/














































































































































































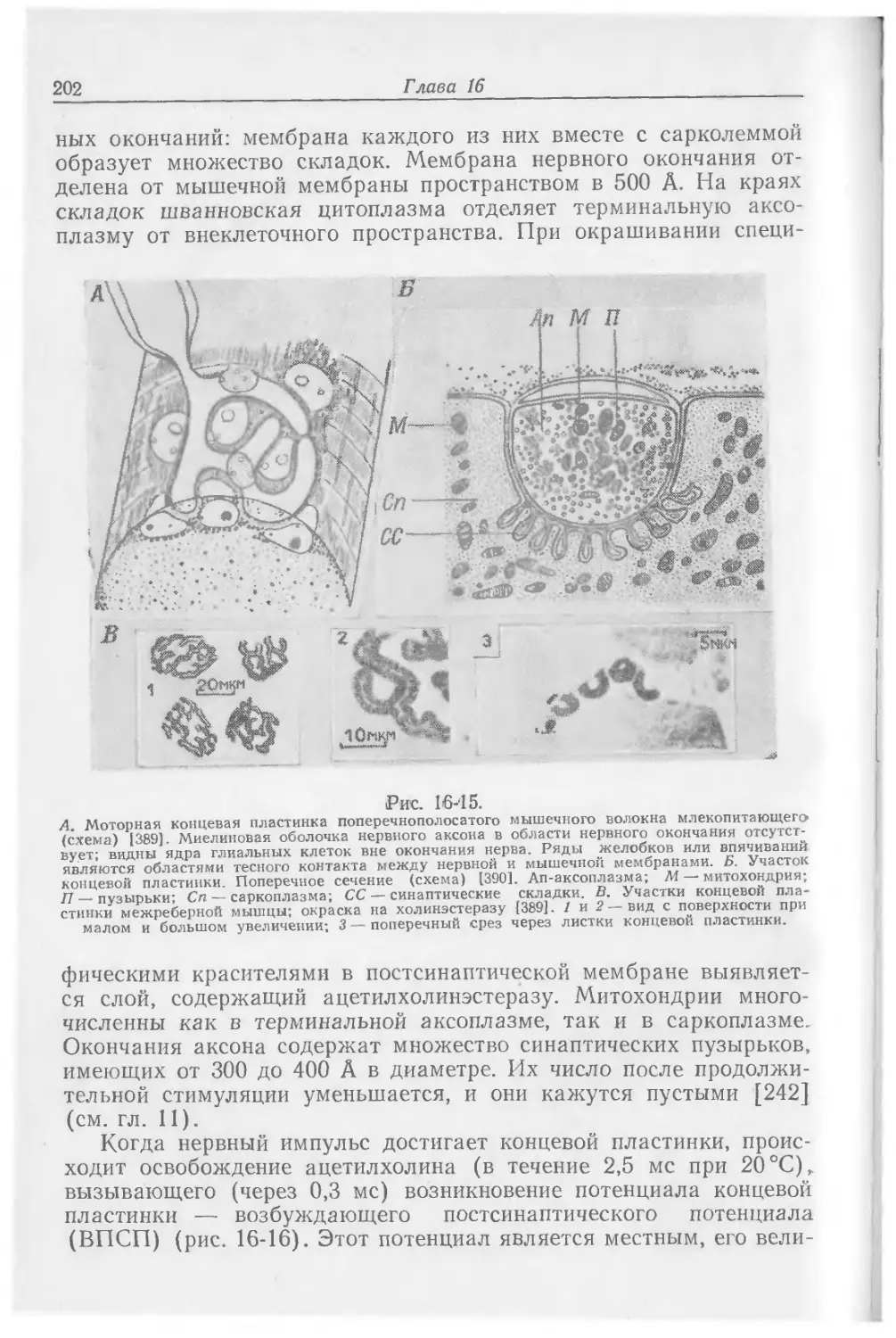
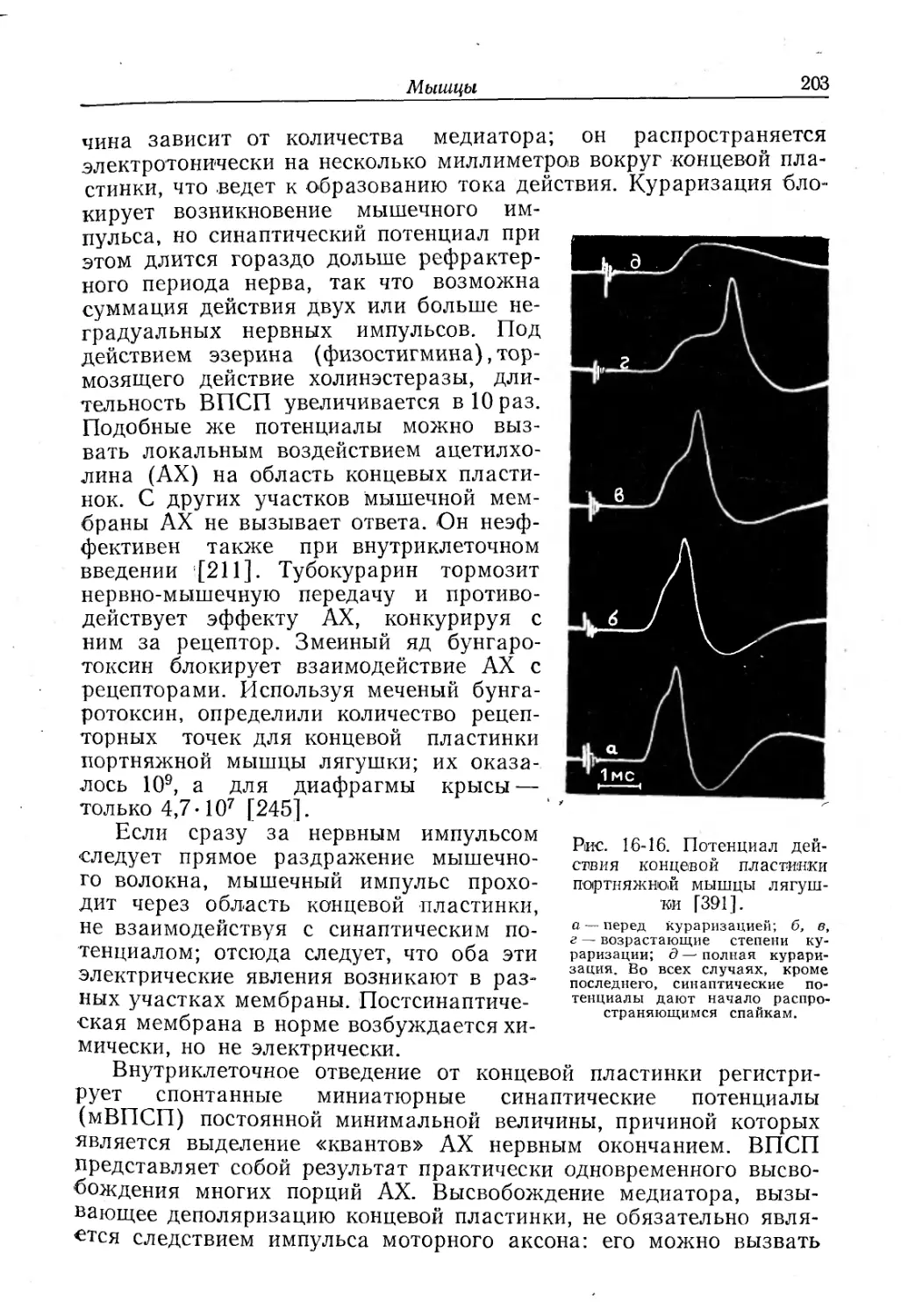



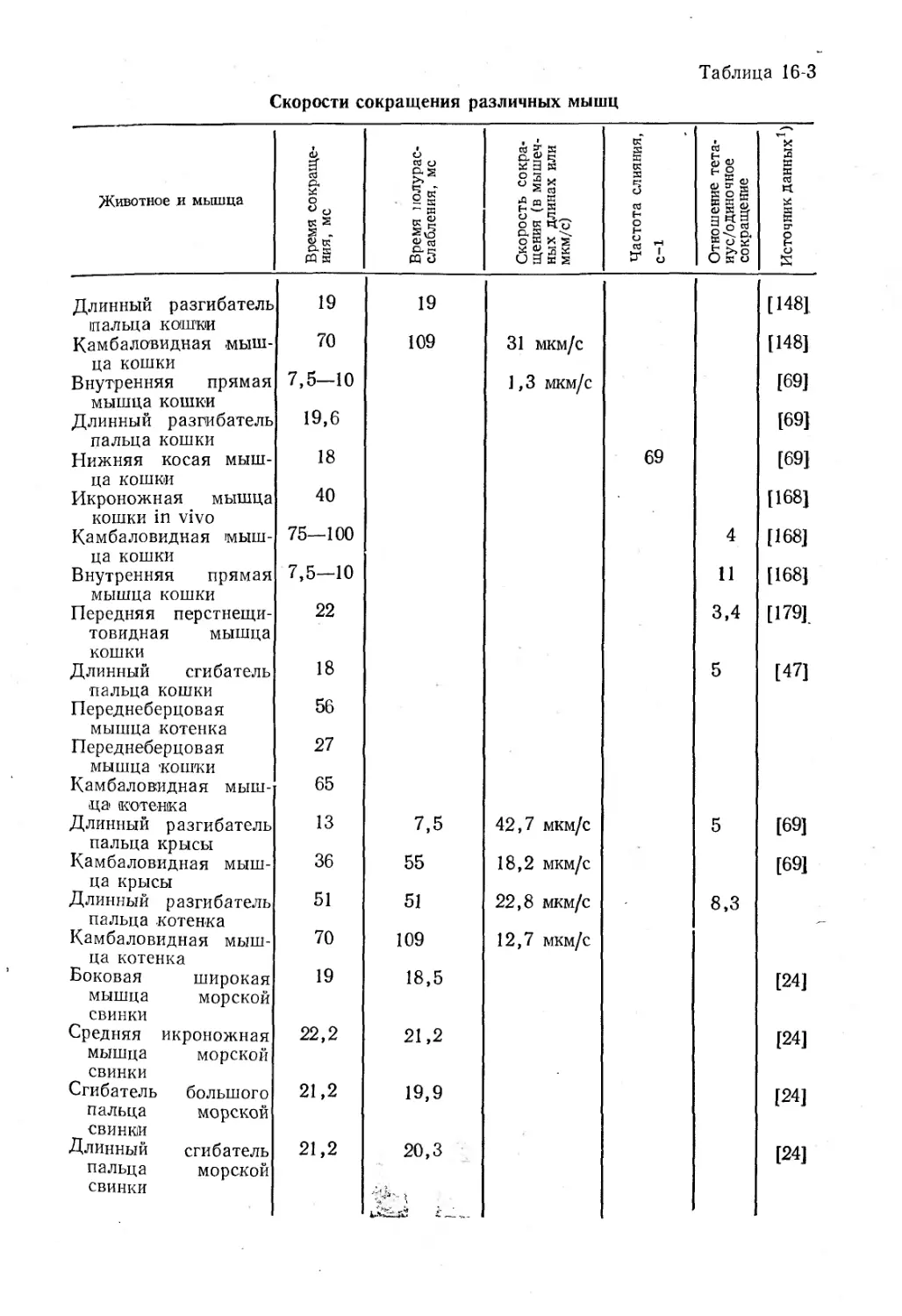

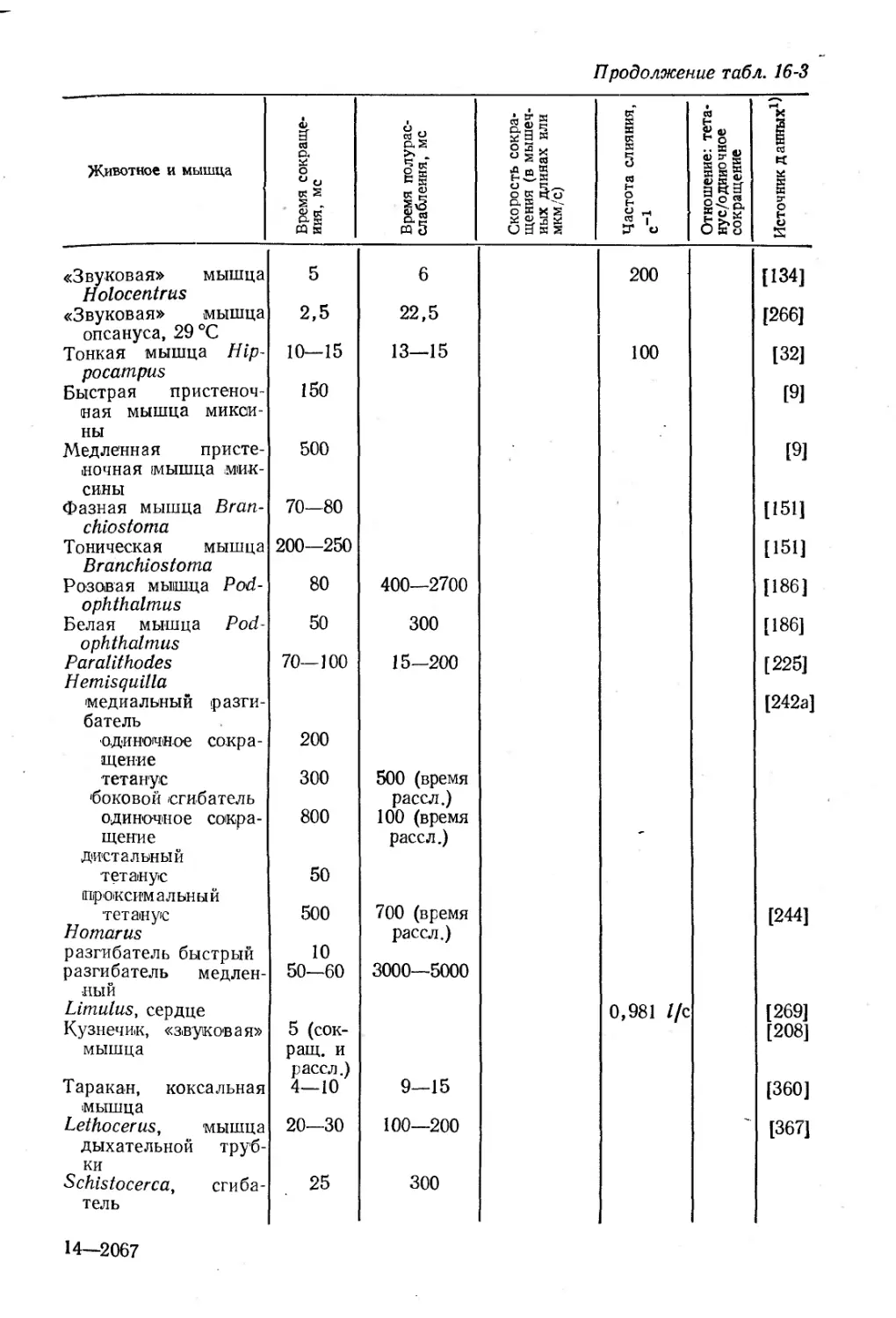

































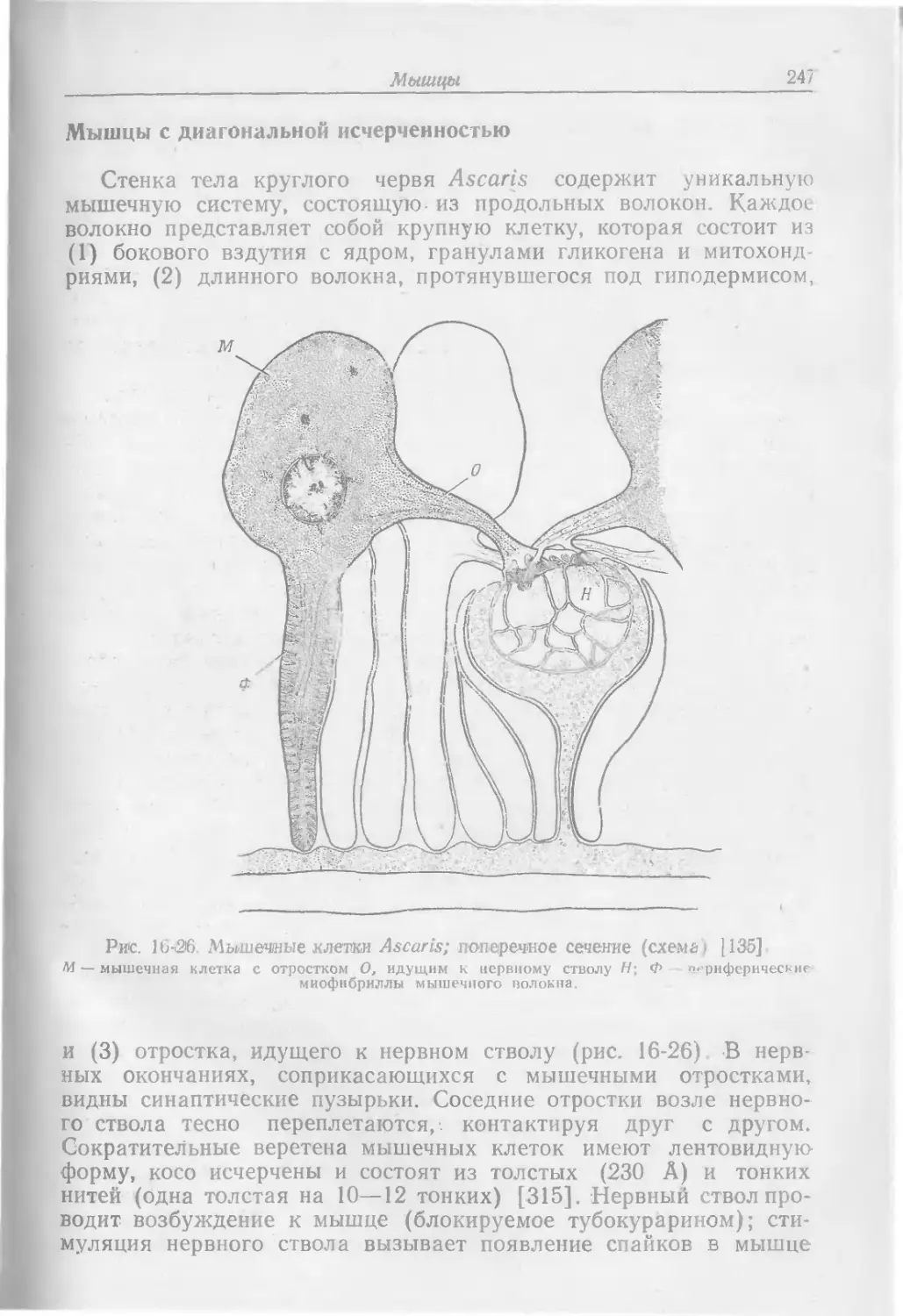

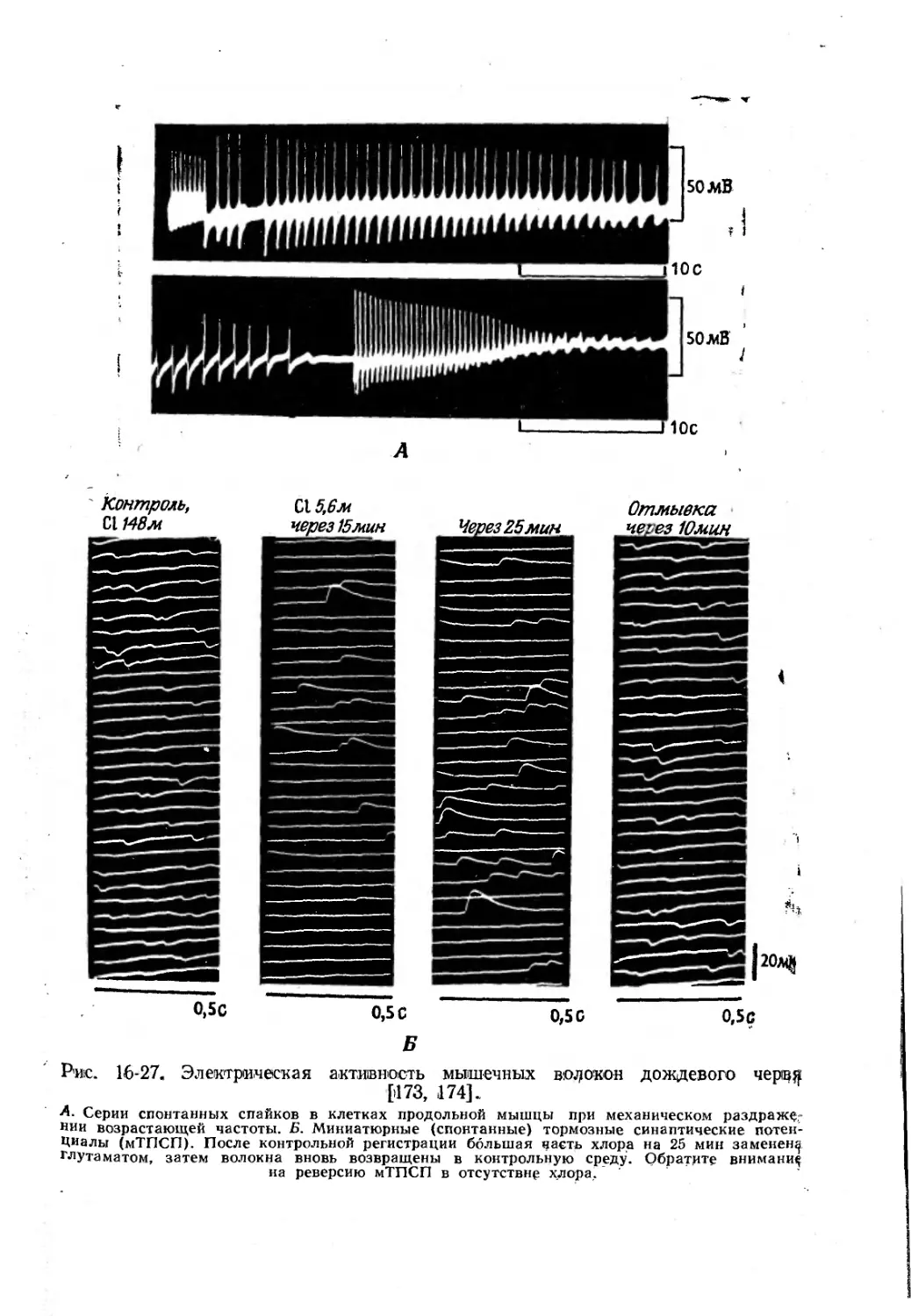






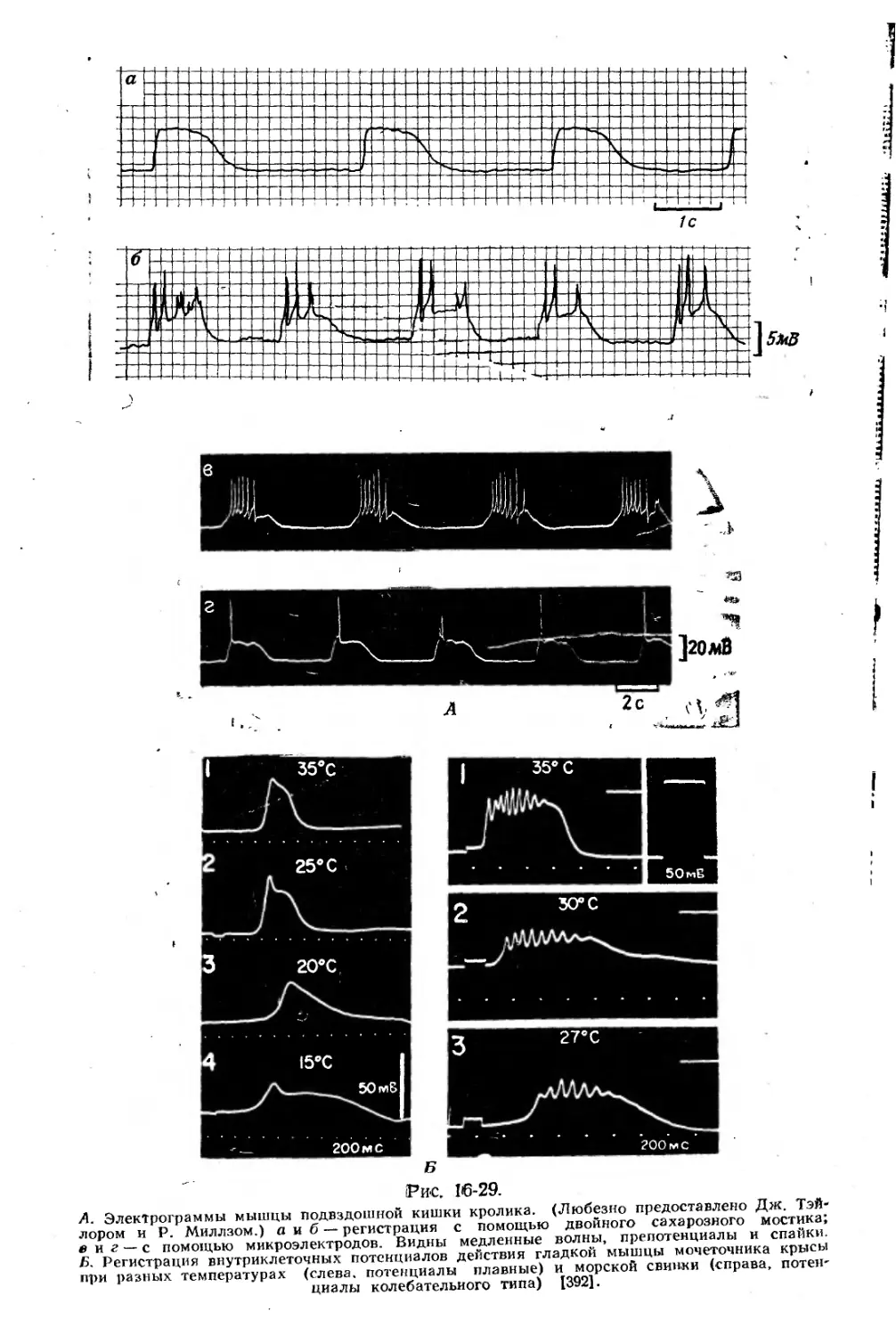








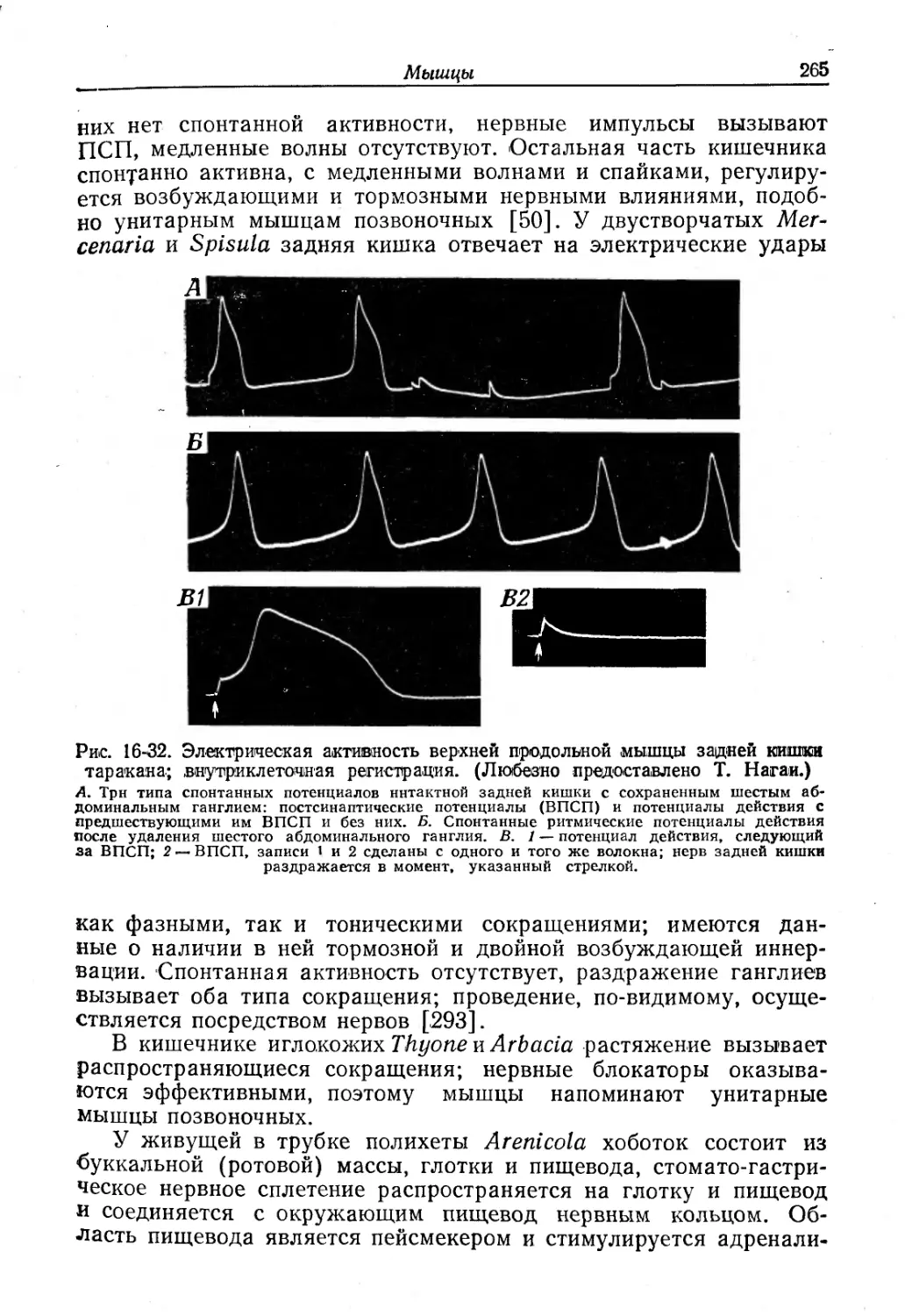




































































































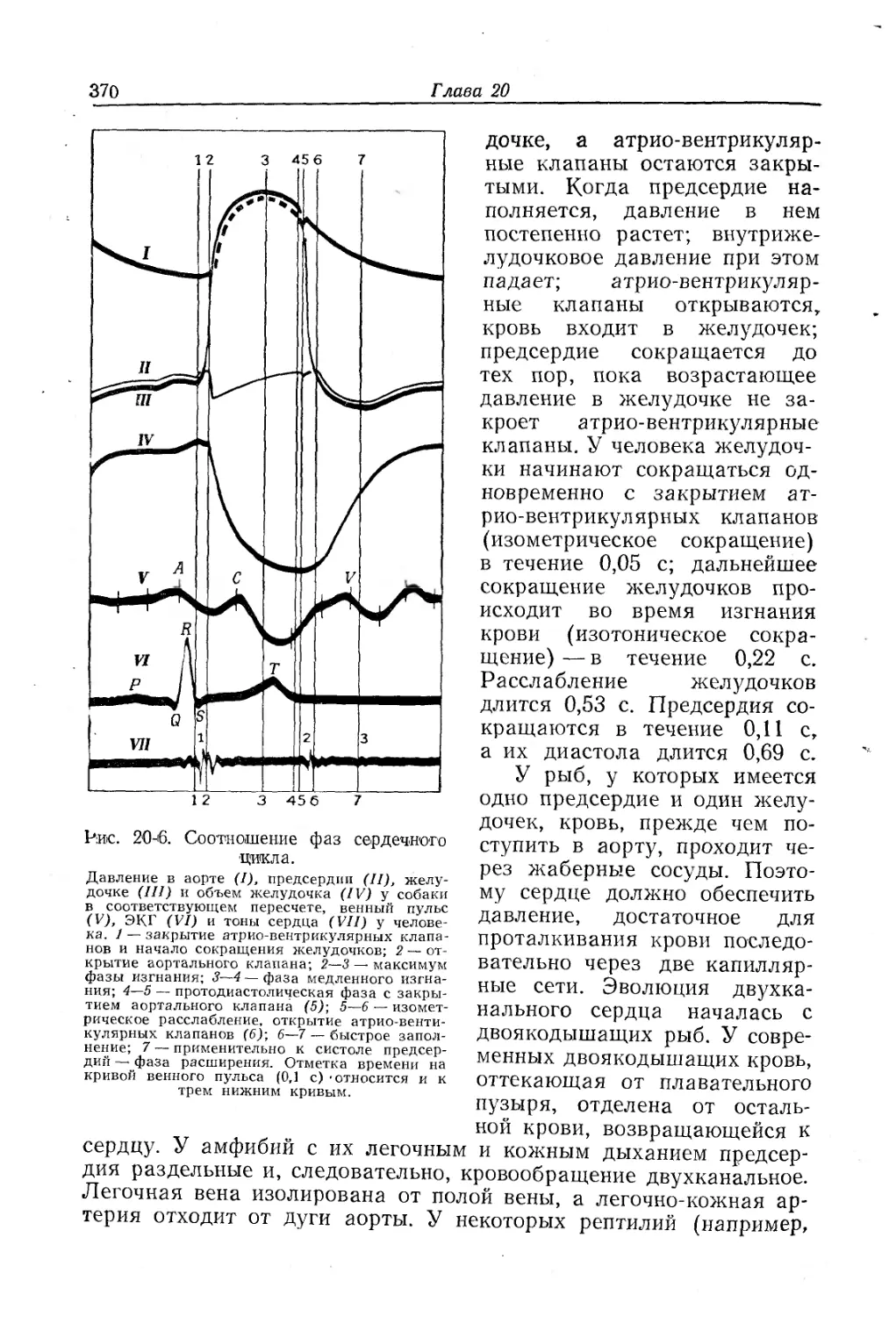





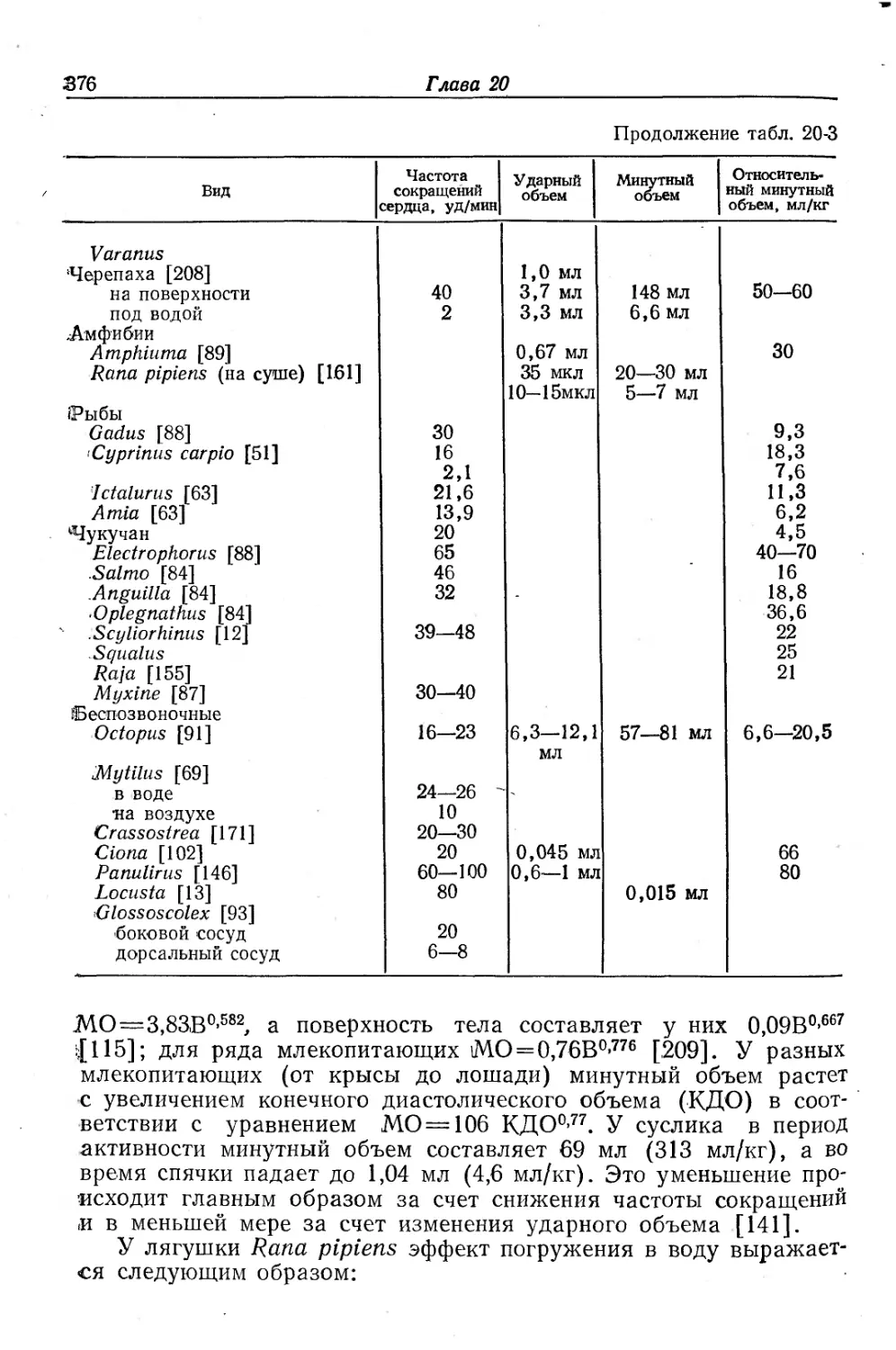









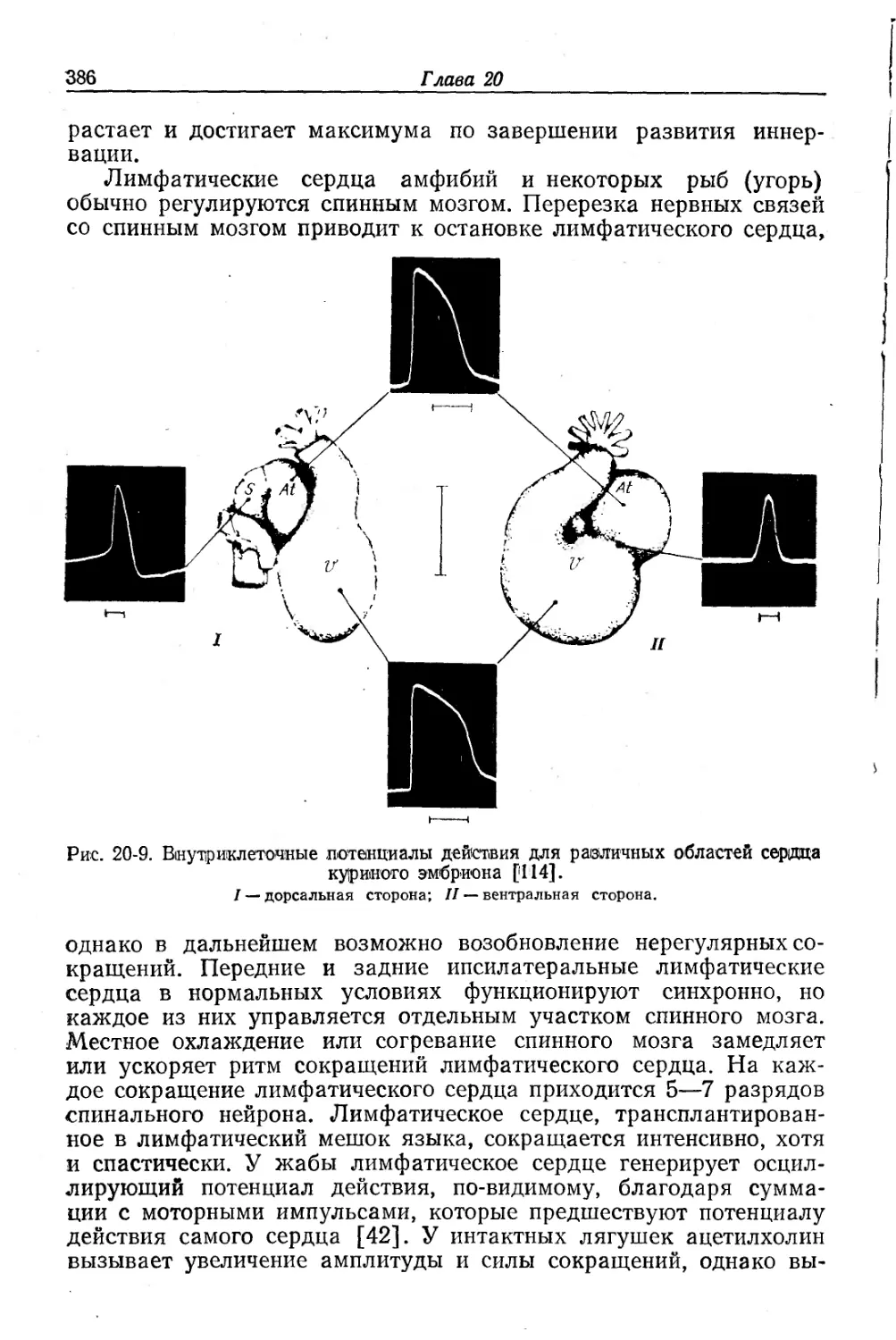








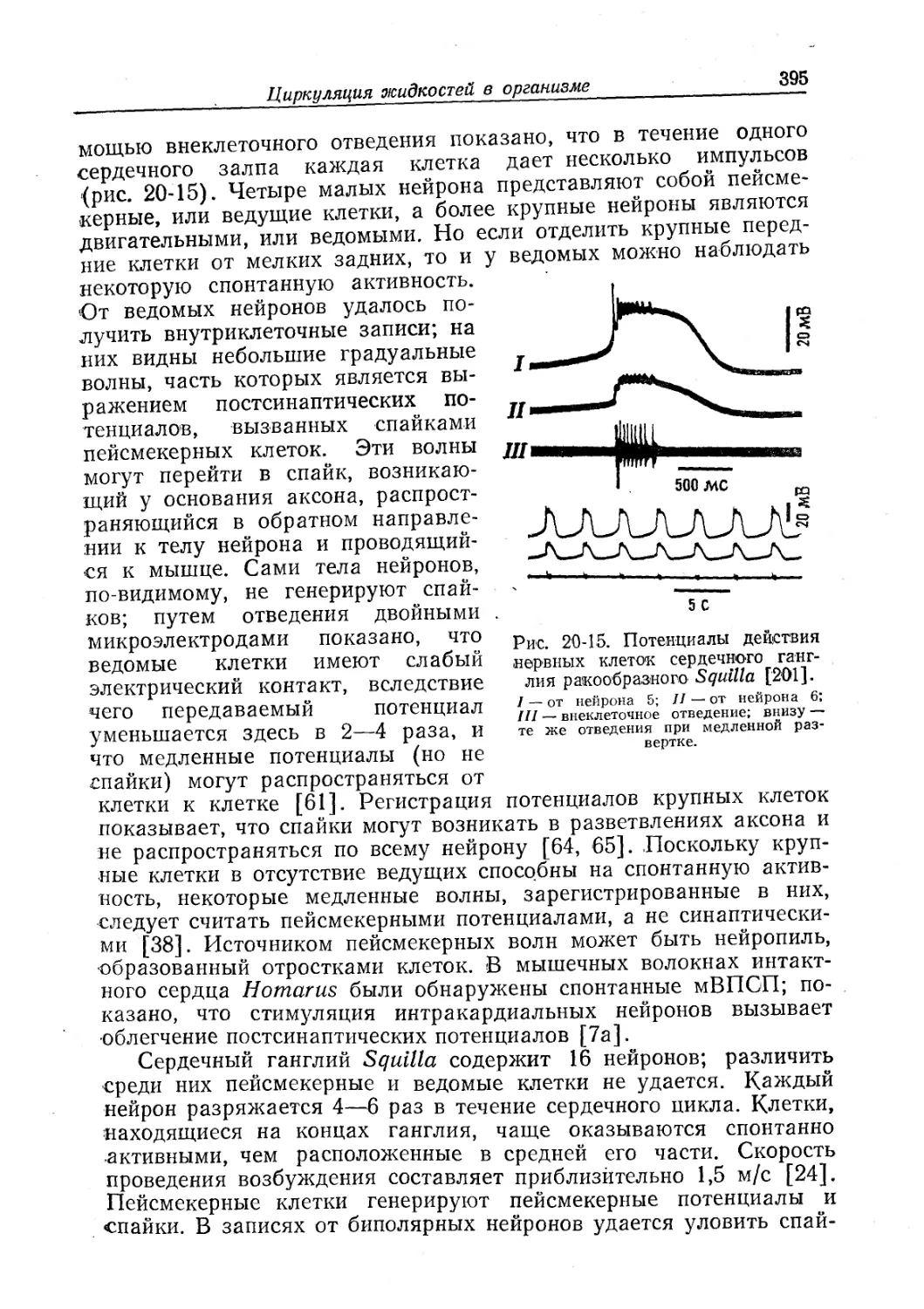




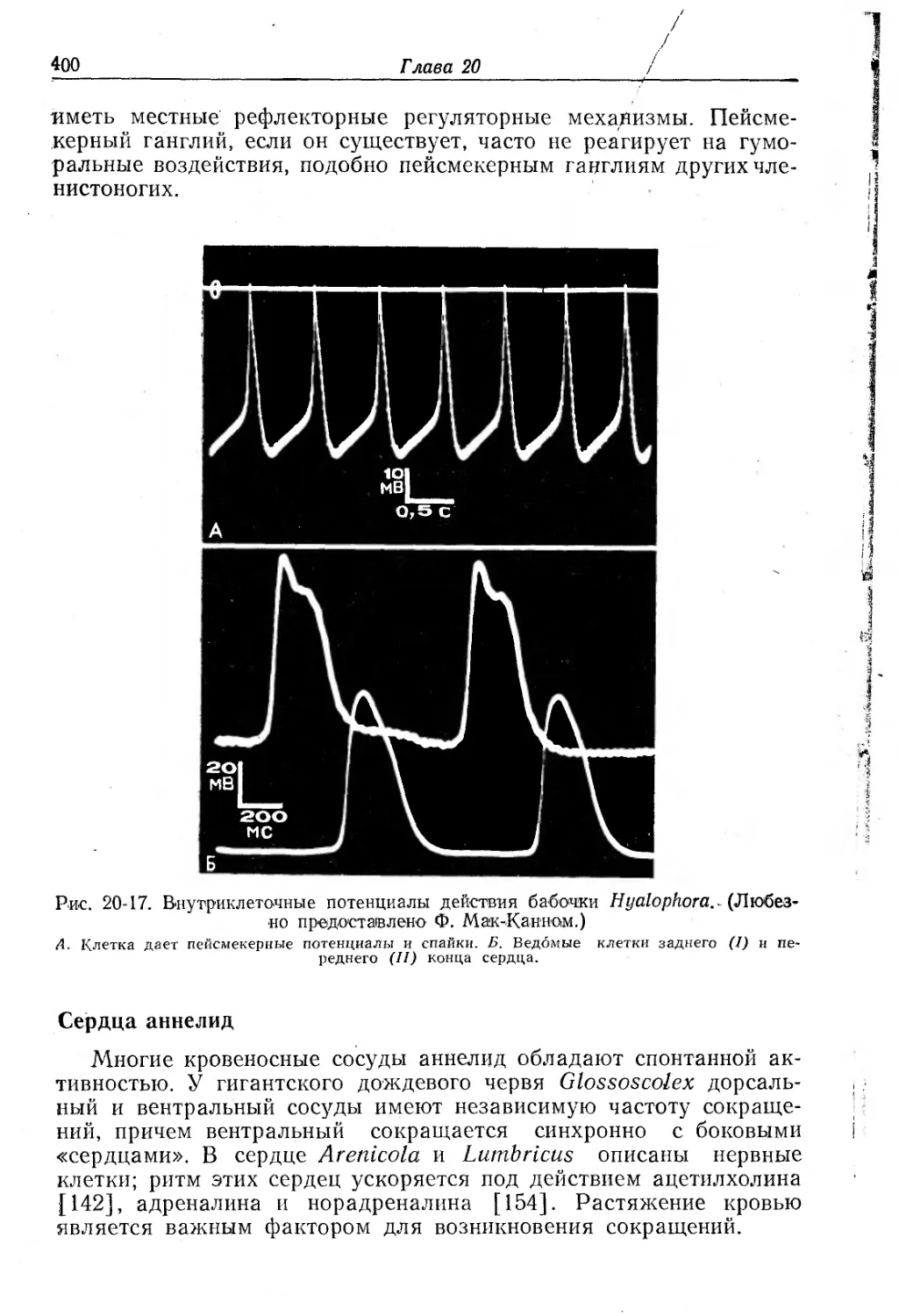
















































































































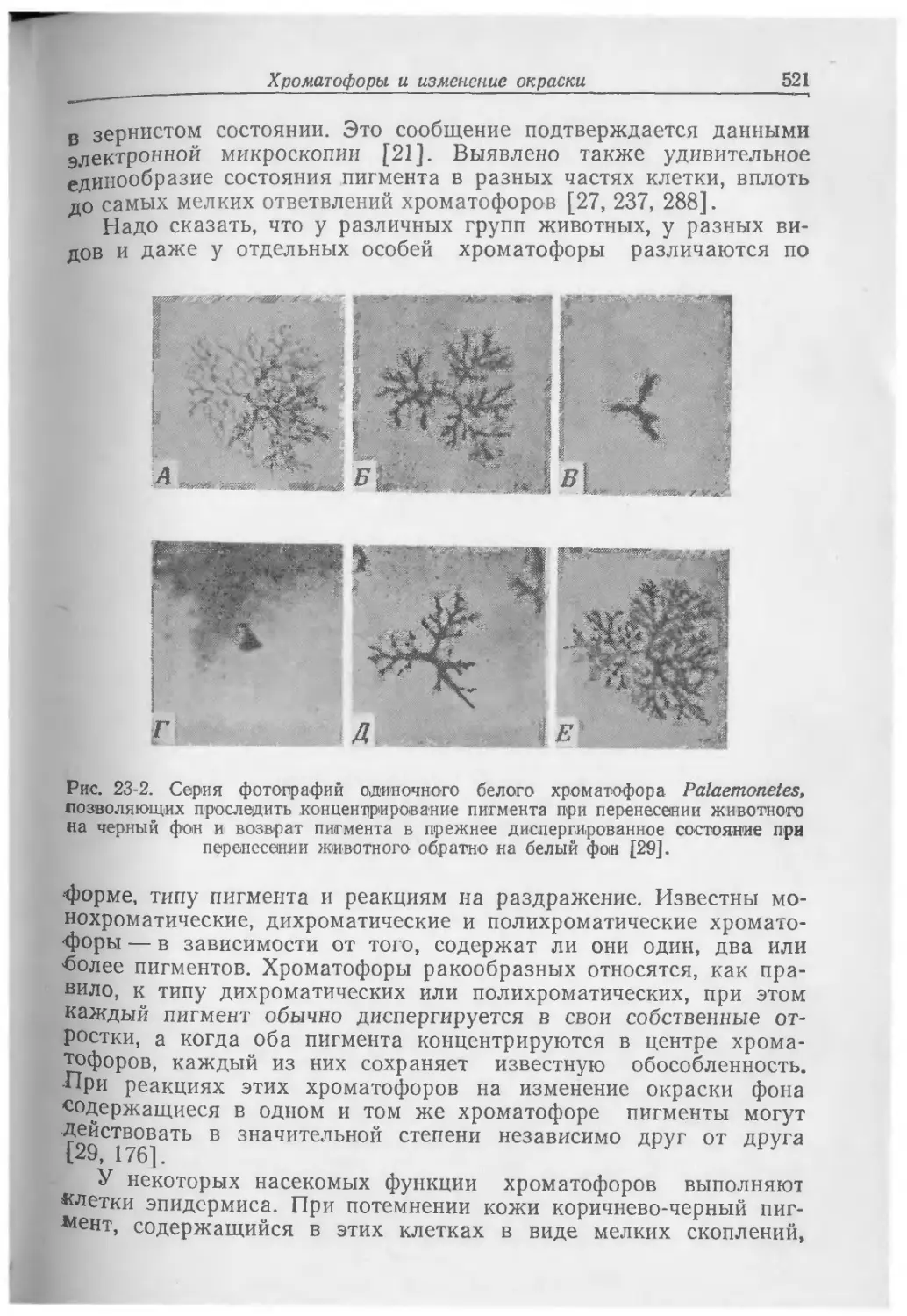


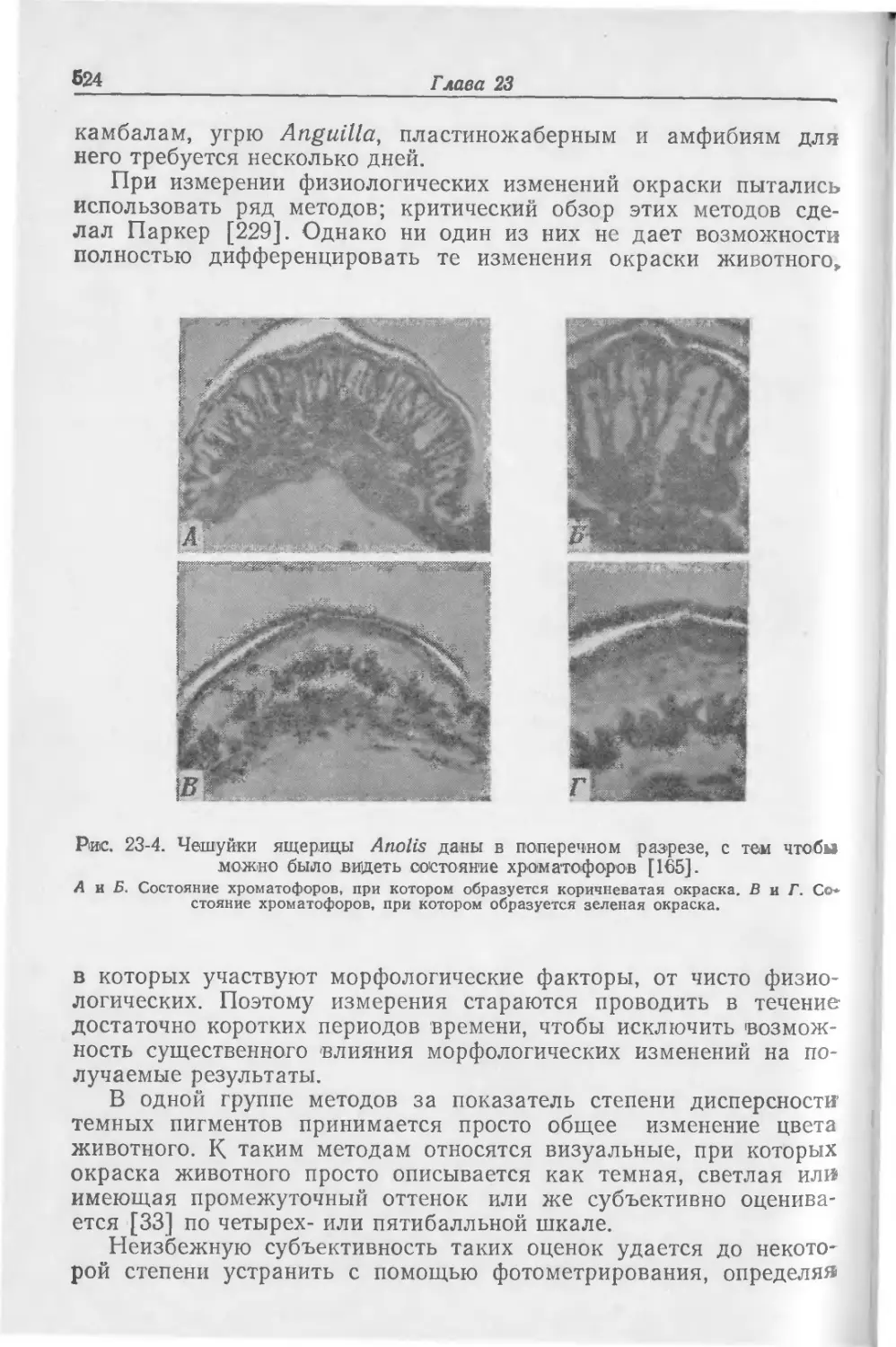























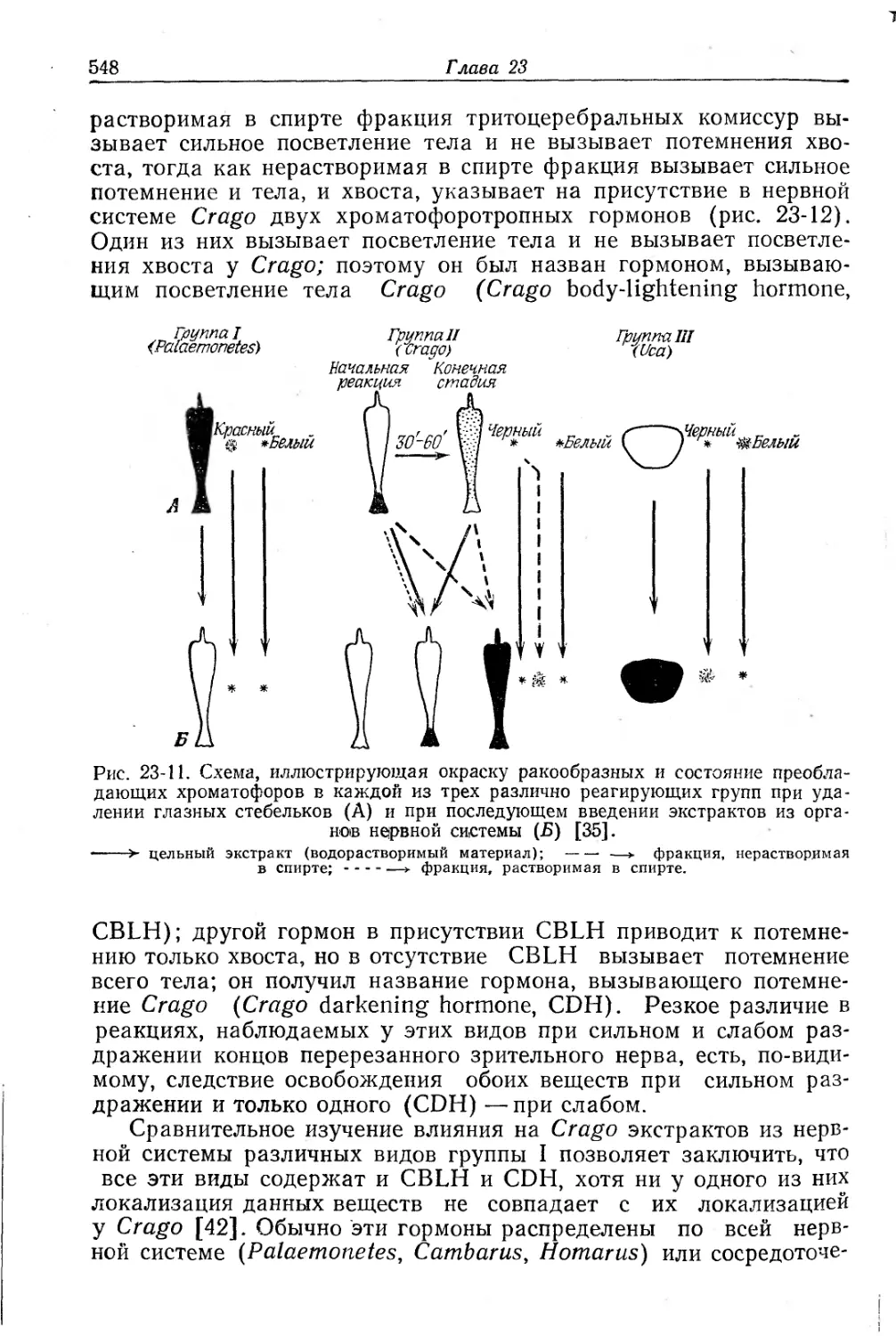

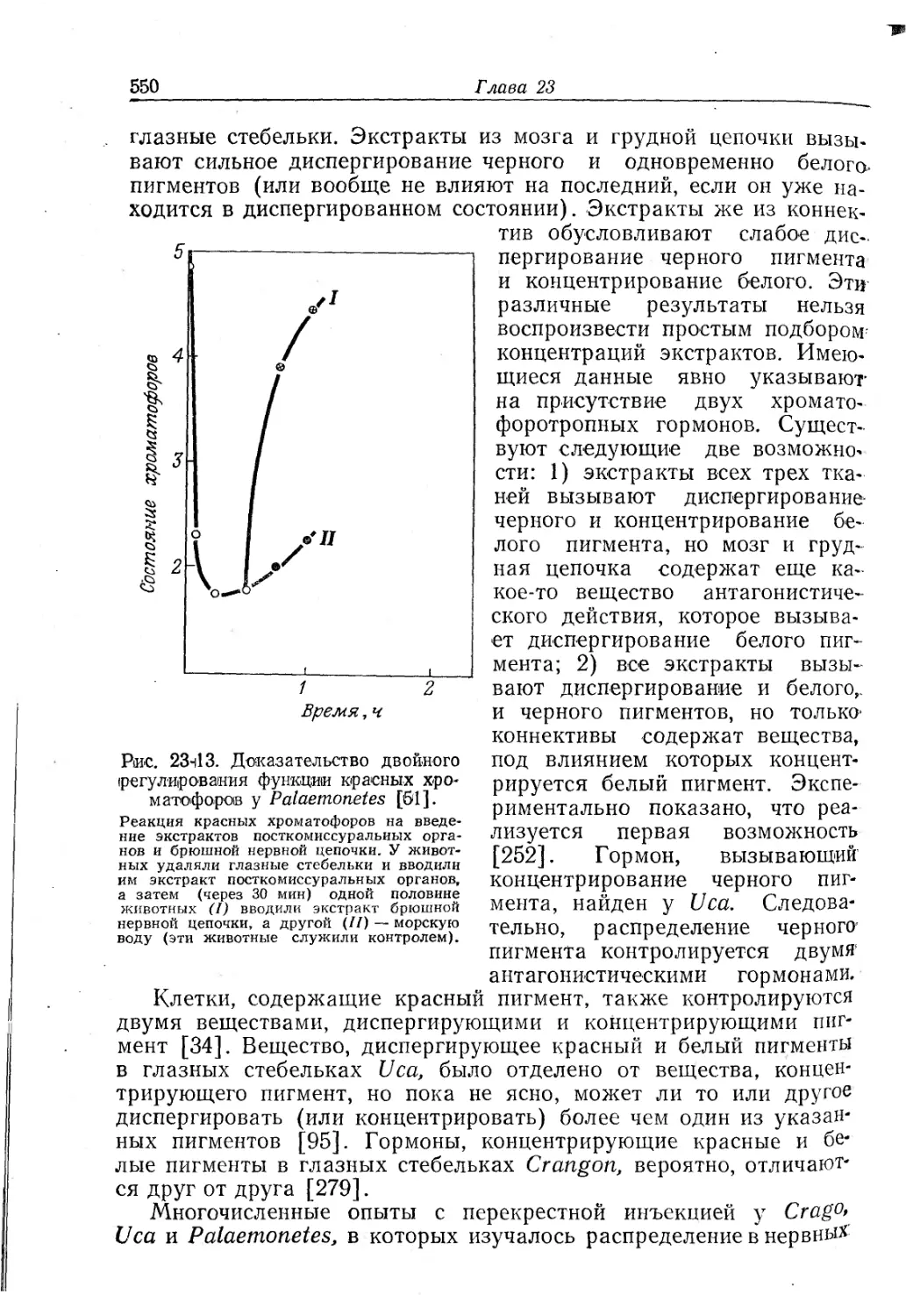


















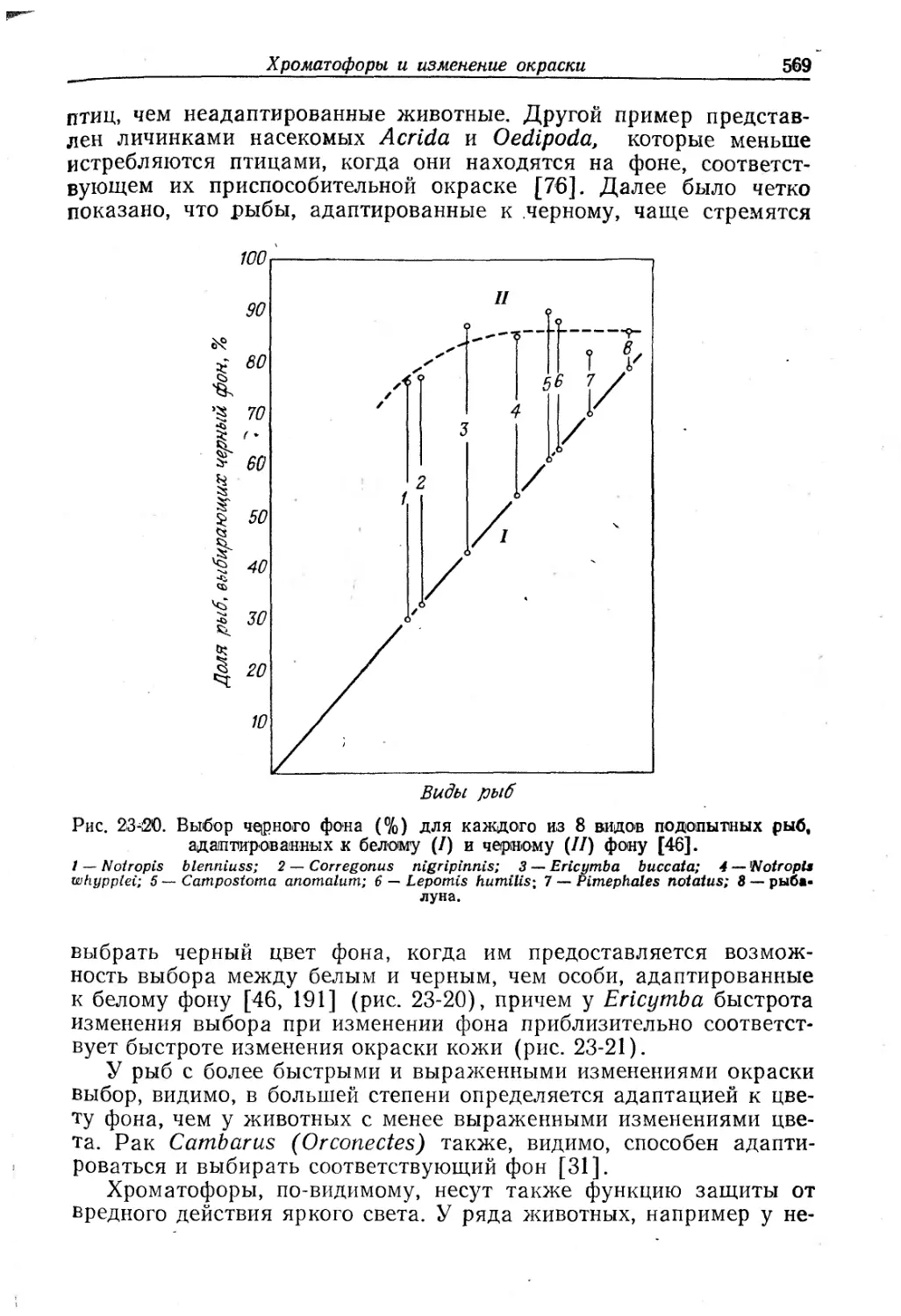
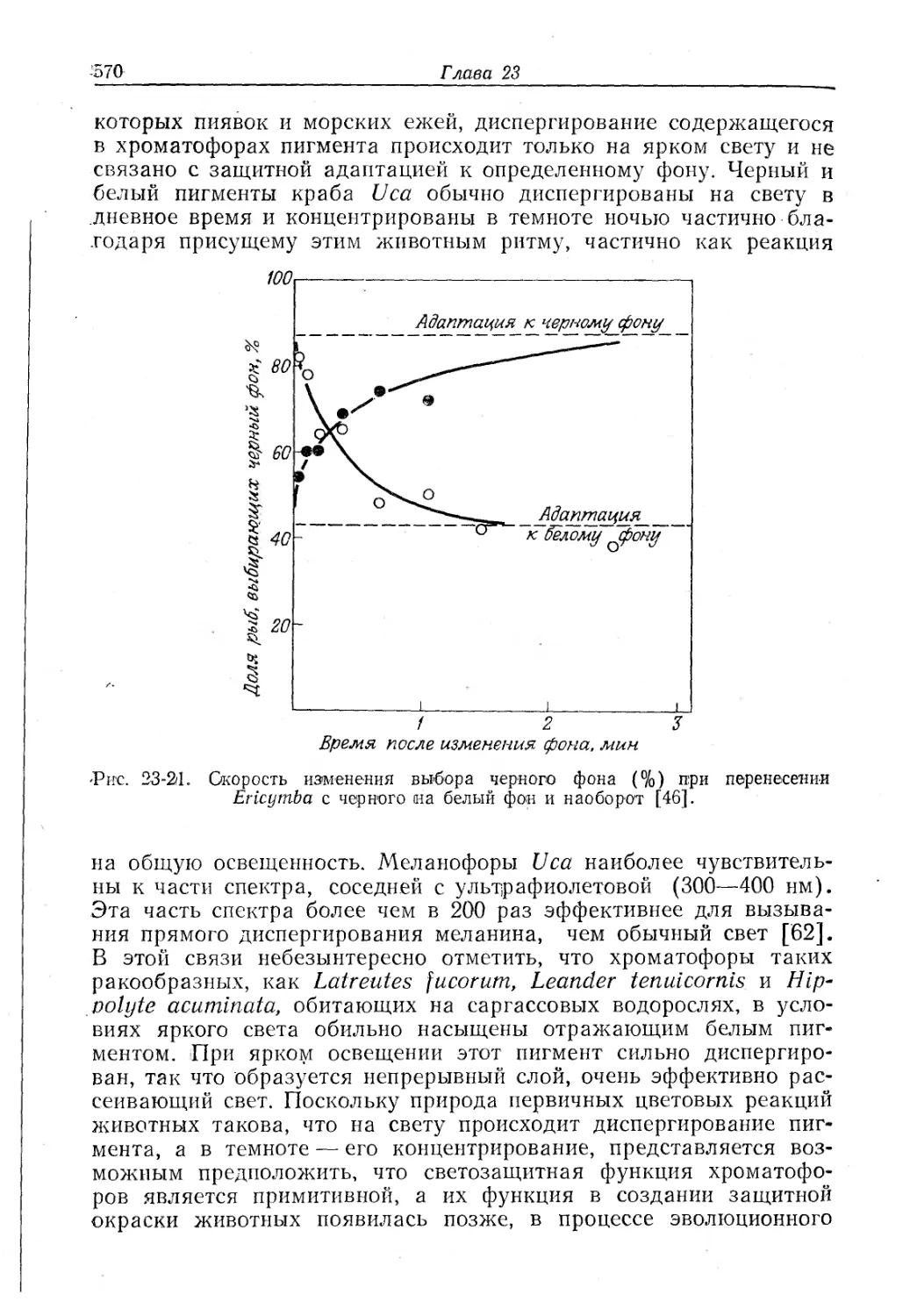
















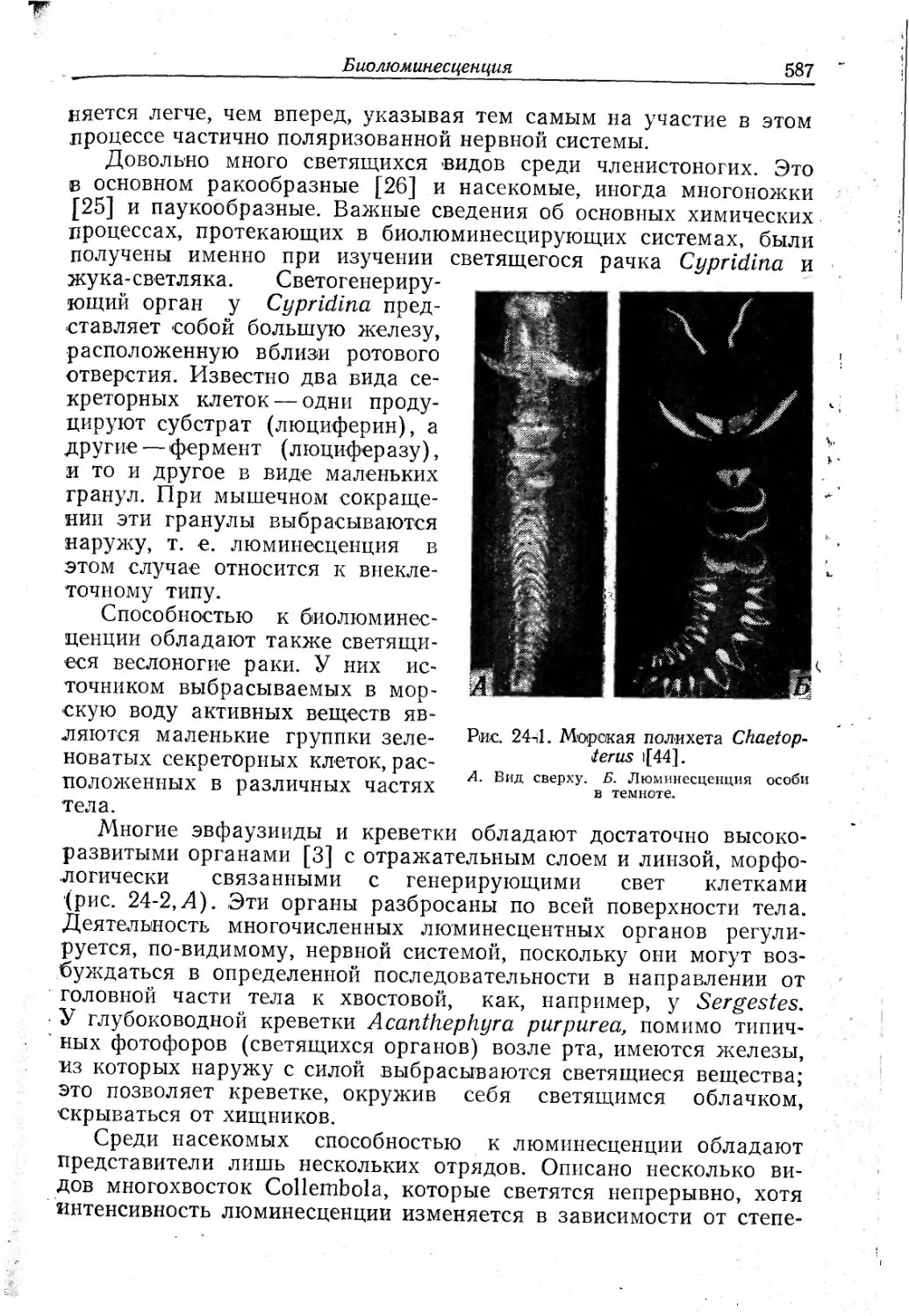
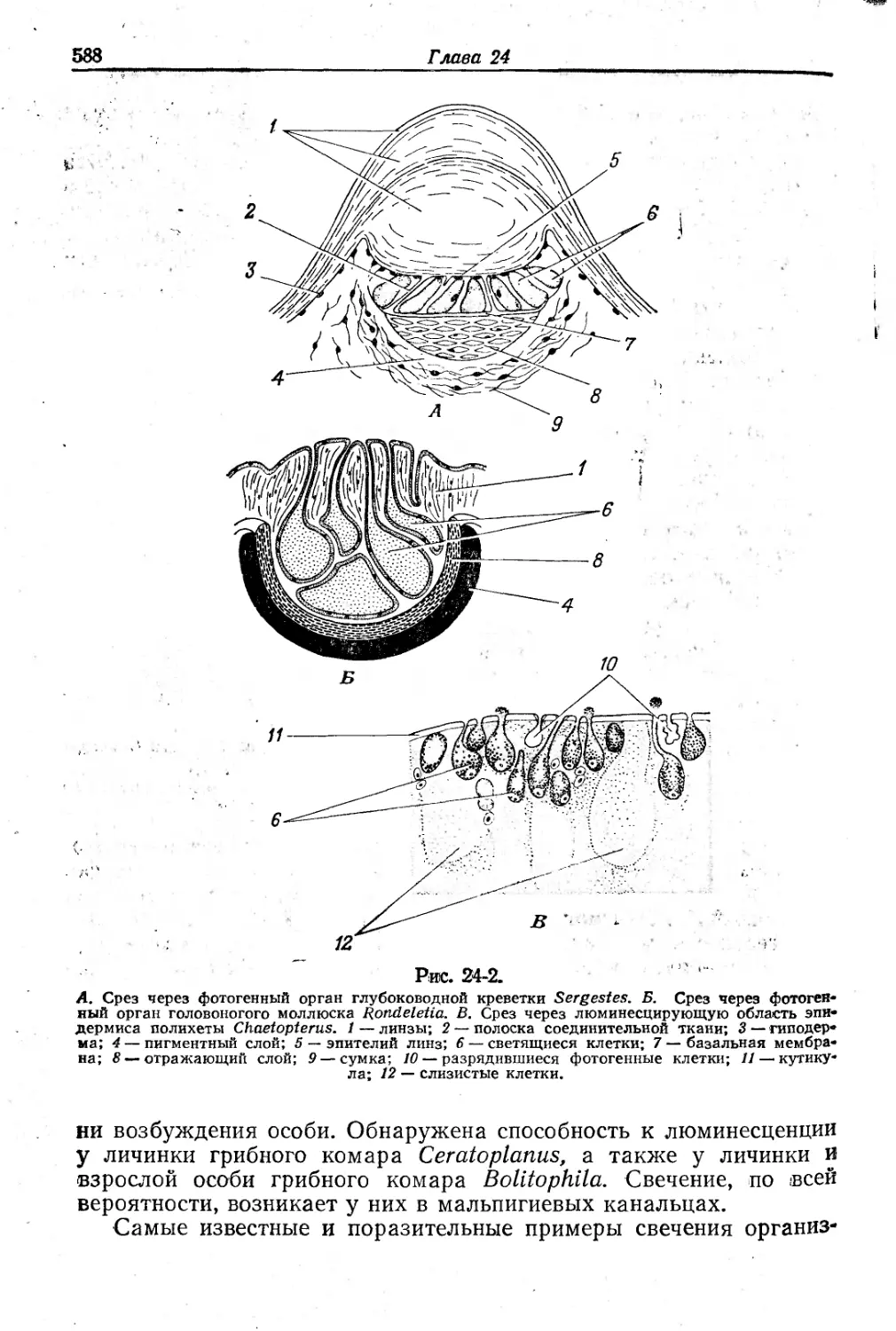

