какие виды вибраций улавливаются вибриссами, до сих пор остается неизвестным, но они играют важную роль в получении информации у некоторых животных, например у кошачьих.
Другие результаты были получены в опытах на общественной (М. socialis) и узкочерепной (М. gregalis) полевках. Так как полученные данные по этим видам были совершенно идентичными, мы будем давать их вместе. Первоначально расстояние между площадкой и диском по вертикали составляло 4 см. Четыре полевки перед началом опытов были энуклеированы, однако процесс обучения в обнаружении площадки протекал у них так же быстро, как и у интактных животных, и уже ко второму опыту площадка обнаруживалась на 80—90% относительно общего количества прыжков с диска. Плавно опуская площадку вниз, мы нашли то критическое расстояние, дальше которого полевки не могли обнаруживать площадку. Величина его оказалась равной 14 см. Увеличение расстояния между диском и платформой не повлияло на число правильных обнаружений ее животными, увеличивалось лишь время поисков и подготовки зверьков к правильному прыжку.
После этого интактные животные были энуклеированы. В первом же опыте после операции полевки показали ухудшение ориентационных способностей. При этом, если площадка устанавливалась в первоначальном положении на расстоянии 4 см от поверхности диска, а затем плавно опускалась, нарушения в поведении животных наблюдались лишь в первом опыте, а уже в следующем обнаружение площадки составляло 90—100% (рис. 107, Б). Если же площадка оставалась на уровне последнего опыта до операции, т. е. на расстоянии 8—10 см от поверхности диска, животные не могли ее обнаружить в течение 6 опытов, несмотря на то что расстояние уменьшалось до 6 см. Лишь при сокращении расстояния до 4 см ослепленные полевки четко находили площадку и затем показывали хорошие результаты при плавном ее опускании. Эти данные показывают, что первоначально интактные полевки при нахождении площадки в основном пользовались зрением, а после энуклеации они должны были заново обучаться находить площадку с использованием других видов сигнализации. Ни одно интактное животное, как и оперированное, так и не смогло в течение нескольких опытов обнаруживать площадку, если расстояние до нее первоначально составляло 8—10 см (рис. 107). Лишь постепенное опускание площадки приносило положительные результаты. Итак, было показано, что
327
функциональная компенсация зрения другими видами рецепций у полевок происходит в течение одного опыта, процесс обучения при поисках площадки и глубинный анализ при замещении осуществляются так же, как и при зрительном восприятии.
По аналогии с опытами на золотистых хомячках у всех энуклеированных полевок были удалены вибриссы, но в противоположность первым в данном случае не произошло нарушений ориентационных способностей у полевок (рис. 107). Возможность использования обонятельной рецепции при обнаружении пло-
А 5
% Ю0 80 60 40 20
0
10108 7 4 4 8'10 8-12 8 8 6 6 44 6'9 см 4 4 7~8 12-13 5~6 101010 44322, 5-8 10-12 10 5-8 10 5~6 10 4 4 7~8 10'12 СМ
Рис. 107. Способность золотистых хомячков (Л) и полевок (Б) обнаруживать площадку после энуклеации и удаления вибрисс.
По оси абсцисс: снизу — расстояние между столиком и площадкой; по оси ординат — положительные ответы.
щадки исключалась экспериментальным путем (после каждого прыжка животного на нее площадка тщательно промывалась). Таким образом, у полевок не оставалось другого способа, кроме акустического, для отыскания местоположения площадки. Выключение слуха путем тампонирования слуховых проходов или перфорации барабанных перепонок подтвердило это предположение.
После выключения слуха (по крайней мере частичного) полевки полностью утратили способность находить площадку даже на минимальном расстоянии 2 см (рис. 107). Количество правильных прыжков не превышало 20—30%, и полевки после долгих безуспешных поисков либо возвращались к выходу на диск, либо прыгали на пол.
Для доказательства эхолокации у полевок оставалось сделать последний шаг — обнаружить издаваемые ими звуки при поисках площадки и проанализировать их. Однако в то время мы не располагали чувствительной аппаратурой в широком ультразвуковом диапазоне. На слух же мы не могли уловить каких-либо звуков, сопровождающих прыжок животных на подставку.
Позднее мы провели опыты на другом виде — рыжих полев
328
ках (Мовчан, 1973). У двух животных вырабатывался прочный пищевой двигательный рефлекс на прямоугольную пластину, вертикально установленную на одной из восьми радиально расходящихся дорожек. Подопытные животные помещались на круглый столик, находящийся в месте пересечения дорожек. Положение вертикальной пластины менялось в процессе опыта в случайном порядке. Эксперименты показали, что рыжие полевки после энуклеации не способны были обнаружить тестовую пластину без участия вибрисс. Сравнение данных, полученных на полевках, показывает, насколько разнообразными могут быть способы ориентации и участие отдельных анализаторов в пространственном анализе даже у близкородственных видов.
Приведенный выше материал представляет собой пеструю и противоречивую картину, не позволяющую сделать окончательный вывод об использовании ультразвуковой или звуковой локации грызунами. По-видимому, необходим более тщательный подбор объектов исследования, основанный на биологии вида и при учете изложенных выше предпосылок к эхолокации. В том, что такое открытие будет сделано, не приходится сомневаться, тем более после представленных доказательств о наличии эхолокации у мелких млекопитающих из отряда насекомоядных (Insectivora), близких по образу жизни к мышевидным грызунам.
Эхолокация у насекомоядных
Этими мелкими насекомоядными, пополнившими список эхолоцирующих животных, оказались землеройки (сем. Soricidae). Уже предварительные сведения давали возможность предполагать наличие у них ультразвуковой локации. Так, многие разрозненные данные в литературе свидетельствуют о том, что землеройки испускают звуки очень высокой частоты при исследовании незнакомой обстановки (Reed, 1944; Pruitt, 1947; Crowcroft, 1957). Хенсон (Henson, 1961), изучая строение среднего уха у примитивных млекопитающих (насекомоядных и рукокрылых) с точки зрения адаптации к ультразвукам, отнес землероек к формам, воспринимающим высокие ультразвуковые колебания свыше 50 кгц. В то же время ежи и кроты были отнесены к формам, не способным улавливать ультразвуки. Правда, последние данные, полученные на ежах методом условных рефлексов, показывают, что диапазон слуха у этих животных простирается от 250 гц до 45 кгц (Richard et al., 1969). Поэтому заключения Хенсона следует понимать лишь относительно: структура среднего уха, например, у землероек и нескольких родов летучих мышей приспособлена для ультразвукового восприятия лучше, чем у других исследованных родов.
Многие морфологи сходятся на том, что зрительный анализатор у землероек развит слабо и, следовательно, функции его ог
329
раниченны (Clark, 1932; Walls, 1942). Исходя из совокупности указанных выше данных, землеройки привлекли внимание американских исследователей, занимающихся изучением эхолокации у разных животных.
В первых предварительных опытах (Gould, 1962) оценивали возможности землероек преодолевать препятствия. В качестве последних использовали стеклянные пузырьки (420 штук), расставленные наугад. Легкое прикосновение к пузырьку вызывало его падение. Число бутылочек, сбитых землеройками при пересечении определенной площади, служило показателем их способности преодолевать препятствия. В результате было показано, что интактные или с удаленными вибриссами землеройки преодолевали препятствия значительно лучше, чем две статистические модели. При закупорке слуховых проходов землеройки бегали медленнее, чем контрольные, и совершали больше ошибок. Однако эта несовершенная методика мало что могла дать для точного доказательства эхолокационных способностей у землероек и в дальнейшем авторы перешли на использование методики круглого столика и площадки, несущественно отличающейся в деталях от описанной нами в опытах на хомячках и полевках (Gould et al., 1964). Землеройка попадала на поверхность диска через хлорвиниловую трубку, достигала края диска, находила платформу, расположенную ниже на несколько сантиметров, прыгала на нее и по наклонной лесенке добиралась до ящика с пищевым подкреплением и водой. Падение с диска на пол служило для них наказанием. Во время первых 3—4 опытов платформу устанавливали на одном уровне с диском, так чтобы животные могли отыскать ее с помощью любого вида рецепции, а затем постепенно опускали, оставляя лишь возможность акустического обнаружения. Влияние обоняния на результаты опытов исключалось экспериментальным путем. Все опыты проводились в полной темноте. Экспериментатор наблюдал за действиями землероек с помощью прибора ночного видения. Возможность использования проприоцептивной сигнализации исключалась путем перемещения площадки в разные положения относительно диска.
Так как, свесившись с диска, землеройки могли достать до площадки на расстоянии 3.5 см, то учитывались лишь те результаты, когда расстояние между диском и площадкой составляло более 4.5 см (обычно 5.5—11 см).
На каком максимальном расстоянии землеройка могла обнаруживать платформу, точно установить не удалось, однако авторы указывают, что даже при расстоянии 24.5 см от поверхности диска поведение животных свидетельствовало об обнаружении площадки, хотя они и не прыгали на нее.
В основном опыты проводились на землеройках из рода Sorex (S. palustris, S. vagranst S. cinereus), но четыре серии были поставлены на землеройках из рода BUwina {В, brevicauda).
330
По расчетам авторов, случайное нахождение платформы зверьком в каждом опыте могло составлять 8%, тогда как возможность случайного обнаружения в 12 поставленных опытах уменьшалась до 0.08%. Животные же показывали 100 % -е обнаружение платформы во всех экспериментах до тех пор, пока они могли использовать слух. После того как в слуховые проходы семи землероек были вставлены миниатюрные хлорвиниловые трубочки и концы их закрыты, землеройки в большинстве случаев не могли обнаруживать платформу и после длительных хождений по диску возвращались на исходное положение в центр его или падали на пол. Контрольные животные с открытыми концами трубочек, вставленными в уши, продолжали находить платформу так же успешно, как и раньше. Результаты этих опытов давали все основания предполагать использование акустической локации землеройками при поисках площадки в темноте. Регистрация ультразвуковых сигналов, которые сопровождали каждый прыжок животных на площадку, развеяла последние сомнения.
На расстоянии 7.5 см от края диска, точно над площадкой, устанавливался микрофон с равномерной ^чувствительностью от 20 до 150 кгц. Принимаемые сигналы усиливались и поступали на вход катодного осциллографа, с экрана которого производилась киносъемка. Иногда записывались звуки во время прохождения животными трубки, ведущей на поверхность диска. Результаты анализа обнаруженных у землероек ультразвуковых сигналов представлены в табл. 23. В темноте животные издавали 2 типа звуков: короткие (0.75—9.5 мсек.) и длинные (11—33 мсек, и более), частота заполнения которых составляла 25—60 кгц. Точно установить продолжительность сигналов второго типа не удалось, так как максимальная развертка осциллографа была 33 мсек, и, следовательно, сигналы большей длительности на ней не умещались. Примерная оценка звукового давления, или интенсивности этих звуков, показала среднюю величину 0.06 дин/см2 (0.02—0.14 дин/см2). Авторы отмечают, что звуковые сигналы излучаются животными довольно редко, но не указывают частоту их повторений во время экспериментов с диском и площадкой. Имеется лишь ссылка на то, что в незнакомой обстановке землеройки издавали 13—16 импульсов в течение 3 мин., при этом они быстро подергивали мордочкой.
Исследователи не исключают возможности использования землеройками в целях эхолокации и других звуков, частоты /которых лежат в звуковом диапазоне. Они отмечали быстрое издавание таких звуков в самом начале опытов при обучении зверьков отыскивать площадку, а также в экспериментах с преодолением препятствий. Частота заполнения их составляла 5— 5.5 кгц, и интервалы между этими звуками были примерно 170 мсек. Эти и ряд других наблюдений (Crowcroft, 1957) дают основание полагать, что в различных ситуациях землеройки
331
Таблица 23
Параметры ультразвуковых ориентационных сигналов землероек, издаваемые ими при выходе на диск (старт) и над площадкой (цель) (Gould et al., 1964)
б ид Ситуация п ^ср Короткие импульсы Длинные импульсы
п | Тор п т ср
Sorex vargans Старт 12 37 16 5.3 5 18
3 33
S. palustris Цель 32 41 — — 1 13.6
Старт 12 36 18 5.1 4 18.7
Blarina hrevicauda Цель 14 41 3 7.0 12 33
» 12 42 8 2.3 4 20
Примечание, п — число исследованных сигналов; стоты, кгц; — среднее значение длительности, мсек.
f - среднее ср
значение ча-
в целях ориентации используют различные звуковые и ультразвуковые сигналы. Однако этот вопрос пока остается неразработанным. Другие звуки слышимого диапазона частот, издаваемые землеройками в неволе, носили, по-видимому, эмоциональный характер и к эхолокации отношения не имели.
Итак, открытие эхолокации у землероек можно считать доказанным, однако интерпретировать биологическое значение ее для животных в естественных условиях представляется затруднительным. Очень слабая интенсивность излучаемых ультразвуковых сигналов, с одной стороны, и неблагоприятные с точки зрения акустики условия обитания этих животных (густая растительность, неровности почвы, узкие ходы земляных нор), где происходит быстрое затухание ультразвука, с другой, казалось бы, .почти полностью исключают возможность акустической локации. Однако поведение этих животных в естественной обстановке и способность ориентации их на местности изучены еще недостаточно хорошо, чтобы судить о том, какую роль может играть эхолокация в их жизни. Можно предполагать, что в отличие от летучих мышей, которые постоянно пользуются эхолокацией, землеройки обращаются к ней лишь в отдельных случаях. Например, при пересечении открытого пространства эхолокация может быть очень эффективным средством для определения ближайших укрытий (пня, кустов). Некоторые авторы (Shillito, 1961) считают, что при неблагоприятных условиях водяные землеройки (Neomys jodiens) совершают миграции по суше. Использование эхолокации в незнакомой местности приобретает особенно важное значение для этих мелких млекопитающих, для которых важно вовремя скрыться в подходящем убежище от разнообразных хищников. А может быть, куторы применяют эхолокацию под водой при ловле мальков рыбы, так как актив
332
ность их проявляется особенно в ночное время. В общем можно найти, очевидно, немало подходящих ситуаций, когда, с человеческой точки зрения, животным выгодно использовать акустическую локацию. Как обстоит дело в действительности — вопрос будущего.
Гульд (Gould, 1965) проводил эксперименты на представителях другого семейства насекомоядных — мадагаскарских щетинистых ежах — тенреках (Tenrecidae) и обнаружил и у них эхолокацию. Им были взяты три различных вида тенреков: Hemicentetes semi-spinosus (20 животных), Echinops telfarii (2 животных) и Microgale (Nosogale) dobsoni (1 животное), отличающихся по многим экологическим показателям. Методика оставалась той же, что
Рис. 108. Осциллограммы импульсов тенрека. (Gould, 1964).
и в экспериментах с землеройками, т. е. круглый столик и платформа под ним. Первоначально платформа находилась на уровне диска, а затем постепенно опускалась. Основные результаты были получены при положении платформы на 11 см ниже уровня диска, но тенреки могли обнаруживать ее и при 14 см. Опыты показали, что без участия зрительной, тактильной и обонятельной рецепции все тенреки обнаруживали платформу и прыгали на нее в 8 случаях из 10 в каждом опыте, т. е. показывали 80% положительных ответов. Однако ориентационные способности их пропадали, когда в слуховые проходы животных были вставлены хлорвиниловые трубочки, наружные концы которых затыкались ватой. Если же трубочки оставались открытыми, животные продолжали успешно обнаруживать платформу. Во время этих экспериментов автор имел возможность регистрировать звуки, издаваемые тенреками на диске и при поисках платформы. В первом случае он отмечал издавание различных звуковых эмиссий животными: высокочастотные скрипы, фырканье, скрежет зубов, щебетание и т. д. Во втором случае все три вида тенреков производили щелчки языком (tongue clicks), особенно когда они находились над площадкой и готовились к прыжку на нее. Echinops при издавании щелчков как бы облизывал губы языком, вытягивая его вбок и вверх над губой, тогда как Hemicentetes и Microgale просто открывали и закрывали рот в момент изда-вания щелчков. Аналогичные щелчки, производимые языком, были обнаружены автором еще у одного рода тенреков — Сеп-tetes. Осциллограммы этих звуков представлены на рис. 108.
333
Как видно на осциллограммах, длительность щелчков тенреков очень мала, составляет всего 0.2—1.2 мсек. Изредка длительность щелчков у Centetes превышала 3 мсек. В щелчках, по крайней мере некоторых, удавалось наблюдать понижение частоты заполнения в конце на несколько сотен герц, т. е. имела место частотная модуляция. Диапазон частот, измеренный лишь на максимальной амплитуде сигнала, составлял в щелчках у Нemicentetes и Centetes 10—16 кгц (в среднем 13 кгц), у двух других видов тенреков — 5—17 кгц. Щелчки издавались либо по одному, либо группами, по 2—3 вместе. Частота их следования была различна у каждого животного. Например, при исследовании пола в незнакомой для них комнате в течение 3 мин. взрослые особи разных видов тенреков издавали от 24 до 64 щелчков.
Доказательством эхолокационного характера щелчков служило то обстоятельство, что животные издавали их всякий раз при поисках площадки либо во время ориентации в новой обстановке. Интенсивность щелчков была различна у разных видов. Так, щелчки Microgale отличались очень малой интенсивностью, тогда как у Centetes их можно было услышать на расстоянии более чем 7 м. К сожалению, автору не удалось снять количественных характеристик величины звукового давления этих звуков.
Особо важное значение этих щелчков в жизни тенреков подтверждается и в исследовании частотно-пороговых характеристик слуха у этих животных (Wever, Herman, 1968). Аудиаграмма одного тенрека (Hemicentetes semispinosus) была получена при отведении микрофонного потенциала улитки. Она показывает, что диапазон воспринимаемых частот у тенрека простирается от 0.2 до 60 кгц, а максимум чувствительности приходится на частоты 10—15 кгц, т. е. точно на ту область частот, которая содержится в ориентационных щелчках этих животных.
Изложенные в этой главе данные об акустической локации грызунов и насекомоядных представляют собой лишь первые шаги в изучении разнообразных средств и способов, используемых этими животными при пространственной ориентации. Однако на основании их возникают оптимистические перспективы дальнейших исследований, которые, по-видимому, с одной стороны, значительно увеличат список животных, обладающих эхолокацией, с другой — расширят наши представления о диапазоне локационных систем этих млекопитающих и позволят окончательно решить вопрос о биологическом значении такого рода рецепции в их повседневной жизни.
ЭХОЛОКАЦИЯ В ОКЕАНЕ
17
ГИДРОЛОКАЦИЯ У КИТООБРАЗНЫХ
Краткая систематическая справка
Китообразные исключительно водные млекопитающие животные. Они живут, рождают детенышей, вскармливают их молоком, дышат атмосферным воздухом и, когда приходит время, умирают в водной стихии. Киты обитают почти во всех морях и океанах Земли и даже в некоторых реках и озерах Азии и Южной Америки.
Размеры этих животных колеблются от 0.9 до 33 м, а вес — от 30 кг до 140 т. Форма их тела вытянутая, хорошо обтекаемая. Задние конечности исчезли и лишь изредка встречаются в виде рудиментарных образований, а передние превратились в веслообразные грудные плавники — рули глубины, поворотов и тормоза. Кроме того, у большинства видов развивается спинной плавник.
Основным органом движения служит хвост, снабженный мощной мускулатурой, слегка уплощенный с боков; стебель его заканчивается хвостовым плавником с горизонтально расположенными лопастями. Киты лишены волосяного покрова, сальных, потовых и слюнных желез.
Окраска тела китообразных варьирует от аспидно-черной до чисто-белой, с различными видовыми вариациями в соотношении темных и светлых участков, образующих пятна, полосы, разводы, что создает характерный для вида рисунок. Видоспецифическая окраска является не только опознавательной, вплоть до семейных группировок у дельфинов, но выполняет защитную, маскирующую функцию и, наоборот, демонстративно-ошеломляющую (белуха, косатка). У некоторых видов окраска меняется с возрастом животного (белуха, нарвал, продельфин и др.). Постоянное обитание в водной среде наложило резкие отпечатки на внешний облик китообразных и повлекло за собой значительные морфофункциональные адаптации.
337
В систематическом отношении современные китообразные делятся на 2 подотряда — усатые киты (Mystacoceti) и зубатые (Odontoceti). Различия между ними довольно отчетливы (ряд черт строения черепа и скелета, белки сыворотки крови, дыхательная, пищеварительная системы и пр.), что дает основания некоторым исследователям считать их произошедшими от разных корней и возводить в ранг самостоятельных отрядов (Клейненберг, 1958; Яблоков, 1964).
Усатые киты характеризуются полным отсутствием зубов (имеются лишь в эмбриональном состоянии) и развитием в ротовой полости (на расширенных небных отростках верхнечелюстных костей) 130—400 роговых пластин с каждой стороны челюсти. Эти пластины свешиваются в ротовую полость, достигая 4.5-метровой длины у гладких китов, и благодаря оторачивающей их по нижнему краю роговой бахроме служат цедильным аппаратом. Этим «ситом» и «языком», действующим как поршень, усатые киты отцеживают из водной массы, попавшей им в рот, различные планктонные организмы, служащие им пищей.
К усатым китам относятся 9 видов, которые объединяются в 3 семейства: полосатики (Balaenoptartdae), гладкие киты (Balaeni-dae) и серые киты (Eschrichtiidae). К полосатикам принадлежат стройные, быстроходные киты (скорость до 37 км/час) длиной от 6 до 33 м. На горле и брюхе у них имеется 12—120 продольных глубоких складок.
Гладкие киты — толстые, медлительные животные (скорость до 17 км/час) длиной 6—17 м, не имеют на теле каких-либо полос и без спинного плавника (за исключением рода Veobalaena),
Семейство серых китов занимает как бы промежуточное положение между двумя предыдущими по своим внешним признакам и включает один вид. Это животные до 14—15 м длиной, буроватосерой с пятнами окраски.
Зубатые киты включают 4 семейства и представлены более чем 50 видами животных, из которых в наших водах обитают 23 вида, относящихся к трем семействам.
Длина тела зубатых китов в целом меньше, чем у усатых, и колеблется у разных видов от 0.9 до 21 м. В их ротовой полости располагается от 2 до 240 монофиодонтных конических зубов. Наружное дыхательное отверстие одно (у усатых китов их два), имеется сложная система воздушных мешков (отсутствующая у усатых китов), связанная с наружным носовым проходом; надгортанники и черпаловидные хрящи удлинены в гортанную трубку, которая входит в хоаны, тем самым разделяя пути пищи и воздуха. На удлиненных межчелюстных и верхнечелюстных костях черепа всегда располагается своеобразная жировая подушка, которая у усатых китов отсутствует.
В этот подотряд входит семейство речных дельфинов (Platanis-tidae), члены которого обитают в реках Ганге и Сузу (платаццста),
338
Амазонке (иния), Ла-Плате (стенодельфис), Янцзы (липотес и белоперый дельфин — буту). Эти обитатели мутных речных вод во многих местах являются священными животными и изучены довольно слабо. Их размеры от 0.9 до 3.5 м, окраска от светло-серой, почти белой до темной, зрение (кроме инии) развито слабо или даже редуцировано (Platanistae). Речные дельфины единственные среди зубатых китов, обладающие во взрослом состоянии вибриссами на голове, которые, по-видимому, играют важную роль в ближней ориентации. Пищей им служат не только различные рыбы, но и плоды, падающие в воду.
Рис. 109. Дельфин афалина Tursiops truncatus.
Второе семейство — кашалотовые (Physeteridae) — включает два монотипических рода: кашалоты (Physeter) и карликовые кашалоты (Kogia). Кашалот — наиболее известный из них когия — в наших водах очень редок. Самый отличительный внешний признак кашалота — огромная голова (до трети длины тела) с огромной жировой подушкой, которая выступает вперед над маленькой и узкой нижней челюстью. Этот «лобный выступ», несколько сжатый в середине с боков, весит до 6 т и покоится на костном ложе из расширенных межчелюстных и верхнечелюстных костей, которые поддерживают его не только снизу, но и с боков. Кашалоты-самцы достигают 21, а самки — 16 м в длину. Размеры когии до 4 м (в среднем самцы — 3 м, самки — 2.5), и пропорции головы (1 : 5) и тела у нее более соразмерны.
339
В самом названии третьего семейства — клюворылые киты (Ziphiidae) — уже указывается, что их рыло вытянуто вперед и напоминает клюв. Длина их тела варьирует у различных видов от 5 до 12.5 м. Все клюворылые киты питаются в основном головоногими моллюсками и являются превосходными ныряльщиками (дыхательная пауза более часа).
Наиболее многочисленное семейство зубатых китов — дельфиновые (Delphinidae) — объединяет 20 родов. Дельфины — это мелкие и средние по своим размерам киты (до 10 м), которые быстро плавают (до 45 км/час), питаются рыбой и головоногими моллюсками и ведут преимущественно стайный образ жизни. Сюда относятся белухи (род Delphinapterus), нарвалы (род Monodon), дельфины белобочки (род Delphinus), продельфины (род Stenella), афалины (род Tursiops), китовидные дельфины (род Lissodel-phis, короткоголовые дельфины (род Lagenorhynchus), серые дельфины (род Grampus), гринды (род Globicephala), косатки (род Orcinus) и малые (черные) косатки (род Pseudorca), бесперые морские свиньи (род Neomeris), белокрылые морские свиньи (род Phocaenoides) и морские свиньи (род Phocaena).
Именно дельфины стали объектами экспериментального изучения явления эхолокации у китообразных, и от таких представителей этого семейства, как афалина (рис. 109), дельфин белобочка, морская свинья (фоцена), продельфины, были получены основные данные на этот счет.
Доказательства наличия эхолокации у дельфинов
Способность дельфинов обнаруживать различные препятствия с помощью эхолокации была продемонстрирована уже в первых экспериментах. Келлог (Kellogg, 1958), используя барьеры из 3-гранных металлических стержней шириной 5 см, подвешенных рядами на расстоянии 2 м 40 см друг от друга в экспериментальном бассейне, где содержались 2 афалины, подсчитывал, сколько раз животные во время плавания заденут за препятствия. В первые 20 мин. испытаний дельфины задели препятствия лишь 4 раза, и то хвостом уже после проплыва. Второй опыт был удачнее, а в последующих испытаниях животные проходили через лабиринт препятствий без единого столкновения. Хотя вода в бассейне была очень мутной и видимость составляла всего около 50 см, Келлог провел аналогичные опыты темной, безлунной ночью, однако и в этом случае дельфины на 100% уклонялись от препятствий.
В других опытах (Norris et al., 1961) дельфин афалина по кличке Кетти должен был по условному сигналу проплыть сквозь барьеры из вертикальных металлических стержней диаметром 2.5 см, расположенных друг от друга на расстоянии 1—2 м, и получить рыбу. Расположение препятствий изменялось в каждом опыте.
340
Зрение исключалось специальными присосками из латекса, надеваемыми на глаза животного. Авторы отмечают, что дельфин плавал по этому лабиринту несколько раз в день в течение 2 месяцев и только один раз коснулся стержня.
На основании этих экспериментов был сделан вывод, что, во-первых, дельфины, как и летучие мыши, используя эхолокацию, способны быстро обнаруживать и своевременно уклоняться от столкновения с многочисленными препятствиями и, во-вторых, применяемые металлические стержни диаметром 5—2.5 см не являются пределом эхолокационного обнаружения для дельфинов.
Попытка оценить порог восприятия сонарной системы китообразных с использованием металлических проволок впервые была предпринята французскими исследователями в 1965 г. (Busnel, Dziedzic, Andersen, 1965). Опыты проводились на одной взрослой самке — морской свинье (Phocaena phocaend) длиной 140 см, весом 35 кг. В бассейне площадью 54 м2 и глубиной 1.2 м устанавливались вертикально натянутые проволоки, диаметр которых от опыта к опыту менялся от 4 до 0.2 мм. Расстояние между проволоками составляло 1 м, и они располагались в шахматном порядке. В каждом опыте в воде находилось 30 препятствий.
Экспериментаторы в каждом опыте наблюдали за плаванием дельфина в бассейне в течение 20 мин. и за это время подсчитывали количество уклонений и встреч с препятствиями. Испытания проводили на животных, лишенных зрения (путем закрывания глаз каучуковыми присосками) и, следовательно, располагающих для обнаружения препятствий только одной сонарной системой.
В результате было показано, что,чем меньше диаметр проволок, тем большее число ошибок (столкновений) совершает животное. Так, проволоки диаметром от 4 до 2.8 мм ослепленная морская свинья преодолевала практически без столкновений (почти в 100%), с проволоками диаметром от 2.8 до 0.5 мм было 90% уклонений, тогда как при проволоках диаметром 0.2 мм число уклонений составляло всего 46 %. Когда животное получало информацию о наличии препятствий по двум каналам одновременно — зрительному и слуховому, способность уклоняться от препятствий оказывалась значительно выше только для критического диаметра 0.2 мм. Таким образом, проволоки диаметром 0.2 мм составляют практически предел эхолокационного обнаружения для морской свиньи.
В этой же работе приводятся некоторые характеристики излучаемых зондирующих сигналов морской свиньи во время обхода препятствий. Отмечается, что интенсивность сигналов составляла примерно 70 дб относительно нулевого уровня, тогда как уровень шума в глубине бассейна из-за всплесков и различных посторонних помех равнялся 35 дб. Продолжительность импульсов дельфина менялась от 3 до 1.2 мсек., а частота повторения импульсов в зависимости от расстояния животного до преграды могла дости
341
гать нескольких сотен герц около цели. Частота заполнения прй максимуме амплитуды сигнала составляла 2 кгц. Следовательно, проволоки диаметром 0.2 мм соответствовали всего 1/1500 длины излучаемой волны. Короткая длительность излучаемых сигналов, их уменьшение во времени по мере приближения животного к препятствию и одновременное нарастание частоты следования импульсов позволили предположить, что принцип определения расстояния до цели у дельфинов, так же как и у летучих мышей семей
ства гладконосых, основан на истинном эхо-принципе.
Изменение частоты следования импульсов у дельфинов описы
вается во многих других экспериментах. Так, например, в опы-
Частота (кгц)
тах Норриса и др. (Norris et al., 1961) по условному сигналу дельфин афалина начинал поиск куска рыбы с поперечным сечением 5 см, опущенного около гидрофона.
Вначале импульсы следовали с частотой 16 гц. Когда дельфин брал рыбу, частота возрастала до 190—416 гц и после этого падала до 26—50 гц.
Рис. НО. Ультразвуковой частотный спектр эхолокационных щелчков у дельфинов афалин. (Kellogg et al., 1953).
В большинстве исследований, посвященных эхолокационному поведению китообразных, отсутствуют точные сведения о дальности действия их гидролокационных систем. Лишь в одной из ранних работ Шевилла и Лоуренс (Schevill, Lawrence, 1956) встречаются указания на то, с какого расстояния дельфин обнаруживал небольших мертвых рыб длиной 8—23 см. Описание этих опытов можно найти в любой сводке на русском языке, посвященной поведению и эхолокации дельфинов (Griffin, 1961; Романенко, 1964; Томилин, 1965), что освобождает нас от необходимости подробно останавливаться на них.
Коротко напомним, что по условному сигналу (всплеску) дельфин, подплывая к лодке, в которой находились экспериментаторы, схватывал рыбу, опущенную в воду. Чтобы точнее определить расстояние, на котором дельфин еще обнаруживал рыбу, от лодки перпендикулярно берегу была протянута сеть. Сеть простиралась от берега на 2.4 м, и дельфин заранее должен был решить, с какой стороны сети ему плыть к лодке, чтобы найти рыбу. Рыба опускалась в воду попеременно то по одну сторону сети, то по другую, но без правильного порядка чередования. В трех случаях из четырех дельфин правильно выбирал сторону сети, хотя одинаково часто получал рыбу с обеих сторон. Однако если рыбы в воде не было, то даже в темную ночь дельфин почти никогда не подплывал к сети.
342
Авторы отмечают, что дельфин обнаруживал рыбу с помощью эхолокации с расстояний, как правило, не превышающих 6 м, хотя изредка приплывал брать рыбу и с 15 м, но считать достоверно установленным, что дельфин обнаруживал рыбу с 15 м, по-видимому, нельзя.
В наших экспериментах, о которых будет подробно сказано далее, дельфин белобочка обнаруживал рыбу (12—15 см длиной) в цементированном бассейне на максимальных расстояниях от 3 до 5 м, что согласуется с результатами Шевилла и Лоуренс.
Однако эти данные, разумеется, не могут служить действительными характеристиками гидролокатора дельфинов, так как сильная реверберация звуковых сигналов в бассейнах, по-видимому, значительно уменьшала дальность обнаружения. Кроме того, рыба не очень хороший объект для подобного рода исследований, так как практически невозможно определить, какова величина отраженного от нее сигнала. Предположение о том, что при обнаружении рыб дельфин ориентируется в основном на эхо-сигналы от плавательного пузыря, не соответствует действительности, так как дельфины с одинаковым успехом вылавливают небольшие куски рыбы и рыб, лишенных плавательных пузырей,
Характеристики локационных сигналов зубатых китов
Древним была хорошо известна способность зубатых китов издавать звуки. Об этом писали еще Аристотель, Плиний Старший и другие философы и естествоиспытатели Древней Греции. Но постепенно их замечательные наблюдения были забыты, и в науке надолго установилось представление, что океан — это «мир безмолвия».
Лишь широкое использование во второй мировой войне акустической аппаратуры позволило вновь открыть богатый мир подводных звуков, издаваемых различными морскими животными, в том числе и китообразными. Нередко зубатые киты оказывались источником серьезных недоразумений, связанных с обнаружением акустической службой непонятных шумов, принимаемых иногда за шум винтов подводной лодки. По-видимому, это и послужило стимулом для планомерного и всестороннего изучения характера звуков, издаваемых китообразными.
Изучение подводных звуков зубатых китов началось лишь с 50-х годов настоящего столетия (Schevill, Lawrence, 1949; Kellogg et al., 1953; McBride, 1956). И сразу же было обнаружено чрезвычайное их разнообразие как у одного и того же животного, так и у представителей различных видов и семейств. Особенно резкие отличия были выявлены между усатыми и зубатыми китами, о чем бодее подробно будет сказано цище,
343
Первоначально давались чисто субъективные описания звуков — щелчков, скрипов, свистов, визгов, трелей, воя, мяуканья, кряканья, мычания, вздохов, скрежета и т. д. В дальнейшем благодаря использованию сложной акустической аппаратуры, позволяющей записывать и снимать физические характеристики сигналов, издаваемых китами, появилась возможность как-то классифицировать эти звуки.
Первая такая попытка была предпринята в 1961 г. Лилли и Миллер, которые разбили все звуки, издаваемые дельфинами, на 3 типа. В первый тип были включены свисты частотой от 4 до 13— 20 кгц, во второй — короткие, широкополосные сигналы, слышимые как щелчки и появляющиеся всякий раз, когда дельфины начинали лоцировать окружающее пространство, и, наконец, в третий тип — комплексные звуковые волны высокой амплитуды, слышимые как кряканье, мяуканье, лай и т. д. (Lilly, Miller, 1961а). Этими же авторами отмечалось, что перечисленные типы сигналов наблюдаются в различных ситуациях в зависимости от конкретных задач животных, но иногда дельфин способен одновременно издавать звуки двух и даже трех типов, что, вероятно, значительно увеличивает возможные комбинации сигналов, но вместе с тем сильно усложняет их изучение и интерпретацию.
По биологическому значению представляется возможным все акустические сигналы зубатых китов условно подразделять на три группы: 1) эхолокационные щелчки — широкополосные высокочастотные импульсы, используемые для ориентации, обнаружения препятствий, поисков пищи, распознавания предметов и т. д.; 2) коммуникационные сигналы, издаваемые дельфинами преимущественно для связи и общения друг с другом, они в основном представлены свистами первого типа, по классификации Лилли и Миллер, и отчасти сигналами третьего типа; 3) сигналы эмоционального и подражательного характера — комплексные звуки третьего типа, биологическое значение которых изучено наиболее слабо (Bateson, 1968; Caldwell, Caldwell, 1968b; Pilleri, Knuckey, 1968; Poulter, 1968; Smith, 1969). Их легче всего вызвать путем электростимуляции отдельных образований головного мозга (Lylly, 1968).
Остановимся на рассмотрении сигналов, используемых дельфинами для локации.
Уже в первых экспериментах по исследованию эхолокационного поведения дельфинов различными авторами было установлено, что, как только дельфин устремлялся на всплеск от брошенной рыбы или других предметов, он начинал излучать серии коротких сигналов, воспринимаемых на слух как щелчки, последовательность которых изменялась по мере сближения животного с объектом. Именно эти наблюдения послужили основанием считать, что с помощью щелчков дельфины получают информацию о местонахождении предметов и об их качественных характеристиках, т. е.
344
щелчки используются животными в целях эхолокации. Способность генерировать локационные сигналы врожденная. Новорожденная амазонская иния уже имела сонар (Caldwell, Caldwell, 1972).
Что же представляют собой эхолокационные сигналы, или щелчки, дельфинов? В целом их можно охарактеризовать как короткие, широкополосные высокочастотные импульсные сигналы, как правило, с крутым передним фронтом, частота следования которых варьирует в зависимости от ситуации. При знакомстве с накопленным экспериментальным материалом нужно иметь в виду, что до настоящего времени вопрос о форме и параметрах локационных сигналов дельфинов не решен. Причина этого в том, что еще не определены требования, которым должна удовлетворять регистрирующая аппаратура, вносящая минимальные искажения в сигналы. Отсюда несоответствие результатов, полученных различными исследователями в отношении формы, длительности и спектрального состава локационных посылок дельфинов (Голубков и др., 1972).
По данным ряда авторов (Kellogg et al., 1953; Norris et al., 1961; Evans, Prescott, 1962; Busnel, Dziedzic, 1966b, 1966c, и др.), длительность локационных сигналов различных китообразных сильно варьирует — от 0.1 мсек, у стено до 10—25 мсек, у косатки. По мере сближения животного с объектом наблюдается тенденция к уменьшению длительности импульсов.
Для некоторых видов описаны щелчки, разделенные одной или большим количеством пауз, т. е. как бы двойные, тройные и т. д. У афалин описаны щелчки, имеющие до 11 пауз (Айрапетьянц и др., 1973). Кашалоты генерируют щелчки пачками из 6— 9 штук, длительностью 0.1—2 мсек, каждый (рис. ИЗ). Косатки генерируют пачки из 10—15 щелчков.
Спектральный анализ, проведенный впервые для сигналов афалин, показал, что, помимо основных частот, представленных в звуковом диапазоне, имеются различные ультразвуковые составляющие, вплоть до 170 кгц. В отношении широкой полосы представленных колебаний локационные щелчки несколько напоминают белый шум (Kellogg et al., 1953). Большая часть энергии концентрируется в диапазоне ниже 30 кгц. Приблизительный спектр ультразвуковых частот локационных щелчков афалин Келлог получил, проведя анализ большого числа сигналов. Этот спектр дает обобщенное представление об относительной интенсивности составляющих в щелчках (рис. 110).
Примерно в это же время Шевилл и Лоуренс (Schevill, Lawrence, 1953а, 1953b) записали на магнитную ленту локационные сигналы, при спектральном анализе которых были обнаружены составляющие с частотой 196 кгц. Так как эта частота была верхним частотным пределом используемой при записи аппаратуры, то нет никаких оснований полагать, что эти составляющие предельны в спектре
345
щелчков дельфина. Спектральный анализ сигналов другого вида дельфинов — гребнезубого дельфина (Steno bredanensis) — показал, что максимум концентрации энергии лежит в пределах 14—25 кгц (Busnel, Dziedzic, 1966а). Имеются указания на то, что в сигналах белухи наибольшую энергию также
несут частоты звукового спектра. В импульсах длинномордого дельфина другими авторами были зарегистрированы частотные составляющие до 208 кгц (Norris, Evans, 1967). У белобочки спектр локационных щелчков широкополосный и прости-. I рается в отдельных случаях до llillllilill , 163 кгц. Интенсивность варьировала от 10 до 130 бар (Титов, Томилин, 1970).
Дальнейшие исследования показали, что энергетический максимум I зондирующих сигналов большинства llllh.Liiil 1 изученных зубатых китообразных лежит в пределах 20—50 кгц. У отдельных видов излучение осуществляется в более низкочастотном диапазоне: у речной соталии —
Рис. 111. Динамика спектра локационного сигнала при дифференцировании сферических тел.
По оси абсцисс — частота; по оси ординат — амплитуда ^гармоник.
Рис. 112. Динамика изменения ширины спектра локационного сигнала для двух случаев дифференцирования сферических тел.
По оси абсцисс — порядковый номер выборки от старта до цели.
свыше 20 кгц (энергетический максимум в районе 8—15 кгц; Caldwell, Caldwell, 1970), у инии — до 16, у малой косатки энергетический максимум лежит в пределах 8— 12 кгц (Busnel, Dziedzic, 1968), у стенеллы — 8—12 (Busnel et al., 1968), у морской свиньи — 0.85—2.5 (в отдельных слу
346
чаях наблюдались сигналы до 100—130 кгц — Титов, Томилин, 1970; Титов, 1971), у гринды — 1.2—1.65, у косатки — 0.25— 0.5 кгц. Причина того, что большинство авторов (Busnel et al., 1965; Busnel, Dziedzic, 1967; Dziedzic, 1968) обнаружило сигналы морской свиньи, лежащие лишь в звуковом диапазоне, объясняется чрезвычайно низким уровнем сигналов, обычно не превышающим 3 бар на расстоянии 1 м от головы животного. Для сравнения напомним, что уровень аналогичных сигналов у афалины и обыкновенного дельфина превышает на том же расстоянии 1000 бар (Дубровский и др., 1970). Н. А. Дубровский и другие наблюдали сигналы до 100 кгц (предел возможности прибора). Высокочастотные импульсы с крутым передним фронтом регистрировались только вместе с низкочастотными, они всегда предшествовали низкочастотным. При этом средний интервал между началом высокочастотного импульса и центром низкочастотного остается практически постоянным.
Большой интерес представляет спектральный анализ сигналов зубатых китообразных. В нашей лаборатории изучалась динамика спектра сигналов, генерируемых в процессе обнаружения и дифференцирования металлических шаров и цилиндров. Для анализа брались отрезки магнитной записи локационных посылок длительностью 400 мсек, в различные моменты поиска и опознания предъявленных мишеней (рис. 111). Наблюдались изменения структуры спектура при соответствующих изменениях временных параметров и стационарность спектрального состава при соответствующей стабильности временных характеристик. Таким образом, очевидно, что в зависимости от локационной ситуации дельфин способен гибко управлять спектральным составом излучения, осуществляя перестройки на протяжении нескольких импульсов при подходе к цели (рис. 112).
Использование дельфинами в эхолокации широкополосных сигналов имеет ряд преимуществ, так как известно, что расширение полосы частот улучшает обнаружение слабого эхо-сигнала на фоне шумов и реверберации, т. е. широкополосные сигналы более помехоустойчивы, чем узкополосные.
Кроме того, использование широкополосных сигналов дает возможность получить информацию не только о расстоянии и местоположении объекта, но и о его природе.
Для дельфинов всех видов характерно увеличение частоты следования локационных сигналов по мере приближения к цели. Афалины, например, изменяли частоту следования сигналов от 16 гц в начале движения к пище по условному сигналу до 190 гц перед взятием рыбы. Когда же животное отплывало от стены бассейна с рыбой в зубах, темп излучения сигналов доходил максимум до 416 гц (Norris et al., 1961).
В экспериментах Эванса и Прескотта (Evans, Prescott, 1962), проведенных также на афалинах, частота следования сигналов
347
менялась от 1—2 до 525 гц, причем при максимальных частотах звук воспринимался как скрип петель открывающейся двери. Белько-вич и Резников (1971) наблюдали у афалины сохранение «спокойного» режима с частотой щелчков в пределах 10—50 гц вплоть до подхода на расстояние 0.4 м к лоцируемому неподвижному объекту. В этот период величина интервалов выдерживалась нестрого, меняясь от интервала к интервалу на 10—20%.
За 0.25 сек. до подхода к лоцируемому объекту животное переходило на «форсированный» режим. Мгновенная частота щелчков резко возрастала с 10—50 до 200 гц, а далее более плавно увеличивалась до 500 гц. В отличие от летучих мышей, у которых увеличение частоты локационных посылок совпадает с моментом обнаружения насекомого, дельфин переходит на «форсированный» режим только вблизи лоцируемого объекта (много позже того, как он его обнаружил и опознал), в процессе подготовки к его захвату.
Проведенные нами наблюдения за локацией афалин подтверждают изложенные выше данные (Айрапетьянц, Воронов и др., 1973). В наших экспериментах по распознаванию сферических и цилиндрических тел частота следования локационных посылок в начале локации, как правило, лежала в пределах 20—40 гц, резко возрастая в момент подхода к облучаемому объекту. Аналогичные изменения в темпе излучения импульсов мы регистрировали у другого вида дельфинов белобочек, у которых максимальное значение во время захвата рыбы превышало 400 гц. Эти дельфины генерируют сигналы сериями продолжительностью 1—3 или 5— 7 сек. (реже 10—20 сек.), с интервалами между сериями 3—5 сек. и более. При этом длительность эхолокационных импульсов изменялась от 0.4 до 7, реже 7—12 мсек, (в среднем 3—4 мсек.) (Титов, Томилин, 1970; Титов, 1971). Для белухи отмечены изменения частоты следования локационных импульсов от 15 до 210 гц (Fisch, Mowbray, 1962) в пределах серии, продолжительность которой могла составлять 1 сек. У морской свиньи по мере приближения к рыбе частота посылок возрастала от 40 до 600 гц, причем длительность импульсов сокращалась с 10 до 0.5 мсек. (Титов, Томилин, 1970).
Морозов и др. (1971) и Акопиан и др. (1971) на большом фактическом материале изучили у афалин изменение величины периода следования и частоты локационных посылок по мере сближения с лоцируемым объектом. Наблюдалось постепенное сокращение периода следования от 34.2 (для расстояния 9—10 м) до 20.7 (для 4— 5 м) и 13.5 мсек, (для 2 м). Авторы приходят к выводу, что каждая очередная локационная посылка генерируется только после прихода эхо-сигнала и обработки содержащейся в нем информации. Поэтому период следования щелчков будет всегда больше некой величины
348
где L — расстояние до цели, а С — скорость распространения звука в воде. На дистанции между 24 и 4 м время задержки генерации импульса, т. е. разница между величиной То и реальной величиной периода следования, составляет 20 мсек. При дальнейшем сближении с целью время задержки сокращается до 3 мсек. Авторы связывают это с улучшением приема эхо-сигнала на близком расстоянии и упрощением локационной задачи.
На основании приведенных выше характеристик локационных сигналов, в частности их краткости и нарастания частоты следования по мере сближения с объектом, можно предполагать, что при
Рис. 113. Осциллограммы одиночных щелчков кашалота. (Bakus, Schewill, 1966).
определении расстояния до цели дельфины используют принцип временной задержки, т. е. время между моментом излучения зондирующего импульса и приходом отраженного сигнала. Однако пока остается неясным, в однозначном или многозначном режиме действует эхолокатор дельфинов. Для ответа на этот вопрос требуются дальнейшие эксперименты.
Большой интерес по исследованию основных акустических характеристик и особенностей щелчков малых косаток и кашалотов, записанных в открытом море, представляют работы американских авторов (Backus, Schevill, 1966; Busnel, Dziedzic, 1968; Сог-cella, Green, 1968). Проводя анализ нескольких сот щелчков кашалота, они обнаружили сосредоточенность большей части энергии щелчков в диапазоне 200—32 гц. Доминирующей частотой щелчков была частота около 5 кгц. Изучение серий, содержащих порядка дюжины щелчков, показало, что частотная модуляция если и есть,
349
то очень незначительная. Однако для обнаружения частотной модуляции следует подвергнуть анализу более длинные серии.
Зарегистрированная частота следования щелчков у кашалотов ниже, чем у остальных зубатых китов. Она меняется от 1 до 50 гц, чаще всего составляя единицы герц.
При более подробном исследовании щелчков оказалось, что в действительности щелчок представляет собой целую серию коротких импульсов длительностью от 2 до 4 мсек. Число импульсов в щелчке меняется от 1 до 9; кроме того, варьируют интервалы между импульсами и их амплитуда (рис. ИЗ). Длительность интервалов между щелчками около 110 мсек.
В рассмотренных записях длительность щелчков менялась от 2 до 24 мсек. Уровень максимального давления составлял 35 дб относительно 1 мкбар на расстоянии 1 ярд (91.44 см).
Отмечалось, что серии щелчков у различных кашалотов отличаются друг от друга. Щелчки, составляющие одну серию, очень похожи друг на друга. Это явно выгодно при использовании их для эхолокации, так как в стае животное должно быть способным различать эхо своих щелчков от эха щелчков других животных.
Сигналы усатых китов
Сведения о том, что усатые киты, подобно зубатым, способны издавать подводные звуки, появились значительно позднее. Совершенно неожиданно на нескольких акустических станциях Тихого и Атлантического океанов были зарегистрированы незнакомые сигналы, похожие на стоны или крики. Крики были низкочастотные, лежащие в инфразвуковом диапазоне. Частота их была всего около 20 гц. Эти звуки слышались минут 15, затем исчезали, а когда их запеленговали, обнаружили, что источник этих звуков передвигается. Долго исследовали участок океана, откуда слышались звуки, использовали даже авиацию, но ничего не нашли. Заметили лишь, что появление звуков совпадало с появлением китовых стад. Но звуки, издаваемые зубатыми китами, уже были известны, тогда как новые звуки не походили на прежние. Лишь в 1961 г. было установлено, что источник звуков — усатые киты финвалы (Schevill, Watkins, 1962). Звуками усатых китов, долгое время считавшихся молчальниками, занялись вплотную.
В настоящее время зарегистрированы звуки 5 видов полосатиков (Томилин, 1955), правда характеристики их не приводятся. Есть сообщения о звуках серого кита Eschrichttus gibbosus (Eberhardt, Evans, 1962), финвала Balaenoptera physalus (Schevill, Watkins, 1966), горбача Megaptera novaeangliae (Griffin, 1955; Schevill, Watkins, 1966), гренландского кита Balaena mysticetus, южного кита Eubalaena glacialis (Schevill, Watkins, 1966). Обычно это низкочастотные (до 1 кгц) сложные «стоны» и «ропли» (крики) 350
(например, у горбачей). С финвалами связывают импульсные сигналы, зарегистрированные многими исследователями в Сев. Атлантике и Тихом океане. Частота сигнала 20 гц, длительность — 1 сек. Они продолжаются с регулярными интервалами несколько раз в минуту в течение 15 мин., затем следует пауза в 2—3 мин., и так в течение нескольких часов. Многие факты: обнаружение сигналов в определенные сезоны, распределение источников сигнала (1 на 300 кв. миль), «бесцельное» перемещение, низкая частота и высокий уровень импульсов (1—8 вт)—говорят о том, что сигналы могут издаваться крупным животным, например китом. Правда, в большинстве случаев авторы не решаются указать определенно вид кита, но Шевилл, Воткинс и Бакус (Backus, Schevill, 1966; Schevill, Watkins, 1966) приводят довольно убедительные доказательства в пользу финвала. Валкер (Walker, 1964) указывает, что скорость перемещения источника обычно 2—3 узла, но может достигать иногда 7—8 узлов. Вследствие большой мощности сигналов (1—25 вт) их можно обнаружить на мелководье на расстоянии 35 миль. Прерывание сигнала может быть связано с выходом кита на поверхность. Отмечена интересная корреляция при сравнении электрокардиограммы кита с временными характеристиками их сигналов.
Перкинс (Perkins, 1966) описывает сигнальные звуки финвала в диапазоне 1500—500 гц, длительностью 50—60 мсек. Ультразвуковые сигналы обнаружены лишь у синих китов (Beamisch, Mitchell, 1971). Возможно, они используются для эхолокации скоплений зоопланктона. Сигналы узкополосные, с частотой 21 и 31 кгц. Мощные низкочастотные звуки, от 12.5 до 200 гц, продолжительностью до 36.6 сек. записаны у берегов Чили (Cummings, Thompson, 1971). Они были самыми сильными из известных звуков, производимых китами. У серого кита Эберхард и Эванс (Eberhardt, Evans, 1962) регистрировали сигналы в диапазоне 40— 700 гц, с максимумом энергии на частотах 80—300 гц. Каждый сигнал длился около 0.1 сек. и состоял из 4—5 посылок; ни свистов, ни щелчков ими обнаружено не было. Шевилл (Schevill, 1964) обнаружил у серого кита звуки во время ухаживания, но не отмечал их во время кормления. Каммингс и др. (Cummings et al., 1968) пришли к выводу, что серые киты производят низкочастотные звуки: «стоны», «вздохи», «бульканье», «стуки». Особенно часты «вздохи». Они составляют 87% всех генерируемых звуков. «Вздохи» издаются ночью и днем, одиночными животными и в группах, но нерегулярно. Из 280 прослушанных животных во время зимовки 2/3 звуков не издавали. Их максимальное количество — 54 «стона» в час. Основная частота «стонов» лежит в пределах 20—200 гц, средняя длительность 154 сек., среднее давление 126 дб (от 85 до 159 дб). «Стуки», или щелчки, имели частоту 350 гц, с уровнем 95—116 дб. Авторы предполагают, что, кроме коммуникационной функции, «стоны» и щелчки используются для локации.
351
Воткинс (Watkins, 1967) описывает необычный звук выдоха с присвистом (wheering brow) у горбача. Этот звук слышим под водой и в воздухе и, видимо, продуцируется во время выдоха; состоит из частот до 2 кгц, длится около 2 сек. и не похож на звук нормального дыхания, который слышен в воздухе и не слышен в воде, непохож па обычные подводные звуки, которые не слышны в воздухе. Нормальный выдох у крупных китов длится 1 сек. Этот необычный звук записывали одновременно через гидрофон и микрофон от 3 горбачей в море. В течение 10 мин. записали 24 свистящих звука, перемежающихся звуками нормального дыхания этих же животных. Видимо, эти звуки издаются произвольно и явно отличаются от звуков при нормальном выдохе. Они много громче даже на воздухе (на 35 дб), чем нормальный звук выдоха. Под водой по частоте он был выше, чем обычные подводные звуки, но того же уровня громкости. В районе Бермудских островов были зарегистрированы продолжительные серии звуков — от 7 до 30 мин. («пение») горбатых китов (Payne, McVay, 1971). Функция их неизвестна.
Пока трудно определить, каков же биологический смысл этих звуков для усатых китов, однако предположение, высказываемое Томилиным (1955) по этому поводу, представляет интерес. Известно, что звуки средних частот под водой распространяются на 15—20 км, а ультразвуки — на 3—5 км. Следовательно, для более дальних локаций желательно использовать звуки более низких частот.
Область миграции усатых китов простирается на тысячи и десятки тысяч километров, причем они из года в год проходят один и тот же путь. Это возможно лишь при наличии эхо-сигнального аппарата, при том, что ориентирами становятся и рельеф дна, и близость берега, и глубина воды. Весьма возможно, что усатые киты, которые кормятся стайной рыбой, на значительном расстоянии лоцируют стаю.
В 1946 г. были открыты подводные звуковые каналы, по которым низкочастотные звуки и инфразвуки проходят тысячи километров. Наличие таких каналов может играть важную роль в ориентации китов, они образуются в связи с увеличением глубины.
При использовании подводных звуковых каналов киты могут ориентироваться по прибою у дальних океанических островов и по этим звуковым маякам определять свое местоположение во время миграций. Возможно, звуковые каналы позволяют китам-одиночкам находить друг друга на весьма значительном расстоянии. Но все эти вопросы, еще ждущие дальнейшего выяснения, лишь нащупывание в проблеме ориентации усатых китов, о которых почти ничего не известно по сравнению с зубатыми китами.
18
МЕХАНИЗМЫ ГЕНЕРИРОВАНИЯ ЗВУКОВ И НАПРАВЛЕННОСТЬ ИЗЛУЧЕНИЯ
Морфологические и физиологические особенности звукоизлучающего аппарата
Еще задолго до того, как были исследованы звуки, издаваемые китами, исследователей занимал вопрос о воздухоносных путях китообразных (Baer, 1826; Murie, 1870; Pouchet, Beauregard, 1885; Rawitz, 1900; Raven, Gregory, 1933, и др.). G разной степенью полноты было сделано описание периферической части дыхательной системы серого дельфина, гринды, обыкновенного дельфина, белухи, нарвала, обыкновенного и карликового кашалота, бутылконоса, фоцены, бесперого дельфина (Kukenthal, 1893; Hawell, 1927). Для понимания механизмов, лежащих в основе генерации звуков китообразных, знание дыхательной системы должно иметь первостепенное значение. Тем не менее противоречивость толкований, отсутствие единообразия терминологии во многом снижали ценность выполненных работ. Более поздние работы Губера (Huber, 1934), Лоуренс и Шевилла (Lawrence, Schevill, 1956), Клейненберга и Яблокова (1958) в известной мере расширили, уточнили указанные исследования и дали возможность с новых позиций рассмотреть функциональные возможности верхних дыхательных путей китообразных. Наиболее полная сводка по этому вопросу приведена в работе Клейненберга и др. (1964).
Как известно, все млекопитающие для производства звуков в той или иной степени используют голосовые связки. Считалось, что китообразные их не имеют. Почти аксиоматическое утверждение, подтвержденное авторитетом анатомов прошлого века (Baer, 1826), совсем неожиданно оказалось заблуждением. В настоящее время убедительно показано (Грачева, 1971) наличие в гортани афалины четко выраженной голосовой связки. Она не рудиментарна, но тем не менее ее функционирование по аналогии с таковой у наземных млекопитающих маловероятно, хотя некоторые исследователи (Гуревич, 1972а, 19726) допускают,
353
что локационные сигналы возникают в гортани и усиливаются акустическими резонаторами — надчерепными воздушными мешками. Выше перечисленное дает основание предполагать, что способы производства акустических сигналов у китообразных существенно отличаются от таковых у наземных млекопитающих.
Строение верхних дыхательных путей китообразных носит четко выраженный адаптивный характер. Это проявляется в том, что они надежно предохраняют дыхательную систему от попадания воды, приспособлены к апериодическому, «взрывному» дыхательному акту (длительностью 0.4—0.7 сек.) и наиболее рациональному использованию запасов воздуха.
Зубатые и усатые китообразные резко различаются по строению дыхательной системы в целом. Единственное сходство сводится к тому, что дыхательное отверстие открывается практически на темени, на самой высокой части головы. Сходство на этом и заканчивается: у зубатых китов дыхательные пути образуют обособленную систему, а у усатых китов, как и у остальных млекопитающих, они пересекаются в глотке с пищеводным путем и их временное разделение осуществляется с помощью надглоточного хряща. Наконец, все зубатые киты имеют сложную систему воздухоносных полостей — «мешков», примыкающих к надчерепным носовым проходам. У усатых китов они отсутствуют, но зато у них имеется своеобразный «гортанный мешок» (Бель-кович, Яблоков, 1963). Столь резкому различию строения дыхательной системы усатых и зубатых китов вполне соответствует и резкое различие в издаваемых ими акустических сигналах (глава 17).