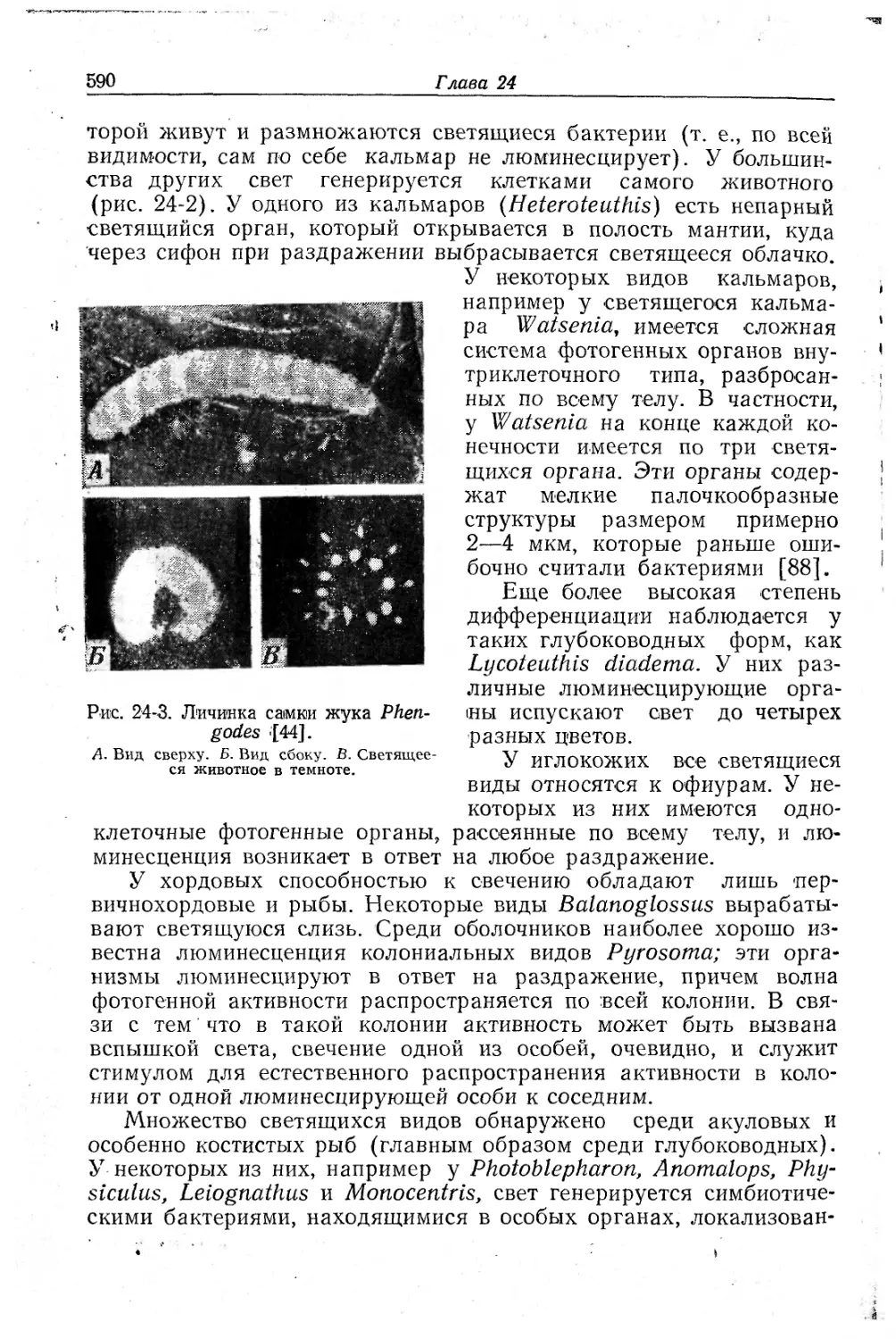













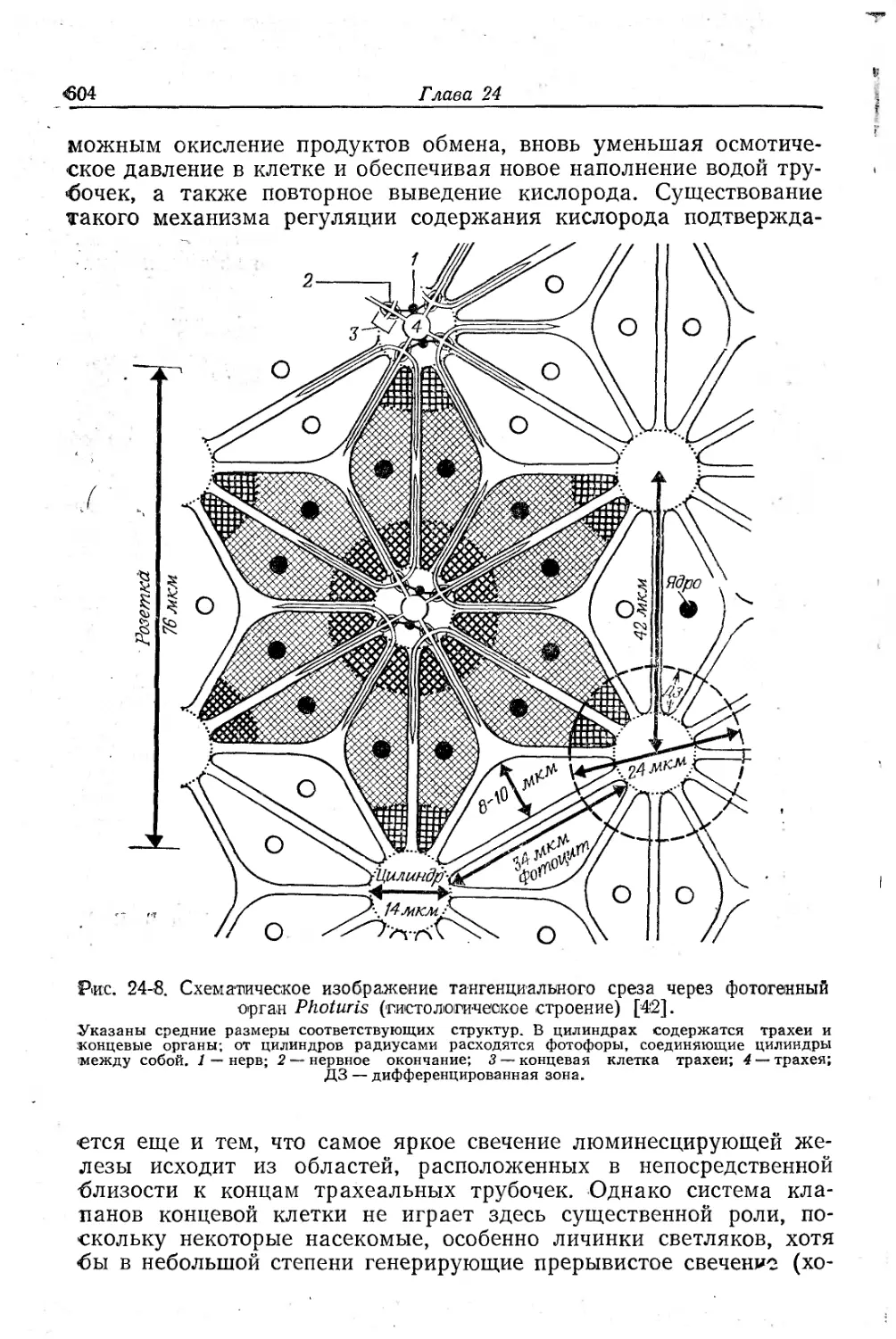
























































Text
СРАВНИТЕЛЬНАЯ ФИЗИОЛОГ ИЯ
ЖИВОТНЫХ
Comparative Animal Physiology
Edited by C. Ladd Prosser
Professor of Physiology and Zoology Department of Physiology and Biophysics University of Illinois at Urbana — Champaign
Third Edition
W. B. SAUNDERS COMPANY PHILADELPHIA LONDON TORONTO 1973
Сравнительная физиология
1
животных том ш
Под редакцией Л. Проссера
Перевод с английского
под редакцией
чл.-корр. АН СССР Т. М. Турпаева
ИЗДАТЕЛЬСТВО «МИР»
МОСКВА
1978
УДК 591.1
,В 1977 г. в издательстве «Мир» вышли первые два тома фундаментального труда по сравни тельной физиологии животных, сочетающего в себе достоинства учебного пособия и монографии, в которой собраны основные факты и наиболее важная литература по этой обширной области науки. В 1-ми 2-м томах приведены материалы по пищеварению, дыханию, 'водному, минеральному и азотистому обмену и т. д.
В третьем томе рассмотрены центральная нервная система, мышцы, электрические органы, амебоидное движение, реснички, циркуляторные системы, эндокринная система, нематоцисты и их аналоги, хроматофоры и изменение окраски, биолюминесценция.
Книга предназначена для биологов всех специальностей, в первую очередь физиологов, биохимиков, экологов, зоологов, для студентов, аспирантов и преподавателей соответствующих кафедр университетов и институтов, а также инженеров, занимающихся проблемами бионики и кибернетики.
Редакция биологической литературы
© 1973 by W. В. Saunders Company 50300—000
С 041(01)—78 Подп- изд-
© Перевод на русский язык, «Мир», 1978
Глава 15
Центральная нервная система
Л. ПРОССЕР
Основной задачей нейробиологии является познание нервных механизмов поведения. За последние годы сравнительная нейрофизиология внесла существенный вклад в выяснение природы нервных импульсов и синаптической передачи, а также механизмов рецепции и мышечного сокращения. Достигнуты также определенные успехи в сравнительно-физиологических исследованиях закономерностей центральной нервной интеграции функции сенсорных и двигательной систем и крупных вставочных нейронов [52, 215].
Методы исследования
Нейробиологи не вполне единодушны в вопросе о наиболее адекватных методах исследования ЦНС. Ниже перечисляются те из методов, с помощью которых были получены новые сведения о функции нервной системы различных животных.
1. Исследования поведения животных как в природных условиях, так и в условиях лабораторных опытов. Очевидно, что ограничение или усложнение движений животных в лабораторных условиях искажает их поведение. Введение телеметрических методик открывает новые возможности для наблюдений в природных условиях.
2. Хирургические и фармакологические воздействия на ЦНС, нарушающие те или иные поведенческие акты (или делающие их более выраженными), служат первым этапом при определении локализации функций. При таких вмешательствах трудно, однако, отделить первичные эффекты от вторичных.
3. Физиологические измерения соотношений между входом и выходом рефлекторных реакций в норме и при наркозе дают ценную информацию для анализа разных типов локомоций, пищевых актов, реакций избегания.
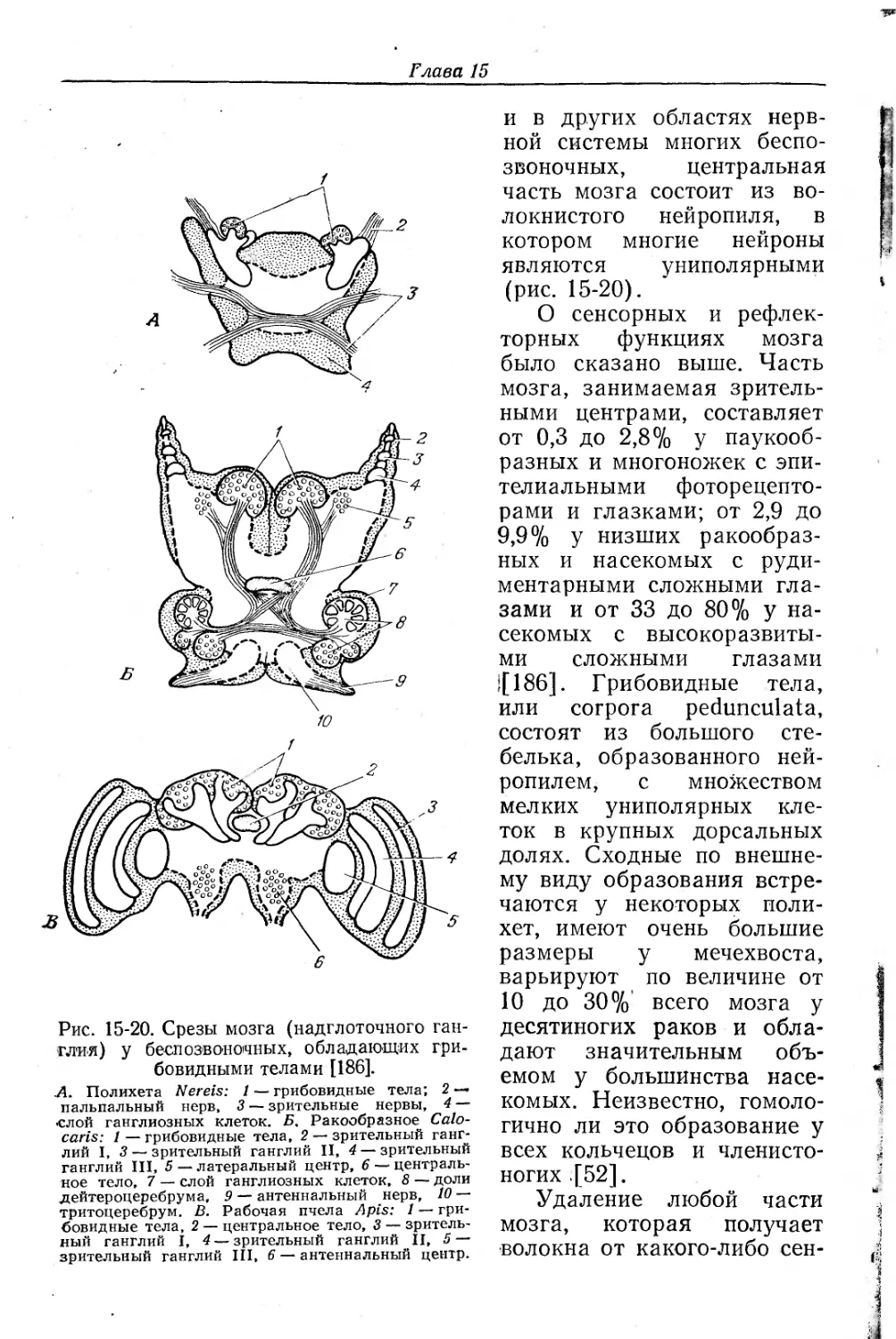
4. Для понимания функции нервной системы необходимо располагать нейроанатомическими сведениями, касающимися всех уровней — макроскопического, микроскопического и ультраструк- г турного. Детали ультраструктуры синапсов были описаны в гл. 11.
6
Глава 15
5. Сравнительное изучение особенностей нейрохимии разных отделов нервной системы способствует пониманию механизма действия синаптических медиаторов и их метаболизма. Современная нейрофармакология создает реальную возможность химического воздействия на разные функции нервной системы.
6. Регистрация электрической активности и измерение различных констант возбудимой мембраны являются основными способами изучения функции нервной системы; электрические свойства аксонов и синапсов описаны в гл. 11. Для анализа интегративной деятельности ЦНС оправдали себя два подхода, а) Регистрация электрической активности одиночных нейронов вне- или внутриклеточными электродами; отведения от отдельных нервных клеток используются при исследовании афферентных, эфферентных и крупных вставочных нейронов; однако еще не существует способов отведения активности от мелких вставочных нейронов нейропиля. б) В нервных системах животных, обладающих корой или послойной нейрональной организацией (например, мозжечок и большой мозг позвоночных), можно записывать суммарные потенциалы, которые представляют собой алгебраическую сумму электрической активности популяции нейронов, и, в частности, суммарные синаптические токи; конфигурация этих суммарных токов дает нам сведения об активации и взаимодействии совокупности нейронов.
7. Создание моделей нервных цепей, в частности машинных моделей, позволило высказать некоторые гипотезы, а также дать рекомендации по проведению экспериментов на живых системах и дальнейшему моделированию функций.
Общие свойства нервной интеграции
Большинство функциональных свойств нервной системы, поддающихся экспериментальному определению, можно обнаружить почти у всех многоклеточных, но некоторые свойства особенно удобно изучать на определенных видах животных.
Распределительные системы: полярность, задержка. Самой простой нервной системой, обладающей свойствами координации, является система, подобная распределительному щиту. Выход ее представляет собой некоторую функцию входа. Преобразование может происходить в отношении один к одному, с суммацией или без суммации конвергирующих входов. Одного входа может быть недостаточно для запуска выхода, и тогда совокупность входов является или аддитивной (суммация), или мультипликативной (облегчение) . Один вход может уменьшать ответ на другой, т. е. быть тормозным. Один вставочный нейрон запускает несколько мотонейронов (дивергенция) или же несколько вставочных нейронов конвергируют на одном мотонейроне.
Центральная нервная система
7
В сложных сетях информация кодируется так, чтобы она избирательно активировала специальные нейроны и хранилась в памяти. Избирательность в центральной нервной системе в значительной мере основана на специфических межнейронных связях и сетях, которые подчас трудно бывает представить в виде схемы. Градация и характер сигналов выражаются амплитудой напряжения при электротонической передаче или же частотой и порядком следования импульсов, подчиняющихся закону «все или ничего». Возможно, существует и химическое кодирование в виде участков специфических мембранных липопротеидов, каждый из которых обусловливает определенное распределение ионной проницаемости.
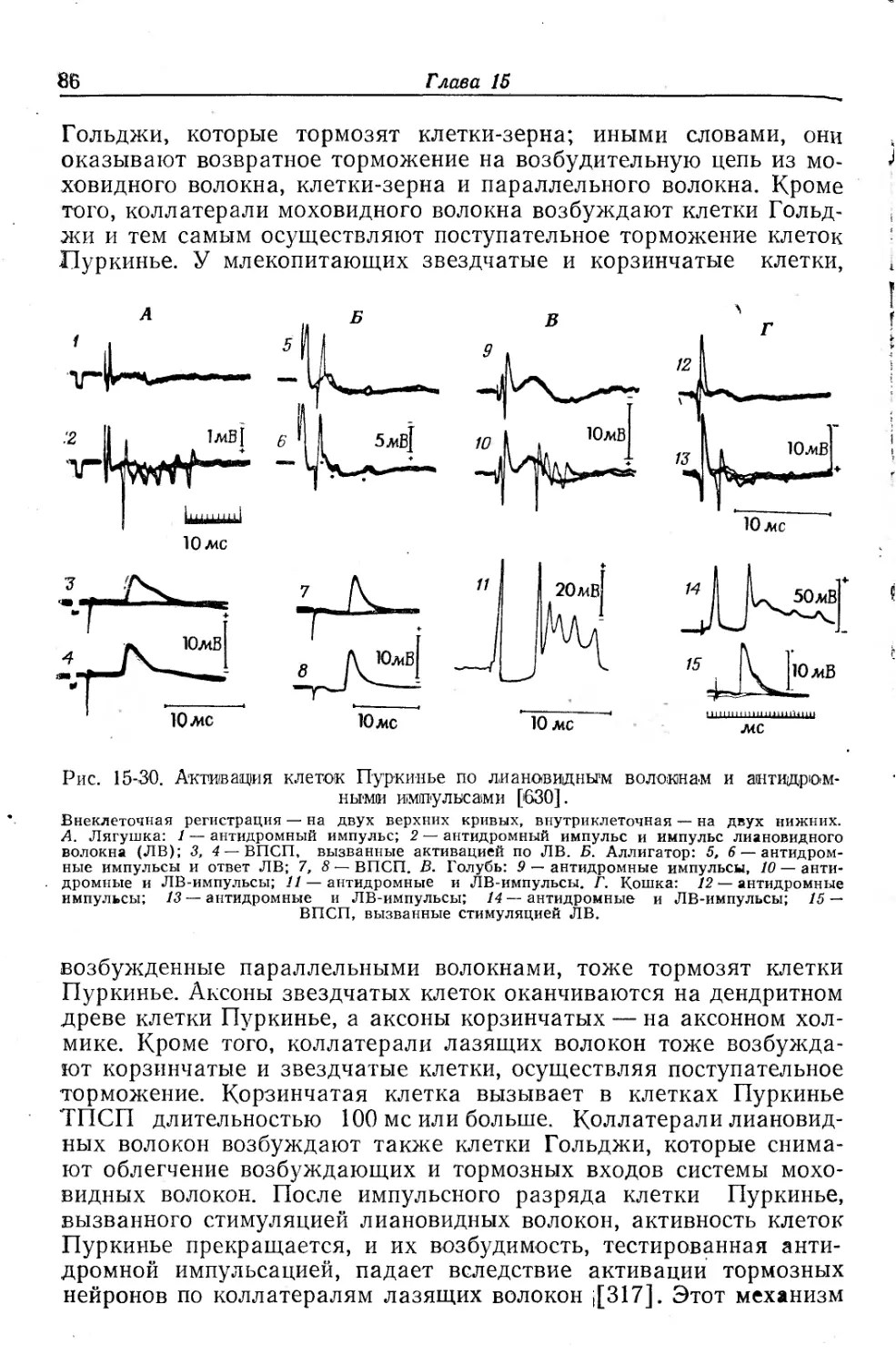
Для того чтобы придать _ смысл рассмотрению центральных уровней нейронной организации у разных групп животных, нужно дать несколько определений, приложимых к установленным реакциям, или рефлексам. В одной из предыдущих глав (гл. 11) дано описание синаптической задержки и полярности. Неполяри-зованные электрические синапсы встречаются сравнительно редко; они обнаружены в перегородчатых гигантских волокнах и в некоторых центральных нервных системах со строгой синхронностью в активности нескольких нейронов. Электрические (поляризованные) синапсы обеспечивают быстроту и синхронность действия, но допускают относительно малую степень интеграции. Поляризация синапсов (химическая или электрическая) гарантирует одностороннее проведение, поскольку она препятствует входу антидромных импульсов в нервный путь. Однако многие нейроны, особенно те, аксоны которых выходят из центра, снабжены возвратными веточками для обратной связи (feed-back) через вставочные нейроны (рис. 15-1). Обычно такая обратная связь осуществляет торможение, блокируя ритмическую импульсацию эфферентного нейрона. Примерами могут служить клетки Реншоу в спинном мозге, клетки Пуркинье в мозжечке млекопитающих и маутнеровские нейроны у рыб. Иногда возвратные коллатерали являются возбуждающими и вновь вызывают ритмическую или круговую импульсацию, как это имеет место в нервных сетях. Иного типа контроль осуществляется аксонами, входящими в центр и отдающими тормозные коллатерали, которые оканчиваются на нейроне, возбуждаемом затем по параллельному пути или же вторым импульсом в первичном пути; таким образом нейрон тормозится или же возбудимость его снижается еще до прихода к нему возбуждения,-Такое торможение называют поступательным (feed-forword inhibition); оно препятствует возникновению импуль-сации вставочного нейрона в ответ на обычные входные сигналы. В качестве примера можно привести ганглии аплизии и параллельные волокна мозжечка, активирующие корзинчатые клетки, которые оказывают тормозное действие на клетки Пуркинье (а эти последние сами возбуждаются параллельными волокнами).
8
Глава 15
В некоторых нервных центрах существует связь между дендритами, как, например, у пейсмекерных нейронов сердечных ганглиев ракообразных. У многих мультиполярных нейронов в центральной нервной системе беспозвоночных дендриты не отли-
Агл чаются от аксонов, и прове-
kJ < дение в одном и том же отростке не обязательно од-
А - — -И.---.—- - ностороннее. Иной меха-
низм кодирования в ветвящихся нейронах основан
на наличии участков волокон разного диаметра и < возникающих вследствие этого различиях в факторе надежности и ограничениях частоты в окончаниях. В об-
щем микроархитектоника нервных центров обеспечивает большое разнообразие сигнализации по поляризо-< ванным межнейронным связям.
Еще один тип локаль-
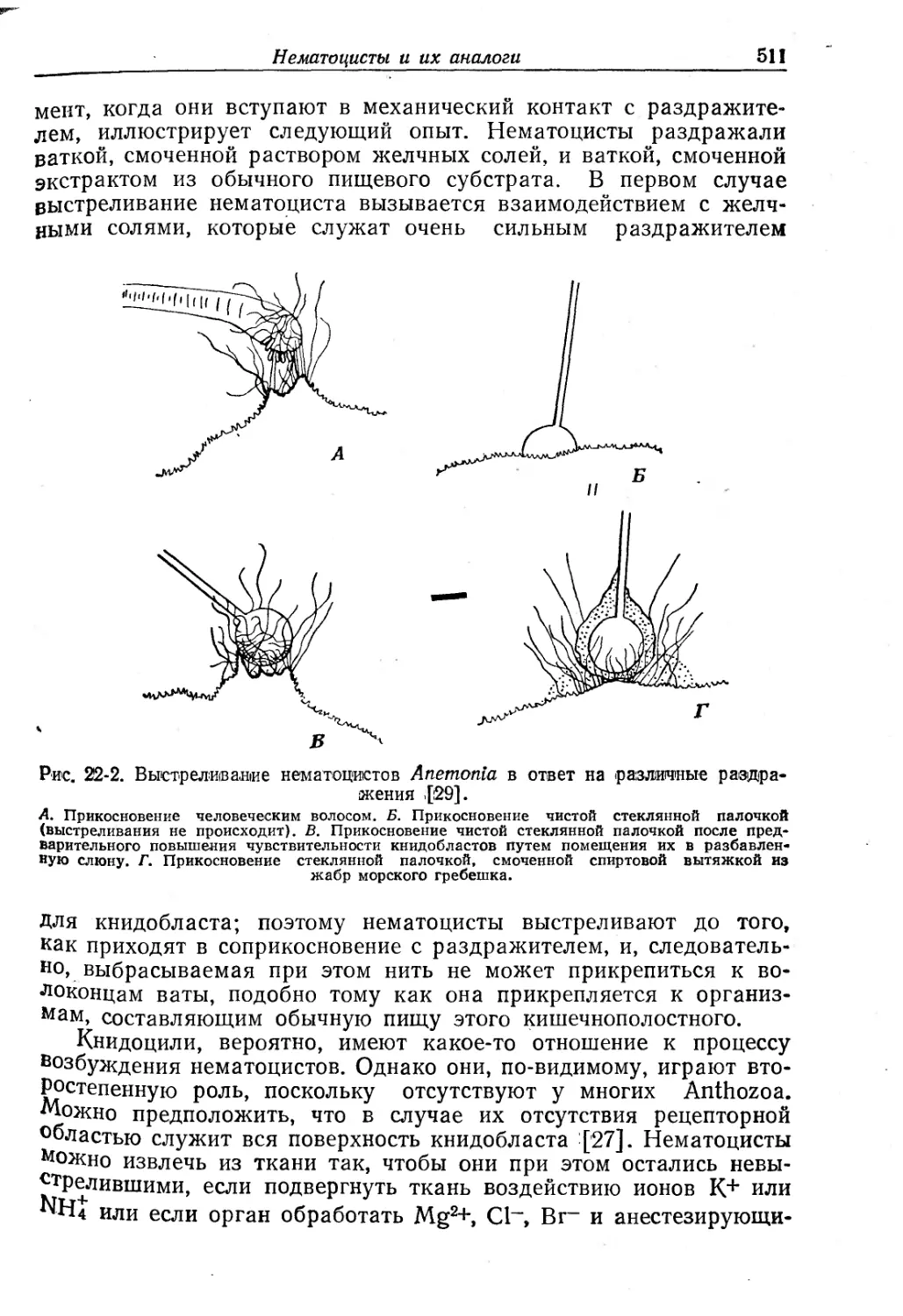
Рис. Г5-'1. Схемы цепей возвратного и поступательного торможения.
А. Непосредственное возвратное торможение. Б. Возвратное торможение через вставочный нейрон. В. Перекрестное пресинаптическое поступательное торможение. Г. Перекрестное постсинаптическое поступательное торможение через вставочные нейроны. Светлые треугольники — возбудительные синапсы, черные треугольники—тормозные синапсы.
ного контроля осуществляется благодаря внешним полям, создаваемым син-< хронизированными импуль-
сами и синаптическими
потенциалами. Так, например, из спинного мозга в передние и задние корешки распространяются медленные потенциалы. У лягушки антидромная стимуляция некоторых аксонов передних к-орешков деполяризует соседние мотонейроны, и
это взаимодействие усиливается ортодромией (сенсорной) импуль-сацией. Таким образом, между перекрывающимися дендритами соседних мотонейронов существует электрическое взаимодействие. Антидромные импульсы в аксонах переднего корешка вызывают
также потенциалы, которые можно зарегистрировать извне в задних корешках, но в отличие от мотонейронных эффектов эти потенциалы уничтожаются холинэргическими блокаторами; следовательно, они обусловлены синаптическими связями возвратных волокон мотонейронов. Вероятно, антидромные импульсы эквивалентны активности возвратных коллатералей ортодромного пути
Центральная нервная система
9
[173]. Ниже описана корреляция между потенциалами поля и вероятностью импульсации корковых нейронов.
Суммация и облегчение. В большинстве случаев при синаптической передаче в интегративных центрах и в некоторых нервно-мышечных соединениях для возникновения спайка недостаточно одного входного импульса. Каждый пресинаптический импульс вызывает синаптический потенциал, однако необходимость большего числа входных импульсов создает фактор надежности против разряда эфферентных нейронов в ответ на однократный афферентный импульс. В релейных, или распределительных, центрах, какими являются некоторые сенсорные ядра, симпатические ганглии или мантийный ганглий кальмара, передача возможна в отношении один к одному. Когда требуется суммация двух входных процессов, то они суммируются либо в пространстве (два или более конвергирующих волокна), либо во времени (два импульса в одном волокне, быстро следующие один за другим). Суммация означает простое сложение потенциалов в синапсе независимо от того, вызваны ли они из одного или из разных источников, а облегчение означает, что два входа вызывают эффект больший, чем их алгебраическая сумма. Так, для двух конвергирующих волокон, или для пространственного облегчения, (a + b)>a + b, а для двух быстро следующих друг за.другом событий в одном волокне, или временного облегчения, (2а)Д>а + а. Эти два вида облегчения могут сочетаться в том смысле, что максимальное облегчение создается при синхронности обоих входных сигналов, т. е. тогда, когда оба синапса находятся на равном расстоянии от места генерации импульса. Облегчение сходит на нет с постоянной времени, близкой к постоянной времени ВПСП. Сгибание одной сенсорной щетинки на хвосте у рака вызывает один импульс или серию импульсов, идущих в шестой брюшной ганглий, но для того, чтобы возник один постсинаптический импульс, требуется стимуляция нескольких (обычно четырех) волосков. В одиночных волокнах, идущих к мышце ракообразного, резко выражено временное облегчение (гл. 16). Центральное облегчение первоначально выражали числом выходных импульсов; по Шеррингтону, этот термин включает пространственную суммацию, которая ведет к облегчению импульсации. Современное применение этого термина допускает различие между суммацией и облегчением на уровне синаптических потенциалов; в препаратах, где нельзя измерить синаптические потенциалы, различие это не всегда ясно.
Другой тип облегчения, измеряемый по рефлекторному выходу, достигает максимума через некоторое время после начального входного сигнала. Так, при рефлекторном сокращении сгибателей у кошки максимальный ответ на второй афферентный залп наступает через 10—20 мс после начального залпа. В некоторых препаратах установлена связь задержанного облегчения с обходной, или параллельной, цепью из нескольких нейронов, по которой им
10
Глава 15
пульсы приходят к постсинаптическому нейрону одновременно со вторым, облегчающим, сигналом. В таком аспекте задержанное облегчение выходной активности в действительности представляет собой пространственную суммацию [ПО]. Явление, обратное облегчению (дисфасилитация), обозначает снижение ПСП при повторении; возможно, оно представляет собой синаптический механизм поведенческого привыкания.
Гетеросинаптическим облегчением называют длительную по-тенциацию синаптического ответа при сочетании слабого тестирующего входного сигнала с более сильным, направляющимся по другому, конвергентному, пути «кондиционирующим» (priming) сигналом. После ряда таких сочетаний один лишь тест-сигнал вызывает сильный ответ. Возможно, что это явление представляет собой пресинаптическое облегчение [281].
Торможение. В большей части интегративных центров торможение является такой же важной синаптической функцией, как и возбуждение. На выходе нейрона создается баланс между этими двумя процессами. Торможение может быть пресинаптическим, и тогда оно снижает выделение возбуждающего медиатора и препятствует разряду постсинаптического нейрона даже в том случае, когда на постсинаптической мембране потенциал не меняется. Торможение может быть по ст синаптическим, меняющим проводимость в постсинаптической мембране, снижая таким образом ответ на возбуждающий медиатор.
Приведем пример пресинаптического торможения в спинном мозге. К окончаниям афферентных волокон, идущих от мышечных веретен, пресинаптическое торможение при быстром сокращении поступает от афферентов, начинающихся в сухожилиях, и в результате мышечные афференты не выделяют медиатора на постсинаптическую мембрану мотонейрона. Примером постсинаптического торможения спинальных мотонейронов служит следующее явление: коллатеральные веточки от аксонов мотонейрона возбуждают вставочные нейроны Реншоу, которые в свою очередь тормозят мотонейрон и препятствуют повторному разряду после начального возбуждения [10]!.
Как правило, постсинаптическое торможение осуществляется через вставочные нейроны, иногда со значительным латентным периодом. Торможение модулирует спонтанную активность и входные сенсорные сигналы. Примеры торможения мышцы и органов чувств даны в гл. 12 и 16. Выходная активность центральных нейронов обычно зависит от алгебраической суммы возбуждения и торможения, и каждый нейрон имеет и возбудительные и тормозные участки рецептивной мембраны. Одним из способов центрального кодирования может служить варьирование относительной величины этих участков.
Ритмические ответы и спонтанная активность. В ответ на длительную деполяризующую стимуляцию многие нервные волокна
Центральная нервная система
И
дают ритмические разряды с частотой более высокой, чем та, которая лимитируется длительностью рефрактерного периода. В некоторых нервных центрах часть рефлекторного ответа нейрона состоит из разряда последействия, или ритмической импульсации. Многие центральные нейроны обладают ритмической активностью и в отсутствие сенсорного или какого-либо другого входа; иными словами, они разряжаются спонтанно. Приходящие сигналы могут модулировать эту спонтанную импульсацию, повышая или снижая ее частоту, что создает большие возможности для градации передачи информации, чем если бы нейрон «молчал», и единственным эффектом входных сигналов был бы запуск его импульсации. В интегративных нервных системах спонтанная нервная активность наблюдается повсюду. В распределительных нервных центрах она встречается реже.
Известны два типа спонтанности в центральной нервной системе. В ганглиях многих беспозвоночных и в большей части интегративных областей головного мозга позвоночных наблюдается спонтанная активность отдельных клеток. Частота импульсации варьирует от клетки к клетке, и синхронизация выражена слабо. Одиночная клетка способна разряжаться регулярно или пачками, разделенными периодами молчания, как в сердечных ганглиях ракообразных. В тот или иной момент времени спонтанно активно лишь небольшое число клеток. Спонтанная импульсация чувствительна к температуре и нуждается в энергии, поставляемой окислительным обменом, но связь между обменом и ритмическими изменениями проводимости не установлена. Второй тип спонтанной активности обнаружен преимущественно в корковых структурах — в коре больших полушарий, мозжечке и тектуме. Здесь синхронизована активность очень многих клеток, и этим создаются медленные ритмы, которые необязательно связаны с импульсацией; это могут быть медленные изменения мембранного потенциала. Синхронизация обеспечивается отчасти реверберационными кругами, а отчасти — электротонической связью между нейронами.
У рака-отшельника и омаров ритм вентиляционных движений регулируется парой нейронов, у которых мембранные потенциалы не генерируют спайков, но вызывают спайки в мотонейронах дыхательных мышц [364]. У аплизии одни нейроны разряжаются равномерно в пределах одной пачки, у вторых частота разряда постепенно снижается, а у третьих она возрастает, а потом падает. При устранении импульсации воздействием тетродотоксина в лишенной кальция среде сохраняются ритмические медленные волны, соответствующие по частоте устраненному импульсному разряду. Эти медленные волны исчезают под действием уабаина или в лишенной натрия среде; возможно, они служат отражением ритмической активности натриевого насоса [519]i. Весьма вероятно, что в создании осциллирующих мембранных потенциалов и
12
Глава 15
ритмической импульсации участвует несколько разных механизмов [15, 63] и что для интегративных нервных центров характерна спонтанная активность.
Ниже мы приведем много примеров поведенческого значения спонтанной активности и запрограммированных последовательностей импульсов в нервных центрах. Наблюдения над животными после деафферентации показывают, что многие координированные движения не требуют обратной сигнализации от органов чувств и придатков. Возможно, что для определения характера ритмической активности сенсорная информация играет большую роль при развитии животного, чем на более поздних стадиях жизненного цикла. После того как структура активности сформировалась, сенсорные сигналы, быть может, лишь запускают запрограммированную последовательность поведенческих актов. Они могут также поддерживать тонический уровень возбудимости в центре. Известны многочисленные примеры эндогенных центральных ритмов ходьбы и полета у насекомых [589]!. Дыхательные нейроны в стволе мозга позвоночных сохраняют активность и после деафферентации. На характер вокализации у птиц не влияет деаффе-рентация, которая устраняет приход звуковых сигналов (см. гл. 12). В картине активности нейронов трудно отделить генетическую детерминированность от онтогенетической. Но так или иначе, ясно, что в интегративных центрах на клеточном уровне существует спонтанная активность и что в нейронных цепях и кругах осуществляется запрограммированная последовательность импульсации.
Системы гигантских волокон
Примерами простых быстродействующих распределительных систем служат гигантские волокна. Они возникли независимо в нескольких группах животных; их следует считать результатом специализации, а вовсе не примитивными структурами.
Существует несколько типов систем гигантских волокон все возрастающей сложности (от волокон с непрерывным проведением до перегородчатых нейронов с электрическими синапсами и до цепей гигантских волокон с химическими синапсами), и в том числе некоторые из самых сложных известных нам одиночных нейронов.
Мы уже упоминали о гигантских волокнах как о приспособлениях, обеспечивающих быстрые реакции, в качестве примеров электронотонических соединений и электрических синапсов (см. гл. 11). Гигантские волокна многоклеточного или функционально синцитиального типа встречаются у кольчатых червей, ракообразных и головоногих моллюсков. Крупные нейроны, называемые гигантскими, встречаются часто, особенно у насекомых и низших позвоночных.
Центральная нервная система
13
Системы гигантских волокон участвуют главным образом в быстрых движениях, в реакциях избегания и хватания, которые начинаются резко и обычно не градуальны. Они эффективны, поскольку один импульс в гигантском волокне активирует мышцы больших участков тела. Скорость проведения в гигантских волокнах в 2—10 раз выше, чем в мелких волокнах у того же животного.
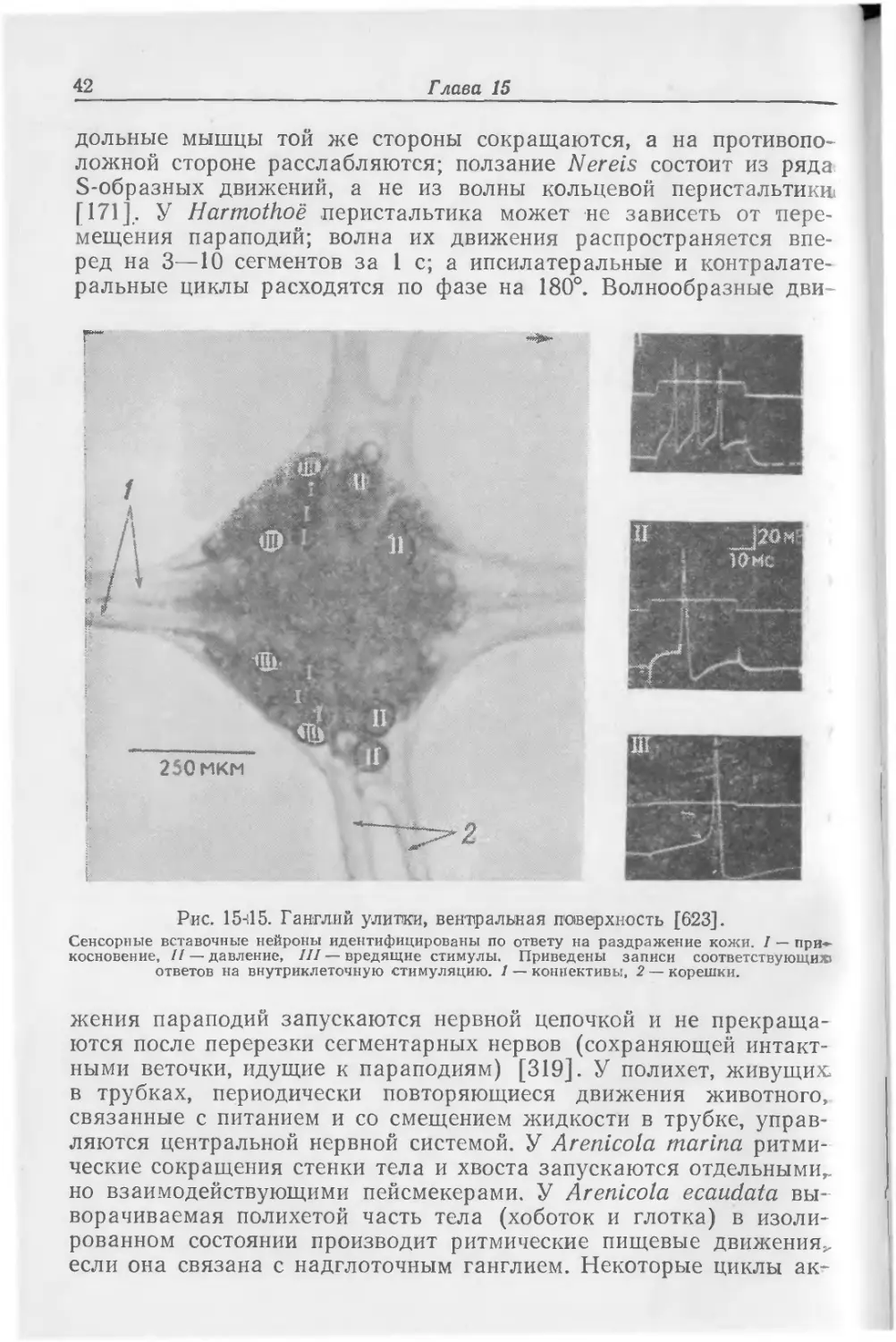
Дождевые черви
У дождевых червей описаны септированные (перегородчатые) гигантские волокна — два латеральных и одно медиальное; каждое из них. соединено с одним телом клетки в каждом сегменте. Оба латеральных волокна соединены между собой перекрестными электрическими синапсами и отдают по 5 веточек в каждый сегмент [384]. По этим гигантским волокнам передаются сигналы быстрого сокращения, распространяющегося из конца в конец туловища. Перерезка отдельных гигантских волокон и последующая тактильная стимуляция показывают, что в медиальном волокне проведение обычно направлено назад, а в латеральных волокнах— вперед. В изолированных цепочках импульсы проводятся в каждом волокне в обоих направлениях. Полярность в проведении возникает потому, что медиальное волокно получает сенсорные связи в переднем конце туловища (в первых 40 сегментах), а латеральный комплекс — в заднем отрезке тела червя [50]. Перегородки (септы) представляют собой остатки эмбриональных границ клеток. Регистрация мышечных сокращений, возникающих при стимуляции гигантских волокон с разной частотой и последовательностью, показывает, что совместное раздражение обоих латеральных и медиального волокон вызывает более сильные быстрые сокращения, чем раздражение только одного волокна. Кроме того, многократно вызываемые ответы подвержены утомлению, которое можно снять облегчающими стимулами, причем изменения возбудимости длятся до 9 с. Эти изменения происходят на уровне соединений гигантских волокон с мотонейронами [440,
Полихеты
У одних полихет нет гигантских волокон (Aphrodite, Chaetop-terus), у других имеется только одно такое волокно (Arenicola, Pista), у третьих их несколько (Neanthes, Glycera) [52];. Myxi-cola обладает одним волокном диаметром 560 мкм. У представи- > телей сабеллид и серпулид гигантские волокна образуют перекресты в середине тела; при утомлении место перехода нисходящих импульсов с одной стороны на другую перемещается к более передним перекрестам. У этих червей можно зарегистрировать си-
Центральная нервная система 15 наптические потенциалы в области связей между двумя волокнами, причем гиперполяризация снижает ВПСП. Электротониче-ских соединений у них мало (1:30), и взаимодействие осуществляется, очевидно, через химические синапсы. Кроме того, в области головы имеются прямые анастомозы [180]. При ритмической низкочастотной стимуляции ответ в синапсе между латеральными гигантскими волокнами и мотонейронами исчезает (наступает утомление); восстановить этот ответ можно высокочастотной стимуляцией (облегчение). Когда синапс между латеральным гигантским волокном и мотонейроном перестает проводить возбуждение (у Nereis или Harmothoe), реактивность синапса можно восстановить стимуляцией медиального волокна [208].
Кальмары
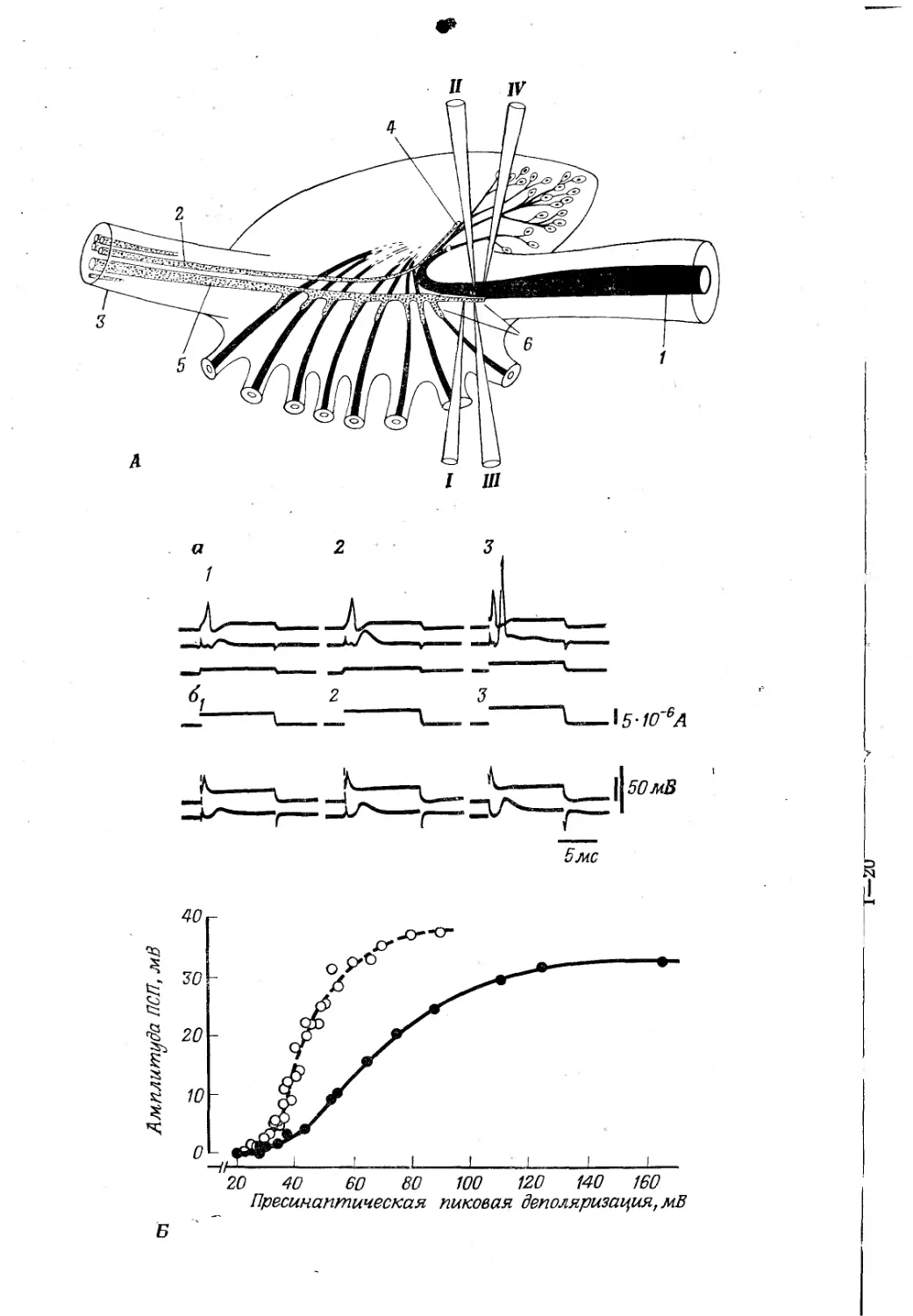
Кальмары имеют два гигантских нейрона в заднем паллиовис-церальном ганглии головного мозга; их аксоны сливаются и перекрещиваются, а затем образуют синапсы в висцеральной части головного мозга со второй группой гигантских нейронов, отростки которых проходят в составе мантийных нервов (рис. 15-2) [602]. Эти гигантские волокна второго порядка образуют синапсы в звездчатом ганглии с третьей группой гигантских нейронов, которые снабжают мантийную мышцу. Гигантские аксоны третьего порядка образованы слиянием отростков многочисленных (300— 1500) нейронов, начинающихся в звездчатом ганглии; каждый гигантский аксон третьего порядка получает ветви от крупного (200 мкм) и мелкого (75 мкм) гигантских нейронов второго порядка. Кроме того, еще одна группа крупных аксонов (добавочных) от головного мозга проходит к звездчатому ганглию, причем некоторые из них образуют синапсы на гигантских нейронах третьего порядка, а другие проходят через ганглий [59],. Синаптическая задержка составляет 0,55 мс; активность двух волокон второго порядка может суммироваться и дать спайк. Синапсы добавочных аксонов с гигантскими нейронами более чувствительны к утомлению и аноксии, чем синапсы гигантских волокон второго
Рис. 15-2.
А. Схема синапсов гигантского волокна мантийного ганглия кальмара (по [46]). Задний гигантский аксон (/), образованный слиянием отростков, идущих от скопления нейронов. Добавочные ^волокна (2) в преганглионарном нерве (3) образуют проксимальные синапсы (4), а крупный преганглионарный аксон (5)—дистальные синапсы (6); проведение через них измеряется посредством раздражающих электродов в пре- и постганглионарном волокнах (соответственно I и II) и отводящих электродов (соответственно III и /V). Б. Соотношение между пресинаптической деполяризацией и постсинаптическими потенциалами [619]. Верхняя запись — пресинаптическое, нижняя — постсинаптическое отведения. 1—3 — возрастающие уровни пресинаптического тока, а — нормальный препарат; ПСП, амплитуда которого возрастет по мере увеличения ответа в пресинаптических терминалях и который генерирует спайк в точке 3. б — препарат после применения тетродотоксина; синаптические токи не блокированы. График выражает отношение между ПСП и пресинаптической деполяризацией в обоих препаратах.
16
Глава 15
порядка. Один из гигантских аксонов третьего порядка крупнее остальных — его диаметр достигает 700—1000 мкм. Каждый гигантский аксон иннервирует большой участок мантии; поскольку к заднему концу идут самые крупные волокна, импульсы приходят ко всей мантии приблизительно одновременно. Скорость проведения (при 10 °C) варьирует от 5 до 25 м/с. Один моторный импульс в гигантском волокне вызывает максимальное сокращение мантийной мышцы. Это приводит к выбросу воды из мантийной полости через воронку, что в свою очередь приводит к движению животного вперед или назад в зависимости от положения воронки.
Рак
Латеральные гигантские волокна рака синаптически связаны: а) с сенсорными элементами и вставочными нейронами в нейропиле, б) с контралатеральным гигантским волокном посредством электрического синапса на месте перекреста в цепочке, в) с латеральным гигантским волокном соседних заднего и переднего сегментов в области перекрытия сегментов и г) с моторными гигантскими волокнами и другими мотонейронами флексоров следующего переднего сегмента [572] (рис. 15-3). Перекрывающиеся соединения латеральных гигантских волокон не поляризованы и обычно проводят в отношении один к одному, но при утомлении иногда отвечают не на каждый, а только на несколько импульсов (создающих облегчение). Тела сегментарных клеток расположены контралатерально и отдают ипсилатерально широкое дендритное ветвление [435]. Латеральные гигантские волокна образуют электрические синапсы с сегментарными гигантскими мотонейронами; на этих нейронах имеются также входы от средних волокон в виде электрических синапсов и от тормозных аксонов нервной цепочки в виде химических синапсов [572, 573]. Медиальные гигантские волокна не образуют синцития; их клетки лежат в надглоточном ганглии, где аксоны перекрещиваются. Импульс в медиальном гигантском волокне вызывает тормозную реакцию (ТПСП) в латеральном гигантском нейроне, которая устраняется пикротоксином. Кроме того, импульс в коллатералях латерального гигантского волокна вызывает возвратное торможение в обоих латеральных волокнах, не чувствительное к пикротоксину. Возвратное торможение в цепочке ограничивает число моторных импульсов, которое может быть вызвано одним афферентным залпом [437]1 Один импульс в гигантском волокне (как в латеральном, так и медиальном) вызывает сокращение брюшных флексоров, что приводит к удару хвостом. При повторных раздражениях наступает очень быстрое привыкание. При стимуляции сегментарных задних корешков в синапсах латеральных гигантских волокон возникают возбудительные потенциалы, состоящие из двух, а иногда из трех компонентов: ранняя волна стабильнее поздней,
Рис. 15-3. [©20].
А. Реконструкция брюшного ганглия рака, показывающая гигантский мотонейрон (отростки, выявленные при помощи инъекции красителя) и его отношение к одному из четырех гигантских волокон. Б. Схема и записи ответов мотонейронов и мышц-сгибателей у рака на стимуляцию различного типа. Стимуляция латерального гигантского волокна (I) вызывает ортодромный ответ в соме нейрона и в корешковых аксонах. Стимуляция сомы нейрона (II) вызывает прямой ответ сомы и синаптический мышечный ответ. Стимуляция корешкового аксона (III) вызывает антидромный ответ сомы нейрона и синаптический мышечный ответ.
18
Глава 15
а эта последняя представляет собой длительную активацию, вызванную вставочными нейронами [302]. Все синапсы на отростках латеральных гигантских нейронов электрические; такой характер передачи, возможно, связан с тем, что эти синапсы находятся на вентральных дендритах, на некотором расстоянии от места возникновения импульса, а также с высоким порогом возбуждения крупного латерального гигантского волокна (рис. 15-4) [617].
Рис. 15-4. Схема нейронов ('брюшко рака), участвующих в реакции избегания [617].
ТР — тактильные рецепторы; А — унисегментарный тактильный вставочный нейрон; Б и В — мультисегментарные тактильные вставочные нейроны; ЛГ — латеральное гигантское волокно; ГМ — гигантский мотонейрон; БМСг — быстрый мотонейрон сгибателя; справа сгибательная мускулатура. Черточками обозначены электрические синапсы; черным кружком — облегчающий химический синапс; светлыми кружками — антиоблегчающие химические синапсы.
Таракан
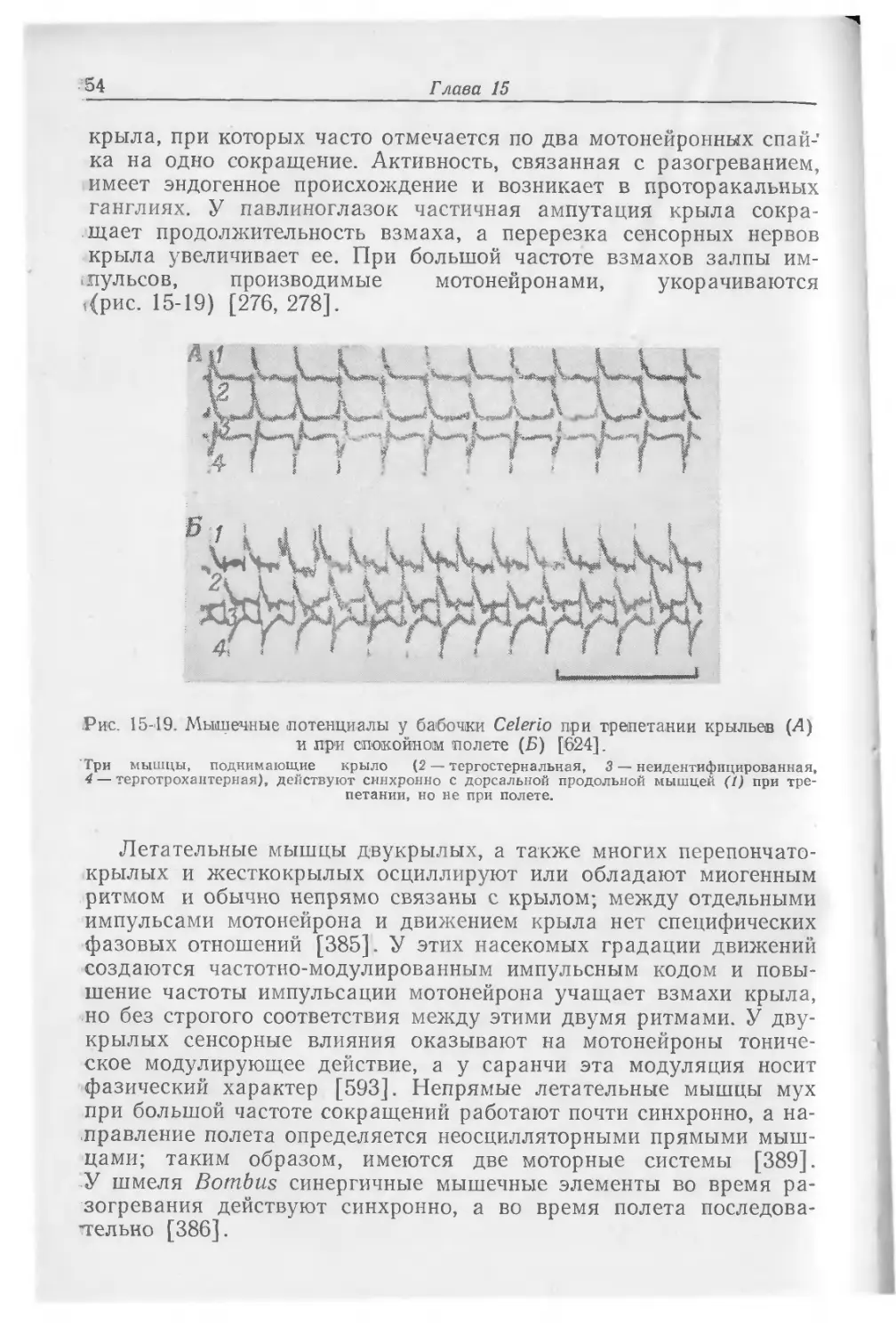
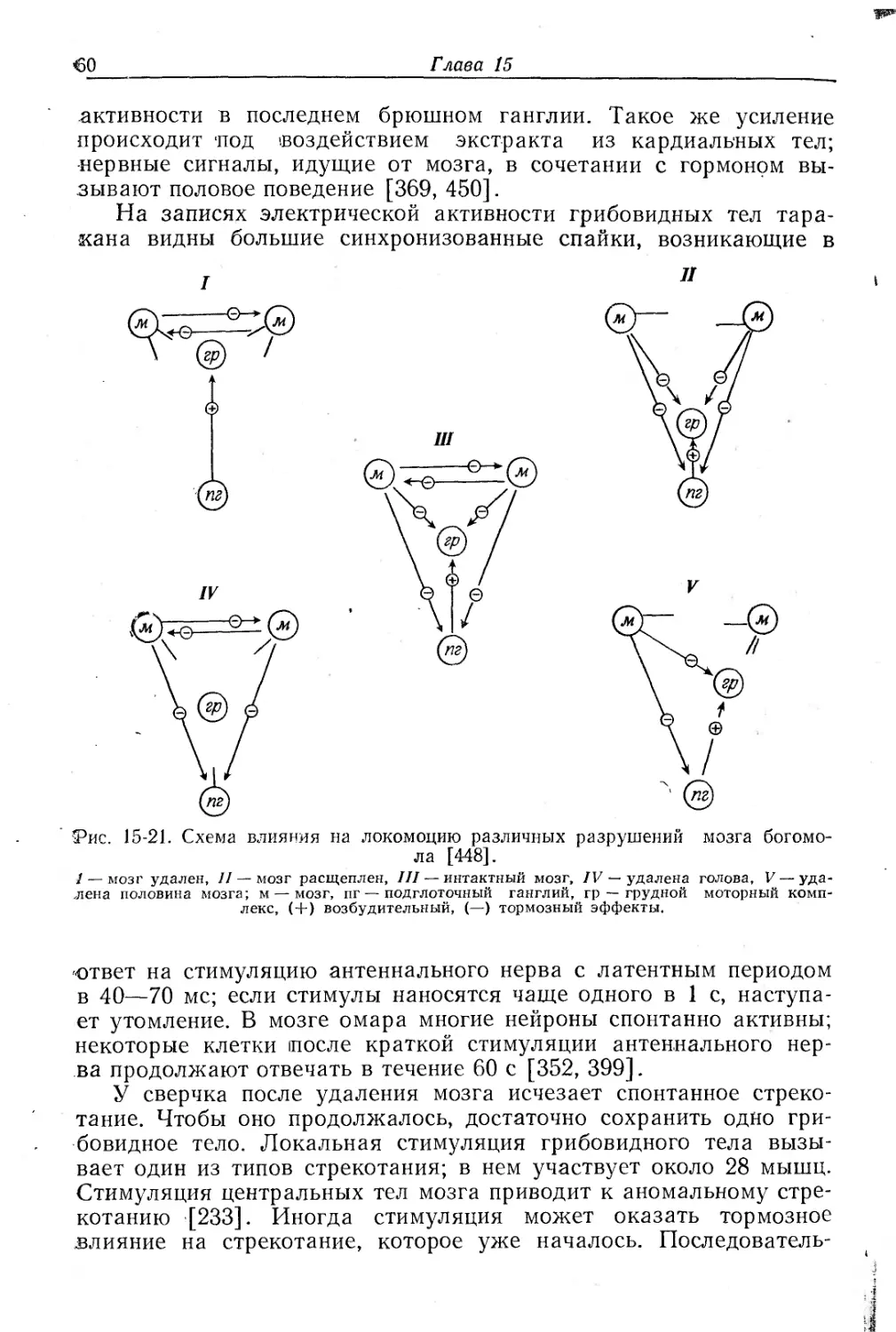
В последнем брюшном ганглии таракана имеются синапсы между сенсорными нейронами, идущими от расположенных на конце брюшка церков, и крупными восходящими несинцитиальными нейронами. Такие же «гигантские» волокна найдены у сверчков и саранчи. Эти синапсы поляризованы — они проводят возбуждение в одну сторону; для них характерна пространственная или временная суммация и некоторая лабильность ответа в зависимости от временной характеристики приходящих импульсов. Для появления спайка в гигантском волокне шестого брюшного ганглия необходимо большое количество пресинаптических импульсов в цервикальных нервных волокнах [64]. У таракана синапсы блокируются при частоте импульсов 40—50 в 1 с, а у сверчка— при 70—100 [448}. Система гигантских волокон принимает участие в прыжке сверчка и вызывается воздушной струей, направленной на церки, расположенные в конце брюшка. Гигантские волокна активируют мотонейроны ног не непосредственно, и латентный период реакции ног на церкальную стимуляцию вдвое длиннее чем если бы импульсы к мотонейронам ног шли по гигантским волокнам. Очевидно, активация мотонейронов осуществляется мелкими волокнами; гигантские же волокна активируют общую систему тревоги, и движение усиков [87]. Гигантские волокна обеспечивают также непрерывную сенсорную сигнализацию, создаваемую движением при реакции избегания. Дорсальные
Центральная нервная система
19
и вентральные гигантские волокна Periplaneta идут непрерывно от последнего брюшного ганглия до подглоточного ганглия, но у них имеются сегментарные сужения, особенно в брюшке, и заметное схождение в грудном отделе. В каждом сегменте происходит задержка по 0,6 мс, и в каждом ганглии проведение может быть блокировано никотином; этот эффект является следствием сегментарных сужений. Несмотря на отсутствие синапсов, проведение не равномерно. Разная его скорость обусловлена различиями в диаметре аксонов [409, 509].
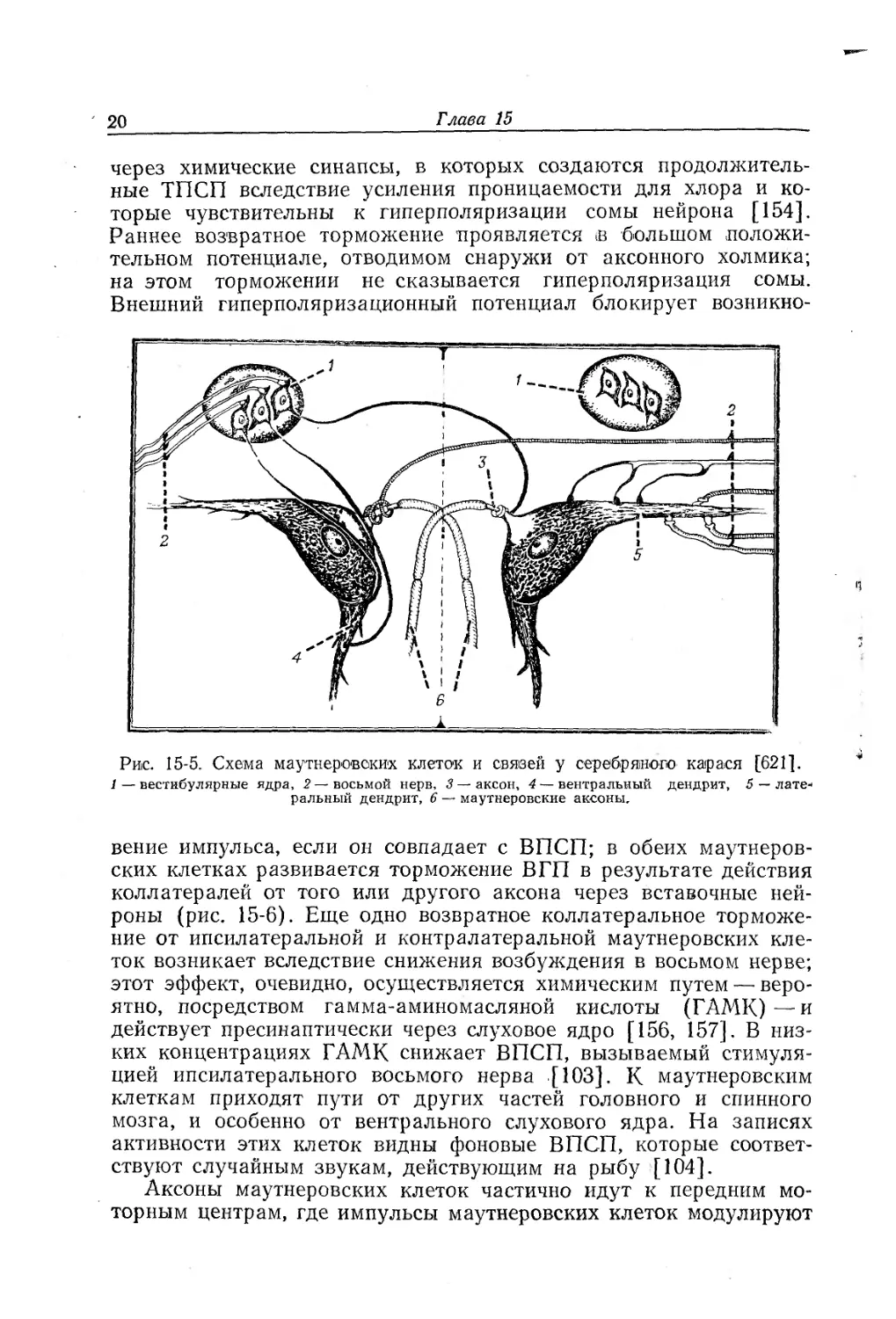
Маутнеровские клетки рыб
Гигантские несинтициальные маутнеровские клетки рыб и хвостатых амфибий обладают множеством входов и, по-видимому, способны к более сложной интеграции входной активности, чем любые другие известные нейроны. У серебряного карася обе маутнеровские клетки имеют по одному крупному дендриту, который идет в каудальном и латеральном направлениях, и по два или более вентральных дендрита. Аксон выходит из большого аксонного холмика, который окружен колпачком или витками окончаний. Аксон образует перекрест, а затем поворачивает в каудальном направлении; от него отходят веточка к преоптическому центру, веточка к контралатеральной маутнеровской клетке и коллатеральная веточка обратно к телу его собственной клетки; затем он идет к мотонейронам спинного мозга (рис. 15-5) [512]. Волокна ипсилатерального восьмого нерва являются возбуждающими и образуют палочковидные окончания (clubs) на дистальной части латерального дендрита и бляшки (boutons) — на его проксимальной части. Ипсилатеральный восьмой нерв генерирует ранний ВПСП с латентным периодом меньше 0,1 мс, вероятно через электрические синапсы палочковидных окончаний, которые построены по типу контактов со щелью; он вызывает также ВПСП с латентным периодом 0,6 мс, вероятно через химические синапсы на окончаниях с бляшками. Контралатеральный восьмой нерв является тормозным и имеет окончания со вздутиями, представляющими собой химические синапсы, на проксимальной части латерального дендрита и на соме. Веточка аксона контралатеральной маутнеровской клетки тоже тормозная и действует через один > или несколько вставочных нейронов.
Кроме того, от каждого аксона идет коллатераль, которая через вставочный нейрон вызывает возвратное торможение в своей собственной маутнеровской клетке, о чем свидетельствует тот факт, что распространение антидромного импульса устраняется предшествующим импульсом; это торможение осуществляется электрическим путем в форме внешнего гиперполяризационного потенциала (ВГП) на аксонном холмике. Кроме того, возникает позднее коллатеральное возвратное торможение, передающееся
2*
20
Глава 15
через химические синапсы, в которых создаются продолжительные ТПСП вследствие усиления проницаемости для хлора и которые чувствительны к гиперполяризации сомы нейрона [154]. Раннее возвратное торможение проявляется в большом положительном потенциале, отводимом снаружи от аксонного холмика; на этом торможении не сказывается гиперполяризация сомы. Внешний гиперполяризационный потенциал блокирует возникно-
Рис. 15-5. Схема маутнеровских клеток и связей у серебряного карася [621].
1 — вестибулярные ядра, 2 — восьмой нерв, 3 — аксон, 4 — вентральный дендрит, 5 — латеральный дендрит, 6 — маутнеровские аксоны.
вение импульса, если он совпадает с ВПСП; в обеих маутнеровских клетках развивается торможение ВГП в результате действия коллатералей от того или другого аксона через вставочные нейроны (рис. 15-6). Еще одно возвратное коллатеральное торможение от ипсилатеральной и контралатеральной маутнеровских клеток возникает вследствие снижения возбуждения в восьмом нерве; этот эффект, очевидно, осуществляется химическим путем — вероятно, посредством гамма-аминомасляной кислоты (ГАМК)—и действует пресинаптически через слуховое ядро [156, 157]. В низких концентрациях ГАМК снижает ВПСП, вызываемый стимуляцией ипсилатерального восьмого нерва [ЮЗ], к маутнеровским клеткам приходят пути от других частей головного и спинного мозга, и особенно от вентрального слухового ядра. На записях активности этих клеток видны фоновые ВПСП, которые соответствуют случайным звукам, действующим на рыбу [104].
Аксоны маутнеровских клеток частично идут к передним моторным центрам, где импульсы маутнеровских клеток модулируют
Центральная нервная система
21
движение челюсти, глазного яблока и жаберной крышки. Основная часть этих аксонных ветвлений спускается по спинному мозгу и активирует мотонейроны, преимущественно на той же стороне, на которой лежит аксон. Скорость проведения по аксону составляет от 50 до 100 м/с (при 15°C), а поведенческий эффект состоит в сильной флексий брюшка, движении плавников и ударе хвостом, что ведет к продвижению вперед. Важно, чтобы нисходящие
Рис. 16-6. Ответы маутнеровской клетки у серебряного карася [>155].
А и Б. Стимуляция ипсилатерального восьмого нерва. А. Отведение от сомы. Б. Отведение от латерального дендрита. В. Антидромная стимуляция. Г. Стимуляция контралатерального восьмого нерва. Д. Стимуляция контралатерального (/) и ипсилатерального (II) восьмых нервов.
импульсы не приходили к мотонейронам одновременно с обеих сторон, так как в противном случае мышечные эффекты нивелируются и удар хвостом не осуществится. Контроль частично осуществляется посредством перечисленных выше тонких коллатеральных тормозных механизмов. Кроме того, реципрокное пере-крестное торможение на уровне спинальных мотонейронов (рис. 15-7) делает возможным разрешение в пределах 0,15— 0,2 мс; два импульса маутнеровских клеток, следующие с меньшим интервалом, не оказывают возбуждающего действия. Ответив на импульс одной маутнеровской клетки, мотонейрон не может ответить на импульс второй маутнеровской клетки в течение примерно 10 мс [104]',.
В общей сложности на одной маутнеровской клетке (рис. 15-8) имеются следующие шесть входов: главный возбуждающий вход
22
Глава 15
от ипсилатерального восьмого нерва через электрические и химические синапсы; пять тормозных входов — от контралатерального восьмого нерва на латеральном дендрите, от него же — на соме, коллатерали маутнеровской клетки через вставочный нейрон посредством ВГП на аксонном холмике, коллатеральный — через химический тормозный синапс и пресинаптическое торможение слухового входа [155—157]. Выход направлен отчасти к голове, но
Рис. 15-7. Схема части спинальных связей маутнероиского рефлекса и его торможения [104].
1 — маутнеровский аксон, 2 — коллатераль, 3 — миелиновая оболочка, 4 — первичный мотонейрон, 5 — вентральный дендрит, 6 — «промежуточный элемент», 7 — перекрестное торможение, 8 — моторный аксон.
преимущественно к спинальным мотонейронам, где благодаря сильной тормозной системе возможна дискриминация активации от обеих маутнеровских клеток с точностью до 0,15 мс. По восьмому нерву приходит как вестибулярная, так и слуховая активность. Значение маутнеровских клеток для поведения состоит в осуществлении быстрых реакций — тревоги и избегания.
В итоге следует сказать, что гигантские волокна возникали в процессе эволюции многократно и делятся на несколько разных типов, но все они служат быстроте и синхронности реакций организма. Это могут быть распределительные системы, сложные одиночные клетки или же функциональные синцитии с относительно фиксированными ответами.
Центральная нервная система
23
Рис. 15-8. Ответы маутнеровокой клетки серебряного карася.
А. Синаптическое взаимодействие [622]. 1 — вестибулярное ядро, 2 — волосковая клетка саккулюса, 3 — VIII нерв, 4—латеральный дендрит, 5—аксон правой М-клетки, 6 — шапочка аксона, 7 — аксон левой М-клетки. В — возбудительные синапсы, Т — тормозные синапсы, Н — вставочный нейрон. Показана блокада утомлением (/), прокаином (II) и стрихнином (III). Б. Схема временных соотношений ответов контралатеральной (I) и ипсилатеральной (II) маутнеровской клетки на стимуляцию VIII нерва [104]. Возбудительные ответы — выше, тормозные — ниже горизонтальной линии.
Нервные сети
Нервная сеть состоит из нейронов, распределенных по плоскости или в трех измерениях и связанных таким образом, чтобы обеспечить диффузное проведение; это придает сети некоторые интегративные свойства. Такие сети составляют главную часть нервной системы у кишечнополостных, гребневиков и кишечноды-щащих; в сочетании со скоплениями нейронов в ганглиях они образуют нервную систему у иглокожих, асцидий и некоторых моллюсков; нервные сети интегрируют движения висцеральной мускулатуры у животных, относящихся ко многим типам. Лучше всего они изучены у кишечнополостных. У сцифоидных медуз и коралловых полипов по нервной сети проводится большая часть сигналов из одной части тела в другую; у гидроидных полипов главную роль играет эпителиальное проведение, но нервные сети имеются у животных также и этого класса [338—340].
24
Глава 15
Кишечнополостные
В ранних гистологических работах нервная сеть полипа или медузы представлялась как некий континуум анастомозирующих волокон от мультиполярных и биполярных нейронов, в котором аксоны не отличимы от дендритов. Однако исследования с помощью поляризационного и электронного микроскопа показали, что отростки обычно не сливаются; в местах их соединения волокна лишь прилегают друг к другу; впрочем, у парусников и, вероятно, у гидр некоторые отростки, возможно, сливаются [337]. В нервной сети медузы крестовичка обнаружены типичные синапсы, по одну сторону которых лежат пузырьки с плотной сердцевиной; синапсы эти, вероятно, поляризованы [568]|. В отличие от них у Aurelia, Phialidium, Cyanea, Cordylophora и сифонофоры Nanomia синапсы симметричные, и проведение через них, по-видимому, двухстороннее [218, 265, 267]. У гидры синапсы имеют 0,8 мкм в длину с синаптической щелью шириной 150 А и пузырьками от 1200 до 2000 А; одни синапсы симметричны, у других пузырьки лежат только с одной стороны [568]. Таким образом, по своей ультраструктуре типы синаптических соединений варьируют от полного слияния мембран через симметричные синапсы и до синапсов с односторонним проведением. Чаще всего встречаются симметричные синапсы, имеющие пузырьки с обеих сторон.
Типичная нервная сеть проводит возбуждение свободно во всех направлениях; импульсы огибают углы, проходят по тканевым мостикам и, миновав узкий участок, расходятся. Общие свойства сети могут быть иными, чем у отдельных нейронов, а приемы исследования свойств отдельных волокон в нервной сети кишечнополостных еще не разработаны. Большинство кишечнополостных обладает двумя или более параллельными сетями [24]; так, например, у сцифомедуз для быстрых плавательных движений служит сеть из крупных волокон, а медленные сокращения при пищевых движениях координируются диффузной сетью из тонких волокон [411]'!. У актиний Calliactis одна медленная система проводит импульсы со скоростью от 4,4 до 14,6 см/с, а другая — со скоростью от 3 до 5,3 см/с, тогда как в быстрой сети мезентерия проведение имеет скорость 120 см/с [358]; еще не известно, из каких проводящих тканей состоят первые две системы.
В пучках волокон в быстро проводящей сети актинии зарегистрированы быстрые потенциалы действия [420]. Различия порогов активации сети в разных ее участках создают полярность особого рода. Ротовой диск актинии в 4000 раз чувствительнее к тактильному раздражению, чем ее стебель [413]. У гребневиков импульсы, вызывающие свечение, идут по нервной сети во всех направлениях, а импульсы, запускающие биение гребня, — от аборального к оральному концу.
Центральная нервная система 25
Гидрозон обладают несколькими проводящими системами, и нельзя сказать с уверенностью, какие из них эпителиальные, а какие — нервные. У Tubularia имеются следующие проводящие системы: 1) система, запускающая электрические процессы в области шейки со скоростью проведения 17 см/с; 2) система дистального открывателя, раскрывающая венчик щупалец со скоростью проведения 15 см/с, и 3) медленная система стебелька со скоростью проведения 6 см/с с высоким порогом и никак не связанная с поведением. Две первые системы, по-видимому, нервные, блокируются Mg2+. Третья, по-видимому, эпителиальная. Система дистального открывателя тормозит шейный пейсмекер [264— 267]. Гидра генерирует связанные с сокращением пачки потенциалов, возникающих в гипостоме и распространяющихся со скоростью 3—5 см/с; число импульсов в пачке варьирует в зависимости от освещения или степени насыщения; эти потенциалы происходят, вероятно, из мышечно-эпителиальных клеток. У гидры отводятся также ритмические потенциалы (1—10 в 1 мин), не связанные с сокращением, которые проводятся со скоростью 5 см/с и возникают у основания стебелька, возможно в клетках гастродермы [268, 412, 474].
Нервные сети кишечнополостных широко варьируют по степени необходимого им облегчения. У актиний и медуз для возникновения максимальной сократительной реакции требуется многократное раздражение, и один залп может вызвать лишь незначительный механический ответ. Опыты с применением блокаторов, например ионов магния, и с регенерацией нервных связей после перерезки показывают, что в большей своей части требуемое облегчение носит нейро-нейрональный характер, а в остальном является нервно-мышечным. Заметный декремент проведения происходит вследствие снижения облегчения в соединениях по мере распространения волны из точки ее возникновения. У люминесци-рующего Renilla неполяризованное, бездекрементное проведение требует облегчения, которое исчезает при интервалах больше 1,5 с. В щупальце Anemonia облегчение сохраняется от 10 до 30 с [91], а у Tealia — в течение 2 мин.
У актинии Calamactis нервные импульсы, распространяющиеся со скоростью 0,9 м/с, оказывают некоторое облегчающее действие в нервно-мышечных соединениях при интервалах в 30 мс, дают максимальное облегчение в интервале 200 мс и небольшой остаточный эффект в интервале от 1 до 4 с [420]. У Metridium быстрее сокращение запускается сетью при частоте стимуляции 1 стимул в 1 с; медленное сокращение той же мышцы вызывается стимуляцией^ частотой 1 стимул каждые 5 с [444]j. Париетальный и кольцевой мышечные слои дают только медленные ответы, а ретрактор и маргинальный сфинктер способны и на быстрый, и на медленный ответы [408]; медленный ответ зависит от облегчения в большей степени, чем быстрый. Рефрактерный период для медленно
26 __________________________Глава 15_________________________
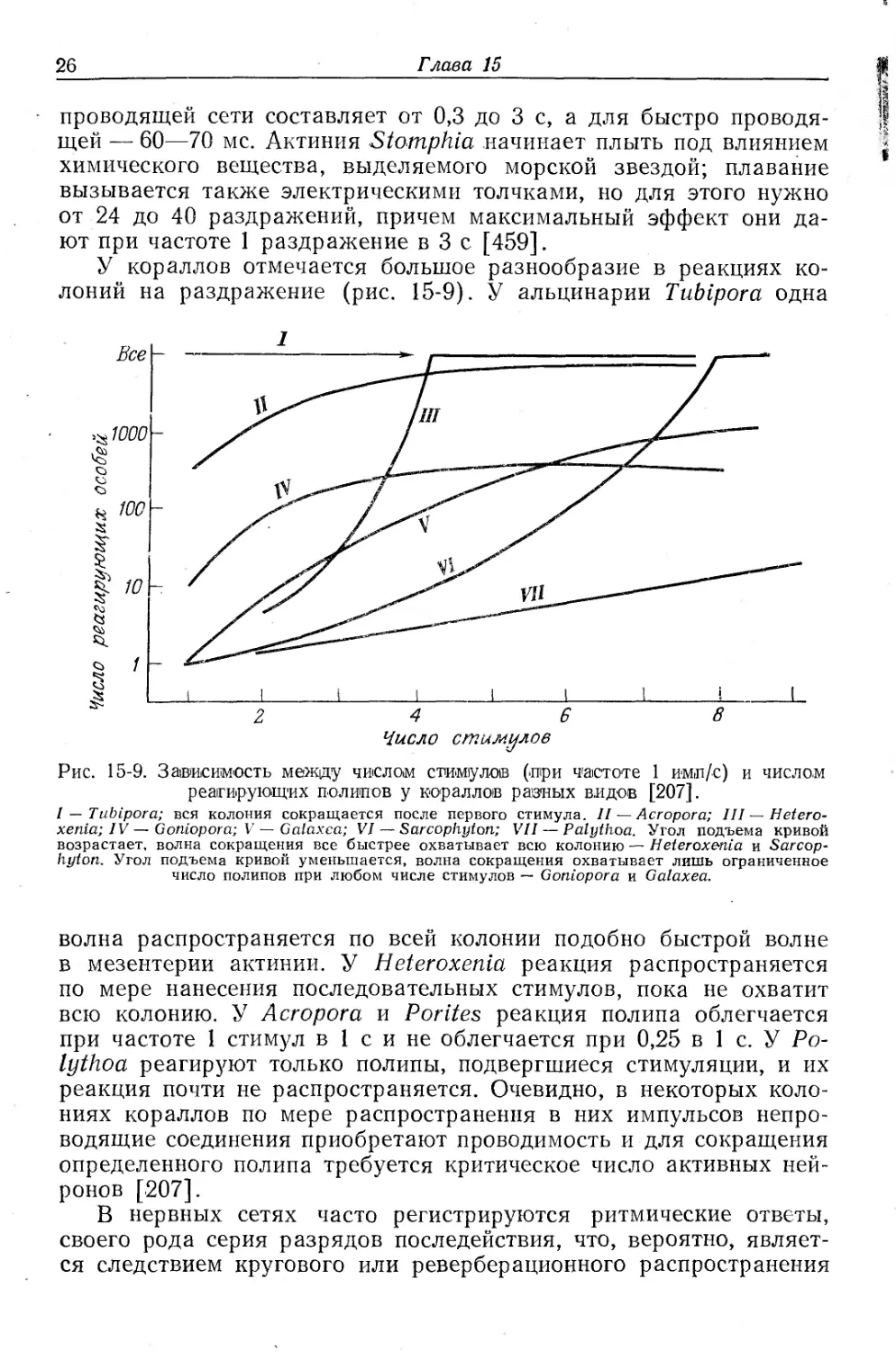
g проводящей сети составляет от 0,3 до 3 с, а для быстро проводя- jl щей — 60—70 мс. Актиния Stomphia начинает плыть под влиянием * химического вещества, выделяемого морской звездой; плавание вызывается также электрическими толчками, но для этого нужно от 24 до 40 раздражений, причем максимальный эффект они дают при частоте 1 раздражение в 3 с [459].
У кораллов отмечается большое разнообразие в реакциях колоний на раздражение (рис. 15-9). У альцинарии Ttibipora одна
Рис. 15-9. Зависимость между числом стимулов (при частоте 1 имп/с) и числом реагирующих полипов у кораллов равных видов [207].
I — Ttibipora; вся колония сокращается после первого стимула. II— Acropora; III—Hetero-xenia; IV — Goniopora; V — Galaxca; VI — Sarcophyton; VII — Palythoa. Угол подъема кривой возрастает, волна сокращения все быстрее охватывает всю колонию — Нeteroxenia и Sarcophyton. Угол подъема кривой уменьшается, волна сокращения охватывает лишь ограниченное число полипов при любом числе стимулов — Goniopora и Galaxea.
волна распространяется по всей колонии подобно быстрой волне в мезентерии актинии. У Heteroxenia реакция распространяется по мере нанесения последовательных стимулов, пока не охватит всю колонию. У Acropora и Porites реакция полипа облегчается при частоте 1 стимул в 1 с и не облегчается при 0,25 в 1 с. У Ро-lythoa реагируют только полипы, подвергшиеся стимуляции, и их реакция почти не распространяется. Очевидно, в некоторых колониях кораллов по мере распространения в них импульсов непроводящие соединения приобретают проводимость и для сокращения определенного полипа требуется критическое число активных нейронов [207].
В нервных сетях часто регистрируются ритмические ответы, своего рода серия разрядов последействия, что, вероятно, является следствием кругового или реверберационного распространения
Центральная нервная система 27
импульсов. Волна сокращения, связанная с циркуляцией возбуждения в нервной сети, может двигаться по вырезанному из колокола медузы кольцу в течение многих часов и дней. У лептомедуз в сети, по которой возбуждение распространяется в радиальном направлении, передача осуществляется медленно, а в сети с кольцевым проведением — быстро; реципрокность в сокращении кольцевых и радиальных мышц свидетельствует о взаимодействии этих сетей, причем радиальная сеть может тормозить кольцевую, но не наоборот [209]j. Сокращения в кольцах, полученных из стебля или педального диска Calliactis, тормозятся при соответствующей стимуляции нервной сети [131].
Вторая функция некоторых нервных сетей состоит в запуске спонтанной ритмической активности. Краевые тельца медузы представляют собой скопления нейронов, которые вызывают ритмические сокращения зонтика. Интактные актинии производят чрезвычайно медленные (от минут до часов) спонтанные ритмические движения [131]. У Tubularia нервная сеть ротового диска представляет собой пейсмекер, но после ее удаления спонтанные ритмы возникают в других участках [264, 266]. Пейсмекером может быть любой участок — шейка гидранта, щупальцы или стебелек [264, 267]>
Нервные сети обеспечивают некоторые очень сложные формы поведения. К ним относятся: отвергание одних пищевых веществ, принятие других и подведение ротового стебелька к пище у гидромедуз; расширение, вытягивание, дефекация и покачивание у актиний. У Stomphia при воздействии пищевым экстрактом прекращается плавание. Calliactis обычно живет на раковинах улиток, в которых поселились раки-отшельники; когда рак переходит в новую раковину, актиния посредством ряда сложных движений перебирается на нее. Рак-отшельник Dardanus помогает актинии в этом перемещении в отличие от рака Pagurus [458].
Если не считать нескольких записей серий нервных импульсов, клеточная активность в сетях кишечнополостных не изучена. У них, вероятно, имеются химические медиаторы, о чем свидетельствует наличие синаптических пузырьков, однако такие подробно изученные у более высокоорганизованных животных медиаторы, как норадреналин и ацетилхолин, оказываются здесь малоэффективными [457]. Обычно для блокирования проведения по сети используется магний, который действует, по-видимому, на синаптические соединения.
Не всегда удается отделить проведение возбуждения по сети от межклеточного проведения по эпителию, особенно у гидрозоев; проведение возбуждения, которое ранее приписывали только нервным сетям, в ряде случаев оказалось эпителиальным.
Системы, которые имеются у кишечнополостных и которые несомненно ’являются нервными сетями примитивного типа, тем не менее обладают многими свойствами нервной системы более вы
28
Глава 15
сокоорганизованных животных. Важную роль в них играет свойство облегчения, благодаря которому животное отвечает не на каждый стимул, а только на значительное их число. Для облегчения требуется также определенная полярность синаптической передачи. Контакты в нервных сетях по своему характеру, вероятно, варьируют от слившихся нейронов до неполяризованных и далее до поляризованных синапсов. Они характеризуются спонтанностью, торможением и реципрокной иннервацией мышц и обеспечивают большой набор поведенческих реакций. Они могут служить моделями сложных нервных систем и никак не могут считаться простыми.
Переход от периферической интеграции к центральной
Координация движений периферическими сетями наблюдается не только у кишечнополостных, но и у некоторых животных, обладающих центральной нервной системой. У брюхоногих и двустворчатых моллюсков по субэпидермальному сплетению в ноге и мантии передаются импульсы, вызывающие сокращение края мантии и околоротовых лопастей. Эта нервная сеть обладает интегративными свойствами. У дождевых червей существует периферическая нервная связь между эпидермальными рецепторами и мышцами. У баланоглосса реакции хоботка могут координироваться периферическим сплетением [421]. У большинства животных, у которых локомоция подчинена центральной регуляции, висцеральные движения координируются или запускаются нервными сплетениями желудочно-кишечного тракта.
Переход от периферического к центральному контролю локомоции хорошо виден у иглокожих. Их нервная система состоит из центрального околоротового кольца, от которого отходят 5 радиальных нервных стволов, содержащих нервные клетки и связанных с периферической сетью в амбулакральных ножках, иглах, педицелляриях и коже. У офиур и морских лилий периферическое сплетение имеется только в амбулакральных ножках. Если у морской звезды перерезать радиальный нерв, то ответ на прикосновение передается по периферическому сплетению к кольцу или к другим лучам. Если удалены все элементы нервного кольца, то изолированный луч движется в сторону своего основания; если же часть кольца сохранена, луч движется по направлению к своему дистальному концу. Если кольцо перерезано в двух местах, морская звезда может разорвать себя надвое. Координированные движения амбулакральных ножек возможны при интактном радиальном нерве, но направление движения зависит от нейронов в местах соединения кольца и радиальных нервов [502]':. Проведение по кольцу идет со скоростью 11 см/с, а по радиальным нервам — со скоростью 17 см/с [424].
У морских ежей в основании каждой иглы лежит по нескольку нейронов, составляющих часть общего сплетения. Проведение
Центральная нервная система
29
возбуждения в сплетении осуществляется со скоростью 6 см/с и не сопровождается движением иглы, т. е. этот процесс не является цепью рефлексов [51]. У ежа Diadema локальное экранирование падающего света вызывает сгибание игл в сторону тени — защитную реакцию, для которой необходим радиальный нерв [601]. Стимуляция радиального элемента вызывает два потенциала действия; один проводится со скоростью 22 см/с и вызывает движение иглы, а второй распространяется со скоростью 6 см/с и тормозит движение иглы [375, 376]1 Реакция выпрямления происходит у морской звезды при участии радиальных нервов и совершается с большей легкостью при наличии нервного кольца.
У плоских червей от головного мозга к концу туловища идут нервные стволы (от 2 до 8), которые содержат нервные клетки, не сгруппированные в ганглии. У планарий и немертин для спонтанных координированных движений, по-видимому, требуется только отрезок одного из этих нервных стволов. У поликлада Planocera возбуждение не переходит с одной стороны тела на другую по периферической сети; для координированной локомоции, нормального выпрямления, ориентации в сторону пищи и питания (но не для глотания и не для откладывания яиц) необходим «головной мозг» [174]. Очевидно, у плоских червей периферическая система не функционирует как независимая нервная сеть.
Функции ганглиев у моллюсков
Брюхоногие и двустворчатые моллюски
У брюхоногих и двустворчатых моллюсков локомоторные и другие рефлексы осуществляются через посредство четырех пар, связанных между собой ганглиев — церебральных, педальных, плевральных и висцеральных. Эти ганглии в разной степени слиты друг с другом и обладают разной степенью симметрии в зависимости от общего строения животного. У улиток, например у Helix, и у голожаберных, например у аплизии, кроме множества мелких нейронов, имеется также небольшое число гигантских вставочных нейронов и крупных мотонейронов, расположенных в определенном порядке [237, 238, 531]. У Helix, Aplysia и Tritonia определены локализация и функция гигантских нейронов. На каждом гигантском вставочном нейроне имеются входы от сенсорных нейронов, иннервирующих ограниченные участки тела; синаптические связи делают возможной фиксированную циркуляцию импульсов; моторные выходы на обеих сторонах нервной системы расположены либо симметрично, либо несимметрично [73, 74, 106, 107]. Синапсы у моллюсков по большей части аксо-аксонные, иногда аксо-соматические, но аксо-дендритных синапсов нет. Ге
30
Глава 15
нерация импульсов происходит на некотором расстоянии от тела клетки независимо от него [528][.
Субэпидермальное нервное сплетение в изолированных около-ротовых лопастях беззубки Anodonta проводит ответы на прикосновение или освещение. У многих моллюсков в периферических нервах лежат тела нервных клеток. У аплизии жабра, отделенная от центральных ганглиев, сокращается при тактильной стимуляции, и для этой реакции, проводимой по периферическому сплетению, может наступить привыкание [414]. Остается неясным, в какой мере периферические нервные сети у брюхоногих и двустворчатых моллюсков взаимодействуют с центральными ганглиями.
Рефлексы у двустворчатых и брюхоногих моллюсков. Как правило, у двустворчатых моллюсков сокращение (фазическое или тоническое) и расслабление аддукторов вызываются разными нейронами. У беззубки импульсы, идущие от висцерального ганглия, вызывают либо тоническое, либо фазическое сокращение в зависимости от того, какой стимул подействовал на ганглий. В церебральном ганглии заложен центр расслабления аддукторов [477, 478]. У литорального моллюска Spisula к специфическим мотонейронам висцерального ганглия приходят импульсы от сенсорных клеток с обеих сторон тела; возбуждение этих нейронов вызывает билатерально симметричные моторные реакции моллюска. Им-пульсация поступает к мотонейронам по церебро-висцеральным коннективам и по нервам от сифона и мантии; в одних мотонейронах возникает деполяризация, за которой следуют гиперполяризационные синаптические потенциалы; в других появляются деполяризационные ответы. От мотонейрона активность поступает на мышцы аддукторы [362]. У морского черенка Ensis сенсорные сигналы от ноги или переднего края мантии идут к одной стороне церебрального ганглия, затем к противоположной стороне, а потом к педальному ганглию и оттуда к ноге; коротколатентный рефлекс проходит только через педальный ганглий [403]i. Плавательные движения у гребешка включают миотатический рефлекс, причем разряд мотонейрона приводит к сокращению, которое в свою очередь вызывает сенсорный разряд, поступающий обратно на мотонейрон [363].
По некоторым данным, у Helix педальный ганглий оказывает на мышцы ноги как возбуждающее, так и расслабляющее действие. Один вставочный нейрон отвечает на раздражение области ноги или головы. У некоторых двустворчатых пути рефлекса и баланс между возбуждением и торможением варьируют в зависимости от интенсивности сенсорной стимуляции. Удаление церебрального ганглия у аплизии усиливает локомоторную активность; следовательно, церебральный ганглий тормозит активность педального, а возбуждение последнего вызывает сокращение мышцы ноги. Прослежены пути рефлексов в центральной нервной
Центральная нервная система
31
системе улитки Buccinium\ значительная сенсорная конвергенция на центральных нейронах в сочетании с определенной эфферентной активностью делает возможными координированные ответы на механическую и химическую стимуляцию рецепторов [20]. Отсюда вывод, что все ганглии моллюсков выполняют ту или иную рефлекторную функцию, но вся нервная система так взаимосвязана, что локализация функций в ганглиях весьма ограничена.
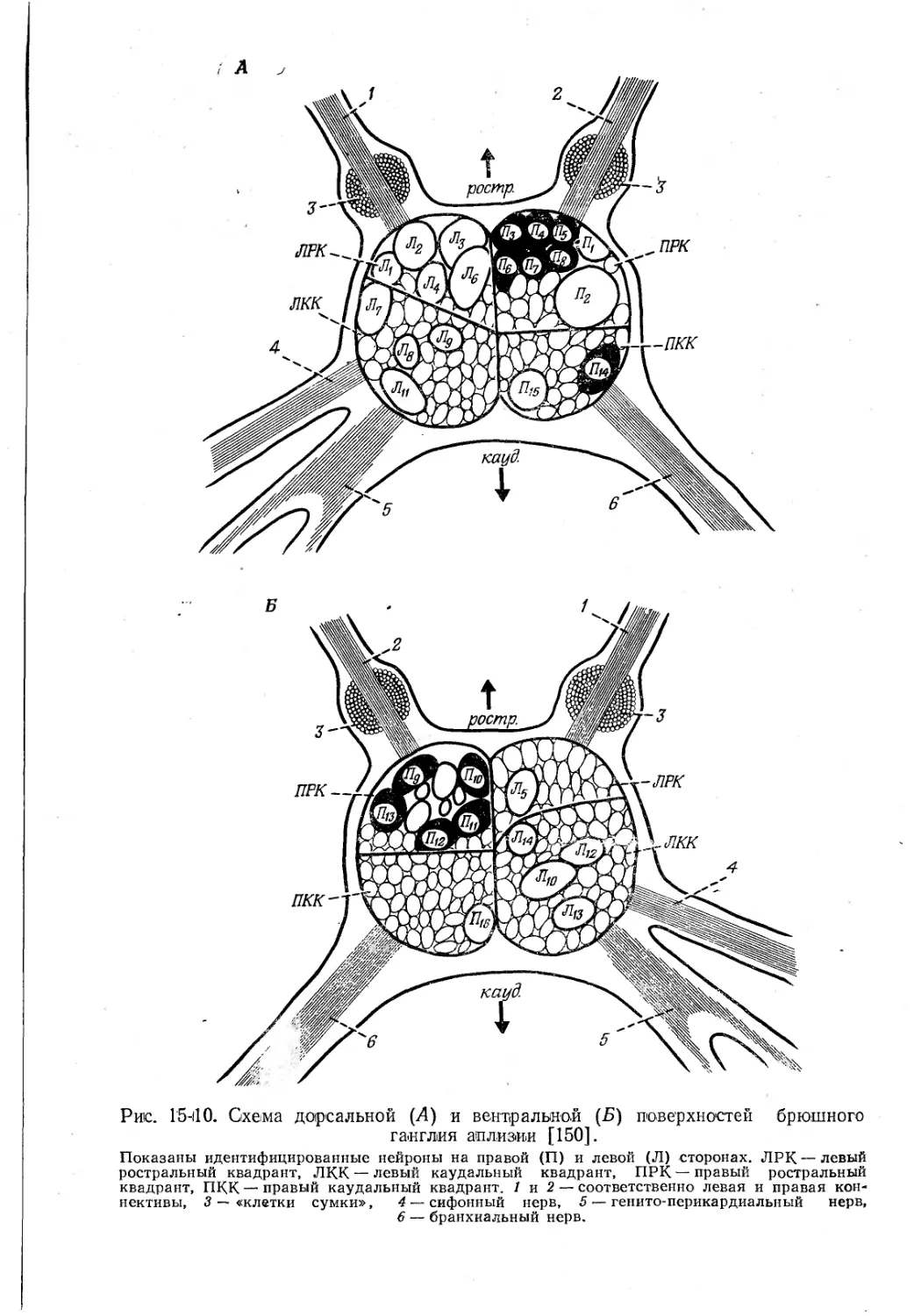
Функция идентифицированных нейронов и скоплений вставочных нейронов. Большие успехи достигнуты в соотнесении специфических функций с отдельными идентифицированными нейронами у брюхоногих моллюсков. Абдоминальный ганглий аплизии содержит от 1000 до 2000 нейронов в зависимости от размеров животного [73], и крупные клетки можно идентифицировать по отдельности на основании их положения в ганглии, пигментации, характера импульсации, синаптических потенциалов, ответов на орто-дромную активность, характера эфферентных путей, связей с другими нейронами и реакций на фармакологические вещества. Удалось идентифицировать 30 крупных клеток и 8 сходных между собой скоплений клеток (рис. 15-10) [150]. Кроме того, имеются нейросекреторные клетки, к которым относятся «клетки сумок» у выхода нервов, а также белые и пигментированные клетки. Некоторые из этих последних реагируют на освещение: наблюдается либо гипер-, либо деполяризация [16]. Все клетки данного скопления ведут себя сходным образом и названы не-идентифицированными вставочными нейронами. Синхронность в активности связанных друг с другом нейронов часто касается пачек импульсов, а не отдельных импульсов. На основании сведенных в таблицы свойств идентифицированных клеток построены модели дивергентных и конвергентных цепей внутри ганглия (рис. 15-11) [283]. Нейроны с одинаковыми функциями локализуются примерно в одной зоне; один разветвленный вставочный нейрон может оказывать множество разных эффектов на следующие клетки, а разные вставочные нейроны вступают друг с другом в разнообразные взаимодействия [280]. Значительная функциональная сложность системы мелких нейронов объясняется резкими индивидуальными различиями ее отдельных элементов. Кодирование осуществляется или модуляцией эндогенных ритмов, или суммацией ПСП, или же комбинацией тормозных и возбудительных эффектов [553]).
Из 30 идентифицированных нейронов абдоминального ганглия у 27 отмечена эндогенная активность, которая обычно выражается в многократных залпах импульсов. Клетка Л15 у аплизии дает ритмические залпы [517]. Эта активность имеет суточный ритм с максимальной частотой залпов после рассвета; такой ритм характерен и для изолированного ганглия [518], но может быть изменен при изменении режима светового дня перед изоляцией ган-
Рис. 15-110. Схема дорсальной (Д) и вентральной (£>) поверхностей брюшного ганглия аплизии [150].
Показаны идентифицированные нейроны на правой (П) и левой (Л) сторонах. ЛРК— левый ростральный квадрант, ЛКК — левый каудальный квадрант, ПРК—правый ростральный квадрант, ПКК — правый каудальный квадрант. 1 и 2 — соответственно левая и правая кон-нективы, 3 — «клетки сумки», 4 — сифонный нерв, 5 — генито-перикардиальный нерв, 6 — бранхиальный нерв.
Центральная нервная система
33
глия [328]. Предполагаемые ионные механизмы ритмичности описаны в гл. 16.
Из 30 идентифицированных нейронов у 28 аксоны входят в состав периферических нервов. Одновременное отведение несколькими электродами показывает, что многие клетки функционально связаны с другими нейронами. Активность большинства нейронов модулируется торможением их эндогенной импульсации, а также
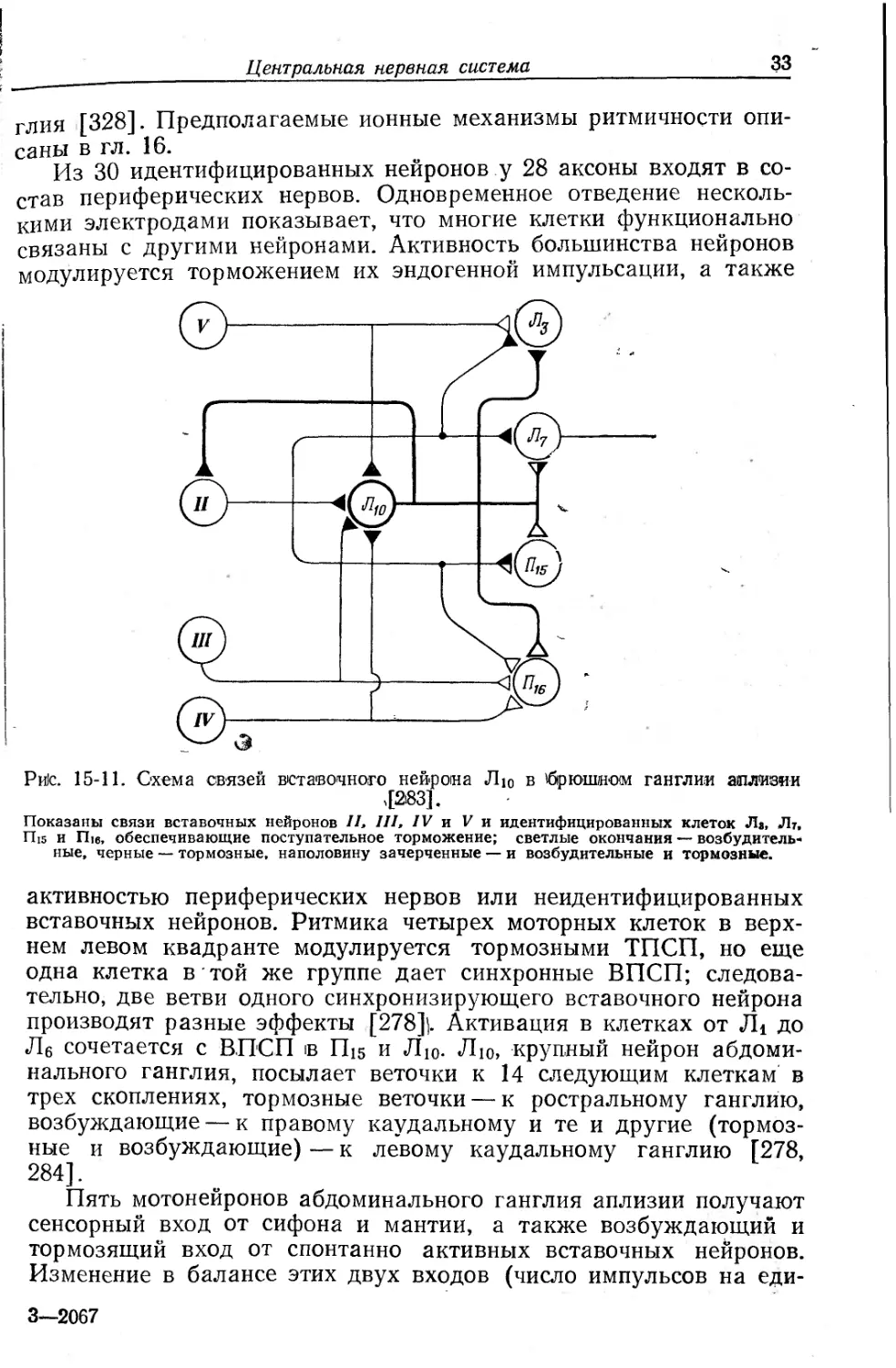
Рйс. 15-11. Схема связей вставочного нейрона Лю в 'брюшном ганглии аплизии ,[283].
Показаны связи вставочных нейронов II, III, IV и V и идентифицированных клеток Л», Лт, Пи и П1в, обеспечивающие поступательное торможение; светлые окончания — возбудительные, черные — тормозные, наполовину зачерченные — и возбудительные и тормозные.
активностью периферических нервов или неидентифицированных вставочных нейронов. Ритмика четырех моторных клеток в верхнем левом квадранте модулируется тормозными ТПСП, но еще одна клетка в той же группе дает синхронные ВПСП; следовательно, две ветви одного синхронизирующего вставочного нейрона производят разные эффекты [278}. Активация в клетках от Л1 до Л6 сочетается с ВПСП в П15 и Лю. Лю, крупный нейрон абдоминального ганглия, посылает веточки к 14 следующим клеткам в трех скоплениях, тормозные веточки — к ростральному ганглию, возбуждающие — к правому каудальному и те и другие (тормозные и возбуждающие) — к левому каудальному ганглию [278, 284].
Пять мотонейронов абдоминального ганглия аплизии получают сенсорный вход от сифона и мантии, а также возбуждающий и тормозящий вход от спонтанно активных вставочных нейронов. Изменение в балансе этих двух входов (число импульсов на еди-
3—2067
34
Глава 15
ницу времени) может превратить осуществляемый через мотонейрон простой рефлекс в сложную реакцию ухода в раковину [307]. Единственная крупная пигментированная клетка в висцеральном ганглии аплизии активируется сенсорными волокнами в каждом из нескольких периферических нервов; при этом импульс в левом висцеральном нерве, предшествующий импульсу в плевральном нерве, эффективнее обратной последовательности. Когда группы из трех сенсорных импульсов активировались при постоянном интервале между Si и S3, но при варьировании момента появления S2, ответ вставочного нейрона был максимальным при S2, близком к S3, но эффект зависел от средней частоты, т. е. от интервала между Si и S3; следовательно, вставочный нейрон опознает характер импульсации на своем входе [492].
Некоторые вставочные нейроны, особенно их группа в правом верхнем квадранте абдоминального ганглия аплизии, отличаются продолжительным гетеросинаптическим облегчением, при котором один вход (primer) облегчает ответ на действие другого (тест-сигнала). Начальный приток активности не изменяет проводимости в гигантском нейроне, следовательно, здесь имеет место пре-синаптическое воздействие на тестируемые аксоны или на цепь вставочного нейрона, включающую эти аксоны [281]!. В другой цепи один вход производит пресинаптическое торможение, которое может длиться целую минуту; ответ клетки абдоминального ганглия на стимуляцию правой коннективы тормозится импульсацией жаберного нерва [529]. Другой тип длительного электрического изменения в мотонейронах коррелирует с поведенческим привыканием (habituation) и отвыканием (dishabituation).
В синаптических ответах гигантских нейронов аплизии и виноградной улитки на медиаторы наблюдаются индивидуальные различия (см. гл. 11). Ацетилхолин (АХ) возбуждает (деполяризует) П15 и в то же время тормозит (гиперполяризует) Л3; две веточки одного и того же вставочного или сенсорного нейрона могут возбуждать одну клетку и тормозить другую. В буккальном ' ганглии аплизии два вставочных нейрона с каждой стороны, во-первых, гиперполяризуют через синапс 6 нейронов, во-вторых, де-: поляризуют один нейрон и, в-третьих, деполяризуют, а затем ги-перполяризуют по одному нейрону с каждой стороны. Ацетилхолин (АХ) оказывает такое же двойное действие, и авторы приходят к заключению, что одна и та же клетка может обладать рецепторами для реакций, связанных с проводимостью и для Na, и. для С1 [158]. Торможение объясняется повышением gci, возбуждение— преимущественно повышением £ка (см. гл. 11). Некоторые клетки возбуждаются АХ и тормозятся дофамином, причем торможение отчасти вызывается усилением g&. Серотонин (5-ок-ситриптамин, 5-ОТ) вызывает деполяризационные ПСП в одних клетках и гиперполяризационные — в других. Таким образом, отдельные нейроны должны обладать разными и иногда нескольки-
Центральная нервная система
35
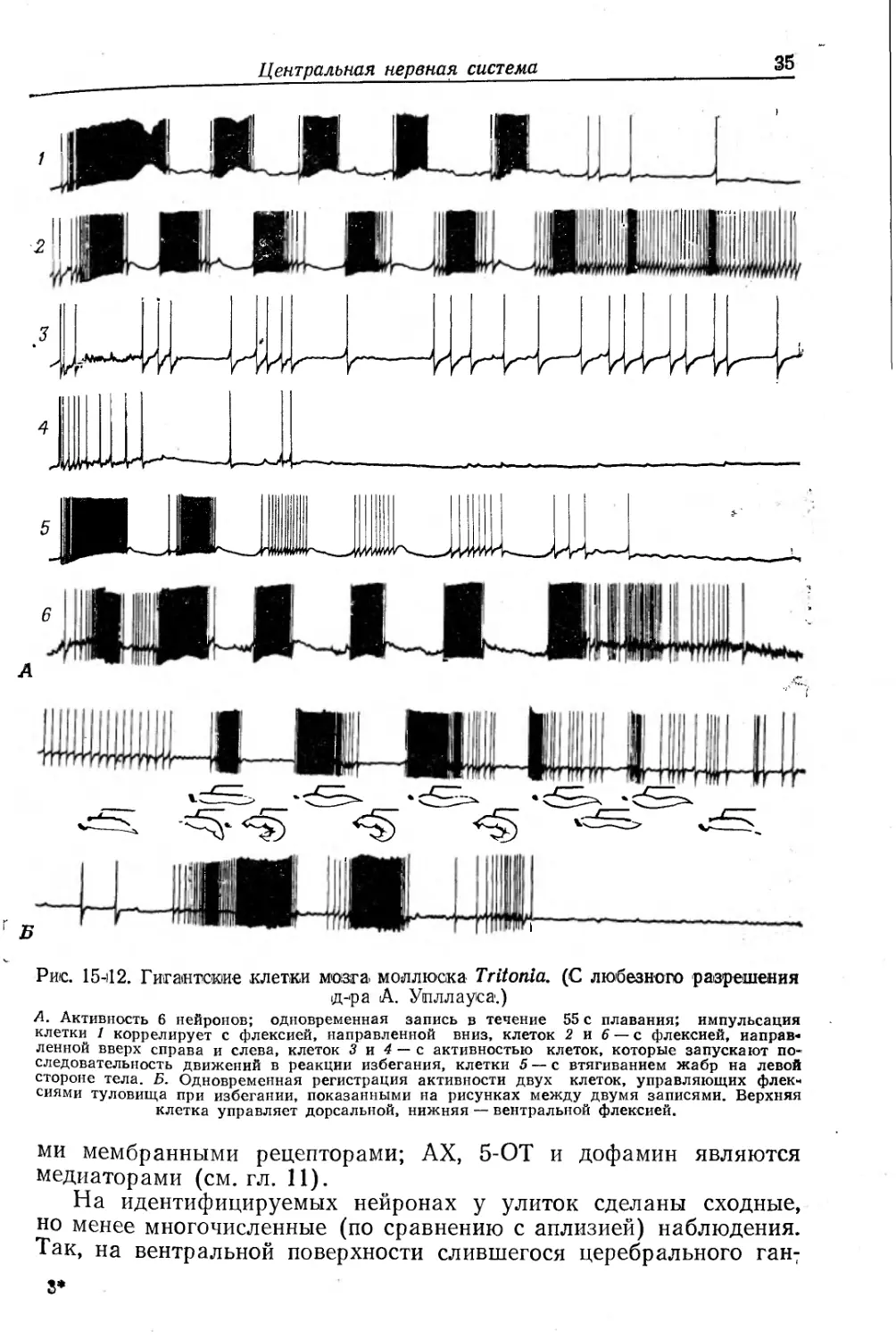
Рис. 15-12. Гигантские клетки мозга моллюска Tritonia. (С любезного разрешения д-ра А. Упллауса.)
А. Активность 6 нейронов; одновременная запись в течение 55 с плавания; импульсация клетки 1 коррелирует с флексией, направленной вниз, клеток 2 и 6 — с флексией, направленной вверх справа и слева, клеток 3 и 4 — с активностью клеток, которые запускают последовательность движений в реакции избегания, клетки 5 — с втягиванием жабр на левой стороне тела. Б. Одновременная регистрация активности двух клеток, управляющих флексиями туловища при избегании, показанными на рисунках между двумя записями. Верхняя клетка управляет дорсальной, нижняя — вентральной флексией.
ми мембранными рецепторами; АХ, 5-ОТ и дофамин являются медиаторами (см. гл. 11).
На идентифицируемых нейронах у улиток сделаны сходные, но менее многочисленные (по сравнению с аплизией) наблюдения. Так, на вентральной поверхности слившегося церебрального ган
8*
36
Глава 15
глия Helix лежат две симметричные клетки, которые посылают отростки к четырем ипси- и четырем контралатеральным нервам, а также к трем ипси- и трем контралатеральным межганглионарным коннективам; на них имеются входы от двух общих вставочных нейронов [282]. В буккальном ганглии Navanax найдены 10 клеток, которые не связаны друг с другом непосредственно через синапсы, но активируют вставочные нейроны, взаимодействующие друг с другом, а затем посылающие обратные связи к 10 крупным клеткам [326]). У виноградной улитки, как и у аплизии, после привыкания к раздражению одного сенсорного участка центральный нейрон способен ответить на сигналы, приходящие из другого участка [563].
У голожаберного моллюска Tritonia в церебральной массе идентифицировано около 50 нейронов. С помощью нескольких электродов, вживленных в головной мозг животного так, что оно могло свободно двигаться, регистрировали и стимулировали отдельные вставочные нейроны [582, 583]. Стимуляция отдельных нейронов вызывала специфическое локальное сокращение, а при стимуляции нескольких вставочных нейронов возникала сложная форма поведения (рис. 15-12). При применении естественных стимулов (например, экстракта морской звезды) и одновременно при отведении активности идентифицированных вставочных нейронов получены 3 типа ответов: 1) импульсация, совпадающая по фазе с дорсальными флексиями латеральных «крыльев» ноги; 2) активность, чередующаяся с ответами (1) и совпадающая по фазе с вентральными флексиями и 3) импульсация, возникающая при обоих плавательных движениях (нейроны общего возбуждения). Очевидно, разряды клеток одной группы синхронизированы, а возбуждение в двух группах флексоров поддерживается положительной обратной связью между нейронами флексоров и нейронами общего возбуждения. Координация попеременных движений основана на реципрокной тормозной связи. В ответ на приход определенной сенсорной активности изолированный мозг может генерировать импульсы по той же программе, что и мозг интактного животного [582а].
Итак, крупные нейроны в ганглиях брюхоногих моллюсков отличаются поразительной индивидуальностью синаптических свойств и соотношения с поведением. Ганглии содержат около 1—2% крупных нейронов, которые удается идентифицировать в каждом животном. Функции остальных 98% нейронов и вообще мелких клеток еще предстоит выяснить. Один центральный нейрон получает волокна от широкого сенсорного поля, и одна клетка может запустить сложную моторную реакцию.
Г оловоногие моллюски
'"У головоногих моллюсков масса мозга представляет собой несколько слившихся ганглиев и обеспечивает сложные формы по
Центральная нервная система
37 -
ведения животного (рис. 15-13, 15-14). Функции разных частей мозга проанализированы в анатомических исследованиях путем разрушений и электрического раздражения. Попытки зарегистрировать электрическую активность в мозге головоногих оказались
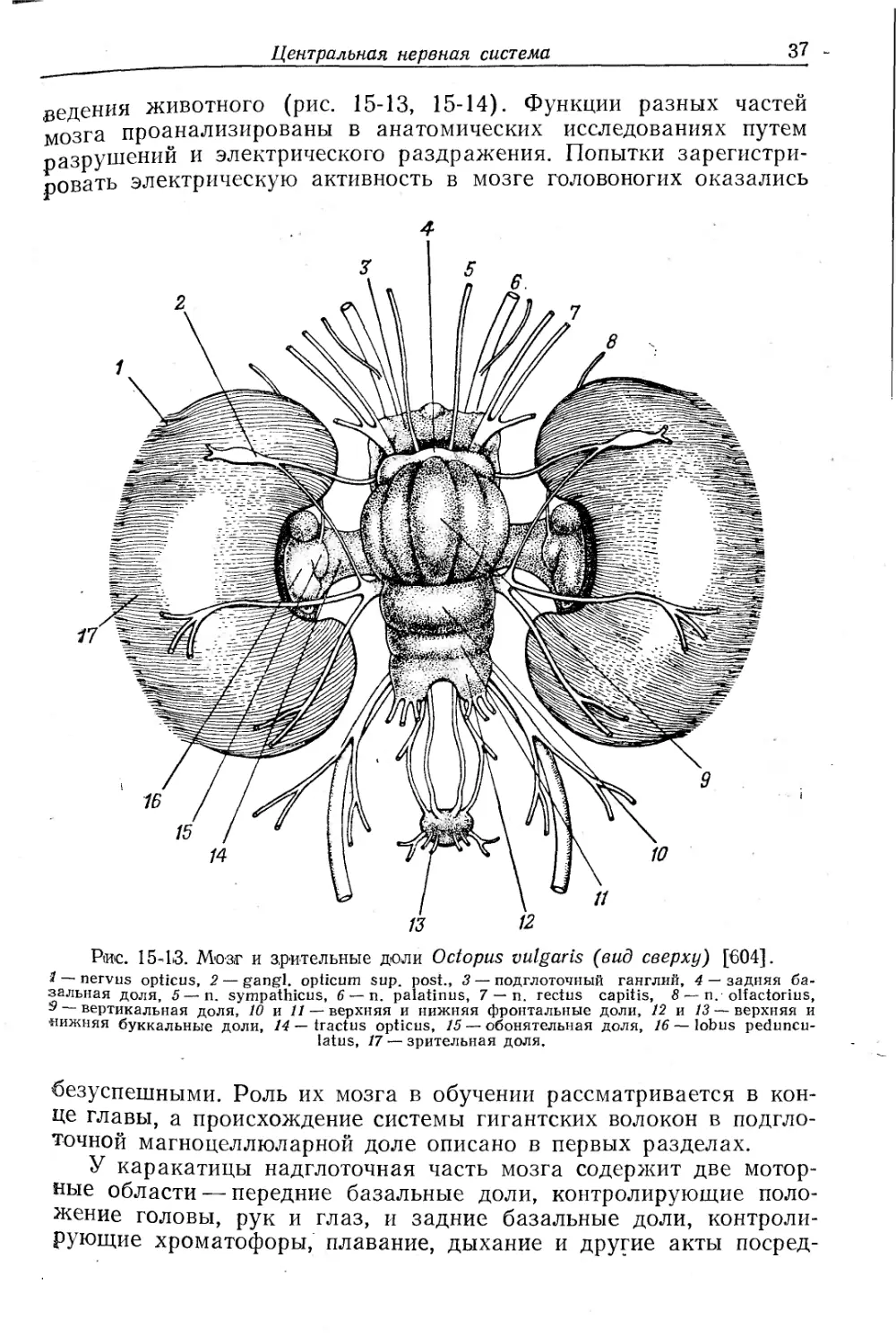
Рис. 15-13. Мозг и зрительные доли Octopus vulgaris (вид сверху) [604].
1 — nervus opticus, 2 — gangl. opticum sup. post., 3 — подглоточный ганглий, 4 — задняя базальная доля, 5—n. sympathicus, 6 — и. palatinus, 7 — и. rectus capitis, 8 — n. olfactorius, Э — вертикальная доля, 10 и 11 — верхняя и нижняя фронтальные доли, 12 и 13 — верхняя и •нижняя буккальные доли, 14 — tractus opticus, 15 — обонятельная доля, 16 — lobus peduncu-latus, 17 — зрительная доля.
безуспешными. Роль их мозга в обучении рассматривается в конце главы, а происхождение системы гигантских волокон в подглоточной магноцеллюларной доле описано в первых разделах.
У каракатицы надглоточная часть мозга содержит две моторные области — передние базальные доли, контролирующие положение головы, рук и глаз, и задние базальные доли, контролирующие хроматофоры, плавание, дыхание и другие акты посред
38
Глава 15
ством шести моторных центров. Над базальными долями лежат нижние и верхние фронтальные доли, а дорсально и в центре — вертикальные и субвертикальные доли. Это ассоциативные области, и их раздражение не вызывает движений; они связаны также с мощным зрительным трактом. Подглоточная часть мозговой
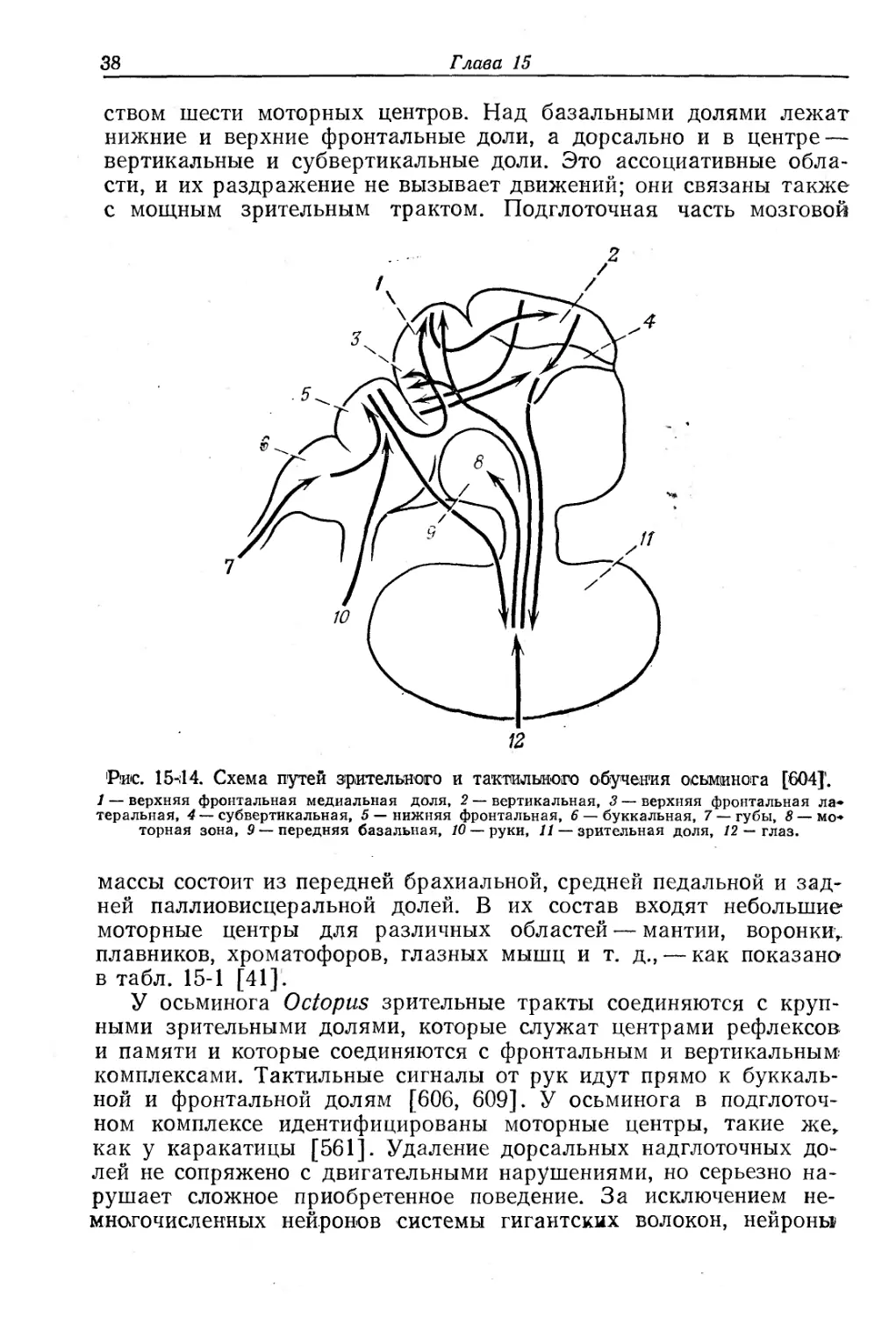
Рис. 15-14. Схема путей зрительного и тактильного обучения осьминога [604]'.
1 — верхняя фронтальная медиальная доля, 2 — вертикальная, 3 — верхняя фронтальная латеральная, 4 — субвертикальная, 5 — нижняя фронтальная, 6 — буккальная, 7 — губы, 8 — моторная зона, 9—передняя базальная, 10 — руки, 11— зрительная доля, 12 — глаз.
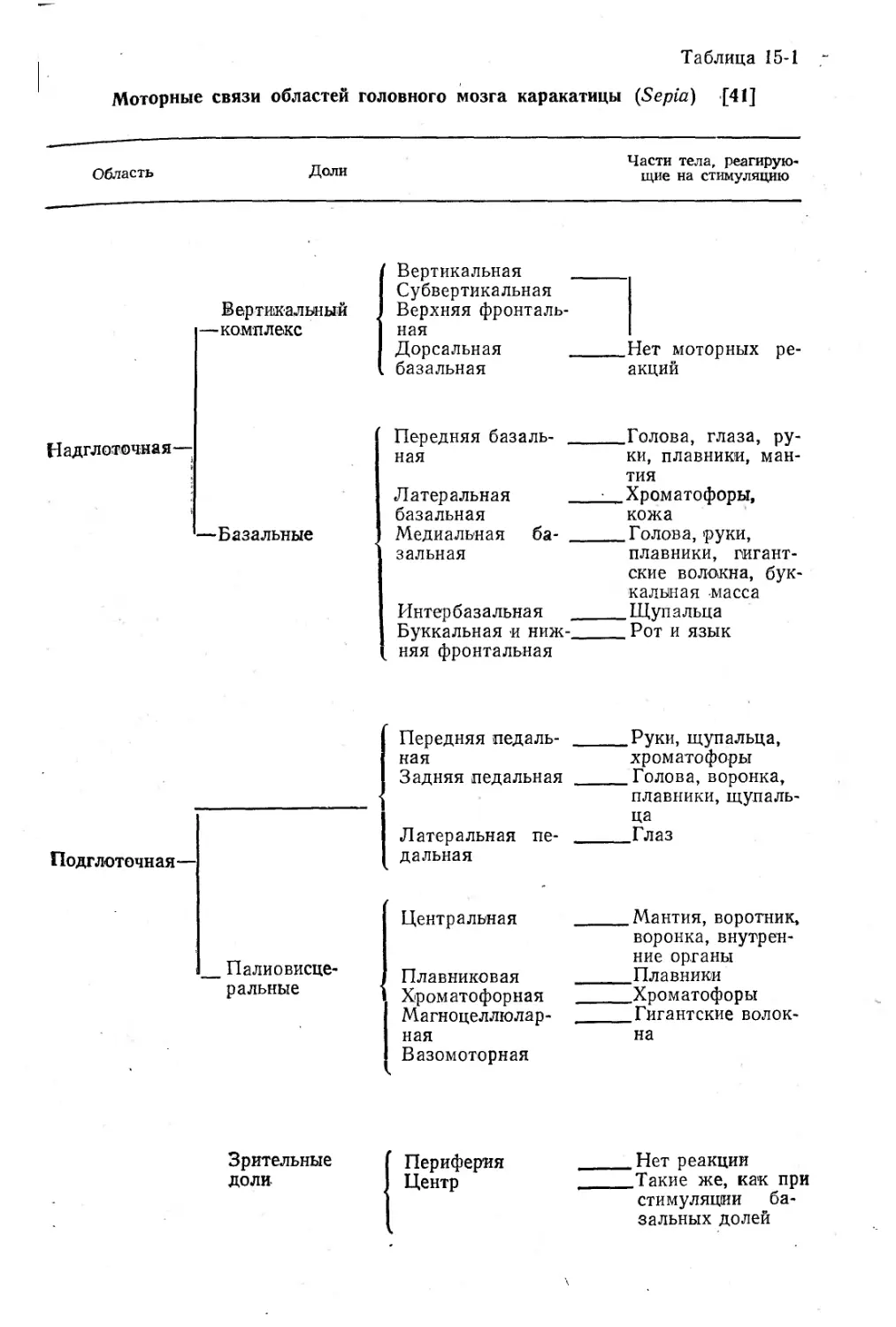
массы состоит из передней брахиальной, средней педальной и задней паллиовисцеральной долей. В их состав входят небольшие моторные центры для различных областей — мантии, воронки,, плавников, хроматофоров, глазных мышц и т. д., — как показано-в табл. 15-1 [41].
У осьминога Octopus зрительные тракты соединяются с крупными зрительными долями, которые служат центрами рефлексов и памяти и которые соединяются с фронтальным и вертикальным комплексами. Тактильные сигналы от рук идут прямо к буккальной и фронтальной долям [606, 609]. У осьминога в подглоточном комплексе идентифицированы моторные центры, такие же, как у каракатицы [561]. Удаление дорсальных надглоточных долей не сопряжено с двигательными нарушениями, но серьезно нарушает сложное приобретенное поведение. За исключением немногочисленных нейронов системы гигантских волокон, нейроны
Таблица 15-1
Моторные связи областей головного мозга каракатицы (Sepia) [41]
Область
Доли
Части тела, реагирующие на стимуляцию
Надглоточная—
Вертикальный
—комплекс
Вертикальная _________
Субвертикальная
Верхняя фронтальная
Дорсальная Нет моторных ре-базальная акций
—Базальные
Передняя базаль- Голова, глаза, ру-ная ки, плавники, ман-
тия
Латеральная Хроматофоры,
базальная кожа
Медиальная ба- Голова, руки,
зальная плавники, гигант-
ские волокна, буккальная -масса
Интербазальная _______Щупальца
Буккальная и ниж-Рот и язык няя фронтальная
Передняя педаль- на я Задняя педальная Руки, щупальца, хроматофоры Голова, воронка, плавники, щупаль-
Подглоточная— Латеральная пе- дальняя ца Глаз
Палиовисце-ральные Центральная Плавниковая Хроматофорная Магноцеллюлар- ная Вазомоторная Мантия, воротник, воронка, внутренние органы Плавники Хроматофоры Гигантские волок-на
Зрительные доли-
Периферия Центр
_____Нет реакции
_____Такие же, как при стимуляции базальных долей
40
Глава 15
мозга головоногих по большей части мелкие (у 90% из них диаметр меньше 5 мкм). Обе зрительные доли содержат около-120-106 нейронов, вертикальная доля 25-106, верхние фронтальные доли 1,8-106. Подсчитано, что мозговая масса осьминога содержит в общем больше 168-106 нейронов [561, 606].
В руках осьминога имеются не только афферентные и эфферентные волокна, но и аксиальные ганглии, которые обеспечивают рефлекторные акты присосок; вставочные нейроны в этих ганглиях дают ответы разной конфигурации на раздражение разных участков тела [461]|.
Физиология ганглионарных нервных систем лестничного типа
Кольчецы
Центральная нервная система кольчатых червей представляет собой цепочки ганглиев по типу лестницы; у некоторых олигохет (дождевых червей) в субэпидермальном сплетении имеются лишь ограниченные связи между эпидермальными рецепторами и сегментарными мышцами, но у других кольчецов (пиявок и полихет} все связи между входом и выходом, очевидно, осуществляются через центральную нервную цепочку. У дождевого червя несколько» сегментов брюшной нервной цепочки, изолированные лигатурой от мышц тела, продолжают проводить перистальтическую волну к области, лежащей за лигатурой. Но в норме эта волна усиливается сегментарными рефлексами. Если отрезок туловища дождевого» червя длиной в 20—40 интактных сегментов подвесить в физиологическом растворе и плавно покачивать, то перистальтики не будет, но легкое растягивание или тактильное раздражение отрезка вызывают сильную перистальтику [79]. Перистальтика, вызванная растяжением, тормозится тактильной стимуляцией. Максимальная частота ритмических сокращений составляет примерно* 1 за 2 с; такие сокращения продолжаются иногда несколько минут после удаления вызвавшего их груза. Кольцевые мышцы сот кращаются во время расслабления продольных, и наоборот, а щетинки управляются координированными рефлексами. Система чередующихся рефлексов осуществляется через посредство центральных волокон с медленным проведением 0,4—0,6 м/с [441]. Перистальтика наблюдается и после перерезки брюшной нервной цепочки или удаления нескольких ее сегментов и даже в том случае, когда червь разрезан надвое, а обе части соединены нитками,,, или же при наркотизации промежуточных сегментов. Но если удалить нервную цепочку червя на протяжении более трех сегментов; и эти сегменты плотно связать, то перистальтическая волна не пройдет через них. Каждый сегментарный нерв обладает сенсорным и моторным полями, охватывающими три сегмента. Отделен
Центральная нервная система
41
ная от ганглия продольная мышца проявляет спонтанную активность (см. гл. 16), а лишенные ганглиев сегменты сокращаются Б ответ на тактильные или световые раздражения. Очевидно, подкожное сплетение обладает какими-то периферическими связями между сенсорными окончаниями и мышцей и обеспечивает конвергенцию многих сенсорных клеток на нескольких нейронах, отростки которых проходят в сегментарных нервах к центральным ганглиям.
У пиявок перерезка нервной цепочки нарушает координированное плавание. Натяжением, прикосновением к рецепторам туловища или стимуляцией присоски можно вызвать рефлекторную перистальтику. В отличие от дождевых червей у пиявок гигантские вставочные нейроны активируются сенсорными окончаниями -с обоих концов тела [318]. В каждом ганглии содержатся 350 нейронов; 14 сенсорных нейронов образуют три группы с каждой -стороны; три отвечают на легкое прикосновение, две на длительное давление и две другие — на повреждающие химические воздействия (рис. 15-15). Сенсорные поля тактильных нейронов перекрываются. Сенсорные нейроны имеют контакты с одной стороной ганглия и с тремя клетками в каждом смежном ганглии через электрические синапсы с двойным выпрямлением (см. гл. 11). Передача от этих тактильных клеток на мотонейроны тоже происходит электрическим путем, но только с одним выпрямлением. От сенсорных клеток, воспринимающих повреждающие воздействия, активность проводится к мотонейронам медленно (с задержкой в 2—4 мс), подавляется ионами магния и усиливается ионами кальция; следовательно, проведение идет через химический синапс. Передача к мотонейронам от клеток, чувствительных к давлению, происходит и химическим и электрическим способом с одним выпрямлением, как у тактильных клеток [392, 393],. Мотонейроны иннервируют в мышце перекрывающиеся поля, причем каждый из них снабжает волокнами один мышечный слой. В каждом ганглии заложены 14 пар возбуждающих и три пары тормозных мотонейронов. Между собой мотонейроны связа-• ны электротонически, но передача активности от них на мышцу осуществляется химическим путем [520]. Связи одного ганглия со следующим не симметричны; связь от заднего к переднему ганглию содержит как тормозные, так и возбуждающие синапсы, :а от переднего к заднему — только возбуждающие. Таким образом, нервная система пиявок характеризуется большой нейронной специфичностью и наличием как электрических, так и химических синапсов.
У большинства полихет перистальтика контролируется сегментарными рефлексами и центральными ритмами. В отличие от дождевых червей у них не найдены периферические связи. У Nereis при локомоции параподии движутся последовательными группами по 4—8. Одновременно с их волнообразными биениями про-
42
Глава 15
дольные мышцы той же стороны сокращаются, а на противоположной стороне расслабляются; ползание Nereis состоит из ряда. S-образных движений, а не из волны кольцевой перистальтика [171]. У Harmothoe .перистальтика может не зависеть от перемещения параподий; волна их движения распространяется вперед на 3—10 сегментов за 1 с; а ипсилатеральные и контралатеральные циклы расходятся по фазе на 180°. Волнообразные дви
Рис. 15-115. Ганглий улитки, вентральная поверхность [623].
Сенсорные вставочные нейроны идентифицированы по ответу на раздражение кожи. I — прикосновение, // — давление, III — вредящие стимулы. Приведены записи соответствующих! ответов на внутриклеточную стимуляцию. 1 — коннективы, 2 — корешки.
жения параподий запускаются нервной цепочкой и не прекращаются после перерезки сегментарных нервов (сохраняющей интактными веточки, идущие к параподиям) [319]. У полихет, живущих, в трубках, периодически повторяющиеся движения животного, связанные с питанием и со смещением жидкости в трубке, управляются центральной нервной системой. У Arenicola marina ритмические сокращения стенки тела и хвоста запускаются отдельными,, но взаимодействующими пейсмекерами. У Arenicola ecaudata выворачиваемая полихетой часть тела (хоботок и глотка) в изолированном состоянии производит ритмические пищевые движения,, если она связана с надглоточным ганглием. Некоторые циклы ак
Центральная нервная система
43
тивности, управляемые из центра, развиваются очень медленно (по одному за 40 мин) [559]j. Nephthys с интактным мозгом роет более сложные норки, чем после децеребрации; обучение в лабиринте после децеребрации замедляется, но если мозг удален после обучения, то приобретенное поведение сохраняется [72, 125, 126].
функции ганглиев у членистоногих
Нервная система членистоногих, как и кольчецов, принадлежит к лестничному типу. Вдоль тела животного идут две симметричные цепочки ганглиев, соединенные коннективами. Парные правый и левый ганглии соединены поперечными коммисурами. В грудном отделе ганглии часто сливаются. Надглоточный ганглий, «мозг», к которому подходят пути от главных органов чувств головы, соединен двумя коннективами с подглоточным ганглием. Число нейронов в нервной системе членистоногих сравнительно невелико: оно не соответствует сложности их поведения.
У рака в каждом брюшном ганглии имеется 500 «крупных» нейронов, в каждом грудном несколько больше, в подглоточном — 6000, в мозге— 10 000, а всего — около 29 000 «крупных» нейронов; «мелких» нейронов насчитывается около 37 000 [574]. Второй брюшной ганглий палочника C.arausius состоит из 600 нейронов, 1000 глиальных и 500 периневральных клеток [25]. Метатора-кальный ганглий таракана содержит 3000 нейронов, из них у 230 диаметр превышает 20 мкм [76]. Кроме обычных нейронов, многие членистоногие обладают системой гигантских волокон; у насекомых и некоторых ракообразных она состоит из крупных, длинных одиночных нейронов, у десятиногих раков — из разделенных перегородками латеральных гигантских волокон (см. рис. 15-3).
У членистоногих меньше половины нейронов сравнительно крупных размеров (5 мкм или больше). Их можно исследовать по ‘отдельности, или расщепив пучок до нескольких волокон, или ,отводя активность внутриклеточно от тела нейрона или крупного аксона. Центральные нейроны большей частью униполярны, и активность может распространяться по ним и по их разветвленным отросткам, не проходя по телу клетки (рис. 15-3, 15-16). Оба синаптических участка и точки генерации импульсов могут лежать в нейропиле на некотором расстоянии от сомы. Нейроны независимы друг от друга в своей спонтанной импульсации и ответах на сенсорные раздражения; здесь нет ни суммарных медленных электрических волн, ни вызванных потенциалов, характерных для позвоночных, быть может, из-за того, что здесь нейроны не расположены слоями, как в коре большого мозга. Поведение членистоногих хотя и стереотипное, но весьма сложное. Очевидно, отдельные нейроны обладают значительными индивидуальными разли-
Рис. 15-16. Нейроны ганглиев таракана, выявленные ионофорезом кобальта. (С любезного разрешения М. Коэна.)
А. Вентральная поверхность метаторакального ганглия; видны гигантские волокна и коллатерали. Б. Дорсальная поверхность шестого брюшного ганглия; видны тело гигантского нейрона, дендриты и часть гигантского аксона. В. Дорсальная поверхность метаторакального ганглия; виден мотонейрон (управляющий быстрой реакцией коксального депрессора).
Центральная нервная система
45
чиями; вставочные нейроны функционируют по иерархии уровней, I а поведение животного определяется простыми нервными сетями.
Локомоция у членистоногих сложнее, чем у кольчецов, из-за наличия членистых конечностей со сложной мускулатурой, эндогенных ритмов в управляющих нервных центрах и модуляции моторной активности проприоцептивными воздействиями и импульсами, идущими от мозга. Членистоногих можно расположить в ряд по возрастающей сложности их локомоции [345]. У Peripattis Наблюдается Основная для членистоногих форма локомоции, I в пределах которой существует несколько «аллюров». При движе-I нии на «первой скорости» парные ноги перемещаются в одной и
| той же фазе и совершают длинное обратное движение, на «сред-
| ней скорости» шаги одинаковые, а на «третьей скорости» шаг короткий, а парные ноги движутся попеременно. У Diplopoda выработалась ходьба по типу «первой скорости», и они отталкиваются ногами; Chilopoda движутся на «третьей скорости» и «тянут ноги».
У тарантула четыре пары ходильных ног; порядок движений при ходьбе непостоянный, попеременные шаги часто сменяются «диагональными». Контралатеральный антагонизм выражен не сильно. Каждая нога со своей половиной ганглия представляет своего рода осциллятор, который соединен отрицательной связью с ногой на противоположной стороне и с соседними ногами. Координация локомоторных движений описана несколько ниже.
Ракообразные. Сенсорные вставочные нейроны. Первичные сенсорные входы нервной системы ракообразных описаны выше для разных органов чувств: рецепторы растяжения, тактильные ре^ цепторы и статоцисты — в гл. 12, глаза — в гл. 14. Аксоны вставочных нейронов, интегрирующих сенсорную информацию, лежат в строго определенных участках нервного ствола и глазного стебелька. Вставочные сенсорные нейроны можно отличить от первичных сенсорных нейронов по большей чувствительности к аноксии, способности усваивать только низкие частоты и широким (часто множественным) периферическим рецептивным полям.
В одной из коннектив между III и IV брюшными ганглиями рака идентифицировано и картировано 75 нейронов. Из 57 вста-> вочных нейронов 30 отвечали на стимуляцию механорецепторов только ипсилатеральную, 10 — только на контралатеральную и 7 — на билатеральную [577]. Самые простые вставочные нейроны ограничены одним сегментом, а другие проводят информацию от нескольких ганглиев. Один вставочный нейрон может иметь входы на нескольких дендритах, у других аксоны образуют перекрест в нервной лестнице. В шестом брюшном ганглии один парный нейрон возбуждается по многим сенсорным корешкам, а на внутриклеточных записях видны единичные ВПСП, возникающие при раздражении отдельных волосков; вставочные ней
46
Глава 15
роны второго порядка в этом и других ганглиях активируются сенсорными вставочными нейронами [291].
В коннективе между грудной и брюшной цепочкой рака най-
дены отличимые друг от друга элементы числом 131, из которых 22 представляют собой первичные сенсорные волокна. Вставочные нейроны распределяются по характеру входов следующим образом (в процентах) [576].
Ипсилатеральные Контралатеральные Билатеральные
От статоцистов 2 1 0
От тактильных волосков 42 8 5
От тактильных волосков и 6 2 2
суставов
Только от суставов 13 4 6
От общей активности 6 5 7
Из этих нейронов 31% направляют активность по нисходящим путям, 51%—по восходящим и 18%—в обоих направлениях. В нескольких клетках зарегистрировано торможение при раздражении головы [576, 579]. Воздействие колебаний воды на антен-нулу вызывало ответ вставочного нейрона в окологлоточной коннективе; ответ был меньше в случае одновременного движения ног или брюшка и при зрительной стимуляции; таким образом, чувствительность вставочного нейрона модулируется разными входами [532].
Глазной стебелек ракообразных имеет три переключения и представляет собой сложную интегративную структуру, которая анатомически представляет собой часть мозга. В глазном стебельке омара или рака идентифицированы 11 вставочных нейронов [13, 575, 580]. По одним волокнам передается информация об интенсивности освещения; вторые реагируют на движение предметов— соответственно на быстрое, умеренное, поступательно-возвратное или очень медленное движение. Третьи вставочные нейроны сигнализируют о движении темных или же светлых предметов. Имеются нейроны с узкими и широкими полями, а также такие, которые реагируют на движение лишь в одном направлении. Мультимодальные вставочные нейроны получают зрительные сигналы от глаз и тактильные сигналы от волосков на теле. Некоторые нейроны с фоновой активностью отвечают на стимуляцию небольшого зрительного поля, окруженного тормозным кольцом с on — off-ответами на границе (см. гл. 14). Волокна, отвечающие на поступательно-возвратное движение, не способны различать направление движения, а волокла с пространственной константностью сигнализируют о константности локализации и размеров поля при изменении положения глаза. В фоновой активности волокон, отводимой вживленными электродами, наблюдаются циркадные ритмы с максимальной реактивностью в период между 22 и 24 ч [13]'. Некоторые вставочные нейроны в мозге находятся
Центральная нервная система 47
под контролем статоцистов, но на них влияет и зрительная активность, и командные мотонейроны, управляющие мышцами глазного стебелька [580].
Итак, многиф вставочные нейроны второго и третьего порядка в нервной системе ракообразных имеют строгую локализацию; на них конвергируют и перекрываются сенсорные входы; они сигнализируют о специфических свойствах сенсорных сигналов и отвечают более чем' на одну модальность; на возбудимость некоторых из них влияют другие вставочные нейроны.
Моторные вставочные нейроны. Функция вставочных нейронов исследовалась также в наблюдениях над моторными ответами на стимуляцию отдельных клеток. Мотонейрон активируется вставочным нейроном, который, возбуждая множество мотонейронов, может вызывать координированные движения нескольких мышц. Один вставочный нейрон возбуждает или тормозит целые мышцы или отдельные пучки мышечных волокон, и эффект его меняется с изменением частоты и порядка следования импульсов. Уронод рака совершает движения в трех плоскостях при помощи 18 разных мышц, которые иннервируются по меньшей мере 55 мотонейронами. Тонические мышцы уропода получают иннервацию от трех (возбуждающих клеток и одной тормозной; фазическими мышцами управляют до 5 возбуждающих нейронов и один (или ни одного) тормозный [313].
Один (командный) вставочный нейрон активирует группу мотонейронов иногда через посредство других вставочных нейронов и, таким образом, вызывает координированное движение. Командные нейроны координируют активность мотонейронов и таким образом управляют степенью и характером сокращений. Командные нейроны вызывают симметричные движения уроподов на обеих сторонах брюшка. Эти движения реципрокны: во время разгибания флексоры получают тормозный сигнал и из центров, и с периферии; при сгибании тормозятся экстензоры [130].
У интактного рака или омара плавательные конечности совершают ритмические биения синхронно в каждом сегменте и последовательно в каждом следующем сегменте. Каждая плавательная конечность приводится в движение 12 мышцами, и рабо-< чие удары и обратные движения взаимосвязаны на обеих сторонах сегмента. В одной коннективе между грудью и брюшком лежит не менее 5 вставочных нейронов, и стимуляция любого из ню; вызывает двустороннее ритмическое движение всех плавательных конечностей брюшка. В каждом сегменте брюшной цепочки с обеих сторон находится по пять командных нейронов; стимуляция любого из них запускает осцилляторный нейрон, который в свою очередь вызывает ритмическую импульсацию группы мотонейронов. Стимуляция с частотой 30 в 1 с вызывает нормальные биения и ритмические разряды частотой один раз в 1 с. Любой из трех вставочных нейронов тормозит ритмические разряды на обе
48
Глава 15
их сторонах [18, 578]!. Каждый из двух нервов плавательной конечности состоит из 6 крупных возбуждающих аксбнов и одного тормозного, а также из многочисленных мелких моторных аксонов; перерезка веточек сенсорных окончаний не i/рерывает ритмическую активность мотонейронов. Мотонейронь^ плавательных j конечностей действуют как одно целое, причем / мелкие клетки -разряжаются раньше крупных. По-видимому, командные нейро- ; ны конвергируют на одном или нескольких еще не идентифицированных осцилляторных нейронах, поочередно возбуждающих и 5 тормозящих группы мотонейронов, которые синергично действуют ' на мышцы и вызывают резкое движение плавательной конечности вверх или вниз [95—97].
Быстрая флексия брюшка и реакция «убегания» запускаются раздражением тактильных волосков; несколько групп продольных мышц активируются мотонейронами, которые управляются или мелкими вставочными нейронами, или же гигантскими волокнами. У рака медиальные или латеральные гигантские волокна действуют посегментно на гигантский мотонейрон и несколько более мелких мотонейронов, иннервирующих фазические флексоры (рис. 15-4). Гигантский мотонейрон получает возбуждение через выпрямляющий электрический синапс от веточки латерального гигантского аксона (см. гл. 11). Моторные синапсы расположены на дендритах, в которых возникают спайки; эти спайки, очевидно, усиливают ВПСП настолько, что за пределами зоны низкого фактора надежности возникает аксонный спайк. Ветвление дендрита создает некоторую лабильность ответов гигантских мотонейронов [617, 618]. Многократные удары хвостом при плавании вызываются или гигантскими волокнами, или менее крупными вставочными нейронами цепочки [488]. Латеральные гигантские волокна управляют также взмахами уроподов [314]!. Ростральная механическая или зрительная стимуляция активирует медиальное гигантское волокно, тело которого лежит в мозге; каудальная стимуляция активирует латеральные гигантские волокна. Обе системы гигантских элементов имеют билатеральные вход и выход [526].
Рефлексы у ракообразных. Рефлексы представляют собой прямые ответы определенной структуры на сенсорный сигнал, которые модифицируются сенсорными и центральными сигналами, конвергирующими на вставочных нейронах. Движение в сочленениях ноги рефлекторно вызывает реципрокное возбуждение и торможение. Так, например, пассивное разгибание дактилоподита возбуждает замыкатель клешни и тормозит размыкатель [60]: стимуляция пары хордотональных органов возбуждает размыкатель клешни и тормозит замыкатель [129].
Между грудью и брюшком, а также между брюшными сегментами лежат по две пары рецепторов растяжения, один из которых тонический, а другой — фазический. Медленно адаптирую
Центральная нервная система 49
щийся, низкоЬороговый рецептор рефлекторно возбуждает один из мотонейронов^, иннервирующих экстензоры брюшка той же стороны; при сокращении экстензора рецептор освобождается от нагрузки. Таким \образом тонический рецептор растяжения участвует в «рефлексе на растяжение», который приводит к стабилизации длины сегмента. Он действует преимущественно ипсилате-рально и IB пределах одного сегмента, функционируя как детектор ошибки в сйстеме сервоконтроля положения сегмента [141, 142].
Примером рефлекса поразительной точности у ракообразных может служить оптокинетическая реакция — движение глазного стебелька, следящее за смещением предмета или маленького светового пятна. Глаз краба может следить за движением солнца даже в том случае, если ориентиры и горизонт заслонены от него. Глаза ракообразных реагируют на очень медленные движения, всего 0,001° в 1 с, следят за движущейся границей на протяжении около 15°, а затем скачком возвращаются обратно—-нистагм. Перемещение изображения по сетчатке преобразуется в возбуждение последовательного ряда дирекционально чувствительных нейронов в мозге, благодаря чему один, ослепленный глаз будет двигаться под влиянием второго, зрячего, глаза, который фиксировали и лишили возможности двигаться; для этого не нужна проприоцептивная обратная связь. Если глаз краба фиксирует ряд определенным образом расположенных полос, а затем свет выключают, передвигают полосы и снова освещают их, то глаз движется так, как будто он .видел, как полосы перемещались. Благодаря памяти в ЦНС коррелируется стимуляция омматидиев до темноты со стимуляцией других омматидиев после темноты. Обычно тремор глаза усиливает восприятие границ [21, 58, 216, 219, 221].
Один из защитных рефлексов у краба состоит во втягивании глазного стебелька, которое вызывается четырьмя мотонейронами в мозге. Каждый из них имеет по два крупных дендрита, которые сильно ветвятся и к которым поступают сигналы от механорецепторов на поверхности тела. При этом регистрируются и ВПСП, однако спайковой активности в крупных дендритах не отмечается. Синаптические потенциалы в двух ипсилатеральных мотонейронах синхронны; между ними имеется слабая электрическая связь, но их синхронность, по-видимому, создается главным образом общими пресинаптическими связями. Активность, приходящая по покровному нерву, оказывает возбуждающее действие, а импуль-сация от окологлоточной коннективы — и возбуждающее, и тормозное [481].
Стоматогастральный ганглий у рака состоит из 25, а у омара из 33 нервных клеток; он иннервирует мышцы передней кишки '[312]. Сенсорные импульсы, возникающие в ответ на механическую стимуляцию стенки желудка, проходят через ганглий по 4—2067
50
Глава 15
стоматогастральному нерву к грудному отделу центральной нервной системы [312]1. Выходящие из нее эфферентные' волокна заканчиваются густым синаптическим нейропилем в соматогастраль-ном ганглии. Нейроны по большей части представляют собой мотонейроны, иннервирующие 25 мышц желудка (рис. 15-17). Одна их группа (9 нейронов) генерирует разряд, приводящий в действие желудочную мельницу; генерация разрядов, по-видимому,
Рис. 15-17. Схема связей стом'атогастральных ’нейронов в пилорическом цикле омара '[351].
Параллельные линии — электротехнические синапсы. Большие светлые кружки — медленные тормозные синапсы; маленькие светлые кружки — быстрые тормозные синапсы; нейроны: ПД — пилорический дилататор; ПЗ — передний нейрон, генерирующий залпы; ВД — вентрикулярный дилататор; ЛП — латеральный пилорический; НК — нижний кардиальный, П — пилорический. Справа — упрощенная схема.
подчинена командным нейронам в центральной нервной системе. Другая группа (14 мотонейронов) активирует пилорический фильтр и связанные с ним мышцы; разряды этих мотонейронов продолжают возникать с той же последовательностью в изолированном ганглии. Их ритмика зависит как от взаимосвязей, так и от мембранных характеристик специфических нейронов. В записях активности идентифицированных отдельных нейронов обнаруживаются быстрые и медленные ТПСП и ВПСП, сильное и слабое электротоническое взаимодействие — результаты многочисленных связей. Нейроны, разряжающиеся с одной и той же временной последовательностью, связаны между собой синапсами, а нейроны, активирующие одну и ту же или синергические мыш
Центральная нервная система
51
цы, обычно связаны электротонически. Химические синапсы по большей части тормозные. На рис. 15-17 дана соответствующая схема [350, 351].
Другим примером программированной активности вставочного нейрона у омара служит контроль дефекации, осуществляемый шестым брюшным ганглием. Моторные импульсы идут к ректальным мышцам по двум задним кишечным нервам и возникают во вставочных нейронах, которые активируются глоточными коннек-тивами или другими отделами нервной цепочки. Вставочные нейроны разряжаются или отдельными импульсами, или пачками или же вовлекаются в ритм стимуляции. Связь между этими клетками варьирует в зависимости от частоты стимуляции нервной. цепочки: одни синапсы устают быстрее других и синхронность работы нейронов меняется с изменением частоты [598].
Насекомые. Ходьба и полет. Локомоция насекомых, очевидно, определяется фиксированными формами активности грудных ганглиев, которая вызывается специфическими стимулами и меняется под действием проприоцептивных сигналов [378]1. Полет обычно запускается прекращением контакта ног с субстратом и поддерживается стимуляцией (движение воздуха) тактильных рецепторов на голове. Точно так же белостома (гигантский водяной клоп) начинает плыть, если подняты его лапки [105]. Ходьба может быть вызвана любым из нескольких конвергирующих сенсорных входов. Часто 6 ног насекомого составляют две попеременно действующие группы по три, и первой совершает шаг та нога, на которую приходится наименьшая нагрузка [236]. Изменение скорости передвижения или же ампутация одной ноги меняет ритм ходьбы. У богомола движения ног при спокойной ходьбе совершаются в такой последовательности: Л (левая) 3, Л2, П (правая) 3, П2; при 'возбуждении и ускорении ходьбы ритм такой: Л2, П2, ЛЗ, ПЗ; когда он ползет — ЛЗ, П1, Л2, ПЗ, Л1, П2 [448]. У некоторых насекомых последовательность движений ног зависит от характера поверхности. У таракана поочередное перемещение ног (по три) сохраняется при любой скорости ходьбы, но по мере ускорения ходьбы возрастает отношение времени вытягивания ноги к времени поджимания [98, 587]. Попеременное сокращение протракторов и ретракторов регулируется центрами, но на скорость этих движений влияют сигналы, идущие по обратным связям от рецепторов [136].
У разных насекомых, возможно, существуют центральные осцилляторы для каждой ноги, связанные друг с другом; сенсорные сигналы от рецепторов на проксимальных частях ноги влияют на активность осциллятора данной ноги; у быстро передвигающегося таракана влияние рецепторов слабее, чем у медленно шагающего палочника (Carausius) [567] (рис. 15-18).
При покалывании брюшка у личинки стрекозы она выбрасывает воду и таким образом движется вперед. Ее брюшная нервная
52
Глава 15
цепочка содержит много клеток со спонтанной активностью, часто идущей регулярными пачками. Одни клетки тормозятся сенсорными раздражениями, у других импульсация учащается, а некоторые молчащие нейроны активируются тактильной стимуляцией. Сенсорное поле вставочного нейрона бывает ипси-, контра- или билатеральным, и на нейроне конвергирует множество волокон [139, 140, 298].
Рис. l<5-il8. Модель управления разгибанием и сгибанием ноги таракана [98]. Активацию осуществляет центральный командный нейрон, который непрерывно посылает сигнальт к элементам А и Б. А преобразует ряды сигналов в синусоидальный сигнал, который затем регулируется порогом Б и выдает периодические пачки импульсов, возбуждающие одну группу мышц. Подобные же, но тормозные пачки импульсов, идущие от Б, взаимодействуют с продолжающейся командой от В, и в результате возникает другая эфферентная активность; в ней пачки точно противоположны по фазе пачкам Б и идут к мышцам-антагонистам. Сенсорные сигналы, приходящие в А, модулируют уровень активности системы.
У саранчи мышцы — закрыватели дыхальца управляются комплексом мотонейронов. Командные вставочные нейроны в метато-рак-альном ганглии вовлекают в свой ритм мотонейроны, которые обладают и собственным ритмом. Они или вовлекают в ритм мотонейроны в отношении 1:1, или облегчают их активность, или же тормозят ее; синаптические участки мотонейрона отделены от пейсмекерных зон [372]'. У богомола механическими или зрительными стимулами или СО2 можно вызвать вентиляцию, причем движения дыхальца сопряжены с локомоцией [374].
В прямых летательных мышцах, например у бабочек и саранчи, разряд мотонейрона и сокращение мышцы находятся в отношении 1 : 1 (см. гл. 16). Привязанная саранча начинает открывать крылья через 45 мс после стимуляции движением воздуха или через 30 мс после нарушения контакта лапок с опорой; непривязанная саранча раскрывает крылья через 15 мс после прыжка, начинающего полет. В полете пронация переднего крыла варьирует в зависимости от скорости ветра, и вставочные нейроны, активируемые рецепторами ветра, участвуют в управлении подъ
Центральная нервная система 53
емом [65]. Боковой ветер вызывает коррекцию направления по-лета. Детекторы ветра могут компенсировать вызываемое повреждением крыла искажение направления, но обычно для сохранения ориентации саранча смотрит на горизонт [167, 589—591]. Вставочные нейроны сигнализируют о направлении и силе ветра и меняют характер импульсации мотонейронов, о чем свидетельствует.' одновременная запись активности нескольких летательных мышц.. Мотонейроны одного пула могут быть связаны друг с другом: электрически, но связь между пулами осуществляется через синапсы [290}. Поочередное сокращение мышц, поднимающих и опускающих крыло, определяется эндогенной ритмикой двух групп мотонейронов. При полете залп импульсов, идущий к мышцам, опускающим крыло, сопровождается паузой в разрядах мышц, поднимающих крыло; иначе говоря, обе группы мотонейронов; связаны по фазе [555, 556, 558]. Полет саранчи менее пластичен,, чем ходьба.
В центральной нервной системе активность определенной! структуры может быть вызвана гормонами. Так, разряд в нерве-фаллуса у таракана, связанный с копуляцией, усиливается введением экстракта из кардиальных тел [368, 369]. Последовательность движений бабочки шелкопряда перед ее выходом из куколки полностью соответствует последовательности электрической активности в нервной цепочке; эта же последовательность регистрируется и в деафферентированной цепочке, обработанной гормоном кардиальных тел, который вызывает превращение куколки [541].
У саранчи ритм полета сохраняется и после удаления головного мозга и подглоточного ганглия; ритмические эфферентные разряды и некоторое движение крыльев возможны и после перерезки идущих от них сенсорных нервов, а также если крылья прижаты и рецепторы, воспринимающие движение воздуха, прикрыты; следовательно, центральный ритм имеет эндогенную природу, но до некоторой степени подвержен рефлекторной модуляции [584, 591]'. Большинство нейронов разряжаются по одному разу на каждый взмах крыла, и учащение их активности соответствует увеличению числа активных клеток и более раннему началу импульсации, так что взмах крыла ограничен [584, 591]. Устранение проприоцептивного входа снижает частоту взмахов; в норме рецепторы дают импульсные разряды один или несколько раз в момент наибольшего подъема крыла и оказывают тонический эффект на эндогенный ритм. Кроме того, ветер также усиливает разряды; Мотонейрона. Растяжение и активность, поступающая к крылу, суммируются в интервале времени, превышающем один цикл движений крыла, и регулируют полет [586].
У ночных бабочек на один взмах крыла приходится один или несколько потенциалов действия. Полет невозможен, пока насекомое не разогреется быстрыми, низкоамплитудными движениями:
54
Глава 15
крыла, при которых часто отмечается по два мотонейронных спайка на одно сокращение. Активность, связанная с разогреванием, имеет эндогенное происхождение и возникает в проторакальных ганглиях. У павлиноглазок частичная ампутация крыла сокращает продолжительность взмаха, а перерезка сенсорных нервов крыла увеличивает ее. При большой частоте взмахов залпы им-. пульсов, производимые мотонейронами, укорачиваются ,(рис. 15-19) [276, 278].
Рис. 15-19. Мышечные .потенциалы у бабочки Celerio при трепетании крыльев (Л) и при спокойном полете (Б) [624].
Три мышцы, поднимающие крыло (2 — тергостернальная, 3 — неидентифицированная, 4 — терготрохантерная), действуют синхронно с дорсальной продольной мышцей (1) при трепетании, но не при полете.
Летательные мышцы двукрылых, а также многих перепончатокрылых и жесткокрылых осциллируют или обладают миогенным ритмом и обычно непрямо связаны с крылом; между отдельными импульсами мотонейрона и движением крыла нет специфических фазовых отношений [385]'. У этих насекомых градации движений создаются частотно-модулированным импульсным кодом и повышение частоты импульсации мотонейрона учащает взмахи крыла, но без строгого соответствия между этими двумя ритмами. У двукрылых сенсорные влияния оказывают на мотонейроны тоническое модулирующее действие, а у саранчи эта модуляция носит фазический характер [593]. Непрямые летательные мышцы мух при большой частоте сокращений работают почти синхронно, а направление полета определяется неосцилляторными прямыми мышцами; таким образом, имеются две моторные системы [389]. У шмеля Bombus синергичные мышечные элементы во время разогревания действуют синхронно, а во время полета последовательно [386].
Центральная нервная система
55
Следовательно, можно заключить, что нейрональный механизм полета насекомых организуется в грудных ганглиях; что у насекомых с прямыми летательными мышцами импульсация более ритмичная, чем у насекомых с осцилляторными мышцами, и у обеих групп сенсорные сигналы оказывают тоническое модулирующее действие. Нейронные цепи ритмически активных центров еще не изучены.
Нелокомоторные рефлексы. Можно привести и другие примеры поведения, которое запускается сенсорным стимулом, а затем следует готовой стереотипной программе. Световые вспышки, испускаемые светляками, по своей длительности, интенсивности и частоте специфичны для данного вида. Самцы тех видов, у которых свечение не является непрерывным, испускают световые вспышки в ответ на краткую световую стимуляцию, и сотни особей одного» вида светятся синхронно. Электрическим раздражением нервной системы можно вызвать вспышку с латентным периодом в несколько сот миллисекунд, тогда как латентный период ответа на прямое раздражение органа свечения составляет от 15 до 75 мс*. [47—49, 67]'|. Нервные пути этой реакции еще не установлены.
О рефлекторном управлении питанием и опорожнением передней кишки у мясной мухи было сказано выше (см. гл. 13). Вкусовые рецепторы на лапках запускают вытягивание хоботка и сосательные движения лабеллы; стимуляция рецепторов растяжения в передней кишке прекращает прием пищи. После перерезки возвратного нерва у мухи формии она проглотила 37,4 мкл сахарного раствора; после перерезки нервной цепочки между мозгом-и грудью — 39,4 мкл, а после «ложной операции» (без перерезки; нервной цепочки)—всего лишь 15—18 мкл. Следовательно, импульсы, возникающие в рецепторах растяжения передней кишки,, идут по возвратному нерву и тормозят двигательный центр питья [99, 166].
Обнаружение звука и центральная реакция на звуковую стимуляцию у бабочек сем. Noctuidae описаны выше (см. гл. 12). Бинауральные первичные слуховые волокна идут ипсилатерально к трем грудным ганглиям; они связаны через релейные нейроны с ипсилатеральными и контралатеральными вставочными нейронами. К вставочным нейронам высших порядков в метаторакаль-ном ганглии относятся: 1) нейроны, отмечающие частоту звуковых стимулов, давая один большой импульс на каждый звуковой толчок, 2) нейроны, отмечающие серии импульсов, т. е. сигнализирующие об их длительности; 3) бинауральные вставочные нейроны, импульсация которых при стимуляции обоих ушей сильнее,, чем при стимуляции одного уха, и 4) спонтанно-активные нейроны,., импульсация которых прекращается при раздражении одного уха-[447—449, 616]. У других вставочных нейронов частота импуль-сации меняется в зависимости от силы звука, а некоторые слуховые клетки в мозгу прекращают свою импульсацию, вызванную»
Глава 15
-56 звуком, при воздействии зрительных стимулов. Центральные связи, управляющие поведением избегания, не прослежены.
У одного из представителей настоящих саранчовых описаны «крупные мезоторакальные слуховые вставочные нейроны двух ти-.пов. 1) Нейрон, получающий и ипсилатеральные, и контралатеральные слуховые волокна и посылающий отросток в головной мозг; многократная стимуляция вызывает в нем привыкание, для ^восстановления ему нужно 25 мин, от мозга к нему приходят тормозные волокна. 2) Вставочный нейрон второго типа со значи-тельной точностью сигнализирует о направлении звука [465]. У кузнечика сем. Tettigonidae один крупный вставочный нейрон (Т) на каждой стороне мезоторакального ганглия возбуждается ^активностью, приходящей от барабанной перепонки; на его реакции воздействует возвратное торможение от заднего ганглия и торможение от контралатеральной барабанной перепонки [521, !522]„ Другой вставочный нейрон, S, отвечает на песню насекомых .данного вида и тормозит нейрон Т через вставочный нейрон D. 8 и Т активируют мотонейроны [360].
Сверчки и некоторые другие прямокрылые издают звуки в .виде специфичной для данного вида стридуляции. Несколько самцов стрекочут попеременно или синхронно, и различные формы стридуляции означают призыв, соперничество или ухаживание (две песни). Если сверчок услышит звуки, характер его песни «изменится [263]. Состоящая из четырех звуков песня создается разными движениями передних крыльев по напильникам и возникает благодаря строгой последовательности точно запрограммированных сокращений ряда тергостернальных, базальных и субаларных мышц. На внутриклеточных записях активности мотонейронов, управляющих складывателем и раскрывателем крыла «сверчка, видны поочередные тормозные и возбудительные синаптические потенциалы. Попеременная импульсация двух групп мотонейронов свидетельствует о наличии обратной связи с осцилля-торным пейсмекером и вставочным нейроном, который поочередно тормозит мотонейроны раскрывателя и складывателя [30— 32]. Подобным же образом последовательные движения ухаживания запрограммированы в грудных ганглиях [123]!.
У «сверхподвижного» мутанта дрозофилы регистрируются быстрые пачки импульсов, идущие от мотонейронов в грудных ганглиях. Структура этой активности не связана с приходом сигналов от органов чувств или от ганглиев более высоких уровней; она представляет собой генетически обусловленный код мотонейронов [240].
У саранчи Schistocerca в метаторакальном ганглии идентифицированы 26 мотонейронов, из которых одни вызывают «медленные», а другие — «быстрые» сокращения определенных мышц. Моторные импульсы возникают на расстоянии 400—500 мкм от тела мотонейрона; дендритные участки, где регистрируются ВПСП
Центральная нервная система
57'
и ТПСП, лежат ближе к телу нейрона; импульсы и синаптические потенциалы пассивно распространяются по телу клетки. Связи: между «быстрыми» и «медленными» мотонейронами частично идут’ через вставочные нейроны; синергизм и распределение во времени активности мотонейронов различных мышечных групп осуществляются премоторными вставочными нейронами. Запись активности вставочных нейронов показывает, что в некоторых из них: происходят выраженные колебания мембранного потенциала, синхронные с залпами импульсов в мотонейронах. В ближайшем будущем, возможно, будут полностью определены нейронные структуры, ответственные за комплекс мышечных сокращений при ходьбе и прыжках, а также механизмы прекращения этих движений [225].
Итак, ганглионарная нервная система ракообразных и насекомых дала возможность подробно изучить интегративные функции одиночных идентифицированных вставочных нейронов. Некоторые из них связаны с сенсорными нейронами; они реагируют-на многочисленные входы, которые состоят из специфических комбинаций сигналов от периферических сенсорных элементов. Другие вставочные нейроны вызывают разряды в соответствующих группах мотонейронов, приводящие к координированным движениям. «Гигантские» волокна представляют собой специальные-мотонейроны крупных размеров, дающие фазические ответы, участвующие в актах «избегания». Многие ритмические движения управляются мотонейронами, которые активируются осциллятор-ными нейронами, а те в свою очередь возбуждаются небольшим числом командных вставочных нейронов. Определенные ритмы импульсации создаются отдельными спонтанно-активными вставочными нейронами или же нервными сетями. Интеграция осуществляется главным образом в нейропиле, но изучение отношения между входом и выходом идентифицированных вставочных и мотонейронов позволяет надеяться, что нам удастся узнать, как программируются сложные движения.
Функция «мозга» членистоногих. Надглоточный ганглий, или мозг, членистоногого животного оказывает тормозное действие на грудные моторные центры; об этом свидетельствует моторная активность, возникающая после разрушения мозга. Степень сложности мозга значительно варьирует у разных членистоногих, но обычно он состоит из нескольких областей: medulla terminalis и добавочные доли являются обонятельными и зрительными центрами; средние и передние области (прото-, дейтеро- и тритоце-ребрум) несут функцию координации и включают крупные дорсальные грибовидные тела (corpora pedunculata), состоящее из нейропиля центральное тело и протоцеребральный мост, а задний дейтеро- и тритоцеребрум получают тактильную иннервацию от-антеннул, содержат моторные центры и дают начало окологлоточным комиссурам и нервам, снабжающим ротовые части. Как
Глава 15
Рис. 15-20. Срезы мозга (надглоточного ганглия) у беспозвоночных, обладающих грибовидными телами [186].
А. Полихета Nereis: 1 — грибовидные тела; 2 — пальпальный нерв, 3 — зрительные нервы, 4 — слой ганглиозных клеток. Б. Ракообразное Calo-caris: 1 — грибовидные тела, 2 — зрительный ганглий 1,3 — зрительный ганглий II, 4 — зрительный ганглий III, 5— латеральный центр, 6 — центральное тело, 7 — слой ганглиозных клеток. 8 — доли дейтероцеребрума, 9 — антеннальный нерв, 10 — тритоцеребрум. В. Рабочая пчела Apis: 1 — грибовидные тела, 2 — центральное тело, 3 — зрительный ганглий I, 4 — зрительный ганглий II, .5— зрительный ганглий III, 6 — антеннальный центр.
и в других областях нервной системы многих беспозвоночных, центральная часть мозга состоит из волокнистого нейропиля, в котором многие нейроны являются униполярными (рис. 15-20).
О сенсорных и рефлекторных функциях мозга было сказано выше. Часть мозга, занимаемая зрительными центрами, составляет от 0,3 до 2,8% У паукообразных и многоножек с эпителиальными фоторецепторами и глазками; от 2,9 до 9,9% у низших ракообразных и насекомых с рудиментарными сложными глазами и от 33 до 80% у насекомых с высокоразвитыми сложными глазами ;[186]. Грибовидные тела, или corpora pedunculata, состоят из большого стебелька, образованного нейропилем, с множеством мелких униполярных клеток в крупных дорсальных долях. Сходные по внешнему виду образования встречаются у некоторых поли-хет, имеют очень большие размеры у мечехвоста, варьируют по величине от 10 до 30% всего мозга у десятиногих раков и обладают значительным объемом у большинства насекомых. Неизвестно, гомологично ли это образование у всех кольчецов и членистоногих [52].
Удаление любой части мозга, которая получает волокна от какого-либо сен
Центральная нервная система
59»
сорного органа, равносильно удалению этого органа. У многих членистоногих после удаления одного глаза или антенны возможна компенсация, а при повреждении мозга дефект поведения оказывается стойким. После перерезки обеих окологлоточных комиссур рефлекторные движения головы сохраняются и возможно-жевание, но не глотание. Сохраняются также рефлексы выпрямления и рефлекторное сокращение ножных мышц, причем могут возникнуть чрезмерные беспорядочные движения ног. После перерезки одной комиссуры или разрушения одной латеральной половины мозга членистоногое животное производит манежные движения в интактную сторону. Удаление подглоточного ганглия или перерезка цепочки позади него может прекратить спонтанную^ активность, но локальные сегментарные рефлексы сохраняются.. Подглоточный ганглий служит для координации ходьбы, плавания или полета, а другие ганглии способны к рефлексам локомоции и автотомии и к значительной спонтанной активности.
Удаление мозга у личинки стрекозы приводит к учащению дыхания, но после удаления подглоточного ганглия частота дыхания снижается. У крабов тормозное действие мозга на брюшные ганглии проявлялось в прекращении круговых движений при элект рической стимуляции перерезанной окологлоточной комиссуры.. После локального разрушения нейронов или стебелька грибовидных тел у гусеницы цекропии она ткет кокон неправильной формы. Повреждение надглоточного ганглия у паука приводит к тому, что он ткет маленькую, неправильной формы паутину [598а].
У сверчка грибовидные тела тормозят локомоторную активность; левая и правая сторона мозга взаимно тормозят друг друга. Центральное тело мозга и подглоточный ганглий возбуждают грудные мотонейроны, но их функция тормозится грибовидными телами [232—235]. У богомола после одностороннего удаления-протоцеребрума ноги противоположной стороны утрачивают то-» нус, а на оперированной стороне становятся более активными;, круговые движения животного направлены в интактную сторону. Обе стороны мозга дают сигналы для поворотов вправо и влево,, а также оказывают тормозное действие на командный центр подглоточного ганглия, который в свою очередь возбуждает мотонейроны (рис. 15-21) [448].
Иногда во время спаривания самка богомола, обхватив самца, отъедает ему голову, но копуляция продолжается. Если у самца Удален только мозг, то половая активность прекращается, но если Удален также и подглоточный ганглий, копуляция может произойти; таким образом, центр копуляции, который находится в последнем брюшном ганглии, в норме тормозится подглоточным ганглием [445, 446, 448]1. После декапитации таракана его ноги совершают координированные движения, а фалломеры сохраняют способность к копуляции. Декапитация таракана или перерезка впереди терминального ганглия приводит к усилению ритмической.*
60
Глава 15
активности в последнем брюшном ганглии. Такое же усиление происходит под 'воздействием экстракта из кардиальных тел; -нервные сигналы, идущие от мозга, в сочетании с гормоном вызывают половое поведение [369, 450].
На записях электрической активности грибовидных тел таракана видны большие синхронизованные спайки, возникающие в
Уис. 15-21. Схема влияния на локомоцию различных разрушений мозга богомола [448].
1 — мозг удален, II — мозг расщеплен, III — интактный мозг, IV — удалена голова, V — удалена половина мозга; м — мозг, пг — подглоточный ганглий, гр — грудной моторный комплекс, (+) возбудительный, (—) тормозный эффекты.
ответ на стимуляцию антеннального нерва с латентным периодом в 40—70 мс; если стимулы наносятся чаще одного в 1 с, наступает утомление. В мозге омара многие нейроны спонтанно активны; некоторые клетки после краткой стимуляции антеннального нерва продолжают отвечать в течение 60 с [352, 399].
У сверчка после удаления мозга исчезает спонтанное стрекотание. Чтобы оно продолжалось, достаточно сохранить одно грибовидное тело. Локальная стимуляция грибовидного тела вызывает один из типов стрекотания; в нем участвует около 28 мышц. Стимуляция центральных тел мозга приводит к аномальному стрекотанию [233]. Иногда стимуляция может оказать тормозное влияние на стрекотание, которое уже началось. Последователь
Центральная нервная система
61
ность работы мышц, по-видимому, запрограммирована в грибовидных телах и преобразуется в команду в центральном теле; вероятно, мышцы активируются мезоторакальными мотонейронами [234]'. Препятствие, создаваемое движению ног и крыльев, не меняет порядка активности мезотор акальных мышц; следовательно, стрекотание запрограммировано в центральной нервной системе, а не вызывается рефлекторно [306]. Активность некоторых центральных нейронов следует циркадному ритму, который проявляется в моторике и стрекотании [19].
Зрительная доля большинства насекомых состоит из периферической ганглиозной пластинки в сетчатке, первой медуллярной пластинки и глубже лежащей второй медуллярной пластинки; все эти три области соединены перекрестами. У бражников и совок найдены сенсорные вставочные нейроны многих типов: монокулярные «оп»-элементы; «оп—оЯ»-нейроны, которые являются детекторами контраста и не зависят от движения; бинокулярные дирекциональные клетки, реагирующие на зрительную стимуляцию, сопровождающую наклон тела и изменение направления движения; нейроны, соединяющие долю одной стороны с противоположной зрительной долей, и небольшое число эфферентных (центробежных) элементов [78]. У саранчи нисходящий нейрон — детектор движения —на противоположной стороне посылает аксон от зрительной доли к грудным ганглиям; многократная сти-’ муляция одной и той же точки сетчатки вызывает привыкание его ответов, которые восстанавливаются при перемещении стимула на другую точку и растормаживаются раздражением цервикальной коннективы или двигательной активностью [463—465];.
Некоторые вставочные нейроны в мозге и подглоточном ганглии мясной мухи не прекращают активности при затемнении сложных глаз, но тормозятся при освещении фасеток; другие пставочные нейроны реагируют на движение антенн, и эта реакция облегчается освещением фасеток [377]. В мозге саранчи некоторые вставочные нейроны отвечают или на зрительные, или на тактильные сигналы; привыкание ответов на стимулы одной модальности не изменяет ответов на другие стимулы [220].
Итак, «мозг» членистоногих состоит из над- и подглоточного «отделов нервной системы. Специфические сенсорные вставочные нейроны осуществляют сенсорную интеграцию. Кроме того, некоторые области «мозга» тормозят вентральные моторные центры и содержат командные вставочные нейроны, управляющие слол$-, ными движениями.
Основные черты морфологии нервной системы позвоночных
В основном у всех позвоночных нервная система организована одинаково: она состоит из нервной трубки, дорсальных сенсорных я вентральных моторных корешков и сходных в общих чертах
62
Глава 15
черепно-мозговых нервов. Детали ее строения можно найти в многочисленных руководствах по сравнительной анатомии [452]. Относительные размеры разных областей мозга показаны на рис. 15-22, а на рис. 15-23 дана сравнительная схема разных областей мозга у млекопитающих и у других видов животных. В то время как основные части нервной системы у позвоночных сходны, гомология их подотделов представляется теперь гораздо менее несомненной, чем несколько лет назад. Важнее разобраться в общности их функций, чем постулировать их гомологию.
Организация центральной нервной системы позвоночных отличается большим количеством нейронов относительно размеров тела, чем у каких-либо беспозвоночных, за исключением головоногих моллюсков. Избыточность выражается в том, что некоторые нейроны могут быть заменены другими, и удаление некоторых; частей мозга не нарушает сколько-нибудь заметно поведения животного; однако более точные измерения показывают, что избыточность эта, пожалуй, кажущаяся. В поясничном сегменте собаки один дорсальный корешок состоит из 12 000 волокон, большей частью афферентных; вентральный корешок содержит 6000 волокон; а в одном поясничном сегменте спинного мозга до-375 000 нейронов, главным образом мелких, и лишь небольшое-число крупных [190, 191]j. Тела сенсорных клеток находятся в; спинальных ганглиях вне спинного мозга, а в самом спинном мозге лежат вставочные и двигательные нейроны (мотонейроны);, и на каждом из последних имеется около 5500 синаптических бля; шек. Таким образом создается потенциальная возможность значительной интеграции в пределах одного сегмента спинного мозга.
У рыб и амфибий главным интегративным отделом мозга считается средний мозг (мезэнцефалон), состоящий из крыши (тек-тум) и покрышки (тегментум); у рептилий большее значение приобретает промежуточный мозг (диэнцефалон); у птиц развиты крупные базальные ядра переднего мозга. Новая кора — шестислойный покров переднего мозга — имеется только у млекопитающих; структурами переднего мозга с такими же связями и функциями обладают рептилии и птицы [102] (рис. 15-23). Иногда встречаются высказывания, что интегративная функция в процессе эволюции переместилась кпереди; но теперь становится ясно, что даже у млекопитающих средний мозг — верхние и нижние бугорки— играет важную роль в простом зрительном опознании предметов и в рефлекторных ответах на эти стимулы, а также в слуховых реакциях. Представление о перемещении функции из «низшего» центра в «высший» сейчас стало не таким незыблемым, как раньше. Кроме того, в пределах отдельных классов (например, млекопитающих) эволюция во многом шла параллельно, и здесь трудно делать обобщения. Примеры развития и переноса функций по мере усиления цефализации будут даны в следующих разделах как для сенсорных, так и для моторных систем.
Рис. 15-22. Сравнительная величина областей головного мозга у некоторых позвоночных [625].
Л. Треска. Б. Аллигатор. В. Кошка. Г. Лягушка. Д. Гусь. Е. Человек. 1 — зрительная доля, •2 — большой мозг, 3 — обонятельная луковица, 4 — мозжечок, 5 — обонятельный тракт, — гипофиз, 7 — нижняя доля, 8 — промежуточный мозг, 9 — воронка, 10 — обонятельная доля, И — зрительный тракт, 12 — эпифиз, 13 — IX и X пары.
Рис. 15-23. Схема головного мозга позвоночных [390].
А. Не-млекопитающие. Б. Млекопитающие; г — гиппокамп, м — миндалина, мж — мозжечек, гип—гипоталамус, тал — таламус, ср. л.—средний мозг, ромб. — ромбэнцефалон, сп. м.— спинной мозг, па — палеостриатум, мкт — медиальное коленчатое тело, лкт — латеральное коленчатое тело, пе — перегородка, вб — верхний бугорок, нб — нижний бугорок; 1 — обонятельная область, 2 — палеостриариый комплекс, 3 — лимбическая система, 4 — наружная стриарная область, 5 — общая кора, 6 — полосатое тело, 7 — новая кора.
Центральная нервная система 65
Одна часть головного мозга осталась сравнительно неизменной в процессе всей эволюции позвоночных — это мозжечок. Возможная причина того, что мозжечок и спинной мозг сохранили одну и ту же структуру во всех классах, состоит в том, что локомоция в этих классах в основном одна и та же, несмотря на различие придатков и способов движения в разных средах.
Как можно видеть у круглоротых и рыб, телэнцефалон развивался вместе с парными обонятельными нервами, и обонятельная луковица, строго говоря, составляет часть переднего мозга. Анатомы не единодушны в своих суждениях о гомологии отделов переднего мозга в разных классах позвоночных. Передний мозг состоит из обонятельной области, лимбической системы, полосатого тела (очень крупного у птиц) и новой коры (хорошо развитой у млекопитающих) [390]. Примитивный плащ (паллиум) выполняет преимущественно обонятельную функцию, и некоторые анатомы считают, что он гомологичен грушевидной доле высших позвоночных (рис. 15-23). Однако передний мозг рыб тоже получает проекции от диэнцефалона и посылает двигательные тракты к стволу мозга и верхнему отделу спинного мозга. Функции необонятельных отделов переднего мозга тесно связаны с функциями диэнцефалона и мало изучены у низших позвоночных. Передний мозг составляет около 60% всей массы головного мозга у акулы и 12% —у серебряного карася; у обоих видов большая его часть не связана с обонянием. В медиальном мозговом плаще амфибий имеются области конвергенции многочисленных сенсорных входов, а дорсолатеральный паллиум представляет собой складку, которая у рептилий превращается в дорсальную стенку желудочка. Неопаллиум рептилий впячивается в желудочек, образуя его дорсальную стенку, средняя часть которой получает слуховые, а латеральная часть — зрительные проекции [396а].
Птицы обладают массивным складчатым плащом; дорсальная стенка желудочка (эктостриатум) получает волокна от круглого ядра промежуточного мозга, каудальный неостриатум — слуховые волокна от медиального коленчатого тела, а дорсомедиальная кора посылает двигательные волокна в спинной мозг. Возможно, что центральные ядра млекопитающих — бледное и чечевицеобразное— гомологичны палеостриатуму пресмыкающихся и птиц. О гомологии говорят высокие концентрации дофамина и ацетил-холинэстеразы в палеостриатуме птиц и в подушке и бледном шаре млекопитающих [390].. Лимбическая система, или необонятельная часть архипаллиума, превращается в гиппокамп и миндалевидный комплекс. У однопроходных и сумчатых животных новая кора имеет слои, но организована иначе, чем у всех других млекопитающих. У высших млекопитающих новая кора состоит из 6 четко очерченных слоев, толщина которых у высших приматов доходит до 100 клеток. Общее число нейронов в коре большого мозга человека составляет 1010, и многие связи между ними 5—2067
66
Глава 15
лежат в самой коре. Подсчитано, что во всей нервной системе на каждый мотонейрон приходится 2000 вставочных нейронов.
Нейрональная организация областей, содержащих множество мелких вставочных нейронов, например центрального ретикулярного вещества в области ствола мозга, остается еще совсем неясной.
Вызванные потенциалы и ритмическая, электрическая активность головного мозга
Внеклеточная и внутриклеточная регистрация импульсной активности отдельных нейронов дала много важных сведений о кодировании информации в нервной системе позвоночных, а также в ганглиях беспозвоночных животных. Однако многие нейроны головного мозга имеют небольшие размеры, а центральный нейропиль представляет собой сложную сеть из тонких отростков; поэтому здесь неприменимы методы, используемые при регистрации активности одиночных клеток. Для изучения притока синхронизированной активности к структурам центральной нервной системы и активности в областях со слоистой, или корковой, структурой применяются три вида суммарной регистрации: запись синхронизированных ответов, вызванных потенциалов в различных областях коры и электроэнцефалография.
В некоторых сенсорных ядрах импульсации, поступающая по группам волокон, так хорошо синхронизирована, что на внеклеточных записях видны воспроизводимые волны. Так, у дельфина Stenella в слуховом ядре продолговатого мозга, в нижнем бугорке и медиальном коленчатом теле сложные вызванные потенциалы, возникающие в ответ на звуковые толчки, состоят из ряда колебаний, каждое из которых соответствует синхронным импульсным разрядам группы нейронов. Изменения амплитуды, формы волны и латентного периода связаны с параметрами звука [54].
В мозжечке, тектуме (особенно у рыб и амфибий), коре больших полушарий и обонятельной луковице специфические типы нейронов образуют слои, причем дендриты их часто ориентированы перпендикулярно поверхности, а аксоны идут в направлении, противоположном дендритам [320]. Число слоев в одной структуре варьирует в зависимости от исследуемого вида да и от усердия гистолога. На записях, полученных от дендритного слоя, видны медленные негативные отклонения, которые представляют собой суммарные синаптические потенциалы, вызываемые синхронизированной входной активностью. По мере погружения электрода в клеточные и более глубокие аксонные слои знак потенциала меняется на обратный, и в типичных случаях имеется последовательность, состоящая из отрицательного (сток) и положительного ^источник) колебаний потенциала (рис. 15-24). Нанесение изопо-
Центральная нервная система
67
тенциальных контуров в коре больших полушарий млекопитающих [539] или тектума рыб [525] показывает форму полей сток — источник (рис. 15-25). Измерения тока (а не напряжения) в разных точках выявляют неоднородность электрического сопротивления в структуре коры.
Рис. 15-24. Суммарные синаптические потенциалы [626].
А. Вызванные потенциалы зрительного нерва лягушки в ответ на его стимуляцию; слева — указана глубина погружения электрода в тектум (в мкм). Б. Микрофотография тектума, видны его слои и темное пятно — место инверсии знака потенциала.
Связанные с этими явлениями регистрируемые внеклеточно потенциалы — электрические волны мозга, или ЭЭГ, — представляют собой суммированные локальные и синаптические потенциалы множества работающих синхронно нейронов данного уровня. В телэнцефалоне трески возникают нерегулярные электрические волны с частотой 7 в 1 с, а в тектуме 8—13 в 1 с [124]. У серебряного карася зарегистрированы волны в мозжечке, идущие с частотой 25—35 в 1 с, в тектуме — с частотой 7—14 в 1 с и в переднем мозгу — 4—8 в 1 с [485, 486]. У каймана волны в перед-5*
68
Глава 15
нем мозге идут с частотой 7—12 в 1 с, а в тектуме — 6—8 в 1 с [419]. У млекопитающих так называемые альфа-волны регистрируются лучше всего (но не исключительно) в затылочных областях; они имеют частоту 7—8 в 1 с у кошки и 10 в 1 су человека. У детей медленные волны (3 в 1 с) появляются в возрасте 3 мес. У щенят отдельные волны появляются на 18—20-й день, стойкие
Время, мс
Рис. 15-25. Профили плотности тока в тектуме серебряного карася при раздражении зрительного нерва в двух препаратах [525].
Линиями обозначены одинаковые токи, заштрихованными участками — движение тока вниз, незаштрихованными — вверх. Во время поверхностно-отрицательного потенциала поля ток течет преимущественно вверх и имеет максимальную плотность в центральной зоне серого вещества тектума. По мере развития ответа пункт максимального направленного вверх тока, смещается на ббльшую глубину.
альфа-волны — к 5-й нед, а характерный для взрослой собаки ритм устанавливается в 5 мес [297]. Исследователи, регистрирующие ЭЭГ, различают медленные дельта-волны (1—3 в 1 с), тета-волны (4—7), отводимые от гиппокампа, альфа-волны (8—12), быстрые бетаг (13—18), бетаг- (19—25) и бета3-волны (26—29) [5]. Корковые ритмы прерываются сенсорной 'стимуляцией — зрительной или звуковой — или же изменением уровня внимания. Во время сна альфа-ритмы заменяются более медленными волнами и иногда — веретенами (см. ниже). Запись множественными электродами выявляет распространение волн по коре со скоростью меньше 1 м в 1 с. Раздражение ретикулярной формации головно
Центральная нервная система 69
го мозга в состоянии покоя может вызвать «реакцию пробуждения»— десинхронизацию альфа-волн и усиленные быстрые коле-, бания. Стимуляция медиальных неспецифических ядер таламуса вовлекает корковые волны в свой ритм (реакция вовлечения) [253, 432]. Перерезка таламических проекций, идущих к коре, приводит к исчезновению синхронных волн. В изолированных полосках коры регистрируется нерегулярная спонтанная активность и горизонтальное проведение. Разные электрические волны мозга слагаются из комбинации спонтанных разрядов нейронов, распространения активности внутри коры и в особенности суммированных В ПСП и ТПСП в .корковых дендритах.
Медленные вызванные потенциалы, отводимые от поверхности коры больших полушарий и тектума, часто используются при исследовании сенсорных входов и их проекций. Так, для соматосенсорной коры составлена карта тактильных и проприоцептивных проекций от различных областей тела (см. ниже). У аллигатора [193], и серебряного карася [431, 489] участки сетчатки проецируются в правильной последовательности на контралатеральном тектуме.
Анализ формы вызванных потенциалов на разной глубине обычно производится при помощи синхронизированной сенсорной стимуляции или электрического раздражения определенных ограниченных трактов. Рассмотрим несколько примеров. Обонятельная луковица представляет собой почти сферическое образование с первичными обонятельными волокнами, которые образуют синапсы на направленных кнаружи дендритах митральных клеток; эти клетки отдают аксоны в обонятельный тракт и, кроме того, посылают коллатерали к клеткам-зернам, которые по обратным связям тормозят митральные клетки (см. гл. 13). Вызванный ответ на антидромное, а также на ортодромное раздражение, отводимый на различной глубине, состоит из трех фаз: 1) начальной негативной на уровне синапсов митральных клеток и позитивной на поверхности; 2) позитивной фазы на уровне клетки в результате реполяризации и второй негативной волны, создаваемой током от клеток-зерен, и 3) завершающей длительной позитивной фазы, обусловленной ТПСП [391, 433].
В мозжечке каждая крупная клетка Пуркинье обладает ветвящимся дендритным древом; синапсы параллельных волокон лежат главным образом на дендритных шипиках близ периферии ветвлении, тогда как синапсы лазящих волокон находятся на центральных веточках. Стимуляция лазящих волокон создает отрицательный вызванный потенциал максимальной амплитуды на уровне тело клеток Пуркинье, не меняющий своей полярности на большой глубине, между тем как в ответ на стимуляцию параллельных волокон у большинства видов (млекопитающие, амфибии, рыбы) возникает вызванный потенциал, отрицательный на поверхности и (положительный у тела клетки. Антидромная стимуляция
70
Глава 15
через аксоны клеток Пуркинье также вызывает ответы, меняющие-свою полярность. Такие ответы трактуют как пассивную дендритную деполяризацию типа сток — источник. В мозжечке аллигатора некоторые ответы на стимуляцию параллельных волокон не-меняют полярности, и в них видят свидетельство возникновения импульсов в дендритах. У всех видов вызванный ответ на стимуляцию лазящих волокон не меняет полярности, потому что их синаптические окончания распределены диффузно в теле и в верхних дендритных слоях клеток Пуркинье [114, 331, 394].
В ответ на стимуляцию сенсорных путей, идущих в кору больших полушарий, также возникают локальные вызванные потенциалы; ответы на стимуляцию ретикулярных и некоторых таламических проекций не столь локальны. Стимуляция таламических входов у кошки вызывает два типа ответов — один с начальным, негативным стоком в зернистом слое и негативностью, перемещающейся к поверхности за 20—30 мс, и другой — с поверхностной негативностью, переходящей в позитивность на глубине ниже 0,3—0,5 мм [505]. В зрительной коре кролика изопотенциальная точка находится на глубине 0,8 мм, а регистрация активности одиночных клеток показывает, что ВПСП длятся около 80 мс [327]'.
В гиппокампе пирамидные клетки ориентированы в направлении, обратном по сравнению с остальной корой, т. е. аксоны выходят с поверхности. При ортодромном возбуждении возникают поверхностно-положительные и отрицательные на глубине вызванные потенциалы [120]. Стимуляция каждого из нескольких входов в гиппокамп вызывает большую положительную волну на глубине 0,5 мм, которая создается ТПСП на уровне тел пирамидных клеток и обусловлена активацией корзинчатых клеток коллатералями от пирамидных нейронов [11]. rt
В тектуме рыбы и лягушки световая стимуляция контралатерального глаза вызывает медленную поверхностно-отрицательную волну, которая меняет свою полярность на средней глубине; отрицательная и положительная волны не строго зеркальны между собой и, возможно, отражают время, которое нужно для элек-тротонического распространения из поверхностных синаптических слоев. Проведение в горизонтальном направлении тоже может сказываться на ответах. В тектуме некоторых рыб на уровне па-равентрикулярных или пириформных нейронов лежит вторая область негативности [429, 431]. При сильном электрическом раздражении зрительного нерва или при обширных полях зрения высокой интенсивности в вызванных потенциалах появляются множественные волны, по-видимому, отражающие синаптические ответы волокон с разными порогами и разной скоростью проведения.
И волны ЭЭГ, и вызванные потенциалы служат показателями состояния головного мозга и синхронной активации большого числа нейронов. Отношений между этими медленными потенциалами
__________________Центральная нервная система______________71 и импульсацией отдельных клеток исследовались при помощи одновременного измерения обоих типов активности, часто одним и тем же электродом, соединенным с усилителями, имеющими малую и большую постоянные времени. В зрительной коре наибольшая вероятность импульсации одиночного нейрона совпадает по времени с самым резким спадом отрицательного отклонения вызванного потенциала, а наименьшая вероятность приходится на самый резкий спад позитивного отклонения; это соотношение сохраняется на любой глубине; следовательно, ответ нейрона определяется местным электрическим полем [547]|. И в зрительной, и в соматосенсорной коре кошки суммирование импульсов усреднителем дает кривую, соответствующую по форме вызванному потенциалу; кроме того, вероятность импульсного разряда соответствует характеру ЭЭГ. Впрочем, волны ЭЭГ появляются и в отсутствие пирамидных импульсов [148]. Когда та или иная область в мозжечке кошки дает вызванные потенциалы в ответ на раздражение гиппокампа и импульсные ответы на звуковые щелчки, то вероятность импульсного ответа хорошо коррелирует с формой и величиной вызванного потенциала [147]. При внутриклеточной регистрации активности пирамидных нейронов в гиппокампе кролика улавливаются импульсы, синхронные с ритмическими тета-^ волнами, причем оба вида активности усиливаются при раздражении седалищного нерва. При помощи методики условных рефлексов (подкрепление вливанием в рот молока) удавалось увеличивать по выбору либо положительную, либо отрицательную фазу вызванного ответа в коре больших полушарий на зрительную стимуляцию [149].
Можно сделать вывод, что вызванные потенциалы и ЭЭГ могут использоваться как показатели региональной активности совокупностей или слоев множества сходных нейронов, но вместе с тем трудно установить точно природу их разных компонентов. Медленные потенциалы могут влиять на порог импульсной активности нейронов, находящихся в поле такого потенциала.
Двигательные системы позвоночных
Спинной мозг и нижние отделы ствола мозга
Первые детальные исследования, установившие принципы рефлекторного поведения, проведены на спинном мозге млекопитающих [83, ПО, 190, 498]. По задним корешкам в спинной мозг идут несколько афферентных входов (см. гл. 12): 1) от рецепторов растяжения, к которым относятся первичные и вторичные волокна мышечных веретен и сухожильные органы Гольджи, реагирующие на напряжение, а не на длину мышцы, и 2) от различных кожных окончаний, которые конвергируют на тех же вставочных нейронах,
72
Глава 15
что и рецепторы растяжения. Одни сенсорные входы ограничены пределами одного сегмента, другие — несколькими сегментами, а иногда происходит переключение на задние столбы, по которым импульсы поднимаются к мозговому стволу. Тракты в задних столбах — спиноталамические и спиноцеребеллярные — являются преимущественно ипсилатеральными. Нисходящие пути — пирамидные, тектоспинальные, руброспинальные, вестибулоспинальные и ретикулоспинальные — конвергируют на мотонейронах всех уровней спинного мозга.
Действие многочисленных входов на мотонейроны одного сегмента спинного мозга подчиняется определенным правилам. Приходящие волокна оказывают возбуждающее действие на один мотонейронный пул и тормозят мотонейроны, иннервирующие антагонистические мышцы; так, моносинаптические рефлекторные сокращения экстензоров, вызываемые афферентами от их веретен, тормозятся афферентами, идущими от флексоров. И наоборот, входы, конвергирующие на мотонейронах нескольких пулов, обеспечивающих одно и то же мышечное движение, обычно оказывают друг на друга облегчающее действие. Торможение может быть постсинаптическим, обычно коротколатентным, или же пресинапти-ческим и достигать пика приблизительно через 15 мс. Стрихнин блокирует йЪстсинаптическое, но не пресинаптическое торможение; это последнее блокируется пикротоксином. Пресинаптическое торможение деполяризует афферентные окончания в спинном мозге; постсинаптическое — повышает проводимость для хлора и обычно гиперполяризует мотонейроны (см. выше) {ПО, 244]. Кроме того, мотонейроны отдают коллатерали, которые образуют возвратные пути и через вставочный нейрон (клетку Реншоу или ее эквивалент) блокируют текущую активность того же и соседних нейронов. Антидромные импульсы тормозят или облегчают другие мотонейроны по коллатералям и могут прервать тоническое торможение, вызываемое вставочными нейронами, запуская множественные разряды >[595, 596]. Вставочные нейроны Реншоу дают высокочастотные залпы в ответ на одиночные антидромные импульсы в мотонейронах; они действуют на мотонейроны постсинаптически.
Самые мелкие мотонейроны (гамма-эфференты) иннервируют интрафузальные мышечные волокна; обычно они спонтанно активны и сильно возбудимы электрически и рефлекторно. Мелкие альфа-эфференты активируются рефлекторно и дают многократные импульсные разряды в ответ на умеренное растяжение, вызывая слабые сокращения экстрафузальных мышечных волокон. Крупные альфа-мотонейроны создают максимальное напряжение, имеют более высокие пороги и весьма чувствительны к торможению. Таким образом, величина мотонейрона обратно пропорциональна его возбудимости и прямо пропорциональна торможению и вызываемому напряжению [190].
Центральная нервная система 73
Управление движением антагонистических мышц осуществляется механизмами обратной связи (см. выше). Первичные мышечные веретена (афференты группы 1а) моносинаптически возбуждают а-мотонейроны, как, например, в экстензорах рефлекса на растяжение, и тормозят антагонистические а-мотонейроны. Вторичные афференты от веретен (группа II) представляют собой волокна с более медленным проведением; они действуют через вставочный нейрон, тормозят а-мотонейроны экстензоров и возбуждают мотонейроны флексоров. Сухожильные рецепторы (афференты группы 1b) тормозят ипсилатеральные а-мотонейроны, особенно те, которые иннервируют экстензоры. Веретена сигнализируют о длине мышцы, а сухожильные рецепторы — о напряжении. Гамма-мотонейроны вызывают сокращение интрафузальных мышечных волокон и тем самым повышают частоту импульсации веретена при данной длине мышцы.
Вставочные нейроны иногда спонтанно активны и обычно генерируют более длительные разряды, чем мотонейроны; самые короткие разряды генерируются альфа-мотонейронами. Вставочные нейроны выполняют функции: 1) распределительных путей для конвергенции и дивергенции; 2) усилителей сенсорной входной активности посредством ряда Т-образных связей или реверберационных кругов с различным числом клеток; 3) своеобразных клапанов между несколькими входами, например в тех случаях, когда нисходящие команды и сенсорная входная активность взаимодействуют, что ведет к облегчению или торможению, и 4) элементов, изменяющих полярность, например когда возбудительный вход меняется на тормозный '[190].
После перерезки спинного мозга у кошки или собаки в верхней грудной области сначала все двигательные рефлексы подавляются вследствие спинального шока. У кошки сгибательные рефлексы восстанавливаются через несколько часов, разгибательные — через несколько дней. У лягушек и рыб спинальный шок очень непродолжителен, т. е. длительность шока обратно пропорциональна степени цефализации. Спинальное млекопитающее не может стоять, но если его подвесить, совершает шагательные движения. Сохраняются рефлексы конечностей, требующие координа-. ции активности многих мышц, как, например, в ритмическом чесательном рефлексе. У спинального млекопитающего можно вызвать висцеральные рефлекторные акты — такие, как мочеиспускание, дефекация и сосудистые реакции. Возможны половые рефлексы. 1акие наблюдения привели к заключению об эндогенной активности и врожденной организации связей в спинном мозге. Впрочем, У Децеребрированных кураризованных кошек зарегистрированы эфферентные импульсы в поясничных передних корешках в ответ на раздражение сенсорных волокон в малоберцовых нервах; при стимуляции нервов на обеих сторонах со слегка различной частотой возникали моторные разряды ритмических шагательных ре
74
Глава 15
акций, если же стимуляцию производили с одинаковой частотой, разряды не возникали ‘[118]. Это подчеркивает важное значение сенсорных входов для временной организации шага у млекопитающих.
У млекопитающих в акте глотания участвует множество мотонейронов продолговатого мозга, иннервирующих около 20 разных мышц. Группа нейронов, лежащих билатерально и дорсолатераль-но по отношению к нижней оливе, координирует последовательность сокращений глотательных мышц. Эта координация смены возбудительных и тормозных реакций осуществляется не рефлекторно, поскольку она не зависит от характера стимула — прикосновения к глотке, быстрого вливания воды в рот или же электрической стимуляции верхнего гортанного нерва; разрыв цепи обратной связи не оказывает существенного влияния на организацию этих движений {109].
Перерезка ствола мозга непосредственно позади среднего мозга или на верхнем уровне моста у млекопитающих приводит к чрезмерной активности экстензоров, которую иногда называют децеребрационной или экстензорной ригидностью. При этом повышаются экстензорные рефлексы, и обычно возникает спастичность. Такой эффект объясняется устранением тормозных влияний, идущих от коры больших полушарий, хвостатого ядра и мозжечка, а также тонической активностью облегчающих путей — ретикулоспинальных й вестибулоспинальных трактов [343]. Приходится заключить, что еще далеко не ясно, каким образом у млекопитающих уравновешиваются эндогенные факторы, формирующие деятельность мотонейронов, и сенсорные и нисходящие влияния, определяющие течение ее во времени.
У амфибий, как и у млекопитающих, входная активность поступает к спинному мозгу от мышечных веретен и сухожильных, кожных и суставных рецепторов. При внутриклеточном отведении активности от мотонейронов в изолированном спинном мозге лягушки в. ответ на раздражение задних корешков регистрируются как возбудительные, так и тормозные синаптические потенциалы... Возбудительные реакции могут быть моносинаптическими (задержка 2,2 мс), а тормозные — включать один вставочный нейрон (задержка 5 мс) [305]. Внеклеточные измерения фокусов синаптических потенциалов на мотонейронах показывают, что импульсы в задних корешках действуют на дистальные зоны дорсально ориентированных дендритов, а нисходящие импульсы нейронов в боковых столбах действуют на тело и близлежащие участки дендритов; импульсация мотонейронов возникает в начальном сегменте аксона, на некотором расстоянии от синапсов [43]. Моторные импульсы могут распространяться электротонически в обратном направлении по телу мотонейрона. В мотонейронах возникают также спонтанные миниатюрные синаптические потенциалы [287].. Антидромная стимуляция через передние корешки также возбуж-
Центральная нервная система 75 дает мотонейроны, вероятно по коллатералям, что приводит к генерации заднекорешкового потенциала. Этот последний подавляется холинолитическими агентами и усиливается нагреванием; возможно, что он возникает в холинэргических синапсах. У лягушек слабая антидромная стимуляция вызывает переднекорешковый потенциал и во многих мотонейронах обнаруживается коротколатентная деполяризация, которая даже генерирует импульсы; переднекорешковый потенциал, вызываемый коротким антидромным залпом, понижается при нагревании и не чувствителен к блокаторам. ^Очевидно, между дендритами многих мотонейронов существует электрическая связь [173, 305].
Имеются данные о наличии у амфибий обоих видов ответов и об их зависимости от сенсорного притока. Перерезка ствола мозга над продолговатым мозгом не приводит к децеребрационной ригидности, наблюдаемой у млекопитающих. При соответствующей стимуляции спинальная лягушка совершает координированные прыжки. Прежние наблюдения над координированными движениями ног показали, что они возможны при сохранении сенсорной и ^моторной сегментарной иннервации [172]. Нормальная последовательность движений ног при ходьбе у жабы Bufo marinus следующая: правая задняя (ПЗ), правая передняя (ПП), левая задняя (ЛЗ), левая передняя (ЛП). После перерезки всех задних корешков спинного мозга нормальные локомоторные движения сохрани-.лись; если в нормальной последовательности совершались три движения, то за ними обычно следовало четвертое >[187]. Головной мозг и черепно-мозговые нервы оставались интактными, но эти .данные показывают, что в передвижении амфибий важную роль играет программа, заложенная в центральной нервной системе. •Эти данные о наличии эндогенного ритма подтверждают наблюдения, касающиеся реиннервации и онтогенеза (см. ниже). У амфибий и рептилий не удается обнаружить различий между интра-фузальными и экстрафузальными мышечными волокнами (см. гл. 12), и для этих классов животных не найдено ничего аналогичного регулирующей системе гамма-петли, свойственной млекопитающим. Возможно, это связано с тем, что у лягушек эндогенные программы эффективнее, чем у млекопитающих.
У рыб, по-видимому, существуют большие видовые различия в степени автономии спинного мозга. Считается, что у них нет мышечных веретен, а кожа так плотно прилегает к мышце, что кожные рецепторы воспринимают движение [439]. У костистых и пластиножаберных рыб рефлекторные движения плавника можно вызвать локальным раздражением кожи. Высокая перерезка спинного мозга у угря почти не нарушает его ритмических волнообразных движений; ритмическая волна проходит вдоль спинного мозга через участок, на котором удалены кожа и мышцы 1[170]. Гольян 'С перерезанным верхним отделом спинного мозга, ослепленный и не соприкасающийся с субстратом, все же способен к спонтанным
76
Глава 15
плавательным движениям. Кошачья акула после перерезки спинного мозга позади головы плавает почти как нормальная рыба, и если ни с чем не соприкасается, то совершает локомоторные движения с частотой 40 волн в 1 мин, которые усиливаются или ослабляются под влиянием тактильной стимуляции. При полной де-афферентации ритмическое движение тела и плавников кошачьей акулы прекращается, но при денервации, охватывающей, скажем, 12 сегментов, программа этого движения может перейти через этот участок на остальную часть спинного мозга |[201]. У серебряного карася перерезка спинного мозга перед спинным плавником приводит к парезу — атонии и исчезновению спонтанных движений, — но при стимуляции возникают координированные движения плавника; через 30 дней спинной мозг регенерирует [34].
На записях электрической активности двигательных нервов кураризованной спинальной кошачьей акулы видны спонтанные залпы импульсов мелких мотонейронов, длящиеся 1—2 ч после перерезки спинного мозга. Однако в отсутствие проприоцептивных входов двигательные разряды больше не возникают. Волны сокращения у кошачьей акулы передаются со скоростью 0,3—0,8 м/с. Нейроны на обеих сторонах спинного мозга организованы так, что разряжаются попеременно, но без фазического сенсорного возбуждения продольная координация локомоторной волны нарушается )[439]. Непарные плавники кошачьей акулы ударяют в ритме, который может не совпадать с ритмом движений туловища; сенсорный приток, вызываемый движением туловища, изменяет ритм плавников [439].
Спинальные круглоротые (миксины) при стимуляции плавают нормально и волна сокращения проходит через область перерезки спинного мозга благодаря локальным рефлексам, возникающим при движениях сегментов за перерезкой. У миноги (Petrotnyzori) тела сенсорных клеток лежат не только в заднекорешковых ганглиях, но и в спинном мозге; они отвечают на прикосновение, давление и вредящее раздражение кожи. Стимуляция сенсорных клеток вызывает ВПСП и ТПСП в мотонейронах, вероятно, через вставочные нейроны. При многократной стимуляции сенсорных клеток или кожных рецепторов в моторном синапсе наступает привыкание [36, 461]. Одни мотонейроны активируют медленные красные мышечные волокна, другие иннервируют быстро сокращающиеся белые мышцы, третьи снабжают ипсилатеральные мышцы, которые вызывают движения плавников. Ни для одного мотонейрона не обнаружено возвратного торможения, но контралатеральные мотонейроны тормозятся при локальном рефлексе [533]. Медленные мотонейроны дают тонические следовые разряды. Сенсорные нейроны имеют Т-образную форму и вызывают как локальные, так и мультисегментные рефлексы.
У миног, кроме дорсальных сенсорных нейронов, в спинном мозге имеются мелкие вставочные нейроны и мотонейроны, а так-
Центральная нервная система 77
;ке несколько исключительно крупных вставочных нейронов. Далее, бульбарная область продолговатого мозга содержит около 8 пар мюллеровских (гигантских) клеток с неперекрещивающимися аксонами и две пары гигантских маутнеровских нейронов с перекрещивающимися аксонами. На мюллеровских и маутнеровских клетках имеются входы от многих источников — от зрительных, тройничных, вестибулярных нервов, от нервов заднего отдела боковой линии и спинного мозга [460]. Многие входы на мюллеровские клетки от спинного мозга являются тормозными; одна пара клеток отвечает при напряжениях ниже тех, какие требуются аксонам дорсальных клеток; возможно, что они возбуждаются восходящими аксонами средней величины. Аксоны мюллеровских клеток образуют синапсы на мотонейронах и вставочных нейронах. Одна пара мюллеровских аксонов вызывает в спинномозговых вставочных нейронах возбуждающие синаптические потенциалы с медленным компонентом, который выражает химическую передачу (последняя подавляется высоким содержанием магния и восстанавливается высоким содержанием кальция), и с быстрым компонентом, для которого характерна электрическая передача (рис. 11-16). ВПСП, вызываемые в мотонейронах мюллеровскими нейронами, имеют небольшую амплитуду и достигают порога только при большой частоте стимуляции [460, 461].
Химические и электрические синапсы на маутнеровских нейронах у рыб были описаны выше (рис. 15-8). У миног входов на маутнеровских клетках больше, чем у костистых рыб. У рыб каждый маутнеровский аксон посылает множество веточек к вставочным нейронам спинного мозга и вызывает ипсилатерально сокращения мышц туловища и хвоста, жаберной крышки, глаз и нижней челюсти; он также тормозит мотонейроны на противоположной стороне. Аддукторы грудного плавника у клинобрюшки Gasteropelecus иннервируются системой гигантского нейрона (рис. 15-26). В продолговатом мозге этой рыбы каждый маутнеровский аксон активирует четыре контралатеральных гигантских волокна через химические синапсы с задержкой 0,4 мс и вызывает ВПСП, который может изменить свой знак при деполяризации гигантского волокна. Гигантские волокна активируют около 40 ипсилатеральных мотонейронов в первых спинальных сегментах через электрические синапсы с задержкой менее 0,05 мс. Система гигантских нейронов позволяет рыбе делать быстрые броски вверх |[18а].
У наземных позвоночных нет образований, которые были бы аналогичны гигантским маутнеровским нейронам. В продолговатом мозге и мосту находится множество функциональных переключений восходящих (сенсорных) и нисходящих (моторных) путей, которые являются центрами вегетативных функций. Выше (гл. 12) были описаны центры слуховых, вестибулярных нервов и нервов боковой линии. У всех позвоночных мозжечок связан многочисленными связями со средним и продолговатым мозгом.
78
Глава 15
Красное ядро пресмыкающихся, птиц и млекопитающих является важным центром, который имеет связи с мозжечком, корой больших полушарий, таламусом и спинным мозгом. Здесь находятся также сосудодвигательный и дыхательный центры. У млекопитающих дыхательный центр моста модулирует (обычно посредством
Рис. 15-26. Схема связей между маутнерювскими волокнами, гигантскими волокнами и мотонейронами аддукторов грудного плавника у Gasteropelecus [18а]. Каждое маутнеровское волокно (2) образует несколько синапсов с ипсилатеральными гигантскими волокнами (1) и один синапс с центральным гигантским волокном. Перекрещивающиеся ветви гигантских волокон идут попарно. Отростки каждого гигантского волокна образуют синапсы с мотонейронами (5). Между маутнеровским и гигантским волокнами имеются химические синапсы (3), а между гигантским волокном и ипсилатеральным мотонейроном— электрические синапсы (4); 6 — нервы, 7—мышца.
торможения) активность инспираторных и экспираторных нейронов. Некоторые нейроны этого центра чувствительны к СО2 или же получают импульсацию от респираторных рецепторов каротидного синуса млекопитающих, а также, возможно, в жабрах и псев-добранхиях у рыб (см. гл. 5). Активность сердечно-сосудистых центров модифицируется другими сенсорными входами, а выходная активность может быть видоизменена условнорефлекторно; примерами могут служить перерыв дыхания и замедление сердечного ритма у рыб [419].
Центральная нервная система
79
Моторные центры в головном мозге, управляющие локомоцией
Локомоторный автоматизм у позвоночных во многом можно объяснить спинальными рефлексами и спонтанными ритмами; роль высших центров в возникновении движений и их контроле изучена хуже. В произвольном движении участвует динамическое кольцо взаимодействия между нейронами сенсомоторной коры, базальных ганглиев, таламуса, мозжечка и сенсорных и моторных ядер в стволе мозга. У обезьян при произвольных движениях глаз, рук и ног [127, 534] меняется частота импульсации в соответствующих областях мозга. Клетки Пуркинье в мозжечке могут давать
разряды при прекращении движения, и некоторые данные говорят о том, что мозжечок оказывает на моторные системы преиму-' щественно тормозное действие [114]. У кошек, у которых вырабатывали мигательный рефлекс на щелчок, вызванный потенциал в двигательной коре возникал с латентным периодом 13 мс и предшествовал мышечной активности на 7 мс [599].
Точечная стимуляция (электрическая или химическая) моторной коры млекопитающих (в области, лежащей ростральнее центральной борозды) вызывает реакции отдельных мышц или отдельных моторных единиц мышцы. Более обширная стимуляция вызывает отдельные движения ног, шеи, туловища или других областей. Карта моторной коры соответствует карте сенсорной коры по другую сторону борозды: нога представлена в дорсальной части, рот и лицо — в вентральной, глаз — спереди, и все это контрала-терально. Величина этих участков у разных видов различна и соответствует размерам конечностей; так, представительство хвоста у паукообразной обезьяны занимает такую же площадь, что и представительства ног (рис. 15-27). Стимуляцией соматосенсорной коры можно вызвать движение по связям с моторной корой. Как правило, стимуляция какой-либо точки моторной коры вызывает
ответ в другом полушарии по связям через • мозолистое тело. У однопроходных и сумчатых животных сенсорные и моторные области перекрываются и накладываются на одну и ту же карту [1, 321—323, 434].
Выходная моторная активность покидает кору млекопитающего животного по аксонам пирамидных нейронов моторной зоны и по соседним экстрапирамидным нейронам. У человека насчитывается 1 млн. пирамидных нейронов. Произвольные выработанные движения запускаются через пирамидные нейроны, но поза и статические реакции контролируются главным образом экстрапира-мидными нервными клетками. Почти все пирамидные пути перекрещены и образуют синапсы на разных уровнях мозгового ствола и спинного мозга; у большей части пирамидных волокон имеются синапсы на вставочных нейронах, но некоторые из них образуют синапсы прямо на мотонейронах. Все экстрапирамидные пути переключаются в базальных ганглиях и других подкорковых ядрах.
80
Глава 15
У ненаркотизированных обезьян с вживленными регистрирующими электродами крупные пирамидные нейроны с максимальной скоростью проведения «молчат» в условиях покоя и фазически активны при движении; пирамидные нейроны, проводящие с мень-
Б
Рис. 15-27. Схема коры большого мозга крысы (Д), кролика (Б), кошки (В) и обезьяны i(jT) [627].
Показана локализация первичных и вторичных моторных (Af) и соматосенсорных (S) областей, а также первичной и вторичной зрительных (V) и слуховых (А) областей. Представительство частей тела показано в виде «гомункулуса». Схемы соответствуют латеральной поверхности полушария; вверху показаны проекции на медиальной поверхности.
шей скоростью, активны в отсутствие движения; во время мышечной активности частота разрядов этих нейронов повышается или понижается [127]. После двусторонней перерезки пирамид животные некоторых видов сохраняют способность к локомоции; животные других видов не способны к координированным движениям. Односторонняя перерезка пирамидного пути сказывается на противоположной стороне — понижаются использование мышц и двигательная инициатива. Относительное значение пирамидных и экс-трапирамидных трактов сильно варьирует у разных видов, а у однопроходных животных вообще нет пирамидного тракта. У сумча
Центральная нервная система
81 -
того Тrichosurus пирамидные аксоны доходят только до середины грудного отдела, где они иннервируют передние конечности; экс-трапирамидные волокна образуют синапсы на вставочных нейронах, которые спускаются по спинному мозгу и снабжают задние конечности i[203].
В переднем мозге низших позвоночных не обнаружено узколокальных истоков двигательных трактов. Удаление переднего мозга у пластиножаберных и костистых рыб не приводит к заметному нарушению позы й локомоции [34]. У лягушек и жаб, лишенных переднего мозга, несколько снижены спонтанные движения. Впрочем, электрическое раздражение переднего мозга не вызывает специфических моторных реакций [2].
Средний мозг осуществляет некоторый моторный контроль, и электрическая стимуляция тектума вызывает движения, вероятно, через посредство тегментума. Так, у серебряного карася при раздражении тектума возникает движение глаза на противоположной стороне; во время движения глаз некоторые нейроны тедтума дают импульсные разряды; эти нейроны не отвечают на зрительные стимулы, но запускают движения, способствующие фиксации изображения [262]. У жабы стимуляция тектума приводит к повороту головы вправо или влево, к схватыванию пищи и глотанию >[132—135]. В тектуме жабы имеется моторное представительство туловища. У лягушки локальная стимуляция тектума вызывает ориентировочное движение и схватывание пищи, а раздражение претектума или таламуса приводит к движению избегания.
У птиц удаление коры большого мозга не нарушает моторики. Управление движениями, по-видимому, локализовано в полосатом теле. Электрическая стимуляция этого отдела вызывает движение конечностей, а его удаление создает тяжелые расстройства моторики и поведения [451].
Таким образом, двигательные системы позвоночных служат хорошей иллюстрацией смещения локализации функций по мере цефализации. У рыб и амфибий локомоция, по-видимому, регулируется преимущественно эндогенными спинальными ритмами, а у млекопитающих контроль осуществляют сенсорные входы путем взаимодействия кожных афферентов, мышечных афферентов трех типов и нисходящих волокон от головного мозга. У круглоротых, и в меньшей степени у рыб и хвостатых амфибий, большие группы мышц управляются специализированными гигантскими нейронами. В ходе эволюции новой коры у млекопитающих появляются пирамидные нейроны — быстродействующие активаторы низших моторных центров. Во всех классах позвоночных некоторые регулирующие влияния на локомоцию оказывает мозжечок.
Мозжечок
Мозжечок представляет собой сложную структуру, входящую в состав нервной системы всех позвоночных. Он не является ни 6—2067
82
Глава 15
сенсорным, ни моторным, ни интегративным образованием в рефлекторном смысле этих понятий; он стоит в стороне от главных, входных и выходных путей и служит органом, направляющим и координирующим нервную деятельность других отделов мозга. Анатомические и электрофизиологические исследования показывают, что принцип нейрональной организации мозжечка один и тот* же во всех классах позвоночных [112, 315, 320]. Детали функциональных связей в мозжечке изучены у американской куньей акулы [115], лягушки [332, 475], ящерицы |[296], аллигатора [331], кошки [ИЗ, 114] и обезьяны'[534].
У круглоротых (миног) мозжечок представляет собой плоскую структуру (архицеребеллум), которая сохраняется в виде флоккуло-нодулярных долек у птиц и млекопитающих. У пластиножаберных и костистых рыб ростральный конец мозжечковой пластинки загибается кверху и образует corpus и vulvuli cerebelli (палеоце-ребеллум) [114]. Электрические рыбы, например мормириды со специализированными электрорецепторными органами, обладают огромным мозжечком, который составляет почти половину головного мозга [29, 396]. У амфибий и рептилий мозжечок состоит из небольшой доли — гомолога палеоцеребеллума птиц и млекопита-щих, передней доли, или червя, и части задней доли. У млекопитающих полушария мозжечка называются неоцеребеллумом.
У рыб и амфибий пути, выходящие из коры мозжечка, направляются как прямо к нижележащим двигательным центрам, так и к глубоким мозжечковым ядрам. У высших позвоночных большая часть выходных путей идет через базальные ядра мозжечка. У рептилий это nuclei fastigiae, emboliformis и globosus; у птиц и млекопитающих к этим трем добавляется зубчатое ядро. От глубоких мозжечковых ядер эфферентные волокна идут к нижележащим двигательным центрам.
Две группы волокон служат входами в кору мозжечка, и обычно каждая сенсорная модальность использует оба пути (рис. 15-28, А, Б). Это, во-первых, лазящие волокна, которые оканчиваются прямо на клетках Пуркинье, и, во-вторых, моховидные волокна, которые оканчиваются на клетках-зернах, по нескольку волокон на одной клетке. Аксоны клеток-зерен поднимаются к поверхностному слою, где они расходятся Т-образно и идут в пучках параллельных волокон, которые активируют клетки Пуркинье. Дендритное древо клетки Пуркинье лежит в одной плоскости с параллельными волокнами и перпендикулярно им (рис. 15-29). В коре мозжечка обнаружены 3 слоя: наружный молекулярный, состоящий из параллельных волокон клеток-зерен и синапсов на дендритах клеток Пуркинье; средний слой, или слой клеток Пуркинье, и более глубокий зернистый слой, состоящий из клеток-зерен. У кошачьей акулы клетки-зерна лежат цепочками по обе стороны от средней линии, а не в зернистом слое.
Рис. 15-28. Основные типы нейронов мозжечка.
Л. Стереограмма [114]. 1 — звездчатая клетка, 2 — параллельные волокна, 3 — корзинчатая клетка, 4~- клетка Пуркинье, 5 — клетка Гольджи, 6 — лиановидные волокна, 7 — аксон, ^ — моховидные волокна. Б. Клетки Пуркинье у различных позвоночных [628]. I—минога, II — акула, III — костистая рыба, IV — черепаха, V — птица, VI — человек.
<5*
84
Глава 15
У высших позвоночных лазящие волокна приходят в мозжечок из переключательных ядер в нижней оливе, а моховидные волокна — из многих ядер восходящих трактов. У млекопитающих моховидные волокна идут из следующих источников: от вестибулярной системы через вестибулярные ядра, от проприоцепторов и соматосенсорных органов через задний спинномозжечковый и клиновидномозжечковый тракты, от сенсомоторной коры больших
Рис. 15-29. Схема связей в мозжечке [629].
Синапсы лиановидного волокна на клетке Пуркинье; синапсы моховидного волокна на гло-меруле мозжечка. Клетки Гольджи, звездчатые и корзинчатые клетки — тормозные; параллельные волокна — возбуждающие. 1 — клетка Пуркинье, 2 — параллельные волокна, 3—звездчатая клетка, 4 — корзинчатая клетка, 5 — клетка Гольджи, 6 — лиановидное волокно, 7 — моховидные волокна, 8 — клетки-зерна, 9 — молекулярный слой, 10 — зернистый слой.
полушарий через ядро переключения в толще варолиева моста и от ретикулярной формации ствола мозга. Информация, поступающая по моховидным волокнам, обычно модально-специфична и тонко градуирована в соответствии с силой стимула [388]. Система лазящих волокон образует поточечную проекцию от нижней оливы на поверхности мозжечка. У кошки стимуляция как разных нервов ноги [388], так и быстро адаптирующихся кожных рецепторов и мышечных афферентов активирует лазящие волокна через нижнюю оливу. Моховидные и лазящие волокна лежат под прямым углом друг к другу, причем моховидные ориентированы в передне-заднем направлении.
Центральная нервная система
85
Активность клеток Пуркинье отчасти эндогенна, но большей частью она вызывается постоянным возбуждающим действием системы моховидных волокон. У кошки нейроны нижней оливы дают ритмические разряды в ответ на различные входные сигналы и могут вовлекать в синфазную активность клетки Пуркинье. Недавно был описан третий вход, состоящий из тонких адренэргических волокон, идущих от голубого ядра и, возможно, оказывающих тормозное действие на клетки Пуркинье [38].
У лягушки раздражение вестибулярного нерва i[332], а также физиологическая стимуляция саккулуса или утрикулуса [330, 332] активируют лазящие волокна, идущие к мозжечку. Сигналы от полукружных каналов проходят преимущественно по моховидным волокнам, а клетки Пуркинье отвечают следующим образом: латеральные клетки отвечают на ипсилатеральное, передние — на контралатеральное вращение; найдены клетки Пуркинье, которые реагируют на вращение в любом направлении, и такие, в которых при любом вращении спонтанная активность прекращается i[332]. Ответы на слуховые и зрительные сигналы варьируют в зависимости от класса животных; у костистых рыб зрительный путь идет через ядро переключения в тектуме [396, 416]. У млекопитающих множество волокон поступает в мозжечок из коры больших полушарий.
Одно лазящее волокно иннервирует одну клетку Пуркинье и образует множество синаптических контактов на базальных дендритах — у кошки около 2000, а у лягушки 300. Параллельные волокна идут пучками вдоль листка перпендикулярно плоскости дендритного сплетения. Их синаптические контакты находятся близ окончаний дендритов клеток Пуркинье и всегда расположены на шипиках; у кошки шипики одной клетки Пуркинье образуют около 200000 синапсов, и на каждую клетку Пуркинье приходят сигналы от нескольких тысяч параллельных волокон.
При нанесении раздражения на ножку мозжечка можно различить 3 возбудительных ответа в клетках Пуркинье. Во-первых, это антидромные импульсы, вызываемые стимуляцией аксонов этих клеток. Во-вторых, это коротколатентные ответы на стимуляцию лазящего волокна, которые регистрируются в виде комплекса «все или ничего» из ВПСП с множественными спайками на них ![27] (рис. 15-30). Число этих импульсов варьирует в зависимости от уровня возбудимости клеток Пуркинье. В-третьих, после длинного латентного периода возникает ответ, который идет по цепи из моховидного волокна, клетки-зерна и параллельного волокна и который состоит из одного или нескольких ВПСП с одним или несколькими импульсами и одного импульса в клетке Пуркинье. Моховидные 'волокна оказывают на клетки-зерна возбуждающее действие, которое, вероятно, передается ацетилхолином [12, 116].
Кроме возбуждающих путей, имеются три типа тормозных вставочных нейронов. Параллельные волокна активируют клетки
86
Глава 15
Гольджи, которые тормозят клетки-зерна; иными словами, они оказывают возвратное торможение на возбудительную цепь из моховидного волокна, клетки-зерна и параллельного волокна. Кроме того, коллатерали моховидного волокна возбуждают клетки Гольджи и тем самым осуществляют поступательное торможение клеток Пуркинье. У млекопитающих звездчатые и корзинчатые клетки,
Рис. 15-30. Активация клеток Пуркинье по лиановидньгм волокнам и антидромными импульсами [630].
Внеклеточная регистрация — на двух верхних кривых, внутриклеточная — на двух нижних. А. Лягушка: 1 — антидромный импульс; 2 — антидромный импульс и импульс лиановидного волокна (ЛВ); 3, 4 — ВПСП, вызванные активацией по ЛВ. Б. Аллигатор: 5, 6 — антидромные импульсы и ответ ЛВ; 7,8— ВПСП. В. Голубь: 9 — антидромные импульсы, 10 — антидромные и ЛВ-импульсы; И — антидромные и ЛВ-импульсы. Г. Кошка: 12 — антидромные импульсы; 13 — антидромные и ЛВ-импульсы; 14 — антидромные и ЛВ-импульсы; 15 — ВПСП, вызванные стимуляцией ЛВ.
возбужденные параллельными волокнами, тоже тормозят клетки Пуркинье. Аксоны звездчатых клеток оканчиваются на дендритном древе клетки Пуркинье, а аксоны корзинчатых — на аксонном холмике. Кроме того, коллатерали лазящих волокон тоже возбуждают корзинчатые и звездчатые клетки, осуществляя поступательное торможение. Корзинчатая клетка вызывает в клетках Пуркинье ТПСП длительностью 100 мс или больше. Коллатерали лиановидных волокон возбуждают также клетки Гольджи, которые снимают облегчение возбуждающих и тормозных входов системы моховидных волокон. После импульсного разряда клетки Пуркинье, вызванного стимуляцией лиановидных волокон, активность клеток Пуркинье прекращается, и их возбудимость, тестированная антидромной импульсацией, падает вследствие активации тормозных нейронов по коллатералям лазящих волокон ,[317]. Этот механизм
Центральная нервная система
87
производит «исправление» или «чистку» и стирает текущую активность или же «освобождает» зоны клеток Пуркинье, дающие-импульсацию, от прихода избыточной возбуждающей или тормозной активности [388].
Торможение звездчатыми клетками показано у кошачьей акулы (оно длится до 125 мс в виде блока ответа лазящих волокон), у лосося (30—150 мс) [554], лягушки [475] и ящерицы {296]. У аллигатора звездчатые клетки блокируют импульсацию клеток. Пуркинье через параллельные волокна, а клетки Гольджи блокируют клетки-зерна {331].
На том основании, что параллельные волокна относительно однородны по своему диаметру и что дендритное древо клеток Пуркинье лежит преимущественно в одной плоскости и под прямыми углами к параллельным волокнам, было выдвинуто предположение, что последовательная активация клеток Пуркинье по пучку параллельных волокон может служить своего рода таймером [42]. В мозжечке лягушки найдена однонаправленная временная корреляция между рядами смежных клеток Пуркинье и определенным пучком параллельных волокон [151]. У лягушки {504] и у рыб-мормирид [274] описано близкое прилегание друг к другу клеток-зерен, иннервируемых одними и теми же моховидными волокнами; это говорит об электрической связи между клетками-зернами и синхронизации активности параллельных волокон одного пучка.
Выходами из коры мозжечка служат только аксоны клеток Пуркинье. У млекопитающих эти клетки во всех областях коры мозжечка (за исключением flocculus, волокна от которого идут к вестибулярным ядрам) дают проекции к глубоким мозжечковым ядрам. Все выходы мозжечка являются тормозными и адресуются к двигательным системам через различные ядра ствола мозга. У низших позвоночных одни аксоны клеток Пуркинье идут прямо-к моторным центрам мозгового ствола, а другие оканчиваются в субцеребеллярных (шатровых) ядрах. У рыбы азиатского змееголова Ophiocephalus некоторые аксоны клеток Пуркинье образуют-синапсы с глазодвигательными нейронами, так что стимуляция мозжечка вызывает в этих нейронах ТПСП и блокирует их ответ-на возбуждающие сигналы от вестибулярных нейронов {294]. Эта система обеспечивает коррекцию ошибок при быстрых и тонких приспособлениях положения и движения глаз [244]. У электрических рыб мормирид в коре мозжечка найдено несколько типов-клеток, соответствующих глубоким ядрам.
Медиатором тормозного действия клеток Пуркинье, вероятно,, является ГАМК {309, 397]. По имеющимся данным, тормозные-эффекты звездчатых [475] и корзинчатых {86] клеток тоже передаются с помощью ГАМК.
У млекопитающих описаны прямые и обратные связи между сенсорной корой и мозжечком. Стимуляция некоторых областей мозжечка вызывает десинхронизацию ритмики коры больших по
88
Глава 15
лушарий и вызывает в сенсомоторной коре серии импульсов более низкой частоты, чем та, с которой импульсы поступали в мозжечок [286]. Раздражением мозжечка можно подавить движения, вызванные раздражением коры.
Повреждения мозжечка нарушают позу и движения в разной степени у разных животных. У кошачьей акулы они не мешают плаванию, хотя часто рыба описывает круги в сторону повреждения. У карпа и серебряного карася возникает мышечная атония на этой стороне, виляние из стороны в сторону, нарушается управление равновесием; у такой рыбы не вырабатываются условные рефлексы на звук и свет, а ранее выработанные рефлексы утрачиваются [286]. Лягушки и ящерицы с удаленным мозжечком способны ходить и прыгать, но у них снижается спонтанность движений, и они вращаются или изгибают тело в сторону повреждения. У птиц и млекопитающих, не относящихся к неприматам, после такой операции создается утрированная поза экстензии, особенно головы и шеи. У приматов возникает флексия конечностей, а также мышечная слабость и ослабление рефлексов на растяжение с мышечных афферентов. При повреждениях мозжечка затруднены начало и координация движения, появляется тремор. Мелкие повреждения флоккуло-нодулярных долек нарушают у птиц и млекопитающих позу и равновесие, а при повреждениях передней доли и полушарий мозжечка страдают двигательные навыки.
Электрическим раздражением мозжечка можно вызвать движения или изменить позу, а также облегчить или подавить двигательную активность, вызываемую из другого источника. Изменение силы и частоты раздражения может превратить облегчающие эффекты в тормозные и наоборот. Как у рыб, так и у млекопитающих короткое раздражение коры мозжечка вызывает движение или определенную позу, за которыми следует постстимульная отдача в пиде длинного ряда других (обычно противоположных) движений или поз.
Организация связей мозжечка позволяет думать, что он способен осуществлять более тонкую функцию, чем описанные двигательные эффекты. В основном эта организация одинакова у всех классов позвоночных; в процессе эволюции произошло лишь совершенствование тормозных элементов и развитие глубоко лежащих ядер. Все классы позвоночных отличаются избыточностью входов (по лазящим и моховидным волокнам), что создает конвергенцию возбуждения за значительный отрезок времени. Торможение действует на одном, двух или трех синаптических уровнях {388’.
Для мозжечка предлагались разные функции (128]: 1) Он получает афферентные сигналы от мышц, активированных большим мозгом, и затем сам посылает модулирующие сигналы, уточняющие движение. 2) Сенсорная активность, направляющаяся к другим отделам мозга, по параллельным путям идет к мозжечку. 3) Моторные сигналы, идущие из коры больших полушарий к мо
Центральная нервная система 89
тонейронам спинного мозга, одновременно следуют в мозжечок, и таким образом выходная активность мозжечка добавляется к выходной активности коры. 4) Импульсный разряд клетки Пуркинье предшествует корковой импульсации и вызывает ее. Тем самым мозжечок контролирует главные входы и выходы. У классов животных, лишенных моторной коры, мозжечок и центры стволовой ретикулярной формации играют особенно важную роль в запуске движений и управлении ими. О значении мозжечка говорит тот факт, что он имеется у всех позвоночных и изменился в эволюции меньше, чем любой другой отдел головного мозга.
Центральные сенсорные пути у позвоночных
В предыдущих главах рассматривались периферические сенсорные структуры и функции и лишь те части центральных путей, которые обеспечивают простые ответы на различные сенсорные стимулы.
Центральные зрительные пути
Свойства той части головного мозга, которая образует сетчатку, описаны в гл. 14, где было показано, что на уровнях биполярных, горизонтальных, амакриновых и ганглиозных клеток происходит значительная переработка зрительной информации. Аксоны ганглиозных клеток образуют зрительный нерв.
Рыбы и амфибии. У костистых рыб и амфибий все зрительные волокна переходят на противоположную сторону и распределяются главным образом в тектуме (дорсальном мезэнцефалоне); кроме того, небольшие зрительные тракты идут в таламус и гипоталамус [66, 169] (рис. 15-31). Посредством повреждений тектума у лягушки установлены пути от него к претектуму и таламусу, а также к спинному мозгу, верхней оливе, ретикулярной формации и зрительному нерву (эфференты) [472]. В тектуме рыб и амфибий многие зрительные волокна образуют синапсы в верхних слоях, а некоторые из них проходят в более глубокие слои. В тектуме вставочные нейроны посылают волокна в мозжечок и спинной мозг по текто-церебеллярным и текто-спинальным трактам; тек-тальные связи с передним мозгом мало изучены. В тектуме содержится несколько типов клеток: пирамидные, некоторое количество горизонтальных и мелкие пириформные клетки в паравентрикуляр-ном слое, к которым относятся 90% нейронов тектума >[320]. У лягушек и ящериц нейроны тектума образуют особенно отчетливые слои, число которых варьирует в зависимости от вида [500],.
В ответ на зрительную стимуляцию в тектуме возникают вызванные потенциалы — отрицательные непосредственно под поверхностью и положительные на большей глубине, на уровне пирамидных нейронов. Посредством локальной стимуляции отдельных
Сетчатка
_____--^-Таламус------------Передний мозг
/х
{ Тектум
/iX |
S' \ Тектум
/ X
< ----*- Таламис-----------Передний мозг
Сетчатка J
Б
Таламус
Доре, отдел ----------
Латеральное коленчатое ' тело
Дорсомедиальная кора
Сетчатка
Сетчатка
Круглое ядро
Круглое ядро
Верхний бугорок
Верхний бугорок
-+ I
. Эктостриатум Дорсальный вентрикулярный край
Эктостриатум Дорсальный вентрикулярный край
Лора отдел ____________► Дорсомедиальная
Латеральное коленчатое тело К0Ра
Рис. 15-31. Схема главных зрительных путей у позвоночных.
Многие связи не показаны, например от среднего мозга к мозжечку. А. Рыбы, амфибии. Б. Птицы. В. Млекопитающие.
•Центральная нервная система 9>.
участков сетчатки установлена ее поточечная проекция на тектум,. в которой назальные квадранты представлены сзади, темпоральные — впереди, нижняя часть сетчатки — медиально (дорсально) и верхняя ее часть латерально (вентрально) у рыб (59, 431, 489] (рис. 15-32), аллигатора [192, 193], лягушки 162 (рис. 15-33), у Xenopus [161] и серебряного карася. В тектуме центр сетчатки имеет более широкое представительство, чем ее периферия. У животных с бинокулярными полями зрения, например у лягушки,, при помощи проецирования света на экран установлено, что центральная часть поля представлена каждым глазом и бинокулярное поле занимает по горизонтальному меридиану 100° [159].
При регистрации импульсов посредством микроэлектродов и высокочастотного усилителя не всегда легко отдифференцировать волокна зрительного нерва от нейронов тектума. В общем по сравнению с клетками тектума для зрительного нерва характерны: более короткий латентный период, меньшего размера рецептивные поля, более постоянные характеристики ответа и более высокая частота вовлечения в ритм световых вспышек; эти различия-легко обнаружить, если раздражать зрительный нерв электрическим током. Многие ганглиозные клетки сетчатки реагируют на концентрические или лежащие рядом возбудительные и тормозные поля; при этом «оп»-клетка усиливает свою импульсацию при-освещении центра поля и ослабляет ее, если освещается внешнее-кольцо; «ofb-клетки дают противоположный ответ. Одна ганглиозная клетка отвечает на неподвижную границу, другая на изменение интенсивности, третья — на движение пятна в одном или нескольких направлениях (см. гл. 14). Классификация ответов ганглиозных клеток зависит от вида животного (лягушка [178, 348], серебряный карась [346, 398]). У лягушки зрительные волокна, идущие от детекторов края, могут оканчиваться в тектуме более поверхностно, чем волокна от детекторов контраста или затенения [164, 176]. У некоторых видов, в частности у рыб, некоторые ганглиозные клетки связаны с колбочками и по-разному отвечают на-цветовой контраст в центре и на периферии.
В средних слоях тектума серебряного карася горизонтальные и пирамидные клетки отвечают с более длинным латентным периодом и обладают более широкими полями, чем ганглиозные клетки, и их ответы зависят от параметров предыдущего раздражения. Для большинства нейронов тектума характерна спонтанная активность и зрительная стимуляция прекращает их импульсацию; У немногих из них зрительные стимулы учащают импульсацию или вызывают ее [398, 525, 615]. Ответы центральных нейронов тектума подвержены привыканию; но при изменении характера раздражения — например, цвета, соотношения освещенных и неосвещенных участков, при движении и ударе по телу рыбы — происходит восстановление ответа [249, 398]. В глубоких слоях тектума серебряного карася пириформные нейроны дают спонтанные-
Дорсальная
Рис. 15-32. Топография поля зрения в левой сетчатке и его проекция к соответствующим положениям электрода на правом тектуме карпа [489].
Л. Сетчатка. Б. Тектум.
Центральная нервная система
93
разряды, которые прекращаются при зрительном раздражении; ответы этих нейронов очень лабильны. Выходы пириформных клеток почти не прослежены; известно, что их аксоны переходят на противоположную сторону тектума и образуют внутритектальные связи. Нейрон в верхнем слое тектума щуки (Esox) чувствителен
Рис. 15-33. Рстинотектальная проекция у лягушки [247].
Монокулярная проекция переходит по зрительному перекресту к противоположному текту-МУ: Б ~ от правого глаза и В—Г — от левого глаза. Бинокулярная проекция Б—В от
каждого глаза к тектуму той же стороны. Интертектальные комиссуры связывают соответствующие точки.
к движению стимулов в одном направлении; нейроны среднего слоя реагируют на небольшие пятна и контраст, а нейроны нижнего слоя — на изменения интенсивности '[615]. Нейроны тектума лягушки делятся на ряд типов. Одни реагируют на край, другие — на движущиеся объекты (светлые или темные) или же на удаляющиеся и приближающиеся предметы. При многократной стимуляции нейронов тектума наблюдается привыкание [177]. В поверхностных слоях тектума лягушки латеральное торможение осу
94
Глава IS
ществляется, по-видимому, по холинэргическим связям (515]'.. Г У саламандры окраска на ацетилхолинэстеразу совпадает с про-екцией зрительных волокон, и некоторые нейроны тектума отве- $ чают или на зрительную, или на контралатеральную тактильную стимуляцию [175].
Зарегистрированы ответы нейронов тектума на вестибулярное ; (у лягушки) [379, 480] и соматосенсорное (у хвостатых амфибий j и серебряного карася) раздражения. Ниже тектума расположен * тегментум, в котором находятся двигательные ядра. В тегменту- ” ме серебряного карася некоторые , нейроны отвечают на зритель- ц ные и слуховые сигналы [407]. У жабы также отмечена конвер- i генция модальностей — зрительной, тактильной и слуховой на суб- I тектальных нейронах {135]. В тектуме лягушки многие элементы, \ отвечающие на ипсилатеральную стимуляцию, получают сигналы, J приходящие через контралатеральный тектум [162, 164]. |
У черепахи для оптокинетической регуляции необходима тек-талъная комиссура [196]. После удаления поверхностной части
тектума у серебряного карася нарушается зрительное обучение ;
(346]. Локальное повреждение одной половины тектума вызывает \ круговые движения и переворачивание. Разряды тектальных ней- t ронов у серебряного карася, возможно, связаны с саккадическими $ движениями глаз. Локальное повреждение тектума у рыбы препятствует выработке уеловнорефлекторного изменения ритма сердечных сокращений на зрительную стимуляцию поля сетчатки, ] посылающего проекции в поврежденный участок; следовательно, для локализации в пределах поля зрения необходима ретинотек-тальная проекция [489]. У лягушки локальная стимуляция тектума вызывает организованные движения. Тектум участвует в движениях, определяемых зрительными стимулами, — поворотах тела, движениях глаз и оптомоторных реакциях. Промежуточный мозг лягушки тормозит или запускает активность идущих от тектума эфферентов, участвующих в ориентировочном и пищевом поведении; иными словами, таламус корректирует тектальные выходы [242].
В заднем отделе таламуса у жабы регистрируются ответы на | зрительные и соматические сенсорные стимулы. Таламические яд- 1 ра посылают проекции к примитивному гиппокампу [134]. Перед тем как напасть на двух мучных червей, жаба медлит дольше,, чем перед тем, как схватить одного, а широкий освещенный фон тормозит нападение; разрушение таламуса и претектума снимает торможение. Некоторые элементы таламуса отвечают не на те зрительные параметры, которые вызывают реакцию тектума. У лягушки аксоны некоторых ганглиозных клеток идут прямо в латеральное коленчатое тело; имеются также проекции от тектума к латеральному коленчатому телу [387]. У лягушек и рыб некоторые волокна проходят от таламуса к переднему мозгу, но эта проекция не изучена физиологически. Судя по последним данным,.
Центральная нервная система
95
можно думать, что повреждение переднего мозга у акулы нарушает различение изображений [169].
У игуаны в таламусе находятся два зрительных ядра; к дорсальному приходит прямая проекция сетчатки, а круглое ядро получает волокна от зрительного тектума [61]. У черепахи зрительные проекции на конечный мозг отвечают приблизительно на 40 мс позднее, чем элементы тектума; но некоторые зрительные волокна минуют тектум и идут через таламус в передний мозг, на что указывают ответы, которые можно зарегистрировать в переднем мозге после удаления тектума [26, 354, 614]. У аллигатора и ящерицы сетчатка проецируется на тектум, а вызванный ответ состоит из одной положительной и двух отрицательных волн, разных на разной глубине [192, 193, 344].
Птицы. У многих птиц зрительная система высокоразвита. Подсчитано, что в сетчатке голубя 20 млн. клеток, из них 2,4 млн. ганглиозных клеток, отростки которых образуют зрительные нервы. Большая доля зрительных волокон переходит к противоположной стороне тектума; из тектума пути направляются к круглому ядру •таламуса, а оттуда к эктостриатуму переднего мозга. Некоторые зрительные волокна идут прямо из сетчатки в таламус и оттуда в передний мозг.
У птиц тектум представляет собой крупное образование; вызванные потенциалы состоят из широкой отрицательной волны под поверхностью (свидетельствующей о двух синаптических стоках) и положительного колебания на более глубоком уровне. Текталь-ные нейроны обычно отвечают всего одним спайком на каждый приходящий к ним импульс и не вовлекаются в частоту стимуляции выше примерно 125 в 1 с. Входные сигналы из сетчатки могут прервать спонтанную активность, и, следовательно, тектальные нейроны у птиц могут тормозиться [200].
У отдельных тектальных нейронов голубя рецептивные поля гораздо больше, чем у ганглиозных клеток сетчатки; в большой их выборке 72% оказались избирательно чувствительными к движению, 21 % — к направлению движения (возбуждались при одном и тормозились при противоположном направлении движения) и 7% были чувствительны к концентрическим и контрастирующим фиксированным полям [254, 255]. У голубей движение и его направление регистрируются также таламическими клетками (соответствующими латеральному коленчатому телу); 75% клеток круглого ядра чувствительны к движению в поле зрения [84]. Значение полосатого тела в зрительной интеграции у птиц выявлено у голубей по нарушению различения яркости и формы (но не цвета) после повреждения гиперстриатума [427].
Млекопитающие. У млекопитающих верхний бугорок (гомолог тектума низших позвоночных) и претектальное ядро участвуют в Движениях наружных глазных мышц и в зрачковых рефлексах. Клетки претектального ядра дают и фазические, и тонические
96
Глава 15
эффекты; по сравнению с бугорком здесь большая их доля относится к тоническим нейронам {499]. Нейроны верхнего бугорка крысы и кошки образуют линейную проекцию от контралатеральной сетчатки, более точную в бугорке, чем в претектуме ,[192]. Нейроны бугорка сравнительно мало чувствительны к стационарным зрительным стимулам, но дают длительные ответы на движущиеся стимулы. Большая часть этих клеток отвечает на бинокулярную стимуляцию, и у каждой клетки есть свой оптимум скорости и направления движения. После удаления или охлаждения зрительной коры у кошки нейроны бугорка становятся малореактивными к движущимся стимулам и утрачивают бинокулярную чувствительность; это показывает, что ответы нейронов бугорка на зрительные стимулы модулируются корой [510, 513, 571]. У денаркотизированных обезьян в поверхностном слое верхнего бугорка обнаружено несколько нейронов, которые отвечали и на плавно движущиеся стимулы и на неподвижные вспышки, а также нейроны, отвечавшие на быстрые, отрывистые движения; в глубоких слоях нейроны разряжались пачками перед началом саккадических движений глаза [487]. Нейроны в тектуме саймири расположены слоями. Ответы на диффузный свет возникают в верхних слоях, на антагонистические поля — в средних и на движущиеся предметы — в глубоких слоях; имеются некоторые признаки ретинотопической проекции [273].
Двигательные реакции, вызываемые у обезьяны раздражением бугорка, хорошо коррелируют с ретинотопическими сенсорными картами на нем. У кролика стимуляция бугорка вызывает повороты головы и глаз в сторону поля зрения, соответствующего месту стимуляции. У крота зрительная система развита слабо, зрительный нерв состоит всего из 200 волокон, проекция на бугорок не обнаружена; в дорсальной части латерального коленчатого тела, по-видимому, осуществляется различение между светом и темнотой [336].
У млекопитающих большая часть зрительных волокон проходит в латеральное коленчатое тело в заднем отделе промежуточного мозга, а оттуда в зрительную кору. Подсчитано, что у макака зрительные нервы состоят из 1 млн. волокон, латеральные коленчатые тела — из 2 млн., а зрительная кора — из 200 млн. нейронов; таким образом, происходит дивергенция зрительных волокон в отношении 1:2: 100 ,[423а] в отличие от периферической части сетчатки, где происходит значительная конвергенция от многих палочек к одной ганглиозной клетке.
У рыб и амфибий в хиазме перекрещиваются все зрительные волокна, а у млекопитающих — лишь некоторая их часть. У приматов волокна от назальной половины сетчатки в хиазме переходят на другую сторону, а волокна от темпоральной части сетчатки идут в ипсилатеральном тракте. Первое переключение происходит в латеральном коленчатом теле, шестислойной структуре, где
Центральная нервная система 97
перекрестные и ипсилатеральные волокна распределены слоями. Одиночные нейроны коленчатого тела дают импульсные ответы, иногда в виде пачек импульсов; они обладают концентрическими । рецептивными полями с возбудительным «оп»-центром и «off»-ne-;риферией или наоборот {226, 227, 230]. В латеральном коленчатом теле кошки большинство клеток обладает рецептивными полями для каждого глаза, причем один глаз доминантный, а второй — недоминантный; он обычно оказывает тормозное действие [483]. От латерального коленчатого тела волокна направляются к затылочной доле коры большого мозга, преимущественно в поле 17 (стриарная кора), а некоторые волокна — в тюля 18 и 19.
Ответы одиночных нейронов в зрительной коре кошки и обезьяны принято делить на простые, сложные и сверхсложные [228]. Работы последних лет, проведенные на разных участках и разными способами зрительной стимуляции, показывают, что эти категории нейронов разграничены не строго и что они составляют некий континуум [70, 71, 359, 508, 516].
Самые простые клетки в зрительной коре имеют рецептивные поля с неконцентрическими возбудительными и тормозными зонами, как и клетки латерального коленчатого тела, но эти зоны находятся по сторонам прямого стационарного края, лежащего под «предпочитаемым» углом. Немного сложнее подобные же клетки (с возбудительной и тормозной зонами), которые дают фазические, ответы на движение края по небольшому полю; у некоторых из них чувствительность одномодальная, или дирекциональная, у других — бимодальная в отношении движения; все они вне периода стимуляции обычно находятся в состоянии покоя. К следующему уровню сложности относятся клетки, у которых нет антагонистических зон, но которые дают тонические высокочастотные ответы на движение по широкому полю в определенном направлении и под определенным углом; эти клетки вне стимуляции проявляют сильную спонтанную активность. Еще более сложные клетки (сверхсложные, лежащие в полях 18 и 19) отвечают на движущиеся ориентированные границы определенной длины, «возбудительную» полоску, по бокам которой находятся участки, вызывающие ответы только при одной ориентации [228] (рис. 15-34). Многие корковые нейроны возбуждаются бинокулярно, причем два рецептивных поля не совпадают; одни нейроны являются детекторами их диспаратности в вертикальном направлении, другие — в горизонтальном {418]. В популяции сверхсложных клеток у кошки 83% были бинокулярными, но для 68% доминировал один глаз, причем большее число нейронов реагировало на контралатеральные стимулы <[228, 229].
Если котенку сшивали веки или закрывали глаз повязкой и через 3 мес регистрировали нейронную активность, то ответы нейронов коленчатого тела характеризовались нормальными концентрическими «оп>- и «ofb-полями. Но у сверхсложных клеток меня-7—2067
Рис. 15-34. Типы клеток в зрительной коре кошки [631].
А. Простые клетки. Возбудительные (светлые) и тормозные (темные) части рецептивного “Поля показаны слева: Клетка а дает «ofb-ответы, клетка б — слабый «оп»-ответ. Б. Сложная клетка обладает спонтарной активностью. Она дает сильный ответ на движение стимула в одном направлении й тормозится его движением в противоположном направлении. чВ. Сверхсложная клетка/ДенДзальнаЯ часть! поля /возбудительная; периферия поля тормозная. Максимальное, цисло рмпульсрр вцзыда^ся,..движением . стимула по центральной части г./.-------------------------* ЧЛОЛЯ.* - ” ,
Центральная нервная система 99
лась бинокулярная доминантность и корковые клетки могли не давать ответа на стимуляцию глаза, который перед тем был выключен; таким образом, в результате неупражнения проводящего пути между коленчатым телом и корой наступало нарушение функции. Число реактивных клеток снижалось и в том случае, когда один глаз был закрыт в течение всего лишь 3—4 дней {231, 581]. Самым критическим для котят периодом являются 4-я и 5-я недели. Эффект незначителен, если односторонняя депривация начинается через 2 или 3 мес после рождения, и он полностью отсутствует при зрительной депривации взрослых животных. Поведенческие проявления вызванного депривацией коркового дефекта состоят в снижении остроты зрения, различения изображений и зрительного контроля движений лапы.
Переработка зрительной информации, наблюдаемая у кошек и обезьян, типична не для всех млекопитающих. В сетчатке у кроликов и бурундуков более выражена интеграция. У бурундука в сетчатке имеются только колбочки, и половина ганглиозных клеток отвечает на контраст (т. е. их рецептивные поля состоят из центрального и периферического участков); 25% ганглиозных клеток отличаются дирекциональностью (т. е. отвечают на движение ориентированного края), а 25% являются рецепторами противоположных цветов [366]. Одна половина волокон зрительного нерва идет к верхнему бугорку, а вторая — к латеральному коленчатому телу. Дирекционально чувствительные элементы направляются только к бугорку, а элементы, реагирующие на контраст и противоположные цвета, — к коленчатому телу. У бурундука нейроны латерального коленчатого тела сходны со сложными и сверхсложными клетками коры кошки [366, 367]. У голубя также имеются сложные ганглиозные клетки.
Итак, у кошек и обезьян ганглиозные клетки обладают концентрическими рецептивными полями; их корковые клетки по степени сложности сходны с ганглиозными клетками лягушки и серебряного карася. А у бурундуков имеются ганглиозные клетки по меньшей мере трех типов и сложные клетки в коленчатом теле. У голубей в стриарной коре находятся сложные клетки разного типа. У лягушки и серебряного карася в тектуме найдены разнообразные интегрирующие клетки.
Слуховые пути
Для большинства позвоночных звук — это важная и сложная сенсорная модальность. В головном мозге происходит переработка следующих ее параметров: временная последовательность интенсивности, частоты и шума; изменения интенсивности при постоянной частоте; модуляция частоты, шума и выделения тона из шума; положение звукового источника. Выше были рассмотрены механизмы звуковой рецепции (гл. 12), а также центральные слуховые
100
Глава 15
пути у рыб, амфибий, птиц и млекопитающих. Кроме того, описаны особые свойства слуховых центров у животных, пользующихся эхолокацией, например у летучих мышей.
У всякого позвоночного животного в продолговатом мозге заложен первичный слуховой центр. У рыб следующим уровнем является torus semicircularis в среднем мозге. Некоторые нейроны слухового ядра посылают аксоны в мозжечок. У элементов torus на аудиограммах одиночных нейронов частотные кривые уже, чем у клеток кохлеарного ядра [406]. На клетки Пуркинье в мозжечке рыб слуховые сигналы могут оказывать либо возбуждающее, либо тормозное действие.
У лягушек звуком активируются три ядра в torus semicircularis тегментума. Ответы состоят из пачек импульсов; тонотопическая организация в torus отсутствует [425]. В своей естественной среде обитания лягушки чрезвычайно чувствительны к комбинациям звуков, издаваемых представителями их вида; но где заложена видоспецифичная «матрица» этих комбинаций, пока неизвестно (см. гл. 12). У рептилий, например у каймана, нейроны кохлеарного ядра дают ответы на широкие полосы звуков, а для нейронов torus звуки, вызывающие ответы, лежат в более узком диапазоне. У этих рептилий <в переднем мозге имеется область — полосатое тело, — которая дает «оп»- и «ofb-ответы на тоны.
Птицы отличаются в этом отношении значительной видовой вариабельностью, но у некоторых из них (например, у сов) в кохлеарном ядре происходит бинауральное взаимодействие, а в мозжечке возникают интенсивные ответы. Нейроны в латеральном ядре среднего мозга (нижнем бугорке) голубя обладают спонтанной активностью; эта активность прерывается в ответ на звуковое раздражение, за исключением узкого возбудительного диапазона. Известно, что птицы каждого вида издают свои специфические звуки, но центральное представительство для них неизвестно.
В кохлеарном ядре млекопитающих (например, кошки) одна и та же клетка может отвечать деполяризацией и связанными с ней импульсами на звуки одного диапазона и гиперполяризацией, сопровождающейся подавлением спонтанной активности, — при другом диапазоне [51 Г]. О тормозных влияниях на кохлеарные ответы, вызываемых стимуляцией эфферентных волокон в оливо-кохлеарном пучке, было сказано выше. Функциональное значение этих эфферентов изучено недостаточно, но известно, что стимуляция эфферентного пучка усиливает кохлеарные микрофонные потенциалы и понижает начальный ответ кохлеарного нерва [300]; перерезка эфферентного пучка усиливает маскировку тонов шумом •[101а]. У серебряного карася стимуляцией среднего мозга можно снизить ответ torus на щелчки, и это возвратное торможение создает быструю (10 мс) модуляцию слуховых сигналов 1[421а].
Значение нижнего бугорка для слуха проявляется в способности животных и человека различать . звуки после удаления слуховой
Центральная нервная система
101
коры. Вызванные потенциалы, отводимые от нижнего бугорка у бурого дельфина, сильно варьируют, причем их различия коррелируют с параметрами звука, а аудиограмма, полученная на основании этих потенциалов, близка по своему диапазону к диапазону поведенческих реакций {54]. У морского льва (отряд китообразных) слуховая система менее специализирована, чем у дельфина; наиболее выраженные ответы нейронов бугорка лежат в пределах от 4 до 6 кГц [53]. У летучих мышей эти нейроны особенно реактивны к частотно-модулированным тонам [546].
Слуховая кора состоит из первичной и вторичной областей в височной доле, а у некоторых видов имеется еще и небольшая третичная область. Обе улитки имеют представительство в коре обоих полушарий с известным тонотопическим распределением; но ко
лонки и пространственное представительство выражены здесь слабее, чем в соматосенсорной и зрительной областях коры [3, 168]. Некоторые корковые нейроны дают нестойкие ответы на тоны, а при многократной стимуляции их активность либо угасает, либо облегчается; небольшое их число отвечает не на чистые тоны, а на час-тотно-модулированные стимулы {545].
Подробнее о центральных механизмах слуховой функции позвоночных сказано в гл. 12.
Соматосенсорные пути
У животных, не относящихся к классу млекопитающих, соматосенсорные проекции мало изучены. У лягушки вызванные потенциалы в ответ на раздражение седалищного нерва регистрируются в таламусе и переднем мозге {550]. У рыб электрические ответы на стимуляцию плавников и туловища возникают в мозжечке и тектуме, но большая часть переработки информации происходит в спинном и продолговатом мозге. В лицевых долях у серебряного карася имеются соматотопические проекции от каждого плавника и от кожи головы и туловища [417]. У тигровой саламандры контралатеральная поверхность тела проецируется на тектум, а окончания восходящих аксонов соматосенсорной системы, по-видимому, соответствуют распределению катехоламиновой флуоресценции f 175]. Таламические ядра у лягушки и черепахи отвечают на соматосенсорную стимуляцию.
У ехидны (Тachy gios sus) кора большого мозга образует складки, которые мало похожи на мозговые извилины высших млекопитающих; обширная область в задней половине коры получает сигналы о тактильном раздражении различных частей тела {322]; она лежит позади моторной области {365]. Подобным же образом насекомоядные, например еж (Erinaceus), обладают неспециализированным передним мозгом с двумя дорсальными областями зрительной проекции, обширной ростральной соматосенсорной проекцией и постеро-вентральной слуховой областью [272]. У сумча
102
Глава 15
тых — опоссума и валлаби — в коре имеются задняя зрительная область, латеральная слуховая область и обширные ростральные соматосенсорные области; сенсорное и моторное представительства частей тела полностью перекрываются и совпадают [323, 342].
В других отрядах млекопитающих соматосенсорная система следует одному общему плану. В спинном мозге волокна от ядер
Рис. 15-35. Схема первичных слуховых, соматосенсорных и зрительных областей,, а также а1ссоциатИ|В1ных областей в коре у кошки [535].
AI — слуховая область 1; АП — слуховая область II, Ер — слуховая область в задней экто-сильвиевой извилине, Ass — слуховая область в передней супрасильвиевой извилине, SI — соматосенсорная область I; SII — соматосенсорная область II, MI — соматомоторная область I, VI — зрительная область I, VII — зрительная область II, V88— зрительная область в супрасильвиевой борозде. I — передняя латеральная ассоциативная область, // — ассоциативная область в передней части средней супрасильвиевой извилины, III — ассоциативная область в задней части средней супрасильвиевой извилины, IV — перикруциатная ассоциативная область. I — слуховая область, 2 — зрительная, 3 — ассоциативная.
заднего столба проводят сигналы от рецепторов прикосновения,, давления и движения к продолговатому мозгу, куда приходит тройничный нерв с такой же информацией от лица и головы. От ядер в продолговатом мозге идут тракты в таламус, где карта проекций в большей степени пропорциональна плотности сенсорной иннервации, чем геометрии тела. Нейроны в таламусе способны различать не только точку тела, от которой приходят сигналы, но также характер и качество механических стимулов, положение частей тела и последовательность отдельных стимулов [382, 383]. Таламические ядра посылают соматосенсорные проекции на три области коры большого мозга, первую — на постцентральную извилину, вторую— на теменную кору и третью — на дорсальную
Центральная нервная система
103
часть передней супрасильвиевой извилины [89, 90] (рис. 15-35). Размеры тактильных областей соответствуют значению сенсорного входа; у обезьяны обширные области соответствуют руке и лицу, У кролика — морде, у кошки — передним конечностям и когтям, у собаки — морде, у кролика и крысы — вибриссам, у овцы — слизистой рта и у енота — передней лапе [535]. У различных обезьян размер представительства хвоста соответствует тому, насколько хвост используется при лазании.
Клетки соматосенсорной коры сгруппированы в функционально раздельные колонки одной и той же сенсорной модальности в соответствии с положением рецептивного поля на периферии. Некоторые клетки в этой области коры не обладают -специфичностью и отвечают не только на тактильную, но и на звуковую стимуляцию |[90]. Корковое представительство уточняет таламическую проекцию и приводит в соответствие сенсорные области с моторными.
Центральные обонятельные и вкусовые пути
У рыб имеется корреляция между крупным передним мозгом я хорошо развитым обонятельным органом (у угря Anguilla), между крупными лицевыми долями продолговатого мозга и вкусовой чувствительностью (у Ameiurus и Nemachilus), между крупным средним мозгом и зрением (у брызгуна Toxotes) [23]. Лосось находит путь, пользуясь обонянием [543]. С поверхности переднего мозга’ рыб отводится спонтанная электрическая активность, на которую накладываются ответы при обонятельной стимуляции. Ответы обонятельной луковицы у лосося изменяются при контралатеральном раздражении; следовательно, сюда приходят эфференты из головного мозга. Удаление обонятельной луковицы у угря влечет за собой дегенерацию в вентролатеральной части переднего мозга и в заднем и вентромедиальном отделах; имеются проекции к паллиуму и субпаллиуму {484]. Вкусовые сигналы приходят от кожи к лицевым долям, а от области рта и жабр — к^вагальным долям; таким образом, вкус и обоняние раздельны
У черепахи каждый обонятельный нерв приходит и к пириформ-ной коре, и к центральным ядрам переднего мозга; обонятельные, слуховые и зрительные пути до известной степени конвергируют в переднем^мозге [404]. У аллигатора обонятельный нерв идет к вентральной части пириформной коры {304].
У большинства млекопитающих обонятельные волокна идут в обонятельную луковицу, обонятельный бугорок, препириформную кору и кортико-амигдалярные ядра [189]. Считается, что обоняние это единственная сенсорная система, не имеющая представительства в таламусе. Вкусовые сигналы передаются через вентральное постеромедиальное ядро таламуса в небольшой участок 'й пресильвисвой борозде. •...-v.. д? з ;,п ц ?.. F
104
Глава 15
Центры эмоций в переднем мозге, участвующие в самосохранении и воспроизведении
Об ассоциативных функциях переднего мозга судят только по поведению. Необонятельная часть древнего переднего мозга получает сенсорный вход из промежуточного мозга и отдает моторные связи в ретикулярную формацию (рис. 15-23). Она тесно связана со средним отделом промежуточного мозга, в частности с гипоталамусом, и, возможно, обладает функциональной константностью по отношению к эмоциям и сложному поведению, связанным с самосохранением и сохранением вида у всех позвоночных [341]. К лимбической системе мозга относятся гиппокамп, септум (перегородка) и миндалины; она не дает дискретных электрических ответов на сенсорное возбуждение и при стимуляции не вызывает движений. Хорошим способом изучения лимбической системы служат наблюдения над поведением животных после повреждения или раздражения этой области. В общем, примитивный передний мозг согласует поведение с окружающей средой, т. е. запускает целые комплексы поведения — как стереотипные, так и нестереотипные.
После удаления переднего мозга у серебряного карася питание и спонтанная локомоция остаются нормальными; у него удается выработать условные рефлексы на свет и темноту; однако исследовательское поведение ограничено, а после одностороннего повреждения рыба плывет по большей части в направлении неповрежденной стороны [182, 250]. У колюшки после удаления необонятельного участка переднего мозга сохраняется нормальное плавание и различение цвета и яркости, но поведение, связанное с заботой о потомстве, и в частности с выбором места для гнезда, оказывается нарушенным. Передний мозг устанавливает баланс между разными формами эмоционального поведения: агрессией, половым влечением и родительским поведением; повреждение ростральных или латеральных областей ослабляет агрессию, сокращает периоды половой активности, но усиливает родительское поведение, а повреждение медиокаудальных областей усиливает агрессию и понижает родительскую активность [182, 490, 491]. Точно так же губаны и лещи с удаленным передним мозгом становятся менее драчливыми; число укусов при встрече с соперником уменьшается с 26 до 8 [138].
Когда рептилии находятся в группе, у них наблюдается сложное ритуальное поведение (демонстрирование определенных частей тела, общая агрессия, движения нападения), которым устанавливается социальная иерархия. После удаления той части мозга, где расположена миндалина (amygdala), ящерицы утрачивают агрессивность и подчиняются другим ящерицам; они могут по-прежнему принимать определенные позы, но не в связи с агрессией. Передняя часть миндалины обеспечивает позы самоутверждения;
Центральная нервная система
105
задняя часть обслуживает функции внимания и реакции на агрессивное поведение других ящериц .[527]. Стимуляция амигдаляр-ного комплекса каймана через вживленные электроды вызывала реакции бегства, а двустороннее повреждение этой области ослабляло реакции нападения и отступления [288].
Раздражая головной мозг кур через глубоко вживленные электроды, можно вызвать сложное поведение: курица кудахчет, садится на яйца, угрожает, нападает, пытается убежать |[202].
У млекопитающих в лимбическую систему входит кора гиппокампа, а также подкорковый комплекс передних септальных и задних амигдалярных ядер. Стимуляция разных частей лимбической системы вызывает вегетативные реакции, как висцеральные, так и сосудистые; раздражение других лимбических областей вызывает пищевые реакции (жевание, облизывание и др.), а некоторые другие раздражения приводят к агрессивному поведению (шипению, вытягиванию когтей), соответствующему данному виду |[295, 443, 542]. Стимуляция септального ядра у кошки вызывает «приятное ощущение», и животные с электродами, вживленными в эту область, получают «удовольствие» от самораздражения. Раздражением переднего гипоталамуса можно вызвать реакцию ярости, половое поведение, а с некоторых областей — пищевые и питьевые реакции. У кошек электрическая стимуляция миндалин подавляет агрессию, вызванную стимуляцией гипоталамуса [119]. У обезьяны нейроны гиппокампа разряжаются при ожидании пищи и перед нажатием на рычаг, в результате которого она эту пищу получает [400]. Удаление миндалин часто увеличивает количество поедаемой пищи (после начального снижения) и иногда создает гиперсексуальность. При удалении миндалин у кошки или собаки растормаживаются реакции ярости, а дикие крысы или макаки, наоборот, становятся при этом спокойными [370, 523]. У больных с повреждением области гиппокампа иногда страдает кратковременная память. Удаление новой коры у кошки делает ее более спокойной. Таким образом, поведенческие проявления повреждений •лимбической системы варьируют в зависимости от вида животного. Новая кора, лимбические центры и гипоталамус образуют систему взаимного контроля и регуляции.
Итак, новая кора служит, особенно у млекопитающих, для связи многочисленных входов с нейронами, запускающими двигательную активность. Старая кора всех классов позвоночных, очевидно, приводит инстинктивное или эмоциональное поведение в соответствие с окружающей средой. Обсуждается вопрос о том, нужны ли для осуществления сложных поведенческих актов нейронные сети, действующие en masse, или же отдельные нейроны обладают на выходе достаточно специфическим кодом, чтобы обеспечивать это сложное поведение. Во всяком случае, в примитивном переднем мозге позвоночных имеются эндогенные системы, имеющие значение для выживания.
106
Глава 15
Сон
В состоянии сна активность во многих областях головного мозга близка к синхронной. У многих млекопитающих (лучше всего это изучено у кошки) обнаружено две фазы сна [270]. В начальной фазе, или фазе медленных волн, волны электрической активности головного мозга, особенно в лобных областях, состоят из высокоамплитудных веретен (11—16 в 1 с), прерываемых медленными (1—4 в 1 с) волнами; движения глаз отсутствуют, но на электромиографических кривых видна постоянная тоническая мышечная активность, особенно в области шеи. Глубокий сон, иногда называемый парадоксальным, или БДГ (быстрые движения глаз — rapid eye movement, REM), отличается низкоамплитудной быстрой активностью, быстрыми движениями глаз и полным отсутствием мышечного тонуса. При глубоком сне крупные пирамидные нейроны моторной коры дают высокочастотные импульсные разряды, и, кроме того, возникает нерегулярная импульсация в латеральном коленчатом теле и ретикулярной формации варолиевою моста. В ретикулярной формации, от продолговатого мозга до среднего, лежат скопления нейронов с высоким содержанием серотонина (5-окситриптамина, 5-ОТ); разрушение этой области или блокирование синтеза 5-ОТ химическим путем сопровождается нарушением сна — наблюдается почти непрерывное бодрствование. Причиной появления характерных для сна медленных волн, или веретен, по-видимому, служит активность серотонинэргической ретикулярной системы [270]. Для глубокого, парадоксального, сна,, очевидно, требуется активность адренэргических нейронов тегмен-тума моста и голубого ядра, хотя в мышечной атонии участвует холинэргическая система. Фаза БДГ, возможно, сопровождается восстановлением медиатора в катехоламиновых системах <[380].
Обе фазы сна наблюдались у крота [9]; у ехидны отсутствовала фаза парадоксального сна [8]. У птиц отмечен медленный сон с короткими периодами парадоксального сна. У черепахи наблюдали медленный сон. Суточная регистрация активности в тектуме и переднем мозге у лягушки выявила и быстрые, и медленные ритмы, но картина сна отсутствовала, и лягушки все время сохраняли реактивность [198]. У рыб наблюдали поведенческие проявления «сна», но никаких его электрических коррелятов, подобных тем, какие обнаружены у млекопитающих, не было [270].
Многие животные — лягушки, жуки и другие — в ответ на угрожающие стимулы впадают в состояние тонической неподвижности, или мнимой смерти. Нейрофизиологические механизмы этой способности еще не установлены.
Развитие нервных функций
Нейробиология развития занимается двумя общими проблемами: 1) корреляцией спонтанных движений у зародышей с ответами
Центральная нервная система
107 "
на стимулы в процессе развития сенсорных, центральных нервных и эффекторных образований; 2) выяснением механизмов роста и дифференциации нервных структур.
Развитие поведенческих актов
Миогенная активность часто предшествует нервному контролю движений у многих низших позвоночных; но у других животных (например, у цыплят) даже самые первые движения управляются нервной системой. Впрочем, мышцы способны сокращаться еще до появления их иннервации.
Разные группы животных заметно отличаются друг от друга по скорости развития и по тому, являются ли самые ранние движения беспорядочными или же интегрированными. Обнаружено несколько градиентов: первичный идет от головного конца назад и вторичный — от проксимальных эффекторов к дистальным.
Переход от миогенной активности к нейрогенной можно проследить на примере дождевого червя Eisenia. После стадии гаст-рулы, снабженной ресничками, зародыш начинает совершать спонтанные сокращения вокруг стомодеума; затем в ответ на механическое раздражение возникают как локальные, так и распространяющиеся сокращения. Когда уже видна ганглионарная организация, передний конец начинает отодвигаться от стимула. Перистальтические реакции постепенно распространяются кзади, пока все животное не становится реактивным по мере развития нервной системы [428].
У зародыша кошачьей акулы Scyliorhinus миотомы ритмически сокращаются независимо на той и другой стороне до того, как возникнут идущие к ним нервные связи, и эти сокращения продолжаются после удаления нервного тяжа; таким образом, миотомы совершают и проводят сокращение. А когда нервный контроль уже установился, стимуляция вызывает сокращения с обеих сторон [188]. Такая же последовательность миогенных сгибаний, нейрогенных движений и рефлексогенной активности наблюдалась у лосося [569]. У Ameiurus развитие нервной регуляции идет в последовательности: спинной мозг, задний мозг и лабиринт; средний мозг и глаз [14].
Зародыши саламандры Ambystomaпроходят следующие стадии: немобильная стадия, когда мышцы сомитов сокращаются в ответ на прямое раздражение; стадия простого изгибания тела в ответ на тактильную стимуляцию; спонтанное сгибание на S-образной •стадии и, наконец, локомоторные волны сокращения. Первыми возникают реакции больших участков — целых частей тела, а локальные рефлексы формируются позднее. Процесс развития свя-с образованием все большего числа центральных синапсов 1[75]. У других амфибий, в частности у Xenopus и Rana, локальные сократительные реакции появляются до иннервации. Если пе
108
Глава 15
ререзка нервного тяжа производится на миогенных стадиях, каудальный стимул вызывает ростральный ответ; но если разрезаны и кожа, и нервный тяж, проведение между двумя концами прекращается. Эпителиальные клетки дают электрические ответы на тактильную и электрическую стимуляцию. Проведение может идти по эпителиальному слою и активировать миотомы [438]. Несколько позднее (на 27-й стадии или позже) для проведения уже требуется нервный тяж. В гл. И были приведены свидетельства электрической связи между клетками зародыша Loligo. У куриных эмбрионов, на стадиях 3—19 сомитов, между клетками одной и той же ткани и между разными тканями (например, между клетками хорды и нервной трубки или между хордой и мезодермой) осуществляется электрическая связь [497]. Но в других группах, помимо амфибий, электрическая связь между эпителиальными клетками и ее возможная роль в проведении на миогенных стадиях не отмечены.
У куриных эмбрионов на стадии закладки конечностей возникают беспорядочные движения, сначала в области шеи, потом туловища и наконец конечностей и хвоста. С этими хаотическими движениями коррелирует спонтанная электрическая активность, регистрируемая в спинном мозге на ранних стадиях развития. Управление координированными рефлексами появляется только после 17 дней инкубации. Проводились опыты по деафферентации путем удаления у двухдневных плодов дорсальной половины поясничного отдела спинного мозга или перерезки в грудном отделе. До 17-го дня периодические движения ног у контрольных и деафференти-рованных плодов были одинаковыми. Обычно ответы на сенсорные стимулы начинаются на 18-й день([183—185]. На электромиограммах видна слабая нерегулярная активность, которая заметно усиливается на 12—15-й день; активность, характерная для взрослого-животного, устанавливается к 17-му дню. По имеющимся данным, у куриного эмбриона и, возможно, также у плодов млекопитающих ранние беспорядочные движения вызываются двигательными разрядами спинного мозга в отсутствие сенсорных сигналов [494]. Впрочем, мышца диафрагмы в культуре ткани обладает миоген-ной ритмикой.
У плода овцы (40 дней беременности) первые движения отрывисты; они вызываются через бульбоспинальную систему. Позднее для продолжительности движений необходимо участие среднего мозга. У плода макака рефлекторное вздрагивание передних конечностей без перекрестного или антагонистического торможения совершается до миелинизации шейных нейронов, но после того, как на дендритах мотонейронов появится некоторое количество синапсов [39, 40].
Делается вывод, что миогенная стадия, наблюдаемая у пла- . стиножаберных и амфибий, возможно, отсутствует у птиц и млекопитающих.
Центральная нервная система
109
Начальное развитие центральных нервных связей происходит и в отсутствие нервных импульсов. У личинок саламандры, которые в течение многих часов находились под наркозом, развитие продолжалось, и после вылупления рефлексы у них были такими же, как если бы животные проделали промежуточные стадии двигательной активности. Эксплантаты спинного мозга крысы в культуре ткани дают спонтанную электрическую активность; добавление ксилокаина или Mg2+ в высокой концентрации, по-видимому,, полностью блокирует электрическую активность, тогда как рост продолжается. После снятия блока сохраняются нормальные свойства полисинаптической сети. Таким образом, для развития синапсов не обязательны эндогенные импульсы [82]. Нейроны развиваются в раннем эмбриогенезе; некоторые из них претерпевают обратное развитие. Самые первые рефлексы проявляются во время замыкания аксо-дендритных связей; образование миелиновых оболочек происходит на поздних стадиях развития, что, возможно, позволяет нейронам «узнать друг друга» на более ранних стадиях [40].
Развитие ритмической активности головного мозга коррелирует со степенью зрелости при рождении или вылуплении (см. выше). У морской свинки электрические волны мозга имеются уже при рождении, но максимальной частоты они достигают через 7 дней; в то же время у крысы первые волны появляются в головном мозге через 4 дня после рождения, .а ритм, характерный для взрослого, возникает через 18 дней. У цыплят некоторые виды волн обнаруживаются на 11—12-й день инкубации, выраженная синхронность — на 17-й день, а активность, свойственная? взрослым, — на 7—9-й день после вылупления. Вместе с тем на энцефалограмме голубей волны отсутствуют до 4-го дня после вылупления [415]. Появление сложных вызванных ответов в коре котят коррелирует с развитием базальных дендритов у пирамидных нейронов [432].
Специализация нейронов в процессе развития
Исследование как регенерации, так и эмбрионального развития показывает, что процессы роста нейронов и периферических структур связаны между собой реципрокно. У упомянутых выше деафферентированных цыплят приблизительно через 17 дней мотонейроны дегенерировали; следовательно, для продолжения их развития требуются сенсорные сигналы [185]. Вкусовые луковицы млекопитающих не развиваются в отсутствие сенсорных нервов. При пересадке метаторакального ганглия таракана в тазик другого таракана иннервацию приобретают только предварительно Денервированные мышцы; в мотонейронах образуется во время процесса активации плотное кольцо РНК {245].
Один из самых интересных вопросов нейробиологии развития — это вопрос о том, каким образом нейронные связи приобретают
по
Глава 15
специализацию. Специализация частично заложена в геноме, а частично создается во время развития. Как правило, сенсорные нейроны специализируются на ранних стадиях развития, а вставочные — несколько позднее. Примеры перехода от отсутствия специализации к точной специализации функции можно найти и среди сенсорных, и среди моторных нейронов.
У головастиков и лягушек пересаживали участки кожи со спины на брюшко или с брюшка на спину; если пересадку производили на поздней личиночной стадии или у взрослого животного, раздражение пересаженного лоскута кожи вызывало рефлексы, нормальные для этой области кожи до пересадки, но если пересадка была совершена у головастика на ранней стадии развития, то направленность возникающих затем рефлексов была не свойственна этому лоскуту кожи; иными словами, специализация сенсорных нейронов изменялась в соответствии с новым положением [247] (рис. 15-36). Отсюда следует вывод, что сенсорные нейроны «инструктируют» центральные связи и что сенсорные нейроны специализируются в соответствии с их положением в теле на поздних стадиях развития.
Аксоны ганглиозных клеток сетчатки у лягушки и рыбы образуют в тектуме проекцию, имеющую специфическое расположение как по поверхности, так и по глубине. Если перерезать зрительный нерв, повернуть глаз и дать возможность зрительным аксонам регенерировать, причем произвести все это у взрослого животного или на поздней стадии головастика, то в зрительных рефлексах и электрических ответах тектума проявляется инверсия нормального расположения проекции; следовательно, ганглиозные клетки специализированы для некоторых областей тектума. Но если эта операция произведена на более ранней стадии, устанавливается нормальная проекция в отношении ориентации глаза, т. е. аксоны ганглиозных клеток тогда еще не были специализированы [248] (рис. 15-37). У Xenopus после операции поворота глаза на стадии 28 проекция сетчатки была нормальной по обеим осям; после поворота на стадии 30 проекция переворачивалась по назотемпораль-ной, но не по дорсовентральной осям; при повороте на стадии 31 возникала инверсия по обеим осям. Таким образом, специализация в одном направлении происходит раньше, чем в другом [246]. У амблистомы нейроны сетчатки специализируются на стадиях 34—36, а у Xenopus — на стадии 30. Между теми областями обеих половин тектума у головастиков Xenopus, которые получают возбуждение, одинаково распределенное в пространстве и времени, образуются связи [163]. Если волокна зрительного нерва у взрослой лягушки переместить в случайном порядке, отдельные волокна будут расти обратно по направлению к месту своего прежнего положения в тектуме [507]. Такое же восстановление прежних проекций наблюдалось у серебряного карася [600].
Центральная нервная система
ш ~
Рис. 15-36. Смещенные рефлексы у лягушки [632].
Показаны области, с которых во все более далекие от операции сроки вызываются смещен* ные рефлексы. Смещенные рефлексы вызываются с зачерненных участков, нормальные — с остальной части трансплантата. А. Трансплантация кожи спины на брюшко проводилась на стадии IV, а тестирование — спустя 31, 48, 51, 58 и 68 дней. 15. Трансплантация на стадии V. а тестирование — спустя 106, 123, 131, 141 и 151 день.
У Цыплят зрительные ответы не нарушаются, если глаз был повернут и реимплантирован в первые 70 ч инкубации. В опытах с зашиванием глазной щели у котят на несколько дней в период от 4-й до 6-й недели после рождения отмечена утрата способности к распознаванию образов, если судить по активности клеток стри-
112
Глава 15
ярной коры [229]. Котят выращивали в клетках, окруженных вертикальными или горизонтальными полосами; когда через несколько месяцев их помещали в сложную среду, они оказывались слепыми по отношению к полосам, перпендикулярным тем, которыми они были раньше окружены. Бинокулярно чувствительные
Вентральная Вентральная
Рис. 15-37. РетиН'ОтекталБная проекция у Xenopus на правую половину тектума (Л) от левого глаза (Г), повернутого на 1'80° на личиночной стадии 28—29, и от правого нормального глаза (В) на левую половину тектума (В) [246].
Проекция от повернутого глаза нормальная. Номерами на тектуме показаны точки, где микроэлектродом регистрировался потенциал действия, который возникал в ответ на маленькое световое пятно, наносимое на точку сетчатки с тем же номером. Стрелкой обозначено рострально-каудальное направление (р—>к); Н — назальная область сетчатки, Т — темпоральная.
нейроны их зрительной коры выказывали предпочтение к направлению этих последних {37, 197]. У крысят, которые получали разнообразные сенсорные раздражения ежедневно, начиная с рождения, к 8-му дню число шипиков на единицу поверхности пирамидных нейронов было значительно выше, чем у крысят, лишенных такой стимуляции; величина клеток не изменилась [493]. Делается вывод, что в процессе развития сенсорные нейроны становятся все более специализированными, что некоторые периоды являются критическими для образования нервных связей и что характер специализации согласуется с их временным и пространственным распределением.
Системы мотонейронов созревают и без сенсорных входов, но в обычных условиях на организацию моторики влияет мускулату-
Центральная нервная система НЗ
ра конечностей. В столбах спинного мозга образуется больше мотонейронов, чем впоследствии используется, и в процессе центральной организации часть из них подвергается дегенерации; мотонейроны дегенерируют, если они не работают, а мышечные волокна дегенерируют, если они не иннервированы. В спинном мозге мотонейроны представлены до некоторой степени топографически; дистальная мускулатура конечностей — в передних столбах дор-сальнее, чем проксимальная, а мышцы экстензоры — латеральнее флексоров [80]. Спинномозговые нервы в течение некоторого времени «ищут» конечность, когда она пересажена не очень далеко от своего нормального положения. Если, например, подсадить саламандре добавочную ногу, сохранив ее правильную ориентацию, неподалеку от нормальной ноги, и если при этом она получит иннервацию, то соответствующие мышцы обеих ног будут сокращаться одновременно. Каждая реиннервированная мышца добавочной конечности сокращается синхронно с соответствующей мышцей нормальной конечности [558]. Если же пересаженная конечность повернута на 180° по сравнению с нормальной, то она движется координированно с последней по времени, но в обратном направлении. Сходные данные получены на цыплятах. Отсюда следует вывод о некоторой миотопической специализации мотонейронов.
Специфичность центральных аппаратов проявляется в реакциях добавочных придатков у ракообразных. Глазной стебелек был заменен у краба наружными сегментами антеннулы. Ее стимуляция вызывала такие же рефлекторные ответы, как и стимуляция нормальной антеннулы. Поскольку при этом использовались новые пути, следует заключить, что центральные связи построены на основе модальности рецепторов [352]. У сверчка брюшные церки регенерируют и их сенсорные волокна образуют нормальные контакты с гигантскими волокнами в терминальном ганглии. При пересадке регенерирующего церка на культю ноги сенсорные волокна контролируют гигантское волокно в груди; следовательно, регенерирующие сенсорные волокна узнают, на каких центральных волокнах им оканчиваться [117].
Пока неизвестно, чем определяется специфичность отростков развивающегося нейрона. Гипотезы о значении тигмо- и гальванотаксиса не оправдывают себя, хотя в культуре тканей на рост отростков нервных клеток можно повлиять ориентированными поверхностями и электрическими полями. Дистантное действие специфических химических веществ мало вероятно >[247]. Наблюдение над нейронами в культуре тканей показывает, что их отростки растут в случайных направлениях и, проходя мимо разных клеток, иногда словно проверяют, годятся ли те для контакта; затем они образуют синапсы на специальных рецептивных клетках, как если бы мембраны этих последних обладали химическим сродством к Данному аксону. Таким образом, химические факторы на близком расстоянии могут играть здесь важную роль.
«—2067
114
Глава 15
На развитие нейронов влияет множество факторов. Скорость развития меняется от упражнения или неупражнения нейрона, и дендриты не развиваются в отсутствие аксонов, которые в норме оканчиваются на них. На нейронах имеются определенные участки для аксонных окончаний данного типа (например, дендриты и тела пирамидных клеток). Зрительная депривация снижает в сетчатке концентрацию РНК, ацетилхолинэстеразы и других веществ, играющих решающую роль. На критических стадиях определенную роль в развитии нейронов играет питание, о чем свидетельствуют случаи умственной отсталости у детей при неполноценном питании; такая умственная отсталость связана с замедлением развития коры больших полушарий и до известного возраста обратима. На созревание нервной системы оказывают влияние гормоны; у головастика центральные связи, обеспечивающие рефлексы закрывания глаза на раздражение роговицы, не развиваются при слишком низком содержании тироксина [299]. Созревание симпатических и некоторых сенсорных нейронов стимулируется фактором роста нервной системы — белком, встречающимся в некоторых опухолях, слюнных железах и сыворотке крови {324, 325]. Введение животному антител, специфичных в отношении этого белкового фактора, препятствует созреванию симпатической системы.
В процессе онтогенеза происходит программирование дифференцировки и дегенерации. Маутнеровские клетки Xenopus образуют отростки и правильно функционируют у головастиков; позднее, в процессе метаморфоза они атрофируются и у взрослых животных отсутствуют. Можно показать не только то, что центральные связи влияют на рост нервных волокон, но и что свойства мотонейронов влияют на дифференцировку иннервируемых ими мышечных волокон. Для маленьких котят характерна средняя скорость сокращения мышц, тогда как у взрослой кошки имеются мышцы, которые сокращаются быстро и бедны некоторыми митохондриальными ферментами (например, икроножная мышца), и мышцы, сокращающиеся медленно и содержащие сравнительно много митохондриальных ферментов (например, камбаловидная мышца). «Перекрест» иннервации этих мышц, произведенный на ранних стадиях онтогенеза, меняет характер развития: у такого животного камбаловидная мышца приобретает способность сокращаться быстрее, а икроножная — медленнее нормальной (см. гл. 16). Двигательные нервы регулируют распределение рецепторов, чувствительных к ацетилхолину, в скелетной мышце позвоночных. В первые 2 нед после перерезки двигательных нервов наблюдается расширение области чувствительности к ацетилхолину, соответствующее классической «денервационной гиперсенситивности» (см. гл. 16). Затем вновь наступает ограничение чувствительности к ацетилхолину, что соответствует образованию новых связей регенерировавших двигательных нервов с мышечными волокнами. В свою очередь мышцы тоже могут влиять на ход диф
Центральная нервная система
115 -
ференцировки иннервирующих их нейронов. Так, в некоторых мышцах ракообразных далеко отстоящие друг от друга терминали одного аксона, находящиеся на одном и том же мышечном волокне, весьма сходны между собой, в то время как рядом расположенные терминали того же аксона, лежащие на другой, непосредственно примыкающей мышце, могут быть совершенно иными.
В общем, нейроны обладают большим диапазоном структурной пластичности. Те животные, у которых деление нейронов продолжается в течение всей жизни, весьма пластичны; у остальных специфичность связей фиксируется на какой-либо определенной стадии развития. Все нейроны способны к некоторым изменениям после повреждения аксона; детали этих процессов в точности не установлены. В процессе развития нейроны подвержены индукции в отношении образования связей; она может осуществляться контактами с сенсорными структурами, с другими нейронами и, возможно, с мышцами. И в свою очередь растущие нейроны действуют на те клетки, с которыми контактируют, и могут влиять на синтез белков в таких иннервируемых ими тканях, как мышцы.
Стойкие изменения функции центральной нервной системы
Процессы синаптической передачи, облегчения и торможения быстротечны — они длятся самое большее несколько сот миллисекунд. Многое в стереотипном поведении можно объяснить, исходя из представления о нервной системе как о распределительном щите, но наиболее адаптивные ее свойства сохраняются долго, и анализ их труден. Одно из самых важных качеств центральной нервной системы — это вариабельность и изменчивость реакций; изменения, которые сохраняются долгое время — секунды, дни или всю жизнь.
К стойким модификациям поведения относятся эффекты упражнения и неупражнения; кратковременное привыкание и отвыкание; выработка условных реакций и обучение, в которые входят краткосрочная и долгосрочная память, и видоизменение врожденных форм поведения. В обзорах и руководствах по психологии и этологии можно найти классификации и теоретические модели стойких изменений поведения [538]. В настоящем разделе кратко рассматриваются: 1) некоторые примеры изменения поведения у разных групп животных, 2) данные о стойких изменениях синаптической функции и 3) возможные объяснения первых,, исходя из вторых.
Классификация стойких изменений
Пластичность, связанная с упражнением и неупражнением, включает в себя утрату навыка или точности реакции после длительного неупражнения двигательной системы (например, ослаб-«*
116
Глава 15
ление спинального рефлекса после перерезки одного заднего корешка и тестирования через другой корешок); компенсаторный перенос функции, в частности от одной конечности на другую (например, изменение походки после ампутации одной ноги у животных, имеющих несколько ног); изменения специализации сенсорного или моторного центра в ходе регенерации или развития {495]. Близкий к этому феномен представляет собой повышенная чувствительность мышцы к нейромедиатору в течение некоторого времени после денервации; он связан с индукцией синтеза рецепторных молекул. Сенсибилизация может играть важную роль в процессе восстановления после повреждения центральных аксонов.
Привыкание. Привыкание характеризуется следующими признаками [537]: снижением ответа при повторении стимула; спонтанным восстановлением после временного прекращения стимуляции; падением реактивности, которая может упасть ниже нулевого уровня. Привыкание сильнее при частых и слабых стимулах. Устранение привыкания вызывают сильные стимулы, обычно другого вида, чем те, которые вызвали привыкание. Привыкание отлично от центрального торможения, утомления и сенсорной адаптации. Поведенческое привыкание можно наблюдать на спинальных рефлексах позвоночных, втягивании жабр и мантии у многих моллюсков, ориентировочных движениях и схватывании пищи у различных животных (например, у паука и жабы), соединении в стаю при появлении хищника (у птиц). Привыкание отдельных компонентов сложного поведения происходит с разной скоростью; например, у хватающего добычу паука ориентация в сторону добычи сохраняется при повторных предъявлениях дольше, чем реакция нападения [426]. В монографии Хорна и Хайнда [204] приведены многочисленные примеры привыкания в поведении.
Выработка условного рефлекса и угашение. Условнорефлекторное поведение, вероятно, может быть разного рода и основано на различных механизмах.
а) При простой выработке рефлекса условный стимул (УС),, за которым следует или с которым совпадает безусловный (БС), приводит при многократных применениях к такому состоянию, когда УС сам по себе вызывает реакцию, по существу такую же, как реакция, вызываемая БС. В отсутствие подкрепления условный рефлекс (УР) исчезает за короткое время. Быстрое угашение достигается также применением общей или неспецифической стимуляции.
б) Павловский условный рефлекс обычно отличается от безусловного тем, что возникает с более длинным латентным периодом, часто является предупреждающим и сохраняется очень долго, если не производится угашение.
в) Основой для обучения в лабиринте, опытов с проблемным ящиком и аналогичных тестов служит инструментальный услов
Центральная нервная система 117~ -
ный рефлекс. Вознаграждение дается после решения задачи или же реакция сама по себе является вознаграждением. Это напоминает обучение методом проб и ошибок.
г) Цепные условные рефлексы допускают переход от одной сенсорной модальности к другой. Число последовательных звеньев, какое может быть добавлено к цепи, варьирует у разных видов.
В качестве контроля образования условного рефлекса надо-применять БС изолированно столько же раз, сколько он применяется с УС; это может привести к сенсибилизации сенсорных путей, которую иногда называют ложным условным рефлексом (pseudoconditioning). Точно так же, если БС применяется до УС, УР может не выработаться. Многие данные показывают, что выработка условного рефлекса, во всяком случае у позвоночных и осьминога, проходит две стадии — раннюю и позднюю [301]. На ранней стадии создается кратковременная память, которую можно разрушить определенными центральными повреждениями или электрошоком. Последовательность событий при выработке УР’ по большей части можно свести к следующей: поступление сенсорных сигналов (УС), классификация или кодирование <в сенсорных ядрах, консолидация классифицированной или кодированной информации, длительное хранение и, наконец, извлечение из памяти, или считывание. Механизмы всех этих операций, вероятно,, различны; отсюда следует, что поиски единого целлюлярного механизма обучения нереальны [251, 252].
Способность к выработке условного рефлекса у разных животных
В поведении многих простейших, особенно Ciliata, проявляются: сложные таксисы: фото-, гео- и гальванотаксисы; такие таксисы способны к видоизменениям. Так, туфельки скапливаются вокруг платиновой проволочки после того, как они регулярно получали корм около таких проволочек [165].
В Т-образном лабиринте, где свет служил УС, а удар электрическим током — БС, планарий обучали выбирать одну ветвь лабиринта. Для «обучения» необходимо наличие церебрального-ганглия, но, по некоторым данным, если червя перерезать после-того, как он уже обучен, регенерировавшая задняя половина способна совершать соответствующий поворот {258, 356, 357]. Име-ются^сообщения о переносе какого-то вещества от обученных планарий к необученным и о соответствующем переносе приобретенного навыка, но эти наблюдения не получили достаточного подтверждения.
Психологи решительно возражают против того, чтобы считать модифицированное поведение туфелек и планарий истинной выработкой условного рефлекса [256]. По мнению Торпа [538], такие
fl 18
Глава 15
изменения представляют собой исследовательское поведение, а не .ассоциативное обучение и животные способны связать одно орто-жинетическое (направленное) движение с другим.
Животные с простыми нервными сетями (кишечнополостные) и те, у которых нервные сети сосредоточены в ограниченных зо-жах (иглокожие), способны 'видоизменять свое поведение. Привыкание гидры к механической вибрации или к освещению сопровождается понижением чувствительности, которое длится несколько часов [473]. Актиний можно до некоторой степени обучить выбору субстрата [458]. Морская звезда перестает пользоваться одним .лучом, если много раз подряд ей не позволяли им двигать. Если морской звезде предъявлять каждые 48 ч свет и пищу в надлежащей последовательности, то она «научается» после нескольких -сочетаний передвигаться по аквариуму в ответ на один только свет [ЗП].
У некоторых кольчецов происходит переделка на обратную реакцию на один сенсорный стимул, если предъявлять его с другим ^стимулом, который обычно вызывает противоположный ответ (например, переделка реакции на свет, сопровождаемый тактильным стимулом). Черви Nereidae, которые на свет и прикосновение обычаю дают отрицательную реакцию, «научаются» вылезать из своих трубочек на свету, если одновременно со светом или прикосновением они получают сок из мидий. Nereis продолжает обучаться в Т-образном лабиринте и после нарушения связи органов чувств переднего конца его тела с головным мозгом (например, если мозг отделен от глаз, пальп и антенн, но сохраняет связь с тен-такулярными органами чувств) [146]. Привыкание к свету происходит и у децеребрированных нереид и, следовательно, является, по-видимому, свойством всех отделов нервной системы {126]. Многие исследователи приучали дождевых червей поворачивать в Т-образном лабиринте в определенном направлении, обычно при .помощи светового или тактильного раздражения в качестве БС. Если у таких червей удалить затем надглоточный ганглий, условная реакция сохраняется. Дождевые черви, которые многократно получали односторонние сильные световые стимулы (отрицательный фототаксис), слабые световые стимулы (положительный фототаксис), или же односторонние электрические или тактильные -стимулы, продолжали и в отсутствие этих стимулов поворачивать в «выученном» направлении, причем это не было результатом изменения порога положительного или отрицательного фототаксиса [303].
Для членистоногих существует много примеров ассоциативного обучения, обучения путем проб и ошибок и обучения в лабиринте [538]. Разнородные стимулы способны вызывать сложные формы врожденного поведения. Свет часто применялся как УС в сочетании с уколом или электрическим стимулом в качестве БС. Так, у мечехвоста был выработан условнорефлекторный удар хвостом
Центральная нервная система119
[501]; сходное условнорефлекторное поведение получено у крабов, креветок, тараканов и других животных. У перепончатокрылых наблюдается сложное поведение, основанное на врожденных стереотипах в сочетании с кратковременной памятью. Пчел приучали летать на определенные цвета, формы или запахи, связанные с местом кормления. Память сохранялась и после того, как пчел подвергали наркозу хлороформом или охлаждению {436]. Приучив пчел прилетать за кормом в определенное время дня, их затем подвергали наркозу в течение 4 ч; на следующий день они прилетали к месту кормления и в тот час, к которому были приучены, и тремя часами позже. Когда к приученным пчелам присоединили пчел из другой колонии, новички вовлекались в выработанное поведение, а первая группа давала третий пик прилетов [329, 361]. Медоносные пчелы сообщают своим товарищам о расстоянии и месте источников питания посредством темпа и направления «танца». Они помнят запахи [152]. Рыжего муравья Formica rufa можно приучить следовать в лабиринте запаховым сигналом.
У рыб вырабатывали пищевые условные рефлексы в челночных ящиках и лабиринтах; освещение разного цвета, форма зрительных стимулов и звуки часто применялись в качестве условных раздражителей. В возвращении мигрирующих рыб к прежнему месту обитания, возможно, важную роль играет обонятельный импринтинг (см. гл. 13). У серебряного карася вырабатывается определенный ритм сердца и частота дыхания в ответ на свет или звущ Если во время выработки рефлекса на свет один глаз рыбки закрыть, то потом условная реакция возникает и при освещении этого глаза. Но если вырабатывается реакция избегания, то перенос-с одного глаза на другой не происходит; иными словами, оба глаза нужны для переноса реакции при зрительно-моторной, но не при сенсорно-вегетативной выработке {355]. У серебряного карася после удаления переднего мозга можно выработать условно-рефлекторное изменение ритма сердца при использовании света в качестве УС, но он не способен научиться различать цвета [33]. Кроме того, удаление у них переднего мозга мешает обучению реакции выбора в челночном ящике и нарушает ранее выработанный навык {181]. Условные рефлексы более чувствительны к охлаждению, чем безусловные плавательные рефлексы [430]. Серебряный карась оказался способным к инструментальному обучению (например, к нажиму на рычаг для получения корма в; двухминутном режиме); охлаждение не нарушало чувства времени [417].
Хвостатых и бесхвостых амфибий обучали в простых лабиринтах и вырабатывали у них условные рефлексы на зрительные ш слуховые стимулы. Способность к образованию ассоциаций сохраняется у них после удаления одной половины переднего мозга, но» утрачивается после удаления обеих половин.
920
Глава 15
Что касается примеров условнорефлекторного поведения птиц и млекопитающих, то можно сослаться на специальные подробные описания [538].
Локализация центров обучения
У многих животных удаление интегративных областей нервной системы приводило к прекращению условнорефлекторной деятельности. В поисках местонахождения «центра обучения» следует исключать сенсорные и моторные нарушения. Прекрасный пример применения метода разрушения определенных структур для анализа сложного обучения можно найти в работе Дж. Янга на головоногом моллюске осьминоге (см. [603—605, 607, 608, 610, 612, €13]).
Осьминог. Осьминог—«разумное» животное, которое обычно устраивает себе жилище под водой в пещере под скалами, откуда выходит для охоты и для исследования новых предметов на своей территории. Его нервная система представляет собой сложную совокупность нервных центров: надглоточный отдел содержит много сенсорных ядер и несколько областей «высшей» интегративной •функции, а подглоточный выполняет главным образом двигательные функции.
Если нормальный осьминог увидит креветку, которая затем «скроется из виду за углом, он поплывет за ней [563, 565]; но если у осьминога удалить вертикальный комплекс головного мозга, он будет гнаться за креветкой, пока видит ее, но не последует за ней, когда она скроется из глаз. В эксперименте у осьминогов вырабатывали условные рефлексы на зрительные и тактильные стимулы. Осьминоги обычно охотятся за движущейся добычей, но их можно приучить нападать и на неподвижные цели. Обычный прием обучения заключается в подкреплении пищей реакции нападения на „данный предмет (например, на пластиковый диск, на котором изображена фигура) при выработке положительной реакции и в наказании электрическим током при выработке отрицательной реакции. Обучение идет быстро, и, комбинируя вознаграждение и наказание, можно обучить осьминога за несколько сочетаний различать две фигуры. Горизонтальные фигуры различаются лучше вертикальных.
Сетчатка осьминога содержит набор рабдомов (зрительных элементов), каждый из которых состоит из четырех клеток, или рабдомеров (гл. 14). Аксоны фоточувствительных клеток обильно ^ветвятся в наружном, или сетевидном, слое зрительной доли. Каждая зрительная доля содержит около 6 -107 нейронов. Дендритные поля в глубоком сетевидном слое доли имеют овальную форму; одни ориентированы вертикально, другие горизонтально. Диаметр одного такого дендритного поля может составлять более 500 мкм. Нейроны, лежащие на большей глубине, имеют широкие кониче
Центральная нервная система
121
ские дендритные поля со входами от наружных клеток. По данным Сатерленда [524], рецепторные клетки сетчатки лежат горизонтальными рядами и вертикальными колонками. Сатерленд предполагает, что зрительная доля кодирует отношение между вертикальными и горизонтальными элементами, которые стимулируются каким-либо изображением; он связывает это кодирование с ориентацией дендритных полей в наружном сетевидном слое. Тестируя способность осьминога к различению разных фигур, Сатерленд нашел, что такое кодирование отношения горизонтальных элементов к вертикальным плюс некоторая способность к восприятию площадей позволяют объяснить способность к различению формы. Далее, если удалены статоцисты, то глаза поворачиваются так, что щелевидный зрачок вместо горизонтального становится вертикальным и реакция на вертикальные предметы оказывается такой, как на горизонтальные. Предполагается, что каждый нейрон в глубоком сетевидном слое зрительных долей отвечает на определенное отношение горизонтальных сигналов к вертикальным и что эта форма ответа устанавливается в процессе развития. Зрительные доли посылают эфферентные волокна прямо к двигательным центрам и таким образом вызывают реакцию нападения.
Тактильное обучение происходит на основе стимуляции сенсорных клеток в руках; этот вид обучения лучше всего изучать в опытах на ослепленных животных. Тактильное различение пластиковых цилиндров с разной шероховатостью или с разными бороздками можно легко продемонстрировать при помощи методики условных рефлексов. Предметы одинаковых размеров и текстуры, но разного веса не различаются. Очевидно, тактильное различение основано на соотношении гладких и шероховатых поверхностей и на величине предмета. Сенсорные нейроны в руках связаны с нижними фронтальными и субфронтальными долями, при удалении которых сразу же утрачивается способность к различению. Химическим чувством осьминог отличает раковину с живым моллюском от раковины с воском.
В верхних фронтальных и вертикальных долях (рис. 15-38), по-видимому, представлены зрительные (глаза) или субфронтальные (руки) доли. Электрическое раздражение верхних и вертикальных долей не вызывает непосредственных двигательных реакций. Аксоны от верхней фронтальной доли проходят к вертикальной доле, которая разделена на 5 долек. Импульсация из вертикальных долей идет через субвертикальную долю к зрительным, верхним и нижним фронтальным долям, т. е. обратно к тем областям, откуда она вышла. Таким образом, имеются две параллельные петли для зрительной и для хемотактильной систем.
Удаление вертикальных долей не приводит к каким-либо явным моторным или сенсорным дефектам. Но осьминог может давать неправильные реакции. Если перед этим он был обучен, то сохраняются некоторые условные реакции. Если он был обучен на
322
Глава 15
падать на предмет, а затем переучен (не нападать), то удаление вертикальной доли возвращает его к первой форме поведения (нападать). Перенос выработанного различения с одного глаза на другой нарушается при удалении вертикальной доли, даже несмотря на наличие волокон, соединяющих обе зрительные доли. После удаления вертикальных долей обучение возможно, но идет медлен-
Медиальный, верхний фронтальный
Латеральный вепхний фронтальный
Вертикалъный
Субвертикальный.
ая
Нападение
Ь®- Отступление
•разаль-Ряные :
Болевые сигналы
А
Рис. 15-38.
Л. Схема центров в мозге Octopus, участвующих в управлении локомоторной активностью (634]. Б. Схема двух комплексов парных центров в системах зрительной и тактильной памяти у
Пищевые и тактильные сигналы
нее и с большим числом ошибок, чем в норме. Способность к обучению сокращается на 75% [566]. Весьма важен тот факт, что реакция, выработанная без вертикальной доли, сохраняется недолго — всего несколько часов, тогда как нормальная память длится дни или недели. Степень нарушения обучения пропорциональна массе ткани, удаленной из вертикальной доли.
От зрительных и нижних фронтальных долей идут пути к верхним фронтальным, оттуда к вертикальным, затем к субвертикальным и обратно к зрительным и нижним фронтальным.
Замкнутые контуры идут от зрительной и нижней фронтальных долей к верхней фронтальной, от нее к вертикальной, затем к субвертикальной и обратно к зрительной и нижней фронтальной
Центральная нервная система 123
долям. Разрыв этого круга сказывается на памяти. Янг предложил следующую гипотезу. В обоих сетевидных слоях происходит извлечение информации и уменьшение избыточности по двоичному коду (вертикальный и горизонтальный ряды). Вероятно, подобным же
/. 2 и 3 — три уровня управления нападением и отступлением в ответ на зрителньый стимул. Octopus [635].
образом нижняя фронтальная доля кодирует тактильные сигналы по признакам гладкий — шероховатый. Затем в верхней фронтальной доле происходит перераспределение и смешивание каналов; в вертикальной доле сильно возрастает число параллельных каналов; от нее передача в зрительную (или субфронтальную) долю идет по меньшему числу каналов, которые оканчиваются в этих Долях двоичным кодом — нападать или не нападать. Предполагается, что первоначальное кодирование идет в зрительной или субфронтальной доле и что там локализуется кратковременная Память; долговременная память сосредоточена в вертикальной до-ле-о Кодирование происходит в ориентированной, тонко расчлененной дендритной системе клеток, в которых каким-то образом осу
Я24 Глава 15
ществляются определенные отношения входа к выходу. Долговременная память локализуется в мощном нейропиле с очень большим числом каналов [611].
Членистоногие. Удаление сенсорных долей показывает, что у многих членистоногих нейропиль играет важную роль в «обучении». Наиболее яркие примеры этого можно найти у насекомых.
После удаления антенн или перерезки между антеннальной долей и альфа-долей грибовидных тел мозга муравей не способен к обучению в лабиринте на основе обонятельных сигналов. Обонятельный центр связан с моторными областями, но для замыкания условной связи, по-видимому, необходима нервная цепь, включающая альфа-долю [552].
В норме таракан очищает свою антенну от нанесенного на нее химического вещества, держа ее контралатеральной передней ногой; если обе передние ноги ампутированы, таракан за неделю научается пользоваться средними ногами, а между окологлоточной коннективой и моторным нервом задней ноги устанавливается путь с короткой синаптической задержкой, которая снижается с 4 до 2 мс [334, 335].
Простым препаратом для изучения процесса выработки «условного рефлекса» избегания электрического тока служит изолированный проторакальный ганглий таракана. Одна лапка получает электрический удар всякий раз, когда нога Р разогнута, а другая нога R получает равное число ударов в случайном порядке. После ряда сочетаний первая нога (Р) удерживается в согнутом положении; этот навык сохраняется до 24 ч. После выработки повышается частота тонических возбудительных импульсов, идущих от ганглия к мышцам ног; выработанная реакция может быть угашена электрической стимуляцией ганглия [121, 122, 211, 224].
Позвоночные. У рыб условные реакции могут замыкаться через средний мозг. Когда у серебряного карася выработали реакцию на зрительную стимуляцию одного глаза при закрытом втором, а затем испытали эту реакцию при открытом втором и закрытом первом глазах, то обнаружили перенос рефлекса на второй глаз; перерезка интертектальной комиссуры нарушила перенос с одного глаза на другой, показав таким образом, что приобретенная энграмма находилась в тектуме [346]. У серебряного карася сначала i был выработан условный рефлекс на зрительный стимул с пище- [ вым подкреплением, а затем в качестве УС второго порядка был Г применен обонятельный стимул (амилацетат) со зрительным сти- > мулом в качестве подкрепления. После удаления части тектума выработка рефлекса второго порядка была нарушена [482]. Пе- | ренос с одного глаза на другой был получен для условнорефлек- 1 торного изменения ритма сердца в ответ на световой УС; после повреждения тектума такой рефлекс не вырабатывался {355]. : Степень участия переднего мозга в условнорефлекторной деятель- | ности, по-видимому, у разных видов разная. У Tilapia удаление •
Центральная нервная система
125
переднего мозга после выработки условного рефлекса избегания на свет привело к замедлению и вариабельности рефлекторного ответа; удаление переднего мозга до выработки снижало способность к обучению [285]. В то же время серебряный карась, лишенный переднего мозга, обучался двухстороннему избеганию так же хорошо, как нормальный '[101]. Отсюда следует, что тектум (вероятно, вместе со связанным с ним тегментумом) способен обеспечить условнорефлекторное поведение.
У птиц кора большого мозга очень слабо развита; она расположена медиальнее и позади большого стриатума, который состоит из гиперстриатума и неостриатума. Птиц можно обучить решению сложных задач, например счету и запоминанию образов. Удаление коры не вносит изменений, но повреждение любой части стриатума заметно нарушает приобретенные навыки [514].
По поводу возможности выработки спинальных условных рефлексов у млекопитающих существуют утвердительные и отрицательные мнения; нет сомнения в том, что и для спинного мозга характерно привыкание и восстановление после него. Некоторые подкорковые образования обеспечивают выработку вегетативных условных рефлексов на звук или свет. После удаления зрительной коры у кошек удавалось выработать рефлексы на общее освещение, но не на фигуры. В последних работах подчеркивается роль префронтальной коры, гиппокам-па и височной доли в процессах памяти [4]. Между гиппокампом и субталамусом существуют реципрокные отношения. У крысы с поврежденным субталамусом нарушается обучение и отсутствуют типичные изменения электрической активности в гиппокампе, возникающие при приближении к цели [4]. Крысы с поврежденным гиппокампом или сводом обучаются в Т-образном лабиринте медленнее, чем нормальные; повреждение гиппокампа после выработки условного рефлекса нарушает его сохранность и возможность повторной выработки {271].
После перерезки мозолистого тела, соединяющего правое и левое полушария, у животного можно выработать зрительный условный рефлекс. Если выработка ведется при стимуляции одного глаза, а испытание рефлекса—-при стимуляции второго, рефлекс сохраняется у нормальных животных (кошек) и нарушается у животных с перерезанным мозолистым телом. Перерезка после выработки показывает, что след памяти переносится из одного полушария в другое только во время обучения [55]. Кроме того, на {85]|Не ПОДКОРКИ ПРОИСХ°ДИТ перекрест соматосенсорных сигналов
Коре больших полушарий свойственна некоторая эквипотенциальность функций. Хотя в зрительной коре имеется поточечная проекция сетчатки, выработанные реакции на зрительные стимулы нарушаются у крыс пропорционально объему удаленной коры и в значительной степени не зависят от локализации дефекта. Слухо
126
Глава 15
вое обучение еще возможно у собак при сохранности любой из трех слуховых областей. Такие наблюдения привели к представлению о «mass action» в коре больших полушарий [316]. Данные последних лет говорят о множестве корково-корковых связей, в особенности в престриарной области, и о том, что некоторые повреждения лобной коры необратимо нарушают ассоциативное обучение, в то время как способность к различению сохраняется. Кошка не научается различать последовательность световых и звуковых стимулов после удаления обеих латеральных фронтальных и префронтальных ассоциативных областей, но способна к такому обучению после повреждения сенсомоторной коры [261, 536]. Следовательно, существует как локализация, так и выполнение одной и той же функции несколькими областями, и некоторые области коры способны замещать другие.
Условными стимулами могут служить не только все сенсорные сигналы, но и электрическое раздражение мозга. У крыс вырабатывали сгибание лапы в ответ на контралатеральную стимуляцию головного мозга [476]. Самостимуляция через электроды, вживленные в гиппокамп, и стимуляция срединного пучка переднего мозга во многих опытах служат даже лучшим подкреплением, чем пища [400, 401]. Стимуляция разных частей коры большого мозга используется как условный стимул [108].
Автономные реакции, в которых обычно не используются корковые пути, но которые модифицируются корой (например, мигательные, зрачковые, кожногальванические рефлексы, ритм сердца и кровяное давление), можно выработать условнорефлекторным путем. Пока не ясно, в какой степени такая выработка происходит в подкорковых структурах. Различные автономные реакции были выработаны с использованием стимуляции медиального пучка переднего мозга в качестве БС (подкрепления) и определенного уровня активности, например частоты сокращений сердца или уровня кровяного давления — в качестве УС [371]. У кураризо-ванных крыс условнорефлекторное изменение этих функций идет так же хорошо или даже лучше, чем у нормальных животных, следовательно, для такой выработки не обязательны произвольные движения.
Стойкие синаптические явления
Посттетаническая потенциация (ПТП). Постсинаптическая реактивность повышается на несколько секунд после ритмической пресинаптической стимуляции; такое повышение наблюдалось во множестве нервно-мышечных и синаптических соединений. Оно возникает в кураризованных мышцах или в симпатических ганглиях, где возможно увеличение амплитуды синаптических потенциалов и квантованное выделение медиатора. В препарате диафрагмальный нерв — диафрагма опустошение депо медиатора и
Центральная нервная система 127
усиленное выделение последнего дают двухфазную кривую реакции. В нервно-мышечном соединении портняжной мышцы лягушки посттетаническая потенциация зависит от общего числа нервных импульсов; при уменьшении количества кальция во внешней среде ПТП наступает раньше и заканчивается скорее [454]. В симпатических ганглиях и при моносинаптических спинальных рефлексах посттетаническая потенциация длится от нескольких секунд до нескольких минут |[110]. в цилиарном ганглии у кур потенциация возникает при химической, но не при электрической передаче в синапсе [347]. В мышце ракообразных ритмическая стимуляция повышает частоту миниатюрных ВПСП.
При моносинаптических рефлексах ВПСП мотонейронов увеличены; полисинаптические рефлексы имеют большую амплитуду при испытании их после тетануса. После ортодромного тетануса рефлекс на растяжение достигает пика за 5—30 с. Тетанизация одного заднего корешка усиливает разряд полисинаптического рефлекса, вызываемого тестирующим залпом в том же самом или соседних задних корешках; антидромная тетанизация не оказывает такого действия. Наблюдается также посттетаническая потенциация торможения <[594].
Согласно одной из гипотез относительно ПТП, во время тета-низации пресинаптические окончания накапливают Са2+ и эффект пропорционален вероятности выхода медиатора [454]. По другому представлению ПТП является следствием гиперполяризации пресинаптических окончаний и тем самым следствием усиления синаптического тока {ПО, 557].
Привыкание и отвыкание; гетеросинаптическое облегчение. Электрофизиологические данные свидетельствуют о том, что во многих нервных центрах повторение сенсорных сигналов приводит к снижению центрального ответа. Сенсорная адаптация при этом исключается, так как привыкание наступает и при низкой частоте сигналов; оно наступает и при прямом раздражении сенсорных нервов. Точно так же исключаются снижение мышечной реактивности и центральное утомление, связанное с метаболической недостаточностью.
Скорость снижения ответа при привыкании является сложной функцией числа стимулов, частоты и интенсивности стимуляции. Если после привыкания прекратить сенсорную стимуляцию, то происходит постепенное спонтанное восстановление реактивности, й какое-нибудь общее раздражение, а также изменение локализации или параметров прежнего стимула (например, перемещение зрительного или тактильного стимула на другую точку) вызовет быстрое отвыкание; такое явление свидетельствует против того, что привыкание обусловлено утомлением.
• Привыкание у позвоночных. В спинном мозге привыкание сопровождается снижением амплитуды как возбудительных, так и тормозных синаптических потенциалов в соответствующих вста-
128
Глава 15
вочных нейронах, а также изменениями частоты импульсации этих нейронов. В полисинаптических рефлекторных дугах амплитуда ВПСП мотонейронов снижается при привыкании и повышается после отвыкания; в моносинаптических рефлексах привыкание почти или совсем отсутствует [506]. В спинном мозге кошки оно достигает максимума при афферентной стимуляции в течение 1 с с частотой 50 в 1 с. Импульсные разряды вставочных нейронов подвержены привыканию, а привыкание полисинаптического рефлекса может вызвать потенциацию моносинаптического рефлекса; следовательно, это не прямое воздействие на мотонейроны '[570]. Восстановление рефлекса после привыкания до половины нормальной амплитуды требует около 6 мин.
У морских миног Petromyzon сенсорные нейроны, связанные с рецепторами, реагирующими на прикосновение, давление или вредоносные стимулы, лежат в дорсальной части спинного мозга. Движение дорсального плавника вызывается рефлекторно раздражением .кожи, при повторении которого наступает привыкание. Соответственно снижаются ВПСП в мотонейронах; привыкание и отвыкание реализуются, очевидно, на уровне синапсов между сенсорными и моторными нейронами Д36].
Привыкание сенсорных вызванных потенциалов наблюдается, например, при тектальных ответах на зрительные стимулы. В одиночных нейронах тектума серебряного карася получено привыкание при многократных раздражениях и восстановление после электрического удара, нанесенного на туловище {398]. Другие нейроны привыкали к стимулу, который двигался в одной части поля зрения, а затем отвечали на тот же стимул в другой его части {525]. Вызванные потенциалы в коре больших полушарий снижаются при повторении; у ненаркотизированных животных вызванные потенциалы снижаются при отвлечении внимания животных. Это значит, что ответ на одну сенсорную модальность может быть подавлен стимулом другой модальности [69, 125]. Для вызванных ответов в первичной слуховой коре на тоны, предъявляемые с двухминутными интервалами, привыкание наступило после 30 предъявлений {293]. В корковой ЭЭГ в ответ на звуковую и световую стимуляцию возникает реакция пробуждения — десинхронизация (см. ранее). Повторение стимулов вызывает привыкание [495].
У высших позвоночных привыкание наступает главным образом в полисинаптических системах. Отвыкание — это не просто исчезновение привыкания, поскольку ответ может превысить исходный уровень; оно представляет собой облегчение, генерализованное по нескольким входным каналам.
Привыкание у беспозвоночных. Синаптическое привыкание наблюдалось в различных распределительных ганглиях. Неясно, идентично ли оно поведенческому привыканию, но по многим па’ раметрам то и другое сходны.
Центральная нервная система 129
В звездчатом ганглии кальмара ВПСП и генерация импульсов ослабляются при повторении 0,5-секундных серий пресинаптичес-ких импульсов через интервалы в 10—15 с; при интервалах в 25— 40 с этот эффект не возникает. После отдыха отвыкание происходит спонтанно [206]. В головном мозге саранчи в одиночных нейронах тритоцеребрума, которые отвечают на движение предметов в контралатеральном поле зрения, при многократной стимуляции наступает привыкание {463, 464]. Оно происходит даже в том случае, когда интервалы между стимулами составляют несколько минут; стимуляция шейных коннектив способна восстановить ответы нейронов тритоцеребрума {467, 468].
Рефлекторная флексия брюшка у рака подвержена привыканию; оно локализуется не в гигантских волокнах и не в мотонейронах, а в сенсорных вставочных нейронах [597, 618]. У рака привыкание — снижение потенциала нервно-мышечного соединения при низкочастотной стимуляции — происходит без снижения проводимости в мышечной мембране; следовательно, это процесс пре-синаптический [44].
Простым препаратом для исследования привыкания и отвыкания может служить нервная система моллюска. У брюхоногих и двустворчатых привыканию подвержено втягивание жабр и мантии в ответ на тактильное раздражение и закрывание створки в ответ на световое раздражение [422]. При внутриклеточном отведении активности крупных нейронов в ганглиях аплизии регистрируется привыкание — ослабление ВПСП при многократной низкочастотной стимуляции соответствующих сенсорных входов {45, 68] (рис. 15-39). Сопротивление постсинаптической мембраны при этом не меняется; следовательно, привыкание происходит, вероятно, на уровне пресинаптической части и выражается в снижении выхода медиатора {307, 308]. Привыкание отличается от длительного торможения (см. гл. 11). После привыкания к ограниченной стимуляции отвыкание происходит при сенсорной стимуляции иного или общего характера. Восстановление может принять форму либо гетеросинаптического облегчения, либо посттетанической по-тенциации [279]. Привыкание можно получить в препарате изолированного ганглия и связанного с ним участка кожи [308]; точно так же, оно возникает в изолированной жабре при многократных прикосновениях к ней; эффект этот длится 2 ч; таким образом, к привыканию способны как центральные ганглии, так и периферии ческие нервные сети [414].
Гетеросинаптическое облегчение. К описанным выше феноменам примыкает гетеросинаптическое облегчение, лучше всего изученное на ганглиях Aplysia. При воздействии на некоторые вставочные нейроны 9—10 сочетаний двух стимулов — одного, вызывающего небольшой ВПСП (тест-бтимул), и другого (priming) , следующего за ним и вызывающего спайки, — ответ на тест становится «условным» — ВПСП делаются больше. Точно так же 9—2067
130
Глава 15
идентифицированные гигантские нейроны усиливают свои ответы на стимуляцию одного входа при сочетании с тетанической стимуляцией разных других входов. Это облегчение длится около 10 мин и, по-видимому, по своей локализации является пресинап-тическим, поскольку такое воздействие на ВПСП может осуществляться и после постсинаптической блокады и поскольку сопротивление постсинаптической мембраны не изменяется. При
Рис. 15-39. Привыкание движения втягивания жабры (жирная линия) и соответствующие импульсы мотонейрона при повторной сенсорной стимуляции, длящей^ ся в течение указанного числа минут [636].
1 — исходные данные, 2 — после отдыха, 3 — после отвыкания.
этом вход может быть или не быть специфическим по отношению ко входу, вызывающему облегчение; эффект длится дольше, чем ПТП, получается с различных входов и не вызывается по колла--теральным обратным связям. Поведенческое гетеросинаптическое облегчение установлено для втягивания жабр и для других рефлексов [278, 281, 308].
Электрические корреляты выработки условного рефлекса
Выше было сказано об увеличении ВПСП в нейронах Aplysia при гетеросинаптическом облегчении и о выработанной десинхронизации альфа-волн у млекопитающих. В зрительной коре кошки после выработки условного рефлекса на зрительный стимул повы
Центральная нервная система 131
шалась амплитуда и укорачивался латентный период вызванных потенциалов [149]. У серебряного карася при выработке реакции на вспышки света появлялись в зрительном тектуме новые длиннолатентные вызванные ответы. Такой выработанный поздний потенциал был более чувствителен к охлаждению, чем первоначальный прямой ответ на свет; это свидетельствует об установлении новых связей посредством выработки [431]. У ненаркотизированных крыс в ответ на звуковые или соместетические стимулы в коре и тегментуме возникают вызванные потенциалы, которые увеличиваются при подкреплении этих стимулов электрическим раздражением коры или тегментума; для этих вызванных потенциалов характерно привыкание i[69]. После выработки УР частота разрядов одних корковых нейронов возрастает, а других — снижается [251, 252]. В некоторых частях головного мозга (главным образом в гиппокампе и, кроме того, в дорсальных и вентральных отделах ретикулярной формации) обнаружены прирост или снижение частоты импульсации нейронов после выработки реакции на тон; эти изменения были связаны с кратковременной памятью [400, 401]. Выработанные изменения импульсных разрядов и поведения происходят без видимых изменений ЭЭГ.
В моторной коре кошки при выработке УР уменьшались импульсные ответы на световые вспышки, тогда как предъявление УС и БС в случайном порядке приводило к небольшому учащению импульсации [399]. Прецентральные (сенсорные) нейроны давали учащение разрядов на световые стимулы при пищевом подкреплении [137]. После выработки УР в некоторых отделах головного мозга крыс возрастала частота импульсации в ответах на свет, применяемый в качестве УС [56]. В разных частях мозга реагирует разное число клеток, но в общем оно составляет меньше половины тестированных нейронов. Некоторые таламические нейроны вначале не отвечали на свет, но после сочетания света с раздражением седалищного нерва в качестве БС появились ответы на световые вспышки. Эти ответы поддаются учащению; у одних нейронов они выражались в ускорении, у других — в замедлении |[275]. Подобные же изменения частоты происходят при выработке реакций мотонейронов в ганглиях таракана [222].