У зубатых китообразных наружное дыхательное отверстие, ноздря, или «дыхало», расположенное в верхней части головы, имеет полулунную форму и замыкается плотным кожно-мышечным клапаном. Эти складки образуют как бы замок, так как выступ одной стенки носового прохода плотно входит в углубление на другой, предохраняя дыхательную систему от попадания воды. Задняя стенка носового прохода примыкает к круто поднимающимся вверх носовым и лобным костям черепа. Спереди она окружена мягкими тканями так называемого лобного выступа, жировой подушки (или «мелона» — «дыни»). Это крайне характерное образование, в большей или меньшей степени развитое у всех зубатых китообразных, действительно состоит в основном из специфической жировой ткани, пронизанной большим числом элементов соединительной ткани, сухожилий, мышц и нервных волокон. Они располагаются на далеко вытянутых вперед парных межчелюстных и верхнечелюстных костях, которые служат для него основанием и вместе с тем образуют столь характерный для зубатых китов рострум.
Почти сразу за клапаном от носового прохода отделяется в толщу лобного выступа первая пара воздушных полостей, или 354
мешков, — дорзальные. Они отходят вперед и вбок от носового прохода, располагаясь в толще жирового слоя, и окружены тонкой сухожильно-мускульной фасцией. С носовым проходом их соединяют довольно широкие отверстия с собственной кольцевой мускулатурой.
Вторая пара мешков — премаксиллярные — отходит в средней части носового прохода. Их вентральная сторона прилежит к костной стенке черепа, а спереди и сверху они окружены тканями лобного выступа.
Рис. 114. Схема строения назальных мешков у некоторых зубатых китообразных. (Белькович, Яблоков, 1963).
1 — белуха; 2 — морская свинья; 3 — кашалот; 4 — берардиус.
Третья пара мешков отходит от носового прохода в том месте, где он делится на два костных канала, проникающих сквозь череп. Это назофронтальные мешки, прилежащие к лобным и носовым костям черепа.
Такова самая общая схема строения воздухоносных полостей, которые окружают наружный носовой проход зубатых китообразных. Строение мешков у различных видов (рис. 114) значительно отличается, но здесь важнее подчеркнуть то, что их объединяет: 1) расположение вокруг носового прохода и сообщение с ним; 2) расположение на разной высоте, поярусно: 3) наличие собственной мускулатуры и эластичности стенок, что в целом позволяет менять размеры и форму полости, полностью ее изолировать от носового прохода или произвольно регулировать степень такого сообщения; 4) расположение между костной стецкой черепа, круто поднимающейся вверх, и лобным выступом»
355
Другой характерной чертой строения воздухоносных путей зубатых китообразных надо считать их разделение на ряд участков, в которых давление воздуха различно. Рассмотренный выше носовой проход с системой воздушных мешков — первый такой участок. Давление воздуха в нем линейно следует за изменением гидростатического давления. Второй изолированный участок — полость костных носовых проходов с небольшой хоанной полостью вплоть до кольцевого сфинктера, удерживающего и замыкающего вершину гортанной трубки. От первого отдела он отделен двумя клапанами — «губами», которые располагаются между второй и третьей парами мешков (Lilly, 1962). Третий участок — полость гортани, трахеи и крупных бронхов вплоть до системы сфинктеров на бронхах 3—4-го порядка. Давление воздуха здесь на 20 мм рт. ст. превышает гидростатическое, если взять за точку отсчета давление на среднем уровне тела животного (Lilly, 1962), в то время как на втором участке оно практически может не отличаться от атмосферного (Яблоков, 1961). В бронхах, а также в альвеолах из-за «проникающего» давления воды поддерживается гидростатическое давление, равное давлению воды.
Отсюда следует, что при выдохе—вдохе весь воздухоносный путь представляет собой единую систему, а при нырянии происходит его разделение мышечными сфинктерами и клапанами на описанные участки с различным давлением воздуха, и таким образом между ними создается перепад давлений. Особенно значителен он между первым и вторым и вторым и третьим участками, так как второй участок — костная несжимаемая трубка. Здесь давление близко к атмосферному, а в первом и третьем участках оно возрастает линейно с глубиной погружения и равно гидростатическому либо выше его, как в третьем участке. Кроме того, разница в давлениях в первом участке может увеличиваться и произвольно за счет активного сокращения мускулатуры наполненных воздухом мешков. Вот, пожалуй, те необходимые анатомические и отчасти функциональные посылки, которые могут нам пригодиться для рассматриваемого вопроса.
Первоначально по аналогии с функционированием голосовых связок наземных млекопитающих предполагалось, что китообразные (дельфины) могут издавать звуки (писк) за счет вибрации наружных краев дыхательного отверстия или перегородок мешков при продувании через них воздуха непосредственно из легких (Томилин, 1955). При этом под водой из дыхала вырывается струйка пузырьков воздуха.
Все остальные типы звуков не сопровождаются выделением воздуха наружу.
До последнего времени механизм генерации звуков остается недостаточно ясным. Достоверно известно лишь, что источник (или источники) звука связан с губами дыхала, диагональной мембраной, двумя внутренними клапанами, тремя парами воз-356
душных мешков и гортанью. Исходя из морфологии этого отдела дыхательной системы зубатых китов, можно представить следующую картину ее функционирования при производстве акустических сигналов. После выдоха-вдоха дыхало закрывается клапаном, описанным выше, в наружном носовом проходе имеются второй клапан и губы почти у границ с началом костных носовых проходов. Эту роль выполняют антеро-латеральные мешки (Huber, 1934; Клейненберг, Яблоков, 1958) и специальная мускульная пробка (Lawrence, Schevill, 1956). Выдох у большинства зубатых китов есть активная часть дыхательного акта и занимает всего 0.3—0.4 сек. Вдох происходит пассивно за счет расслабления мышц грудной клетки и диафрагмы. Наблюдения В. М. Белько-вича над дыхательным актом дельфинов показали, что, видимо, первым закрывается только наружный клапан. При этом сразу же происходит некоторое сокращение грудных мышц и диафрагмы, которое повышает давление воздуха во всей системе. По измерениям в гортани, оно составляет примерно 20 мм рт. ст. (Lilly, 1962). Это повышение давления воздуха создает условия для наполнения и расправления всех мешков наружного носового прохода. Лишь после этого может происходить перекрывание воздухоносных путей на ряд участков. Рентгенологический анализ дыхательного акта дельфина белобочки (Гуревич и др., 1972) показал, что в воздушных мешках никогда не бывает значительных объемов воздуха. Необходимо заметить, что представление о постоянном разделении воздухоносных путей на ряд участков (Яблоков, 1961; Белькович, Яблоков, 1963) с разным давлением воздуха в них как о безусловнорефлекторном акте, обязательно сопутствующем погружению животного и вызываемом им, оказалось ошибочным (Белькович и др., 1967). Об этом свидетельствует то, что животные под водой могут произвольно выпускать из дыхала огромные пузыри воздуха (примерно 0.2 объема легких). Они прибегают к этому для выражения эмоционального состояния недовольства (белобочка) и для резкого смещения центра тяжести тела (белобочка и афалина), например, чтобы с разгона встать в воде вертикально. Эти наблюдения показали, что дыхательный акт в целом условнорефлекторный и более сложный процесс, чем это представлялось ранее, процесс, произвольно управляемый самим животным.
Для решения вопроса о механизмах производства звуков рассмотрим гипотезу о временном и произвольном разделении дыхательного тракта на ряд участков. При этом перепад давлений между первым и вторым, вторым и третьим участками будет реально иметь место, как показано выше. Кроме того, повышение давления внутри мешков за счет сокращения собственных мышц создает предпосылки для создания нужного давления при той или иной глубине погружения и для тонкой регулировки перепускания воздуха с тем, чтобы создавать все много-
357
образие эхолокационных и сигнальных звуков, менять их интенсивность, частоту и т. д. за счет струйных и вибрационных эффектов.
Один из способов производства свистовых сигналов с выходом из дыхала гирлянд пузырьков был описан. Другой вероятный способ состоит в пережимании воздуха из одного мешка в другой в замкнутой системе наружных дыхательных путей без потерь запаса воздуха. При постоянном объеме носового канала выжимание воздуха из одного или нескольких мешков приведет к повышению давления в нем, что может быть использовано для увеличения давления в другцх мешках. Кроме того, расслабление мышц «выжатого» мешка позволит ему вновь наполниться воздухом. В итоге представляется, что при замкнутой системе генерация сигнала может продолжаться дольше, чем длится дыхательная пауза. На основании морфологического изучения системы наружного носового прохода афалины и обыкновенного дельфина Белькович и Резников (1971) считают, что локационные сигналы генерируются при прохождении воздуха между стенками наружного носового прохода и внутренними носовыми пробками. Направление и скорость потока воздуха могут обеспечиваться двумя парами мешков, вестибуляторными наверху и премаксил-лярными снизу. При этом предполагается, что тубулярный мешок, охватывающий наружный носовой проход в его средней части может менять натяжение стенки, противолежащей внутренней носовой пробке, и тем самым регулировать частоту колебаний при прохождении воздушного потока. Каждая из пробок способна функционировать независимо от другой. Кости черепа, вероятно, отражают вперед задний лепесток восьмеркообразной мгновенной диаграммы направленности, а жировая подушка ее сужает или расширяет.
Изложенные выше новые представления о произвольном управлении животным разделением дыхательного тракта на участки указывают и второй способ для создания разницы давлений в первом участке. После вдоха открытие клапанов всего дыхательного тракта под водой, кроме клапана дыхала (назального), наполнит все мешки воздухом под гидростатическим давлением. Затем тракт вновь перекрывается на участки и воздух (весь или часть) из первого участка удаляется в воду. Тем самым в первом участке создаются регулируемое разрежение и благоприятные условия для генерации сигналов по второму способу.
Предложенные выше предположительные схемы генерации звуков предполагают достаточно сложный механизм с точной регулировкой всех фаз его деятельности. Между тем односторонняя перерезка лицевого нерва у морской свиньи (Агарков, 1972) не вызывала заметных изменений спектра локационных посылок, сказавшись лишь в нестационарное™ частоты повторения звуковых посылок, 35g
Мы рассмотрели Принципиальные возможности производств^ Сигналов главным образом в первом участке — наружном носовом проходе. Сказать же с уверенностью, какая из описанных выше структур производит те или иные звуки, в настоящее время не представляется возможным.
Исследуя сигналы дельфина белобочки, Вюснель и Дзедзик (Busnel, Dziedzic, 1966а) отметили, что свист длительностью 1.1 сек; может прерываться одним или серией щелчков, а затем вновь продолжаться. Эти же авторы показали, что свист и щелчки могут производиться одновременно как дельфином белобочкой, так и гриндой. У белобочки свист в этом случае состоял из одной части длительностью 250 мсек., с падением частоты до 6 кгц и другой части длительностью 350 мсек., со средней постоянной частотой около 10 кгц. На этом фоне продуцируются щелчки с интервалом 0.5 мсек. У гринды зарегистрированы свисты с одновременным скрипом (щелчки с высокой частотой повторения). Один тип таких сигналов характеризуется основной гармоникой и 3— 4 гармониками, уменьшающимися по интенсивности либо сразу после подъема частоты, либо после следующего за ним понижения частоты. Начальная частота 4—8, конечная — 1—4 кгц. В другом двойном сигнале, длительностью 1.4—2.7 сек., на фоне также частотно-модулированного свиста продуцировались скрипы значительной интенсивности, но в 2—3 раза короче. В третьем двойном сигнале, при частоте порядка 3 кгц и длительности 550 мсек., щелчки длились 180—270 мсек., а свисты — 160—400 мсек. Все эти примеры показывают, насколько животное может быстро и произвольно менять характер генерирования сигналов, а также говорят о наличии двух источников, работающих одновременно. Кроме того, анализ длительных свистов гринды, в 700—800 мсек., позволил авторам предполагать наличие одного источника для свистов частотой 3 кгц и второго — для свиста частотой порядка 11 кгц. Ими же отмечен специфический резкий свист: серии по 5 и более почти синусоидальных колебаний без гармоник, с непостоянной частотой модуляции, с периодом колебаний 0.2 сек. Имеется ли для его производства специфический источник, или вся система достаточно полифункциональна, пока остается невыясненным.
В условиях экспериментального изучения поведения афалины также многократно отмечали (Lilly, Miller, 1961а, 1961b) одновременное звучание свистов и щелчков, свистов и визга, лая и других сложных звуков. Кроме того, эти авторы полагают, что щелчки могут иметь не только локационный, но и связной характер. Первые (локационные) имеют частоты свыше 20 кгц при длительности 0.1 мсек., вторые (связные) имеют частоты 1—20 кгц и длятся до 5 мсек. Щелчки обоих типов могут издаваться одновременно и раздельно, что дает авторам основание считать, что имеется по крайней мере два источника звучания.
359
О совершенстве и сложности системы генерации Сигналов свидетельствует сильно варьирующая частота повторения щелчков у афалины, хотя возможно, что это связано не столько с локационными задачами, которые при этом решались животным, сколько с его индивидуальными особенностями, или же вообще носит ошибочный характер в случае крайне высоких значений. То обстоятельство, что локационные щелчки афалины часто парные, с интервалом в 0.4—0.7 мсек. (Norris et al., 1966), говорит об очень стабильной работе системы генерации сигналов.
Многие исследователи полагают (Schevill, 1964), что свисты возникают в удлиненных хрящах гортани, хотя есть мнение об их продуцировании носовыми мешками (Томилин, 1955; Lilly, 1962). Относительно щелчков мнение большинства исследователей совпадает, и считается, что их производят мешки наружного носового прохода (Wood, 1954; Evans, Prescott, 1962, и др.). Некоторое экспериментальное подтверждение важности гортани, костных и наружных носовых проходов для генерации звуков было получено при пропускании через эти отделы воздуха под давлением 0.07—0.14 атм. в голове мертвого дельфина. Полученные при этом звуки оказались сходными с естественными (Evans, Prescott, 1962).
В производстве звуков у усатых китов, очевидно, главную роль играет гортань, к которой прилежит своеобразный, отсутствующий у других млекопитающих гортанный мешок. Функция последнего еще неясна: возможно, он служит своеобразным резонатором или усиливает инфразвуки (Белькович, Яблоков, 1963). Существует мнение, что легкие как таковые ответственны за низкочастотные звуки у усатых китов, но вряд ли это так. Во всяком случае необходимо показать, что частота сигналов у усатых китов систематически меняется с глубиной погружения и связана с размерами животного.
У горбача (Watkins, 1967) свистящий звук начинается увеличением амплитуды и постепенным поднятием частотных компонентов, что совпадает со сжатием губ дыхала. Во время остального выдоха частота сигнала почти постоянна, но падает к концу, по мере того как падает давление воздуха или ослабевает напряжение дыхала. Явное частичное сжатие губ дыхала и их вибрация во время звучания, видимо, указывают на то, что звук производится дыхалом во время выдоха (звук издается на поверхности, но слышен и под водой). Возможно, наоборот, поток воздуха заставляет вибрировать губы дыхала. Анализ одного и того же звука, записанного через микрофон (т. е. на воздухе) и через гидрофон (подводный звук), позволил обнаружить разницу между ними. В звуке на воздухе преобладают сильные низкочастотные звуки и непрерывность спектра. В подводном сигнале преобладают дискретные частоты. Это дает основание полагать, что в воздухе звук продуцируется вибрацией и завихрениями выдыхаемого воздуха,
360
а в воде — только вибрацией воздуха в респираторном тракте. Этот звук в воздухе в какой-то мере аналогичен звукам (некоторым) дельфинов, поощряемых тренером. Он производится с открытым дыхалом вне воды, так что звук хорошо излучается в воздухе, но не слышен под водой. Лай частично погруженных тюленей и морских львов относится к этой же категории. Он отличен от истинных подводных звуков этих животных, которые издаются с закрытым ртом и ноздрями без выпускания воздуха.
Направленность излучения локационных сигналов
Вся система, генерирующая звуки у зубатых китов, как уже отмечалось выше, располагается на костном ложе, образованном костями черепа. Перед ними располагается жировая подушка, имеющая различную форму у разных видов, но обязательно присутствующая у всех. Было высказано предположение, что стенки черепа служат как бы рефлектором (Белькович, Яблоков, 1963), лобный выступ — своеобразной «линзой» (Evans, Prescott, 1962; Белькович, 1972), концентрирующей, направляющей акустический пучок, т. е. голову зубатых китообразных можно в известной мере сравнить с «акустическим прожектором» (Белькович, Яблоков, 1963).
Однако надо иметь в виду, что кости в воде являются для. звуковых волн «полупрозрачным» материалом (Белькович, Нестеренко, 1971). Даже весьма плотная кость слонового бивня отражает лишь 35% энергии нормально падающей звуковой волны. По данным Чевненко (1969) и Мастрюкова (1971), коэффициент отражения ультразвука на границе воды с жиром и костью черепа соответственно равняется 0.37 и 37 %, а на границе жир—мышца — 12%. Поэтому мышечный или костный рефлектор следует рассматривать как ненаправленный отражатель с плохим кпд.
Использование костного рефлектора находит известное подтверждение при анализе строения надчерепных носовых проходов и конфигурации черепа у зубатых китообразных с различной экологией. Оказалось, что наиболее совершенными отражательными поверхностями обладают черепа речных дельфинов, кашалотов и клюворылых китов (Белькович, Яблоков, 1963; Norris, 1964). Первые живут постоянно в условиях очень мутной воды (Амазонка, Ганг), и некоторые из них даже теряют зрение, другие — кашалоты и клюворылы — питаются на таких глубинах, куда не проникает свет. Интересно, что кашалоты и клюворылы обладают самыми сложными по строению системами воздухоносных проходов и мешков.
Концентрирование акустических волн жировой подушкой связано с ее преломляющим действием, основанным на изменении фазовой скорости волны в жировом веществе линзы по сравнению
361
со скоростью в окружающей среде. Сферическая форма жировой подушки позволяет ее рассматривать как замедляющую линзу с двумя преломляющими плоскостями, способную благодаря этому создавать на своей наружной поверхности то или иное фазовое и амплитудное распределение, а значит, преобразовывать сферический фронт волны в плоский и формировать диаграмму направленности.
Жировая подушка нерезко отграничена от окружающих тканей. Лишь ее центральные районы состоят почти исключительно из жировых клеток, чем снижается отражение от пограничного слоя и увеличивается коэффициент преломления центральных отделов. Неоднородность вещества жировой подушки создает дополнительные возможности для управления фазовым и амплитудным полем (Cummings et aL, 1968; Lord, 1969; Каневский, Суриков, 1970, 1971). Перемещая источник излучения в направлении, перпендикулярном оптической оси линзы, или линзу относительно источника излучения, можно осуществлять сканирование диаграммой направленности. Изменяя форму лобного выступа с помощью окружающих мышц, дельфин, очевидно, способен фокусировать акустический пучок. Кроме того, путем изменения конфигурации осуществляется определенная корреляция направленности излучения при значительных изменениях среды: температуры, солености, гидростатического давления.
Акустическая линза дельфина имеет небольшой размер, поэтому на ее долю падает лишь незначительная часть энергии излучателя. Остальная часть акустических лучей или гасится окружающими тканями, главным образом мышцами, или попадает в линзу вторично, после отражения от них, что увеличивает сложность структуры излучаемых сигналов. Как указывалось выше, кости черепа, вероятно, отражают вперед задний лепесток восьмеркообразной мгновенной диаграммы направленности, а жировая подушка ее сужает, расширяет или меняет направленность.
Фокусирующее действие жировой подушки не имеет пока бесспорных доказательств. Модельные эксперименты, проведенные на свежем трупе дельфина афалины, как будто не подтверждают предположение о том, что жировая подушка выполняет роль акустической линзы. Как сигналы разной частоты, излученные при одном и том же положении гидрофона, так и сигналы одной частоты, излученные из разных точек наружного надчерепного прохода, могли иметь различную направленность.
Особенности распространения излучаемых звуков впервые исследовались Норрисом с соавторами (Norris et al., 1961). Эти эксперименты свидетельствуют в пользу направленности звуко-излучения. Звуки афалины регистрировались с помощью гидрофона, чувствительного к колебаниям в узком диапазоне, в области 100 кгц. Наиболее эффективно гидрофон воспринимал сигнал животного при строгой направленности головы дельфина
362
на него. Эти же исследователи выявили способность дельфина обнаруживать куски рыбы в различных местах около головы. Во время этого опыта глаза животного закрывались специальными присосками. Оказалось, что быстрее всего рыба обнаруживалась над челюстями и не обнаруживалась совсем ниже уровня нижней челюсти.
Экспериментальные данные о направленности излучения у дельфинов были получены Эвансом и Прескоттом (Evans, Prescott, 1962) при продувании воздуха через дыхательный тракт (включая гортань) головы мертвой афалины и продельфина. Полученные ими диаграммы распространения сигналов в вертикальной и горизонтальной плоскостях указывали на слабую направленность их. Диаграммы строились по данным измерений общего уровня давления на расстоянии 38 см от головы с помощью конденсаторного микрофона в воздушной среде.
Эванс и др. (Evans et al., 1964) приводят данные по измерению звукового давления от миниатюрного источника, помещавшегося в различных местах туши над черепом и в области гортани неполовозрелого продельфина (длиной 78 см) и черепе взрослой афалины. Измерения проводили под водой как в горизонтальной, так и в вертикальной плоскостях через каждые 15°. Хотя измерения делали при частотах 20—70 кгц, приводятся результаты только для низших, средних и высших частот этого диапазона (для продельфина — 20, 40 и 60 кгц, для афалины — 40, 50 и 70 кгц).
Полученные данные показывают, что звук, производимый над черепом в области носовых мешков, отражается вверх и наружу определенным лучом. Угол максимального сигнала вверх составляет 15° выше рострума, а в горизонтальной плоскости равен 30—40° вправо и влево от рострума. Когда источник помещен ниже уровня межчелюстных костей, недалеко от гортани, область максимального звучания располагается под углом 90° вниз от линии рострума, как раз под животными, и очень мало энергии звука идет вперед. Во всех экспериментах ширина луча строго зависела от частоты сигнала. В случае продельфина ширина луча (вертикального) для падения уровня сигнала на 10 дб была равна 170° для 20 кгц, 100° — для 40 и 45° — для 60 кгц. Точка максимального уровня излучения, определенная графически, зависит от положения источника. Данные по направленности на черепе афалины, полученные экспериментально и графическим методом, до некоторой степени согласуются.
Романенко (1964), Романенко и др. (1965) приводят диаграммы направленности излучения у дельфина белобочки, полученные следующим образом.
Излучатель-сферу (диаметр 1.6 см) из керамики титаната бария помещали в отрезанной голове в области воздушных полостей. Приемник-сферу из титаната бария (диаметром 3 см) помещали на расстоянии 1.5 м от излу-
363
чателя. Отсчеты производились при погружении системы в воду на глубину 1 м. Голову поворачивали относительно вертикальной оси, проходящей через излучатель. Режим работы излучателя был импульсный. Длительность импульса 1 мсек. Скважность импульсов обеспечивала полное затухание сигналов, отраженных посторонними предметами. Отсчеты снимались при частотах 10—40, 60, 80, 100, 110, 140, 170 и 180 кгц.
На частотах 30 кгц диаграмма направленности имела ширину около 90°, а на частоте 180 кгц — 17°. На частотах 80 кгц и выше диаграмма направленности была вдвое шире, если голову перед опытом освобождали от мягких тканей.
Аналогичные модельные эксперименты были поставлены В. И. Марковым на свежем трупе афалины.
Опыты проводились в ванне, стенки которой для уменьшения отражения экранировались рупорной резиной. Сферический излучатель из пьезокерамики ЦТС-19 диаметром 10 мм вводился последовательно через дыхало в 9 точек наружного надчерепного хода: 1-я — на глубине 3.5 см, у устья вестибулярных мешков; 2—3-я — на глубине 9 см, в левом или правом носовых проходах, у устья премаксиллярных мешков; 4-я — на глубине 25 см, у гортани; 5-я—на глубине 10 см, у дна левого вестибулярного мешка; 6-я— на глубине 7 см, у дна правого (меньшего по размерам) вестибулярного мешка; 7-я — на глубине 5 см, в правом носовом проходе, на уровне устья тубулярного мешка; 8-я и 9-я — на глубине 15 см, в правом или левом носовом проходе.
При каждом из 9 положений излучателя подавались короткие посылки тональных сигналов с частотами 20, 40, 60, 80, 100, 120, 140, 160 кгц. Излучаемые сигналы контролировались путем последовательного перемещения приемного гидрофона в 7 точек пространства, расположенных в непосредственной близости от головы трупа. Три из них располагались в сагиттальной плоскости, 5 — в горизонтальной. Результаты измерений для каждой частоты брались отдельно и пересчитывались относительно соответствующих значений звукового давления в точке 1, лежащей на продольной оси тела животного в месте пересечения сагиттальной и горизонтальной плоскостей.
Результаты исследования показали, что в сагиттальной плоскости при всех положениях излучателя наблюдается обострение диаграммы в направлении продольной оси тела, особенно значительное при генерации сигнала у гортани, на уровне сфинктеров премаксиллярных и тубулярных мешков.
В горизонтальной плоскости диаграмма направленности имела пик в направлении оси тела для всех положений излучателя только на частотах 20 и 40 кгц. Начиная с 60 кгц при некоторых положениях излучателя возникала асимметрия диаграммы или появлялись боковые лепестки. Наиболее симметричной диаграмма была при нахождении излучателя у гортани, у устья премаксиллярных мешков, в глубине правого и левого носовых проходов. Наиболее асимметричны диаграммы при неглубоком погружении излучателя. Важно отметить, что и для сагиттальной плоскости при одной и той же частоте, но при различных положениях излучателя пики диаграммы могут иметь различное направление. Таким образом, модельные опыты показывают, что при излучении 364
сигналов в некоторых точках наружного носового прохода, которые предположительно можно оценивать как места генерации звука, направленность излучения может не совпадать с направлением оси тела животного. Как мы увидим из дальнейшего, данные модельных экспериментов получили подтверждение при изучении реальной направленности зондирующих сигналов дельфина, излучаемых в процессе поиска рыбы.
Еще один модельный эксперимент, в ходе которого регистрация звукового давления осуществлялась в горизонтальной плоскости для трех точек, расположенных на линии, перпендикулярной оси тела, вынесенной на 20 см вперед от кончика рострума, показал, как тонко изменяется направленность излучения при относительно незначительных смещениях излучателя. Регистрирующие гидрофоны помещались: один — на линии, продолжающей ось тела, два других—в 20 см вправо и влево от него. Излучатель устанавливался на уровне сфинктеров правого или левого премаксиллярных мешков, т. е. на 2 см в сторону от продольной оси тела. При этом наблюдалось значительное превалирование уровня звукового давления, регистрируемого гидрофоном, смещенным в ту же сторону от средней линии. Этот эксперимент свидетельствует о том, что, если животные в состоянии изменять местоположение генерации локационных посылок, они имеют возможность осуществлять тонкое сканирование диаграммой направленности.
Полученные диаграммы нельзя, конечно, полностью отождествлять с диаграммами направленности живого дельфина: излучатель, использованный в опытах, существенно отличается от естественного излучателя животного. Кроме того, посмертные изменения тканей тела несомненно влияют на акустические свойства лобного выступа. Впервые эксперименты на живых объектах были проведены Норрисом и Эвансом (Evans et al., 1967). В круглом бассейне гребнезубого дельфина (Steno bredanensis), «ослепленного» с помощью присосок, заставляли двигаться по диаметру между 2 натянутыми тросами. Звуки животного записывали с помощью одного гидрофона, погруженного на 1 м в воду, с последующим анализом частотного состава и звукового давления в различных точках звукового поля (гидрофон перемещался по периметру бассейна). Исследователями отбирались записи, соответствующие одному и тому же положению головы дельфина относительно пути следования его к цели (т. е. когда звук издавался в одной постоянной точке). Анализ показал, что звуковое поле — узкое и направлено вперед. Высокочастотные компоненты выявлялись только впереди животного и очень резко падали по обе стороны от оси тела. Полученные данные не позволяли судить о том, направлены ли высокочастотные компоненты по средней линии или излучались асимметрично. Звуковое давление было много выше во фронтальном спектре, чем с боков, и имела
365
Место асимметрия (рис. 115). Ёысшая Частота, порядка 208 кЩ, зарегистрирована только при 0° и в секторе, ограниченном 5°. По обе стороны от средней линии частота стремительно падала до 150 кгц; при 12° — до 100, при 17° — до 50 кгц с каждой стороны. Значительная часть звуковой энергии обнаружена на частотах порядка 20 кгц (высшая частота при углах 20—25° с каждой стороны от средней линии). Межщелчковый интервал при 0° составил в среднем 1.8 мсек, и увеличивался до 4.5 мсек, при 40° (в среднем). Остронаправленное поле было несколько шире в от-
Рис. 115. Диаграммы направленности излучения зондирующих сигналов, измеренные у гребнезубого дельфина. (Norris, Evans, 1967).
А — диаграмма изменения интенсивности; Б — диаграмма изменения частоты.
ношении звукового давления, чем в отношении частоты. Пик и близкие к нему значения давлений менее концентрированы в наружных долях, но еще хорошо фокусированы. Звуковое давление падало на 1/2 (от уровня при 0°) при отклонении на 10— 12° в стороны (—6 дб). Оно уменьшалось еще вдвое при угле 24—26° и было меньше на 20 дб на траверзе животного (90°). Сзади животного регистрировали только низкие частоты и низкое звуковое давление.
Карадагским отделением Института биологии южных морей изучалась направленность излучения в горизонтальной плоскости у морских свиней. Опыты ставились на животном, свободно передвигающемся в вольере размером 18x13x2 м. Регистрация звукового давления велась на двухдорожечном магнитофоне с двух приемных гидрофонов, разнесенных друг от друга на 5 м. Регистрация местоположения и ориентации животного в простран
366
стве осуществлялась визуально по отметкам, делящим вольер на квадраты со стороной, равной 1 м. Отсчет звукового давления начинался, когда голова животного находилась на расстоянии 2.5 м от обоих гидрофонов, а продольная ось тела была направлена на один из них. Таким образом, второй гидрофон в момент записи всегда находился под некоторым углом к оси тела. Момент нахождения дельфина в точке, равноудаленной от обоих гидрофонов, определялся путем анализа времени достижения звуковой волны обоих приемников. Анализ экспериментального материала показал, что в диапазоне 0.8—4 кгц диаграмма направленности излучения имеет круговую форму, т. е. излучение одинаково во всех направлениях.
Заславский (1972) изучал направленность излучения у афалины в бетонированном бассейне с помощью двух гидрофонов и двухканальной магнитофонной регистрации сигнала. Положение животного определялось на основе разности во времени прихода локационной посылки на каждый из гидрофонов. Максимум излучения в горизонтальной плоскости находился в секторе 14°. Ширина сектора направленности, измеренная по уровню 3 дб от максимума, составляла приблизительно 30°, ширина раствора лепестка — около 160°. Излучения позади дельфина в секторе от 90 до 270° вообще не наблюдалось. Ширина диаграммы в вертикальной плоскости оценена в 70°. Направленность излучения в горизонтальной плоскости на разных частотах в пределах спектра импульса была одинаковой.
Яблоков и др. (1972) приводят данные о диаграмме направленности излучения дельфина афалины, полученные с помощью одновременной регистрации локационных сигналов на четырех каналах и одновременной киносъемки двумя кинокамерами, позволявшими определять положение животного в пространстве в системе трех координат. Проведенные эксперименты позволили прийти к заключению, что в среднем максимум акустического излучения по уровню давления 0.5 в горизонтальной плоскости сосредоточен в углу +20° от оси головы (рис. 116) и слегка смещен вправо. В полупространстве ниже уровня рта прямой сигнал обнаружен лишь до углов 30—40° с амплитудой 0.2—0.3 от уровня максимума. Значение мощности в импульсе первичного акустического излучения (приведенное к эквивалентному направленному излучателю) оказалось порядка 0.1 вт, а результирующая амплитуда давления на оси излучения (приведенная к расстоянию 1 м от источника) — порядка 103 бар. В условиях уверенной локации амплитуда зондирующего сигнала остается приблизительно на одном уровне и не зависит от расстояния до объекта.
Несомненно, что узкая направленность звукоизлучения выгодна для животного, так как не требует значительных энергетических затрат и может обеспечить высокую разрешающую способность.
367
Однако предположение о постоянной направленности излучения по оси тела животного не могло быть принято безоговорочно, так как в этом случае был бы сильно затруднен широкий обзор
животным окружающего пространства, что явно не соответствует действительности. В связи с этим возникло представление, что дельфины сканируют звуковым лучом, поворачивая голову. Действительно, большинство исследователей отмечает, что во время движения, особенно в момент локации, животные покачи
вают из стороны в сторону головой, поворачивая ее на 10—13°. Однако есть основание считать, что дельфины обладают более совершенным механизмом сканирования и могут произвольно менять ширину луча (зону облучения), то делая его узконаправленным, то достаточно широким.
Первые исследования по изучению сканирования осуществлены Бельковичем и Резниковым (1971) на афалинах. Локационные сигналы животного регистрировали с помощью четырех гидрофонов, расположенных на равных расстояниях друг от друга на линии, перпенди-
Рис. 116. Диаграмма направленности амплитуд давления дельфина афалины. (Яблоков и др., 1972).
Звуковое давление уменьшается наполовину при отклонении на 10—15° от осевой линии.
кулярной направлению движения животного. Положение животного регистрировалось с помощью двух кинокамер. Таким образом, имелась возможность определить положение животного в пространстве в момент генерации каждого локационного сигнала. Если бы направленность излучения была всегда постоянной, то время достижения звуковой волной каждого из гидрофонов должно было бы строго зависеть от расстояния между ним и головой дельфина. При сопоставлении записи, сделанной по всем четырем каналам, такая зависимость не обнаружилась. Зная время достижения звуковой волной трех и более точек пространства, нетрудно определить местоположение источника звука (если он точечный и расположен в одной плоскости
368
с приемными гидрофонами). Обработка сделанных записей с целью определения координат эффективного центра излучения показала, что он всегда расположен справа от животного, вне его головы. Естественно, что центр звукообразования располагаться вне тела животного не может. Авторы исследования объяснили расхождение наблюдаемых в эксперименте фактов с твердо установленными законами распространения звуковых волн способностью дельфина менять направление излучения локационных сигналов. В этом случае запаздывание прихода звука на второй гидрофон (по очередности прихода к гидрофону звуковой волны) будет зависеть от разности расстояний каждого из них до центра звукообразования и времени, необходимого для изменения направления звукового луча. Произведя соответствующие расчеты, авторы пришли к выводу, что сканирование происходит справа налево (этим объясняется мнимое смещение эффекторного центра вправо) с огромной скоростью, порядка 1000 оборотов звукового луча в секунду.
Способность изменять направленность излучения может быть удовлетворительно объяснена наличием у дельфина нескольких источников звукообразования. Система из двух разнесенных на некоторое расстояние точечных источников модулированного излучения при определенных условиях создает вращение диаграммы направленности. Агарков и др. (1971) считают, что характеристики направленности формируются в результате совместной работы 4 воздушных мешков (тубулярных и премаксиллярных) и назального хода. При этом высказывается предположение, что изменение направленности осуществляется путем изменения фазовых соотношений между колебаниями, порожденными в изолированных источниках излучения: в горизонтальной плоскости между колебаниями воздушных мешков, а в вертикальной — между частями назального хода. Далее предполагается, что воздушные мешки одной стороны работают в статическом режиме, не меняя ни амплитуды, ни фазы генерируемых колебаний, а мешки противоположной стороны непрерывно меняют характеристики колебаний. Кстати, необходимостью иметь для этого на одной стороне головы более развитую мускулатуру указанные выше авторы пытаются объяснить асимметрию черепа дельфинов и вторично возникшую асимметрию их мозга.
Вращение диаграммы направленности объясняет ряд особенностей локационных сигналов дельфинов, их кратковременность и крутизну фронтов. Наличие локационных щелчков, разделенных паузами, может быть связано с повторным многократным прохождением звукового луча через точку пространства, где расположен приемный гидрофон. Появление перед импульсом сопутствующих колебательных процессов может быть вызвано прохождением боковых лепестков вращающейся мгновенной диаграммы направленности.
369
На возможность сканирования диаграммой направленности указывают наши эксперименты (Айрапетьянц и др., 1972а; Айра-петьянц, Воронов и др., 1973).
Исследование также осуществлялось на афалинах. Локационные сигналы регистрировались с помощью приемных гидрофонов, расположенных перпендикулярно или параллельно направлению движения животного (в процессе поиска рыбы). Помещая рыбу в определенные точки вольера, экспериментаторы пытались создать ситуации, наиболее благоприятные для построения диаграммы направленности. В исследовании использовался принцип гетеродинирования. Последовательно перестраивая гетеродин, можно было производить отсчет амплитуды одной из следующих гармонических составляющих: 10, 30, 40, 50, 60, 70 и 90 кгц. Положение животного в пространстве регистрировалось электрофотокамерой со скоростью до 15 кадров в секунду. Огибающие амплитуд звукового давления и метки кадров фотокамеры совмещались на шлейфном осциллографе.
Анализ сделанных записей показал, что мгновенные диаграммы направленности при одних и тех же положениях в пространстве головы животного очень редко повторяются. Это свидетельство-
Рис. 117. Направленность излучения в горизонтальной плоскости для 90 кгц.
Справа — три положения головы животного (номера кадров 13, 17 и 21) в момент генерации локационных посылок. Ш и ПИ — приемные гидрофоны.
вало о том, что направленность излучения постоянно меняется. Количество случаев, когда уровень звукового давления, зарегистрированный с помощью гидрофона, на который был направлен рострум животного, значительно превышал амплитуду звукового давления, зарегистрированную вторым гидрофоном, было относительно невелико. Это означало, что направленность излучения может не совпадать с направлением оси тела животного. Последнее положение хорошо иллюстрирует рис. 117, на котором
370
изображены 3 положения головы животного относительно приемных гидрофонов и векторным способом показан размах изменения амплитуд звукового давления, происходящего за время экспозиции данных кадров. Излучение направлено почти перпендикулярно к оси тела животного. Кроме того, представлены пределы (разность длины сплошных и штриховых линий), в которых может изменяться звуковое давление на протяжении одного кадра. Запись огибающей амплитуд звукового давления (рис. 118)
показывает, что в отдельные моменты времени амплитуда звуко
вого давления, регистрируемая каждым из гидрофонов, изменяется или однонаправленно (на рисунке эти участки обозначены
знаком плюс), или в противофазе (отмечено знаком минус). Изменения звукового давления достаточно велики и происходят от кадра к кадру и даже на протяжении одного кадра, т. е. без изменения положения в пространстве головы животного или при столь незначительных смещениях, что ими можно пренебречь. Время
Рис. 118. Диаграмма изменения амплитуд гармонических составляющих / = 70 кгц локационных посылок.
По оси абсцисс — номера импульсов; по оси ординат — амплитуда, в делениях шкалы осциллографа. 1 и 2 — огибающая амплитуд по приемным гидрофонам П1 и ПИ (см. рис. 117). Черные квадраты — моменты смены кадра.
экспозиции одного кадра составляло 0.02 сек. За это время положение головы животного практически не меняется. Время перехода к следующему кадру 0.06—0.1 сек. позволяет голове животного сместиться максимум на 1—2°. Падение звукового давления, регистрируемое одним гидрофоном, и одновременное повышение амплитуды звукового давления, регистрируемое другим, при неизменном положении головы животного свидетельствует о возможности изменять направленность излучения, не меняя в пространстве положения головы. Все это говорит в пользу сканирования диаграммой направленности излучения.
Его целесообразность не подлежит сомнению. Благодаря сканированию достигаются кратковременность облучения по сравнению с длительностью локационного импульса и крутизной фронта импульса, что повышает разрешающую способность. С другой стороны, сканирование импульсом достаточно большой длительности обеспечивает значительную ширину зоны облучения.
Возможность подобного характера сканирования диаграммой направленности получила косвенное подтверждение в исследованиях Маркова (1972).
371
На основании модельных опытов на трупе афалины он отрицает обязательную направленность звукоизлучения вдоль оси тела животного. Сферический излучатель из пьезокерамики ЦТС-19 диаметром 10 мм вводился через дыхало к гортани, в правый и левый носовые проходы на уровень сфинктеров премаксиллярных мешков. Звуковое давление для тональных сигналов с частотой 20, 40, 60, 80, 100, 120, 140 и 160 кгц изучалось в трех точках, лежащих на горизонтальной плоскости, проходящей через среднюю ось тела (линию, соединяющую кончик рост-рума и гортань): 1-я точка лежит на оси тела, в 10 см от кончика рострума; 2-я и 3-я находятся в 30 см от кончика рострума и в 20 см вправо и влево от оси тела. Только при нахождении излучателя у гортани диаграмма направленности имела симметричный вид. Смещение излучателя от оси тела на 1.5—2 см к сфинктеру премаксиллярного мешка приводило к нарушению симметрии в сторону больших значений звукового давления на одноименной стороне. Наличие асимметрии автор объясняет возможностью раздельного проведения звука костями рострума, изолированными друг от друга хрящевой прослойкой. Сделанные наблюдения дали основание высказать предположение о возможности тонкого сканирования диаграммой направленности зондирующего луча путем последовательного переключения работы генераторов звука без изменения положения в пространстве головы животного.
Наши эксперименты (Айрапетьянц, Воронов и др., 1973), описанные выше, не подтвердили предположение В. М. Бель-ковичаи А. Е. Резникова о вращении диаграммой направленности в одном определенном векторе и заставляют с осторожностью отнестись к выводу о постоянной и весьма высокой скорости сканирования.
В настоящее время не представляется возможным объяснить причину того, что ряд исследователей не сумел заметить способность дельфинов в широких пределах изменять направленность звукового излучения. Не исключено, что запись локационных сигналов производилась в момент непосредственной близости животного от лоцируемого объекта или препятствия. В этом случае внимание животного сосредоточено на держании в поле зрения или захвате интересующего объекта, тогда как на более отдаленных участках траектории животное активно исследует все лежащее перед ним пространство. Использование в ряде исследований лишь одного гидрофона, естественно, не позволило изучить изменения направленности излучения.
Механизм сканирования остается пока недостаточно ясным. Из наиболее вероятных следует упомянуть раздельное использование нескольких источников звука и звуководов, использование звукофокусирующих устройств, сложное взаимодействие нескольких одновременно работающих источников звука.
372
Очень важным представляется вопрос о формировании звукового поля дельфина. Дело в том, что дальнее звуковое поле формируется не только самим дельфином, но и окружающей средой. Для выяснения механизма излучения и формирования звуковых сигналов необходимо изучать звуковое поле в непосредственной близости от головы дельфина.
Романенко (1972) подверг изучению ближнее поле излучения. Это позволило ему избегнуть искажений, вносимых близостью свободной границы раздела воздух—вода, интерференцией прямых и отраженных сигналов, дифракцией и дисперсией скорости звука. Исследование осуществлено с помощью трех миниатюрных цилиндрических гидрофонов (диаметром 2 мм, высотой 3 мм), закреплявшихся с помощью присосок на голове афалины, и трехканального, носимого животным магнитофона, позволявшего регистрировать звуковые сигналы в полосе частот от 500 гц до 50—80 кгц. Было установлено, что свисты могут излучаться односторонне (разность уровней звукового давления между правой и левой стороной достигает 12—15 дб), но принимают на звуковой подушке среднее значение. Импульсные акустические сигналы в ближнем поле более богаты низкими частотами, чем сигналы в дальнем поле. Локационные импульсы по форме очень напоминают электрические разряды, что свидетельствует о нерезонансном механизме их излучения. Однако, проходя через жировую подушку, локационные сигналы принимают более сложную форму.
Подытоживая эту краткую сводку, можно сделать следующее заключение. Органы, создающие условия для генерации сигналов у зубатых и усатых китов, резко отличаются по сложности своего строения, что находит выражение в сравнительно немногочисленных и простых сигнальных звуках усатых китов и в сложных и многообразных звуках зубатых китов.
Система генерации сигналов зубатых китообразных конструктивно состоит не только из нескольких источников звуков в диапазоне от 150 гц до 208 кгц, которые тонко и произвольно регулируются животным, но также из костного рефлектора и акустической линзы, формирующих звуковое поле. Фазовое манипулирование несколькими источниками звукообразования, использование жировой линзы и мышечно-костного рефлектора, а также, возможно, и другие, еще в настоящее время неизвестные механизмы обеспечивают узкую направленность излучения и сканирование диаграммой направленности без изменения положения в пространстве головы животного. Таким образом, передающий акустический тракт зубатых китов как высокосовершенная часть телерецептора обеспечивает навигацию, коммуникацию и тонкое локационное дифференцирование.
19
СЛУХОВОЙ АНАЛИЗАТОР
Строение анализатора
В настоящее время принято считать, что китообразные произошли от древних хищных млекопитающих, которые в процессе эволюции перешли к водному образу жизни. Организм этих животных подвергся вторичной адаптации к условиям обитания в водной среде. Соответствующие изменения произошли и в строении органов слуха. Ухо наземных млекопитающих приспособлено к восприятию звуков в воздухе и почти совершенно непригодно для подводного слушания. Вибрирующий под воздействием звуковых волн череп оказывается не способным осуществлять под водой полную изоляцию ушей друг от друга, что приводит к утрате направленного слуха (Reysenbach de Haan, 1957b). Поэтому у китообразных в процессе эволюции произошли существенные перестройки. Необходимость иметь в своем распоряжении два не зависящих друг от друга приемника звука реализована у зубатых китов следующим образом. Органы слуха не сращены с черепом, как у наземных млекопитающих, а подвешены на короткой сухожильной связке. Удивительная особенность слухового анализатора китообразных заключается в том, что среднее и внутреннее ухо располагаются в одном костном образовании, называемом барабанно-околоушной костью (os petrotympanicum). Кость эта чрезвычайно твердая, и вещество, из которого она состоит, напоминает зубную эмаль. Такие «ушные кости», известные под названием «цитолиты», не разрушаются со временем, как другие кости скелета этих животных, и иногда извлекаются со дна океана драгами и тралами. Расчеты Рейшенбаха де Хаана показывают, что орган слуха надежно изолирован от колебаний черепа (рис. 119).
Свойства подобной «маятниковой» системы зависят от величины подвешенной массы os petrotympanicum и от жесткости самого подвеса, т. е. связки. Чем больше подвешенная масса при данной 374
жесткости подвеса, тем ниже резонансная частота самой системы, и наоборот, чем больше жесткость подвеса при данной массе, тем выше специфичная для нее частота. Соотношение между частотами, воздействующими на эту систему, и ее собственной, специфичной для нее частотой у китообразных таково, что весь костный барабанно-околоушной комплекс находится в покое относительно биологически значимых звуковых частот (Reysen-bach de Haan, 1957b, 1966).
Второе важное приспособление: среднее и внутреннее ухо окружены со всех сторон специальными синусами, заполненными
Рис. 119. Схема поперечного разреза головы зубатого кита. (Reysenbach de Haan, 1957b).
н. с. п. — наружный слуховой проход.
воздухом и пеной из белковой эмульсии. Пена, содержащая множество воздушных пузырьков, поглощает все звуковые колебания. Благодаря такой изоляции для звука остается единственный путь — попасть во внутреннее ухо через систему косточек среднего уха (Boenninghans, 1903; Fraser, Purves, 1954; Reysenbach de Haan, 1957b). В системе синусов располагаются сосудистые сплетения, за счет их большего или меньшего наполнения кровью («набухания») в среднем ухе происходит выравнивание давления с наружным. Этим обеспечивается нормальная работа органа слуха при быстро меняющихся давлениях, которые постоянно испытывают китообразные во время всплытия и погружения.
375
Рассмотрим более подробно каждый из отделов слухового анализатора китообразных (рис. 120).
Наружное ухо, или, вернее, то, что от него осталось, представлено узким слуховым проходом, который открывается маленьким отверстием (у афалины диаметр его около 1.5—2 мм) сбоку головы, позади глаза. Внешние ушные раковины у китообразных отсутствуют. Почти сразу за наружным отверстием
Рис. 120. Схематический дорзо-вентральный разрез через слуховой аппарат зубатого кита. (Reysenbach de Haan, 1957b).
1 — барабанный конус; 2 — молоточек; 2а — тонкий отросток; 3 — барабанный тензор; 4 — наковальня; <5 — жир; 6 — перилимфатическая часть лабиринта; 7 — эндолимфатическое пространство в канале улитки; 8 — латеральный полукружный канал; 9 — круглая ямка, прикрытая односторонней мембраной; 10 — каменнососцевидный или околоушной отросток; 11 — тимпаникум; 12 — воздушные мешки в барабанной полости; 12а — воздушные мешки вокруг периотикума; 13 — лигаментозная суспензия периотикума; 14 — соединительная ткань; 15 — наружный слуховой проход; 16 — кости черепа.
слуховой проход становится узкой щелью (360 X 36 мкм у афалины и 330x32 мкм у белобочки), которая резко сужается, и в жировом слое просвет полностью зарастает. Пройдя жировой слой, слуховой проход делает S-образный изгиб, и в нем снова появляется просвет, который начинает резко увеличиваться в размерах, постепенно приобретая овальную форму (2250x1305 мкм у афалины и 1620x810 мкм у белобочки) у входа в полость среднего уха, где заканчивается видоизмененной барабанной перепонкой (Белькович, Солнцева, 1970; Солнцева, 1971).
Тщательное морфологическое изучение строения наружного
376
слухового прохода, проведенное в Последние годы ПурвеСоМ (Purves, 1966), показало, что отверстие прохода, фиброэластичный конус, мышцы и хрящи, окружающие слуховой проход, можно рассматривать как единый комплекс и считать гомологичным наружному уху наземных млекопитающих.
Среднее ухо у китообразных в целом очень массивно по сравнению со средним ухом наземных млекопитающих, а барабанная кость жестко связана с околоушной костью, которая заключает в себе систему внутреннего уха. Наружный слуховой проход заканчивается у входа в среднее ухо барабанной перепонкой, которая у китообразных имеет весьма своеобразное строение, напоминая сложенный (а не раскрытый, как у наземных млекопитающих) зонтик. Вершина ее соединена под углом с укороченной рукояткой молоточка, который в свою очередь связан тонким отростком с барабанной костью. Отношение площади барабанной перепонки к площади подножной пластинки стремени равно 90 (Липатов, Солнцева, 1972)» Все три слуховые косточки — молоточек, наковальня и стремечко — плотные, небольшой величины, жестко соединены между собой, специализированы для передачи звуков широкого диапазона частот, включая и ультразвуки. За счет изменения соотношения своих элементов (длины pr. gracilis, площади стремечка и т. д.) косточки среднего уха приспособлены к передаче на внутреннее ухо колебаний из водной среды с уменьшенной в 60 раз амплитудой и увеличенной интенсивностью (Reysenbach de Haan, 1957b). В общей сложности среднее ухо обеспечивает усиление передачи в 105—158 раз (Липатов, Солнцева, 1972; Солнцева, 1972).
Внутреннее ухо китообразных по своей внешней структуре^соответствует таковому у наземных млекопитающих. В состав околоушной кости входят преддверие, улитка и полукружные каналы. Однако при тщательном рассмотрении внутреннего уха китообразных обнаруживаются существенные отличия, указывающие на способность этих животных к восприятию звуков очень высокой частоты.
У китообразных, и в особенности у зубатых китов, улитка сильно увеличена по сравнению с вестибулярным аппаратом. Костный канал улитки делает 1—2 оборота вокруг костного стержня (modiolus), причем базальный виток, достигающий больших размеров, почти целиком охватывает последующий. Базальный, или основной, виток улитки по всей длине канала перегорожен первичной спиральной пластинкой (ossea primaria), отходящей от осевого столбика (modiolus), и вторичной (ossea secundaria), идущей от наружной стенки. Между обеими пластинками имеется узкий просвет, закрытый базилярной мембраной (memb-rana basilaris), на поверхности которой располагается кортиев орган. Просвет расширяется от основания улитки кверху за счет
377
Постепенного уменьшения вторичной пластинки в том же направлении.
Весьма примечательно, что такая же конструкция улитки, но менее развитая, отмечена у летучих мышей и у некоторых грызунов, воспринимающих ультразвуки.
Базилярная мембрана у основания улитки очень узкая и твердая, жестко прикрепляется к хорошо развитой спиральной связке. К вершине улитки мембрана становится гораздо шире. У афалины ширина базального конца базилярной мембраны 25 мкм, а у вершины — 350 мкм. Увеличение в 14 раз соответствует высокой чувствительности уха дельфина в области высоких частот (Wever et al., 1971а, 1971b, 1971с), а расположенный вдоль длины мембраны кортиев орган постепенно увеличивается к вершине улитки, тогда как соседствующие с ним опорные клетки Клау-диса пропорционально понижаются.
Рецепторными элементами кортиева органа служат наружные и внутренние волосковые клетки. Внутренние располагаются в один ряд и имеют кувшинообразную форму. Крупное ядро находится у основания клетки. Наружные волосковые клетки цилиндрической формы также с крупным ядром у основания, расположены в три ряда.
У афалины в кортиевом органе обнаружено 17 400 волосковых клеток, т. е. столько же, сколько и у человека. Ганглиозных клеток 95 000, или в 3 раза больше, чем у человека (Wever et al., 1971а, 1971b, 1971с). У тихоокеанского короткоголового дельфина волосковых клеток 12 900, а ганглиозных — 60 000— 70 000 (Wever et al., 1972). Большое число волосковых клеток свидетельствует о хороших способностях к тональному различению, а ганглиозных — к переработке акустической информации уже на периферии анализаторной системы.
Волосковые клетки окружены опорными, представленными наружными и внутренними клетками, столбами кортиева органа, располагающимися здесь же, на базилярной мембране.
Уже в самом рецепторном органе принцип соотношения между жесткостью подвеса и подвешенной массой у китообразных также используется с максимальным эффектом. Сравнение кортиева органа и способа его подвеса у человека (как типичного млекопитающего с низкочастотным слухом) с таковыми у других животных (грызунов, летучих мышей, зубатых китов), имеющих «высокочастотный» слух, показывает, что наименьший по своей массе и самый жесткий по способу крепления (подвеса) кортиев орган у зубатых китов. А это означает, как указывает автор, что зубатые киты обладают всеми данными для оптимального подводного слуха и восприятия ультразвуковых частот, включая акустическую изоляцию органов слуха друг от друга, способ жесткого подвеса к черепу и особенности структур среднего и внутреннего уха.
378
Слуховые центры. Нервная система китообразных изучена пока слабо. Особый интерес представляет высоко развитый конечный мозг — telencephalon. Коэффициент цефализации у дельфина равен 75.6 (Никитенко, 1965), он соседствует с таковыми у приматов (80).
Вес мозга дельфинов колеблется в зависимости от вида: у Phocaena phocaena — от 380 до 500 г при весе тела от 23 до 51 кг; у Tursiops truncatus — 1140 г (Andersen, 1967). По данным Кесарева (1970), относительная величина неокортекса наиболее высока у дельфина, составляет 97.8% поверхности всей коры.
Наиболее полное исследование мозга афалины, включая слуховую систему, было сделано Лангворси (Langworthy, 1932), который пришел к выводу, что последняя по сравнению с таковой у других млекопитающих достигает у этого вида высшей стадии развития.
При сравнительном изучении слуховой системы мозга у зубатых и усатых китообразных выяснилось, что у дельфинов дорзальные структуры среднего мозга, моста и продолговатого мозга заметно сдвинуты каудально (Ogawa, Arifuku, 1948). Было также показано, что мозговой ствол дельфинов сужен трансверсально и расширен дорзо-вентрально, отмечено наибольшее развитие вентральных кохлеарных ядер, ядер верхних олив, трапециевидного тела и латеральной петли.
Несмотря на принципиальную общность отдельных звеньев слухового анализатора дельфина отмечается их структурное отличие от аналогичных образований у представителей своего класса: во-первых, гипертрофия всех подкорковых центров слуха; во-вторых, обилие крупноклеточных нейронов в них; в-третьих, плотность клеточных элементов.
На основании широкого количественного сопоставления величин слуховой подкорки у различных млекопитающих (грызунов, хищных, китообразных и приматов) Зворыкин (19636) высказывает предположение о том, что необыкновенное разрастание слуховых подкорковых центров у китообразных и летучих мышей может быть объяснено способностью этих животных воспринимать звуки высокой частоты, лежащие над верхним порогом слуха человека (табл. 24). Зворыкин установил, а позднее Богословская (1968, 1972) подтвердила, что у дельфинов преимущественное развитие получили переднее и заднее вентральные кохлеарные ядра. Особенно сильно развитым оказалось переднее вентральное ядро, которое связано с верхними оливами, ответственными за бинауральный эффект (Katsuki, 1961).
Кроме различий в величине, на основании подсчета числа клеток было установлено, что большие размеры тех или иных звеньев слуховой подкорки сочетаются и с большим числом составляющих их клеточных элементов. Так, например, на срезе центрального ядра латеральной петли у дельфина было под-
379
Таблица 24
Объем (мм3) системы подкорковых образований слухового анализатора и вес мозга (г) дельфина и человека
(Зворыкин, 19636)
Дельфин Человек Слуховые подкорковые центры Bee мозга
ta nvc os ntr nllv nllim nlld ci cgm
7.0 1.2 160.0 10.3 150.0 0.96 85.7 217.0 1.1 160 40.0 0.43 470 38.0 340 48.5 780 1250
Примечание, ta — слуховой бугорок; nvc — вентральное слуховое ядро; os — верхняя олива; ntr — ядро трапециевидного тела; nllv — вентральное ядро латеральной петли; nllim — интермедиальное ядро латеральной петли; nlld — дорзальное ядро латеральной петли; ci — заднее двухолмие; cgm — внутреннее коленчатое тело.
считано 1380 нейронов, а у человека — всего 25, в верхней оливе дельфина 510 нейронов, а у человека — 57, во внутреннем коленчатом теле соответственно 11 450 и 2840. Видно, что величина различных звеньев слуховой подкорковой области у дельфина от 6 до 640 раз превышает таковую у человека (Зворыкин, 19636).
Интересной особенностью вентрального кохлеарного ядра является наличие гигантских клеток, отчетливо видимых в его дорзальной части, которые считаются чувствительными к высоким тонам (Galambos, 1954; Rose et al., 1959; Osen, Jansen, 1965).
У дельфина и человека измерялся объем кохлеарных ядер и подсчитывалось относительное количество нейронов и волокон в них (Hall, 1967). У человека отношение количества нервных волокон, несущих импульсы к нервным клеткам, к количеству нейронов равно 1/4, тогда как у дельфина — 1/6.
Подробно нейронный состав кохлеарных ядер дельфинов представлен Богословской (1968, 1972). Автор считает, что дорзальные области переднего и заднего вентральных ядер, куда, по-видимому, проецируются высокие частоты, без перерыва продолжают друг друга и обладают сходным набором нервных элементов. Помимо этого, Л. С. Богословской в обоих вентральных ядрах выделяется поверхностная зона, наиболее разнообразная по клеточному составу, куда коллатерали слуховых афферентов проецируют средние и высокие частоты и которая обладает своей внутренней системой тангенциальных волокон.
Изучению верхнеоливарного комплекса дельфинов посвящен ряд исследований (Hatschek, Schlesinger, 1902; Hofmann, 1908; Jelgersma, 1934; Зворыкин, 1960, 19636; Петелина, 1972). В составе верхнеоливарного комплекса медио-вентральное ядро наиболее развито. Предполагается, что в слуховом пути дельфиноц
380
ядра трапециевидного тела имеют бблыпую функциональную нагрузку, чем у других животных с таким же типом слуховых связей, так как все развитие верхнеоливарного комплекса дельфинов в значительной степени обусловлено представительством ядер именно этого образования.
Ядра латеральной петли у дельфина значительны по своим размерам и имеют чрезвычайно полиморфный клеточный состав (Breatnach, 1960; Зворыкин, 1963в).