Ранние стадии зрительного и слухового обучения, по-видимому, объясняются реакциями активации и десинхронизации альфа-ритма, вероятно связанными с неспецифическими путями главным образом в ретикулярной системе [259, 260, 381]. Можно вызвать снижение амплитуды альфа-волн, десинхронизацию и замену быстрыми волнами; сначала эти эффекты широко распространяются по коре, -а затем становятся более локальными. Условнорефлекторные изменения ЭЭГ не обязательно сопровождаются поведенческими коррелятами.
Помимо изменений вызванных потенциалов, ЭЭГ и частоты импульсации некоторых нейронов, зарегистрированы изменения по-9*
132
Глава 15
стоянных, или медленных, потенциалов (ПП), охватывающих широкие области коры больших полушарий. Эти очень медленные потенциалы имеют либо положительный, либо отрицательный знак и меняются в зависимости от состояния коры (например, при распространяющейся депрессии, создаваемой локальным нанесением КС1). На корковые медленные потенциалы можно воздействовать раздражением ретикулярной формации; сочетания звукового или светового стимула с электрическим раздражением ретикулярной формации приводят к медленным изменениям ответов на изолированный звук или свет [70, 168, 381, 469, 470]. Реактивность корковых нейронов меняется также от поляризации коры постоянным током [56]. Происхождение очень медленных потенциалов коры еще не ясно, но предположительно они представляют собой активность дендритов.
Многие электрические корреляты выработки УР идентифицированы с полной очевидностью. Являются ли даже изменения синаптических потенциалов прямыми показателями консолидации следов памяти — вопрос спорный.
Химические сдвиги при изменениях нейронной активности
Синтез специфической РНК и белка; чувствительность к медиаторам. По предположению Хидена [239], нейроны кодируют энграммы посредством специфических видов мРНК. Разработаны методы выделения и анализа отдельных нейронов из наиболее важных областей головного мозга крыс. Проведен анализ нейронов из ядра Дейтерса у крыс, обученных балансированию, и установлено, что отношение аденин/урацил в них повышено по сравнению с контрольными. В корковых нейронах было определено содержание РНК в связи с предпочитаемой лапой. Когда крыс заставляли действовать непредпочитаемой лапой, содержание РНК в корковых нейронах повышалось с 22 до 27 мкм/кг, а отношение Г+Ц/А+У снижалось. Отмечалось также повышение концентрации РНК в глии. У крыс, получивших 8-азагуанин, который блокирует синтез мРНК, обучение шло хуже. У серебряного карася, обученного плавать в ненормальной позе с привязанным поплавком, изменялось отношение У/Ц в РНК ткани мозга [496]. Был сделан вывод, что во время приобретения навыка образуется новая РНК, которая, возможно, кодирует определенные энграммы.
Путем введения блокирующих агентов были получены данные об участии синтеза белка в обучении. Введенный в гиппокамп мышей пуромицин препятствовал развитию краткосрочной памяти (меньше 9 ч) в Y-образном лабиринте, а при введении в новую кору — консолидации следов долговременной памяти {22, 143— 145, 442]. Ацетоксициклогексимид блокирует синтез белка эффективнее пуромицина, однако по имеющимся данным он мало влияет на память у крыс; в опытах на серебряном карасе этот инги
Центральная нервная система
133
битор оказался эффективным. Проведенные недавно исследования показывают, что инъекция ацетоксициклогексимида в гиппокамп крыс незадолго перед обучением в Y-образном лабиринте не вызывает никаких нарушений через 3 ч после обучения, но приводит к заметным нарушениям памяти через 1 и 7 дней. Введение препарата после обучения вообще не давало эффекта. Отсюда был сделан вывод, что кратковременная память не требует нового белка, а долговременная нуждается в нем [88].
У серебряного карася, обучаемого в челночном ящике, электрошок (ЭШ), вызываемый через 90 мин после обучения (или пу-ромицин, введенный через 30 мин), нарушал обучение; при охлаждении время, в течение которого ЭШ оказывался эффективным, увеличивалось [6, 92—94]. Действие пуромицина сказывается лишь при введении его не позднее чем через 3 ч после обучения; надо думать, следовательно, что именно на протяжении этого времени происходит консолидация. У серебряного карася ацетокси-циклогексимид, введенный немедленно после тренировки в ящике, блокирует и кратковременную, и долговременную память, а при введении через 6 ч или более подобного влияния не оказывает.
В приведенных выше опытах для контроля побочных эффектов производили инъекцию этого вещества в другие области тела. Эти и другие опыты приводят к заключению, что синтез РНК и белка необходим, но недостаточен для обучения и что синтез этих веществ зависит от состояния организма [453].
Другие химические изменения связаны с медиаторами, в частности с ацетилхолином. Крысам, обучавшимся в Y-образном лабиринте, вводили в разные сроки после обучения антихолинэсте-разы — ДФФ или физостигмин; такие крысы в течение 14 дней выполняли задачи лучше контрольных. Кроме того, инъекция ацетилхолиноподобного вещества карбахола блокировала навык, приобретенный за 7 дней до этого. Предполагается, что на определенный период консолидации постсинаптическая мембрана становится более чувствительной к ацетилхолину, после чего ее чувствительность снижается [100]. В ганглиях таракана после обучения можно обнаружить снижение содержания ацетилхолинэстеразы и глутаматдекарбоксилазы {402].
Все указанные химические изменения сопутствуют процессу обучения; однако участие их в механизмах хранения следов памяти не доказано. Эти изменения, возможно, отражают воздействие на активацию, внимание и двигательные акты. Химические преобразования, быть может, лежат также в основне изменения свойств синаптической мембраны.
Структурные изменения
Одна из гипотез о механизме обучения была предложена давно, но не получила твердого обоснования. Согласно этой гипотезе, между нейронами образуются новые связи. В начале раз
134
Глава 15
вития нейронов направление их роста точно определено, но на более поздних стадиях это почти или совсем (в зависимости от вида животного) не так (см. выше). Перерезка аксона стимулирует синтез белка в теле клетки, образование большого количества новой РНК и некоторую реорганизацию отростков. В ганглиях таракана каждый нейрон обладает своей характерной формой дендритного древа; после перерезки аксона или введения колхицина кольцо РНК вокруг ядра увеличивается до максимума, а затем на 10-й день исчезает *[77]. Тело нейрона в норме не генерирует импульсов при деполяризации или антидромной стимуляции, но в стадии кольца РНК оно продуцирует высокие спайки. Таким образом, между состоянием регенерации и возбудимостью существует тесная связь [423].
Если детенышей млекопитающих выращивают в «обогащенной» среде — среди освещенных фигур, игрушек, звуков, в обществе других особей, — у них заметно увеличиваются толщина и вес коры большого мозга (особенно зрительной коры) по сравнению с животными, выращенными в «обедненной» среде. На дендритах пирамидных клеток больше шипиков, поперечное сечение синапсов увеличено на 50%, а число их невелико; общее содержание ацетилхолинэстеразы повышается, но ее концентрация на 1 г коры падает. У крыс, выращенных под открытым небом, в условиях, близких к естественным, кора-была развита даже сильнее, чем у крыс, содержавшихся в клетках, в «обогащенной» среде [28, 455, 456]. Выше упоминалось об опытах с выключением зрения, а зрительная стимуляция сама по себе является частью обогащенной среды.
Упражнение и неупражнение нейронов приводят к изменениям в количестве синаптических пузырьков. Возможно, что синапсы сохраняют способность к структурным изменениям, несмотря даже на то, что нейрон как целое относительно стабилен [81]. Выше было описано, как меняются свойства зрительных нейронов в коре кошки в зависимости от опыта, приобретаемого в процессе развития.
Итак, в различных условиях деятельности в нейронах могут происходить стойкие структурные изменения. Связаны ли они с обучением, пока не ясно.
Обзор данных о развитии и стойких изменениях
Сравнительное изучение нервной системы показывает несостоятельность попыток распространить одну какую-нибудь гипотезу о нервной пластичности на все случаи обучения. Ниже перечисляются некоторые установленные стабильные нейрональные феномены.
Нейроны представляют собой клетки, химически весьма активные. Химические поля так направляют растущие нейроны, чтобы
Центральная нервная система 135
они могли образовать генетически детерминированные связи. Во всех нейронах происходит активный синтез липопротеидов мембран, а также медиаторов и перенос сложных молекул, в частности белков, током протоплазмы от тела клетки к терминалям отростков; вся эта метаболическая активность регулируется ядром. Химическая активность нервных клеток изменяется под влиянием перерезки отростков, а также при «упражнении и неупражне-нии» функции проведения импульса. Кроме того, при «упражнении» у некоторых нейронов увеличиваются их дендритное ветвление и синаптическая структура. Таким образом, рост, разного рода биохимические синтетические процессы и структурные связи изменяются под влиянием опыта.
В пределах более сжатой временной шкалы изменениям подвержена синаптическая передача. Посттетаническая потенциация, возможно, отражает поглощение кальция или пресинаптическую гиперполяризацию и тем самым повышенную пресинаптическую дееспособность. Привыкание и отвыкание, по-видимому, представляют собой пресинаптические процессы, происходящие во вставочных нейронах. Гетеросинаптическое облегчение может быть следствием измененного баланса между тормозными и облегчающими окончаниями. Выработка условного рефлекса, возможно, происходит отчасти посредством модуляции врожденной ритмической активности. Эти изменения возникают до некоторой степени и в относительно простых нервных системах. Очевидно, любая нервная система (нервные сети, ганглии и корковые структуры) способна видоизменяться под влиянием опыта.
Но все же представляется сомнительным, чтобы процессами, возникающими в результате роста и дегенерации или в результате изменений синаптической передачи в простых моносинаптических или полисинаптических цепях, можно было бы объяснить обучение в сложных нервных системах головоногих моллюсков и позвоночных. В таких нервных системах удается различить кратковременное и долговременное обучение. Кроме того, они обладают иерархией параллельных путей, ib которой те пути, что лежат ближе всего к сенсорным входам, возможно, служат для кратковременного обучения, а те, что лежат дальше — для долговременной памяти и команд. Аналогия между обучением, и кодированием сенсорных сигналов, хранением кода, как на магнитной пленке, и его извлечением по мере надобности представляет собой чрезмерное упрощение. Ведется живая полемика по вопросу о том, осуществляются ли «высшие» функции в головном мозге позвоночных целиком посредством нервных импульсов и синаптической передачи или же в интегративной деятельности играют роль также и эффекты поля. Существуют некоторая эквипотенциальность между отдельными областями мозга и значительная синхронизация электрических ответов разных зон; имеются также данные о коммуникациях между дендритами в корковых структурах.
136
Глава 15
Установлено много явлений, сопутствующих выработке УР„ Некоторые его типы, по-видимому, связаны с синтезом белка и кодируются РНК. Обнаружены изменения чувствительности к медиаторам. Некоторые постсинаптические мембраны неоднородны, во всяком случае в отношении возбуждения и торможения, и один из типов кодирования, возможно, осуществляется посредством изменения на мембранах относительного числа участков с разной постсинаптической чувствительностью. Для сложных нервных систем статистический подход представляется более перспективным^ чем анализ, основанный целиком на специфических свойствах отдельных нейронов.
Заключение
За последние годы достигнуты серьезные успехи в понимании механизмов проведения нервных импульсов и синаптической передачи (гл. 11). В какой мере нейробиология достигает своей главной цели — познания механизмов поведения животных? Нервные системы и субсистемы образуют ряд уровней интеграции, непрерывный переход от простого к сложному, в котором, однако, можно различить эти разные уровни.
Наиболее простые нервные центры характеризуются отношениями один к одному обычно с конвергенцией или дивергенцией входов и выходов. В одних таких соединениях осуществляется электрическая передача, в других — химическая. Примером служат системы гигантских волокон, которые обеспечивают быстрые поведенческие реакции, часто сопровождающиеся синхронной активностью обширных мышечных зон. Передача в отношении один к одному происходит от сенсорного нейрона- к гигантскому волокну, между гигантскими волокнами и от гигантского волокна к мотонейронам. В других нервных центрах электрические соединения между вставочными нейронами создают значительную синхронность двигательной активности (например, в центрах, управляющих разрядом электрических органов). В химических синапсах распределительных центров устойчивые эффекты могут возникать в форме посттетанической потенциации. Выделение медиатора запускается или дискретными импульсами, или градуальными потенциалами, и постсинаптический эффект обычно является деполяризационным (но может быть и гиперполяризационным).
Простой уровень интеграции в противоположность передаче в отношении один к одному представляет собой такую активность центральной нервной системы, при которой выход не равен и не прямо пропорционален входу, а дискретные входные импульсы (подчиняющиеся закону «все или ничего») преобразуются в аналоговые (градуальные) сигналы, сортируются и преобразуются обратно в импульсные сигналы. Выход может быть усилен облегчением или ослаблен торможением; иными словами, возбуждающие
Центральная нервная система
137
входы суммируются или умножаются, а тормозные входы либо ослабляют действие возбуждающего (пресинаптическое торможение), либо снижают реактивность нейронов на выходе (постсинап--гическое торможение). Кроме того, возможны разряд последействия, модуляция спонтанной активности и послетормозная отдача. Соединения, лежащие на облегчающе-тормозном уровне интеграции, подвержены некоторым длительным изменениям, например привыканию и отвыканию. Привыкание наступает в моносинапти-ческих путях в некоторых нейронах аплизии, где это явление, по всей видимости, объясняется снижением пресинаптической активности. Привыкание происходит также во вставочных нейронах по-лисинаптических путей спинного мозга позвоночных. Этот облег-чающе-тормозный уровень интеграции наблюдается в самых примитивных нервных системах — нервных сетях — и сохраняется <в субсистемах любой центральной нервной системы. Он подробно описан для спинальных рефлексов позвоночных и для локомоторных ответов ганглиозных цепочек многих беспозвоночных. Обычно этот уровень интеграции осуществляется на основе химических синапсов.
Следующий уровень интегративного контроля принимает форму выбора из множества возможных (входов и выходов и их включения. Это происходит или а) по запрограммированным связям в центре, или б) посредством команд со стороны «высших», или специализированных, командных нейронов. Нейрональные программы определяются генетически и в процессе развития, и если какая-то последовательность запущена соответствующим входом, она считывается до конца. Примерами моторных программ могут служить совокупности движений при полете у насекомых или плавании у рыб. Близкий к этому способ кодирования осуществляется в сенсорных вставочных нейронах, дающих специализированные ответы (например, слуховые вставочные нейроны некоторых насекомых), или в нейронах, отвечающих на специфические компоненты сенсорного входа (например, в зрительных вставочных нейронах рака, или вставочных нейронах тектума у рыб, или латерального коленчатого тела у млекопитающих). Моторный выход большинства таких программированных систем состоит из ритмических разрядов; временные параметры этих колебаний заложены или 1) (в сети, как, например, в дыхательном центре, или 2) в одном ритмически активном нейроне, как при ритмической регуляции движений некоторых придатков у ракообразных (плавательных конечностей). Командные нейроны могут находиться в центре с запрограммированной активностью и подвергаться воздействию разнообразных входов. Еще не ясно, каким образом эти нейроны принимают «решения». Однако именно в отношении этого уровня нейрофизиология подошла ближе всего к полному описанию последовательности некоторых поведенческих актов (например, начала, продолжения и окончания движения плавательных конеч
138
Глава 15
ностей у десятиногих раков, полета саранчи и ходьбы у тараканов). В этих процессах может быть определена роль каждого нейрона.
Следующий уровень интеграции состоит в модуляции активности рефлекторного центра со стороны уровней «высшего контроля». Механизмы такого рода контроля изучены плохо, но обычно он осуществляется через те области, стимуляция которых не вызывает никакой видимой реакции. Так, например, области надглоточного ганглия у членистоногих оказывают тормозное действие на грудные двигательные центры. У позвоночных мозжечок контролирует сообщения, идущие от органов чувств в средний и передний мозг, а также выходные сигналы, идущие в двигательные области ствола мозга и спинного мозга. В мозге осьминога несколько уровней контроля организованы по иерархии, от простых реакций до сложных решений о нападении или убегании. С регулирующими функциями связаны те области нервной системы, которые задают программы для низших центров. У сверчка определенные участки грибовидных тел в «головном мозге» запускают специфические последовательности звуковых сигналов, составляющие их песню. Моторная кора у млекопитающих и моторные ядра ретикулярной формации каким-то неизвестным образом запускают локомоцию.
На уровне, который, возможно, является высшим уровнем интеграции, можно выделить функции двух видов, хотя ни один из них еще не изучен. Функции первого вида связаны с поведением, которое служит для сохранения особи, когда она подвергается вредному воздействию или стрессу, а также для продолжения рода. У позвоночных эти функции локализуются в значительной степени во всей лимбической системе — в комплексе, состоящем из ретикулярной формации, гипоталамуса и несенсорного примитивного переднего мозга, иногда называемого «эмоциональным мозгом». Второе свойство высшего уровня нейрональной организации — это изменчивость, связывание одного входа с другим, как при выработке условного рефлекса, сохранение приобретенной системы. и извлечение ее из памяти в случае надобности. Связывание нескольких входов, которое создает длительно сохраняющуюся способность к реакции, может происходить в некоторых ганглионарных системах (например, у головоногих моллюсков), а выработка условного рефлекса возможна у позвоночных в среднем мозге и на уровне промежуточного мозга. Вероятно, существует множество разных механизмов условнорефлекторной деятельности, но свойства инструментального обучения и извлечения следов долговременной памяти еще не описаны в строгих нейрофизиологических понятиях. Электрические явления, сопутствующие выработке условного рефлекса, выражаются в понижении порога импульсации, пресинаптическом привыкании и изменениях эндогенной ритмической активности. Химические и структурные процессы, сопут
Центральная нервная система
139
ствующие выработке, состоят в изменениях дендритного древа по мере приобретения опыта и в стимулировании обмена белков и РНК в связи с упражнением и неупражнением (и, возможно, в связи с выработкой условного рефлекса). Однако эти сопутствующие явления не объясняют возникновения измененных систем или энграмм.
Помимо установления иерархии и описания поведения на основе функции отдельных нейронов на промежуточных уровнях организации центральной нервной системы, сравнительная нейрофизиология помогает разрешить противоречие между унитарным и холистическим характером нервной интеграции [52]. Содержание информации в идентифицированных нейронах поразительно велико. В ганглиях некоторых брюхоногих моллюсков каждый из 30 известных нейронов характеризуется своими собственными соотношениями входов и выходов. Одни нейроны получают сенсорный вход от обширных рецептивных зон; другие, вставочные, нейроны тормозят одни мотонейроны и возбуждают другие или же обладают двояким действием; третьи клетки при стимуляции вызывают -сложные движения с участием многих мышц. Индивидуальные функции установлены также для сенсорных и моторных вставочных нейронов членистоногих. У позвоночных благодаря интенсивному исследованию отдельных нейронов, особенно в сенсорных системах, становится очевидной неповторимость функций отдельных клеток независимо от того, производится ли передача импульсами по закону «все или ничего» или же градуальными потенциалами. Согласно одной точке зрения, если бы было известно информационное значение каждого нейрона, нервная интеграция была бы понята. Пока еще нет возможности распространить это представление на области с мелкими нервными клетками; даже в ганглиях брюхоногих моллюсков в настоящее время удается идентифицировать меньше 1 % нейронов. В корковых структурах позвоночных — тектуме, мозжечке, коре большого мозга — синаптические потенциалы и мембранные осцилляции часто синхронизованы на больших участках. Эти «потенциалы поля» отражают входную активность корковых областей; их можно модифицировать и связать -с вероятностью импульсации отдельных нейронов. Таким образом, согласно второй точке зрения, нервная интеграция в «высших» субсистемах позвоночных осуществляется частично посредством потенциалов поля. Согласно третьей точке зрения, интегративные свойства создаются цепями, сетями нейронных связей. Получено много данных в пользу информационного значения соответствующих связей и сведений о том, что на связи, образуемые растущим нейроном, влияют его входы и выходы. Математические модели показывают, что сети обладают свойствами, которых лишены составляющие их компоненты. Следует заключить, что некоторые Целлюлярные механизмы являются общими для всех нейронов, что реакции отдельных клеток и групп нейронов обладают специфич
140
Глава 15
ностью и что каждый уровень организации построен на предыдущих уровнях, и в то же время на нем добавляются новые свойства, присущие только ему.
СПИСОК ЛИТЕРАТУРЫ
1. Abbie A. A., J. Comp. Neurol. 72: 469—488, 1940. Excitable cortex in marsupials.
2. Abbie A. A., Adey W. R., J. Comp. Neurol. 92: 241—292, 1950. Motor mechanisms of anuran brain.
3. Abeles M., Goldstein M. H., J. Neurophysiol. 33: 172—197, 1970. Organization of cat auditory cortex.
4. Adey W. R., pp. 233—276 in Brain Function II, edited by M. Brazier, University of California Press, 1964. Hippocampal mechanisms in processes of memory.
5. Adey W. R., pp. 224—243 in The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. Electrical brain rhythms accompanying learned responses.
6. Agranoff B. W., Davis R. E., Brink J. J., Proc. Nat. Acad. Sci. 54: 788—793,. 1965; also Brain Res. 1: 303—309, 1966. Protein synthesis and learning.
7. Akert R., Helvet. Physiol. Acta 7: 112—134, 1949. Motor representation on tectum, fish.
8. Allison T., Groff W. R., Psychophysiology 5: 200—201, 1968. Sleep in
echidna.
9. Allison T., Van Twyver H., Exp. Neurol. 27: 564—578, 1970. Sleep in moles.
10. Andersen P., Eccles J. C., Nature 196: 645—647, 1963. Inhibitory phasing of neuronal discharge.
11. Andersen P., Eccles J. C., Loyning Y., Nature 198: 541—542, 1963. Recurrent inhibition in hippocampus.
12. Andersen P., Eccles J. C„ Voorhoeve P. E., J. Neurophysiol. 27: 1138—1153^ 1964. Postsynaptic inhibition of Purkinje cells.
13. Arechiga H., Wiersma C. A. G., J. Neurobiol. 1: 71—85, 1969. Circadian rhythm of crayfish visual units.
14. Armstrong P. B., Higgins D. C., J. Comp. Neurol. 143: 371—384, 1971. Behavior in bullhead embryo.
15. Arvanitaki A., Chalazonitis N., Bull. Inst. Oceanogr. Monaco, No. 1164: 1—83,. 1960; No. 1224: 1—15, 1961. Rhythmic synaptic potentials, Aplysia neurons.
16. Arvanitaki A., Chalazonitis N., pp. 194—231 in Nervous Inhibition, edited by E. Florey. Pergamon Press, New York, 1961. Photostimulation of Aplysia neurons.
17. Atema J., Brain Behav. Evol. 4: 273—294, 1971. Sense of taste in cat-fish ictalurus.
18. Atwood H. L., Wiersma C. A. G., J. Exp. Biol. 46: 249—261, 1967. Command interneurons in the crayfish central nervous system.
18a. Auerbach A. A., Bennett M. J. V., J. Gen. Physiol. 53: 183—239, 1969. Mauth-ner cell giant neuron system in hatchet fish.
19. Азарян А. Г., Тищенко В. П., Роль нейронов мозга в регуляции циркадного» ритма поведения сверчка Gryllus domesticus L. ДАН СССР, 186, 464—466,. 1969.
20. Bailey D. Т., Laverack М. S., J. Exp. Biol. 44: 131—148, 1966. Neurophysiology of Buccinium.
21. Barnes W. J. P., Horridge G. A., J. Exp. Biol. 50: 651—671, 1969. Eyecup movements in crab.
22. Barondes S. H., Cohen H. D., Brain Res., 4: 44—51, 1967. Effects of cycloheximide and puromycin on cerebral protein synthesis and consolidation of memory.
Центральная нервная система
141
23. Barth Н., Z. wiss. Zool. 167: 238—290, 1962. Comparative anatomy of sensory centers in fishes.
24. Batham E. J., Pantin C. F., Robson E. A., Quart. J. Mier. Sei. 101: 487—510, 1960. Nerve net of sea anemone Metridium, mesenteries and the column.
25. Becker H. W., Experientia 21: 719, 1965. The number of neurons in an insect ganglion.
26. Belekhova M. G., Kosareva A. A., Brain Behav. Evol. 4: 337—375, 1971. Organization of the turtle thalamus.
27. Bell С. C., Kawasaki T., J. Neurophysiol. 35: 155—169, 1972. Responses of Purkinje cells to climbing fiber input.
28. Bennett E. L. et al., Science 146: 610—617, 1964. Chemical and anatomical plasticity of brain.
29. Bennett M. J. V., Steinbach A., pp. 207—214 in Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association, Chicago, 1969. Cerebellum of electric fish.
30. Bentley D. R., Z. vergl. Physiol. 62: 267—283, 1969. Neuronal activity and song patterns, cricket.
31. Bentley D. R., J. Insect Physiol. 15: 677—699, 1969. Activity of cricket neurons during behavior patterns.
32. Bentley D. R., Kutsch W., J. Exp. Biol. 45: 151—164, 1966. Neuromuscular mechanism of stridulation in crickets.
33. Bernstein J. J., Exp. Neurol. 3: 1—17, 1961. Loss of hue discrimination in forebrain-ablated fish.
34. Bernstein J. J., pp. 2—90 in Fish Physiology, vol. 4, edited by W. S. Hoar and D. J. Randall Academic Press, New York, 1970. Review of fish central nervous systems.
35. Binstock L., Goldman L., J. Gen. Physiol. 54 : 730—764, 1969. Myxicola giant axons.
36. Birnberger K. R-, Rovainen С. M., J. Neurophysiol. 34: 983—989, 1971. Habituating fin reflex in sea lamprey.
37. Blakemore C., Pettigrew J. D., Nature 225: 426—429, 1970; also Blakemore С.» Cooper G. F., Nature 228: 477—478, 1970. Eye dominance in visual cor-tex.
38. Bloom T. E., Hoffer B. J., Siggins G. A., Brain Res. 25: 501—521; 523—534; 535—553, 1971. Norepinephrine-containing afferents to Purkinje cells.
39. Bodian D., Bull. Johns Hopkins Hosp. 119: 129—149, 1966. Embryonic development of spinal cord in monkey.
40. Bodian D., pp. 129—140 in The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. A model of synaptic and behavioral ontogeny.
41. Boycott В. B„ Proc. Roy. Soc. Lond. B„ 153: 503—534, 1961. Functional organization of brain of cuttlefish.
42. Braitenberg V., Kybernetik 2: 248—287, 1965. Perception of movement by' the frog.
43. Brookhart J. M., Fadiga E., J. Physiol. 150: 633—635, 1960. Potential fields in frog spinal cord.
44. Bruner J., Kennedy D., Science 169: 92—94, 1970. Habituation in crayfish neuromuscular junction.
45. Bruner J., Tauc L„ J. Physiol. Paris 57: 230—231, 1965; also Nature 210: 37—39, 1966. Habituation at synaptic level, Aplysia.
46. Bryant S. H., J. Gen. Physiol. 41: 473—484, 1958; 42: 609—619, 1959. Transmission in squid giant synapses.
47. Buck J., Case J. F., Biol. Bull., 121: 234—256, 1961. Control of flashing in fireflies.
48. Buck J., Case J. F„ Hanson J. F., Biol. Bull. 125: 251—269, 1963. Peripheral excitation of photic organ.
49. Buck J.., Buck E., Science 159: 1319—1327, 1968. Rhythmic synchronous flashing of fireflies.
142
Глава 15
50. Bullock Т. Н., J. Neurophysiol. 8: 55—72, 1945. Function of giant fibers in Lutnbricus.
51. Bullock T. H., Amer. Zool. 5: 545—562, 1965. Conduction systems in echinoids and asteroids.
52. Bullock T. H., Horridge G. A., Structure and Function in the Nervous Systems of Invertebrates. W. H. Freeman, San Francisco, 1965, 798 p.
53. Bullock T. H., Ridgway S. H., Suga N., Z. vergl. Physiol. 74: 372—387, 1971. Midbrain responses to sound in sea lion.
54. Bullock T. H. et al., Z. vergl. Physiol. 59: 117—156, 1968. Central auditory responses in cetaceans.
55. Bures J., Buresova O., pp. 211—238 in Neural Control of Behavior, edited by R. E. Whalen et al. Academic Press, New York, 1970. The reunified split brain.
56. Bures J., Buresova O., pp. 363—403 in Short-Term Changes in Neural Activity and Behaviour, edited by G. Horn and R. A. Hinde, Cambridge University Press, 1970. Plasticity in single neurons.
57. Burns B. D., Salmoiraghi G. C., J. Neurophysiol. 23: 27—46, 1960. Patterns of rhythmicity in single respiratory neurons of cat.
58. Burrows M., Horridge G. A., J. Exp. Biol. 49: 223—250; 285—297, 1968. Optokinetic movements in Carcinus.
59. Buser P., Arch. Sci. Physiol. 3: 471—487, 1949; also C. R. Soc. Biol. (Paris) 143: 817—819, 1949; also J. Physiol. (Paris) 43: 673—677, 1951, Responses of optic tectum, catfish.
60. Bush B. M. H., Comp. Biochem. Physiol. 10: 273—290, 1963; 15: 567—587, 1965. Reflexes elicited by chordotonal organs in crabs.
61. Butler A. B„ Northcutt R. G., Brain Res. 26: 1—13, 1971. Retinal projections in Iguana and Anolis.
62. Cabral R. J., Johnson J. I., J. Comp. Neurol. 141: 17—36, 1971. Mechanore-ceptive projections in thalamus of sheep.
63. Callee J. J., Boistel J., C. R. Soc. Biol. (Paris) 160: 1943—1947, 2418—2424, 1966. Rhythmic synaptic potentials, abdominal ganglia, Periplaneta.
64. Callee J. J. et al., J. Exp. Biol. 55: 123—149, 1971. Synaptic transmission in cockroach.
65. Camhi J. M., J. Exp. Biol. 50: 335—348, 349—362, 363—373, 1969. Locust wind receptors.
66. Campbell С. B. G., Brain Behav. Evol. 2: 415—430, 1969. Optic systems of teleost.
67. Case J. F., Buck J., Biol. Bull. 125: 234—250, 1963. Central nervous aspects of flashing in fireflies.
68. Castellucci V. et al., Science 167: 1745—1748, 1970. Synaptic habituation in Aplysia.
69. Caviedes E., Bures J., Brain Res. 19: 249—621, 1970. Habituation of evoked responses in rat cortex and tegmentum.
70. Chow K. L., J. Neurophysiol. 24: 377—390, 1961, Brain electropotentials during visual discrimination learning in monkey.
71. Chow K. L. et al., Brain Res. 33: 337—357, 1971. Receptive field characteristics of striate cortical neurons in rabbit.
72. Clark R. B., Soc. Exp. Biol. Symp. 20: 345—379, 1966. Integrative action, worm brain.
73. Coggeshall R. E., J. Neurophysiol. 30: 1263—1287, 1967. Light and electron microscope study of abdominal ganglion, Aplysia.
74. Coggeshall R. E. et al., J. Cell Biol. 31: 363—368, 1966. Neuronal map, Aplysia ganglion.
75. Coghill G. E., Anatomy and the Problem of Behavior. Macmillan Co., New York, 1929, 113 p.
76. Cohen M. J., Jacklet J. W., Phil. Trans. Roy. Soc. Lond. В 252: 561—579, 1967. Organization of motorneurons in insect ganglia.
77. Cohen M. J., pp. 798—812 in the Neurosciences: Second Study Program, edited by F. O. Schmitt, Rockefeller University Press, New York, 1970. A comparison of invertebrate and vertebrate central neurons.
Центральная нервная система 143
78. Collett Т„ J. Neurophysiol. 33: 239—256, 1970, Visual cells, medulla of sphinx and noctuid insects.
79. Collier H. O., J. Exp. Biol. 16: 286—299, 300—312, 1939. Central reflexes of earthworms.
80. Coulombre A. J., pp. 108—116 in the The Neurosciences: Second Study Program, edites by F. O. Schmitt. Rockefeller University Press, New York,. 1970. Development of the vertebrate motor system.
81. Cragg B. G., pp. 1—60 in Structure and Function of Nervous Tissue, vol. 4, edited by G. Bourne. Academic Press, New York, 1972. Plasticity of synapses..
82. Crain S. M., Bornstein M. B., Peterson E. R., Brain Res. 8: 363—372, 1968. Tissue cultures of spinal cord.
83. Creed R. S. et al., Reflex Activity of the Spinal Cord. Oxford University Press, 1938. 183 p.
84. Crossland W., Ph. D. Thesis, University of Illinois, 1971. Visual responses in brain of pigeon.
85. Cuenod M„ pp. 455—506 in Structure and Function of Nervous Tissue, vol. 5, edited by G. Bourne. Academic Press, New York, 1972. Split-brain studies.
86. Curtis D. R., Felix D., Brain Res. 34: 301—321, 1971. Synaptic inhibition in cerebral and cerebellar cortex.
87. Dagan D., Parnas I., J. Exp. Biol., 52: 313—324, 1970. Giant fiber and small fiber pathways involved in the evasive response of the cockroach.
88. Daniels D., J. Comp. Physiol. Psychol. 76: 110—118, 1971. Effects of aceto* xycycloheximide on conditioning in rats.
89. Darian-Stnith I. et al., J. Physiol. 182: 671—689, 1966. Somatic sensory cortical projection areas excited by tactile stimulation of the cat.
90. Darian-Stnith I. et al., Science 160: 791—794, 1968. Somatosensory pathways in mammalian brain.
91. Davenport D., Bull. Inst. Oceanogr. Monaco, No. 1236: 1—24, 1962. Responses of tentacles of actinians to electrical stimulation.
92. Davis R. E., J. Comp. Physiol. Psychol. 65: 72—78, 1968. Environmental control of memory fixation goldfish.
93. Davis R. E., Agranoff B. W., Proc. Nat. Acad. Sci. 55: 555—589, 1966. Stages of memory formation in goldfish: evidence for an environmental trigger.
94. Davis R. E., Bright P. J., Agranoff B. W., J. Comp. Physiol. Psychol. 60: 162—166, 1965. Effect of ECS and puromycin on memory in fish.
95. Davis W. J., J. Exp. Zool. 168: 363—378, 1968. Neuromuscular basis of lobster swimmeret beating.
96. Davis W. J., J. Exp. Biol. 50: 99—117, 1969; 51: 547—574, 1969. Reflex organization, lobster swimmeret.
97. Davis W. J., Kennedy D., J. Neurophysiol. 35: 1—12, 1972. Command interneurons controlling swimmerets of lobster.
98. Delcomyn F., J. Exp. Biol. 54: 443—452, 453—469, 1971. Cockroach locomotion.
99. Dethier V. G., Gelperin A., J. Exp. Biol. 47: 191—200, 1967. Hyperphagia in the blowfly.
100. Deutsch J. A., Science 128: 288—294, 1971. Cholinergic synapse and the site of memory.
101. Dewsbury D. A., Bernstein J. J., Exp. Neurol. 23: 445—456, 1967. Effects of forebrain lesions in goldfish.
101a. Dewson J. H„ J. Neurophysiol. 30: 817—832, 1967; 31: 122—130, 1967. Effects of efferent fibers on auditory responses, monkey.
102. Diamond I. T., Hall W. C., Science 164: 251—262, 1969. Evolution of neocortex.
103. Diamond J., Nature 199: 773—775, 1963; also J. Physiol. 194 : 669—723, 1968. Sensitivity to gamma-aminobutyric acid of different regions of Mauthner neuron.
104. Diamond I., pp. 265—346 in Fish Physiology, vol. 5, edited by W. S. Hoar, Randall D. J., Academic Press, New York, 1970, The Mauthner cell.
144
Глава 15
105. Dingle Н., Biol. Bull. 121: 117—128, 1961. Flight and swimming reflexes in giant water bugs. Lethocerus and Benacus.
106. Dorsett D. A., J. Exp. Biol. 46: 137—151, 1967. Giant neurons and axon pathways in brain of Tritonia.
107. Dorsett D. A., J. Exp. Biol. 48: 127—140, 1968. Pedal neurons of Aplysia punctata.
108. Doty R. IF., The Physiologist 5: 270—284, 1962. Behavioral aspects of neurophysiology.
109. Doty R. W, pp. 1861—1902 in Handbook of Physiology, Vol. 4, Sec. 6, edited by C. F. Code. Amer. Physiol. Soc., Washington, D. C., 1968. Neuronal organization of deglutition.
110. Eccles J. C., The Physiology of Synapses. Springer-Verlag, Berlin, 1964. 316 p. (Экклс Дж., Физиология синапсов, Изд-во «Мир», 1966.)
ill. Eccles J. С., рр. 408—427 in The Neurosciences: A Study Program, edited by G. C. Quarton et al. Rockefeller University Press, New York, 1967. Post-synaptic inhibition.
112. Eccles J. C., Naturwissenschaften 56: 525—534, 1969. Comparative physiology of cerebellum.
113. Eccles J. C., Brain Res. 15 : 267—271, 1969. Cerebro-cerebellar connections in mammals.
114. Eccles J. C., Ito M., Szentagothai J., The Cerebellum as a Neuronal Machine. Springer-Verlag, Berlin, 1967, 335 p.
115. Eccles J. C., Taborekova H., Tsukahara N., Brain Res. 17: 87—102, 1970. Responses of granule cells of elasmobranch cerebellum.
116. Eccles J. C. et al., Exp. Brain Res. 13: 15—35, 1971. Mossy fiber input to granule cells.
117. Edwards J. S., Sahota T. S., J. Exp. Zool. 166: 387—396, 1967. Regeneration in sensory system of cricket Acheta.
118. Egger M. D., Wyman R. J., J. Physiol. 202: 501—516, 1969. Reflex stepping in cat.
119. Egger M. D., Flynn J. P., Science 136: 43—44, 1962. Effect of amygdaloid on hypothalamus stimulation.
120. Eidelberg E., J. Neurophysiol. 24: 521—533, 1961. Hippocampal dendritic responses in rabbits.
121. Eisenstein E. M., Cohen M. J., Anim. Behav. 13: 104—108, 1965. Learning in an isolated prothoracic insect ganglion.
122. Eisenstein E. M., Rrasilovsky G. H., pp. 329—332 in Invertebrate Nervous Systems, edited by C. A. G. Wiersma, University of Chicago Press, Chicago, 1967. Learning in isolated insect ganglia.
123. Elsner N., Huber F„ Z. vergl. Physiol. 65: 389—423, 1969. Central nervous basis for song patterns in crickets.
124. Enger P. S., Acta Physiol. Scand. 39: 55—72, 1957. The electroencephalogram of the codfish.
125. Evans S. №., Anim. Behav. 11: 172—178, 1963. Learning in polychaetes.
126. Evans S. M„ Biol. Bull. 137: 95—104, 1969. Habituation of withdrawal response in nereid polychaetes.
127. Evarts E. V., J. Neurophysiol. 31: 14—27, 1968. Motorneuron firing in pyramidal tract correlated with movement.
128. Evarts E. V., Thach W. T., Ann. Rev. Physiol. 31: 451—498, 1969. Motor mechanisms of the CNS: cerebrocerebellar interrelations.
129. Evoy W. H., Cohen M. J., J. Exp. Biol. 51: 151—169, 1969. Sensory and motor interaction in locomotor reflexes of crabs.
130. Evoy W. H., Kennedy D., J. Exp. Zool. 165: 223—238, 1967. Command neurons controlling muscle groups in crayfish.
131. Ewer D. W., J. Exp. Biol. 37: 812—831, 1960. Inhibition and rhythmic activity of circular muscles of Calliactis.
132. Ewert J. P., Z. vergl. Physiol. 54: 455—480, 1967. Behavior elicited by midbrain stimulation. Bufo.
Центральная нервная система
145
133. Ewert J. Р., Pfliigers Arch. 306: 210—218, 1969; also Brain Behav. Evol. 3: 36—56, 1970; also Z. vergl. Physiol. 68: 84—110, 1970. Prey catching behavior by Bufo.
134. Ewert J. P., 7. vergl. Physiol. 74: 81—102, 1971. Toad caudal thalamus.
135. Ewert J. P., Borchers H. W., Z. vergl. Physiol. 71: 165—189, 1970. Tectum of toad.
136. Ewing A. W., Manning A., J. Insect Physiol. 12: 1115—1118, 1966. Control of walking in the cockroach.
137. Fetz E. E., Science 163: 955—958, 1969. Conditioning of cortical units.
138. Fiedler K., J. Hirnforsch. 9: 480—563, 1967. Forebrain influences on attack behavior in fish.
139. Fielden A., J. Exp. Biol. 40: 541—552, 1963, Interneurons, abdominal nerve cord of dragonfly nymph.
140. Fielden A., Hughes G. M., J. Exp. Biol. 39: 31—44, 1962. Unit, activity in abdominal nerve nord of dragonfly nymph.
141. Fields H. L., J. Exp. Biol. 44: 455—468, 1966. Proprioceptive control of posture in crayfish abdomen.
142. Fields H. L., Kennedy D., Nature 206: 1232—1237, 1965. Muscle receptor organs in crayfish.
143. Flexner L. B., Flexner J. B., Roberts R. B., Science 155: 1377—1383, 1967. Memory in mice.
144. Flexner L. B„ Flexner J. B., Proc. Nat. Acad. Sci. 55: 369—374, 1966; also Science 159: 330—331, 1968. Protein synthesis and memory in mice.
145. Flexner J. B. et al., Science 141: 57—59, 1963. Effects of puromycin on learning.
146. Flint P., Anim. Behav. 13: 187—193, 1965. Sensory deprivation and behavior of the polychaete Nereis in T-mazes.
147. Fox S. S., pp. 243—259 in The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. Evoked potential, coding, and behavior.
148. Fox S. S., O’Brien J. H., Science 147: 889—890, 1965. Duplication of evoked potential waveform by curve of probability of firing of a single cell.
149. Fox S. S., Rudell A. P., J. Neurophysiol. 33: 548—561, 1970. Cortical potentials and learning.
150. Frazier W. T. et al., J. Neurophysiol. 30: 1288—1351, 1967. Properties of identified neurons in Aplysia.
151. Freeman J. A., pp. 397—420 in Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association. Chicago, 1969. The cerebellum as a timing device in frog.
152. von Frisch K., Kratky A., Naturwissenschaften 49: 409—417, 1962. Flight distance and dance pattern, bees.
153. von Frisch K-, Kosmos (Stuttgart) 59: 279—283, 1963. Olfactory cues in bees.
154. Fukatni Y. et al., J. Gen. Physiol. 48: 581—600, 1965. Excitability, Mauthner cell; collateral inhibition.
155. Furshpan E. J., Furukawa T„ J. Neurophysiol. 25: 732—771, 1962. Synaptic excitation of Mauthner neurons.
156. Furukawa T., Prog. Brain Res. 21A: 44—70, 1966. Synaptic interaction, Mauthner cell of goldfish.
157. Furukawa T. et al., J. Neurophysiol. 26: 140—176, 759—774, 1963, Inhibition in Mauthner cell of goldfish.
158. Gardner D., Kandel E. R., Science 176: 675—678, 1972. Synaptic transmission in Aplysia.
159. Gaze R. M., Jacobson M., Proc. Roy. Soc. bond. В 157: 420—448, 1963. Re-tinotectal projection in frog.
160. Gaze R. M., Jacobson M., J. Physiol. 169: 92P — 93P, 1963. Single-unit responses, different depths in optic tectum, goldfish.
161. Gaze R. M., Jacobson M., Szekely G., J. Physiol. 165: 484—499, 1963. Retino-tectal projections in Xenopus.
Ю—2067
146
Глава 15
162. Gaze R. M., Keating M. J., Quart. J. Exp. Physiol. 35: 143—152, 1970. Reti-notectal projection in frog.
163. Gaze R. M., Keating M. J., Szekely G., Beazley L., Proc. Roy. Soc. bond. В 175: 107—147, 1970. Binocular interaction in the formation of intertectal neuronal connections, Xenopus.
164. Gaze R. M. et al., Quart. J. Exp. Physiol. 47: 273—280, 1970; 55: 143—152, 1970; also J. Physiol. 200: 128P—129P, 1969. Projection of binocular visual field on optic tecta of frog.
165. Gelver B., Anim. Behav. 13 (Suppl.): 21—29, 1965. Learning behavior, Paramecium.
166. Gelperin A., Z. vergl. Physiol. 72: 17—31, 1971. Negative feedback and feeding behavior of the blowfly.
167. Gettrup E., Wilson D. M„ J. Exp. Biol. 41: 183—190, 1964. Liftcontrol reaction of flying locusts.
168. Goldstein M. H. et al., J. Neurophysiol. 33: 188—197, 1970. Functional organization, cat primary auditory cortex.
169. Graeber R. C., Ebbesson S. О. E., Comp. Biochem. Physiol. 42A: 131—139, 1972. Visual learning in normal and tectum-ablated nurse sharks.
170. Gray J., J. Exp. Biol. 13: 181—191, 200—209, 210—218, 1936. Locomotor reflexes in fishes.
171. Gray J., J. Exp. Biol. 16: 9—17, 1939. Locomotor reflexes in Nereis.
172. Gray J., Lissmann H., J. Exp. Biol. 17: 227—236, 237—251, 1940; 23: 121— 132, 133—142, 1946. Locomotor reflexes in amphibians.
173. Grinnell A. D., J. Physiol. 182: 612—648, 1966. Motorneurons in frog spinal cord.
174. Gruber S. A., Ewer D. W., J. Exp. Biol. 39: 459—477, 1962. Myoneural physiology of polyclad. Planocera.
175. Gruberg E. R., Ph. D. Thesis, University of Illinois, 1969. Organization of tectum of salamander Ambystoma.
176. Grusser-Cornehls U., Pfluger. Arch. 303: 1—13, 1968. Movementdetecting neurons of frog’s retina.
177. Grusser-Cornehls U., Grilsser O. J., pp. 275—286 in: Neurophysiologie and Psychophysiologie des visuellen Systems, edited by R. Jung and H. Kornhu-ber. 1960. Reaction model of neuron in central visual system of fish, rabbits and cats with monocular and binocular light stimulation.
178. Grusser-Cornehls U., Grilsser O. J., Bullock T. H., Science 141: 820—822, 1963. Visual responses in frog tectum.
179. Guthrie D. M., Banks J. R., J. Exp. Biol. 50: 255—273, 1969. Development of patterned activity by implanted ganglia and their peripheral connections in Periplaneta.
180. Hagiwara S. et al., Comp. Biochem. Physiol. 13: 453—460, 1964. Transmission in giant axons, sabellid worm.
181. Hainsworth F. R., J. Comp. Physiol. Psychol. 63: 111—116, 1967. Learning in goldfish.
182. Hale E. B., Physiol. Zool. 29: 107—127, 1956. Effects of forebrain lesions on the aggressive behavior of green sunfish.
183. Hamburger V., 27th Symp. Soc. Devel. Biol., Suppl. 2, 251—271, 1968. Emergence of nervous coordination.
184. Hamburger V., pp. 141—151 in The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. Embryonic motility in vertebrates.
185. Hamburger V., Wenger E., Oppenheim R., J. Exp. Zool. 162: 133—160, 1966. Motility in chick embryo in absence of sensory input.
186. Hanstrom B., Vergleichende Anatomie des Nervensystems der wirbellosen Tiere. Springer-Verlag, Berlin, 1928, 628 pp.
187. Harcombe E. S., Wyman R. J., J. Exp. Biol. 53: 255—263, 1970. Diagonal locomotion in de-afferented toads.
'Центральная нервная система 147
188. Harris J. Е., Whiting Н. Р., J. Exp. Biol. 31: 501—524, 1954. Function in the locomotory system of the dogfish embryo.
189. Heimer L., Ann. N. Y. Acad. Sci. 167: 129—146, 1969. Olfactory connections in vertebrate brain.
190. Henneman E., pp. 1717—1732 in: Medical Physiology, edited by V. B. Mountcastle С. V. Mosby, St. Louis, 1968. Organization of the spinal cord.
191. Henneman E. et al., J. Neurophysiol. 28: 560—580, 1965. Significance of cell size in spinal motorneurons.
192. Heric T. M„ Kruger L., J. Comp. Neurol. 124: 101—112, 1965. Organization of the visual projection upon the optic tectum of reptile (Alligator).
193. Heric T. M., Kruger L., Brain Res. 2: 187—199, 1966. Electrical response evoked in reptilian optic tectum by afferent stimulation.
194. Hermann H. T., Brain Res. 26: 293—304, 1971. Saccade correlated potentials in optic tectum and cerebellum of Carassius.
195. Hernandez-Peon R., Electroenceph. Clin. Neurophysiol. Suppl. 13: 101—114, 1960. Neurophysiological correlates of habituation.
196. Hertzler D. R., Hayes W. N., J. Comp. Physiol. Psychol. 63: 444—447, 1967. Cortical and tectal function, visually guided behavior, turtles.
197. Hirsch H. B., Spinelli D. N., Science 168: 869—871, 1970. Effect of visual experience on visual cortex, kittens.
198. Hobson J. A., Electroenceph. Clin. Neurophysiol. 22: 113—121, 1967. Frog EEG with reference to sleep.
199. Hoffmann К.-P., Z. vergl. Physiol. 67: 26—57, 1970. Retinal connection to receptive fields in optic tectum and pretectum of cat.
200. Holden A. L., J. Physiol. 194: 75—90, 91—104, 1968. Field potentials and unit responses in optic tectum, pigeon.
201. von Holst E., Z. vergl. Physiol. 20: 582—599, 1934; 26: 481—528, 1939; also Pfliiger. Arch. 236: 149—158, 1935. Reflexes and locomotor rhythms in fish.
202. von Holst E., St. Paul U., Naturwissenschaften 45: 579, 1960; 47: 409—422, 1960. Brain stimulation, chickens.
203. Ноге I., Porter R., Brain Res. 30: 232—234, 1971. Pyramidal neurons in a marsupial.
204. Horn G., Hinde R. A., editors, Short-Term Changes in Neural Activity and Behavior. Cambridge University Press, 1970, 628pp.
205. Horn G., Rowell С. H. F., J. Exp. Biol. 49: 143—169, 1968. Long-term changes in behavior of visual neurons in tritocerebrum of locust.
206. Horn G., Wright M. J., J. Exp. Biol. 52: 217—231, 1970. Habituation in squid stellate ganglion.
207. Horridge G. A., Phil. Trans. Roy. Soc. Lond. В 240: 495—528, 1957. Inter-neural ficilitation, probability basis for transmission.
208. Horridge G. A., Proc. Roy. Soc. Lond. В 150: 245—262, 1959. Analysis of rapid responses in Nereis and Harmothde.
209. Horridge G. A., J. Exp. Biol. 36: 72—91, 1959. Properties of fast and slow nerve nets of medusae.
210. Horridge G. A., J. Physiol. 155: 320—336, 1961. Sequence of impulses initiated from ganglia of clam.
211. Horridge G. A., Proc. Roy. Soc. Lond. В 157: 33—52, 1963. Learning of leg position, ventral nerve cord in insects.
212. Horridge G. A., Proc. Roy. Soc. Lond. В 157: 199—222, 1963. Proprioceptors, bristle receptors, efferent sensory impulses.
213. Horridge G. A., J. Exp. Biol. 44: 233—245, 1966. Optokinetic memory in crab.
214. Horridge G. A., Anim. Behav. Suppl. 1: 163—182, 1966. The electrophysiological approach to learning in isolatable ganglia.
215. Horridge G. A., Interneurons. W. H. Freeman. San Francisco, 1968. 436 p.
216. Horridge G. A., Burrows M. J. Exp. Biol. 49: 269—284, 315—324, 1968. Eyecup movement, crab Carcinus.
217. Horridge G. A., Chapman D. M., MacKay B., Nature 193: 889—890, 1962. Electron microscopy of Aurelia and Cyanea.
10*
148
Глава 15
218. Horridge G. A., Mackie G. O., Quart. J. Mier. Sci. 103: 531—542, 1962. Electron microscopy of synapses, jellyfish.
219. Horridge G. A., Sandeman D. C., Proc. Roy. Soc. bond. В 161: 216—246, 1964. Nervous control of optokinetic responses in crab Carcinus.
220. Horridge G. A., Scholes J. H., Shaw S., Tunstall J., pp. 165—202 in: Physiology of the Insect Central Nervous System, edited by J. E. Treherne and J. W. Beament. Academic Press, New York, 1965.
221. Horridge G. A., Shepheard P. R. B., Nature 209: 267—269, 1966. Perception of movement by a crab.
222. Hoyle G., pp. 203—232 in Physiology of the Insect Central Nervous System, edited by J. E. Treherne and J. W. Beament. Academic Press, New York, 1965. Learning in headless insects.
223. Hoyle G., pp. 349—444 in Advances in Insect Physiology, vol. 7, edited by J. W. Beament. Academic Press, New York, 1970.
224. Hoyle G., J. Exp. Biol. 44: 413—427, 1966. Conditioning of locust ganglion.
225. Hoyle G., Burrows M., personal communication. Neural mechanisms of behavior in Schistocerca.
226. Hubei D. H., J. Physiol. 147: 226—238, 1959; 150: 91—104, 1960. Single unit activity in geniculate and optic cortex, cat.
227. Hubei D. H., Wiesel T. N., J. Physiol. 155: 385—398, 1961. Integrative action in cat’s lateral geniculate body.
228. Hubei D. H., Wiesel T. N., J. Physiol. 160: 106—154, 1962; 164: 559—568, 1963; also J. Neurophysiol. 28: 229—289, 1965. Visual fields of cortical neurons.
229. Hubei D. T., Wiesel T. N., J. Neurophysiol. 26: 998—1002, 1965. Receptive fields in striate cortex, young kittens.
230. Hubei D. T., Wiesel T. N., J. Physiol. 195: 215—243, 1968. Receptive fields and functional architecture of monkey striate cortex.
231. Hubei D. H., Wiesel T. N., J. Physiol. 206: 419—436, 1970. The period of susceptibility to physiological effects of unilateral eye closure in kittens.
232. Huber F., Naturwissenschaften 43: 317—321, 1956. Sound patterns, grasshoppers and crickets.
233. Huber F., Z. vergl. Physiol. 43: 359—391; 44: 60—132, 1960. Respiration, locomotion, sound production, Orthoptera.
234. Huber F., pp. 334—406 in The Physiology of Insecta, vol. 2, edited by M. Rock-stein, Academic Press, New York, 1965. Neural integration (central nervous system).
235- Huber F., pp. 333—351 in: Invertebrate Nervous Systems, edited by C.A.G. Wiersma. University of Chicago Press, 1967. Central control of movements and behavior of insects.
236. Hughes G. M., J. Exp. Biol. 29: 267—284, 1952; 34: 306—333, 1957. 35: 567—583, 1958. Neural coordination of insect movements.
237. Hughes G. M., J. Exp. Biol. 46: 169—193, 1967. Physiological anatomy of left and right giant cells in Aplysia.
238. Hughes G. M„ Tauc L., J. Exp. Biol. 39: 45—69, 1962; 40: 469—486, 1963. Electrophysiological evidence for central nervous pathways in Aplysia.
239. Hyden H., Egyhazi E., Proc. Nat. Acad. Sci. 49: 618—624, 1963; 52: 1030— 1035, 1964. Changes in RNA in cortical neurons of rats in learning.
240. Ikeda K., Kaplan W. D., Proc. Nat. Acad. Sci. 66: 765—772, 1970. Electrical activity in Drosophila neurons.
241. Ikeda K„ Wiersma C. A. G., Comp. Biochem. Physiol. 12: 107—115, 1964. Rhythmicity in abdominal ganglia of crayfish.
242. Ingle D., Brain Behav. Evol. 3: 57—71, 1970. Visual-motor functions of frog optic tectum.
243. Ingle D., Exp. Neurol. 33: 329—342, 1971. Potentials in goldfish optic tectum.
244. Ito M. et al., J. Physiol. 164: 150—156, 1962; also Proc. Roy. Soc. bond. В 161:
Центральная нервная система 149’
92—108, 132—141, 964. Anion permeability of inhibitory postsynaptic membrance of cat motorneurons.
245. Jacklet J. W., Cohen M. J., Science 156: 1638—1640, 1967. Synaptic connection of transplanted insect ganglion and muscles of the foot.
246. Jacobson M., Devel. Biol. 17: 202—218, 1968; also Science 163: 543—547, 1969. Neuronal specificity in retinal ganglion cells, Xenopus.
247. Jacobson M., Developmental Neurobiology. Holt, Rinehart and Winston, New York, 1970, 465 p.
248. Jacobson M., pp. 116—129 in The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. Development, specification, and diversification of neuronal connections.
249. Jacobson M., Gaze R. M., Quart. J. Exp. Physiol. 49: 199—209, 1964. Response of units in optic tectum of goldfish.
250. Janzen W., Zool. Jahrb. 52: 591—628, 1933. Goldfish forebrain function.
251. Jasper H. H., Doane B., pp. 79—117 in Progress in Physiological Psychology, vol 2, edited by E. Stellar and J. M. Sprague. Academic Press, New York,. 1968. Neurophysiologic-al mechanisms in learning.
252. Jasper H. H., Ricci G., Doane B., Electroenceph. Clin. Neurophysiol. (Suppl.) 13: 137—155, 1960. Cortical cell discharges during avoidance conditioning,, monkey.
253. Jasper H. H., Siefanis C., Electroenceph. Clin. Neurophysiol. 18: 541—553,. 1965. Oscillatory rhythms in pyramidal neurons, cat.
254. Jassik-Gerschenfeld D., Brain Res. 24: 407—421, 1970. Pigeon optic tectum.
255. Jassik-Gerschenfeld D., Guichard J., Brain Res. 40: 303—317, 1972. Visual receptive fields of single cells in pigeon’s optic tectum.
256. Jensen D. D., Anim. Behav. 13 (Suppl. 1): 9—20, 1965. Pseudo-learning in paramecia and planaria.
257. Jha R. X-, Mackie G. O., J. Morph. 123: 43—61, 1967. Nerve net structure in. Cordylophora and siphonophore Sarsia.
258. John E. R., pp. 161—182 in Brain Function, vol. 2, edited by M. A. Brazier. University of California Press, 1965. Studies on learning and retention in Planaria.
259. John E. R., pp. 690—704 in The Neurosciences: A Study Course, edited by G. C. Quarton et al., Rockefeller University Press, New York, 1967. Electrophysiological studies of conditioning.
260. John E. R., Ruchkin D. S., Villegas J., Ann. N. Y. Acad. Sci. 112: 362—420,.
1964. Behavioral correlates of evoked potential configurations in cats.
261. Johnson R. H., Thompson R. F., J. Comp. Physiol. Psychol. 69: 485—491, 1969. Association cortex in auditory-visual conditional learning in cat.
262. Johnstone J. R., Mark R. F., J. Exp. Biol. 54: 403—414, 1971. Efference copy neuron in goldfish.
263. Jones M. D. R., J. Exp. Biol. 45: 15—30, 1966. Acoustic behavior of bush cricket.
264. Josephson R., Comp. Biochem. Physiol. 5: 45—58, 1962. Spontaneous electrical activity in a hydroid polyp.
265. Josephson R., J. Exp. Biol. 42: 139—152, 1965. Conducting Systems in stalk of hydroid.
266. Josephson R. X-, Cnidarian neurobiology in Perspectives in Coelenterate Biology, edited by L. Muscatine and H. M. Lenhoff. Academic Press, New York,. 1973.
267. Josephson R. X., Mackie G. O., J. Exp. Biol. 43: 293—332, 1965. Multiple pacemakers in hydroid Tubularia.
268. Josephson R. X., Macklin M., J. Gen. Physiol. 53: 638—665, 1969. Electrical’ properties, body wall of Hydra.
269. Josephson R. Х-, Uhrich J., J. Exp. Biol. 50: 1—14, 1969. Inhibition of pacemaker systems in a hydroid.
270. Jouvet M., Physiol. Rev. 47: 117—177, 1967; also pp. 529—544 in: The Neuro-sciences: A Study Program, edited by G. C. Quarton et al., Rockefeller Uni
150
Глава 15
versify Press, New York, 1967; also et al., Science 159: 112—114, 1968; 163: 32—41, 1969. Physiology of states of sleep.
271. Kaada B. R., Rasmussen E. W., Kriem A., Exp. Neurol. 3: 333—355, 1961. Hippocampus in memory storage in rodents.
272. Kaas J., Hall W. C., Diamond I. T., J. Neurophysiol. 33: 595—614, 1970. Erinaceus (hedgehog) cortical sensory areas.
273. Kadoya S., Wolin L. R., Massopust L. C., Jr., J. Comp. Neurol. 142: 495—508, 1971. Optic tectum of squirrel monkey.
274. Kaiserman-Abramof I. R., Palay S. L„ pp. 171—205 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association, Chicago, 1969. Fine structural studies of the cerebellar cortex in mormvrid fish.
275. Kamikawa K„ McIlwain J. T., Adey W. R., Electroenceph. Clin. Neurophysiol.
17: 485—496, 1964. Response patterns of thalamic neurons during classic conditioning.
276. Kammer A. E., J. Exp. Biol. 47: 2772—95, 1967. Muscle activity during flight | in some large Lepidoptera.
277. Kammer A. E., J. Exp. Biol. 48: 89—109, 1968. Motor warmup in Lepidoptera. I
278. Kandel E. R., pp. 666—689 in: The Neurosciences: A Study Program, edited i
by G. C. Quarton et al., Rockefeller University Press, New York, 1967.
279. Kandel E., Castellucci V., Pinsker H., Kupferman I., pp. 281—322 in: Short-Term Changes in Neural Activity and Behaviour, edited by G. Horn and $ R. A. Hinde, Cambridge University Press, 1970. Synaptic plasticity in short-term modification of behavior.
280. Kandel E., Frazier W. T„ Coggeshall R. E., Science 155: 346—349, 1967.
Synaptic actions mediated by different branches of an identifiable interneuron Г in Aplysia. r
281. Kandel E., Tauc L„ J. Physiol. 181: 1—27, 28—47, 1965. Heterosynaptic facilitation in neurons of abdominal ganglion of Aplysia.
282. Kandel E., Tauc L., J. Physiol. 183: 269—286, 1966. Input organization of two symmetrical giant cells in the snail brain.
283. Kandel E., Wachtel H., pp. 17—65 in: Physiological and Biochemical Aspects of Nervous Integration, edited by F. D. Carlson. Prentice-Hall, 1967. Functional organization of neural aggregates in Aplysia.
284. Kandel E. et al., J. Neurophysiol. 30: 1352—1376, 1967. Interactions between neurons in Aplysia.
285. Kaplan H., Aronson L. R., Anim. Behav. 15: 438—448, 1967. Conditioned behavior in Tilapia, fish.
286. Karamian A. I., Fanardjian V. V., Kosareva A. A., pp. 639—671 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association, Chicago, 1969. Functional and morphological evolution of the cerebellum.
287. Katz B., Miledi R., J. Physiol. 168: 389—422, 1963. Spontaneous miniature potentials in spinal motorneurons.
288. Keating E. G:, Kormann L. A., Horel J. A., Physiol, and Behav. 5: 55—59, 1970. Stimulation of amygdala of caiman.
289. Keating M. J., Gaze R. M., Quart. J. Exp. Physiol. 55: 129—142, 1970. Frog retinal ganglion cell units.
290. Kendig J., J. Exp. Biol. 48: 389—404, 1968. Motorneuron coupling in locust flight.
291. Kennedy D„ The Physiologist 14: 5—30, 1971. Crayfish interneurons.
292. Kennedy D., Mellon D., Comp. Biochem. Physiol. 3: 275—300, 1964. Receptive fields in crayfish interneurons.
293. Key B. J., Nature 207: 441—442, 1965. Correlation of behavior with changes in amplitude of cortical potentials evoked during habituation.
294. Kodokoro Y., pp. 257—272 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association. Chicago, 1969. Cerebellar and vestibular control of fish oculomotor neurons.
Центральная нервная система
151
295. King F. A., Meyer Р. М., Science 128: 655—656, 1958. Amygdalectomy in с st
296. Kitai S. et al., Brain Res. 22: 381—385, 1970; also pp. 481—489 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas, American Medical Association, Chicago, 1969. Cerebellum of reptiles.
297. Klemm W. R„ Animal Electroencephalography, Academic Press, New York, 1969. 292 p.
298. Knights A., J. Exp. Biol. 42: 447—461, 1965. Activity of single motor fibers in arthropods.
299. Kollross J. J., pp. 179—199 in: Growth of the Nervous System, edited by G. E. W. Wolstenhome and M. O’Connor. Little, Brown Co., Boston, 1968. Endocrine influences in neural development.
300. Konishi R., Slepian J. Z., J. Acoust. Soc. Amer. 49: 1762—1769, 1971. Effect of efferent impulses on cochlear responses, guinea pig.
301. Konorski J., pp. 115—130 in: Brain Mechanisms and Learning, edited by J. F. Delafresnaye. Blackwell, Oxford; 1961. The physiological approach to the problem of recent memory.
302. Krasne F. B., J. Exp. Biol. 50: 29—46, 1969. Excitation and habituation of crayfish escape reflex: depolarizing response in lateral giant fibers.
303. Krivanek J. O., Physiol. Zool. 29: 241—250, 1956. Habit formation in the earthworm.
304. Kruger L„ Berkowitz E. C., J. Comp. Neurol. 115: 125—140, 1960. Afferent connections, reptile telencephalon.
305. Kubota K., Brookhart J. M., Amer. J. Physiol. 204: 660—666, 1963. Inhibitory synaptic potential of frog motorneurons.
306. Kubota K-, Brookhart J. M., J. Neurophysiol. 26: 877—893, 1963. Recurrent facilitation of frog muscle.
307. Kulsch E., Huber F., Z. vergl. Physiol. 67: 140—159, 1970. Central vs. peripheral control of cricket singing.
308. Kupfermann I. et al., Science 164: 847—850, 1969; 167: 1743—1745, 1970. Behavioral habituation, Aplysia.
309. Kupfermann I. et al., Science 174: 1252—1255, 1971. Central and peripheral control of gill movements, Aplysia.
310. Kuriyama K. et al., Proc. Nat. Acad. Sci. 55: 846—852, 1966. GABA as transmitter in cerebellum.
311. Landenberger D. E., Anim. Behav. 14: 414—418, 1966. Conditioning of starfish.
312. Larimer J. L., Kennedy D., J. Exp. Biol. 44: 345—354, 1966. Afferent signals in crayfish stomatogastric ganglion.
313. Larimer J. L., Kennedy D., J. Exp. Biol. 51: 119—133, 135—150, 1969. Innervation patterns of muscles in responses of crayfish.
314. Larimer J. L. et al., J. Exp. Biol. 54: 391—402, 1971. Connections of lateral and medial giant fibers of crayfish.
315. Lar sell O., The Comparative Anatomy and Histology of the Cerebellum. University of Minnesota Press, Minneapolis, 1970. 269 p.
316. Lashley K. S., Comp. Psychol. Monogr. 11: 3—79, 1935; also Quart. Rev. Biol. 24: 28—42, 1949. Neural basis of learning in rats.
317. Latham A., Paul D. H., J. Physiol. 213: 135—156, 1971. Activity of cerebellar Purkinje cells.
318. Laverack M. S., J. Exp. Biol. 50: 129—140, 1969. Mechanoreceptors, photoreceptors and rapid conduction pathways in leech Hirudo.
319. Lawry J. V., Comp. Biochem. Physiol. 37: 167—179, 1970. Locomotion, po-lychaete.
320. Leghissa S„ Z. Anat. EntwickL 118: 427—463, 1955. Structure of optic tectum,
321. Lende R. A., J. Comp. Neurol. 121: 395—403, 405—415, 1963; also Science
141: 730—732, 1963. Sensory and motor representation, cerebral cortex, opossum.
322. Lende R. A., J. Neurophysiol. 27: 37—48, 1964. Cerebral cortex of echidna.
352
Глава 15
323. Lende R. A., Ann. N. Y. Acad. Sci. 167: 262—275, 1969. Neocortex of mo-notremes, marsupials, and insectivores.
324. Levi-Montalcini R„ Angeletti P. U., Physiol. Rev. 48: 534—569, 1968. Nerve growth factor.
325. Levi-Montalcini R., Angeletti R., Angeletti P. U., pp. 1—38 in: Structure and Function of Nervous Tissue, vol. 5, edited by G. H. Bourne. Academic Press, New York, 1972. Nerve growth factor.
.326 . Levitan H., Tauc L., Segundo J. P., J. Gen. Physiol. 55: 484—496, 1970. Neuronal interactions in buccal ganglion of mollusc Navanax.
327. Li C., Chou S. N., J. Cell. Comp. Physiol. 60: 1—16, 1962. Cortical intracellular synaptic potentials.
328. Lickey M. E., J. Comp. Physiol. Psychol. 68: 9—17, 1969. Entrainment of circadian rhythm in Aplysia neuron.
329. Lindauer M., Medugorac 1., Colloq. Internat. Centr. Nat. Recher. Sci. 15—25, 1968. Retention of learned response after narcosis, bees.
330. Llinas R„ Hillman D. E., pp. 43—73 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas, American Medical Association, Chicago, 1969. Comparative morphology of the cerebellum.
331. Llinas R., Nicholson C., J. Neurophysiol. 34: 532—551, 1971; also pp. 431— 464 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas, American Medical Association, Chicago, 1969. Properties of dendrites and somata in alligator Purkinje cells.
332. Llinas R. et al., Brain Res. 6: 371—378, 1967; also Exp. Brain Res. 13: 408— 431, 1971. Vestibular input to cerebellum, frog.
333. Luco J. V., pp. 135—159 in: Brain Function, vol. 2, edited by M. A. Brazier. University of California Press, 1965. Plasticity of neural function in learning and retention.
334. Luco J. V., Aranda L. C., Nature 201: 1330—1331, 1964. Electrical correlate of learning.
335. Luco J. V., Aranda L. C., Nature 209: 205—206, 1977. Reversibility of electrical . correlate to process of learning.
336. Lund R. D., Lund J. S., Exp. Neurol. 13: 302—316, 1965. Vision in mole.
337. Mackie G. O., Quart. J. Mier. Sci. 101: 119—131, 1960. Velella — dual nerve net.
338. Mackie G. O., Proc. Roy. Soc. Lond. В 159: 366—391, 1964. Locomotion in a siphonophore colony.
339. Mackie G. 0., Amer. Zool. 5: 439—453, 1965. Non-nervous (epithelial) conduction in a cnidarian.
340. Mackie G. O., Passano L. M., J. Gen. Physiol. 52: 600—621, 1968. Epithelial conduction in coelenterates.
341. MacLean P. D., pp. 336—349 in: The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. Emotion brain and subjective brain.
342. Magalhaes С. B., Saraiva P. E. S., Brain Res. 34: 291—299, 1971. Sensory and motor representation in cerebral cortex of marsupial Didelphis.
343. Magoun H. W., The Waking Brain. Charles C. Thomas, Springfield, I1L, 1958, 188 pp The reticular system in the brain. (Мэгун Г., Бодрствующий мозг, ИЛ, 1960.)
344. Мантейфель Ю. Б., Вызванные потенциалы в тектуме ящерицы, ДАН СССР, 168, 414 (1966) - п
345. Manton S. М., J. Linn. Soc. Lond. 41: 529—570, 1950; 42: 93—166, 299—368, 1954. Locomotor rhythms in arthropods.
346. Mark R. F„ Exp. Neurol. 16: 215—225, 1966; 21: 92—104, 1968. Intertectal connections, goldfish.
347. Martin A. R., Pilar G„ J. Physiol. 168: 443—475, 1963. Synaptic transmission, chick ciliary ganglion.
348. Maturana H. et al., J. Gen. Physiol. 43: 129—176, 1960. Vision in frog.
349. Maynard D. M., Symp. Soc. Exp. Biol. 20: 111—150, 1966. Integration in lobster brain.
Центральная нервная система
153
350. Maynard D. М., Science 158: 531—532, 1967. Stomatogastric ganglion of mangrove crab.
351. Maynard D. M., Ann. N. Y. Acad. Sci. 193: 59—72, 1972. Stomatogastric ganglion in crustaceans.
352. Maynard D. M„ Cohen M. J., J. Exp. Biol. 43: 55—78, 1965. Heteromorph an-tennule in lobster.
353. Maynard D. M., Yager J. G., Z. vergl. Physiol. 59: 241—249, 1968. Function of an eyestalk ganglion.
354. Мазурская П. 3., Смирнов Г. Д., Зрительные проекции в теленцефалоне черепахи, ДАН СССР, 167, 285 (1966).
355. McCleary R. A., J. Comp. Physiol. Psychol. 53: 311—321, 1960. Interocular transfer in fish.
356. McConnell J. V., Ann. Rev. Physiol. 28: 107—136, 1966. Learning in invertebrates.
357. McConnell J. V. et al., J. Comp. Physiol. Psychol. 52: 1—5, 1959; also Amer. J. Psychol. 73: 618—622, 1960. Conditioned response in planarian.
358. McFarlane I. D., J. Exp. Biol. 51: 377—385, 1969. Slow condition systems in sea anemone Calliactis.
359. McIlwain J. T., Ann. Rev. Physiol. 34: 291—314, 1972. Visual cortex and superior colliculus.
360. McKay J. M., J. Exp. Biol. 53: 137—145, 1970. Cerebral control of an insect sensory interneuron.
361. Medugorac I., Lindauer M., Z. vergl. Physiol. 55: 450—474, 1967. Memory in bees after narcosis.
362. Mellon D., J. Exp. Biol. 43: 455—472, 1965; 46: 585—597, 1967; 53: 711—725, 1970. Reflexes in surf clam, Spisula.
363. Mellon D., Z. vergl. Physiol. 62: 318—336, 1969. Reflex control of rhythmic motor output during swimming in scallop.
364. Mendelson M., Science 171: 1170—1173, 1971. Oscillator neurons in crustacean ganglia.
365. Meulders M., et al., J. Comp. Neurol. 126: 535—545, 1966. Sensory projections in sloth.
366. Michael C. R., J. Nerophysiol. 31: 249—282, 1968. Receptive fields of optic nerve fibers in ground squirrel all-cone retina.
367. Michael C. R., Brain Behav. Evol. 3: 205—209, 1970; also Vision Res. Suppl. 3: 299—308, 1971. Superior colliculus of ground squirrel.
368. Milburn N. S., Roeder K- D., Gen. Comp. Endocr. 2: 70—76, 1962. Control of efferent activity in cockroach.
369. Milburn N. S., Weiant E. A., Roeder K. D., Biol. Bull. 118: 111—119, 1960. Release of efferent nerve activity in Periplaneta in extracts of corpus car-diacum.
370. Miller N. E., Science 148: 328—338, 1965. Chemical coding of behavior in the brain.
371. Miller N. E., Science 168: 434—445, 1969. Learning of visceral and glandular responses.
372. Miller P. L., J. Exp. Biol. 46: 349—371, 1967. Derivation of motor command to spiracles in locust.
373. Miller P. L., pp. 475—498 in: Short-Term Changes in Neural Activity and Behaviour, edited by G. Horn and R. A. Hinde. Cambridge University Press, 1970. Learning in insect central nervous system.
374. Miller P. L., J. Exp. Biol. 54: 587—608, 1971. Rhythmic activity in insect nervous system.
375. Millott N., pp. 465—485 in: Physiology of Echinodermata, edited by R. Bo-lootian. Interscience, New York, 1966. Coordination of spine movement in echinoids.
376. Millott N., Okumura H., J. Exp. Biol. 48: 279—287, 1968. Electrical activity of the radial nerve in Diadema and certain other echinoids.
377. Mimura K. et al., Z. vergl. Physiol. 62: 382—394, 1969. Unit recording, brain of fleshly.
154
Глава 15
378. Mittelstaedt Н., Ann. Rev. Entomol. 7: 177—198, 1962. Control systems of orientation in insects.
379. Мкртичева Л. И., Элементы функциональной организации зрительной системы у лягушки, Ж. высш, нервн. деят., 5, 513 (1966).
380. Morgane Р. J., Stern W. С., Ann. N. Y. Acad. Sci. 193: 95—ПО, 1972. Relation of sleep to neuroanatomical circuits.
381. Morrell F., Electroenceph. Clin. Neurophysiol. Suppl. 13: 65—79, 1960. Dendritic locus of closure.
382. Mountcastle V. B., pp. 1345—1371 in: Medical Physiology, vol. 2, edited by V. B. Mountcastle. С. V. Mosby, St. Louis, 1968. Physiology of sensory receptors: introduction to sensory processes.
383. Mountcastle V. B., Davies P. W., Bermann A. L., J. Neurophysiol. 20: 374— 407, 1957. Responses of somatic sensory cortex, cat.
384. Mulloney B., Science 168: 994—998, 1970. Structure of the giant fibers of earthworms.
385. Mulloney B., J. Neurophysiol. 33: 86—95, 1970. Organization of flight motorneurons of flies.
386. Mulloney B., J. Exp. Biol. 52: 59—78, 1970. Impulse patterns in flight motor-netirons of Bombus.
387. Muntz W. R. A., J. Neurophysiol. 25: 699—711, 1962. Diencephalon of frog.
388. Murphy J. T., Sabah N. H., Brain Res. 25: 449—467, 1971. Climbing fiber activation of Purkinje neurons.
389. Nachtigall W., Z. vergl. Physiol. 61: 1—20, 1968. Flight patterns in flesh flies.
390. Nauta W. J. H., Karten H. J., pp. 7—26 in: The Neurosciences: Second Study Program, edited by F. O. Schmitt. Rockefeller University Press, New York, 1970. General profile of the vertebrate brain, with sidelights on the ancestry of the cerebral cortex.
391. Nichol R. F., Exp. Brain. Res. 14: 185—197, 1972. Excitation in olfactory bulb.
392. Nicholls J. G., Baylor D. A., J. Neurophysiol. 31: 740—756, 1968. Sensory neurons of leech.
393. Nicholls J. G., Purves D., J. Physiol. 209: 647—668, 1970. Connections between sensory and motor cells in leech.
394. Nicholson C., Llinas R., J. Neurophysiol. 34: 509—531, 1971. Field potentials in alligator cerebellum.
395. Nicol J. A. C., Quart. J. Mier. Sci. 89: 1—45, 1968. Giant fiber system of My-xicola.
396. Nieuwenhuys R., Nicholson C., pp. 107—170 in: Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association, Chicago, 1969. Cerebellum of mormyrid fishes.
396a. Northcutt R. G., Ann. N. Y. Acad. Sci. 167: 180—185, 1969. Homologies in vertebrate brain.
397. Obata K. et al., Exp. Brain Res. 4: 43—57, 1967. GABA as cerebellar transmitter.
398. O’Benar J. D., Ph. D. Thesis, University of Illinois, 1971. Electrophysiology of goldfish optic tectum.
399. O’Brien J. H., Fox S. S., J. Neurophysiol. 32: 267—296, 1969. Single cell activity in cat motor cortex.
400. Olds J., pp. 257—293 in Neural Control of Behavior, edited by R. E. Whalen et al. Academic Press, New York, 1970. Behavior of hippocampal neurons during conditioning experiments.
401. Olds J., Hirano T„ Electroenceph. Clin. Neurophysiol. 26: 159—166, 1969. Conditioned responses of hippocampal and other neurons.
402. Oliver G. W. O. et al., Comp. Biochem. Physiol. 38B: 529—535, 1971. Changes in GABA level, GAD, AChE activity in CNS of an insect during learning.
403. Olivo R. F., Comp. Biochem. Physiol. 35: 787—807, 1970. Relfexes in razor clam.
Центральная нервная система
155
404. Orrego F„ Arch. Ital. Biol. 99: 425—445, 1961. Olfactory pathways and cortical areas in turtle.
405. Packard A., Nature 221: 875—877, 1969. Jet propulsion and giant fiber response of Loligo.
406. ’ Page С. H., J. Neurophysiol. 33: 116—128, 1970. Electrophysiology, auditory responses, goldfish brain.
407. Page С. H., Sutterlin A. M., J. Neurophysiol. 33: 129—136, 1970. Visual-auditory unit responses in goldfish tegmentum.
408. Pantin C. F. A., Amer. Zool. 5: 581—589, 1965. Capabilities of coelenterate behavior.
409. Parnas I. et al., J. Exp. Biol. 50: 635—649, 1969. Conduction in giant axons of Periplaneta.
410. Parsons L. C., Huggins S. E., Proc. Soc. Exp. Biol. Med. 119: 397—400, 1965. Electrical activity in brain of Caiman.
411. Passano L. M., Amer. Zool. 5: 465—481, 1965. Recordings from Scyphome-dusae: giant fiber net for swimming beat and diffuse nerve net.
412. Passano L. M., McCullough С. B., Nature 199: 1174—1175, 1963; also J. Exp. Biol. 41: 643—664, 1965; 42: 205—231, 1965. Coordinating system and behavior, Hydra.
413. Passano L. M., Pantin C. F. A., Proc. Roy. Soc. London. В 143: 226—238, 1955. Responses to mechanical stimulation in Calliactis.
414. Peretz B., Science 166: 1167—1172, 1969; 169: 379—381, 1970. Control of gill movements in Aplysia.
415. Peters J., Vonderahe A., Schmid D., J. Exp. Zool. 60: 255—262, 1966. Onset of cerebral electrical activity associated with sleep and attention in developing chick.
416. Peterson R. H„ Brain Res. 41: 67—79, 1972. Electrical responses of goldfish cerebellum.
417. Peterson R. H., Copeia 816—819, 1972. Somatosensory projection to facial lobe in goldfish.
418. Pettigrew J., Nikara T., Bishop P. O., Exp. Brain Res. 6: 373—390, 1968. Cat. striate cortex.
419. Peyraud C., These, University of Toulouse, 1965. 234 p. Respiratory movements in carp.
420. Pickens P. E., J. Exp. Biol. 51: 513—528, 1969. Rapid contractions and associated potentials in a sand dwelling anemone.
421. Pickens P. E„ J. Exp. Biol. 53: 515—528, 1970. Conduction along ventral nerve cord of a hemichordate worm.
421a. Piddington R. W., J. Exp. Biol. 55: 569—584, 585—610, 1971. Central control of auditory input in goldfish.
422. Pinsker K. et al., Science 167: 1740—1742, 1970. Habituation and dishabituati-on, Aplysia.
423. Pitman R. M., Tweedie C. D„ Cohen M. J., Science 178: 507—509, 1972. Responses of insect central neurons.
423a. Poggio G. F., pp. 1592—1625 in Medical Physiology, edited by V. B. Mountcastle, С. V. Mosby, St. Louis, 1968. Central visual pathways, mammals.
424. Pople W., Ewer D. W., J. Exp. Biol. 32: 59—69, 1955. Nervous system of holothurian Cucumaria.
425. Potter H. D., J. Neurophysiol. 28: 1132—1152, 1965. Mesencephalic auditory region of the bullfrog.
426. Precht H., Freytag G., Behaviour 13: 143—211, 1958. Habituation of prey capture in spiders.
427. Pritz M. B., Mead W. R., Northcutt R. G., J. Comp. Neurol. 140: 81—100, 1970. Effects of Wulst ablation in pigeons.
428. Prosser C. L., J. Comp. Neurol. 58: 603—630, 1934. Development of behavior in earthworm embryos.
429. Prosser C. L., Z. vergl. Physiol. 50: 102—118, 1965. Electrical responses, fish optic tectum.
256
Центральная нервная система
430. Prosser С. L., Farhi Е., Z. vergl. Physiol. 50: 91—101, 1965. Temperature effects on conditioned reflexes, goldfish.
431. Prosser C. L., Nagai T., pp. 171—180 in The Central Nervous System and Fish Behavior, edited by D. Ingle, University of Chicago Press, 1968. Effect of low temperature on conditioning in goldfish.
432. Purpura D. P., Int. Rev. Neurobiol. 1: 47—163, 1959. Nature of electrocortical potentials and synaptic organizations in cerebral and cerebellar cortex.
433. Rail IF., Shepherd G. M„ J. Neurophysiol. 30: 1138—1167, 1968; 31: 884—915, 1968. Evoked potentials in olfactory bulb.
434. Rees S„ Ноге J., Brain Res. 20: 439—451, 1970. Motor cortex of brush-tailed possum.
435. Render M., Selverston A., Kennedy D„ Science 162: 281—283, 1968. Lateral giant fibers of crayfish.
436. Ribbands C. R., J. Exp. Biol. 27: 302—310, 1950. Memory in bees after anesthesia.
437. Roberts A., J. Exp. Biol. 48: 545—567, 49: 645—656, 1968. Giant fibers and escape reflexes in crayfish.
438. Roberts A., Z. vergl. Physiol. 71: 295—310, 1971; 75: 388—401, 1971. Skin impulses in sensory system of young tadpoles.
439. Roberts B. L., J. Marine Biol. Assoc. U.K. 49: 33—49, 51—75, 1969. Function in spinal cord of dogfish.
440. Roberts M. В. V., J. Exp. Biol. 39: 219—227, 229—237, 1962. Giant fiber reflexes in earthworm.
441. Roberts M. В. V., J. Exp. Biol. 45: 141—150, 1966; 46: 571—583, 1967. Fast and slow reflexes, earthworm.
442. Roberts R. B., Flexner L. B., Quart. Rev. Biochem. Biophys. 2: 135—173, 1969. Biochemistry of long-term memory.
443. Roberts IF. IF., pp. 175—206 in Neural Control of Behavior, edited by R. E. Whalen et al., Academic Press, New York, 1970. Hypothalamic mechanisms for motivational and species-typical hebavior (cat, opossum).
444. Robson E. A., Jesephson R. K-> J. Exp. Biol. 50: 151—168, 1969. Neuromuscular properties of mesenteries from sea anemone.
445. Roeder K. D., Biol. Bull. 69: 203—220, 1935; also J. Exp. Zool. 76: 353—374, 1937. Nervous control of sexual behavior in mantids.
446. Roeder K. D., pp. 423—487 in Insect Physiology, edited K- D. Roeder. John Wiley and Sons, New York, 1953, Rhythms in insect nervous systems.
447. Roeder K. D., Science 154: 1515—1521, 1966. Auditory system of noctuid moths.
448. Roeder K. D., Nerve Cells and Insect Behavior. Harvard University Press, 1967. 238 pp.
449. Roeder K. D„ J. Insect Physiol. 12: 1227—1244, 1966; 15: 825—838, 1713— 1718, 1969. Acoustic interneurons, noctuid moths.
450. Roeder K. D., Tozian L., Weiant E. A., J. Insect Physiol. 4: 45—62, 1960. Nerve activity and behavior in mantis and cockroach.
451. Rogers F. T., J. Comp. Neurol. 35: 61—65, 1922; also Amer. J. Physiol. 86: 639—650, 1928. Localization of function in pigeon brain.
452. Romer A. S., The Vertebrate Body, 4th edition. W. B. Saunders Co., Philadelphia, 1970. 601 p.
453. Rose S. P. R., pp. 517—551 in Short-Term Changes in Neural Activity and Behaviour, edited by G. Horn and R. A. Hinde. Cambridge University Press, 1970. Neurochemical correlates of learning.
454. Rosenthal J., J. Physiol. 203: 121—133, 1969. PTP at neuromuscular junctions of frog.
455. Rosenzweig M. R., Bennett E. L., Diamond M. C., Sci. Amer. 226: 22—29, 1972. Brain changes in response to experience.
456. Rosenzweig M. R., Bennett E. £., Diamond M. C. et al., Brain Res. 14: 427— 445, 1969. Effect of environmental complexity and visual stimulation on occipital cortex in rat.
Центральная нервная система 157
457. Ross D. М., рр. 253—261 in Essays on Physiological Evolution, edited by I. Turpayev and J. Pringle. Pergamon Press, New York, 1964. Problems of neuromuscular activity and behavior.
458. Ross D. M., Sutton L., Proc. Roy. Soc. bond. В 155: 266—281, 1961. Response of sea anemone Calliactis to shells of hermit crab.
459. Ross D. M., Sutton L., J. Exp. Biol. 41: 735—749, 1964. Swimming response of sea anemone Stomphia to electrical stimulation.
460. Rovainen C., J. Neurophysiol. 30: 1000—1023, 1024—1042, 1967. Sea lamprey, functions of Muller and Mauthner cells; giant cells of spinal cord.
461. Rovainen C., Birnberger K. J., J. Neurophysiol. 34: 974—982, 1971. Motorneuron function, sea lamprey.
462. Rowell С. H. F., J. Exp. Biol. 40: 257—270, 1963; 44: 589—601, 1966. Arm of Octopus, reflexes.
463. Rowell С. H. F., J. Exp. Biol. 41: 559—573, 1964. Central control of insect segmental reflex.
464. Rowell С. H. F., pp. 237—280 in Short-Term Changes in Neural Activity and Behaviour, edited by G. Horn and R. A. Hinde. Cambridge University Press, 1970. Incremental and decremental processes in insect central nervous system.
465. Rowell С. H. F., J. Exp. Biol. 55: 727—747, 749—761, 1971. Visual interneuron in locust.
466. Rowell С. H. F., McKay J. M., J. Exp. Biol. 51: 231—245, 1969. An acridid grasshopper auditory interneuron.
467. Rowell С. H. F., Horn G., J. Exp. Biol. 49: 143—169, 171—184, 1968. Longterm changes in neurons in locust.
468. Rowell С. H. F., Horn G., J. Exp. Biol. 49: 176—183, 1968. Dishabituation and arousal in response of single nerve cell.
469. Rowland V., pp. 482—495 in The Neurosciences: A Study Program, edited by G. C. Quarton. Rockefeller University Press, New York, 1967. Steady phenomena of cortex.
470. Rowland V., Goldstone M., Electroenceph. Clin. Neurophysiol. 15 : 474—485, 1963. Conditioned and drive-related baseline shift in cat cortex.
471. Rozin P. N., Mayer J., Science 134: 942—943, 1961. Thermal reinforcement and thermoregulatory behavior in goldfish, Carassius.
472. Rubinson K-, Brain Behav. Evol. 1: 529—561, 1968. Projections of optic tectum of frog.
473. Rushforth N. B., Animal Behavior: Suppl. 1: 30—42, 1965. Habituation in Hydra.
474. Rushforth N. B„ Biol. Bulk 140: 255—273; 502—519, 1971. Behavioral and electrophysiological studies of Hydra. I. Analysis of contraction pulse patterns.
475. Rushmer D. S., Woodward D. J., Brain Res. 33: 83—90, 315—335, 1971. Responses of Purkinje cell in frog cerebellum.
476. Rutledge L., J. Science 148: 1246—1248, 1965. Response enhanced by conditional excitation of cerebral cortex.
477. Salanki J., Symp. Neurobiol. Invert. 493—501, 1967. Cerebral ganglia in regulation of activity in freshwater mussel Anodonta. .
478. Salanki J., Pecsi T., Labos E., Acta Biol. Acad. Sci. Hung. 19: 391—406, 1968. Ganglionic regulation of tonic muscle, mollusc.
479. Salmoiraghi G. C., Burns B. D„ J. Neurophysiol. 23: 2—13, 14—26, 1960. Rhythmicity in respiratory neurons, cat.
480. Самсонова В. Г., Функциональная организация нейронов зрительного центра у лягушки. Ж. высш, нервн. деят., 16, 125 (1966).
481. Sandeman D. С., J. Exp. Biol. 50: 771—784, 1969; also Z. vergl. Physiol. 64: 450—464, 1969; 72: 111—130, 1971. Function of identified motorneurons in crab brain.
482. Sanders F. K., J. Exp. Biol. 17: 416—434, 1940. Second order learning, tectum of goldfish.
158
Глава 15
483. Sanderson К. J., Bishop P. 0., Darian-Smith I., Exp. Brain Res. 13: 178—207, 1971. Binocular receptive fields of lateral geniculate neurons.
484. Scalia F., Brain Behav. Evol. 4: 376—399, 1971. Central projections of olfacto-ry bulb in Gymnothorax.
485. Schade J. P., Arch. Neerl. Zool. 14: 604—605, 1962. Visual responses of midbrain in fish.
486. Schade J. P., Beiler I. J., J. Exp. Biol. 36: 435—452, 1959. Electroencephalographic patterns of the goldfish (Carassius).
487. Schiller P. H., Koerner F., J. Neurophysiol. 34: 920—936, 1971. Unit activity in superior colliculus of rhesus monkey.
488. Schrameck J. E., Science 169: 698—700, 1970. Crayfish swimming: motor output and giant fiber activity.
489. Schwassman H. O., Kruger L., J. Comp. Neurol. 124: 113—126, 1965. Visual projection to optic tectum in fish.
490. Seegar J., Progr. Brain Res. 14: 143—231, 1965. Telencephalon of teleosts in relation to behavior.
491. Seegar J., Nieuwenhuys, Anim. Behav. 11: 331—344, 1963. Etho-physiological experiments with Gasterosteus.
492. Segundo J. P., Moore G. P., Stensaas L. J., Bullock T. H., J. Exp. Biol. 40: 643—667, 1963. Sensitivity of neurons in Aplysia to input patterns.
493. Shapiro S., Vukovish K- R., Science 167: 292—294, 1970. Developmental effects on cortical dendrites.
494. Sharma S. C., Provine R. R., Hamburger V., Sardel T. T., Proc. Nat. Acad. Sci. 66: 40—47, 1970. Unit activity in isolated spinal cord of chick embryo.
495. Sharpless S. K, Ann. Rev. Physiol. 26: 357—388, 1964. Reorganization of brain in use and disuse.
496. Shashona V. E., Nature 217: 238—240, 1968. RNA in goldfish brain.
497. Sherman J. D., J. Cell Biol. 37: 650—659, 1968. Electrical coupling between cells of chick embryo.
498. Sherrington C. S., The Integrative Action of the Nervous System. Scribner’s, New York, 1906.
499. Siminoff R., Schwassmann H. O., Kruger L., J. Comp. Neurol. 127: 435—444,
1966; 130: 329—342, 1967. Visual projection to superior colliculus, rat.
500. Смирнов Г. Д., pp. 263—298 in: Brain and Behavior, vol. 7, edited by
M. A. B. Brazier. Amer. Inst. Biol. Sci., Washington, D. C., 1961. Neurophy-
siology of vision.
501. Smith J. C., Baker H. D., J. Comp. Physiol. Psychol. 53: 279—281, 1960. Conditioning in Limulus.
502. Smith J. E„ pp. 503—511 in: Physiology of Echinodermata, edited by R. Bo-lootian. Interscience, New York, 1966.
503. Соколов E. H., Дуленко В. П., Электрическая активность виноградной улитки при тактильном раздражении. Ж. высш. нерв, деят., 18, ИЗ (1968).
504. Sotelo С., рр. 327—371 in Neurobiology of Cerebellar Evolution and Development, edited by R. Llinas. American Medical Association, Chicago, 1969. Ultrastructural aspects of cerebellar cortex of frog.
505. Spencer IF. A., Brookhart J. M., J. Neurophysiol. 24: 26—49, 50—65, 1961. Sensorimotor cortex of cat.
506. Spencer W. A., Thompson R. F., Neilson D. R., J. Neurophysiol. 29: 221—274, 1969. Conditioning in spinal cord.
507. Sperry R. IF., Proc. Nat. Acad. Sci. 50: 703—709, 1963. Chemoaffinity in orderly growth of nerve fiber patterns and connections.
508. Spinelli D. N., Barrett T. W„ Exp. Neurol. 24: 76—87, 1969. Visual cortex, cat
509. Spira M. E., Parnas I., Bergmann F., J. Exp. Biol. 50: 615—627, 1969. Organization of giant axons of cockroach Periplaneta.
510. Sprague J. M., Science 153: 1544—1546, 1966. Interaction of cortex and superior colliculus.
511. Starr A., Britt R., J. Neurophysiol. 33: 137—147, 1970. Intracellular recording from cat cochlear nucleus during tone stimulation.
Центральная нервная система
159
512. Stefanelli A., Quart. Rev. Biol. 6: 17—34, 1951. Synaptic structure and Mauthner cells.
513. Sterling P., Wickelgren B. G., J. Neurophysiol. 32: 1—16, 1969. Visual responses of superior colliculus, cat.
514. Stettner L. J., Schulz IF. /., Science 155: 1689—1692, 1967. Effects of brain lesions on learning in birds.
515. Stevens R. J., Ph. D. Thesis, University of Illinois, 1969. Inhibition in frog optic tectum.
516. Stewart D. L., Chow K. L., Masland R. H., Jr., J. Neurophysiol. 34: 139—147, 1971. Lateral geniculate of rabbit.
517. Strumwasser F., in Physiological and Biochemical Aspects of Nervous Integration, edited by F. D. Carlson. Prentice-Hall, 1968. Membrane and intracellular mechanisms of endogenous activity in neurons.
518. Strumwasser F., pp. 442—462 in Circadian Clocks, edited by J. Aschoff, North-Holland Publ. Co., Amsterdam, 1965; also pp. 516—528 in The Neurosciences: A Study Program, edited by G. C. Quarton et al., Rockefeller University Press, New York, 1967; also pp. 291—320 in Invertebrate Nervous Systems, edited by C. A. G. Wiersma. University of Chicago Press, 1967. Circadian rhythm in a single neuron.
519. Strumwasser F., J. Psychiatr. Res. 8: 237—257, 1971. Rhythmic activity in molluscan neurons in culture.
520. Stuart A. E., Science 165: 817—819, 1969; also J. Physiol. 209: 627—646, 1970. Motorneurons in leech.
521. Suga N., Japan. J. Physiol. 11: 666—677, 1961. Tympanic neurons in noctuid moths.
522. Suga N., J. Insect Physiol. 12: 1039—1050, 1966. Ultrasonic production and its reception in some neotropical Tettigoniidae.
523. Summers T. B., Kaelber IF. M, Amer. J. Physiol. 203: 1117—1119, 1962. Amyg-dalectomy in cats.
524. Sutherland N. S., J. Comp. Physiol. Psychol. 54: 43—48, 1961. Discrimination, horizontal and vertical, octopus.
525. Sutterlin A. M., Prosser C. L., J. Neurophysiol. 33: 36—45, 1970. Electrical responses, optic tectum, goldfish.
526. Takeda K., Kennedy D., J. Cell. Comp. Physiol. 64: 165—182, 1964. Modes of activation of crayfish motorneurons.
527. Tarr R. S., Ph. D. Thesis, University of Illinois, 1971. Neuro-ethological study of ritualized aggressive behavior in the western fence lizard, Sceloporus.
528. Tauc L., J. Gen. Physiol. 45: 1077—1097, 1099—1115, 1962. Active membrane areas in giant neuron of Aplysia.
529. Tauc L., C. R. Acad. Sci. Paris 259: 885—888, 1964. Presynaptic inhibition, Aplysia neurons.
530. Tauc L., pp. 247—257 in Mechanisms of Synaptic Transmission, edited by K. Akert and P. G. Waser. Elsevier, Amsterdam, 1969, Polyphasic synaptic activity.
531. Tauc L., Hughes G. M., J. Gen. Physiol. 46: 533—549, 1963. Spikes in mollusc central neurons.
532. Taylor R. C., Comp. Biochem. Physiol. 33: 911—921, 1970. Contral of sensitivity of single crayfish interneuron.
533. Teravainen H„ Rovainen С. M., J. Neurophysiol. 34: 990—998, 999—1009, 1971. Motorneurons, sea lamprey.
534. Thach W. T„ J. Neurophysiol. 33: 527—536, 537—547, 1970. Discharge of cerebellar neurons related to two maintained postures and two prompt movements.
535. Thompson R. F., Johnson R. H., Hooper J. J., J. Neurophysiol. 26: 343—364, 1963. Organization of auditory, somatic sensory, and visual projection to association fields of cerebral cortex in cat.
536. Thompson R. F., Shaw J. A., J. Comp. Physiol. Psychol. 60: 329—339, 1965; 69: 485—491, 1969. Association cortex, cat.
537. Thompson R. F., Spencer IF. A., Psychol. Rev. 73: 16—43, 1966. Habituation.
160
Глава 15
538. Thorpe W. H., Learning and Instinct in Animals. Methuen, London, 1963. 558 p.
539. Towe A. L., Exp. Neurol. 15: 113—139, 1966. On nature of primary evoked response.
540. Treherne J. E., Beament I. W., editors, Physiology of the Insect Central Nervous System. Academic Press, New York, 1965, 277 p.
541. Truman J. W., Sokolove P. G., Science 175: 1491—1493, 1972. Silk moth eclosion.
542. Turner B., J. Comp. Physiol. Psychol. 71: 103—113, 1970. Rage syndrome of rat.
543. Ueda К. T., Hara J., Gorbman A., Comp. Biochem. Physiol. 21: 133—143, 1967. Olfactory discrimination in spawning salmon.
544. Van der Kloot W. G., Williams С. M., Behavior 5: 141—174, 1953; 6: 233—255, 1954. Hormonal and neural control of construction in silkmoths.
545. Вардапетян Г. А., Ответы одиночных нейронов в слуховой коре крыс, Ж. высш, нервн. деят., 17, 95 (1967).
546. Васильев А. Г., Электрическая активность заднего двухолмия летучей мыши, раздражаемой ультразвуком. ДАН СССР, 175, 541 (1967).
547. Verzeano №., рр. 27—54 in: Neuronal Control of Behavior, edited by R. E. Whalen et al. Academic Press, New York, 1970. Evoked responses and network dynamics.
548. Verzeano №.. Dill R. C. et al., Experientia 24: 696—698, 1968. Evoked responses, lateral geniculate.
549. Веселкин H. П., Электрические реакции в среднем, продолговатом и спинном мозге миноги при зрительной стимуляции. Физиол. ж. СССР, 52, 131 (1966).
550. Веселкин Н. П. и др., Brain Behav. Evol. 4: 295—306, 1971. Thalamotelen-cephalic afferent system in frogs.
551. Воронин Л. Л., Скребицкий В. Г., Вне- и внутриклеточное исследование ответов нейронов двигательной зоны коры бодрствующего кролика на звуковые и световые стимулы. Ж. высш, нервн. деят., 17, 523 (1967).
552. Vowles D. М., J. Comp. Physiol. Psychol. 58: 105—111, 1964. Offactory learning and brain lesions in wood ant Formica.
553. Wachtel H., Handel E. R„ Science 158: 1206—1208, 1967. Synaptic connection mediating both excitation and inhibition.
554. Waks M. D., Westerman R. D., Comp. Biochem. Physiol. 33: 465—469, 1970. Purkinje cells of Salmo.
555. Waldron J., J. Exp. Biol. 47: 201—212, 1967. Motor output pattern in flying locusts.
556. Waldron J., Z. vergl. Physiol. 57: 331—347, 1968. Mechanism'of coupling of locust flight oscillator to escillatory inputs.
557. Wall P. D., Johnson A. R., J. Neurophysiol. 21: 148—158, 1958. Post-tetanic potentiation of a monosynaptic reflex.
558. Weiss P., J. Comp. Neurol. 67: 269—315, 1937; also Comp. Psychol. Monogr.
17: 1—96, 1941. Coordination of transplanted amphibian limbs.
559. Wells G. P., J. Marine Biol. Assoc. U. K. 28: 447—464, 1949; also J. Exp. Biol. 28: 41—56, 1951. Feeding rhythms in Arenicola.
560. Wells M. J., J. Exp. Biol. 38: 811—826, 1961. Tactile and visual learning in brain of Octopus.
561. Wells M. J., Brain and Behavior in Cephalopods. Stanford University Press, 1962. 171 p.
562. Wells M. J., Ergebn. Biol. 26: 40—84, 1963. Orientation of Octopus.
563. Wells M. J., J. Exp. Biol. 41: 621—642, 1964. Detour experiments with octopuses.
564. Wells M. J., J. Exp. Biol. 42: 233—255, 1965. Orbital lobe and touch learning in Octopus.
565. Wells M. I., J. Exp. Biol., 47: 393—408, 1967. Short-term learning and interocular transfer in detour experiments with octopuses.
Центральная нервная система 161
566. Wells М. J., Young J. Z„ J. Exp. Biol. 50: 515—526, 1969. Brain lesions and effect of splitting part of brain or removal of median inferior frontal lobe on touch learning in Octopus.
567. Wendler G., Soc. Exp. Biol. Symp. 20: 229—249, 1966. Coordination of walking movements in arthropods.
568. Westfall J. A., J. Ultrastruct. Res. 32: 237—246, 1970; also J. Cell Biol. 51: 318—323, 1971. Synapses in a primitive coelenterate.
569. Whiting H. P., pp. 85—103 in International Neurochemical Symposium, 1st, Biochemistry of the Developing Nervous System: Proceedings, edited by H. Waelsch. Academic Press, New York, 1955. Functional development in the nervous system.
570. Wickelgren B. G., J. Neurophysiol. 30: 1404—1423, 1967. Habituation of spinal motorneurons.
571. Wickelgren B. G., Sterling P., J. Neurophysiol. 32: 16—23, 1969. Influence of visual cortex on receptive fields in superior colliculus.
572. Wiersma C. A. G„ J. Neurophysiol. 10: 23—38, 1947. Giant nerve fiber system of the crayfish.
573. Wiersma C. A. G., J. Cell. Comp. Physiol. 40: 399—419, 1952; also Physiol. Rev. 33: 326—355, 1953. Transmission in giant fiber systems, crayfish.
574. Wiersma C. A. G., pp. 241—279 in Physiology of Crustaceans, vol. 2, edited by T. Waterman. Academic Press, New York, 1961. Central nervous systems, crustaceans.
575. Wiersma C. A. G., pp. 269—284 in: Invertebrate Nervous Systems, edited by C. A. G. Wiersma. University of Chicago Press, 1967. Visual interneurons, crayfish.
576. Wiersma C. A. G., Bush В. M. H., J. Comp. Neurol. 121: 207—235, 1963. Neuronal connections between thoracic and abdominal cords of crayfish.
577. Wiersma C. A. G., Hughes G. M., J. Comp. Neurol. 116: 209—228, 1961. Functional anatomy of neuronal units in abdominal cord of crayfish.
578. Wiersma C. A. G„ Ikeda Ц., Comp. Biochem. Physiol. 12: 509—525, 1964. Interneurons commanding swimmeret movements in crayfish.
579. Wiersma C. A. G., Mill P. S., J. Comp. Neurol. 125: 67—94, 1965. Descending neuronal units in commissure of crayfish CNS.
580. Wiersma C. A. G., Yamaguchi T., J. Exp. Biol. 47: 409—431, 1967. Integration of visual stimuli by crayfish central nervous system.
581. Wiesel T. N., Hubei D. H., J. Neurophysiol. 26: 1003—1017, 1965; 28: 1060— 1072, 1965. Single-cell responses in striate cortex of kittens deprived of vision in one eye.
582. Willows A. O. D., in: Physiological and Biochemical Aspects of Nervous Integration, edited by F. D. Carlson, Prentice-Hall, 1967; also Science 157: 570—574, 1967. Behavioral acts elicited by stimulation of single identifiable nerve cells, Tritonia.
582a. Willows A. O. D., Sci. Amer. 224: 68—75, 1971. Behavioral function, single neurons, Tritonia.
583. Willows A. O. D., Hoyle G., Science 166: 1549—1551, 1969. Neuronal network triggering a fixed action pattern.
584. Wilson D. M., J. Exp. Biol. 38: 471—490, 1961; 41: 191—205, 1964. Locust flight, nervous control.
585. Wilson D. M., J. Exp. Biol. 47: 133—151, 1967. Stepping patterns in tarantulas.
586. Wilson D. M., in: Neural Theory and Modeling, edited by R. F. Reis. Stanford University Press, 1964. The origin of flight-motor command in grasshoppers.
587. Wilson D. M., J. Exp. Biol. 43: 397—409, 1965. Proprioceptive leg reflexes in cockroaches
588. Wilson D. M., J. Exp. Biol. 39: 669—677, 1962. Bifunctional muscles in thorax of grasshoppers.
589. Wilson D. M., J. Exp. Biol. 48: 631—641, 1968. Reflex modulation of locust flight.
11—2067
162
Глава 15
590. Wilson D. M., Adv. Insect Physiol. 5: 289—338, 1968. The nervous control of insect flight.
591. Wilson D. M., Gettrup E., J. Exp. Biol. 40: 171—185, 1963. Stretch reflex controlling wingbeat frequency in grasshopper.
592. Wilson D. M., Weis-Fogh T., J. Exp. Biol. 39: 643—667, 1962. Coordinated motor units in flying locusts.
593. Wilson D. M., Wyman R. I., J. Insect Physiol. 9: 857—865, 1963. Phasically unpatterned nervous control of dipteran flight.
594. Wilson V. J., J. Gen. Physiol. 39: 197—206, 1955; 41: 1005—1018, 1958. Post-tetanic potentiation of polysynaptic reflexes in spinal cord.
595. Wilson V. J., Burgess P. R., J. Neurophysiol. 25: 636—650, 1962. Antidromic conditioning of some motorneurons and interneurons.
596. Wilson V. J. et al., J. Neurophysiol. 27: 1063—1079, 1964. Inhibitory convergence on Renshaw cells.
597. Wine J. J., Krasne F. B., J. Exp. Biol. 56: 1—18, 1972. Escape behavior in crayfish.
598. Winlow W., Laverock M. S., Marine Behav. Physiol. 1: 1—27, 29—47, 93—121, 1972. Control of hindgut motility in lobster.
598a. Witt P. N., Amer. Zool. 9: 121—131, 1969. Effects of brain lesions in spider Araneus.
599. Woody C. D„ J. Neurophysiol. 33: 838—850, 1970. Conditioned eye blink.
600. Yoon M., Exp. Neurol. 33: 395—411, 1972. Connections of retina to tectum, goldfish.
601. Yoshida M., pp. 435—465 in: Physiology of Echinodermata, edited by R. Bo-lootian. Interscience, New York, 1966. Photosensitivity of echinoids.'
602. Young J. Z., Quart. J. Mier. Sci. 78: 367—386, 1936; also Phil. Trans. Roy. Soc. Lond. В 229: 465—501, 1939. Giant fiber system of cephalopods.
603. Young J. Z., Proc. Roy. Soc. Lond. В 149: 463—483, 1958. Responses of untrained octopuses to various figures and the effect of removal of the vertical lobe.
604. Young J. Z., Proc. Roy. Soc. Lond. В 153: 18—46, 1960. Failure of discrimination learning following removal of vertical lobe of Octopus.
605. Young J. Z., Biol. Rev. 36: 32—96, 1961. Learning and discrimination in Octopus.
606. Young J. Z., Phil. Trans. Roy. Soc. Lond. В 245: 1—58, 1962. Retina and optic lobes of cephalopods.
607. Young J. Z. Quart. J. Exp. Psychol. 14: 193—205, 1962. Revers?.! of learning in Octopus and effect of removal of vertical lobe.
608. Young J. Z., Nature 198: 626—630, 1963. Essentials of neural memory systems.
609. Young J. Z., Phil. Trans. Roy. Soc. Lond. В 249: 27—44, 45—67, 1965. Octopus, buccal nerve system; centers for touch discrimination.
610. Young J. Z., J. Exp. Biol. 43: 595—603, 1965. Influence of previous preferences on memory of Octopus.
611. Young J. Z., pp. 353—362 in: Invertebrate Nervous Systems, edited by C. A. G. Wiersma. University of Chicago Press, 1967. Cephalopod brain.
612. Young J. Z., J. Exp. Biol. 49: 413—419, 1967. Reversal of visual preference in Octopus after removal of vertical lobe.
613. Young J. Z„ J. Exp. Biol. 52: 385—393, 1970. Short and long memories in Octopus and the influence of the vertical lobe system.
614. Загорулько T. M„ Вызванные ответы общей коры и tectum opticum у черепах (1969).
615. Зенкин Г. М., Пигарев И. Н„ Детекторные свойства ганглиозных клеток сетчатки щуки. Биофизика, 14, 722 (1969).
616. Жантиев Р. Д., Функциональные свойства нейронов в центральной нервной системе прямокрылых насекомых. ДАН СССР, 181, 477 (1968).
,617. Zucker R. S., J. Neurophysiol. 35: 599—620, 621—637, 1972. Crayfish escape behavior and central synapses.
Центральная нервная система
163
618. Zucker R. S., J. Neurophysiol. 35: 638—651, 1972. Excitation of fast flexor motorneurons.
619. Kusano K-, J. Gen. Physiol. 52: 326—345, 1968.
620. Kennedy L)., Selverston A. I., Rentier M. P., Science 164: 1488—1496, 1969.
621. Retzlaff E., J. Comp. Neurol. 101: 407—445, 1954.
622. Furukawa T., in: Tokirane T. a. Schade J. P. (Eds.), Correlative Neurosciences, Progr. Brain Res. 21A, 44—70, 1966.
623. Baylor D. A., Nicholls J. G., in: Carlson F. D., ed. Physiological and biochemical aspects of nervous integration. Prentice-Hall, 3—16, 1967.
624. Kammer A. E., Z. vergl. Physiol. 70: 45—56, 1970.
625. Truex R. C., Carpenter M. B., eds., Strong and Elwyn’s human neuroanatomy, 5th ed. Williams, Wilkins, Baltimore, 1964.
626. Vanegas EE, Essayag-Millan E., Laufer M., Brain Res. 31: 107—118, 1971.
627. Woolsey C. N., in: Harlow H. F., Woolsey C. N., eds., Biological and biochemical basis of behavior, University of Wisconsin Press.
628. Nieuwenhuys R., in: Fox C. A., Snider R. S., eds., The Cerebellum, Progr. Brain Res. 25: 1—93, Elsevier, Amsterdam, 1967.
629. Eccles J. C., Llinas R., Sasaki K. et al., Exp. Brain Res. 1: 161—183, 1966.
630. Llinas R. R. et al., in: Fields W. S., Willis W. D., eds., The Cerebellum in health and disease. Warren H. Green Publishers, St. Louis, 1969.
631. Michael C. R„ Retinal processing of visual images, Scientific American, May 1969, 109.
632. Jacobson M., Baker R. E., J. Comp. Neurol. 137: 121—141, 1969.
633. Young J. Z., Proc. Roy. Soc. Lond. В 159: 565—588, 1964.
634. Young J. Z., Proc. Roy. Soc. Lond. В 163: 285—320, 1965.
635. Kupfermann I., Castelucci V., Pinsker H., Kandel E., Science 167: 1743—1745,
1970.
Глава 16
Мышцы
Л. ПРОССЕР
Скорость передвижения животного и, следовательно, возможность добывать пищу или избегать нападения, зависит от скорости реакции его мышц. Помимо двигательной функции мышцы выполняют также функции, связанные с пищеварением, выделением, размножением и вокализацией; у большинства животных имеется несколько типов мышц, обладающих различными временными характеристиками.
Сократительные фибриллы встречаются уже у простейших (например, мионемы Vorticella и Stentor); сократительные миоциты могут закрывать поры у губок.
Клеточный механизм сокращения и расслабления обеспечивается особыми, весьма своеобразными белками. Один и тот же сократительный механизм лежит, по-видимому, в основе как быстрых движений глаз млекопитающих или комариного крыла, так и вялых движений актинии или кишечника позвоночных.
Мышечная ткань представляет собой наиболее яркий пример приспособления одного и того же механизма к выполнению самых разнообразных функций. Гистологические, биохимические и механические особенности мышц весьма разнообразны. Сравнительное изучение выявило следующие коррелирующие между собой изменения свойств мышц: 1) структурную организацию сократительных элементов, 2) природу сократительных белков, 3) обеспечение энергией процессов сокращения и расслабления, 4) временные характеристики сократительного акта, 5) особенности нервно-мышечной передачи и ее изменение при облегчении, 6) связь функции мембран с сокращением, 7) влияние механических (вязкоэластических) свойств при длительном поддержании напряжения, тонуса и при быстром растяжении и расслаблении, 8) спонтанную ритмическую активность и ее нервную и механическую регуляцию.
Одни мышцы — позные—по происхождению и месту прикрепления связаны со скелетом, внутренним или внешним, а также с кожей. Примерами могут служить мышцы, приводящие в движение ноги, крылья и ротовые части; мышцы, выдвигающие хоботки
Мышцы
165
и щупальца, или мышцы, закрывающие створки раковин моллюсков. Такие мышцы часто образуют пары антагонистов: сокращение одной мышцы рефлекторно тормозит сокращение другой, выполняющей противоположную функцию. По отношению к некоторым мышцам, например аддуктору моллюсков, антагонистическую функцию выполняют эластические связки. Обычно это сравнительно быстрые или фазные мышцы; они образуют часть рычажной системы и работают либо изотонически (укорачиваясь во время сокращения), либо изометрически (развивая напряжение при постоянной длине). Строго говоря, ни одна мышца не работает только изотонически или только изометрически — она может только приближаться к одному из этих режимов.
Другие мышцы располагаются в стенках полых органов; они составляют единую систему, сокращение одной ее части растягивает соседние. Примерами могут служить мышцы мочевого пузыря, мочеточника, матки, пищеварительного тракта, сердца, а также мышцы стенок тела кольчатых червей, голотурий и кишечнополостных. Как правило, мышцы внутренних органов сокращаются медленно (их называют тоническими); они могут быть парными (например, кольцевые и продольные), а в других случаях могут сокращаться одновременно, например при сжимании полостей, наполненных жидкостью.
Регуляция как фазных (быстрых), так и тонических (медленных) мышц осуществляется рефлекторным путем; однако многие тонические мышцы обладают спонтанной ритмической активностью, которая может контролироваться нервной системой. Фазные мышцы часто образуют двигательные (моторные) единицы, т. е. группы волокон, иннервируемых одним мотонейроном. Сила и скорость сокращения могут определяться числом возбужденных моторных единиц, частотой двигательных импульсов, соотношением между количеством участвующих в сокращении быстрых и медленных, возбуждающих и тормозных нейронов и, наконец, механическими свойствами соединительной ткани и сократительных волокон.
Гистологические типы мышц
Основным структурным элементом мышцы является мышечная нить, или протофибрилла, состоящая из сократительного белка.. Толстые нити (диаметром 150—250 А) содержат миозин, а тонкие (45—65 А) — актин. Нити иногда собраны в пучки — миофибриллы. Основной клеточной единицей мышцы является мышечное волокно, которое может быть одноядерным или многоядерным. Волокна могут образовывать пучки. Количество соединительной ткани между волокнами сильно колеблется в разных типах мышц. Наружный слой мембраны волокна (наружная сарколемма) связан, по-видимому, с соединительной тканью, в то время как внут
166
Глава 16
ренний слой мембраны представляет собой физиологическую плазматическую мембрану.
Мышцы можно грубо разделить на поперечнополосатые и гладкие, однако существует много промежуточных типов, например «спиральноисчерченные» мышцы круглых червей.
Поперечнополосатые мышцы. Для поперечнополосатых мышц характерно упорядоченное расположение толстых и тонких нитей
I а I
z
Z
Н
I I ’
iiiiiiii iinTffi»*
HIIIIII ПНИ»*
illlllll Н1Ж#н - НИН
illlllll liTW*
Iilllllll 11Ж1МН
п||)1111 illliyi'
IIHHII liTW
• • ® ® ®
• ее
® • о ®
• ® • •
• • • •
• • ®
Рис. 164. Схема строения поперечнополосатой мышцы с перекрывающими друг друга рядами тонких и толстых нитей, поперечными мостиками и Z-пластинками в середине 1-дисков [1196].
Вверху — продольный разрез, внизу — поперечный, через разные участки саркомера.
и наличие системы продольных и поперечных канальцев (трубочек). Поперечная исчерченность возникла на ранних этапах эволюции: уже многие волокна кишечнополостных являются поперечнополосатыми. Поперечная исчерченность есть результат чередования участков с сильным и слабым лучепреломлением — анизотропных (А) и изотропных (I) дисков. 1-диски делятся пополам Z-пластинками (рис. 16-1), которые представляют собой поперечные границы между структурными единицами волокна — саркомерами. 1-диск состоит из тонких актиновых нитей, а А-диск — из толстых миозиновых нитей и заходящих между ними концов тонких нитей, однако в середине диска может быть более светлая Н-зона, в которой тонких нитей нет; в центре ее иногда заметна линия М. Тонкие и толстые нити в области взаимного перекрытия соединены поперечными мостиками. Тонкие нити прикреплены к Z-пластинкам.
Тубулярная система, или система канальцев, состоит из 1) впя-чиваний сарколеммы, проникающих в волокно на различные расстояния в области Z-пластинки или на границе А- и 1-дисков и называемых поперечными канальцами (или Т-канальцами) и 2) сети продольных канальцев (саркоплазматический ретикулум,
Мышцы
167
СР) и пузырьков между миофибриллами, образующих ячеистую «оплётку» вокруг саркомера (рис. 16-3), с вздутиями (цистернами), прилегающими к Т-канальцам. По Т-канальцам мембрана сарколеммы впячивается внутрь волокна, и по ней возбуждение передается с наружной мембраны саркоплазматическому ретикулуму, обеспечивая переход от возбуждения к сокращению (exita-tion-contraction coupling).
Скорость сокращения мышцы коррелирует со следующими факторами: длиной саркомера, упорядоченностью и плотностью упаковки протофибрилл, количеством митохондрий и липидных капелек, присутствием или отсутствием миоглобина в саркоплазме. Быстрые мышцы с так называемой фибриллярной структурой (Fibrillenstruktur) бедны саркоплазмой, содержат много правильно расположенных нитей (обычно вокруг одной толстой нити имеется шесть тонких); их тубулярная система имеет постоянную структуру; Z-пластинки проходят точно поперек волокна (рис. 16-2, 16-4А). Другие, более медленные мышцы богаты саркоплазмой (имеют Felderstruktur—структуру «полей»), с менее правильным расположением нитей и канальцев, с большим количеством митохондрий и зигзагообразной Z-пластинкой (рис. 16-4,Б). Напряжение, развиваемое мышцей на единицу поперечного сечения, у мышц с длинными саркомерами, как правило, больше, так как при одинаковом расстоянии между расположенными параллельно мостиками их общее количество, приходящееся на одну нить, будет большим (что позволит развить большее напряжение). С другой стороны, мышцы, имеющие короткие саркомеры, способны к быстрым сокращениям. Обычно мышцы с фибриллярной структурой имеют одну зону иннервации, в которой каждое волокно получает одно пластинчатое (en plaque) нервное окончание, между тем как волокна мышцы со структурой «полей» снабжены несколькими гроздевидными (en grappe) окончаниями или многочисленными нервными бляшками. Волокна поперечнополосатых мышц многоядерные и имеют большой диаметр (около 100 мкм). На рис. 16-3 и 16-4, А показана структура быстрой поперечнополосатой мышцы. В портняжной мышце лягушки миофибриллы или пучки нитей малы, а Т-канальцы, пронизывающие волокно на уровне Z-пластинки, образуют плоские дискообразные полости, или цистерны. К ним непосредственно примыкают концевые цистерны продольной ретикулярной системы, образуя триады — две цистерны с Т-канальцем между ними. Концевые канальцы (цистерны) соединены с продольными канальцами саркоплазматического ретикулума (СР-система). Поверхность поперечных канальцев приблизительно в семь раз превышает наружную поверхность волокна [276]. В «медленных» мышцах млекопитающих объем поперечных канальцев составляет 0,18% объема волокна, а в «быстрых» — 0,39%; площадь соприкосновения диад у «быстрых» волокон в, 5—10 раз больше, чем у «медленных».
168
Глава 16
У змеи Thamnophis быстрые волокна имеют поперечные канальцы на уровне границы А- и 1-дисков (т. е. имеют по две Т-системы в каждом саркомере); их связь с внеклеточным пространством была выявлена методом импрегнации зернами ферритина [286]. В мышцах рыб триады также могут быть на уровне
Рис. 16-2. Ультраструктура «быстрой» поперечнополосатой мышцы [304].
Электронная микрофотография крикотиреоидной мышцы летучей мыши. Видно взаимное расположение T-каналъцев, миофибрилл и митохондрий.
границы А- и 1-дисков, как у экстраокулярной мышцы Fundulus или на уровне Z-пластинки, как в тонкой спинной мышце Hipp0' campus [32]. В очень быстрой «звуковой» мышце плавательного пузыря опсануса саркомеры имеют длину всего 2 мкм, нити упорядочены, особенно в центральной части волокна, а поперечные канальцы проникают в волокно на уровне границы А- и 1-дисков
Мышцы
169
(т. е. на каждый саркомер приходится по два Т-канальца) [ИЗ, 304]. В мышцах ноги речного рака имеются большие впячивания сарколеммы во многих участках саркомера, которые разветвляются на канальцы диаметром 300 А, контактирующие с канальцами саркоплазматического ретикулума на границе А- и 1-дисков, образуя диады [44] (рис. 16-5). Расстояние от Т-канальцев до
Р.ис. 16-3. Модель папиллярной мышцы желудочка сердца кошки [114].
А — субсарколемные цистерны, 2 — митохондрия, 3 — Т-каналец, 4 — саркоплазматический ретикулум.
центра фибриллы не превышает 0,4 мкм [202]. Подсчитано, что общая поверхность мембран тубулярной системы, включая поперечные канальцы, приблизительно в 10 раз превышает площадь наружной мембраны волокна [303]. Сходные впячивания или щели встречаются и в мышечных волокнах краба, где на уровне Z-пластинки они дают начало слепым канальцам, а на уровне границы А- и 1-дисков — Т-канальцам, образующим диады с СР-элементами [275]. В быстрых мышцах веслоногих рачков (Copepoda), как и у речного рака, канальцы проникают на уровне Z-пластинки и дают ответвления к цистернам [НО]. На рис. 16-6 показана структура быстрых и медленных мышц скорпиона.
Соотношение количества тонких и толстых нитей варьирует в Разных мышцах и даже в различных частях одного и того же волокна [333, 334]. В мышцах мечехвоста (Litnulus), где длина СаРкомера достигает 8 мкм, а толстые нити имеют плотную сердцевину и менее плотную оболочку, на одну толстую нить в центре
170
Глава 16
A-диска приходится 12 тонких, а на концах диска — еще больше [197]. В некоторых мышцах ног речных раков и тараканов отношение числа тонких нитей к числу толстых составляет 6:1, в грудных мышцах таракана и бабочек —4: 1, в летательных мышцах стрекозы и падальной мухи — 3:1, а у большинства позво-
Рис. 16-4. Сравнение фазных и тонических мышечных волокон [265].
Электронные микрофотографии. А. Фазное волокно. Б. Тоническое волокно; продольное сечение.
ночных в поперечнополосатых мышцах и в сердечной мышце — 2:1 [153Ь].
Поперечные мостики между толстыми и тонкими нитями образуют левую спираль с расстоянием между мостиками, равным 146 А; в состоянии окоченения (ритора) они образуют шевроноподобный узор [229]. У насекомых трахеолы могут проникать внутрь крупных волокон, а митохондрии (саркосомы) могут составлять до 40% общего объема волокна [333].
Между диаметром волокна и количеством канальцев ретикулума существует прямая зависимость. В волокнах папиллярной мышцы сердца кошки клетки размером 10—12 мкм имеют большие Т-канальцы, развитую сеть саркоплазматического ретикулума; в то же время в клетках предсердия, имеющих диаметр 5—6 мкм, отмечается лишь небольшое количество разбросанных Т-каналь-
Мышцы
171
цев, причем цистерны CP-системы в них направлены скорее к периферии клетки, чем к Т-системе. Миотомы волокон Branchiosto-та представляют собой плоские пластинки толщиной 8 мкм, шириной 100 мкм и длиной 600 мкм с небольшим количеством Т-ка-нальцев [274]. Крупные волокна насекомых имеют столь богатую
Рис. 16-5. Модель мышечного волокна речного рака [303].
Видно впячивание сарколеммы (С) и Т-канальцы (Т), которые с примыкающими к ним CP-канальцами (СР) образуют диады (Д).
систему канальцев, что любая часть мышечного волокна отстоит ют канальца не дальше чем на 2 мкм [333].
Резюмируя, можно сказать, что в медленных исчерченных мышцах нити образуют преимущественно нерегулярную, «полевую» структуру (Felderstruktur), Z-пластинки расположены беспорядочно, триады редки или отсутствуют, канальцы немногочисленны, волокна могут иметь множественную иннервацию с гроздевидными (en grappe) окончаниями. Медленные мышцы имеют Длинные саркомеры и содержат обычно больше митохондрий и жировых капелек, чем фазные. Медленно утомляющиеся тонические мышцы часто содержат миоглобин, придающий им красный Цвет, в отличие от быстро утомляющихся фазных мышц, которые называются белыми. Исключения из этих обобщений будут приведены позже.
Мышцы
173
Промежуточные типы мышц. Некоторые мышцы имеют крайне своеобразную исчерченность. Так, у некоторых беспозвоночных, особенно трематод и асцидий, мышечные волокна имеют типичную исчерченность лишь на одной стороне клетки; на другой стороне расположены митохондрии. Другие мышцы беспозвоночных имеют косую исчерченность. Z-пластинки могут быть расположены беспорядочно или уступами по диагонали '[248]. Такое расположение видно на исчерченных полупрозрачных мышцах двустворчатых моллюсков, например у гребешка (Pecten) и в мышцах хроматофоров кальмара [123]. У дождевых червей мышечные волокна имеют ленточную форму с пучками нитей в кортексе, рас-
Рис. Ь6-6. Волокна мышцы скорпиона. (Любезно .предоставлено А. Джилаи и И. Парнасом.)
Л. Поперечное сечение; вверху — быстрое волокно, в середине—медленное и внизу — промежуточное; обратите внимание на разницу в упорядоченности взаимного расположения нитей и на Т- и С-канальцы. Б. Модель части волокна. Видны Т-канальцы, цистерны против A-дисков и С-канальцы между цистернами.
174
Глава 16
положенными по спирали; такая спиралевидная структура особенно отчетливо выявляется в поляризованном свете.
В мышцах стенки тела Ascaris и в миотомах Branchiostoma от одной из сторон волокна отходят отростки по направлению к нервному стволу, где они получают синаптическую иннервацию. Мышца как бы сама направляется к нервной системе.
Все промежуточные, или полуисчерченные, мышцы имеют ограниченную тубулярную систему, менее развитую в сильно уплощенных волокнах по сравнению с нормальными (толстыми).
Неисчерченные мышцы. Волокна неисчерченных мышц могут быть длинными (до 2 см) или короткими (100—300 мкм) и веретенообразными. Все волокна имеют небольшую толщину, обычно от 2 до 5 мкм в поперечнике. Т-канальцев нет, саркотубулярная система либо отсутствует, либо слабо выражена, но в некоторых случаях отмечается наличие большого количества пиноцитозных пузырьков. Это наблюдение согласуется с представлением о том, что Т-канальцы необходимы для проведения возбуждения от мембраны внутрь волокна и что при отсутствии Т-канальцев в волокнах малого диаметра имеется прямое взаимодействие между мембраной и сократительной системой.
В неисчерченных мышцах отсутствует поперечное ориентирование толстых и тонких нитей; тем не менее во многих неисчерченных мышцах толстые и тонкие (миозиновые и актиновые) нити образуют подобие правильного узора (рис. 16-7). Среди нитей разбросаны темные тельца, которые являются, по-видимому, гомологами Z-пластинок. Толстые и тонкие нити особенно хорошо различаются в неисчерченных мышцах беспозвоночных. Даже в миоцитах губок можно видеть нити двух размеров. В висцеральных гладких мышцах позвоночных толстые нити часто бывают не видны, однако из них экстрагируется миозин. В некоторых сокращенных гладких мышцах (мускульный желудок цыпленка, taenia coli морской свинки и некоторые мышцы сосудов) толстые нити видны так же хорошо, как и тонкие [92, 305, 328]. Возможно, что в расслабленном состоянии миозин присутствует в виде димеров, которые при сокращении полимеризуются. Актин тонких нитей с помощью боковых отростков прикрепляется к темным тельцам и к мембранным бляшкам [267].
В некоторых неисчерченных мышцах, имеющих как короткие, так и длинные волокна, видны очень толстые нити (1000—1500 А) с отчетливыми полосами шириной 145 А, повторяющиеся через каждые 760 А. Эти очень толстые нити содержат парамиозин (тропомиозин А), миозин же расположен, по-видимому, на их поверхности. Парамиозиновые нити видны в различных запирательных мышцах, например в аддукторах моллюсков, в биссусном ретракторе Mytilus, ретракторе хоботка сипункулиды (Golfingia) в мышцах стенки тела круглого червя волосатика (Paragordius) и в хорде Brdnchiostotna.
Рис. 16-7. Гладкомышечное волокно воротной вены кролика; электронные микрофотографии. (Любезно предоставлено К. Девайном и А. Сомлио.)
А. Продольное сечение периферической части волокна; видны миофибриллы и прилегающие непосредственно к ним пузырьки и С-канальцы. Б. Поперечное сечение; видны толстые нити, рассеянные среди множества тонких, митохондрии и периферические пузырьки; некоторые из них соединяются с С-канальцами.
176
Глава 16
Некоторые гладкие мышцы (как с короткими, так и с длинными волокнами) богато иннервированы, причем в их иннервации зачастую участвуют два нейрона. В других, например в висцеральных мышцах позвоночных, нервные волокна расположены далеко друг от друга и мышечная клетка может быть отделена от нервного окончания пятью и даже десятью другими клетками. В этих условиях медиатор, по-видимому, диффундирует в межклеточное пространство и его действие передается по группе клеток, поскольку функциональной единицей здесь является группа электрически взаимодействующих клеток [50а].
Неисчерченные мышцы с короткими волокнами подразделяются на 1) мультиунитарные, в которых активация и проведение обеспечиваются нервным путем, и 2) унитарные, в которых возбуждение возникает спонтанно (миогенный ритм) и передается от волокна к волокну через контакты с малым электрическим сопротивлением. Нервная система лишь регулирует подобные сокращения. Унитарные мышцы более чувствительны к растяжению, чем мультиунитарные. Большинство гладких позных мышц беспозвоночных относится к мультиунитарным, например ретракторы хоботка сипункулиды Golfingia, ретракторы радулы улитки и различных иглокожих и некоторые мышцы кишечнополостных; мультиунитарные мышцы у позвоночных найдены в мигательной перепонке, пиломоторах и в некоторых кровеносных сосудах; унитарные мышцы встречаются главным образом в висцеральных оога-нах позвоночных, у некоторых моллюсков и иглокожих..
Сократительные белки и теории сокращения
Обычная последовательность событий, наблюдаемых при мышечном сокращении, такова: электрический сигнал в мембране волокна передается внутрь по Т-канальцам, которые каким-то образом активируют саркоплазматический ретикулум. В мышцах, лишенных тубулярной системы, поверхностная мембрана непосредственно взаимодействует с сократительными элементами. В любом случае происходит высвобождение ионов Са2+ либо в пузырьках CP-системы, либо в мембране волокна; благодаря кальцию в присутствии АТФ возникает взаимодействие между актином и миозином. Гидролиз АТФ вызывает в комплексе актин—миозин конформационные изменения, природа которых неясна; в исчерченной мышце толстые и тонкие нити начинают двигаться (скользить) друг, относительно друга, возможно, из-за изменения угла наклона поперечных мостиков (рис. 16-8). Затем кальций активно связывается CP-мембранами и наступает расслабление. Удаление Са2+ тормозит действие АТФазы актомиозина (AM); при этом поглощение Са2+ саркоплазматическим ретикулумом связано с
Мышцы
177
ДТФазой CP-пузырьков. Основная реакция такова: расслабление
AM " .......*• А + М;
сокращение
Са2+ способствует сдвигу реакции влево.
В задачу настоящей главы не входит детальное обсуждение физической химии миофибриллярных белков, хорошо изученных на поперечнополосатых мышцах млекопитающих; необходимо, однако, привести здесь некоторые общие сведения, полезные для сравнительно-физиологического подхода.
Миозин. Миозин получают экстрагированием мышцы в растворе КС1 высокой ионной силы, причем в такой экстракт переходит 50—55% структурного белка миофибрилл и весь или почти весь белок толстых нитей. Миозин состоит из крупных и сложных молекул с мол. весом от 470 000 до 490 000 >[96—98]. Миозиновые молекулы настолько велики, что их можно видеть с помощью электронного микроскопа. Они состоят из длинной хвостовой части (ее длина приблизительно равна 1400 А, а диаметр — 20 А) и глобулярной головки (длина около 200 А, диаметр 50—70 А). Глобулярная головка состоит из двух долей [228]. Были проведены физико-химические исследования фрагментов миозиновой молекулы, получающихся при обработке миозина веществами, нарушающими его четвертичную структуру, но не разрушающими ковалентных связей; они позволили установить, что миозин состоит из двух больших полипептидных субъединиц или цепей с мол. весом от 200 000 до 212 000, образующих в хвостовой части две перевитые ia-спирали, а в головной — две глобулярные доли. Кроме того, три или четыре полипептида (легкие цепи или легкие компоненты) локализованы в головной части и составляют 12— 15% всей массы миозиновой молекулы [84а, 228а, 325].
Состав легких и тяжелых цепей миозина зависит от мышц, из которых они экстрагированы. Белые скелетные мышцы млекопитающих по числу компонентов их легких и тяжелых цепей отличаются от красных и сердечной мышц [325]. Белые скелетные мышцы взрослого кролика дают при электрофорезе четыре фракции, в то время как в красной и сердечной мышцах и в белой мышце плода — две более подвижные фракции неотчетливы или отсутствуют [279а]. Миозин белых скелетных мышц кролика можно расщепить на три декапептида, которые содержат 3-метилгис-тидин; в гомологичных цепях сердечной мышцы гистидин не метилирован. Различия в последовательности аминокислот указывают на то, что структура миозина сердечной и скелетной мышц закодирована в различных генах. В сердечной мышце, по-видимому, отсутствует фермент для метилирования гистидина [191а]. Путем соответствующей обработки легкие цепи миозина можно отделить от тяжелых. Удаление всех легких цепей ведет к полной 12—2067
Рис. 16-8.
А. Продольное сечение быстрой скелетной мышцы [193J.
Видны Z-пластинки (вверху и внизу), толстые нити в A-диске и тонкие нити в 1-диске, рас» положенные между толстыми и соединенные с ними поперечными мостиками. Б. Электронная микрофотография и схема исчерченной мышцы насекомого. Показано положение поперечных мостиков при расслабленном состоянии мышцы и при ее окоченении 1299]. I — око-о
ченение; II— расслабление; 1 — по данным электронной микроскопии — ЗВО А; по данным о о
рентгеноструктуриого анализа — 388 А; 2 — по данным электронной микроскопии — 143 А, О
по данным рентгеноструктурного анализа — 146 А.
Мышцы
179
.неспособности миозина взаимодействовать с АТФ и к потере его ДТФазной активности.
С помощью гидролиза трипсином молекулу миозина можно расщепить на два фрагмента: а) большая часть хвоста (легкий
Рис. '16-9.
А. Схематическое изображение различных частей миозиновой молекулы, показывающее воз* можное расположение фрагментов (по результатам опытов с расщеплением) [196]. 1 — точки действия трипсина и папаина; 2 — «головка» или Si-субъединицы, обладающие АТФаз-ной активностью и взаимодействующие с актином. Б. Расположение миозиновых молекул в толстой нити. Основа состоит из хвостовых частей молекул, образующих два антипарал-лельных пучка [196]. В. Возможные механические изменения, возникающие в поперечных мостиках при сокращении [192]. Головка миозина (Г), соединенная с толстой нитью эла* стичным компонентом, который может изменять положение от В2 до Bi, а с тонкой нитью иосредством элемента, который может менять место прикрепления от Mi до М<.
меромиозин, ЛМ.М.) с мол. весом 120 000—150000 и длиной 800— 900 А; из него состоит сердцевина толстых нитей (рис. 16-9); б) головная часть с остатком хвоста (тяжелый меромиозин, ТММ)
12*
180
Глава 16
с мол. весом около 340 000; ТММ сохраняет АТФазную активность, способность соединяться с актином и образует поперечные мостики. При дальнейшем «переваривании» трипсином тяжелый меромиозин можно расщепить еще на три части. Две из них соответствуют двум глобулярным долям головки миозиновой молекулы, каждая с мол. весом в 110 000, сохраняют АТФазную активность и способность взаимодействия с актином; одна соответствует остатку хвоста и имеет мол. вес 60 000 и длину от 400 до 500 А [136] (рис. 16-9).
Рентгенографические и электронно-микроскопические исследования натуральных и искусственных нитей, а также физические измерения формы и размера позволили заключить, что стержневые части миозиновых молекул частично перекрывают друг друга в середине толстой нити, а головки каждой пары молекул направлены в противоположные стороны [196] (рис. 16-9). Результатом такого расположения является «ложная» Н-зона (обнаженная часть толстой нити), где нет выступающих поперечных мостиков. В каждой половине толстой нити молекулы миозина упакованы так, что их головки ориентированы в одном направлении. Поперечные мостики выступают, образуя подобие ступенчатой спирали, причем спирали двух половин A-диска закручены в противоположные стороны [193, 195]. Поперечные мостики выступают попарно на противоположных сторонах нити, каждая пара на расстоянии 143 А от другой с поворотом на 120°. Таким образом, на один оборот приходится три пары мостиков, занимающих 429 А по длине нити {196]. В быстрых мышцах насекомых угол между соседними мостиками составляет 67,5°, что дает поворот в 190° на каждые 380 А длины нити [299].
Актин. Актин может быть выделен в виде мономера, имеющего мол. вес около 46000 и содержащего только одну полипептидную цепь; он составляет 20—25% структурных белков миофибрилл и представляет собой главную составную часть тонких нитей. Актин может в соответствующих солевых растворах переходить из глобулярной формы (G-актин) в фибриллярную (F-актин), структура которой напоминает тонкие нити. Полимеризованный F-актин состоит из линейной цепи бусинок G-актина. Актиновые нити в мышцах во всех случаях имеют диаметр 60—90 А и состоят из двух перевитых правозакрученных спиралей с повторяющимися участками по 360 А, в каждом из которых имеется 13 бусинок (диаметром 56,5 А) G-актина (рис. 16-10,А) [158]. Актин, выделенный из поясничной мышцы кролика, аддукторов морского гребешка Pecten и Crassostrea, мантии Loligo, продольной мышцы Lnmbricus и толстой кишки морской свинки, состоит из совершенно одинаковых нитей [158].
Разделить актин и миозин в мышцах лягушки, рыбы, мечехвоста и некоторых насекомых не так легко, как в мышцах млекопитающих. Экстрагируемая смесь актомиозина называется мио-
Тропонин Актин
Рис. 16-10.
А. Спиральная форма молекул F-актина [158]. Б. Схема строения тонкой нити, показывающая предполагаемое взаиморасположение молекул актина и тропонина вдоль тропомиозиновой молекулы [105].
182
Глава 16
зином В; кроме миозина и актина он содержит некоторые другие белки миофибрилл. Актомиозин был выделен также из гладких мышц моллюсков [218] и рыб [77, 78] и по своим свойствам очень близок к миозину В млекопитающих. Миозин форели менее термостоек, чем миозин млекопитающих [53]. В период развития головастика свойства его актомиозина меняются, указывая на то, что вместе с увеличением количества актомиозина происходят и его химические перестройки [260]. В миобластах эмбрионов, пока псчерченность еще не видна в микроскоп, наличие миозина может -быть продемонстрировано иммунологическими методами. В курином эмбрионе до 12-го дня инкубации содержание актомиозина •составляет менее 1 мг/г; затем оно быстро возрастает и к моменту вылупления доходит до 50 мг/г. В гладкой мышце матки •содержание актомиозина под действием эстрогена и при беременности возрастает в несколько раз. В 0,6 М растворе КС1 актомиозин содержит миозин и F-актин в весовом соотношении 3,7 : 1, или одну молекулу миозина на 2,7 мономера актина '[105]. При низкой ионной силе раствора тяжелый меромиозин соединяется с актином в молярных отношениях. Возможно, что контакт одной головки ТММ (или нативного миозина) с актином влияет на связывание другой головки.
Тропомиозин и тропонин. Из миофибрилл может быть экстрагирован белковый комплекс, называемый тропомиозином, который, как теперь известно, состоит из двух белков: тропомиозина В и тропонина. Поперечнополосатая мышца кролика содержит 54% миозина, 21% актина и 15% нативного тропомиозина. Мышцы насекомых— 55,2% миозина, 21% актина и 9% тропомиозина [62]. Тропомиозин В и тропонин в пропорции 1 : 1,3 найдены в мышце кролика [104]. Молекулярный вес тропомиозина В равен 70 000, а его константа седиментации (S20) равна 2,7; молекулярный вес тропонина оценивается в 80 000—90 000, а его S20 равна 3,8. По современным представлениям тропомиозин состоит из трех компонентов; один из них — тропонин А (фракция, связывающая Са2^) имеет мол. вес от 18000 до 20 000, несет большой отрицательный заряд и имеет очень большое сродство к ионам кальция — четыре иона Са2+ на одну молекулу или 22 мкМ Са2+ на 1 г белка.
Тропонин В имеет мол. вес 60 000 и состоит из двух субъединиц, имеющих мол. вес 23 000 и 37 000 [104]. Молекула тропомиозина имеет палочковидную форму и следующие размеры: длина — 400 А, ширина 20—ЗОА; они располагаются по всей длине тонкой нити (рис. 16-10, Б) [105, 106]. Тропонин оказывается связанным с тропомиозином через каждые 400 А по длине нити, что показано методом меченых антител; по-видимому, он прикрепляется в определенных пунктах к молекулам тропомиозина.
Одним из важнейших событий при сокращении является образование и разрушение поперечных мостиков между миозином и .актином. Этот процесс происходит и in vitro при суперпреципита
Мышцы 183
ции актомиозина, сопряженной с гидролизом АТФ; in vivo АТФаза миозина активируется актином в присутствии ионов магния. Энергия для мышечного сокращения освобождается в этом случае Mg-АТФазой актомиозина. В отсутствие ионов кальция тропонин, по-видимому, подавляет взаимодействие актина с миозином, и это тормозящее действие, вероятно, оказывает тропонин А через тропомиозин В. Добавление Са2+ снимает тормозящее действие тро-понин-тропомиозина, актомиозиновая АТФаза освобождается, и происходит гидролиз Mg-АТФ.
In vitro тропонин В может тормозить Mg-АТФазу и без тропомиозина, но он не чувствителен к Са2+; добавление тропомиозина В ведет к еще большему торможению АТФазы. Тропонин А оказывает незначительное влияние на Mg-АТФазу, но энергично связывает кальций. Добавление тропонина А к тропонин-тропомиозин-актомиозину in vitro уменьшает степень торможения АТФазы в отсутствие Са2+ и активирует фермент в его присутствии [163, 164]. Следует заключить поэтому, что in vivo тропонин-тропомио-зиновая система является предохранительным механизмом, который в отсутствие Са2+ препятствует взаимодействию миозина с актином и активации Mg-миозиновой АТФазы. Когда ионы Са2+ освобождаются из саркоплазматического ретикулума, они связываются с тропонином А, в результате чего устраняется тормозящее действие тропонина А и тропомиозина на АТФазу и начинается циклическое взаимодействие актина с миозином, ведущее к. сокращению. Таким образом, роль Са2+ состоит скорее в устранении торможения актомиозиновой АТФазы, чем в активации этого фермента.
Иной способ кальциевого контроля найден у моллюсков. Тонкие нити мышц моллюсков содержат только актин и тропомиозин (тропонин отсутствует). Однако в противоположность миозину позвоночных миозин моллюсков связывает Са2+, миозиновая АТФаза зависит от Са2+, а актин не активирует актомиозиновую АТФазу без Са2+. Регулирующие участки, связывающие Са2+, расположены на легкой цепи миозина моллюсков. Сравнительные исследования показали, что такой миозиновый тип регуляции существует также-у брахиопод, немертин и сипункулид. Функция тропомиозина у них не установлена. У членистоногих тропонин в отличие от тропонина позвоночных имеет другие электрофоретические свойства, и связывает меньше Са2+.
У кольчатых червей имеются как миозиновый, так и тропониновый механизмы кальциевой регуляции. Тропонин у иглокожих напоминает тропонин позвоночных. Полагают, что регуляция,, связанная с миозином, более примитивна и что тропонин в процессе эволюции возникал дважды [214а, 227а].
Имеется очень мало сравнительных данных, касающихся тро-помиозин-тропонина. Константа связывания (величина, обратная: константе диссоциации) Са2+ с тропонином белых скелетных мышц.
184
Глава 16
кролика в два раза выше, чем константа связывания этих ионов с тропонином красных мышц; в случае тропонина сердечной мышцы она выше втрое {104]. Однако АТФаза сердечной мышцы по сравнению с АТФазой белой мышцы гораздо более чувствительна к стронцию; путем различного комбинирования белков было найдено, что стронциевый эффект связан с тропонином, поэтому тропонин сердечной и белой мышц должны отличаться друг от друга. «Быстрая» белая мышца содержит гораздо больше тропонина, чем «медленная» красная; белая часть широкой латеральной мышцы кролика содержит в три раза больше тропонина, чем красная ее часть (при сравнении количества Са2+-связей на единицу миозина) [130]. Гладкая мышца артерии млекопитающих содержит во много раз меньше актомиозина (2—10 мг/г) по сравнению со скелетной (50—100 мг/г) [321]. Однако у свиньи в мышце сонной артерии содержание тропомиозина выше (8,2 мг/г), чем в поперечнополосатой мышце (5,8 мг/г) [334b]. Тропомиозин из некоторых мышц позвоночных и беспозвоночных образует субъединицы с мол. весом 35 000, образующие двухцепочечную спираль, характерную для тонких нитей [252]. Различия ® аминокислотном составе тропомиозинов таких животных, как, например, кролик, корова, треска, минога, кальмар, моллюск, омар.незначительны [183].
а-Актинин. Молекула а-актинина имеет мол. вес около 180 000 и состоит из двух полипептидных цепей одинакового размера (с мол. весом по 90 000) и имеет значение S2o, равное 6,2. «-Актинии ускоряет суперпреципитацию и усиливает АТФазную активность миозина in vitro. «-Актинин проявляет это действие через актин, с которым он связывается, конкурируя с тропомиозином. Оказалось, что а-актинин локализован в Z-пластинке. Недавно его удалось получить в чистом виде; при этом выяснилось, что он составляет около 1 % всех миофибриллярных белков. Поскольку Z-пластинка составляет около 6% сухого веса миофибрилл, она должна содержать помимо а-актинина и другие белки {310].
Состав а-актинина быстрых (белых скелетных) мышц млекопитающих несколько отличается от состава а-актинина медленных красных и сердечной мышц. Подобного различия не найдено между белыми, красными, сердечной и гладкой мышцами цыпленка [310].
Другие миофибриллярные белки. Из мышечных экстрактов было выделено еще несколько белков (некоторые из них являются, вероятно, продуктами загрязнения или денатурации). Два таких белка (р-актинин и М-актинин), по-видимому, имеют какое-то функциональное значение. р-Актинин был выделен из неочищенного препарата актина; он, вероятно, тормозит перекрестные связи цепочек F-актина. М-белок, полученный иммунохимическим методом, вероятно, образует, по крайней мере частично, М-линию, в которой соседние толстые нити связаны между собой, образуя упорядоченную структуру [220а].
Мышцы
185
Теории сокращения. Существует много теорий сокращения, но большинство из них основано на развитой Хаксли идее взаимного скольжения толстых и тонких нитей. 'В исчерченных мышцах максимальное напряжение развивается при полном перекрытии миозиновых и актиновых нитей друг другом в области образования мостиков. Напряжение падает до нуля, когда мышца растянута до такой степени, что перекрытие отсутствует; при меньших (чем в случае максимального напряжения) длинах напряжение падает, так как нити «налезают» друг на друга.
Чтобы возникло скольжение толстых нитей вдоль тонких, ведущее к сокращению или развитию напряжения без изменения длины нитей, должны произойти какие-то конфигурационные изменения миозина, кроме образования мостиков и гидролиза АТФ. Согласно одной из гипотез, при расщеплении АТФ изменяется угол наклона мостиков; это может вызвать вращение головок миозина [195] или скручивание миозиновой цепочки [279]. Изменение угла наклона поперечных мостиков было отмечено в мышце насекомого, находящегося в состоянии окоченения [299]. Другая гипотеза состоит в том, что последовательные взаимодействия мостиков создают торсионное усилие и вызывают скручивание нитей [96, 98]. Современные представления об изменении напряжения без изменения длины основаны на следующем наблюдении: если сокращенную мышцу внезапно расслабить, ее напряжение восстанавливается в два этапа. Предполагается, что каждый мостик состоит из эластичного плеча, которое вступает в контакт с актиновой нитью, находясь в одном из нескольких энергетически стабильных положений. Причина восстановления напряжения может лежать, во-первых, в эластичности основания мостика и, во-вторых, в повороте головки из одного положения в другое, имеющее более низкий уровень потенциальной энергии.
Роль кальция. «Спусковым крючком» сокращения как в гладких, так и в исчерченных мышцах является Са2+. При микроинъецировании различных ионов внутрь волокон исчерченных мышц только кальций (в низких концентрациях) вызывал сокращение. В поперечнополосатых мышцах теплокровных пороговая концентрация Са2+, вызывающая сокращение, равна 10-6 М, а концентрация, вызывающая максимальный эффект, равна примерно 10-5 М. Мышечные волокна лягушки со снятой сарколеммой реагируют на 5-10~7 М и дают максимальный ответ при 1(Н М Са2+ [105]. В глицеринизированных мышцах насекомых актомиозиновая АТФаза активируется при 10~8 М, а 7,5-10-7 М Са2+ соответствует максимальной активации. Актомиозиновая АТФаза млекопитающих активна при 10~9 М Са2+. Содержание кальция на 1 мг актомиозина в разных мышцах, несмотря на различия в весовом содержании актомиозина, сходно [33].
386
Глава 16
Актомиозин (AM) мг/г Са, мкмоль/г(АМ) сырого веса Са, мкмоль/мг AM
Портняжная мышца лягуш- 70 90 1,3
ки
•Сердце лягушки 38 50 1,3
Толстая кишка .морской 10 13 1,3
свинки
Универсальность значения Са2+ для мышечного сокращения .демонстрируется и тем, что миоциты губок могут сокращаться только в присутствии ионов кальция (или заменяющих их ионов стронция или магния) [288].
Большая часть (90%) кальция в покоящейся мышце связана внутриклеточными мембранами (главным образом системой саркоплазматического ретикулума). Потенциал действия приводит к высвобождению кальция; это подтверждается появлением внезапного пика в абсорбции мурексида (красителя, имеющего сродство к Са) [206] и излучением света во время сокращения мышцы, в которую был введен экворин (белок, для люминесцентного свечения которого необходимы ионы Са2+). Количество высвобождаемого кальция пропорционально развиваемому напряжению и соответствует по времени начальной теплопродукции [61].