Сведения о строении заднего двухолмия дельфина содержатся в работах Лангворси (Langworthy, 1932) и Зворыкина (1966). Но наиболее полно оно было изучено Суховской (1968, 1972). Ею было показано, что у данного образования наиболее выражены латеральная и центральная клеточные группы. Анализ нейронной организации заднего двухолмия выявил преобладание в латеральной группе мелких афферентных нейронов, которые, как считается, выполняют прежде всего переключательную и рецепторную функции. Нейроны этой группы воспринимают специфическую импульсацию и передают ее в слуховую кору.
Центральная клеточная группа содержит крупные мультипо-лярные нейроны, по-видимому, являющиеся двигательными нервными клетками, которые переключают специфическую импульсацию на различные моторные образования ствола.
Наименее изученным отделом слухового анализатора дельфина являются внутренние коленчатые тела (Kruger, 1959). Зворыкин (1963а) описал их как ушковидные выпячивания на наружной стенке промежуточного мозга, чрезвычайно богатые клеточными элементами. Автор подсчитал количество нейронов в медиальном коленчатом теле и выяснил, что у дельфина их плотность в 5 раз выше, чем у человека. В настоящее время изучение нейронного состава и межнейронных связей этого образования изучается в нашей лаборатории.
Данные, касающиеся слуховой коры дельфинов, представлены в литературе достаточно широко, но они крайне противоречивы (Rose, 1926; Riese, 1927; Langworthy, 1932; Kojima, 1951; Зворыкин, 1963a; Morgane, 1965; Hall, 1967; Pilleri, 1967a, 1967b; Краснощекова, 1969, 1972; Ладыгина, Супин, 1970).
Микроскопическое строение височной доли больших полушарий дельфина достаточно сложно. Создается впечатление о большей сложности, чем имеют аналогичные образования человека. Подобные предположения вряд ли обоснованы. У человека височная кора характеризуется большей шириной в поперечнике, а также лучшей дифференцировкой формы составляющих ее нейронов. Вместе с тем по сравнению с другими животными височная кора дельфина организована более сложно и превосходит обилием клеточных элементов.
Большинство авторов (Morgane, 1965; Краснощекова, 1969, 1972; Кесарев, 1970, и др.) склонны выделять в височной области
381
коры дельфинов 6 слоев с нечеткой границей между III и IV слоями. Височная кора обладает пирамидными нейронами меньшего калибра, чем, например, двигательная кора, и большим коли-чеством звездчатых нейронов, среди которых преобладают короткоаксонные. В нашей лаборатории на нейронах слуховой области коры были выявлены аксо-соматические и аксо-дендритические синапсы, преимущественно петлевидной формы, а на препаратах, обработанных по методу Гольджи, найдены дендритные разветвления, густо покрытые шипиками.
Сопоставление височной коры дельфина с другими областями его мозга показывает, что она не отличается какими-либо более прогрессивными чертами строения. То же самое отмечалось и для летучих мышей в соответствующей главе. На основании проведенных исследований нам кажется правомерным сделать следующий основной вывод. Как и у летучих мышей, у дельфинов координация деятельности эхолокационной системы в значительно большей степени осуществляется на уровне подкорковых центров, чем за счет слуховой коры мозга, что подтверждают электрофизиологические исследования (Bullock et al., 1968, 1972).
Звуковая рецепция
Как ни банальна ссылка на Аристотеля, но ведь на самом деле великий энциклопедический «ясновидец» успел осуществить наблюдение и заметить, что хотя у дельфинов отсутствуют наружные ушные раковины, однако они слышат подводные звуки. В XX в. первые публикации, посвященные слуху дельфинов, появились около 40 лет назад. В частности, Хоуэлл (Hawell, 1933) сообщал, что при включении ультразвукового глубиномера (эхолот) судна вблизи стаи дельфинов те стремительно уплывают прочь.
Отсутствие ушных раковин и узость слухового прохода заставляли задуматься над тем, каким образом звуковые волны доходят до внутреннего уха. У исследователей не было уверенности в том, что среднее и внутреннее ухо, окруженные слоем жира и мышц, способны участвовать в приеме звуков. Последнее обстоятельство, а также особые требования к звуковоспри-ятию в процессе активной локации невольно наталкивали на поиски иных рецепторных образований для анализа эхо-сигналов.
В вопросе о функции наружного слухового прохода у китообразных мнения исследователей расходятся. Первоначально (Camper, 1777) считалось, что киты слышат лишь в мгновения, когда выставляют голову из воды. Затем стали полагать (Hyrtl, 1845), что они могут слышать только под водой и 'совершенно не слышат на воздухе. Когда от этого пришлось отказаться, была выдвинута (Lillie, 1910) теория костной проводимости.
382
До недавнего времени существовало представление, что слуховой проход и барабанная перепонка не имеют отношения к проведению звуковых колебаний (Boenninghans, 1903; Lillie, 1910; Turner, 1913; Kellogg, 1938, и др.). Звук проходит мягкие ткани головы, поступает непосредственно в среднее ухо и затем благодаря системе слуховых косточек проводится во внутреннее ухо. Выдвигалось даже предположение, что слуховой проход и окружающие его мышцы выполняют лишь функцию «манометра» (Yamada, 1953), измеряющего величину давления при нырянии китообразных. Однако уже в конце 50-х годов настоящего столетия на основании экспериментальных исследований некоторые авторы пришли к заключению, что в проведении звука в среднее ухо участвуют проксимальный отдел наружного слухового прохода и барабанная перепонка, тогда как дистальный конец прохода и мышцы, окружающие его, не функционируют (Reysenbach de Haan, 1957b; Dudok van Heel, 1962). Последний вывод подтверждается тем обстоятельством, что наружный проход полностью закрывается. И наконец, по еще одной гипотезе, подтвержденной в известной мере экспериментом, через наружный слуховой проход продольные звуковые волны проходят к барабанной перепонке и системе слуховых косточек среднего уха с наименьшими потерями интенсивности (Fraser, Purves, 1954, 1960; Purves, 1966). Как было показано выше, изоляция среднего и внутреннего уха синусами не оставляет других путей для звука, кроме слухового прохода. По мнению авторов, этим обеспечивается направленность восприятия и лучшая акустическая передача при проведении звуков. Была предпринята попытка проверить значение наружного слухового прохода для проведения звуковых волн на живом дельфине. Для этого афалине прикрепляли небольшие чашечки — присоски из латекса, закрывающие отверстия слуховых проходов с обеих сторон; это не помешало дельфину хорошо ориентироваться в бассейне (Norris et al., 1961).
Мак-Кормик и др. (McCormic et al., 1970) исследовали прохождение звука к внутреннему уху у двух видов дельфинов — у атлантической афалины и беломордого (Lagenorhynchus obligui-dens) — методом отведения микрофонных потенциалов.
Данные экспериментов свидетельствуют о том, что звуковые волны проходят через ткани головы, а редуцированный слуховой проход для звукопроведения не используется.
। Ряд исследователей (Norris, 1964; Bullock et al., 1968) высказал предположение, что волноводом служит нижняя челюсть. Действительно, звуковые колебания могут проходить через 3—4 отверстия (foramina mentales), расположенные с каждой стороны назального отрезка челюсти и по каналу нижней челюсти (сапа-lis mandibularis), заполненному жиром, хорошо проводящим звук, поступать в район сустава челюсти, т. е. в область расположения среднего и внутреннего уха. Солуха и Валиулина (1972)
383
построили математическую модель звукового приема (сознательно допустив значительные упрощения). Так как отверстия в кости челюсти гораздо меньше минимальной длины волны эхо-сигнала, данные приемники являются ненаправленными. По построенной модели, первичные слуховые центры мозга должны получать информацию, включающую постоянную и переменную составляющие. Благодаря этому, сканируя движением головы, животное может определять пеленг цели, так как при движении прямо на лоцируемый объект переменная составляющая должна исчезать. По изменению тона переменной составляющей дельфин может судить об изменении расстояния до объекта.
В нашей лаборатории (Айрапетьянц, Воронов и др., 1973) на морских свиньях была предпринята попытка экспериментально оценить значение каждого из перечисленных выше каналов, подтвердить роль внутреннего уха для звуковосприятия и выяснить долю участия в нем лобного жирового выступа.
Животные во время опыта помещались в звукозаглушенную ванну и жестко фиксировались. У каждого из 5 дельфинов определялись пороги слышимости тональных сигналов с частотой от 5 до 100 кгц при трех положениях излучателя: прямо против рострума, против слуховых проходов и позади них под углом в 135° к оси тела животного. Каждая из указанных точек находилась в 20 см от животного. Излучатели и линия рострума находились на 12—20 см ниже поверхности. О восприятии звукового сигнала судили по выраженности сердечного компонента электрокожного оборонительного условного рефлекса. Аудиограмма интактных животных сравнивалась с аудиограммой, полученной после акустического экранирования двух участков головы дельфина: нижней челюсти и районов ушных отверстий.
Было обнаружено (рис. 121), что для сигналов с частотой выше 30 кгц имеются заметные различия в восприятии звуков, идущих под углом 135° к оси тела и двух других положений излучателя. При частоте 50 кгц разница достигала 8 дб. Дальнейшее повышение частоты тонального сигнала выявило разницу в восприятии звука для всех трех положений излучателя. Наибольшая чувствительность обнаружена для сигналов, идущих спереди вдоль оси тела. Для тональных сигналов частотой 90— 100 кгц, поступающих от излучателя, расположенного против ушных отверстий, характерно понижение чувствительности на 7, а для поступающих под углом 135° — на 13—15 дб. Понижение чувствительности к сигналам, идущим через ушные отверстия, вероятно, связано с физическими условиями прохождения звука через узкие каналы.
Восприятие тональных сигналов, следующих со стороны рострума, после экранирования ушных отверстий ухудшилось лишь для частот 10 и 30 кгц примерно на 5 дб. Для звуков, идущих со стороны ушных отверстий, чувствительность упала значительнее, до 20 дб по отношению к сигналам всех использованных частот.
384
дб
Экранирование нижней челюсти привело к резкому повышению порога чувствительности для всех сигналов, поступающих со стороны рострума, но не отражалось на чувствительности к звукам, идущим из двух других точек.
Таким образом, анализ полученных нами материалов дает основание считать, что звуки частотой ниже 30 кгц поступают к кохлеарному аппарату по двум каналам: через область слуховых проходов и нижнюю челюсть. Учитывая некоторое снижение чувствительности к звукам, идущим вдоль оси тела при экранировании ушных отверстий, можно предполагать, что слуховые проходы для данной категории звуков служат ведущим волноводом. Для частот свыше 30 кгц главный путь достижения внутреннего уха — это нижняя челюсть.
Д анный эксперимент подтверждает предположение, что единственным звукоприемником служит кохлеарный аппарат. Верхняя челюсть и лобный жировой выступ не принимают участия в звуко-проведении, так как ока-
го
ю
о
-10
135'
f. 90
0°
60 80 100
-20-~30--90-~50-
5 10 20 90
Рис. 121. Аудиограммы морской свиньи, полученные при трех положениях излучателя: 0° — против рострума, 90° — против слухового отверстия, 135° — позади слухового отверстия.
зались не в состоянии компенсировать потерю обычных путей звукопроведения при их экранизации.
Изучение морфологии возможных путей, по которым звуковые волны могли бы достигать среднего и внутреннего уха (Fraser, Purves, 1960; Purves, Utrecht, 1964), показало, что булля в значительной мере экранирована за счет ослабления звука при переходе из жировой ткани в мышечную изменением напряженности окружающих мышц, наличием белково-воздушной эмульсии и других образований перибульбарной области. Эти наблюдения заставляют предполагать существование направленности слухового приема.
Таким образом, в проведении звуковых колебаний принимают участие наружный слуховой проход и нижняя челюсть, барабанная перепонка, слуховые косточки, кольцевая связка овального окна, мембрана круглого окна, жидкость лабиринта (перилимфа), базилярная мембрана. Барабанная перепонка и слуховые косточки трансформируют воздушные колебания с большой амплитудой
385
И малым давлёнйём в колебания перилимфы с малой амплитудой и большим давлением. Колебания перепонки овального окна передаются перилимфе, наполняющей улитку, вызывая движение волоконец базилярной и тенториальной мембран, колебания которых приводят к раздражению рецепторных клеток кортиева органа, вследствие чего в них возникает процесс возбуждения, т. е. происходит трансформация физической энергии звуковых колебаний в физиологический процесс. Далее информация об эхо-сигналах поступает в продолговатый и средний мозг, где и осуществляется их анализ, а о коммуникативных — в височные отделы коры (Bullock, Ridgway, 1972).
Помимо кохлеарной теории звуковосприятия, в мировой литературе высказано немало предположений о наличии и других видов звуковых рецепторов. Большое значение придается лобному жировому выросту. Дреер (Dreher, 1967) обнаружил в коже, покрывающей жировую подушку, трубочки дермальных сосочков, которым он приписал роль голографической решетки. Плотность их такова, что могла бы обеспечить разрешение около 100 линий на 1 мм (если описанные структуры действительно обладают рецепторными функциями). Экспериментального подтверждения эта теория пока не имеет.
Некоторые исследователи (Агарков, Хоменко, 1968, 1969; Хоменко, 1969, 1970а, 19706; Резников, 19706, 1971а; Белько-вич, Резников, 1971; Агарков, Солуха, Хоменко, 1971; Хоменко, Нечаева, 1971; Агарков, Хоменко, Слезин, 1972; Хоменко, Солуха, Нечаева, 1972) предполагают участие рецепторных аппаратов мягких тканей головы в восприятии звуковых колебаний. Резников (19706, 1971а) и Белькович и Резников (1971) допускают существование у дельфит ов «акустических глаз». В этом случае жировой подушке приписывается роль хрусталика, формирующего акустическое изображение лоцируемых объектов, а воздушным мешкам, выстланным плоским эпителием, — роль воспринимающей части рецептора. Резников (1970а) предполагает, что акустическое излучение, приходящее на линзообразную лобную жировую подушку из верхнего полупространства, фокусируется на воздухоносных мешках и создает на них действительные акустические изображения активных или пассивных источников звука. Он подтверждает свое предположение построением лучевой картины, расчетом размеров фокальных пятен и амплитуды смещений и натяжений, возникающих под действием звука на граничных поверхностях воздушных мешков.
Расчеты показывают, что осязательными (или иными) рецепторными образованиями стенок мешков могут быть восприняты эффекты звуковых воздействий. Лучевая картина показывает, что при падении звука спереди он должен фокусироваться «сферическими» участками жировой подушки, при падении сверху — только «цилиндрическим» участком. Поэтому спереди над линией
386
рта должна находиться зона относительно четкого изображения, а над верхней частью жировой подушки — зона нечеткого изображения с анализом объектов лишь в плоскости, перпендикулярной оси тела. Предполагается, что зона четкого звуковидения позволяет осуществлять угловую ориентацию и одновременное опознание многих мишеней. Зона нечеткого звуковидения способна лишь обеспечить информацию о наличии их над дельфином и привязку к поверхности. Если бы подобное устройство действительно имело место, то оно оказалось бы способным обеспечить высокую помехозащищенность путем пространственного разделения акустических пятен. По автору, реверберация и зондирование того же предмета другими животными должны в этом случае улучшать распознавание, так как увеличивают суммарную энергию сигнала, отраженного от цели. Гипотеза звуковидения, по мнению А. Е. Резникова, убедительно подтверждается неспособностью дельфинов обнаруживать цели в нижнем полупространстве, наличием в верхнем полупространстве зон четкого и нечеткого обзора, уверенной локацией с заглушками на ушах и нарушением локации при экранизации подушки. Таким образом, в соответствии с изложенной выше гипотезой изображения объектов должны восприниматься животными как некие образы «пространственного осязания» и значительно дополнять информацию об окружающей среде, получаемую кохлеарным аппаратом о тех же эхо-сигналах. Никаких прямых подтверждений данная теория не имеет. Трудно себе представить одновременное использование одного и того же инструмента для генерации (и, естественно, для осуществления контроля над качеством зондирующих сигналов) (Хоменко, 1970а, 19706; Агарков, Хоменко, Слезин, 1972) и восприятия локационных сигналов.
В последние годы выполнена серия работ по изучению иннервации головы дельфинов (Агарков и др., 1968, 1969, 1971, 1972; Хоменко, 1969, 1970а, 19706; Хоменко и др., 1971, 1972). Эти исследователи на большом фактическом материале показали наличие на голове дельфинов вокруг ротовой щели, дыхала и по бокам лобно-жировой подушки нескольких зон с высокой концентрацией кожных рецепторов. Интерарганная нервная периферия мягких тканей головы дельфина весьма разнообразна. Она представлена обширными арборизациями и компактными кустиками с фибриллярными пластинками, пуговками, петельками и сложноустроенными инкапсулированными приборами типа колб Крузе и фатер-пачаниевых телец. Авторы относят их к ангио-, трофо-, баро- и механорецепторам, а также к своеобразным рецепторным аппаратам, образующим «сетчатку» лобного выступа. Перечисленные рецепторы являются образованиями глазничной и верхнечелюстной порций тройничного и дорзальной щечно-губной ветви лицевого нерва. Указанные выше авторы отмечают, что рецепторный аппарат надкостницы максиллярной и фронталь
387
ной костей развит значительно сильнее, чем надкостницы области затылка.
Механизм работы «сетчатки» лобного выступа представляется следующим образом. В звуковой посылке, отраженной от объекта локации, происходит изменение амплитуды и фазы отдельных спектральных компонентов. Принимая отраженный сигнал с помощью системы, состоящей из достаточно большого количества отдельных приемников, и обрабатывая его определенным образом, можно получить представление о внешнем виде объекта. Предполагается наличие у дельфина голографического метода обработки эхо-сигнала. Если подобное рецепторное устройство действительно существует, оно может обеспечить детальное изучение подводного объекта по величине, не превышающей размер приемника, при условии, что объект будет находиться не далее 1 м от животного и полностью уместится внутри конуса лоцирующего луча, а частота излучения будет достаточно высокой.
Неясно функциональное значение обнаруженных рецепторов. В виде предположения допускается, что они принимают участие в регуляции дыхательного акта, в контроле за давлением в воздухоносных полостях и за генерацией различного рода сигналов, в оценке сопротивления воды при различных режимах плавания и, наконец, для гидроэхолокации.
Кроме вышеперечисленных, в научной литературе появились суждения относительно физиологического механизма локации. К их числу относится соображение о голографическом видении с использованием мозга в качестве приемного устройства (Кузин, Слезин, 1972). Основанное на ряде допущений, строится предположение, что дельфин излучает когерентные ультразвуковые сигналы. Отраженные от различных объектов, они могут собираться жировой подушкой и фокусироваться в некоей зоне головного мозга. Авторы полагают, что ультразвук вызывает в нервных клетках какие-то физико-химические сдвиги, носящие обратимый характер, однако эти соображения пока не подкреплены экспериментальными исследованиями.
Функциональные характеристики центрального звена анализатора
Изучение морфологии центральных аппаратов слухового анализатора пока не дало материала для понимания его функциональной организации. Некоторые сведения дают об этом немногочисленные электрофизиологические исследования. МакКормик и др. (McCormick et al., 1970) методом регистрации микрофонных потенциалов пытались составить представление о том, какие изменения претерпевает звуковой сигнал на пути
388
к внутреннему уху и с каким исходным материалом имеют дело слуховые центры мозга.
Первые записи биопотенциалов мозга дельфинов были осуществлены группой японских и американских исследователей, работавших в период с 1965 по 1966 г. в морских океанариумах в Ито (Япония). Буллок и др. (Bullock et al., 1968) на 24 дельфинах четырех видов (Tursiops truncatus и три вида Stenella, главным образом на полосатом дельфине — Stenella caeruleoalbus) изучали вызванные потенциалы, отводимые от слуховых центров продолговатого мозга, заднего двухолмия и внутреннего коленчатого тела. Эксперименты проводились на наркотизированных закисью азота и галотаном и частично кураризированных с применением искусственного дыхания животных.
Звуковые стимулы подавались в воду, в воздух или через гидрофоны, прижатые к голове животного.
Вызванные потенциалы имели сложную форму. Их латентный период обычно равнялся 2—4 мсек. Наибольшее отклонение потенциала очень коротко, его продолжительность менее 1 мсек. Типичные вызванные ответы среднего мозга возникали только на ультразвуковые, короткие, быстро затухающие сигналы.
Аудиограммы, снятые по вызванным потенциалам задних холмов, совпадали с полученными в поведенческих опытах. Наибольшая чувствительность обнаружена к сигналам с частотой, лежащей в пределах от 20 до 70 кгц. Максимальной была чувствительность к сигналам 60 кгц. В интервалах между 20 и 10 кгц и 60—100 кгц чувствительность понижалась на 25 дб. Наибольшей частотой акустического сигнала, на которую еще возникали вызванные потенциалы, была 120—140 кгц, наименьшей — 5 кгц.
Различия в восприятии звуковых сигналов у разных видов дельфинов были незначительными. Форма и амплитуда вызванных потенциалов даже при незначительных изменениях частоты стимула изменялись гораздо больше, чем пороги.
Наибольший эффект вызывали ЧМ-импульсы. Ответ на них был гораздо большим по сравнению с любыми стимулами, имеющими постоянную частоту в том же диапазоне. Интересно отметить, что обнаружена зависимость амплитуды вызванных потенциалов от направления частотной модуляции стимула. В отдельных случаях ЧМ-импульсы вызывали больший эффект примерно на 10 дб при изменении их частот от высоких к низким, чем при изменении частот от низких к высоким. Таким образом, установлено наличие детекторов, выявляющих изменение частоты определенного типа. Это дает основание предположить, что в слуховых центрах дельфинов существует значительное число нейронов, специально приспособленных для восприятия ЧМ-звуков и способных реагировать на определенные комбинации частот в импульсах.
Слуховые центры дельфинов способны производить очень
389
быстрый временной анализ. Так, полное восстановление ответов на второй стимул наблюдалось при интервале 0.8—1 мсек., между первым и вторым стимулами, в отдельных случаях при 0.5 мсек. Максимальная частота повторения импульсов, на которую еще реагировали слуховые центры, составляла более 2000 импульсов в секунду. Однако наиболее типичные записи показывали, что для полного восстановления ответа на второй стимул требуется 3—5 мсек.
Вызванные потенциалы не возникали, если время нарастания звука превышало 5 мсек. В этом отношении дельфины резко отличаются от калифорнийских морских львов (Zalofus kalifornianus)* также, по-видимому, способных к локации (Bullok et al., 1971).
Анализ биоэлектрических реакций позволил оценить диаграмму направленности звукового приема. Оказалось, что конус оптимальной рецепции направлен вперед и вниз.
Буллок и Ридгвей (Bullok, Ridgway, 1971) провели исследование в условиях хронического эксперимента на ненаркотизированных дельфинах, специально обученных генерировать по сигналу звуковые посылки типа локационных сигналов. Во время опыта движения животного были ограничены или лямками, или размерами ванны, куда оно помещалось. Электроды диаметром 75—150 мкм в количестве 7—18 вводились с помощью шприцевых игл веером на расстоянии 5—10 мм друг от друга. Ввиду того что стереотакси-чески координаты слуховых центров не были определены, точность попадания в задние холмы, имеющие диаметр около 10 мм, была невелика. Из 137 электродов только 30 попало в слуховые системы среднего мозга и 7 — над тенториумом. Излучающий и контрольный (приемный) гидрофон крепились на голове животного. Раздражением служили собственные «щелчки» дельфина, генерируемые им по команде тренера. Тоновый генератор через заданный интервал генерировал искусственное «эхо». Этот прием обеспечивал неизменность эха. Запись биоэлектрических реакций осуществлялась с помощью магнитофона и обрабатывалась электронно-вычислительным устройством.
В ходе эксперимента регистрировались два типа ответов — низкочастотные потенциалы и высокочастотные. Кратковременные, низкочастотные, медленно развивающиеся потенциалы возникают с латентным периодом ~ 22 мсек. Продолжительность основного отклонения такого потенциала 20 мсек. Эти потенциалы вызывались тонами 5—10 кгц. Короткие, быстрые, характерны главным образом для среднего мозга и лучше всего вызываются частотами выше 30 кгц при условии, что нарастание тона короче 4 мсек. Если нарастание было большим, то даже звуковые посылки на уровне 50 дб не были способны вызвать потенциал. При времени нарастания сигнала порядка 2 мсек, ответ был слабым. Максимальный ответ возникал, если время нарастания сигнала было короче 30—50 мксек. Удалось найти
390
участки, которые не отвечали на голос и свисты, прикосновение к коже под водой и на укол иглой, но отвечали на падение в воду или на кожу нитки или прикосновение пальцем к коже извлеченного из воды животного и на охлаждение его хвоста. Очень четкие вызванные потенциалы возникают на локационные щелчки. Амплитуда вызванных потенциалов, которые возникают вслед за каждым собственным щелчком дельфина, чаще не коррелирует с амплитудой щелчка. Звуки, которые сами не были способны вызвать ответ, хорошо маскировали вызванный потенциал на щелчок. Особенно хороший маскирующий эффект давали шипение и почесывание кожи дельфина. Чистые тоны никогда не могли подавить вызванные потенциалы.
Интересные результаты получены относительно продолжительности цикла восстановления. Кортикальные низкочастотные потенциалы на свисты развиваются в течение 50 мсек. Цикл восстановления для щелчков мог быть короче 1 мсек.
Дельфины в отличие от рыб оказались не способны различать фазу сигнала. Характер вызванных потенциалов не менялся, когда дельфину предъявляли сигналы 20, 50 и 70 кгц с нулевой фазы, за которой следовала компрессия, или разрежение.
Большой интерес представляют эксперименты, имевшие целью получить вызванный ответ от эхо-сигнала, отраженного диском диаметром 15 см с расстояния 1—3 м. Однако положительных результатов добиться не удалось. Щелчки не вызывали облегчения для вызванных потенциалов от эха, но восстановление было более быстрым, чем после обычных звуковых посылок.
Проведенные эксперименты позволяют считать, что звуки типа локационных щелчков анализируются в заднем четверохолмии и в прилегающих областях, а коммуникационные сигналы типа свистов — в коре головного мозга.
Ладыгина и Супин (1970) изучали вызванные потенциалы коры головного мозга морской свиньи. Они обнаружили, что фокус вызванных ответов на звуки лежит на 1.5—3 см латераль-нее сагиттального шва и на 3.5—6 см каудальнее коронарного шва черепа дельфина, т. е. данная часть акустической зоны находится в теменной коре. Первичный ответ негативный, с латентным периодом 6 мсек. Время до максимального отклонения 15— 16 мсек. Потенциалы имели истинное корковое происхождение, так как на глубине 8—9 мм наблюдалась их реверсия, а при дальнейшем погружении электрода ответы исчезали. Вторичный ответ был более медленным, с латентным периодом 15—20 мсек, и не коркового происхождения. Данная работа не дает возможности сделать заключение о точной локализации акустической зоны.
20
ХАРАКТЕРИСТИКИ ПРИЕМНОЙ СИСТЕМЫ
Диапазон слухового восприятия
Почти одновременно и независимо друг от друга, используя разные методики и различную аппаратуру, две группы исследователей попытались определить верхний и нижний частотные пороги слухового восприятия у дельфинов, содержащихся в морской студии Мэриленда, во Флориде и Брайтоне на Бимини. Одна из них, под руководством Келлога (Kellogg, Kohler, 1952), проводила опыты непосредственно в бассейне, где постоянно содержались 12 дельфинов (10 афалин и 2 длиннорылых дельфина). Комплекс аппаратуры состоял из генератора с частотным диапазоном от 20 гц до 200 кгц, сигнал с которого поступал на усилитель мощностью 20 вт, с линейной частотной характеристикой до 100 кгц и подавался под воду с помощью излучателя. Данный комплекс позволял излучать звуковые колебания вплоть до 200 кгц. При проведении экспериментов звук подавался в течение 2—3 сек., с интервалами от 30 сек. до нескольких минут. Минимальное звуковое давление в воде на расстоянии 4 м от излучателя составляло 4.03 дин/см2. О восприятии звуков судили по изменению поведения животных. Результаты этих экспериментов показали, что дельфины реагировали на звуки от 100 гц до 80 кгц. Более подробно они были описаны в 1953 г. (Kellogg, 1953).
Другая группа исследователей проводила опыты в небольшом мутном заливчике с илистым дном на одной взрослой самке афалины (Schevill, Lawrence, 1953а). В опытах была использована пищевая условнорефлекторная методика. Если дельфин слышал звуковой сигнал, он подплывал к экспериментатору и получал вознаграждение в виде рыбки. Авторы, используя генератор, усилитель и несколько гидрофонов различного частотного диапазона, проверяли способность дельфина воспринимать звуки частотой от 150 гц до 153 кгц. Длительность звукового раздражителя составляла 3—5 сек.
392
В результате проведенных экспериментов выяснилось, что дельфины на звуки частотой от 150 гц до 120 кгц давали около 100% положительных ответов. При подаче сигналов частотой свыше 120 кгц наблюдалось заметное падение процента положительных ответов. Уже на частоте 126 кгц число положительных реакций составляло 50%, а на частотах 151—153 кгц — всего 13%.
Шевилл и Лоуренс попытались объяснить, почему в опыте, описанном выше (Kellogg, Kohler, 1952), дельфины воспринимали звуки лишь до 50 или 80 кгц. Как они сообщают, излучатель, которым пользовались предыдущие исследователи, был предназначен для частот от 0.1 до 10 кгц, а на более высоких частотах выходной сигнал настолько ослаблялся, что его весьма трудно было использовать. Позднее Келлог (Kellogg, 1961) в свою очередь подверг критике опыты Шевилла и Лоуренс, указывая, что дельфин мог обнаруживать пищевое подкрепление в воде и именно оно служило сигналом независимо от частоты подаваемого раздражителя. Однако Норрис (Norris, 1964) отмечал, что в обоих случаях опыты ставились вполне корректно и результатам можно верить. Эти начальные сведения о диапазоне слухового восприятия у дельфинов получили дальнейшее развитие лишь в последние годы.
Джонсон (Johnson, 1966), применив метод условных рефлексов, исследовал частотно-пороговые характеристики слуховой системы афалины в частотном диапазоне 0.04—200 кгц. В результате этих исследований было установлено, что на частотах 20 — 100 кгц пороги были наименьшими, особенно около 50 кгц, и дельфин реагировал на сигналы, интенсивность которых составляла всего 10~16—10 ~17 вт/см2, т. е. примерно —40 дб относительно нулевого уровня 1 мкбар. В то же время реакция на ультразвук с частотой 150 кгц отмечалась лишь при интенсивности 10"10 вт/см2.
Кроме того, выяснилась зависимость абсолютного порога от длительности сигнала. В этой серии опытов использовались сигналы с частотами 0.25, 1.4, 20, 45 и 100 кгц, длительность которых по ходу опыта уменьшалась от 1 до 10 ~5 сек. Если длительность сигнала составляла 10сек. и меньше, наблюдалось резкое повышение порога (примерно вдвое), тогда как при длительностях сигналов 1—10~3 сек. абсолютные пороги слуховой системы у дельфина почти не менялись (10 кгц — 10 дб, 20 кгц — 35, 45 кгц — 25 дб).
Результаты поведенческих опытов Джонсона и снятая им аудиограмма слуха афалины почти полностью подтвердились в электрофизиологических исследованиях (Bullock et aL, 1968).
Для обыкновенного дельфина границы слухового восприятия были определены Бельковичем и Солнцевой (1970) методикой условных рефлексов. В начале эксперимента вырабатывался условный рефлекс на конкретный звуковой раздражитель. В дальнейшем
393
подкреплялся любой акустический сигнал. Условнорефлекторная реакция возникала в диапазоне частот от 10 гц до 280 кгц. Авторы определяют указанный частотный интервал как диапазон восприятия, хотя отмечают (Яблоков и др., 1972), что на сигнал 320 кгц животные или отвечали бурной двигательной реакцией, или «дергались», оставаясь на стартовой позиции. Совершенно очевидно, что акустический раздражитель ими воспринимался.
Диапазон слухового восприятия морской свиньи изучался с использованием метода кожно-гальванической реакции на фиксированном животном (Супин, Сухорученко, 1970; Сухорученко, 1971). Слуховой порог определялся постепенным ступенчатым
FjKzy
снижением уровня звука на 10—5 дб в диапазоне 4— 180 кгц через каждые пол-октавы (рис. 122). Как видно из рисунка, максимум чувствительности приходится на частоту 128 кгц. При дальнейшем увеличении частоты чувствительность резко падает. С понижением частоты чувствительность меняется более медленно: 6—7 дб/октава.
Рис. 122. Пороговая кривая морской свиньи для тональных сигналов. (Супин, Сухорученко, 1970).
I — уровень звукового давления относительно 1 бара; L — интенсивность предъявляемых стимулов.
Самые низкие пороги имели величину 10 ~18 вт/см2, самые высокие, на максимуме чувствительности — менее 10 "16 вт/см2. Кривая порога на шумы повторяла форму аудиограммы, проходя несколько ниже последней. В исследовании использовался полосовой шум, в совокупности перекрывающий диапазон частот 3—100 кгц. Каждая из полос имела ширину порядка октавы.
Для амазонской инии (Inia geoffrensis) диапазон слухового восприятия сигналов длительностью 5 сек. лежит в пределах 1—105 кгц. Максимальная чувствительность в области .75—90 кгц для величины звукового давления равна —50 дбЪтносительно уровня 1 дин/см2 (Jacobs, Hall, 1972). Диапазон звуков, воспринимаемых косаткой {Orcinus orca), значительно уже — 15—32 кгц (Hall, Johnson, 1972).
Указанными работами по существу и ограничиваются сведения о частотном и временном диапазонах слухового восприятия у китообразных. О способности дельфинов определять направление на источник звука упоминается в ряде сообщений (Schevill, Lawrence, 1956; Kellogg, 1958, 1959а, 1959b, 1961;' Norris et al., 1961), где отмечается, что дельфин точно цдет к .месту всплеска,,
ч394
создаваемого падающими в воду предметами с расстояний 18— 26 м и более. При этом даже такие слабые звуки, как всплеёк падающей дробинки или нескольких капель воды, вызывали целенаправленную реакцию животного на источник звука.
Способность дельфина афалины определять направление на источник шумового сигнала изучали Дьяченко и др. (1971). Животное' должно было по длительному сигналу определить, с какого из двух гидрофонов подавался сигнал, и подойти к нему. Дифференцирование начинали вырабатывать при расстоянии между излучателями в 1 м (что соответствовало углу 5°). Звуковое давление в конце разделительной сети (минимальное расстояние, на котором должно произойти дифференцирование) составляло 300 бар. Животное уверенно определяло направление на источник звука на уровне не ниже 80% правильных ответов до углового расстояния между гидрофонами, равного 1?5. При угле 1° наблюдалось резкое ухудшение дифференцировки (57 % правильных реакций). Дальнейшее сближение излучателей исключало возможность правильного выбора направления на источник звука. Снижение уровня сигнала на 40 дб для угла 1?5 не привело к существенному ухудшению дифференцирования.
Дифференциальные пороги слуха по интенсивности
Важной характеристикой слуха является дифференциальный порог по интенсивности, т. е. та минимальная величина приращения интенсивности сигнала, которую еще способен уловить слуховой анализатор. Было проведено исследование по определению дифференциального порога по интенсивности для белого шума (Бурдин и др., 1971а, 19716).
Как известно, при восприятии амплитудно-модулированных шумов с низкой частотой модуляции амплитудная модуляция воспринимается как изменение интенсивности (Цвикер, Фельд-келлер, 1971), а пороговая разность между максимальным и минимальным уровнями шума близка к величине дифференциального порога по интенсивности для белого шума (Дубровский, Тумар-кина, 1967).
В связи с этим у дельфинов определялась величина минимальной глубины модуляции белого шума, еще различимая животным. Для этого вырабатывался двигательно-пищевой условный рефлекс на звуковую посылку длительностью 30 сек. широкополосного (с полосой от 0.5—до 100 кгц) белого шума. Дифференциро-вочным раздражителем был аналогичный сигнал, модулированный 'частотой 3 кгц, при глубине модуляции, равной 20%. После упрочения дифференцировки к модулированному белому шуму подмешивался немодулированный шум, чем вызывалось уменьшение глубины модуляции, и смесь предъявляли животному. Посте
395
пенно, увеличивая долю немодулированного, определяли момент, когда дельфин переставал воспринимать «смесь» как модулирован-
ный шум. Уровень подаваемых сигналов был постоянным и равнялся 50 бар. Анализ полученных результатов свидетельствует о том, что вплоть до глубины модуляции, равной 3.3%, животные еще хорошо отличали данный сигнал от немодулированного
Рис. 123. Величина дифференциального порога по интенсивности для белого шума.
По оси абсцисс — Мэкв (отношение модулированного к немодулированному шуму, %); по оси ординат — вероятность правильного различения. Сплошная линия — смесь шумов; штриховая — смодулированный шум.
шума. При глубине модуляции, равной 4—7 %, распознавание осуществлялось на уровне 85—95% правильных реакций (рис. 123),
Дифференциальные пороги слуха по частоте
Определение диапазона воспринимаемых дельфином частот недостаточно для характеристики его сонарной системы. Очень важно знать дифференцированные пороги по частоте. Подобные исследования впервые осуществлены Абрамовым и др. (1972) на афалинах. Опыты проводились по методу условных рефлексов. В качестве положительных условных раздражителей использовались тональные посылки частотой 12.5, 25, 50 и 100 кгц, что перекрывало весь диапазон, в котором чувствительность слухового анализатора дельфина близка к максимальной. Все тональные сигналы других частот были отрицательными раздражителями. Максимальное отклонение частоты отрицательных сигналов составляло половину от частот положительного сигнала. Минимальное изменение частоты сигнала, воспринимаемого дельфином по уровню 70% правильных ответов для частот 12.5—25кгц, при уровне сигналов +35 дб относительно 1 бара на расстоянии 1 м, составляет 14+7%, а для частот выше 25 кгц — 7+1%. Таким образом, частотная избирательность слухового анализатора дельфина растет по мере увеличения частоты анализируемых сигналов.
Сходные результаты получены для морских свиней (Сухору-ченко, 1971, 1972). По показаниям условных дыхательных и кожно-гальванических рефлексов, у фиксированных животных для частот в диапазоне от 3 до 190 кгц, при уровне звука 40—60 дб над средним порогом, дифференциальные пороги по частоте оставались в пределах 0.02—0.2%.
396
Наши исследования, проведенные в аналогичных условиях на 2 афалинах, включали три этапа: 1) образование двигательнопищевого условного рефлекса на тон 30 или 70 кгц; 2) выработку грубой дифференцировки на тон 60—70 или 30 кгц; 3) определение дифференциального порога методом постепенного сближения частоты дифференцируемого раздражителя с частотой положительного. Минимальное изменение частоты, воспринимаемое дельфином, равнялось 4 кгц, что составляло 13.3% от несущей частоты положительного сигнала.
Теоретический анализ позволяет предполагать, что дифференцирование длительных сигналов будет существенно отличаться от различения импульсных, поскольку сложная спектральная структура последних может накладывать определенные ограничения на частотное разрешение. Между тем для локации животные используют серии коротких сигналов. В связи с этим было интересно выяснить, произойдут ли изменения в тонкости дифференцирования по частоте акустических сигналов при предъявлении дельфинам последовательности импульсов длительностью 200 мксек, с различной частотой следования. В первой серии частота следования сигнала составляла 3.3 кгц. Положительный условный раздражитель, как и при дифференцировании тональных сигналов, имел частоту заполнения 30 кгц. При такой последовательности импульсных сигналов большая часть энергии импульса содержится вглавном лепестке спектра, имеющем 3 основные спектральные составляющие (рис. 124).
Рис. 124. Спектры радиоимпульсов с частотой заполнения 30, 32 и 34 кгц {сверху вниз) и частотой следования 3.3 кгц.
Вновь выработать условный рефлекс на импульсный сигнал не пришлось, дельфин отвечал на него положительной условной реакцией, как на тональный сигнал с той же частотой заполнения— 30 кгц. Однако отдифференцировать импульсные сигналы с частотой заполнения 30 и 50 кгц без специальной тренировки животное оказалось не в состоянии. Применение в качестве дифферен-цировочного раздражителя импульсного сигнала вызвало угаше-ние условного рефлекса на положительный раздражитель, и для его восстановления потребовалось 85 сочетаний. Это свидетельствует о том, что для животного моночастотный длительный и импульсный сигналы являются самостоятельными раздражителями. Их обобщение, происходящее в момент перехода к импульсным
397
сигналам, оказалось непрочным. В конечном итоге было достигнуто дифференцирование сигнала с частотой заполнения 34 кгц. Более тонкую дифференцировку на импульсный сигнал частотой 32 кгц выработать не удалось. Таким образом, предел дифференцирования импульсных сигналов, следующих с достаточно высокой частотой — 3.3 кгц, практически не отличается от дифференцирования тональных сигналов. Минимальное отличие частотных характеристик сигнала, воспринимаемое животным в обоих случаях, составляет 4 кгц, т. е. 13.3% от частоты положительного сигнала.
Дифференцирование импульсных сигналов, следующих с частотой 0.9 кгц, оказалось более трудной задачей. Дифференцировка выработалась на сигнал 40 кгц, более тонкого различения сигнала 36 кгц осуществить не удалось. Следовательно, в этом случае минимальное изменение частоты, воспринимаемое животным, равняется 33.3% частоты положительного сигнала. Увеличения дифференциального порога при уменьшении частоты следования раздражителя можно было ожидать, поскольку в этом случае «вес» основной гармоники (несущей частоты) уменьшается и распознавание должно осуществляться как бы в условиях «аддитивных помех», роль которых играют боковые гармонические составляющие. В данном случае на интервал 10 кгц приходится 11 гармонических составляющих, тогда как сигналы, следующие с частотой 3.3 кгц, имели всего 3 гармоники.
Таким образом, проведенное исследование показало, что дифференцирование моночастотных сигналов существенно отличается от дифференцирования импульсных. Такое различение частот тонального сигнала от аналогичного раздражителя, но
Таблица 25
Пороговое различение частоты афалиной (Jacobs, 1972)
Основная частота (F), кгц Минимальное изменение основной частоты, различаемое животным (ДГ), гц Отношение / др \ Вебера 1 —)
0.9 7.9 0.0088
10.9 0.0121
2.0 6.1 0.0031
19.5 0.0098
5.0 14.1 0.0028
9.0 31.7 0.0032
20.0 90.0 0.0045
87.0 0.0045
50.0 532.0 0.0107
368.0 0.0074
90.0 1038.0 0.0116
398
модулированного частотой 2 гц, проведено на атлантической афалине (Jacobs, 1972). Амплитуду модулирующего сигнала уменьшали ступенчато через 3 дб. Минимальную девиацию модулированного сигнала, при котором животное было еще способно отличать один сигнал от другого, считали за пороговое различение частоты в данном диапазоне (табл. 25).
В противоположность человеку, у которого область наилучшего различения сигнала по частоте совпадает с областью наиболее хорошо слышимых частот, у дельфинов эти области лежат в разных частотных диапазонах. У афалин наилучшее различение сигналов по частоте приходится на диапазон 2—20 кгц, а область высокой чувствительности слуха — 50—70 кгц.
Пределы дифференцирования временных характеристик сигнала
Способность животного различать сдвиг эхо-сигнала по оси времени позволяет ему определять расстояние до лоцируемых объектов и их параметры. Способность различать временные интервалы важна еще и потому, что позволяет определять частоту воспринимаемого сигнала не только с помощью набора резонаторов или фильтров, но и анализом периодичности звукового сигнала, т. е. измерения длительности интервалов между максимумами (или минимумами) амплитуды колебаний (Schouten et al., 1962). Такой способ обработки сигнала необходим при восприятии частоты сложных звуков (Bekesy, 1963).
В опытах на дельфинах белобочках по дифференцированию трехступенчатых пирамид из пенопласта (Белькович и др., 1969) было показано, что животные способны обнаружить уменьшение толщины среднего сегмента площадью 49 см2 с 12 до 6 мм и значительно труднее реагируют на такое же уменьшение толщины верхнего сегмента площадью 9 см2, что должно соответствовать задержке эхо-сигнала порядка 4х10-6 сек.
Проведенные нами исследования на афалинах по дифференцированию расстояния между двумя пластинами, одна из которых была изготовлена из гетинакса, а другая — из дюралюминия, и между торцевыми стенками полых латунных цилиндров, заполненных водой, выявили весьма высокую разрешающую способность. При максимальном расстоянии между отражающими плоскостями 92 мм, принятом в качестве положительного раздражителя, животные с 3 м дифференцировали данную мишень от аналогичной, имеющей расстояние между отражающими плоскостями на 1.5 мм меньше. Если предположить, что животные дифференцировали расстояние между двумя отражающими звук плоскостями ца основании разницы во времени прихода первого и второго эхо-
•399
сигналов, придется признать, что дельфины способны дифференцировать временные интервалы, отличающиеся всего на 2 X10 "6 сек.
Хотя несомненно, что измерение временной задержки эхо-сигнала позволяет лоцирующим животным определять расстояние до лоцируемого объекта, разрешающая способность по дифференцированию расстояния не может явиться безоговорочным критерием для определения дифференциальных порогов по времени. Дело в том, что в большинстве экспериментов животному предлагается дифференцировать или расстояние от передней до задней поверхности мишени, или расстояние между двумя близко расположенными объектами. В этом случае объекты локации представляют собой сложную колебательную систему, создающую частотно окрашенное звуковое поле, спектральный состав которого будет меняться при изменении расстояния между формирующими поле объектами. Экспериментатор не имеет возможности судить, пользуется ли в данном случае животное временной задержкой или использует для дифференцирования другие показатели эхо-сигнала.
Из тех же соображений нельзя безоговорочно принять выводы Абрамова и др. (1972). В проведенном ими исследовании дельфину предъявлялась задача дифференцировать последовательности пар сдвоенных звуковых импульсов по временному интервалу между сигналами в парах. За положительный раздражитель были приняты сдвоенные импульсы, следующие друг за другом с интервалом 50 мксек. На основании полученных результатов авторы делают вывод, что разрешающая способность афалины по времени на уровне 70% различения равняется 17 мсек. Однако в работе отсутствует оценка спектрального состава предъявляемых сигналов. Не исключено, что животные помимо временных характеристик использовали при дифференцировании изменения частотных параметров сравниваемых раздражителей.
Более объективные результаты можно получить методом изучения электрической активности мозга. В опытах Буллока и др. (Bullock et al., 1968) первый признак наличия реакции на второй из двух одинаковых сигналов возникает при интервале между ними, равном 0.5 мсек.
В наших последних исследованиях на афалинах мы предъявляли животным для дифференцирования такие раздражители, которые могли бы различаться лишь по временным характеристикам. Теоретически любые временные изменения в сигнале неизбежно приводят к изменению его спектрального состава. Однако путем соответствующего выбора параметров сигнала можно избежать их влияния на результаты эксперимента. У животных вырабатывалось дифференцирование величины интервала между двумя звуковыми посылками. Положительная и дифференцировоч-ная серии пар звуковых посылок, состоящие из прямоугольных сигналов длительностью 50 мксек., заполненных белым шу
400
мом, с полосой А/, равной 50 кгц, с частотой повторения 2 гц, предъявлялись животному одновременно на 2 разнесенных на 3 м гидрофона. За положительный условный раздражитель была выбрана последовательность звуковых посылок, следовавших друг за другом, с величиной интервала в парах между концом одного импульса и началом следующего, равной 1000 мксек. Дифферен-цировочным раздражителем была последовательность звуковых посылок, подававшихся с интервалом в парах 100 мксек. Проведенное исследование (30 опытов) показало, что дельфин не способен дифференцировать такие раздражители. Дифференцирование второй пары серии звуковых посылок с интервалами между импульсами 2000 мксек, (положительный раздражитель) и 250 мксек, (отрицательный раздражитель) также не увенчалось успехом.
В другой серии опытов дельфину предлагалось дифференцировать длительность импульсов (или пауз между ними), следующих с частотой 5 гц. Длительность импульсов положительной серии 190 мсек, (интервал между импульсами 10 мсек.), дифферен-цировочной — 199 мсек, (интервал 1 мсек.).
Такие параметры импульсов не соответствуют длительностям локационных импульсов дельфинов, но исключают разрешение их по частотному параметру. Дифференцирование выработать не удалось. Различение оказалось возможным, когда длительность импульсов сократили до 100 мсек, (интервал 100 мсек.). Однако в данном случае, помимо различий во временном интервале, раздражители значительно отличались в энергетическом отношении.
Таким образом, дифференцирование дельфином величины двух интервалов, отличающихся на 900 и 1750 мксек., или длительности двух сигналов, отличающихся на 9 мсек., оказалось невозможным. Полученные результаты резко отличаются от данных по дифференцированию расстояния между подводными объектами. При локации использованные интервалы должны были бы соответствовать дальности до лоцируемых объектов в 0.67 и 1.3 м. Способность дельфинов определять расстояния намного превышает указанные величины.
Возможность использования фазы сигнала
Анализ характера локационных сигналов дельфинов делает допустимым предположение о принципиальной возможности использования ими фазы сигнала для наиболее уверенного выделения его из помех и для определения расстояния до облучаемого тела, размеров и угловых координат подводных объектов. На основе современной теории связи, в частности теории приема радиолокационных эхо-сигналов в условиях помех, известно, что использование фазы сигнала позволяет получить наиболее высокую точность определения пространственного положения
401
объекта. Существующие в настоящее время фазовые пеленгаторы дают возможность осуществлять измерения углов с предельно высокой точностью.
Использование фазы сигнала способно обеспечить наибольшую достоверность передачи информации, так как она является и информативным признаком, и параметром, обеспечивающим устойчивое выделение сигнала из помех. Особенно широкие возможности использования фазы открываются при применении шумоподобных сигналов. Несколько ограниченное применение фазовых систем в технике в настоящее время объясняется трудностями, связанными с их технической реализацией, сложностью физического истолкования и математических методов анализа процессов, протекающих в этих системах (Williamson, 1969). В связи с этим изучение возможности распознавания дельфином сигналов по их начальным фазам, а также использование животным фазо-манипулированных сигналов при ориентации, распознавании объектов, измерении их размеров и при определении пространственных координат представляют значительный интерес.
Наиболее вероятным кажется использование дельфинами фазы сигнала для повышения его помехозащищенности. При определенных способах модуляции по фазе импульсных сигналов (например, кодами Баркера, Хафмана, Диджилок и др.) амплитудно-частотный спектр сигнала оказывается почти равномерным, как и у шума. В этом случае все особенности сигнала отражаются в фазово-частотном спектре. При приеме такого сигнала с помощью согласованного с ним фильтра (а далее будет показана правомерность предположения о существовании у дельфинов некоторых типов согласованных фильтров) на выходе системы происходит «сжатие» сигнала во времени, вплоть до получения одного короткого выброса. Следовательно, используя модуляцию по фазе, можно получить сигналы с автокорреляционной функцией, имеющей один основной узкий выброс при малых уровнях боковых. Чем шире спектр сигнала, тем ^же выброс во времени. Это обеспечивает уверенное разделение даже весьма длинных сигналов и высокую помехоустойчивость системы по отношению к помехам, носящим стационарный характер.
Способность дифференцирования фазы сигнала показана для человека и ряда животных (Темчин, 1972). Ее механизм связан с неравноценностью фаз микрофонного потенциала. Отрицательная фаза сигнала связана с гиперполяризацией волосковых клеток, а положительная — с их деполяризацией, приводящей к возбуждению кохлеарных нейронов. Информация о текущей фазе сигнала передается в виде «микрофонической» реакции ядер продолговатого мозга.
В связи с вышеизложенным нами было предпринято исследование с целью установить, способны ли дельфины дифференцировать сигналы по их начальной фазе. Если бы такая способность
402
у зубатых китообразных была доказана, следовало предпринять попытку выяснить, является ли способность дифференцировать фазу сигнала необходимым условием для локационных измерений.
Чтобы осуществить подобное исследование, нам необходимо было иметь возможность генерировать сигналы, совпадающие по всем параметрам, кроме начальной фазы. Частоту заполнения сигнала, его амплитуду, длительность и величину периода повторения нетрудно сделать постоянными. Однако при изменении начальной фазы высокочастотного сигнала его спектр будет неизбежно изменяться строго определенным образом. Выходом из создавшегося затруднения был такой подбор параметров дифференцируемых сигналов, при котором их спектры отличались бы на величину, меньшую разрешающей способности дельфина по частоте. В наших экспериментах мы ее оценили равной 1%, что позволило допустить различия в спектре дифференцируемых сигналов 10—15%.
Предварительно проведенные расчеты показали, что для уменьшения различия в спектрах двух импульсов с различными начальными фазами необходимо увеличивать длительность импульсов. В качестве критерия, по которому производился расчет минимально допустимой длительности импульса, была выбрана относительная ширина полосы частот, необходимая для передачи импульса.
В экспериментах, проведенных на двух афалинах, была сделана попытка выработать дифференцирование двух импульсных сигналов длительностью 250 мксек., с частотой заполнения 30 кгц, следующих с частотой 1 кгц, имевших начальную фазу 0 или 90°, подаваемых на излучатель. Было произведено раздельно 32 и 40 сочетаний — дифференцировка не образовалась.
Направленность звукового приема
Эффективность приема любой сонарной системы во многом зависит от того, насколько направленно она способна осуществлять прием. Направленный прием позволяет значительно увеличить дальность действия системы и ее разрешающую способность по угловым координатам. Сужая диаграмму направленности, можно повысить помехоустойчивость системы.
Буллок и др. (Bullock et al., 1968) в электрофизиологических исследованиях изучали особенности звукового приема, показав на полосатом и гребнезубом дельфине, дельфине Билля и карликовой косатке, что максимальная чувствительность к звукам имеет место при их направленности в области, отстоящей на 5— 30° от средней линии тела животных.
Проведенные нами исследования (Айрапетьянц, Воронов и др., 1972а, 1973; Иваненко, Чилингирис, 1973; Чилингирис, Иваненко, 1973) подтвердили эти наблюдения.
403
Изучалась диаграмма направленности звукового приема тональных и импульсных сигналов на черноморских афалинах и морских свиньях. Эксперименты проводились методом условных рефлексов на афалинах по двигательно-пищевой, а на морских свиньях — по электрооборонительной методикам. Условный рефлекс вырабатывался на звуковые сигналы в диапазоне 30—120 кгц. Во время эксперимента морские свиньи фиксировались, положение афалин в момент восприятия звукового сигнала регистрировалось
с помощью скоростной фотокамеры. В этом случае характеристики направленности для каждой из частот (30, 50, 80 и 120 кгц) находились путем последовательного перемещения излучателя в различные точки вольера. Пороговое значение излучаемой мощности определялось последовательным уменыие-
0 Ю 20 30 40 50 60
Рис. 125. Диаграмма направленности дельфина афалины при приеме тональных сигналов (в пределах 180—360° положение животного определялось визуально).
ft кг и,
Рис. 126. Главный лепесток спектра импульсного сигнала частотой 30 кгц и длительностью 40 мксек, на входе {сплошная линия) и выходе {штриховая) пространственно-частотного фильтра.
По оси абсцисс — часто.та; по оси ординат — амплитуда огибающей спектра.
нием сигнала дискретно, с шагом в 2 дб. Продолжительность тональных сигналов составляла 3—5 сек., импульсных — 40—1100 мксек. Диаграммы построены на основе 4000 кинокадров.
Изучение направленности звукового приема в горизонтальной плоскости для тональных сигналов у афалин показало, что с повышением частоты принимаемого сигнала диаграмма направленности сужается (рис. 125).
Исследование направленности звукового приема у морской свиньи дало сходные результаты, показав, что наименьшие пороги слуховой чувствительности обнаруживаются к звукам, источник которых расположен на продолжении оси тела. С повы
404
Шением частоты принимаемого сигнала диаграмма направленности обостряется.
Из полученных диаграмм направленности для тональных сигналов можно предположить, что при переходе на импульсный режим работы должна наблюдаться «деформация» диаграммы из-за наличия ряда гармоник в спектре излучаемого сигнала. Так, для сигналов длительностью 40 мксек, помимо основной несущей частоты 30 кгц будут присутствовать более высокочастотные гармоники, вплоть до 55 кгц (рис. 126). Именно в области 60—70 кгц лежит наибольшая чувствительность слуха афалины. Поэтому животные в этом случае могут иметь возможность реагировать не на несущую частоту, а на более высокочастотные составляющие, лежащие в области максимальной чувствительности, и, следовательно, диаграмма направленности будет строиться соответственно чувствительности к высокочастотным гармоникам.
Проведенные исследования подтвердили это предположение. Изучение направленности приема сигналов длительностью 40 и 1100 мксек, с несущей частотой 30 кгц и частотой повторения 330 гц показало, что уменьшение длительности излучаемого сигнала, а следовательно, расширение его спектрального состава вызывает уменьшение ширины диаграммы чувствительности. По форме и ширине диаграмма направленности приема для импульсного сигнала с несущей частотой 30 кгц приближалась к диаграмме направленности для тонального сигнала 50 кгц. Увеличение длительности сигнала до 1100 мксек, приводило к существенному расширению диаграммы направленности (рис. 127).
Диаграмма направленности, показанная на рис. 128, дает более наглядное представление об особенности восприятия звуков, если ее изобразить в системе трех координат с учетом ЧПК слухового анализатора (Bullock et al., 1968) и величины угла (относительно продольной оси тела животного), под которым сигнал принимается, т. е. представить некоторым объемом V=/ (F, Q, f). Рассекая «тело направленности» плоскостями, параллельными соответствующим координатным плоскостям, мы можем проследить, как принимается сигнал при постоянстве одного из двух параметров. Диаграмма направленности при фиксированных углах приема существенно зависит от частоты. Она позволяет оценить особенности приема сигналов дельфинами со спектральной точки зрения. Если рассматривать сечение F (Q3 f) как амплитудно-частотную характеристику некоторого приемного фильтра, легко оценить влияние диаграммы направленности на спектр сигнала, пропускаемого через этот фильтр. На рис. 129, 130 представлены спектры одиночного импульса на входе и выходе подобного фильтра. Спектр на выходе претерпевает определенную деформацию: максимум спектра сдвигается в область более высоких частот, а в огибающей появляется
405
асимметрия. Сопоставление рисунков показывает, что при уменьшении длительности сигнала частотный сдвиг максимума ‘ увели-
чивается. При длительности импульса 100 мксек, сдвиг равен 2.5 кгц, т. е. 6.25% от несущей, а при сокращении длительности импульса до 50 мксек, этот сдвиг составляет уже 6 кгц, т. е. 15% от несущей.
Аналогичная деформация спектра на выходе пространственно -частотного фильтра будет происходить и при изменении направления приема (рис. 131). Величина частотного сдвига зависит от угла, под которым принят сиг-
Рис. 127. Диаграмма направленности (в горизонтальной плоскости) дельфина афалины при приеме тонального сигнала 39 кгц (7) и импульсных сигналов длительностью 1100 мксек. (2) и 40 (3) с частотой заполнения 30 кгц.
Рис. 128. Диаграмма направленности слухового приема как функция частоты и направления.
По оси / — частота в кгц; по оси 9 — направление источника звука; по оси S — значения слухового порога относительно пороговых значений для 80 кгц.
нал. Асимметрия спектра и смещение энергетического максимума уменьшаются при увеличении угла приема до 90°, так как в этом случае амплитудно-частотная характеристика фильтра ста
406
новится более равномерной во всем диапазоне частот. Проведен-
ные теоретические расчеты хорошо совпадают с экспериментальными данными. Как выше было показано, при прохождении сигнала частотой 30 ’ кгц и
длительностью 40 мксек, через пространственно-частотный фильтр его энергетический максимум на 11.5 кгц сдвигается в область более высоких частот, наблюдается асимметрия спектра (рис. 126). Это должно означать, что подобный сигнал дельфин будет воспринимать как колебания с частотой порядка 41—42 кгц и большей длительности. Действительно,
диаграмма направленности
Рис. 129. Спектр импульсного сигнала частотой 40 кгц и длительностью 100 мксек, на входе (сплошная линия) и выходе (штриховая) пространственночастотного фильтра.
По оси ординат —'амплитуды гармоник спектра, нормированные относительно несущей.
Рис. 130. Главный лепесток спектра импульсного сигнала частотой 40 кгц и длительностью 50 мксек, на входе (сплошная линия) и выходе (штриховая) пространственно-частотного фильтра.
Остальные обозначения те же, что и на рис. 129.
Рис. 131. Главный лепесток спектра импульсного сигнала частотой 60 кгц и длительностью 50 мксек, на входе (сплошная линия) пространственно-частотного фильтра и выходе в случае приема его под углом 0° (штрихпунктирная линия) и 10° (штриховая).
Остальные обозначения те же, что и на рис. 126.
моночастотных сигналов по мере сокращения их длительности сужается и для импульсного сигнала частотой 30 кгц и длитель
407
ностью 40 мксек, приближается по своей форме к диаграмме приема тонального сигнала частотой 50 кгц (рис. 127).
Таким образом, очевидно, что дельфины способны осуществлять пространственно-частотную фильтрацию. Изменяя положение головы относительно направления приема, они весьма значительно изменяют характеристики своего пространственно-частотного фильтра, согласуй их с параметрами принимаемых сигналов, что создает предпосылки для оптимальной фильтрации. Метод согласованной фильтрации осуществляет оптимальную линейную обработку сигнала. При такой обработке исходная локационная информация, поступающая в приемный тракт, преобразуется к виду, удобному для принятия животным оптимального решения об обнаружении и определении параметров цели. Если учесть, что сканирование приема путем поворота головы может осуществляться как в горизонтальной, так и вертикальной плоскости, вариабельность характеристик фильтра должна быть весьма высокой и обеспечивать «согласованный» прием эхо-сигнала. Направленный и тем более «согласованный» прием значительно увеличивает дальность действия системы, повышает ее разрешающую способность и помехоустойчивость.
21
ДИАПАЗОН ДЕЙСТВИЯ, ПОМЕХОУСТОЙЧИВОСТЬ И ПРЕДЕЛЫ РАСПОЗНАВАНИЯ
Дальность действия эхолокатора при обнаружении искусственных объектов
Одна из первых работ по изучению дальности действия эхолокатора выполнена на морской свинье (Заславский и др., 1969). Животное должно было обнаружить металлические цилиндры (диаметром и высотой 75 мм) и цилиндры из дерева и пластмассы (высотой 115 мм). В опыте отчетливо проявилась зависимость дальности обнаружения от материала мишени. В 90% испытаний животное правильно обнаруживало металлические цилиндры с расстояния в И м, пластмассовый — 7.3, деревянный — 6.8 м. Дальность обнаружения афалиной шаровых мишеней, изготовленных из различных материалов, определялась Титовым (1972). Шары диаметром 50 мм из резины, воска, свинца и стали в начале эксперимента обнаруживались с расстояния в 5.9, 7.8, 11.2 и 12.4 м. Через месяц дальность действия настолько возросла, что все мишени обнаруживались с максимального для данного бассейна расстояния (около 13). При переходе к мишеням меньшего размера оказалось, что пенопластовый цилиндр (10x10 мм) и шары (диаметром 10 мм) из свинца, стали и дюраля дельфин безошибочно находил с расстояния 11 м. В наших экспериментах (Айрапетьянц, Воронов и др., 1973) дальность различения металлических шаров и цилиндров диаметром и высотой 10 мм и больших размеров превышала 22—24 м.
Зависимость порога обнаружения препятствий — нитей различного диаметра — от материала наблюдалась и в опытах группы Бюснеля (Dziedzic, 1968). Морская свинья могла обнаружить металлическую нить (диаметром 0.2 мм), крученую нить из перлона (диаметром 0.8 мм) и нейлоновую нить (диаметром 1 мм). В этом исследовании и во всех ранних работах по эхолокации дельфина дальность обнаружения препятствий не определялась (Kellogg, 1958; Norris et al., 1961; Busnel et al., 1965). Авторы ограничивались лишь указаниями параметров препятствия, чаще
409
fecero барьеров из вертикально установленных стержней или проволок (нитей). Первые достаточно строгие измерения дальности действия эхолокатора были проведены в работе Бабкина и Дубровского (1971). Дельфину афалине предъявлялись для обнаружения стальные и свинцовые шары, подвешенные в воде на капроновых нитях (диаметром 0.1 мм) на глубине 25—30 см (рис. 132). Свинцовый шарик диаметром 3 мм обнаруживается дельфином с расстояния в 4.5 м. Пороговое обнаружение каждого вида шариков происходило при одном и том же угловом размере мишени. В обнаружении
стальных и свинцовых шариков имеются некоторые различия, однако нет уверенности, что оно зависит только от материала мишени. Существенную роль могла также играть различная степень натренированности животного.
Постоянство отношения диаметра мишени к дальности обнаружения может сви-
Рис. 132. Дальность обнаружения шаров дельфином афалиной. (Бабкин, Дубровский, 1971).
По оси абсцисс — расстояние; по оси ординат — диаметр мишени. Штриховая линия — постоянный угловой размер мишени 2.5', пунктирная — 5'. Кружками отмечены данные по обнаружению свинцовых (для двух животных), крестиками — стальных мишеней.
детельствовать о том, что дельфин обнаруживал мишени при достижении эхо-сигналом некоторого фиксированного порогового уровня. Так как длины волн основного рабочего диапазона (15—50 кгц) значительно превосходили диаметр мишеней (d; область релеевского расстояния), то по крайней мере для стальных мишеней амплитуда эха уменьшалась как i/d2. С другой стороны, эта амплитуда уменьшалась пропорционально г2 (г — расстояние между дельфином и мишенью). Отсюда следует, что при одновременном изменении d nr постоянное значение амплитуды эха, а следовательно, и пороговое отношение сигнал/помеха достигались при постоянном отношении d/r, т. е. при постоянном угловом размере мишени. Отметим, что, по данным Хиклинга (Hickling, 1962), для мягких мишеней отсутствует область релеевского рассеяния звука, поэтому приведенное выше рассуждение не позволяет объяснить постоянство r/d для свинцовых мишеней.
Исходя из особенностей рассеяния звука свинцовыми шариками (при ко, < 1.2 рассеяние пренебрежимо мало; здесь к — волновое число, а — радиус шара), можно показать, что дельфин при обнаружении мишени эффективно использовал участок спектра 410
зондирующего импульсного сигнала на частотах выше 100 кгц. Имеющиеся данные, однако, не позволяют определить верхнюю границу диапазона частот, эффективно используемого дельфином при эхолокации.
Дальность действия эхолокатора дельфина зависит от интенсивности излучения зондирующего сигнала, направленности звукоизлучающей и звукоприемной систем, чувствительности последней, а также от уровня шумовых и реверберационных помех. Определенное влияние могут оказывать также и условия распространения сигнала. Например, волнующаяся поверхность моря может сильно рассеивать сигнал. Пузырьки воздуха, насыщающие при обрушении волн поверхностные слои, увеличивают затухание сигнала.
В нормальном режиме излучения звуковое давление зондирующих сигналов дельфина составляет 2*103 дин/см2 на расстоянии 1 м. В экстремальных условиях (например, при действии интенсивных помех) оно может возрастать в среднем до 4-104 дин/см2 (Бабкин, Дубровский, 1971; Титов, 1972). Чувствительность эхолокационного аппарата к сигналам, имитирующим зондирующие импульсы дельфина (длительностью около 150 мксек., на частоте 60 кгц, повторяющимся 30 раз в секунду), оказалась равной (5.1 + +0.3)-10"3 дин/см2 (Вельмин и др., 1973). Дальность действия при отсутствии помех можно определить из следующего уравнения:
E = S + T-201g [r(r-r0)J,
где S — уровень зондирующих сигналов, дб; Т — сила цели, дб; Е — уровень чувствительности эхолокатора дельфина; г — расстояние дельфин—мишень, г0 — расстояние от дельфина, на котором производилось измерение S. Если принять за уровень отсчета Ро=1О дин/см2, то 5=46 дб для нормального и 72 дб для экстремального режима излучения, а £=—66 дб. Тогда для нормального режима
20 1g [г (г - г0)] = 112 + Т
и для экстремального режима
201g [г (г-г0)] = 138 +Г.
При г >* г0 вышеприведенные выражения упрощаются и принимают вид
401g г =112 +Г и
401g г = 138 + Г.
Значения Т должны определяться экспериментально для сигнала, имитирующего эхолокационный сигнал дельфина. Отметим, что сила цели может быть выражена через сечение рассеяния препятствия:
а
r = 101g??
411
Значения о для моночастотных сигналов измерены для многих видов морских обитателей и для различных искусственных мишеней.
Дальность обнаружения рыбы
Вопрос о дальности обнаружения рыбы дельфином неоднократно обсуждался в литературе. Шевилл и Лоуренс (Schevill, Lawrence, 1956) нашли, что дельфин в 75% случаев обнаруживал погруженную в воду мертвую рыбу длиной 8—23 см с расстояния, не меньшего 2.5 м. В опытах Константинова с соавторами (1968) дельфин белобочка обнаруживал рыбу длиной 12— 15 см на максимальных расстояниях от 3 до 5 м.
Бабкин и Дубровский (1971) поставили своей целью определить у афалин в контролируемых условиях дальность обнаружения рыб одного и того же размера (ставрида длиной 13.5 + 1 см вместе с хвостовым плавником, шириной 3.8 +0.5 и толщиной 1.8+0.3 см) с удаленным плавательным пузырем.
Было проведено несколько серий опытов, в продолжение которых оставляли неизменным угол, составленный линией, проведенной вдоль рыбы, и локационным направлением (азимут). Этот угол мог принимать следующие значения: 0 + 10°, 45 + 10°, 90 + 10°, 180 + 10°, если принять за начало отсчета положение, при котором рыба направлена параллельно длинной стороне бассейна (разброс азимутных углов обусловлен тем, что при различных траекториях движения дельфина локационное направление слегка варьировало).
В двух сериях опытов рыба предъявлялась в вертикальном положении головой вниз и боком по отношению к дельфину. В этом случае пороговая дальность обнаружения (75% правильных ответов) составила 9.1 м. Легко видеть, что эта дальность почти в четыре раза превышает величину, приведенную Шевиллом и Лоуренс (Schevill, Lawrence, 1956), и в 2—3 раза — величину, называемую Константиновым с соавторами (1968). Через месяц количество случаев уверенного обнаружения рыбы дельфином
Таблица 26
Обнаружение дельфином горизонтально расположенных рыб
Дальность, м Число предъявлений .... Азимут, град.
0 45 90 180
3.8 40 9.0 120 9.0 100 3.4 75
412
возросло до 80%, однако на расстоянии 10 м оно осталось практически тем же (72% в начальной серии и 73% — в повторной). При этом пороговая дальность обнаружения возросла, достигнув 9.8 м.
Результаты опытов по обнаружению дельфином горизонтально расположенных рыб сведены в табл. 26.
Максимальная дальность наблюдается при боковой локации рыбы (9 м), а минимальная — при локации либо с хвоста (3.4 м), либо с головы (3.8 м). Таким образом, угловое расположение рыбы-мишени сильно влияет на ее обнаруживаемость дельфином.Этим можно объяснить большой разброс оценок дальностей, наблюдаемых у различных авторов.
По данным Колобаева (1958), у ставриды (размером 15x2x1 см) после удаления плавательного пузыря сечение рассеяния в диапазоне частот 20—70 кгц в среднем в два раза меньше, чем у живой. Исходя из этого, можно ожидать, что дальность обнаружения у живой ставриды данного размера возрастает в 2 раза и составит приблизительно 12 м.
В работах Колобаева (1958, 1970) была измерена полярная диаграмма рассеяния различных рыб, в том числе ставриды. Оказалось, что интенсивность рассеяния звука от рыб сильно зависит от их ориентации. Так, например, на частотах 20 и 50 кгц отношение интенсивности звука, рассеянного при падении звуковой волны перпендикулярно оси рыбы, к интенсивности звука, рассеянного при падении волны вдоль продольной оси рыбы, составляет величину около 5. Следовательно, дальность обнаружения во втором случае будет в 5 = 1.5 раза меньшей, чем в первом. Действительно, в этом опыте отношение дальности при двух указанных аспектах локации составляло величину — 2.5.
Данные по обнаружению одиночной рыбы могут быть использованы для оценки дальности обнаружения стаи ставриды в естественных условиях (Титов, 1972). Экспериментально установлено, что в первом приближении величина эхо-сигнала от стаи увеличивается в \/п раз (п — количество особей в объеме пространства, охватываемом локационным импульсом). При п свыше 4000 величина сигнала перестает увеличиваться и остается практически постоянной. Расчет показывает, что дальность обнаружения стаи при п 4000 должна быть в восемь раз больше дальности обнаружения одиночной рыбы, и для принятой нами средней дальности обнаружения ставриды — 13 м — она составит 100 м. Для стай с меньшей концентрацией особей (п < 4000) было получено выражение
tfCT=^-7.5rt.tga,
где Дст и Дг — дальности обнаружения стаи и одиночной рыбы; а — ширина диаграммы направленности дельфина по уровню
413
половинной мощности. Следует учесть, что дальность обнаружения может возрасти за счет увеличения амплитуды локационных сигналов. Так, при максимальном звуковом давлении сигналов 4-10"4 дин/см2, отмеченном в опытах Дубровского и др. (1971), дальность обнаружения должна увеличиться в 5 раз. В реальных условиях при частоте заполнения локационных сигналов 25~ 30 кгц поглощение сигналов составит 6 дб/км и, следовательно, дальность обнаружения стаи будет равна ~ 350 м.
Так как эффективное сечение рассеяния плотной стаи ставриды больше, чем других съедобных объектов, дальность эхолокационного обнаружения объектов питания черноморских дельфинов не должна превышать полукилометра.
Одними из важнейших вопросов в экологии дельфинов являются ориентация в стае и дальность взаимного эхолокационного обнаружения особей. Зная объем легких дельфинов (Яблоков и др., 1972) и исходя из полученного в экспериментах постоянства углового размера обнаруживаемых объектов и его минимальной величины, нетрудно рассчитать, что дальность обнаружения взрослых афалин с учетом затухания должна составить ~100— 130 м. Из вышесказанного следует, что эхолокационный анализатор используется дельфинами для обнаружения пищи и ориентации на расстояниях не более нескольких сот метров и является, таким образом, анализатором ближнего действия.
Помехоустойчивость эхолокационного аппарата
Дальность действия эхолокатора, его чувствительность к слабым эхо-сигналам, как уже указывалось, существенно зависит от уровня маскирующих помех. Поскольку в естественных условиях обитания дельфин постоянно находится как под воздействием аддитивных акустических помех (шумов ветрового волнения, шумов биологического происхождения и т. п.), так и помех, причинно связанных с излученными сигналами (собственные зондирующие сигналы, реверберационные помехи, т. е. отражения от препятствий, не являющихся в данный момент объектами эхолокации), то естественно ожидать наличия хороших приспособительных способностей гидролокационного аппарата дельфина по отношению к указанным помехам.
Несмотря на очевидную значимость данного вопроса для оценки возможностей эхолокатора дельфина, имеющиеся экспериментальные данные весьма скудны. Гриннел (Grinnell, 1967) считает, что устойчивость к любым видам помех зависит в основном от исключительно тонкого временного различения и от высокой точности пространственного разрешения при различении собственного эха от посторонних звуков.
Возможно, что нейрофизиологические механизмы слухового анализатора у дельфина обладают свойствами, сходными со своц-
414
ствами аналогичных механизмов летучих мышей. Так, по даннЬШ Буллока и др. (Bullock et al., 1968), способность реагировать на последовательные стимулы восстанавливается в течение 0.5— 1 мсек., что соответствует возможности реагировать на последовательность стимулов с частотой до 2000 в 1 сек. В работе этих авторов указывается на способность слуховой системы дельфина к избирательной реакции вызванного потенциала на изменение параметров ЧМ-стимулов.
Одним из направлений изучения помехозащищенности являются эксперименты по выяснению способности животных обнаруживать подводные объекты в условиях акустических помех. В экспериментах Бабкина и Дубровского (1971) изучалась способность дельфина афалины обнаруживать стальной шарик (диаметром 8.7 мм) с расстояния 8 м при наличии широкополосных аддитивных шумовых помех с различными звуковыми давлениями. Помеха излучалась со стороны мишени. Звуковое давление шума в полосе 10—23 кгц; (по уровню —3 дб от максимума), развиваемое излучателем помехи на расстоянии 8 м от мишени измерялось при помощи того же измерительного гидрофона, которым измерялись и зондирующие сигналы афалины.
С ростом звукового давления помехи (Рп) от 0 до 140 дин/см2 звуковое давление локационных сигналов дельфина (Рс) возрастает нелинейно, но монотонно, в среднем от 2-103 до 4-104 дин/см2. Особенно резкое нарастание Р — при начальном введении помех (до 30 дин/см2). При дальнейшем увеличении помех сигнал растет почти линейно. Это означает, что отношение сигнал/помеха, а также отношение звукового давления эхо-сигнала к звуковому давлению помехи сохраняются постоянными. Уровень локационных сигналов был настолько велик, что их низкочастотные компоненты свободно воспринимались на слух экспериментатором, стоящим на мостках, когда дельфин находился от него на расстоянии 3—5 м и на глубине около 0.5 м.
Вводимые помехи не мешали сколько-нибудь заметно правильному обнаружению мишени. Это говорит о скрытых резервах эхолокатора, которые при проведении опытов в отсутствие помех не обнаруживаются экспериментатором.
Приведенные данные подтверждают в первую очередь наличие у дельфина простого и эффективного механизма эхолокации в условиях аддитивных помех. Работа этого адаптивного механизма проявляется прежде всего в резком повышении уровня зондирующих сигналов при появлении помехи. При этом максимальные значения звукового давления достигали в ряде случаев 5-104 дин/см2 на расстоянии 1 м от головы животного. Любопытно отметить, что аналогичные эффекты наблюдались также и у человека. Известно, например, что на* введение помех человек реагирует увеличением громкости голоса и растягиванием гласных звуков. Анализ длительностей зондирующих сигналов в на
415
ших опытах не показал, однако, заметного изменения длительности импульсов с ростом звукового давления помехи.
К перечисленным выше особенностям эхолокации в условиях помех можно добавить и изменение спектра зондирующего сигнала (рис. 133).
Рис. 133. Влияние помехи на спектр сигнала. (Бабкин, Дубровский, 1971).
Спектр: а — помехи, б — сигнала, в — сигнала при наличии помехи, г —при отсутствии помехи (штриховая линия) и при наличии помехи (сплошная линия). Горизонтальными черточками отмечены предельные значения разброса пиковых напряжений на выходе фильтров спектроанализатора.
На рис. 133 представлены спектр шумовой помехи, записанной и проанализированной теми же измерительными трактами, что и локационные сигналы (а), типичный спектр щелчков афалины при обнаружении стального шарика диаметром 8.7 мм в отсутствие искусственных помех (б, а), а также спектр зондирующего сигнала при наличии помехи (в, а). Появление помехи изменяет сигнал дельфина. Максимум этого спектра смещается в область максимума спектра помехи, что способствует поддержанию на постоян
416
ном уровне не только интегрального отношения РСЛРП, но и отношения спектральных плотностей сигнала и помехи во всем исследованном диапазоне частот.
Иную картину наблюдали Абрамов и др. (1971). В опыте изучалось опознание дельфинами подводных объектов на фоне акустических помех: шума в полосе частот 1—150 кгц (с уровнем 9000 дин/см2), тона 80 кгц (уровень 5400 дин/см2) или импульса с частотой заполнения 80 кгц. При этом энергетический максимум в спектре локационных сигналов дельфина смещался в область более низких частот (35—50 кгц). Наличие в фоне тонального сигнала частотой 30 кгц или тонального импульса с частотой заполнения 30 кгц и звуковым давлением 1.200 дин/см2 вызывало сдвиг спектрального максимума в область высоких частот (65— 85 кгц). Таким образом, дельфин смещал спектр сигнала в область частот, на которых интенсивность помех значительно меньше интенсивности локационного сигнала, т. е. отстраивался от излучаемой помехи по частоте. Несмотря на это, помехи приводили к нарушению дифференцировки, за которую авторы принимали снижение правильных решений со 100 до 85%. Лишь радиоимпульсы не ухудшили распознавания. Это, вероятно, связано с тем, что в данном случае дельфин использовал для генерации своих сигналов временные паузы между импульсами помех. Для подавления полезного сигнала уровень маскирующего сигнала должен превышать его на 30 дб (Абрамов и др., 1972).
Описанные выше опыты проводились с использованием аддитивных (шумовых, тональных или импульсных) помех, т. е. помех, не связанных причинно с зондирующим сигналом дельфина. Между тем в жизни дельфина (особенно прибрежных видов) не меньшее значение имеют реверберационные помехи. Для исследования такой помехи была искусственно создана эхолокационная ситуация, типичная для обнаружения придонных объектов питания на каменистом дне (Титов, 1,972). Были отобраны гладкие круглые камни размерами 5, 10, 15, 20, 25, 30 мм, по сто штук каждого размера. Камни равномерно рассыпали на дне в виде двух окружностей (каждая диаметром 40 см) по обе стороны разделительной сети. Мишень опускалась над центром окружности, вначале на дно среди камней, а затем поднималась на высоту, при которой происходило ее пороговое обнаружение.
Дельфин обнаруживал в 75% испытаний с расстояния 5 м свинцовый шар (диаметром 50 мм), лежащий на дне. Стальной шар (диаметром 33 мм) животное обнаруживало с 5 м с той же вероятностью в том случае, когда расстояние между нижним краем шара и дном составляло 1.7 см. Измеренная в этот же период дальность обнаружения обеих мишеней на глубине 30 см от поверхности была больше 11 м. Таким образом, животное могло обнаружить предъявляемые мишени с расстояния 5 м только в том случае, если они выступали над самыми крупными по размеру камнями (шероховатостями дна)
417
нё мёнёе чем на 2.0 см. В обоих последних опытах дельфин сразу со старта уходил на глубину и двигался к мишеням очень медленно с тем, чтобы уменьшить донную реверберацию, лоцируя мишень не сверху вниз, а под малым углом ко дну.
Эксперименты показали принципиальную возможность обнаружения дельфином среди камней и различных препятствий па дне как донных, так и придонных объектов питания при условии, что размер их превышает средний размер препятствий или неровностей дна. В противном случае добыча должна располагаться на некотором расстоянии от дна. Таким образом, можно предположить, что дельфину легче обнаружить добычу, если он плывет, лоцируя, вдоль днав
Помехоустойчивость эхолокатора исследуется также на уровне приемной (слуховой) системы дельфина (Johnson, 1968). Способность слуховой системы обнаруживать тональные сигналы на фоне помех характеризуется критической полосой слуха (КП), которая служит одновременно и мерой разрешающей способности слуха по частоте (Фрейдин, 1968).
Оценки КП—у афалины были сделаны по методу Флетчера (метод критического отношения — Johnson, 1968). Значение КП по этому методу определяется из соотношения
I
Ло-Д/ -1,
где I — интенсивность сигнала на пороге маскировки; 7У0 — спектральная плотность интенсивности; А/ — ширина полосы шумов, эффективно маскирующих сигнал. Опытные данные для дельфина хорошо экстраполируют зависимость КП от частоты для человека. Заметим, что оценки КП по методу Флетчера дают приблизительно в 2.0—2.5 раза меньшие значения, чем более точные методы (Фрейдин, 1968).
Важный вывод из этого эксперимента заключается в том, что слуховая система афалины работает очень сходно со слуховой системой человека.
Дифференцирование величины подводных объектов
Первое сообщение о способности афалины с помощью эхолокации оценивать размер подводных объектов принадлежит Келлогу (Kellogg, 1959а). В его экспериментах дельфин с расстояния в 1—3 м безошибочно дифференцировал рыб длиной 10 и 20 см. Константинов и др. (1968) подтвердили способность дельфинов различать размер рыб. Белобочки с расстояния 2.6 ± +0.4 м в 87% случаев правильно дифференцировали рыб длиной 13 и 26 см.
Американские ученые (Turner, Norris, 1966; Norris et al., 1967) изучали способность афалин дифференцировать стальные 418
шары диаметром 6.35 см и диаметром от 2.5 до 5.71 см с расстояния 105—120 см. Животные в 100% случаев правильно дифференцировали мишени, если разница их диаметров равнялась 0.96 см, и в 77% при разнице 0.64 см. Следует иметь в виду, что эксперимент не был достаточно корректным, так как шары были связаны между собой металлическим стержнем, а расстояние между ними всего в два раза превышало диаметр большего шара, что не исключало акустического взаимодействия через воду. Таким образом, правильнее говорить не о дифференцировании шаров, а о различении асимметрии звукового поля, рассеянного системой из двух шаров и соединительного стержня.
Способность морских свиней различать по высоте одновременно предъявляемые стальные цилиндры (диаметрами 75 мм) изучали Заславский и др. (1969). С расстояния в 2.5 м цилиндр высотой 75 мм правильно отличался от цилиндра высотой 85 мм лишь в 50% случаев. При увеличении разницы в высоте до 20 мм правильное дифференцирование осуществлялось в 80%. При дальнейшем увеличении разницы высот цилиндров различение медленно приближалось к 100%.
В исследованиях Айрапетьянца, Голубкова и др. (1969), а также в работе Голубкова и др. (1969) у афалин определялся предел дифференцирования стальных цилиндров (диаметром 110 мм) по высоте при последовательном предъявлении их животным. В этом случае для идентификации цилиндра высотой 25 мм животным приходилось использовать долговременную память, поскольку интервал между последовательными предъявлениями был не меньше 1 мин. Естественно, что при одновременном предъявлении дифференцируемых мишеней различение должно быть более точным. Известно, что для сенсорных систем человека удлинение интервала между парой сравниваемых стимулов 1 до 60 сек. может приводить к 10-кратному увеличению предела различения. Эксперименты показали, что дельфины могут различать пару цилиндров с высотами 35 и 25 мм со 100 %-й надежностью и пару с высотами 35 и 25 мм с 70%-й надежностью. Голубков и др. (1969) приводят цифры 47 и 60%. Цилиндры с высотами 28 и 25 мм дельфины не различали. Титов (1972) нашел, что для афалины пороговое отношение высот различаемых цилиндров, соответствующих 75% правильных реакций животного, составило 1.3. Таким образом, данные различных авторов по различению дельфинами высоты цилиндров удовлетворительно согласуются между собой. Пороговые отношения высот оказываются в пределах 1.1—1.3.
Титовым (1972) были проведены также опыты по различению толщины цилиндрических оболочек. Афалине с расстояния в 5 м предъявлялись стальные цилиндрические оболочки высотой 50 мм и внешним диаметром 50 мм. Толщина оболочки изменялась от 0.1 до 2 мм. Пороговая толщина оболочки
419
(75% правильных реакций) была равна 0.2 мм. 100% правильных реакций афалины наблюдалось при толщине оболочки 1 мм.
Исследование способности белобочки распознавать размеры пенопластовых квадратов было выполнено в работе Бельковича и др. (1969а). Порог различения составил 9.7% при 77% правильных реакций животного. Дальность различения составила в среднем 5 м.
Исследования по различению размеров шаровых мишеней были выполнены на Карадагской биостанции (Дубровский и др., 1970, 1971; Дубровский, Краснов, 1971; Краснов, 1972; Дубровский, 1972, 1973; Титов, 1972; Фадеева, 1973). Они
проводились в основном в условиях, исключающих зрительное восприятие дифференцируемых мишеней. В первой же серии у дельфинов вырабатывалось дифференцирование свинцового шара диаметром 5 см от шаров с диаметрами 10, 6 и 5.5 см. Точность дифференцирования почти всюду монотонно падала по мере увеличения расстояния до мишеней. Для шаров 5.5 и
Диаметр шара, мм
Таблица 27
Дифференцирование дельфинами стальных шаров различного диаметра
положительного дифференциро-вочного дифференцирования, %
16 11.3 90
17.3 11.3 93
18.5 11.3 100
24 11.3 100
18.5 16 46
17.3 16 50
5.0 см надежность различения с расстояния 8 м составляла 78%.
Результаты второй серии экспериментов по дифференцированию стальных шаров представлены в табл. 27. Опознание мишеней осуществлялось с расстояния от 2 до 6 м.
В третьей серии исследовалась различимость двух свинцовых шаров с диаметрами 16 (положительная мишень) и 14 мм (отрицательная мишень). С расстояний 2.4 и 5 м была достигнута надежность различения 87 и 64% соответственно.
Наиболее важным из приведенных выше результатов мы счи
таем то, что на надежность различения размеров существенно влияет материал мишеней. Так, например, свинцовые шары с диаметрами 14 и 16 мм распознавались дельфином в 87% случаев, в то время как стальные с диаметрами 18.5 и 16 мм совершенно не распознавались.
Обращает на себя внимание тот факт, что пределы различения размеров определенно зависят от удаления животного от мишеней. Для афалин максимальная дальность дифференцирования сферических мишеней с диаметром 50 мм (положительная) от шаров с диаметрами 5, 20 ,100 и 150 мм составляла 17 м, а для цилиндров с аналогичными величинами высоты и диаметра — 18.5 м (Айрапетьянц, Воронов и др., 1973). Было замечено, что если
420
разница в размерах мишенеи намного превышает предельную, то максимальная дальность надежного различения остается практически постоянной. Напротив, с приближением разницы размеров к пороговой дальности различение начинает резко ухудшаться. Например, для всех пар свинцовых шаров (с диаметром больше 5 см) процент различения остается высоким на дальностях около 6—8 м, а для восковых шаров он мало зависит от дальности для пар с диаметрами 5 и 10 см, 5 и 9, 5 и 8 и 5 и 7 см, однако для пары с диаметрами 5 и 6 см резко падает на дальностях, больших 3 м.
Указанная выше независимость предельной дальности различения от разницы в размерах мишеней обусловлена все возрастающей натренированностью животного по мере перехода к различению все уменьшающейся разницы в размерах мишеней. Особенно ярко это проявилось при различении пары восковых шаров с диаметрами 5 и 7 см. На большом интервале дистанций различение этой пары оставалось на уровне, близком к 70%. Тренировка по дифференцированию пары свинцовых шаров с диаметрами 5 и 6 см в течение приблизительно двух недель привела к увеличению дальности различения с 2 до 6 м (считая
Таблица 28
Дифференцирование дельфином афалиной № 1 сферических тел
Материал шара Диаметр шара, мм Разность диаметров дифференцируемых шаров Число предъявлений Правильность дифференцирования, %
положительного дифференци-ровочного
Свинец 10.2 10.5 0.3 37 52
10.2 10.8 0.6 112 55
12.0 10.8 1.2 40 67
11.7 10.2 1.5 128 75
12.7 10.2 2.5 60 88
14.3 10.2 4.1 40 93
50.0 52.5 2.5 80 52
50.0 54.0 4.0 78 65
50.0 55.0 5.0 177 70
50.0 60.0 10.0 158 80
Сталь 11.5 12.7 1.2 34 56
10.0 13.5 1.5 73 58
10.4 12.7 2.3 81 62
10.0 12.7 2.7 65 71
10.4 14.3 3.9 94 75
10.0 14.3 4.3 100 73
11.5 20.7 9.2 50 88
50.8 50.8 0.0 36 51
50.8 54.0 3.2 81 68
54.0 60.0 6.0 29 90
50.8 60.0 9.2 15 94
57.1 63.5 6.4 — 77
421
Таблица 29
Дифференцирование дельфином афалиной № 2 сферических тел
Материал шара Диаметр шара, мм Разность диаметров дифференцируемых шаров Правильность дифференцирования,. %
положительного дифференци-ровочного
Свинец 14.0 16.0 2.0 87*
50.0 55.0 5.0 78
50.0 60.0 10.0 100
50.0 70.0 20.0 87
50.0 80.0 30.0 93
50.0 100.0 50.0 100
Сталь 11.3 16.0 4.7 90
11.3 17.3 6.0 93
11.3 18.5 7.2 100
11.3 24.0 12.7 100
16 18.5 2.5 46
16 17.3 1.3 50
по уровню 80%). Следовательно, и оценка пределов разрешающей способности дельфина существенно зависит от фактора тренировки, что полностью согласуется с наблюдениями американских авторов (Turner, Norris, 1966; Norris, Evans, 1967).
Эксперименты, проведенные на двух афалинах (Дубровский, Краснов, 1971; Титов, 1972), показали, что дельфины могут с высокой точностью улавливать различия в размерах и площади дифференцируемых объектов, имеющих одинаковую форму и структуру материала (табл. 28, 29). Это установлено для водоподобных (воск), жестких (сталь, свинец) и акустически мягких материалов (пенопласт) и для естественных объектов питания — рыб.
Механизм дифференцирования подводных объектов по размеру и материалу
К настоящему времени накоплен довольно большой экспериментальный материал, характеризующий способность дельфинов различать материал мишеней. Первый эксперимент по различению материалов мишеней проделали Эванс и Пауэлл (Evans, Powell, 1967), которые определили надежность распознавания афалиной дисков диаметром 20 см разной толщины из меди, латуни и алюминия и медного диска толщиной 2.2 мм, принятого в качестве стандарта. Был выполнен расчет отражательной способности дисков в диапазоне частот от 2 до 200 кгц. Использованные мишени имели различную отражательную способность. Авторы нашли, что распознаваемость дисков нельзя объяснить только разностью интенсивностей эха. В ходе дискуссии цо данной работе они, а также Джонсон (Johpsop, 422
1967) предположили, что эхолокационное различение дисков могло определяться изменением фазовых соотношений между компонентами зондирующего сигнала дельфина при отражении. О способности дифференцировать материал мишени морскими свиньями свидетельствует работа Заславского с соавт. (1969). Животному предъявлялись цилиндры диаметром 75 и высотой 115 см, выполненные из разных материалов. В 75% случаев животные правильно дифференцировали пары металл—дерево на расстоянии 6.3 м, металл—пластмасса — 6, дерево—пластмасса — 5 и металл—стекло — на расстоянии 4 м.
В работах Абрамова и др. (Абрамов, Бурдин, Ершова и др., 1970; Абрамов, Айрапетьянц и др., 1971; Абрамов, Голубков, Ершова, 1972) сообщается о результатах опытов по распознаванию дельфинами материалов сферических и цилиндрических тел. Применялись латунь, сталь, эбонит, дюралюминий и другие материалы (табл. 30). Было обнаружено, что латунные мишени дельфины отличают от мишеней, изготовленных из других материалов, с надежностью от 76 до 100%.
Авторы вышеупомянутых работ ограничиваются, как правило, лишь констатацией достигнутых пределов различения и не обсуждают вопроса о физиологическом механизме, лежащем в основе различения. Чтобы понять механизм различения материала, необходимо знать физические характеристики эха зондирующих сигналов дельфинов от применяемых в опытах мишеней. К сожалению, выбор мишеней в настоящее время ограничен телами простой формы, такими как шар, цилиндр бесконечной длины или эллипсоид, для которых известно достаточно строгое решение задачи о нахождении эхо-сигнала. Поэтому в первую очередь изучалось различение материала сферических мишеней (Дубровский и др., 1970; Дубровский, Краснов, 1971).
Таблица 30
Дифференцирование дельфинами мишеней (шаров диаметром 20 мм и цилиндров высотой 50 и диаметром 52 мм) по материалу
Шар Цилиндр
дифференцировоч-ный раздражитель правильность дифференцирования, % дифференцировоч-ный раздражитель правильность дифференцирования, %
Сталь 76 Сталь 87
Дюраль 90 Дюраль 94
Фторопласт 87 Тефлон 98
Эбонит 91 Эбонит 100
Текстолит 100
Примечание. Положительными мишенями служили шар и цилиндр из латуни.
423
Хиклинг (Hickling, 1962) показал, что для тонального зондирующего сигнала амплитуда эха от шара радиусом а может быть представлена в виде
^эха = 2r2 I I’
где PQ — амплитуда звукового давления зондирующего сигнала
в точке излучения; г — расстояние от точки излучения до шара;
й=2ти/Х — волновое число (X — длина волны), а До (ка) — безразмерная функция частоты ка. Эта функция представляет со-
бой кривую, состоящую из последовательно расположенных
максимумов и минимумов (рис. 134), средний интервал между которыми Д монотонно связан со скоростью сдвиговых волн С2 в упругом материале, причем чем больше Д, тем больше С2 (рис. 135). На примере зондирующего сигнала с прямоугольной огибающей
/с/
О 4 8 12 16 20 ка
Рис. 134. Амплитуда давления синусоидального сигнала, отраженного от шара из латуни, в зависимости от ка. (Дубровский и др., 1973).
Рис. 135. Величина среднего периода осцилляций частотных характеристик спектра отражения шаровых мишеней А и ее зависимость от скорости сдвиговых волн в материале С2 (Бабкин, Фадеева, 1971).
было показано, что эхо от шара состоит из двух частей: начального, или «первичного», и более позднего во времени, «вторичного». Первичное эхо соответствует отражению от упругого шара как от абсолютно жесткого тела. Вторичное эхо зависит как от размера, так и от материала шара и содержит информацию о характере собственных колебаний мишени.
В эксперименте использовались шары, имеющие одинаковые диаметры и одинаковые акустические сопротивления, но изготовленные из более мягкого и более жесткого материалов. Следовало ожидать, что различение такого рода мишеней не представит для животного большого труда. Для сравнения изучалась различимость двух твердых материалов, сильно отличающихся своими волновыми сопротивлениями, но имеющих близкие значения скорости сдвиговых волн, т. е. близкие значения интервала Д
424
между максимумами в функции \fm(kd)\. Такими материалами могли служить сталь и дюраль (табл. 31).
Таблица 31
Физические константы мишеней
Материал мишени Плотность, г/см3 Скорость продольных волн, м/сек. Скорость сдвиговых волн, м/сек. Волновое сопротивление, кг/м2«сек. д
Свинец . . 11.3 1960 700 2.22-10® 0.30
Сталь . . . 7.70 5960 3240 45.7 -10® 2.40
Дюраль . . 2.79 6320 3130 17.7 -10® 2.30
Таблица 32
Дифференцирование афалиной материала шаровых мишеней диаметром 50 мм
Материал мишени Правильность дифференцирования (%) для расстояний (м)
3 5 6 8 11
Свинец—дюраль 100 87 100 94 75
Свинец—сталь — 100 — 100 100
Сталь—дюраль 46 50 67 65 67
Различение свинец—сталь абсолютно надежно на расстоянии 11 м, а свинца и дюраля — до 8 м, несмотря на близость волновых сопротивлений (табл. 32).
С другой стороны, несмотря на трехкратное различие в волновых сопротивлениях, мишени из стали и дюраля совершенно не различаются даже на минимальном удалении животного от них. Таким образом, именно скорость сдвиговых волн в материале, а не акустическое сопротивление последнего позволяет дельфинам различать материал мишеней.
Так как скорость сдвиговых волн монотонно зависела от величины Д, т. е. среднего интервала между максимумами функции |/оо(А:а)|, то следовало ожидать, что процент различения шаровых мишеней, отличающихся материалом, будет зависеть только от различий в величинах Д для распознаваемых мишеней, а не от абсолютных значений этих величин. Данная гипотеза была подвергнута экспериментальной проверке (Дубровский, Краснов, 1971; Дубровский и др., 1971). Дельфинам предъявлялись мишени из следующих материалов: свинец, сталь, дюраль, эбонит,
425
оргстекло, латунь. Было показано, что надежность различения материалов е шаровых мишеней одинакового размера (50 мм) увеличивается с ростом разности 8Д и не зависит от абсолютных значений Дх и Д2. При этом пороговое значение, определяемое по уровню 75 %-й надежности различения, составило для двух афалин и одной азовки около 0.2.
Обсуждение гипотезы о механизме различения мишеней по их спектрам и пределах ее применимости для анализа экспериментальных материалов дано в работе Дубровского и Краснова (1971). В ней рассмотрен вопрос об эквивалентности различий спектра мишеней, обусловленных различиями либо в материале, либо в размере сравниваемых мишеней. Различия в спектре мишеней будут по-прежнему характеризоваться при помощи среднего периода осцилляций Д. По определению,
2 тс- bf-a
Д= ,
где С — скорость звука в воде; а — радиус шара; Д/ — диапазон частот (гц) между соседними максимумами в спектре мишени, т. е. средний интервал осцилляций данного спектра.
Различию в величинах Д для всех сравниваемых материалов 8Д соответствует различие в величине Д/. Обозначим последнее через
ЪЬ-С
= . (1)
J 2тса v ’
Нетрудно показать, что изменение спектра, обусловленное изменением радиуса шаровой мишени от аг до а2, равно
_ Д-С Да 2тс ’а^’
где Да=а2—(а2 > aj.
Теперь можно сформулировать правило распознавания, основываясь на выражениях (1) и (2).
Если 8/ > 8/0 г(где 8/0 — некоторое пороговое изменение спектра, зависящее от выбранной для определения порога надежности распознавания, а также от индивидуальных особенностей животного), то принимается решение о том, что мишени различны; если 8/ < 8/0 —- мишени одинаковы.
Из соотношения (1) следует, что пара шаровых мишеней, отличающихся одновременно по размеру и материалу, будет неразличима, если
Формулы (1) и (2) дают возможность сопоставить экспериментальные данные цо различению размеров и материалов (табл. 33).
426
Таблица 33
Дифференцирование афалинами сферических тел с различными акустическими характеристиками
Материал мишени зд 3/, кгц Правильность дифференцирования, °/0
Сталь—дюраль .... 0.05 0.25 58 и 65
Оргстекло—эбонит . . 0.29 1.39 80
Оргстекло—свинец . . 0.45 2.15 100
Латунь—эбонит . . . 0.60 2.87 91 и 95.5
Латунь—сталь .... 0.75 3.58 100
Латунь—алюминий . . 0.80 3.82 92
Латунь—дюраль . . . 0.80 3.82 100
Примечание. В таблице объединены результаты экспериментов Дубровского и Краснова (1971), выполненные на 2 животных, и Абрамова и др. (1970, 1971). Значения 8д получены на основе модельных опытов (Бабкин, Фадеева, 1941).
Анализируя эти данные, можно отметить, что наблюдается удовлетворительное совпадение зависимости по материалам и размерам для малых свинцовых и больших стальных шаров от величины 8/. В то же время имеется резкое несоответствие этих данных с результатами опытов по различению дельфинами размеров малых стальных шаров. Так, если 75%-я надежность различения материалов и размеров малых свинцовых и больших стальных шаров достигается одним из дельфинов при 8/0=1.1-Ь 4-1.8 кгц, то различение малых стальных шаров происходит при 8/0=31+73 кгц и 8/0=24 кгц. Таким образом, наше предположение об общности механизма распознавания дельфинами материалов и размеров по изменению среднего периода осцилляций спектра мишени достаточно хорошо выполняется только для малых свинцовых и больших стальных шаров и совершенно не выполняется для малых стальных шаров. Особо следует сказать о различении дельфинами размеров свинцовых шаров с минимальным диаметром 50 мм. Пороговая величина для одного дельфина равна 0.37 кгц и для другого — 0.25 кгц (по уровню 78%), т. е. превышает в 3—5^раз величину 8/0, необходимую для 75%-го различения материалов. Вероятно, и различение размеров больших свинцовых мишеней происходит на основе других признаков, нежели различение материалов и размеров малых свинцовых и больших стальных мишеней.
Для выяснения причин указанного выше несоответствия в величинах 8/0 рассмотрим особенности спектра малых стальных и больших свинцовых мишеней (табл. 34).
При получении данных, представленных в табл. 34, учитывался диапазон волновых размеров ка > 1, так как регулярные
427
Таблица 34
Число периодов осцилляций в спектрах различных мишеней для диапазона частот от 1 до 100 кгц
Размер, мм Материал
свинец сталь дюраль латунь эбонит оргстекло
10 13 1.5 1.7 2.5 4.0 5.3
50 70 8 8.4 12.3 19.2 26.2
осцилляции в спектрах мишеней начинаются именно с частоты, приближенно определяемой из условия каЛ (Hickling, 1962).
Из таблицы видно, что число периодов для малых стальных шаров составляет величину порядка 1, т. е. говорить об изменении степени изрезанности спектра или об изменении его периодичности уже невозможно. С другой стороны, число периодов осцилляций спектра свинцовой мишени с минимальным диаметром 50 мм составляет величину порядка 100. В то же время малые свинцовые и большие шары из всех остальных материалов имеют число периодов осцилляций в своем спектре порядка 10. Сопоставление этих величин с опытными данными, приведенными в табл. 30—31 и 35, позволяет прийти к заключению, что слуховая система дельфина, которая производит частотный анализ принятого эхо-сигнала, обладает достаточной чувствительностью к изрезанности спектра только в ограниченном диапазоне чисел периодов осцилляций. Если число периодов спадает до 1 или стремится к 100, то чувствительность к изменению периодичности спектра или, точнее, его изрезанности резко снижается. При этом дельфин может использовать для распознавания другие признаки.
Предложенная нами модель механизма распознавания шаровых мишеней может быть выражена эквивалентным образом на временном языке. Если предположить на момент, что первичное и вторичное эхо от шара имеют одну и ту же форму, отличаясь лишь амплитудой Л, то полное эхо можно представить в виде
Е (Z) =/(0 + — т),
где t — задержка вторичного эха относительно первичного. Если спектр первичного эха равен S (со), то спектр вторичного (в соответствии с теоремой о сдвиге) равен AS (м) cos сот. Тогда спектр всего эха будет иметь вид:
SЕ (со) = S (со) (1 + A COS сот),
428
т. е. он будет состоять из последовательности максимумов и минимумов, средний интервал между которыми А/ будет связан с задержкой т следующим соотношением:
Если принять
т=1/Д/.
Т1Т2 от =-------
Т2 —Т1
то правило распознавания можно сформулировать при помощи временного описания следующим образом: при > 8т2 принимается решение о различии мишеней. Вопрос об эквивалентности описания механизма распознавания мишени на спектраль
Рис. 136. Спектры эхо-сигналов от стальных шаров.
1 — шар диаметром 10 мм; 2 — шар диаметром 12.7 мм.
Рис. 137. Спектр эха от шара диаметром 15 мм из оргстекла.
ном и временном языках обсуждается в работах Дубровского (1973), Дубровского и Фадеевой (1973), Фадеевой (1973).
Дальнейшее развитие модели механизма распознавания получено при анализе результатов экспериментов по различению размеров мишеней малого размера из стали, свинца и оргстекла (Фадеева, 1973). Анализ экспериментов был проведен с использованием результатов расчета на ЭВМ спектров и осциллограмм эхо-сигнала дельфина от примененных в опытах мишеней. На рис. 136 приведены спектры эхо-сигналов от стальных шаров с диаметрами 10 и 12.7 мм. Различение этих шаров дельфинами происходило в 71% испытаний (Дубровский, Краснов, 1971). Небольшие осцилляции в спектре связаны с дифракцией звука на шаре, а изрезанность, обусловленная присутствием вторичного эха, отсутствует. Очевидно, что в этом случае разность средних периодов осцилляций в спектре не может служить признаком различения.
На рис. 137 показаны спектры эхо-сигнала для шара диаметром 15 мм из оргстекла. В спектре эха от акустически более
429
мягких материалов малого размера (порядка 10 мм) содержится 5—7 периодов осцилляций, и в этом случае разность средних периодов осцилляций вполне может быть признаком различения. Экспериментальные данные по распознаванию дельфином шаров
Рис. 138. Форма зондирующего импульса дельфина, принятая в качестве стандарта (а), и эхо импульса (б) от шара из оргстекла диаметром 15 мм.
Рис. 139. Эхо стандартного импульса от стального шара диаметром 10 мм.
из оргстекла и свинца малых размеров (табл. 35) демонстрируют наличие монотонной зависимости между надежностью различения и разностью средних периодов осцилляций в спектрах эха.
Аналогичные заключения можно сделать на основании сравнения формы эхо-сигналов мишеней (рис. 138, 139). Эхо от шара
Таблица 35
Дифференцирование афалиной шаров'малого размера из оргстекла и свинца при различных соотношениях диаметров (мм)
Оргстекло Свинец
14.5/12 14.5/13 14.5/13.5 14.5/14 14,16 10/10.7
А«, мм 1.25 0.75 0.50 0.25 1 0.35
&/, кгц 5.5 3.1 1.95 0.94 1.28 0.93
Правильность дифференцирования, % . . . 100 88 90 80 87 75
430
из оргстекла представляет собой последовательность затухающих импульсов, разницу интервалов между которыми в эхе от шаров с близкими по величине радиусами составляют величины 1.5—2 мксек. Эхо от стального шара, напротив, представляет собой единичный импульс, амплитуда которого пропорциональна радиусу шара. Таким образом, признаком различения стальных шаров малого размера будет разность амплитуд первичного эха, причем порог распознавания равен ^2—3 дб. Это значение совпадает с дифференциальным порогом человека для коротких импульсов.
Рассмотрим теперь, обладают ли спектры эха от естественных объектов периодичностью и возможно ли распознавание этих объектов по различию в изрезанности спектров. Как показали результаты модельных экспериментов на рыбах и других морских животных, служащих объектами питания дельфинов, в диапазоне частот 10—100 кгц амплитуда рассеяния в зависимости от частоты подвержена большим осцилляциям (Колобаев, 1970). Появление осцилляций есть результат интерференции эха от передней и задней границ объекта, и в частности от боковых или спинной и брюшной поверхностей рыбы.
Дифференцирование мишеней сложной конфигурации
Для выявления физиологических механизмов эхолокационного распознавания весьма существенны опыты с мишенями сложной конфигурации, так как они позволяют приблизиться к пониманию эхолокационных возможностей дельфина в естественных условиях. Различными авторами был проведен ряд опытов со сложными мишенями, составленными из пар — шаров, цилиндров, пластин разного размера (Абрамов и др., 1970; Белькович и др., 1970; Титов, Фадеева, 1971; Абрамов и др., 1972; Титов, 1972; Резвов и др., 1973).
Задача опыта состояла в том, чтобы определить, при каком минимальном разнесении элементов сложной мишени по дальности и угловым координатам (углу места Дер и азимуту А Я) дельфин еще в состоянии различить мишени при помощи эхолокатора. Отметим, что в ряде работ связывали проведение этих экспериментов с решением задачи о разрешающей способности эхолокатора по дальности и угловым координатам. Используемые мишени и способы их крепления у разных авторов различны. В работе Титова и Фадеевой (1971) сложные мишени составлялись из пар шаров, подвешиваемых на тонких капроновых лесках. В других работах цилиндры перемещались один относительно другого, причем их оси оставались параллельными (Белькович и др., 1970; Абрамов и др., 1972; Резвов и др., 1973).
431
По-видимому, эти методические особенности повлияли на полученные результаты. Значительное влияние оказывало исходное расстояние между элементами сложной мишени (Титов, Фадеева, 1971; Титов, 1972). По данным работ Бельковича и др. (1970), Резвова и др. (1973), порог различения разницы в разнесении цилиндров с расстояния от 9 до 15 м по горизонтали оказался равным 2—3 мм, а по вертикали — 3—5 мм. Пороговая разница в угловых размерах мишеней ~Г. В этих опытах цилиндры в одной из мишеней были сдвинуты вплотную.
В работе Титова, Фадеевой (1971) использовались пары стальных и свинцовых шаров различного диаметра, которые разносились один от другого по горизонтали (азимуту), вертикали (углу места) или дальности. В положительной мишени расстояние между центрами шаров (база мишени) в течение опыта оставалось постоянным, в отрицательной расстояние между шарами менялось.
Результаты опытов сведены в табл. 36, 37 и 38.
Таблица 36
Пороги распознавания при разнесении шаров по азимуту
Материал и диаметр шаров, мм Расстояние до мишеней, м База Число испытаний Пороги по азимуту
угл. ед. см
Свинец 50 5 4Г 6 669 31'
8 26' 6 48 39'
5 5°43' 50 239 2°36'
Сталь 17.3 5 17' — 320 21'
8 И' 2.5 248 18'
5 2°17' 20 118 1°5'
5 4°17' 39 39 Г58'
Таблица 37
Пороги распознавания при разнесении шаров по углу места
Материал и диаметр шаров, мм Расстояние до мишеней, м База Число испытаний Пороги по углу
угл. ед. см
Свинец 50 5 48' 7 233 17'
Сталь 17.3 3 34' 3 37 12'
5 20' 3 446 9'
Из полученных данных следует, что порог по ДЯ для свинцовых шаров почти в 2 раза выше, чем для стальных;, а по углу места пороги в 2 раза ниже, дем по азимуту. Пороги для свиц-
432
цоцых шаров в этих экспериментах также оказались значительно выше, чем для стальных.
Таблица 38
Пороги распознавания при разнесении шаров по дальности
Материал и диаметр шаров, мм База, см Число испытаний Пороги по дальности, см
Свинец 50 5 205 4.4
Сталь 33 8 67 1.0
5 62 1.2
Сталь 17.3 3.5 241 0.9
7 67 1.5
Имеется значительное различие в порогах по азимуту для мишеней с малыми и большими угловыми размерами. Так, для свинцовых мишеней с угловыми размерами 4Г и 5°43' пороги отличаются в 5 раз, а для стальных с размерами 17' и 4°17' — в 6.6 раза. Поскольку увеличение порогов характерно для обоих материалов, можно предположить, что в случае малых и больших баз дельфин использовал при распознавании различные информативные признаки. Так как рассеянное звуковое поле от пары шаров будет отличаться от поля одиночного шара наличием интерференции полей от каждого из шаров, то с увеличением расстояния (базы) между шарами интерференция рассеянных полей одиночных шаров должна ослабевать. Проведенный анализ показал, что парные мишени с малым расстоянием между центрами дельфин должен воспринимать как одну мишень сложной формы, не выделяя каждого шара в отдельности. Следовательно, и распознавание мишеней с малыми базами животное могло производить по различиям в спектрах результирующих эхо-сигналов от каждой пары шаров. Этим и объясняется малое значение порогов, полученное в данной работе (за исключением мишеней с большими базами) и в аналогичных по методике работах других авторов (Белькович и др., 1970, и др.).
При больших расстояниях между центрами шаров дельфин не может использовать различия в интерференционных картинах из-за слабого взаимодействия полей от каждого из шаров и распознает мишени по разности расстояний между шарами, воспринимая их по отдельности. Именно в этом случае порог углового различения может служить мерой угловой разрешающей способности эхолокатора дельфина, которая по порядку величины должна совпадать с шириной диаграммы направленности эхолокатора дельфина.
Пороги распознавания мишеней, полученные другими исследователями (Белькович и др., 1969а, 19696, 1970), оказались
433
в несколько раз ниже, чем в наших экспериментах. Это объяс
няется тем, что жесткое крепление цилиндров, расположение их в положительной мишени вплотную друг к другу, а также использование в качестве мишеней ступенчатых пирамид обеспечили более четкую и устойчивую интерференционную картину, позволившую получить более низкие пороги.
Пороги распознавания мишеней, отличающихся разнесением элементов по дальности, оказались порядка 1 см. В соответствии с современными физическими представлениями разрешающая способность по порядку величины должна быть равна пространственной протяженности сигнала. Приняв минимальную длительность зондирующего сигнала дельфина равной 30 мксек., легко рассчитать, что разрешающая способность эхолокатора дельфина должна быть около 3 см, что соответствует пороговой разнице в разнесении шаров по дальности в 1.5 см. Таким образом, можно допустить, что и в случае различения мишеней с элементами, разнесенными по дальности, дельфин использовал изменение в изре-
занности спектра эхо-сигнала в качестве признака распознавания.
В работе Бельковича и Борисова (1971) обсуждается вопрос о распознавании мишеней сложной формы. Тест-объектами служили квадраты из плексигласа толщиной 3 мм, обклеенные снаружи слоем пористой резины толщиной 3.5 мм, которая представляла собой поверхность, хорошо отражающую звуковую волну. Положительной фигурой был сплошной квадрат размером 10x10 см, отрицательной — сплошной квадрат 7x7 см, контрольными мишенями служили квадраты размером 10x10 см с квадратными отверстиями в центре, площадью от 1 до 86.5 см2.
В контрольных опытах дельфину одновременно предъявляли меньший отрицательный квадрат и квадрат с вырезом в центре. Животное в 89.5% случаев выбирало фигуру с большей площадью отражающей поверхности, несмотря на то что при обучении она была отрицательной.
Дальнейшая работа сводилась к определению минимальной величины отверстия, при котором «контур» еще дифференцировался дельфином от квадрата тех же размеров. Фигура с отверстием площадью 25 см2 дифференцировалась в 80% случаев, с отверстием 6.2 см2 — в 72.6%, а с отверстием 1 см2 практически не различалась (50% правильных реакций).
Представлялось, что при распознавании квадратов с отверстиями дельфин получает информацию по двум контролируемым
параметрам: интенсивности эха и спектральным характеристикам. Искажение спектров сигнала при его отражении от сплошных фигур будет происходить в более низкочастотной области, чем от контуров, так как область максимальных искажений зависит от отношения длины звуковой волны X к стороне квадрата, или
к толщине «контура»
Zi — *2 2
(Zj и Z2 — наружная и внутренняя
434
стороны «контура»). Поскольку ранее было показано, что ин-тенсивность эха анализируется с ошибкой в 10%, то можно полагать, что введение вторичного отличительного параметра по спектру позволит животному различать фигуры с большей точностью. Полученные экспериментальные данные подтвердили важность для дельфина анализа интенсивности эхо-сигнала. Однако различение сплошной фигуры от контура, отличающегося по площади всего на 6.2%, осуществляется лучше, чем распознавание однотипных плоских фигур, и это дает основания полагать, что животное могло использовать, кроме интенсивности, также и отличия по спектру.
Бельковичу и др. (1969а) при работе с обыкновенным дельфином не удалось добиться дифференцирования квадрата и треугольника, равных по площади. Однако в работе Дмитриевой с соавт. (1972) проведен более детальный анализ. Дифференцировка вырабатывалась между квадратом со стороной 10 см и равновеликими ему треугольниками со сторонами 5x20, 15.4x6.5 и 12.5x8 см, ромбами размером 22x9 и 17x12 см и кругом диаметром 11.3 см. Всего было испытано 7 пар фигур, изготовленных из плексигласа толщиной 95 мм, покрытых слоем пористой резины 5 мм толщиной. Дифференцировка осуществлялась с расстояния 10—18 м. Предварительно животное обучали отличать от квадрата горизонтально ориентированные прямоугольники с меньшей площадью (65 и 85 см2), и лишь затем ставились пробы по дифференцировке фигур с площадью 100 см2, отличающихся только формой. Все дифференцировки удалось выработать.
Адаптация сонара к параметрам лоцируемых объектов
Одно из замечательных свойств эхолокационного анализатора — это его адаптивность, которая особенно ярко проявляется в экстремальных условиях работы эхолокационного анализатора. Так, например, при увеличении уровня аддитивных помех дельфин увеличивал в среднем на 26 дб уровень зондирующих импульсов (Бабкин, Дубровский, 1971). Характерно, что надежность обнаружения мишени при этом оставалась на постоянном уровне, близком к 90%. Одновременно наблюдалось повышение спектральной плотности зондирующего сигнала в области максимума спектра помехи.