В мышцах позвоночных высвобождаемые ионы Са2+ немедленно связываются тропонином А, который после этого приобретает способность снимать тропонин-тропомиозиновое торможение взаимодействия актина с миозином, о чем говорилось выше. В мышцах моллюсков ионы Са2+ присоединяются к активным центрам легких цепей миозина, после чего становится возможным взаимодействие других активных центров миозина с актином. Перед расслаблением кальций снова должен быть удален. Это осуществляется активным поглощением кальция мембранами саркоплазматического ретикулума (СР). В экстрактах из гомогената мышцы частицы СР содержатся в виде мелких пузырьков или микросом, которые связывают Са и образуют расслабляющую систему; эта система может быть экстрагирована [370]. Для активного связывания кальция мембраной СР необходима энергия, которую дает расщепление АТФ. Этот процесс совпадает во времени с теплопродукцией расслабления. Необходимая для гидролиза АТФ АТФаза входит в состав самой мембраны СР; она активируется ионами магния и кальция и имеет мол. вес около 147 000 (102 000 — белки и 45 000 — липиды). Функцией второго белка мембраны СР, который содержится в меньшем количестве, является связывание кальция в количестве до 43 молей ионов кальция на моль белка [231а]. Радиоавтографическое исследование мышцы лягушки, меченной 45Са, показало, что сразу после тетануса кальций накачивается обратно в продольные и промежуточные канальцы СР-си-стемы и при полном восстановлении его количество в терминаль
М ышцы 187'
ных канальцах или цистернах возрастает; при возбуждении он высвобождается снова [380].
Частицы саркоплазматического ретикулума (микросомная фракция) из различных мышц позвоночных и беспозвоночных обладают одинаковыми расслабляющими свойствами и способностью связывать кальций. Скорость его поглощения частицами иа красных мышц составляет лишь 16% скорости поглощения в белых мышцах млекопитающих [336]. Пузырьки СР из мышцы омара могут убирать стронций так же успешно, как и кальций; пузырьки крыс стронций не связывают. У омара пузырьки поглощают кальций со скоростью 0,6-10~2 мкМ/мг белка, что достаточно для обеспечения расслабления [364]. Вещества, которые, подобно NO3, увеличивают продолжительность сокращения, подавляют связывание кальция пузырьками; другие вещества, как, например,. Zn, удлиняют сокращение, тормозя АТФазу саркоплазматического ретикулума [58].
В гладких мышцах млекопитающих цитоплазматические гранулы связывают Са2+ так же, как и в поперечнополосатых мышцах, но связывание Са2+ и АТФазная активность частиц мышцы-матки в 10 раз ниже, чем эти же показатели, полученные на сердечной мышце [57]. Микросомные частицы из матки связывают Са2+ из раствора с концентрацией не ниже 6-10-7 М '[26]. В гли-церинизированной гладкой мышце содержится меньше кальция, чем в поперечнополосатой, и связывание прекращается при более низкой его концентрации [165, 166].
Переход от возбуждения к сокращению (электромеханическая связь). Чтобы процесс возбуждения вызвал сокращение, электрический сигнал должен перейти с мембраны внутрь клетки. В быстрых мышцах эту функцию выполняет система поперечных канальцев (Т-система). Это впервые было показано методом локальной стимуляции различных участков саркомера с помощью электрода, диаметр которого меньше длины саркомера. В волокнах лягушки минимальный порог сокращения был на уровне Z-мембраны, а у волокон краба — на границе А- и 1-дисков; эти точки соответствуют расположению Т-канальцев. При фиксации напряжения на внешней мембране внеклеточным электродом внутриклеточный микроэлектрод регистрирует во время возбуждения мышцы небольшие спайки, которые, вероятно, идут от Т- и -СР-канальцев. По последовательности активации миофибрилл от периферии к центру определили скорость проведения возбуждения внутри волокна; она оказалась равной 7 см/с (при 20 °C) [145].
Каким образом электрический сигнал в Т-канальцах вызывает высвобождение Са2+ из CP-системы, неясно. Можно наблюдать, что миофибриллы на периферии волокна начинают сокращаться раньше, чем в центре [5]. После удаления сарколеммы с поперечнополосатого мышечного волокна лягушки локальное воздействие кальцием вызывает его сокращение, величина которого пропор
188
Глава 16
циональна концентрации Са2+. Радиальное распространение возбуждения уменьшается при воздействии тетродотоксином или при недостатке Na; отсюда по аналогии с поверхностной мембраной может следовать, что потенциал действия активно распространяется вдоль Т-канальцев [81]. При обработке мышечных волокон гипертоническим раствором Т-канальцы благодаря добавлению глицерина разбухают и даже разрываются; такие волокна еще проводят во внешней мембране нормальные потенциалы действия, но они не вызывают сокращения и не имеют негативных следовых потенциалов [132]. Наружная мембрана в покое обладает К- и Cl-проводимостью. В Т-канальцах мышцы лягушки Cl-проводимость отсутствует [133]. В мышце ракообразных во время возбуждения ионы С1~, по-видимому, проходят через Т-мембраны [44]. Очевидно, что электрическое проведение сигнала от наружной мембраны внутрь волокна осуществляется через Т-канальцы, однако каким образом он переходит на CP-канальцы, пока неизвестно.
Механические свойства, «активное состояние» и теплопродукция
Механика. Мышцы обладают определенными механическими свойствами, влияющими на их сокращение. Схематически мышцу можно изобразить в виде системы, состоящей из сократительного элемента, соединенного с эластическими элементами, которые в соответствии с их расположением по отношению к сократительному элементу делятся на последовательные и параллельные.
В скелетных мышцах последовательными эластическими элементами являются главным образом сухожилия и другие виды соединительной ткани на концах мышцы. В мышцах с короткими волокнами различия между последовательными и параллельными эластическими элементами не столь отчетливы. Напряжение последовательных эластических элементов может быть измерено, если изометрически сокращенной мышце позволить укорачиваться при различных нагрузках или если напряжение параллельных элементов исключается при укорочении. Податливость (определяемая как смещение на единицу приложенной силы) последовательных эластичных элементов может быть увеличена, если к концам мышцы прикрепить пружины; скорость развития напряжения в этом случае уменьшается, высота одиночного сокращения понижается, а отношение величины тетануса к одиночному сокращению увеличивается.
Существование параллельной эластичности демонстрируется пассивной кривой напряжение — длина. Когда невозбужденная мышца растягивается, в ней развивается эластичное напряжение, непропорциональное степени растяжения: в начале растяжения
Напряжение, % от максимума
--------------3.65 мкм (а+б)----------*
1
-*—1,85-1,90мкм(б-в)-^
Рис. 1641/1. Зависим ость активного напряжения от длины саркомера [376а].
Длина саркомера и относительное расположение толстых и тонких нитей в нижней части рисунка соответствуют пронумерованным точкам на графике длина — напряжение (вверху).
190
Глава 16
напряжение растет незначительно, но, по мере того как растяжение приближается к точке разрушения, оно увеличивается со все возрастающей скоростью (рис. 16-11). Напряжение в параллельных эластичных элементах сильно возрастает, когда длина выходит за пределы длины покоя (/0) (рис. 16-12,Л). При меньшей, чем в состоянии покоя, длине механическая система состоит только из сократительных и последовательных эластичных элементов. Сопротивление растяжению параллельных элементов возни-
Рис. 16-12. Кривые
А. Мышцы в состоянии покоя [156]. 1 — летательная мышца шмеля; 2 — летательная мышца сусный ретрактор Mytilus (14 °C); 5 — ретрактор глотки улитки Helix (14 °C). Б. Мышцы в тор Mytilus-, 3 — летательная мышца саранчи; 4 — ретрактор глотки улитки Не О активное напряжение, * разница между
Мышцы
191
кает в соединительной ткани между волокнами, в сарколемме и саркоплазме. Летательные мышцы насекомых, в которых количество соединительной ткани невелико, менее растяжимы, чем мышцы позвоночных. Многие гладкие мышцы с короткими волокнами более растяжимы, чем поперечнополосатые. Ретрактор хоботка сипункулиды Golfingia легко растягивается более чем в 10 раз по отношению к длине покоя за счет расправления поперечных складок, в которые собираются волокна (см. ниже). Для многих гладких мышц /0 или длину покоя in vivo трудно определить, так как она может меняться под влиянием гормональных и других факторов.
В расслабленном состоянии сократительные элементы очень пластичны. В возбужденной мышце их жесткость резко возрастает и может сохраняться после окончания активного состояния. О свойствах сократительных элементов можно судить по скорости укорочения (в изотоническом режиме сокращения), по эффекту
длина —напряжение.
саранчи (Schistocerca, при 11 °C); 3 — портняжная мышца лягушки (0°С); 4— передний биоактивном состоянии [156]. / — портняжная мышца лягушки; 2 — передний биссусный ретрак-«х; 5 — летательная мышца шмеля. В. Кольцевая мышца кишечника кошки [243]. А покой, активным и пассивным напряжением.
192
Глава 16
растяжения или ослабления во время сокращения и по максимальному напряжению при изометрическом сокращении.
Изотоническим сокращением называется такое сокращение, при котором укорочение мышцы возникает при постоянной нагрузке;
Рис. 16-13.
А. Кривые сила — скорость для различных мышц [1561. 1 — передний биссусный ретрактор Mytilus; 2 — ретрактор глотки улитки Helix; 3 — исчерченная часть аддуктора Pecten; 4 — летательная мышца саранчи. Б. Скорость укорочения мышцы матки кролика (5) и портняжной мышцы лягушки (6) при различных нагрузках [388]. Гиперболическая кривая относится к Рп— р
скорости (см/с), прямая линия выражает зависимость величины ------от нагрузки.
изучение этого вида сокращения позволяет получать данные о динамике сократительного процесса. Скорость сокращения является функцией нагрузки (зависимость сила — скорость, рис. 16-13,Л) и определяется известным уравнением А. Хилла:
Ь(Р0-Р) р Ь(Р0—Р)
V— (P-f-a) ’ ИЛИ Р-------------V---
а,
где V — скорость сокращения, Ро—максимальное напряжение,
Мышцы
193
развиваемое мышцей, сокращающейся без укорочения, т. е. максимальное тетаническое напряжение, Р — нагрузка, а — константа с размерностью силы, b — константа с размерностью скорости. Vo — максимальная скорость сокращения, или получаемая путем экстра-а Г? Л)-?
полиции скорость укорочения без нагрузки. Если величину — изобразить как функцию Р, получим прямую линию с наклоном Ь, пересекающую ось абсцисс в точке —а (рис. 16-13,Б).
Выходная мощность мышцы (произведение нагрузки на скорость) достигает максимума при определенных промежуточных значениях силы и скорости.
Как видно из графиков, кривые сила — скорость для разных мышц одинаковы по форме, но имеют различные константы. Если мы Vo выразим не в мм/с, а в длинах мышцы в секунду, то получим присущую данной мышце величину, которую называют характеристической скоростью (intrinsic velocity или intrinsic speed) . Эта скорость равна Vo//, где I — длина мышцы, а ее размерность— с-1.
У наружной мышцы глаза котенка, например, VQ/l равно 70 с-1, у портняжной мышцы лягушки (при 0°С)—1,3 с-1, а у мышцы черепахи — 0,1 с^1 [177, 376]. Характеристическая скорость в более строгом понимании есть скорость перемещения актиновых нитей по миозиновым во время сокращения. У мелких животных (например, мыши) длина саркомера не слишком отличается от длины саркомера крупных животных (например, лошади); однако, поскольку длина мышц разнится очень сильно, характеристическая скорость V0/l, необходимая для перемещения в пространстве с данной скоростью, много больше у мелких животных по сравнению с крупными. Например, колибри может делать своими крыльями до 100 взмахов в*секунду, а пеликану достаточно всего одного взмаха за несколько секунд, чтобы поддержать ту же скорость. Характеристическая скорость мышц крупных животных, таких, как кит или слон, составляет лишь малую долю той, которая необходима дельфину или тушканчику, чтобы передвигаться с такой же скоростью.
Характеристическая скорость мышц выражается показательной функцией от веса тела и обратно пропорциональна длине конечности. Мощность, развиваемая мышцей, определяется как произведение силы на скорость и обычно максимальна при 0,3 Vo и 0,3 Ро. Коэффициент полезного действия (КПД)—отношение механической работы к затраченной энергии — максимален при 0,2 Vo. и 0,5 Ро. Таким образом, максимальная мощность и максимальный КПД достигаются при различных значениях скорости и нагрузки. В прекрасной статье Хилл [177] показал, что характеристическая скорость мышц адаптирована к размерам тела. Ниже будет показано, что характеристическая скорость определяется-специфичностью миозина, т. е. может оказаться, что миозины жи*-13—2067
194
Глава 16
вотных адаптированы |к размерам тела и скорости мышечного сокращения, так же как гемоглобин различных животных адаптирован к потреблению кислорода и его доступности.
При изометрическом сокращении длина мышцы остается постоянной, так как оба ее конца закреплены и укорочение невозможно. Сокращение такого рода 'можно получить при любой длине мышцы; для сократительных элементов оно не является строго изометрическим, так как они могут сокращаться за счет последовательных эластичных элементов. Когда мышца подвергается раздражению при различных длинах, изометрическое напряжение достигает максимума при длине, близкой длине покоя. По мере растягивания мышцы общее напряжение приближается к максимальному пассивному напряжению (рис. 16-12). По разности между общим и пассивным напряжением можно построить зависимость активного напряжения от длины. Активное напряжение достигает максимума при разных длинах: для большинства исчерченных мышц оно максимально приблизительно при /0, когда Р=Ро (Io обычно совпадает с нормальной длиной мышцы в организме), а для сердечной и гладкой мышц активное напряжение максимально при несколько больших длинах. В taenia coli максимальное напряжение развивается при длине, равной 130— 180% 1о.
Максимальное напряжение или развиваемую силу определяют при тетанусе и выражают в граммах на единицу поперечного сечения (г/см2). Некоторые значения этой величины даны в табл. 16-1. Как правило, мышцы с высокой скоростью укорочения развивают небольшое напряжение, а мышцы, выполняющие функцию поддержания, развивают наиболее высокое напряжение.
Возрастание активного напряжения до максимума при растяжении сменяется падением при больших длинах. Это можно объяснить с помощью представления о взаимном перекрывании нитей. При растяжении поперечнополосатой мышцы длина саркомеров увеличивается. При максимальной длине, которую мышца еще может выдержать без разрушения, взаимного перекрывания между толстыми и тонкими нитями уже нет. При длине максимального напряжения перекрывание в области поперечных мостиков также максимально и тонкие нити подходят вплотную друг к другу в середине А-диска. При меньшей длине возможно перекрывание или даже сминание тонких нитей. Таким образом, максимальное количество поперечных мостиков образуется при длине, соответствующей полному перекрыванию [147]. Эта упрощенная модель приложима только к поперечнополосатым мышцам [385], хотя взаимное перекрывание тонких нитей недавно было подтверждено на летательных мышцах насекомых. В гладких и сердечной мышцах (а возможно, и в некоторых поперечнополосатых) взаимное расположение молекул внутри нитей также может влиять на развитие напряжения.
Мышцы 195
Таблица 16-1
Максимальная сила, развиваемая мышцами
Мышцы Максимальная сила, кг/см 2 Источник данных
М. tenuissimus кошки Длинный разгибатель пальца крысы 1,4 3,0
Икроножная крысы Камбаловидная крысы 1,8 0,28; 0,22 [330]
Сердечная крысы 0,016 [269]
Икроножная ленивца Диафрагма ленивца 1,62 2,1 [232]
Папиллярная кошки 0,8 [339]
Гладкая мышца трахеи собаки 0,8 [337]
Матка кролика 0,13; 0,7 [339]
Taenia coli морской свинки 1,5 [339]
Taenia coli кролика 0,89 [339]
Двенадцатиперстная кишка (кольцевая мышца) 0,42 [339]
Портняжная лягушки 2,0 [339]
«Звуковая» мышца опсануса 0,1 [266]
Кожнореберная змеи 0,7—1,0 [306]
Разгибатель омара, медленная часть Разгибатель омара, быстрая часть Коксиальная мышца таракана 2,8 0,025 0,8 [244]
Ретрактор дыхательной трубки Lethocerus 1,25 [367]
Аддуктор устрицы 12 (кратковременно) 0,6 (длительно) [250]
Большинство мышц при повторяющихся раздражениях развивает большее напряжение, чем при одиночном сокращении. Стимуляция in vivo осуществляется сериями нервных импульсов, и более высокое напряжение при тетанусе может частично объясняться возникновением явления облегчения в нервно-мышечных синапсах и вовлечением в действие большего количества волокон. Кроме облегчения возбуждения, возможно механическое суммирование, в случае если максимальные стимулы следуют с интервалами, исключающими полное расслабление. Мышцы, дающие слабый ответ на одиночный импульс и требующие ритмического раздражения или облегчения, называются итеративными. Те же, которые на одиночное раздражение дают ответ, близкий к максимальному, и в которых при ритмической стимуляции наблюдается лишь суммация, называются неитеративными. В условиях максимальной стимуляции или прямого раздражения мышечных волокон напряжение при тетанусе выше, чем при одиночном сокращении. Это объясняется тем, что в процессе одиночного сокращения энергия тратится главным образом на растяжение последовательных эластичных элементов и лишь при тетанусе после полного их растяжения достигается максимальное напряжение; в дальнейшем вся энергия сокращения идет на его поддержание. Частота, при кото-13*
196
Глава 16
рой происходит полное слияние отдельных сокращений (гладкий тетанус), является мерой максимальной скорости расслабления.
Цепь событий между возбуждением мембраны и началом сокращения составляет содержание упоминавшегося выше понятия «exitation-contraction coupling». Раньше эта цепь не расчленялась и рассматривалась целиком как причина, вызывающая «активное состояние». Перед развитием изометрического сокращения при длинах, больших чем 10, может наблюдаться латентное расслабление (сопровождающееся кратковременным и незначительным падением напряжения). Его происхождение еще не установлено.
Активное состояние. Уменьшение растяжимости, т. е. увеличение жесткости, предшествующее развитию напряжения, является признаком активации сократительных элементов. Кривая развития активного состояния получена с помощью комбинированных механических измерений. Если растянуть мышцу во время латентного периода одиночного сокращения, развивающееся напряжение может быть таким же, как при тетанусе, поскольку растяжение эластичных элементов уже произведено. Если мышцу растянуть во время пика одиночного сокращения, дополнительное напряжение не развивается. Точка нулевого прироста напряжения (соответствующая пику одиночного сокращения) лежит на кривой активного состояния. Если мышцу внезапно осл,абить во время максимума сокращения, напряжение резко падает, но затем восстанавливается до уровня, несколько меньшего, чем в соответствующей фазе одиночного сокращения. Серия таких быстрых ослаблений с последующим восстановлением позволяет проследить угасание активного состояния. Кривая напряжения, полученная методом быстрых растяжений мышцы до пика и последующих быстрых ослаблений напряжений, дает временную характеристику активного состояния сократительных элементов [70]. Начало возрастания напряжения в ответ на второй из двух спаренных стимулов и момент начала падения напряжения после тетануса могут быть использованы для определения начала фазы падения активного состояния. Как правило, активное состояние возрастает быстро (в течение 40 мс для портняжной мышцы лягушки при 0°С). Падение до нуля занимает половину общей продолжительности одиночного сокращения. В некоторых гладких мышцах восстановление напряжения при ослаблении после пика незначительно; отсюда следует, что активное состояние угасает быстрее, чем напряжение. Угасание активного состояния имеет больший температурный коэффициент по сравнению со скоростью укорочения. Напряжение одиночного сокращения у зимних лягушек имее.т максимум при более низкой температуре, чем у летних. Сезонные изменения сказываются и на многих других механических свойствах мышцы лягушки. Течение активного состояния во времени в мышцах лягушки изменяется многими агентами—нитратами, кофеином, гипертоническими растворами; теперь известно, что они действуют
Мышцы
197
на определенные звенья процессов, связанных с развитием сокращения.
Некоторые гладкие мышцы сохраняют остаточную жесткость длительное время после расслабления. Это указывает на устойчивые структурные изменения, остающиеся после нормального возбуждения, но не связанные с активным поддержанием напряжения [243].
Энергетика и теплопродукция. КПД мышц меньше 25%, т. е. большая часть энергии химических превращений идет не на механическую работу, а превращается в тепло. В папиллярной мышце кролика лишь 11,6% химической энергии превращается в механическую {137]. Обмен в мышечных клетках, находящихся в состоянии покоя, сопровождается низкой теплопродукцией. Теплопродукция покоящейся сердечной мышцы резко возрастает при ее растяжении. При одиночном сокращении или тетанусе теплопродукция стремительно возрастает. Большая часть серьезных работ по изучению этих явлений проведена на портняжной мышце лягушки при О °C.
В коротком тетанусе быстрой поперечнополосатой мышцы лягушки при О °C одновременно с подъемом напряжения освобождается большое количество тепла. Это — начальная теплопродукция, и при 10 ее скорость приблизительно равна 30 мкал/с на 1 г. В начале расслабления тепло продолжает освобождаться, но с меньшей скоростью. Это — задержанная теплопродукция, сопровождающая задержанные окислительные процессы; она может длиться несколько минут. При длительных тетанусах тепло выделяется с постоянной скоростью до тех пор, пока поддерживается напряжение; такое тепло называется теплом поддержания. Если мышца постепенно растягивается (от длины /0) и стимулируется при разных длинах, изометрическая теплопродукция и напряжение по мере уменьшения взаимного перекрытия протофибрилл закономерно уменьшаются. Таким образом, начальное тепло зависит от длины и максимально при /0. Его можно расценить как результат быстрого распада креатинфосфата для ресинтеза АТФ; само по себе сокращение поперечнополосатой мышцы не требует кислорода. Мышца сердца, наоборот, в отсутствие кислорода теряет способность к сокращению. Если поперечнополосатая мышца растянута до длины, при которой протофибриллы уже не перекрывают друг друга, при раздражении продолжает выделяться часть начального тепла в количестве 0,5—1,0 мкал/г; такое тепло называется теплом активации. Это сумма термохимических процессов, связывающих возбуждение с сокращением, — движение Са24', распад АТФ и другие процессы. Если мышца сокращается под нагрузкой, производя работу, то выделяется тепло укорочения, пропорциональное произведению укорочения на коэффициент, являющийся функцией силы. Тепло укорочения выделяется как дополнительное, когда мышца во время тетануса получает возмож
198
Глава 16
ность укорачиваться. При длинах, меньших 10, тепло укорочения возрастает, а тепло поддержания напряжения уменьшается. Работа укорочения совершается со скоростью , где Р — нагрузка, а I — величина укорочения. При 10 отдельные компоненты теплопродукции трудно разделить. При возрастании напряжения эла-
Рис. 1'6-44. Зависимость .количества энергии, освобождающейся при одиночном сокращении мышцы лягушки, от величины нагрузки. (Любезно предоставлено К. Гиббсом.)
Тепло укорочения максимально при нулевой нагрузке (Р/Ро=0) и минимально при максимальной (Г’/Г’о=1). 1 — работа; 2 — тепло укорочения; 3 — тепло поддержания; 4 — тепло активации.
стичные элементы поглощают тепло, которое может освобождаться при расслаблении. Тепло укорочения и тепло поддержания суммируются.
На рис. 16-14 дана упрощенная схема энергетического баланса для портняжной мышцы лягушки при О °C. Тепло укорочения и тепло поддержания находятся в обратных отношениях, причем первое максимально при нулевой нагрузке. В других мышцах или при других температурах количество тепла укорочения может меняться, а наклон кривой тепла поддержания значительно варьирует. В скелетных мышцах количество тепла активации явно зависит от длины, а в сердечной мышце — от степени сократимости.
Мышцы
199
У всех мышц кривая работы имеет колоколообразную форму, а величина работы, вероятно, зависит от тепла активации.
Портняжная мышца лягушки содержит около 24 мкМ креатинфосфата на грамм; при одиночном сокращении освобождается около 3 кал/г, а при 10-секундном тетанусе — около 40 мкал/г [56]. Сумма начальной теплопродукции и механической работы хорошо коррелирует со свободной энергией расщепления креатинфосфата, при котором освобождается 11 ккал/моль.
Фосфагены и химические источники энергии. Из анализа теорий мышечного сокращения следует, что непосредственным источником энергии для пространственной перестройки сократительных белков является аденозинтрифосфорная кислота (АТФ). Однако количества АТФ в покоящейся мышце (приблизительно 5-Ю-6 моль/г) достаточно лишь для нескольких одиночных сокращений, а поддержание сокращения требует 10-4—10-3 моль/мин на 1 г [27, 376]. Таким образом, синтез АТФ должен идти непрерывно; в этом синтезе принимают участие фосфагены, которые у позвоночных представлены креатинфосфатом, а у многих беспозвоночных — аргининфосфатом. Фермент креатинкиназа катализирует перенос богатого энергией фосфата с креатинфосфата на АДФ, образуя АТФ. Если креатинкиназу блокировать динитрофторбензолом (ДНФБ) и мышцу в состоянии сокращения быстро заморозить, то, как показывает последующий анализ, расщепление АТФ происходит в фазе сокращения [254]; при 0,2-секундном тетанусе при 0 °C АТФ расходуется <в количестве 0,34 мкмолей. Фосфорилирование АДФ происходит при расслаблении или во время поддержания сокращения [56].
Креатин рефосфорилируется либо окислительным путем, либо в гликолитическом цикле с распадом гликогена и образованием молочной кислоты. В аэробных условиях in vitro часть молочной кислоты окисляется, давая энергию для ресинтеза остальной части молочной кислоты в гликоген. In vivo молочная кислота поступает из мышц в кровь, а оттуда в печень, где превращается в гликоген. Отсюда по мере надобности в кровь поступает глюкоза, которая в мышцах превращается снова в гликоген, пополняя его запасы. Ликвидация кислородного долга, образующегося в результате физической работы, обеспечивает восстановление мышечного гликогена. В анаэробных условиях молочная кислота накапливается в крови, а креатинфосфат ресинтезируется, пока в мышцах не иссякает запас гликогена. В мышце, лишенной доступа О2 и отравленной иодацетатом, блокирующим фосфоглицеральдегиддегидрогеназу, гликолиз останавливается на стадии триозофосфата и сокращение продолжается, пока не истощится запас креатинфосфата, необходимого для ресинтеза АТФ. Количество расщепляющегося креатинфосфата пропорционально количеству одиночных сокращений, иными словами сумме тепла активации и работы. Отравленная иодацетатом портняжная мышца лягушки
200
Глава 16
в атмосфере азота расщепляет 0,286 мкмоля креатинфосфата на 1 г мышцы за одно сокращение [56]. Сердечная мышца больше зависит от окислительного восстановления гликогена, чем поперечнополосатая, поэтому она, как и гладкие мышцы, содержит гораздо меньше креатинфосфата. Изофермент лактатдегидрогеназы в сердце лучше приспособлен к аэробному обмену, чем изофермент того же фермента в поперечнополосатой мышце (см. гл. 6).
Как правило, быстрые мышцы менее экономичны при поддержании напряжения, чем медленные. Например, задняя широчайшая мышца спины цыпленка в 15 раз «быстрее», чем передняя широчайшая мышца спины, однако передняя может поддерживать напряжение в 13 раз дольше, чем задняя. Содержание АТФ в передней мышце составляет 1 мкмоль/г, а в задней 3 мкмоль/г; при отравлении ДНФБ передняя (но не задняя) мышца может поддерживать активное состояние многие минуты. Ниже приведены данные, позволяющие сравнить экономичность некоторых мышц [143, 144].
Суммарное изометрическое напряжение в 103 г • с/г мышцы на 1 мкМАТФ
Хомячок
m. biceps brachii 30,8
диафрагма 36,3
длинный разгибатель пальца 103
камбаловидая мышца 191
Цыпленок
передняя широчайшая мышца спи- 540
ны
Мышца цыпленка оказалась в 2,5 раза более экономичной, чем медленная камбаловидная мышца хомячка.
Фосфагены содержат различные гуанидиновые основания. Многие из них образуются трансаминированием из аргинина (см. гл. 7), но только некоторые из гуанидиновых оснований имеют соответствующие киназы для фосфорилирования и дефосфорили-рования и могут служить фосфагенами. Например, октопин, яв- 1 ляющийся продуктом соединения аргинина и пирувата, найден во I всех классах моллюсков, однако фосфагеном у всех моллюсков | служит фосфоаргинин. |
Мышцы всех хордовых содержат креатинфосфат; мышцы чле- 1 нистоногих, моллюсков, кишечнополостных и некоторых низших 1 червей — аргининфосфат (табл. 16-2). Из простейших Stentor содержит аргининкиназу как единственную фосфагенкиназу [369]. }
Среди иглокожих голотурии и морские звезды содержат только аргининфосфат, а офиуры — только креатинфосфат; некоторые J
роды морских ежей имеют как аргининфосфат, так и креатинфос- ;
фат, другие роды — только аргининфосфат. Полухордовые имеют
Мышцы
201
оба соединения; бесчерепные, как и позвоночные, только креатинфосфат [18, 352].
Кольчатые черви и сипункулиды отличаются большим разнообразием гуанидиновых оснований; некоторые из них служат мышечными фосфагенами; найдены также соответствующие киназы.
Таблица 16-2
Распределение фосфагенов у некоторых червей и родственных им животных1) [352]
Polychaeta
Nereidae
Nephthydae
Glyceridae
Eunicidae
Capitellidae
Terebellidae
Serpulidae Sabellidae Arenicolidae
Ophelidae
Oligochaeta Lumbricidae
Phoronida
Echiurida:
Urechis
Nemertea
Sipunculida
(за исключением Sipunculus)
Ч Обозначения:
(КФ) ГлФ ГлФ
КФ
КФ
АФ ’ КФ
АФ ? КФ ТФ
АФ ’ КФ ТФ
АФ £ КФ ТФ
ГиФ
АФ ЛФ ОФ
ЛФ
КФ
ЛФ
АФ
ГиФ ТФ
ГлФ — гликоциаминфосфат, ОФ — офелин-
АФ — аргининфосфат, КФ — креатинфосфат, ГиФ — гипотауроциаминфосфат, ЛФ — ломбрицинфосфат, фосфат, ТФ — тауроциаминфосфат.
В табл. 16-2 приводится несколько характерных примеров. Как правило, у свободноплавающих полихет отсутствует аргининфосфат; сидячие полихеты имеют большой набор оснований, но у них нет гликоциаминфосфата. Нереиды используют креатинфосфат и гликоциаминфосфат. Сперматозоиды кольчатых червей большей частью содержат креатинфосфат, независимо от вида мышечного фосфагена (у взрослых). Пиявка Hirudo содержит большое количество основания гирудина, a Bonella имеет другое основание — бонеллидин, но эти основания не фосфорилируются, и фосфагены для этих животных неизвестны. Сипункулиды (за исключением рода Sipunculus) содержат много фосфогипотауроциамина и некоторое количество фосфотауроциамина. Объяснения большого биохимического разнообразия фосфагенов, наблюдаемого у морских червей, не найдено.
Нервная активация быстрых скелетных мышц позвоночных
Моторные концевые пластинки позвоночных изображены на рис. 16-15. Вблизи концевой пластинки моторный аксон теряет -Миелиновую оболочку и разветвляется, образуя несколько нерв-
202
Глава 16
них окончаний: мембрана каждого из них вместе с сарколеммой образует множество складок. Мембрана нервного окончания отделена от мышечной мембраны пространством в 500 А. На краях складок шванновская цитоплазма отделяет терминальную аксоплазму от внеклеточного пространства. При окрашивании специ-
Рис. 16-15.
А. Моторная концевая пластинка поперечнополосатого мышечного волокна млекопитающего (схема) [389]. Миелиновая оболочка нервного аксона в области нервного окончания отсутствует; видны ядра глиальных клеток вне окончания нерва. Ряды желобков или впячиваний являются областями тесного контакта между нервной и мышечной мембранами. Б. Участок концевой пластинки. Поперечное сечение (схема) [390]. Ап-аксоплазма; М — митохондрия; П _ пузырьки; Сп — саркоплазма; СС — синаптические складки. В. Участки концевой пластинки межреберной мышцы; окраска на холинэстеразу [389]. 1 и 2—вид с поверхности при малом и большом увеличении; 3 — поперечный срез через листки концевой пластинки.
фическими красителями в постсинаптической мембране выявляется слой, содержащий ацетилхолинэстеразу. Митохондрии многочисленны как в терминальной аксоплазме, так и в саркоплазме. Окончания аксона содержат множество синаптических пузырьков, имеющих от 300 до 400 А в диаметре. Их число после продолжительной стимуляции уменьшается, и они кажутся пустыми [242] (см. гл. 11).
Когда нервный импульс достигает концевой пластинки, происходит освобождение ацетилхолина (в течение 2,5 мс при 20°С)г вызывающего (через 0,3 мс) возникновение потенциала концевой пластинки — возбуждающего постсинаптического потенциала (ВПСП) (рис. 16-16). Этот потенциал является местным, его вели
Мышцы 203
чина зависит от количества медиатора; он распространяется электротонически на несколько миллиметров вокруг концевой пластинки, что ведет к образованию тока действия. Кураризация бло
кирует возникновение мышечного импульса, но синаптический потенциал при этом длится гораздо дольше рефрактерного периода нерва, так что возможна суммация действия двух или больше неградуальных нервных импульсов. Под действием эзерина (физостигмина), тормозящего действие холинэстеразы, длительность ВПСП увеличивается в 10 раз. Подобные же потенциалы можно вызвать локальным воздействием ацетилхолина (АХ) на область концевых пластинок. С других участков мышечной мембраны АХ не вызывает ответа. Он неэффективен также при внутриклеточном введении [211]. Тубокурарин тормозит нервно-мышечную передачу и противодействует эффекту АХ, конкурируя с ним за рецептор. Змеиный яд бунгаротоксин блокирует взаимодействие АХ с рецепторами. Используя меченый бунгаротоксин, определили количество рецепторных точек для концевой пластинки портняжной мышцы лягушки; их оказалось 109, а для диафрагмы крысы — только 4,7-107 [245].
Если сразу за нервным импульсом следует прямое раздражение мышечного волокна, мышечный импульс проходит через область концевой пластинки,
Рис. 16-16. Потенциал действия концевой пластинки портняжной мышцы лягушки [391].
а — перед кураризацией; б, в, г — возрастающие степени ку-раризации; д — полная кураризация. Во всех случаях, кроме последнего, синаптические потенциалы дают начало распространяющимся спайкам.
не взаимодействуя с синаптическим потенциалом; отсюда следует, что оба эти электрические явления возникают в разных участках мембраны. Постсинаптическая мембрана в норме возбуждается хи-
мически, но не электрически.
Внутриклеточное отведение от концевой пластинки регистрирует спонтанные миниатюрные синаптические потенциалы (мВПСП) постоянной минимальной величины, причиной которых является выделение «квантов» АХ нервным окончанием. ВПСП представляет собой результат практически одновременного высвобождения многих порций АХ. Высвобождение медиатора, вызывающее деполяризацию концевой пластинки, не обязательно является следствием импульса моторного аксона: его можно вызвать
204
Глава 16
катодной деполяризацией нервных окончаний; частота (но не величина) мВПСП возрастает при искусственной деполяризации [212]. Для высвобождения АХ необходимо наличие ионов Са2+, которые воздействуют на пресинаптическую мембрану (даже в отсутствие Na). Амплитуда ВПСП увеличивается до максимума при увеличении внешней концентрации кальция; в портняжной мышце лягушки на одну молекулу высвобождающегося АХ требуется четыре иона Са24’, а в диафрагме крысы — три. Mg2+ конкурентно тормозит высвобождение АХ кальцием, поэтому для увеличения ВПСП в присутствии Mg2+ требуется больше Са2+. Лантан и марганец блокируют высвобождение медиатора [132]. Из кинетического анализа следует, что у лягушки четыре иона Са2* связываются одним участком X рецептора и что количество освобожденных квантов АХ пропорционально СаХ [4]. Частота спонтанных миниатюрных потенциалов растет с увеличением наружной концентрации Са, но они все же могут возникать и в бескальциевом растворе; таким образом, спонтанное высвобождение АХ в небольшом количестве происходит и при полном отсутствии наружного Са. Однако кальциевый комплекс необходим для высвобождения АХ нервным импульсом [95, 189, 296].
Величина синаптического тока, генерируемого постсинаптической мембраной, пропорциональна количеству высвобождаемого АХ. Сопротивление мембраны концевой пластинки уменьшается в результате одновременного увеличения проводимости для Na+ и К+. Если повысить потенциал покоя путем поляризации вторым микроэлектродом, ВПСП в отличие от потенциала действия возрастет, а при деполяризации мембраны уменьшится. Для портняжной мышцы лягушки потенциал равновесия равен 15—20 мВ. ВПСП вызывает изменения проводимости, пропорциональные количеству АХ и независимые от мембранного потенциала, в то время как изменения проводимости, вызываемые потенциалом действия, зависят от его вольтажа. Мышечный импульс, как и нервный, вызывается повышением натриевой проводимости во время фазы подъема тока действия, за которой следует повышение калиевой проводимости. Тетродотоксин, блокирующий распространение импульса по нервному волокну, не действует на синаптическую мембрану, но препятствует изменению натриевой проводимости во время мышечного спайка. Разрушение Т-канальцев при кратковременном действии гипертонического раствора блокирует активацию сокращения, но не затрагивает ВПСП и потенциала действия.
После денервации миниатюрные потенциалы исчезают, а через несколько дней вся мышечная мембрана становится чувствительной к АХ. Потенциалы действия в ответ на прямое раздражение после денервации или кураризации сохраняются, но становятся нечувствительными к тетродотоксину. В диафрагме крысы максимум увеличения чувствительности к АХ достигается на 15-й день
Мышцы
205
после денервации. Возрастание чувствительности можно предотвратить веществами, блокирующими синтез белков. Распределение радиоактивного бунгаротоксина, который связывается с контактными участками АХ-рецепторов (например, в концевых пластинках), показывает, что после денервации на поверхности мышцы образуются новые рецепторы [164а].
Предыдущее описание нервно-мышечной передачи в наибольшей мере относится к портняжной мышце лягушки. Однако главные его черты наблюдались у ящериц, птиц и млекопитающих. В диафрагме крысы максимальная частота мВПСП наблюдается при 10 мМ Са2+. Декаметоний, блокирующий нервно-мышечную передачу, в отличие от тубокурарина действует как деполяризующий агент; ботулотоксин нарушает высвобождение АХ. Нервно-мышечные синапсы курицы, кошки и человека более чувствительны к декаметонию, чем к тубокурарину, а у собаки и кролика чувствительность синапсов к обоим 'веществам одинакова. Ионы тетраэтиламмония и прокаина уменьшают ток калия из клетки и 'могут блокировать постсинаптические ответы.
В мышечных волокнах миотомов Branchiostoma, когда натриевые спайки блокированы тетродотоксином или погружением в без-натриевый раствор, а ток калия блокирован прокаином, наблюдаются большие спайки, причиной которых является ток кальция внутрь клетки. В обычных условиях кальций засасывается внутрь током натрия. Таким образом, мышца способна давать как тот, так и другой типы спайков [ 152b].
Скорость сокращения мышц
Исчерченные мышцы обладают широким диапазоном скоростей сокращения в соответствии с различными специфическими функциями. Многие попытки дифференцировать поперечнополосатые мышцы позвоночных на быстрые и медленные, фазные и тони-, ческие основываются обычно на выделении одного какого-либо свойства. На самом деле скорость определяется совокупностью факторов. Ленивец ползет со скоростью 50 м/ч, но его можно заставить увеличить ее до 250 м/ч, в то время как максимальная скорость передвижения кошки составляет 40 км/ч, а гепарда — 104 км/ч. Эта разница в скоростях, очевидно, зависит от сократительных свойств мышц конечностей [140]. Многие качества, определяющие скорость, регулируются трофическим влиянием моторных нервов, что предполагает наличие химического воздействия на мышцы независимо от нервных импульсов.
Для характеристики скорости сокращения и расслабления используются различные показатели: время изометрического сокращения, время расслабления до половины максимального напряжения, скорость укорочения при изотоническом сокращении, выражаемая в изменении в длине мышцы или саркомера в секунду,
206
Глава 16
частота слияния при тетанусе. Скорость сокращения хорошо определяется с помощью кривой зависимости скорости укорочения от нагрузки. I
Скорость сокращения зависит от количества последовательных и параллельных вязкоэластичных элементов, от длины волокон и саркомеров, от кинетики и активности миозиновой АТФазы, от характеристических свойств сократительных белков, от особенностей обмена в различных мышцах. Скорость зависит также от того, активирована ли мышца мышечными импульсами (спайками) или синаптическими потенциалами, от количества Т-канальцев и т. д. Скорость расслабления зависит от упомянутых механических факторов и от скорости связывания кальция пузырьками СР. Характеристическая скорость, выраженная в длине саркомера в секунду, характеризует скоростно-силовые качества мышцы, но не зависит от активного состояния. Скорость для одного или многих параллельных актино-миозиновых мостиков одинакова при строго нулевой нагрузке и незначительном внутреннем трении. Несколько миофибрилл в этих условиях сокращаются с той же скоростью, что и большое их количество.
В табл. 16-3 приведены величины скоростей сокращения и максимального напряжения. Как правило, очень быстрые мышцы развивают меньшее напряжение, чем медленные. Скорость расслабления колеблется в гораздо больших пределах, чем скорость сокращения; некоторые запирательные мышцы могут поддерживать напряжение в течение многих часов. Быстрые мышцы скорее утомляются при повторяющихся сокращениях, чем медленные. Мышцы, связанные с генерацией звука, обладают очень большой частотой сокращения: например, крикотиреоидная мышца летучей мыши сокращается 200 раз в секунду, звуковая мышца плавательного пузыря опсануса 300 раз в секунду (сокращение и расслабление длятся 1—1,5 мс [332]), разгибатель омара — от 100 до 200 раз в секунду [244]. При обычном и неподвижном (висячем) полете у колибри частота взмахов крыльев равна 35—45, у зяблика — 22—25 раз в секунду {152]. Самыми быстрыми мышцами млекопитающих являются глазодвигательные. Нижняя прямая мышца глаза крысы (при 35 °C) сокращается за 5—6 мс со скоростью укорочения 65 мкм/с [69а]. М. gracilis крысы проводит возбуждение со скоростью 3,5—5,0 м/с (при 35°C), а портняжная мышца лягушки —со скоростью около 1 м/с (при 20 °C).
Иннервация, мембрана, механические свойства мышцы и скорость сокращения. Очень немногие мышцы являются только быстрыми или только медленными: большинство мышц — смешанные, имеющие как быстрые, так и медленные волокна. В мышцах лягушки моторные нервные волокна, вызывающие быстрые сокращения, имеют диаметр 10—12 мкм и скорость проведения 20— 35 м/с. Многие мышцы лягушки, кроме толстых аксонов, иннервируются еще и более тонкими (5—8 мкм) со скоростью проведе-
Таблица 16-3
Скорости сокращения различных мышц
Животное и мышца Время сокращения, мс Время иолурасслабления, мс Скорость сокращения (в мышечных длинах или мкм/с) Частота слияния, С-1 Отношение тета-иус/одиночное сокращение Источник данных1)
Длинный разгибатель пальца кошки Камбаловидная мыш- 19 70 19 109 31 мкм/с [1481 [148]
ца кошки Внутренняя прямая мышца кошки Длинный разгибатель пальца кошки Нижняя косая мышца кошки Икроножная мышца кошки in vivo Камбаловидная мышца кошки Внутренняя прямая мышца кошки Передняя перстнещитовидная мышца кошки Длинный сгибатель пальца кошки Переднеберцовая мышца котенка Переднеберцовая мышца кошки Камбаловидная мышца котенка Длинный разгибатель 7,5—10 19,6 18 40 75—100 7,5—10 22 18 56 27 65 13 7,5 1,3 мкм/с 42,7 мкм/с 69 4 11 3,4 5 5 [69] [69] [69] [168] [168] [168] [179]. [47] [69]
пальца крысы Камбаловидная мыш- 36 55 18,2 мкм/с [69]
ца крысы Длинный разгибатель 51 51 22,8 мкм/с 8,3
пальца котенка Камбаловидная мышца котенка Боковая широкая мышца морской свинки Средняя икроножная мышца морской свинки Сгибатель большого пальца морской свинки Длинный сгибатель пальца морской свинки 70 19 22,2 21,2 21,2 109 18,5 21,2 19,9 20,3 12,7 мкм/с [24] [24] [24] [24]
Продолжение табл. 16-3
Животное и мышца Время сокращения, мс Время полурасслабления, мс Скорость сокращения (в мышечных длинах или мкм/с) Частота слияния, с-1 Отношение: тета-нус/одиночиое сокращение Источник данных!)
Камбаловидная мыш- ’182,3 113,8 [24]
ца морской свинки Длинный разгибатель 65—98 169—190 [142]
пальца ленивца 4—5,8
Икроножная мышца 174 111—245 [142]
ленивца Сгибатель когтей 140—300 240—300 15—20 [140]
Bradypus Кржотир ео идн а я 4 200 [304]
мышца летучей мы-
ши
Длинный разгибатель 56,5 17,3 мкм/с 2 [67]
пальца новорож- денной крысы 19,3 мкм/с 1,4
Камбаловидная мыш- 65 [67]
ца крысы Щитовидно-черпало- 6,5 [154]
видная мышца кролика
Крикотиреоидная 24—30 [157]
мышца кролика Большеберцовая 24—28 [154]
мышца кролика Летательная мышца 8 250—300 [152]
колибри Летательная мышца 14 100—150 [152]
зяблика Полусухожильная 800 900
мышца жабы (0°С) Фазное волокно мыш- *22 (85%
цы лягушки 'от макс.) [4]
Тоническое волокно 46 (85%
мышцы лягушки Быстрое волокно без от макс.) 6—12 мкм/с [4]
сарколеммы (лягушка) 1 мкм/с [81]
Медленное волокно [81]
без сарколеммы (лягушка) 6 1/с
Портняжная мышца 75 НО 16—20 [148]
лягушки (16 °C) Лестничная мышца 40 30 1,3 Z/c
змеи Thamnophis Ретрактор penis че- 400 1000 0,36 1/с [168]
репахи
Кожнореберная мыш- 37—54 [306]
ца змеи
«Звуковая» мышца 5 8 6 1/с [332]
опсануса
14—2067
v-ottiotuCC/ UU, UHUd- тель 1 VI V41 1 'боковой сгибатель одиночное сокращение дистальный тетанус проксимальный тетанус Homarus разгибатель быстрый разгибатель медленный Limulus, сердце Кузнечик, «звуковая» мышца Таракан, коксальная мышца Lethocerus, мышца дыхательной трубки ophthalmus Белая мышца Pod-ophthalmus Paralithodes Hemisquilla медиальный разгибатель одиночное сокращение ночная мышца миксины Фазная мышца Вгап-chiostoma Тоническая мышца Branchiostoma Ртпияя мыпп.пя Pnd- Ьыстрая пристеночная мышца миксины Медленная поисте- «Звуковая» мышца Holocentrus «Звуковая» мышца опсануса, 29 °C Тонкая мышца Hippocampus Животное и мышца
t с to сл с с uu—UU 5 (сок-ращ. и рассл.) 4—10 90 ЧП 50 500 10 Кп сл ю о с D to х> to ? ? ? DO 1 СЛ 00 DO “ О О <io О 8 ° сл ~ £ to о сл » сл о о сл сл Время сокращения, мс
с с о S чз 2 тз ® о <о о ° К ° с Э II 1 р ~ g "o' р 'и' 1 D to >— слЬтз я чз «а -о к о сл о 2 S о о S S S с о » йз м 300 4*-О О -о D D 6 22,5 13—15 Время полурас-слаблеиня, мс
Скорость сокращения (в мышечных длинах или мкм/с)
0,981 Z/c 200 100 Частота слияния, с-1
I Отношение: тета-нус/одииочное сокращение
—t—Ч г—-!Г— СО СО tO tO to о 05 О 03 ►£- -q О 00 СО фь %, ^-и 1 U-J I—Ч t—! Iя—Ч Г““Ч I"—Ч 1—*4 ЬО КЭ * ф- to 00 00 сл сл _ г- tO СЛ .03 03 »— 1—( с£> С£ Е0 Cz-I |—1 i_, Ь—' —‘ (-L >— 1—"Ч F«4 >—1 to >— СО 03 со to оз к— 1——1 Источник данных1^
Продолжение табл. 16-3
Продолжение табл. 16-3
Животное и мышца Limulus, сгибатель брюшка Улитка (Helix) ретрактор щупальца ретрактор глотки У стр ица, непрозр ач- ная мышца Mytilus, передний бис-сусный ретрактор (быстрая) Lumbricus продольная (фазная) продольная (тоническая) кольцевая Pecten исчерченная неисчерченная Aurelia Metridium Golfingia ретрактор хоботка (фазный) ретрактор хоботка (тонический) веретенообразная мышца Thyone, мышца-ретрактор Головоногие, мышца хроматофора Кальмар, мантия Трахейная гладкая мышца (37 °C) Матка (27 °C) Папиллярная мышца (27 °C) Taenia -coli Черепаха, кишка Кошачая акула, -брыжейка (фазная) S’ <0 а я о О у к S S . « к Q< а pq а 195 2500 200—250 1000 70—80 300—500 46 2280 500—1000 500— 30 000 (1-3 мин) 87 540 1800 3900 НО 68 16,6 30000 1800 CJ Я о й2 5 § С я к « и 3 435 25 000 1300—1500 4000—17000 200 800—1500 40 5140 600—1000 1—2 мин (2—6 мин) 95 2700 3500 5700 300 1060 36 000 2500 .° _ О О О — е-> ю « - - - - _ Скорость сокра- ьэ оо ^4 сл S2 со щения (в мышеч- g. ►— — ных Длинах или g -- -г-* > мкм/с) 7? ° 77 77 77 „ К1 частота слияния, со 04 00 _! С с ишишепис. тета-нус/одиночное сокращение § tt ‘я' S я СТ о н о S [250] [250] [351а, Ь] [64] [64] [292] [287] [339] [3]
’) Отсутствующие ссылки можно найти в табл. 50, стр. 479, I издания этой книги, «Мир», 1967.
Мышцы
211
ния от 2 до 8 м/с. Если проведение в толстых аксонах блокировано или раздражаются только отпрепарованные пучки тонких аксонов, мышца в ответ на одиночный импульс дает незначительное сокращение, возрастающее при повторных импульсах; такое сокращение много медленнее, чем наблюдаемое при стимуляции крупных аксонов. При стимуляции тонких нервных волокон в мышцах возникают не распространяющиеся по закону «все или
Юме 100 мс
Рис. 16-17. Регистрация изометрических одиночных сокращений и тетанусов моторных единиц крысы [i67 b].
А. Медленная мышца (камбаловидная). Б. Моторная единица промежуточного типа (из камбаловидной мышцы). В. Быстрая мышца.
ничего» импульсы, а только синаптические потенциалы, более слабые (от 7 до 15 мВ), чем ВПСП системы «быстрых» аксонов. Длительность ВПСП медленных мышц значительно больше, чем ВПСП быстрых мышц, а при повторяющихся раздражениях возникает облегчение с образованием плато на уровне деполяризации в 20—50 мВ.
Потенциал покоя медленных мышечных волокон лягушки ниже (около 60 мВ), чем у быстрых (около 90 мВ). У медленных волокон лягушки емкость мембраны в три раза больше, а удельное сопротивление в 10 раз больше, чем у быстрых волокон [4] (табл. 16-4). Максимальная скорость укорочения обратно пропорциональна времени сокращения, однако отношение величин одиночного сокращения и тетануса не зависит от времени сокращения (рис. 16-17). Мышечные волокна медленного типа имеют много двигательных нервных окончаний, так что возбуждение всего волокна обеспечивается нервным путем и нет необходимости в распространяющихся мышечных импульсах. Медленные мышечные 14*
212
Глава 16
волокна, сокращающиеся градуально, примерно в 100 раз менее утомляемы, чем быстрые фазные волокна. АХ или прямое электрическое раздражение вызывают в медленных волокнах продолжительное сокращение; это свойство позволило использовать прямую мышцу живота лягушки в качестве тест-объекта для определения концентрации ацетилхолина. Портняжная мышца или длинная приводящая мышца лягушки дают только быстрые ответы, а подвздошно-берцовая — преимущественно медленные. В прямой мышце живота лягушки вентральные поверхностные волокна имеют низкий потенциал покоя, иннервируются аксонами с высоким порогом, большим латентным периодом, небольшими ВПСП с облегчением. Глубже расположенные волокна имеют высокий потенциал покоя, низкий порог, короткий латентный период, ВПСП, вызывающие потенциал действия и фазные сокращения [125].
У млекопитающих большинство поперечнополосатых мышечных волокон имеет, по-видимому, по одному нервному окончанию, поэтому различия в скорости их сокращения определяются не характером иннервации, а другими причинами. Однако в некоторых волокнах верхней косой мышцы глаза найдены множественные гроздевидные (en grappe) нервные окончания [171]. Различия в скорости отражают скорее различия в сократительных белках, чем в активации мембран.
Птицы имеют две широкие спинные мышцы; одна из них, задняя, является фазной, а передняя — тонической. Электрические свойства мембран у них различны: мембрана задней имеет меньшую временную константу (т)1 и меньшую пространственную константу (Tv)2, [116, 180] (табл. 16-4). Ее волокна получают по одному нервному окончанию пластинчатого типа (en plaque). При ионофоретическом исследовании действия АХ на каждом волокне обнаруживается лишь одно чувствительное место с относительно высоким порогом чувствительности к АХ. Передняя мышца, наоборот, имеет множественную иннервацию; точки, высокочувствительные к АХ, расположены через каждые 740 мкм.
У подвязковой змеи Thamnophis короткие 'подкожные мышцы между чешуйками имеют время сокращения 40 мс и период полурасслабления 30 мс; пороговая деполяризация равна —40 мВ, максимальное сокращение при деполяризации — между —10 и 0 мВ [167]. Фазные волокна с одиночной иннервацией и потен-
1 Временная константа г характеризует скорость нарастания или спада потенциала мембраны при раздражении ее прямоугольным импульсом тока. Она равна произведению сопротивления мембраны на ее емкость, измеряется временем (в секундах), в течение которого потенциал падает до 0,37 от исходной величины, т. е. уменьшается в «е» раз (е— основание натуральных логарифмов, равно 2,7,1). — Прим, перев.
2 Пространственная константа % характеризует декремент потенциала импульса вдоль волокна и измеряется расстоянием (в см), на котором потенциал уменьшен ,в «е» раз. — Прим, перев.
Таблица 16-4
Константы мышечной мембраны
Мышца X, мм т, мс R, Ом/см2 с, мкФ/см2 Источник Данных ;
Лягушка портняжая мышца фазная мышца портняжная мышца разгибатель пальца разгибатель пальца портняжная мышца Крыса, диафрагма Мышь камбаловидная мышца длинный разгибатель пальца Лягушка тоническая мышца Цыпленок передняя широчайшая спины задняя широчайшая спины Змея, кожнореберная мышца Ящерица быстрая мышца медленная мышца Минога париетальные волокна центральные волокна Краб Carcinus, мышцы ног волокна 500 мкм 210 мкм 105 мкм Скорпион, закрыватель клешни Таракан, прямая кишка Краб, мышца ноги Portunus Морская свинка, taenia coli Кошка, кишка кольцевая мышца продольная мышца 2,2 1,1 0,5—0,8 1,1 0,5—0,8 0,42—0,53 0,92 0,55 1,87 0,68 4,4 4 0,4 1,5 2,3 0,6 1,5—4 2 0,9 1 1—1,5 15 18 10,8 18 10,8 4,4 2,8 35 3,7 37,2 140 5,2 4 15 41,4 3 70 5 4,6 133 (вычисл.) 52—105 (вычисл.) 3140 4000 2080 4000 2080 360—400 2170 708 29 000 4388 561 38280 4000 31 000 36 000 88,5 282 1170 830 2-104—1,6- 10б 5000 116 20-103—30-103 1000 780 6,1 6,8 6 5,2 4,8 5,2 3,4—3,8 1,6 8,2 7,0 1,04 7 3 4 45 54,6 36 3,6 0,4—3,2 1,1 4,2 3 30 12,7 [132] [41 [229а] [H6J [306] [286] 1286] [351с] [13] [138] [26b] [П2] [353] [220] [220]
214
Глава 16
циалами действия перемешаны с тоническими волокнами, имеющими множественную иннервацию, которая обеспечивает лишь синаптические потенциалы.
В мышцах рыб встречаются оба типа иннервации — и одиночная, и множественная. Звуковая мышца рыбы-белки Holocentrus при производстве звука сокращается с частотой до 100 с-1 и дает гладкий тетанус при частоте сокращений 200 с-1. Она имеет множественную иннервацию; нервные окончания располагаются на расстоянии 10 мкм друг от друга по длине волокна. В ней были зарегистрированы множественные ПСП и потенциалы действия. Таким образом, эта мышца иннервирована по типу медленных мышц, в то время как ее мембрана имеет свойства быстрых [134]. У змееголова Ophiocephalus и красные, и белые мышцы имеют М-полосу, триады и множественные гроздевидные окончания; иными словами, белые волокна своими функциональными особенностями напоминают быстрее волокна лягушки, а характером иннервации— медленные {259]. Красные волокна — тонкие и дают градуальные суммирующиеся потенциалы, в то время как белые волокна толще и дают ВПСП со спайками [347].
Брюшные мышцы рыбы Cottas иннервированы аксонами диаметром 10—14 мкм, сокращаются быстро, дают спайки, но их иннервация — множественная: одно волокно может иметь от 8 до 22 окончаний, что показано специфическим окрашиванием, выявляющим ацетилхолинэстеразу [190]. Париетальная мышца миксины имеет волокна двух типов. Толстые более глубоко расположенные волокна дают сравнительно быстрые отдельные сокращения продолжительностью 150 мс; их потенциал покоя равен —75 мВ; иннервация одиночная. У поверхностных тонких волокон имеется по два моторных аксона с распределенными по всей длине волокна окончаниями, отдельное сокращение длится более 500 мс, а потенциал покоя составляет около —46 мВ [9, 203]. Сегментные миотомы миноги имеют центральные волокна и париетальные волокна на каждой стороне миотома; центральные волокна дают отдельные сокращения и спайки с «овершутом» и электротонически взаимодействуют между собой, тогда как париетальные волокна дают медленные ответы без спайков, снабжены множественными концевыми пластинками и в них регистрируются мВПСП (351с).
В миотомах ланцетника (Branchiostomd) мышечные волокна представляют собой лентовидные клетки толщиной в 1—2 мкм и длиной 600 мкм; Т-канальцы отсутствуют; изменения натриевой и кальциевой проводимости могут происходить параллельно; кальций может участвовать в кофеиновой контрактуре [152а]. Поверхностные волокна — медленные (одно сокращение длится 200—250 мс), внутренние — быстрые (70—80 мс), дают спайки [151].
Мышцы
215
Структура мышцы и скорость сокращения. Ультраструктура быстрых и медленных мышц была описана выше. Здесь мы сравним фазные и тонические позные мышцы лягушки.
Быстрые мышцы Медленные мышцы
Примеры Полусухожильная Прямая живота, часть волокон Портняжная т. iliofibularis
Миофибриллы 1 мкм и меньше, цилиндриче- Лентовидные, полевидной ские, фибриллярной струк- структуры (Felderstrukture) туры
Окончания Z-пластинки М-линия Канальцы Пластинчатые (ей plaque) Гроздевидные (еп grap.pe) Поперечные, прямые Зигзагообразные Ясно обозначена Отсутствует Развиты, с триадами Неразвиты (редки), без выра- женных триад
М. stapedius кошки относится к быстрым мышцам, мышца, натягивающая барабанную перепонку, — к смешанным; она имеет примесь медленных волокон без М-линий и зигзагообразную Z-пластинку {П7].
Корреляцию между скоростью и длиной саркомера можно проследить на разных волокнах сгибателя брюшка омара [1, 200, 201].
Длина саркомера, мкм Число толстых нитей на 1 мкм2 Число диад на 1 мм волокна Продолжительность сокращения, мс
Поверхностные волокна Глубоко расположенные волокна 6—8 2—4 350 450 600 2500 500—2000 30—50
Из этого сопоставления видно, что быстрые сокращения коррелируют с короткими саркомерами, правильными Т-канальцами, развитой 'CP-системой и упорядоченным расположением фибрилл.
Миоглобин, метаболические ферменты мышцы и скорость сокращения. Поперечнополосатые мышцы позвоночных бывают красными (богатыми миоглобином), белыми (лишенными миоглобина) и промежуточных типов. Как правило, красные мышцы сокращаются более медленно и не так быстро утомляются, как белые. Например, красная камбаловидная мышца кошки сокращается вдвое медленнее белой икроножной мышцы, однако они развивают примерно равное напряжение на единицу поперечного сечения. Кровоток в красных мышцах в покое может быть в три раза интенсивнее, чем в белых, но при работе возрастает сильнее в белых, чем в красных. По составу ферментов белые мышцы лучше приспособлены к анаэробному гликолизу, а красные — к окислительному обмену. Результаты гистохимического исследования ферментов показывают, что во многих мышцах среди красных волокон
216
Глава 16
разбросаны белые, встречаются волокна и с промежуточными свойствами. Сравнение двух типов мышц дает следующую картину:
Красные мышцы крысы
М. soleus
Много крупных митохондрий
Имеется запас липидов
Высокая активность фосфорилазы
Высокая активность цитохромок-сидазы и большое количество НАД-Н
Высокая активность сукцинатде-гидрогеназы
Окислительный обмен
Утомляется медленно
Белые мышцы крысы
М. gastrociiemius
(Мало митохондрий
Нет запаса липидов, но много гликогена
Низкая активность фосфорилазы
Высокая активность мышечной лактатдегидрогеназы
Низкая активность сукцинат-дегидрогеназы
Анаэробный гликолиз
Утомляется быстро
Мышца сердца — красная, в ней мало гликогена, но много ферментов аэробного обмена '[99]. Известны исключения из приведенного выше правила. Так, крикотиреоидная мышца летучей мыши и грудная мышца колибри содержат много митохондрий и липидных капелек, но относятся тем не менее к быстрым мышцам. Лактатдегидрогеназа содержится в мышцах в двух формах, что можно определить с помощью электрофореза; одна из них характерна для сердца и адаптирована к высокому уровню кислорода и к окислительному обмену, другая характерна для скелетных мышц и адаптирована к низкому содержанию кислорода (см. гл. 6). Существуют разные объяснения наличия двух дегидрогеназ, но все они сходятся на признании этой основной корреляции.
Фазные мышечные волокна лягушки содержат много митохондрий, и их ядра располагаются ближе к центру волокна; медленные волокна содержат меньше митохондрий, а их ядра расположены под сарколеммой [108]. Гистохимические различия волокон видны из следующего сопоставления:
Волокна Диаметр Активность АТФазы Активность НАД Н-дегидрогеназы Активность фосфорилазы
Тонические Фазные » Малый » Большой Очень низкая Высокая » Низкая Высокая Низкая Очень низкая Высокая Средняя
Содержание РНК в медленной передней широчайшей мышце спины цыпленка на 40% больше (а протеолитическая активность выше на 200%), чем в быстрой задней; это указывает на более интенсивный обмен белков в медленной мышце [343].
Мышцы
217
В радиальной мышце непарного плавника кошачьей акулы имеется наружный слой красных волокон, которые характеризуются наличием ВПСП, но являются тоническими. Внутренний слой этой мышцы состоит из крупных белых волокон фазного типа, образующих спайки [307а]. Поперечнополосатые мышцы позвоночных могут быть самых разнообразных видов — от чисто аэробных красных мышц до чисто гликолитически белых, включая сюда ряд определяемых гистохимически промежуточных типов. Их ферментативные различия коррелируют со скоростью расслабления и развиваемой силой, но не обязательно с характером иннервации.
Корреляция между активностью миозиновой АТФазы и сокращением, а также между связыванием кальция и расслаблением. О некоторых свойствах миозиновой АТФазы уже говорилось в разделе, посвященном тропомиозину и тропонину. Удельная активность этого фермента, оцениваемая по активации М§2+-зави-симым актином (in vivo) или ионами кальция (in vitro), у многих животных исключительно хорошо коррелирует со скоростью сокращения. В задней широчайшей мышце спины цыпленка активность миозиновой АТФазы в три раза выше, чем в передней [143]. В красных мышцах крысы активность миозиновой АТФазы низка, а в белой — высока [99]; в быстрой исчерченной части аддуктора раковины гребешка она в 5 раз выше, чем в медленной запирательной части [20]. Температура оказывает на миозиновую АТФазу такое же действие, как на укорочение мышцы [19].
Табл. 16-5 демонстрирует корреляцию между активностью миозиновой АТФазы и скоростью сокращения [19].
Для мышцы моллюска Pecten имеются следующие данные [340]:
Mg-АТФаза, в мкмолях
Рнеорг/Мин на 1 мг белка
Исчерченный аддуктор (быстрый) 1,75
Прозрачная гладкая мышца (медленная) 1,21
Непрозрачная гладкая мышца (запирательная) 0,36
Активность миозиновой АТФазы в камбаловидной мышце млекопитающих низка, а в медиальной головке икроножной — высока. По активности этого фермента мышцы кошки в 2—4 раза превосходят соответствующие мышцы ленивца [19]. В глицеринизи-рованных мышечных волокнах насекомых величина напряжения примерно пропорциональна уровню распада АТФ, отмечаемому при повышении концентрации кальция от 10~8 до 10~7 М [60]. Таким образом, скорость сокращения хорошо коррелирует с уровнем АТФазной активности.
Выше уже говорилось что при расслаблении мышц ионы кальция активно связываются пузырьками саркоплазматического ретикулума, которые содержат активируемую магнием АТФазу.
218
Глава 16
Таблица 16-5
Корреляция между активностью миозиновой АТФазы и скоростью сокращения [19]
Скорость укорочения мышцы, мм/d) Активируемая актином АТФаза2)
Мышь, длинный разгибатель пальца 24 30,03
Крыса, длинный разгибатель пальца 17,2 27,95
Мышь, камбаловидая мышца 12,8 14,38
Крыса, камбаловидная мышца 7,2 10,74
Моллюск Pecten, исчерченная мышца 3 4,03
Лягушка, портняжная мышца 2 2,04
Кролик, мышца матки 0,2 0,23
Черепаха, подвздошно-берцовая мышца 0,1 0,13
Моллюск Mytilus, задний аддуктор 0,1 0,15
') При 37—38 °C для млекопитающих и при 18 °C — для других животных.
2) В микромолях неорганического фосфата на 1 мг миозина в 1 мин.
В большинстве случаев скорость связывания Са2+ (in vitro) изолированными пузырьками (микросомами) соответствует главным образом Скорости расслабления. Связывание кальция частицами гладкой мышцы млекопитающих в 10 раз меньше, чем частицами сердца {57]. Скорость связывания Са2+ микросомами белой мышцы кролика выше, чем скорость связывания микросомами красной. Вот некоторые данные, характеризующие связывание кальция [340]:
Саранча, мышцы крыльев мышцы задних ног
Мышь, ягодичная мышца
Lumbricus, мышца стенки тела
Mytilus, передний биссусный ретрактор
Phormia, синхронные летательные мышцы
Связывание Са, мкмоль/г за 10 мин при 30°С
111
53
88
10
10
14
В лишенных сарколеммы волокнах тонического пучка подвздошно-берцовой мышцы лягушки сокращение при микроперфузии кальция распространяется лучше в медленных, чем в быстрых волокнах. Причина этого, вероятно, заключается в более эффективном связывании Са быстрыми волокнами [82].
Трофическое влияние нервной системы на сократительные свойства мышц; корреляция с АТФазой. Из многих наблюдений над мышцами млекопитающих следует, что характер их иннервации влияет на некоторые свойства ферментов, мембран и структурных белков; соответственно различаются и скорости сокращения. Разительные перемены происходят во время развития и рос
Мышцы
219
та, а также в тех случаях, когда нервы «быстрых» мышц врастают в «медленные», и наоборот. Например:
Длинныг i сгибатель i (быстрая шца) Камбаловидная мышца (медленная) Источник данных
пальце!
мы
перекрестная иннервация норма перекрестная иннервация
норма
Кошка
время сокращения (мс) 25,5 59 75 33,3 [48]
время сокращения 18—20 60 70—80 35—40
Отношение тетанус/одиночное 7,1 4,3 5,6 11,7 [47]
сокращение
Кошка, время сокращения (мс) 32,8 48,8 69,7 47
Крыса, время сокращения (мс) 23,2 32,2 45,8 28,7 [314]
Длинный разгибатель пальцев Камбаловидная мышца
Крыса
время сокращения (мс) 13 25 34 15
скорость сокращения (мкм/с) 45,1 22,5 19,8 33,8 [68]
Кривые сила — скорость представлены на рис. 16-18.
Приведенные выше данные показывают, что путем перекрестной иннервации быстрые мышцы можно сделать более медленными, а медленные — более быстрыми; при этом, в частности, изменяется скорость сокращения, что означает возникновение изменения в контрактильных белках и в продолжительности активного состояния. В норме волокна камбаловидной мышцы чувствительны к АХ на большей части своей длины, между тем как длинный разгибатель пальца — только в области концевых пластинок. В результате иннервации смешанным седалищным нервом чувствительность некоторых волокон камбаловидной мышцы к АХ уменьшается [247]. Подобным же образом перекрестная иннервация увеличивает размер чувствительной к АХ области в задней широчайшей мышце спины цыпленка и уменьшает в передней.
Миозиновая АТФаза была выделена из медленной мышцы (камбаловидной) и быстрой (длинного сгибателя или разгибателя пальцев) и из тех же мышц после взаимной перекрестной иннервации. Активность актин- и Mg-активируемой АТФазы из камбаловидной мышцы крысы и длинного разгибателя пальцев пропорциональна скорости их укорочения. АТФаза камбаловидной мышцы более термостабильна, менее устойчива к воздействию Щелочей и имеет другую электрофоретическую подвижность [135]. После перекрестной иннервации миозин быстрой мышцы уменьшает свою АТФазную активность и увеличивает устойчивость по отношению к щелочам, в то время как в камбаловидной мышце АТФазная активность и чувствительность к кислотам возрастают [324]. Зависимость активности миозиновой АТФазы от pH показывает, что перекрестная иннервация изменяет миозин. В медлен-
Рис. 16-18.
А. Изометрические одиночные сокращения камбаловидной мышцы (1), длинного сгибателя большого пальца (2) и длинного разгибателя пальца (3) трех млекопитающих [67а]. Б. Кривые сила — скорость для двух мышц крысы: медленной — камбаловидной (1) и быстрой — длинного разгибателя пальца (3) при нормальной (а), после естественной (б) и после перекрестной (в) иннервации [68].
Мышцы
221
пых мышцах уменьшается динитрофенилирование, а миозин становится более устойчивым к кислотам [217]. Таким образом, миозин быстрых и медленных мышц качественно различен и его форма, синтезируемая в мышце, зависит от характера иннервации. До сих пор неизвестно, каким способом осуществляется это влияние— особой последовательностью нервных импульсов или какими-либо «трофическими» веществами.
Результаты опытов по перерезке сухожилий и длительной стимуляции мышц у кролика свидетельствуют о влиянии нервных импульсов на сократительные белки. В норме камбаловидная мышца постоянно получает тонические моторные импульсы от спинного мозга; в быстрой мышце постоянная активация отсутствует. После тендотомии время сокращения камбаловидной мышцы уменьшается от 62 до 20 мс, но непрерывная длительная стимуляция (до 20 дней) предотвращает этот эффект [323, 366].
В период индивидуального развития у млекопитающих происходят изменения скорости сокращения мышц, сходные с теми, которые наблюдаются при перекрестной иннервации. Об этом свидетельствуют следующие данные {70, 71]:
Длинный разгибатель пальца Камбалови дная мышца котенок кошка
котенок кошка
Время сокращения (мс) 51 30 70 70—80
Время полурасслабления (мс) Скорость укорочения (мкм/с) 51 22,8 31 109 12,7 13
Таким образом, в процессе развития животного скорость сокращения длинного разгибателя увеличивается, а камбаловидной мышцы — нет. Подобным же образом у новорожденных крыс кривые сила — скорость и скорости сокращения для разгибателя пальцев и камбаловидной мышцы одинаковы, но в процессе развития эти показатели для длинного разгибателя пальца возрастают в 2,5 раза, а для камбаловидной мышцы остаются без изменений [67].
Потенциал покоя мышечной мембраны с возрастом увеличивается, что видно из следующих данных:
Мышь, потенциал покоя (мВ)
Длинный разгибатель пальца
1 нед 8 нед
—57,8 —79,5
Камбаловидная мышца
1 нед 8 нед
—67,4 —77,2
Чувствительность к локальному действию АХ и реакция на кофеин также зависят от иннервации, следовательно, СР-система (саркоплазматический ретикулум) и распределение рецепторов АХ находятся под нервным контролем {247].
222
Глава 16
Подытоживая, можно сказать, что быстро сокращающиеся поперечнополосатые мышцы отличаются от более медленных целым рядом признаков: ультраструктурой, распределением нервных окончаний и областей, чувствительных к АХ, окислительными и гликолитическими ферментами, механическими свойствами (связанными с миозином и миозиновой АТФазой). Некоторые черты, важные для одной мышцы, могут не быть таковыми для другой, имеющей сходную скорость сокращения, особенно если речь идет о разных видах животных. Многие мышцы являются смешанными, т. е. содержат волокна двух или трех типов, различающихся главным образом распределением ферментов.
Скорость сокращения коррелирует со скоростью гидролиза АТФ, а скорость расслабления — со связыванием кальция. Многие биохимические свойства мышц определяются влиянием двигательных нервов. Мышцы в свою очередь влияют на распределение нервных окончаний врастающих в них нервов. Влияние иннервации определенно сказывается на синтезе мышечных белков и, следовательно, может осуществляться посредством генетического кодирования.
Какова относительная значимость влияния трофических химических агентов и моторных импульсов, предстоит еще выяснить.
Множественная иннервация мышц у членистоногих, особенно у ракообразных
Некоторые гистологические и химические свойства мышц членистоногих коррелируют со скоростью сокращения так же, как и свойства мышц позвоночных. Короткие саркомеры, упорядоченное распределение волокон и развитая CP-система коррелируют с быстрым сокращением.
Различия в длине саркомеров между быстрыми и медленными волокнами позвоночных гораздо меньше (от 2 до 3,5 мкм), чем между волокнами членистоногих (от 2 до 14 мкм). Медленные волокна — тонкие, с длинными саркомерами и глубокими складками сарколеммы [14, 15]. Длина саркомеров фазных волокон краба Cancer равна 4—5 мкм; одна толстая нить у них приходится на шесть тонких. Тонические же волокна имеют более крупные миофибриллы с саркомерами в 12 мкм, одна толстая нить приходится на 10—12 тонких [ИО]. Скорость сокращения коррелирует не только с длиной саркомера, но также и с развитием СР-системы. Время сокращения дистальной части бокового сгибателя Не-misquilla, иннервируемой «быстрым» моторным аксоном, равно 50 мс, а проксимальной части той же мышцы, иннервируемой «медленным» аксоном, — 500 мс; в обеих частях длина саркомера равна 4,4 мкм, но дистальная содержит 24 диады, а проксимальная на той же площади сечения— 12 [242а].
Мышцы
223
Большая часть волокон мышц ракообразных имеет очень большой размер; это связано, возможно, с периферическим контролем сокращения и с малым количеством нейронов, более крупных, чем у позвоночных. Мышечные волокна морского желудя (бала-нуса) могут иметь диаметр до 2 мм, а волокна ножных мышц камчатского краба —до 4 мм. Средняя величина саркомеров бала-нуса — 9,2 мкм, и его мышца может сократиться до 30% длины покоя (т. е. более чем в 3 раза); в этом состоянии толстые нити проникают сквозь Z-пластинку и перекрывают нити соседнего саркомера [187].
Характер иннервации. Нервные окончания у членистоногих не образуют отдельных концевых пластинок; медиатор выделяется там, где аксон лишен шванновских клеток в любом месте его контакта с мышечными волокнами; нервные волокна могут опутывать мышечные клетки плотной сетью (рис. 16-19).
Целые мышцы, состоящие из многих волокон, могут у ракообразных и насекомых иннервироваться лишь двумя (или несколькими) аксонами, а каждое мышечное волокно может иметь поли-нейронную иннервацию, т. е. получать иннервацию от двух и более (до пяти) аксонов. Нервные окончания густо покрывают поверхность мышечного волокна, и, несмотря на то что большинство волокон иннервировано определенным аксоном, число возбужденных волокон может варьировать в зависимости от частоты нервной импульсации. Этот факт, по-видимому, отражает различия в нервно-мышечном облегчении и проводимости в различных точках аксона. Обычно один из аксонов является тормозным, другие— возбуждающими и могут вызывать сокращения различной силы и необходимую степень облегчения. В тех случаях, когда тормозные и возбуждающие аксоны могли быть идентифицированы, было найдено, что синаптические пузырьки возбуждающих аксонов имеют сферическую форму, а более мелкие тормозные— овальную или неправильную форму [224а, 357а].
Не все волокна мышцы иннервируются всеми входящими в нее аксонами. Исследование большого числа волокон флексора клешни Panulirus показало, что 7% волокон получают по четыре возбуждающих аксона, 27% —по три, 26% —по два и 38% —только один аксон [374, 375]. Внешняя часть мышцы, поднимающей глазной стебелек краба Podophthalmus, состоит из быстрых белых волокон, а внутренняя — из розовых тонических; белая часть получает два быстрых аксона, а розовая — два возбуждающих, быстрый и медленный, и один тормозной [186]. У речного рака Не-misquilla средний разгибатель мероподита получает один быстрый возбуждающий (без облегчения) и один тормозной аксон; боковой разгибатель получает один быстрый и два медленных (облегчающих) аксона, а срединный сгибатель — два медленных моторных аксона {51а].
224
Глава 16
У ракообразных имеется большое разнообразие в характере иннервации разных мышц одного животного и одних и тех же мышц разных животных (рис. 16-20). Часто один аксон иннервирует несколько мышц. У рака такие мышцы, как открыватель клешни и вытягиватель карпоподита, имеют один возбуждающий аксон. Закрыватель, сгибатель и разгибатель могут иметь по два
Мышцы
225
аксона каждый, а флексор имеет даже четыре (рис. 16-20) [375]_ Закрыватель, сгибатель и разгибатель клешни обычно имеют по одному быстрому и одному медленному аксону. Когда один аксон, возбуждающий или тормозной, иннервирует две мышцы, сокращение одной или другой может определяться частотой стимуляции.
Периферическое торможение бывает двух родов: пре- и постсинаптическое. В норме при постсинаптическом торможении имеет место гиперполяризация мембраны мышечного волокна. При поляризации клетки с помощью внутриклеточного микроэлектрода мембрана становится более отрицательно заряженной, в этом случае тормозные влияния увеличивают проводимость (уменьшают сопротивление) малых анионов (особенно С1_), что приводит к деполяризации. В брюшном сгибателе рака торможение также постсинаптическое {215]. При пресинаптическом торможении
Рис. 16-19. Микрофотографии, полученные при помощи сканирующего1 электронного микроскопа; мышцы ракообразных. (Любезно предоставлено Ф. Лангом.) А. Видно, как нервное волокно, переходя от одного мышечного волокна к другому, образует попутно (en passant) синапсы и веточки и затем возвращается к первому волокну. / нерв, 2 — мышечное волокно; 3 — синапс. Б. Виден диффузный характер нервных окончаний на мышечном волокне. 4 — нерв; 5 — тонкие веточки. В. Нервно-мышечные и аксо-аксонные синапсы в открывающей клешню мышце рака. Возбуждающее окончание (ВО) содержит круглые синаптические пузырьки и образует на мышце две синаптические мембраны; тормозное окончание (ТО) содержит более мелкие разбросанные пузырьки и образует пре паптнческий тормозной контакт с возбуждающим нервным окончанием 1224а)..
15—2067
в
Рис. 16-20.
А. Схемы иннервации различных мышц в нескольких группах десятиногих рг: 1375J п открыватель и 3 — закрыватель клешни; В — вытягиватель; С — сгибатель кар флексор; А — аддуктор и ₽ — разгибатель мероподита. Б. Схема синаптичес, зей возбуждающего (В), специфического тормозного (СТ) и общего тормозн нов вытягивающей мышцы краба Grapsus [14aJ. Мышечное волокно 1 —медле?
ми саркомерами; мышечное волокно 2— быстрое, с короткими саркомерам, н, , слабым облегчением имеют длинные сплошные синаптические мембраны, а сиил1н„ы с ш мальным облегчением — короткие синаптические .участки.
Мышцы
227
уменьшается выделение возбуждающего медиатора и отсутствует прямое влияние на сопротивление мышечной мембраны. В открывателе дактилоподита рака тормозной импульс опережает возбуждающий на 1—6 мс, что приводит к уменьшению ВПСП в четыре раза ([100, 101]. В ряде случаев (например, в случае закры-вателя Pachygrapsus) действие медленного возбуждающего аксона ослабляется пресинаптическим торможением в большей степени, чем действие быстрого [15]. Пресинаптическое торможение уменьшает или блокирует также спонтанные мВПСП. Торможение открывателя клешни рака при частоте раздражения 10 Гц является на 90% пресинаптическим, а при 40 Гц — лишь на 50%. Таким образом, действие одного и того же аксона зависит .от частоты его импульсов. Сила пре- и постсинаптического торможения изменяется от волокна к волокну, а облегчение торможения зависит от количества выделяющегося медиатора [14а].
Принято считать, что у ракообразных распространяющиеся токи действия мышечных волокон не являются необходимыми, так как скорость проведения по аксону высока- а количество нервных окончаний вдоль мышечных волокон, достаточно велико; в результате этого медиатор выделяется практически одновременно во всех участках мышечного волокна. Синаптические потенциалы, являющиеся следствием повышения , натриевой проводимости, достаточны для того, чтобы вызвать сркращение [264]. Синаптические потенциалы могут быть быстрыми, с небольшим облегчением или без него, и могут достигать амплитуды, которая вызывает спайкоподобную деполяризацию. В других случаях они могут; быть медленными и облегченными,; иногда эти ПСП достигают уровня деполяризации, дающей начало «градуальным» спайкам [14а]. У Hemisquilla медиальный сгибатель меропбдита получает два медленных моторных аксона; в нем наблюдаются серий: облегченных ВПСП с возникающими время"ОТ-’ времени-быстрыми спайками [51а]. При прямом раздражении" ВйутрйклёТЬчйыми электродами могут возникать потенциалы действий;‘йрёвышающие потенциал покоя (т. е. «с овершутом»), в йе^ОТб^ЫХ МЁГГлечных волокнах в о.твет на стимуляцию моторных- айёбйбн вбзнйкают распространяющиеся по закону «все
Спайки, образующиеся у ракообразныхответ! ца-Тр^1мёй-бранную стимуляцию1 мыщц, обусловлен.^;. возросшей- ;й|ю$одй-мостью к ионам кальция. В, некоторых мый^аХ^Ййа^алЙЦйЙ может быть заменен стронцием; при этом, у'даё^я .наблюдавьИокй действия, но не сокращения [111].'
-ризация при трансмембранной стимуляции •Щ^др^'иЬйаЛьНа.’.-тра-диенту Са. Она избирательно блокируемся ирными 'Mfi2* или’\С<Я+ в малых концентрациях *[153].
Скорость сокращения и расслабления^. а тдкже; й облегчение зависят как от свойств мышечных волокон —фазных или тони
15*
228
Глава 16
ческих, — так и от свойств возбуждающих моторных аксонов — быстрых или медленных. Некоторые мышцы получают по одному возбуждающему и одному тормозному аксону. В некоторых из них, например в добавочном флексоре краба, проксимальные волокна имеют длинные саркомеры и являются тоническими, дистальные же имеют короткие саркомеры и являются быстрыми. В открывателе клешни рака волокна центрального участка мышцы дают небольшие ВПСП с облегчением, а волокна дистального участка — более значительные, со слабым облегчением, хотя обе части иннервируются одним и тем же аксоном {14а]. В фазных волокнах вытягивателя карподита Cancer возникают короткие ВПСП с облегчением, а в тонических волокнах — большие и длительные ВПСП, и в этом случае оба типа ответов также вызываются одним и тем же аксоном ’[15]. Вообще в тех случаях, когда мышца с одним моторным аксоном дает как быстрые, так и медленные сокращения, в медленных волокнах отмечаются большие ВПСП со слабым облегчением, а в быстрых — небольшие ВПСП с сильным облегчением, которые при большой частоте раздражения часто вызывают спайки. Таким образом, для сокращения медленных волокон необходима низкая частота раздражающих импульсов, а для быстрых — высокая. Облегчение и скорость нарастания ВПСП в разных синапсах одного и того же аксона различны и зависят от количества выделяемого медиатора. Скорость сокращения и его амплитуда коррелируют также с мембранным потенциалом; в некоторых мышцах ног омара скорость развития напряжения возрастает с увеличением деполяризации мембраны, вызванным внутриклеточными электродами [202а].
В мышцах, имеющих как быстрые, так и медленные возбуждающие аксоны, стимуляция крупного моторного аксона вызывает сокращение с коротким латентным периодом, быстрым укорочением и высоким напряжением, почти одинаковым для одиночного сокращения и для тетануса; ВПСП велики, но обычно без «овер-шута» и с незначительным облегчением. Раздражение второго или среднего по величине аксона дает медленно нарастающее сокращение без отчетливого латентного периода, с заметным облегчением и низким напряжением. Мышечные потенциалы вначале малы, но благодаря облегчению при повторном- раздражении сильно увеличиваются. При раздражении всего нерва с возрастающей частотой быстрое сокращение переходит в медленное. Стимуляция третьего, обычно самого тонкого аксона тормозит действие возбуждающих аксонов. В период торможения сокращение может быть подавлено, но облегчение может продолжаться, что обнаруживается после прекращения торможения.
Таким образом, в мышцах с двойной или множественной иннервацией ответ может определяться типом и местоположением мышечных волокон и свойствами активного в данный момент аксона. В некоторых волокнах закрывателя отдельных видов крабов
Мышцы
229
в ответ на раздражение любого возбуждающего аксона возникает значительное облегчение; в других — при раздражении быстрых аксонов отмечается появление крупных ВПСП, а при раздражении медленных аксонов — появление небольших ВПСП. Эти различия в ответах более выражены при низкой частоте стимуляции, чем при высокой.
Мышца-разгибатель второй антенны омара состоит из двух частей, каждая из которых снабжена большим количеством нервных окончаний и полинейронной иннервацией. Быстрая часть, содержащая большое количество саркоплазматических канальцев, дает одиночные сокращения без облегчения продолжительностью менее 10 мс, и развивает напряжение в 25 г/см2. Время сокращения медленной части составляет 50—60 мс и сопровождается облегчением ВПСП; она расслабляется медленно (время полурасслабления 3—5 с), развивая напряжение, равное 2800 г/см2 {244]. В закрывателе клешни Carcinus некоторые волокна дают крупные ВПСП, в них возникают одиночные сокращения, они электрически невозбудимы, относительно нечувствительны к торможению, имеют густую иннервацию от быстрого аксона, небольшое сопротивление мембраны и временную константу, равную 5 мс. Во второй группе волокон в ответ на раздражение медленного аксона появляются крупные и длительные ВПСП, а в ответ на раздражение быстрого аксона — небольшие (в последнем случае реакции может не быть совсем); эти волокна электрически невозбудимы, чувствительны к торможению, временная константа их мембраны составляет 30—60 мс. Третья группа волокон дает как быстрые, так и медленные ВПСП с облегчением в обоих случаях '[13]. У открывателя клешни рака ВПСП, наблюдаемые в дистальных волокнах, примерно в 5 раз превышают ВПСП центральных волокон; вероятность высвобождения медиатора больше в дистальных волокнах [34]. У камчатского краба Paralith&des закрыватель имеет два возбуждающих аксона; один (быстрый) в проксимальной части вызывает крупные, слабо облегчающиеся ВПСП, в дистальной — мелкие ВПСП, в центральной — ВПСП среднего размера, с сильным облегчением; другой (медленный) аксон вызывает в дистальной части возникновение крупных ВПСП с умеренным облегчением и мелких, с очень сильным облегчением — в центральной части [225] (рис. 16-21).
В брюшке речного рака и омара глубокий или медиальный сгибатель является фазной мышцей; некоторые ее волокна имеют по три возбуждающих и по два тормозных аксона, другие — соответственно два и один. Поверхностная или боковая мышца — тоническая, медленно утомляемая, получает пять возбуждающих и один тормозной аксоны; ее иннервация в разных волокнах сильно варьирует t[l, 215]. Считается, что различные аксоны дают различные эффекты, особенно при низких частотах, что мембраны отдельных мышечных волокон имеют различные свойства, что раз-
4
Мышцы 231
ные волокна одной мышцы имеют разную иннервацию и что все эти различия могут зависеть от локализации волокна в мышце.
Еще один вид. регуляции сокращения осуществляется распределением импульсов во времени. Например, открыватель клешни Eupagurus, имеющий один моторный аксон, при стимуляции с низкой частотой дает медленно усиливающееся сокращение, в то же время дополнительные раздражения вызывают в нем быстрые, сильные сокращения. Таким образом, один и тот же аксон способен в зависимости от характера раздражения вызывать либо медленные, либо быстрые сокращения. Некоторые возбуждающие аксоны реагируют на один короткий стимул серией импульсов; они имеют длительную аккомодацию и короткие константы возбуждения. Аксон быстрого закрывателя клешни рака, не дающий разряда импульсов, имеет высокий порог и константу аккомодации 717 мс, а аксон одиночного открывателя клешни дает серии разрядов, имеет константу аккомодации 48,2 мс, и меньшую временную константу возбуждения [384а]. У Uca общий возбуждающий аксон обслуживает две мышцы — растягиватель и открыватель,— но тормозные нервы у них разделены; торможение возникает рефлекторно: тормозной аксон открывателя дает залп импульсов вслед за разрядами возбуждающего аксона, а залп тормозного аксона растягивателя предшествует им. Это коррелирует с тем, что открыватель тормозится преимущественно постсинаптически, а растягиватель — пресинаптически [335].
У некоторых членистоногих флексоры и экстензоры ног не являются антагонистами, так как разгибание осуществляется гидростатическим способом. Предполагается, что торможение имеет место только при наличии антагонистов. В клешне скорпиона закрывание представляет собой активный процесс, а открывание— пассивный, волокна флексора имеют по два возбуждающих аксона, один вызывает ПСП, облегчающиеся до спайков, а другой вызывает быстрые ПСП без облегчения. Каждая из двух или трех моторных единиц закрывателя имеет свою собственную пару аксонов, но при этом периферическое торможение не обнаруживается [139]. Флексор ноги паука получает три моторных аксона и ни одного тормозного. Одиночное мышечное волокно дает три
Р,ис. 16-21.
/. Электрическая (вверху) и механическая (внизу) регистрация активности волокон мышцы камчатского краба при раздражении медленных (А, Б, В, Г) и быстрых (Д, Е, Ж) моторных аксонов [225]. II. Электрические ответы волокон вытягивающей мышцы краба Grapsus на стимуляцию, производимую с различной частотой (мышца имеет только один возбуждающий аксон) [14а]. А. ВПСП крупные, со слабым облегчением (в медленных волокнах с длинными саркомерами). Б. ВПСП вначале малы, ио затем увеличиваются в результате сильного облегчения (в быстрых волокнах с короткими саркомерами). В. ВПСП умеренной величины; при низкой частоте переходят в спайкообразиые ответы. Калибровка: А, Б. 20 мВ и 1 с. В. 20 мВ и 0,5 с. III. Развитие напряжения в разгибателе омара в зависимости от величины Деполяризации мембраны [202а]. Верхние записи — изменение мембранного потенциала в ответ иа пропускаемый ток, нижние записи — напряжение. А. В быстром волокне напряжение пропорционально деполяризации. Б. В медленном волокне напряжение медленно растет в ответ на постоянную деполяризацию. Калибровка: 0,2 с, 10 мВ, 20 мг (напряжение).
232
Глава 16
различных ответа: один аксон вызывает быстрое сокращение, другой— медленное и третий — быстрое, но с высоким порогом {298]. Крупные волокна закрывателя у балануса также не имеют антагонистов; они иннервируются двумя или тремя возбуждающими аксонами и не получают тормозных; они дают нераспространяю-щиеся ответы, и, чтобы вызвать сокращение, деполяризация при ПСП должна превысить 18 мВ [187]. Закрывающая мышца мечехвоста (Limulus) иннервирована шестью возбуждающими аксонами, а каждое волокно может получать до трех аксонов и давать градуальные облегченные ответы. Тормозной аксон трудно выделить, и тормозные ПСП (ТПСП) не были обнаружены, однако механическая активность уменьшалась при высокой интенсивности стимуляции или при воздействии гамма-аминомасляной кислотой [270].
Медиаторы. Фармакологические вещества, блокирующие или усиливающие нервно-мышечную передачу у позвоночных, неэффективны у ракообразных (в обычных концентрациях). Очевидно при этом, что ни АХ, ни норадреналин у них в нервно-мышечной передаче не участвуют. Наиболее вероятным медиатором является L-глутамат: он деполяризует мышцу, вызывает сокращение, действует синэргично с возбуждающими нервными импульсами и, по-видимому, воздействует на те же рецепторы, что и возбуждающие медиаторы [349—351]. Высокая концентрация магния или пониженная концентрация кальция блокируют передачу возбуждения (в закрывателе клешни Pachygrapsus); при этом в облегчающих окончаниях возбуждающего аксона имеется много синаптических пузырьков [15]. Количество участков, чувствительных к глутамату, пропорционально числу нервных окончаний [349]. Синаптические потенциалы и ответы на вводимый с помощью электрофореза глутамат исчезают при замене натрия ионами трис-буфера или лития [264]. Потенциал равновесия для ВПСП равен — 20 мВ (у рака); во время ВПСП сопротивление мембраны уменьшается по сравнению с состоянием покоя втрое [303]. В открывателе клешни рака (абдукторе) наблюдаются спонтанные мВПСП, увеличивающиеся при деполяризации нервных окончаний возбуждающего аксона. Подсчитано, что при частоте 5 с-1 каждый импульс освобождает в среднем 2,2 «кванта» медиатора (один квант равен количеству, необходимому для одного мВПСП). Облегчение увеличивает вероятность освобождения квантов возбуждающего медиатора [103]. В отдельных окончаниях при низкочастотной стимуляции медиатор в дистальных волокнах высвобождается с большей вероятностью, чем в центральных, т. е. освобождение медиатора отклоняется от кривой распределения Пуассона [35].
Гамма-аминомасляная кислота (ГАМК) имитирует действие тормозного медиатора, вызывая гиперполяризацию мембраны посредством увеличения проводимости для С1~; она является антагонистом пикротоксина при низких концентрациях (10~7 М) и
Мышцы,
233
тормозит сокращение мышц рака. У открывателя клешни омара реверсивный потенциал ТПСП равен —75 мВ, у рака —87 мВ [255], или на 5—10 мВ ниже потенциала покоя. ГАМК уменьшает величину ВПСП пропорционально концентрации ионов хлора. Постсинаптическая мембрана тормозных окончаний проницаема для мелких анионов (NO3, SCN, Cl), но не для крупных (изотионат, метилсульфат), и эта проницаемость под влиянием ГАМК возрастает [255]. Пресинаптическое торможение является следствием увеличенной проницаемости для С1~; оно менее эффективно при уменьшении внешней концентрации хлора {101]. Пикротоксин уменьшает ТПСП и гиперполяризацию, вызванную ГАМК; на ВПСП он не влияет. В перфузате мышцы ГАМК может при торможении быть выделена в количествах, пропорциональных тормозной стимуляции. Таким образом, можно считать установленным, что ГАМК является медиатором как для пресинаптического, так и для постсинаптического торможения у ракообразных.
В тибиальной мышце скорпиона один аксон вызывает ВПСП, другой — спайкоподобные ответы. Ни ГАМК, ни глутамат не оказывают никакого влияния на мышцу, из чего следует, что у скорпиона медиатором служит другое вещество Ц38].
В синапсах гигантского моторного аксона в брюшном флексоре рака наблюдались длительные изменения возбудимости: стимуляция аксона с частотой 1 имп/мин ведет к уменьшению ВПСП (привыкание); внезапное увеличение частоты снимает этот эффект /[45]. В мышцах рака и краба стимуляция с частотой свыше 5 имп/с ведет к длительному увеличению количества выделяемого медиатора: ВПСП возрастают в 2—5 раз, вероятно, вследствие накопления натрия в нервных окончаниях [331а].
Суммируя, можно сказать следующее. Полинейронная, с множеством нервных окончаний иннервация мышц ракообразных создает возможность широкой моторной интеграции, возникающей на периферии. Существуют различные схемы иннервации. Один и тот же аксон может оказывать различное действие в зависимости от частоты импульсов. Некоторые мышечные волокна дают тонические, а другие — фазные сокращения. Медиатор возбуждения — глутамат — может вызывать как быстрые, так и медленные сокращения, в зависимости от распределения нервных окончаний. Тормозной медиатор — гамма-аминомасляная кислота — может действовать пре- и постсинаптически, повышая проницаемость для ионов хлора. Очевидно, что многие регуляторные функции, осуществляемые у позвоночных на уровне спинного мозга, у ракообразных перенесены в нервно-мышечные синапсы.
Неосциллирующие мышцы насекомых
Ножные и брюшные мышцы насекомых, так же как и их летательные мышцы с низкой частотой движений крыла, имеют Двойную, а иногда тройную иннервацию и в этом отношении на
234
Глава 16
поминают мышцы других членистоногих. Нервные окончания с синаптическими пузырьками равномерно распределены вдоль мышечных волокон. Один аксон бывает обычно быстрого типа, дру гой — медленного.
Раздражение быстрого аксона мышцы ноги насекомого визы вает в отдельном волокне значительные электрические ответы, которые могут быть градуальными или спайкоподобными, переходящими нулевой потенциал мембраны («овершут»). Быстрые4от веты не облегчаются. При охлаждении препарата или увеличении содержания магния в растворе ответ уменьшается, и синаптические потенциалы и спайки разделяются. Реакция оказывается одинаковой во всех точках волокна, т. е. спайк по волокну не распространяется. Трансмембранная стимуляция вызывает нераспро-страняющиеся градуальные спайки, поэтому они могут быть названы «локальными спайками». Как быстрые, так и медленные синаптические потенциалы отличаются От нераспростра'няЮгЦихся спайков; синаптические потенциалы увеличиваются при гипергюля-ризации и уменьшаются при деполяризации мышечной мембраны; спайки же при деполяризации превращаются в залпы импульсов [184].
У саранчи стимуляция медленного аксона с частотой ниже 10 имп/с вызывает незначительное сокращение мышц ноги, но с повышением частоты до 150 имп/с сокращение усиливается до максимума. Только» от 30 до 50% волокон получает медленные аксоны.
При внутриклеточном отведении обнаруживается заметная рг ница между волокнами: некоторые дают ПСП с облегчением, др гие — большие потенциалы с «овершутом» по отношению к т тенциалу покоя, равному 60 мВ. Величина ответов при облегчение пропорциональна мембранному потенциалу покоя [184]. Базилярные мышцы пластинчатоусых жуков иннервируются только медленными волокнами, мышечные потенциалы облегчаются.
В проторакальной мышце Carausius в ответ на раздражение ее быстрых нервных волокон возникают синаптические потенциалы, которые могут суммироваться и спайки, имеющие рефрактерный период; в ответ на раздражение медленных нервных волокон в этой мышце возникают синаптические потенциалы, облегчающиеся при повторении [381]. Не все мышцы ног насекомых имеют двойную иннервацию; так, разгибатель голени метаторакальной ноги саранчи получает лишь один быстрый аксон. Вертлужная мышца таракана. (Periplaneta) дает реакцию типа синаптического потенциала.
В депрессоре тазика таракана самьщ крупные моторные аксоны вызывают быстрые сокращения, более тонкие — медленные сокращения, а самые тонкие являются тормозными. Некоторые мышцы имеют три тормозных аксона; в то же время одни мышцы могут иметь два возбуждающих аксона, а другие — один [278]..
Мышцы
235
Мышца j закрывающая дыхальце у саранчи, .имеет двойную иннервацию, а у павлиноглазки та же мышца получает четыре аксона, один из которых более «быстрый», чем остальные. При повышении концентрации СО2 снижается поток моторных импульсов от нервной системы, мышца расслабляется и дыхальце открывается. В мышце обнаружены спонтанные пейсмекерные потенциалы и потенциалы действия, уменьшающиеся при повышении концентрации Ср2 [363]. У таракана Blaberus правое и левое дыхальца имеют отдельные тормозные аксоны, но один общий возбуждающий '[249].
У некоторых ночных бабочек с хорошей теплоизоляцией в период предполетного разогрева грудные мышцы, ритмически сокращаясь, повышают температуру груди и особенно нервного ствола (см. гл. 9). У,сатурнид во время разогревания поднимающие и опускающие крылья мышцы могут сокращаться синхронно, но при полете их сокращения происходят поочередно (209].
У кузнечика Neoconocephalus стрекотанье производится надкрыльями, движения которых вызываются синхронными сокращениями мышц с частотой от 145 до 212 с~к (в зависимости от температуры); на каждое сокращение приходится один потенциал действия [208а]. Более высокая частота обнаружена у стридуля-ционной мышцы сверчка Orocharis — до 280 с-1’*йри 35 °C [367а].
Ранее считалось, что мышцы насекомых имеют по 2—3 возбуждающих аксона, но что тормозные аксоны у них отсутствуют; однако теперь тормозные аксоны обнаружены у таракана и саранчи. Метаторакальный задний леватор тазика таракана Periplaneta получает четыре возбуждающих и два тормозных аксона. Три возбуждающих аксона имеют малый диаметр и вызывают медленные ответы, а четвертый значительно крупнее и вызывает быстрые ответы; общий тормозной аксон обслуживает как синергические, так и антагонистические мышцы ноги. У саранчи (£о-eusta) также найден общий тормозной аксон, вероятно регулирующий положение ноги и облегчающий расслабление [277]. Саранча (Schistocercd) для управления движениями каждого крыла имеет четыре мышцы — три‘ леватора и один депрессор. Депрессор получает два возбуждающих быстрых аксона, один леватор имеет три быстрых возбуждающих аксона, а второй и третий получают по одному медленному возбуждающему и одному тормозному. Так же как и у ракообразных, не все волокна в мышце одинаково иннервированы [277]. У пустынной саранчи тормозной аксон метаторакального экстензора голени дает разряд импульсов непосредственно перед сгибанием; это значит, что сенсорная сигнализация контролирует разряды центральной нервной системы и торможение ускоряет расслабление и устанавливает исходное тоническое напряжение [362]. В коксальном аддукторе Schistocerca и в мышцах других насекомых каждое волокно имеет по два возбуждающих и по одному тормозному аксону [280].
236
Глава 16
У насекомых, как и у ракообразных, действие возбуждающих окончаний не блокируется холи политика ми и не воспроизводится холиномиметиками; у них не обнаружено также чувствительности к ацетилхолину ни в процессе развития, ни при дегенерации [217]. Наиболее эффективным веществом, имитирующим действие нерва при перфузии через мышцы насекомых, оказался /-глутамат [361]. В ноге таракана в нервно-мышечном соединении глутамат вызывает миниатюрные синаптические потенциалы и сокращения при концентрации 10~6 М [217]. Чувствительность к локальному действию глутамата уменьшается с увеличением расстояния от нервных окончаний, но после дегенерации <нерва зона чувствительности к глутамату расширяется [359]. В концентрации Ю '1 М глутамат вызывает контрактуру в мышцах таракана. Гемолимфа содержит глутамат, который в норме связан с белками плазмы [361].
Гамма-аминомасляная кислота (ГАМК) — наиболее вероятный медиатор торможения, так как она уменьшает действие возбуждающих нервов. Пикротоксин блокирует как нервное торможение, так и действие ГАМК. Вероятно, торможение является следствием возрастания проницаемости мембраны для ионов хлора, с реверсивным потенциалом —70 мВ (в мышце саранчи) .[360]. Потенциал покоя мышечной мембраны личинки мучного хрущака Tenebrio сравнительно малочувствителен к наружной концентрации калия и в норме положителен по отношению к потенциалу равновесия калия; при активации возросшая калиевая проводимость может вызвать деполяризацию, если мышца находится в среде с высокой концентрацией калия [26b].
Резонирующие летательные и тимпанальные мышцы насекомых
Летательные мышцы перепончатокрылых и двукрылых и мышцы, производящие звук у равнокрылых, сокращаются с частотой, превышающей скорость восстановления возбудимых мембран. У стрекоз, бабочек и прямокрылых летательные мышцы прямые: они прикрепляются непосредственно к суставам крыла и получают один моторный импульс на каждое сокращение. Летательные мышцы более быстрых насекомых — непрямые: они соединены со сложной системой рычагов, имеют очень крупные волокна и упорядоченное расположение протофибрилл.
Частота движений крыла, измеренная стробоскопическим методом или по высоте звука, издаваемого в свободном полете, соответствует мышечным сокращениям и потенциалам действия, зарегистрированным в условиях прикрепленного полета. У двукрылых частота взмахов при подрезании крыльев или при повышении температуры возрастает. Частота движения крыла обратно пропорциональна корню кубическому из момента инерции крыла, а также обратно пропорциональна атмосферному давлению. Макси
Мышцы
237
мальная частота в 2218 взмахов в секунду была зарегистрирована при высокой температуре среды у комара-звонца Forcips, крылья которого были подрезаны. Высота звука при полете Chironomus равна 600—650 Гц. Движения груди у прикрепленного насекомого имеют несколько более низкую частоту. Его непрямые летательные мышцы должны обладать способностью сокращаться и расслабляться в пределах 0,45 мс [334а]. У более крупных насекомых частота взмахов крыльев не зависит от подрезания и груза.
Было проведено много исследований по аэродинамике и особенно энергетике полета насекомых. У пустынной саранчи (Lo-custa) выходная мощность высока — около 5 ккал/ч-кг; скорость полета зависит от частоты взмахов и инерции крыла, формы тела и некоторых внешних факторов среды. У некоторых мух скорость полета достигает 150—375 см/с [179а]. Подъемная сила и сила тяги в какой-то степени независимы. На подъемную силу больше влияет наклон крыла, чем угол наклона корпуса. У саранчи, как и у многих других насекомых, полет рефлекторно вызывается прерыванием контакта лапок с опорой, общим неспецифическим раздражением и обдуванием чувствительных окончаний, расположенных на голове; для поддержания полета требуется обдувание крыльев или головы.
У насекомых с прямыми летательными мышцами частота взмахов достигает 20—40 с-1, сокращения мышц синхронны с токами действия, ампутация крыла мало влияет на частоту: она запрограммирована в грудных ганглиях. У бабочки Agrotis мышечные потенциалы могут повторяться с интервалами в 20 мс при отсутствии облегчения [311]. У мух и перепончатокрылых, имеющих непрямые летательные мышцы, синхронность между мышечными сокращениями и токами действия отсутствует; у интактного насекомого один мышечный спайк может приходиться на 5—20 взмахов крыла. Ампутация крыльев может почти удвоить частоту сокращений без изменения частоты спайков. Изолированные непрямые мышцы нельзя заставить сокращаться под влиянием нервной стимуляции с той же частотой, с которой они сокращаются при нормальном полете. Летательные мышцы шмеля Дают тетанус при частоте 40 с-1; у мух сокращение прикрепленной к рычагу мышцы не зависит от частоты раздражения, если она выше 40 с-1. Таким образом, миогенный ритм превышает частоту стимуляции через нерв.
У некоторых видов цикад (например, Platypleura) тимпанальный орган производит звук частотой 200 Гц (а максимально — до 5400—7000 Гц); частота токов действия в мышце равна 50 с-1, но при этом мышца может сокращаться несколько раз в ответ на один потенциал действия. Один аксон иннервирует всю мышцу. После разрушения тимпанальной мембраны отношение между мышечными потенциалами и сокращениями становится равным 1:1; если тимпанальную мембрану нагрузить, частота биений
238
Глава 16
•уменцшдется [284]. У других вцдов насекомых, например Meimura,. частота сокращений тимпанальной мышцы (как и в летательных мышцах саранчи) совпадает с частотой нервных импульсов; слияние электрической реакции происходит при частоте сокращений, примерно равной 120 су1. Как и в летательных мышцах при отношении: частоты сокращений мышцы тимпанального органа и потенциалов действия нерва, равном 1:1, ампутация тимпанальной мембраны дает малый эффект, тогда как в тех тимпанальных органах,: где частота звука превышает частоту импульсов, частота биений при ампутации возрастает.
То,, что одному импульсу соответствует несколько сокращений, а частруа движений крыла превышает частоту слияния при тетанусе, оказывается возможным благодаря особым механическим свойствам мышцы. У двукрылых непрямые летательные мышцы связащя со скелетными элементами, включающими щитковый рычаг, имеющий два стабильных положения, — верхнее и нижнее, Когда, под влиянием сокращения мышцы один конец этого рычага перемещается вниз за его среднюю точку, скелетные элементы быстро переходят* в стабильное положение, соответствующее меньшей длине мышцы и таким образом ослабляют (разгружают) ее.. Затем' вследствие эластичности скелетных элементов (в тимпанальном органе) или под влиянием сокращения антагониста (непрямой летательной мышцы) рычаг восстанавливается в своей верхней позиции и растягивает мышцу (рис. 16-22). Быстрое растяжение вновь вызывает развитие напряжения, и сокращение снова переводит рычаг вниз. Подобным же образом и в тимпанальном органе мышца перетягивает мембрану через точку неустойчивого равновесия и она изгибается внутрь, ослабляя напряжение мышцы; затем мембрана восстанавливает положение за счет собственной эластичности и растягивает мышцу, вызывая новое сокращение. Этот цикл может повторяться до тех пор, пока сохраняется «активное состояние» мышцы; для поддержания последнего требуется лишь достаточное количество нервных импульсов [284]..
Колебательный характер сокращений проявляется не только в интактных мышцах вне организма, но; и в пучках глицеринизиро-ванных волокон. Если сокращения записываются в изометрическом режиме (или изотоническом, когда нагрузка мышцы превышает критическую демпфирующую), напряжение растет по типичной активной кривой длина—напряжение (рис. 16-22,Б). Если, однако, нагрузка мышцы ниже критической демпфирующей, напряжение отстает от длины, так что при критической длине активное ; напряжение развивается медленно, вызывая небольшое укорочение с последующим падением .напряжения; однако продол, жающееся растяжение • ведет к новому подъему напряжения, и возникает колебательный режим. Запаздывающий подъем напряжения, при растяжении превышает падение напряжения при рас-
Рис. 16-22. ‘
Колебательные сокращения глицеринизированнбГо волокна .непрямой летательной ллышцы J.ethocerus [285]. Толщина волокна 70 мкм, длина—0,5 .см, частота .синусоидального растяжения 2 с—’, амплитуда — 36% первоначальной длины. Б. За^ибимоеть напряжения от длины глицеринизнрованного волокна гигантского водяного клопа [320]. Вынужденные осцилляции и петля, образующаяся при записи напряжения как функций- длины pH 6,5; 5 мМ Mg-АТФ; 20 мМ гистидин, 70 мМ КСГ.?
240 Глава 16
сл’аблении. Кривая, описываемая при этих колебаниях, лежит между активной и пассивной кривыми длина — напряжение: ее положение определяется нагрузкой и характерными свойствами мышцы. Глицеринизированные волокна могут расслабляться действием ЭГТА (этиленгликоль-бнс-тетраацетат); если их затем погрузить в раствор слабой ионной силы, содержащий АТФ и Са2* (10 s мэкв), и растянуть, волокна начнут колебаться с частотой, зависящей от нагрузки [285]. При более высоком содержании Са2+ (10~7 мэкв) в мышце развивается более значительное напряжение, но колебания отсутствуют. Глицеринизированные волокна гигантского водяного клопа колеблются при 8-10~8 М Са, но сокращаются без колебаний при 10~7 М Са [23]. Глицеринизированные волокна тимпанальной мышцы цикады начинают колебаться при синусоидальном растяжении в среде, содержащей АТФ и Са2+ [7]. В растворе 5 мМ АТФ синусоидальное напряжение отстает от длины и колебания возникают при Ю-6 М Са2+ [320]. Миозиновая АТФаза расщепляет АТФ во время активных колебаний, но не при пассивном растяжении и ослаблении мышцы [320]. Однако растяжение увеличивает активность активируемой кальцием миозиновой АТФазы, т. е. растяжение действует подобно возрастанию содержания кальция [285].
В летательных мышцах кузнечика на кривой напряжение — длина наблюдается гистерезис; если они растягиваются синусоидально, изменения напряжения отстают от изменения длины. При уменьшении частоты кривая падения напряжения ложится над кривой повышения частоты, так что напряжение при промежуточной частоте во время падения частоты выше, чем во время ее подъема [379].
Запирательные мышцы и задержанное расслабление
Неисчерченные непрозрачные мышцы аддукторов двустворчатых моллюсков, сократившись, способны поддерживать напряжение в течение многих часов. Это может засвидетельствовать каждый, кто пытался открыть устрицу. Аддуктор устрицы способен поддерживать напряжение в 0,56 кг/см2 в течение 20—30 дней ,[250]. Максимальное начальное напряжение, развиваемое различными мышцами при минимальной длине («длина покоя»), выражается в следующих величинах (кг/см2): аддуктор Venus—12,8; аддуктор тридакны—6,7; передний биссусный ретрактор Myti-lus—10—12; аддуктор Unto — 3,1; гребешок — 2—8 [226, 250].
У интактного гребешка (Pecten) в тоническом состоянии можно зарегистрировать незначительную электрическую активность, а редкая стимуляция висцерального нерва поддерживает мышцу в сокращенном состоянии. Отсюда можно было бы сделать вывод о том, что задержка расслабления является, хотя бы частично, тетаническим ответом на редкую стимуляцию. Однако если при
Мышцы
241
сокращенном аддукторе висцеральный нерв перерезать, то мышца останется сокращенной.
Биссусный ретрактор Mytilus состоит из тонких (5 мкм) неисчерченных волокон [356] длиной 1 —1,5 мм [354]. При стимуляции импульсами постоянного тока продолжительностью 1 с или больше мышца сокращается и может не расслабляться в течение многих минут. При стимуляции в течение нескольких секунд переменным током или сериями коротких (5—20 мс) импульсов постоянного тока частотой от 10 до 60 с-1 мышца отвечает быстрыми сокращениями и расслаблениями. Если мышца находится в состоянии тонического сокращения после стимуляции постоянным током, то короткие серии импульсов ведут к немедленному расслаблению [207]. Нервные волокна, вызывающие сокращение, и волокна, вызывающие расслабление, удалось разделить. Напряжение при фазных сокращениях увеличивается с повышением частоты стимуляции до нескольких импульсов в секунду; при этом амплитуда потенциалов действия увеличивается при частоте 0,5 с-1 и при частоте от 5 до 7 с-1 они сливаются [119]. Скорость проведения нервных импульсов составляет 25—50 см/с. После обработки веществами, блокирующими нервное проведение, распространяющихся импульсов не наблюдалось.
Потенциал покоя волокон переднего биссусного ретрактора равен 65 мВ, а синаптические потенциалы достигают 25 мВ; они максимально облегчаются при стимуляции нерва 2—3 раза в секунду. Фазным сокращениям предшествуют мышечные спайки продолжительностью 50 мс и амплитудой, равной примерно 50 мВ [355].
АХ может вызвать сокращение, которое удерживается длительное время после деполяризации. Локальное воздействие АХ вызывает деполяризацию синаптического типа с таким же реверсивным потенциалом, какой наблюдается у синаптического потенциала, вызванного нервным импульсом. Ацетилхолиновое сокращение блокируется бантином (метантелином), который, очевидно, блокирует рецепторы АХ. d-Тубокурарин, бензохинон, атропин, гекса-метоний и декаметоний также обладают эффективным блокирующим действием. Серотонин (5-окситриптамин, 5-ОТ) снимает тоническое сокращение независимо от того, вызвано оно раздражением нерва или ацетилхолином. Считается, что возбуждающие нервы являются холинэргическими, а расслабляющие (выклю-щающие запирательное действие) — триптаминэргическими. Кривые сила — длительность для сокращений и расслаблений, возникающих в ответ на стимуляцию висцерального нерва, показывают, что расслабляющие волокна имеют при любой длительности стимуляции более высокий порог, чем возбуждающие нервы [46]. Концентрация 5-ОТ, необходимая для расслабления запирательных мышц, достаточно низка —от 10~8 до 10~7 М. Педальный ганглий содержит значительное количество 5-ОТ [322] и дофа-16—2067
л
10В 20В ЗОВ 40В 40В (1/2,5 с ) ЮВ 15В 25В 25В (1/15 с )
!-•-----Переменный ток (0,5 с)----►Н---------Постоянный ток (1 с) ----------
Рис. 16-Q3. Запирательные свойства мышцы червя Paragoning [342J-
Мышцы
243
мина. 5-ОТ не оказывает влияния на потенциал покоя, но он снижает сопротивление мембраны, понижает порог возникновения спайков и увеличивает Их амплитуду.
Высота спайков в переднем биссусном ретракторе Mytilus пропорциональна логарифму концентрации Са2+ во внешней среде
Ри» 16 2'4. Прюгофибрмлла ретрактора хоботка Golfingia, содержащая парам'иню-зин (Любезно предоставлено В. Эрнстом.)
(когда мышца находится в растворе, лишенном натрия); спайки блокируются антагонистами кальция, например лантаном, но тет-родотоксин на них не действует. Отсюда следует, что мышечные спайки являются результатом повышения проводимости для Са2+ Есть сведения, что 5-ОТ способствует связыванию кальция внутриклеточными частицами; таким образом, возрастание высоты спайков может быть следствием уменьшения количества ионов Са2+ внутри клетки [357]. Активное напряжение при данной длине уменьшается пропорционально концентрации 5-ОТ [355].
ь
244
Глава 16
Состояние запирательного тонуса (рис. 16-23) есть определенное физическое состояние мышцы, при котором она, в частности, приобретает устойчивость к растяжению, расслабленная мышца пластична, мышца, находящаяся в состоянии запирательного тонуса, ригидна. Сопротивление растяжению обратно пропорционально концентрации 5-ОТ; это означает, что 5-ОТ делает мышцу более растяжимой [355]. Если мышце в состоянии запирательного
гонуса дать возможность укоротиться, напряжение в ней не восстановится. Активное состояние, оцениваемое по восстановлению напряжения, продолжается только 1—7 с, а между тем запирательный тонус поддерживается в течение длительного времени после угасания активного состояния.
На электронных микрофотографиях мышц двустворчатых моллюсков, способных к запирательному тонусу, видны толстые нити (диаметром 300—500 А), окруженные тонкими (55—60 А) (рис. 16-24). Толстые нити состоят из белка парамиозина (тропомиозина А), который можно отделить от других белков, используя неодинаковую растворимость в спирте или в растворах разной ионной силы. Тонкие нити состоят из актина, миозин же образует оболочку вокруг парамиозиновых нитей [327, 344]. Эти парамиозиновые нити (и выделенный из раствора парамиозин)
имеют характерную периодичность в 145 и 725 А. Парамиозин образует тактоиды через 725 А с перекрывающими друг друга молекулами (с мол. весом 200 000) [75] (рис. 16-25). На каждую толстую нить приходится 12 или более тонких [318]. Среди нитей разбросаны темные тельца, которые считают гомологами Z-пла-стинок. Удаление тонких (актиновых) нитей выявляет стреловидную структуру мостиков толстых нитей, обусловленную, вероятно,
Мышцы
245
миозином, окружающим парамиозин.
Согласно одному предположению, сокращение, являющееся следствием взаимодействия актина и миозина, создает трудноизменяемую конфигурацию парамиозина (исключением является расслабляющее действие 5-ОТ). Это объяснение подтверждается сходством между запирательным тонусом интактной мышцы и изменением состояния глицеринизированных волокон в их зависимости от pH среды и от АТФ. Запирательный тонус нельзя вызвать при температуре выше 30 °C [355]. Отсутствие видимых различий в дифракционной картине при рентгеноструктурном анализе свидетельствует против трактовки явления запирательного тонуса как следствия конфигурационных изменений [250, 251].
Согласно другой гипотезе, при запирательном тонусе парамиозин стабилизирует актнно-миозиновые мостики и они разрушаются с трудом. Вариантом этой гипотезы является идея о том, что парамиозин препятствует соединению кальция с расслабляющей системой [261, 355]. 5-ОТ может удалить кальций из поперечных мостиков, т. е. он способствует связыванию кальция. Запирательный тонус (остаточное напряжение) слегка уменьшается с увеличением концентрации Са2+, в то время как максимальное напряжение и Са-АТФазная активность возрастают [227]. Парамиозин
Рис. 16-25.
Л. Паракристалл парамиозина, полученный с помощью преципитации из аддуктора Мегсепа-о
ria Периодичность — приблизительно 725 А. Б. Парамиозин из мышцы Airina, полученный преципитацией ионами бария из раствора тиоцианата. [75]
246
Глава 16
может подвергаться фазовым изменениям, сопряженным с движением поперечных мостиков, а после образования мостиков парамиозин может препятствовать их разрушению [75].
Одно время думали, что запирательный тонус поддерживается без энергетических затрат; аргининфосфат распадается лишь , во время фазного сокращения и расслабления; в,процессе же поддержания запирательного тонуса не было обнаружено заметного возрастания неорганического фосфата или убыли аргининфосфа та. Количество аргининфосфата, расходуемого при фазном и обычном тоническом сокращениях, одинаково, но в случае запирательного тонуса его расход может маскироваться ресинтезом [261]. В покое при 20 °C передний биссусный ретрактор потребляет 78 мкмолей Ог/мин-г; если мышцу отпустить до 75% длины покоя, потребление ее О2 возрастет, а при растяжении уменьшится. Во время фазных сокращений потребление Ог возрастает на 83 мкмоля О2/мин-г (при напряжении 5 мг/см2), и этот дополнительный расход энергии восстанавливается за 20 мин на •80%. Энергетические затраты переднего биссуСного ретрактора при фазных сокращениях в 250 раз ниже, чем затраты, обнаруженные в портняжной .мышце лягушки. Мышцы в состоянии запи радельного тонуса вначале потребляют повышенное количество О2, однако через 20 мин их избыточный обмен 'снижается до 26 мкмолей О2/мин-г при напряжении 5 кг/см2. Если мышцу при запирательном тонусе ослабить, потребление О2 упадет с 26 до 13 мкмолей О2/мин-г, а если снова растянуть, оно возрастает до .27 мкмолей О2/мин-г. Серотонин увеличивает потребление О2.
Парамиозиновые нити найдены не только у двустворчатых моллюсков, но и в мышцах других систематических групп. Толстые парамиозиновые нити обнаружены в ретракторе хоботка Golfingia (рис. 16-24), в мышцах стенки тела круглого червя волосатика (Aschelminthes) [342] и в хорде ланцетника ![122]; у .ланцетника сокращение хорды создает гидростатическое давление для поддержания формы тела [151]. Следующие данные, полученные для запирательного тонуса червя Paragordius, демонстрируют его сходство с запирательным тонусом моллюсков:
Раздражение Время сокращения, с Время полурасслабления Максимальное напряжение, мг
-Переменный ток 0,8—1,6 0,3—08 с 450—600
Постоянный ток, I с 14 18 с 840
Постоянный ток, 5 с 6,4 42 мин 6853
АХ 180—240 58 мин 8328
Менее выраженный запирательный тонус часто встречается в неисчерченных мышцах. Роль парамиозина, так же как и меха низм задержки расслабления при возрастании жесткости, оста ется невыясненной. В мышцах моллюсков расслабление вызывается серотонином, возможно, посредством перераспределения кальция.
Мышцы
247
Мышцы с диагональной исчерченностью
Стенка тела круглого червя Ascaris содержит уникальную мышечную систему, состоящую- из продольных волокон. Каждое волокно представляет собой крупную клетку, которая состоит из (1) бокового вздутия с ядром, гранулами гликогена и митохондриями, (2) длинного волокна, протянувшегося под гиподермисом,
Рис. 16-.26 Мьшючиые клетки Ascaris; поперечное сечение (схема» J135] М— мышечная клетка с отростком О, идущим к нервному стволу Н; Ф периферические миофнбриллы мышечного волокна.
и (3) отростка, идущего к нервном стволу (рис. 16-26) В нервных окончаниях, соприкасающихся с мышечными отростками, видны синаптические пузырьки. Соседние отростки возле нервного ствола тесно переплетаются,, контактируя друг с другом. Сократительные веретена мышечных клеток имеют лентовидную форму, косо исчерчены и состоят из толстых (230 А) и тонких нитей (одна толстая на 10—12 тонких) [315]. Нервный ствол проводит возбуждение к мышце (блокируемое тубокурарином); стимуляция нервного ствола вызывает появление спайков в мышце
248
Глава 16
[86]. ГАМК гиперполяризует мышцу, а АХ деполяризует. В мышечных волокнах наблюдаются спонтанные синаптические потенциалы с частотой от 1,5 до 7 с-1, иногда переходящие в спайки. Эти спонтанные спайки усиливаются с повышением температуры; местом их возникновения является область синцития. Потенциал покоя мышечных волокон необычен тем, что уменьшается с возрастанием концентрации наружного CI. Гиперполяризационная фаза спайков является следствием усиления калиевого тока. Электрические стимулы, приложенные к одному волокну, распространяются на соседние, что указывает на тесное электрическое взаимодействие [88, 89, 149].
Еще один вид мышц, отростки волокон которых подходят к нервному стволу (а не наоборот), представляет собой миотомы ланцетника [121]. Волокна с диагональной исчерченностью найдены также в прозрачной (или быстрой) части аддуктора двустворчатых моллюсков. У гребешка (Pecten) и устрицы (Grasso-strea) тонкие и толстые нити переплетаются, образуя спиральную •структуру (более «поперечную» у гребешка, с саркомерами от 2 до 3 мкм) [41]. Эти мышцы сокращаются быстро (продолжительность одиночного сокращения равна 100 мс). У гребешка замок раковины эластичен, что позволяет ему резкими сокращениями выбрасывать воду и продвигаться вперед толчками.
Мышцы дождевого червя имеют толстые и тонкие нити, образующие спираль по краям лентообразных волокон. Толстые нити имеют диаметр 200 А, тонкие — 50 А; на одну толстую нить приходится четыре и больше тонких [197]. Проникающие в волокно Т-канальцы идут от сарколеммы {172]. Внутриклеточное отведение от волокон продольной мышцы выявляет спонтанные ПСП, главным образом гиперполяризационные (рис. 16-27). При деполяризации клетки внутриклеточными электродами величина тормозных миниатюрных синаптических потенциалов (мТПСП) возрастает, а при гиперполяризации — уменьшается. Реверсивный потенциал составляет —58 мВ при потенциале покоя —35 мВ. ГАМК также вызывает гиперполяризацию клетки и уменьшает сопротивление мембраны. мТПСП чувствительны к хлору и блокируются пикротоксином; эти факторы изменяют также характер влияния ГАМК- Наблюдались и возбуждающие миниатюрные синаптические потенциалы (мВПСП); они блокируются тубокура-рином. В продольной мышце дождевого червя наблюдаются также спонтанные спайки, которые могут превосходить мембранный нулевой потенциал на 18 мВ («овершут»); за спайками следует гиперполяризация (или «андершут»); внутриклеточная деполяризация увеличивает частоту спонтанных спайков. Амплитуда спайков и скорость их подъема пропорциональны концентрации наружного кальция; марганец блокирует спайки; по-видимому, они зависят от тока кальция внутрь клетки [199]. Миниатюрные ги-перполяризующие ПСП обусловлены уровнем проницаемости для
' Контроль, Cl 148м
Cl 5,6м
через 15мин
O,5C
Отмывка
0,5 С
Б
0,5 С
0,5с
A
Рис. 16-27. Электрическая активность мышечных волокон дождевого червя [173, 174].
А. Серии спонтанных спайков в клетках продольной мышцы при механическом раздражу нии возрастающей частоты. Б. Миниатюрные (спонтанные) тормозные синаптические потенциалы (мТПСП). После контрольной регистрации большая насть хлора на 25 мин заменен^ глутаматом, затем волокна вновь возвращены в контрольную среду. Обратите внимание на реверсию мТПСП в отсутствие хлора.
250
Глава 16
ионов хлора и могут быть вызваны действием ГАМК, являющейся тормозным медиатором [198].
Продольная мышца дождевого червя (Lumbricus) дает гладкий тетанус при стимуляции (предпочтительно через собственные нервы) с частотой 0,4 имп/с. Облегчающее действие длится 0,5 с после одиночного стимула, 30 с — после тетануса и еще дольше — после эзеринизации.
В периферической части мышечных волокон пиявки миофибриллы ориентированы радиально и имеют миофиламенты, длинные трубочки саркоплазматического ретикулума и некоторые филаменты, ориентированные поперек волокна от сарколеммы к осевой саркоплазме [312]. В спинной мышце пиявки, как и у дождевого червя, возникают спонтанные спайки, за которыми следует волна гиперполяризации. Между волокнами имеется непосредственная электрическая связь [368].
У нереид волокна продольной мышцы иннервируются несколькими нейронами; в них обнаружены кальциевые спайки, аналогичные наблюдаемым у дождевого червя [199].
Соматические мышцы с неисчерченными короткими волокнами
Основные соматические мышцы многих беспозвоночные меют неисчерченные веретенообразные волокна длиной от 0 до 500 мкм и максимальным диаметром 5 мкм, часто с тот и тонкими нитями, не образующими упорядоченной систем
Мантия кальмара или осьминога, чтобы создать рег ю силу для атаки или бегства, должна сокращаться быстра 'я одновременно. Мантия Loligo иннервируется звездчатым или я-тийным ганглием с помощью пучков гигантских аксонов то
10 аксонов в пучке) с каждой стороны; один из них, идуп к
задней части тела, особенно велик. Один импульс каждой гантского аксона вызывает сокращение (по закону «вес ничего») тысяч мышечных волокон мантии. Повторная сти ция не усиливает ответа. Время сокращения равно 60 мс, a i расслабления — 200 мс. Мантия получает также много тонк локон, идущих вдоль гигантских аксонов; они вызывают ное сокращение с облегчением.^ Электрическая регистра тверждает двойную («быструю» и «медленную») иннервг о м; шечных волокон мантии кальмара и осьминога.
Хроматофоры головоногих представляют собой одноклеточны пигментированные мешочки, растягиваемые сокращением ради ально расположенных мышечных волокон. Их протофибриллы сосредоточены по периферии волокна и отсутствуют в середине. i Волокна получают полинейронную иннервацию; стимуляция нер- .. вов вызывает ВПСП. АХ увеличивает частоту мПСП, а 5-ОТ ее ? уменьшает. Тормозных нервов не найдено [123]. te
Мышцы
251:
Потенциалы действия ретрактора глотки улитки Helix (при* внеклеточной регистрации) состоят из двух волн —быстрой и медленной. Внутриклеточная регистрация обнаруживает суммирующиеся ПСП и спайки [326]. Ритмическая стимуляция усиливает сокращение мышцы до максимума при интервалах в 20 мс. Z-Глутамат в низких концентрациях действует на мышцу возбуждающе [216]. Протрактор (вытягивающая мышца) радулы Busycon состоит из множества моторных единиц; при помещении в сосуд со смесью ацетилхолина (возбуждающее действие) и триптамина (расслабляющее действие) мышца начинает ритмически сокращаться [178].
У двустворчатого моллюска Spisula край мантии снабжен мышечной полоской, способной как к фазным, так и к тоническим? сокращениям. Ганглиозные клетки, заключенные в мантии, проявляют спонтанную активность, вызывающую тоническое сокращение. Тоническая активность уменьшается под действием дофамина и 5-ОТ. Фазные сокращения, возникающие в ответ на электрическое раздражение, блокируются трикаином и усиливаются ме-тисергидом [377]. У гребешка волокна быстрого аддуктора имеют множественную иннервацию; все синапсы принадлежат к «быстрому» типу [90].
У иглокожих ретракторы глотки или аристотелева фонаря представляют собой полоски неисчерченных мышц с короткими-волокнами. У морского ежа (Echinus) фонарь состоит из 40 известковых зубов, движение которых осуществляется шестью группами мышц. Отдельные мышечные волокна имеют диаметр от & до 12 мкм и длину до 1 мм [74]. Скорость проведения в мышце составляет 4 см/с; стимуляция вызывает как быстрые, так и медленные электрические и механические ответы ![72]. Ретрактор-глотки голотурии Cucutnaria получает иннервацию от околородового кольца; раздражение нерва вызывает в нем фазные и тонические сокращения с быстрыми и медленными потенциалами действия. Слияние сокращений происходит при частоте 2,5 т1, а максимальное напряжение достигается при частоте 5 с-1. -Облегчение ответа усиливается эзерином; тубокурарин блокирует ответ. Мышца очень чувствительна к АХ. В длинных ретракторах стенки тела голотурии Thybne напряжение увеличивается с увеличением частоты импульсации до 1 с-1 и поддерживается в течение 10 с. Мышцы иннервируются веточками радиальных ’ Нервов с промежутками между синапсами менее Lmm; скорость проведения на коротких расстояниях равна примерно 17 см/с-![287].
Укорочение неисчерченных волокон путем образования складок^ Ретракторы хоботка сипункулиды Golfingia состоят из нейСчерЧен-ных волокон диаметром 5 мкм и^длиной 0,5—1 мм. Эти мышцы? втягивают хоботок, преодолевая высокое гидростатическое давление в полости тела, и могут вытягиваться вместе с хоботком до Длины, в 10 раз превышающей длину сокращенной мышцы. Ре
252
Глава 16
тракторы богато иннервируются аксонами двух видов, идущими от церебрального ганглия. Мышца дает фазные сокращения (время укорочения — 87 мс, время полурасслабления — 950 мс) и тонические (время полурасслабления — 2700 мс). Внеклеточная регистрация выявила два типа потенциалов действия — спайки со скоростью проведения 1,3 м/с, уменьшающиеся при ритмической стимуляции, и медленные волны, распространяющиеся со скоростью 0,3 м/с, с облегчением при повторном раздражении. Быстрые потенциалы вызывают быстрые сокращения, медленные — тонические. Мышца чувствительна к АХ; эзерин усиливает медленные волны, а тубокурарин блокирует оба типа ответов. Нервные импульсы могут быть зарегистрированы до появления быстрых и медленных мышечных потенциалов; после дегенерации нервов проводимость исчезает. Внутриклеточная регистрация показывает, что в некоторых волокнах возникают только быстрые или только медленные потенциалы, а в некоторых — и те и другие. Установлено наличие двойной иннервации [292].
В мышечных волокнах Golfingia при сокращении возникают складки, располагающиеся через равные промежутки (в 10 мкм) по всей длине волокна.
Складки, повторяясь в сотнях волокон и совпадая друг с другом, создают своеобразный рисунок (рис. 16-28); в поляризован^ ном свете можно видеть чередование двоякопреломляющих полос. Такое регулярное расположение складок свидетельствует о наличии плотной упаковки и связей между волокнами. Кривые длина — напряжение имеют два пика ([237]. Первый отражает небольшое увеличение напряжения, соответствующее длине мышцы, при которой напряжение в покое не возрастает (для невозбужденной мышцы); второй соответствует длине, при которой напряжение покоя возрастает. Наблюдения, проведенные с помощью микроскопа, показывают, что первый пик связан с растягиванием складок, а второй является результатом растяжения протофибрилл, не вполне обратимого. Благодаря способности к образованию складок мышца может изотонически сокращаться в большом диапазоне длин. В другой очень растяжимой мышце Golfingia возможность механического растяжения одного волокна другим используется для проведения возбуждения. Это веретенообразная мышца с кольцами кишок вокруг нее. Небольшое растяжение побуждает ее к сокращению, но при механической иммобилизации средней части проведение блокируется. По-видимому, нервное проведение, обнаруживаемое в ретракторе хоботка, здесь отсутствует.
Сократительные клетки кишечнополостных и губок. Исчерченные и неисчерченные сократительные клетки могут функционировать одновременно. У некоторых медузоидных кишечнополостных верхняя поверхность зонтика снабжена неисчерченными волокнами, а нижняя поверхность и парус имеют исчерченные волокна.
Мышцы
253
В щупальцах находятся как исчерченные, так и неисчерченные волокна, иногда перекрывающие друг друга; наличие толстых и тонких нитей свидетельствует о присутствии миозина и актина [64]. Щупальца способны к быстрым и медленным сокращениям;
Рис. 16-28. Ретрактор хоботка Golfingia; поперечный срез [291]., Видна упорядоченная складчатость волокон.
все области тела медузы имеют обильную иннервацию. Субум-брелла медузы Obelia в срединной своей части имеет слой миоцитов с исчерченными фибриллами; эти фибриллы образуют два слоя, расположенные под прямым углом друг к другу, и имеют саркомеры с диаметром 1,4 мкм. Во внешней части субумбреллы находятся спиральные пучки фибрилл с толстыми и тонкими нитями [63].
254
Глава 16
Актинии способны к быстрым и медленным сокращениям; их сфинктеры сокращаются более быстро, мышцы тела — более медленно. Латентный период быстрой реакции может быть равен 0,1 с, а медленной —30 с и более. Мышцы тела сокращаются спонтанно (но, вероятно, под контролем центральной нервной системы) с интервалами, измеряемыми многими минутами {317]. Химические вещества, активно действующие на мышцы животных других филогенетических групп, в случае кишечнополостных малоэффективны. Из актиний были экстрагированы сократительные белки, напоминающие актин и миозин, а также АТФаза, активируемая магнием и кальцием {234, 235]. Вероятно, многие мышечные волокна кишечнополостных имеют двойную возбуждающую иннервацию.
У губок контрактильные клетки — миоциты — расположены в основном вокруг пор (где они могут регулировать размер отверстия). Миоциты напоминают неисчерченные мышечные волокна и могут содержать толстые и тонкие нити; размеры этих нитей свидетельствуют о том, что они содержат миозин и актин 1[16]«Нервные волокна отсутствуют, но миоциты активно реагируют на механические раздражения. Электрическая стимуляция не вызывает сокращения, потенциалов действия не обнаружено. Вызванное механическим раздражением сокращение сохраняется при замене натрия в среде на калий (или литий); таким образом, обычное взаимодействие между мембраной и сократительными белками здесь, по-видимому, необязательно. Для сокращения требуется кальций (или магний, а также стронций); обнаружена АТФаза актомиозинового типа [288]. Возможно, что механическая стимуляция непосредственно вызывает взаимодействие кальция с сократительными белками.
Гладкие мышцы позвоночных
Гладкие мышцы позвоночных бывают двух типов — унитарные и мультиунитарные. Имеются также промежуточные типы. Унитарные мышцы включают мышцы внутренних органов — матки, мочеточника, пищеварительного тракта; эти мышцы способны к спонтанной ритмике и характеризуются электрическим проведением возбуждения от волокна к волокну. Растяжение унитарных мышц вызывает в них деполяризацию и сокращение; их активность может регулироваться нервной системой. Мультиунитарные мышцы включают мигательную перепонку, пиломоторные, цилиарные мышцы и мышцы радужной оболочки. В норме они возбуждаются нервными импульсами или гормонами, сокращаются сравнительно быстро; проведение возбуждения в пределах мышцы осуществляется через нервы; многие из них быстрым растяжением не возбуждаются. Между некоторыми клетками мультиунитарных гладких мышц, возможно, существуют нексусы (электрические
Мышцы
255
соединения); однако группы клеток не составляют функционального синцития,, как это происходит в унитарных мышцах. Некоторые гладкие мышцы имеют свойства как унитарных, так и мультиуни-тарных мышц: стенки семявыносящего протока и крупных артерий своей зависимостью от нервной системы больше напоминают мультиунитарные мышцы, а мышцы мочевого пузыря и воротной вены благодаря чувствительности к растяжению и способности к миогенному ритму больше сходны с унитарными.
Мышцы, активируемые нервами. Мигательная перепонка кошки отвечает на импульсы симпатической нервной системы потенциалами действия (определяемыми с помощью внеклеточной регистрации); их амплитуда по мере увеличения числа раздражаемых аксонов увеличивается, а форма усложняется. Постганглио-нарные волокна, активирующие мигательную перепонку, — в большинстве своем адренэргические (некоторые могут быть холинэрги-ческими). Иннервация радужной оболочки — холинэргическая; у некоторых амфибий и рыб гладкая мышца радужной оболочки реагирует непосредственно на освещение. В таких мышцах, как мигательная перепонка, напряжение развивается медленно, увеличиваясь с увеличением количества раздражающих импульсов, но до ойределенного предела. Не все мышечные волокна иннервированы; они возбуждаются, вероятно, диффундирующим к ним медиатором, выделяемым терминальными веточками аксонов.
Семявыносящие протоки млекопитающих иннервируются подчревным нервом; у морской свинки были зарегистрированы облегчающиеся ВПСП. Норадреналин деполяризует клетки; мето-г дом градуальной стимуляции было выявлено, что отдельные мы- щечные волокна возбуждаются несколькими аксонами [129,182]. , ВПСП могут суммироваться, рождая спайк [30, 182]. Установле-? но, что 25% мышечных волокон в семявыносящем протоке мыши находятся на расстоянии 0,25 мкм от аксона, а 1 % — в пределах 200 А (0,02 мкм), следовательно, возбуждение распространяется преимущественно от волокна к волокну [50а].
Висцеральные мышцы с миогенным ритмом. Свойства гладких, мышц позвоночных. Висцеральные гладкие мышцы исключительно разнообразны, особенно это относится к их электрическим < свойствам и к действию на них нейромедиаторов и некоторых дру-? гих веществ; различия существуют не только между мышцами | разных органов, но и между одноименными мышцами разных ви-^дов животных.
В мышце матки возникают ритмические спайки, каждый из | которых может вызвать сокращение; серии спайков могут поддер-йЖивать сокращение. В некоторых клетках препотенциал или пей-|смекерный потенциал предшествуют каждому спайку, в то время & как в соседних клетках наблюдаются только спайки. Спайки
A.
Рис. 16-29.
... Электрограммы мышцы подвздошной кишки кролика. <Л(обезно пР^тавлено Дж Тэй-
Ж1-". 5
"*"?>»<>» — <«"»“• ’°™' циалы колебательного типа) [оУ2].
Мышцы
257
пропорциональны наружной концентрации натрия [10, 233] и этим напоминают спайки поперечнополосатых мышц. Активность мышцы матки усиливается эстрогеном, подавляется прогестероном и изменяется в зависимости от репродуктивного цикла.
В taenia coli морской свинки также образуются спонтанные пейсмекерные потенциалы, каждый из которых может генерировать один или несколько спайков. Каждый спайк или разряд импульсов вызывает короткое сокращение. Скорость проведения от волокна к волокну равна примерно 1 см/с [30]. В отличие от мышцы матки спайки в taenia coli мало зависят от наружной концентрации натрия и вызываются, по-видимому, током ионов кальция внутрь клетки [222, 223]. Как и в большинстве гладких висцеральных мышц, проведение импульса обеспечивается большим количеством параллельно расположенных клеток: отдельная клетка не является электрической единицей. Пространственная константа во много раз больше длины одной клетки (1,5 мм при длине клеток, равной 100—200 мкм, и при условии, что клетки перекрывают друг друга на 50%); измеренная временная константа оказалась продолжительнее (100— 300 мс), чем расчетная (30 мс), полученная на основании значений сопротивления’и емкости отдельной клетки [353]. Таким образом, теория кабеля, описывающая проведение в длинных нервных или поперечнополосатых волокнах, неприменима к отдельной мышечной клетке. Гладкая мышца по своим свойствам скорее напоминает «загрязненный» кабель.
Мочеточник отличается от других гладких мышц спайкоподобными потенциалами действия, за которыми следует плато, напоминая этим сердечную мышцу (рис. 16-29, Б). У некоторых видов (в частности, у морской свинки) на плато возникают осцилляции; ряд веществ, например прокаин, могут удлинить плато. Медленное проведение соответствует скорости распространения волны перистальтики по мочеточнику.
Амплитуда спайков зависит от концентрации наружного кальция; плато чувствительно к натрию. При нулевой концентрации натрия или высокой концентрации кальция вместо плато может появиться серия спайков. Влияние кальция на высоту спайков лучше выражено в присутствии натрия, чем ib его отсутствие, поэтому предполагается существование параллельных каналов’ для кальция и натрия [219].
Электрическая активность обоих слоев тонкой кишки млекопитающих очень различна; имеются различия между разными ее участками; видовые отличия для одной и той же области кишки весьма велики. Кольцевой слой тонкой кишки млекопитающих толще и производит большую часть механической работы. В нем возникают спайки, которые in vitro могут быть спонтанными и редкими, a in vivo стимулируются продольным слоем. В мышеч-17—2067
258
Глава 16