В ходе опытов по различению размеров шаровых мишеней (Абрамов и др., 1971) в условиях действия помех отмечалось смещение максимума спектра зондирующего сигнала в сторону от максимума спектра помехи. При предъявлении импульсных помех частота следования эхолокационных импульсов синхронизировалась с частотой излучения импульсов помехи, что приво-
435
дило к лучшей временной селекции эхо-СиГналов от импульсных помех.
Адаптивные свойства эхолокационного анализатора дельфина проявлялись не только в изменении параметров эхолокационных сигналов, но и в целесообразном изменении поведения, например траектории движения животного. При обнаружении металлических шаров с диаметрами от 20 до 50 мм вблизи дна бассейна, специально выложенного галькой с характерными размерами от 5 до 30 мм, дельфин стремился уменьшить реверберационную помеху, лоцируя мишени под скользящими углами и двигаясь к мишени не вблизи поверхности, как обычно, а вблизи дна (Титов, 1972).
В работе Норриса (Norris, 1967) основное внимание было уделено изменению продолжительности серии, частоте повторения локационных импульсов и скорости движения животного к цели. Авторы нашли, что длительность серии локационных импульсов возрастает при уменьшении различий в диаметрах шаров, а скорость движения животного уменьшается в среднем от 111 до 86 см/сек. Средняя частота повторения импульсов увеличивалась с усложнением эхолокационной задачи от 65 до 78 имп./сек. Приведенные авторами значения средних (и даже максимальных) частот повторения импульсов много ниже отмеченных другими авторами.
В работе Гиро и Дубровского (1972) подробно исследована подстройка частоты следования зондирующих импульсов (табл. 39, 40)
1 F = ~f’
где Т — межимпульсный интервал.
Рассматривалась корреляция межимпульсного интервала со степенью сложности задачи: 1) обнаружение металлических шаров разной величины при фиксированном расстоянии; 2) обнаружение металлического шара при разных расстояниях до него; 3) дифференцирование величины шаров, изготовленных из одинакового материала; 4) дифференцирование материала шаров стандартной величины.
Были вычислены кривые точной и прямые приближенной регрессии, коэффициенты корреляции между межимпульсными интервалами и значениями параметра сложности. Было выявлено, что рассматриваемое распределение есть двумерное нормальное распределение. Основываясь на этом, были произведены оценки достоверности и значимости полученных коэффициентов корреляции.
Сопоставление данных показывает, что при правильности обнаружения, близкой к 100%, статистическая взаимосвязь между расстоянием и величиной межимпульсного интервала 436
Таблица 39
Зависимость величины межимпульсного интервала от сложности локационной задачи при обнаружении сферических мишеней
Локационная задача Материал мишени Параметр сложности Средние значения параметра сложности ё, °/о р Pi ?2 1 Ро 1 Объем выборки межимпульсных интервалов N
Обнаружение шара Свинец 2.25, 3.75, 5.25, 100 0.07 0.00 0.14 0.07 804
диаметром 8.5 мм Z, м 6.75
То же 11 мм Сталь 4, 5, 6 96 —0.08 —0.16 —0.03 0.08 599
» » 6 мм » 4, 5, 6 72 -0.63 —0.69 -0.58 -0.06 446
Обнаружение ша- 9, И, 13 92 0.31 0.23 0.38 0.08 560
ров при длине d, мм
сети 3 м »
То же 6 м » 1 9, И, 13 73 0.64 0.58 0.71 0.08 284
Примечание. £ — процент правильного обнаружения, усредненный по параметру сложности; р — коэффициент корреляции между параметром сложности и межимпульсным интервалом; р! —нижняя граница 95%-го доверительного интервала для р; р2 — верхняя граница 95°/0-го доверительного интервала для р; р0 — граница критической области, соответствующей выполнению гипотезы р=0.
Таблица 40
Зависимость величины межимпульсного интервала от сложности локационной задачи при дифференцировании сферических мишеней *
Локационная задача Материал мишени Средние значения параметра сложности 8/, гц р Pi ?2 1 Ро 1 t, “Io Объем выборки J межимпульсных интервалов N
Различение шаров по диаметру при длине сети 3 м Сталь 400, 1200, 2000, 2800, 3000 0.53 0.49 0.56 0.03 76 1969
То же Свинец 45, 135, 225, 315 0.65 0.59 0.72 0.06 59 309
» » » 225, 675, 1125, 1575, 2025, 2475, 2925, 3395 0.71 0.69 0.73 0.02 74 2540
Различение шаров (диаметром 50 мм) по материалу при длине сети 8 м Сталь, свинец 400, 1200, 2000 0.92 0.90 0.93 0.01 76 484
* См. прим, к табл. 39.
отсутствует. При усложнении задачи — переход с обнаружений шара диаметром И мм на обнаружение шара диаметром 6 мм — средний процент правильного обнаружения падает до 72. Кривые регрессии в этом случае указывают на тесную связь этих параметров: при увеличении расстояния от 4 до 6 м межимпульсный интервал уменьшается приблизительно с 50 до 40 мсек. Коэффициент корреляции —0.63; это значение является достоверным и значимым. Таким образом, переход от надежного обнаружения к пороговому сопровождается появлением зависимости частоты следования импульса от степени сложности эхолокационной задачи. Сходная ситуация наблюдается и при обнаружении шаров различных диаметров с расстояния 3 и 6 м (правильность обнаружения соответственно 93 и 73%). Коэффициент корреляции между диаметром шара и межимпульсным интервалом возрастает от 0.31 до 0.64. При изменении диаметра от 9 до 13 мм среднее значение межимпульсного интервала изменилось от 41 до 43 мсек, в одном случае и от 42 до 51 мсек. — в другом.
Аналогичная зависимость обнаружилась при дифференцировании размера и материала сферических мишеней. Во всех случаях различение было припороговым на уровне 59—76% правильных реакций, а коэффициент корреляции находился в пределах 0.53—0.92. Особенно высок уровень корреляции для задач по различению материалов мишеней.
Таким образом, частота следования импульсов увеличивалась с усложнением эхолокационной задачи лишь в том случае, если задача ставилась как пороговая и, решая ее, дельфин был вынужден осуществлять подстройку эхолокатора применительно к эхолокационной ситуации.
Следует заметить, что данные Акопиана и др. (1973), обнаруживших уменьшение частоты следования локационных импульсов при поисках рыбы, не подтверждают этот вывод.
Каковы же причины увеличения частоты следования импульсов по мере усложнения эхолокационной задачи. Естественно допустить, что дельфин способен накапливать во времени последовательные эхо-сигналы. Предположим, что дельфин осуществляет когерентное накопление эхо-сигналов. Тогда увеличение отношения сигнал/помеха при возрастании частоты следования от FT до F2 будет равно
При увеличении расстояния до мишеней с 4 до 6 м период следования импульсов уменьшился от 50 до 40 мсек., что соответствует увеличению отношения сигнал/помеха приблизительно на 1.8 дб. В то же время уменьшение отношения сигнал/помеха, обусловленное увеличением расстояния с 4 до 6 м, составит более 6 дб. Таким образом, путем увеличения частоты следования им-438
пульсов дельфин даже при когерентной обработке эхо-сигнала не компенсирует уменьшения отношения сигнал/помеха. Учитывая, что эхо-сигнал составляет по времени приблизительно 0.001-ю долю межимпульсного интервала, следует признать, что любое накопление последовательных эхо-сигналов, будь то когерентное или некогерентное, невозможно без временного стробирования.1
В данном случае оно может выражаться в периодическом блокировании слуховой системы в моменты как излучения зондирующего сигнала, так и прихода мощных импульсов реверберации и в повышении чувствительности слуха в моменты прихода эха. Наличие блокировки слуховой системы летучей мыши за счет снижения коэффициента передачи среднего уха при излучении зондирующего сигнала было ранее показано Хенсоном (Henson, 1967). Физиологические доказательства существования эффекта стробирования у дельфинов пока отсутствуют.
Не вполне ясно, почему дельфин не увеличивает частоту следования в гораздо больших пределах, хотя имеется возможность такого увеличения без наложения последующего зондирующего импульса на эхо, вызванное предыдущим импульсом. В вышеприведенном примере минимально допустимый межимпульсный интервал равен приблизительно 7 мсек. По-видимому, такое «самоограничение» определяется акустическими условиями бассейна. Похоже, что дельфин излучает последующий импульс лишь после того, как затухнут до допустимого уровня последовательные отражения от стенок бассейна, вызванные предыдущим зондирующим сигналом.
Приведенные выше соображения в равной мере могут быть применены и к обсуждению причин увеличения частоты следования сигналов при усложнении задачи различения мишени. Использование последовательных эхо-сигналов может привести к более точной оценке характеристик, определяющих различие мишеней.
Другие исследования (Айрапетьянц и др., 19726; Айрапетьянц, Воронов, Иваненко и др., 1973; Айрапетьянц, Воронов, Ордовский и др., 1973) были сосредоточены на изучении особенностей изменения локационных посылок, генерируемых афалиной, при дифференцировании величины сферических и цилиндрических мишеней. Дифференцируемый ряд состоял из 5 шаров и 5 цилиндров диаметром и высотой от 5 до 150’мм/Было установлено, что при повторных предъявлениях одной и той же пары дифференцируемых объектов и сохранении неизменными всех остальных условий опыта падали продолжительность и частота щелчков в первой и второй сериях (опознание мишени осуществлялось в процессе
1 Восприятие дискретных эхо-ответов как процесс непрерывного изменения сщнада.
439
генерации первой серии локационных посылок), а интервал между сериями увеличивался. Средняя и доминирующая частоты заполнения щелчков сдвигались в область более низких частот.
При любых изменениях условий опыта, особенно ведущих к усложнению задачи (увеличение расстояния до лоцируемых мишеней, предъявление более трудной для дифференцирования пары объектов), доминирующая и средняя частоты заполнения сдвигались в область более высоких частот. Подобный характер изменения локационных сигналов в одинаковой степени характерен для задач по опознанию и шаров, и цилиндров. Разница состояла лишь в том, что при опознании сферических тел энергетический максимум более значительно сдвигался в высокочастотную часть спектра, чем при опознании цилиндров. Таким образом, установлен факт адаптации локационных сигналов дельфина к параметрам лоцируемых объектов. Однако, учитывая, что эксперимент проведен только на одном животном, его результаты следует оценивать как предварительные.
Большинство исследователей отмечает значительную вариабельность траектории движения животного при подходе к дифференцируемым объектам. Ивановым и Курганским (1973) изучалась траектория движения при распознавании шаров и цилиндров. Расстояние до мишеней около 24 м. Расстояние, с которого животное должно было осуществить дифференцирование, было короче, оно задавалось экспериментатором. Анализ траектории осуществлялся путем измерения величины отклонения от «оптимальной» траектории в фиксированных точках с дискретностью 2 м. За «оптимальную» траекторию принималось кратчайшее расстояние между местом старта и положительной мишенью. На основании проведенных измерений вычислялись среднее отклонение и дисперсия, характеризующие вариабельность траектории.
Произведенный анализ показал, что траектория подхода животных к дифференцируемым мишеням одинакова для обоих классов объектов. Следовательно, ее характер не был связан с задачами по распознаванию. Момент опознания объектов всегда соответствовал моменту наибольшей вариабельности траектории. Это говорит о том, что опознание использованных мишеней одинаково легко осуществлялось при любом характере траектории движения дельфина.
Приведенный в этой главе материал свидетельствует о том, что дельфины способны, меняя различные параметры сигнала, добиваться оптимальных условий, необходимых для получения исчерпывающей информации о подводных объектах,
22
ЭХОЛОКАЦИЯ У ЛАСТОНОГИХ
Вслед за открытием акустической локации у китообразных внимание исследователей обратилось к представителям другого отряда млекопитающих — ластоногим (Pinnipedia). Ластоногие, подобно китообразным, полностью приспособились к водному образу жизни, но в отличие от последних не утратили связи с сушей. Это хищные млекопитающие с пятипалыми конечностями, превращенными в плавники (ласты). Задние конечности, обращенные назад, представляют основной орган движения в воде.
Отряд ластоногих подразделяется на 3 семейства: 1) ушастые тюлени (Otariidae), к которым относятся морской котик (Callor-hinus ursinus), сивуч (Eumetopias jubatus) и калифорнийский морской лев (Zalophus californianus); 2) моржевые (Odobenidae) с одним видом — морж (Qdobenus rosmarus), и 3) безухие тюлени (Phocidae) с многочисленными представителями, например обыкновенный тюлень (Phoca vitulina), нерпа (Phoca faetida), хохлач (Cystophora cristata) и др.
Многие исследователи, которые изучали ластоногих, полагают, что звук должен играть важную роль в их жизни, поскольку эти животные с успехом обитают в темных и мутных водах. Более того, некоторые авторы (Poulter, 1963а) в течение долгого времени вели наблюдения за полностью слепыми экземплярами ластоногих (морских львов) и показали, что их поведение и физиологическое состояние почти не отличаются от животных с нормальным зрением. Уже эти отрывочные сведения наводят на мысль, что по крайней мере некоторые ластоногие могут использовать звук в целях ориентации в окружающем подводном мире. Однако долгое время не удавалось обнаружить никаких подводных звуков, издаваемых этими животными, хотя, как сообщают Шевилл и др. (Schevill et al., 1963), ряд исследователей проводил специальные прослушивания под водой. Полтер из Стенфордского института (Калифор-
441
йия) первым записал подводные звуки одного из представителей ластоногих — калифорнийского морского льва и дал их характеристику (Poulter, 1963а, 1963b). Как отмечает автор, все записанные сигналы могли быть использованы животными для целей эхолокации.
В бассейн, где содержались морские львы, бросали куски рыбы и куски конского мяса на расстоянии 2.5—3 м от ближайшего животного. Обычно куски рыбы морские львы находили и схватывали быстро, тогда как куски конины ни одно животное не трогало. Они часто начинали движение к несъедобному объекту, но в каждом случае сворачивали в сторону с расстояния 45—60 см. Эксперименты проводили в ночное время, и различение объектов с помощью зрения было затруднено. Морские львы быстро обучились различать даже виды рыб и всегда устремлялись к той, которую они «предпочитали», тогда как от «нелюбимой» рыбы сворачивали уже за метр.
Эти первые простые эксперименты давали основания предполагать, что при нахождении пищи морские львы, возможно, используют эхолокацию. Сразу отметим, что обоняние плохо развито у этих животных (Scheffer, 1962) и трудно допустить, чтобы оно могло значительно облегчить им поиски добычи под водой.
Установив гидрофоны около места подачи рыбы, Полтер обнаружил, что каждый подход животного к съедобному или несъедобному объекту сопровождается излучением серий коротких звуковых импульсов, частота следования и другие параметры которых изменяются по мере приближения животного к цели.
Когда животные производили поиск рыбы в воде, они часто повторяли короткий импульс или группу импульсов. Анализ 250 таких групп позволил рассчитать следующие их параметры:
1) длительность каждого импульса составляла 3—5 мсек.;
2) основная частота заполнения импульса изменялась от 3 до 13 кгц; средняя частота приблизительно равнялась 7.5 кгц;
3) частота заполнения импульсов в группе менялась следующим образом: вначале частота импульсов составляла 5 кгц, затем повышалась до 8—10 кгц и плавно понижалась к концу примерно до 4—5 кгц;
4) число импульсов в группе в среднем было около 11; промежутки между импульсами составляли в среднем 33.9 мсек.; средняя частота следования импульсов равнялась 30 гц (при минимальной частоте следования 15 гц и максимальной 80 гц);
5) в начале импульса амплитуда колебаний была максимальной, а затем постепенно уменьшалась.
Когда же животное слышит всплеск брошенного куска рыбы, оно немедленно переключает локационный аппарат на иной режим, при котором издаются серии импульсов, имеющих одну и ту же частоту, и морской лев начинает приближаться к добыче.
На последней стадии подхода к цели вместо одиночных импуль
442
сов морской лев переходит на режим излучения двойных импульсов. Каждый двойной импульс состоит из предварительного импульса весьма малой длительности и основного импульса, имеющего гораздо большую длительность. Так, длительность предва рительного импульса в среднем составляла 0.7 мсек. (0.3—1 мсек.), тогда как длительность основного импульса примерно в 8 раз превышала длительность предварительного. Частота заполнения предварительного импульса оказывалась всегда выше основного и в среднем составляла 7.23 кгц (4.5—13.4 кгц), тогда как среднее значение частоты основного импульса равнялось 4.8 кгц (2—11 кгц).
Длительность периода между предварительным и основным импульсами изменялась от 0.1 до 1 мсек. Амплитуда предварительного и основного импульсов могла меняться в широком диапазоне.
Следует подчеркнуть, что и длительность, и амплитуда основных импульсов нарастают по мере приближения животного к цели, но в самом конце длительность основных импульсов уменьшается, как предполагает автор, чтобы предотвратить наложение его на последующий предварительный импульс.
На основании анализа этой и многих аналогичных серий импульсов принцип действия эхолокации ластоногих объясняется следующим образом.
Посылая короткий предварительный импульс, морской лев затем слушает отраженный эхо-сигнал, изменяя при этом интервал между основным и предварительным импульсами до тех пор, пока эхо-сигнал не попадает в основной импульс, и тогда промежуток времени между двумя “сильными импульсами может служить мерой расстояния до цели.
Промежутки между двойными импульсами в начале серии были довольно большими, а затем по мере приближения животного к куску рыбы частота следования импульсов увеличивалась.
По всей вероятности, эхолокатор ластоногих обладает высокой помехозащищенностью, как, впрочем, и эхолокаторы всех других животных, о которых речь шла выше. Несмотря на то что в бассейне, где проводились эксперименты, была значительная реверберация и, кроме того, другие морские львы, плавающие тут же, могли облучать своими сигналами тот же кусок рыбы, животное всегда без колебаний следовало по наиболее прямому пути к цели. Даже при возникновении дополнительных акустических помех морские львы быстро перестраивали режим своего эхолокатора и таким путем избегали заглушения отраженных сигналов. Подтер описывает один такой случай. В соседнем здании работал насос, который качал воду в бассейн, где находились экспериментальные животные. При работе насоса в бассейне возникал быстрый пульсирующий скрип, который создавал дополнительные трудности при обнаружении животным пищи акустическим путем. Морской лев сначала использовал случайную серию импульсов
443
с различными частотами, которая накладывалась на этот шумовой фон, но затем он быстро перестроился и начал работать синхронно с насосом, подавая импульсы между скрипами насоса. Морской лев излучал короткие сигналы, состоящие из 5—8 импульсов, с изменением частоты от 5 до 7 кгц вместо обычного при поиске диапазона 3—13 кгц.
Эти первые исследования, которые затем были расширены, и произведенный математический анализ различных сигналов морского льва (Poulter, 1966, 1968; Schaver, Poulter, 1967) остаются пока единственными, подтверждающими наличие эхолокации у ластоногих.
Уже в этих работах Полтер упоминает о том, что ему удалось записать импульсные сигналы и у других представителей ластоногих: обыкновенного тюленя, морского слона и сивуча, но количественные характеристики этих сигналов он не приводит. Этот пробел в какой-то степени восполняет работа других исследователей (Schevill et al., 1963), которые записали подводные звуки у нескольких видов ластоногих, содержащихся в аквариумах и Зоологическом парке Нью-Йоркского зоологического общества. Авторы признают, что, прослушав в течение лишь нескольких часов каждого из ластоногих, содержащихся в неволе, они сделали только выборки из их вокального репертуара, которые не подтверждают использования животными этих звуков для эхолокации. Однако авторы отмечают, что эти импульсные звуки всегда были связаны с поисками пищи и обычно отсутствовали, когда в дневное время тюлень спокойно проплывал около гидрофонов.
Сравнивая по интенсивности импульсы тюленей и дельфинов, Шевилл и другие указывают, что подводные звуки (щелчки) всех исследованных ластоногих очень слабы и, вероятно, могут использоваться животными в целях эхолокации лишь на небольших расстояниях, на последних критических стадиях поимки добычи, тогда как для предварительного определения места пищи на большом расстоянии они малопригодны. Как низкий уровень, так и довольно низкая частота заполнения особенно затрудняют улавливание звуков ластоногих на фоне шумов. Возможно, низкий уровень звука был следствием содержания животных в неволе в реверберирующем бетонном бассейне, или это объяснялось тем, что записи производились в дневное время. Во всяком случае авторы могли улавливать звуки некоторых ластоногих лишь тогда, когда животные находились на расстоянии не более 3 м от гидрофона. По этой причине экспериментаторы не смогли записать сигналы сивучей, морских котиков и моржа, которые избегали приближаться к гидрофонам.
В работе Шевилла и других приводятся краткие сведения о характере подводных звуков калифорнийских морских львов и 5 видов тюленей. Отмечается, что морские львы издают одинаковые щелчки как под водой, так и в воздухе, причем в воздухе
444
щелчки и лай производились животными через рот одновременно. Частота заполнения импульсов морских львов в записях Шевилла и других была примерно в 4 раза ниже, чем в экспериментах Пол-тера. Чем вызваны такие различия, остается неясным.
Щелчки, издаваемые обыкновенными тюленями, имели основную частоту в районе 12 кгц. Нерпа кольчатая — Phoca hispida — издавала щелчки, которые часто шли парами. Промежутки между щелчками составляли от 2 до 20 мсек., а основная частота заполнения каждого щелчка лежала около 4 кгц и сопровождалась гармониками вплоть до 6-й с различными интенсивностями. Структура сигнала гренландского тюленя {Phoca groenlandicd) в общем напоминала структуру щелчка нерпы кольчатой, но основная частота составляла всего 2 кгц. Частота следования сигналов достигала 130 гц. Серые тюлени {Halichoerus ^rypus) издавали щелчки либо одиночные, либо парные, либо сериями с частотой заполнения от 6 до 12 кгц, в других случаях она достигала 30 кгц. Один молодой тюлень хохлач издавал 2 типа щелчков. Щелчки первого типа характеризовались довольно узкой полосой частот в районе либо 4, либо 16 кгц. Другой тип щелчков отличался от сигналов всех других ластоногих тем, что эти щелчки были широкополосными — от 100 гц до 3 кгц, а иногда и до 16 кгц. Наиболее высокая частота следования щелчков, зарегистрированная у хохлача, составляла 20 гц.
Из приведенного обзора видно, что исследования эхолокации ластоногих только начинаются и все основные эксперименты еще впереди.
ЗАКЛЮЧЕНИЕ
Как мог увидеть читатель, давние истоки исследований эхолокации животных лишь в наше время превратились в широкую проблему и теперь стали интересны для многих научных дисциплин. Нельзя не отметить поучительный пример эволюции подлинного научного открытия и то, насколько же тернист и долог путь идеи от догадки до доказательства. Потребовалось 150 лет, чтобы тайна ночных полетов летучей мыши от старта спалланцаниевой гипотезы дошла до гриффиновских открытий. А далее, через 2—3 десятилетия, она была доведена до той степени расшифровки, когда начинается подлинное знание с всесторонним пониманием сущности явления. Разгадка необыкновенных акустических дарований рукокрылых может стать еще одной общебиологической призмой в освещении образа жизни ряда животных. Более того, опознанный принцип эхолокации животных не только блестящая модель в инженерной науке, но и оптимистический вестник для человека, потерявшего зрение, — он обретет свободу самостоятельного передвижения и более полное восприятие жизни.
Книга наша биологического содержания, и, естественно, в ней представлен материал, который в целом рисует зоологические и физиологические аспекты эхолокационной функции животных. Становится очевидным, что способность к эхолокации возникла независимо у разных отрядов, у отдельных представителей животных, как наземных, так и водных позвоночных (есть много оснований, чтобы сказать также — и у ряда беспозвоночных). Здесь основным преобразователем поведения выступает экологический фактор. На протяжении длительного исторического периода в относительно схожих условиях обитания все более совершенствовалась эта приспособительная узкоспециализированная функция. Несомненно, что в эволюционном плане эхолокация развивалась по тем путям прогресса, который в терминах А. Н. Северцова обозначен как идиоадаптация. Это действительно своеобразное частное приспособление в системе анализаторной функции,
446
Важно подчеркнуть, что животных, Пользующихся, ТоЧнёб сказать, вынужденных пользоваться этим специфическим акустическим способом ориентации в пространстве, объединяет одна непременная зависимость. Все они, или по крайней мере большинство, проводят основную часть своей активной жизни в условиях, где зрительный анализ так или иначе либо ограничен, либо в определенные периоды выключен. Но роль эхолокации в жизни указанных животных оказывается отнюдь не тождественной и даже отчетливо различной. Только у летучих мышей эхолокация есть основной, а у некоторых из этих видов почти единственный способ ориентации. У других наземных эхолоцирующих животных она по сравнению, например, со зрительной информацией служит вспомогательным средством ориентации. Различный уровень эхолокации в комплексе анализаторов, обеспечивающих пространственный анализ, доказан точными экспериментами. Отсюда возникает очередная задача — вскрыть механизмы становления особой системы слуховой информации о среде — эха от собственных звуковых излучений.
В предисловии мы коснулись некоторых результатов исследования нашей лаборатории о перестройке (реинтеграции) в деятельности анализаторов при выключении одного или группы дистантных рецепторов. Опыты показали, что в подобной ситуации мозг вводит в действие виртуальные механизмы. Иначе говоря, в данных, конкретных критических обстоятельствах формируются возможные комбинации нейронных коммуникаций с повышенной их активностью. Оставшиеся в целости анализаторы не только обостренной рецепцией восполняют дефицит информации, но одновременно обогащаются новой функцией: в пространственном восприятии наступает викарирование того приемника, который оказался недействующим. В естественных условиях жизни при экстренной или длительной во времени смене экологических факторов разворачивается подобного типа функциональная реконструкция в системе межанализаторных взаимоотношений.
Нам представляется, что история становления эхолокации полностью укладывается в описанную экспериментальную модель. Дело, конечно, в определенной последовательности замещения, специфичности, объеме новообразования дополнительных средств освоения пространства и пространственных отношений.
Может быть, для наглядности допустимо обратиться к формированию относительно грубой, примитивной формы эхолокационных надстроек у слепого (ослепшего) человека. Специально поставленные опыты показали, что утерянное зрительное восприятие постепенно замещается преимущественно акустической ориентировкой. Различные звуковые шумы, сопровождающие движение зрячего человека (например, шум шагов), и некоторые акустические отражения от предметов в известной степени способствуют обнаружению препятствий на пути слепого человека. Контрольные
447
наблюдения над слепыми с закрытыми ушами или при сильном постороннем шуме (помехе) исключают викарирование слухом. Достоверные факты указывают, что некоторые слепые, тренируя слух, могут достигнуть существенной репродукции «видения» слуховым анализатором.
У животных разных биотопов, ночного или ограниченного светового режима существования эхолокационное замещение зрения развивалось поэтапно. Вначале наступало лишь усиление функций приемного (рецепторного) звукового аппарата. Далее слуховой диапазон расширялся в область ультразвука (или, наоборот, специализация направилась в область инфразвука). Стало возможным улавливание высокочастотных звучаний, сопровождающих передвижение жертвы или агрессора. Подобный слуховой анализ, например у летучих мышей в форме «подслушивания», обозначают термином «пассивный», желая подчеркнуть решающую роль высокочувствительного приемника по типу звукового пеленгатора. Но следует заметить, что улавливание любого звука, в том числе избирательное подслушивание в ультра- или инфразвуковых диапазонах, есть результат регулируемой, целенаправленной настройки приемника, осуществляемой центрами слухового и двигательного анализаторов. У некоторых животных указанным механизмом завершается специализированная прибавка в пространственной ориентации.
Истинная эхолокация возникает, когда к аппарату восприятия ультразвука добавляется ультразвуковая излучающая система с инициативой запуска особых «щупалов» — зондирующих дискретных импульсов. В микроинтервалах времени мозг успевает последовательно обрабатывать каждый возвратившийся импульс, несущий в себе информацию — показания эха, которое служит тончайшим средством ориентации в пространстве, например в пищевом поведении. Хотя здесь в оптимальных возможностях осуществляется викарирование зрения и слуховая эхолокационная система приобретает господствующее значение, однако и в этой стадии развития эхолокации обнаруживается комплексность в деятельности анализаторов. Так, в наших опытах показаны взаимосвязь между эхолокацией и двигательным анализатором и сателлитное участие зрительной функции в эхолоци-ровании. Общепринятое утверждение о полном редуцировании зрительной информации у летучих мышей не подтвердилось. После выключения зрения (временной повязки на глазах, постоянной — энуклеации) ориентация в пространстве, как и следовало ожидать, не нарушается, но происходит за счет расширения механизма викарирования: резко усиливается активность эхолокации, проявляется она в увеличенной в несколько раз частоте зондирующих импульсов. Сохранение функции зрения в роли спутника эхолокации в наибольшей степени выражено у некоторых крыланов и птиц, гнездящихся в пещерах. Но вместе с этим летучие мыши
448
и зубатые киты даже в условиях достаточной освещенности продолжают стереотипно пользоваться акустической информацией, можно сказать, «не доверяя» только показаниям зрения.
Привлекает к себе внимание еще один аспект деятельности эхолокационного аппарата летучих мышей как органа пространственного анализа. Заметим, что у всех других млекопитающих морфофизиологические сложные структуры головного мозга предназначены для управления множеством каналов связи организма со средой и, следовательно, для обеспечения в широком диапазоне и в разнообразнейшей модальности анализа пространственных факторов, иначе говоря, поведение «обычных» животных основывается на разном объеме и разном уровне энциклопедической информации о среде. У летучих мышей все системы мозга конвергируются преимущественно на одном афферентно-эфферентном канале связи со средой — крайне узкоспециализированном аппарате слухового анализа. Естественно, что исключительное значение приобретает многообразие параметров ультразвука как при пеленгации, так и в особенности при зондировании. Поведение животного при восприятии озвученного мира основывается на «уточненной и утонченной» (И. П. Павлов), приобретенной и врожденной звуковой сигнализации. Результаты физиологических исследований обнаружили у летучих мышей в области заднего двухолмия среднего мозга один из центров эхолокации и особый режим функционирования в этом районе отдельного нейрона. По сравнению с другими млекопитающими у летучих мышей существуют иные соотношения между корковыми и подкорковыми этажами головного мозга. Состояние пищевой доминанты у рукокрылых с их разрешающей эхолокационной системой характеризуется необычными чертами. Доминанта реализуется высокоподвижными согласованными механизмами координаций акустических средств и вектором полета с быстрой их сменой, с мгновенно действующими преобразователями частоты и структуры зондирующих импульсов, с усилением мощности генерации излучения для перекрытия внешних звуковых помех и т. д. Таким образом, молниеносно комбинируется множество акустических вариантов и слуховых интеграций, чтобы с наибольшей достоверностью животное могло завершить решение биологической задачи.
Для иллюстрации сказанного напомним лишь два факта, полученных в наших опытах на подковоносых летучих мышах. У подковоноса в состоянии пищевой доминанты, но при спокойном курсирующем полете зондирование производится одиночными моно-частотными длительными импульсами (60—100 мсек., с частотой излучения 4—6 в секунду). При трудной же задаче, требующей тонкого анализа или четкого опознавания нового предмета, происходит мгновенное дробление одного обычного длинного импульса на ряд коротких. Образуются пачки импульсов, следующих друг за другом, каждая из которых теперь состоит из 2—5—8 и даже
449
20 импульсов. В результате возрастает эффективность зондирования, увеличиваются за одно и то же время количество эхо-сигналов и соответственно объем информации. Так удалось обнаружить два режима эхолокации у одного и того же животного.
Другая иллюстрация. Остроухие ночницы и подковоносы на каком-то расстоянии вначале средствами пеленгации обнаруживают насекомое по тем звукам, которые оно производит в полете. А вслед за этим у каждого животного в разном режиме завершается добыча пищи. Остроухие ночницы сейчас же запускают зондирующие импульсы — истинную эхолокацию. В преследовании жертвы все более учащается ритм излучения, сокращается длительность импульсов, хотя частотное заполнение их остается почти неизменным. Перед самой атакой зондаж напоминает «пулеметную очередь». Совершенно иначе решают задачу захвата жужжащего насекомого мелкие подковоносы. Они не пускают в ход эхолокацию, а лишь усиливают пеленгацию: ориентируясь на источник звучания, подковоносы стремительно, но молча подлетают и захватывают жертву. Если же насекомое по тем или иным причинам замолкнет (перестанет жужжать), то подковонос теряет мишень, пролетает мимо. Исчез адекватный звук — исчезает пространственная информация о пище.
Хотя уже накоплен в ряде лабораторий большой экспериментальный материал и расширяется отряд исследователей, все же надо признать, что исходные знания об эхолокации животных недостаточны, в первую очередь пока еще мало изучены ее биологические основы. Например, в только что приведенном опыте о звуковой тактике подковоноса при ловле насекомых можно полагать, что и насекомое в борьбе с подковоносом и ему подобным своим врагом выработало защитные средства — оно тоже пеленгирует. Какова физическая природа подобного превентивного звукоулавливания у насекомых, в нашем опыте — у бабочек? Только ли звуки агрессора? И далее. Что вынудило одних летучих мышей при ловле насекомых пользоваться зондированием, а других лишь пеленгацией? В чем заключается биологическая оправданность использования различных эхолокационных систем, в том числе структуры импульсов (частотно-модулированных и моночастот-ных) у гладконосых и подковоносых летучих мышей, которые в экологическом отношении почти не отличаются? Для чего применяются высокочастотные сложные, обладающие очень слабой интенсивностью импульсы у «шепчущих» неотропических рукокрылых? Совершенно неясно, используют ли и как летучие мыши эхолокацию при возвращении в свои дневные убежища после нескольких ночных часов охоты за насекомыми.
Определенный интерес в бионическом аспекте представляет изучение надежности пространственного анализа у эхолоцирующих животных. С этой целью в нашей лаборатории исследовалось влияние длительных гравитационных перегрузок (гипервесомо
450
сти) на пространственную ориентацию остроухих ночниц, больших подковоносов и нетопырей-карликов. При этом установлена исключительная выносливость летучих мышей к гипергравитации, превышающая таковую у всех млекопитающих. Так, летучие мыши оказались способными перенести 60 сек. перегрузки величиной 25 G; 30 сек. перегрузки — 50; 15 сек. перегрузки — 70 и даже 120 G. Наиболее существенным параметром перегрузок в отношении влияния на летучих мышей оказалась длительность воздействия.
В отличие от других животных поперечные перегрузки (спина— грудь, грудь—спина) влияли на летучих мышей сильнее, чем продольная перегрузка голова—таз. Перегрузка таз—голова при меньших ее величинах и длительности влияет на летучих мышей аналогично воздействию голова—таз, а при более значительных параметрах — тяжелее.
Сопоставление реакций летучих мышей на гравитационные перегрузки, подъемы в барокамере и общее рентгеновское облучение выявило специфичность выносливости этих животных к гипергравитации, что, надо думать, зависит прежде всего от экологической адекватности ускорений в полете для этих летающих млекопитающих.
Установленная высокая надежность их пространственного анализа в этих условиях определяется, очевидно, в значительной степени компенсаторными и викарирующими способностями центральных звеньев аппарата эхолокации (Айрапетьянц, Зворыкин, Савин, 1972, 1973; Айрапетьянц, Зворыкин, Петрова, 1973).
Но еще больше вопросов возникает при анализе результатов исследований морских млекопитающих. Практически ничего не известно о способах использования эхолокации китами, дельфинами и ластоногими в естественных условиях. Имеются лишь предположения, основанные на экстраполяции результатов опытов, проведенных в небольших вольерах и бассейнах. Содержание в неволе, особенно крупных животных, неизбежно накладывает отпечаток на их поведение. Достаточно упомянуть, что дельфины, которых даже непродолжительное время содержат в вольере, перестают охотиться за живой добычей и быстро приучаются получать ее из рук экспериментатора.
Животные морских глубин пока еще недоступны для знания. Можно уверенно сказать, что китообразные и ластоногие не единственные, применяющие эхолокацию в водной среде. В нашей морской лаборатории (Дальние Зеленцы) описана своеобразная форма гидролокации у морских ежей, которые на некотором расстоянии точно определяют местонахождение камня, направляются к нему и прикрепляются амбулакральными ножками. Что же касается рыб и других животных, обитающих на большой глубине, куда свет практически не проникает, то они несомненно используют звуковую и электрическую локацию в целях ориентировки
451
и при добывании пищи. Пока нет еще экспериментальных доказательств существования эхолокации у рыб, однако предположения на этот счет высказываются некоторыми исследователями.
Какие же перспективы таит в себе изучение эхолокации у различных животных?
Открытие акустического способа ориентации в живой природе не явилось принципиально новым с точки зрения технического прогресса. К тому времени человек уже имел ряд приборов — эхолот, гидролокатор, радиолокатор, работающих по аналогичному принципу. Скорее наоборот, это открытие смогло произойти именно потому, что сознание исследователей было уже подготовлено предшествующими техническими достижениями. Тем не менее интерес со стороны физиков и инженеров к «живым эхолокаторам» увеличивается с каждым годом. Что же ожидается получить в результате расшифровки конструкций биологической модели эхолокации?
Не требуется большого воображения для того, чтобы заметить принципиальное сходство между эхолокаторами, или сонарами, животных и радио- и гидролокационными системами, созданными человеком. Те и другие посылают в пространство кванты энергии и по эху определяют наличие предметов и расстояние до них. Каждая из этих систем в своем диапазоне использует преимущественно высокие частоты и относительно короткие сигналы. Наконец, каждая система вполне оправдывается теми целями, для которых она предназначается. Не в пользу природы говорит сравнение дальности действия этих систем. Животные получают необходимую информацию на расстояниях, исчисляющихся метрами, и для их жизненных потребностей этого вполне достаточно. В то же время радиолокаторы зондируют пространство на сотни, тысячи и даже миллионы километров, как это доказано для космических сфер. Однако по многим другим показателям «живые локаторы» вызывают зависть инженеров.
Помимо основного, главного преимущества сверхминиатюрных конструкций весом всего-навсего несколько граммов эхолокационные системы летучих мышей обладают высокой разрешающей способностью. Используя широкополосные ультразвуковые сигналы, эти животные могут надежно различать цели, отличающиеся по размерам всего на 5—6 мм, и обнаруживать и избегать препятствия (тонкие проволоки), диаметр которых почти в 50 раз меньше длины излучаемой волны ультразвука. Далее, эхолокационная система содержит механизмы, обеспечивающие высокую помехоустойчивость, продемонстрированные как в опытах с летучими мышами при использовании низкочастотных и ультразвуковых акустических помех, так и в экспериментах с дельфинами в условиях сильной реверберации бассейна. И наконец, эхолокатор этих животных есть специализированный орган мозга, неотъемлемая его часть. Мозг в целом осуществляет эхолокацию, или, иначе говоря, в распоряжении эхолокации — весь мозг со всеми его регу
452
ляторными, активизирующими аппаратами. Отсюда и принци’ пиально особое качество «эхолокационного мозга»—неограниченность свободы действий. Перечисленными возможностями пока не обладает ни одна из современных радиолокационных или ультразвуковых систем, созданных человеком.
Однако в соревновании творчества природы и творческой мысли человека в итоге победу, конечно, одержит человек, как только будут описаны законы, лежащие в основе достижений природы. Совместные усилия биологов и инженеров, работающих в области бионики, и должны ускорить приобретение этих знаний. Несомненно будут созданы более универсальные и менее громоздкие радио- и гидролокационные установки, они будут более совершенны, чем у любого животного, вызывающего ныне удивление.
Уже сейчас в различных странах мира, в том числе и в Советском Союзе, создаются различные модели ультразвуковых приборов-поводырей для слепых людей, основанные на принципах эхолокации летучих мышей. Примером может служить ультразвуковой прибор «Ориентир», созданный советскими инженерами, который в настоящее время успешно проходит испытания. Дальность действия этого прибора порядка 10 м. Собран он на полупроводниках, вес его всего 230 г. Небольшой передатчик ультразвуковых колебаний слепой несет в руке и пользуется им как карманным фонариком. Отражения ультразвукового луча от встречных предметов усиливаются, преобразуются в звуковые колебания и вызывают изменение тона звука в микронаушниках. При этом по высоте тона слепой может судить о расстоянии до предмета, а по тембру звука — о характере предмета: изменение тона сигнала указывает на то, что предмет движется — удаляется или приближается. Принцип, заложенный в этом приборе, очень напоминает тот, который используется подковоносыми летучими мышами.
Наконец, недолго придется ждать, когда завершится успехом попытка создания миниатюрного ультразвукового прибора, предназначенного для ориентации аквалангистов. В основу этого прибора положен принцип эхолокации, заимствованный у дельфинов.
Неисчерпаемы перспективы применения эхолокации животных в области техники, инженерного творчества и всех тех задач, которые выдвигаются бионикой. В этом направлении написано много статей, книг, проведены разносторонние обсуждения на специальных конференциях, симпозиумах. И пусть в них пока что более отражены гипотезы, поиски, желания, мечты, чем конкретные результаты. Уместно вспомнить мудрые слова профессора Ленинградского университета А. А. Ухтомского: «Человек очень сильное существо: если оно начинает серьезно мечтать, то это значит, что рано или поздно мечта сбудется».
ЛИТЕРАТУРА
Абрамов А. П., Айрапетьянц Э. Ш., Бурдин В. И., Голубков А. Г., Е р ш о в а И. В., Ж е ж е р и н А. Р., Королев В. И., Малышев Ю. А., Ульянов Г. К., Фрадкин В. Б. 1971. Исследование способностей дельфинов к дифференцировке объемных предметов по линейным размерам и материалам. Тез. докл. 7-й Всес. акуст. конф., Л. : 3.
Абрамов А. П., Б у р д и н В. И., Е р ш о в а И. В., Королев В. И., М а л ы ш е в Ю. А. 1970. Методика и аппаратура проведения экспериментов по дифференцировке объемных мишеней дельфинами. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 59.
Абрамов А. П., Бурдин В. И., Иваненко Ю. В., Королев В. И., Малышев Ю. А. 1970. Определение разрешающей способности дельфинов по углу. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 65.
Абрамов А. П., Голубков А. Г., Е р ш о в а И. В., Жеже-рин А. Р., Малышев Ю. А., Фрадкин В. Б. 1971. Некоторые характеристики гидролокационных сигналов дельфинов. Тез. докл. 7-й Всес. акуст. конф., Л. : 3.
Абрамов А. П., Голубков А. Г., Е р ш о в а И. В., Ф р а д-к и н В. Б., К о р о л е в В. И. 1972. Исследование способности дельфина к дифференцировке объемных предметов по линейным размерам и материалам. В сб.: Некоторые проблемы биологической кибернетики, Л. : 101—103.
Абрамов А. П., Голубков А. Г., Игнатьева Е. А., Королев В. И., Фрадкин В. Б. 1972. Дифференциальные пороги слухового анализатора дельфина по частоте. Тез. докл. 25-й научнотехн. конф. ЛИАП, Л. : 73.
Абрамов А. П., Голубков А. Г., Королев В. И., Фрадкин В. Б. 1971. О помехозащищенности гидролокатора дельфина. Тр. Акуст. инет., вып. 17 : 24—28.
Абрамов А. П., Голубков А. Г., Фрадкин В. Б. 1972. Аппаратура для исследования адаптации гидролокатора дельфина к различным помехам. В сб.: Некоторые проблемы биологической кибернетики, Л. : 250.
Абрамов А. П., Жежерин А. Р., Королев В. И., Малышев Ю. А. 1970. Исследование возможностей дельфинов (Tursiops truncatus) различать объемные предметы, изготовленные из различных материалов. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 63.
Абрамов А. П., Жежерин А. Р., Королев В. И., Ульянов Г. К. 1970. Исследование помехозащищенности гидролокационных сигналов дельфина. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 64—65.
Абрамов А. П., Игнатьева Е. А., Королев В. И., Фрадкин В, Б, 1972. Определение дифференциального порога и пре
454
дельного соотношения интенсивностей двух сигналов для слухового анализатора дельфина по времени задержки импульсов. Тез. докл. 25-й научно-техн. конф. ЛИАП, Л. : 76—77.
Абрамов А. П., Игнатьева Е. А., Малышев Ю. А., Фрадкин В. Б. 1972. Исследование разрешающей способности эхолокационного аппарата дельфина по угловым координатам. Тр. ЛИАП, вып. 76 : 3—8.
Абрамов А. П., Королев В. И., Фрадкин В. Б. 1972. О многоканальное™ слухового анализатора морских млекопитающих. Тез. докл. 25-й научно-техн. конф. ЛИАП, Л. : 74.
Агарков Г. Б. 1972. Морфофункциональный анализ адаптации черноморских дельфинов к водной среде обитания. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 5—8.
Агарков Г. Б., С о л у х а Б. В., Хоменко Б. Г. 1971. О гидролокационной способности дельфинов. В сб.: Бионика, вып. 5, изд. «Наукова думка», Киев : 52—57.
Агарков Г. Б., Хоменко Б. Г., С л е з и н В. Б. 1972. Нейро-морфологические основы эхолокации и поведения некоторых китообразных. Тр. Северо-Западного заочного политехи, инет., № 22, Л. : 8—12.
Айрапетьянц Э. Ш. 1960. Опыт сравнительного изучения принципа замещаемости и межанализаторной интеграции. В сб.: Вопр. сравн. физиол. анализаторов, вып. 1, Изд. ЛГУ : 9.
Айрапетьянц Э. III. 1961а. К вопросу о функциональной структуре пространственного анализа. В сб.: Пробл. восприятия пространства и пространств, представлений, Изд. АПН РСФСР, М. : 11.
Айрапетьянц Э. Ш. 19616. Целостность мозга и физиология пространственного анализа. В сб.: Пробл. восприятия пространства и времени, Изд. ЛГУ : 16.
Айрапетьянц Э. Ш. 1967а. Бионический аспект исследования механизмов информации о пространстве — об эхолокации летучих мышей. Физиол. журн. СССР, т. 53, № 4 : 368.
Айрапетьянц Э. Ш. 19676. Механизмы мозга и пространственный анализ. В кн.: Исследования аппаратов сигнализации мозга. Изд. «Наука», Л. : 11.
Айрапетьянц Э. Ш. 1970. К физиологической теории пространственного анализа. IX Всес. съезд физиол., т. 1, изд. «Наука», Л. : 153— 158. То же, 1973. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 9—20.
Айрапетьянц Э. Ш., Бурикова Н. В., Константинов А. И., Котеленко Л. М., Куликов Г. А., С о к о-л о в а Н. Н. 1972. Организация слуховой коры рукокрылых в ряду млекопитающих. Тез. докл. XXIII совещ. по пробл. высш, нервн. деят., т. -2, Горький : 219—220.
Айрапетьянц Э. Ш., Бурикова Н. В., Константинов А. И., Котеленко Л. М., Куликов Г. А., Соколова Н. Н. 1973. Организация слуховой коры рукокрылых в ряду млекопитающих. Журн. высш, нервн. деят., т. 23, вып. 2 : 392—402.
Айрапетьянц Э. Ш., Васильев А. Г. 1970а. Характеристика вызванных потенциалов слухового анализатора летучих мышей на стимулы с разной частотой заполнения. Физиол. журн. СССР, т. 56, № 12 : 1721-1730.
Айрапетьянц Э. Ш., Васильев А. Г. 19706. Характеристика вызванных потенциалов слухового анализатора летучих мышей на стимулы с разной частотой заполнения. Реф. докл. IV Всес. конф, по нейрокибернетике, Ростов-на-Дону : 5.
(Айрапетьянц Э. Ш., Васильев А. Г.) A i г а р е t i a n z Е. Sh., Vasilyev A. G. 1971. On neurophysiological mechanisms of the echolocating apparatus in bats (frequency parameters). Intern. J. Neuroscience, v. 1 : 279—286.
455
А й
А й
А й
А й
А й
А й
А й А й А й
А й
рапетьянц Э. I1L, Воронов В. А., Иваненко 10. В., Иванов М. П., Орловский Д. Л., Попов В. В., Сергеев Б. Ф., Чилингирис В. И. 1972а. Физиологические механизмы гидролокатора дельфина. Тез. докл. 6-го научн. совещ. и симпоз. по эволюц. физио л., Л. : 6.
рапетьянц Э. Ш., Воронов В. А., Иваненко Ю. В., Иванов М. П., Орловский Д. Л., П о п о в В. В., Сергеев Б. Ф., Чилингирис В. И. 19726. Изменение параметров локационных сигналов дельфинов в процессе дифференцировки сфе-
рических и цилиндрических мишеней. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 14.
рапетьянц Э. Ш., Воронов В. А., Иваненко Ю. В., Иванов М. П., Ордовский Д. Л., Попов В. В., Сергеев Б. Ф., Чилингирис В. И. 1973. К физиологии сонарной системы черноморских дельфинов. Ж урн. эволюц. биох. и физиол., № 4:9.
рапетьянц Э. Ш., Воронов В. А., Ордовский Д. Л., Сергеев Б. Ф., Чилингирис В. И. 1973. Дифференцирование дельфином афалиной сферических и цилиндрических объектов. Реф. докл. IV Всес. конф, по бионике, т. II, М. : 1—5.
рапетьянц Э. Ш., Голубков А. Г., Ершова И. В., Ж е-жерин А. Р., Зворыкин В. Н., Королев В. И. 1969. Эхолокационное дифференцирование и характеристика излучаемых импульсов у дельфинов. ДАН СССР, т. 188, № 5 : 1197—1199.
рапетьянц Э. Ш., Зворыкин В. Н., Петрова Л. М. 1973. Пространственный анализ у больших подковоносов после действия гравитационных перегрузок при ускорениях. Вести. ЛГУ, вып. 3, № 15 : 51—60.
рапетьянц Э. Ш., Зворыкин В. Н., Савин Б. М. 1972. Пространственная ориентация летучих мышей под влиянием повышенной гравитации. ДАН СССР, т. 203, № 3 : 723—725.
рапетьянц Э. Ш., Зворыкин В. Н., Савин Б. М. 1973. Материалы о влиянии гипервесомости на летучих мышей. В сб.: Сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 87—93.
рапетьянц Э. Ш., Константинов А. И. 1965. К вопросу о роли эхолокации в анализе пространства летучими мышами. В сб.: Бионика, изд. «Наука», М. : 334.
рапетьянц Э. Ш., Константинов А. И. 1967а. О взаимодействии анализаторов в эхо локационной активности у летучих мышей. В сб.: Исследование аппаратов сигнализации мозга, изд. «Наука», Л. : 22.
Айрапетьянц Э. Ш., Константинов А. И. 19676. Исследование нервных механизмов эхолокации у летучих мышей. В сб.: Вопр. бионики, изд. «Наука», М. : 445.
(Айрапетьянц Э. Ш., Константинов А. И.) Airapetian z Е. Sh., Konstantinov А. I. 1968. Echo-locating space analysis of animals. XXIV Intern, congr. physiol, sci., v. XII, Waching-ton : 5.
Айрапетьянц Э. Ш., Константинов А. И. 1970a. Эхолокация в природе. Изд. 1-е. Изд. «Наука», Л.
Айрапетьянц Э. Ш., Константинов А. И. 19706. Эхолокационный пространственный анализ у животных. В сб.: Пространственная ориентация животных, под ред. Н. П. Наумова, Изд. МГУ : 50—63.
Айрапетьянц Э. Ш., Константинов А. И. 1973. Физиологические исследования ультразвуковой эхолокации животных. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 20—28.
Айрапетьянц Э. Ш., Константинов А. И., Матюшкин Д. П. 1968. Мозговые механизмы эхолокации и бионика. Вести. ЛГУ, вып. 2 : 57.
456
(Айрапетьянц Э. Ш., Константинов А. И., Матюшкин Д. П.) Airapetianz Е. Sh., Konstantinov A. I., Matjushkin D. P. 1969. Brain echolocation mechanisms and bionics. Acta Physiol. Acad. Hung., v. 35 (1) : 1—17.
Айрапетьянц Э. Ш., Константинов А. И., Матюш-к и н Д. П. 1973. К теории эхолокации летучих мышей. В сб.: Проблемы бионики, изд. «Наука», М. : 125—135.
Акопиан А. Н., Бурдин В. И., Зайцева К. А., Морозов В. П., С о к о в ы х Ю. А. 1971. Некоторые особенности локационных сигналов дельфинов при поиске и обнаружении пищи. Тез. докл. VII Всес. акуст. конф., Л. : 4.
Акопиан А. Н., Бурдин В. И., Зайцева К. А., Морозов В. П., Соковых Ю. А. 1973. Влияние шума на частоту следования эхолокационных сигналов дельфинов. Реф. докл. VIII Всес. акуст. конф., т. I, М. : 135.
Алексеева Т. В., Голубков А. Г., Григорьев В. П., И г-натьева Е. А., Малышев Ю. А. 1972. Некоторые вопросы аппаратурного моделирования гидролокатора дельфина. Тез. докл. 25-й научно-техн. конф. ЛИАП, Л. : 81.
Алексеева Т. В., Голубков А. Г., Ершова И. В. 1971. К вопросу об активной ширине спектра локационных сигналов дельфинов. Тр. Акуст. инет., вып. 17 : 99—103.
Альтман Я. А. 1967. Характеристика импульсной активности нейронов заднего двухолмия кошки при действии моноуральных и бинауральных раздражений. В сб.: Пробл. физиол. акустики, т. 6, изд. «Наука», Л. : 158.
Альтман Я. А. 1972. Локализация звука. Изд. «Наука», Л.
Альтман Я. А., Дубровский Н. А. 1972. Пространственный слух. В кн.: Физиология сенсорных систем, ч. II, Л. : 398—426.
Андреева Н. Г. 1973. Некоторые механизмы распознавания сигналов на уровне медиальных коленчатых тел гладконосых и подковоносых летучих мышей. Тез. докл. IV Всес. конф, по бионике, т. II, М. : 6—9.
Аннарауд Д. К., Беляев В. В., Протасов В. А., Усов В. В., Ч е р е п а н о в И. М. 1972. Методика и программное обеспечение для сравнительного изучения строения мозга. Тез. докл. 6-го научн. совещ. и симпоз. по эволюц. физиол., Л. : 9.
Афанасьев М. Е. 1946. Новые данные о структуре и функции четверохолмия млекопитающих. Автореф. канд. дисс. М.
Бабкин В. П., Дубровский Н. А. 1971. О дальности действия и помехоустойчивости эхолокационного аппарата дельфина афалины при обнаружении различных мишеней. Тр. Акуст. инет., вып. 17 : 29—42.
Бабкин В. П., Дубровский Н. А., Краснов П. С., Т и-т о в А. А. 1971. Различение материалов шаровых мишеней дельфином афалиной. Тез. докл. VII Всес. акуст. конф., Л. : 5.
Баб к'и н В. П., Ф а д е е в а Л. М. 1971. Модельные эксперименты по аттестации шаровых мишеней. Тр. Акуст. инет., вып. 17 : 80—97. Багданас А., Белькович В. М., Крушинская Н. Л.
1970. Взаимодействие анализаторов у дельфина белобочки при различении геометрических фигур под водой. Журн. высш, нервн. деят., т. 20, вып. 5 : 1070—1074.
Бару А. В. 1966. О роли височных отделов коры больших полушарий в обнаружении звуковых сигналов различной длительности. Журн. высш, нервн. деят., т. 16 : 655—665.
Б а р у А. В. 1967. Дифференциальные пороги по частоте в зависимости от длительности тональных посылок у животных после удаления слуховой коры. В сб.: Механизмы слуха, Л. : 121.
Бару А. В. 1972. Роль различных отделов слуховой системы в восприятии звуков. В кн.: Физиология сенсорных систем, ч. II. Изд. «Наука», Л. : 261-279.
457
Б а р у А. В., К а р а с е в а Т. А. 1971. Мозг и слух. Изд. МГУ. Батуев А. С., Сытинский И. А. 1962. Влияние гамма-аминомасляной кислоты на стрихнинный потенциал коры головного мозга. ДАН СССР, т. 147, № 5 : 1242.
Беленков Н. Ю. 1965. Условный рефлекс и подкорковые образования мозга. Изд. «Медицина», М.
Беленков Н. Ю. 1969. Старые и новые проблемы взаимоотношений коры и подкорковых образований. Журн. высш, нервн. деят., т. 19, вып. 5 : 752.
Беленков Н. Ю., Горева О. А. 1969. Роль задних бугров четверохолмия в осуществлении ориентировочного рефлекса. Журн. высш, нервн. Деят., т. 19, вып. 3 : 453—461.
Беленков Н. Ю., Горева О. А., Калинина Т. Е. 1969. Исследование роли заднего двухолмия в анализе звуковых раздражений. В сб.: Зрительный и слуховой анализаторы, М. : 12—18.
Белькович В. М. 1962. Приспособительные особенности строения кожного покрова водных млекопитающих. Автореф. канд. дисс. М.
Белькович В. М. 1970. Некоторые новые представления о локализационных возможностях дельфинов. Экология, № 6 : 89—93.
Белькович В. М. 1972. Взаимодействие органов чувств китообразных при ориентации. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 38—42.
Белькович В. М., Борисов В. И. 1971. Локационное распознавание дельфинами фигур сложной конфигурации. Тр. Акуст. инет., вып. 17 : 19—23.
Белькович В. М., Борисов В. И., Гуревич В. С. 1970. Возможности углового разрешения локационного аппарата дельфина белобочки. Докл. 23-й научно-техн. конф. ЛИАП, Л. : 66—67.
Белькович В. М., Борисов В. И., Доронин А. Я. 1971. Улыбка дельфина. «Природа», № 3 : 58—61.
Белькович В. М., Борисов В. И., Гуревич В. С., Круши нс к а я Н. Л. 1969а. Эхолокационные возможности дельфина белобочки. Зоол. журн., т. 48, вып. 6 : 876—883.
Белькович В. М., Борисов В. И., Гуревич В. С., Круши нс к а я Н. Л. 19696. Эхолокация у зубчатых китообразных. Тез. докл. конф. МГУ по проблеме «Океан», М. : 3—7.
Белькович В. М., Борисов В. И., Гуревич В. С., Кру-шинская Н. Л., Дмитриева И. Л. 1972.Локационное распознавание расстояния между предметами (угловое различение). Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 30—33.
Белькович В. М., Клейненберг С. Е., Яблоков А. В. 1965. Загадка океана. Изд. «Молодая гвардия», М.
Белькович В. М., Клейненберг С. Е., Яблоков А. В. 1967. Наш Друг — дельфин. Изд. «Молодая гвардия», М.
Белькович В. М., Крушинская Н. Л., Гуревич В. С. 1969. Поведение дельфинов в неволе. Природа, № 11 : 18—28.
Белькович В. М., Нестеренко Ю. И. 1971. Как работает локатор дельфина. Природа, № 7 : 71—75.
Белькович В. М., Резников А. Е. 1971. Новое в гидролокации у дельфинов. Природа, № 11 : 84—90.
Б е "л ь к о в и ч В. М., Солнцева Г. Н. 1970. Морфофункциональные особенности органа слуха дельфина. Зоол. журн., т. 49, вып. 2 : 275-282.
Белькович В. М., Яблоков А. В. 1963. Обитатели моря «делятся опытом» с конструкторами. Наука и жизнь, № 5 : 61—64.
Берг А. И. 1965. Бионика и ее значение для развития техники. В кн.: Бионика. Изд. «Наука», М.
Бергман Л. 1957. Ультразвук и его применение в науке и технике. ИЛ*
458
Бехтеревым. 1895. Заднее двухолмйе как цен!р слуха, голоса и отраженных движений. Неврол. вести., т. 3:2.
Бехтерев В. М. 1906. Основы учения о функциях мозга. Вып. 5. СПб.
Бибиков Н. Г., Сухорученко М. Н. 1966. Исследование динамики маскировки шумом суммарного ответа улитки на щелчок. Журн. высш, нервн. деят., т. 16, вып. 5 : 924.
Блохин С. А. 1972. Топография костей черепа, спермацетового органа и носовых проходов на спилах головы эмбриона кашалота. Изв. Тихо-океанск. НИИ рыбн. хоз. и океаногр., 81 : 253—257.
Богословская Л. С. 1968. Нейроны и межнейронные связи кохлеарных ядер млекопитающих. Автореф. канд. дисс. М.
Богословская Л. С. 1972. Морфология первичных акустических центров мозга млекопитающих. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 48—54.
Борисов Ю. Я., Марков Л. О. 1960. Ультразвук в технике настоящего и будущего. М.
Бурдин В. И., Марков В. И., Резник А. М., Скорняков В. М., Ч у п а к о в А. Г. 1971а. О способности черноморской афалины (Tursiops truncatus Barabasch) различать полезный сигнал на фоне шума. В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 106—112.
Бурдин В. И., Марков В. И., Резник М. М., Скорняков В. М., Чупаков А. Г. 19716. Определение величины дифференциального порога по интенсивности для белого шума у черноморской афалины (Tursiops truncatus Barabasch). В сб.: Морфология и экология морских млекопитающих, М. : 112.
Буреш Я., Петрань М., Захар И. 1962. Электрофизиологические исследования. ИЛ.
Бурикова Н. В. 1971. Эфферентные связи слуховой коры головного мозга летучей мыши. Журн. эволюц. биохим. и физиол., т. 7, № 5 : 529-533.
Бурикова Н. В. 1973. Эфферентные связи нижнего двухолмия летучей мыши. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 13—17.
Вартанян И. А. 1966. Электрофизиологическое исследование слуховых отделов среднего мозга. Автореф. канд. дисс. Л.
Васильев А. Г. 1967а. Сравнительная характеристика слуховой системы гладконосых и подковоносых летучих мышей (электрофизиологические данные). ДАН СССР, т. 175, № 6 : 1414.
Васильев А. Г. 19676. Электрическая активность заднего двухолмия летучей мыши, раздражаемой ультразвуком. ДАН СССР, т. 175, № 3 : 741.
Васильев А. Г. 1968а. Электрическая активность заднего двухолмия гладконосых и подковоносых летучих мышей при раздражении ультразвуком. Автореф. канд. дисс. Л.
Васильев А. Г. 19686. Вызванные потенциалы и реакции отдельных нейронов заднего двухолмия летучих мышей на ультразвуковые стимулы с разной длительностью их фронтов. Вести. ЛГУ, сер. биол., вып. 1 : 167.
Васильев А. Г. 1969а. Характеристика электрических ответов задних холмов четверохолмия гладконосых и подковоносых летучих мышей при изменении силы ультразвука. В сб.: Механизмы нервной деятельности, Л. : 180—187.
Васильев А. Г. 19696. Характеристика вызванных потенциалов заднего двухолмия летучих мышей при изменении направления действия раздражителя. Биол. науки, № 11 : 57—62.
Васильев А. Г. 1973а. Характеристика реакций отдельных нейронов заднего двухолмия летучих мышей при изменении некоторых пара
459
метров ультразвуковых стимулов. В сб.: Проблемы бионики, изд» «Наука», М. : 324—333.
Васильев А. Г. 19736. Некоторые особенности слуховой системы гладконосых и подковоносых летучих мышей (электрофизиологические данные). В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 94—110.
Васильев А. Г., Андреева Н. Г. 1971. Характеристика электрических реакций внутренних коленчатых тел гладконосых и подковоносых летучих мышей на ультразвуковые стимулы с разной частотой заполнения. Нейрофизиология, т. 3, № 2 : 138—144.
Васильев А. Г., Матюшкин Д. П. 1967. К физиологической характеристике слуховой системы летучих мышей по реакциям на ультразвуковые стимулы. Физиол. жури. СССР, т. 53, № 12 : 1407.
Васильев А. Г., Стосман И. М. 1970. Характеристика потенциалов действия нейронов кохлеарного ядра летучих мышей при раздражении ультразвуком с разных направлений. Журн. эволюц. биохим. и физиол., т. 6, № 6 : 642—644.
Васильев А. Г., Стосман И. М., Куликов Г. А. 1973. Механизмы определения некоторых параметров ультразвуковых стимулов гладконосых и подковоносых летучих мышей. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 22—26.
Васильева Т. Е. 1973. Реакция нейронов верхнеоливарного комплекса гладконосых и подковоносых летучих мышей на ультразвуковые стимулы. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 18—21.
Великий Н. 1872. Об окончании нервов в волосковых сумках млекопитающих. Тр. СПб. общ. естествоиспыт., т. 3 : 421.
Вельмин В. А., Дубровский Н. А., Юркевич Л. И. 1973. Об исследовании слуха дельфинов поведенческим методом. Реф. докл. VIII Всес. акуст. конф., т. I, М. : 125.
Вербицкая Л. Б. 1970. Строение ядер мозжечка дельфина (Delphinus delphis). Архив анат., гистол. и эмбриол., т. 59, № 10 : 26—33.
Винников Я. А., Титова Л. К. 1961. Кортиев орган. Изд. АН СССР, М.-Л.
Воронин В. Р., Голубков А. Г., Малышев Ю. В. 1972. Исследование диаграммы направленности излучающего аппарата дельфина по частотам. Тез. докл. 25-й научно-техн. конф. Л И АП, Л. : 81.
Воронин Л. Г. 1970. Поведение «примата» моря — дельфина афалины (Tursiops truncatus). Усп. совр. биол., т. 69, вып. 2 : 191—207.
Воронин Л. Г., Козаровицкий Л. Б. 1969. Некоторые особенности высшей нервной деятельности черноморского дельфина афалины (Tursiops truncatus). Журн. высш, нервн. деят., т. 19, вып. 2 : 233.
Воронин Л. Г., Козаровицкий Л. Б., Стародубцев Ю. Д. 1971. О групповом поведении черноморских дельфинов афалин (Tursiops truncatus Pont.). В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 88—93.
Воронин Л. Н., Соколов В. Е. 1971. Обзор материалов по изучению дельфинов. В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 3—26.
В оро пин Л. Г.,Стародубцев 10. Д., Козаровицкий Л. Б. 1971. Динамика тренировки поведенческих навыков у черноморского дельфина афалины (Tursiops truncatus Pont.). В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 73—87.
Г а п и ч Л. И., Ладыгина Т. Ф., С у и и н А. Я., Сухорученко М. Н. 1971. Техника электрофизиологических исследований на мозге дельфина. В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 98—105.
Гер ш у н и Г. В. 1963. Вызванные потенциалы и механизм различения внешнего сигнала. Журн. высш, нервн. деят., т. 13 : 882—890-
460
Гершуни Г. В. 1964. Электрофизиологические показатели функций слуховой системы. В сб.: Современные проблемы электрофизиологических исследований нервной системы, изд. «Медицина», М. : 361.
Г е р ш у н й Г. В. 1967. О механизмах слуха (в связи с исследованием временных и временно-частотных характеристик слуховой системы). В сб.: Механизмы слуха, Л. : 3—32.
Гершуни Г. В., Вартанян И. А. 1972. Реакции центральных нейронов на звуки с различной временной структурой. Физиол. жури. СССР, т. 58, № 7 : 988—1000.
Гиро Л. Р., Дубровский Н. А. 1972. Зависимость частоты следования эхолокационных сигналов дельфина от сложности эхолокационной задачи. Морское приборостроение, сер. «Акустика», № 2 : 271 — 290.
Голубков А. Г., Ершова И. В., Малышев Ю. А. 1970. Дальность связи и дальность действия эхолокатора черноморских афалин (Tursiops truncatus). Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 62—63.
Голубков А. Г., Ершова И. В., Розет Т. А. 1972. Восстановление формы зондирующих сигналов дельфинов. Тр. ЛИАП, вып. 75 : 151—156.
Голубков А. Г., Ершова И. В., Ульянов Г. К. 1970. Некоторые характеристики гидролокационных сигналов дельфинов при различении ими объемных предметов по линейному размеру. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 61.
Голубков А. Г., Зворыкин В. Н., Ершова И. В., Королев В. И., Бурдин В. И., М а л ы ш е в Ю. А. 1969. Некоторые экспериментальные данные и перспективы изучения анализаторной способности дельфина. Тр. ЛИАП, вып. 64 : 128—132.
Голубков А. Г., Иваненко Ю. В. 1970. Исследование разрешающей способности по дальности эхолокатора дельфина. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 65.
Голубков А. Г., Константинов А. И., Макаров А. К. 1969. Дальность действия и чувствительность эхолокатора летучих мышей. Тр. ЛИАП, вып. 64 : 117—127.
Гольдбурт С. Н. 1964. Нейродинамика слуховой системы человека. Изд. ЛГУ.
Горлинский И. А. 1972. О локализации ультразвуковых сигналов летучими мышами. Тез. докл. VI научи, совещ. и симпоз. по эволюц. физиол., изд. «Наука», Л. : 53.
Горшков С. И., Горбунов О. Н., Антропов Г. А. 1965. Биологическое действие ультразвука. Изд. «Медицина», М.
Грачева М. С. 1971. Некоторые особенности строения гортани афалины. Зоол. журн., т. 50, № 10 : 1539—1545.
Гришин В. Г. 1971. Аппаратура для спектрального анализа акустических сигналов морских животных. В сб.: Морфология и экология морских млекопитающих, М. : 120.
Громов И. М., Гуреев А. А., Новиков Г. А., С о к о-л о в И. И., Стрелков П. П., Чапский К. К. 1963. Млекопитающие фауны СССР, Ч. 1. М.—Л.
Гуревич В. С. 1969. О способности дельфина Delphinus delphis к эхолокационному распознаванию геометрических фигур. Вести. МГУ, сер. биол., № 3 : 109—112.
Гуревич В. С. 1972а. Морфофункциональное исследование надчерепных дыхательных путей обыкновенного дельфина. В сб.: Применение математических методов и вычислительной техники в медицине и биологии, Л. : 19—20.
Гуревич В. С. 19726. О возможном взаимодействии гортани и надчерепных носовых структур в звукообразовании у зубатых китообразных.
461
Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 66—68.
Гуревич В. С., Евдокимов В. Н., Корольков Ю. И. 1972. Использование рентгенологического метода для исследования надчерепных дыхательных путей обыкновенного дельфина. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 70—72.
Д а н и л о в В. Н. 1972. Об одном методе математического моделирования слухового анализатора. В сб.: Некоторые проблемы биологической кибернетики, Л. : 289.
Дмитриева И. М., Круши некая Н. Л., Белькович В. М., Ш у р х а л А. В. 1972. Локационное распознавание формы предметов дельфином. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 80—83.
Добрачев Ю. П., Масленникова М. П., П анютин К. К., Патлякевич Н. Д., Симкин Г. Н., Федорова Е. С. 1968. Локационные сигналы отечественных летучих мышей. Тез. докл. VI Всес. акуст. конф., М.
Добрачев Ю. П., Симкин Г. Н. 1973. Математическое представление некоторых характеристик зондирующего локационного сигнала и анализ отраженного импульса у гладконосых летучих мышей. В сб.: Проблемы бионики, изд. «Наука», М. : 155—162.
Добрякова О. А. 1968. К вопросу о цитоархитектонике слуховой области коры больших полушарий головного мозга летучей мыши. Тез. докл. V научи, совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 92.
Дубровский Н. А. 1972. Эхолокационное различие объектов у дельфинов. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 83—86.
Дубровский Н. А. 1973. Проблемы исследования и моделирования эхолокатора у дельфина. Реф. докл. IV Всес. конф, по бионике, т. 1, М. : 7—13.
Дубровский Н. А., Краснов П. С. 1971. Распознавание дельфином афалиной упругих шаров по материалу и размеру. Тр. Акуст. инет., вып. 17 : 9—17.
Дубровский Н. А., Краснов 11. С., Титов А. А. 1970. К вопросу об излучении дельфином азовкой ультразвуковых локационных сигналов. Акуст. журн., т. 16, вып. 4 : 521—525.
(Дубровский Н. А., Краснов П. С., ТитовА. A.) Dubrovsky N. A., Krasnov Р. S., Titov А. А. 1971. Discrimination of solid elastic spheres by an echolocating porpoise Tursiops truncatus. VII Intern, congr. Acoust., Budapest: 533—536.
Дубровский H. А., Титов А. А., Краснов П. G., Бабкин В. П., Лекомцев В. М., Николенко Г. В. 1970. Исследование разрешающей способности эхолокационного аппарата черноморской афалины. Тр. Акуст. инет., вып. 10 : 163—170.
Дубровский Н. А., Тумаркина Л. Н. 1967. Исследование восприятия человеком амплитудно-модулированных шумов. Акуст. журн., т. 13, вып. 1 : 76—79.
Дубровский Н. А., Фадеева Л. М. 1973. Распознавание шаровых мишеней дельфинами. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 29-34.
Дьяченко С. М., Калита Е. В. 1970. О спектральном анализе сигналов, издаваемых животными. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 63.
Дьяченко С. М., Королев Л. Д., Р е з в о в Р. Н., Чемоданов Б. К. 1971. Исследование способности дельфина афалины определять направление на источник шумового сигнала. Тр. Акуст. пнет., вып. 12 : 43—46.
462
Ермолин С. И. 1973. Изучение механизмов обнаружения эха на уровне верхнеоливарного комплекса эхолокатора гладконосых летучих мышей. Тез. докл. IV Всес. конф, по бионике, т. II, М. : 94—97.
Ершова И. В. 1972. Влияние полосы пропускания регистрирующей аппаратуры на искажения формы локационных сигналов дельфинов. Тез. докл. 25-й научно-техн. конф. ЛИАП, Л. : 78.
ЖежеринА. Р. 1972. Исследование временной функции сигнала, отраженного от упругой сферы, погруженной в воду. Тез. докл. 25-й научно-техн. конф. ЛИАП, Л. : 79.
Забусов Н. П. 1910. Исследование иннервации летательной перепонки летучих мышей. Тр. Общ. естествоисп. Казанского унив., т. XLIII, вып. 1:1.
Замбржицкий И. А. 1956. Цитоархитектоника и нейронное строение лимбической области у некоторых млекопитающих. Архив анат., гистол. и эмбриол., т. 33, вып. 4 : 41.
Заславский Г. Л. 1971. О направленности звукоизлучения у черноморского дельфина афалины. Тр. Акуст. инет., вып. 17 : 60-70.
Заславский Г. Л. 1972. Исследование локационных сигналов дельфина афалины при помощи двухканальной системы регистрации. Биофизика, т. 17, № 4 : 717—720.
Заславский Г. Л., Титов А. А., Лекомцев В. М. 1969. Исследование гидролокационных способностей дельфина азовки. Тр. Акуст. инет., вып. 8 : 134—138.
Зворыкин В. П. 1959. Морфологические основы локационных и ультразвуковых свойств летучих мышей. Архив анат., гистол. и эмбриол., т. 36, вып. 5 : 19.
Зворыкин В. П. 1961. Морфологические основы ультразвукового восприятия млекопитающих. В сб.: Структура и функции нервной системы, М. : 284—295.
Зворыкин В. П. 1963а. Морфологические основы ультразвуковых и локационных качеств дельфина. Архив анат., гистол. и эмбриол., вып. 7 : 3.
Зворыкин В. П. 19636. Некоторые цитоархитектопические и количественные закономерности системы подкорковых образований слухового анализатора в сравнительном ряду млекопитающих. Автореф. докт. дисс. М.
Земский В. А. 1970. Киты Южного полушария (биология и морфология). Тр. Атлантич. НИИ рыбн. хоз. и океаногр., вып. 29, Калининград : 234.
Златоустова Л. В., Низова А. Б. 1971. Опыт акустического анализа некоторых свойств афалины (Tursiops truncatus Pont.). В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 116-119.
Иваненко Ю. В. 1973. Разрешающая способность сонара дельфина по дальности. Тез. докл. IV Всес. конф, по бионике, т. II, М.: 102— 105.
Иваненко Ю. В., Чилингирис В. И. 1973. Характеристики приема сонара дельфина. Реф. докл. VIII Всес. акуст. конф., т. I, М. : 126.
Иванов М. П., Курганский Н. А. 1973. Траектория движения дельфина в процессе дифференцировки сферических и цилиндрических тел. Тез. докл. IV Всес. конф, по бионике, т. II, М. : 106-109.
Ильичев В. Д. 1970. Пассивная локация сов. Усп. совр. биол., т. 70, вып. 1 (4) : 120-136.
Ильичев В. Д. 1972. Биоакустика птиц. Изд. МГУ.
Ильичев В. Д., Голубева Т. Б., Кравцов Б. Г., Чер-л ы й А. Г. 1973. Электрофизиологические характеристики рецептор
463
ного отдела слухового анализатора летучих мышей и сов в связи с особенностями пространственного слуха. В сб.: Проблемы бионики, изд. «Наука», М. : 169—181.
Казнадзей В. В. 1971. Поведение дельфина афалины при обучении решению локационных задач. Тр. Акуст. инет., вып. 17 : 71—77.
Калинина Т. Е. 1962. Влияние удаления слуховой и смежных областей коры на звуковые условные рефлексы. Журн. высш, нервн. деят., т. 12, № 4 : 720—725.
Калинина Т. Е. 1971. О структурно-функциональной организации слуховой системы. Автореф. докт. дисс. М.
Каневский И. Н., Суриков Б. С. 1970. Об одном варианте акустической линзы Люнеберга. Акуст. журн., т. 16, вып. 4 : 536-541.
Каневский И. Н., Суриков Б. С. 1971. Однородные сферические и цилиндрические линзы для фокусирования звуковых волн. Акуст. журн., т. 17, вып. 1 : 55—60.
Карандеева О. Г. 1969. Особенности поведения в неволе и дрессировка дельфинов. Журн. высш, нервн. деят., т. 19, № 1 : 55—58.
Карасева Т. А. 1967. Особенности обнаружения звуковых сигналов малой длительности при очаговом поражении височной доли головного мозга. В сб.: Механизмы слуха, изд. «Наука», Л. : 135—143.
Кесарев В. С. 1970. Некоторые данные о нейронной организации новой коры дельфина. Архив анат., гистол. и эмбриол., вып. 8 : 71—77.
Кесарев В. С., Малофеева Л. И. 1969. Структурная организация двигательной зоны коры большого мозга дельфина. Архив анат., гистол. и эмбриол., т. 56, вып. 4 : 48—55.
Кибернетические проблемы бионики. 1971. М.
Клейненберг С. Е. 1958. К вопросу о происхождении китообразных. ДАН СССР, т. 122, Яз 5 : 950.
Клейненберг С. Е., Белькович В. М. 1967. Дельфины-мифы и действительность. Изд. «Знание», М.
Клейненберг С. Е., Яблоков А. В. 1958. О морфологии верхних дыхательных путей китообразных. Зоол. журн., т. 37, № 7 : 1091.
Клейненберг С. Е., Яблоков А. В., Белькович В. М., Тарасевич М. Н. 1964. Белуха. Опыт монографического описания вида. Изд. «Наука», М.
Клосовский В. Н. 1946. Строение и функция среднего мозга. 1. О прямой проводниковой связи ядра заднего двухолмия с красным ядром (Fasciculus colliculo-rubralis). Бюлл. эксп. биол. и мед., т. 21, вып. 5 : 3—4.
Клявина М. П., Марусева А. М. 1963. Электрические ответы улитки новорожденных. ДАН СССР, т. 149, № 5 : 1221—1224.
Кожин В. Н. 1970а. Рассеяние и поглощение плоской звуковой волны цилиндром произвольного радиуса в вязкой среде. Акуст. журн., т. 16, вып. 3 : 403—408.
Кожин В. Н. 19706. Излучение и рассеяние звука цилиндром в вязкой среде. Акуст. журн., т. 16, вып. 2 : 269—274.
Козлова Н. А., Малашкин В. И., Минайкин Ю. И., П е-р е с а д а В. П. 1970а. Анализ методов обработки широкополосных сигналов в приемной системе эхолокатора летучей мыши. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 56—57.
Козлова Н. А., Малашкин В. И., Минайкин Ю. И., Переезда В. П. 19706. Экспериментальные исследования помехоустойчивости эхолокатора летучей мыши. Тез. докл. 23-й научнотехн. конф. ЛИАП, Л. : 55—56.
Козлова Н. А., Малашкин В. И., Минайкин Ю. И., П е-ресада В. П. 1970в. Исследование характеристик эхолокатора
464
летучей мыши. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 54—55.
Колчин С. П., Белькович В. М. 1970. Некоторые особенности функции сердца у дельфинов. Журн. эвол. биохим. и физиол., т. 6, № 4 : 411-417.
Комаров В. Т. 1972. Акустикоизлучающая система зубатых китов. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 104—107.
Константинов А. И. 1964а. О роли условной сигнализации в пространственном анализе у летучих мышей. Журн. высш, нервн. деят., т. 14, вып. 4 : 701.
Константинов А. И. 19646. Влияние выключения зрительной рецепции на направленное поведение у летучих мышей. Вести. ЛГУ, № 15, сер. биол., вып. 8 : 72.
Константинов А. И. 1965а. Влияние частичного и полного выключения коры головного мозга на эхолокацию у летучих мышей. ДАН СССР, т. 161, № 4 : 989.
Константинов А. И. 19656. Влияние выключения заднего двухолмия на локационно-двигательную активность у летучих мышей. Тез. докл. научи, совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 153.
Константинов А. И. 1965в. Материалы по физиологии пространственного анализа летучих мышей (опыт физиологического исследования эхолокации). Автореф. канд. дисс. Л.
Константинов А. И. 1966а. Основы ультразвуковой пространственной ориентации летучих мышей. В сб.: Вопр. сравн. физиол. анализаторов, вып. 2, Изд. ЛГУ : 93.
Константинов А. И. 19666. Распознавание геометрических фигур летучими мышами с помощью ультразвуковой локации. Тез. докл. XXI совещ. по пробл. высш, нервн. деят., изд. «Наука», М. — Л. : 165.
Константинов А. И. 1968. Эхолокационное распознавание целей летучими мышами. Тез. докл. V научи, совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 159.
Константинов А. И. 1969. Связь слухового восприятия и эхолокации при обнаружении и ловле насекомых гладконосыми летучими мышами (Vespertilionidae). Журн. эволюц. физиол. и биохим., т. 5, № 6 : 566-572.
Константинов А. И. 1970. Характеристика ультразвукового локатора рыбоядных летучих мышей. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 51-52.
Константинов А. И. 1972а. Особенности постэмбрионального развития и пути формирования эхолокационного сигнала у летучих мышей. Тр. IV Всес. конф, по экол. физиол., Краснодар : 133.
Константинов А. И. 19726. Развитие функций ультразвуковых локационных систем у летучих мышей в постнатальном онтогенезе. Тр. VI научи, совещ. по эволюц. физиол., посвящ. памяти акад. Л. А. Орбели, Л : 103.
Константинов А. И. 1973а. Эхолокационные системы летучих мышей подотряда Microchiroptera. Автореф. докт. дисс. Л.
Константинов А. И. 19736. Развитие эхолокации у летучих мышей в постнатальном онтогенезе. Вести. ЛГУ, № 3 : 67—77.
(Константинов А. И.) Konstantinov А. I. 1973в. Development of echolocation in bats in postnatal ontogenesis. Period, biologorum, v. 75 : 13—17.
Константинов А. И. 1973г. Основные показатели эхолокационных систем летучих мышей. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 45-50.
465
Константинов А. И., Ахмарова Н. И. 1968. Распознавание (анализ) предметов при помощи эхолокации остроухой ночницей (Myotis oxygnathus). Биол. науки, т. 4 : 22.
Константинов А. И., Ахмарова Н. И., Головина С. С. 1966. О возможности использования ультразвуковой локации мелкими грызунами. Тез. докл. XXI совещ. по пробл. высш, нервн. деят., изд. «Наука», М.—Л. : 155.
Константинов А. И., Макаров А. К. 1973. Эхолокационные сигналы некоторых отечественных видов гладконосых летучих мышей (семейство Vespertilionidae). В сб.: Вопр. сравн. физиол. анализаторов, вып. 8. Изд. ЛГУ : 29—44.
Константинов А. И., Малашин В. И., Макаров А. К., Козлова Н. А., Соколова Н. Н., Чилингирис В. И. 1973. Диапазон и принципы эхолокационного распознавания у летучих мышей. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 57-77.
Константинов А. И., Макаров А. К., Соколов Б. В., Соколова Н. Н. 1973. Биоакустические особенности и принципы функционирования эхолокационной системы подковоносых летучих мышей. Реф. докл. VIII Всес. акуст. конф., т. 1, М. : 136.
Константинов А. И., Мельников Н. Ф., Титов А. А. 1968. О способности дельфинов распознавать предметы. Тез. докл. II республ. конф, по бионике, Киев : 57.
(Константинов А. И., Мовчан В. Н., Макаров А. К.) Konstantinov А. I., М о vtchan V. N., Makarov А. К. 1973. Influence of band limited noises on the efficiency of echolocation detection of targets by Rhinolophus ferrum-equinum. Period, biologorum, v. 75:7-11.
Константинов А. И., Рахматулина И. К. 1968. Исследование хоминга у летучих мышей. Тез. докл. V научи, совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 112.
Константинов А. И., Саноцкая Н. Н., Соколова Н. Н. 1971. Частотно-пороговые характеристики слуховой системы у летучих мышей, измеренные методом условных рефлексов. Журн. высш, нервн. деят., т. 21 : 535—541.
Константинов А. И., Соколов Б. В. 1967. Активная и пассивная локация при ловле насекомых у летучих мышей. Тез. докл. III конф, по экол. физиол., биохим. и морфол., Новосибирск : 68.
Константинов А. И., Соколов Б. В. 1969. Характеристика ультразвуковых ориентационных сигналов подковоносых летучих мышей (Rhinolophidae). Журн. эволюц. физиол. и биохим., т. 5, № 1 : 90—97.
Константинов А. И., Соколов Б. В. 1973. Роль и взаимодействие эхолокации и слухового восприятия при ловле насекомых у летучих мышей. Веб.: Проблемы бионики, изд. «Наука» М. : 162—169.
Константинов А. И., Соколов Б. В., Стосман И. М. 1967а. Сравнительное исследование чувствительности эхолокации летучих мышей. ДАН СССР, т. 175, № 6 : 1418.
Константинов А. И., Соколов Б. В., Стосман И. М. 19676. Чувствительность эхолокации у гладконосых и подковоносых летучих мышей. Тез. докл. III конф, по экол., физиол., биохим. и морфол., Новосибирск : 66.
Константинов А. И., Соколова Н. Н. 1970. О возможности функционирования эхолокационной системы летучих мышей без участия слуховой коры. Журн. эволюц. биохим. и физиол., т. 6, № 3 : 347—350.
Константинов А. И., Стосман И. М. 1972. Электрическая активность заднего двухолмия при действии ультразвуковых стиму-
466
лов в онтогенезе летучей мыши Myotis oxygnathus. Журн. эволюц. биохим. и физиол., т. VIII, № 2 : 182—187.
Константинов А. И., Стосман И. М. 1973. Сравнительная характеристика электрических ответов заднего двухолмия гладконосых и подковоносых летучих мышей в онтогенезе. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 137—146.
Константинов А. И., Стосман И. М., Горлинский И. А. 1973. Характеристики направленности приема и точность локализации ультразвука у эхолоцирующих летучих мышей. Реф. докл. IV Всес. конф, по бионике, т. IV, М. : 51—56.
Котеленко Л. М. 1968. Некоторые свойства слуховой области коры летучей мыши. Тез. докл. V научн. совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 135.
Котеленко Л. М. 1973. Реакции слуховой области коры у гладконосых летучих мышей на ультразвуковые раздражители. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 111—118.
Котеленко Л. М., Васильев А. Г. 1971. Электрические реакции слуховой области коры летучих мышей на ультразвуковые стимулы с разной частотой заполнения. Нейрофизиология, т. 3, № 5 : 526-532.
Котеленко Л. М., Матюшкин Д. П. 1969. Свойства нейронов слуховой коры летучей мыши (реакции на ультразвуки разной высоты и интенсивности). Журн. высш, нервн. деят., т. 19, вып. 3 : 502.
Кравцов Б. Г., Звонов Б. М. 1969. Зависимость суммарной реакции слухового нерва ночницы от пространственного расположения источника звука. Вести. МГУ, № 5 : 99—101.
Кравцов Б. Г., Черный А. Г., Ильичев В. Д. 1969. Зависимость суммарной реакции слухового нерва ночницы от пространственного расположения источника звука. Биол. науки, № 6 : 33—37.
Краснов П. С. 1972. Различение дельфином белобочкой шаровых мишеней по материалу и размеру. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 110—111.
Краснощекова Е. И. 1969. К морфологии слуховой области коры мозга дельфина и летучей мыши. Матер. XII конф, студентов и аспирантов морф, кафедр, Л. : 22—23.
Краснощекова Е. И. 1972. Некоторые данные по нейронной организации слуховой области коры мозга дельфина Delphinus delphis (поле Т4). Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 1, Махачкала : 238—239.
Кузин В. С., С л е з и н В. Б. 1972. Гипотеза голографического звуко-видения у китообразных. В сб.: Некоторые проблемы биологической кибернетики, Л. : 104—109.
Кузякин А. П. 1950. Летучие мыши. Изд. «Советская наука», М.
Кузякин А. П. 1958. К теории вида и видообразования. Уч. зап. Моск, обл. пед. инет., т. 65, Тр. кафедры зоол. : 21—99.
Куликов Г. А. 1968. О некоторых кортико-фугальных влияниях в слуховом анализаторе у летучих мышей и белых крыс. Тез. докл. Обще-городск. научн. конф, по биол., Л. : 40.
Куликов Г. А. 1970а. Кортикальная регуляция некоторых образований в слуховом анализаторе у летучих мышей и белых крыс. Биол. науки, № 3 : 57-60.
Куликов Г. А. 19706. К проблеме обратных связей в системе слухового анализатора. Тез. докл. IV Всес. конф, по нейрокибернетике, Ростов-на-Дону : 79.
Куликов Г. А. 1972а. Влияние коры больших полушарий на частотнопороговые характеристики электрических реакций заднего двухолмия летучих мышей. Журн. эволюц. биохим. и физиол., т. 8, № 5 : 563— 564.
467
Куликов Г. А. 19726. Реакции нейронов вентрального кохлеарного ядра подковоносых летучих мышей на ультразвуковые стимулы при удалении ипсилатеральной улитки. Нейрофизиология, т. 4, № 1 : 32—40.
Куликов Г. А. 1973а. О возможности кортикальной регуляции сенсорного потока в слуховом анализаторе летучих мышей. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 77—86.
Куликов Г. А. 19736. Физиологическая характеристика нисходящих влияний в слуховом анализаторе летучих мышей. Автореф. канд. дисс. Л.
Куликов Г. А., Самойлова Л. А. 1968. К вопросу о кортикальной регуляции в слуховом анализаторе у летучих мышей и белых крыс. Тез. докл. V научи, совещ. по эволюц. физиол., посвящ. памяти Л. А. Орбели, Л. : 142.
(Курепина М.) Kurepina М. 1967. Cytoarchitektonic des Seh-hiigels (Thalamus Dorsalis) der Chiroptera. In: Evolution of the Forebrain. Eds. R. Hassler a. H. Stephan. Plenum Press, N. Y. : 356—364.
Курепина M. M., Хачатурян E. A. 1970. Сравнительная цитоархитектоника коры кожно-двигательного анализатора летучих мышей. Уч. зап. МГПИ им. Ленина, Вопр. нервной регуляции функций, № 360 : 221—238.
Курское А. Н. 1966а. Живые радары (летучие мыши). Изд. «Наука и техника», Минск.
Ладыгина Т. Ф., Супин А. Я. 1970. Об акустической проекции в коре головного мозга дельфина. Физиол. журн. СССР, т. 56, № И : 1554-1560.
Лившиц М. С. 1973. Корреляционная гипотеза эхолокации летучих мышей. Реф. докл. VI Всес. конф, по бионике, т. IV, М. : 57—60.
Липатов Н. В. 1972. Функциональные основы подводного слуха млекопитающих. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 137—140.
Липатов Н. В., Солнцева Г. Н. 1972. Некоторые особенности биомеханики среднего уха дельфина. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 140—143.
Макаров А. К. 1973а. Эффективность эхолокационного обнаружения мишеней в присутствии шумов у летучих мышей. Тез. докл. IV Всес. конф, по бионике, т. IV, М. : 70—75.
Макаров А. К. 19736. Акустические методы и аппаратура для исследования ультразвуковой эхолокации у животных. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 77—86.
Макаров А. К., Соколов Б. В. 1970. О роли интерференции в формировании направленности эхолокационного излучения летучих мышей семейства подковоносых. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 52-53.
Малашин В. И. 1970а. Экспериментальные исследования предельных возможностей распознающей эхо локационной системы летучей мыши. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 53—54.
Малашин В. И. 19706. Теоретические и экспериментальные исследования частотно-модулированных сигналов, отраженных от объектов, распознаваемых летучей мышью. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 57.
М а н г е р А. П. 1972. Особенности строения и иннервации гортани и трахеи у некоторых китообразных. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 156—157.
Марков В. И. 1972. О тонком сканировании диаграммой направленности звукового луча при локационном зондировании подводных предметов у дельфина афалины. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 148.
468
Мастрюков В. А. 1971. Ультразвук и возможности использования его в физиологических исследованиях. Усп. физиол. наук, т. 2, № 2 : 105—125.
Марусева А. М. 1967. О временных характеристиках нейронов заднего двухолмия крыс с различным типом ответов на звуковые сигналы. В сб.: Механизмы слуха, Л. : 50—62.
Марусева А. М., Малышева В. И. 1970. Импульсная активность нейронов заднего двухолмия котят при действии звуковых сигналов. Журн. эволюц. биохим. и физиол., т. 4, № 2 : 225—234.
Матюшкин Д. П. 1969. Некоторые характеристики входа слуховой системы летучей мыши (о торможении кохлеарных нейронов сильными адекватными стимулами). Биол. науки, № 3 : 28.
Матюшкин Д. П. 1973. Электрические реакции слухового нерва летучей мыши на ультразвуки разной высоты и интенсивности. В сб.: Проблемы бионики, изд. «Наука», М. : 112—115.
Матюшкин Д. П., Васильев А. Г. 1966. Вызванные потенциалы и разряды одиночных нейронов задних холмов летучей мыши при ультразвуковой стимуляции. Матер. V Всес. конф. «Электрофизиология центральной нервной системы», Ростов-на-Дону : 25.
Матюшкин Д. П., Васильев А. Г. 1968. Биоэлектрические реакции на ультразвуки и их маскировка белым шумом в слуховой системе летучих мышей. Вести. ЛГУ, вып. 3, № 15 : 94.
Матюшкин Д. П., Котеленко Л. М. 1973. Характеристика тональных настроек слуховой коры летучей мыши. В сб.: Проблемы бионики, изд. «Наука», М. : 116—119.
Матюшкин Д. П., Стосман И. М. 1967. К характеристике нейронных сетей слухового анализатора, участвующих в определении направления приходящего сигнала. В сб.: Пробл. нейрокибернетики, Изд. Рост. унив. : 92.
Матюшкин Д. П., Стосман И. М. 1968. Чувствительность заднехолмовых нейронов летучей мыши к ультразвуковым сигналам, приходящим с разных направлений (в пространственном угле 180°). ДАН СССР, т. 178, № 4 : 973.
Матюшкин Д. П., Стосман И. М. 1969. К анализу нервного механизма, определяющего направление акустического сигнала у летучей мыши. В сб.: Механизмы нервной деятельности, Л. : 200—208.
Меринг Т. А. 1963. Роль различных звеньев слухового анализатора в замыкании двигательных пищевых условных рефлексов. Автореф. докт. дисс. М.
Меринг Т. А. 1964. О роли заднего двухолмия в условнорефлекторной деятельности. Журн. высш, нервн. деят., т. 14 : 799.
Меринг Т. А. 1967. Особенности замыкания условнорефлекторной связи. Изд. «Медицина», М.
Метелкина Р. В., Суриков Б. С. 1971. Модификация акустической линзы Люнеберга. Акуст. журн., т. 17, № 1 : 161—163.
Мовчан В. Н. 1973. Электрофизиологические исследования частотных характеристик слухового восприятия некоторых видов сонь (сем. Муо-xidae). Реф. докл. VIII Всес. акуст. конф., М., т. I : 49.
М о р з П. 1949. Колебания и звук. М.—Л.
Морозов В. П., Акопиан А. И., Бурдин В. И., Донсков А. А., Зайцева К. А., Соковых Ю. А. 1971. Аудиограмма дельфина Tursiops truncatus. Физиол. журн. СССР, т. 57, № 6 : 843-848.
Морозов В. П., Акопиан А. И., Бурдин В. И., Зайцева К. А., Соковых Ю. А. 1972. Частота следования локационных сигналов как функция расстояния до цели. Биофизика, т. 17, № 1 : 139-145.
Морозов Д. А. 1972. Принципы проектирования морских вольеров для содержания дельфинов. Тр. молодых ученых ВНИИ морск. рыб. хоз. и океаногр., вып. 5 : 81—88.
469
Мясников Б. С. 1972. Звуковые сигналы молодых белух. В сб.г Вопр. гидробиол. некоторых районов Тихого океана, Владивосток : 100—107.
Наумов Н. П. 1948. Очерки сравнительной экологии мышевидных грызунов. М.
Наумов Н. П., Симкин Г. Н. 1973. Сравнительное исследование эхолокации у летучих мышей. В сб.: Проблемы бионики, изд. «Наука», М. : 136-146.
Никитенко М. Ф. 1969. Эколого-морфологическая характеристика головного мозга рукокрылых. В сб.: Фауна и экология животных Белоруссии, изд. «Наука и техника», Минск : 32—74.
Никитенко М. Ф. 1971. Сравнительная морфология головного мозга млекопитающих в связи с их эволюцией и образом жизни. Автореф. докт. дисс. Свердловск.
Новиков Г. А., Айрапетьянц А. Э., Пукинский Ю. Б., Стрелков П. П. , Тимофеев Е. К. 1970. Звери Ленинградской области (фауна, экология и практическое значение). Изд. ЛГУ.
Огнев С. И. 1928. Звери Восточной Европы и Северной Азии. Т. 1. Насекомоядные и летучие мыши. Изд. АН СССР, М.—Л.
Павлов И. П. 1951. Поли. собр. соч., т. III, кн. 1 и 2, Изд. АН СССР, М.—Л.
Петелина Е. В. 1972. Гистологические исследования верхнеоливарного комплекса дельфина. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 169—171.
П у м п е р Е. Я. 1946. Ультразвуковая локация у летучих мышей. Наука и жизнь, № 5—6 : 9.
Радионова Е. А. 1966. Исследование временных характеристик реакций нейронов кохлеарного ядра. Тез. докл. XVIII Междунар. психол. конгр., № 15, М. : 152.
Резвов Р. Н., Савельев М. А., Ф л е н о в А. Б. 1973. Исследование возможности локатора дельфина при различении целей, отличающихся сдвигами элементов по дальности и углу в вертикальной и горизонтальной плоскостях. Реф. докл. VIII Всес. акуст. конф., т. 1, М. : 135.
Резник А. М., Скорняков В. М., Чернышов О. Б., Чу-паков А. Г. 1968. О некоторых особенностях коммуникационных сигналов дельфинов. Тез. докл. VI Всес. акуст. конф., М. : 118—121.
Резник А. М., Скорняков В. М., Чупаков А. С. 1970. Локационная активность черноморской афалины при предъявлении ей мишеней. Тр. Акуст. инет., вып. 12 : 116—120.
Резников А. Е. 1970а. О механизмах гидроакустической локации дельфина. Природа, № 9 : 103—104.
Резников А. Е. 19706. О звуковидении у дельфинов. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 67.
Резников А. Е. 1971а. «Акустические глаза» дельфина. Природа, № 10 : 56—61.
Резников А. Е. 19716. О механизмах гидроакустической локации дельфина. Природа, № 9 : 103—104.
Резников А. Е. 1971в. «Звуковидение» у Дельфина. Тез. докл. VII Всес. акуст. конф., Л. : 13.
Розенберг Л. Д. 1957. Применение ультразвука. М.
Ройтбак А. И. 1954. Биоэлектрические явления в коре больших полушарий. Сообщ. АН Груз. СССР, т. 15, № 1 : 41.
Романенко Е. В. 1964. О гидролокационной способности дельфинов. (Обзор). Акуст. журн., т. 10, вып. 4 : 385.
Романенко Е. В. 1972. Ближнее акустическое поле дельфина афалины. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 200.
470
Романенко Е. В. 1973. О механизмах излучения и формирования эхолокационных сигналов афалиной. Реф. докл. VIII Всес. акуст. конф., т. 1, М. : 131.
Романенко Е. В., Томилин А. Г., Артеменко В. А. 1965. К вопросу о звукообразовании и направленности звуков у дельфинов. В сб.: Бионика, изд. «Наука», М. : 269.
Сиверс A. IL, Суслов Н. А., Метельский В. И. 1959. Основы радиолокации. Судпромгиз, Л.
Симкин Г. Н. 1970а. Некоторые результаты исследования эхолокации у летучих мышей в Московском университете. В сб.: Пространственная ориентация животных, М. : 64—77.
Симкин Г. Н. 19706. О некоторых закономерностях развития локационного процесса у остроухих ночниц при пеленгации, распознавании и выборе объемных мишеней сложной конфигурации в условиях свободного полета. Биол. науки, № 3 : 61—71.
Симкин Г. Н. 1970в. Эхолокация, приемы распознавания и пеленгации цели у гладконосых летучих мышей. Вести. МГУ, № 1 : 111—113.
Симкин Г. Н. 1971а. Главные направления эволюции локационных систем летучих мышей. Усп. совр. биол., т. 71, вып. 1 : 123—136.
Симкин Г. Н. 19716. Приемы распознавания и пеленгации цели у гладконосых летучих мышей. Вести. МГУ, № 2 : 17—24.
Симкин Г. Н., Макаревич А. С. 1972. Эхолокация, приемы распознавания и пеленгации цели у большой ночницы. Биол. науки, № 1 : 45—56.
Симкин Г. Н., Патлякевич Н. Д. 1969а. Локационные системы летучих мышей фауны СССР. Сообщение 1. Род Myotis. Ночницы. Зоол. журн., т. 48, вып. 10 : 1526—1542.
Симкин Г. Н., Патлякевич Н. Д. 19696. Движение летучих мышей на плоские мишени и характер изменения локационных сигналов в процессе выбора и распознавания цели. Биол. науки, № 3 : 48—57.
Симкин Г. Н., Патлякевич Н. Д. 1970. Локационные системы летучих мышей фауны СССР. Сообщение 2. Род Vespertilio. Кожаны и нетопыри. Зоол. журн., т. 49, вып. 8 : 1189—1207.
Симкин Г. Н., Патлякевич Н. Д., Добрачев Ю. П. 1973. К характеристике локационных систем летучих мышей семейства подковоносов. В сб.: Проблемы бионики, изд. «Наука», М. : 146—154.
С лезин В. Б., Иваненко Ю. В., Краснощекова Е. И. 1972. Метод электростимуляции при изучении функций подкорковых структур китообразных. Краткие тез. докл. конф, молод, ученых «Применение математических методов и вычислительной техники в медицине и биологии», Л.
Слепцов М. М. 1953. Китообразные дальневосточных морей. Т. 38. Изд. ТИНРО, М.
Соколов Б. В. 1972а. Эхолокационный пространственный анализ у подковоносых летучих мышей (Rhinolophidae). Автореф. канд. дисс. Л.
Соколов Б. В. 19726. О взаимодействии слухового восприятия и эхолокации у летучих мышей семейства Rhinolophidae в процессе охоты за насекомыми. Вести. ЛГУ, сер. биол., вып. 1 : 96—104.
Соколов Б. В. 1973. Эхолокационные сигналы подковоносых летучих мышей и некоторые особенности их излучения. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 44—56.
Соколов Б. В., Макаров А. К. 1971. Направленность ультразвукового ориентационного излучения большого подковоноса и роль носовых выростов в ее формировании. Биол. науки, №’7 : 37—44.
Соколов Б. В., Соколова Н. Н. 1972. Особенности слухового восприятия у подковоносых летучих мышей (Rhinolophus ferrum-equi-num). Тез. и реф. докл. VI научи, совещ. по эволюц. физиол., Л. : 200.
471
Соколов В. Е. 1973. Систематика млекопитающих. Изд. «Высшая школа», М.
Соколов В. Е., Волкова О. В. 1971. Строение языка дельфинов. В сб.: Морфология и экология морских млекопитающих, изд. «Наука», М. : 64-72.
Соколов В. Е., Ладыгина Т. Ф., Супин А. Я. 1972. Локализация сенсорных зон в коре головного мозга дельфина. ДАН СССР, т. 202, № 2 : 490-493.
Соколова Н. Н. 1968. Влияние выключения слуховой коры мозга летучих мышей на эхолокационное поведение. Тез. докл. Общегородек. научн. конф, по биологии, Л. ; 71.
Соколова Н. Н. 1973а. Восприятие и анализ отраженных ультразвуковых сигналов малой длительности летучими мышами после удаления слуховой коры. В сб.: Вопр. сравн. физиол. анализаторов, вып. 3, Изд. ЛГУ : 154—165.
Соколова Н. Н. 19736. Измерение некоторых параметров приемной системы эхолокатора летучих мышей. Тез. докл. IV Всес. конф, по бионике, т. IV, М. : 104—109.
Солнцева Г. Н. 1971. Сравнительно-анатомические и гистологические особенности строения наружного и внутреннего уха некоторых дельфинов. В сб.: Исследования морских млекопитающих, Калининград : 226—237.
Солнцева Г. Н. 1972. Сравнительно-анатомические особенности строения среднего уха наземных, по л у водных и водных млекопитающих. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 216—220.
Солнцева Г. Н. 1973. Морфофункциональные особенности периферического отдела слухового анализатора наземных, полуводных и водных млекопитающих. Автореф. канд. дисс. М.
Солуха Б. В., Валиулина Ф. Г. 1972. К гипотезе Норриса о приеме звуковых сигналов нижней челюстью у Cetacea. В сб.: Некоторые проблемы биологической кибернетики, Л. : 109.
Стосман И. М. 1973а. Характеристика диаграммы направленности приема на различных уровнях слухового анализатора летучих мышей (слуховой нерв, кохлеарные ядра, задние холмы). В сб.: Вопросы сравн. физиол. анализаторов, вып. 3. Изд. ЛГУ : 118—129.
Стосман И. М. 19736. Пространственная чувствительность слуховой системы летучих мышей по порогам электрических реакций заднего двухолмия. В сб.: Проблемы бионики, изд. «Наука», М. : 119—122.
Стосман И. М., Константинов А. И. 1972. Характеристика вызванных потенциалов заднего двухолмия у детенышей подковоносых летучих мышей (Rhinolophidae) при действии ультразвука. Журн. эволюц. биохим. и физиол., т. 6, № 2 : 129—135.
Стосман И. М., Этманова Н. А. 1971. Характеристика электрических реакций слухового нерва и кохлеарных ядер гладконосых и подковоносых летучих мышей при изменении угла предъявления раздражителя. Физиол. журн. СССР, т. 57, № 4 : 525—531.
Супин А. Я., Сухорученко М. Н. 1970. Определение слуховых порогов дельфинов методом условной кожно-гальванической реакции (Phocaena phocaena). Тр. Акуст. инет., вып. 12 : 194—199.
Супин А. Я., Сухорученко М. Н. 1971. Исследования слуховой чувствительности дельфинов. Тез. докл. VII Всес. акуст. конф., Л. : 14.
Суховская Л. И. 1965. Нейрогистологическое исследование заднего двухолмия мозга летучей мыши. Тр. IV совещ. по эволюц. физиол., Л.
Суховская Л. И. 1966. Нейронный состав и межнейронные связи заднего двухолмия мозга летучей мыщи. Архив анат., гистол. и эмбриол., т. 51, № 12 : 33,
472
Суховская Л. И. 1968. МатериалЫ к морфологии заднего двухолмия мозга дельфина. Архив анат., гистол. и эмбриол., т. 4, № 10 : 12—16.
Суховская* Л. И. 1972. Особенности цитоархитектоники и нейронной организации заднего двухолмия мозга некоторых морских млекопитающих. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 244.
Сухорученко М. Н. 1971. Верхний предел слуха дельфинов по частоте. Тр. Акуст. инет., вып. 17 : 54—59.
Сухорученко М. Н. 1972. Условные вегетативные рефлексы дельфинов на частотно-модулированные звуковые и ультразвуковые сигналы. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 242—243.
Теоретические основы радиолокации. 1970. Ред. Я. Д. Ширман. Изд. «Сов. радио», М.
Титов А. А. 1971. Особенности звуковой сигнализации дельфинов белобочек в новых условиях. В сб.: Бионика, вып. 5, Киев : 62—67.
Титов А. А. 1972. Исследование звуковой активности и феноменологических характеристик эхолокационного анализатора черноморских дельфинов. Автореф. канд. дисс. Л.
Титов А. А., Томилин А. Г. 1970. О звуковой активности дельфинов белобочки и морской свиньи в различных ситуациях. В сб.: Бионика, вып. 4, Киев : 88—94.
Титов А. А., Томилин А. Г., Барышников Н. С., Юрке-вич Л. И., Лекомцев В. М. 1971. Коммуникационно-эмоциональные сигналы черноморских дельфинов. В сб.: Бионика, вып. 5, Киев : 67—72.
Титов А. А., Фадеева Л. М. 1971. О пространственной разрешающей способности эхолокатора дельфина. Тр. Акуст. инет., вып. 17 : 47—53.
Титов А. А., Юркевич Л. И. 1971. Физические характеристики нелокационных звуков черноморских дельфинов. В сб.: Бионика, вып. 5, Киев : 57—62.
Томилин А. Г. 1955. О поведении и звуковой сигнализации китообразных. Тр. Инет, океаногр. АН СССР, т. 18 : 45.
Томилин А. Г. 1957. Звери СССР и прилежащих стран. Китообразные. Изд. АН СССР, М.
ТомилинА. Г. 1962. Китообразные фауны морей СССР. Изд. «Наука», М.
ТомилинА. Г. 1964. Ультразвуковой прожектор и сверхдальние сигналы китов. Тр. Всес. с.-х. инет, заочного образования, вып. 17, ч. 2 : 102-108.
Томилин А. Г. 1965. История слепого кашалота. М.
Туссенель А. 1874. Нравы животных. Природа, кн. 1.
Уфимцев П. Я. 1962. Метод краевых волн в физической теории дифракции. Изд. «Сов. радио», М.
Ухтомский А. А. 1954. Собр. соч., т. 4, Изд. ЛГУ.
Ухтомский А. А. 1966. Доминанта. Изд. «Наука», М.—Л.
Фадеева Л. М. 1973. Распознавание дельфином шаровых мишеней с различнойхструктурой эхо-сигнала. Реф. докл. VIII Всес. акуст. конф., М., т. 1 : 134.
Хананашвили М. М. 1965. Дифференцирование звуковых условных раздражителей по признаку времени нарастания интенсивности и значение височной области коры полушарий в этой функции. Журн. высш, нервн. деят., т. 15, вып. 1 : 5.
Хачатурян Е. А. 1969. Сравнительная морфология кожно-двигательного анализатора летучей мыши. Автореф. канд. дисс. М.
Хоменко Б. Г. 1969. Особенности строения периферической'нервной системы жировой подушки дельфина афалины. В сб.: Вопросы морской биологии, Севастополь : 137—139.
473
Хоменко Б. Г. 1970а. Некоторые особенности гидроСтрукТуры и иннервации лобного выступа черноморских дельфинов. Бионика, вып. 4, Киев : 70.
Хоменко Б. Г. 19706. Сравнительно-морфологический анализ строения и иннервации лобного выступа дельфинов. Автореф. канд. дисс. Киев.
Хоменко Б. Г. 1972. К вопросу о морфологии некоторых черепномозговых нервов у дельфинов. Тез. докл. V Всес. совещ. по изуч. морских млекопитающих, ч. 2, Махачкала : 267—271.
Хоменко Б. Г., Нечаева О. В. 1971. Нейроморфологические основы поведения и звуковой ориентации дельфинов. В сб.: Повышение продуктивности сельскохозяйственных животных, т. 2, вып. 2, Ульяновск : 43—46.
Хоменко Б. Г., Солуха Б. В., Нечаева О. В. 1972. К вопросу о моделировании приемной системы эхолокатора дельфинов. Краткие тез. докл. к конф, молодых ученых «Применение математических методов и вычислительной техники в медицине и биологии», Л. : 74—75.
Чевненко А. А. 1969. Физические основы диагностики с помощью ультразвука. В кн.: Руководство по ультразвуковой диагностике. Ташкент.
Чемоданов Б. К., Калита Е. В., Кедров Б. И., Р е з-вов Р. Н. 1970. Аттестация целей, используемых при исследовании локационных способностей дельфинов, методами гидроакустического моделирования. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 60.
Чемоданов Б. К., Лебедев В. И., Савельев М. А., С е-ливанов Ю. И. 1970. Исследование зондирующего сигнала дельфина афалины на траектории движения к цели. Тез. докл. 23-й научно-техн. конф. ЛИАП, Л. : 61—62.
Чепурнов А. В. 1971. Биоакустика дельфинов и электросигналы рыб Черного моря. В сб.: Вопросы продукц. сан. и техн, гидробиол. южн. морей, Киев : 146—152.
Чилингирис В. И., Иваненко Ю. В. 1973. Направленность слухового приема черноморских дельфинов. Реф. докл. IV Всес. конф, по бионике, т. II, М. : 247—250.
Шелепа Д. Д. 1970. К вопросу о морфологии нервного аппарата мягкой оболочки головного мозга дельфинов. Тр. Крымск, мед. инет., т. 44 : 50-54.
Шендеров Е. А. 1972. Волновые задачи гидроакустики. Изд. «Судостроение», Л.
Ш ишкова Е. В. 1965а. Звуки черноморских дельфинов. Рыбное хоз., № 1 : 25—28.
Шишкова Е. В. 19656. Аналитический метод определения звукового давления сигналов дельфина в точке приема. Рыбное хоз., № 2 : 25—27.
Шишкова Е. В. 1967. Об акустической активности у дельфинов. Рыбное хоз., № 2 : 25.
Яблоков А. В. 1961. Функциональная морфология органов дыхания зубатых китообразных. Тр. I Всес. совещ. по изучению морских млекопитающих, Изд. АН СССР, М.
Яблоков А. В., Бель кович В. М., Борисов В. И. 1972. Киты и дельфины. Изд. «Наука», М.
Adams J. L., Burton J. A., Sitwell N. 1972. Dolphinaria in Britain. Animals, v. 14, № 1 : 10—14.
Ades H. W. 1959. Central auditory mechanisms. In: Handbook of physiology, sect. I. Neurophysiol., v. 1, Washington : 585—613.
Ades H. W., Brookhart A. 1950. The central auditory pass way. J. Neuroph., v. 13. M 3 : 189—205.
474
Alford В. R., Ruben R. I. 1963. Physiological, behavioral and anatomical correlates of the development of hearing in the mouse. Ann. OtoL, v. 72 : 237—247.
Allen G. M. 1939. Bats. Mass., Harvard Univ. Press. Cambridge.
Aites R. A. 1971. Computer derivation of some dolphin echolocation signals. Science, v. 173, № 4000 : 912—914.
Aites R. A., T i 11 e b a u m E. L. 1970. Bat signals as optimally Doppler tolerant waveforms. J. Acoust. Soc. Amer., v. 48 : 1014— 1020.
Altman I., Carpenter M. B. 1961. J. Comp. Neurol., v. 116, № 2 : 157—177.
Andersen K. 1905. On some bats of the genus Rhinolophus, with Remarks on their matual affinities, and descriptions of twentysix new forms. Proc. Zool. Soc., v. II : 75—145.
Andersen K. 1918. Diagnoses of new bats of the families Rhino-lophidae and Megadermatidae. Ann. Mag. Nat. Hist., v. II, № 10 : 374—383.
Andersen H. T. 1967. Cardiovascular adaptations in diving mammals. Amer. Heart. J., v. 74, № 3 : 295—298.
Andersen H. T. (Ed.). 1969. The biology of marine mammals. Acad. Press, N. Y.—London.
Andersen S. 1971. Orientiring hos delphiner. Nat. verden, v. 5 : 181—187.
Anderson J. W. 1954. The production of ultrasonic sounds by laboratory rats and other mammals. Science, № 119 : 108—109.
Anggard L. 1964. On the development of cochlear function in rabbit. Acta Physiol. Scand., v. 60 : 383—384.
Backus R. H., Schevill W. E. 1966. Physeter clicks. In: Whales, dolphins, and porpoises. Univ. Calif. Press, Berkeley—Los Angeles : 510-527.
Baer К. E. 1826. Die Nase der Cetaceen erlautert durch Untersuchungen der Nase des Braunfisches. Bull. Sci. Acad. St.-Petersb., Bd. 1 : 811 — 847.
Bastian I. 1967. The transmission of arbitrary environmental information between bottlenose dolphins. Animal sonar systems. Biology and bionics. T. 2. Ed. R. G. Busnel, INRA—CNRZ : 803-873.
Bateson G. 1966. Problems in cetacean and other mammalian communication. In: Whales, dolphins, and porpoises. Univ. Calif. Press, Berkeley— Los Angeles : 569—579.
Beamish P., Mitchell E. 1971. Ultrasonic sounds recorded in the presence of a blue whale Balaenoptera musculus. Deep-sea Res., v. 18, № 8 : 803—809.
Beauregard H. 1894a. Recherches sur Pappareil antitif chez les mam-miferes. J. Anat. Paris, № 30 : 367—413.
Beauregard H. 1894b. Recherches sur Uoreille interne de la rousset de 1’Inde (Pteropus medius). C. R. Acad, sci., t. 118, № 24 : 1351—1353.
Beddard F. E. 1900. A book of whales. London.
Bekesy G. J. 1951. DC potentials and energy balance of the cochlear partition. J. Acoust. Soc. Amer., № 23 : 576.
Bekesy G. J. 1953. Shearing microphonics producted by vibrations near the inner and outer hair cells. J. Acoust. Soc. Amer., № 25 : 786.
Bekesy G. J. 1956. Current status of theories of hearing. Science, № 123 : 779.
Bekesy G. J. 1963. Hearing theories and complex sounds. J. Acoust. Soc. Amer., № 35 : 588—601.
(Bekesy G., Rosenblith V. А.) Бекеши Г., Розен-б л и т В. А. 1963. Механические свойства уха. В сб.: Экспериментальная психология, т. 1, ред.-сост, Е, И» Стивенс, ИЛ : 682.
475
Birch L. М.» Dickens W., Roberts R., Diran M. 1968. Behavioral measuarements of pure tone threshold in normal. GBA— Iticc. J.‘ And. Res., v. 8, № 4 : 459—468.
Blair S. F. 1964. Perceptual theories of middle ear muscle function. Ann. Otol. Rhinol. LaryngoL, v. 73, № 3 : 724.
Boenninghans G. 1903. Das Ohr des Zahn wales, zugleich ein Beitrag zur Theorie der Schalleitung. Zool. Jahrb., Bd. 19 : 189—360.
Bondy G. 1907. Beitrage zur vergleichenden Anatomie des Gehdrorgans der Sauger. (Tympanicum, Membrana Shrapnelli und Ghordaverlauf). Ergeb. Anat. Entwicklungsgesch., Bd. 35 : 293—408.
Bourliere F. 1955. In: Grasse P. (Ed.). Traite de Zoologie. Mammi-feres, v. 17 (1 et 2 fasc.). Paris.