ных клетках продольного слоя возникают регулярные, почти синусоидальные медленные электрические волны, частота которых максимальна в двенадцатиперстной кишке (у собаки 14 в 1 мин) и уменьшается в каудальном направлении (рис. 16-29,Л). Эти волны в пределах кишечного сегмента по существу синхронны; их ‘Проведение ограничено коротким расстоянием в направлении длинных осей волокон; через соединительные мышечные мостики волны распространяются на кольцевой слой. На медленных волнах могут быть спайки, в начале каждого из них имеется пей-смекерный потенциал. Такие же потенциалы могут предшествовать спайкам и в кольцевых волокнах. Медленные волны чувствительны к концентрации натрия, к действию метаболических ингибиторов и к температуре; они исчезают под действием уабаина и связаны с током натрия из клетки [205а, 227а]. Они, по-ви-димому, являются результатом действия натриевого насоса, подчиняющегося ритмам окислительных циклов. Спайки и пейсме-керные потенциалы относительно нечувствительны к натрию, но их амплитуда пропорциональна концентрации кальция вплоть до предельной. Поскольку кальций необходим для сокращения, а гладкая мышца лишена Т-канальцев и развитой CP-системы, возможно, что ток кальция, с которым связано возникновениеспайков, активирует также и сократительную систему [227b].
Ритм медленных волн и серий спайков соответствует ритму движений сегментов кишки. Ритмическое удлинение и укорочение кишечных сегментов может быть вызвано сокращением и расслаблением кольцевых мышц при незначительной активности продольных мышечных слоев [383]. Спайки в слоях кольцевой мышцы распространяются от клетки к клетке по областям, имеющим низкое электрическое сопротивление, через соединения, разделенные щелью, или по нексусам [93, 94, '220, 258].
Обработка гипертоническими растворами (в частности, сахарозой, но не мочевиной, проникающей в клетку) приводит к возрастанию электрического сопротивления, прекращению распространения спайков, а в некоторых препаратах — к исчезновению нексусов. Однако электротоническое распространение может продолжаться, и эффект сахарозы, возможно, связан со сморщиванием клеток и частично — с гиперполяризацией. В норме медленные волны проводятся на короткие расстояния, определяемые перекрывающими друг друга пейсмекерами, и в данной точке можно зарегистрировать как пришедшие сюда волны, так и волны местного происхождения; таким образом, in vivo можно наблюдать различные комбинации этих явлений.
В расположении продольных и диагональных пучков мышечных волокон желудка отмечаются значительные видовые различия. Кроме спайков, обусловленных, по-видимому, кальцием, имеются еще два вида медленных электрических процессов. Один да них представляет собой спайкоподобные колебания; они могут
Мышцы
259
вызывать мышечные сокращения и зависят от концентрации кальция; другой вид соответствует медленным натриевым волнам тонкого кишечника [83, 2681.
В толстой кишке кольцевой слой является пейсмекером; в нем, как и в продольном слое тонкого кишечника, обнаруживаются медленные волны [65].
В пищеводе имеются как исчерченные, так и гладкие мышцы, различно локализованные: у человека и опоссума верхняя его четверть состоит из исчерченных волокон; у крысы, овцы и коровы исчерченные волокна расположены по всей длине пищевода; у собаки и свиньи продольный слой — исчерченный, а кольцевой, вблизи кардиальной части желудка, — гладкий, а в других областях— исчерченный. Мышцы пищевода не проявляют спонтанной активности, а возбуждаются нервами [66] и поэтому не являются мультиунитарными. Часть гладких мышечных волокон возбуждается непосредственно из центральной нервной системы, часть— через собственные ганглии [230].
Многие гладкие висцеральные мышцы возбуждаются адекватным механическим раздражением даже при отсутствии местных рефлексов. Увеличение натяжения полости толстой кишки морской свинки понижает мембранный потенциал и усиливает сокращение. Амнион цыпленка, в котором отсутствуют нервы, легко стимулируется растяжением. В мышцах мочеточника и мочевого пузыря растяжение вызывает сокращение даже после блокирования нервов. Следует отметить, что в этих мышцах механическая иммобилизация не препятствует проведению, т. е. оно не зависит от натяжения, передаваемого от одной клетки к другой.
Нервная, регуляция. Гладкие висцеральные мышцы иннервируются симпатической и парасимпатической нервными ^системами, причем многие гладкие мышцы имеют собственные нервные сплетения, которые регулируют активность и служат релейными станциями для афферентных и эфферентных импульсов. По классическим представлениям, парасимпатические волокна выделяют в качестве медиатора ацетилхолин, а симпатические волокна — норадреналин. Однако с помощью блокирующих веществ было обнаружено, что в периферических стволах обоих видов автономной системы представлены как холинэргические, так и адренэргиче-ские волокна [29, 48а]. Для каждого медиатора имеется по два вида рецепторов: мускариновые и ’никотиновые рецепторы для ацетилхолина и а- и p-рецепторы—для норадреналина. Парасимпатическая иннервация у млекопитающих — холинэргическая и является возбуждающей для желудка и кишечника; ее действие блокируется скополамином (или атропином).
Высвобождение АХ вызывает возбуждающие миниатюрные синаптичесие потенциалы (рис. 16-30). Иногда ответную сократи-j тельную реакцию на прекращение торможения (сокращение «от-
17*
a
2a
Б
Рис. 16-30. Типы синаптических потенциалов в клетках гладкой мышцы кишки морской свинки [128].
А. Возбуждение (а), торможение (б), двухфазный потенциал (в) и ответ на раздражение собственных нервных волокон (г). Б. Спонтанные возбуждающие миниатюрные синаптические потенциалы (а), спонтанный и вызванный тормозные потенциалы (б).
Мышцы
261
дачи» _ rebound contraction) принимают за обычное сокращение, возникающее в ответ на возбуждающий стимул.
Много путаницы, связанной с тормозными и возбуждающими эффектами катехоламинов, обусловлено тем, что различные органы или одни и те же органы у различных видов животных могут иметь а-рецепторы (блокируемые феноксибензамином или фен-толамином) или p-рецепторы (блокируемые пропранололом). Норадреналин может возбуждать одни гладкие мышцы и тормозить другие. Свойства а- и p-рецепторов приведены в табл. 11-3. Материал, касающийся влияния катехоламинов на клетки, помещен в гл. 11.
Трансмуральная стимуляция кишки млекопитающих активирует тормозные нейроны, действующие на мышечные слои с помощью медиатора, который не блокируется веществами, блокирующими катехоламины. Имеются данные о том, что таким нека-техоламиновым медиатором может быть АТФ (в случае пуринэр-гических нервов). АТФ тормозит кишечные мышцы и возбуждает мышцы мочевого пузыря. Пуринэргическая система может быть более важной для торможения кишечных мышц рыб и амфибий, чем катехоламины [48а, 51]. Тормозные (гиперполяризующие) синаптические потенциалы кишечника млекопитающих нечувствительны к адренэргическим блокаторам. Почти в каждой исследованной мышечной клетке толстой кишки кролика и морской свинки при трансмуральном раздражении появлялись ТПСП, представляющие собой, по-видимому, результат деятельности пу-ринэргических нейронов [128].
Предполагалось, что такие вещества, как серотонин (5-ОТ), простагландины, гистамин, являются нервными медиаторами гладкой мышцы, но теперь это считают маловероятным.
По данным флуоресцентной микроскопии, синапсы нервных сплетений, расположенных между мышечными элементами и под слизистой оболочкой кишечника, являются адренэргическими. Норадреналин стимулирует в сплетениях некоторые нейроны, которые, .вероятно, тормозят спонтанную спайковую активность мышечных волокон. Многие нейроны сплетений проявляют спонтанную активность, другие стимулируются механическим растяжением [382а]. Разряды нейронов не имеют прямого отношения к миогенной активности мышечных слоев; однако блокирование спонтанной активности нейронов снимает торможение с кишечных мышц, и их ритмическая активность усиливается. Нейроны механорецепторов участвуют, вероятно, в локальных рефлексах на растяжение. Автономный контроль гладких мышц сосудов обсуждается в гл. 20.
Изучение висцеральных мышц низших позвоночных подтверждает полученные на млекопитающих данные, свидетельствующие с разнородности гладких мышц и их автономной регуляции.
Иннервация желудка
Рис. 16-31. Схематическое изображение автономной иннервации мышцы желудка [48а].
Вагусная ветвь у рыб и амфибий является тормозной, а симпатические холинэргические волокна — возбуждающими. У рептилий и млекопитающих холинэргические возбуждающие волокна идут в парасимпатическом стволе, а адренэргические тормозные — в симпатическом. 1 — рыбы; 11 — амфибии; Ill — рептилии; IV — млекопитающие. — холинэргические возбуждающие нервы; ------- адренэргические нервы; 1----------------• неадренэргические тормозные
нервы. Жирными линиями обозначено большое количество волокон, тонкими — малое.
Мышцы пищеварительного тракта у рептилий, как и у млекопитающих, снабжаются вагусными холинэргическими возбуждающими волокнами, преганглионарные вагусные волокна активируют тормозные неадренэргические нейроны, а также симпатические холинэргические возбуждающие и адренэргические тормозные волокна [506].
У рыб и амфибий ветви черепных парасимпатических и передних симпатических нервов идут вместе в составе ва-госимпатического нерва (рис. 16-31), поэтому выяснение происхождения отдельных волокон весьма затруднительно. В желудке лягушки наблюдаются ритмические электрические волны, возникающие с частотой от 1 до 4 в 1 мин и длящиеся 10—15 с; они распространяются со скоростью от 0,3 до 1,4 мм/с [365]. Эти волны вызывают сокращения мышц желудка и, по-видимому, аналогичны «медленным спайкам» желудка млекопитающих. Вагосим-патические нервы лягушки содержат холинэргические возбуждающие волокна симпатического происхождения и вагусные преганглионарные волокна, которые тормозят сокращения желудка через не-катехоламиновые нейроны [51]. У бесхвостых амфибий желудок снабжается адренэр-гическими тормозными веточками чревного нервн [54].
У рыб влияние вагуса распространяется немногим далее
Мышцы
263
желудка. У форели, например, вагусные волокна желудка являются тормозными (через неадренэргические 'нейроны); за торможением у них следует сокращение «отдачи» (rebound сопгас-tion); вагус содержит также некоторое количество холинэргиче-оких возбуждающих волокон симпатического происхождения. Желудок получает и возбуждающие адренэргические волокна от чревного нерва [486]. На кишечник форели в противоположность желудку адреналин оказывает тормозящее действие, так же как и высокочастотная стимуляция спинного мозга, сопровождающаяся послетормозной «одчачей» [54b]. АХ возбуждает все отделы кишечника рыб; адреналин возбуждает желудок и тормозит кишечник. В кишечнике линя (Tinea) слой исчерченных волокон покрывает «чехлом» слой гладких; вследствие этого при электрической стимуляции возникают как быстрые (в исчерченных волокнах), так и медленные (в гладких) сокращения [263]. В мышечных стенках кишечника и желудка рыб имеется много нейронов.
Нервная регуляция деятельности мускулатуры желудка у пластиножаберных неясна. Действие вагуса имеет, по-видимому, тормозной характер и сопровождается послетормозным возбуждением; спинальный отдел автономной системы имеет как тормозные, так и возбуждающие волокна [54b]. Мышца брыжейки, образующая у акулы спиральный клапан, стимулируется легким растяжением; она нечувствительна к АХ, но возбуждается адреналином, действие которого блокируется (пропранололом. У .миксин вагус тормозит верхнюю кишку.
Гладкие мышцы легких также регулируются автономной нервной системой [48а, 54а], что видно из следующего сопоставления:
Возбуждающие нервы
Амфибии Симпатические холинэргические, некоторые адрен-
эргические
Рептилии и млекопитающие Парасимпатические холинэргические
Тормозные нервы
Амфибии Вагусные неадренэргические
Рептилии Вагусные неадренэргические, симпатические ад-
ренэргические
Млекопитающие Симпатические адренэргические
Таким образом, характер иннервации легких изменяется при переходе от амфибий к рептилиям.
Гладкие мышцы мочевого пузыря получают холинэргическую возбуждающую иннервацию во всех классах позвоночных, в которых она имеется. В стенках пузыря амфибий, рептилий и млекопитающих много интрамуральных нейронов. Адренэргические тормозные нервы имеются у рептилий и млекопитающих. Роль нервов, содержащих адреналин, у лягушек неясна, поскольку адреналин вызывает у них сокращение пузыря [48с].
264
Глава 16
Морфологически обособленная симпатическая система впервые возникает у пластиножаберных, где она является холинэргической возбуждающей, у других рыб, амфибий, рептилий и птиц — смешанной (холинэргической и адренэргической), у млекопитающих— преимущественно адренэргической тормозной. Примитивная вагусная иннервация гладких висцеральных мышц была первоначально неадренэргической тормозной, с послетормозным рефлекторным возбуждением (rebound exitation). На поздних стадиях эволюции вагусная иннервация мышц пищеварительного тракта приобрела холинэргическую возбудительную функцию.
Висцеральные мышцы беспозвоночных
Исчерченная мускулатура кишечника насекомых и большинства ракообразных весьма непохожа на висцеральную мускулатуру позвоночных. В некоторых висцеральных мышцах насекомых каждая миозиновая нить окружена двенадцатью тонкими нитями (по сравнению с шестью в локомоторных мышцах) [334]. В волокнах исчерченных висцеральных мышц задней кишки таракана наблюдаются спонтанные флуктуации потенциала покоя с препотенциалами, которые могут вызвать спайк, особенно если мышца растягивается; нервы шестого брюшного ганглия также могут вызывать ВПСП, переходящие в спайки (рис. 16-32). Наблюдаются и миниатюрные ВПСП [256, 257]. К мышечным волокнам подходит множество нервных окончаний от разных нейронов. Таким образом, задняя кишка обладает как миогенной, так и нервной активацией. У саранчи (Locusta) спонтанная активность в передней кишке выше, чем в задней. Передняя кишка возбуждается серотонином в концентрации 0,2 мкг/мл, а для задней требуется 20 мкг/мл [126].
Кишечник речного рака имеет толстый внутренний слой продольных волокон и наружный слой кольцевых. Для всех этих волокон характерно наличие длинных саркомеров и исчерченносты Имеется хорошо развитое нервное сплетение, и сокращения, по-видимому, имеют нейрогенную природу.
Кишечник моллюсков содержит неисчерченные мышечные волокна. В желудке кальмара перистальтика возбуждается растяжением и тормозится ацетилхолином, спайки тетродотоксином не блокируются [382]. Особенности деятельности этих мышц напоминают особенности деятельности унитарных мышц позвоночных/ У слизня Arion характер сокращения в различных областях тела различен, движения регулируются, по-видимому, центральной нервной системой [307]. У хитона Poneroplax иннервация богаче в области прямой кишки и менее выражена в области задней кишки. Мышцы задней кишки обладают спонтанной активностью, проявляющейся в виде медленных волн и спайков. Мышцы прямой кишки напоминают мультиунитарные мышцы позвоночных: в
Мышцы
265
них нет спонтанной активности, нервные импульсы вызывают ПСП, медленные волны отсутствуют. Остальная часть кишечника спонтанно активна, с медленными волнами и спайками, регулируется возбуждающими и тормозными нервными влияниями, подобно унитарным мышцам позвоночных [50]. У двустворчатых Мег-cenaria и Spisula задняя кишка отвечает на электрические удары
Рис. 16-32. Электрическая активность верхней продольной мышцы задней кишки таракана; внутриклеточная регистрация. (Любезно предоставлено Т. Наган.)
А. Три типа спонтанных потенциалов интактной задней кишки с сохраненным шестым абдоминальным ганглием: постсинаптические потенциалы (ВПСП) и потенциалы действия с предшествующими им ВПСП и без них. Б. Спонтанные ритмические потенциалы действия после удаления шестого абдоминального ганглия. В. 1 — потенциал действия, следующий за ВПСП; 2— ВПСП, записи 1 и 2 сделаны с одного и того же волокна; нерв задней кишки раздражается в момент, указанный стрелкой.
как фазными, так и тоническими сокращениями; имеются данные о наличии в ней тормозной и двойной возбуждающей иннервации. Спонтанная активность отсутствует, раздражение ганглиев вызывает оба типа сокращения; проведение, по-видимому, осуществляется посредством нервов [293].
В кишечнике иглокожих Thyone и Arbacia растяжение вызывает распространяющиеся сокращения; нервные блокаторы оказываются эффективными, поэтому мышцы напоминают унитарные мышцы позвоночных.
У живущей в трубке полихеты Arenicola хоботок состоит из буккальной (ротовой) массы, глотки и пищевода, стомато-гастри-ческое нервное сплетение распространяется на глотку и пищевод и соединяется с окружающим пищевод нервным кольцом. Область пищевода является пейсмекером и стимулируется адренали
266
Глава 16
ном и ацетилхолином. У Glycera активность глотки и пищевода также, по-видимому, стимулируется стомато-гастрическим сплетением. Изолированный зоб и задний желудок дождевого червя спонтанно активны при растяжении и возбуждаются ацетилхолином при концентрации 10~10 (г/мл); этот эффект блокируется атропином. Торможение адреналином ритмических сокращений зоба и желудка может быть прекращено [3-блокаторами, однако адреналин действует на а-рецепторы мышц стенки тела.
Заключение f
•Скорость движения, развиваемая мощность, напряжение, степень укорочения подвижных структур, необходимые для существования животного, обеспечиваются мышцами. Скорости сокращения мышц от самых быстрых до самых медленных различаются в 10000 раз. Нижеследующая классификация подытоживает основные свойства мышц, имеющие отношение к различиям в скорости сокращения и расслабления. Исключения и промежуточные типы были упомянуты выше.
I. Поперечнополосатые мышцы. Имеют развитую систему саркоплазматического ретикулума и поперечных канальцев, длинные, прямые волокна. Представлены скелетными мышцами.
А. Одно (или два) пластинчатое (en plaque) нервное окончание на волокне, мононейронная иннервация; «фибриллярная структура» и упорядоченная Т-система; проведение мышечными потенциалами действия.
1. Белые фазные мышцы: гликолитический цикл; специфический состав миозина. (Примеры: разгибатель пальца млекопитающих, задняя широчайшая мышца спины цыпленка, портняжная мышца лягушки.)
2. Красные тонические мышцы: длительное поддержание напряжения; окислительный цикл; более слабая, чем у фазных, миозин-АТФаза. (Камбаловидная и поясничная мышцы млекопитающих.)
Б. Множественные гроздевидные (en grappe) моторные окон-; чания; градуальные синаптические потенциалы; редкие распространяющиеся мышечные спайки; полевидная структура (Felderstruktur); неупорядоченная Т-система.
1. Мононейронная иннервация; обычно тонические или медленные мышцы (передняя широчайшая мышца спины цыпленка, прямая брюшная мышца лягушки, многие мышцы рыб).
2. Полинейронная иннервация; быстрые и медленные мышцы; различные комбинации возбудительной и тормозной иннервации; возбуждаются медленно (с облегчением) или быстро (по закону «все или ничего»); мы
Мышцы
267
шечные потенциалы в большинстве своем синаптические, изредка возникают потенциалы действия.
а. Синхронные: одно сокращение на один моторный импульс (многие позные мышцы насекомых и ракообразных).
б. Асинхронные: несколько сокращений на один моторный импульс (непрямые летательные мышцы двукрылых и перепончатокрылых).
II. Исчерченные сердечные мышцы; волокна часто образуют синцитий; умеренная скорость сокращения; окислительный цикл (сердца позвоночных и большинства членистоногих). III. Промежуточные типы.
А. Исчерченность только на одной стороне или одном отростке волокна (некоторые трематоды, Ascaris, оболочники).
Б. Миофибриллы располагаются по спирали вокруг волокна (прозрачная мышца устрицы, мантия кальмара, стенка тела дождевого червя).
IV. Неисчерченные мышцы; отсутствуют Т-канальцы, слабо выражен саркоплазматический ретикулум, тонкие волокна.
А. Длинные волокна, содержащие парамиозин; множественная иннервация (запирательные мышцы моллюсков).
Б. Короткие, веретенообразные тонкие волокна.
1. Мультиунитарные мышцы, активация и проведение нервное (ретракторы хоботка у сипункулид, ретракторы фонаря у иглокожих, ретрактор глотки улитки Helix, пиломоторы млекопитающих, мигательная перепонка).
2. Унитарные мышцы; активность спонтанная, проведение от волокна к волокну, часто возбуждаются растяжением, активность может регулироваться нервной системой (мышцы внутренних органов позвоночных).
"Самые быстрые сокращения наблюдаются .у летательных мышц двукрылых, но частота сокращений в этом случае определяется механическими резонансными свойствами сократительных ‘белков; полной активации при каждом сокращении не происходит. Самыми быстрыми мышцами, характеризующимися полным возбуждением, сокращением и расслаблением, являются экстра-окулярные мышцы млекопитающих и «звуковые» мышцы рыб (время сокращения 5—10 мс). Позные мышцы позвоночных и многих беспозвоночных сокращаются с умеренными скоростями (40—100 мс). Запирательные мышцы моллюсков сокращаются быстро, но могут поддерживать напряжение в течение многих ча-•сов или дней. Сокращения и расслабления висцеральных мышц позвоночных длятся секунды, а у некоторых актиний периоды сокращения измеряются минутами.
Скорость сокращения коррелирует со структурными и ферментативными свойствами мышц. Быстрые мышечные волокна
268
Глава 16
исчерчены и имеют короткие саркомеры, много миофибрилл, богатую саркотубулярную систему и одно или два нервных окончания; они часто содержат больше гликолитических ферментов, чем окислительных. Медленные исчерченные мышцы имеют обычно большие резервы гликогена и жира, богаты окислительными ферментами, могут быть снабжены большим количеством нервных окончаний и иметь неупорядоченную систему Т-канальцев. Наиболее медленные мышцы характеризуются тонкими волокнами^ отсутствием исчерченности и выраженной тубулярной системы. Некоторые мышцы проявляют свойства как исчерченных, так и неисчерченных типов мышц, имеют тонкие волокна с редкими канальцами, ступенчатое или диагональное взаиморасположение саркомеров соседних миофибрилл. Скорость сокращения описывается кривой сила — скорость и измеряется изменением длины мышцы в секунду. Эта мера хорошо коррелирует с активностью^ миозиновой АТФазы; свойства миозина исчерченных волокон могут определяться характером иннервации. Скорость расслабления хорошо коррелирует со скоростью связывания цитоплазматическими гранулами высвобождаемого кальция. Запирательные мышцы беспозвоночных содержат крупные нити парамиозина — белка, который сам не способен к развитию напряжения, но каким-то образом задерживает расслабление.
Все мышцы содержат актин, образующий тонкие нити, к которым присоединяются миозиновые мостики. Молекула миозина очень велика и сложна и существует, по-видимому, во многих модификациях. Она состоит из нескольких частей — нитевидного хвоста, шейки и мостиков (к актину) и центров с АТФазной активностью. Энергия для сокращения поступает от АТФ при его гидролизе АТФазой миозина (in vivo) в присутствии магния и актина.’Взаимодействие между актином и миозином, а также АТФаза активируются ионами Са2+, которые либо устраняют тормозящее действие тропомиозинтропонина, либо присоединяются к миозину. Освобождение Са2+ из пузырьков саркоплазматического ретикулума обеспечивает связь мембраны с сократительными белками. Для расслабления необходимо, чтобы кальций снова был1 связан в пузырьках. Мышца содержит также несколько актини-нов с неизвестными функциями, один из которых найден в Z-пластинке. АТФ ресинтезируется фосфогеном (креатинфосфатом или аргининфосфатом). Креатин или аргинин рефосфорилиру-ются в гликолитическом цикле. Продукты этого распада восстанавливаются в процессе окисления или восполняются из других источников. Только часть энергии АТФ (около 25%) превращается в механическую работу, что подтверждается измерениями' теплопродукции.
Разнообразие способов активации мышц так же велико, как диапазон скоростей сокращения. Наиболее быстрые мышцы активируются одним нервным импульсом, который с помощью медиа
Мышцы
269
тора (ацетилхолина у позвоночных) вызывает синаптический потенциал одной (или двух) концевой пластинки. Освобождение медиатора требует деполяризации моторных нервных окончаний; это в свою очередь требует присутствия кальция, который, по-видимому, соединяется с молекулами-переносчиками, участвующими в освобождении медиатора. Свободный АХ немедленно гидролизуется АХ-эстеразой субсинаптической мембраны.
Величина синаптического потенциала зависит от количества выделившегося медиатора и является следствием возросшей натриевой и калиевой проводимости; достигнув критической величины, он вызывает потенциал действия мышцы (дальнейшее увеличение Na- и К.-проводимости), который распространяется внутрь клетки по Т-канальцам. Сигналы с Т-канальцев каким-то образом активируют саркотубулярную систему, которая освобождает ионы кальция, необходимые для активации актомиозина. Потенциалы действия мышцы активируют волокна по закону «все или ничего», но синаптические потенциалы могут вызывать и градуальные сокращения.
Медленные исчерченные мышечные волокна (как у позвоночных, так и у членистоногих) имеют много нервных окончаний и для активации нуждаются в синаптических потенциалах, распределенных по всей мышечной мембране. Отдельный моторный импульс редко вызывает максимальный ответ: обычно необходимо облегчение. Многие мышцы беспозвоночных, особенно у членистоногих, имеют по нескольку возбуждающих аксонов, использующих, по-видимому, один и тот же медиатор (например, глутамат), и каждый из них вызывает ответ с определенной скоростью и степенью облегчения. Некоторые мышцы ракообразных, кроме возбудительной, получают и тормозную иннервацию, медиатором которой является гамма-аминомасляная кислота (ГАМК). Действие ГАМК может быть пресинаптическим (на моторные нервные окончания) или постсинаптическим (на мышечную мембрану). Торможение в данном случае является результатом увеличения анионной проводимости.
Неисчерченные мышцы могут активироваться моторными аксонами (например, позные мышцы моллюсков, сипункулид, кишечнополостных и иглокожих и некоторые мультиунитарные мышцы позвоночных). Другие неисчерченные мышцы способны к спонтанной (миогенной) активности, которая может регулироваться нервной системой; это в большинстве своем мышцы внутренних органов (например, у позвоночных, моллюсков и, вероятно, иглокожих). Многие неисчерченные мышцы, в особенности унитарные, возбуждаются растяжением. У различных висцеральных мышц позвоночных медиатором могут служить АХ и норадреналин; их влияние — возбуждающее или тормозящее — находится в зависимости от особенностей мышцы. Между многими гладкими мышечными волокнами существует прямое электрическое взаимодейст
270
Глава 16
вие, так же как и между пучками волокон унитарных висцеральных мышц. Нервная регуляция может быть как возбуждающей, так и тормозной; часто она дополняется гормональной регуляцией.
Градуальный характер движений у позвоночных обеспечивается путем изменения числа активируемых моторных единиц спинного мозга. В медленных мышцах градуальность достигается изменением частоты импульсации и изменением соотношения быстрых и медленных сократительных систем — в пределах одного и того же волокна (у членистоногих) или между различными волокнами мышцы (у низших позвоночных). У членистоногих в осуществлении градации принимают участие также тормозные аксоны. Система взаимодействия актина с миозином, используемая во всех мышцах, сходна, и фактически все мышцы активируются электрическими сигналами клеточной мембраны; их связь с сократительными элементами осуществляется высвобождающимися ионами кальция. Широкий диапазон скоростей сокращения, расслабления, величин напряжения коррелирует с различиями, имеющимися в макро- и микроструктуре мышц, с разновидностями миозинов и особенно с характером нервной активации.
СПИСОК ЛИТЕРАТУРЫ
1. Abbott В. С., Parnas J. Gen. Physiol. 48: 919—931, 1965. Electrical and mechanical properties of crustacean muscle.
2. Abe Y., Tomita T., J. Physiol. 196: 89—100, 1968. Cable properties of smooth muscle.
3. Aberg A. K. G., Axelson J., Acta Physiol. Scand. 64: 15—27, 1965. Mechanical properties of intestinal smooth muscle.
4. Adrian R. H., Peachey L. D., J. Physiol. 181: 324—336, 1965. Membrane capacity, frog muscle fibers.
5. Adrian R. H. et al., J. Physiol. 204: 231—257, 1967. Radial spread of contraction in frog fiber.
6. Ahlquist R. P., Ann. Rev. Pharmacol. 8: 259—272, 1968. Agents blocking adrenergic P-receptors.
7. Aidley D. J., White D. C. S., J. Physiol. 205: 179—192, 1969. Mechanical properties of tymbal muscles in cicada.
8. Ambache N., Dixon A. S., Wright E. A., J. Exp. Biol. 21: 46—57, 1945. Properties of intestinal muscles in earthworm.
9. Andersen P., Jensen J. K. S., Loyning Y., Acta Physiol. Scand. 57: 167—179, 1963. Slow and fast muscle fibers of hagfish.
10. Anderson N. C., J. Gen. Physiol. 54: 145—165, 1969; 58: 322—339, 1971. Voltage clamp on uterine smooth muscle.
11. Anderson R., Fange R., Arch. Int. Physiol. Biochem. 75: 461—468, 1967. Pharmacologic receptors in Lumbricus.
12. Ashley С. C., Ridgway E. B.t Nature 219: 1168—1169, 1968. Calcium transport in barnacle fibers.
13. Atwood H. L., Comp. Biochem. Physiol. 10: 17—32, 77—81, 1963. Fast and slow contractions in Carcinus and Nephrops.
14. Atwood H. L., Comp. Biochem. Physiol. 16: 409—426, 1965. Excitation and inhibition in crab muscle fibers.
Мышцы
271
14а. Atwood Н. L., Bittner G„ J. Neurophysiol. 34: 157—170, 1971. Matching of excitatory and inhibitory inputs to crustacean muscle fibers.
15 Atwood H. L., Johnston H. S., J. Exp. Zool. 167: 456—470, 1968; also Atwood H. L., Morin W. A., J. Ultrastruc. Res. 32: 351—369, 1970. Neuromuscular synapses in crab and crayfish.
15a. Aubert X., Lebacq J., J. Physiol. 216: 181—200, 1971. Heat of shortening during plateau of tetanic contraction and at end of relaxation.
16. Bagby R. M., J. Morph. 118: 167—182, 1966. Fine structure, sponge myocy-
17. ^Baguet F., Gillis J. M., J. Physiol. 188: 67—82, 1967; 198: 127—143, 1968; also Baguet F., Gillis F. J., Dainoff G., Arch. Int. Physiol. Biochem. 75: 523—527, 1967; also Baguet F., Marechai G., Aubert X., Arch. Int. Physiol. Biochem. 70: 416—417, 1962. Energy cost of tonic contraction in bivalve muscles.
18. Baldwin E., Yudkin W. H., Proc. Roy. Soc. Lond. В 136: 614—631, 1950. Phosphagens in invertebrates.
19. Bdrany M., J. Gen. Physiol. 50 (Suppl.): 197—218, 1967. ATPase activity of myosin in relation to contraction speed.
20. Bdrany M., Bdrany K., Biochem. Zeitschr. 345: 37—56, 1966. Myosin from adductor of Pecten.
21. Bdrany M., Close R. I., J. Physiol. 213: 455—474, 1971. Changes ofv myosin in cross-innervated rat muscles.
22. Bdrany M. et al., Europ. J. Biochem. 2: 156—164, 1967. Myosin and contraction in muscle of cat and sloth.
23- Barber S. B., Pringle J. W. S., Proc. Roy. Soc. Lond. В 164: 21—39, 1966. Flight muscles of belostomatid bug.
24. Barnard R. et al., Amer. J. Physiol. 220: 410—414, 1971. Hystochemistry of red and white muscle fibers.
25. Bastian J., Esch H., Z. vergl. Physiol. 67: 307—324, 1970. Control of flight muscles of honeybee.
26. Batra S. C., Daniel E. E., Comp. Biochem. Physiol. 38A: 369—385, 1971. Calcium uptake by particulates of smooth muscle.
26a. Belton P., Brown В. E., Comp. Biochem. Physiol. 28: 853—863, 1969. Electrical constants, cockroach visceral muscle.
26b. Belton P., Grundfest H., Amer. J. Physiol. 203: 588—594, 1962. K-spikes in muscle of mealworm larvae.
27. Bendall J. R., Muscles, Molecules and Movement. Heinemann, London, 1969. 219 pp.
28. Bennett M. R., J. Physiol. 185: 132—147, 1966. Nervous transmission to taenia coli.
29. Bennett M. R., Burnstock G., pp. 1709—1732. In Handbook of Physiology, Sec. 6, Vol. IV, edited by C. F. Code. Amer. Physiol. Soc., Washington, D. C., 1968. Effects of extrinsic nerves on smooth muscle.
30. Bennett M. R., Burnstock G., Holman M. E., J. Physiol. 182: 527—540, 1966. Neural inhibition of smooth muscle.
31. Bennett T., Comp. Biochem. Physiol. 32: 669—680, 1970. Excitation and inhibition of bird gizzard.
32. Bergman R. A., Bull. Johns Hopkins Hosp. 114: 325—343, 344—353, 1964. Dorsal fin muscles of teleosts.
33. Bianchi С. P., pp. Ill—120. In Protein Metabolism and Biological Function, edited by С. P. Bianchi and R. Hilf. Rutgers University Press, 1970. Calcium regulation of muscle contraction.
34. Bittner G. D., J. Gen. Physiol. 51: 731—758, 1968. Differentiation of nerve terminals in crayfish muscle.
35. Bittner G. D., Harrison J., J. Physiol. 206: 1—23, 1970.. Crustacean transmitter release.
36. Bolton T. B., J. Physiol. 216: 403—418, 1971. Intestinal smooth muscle, slow waves, ACh or carbachol.
37. Boltt R. E., Ewer D. W., J. Exp. Biol. 40: 713—726, 1963. Neural transmission to muscles of echinoderms.
272
Глава 16
38. Bortoff A., Amer. J. Physiol. 201: 203—208, 1961. Slow potentials of small intestine.
39. Bortoff A., Amer. J. Physiol. 209: 1254—1260, 1965. Electrical transmission from longitudinal to circular intestinal muscle.
40. Boyd H., Burnstock G„ Rogers D., Brit. J. Pharm. Chemother. 23: 151—163, 1964. Innervation of large intestine of Bufo.
41. Bowden J., Int. Rev. Cytol. 7: 295—335, 1958, Structure and innervation of clam muscle.
42. Bozler E., Amer. J. Physiol. 207: 701—704, 1964. Mechanical properties and excitation of smooth and cardiac muscle.
43. Brady A. J., The Physiologist 10: 75—86, 1967. Mechanical model of muscle.
44. Brandt P. W., Reuben J. P„ Grundfest H„ J. Cell. Biol. 38: 115—129, 1968; also Brandt P. W., Reuben J. P., Girardier L., Grundfest H., J. Cell Biol. 25: 233—260, 1965. Structure and function in single muscle fibers of crayfish.
45. Bruner J., Kennedy D., Science 169: 92—94, 1970. Habituation at a neuromuscular junction.
46. Bullard B., Comp. Biochem. Physiol. 23: 749—759, 1967. Nervous control of byssus retractor of Mytilus.
47. Buller A. J., Lewis D. M., J. Physiol. 178: 343—358, 1965. Crossinnervation of skeletal muscles.
48. Buller A. J., Mommaerts W. F. H. M., Seraydarian K-, J. Physiol. 205: 581— 597, 1969. Properties of myosin in fast and slow muscles of cat.
48a. Burnstock G., Pharmacol. Res. 21: 247—324, 1969. Autonomic innervation of visceral and vascular muscles of vertebrates. Washington, D. C., 1968. Comparative physiology of gastrointestinal motility.
48b. Burnstock G., Quart. J. Microscope Sci. 100: 199—219, 1959. The innervation of the gut of the brown trout.
48c. Burnstock G., Campbell G., J. Exp. Biol. 40: 421—437, 1963. Innervation of ringtail possum bladder.
48d. Burnstock G., Pharmacol. Rev. 24: 509—581, 1972. Purinergic nerves.
49. Burnstock G., Campbell G., Rand M. J., J. Physiol. 182: 504—526, 1966. Inhibitory innervation of taenia.
50. Burnstock G., Greenberg M. J., Kirby S., Willis A. G., Comp. Biochem. Physiol. 23: 407—429, 1967. Pharmacology of visceral muscle of mollusc.
50a. Burnstock G., Iwayama T„ Prog. Brain Res. 34: 389—404, 1971. Fine-structural identification of autonomic nerves and their relation to smooth muscle.
50b. Burnstock G., O’Shea J., Wood M., J. Exp. Biol. 40: 403—420, 1963. Innervation of toad bladder.
50c. Burnstock G., Wood M. E., Comp. Biochem. Physiol. 20: 675—690, 1967; 22: 815—831, 1967. Innervation of lungs of sleepy lizard.
51. Burnstock G. et al., Brit. J. Pharm. Chemother. 44: 668—688, 1970. ATP as an inhibitory transmitter.
51a. Burrows M., Hoyle G., J. Exp. Zool. 179: 379—394, 1972. Neuromuscular transmission in mantis shrimp Hemisquilla.
52. Bush В. M. H., J. Exp. Biol. 39: 71—88, 1962. Reflex inhibition in claw of Carcinus.
53. Buttkus H., J. Fish. Res. Bd. Canad. 24: 1607—1612, 1967. Amino acid composition of myosin, trout muscle.
54. Campbell G., Comp. Biochem. 31: 693—706, 1969. Autonomic innervation of stomach of toad.
54a. Campbell G., Comp. Gen. Pharmacol. 2: 281—286, 1971. Autonomic innervation of lung musculature, toad.
54b. Campbell G., Burnstock G., pp. 2213—2266. In Handbook of Physiology, Sec. 6, Vol. IX, edited by C. F. Code. Amer. Physiol. Soc.,
55. Carlson F. D., Prog. Biophys. Molec. Biol., 13: 262—314, 1963. Mechanochemistry of muscular contraction.
56. Carlson F. D., Hardy D. J., Wilkie D. R., J. Gen. Physiol. 46: 851—882, 1963; J. Physiol. 189: 209—235, 1967. Energetics of muscle contraction.
Мышцы
273
57. Carsten М. Е„ J. Gen. Physiol. 53: 414—426, 1969. Calcium binding in uterine smooth muscle.
58. Carvalho A. P., J. Gen. Physiol. 52: 622—642, 1968. Calcium binding by SR granules.
59. Cerf J. A., Grundfest H., Hoyle G., McCann F. V., J. Gen. Physiol. 43: 377— 395, 1959. Dual responses in grasshopper muscle.
60. Chaplain R. A., Arch. Biochem. Biophys. 115: 450—461, 1966; 121: 154—168, 1967. Insect actomyosin ATPase.
61. Chaplain R. A., Pfister E., Experientia 26: 505—506, 1970. Heat and calcium transference in frog sartorius.
62. Chaplain R. A., Tregear R. T„ J. Molec. Biol. 21: 275—280, 1966. Distribution of myosin and insect fibrillar muscle.
63. Chapman D. M., J. Marine Biol. Assoc. U. K. 48: 667—688, 1968. Muscle cells in medusa of Obelia.
64. Chapman D. M., Pantin C. F. A., Robson E. A., Rev. Canad. Biol. 21: 267— 278, 1962. Muscles in coelenterates.
<65- Christensen J., Caprilli R., Lund G. F., Amer. J. Physiol. 217: 771—776, 1969. Slow waves in cat colon.
66. Christensen J., Daniel E. E., Amer. J. Physiol. 211: 387—394, 1966. Pharmacology of esophageal smooth muscle.
67. Close R., J. Physiol. 173: 74—95, 1964. Dynamic properties of fast and slow muscle in developing rats.
68. Close R., J. Physiol. 204: 331—346, 1969. Fast and slow muscle of rat after cross-innervation.
69. Close R., pp. 142—149. In Exploratory Concepts in Muscular Dystrophy and Related Disorders, Proc. Int. Conf. Harriman, New York, 1966. Dynamic properties of fast and slow skeletal muscle.
69a. Close R., Physiol. Rev., 52: 129—197, 1972. Dynamic properties of mammalian skeletal muscles
70. Close R., Hoh J. F. Y., J. Physiol. 192: 815—822, 1967. Forcevelocity proper-ties of kitten muscle
71. Close R., Hoh J. F. У., J. Physiol. 198: 103—125, 1968. Crossinnervation of fast and slow muscles in toad.
72. Cobb J. L. S., Comp. Biochem. Physiol. 24: 311—315, 1968. Electrical activity of lantern retractor, Echinus.
73. Cobb J. L. S., Bennett T., J. Cell Biol. 41: 287—297, 1969. Nexuses in visceral smooth muscle.
74. Cobb J. L. S., Laverack M. S., Proc. Roy. Soc. Lond. В 164: 624—640, 641— 650, 1966. Function and structure in lantern muscle of Echinus.
75. Cohen C. et al., J. Molec. Biol. 56: 223—237, 239—258, 1971. Paramyosin, organization of filaments.
76. Colano F., Rahamimoff R., J. Physiol. 198: 203—218, 1968. Frog neuromuscular junction, Ca and Na effects.
77. Connell J. J., Biochem. J., 80: 503—509, 1961. Muscle proteins of various animals.
78. Connell J. J., Mackie I. M., Nature 201: 78—79, 1964. Molecular weight of rabbit and cod myosin.
79. Cooke I. M., Grinnell A. D., J. Physiol. 175: 203—210, 1964. Drug effects on denervated skeletal muscle.
80. Cooke P. H„ Chase R. H., Exp. Cell. Res. 66: 417—425, 1971. Myosin and actin filaments from vertebrate gizzard.
81. Costantin L. L., J. Gen. Physiol. 55: 703—715, 1970. Role of sodium in inward spread of contraction in frog muscle.
82. Costantin L. L., Podolsky R. J., Tice L. W„ J. Physiol. 188: 261—271, 1967. Calcium activation of frog slow muscle.
82a. Dancker P., Pfliiger. Arch. 315: 187—197, 1970. Effect of tropomyosin on Ca-activation of actomyosin ATPase.
83. Daniel E. E., Gasrtoenterology 49: 403—418, 1965. Properties of stomach muscle.
18—2067
274
Глава 16
84. Daniel Е. Е., Chapman К. М., Amer. J. Digest. Dis. 8: 54—102, 1963. Electrophysiology of gastrointestinal tract.
84a. Daw J., Stretcher A., Proc. Nat. Acad. Sci. 68: 1107—1110, 1971. Light chains of myosin in chicken.
85. DeBell J. T., Quart. Rev. Biol. 40; 233—251, 1965. Neuromuscular junctions in nematodes.
86. DeBell J. T., Castillo J. del, Sanchez V., J. Cell. Comp. Physiol. 62: 159—177, 1963. Electrophysiology of muscle of Ascaris.
87. DeBell J. T., Sanchez V., J. Exp. Biol. 48: 405—410, 1968. Effect of temperature on Ascaris muscle fibers.
88. del Castillo J., de Mello W. C., Morales T., J. Exp. Biol. 46: 263—279, 1967. Action potentials in Ascaris muscle.
89. del Castillo J., de Mello W. C., Morales T., J. Gen. Physiol. 48: 129—140, 1964. Effects of ions on Ascaris muscles.
90. de Mellon F., Science 160: 1018—1020, 1968. Neuromuscular junctions in scallop.
91. de Villafranca G. W., Campbell L. K., Comp. Biochem. Physiol. 29: 775—783, 1969; also Biochem. J. 104: 263—269, 1967. Activation of actomyosin ATPase, Limulus.
92. Devine C., Somlyo A. V., J. Cell Biol. 49: 636—649, 1971. Thick filaments in smooth muscle.
93. Dewey M. M., Barr L., Science 137: 670—672, 1962. Nexuses between smooth muscle fibers.
94. Dewey M. M., Barr L., pp. 1629—1654. In Handbook of Physiology, Sec. 6, Vol. IV, edited by C. F. Code. Amer. Physiol. Soc., Washington, D. C., 1968; also J. Cell Biol. 23: 553—585, 1964. Ultrastructure of intestinal smooth muscle.
95. Dodge F. A., Rahamimoff R., J. Physiol. 193: 419—432, 1967. Effect of calcium on neuromuscular transmission.
96. Dreizen P., Ann. Rev. Med. 22: 365—390, 1971. Structure and function of myofibrillar proteins.
97. Dreizen P., Gershman L. C. et al., J. Gen. Physiol. 50: 85—110, 1967. Myosin subunits and their interactions.
98. Dreisen P., Gershman L. C., Trans. N. Y. Acad. Sci. Ser. II 32: 170—203, 1970. Properties of myosin.
99. Dubrowitz V., Neuman D. L., Nature 214: 840—841, 1967. Enzyme changes in cross-innervated muscle.
100. Dubel J., Kuffer S. W., J. Physiol. 155: 514—529, 1961. Neuromuscular transmission in crayfish.
101. Dudel J., Kuffler S. W., pp. Ill—113. In Nervous Inhibition, edited by E. Florey, Academic Press, New York, 1961; also J. Physiol. 155: 543—562, 1961. Presynaptic inhibition in crayfish muscle junctions.
102. Dudel J., Rildel R., Pfliiger. Arch. 301: 16—30, 1968. Electromechanical coupling in crayfish muscle.
103. Ebashi S., Ebashi F., Kodama A., J. Biochem. (Tokyo) 62: 137—138, 1967. Troponin as calcium receptor protein.
104. Ebashi S., Endo M., Prog. Biophys. Molec. Biol. 18: .125—183, 1969. Muscle proteins, especially troponin.
105. Ebashi S., Endo M., Ohtsuki 1., Quart. Rev. Biophys. 2: 351—384, 1969. Control of muscle contraction.
106. Ebashi S„ Kodama A., J. Biochem. 58: 7—12, 13—17, 1965; 59: 422—424, 1966; 60: 733—734, 1966. Alpha-actinin.
107. Edwards C., Lorkovic H., Amer. J. Zool. 7: 615—622, 1967. Calcium in excitation contraction coupling.
108. Engel W. K., Irwin R. L., Amer. J. Physiol. 213: 511—518, 1967. Histochemistry of muscle enzymes.
109. Feeder I. R., O'Brien R. D., Saltpeter M. M., J. Exp. Zool. 173: 187—202, 1970; also Faeder I. R„ O’Brien R. D., J. Exp. Zool. 173: 203—214, 1970. Pharmacology of cockroach neuromuscular transmission.
Мышцы
275
ПО. Fahrenbach W. Н., J. Cell Biol. 17: 629—640, 1963; 35: 69—80, 1967. Fine structure of crustacean muscles.
111. Fatt P., Ginsborg B. L„ J. Physiol. 142: 516—543, 1958. Ionic basis for action potentials in crustacean muscle.
112. Fatt P., Katz B„ J. Physiol. 120: 171—204, 1953; 121: 374—389, 1953. Properties of crustacean muscle fibers.
113. Fawcett D. W., Circulation 24: 336—348, 1961. Sarcoplasmic reticulum of ske-1РГЯ1 ЯПП СЯГЛ1ЯС TTHIQCiP
114. Fawcett D. W., McNutt'N. S„ J. Cell Biol. 42: 1—45, 46—67, 1969. Ultrastructure of cardiac muscle.
115. Fawcett D. W., Revel J. P., J. Biophys. Biochem. Cytol. (Suppl.) 10: 89—103, 1961. Sarcoplasmic reticulum in fast muscles of fish.
116. Fedde M. R., J. Gen. Physiol. 53: 624—637, 1969. Electrical properties of avian muscle.
117. Fernard V. S. V., Hess A., J. Physiol. 200: 547—554, 1969. Slow and fast muscle fibers in muscles of ear.
118. Fleckenstein A., pp. 71—92. In Cellular Functions of Membrane Transport, edited by J. F. Hoffman. Prentice-Hall, New York, 1964. Excitation-contraction coupling.
119. Fletcher С. M., J. Physiol. 90: 233—253, 415—428, 1937. Responses of Mytilus byssus retractor.
120. Flitney F. W., J. Physiol. 217: 243—257, 1971. T system of slow muscle.
121. Flood P„ J. Comp. Neur. 126: 181—217, 1966. Innervation of muscle in Amphioxus.
122. Flood P„ Guthrie D. M., Banks J. R., Nature 222: 87—89, 1969. Paramyosin in notochord of Amphioxus.
123. Florey E., Kriebel M. E., Z. vergl. Physiol. 65: 98—130, 1969. Chromatophore muscles of squid.
124- Florey E., woodcock B., Comp. Biochem. Physiol. 26: 651—661, 1968. Presy -naptic actions of glutamate on crab muscles.
125. Forrester T., Schmidt H., J. Physiol. 207: 477—491, 1970. Electrophysiology of slow fibers in frog muscle.
126. Freeman M. A., Comp. Biochem. Physiol. 17: 755—764, 1966. Pharmacology of alimentary tract of locust.
127. Fuchs F., Briggs I. N.t J. Gen. Physiol. 51: 655—676, 1968; 52: 955—968, 1968. Calcium and troponin in the control of contractions.
128. Furness J. B., J. Physiol. 205: 549—562, 1969. Innervation of the colon.
129. Furness J. B., Pfliiger. Arch. 314: 1—13, 1970. Neural control of smooth muscle cells.
130. Furukawa T„ Peter J. B., Exp. Neurol. 31: 214—222, 1971. Troponin in different fiber types.
131. Gage P. IF., Fed. Proc. 26: 1627—1632, 1969. Excitation-secretion coupling in presynaptic terminals.
132. Gage P. IF., Eisenberg R. S., J. Gen. Physiol. 53: 265—278, 298—310, 1969. Electrical properties of transverse tubules in frog sartorius.
133. Gage P. IF., Eisenberg R. S., Science 158: 1700—1701, 1702—1703, 1967. Action potentials without contractions.
134. Gainer H., Klancher J. E., Comp. Biochem. Physiol. 15: 159—165, 1965. Neuromuscular junctions in a fast fish muscle.
135. Gergely J., Ann. Rev. Biochem. 35: 691—722, 1966. Contractile proteins.
136. Gershman L. C., Stracher A., Dreizen P., J. Biol. Chem. 244: 2726—2736, 1969. Subunits structure of myosin.
137. Gibbs C. L., Mommaerts IF. F. H. M., Ricchiuti N. V., J. Physiol. 191: 25—46, 1967. Heat production , in cardiac muscle.
138. Gilai A., Parnas I., J. Exp. Biol. 52: 325—344, 1970. Neuromuscular transmission in scorpion.
139. Gillespie C. A., Simpson D. R., Edgerton V. R., J. Histochem. Cytochem. 18: 552—558, 1970. Glycogen content in red and white muscle.
18*
276
Глава 16
140. Goffart М., Function and Form in the Sloth. Pergamon Press, New York, 1971. 225 pp.
141. Goffart M., Electromyography 8: 245—251, 1968. Sloth muscles.
142. Goffart M., Holmes O., Bacq Z. M., Arch. Int. Physiol. Biochem. 70: 103—106» 1962. Mechanical properties of muscle in the sloth.
143. Goldspink G„ Larson R. E., Davies R. E„ Z. vergl. Physiol. 66: 379—388, 19701 Thermodynamic efficiency of chicken ALD muscle.
144. Goldspink G., Larson R. E., Davies R. E., Z. vergl. Physiol. 66: 389—397» 1970. Energetics of contraction in hamster muscles.
145. Gonzalez-Serratos H., J. Physiol. 212: 777—799, 1971. Inward spread of activation in vertebrate muscle fibers.
146. Goodwin L. G., Vaughan Williams E. M., J. Physiol. 168: 857—871, 1963. Neuromuscular transmission in Ascaris.
147. Gordon G., Huxley A. F., Julian F. J., J. Physiol. 171: 28P — 30P, 1964. Length-tension diagram of single vertebrate striated muscle fibers.
148. Gordon G., Phillips C. G., Quart. J. Exp. Physiol. 38: 35—45, 1953. Slow and fast components in a flexor muscle.
149. Grundfest H., J. Gen. Physiol. 50: 1955—1958, 1967. Anomalous spikes of Ascaris esophageal cells.
150. Guth L., Samoha F. J., Exp. Neurol. 25: 138—152, 1969. Actomyosin ATPase if slow and fast muscles.
151. Guthrie D. M., Banks J. R., J. Exp. Biol. 52: 125—138, 1970. Properties of paramyosin in notochord of Amphioxus.
152. Hagiwara S., Chichibu S., Simpson N., Z. vergl. Physiol. 60: 209—218, 1968. Neuromuscular mechanisms of wingbeat in hummingbirds.
152a. Hagiwara S., Henkart M. P., Ridokoro Y., J. Physiol. 219: 233—251, 1971. Excitation-contraction coupling in Amphioxus muscle.
152b. Hagiwara S., Ridokoro Y., J. Physiol. 219: 217—232, 1971. Ionic components of action potentials in Amphioxus muscle.
152c. Hagiwara S., Takahashi R., J. Physiol. 190: 499—518, 1967. Electrical activity in muscles of fish.
153. Hagiwara S., Takahashi R., Junge D., J. Gen. Physiol. 51: 157—175, 1968. Excitation-contraction coupling in barnacle muscle.
153a. Hagiwara S. et al., J. Physiol. 219: 233—251, 1971. EC coupling in Amphioxus muscle myotomes.
154. Hall-Craggs E. С. B., J. Anat. 102: 241—255, 1968. Speeds of rabbit laryngeal muscles.
155. Hanson J., J. Biophys. Biochem. Cytol. 3: 111—122, 1957. Structure of body wall muscle of earthworm.
156. Hanson J., Lowy J., pp. 265—335. In The Structure and Function of Muscle, Vol. I, edited by G. Bourne. Academic Press, New York, 1960. Structure and function of invertebrate muscle.
157. Hanson J., Lowy J., Proc. Roy. Soc. Lond. В 154: 173—196, 1961. Structure of muscle fibers in adductor of oyster.
158. Hanson J., Lowy J., J. Molec. Biol. 6: 46—60, 1963. Structure of actin.
159. Hanson J., Lowy J., Proc. Roy. Soc., Lond. В 160: 449—460, 1964. Locations of proteins in smooth muscles.
160. Harris A. J., Miledi R., J. Physiol. 217: 497—515, 1971. Botulinum toxin and neuromuscular transmission block in frogs.
161. Harris J. B., Luff A. R., Comp. Biochem. Physiol. 33: 923—931, 1970. Resting potentials in fast and slow muscles.
162. Harrison R. G., Lowey S., Cohen C., J. Molec. Biol. 59: 531—535, 1971. Paramyosin in muscle.
163. Hartshorne D. J., J. Gen. Physiol. 55: 585—601, 1970. Interactions of muscle proteins.
164. Hartshorne D. J. et al., Biochim. Biophys. Acta 175: 320—330, 1969; 229: 698—711, 1971. Properties of troponin-tropomyosin.
164a. Hartzell H. C., Fambrough D. M., J. Gen. Physiol. 60: 248—262, 1972. Extra-junctional ACh receptors after denervation, rat diaphragm.
Мышцы
277
165. Hasselbach W., Ann. N. Y. Acad. Sci. 137: 1041—1048, 1966. Calcium transport by membranes of muscle reticulum.
166. Hasselbach W., Prog. Biophys. 14: 167—222, 1964. Relaxation of muscle.
167. Heistracher P., Hunt С. C., J. Physiol. 201: 589—611, 1969. Membrane properties in relation to contraction, snake muscle.
168. Henneman E. C., Olsen B., J. Neurophysiol. 28: 581—598, 1965. Structure and . function in skeletal muscle.
169. Hess A., J. Physiol. 157: 221—231, 1961. Fast and slow extrafusal fibers* chickens.
170. Hess A., J. Cell Biol. 26: 467—476, 1965. Motor terminals of slow and twitch’ muscle fibers in snake.
171. Hess A., Pilar G., J. Physiol. 169: 780—798, 1964. Slow fibers in extraocular muscles of cat.
172. Heumann H. G., Zebe E., Z. Zellforsch. 78: 131—150, 1967. Fine structure and function of outer muscle layer in earthworm.
173. Hidaka T., Ito У., Kuriyama H., J. Exp. Biol. 50: 387—403, 1969. Membrane properties of earthworm muscle.
174. Hidaka T„ Ito Y„ Toshiro N., J. Exp. Biol. 50: 405—415, 417—430, 1969. Effects of ions and drugs on excitation and transmission in earthworm: muscle.
175. Hidaka T., Kuriyama H., Yamamoto T., J. Exp. Biol. 50: 431—443, 1969. Mechanical properties of earthworm muscle.
176. Hidaka T., Osa T., Twarog В. M., J. Physiol. 192: 869—877, 1967. Action of 5-HT on Mytilus muscle.
177. Hill A. V., Sci. Prog. 38: 209—230, 1950. Dimensions of animals and muscledynamics.
178. Hill R. B., Greenberg M. J., Irisawa H., Nomura H., J. Exp. Zool. 174: 331— 348, 1970. Electromechanical coupling in muscle of snail.
179. Hirose H. et al., Ann. Otol. 78: 297—306, 1969. Time constants of laryngeal’ muscles of cat.
179a. Hocking B., Trans. Roy. Entomol. Soc. Lond. 104: 223—345, 1953. Intrinsic range and speed of flight of insects.
180. Hoekman T. H., Thesis Ph. D., University of Illinois, 1968. Mechanical properties of muscles of chicken.
181. Hoffman-Berling H., Biochim. Biophys. Acta 27: 247—255, 1958. Contraction-properties of Vorticella.
182. Holman M. E., pp. 1665—1708. In Handbook of Physiology, Sec. 6, Vol. IV,. edited by C. F. Code. Amer. Physiol. Soc., Washington, D. C., 1968. Electrophysiology of visceral muscle.
183. Hoogland P. L., Freeman H. C., Truscote B., Waddell F. E., J. Fish. Res^ Bd. Canad. 18: 501—512, 1961. Amino acid composition of cod tropomyosin.
184. Hoyle G., Comparative Physiology of Nervous Control of Muscular Contractions. Cambridge University Press, Cambridge, 1957. 147 pp.
185. Hoyle G., J. Exp. Zool. 167: 471—486, 1968. Neuromuscular physiology of eyestalk muscle of a crab.
186. Hoyle G., McNeill P. A., J. Exp. Zool. 167: 487—522, 523—550, 1968. Ultrastructure of eyestalk muscles in a crab.
187. Hoyle G., Smyth T., Comp. Biochem. Physiol. 10: 291—314, 1963. Neuromuscular physiology of a barnacle.
188. Hubbard J. I., Jones S. F., Landau E. M., J. Physiol. 196: 75—86, 1968. Endplate potentials in rat diaphragm.
189. Hubbard J. I. et al., J. Physiol. 194: 355—380, 1968. Effects of varying calcium and magnesium on spontaneous release of transmitter from motor nerve terminals.
190. Hudson R. C. L., J. Exp. Biol. 50: 47—67, 1969.xPolyneuronal innervation of fast muscles in fish Cottus. J 1
191. Huik P., Jirsmanova I., Vyklicky L., Zelena J., J. Physiol. 193: 309—325, 1967. Cross-innervation of chick muscles.
278
Глава 16
191а. Huszar G., Elzinga M., J. Biol. Chem. 247: 745—753, 1972. Methylated and non-methylated histidines in myosins of skeletal and cardiac muscle.
192. Huxley A. F., Simmons R. M., Nature 233: 533—538, 1971. Quick changes in length, tension redevelopment.
Л92а. Huxley A. F., Taylor R., J. Physiol. 144: 426—441, 1958. Inward spread of excitation in striated muscle fibers.
193. Huxley H. E., J. Molec. Biol. 7: 281—308, 1963. Electron microscopy of protein filaments from muscle.
194. Huxley H. E., Circulation 24: 328—335, 1961. Contractile structure of cardiac and skeletal muscle.
195. Huxley H. E., Science 164: 1356—1366, 1969. Theory of muscle contraction.
196. Huxley H. E., Proc. Roy. Soc. Lond. В 178: 131—149, 1971. Structural basis of muscular contraction.
,197. Ikemoto N., Kawaguti S., Biol. J. Okayama Univ. 9: 81—126, 1963; 14: 21—33, 1968. Electron microscopy of muscles of earthworm and horseshoe crab.
198. Ito Y., Tashiro N., J. Exp. Biol. 53: 597—609, 1970. Calcium spikes in a po-lychaete muscle.
.199. Ito Y., Ruriyama H., Tashiro N., J. Exp. Biol. 51: 363—375, 1969; 54: 167— 186, 1971. Pharmacology of earthworm muscle.
200. Jahromi S. S., Atwood H. L., Canad. J. Zool. 45: 601—606, 1967. Ultrastructure of crayfish muscle.
201. Jahromi S. S., Atwood H. L., Experientia 25: 1046, 1969. Tension and structure in crayfish muscle.
202. Jahromi S. S., Atwood H. L., J. Exp. Zool. 171: 25—37, 1964. Speeds of various crayfish muscles.
202a. Jahromi S. S., Atwood H. L., J. Exp. Zool. 176: 475—486, 1971. Structure and function of muscle fibers in lobster leg.
203. Jansen P., Andersen J. K- S., Loyning Y., Acta Physiol. Scand. 57: 167—179, 1963. Fast and slow muscles in hagfish.
204. Jewell B. R., J. Physiol. 149: 154—177, 1959. Phasic and tonic responses of muscle of Mytilus.
205. Jewell B. R., Ruegg J. C., Proc. Roy. Soc. Lond. В 164: 428—459, 1966. Oscillations of insect fibrillar muscles.
205a. Job D. D., Amer. J. Physiol. 217: 1534—1541, 1969. Ionic fluxes in slow potentials of intestine.
206. Jobsis I. F., O'Connor M. J., Biochem. Biophys. Res. Comm. 25: 246—252, 1966. Calcium release and reabsorption in sartorius muscle.
207. Johnson W. H., Rahn J. S., Szent-Gyorgyi A. G., Science 130: 160—161, 1959. Paramyosin and contraction of catch muscles.
208. Josephson R. K-, Elder H. У., Biol. Bull. 135: 409, 1968. Muscle of sound production in katydid.
208a. Josephson R. K., Halverson R. C., Biol. Bull. 141: 411—433, 1971. High frequency muscles of sound production in katydid.
209. Kammer A. E., J. Exp. Biol. 48: 89—109, 1968. Motor patterns in Lepidoptera.
210. Katz B., Biol. Rev. 24: 1—20, 1949. Neuromuscular transmission in invertebrates.
2I0a. Katz B., Nerve, Muscle and Synapse. McGraw-Hill Book Co., New York, 1966. 193 pp.
211. Katz B., Miledi R., J. Physiol. 203: 689—706, 1969; also Nature 207: 1096— 1097, 1965; 215; 651, 1967. Spontaneous and evoked activity at motor nerve endings.
212. Katz B., Miledi R., Proc. Roy. Soc. Lond. В 161: 453—482, 483—495, 496'—503, 1971. Miniature junction potentials.
213. Kendrick-Jones J., Cohen C., Szent-Gyorgyi A. G., Longley W., Science 163: 1196—1198, 1969. Biophysics of paramyosin.
214. Kendrick-Jones J., Lehman W., Szent-Gyorgyi A. G., J. Molec. Biol. 54: 313— 326, 1971. Ultrastructure of paramyosin.
Мышцы
279
214а. Kendrick-Jones J. et al., J. Molec. BioL, 1972 (in press). Myosin-linked regulation of calcium, molluscan muscle.
215. Kennedy D., Evoy W. H., J. Gen. Physiol. 49: 457—468, 1966. Pre- and post-synaptic inhibition in crustaceans.
216. Kerkut G. A., Leake L. D., Comp. Biochem. Physiol. 17: 623—633, 1966. Effect of drugs on snail muscle.
217. Kerkut G. A., Walker R. J., Comp. Biochem. Physiol. 17: 435—454, 1966. Pharmacology of leg muscles of cockroach.
218. Kishimoto U., Comp. Biochem. Physiol. 2: 81—89, 1961. ATPase activity of myosin.
219. Kobayashi M., Amer. J. Physiol. 206: 205—210, 1964; 208: 715—719, 1965;
216: 1279—1285, 1969. Electrophysiology of cat ureter.
220. Kobayashi M., Prosser C. L., Nagai T., Amer. J. Physiol. 213: 275—286, 1967. Electrical properties of intestinal muscle, measured intracellularly and extra-cellularly.
220a. Kundrat E., Pepe F. A., J. Cell Biol. 48: 340—347, 1971. M-protein.
221. Kuriyama H., Mekata F., J. Physiol. 212: 667—683, 1971. Longitudinal muscle of guinea pig rectum.
222. Kuriyama H., Osa T., Tasaki H., J. Gen. Physiol. 55: 48—62, 1970. Electrophysiology of stomach.
223. Kuriyama H., Osa T., Toida N., J. Physiol. 191: 239—255, 1967. Electrophysiology of intestinal muscle.
224. Kuriyama H., Tomita T., J. Gen. Physiol. 55: 147—162, 1970; J. Physiol. 178: 270—289, 1965. Response of ureter and of taenia coli to applied currents.
224a. Lang F., Atwood H. L., Morin W. A., Z. Zellforsch. 127: 189—200, 1972. Electron microscopy of nerve supply to crayfish opener muscle.
225. Lang F., Sutterlin A., Prosser C. L., Comp. Biochem. Physiol. 32: 615—628, 1970. Physiology of leg muscle in Paralithodes.
226. Leenders H. J., Pfliiger. Arch. 295: 127—135, 1967. Mechanical properties of Mytilus catch muscle.
227. Leenders H. J., J. Physiol. 192: 681—693, 1967; also Comp. Biochem. Physiol.
31: 187—196, 1969. Calcium coupling and ATPase activity in catch muscles.
227a. Lehman W., Kendrick-Jones J., Szent-Gyorgyi A. G., J. Molec. Biol., 1972 (in press). Comparative studies of Ca regulation in muscles.
227b. Liu J., Prosser C. L., Job D. D., Amer. J. Physiol. 217: 1542—1547, 1969. Ionic dependence of slow waves and spikes in intestinal muscle.
228. Lowey S. et al., J. Molec. Biol. 4: 293—308, 1962; 42: 1—29, 1969. Structure of myosin.
228a. Lowey S., Risby D., Nature 234: 81—85, 1971. Light myosin chains from fast and slow muscles.
229. Lowy J., Millman В. M., Trans. Roy. Soc. 246: 105—148, 1963. Contractile mechanism of Mytilus muscle.
229a. Luff A. R., Atwood H. L., Amer. J. Physiol. 222: 1435—1440, 1972. Mouse muscle speeds.
230. Lund G. F„ Christensen I., Amer. J. Physiol. 217: 1369—1374, 1969. Electrical stimulation of the esophagus.
231. Lymn R. W., Taylor E. W., Biochemsitry 9: 2975—2983, 1970. ATPase activity of myosin.
231a. MacLennan D. H. et al., J. Biol. Chem. 246: 21(02—2710, 1971; also Proc. Nat. Acad. Sci. 68: 1231—1235, 1971. Calcium binding and Ca-ATPase of sarcoplasmic reticulum.
232. Marechai G., Goffart M., Aubert X., Arch. Int. Physiol. Biochem. 71: 236—240, 1963. Muscle speeds in sloths.
233. Marshall J. M„ J. Physiol. 197: 935—942, 1959; 204: 732—738, 1963. Electrical, activity of uterine smooth muscle.
234. Maruyama K-, Biochim. Biophys. Acta 102: 542—548, 1965. Actinin.
235. Maruyama K., Sci. Papers Coll. Gen. Ed., Univ, of Tokyo 6: 95—111, 1956;. Properties of contractile proteins from a sea anemone.
280
Глава 16
236. Mashima Н., Yoshida Т., Japan. J. Physiol. 15: 463—477, 1965. Tension-length curves in taenia coli.
•237. Matsumoto Y., Abbott В. C., Comp. Biochem. Physiol. 26: 927—936, 1968. Folding muscle fibers of Golfingia.
'238. Matyushkin D. P., Fed. Proc. TransL Suppl. 22: Т728 — T731, 1963. Innervation of tonic muscle fibers in oculomotor system.
239. Maynard D. M., Burke W., Comp. Biochem. Physiol. 38A: 339—350, 1971. Tension development by adductor of Tridacna.
'2 40. McCann F. V., Boettiger E. G., J. Gen. Physiol. 45: 125—142, 1961. Electrophysiology of fibrillar flight muscles.
241. McCoy E. J., Baker R. D„ Proc. Soc. Exp. Biol. Med. 127: 562—563, 1968. Intestinal slow waves.
242. McLennan FL, Synaptic Transmission, 2nd Ed. W. B. Saunders Co., Philadelphia, 1970. 178 p.
242a. McNeill P. et al., J. Exp. Zool. 179: 395—416, 1972. Fine structure of muscle of mantis shrimp.
243. Meiss R. A., Amer. J. Physiol. 220: 2000—2007, 1971. Some mechanical properties of cat intestinal muscle.
244. Mendelson M., J. Cell Biol. 42: 548—563, 1969. Properties of a very fast lobster muscle.
.245. Miledi R., Potter L. T., Nature 233: 599—603, 1971. Acetylcholine receptors in muscle fibers.
246. Miledi R., Slater C. R., Proc. Roy. Soc. Lond. В 174: 253—269, 1969. Ultrastructure of denervated skeletal muscle.
247. Miledi R., Stefani E., Nature 222: 569—571, 1969. Non-selective re-innervation of slow and fast muscle fibers in rat.
.248. Mill P. L, Frapp M. F„ J. Cell. Sci. 7: 233—262, 1970. Fine structure of earthworm muscle.
248a. Miller M. D., Marshall I. M., J. Physiol. 209: 859—865, 1965. Catecholamine receptors in uterus.
249. Miller P. L., Nature 221: 171—173, 1969. Inhibitory innervation of insect spi-r acles
250. Millman В. M., J. Physiol. 173: 238—262 , 1964; also Proc. Roy. Soc. Lond В 160: 525—536, 1964. Contraction fo opaque adductor of oyster.
251. Millman В. M., Elliott G. E., Nature 206: 824—825, 1965. X-ray diffraction of molluscan muscle.
252. Millward G. R., Woods E. F„ J. Molec. Biol. 52: 585—588, 1970. Crystals of tropomyosin.
253. Mommaerts W. F. FL M., Physiol. Rev. 49: 427—508, 1969. Energetics of muscular contraction.
254. Mommaerts W. F. H. M., Wallner A., J. Physiol. 193: 343—357, 1967. Energetics in frog sartorius.
255. Motdkisawa F., Reuben J. P., Grundfest FL, J. Gen. Physiol. 54: 437—461, 1969. Ion properties of inhibitory receptors in lobster muscle.
256. Nagai T., J. Insect Physiol. 16: 437—448, 1970. Properties of insect visceral muscle.
257. Nagai T., Brown В. E., J. Insect Physiol. 15: 2151—2167, 1969. Properties of cockroach visceral muscle.
258. Nagai T., Prosser C. L., Amer. J. Physiol. 204: 915—924, 1963. Electrical parameters of smooth muscles.
259- Nakajima Y., Tissue and Cell 1: 229—246, 1969. Fine structure of red and white muscle in snake fish.
260. Nass M. M. K„ Develop. Biol. 4: 289—320, 1962. Developmental changes in actomyosin.
261. Nauss К. M„ Davies R. E„ Biochem. Zeitschr. 345: 173—187, 1966. Energetics of catch muscle of Mytilus.
262. Nilsson S., Fange R., Comp. Biochem. Physiol. 30: 691—694, 1969. Neural control of stomach of cod.
Мышцы
281
263. Ohnesorge F. К., Rauch R., Z. vergl. Physiol. 58: 153—170, 1968. Peristalsis in intestine of Tinea.
264. Ozeki M„ Grundfest H., Science 155: 478—481, 1967. Ionic mechanisms incrayfish muscle.
265. Page S. G., J. Cell Biol. 26: 477—497, 1965. Ultrastructure of frog fast and slow fibers.
266. Pak M. J., Abbott В. C., personal communication. Electromechanical properties of toadfish sonic muscle.
267. Panner B. J., Honig C. R., J. Cell Biol. 35: 303—321, 1967. Filament ultrastructure of smooth muscle.
268. Papasova M. P., Nagai T., Prosser C. L., Amer. J. Physiol. 214: 695—702;. 1968. Two component slow waves in cat stomach.
269. Parmley W. W., Yeatman L. A., Sonnenblick E. H., Amer. J. Physiol. 220: 546—550, 1970. Forse-velosity relations, striated muscle.
270. Parnas I., Abbott В. C., Lang F., Comp. Biochem. Physiol. 26: 467—478, 1968; Neuromuscular system of Limulus leg muscle.
271. Parnas I., Atwood H. L., Comp. Biochem. Physiol. 18: 701—723, 1966. Neuromuscular system of abdominal muscles of crayfish.
272. Parnas I., Dagan D„ Comp. Biochem. Physiol. 28: 359—369, 1969. Abdominal muscles of prawn Palaemon.
273. Paul W. M., Daniel E. E., Kay С. M., Monckton G„ editors, University of Alberta Medical School Symposium: Muscle. Pergamon Press, New York, 1965. 584 p.
274. Peachey L. D., Biochem. Biophys. CytoL Suppl. 10: 159—176, 1961. Ultrastructure of muscle of Amphioxus.
275. Peachey L. D., Amer. Zool. 7: 505, 1967. Membrane system in crab muscle.
275a. Peachey L. D., Ann. Rev. Physiol. 30: 401—440, 1968. Ultrastructure of muscle, correlation with contraction.
276. Peachey L. D., Huxley A. F., J. Cell Biol. 13: 177—180, 1962. Structural identification of fast and slow fibers in frog.
277. Pearson K. G., Bergman S. J., J. Exp. Biol. 50: 445—471, 1969. Neuromuscular inhibition in insects.
278. Pearson K- G., Iles J. F., J. Exp. Biol. 54: 215—232, 1971. Neuromuscular transmission in cockroach.
279. Pepe F. A., J. Cell Biol. 28: 505—525, 1966; also J. Molec. Biol. 27: 203—225, 1967; also pp. 75—96 in Progress in Biophysics and Molecular Biology, edited by J. A. V. Butler and D. Noble, Pergamon Press, Oxford, 1970; also pp. 323—353 in Fine Structure of Proteins and Nucleic Acids (Biological Macromolecular Ser. Vol. 4), edited by S. Timasheff and G. Fasman, Marcel Dekker, Inc., New York, 1970. Structural components of the striated muscle fibers.
279a. Perrie W. T., Perry S. V., Biochem. J. 119: 31—38, 1970. Low molecular weight components of myosin.
280. Piek T., Mantel P., Comp. Biochem. Physiol!. 34: 935—951, 1970. Electrical activity in insect muscle. V
281. Pilar G., J. Gen. Physiol. 50: 2289—2300, 1967. Electromechanical properties of slow fibers in extraocular muscles.
282. Podolsky R. D., Teickholz L. E., J. Physiol. 211: 19—35, 1970. Ca++ and contractions in skinned fibers.
283. Prewitt M. A., Salafsky B., Amer. J. Physiol. 213: 295—300, 1967; 218: 69—74, 1970. Changes in fast and slow muscles after cross-innervation.
284. Pringle J. W. S„ Insect Flight. Cambridge University Press, Cambride, 1957. 133 p.
285. Pringle J. W. S., Prog. Biophys. 17: 1—60, 1967; also J. Gen. Physiol. 50: 139—156, 1967. Contractile mechanisms in insect fibrillar muscle.
286. Proske U., Vaughan P., J. Physiol. 199: 495—510, 1968. Histological correla- tions with functions, lizard skeletal muscle.
287. Prosser C. L., J. Cell. Comp. Physiol. 44: 247—254. Activation of muscle of Thy one.
282
Глава IS
288. Prosser С. L., Comp. Biochem. Physiol. 6: 69—74, 1962; Z. vergl. Physiol. 54: 109—120, 1967. Contractions in sponge.
289. Prosser C. L., Bortoff A., pp. 2025—2050. In Handbook of Physiology, Sec. 6. Vol. IV, edited by C. F. Code. Amer. Physiol. Soc., Washington, D. C 1968.
290. Prosser C. L., Burnstock G., Holman M., Physiol. Rev. (Suppl. 5) 42: 193— 212, 1962. Conduction in non-striated muscles.
291. Prosser C. L., pp. 133—149. In Invertebrate Nervous Systems, edited by C. A. G. Wiersma. University of Chicago Press, Chicago, 1969. Comparative physiology of non-striated muscle.
292. Prosser C. L., Curtis H. J., Travis D., J. Cell. Comp. Physiol. 38: 299—319,
1951; also Prosser C. L., Melton С. E., J. Cell. Comp. Physiol. 44: 255—275,
1954; also Prosser C. L., Sperelakis N., J. Cell. Comp. Physiol. 54: 129—133,
1959. Nervous conduction in smooth muscle of Phascolosoma (Golfingia).
293. Prosser C. L., Nystrom R., Nagai T„ Comp. Biochem. Physiol. 14: 53—70, 1965. Intestinal muscle of invertebrates.
294. Prosser C. L., Rafferty N. S., Amer. J. Physiol. 187: 546—548, 1956. Electrical activity in chick amnion.
295. Prosser C. L., Ralph C. L., Steinberger W. W., J. Cell. Comp. Physiol. 54: 135—146, 1959. Responses to stretch in invertebrate muscles.
296. Rahamimoff R., J. Physiol. 195: 471—480, 1968. Frog neuromuscular junction; ions, glycerol effects.
297. Rai B. S. D., Cohen M. J., Naturwissenschaften 51: 224—225, 1964. Structurefunction correlation in crab muscle.
298. Rathmayer W., Comp. Biochem. Physiol. 14: 673—687, 1965. Neuromuscular transmission in spider.
299. Reedy M. R., Amer. Zool. 7: 465—481, 1967. Ultrastructure of insect flight muscles.
300. Rees M. R„ Young M„ J. Biol. Chem. 242: 4449—4458, 1967. Properties of actin.
301. Reger J. F., Biochem. Biophys. Cytol. Suppl. 10: 111—121, 1961. Fine structure of ocular muscles of Fundulus.
302. Resis D. J., Wooten G. F., Hollenberg M., Amer, J. Physiol. 213: 592—596, 1967. Enzymes in red and white skeletal muscle.
303. Reuben J. P., Brandt P. W., Garcia H., Grundfest H., Amer. Zool. 7: 623—645, 1967. Excitation-contraction coupling in crayfish.
304. Revel J. P., J. Cell Biol. 12: 571—588, 1962. Sarcoplasmic reticulum of fast muscle in bat.
305. Rice R. V. et al., Nature 231: 242—246, 1970. Thick filaments in mammalian smooth muscle.
306. Ridge R. M., J. Physiol. 217: 393—418, 1971. Contraction times of snake muscle fibers.
307. Roach D. R., Comp. Biochem. Physiol. 24: 865—878, 1968. Rhythmic activity in digestive tract of snail.
307a. Roberts B. L., J. Marine Biol. Assoc. U. K. 49: 357—378, 1969. Properties of dogfish muscles.
308. Robison G. A., Butcher R. W., Sutherland E. W„ Ann. N. Y. Acad. Sci. 139: 703—723, 1966. Adenyl cyclase as an adrenergic receptor.
309. Robison G. A., Sutherland E. W., Circ. Res. 26 (Suppl.): 147—161, 1970. Role of cyclic AMP in adrenergic responses.
310. Robson R. M., Goll D. E., Arakawa N., Stromer M. H., Biochim. Biophys. Acta 200: 296—318, 1970. Properties of alpha-actinin.
311. Roeder R. D., Biol. Bull. 100: 95—106, 1951. Potentials in flight muscles of insects.
312. Rohlich P., J. Ultrastruct. Res. 7: 399—408, 1962. Fine structure of muscle of leech.
313. Romanul F. C. A., Arch. Neurol. 11: 355—368, 1964. Enzymes in striated ; muscle. h
Мышцы
283-
314. Romanul F. C. A., Van der Meulen J. P., Arch. Neurol. 17: 387—402, 1967. Slow and fast muscles after cross-innervation.
315. Rosenbluth J., J. Cell Biol. 25: 495—515, 1965; 26: 579—591, 1965. Ultrastructure of muscle of Ascaris.
316. Rosenbluth J., J. Cell Biol. 42: 534—547, 1969. Ultrastructure of a fast crustacean muscle.
317. Ross D. M., J. Exp. Biol. 37: 732—752, 753—774, 1960. Effects of ions and drugs on muscle of a sea anemone.
318. Ruegg J. C., Biochem. Biophys. Res. Somm. 6: 24—28, 1961; also Proc. Roy. Soc. Lond. В 154: 209—249, 1961. Proteins in tonic muscles of lamellibranchs.
319. Ruegg J. C. et al., Proc. Roy. Soc. Lond. В 158: 156—176, 177—195, 1963. Contractile properties of tonic molluscan muscles.
320. Ruegg J. C., Amer. Zool. 7: 457—464, 1967; also Experientia 24: 529—536, 1968; also Ruegg J. C., Tregear R. T., Proc. Roy. Soc. Lond. В 165: 497—512, 1966. Oscillatory flight muscles of insects.
321. Ruegg J. C., Physiol. Rev. 51: 201—248, 1971. Smooth muscle tone.
322- Salanki J., Hiripi L., Comp. Biochem. Physiol. 32: 629—636, 1970. Transmission in adductor muscles of Anodonta.
323. Salmons S., Vrbova G., J. Physiol. 201: 535—549, 1969. Effect of exercise on mammalian fast and slow muscles.
324. Samaha F. J., Guth L., Albers R. W., Exp. Neurol. 27: 276=—282, 1970. Neural regulation of chemical development in muscles.
325. Sarkar S., Sreter F. A., Gergely J., Proc. Nat. Acad. Sci. 68: 946—950, 1971. White, red, and cardiac myosin of rabbit.
326. Sato M„ Tamasige M., Ozeki M., Japan J. Physiol. 10: 85—98, 1960. Electrical activity of pharynx of snail.
327. Schmit F. O., Bear R. S., Hall С. E., Jakus M. A., Ann. N. Y. Acad. Sci. 47: 799—812, 1947. Ultrastructure of paramyosin.
328. Schoenberg C. F., Tissue and Cell 1: 83—96, 1969. Electron microscopy of myosin filaments in chicken gizzard.
329. Scopes R. K-, Perry I. F., Biochim. Biophys. Acta 236: 409—415, 1971. Subunits of muscle proteins.
330. Sexton A. W., Gersten J. W., Science 157: 199, 1967. Mechanical properties of red and white muscle.
331. Sherman R. G., Atwood H. L., J. Exp. Zool. 176: 461—474, 1971. Neuromuscular physiology of lobster.
331a. Sherman R. G., Atwood H. L., Science 171: 1248—1250, 1971. Long-term neuromuscular facilitation in crustaceans.
332. Skoglund C. R., Biochem. Biophys. Cytol. Suppl. 10: 187—200, 1961. Sound producing muscles in toadfish.
333. Smith D. S., Rev. Ganad. Biol. 21: 279V-301, 1962. Ultrastructure of insect flight muscle. /
334. Smith D. S., Gupta B. L., Smith N., J. Cell. Sci. 1: 49—57, 1966. Organization of myofilaments in insect visceral muscle.
334a. Sotovalta O., Biol. Bull. 104: 439—444, 1953. Flight tone and thoracic vibration frequency in midges.
334b. Sparro w M., personal communication.
335. Spirito С. P., Z. vergl. Physiol. 68: 211—228, 1970. Control of leg muscles in Uca.
336. Sreter F. A., Arch. Biochem. 134: 25—33, 1969. ATPase in red and whitemuscle.
337. Stephens N. L., Chiu B. S., J. Physiol. 219: 1001—1008, 1970. Mechanical properties of tracheal muscle.
338. Stephens N. L., Kramer U., Amer. J. Physiol. 220: 1890—1895, 1971. Series elastic component of tracheal smooth muscle.
339. Stephens N. L., Kroeger E., Mehta J. A., J. Appl. Physiol. 26: 685—692, 1969. Force-velosity properties of tracheal smooth muscle.
-340. Stossel W., Zebe E., Pfliiger. Arch. 302: 38—56, 1968. Control of relaxation; in skeletal muscle.
т
*284 Глава 16
-340а. Stromer М., Hartshorne D„ Mueller Н., Rice R. V., J. Cell Biol. 40: 167—178, 1969. Effects of protein fractions on Z-and M-line reconstitution.
341. Sugi H., О chi R., J. Gen. Physiol. 50: 2145—2176, 1967. Spread of contraction in crayfish fibers.
342. Swanson C. J., Z. vergl. Physiol. 74: 403—410, 1971; also Nature 232: 122, 1971. Paramyosin in Nematomorpha.
•343. Syrovy I., Gutmann E., Nature 213: 937—938, 1967. Biochemical differentiation in fast and slow muscles of chick.
.344. Szent-Gyorgyi A. G., Cohen C., Kendrick-J ones J., J. Molec. Biol. 56: 239—258, 1971. Ultrastructure of paramyosin.
345. Szurszewski J. H., Elveback L. R., Code C. F., Amer. J. Physiol. 218: 1468— 1473, 1970. Electrical siwo waves in dog intestine.
•346. Takahashi K-, Annot. Zool. Japan. 33: 67—84, 1960. Nervous control in catch muscle of Mytilus.
347. Takeuchi A., J. Cell. Comp. Physiol. 54: 211—220, 1959. Neuromuscular transmission in fish.
348. Takeuchi N., J. Physiol. 167: 128—155, 1963. Conductance changes in motor endplates.
349. Takeuchi A., Takeuchi N., Fed. Proc. 26: 1633—1638, 1967. Effect of GABA on synapses.
350. Takeuchi A., Takeuchi N., J. Physiol. 170: 296—317, 1964. Effect of glutamate on crayfish muscle.
351. Takeuchi A., Takeuchi N., J. Physiol. 217: 341—358, 1971. Crayfish muscle conductance.
351a. Tashiro N., J. Exp. Biol. 55: 101—110, 1971. Mechanical properties of earthworm muscles.
351b. Tashiro H., Yamamoto T., J. Exp. Biol. 55: 111—122, 1971. Contraction in earthworm muscle.
351c. Teravainen H., J. Neurophysiol. 34: 954—973, 1971. Structure and function of lamprey muscles.
352. Thoai N., Robin Y., pp. 163—203. In Chemical Zoology, Vol. 4, edited by M. Florkin and В. T. Scheer. Academic Press, New York 1969. Guanidine compounds and phosphagens.
353. Tomita T. J., J. Theor. Biol. 12: 216—227, 1966. Capacity and resistance in mammalian smooth muscle.
354. Twarog В. M., J. Physiol. 152: 220—235, 236—242, 1960. Innervation and contraction in molluscan smooth muscle.
355. Twarog В. M., Life Sci. 5: 1201—1213, 1966; also J. Physiol. 192: 847—856, 857—868, 1967. Effects of calcium and 5-HT on molluscan muscles.
.356. Twarog В. M., J. Gen. Physiol. 50: 157—169, 1967. Review of regulation of catch in molluscan muscles.
357. Twarog В. M., Hidaka T., J. Gen. Physiol. 57: 252, 1971. Effect of 5-HT on electrical properties of molluscan muscle fibers.
357a. Uchizono K., Nature 214: 833—834, 1967. Crayfish vesicles.
358. Usherwood P. H. R., J. Exp. Biol. 49: 201—222, 1968. Inhibitory innervation of muscle in insects.
359. Usherwood P. N. R., Nature 223: 411—413, 1969. Glutamate stimulation of insect muscle.
360. Usherwood P. N. R., Grundfest H., Science 143: 817—818, 1964; also J. Neuro-physiol. 28: 497—518, 1965. Inhibition in grasshopper muscle.
361. Usherwood P. N. R., Machili P., J. Exp. Biol. 49: 341—361, 1968. Pharmacology of excitatory junctions in locust muscle.
362. Usherwood P. H. R., Runion H. I., J. Exp, Biol. 52: 39—58, 1970. Mechanical properties of tibial muscle of locust.
363. Van der Kloot W. S., Comp. Biochem. Physiol. 9: 317—334, 1963. Control of spiracular muscles in silk moth.
364. Van der Kloot W. S., Comp. Biochem. Physiol. 15: 547—565, 1965. Uptake of calcium by vesicles of lobster muscles.
Мышцы
285
365. Van Harn G. L., Amer. J. Physiol. 215: 1351—1358, 1968. Frog stomach smooth muscle.
366. Vrbova G., J. Physiol. 169: 513—526, 1963. Activation of striated muscle by motorneurones.
367. Walcott B., Burrows M., J. Insect Physiol. 15: 1855—1872, 1969. Abdominal air-guide retractor muscles, giant bug Lethocerus.
367a. Walker T. J., Ann. Entom. Soc. Amer. 62: 752—762, 1969. Acoustical behavior of cricket Orocharis.
368. Washizu Y., Comp. Biochem. Physiol. 20: 641—646, 1967. Electrical properties of leech muscle.
369. Watts D. C„ Moreland B., Tatshell E. C„ Bannister L. H., Comp. Biochem. Physiol. 25: 553—558, 1968. Phosphoarginine in protozoan Stentor.
370. Weber A., pp. 203—254. In Current Topics in Bioenergetics, edited by D. R. Sandi. Academic Press, New York, 1966. Calcium transport and relaxation.
371. Weber A., Herz R., Reiss I., J. Gen. Physiol. 46: 679—702, 1963. Relaxing factor in sarcoplasmic reticulum.
372. Weis-Fogh T., J. Exp. Biol. 33: 668—684, 1956; also Phil. Trans. Roy. Soc. Lond. В 239: 415—510, 553—584, 1956. Aerodynamics of insect flight.
373. Welsh J., Moorhead M., J. Neurochem. 6: 146—169, 1960. 5-Hydroxytryptamine in molluscan tissues.
374. Wiersma C. A. G., pp. 143—159. In Recent Advances in Invertebrate Physiology» edited by B. Scheer. University of Oregon Press, 1957. Neuromuscular transmission in crustaceans.
375. Wiersma C. A. G., pp. 191—240. In The Physiology of Crustacea, Vol. II, edited by T. Waterman. Academic Press, New York, 1961. Neuromuscular system of crustaceans
376. Wilkie D. R., Prog. Biophys. 4: 288—324, 1954; also J. Physiol. 134: 527—530, 1956; 195: 157—183, 1968. Mechanical properties of frog muscle.
376a. Wilkie D. R., Muscle. St. Martin’s Press, Inc., New York, 1968, 63 pp.
377. Wilson D. F., Comp. Biochem. Physiol. 29: 703—715, 1969. Fast and slow contractions in pelecypod mantle.
378. Wilson D. M., J. Exp. Biol. 37: 57—72, 1960. Nervous control of muscle in cephalopods.
379. Wilson D. M., Smith D. O., Dempster P., Amer. J. Physiol. 218: 916—922, 1970. Length-tension hysteresis in arthropod muscle.
380. Winegrad S., J. Gen. Physiol. 51: 65^-83, 1968; 55: 77—88, 1970. Intracellular localization of calcium in contract ign..'
381. Wood D. W., J. Exp. Biol. 35: 850—861, 1958. Fast and slow muscles in stick insect Carausius.
382. Wood J. D., Comp. Biochem. Physiol. 30: 813—824, 1969. Electrical and mechanical activity of stomach of squid.
382a. Wood J. D., Amer. J. Physiol. 219: 159—169, 1970. Electrical activity from neurons in Auerbach’s plexus.
383. Wood J. D., Perkins W. E., Amer. J. Physiol. 218: 762—768, 1970. Interaction between longitudinal and circular axes of the intestine.
384. Wood M. E„ Burnstock G., Comp. Biochem. Physiol. 22: 755—766, 1967. Innervation of lung of toad.
384a. Wright E. B., Coleman P. D., J. Cell. Comp. Physiol. 43: 133—164, 1954. Excitation of motor axons of crayfish.
384b. Yasui V., Fuchs F., Briggs F. M., J. Biol. Chem. 243: 735—742, 1968. Calcium and troponin control of contractions.
385. Zebe E., Meinrenken W., Ruegg I. C., Z. Zellforsch. 87: 603—621, 1968. Contractions of glycerinated muscle.
386. Zolovick A. J„ Norman R. L., Fedde M. R., Amer. J. Physiol. 219: 654—657, 1970. Membrane consants of rat diaphragm.
387. Zs-Nagy I., Labos E., Ann. Biol. Tihany 36: 123—133, 1969. Ultrastructure of adductor muscle in Anodonta.
286
Глава 16
388. Csapo A., in: The Structure and Function of Muscle, vol. 1, ed. by G. Bourne. Ac. Press, N. Y., 1960.
389. Couteaux R., in: Problemes de structure, d’ultrastructure et de fanctions cellu-laires, ed. by J. A. Thomas, Masson et Cie, 1955.
390. Couteaux R., in: The Structure and Function of Muscle, ed. by G. Bouraer Ac. Press, N. Y., 1960.
391. Kuffler S. W„ J. Neurophysiol. 5: 18—26, 1942.
392. Irisawa H., Kobayashi M„ Japan J. Physiol. 13: 421—430, 1963.
Глава 17
Электрические органы и электрорецепторы
Л. ПРОССЕР
Первыми обнаруженными человеком биоэлектрическими потенциалами были разряды электрических рыб; в настоящее время большие плоские клетки их электрических органов являются весьма ценным объектом для изучения природы биопотенциалов. Электрического сома Malapterurus изображали на гробницах в древнем Египте [23], у римлян встречаются упоминания об электрическом скате Torpedo. «Электрическую терапию» с помощью электрических рыб рекомендовал Гален; ее широко использовали европейские врачи восемнадцатого столетия. Большой электрический угорь Electrophorus был привезен в Европу исследователями Южной Америки, и появились 'первые предположения о природе его удара. В конце XVIII в. некоторые естествоиспытатели предполагали, что удар электрического угря сходен с молнией или электростатическим разрядом лейденской банки. В 1773 г. Дж. Уолш выяснил, что электрический удар проводится через металл и не проходит через стекло и воздух. Физик из Кембриджа Кавендиш создал модель Torpedo, на основе которой установил различие между понятиями разности электрических потенциалов и количеством электричества. Фарадей измерял ток разряда Electrophorus с помощью гальванометра. Он пророчески заметил, что, если бы удалось понять природу электрического разряда рыб, можно было бы «превращать электрическую силу в нервную».
Электрические органы представляют собою прекрасный пример эволюционной конвергенции — мышцы, а в двух случаях нервные клетки превратились в электрические органы. По-видимому, электрические органы возникали в процессе эволюции животных шесть раз, и их функциональные свойства приспособлены к конкретным особенностям экологии этих животных [4, 8]. Высоковольтными электрическими органами обладают лишь несколько семейств рыб: к ним относятся морские электрические скаты (и, возможно, также звездочеты) и пресноводные электрические угри и сомы; для них характерен рефлекторный высоковольтный разряд. Морские скаты, пресноводные клюворылые, гимнарховые,
288
Глава 17
Рис. 17-1. Схемы эквивалентных электрических цепей электрощитов различных типов и запись потенциалов иннервированной поверхности Ри, неиннервированной поверхности VH и целой клетки (Ркл) [8].
А. Электроцит сильноэлектрической морской рыбы; иннервированная поверхность генерирует ВПСП. Б. Электроцит ската из сем. Rajidae; иннервированная поверхность генерирует ВПСП, а неиннервированная обнаруживает задержанное выпрямление. В. Электроцит электрического угря и некоторых гимнотид; иннервированная поверхность генерирует спайк с овершутом. Г. Электроциты клюворылых и некоторых гимнотид; обе поверхности генерируют спайки, потенциал V КЛ двухфазный. Д. Электроцит электрического сома; стебель имеет высокий порог и генерирует небольшой спайк, значение V Кл отрицательно на поверхности, не имеющей стебля. Е. Электроцит Gumnarchus; неинервированная поверхность функционирует подобно последовательно присоединенной емкости. Цифрами на эквивалентных схемах и кривых указаны переходы от состояния покоя к состоянию активности в различные моменты времени.
гимнотиды и некоторые другие рыбы обладают небольшой электрической мощностью; за исключением скатов, они почти непрерывно с переменной частотой и регулярностью генерируют разряды импульсов.
Генерация электрических разрядов
Электрические органы представлены множеством типов, поэтому для обозначения любых клеток, способных к генерации
Электрические органы и электрорецепторы 289
электрических разрядов (независимо от их эмбрионального происхождения), удобно применить общий термин электроцит. Большие плоские электроциты называются электропластинками.
Свойства некоторых электрических органов различных типов приведены в сжатой форме в нижеследующем перечне, составленном в основном по данным, опубликованным Беннетом [4, 8] (рис. 17-1).
У сильноэлектрического ската Torpedo (из морских пластиножаберных) каждый орган содержит примерно 45 дорсовент-ральных столбиков из 700 электроцитов (или электропластинок) в каждом. Электропластинки в столбике соединены последовательно, а сами столбики — параллельно. Разряд Torpedo обычно составляет 20—30 В, иногда 50 В и создает в морской воде ток в несколько ампер. По некоторым данным мощность электрического органа Torpedo occidentalis доходит до 1000 Вт. Все пластинки в столбике обильно иннервируются с вентральной стороны, на которой генерируются нераспространяющиеся градуальные постсинаптические потенциалы ^(ВПСП); при этом вентральная сторона пластинки отрицательна. Мембрана иннервированной поверхности клетки является холинорецептивной; ответы блокируются тубокурарином и увеличиваются по длительности эзерином. Не-иннервированная мембрана обладает более низким сопротивлением по сравнению с иннервированной; кожа электрических органов также имеет низкое сопротивление, поэтому ток может протекать даже в том случае, если деполяризуется только одна поверхность клетки (рис. 17-1, Л) [3, 8, 10].
У другой группы морских пл-астиножаберных — Rajidae — электрические органы расположены у основания хвоста; они образованы чашеобразными или дисковидными электроцитами. Иннервированная поверхность каждого электроцита генерирует ВПСП длительностью 50 мс или более. В дисковидных электро-цитах неиннервированная поверхность в течение фазы спада ВПСП уменьшает свое сопротивление от 5—30 Ом-см2 до 1,5 Ом-см2 (задержанное выпрямление), создавая таким образом некоторое увеличение тока во внешней цепи (рис. 17-1,Б) [4].
Морской звездочет Astroscopus имеет расположенные кнаружи от глаза мышцы, видоизмененные в электрические органы, которыми он пользуется для того, чтобы парализовать свою добычу. Как и у Торпедо и скатов, иннервированная поверхность каждого электроцита у звездочета дает отрицательный ВПСП (10— 20 мВ), который является холинэргическим по своей природе. Неиннервированная поверхность обладает небольшим электрическим сопротивлением [9].
Таким образом, у всех морских электрических рыб разряд представляет собою постсинаптические потенциалы, которые арифметически складываются при последовательном соединении электроцитов, Мембраны электроцитов электрически невозбудимы, но
19—2067
290
Глава 17
иннервированная сторона мембраны стимулируется ацетилхолином; неиннервированная сторона обладает небольшим сопротивлением или способностью к задержанному выпрямлению. Внешний ток, в особенности создаваемый Torpedo и Astroscopus, является очень сильным в морской воде, имеющей низкое сопротивление.
Сильноэлектрический уголь Electrophorus, обитающий в Амазонке, имеет несколько электрических органов; основной из них содержит в каждом столбике около 1000 продольно ориентированных пластинок. Отдельные импульсы достигают в разомкнутой цепи 400 В, а при коротком замыкании сила тока доходит до 1 А. Каждый разряд представляет обычно серию из 3—5, иногда 20— 30 импульсов. Одиночная электропластинка имеет потенциал покоя 90 мВ (отрицательный с внутренней стороны) при измерении на любой из поверхностей. Стимуляция нерва дает градуальный ВПСП, который может вызвать спайк на иннервированной стороне. Полярность этой (задней) поверхности изменяется на противоположную, достигая примерно 60 мВ, но неиннервированная сторона имеет низкое сопротивление, электрически не возбудима и не деполяризуется; следовательно, две поверхности, соединенные последовательно, дают суммарный клеточный потенциал в 150 мВ (рис. 17-1, В) [1, 24]. Спайк обусловлен активацией натриевого канала и блокируется тетродотоксином. В отличие от спайков, возникающих в нервах или скелетных мышцах, увеличения проводимости ионов калия в этом случае не происходит; восстановление потенциала покоя обусловлено скорее всего увеличением сопротивления (аномальное выпрямление) при натриевом токе в фазе возрастания ВПСП [8].
ВПСП снижается тубокурарином; ритмические нервные импульсы вызывают облегчение, которое усиливается антихолин-эстеразами. Электрические органы богаты ацетилхолином (АХ) и ацетилхолинэстеразой (АХЭ), и, вероятно, АХ является медиатором. Спайки обусловлены увеличением входящего тока натрия и блокируются тетродотоксином. Сопротивление пассивной неиннер-вированной стороны меньше, чем сопротивление иннервированной мембраны. Так, иннервированная мембрана состоит из участков синаптического типа, которые чувствительны к АХ и электрически невозбудимы, и чередующихся с ними электрически возбудимых участков, в которых возникают спайки. АТФ гидролизуется при разряде и регенерируется из креатинфосфата (гл. 16); электроциты обладают высокой активностью Па+-К+-АТФазы, которая может участвовать в выведении натрия после периода активности.
Слабую электрическую активность проявляют некоторые гим-нотиды и клюворылые пресноводные рыбы. Их электроциты генерируют спайки, причем у разных видов свойства двух поверхностей могут различаться, так что спайки бывают двух- и трехфазными. У Gymnotus электрически возбудимы обе поверхности
Электрические органы и электрорецепторы 291
электроцита, и каждая дает ответ в виде спайка (рис. 17-1, Г). Однако нижняя, неиннервированная сторона возбуждается первой, так что суммарный ответ является двухфазным. У Hypopomus спайк, возникающий на каудальной стороне, менее длителен, чем на ростральной. У Sternopygus и Eigenmannia на медленный или постоянный потенциал накладывается ряд ВПСП и спайков; голова рыбы оказывается эдектроотрицательной. Измерение сопротивления электроцитов показало обычное снижение сопротивления в период нарастания спайка, а затем увеличение сопротивления мембраны во время максимума потенциала действия (аномальное выпрямление)^ с последующим уменьшением (задержанное выпрямление) в период его спада [9].
У некоторых клюворылых рыб электроцит имеет вид стебля, который разветвляется и обильно иннервируется. Потенциал действия неиннервированной стороны обладает большей амплитудой и длительностью, чем иннервированной, так что преобладает вторая фаза спайка [38].
У сильноэлектрического сома Malapterurus, обитающего в реках африканского континента, один гигантский нейрон иннервирует несколько миллионов электроцитов на одной стороне тела. Иннервированная сторона каждого электроцита генерирует небольшой кратковременный спайк (рис. 17-1, Д); за ним следует большой спайк на неиннервированной поверхности, иногда с двумя максимумами. Таким образом, иннервированная сторона дает ВПСП и короткий спайк, который распространяется на неиннер-вированную сторону, где длительность спайка больше.
У другого пресноводного обитателя Африки — Gymnarchus — иннервированная сторона электроцита также генерирует спайк; неиннервированная сторона электрически невозбудима и обладает большей емкостью и сопротивлением. Ток от иннервированной стороны заряжает последовательно соединенные емкости неиннервированной мембраны (рис. 17-1, Е), которые затем пассивно разряжаются; максимальное значение разрядного тока у них достигается позже, чем максимальное значение тока от иннервированной стороны.
По меньшей мере у двух слабоэлектрических рыб миогенногс компонента нет, и электрические органы происходят из моторных нервов. У Sternarchus спинномозговые нервы образуют орган, расположенный вентрально по отношению к позвоночнику; множество нервных волокон направляется вперед, затем, делая петлю, идут назад и слепо заканчиваются. В средней части пути вперед и назад нервные волокна имеют увеличенный диаметр. Разряд у Sternarchus двухфазный (голова вначале положитель-на), он является результатом синхронизации спайков нервных волокон в обеих частях петли. Протяженность перехватов Ранвье в проксимальных и центральных участках обеих ветвей петли составляет 1 мкм; в дистальной части имеется 3—5 перехватов по 19*
292
Глава 17
50 мкм каждый, а мембрана в этом месте имеет отростки. На основе электрических данных предполагают, что участки с перехватами в 1 мкм генерируют спайки, а участки поверхности больших перехватов играют роль последовательных емкостей и обладают лишь электротонической проводимостью. Таким образом, начальная, положительная по отношению к голове, фаза генерируется .в (волокне, идущем вперед, а отрицательная — в волокне, идущем назад [41]. У Andontosternarchus добавочный орган под нижней челюстью является производной сенсорных нейронов [37].
Неиннервированная поверхность большинства электрощитов, как правило, складчата, и это увеличение площади мембраны обеспечивает большую ее проводимость или емкость. Все сильноэлектрические и некоторые слабоэлектрические рыбы генерируют непрерывный поток импульсов, создавая постоянное электрическое поле. Электроциты пресноводных рыб имеют мембраны, генерирующие спайки, а электроциты морских видов таковыми не обладают. Сопротивление мембран, в особенности на неиннерви-рованной стороне, невелико, что способствует распространению тока.
Электрорецепция
Все животные при асимметричной секреторной и нервно-мышечной активности генерируют низкочастотные электрические поля. Низкочастотные поля, генерируемые многими нервными системами, могут быть относительно сильными, и спайковая активность некоторых нейронов в этих полях изменяется в соответствии с изменениями напряженности поля (гл. 16). Органы чувств, способные обнаруживать малые электрические поля, известны только у рыб и обычно являются видоизменением боковой линии, если считать, что она включает эпидермальные рецепторы, как, например, ампулы Лоренцини. Электрорецепторы весьма разнообразны, однако их можно разбить на две основные группы [6, 8] (рис. 17-2). 1) Тонические рецепторы, для которых характерна спонтанная ритмическая импульсация, дают продолжительные ответы на стимуляцию низкочастотным или постоянным током. В каждом органе в ампуле у основания канала, который открывается наружу, находится по нескольку рецепторных клеток (рис. 17-3). 2) Фазные рецепторы — быстро адаптируемые, чувствительные к высокочастотной стимуляции, на постоянный ток дают быстро угасающие ответы; эти органы бугристы и имеют эпителиальные клетки, расположенные между рецепторами и наружной частью тела [39]. Часто у одной рыбы имеются электрорецепторы обоих типов. Они реагируют на изменения электрического поля и возбуждают нервные окончания у основания рецепторной клетки.
Электрические органы и электрорецепторы
293
Тонические электрор^ецепторы, например, такие, как в боковой линии у сома и ъ ампулах Лоренцини у пластиножаберных, имеются у многих видов рыб, не обладающих электрическими органами. Они обнаруживают потенциалы действия дыхательных и плавательных мышц. Частота их спонтанной спайковой активности может изменяться под действием слабых токов.
Б
Рис. 17-2. Схематическое изображение структуры и эквивалентные электрические схемы электрорецепторов пресноводных рыб [6].
А. Тонический ампулярный рецептор. Б. Фазный бугристый рецептор. гср — сопротивление внешней среды; г,,— сопротивление кожи; г — сопротивление внутренней части клетки. Электроды для стимуляции и отведения потенциалов расположены у входа в рецептор. Иннервация указана под рецепторными клетками.
На рис. 17-4 приведены ответы тонического рецептора у Gym-notus, измеренные in vitro. Рецептор разряжается спонтанно с частотой около 100 импульсов в 1 с; анодный ток у входа в канал увеличивает частоту, а по прекращении его пропускания наступает период молчания. Катодный ток (гиперполяризующий внутреннюю поверхность рецепторной клетки) уменьшает частоту потенциалов действия [7]. В клетках фазного рецептора возникает рецепторный потенциал, иногда осцилляторного типа, частота осцилляций которого может отличаться от частоты сенсорных спайков (в нерве). Осциллирующий рецепторный потенциал может генерироваться непрерывно или затухать по окончании стимула [7].
294
Глава 17
Возбуждение нервных волокон в большинстве случаев осуществляется в химических синапсах. На это указывает наличие в пресинаптических окончаниях множества пузырьков. Минимальный период задержки при стимуляции рецептора составляет
Рис. 17-3. Микроскопический срез тонического электрорецептора Eigenmannia (гимнотиды) [39].
Полость рецептора (1) открыта наружу. На рецепторных клетках (2) показаны нервные терминальные участки в виде темных пятен у основания клетки.
1,6 мс; ответ нерва продолжается в течение некоторого времени после окончания стимуляции. Природа медиатора неизвестна, у некоторых гимнотид его роль выполняет, по-видимому, глутамат.
У клюворылых размеры электрорецепторов меняются в широких пределах. В рецепторах больших размеров имеется несколько клеток (от 1 до 8), которые могут быть иннервированы одним
Электрические органы и электрорецепторы
295
нервным волокном. Они имеют короткий период задержки (0 2 мс), поэтому передача вполне может быть электрической [20, 21].
Ампулы Лоренцини у пластиножаберных представляют собою луковицеобразные рецепторы, особенно многочисленные на голове.
300 н
200
а) .lIllIlllUIJIlilLLmiL
uillillllllHIlllHI—Li
100
Частота разрядов е состоянии покоя
Напряжение стимуляции, мВ
Рис. 17-4. Реакции тонического рецептора Gymnoius [7].
На вставке верхняя запись представляет афферентные импульсы, нижняя — стимулирующий потенциал у входа в рецептор; а — спонтанный разряд; б — анодная стимуляция; в — катодная стимуляция. На диаграмме изображена зависимость средней частоты импульсов от напряжения стимуляции, справа положительного и слева отрицательного.
Разряжаются они спонтанно, чувствительны к температуре, тактильным раздражениям, СОз и содержанию солей, но их первичной функцией является, по-видимому, электрорецепция. Например, у Scyliorhinus и Raja в ответ на мышечные потенциалы камбалы, находящейся на расстоянии 5—16 см, могут быть получены условнорефлекторные изменения кровообращения. Судя по поведению акулы, она реагирует на столь малые продольные градиенты поля, как 0,02 мкВ-см-1 [17, 22]. Частота сокращений сердца Raja повышается, если в воде создано электрическое поле, составляющее всего лишь 0,01 мкВ-см-1; выключение других рецепторов показало, что эта реакция вызывается через ампулы Лорен-Цини [17]; порог для боковой линии составляет 3 мкВ-см-1. У ската Raja ампулы разряжаются спонтанно с частотой около 40 имп-с-1; если по студенистой трубке ампулы пропускать ток, ‘При катодном токе ответы увеличиваются, а при анодном уменьшаются [31]. Вычислено, что порог составляет 5-10-11 А, т. е. чувствительность ампулярного рецептора в 103—104 раз выше чувствительности боковой линии [31]. Изменение частоты серд
296
Глава 17
цебиений можно вызвать изменением напряженности внешнего поля на 0,01—0,02 мкВ-см-1 [22].
У интактной электрической рыбы (Gymnotus или Eigenman-nia} нервные волокна боковой линии в ответ на отдельный разряд электрического органа дают несколько спайков — до 200— 750 в 1 с в зависимости от особи. Если внешнее поле изменить, например, введением куска металла, то изменится и частота сенсорного разряда. Градация ответа осуществляется изменением числа спайков на электрический импульс или добавлением или выпадением нескольких спайков в серии [18, 19].
Сигналы, поступающие на вход электрорецепторов, преобразуются в импульсы волокон боковой линии электрических рыб несколькими способами. Некоторые из них посылают афферентные импульсы непрерывно, безотносительно к электрическому разряду. Другие являются несинхронными фазными единицами, которые сигнализируют о присутствии вблизи рыбы объектов различной электрической проводимости. Есть волокна, разряжающиеся синхронно, с частотой разряда электрического органа данной рыбы, но их сигнал либо изменяется по фазе по отношению к разряду, либо кодирует вероятность разряда, пропуск его или изменение частоты [15].
У электрического сома ямковые рецепторы в коже восприимчивы, судя по поведению рыбы, к постоянному электрическому полю в 0,3 мВ-см-1 или току в 0,005 мкА-мм-2 [35, 36]. Электрические рыбы способны различать направление и полярность поля, изменения его формы под действием проводников или непроводников, например проволочной сетки.
Устройство ампулы таково, что рецептор способен воспринимать небольшие электрические токи. Кожа клюворылых имеет сопротивление 50 кОм-см2 — почти в 100 раз выше, чем у серебряного карася; сопротивление кожи Gymnotus составляет 1—3 кОм-см2. Продольные стимулы наиболее эффективно действуют на рецепторы головы, менее эффективно — на хвостовые и минимально — на рецепторы средней части тела рыбы.
Изучение поведения животных свидетельствует о том, что многие из них способны реагировать на магнитные поля, близкие по величине к магнитному полю Земли (несколько меньше 1 Гс). Например, планария Dugesia, удаляясь от источника света, отклоняется от прямолинейного пути, сворачивая вправо, если ее путь ориентирован к северному или южному полюсу магнитного поля, индукция которого менее 5 Гс, и влево — когда путь ориентирован на восток или запад. В более сильном поле (10 Гс) ориентировка север — юг и восток — запад изменяется на противоположную. Траектория перемещения ильной улитки Nassarius, определяемая фототаксисом, в слабом магнитном поле отклоняется от прямолинейной; моллюск способен отличить северный полюс магнита от южного. Реакция на слабое магнитное поле
Электрические органы и электрорецепторы 297
обнаруживает периодичность, равную солнечным и лунным суткам и двум неделям.
у медоносных пчел, возвратившихся со взяткой из полета, наклон оси восьмерки, описываемой при виляющем танце на вертикальной поверхности сот, уменьшается, если магнитное поле Земли скомпенсировать, и увеличивается при увеличении магнитного поля [27]. У Drosophila при вылете из темной трубки изменяется реакция поворота при изменении магнитного поля [33]. У почтового голубя вырабатывается условнорефлекторное изменение пульса в магнитном поле в 0,8 Гс, ориентированном под углом 180° по отношению к полю Земли [34].
Существует немало разнообразных видов животных, чувствительных к слабым электростатическим полям. Механизм обнаружения животными слабых электромагнитных полей пока не выяснен. Известно, что порог чувствительности ампул Лоренцини у ската ниже амплитуды поля, вызываемого перемещением морской воды в магнитном поле Земли; следовательно, движущийся скат обнаруживает магнитные поля своей ампулярной системой [22]. Разумно предположить, что одним из механизмов обнаружения магнитного поля является индукция электрического поля в чувствительных к нему клетках.
Поведенческие реакции и нервная регуляция
Электрические органы могут выполнять две функции — оглушение жертвы и электролокацию. Большинство слабоэлектрических рыб обитают в мутной воде или активны ночью. Они посылают постоянные серии импульсов и обнаруживают небольшие изменения электрического поля окружающей среды; некоторые из них для более тщательного зондирования среды изменяют частоту посылаемых импульсов [28]. Сильноэлектрические рыбы дают мощные разряды, которые оглушают жертву или хищника. Некоторые рыбы, например Electrophorus, помимо основного органа, испускающего высоковольтные импульсы, имеют органы, непрерывно генерирующие низковольтные сигналы. Среди слабоэлектрических рыб Амазонки у «низкочастотных» видов электрические органы произошли из мышц, а органы «|Высокочастотных» видов преобразованы из нервов [37]. Sternarchella разряжается с частотой 1200 имп-с-1, a Eigenmannia— 320—750 ими-с-1.
Характер разряда различен у разных видов и меняется в зависимости от условий. У некоторых слабоэлектрических рыб, например у клюворылых, частота разрядов обычно составляет * 15 имп-с-1, но, когда они преследуют жертву, частота увеличивается до 130 имп*с~1 [28—30]. Частота локаций у Gymnarchus Достигает 250 имп-с-1. Звездочет Astroscopus погружается в донный песок и выставляет наружу только глаза; при приближении небольших рыб, перед тем как раскрыть рот или одновременно с
298
Глава 17
этим, он генерирует высокочастотный импульс, а через 100 мс — серию импульсов длительностью в несколько секунд [32].
У Torpedo электрический разряд начинается через 80 мс после первого движения ската по направлению к жертве, которую он
Пейсмекерные нейроны: доминирующий подчиненный
/] п Переключатель пподол-/1 / \ гоеатого мозга '
Переключатель спинного мозга
Синапсы;
Орган * 2мс
X Дендродендритный ч
уэлектрические
Аксосоматический
А Аксосоматический-химический
Рис. 17-5. Схема нейронного контроля электрических органов у некоторых, электрических рыб [6].
У Malapterurus, гимнотид и Astroscopus каждому разряду органов предшествует одиночный командный сигнал на каждом уровне, тогда как у клюворылых активность нейронов изменяется по сложности на разных уровнях.
обнаруживает с помощью механорецепторов; перед самым захватом добычи частота разрядов составляет 140—290 имп-с-1,. а затем разряды становятся нерегулярными и их частота уменьшается. Жертва может быть парализована с расстояния 15 см [3]..
Электрические органы и электрорецепторы 299
Контроль деятельности мощных электрических органов со стороны центральной нервной системы позволяет строго синхронизовать разряд тысяч электроцитов. Два органа Torpedo разряжаются в пределах нескольких десятых долей миллисекунды, а все электропластинки полутораметрового Electrophorus активируются за 1,5 мс. У слабоэлектрических рыб сходная картина наблюдается при разряде всего органа.
У электрического сома два нейрона в первом спинномозговом сегменте возбуждают электрические органы на соответствующих им сторонах тела; эти два командных нейрона электротонически сопряжены, что обеспечивает синхронность их активности. У гим-нотид группа пейсмекерных нейронов в дорсальной части продолговатого мозга активирует вентральные ядра переключения в продолговатом мозге, которые в свою очередь активируют нейроны •спинного мозга; электротоническое сопряжение осуществляется и на уровне продолговатого, и на уровне спинного мозга [5, 11] (рис. 17-5). Клювокрылые (Mormyridae) имеют симметрично расположенные пейсмекерные нейроны в среднем мозге, которые активируют ядра второго порядка в продолговатом мозге; последние в свою очередь управляют нейронами спинного мозга. Клетки переключения в продолговатом мозге возбуждаются двойными импульсами, а из клеток переключения спинного мозга могут исходить тройки импульсов [8, 11]. Тесную взаимосвязь между нейронами осуществляет обратная связь, обеспечиваемая электро-тоническим сопряжением между ними на всех трех уровнях.
Электроциты Astroscopus, у которого электрический орган является дериватом гладких мышц, иннервируются от глазодвигательных ядер. У электрического угря Electrophorus единственный командный центр в продолговатом мозге активирует как орган электролокации Сакса, так и основной орган электрического разряда; основной орган реагирует только на облегчающие, с небольшими интервалами моторные импульсы, в то время как орган Сакса возбуждается в ответ на одиночные импульсы. Синхронность у этой рыбы, имеющей удлиненную форму тела, обеспечивается за счет более длительной задержки в переключателях передней части, чем в переключателях задней части спинного мозга, а также за счет относительно более низкой проводимости в ростральном направлении, так что электроциты в передней и задней частях тела рыбы активируются почти одновременно.
иКак правило, небольшое число командных и переключающих нейронов активирует большее число нейронов спинного мозга, которые в свою очередь активируют еще большее количество электроцитов. Командные ядра находятся под контролем по типу обратной связи, а нейроны переключения импульсов часто сопряжены электротонически. Таким образом, высокая синхронизация электрических разрядов обеспечивается комбинацией электрического сопряжения, использованием небольшого числа пей-
300
Глава 17
смекерных нейронов в командном центре, взаимодействием по типу обратной связи, соответствующей скоростью проведения и синаптической задержкой.
Помехозащита
Слабоэлектрические рыбы реагируют на приложенное поле большей или меньшей частоты, чем частота генерируемых ими им-пульсрв, помехозащитной реакцией. Когда Eigenmannia раздражают повторяющимися стимулами, например переменным током различной частоты, происходит рефлекторное изменение частоты собственных разрядов электрического органа рыбы, но это имеет место только в том случае, если частота стимуляции отличается от нормальной частоты разрядов органа не более чем на 10 Гц. При отклонении частоты стимуляции в сторону увеличения частота разрядов у рыбы уменьшается, а в обратном случае — увеличивается, т. е. происходит как бы защита от путаницы при сенсорном восприятии. Чувствительность к полю, ориентированному вдоль тела рыбы, не превышает 70 мкВ-см-1 [40]. У Нуроро-mus при стимуляции в перерывах между разрядами органа порог чувствительности ниже, чем при совпадении ее с собственными разрядами [26].
Частота разрядов Eigenmannia увеличивается при уменьшении частоты раздражения и уменьшается при ее увеличении по сравнению с частотой разрядов рыбы; рыба лучше реагирует на колебание частоты, чем на стимуляцию фиксированной частотой. Разряды отдельных рыб незначительно отличаются по частоте, и одна рыба реагирует на другую таким образом, что разница между частотами их' разрядов увеличивается, т.е. каждая рыба как бы стремится сохранить свою собственную частоту [15, 16] (рис. 17-6).
Способ проведения сигналов в головной мозг к командным центрам изучен недостаточно, но были обнаружены проекции в латеральную слуховую долю. Интересно отметить, что у большинства электрических рыб мозжечок имеет необычно большие размеры. Сильноэлектрические рыбы защищены от своих собственных разрядов кожей с высоким сопротивлением, прослойками жировой изоляции вокруг головного мозга и толстыми миелиновыми оболочками на многочисленных нервных волокнах; несмотря на это, они, несомненно, ощущают свои разряды [8].
У некоторых видов порог чувствительности, определяемый по поведению, составляет около 1/100 электрофизиологического порога. Причина этого несоответствия неясна, для его объяснения ссылаются на такие факторы, как экстраполяция тангенциальных полей на боковые стороны и концы (головной и хвостовой) рыб, плохое состояние рыб в условиях регистрации, неуверенность в том, что регистрируются сигналы наиболее чувствитель
Электрические органы и электрорецепторы
301
ных рецепторов, облегчение в центральной нервной системе. После введения ряда поправок Беннет [4, 8] сделал вывод, что в действительности различие порога восприимчивости у рыб, определяемого тем и другим методами, может достигать величины только одного порядка. В любом случае рыбы, обладающие
Рис. 17-6. Схема строения нервных цепей, осуществляющих помехозащитную реакцию [16].
Пунктирные линии представляют вероятные, но еще не установленные связи.
электрорецепторами, в 105 раз более чувствительны к электрическим токам, чем рыбы, такими специализированными рецепторами не обладающие [4].
Заключение
В процессе эволюции рыб мышечные клетки видоизменялись, приобретая способность к генерации электрического тока; постепенно были утеряны контрактильные элементы, волокна становились уплощенными, а иннервированные и неиннервированные мембраны приобрели различные свойства. У некоторых рыб элек-трогенный «столб» соединен с нервными волокнами, а у других электрический орган образуется из самих моторных нервных волокон.
Часто электроциты имеют асимметричные потенциалы, так что электрические потенциалы двух поверхностей клетки складываются; множество клеток соединяется последовательно. Все клет
302
Глава 17
ки активируются холинергическими веществами и в высокой степени синхронизованы нервной системой.
Процесс генерации импульсов включает: генерацию постсинаптических потенциалов (ВПСП) на одной поверхности электрощита при низком сопротивлении противоположной поверхности с дальнейшим снижением ее сопротивления во время импульса; генерацию спайков только на иннервированной поверхности; генерацию спайков на каждой поверхности с проведением от одной поверхности к другой; двухступенчатое снижение сопротивления. Таким образом, в электроцитах осуществляется клеточный электрогенез многих известных типов.
У слабоэлектрических и некоторых неэлектрических рыб имеются электрорецепторы, которые являются модифицированными рецепторами боковой линии. Они способны обнаруживать ничтожные продольные поля, напряженность которых составляет всего 10~7—10~8 В-см-1. Рецепторные клетки передают сигналы окончаниям сенсорного нерва обычно через химический синапс, а сенсорное кодирование осуществляется главным образом с помощью изменения характера последовательности спонтанных импульсов. Таким образом, существовавшие ранее структуры — мышцы и рецепторы боковой линии — преобразовались в органы генерации и обнаружения электрических импульсов.
Электрические разряды слабоэлектрических рыб используются ими для локации в темноте или в мутной воде, тогда как сильноэлектрические рыбы своими разрядами парализуют добычу. Синхронизация разрядов очень большого количества клеток (до нескольких миллионов) осуществляется небольшим числом электротонически сопряженных нейронов центральной нервной системы.
СПИСОК ЛИТЕРАТУРЫ
1. Altamirano М. et al., J. Gen. Physiol. 38: 319—360, 1955. Direct and indirect excitation of electroplaques of Electrophorus.
2. Bauer R., Z. vergl. Physiol. 59: 371—402, 1968. Electric discharges in Ma-lapterurus.
3. Belbenoit P., Z. vergl. Physiol. 67: 205—216, 1970. Electric discharge in Torpedo.
4. Bennett M. V. L., Ann. N. Y. Acad. Sci. 94: 458—509, 1961; Ann. Rev. Physiol. 32: 471—528, 1970. Modes of operation of electric organs.
5. Bennett M. V. L., Ann. N. Y. Acad. Sci. 137: 509—539, 1966. Electrotonic junctions.
6. Bennett M. V. L., pp. 313—393. In: Lateral Line Detectors, edited by P. Cahn, Indiana University Press, 1967; also pp. 147—169 in: The Central Nervous System and Fish Behavior, edited by D. Ingle, University of Chicago, 1968. Electroreception and neural control.
7. Bennett M. V. L., pp. 73—128. In: Physiological and Biophysical Aspects of Nervous Integration, edited by F. D. Carlson. Prentice-Hall, Englewood Cliffs, N. J., 1968. Chemical and electrical synapses.
;8. Bennett M. V. L., pp. 347—491. In: Fish Physiology, Vol. 5, edited by W. S. Hoar and D. J. Randall. Academic Press, New York, 1971. Electric organs and electroreception.
Электрические органы и электрорецепторы 30?
9. Bennett М. V. L., Grundfest Н., J. Gen. Physiol. 42: 1067—1104, 1959; 44: 805—843, 1961. Electric organs of Gymnotus, N ar cine, and Astroscopus.
10. Bennett M. V. L. et al., J. Gen. Physiol. 44: 757—804, 1961. Electric organs of Torpedo.
11. Bennett M. V. L. et al., J. Neurophysiol. 30: 180—300, 1967. Central nervous control of electric organ discharge.
12. Brown F. A., Biol. Bull. 123: 264—281, 282—294, 1962. Responses of planarian Dugesia to very weak horizontal magnetic and electrostatic fields.
13. Brown F. A. et al., Biol. Bull. 127: 221—231, 1964. Rhythmic modifications of responses of mud snails to geomagnetic fields.
14. Brown F. A., Ann. N. Y. Acad. Sci. 188: 224—241, 1971. Orientational influences of electromagnetic fields.
15. Bullock T. H., Chichibu S., Proc. Nat. Acad. Sci. 54: 422—429, 1965. Analysis of sensory coding in electroreceptors of electric fish.
16. Bullock T. H. et al., J. Comp. Physiol. 77: 1—22, 23—48, 1972. Jamming avoidance response of high frequency electric fish.
17. Dijkgraaf S., Kalmijn A. J., Z. vergl. Physiol. 53: 187—194, 1966. Function of ampullae of Lorenzini.
18. Hagiwara S., Morita H., J. Neurophysiol. 26: 551—567, 1963. Coding of electro-receptive fibers.
19. Hagiwara S., Szabo T., Enger P. S., J. Neurophysiol. 28: 775—799, 1965. Properties of electroreceptors, Electrophorus and Sternarchus.
20. Harder W., Z. vergl. Physiol. 59: 272—318, 1968. Electroreception in mormy-rids.
21. Harder W. et al., Z. vergl. Physiol. 54: 89—108, 1967. Sensitivity of some weakly electric fish to electrical stimulation.
21a. Hopkins C. D., Science 176: 1035—1037, 1972. Sex differences in electric signaling in an electric fish.
22. Kalmijn A. J., J. Exp. Biol. 55: 371—383, 1971. Electric sense of sharks and rays.
23. Kellaway P., Bull. Hist. Med. 20: 112—137, 1946. Electric fish in history of electrobiology and medicine.
24. Keynes R. D., Martins-Ferreira H., J. Physiol. 119: 315—351, 1953. Membrane potentials in electroplaques of Electrophorus.
25. Keynes R. D. et al., pp. 102—112. In: Bioelectrogenesis, edited by C. Chagas and A. P. de Carvalho. Elsevier, Amsterdam, 1961. Electrophysiology of electric organ of Malapterurus.
26. Larimer J. L., MacDonald J. A., Amer. J. Physiol. 214: 1253—1261, 1968. Sensory feedback from electroreceptors to electropacemakers in gymnotids.
27. Lindauer M., Martin H., Z. vergl. Physiol. 60: 219—243, 1968. Gravity orientation of bees in earth’s magnetic field.
28. Lissman H. W., J. Exp. Biol. 35: 156—191, 1958. Evolution and function of electric organs.
29. Lissman H. W., Machin К. E., J. Exp. Biol. 35: 451—486, 1958. Electrolocation of objects by Gymnarhus.
30. Machin К. E., Lissman H. W., J. Exp. Biol. 37: 801—811, 1960. Electroreceptors in Gymnarchus.
31. Murray R. W., J. Physiol. 180: 592—606, 1965. Sensory function of ampullae of Lorenzini.
32. Pickens P. E., MacFarland W. N., Anim. Behav. 12: 362—367, 1964. Electric discharge and behavior in stargazer.
33. Picton H. D., Nature 211: 303—304, 1966. Orienting response of Drosophila to electric fields.
34. Reille A., J. Physiol. Paris 60: 85—92, 1968. Conditioning of homing pigeons to a magnetic field.
35. Roth A., Z. vergl. Physiol. 61: 196—202, 1968; 65: 368—388, 1969. Electroreception in catfish Ictalurus.
36. Roth A., Szabo T., Z. vergl. Physiol. 62: 395—410, 1969. Receptor function in tuberous organs of mormyrid Gnathonemus.
304
Глава 17
37. Steinbach А. В., Biol. Bull. 138: 200—210, 1970. Survey of electric fishes in Rio Negro, Brasil.
38. Szabo 7L pp. 20—23 in: Bioelectrogenesis, edited by C. Chagas and A. P. de Carvalho, Elsevier, Amsterdam, 1961; also pp. 295—312 in: Lateral Line Detectors, edited by P. Cahn, Indiana University Press, 1967. Peripheral and central components in electroreception.
39. Szamier R. B„ Wachtel A. W., J. Ultrastruc. Res. 30: 450—471, 1970. UltrastrU’ cture of electroreceptor organs.
40. Watanabe A., Takeda K., J. Exp. Biol. 40: 57—66, 1963. Change in discharge frequency by ac stimulation in weakly electric fish.
41. Waxman S. G., Pappas G. D., Bennett M. V. L., J. Cell Biol. 53: 210—224, 1972. Differentiation of nodes along fibers of neurogenic electric organ, Ster-narchus.
Глава 18
\
\ \
Амебоидное движение
Дж. АНДЕРСОН
\
Подвижность в биологических системах не ограничивается мышечным сокращением. С тех пор как был изобретен микроскоп, каждое последующее поколение биологов наблюдало движение цитоплазмы, скольжение клеток, образование псевдоподий, токи протоплазмы, движение хромосом при клеточном делении, биения ресничек и жгутиков и множество других немышечных видов движения. Важность таких движений понимали давно. Фагоцитарная функция лейкоцитов, миграция клеток нервного гребня в эмбриогенезе, строго упорядоченная последовательность событий митоза, удаление слизи из дыхательных путей и движение сперматозоидов— это лишь несколько примеров из множества важнейших физиологических процессов, зависящих от немышечных форм двйжения.
Движение протоплазмы наблюдается во многих живых тетках, а может быть и во всех. Диффузия представляет собой сл. сом медленный процесс, чтобы обеспечить доставку веществ из ( ной части клетки в другую, тогда как активное движение цитопл; мы служит средством быстрого транспорта. Движение может п исходить по фиксированному пути и быть достаточно быстрых гак что его легко непосредственно наблюдать под микроскоп' : таково, например, круговое движение протоплазмы (циклоз) округ вакуолей в некоторых растительных клетках или тран орт пищеварительных вакуолей и гранул у ресничных инфу: >ий. В других клетках течение может быть медленным и состогг :ак бы в перемешивании протоплазмы, без образования посте' ?ых токов; такое движение цитоплазмы лучше всего выявляется * это-дом цейтраферной съемки, например в культуре фибробластов или в кончиках растущих нервных волокон. Хотя влияние гчда внешних факторов на движение протоплазмы изучалось довольно подробно, точный молекулярный механизм явления остается невыясненным.
Типы амебоидных клеток и псевдоподий
Амебоидное движение имеет много общего с токами протоплазмы, так что выяснение закономерностей процесса превращения химической энергии в механическую работу в одном случае 20—2067
306 Глава 18 /
должно облегчить понимание, этого процесса в другом. Амебоидное движение сопровождается изменением формы клетки,/появлением псевдоподий, а часто поступательным движение^. Оно может носить характер направленного перемещения, как у корненожек и плазмодия миксомицетов, а также у подвижных лейкоцитов, амебоцитов или блуждающих клеток многих жйвотных; к амебоидному движению относятся также вытягивание/ втягивание и изменение формы отростков (псевдоподий), связайные преимущественно с питанием, как у большинства фораминифер, солнечников и радиолярий, макрофагов позвоночных и ретикулоэндотелиальных клеток. Локомоторное амебоидное движение возможно лишь при наличии сцепления с каким-либо субстратом; ненаправленное амебоидное движение присуще свободным псевдоподиям.
У свободноживущих амебоподобных животных способ перемещения несколько различается в зависимости от формы клетки и типа псевдоподий [82]. Псевдоподии могут быть широкими или цилиндрическими, округлыми на конце (лобоподии); они могут быть нитевидными (филоподии), тонкими и заостренными; могут быть сетчатыми (ретикулоподии), разветвляющимися и анастомозирующими, как у фораминифер; наконец, это могут быть аксо-подии — с центральным стержнем, как у солнечников и радиолярий (рис. 18-1). Перемещение лобоподий в значительной мере изучено; в филоподиях, ретикулоподиях и аксоподиях наблюдаются токи протоплазмы, не сопровождающиеся изменениями длины; для аксоподий характерна сократимость.
Амебы сильно различаются по форме, скорости течения цитоплазмы и передвижения клетки в целом [25]. Amoeba Umax образует одну лопастевидную псевдоподию; у амеб звездчатой формы много свободных псевдоподий. A. proteas и A. dubia имеют также по многу псевдоподий; некоторые из них прикрепляются к субстрату, причем подвижность протоплазмы в них различна. У некоторых амеб бывают поверхностные складки, например у A. verrucosa, у некоторых — бесформенный «хвост», или уроид, как у Pelomyxa palustris (рис. 18-1). Разнообразна и форма лимфоцитов: для одних характерна широкая, вытянутая псевдоподия и небольшой «хвост», для других — червеобразная форма. Отдельные амебоидные клетки могут принимать различную форму в разных условиях. Например, Amoeba proteus в дистиллированной воде становится звездчатой, лучистой, а в разбавленном солевом растворе у нее лишь одна псевдоподия. Перед делением она также становится звездчатой. Амеба состоит из центральной жидкой эндоплазмы (плазмазоль) и наружной вязкой эктоплазмы (плаз-магель). Отношение массы эктоплазмы к массе эндоплазмы велико у A, proteus, A. verrucosa, Chaos carolinensis и Pelomyxa palustris, но мало у A. dubia и у паразитической A. blattae.
Рис. 18-/1. Схематические рисунки амеб с различными типами псевдоподий.
A. Amoeba proteus; Б. A. discoides; В. A. dubia; Г. А. Птах-, Д. A. verrucosa; Е. Pelomyxa palustris; Ж. Aciinosphaerium eichhorni.
* — ядро, 2 — пищеварительная вакуоль, 3 — сократительная вакуоль, 4 — плазмалемма, 5 — плазмазоль, 6 — гиалиновый слой, 7 — уроид, 8 — псевдоподия.
20*
308
Глава 18
Наиболее подвижные и сложные псевдоподии образуют/фора-миниферы и радиолярии. Нитевидные псевдоподии часто разветвляются и образуют сетки [5, 6, 67]. Течение протоплазмы в этих нитчатых псевдоподиях происходит в двух направлениях — наружу на одной стороне и внутрь, на противоположной; оно так сильно отличается от движения протоплазмы у обычных амеб, что была предложена новая классификация саркодовых, основанная на различиях в движении протоплазмы [65].
Общая картина амебоидного движения
«Типичная» амеба имеет наружный слой — плазмалемму; этот слой липкий, не смачивается водой и легко скользит по прилегающему к нему внутреннему слою. С помощью электронного микроскопа установлено, что плазмалемма состоит из наружной волокнистой оболочки и внутренней мембраны [100]. Волокна имеют диаметр около 80 А и длину 0,1—1 мкм. Оболочка содержит 35% липидов, 26% белка и 16% полисахаридов [98]. Под плазмалеммой лежит гиалиновый слой, который, судя по броуновскому движению попавших в него частиц, жидкий. В точках прикрепления амебы к субстрату этот слой очень тонкий, но в передней части вытягивающейся псевдоподии он часто утолщается, образуя широкий гиалиновый колпачок. Далее следует эктоплазма, или цилиндр из плазмагеля, довольно вязкий. У амеб с одной псевдоподией эктоплазма постепенно утолщается от переднего конца к заднему. У многих видов она вытянута в виде тонкого слоя плазматического геля под передним гиалиновым колпачком, слоя, который часто прорывается и позволяет гранулам проникать в гиалиновый колпачок. Частота прорывов в разных вытягивающихся псевдоподиях может быть различной. Эндоплазматической основой амебы является плазмазоль со свободно плавающими в нем гранулами. При реакциях на нейтральные полисахариды он окрашивается более интенсивно, чем эктоплазма [112]. Ядро обычно находится в плазмазоле; у некоторых видов оно занимает центральное положение, несмотря на интенсивные токи протоплазмы вокруг него [45]. Эндоплазма и эктоплазма содержат разнообразные гранулы, пищеварительные вакуоли и кристаллические включения различного типа. Протоплазма гиперосмотична у пресноводных амеб и примерно изоос-мотична у паразитических и морских амеб. Эктоплазма обладает тургесцентностью, поэтому клетки обычно не имеют сферической формы.
Судя по данным микроскопии, перемещение амебы зависит от трех основных факторов.
1. Прикрепление к субстрату, которое облегчается, если среда содержит следы солей, в частности солей кальция; действие Са2+, Mg2+ и К+ аддитивно в отношении прикрепления, но не переме
Амебоидное движение
309
щения. Наиболее прочно прикрепляются к субстрату кончики псевдоподий; новые псевдоподии прикрепляются лучше, чем старые.
2. На заднем конце клетки или в какой-либо другой определенной области эктоплазма непрерывно превращается в эндоплазму, а в передней части клетки или в любой вытягивающейся псевдоподии эндоплазма превращается в эктоплазму. Во время движения эндоплазмы вперед гранулы либо отклоняются в сторону и превращаются в эктоплазму, либо внедряются в гиалиновый колпачок и желатинизируются по мере образования нового колпачка.
3. Движение эндоплазмы вперед происходит под действием эластических сил, давления и сократительной силы.
Имеется несколько теорий относительно природы сил в амебоидном движении, но большинство из них предполагает наличие сократительных белков, обеспечивающих процессы «сдавливания» и «растягивания». Наблюдая сбоку за гранулами амеб и лимфоцитов и за частицами, прилипшими к плазмалемме, можно заметить, что в перемещающихся клетках гранулы в эктоплазме остаются фиксированными до тех пор, пока они не окажутся у заднего конца тела, где переходят в эндоплазму. Частицы на участке прикрепленной к субстрату плазмалеммы также неподвижны, но частицы на других участках плазмалеммы, дорсальных и вентральных, движутся вперед. У A. verrucosa описано «вращательное» движение плазмалеммы [69, .87]; у других видов, например у Difflugia, можно наблюдать «шагающее» движение: вытягивающийся кончик прикрепляется к субстрату, а затем подтягивается остальная часть. В ретикулоподиях фораминифер вместо наружной оболочки эктоплазмы имеется что-то вроде тяжей геля; по одному из них гранулы движутся вперед, а по другому — назад [67].
Скорости движения весьма различны — от едва заметной до 1350 мкм/с в потоке цитоплазмы у плазмодия бесклеточного слизистого гриба Physarum polycephalum [75]. Имеются наблюдения о перемещении мигрирующих плазмодиев со скоростью 5—6 см/ч .[*15]. Свободно ползающие амебы движутся со скоростью от 0,5 до 4,5 мкм/с, большая часть из них — со скоростью около 1 мкм/с. Особи с одной псевдоподией перемещаются быстрее .4,6 мкм/с), чем имеющие много псевдоподий (2,1 мкм/с) 101—103]. Лимфоциты в культуре перемещаются со средней скоростью 0,55 мкм/с, а неполяризованные макрофаги — со скоростью 0,005 мкм/с или меньше [35—37].
В процессе питания амебы, которые перемещаются с помощью небольших псевдоподий (например, A. proteus), образуют подобие чашечек, состоящих из латеральных и дорсальных псевдоподий, обволакивающих пищевые частицы, которые и сами могут быть подвижными. Псевдоподии чашечки смыкаются, и пищевая ч.а-
310
Глава 18
стица оказывается заключенной в пищеварительную вакуоль. У амеб с одной широкой псевдоподией, как у A. Umax, пища, по-видимому, прилипает к плазмалемме. Ретикулоподии и аксоподии также имеют липкую поверхность, и пищевые частицы прилипают к ней [77].
Эндоплазма течет как бы по каналам между островками эктоплазмы. Отдельные гранулы миксомицета Physarum polycepha-lum, находящиеся друг от друга лишь в нескольких микронах, могут перемещаться в противоположных направлениях; поток, образующийся в данной точке, время от времени меняет направление; строгой периодичности при этом не наблюдается [115].
Теории амебоидного движения
В сократимых системах, таких, например, как поперечнополосатые мышцы, реснички и жгутики, движение связано с определенной структурой. Такую связь трудно установить уже для клеток гладких мышц и тем более для амебоидных клеток. Отсутствие структурных критериев не позволяет решить, существует ли обособленная механо-химическая система, обеспечивающая амебоидное движение [129].
Для объяснения различных проявлений амебоидного движения предложен ряд теорий. Эти теории не исключают друг друга, а иногда даже дополняют одна другую. Очень мало вероятно, что какая-либо из этих теорий применима ко всем видам амеб ,[6], но существует предположение, что у всех теорий, несмотря на их кажущееся различие, имеется общая основа, возможно на макромолекулярном уровне [41].
В середине XIX в. цитологические исследования выявили сетчатую структуру цитоплазмы [124]. Неудивительно, что в самых ранних теориях амебоидного движения предполагалось наличие сократительных тяжей в различных участках клетки, в частности в вытягивающихся псевдоподиях, которые тянут массу протоплазмы вперед [38]. Эта теория в видоизмененной форме возродилась в 60-х годах [4]. В связи с успехами физики и химии в изучении поверхностного натяжения появились теории, рассматривавшие изменения поверхностного натяжения как источник силы для амебоидного движения. От этих теорий в общем пришлось отказаться [38], хотя поверхностные силы, вероятно, в какой-то степени определяют форму клетки. Изучение коллоидов и их свойств в более позднее время привело к «гель — золь»-концепции амебоидного движения; широкое признание получили модели, основанные на предполагаемой сократимости эктоплазмы (геля) [25].
Опубликованы критические обзоры с анализом разнообразных гипотез [3, 38, 66, 75, 79, 129]. Для тех организмов, которые обладают хорошо выраженными эктоплазмой и эндоплазмой, все тео
Амебоидное движение
311
рии могут быть сведены к двум основным моделям, включающим в себя часть главных положений многих предложенных гипотез и моделей.
1. Гидравлическая модель. Пассивное или активное сокращение эктоплазмы и (или) активно сдвиговые силы, возникающие в эктоплазме либо на границе раздела эктоплазмы с эндоплазмой, либо между микрофибриллами, создают давление на более жидкую эндоплазму, вызывая ее движение. Эктоплазма превращается в золь иногда лишь в определенных участках, образуя эндоплазму, которая затем течет от участков превращения в золь и желатинизируется в других участках, вновь превращаясь в эктоплазму (см. [42, 43, 46, 48, 63—66, 68, 80, 86, 87, 92, 99, 107, 108, 114, 117]).
2. Модель фронтального сокращения. Сократительная сила возникает в передней части псевдоподии как результат вязкости и эластичности эндоплазмы и достаточна для ее подтягивания вперед. У кончика псевдоподии эндоплазма, сокращаясь, превращается в эктоплазму. Считается, что самая передняя часть эктоплазматической оболочки представляет собой только что сокращенную цитоплазму [2—4]. Если крупную Chaos carolinetisis поместить в тонкую кварцевую трубочку и «переломить» пополам под слоем масла, то в изолированных ее фрагментах можно наблюдать токи — вначале осевая эндоплазма течет вперед, а эктоплазма назад, затем масса делится на несколько каналов и возникают U-образные изгибы у одного конца. Скорость потока по направлению к изгибу выше, а канал более узок, чем после изгиба в геле. В нормально вытягивающейся псевдоподии цилиндр эндоплазмы в зоне «фонтана» укорачивается и утолщается. Полагают, что тяжи геля, прикрепленные к краю трубки, сокращаются и тянут переднюю часть золя вперед; вода, выделяемая под гиалиновым колпачком при синерезисе, идет назад подплаз-малеммой, возвращаясь в заднюю зону восстановления эндоплазмы [7—9]. Течение происходит в основном в пограничной зоне или в зоне скольжения между эндоплазмой и эктоплазмой. Считается, что сокращение у U-образного изгиба между эндоплазмой и эктоплазмой может одинаково хорошо объяснить амебоидное движение и циклоз. Эта модель получила подтверждение после обнаружения упругого эффекта вблизи кончиков псевдоподий [10]. Дальнейшие данные в пользу гипотезы фронтального сокращения и против гидравлической модели образования псевдоподий У Chaos carolinetisis были получены в опытах, в которых одна псевдоподия всасывалась в капилляр; при этом уменьшение давления на заключенную в капилляр шсевдоподию до 30—35 см вод. ст. не предотвращало вытягивания других псевдоподий при нормальном атмосферном давлении [11].
Получены многочисленные данные, свидетельствующие о способности эктоплазмы к сокращению. Ее толщина увеличивается
312
Глава 18
в направлении назад [88]; с помощью электронной микроскопии обнаружены нити в участках, в которых следует ожидать развития напряжения (см. [111, 127]). Эластичностью и сократимостью можно объяснить течение в двух противоположных направлениях от сокращенного участка эктоплазмы в область меньшей эластичности [63, 92, 114]. Если компонентом аппарата сокращения служат нити, то возможно, что этот аппарат функционально представляет собой разновидность механизма скольжения нитей, лежащего в основе мышечного сокращения (см. гл. 16). Было высказано предположение о скольжении нитей в эктоплазме [97] и в тонких сетчатых псевдоподиях фораминиферы Allogromia [67]. Эти ретикулоподии толщиной 5 мкм лишены центральной эндоплазмы и наружного гиалинового слоя; они, по-видимому, состоят из двух полукруглых нитей геля, одна из которых движется вперед, а другая — назад; при этом одна нить геля скользит по другой. Предполагают также наличие многочисленных сократительных участков вдоль нити [5].
Несколько иную роль приписывают эктоплазме на основании ее коллоидных свойств. При повышении гидростатического давления и при понижении температуры вязкость эктоплазмы амебы уменьшается. Точно так же ведет себя актомиозин. Гели такого рода при желатинизации увеличиваются в объеме (+AV) и поглощают тепло [80, 86]. На плазмодиях миксомицетов установлено, что в области, где эктоплазма переходит в эндоплазму, температура выше, чем в участках, в которые эндоплазма втекает [12].
Если живую амебу подвергнуть высокому гидростатическому давлению, ее вязкость уменьшается, эктоплазма превращается в золь, и перемещение прекращается. При давлении 15 атм псевдоподии удлиняются, приобретают цилиндрическую форму; при 45 атм новые псевдоподии не образуются; приблизительно при 50 атм на концах псевдоподий появляются сферические образования и вся клетка превращается в жидкий шар. При высоком давлении, когда жесткость протоплазмы снижается, прекращается ряд функций: амебоидное движение, расширение хроматофоров, циклоз у Elodea, дробление яиц Arbacia [30, 53, 81, 85]. Вместе с тем другие функции — биолюминесценция, биение ресничек, мышечное сокращение и нервное проведение — при повышении давления усиливаются, вероятно, в связи с мембранными эффектами [29]. Критическое давление, при котором псевдоподии исчезают, а клетка принимает сферическую форму, при низких температурах меньше, чем при высоких; прочность геля, измеряемая методом центрифугирования в условиях высокого давления, уменьшается быстрее при 15 °C, чем при 25° [85]. Если к среде добавить АТФ, давление, необходимое для придания амебе сферической формы, оказывается более высоким; это согласуется с началь
__________________Амебоидное движение___________________313
ным повышением вязкости белков геля при его обработке АТФ и заметным начальным сокращением живой амебы при инъекции АТФ [46].
Сократительные белки
В 1952 г. Леви впервые показал, что немышечная сократимая система содержит сократительные белки. Из плазмодия бесклеточного миксомицета Physarum polycephalum он экстрагировал белок, который обладал многими свойствами актомиозина: растворялся в 1 М КС1 при pH 8,2 с образованием вязкого раствора, причем вязкость эта уменьшалась при добавлении АТФ; по-видимому, увеличение вязкости сопровождалось дефосфорилированием АТФ с образованием АМФ [83]. Этот белок, названный миксо-миозином, был в дальнейшем очищен, а Тс’о с сотрудниками определил его физические свойства. Мол. вес этого белка около 6-106, молекулы имеют форму стержня длиной 4000—5000 А и диаметром 70 А [120—122]. В пользу существования сократимой системы свидетельствовали данные о том, что АТФ ускоряет течение протоплазмы миксомицетов [75] и вызывает токи в изолированной цитоплазме Amoeba proteus [131]. Несколько позже Накаяма [95] показал, что очищенный миксомиозин, выделенный из миксомицета, обладает рядом свойств мышечного миозина В. Оказалось, что миозин В из плазмодия обладает такой же чувствительностью к АТФ, как и миозин В из мышц кролика; ионы Mg2+ увеличивают скорость его преципитации, а ионы Са2+ ее снижают [54, 58].
Актиноподобный белок, выделенный из миксомицета, при добавлении к миозину В из мышц кролика образует актино-миози-новый комплекс. И наоборот, миозин В из миксомицета реагирует с актином млекопитающих [56, 57, 59]. Актин был выделен также из яиц морского ежа [55] и из амеб .['123]. Между актином, выделенным из этих объектов, и актином, выделенным из мышц, много общего: 1) в обоих случаях на 1 моль белка приходится 1 моль 3-метилгистидина; 2) молекулярные веса этих белков очень близки; 3) оба осаждаются миозином в растворах небольшой ионной силы; 4) увеличение вязкости после добавления миозина при высокой ионной силе обратимо под действием АТФ; 5) оба способны к обратимому G-F-превращению [59, 123]. Сходство этих актинов позволяет предположить наличие общей эволюционной основы подвижности. Различные формы движения клеток связаны с самой разнообразной структурной организацией — от макроскопического до электронно-микроскопического уровня. Однако сходство сократительных белков предполагает существование единого принципа превращения химической энергии в механическую [41].
314
Глава 18
Фибриллы, микрофиламенты и микротрубочки
Вольфарт-Боттерман с сотрудниками (см. [111, 125—127]) первым описал у Amoeba proteus и миксомицета Physarum polyce-phalum микрофиламенты диаметром около 50 А. Их количество оказалось небольшим по сравнению с общим количеством основного вещества. Они были найдены в областях, где можно было предполагать развитие напряжений, а именно вокруг вакуолей, в псевдоподиях и эктоплазме каналов миксомицетов. Этим структурам приписали сократительную функцию, но небольшое их число и отсутствие прикрепления к мембранам породили сомнения относительно их роли в движении цитоплазмы.
Наличие микрофиламентов было подтверждено рядом работ [89, 90, 94]. Кроме их возможного участия в движении цитоплазмы, предполагалось, что они удерживают при движении органеллы [60] и что они связаны с сокращением вакуолей [106].
Вольперт и сотр. [130, 131] впервые продемонстрировали движение и сокращение изолированной цитоплазмы Amoeba proteus при добавлении АТФ в присутствии ЭДТА. При электронной микроскопии сокращенного геля обнаружено множество филаментов диаметром 70 А. Сообщалось о двух фазах сокращения и перемещения в таких препаратах [104]. В первой фазе происходит увеличение вязкости и появляются нити диаметром 50—70 А, затем нити диаметром 160 А, из которых создаются волокноподобные структуры диаметром 0,1—1,0 мкм. Сообщалось, что описанные нити связывают тяжелый меромиозин [105]. Образование этих ’нитей подавляется детергентом салирганом [1], но не колхицином [93]. Микрофиламенты найдены в глицеринизирован-ных препаратах амеб и миксомицетов; добавление АТФ сопровождается их сокращением [61, 78]. Актомиозиновые нити, полученные из миксомицетов и амеб, укорачиваются и утолщаются под действием АТФ [16—18].
Микротрубочки и микрофиламенты обнаружены в изобилии в аксоподиях солнечников [109], но не найдены в плазмодии миксомицетов и у больших амеб в нормальных условиях. Сообщалось о наличии таких структур у Amoeba proteus, голодавших в течение 16 дней [33], и у небольшой амебы Нartmannella sp. [60]. Микротрубочки необратимо разрушаются под действием ионов меди и никеля [ПО] и обратимо — при обработке разбавленной мочевиной [113]. В культуре макрофагов, обработанной колхицином или винбластином, скользящая форма перемещения заменяется амебоидным движением. В обработанных клетках не найдено микротрубочек, но имеются микрофиламенты [21]. Обработка колхицином изменяет форму фибробластов — они становятся эпителиоидными; при этом перемещение клеток замедляется [47].
Возникает вопрос, одинаковы ли химический состав и структура субъединиц микрофиламентов и микротрубочек. Колхицин
Амебоидное движение
315
мик-
и в аксоподиях [5], свидетельствует об макромолекулярных структур. Обнаружение мик-микротрубочек свидетельствует в пользу этого Полагают, что нити и микротрубочки играют перемещении — участвуя либо в процессах, сход-
разрушает микротрубочки и замедляет образование новых 47], но не препятствует образованию пучков микрофиламентов [93]. Микрофиламенты и микротрубочки из аксона кальмара различаются по молекулярному весу, аминокислотному составу, иммунологическим реакциям и электрофоретической подвижности [34, 62].
Двойное лучепреломление, наблюдаемое у амеб [10], сомицетов [96] упорядоченности рофиламентов и предположения, какую-то роль в ных с известными по модели скользящих нитей при мышечном сокращении [Ю7], либо в развитии сдвиговых сил [И7]. Данные о наличии у миксомицетов пузырьков, накапливающих Са2+, подтверждают аналогию с мышечным сокращением [26]. Однако пока отсутствуют экспериментальные данные, которые позволили бы объяснить быстрые, иногда циклические изменения двойного лучепреломления, наблюдаемые у амеб и миксомицетов, или периодичность челночных токов в плазмодии миксомицетов.
Роль ядра
Для исследования ядерно-цитоплазматического взаимодействия были поставлены многочисленные опыты на Amoeba proteus [27, 28]. Ядро может быть пересажено от одной особи к другой; ядро и практически всю цитоплазму можно удалить из одной амебы, а затем ввести в нее цитоплазму из другой амебы, а ядро — из третьей. Такая «комбинированная» амеба оказывается жизнеспособной [73]. Лишь при удалении ядра выяснилась его роль в амебоидном движении. У лишенной ядра амебы вскоре прекращается упорядоченный направленный ток цитоплазмы, вызывающий движение. Циклоз сохраняется, псевдоподии же либо не образуются, либо приобретают аномальную форму. Амеба утрачивает способность прикрепляться к субстрату и питаться. Введение ядра быстро восстанавливает нормальную деятельность амебы — происходит реактивация. Ядро необходимо для образования и сохранения сократительных вакуолей и телец Гольджи [39, 40]. Ядра, взятые из клеток одного штамма и пересаженные в цитоплазму другого, создают новый штамм со свойствами, отличными от свойств любого из исходных штаммов [84]. Ядра, выдержанные в солевом растворе в течение 20 мин, теряют свою генетическую функцию, но сохраняют способность реактивировать клетку [70]. Ясно, по-видимому, что движение протоплазмы у Physarum polycephalum зависит от наличия неповрежденного ядра в интерфазе, так как в период деления ядра быстрые челночные токи прекращаются [15, 50].
316
Глава 18
Ядерно-цитоплазматические взаимодействия, которые обеспечивают контроль и регуляцию тока цитоплазмы и амебоидного движения, не выяснены, но открытие синтеза актиноподобного белка в ядрах миксомицетов должно стимулировать исследования, которые позволят выяснить роль ядра в амебоидном движении [74].
Роль мембраны
Очевидно, что при изменении формы амебоидной клетки от сферической до обычной с псевдоподиями общая площадь поверхности сильно увеличивается; при втягивании псевдоподий площадь снова уменьшается. Теории механизма этих изменений распадаются на'три группы: 1) мембрана представляет собой жидкость или пластичную поверхность, которая свободно скользит по эктоплазме; 2) новая мембрана быстро образуется при необходимости и резорбируется при уменьшении площади поверхности; 3) общее количество вещества мембраны относительно неизменно, но она складчата и может расправляться.
Несколько работ по изучению движения прикрепленных частиц на движущейся амебе подтверждают концепцию жидких или гибких мембран. Частицы движутся вперед мимо фиксированной точки и по направлению к кончику псевдоподии [49] или могут огибать его и течь назад, создавая впечатление движения плаз-малеммы [69, 87].
Имеются наблюдения, что частицы, прикрепленные к задней части клетки, перемещаются назад и что на заднем конце мембрана сильно сморщена. Это позволило предположить, что на заднем конце происходит всасывание мембраны и что клеточная поверхность обновляется всякий раз по прохождении клеткой расстояния, равного собственной длине [44]. Сморщивание мембраны около уроида Amoeba proteus происходит циклически. Во время быстрого образования псевдоподий сморщивание прекращается, а затем по окончании движения вперед усиливается [128].
Вольперт и О’Нейл [129—131] пометили поверхность Amoeba proteus флуоресцирующими антителами и обнаружили, что период полусуществования метки составляет около 5 ч, что указывает на скорость обновления 0,2% в 1 мин. Эта величина в 100 раз меньше скорости, которой следовало бы ожидать, если бы мембрана полностью обновлялась при перемещении клетки на расстояние, равное ее длине. Однако указанные антитела были связаны лишь с поверхностной оболочкой, и если этот слой механически удерживается сзади во время вытягивания псевдоподий, мембрана, покрывающая новую псевдоподию, оказывалась немеченой [72], так что вопрос о скорости обновления мембран остается открытым.
Амебоидное движение
317
Некоторые электронно-микроскопические и кинематографические исследования поддерживают концепцию образования складок и расправления мембран и свидетельствуют о том, что площадь мембраны остается относительно постоянной [32, 51, 116].
Хотя не существует данных о том, что в мембране возникают силы, достаточные для амебоидного движения, ее механические и физиологические свойства могут оказывать влияние на форму клетки и на движение. В точке прикрепления расстояние между плазмалеммой и эктоплазмой меньше, чем в других участках [20]. Высказывалось предположение, что контакт между плазмалеммой и эктоплазмой у A. proteus индуцирует сокращение [44, 45].
Амебоидные клетки реагируют на электрическое поле и создают биопотенциалы. В электрическом поле у A. proteus обнаруживается образование золя на стороне, обращенной к катоду; в этом же направлении вытягиваются псевдоподии [52]. Миграция плазмодия Physarum по направлению к катоду объясняется торможением на стороне анода, при этом содержание К+ в задней части плазмодия на 15—20% меньше [13, 14].
У миксомицетов механический стимул вызывает электрическую реакцию длительностью в несколько секунд [118]; сообщалось, что после электрической стимуляции удается зарегистрировать потенциалы действия, но, так как они не распространяются, вряд ли правильно их так называть.
Миксомицет можно поместить в сосуд из двух отделений, соединенных каналом (по типу песочных часов). Избыточное гидростатическое давление в одном из отделений останавливает поток протоплазмы в этом направлении; величина уравновешивающего давления, необходимого для остановки потока, изменяется периодически, в пределах 12,5—25 см вод. ст. [75]. Разность потенциалов между двумя отделениями камеры также изменяется периодически, но эти изменения отстают от изменений давления. Следовательно, движение протоплазмы не связано с изменением разности потенциалов.
У амеб и миксомицетов регистрируются трансмембранные потенциалы в 40—70 мВ [22—,24, 91, 119]. У амеб кончик вытягивающейся псевдоподии заряжен отрицательно по отношению к внутренней и положительно по отношению к наружной поверхности заднего конца [24]; химические агенты, которые вызывают образование псевдоподий, снижают трансмембранный потенциал [19, 71]. Регистрация изменений потенциалов у амеб [119] и миксомицетов [91] выявила быстрые изменения в 6—10 мВ, которые не коррелируют с направлением движения. Предполагается, что они отражают изменения в мембранах, но их природа неизвестна. Поэтому, хотя плазмалемма обладает некоторыми электрическими свойствами, сходными со свойствами мембран других клеток, ее роль в амебоидном движении пока не выяснена.
318
Глава 18
СПИСОК ЛИТЕРАТУРЫ
1. Achterrath М., Cytobiologie 1: 159—168, 1969. The inhibition of formation of fibrillar pattern in slime mold plasmodia by Salyrgan.
2. Allen R. D., J. Biophys. Biochem. Cytol. 8: 379—397, 1960. The consistency of ameba cytoplasm and its bearing on the mechanism of ameboid movement.
3. Allen R. D., In: The Cell, Vol. II, edited by J. Brachet and A. E. Mirsky, Academic Press, New York, pp. 135—216, 1961. Ameboid movement.
4. Allen R. D., Exp. Cell Res., Suppl. 8: 17—31, 1961. A new theory of amoeboid movement and protoplasmic streaming.
5. Allen R. D., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 407—432, 1964. Cytoplasmic streaming and locomotion in marine Foraminifera.
6. Allen R. D., Symp. Soc. Exp. Biol. 22: 151—168, 1968. Differences of a fundamental nature among several types of amoeboid movement.
7. Allen R. D., Cooledge J. W., Hall P. J., Nature 187: 896—899, 1960. Streaming in cytoplasm dissociated from the giant amoeba, Chaos chaos.
8. Allen R. D., Cowden R. R., Hall P. J., J. Cell Biol. 12: 185—189, 1962. Syneresis in ameboid movement.
9. Allen R. D., Francis D. W., Symp. Soc. Exp. Biol. 19: 259—271, 1965. Cytoplasmic contraction and the distribution of water in the amoeba.
10. Allen R. D., Francis D. W., Nakajima H., Proc. Nat. Acad. Sci. 54: 1153—1161, 1965. Cyclic birefringence changes in pseudopods of Chaos carolinensis revealing the localization of the motive force in pseudopod extension.
11. Allen R. D., Francis D., Zeh R., Science 174: 1237—1240, 1971. Direct test of the positive pressure gradient theory of pseudopod extension and retraction in йтобЬй c
12. Allen R. D., Pitts W. R., Jr., Speir D., Brault J., Science 142: 1485—1487, 1963. Shuttle-streaming: synchronization with heat production in slime mold.
13. Anderson J. D., J. Gen. Physiol. 35: 1—16, 1951. Galvanotaxis of slime mold.
14. Anderson J. D., J. Gen. Physiol. 45: 567—574, 1962. Potassium loss during galvanotaxis of slime mold.
15. Anderson J. D., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press New York, pp. 125—136, 1964. Regional differences in ion concentration in migrating plasmodia.
16. Beck R., Hinssen H., Komnick H., Stockem W., Wohlfarth-Botterman К- E., Cytobiologie 2: 259—274, 1970. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. V. Contraction, ATPase-acti-vity and fine structure of actomyosinthreads from Physarum polycephalum.
17. Beck R., Komnick H., Stockem W., Wohlfarth-Botterman К- E., Cytobiologie 1: 99—114, 1969. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. IV. Comparative studies on actomyosinthreads and glycerinated cells.
18. Beck R., Komnick H., Stockem W., Wohlfarth-Botterman К- E., Cytobiologie 2: 413—428, 1970. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. VI. Comparative studies on actomyosinthreads isolated from obliquely striated and smooth muscles.
19. Bell L. G. E., J. Theoret. Biol. 1: 104—106, 1961. Surface extension as the mechanism of cellular movement and cell division.
20. Bell L. G. E„ Jeon K. W., Nature 198: 675—676, 1963. Locomotion of Amoeba proteus.
21. Bhisey A. N., Freed J. J., Exp. Cell Res. 64: 419—429, 1971. Ameboid movement induced in cultured macrophages by colchicine or vinblastine.
22. Bingley M. S„ J. Exp. Biol. 45: 251—267, 1966. Membrane potentials in Amoeba proteus.
23. Bingley M. S., Bell L. G. E., Jeon K. W., Exp. Cell Res. 28: 208—209, 1962. Pseudopod initiation and membrane depolarization in Amoeba proteus.
24. Bingley M. S., Thompson С. M., J. Theoret. Biol. 2: 16—32, 1962. Bioelectric potentials in relation to movement in amoebae.
Амебоидное движение
319
25. Bovee Е. С., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 189—219, 1964. Morphological differences among pseudopodia of various small amebae and their functional significance.
26. Braatz R., Komnick H., Cytobiologie 2: 457—463, 1970. Histochemical demonstration of a calcium accumulating system in myxomycete plasmodia.
27. Bracket J., Ann. N. Y. Acad. Sci. 78: 688—695. Cytoplasmic dependence in amoeba.
28. Bracket J., In: The Cell, Vol. II, edited by J. Brachet and A. E. Mirsky, Academic Press, New York, pp. 771—841, 1961. Nucleocytoplasmic interactions in unicellular organisms.
.29 . Brown D. E. S., J. Cell. Comp. Physiol. 8: 141—157, 1936. The effects of rapid compression upon events in the isometric contraction of skeletal muscle.
30. Brown D. E. S., Marsland D. A., J. Cell. Comp. Physiol. 8: 159—165, 1936. The viscosity of amoeba at high hydrostatic pressure.
31. Burr H. S., J. Exp. Zool. 129: 327—341, 1955. Certain electrical properties of the slime mold.
32. Czarska L., Grebecki A., Acta Protozool. 4: 201—239, 1966. Membrane folding and plasma-membrane ratio in the movement and shape transformation in Amoeba proteus.
33. Daniels E. W., Breyer E. P., Z. Zellforsch. 91: 159—169, 1968. Starvation effects on the ultrastructure of amoeba mitochondria.
34. Davison P. E., Hunneus F. C., J. Molec. Res. 52: 429—439, 1970. Fibrillar proteins from squid axons: microtubule protein.
35. DeBruyn P. P. H., Anat. Rec. 89: 43—63, 1944. Locomotion of blood cells in tissue cultures.
36. DeBruyn P. P. H., Anat. Rec. 93: 295—315, 1945. The motion of the migrating cells in tissue cultures of lymph nodes.
37. DeBuyn P. P. H., Anat. Rec. 95: 177—191, 1946. The amoeboid movement of the mammalian leukocyte in tissue culture.
38. DeBruyn P. P. H., Quart. Rev. Biol. 22: 1—24, 1947. Theories of amoeboid movement.
39. Flickinger C. J., J. Cell Biol. 49: 221—226, 1971. Decreased formation of Golgi bodies in amebae in the presence of RNA and protein synthesis inhibitors.
40. Flickinger C. J., Coss R. A., Exp. Cell Res. 62: 326—330, 1970. The role of the nucleus in the formation and maintenance of the contractile vacuole in Amoeba proteus.
41. Gibbons I. R., Ann. Rev. Biochem. 37: 521—546, 1968. The biochemistry of motility.
42. Goldacre R. J., Rev. Cytol. 1: 135—164, 1952. The folding and unfolding of protein molecules as a basis of osmotic work.
43. Goldacre R. J., Symp. Soc. Exp. Biol. 6: 128—144, 1952. The action of general anaesthetics on amoebae and the mechanism of the responses to touch.
44. Goldacre R. J., Exp. Cell Res. Suppl. 8: 1—16, 1961. The role of the cell membrane in the locomotion of amoebae and the source of the motive force and its control by feedback.
45. Goldacre R. In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 237—255, 1964. On the mechanism and control of amoeboid movement.
46. Goldacre R. J., Lorch I. J., Nature 166: 497—500, 1950. Folding and unfolding of protein molecules in relation to cytoplasmic streaming, amoeboid movement and osmotic work.
47. Goldman R. D., J. Cell Biol. 51: 752—762, 1971. The role of three cytoplasmic fibers in BHK-21 cell motility: microtubules and the effects of colchicine.
48. Griffin J. L., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 303—327, 1964. The comparative physiology of movement in the giant, multinucleate amebae.
320
Глава 18
49. Griffin J. L., Allen R. D., Exp. Cell Res. 20: 619—622, 1960. Movement of particles attached to the surface of amebae in relation to current theories of ameboid movement.
50. Guttes E., Guttes S., Exp. Cell Res. 30: 242—244, 1963. Arrest of plasmodial motility during mitosis in Physarum polycephalum.
51. Haberey M., Wohlfarth-Botterman К. E., Stockem W., Cytobiologie 1: 70—84, 1969. Pinocytosis and locomotion of amoebae. VI. Kinematographic studies on the behavior of the cell surface of moving Amoeba proteus.
52. Hahnert W. F., Physiol. Zool. 5: 491—526, 1932. A quantitative study of reactions to electricity in Amoeba proteus.
53. Harvey E. N., Marsland D. A., J. Cell. Comp. Physiol. 2: 75—97, 1932. The tension at the surface of Amoeba dubia with direct observations on the movement of cytoplasmic particles at high centrifugal speeds.
54. Hatano S., Exp. Cell Res. 61: 199—203, 1970. Specific effect of Ca++ on movement of plasmodial fragment obtained by caffeine treatment.
55. Hatano S., Rondo H., Miki-Noumura T., Exp. Cell Res. 55: 275—277, 1969. Purification of sea urchin egg actin.
56. Hatano S., Oosawa F., J. Cell Physiol. 68: 197—202, 1966. Extraction of an actin-like protein from the plasmodium of a myxomycete and its interaction with myosin A from rabbit striated muscle.
57. Hatano S., Oosawa F., Biochim. Biophys. Acta 127: 488—498, 1966. Isolation and characterization of plasmodium actin.
58. Hatano S., Tazawa M., Biochim. Biophys. Acta 154: 507—519, 1968. Isolation, purification and characterization of myosin В from myxomycete plasmodium.
59. Hatano S., Totsuka T., Oosawa F., Biochim. Biophys. Acta 140: 109—122, 1967. Polymerization of plasmodium actin.
60. Holberton D., Nature 222: 680—681, 1969. Microtubules in the cytoplasm of an amoeba.
61. Holberton D. V., Preston T. M., Exp. Cell Res. 62: 473—477, 1970. Arrays of thick filaments in ATP-activated Amoeba model cells.
62. Huneeus F. C., Davison P. F., J. Molec. Biol. 52: 415—428, 1970. Fibrillar proteins from azons: neurofilament protein.
63. Jahn T. L., Biorheology 2: 133—152, 1964. Protoplasmic flow in the myceto-zoan, Physarum: The mechanism of flow; a re-evaluation of the contraction-hydraulic theory and of the diffusion drag hypothesis.
64. Jahn T. L., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press,. New York, pp. 279—302, 1964. Relative motion in Amoeba proteus.
65. Jahn T. L., Bovee E. C., Amer. Midi. Nat. 73: 30—40, 1965. Mechanisms of movement in taxonomy of Sarcodina as a basis for a new major dichotomy into two classes, Autotractea and Hydraulea.
66. Jahn T. L., Bovee E. C., Physiol. Rev. 49: 793—862, 1969. Protoplasmic movements within cells.
67. Jahn T. L., Rinaldi R. A., Biol. Bull. 117: 100—118, 1959. Protoplasmic movement in the foraminiferan, Allogromia, Lati collaris, and theory of its mechanism.
68. Jahn T. L., Rinaldi R. A., Brown M., Biorheology 2: 123—131, 1964. Protoplasmic flow in the mycetozoan, Physarum: Geometry of the plasmodium and the observable facts of flow.
69. Jennings H. S., Carnegie Inst. Wash. Publ. No. 16, 129—234, 1904. The movements and reactions of amoeba.
70. Jeon R. W., Exp, Cell Res. 50: 467—471, 1968. Nuclear control of cell movement in amoebae: nuclear transplantation study.
71. Jeon K. W., Bell L. G. E., Exp. Cell Res. 27: 350—352, 1962. Pseudopod and foodcup formation in Amoeba proteus.
72. Jeon K. W., Bell L. G. E., Exp. Cell Res. 33: 531—539, 1964. Behavior of cell membrane in relation to locomotion in Amoeba proteus.
73. Jeon K. W., Lorch I. J., Danielli J. F., Science 167: 1626—1627, 1970. Reassembly of living cells from dissociated components.
Амебоидное движение
321
74. Jockusch В. М., Brown D. F., Rusch FL Р., J. Bact. 108: 705—714, 1971. Synthesis and some properties of an actin-like nuclear protein in the slime mold Physarum polycephalum.
75. Kamiya N., Protoplasmatologia 8: 1—199, 1959. Protoplasmic streaming. (Камия H., Движение протоплазмы, ИЛ, 1962.)
76. Kamiya N., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 257—277, 1964. The motive force of endoplasmic streaming in the ameba.
77. Kitching J. A., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 445—455, 1964. The axopods of the sun animalcule Actinophrys sol (Heliozoa).
78. Komnick H„ Stockem W., Wohlfarth-Botterman К. E., Z. Zellforsch. 109: 420— 430, 1970. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. VII. Experimental induction, contraction and extraction of the protoplasmic fibrils of Physarum polycephalum.
79. Komnick H., Wohlfarth-Botterman К. E., Fortschr. Zool. 17: 1—154, 1966. Morphology of cytoplasms.
80. Landau J. V., Ann. N. Y. Acad. Sci. 78: 487—500, 1959. Gel-sol transformations in amoebae.
81. Landau J. V., Zimmerman A. M., Marsland D. A., J. Cell Comp. Physiol. 44: 211—232, 1954. Temperature-pressure experiments on Amoeba proteus; plasmagel structure in relation to form and movement.
82. Leidy J., Freesh-Water Rhizipods of North America. Government Printing Office, Washington, D. C., 1879, 324 p.
83. Loewy A. G„ J. Gen. Physiol. 40: 127—156, 1952. An actomyosin-like substance from the plasmodium of a myxomycete.
84. Lorch I. J., Jeon K. W„ Exp. Cell Res. 57: 223—229, 1969. Character changes induced by heterologous nuclei in amoeba heterokaryons.
85. Marsland D. A., In: A Symposium on the Structure of Protoplasm; A Monograph of the American Society of Plant Physiologists, edited by W. Seifriz, Iowa State College Press, Ames, Iowa, pp. 127—161, 1942. Protoplasmic streaming in relation to gel structure in the cytoplasm.
86. Marsland D. A., Int. Rev. Cytol. 5: 199—227, 1956. Protoplasmic contractility in relation to gel structure: Temperature-pressure experiments on cytokinesis and amoeboid movement.
87. Mast S. O., J. Morphol. 41: 347—425, 1926. Structure, movement, locomotion, and stimulation in Amoeba.
88. Mast S. O., Prosser C. L., J. Cell. Comp. Physiol, 1: 333—354, 1932. Effects of temperature, salts and hydrogen-ion concentration on rupture of the plasmagel sheet, rate of locomotion, and gel/sol ratio in Amoeba proteus.
89. McManus S. M. A., Amer. J. Bot. 52: 15—25, 1965. Ultrastructure of myxomycete plasmodia of various types.
90. McManus S. M. A., Roth L. E., J. Cell Biol. 25: 305—318, 1965. Fibrillar differentiation in myxomycete plasmodia.
91. Miller D. M., Anderson J. D., Abbott В. C., Comp. Biochem. Physiol. 27: 633— 646, 1968. Potentials and ionic exchange in slime mold plasmodia.
82. Miller D. M., Reed X. B., Acta Protozool. 8: 119—128, 1970. The distributed solation hypothesis: an explanation of protoplasmic movement in slime mold plasmodia.
93. Morgan J., Exp. Cell Res. 65: 7—16, 1971. Microfilaments, from Amoeba proteus.
94. Nagai R., Kamiya N„ Proc. Japan Acad. 42: 934—939, 1966. Movement of the myxomycete plasmodium: electron microscopic studies on fibrillar structures in the plasmodium.
95. Nakajima H., In: Primitive Mothile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 111—123, 1964. The mechanochemical system behind streaming in Physarum.
21—2067
322
Глава 18
96. Nakajima Н., Allen R. D., J. Cell Biol. 25: 361—374, 1965. The changing pattern of birefringence in plasmodia of the slime mold, Physarum polycepha-lum.
97. Noland L. E., J. Protozool. 4: 1—6, 1957. Protoplasmic streaming: a perennial puzzle.
98. O’Neill С. H., Exp. Cell Res. 35: 477—496, 1964. Isolation properties of the cell surface membrane of Amoeba proteus.
99. Pantin C. F. A., J. Marine Biol. Assoc. U. K. 13: 24—69, 1923. On the physiology of amoeboid movement.
100. Pappas G. D., Ann. N. Y. Acad. Sci. 78: 448—473, 1959. Electron microscope studies on amoeba.
101. Pitts R. F., Mast S. 0., J. Cell. Comp. Physiol. 3: 449—462, 1933. The relation between inorganic salt concentration, hydrogen ion concentration and physiological processes in Amoeba proteus. I. Rate of locomotion, gel/sol ratio, and hydrogen in balanced salt solutions.
102. Pitts R. F., Mast S. O., J. Cell. Comp. Physiol. 4: 237—256, 1934. The relation between inorganic salt concentration, hydrogen ion concentration and physiological processes in Amoeba proteus. II. Rate of locomotion, gel/sol ratio, and hydrogen ion concentration in solutions of single salts.
103. Pitts R. F., Mast S. O., J. Cell. Comp. Physiol. 4: 435—455, 1934. The relation between inorganic salt concentration, hydrogen ion concentration and physiological process in Amoeba proteus. III. The interaction between salts (antagonism) in relation to hydroge ion concentration and salt concentration.
104. Pollard T. D., Ito S., J. Cell Biol. 46: 267—289, 1970. Cytoplasmic filaments of Amoeba proteus: the role of filaments in consistency of changes and movement.
105. Pollard T, D., Korn E. D., J. Cell Biol. 48: 216—219, 1971. Filaments of Amoeba proteus: binding of heavy meromyosin by thin filaments in motile cytoplasmic extracts.
106. Prusch R. D., Dunham P. B., J. Cell Biol. 46: 431—434, 1970. Contraction of isolated contractile vacuoles from Amoeba proteus.
107. Rinaldi R. A., Baker W. R., J. Theoret. Biol. 23: 463—474, 1969. A sliding filament model of amoeboid motion.
108. Rinaldi R. A., Jahn T. L., J. Protozool. 10: 344—357, 1969. On the mechanism of ameboid movement.
109. Roth L. E., Pihlaja D. J., Shigenaka Y., J. Ultrastruct. Res. 30: 7—37, 1970. Microtubules in the heliozoan axopidium. I. The gradion hypothesis of allosterism in structural proteins.
110. Roth L. E., Shigenaka Y., J. Ultrastruct. Res., 31: 356—374, 1970. Microtubules in the heliozoan axopodium. II. Rapid degradation by cupric and nickelous ions.
111. Schneider L., Wohlfarth-Botterman К. E., Protoplasms 51: 377—389, 1959. Unicellular animal studies. IX. Electron microscopic investigation of Amoeba, with particular consideration of the fine structure of cytoplasms.
112. Sheen S. J., Gailey F. B., Miller D. M., Anderson J. D., Bargman T. J., Carter D. A., Bioscience 19: 1003—1005, 1969. Sol-gel differences in plasmodia of the acellular slime mold, Physarum polycephalum.
113. Shigenaka Y., Roth L. E., Pihlaja D. J., J. Cell Sci. 8: 127—151, 1971. Microtubules in the heliozoan axopodium. III. Degradation and reformation after dilute urea treatment.
114. Stewart P. A., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya. Academic Press, New York, pp. 69—78, 1964. The organization of movement in slime mold plasmodia.
115. Stewart P. A., Stewart В. T., Exp. Cell Res. 17: 44—58, 1959. Protoplasmic movement in slime mold plasmodia: the diffusion drag force hypothesis.
116. Stockem W„ Wohlfarth-Botterman К. E., Haberey M., Cytobiologie 1: 37—57, 1969. Pinocytosis and locomotion of amoebae. V. Outline-changes and foldingdegree of the cell surface of Amoeba proteus.
Амебоидное движение
323
117. Subirana J. A., J. Theor. Biol. 28: 111—120, 1970. Hydrodynamic model of amoeboid movement.
118. Tasaki I., Kamiya N., Protoplasma 39: 333—343, 1950. Electrical responce of a slime mold to mechanical and electrical stimuli.
119. Tasaki 1., Kamiya N., J. Cell. Comp. Physiol. 63: 365—380, 1964. A study on electrophysiological properties of canivorous amoeba.
120. Ts’o P. О. P., Bonner J., Eggman L., Vinograd J., J. Gen. Physiol. 29: 325— 347, 1956. Observations on an ATP-sensitive protein system from the plasmo-dia of a myxomycete.
121. Ts’o P. О. P., Eggman L„ Vinograd J., J. Gen. Physiol. 39: 801—812, 1956. The isolation of myxomyosin, an ATP-sensitive protein from the plasmodium of a myxomycete.
122. Ts’o P. О. P., Eggman L., Vinograd J., Biochim. Biophys. Acta 25: 532—542, 1957. Physical and chemical studies of myxomyosin, an ATP-sensitive protein in cytoplasm.
123. Weihing R. R., Korn E. D., Biochem. Biophys. Res. Comm. 35: 906—912, 1969. Ameba actin: the presence of 3-methylhistidine.
124. Wilson E. B., The Cell in Development and Heredity, 3rd Ed., MacMillan Co., New York, pp. 63—65, 1934.
125. Wohlfarth-Botterman К. E., Protoplasma 54: 514—539, 1962. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. I. Electron microscopic evidence and fine structure.
126. Wohlfarth-Botterman К. E„ Protoplasma 57: 747—761, 1963. Extensive fibrillar protoplasmic differentiations and their significance for protoplasmic streaming. II. Light microscopic description.
127. Wohlfarth-Botterman К. E., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 79—109, 1964. Differentiations of the ground cytoplasm and their significance for the generation of the motive force of ameboid movement.
128. Wohlfarth-Botterman К. E., Stockem W., Z. Zellforsch. 73: 444—474, 1966. Pinocytosis and locomotion of amoebae. II. Information on permanent and induced pinocytosis by Amoeba proteus.
129. Wolpert L., Symp. Soc. Gen. Microbiol. 15: 270—293, 1965. Cytoplasmic , streaming and amoeboid movement.
130. Wolpert L„ O’Neill С. EL, Nature 196: 1261—1266, 1962. Dynamics of the membrane of Amoeba proteus studied with labelled specific antibody.
131. Wolpert L., Thompson С. M., O’Neill С. H., In: Primitive Motile Systems in Cell Biology, edited by R. D. Allen and N. Kamiya, Academic Press, New York, pp. 143—168, 1964. Studies on the isolated membrane and cytoplasm of Amoeba proteus in relation to ameboid movement.
Глава 19
Реснички
Ф. БРАУН
Реснички — это органеллы с весьма характерной внутренней структурой, расположенные на поверхности клетки [19, 20, 38, 60, 64, 69, 76]. Их основная функция заключается в перемещении жидкости над поверхностями посредством колебательных движений. Реснички принято подразделять на 2 типа: 1) относительно длинные жгутики (обычно в клетке один или несколько жгутиков) и 2) короткие собственно реснички (в клетке их много). Жгутики типичны для Mastigophora— одного из классов простейших. Ими снабжены также хоаноциты губок, гастродерм многих кишечнополостных, мерцательные терминальные клетки протонефридиев у ряда коловраток, соленоциты аннелид и сперматозоиды большинства животных. Собственно реснички характерны для инфузорий и обычно встречаются на поверхности тела кишечнополостных, турбеллярий и немертин. Реснички имеются у представителей всех типов животных, за исключением нематод и членистоногих; они располагаются на определенных участках наружной или внутренней поверхности тела1.
Реснички проявляют активность только в водной среде и поэтому обнаруживаются лишь на тех поверхностях, которые погружены в жидкость или по крайней мере покрыты тонким слоем жидкости. Типичные движения ресничек могут иметь различный результат в зависимости от инерции снабженной ресничками поверхности. Если инерция мала, то покрытое ресничками тело будет перемещаться в водной среде. Если же инерция значительна или если снабженная ресничками структура неподвижна, то будет перемещаться внешняя среда. Следовательно, успешно использовать реснички для быстрого передвижения могут только такие мелкие организмы, как простейшие, коловратки и покрытые ресничками личинки. Эти формы очень быстро развивают максимальную скорость и очень быстро останавливаются после прекращения активности ресничек.
1 Недавно у рада свободноживущих нематод найдены неподвижные реснички в амфидах (органах химического чувства) и подвижные — в кишечнике. — Прим. ред.
Реснички§25
При диаметре тела свыше 1 мм организмы способны перемещаться с помощью ресничек, но более медленно. Свободно плавающие организмы, такие, как гребневики, должны обладать для этого плотностью, очень близкой к плотности окружающей среды. Гребневик Pleurobrachia, например, содержит 94,73% воды и имеет плотность тела 1,02741—действительно весьма близкую к плотности морской воды.
Многие плоские черви и моллюски, такие, как Nassarius, используют реснички лишь для ползания по водной или увлажненной поверхности в слое секретируемой ими слизи.
Поскольку большинство снабженных ресничками (мерцательных) поверхностей сохраняет активность на протяжении всей жизни животного, имеет значение, по-видимому, то обстоятельство, что реснички, основная функция которых заключается в локомоции, обычно регулируются координирующими системами организма. Это дает возможность животному адаптироваться, меняя направление и скорость своих движений в ответ на внешнее раздражение. Однако подобная регуляция локомоторной деятельности ресничек накладывается, по-видимому, на присущий им автоматизм [28].
В тех случаях, когда движется не сама снабженная ресничками клетка, а окружающая среда, реснички участвуют в выполнении одной или нескольких функций: питание, циркуляция жидких и твердых веществ, .выделение, дыхание. Движения ресничек обеспечивают создание токов питательных веществ у многих простейших, у сидячих или малоподвижных животных, принадлежащих к различным систематическим группам. Особенно хорошо заметны эти токи у коловраток. У многочисленных •актиний и кораллов реснички щупалец и оральной области обеспечивают не только питание, но и удаление несъедобных частиц изо рта и с вершин щупалец [7, 87]. При контакте же с пищей •Щупальца наклоняются к ротовому отверстию и таким образом проталкивают пищу в гастроваскулярную полость. Известно, что у Metridium marginatum и у других видов актиний направление эффективного биения ресничек в стомодеуме под влиянием пищевых раздражителей меняется на противоположное [61]. В норме в отсутствие раздражений биение ресничек в стомодеуме направлено наружу, но в присутствии пищи, например мяса краба, оно направлено внутрь. Сходную реакцию вызывает и КС1. В стомодеуме Actinolob а имеются продольные бороздки. Биение ресничек, расположенных в бороздках, направлено внутрь, а расположенных на гребнях наружу. Поэтому основное направление потоков в этом органе в значительной мере зависит от степени мышечного сокращения, изменяющего складчатость стомодеума.
Вероятно, ни у одного другого животного мерцательный механизм не получил такого развития, как у форм, фильтрующих питательные вещества с помощью ресничек. К таким формам от
326
Глава 19
носятся главным образом пластинчатожаберные и некоторые брюхоногие моллюски, а также полухордовые [5, 65]. У этих животных система ресничек организована так, чтобы создавать токи пищи, отфильтровывать взвешенные частицы, собирать их в особые мерцательные тракты и доставлять к ротовому отверстию. На этом пути часто встречаются специальные мерцательные структуры для сортировки частиц по их величине и для удаления более крупных из них.
Благодаря ресничкам происходит перемещение пищевых частиц в пищеварительной системе у самых различных беспозвоночных. У Aurelia и Pleurobrachia каналы гастроваскулярной системы представляют собой пути прохождения пищи. Движения ресничек играют важную роль в транспорте пищи в пищеварительной системе некоторых иглокожих, многочисленных моллюсков [11] и многих других животных.
У сидячих и малоподвижных форм реснички часто служат для удаления инородных веществ. У многих морских звезд биение ресничек происходит в направлении от ротового отверстия к анальному по всей поверхности тела, тем самым обеспечивая удаление отбросов. Реснички эпителия ротовой полости у лягушек и респираторного эпителия у млекопитающих, по-видимому, играют такую же роль [35].
Само собой разумеется, что движения ресничек на поверхности тела у водных животных, дышащих кожей, или на специальных дифференцированных респираторных поверхностях должны благодаря перемещению компонентов окружающей среды облегчать газообмен.
Реснички необходимы для обеспечения нормальной циркуляции жидкости в теле некоторых аннелид, таких, как пелагическая полихета Tomopteris, а также сипункулид и эхиурид [49, 50], у которых истинная кровеносная система рудиментарна или отсутствует. У этих червей специальная система мерцательных трактов обеспечивает эффективную циркуляцию целомической жидкости. С помощью такого же механизма осуществляется перемещение целомической жидкости и у морских звезд, у которых реснички обеспечивают эффективную циркуляцию даже в амбулакральных ножках. У лягушек спинномозговая жидкость циркулирует за счет активности мерцательного эпителия, выстилающего желудочки мозга [14].
Очень часто реснички облегчают перемещение веществ внутри канальцев, в частности в канальцах нефридиев и почек и в протоках гонад.
Приведенные выше сведения о ресничках объясняют широкое распространение этих органелл и их различную функциональную роль. Хотя реснички не в состоянии производить движения, столь же заметные и сильные, как движения, совершаемые при мышечном сокращении, однако можно привести многочисленные приме-
I
Реснички
327
ры, когда они являются незаменимыми функциональными компонентами организма. Во многих этих случаях реснички выполняют •свою роль более эффективно, чем любой типичный мышечный механизм.
Строение ресничек
Реснички разных клеток построены по единому типу [2, 3, 13, 20, 32, 33, 85]. При рассмотрении в оптический микроскоп они кажутся совершенно гомогенными.
У некоторых жгутиковых, например у Euglena, жгутик прикреплен к футляру группами диагонально ориентированных выростов, или мастигонем, которые придают ему вид пера [7, 23, 62]. Подобные жгутики в отличие от более простых называют «перообразными», «реснитчатыми» или «стихонематическими». Проксимальный конец осевой нити обязательно связан с базальным тельцем, которое, как полагают, происходит из центросомы клетки. Многие цитологи описывали системы фибрилл, идущих от базальных телец к ядру клетки.
С помощью электронной микроскопии была изучена ультра-структурная организация ресничек у многочисленных одноклеточных и многоклеточных организмов.
Диаметр ресничек и жгутиков, как правило, колеблется в пределах от 0,20 до 0,25 мкм. Длина ресничек обычно составляет 10—20 мкм, жгутики же могут иметь большие размеры и достигают в длину от 20 мкм до нескольких миллиметров [76]. К наиболее длинным жгутикам относятся хвосты сперматозоидов.
Как правило, ресничка состоит из пучка продольных фибрилл [1, 20, 27], аксонемы (рис. 19-1, А), ограниченной мембраной, которая служит продолжением плазматической мембраны клетки. Аксонема состоит из наружного цилиндра, образуемого 9 парами фибрилл, окружающих центральную пару. Все эти фибриллы представляют собой микротрубочки [1, 20, 27]. Центральные микротрубочки имеют диаметр около 20 нм при толщине стенки около 4 нм. Стенки как у двух центральных, так и у 9 двойных периферических микротрубочек образованы приблизительно Грядами глобулярного белка тубулина, который по аминокислотному составу подобен мышечному белку актину [25, 66, 68]. (Микротрубочки каждой периферической пары имеют общую стенку, состоящую из трех рядов тубулина; в результате каждая такая пара образована 23 рядами тубулина.
При входе в клетку аксонема меняет свою структуру. Центральные фибриллы оканчиваются у поверхности базального тельца реснички, а двойные периферические становятся тройными в результате добавления третьей микротрубочки. Базальные тельца, имеющие длину около 0,5 мкм, снабжены волокнистыми образованиями, с помощью которых реснички прикрепляются к поверхно-
328
Глава 19
сти клетки. Периферические фибриллы аксонемы снабжены, за> исключением их основания и вершины, ручками (рис. 19-1,Б). На поперечном срезе реснички можно видеть, что одна из фибрилл каждой пары несет 2 ряда таких ручек, которые направлен
Рис. 19-1.
А. Схематическая реконструкция жгутика и базального тельца переднего отдела тела Pseudotrichonympha [26.] Б—Ж. Поперечные срезы на указанных уровнях. 1 — ручки, 2 — фиксирующие гранулы, 3 — базальная пластинка, 4 — центральная фибрилла, 5 — тельце в форме полумесяца, 6 — клеточная мембрана, 7 — спицеобразные структуры, 8 — цилиндр, 9 — дистальный участок базального тельца, 10— мембрана жгутика, 11— наружные фибриллы. 12 — проксимальный участок базального тельца, /3 — центральный футляр, 14 — дистальный конец субфибриллы, 15 — вторичная фибрилла, 16 — переходная фибрилла. 3 — схема поперечного среза жгутика. Прямая х—х указывает направление аксиальной плоскости наружной фибриллы.
ны по ходу часовой стрелки в сторону не имеющей ручек фибриллы соседней пары. Ручки длиной около 15 нм, расположенные с интервалом около 17 нм, состоят из молекул белка, называемого динеином. В присутствии двухвалентных катионов — кальция и магния — в соответствующих концентрациях динеин играет роль фермента, расщепляющего молекулы АТФ с освобождением энергии.
В аксонеме описаны также и другие структуры. Палочки, отходящие радиально внутрь от периферической пары фибрилл и
Реснички
329
•оканчивающиеся маленькими булавовидными утолщениями на расстоянии около половины пути к центральным фибриллам, считаются либо вторым рядом более коротких продольных фибрилл, .лежащих в наружном кольце аксонемы, либо фибриллярной системой, связывающей периферические и центральные фибриллы. Описана также оболочка, окружающая две центральные фибриллы. Однако, образована ли она спиральными нитями или мембраной, остается неясным.
Несмотря на то что общая структура ресничек едина, известны некоторые ее модификации. Для специализированных жгутиков и ресничек, функция которых не связана с движением, характерно упрощение строения, такое, как утрата центральных фибрилл. Усложнения структуры, включающие добавочные компоненты к 9 типичным периферическим микротрубочкам, особенно выражены в некоторых жгутиках, например у сперматозоидов.
Реснички часто объединяются, образуя сложные органеллы. Цирры Euplotes и крупные реснички жабр Nephthys можно рассматривать как сложные образования, состоящие из множества ресничек, отходящих от ряда базальных телец, которые расположены в круглом или овальном поде. В подобных циррах отдельные реснички тесно прилегают друг к другу, будучи скреплены вязким матриксом, и сгибаются одновременно. Мембранеллы адоральной области многих инфузорий из отряда Peritricha представляют собой небольшие пластинки из спаянных и синхронно колеблющихся ресничек, а ундулирующие мембраны таких инфузорий, как Blepharisma,—длинные ряды ресничек, каждая из которых вплотную прилежит к соседней. Сложный характер этих мембран часто можно выявить, расщипывая их с помощью мик-ропрепаровальных игл. Составляющие мембрану реснички, отде-ленные друг от друга, начинают двигаться совершенно независимо; если же их снова объединить, то присущая им характерная координированная активность вызывает восстановление ундули-рующего движения.
Характеристика движения ресничек
Ритм работы ресничек обычно находится в пределах от 10 до 40 ударов в 1 с. Активность любой реснички можно отнести к «одному из четырех основных типов движения или их комбинации 1'29]. Одним из них является маятникообразное движение, при котором ресничка перемещается назад и вперед, сгибаясь только у основания [37]. Такое движение характерно для ресничек эпи-телия глотки лягушки или цирр брюхоресничных инфузорий. Рабочая и возвратная фазы движения отличаются только по скорости. Второй тип движения — сгибание. Изгиб появляется сначала на вершине реснички 'и перемещается к ее основанию; в возвратную фазу ресничка постепенно выпрямляется от основания к вер
830
Глава 19
шине. Такое сгибание в форме крючка встречается у ресничек, расположенных на передне-боковых поверхностях жабр пластинчатожаберных моллюсков. Третий тип движения представляет ундулирующее движение; такой тип характерен для жгутиков. Волны обычно проходят вдоль жгутика от основания к кончику; реже — в обратном направлении [31].
Пожалуй, наиболее общим способом движения ресничек является сочетание маятникообразного типа движения и сгибания.
Рис. 19-2. Стадии движения реснички из жабр полихеты [80].
Указано время (мс) каждой стадии от начала эффективного удара.
У Mytilus и Sabellaria реснички, покрывающие переднюю поверхность жабр, осуществляют рабочий удар за счет быстрого, резко затухающего маятникообразного движения (рис. 19-2). Возвратная фаза включает наклон в обратном направлении, начинающийся у основания реснички [75, 78] и быстро перемещающийся к вершине. Эта фаза движения реснички начинается до того, как кончается рабочий удар.
В мерцательном эпителии, где направление биения ресничек является постоянным, плоскость биения перпендикулярна оси, проходящей через две центральные фибриллы [20, 26, 82]. У сперматозоидов морской свинки плоскость биения реснички отклоняется от перпендикулярной на 20—30° [21].
Как рабочая, так и возвратная фазы биения, а также и унду-ляции могут совершаться либо в одной плоскости, либо в трех измерениях [57, 70]. Последний тип движения характерен для локомоторных ресничек некоторых простейших (Paramecium и Col-poda), у которых основание реснички в течение возвратной фазы, по-видимому, вращается против часовой стрелки, тогда как рабочая фаза удара совершается в одной плоскости. Ундулирую-щие движения жгутиков могут быть винтообразными, а не лежащими в одной плоскости. Движение в трех измерениях свойствен
Реснички
331
но ресничкам эпидидимиса млекопитающих и жгутикам некоторых простейших, например Trypanosoma. У последних ундулирую-щие и маятникообразные колебания не обязательно совершаются в одной плоскости и не обязательно совпадают во времени. Таким образом, вершина движущегося жгутика описывает эллипс, восьмерку или более сложную фигуру.
Как было показано, единственный жгутик Monas обладает способностью выполнять многочисленные сложные движения [40]. Перемещение вперед сопровождается быстрым маятникообразным
Рис. 19-3. Некоторые виды движений жгутика при перемещении Monas.
А. Эффективная (I) и возвратная (II) фазы при быстром движении вперед. Б. Те же фазы при медленном движении. В. Движение назад. Г. Движение вбок.
движением жгутика из положения, совпадающего с направлением движения животного, в положение, составляющее почти прямой угол с первоначальным направлением (рис. 19-3, Л). Возвратная фаза представляет собой типичное «крючкообразное» сгибание. Рабочий удар может начинаться у основания и быстро распространяться к кончику. При медленном движении животного вперед полная амплитуда колебания жгутика может уменьшаться почти вдвое (рис. 19-3, Б) без изменения формы колебания, характерного для быстрого движения. Движение животного назад осуществляется за счет ундулирующего колебания жгутика, когда волна проходит от его основания к кончику (рис. 19-3, В). Боковые движения животного являются результатом ундулирующих колебаний жгутика, согнутого приблизительно на 90° (рис. 19-3,Г). Колебания согнутого назад жгутика вызывают движение животного вперед. Ундулирующее движение может охватывать весь жгутик или только его кончик.
Колебание жгутика было изучено при помощи скоростной микрокиносъемки J43]. Особенно полезным для изучения последовательных стадий колебания жгутика в течение всего цикла движения оказался метод мгновенной фиксации, при котором реснички внезапно останавливаются и фиксируются на любой стадии их Движения. При этом было показано, что ундуляторная активность в большинстве случаев распространяется от основания к вершине жгутика как по окружности, так и вдоль него. Такое колебание
32
Глава 19
сообщает организму вращательное и спиралеобразное движение наряду с толчком назад. Возникающее в результате этого вращение тела животного вокруг оси, которая совпадает с направлением его перемещения, создает, как предполагают, важное, если не основное усилие, двигающее организм вперед. Тело животного действует в этих условиях как гребной винт [42].
Из приведенных выше данных видно, что характер колебания даже отдельной реснички может в значительной степени варьировать; по всей вероятности, активность ресничек находится под контролем механизма, обусловливающего общую реакцию организма.
Посредством активных колебаний реснички в состоянии перемещать организм в воде или, напротив, создавать ток жидкости над неподвижной клеткой, снабженной ресничками. При этом: большая часть работы должна совершаться во время рабочей, а не возвратной фазы колебания реснички. Механизм этого явления очевиден в тех случаях, когда рабочий удар является маятникообразным, а возвратный — крючкообразным. Простые маятникообразные колебания реснички, как и следовало ожидать, оказываются непригодными для направленного движения.
Хотя по визуальным наблюдениям создается впечатление, что» реснички движутся в жидкой среде с очень большой скоростью, простой расчет показывает, что на самом деле они движутся очень медленно. Угловая скорость ресничек достаточно велика (это* следует из того факта, что они могут совершать много биений в секунду), но их вершина в действительности движется с максимальной скоростью, не превышающей нескольких десятков сантиметров в 1 ч. Эта скорость мала по сравнению с обычными скоростями гидродинамических механизмов, созданных человеком.. Скорость движения ресничек при рабочем ударе, как правило, в 2—10 раз выше, чем при возвратном.
Известно, что реснички могут изменять направление удара на противоположное (явление реверсии). У ресничных инфузорий, мелких турбеллярий, таких, как Stenostomum, и у снабженных ресничками личинок изменение в направлении движения связано»' с реверсией колебания ресничек. Однако некоторые ранее опубликованные сообщения о реверсии колебаний ресничек у многоклеточных могут иметь и другое объяснение. Так, кажущаяся реверсия колебаний ресничек на околоротовых щупальцах Ostrea и в глотке кишечнополостного Actinoloba объясняется,, по-видимому, тем, что в продольных бороздках и на гребешках, этих органов реснички движутся в противоположных направлениях, и в зависимости от степени мышечного сокращения стенок: становится заметной деятельность какого-нибудь одного из этих мерцательных трактов.
У актиний Metridium, а также у ряда других коралловых полипов, у которых короткие щупальца не могут успешно пере
Реснички
333
носить пищу в рот, реснички на губах и в глотке действительно способны менять направление своего движения в ответ на непосредственное раздражение химическими веществами. Так, в норме у невозбужденных актиний эти реснички сгибаются наружу. Если, однако, к мерцательному ресничному эпителию поднести кусочек мяса краба, то находящиеся в непосредственной близости от пищи реснички изменят направление своих колебаний и перенесут ее в глотку. Сразу же после прохождения кусочков пищи восстанавливается прежнее направление колебаний ресничек.
Такую реакцию ресничек Metridium на мясо краба можно имитировать, нанося 2,5%-ный раствор КС1 в морской воде или гликоген. После подобных воздействий направление рабочего колебания ресничек и метахронной волны меняется на противоположное. Нанесение этих веществ на щупальца и сифоноглифы не-, приводит, однако, к реверсии колебаний ресничек.
КС1 и соли других одновалентных катионов вызывают реверсию движения ресничек у Paramecium [8, 56]. Если животное поместить в слабое электрическое поле, возникнет реверсия биения ресничек на том полюсе животного, который ближе к катоду. Наблюдаемая иногда реверсия винтообразного движения может быть обусловлена рядом факторов, в том числе и вязкостью-среды [74].
Реверсию колебаний ресничек можно наблюдать на ресничках эктодермы личинок амфибий [84]; локальное изменение реверсии происходит в ответ на механическое раздражение. У более высоко организованных многоклеточных реверсия колебаний ресничек становится невозможной.
Регуляция и координация активности ресничек
Реснички, по-видимому, обладают хорошо развитым автоматизмом. Они продолжают двигаться даже в изолированных кусочках эпителиальной ткани. У простейших активность ресничек сохраняется в течение некоторого времени и в безъядерных фрагментах тела. Сперматозоиды после удаления головки проявляют* активную локомоцию. Следовательно, автоматизм присущ самой: ресничке.
Поверхности, покрытые ресничками, обычно обнаруживают метахронизм, указывающий на наличие общего координирующего механизма. Термином метахронизм обозначают определенную последовательность возникновения колебаний ресничек, в расположении которых на поверхности заметна какая-то пространственная упорядоченность (рис. 19-4). Колебания соседних ресничек вдоль оси,, совпадающей с направлением распространения мета-хрональной волны, очень незначительно сдвинуты по фазе., еснички, расположенные под прямым углом к направлению распространения метахрональной волны, колеблются изохронно, т. е.
Глава 19
в одной фазе друг с другом, создавая картину волн, проходящих по поверхности эпителия. Гребни волн создаются ресничками в момент пика их рабочего удара; впадины включают в себя рес
I П
—<
Рис. 19-4. Схематическое изображение двух видов метахронизма [60]. /«-отдельные стадии эффективного удара вправо, II— стадии возвратного удара влево. А и Б. Метахрональный ритм ресничек, при котором стадии следуют друг за другом справа налево (симплектический метахронизм. А) или слева направо (антиплектический метахронизм, Б). Схема метахрональных волн на поверхности тела Paramecium multimicronucleatum, стрелкой обозначено направление плавания. Г. Схема цикла би»ния, соответствующего области, отмеченной пунктиром на В; расстояние между ресничками увеличено в обоих направлениях для ясности изображения.
А
нички, находящиеся в конце их рабочего удара и в начале возвратного удара.
Определенное направление метахрональной волны, так же как и направление колебаний отдельных ресничек, по-видимому,
I
\ Реснички 335
I ________________________________________________________________
-—,...^ .....--
1
обусловлено основными свойствами поверхности, покрытой ресничками. Даже небольшие изолированные участки такой поверхности могут сохранять присущее им направление метахрональной волны. Это было показано, в частности, на кусочке мерцательного эпителия из ротовой полости лягушки, который выделяли, а затем
вновь трансплантировали,, повернув на 180°. При этом направление метахрональной волны в трансплантирован ном участке и в окружающем эпителии оказалось противоположным.
В тканях различного происхождения могут наблюдаться разные соотношения между направлением метахрональной волны и направлением колебания ресничек в рабочей фазе. Была предложена общая терминология для описания основных взаимоотношений направления (рис. 19-5) биений ресничек с направленнием распространения метахрональной волны-.1 симплектическое движение — если оба направления совпадают, анти-плектическое — если они противоположны, и леоплектическое или дексио-плектическое движения, когда направления составляют угол около 90° соответственно влево или вправо [39]. По-видимому, имеет место корреляция между типом волнообразного движения и функцией эпителия. Так, с
Рис. 19-5. Номенклатура видов метахронного движения ресничек, предложенная Найт-Джонсом (Knight-Jones, 1954) (см. [78]).
Стрелки указывают направление метахронных волн. / — антиплекти-ческий метахронизм, 2 — дпаплектп-ческие ряды, 3 — ортоплектические ряды, 4 — симплектнческпй метахронизм.
ортоплектическими типами
связывают перемещение твердых частиц и слизи, а с диаплектиче-скими — перемещение воды.
В эпителии рта лягушки метахрональная волна и движение реснички в рабочей фазе осуществляются в одном и том же направлении. На гребных пластинках гребневиков они направлены противоположно [15]. В жабрах аннелиды Nephthys [16] и на боковых поверхностях жабр Mytilus [29, 37] метахрональная волна всегда распространяется под прямым углом к направлению удара ресничек. У Paramecium и Colpoda быстрая метахрональная волна следует от заднего полюса животного к переднему, а удар реснички направлен по отношению к ней под углом [57]. У Opalina метахрональная волна может легко изменять свое направление назад, направо или налево в зависимости от направления движения животного (рис. 19-6) [75]. Было высказано предположение, что при определенных обстоятельствах, например при высокой вязкости среды, движущей силой служит метахрональная волна в целом, а не колебания отдельных ресничек.
336
Глава 19
Жгутики, плотно прилегающие друг к другу, независимо от того, принадлежат ли они одной клетке или нескольким соседним, как, например, жгутики сперматозоидов, стремятся при ундули-рующем движении сохранять примерно одну и ту же длину волны
Рис. 19-6.
Z1. Фиксированные метахрональные волны на участках тела Opalina: животное было зафиксировано в растворе Рингера. Б. Увеличенное изображение участка в рамке с ресничками в эффективной (1) и возвратной (2) фазах. Волны просматриваются сзади—сверху. П — переднее направление, з — заднее, пр — правое, л - левое [82].
и частоту [7]. Функциональное значение такой синхронности очевидно. Хотя плавно распространяющееся непрерывное усилие может генерироваться и единичным жгутиком, однако при их согласованной работе достигается максимальная эффективность за счет уменьшения до минимума взаимодействия между ними, являющегося следствием вязкости среды. Для ресничек, движущихся периодически, метахронизм должен был бы сглаживать их
Реснички
337
собственное движение, а также повышать эффективность их движения путем синхронизации деятельности соседних ресничек и уменьшать их взаимные помехи, являющиеся следствием вязкости среды. Эксцентрическое биение ресничек вызывает генерацию метахронизма, а также приводит к возникновению временных различий, соответствующих максимуму гидродинамической интерференции для рабочей и возвратной фаз движения [79]. Явление леоплектического и дексиоплектического метахронизма наиболее характерно для тех случаев, когда имеется узкая полоска ресничек и плоскость биения ресничек наклонена к плоскости распространения метахрональной волны.
Теории механизма координации движения ресничек [38, 58,60, 76, 77] учитывают: а) нейроподобный тип передачи волны возбуждения вдоль клеточной поверхности, б) триггерный способ биения соседних ресничек, возникающий при механическом контакте ресничек друг с другом или же при какой-либо передаче движения от основания одной реснички к следующей, и в) просто процесс взаимодействия в вязкой среде. Последнее положение, по-видимому, более точно объясняет большинство наблюдений и экспериментальных данных.
Хотя колебания ресничек в основном осуществляются автономными механизмами, во всех случаях они регулируются механизмами, ответственными за общие реакции организма. Этот контроль включает простое торможение, регуляцию скорости биения, реверсию биения [4], а также дальнейшую регуляцию направления движения у некоторых простейших — Opalina и Paramecium [59, 60].
У брюхоресничных инфузорий Uronychia и Euplotes отмечено несколько типов локомоторной активности. Экспериментальное повреждение участков нейромоторной системы выявило ее участие в управлении движением у Euplotes. Имеются прямые и косвенные доказательства способности нервной системы изменять характер движения ресничек у некоторых многоклеточных
У гребневиков колебания ресничек регулируются импульсами, идущими от аборального органа по гребным пластинкам [5]. Известно, что нервная система регулирует движение ресничек у 'турбеллярии Stenostomum, а также у личинок многих моллюсков и аннелид. Реснички у этих видов могут быть подвижными, а у Stenostomum могут изменять направление своих колебаний. На •свежеизолированных кусочках губ брюхоногого моллюска Physa реснички начинают двигаться только после раздражения волокон, иннервирующих эту область. В то же время движения ресничек на ноге Alectrion, по-видимому, тормозятся нервными импульсами. На отрезанной ноге реснички временно перестают двигаться, и затем возобновляют свои колебания и продолжают их все время, пока эпителий не погибнет.
22—2067
338
Глава 19
Электрическое раздражение черепно-мозговых нервов увели-чивает амплитуду и скорость колебаний ресничек эпителия в глотке лягушки [45, 73]. Реакция, имеющая латентный период около 1 с, продолжается в течение 10 мин после прекращения раздражения. Не известно, относятся ли регулирующие нервные волокна к симпатической или парасимпатической системе, хотя большая часть данных свидетельствует об их принадлежности к лицевому нерву. Как сообщают, ацетилхолин усиливает активность ресничек [9, 10, 48, 72]. Наличие нервной регуляции подтверждается также данными о том, что неподвижные в норме реснички в глотке лягушки начинают колебаться в ответ на раздражение, например, инородными частицами и прекращают свои колебания после удаления этих частиц [46].
Механизмы координации и регуляции активности ресничек остаются еще в значительной степени невыясненными. Есть основания предполагать, что у простейших, у которых наблюдается самая разнообразная локомоторная активность, эти механизмы сложнее, чем в мерцательном эпителии у более крупных многоклеточных беспозвоночных и позвоночных. Последние имеют, по-видимому, единый способ координации активности ресничек.
Вызывая у простейших состояние наркоза, установили, что реснички прежде всего утрачивают способность к реверсии. Затем исчезает метахрональная координация, и реснички начинают колебаться независимо друг от друга. В последнюю очередь прекращается двигательная активность ресничек. На оснований этих данных было выдвинуто предположение, что колебания ресничек, метахронизм и реверсия регулируются тремя отдельными механизмами. Реверсия должна управляться механизмом с относительно быстрой передачей так, чтобы процесс возбуждения охватывал все реснички организма почти одновременно и распространялся во всех направлениях. У изучавшихся с этой целью простейших глубокие поперечные надрезы тела не влияли заметным образом на распространение возбуждения в ответ на раздражения, вызывающие реверсию колебаний ресничек. У инфузорий изменения колебаний ресничек сопровождаются изменениями мембранного потенциала, который в свою очередь связан с потоком ионов, в частности ионов кальция [18, 18а, 34, 50а, 51, 52, 53, 55].
Теории движения ресничек
Исторически все выдвигавшиеся теории можно разделить на две основные группы: 1) теории, которые предполагали, что движущаяся сила возникает в теле клетки, тогда как реснички колеблются лишь пассивно, 2) теории, которые предполагали, что движущая сила возникает в самой ресничке, обладающей самостоятельной способностью к сокращению. Шефер в 1891 г. пред-
Реснички 339
положил, что реснички представляют собой пассивные полые •структуры с различной степенью эластичности стенок на разных участках, а колебания ресничек происходят вследствие ритмического притока и оттока жидкости в этих отростках.
Точка зрения, более совместимая с новыми теориями механизма действия, была высказана Гейденгайном в 1911 г. Он предположил, что реснички сами по себе содержат активные сократимые элементы, обеспечивающие их колебания. Многочисленные наблюдения подтверждают эту точку зрения. Амплитуда волн, проходящих вдоль по жгутику от основания к вершине, как правило, не уменьшается; в ряде случаев наблюдается даже ее увеличение. Уменьшения амплитуды следовало бы ожидать, если бы кинетическая энергия генерировалась только в теле клетки у основания жгутика. У таких 'видов, как Регапета, ундулирую-щие колебания могут захватывать только кончик жгутика, тогда как основная часть остается относительно неподвижной. У трипаносом волна биения жгутика проходит от кончика к основанию.
Современные теории механизма движения ресничек основываются на предположении, что сложная структурная организация ресничек, весьма сходная у всех организмов, тесно связана с процессом движения. Исследователи пытаются отыскать в структуре реснички принципиальные функциональные свойства, которые выявлялись бы при движении реснички. Одним из таких свойств является способность к сгибанию, причем в двух или нескольких направлениях. Другим свойством является способность передавать волну вдоль оси и третьим — способность сохранять в соответствующее время высокую степень жесткости. Почти все участки реснички по ее оси могут проявлять все эти свойства в одно и то же или же в разное время в каждой фазе биения. Ясно, что эти свойства присущи лишь активной клеточной органелле. Очевидно, что система продольных фибрилл играет роль в создании упругости реснички, особенно если между фибриллами имеются поперечные связи. Менее ясны свойства механизмов изгиба и его распространение.
Для объяснения способности к сгибанию ресничек выдвигаются две группы гипотез. Согласно одной из них, предполагают, что фибриллы обладают способностью сокращаться и что сгибание может быть вызвано различным сокращением периферических фибрилл. Изгиб, таким образом, может осуществляться в любой плоскости, распространяясь или непосредственно вдоль реснички, или же спирально, вызывая круговое или спиральное Движение. Данных, подтверждающих справедливость этой гипотезы, в настоящее время недостаточно. Другая группа гипотез, основывающаяся на фактах, используемых для объяснения работы мышечных волокон, учитывает то обстоятельство, что периферические фибриллы не сокращаются, но активно скользят относи-22*
340
Глава 19
тельно друг Друга, относительно центральных фибрилл или же относительно матрикса реснички и тем самым вызывают изгиб, так как все фибриллы прочно закреплены корешком у основания реснички. Результатом действия такого механизма для фибрилл неизменяющейся длины должно быть то, что периферические двойные фибриллы на вогнутой стороне изогнутой реснички, независимо от направления ее изгиба, должны подходить ближе к кончику реснички, чем двойные периферические фибриллы на ее выпуклой стороне. Приводятся доказательства такого механизма работы ресничек [71].
Хотя можно думать, что сила, побуждающая фибриллу скользить, действует между периферическим набором двойных фибрилл и непосредственно ее окружающим матриксом или даже между двумя центральными фибриллами, ориентация АТФ-содержащих динеиновых выростов («ручек»), направленных к соседней паре периферических фибрилл, дает возможность предположить, что между соседними парами действует сила скольжения. Следовательно, можно допустить, что механизм движения ресничек включает динеиновые связи между молекулами тубулина соседних пар* фибрилл, лежащими на одном уровне.
Можно также предположить, что в ресничке, выпрямленной в. начале эффективной фазы движения, все фибриллы оканчиваются на одном уровне по отношению к ее кончику [80]. Перед началом движения вдоль всей реснички, за исключением ее основания и кончика, где «ручки» отсутствуют, по-видимому, существуют связи между соседними дублетами. Эти связи вызывают силу скольжения, результатом действия которой будет изгиб в области жесткого, лишенного связей основания реснички, причем сама ресничка останется относительно жесткой. Ресничка сохраняет жесткость за счет жесткости, присущей самим фибриллам и увеличенной связями между фибриллами. Необходимая скорость скольжения, изменяющаяся вместе со скоростью движения реснички,, составляет всего несколько микрометров в 1 с. Как только эффективная фаза движения подходит к концу, основание реснички начинает возвращаться в вертикальное положение за счет' реверсии направления скольжения фибрилл в месте сгиба у основания. Затем локальная зона обратного скольжения распространяется вдоль реснички по направлению к вершине, пока она вся: не возвратится в первоначальное состояние, будучи готова к следующему активному удару.
Скорость распространения волны биения в течение возвратной’ фазы, по-видимому, соответствует скорости скольжения фибрилл. При этом предполагается, что скорость распространения сама по1 себе должна зависеть от скорости, при которой во время скольжения происходит разрыв старых и формирование новых свя* зей [6].
Реснички 34 Г
Та же самая теория может быть применена к ундулирующе-»’ му движению жгутиков, если предположить, что определенные-участки биения распространяются по оси реснички в противоположных направлениях.
Энергия движения ресничек
Сила удара ресничек у Paramecium, по-видимому, остается постоянной; в растворах гуммиарабика скорость движения реснички в рабочей фазе обратно пропорциональна вязкости-среды [63].
Активность ресничек, обычно связана с использованием кислорода, интенсивность потребления которого зависит от скорости-биения ресничек [29]. После обработки 2,4-динитрофенолом потребление Ог возрастает, вероятно, из-за разобщения стадий использования энергии [86]. Однако, подобно мышце, реснички могут в течение некоторого времени функционировать в анаэробных условиях. После нанесения 0,1°/о-ного раствора NaCN на.1 кусочек жабры Mytilus потребление кислорода резко уменьшается, а биения ресничек постепенно замедляются. Биения ресничек в изолированных кусочках жабры Mytilus, помещенных в слабый раствор гемоглобина в камере, перфузируемой увлажненным водородом, продолжаются в течение некоторого времени после-полного восстановления гемоглобина. В таких анаэробных условиях биения ресничек становятся очень слабыми, но сразу же ускоряются после возобновления снабжения кислородом и окисления гемоглобина. В то же время сообщалось, что реснички жабр Pecten немедленно прекращают биения после уменьшения содержания кислорода до уровня, при котором восстанавливаются такие красители, как янус зеленый и нейтральный красный, но еще не восстанавливается нильский голубой. В среде, не содержащей кислорода, реснички Paramecium перестают двигаться в. течени^нескольких секунд [24]. Тем не менее известно, что у некоторых инфузорий нормальное движение ресничек осуществляется: в среде, практически лишенной О2. Пока еще не ясно, существуют ли между различными видами животных только количественные или также и качественные различия в потреблении Ог.
Работа, выполняемая при движении ресничек или жгутиков, главным образом направлена на преодоление вязкости среды и: эластического сопротивления реснички сгибанию [67, 81]. Силы инерции составных элементов реснички не известны. Сила, необходимая для активного биения реснички, пропорциональна угловой скорости, длине и диаметру реснички, а также величине вязкости среды. Так, для реснички длиной 32 мкм при 20 °C работа эффективного удара исчисляется величиной около 4-10_9 эрг, что> в 4 раза больше работы в возвратной фазе. Вычислено, что выделяющаяся энергия будет около 4 • 10-7 эрг/с, если в единичном^
342
Глава 19
цикле каждой динеиновой молекулы расщепляется одна молекула АТФ. Рабочая мощность одного жгутика, как было найдено, измеряется величиной порядка от 1 до 4-10—7 эрг/с.
Влияние факторов окружающей среды на активность ресничек
Большое влияние на двигательную активность ресничек оказывают температура и концентрация водородных ионов. Максимальная частота биений ресничек в жабрах Mytilus наблюдается при .34 °C, а в эпителии лягушки — при 35° [29, 45, 61]. Qio варьирует в зависимости от температуры. При низких температурах Qio для активности ресничек Mytilus приблизительно равен 3, но у верхнего предела физиологического диапазона температур он несколько меньше двух. При температуре выше критической биения ресничек быстро замедляются; эта критическая температура, как можно ожидать, зависит от средней температуры местообитания животного.
Концентрация водородных ионов, весьма изменчивая в природных условиях, влияет на частоту биений ресничек в большей степени, чем концентрация любых других ионов [12, 54, 83]. Кислоты типа угольной, способные быстро проникать в клетку, оказывают больший эффект, чем медленно проникающие. При высокой концентрации водородных ионов колебания ресничек могут полностью прекратиться. Биологическое значение подобной реакции ресничек на повышение кислотности четко проявляется у пластинчатожаберных моллюсков. Спустя 2—3 ч после закрывания их створок концентрация СО2 повышается настолько, что вызывает торможение активности ресничек. Адаптивное значение тормозного влияния СО2 очевидно, поскольку скорости биения ресничек прямо пропорциональны потреблению О2 мерцательным эпителием. Минимальная концентрация водородных ионов, прекращающая колебания ресничек жабр, различна у разных видов моллюсков. Например, у моллюска Муа, обычно живущего в хорошо аэрируемой воде, эта концентрация ниже, чем у Ostrea .или Mytilus.
Поразительный пример адаптивного значения минимальной концентрации водородных ионов, необходимой для торможения активности ресничек, был установлен для различных отделов пищеварительной системы Муа. У этого моллюска выявилось определенное соответствие между нормальной величиной pH каждого отдела пищеварительной системы и величиной pH, при которой реснички данного отдела прекращают движение.
Другие катионы и анионы также оказывают влияние на активность ресничек [29]. Калий вызывает кратковременное усиление биений ресничек у Mytilus [36]. Вообще концентрация этих ионов .в любой естественной среде относительно постоянна, и ее изме-
Реснички
343
некие, по-видимому, имеет для ресничек не меньшее значение, чем для клетки в целом. Как и в случае некоторых других клеточных процессов, катионы оказывают большее влияние на биение ресничек, чем анионы.
СПИСОК ЛИТЕРАТУРЫ
1. Afzelius В. A., J. Biophys. Biochem. Cytol. 5: 269—278, 1959. Ultrastructure of cilium.
2. Afzelius B. A., J. Biophys. Biochem. Cytol. 9: 383—394, 1961. Fine structure of ctenophore ciliary plates.
3. Allen R. D., J. Cell Biol. 37: 825—831, 1968. Electron micrograph cross sections-of Tetrahymena cilia.
4. Andrivon C., L’Annee Biol. Ser. 4, 8: 99—114, 1969. The phenomenon of ciliary reversal in protozoans.
5. Atkins D„ Quart. J. Mier. Sci. 80: 321—435, 1938. Ciliary mechanisms, lamel-libranchs.
6. Brokaw C. J., Symp. Soc. Exp. Biol. 22: 101—116, 1965. ATP in ciliary mo-, vement.
7. Brown H. P., Ohio J. Sci. 45: 247—301, 1945. Structure of protozoan flagellum.
8. Brutkowska M., Acta Protozool. 4: 353—364, 1967. Ciliary reversal.
9. Bulbring E., Burn J. FL, Shelley EL J., Proc. Roy. Soc. Lond. В 141: 445—466,. 1953. Acetylcholine and ciliary movement.
10. Burn J. H., Pharmacol. Rev. 6: 107—112, 1954. Acetylcholine and ciliary movement.
11. Carriker M. R., Biol. Bull. 91: 88—111, 1946. Ciliation in gut of Lymnaea.
12. Chase A. M., Glaser O., J. Gen. Physiol. 13: 627—636, 1930. H+ ion concentration and ciliary activity.
13. Chasey D., J. Cell. Sci. 5: 453—458, 1969. Fine structure of Tetrahymena. cilia.
14. Chu Hsiang-Vao, Amer. J. Physiol. 136: 223—228, 1942. Ciliary circulation of anuran cerebrospinal fluid.
15. Coonfield B. R., Biol. Bull. 66: 10—21, 1934. Coordination of swimming combs-of ctenophores.
16. Coonfield B. R., Biol. Bull. 67: 399—409, 1934. Ciliary movement on Nephthys gills.
17. Drai A. D. G., Neth. J. Sea Res. 3: 391—422, 1967. Activity and roles of latero-frontal cilia of Mytilus.
18. Dryl S., Acta Protozool. 7: 325—333, 1970. Excitation in ciliary control.
18a. Eckert R., Science 176: 473—481, 1972. Control of ciliary action.
19. Faure-Fremiet E., Biol. Rev. 36: 464—536, 1961. General review ciliary movement.
20. Fawcett D. W., pp. 217—297. In the Cell, Vol. 2, edited by J. Brachet and* A. A. Mirsky, Academic Press, New York, 1961. General treatment of cilia and flagella.
21. Fawcett D. W., J. Cell. Sci. 3: 187—198, 1968. Beat direction in relation to-central tubules.
22. Fawcett D. W., Porter K. R., J. Morphol. 94: 221—281, 1954. Fine structure of cilia.
23. Foster E., Baylor M. B., Meinkoth N. A., Clark G. I., Biol. Bull. 93: 114—121, 1947. Structure of protozoan flagellum.
24. Gersch M., Protoplasma 27: 412—441, 1937. O2 and ciliary activity in Paramecium.
25. Gibbons I. R., Symp. Internat. Soc. Cell Biol. 6: 99—113, 1968. Chemical composition of cilia.
3344
Глава 19
<26. Gibbons I. R., J. Biophys. Biochem. Cytol. 11: 179—205, 1961. Relation of fine structure to beat direction in lamellibranch.
-27. Gibbons I. R., Grimstone A. V., J. Biophys. Biochem. Cytol. 7: 697—716, 1960. Ciliary ultrastructure.
.28. Gothlin G. L., Skand. Arch. Physiol. 58: 11—32, 1930. Influence of nerves on ciliary activity.
29. Gray J., Ciliary Movement. Cambridge University Press, 1928.
- 30. Grim J. N., J. Protozool. 14: 625—634, 1967. Ultrastructure of Euplotes ciliary structure.
;31. Holiwell M. E. J., Physiol. Rev. 46: 696—785, 1966. General review of cilia.
• 32. Horridge G. A., Proc. Roy. Soc. Lond. В 162: 351—364, 1965. Macrocilia of ctenophore (Beroe) lips.
• 33. Horridge G. A., Tamm S. L., Science 163: 817—818, 1969. Scanning electron micrographic studies of Opalina cilia.
34. John T. L., J. Cell. Physiol. 70: 79—90, 1967. Mechanism of ciliary movement.
.’35. Rilburn К. H., Ed. Suppl. Amer. Rev. Resp. Dis. 93: 1966. Respiratory cilia.
36. Kinosita H., Annot. Zool. Japan. 25: 8—14, 1952. Effect of KC1 on cilia.
37. Kinosita H., Kamada T., Japan. J. Zool. 8: 291—310, 1939. Ciliary movement in Mytilus.
38. Kinosita H., Murakami A., Physiol. Rev. 47: 53—82, 1967. General review of ciliary movement.
:39. Knight-Jones E. W., Quart. J. Mier. Sci. 95: 503—521, 1954. Ciliary metachro-nism.
40. Krijgsman B. J., Arch. Protist. 52: 478—488, 1925. Flagellar movement in Monas.
•41. Lowndes A. G., Nature 150: 579—580, 1942. Ciliary movement and density, Pleurobrachia.
42. Lowndes A. G., Proc. Zool. Soc. Lond. 114: 325—338, 1944. Flagellar activity, Monas and Peranema.
43. Lowndes A. G., School Sci. Rev. 100: 319—332, 1945. Locomotion in Euglena.
44. Lucas A. M., J. Morphol. 53: 243—262, 1932. Influence of nerves in ciliary activity.
- 45. Lucas A. M., J. Morphol. 53: 265—276, 1932. Influence of temperature on ciliary beat.
- 46. Lucas A. M., Amer. J. Physiol. 112: 468—476, 1935. Control of cilia of frog mouth.
- 47. Machin К- E., Proc. Roy. Soc. Lond. В 158: 88—104, 1963. Ciliary coordination.
• 48. Maroney S. P. J., Ronkin R. R., Biol. Bull. 105: 378, 1953. Cholinesterase and ciliary movement.
• 49. Meyer A., Z. wiss. Zool. 135: 495—538, 1929. Coelomic ciliation of annelids.
50. Meyer A., Zool. Jahrb. Abt. und Ont. Tiere 64: 371—436, 1938. Ciliated coelom in Tomopteris.
- '50a. Murakami A., Eckert R., Science 175: 1375—1377, 1972. Ca ions in ciliary regulation.
.51. Naitoh Y., Science 154: 660—662. Ciliary reversal in Paramecium.
52. Naitoh Y., J. Gen. Physiol. 51: 85—103, 1968. Mechanism of ciliary reversal, «a calcium hypothesis».
53. Naitoh Y., J. Gen. Physiol. 53: 517—529, 1969. Regulation of cilia in Paramecium.
-54. Nomura S., Protoplasma 20: 85—89, 1933. H+ ions and ciliary movement; O2 consumption of cilia.
55. Okaljima A., Japan. J. Zool. 11: 87—100, 1953. Membrane potential and beat direction.
-56. Oliphant J. F., Physiol. Zool. 15: 443—452, 1942. Influence of chemicals on ciliary beat in Paramecium.
Реснички
345»
57. Parducz В., Acta Biol. Acad. Sci. Hungaricae 5: 169—212, 1954. Ciliary activity-in infusoria.
58. Parducz B., J. Protozool. 6: Suppl. 27, 1962. Ciliary coordination.
59. Parducz B., Acta Protozool. 2: 367—374, 1964. Environmental regulation oF ciliary movement.
60- Parducz B., Int. Rev. Cytol. 21: 91—128, 1967. General review of ciliary movement.
61. Parker G. H., Marks A. P., J. Exp. Zool. 52: 1—7, 1928. Ciliary reversal,. Metridium.
62. Peterson J. B., Bot. Tidskr. 40: 373—389, 1929. Structure of feathered flagella.
63. Pigon A., Szarski H., Bull. Acad. Polonaise Sci. 3: 99—102, 1955. Velocity and* force of ciliary beat.
64. Pitelka D. R., Child F. M., pp. 131—198. In Biochemistry and Physiology of Protozoa, Vol. 3, edited by S. H. Hutner. Academic Press, New York, 1964. Review of ciliary structure and function.
65. Purchon R. D., Proc. Zool. Soc. Lond. 124: 859—911, 1955. Ciliation of Pho-ladidae.
66. Renaud F. L., Rowe A. J., Gibbons I. R„ J. Cell. Biol. 36: 79—90, 1968. Proteins of cilia.
67. Rikmenspoel R., Sleigh M. A., J. Theoret. Biol. 28: 81—100, 1970. Ciliary-work.
68. Ringo D. L., J. Ultrastruct. Res. 17: 266—277, 1967. Ciliary fine structure.
69. Rivera J. A., Cilia, Ciliated Epithelium and Ciliary Activity. Macmillan Co., New York, 1962. General review of cilia.
70. Satir P., J. Cell. Biol. 18: 345—365, 1963. Form of ciliary movement.
71. Satir P., J. Cell. Biol. 39: 77—94, 1968. Mechanism of ciliary bending.
72. Seaman J. R., Biol. Bull. 99: 347, 1950. Acetylcholine and ciliary movement.
73. Seo A., Japan. J. Med. Sci. Ill Biophysics 2: 47—75, 1931. Nervous control of frog mouth.
74. Seravin L. N„ Acta Protozool. 7: 313—323, 1970. Regulation of protozoan-spiralling.
75. Sleigh M. A., J. Exp. Biol. 37: 1—10, 1960. Ciliary beat, Opalina and Sten-tor.
76. Sleigh M. A., The Biology of Cilia and Flagella. Pergamon Press, Oxford, 1962. General account of cilia.
77. Sleigh M. A., Symp. Soc. Exp. Biol. 20: 11—31, 1966. General review of ciliary coordination.
78. Sleigh M. A., Symp. Soc. Exp. Biol. 22: 131 —150, 1968. Form of ciliary beat.
79. Sleigh M. A., Int. Rev. Cytol. 25: 31—54, 1969. Mechanism of metachronal coordination.
80. Sleigh M. A., Endeavour 30: 11—17, 1971. Review of ciliary movement.
81. Sleigh M. A., Holiwell M. E. J., J. Exp. Biol. 50: 733—743, 1969. Ciliary work.
82. Tamm S. L., Horridge G. A., Proc. Roy. Soc. Lond. В 175: 219—333, 1970. Central fibrils and ciliary beat in Opalina.
83. Tomita G., J. Shanghai Sci. Inst. 1: Sec. IV, 69—76, 77—84, 1934. H+ ions and, ciliary movement.
84. Twitty V. E., J. Exp. Zool. 50: 319—344, 1928. Ciliary reversal in Amphibia.
85. Warner F. D., J. Cell. Biol. 47: 159—182, 1970. Flagella fine structure.
86. Weller H., Ronkin R. R., Proc. Soc. Exp. Biol. Med. 81: 65—66, 1952. 2,4-Di-nitrophenol on cilia.
87. Yonge С. M., Great Barrier Reef Expedition Sci. Rep. 1: 13—58, 1930. Ciliary feeding in corals.
Глава 20
Циркуляция жидкостей в организме
/
Л. ПРОССЕР
Кроме механизмов диффузии, у всех животных существуют особые механизмы, обеспечивающие перенос определенных газов, питательных веществ и продуктов метаболизма из одной части организма в другие. У всех животных, включая одноклеточных, имеется несколько жидкостных «отсеков» (compartments).
Типы циркуляторных систем
Существует несколько типов циркуляторных систем, выполняющих транспортную функцию (рис. 20-1).
Внутриклеточный транспорт. У простейших диффузия обычно дополняется интенсивным движением протоплазмы. Токи протоплазмы могут иметь определенное направление, как, например, при движении пищеварительных вакуолей у парамеции. У многоклеточных токи протоплазмы наблюдаются почти во всех клетках; их легко обнаружить с помощью цейтраферной съемки.
Перенос веществ с использованием компонентов внешней среды. Некоторые животные, в частности губки и кишечнополостные, используют для переноса различных веществ воду, в которой обитают. Она протекает по определенным каналам и приводится в движение ресничками, жгутиками или сокращением мышц.
Движение жидкости, поддерживаемое соматическими мышцами. У нематод, мшанок и коловраток наблюдается циркуляция жидкости в псевдоцеломической полости. У иглокожих, кольчецов, сипункулид и хордовых перенос веществ осуществляется через целомическую полость, выстланную клетками мезодермального происхождения. У некоторых моллюсков целомическая полость редуцирована и превратилась в полость перикарда, полости половых желез и почек. Последнее произошло у ракообразных. У моллюсков и членистоногих, имеющих «открытый» гемоцель, проталкивание жидкости достигается ib основном благодаря сокращению соматической мускулатуры. У позвоночных дериватами целома являются полости перикарда, брюшины и плевры.
Движение гемолимфы в незамкнутой циркуляторной системе. Большинство членистоногих, многие моллюски и асцидии имеют
Циркуляция жидкостей в организме-
34?
циркуляторную систему, в которой сердце нагнетает гемолимфу через сосуды и гемоцель, являющийся дериватом первичной поло-сти (бластоцель). Гемолимфа проходит из сосудов в тканевые-пространства и возвращается через синусы к сердцу. Протяженность сосудов у животных различных классов сильно варьирует; у иглокожих целомическая жидкость, гемолимфа и межклеточная
Рис. 20-1. Схема различных транспортных механизмов.
А. Губки. Б. Кишечнополостные. В. Нематоды; имеется первичная псевдоцеломическая полость. Г. Олигохеты; в переносе веществ участвуют целомическая полость и замкнутая кровеносная система. Д. Ракообразные и моллюски; кровеносная система сообщается с гемо-Целем, который развивается из первичной полости или из бластоцеля. Е. Позвоночные; целомическая полость, замкнутая кровеносная система и лимфатическая система. 1 — ротовое отверстие; 2—ресничные каналы; 3— полость тела; 4 — псевдоцель; 5 — пищеварительная трубка; 6 — дорсальный кровеносный сосуд; 7 — капиллярная сеть; 8 — целом; 9 — боковые *сердца»; 10— артерия; 11— сердце; 12 — остие; 13— гемоцель; 14— дорсальная артерия;
— капиллярная сеть; 16 — вена; 17 — лимфатический сосуд; 18 — лимфатический проток; 19 — предсердие; 20 — желудочек.
348
Глава 20
жидкость сообщаются между собой в неодинаковой степени; амбулакральная система изолирована от них и может фактически содержать морскую воду.
Движение крови в замкнутой сосудистой системе. Замкнутая '.система трубок, часто с одним или несколькими нагнетательными устройствами, обнаружена у олигохет, у многих полихет, пиявок, •форонид, немертин, головоногих моллюсков, голотурий, иглокожих и позвоночных. Кровь вступает в тесный контакт с тканями в капиллярах, иногда образующих синусы. У некоторых позвоночных кровь совершает один-единственный цикл — от сердца через органы дыхания к тканям; у других кровь дважды прохо-.дит через сердце.
Лимфатические пути. У позвоночных межклеточные пространства (первичная полость тела) соединяются с кровеносной системой через лимфатические пути, впадающие в венозную систему. Лимфатические сосуды образуют обширную сеть, иногда не усту^ пающую по протяженности капиллярной сети. У некоторых животных имеются лимфатические сердца.
Циркуляторная система (как незамкнутая, так и замкнутая) состоит из двух частей: периферических сосудов и нагнетающего сргана, т. е. сердца.
Жидкостные отсеки
Приведем перечень, в котором сопоставляются жидкостные отсеки у различных групп животных.
Позвоночные
Клетки тела } D
( Форменные элементы Внутриклеточное Кровь 1 Плазма в замкнутой со- ПР стРанств0
( судистой системе
Лимфа (исключая клетки)
Целом (полости перикарда, брюшины Внеклеточное про-и плевры) странство
Первичное межклеточное пространство (бластоцель)
Членистоногие, моллюски
Клетки тела
Первичное пространство гемолимфа
’ Клетки I
Плазма в не- замкнутой системе (со- • суды и синусы)
Внутриклеточное пространство
Внеклеточное пространство
Иглокожие
Целом (полости гонад, почек)
Целом
Кровяные пространства (лакуны)
Амбулакральная система Межклеточное пространство
Внеклеточное пространство
Циркуляция жидкостей в организме 349
Суммарное количество воды, содержащееся в организме, можно измерить несколькими способами. Наиболее точный из них — высушивание; у 'интактных животных .можно использовать в качестве метки D2O или 3Н2О. Содержание воды в организме варьирует в пределах 60—80% веса тела (см. гл. 1). Время полного кругооборота воды в организме зависит от осмотических процессов; период полуобновления всей воды организма составляет у лабораторных мышей 1,1 дня, у человека —9,5 дня, у Dipodomys — 12 дней [149].
Определить объем внеклеточной жидкости можно, измеряя степень разведения таких веществ, которые не способны проникать в клетки тканей, не слишком быстро выводятся и не подвергаются фагоцитозу. Необходимо определить время полного перемешивания их с кровью. Универсального вещества, пригодного для исследования всех организмов, не существует; вещество выбирают в соответствии с объектом исследования. Наиболее употребительны инулин, ион тиоцианата, меченая сахароза, а в отдельных случаях — хлор и натрий. Для большей точности следует говорить не об объеме внеклеточной жидкости вообще, а об объеме жидкости, в которой растворилось введенное вещество. Объем плазмы измеряют посредством введения вещества метки, которое не должно проникать в межклеточные пространства тканей; добавляя к полученной величине объем форменных элементов, получают значение объема крови. Объем плазмы обычно вычисляют по степени разведения красителя (синий Эванса) или белка, меченного 1311. Объем форменных элементов крови определяют по показателю гематокрита или вводя эритроциты, меченные 32Р [58]. Концентрация форменных элементов в крови, взятой из крупных сосудов, может быть меньше, чем средняя во всей сосудистой системе, так как существует их запас в капиллярах. Как объем внеклеточной жидкости, так и объем крови обычно выражают в процентах от веса тела, принимая удельный вес последнего за 1,0. В случае отклонения удельного веса от единицы целесообразно выражать объем крови в процентах к объему, а не весу тела.
Объем плазмы и внеклеточной жидкости у некоторых животных приведен в табл. 20-1.
У млекопитающих и птиц объем крови составляет 7—10% веса тела, а объем внеклеточной жидкости—18—25%. У женщин объем крови несколько меньше, чем у мужчин. Абсолютная величина объема крови у взрослых людей довольно постоянна, но с возрастом ее отношение к весу тела часто снижается. У крупных Животных объем крови относительно меньше, чем у мелких {табл. 20-1). Во время зимней спячки объем крови уменьшается вчетверо. Его величины у амфибий и млекопитающих близки: содержание воды в организме Rana catesbiana составляет 79%, а объем внеклеточной жидкости — 22 %. У Bufo содержание воды в
Таблица 20-1
Объем жидкости у разных видов »
(Использованы данные из источников, указанных в скобках, а также из второго издания этой книги и из Биологического справочника)
Вид Объем плазмы Объем крови и метод определения Объем внеклеточной жидкости и метод определения
Человек, мужчина 4,3—4,8 7,1 — 7,8 17,5
женщина 4,1—4,6 6,5—7,1
(на высоте 4500 м) Собака 5 8,3 8,6 (краситель)
Коза Корова 3,9 7,3 (краситель) 6,1 (5,Сг) 5,2—5,7 (меченые
Лошадь 4,2—6,2 клетки) 7,6; 10,3
Кошка 4,1 5,6 (SICr)
Лабораторная крыса 4,0 8,5 (краситель, 32Р)
(137] Морская свинка Летучая мышь Myotis 6,5 7,5 7,5 (13Ч) 13,0 (краситель)
Marmota 5,1 10,0 (краситель)
Еж летом [45] (В спячке Опоссум Кролик Птицы Голубь Курица Петух Утка 6,5 8 (краситель) 2—3 5,8 (краситель) 5,5 (клетки, меченные 32р) 5,7 (краситель) 7,8 9 7 10,2 (13Ч)
Голубь [19а] 4,4 9,2
Рептилии и амфибии Pseudemys [77] Черепаха [184] 7,4 12 (черепаха весом 400 г) 4 (черепаха весом 1600 г) 9,1 (краситель)
Аллигатор [184] 6,0 7,3 (,3Ч)
Аллигатор 5,8 13,2
Крокодил [184] 3,5 15,4 *
Игуана 4,2 16,8
Rana pipiens [183] 8 9,5 22
Necturus [183] Cryptobranchus [183] Рыбы Ictalurus 3,5 24 22
1,2 1,8 (краситель)
Salmo 1,6 2,8 (,3Ч)
Anguilla Anguilla [1'23 a] Некоторые костистые ricol 1,8 2,9 12,2 (инулин) 13,9 2,2—3,6 (краситель) И—16,6 (сахаро-
LlozJ Костные галоиды [182] 2,1 3,1 16 (сахароза)
Продолж. табл. 20-1
Вид Объем плазмы Объем крови и метод определения Объем внеклеточной жидкости и метод определения
Хрящевые [182] Salmo [39] Акула [181] Морская акула [181] Акула [181] Лосось красный [170] Горбуша [170] Petromyzon [181] Lampetra, взрослая особь личинка Carcharhinus Negaprion [181] Chaenocephalus [70] ’Черви Sipunculus Dendrostomum ’Членистоногие Limulus [151] Carcinus [151] Maj а (после линьки) Homarus Eriocheir Cambarus Locusta [13] Apis Periplaneta [113] Schistocerca Carausius [189] Скорпион Тарантул [175], самец самка Моллюски Астаеа [202] Strombus [117а] Viviparus [lil7a] (в пресной воде) (в соленой воде) Margaritifera Mytilus Aplysia Octopus Sepia [151] Eledone Bullia 2,5 5,1 5,4 5,5 3,3 7,8 5,5 5,5 4,8 5,1 5,4 3,5 3,5 (краситель) 6,1 7,0 6,8 4,8 (краситель) 9,8 6,2 4,5 (кровь донора) 8,5 8,5 7,9 6,8 7,0 7,6 5,9 (краситель) 8,5 (инулин) 14,7 18,4 (сахароза) 19,7 (сахароза) 21,2 21,2 23,9 19,7 21,2 50 47 51,6 (инулин) 32,6 (инулин) 29 17 (краситель) 33 (SCN) 25,1 (краситель) 25,6 (SCN) 18,1 25—30 15,7—17,5 14,4 (краситель) 10,4-45,2 34 (краситель) 19,6 18,1 25,9 64,6 23 (инулин) 33 (инулин) 49 51 (инулин) 79 (инулин) 28 (инулин) 14,5 (сахароза) 21,8 61—64 (краси- тель)
') Указан в процентах от веса тела (уд. вес принят за 1,0).
352
Глава 20
организме равно 74% при объеме внеклеточной жидкости 25% [183].
У пластиножаберных рыб объем крови составляет около 5%’ веса тела, а у костистых — меньше, чем у всех позвоночных, обычно 1,5—3%. Объем крови лосося, измеренный методом разведения красителя, составляет 6,2% веса тела, однако измеренный более точный метод разведения меченой крови, взятой от рыбы-донора, показал, что он составляет 4,5% [169]. У осьминога (моллюск с замкнутой циркуляторной системой) кровь составляет 5,8%, а внеклеточная жидкость 28% веса тела [122].
У животных с незамкнутой циркуляторной системой объем крови соответствует объему внеклеточной жидкости животных с замкнутой системой циркуляции (табл. 20-1). Его величина у ракообразных составляет 25—30% объема тела; отнесенный к весу тела, он может составлять 30% сразу после линьки и 10% после затвердевания панциря, и поэтому отношение объема крови к весу тела нельзя считать корректной характеристикой. У большинства взрослых форм насекомых объем крови меньше, чем в личиночной стадии. У личинки мухи Celerio он составляет 18,6%, тогда как у. взрослого насекомого только 7,2—7,8% общего веса тела. У личинки мухи Sarcophaga вес гемолимфы достигает 35—42 %‘ общего веса, а у куколки — только 24—33%.
Объем гемоцеля у брюхоногих и двустворчатых моллюсков больше, чем у каких-либо других животных. У Chiton объем гемолимфы 90% объема тела, а у сухопутной улитки Achatina он составляет 40% [122].
Объем циркулирующей крови может изменяться под влиянием стресса. На больших высотах объем плазмы у млекопитающих уменьшается, а объем эритроцитов увеличивается и объем крови может несколько возрасти (до 8,3% веса тела на высоте 4500 м против 7% на уровне моря) [23]. Объем крови играет определенную роль в поддержании постоянного артериального давления; при массивной кровопотере плазма и форменные элементы поступают в кровоток из депо, например из селезенки. Объем крови млекопитающих в некоторой степени регулируется благодаря интероцептивным рефлекторным механизмам (см. гл. 12). Внезапная кровопотеря, превышающая 30% общего объема крови, для человека смертельна. Японский жук, напротив, может перенести потерю 50% гемолимфы. Небольшой объем циркулирующей крови (например, у костистых рыб) достаточно эффективен благодаря тому, что ее полный оборот совершается достаточно быстро. Однако большой объем крови представляет собой резерв жидкости и растворенных в ней веществ; у некоторых животных, кроме того, циркуляторная система выполняет функции гидростатического скелета (см. ниже).
Циркуляция жидкостей в организме________ 353
Давление и ток жидкости в циркуляторных системах позвоночных
Млекопитающие. В любой замкнутой системе труб, снабженной нагнетающим устройством типа насоса, максимум давления приходится на область вблизи насоса. По мере продвижения по сосудам давление расходуется на преодоление трения. Падение давления уменьшается, если трубки растяжимы. Основы гемодинамики изучены довольно подробно [29]. Суммарная энергия единицы объема крови может быть определена из следующего уравнения: 1
E=P+pgh+-±-pV\ £
где Р — давление (в динах на квадратный сантиметр), р — плотность крови (в г/см3), g— ускорение силы тяжести (980 см/с2), h — высота столба крови в см, и V — скорость тока крови (в см/с).
Средние величины давления в крупных артериях (например, в аорте или сонной артерии) приведены в табл. 20-2. У молодых мужчин давление во время сокращения сердца (систолическое) около 120 мм рт. ст., а во время расслабления сердечной мышцы (диастолическое) — 80 мм рт. ст. Это принято обозначать в виде дроби: 120/80. Разность между этими двумя цифрами, или пульсовое давление, уменьшается по направлению к мелким, периферическим сосудам. Давление в легочной артерии составляет около 25/10 мм рт. ст. [172].
У дельфина давление крови в аорте колеблется от 142/111 до 160/130 мм рт. ст., а давление в легочной артерии — от 55/35 до 76/50 мм рт. ст. [172]. Скорость кровотока в обширной капиллярной сети невелика; в венах она слегка возрастает, хотя давление продолжает падать. У млекопитающих суммарная площадь поперечного сечения капилляров приблизительно в 800 раз больше, чем площадь поперечного сечения аорты. Если две капиллярных сети расположены последовательно, то давление во второй из них очень мало; так, в печеночной артерии давление может достигать 120 мм рт. ст., а в воротной вене, которая собирает кровь из капилляров кишечника, всего 5 мм рт. ст. У ламы, обитающей в низине (на уровне моря), давление в стволе легочной артерии составляет 14 мм рт. ст., тогда как на высоте 3400 м над уровнем моря оно повышается до 23 мм рт. ст. вследствие повышения сопротивления сосудов кровотоку [10].
Кровяное давление обычно зависит от: 1) периферического сопротивления, 2) нагнетающей силы сердца и 3) объема крови.
Периферическое сопротивление изменяется в зависимости от сужения или расширения артериол и капилляров; возможно и пассивное изменение их просвета благодаря растяжимости колла-23—2067
Таблица 20-2
Артериальное давление у разных видов
(Использованы данные из источников, указанных в скобках, а также из второго издания этой книги
и Биологического справочника)
Вид Сосуд Артериальное давление, мм рт. ст.
Человек (в 'возрасте 20 лет) Лучевая артерия 120/75
(40 лет) 138/84
(60 лет) 140/85
Человек Легочная артерия 25/10
Лошадь 171/103
Собака 112/56
Крыса 130/91
Морская свинка 77/47
Мышь 113/81 1
Жираф (под наркозом) [53] 240—315/ /185—240
(стоя, без наркоза) [37] 120/75
Овца (без наркоза) [120] 135/112
Морской слон (без наркоза) [36] 120/90
Т иг stops [172] Сонная артерия 142/111—
Легочная артерия 160/130 55/35—76/50
Утка (без наркоза) 100—175
Курица (14 мес) 149/43
Голубь (местная анестезия) 135/105
Ящерица Tiliqua [26b] 14/10
Sauromalus [180] 80/60
Pseudemys 42/32
Xenopus [161] 38/34
Rana temporaria [161] 32/19
Rana pipiens [161] 30/20
Bufo [161] 32/19
Salamandra [161] 22/12
Amphiuma [161] 30/25
Bufo marinus [26b] 25/16
Sa.lmo (без наркоза) [174] Спинная аорта 29/25
Брюшная аорта 40/32
Oncorhynchus (без наркоза) [20a] Спинная аорта 44/38
Gadus (без наркоза) Bulbus arteriosus 29/18
Mustelus Брюшная аорта 26/19
Squalus [155] Спинная аорта 17/16
Брюшная аорта 30/24
Raja То же 16/14
Myxine Жаберная артерия [33] 17/7 f.
Спинная аорта [87] 5/3 V
Брюшная аорта [87] 9 5-
Octopus [90] Головная аорта 44/22 s’
Афферентная жаберная 18—37/11 1
артерия 7,5—18/3,7—11
Эфферентная жаберная F
артерия 0—12,5 g
Полая вена I J и
Циркуляция жидкостей в организме
355
Продолж. табл. 2
Вид Сосуд Артериальное давление, мм рт. ст.
Glossoscolex [93] Дорсальный сосуд 17,6/10,3
Вентральный сосуд 48/34,5
Locusta [13] Homarus Спинная аорта Желудочек в покое в движении 6,3/2,3 13/1 27/13
Maja Сердце Грудной синус 4/3,3 1,8/2,5
Рак Mytilus [171] Pecten [171] Crassostrea [171] Macoma [171] Mya [171] Ножной синус 7,4 1,9/0 0,5/0 1,4/0 0,8/0 0,6/0
Helix [95] Arenicola Neanthes Lumbricus Golfingia Сердце Полость тела в покое в движении Полость тела в покое в движении Полость тела в покое в движении Полость тела в покое в движении 24/5 0,73 11 1,1—7,2 17,6 1,5 10 2,2—2,9 14—79
гена и эластина; активное изменение сопротивления происходит при сокращении гладкой мускулатуры сосудов и эндотелиальных клеток капилляров. У самцов птиц и млекопитающих среднее давление обычно выше, нежели у самок, и с возрастом оно повышается. Как правило, в условиях покоя артериальное давление у крупных млекопитающих выше, чем у мелких. Давление в сонных артериях лошади может достигать 190 мм рт. ст. У крысы в течение первых трех недель жизни артериальное давление повышается от 20 до 75 мм рт. ст. [117]. Под влиянием силы тяжести наблюдается повышение давления в сосудах ног (у стоящего мужчины ростом 180 см — на 60 мм рт. ст.) и снижение давления в сосудах головного мозга приблизительно на такую же величину. Некоторое противодействие силе тяжести оказывают венозные клапаны, сокращение скелетных мышц, а также рефлекторные вазомоторные механизмы. Влияние вертикального положения тела лучше компенсируется у таких животных, для которых вертикальное положение является обычным, и хуже у тех, которым свойственно горизонтальное положение. Сонные артерии жирафа достигают в длину почти двух метров; когда животное 23*
356
Глава 20
лежит, давление в сонных артериях составляет 260/160 мм. рт. ст.; у стоящего жирафа — оно равно 120/75 мм. рт. ст. Скорость кровотока в сонной артерии у лежащего животного равна 50 мл/с, а у стоящего — 35 мл/с. Аналогичным образом, давление в артериях головного мозга у стоящего человека меньше, чем у лежащего [36].
Физическая нагрузка обычно сопровождается повышением артериального давления и увеличением минутного объема. У человека статическое изометрическое усилие, например длительное сжатие кисти, вызывает заметное повышение артериального давления и умеренное ускорение ритма сердца, тогда как динамическая работа, например бег на третбане, ведет к значительному повышению частоты сердечных сокращений и небольшому изменению артериального давления [116].
Работа, совершаемая сердцем, определяется уравнением
W=PxSV,
где Р — систолическое давление, a SV— ударный объем сердца. Увеличение минутного объема является результатов ускорения ритма сердца, с одной стороны, и следствием увеличения ударного объема — с другой. При небольших нагрузках доминирует повышение частоты сокращений сердца, но у тренированных спортсменов основным фактором повышения минутного объема может быть увеличение ударного выброса.
Венозное давление и кровоток, особенно в сосудах конечностей, при сокращении мышц повышаются.
В крыле летучей мыши имеются вены, ритмично сокращающиеся через каждые 10 с, причем волна каждого сокращения распространяется со скоростью от 0,33 до 3,5 мм/с [40].
Регуляция просвета сосудов и сокращений сердца осуществляется с помощью рефлекторных и гормональных механизмов. Местные реакции капилляров на раздражение могут быть обусловлены прямым действием раздражителя (например, гистамина) на сосудистую стенку или аксон-рефлексом (рефлекс по эфферентным веточкам чувствительных нейронов). Барорецепторы располагаются в стенке каротидного синуса, дуги аорты, легочной артерии и сердца (главным образом предсердий), а также в стенке сосудов надпочечников.
Чувствительные нервные окончания в стенке каротидного синуса дают разряды в соответствии с частотой пульсации артерии. У кошки частота импульсации рецепторов предсердий выше, чем аортальных; ее регистрируют путем отведения потенциалов от афферентных волокон блуждающего нерва [74]. Частота афферентных импульсов в нерве, идущем от надпочечников, пропорциональна уровню системного артериального давления [135]. Афферентные импульсы идут к вазомоторному центру продолговатого мозга, откуда выходят сосудодвигательные волокна веге
Циркуляция жидкостей в организме
357
тативной нервной системы. В составе симпатических нервов кровеносных сосудов у млекопитающих имеются холинэргические волокна, которые обеспечивают расширение сосудов некоторых мышц, кожи шеи и лицевой части головы; холинэргические волокна встречаются в парасимпатических нервах как краниального, так и сакрального отделов. Хорошо известны волокна, расширяющие сосуды половых органов. Сужение сосудов осуществляется симпатическими волокнами, которые являются адренэргическими; им соответствуют альфа-рецепторы, тогда как тормозным (сосудорасширяющим) нервным волокнам принадлежат бета-рецепторы (см. гл. 11).
Норадреналин (выделяемый окончаниями симпатических нервов, а также составляющий от 20 до 50% катехоламинов, продуцируемых мозговым веществом надпочечников) обладает сосудосуживающим действием и повышает артериальное давление. Адреналин (выделяемый надпочечниками) вызывает сужение сосудов кожи и почек, но расширение коронарных сосудов, а также сосудов мышц и печени. В высоких концентрациях адреналин
вызывает сужение сосудов, питающих скелетные мышцы; возможно, этот эффект опосредован через метаболические процессы.
Ацетилхолин, как правило, вызывает расширение сосудов и снижение артериального давления. Гистамин также обладает сосудорасширяющим действием. Серотонин (5-окситриптамин) в соответствующих дозах повышает артериальное давление у собак; у кошек он вызывает понижение давления. Вазопрессин, входящий в состав экстракта задней доли гипофиза у млекопитающих,
повышает артериальное давление; окситоцин оказывает на сосуды слабое действие. У птиц, напротив, преобладает депрессорный эффект окситоцина. В опытах in vitro у собаки ацетилхолин вызывал сужение легочных артерий; на почечные, брыжеечные и
мозговые артерии он никакого влияния не оказывал. При действии адреналина наблюдалось сужение аорты, а также почечных и брыжеечных артерий; легочные и мозговые артерии на адреналин не реагируют [19].
Сосуды млекопитающих существенно отличаются спонтанной
ритмической активностью их гладкой мускулатуры. Например, у крыс сосуды составляют следующий ряд по убыванию спонтанной ритмической активности; воротная вена > пупочная артерия> >аорта>мозговые сосуды [18]. Изолированная бедренная артерия кролика очень чувствительна к охлаждению: ее реакция на норадреналин исчезает при охлаждении до 13 °C, хотя реакция ушной артерии максимальна при 24 °C и не исчезает даже при 7 С; следовательно, гладкие мышцы этих двух сосудов различны по температурной чувствительности [52а].
При снижении давления в почечной артерии изменяется активность барорецепторов и из юкстагломерулярных клеток выделяется ренин; выделение ренина стимулируется также симпатической
358
Глава 20
импульсацией. Ренин в свою очередь активирует содержащийся в плазме ангиотензин, который вызывает компенсаторное повышение артериального давления путем сужения сосудов. Выделение ренина происходит также при гипонатриемии [195].
Ренин образуется в почках рыб, причем у пресноводных видов его больше, чем у морских. Его роль в поддержании ионного баланса точно установлена, участие же в регуляции давления не доказано.
Под вазомоторной ауторегуляцией подразумевают регуляцию периферического кровообращения путем сужения и расширения сосудов в соответствии с метаболическими потребностями. Так, коронарные сосуды реагируют расширением на местную гипоксию, поддерживая на необходимом уровне снабжение сердечной мышцы кислородом. Точно так же почечный кровоток линейно возрастает при повышении давления примерно на 100 мм рт. ст., однако дальнейшее повышение артериального давления почти не сказывается на почечном кровотоке, иначе говоря, почечные артерии способны к ауторегуляции. В мышцах конечностей в случаях обтурации артерий или после физической нагрузки происходит значительное усиление кровотока, компенсирующее кислородную задолженность. Можно полагать, что кроме общих нейро-гормональных механизмов вазомоторной регуляции существуют механизмы местной регуляции — непосредственное действие на сосудистую стенку метаболитов, например молочной кислоты.
Реакции разных сосудов на раздражения такого рода неодинаковы, и даже одни и те же сосуды у животных разных видов ведут себя по-разному.
Важным следствием изменений потребности в кислороде и снабжения им тканей являются изменения степени раскрытия соответствующих капилляров и артерио-венозных анастомозов (шунтов). У крыс при Ро2 =100 мм рт. ст. среднее межкапиллярное расстояние составляет 20 мкм и функционируют лишь 50% всех капилляров мышцы. В условиях гипоксии (Ро2 = = 32,2 мм рт. ст.) раскрываются все капилляры • и среднее расстояние между ними уменьшается до 18 мкм [123].
Если в интерстициальную жидкость подкожной клетчатки или мышцы ввести капиллярную трубку, то можно наблюдать в ней возникновение отрицательного давления, колеблющегося в пределах от •—0,7 до —6,8 см вод. ст. У всех наземных животных (млекопитающие и рептилии) давление в интерстиции составляет в среднем —5,6 см вод. ст., а у лягушек (исследованы только два вида) — 1 см вод. ст. Эти отрицательные величины становятся выше в условиях дегидратации и являются результатом взаимодействия капиллярного всасывания и коллоидно-осмотического давления белков плазмы, вследствие чего повышается сопротивление выходу жидкости из сосудов [155Ь].
Циркуляция жидкостей в организме
359
Регуляция кровообращения является в конечном счете одним из механизмов гомеостаза. Кровоснабжение тканей должно соответствовать постоянно изменяющимся местным условиям, однако не должно превышать потребностей. Артериальное давление и кровоток регулируются в широких пределах путем сужения и расширения периферических сосудов с включением шунтов между артериями и венами. Гладкая мускулатура периферических сосудов служит тем эффектором, который осуществляет реакции сосудов на локальные химические, термические и механические воздействия и обеспечивает рефлекторные ответы на раздражение рецепторов, воспринимающих изменения давления и объема, а также на центральные влияния. Различные сосудистые системы— коронарная, легочная, почечная и мышечная—способны по-разному реагировать на одинаковые стимулы. Кибернетическое моделирование взаимоотношений между стимулом и реакцией в различных областях сердечно-сосудистой системы позволяет предсказывать изменения различных переменных.
Низшие позвоночные. Амфибии и рептилии имеют два круга кровообращения, хотя желудочек у них не разделен продольной перегородкой. Сердце у этих животных не только обеспечивает разделение оксигенированной и неоксигенированной крови, но и позволяет создавать более высокое давление в артериальной системе по сравнению с давлением <в жаберных капиллярах. У лягушки давление в артериях легких и кожи ниже, чем в системных артериях. Эта разность уменьшается во время систолы; когда сосудистое сопротивление в легких и коже относительно выше, давление там падает более значительно, чем в системных артериях (рис. 20-2) [161]. У лягушек и других амфибий адренэргические рецепторы принадлежат к альфа-типу, и в ряде сосудистых областей участвуют в сосудосуживающих реакциях. У жабы блуждающий нерв повышает давление в легочных сосудах, а симпатические волокна вызывают небольшое его понижение.
Кровеносные сосуды рептилий снабжены как холинэргической, так и адренэргической иннервацией. У черепахи адреналин вызывает сужение, а ацетилхолин — расширение коронарных сосудов (в противоположность млекопитающим, у которых коронарные сосуды под влиянием адреналина расширяются) [26а].
У рыб и круглоротых сердце является по сути жаберным, так как оно прогоняет кровь через сосуды жабр, после чего она по спинной аорте направляется к периферии.
У радужной форели в покое давление в брюшной аорте составляет 40/32 мм рт. ст., а в спинной аорте — 29/25 мм рт. ст. Во время движения венозное давление достигает 9 мм рт. ст., Давление в брюшной аорте возрастает на 40%, а в спинной аорте на 16% [174]. У акулы брюшная аорта содержит втрое ©ольше эластина, чем спинная. Эластин обладает гораздо более
360
Глава 20
низким модулем упругости, чем коллаген; поэтому брюшная аорта сравнительно ригидна, и при повышении давления поток крови через жабры минимален [155а]. У форели во время движения ритм сердца ускоряется; возврату крови способствует насосная функция хвостовой мускулатуры [177].
Систола
Рис. 20-2. Различия в гемодинамике между респираторным (/) и системным (II) кругами циркуляции у лягушки [Г61].
Вверху — пульсовые кривые. Внизу — изменения давления в сосудах того и другого круга во время систолы и диастолы. В фазе систолы разность между давлением в этих системах уменьшается, а в фазе диастолы — возрастает. Остальные объяснения — в тексте.
Давление в полости перикарда у акул колеблется от —2 см до —5,6 см вод. ст. во время систолы. Благодаря отрицательному давлению возникает присасывающий эффект, облегчающий заполнение сердца и наблюдающийся у пластиножаберных рыб и круглоротых; у костистых рыб его нет [155а].
У круглоротых имеются добавочные сердца, способствующие поддержанию кровотока в системе низкого давления с ложными синусами. У миксины, например, кровь проходит через системное сердце под давлением 14—16/5—8 мм рт. ст. и поступает в мышечные жабры, а затем под давлением около 5 мм рт. ст. — в синусы. Отсюда часть крови направляется в каудальные добавочные сердца, а часть — через сосуды кишечника в портальное до-
Циркуляция жидкостей в организме 36!
б явочное сердце под давлением 3—6 мм рт. ст. и наконец возвращается в системное сердце [33, 87].
У рыб сердце иннервируется блуждающим нервом, а периферические сосуды имеют симпатическую иннервацию. Если ток воды через жабры ската или акулы прекращается, то сердце останавливается и давление падает. Если же рыба атропинизирована, то прекращение водного потока через жабры не сказывается ни на работе сердца, ни на давлении. Повышение давления в перфузируемых жаберных артериях усиливает афферентную импульсацию жаберных нервов, и их афферентные залпы возникают при каждом сокращении сердца [83]. Псевдобранхиальные сосуды гомологичны каротидным синусам и каротидным тельцам млекопитающих и птиц. Рецепторы жабр способны реагировать на такие раздражения, как кратковременное прикосновение или длительное надавливание. Псевдобранхии некоторых костистых рыб снабжены барорецепторами, афференты которых идут в составе языкоглоточных нервов. Инъекция адреналина вызывает у акулы прессорную реакцию; жаберные сосуды в ответ на адреналин расширяются подобно коронарным артериям млекопитающих и в противоположность висцеральным артериям, которые отвечают на введение адреналина сужением [32]. У угря ацетилхолин вызывает сужение жаберных сосудов, адреналин их расширяет, но вызывают сужение системных артериол; в жабрах угря кровь может протекать по пластинкам, но может и шунтироваться, проходя по оси листочков и минуя респираторные поверхности. При высокой потребности в кислороде каналы пластинок под действием норадреналина раскрываются и кровоток через жабры усиливается; если же такой повышенной потребности нет, то под действием ацетилхолина начинают функционировать шунты и жаберный кровоток снижается [100].
У костистых рыб в регуляции периферических сосудов участвуют и холинэргические и адренэргические механизмы. Ацетилхолин вызывает у костистых рыб сужение жаберных сосудов, адреналин— их расширение [138]. У земноводных адреналин вызывает расширение легочных сосудов [96а]. Сопоставление реакций крупных артерий на катехоламины у рыб, амфибий и рептилий позволяет думать, что в процессе эволюции значение адренэрги-ческой регуляции прогрессивно возрастало [26b]. Установлено, что у рыб, амфибий и рептилий в противоположность млекопитающим катехоламины не вызывают расширения системных сосудов [26а]. У млекопитающих многие сосуды находятся лишь под адренэргическим контролем. У рыб и амфибий лишь немногие вены имеют адренэргическую иннервацию, тогда как у рептилий, птиц и млекопитающих такая иннервация вен широко распространена. Расширяющее действие норадреналина на крупные сосуды наблюдается только у птиц и млекопитающих i[26a, 49а]« У рыб ацетилхолин, как правило, вызывает сужение болыпинст-
362
Глава 20
ва сосудов и повышает давление, у млекопитающих же он оказывает сосудорасширяющее действие и понижает давление. Адреналин вызывает сужение большинства системных сосудов и у млекопитающих и у рыб, расширяя коронарные сосуды у первых и жаберные — у вторых.
У черепах повышение температуры переднего гипоталамуса даже на 1 °C сопровождается повышением артериального давления на 10—20%, а охлаждение на 1 °C ведет к неуклонному падению давления, продолжающемуся до тех пор, пока температура тела не достигнет 20 °C [67].
Таким образом, регуляция кровяного давления и кровотока у рыб, у которых кровь проходит через сердце только один раз за весь кругооборот, слагается из регуляции кровообращения в двух включенных последовательно капиллярных сетях — жаберной и периферической. Между ними, несомненно, функционируют шунты; объем циркулирующей крови невелик. У амфибий и рептилий кровь на протяжении каждого кругооборота дважды протекает через сердце; хотя оно имеет неполную перегородку, кровь, идущая через правую половину сердца, достаточно надежно изолирована от крови, проходящей по левой половине.
Ныряющие животные; реакции на гипоксию. Универсальной реакцией сердечно-сосудистой системы при нырянии животных, которые дышат воздухом, является выраженная брадикардия. У морского слона (Mirounga) частота сокращений сердца в покое составляет 60 ударов в 1 мин, а когда животное погружено в воду — падает до 4. В покое кровоток в сонной артерии достигает 1200 мл/мин, а в подвздошной артерии— 1600 мл/мин. Под водой кровоток в подвздошной артерии совершенно прекращается, а в сонных артериях остается неизменным, так же как и давление в этих сосудах [36]. После ныряния наблюдается тахикардия и расширение сосудов. У тюленя при погружении подводу происходит сужение всех артерий, кроме коронарных и мозговых; во всех органах, кроме сердца и мозга, кровоток практически прекращается [21]. Точно так же падает кровоток у ныряющей утки: под водой он составляет в сосудах зоба 4%, в коже — 29%, а в икроножной мышце—11%. Мозговой кровоток, напротив, возрастает, а коронарный увеличивается вчетверо [90]. Давление в сонных артериях ныряющей утки остается неизменным, хотя центральное венозное давление и диастолическое давление в левом желудочке возрастают [90]. У хороших ныряльщиков, например у дельфинов и китов, мозговое кровообращение отделено от системного, и сужение системных сосудов создает условия для сохранения кровоснабжения головного мозга [132].
Рептилии и амфибии обладают рефлекторными механизмами, замедляющими ритм сердца при погружении в воду. У игуаны частота сокращений сердца может падать до 1% по сравнению с состоянием покоя; при этом значительно удлиняется интервал
Циркуляция жидкостей в организме
363
между зубцами Т и Р на ЭКГ. Атропин снимает эту реакцию; следовательно, речь идет о вагусном рефлексе [208]. У черепахи Pseudemys наблюдается брадикардия (41% от обычной частоты), которая также снимается атропинизацией. У этой черепахи имеется запас кислорода, достаточный для пребывания под водой в течение 2—3 ч [14]. Резко выраженная брадикардия отмечается и у морской черепахи Chelonia, если во время погружения повышается гидростатическое давление: при 19 атм сердце животного сокращается один раз через каждые 9 мин [15]. У Rana погружение вызывает быстро наступающую брадикардию, опосредованную вагусным рефлексом. В то же время при асфиксии (помещение в атмосферу азота) брадикардия сильно запаздывает; в этом случае она уже не связана с блуждающим нервом [119]. Системное артериальное давление у Rana на поверхности воды — 32/21 мм рт. ст.; при погружении оно падает до 23/16 мм рт. ст. Давление в артериях легких и кожи на поверхности воды составляет 31/16, а под водой — 26/15 мм рт. ст. Таким образом, систолическое давление во всех сосудистых областях при погружении снижается, а диастолическое остается относительно постоянным, несмотря на уменьшение частоты сокращений и ударного объема сердца [161].
У рыбы, извлеченной из воды, наступает брадикардия, подобно тому как это происходит с наземными животными при погружении в воду. У литоральных прилипал Sicyases в естественных условиях частота сокращений сердца составляет 70—80 в 1 мин; по извлечении животного из воды она снижается до 40 [54]. У карпа при снижении содержания кислорода в воде до 2,5 мл/л ритм сердца остается неизменным, но при дальнейшем нарастании гипоксии он несколько учащается; если же содержание кислорода в воде ниже 1,8 мл/л, то частота сокращений сразу падает. В случае предварительной ваготомии гипоксическая брадикардия запаздывает [158]. Аналогичным образом у Salmo gairdneri снижение парциального давления О2 в воде с 155 до 60 мм рт. ст. не влияет на устойчивую частоту сокращений сердца (65 в 1 мин), но когда Ло2 достигает 40 мм рт. ст., наступает брадикардия (22 в 1 мин). Этого не происходит, если рыба предварительно атропинизирована. Рецепторы, реагирующие на кислород, находятся в жабрах или в слизистой полости рта [145].
Гипоксия, как правило, вызывает у рыб рефлекторную брадикардию, увеличение ударного объема, повышение сосудистого сопротивления в жаберном русле и на периферии, а также усиление гликолиза [155а].
У рыб во время двигательной активности значительно снижается сопротивление в периферической сосудистой сети; это объясняется расширением жаберных и мышечных сосудов, хотя сосуды внутренних органов при этом сужаются [155а].
1
i
364 Глава 20
Периферическая циркуляция у беспозвоночных
Замкнутые циркуляторные системы. Головоногие моллюски обладают замкнутой циркуляторной системой с крупным системным сердцем, посылающим кровь на периферию, и с добавочными жаберными сердцами, проталкивающими кровь по капиллярам, жабр. У крупного осьминога системное давление крови может достигать 60/30 мм рт. ст., а давление в жаберных сосудах—> 10—25/7—15 см. вод. ст. [91]. У него, как и у других моллюсков,, через стенку сердца происходит фильтрация не содержащей белков плазмы в полость перикарда, после чего жидкость напраа-ляется к почкам.
У олигохет и пиявок кровь циркулирует в замкнутой системе. У гигантского дождевого червя Glossoscolex давление в спинном, сосуде достигает 24/14 см вод. ст. и сосуд сокращается 6—8 раз в 1 мин, при 18 °C волна сокращения распространяется по сосуду со скоростью 12 сегментов в 1 с. Спинной сосуд наполняет латеральные сердца, которые сокращаются 20 раз в 1 с, нагнетая кровь в брюшной сосуд, где во время систолы сердца может создаваться давление 100 см вод. ст. [93].
Незамкнутые циркуляторные системы. Различия между замкнутой и незамкнутой циркуляторными системами не столь значительны, как принято думать. Сосуды некоторых ракообразных у места впадения в тканевые синусы по размерам приближаются к капиллярам позвоночных. Однако в незамкнутых системах давление обычно низкое и крайне непостоянное, а объем гемолимфы велик. У крупных ракообразных, например у омара, систолическое давление в покое составляет 12—17 см вод. ст., а диастолическое — меньше 1 см вод. ст. [26]. При движении внутрижелудочковое давление у омара может возрасти до 37 см вод. ст. во время систолы и до 17 см вод. ст. — во время диастолы (рис. 20-3). Таким образом, в условиях покоя разность между систолическим и диастолическим давлением, так же как амплитуда их изменения в условиях двигательной активности, при незамкнутой системе гораздо больше, чем при замкнутой. Одновременное измерение давления у речного рака в разных сосудистых областях показывает, что колебания давления неодинаковы по всей системе—• в активной конечности оно выше, чем в неподвижной, в дистальной части конечности иногда выше, чем в проксимальной. Кровоток в синусах регулируется, по-видимому, благодаря сокращениям соматических мышц, а не под действием сердца и сосудов; у рака и омара афферентные и эфферентные каналы жабр разделены перегородкой. Аналогичное разделение имеется в конечностях личинок стрекозы. У двустворчатых моллюсков систолическое давление в сердце достигает лишь 7—26 мм вод. ст. Давление во время диастолы равно нулю. У Anodonta зарегистриро* вано отрицательное давление в перикарде во время систолы.
«А
Циркуляция жидкостей в организме
365
Таким образом, наполнение предсердия облегчено благодаря присасывающему эффекту. Мышечное сокращение, замыкающее створки моллюска, может повысить давление в сердце до 170 мм вод. ст. Моча выводится под давлением [69]. У Locusta аортальное давление составляет 8,6/3,2 см вод. ст. при частоте
Рис. 20-3. Давление в разных областях кровеносной системы краба Carcinus на протяжении двух сердечных циклов [16].
I — дорсальный перикард; II — ножной синус; III — жаберный синус; IV—жаберная вена; V—латеральный перикард; VI — желудочек; VII — жаберная камера.
сокращений сердца 79 в 1 мин. Движения туловища и ног повышают давление в гемоцеле, а в некоторых синусах в это же время зарегистрировано отрицательное давление, от —2,4 до —3,6 см вод. ст. [13]. Вычисленный ударный объем достигает 0,17 мкл.
У моллюсков (кроме головоногих) и у ракообразных величины, характеризующие объем крови, сопоставимы с величинами, известными для внеклеточной жидкости позвоночных (см. табл. 20-1), хотя кровяное давление рака весом 20 г составляет всего 30% величины артериального давления у мыши того же
366
Глава 20
веса. Поэтому скорость циркуляции у рака очень низка, у омара время кругооборота крови колеблется от 3 до 8 мин [26], (у человека— всего 21 с). Такое медленное кровообращение могло послужить фактором, ограничивающим размеры и активность животных с незамкнутой циркуляторной системой; их дыхательный газообмен осуществляется в большей степени через трахеи, нежели с помощью крови.
Гидростатический скелет. У многих животных, у которых давление в гемоцеле или целомической полости зависит от двигательной активности, заполненная жидкостью система обладает ригидностью, выполняя опорную роль. Гидростатические скелеты имеют значение для роющих мягкотелых животных. Кроме того, гидростатическое давление в синусах может играть важную роль и для движения конечностей у животных с незамкнутой циркуляторной системой. Так, у пауков сгибание конечностей осуществляют мышцы, разгибание же происходит за счет повышения гидростатического давления в конечности. Если объем крови почему-либо уменьшился, то конечности остаются в согнутом положении; при избыточном объеме жидкости сохраняется разогнутое положение [46].
Гидравлические механизмы иглокожих играют важную роль в передвижении этих животных, происходящем с помощью амбулакральной системы. Вытягивание амбулакральной ножки морской звезды происходит в результате перехода жидкости из ампулы в ножку. Обратный результат достигается при сокращении продольных мышц ножки [168]. Благодаря значительному отрицательному давлению в присосках луч морской везды может развивать усилие около 3000 г, и несколькими толчками животное в состоянии раскрыть раковину двустворчатого моллюска [111].
Передняя амбулакральная система голотурий состоит из двух частей, разделенных клапаном: тентакулярных каналов, подающих жидкость в щупальца, и полиева комплекса, состоящего в свою очередь из радиальных каналов, амбулакрального кольца и полиевых пузырей. Этот комплекс служит резервуаром жидкости для тентакулярной системы. У Thyone давление в каналах щупалец обычно выше (10—25 см вод. ст.), чем в полиевом комплексе (3—3,5 см вод. ст.). Если животное находится в расслабленном состоянии, то давление в полости его тела низкое (0— 2,5 см вод. ст.); во время сокращения мышц оно может достичь 35 см вод. ст. [211]. У голотурии целомическое давление в покое составляет 1,7 см вод. ст., а при сокращениях—19 см вод. ст.
Двустворчатые моллюски, роющие норы, делают это с помощью гидравлического скелета ноги. Teredo, находясь в своей норке, раздувается, и тогда давление в его мантии достигает 5— 17 мм вод. ст.; когда же моллюски вылезают наружу, давление снижается до 2—5 мм вод. ст. Во время рытья норы у Ensis зарегистрировано повышение давления до 50 см вод. ст., а у Апо-
\
х Циркуляция жидкостей в организме
—----------------------— - —
dontaS.^Q 40 см вод. ст. (рис. 20-4) [193]. Вода, изгоняемая из мантийной полости, разрыхляет песок; затем под повышенным давлением вытягивается нога, ее кончик раздувается, заякоривая, животное, и сифоны закрываются. После этого створки раковины частично или полностью смыкаются, и мышца-ретрактор ноги
Рис. 20-4. Схема двух стадий зарывания двустворчатого моллюска [190].
Фиксация ноги и фиксация раковины показаны стрелками. А. Створки сближены мышцей-аддуктором (1), что вызывает расширение ноги и освобождает пространство в грунте вокруг раковины (2). Б. Створки разведены благодаря эластичности связки, фиксирующей раковину, пока сокращение tn. protractor (3) и поперечных мышц (5) не выдвинет ногу. 4 — мантийная полость; 6 — гемоцель.
втягивает животное в грунт; теперь створки расходятся, заклинивая раковины в грунте, и нога вновь сокращается. Раскрытие створок раковины происходит у одних видов под давлением ноги и сифона, а у других — при помощи эластической связки. Муа может поочередно сокращать переднюю и заднюю мышцу, сжимающую створки [8, 193]. У некоторых двустворчатых моллюсков полость ноги изолирована от основного гемоцеля.
У роющих норы червей возможно изменение давления целомической жидкости, что ведет к изменению поперечника их тела при внедрении в грунт. У Arenicola во время рытья норы целомическое давление ритмично колеблется. При его повышении происходит раздувание хоботка; его кончик расширяется наподобие воротничка и служит якорем, способствуя втягиванию тела в грунт.
/ i /
368 Глава 20 >
-------— .— -------------------------------------------4--------
Давление в целомической полости в покое у Arenicalk около 2 см вод. ст., но во время рытья норы оно достигает ПО см вод. ст. Сокращение кольцевых мышц создает волну высокого давления каждые 5—7 с, причем расчетное натяжение этих мышц достигает 3 кг/см2 (рис. 20-5, Б) [190, 191]. Если удалить целомическую жидкость, Arenicola утрачивает способность к рытью норы.
Рис. 20-5.
А. Колебания давления в целомической полости дождевого червя Lumbricus при прохождении перистальтической волны; горизонтальной штриховкой показана фаза уменьшения поперечника и увеличения длины тела при сокращении кольцевой мускулатуры; вертикальная штриховка соответствует раздуванию и укорочению тела; черная полоса — выдвижение щетинок [160]. Б. Давление, оказываемое на грунт червем Arenicola во время зарывания.
/ — расширение; П — выворачивание хоботка [190].
У свободно движущихся Glycera и Nereis давление возрастает до 15—18 см вод. ст., а в покое оно не превышает 1—2 см вод. ст. У дождевого червя <в покое давление всего 0,26 см вод. ст., а при сокращении продольной и круговой мускулатуры — 7— 12 см вод. ст. (рис. 20-5, Д) [160]. При диаметре тела 0,6 см повышение давления от 2 до 13,5 см вод. ст. обеспечивает животному внедрение в грунт с силой около 8,5 г. Во время рытья норы передний конец тела внедряется в почву и раздувается, а затем перисталическими движениями животное продвигается дальше: этому способствуют щетинки. Давление в передней и задней частях тела может быть неодинаково, и гибкие перегородки передают изменения давления всей системе.
У сипункулид и эхиурид способность сильно увеличивать диаметр тела является специальным приспособлением для рытья нор. В полости тела у крупных представителей сипункулид заре-
Циркуляция жидкостей в организме
369
гистрировано давление до 600 см вод. ст., а у мелких Golfingia— нарастание давления от 2,8 см вод. ст. в момент расслабления до 108 см вод. ст. при сокращении [211]. Sipunculus утрачивает способность рыть норы, если вызвать паралич каудальной части •его тела, удалив ганглий; но если впереди парализованной части наложить лигатуру, то передняя часть быстро вбуравливается в песок. У червей, обитающих в трубках, нагнетание воды совершается отчасти при помощи ресничек, но основная роль в этом принадлежит перистальтическим сокращениям мышц. Перистальтические волны сокращений у Urechis следуют с интервалами в 25 с. Их пейсмекер расположен в нервном кольце, охватывающем пищевод, причем перистальтические волны имеют рефлекторную природу. Один такой червь способен перекачивать за сутки 29 л воды [НО]. Каждая перистальтическая волна перемещает 20— 30 мл воды под давлением 3—4 см вод. ст., т. е. гораздо меньшим, чем во время рытья нор (100 см вод. ст.) [34].
При сокращении тела актинии давление во внутренней полости повышается от 1—2 до 10—15 см вод. ст.
Гидростатический скелет, придающий в надлежащий момент жесткость телу, не может вместе с тем обеспечить такую независимость его отдельных частей, какую дает сегментированный наружный или внутренний скелет. Однако жесткость гидростатического скелета может увеличиваться и уменьшаться в зависимости от потребностей животного при соответствующих изменениях внутреннего давления, придавая телу различную степень мягкости или ригидности. Следовательно, у многих животных жидкостная система функционирует не только в качестве циркуляторной, но и выполняет опорную функцию.
Типы сердец
Любая система, предназначенная для циркуляции жидкости, должна иметь насос. Чтобы обеспечить движение жидкости в определенном направлении, такой насос либо должен иметь клапаны, препятствующие обратному току, либо должен нагнетать жидкость непрерывно. Морфологически различают следующие типы сердец: 1) камерные сердца, 2) трубчатые сердца, 3) пульсирующие сосуды и 4) ампулярные добавочные сердца.
Камерные сердца. Общеизвестны камерные сердца позвоночных и моллюсков. Последовательность сокращений сердца у позвоночных по существу одинакова, независимо от числа его камер, о время сокращения желудочка, когда аортальные (полулунные) клапаны открыты, аорта наполняется кровью, а сердце опорожняется. Систола желудочка (сокращение) заканчивается, гда давление в желудочке падает ниже уровня давления в аорте, и аортальные клапаны закрываются (рис. 20-6). Во время кращения желудочка давление в предсердии ниже, чем в желу-24—2067
370
Глава 20
Рис. 20-6. Соотношение фаз сердечного цикла.
Давление в аорте (I), предсердии (II), желудочке (III) и объем желудочка (IV) у собаки в соответствующем пересчете, венный пульс (V), ЭКГ (VI) и тоны сердца (VII) у человека. 1 — закрытие атрио-вентрикулярных клапанов и начало сокращения желудочков; 2 — открытие аортального клапана; 2—3 — максимум фазы изгнания; 3—4 — фаза медленного изгнания; 4—5 — протодиастолическая фаза с закрытием аортального клапана (5); 5—6 — изометрическое расслабление, открытие атрио-венти-кулярных клапанов (6); 6—7 — быстрое заполнение; 7 — применительно к систоле предсердий—фаза расширения. Отметка времени на кривой венного пульса (0,1 с) 'относится и к трем нижним кривым.
дочке, а атрио-вентрикуляр-ные клапаны остаются закрытыми. Когда предсердие наполняется, давление в нем постепенно растет; внутрижелудочковое давление при этом падает; атрио-вентрикуляр-ные клапаны открываются, кровь входит в желудочек; предсердие сокращается до тех пор, пока возрастающее давление в желудочке не закроет атрио-вентрикулярные клапаны. У человека желудочки начинают сокращаться одновременно с закрытием ат-рио-вентрикулярных клапанов (изометрическое сокращение) в течение 0,05 с; дальнейшее сокращение желудочков происходит во время изгнания крови (изотоническое сокращение) — в течение 0,22 с. Расслабление желудочков длится 0,53 с. Предсердия сокращаются в течение 0,11 с, а их диастола длится 0,69 с.
У рыб, у которых имеется одно предсердие и один желудочек, кровь, прежде чем поступить в аорту, проходит через жаберные сосуды. Поэтому сердце должно обеспечить давление, достаточное для проталкивания крови последовательно через две капиллярные сети. Эволюция двухканального сердца началась с двоякодышащих рыб. У современных двоякодышащих кровь, оттекающая от плавательного пузыря, отделена от осталь-
ной крови, возвращающейся к сердцу. У амфибий с их легочным и кожным дыханием предсердия раздельные и, следовательно, кровообращение двухканальное. Легочная вена изолирована от полой вены, а легочно-кожная артерия отходит от дуги аорты. У некоторых рептилий (например,
Циркуляция жидкостей в организме
371
у крокодилов) межжелудочковая перегородка сформировалась полностью. У ящериц, черепах и змей она неполная. При помощи радиоактивной метки, методом разведения красителя, а также путем определения содержания кислорода было показано, что в желудочках ящерицы, каймана и черепахи существует достаточно полное разделение потоков оксигенированной и неоксигенированной крови; однако, несмотря на это, при пониженной потребности в кислороде может происходить сброс некоторого количества неоксигенированной крови из правого сердца в левое; в зависимости от потребности в кислороде это количество варьирует. У гигантской жабы кровь легочной вены оксигенирована на 96%, а кровь левой половины желудочка — на 87,6%; в то же время в правой половине желудочка она оксигенирована всего на 8,9%, следовательно, степень перемешивания крови крайне незначительна [92]. У Ат-phiuma степень перемешивания крови в сердце изменяется в зависимости от обстоятельств [92]. У аллигатора во время пребывания под водой возникает брадикардия (см. выше), и при этом кровь из правой половины желудочка направляется в левую. Таким образом достигается преимущественное снабжение кислородом головного мозга — механизм адаптации, которым не располагают млекопитающие и птицы [207, 208]. У игуаны в условиях высокой температуры сброс крови в левую половину сердца составляет 20—25%. Благодаря этому больше крови направляется в сосуды кожи и меньше—в легочные сосуды .[9].
Камерные сердца моллюсков состоят из двух предсердий (у четырехжаберных головоногих — из четырех) и одного желудочка. У головоногих моллюсков кровь из всех областей тела идет к жаберным сердцам у основания жабр, а из жабр направляется по венам в системные предсердия. У других моллюсков кровь покидает артериолы, поступает в тканевые синусы, после чего кровоток в различных соотношениях разделяется между жабрами и почками, а затем кровь по венам возвращается в предсредия..
Трубкообразные сердца. Системные сердца большинства членистоногих представляют собой сократимые трубки. У некоторых, например у Limultis, имеется тонкостенная приемная камера (предсердие), частично охватывающая собственно сердце. У других сердце свободно лежит в широкой полости перикарда (синуса). Оно обычно прикреплено в нескольких точках и принимает кровь через парные отверстия с клапанами. У ракообразных кровь из сердца выбрасывается в артерии, проходит через мелкие артериолы в тканевые синусы, откуда в конце концов попадает в жабры; по жаберным венам кровь возвращается к перикарду. У многих насекомых сердце «подвешено» при помощи крыловидных мышц, имеющих различное строение. Эти мышцы растягивают сердце и, сокращаясь, могут создавать в нем отрицательное давление. Кровь оттекает от сердца по одной или нескольким артериям. Последние всегда начинаются от передней 24*
372
Глава 20
части сердца, а иногда, кроме того, еще от латеральной и задней. Сокращение либо сразу охватывает все сердце, либо распространяется по нему волнообразно.
У оболочников сердце представляет собой тонкостенную трубку, лишенную клапанов и лежащую в перикарде. Оно гонит кровь то в одном направлении, то в противоположном, изменяя характер сокращения.
Пульсирующие сосуды. Кровеносные сосуды, способные перистальтически сокращаться, встречаются у многих животных. У кольчецов ритмичной перистальтикой обладают многие сосуды. У дождевого червя волны сокращений дорсального сосуда распространяются от его заднего конца к переднему. Отсюда кровь проходит через несколько парных латеральных сосудов — так называемых «боковых сердец»—в сокращающейся вентральный сосуд. У пиявок существуют два боковых сердца, которые сокращаются поочередно.
У Arenicota кровь движется по дорсальному сосуду, проходя в боковые гастральные сосуды, лежащие по обеим сторонам гастрального сплетения, а оттуда — к боковым сердцам, которые перегоняют кровь в брюшной сосуд. Собственной перестальтикой обладают дорсальный, латеральный и пищеводный сосуды, а также некоторые нефридиальные.
У ряда голотурий сокращения кровеносных сосудов носят перистальтический характер. У многих насекомых трудно провести грань между пульсирующими сосудами и трубкообразными сердцами. У ланцетника многие сосуды способны сокращаться, а сердце представляет собой нечто вроде sinus venosus или conus arteriosus. У более высоко организованных позвоночных ветре- , чаются пульсирующие вены, например в крыле летучей мыши. < Сокращения вен способствуют движению крови к сердцу. '
Ампулярные сердца. Добавочные сердца прогоняют кровь по периферическим каналам. Жаберные сердца головоногих моллюс- ‘
ков состоят из губчатой эндотелиальной ткани, окружающей ;
множество мелких сосудов. У насекомых добавочные сердца обычно находятся в основании антенн, в области прикрепления крыльев и в конечностях.
У миксины три добавочных сердца: в воротной, главной и хвостовой венах. Поперечнополосатая мышца также образует добавочные сердца в жабрах; эти сердца гонят кровь к переднему концу тела [87].
У рыб, амфибий и рептилий имеются лимфатические «сердца»; эти способные сокращаться расширения лимфатических сосудов гонят лимфу в вены. У таких животных лимфа поступает в вены не только через грудной проток, как у птиц и млекопитающих. । Лимфатические сердца состоят из исчерченных анастомозирующих волокон, у некоторых животных эти сердца снабжены клапанами, препятствующими обратному току лимфы. ?
Циркуляция жидкостей в организме
313
Ритм сердечной деятельности и минутный объем
Определение частоты сокращений сердца, вероятно, самый» распространенный в сравнительной физиологии вид измерения», хотя этот физиологический параметр относится к наиболее изменчивым. В табл. 20-3 представлен ряд цифр, полученных преимущественно при исследовании животных без наркоза, а зачастую— при свободном движении. Применение телеметрических; методов в естественных условиях дает несколько меньшую, нсу более стабильную величину.
Частота сердечных сокращений у мелких животных, как правило, выше, чем у крупных. У слона и у лошади частота сокращений сердца составляет 25 — 40 ударов в 1 мин, у собаки — 80» у кошки — 125, у кролика — 200, у мыши — от 300 до 500. У мелких птиц сердце сокращается 500—600 раз в 1 мин, у домашней птицы— 150—300. Из ракообразных можно назвать Daphnia-,. у которой сердце бьется с частотой от 250 до 450 ударов в 1 мию (при 20°C); частота сокращений у Asellus— 180—200, у речного* рака — 30—60 в 1 мин. В ряду крабов частота сердечных сокращений с увеличением размеров тела уменьшается экспоненциально, но несколько менее круто, чем интенсивность обменных процессов. Частота сокращений сердца весьма значительно меняется' при физической нагрузке, в покое, при нервном возбуждении; У малоподвижных животных она обычно ниже, чем у более активных. У двустворчатых моллюсков частота сокращений сердца варьирует в пределах от 0,2 до 22 ударов в 1. мин, у кальмара и осьминога — от 40 до 80, у быстро плавающих частота сокращений сердца выше, чем у медленно передвигающихся. У бабочки сфинкса сердце сокращается в условиях покоя 40—45 раз в 1 мин, а во время полета — ПО—140 раз. Частота сокращений сердца, особенно у животных с незамкнутой циркуляторной системой, зависит от давления в полостях. Так, у двустворчатых моллюсков втягивание ноги сопровождается повышением давления гемолимфы и одновременно учащением сокращений сердца. У бодрствующей, но находящейся в покое летучей мыши частота сокращений сердца составляет 250—450 в 1 мин, в состоянии возбуждения — 880, во время дневного сна — 120—180, и во время зимней спячки— 18 ударов в 1 мин [105].
Ритм сердца весьма чувствителен к изменениям температуры тела и потребности в кислороде. У большинства пойкилотермных животных он ускоряется в 2—3 раза при повышении температуры на 10°C (в пределах физиологического диапазона). Подобное ускорение ритма сердца с повышением температуры происходит и у гомойотермных животных, но обычно средний ритм сердца у них выше, чем у пойкилотермных при соответствующей температуре. У ныряющих млекопитающих, птиц и черепах при погружении под воду наблюдается брадикардия. Замедление ритма, от
374
Глава 20
мечено у многих рыб при извлечении из воды, причем оно выражено значительнее, чем бывает при обеднении воды кислородом, которое, как правило, ведет к брадикардии. При возвращении к нормальным условиям у ныряющих животных, дышащих воздухом, и у рыб, извлеченных из воды, брадикардия сменяется тахикардией. У моллюсков, например у Mytilus, частота сокращений сердца в воде, бедной кислородом, или в воздухе ниже, чем в аэрированной воде.
Некоторые изменения ритма сердца являются результатом прямого действия внешних факторов. Большинство таких реакций имеет сложный рефлекторный механизм, расшифрованный лишь частично.
Минутный объем сердца — это объем крови, выбрасываемой за 1 мин каждым желудочком двухканального сердца или единственным желудочком одноканального. Один из способов измерения минутного объема — определение поглощения кислорода по разности в его содержании между артериальной кровью и кровью, направляющейся к дыхательным поверхностям (метод Фика).
дс „ - , , ч Количество поглощенного О2 в 1 мин
Минутный объем (л/мин) =—«---------------------------.