Bradbury J. W. 1968. Mechanisms of target discrimination by the echolocating bat Vampyrum spectrum. Ph. D. diss. The Rockefeller Univ. N. Y.
Bradbury J. W. 1970. Target discrimination by the echolocating bat Vampyrum spectrum. J. Exp. Zool., № 173 : 23—46.
Breathnach A. S. 1960. The cetacean central nervous system. Biol. Rev., v. 35, № 2 : 187—230.
Brodmann K. 1909. Vergleichende Lokalisationslehre der Grosshirn-rinde. Barth. Leipzig.
Brown A. M. 1970. Bimodal cochlear response curves in rodents. Nature (Gr. Brit), v. 228, № 5271 : 576—577.
Bullock T. H., Grinnell A. D., I к e z о n о E., К a m e d a K., Katsuki Y., Nomoto M., Sato O., Suga N., Y a n a-gisawa K. 1968. Electrophysiological studies of central auditory mechanisms in Cetaceans. Z. vergl. Physiol., Bd. 59, № 2 : 117—156.
Bullock T. H., Ridgway S. H. 1972. Evoked potentials in the central auditory system of alert porpoises to their sounds. Neurobiol., v. 3, № 1 : 79—99.
Bullock T. H., Ridgway S. H., Suga N. 1971. Acoustically evoked potentials in midbrain auditory structures in sea lions (Pinnipedia). Z. vergl. Physiol., Bd. 74, № 4 : 372—387.
Busnel R. G., Dziedzic A. 1966a. Acoustic signals of the pilot whale Globicephala malaena and of the porpoises Delphinus delphis and Phocaena phocaena. In: Whales, dolphins, and porpoises. Univ. Calif. Press, Berkeley : 607—646.
Busnel R. G., Dziedzic A. 1966b. Acoustique physiologique— caracteristiques physiques de certains signaux acoustiques du Delphi-nide Steno bredansis. C. R. Acad, sci., t. 262 : 143—146.
Busnel R. G., Dziedzic A. 1967. Resultats metrologiques experi-mentaux de UEcholocation chez le Phocaena phocaena, et leur comparison avec ceux de certaines Chauves-Souris. Animal sonar systems. Biology and bionics. Ed. R. G. Busnel. INRA-GNRZ : 307-334.
Busnel R. G., Dziedzic A. 1968. Caracteristiques physiques des signaux acoustiques de Pseudorca crassidens Owen (Cetacea, Olonto-ceti). J. Mamm., v. 32, № 1 : 1—5.
Busnel R. G., Dziedzic A., Andersen S. 1965. Seuils de perception du systeme sonar du marsouin Phocaena phocaena, et function du diametre d’un obstacle filiforme. G. R. Acad, sci., t. 260, № 1 : 295— 297.
Busnel R. G., Pilleri G., Fraser F. G. 1968. Notes concerant le dauphin Stenella styx Gray 1846. Extr. Mammalia, v. 32, № 2:192— 203.
Cahlander D. A. 1967. Discussion in «Animal sonar systems». Biology and bionics. V. 2. Ed. R. G, Busnel. INRA—GNRZ : 1052-* 1081.
475
Cahlander D. A., McCue J. J. G., Webster F. A. 1964. The determination of distance by echolocating bats. Nature, v. 201 : 544—546.
Caldwell D. K., Caldwell M. C. 1968a. The dolphin observed. Nat. Hist., v. 77, № 8 : 58—63.
Caldwell M. C., Caldwell D. K. 1968b. Vocalization of naive captive dolphins in small groups. Science, v. 159, № 3819 : 1121—1123.
Caldwell M. C., Caldwell D. K. 1970a. Echolocation-type signals by dolphins genus Inia. Sea Frontiers, v. 15, № 6 : 343—355.
Caldwell D. K., Caldwell M. C. 1970b. Echolocation-type signal by two dolphins. Genus Sotalia Quart. J. Fla Acad. Sci., v. 33, № 2 : 124-131.
Caldwell M. C., Caldwell D. K. 1972. The littlest ugly dolphin. Sea Front, v. 18, № 1 : 24—29.
Caldwell M. C., Haugen R. M., Caldwell D. K. 1962. High-energy sound associated with fright in the dolphin. Science, v. 138, № 3543 : 907—908.
Camper P. 1777. Verhandeling over de Zitplaats van det Beening Gehoor-ting. . . in de Walvischen. Verh. Holland. Maatsch., v. 17, pt. 2 : 157— 200.
Chaloupka Z., Zahlava J., Myslivecek J., Hasman-nova J. 1968. Development of functional abilities of the auditory system. In: Ontogenesis of the brain, Praha : 387—393.
Chase J. 1969. The role of vision in echolocating bats. Ph. D. Thesis, Indiana University. Bloomington, Indiana.
Chase J., Suthers R. A. 1969. Visual obstecle avoidance by echolocating bats. Anim. Behav., v. 17 : 201—207.
Clark W. E. 1932. The brain of the Insectivora. Proc. Zool. Soc. London, v. 11, № 102 : 975—1013.
Corcell a A. T., Green M. 1968. Investigation of impulsive deep-sea noise resembling sounds produced by a whale. J. Acoust. Soc. Amer., v. 44, № 2 : 483-494.
Cranbrook E., Medway L. 1965. Lack of ultrasonic frequencies in the calls of swiftlets (Collocalia spp.) Ibis, v. 107, № 2.
Crowcroft P. 1957. The life of the Shrew. London.
Crowley D. E., Hepp-Reumond M. C. 1966. Development of cochlear function in the ear of the infant rat. J. Compar. a. Physiol. Psychol., v. 62 : 427.
Cummings W. C., Fish J. F., Thompson R. O., Jehl J. R. 1971. Bioacoustics of marine mammals off Argentina. R/V Hero Cruise 71—3. Antarct. G. U. S., v. 6, № 6 : 266—268.
Cummings W. C., Thompson P. O. 1971. Underwater sounds from the blue whale, Balaenoptera musculus. J. Acoust. Soc. Amer., v. 50, № 4, pt. 2 : 1193—1198.
Cummings W. C., Thompson P. O., Cook R. 1968. Underwater sounds of migrating gray whales, Eschrichtius glaucus (Cope). J. Acoust. Soc. Amer., v. 44, № 5 : 1278—1281.
Curtis W. E. 1952. Quantitative studies of echolocation in bats (Myotis 1. lucifugus): studies of vision of bats (Myotis and Eptesicus) and quantitative studies of vision of owls. Thesis deposited in Library of Coronell Univ. Ithaca, N. Y.
Cuvier G. 1795. Conjectures sur le sixieme sens qu’on a cru remarquer dans les chauves-sourism lues a la Societe d’Historie Naturelie le 17 Ventose. Mag. encyclop., t. 6 : 297—301.
Cuvier G. 1800. Lecons d’anatomie comparee. T. 2. Paris : 581.
Cuvier G. 1805. Lecons d’anatomie comparee. T. 5. Paris.
Cuvier G. 1836, Le renge animal, T. 20. Paris,
477
Dalland J. I. 1965a. Auditory thresholds in the bat: a behavioral technique. J. Audit. Res., v. 5 : 95—108.
Dalland J. I. 1965b. Hearing sensitivity in bats. Science, v. 150 : 1185— 1186.
Dalland J. I., Vernon J. A., Peterson R. A. 1967. Hearing and cochlear microphonic potentials in the bat Eptesicus fuscus. J. Neu-rophys., v. 30, № 4 : 697—709.
Dalquest W. W., Werner H. J. 1954. Histological aspects of the faces of North American bats. J. Mamm., v. 35 : 147—160.
(Davis G.) Дэвис Г. 1963. Психофизиология слуха и его нарушений. В сб.: Экспериментальная психология, ИЛ : 615.
(Davis G.) Дэвис Г. 1964. Периферическое кодирование слуховой информации. В сб.: Теория связи в сенсорных системах, изд. «Мир», М. : 232.
Davis W. Н., В ar houг R. W., Hassell М. D. 1968. Colony behavior of Eptesicus fuscus. J. Mamm., v. 49 : 44—50.
Dice L. R., Barto E. 1952. Ability of mice of the genus Peromyscus to hear ultrasonic sounds. Science, v. 116 : 110—111.
Diercks K. J., Trochta R. T., Greenlaw G. F., Evans W. E. 1971. Recording and analysis of dolphin echolocation signals. J. Acoust. Soc. Amer., v. 49, № 6, pt. I : 1729—1732.
D i j k g r a a f S. 1943. Over een merkwaardige functie van den gehoorzin bij vleermuized. Versl. Nederland, Akad. van Wetensch. Afd. Natur., Bd. 52, № 9 : 622—627.
D i j k g r a a f S. 1946. Die Sinneswelt der Fledermause. Experientia, v. 2 : 438-448.
Dijkgraaf S. 1949a. Spallanzani und die FledermSuse. Experientia, Bd. 5: 90.
Dijkgraaf S. 1949b. Nieuws omtrent Spallanzanis vleermuizen-proef. Vakblad voor Biologen, Bd. 29 : 21—24.
Dijkgraaf S. 1957. Sinnesphysiologische Beobachtungen an Fledermau-sen. Acta physiol, et pharmacol. Neerlandica, v. 6 : 675—684.
Dobson G. E. 1878. Catalogue of the Ghiroptera in the Collection of the British Museum. Brit. Mus. (Nat. Hist.). London.
Doran A. M. G. 1878. Morphology of the mammalian ossicula auditus. Trans. Linn. Soc. London, ZooL, v. 1 : 371—498.
D r a 1 A. D. G. 1972. Aquatic and aerial vision in the bottle-nosed dolphin. Neth. J. Sea Res., v. 5, № 4 : 510—513.
Dreher J. J. 1967. Acoustic holographic model of Cetacean echolocation. In: Acoustical holography, v. I, Eds. A. E. Metherell, M. A. El-Sum, L. Larmore, Plenum Press, N. Y. : 127—137.
Dreher J. J., Evans W. E. 1964. Cetacean Communication. In: Marine Bio-Acoustic. Ed. W. Tavolga. Pergamon Press : 373—394.
Dudok van Heel W. H. 1959. Audio-direction finding in the porpoise (Phocaena phocaena). Nature, v. 183, № 4667 : 1063.
Dudok van Heel W. H. 1962. Sound and Cetacea. Netherlands J. Sea Res., № 1 : 407—507.
Dudok van Heel W. H. 1966. Navigation in Cetacea. In: Whales, dolphins, and porpoises. Berkeley—Los Angeles, Univ. Calif. Press : 597—606.
Dziedzic A. 1968. Quelgues performances des systemes de detection par echos des chauves-souris et des delphinidae. Rev. Acoust., № 1 : 23—28.
Eberhardt R. F., Evans W. E. 1962. Sound activity of the California gray whale (Eschrichtius glaucus). J. Aud. Eng. Soc., v. 10 : 324-328.
Eisentraut M. 1957. Aus demJLeben der Fledermause und Flughunde. Veb. G. Fischer Verlag, Jena.
Elias M. 1907. Zur Anatomie des Kehlkopfes der Microchiropteren, Morphol, Jahrb., v. 37 : 70-118,
478
Erulkar S. D. 1959. The responses of single units of the inferior colliculus of the cat to acoustic stimulation. Proc. Roy. Soc., v. 150, № 940 : 336.
Evans W. E., Powell B. A. 1967. Dicrimination of different metallic plates by on echolocating delphinid. In: Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 363—383.
Evans W. E., Prescott J. H. 1962. Observation of the sound production capabilities of the bottlenose porpoise: a study of whistles and cliks. Zoologica, v. 47, № 3 : 121 — 128.
Evans W. E., Sutherland W. W., Beil R. G. 1964. The directional characteristics of delphinid sounds. In: Marine Bio-Acoust. Ed. W. Tavolga. Pergamon Press, N. Y. : 353—372.
Ewer D. W. 1945. Acoustical control in the flight of bats. Nature, v. 156 : 692.
Feldman H. R. 1969. Nearly spherical acoustic lenses. J. Acoust. Soc. Amer., v. 45, № 4 : 868—871.
F i r b a s W., Welleschik B. 1973. A quantitative study on the spiral ganglion of the chiroptera. Period, biol., v. 75 : 67—70.
Fischer H. 1962. Der Kehlkopf der Fledermaus Myotis myotis, ein primiti-ver Saugerlarynx mit der Sonderleistung der Ultraschallerzeugung. Orch. Ohren-Nasen u. Kehlkopfheilkunde, Bd. 178, № 2 : 315—316.
Fischer M., V о m e 1 H. J. 1961. Der Ultraschallapparat des Larynx von Myotis myotis. Eine morphologische Studie uber einen primitiven Saugerkehlkopf. Gegenbaurs Jahrb. Morphol. u. mikroskop. Anat., Abt. 1, Bd. 102, № 2 : 200—226.
Fish M. P., Mowbray W. H. 1962. Production of underwater sound by the white whale or beluga, Delfinopterus leucas (Pallas). J. Mar. Res., v. 20, № 2 : 149—162.'
Fletcher H. 1940. Auditory Patterns. Rev. Mod. Phys., № 12 : 47—65.
FliegerE., Schnitzler H.-U. 1973. Ortungaleitungen der Fledermaus Rhinolophus ferrumequinum bei ein- und beidseitiger Ohrverstopfung. J. Comp. Physiol., v. 82 : 93—102.
Fraser F. G., Purves R. E. 1954. Hearing in cetaceans. Bull. Brit. Mus. (Nat. Hist), v. 2, № 5 : 103—116.
Fraser F. G., Purves R. E. 1959. Hearing in whales. Endeavour, v. 18 : 94-98.
Fraser F. G., Purves R. E. 1960. Hearing in cetaceans. Bull. Brit. Mus. (Nat. Hist.), v. 7 : 1—140.
(Freedman А.) Фридман A. 1963. Изображение формы тела с помощью звуколокационной или радиолокационной системы. Зарубежн. радиоэлектроника, № 8.
Freeman W. 1950. The curlew’s echo system. Field.-Naturalist, v. 202. Freeman W., Williams J. M. 1952. Human sonar: The amygdaloid nucleus in relation to auditory hallucinations. J. Nerv. Ment. Dis., v. 116 : 456—462.
Friend J. H., Suga N., Suthers R. A. 1966. Neural responses in the inferior colliculus of echolocating bats to artificial orientation sounds and echoes. J. Cell. Physiol., v. 67 : 319—332.
Frishkopf S. 1964. Excitation and inhibition of primary auditory neurons in little brown bat. J. Acoust. Soc. Amer., v. 36, № 5 : 1016.
F u 1 1 i s J. 1970. Reffered orientation in rocks produced by dolphine. Science, v. 168, № 3937 : 1344—1347.
Fuse G. 1920. Ein Beitrag zur Anatomie der centralen acustischen Bahnen beim fliegenden Hunde (Pteropus pselaphon). Arb. Anat. Inst. Kaiser.-Jap. Univ. Sendai, Bd. 5 : 71—74.
Galambos R. 1941a. The production and reception of supersonic sounds by flying bats. Summ. Theses Harvard Univ., Cambridge Mass. : 42—44. Galambos R. 1941b. Cochlear potentials from the bat. Science, v. 93 : 215.
479
Gal am bo s R. 1942a. Cochlear potentials elicited from bats by supersonic sounds. J. Acoust. Soc. Amer., v. 14, № 1 : 41—49.
Galambos R. 1942b. The avoidance of obstacles by flying bats: Spallanzanis ideas (1794) and later theories. Isis, v. 34 : 132—140.
Galambos R. 1952. Microelectrode studies on Medial geniculate body of cat. Ill Response to pure tones. J. Neuroph., v. 15, № 5 : 381.
Galambos R., Griffin D. R. 1940. The supersonic cries of bats. Anat. Rec., v. 78 : 95.
Galambos R., Griffin D. R. 1942. Obstacle avoidance by flying bats, the cries of bats. J. Exp. Zool., v. 89 : 475—490.
Gehuchten A., van. 1902. Recherches sur la voie acoustique centrale. Mem. cor. J. Acad. Roy. Relgique, № 15.
Gould E. 1955. Feeding efficiency of insectivorous.bats. J. Mamm., v. 36 : 399—407.
Gould E. 1959. Further studies on the feeding efficiency of bats. J. Mamm., v. 40, № 1 : 149—150.
Gould E. 1962. Evidence for echolocation in Shrews. Ph. D. thesis. Tulane Univ.
GouldME. Ц965. Evidence for echolocation in the Tenrecida of Madagascar. Proc. Amer. Philosoph. Soc., v. 109, № 6 : 352—360.
Gould E. 1970. Echolocation and communication in bats. In: About Bats. Eds. Slanghter a. Walton. Southern Method. Univ. Press, Dallas, Texas : 144—161.
Gould E. 1971. Studies of maternal-infant communication and development of vocalization in the bats Myotis and Eptesicus. In: Communications in behavioral biology, v. 5 : 263—313.
Gould E. N., Negus C., Novick A. 1964. Evidence for echolocation in Shrews. J. Exp. Zool., v. 156, № 1 : 19—38.
Gould J., Morgan С. T. 1941. Hearing in the rat at high frequencies. Science, v. 94 : 168.
Grasse P. P. (Ed.). 1955. Traite de zoologie, anatomie, systematique, biologie. T. XVII. Mammiferes, second fasc. Paris.
Gray A. A. 1907. The labyrinth of animals. V. 1. Churchill. London.
Griffin D. R. 1944. Echolocation in blind men, bats and radar. Science, v. 100, № 2609 : 589-590.
Griffin D. R. 1946. The mechanism by which bats produce supersonic sounds. Anat. Rec., v. 96 : 519.
Griffin D. R. 1950. Measurements of the ultrasonic cries of bats. J. Acoust. Soc. Amer., v. 22 : 147—255.
Griffin D. R. 1951. Audible and ultrasonic sounds of bats. Experientia, v. 7 : 448—453.
Griffin D. R. 1952. Mechanisms in the bat larynx for production of ultrasonic sounds. Fed. Proc., v. 11 : 59.
Griffin D. R. 1953a. Acoustic orientation in the oil bird, Steatornis. Proc. Nat. Acad. Sci., v. 39 : 884—893.
Gkr i f f i n D. R. 1953b. Bat sounds under natural conditions with evidence for the echolocation of insect prey. J. Exp. Zool., v. 123 : 456—466.
Griffin D. R. 1958. Listening in the dark. Yale Univ. Press, New Haven.
(Griffin D. R.) Гриффин Д. P. 1961. Эхо в жизни людей и животных. Изд. физ .-мат. литер., М.
G г i f f i n D. R. 1962. Comparative studies of the orientation sounds of bats. Symp. Zool. Soc. Lond., v. 7 : 61—72.
GY i f f i n D. R. 1967. Discriminative echolocation in bats. In: Animal sonar systems, v. 1. Ed. R. G. Busnel. INRA—CNRZ : 273—299.
Griffin D. R. 1971. The importance of atmospheric attenuation for the echolocation of bats (Chiroptera). Anim. Behav., v. 19 : 55—61.
Griffin D. R., Dunning D. С., C ah lan der D. A., Webster F. A. 1962. Correlated orientation sounds and ear movements of horseshoe bats. Nature, v. 186 : 1185—1186.
480
Griffin D. R., Friend I. H., Webster F. A. 1965. Target discrimination by the echolocation of bats. J. Exp. Zool., v. 158, № 2 : 155-168.
Griffin D. R., Galambos R. 1941. The sensory basis of obstacle avoidance by flying bats. J. Exp. Zool., v. 86 : 481—506.
Griffin D. R., Grinnell A. D. 1958. Ability of bats to discriminate echoes from louder noise. Science, v. 128, № 3316 : 145—147.
Griffin D. R., Hollander P. 1973. Directional patterns of bats orientation sounds. Period, biol., v. 75 : 3—6.
Griffin D. R., McCue J. J. G., Grinnell A. D. 1963. The resistance of bats to jamming. J. Exp. Zool., v. 152, № 3 : 223—250.
Griffin D. R., Novick A. 1955. Acoustic orientation in neotropical bats. J. Exp. Zool., v. 130 : 251—300.
Griffin D. R., Novick A., Kornfield M. 1958. The sensitivity of echolocation in the fruit bat, Rousettus. Biol. Bull., v. 115, № 1 : 107—113.
Griffin D. R., Suthers R. A. 1970. Sensitivity of echolocation in cave swiftlets. Biol. Bull., v. 139 : 495—501.
Griffin D. R., Webster F. A., Michael C. R. 1960. The echolocation of flying insects by bats. Anim. Behav., v. 8, № 3—4 : 141—154.
Grinnell A. D. 1963a. The neurophysiology of audition in bats: intensity and frequency parameters. J. Physiol., v. 167 : 38—66.
Grinnell A. D. 1963b. The neurophysiology of audition in bats: temporal parameters. J. Physiol., v. 167 : 67—96.
Grinnell A. D. 1963c. The neurophysiology of audition in bats: directional localization and binaural interaction. J. Physiol., v. 167 : 97—113.
Grinnell A. D. 1963d. The neurophysiology of audition in bats: resistance to interference. J. Physiol., v. 167 : 114—127.
Grinnell A. D. 1967. Mechanisms of overcoming interference in echolocating animals. Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA-CNRZ : 451—481.
Grinnell A. D. 1970. Comparative auditory neurophysiology of neotropical bats employing different echolocation signals. Z. vergl. Physiol., v. 68, № 2 : 117-153.
Grinnell A. D. 1973. Rebound Excitation (off-responses) following non-neural suppression in the cochleas of echolocating bats. J. comp, physiol., v. 82 : 179—194.
Grinnell A. D., Griffin D. R. 1958. Sensitivity of echolocation in bats. Biol. Bull., v. 114, № 1 : 10—22.
Grinnell A. D., Grinnell V. S. 1965. Neural correlates of vertical localization by echolocating bats. J. Physiol., v. 181 : 830—851.
Grinnell A. D., Hagiwara S. 1972a. Adaptations of the auditory nervous system for echolocation. Z. vergl. Physiol., v. 76 : 41—81.
Grinnell A. D., Hagiwara S. 1972b. Studies of auditory neurophysiology in non-echolocating bats, and adaptations for echdYocation in one genus, Rousettus. Z. vergl. Physiol., v. 76 : 82—96.
Grinnell A. D., McCue J. J. G. 1963. Neurophysiological investigation of the bat Myotis lucifugus. Stimulation by frequency modulated acoustical pulses. Nature, v. 198, № 4878 : 453—455.
Grosser O. 1900. Zur Anatomie der Nasenhole und des Rachens der ein-heimischen Chiropteren. Gegenbaur’s Morphol. Garb., Bd. XXIX, pt. 1.
Gross N. B., Thurlow N. R. 1951. Microelectrode studies of neural auditory activity of cat. II Med. geniculate body. J. Neuroph., v. 14, № 5 : 409.
Grummon R. A., Novick A. 1963. Obstacle avoidance in the bat, Macrotus mexicanus. Physiol. Zool. (Chikago), v. 36 : 361—369.
Hahn W. L. 1908. Some habits and sensory adaptations of cave inhabiting bats. Biol. Bull,, v. 15 : 165—193.
481
Hall J. G. 1967. Hearing and primary auditory centres of the whales. Acta Oto-Laryngol., suppl. № 224 : 244—250.
Hall J. G., J ohnsonC. S. 1972. Auditory thresholds of a killer whale Orcinus orca Lin. J. Acoust. Soc. Amer., v. 51, № 2, pt. 2 : 515— 517.
Hammelbo T. 1972. On the development of the cerebral fissures in Cetacea. Acta Anat., v. 82, № 4 : 606—618.
Harrison J. B. 1965. Temperature effects on responses in the little brown bat Myotis lucifugus. Physiol. Zool., v. 38, № 1 : 34—48.
Harrison T. 1966. Onset of echolocation in Collocalia swiftlets. Nature, v. 212, № 5061 : 530-531.
Harrison J. M., Irving R. 1966. Visual and nonvisual auditory systems in mammals. Science, № 154 : 738—742.
Harrison J. M., Warr B. 1962. A study of the cochlear nuclei and ascending auditory pathways of the medulla. J. Comp. Neurol., v. 119 : 341-379.
Hartridge H. 1920. The avoidance of objects by bats in their flight. J. Physiol., v. 54 : 54—57.
(Hartridge H.) Хартридж Г. 1946. Слуховое управление полетом у летучих мышей. Усп. совр. биол., т. 22, вып. 1 : 125.
Hatschek R., Schlesinger Н. 1902. Der Hirnstamm des Desep-hins. Arb. Neural. Just. Wiener Univ., Bd. 9 : 1—117.
H a w e 1 1 B. 1927. Contribution to the anatomy of the Chinese Tialess Por-pose, Neomeris phocaenoides. Proc. Un. St. Nation. Mus., v. 70, art. 13 : 1.
H a w e 1 1 B. 1933. Aquatic mammals. Their adaptations to life in the water. Springfield-Baltimore.
Hawen H., Gregory W. 1933. The spermaceti organ and nosal passages of the sperm whale (Physeter catodon) and other Odontoceti. Amer. Mus. Novitates, № 667.
Hawkins J. E., Stevens S. S. 1950. The masking of pure tones and speech by white noise. J. Acoust. Soc. Amer., v. 22, № 1 : 6—13.
Heel D. 1970. Dolfijn, hoe doe je het. Eds. P. N. van Kampen a. N. V. Toon. Amsterdam : 5—158.
Henson O. W., Jr. 1961. Some morphological and functional aspects of certain structures of the middle ear in bats and of insectivores. Univ. Kansas Science Bull., v. 62, № 3 : 151—255.
Henson O. W., Jr. 1964. Echolocation and hearing in bats with special reference to the function of the middle ear muscles. Ph. D. Thesis. Yale Univ.
Henson O. W., Jr. 1965. The activity and function of the middle ear muscles in echolocation bats. J. Physiol., v. 180, № 49 : 871—887.
Henson O. W., Jr. 1966. Comparative physiology of middle ear muscle activity during echolocation in bats. Amer. Zool., № 6 : 603.
Henson O. W., Jr. 1967. The perception and analysis of biosonar signals by bats. Animal sonar systems. Biology and bionics. V. 2. Ed. R. G. Busnel. INRA—CNRZ : 949—1003.
Henson O. W., Jr. 1970a. The central nervous system. In: Biology of bats. V. II. Ed. W. A. Wimsatt. Acad. Press, N. Y. : 57—152.
Henson O. W., Jr. 1970b. The car and audition. In: Biology of bats. V. II. Ed. W. A. Wimsatt. Acad. Press, N. Y. : 181—263.
Henson O. W., Eager R. P., Henson A. M. 1968. Tecto-cerebellar projections in the bat, Tadarida brasiliensis murina. Anat. Res., v. 160 : 473.
Herman P. N., Clark T. D. 1963. Post-stimulatory shift of the Preyer reflex threshold in the white rat. J. Anat. Res., v. 3 : 189—204.
H i с к 1 i n g R. 1962. Analysis of echoes from a solid elastic sphere in water. J. Acoust. Soc. Amer., v. 34, № 20 : 1582—1592.
482
Hoffmann F. 1908. Die obere Olive der Saugetiere nebst Bemerkungen uber die Lage der Cochlearisenkerne. Arb. Neurol. Inst., Wien. Univ., Bd. 14 : 76-328.
Hollmann H. E. 1952. Die Orientierungsmusik der Fledermause. Natur-wiss., Bd. 39 : 384.
Hornet Th. 1933. Vergleichend-anatomische Untersuchungen fiber das Corpus geniculatum mediale. Arb. Neurol. Inst., Wien. Univ., Bd. 35 : 76—92.
Houseknecht C. 1968. Sonographic analysis of vocalizations of three species of mice. J. Mammal., v. 49, № 3 : 555.
Huber E. 1934. Anatomical notes on pinnipedia and Cetacea. Publ. Carnegie Inst. Wash., № 447 : 105-136.
Huberg-H ansen R. 1964. Exp. Neurol., v. 9, № 3 : 212—227.
H yr tie J. 1845. Vergleichend-anatomische Untersuchungen fiber das innere Gehororgan des Menschen und der Saugetiere. F. Ehrlich. Prag.
Ikeda Y., Yokote T. 1939. Uber einige, teils bischer noch unbekan-nte Eigentiimlichkeiten in der Schnecke einer Art von Fledermaus (Rhinolophus nippon Temminck). Acta Med. Nagasak, v. 1 : 1—20.
I n u m a Y. 1958. Comparative anatomy of the body labyrinth of bats. Hirosaki Med. J., v. 9 : 106—107.
Irvine B. 1972. Behavioral changes in dolphins in a strange environment. Quart. J. Ela Acad. Sci., v. 34, № 3-4 : 206—212.
Irving R., Harrison J. M. 1967. The superior olivary complex and audition: a comparative study. J. Comp. Neurol., v. 130 : 77—86.
Iskii H., Rimura K., Harada O. 1965. Stress and auditory characteristics of the rat. Ann. Otol., Rhinol., Lar., v. 73 : 948—956.
I w a te N. 1924. Uber das Labyrinth der Fledermaus mit besonderer Beriick-sichingung des statischen Apparates. Aichi J. Exp. Med. Nagoya, v. 1 : 41—173.
Jacobs D. W. 1972. Auditory frequency discrimination in the Atlantic bottlenose dolphin Tursiops truncatus Montague. J. Acoust. Soc. Amer., v. 52, № 2, pt. 2 : 696—698.
Jacobs D. W., Hall J. D. 1972. Auditory thresholds of a fresh water dolphin Inia geoffrensis Blainville. J. Acoust. Soc. Amer., v. 51, № 2, pt. 2 : 530—533.
Jacobs M. S., Morgane P. J., McFarland W. L. 1971. The anatomy of the brain of the bottlenose dolphin (Tursiops truncatus). I. The paleocortex. J. Comp. Neurol., v. 141, № 2 : 205—272.
Jansen J. 1950. Studies on the cetacean brain. Hvalradets Scrft, № 37. Jelgersma G. 1934. Das Genirn der Wassersaugetiere. L. A. Barth. Leipzig.
J e p s en G. L. 1970. Bat origins and evolution. In: Biology of bats. V. I. Ed. W. A. Wimsatt. Acad. Press., N. Y.—London.
J о b e r t M. 1872. Etude d’anatomie comparee sur les organes due touches. Annales des sciences naturelies. Zoologie (цит. по: 3 а б у с о в, 1910). Johnson C. S. 1966. Auditory thresholds of the bottlenosed porpoise (Tursiops truncatus, Montagu). U. S. Naval Ordnance Test Station Rep (NOTS T. P. 4178).
Johnson C. S. 1967. Discussion in Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. IRNA—CNRZ : 384—386.
Johnson C. S. 1968a. Masked tonal thresholds in the Bottlenosed porpoise. J. Acoust. Soc. Amer., v. 44, № 4 : 965—967.
Johnson C. S. 1968b. Relation between absolute threshold and duration of tone pulses in the bottlenosed porpoise. J. Acoust. Soc. Amer., v. 43 : 757—763.
Johnson T. N. 1954. The superior and inferior colliculi of the mole (Scalo-pus aquaticus mechrinus). J. Comp. Neurol., v. 101, № 3 :765—799.
Jones C. 1967. Growth development and wing loading in the evening bat Nycticeus humeralis (Rafinesque). J. Mamin., v. 48 : 4—19.
483
J urine L. 1798. Experiments on bats deprived of sight by M. de Jurine. Philos. Mag., v. 1 : 136—140.
Kahmann H., Ostermann K. 1951. Wahrnehmen und Hervorbringen hoher Tone bei Kleinen Saugetieren. Experientia, v. 7 : 268—269.
Katsuki Y. 1961. Neural mechanism of auditory sensation in cats. Sensory communication, № 4. Ed. V. A. Rosenblith. London : 561.
(Katsuki Y.) Кацу к и Я. 1964. Нервные механизмы слуховых ощущений у кошки. В сб.: Теория связи в сенсорных системах, изд. «Мир», М. : 288—306.
Katsuki Y., Sumi Т., Uchiyama Н., Watanabe Т. 1958. Electric responses of auditory neurons in cat to sound stimulation. J. Neurophysiol., v. 21, № 6 : 569.
Katsuki Y., Watanabe T., Maruyama N. 1959. Activity of auditory neurons in upper levels of brain of cat. J. Neurophysiol., v. 22, № 4 : 343.
Kay L. 1961a. The orientation of bats and men by ultrasonic echolocation. Brit. Comm. Electronics, v. 8, № 8 : 582—586.
К a у L. 1961b. Reception of distance in animal echolocation. Nature, v. 190, № 4773 : 361.
Kay L. 1962. A plausible explanation of bats echolocating acuity. Anim. Behaviour., v. 10 : 34—41.
Kay L. 1967. Enhanced environmental sensing by ultrasonic waves. Discussion of Henson’s paper. Animal sonar systems. Biology and bionics. V. 2. Ed. R. G. Busnel. INRA—CNRZ : 1022—1025.
Kellogg R. 1938. Adaptation of structure to function in whales. Publ. Carneg. Inst. Wash., № 501 : 649—682.
Kellogg W. N. 1953. Ultrasonic hearing in the porpoise. J. Comp. Physiol. Psych., v. 46 : 446—450.
Kellogg W. N. 1958. Echo-ranging in porpoise. (Perception of objects by reflected sound is demonstrated for the first time in marine animals). Science, v. 128, № 3330 : 981—988.
Kellogg W. N. 1959a. Auditory perception of submarged objects by porpoise. J. Acoust. Soc. Amer., v. 31, № 1 : 1—6.
Kellogg W. N. 1959b. Sire discrimination by reflected sound in a bottlenoise porpoise. J. Comp. Physiol. Psych., № 52 : 509—514.
Kellogg W. N. 1961. Porpoises and sonar. Univ. Chicago Press.
Kellogg W. N., Kohler R. 1952. Responses of the porpoise to the ultrasonic frequencies. Science, v. 116 : 250—252.
Kellogg W. N., Kohler R., Morris H. N. 1953. Porpoise sounds as sonar signals. Science, v. 117 : 239—243.
Kellogg W. N., Raice Ch. 1966. Visual discrimination and problem solving in a bottlenose dolphin. In: Whales, dolphins, and porpoises. Ed. K. Norris. Univ. Calif. Press. : 731.
Kellogg W. N., Winthrop N. 1962. Dolphins and hearing. Nat. His., v. 71, № 2 : 30—39.
Kleesattel C. 1952. Die Ultraschallorientierung der Fledermause. Na-turwiss., Bd. 39 : 574.
Kleiman D. G. 1969. Maternal care growth rate and development in the noctule (Nyctalus noctula) pipistrelle (Pipistrellus pipistrellus) and serotine (Eptesicus serotinus) bats. J. Zool. (London), v. 157 : 187—211.
К о j i m a T. 1951. On the brain of the sperm whale (Physeter catodon L.). Sci. Rep. whales Res. Inst. Tokyo, v. 6 : 49—72.
Kolb A. 1959. Uber die Nahrungsaufnahme einheimischer Fledermause vom Boden. Zool. Anz., Bd. 22 : 162—168.
Kolb A. 1961. Sinnesleistungen einheimischer Fledermause bei der Nahrung-ssuche. Z. vergl. Physiol., Bd. 44, № 5 : 550.
Kraus C., Gihr M. 1967. Architectonical [and volumetrical investigations on the thalamus of the Cetacea in comparison with the human brain, Experientia, v. 23 ; 973—976,
484
Kraus C., Gihr M. 1968. Vergleichendmorphometrische Untersuchungen uber den Thalamus der Cetacea. J. Hirnforsch., v. 10 : 441—449.
Kraus C., Pilleri G. 1969a. Quantitative Untersuchungen fiber die Grophirnrinde der Cetaceen. In: Investigations on Cetacea. V. I. Ed. G. Pilleri, Bern : 127—150.
Kraus C., Pilleri G. 1969b. Zur Histologie der Grophirurinde von Balaenoptera borealis. In: Investigations on Cetacea. V. I. Ed. G. Pilleri. Bern : 151 — 172.
К r i g h A. 1934. Physiology of the blue whale. Nature, v. 133.
Kruger L. 1959. The thalamus of the dolphin (Tursiops truncatus) and comparison with other mammals. J. Compar. Neurol., v. Ill, № 1 : 133— 194.
Kruger L. 1966. Specialized features of the cetacean brain. In: Whales, dolphins and porpoises. Univ. Calif. Press, Berkeley: 232—254.
Kiikenthal W. 1893. Vergleichend anatomische und Entwicklungsge-schichtliche Untersuchungen an Waltiere. T. II. Jena.
Kulzer E. 1956. Flughunde erzeugen Orientierungslaute durch Zungen-schlag, Naturwiss., Bd. 43, № 5 : 117—118.
Kulzer E. 1960. Physiologische und morphologische Untersuchungen fiber die Erzeugung der Orientierungslaute von Flughunden der Gattung Rousettus. Z. vergl. Physiol., Bd. 43, № 3 : 231—268.
Kulzer E. 1962. Uber die Jugendentwicklung der Angola—Bulldogfleder-maus Tadarida (Mops) condylura (A. Smith, 1833) (Molossidae). Sau-getierkunde Mitt., Bd. 10 : 116—124.
Langworthy 0. R. 1932. A description of the central nervous system of the porpoise (Tursiops truncatus). J. Compar. Neurol., v. 54, № 2 : 537—588.
Lawrence B., Schevill W. E. 1956. The functional anatomy of the dolphinid nose..Bull. Mus. Comp. Anat., v. 114, № 4 : 103—151.
L e i d i g F. 1869. Uber die ausserem Bedeckungen der Saugetiere. Arch. Anat. Physiol, u. wiss. Mag., H. 1—6 : 677—747.
Lillie D. G. 1910. Observations on the anatomy and general biology of some members of the larger Cetacea. Proc. Zool. London : 769— 792.
Lilly J. C. 1962. Vocal behaviour of the bottlenose dolphin. Proc. Amer. Philos. Soc., v. 106, № 6 : 520-529.
Lilly J. C. 1963. Distress call of the bottlenose dolphin: stimuli and evoked behavioral responses. Science, v. 139, № 3550 : 116—118.
Lilly J. C. 1966. Sonic-Ultrasonic emissions of the bottlenose dolphin.
In: Whales, dolphins, and porposes. Berkeley—Los Angeles : 503—508.
Lilly J. C. 1968. Sound production in Tursiops truncatus (Bottlenose dolphin). Ann. N. G. Acad. Sci., v. 155, № 1 : 320—341.
Lilly J. C., Miller A. M. 1961a. Sounds emitted by the bottlenose dolphin. Science, v. 133, № 3465 : 1689—1693.
Lilly J. С., M i 1 1 er A. M. 1961b. Vocal exchanges between dolphins. Science, v. 134, № 3493 : 1873.
Lilly J. C., Miller A. M., T r u b у U. M. 1968. Reprogramming of the sonic output of the dolphin sonic Burst count matching. J. Acoust. Soc. Amer., v. 43, № 6 : 1412—1424.
Lord G. 1969. Wave analysis of a Luneberg-Gutmn fluid acoustic lens. J. Acoust. Soc. Amer., v. 45, № 4 : 885—891.
Lfitgemeier F. 1962. Histologische Sonderheiten der Gehirne von Chi’ ropteren Korpergrosse und verschiedener Orientierungsweise. Z. Mor-phol. Oekol., Tiere, v. 50 : 687—725.
Macalister A. 1872. The myology of the Chiroptera. Phil. Trans. Roy. Soc. London, Ser. B, v. 162 : 125—171.
M ai wal d D. 1967. Beziehungen zwischen Sohallspktrum,. Mithorschwell Und der Erregung des Gehore, Acustica, v. 18, № 2 : 69—80.
485
Mangold-Wirz К. 1966. Cerebralisation und Ontogenesemodus bei I Eutherien. Acta Anat., v. 63 : 449—508.
Mann G. 1960. Neurobiologia de Desmodus rotundus. Inv. Zool. Chil.r. v. 6 : 79-99.
Maruyaifla N., Kanno J. 1961. Experimental study on functionals compensation after bilateral removal of auditory cortex in cat. J. Neuro--physiol., v. 24, № 2 : 193-202.
Maxim H. 1912. The sixth sense of the bat. Sir Hiram’s contention. The possible prevention of sea collisions. Sci. Amer., v. 7 suppl. : 148—150.
McBride 1956. Evidence for echolocation by cetaceans. Deep-sea Research., v. 3 : 153—154.
McCormick J. G. 1968. Theory of hearing for delphinids. Diss. Princeton Univ.
McCormick J.G., Wever E.G., Palin J., Ridgway S. H. 1970. Sound conduction in the dolphin ear. J. Acoust. Soc. Amer., v. 48, № 6, pt. 2 : 1418—1428.
McCormick J. G., Wever E. G., Ridgway S., Palin J. 1969. Function of the porpoise ear as shown by its electrical potentials. J. Acoust. Soc. Amer., v. 47, № 1, pt. 1 : 67—68.
McCue J. J. G. 1966. Aural pulse compression by bats and humans. J. Acoust. Soc. Amer., v. 40 : 445—448.
McCue J. J. G. 1969. Signal processing by the bat, Myotis lucifugus. J. Audit. Res., v. 9 : 100—107.
Medway L. 1959. Echo-location among Collocalia. Nature, v. 184 : 1352.
Medway J. 1962. The swiftlets (Collocalia) of Nian Cave Sarawan. J. Ornit-hol., v. 104, № 2 : 227.
Medway L. 1967. The function of echonavigation among swiftlets. Anim. Behav., v. 15 : 416—420.
Miller G. 1907. The families and genera of bats. Bull. U. S. Nation.. Mus., v. 57 : 1—282.
Mizue K., Nishiwki M., Takemura A. 1971. The underwater sound of Ganges river dolphins (Platanista gangetica). Sci. Repts Whales Res Inst v 23 * 123___________128
Moffett M., Moffett R. 1971. Dolphin. Brit. Nat. Bibliogr., № 1147:: 23.
M 6 h r e s F. P. 1950. Zur Funktion der Nasenaufsatze bei Fledermausen.. Naturwiss., Bd. 37, № 22 : 526.
M 6 h r e s F. P. 1951. Uber eine neue Art von Ultraschall-Orientierung bei Fledermausen. Verh. Dtsch. Zool. Ges. : 179—186.
M 6 h r e s F. P. 1952. Die Ultraschall orientierung des Fledermause. Naturwiss., Bd. 39, № .12 : 273—279.
M 6 h r e s F. P. 1953. Uber die Ultraschallorientierung der Hufeisennasen (Chiroptera—Rhinolophinae). Z. vergl. Physiol., Bd. 34 : 547—588.
M 6 h r e s F. P. 1967a. Ultrasonic orientation in Megadermatid bats. Animal sonar systems. Biology and bionics. V. 2. Ed. R. G. Busnel. INRA — CNRZ : 115-127.
M 6 h r e s F. P. 1967b. General characters of acoustic orientation signals and performances of sonar in the order of Chiroptera. Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 401-408.
M 6 h r e s F. P. 1967c. Communicative character of sonar signals in bats. Animal sonar systems. Biology and bionics. V. 2. Ed. R. G. Busnel. INRA—CNRZ : 939—948.
Mohres F. P., Kulzer E. 1955a. Ein neue kombinierter Typ der Ultraschallorientierung bei Fledermausen. Naturwiss., Bd. 42, № 5 : 131-132.
Mohres F. P., Kulzer E. 1955b. Untersuchungen uber die Ultraschallorientierung von vier afrikanischen Fledermausfamilien. Verh. Dtsch» Zool. Ges. : 59—65.
486
Mohres F. P., Kulzer E. 1956. Uber die orientierung der Flughunde (Chiroptera—Pteropodidae). Z. vergl. Physiol., Bd. 38 : 1—29.
Mohres F. P., Kulzer E. 1957. Megaderma — ein Konvergenter Zwi-schentyp der Ultraschallpeilung bei Fledermausen. Naturwiss., Bd. 44, № 1 : 21—22.
Mohres F. P., Neuweiler G. 1966. Die Ultraschallorientierung der Grobblatt—Fledermause (Chiroptera—Megadermatidae). Z. vergl. Physiol., Bd. 53 : 195-227.
Morgane P. J. 1965. Lamination characteristics and areal differentiation in the cerebral cortex of bottlenose dolphin (Tursiops truncatus). Anat. Rec., v. 151, № 3 : 390-391.
Morgane P. J., Jacobs M. S., Jakovlev P. J., McFarland W. Z., Pijero S. J. 1969. Surface features of the brain of the bottlenose dolphin in comparison to that of five other cetacean species. J. Comp. Neurol., v. 38, № 1 : 111—121.
Morgane P. J., McFarland W. Z., Jacobs M. S., Pijero S J., Jakovlev P. J. 1966. The gross and microscope structure of the brain of the dolphin (Tursiops truncatus). Anat. Rec., v. 151, № 3 : 491.
Moris F. 1969. Etude anatomique de la region cephalique du marsouin, Phocoena phocoena. J. Mamm., v. 4 : 667—705.
M о t t a M. G. 1956. II volo cieco delo pipistrelli e le pieghe ari-epiglottiche quale organo produttore degli ultrasuoni. Atti Accad. naz. Lincei, Mem. Cl. Sci. fis.-mat. et nat., Ser. XIII, v. 5, № 3 : 83—133.
M u r i e J. 1870. Lagenorynchus albirostris. J. Linnean Soc. London.
Myrberg A. A. J. 1972. Using sound to influence the behavior of free-ranging marine animals. In: Behavior of marine animals. V. 2. N. Y.— London : 435—468.
Myslivecek J., Hasmannova J. 1963. Quelques caracteristiques des correlation diencephalocorticales ches les mammiferes nouveaunes. J. Physiol. (Paris), t. 55 : 306—307.
Negus V. E. 1929. The mechanisms of the larynx. London, Heinemann.
Ness A. R. 1967. A measure of asymmetry of the skulls of odontocete whales. J. Zool. London, v. 153 : 209—221.
Neuweiler G. 1962. Bau und Leistung des Flughundauges (Pteropus giganteus Brunn). Z. vergl. Physiol., Bd. 46 : 13—56.
Neuweiler G. 1967. Interaction of other sensory systems with the sonar system. In: Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 509—533.
Neuweiler G. 1970. Neurophysiologische untersuchungen zum Echo-ortungssystem, der Groben Hufeisennase Rhinolophus ferrum-equinum. Z. vergl. Physiol., Bd. 67 : 273—306.
Neuweiler G., Schuller G., Schnitzler H.-U. 1971. On-and Off-responses in the inferior Colliculus of the greater horseshoe bat to pure tones. Z. vergl. Physiol., Bd. 74 : 57—63.
N о i г о t E. 1965. Ultrasons et comportements chez les petits rongeurs. Ann. Soc. Roy. Zool. Belg., t. 95, № 2 : 47—56.
Nordmark J. 1960. Perception of distance in animal echolocation. Nature, v. 188, № 4755 : 1009—1010.
Nordmark J. 1961. Reception of distance in animal echolocation. Nature, v. 190, № 4773 : 363-364.
Norris K. S. 1964. Some problems of echolocation in cetaceans. Marine Bio-Acoust. Pergamon Press. Ed. W. Tavolga. N. Y. -.317—336.
Norris K. S. 1968. The evolution of acoustic mechanisms in odontocete cetaceans. In: Evolution and environment Yale Univ. Press. New Haven : 297—324.
Norris K. S. 1969. The echolocation of marine mammals. In: The biology of marine mammals. Ed. H. T. Andersen. Acad. Press. N. Y.—London : 391—423.
487
N о r r i s К. S., Ё vans W. Е. 1967. Directionality of echolocation click in the rough-tooth porpoise. Steno bredanensis. (Lesson). Marine BioAcoustic. V. 2. Ed. W. N. Tawolga. Proc. Perg. Press : 305—316.
Norris K. S., Evans W. E., Turner R. N. 1967. Echolocation in an atlantic bottlenose porpoise during discrimination. Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 409-437.
Norris K. S., Prescott J. H., Asa-Dorian P. V., Perkins P. 1961. An experimental demonstration of echolocation behavior in the porpoise, Tursiops truncatus (Montagu). Biol. Bull., v. 120, № 2 : 163—176.
Novick A. 1955. Laryngeal muscles of the bat and production of ultrasonic sounds. Amer. J. Physiol., v. 183, № 3 : 648.
Novick A. 1958a. Orientation in paleotropical bats. I. Microchiroptera. Exp. Zool., v. 138 : 81—154.
Novick A. 1958b. Orientation in paleotropical bats. II. Megachiroptera. J. Exp. Zool., v. 137, № 3 : 443—462.
Novick A. 1959. Acoustic orientation in the cave swiftlet. Biol. Bull., v. 117, № 3 : 497—503.
Novick A. 1962. Orientation in neotropical bats. I. Natalidae and Emballonuridae. J. Mamm., v. 43 : 449—455.
Novick A. 1963a. Orientation in neotropical bats. II. Phyllostomatidae and Desmodontidae. J. Mamm., v. 44, № 1 : 44—56.
Novick A. 1963b. Pulse duration in the echolocation of insects by the bat, Pteronotus. Ergebn. Biol., v. 26 : 21—26.
Novick A. 1965. Echolocation of flying insects by the bat Chilonycteris psilotis. Biol. Bull., v. 128, № 2 : 297—314.
Novick A. 1971. Echolocation in bats. Some aspects of pulse design. Amer. Sci., v. 59 : 198—209.
Novick A., Griffin D. R. 1961. Laryngeal mechanisms in bats for the production of orientation sounds. J. Exp. Zool., v. 148, № 2 : 125— 146.
Novick A., Leen N. 1970. The world of bats. New York.
Novick A., Vaisnys D. 1964. Echolocation of flying insects by the bat Chilonycteris parnellii. Biol. Bull., v. 127, № 3 : 478— 487.
Ogawa T., Arifuku S. 1948. On the acoustic system in the cetacean brains. Sci. Repts Whales Res., Inst. Tokyo, v. 2 : 1—20.
О r r R. T. 1954. Natural history of the pallid bat, Antrozous pallidus. Proc. Calif. Acad. Sci., v. 28 : 165-248.
О rr R. T. 1970. Development: prenatal and postnatal. In: Biology of bats. V. I. Ed. W. A. Wimsatt. Acad. Press, N. Y. : 217-234.
Osen K.K., Jansen J. 1965. The cochlear nuclei in the common porpoise Phocaena phocaena. J. Compar. Neurol., v. 125 : 223—258.
Paul L. J. 1969. Dolphin noises recorded by echo-sounder. New Zealand J. Marine a. Freshwater Res., v. 3, № 2 : 343—348.
Payne R. 1962. Prey location in owls. In: Biology prototypes and synthetic systems. Cornell. Univ.
(Payne R.) Пэйн P. 1965. Локация жертвы у сипухи. В сб.: Проблемы бионики, изд. «Мир», М. : 69.
Payne R. S. 1971. Acoustic location of prey by the barn owl, Tyto alba. J. Expt. Biol., v. 54, № 3 : 535—573.
Payne R. S., McVay S. 1971. Songs of Humpback whales. Science, v. 173, № 3997 : 585—597.
Peff T. C., Simmons J. A. 1972. Horizontal-angle resolution by Echolocating bats. J. Acoust. Soc. Amer., v. 51, № 6 : 2063—2065.
Perkins P. J. 1966. Communication sounds of finback whales. Norsk Hval-fangst-Tidend., v. 55, № 10 : 199-200.
488
Perkins P. J., Fish M. P., Mowbray W. H. 1966. Underwater communication sounds of sperm whale Physeter catodon. Norsk Hval-fangst-Tidende, v. 55, № 12 : 225—228.
Pierce G. W., Griffin D. R. 1938. Experimental determination of supersonic notes emitted by bats. J. Mamm., v. 19 : 454—455.
Pilleri G. 1966. Uber die Anatomie des Gehirns der Ganges Delphins, Platanista gangetica. Rev. suisse Zool., v. 73 : 113—118.
Pilleri G. 1967a. Consideration sur le cerveau et le comportement du Delphinus delphis. Rev. suisse Zool., v. 74 : 665—677.
Pilleri G. 1967b. Rehaviour of the Pseudorca crassidens (Owen) off the Spanish Medeterranean coast. Rev. suisse Zool., v. 74 : 679—683.
Pilleri G. 1970a. Investigations on cetacea brain. Anat. inst. Univ. Hern. Switzerland.
Pilleri G. 1970b. Wissenschaftliche Expedition des Berner Hirnanatomi-schen Institutes nach Westpakistan und Assam im Jahre 1969 zur Er-forschung des Gangesdelphins (Platanista gangetica). Vierteljahr. der Naturforsch. Ges. Zurich, Bd. 115, № 3 : 281—332.
Pilleri G. 1972. Einige Beobacktungen fiber Habitat und Verhalten von Platanista gangetica in Ostbengalen und Indien. Rev. suisse Zool., v. 97, № 1 : 227—233.
Pilleri G., Gihr M. 1968. On the brain of Amazon dolphin Inia geof-frensis de Blainville. 1817 (Cetacea, Susidae). Experientia, v. 124, № 2 : 932—933.
Pilleri G., Knuckey J. 1968. The distribution, navigation and orientation by the Delphinus delphis L. in the Western Mediterranean. Experimentia, v. 24, № 4 : 394—396.
Pilleri G., Knuckey J. 1969. Behaviour patterns of some delphini-dae observed in the Western Mediterranean. Z. Tierpsychol., v. 26, № 1 : 8-59.
Pollak G., Henson 0. W., Novick A. 1972. Cochlear microphonic audiograms in the «pure tone» bat, Chilonycteris parnellii parnellii. Science, v. 176 : 66—68.
Polyak S. 1926a. Untersuchungen am Octavussystem der Saugetiere and dem mit diesem Koordinierten motorischen Apparate des Hirnstammes. Z. Psychol, u. Neurol., Bd. 32 : 170—231.
Polyak S. 1926b. The connections of the acoustic nerve. J. Anatomy, v. 60 : 465—469.
Pouchet L., Beauregard N. 1885. Note sur 1’organe du spermaceti. C. r. Soc. biol., t. 2 : 342-344.
Poulter T. C. 1963a. The sonar of the Sea Lion. IEEE Trans. Ultrasonic Eng., v. 10 : 109—111.
Poulter T. C. 1963b. Sonar signals of the Sea Lion. Science, v. 139 : 753 — 755.
Poulter T. C. 1966. The use of active sonar by the California Sea Lion. J. Audit. Res., v. 6:165—173.
Poulter T. C. 1968. Marine mammals. In: Animals communication; techniques of study and results of research. Indiana Univ. Press. Bloomington : 405—465.
Poulter T. 1969. Sonar of penguins and fur seals. Proc, calif. Acad. Sci., v. 36, № 13 : 363—380.
Pruitt W., Jr. 1947. A survey of the mammalian family Soricidae (Shrews). Sonderdruck aus Saudetierkundliche Mitteilungen, Bd. 5 : 18—27.
P r u s I. 1899. Untersuchungen fiber electrische Reizunr der Vierhiidel. Wien. Klin. Wschr., Bd. 12, № 45 : 1124—1130.
Pujol R., Marty R. 1968. Structural and physiological relationships of the maturing auditory system. In: Ontogenesis of the brain. Praha : 377—385,
489
Purves P. E. 1966. Anatomy and physiology of the outer and middle ear in cetaceans. In: Whales, dolphins, and porpoises. Univ. Calif. Press. Berkeley—Los Angeles : 320—376.
Purves P. E. 1967. Anatomical and experimental observations on the cetacean sonar system. Animal sonar systems. Biology and bionics, v. 1. Ed. R. G. Busnel. INRA-CNRZ : 197-270.
Purves P. E., U n t r e c h t W. L., van. 1964. The anatomy and function of the ear of the bottlenose dolphin, Tursiops truncatus. Beaufortia,v. 3, № 9 : 241—255.
Pye A. 1964. A comparative anatomical study of the auditory apparatus of selected members from the orders Chiroptera and Rodentia. Ph. D. Thesis, London Univ.
Pye A. 1966a. The structure of the cochlea in Chiroptera. I. Microchiroptera. Emballonuroidea and Rhinolophidae. J. Morphol., v. 118, № 4 : 495— 511.
P у e A. 1966b. The structure of the cochlea in Chiroptera. II. Megachiroptera and Vespertillionoidea of the Microchiroptera. J. Morphol., v. 119 : 101 — 109.
Pye A. 1967. The structure of the cochlea in Chiroptera. III. Microchiroptera. Phyllostomatoidea. J. Morphol., v. 121 : 241—254.
P у e A. 1970. The structure of the cochlea in Chiroptera. A selection of Microchiroptera from Africa. J. Zool. Lond., v. 162 : 335—343.
Pye A. 1973. The structure of the cochlea in chiroptera from Africa. Period, biol., v. 75 : 83-87.
Pye J. D. 1960. A theory of echolocation by bats. J. Laryngol. Otol., v. 74 : 718—729.
Pye J. D. 1961a. Echolocation by bats. Endeavour, v. 20, № 78 : 101—111.
Pye J. D. 1961b. Reception of distance in animal echolocation. Nature, v. 190, № 4773 : 362—363.
Pye J. D. 1963. Mechanisms of echolocation. Ergebn. Biol., v. 26 : 12—20.
Pye J. D. 1967. Synthesizing the waveforms of bat’s pulses. Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 43-64.
Pye J. D. 1968a. Hearing in bats. In: Hearing mechanisms in vertebrates. Ed. A. V. S. de Reuck a. J. Knight. Churchill Ltd. London : 66—84.
Pye J. D. 1968b. Animal sonar in air. Ultrasonics., v. 6 : 32—38.
Pye J. D. 1971. Bats and fog. Nature, v. 229 : 572—574.
Pye J. D. 1972. Bimodal distribution of constant-frequencies in some Hippo-siderid bats (Mammalia : Hipposideridae). J. Zool. Lond., v. 166 : 323—335.
Pye J. D. 1973. Echolocation by constant frequency in bats. Period, biol., v. 75 : 21—26.
Pye J. D., Flinn M., Pye A. 1962. Correlated orientation sounds and ear movements of horseshoe bats. Nature, v. 186 : 1186—1188.
Pye J. D., Roberts L. H. 1970. Ear movements in a Hipposiderid bat. Nature, v. 225 : 285—286.
Quay W. B. 1970. Peripheral nervous system. In: Biology of bats. V. II. Ed. W. Wimsatt. Acad. Press. N. Y. : 153—179.
Ralls K. 1967. Auditory sensitivity in mice: Peromyscus and Mus musculus. Anim. Behav., v. 15, № 1 : 123—128.
Ramon у Cajal. 1902. Textura del systema nervoso del hombre у dex los vertebrates. Fasc. 6. Madrid.
Ramon у Cajal. 1909—1911. Histologie du systeme nerveux de 1’homme et des vertebres. Paris.
R a witz B. 1900. Die Anatomie des Kehlkopes und der Naze von Phocaena commons. Intern. Monatsschr. Anat. Phys., Bd. 14 : 468.
R e d t e 1 H. S. 1873. Der Nasenaufsats des Rhinolophus hippocrepis. Z. wiss, Zool., Bd. 23 : 254—288,
490
Reed С. A. 1944. Behaviorof a shrew note in captivity. J. Mammal., v. 25* : 186—198.
Rehman L R. 1964. Porpoise aids research. Undersea Techn. Mag., v. 2 : 36—39..
Revel I. P. 1962. The sarcoplasmic reticulum of the bat cricothyroid muscle. J. Cell. BioL„ v. 12: 571-580.
Reysenbach de Haan F. W. 1957a. De gehoorzin van Cetacea. Vakbl. biol., t. 37, № 8 : 117—127.
Reysenbach de Haan F. W. 1957b. Hearing in whales. Acta Otolaryngol., suppl. № 134, Stockholm : 1—114.
Richard R. J., Henry H. E., Bruce M. 1969. Hearing in primitive> mammals. Il Hedgehog (Hemiechinus auritus). J. Aud. Res., v. 9, № 1 : 8-11.
Ridgway S. H., McCormick J.G. 1967. Anesthetization of porpoises for major surgery. Science, v. 158, № 3800 : 510—512.
R i e s e W. 1927. Uber den Ban und die Leistungen des akustischen Systems der Whale. J. Psychol. Neurol., v. 34 : 194—201.
Riley D. A., Rosenzweig M. 1957. Echolocation in rats. J. Comp. Physiol. Psych., v. 50 : 323—328.
Roberts L. H. 1972. Variable resonance in constant frequency bats. J. Zool. bond., v. 166 : 337-348.
Roberts L. H. 1973. Cavity resonances in the production of orientation cries. Period biol., v. 75 : 27—32.
Robin H. 1881. Recherches anatomiques sur les mamiferes de 1’ordre des chiropteres. Ann. Sci. Nat., v. 12 : 110.
Roeder K. 1963. Echoes of ultrasonic pulses from flying moths. Biol. Bull., v. 124 : 200-210.
(Roeder К.) Редер К. 1965. Взаимодействие между летучими мышами и ночными бабочками при помощи ультразвука. В сб.: Проблемы бионики, изд. «Мир», М. : 75.
Roeder К. D., Treat А. Е. 1961. The detection of bat cries by moths. In: Sensory communication. Ed. W. A. Rosenblith. N. Y.—London. M. I. T., Techn. Press.
(Roeder K., Treat А.) Редер К., Трит A. 1964. Восприятие звуков, издаваемых летучими мышами, тимпанальным органом ночных бабочек. В сб.: Теория связи в сенсорных системах, изд. «Мир», М. : 308.
Rollinat R., Trouessart Е. 1900. Sur le sens de la direction chez chauves-souris. C. r. Soc. Biol., t. 52, Paris : 604—607.
Rose M. 1912. Histologische localisation der grosshirnrinde bei kleinen Saugel-tieren (Rodentia, Insectivora, Chiroptera). J. Psychol., Neurol., v. 19 : 1-119.
Rose M. 1926. Der Grundplan der Cortextektonik beim Delphin. J. Psychol. Neurol., v. 32.
Rose J. E., Galambos R., H u g e s J. R. 1959. Microelectrode studies of the cochlear nuclei of the cat. Jonhs Hopk. Ins. Hosp. Bull., v. 104, № 5 : 211.
Rose J. E., Greenwood D. D., Goldberg I. M., Hind . E., 1963. Some discharge characteristics of single neurons in the inferior colliculus of the cat. J. Neurophysiol., v. 26 : 294.
Rosenblith W. A., Kahana L., Galambos R. 1950. Effect of temperature-change upon round window response in hamster and bat. Fed. Proc., v. 9 : 109.
Rosenzweig M. R., Riley D. А., К r e c h K. 1955. Evidence for echolocation in the rat. Science, v. 121 : 600.
Rosevear D. R. 1965. The bats of West Africa. Brit. Mus. (Nat. Hist.) London.
491
S c h arv e г H. N., Poulter T. C. 1967. Sea Lion echo ranging. J. Acoust. Soc. Amer., v. 42, № 2 : 428—437.
Scheffer V. B. 1962. Pelage and surface topography of the northern pur seal. N. Amer. Fauna, v. 64 : 1.
Scheibel I., Scheibel A., Mollica A., Moruzzi G. 1955. Convergence and interaction of offerent impulses on single units of reticular formation. J. Neurophysiol., v. 18, № 4 : 309—331.
Schevill W. E. 1964. Underwater sounds of cetaceans. In: Marine Bio-Acoust. Pergamon Press, N. Y. : 307—316.
Schevill W. E., Lawrence B. 1949. Underwater listening to the white porpoise (Delphinapterus leucas). Science, v. 109 : 143—144.
Schevill W. E., Lawrence B. 1953a. High frequency auditory response of a bottlenosed porpoise. J. Acoust. Soc. Amer., v. 25, № 5 : 1016-1017.
Schevill W. E., Lawrence B. 1953b. Auditory response of a bottlenosed porpoise, Tursiops truncatus, to frequencies above 100 kc. J. Exp. Zool., v. 124 : 147—165.
Schevill W. E., Lawrence B. 1956. Food-finding by a captive porpoise (Tursiops truncatus). Breviora, Mus. Comp. Zool., Harvard, v. 52 : 1-15.
Schevill W. E., Watkins W. A. 1962. Whale and porpoise voices, a phonograph record. Woods Hole Oceanogr. Inst.
Schevill W. E., Watkins W. A. 1966. Sound structure and directionality in Orcinus (Killer whale). Zoologica, v. 51, № 2 : 71—76.
Schevill W. E., Watkins W. A., R а у C. 1963. Underwater sounds of Pinnipeds. Science, v. 141 : 50—53.
Schleidt W. 1948. Tone hoher Frequenz bei Mausen. Experientia, Bd. 4 : 145—146.
Schleidt W. 1951. Tone hoher Frequenz bei Mausen (Zweite Mitteilung). Experientia, Bd. 7 : 65—66.
Schleidt W. 1952. Reaktionen auf Tone hoher Frequenz bei Nagern. Naturwiss., Bd. 39 : 60—70.
Schleidt W. 1965. Personal communication.
Schneider H. 1957. Morphologische Untersuchungen am Gehirn der Chiroptera. (Mammalia). Abhandl. Senckenberg, Nat. Ges., № 495 : 92.
Schneider H. 1961. Die Ohrmuskulature von Asellia tridens Geoffr. (Hipposideridae) und Myotis myotis Borkh. (Vcspertilionidae). Zool. Jahresber., Bd. 79 : 93—122.
Schneider H., Mohres F. P. 1960. Die Ohrbewegungen der Hufeisen fledermause (Chiroptera, Rhinolophidae) und der Mechanismus des Bild-horens. Z. vergl. Physiol., Bd. 44 : 1—40.
Schnitzler H. U. 1967a. Discrimination of thin wires flying Horseshoe bats (Rhinolophidae). Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA-CNRZ : 69-87.
Schnitzler H. U. 1967b. Kompensation von Dopplereffekten bei Hufei-sen-Fledermausen. Naturwiss., Bd. 54 : 523.
Schnitzler H. U. 1968. Die Ultraschall-Ortungslaute der Hufeisen-Fledermause (Chiroptera—Rhinolophidae) in verschiedenen Orientierungs-situationen. Z. vergl. Physiol., Bd. 57 : 376—408.
Schnitzler H. U. 1970a. Echoortung bei der Fledermaus Chilonycteris rubiginosa. Z. vergl. Physiol., Bd. 68 : 25—39.
Schnitzler H. U. 1970b. Comparison of the echolocation behavior in Rhinolophus ferrum-equinum and Chilonycteris rubiginosa. Bijdr. Dierk., Bd. 40 : 77-80.
Schnitzler H. U. 1971. Fledermause in Windkanal. Z. vergl. Physiol., Bd. 73 : 209—221.
Schnitzler H. U. 1973a. Control of Doppler shift compensation in the greater horseshoe bat, Rhinolophus ferrum-equinum. J. Comp. Physiol., v. 82 : 79-92.
492
Schnitzler H. U. 1973b. Die Echoortung der Fledermause und ihre horphysiologischen Grundlagen. Sonderdruck aus «Fortschritte der Zo-ologie». Bd. 21, Ht. 2/3. Gustav Fischer. Verlag Stuttgart : 136—189.
Schnitzler H. U., Schuller G., Neuweiler G. 1971. Ant-worten des Colliculus inferior der Fledermaus Rhinolophus euryale auf tonale Reizung. Naturwiss., Bd. 58 : 627.
S c h о b e r W. 1959—1960. Zur Cytoarchitektonik und Cytologie der Medulla oblongata der Mausohr-Fledermaus, Myotis myotis. (Borkhausen) Wiss. Z. Karl-Marx-Univ. Leipzig, Math.-Naturwiss., Reihe 9 : 745—771.
S c h 6 b 1 J. 1871. Die Flughaut der Fledermause namentlich die Endigung ihrer. Nerven Arch. Mier. Anat., Bd. 7 : 1—31.
S c h о u t e n J. F., R i t s m a R. J., Cardoso B. G. 1962. J. Acoust. Soc. Amer., v. 34 : 1418.
Schuller G. 1972. Echoortung bei Rhinolophus ferrum-equinum mit frequenz-modulierten Lauten. Evoked Potentials in Colliculus inferior. J. Com. Physiol., v. 77 : 306—331.
Schuller G., Neuweiler G., Schnitzler H.-U. 1971. Col-licular responses to the frequency modulated final part of echolocation sounds in Rhinolophus ferrum-equinum. Z. vergl. Physiol., Bd. 74 : 153—155.
Schusterman R. J. 1965. Visibility and reineorcement as variables ine-luecing the underwater click vocalizations of a California sea lion. Amer. Zool., v. 5 : 329.
Schusterman R. J. 1966. Underwater click vocalizations by a California sea lion: effects of visibility. Psychol. Rec., v. 16 : 129—136.
Schusterman R. J. 1967. Perception and determinants of underwater vocalization in the California sea lion. Animal sonar systems. Biology and bionics, v. 1 : 535—617.
Schusterman R. J., R. Gentry, J. Sc h moo k. 1966. Underwater vocalization by sea lions: social and mirror stimuli. Science, v. 154 : 540-542.
Schusterman R. J., R. Gentry, J. Schmook. 1967. Underwater sound production by captive California sea lions, Zalophus cali-fornianus. Zoologica, v. 52 : 21—24.
Schwartzkopff J. 1955. On the hearing of birds. Auk, v. 72.
Schwartzkopff J. 1962. Zur Frage des Richtungshorens von Eulen (Striges). Z. vergl. Physiol., Bd. 45.
Sewell G. D. 1970. Ultrasonic signals from rodents. Ultrasonics, v. 8, № 1 : 26—30.
Shaver H. N., Poulter T. C. 1967. See lion echo ranging. J. Acoust. Soc. Amer., v. 42, № 2 : 428—437.
Shillito J. 1961. Observations on range and movements of shrews. Animal. Behav., v. 9 : 237—238.
Sgonina K. 1935. Der Spallanzanische Fledermausversuch. Zool. Anz., v. 109 : 325-327.
Simmons J. A. 1968a. Depth perception by sonar in the bat Eptesicus fuscus. Ph. D. diss. Princeton Univ.
Simmons J. A. 1968b. Echolocation: auditory cues for range perception in bats. Proc. 76th Ann. Convention, APA : 301—302.
Simmons J. A. 1968c. The sonar sight of bats. Psychol, today. November : 50-57.
Simmons J. A. 1969. Acoustic radiation patterns for the echolocating bats Chilonycteris rubiginosa and Eptesicus fuscus. J. Acoust. Soc. Amer., v. 46 : 1054—1056.
Simmons J.A. 1971a. Echolocation in bats: signal processing of echoes for target range. Science, v. 171, № 3974 : 925—928.
Simmons J.A. 1971b. Narrow band cw/fm echolocation in bats. J. Acoust. Soc. Amer., v. 50 : 148.
493
Simmons J. A. 1973. Response of the Doppler sonar system in the bat, Rhinolophus ferrum-equinum. J. Acoust. Soc. Amer., v. 53.
Simmons J. A., Howell D. J. 1973. Multiple sonar systems in bats. Period, biol., v. 75 : 33—35.
Simmons J. A., Vernon J. A. 1971. Echolocation: discrimination of target bv the bat, Eptesicus fuscus. J. Exp. Zool., v. 176, № 3:315— 328.
Simpson G. G. 1945. The principles of classification and classification of mammals. Bull. Amer. Mus. Nat. Hist., v. 85 : 1—350.
S 1 i j p e r E. J. 1962. The whales. London.
Smith G. 1902. Elliot notes on the brain of Macrocelides and other Insecti-vora. J. Linn. Soc. Zool., v. *28.
Smith J. D. 1972. Systematics of the chiropteran family Mormoopidae. Univ. Kansas Mus. Nat. Hist., № 56 : 3—139.
Smith W. J. 1969. Messages of vertebrate communication. Science, v. 165, № 3889 : 145-150.
Spallanzani L. 1932. Lettere sopra il sospetto d’un nuovo senso nei Pipistrelli di Lazzaro Spallanzani. 3. Millan, 1793.
S p о n g P., White D. 1971. Visual acuity and diseri mination learning in the dolphin (Lagenorhynchus obliquidens). Exp. Neurol., v. 31, № 3 : 431-436.
Spotte S. H. 1967. Intergeneric behavior between captive Amazon river dolphins Inia and Sotalia. Underwater Natur., v. 4, № 2 : 9—13.
Stanek V. J. 1933. К Topograficke a Srovnavaci Anatomii Sluchoveho Organu Nasich Chiroptar. Nakladem Ceske Akad. Ved. a umeni. Prague.
Stef anelli A. 1944a. La morphologia dell’apparato Statico e dell’apparato acoustico del Chirotteri in relazione allo condizioni di vollo. Pontif. Acad. Sci., Acta, t. 8 : 1—5.
Stefanelli A. 1944b. Considerazioni morfo-ecologiche sull’apparato acu-stico e statico del Microchirotteri. Mem. Acad. Italy, t. 14 : 805—816.
Stieda L. 1872. Die angeblichen Terminal-Korperchen an den Haaren einiger Saugetiere. Arch. mikr. Anat., Bd. 8 : 274—278.
Strother G. K. 1961. Note on the possible use of ultrasonic pulse compression by bats. J. Acoust. Soc. Amer., v. 33, № 5 : 696—697.
Strother G. K. 1967. Comments on aural pulse compression in bats and humans. J. Acoust. Soc. Amer., v. 41 : 529.
Strother G. К., M о g u s M. 1970. Acoustical beam patterns for bats: some theoretical considerations. J. Acoust. Soc. Amer., v. 48, № 6, pt. 1 : 1430-1432.
Suga N. 1964a. Recovery cycles and responses to frequency modulated tone pulses in auditory neurons of echo-locating bats. J. Physiol., v. 175, № 1 : 50-80.
Suga N. 1964b. Single unit activity in cochlear nucleus and inferior colliculus of echolocating bats. J. Physiol., v. 172, № 3 : 449—474.
Suga N. 1965a. Functional properties of auditory neurones in the cortex of echolocating bats. J. Physiol., v. 181 : 671—700.
Suga N. 1965b. Analyses of frequency-modulated sounds by auditory neurones of echolocating bats. J. Physiol., v. 179, № 1 : 26—53.
Suga N. 1965c. Responses of cortical auditory neurons to frequency modulated sounds in echo-locating bats. Nature, v. 206 : 890—891.
Suga N. 1967. Discussion of Henson’s paper (1967a). In: Animal sonar systems. V. 2. Ed. R. G. Busnel. INRA-CNRZ : 1004—1020.
Suga N. 1968. Analysis of frequency-modulated and complex sounds by single auditory neurons of bats. J. Physiol., v. 198 : 51—80.
Suga N. 1969a. Echo-location of bats after ablation of auditory cortex. J. Physiol., v. 203 : 729-739.
Suga N. 1969b. Echo-location and evoked potentials of bats after ablation of inferior colliculus. J. Physiol., v. 203, № 3 : 707—728.
494
Suga N. 1973. Neurophysiological analysis of echolocation in bats. In: Animal orientation and navigation : 341—353.
Suga N., Griffin D. R. 1964. Single unit activities in the cochlear nucleus and inferior colliculus of echolocating bats. Fed. Proc., v. 23, pt. 2, № 1 : 414.
Suga N., Schlegel P. 1972a. Neural attenuation of responses to emitted sounds in echolocating bats. Science, v. 177 : 82—84.
Suga H., Schlegel P. 1972b. Echolocating bats: vocalization evoked by electrical stimul and responses of inferior collicular neurons to emitted sounds. Fed. Proc., v. 31, № 2 : 359.
Suthers R.A. 1965. Acoustic orientation by fish catching bats. J. Exp. Zool., v. 158, № 3 : 319-348.
Suthers R. A. 1967. Comparative echolocation by fishing bats. J. Mamm., v. 48, № 1 : 79—87.
Suthers R.A. 1970a. Vision, Olfaction, Taste. In: Biology of bats. v. II. Ed. W. A. Wimsatt. Acad. Press, N. Y. : 265—309.
Suthers R.A. 1970b. A comment on the role of choroidal papillae in the fruit bat retina. Vision Res., v. 10 : 921—922.
Suthers R. A., Chase J., Braford B. 1969. Visual form discrimination by echolocating bats. Biol. Bull., v. 137 : 535—546.
Suthers R. A., Thomas S. P., Suthers В. J. 1972. Respiration, wing-beat and ultrasonic pulse emission in an echo-locating bat. J. Exp. Biol., v. 56 : 37-48.
Tamura I. 1950. Comparative anatomical studies on the brain stem, with special references to the reticular formation and its relating nuclei of Chiroptera. Acta Anat. Niigata, Ensia Niigata, v. 50 : 65—98.
Taniguchi H. 1959. A comparative anatomical study on the cochlear nucleus in the rodent and the Chiroptera. Bull. Kobe Med. Coll., v. 17 : 483-514.
Tepaske E. R. 1964. Some morphological aspects and taxonomic relationships of the middle ear of bats. Ph. D. Thesis. Oklahoma State Univ.
Thompson R. F., Sindberg R. M. 1960. Auditory response fields in association and motor cortex of cat. Neurophysiol., v. 13, № 1 : 87— 105.
Thorpe W. H. 1961. Bird-song. Cambridge.
Treat A. E. 1955. The response to sound of certain Lepidoptera. Ann. Ent. Soc. Amer., v. 48 : 272—284.
Turkewitsh B. G. 1933a. Vergleichend-anatomische Untersuchungen uber das Rnoeherne Labvrinth der Saugetiere (Ampullae). Anat. Anz., Bd. 76 : 206-214.
Turkewitsch B. G. 1933b. Zur Anatomic des Gehororganes der Saugetiere (Canales semicirculares). Anat. Anz., Bd. 76 : 369—408.
Turkewitsch B.G. 1935. Beitrage zur vergleichenden Anatomie der Cochlea der Saugetiere. (Aquaeductus cochlea et Fenestra cochleae). Anat. Anz., Bd. 78 : 155-166.
Turkewitsch B. G. 1937. Charakteristische Eigentiimlichkeiten der anatomischen Struktur des Knochernen inneren Ohres bei den Vertratern der Familie Rhinolophidae. Zool. Jahresb., Bd. 63 : 471—482.
Turner W. 1913. Observations on the auditory organ in Cetacea. Proc. Roy. Sci. Edinburgh, v. 34 : 10—26.
(Turner R.) T у p н e p P. 1962. Методы изучения китообразных. В сб.: Морская биоакустика, изд. «Судостроение», Л. : 362—379.
Turner R. N., Norris К. S. 1966. Discriminative echolocation in a porpoise. J. Exp. Analysis Behav., v. 9, № 5 : 535—544.
Thurlow W. K., Small A. H. 1955. Pitch perception for certain periodic auditory stimuli. J. Acoust. Soc. Amer., v. 27 : 132.
Vale ton M. T. 1908. Beitrag zur vergleichenden Anatomie des hinteren Vierhugels des Menschen und einigen Saugetiere. Arb, Neurol. Inst. Wiener Uniy., Bd. 14 : 29—75r
495
Vernon J. A., Dalland J. I., Wever E. G. 1966. Further studies of hearing in the bat Myotis lucifugus, by means of cochlear potentials. J. Audit. Res., v. 6 : 153—163.
Vernon J. A., Peterson E. A. 1965. Echolocation signals in the freetailed bat, Tadarida mexicana. J. Audit. Res., № 5 : 317—330.
Vernon J. A., Peterson E. A. 1966. Hearing in the vampire bat, Desmodus rotundus murinus, as shown by cochlear potentials. J. And. Res., v. 6 : 181—187.
Vincent F. 1963. Acoustic signals for auto-information or echolocation. In: Acoustic behavior of animals. Ed. R. G. Busnel. Amsterdam : 183— 227, 789-793.
Walker E. 1964. Mammals of the World. V. I. Johns Hopkins Press. Baltimore, Maryland.
Walker R. A. 1964. Some widespread high level underwater noise pulses of apparent biological origin opp Gape God. Marine Bio-Acoust. Perg. Press, N. Y. Ed. W. Tavolga : 121—124.
Walls G. L. 1942. The vertebrate eye. Michigan.
W a s s i f K. 1946. The processus muscularis and the tensor Tympani muscle of bats. Nature, v. 157 : 877—878.
W a s s i f K. 1948. Studies on the structure of the auditory ossicles and tympanic bone in Egyptian insectivora. Chiroptera and Rodentia, № 27 : 177—213.
W a s s i f K. 1950. The tensor Tympani Muscle of bats. Ann. Mag. Nat. Hist., v. 12 : 811—812.
Watkins W. A. 1967. Air-bone sounds of the Hympback whale Megaptera novaengliae. J. Mamm., v. 48, № 4 : 573.
Watkins W. A., S c h e v i 11 W. E., Ray G. 1971. Underwater sounds of Monodon (Narwhal). J. Acoust. Soc. Amer., v. 49, № 2, pt. 2 : 595—599.
Weber M. 1928. Die Saugetiere Gustav Fischer. 2. Jena.
Webster D. B. 1966. Ear structure and function in modern mammals. Amer. Zool., v. 6, № 3 : 451—466.
Webster F. A. 1967. Interception performance of echolocating bats in the presence of interference. Animal sonar systems. Biology and bionics. V. 1. Ed. R. G. Busnel. INRA—CNRZ : 673—712.
Webster F. A. 1968. Some acoustical deferences between bats and men. Proc. Int. Conf. Sensory devices for the blind. London : 63—87.
Wever E. G. 1966. Electrical potentials of the Cochlea. Physiol. Rev., v. 46, № 1 : 102.
Wever E. G., McCormick J. G., Palin J., Ridgway S. H. 1971a. The cochlea of the dolphin, Tursiops truncatus: hair cells and ganglion cells. Proc. Nation. Acad. Sci. USA, v. 68, № 12 : 2908—2912.
Wever E. G., McCormick J. G., Palin J., RidgwayS. H. 1971b. The cochlea of the dolphin Tursiops truncatus: general morphology. Proc. Nation. Acad. Sci. USA, v. 68, № 10 : 2381—2385.
Wever E. G., McCormick J. G., Palin J., Ridgway S. H. 1971c. Cochlea of the dolphin Tursiops truncatus: the basilar membrane. Proc. Nation. Acad. Sci. USA, v. 88, № 11 : 2708—2711.
Wever E. G., McCormick J. G., Palin J., Ridgway S. H. 1972. Cochlear structure in the dolphin Lagenorhynchus obliquidens. Proc. Nation. Acad. Sci. USA, v. 69, № 3 : 657—661.
Wever E. G., Vernon J. A. 1961a. The protective mechanisms of the bats ear. Ann. OtoL, et St. Louis, v. 70 : 1—13.
Wever E. G., Vernon J. A. 1961b. Hearing in the bat, Myotis lucifugus as shown by the cochlear potentials. J. Audit. Res., v. 2 : 158—175.
Wever E. G., Vernon J. A., Peterson E. A. 1963. The high-frequency sensitivity of the Guinea pig ear. Proc. Nat. Acad. Sci., v. 49, № 3 : 319-322.
Whitaker A. 1906. The development of the sence in bats. Naturalist (London) : 145-151.
496
Williamson R. G. 1969. Echo phase-comparison technique and measurement of sound velocity in water. J. Acoust. Soc. Amer., v. 45, № 5 : 1251_________1257.
Winn H. E., Olla B. L. (Eds). 1972. Behavior of Marine animals. V. 2. Vertebrate. Plenum Press. N. Y.—London.
W i r z K. 1954. Ontogenese and Cerebralisation bei Eutheria. Acta Anat., v. 20 : 318—329.
Yamada M. 1953. Contribution to the anatomy of the organ of hearing of whales. Sci. Rep. Whales Res. Inst., № 8 : 1—79.
Zimmerman K. W. 1926. Wie werden die Harchen der Horzellen erregt. Schweiz, Med Wochenschr., v. 56 : 566—569.
Zippelius H. M., Schleidt W. M. 1956. Ultraschall-laute beu jungen Mausen. Naturwiss., Bd. 43 : 502.
Zunino D. G. 1911. Sulla citoarchitectonica della corteccia cerebrale dei Microchirotteri. Arch. I tai. Anat. Embriol., t. 10 : 145—167.
(Z w i с к e r E., Feldtkeller R.) Цвикер Э., Фе л ьдке л лер Р. 1971. Ухо как приемник информации. Изд. «Связь», М.
Z w i с k е г Е., Flottorp S., Stevens S. S. Critical band width in loudness summation. J. Acoust. Soc. Amer., v. 29, № 2 : 548—557
ПРИЛОЖЕНИЕ
Работы по эхолокации животных, опубликованные сотрудниками кафедры физиологии высшей нервной деятельности Ленинградского университета (1964—1973 гг.)
19 6 4
Влияние выключения зрительной рецепции на направленное поведение у летучих мышей. (А. И. Константинов).
О роли условной сигнализации в пространственном анализе у летучих мышей. (А. И. Константинов).
19 6 5
Влияние выключения заднего двухолмия на локационно-двигательную активность у летучих мышей. (4. И. Константинов).
Влияние частичного и полного выключения коры головного мозга на эхолокацию у летучих мышей. (4. И. Константинов).
К вопросу о роли эхолокации в анализе пространства летучими мышами. (Э. III. Айрапетьянц, А. И. Константинов).
Материалы по физиологии пространственного анализа летучих мышей (опыт физиологического исследования эхолокации). (4. И. Константинов).
19 6 6
Бионический аспект исследования механизмов информации о пространстве — об эхолокации летучих мышей. (Э. Ш. Айрапетьянц). Вызванные потенциалы и разряды одиночных нейронов задних холмов летучей мыши при ультразвуковой стимуляции. (4. Г. Васильев, Д. П. Матюшкин).
Нейронный состав и межнейронные связи заднего двухолмия мозга летучей мыши. {Л. И. Суховская).
О возможности использования ультразвуковой локации мелкими грызунами. (4. И. Константинов, Н. И. Ахмарова, С. С. Головина).
Основы ультразвуковой пространственной ориентации летучих мышей. (4. И. Константинов).
Распознавание геометрических фигур летучими мышами с помощью ультразвуковой локации. (4. И. Константинов).
19 6 7
Активная и пассивная локация при ловле насекомых у летучих мышей. (4. И. Константинов, В. В. Соколов).
Исследование нервных механизмов эхолокации у летучих мышей, (Э. III. Айрапетьянц, А. И. Константинов).
498
К физиологической характеристике слуховой системы летучих мышей по ее реакциям на ультразвуковые стимулы. (А.. Г. Васильев, Д. П. Матюшкин).
К характеристике нейронных сетей слухового анализатора, участвующих в определении направления приходящего сигнала. (Д. П. Матюшкин, И. М. Стосман).
Механизмы мозга и пространственный анализ. (Э. Ш. Айрапетьянц).
О взаимодействии анализаторов в эхолокационной активности у летучих мышей. (Э. Ш. Айрапетьянц, А. И. Константинов).
Сравнительная характеристика слуховой системы гладконосых и подковоносых летучих мышей (электрофизиологические данные). (Л. Г. Васильев).
Сравнительное исследование чувствительности эхолокации летучих мышей. (А. И. Константинов, Б. В. Соколов, И. М. Стосман).
Чувствительность эхолокации у гладконосых и подковоносых летучих мышей. (Л. И. Константинов, Б. В. Соколов, И. М. Стосман).
Электрическая активность заднего двухолмия летучей мыши, раздражаемой ультразвуком. (4. Г. Васильев).
196 8
Биоэлектрические реакции на ультразвуки и их маскировка белым шумом в слуховой системе летучих мышей. (Д. П. Матюшкин, А. Г. Васильев).
Влияние выключения слуховой коры летучих мышей на эхолокационное поведение. (Я. Н. Соколова).
Вызванные потенциалы и разряды отдельных нейронов заднего двухолмия летучих мышей при изменении некоторых параметров ультразвуковых стимулов. (4. Г. Васильев).
Вызванные потенциалы и реакции отдельных нейронов заднего двухолмия летучих мышей на ультразвуковые стимулы с разной длительностью их фронтов. (4. Г. Васильев).
Исследование хоминга у летучих мышей. (4. И. Константинов, И. К. Рахматулина).
К вопросу о кортикальной регуляции в слуховом анализаторе у летучих мышей и белых крыс. (Г. А. Куликов, Л. А. Самойлова).
К вопросу о цитоархитектонике слуховой области коры больших полушарий головного мозга летучей мыши. (О. А. Добрякова).
Мозговые механизмы эхолокации и бионика. (Э. Ш. Айрапетьянц, А. И. Константинов, Д. П. Матюшкин).
Некоторые свойства слуховой области коры летучей мыши. (Л. М. Котеленко).
О некоторых кортико-фугальных влияниях в слуховом анализаторе у летучих мышей и белых крыс. (Г. А. Куликов).
О нисходящих связях слуховой области коры летучей мыши. (Н. В. Бурикова).
Особенности проекции медиального коленчатого тела на слуховую кору у летучих мышей. (Я. В. Бурикова).
О способности дельфинов распознавать предметы. (4. И. Константинов, Н. Ф. Мельников, А. А. Титов).
Пространственная чувствительность слуховой системы летучих мышей по порогам электрических реакций заднего двухолмия на ультразвук. (Я. М. Стосман).
Распознавание (анализ) предметов при помощи эхолокации остроухой ночницей (Myotis oxygnathus). (4. И. Константинов, Н. И. Ахма-рова).
Сравнительная характеристика слуховой системы гладконосых и подковоносых летучих мышей. (4. Г. Васильев).
499
Сравнительная характеристика электрической активности заднего двухолмия летучих мышей при изменении длительности ультразвукового стимула. (А. Г. Васильев).
Характеристика тональных настроек нейронов слуховой коры летучих мышей. (Д. П. Матюшкин, Л. М. Котеленко).
Чувствительность заднехолмовых нейронов летучей мыши к ультразвуковым сигналам, приходящим с разных направлений (в пространственном угле 180°). (Д. П. Матюшкин, И. М. Стосман).
Электрическая активность заднего двухолмия гладконосых и подковоносых летучих мышей при раздражении ультразвуком. (Л. Г. Васильев).
Электрические реакции слухового нерва летучей мыши на ультразвуки разной высоты и интенсивности. (Д. П. Матюшкин).
Эхолокационное распознавание целей летучими мышами. (А. И. Константинов).
Echolocating space analysis of animals. (E. Sh. Airapetiantz, A. I. Konstantinov).
196 9
Brain echolocation mechanisms and bionics. (E. Sh. Airapetiantz, A. I. Konstantinov, D. P. Matjushkin).
Влияние коры больших полушарий на частотно-пороговые характеристики заднего двухолмия летучих мышей. (Г. А. Куликов).
Дальность действия и чувствительность эхолокатора летучих мышей. (Л. Г. Голубков, А. И. Константинов, А. К. Макаров).
К анализу нервного механизма, определяющего направление акустического сигнала у летучих мышей. (Д. П. Матюшкин, И. М. Стосман).
Некоторые характеристики входа слуховой системы летучей мыши (о торможении кохлеарных нейронов сильными адекватными стимулами). (Д. П. Матюшкин).
Некоторые экспериментальные данные и перспективы изучения анализаторной способности дельфина. (А. Г. Голубков, В. Н. Зворыкин, И. В. Ершова, В. М. Королев, В. И. Бурдин, Ю. А. Малышев).
Свойства нейронов слуховой коры летучей мыши (реакция на ультразвуки разной высоты и интенсивности). (Л. М. Котеленко, Д. П. Матюшкин).
Связь слухового восприятия и эхолокации при обнаружении и ловле насекомых гладконосыми летучими мышами (Vespertilionidae). (Л. И. Константинов).
Характеристика вызванных потенциалов заднего двухолмия летучих мышей при изменении направления действия раздражителя. (Л. Г. Васильев).
Характеристика вызванных потенциалов и реакций отдельных нейронов заднего двухолмия летучих мышей при раздражении парными ультразвуковыми стимулами. (Л. Г. Васильев).
Характеристика областей ответов отдельных нейронов заднего двухолмия гладконосых и подковоносых летучих мышей при ультразвуковой стимуляции. (Л. Г. Васильев).
Характеристика ультразвуковых ориентационных сигналов подковоносых летучих мышей. (Л. И. Константинов, Б. В. Соколов).
Характеристика электрических ответов задних холмов четверохолмия гладконосых и подковоносых летучих мышей при изменении силы ультразвука. (Л. Г. Васильев).
1970
Исследование разрешающей способности по дальности эхолокатора дельфина. (Л. Г. Голубков, Ю. В. Иваненко).
Кортикальная регуляция некоторых образований в слуховом анализаторе у летучих мышей и белых крыс. (Г. А. Куликов).
500
К проблеме обратных связей в системе слухового анализатора. (Г. А. Куликов).
К физиологической теории пространственного анализа. (Э. Ш. Айрапетьянц).
О возможности функционирования эхо локационной системы летучих мышей без участия слуховой коры. (4. И. Константинов, Н. Н. Соколова).
Определение разрешающей способности дельфинов по углу. (4. П. Абрамов, В. И. Бурдин, Ю. В. Иваненко, В. И. Королев, Ю. 4. Малышев).
О роли интерференции в формировании направленности эхолокационного излучения летучих мышей семейства подковоносых. (4. К. Макаров, Б. В. Соколов).
Соотношение физических и нервных факторов в бинауральном слухе летучих мышей. (И. М. Стосман, Н. А. Этманова).
Ультразвуковые эхолокационные сигналы животных и проектирование радио- и гидросистем. (Э. Ш. Айрапетьянц, 4. Г. Голубков, И. В. Ершова, А. И. Константинов, В. П. Пересади, Г. К. Ульянов).
Характеристика вызванных потенциалов слухового анализатора летучих мышей на стимулы с разной частотой заполнения. (Э. HI. Айрапетьянц, А. Г. Васильев).
Характеристика потенциалов действия нейронов кохлеарного ядра летучих мышей при раздражении ультразвуком с разных направлений. (4. Г. Васильев, И. М. Стосман).
Характеристика ультразвукового локатора рыбоядных летучих мышей. (4. И. Константинов).
Электрические реакции слуховой области коры летучих мышей на ультразвуковые стимулы. (4. Г. Васильев, Л. М. Котеленко).
Электрические реакции слуховой области коры летучих мышей на ультразвуковые стимулы. (Л. М. Котеленко).
Эхолокация в природе. (Э. Ш. Айрапетьянц, А. И. Константинов). Эхолокационный пространственный анализ у животных. (Э. Ш. Айрапетьянц, А. И. Константинов).
19 7 1
Исследование способности дельфинов к дифференцировке объемных предметов по линейным размерам и материалам. (Э. Ш. Айрапетьянц, А. П. Абрамов, В. И. Бурдин, А. Г. Голубков, И. В. Ершова,
А. Р. Жежерин, В. И. Королев, Ю. А. Малышев, Г. К. Ульянов, В. Б. Фрадкин).
Исследование частотно-пороговых характеристик слуховой системы остроухой ночницы методом условных рефлексов. (4. И. Константинов, Н. Н. Саноцкая, Н. Н. Соколова).
Метод электростимуляции при изучении функций подкорковых структур китообразных. (Ю. В. Иваненко, Е. Д. Краснощекова, В. Б. Слезин).
Направленность ультразвукового ориентационного излучения большого подковоноса и роль носовых выростов в ее формировании. (Б. В. Соколов, А. К. Макаров).
On neurophysiological mechanisms of the echolocating apparatus in bats (frequency parameters). (E. Sh. Airapetiantz, A. G. Vasilyev). Physi ological investigations of ultrasonic echolocation in animals. (E. Sh. Airapetiantz, A. I. Konstantinov).
Характеристика электрических реакций внутренних коленчатых тел гладконосых и подковоносых летучих мышей на ультразвуковые стимулы с разной частотой заполнения. (4. Г. Васильев, И. Г. А ндреева).
Характеристика электрических реакций кохлеарных ядер гладконосых и подковоносых летучих мышей на ультразвуковые стимулы с разной частотой заполнения. (4. Г. Васильев).
501
Характеристика электрических реакций слухового нерва и кохлеарных ядер гладконосых и подковоносых летучих мышей при изменении угла предъявления раздражителя. (И. М. Стосман, Н. А. Этманова).
Электрические реакции слуховой области коры летучих мышей на ультразвуковые стимулы с разной частотой заполнения. (Л. М. Котеленко, А. Г. Васильев).
Эфферентные связи слуховой коры головного мозга летучей мыши. (Н. В. Бурикова).
19 72
Влияние коры больших полушарий на частотно-пороговые характери стики электрических реакций заднего двухолмия летучих мышей (Г. А. Куликов).
Изменение параметров локационных сигналов дельфинов в процессе дифференцирования сферических и цилиндрических мишеней. (Э. Ш. Айрапетьянц, В. А. Воронов, Ю. А. Иваненко, М. П. Иванов, Д. Л. Ордовский, В. В. Попов, Б. Ф. Сергеев, В. И. Чилингирис).
Некоторые данные о нейронной организации слуховой области коры мозга дельфинов. (Е. И. Краснощекова).
О взаимодействии слухового восприятия и эхолокации у летучих мышей семейства Rhinolophidae в процессе охоты за насекомыми. (Б. В. Соколов).
О локализации ультразвуковых сигналов летучими мышами. (И. А. Горлинский).
Организация слуховой коры рукокрылых в ряду млекопитающих. (Э. Ш. Айрапетьянц, И. В. Бурикова, А. И. Константинов, Л. М. Котеленко, Г. А. Куликов, И. И. Соколова).
Особенности слухового восприятия у подковоносых летучих мышей (Rhinolophus ferrum-equinum Schreb.). (Б. В. Соколов, И. И. Соколова).
Особенности постэмбрионального развития и пути формирования эхолокационного сигнала у летучих мышей. (Л. И. Константинов). Пространственная ориентация летучих мышей под влиянием повышенной гравитации. (Э. Ш. Айрапетьянц, В. И. Зворыкин, Б. М. Савин).
Развитие функций ультразвуковых локационных систем у летучих мышей в постнатальном онтогенезе. (А. И. Константинов).
Реакции нейронов вентрального кохлеарного ядра подковоносых летучих мышей на ультразвуковые стимулы при удалении ипсилатеральной улитки. (Г. А. Куликов).
Физио логические механизмы гидролокатора дельфина. (Э. Ш. Айрапетьянц, В. А. Воронов, Ю. В. Иваненко, Л. В. Иванов, Д. Л. Ордовский, В. В. Попов, Б. Ф. Сергеев, В. И. Чилингирис).
X арактеристика вызванных потенциалов заднего двухолмия у детенышей подковоносых летучих мышей (Rhinolophidae) при действии ультразвука. (И. М. Стосман, А. И. Константинов).
Электрическая активность заднего двухолмия у детенышей гладконосых летучих мышей при действии ультразвуковых тональных сигналов. (А. И. Константинов, И. М. Стосман).
Эхолокационный пространственный анализ у подковоносых летучих мышей (Rhinolophidae). (Б. В. Соколов).
1973
Акустические методы и аппаратура для исследования ультразвуковой эхолокации у животных. (А. К. Макаров).
Б иоакустические особенности и принципы функционирования эхолокационной системы подковоносых летучих мышей. (А. И. Константинов, А. К. Макаров, Б. В. Соколов, Н. Н. Соколова).
502
Временная суммация в слуховой области коры летучих мышей. (Л. М. Котеленко).
Диапазон эхолокационного распознавания у летучих мышей. (4. И. Константинов, В. И. Малашин, А. К. Макаров, Н. Н. Соколова, В. И. Чилингирис, Н. А. Козлова).
Development of echolocation in bats in postnatal ontogenesis. (A. I. Konstantinov).
Дифференцирование дельфином афалиной сферических и цилиндрических объектов. (5. Ш. Айрапетьянц, В. А. Воронов, Д. Л. Ордовский, Б. Ф. Сергеев, В. И. Чилингирис).
Измерение некоторых параметров приемной системы эхолокатора летучих мышей. (Я. Я. Соколова).
Изучение механизмов обнаружения эха на уровне верхнеоливарного комплекса эхолокатора гладконосых летучих мышей. (С. И. Ермолин).
Influence of band limited noises on the efficiency of echolocation detection of targets by Rhinolophus ferrum-equinum. (4. I. Konstantinov, V. N. Movtchan, A. K. Makarov).
К теории эхолокации у летучих мышей. (Э. Ш. Айрапетьянц, А. И. Константинов, Д. П. Матюшкин).
К физиологии сонарной системы черноморских дельфинов.
(Э. Ш. Айрапетьянц, В. А. Воронов, Ю. В. Иваненко, М. П. Иванов, Д. Л. Ордовский, Б. Ф. Сергеев, В. И. Чилингирис).
К физиологической теории пространственного анализа. (Э. Ш. Айрапетьянц).
Материалы о влиянии гипервесомости на летучих мышей. (Э. Ш. Айрапетьянц, В. Н. Зворыкин, Б. М. Савин).
Механизмы определения некоторых параметров стимулов гладконосых и подковоносых летучих мышей. (4. Г. Васильев, И. М. Стосман, Г. А. Куликов).
Направленность слухового приема черноморских дельфинов. (В. И. Чилингирис, Ю. В. Иваненко).
Некоторые механизмы распознавания сигналов на уровне медиальных коленчатых тел гладконосых и подковоносых летучих мышей. (Я. Г. Андреева).
Некоторые особенности слуховой системы гладконосых и подковоносых летучих мышей (электрофизиологические данные). (4. Г. Васильев).
О возможности кортикальной регуляции сенсорного потока в слуховом анализаторе летучих мышей. (Г. А. Куликов).
О нисходящих связях слуховой области коры (area temporalis communis) летучей мыши. (Я. В. Бурикова).
Определение возможности различения дельфином сложных модулированных шумов. (Ю. В. Иваненко, В. Б. Слезин).
Эхолокационные системы летучих мышей подотряда Microchiroptera. (4. И. Константинов).
Организация слуховой коры летучих мышей в сравнительном ряду млекопитающих. (Э. Ш. Айрапетьянц, Н. В. Бурикова, А. И. Константинов, Л. М. Котеленко, Г. А. Куликов, Н. Н. Соколова).
О реакциях слуховой области коры гладконосых летучих мышей на ультразвуковые раздражения. (Л. М. Котеленко).
Основные показатели эхолокационных систем гладконосых и подковоносых летучих мышей. (4. И. Константинов).
Особенности нисходящих путей слуховой коры летучей мыши. (Я. В. Бурикова).
Пространственная чувствительность слуховой системы летучих мышей по порогам электрических реакций заднего двухолмия на ультразвук. (Я. М. Стосман).
Пространственный анализ у больших подковоносов после действия гравитационных перегрузок при ускорениях. (Э. Ш. Айрапетьянц, р. Н, Зворыкин, Л. И. Петрова).
503
Развитие эхолокации у летучих мышей в постнатальном онтогенезе. (4. И. Константинов),
Разрешающая способность сонара дельфина по дальности. (Ю. В, Иваненко).
Реакция нейронов верхнеоливарного комплекса гладконосых и подковоносых летучих мышей на ультразвуковые стимулы (параметр частоты). (Т. Е, Васильева).
Реакции слуховой области коры у гладконосых летучих мышей на ультразвуковые раздражители. (Л. М. Котеленко).
Роль и взаимодействие эхолокации и слухового восприятия у летучих мышей при ловле насекомых. (4. И. Константинов, Б. В. Соколов).
Сравнительная характеристика электрических ответов заднего двухолмия гладконосых и подковоносых летучих мышей в онтогенезе. (4. И. Константинов, И. М. Стосман).
Траектория движения дельфина в процессе дифференцирования сферических и цилиндрических тел. (М. П. Иванов, Н. А. Курганский).
Физиологическая характеристика нисходящих влияний в слуховом анализаторе летучих мышей. (Г. А. Куликов).
Физиологические исследования ультразвуковой локации у животных. (Э. Ш. Айрапетьянц, А. И. Константинов).
Характеристика вызванных потенциалов заднего двухолмия летучих мышей на разные ультразвуковые стимулы. (4. Г. Васильев, И. М. Стосман, В. Б. Слезин).
Характеристика диаграммы направленности приема на различных уровнях слухового анализатора летучих мышей (слуховой нерв, кохлеарные ядра, задние холмы). (И. М. Стосман).
Характеристики направленности приема и точность локализации ультразвука у эхолоцирующих летучих мышей. (4. И. Константинов, И. М. Стосман, И. А. Горлинский).
Характеристики приема сонара дельфина. (В. И. Чилингирис, Ю. В. Иваненко).
Характеристика реакций отдельных нейронов задних холмов летучих мышей на ультразвуковые стимулы при изменении их частоты и силы. (4. Г. Васильев).
Характеристика тональных настроек нейронов слуховой коры летучих мышей. (Д. И. Матюшкин, Л. М. Котеленко).
Характеристика электрической активности верхних олив гладконосых и подковоносых летучих мышей на ультразвуковые стимулы с разной частотой заполнения. (А. Г. Васильев).
Электрические реакции слухового нерва летучей мыши на ультразвуки разной высоты и интенсивности. (Д. И. Матюшкин).
Эффективность эхолокационного обнаружения мишеней в присутствии шумов у летучих мышей. (А. К. Макаров).
Эфферентные связи нижнего двухолмия летучей мыши. (Я. В. Бурикова).
Эхолокационные сигналы некоторых отечественных видов гладконосых летучих мышей (семейство Vespertilionidae). (А. И. Константинов, А. К. Макаров).
Эхолокационные сигналы подковоносых летучих мышей и некоторые особенности их излучения. (В. В, Соколов),
ОГЛАВЛЕНИЕ
Стр.
Предисловие ко второму изданию ............................... 3
Предисловие к первому изданию................................. 6
Введение..................................................... 13
Сущность эхолокации..................................... 13
Природа звуковых волн................................... 15
Ультразвук и его особенности............................ 19
История открытия эхолокации............................. 21
ЭХОЛОКАЦИЯ В ВОЗДУХЕ
Глава 1. Типы локационных систем у летучих мышей............. 33
Основные систематические группы рукокрылых.............. 34
Методы регистрации и измерения основных параметров ультразвуковых сигналов.................................. 36
Эхолокация у примитивных летучих мышей.................. 39
Особенности эхолокации у щелегубов и подковоносов....... 46
Локационные системы летучих мышей Нового Света.......... 54
Локационные системы бульдоговых и гладконосых летучих мышей ............................................. 68
Пути развития эхо локационных систем летучих мышей .... 77
Глава 2. Звукоизлучающая система и характеристики направленности излучения...................................... 85
Особенности строения гортани^........................... 85
Нервная регуляция голосового аппарата................... 87
Механизмы излучения ультразвука......................... 93
Характеристики направленности излучения локационных сигналов .......................................... 96
Глава 3. Структурно-функциональные особенности слухового анализатора летучих мышей.............................. 104
Наружное ухо........................................... 104
Среднее ухо............................................ 106
Внутреннее ухо. Микрофонные потенциалы улитки.......... 109
Проводниковый отдел..................................... ИЗ
Слуховая область коры.................................. 117
505
Глава 4. Общая функциональная схема нервной регуляции эхолокатора ............................................. 121
О функциональной роли слуховой области коры............. 121
О функциональной роли заднего двухолмия................. 126
Схема регуляции эхолокатора как целостной системы .... 130
Глава 5. Особенности слухового восприятия акустических сигналов по параметрам частоты и длительности...................... 135
Границы частотного диапазона и зоны оптимального слуха . 135
Зависимость порогов слуха от длительности сигнала....... 142
Глава 6. Электрофизиологические характеристики нервных^аппара-тов слуховой системы летучих мышей...................... 145
Потенциалы действия слухового нерва, свойства первичных слуховых нейронов................................... 145
Электрические реакции и свойства элементов кохлеарных ядер 151
Электрические явления в задних холмах четверохолмия и свойства нейронов задних холмов..................... 155
Электрические явления во внутренних коленчатых телах ... 169
Электрические явления в слуховой корковой области .... 172
Некоторые соображения о принципах анализа ультразвуковых сигналов в слуховой системе......................... 180
Глава 7. Пространственный слух и точность локализации ультразвука ........................................................ 185
Формирование направленности приема на разных уровнях слуховой системы ............................................. 185
Точность локализации источника ультразвука.............. 190
Глава 8. Развитие эхолокации у летучих мышей в онтогенезе 195
Развитие детенышей у ночниц и подковоносов.............. 196
Характер призывных криков и становление локационного сигнала............................................. 197
Развитие функций слуховой системы..................... 205
Глава 9. Особенности функционирования эхолокационных систем летучих мышей при преследовании и ловле добычи........... 209
Возможность пассивного (слухового) обнаружения насекомых 211
Деятельность эхолокационных систем у гладконосых летучих мышей на разных этапах ловли насекомых.............. 213
Особенности эхолокации подковоносов при ловле насекомых . 220
Глава 10. Пределы обнаружения, дальность действия и чувствительность эхолокационных систем............................... 226
Пределы обнаружения..................................... 226
Дальность действия...................................... 230
Пороговая чувствительность эхолокатора.................. 238
Глава 11. Точность измерения дальности и разрешающая способность эхолокатора........................................ 241
Точность определения расстояния......................... 241
О разрешающей способности эхолокатора по дальности .... 247
Разрешающая способность эхолокатора по угловым координатам ................................................... 250
506
Глава 12. Диапазон и принципы эхо локационного распознавания предметов..................................................... 254
Пределы эхолокационного распознавания мишеней, отличающихся по размерам.................................. 255
Распознавание мишеней, отличающихся по форме и материалу 262
Пределы распознавания регулярных решеток................ 265
Исследование параметров локационных сигналов летучих мышей при распознавании.................................. 267
Механизмы формирования отраженного сигнала.............. 273
Глава 13. Помехозащищенность эхолокаторов..................... 279
Особенности приема ультразвука в присутствии шумов .... 280
Оценка помехозащищенности эхолокатора................... 286
Глава 14. Сравнение возможностей эхолокационных систем и принципов их действия у гладконосых и подковоносых летучих мышей........................................................ 293
Глава 15. Акустическая локация у пещерных птиц и крыланов 305
«Жирная птица из Карипе»................................ 306
Эхолокация у стрижей-саланган........................... 309
Способы ориентации у крыланов........................... 313
Глава 16. Эхолокация у грызунов и насекомоядных........... 321
Эхолокация у грызунов................................... 321
Эхолокация у насекомоядных.............................. 329
ЭХОЛОКАЦИЯ В ОКЕАНЕ
Глава 17. Гидролокация у китообразных......................... 337
Краткая систематическая справка......................... 337
Доказательства наличия эхолокации у дельфинов........... 340
Характеристики локационных сигналов зубатых китов .... 343
Сигналы усатых китов.................................... 350
Глава 18. Механизмы генерирования звуков и направленность излучения.................................................... 353
Морфологические и физиологические особенности звукоизлучающего аппарата........................................ 353
Направленность излучения локационных сигналов........... 361
Глава 19. Слуховой анализатор................................. 374
Строение анализатора.................................. 374
Звуковая рецепция....................................... 382
Функциональные характеристики центрального звена анализатора ................................................... 388
Глава 20. Характеристики приемной системы..................... 392
Диапазон слухового восприятия........................... 392
Дифференциальные пороги слуха по интенсивности.......... 395
Дифференциальные пороги слуха по частоте................ 396
Пределы дифференцирования временных характеристик сигнала ................................................... 399
Возможность использования фазы сигнала.................. 401
Направленность звукового приема........................ 403
507
Глава 21. Диапазон действия, помехоустойчивость и пределы распознавания . •........................................... 409
Дальность действия эхолокатора при обнаружении искусственных объектов.......................................... 409
Дальность обнаружения рыбы............................ 412
Помехоустойчивость эхолокационного аппарата........... 414
Дифференцирование величины подводных объектов......... 418
Механизм дифференцирования подводных объектов по размеру и материалу ............................................ 422
Дифференцирование мишеней сложной конфигурации .... 431
Адаптация сонара к параметрам лоцируемых объектов .... 435
Глава 22. Эхолокация у ластоногих........................... 441
Заключение ................................................. 446
Литература.................................................. 454
Приложение, Работы по эхолокации животных, опубликованные сотрудниками кафедры физиологии высшей нервной деятельности Ленинградского университета (1964—1973 гг.)...... 498
CONTENTS
Page
Preface to the second edition................................... 3
Preface to the first edition...................................... 6
Introduction..................................................... 13
The essence of echolocation................................ 13
The nature of acoustic waves............................... 15
Ultra-sound and its features............................... 19
History of echo location discovery......................... 21
ECHOLOCATION IN THE AIR
Chapter 1. Sonar types in bats................................. 33
The main systematic groups of the Chiroptera.................. 34
Methods of registration and determination of pulse parameters in bats....................................................... 36
Echolocation in primitive bats............................... 39
Echolocation peculiarities in Hipposideridae and Rhinolophidae bats ......................................................... 46
Sonar systems in bats of the New World........................ 54
Sonar systems in Molossidae and Vespertilionidae bats......... 68
The ways of bats’ sonar systems development................... 77
Chapter 2. Sound emission systems and directional beaming ... 85
Anatomical specialization of the bat larynx................... 85
Nervous regulation of laryngeal apparatus..................... 87
Mechanisms of ultra-sound emission............................ 93
Directional beaming patterns.................................. 96
Chapter 3. Structure and function of auditory system in bats . . . 104
The external ear............................................... 104
The middle ear................................................. 106
The inner ear. Cochlear microphonic potentials................. 109
Auditory centers............................................... 113
The auditory cortex , , . , , , , t t . 117
509
Chapter 4. Common functional regulation of sonar system .... 121
On the functional role of the auditory cortex...................... 121
On the functional role of inferior colliculi....................... 126
Nervous regulation of sonar system................................. 130
Chapter 5. The peculiarities in auditory perception of acoustic signals (frequency and duration)............................................ 135
Frequency range and areas of optimum audition...................... 135
The dependence of auditory thresholds on stimulus duration 142
Chapter 6. Electrophysiological characteristics of the nervous apparatuses of the auditory system in bats............................... 145
Action potentials of auditory nerve, properties of primary auditory neurons ........................................................... 145
Electrical reactions and properties of the elements of cochlear nuclei ............................................................ 151
Electrical phenomena and properties of neurons in the inferior colliculi ............................................................ 155
Electrical phenomena in medial geniculate bodies................ 169
Electrical phenomena in the auditory cortex..................... 172
Some ideas of the principles of analysis of ultra-sonic signals in the auditory system of bats..................................... 180
Chapter 7. Spatial hearing and the accuracy of ultra-sound localization 185
Directional sensitivity on different levels of auditory system 185
The precision of the localization of ultra-sound source............ 190
Chapter 8. Development of echolocation in bats in ontogenesis 195
Development of Myotis and Rhinolophus infants...................... 196
Pattern of attractive calls and development of the locational signal ............................................................ 197
Development of the auditory system functioning.................. 205
Chapter 9. Functioning of bat sonar system in pursuit and catching of insect prey........................................................... 209
Ability of passive (auditory) detection of insects............... 211
The function of sonar system in Vespertilionidae bats at different stages of insects catching......................................... 213
Insects catching by horseshoe bats................................. 220
Chapter 10. Limits of detection, range of action and sensitivity of sonar systems in bats................................................. 226
Limits of detection................................................ 226
Range of action.................................................... 230
Threshold sensitivity.............................................. 238
Chapter 11. The accuracy of distance determination and sonar resolution 241
The accuracy of distance determination............................. 241
On distance resolution of sonar system......................... 247
Sonar angle resolution ................................, . . . . 250
510
Chapter 12. Range and principles of discriminative echolocation by bats ................................................................ 254
Limits of discrimination of targets different in size . .......... 255
Discrimination of targets different in shape and material .... 262
Limits of discrimination of regular arrays .......... 265
Parameters of bats locational signals during discrimination . . . 267
The mechanism of echo signals formation........................... 273
Chapter 13. Resistance of bat sonar system....................... 279
Specifities of ultra-sonic signals reception in noise............. 280
Estimation of sonar systems noise screening....................... 286
Chapter 14. Comparison of possibilities and principles of activity in Vespertilionidae and Rhinolophidae sonar systems..................... 293
Chapter 15. Acoustic location in cave—birds and flying foxes . . . 305
«The Oil bird of Caripe».......................................... 306
Echolocation in swiftlets......................................... 309
Orientation of Megachiroptera bats.............................. 313
Chapter 16. Echolocation in rodents and insectivora .............. 321
Echolocation in rodents........................................... 321
Echolocation in shrews and tenrecs................................ 329
ECHOLOCATION IN THE OCEAN
Chapter 17. Hydrolocation in Cetacea............................. 337
Short information on taxonomy..................................... 337
Evidence of echolocation in dolphins.............................. 340
Characteristics of locational signals in Odontoceti .............. 343
Signals of Whales................................................. 350
Chapter 18. Mechanism of emission and directional beaming .... 353
Morphological and physiological specificities of sound emissional apparatus.................................................. 353
Emission direction of orientation signals............... 361
Chapter 19. Auditory system...................................... 374
Structure of auditory system...................................... 374
Sound reception................................................... 382
Functional characteristics of auditory centers.................... 388
Chapter 20. Characteristics of reception (auditory) system .... 392
Range of auditory reception....................................... 392
Differential thresholds of intensity.............................. 395
Differential thresholds of frequency.............................. 396
The limits of signal temporal characteristics discrimination 399
Possibility of phase use.......................................... 401
The auditory receptive direction.................................. 403
Chapter 21. Range of action, resistance and discrimination limits of sonar system......................................................... 409
Range of action at detection of artificial objects................ 409
Range of fish detection........................................... 412
511
Resistance of sonar system .... *.............................. 414
Discrimination of underwater objects size...................... 418
Mechanism of differentiation in size and material of underwater objects ................................................ 422
Discrimination of complex-shape objects........................ 431
Sonar adaptation to parameters of located objects.............. 435
Chapter 22. Echolocation in fin-legged animals................. 441
Conclusion .......................................................... 446
Literature .......................................................... 454
Supplement, List of works on echolocation of animals published by members of the Chair of Higher Nervous Activity, Leningrad State University (1964—1973).................................... 498
Эрванд Шамирович Айрапетьянц Алексей Иванович Константинов
ЭХОЛОКАЦИЯ В ПРИРОДЕ
Издание второе, переработанное и дополненное
Утверждено к печати
Отделением физиологии Академии наук СССР
Редактор издательства Г. Т. Никифорова
Художник Л. А. Яценко
Технический редактор Г. А. Бессонова
Корректоры Л. М. Азад ж ано в а и Э. Н. Липпа
Сдано в набор 12/XII 1973 г.
Подписано к печати 29/IV 1974 г.
Формат бумаги бОхЭО1/^- Бумага №2.
Печ. л. 32=32 усл. печ. л.
Уч.-изд. л. 35.89. Изд. Ка 5458.
Тип. зак. К 795. М-03449.
Тираж 3600. Цена 2 р. 39 к.
Ленинградское отделение издательства «Наука». 199164, Ленинград, В-164,
Менделеевская линия, д. 1
1-я тип. издательства «Наука». 199034, Ленинград, В-34, 9 линия, д. 12
ИСПРАВЛЕНИЯ И ОПЕЧАТКИ
Страница Строка Напечатано Должно бытъ
33 10 сверху Щелегубы Подковогубы
92 И снизу 30 20
92 8 » под над
125 Табл. 8, графа 1 справа, 83.9 ± 0 83.9 ± 0.7
4 снизу
Э. III. Айрапетьянц и А. И. Константинов