J \ / Артерио-венозная разница по О2
Другие методы сводятся к аналогичным измерениям с использованием инородных для организма газов или красителей. Сердечный индекс — отношение минутного объема к площади поверхности тела или к весу (в степени 2/3). Наиболее приемлемым показателем для сравнения разных животных следует считать минутный объем, который выражают в литрах крови в 1 мин и <i
относят к весу тела в килограммах. Минутный объем может i
измениться вследствие изменения ударного объема сердца или ча- i ътоты его сокращений. '
В табл. 20-3 приведены величины минутного объема сердца у разных животных. У человека минутный объем с момента ' рождения возрастает, достигая максимальной величины в возрасте 3—5 лет, а затем в течение всей последующей жизни несколько уменьшается.
Средняя величина- минутного объема сердца у 45-летнего мужчины составляет 5,2 л, что соответствует 3 л в 1 мин на 1 м2 поверхности тела при 68 ударах в 1 мин, или ударном объеме 74 мл [59]. При физической нагрузке минутный объем у человека возрастает (иногда в 6 раз) в основном за счет ускорения ритма; при переходе из горизонтального положения в вертикальное минутный объем падает; повышение температуры тела сопровождается увеличением минутного объема. У млекопитающих минутный объем находится в прямой зависимости от веса (В) тела животного, но отношение минутного объема (МО) к поверхности тела остается почти постоянным (рис. 20-7) [73]. Для большинства млекопитающих справедливо равенство МО = В0'776; у крыс i
Таблица 20-3
Частота сокращений сердца, ударный, минутный и относительный объемы в условиях покоя
(Данные из источников, указанных в скобках, и из Биологического справочника)
Вид Частота сокращений сердца, уд/мин Ударный объем Минутный объем Относитель- * ный минутный] объем, мл/кг]
Млекопитающие
Человек [59] 75 77 мл 5,5—6 л 80—90
(2,1—4,9л/м2)
Tursiops [172] 84—140 12,2 л 47—105
Phoca 5,8 л
Phoca [131] 5,2 л
. на поверхности (при 10 °C) 0,62 л
под водой 8,3 л
после ныряния
Морской слон [36, 37]
на поверхности 60
под водой 4
Жираф [53] 90 100 мл 20—40 л
Слон 30—40
Корова 45—60 45,8 л 113
Свинья 4,5 л 146
Лошадь 35—40 18—24 л
Лама (на уровне моря) 118
Собака 90—100 2-3 л
Крыса [141] 350 286
Мышь 498
Citellus [141]
|бО!Др1СТ1ВуЮЩИЙ 276 69 л 313
в спячке 2—7 1,04 л 4,6
Летучая мышь [105]
бодрствующая, в покое 250—450
возбужденная 880
дневной сон 120—180
глубокая спячка 18
Птицы
Утка [176]
самец 175 287
самка 185 253
Курица 178—460 400 мл 262
Воробей 460
Troglodytes 450
Рептилии
Iguana [194] (при 30 °C) 38,2 1,5 мл 58
Iguana [207] 60
Iguana [9]
при нормальной темперагу- 44
ре 80
при обогреве
Tiliqua [26b]
зимой 26
летом 38
376
Глава 20
Продолжение табл. 20-3
Вид Частота сокращений сердца, уд/мин Ударный объем Минутный объем Относительный минутный объем, мл/кг
Varanus
‘Черепаха [208] 1,0 МЛ
на поверхности 40 3,7 мл 148 мл 50—60
под водой 2 3,3 мл 6,6 мл
.Амфибии
Amphiuma [89] 0,67 мл 30
Rana pipiens (на суше) [161] 35 мкл 20—30 мл
10—15мкл 5—7 мл
(Рыбы
Gadus [88] 30 9,3
• Cyprinus carplo [51] 16 18,3
2,1 7,6
'Ictalurus [63] 21,6 11,3
Amia [63] 13,9 6,2
'Чукучан 20 4,5
Electrophorus [88] 65 40—70
Salmo [84] 46 16
.Anguilla [84] 32 - 18,8
Oplegnathus [84] 36,6
.Scyliorhinus [12] 39—48 22
Squalus 25
Raja [155] 21
Myxine [87] 30—40
Беспозвоночные
Octopus [91] 16—23 6,3—12,1 57—81 мл 6,6—20,5
мл
Mytilus [69]
в воде 24—26 ' -
•на воздухе 10
Crassostrea [171] 20—30
С io па [102] 20 0,045 мл 66
Panulirus [146] 60—100 0,6—1 мл 80
Locusta [13] 80 0,015 мл
iGlossoscolex [93]
боковой сосуд 20
дорсальный сосуд 6—8
МО=3,83В0’582, а поверхность тела составляет у них 0,09В°>667 {115]; для ряда млекопитающих iMO = 0,76B°>776 [209]. У разных млекопитающих (от крысы до лошади) минутный объем растет с увеличением конечного диастолического объема (КДО) в соответствии с уравнением МО = 106 КДО°>77. У суслика в период активности минутный объем составляет 69 мл (313 мл/кг), а во время спячки падает до 1,04 мл (4,6 мл/кг). Это уменьшение происходит главным образом за счет снижения частоты сокращений и в меньшей мере за счет изменения ударного объема [141].
У лягушки Rana pipiens эффект погружения в воду выражается следующим образом:
Циркуляция жидкостей в организме
37Т
На воздухе Под водой
Число сокращений в 1 мин 40 25—32
Ударный объем, мкл 35 10—15
Минутный объем, мл 20—23 5—7
Такое же уменьшение минутного объема, связанное с выражеш-ной брадикардией, наблюдается и у черепах.
W00
750
500
250
500 1000 мл
Рис. 20-7. Соотношения между объемом в конце диастолы и периферическим сопротивлением (Л), а также между объемом в конце систолы и ударным объемом' (Б) у млекопитающих разного размера [73].
X собака, А свинья, ошадь, □ бык.
В исследованиях, проведенных на обездвиженных рыбахг 5ыло показано, что минутный объем сердца у пластиножаберных? дольше, чем у костистых. У карпа (без наркоза) минутный эбъем составлял 18,3 мл/кг, а у акулы он достигал 22 мл/кг [12]. Работа сердца определяется из уравнения.
Работа = Давление в желудочке хОбъем/с.
378
Глава 20
У человека работа, производимая левым желудочком (Р = = 100 мм рт. ст.), составляет 1,1 -107 эрг/с; работа правого желудочка (Р — 20 мм рт. ст.) равна 0,2-107 эрг/с [29].
Системные сердца позвоночных
Возникновение и проведение возбуждения. У позвоночных, оболочников и моллюсков сокращение сердца имеет миогенную природу. У взрослой лягушки или рыбы импульс к нормальному сокращению сердца возникает в венозном синусе, а у птиц и млекопитающих в синоаурикулярном узле. Волокна сердечной мышцы эмбриона в культуре ткани ритмично сокращаются. У куриного эмбриона координированные сокращения появляются у аортального1 конца желудочка лишь на 29-м часу инкубации; по мере дальнейшего развития пейсмекер перемещается по направлению к синусу, где восстановительные процессы в интервалах между сокращениями протекают быстрее. Если у амфибии удален венозный синус, то ритм сердца нарушается — сокращения начинают генерировать другие области сердца. Эти сокращения ритмичны, но частота их ниже. Частота сокращений синоаурикулярной полоски из сердца крысиного эмбриона вдвое выше, чем наблюдаемая в вентрикулобульбарной полоске [62]. Локальное охлаждение или согревание сердца взрослого млекопитающего in situ вызывает особенно резкое изменение частоты сокращений, если термический раздражитель приложен к синоаурикулярной области. При локальном обогревании разных областей сердца рыбы пейсмекером может стать синус, нижний отдел предсердия или атрио-вентрикулярная область.
Клетки синусного узла лягушки характеризуются градуальным медленным нарастанием потенциала, который, достигнув уровня деполяризации около 15 мВ, переходит в спайк (до 70 мВ), проводящийся по предсердиям [76]. Пейсмекерный потенциал в синусе лягушки и черепахи снижается под влиянием импульсов блуждающего нерва [76]. Импульсы блуждающего нерва вызывают гиперполяризацию клеток в синоаурикулярном узле кролика и уменьшают скорость нарастания пейсмекерного потенциала i[186]. У черепахи, и в лишенном иннервации сердце миксины повышение внутреннего давления ведет к усилению пейсмекерной активности: частота сокращений может возрасти на 75 % . [85].
Волна возбуждения, которая возникает в синусе, проводится по миокарду предсердий (у млекопитающих — со скоростью 1 м/с) и затем, после атрио-вентикулярной задержки, охватывает мышцу желудочков (у пойкилотермных) или со скоростью 5—6 м/с распространяется по специализированным проводящим волокнам Пуркинье (у птиц и млекопитающих). Фронт электрической волны, проходящей по сердцу, имеет достаточную амплитуду и крутизну, так что ее можно зарегистрировать в виде
Циркуляция жидкостей в организме
379
электрокардиограммы (ЭКГ) на некотором удалении от сердца. Типичная ЭКГ у всех позвоночных, записанная методом внеклеточного отведения, состоит из нескольких зубцов — медленных, направленных вверх, обозначаемых Р, R и Т, и направленных
Б
Рис. 20-8.
А. Потенциалы действия при внутриклеточном отведении и кривые сокращения сердца у разных млекопитающих. I — человек (предсердие); II — шимпанзе (видны двойные спайки); III — обезьяна; IV— крыса (две кривые при разных скоростях развертки); V — собака; VI — кролик (вторые записи потенциалов предсердий у собаки и кролика сделаны через 10 и 5 с после первых); VII — мышь; а — предсердие, б — желудочек [166]. Б. Влияние адреналина (I) и ацетилхолина (II) на сокращения (верхние кривые) и на потенциалы (нижние кривые) предсердия морской свинки, / — контроль; 2 — адреналин в разведении 1,2-10—6 и ацетилхолин в разведении 4,4-10~8 М. (Любезно предоставлено У. Слейтором.)
вниз — зубцов Q и S. Зубец Р соответствует проведению возбуждения по предсердиям, интервал Р—Q — атрио-вентрикулярной задержке проведения, а комплекс QRS — проведению в желудочках. Зубец Т отражает реполяризацию поверхности желудочков; направленный вверх, он указывает на более раннюю реполяризацию левой стороны сердца, а направленный вниз (отрицательный) Т говорит о более ранней реполяризации правой стороны сердца. У амфибий и рыб зубцу Р предшествует формирующийся в области венозного синуса зубец V. У пластиножаберных на ЭКГ имеется зубец В, отражающий процесс деполяризации в conus arteriosus и возникающий между зубцами S и Т. Кривые, полу-
380
Глава 20
ценные с помощью внутриклеточных электродов, свидетельствуют о том, что каждое волокно деполяризуется с большой скоростью; затем, после начальной реполяризации, оно остается некоторое время частично деполяризованным, что выражается на кривой в виде плато, котброе может иметь протяженность 0,1—1,5 с в зависимости от типа сердца (рис. 20-8, А). Волокна предсердия характеризуются менее выраженным плато, чем желудочковые. Волна возбуждения распространяется сначала вниз по проводящим пучкам, а затем к наружной поверхности сердца, сквозь толщу миокарда. Суммарные электрические диполи правого и левого желудочков или правой и левой половин единственного желудочка трехкамерного сердца направлены противоположно. Алгебраическая сумма потенциалов действия желудочков, зарегистрированная при отведении от обеих сторон тела, показывает, что считающийся «изоэлектрическим» интервал S—Т является на самом деле результатом взаимной нейтрализации двух плато, предшествующих реполяризации, и что зубец Т возникает благодаря тому, что левый желудочек реполяризуется раньше, чем правый [29].
Гистологически сердце позвоночного состоит из ветвистых по-перечноисчерченных волокон. Волокна Пуркинье в желудочках млекопитающих и птиц представляют собой мышечные волокна, приспособленные для быстрого проведения возбуждения (2 м/с). Часто волокна сердечной мышцы соединены друг с другом с помощью поперечных вставочных дисков, которые имеют десмосом-•ную область (видимо, для механического соединения) и нексусы, или «контакты со щелью», которые, как выяснено, служат областью электрического проведения импульса из одной клетки в другую (см. гл. 11) [11]. Постоянная расстояния (space constant) — т. е. расстояние, на котором приложенный потенциал падает до 1/е исходной величины, — в предсердии крысы составляет 130 мкм вдоль оси волокна и 65 мкм перпендикулярно оси. -Сопротивление в области контакта волокон, как показывают вычисления, составляет около 1 Ом/см2 |[203, 204, 210]. В волокнах Пуркинье (у овцы) постоянная расстояния в 2—4 раза больше, чем в поперечнополосатых мышцах, и, по-видимому, вставочные диски фактически не создают сопротивления электрическому проведению [76, 188]. Постоянная расстояния одиночных волокон правого желудочка овцы или теленка.—880 мкм, а постоянная времени (time constant) составляет 4,4 мс при /?т=9100 Ом/см2, а Ст = 0,81 мкФ/см2. Несомненно, электрическая емкость волокна Пуркинье гораздо выше; по результатам измерений на пучках электрически связанных клеток она составляет 10 мкФ/см2; учитывая их поверхность, можно считать, что электрическая -емкость, приходящаяся на единицу площади, несколько меньше [204]. Парасимпатический медиатор ацетилхолин вызывает уменьшение постоянной расстояния и постоянной времени сердеч
Циркуляция жидкостей в организме 381
ной мышцы и уменьшает протяженность электрического плато. Имеются данные о том, что в культуре ткани сердечной мышцы существует проведение от клетки к клетке, несмотря на высокое сопротивление межклеточных соединений [173].
Длительность интервала Р—R на ЭКГ составляет 30—50%; всего сердечного цикла независимо от того, какова частота сокращений сердца —- 36 ударов в 1 мин (крокодил, лошадь) или свыше 600 (мышь). Однако у млекопитающих во время спячки интервал P—R удлиняется больше, чем другие компоненты ЭКГ. Иначе говоря, атрио-вентрикулярная задержка увеличивается.
Если ткань миокарда млекопитающего погрузить в раствор Тироде и поместить в холодильник, то в ней повышается содержание натрия, но понижается содержание калия; при последующем согревании потенциал покоя может возрасти по сравнению с нормальным уровнем (70 мВ) более чем вдвое. Такое возрастание электроотрицательности связано с кратковременным усилием введения натрия; это можно предотвратить, блокируя натриевый насос уабаином [178]. Для волокон Пуркинье в сердце характерен потенциал покоя, равный 90 мВ, и потенциал действия — 120 мВ; длительность последнего достигает 300—500 мс. Снижение содержания Na в питательной среде ведет к падению овершута, а при его снижении на 70% проведение возбуждения прекращается. В опытах на волокнах Пуркинье кошки установлено, что при замещении натрия литием спайковая активность сохраняется, но после замещения холином спайки не возникают [31]. Точно так же для сердца лягушки установлена прямая зависимость овершута от концентрации Na во внешней среде ![20].
Во время восходящей фазы спайка сопротивление мембраны резко падает, а затем, на протяжении плато, повышается до более высокого уровня по сравнению с сопротивлением покоя; во время реполяризации сопротивление мембраны вновь падает. Под влиянием бария плато удлиняется, под влиянием калия внешней среды и ацетилхолина укорачивается. Методом фиксации напряжения в сочетании с замещением ионов установлено, что в норме началу спайка соответствует повышение натриевой проводимости, за которым следует понижение калиевой проводимости (во время плато); затем, во время реполяризации, происходит медленное восстановление калиевой проводимости [48, 136]. Для пейсмекер-ных клеток в покое характерен значительный ток натрия внутрь; это в сочетании с зависимостью калиевой проводимости от потенциала и от времени делает возможной ритмическую активность (см. гл. 11) [66, 188]. Адреналин усиливает калиевый ток при более низком, чем обычно, мембранном потенциале, и поэтому частота импульсов пейсмекера возрастает. В желудочке лягушки сокращение сопровождается поглощением кальция. Ток кальция внутрь клетки растет также при повышении его внешней концентрации или понижении концентрации натрия [134]. В предсер
382
Глава 20
дии кролика высокая концентрация внеклеточного кальция повышает крутизну нарастания пейсмекерного потенциала [157]. В волокнах Пуркинье, находящихся в состоянии частичной деполяризации, медленный ток Са зависит от Na: при нормальной концентрации Na ток кальция невелик, если же концентрация внеклеточного натрия низка, то ток Са увеличивается [147]. В сердечной мышце млекопитающих направленный внутрь клетки спайковый ток в норме представлен током натрия, но до некоторой степени в этом участвует и кальций. В сердце лягушки основная часть тока возникает за счет кальция. Кроме того, кальций изменяет проницаемость мембраны для натрия и калия. У морской свинки удлинение плато происходит при полном отсутствии кальция и магния. Высокая концентрация кальция укорачивает плато, ускоряя таким образом реполяризацию, обусловленную выходом калия [52]. Ацетилхолин укорачивает плато,, регистрируемое в клетках предсердия и пейсмекера (рис.20-8,Б). У человека и шимпанзе при низкой температуре потенциал предсердий в фазе плато понижен и протяженность плато увеличена. Одиночный стимул может вызвать два спайка, второй ниже первого, и это также зависит от натриевой проводимости, которая связана с внеклеточной концентрацией кальция [166, 167].
Сокращение и регуляция. Благодаря электрическому распространению потенциалов по всей сердечной мышце сокращение сердца у позвоночных происходит по закону «все или ничего». При тетаническом раздражении сердце совершает спазматические беспорядочные сокращения — состояние delirium cordis. Сердце абсолютно рефрактерно по отношению к электрической стимуляции на протяжении большей части систолы; его удается стимулировать в период относительной рефрактерности — в конце систолы и во время диастолы,— но внеочередное сокращение, вызванное таким раздражением, достигает лишь субмаксимальной силы. Вслед за экстрасистолой, возникшей в ряду нормальных сокращений, можно наблюдать компенсаторную паузу большей длительности, чем нормальная диастола; длительность ее зависит от того, в какой момент произошла экстрасистола.
Сердечные сокращения происходят по закону «все или ничего» применительно к определенной степени растяжения полостей сердца, т. е. применительно к его объему. Если полоска сердечной мышцы растянута, то сила ее сокращения возрастает до максимума. Соответственно, если желудочек растянут повышенным давлением, то сила его сокращения увеличивается. Иначе говоря, сердце автоматически компенсирует увеличение нагрузки. При растяжении сверх определенного предельного объема (радиуса кривизны) давление, необходимое для поддержания этого чрезмерного растяжения, сильно падает и ударный объем сердца уменьшается; это называют фазой декомпенсации [29]. Под влиянием нагрузки или раздражения симпатического нерва энергия сокра
Циркуляция жидкостей в организме
383
щения может возрасти и при неизмененном давлении в полости сердца; сократительная способность сердца повышается. У разных видов животных скорость укорочения сердечной мышцы при сокращении находится в соответствии с присущей им нормальной частотой сокращений. У собаки с длительностью систолы около 161 мс полоски миокарда левого желудочка, сокращаясь, укорачиваются за 1 с примерно в 3 раза, а у крысы (продолжительность систолы—50 мс) — в 6,5 раз [71].
Рефлекторная регуляция ритма сердца согласуется с периферической регуляцией, причем частота сокращений сердца, возрастая, компенсирует снижение артериального давления; при повышении артериального давления ритм замедляется. Под влиянием физической нагрузки возрастает и частота, и амплитуда сокращений, но изменения частоты имеют большее значение для регуляции гемодинамики. Блуждающие нервы замедляют ритм сердца и уменьшают амплитуду сокращений. При этом тормозное влияние правого вагуса выражено сильнее, чем левого. Первые нейроны блуждающих нервов оканчиваются в ганглиях, расположенных в синоаурикулярном узле и стенке предсердий, но не в желудочках. Ацетилхолин, выделяемый окончаниями блуждающего нерва, оказывает отрицательное инотропное и хронотропное действие. Атропин блокирует тормозное влияние блуждающих нервов и прямое действие ацетилхолина, а физостигмин (эзерин), напротив, усиливает эти тормозные влияния, блокируя холинэстеразу. Атропин действует на уровне рецепторов миокарда, тогда как никотин блокирует ганглии.
Симпатические нервные волокна, идущие от звездчатого узла, иннервируют синоаурикулярный узел, предсердия и желудочки; кроме того, клетки, содержащие адреналин, обнаружены в самой стенке сердца. Постганглионарные симпатические волокна активируют сердечную деятельность, выделяя норадреналин. Симпатические волокна стимулируют выделение адреналина надпочечниками. Симпатический эффект выражается в увеличении как частоты сокращений сердца (влияние на пейсмекер), так и их амплитуды. Влияние адренэргических нервных окончаний реализуется через бета-рецепторы; чувствительность к катехоламинам убывает в следующем порядке: изопротеренол>адреналин>но-радреналин. Это подтверждается также действием бета-блокаторов (см. гл. 11).
Регуляция работы сердца у млекопитающих, птиц, рептилий и амфибий в основном сходна. У всех этих животных ваготомия приводит к учащению сокращений сердца и повышению артериального давления. Возможно, что тоническое действие блуждающих нервов в обычных условиях слегка подавляет активность сердца. Раздражение блуждающих нервов вызывает брадикардию, а стимуляция центрального конца одного из них тормозит сердечную деятельность и снижает артериальное давление. В со-
384
Глава 20
ставе блуждающих и синокаротидных нервов обнаружены аффе* рентные волокна ['89]. Атропинизация или перерезка блуждающих нервов у ныряющих животных снимает характерную для них брадикардию, возникающую при погружении под воду (см.выше). Раздражение блуждающих нервов и ацетилхолин изменяют потенциалы действия миокарда, укорачивая плато и снижая овершут. Во время впадения в оцепенение ритм сердца у белоногого хомячка (Peromyscus) падает быстрее и восстанавливается также быстрее, чем температура тела. По-видимому, блуждающие нервы способствуют впадению в оцепенение, а симпатические — пробуждению [121].
У амфибий и рыб симпатические нервы сердца часто проходят в том же стволе, что и парасимпатические. У бесхвостых амфибий все симпатические волокна к сердцу идут вместе с вагусом, а у хвостатых обнаружены и вагосимпатические, и чисто симпатические нервные стволы.
В сердце лягушки преганглионарные вагусные парасимпатические волокна могут оканчиваться во внутрисердечных ганглиях ;[187]. Отдельные парасимпатические нейроны можно обнаружить в межпредсердной перегородке. Каждый пресинаптический аксон образует около 25 синаптических бляшек, возбуждающий постсинаптический потенциал (ВПСП) возникает с задержкой около 40 мс. Потенциал реверсии в синапсе составляет 0—12 мВ; ионы кальция повышают частоту миниатюрных синаптических потенциалов. Чувствительность клетки к ацетилхолину обычно ограничена областью синапса, однако после денервации чувствительной становится вся клетка [Ю4].
У опистогнатовых рыб ритм сокращений сердца под действием ацетилхолина замедляется; у этих рыб обнаружена тормозная холинергическая вагусная иннервация венозного синуса и предсердия (но не желудочка, который не реагирует на ацетилхолин). Вагусное торможение сердца у пластиножаберных (Scyllium) может быть достигнуто рефлекторно при афферентном раздражении блуждающего и поджаберного нерва и нерва боковой линии, а у ганоидных (Acipenser) раздражение кожи, особенно на голове, плавниках и усиках, приводит к остановке сердца — эффект, противоположный атропиновому. У Tinea гипоксия сначала усиливает дыхание, а затем вызывает урежение ритма сердца [144]. У форели электрическое раздражение блуждающего нерва оказывает тормозное действие, но если это торможение блокировать атропином, то дальнейшая стимуляция приводит к ускорению ритма, и этот эффект в свою очередь блокируется гуанетидином и пронетало-лом. По-видимому, у рыбы в составе блуждающего нерва наряду с холинэргическими тормозными волокнами идут адренэргические стимулирующие [50]. Флуоресцентная микроскопия позволила выявить в сердце форели множество нервных окончаний, содержащих катехоламины, и очень немного настоящих адренэргиче-
I
i
Циркуляция жидкостей в организме______________________ 385
ских ганглиозных клеток. Адреналин и норадреналин оказывают на сердце этих рыб положительное инотропное и хронотропное действие [50].
Известны три способа адреэнергического возбуждения сердца; 1) через симпатические нервные волокна, либо идущие в составе блуждающих нервов (у форели — ко всем отделам сердца, у линя — только к венозному синусу), либо отходящие к сердцу непосредственно от симпатической цепочки (у акулы Heterodon-tus — к синусу); 2) через хромаффинные клетки, рассеянные по ходу вен (у форели), идущих к сердцу или находящиеся в заднем каротидном синусе (у Heterodontus); 3) через хромаффинные клетки, лежащие в толще миокарда желудочка и предсердия (у ската и двоякодышащих рыб) [50].
Что касается круглоротых, то миксины отличаются от миног; у миксин не обнаружено нервов, идущих к сердцу, которое у этих животных лишено чувствительности к ацетилхолину даже в высоких концентрациях (10~2 г/мл); однако хромаффинные клетки у миксин имеются; клетки эти опорожняются под действием резерпина [87]. У миног нет адренэргических нервов, но холинэргические волокна блуждающего нерва (как и введение ацетилхолина) повышают частоту и понижают амплитуду сокращений сердца. В сердце имеются никотиновые холинэргические рецепторы (которые блокируются тубокурарином). Хромаффинные клетки описаны также у миног; адреналин стимулирует у них сердечную деятельность [85].
Из сказанного можно сделать следующие выводы.
1. У круглоротых иннервация сердца либо отсутствует, либо представлена только возбуждающими холинэргическими нервами.
2. Для всех остальных позвоночных характерно наличие тормозной вагусной холинэргической иннервации мускаринового типа (блокируется атропином).
: 3. У некоторых рыб и амфибий возбуждающие адренэргиче-
ские волокна идут в составе блуждающих нервов, а у млекопитающих, птиц и рептилий — в составе обособленного симпатического ствола. Хромаффинные клетки, содержащие катехоламины, обнаружены в сердце всех позвоночных, но функция этих клеток неясна.
' Ритмические потенциалы действия в сердце куриного эмбриона появляются на стадии 9 сомитов, а спайки с овершутами на стадии 20 сомитов [101].
Потенциалы сердечной мышцы эмбриона имеют плато и сход--аы с потенциалами сердца взрослой особи (рис. 20-9) [114]. До-гех пор, пока не сформировалась вагусная иннервация, сердце-эмбриона либо совершенно нечувствительно к ацетилхолину (например, у Fundtdus), либо отвечает торможением лишь на действие высоких концентраций ацетилхолина (например, у цыпленка; пли крысы). В онтогенезе активность ацетилхолинэстеразьг воз* >5—2067
386
Глава 20
растает и достигает максимума по завершении развития иннервации.
Лимфатические сердца амфибий и некоторых рыб (угорь) обычно регулируются спинным мозгом. Перерезка нервных связей со спинным мозгом приводит к остановке лимфатического сердца,
Рис. 20-9. Внутриклеточные .потенциалы действия для различных областей сердца куриного эмбриона [114].
I — дорсальная сторона; II — вентральная сторона.
однако в дальнейшем возможно возобновление нерегулярных сокращений. Передние и задние ипсилатеральные лимфатические сердца в нормальных условиях функционируют синхронно, но каждое из них управляется отдельным участком спинного мозга. Местное охлаждение или согревание спинного мозга замедляет или ускоряет ритм сокращений лимфатического сердца. На каждое сокращение лимфатического сердца приходится 5—7 разрядов спинального нейрона. Лимфатическое сердце, трансплантированное в лимфатический мешок языка, сокращается интенсивно, хотя и спастически. У жабы лимфатическое сердце генерирует осциллирующий потенциал действия, по-видимому, благодаря суммации с моторными импульсами, которые предшествуют потенциалу действия самого сердца [42]. У интактных лягушек ацетилхолин вызывает увеличение амплитуды и силы сокращений, однако вы
Циркуляция жидкостей в организме 387
сокие концентрации ацетилхолина приводят к остановке лимфатического сердца [41].
Таким образом, для системного сердца позвоночных характерно поддержание постоянной активности с адаптацией к изменяющейся потребности в кровоснабжении. Миокард как целое, обладает миогенной, или спонтанной, активностью и проводит возбуждение как функциональный синцитий. Его потенциал действия имеет плато, так как фаза деполяризации при каждом сокращении достаточно продолжительна. Сила сокращения сердца тем больше, чем сильнее исходное растяжение. Отдельные области миокарда специализируются, превращаясь в пейсмекер, проводящую систему, сократительные элементы. Регуляция работы сердечной мышцы осуществляется как благодаря прямым реакциям мышечных волокон на растяжение и на местные химические воздействия (ионы, катехоламины), так и через гормональные и рефлекторные механизмы.
Сердца моллюсков
У моллюсков сердца относятся к миогенным. У головоногих в сердце или вблизи него встречаются нервные клетки, которые являются вторыми нейронами регуляторных нервов. У мидии или улитки любой участок сердца способен генерировать сокращения, как распространяющиеся на все сердце, так и локальные. У двустворчатых моллюсков сердце состоит из двух тонкостенных предсердий и более мускулистого желудочка, через который проходит кишка. У головоногих есть системное сердце, которое состоит из двух предсердий и желудочка, а также добавочные жаберные сердца.
Сердца моллюсков отличаются высокой чувствительностью к давлению. Они неспособны развить максимальное сокращение без предварительного растяжения; при повышении внутрисердечного давления сокращения сердца становятся сильнее и чаще. У Octopus при внутрисердечном давлении ниже 2 см вод. ст. зарегистрировать сокращения сердца не удается, при 8,5 см вод. ст. частота сокращений составляет 69 в 1 мин, а при 4,5 см вод. ст.—52 [49]. Сокращение сердца у улитки Rapana происходит при давлении 40 см вод. ст., а у Aplysia сердце сокращается уже при 2 см вод. ст. [72].
Электрокардиограмма моллюска состоит из нескольких медленных волн, часто с более быстрым двухфазным компонентом в самом начале сокращения и более медленным компонентом, совпадающим с последующей частью сокращения. При внутриклеточном микроэлектродном отведении у устрицы в некоторых волокнах зарегистрированы пейсмекерные потенциалы, предшествующие спайку. В сердце улитки зарегистрированы пейсмекерные 25*
388
Глава 20
потенциалы, спайки и плато [72]. .Плато иногда следует за спай-ком аналогично тому, что наблюдается при отведении от желудочка у позвоночных. Амплитуда спайка у устрицы достигает 53 мВ при потенциале покоя 57 мВ [78, 80].
Отводя потенциалы от двух участков сердца улитки, соединен-, ных мышечным мостиком, можно наблюдать распространение возбуждения [43, 44]. Для возникновения нормального спайка необходимы натрий и кальций. При отношении Ca/Na, равном 4,2-10-5, амплитуда спайка пропорциональна Na0, тогда как в среде, лишенной натрия, высота спайка пропорциональна Са0; приращение амплитуды составляет 22 мВ у Mytilus и 19 мВ у устрицы (на единицу логарифма концентрации Са2+). Мп снижает спайки точно так же, как и в других системах [82]. Анодная поляризация волокон в сердце устрицы устраняет плато. Гиперполя-ризованные волокна (большое значение £m) дают высокие спайки, возникающие благодаря притоку натрия; при низком потенциале покоя возникают небольшие спайки под влиянием кальция [78, 81].
Сердечные волокна устрицы содержат толстые и тонкие мышечные нити. Т-системы у этих животных отсутствуют, а контакты по типу нексусов обнаруживаются; проведение возбуждения имеет, по-видимому, электрическую природу, как и в сердечной мышце позвоночных [78, 81].
У большинства моллюсков сердце снабжено как тормозными, так и ускоряющими нервами; у двустворчатых моллюсков и брюхоногих оба типа волокон начинаются в висцеральных ганглиях. У некоторых животных (например, Муа, Anodonta и Мегсе-naria) преобладает тормозная иннервация; у других (например, Chiton, Aplysia, Haliotis и Ariolimax) при раздражении висцеральных ганглиев чаще возникает ускорение ритма. У Helix, Limax и Triopha тормозные и ускоряющие волокна берут начало в плевральном ганглии. У Mercenaria за ускорением часто следует нервное торможение, и если тормозные нервные окончания блокировать митолоном (бензохиноном), то в результате стимуляции нерва ритм ускоряется [205]. У Dolabella задняя часть висцерального ганглия замедляет, а передняя — ускоряет ритм сердца. Висцеральные нервы Octopus снижают частоту сокращений; в этих стволах содержатся, однако, и ускоряющие волокна с более высоким порогом раздражения, чем у тормозных. У головоногих сокращение жаберных сердец регулируется с помощью висцеральных ганглиев. У двустворчатого моллюска Clinocardium источником ускоряющих импульсов служит церебральный ганглий, а источником тормозных — висцеральный. Вытяжению ноги предшествует стимуляция сердца, а сокращению ноги — торможение сердца [165]. У Aplysia стимуляция висцерального нерва, идущего к сердцу, может вызвать как возбуждающие синаптические потенциалы с длительностью фазы нарастания 35 мс, так и тормоз
Циркуляция жидкостей в организме
389
ные синаптические потенциалы с длительностью фазы нарастания 27 мс [106].
Ацетилхолин тормозит сердечную деятельность у большинства моллюсков. Эзерин усиливает нейрогенное торможение и действие ацетилхолина у таких моллюсков, как Mercenarla и Dolabefla. Бензохинон является антагонистом как ацетилхолина, так и нервного торможения у Mercenaria (но не у Helix). Во время нервного торможения у Mercenaria и Dolabella в оттекающем от сердца перфузате появляется тормозный агент, по всей вероятности ацетилхолин. Однако перфузат, оттекающий от сердца во время стимуляции висцеральных нервов у Octopus и Eledone, ацетилхолина не содержит [49]. У Helix стимуляция висцеральных нервов ведет к выделению агента, который оказывает тормозное действие на сердце другой улитки, но это вещество, по-видимому, не ацетилхолин [96]. У некоторых двустворчатых моллюсков тормозным действием обладают чрезвычайно малые концентрации ацетилхолина (10~12—10~10 iM у Mercenaria); это торможение можно блокировать тетраэтиламмонием или митолоном, но атропин на него не влияет. Таким образом, молекулы, служащие рецепторами ацетилхолина, у моллюсков и позвоночных различны. Так, у Lymnaea ацетилхолин в концентрации 10~12 М тормозит сокращение сердца, а бензохинон блокирует этот эффект [152, 153]. У большинства видов, имеющих низкий порог торможения частоты и амплитуды сокращений сердца, при действии высоких концентраций ацетилхолина может возникнуть противоположный эффект — тоническое возбуждение сердца. Так, ацетилхолин в высокой концентрации может вызвать сокращение неподвижного сердца. У других видов двустворчатых моллюсков, например Mytilus, Gryphaea и Modiolus, ацетилхолин в низких концентрациях на работу сердца не влияет, а в концентрации 10-5—10~4М оказывает стимулирующее действие (рис. 20-10). Сопоставляя характер ацетилхолинового эффекта (тормозный или стимулирующий) у 40 разных видов моллюсков, никакой корреляции с экологией или систематическим положением этих животных отметить не удается [56]. Ацетилхолин снижает частоту и амплитуду сокращений сердца устрицы и вызывает гиперполяризацию мышечных волокон — эффект, который не возникает в отсутствие С1. Таким образом, тормозное действие ацетилхолина состоит в повышении хлорной проводимости при потенциале реверсии 68мВ [464]. У Mytilus ацетилхолин повышает частоту сокращений сердца и деполяризует мышечные волокна; в среде, лишенной Na, -подобный ответ не возникает. По-видимому, возбуждающее действие ацетилхолина реализуется путем повышения натриевой проводимости [164]. Эффект ацетилхолина — тормозный или возбуждающий — зависит от молекулярной организации мембран.
Возбуждающим медиатором висцеральных нервных волокон, ^возможно, служит катехоламин. У Mercenaria 5-окситриптамин
390
Глава 20
(5-ОТ) в концентрации 10-7 М оказывает возбуждающее действие на сердце, а метил-альфа-лизергиновая кислота блокирует возбуждающий эффект как 5-ОТ, так и стимуляции нерва [55]. Истощение запасов 5-ОТ в нервных волокнах под влиянием резерпина прекращает процесс возбуждения в них [118]. Сходным образом 5-ОТ в концентрации 10-10 М оказывает возбуждающее действие на сердце Lytnnaea; лизергиновая кислота блокирует этот эффект. Дофамин в концентрации 10-10 М и адреналин в
Рис. 20-10. Действие ацетилхолина (момент нанесения отмечен стрелкой) на электрическую активность сердец Mytilus (I) и устрицы (//) [>164].
концентрации 10~9 М вызывают увеличение амплитуды и частоты сердечных сокращений [153]. В экспериментах с перфузией установлено, что для сердца Helix 5-ОТ служит возбуждающим медиатором [152].
Как системные, так и жаберные сердца головоногих моллюсков богато иннервированы [5]; однако нейромедиаторы у них пока не найдены.
Сердца оболочников
У оболочников сердце представляет собой тонкостенную трубку, лишенную клапанов. Такое сердце гонит кровь то в одном направлении, то в противоположном, изменяя характер сокращений. Пейсмекеры имеются у обоих концов такого сердца. Частота сокращений сердца у оболочников достигает максимума, когда давление в нем составляет 5 мм вод. ст. [102, 103]. Изменение направления потока в изолированном сердце происходит в отсутствие изменений давления, и если на сердце наложить лигатуру у его середины, то оба конца будут сокращаться независимо друг от друга. Таким образом, перемена направления потока не является следствием создаваемого давления. Электрическая стимуляция с интервалами около 2,6 с может вызывать сокращения
Циркуляция жидкостей в организме 391
сердца; наименьший порог возбуждения обнаружен на концах трубки. По-видимому, группы клеток на концах сердечной трубки, связанные электрическими контактами, отличаются периодическими колебаниями возбудимости [7]. У Ciona нормальная частота сокращений сердца составляет около 50 в 1 мин. Порог эффективного раздражения сердца максимален в средней его
Рис. 20-111. Распространение фронта волны сокращения вдоль сердца асцидии Ciona [1102].
Вверху — эффективные потенциалы действия, отведенные от 15 точек, расположенных вдоль сердца соответственно цифрам на нижнем рисунке. Фронт волны штрих-пунктирными линиями (интервал 0,2 с).
части и минимален вблизи концевых пейсмекеров [103]. Регистрация внутриклеточных потенциалов обнаруживает спайки высотой 75 мВ, потенциал покоя—71 мВ при длительности около 1,2 с [102] (рис. 20-11). Мышечные волокна однослойной стенки сердца имеют размер около 5-100 мкм и ориентированы под углом 70° к продольной оси органа. Фронт волны возбуждения параллелен оси клетки, а скорость проведения составляет 7,7 см/с при 10 °C. Кроме того, возбуждение распространяется из точки стимуляции в направлении, перпендикулярном оси клетки, со скоростью 0,87 см/с [103]. Распространение возбуждения, как показывает регистрация внутриклеточными электродами, происходит посредством электрической передачи в 25—50% соприкасающихся друг с другом волокон, причем сопротивление области кон
392
Глава 20
такта между клетками составляет 0,2 Ом/см2. Скорость провед е-> ния возбуждения в концевых областях сердечной трубки выше, чем в средней ее части [103].
Удаление центрального ганглия не влияет на активность серд* ца — рефлекторной регуляции оно, по-видимому, не имеет. Указывают, что ацетилхолин в концентрации 10-6 iM, а также эзерин не влияют на сердце Ciona [156]. По другим данным вскрытое сердце может прекратить сокращения на 1 мин при действии ацетилхолина в концентрации 10~8 М; эффект этот предотвращается атропином [102]. Адреналин в разведении 10~5 IM сначала ускоряет ритм сердца, а затем приводит к его остановке. Данных об опытах со стимуляцией ганглия в литературе не приведено.
Сердца ракообразных и Limulus
У большинства ракообразных, а также у мечехвоста (Limulus) в ганглиях дорсальной стороны сердца имеются нервные клетки, генерирующие волну возбуждения, которая вызывает сокращения сердца. У Ligia такой сердечный ганглий состоит из шести нейронов, у Astacus — из 15, у Squilla— из 16. Многие десятиногие из подотрядов длиннохвостых, мягкохвостых и короткохвостых имеют по 9 нейронов [4]. Омары Panulirus и Homarus имеют в ганглии 5 больших и 4 малых нейрона. Дорсальный сердечный ганглий Limulus содержит множество мелких мультиполяр-ных нейронов, перемежающихся с большими униполярными клетками.
В сердце Limulus имеется 8 пар отверстий, которые делят его на 9 сегментов. Кровь поступает в сердце из окружающего его перикарда через эти отверстия, снабженные клапанами. От передней половины сердца отходят 5 парных артерий и одна передняя срединная артерия. По обе стороны сердца лежат два латеральных нерва, связанные боковыми ветвями со срединным ганглием — пейсмекером. Локальное согревание сердца ускоряет ритм его сокращений. Особенно сильно действует тепловое раздражение области 4-го и 5-го сегментов. Удаление пейсмекерного ганглия ведет к остановке сердца, но если после этого сердце растягивается кровью, то могут появиться перистальтические сокращения. При помещении сердца в раствор, лишенный кальция, возникают локальные сокращения. Сокращения сердца, вызванные растяжением и отсутствием кальция, отличаются от нормальных синхронизированных сокращений [107]. У эмбриона Limulus миогенные перистальтические сокращения сердца начинаются с 22-го дня; нервы до 28-го дня не обнаруживаются [30].
Электрокардиограммы ракообразных и Limulus отражают быструю начальную деполяризацию, за которой следует непостоянное число осцилляций. Такая картина типична для тетануса (рис. 20-12). Каждое отдельное мышечное волокно генерирует ряд
Циркуляция жидкостей в организме
393
сливающихся синаптических потенциалов, из которых каждый соответствует одному нервному импульсу (рис. 20-13). У рака-отшельника два таких синаптических потенциала могут, сливаясь, вызвать овершут и спайк. Потенциалы сердечной мышцы обусловлены нарастанием кальциевой проводимости [197]. В сердечной
WJJ-i
II L—-1
Рис. 20-112. Внутриклеточные потенциалы действия сердца ракообразных.
I — Maja; II — Carcinus; III — Palaemon.
мышце Squi/la слияние постсинаптических потенциалов облегчается, когда они следуют друг за другом с интервалами меньше 630 мс [24]. У Limulus ПСП могут вызвать длительную деполяризацию; опыты по стимуляции боковых нервов показывают, что одно мышечное волокно могут иннервировать до 6 разных аксонов, принадлежащих разным сегментам [1]. ПСП носят градуальный характер; у Limulus максимум деполяризации может достигать 35 мВ при потенциале покоя —45 мВ [150]. Гиперполяризация мышечных волокон увеличивает амплитуду ПСП, а деполяризация ее снижает [179]. При множественной иннервации .мышечного волокна приближение к синхронности сокращений сердца достигается за счет проведения возбуждения по сердечным нейронам.
Сердечный ганглий омара состоит из пяти крупных передних я четырех более мелких задних нейронов (рис. 20-14). С по-
Рис. 20-16. Электрокардиограмма Limulus (при разных скоростях развертки) [ilO7a].
Верхние кривые внутриклеточные потенциалы миокарда, нижние — внеклеточные записи разрядов ганглия. Фаза нарастания начальной деполяризации мышцы имеет волны, соответствующие синаптическим потенциалам. Калибровка верхних записей — 10 мВ, нижних — 40 мкВ. /— 500 мс, //—200 мс, ///—100 мс.
Рис. 20-14. Схема нервов и расположение нервных клеток в сердечном гангли» омара [64].
Пунктиром обозначены остин, цифрами — нейроны.
Циркуляция жидкостей в организме
395
мощью внеклеточного отведения показано, что в течение одного •сердечного залпа каждая клетка дает несколько импульсов (рис. 20-15). Четыре малых нейрона представляют собой пейсмекерные, или ведущие клетки, а более крупные нейроны являются двигательными, или ведомыми. Но если отделить крупные передние клетки от мелких задних, то и у ведомых можно наблюдать
некоторую спонтанную активность. От ведомых нейронов удалось получить внутриклеточные записи; на них видны небольшие градуальные волны, часть которых является выражением постсинаптических потенциалов, вызванных спайками пейсмекерных клеток. Эти волны могут перейти в спайк, возникающий у основания аксона, распространяющийся в обратном направлении к телу нейрона и проводящийся к мышце. Сами тела нейронов, по-видимому, не генерируют спайков; путем отведения двойными
микроэлектродами показано, что ведомые клетки имеют слабый электрический контакт, вследствие чего передаваемый потенциал уменьшается здесь в 2—4 раза, и что медленные потенциалы (но не спайки) могут распространяться от
клетки к клетке [61]. Регистрация
Рис. 20-15. Потенциалы действия нервных клеток сердечного ганглия ракообразного Squilla [201].
I — от нейрона 5; II — от нейрона 6: III — внеклеточное отведение; внизу — те же отведения при медленной развертке.
потенциалов крупных клеток
показывает, что спайки могут возникать в разветвлениях аксона и не распространяться по всему нейрону [64, 65]. Поскольку крупные клетки в отсутствие ведущих способны на спонтанную актив-
ность, некоторые медленные волны, зарегистрированные в них, следует считать пейсмекерными потенциалами, а не синаптическими [38]. Источником пейсмекерных волн может быть нейропиль, •образованный отростками клеток. В мышечных волокнах интактного сердца Homarus были обнаружены спонтанные мВПСП; показано, что стимуляция интракардиальных нейронов вызывает облегчение постсинаптических потенциалов [7а].
Сердечный^ ганглий Squilla содержит 16 нейронов; различить среди них пейсмекерные и ведомые клетки не удается. Каждый нейрон разряжается 4—6 раз в течение сердечного цикла. Клетки, находящиеся на концах ганглия, чаще оказываются спонтанно •активными, чем расположенные в средней его части. Скорость проведения возбуждения составляет приблизительно 1,5 м/с [24]. Пейсмекерные клетки генерируют пейсмекерные потенциалы и спайки. В записях от биполярных нейронов удается уловить спай-
396
Глава 20
ковые компоненты каждого аксона и собственный потенциал тела клетки. Таким образом, возбуждение может распространяться по телу такого нейрона в двух направлениях. Нейроны связаны друг с другом электрическим контактом [201]. Спайки возникают на некотором расстоянии от тела нейрона, причем в униполярных клетках дальше, чем в биполярных [3].
У Limulus на записях, полученных путем отведения от тел больших нейронов, выявляются серии спайков, соответствующие суммарному разряду ганглия. Иногда спайки возникают между залпами, а иногда первому спайку предшествует медленный потенциал, по-видимому, ВПСП (рис. .20-16). Спайки в залпе варьируют по амплитуде и предположительно происходят из разных, ветвей аксона. На некоторых записях видны два типа спайков в одной и той же клетке, что указывает на существование двух независимых участков возбуждения. Редкие отведения от одиночных малых нейронов выявляют пейсмекерные потенциалы, дающие-начало одному или двум спайкам большой амплитуды. По-видимому, один или два пейсмекерных спайка служат триггером, запускающим серии спайков в ведомых клетках [Ю7].
По механическим свойствам сердечная мышца ракообразных и моллюсков отличается от сердечной мышцы позвоночных. Сердца моллюсков и ракообразных способны к градуальным, а также* к тетаническим сокращениям. После ганглионэктомии сердце-Limulus способно к спонтанным сокращениям, которые сопровождаются большими спайками, зависящими от концентрации натрия-в окружающей среде, если последняя лишена кальция и если сердце растянуто до двойного диаметра по сравнению с нормой. Кроме того, сердце Limulus проявляет способность к перистальтическим сокращениям, сопровождающимся выраженными спайками. В сердце эмбриона Limulus обнаруживаются локальные сокращения еще до появления ганглиозных клеток. Следовательно^ сердечная мышца способна к миогенной активности, но в обычных условиях сокращается благодаря импульсам пейсмекерного ганглия [107].
У ракообразных и Limulus к сердцу подходят несколько регуляторных нервов, окончания которых распределяются в пейсме-керном ганглии и сердечной мышце. Сердце Callinectes получает две пары ускоряющих и одну пару тормозных нервов. У Limulus тормозные нервы выходят из задней части мозга; у ракообразных и тормозные, и ускоряющие нервы выходят из околопищеводного-ганглия, причем тормозные расположены впереди ускоряющих.
У Squilla тормозные импульсы гиперполяризуют сердечные нейроны. Порог для образования спайка обычно соответствует деполяризации 7 мВ, а на фоне торможения—15 мВ. Ускоряющие нервы действуют на мембрану пейсмекерных клеток и увеличивают крутизну нарастания пейсмекерных потенциалов [201]. У Limulus 7-й и 8-й нервы, идущие от центрального кольца, пре
Циркуляция жидкостей в организме
397
имущественно тормозные, а 9—13-й — возбуждающие [27]. Стимуляция последних наиболее эффективна при частоте 5 импульсов в 1 с, а латентный период может достигать 30—40 с. Тормозные ответы наиболее выражены при стимуляции с частотой 40 имп/с [27, 40].
У ракообразных и Limulus не удалось точно определить ни возбуждающий, ни тормозный медиаторы. Ацетилхолин вызывает возбуждение клеток ганглия, но лишь в сравнительно высоких
Рис. 20-il6. Потенциалы действия нейронов сердечного ганглия Limulus. (Любезно предоставлено Ф. Лангом.)
I — внутриклеточное отведение; II — внеклеточное отведение. А. Залпы больших ведомых нейронов. Б. Одиночные спайки малого пейсмекерного нейрона; каждый ведомый нейрон отвечает длительной деполяризацией и множественными спайками на одиночный спайк ведущей (пейсмекерной) клетки. В. Развернутая запись потенциалов пейсмекерной клетки, показывающая, что пейсмекерный потенциал предшествует спайку. Калибровка: А. I — 10 мВ, II — 20 мкВ, 500 мс. Б. I — 20 мВ, II — 20 мкВ, 500 мс. В. 20 мВ, 40 мс.
Концентрациях; применение блокаторов и потенцирующих агентов не вызывает эффектов, характерных для холинэргической иннервации. Глутамат, который известен как возбуждающий медиатор у некоторых членистоногих, вызывает сокращение сердца Limulus после ганглионэктомии в разведении 10-6 М. При интактной иннервации глутамат оказывает стимулирующее действие лишь в концентрации 10~4 М [1]. Гамма-аминомасляная кислота (ГАМК) — ингибитор нервно-мышечных синапсов у ракообразных— уменьшает длительность и частоту разрядов ганглия в концентрации 10~4 М [1]. Возбудителем клеток ганглия у Squill а служит 5-ОТ [24]. У некоторых короткохвостых крабов группа нейросекреторных клеток, входящая в состав перикардиальной железы, возбуждает активность сердца при посредстве вещества, близкого к 5-ОТ, но не самого 5-ОТ [6]. Вещество, экстрагируемое из сердца Carcinus, ускоряет ритм сердца у других животных GOT^^rS]*0’ является производным триптамина (возможно, Ацетилхолин не оказывает влияния на сердце Daphnia, электрокардиограмма которой не имеет осцилляторного характера.
Сердечные ганглии ракообразных и Limulus могут служить моделью нервной интеграции. Здесь немногочисленные нейроны взаимодействуют чрезвычайно точно и согласованно во времени. Есть основания надеяться, что с помощью этой модели удастся
398
Глава 20
получить интересные данные относительно природы спонтанного ритма нейронов. Сердечная мышца этих животных, хотя и способная к миогенной активности, находится под нервным контролем и напоминает в большей части своих проявлений поперечнополосатые соматические мышцы.
Сердца пауков, скорпионов и насекомых
У тарантула Eurypelma сердце имеет ганглий с униполярными, биполярными и мультиполярными нейронами; ЭКГ тарантула носит осцилляторный характер, в основном такой же, как у Limulus. Следовательно, сердце этого животного можно признать нейрогенным. Ганглий возбуждается ацетилхолином и адреналином, а также норадреналином в концентрациях 10-6 М, а глутаматом и 5-ОТ — в концентрациях 10-4 М. Гамма-аминомасляная кислота оказывает тормозное действие в концентрации 10-5 М [162, 163].
У скорпиона Urodacus сердце имеет пейсмекерный ганглий, удаление которого ведет к остановке сердца; даже будучи изолированным, этот ганглий генерирует ритмичные залпы. Сердце получает стимулирующую и тормозную иннервацию от центральной нервной системы. Ацетилхолин не вызывает ускорения ритма; в концентрации 10~4 М он замедляет ритм. Однако эксперименты с применением различных фармакологических агентов не дают оснований считать ацетилхолин тормозным медиатором для сердца Urodacus [212].
Поскольку у насекомых сердце не должно обеспечивать респираторный транспорт и поскольку развитие разных насекомых происходит по-разному, не удивительно, что их сердца отличаются значительным разнообразием. Одни имеют нейрогенную природу, другие — миогенную. Методом внутриклеточного отведения сердечной мышце мухи Sarcophaga выявлены суммарные осцилляции, сходные с обнаруженными у Limulus [25]. Осцилляторная ЭКГ встречается у Melanoplus, Galleria и Dytiscus. У цикад пей--смекерные ганглии расположены во 2—7-м сегментах [79]. У таракана, кузнечика, сверчка, медоносной пчелы отмечено повышение частоты сокращений под действием ацетилхолина; у некоторых из них обнаружена высокая чувствительность к этиловому эфиру, характерная для нейрогенных сердец ракообразных. Сердца многих насекомых обычно находятся в состоянии растяжения благодаря дорсальным связкам и крыловидным мышцам. У некоторых видов (но не у всех) перерезка этих мышц и связок ведет к остановке сердца. У цикады Cryptotympana ЭКГ после перерезки крыловидных мышц упрощается, но все же сохраняет осцилляторный характер [79].
Отсутствие нейронов в сердце отмечали у многих насекомых, особенно в личиночной стадии. В сердце личинки А пах и взрослой
\ Циркуляция жидкостей в организме 399
-------------
\
формы Belostdma нет нервных клеток; такое сердце сокращается при сохранности дорсальных связок. Сердца личинок Galleria и Anopheles нечувствительны к ацетилхолину.
Сердце взрослого таракана Periplaneta имеет ганглий, состоящий из 6 нейронов, разряды которых координированы с сердечными сокращениями. Однако ритмичные сокращения не прекращаются и после удаления ганглия. Это позволяет считать, что* сердечный ганглий стимулируется растяжением, а сам в свою очередь повышает миогенную возбудимость. Ацетилхолин оказывает возбуждающее действие, возможно, путем усиления регуляторных влияний со стороны ганглия [129]. Перистальтические сокращения продолжаются и после блокады активности ганглия тетродотоксином. Изолированный сердечный ганглий не дает спайковых разрядов, а в ганглии in situ спайковая активность при растяжении сердца возрастает [172а]. Мышечные волокна получают полинейронную иннервацию через многочисленные окончания [130]. Нервные волокна, идущие из центральной нервной системы, способны модулировать активность как сердечного ганглия, так и самой мышцы.
Сердце бабочки Hyalophora cecropia может сокращаться в ответ на растяжение крыловидных мышц, достаточно интенсивное, чтобы вызвать их сокращение. Сердце не содержит нервных клеток и не имеет локализованной пейсмекерной зоны. На его сокращения не влияют ни ацетилхолин, ни адреналин, ни эфир, стало быть, оно относится к миогенным. Внутриклеточные потенциалы представляют собой спайк с овершутом и задержкой реполяризации— нечто напоминающее плато, характерное для сердца позвоночных (рис. 20-17). Кровь этой бабочки (как это характерно для многих насекомых; см. гл. 2) богата калием и магнием и бедна натрием. Потенциал покоя сердечной мышцы составляет —47 мВ, тогда как потенциал равновесия для калия равен —11,6 мВ. Следовательно, здесь потенциал покоя определяется не ионами калия, а другими ионами. Большие спайкообразные потенциалы действия продолжаются и в отсутствие натрия в среде, но если ее лишить кальция, они исчезают, и сердце останавливается. Миогенная активность зависит от кальция, и спайки отражают изменения кальциевой проводимости [124—128].
У насекомых регуляция работы сердца со стороны центральной нервной системы исследована недостаточно. У бабочки Sphinx сердце (в брюшке) изменяет частоту и амплитуду сокращений в соответствии с температурой области груди. Это происходит главным образом благодаря участию тормозных и возбудительных нервов [68].
Из сказанного ясно, что сердца скорпионов, пауков и насекомых весьма разнообразны в смысле источника сократительной деятельности: они могут быть миогенными, нейрогенными, могут
400
Глава 20
иметь местные рефлекторные регуляторные механизмы. Пейсме-керный ганглий, если он существует, часто не реагирует на гуморальные воздействия, подобно пейсмекерным ганглиям других членистоногих.
Рис. 20-17. Внутриклеточные потенциалы действия бабочки Hyalophora. - (Любезно предоставлено Ф. Мак-Канном.)
А. Клетка дает пейсмекерные потенциалы и спайки. Б. Ведомые клетки заднего (I) и переднего (II) конца сердца.
Сердца аннелид
Многие кровеносные сосуды аннелид обладают спонтанной активностью. У гигантского дождевого червя Glossoscolex дорсальный и вентральный сосуды имеют независимую частоту сокращений, причем вентральный сокращается синхронно с боковыми «сердцами». В сердце Arenicola и Lumbricus описаны нервные клетки; ритм этих сердец ускоряется под действием ацетилхолина [142], адреналина и норадреналина [154]. Растяжение кровью является важным фактором для возникновения сокращений.
Циркуляция жидкостей в организме 401
Сердце пиявки лишено нервных клеток и реагирует на адреналин ускорением, а на мускарин — замедлением ритма; у этих животных передние корешки сегментарных нервов являются возбуждающими, а задние — тормозными [196].
Заключение
Чтобы обеспечить адаптацию организма к изменяющимся условиям, циркуляторная система должна реагировать на стресс; она должна обеспечивать доставку крови туда, где это в данный момент необходимо, и в таком количестве, какое требуется. Лишенные сердца незамкнутые циркуляторные системы (целом, псевдоцелом или гемоцель) оказываются достаточными в условиях низких респираторных и алиментарных потребностей. Однако увеличение размеров тела и повышение двигательной активности в процессе эволюции привело к появлению сердца в некоторых незамкнутых системах, например у ракообразных и моллюсков. Правда, в таких системах сердце еще неспособно создавать высокое давление и циркуляция осуществляется отчасти благодаря сокращениям соматической мускулатуры. Объем крови в таких системах велик, и перенос кислорода от жабр к мышцам происходит сравнительно медленно. У кольчецов имеются замкнутые системы кровеносных сосудов, которые служат для переноса кислорода, но главным циркуляторным механизмом, обеспечивающим питание и выделение, остается целомическая полость. Незамкнутая система имеется и у насекомых, но у них гемоцель выполняет лишь меньшую часть этих функций, а воздух доставляется непосредственно к мышцам по трахеям. У некоторых ракообразных артериальные сосуды перед открытием в тканевые пространства интенсивно ветвятся. У головоногих моллюсков и позвоночных циркуляторная система замкнутая; для нее характерны высокое системное давление и большая скорость кровотока при малом объеме крови. Гладкие мышцы кровеносных сосудов в разных органах и у разных видов варьируют по характеру адаптивных реакций на такие медиаторы, как ацетилхолин и катехоламины. У низших позвоночных имеется лимфатическая система с добавочными лимфатическими сердцами. У этих животных лимфатическая система играет более важную роль, чем у высших позвоночных.
У позвоночных существуют сложные рефлекторные и гормональные механизмы, а также механизмы местной сосудистой регуляции. Все это обеспечивает поддержание кровяного давления. Природа механизмов, регулирующих давление у беспозвоночных, неизвестна. Часто заполненные жидкостью полости выполняют не столько транспортную, сколько опорную функцию, создавая гидростатический скелет. Такой скелет способен приобретать жесткость или гибкость в зависимости от функциональных потребно-26—2067
402 Глава 20
стей животного; он особенно важен для роющих животных. Однако подобный скелет не обеспечивает локальных движений и ; не выполняет защитных функций, чем невыгодно отличается от I сегментарного и наружного скелетов. |
Сердечная мышца у всех видов животных сочетает в себе не- |
которые свойства висцеральной и соматической мускулатуры. 1
Сердце позвоночных сокращается по закону «все или ничего». I
У моллюсков и членистоногих это наблюдается не всегда. Мио- |
генный автоматизм сердца (более примитивный) сохраняется у |
взрослых позвоночных и моллюсков, но у многих зрелых форм ’
членистоногих этот механизм заменен нейрогенным. Небольшое число нейронов в пейсмекерных ганглиях ракообразных составляет простую интегративную систему. Для сердца характерны длительные потенциалы действия, различные модификации пейсмекеров, проводящая система и изменчивая длительность деполяризации. Здесь встречаются разные виды клеточных мембран, одним из которых (таких большинство) свойственна натриевая проводимость, другим — кальциевая.
Большинству животных присуща нервная регуляция сердечной J деятельности. Этот механизм чаще представлен тормозными нер-вами, иногда — тормозными и ускоряющими. У эмбрионов сердце I обычно начинает сокращаться еще до формирования иннерва- -ции, а у некоторых животных (у Artemia, у многих насекомых, например у личинки Anopheles, и у миксин) сердце обладает мио- j генным автоматизмом и лишено иннервации. Сердечные нервы N осуществляют свою функцию путем выделения химических медиа- j торов. У позвоночных и некоторых моллюсков тормозным медиа-тором служит ацетилхолин. К ускоряющим медиаторам относятся . адреналин и норадреналин (у позвоночных) и серотонин (у неко- ’ торых моллюсков). Нейромедиаторы способны изменять времен- 1 ной ход мембранной проводимости, они влияют также на функ- 1 ции внутриклеточных структур. d
Эволюция циркуляторных механизмов шла по нескольким параллельным путям, но все они вели к тому, чтобы обеспечить J более эффективное выполнение транспортных функций данным объемом жидкости. Различные типы периферической циркуляции представляют собой системы, удобные для кибернетического изучения и моделирования механизмов гомеостаза.
СПИСОК ЛИТЕРАТУРЫ
1. Abbott В. С., Lang F., Parnas I., Comp. Biochem. Physiol. 28: 149—158; also pp. 232—243 in: Comparative Physiology of the Heart, edited by F. McCann, Birkhauser, Basel, 1969. Muscle and ganglion of heart of Limulus.
2. Ai N., Sci. Rep. Tokyo Kyoiku Daigaku В 12: 131—149, 1966. Pacemaker of heart of Ligia.
3. Akiyama T., Comp. Biochem. Physiol. 27: 197—211, 1968. Pacemaker ganglion of Squilla.
Циркуляция жидкостей в организме
403
4. Alexandrowicz J. S., J. Marine Biol..Assoc. U.K. 31: 85—96, 1952; 33: 709— 719, 1954; 34: 47—53, 1955; also Quart. J. Mier. Sci. 75: 291—309, 1932. Innervation of crustacean hearts.
5. Alexandrowicz J. S., Acta Zool. 41: 65—100, 1960. Innervation of heart of Sepia.
6. Alexandrowicz J. S„ Carlisle D. B., J. Marine Biol. Assoc. U. K. 32: 175—192, 1953. Pericardial organs in Crustacea.
7. Anderson M., J. Exp. Biol. 49: 363—385, 1968. Initiation and reversal of heart beat in Ciona.
7a. Anderson M., Cooke I. M„ J. Exp. Biol. 55: 449—468, 1971. Activation of heart muscle in lobster.
8. Ansell A. D., Trueman E. A., J. Exp. Biol. 46: 105—115, 1967. Burrowing by M.er cetiaria.
9. Baker L. A., White F. N., Comp. Biochem. Physiol. 35: 253—262, 1970. Cardiac output in Iguana.
10. Banchero N., Grover R. F., Will I. A., Amer. J. Physiol. 220: 422—427, 1971. Pulmonary hypertension in llama at high altitude.
11. Barr L., Berger W., Pfliiger. Arch. 279: 192—194, 1964; also Barr L., Dewey M. M., Berger W., J. Gen. Physiol. 48: 797—823, 1965. Conduction and structure in cardiac muscle.
12. Baumgarten-Schulmann D., Piiper J., Respir. Physiol. 5: 317—325, 1968. Cardiac output in dogfish.
13. Bayer R., Z. vergl. Physiol. 58: 76—135, 1968. Circulation in Locusta.
14. Belkin D. A., Copeia 321—330, 1964. Changes in heart rate during diving in Pseudemys.
15. Berkson H., Comp. Biochem. Physiol. 21: 507—524, 1967. Physiology of diving in Chelonia.
16. Blatchford J. Q., Comp. Biochem. Physiol. 39A: 193—202, 1971. Hemodynamics in Carcinus.
17. Bloom G. et al., Acta Physiol. Scand. 53 (Suppl. 185); 1—38, 1961. Catecholamines in hearts of cyclostomes.
18. Bohr D. F., pp. 342—355. In: Electrolytes and Cardiovascular Diseases, edited by E. Basjusz, Karger, Basel, 1965. Differences among vascular smooth muscles.
19. Bohr D. F. et al., Angiology 12: 478—485, 1961. Mechanical properties of resistance vessels
19a. Bond C. F., Gilbert P. W., Amer. J. Physiol. 194: 519—521, 1958. Blood volume of pigeon.
20. Brady A. J., Woodbury J. W., J. Physiol. 154: 385—407, 1960. Bioelectrics of frog ventricle.
20a. Brett J. R., Davis J. C., J. Fish. Res. Bd. Canad. 24: 1775—1790, 1967. Cardiovascular dynamics of salmon.
21. Bron К. M. et al., Science 152: 540—542, 1966. Vasomotor responses in diving mammals.
22. Brown A. C., J. Exp. Biol. 41: 837—854, 1964. Blood distribution in sinuses of snail Bullia.
23. Brown E., Hoffer J., Wennesland R., Ann. Rev. Physiol. 19: 231—254, 1957. Blood volume and its regulation.
24. Brown H. F., J. Exp. Biol. 41: 689—700, 701—734, 1964. Electrophysiology of heart of Squilla.
25. Bruen J. P., Ballard R. C., Comp. Biochem. Physiol. 32: 227—236, 1970. Action potentials in heart of fly Sarcophaga.
25a. Bull J. M., Morris R., J. Exp. Biol. 471: 485—494, 1967. Blood volume in lamprey.
26. Burger J. W., Smythe С. M„ J. Cell. Comp. Physiol. 42: 369—383, 1953. Pattern of circulation in Homarus.
26a. Burnstock G., Pharmacol. Rev. 21: 247—313, 1969. Evolution of autonomic innervation of visceral and cardiovascular systems in vertebrates.
26*
404
Глава 20
26b. Burnstock G., Kirby S., J. Pharm. Pharmacol. 20: 404—406, 1968. Absence of inhibitory effects of catecholamines on lower vertebrate arterial strip preparations.
27. Bursey C. R., Pax R. A., Comp. Biochem. Physiol. 35: 41—48, 1970. Cardioregulation in Limulus.
28. Bursey C. R., Sherman R. G., Comp. Gen. Pharm. 1: 160—170, 1970. Spider cardiac physiology.
29. Burton A. C., Physiology and Biophysics of Circulation. Yearbook Medical Publishers, Chicago, 1968, 217 pp.
29a. Campbell G., Comp. Gen. Pharm. 2: 287—294, 1971. Pulmonary vascular bed in toad Bufo marinus.
30. Carlson A. J., Amer. J. Physiol. 12: 55—66, 67—74, 1904; 14: 16—53, 1965;
15: 207—234, 1906; 17: 179—210, 1906. Invertebrate hearts.
31. Carmeliet E. E., J. Gen. Physiol. 47: 501—530, 1964. Effects of ions on cardiac muscle.
32. Carvalho O. et al., Amer. J. Physiol. 196: 483—488, 1959. Electrical properties of rabbit atrium.
33. Chapman С. B., Jensen D., Widenthal K, Circ. Res. 12: 427—440, 1963. Circulatory control of hagfish.
34. Chapman G., J. Exp. BioL 49: 657—667, 1968. Hydraulic system of Urechis.
35. Chapman G., Newell G. E., Proc. Roy. Soc. Lond. В 145: 564—580, 1956. Hydraulics in molluscs.
36. van Citters R. L. et al., Comp. Biochem. Physiol. 16: 267—276, 1965. Cardiovascular adaptations to diving in elephant seal.
37. van Citters R. L., Kemper W. S., Franklin D. L., Comp. Biochem. Physiol. 24: 1035—1042, 1968. Science 152: 384—386, 1966. Blood pressure in giraffe.
38. Connor J. A., J. Exp. Biol. 50: 275—295, 1969. Pacemaker ganglion of lobster heart.
39. Conte F. P., Wagner H. H., Harris T. O., Amer. J. Physiol. 205: 533—540, 1963. Blood volume in Salmo.
40. D’Agrossa L. S., Amer. J. Physiol. 218: 530—535, 1970. Venous vasomotion in bat wing.
41. Day J. B., Rech R. H., Robb J. S., J. Cell. Comp. Physiol. 62: 33—42, 1963. Electrophysiology of frog lymph heart.
42. Del Castillo J., Sanchez V., J. Cell. Comp. Physiol. 57: 29—46, 196 L Bioelectrics of amphibian lymph heart.
43. Ebara A., Japan J. Physiol. 16: 371—379, 1966. Plateau potential of oyster heart.
44. Ebara A., Sci. Rep. Tokyo Kyoiku Duigaku В 13: 129—137, 1967. Conduction in heart of oyster.
45. Eliassen E., Nature 192: 1047—1049, 1961. Blood volume in hibernating hedgehog.
46. Ellis С. H„ Biol. Bull. 86: 41—50, 1944. Hydraulics of leg of spider.
47. Folkow B. et al., Acta Physiol. Scand. 70: 347—361, 1967. Cardiac output of duck.
48. Fozzard H. O., Sleator W., Amer. J. Physiol. 212: 945—951, 1967. Ionic basis of conductance in guinea pig atrial muscle.
49. Fredericq H., Bacq Z., Arch. Int. Physiol. Biochem. 49: 490—496, 1939; 50: 169—184, 1940. Effects of drugs on cephalopod hearts.
49a. Furness J. B., Moore J., Z. Zellforsch. 108: 150—176, 1970. Adrenergic innervation of cardiovascular system in lizard Trahysaurus.
50. Gannon B. J., Burnstock G., Comp. Biochem. Physiol. 29: 765—773, 1969. Excitatory innervation of fish heart.
51. Garey W., Comp. Biochem. Physiol. 33: 181—189, 1970. Blood volume and cardiac output of fishes.
52. Garnier D„ Coraboeuf E., Paille A., Compt. Rend. Soc. Biol. 155: 2430, 1961; 155: 1349—1357, 1962. Effects of ions on potentials in heart of guinea pig and rat.
Циркуляция жидкостей в организме 405»
52а. Glover W. Е. et al., J. Physiol. 194: 78—79P, 1967. Cooling of rabbit arteries.
53. Goetz R. H. et al., Circ. Res. 8: 1049—1059, 1960. Circulation of giraffe.
54. Gordon M. S., Fischer S., Tarifeno E., J. Exp. Biol. 53: 559—572, 1970. Physiology of heart of amphibious fish.
55. Greenberg M. J., Brit. J. Pharmacol. Chemother. 15: 365—374, 375—388, 1960. Pharmacology of heart of Venus.
56. Greenberg M. J., Comp. Biochem. Physiol. 14: 513—539, 1965. Responses of bivalve hearts to acetylcholine.
57. Greenberg M. J., Comp. Biochem. Physiol. 33: 259—295, 1970. Effect of acetylcholine on hearts of various molluscs.
58. Gregersen M. I., Shu C., pp. 244—261. In: Medical Physiology, edited by: В. B. Mountcastle, С. V. Mosby, St. Louis, 1968. Blood volume.
59. Guyton A. C., Cardiac Physiology. W. B. Saunders Co., Philadelphia, 1963,, 468 pp. Cardiac output and its regulation.
60. Hagiwara S., Ergebn. Biol. 24: 287—311, 1961. Electrophysiology of heart; in crustaceans.
61. Hagiwara S., Watanabe A., Saito N., J. Neurophysiol. 22: 554—572, 1959. Cardiac ganglion of lobster.
62. Hall E. K., Anat. Rec. Ill: 381—400, 1951; 118: 175—184, 1954; also J. Cell..
Comp. Physiol. 49: 187—200, 1957. Properties of embryonic rat heart.
63. Hart J. S., Canad. J. Res. D 21: 77—84, 1943. Cardiac output of fish.
64. Hartline D. K, J. Exp. Biol. 47: 327—340, 1967. Mapping of cardiac ganglion” of lobster.
65. Hartline D. K-> Cooke I. M., Science 164: 1080—1082, 1969. Control of cardiac ganglion in lobster.
66; Hauswirth O., Noble D., Tsien R. W., Science 162: 916—917, 1968. Action off epinephrine on cardiac muscle.
67. Heath J. E., Gasdorf E., Northcutt R. G., Comp. Biochem. Physiol. 26: 509’— 518, 1968. Blood pressure of turtle.
68. Heinrich B., Science 169: 606—607, 1970. Nervous control of heart of sphinx' moth.
69. Helm M. M., Trueman E. R., Comp. Biochem. Physiol. 21: 171—177, 1967. Heart rate in Mytilus.
70. Hemmingsen E. A., Douglas E. L., Comp. Biochem. Physiol. 33: 733—744, 1970. Blood volume in Antarctic fish.
71. Henderson A. H. et al., Proc. Sci. Exp. Biol. Med. 134: 930—932, 1970. Spe--cies differences in cardiac contractility.
72. Hill R. B., Irisawa H., Life Sci. 6: 1691—1696, 1967. Pressure relations inn heart of marine gastropod. .
73. Holt J. P., Rhode E. A., Peoples S. A., Nines H., Circ. Res. 10: 798—806,. 1962. Cardiac output in animals of different sizes.
74. Momma S., Suzuki S., Japan. J. Physiol. 16: 31—41, 1966. Sensory discharge-from aortic and atrial receptors.
75. Huggins S. E., Percoco R. A., Proc. Soc. Exp. Biol. Med. 119: 678—682, 1965.. Blood volume in alligators.
76. Hutter O. F., Trautwein W., J. Gen. Physiol. 39: 715—733, 1956. Bioelectrics-of the sinus venosus.
77. Hutton К. E., Amer. J. Physiol. 200: 1004—1006, 1961. Blood volume inturtles.
78. Irisawa H., Kobayashi M„ Japan. J. Physiol. 13: 421—430, 1963. Molluscam heart.
79. Irisawa H. et al., Japan. J. Physiol. 6: 150—161, 1956. Electrocardiogram of cicada.
80. Irisawa H. et al., Japan.'J. Physiol. 11: 162—168, 1961. Action potentials of oyster heart.
81. Irisawa H. et al., pp. 176—191. In: Comparative Physiology of the Heart,, edited by F. McCann. Birkhauser, Basel, 1969. Ion effects on bivalve heart.
406
Глава 20
82. Irisawa Н. et al., Comp. Biochem. Physiol. 23: 199—212, 1969; Japan J. Physiol. 18: 157—168, 1968. Effects of calcium and sodium on oyster and Mytilus heart.
83. Irving L. et al., J. Physiol. 84: 187—190, 1935. Branchial pressure receptors in dogfish.
84. Itazawa V., Bull. Japan. Soc. Sci. Fish. 36: 926—931, 1970. Cardiac output of fish.
85. Jensen D., Comp. Biochem. Physiol. 2: 181—201, 1961. Cardiac regulation in aneural heart of hagfish.
86. Jensen D., C©mp. Biochem. Physiol. 30: 685—690, 1969. Cardiac regulation in lamprey and trout.
87. Johansen K-, Biol. Bull. 118: 289—295, 1960. Circulation in hagfish My-xine.
88. Johansen K-, Comp. Biochem. Physiol. 7: 169—174, 1962. Cardiac output and aortic flow in Gadus.
89. Johansen K-> Acta Physiol. Scand. 60 (Suppl. 217): 1—82, 1963. Cardiovascular dynamics in Amphiuma.
90. Johansen K-, Acta Physiol. Scand. 62: 1—17, 1964; also Johansen K-, Aarhus T., Amer. J. Physiol. 205: 1167—1171, 1963. Blood flow during submersion in duck.
91. Johansen K-, J. Exp. Biol. 42: 475—480, 1965; also Comp. Biochem. Physiol. 5: 161—176, 1962. Cardiac output in Octopus.
92. Johansen K-, Ditadi A. S. F., Physiol. Zool. 39: 140—150, 1966. Double circulation in giant toad.
93. Johansen K-, Martin A. W., J. Exp. Biol. 43: 337—347, 1965. Circulation of giant earthworm Glossoscolex.
94. Johansen K-, Reite О. B., Comp. Biochem. Physiol. 12: 479—487, 1964. Cardioregulation in birds.
95. Jones H. D., Comp. Biochem. Physiol. 39A: 289—295, 1971. Circulatory pres-stires in Helix.
96. Jullien A., Ripplinger J., C. R. Soc. Biol. 144: 544—545, 1950; 146: 1326—1329, 1952. Cardiac regulation in snails and fishes.
96a. Kadatz R., Pfliiger. Arch. 252: 1—16, 1949. Effects of acetylcholine and noradrenaline on lung vessels in amphibians.
97. Kawakami У., Natelson В. H., DuBois A. B., J. Appl. Physiol. 23: 964—970, 1967. Cardiovascular reflexes of diving in man.
98. Kerkut G. A., Price M. A., Comp. Biochem. Physiol. 11: 45—52, 1964. Cardioaccelerators of Carcinus.
99. Keyes A., Bateman B., Biol. Bull. 63: 327—336, 1932. Branchial responses to epinephrine in eel.
100. Kirschner L. B., Amer. J. Physiol. 217: 596—604, 1969. Circulation in gills of eel.
101. Kreski V., Sleator W. W„ Life Sci. 5: 1441—1446. 1966. Development of electrical activity in chick embryo hearts.
102. Kriebel M. E., J. Gen. Physiol. 50: 2097—2107, 1967; 52: 46—59, 1968; also Biol. Bull. 134: 434—455, 1968; also Life Sci. 7: 181—186, 1968; also Amer. J. Physiol. 218: 1194—1200, 1970. Action potentials of pacemakers and conduction in tunicate heart.
103. Kriebel M. E., Biol. Bull. 135: 166—173, 1968. Pacemaker properties of tuni-
104. Kuffler S. W„ Dennis M. Harris A. J., Proc. Roy. Soc. Lond. В 177:
488—508, 509—539, 541—553, 555—563, 1971. Properties of synapses in vagal secondary neurons of frog heart.
105. Kulzer E., Z. vergl. Physiol. 56: 63—94, 1967. Heart beat in hibernating bats.
106. Kuwasawa K., Sci. Rep. Tokyo Kyoiku Diagaku В 13: 111—128, 1967. Nervous regulation in molluscan heart.
107. Lang F., J. Exp. Biol. 54: 815—826, 1971. Intracellular studies on pacemaker and follower neurons in Limulus cardiac ganglion.
Циркуляция жидкостей в организме
407
107а. Lang F., Biol. Bull. 141; 269—277, 1971. Myogenic activity in Limulus heart.
108. Laplaud J. et al., C. R. Soc. Biol. 155: 1990—1993, 1961. Electrocardiogram of Carcinus.
109. Larimer J. L., Tindel J. T., Animal Behav. 14: 239—245, 1966. Reflex regulation of heart in crayfish.
110. Laury J. V., J. Exp. Biol. 45: 343—356, 1966. Hydraulics of burrowing Urechis.
111. Lavoie M. E., Biol. Bull. Ill: 114—122, 1956. Hydrostatic movement in surffish.
112. Leblanc J., Ann. N. Y. Acad. Sci. 134: 721—732, 1966. Cardiovascular adaptations to cold in man.
113. Lee R. M., J. Insect Physiol. 6: 36—51, 1961. Blood volume in locust.
114. Lieberman M., de Carvalho A. P., J. Gen. Physiol. 49: 351—363, 1965. Electrocardiogram of chick embryo.
115. Lin V., Dawson C. A., Horvath S. M., Comp. Biochem. Physiol. 33: 901—909, 1970. Cardiac output in rat.
116. Lind A. R., Circulation 41: 173—176, 1970. Cardiovascular response to exercise in man.
117. Litchfield J. B„ Physiol. Zool. 31: 1—6, 1958. Blood pressure in infant rats.
117a. Little C., J. Exp. Biol. 46: 459—474, 1967. Hemolymph volume in snail Strombus.
118. Loveland R. E., Comp. Biochem. Physiol. 9: 95—104, 1963. Serotonin as excitatory transmitter to heart of Mercenaria.
119. Lund G. F., Dingle H., J. Exp. Biol. 48: 265—277, 1968. Vagal control of diving bradycardia in frog.
120. Magarey F. R., Stebbens W. E., J. Exp. Biol. Med. Sci. 35: 347—351, 1957. Blood pressure in sheep.
121. Markardt J. E., Comp. Biochem. Physiol. 33: 441—457, 1970. Regulation of heart rate in Peromyscus.
122. Martin A. W., J. Exp. Biol. 35: 260—279, 1958. Blood volume in molluscs.
123. Martini J., Honig C. R., Microvasc. Res. 1: 244—256, 1969. Capillary distribution in rat heart.
123a. Mayer N., Nibelle N., Comp. Biochem. Physiol. 31: 589—597, 1969. Blood volume in Anguilla.
124. McCann F. V., J. Gen. Physiol. 46: 803—821, 1963. Electrophysiology of insect heart
125. McCann F. V., Ann. N. Y. Acad. Sci. 127: 84—99, 1965; also Comp. Biochem. Physiol. 17: 599—608, 1966. Electrical properties of moth myocardium.
126. McCann F. V., Comp. Biochem. Physiol. 40A: 353—357, 1971. Ionic basis of moth heart potentials.
127. McCann F. V., Wira C. R., Comp. Biochem. Physiol. 22: 611—615, 1967. Ionic gradients in lepidopteran hearts.
128. McCann F. V., ed., Comparative Physiology of the Heart. Birkhauser, Basel, 1969.
129. Miller T„ J. Insect. Physiol. 14: 1265—1275, 1968. Neuronal control of cockroach heart.
130. Miller T., Usherwood P. N. R., J. Exp. Biol. 54: 329—348, 1971. Cardioregulation in cockroach.
131. Murdaugh H. V. et al., Amer. J. Physiol. 210: 176—180, 1966. Cardiac output in seal.
132. Nagel E. L. et al., Science 161: 898—900, 1968. Cerebral circulation in dolphin.
133. Nayler W. G., Chipperfield D., Amer. J. Physiol. 217: 609—614, 1969. Calcium binding by cardiac sarcoplasmic reticulum.
134. Niedergerke R., J. Physiol. 143: 486—503, 1958; 167: 515—550, 551—586,
1963. Movements of calcium ions in frog heart.
-408
Глава 20
135. Nijima A., Winter D. L., Science 159: 434—435, 1968. Baroreceptors in adrenal gland.
136. Noble P., Tsien R. W., J. Physiol. 195: 185—214, 1968. Membrane properties of cardiac muscle.
137. Ormond A. P., Rivera-Velez J. M., Proc. Soc. Exp. Biol. Med. 118: 600—602, 1965. Blood volume in rat.
138. Ostlund E., Fange R., Comp. Biochem. Physiol. 5: 307—309, 1962. Circulation in gill of fish.
139. Parer J. T., Metcalfe J., Resp. Physiol. 3: 151—159, 1967. Cardiac output mo-notremes.
140. Pax R. A., Comp. Biochem. Physiol. 28: 293—305, 1969. Cardioregulation in Limulus.
141. Popovic V., Amer. J. Physiol. 207: 1345—1348, 1964. Cardiac output of hibernating ground squirrels.
142. Prosser C. L., J. Cell. Comp. Physiol. 21: 295—305, 1943; also Biol. Bull.
98: 254—257, 1950. Pacemaker activity in Limulus and Arenicola.
143. Prosser C. L., Weinstein S. J., Physiol. Zool. 23: 113—124, 1950. Blood volume of catfish and crayfish.
144. Randall D. J., Shelton G., Comp. Biochem. Physiol. 9: 229—239, 1963. Respiratory and cardiovascular reflexes in fish.
145. Randall D. J., Smith J. C. Physiol. Zool. 40: 104—113, 1967. Cardiac regulation in hypoxia fish.
146. Redmond J. R„ J. Cell. Comp. Physiol. 46: 209—247, 1955. Cardiac output in crus taceans
147. Reuter H., J. Physiol. 192: 479—492, 1967; 197: 233—253, 1968. Membrane properties of Purkinje fibers.
148. Rhodin J. A., Missier A. G. P., Reid L. C. Circulation 24: 349—367, 1961. Conduction in beef heart.
149. Richmond C. R., Langham W. H., Trujillo T. T., J. Cell. Comp. Physiol. 59: 45—53, 1962. Body water in mammals.
150. Robb J. S., Rech R. H., J. Cell. Comp. Physiol. 63: 299—307, 1964. Electrocardiogram of Limulus.
.151 . Robertson J. D., Biol. Bull. 138: 157—183, 1970. Inulin Space in arthropods.
152. Rozsa K., Nagy I. Z., Comp. Biochem. Physiol. 23: 373—382, 1967. Neuro-endocrines of heart of snail.
153. Rozsa R., Perenyi L., Comp. Biochem. Physiol. 19: 105—113, 1966. Serotonin as excitor in Helix heart.
154. Rozsa R., Szoke L, Ann. Biol. Tihany 37: 85—97, 1970. Pharmacology of heart of Limulus.
1155. Satchell G. H„ Fed. Proc. 29: 1120—1123, 1970; also Satchell G. H., Hanson D., Johansen R., J. Exp. Biol. 52: 721—726, 1970. Regulation of peripheral circulation in fish.
155a. Satchell G. H., Circulation in Fishes. Cambridge University Press, 1971, 131 pp.
155b. Scholander P. F. et al., Science 161: 321—328, 1968; also Stromme S. B. et al., J. Appl. Physiol. 27: 123—126, 1969. Interstitial fluid pressures in vertebrates.
156. Scudder C. L., Akers T. R., Rarczmar A. G., Comp. Biochem. Physiol. 9: 307—312, 1963. Electrocardiogram of tunicate.
157. Seifen E., Schoer H„ Marshall J. M., Nature 202: 1223—1224, 1964. Membrane properties of rabbit atrium.
158. Serfaty A., Labat R., Berant A., Rev. Canad. Biol. 24: 1—5, 1965. Cardiac reaction to anoxia in carp.
159. Seyama L, Amer. J. Physiol. 216: 687—692, 1969; also Seyama L, Irisawa H., J. Gen. Physiol. 50: 505—518, 1967. Effect of sodium and calcium on heart of skate.
160. Seymour M. R., J. Exp. Biol. 51: 47—58, 1969. Hydrodynamics of Lumbri-cus.
Циркуляция жидкостей в организме
409»
161. Shelton G., Jones D. R., J. Exp. Biol. 42: 339—357, 1965; 43: 479—488, 1965;
49: 631—643, 1968. Blood pressure, volume regulation, and cardiac output in lower vertebrates.
162. Sherman R. D., Pax R. A., Comp. Biochem. Physiol. 26: 529—536, 1968; also-Comp. Gen. Pharm. 1: 171—184, 185—195, 1970. Spider cardiac physiology.
163. Sherman R. G., Pax R. A., Comp. Biochem. Physiol. 28: 487—489, 1969. Electrical activity in spider hearts.
164. Shigeto N., Amer. J. Physiol. 218: 1773—1779, 1970. Acetylcholine excitation and inhibition in molluscan heart.
165. Silvey G. E., Comp. Biochem. Physiol. 25: 257—269, 1968. Nervous regulation of heart in cockle.
166. Sleator W., de Gubareff T., Amer. J. Physiol. 206: 1000—1014, 1964. Electrical activity of human atrium.
167. Sleator W., de Gubareff T., pp. 107—119. In Paired Pulse Stimulation of the Heart, edited by P. F. Cranefield and B. F. Hoffman, Rockefeller University Press, New York, 1969. Action potentials of the human heart.
168. Smith J. E., Phil. Trans. Roy. Soc. B. 232: 279—310, 1946. Hydraulics of starfish tube feet.
1’69. Smith L. S., J. Fish. Res. Bd. Canad. 23: 1439—1446, 1966; also Smith L. S.„ Bell G., J. Fish. Res. Bd. Canad. 21: 711—717, 1964. Blood volume in salmon.
170. Smith L. S., Brett J. R., Davis J. C., J. Fish. Res. Bd. Canad. 24: 1775—1790» 1967. Cardiovascular dynamics in salmon.
171. Smith L. S., Davis J. C., J. Exp. Biol. 43: 171—180, 1965. Hemodynamics in molluscs.
171a. Smith N. A., Experientia 15: 200—205, 1969. Rhythmicity in cardiac ganglion of cockroach.
172. Sommer L. S. et al., Amer. J. Physiol. 215: 1498—1505, 1968. Hemodynamics and coronary flow in dolphin Tursiops.
173. Sperelakis N., Lehmkuhl D., J. Gen. Physiol. 47: 895—927, 1964. Action potentials in cultured heart cells.
174. Stevens E. D., Randall D. J., J. Exp. Biol. 46: 307—315, 1967. Cardiovascular changes in swimming in trout.
175. Stewart D. M„ Martin A. W., Z. vergl. Physiol. 70: 223—246, 1970. Blood volume in tarantula.
176. Sturkie P. D., Proc. Soc. Exp. Biol. Med. 123: 487—488, 1966. Cardiac output in ducks.
177. Sutterlin A. M., Physiol. Zool. 42: 36—52, 1969. Effect of exercise on heart of teleosts.
178. Tamai T., Kagiyama S., Circ. Res. 22: 423—433, 1968. Effect of hypothermia on cardiac muscle.
179. Tanaka I., Sasaki У., Shin-mura H., Japan. J. Physiol. 16: 142—153, 1966. Electrical activity of heart of horseshoe crab.
180. Templeton J. R., Physiol. Zool. 37: 300—306, 1964. Cardiovascular response in lizard.
181. Thorson T. B., Science 138: 99—100, 688—690, 1959. Partitioning of body I fluids in sea lamprey, marine sharks, and freshwater shark.
182. Thorson T. B., Biol. Bull. 120: 234—254, 1961. Partitioning of body water in fishes.
183. Thorson T. B., Physiol. Zool. 37: 395—399, 1964. Partitioning of body water in amphibians.
184. Thorson T. B., Copeia 592—601, 1968. Body fluid partitioning in reptiles.
185. Tille J., J. Gen. Physiol. 50: 189—202, 1966. Electrotonic interaction between muscle fibers in ventricle.
186. Toda N., West T. C., Nature 205: 808—809, 1965. Effect of vagal stimulation on rabbit atrium.
187. Tokchieva E. P., Fed. Proc. Transl. Suppl. 25: T739 —T742, 1966. Responses of neurons in frog heart.
410
Глава 20
187а. Trautwein W., Pharmacol. Rev. 15: 277—332, 1963. Ionic mechanisms in heart conduction.
188. Trautwein W., Kassebaum D. G., J. Gen. Physiol. 45: 317—330, 1961. Pacemaker activity of heart.
189. Treherne J. E., J. Exp. Biol. 42: 7—27, 1965. Blood volume in stick insect, Carausius.
190. Trueman E. R., Biol. Bull. 131: 369—377, 1966. Hydrodynamics of burrowing in Arenicola.
191. Trueman E. R., J. Exp. Biol. 44: 93—118, 1966. Burrowing reactions in Arenicola.
192. Trueman E. R. Science 152: 523—525, 1966. Fluid dynamics of burrowing.
193. Trueman E. R., Brand A. R., Davis P., J. Exp. Biol. 44: 469—492, 1966. Burrowing in bivalves.
194. Tucker V. A., J. Exp. Biol. 44: 77—92, 1966. Circulation in Iguana.
195. Vander A. J., Physiol. Rev. 47: 359—382, 1967. Control of renin release.
196. Van der Kloot W. G., Fed. Proc. 26: 975—980, 1967. Regulation of heart of leech.
197. Van der Kloot W. G., J. Exp. Zool. 174: 367—380, 1970. Electrophysiology of cardiac muscle of crustaceans.
198. Van der Kloot W. G., Dane B., Science 146: 74—75, 1964. Conduction in frog ventricle.
199. Vogel J. A., Sturkie P. D., Science 140: 1404—1406, 1963. Cardiac output in hen.
200. Watanabe A., Japan. J. Physiol. 8: 305—318, 1958. Interaction between neurons and lobster cardiac ganglion.
201. Watanabe A., Obara S., Akiyama T., J. Gen. Physiol. 50: 813—838, 839—862, 1967; 52: 908, 1968; 54: 212—231, 1969. Cardiac ganglion of Squilla.
202. Webber H. H., Dehnel P. A., J. Exp. Zool. 168: 327—336, 1968. Water compartments in snail Acmaea.
203. Weidman S., J. Physiol. 118: 348—360, 1952. Electrical constants of Purkinje fibers.
204. Weidmann S., J. Physiol. 210: 1041—1054, 1970. Electrical constants of mammalian heart.
206. Whartun D., Whartun M. L., Lola J., J. Insect Physiol. 11, 391—404, 1965. Blood volume in Periplaneta.
207. White F. N„ Fed. Proc. 29: 1149—1153, 1970. Vascular shunt in reptiles.
208. White F. N., Ross G., Amer. J. Physiol. 211: 15—18, 1966. Circulatory changes in diving in turtle.
209. White L. et al., Comp. Biochem. Physiol. 27: 559—566, 1968. Cardiac output of various mammals.
210. Woodbury J. W„ Crill W. E., pp. 124—435. In: Nervous Inhibition. Proceedings of the International Symposium, edited by E. Florey. Pergamon Press, New York, 1961. Conduction in the atrium.
211. Zuckerkandl E., Biol. Bull. 98: 161—173, 1950. Pressure in holothurians and sipunculids.
212. Zwicky К. T., Nature 207: 778—779, 1965; Comp. Biochem. Physiol. 24: 799— 808, 1968. Pharmacology of scorpion heart.
Глава 21
Эндокринные механизмы
р. И ОВАЛ ЕС, Л. ГИЛБЕРТ, Ф. БРАУН
В любом живом организме вырабатываются химические вещества, которые разносятся по всему телу и играют определенную интегрирующую и координирующую роль, дополняя тем самым деятельность нервных элементов. Такие вещества можно назвать описательно «химическими координаторами». В самом широком смысле химическим координатором является любое вещество, которое попадает в жидкости тела из окружающей среды или из клеток, составляющих высший организм, и участвует таким образом в поддержании нормального состава внутренней среды. Например, важную роль в координации функций организма играют Ог и СО2. Другие вещества, такие, как витамин D, могут попадать в организм извне или в определенных условиях синтезироваться некоторыми клетками организма и переходить из них в кровь. Есть также вещества, которые образуются в ограниченных участках тела и служат факторами адаптивной регуляции каких-то специальных функций. Например, панкреозимин (холецистокинин) и секретин, выделяемые слизистой двенадцатиперстной кишки в ответ на попадание в нее пищи, стимулируют выделение панкреатического сока. У многих высших животных имеются специализированные железистые клетки, ткани или органы, которые вырабатывают вещества, влияющие на организм в целом. Развитие этих специфических структур, по-видимому, шло параллельно со специализацией функций многих других органов и систем; они могли возникнуть только при наличии эффективных средств переноса веществ внутри организма, т. е. тогда, когда появилась циркуляторная система. В соответствии с этим мы находим специализированные эндокринные органы главным образом у кольчатых червей, моллюсков и ракообразных [78, 201], а также у насекомых [166], других членистоногих и, наконец, у позвоночных [409].
Гормонами называют особые химические координаторы, которые вырабатываются в определенном, более или менее ограниченном участке (или участках) организма и оказывают специфическое действие обычно где-то в других частях тела. Как правило (но не всегда), они образуются в хорошо обособленных желези-
412
Глава 21
«стых органах. Среди эндокринологов нет единого мнения о том, насколько широко следует понимать термин «гормон». Поскольку химические координаторы всех типов теснейшим образом связаны между собой, очевидно, что нельзя провести резкую границу между гормонами, с одной стороны, и прочими химическими координаторами— с другой. Аскорбиновая кислота, например, выполняет по существу роль гормона в организме крысы, но в то же время является витамином для человека.
В выяснении механизма действия гормонов достигнут значительный прогресс. Некоторые из них (например, адреналин, глюкагон, паратгормон, антидиуретический и тиреотропный гормоны, кортикотропин, лютеинизирующий гормон, гипоталамические высвобождающие факторы и меланоцитстимулирующий гормон)1, по-видимому, действуют, стимулируя в клетках-мишенях фермент, связанный с мембраной, —аденилатциклазу. В результате этого возрастает внутриклеточное содержание циклического 3',5'-аде-нозинмонофосфата (цАМФ). Согласно концепции Сьюзерленда и его сотрудников (рис. 21-1) [346, 347], образующийся в клетках цАМФ вызывает гормональные эффекты, действуя в качестве второго посредника; сейчас затрачивается много усилий на то, чтобы выяснить конкретные механизмы осуществления этих эффектов.
Другие гормоны (например, гормоны щитовидной железы, эстрогены, андрогены, прогестерон, кортикостероиды, экдизон и гормон роста), по-видимому, стимулируют образование специфических белков, воздействуя на синтез белка или РНК- «Точка приложения» зависит от типа гормона и его клеток-мишеней [405]; это может быть ядро, как в случае влияния эстрадиола на матку [175], или цитоплазма, как при действии гормона роста на полисомы в клетках печени [240]. Согласно одной из общих концепций эндокринологии, гормоны присоединяются в клетках-мишенях к специфическим рецепторам. Эта концепция особенно хорошо объясняет влияние эстрадиола на матку. По-видимому, этот гормон взаимодействует в цитоплазме со специфическим рецепторным белком, который при этом изменяет свою конформацию, а затем образовавшийся комплекс переходит в ядро [175] и оказывает здесь действие на синтез РНК- Другие гормоны, например инсулин, влияют главным образом на плазматическую мембрану, регулируя ее проницаемость для специфических метаболитов [344]. Все три указанных механизма могут сочетаться между собой; известно, что такое комбинированное действие имеет место при осуществлении некоторых гормональных эффектов.
Определить понятие «гормон» в физиологическом плане затруднительно, но можно дать такое определение, которое будет по
1 В настоящее время Комиссией .по биохимической номенклатуре Международного общества чистой и прикладной химии и Международного биохимического общества принята новая номенклатура пептидных гормонов (ом. Реферативный журнал «'Биология», 1976, 1ФЗ). — Прим. ред.
Эндокринные механизмы
413
меньшей мере полезно в качестве основы для дискуссии: гормон— это специфическое химическое вещество, выделяемое особыми клетками в определенном участке тела (эндокринной железе), которое поступает в кровь и затем оказывает специфическое действие на определенные клетки или органы-мишени, расположенные
Эндокринная железа
^стимуляция Блокатор^
Аденилат\ циклаза^ Ixcucmeni-мишень
Гормон (1-й посредник)
т~* ♦
Модулятора
АТФ
32+ метил-ксаптины
Аденин
Мд'
5-АМФ
Фосфодиэстераза (модулятор)
Инактивированный гормон
С^,—У/й Н0. Р +
ОН ^5
Циклический 3,5-АМФ (2-й посредник)
2-ясистема- , „ * „Физиологиче-мишень ская“реакция)
Зффекторная клетка
Рис. 21-1. Система двух передатчиков, включающая гормон и аденилатциклазу (Sutherland Е. W., Dye I., Butcher R. W., Rec. Progr. Horm. Res. 21: 623—646, 1965).
s другой области тела, что приводит к координации функций организма в целом. В настоящей главе речь будет идти в основном о веществах, соответствующих этому определению.
С точки зрения сравнительной физиологии важно подчеркнуть, что у представителей некоторых типов и классов животных образуются особые химические вещества, необходимые для нормального развития и функциональной целостности организма. Место образования гормонов в организме, их химическая природа и способы их переноса имеют второстепенное значение. Характер вызываемых эффектов зависит как от природы реагирующих тканей, так и от химических свойств циркулирующего гормона. Очевидно, что гормоны, распределяющиеся в организме случайным образом, не могли бы вызывать дифференцировку тканей и органов или какую-либо направленную и упорядоченную активность при отсутствии генетически предопределенной дифференциации. Таким образом, влияние гормонов в известном смысле осуществляется на фоне некоторой уже имеющейся структуры организма и способствует полному функциональному проявлению многочисленных и сложных «скрытых» дифференцировок. Поэтому
414
Глава 21
при исследовании эндокринных механизмов изучить фило- и онтогенетическое развитие способности тканей реагировать на гормо- г ны не менее важно, чем проследить возникновение самих гор- , МОНОВ. i
Две главные интегративные системы организма — нервная и эндокринная—функционально тесно связаны друг с другом [276, 367]. Нервные клетки обычно выполняют двоякую функцию: они, । во-первых, проводят возбуждение и, во-вторых, секретируют такие нейроэндокринные вещества, как ацетилхолин [280], норадреналин [423] и 5-окситриптамин [260, 310, 432], которые играют важную роль в межнейронных синапсах и нервно-мышечных соединениях. Кроме того, особые группы нервных клеток специализировались в качестве источников нейросекреторных веществ, которые переносятся кровью и функционируют совершенно таким же образом, как прочие гормоны — продукты эндокринных клеток, не имеющих отношения к нервам. Примерами таких нейрогормонов служат окситоцин и вазопрессин, вырабатываемые клетками гипоталамуса позвоночных, активные вещества (уротензины), выделяемые группой клеток в каудальной области спинного мозга рыб — урогипофизом [150], и гормоны насекомых, образующиеся в клетках мозга и нервной цепочки. Само мозговое вещество надпочечников имеет нейрогенное происхождение; оно возникает в эмбриогенезе из мигрировавших на периферию клеток, гомологичных симпатическим постганглионарным нервным элементам.
Нейросекреция — один из наиболее важных эндокринных механизмов [158]. Строго говоря, это секреция нейроном любого вещества, например синаптического медиатора. Однако обычно этот термин употребляют только для обозначения секреции истинных гормонов нервами, которые содержат гранулы, видимые при специальной окраске в обычный микроскоп. Находящиеся в этих гранулах гормоны переносятся по аксону и затем выделяются нервными окончаниями, образующими иногда нейрогемальнЫе органы, специально приспособленные для этой цели. Есть также данные о том, что гормоны нейрогипофиза позвоночных присоединяются к особому транспортному белку — нейрофизину.
Существуют и иные связи между нервной и эндокринной системами. Секреция тропных гормонов передней долей гипофиза регулируется другими гормонами — стимулирующими или тормо*-зящими факторами, которые образуются в гипоталамусе и попадают в переднюю долю гипофиза по высокоспециализированным сосудистым путям — так называемой гипоталамо-гипофизарной портальной системе (рис. 21-2). Возбуждение мозгового вещества надпочечников и, возможно, pars intermedia гипофиза у некоторых животных вызывается непосредственно нервами, а остальные железы могут находиться под непрямым нервным контролем. Короче говоря, очевидно, что эндокринная система в значительной мере служит «орудием» нервной системы и частично развивается
Эндокринные механизмы
415
из нее. Такое тесное соподчинение двух систем лежит в основе высокоэффективных механизмов биологической координации.
У трех групп животных, эндокринная система которых исследована наиболее полно,— у позвоночных, насекомых и ракообразных— обнаруживаются черты удивительного сходства функций и внешнего плана организации этой системы, но здесь нет истинных
Рис. 21-2. Схема анатомических связей между гипоталамусом и гипофизом млекопитающего [409].
Происхождение волокон, направляющихся к серединному возвышению (eminentia mediana), не так определенно, как это обозначено на рисунке. 1 — нейросекреторные клетки в ядрах гипоталамуса; 2 — первичное сплетение (в eminentia mediana); 3 — третий желудочек; 4 — перекрест зрительных нервов; 5 — артерия, идущая к eminentia mediana; 6 — артерия, идущая к pars distalis; 7 — портальные вены гипофиза; 8 — артерия, идущая к pars nervosa; S — pars nervosa (нейрогемальный орган); 10 — отводящая вена; 11 — синусоиды (вторичное сплетение).
гомологий. В пределах каждой из этих трех групп гормоны более или менее сходны по своей химической природе, но они обычно отличаются от гормонов, свойственных другим группам. Поэтому лучше всего будет рассматривать интересующий нас материал в сравнительном аспекте по каждой группе в отдельности.
Позвоночные
Эндокринные железы
Щитовидная железа. Эта железа, филогенетически связанная с эндостилем первичнохордовых [20], возникает у млекопитающих как непарное эктодермальное выпячивание вентральной части глотки. Она теряет связь с кишкой и превращается в обособленный орган, состоящий из двух долей. Железа построена из много-
416
Глава 21
численных фолликулов, окаймленных секреторным эпителием и содержащих белковый «коллоид», богатый иодом. Высота эпителиальных клеток изменяется в прямой зависимости от активности железы. Тиреоидный гормон резорбируется из коллоида фолликулярным эпителием и затем выделяется в кровь. Коллоид содержит белок с большим молекулярным весом (680 000) — иодтирео-глобулин.
Щитовидная железа поглощает неорганические иодиды, поступающие в организм с пищей. Эпителий железы обладает наибольшей из всех тканей тела способностью концентрировать иодид. Накопленный иодид подвергается ферментативному окислению до свободного иода или Ю~. Образовавшийся иод присоединяется к остаткам тирозина, содержащимся в белках фолликулов. Это приводит к образованию ряда тиронинов — дииодтирозина, трииодти-ронина (Тз) и тетраиодтиронина, или тироксина (Т4, см. ниже). Для осуществления секреции гормона необходимо, чтобы Т3 и Т4 высвободились из тиреоглобулина в результате расщепления его протеазой щитовидной железы. Гормоны, большей частью связанные со специфическими а-глобулинами, циркулируют в крови. Катаболизм тиреоидных гормонов осуществляется главным образом в печени и почках. Т3 и Т4 оказывают на различные процессы неодинаковое влияние, но гормоном щитовидной железы обычно считают тироксин (Т4).
Тироксин (тетраиодтиронин)
ZNH2 но" р—\ р—СН2—СНХ
у=/ \соон
Наиболее характерным эффектом тироксина у гомойотермных животных является калоригенное действие, связанное с усиленным потреблением кислорода большинством тканей организма. У пойкилотермных позвоночных этот эффект не был обнаружен. У рыб такого рода исследования сложны еще и в техническом отношении: необходимо, например, предотвратить избыточные движения после инъекции тиреоидных гормонов. В одном исследовании на мальках акулы Squalus suckleyi, в котором это условие удалось выполнить, тироксин не давал регулярного эффекта в отношении потребления О2, тогда как введение аналога тироксина — трииод-тироуксусной кислоты — определенно повышало интенсивность обмена по сравнению с контролем [335]. Возможно, что у ряда холоднокровных животных отсутствие эффекта бывает связано с низкой температурой обитания. Если ящериц Anolis carolinensis содержать при температуре значительно выше комнатной, то можно обнаружить существенное влияние Т4 и Т3 на окислительный метаболизм этих животных [273]. Однако эти результаты не
Эндокринные механизмы
417
изменяют основного вывода о том, что холоднокровные животные по существу нечувствительны к калоригенному действию тиреоидных гормонов [172].
Хотя молекулярные механизмы калоригенного эффекта пока неизвестны, получены веские данные в пользу того, что на первых этапах происходит стимуляция синтеза РНК в ядрах клеток. Актиномицин D — мощный ингибитор ДНК-зависимого синтеза РНК— блокирует калоригенный эффект тироксина у тиреоидэктомиро-ванных крыс [403]. Сейчас уже очевидно, что калоригенный эффект определяется рядом последовательных событий [404]. Сначала возрастает включение предшественников РНК, затем в ядрах повышается уровень РНК-полимеразы, активируемой Mg2+, усиливается синтез рибосомной РНК и, наконец, ускоряется включение аминокислот в белки. Поскольку ранее было показано,что после введения тироксина возрастает синтез окислительных ферментов, можно предположить, что именно этот синтез и приводит к калоригенному эффекту.
Гормон щитовидной железы необходим для нормального роста и дифференцировки, особенно для образования таких твердых или ороговевающих структур, как кости и волосы; он регулирует синтез межклеточного вещества в тканях и способствует использованию углеводов, поддерживает нормальную возбудимость ряда функциональных систем, например нервных центров и сердечной мышцы.
Щитовидная железа оказывает также влияние на другие эндокринные железы. Она стимулирует кору надпочечников, от ее активности зависит нормальная функция половых желез. Тироксин, по-видимому, каким-то образом регулирует водный баланс совместно с антидиуретическим гормоном. И наконец, он участвует в регуляции своего собственного синтеза, подавляя выделение тиреотропного гормона передней долей гипофиза, а может быть, и непосредственно воздействуя на щитовидную железу.
Функция щитовидной железы регулируется главным образом тиреотропным гормоном гипофиза, который стимулирует все фазы ьсинтеза и выделения тиреоидного гормона.
| Секреция тиреотропного гормона (ТТГ) находится под кон-тролем гипоталамического ТТГ-высвобождающего фактора, ^структура которого теперь известна [48]: это (пиро) Глу-Гис-Про-NH2; он стимулирует выделение ТТГ из передней доли гипофиза крыс как in vitro, так и in vivo. Еще раньше было показано, что секреция ТТГ возможна, по-видимому, только при неповрежденном гипоталамусе [100]. Главным внешним фактором, активирующим функцию щитовидной железы, является низкая температура [58], которая способствует усиленному выделению ТТГ, а тем самым и секреции тиреоидного гормона у большинства видов животных. У ящериц, однако, этот эффект может отсутствовать: было найдено, что у Sceloporus occidentalis температура в пределах от 27—2067
418
Глава 21
21 до 38 °C не оказывает существенного влияния на секрецию щитовидной железы [84]; таким образом, у этого вида роль температуры в активации щитовидной железы может быть лишь «разрешающей».
Теперь известно, что ТТГ влияет на щитовидную железу, повышая образование в ней циклического АМФ в результате стимуляции аденилатциклазы [371]. Концентрации циклического АМФ в железе под действием ТТГ возрастают, и получены убедительные данные о том, что циклический АМФ вызывает в железе следующие эффекты: он стимулирует связывание иодида и перевод его в органическую форму, стимулирует синтез иодтиронинов, способствует выделению иода и обмену фосфолипидов [371].
В настоящее время изучается механизм стимуляции аденилат-циклазы тиреотропным гормоном и роль циклического АМФ в этом процессе.
Еще много лет назад было обнаружено [479], что скармливание ткани щитовидной железы ускоряет метаморфоз амфибий, а удаление зачатка этой железы у личиночных форм препятствует метаморфозу. У тиреоидэктомированных особей можно вызвать метаморфоз, вводя им тиреоглобулин или тироксин. Менее эффективен дииодтирозин и еще менее — неорганические соли иода или свободный иод, которые только в больших количествах вызывают метаморфоз [6]. Эта реакция на тироксин совершенно не связана с повышением общей интенсивности метаболизма. Динитрофенол — мощный стимулятор обмена веществ —не оказывает никакого влияния на метаморфоз. В то же время ацетилированный тироксин сохраняет способность вызывать метаморфоз, хотя полностью утрачивает стимулирующую активность в отношении метаболизма.
Разные виды амфибий сильно различаются между собой по способности реагировать на тироксин. На ранних этапах онтоге-* неза ни один из них не реагирует на этот гормон, но у большинства амфибий эта способность появляется на каком-то определенном этапе. У некоторых видов, например у мексиканского аксолотля, никогда не развивается достаточно высокая чувствительность к этому фактору, и поэтому метаморфоза у них обычно не происходит. Однако большими дозами тироксина можно вызвать метаморфоз и у аксолотля; у некоторых других видов, например Necturus, по-видимому, вообще не развивается чувствительность к этому гормону, и у них никогда не бывает метаморфоза. Отсутствие метаморфоза у Necturus связано именно с неспособностью его тканей реагировать на гормон, а не отсутствием соответствующего фактора в щитовидной железе; это видно из того, что щитовидная железа Necturus способна ускорять метаморфоз у Rana clamitans [398].
Развитие амфибий, включая метаморфоз, представляет собой упорядоченную последовательность событий. По-видимому, эта
Эндокринные механизмы
419
последовательность, по крайней мере отчасти, обусловлена постепенно возрастающей активностью щитовидной железы и развивающейся способностью различных тканей реагировать на ее гормон [25, 124, 238, 272, 320].
После гипофизэктомии амфибии утрачивают способность к метаморфозу [387]. Гипофизарный фактор, участвующий в метаморфозе, выделяется передней долей гипофиза, он перестает оказывать свое действие после полной тиреоидэктомии. Известно также, что у гипофизэктомированных животных щитовидная железа не накапливает коллоида. Все это показывает, что у амфибий выработка и выделение гормона щитовидной железой, а значит, и метаморфоз регулируются гипофизом. Существенную роль в регуляции метаморфоза может также играть пролактин [125].
Известно, что у некоторых костистых рыб щитовидная железа регулирует метаморфоз, а у других рыб изменение количества свободного тироксина приводит к изменениям скорости роста и формы тела [12, 203, 205]. У низших позвоночных, так же как и у высших, гипофиз вырабатывает тиреотропный гормон, от активности которого зависят развитие и секреторная функция щитовидной железы [169].
Влияние тироксина на развитие опосредовано через действие его на синтез РНК и белков. Инъекция актиномицина подавляет атрофию хвоста, которая в норме происходит у головастиков Хе-nopus во время метаморфоза [427]. При этом повышение количества катепсина, обычно наблюдаемое при атрофии хвоста, бывает значительно менее выражено; это указывает на то, что тироксин, вероятно, стимулирует синтез катепсина, а не его высвобождение. Ряд других онтогенетических изменений, вызываемых тироксином, тоже связан со стимуляцией синтеза РНК [151], как и в случае влияния на потребление кислорода.
Интересно то, что хотя у костистых рыб щитовидная железа определенно воздействует на рост и развитие, влияние ее на общую интенсивность метаболизма у рыб до сих пор окончательно не доказано [126]. Вообще на разных видах пойкилотермных животных результаты получаются весьма различными.
Как показывают эксперименты с введением гормона щитовидной железы, у гуппи и серебряных карасей этот гормон не влияет на потребление Ог [126]. Обработка тиомочевиной не ведет к существенному изменению потребления О2 у Fundulus. Введение экстрактов щитовидной железы бермудской рыбы-попугая увеличивает потребление О2 у рыб Haemulon, но только у особей весом оолее 15 г [385]. Введение гормона щитовидной железы головастикам капа pipiens до метаморфоза не вызывало у них (также как и у большинства рыб) изменений в потреблении Ог. Между тем у взрослой R. pipiens потребление Ог при этом существенно возрастало [425], а если животных содержали при температуре ыше 13 С, то наблюдалось снижение веса тела. У рептилий и
27*
420
Глава 21
птиц влияние гормона щитовидной железы на интенсивность основного обмена, по-видимому, очень сходно с наблюдаемым у млекопитающих. У голубей после полной тиреоидэктомии отмечается заметное снижение теплопродукции в условиях покоя. У молодых питонов длительное кормление щитовидной железой приводит к значительному повышению возбудимости и потере веса.
Имеются данные, что у рыб щитовидная железа выполняет и другие функции. Она участвует в регуляции содержания гликогена в печени и в осморегуляции [386]. У эвригалинных рыб активность щитовидной железы возрастает при повышенной солености воды [199]. У колюшки (Gasterosteus) именно увеличение активности щитовидной железы стимулирует миграцию из соленой воды в пресную в период размножения; это изменение активности железы в свою очередь регулируется фотопериодом [15].
Тироксин оказывает влияние на головной мозг серебряного карася. Он изменяет, например, электрические ответы, возникающие в обонятельном тракте при вливании в обонятельный мешочек слабого солевого раствора. При воздействии тироксином наблюдали также локальное облегчение ответов в обонятельной луковице и в то же время торможение центробежных влияний конечного мозга на реакцию, вызываемую NaCl [307]. Дальнейшие исследования показали, что в одиночных клетках, расположенных в различных слоях обонятельных луковиц, наблюдаются разряды трех типов; тироксин повышает частоту «спонтанных» разрядов типа I [308]. Кроме того, тироксин играет важную роль в определенном этапе развития лососей — в превращении пестрятки в серебрянку (смолтификация), сопровождающемся, помимо других изменений, отложением гуанина в коже [172].
Реакция щитовидной железы при адаптации к изменениям температуры у разных рыб различна. В то время как у большинства рыб активность железы возрастает при более высоких температурах, у Salmo и Umbra она повышается при низкой температуре, как у гомойотермных животных [171].
Щитовидная железа — высокоспециализированный орган, приспособленный для связывания иода. Эта способность присуща не только эндостилю ланцетника [19] и оболочников, например Cio-па [18], но и кожным железам полухордовых. Связывание иода склеропротеинами наружного скелета наблюдается у многих беспозвоночных [30, 170, 173].
Паращитовидные железы. Эти железы отсутствуют у рыб, но имеются у амфибий и высших позвоночных. У млекопитающих две пары этих желез вкраплены в ткань щитовидной железы и образуются из энтодермы третьего и четвертого глоточных карманов. Железы содержат клетки двух типов: главные, в которых образуется паратгормон, и богатые митохондриями оксифильные клетки, функция которых неизвестна. Бычий паратгормон — это одноце
Эндокринные механизмы
421
почечный полипептид с мол. весом 9500, состоящий из 83 аминокислотных остатков [333].
Главная функция этого гормона — регуляция обмена кальция и фосфатов. Он понижает содержание фосфата в крови, повышает содержание в ней кальция и способствует выделению фосфатов и кальция почками. Влияние гормона затрагивает только соответствующие ионы и не распространяется на кальций и фосфат, связанные с белками или находящиеся в жидкостях тела в неио-
низированном состоянии.
Секреция паратгормона регулируется содержанием кальция в сыворотке. Падение уровня кальция активирует железу. Повышение концентрации фосфата в сыворотке оказывает аналогичное действие, но только косвенным путем — через соответствующие изменения в содержании кальция.
Действие паратгормона как на почечную, так и на костную ткань можно объяснить влиянием его на аденилатциклазу. Физио-
логические количества гормона повышают выделение циклического АМФ с мочой [79], a in vitro гормон стимулирует аденилатциклазу в кости и в почечной ткани [81]. Дибутирилпроизводное циклического АМФ действует на эксплантаты костей подобно паратгормону [412]. В настоящее время ведется изучение механизма, с помощью которого циклический АМФ вызывает переход кальция из костей в кровь. Если мы вспомним о многообразной роли кальция в организме (поддержание возбудимости нервов и мышц, проницаемости мембран для различных ионов, сопряжение между возбуждением мышцы и ее сокращением) и об участии фосфата в буферных системах, углеводном обмене и построении
нуклеиновых кислот, то станет ясно, насколько важна регуляция обмена этих двух веществ.
Ультимобранхиальные железы. Особые клетки, происходящие из ультимобранхиальных телец эмбриона, секретируют гормон кальцитонин. У взрослых позвоночных (кроме млекопитающих) эти клетки образуют обособленные железы, а у млекопитающих они врастают в щитовидную железу [91]. Кальцитонин понижает концентрацию кальция в крови, подавляя резорбцию его из костей [292]. В щитовидной железе млекопитающих эти клетки называют кальцитониновыми (С-клетками) или парафолликулярными, так как они часто бывают расположены между фолликула-П_”ИТОНИНЫ свиньи и лосося состоят из 32 аминокислотных я^™ПрМ\т^ЛаЗЛИЧаются 110 своей структуре. Гормон лосося более npw ’ а свинои> при испытании на млекопитающих (судя по оазггиптяртся стандаРТН0Г0 теста) [91]; кроме того, он медленнее яктиппогтт. Си Рганизме [181] и поэтому дольше сохраняет свою жит гипрп^я nJr алом’ вызывающим секрецию кальцитонина, слу-тии гопмпия Г9091ИЯ’ тт ЭТ0Т Факт сыгРа^ важную роль в откры-нопмяшлв.п- J 9 1* Доказать, что кальцитонин выделяется в
Р условиях в ответ на изменение концентрации Са в
422
Глава 21
плазме, было не так просто, и поэтому положение кальцитонина среди гормонов остается несколько спорным [292].
Поджелудочная железа. Эндокринные участки поджелудочной» железы, так называемые островки Лангерганса, имеют эктодермальное происхождение — они отпочковываются от протоков^ Островки содержат клетки трех типов. Альфа-клетки с ацидофильными гранулами, более многочисленные в периферических зонах, островков, считаются источником глюкагона [208]. Бета-клетки, составляющие большинство в островковой ткани, вырабатывают инсулин. Кроме того, есть еще элементы третьего типа, не содержащие гранул, —дельта-клетки, функция которых неизвестна; возможно, что они секретируют гастрин1. Недавно Эппл [123] описал в островковой ткани низших позвоночных амфифильные клетки,, которые окрашиваются и по типу бета-клеток, и по типу дельтаклеток. У птиц и млекопитающих таких клеток нет. У жабы Bufo bufo эти клетки могут быть обнаружены только весной. У акулы Scyliorhinus canicula найдены другие клетки, промежуточные по некоторым свойствам между альфа- и бета-клетками.
Оба гормона — инсулин и глюкагон—представляют собой белки. Молекула инсулина (мол. вес 6000, изоэлектрическая точка при pH 5,4) состоит из 51 аминокислоты, взаимное расположение которых точно установлено. Молекула построена из двух пептидных цепей, одна из которых носит кислый, а другая — основной характер; они соединены между собой двумя дисульфидными связями. Обнаружены видовые различия, которые касаются трех аминокислот, находящихся в определенных участках молекулы [191]. Инсулин образуется из проинсулина [391] —белка, состоящего из двух цепей инсулина, соединенных вставочным пептидом. Ферментативное отщепление этого пептида приводит к образованию инсулина, две цепи которого соединены дисульфидными мостиками. Хотя инсулины разных животных иммунологически сходны, это сходство не абсолютно, о чем свидетельствует выработка антител [288]. Молекула глюкагона представляет собой однупо-липептидную цепь, состоящую из 29 аминокислотных остатков [27, 143].
Инсулин играет важную роль в регуляции углеводного обмена. Он стимулирует процесс превращения глюкозы ib гликоген в мышцах и печени, облегчает образование жира из углеводов и способствует синтезу белков. Кроме того, инсулин ускоряет окисление глюкозы в мышцах и в других тканях. Механизм действия инсулина до сих пор с достоверностью не известен. Согласно гипоте-
1 В самое последнее время получены доказательства, что так называемые D- или дельта-клетки поджелудочной железы в нормальных условиях вырабатывают соматостатин — пептид, обнаруженный впервые в гипоталамусе и угнетающий секрецию как гормона роста, так и ряда других гормонов, в том числе инсулина и глюкагона (ом. Arimura et al., Science, 1975, 189, 1007). — Прим, перев,.
Эндокринные механизмы 423
зе, в пользу которой накопилось больше всего данных, инсулин облегчает перенос глюкозы и аминокислот через клеточные мембраны [344].
Когда концентрация инсулина падает ниже определенного уровня, содержание глюкозы в крови возрастает вследствие подавления механизмов поглощения ее из крови и утилизации в тканях; при этом часть глюкозы выводится с мочой. При недостатке инсулина окисление жиров усиливается, и это ведет к накоплению кислых кетоновых тел, таких, как ацетон, ацетоуксусная и оксимасляная кислоты. Кетоновые тела выводятся в виде натриевых и калиевых солей; в результате уменьшается щелочной резерв организма и снижается pH крови. Происходит расщепление белков, и азот выделяется из организма.
Второй гормон островковой ткани, глюкагон, действует как антагонист инсулина — он стимулирует гликогенолиз в печени. Этот эффект развивается по «каскадному» типу, глюкагон активирует аденилатциклазу, в результате чего из АТФ образуется больше циклического АМФ [346], а последний в свою очередь увеличивает содержание в печени активной фосфорилазы, катализирующей образование глюкозо-1-фосфата из гликогена. Образующийся в результате изомеризации глюкозо-6-фосфат под действием глюкозо-6-фосфатазы превращается в глюкозу, которая выделяется в кровь, и это приводит к глюкагоновой гипергликемии. По-видимому, секреция обоих гормонов поджелудочной железы регулируется главным образом самим содержанием сахара в крови: повышение его стимулирует секрецию инсулина, а падение — секрецию глюкагона, т. е. по сути дела глюкагон стимулирует выделение инсулина, и наоборот. Соматотропный гормон аденогипо; физа усиливает секрецию инсулина, а также, вероятно, глюкагона, что приводит к гипергликемическому эффекту.
Относительное значение двух гормонов поджелудочной железы у разных позвоночных различно. Среди млекопитающих высокая чувствительность к недостатку инсулина свойственна собакам, кошкам и крысам, а у травоядных она обычно выражена в меньшей степени. У рептилий и птиц ткани менее чувствительны к инсулину [396]; у птиц недостаток этого гормона вызывает лишь преходящий эффект [284]. У черепах после удаления поджелудочной железы развивается гипергликемия [144]. Вместе с тем рептилии и птицы весьма чувствительны к глюкагону, и у рептилий поджелудочная железа особенно богата альфа-клетками. Амфибии в отличие от пресмыкающихся и птиц обычно очень чувствительны к инсулину или его недостатку; при этом у них часто отсутствуют альфа-клетки [286, 287, 456].
Мозговое вещество надпочечников. Ткань мозгового вещества надпочечников имеет эктодермальное происхождение; она возникает из нервных гребешков, т. е. тех же эмбриональных зачатков, из которых образуются симпатические ганглий. Секреторные
424
Глава 21
клетки по сути дела представляют собой постганглионарные элементы симпатической системы и сохраняют связь при помощи синапсов с холинэргическими преганглионарными волокнами. Мозговое вещество надпочечников богато васкуляризировано. Известно, что у млекопитающих оно выделяет два родственных друг другу гормона: первичный амин норадреналин и вторичный амин адреналин .[423]. Физиологической активностью обладают только их природные левовращающие изомеры. По структуре эти гормоны связаны между собой следующим образом:
Норадреналин
Перенос метильной группы (а метионина)
+СНд
Адреналин
>Hof3 —CH—СН2—NH—СН, НО' ' он
Относительное содержание этих двух гормонов у разных животных различно. Соотношение выделяемых гормонов зависит от иннервации. Судя по некоторым данным, эти два гормона секретируются в мозговом веществе клетками двух разных типов, но окончательно это еще не доказано. В клетках мозгового вещества гормоны содержатся в особых пузырьках [36].
Адреналин и норадреналин оказывают в общем качественно сходное действие на сердце, кровеносные сосуды и гладкие мышцы, а также на углеводный обмен. Имеются, однако, важные количественные различия. Действуя на сосудистую систему, оба гормона увеличивают возбудимость и силу сокращения сердечной мышцы, что ведет к повышению кровяного давления. Оба гормона вызывают сужение сосудов внутренних органов, кожи и слизистых оболочек и повышают давление в сосудах легких; однако адреналин усиливает ток крови через мышцы, сердце, печень и головной мозг, так что общее сопротивление периферических сосудов падает, а норадреналин в различной степени вызывает сокращение всех периферических сосудов, за исключением сосудов скелетных мышц и печени, что ведет к некоторому повышению периферического сопротивления. После введения норадреналина минутный объем сердца рефлекторно (в результате раздражения прессоре-цепторов) уменьшается.
Адреналин вызывает быстрое повышение содержания сахара в крови; норадреналин в этом отношении примерно вчетверо менее эффективен. Так же как и при действии глюкагона, выделяю
Эндокринные механизмы
425
щийся в кровь сахар образуется из гликогена печени, причем вначале получается глюкозо-6-фосфат, который затем расщепляется глюкозо-6-фосфатазой. Циклический АМФ в биологическом материале открыли Сьюзерленд и Ролл, изучавшие механизм действия адреналина и глюкагона на гликогенолиз в печени. Во фракции субклеточных частиц, содержавшей фрагменты мембран, при добавлении адреналина или глюкагона образовывался термостабильный аденинрибонуклеотид [397],' который и оказался циклическим АМФ. Адреналин и глюкагон влияют, очевидно, на разные рецепторы, регулирующие образование аденилатциклазы, поскольку действие адреналина можно специфически блокировать эрготамином; однако в обоих случаях происходит стимуляция аденилатциклазы, а затем фосфорилазы. В мышцах, в которых нет надлежащей фосфатазы, образующийся глюкозо-6-фосфат окисляется до молочной кислоты, и этот конечный продукт выделяется в кровь. Большая часть молочной кислоты превращается затем в гликоген, главным образом в печени. Между мышечной фосфорилазой и фосфорилазой печени также есть некоторые различия.
Гормоны мозгового вещества надпочечников воздействуют на рецепторы двух различных типов, обозначенные Алквистом как альфа- и бета-рецепторы. Хотя иногда трудно с точностью определить, какой именно тип рецепторов участвует в осуществлении того или иного эффекта, обычно используют три критерия: 1) относительную эффективность ряда катехоламинов [3], 2) влияние блокирующих агентов [4, 300] и 3) характер изменений концентрации циклического АМФ в тканях [411]. Например, с участием альфа-рецепторов связывают три реакции: сокращение артериол, селезенки и сфинктеров желудочно-кишечного тракта [299]; с помощью бета-рецепторов осуществляются расслабление артериол, желудка и небеременной матки кошки [299]. Можно привести много других примеров, которые показывают, что изучение адрен-эргических рецепторов является очень важным моментом при выяснении механизма любой реакции, связанной с мозговым веществом надпочечников.
Адреналин вызывает гораздо более сильное расслабление мускулатуры бронхов, чем норадреналин. Этим различием, а также существенно разным действием обоих гормонов на сердечно-сосудистую систему, вероятно, можно в значительной мере объяснить возникновение реакции тревоги и других нервных ответов при повышении^ концентрации адреналина и отсутствие тех же реакций при действии норадреналина.
Адреналин вызывает и многочисленные другие реакции. К ним относятся: расширение зрачков, пиломоторная реакция, торможение потоотделения, расслабление мышц кишечника и мочевого пузыря (кроме сфинктеров, действие которых, наоборот, усиливается), сокращение капсулы селезенки, усиленное слюноотделение и повышение свертываемости крови.
426
Глава 21
Функция мозгового вещества надпочечников регулируется исключительно симпатической нервной системой. Степень ее тонической активности связана, как полагают, с общей афферентной нервной активностью и, вероятно, колеблется вместе с последней. Организм, по-видимому, может обойтись без этой ткани: ее отсутствие полностью компенсируется образованием норадреналина в-адренэргических нервных волокнах. Однако в обычных условиях любой стресс или резкое падение содержания сахара в крови стимулирует секрецию адреналина, который способствует быстрому развитию компенсаторной реакции. Какого рода раздражение вызывает обычно специфическую секрецию норадреналина, не вполне ясно.
Кора надпочечников. Стероидные гормоны вырабатывают четыре железы: кора надпочечников, семенники, яичники и плацента. Все стероиды содержат в своей структуре циклопентанпергидрофенантреновое ядро, и для того, чтобы понять их биологическое действие, необходимо знать их химические особенности [409].
У разных позвоночных встречаются различные взаимоотношения между корковым и мозговым веществом надпочечников '[172]. Расположение коры в виде слоя, окружающего мозговое вещество, свойственно почти исключительно млекопитающим. В тех случаях, когда кортикальная ткань внедрена в почки (у костистых рыб), ее называют интерренальной тканью. Корковое вещество надпочечников образуется из мезодермы по обе стороны от основания спинной брыжейки на уровне первичной мочеполовой закладки. У млекопитающих кора надпочечников, как это следует из ее названия, полностью окружает элементы мозгового слоя. Она относительно толстая, желтого цвета и разделена на зоны: тонкую, самую наружную — клубочковую, среднюю — пучковую и внутреннюю — сетчатую.
В экстрактах железы обнаружено более пятидесяти различных стероидов, хотя лишь относительно немногие из них попадают в кровяное русло. Часть из них служит предшественниками при биосинтезе гормонов.
Кортикальные гормоны можно разделить на три категории: глюкокортикоиды, минералокортикоиды и половые гормоны. Вопрос о месте их образования в коре выяснен еще не до конца; однако, по общему мнению, глюкокортикоиды образуются в пучковой и сетчатой зонах, а минералокортикоиды-—в клубочковой: зоне. Кора надпочечников выделяет половые гормоны трех типов: эстрогены, андрогены и (Прогестерон; у млекопитающих, по-види-мому, здесь преобладает выработка андрогенов. Как у генетических самцов, так и у генетических самок эти гормоны, по-видимому, участвуют в регуляции нормального развития мышц и: скелета, дифференцировки наружных половых органов, роста волос по определенному типу, а также полового влечения и поведения.
Эндокринные механизмы
427
Глюкокортикоиды оказывают большое влияние на метаболические превращения в организме, которые в свою очередь лежат в основе изменений поведения. Таким образом, эта группа стероидов чрезвычайно важна для поддержания жизнедеятельности взрослого организма. Введение одних только глюкокортикоидов обеспечивает выживание особей с удаленными надпочечниками. Приводим структуру наиболее изученных стероидов этой группы:
Кортизон (соединение Е)
В результате обширных исследований была обнаружена довольно сходная картина синтеза кортикостероидов у различных позвоночных [357]. Круглоротые синтезируют кортизол и кортикостерон, хрящевые рыбы—1-а-гидрокортизон [213], кортизол и кортикостерон, костистые рыбы — кортизол, кортизон и И-дезоксикортизол; у амфибий, рептилий и птиц образуются кортикостерон, альдостерон и 18-гидрокортикостерон, а у большинства млекопитающих—кортизол, кортизон, кортикостерон, 18-оксикортикосте-рон и альдостерон. Таким образом, либо кортикостерон, либо кортизол, либо оба эти гормона синтезируются у представителей всех классов позвоночных, что говорит о древнем происхождении глюкокортикоидов и их неизменном присутствии. Однако альдостерон, мощный минералокортикоид, не образуется у рыб и впервые появляется только у четвероногих, а кортизон 'встречается -лишь в немногих классах позвоночных.
Для глюко- и минералокортикоидов характерно наличие кетогрупп в положениях 3 и 20 и гидроксильной группы в положении 21. Глюкокортикоиды содержат кислород в положении И, а в минералокортикоидах его обычно здесь нет.
428
Глава 21
Глюкокортикоиды оказывают большое и разностороннее влияние на обмен белков. Они способствуют увеличению количества углеводов, стимулируя процессы глюконеогенеза; содержание гликогена в печени возрастает. В то же время эти гормоны подавляют обратное всасывание глюкозы в почечных канальцах, что приводит к глюкозурии. Изменившийся под влиянием глюкокортикоидов обмен веществ благоприятствует распаду белков и таким образом усиливает выделение азота. Морфологически действие глюкокортикоидов проявляется в уменьшении массы соединительной, лимфатической и мышечной тканей, в изменении структуры Гостей и, кроме того, в подавлении различных процессов, участвующих обычно в реакции организма на раздражение, повреждение и аллергены. Глюкокортикоиды (кроме кортизола) влияют также и на другие процессы, например на распределение воды и электролитов в организме. Участвуя в регуляции водно-солевого равновесия, они действуют таким же образом, как и минералокортикоиды (способствуют быстрой реакции на избыток воды), однако в этом отношении они менее активны. Глюкокортикоиды поддерживают проницаемость мембран, в том числе синовиальных, на нормальном низком уровне; они стимулируют желудочную секрецию и поэтому способствуют образованию язвы желудка; и наконец, исходя из метаболических функций этих гормонов, можно предположить, что с увеличением их количества могут быть также связаны психические расстройства.
Из минералокортикоидов наиболее изучены 11-дезоксикортикостерон и альдостерон:
[ 11 -дезоксикортикостерон
СН2ОН
С=О
Как и следовало ожидать, исходя из наличия гидроксильной группы в положении 11, альдостерон в соответствующих количествах обладает также и глюкокортикоидной активностью.
Функция минералокортикоидов состоит главным образом в регуляции содержания электролитов и воды в организме. 11-дезок-сикортикостерон, который полностью лишен глюкокортикоидной активности, усиливает выведение калия и фосфатов с мочой и снижает выведение натрия, хлоридов и воды. Полагают, что он действует в основном на почечные канальцы, в частности на вое-
Эндокринные механизмы
429
ходящее колено петли Генле и дистальные извитые канальцы, стимулируя обратное всасывание натрия в обмен на выведение ионов водорода и калия [289].
Интенсивность секреции гормонов корой надпочечников меняется с изменением физиологических потребностей организма под влиянием внешних факторов. Самый важный из факторов, регулирующих функцию надпочечников, — это, несомненно, адренокортикотропный гормон аденогипофиза (АКТГ), который, по-видимому, стимулирует выработку глюкокортикоидов.
Реакция организма на любой стрессорный фактор связана с выделением АКТГ аденогипофизом. Происходящее при этом повышение секреции адренокортикальных гормонов удовлетворяет возросшие потребности организма. Для объяснения механизма секреции АКТГ при стрессе выдвинуты три теории. Согласно первой из них, основным стимулом служит нехватка кортикальных гормонов в результате их использования. По второй теории на гипофиз воздействует адреналин, выделяющийся при стрессе. В третьей теории предполагается, что выработка АКТГ находится под контролем гипоталамуса, а при стрессе возбудимость последнего возрастает. Судя по некоторым данным, секреция минералокортикоидов корой надпочечников прямо регулируется уровнем калия в крови. По-видимому, существенную роль в стимуляции выделения альдостерона может играть также ангиотензин II. Этот пептид образуется в крови из ангиотензина I под действием ренина — вещества, вырабатываемого в почках. Ангиотензин II может оказывать влияние на клубочковую зону, способствуя выделению альдостерона.
Секрецию адренокортикотропного гормона регулирует гипоталамус, в котором образуется фактор высвобождения АКТГ; нарушение гуморальной и нервной связи между промежуточным мозгом и гипофизом блокирует ответную реакцию на стресс.
Есть достаточные основания предполагать сходство общей схемы регуляции водно-солевого и углеводного обмена у всех позвоночных животных. Интересно, что кортизол, относимый обычно к глюкокортикоидам, у костистых рыб оказывает существенный осморегуляторный эффект; функция кортизола отличается здесь от той преимущественно метаболической роли, которую этот гормон играет у млекопитающих. Это служит примером того, как в результате эволюции может изменяться функция гормона. Таким образом, эволюционировать могли не сами гормоны, а способы их использования. Еще одним примером может служить рассмотренный выше гормон щитовидной железы.
У лягушек отмечены интересные сезонные различия в реакции на полную адреналэктомию. Зимой лягушки переносят ее, а летом, если оперированных лягушек не содержат в изотоническом солевом растворе, в их организме накапливаются вода и К+, животные теряют Na+ и погибают в течение одного-двух дней [219, 220].
430
Глава 21
Кортикостероиды, очевидно, стимулируют овуляцию как у млекопитающих [400], так и у амфибий [72, 451, 453], даже in vitro [457]. Кроме того, альдостерон обладает способностью усиливать перенос натрия в мочевом пузыре лягушки. По-видимому, это связано с ускорением синтеза какого-то белка, который затем стимулирует перенос натрия [329].
Яичник. Этот орган выделяет стероидные гормоны двух типов. Первый тип — эстрогены. До овуляции они секретируются клетками theca interna развивающегося фолликула, а после овуляции—клетками membrana granulosa и theca lutea. Эстрогены выделяются также плацентой. Приводим структурные формулы двух эстрогенов, относящихся к числу наиболее распространенных (хотя описаны и другие гормоны этого типа).
Эстрон
Как показали многочисленные исследования [417], у большинства низших позвоночных в яичнике образуется 17|3-эстрадиол или эстрон. Таким образом, в этом отношении различия между разными группами животных даже меньше, чем в случае адренокортикальных стероидов.
Эстрогены переносятся кровью преимущественно в связанной форме (в соединении с белком). Наиболее сильным действием обладает эстрадиол. Эстрогены необходимы для развития вторичных половых признаков и придаточных структур половой системы у самок млекопитающих. Они стимулируют рост эндометрия и желез матки, ее васкуляризацию и активность миометрия. Они вызывают ороговение эпителия влагалища и обусловливают завершение развития яйцеводов. В низкой концентрации эстрогены обычно способствуют дифференцировке фолликулов, но в более высоких концентрациях подавляют этот процесс, снижая секрецию фолликулостимулирующего гормона. Они способствуют также нормальному росту и дифференцировке молочных желез.
Эстрогены влияют на рост костей, определяя телосложение женского типа. Эти гормоны подавляют рост длинных трубчатых костей и способствуют окостенению эпифизарных хрящей. Они стимулируют синтез белка и вызывают задержку в организме натрия, кальция, фосфата и воды; кроме того, они снижают активность сальных желез.
Эндокринные механизмы
431
Второй гормон яичников — прогестерон:
У млекопитающих образуются и другие гормоны, сходные по своему действию с прогестероном:
20р-оксипрегн-4-ен-3-он 20а-оксипрегн-4-ен-3-он
Они вырабатываются клетками theca и granulosa желтого тела. Прогестерон секретируют также плацента и кора надпочечников. Этот гормон ответствен за активацию секреторной фазы эндометрия и дополняет тем самым действие эстрогенов; однако в отличие от последних он подавляет функцию миометрия. Прогестерон усиливает выделение секрета слизистой влагалища. В низких концентрациях он совместно с эстрогенами стимулирует выработку лютеинизирующего гормона, а в более высоких концентрациях подавляет ее. Прогестерон дополняет действие эстрогенов, стимулируя развитие молочных желез.
Секреторная функция яичников [17] регулируется тремя гормонами аденогипофиза (рис. 21-3). Фолликулостимулирующий гормон (ФСГ) способствует развитию фолликулов и усиливает секрецию эстрогенов. Его синергистом служит лютеинизирующий гормон (ЛГ), который, кроме того, вызывает овуляцию и развитие желтого тела. Третий гормон, пролактин (а у некоторых видов — сам ЛГ), стимулирует секрецию прогестерона желтым телом.
Эстрогены подавляют секрецию ФСГ и стимулируют выработку ЛГ и пролактина. Прогестерон подавляет секрецию ЛГ и способствует выработке ФСГ аденогипофизом. Согласованное взаимодействие яичника и гипофиза при участии этих пяти гормонов лежит в основе регуляции менструального и астрального циклов у млекопитающих.
432
Глава 21
Во время беременности плацента выполняет роль эндокринного органа, вырабатывая гормоны, которые в той или иной мере обеспечивают ее собственное существование до самых родов. Внутренний цитотрофобласт плаценты секретирует хорионический гонадотропин— гормон, сходный по физиологическому действию с ЛГ или пролактином. Недавно был очищен плацентарный лактоген человека и расшифрована его структура [380]. Он обнаруживает удивительное сходство с человеческим гормоном роста, будучи идентичен ему в 85% соответствующих аминокислотных позиций. Наружный синцитиальный трофобласт выделяет эстрогены и прогестерон; по существу это те же самые гормоны, которые обычно образуются в яичниках.
Хотя толчок к развитию молочных желез дают эстрогены и действие последних дополняется влиянием прогестерона, относительное значение этих двух гормонов у разных видов различно. Пролактин, стимулирующий секрецию прогестерона, кроме того, непосредственно способствует развитию молочных желез и секреции ими молока. Существенную роль в стимуляции выработки молока играют также стероиды коры надпочечников. Нейрогипофизарный гормон окситоцин способствует активному выведению молока, воздействуя на сократимые элементы протоков железы.
Семенник [267]. Семенник вырабатывает андрогены. Наиболее важный и самый активный из них—тестостерон, хотя существует и ряд других андрогенов.
Тестостерон
Имеется еще один важный андроген, циркулирующий в крови,— андростендион:
О
Кроме того, получены данные о том, что in vivo активным гормоном может служить также дигидротестостерон, образующийся в тканях, чувствительных к андрогенам [409].
Андрогены образуются в интерстициальных клетках Лейдига, а возможно, и в других участках семенника. 'Функции андрогенов многочисленны. Эти гормоны необходимы для развития придаточ-
Эндокринные механизмы
433
них желез мужской половой системы и вторичных мужских половых признаков, а также нормального полового поведения. В своем действии на общий метаболизм андрогены дополняют гормон роста, ускоряя рост тканей и синтез белков. Они стимулируют рост костей и окончательное окостенение эпифизарных хрящей, повышают интенсивность эритропоэза и усиливают кровоток в тканях; кроме того, они стимулируют активность кожных желез и образование меланина.
[окружающая среда
Рис. 21-3. Схема эндокринной регуляции обычного полового цикла у половозрелых самок млекопитающих.
Сплошными линиями показаны стимулирующие влияния, пунктирными — подавляющие.
Регуляция функций семенников осуществляется в первую очередь аденогипофизом, хотя на их активность влияют также щитовидная железа и кора надпочечников (рис. 21-4). Главную роль в этой регуляции играют гормоны гипофиза — ФСГ и Л Г, секрецию которых в свою очередь регулируют гипоталамические высвобождающие факторы. ФСГ стимулирует зачатковый эпителий и вызывает сперматогенез, а ЛГ побуждает семенники к выработке тестостерона и таким образом регулирует развитие половых признаков. Семенники в свою очередь также влияют на аденогипофиз. После кастрации бета-клетки гипофиза увеличиваются в размерах и секреция ФСГ усиливается; это, очевидно, бывает вызвано отсутствием тестостерона и, возможно, эстрогенов, которые в норме подавляют секрецию ФСГ.
В низкой концентрации андрогены, по-видимому, необходимы для сперматогенеза, хотя в более высокой концентрации они оказывают на него тормозящее влияние — как косвенно, через ФСГ, так и прямым путем, воздействуя на зачатковый эпителий.
Аденогипофиз. Эта железа имеет эктодермальное происхождение и образуется как выпячивание верхней стенки глотки. Это выпячивание, или карман Ратке, приходит в тесное соприкоснове-28—2067
434
Глава 21
ние с выпячиванием воронки, которая позже превратится в нейро- ;
гипофиз. Карман Ратке дает начало трем главным частям аде- j
ногипофиза: задняя стенка остается тонкой, превращаясь в промежуточную часть, pars intermedia (у тех животных, у которых она I есть); передняя стенка сильно утолщается и образует дистальную I часть, pars distalis, а дорсальный участок кармана вытягивается | по направлению к воронке, образуя бугровую часть, pars tubera- • lis. В аденогипофизе обнаружено 7 гормонов. Поскольку по край- j ней мере 5 из них оказывают важное регуляторное влияние на
Рис. 21-4. Схема эндокринной регуляции обычного полового цикла у половозрелых самцов млекопитающих.
Сплошными линиями показаны стимулирующие влияния, пунктирными — подавляющие.
другие эндокринные железы, аденогипофиз можно с полным основанием рассматривать как центральную железу эндокринной системы.
Гормоны роста, или соматотропные гормоны. Это белки смол, весом около 22 000, состоящие из длинных разветвленных или коротких неразветвленных пептидных цепей [263, 264, 439]. Каждый такой белок наиболее активен по отношению к тому виду животных, от которого он получен. Однако было высказано предположение, что все они обладают общей центральной структурой. Судя по некоторым данным, эти гормоны образуются в дистальной части аденогипофиза, в ацидофильных клетках, окрашивающихся в оранжевый цвет.
Гормон роста в нормальных условиях стимулирует рост молодых животных, ускоряя синтез белка и подавляя распад аминокислот. В результате в организме задерживается азот, повышается содержание воды и солей и уменьшается количество жиров. Этот гормон способствует также усиленному росту костей: он стимулирует периостальное окостенение, пролиферацию хряща и образование кости в зоне эпифизарного хряща, т. е. рост костей в; длину.
У взрослых плотоядных животных гормон роста оказывает диабетогенное влияние, нейтрализуя действие инсулина. При введении этого гормона бета-клетки островков Лангерганса сна
Эндокринные механизмы
435
чала пролиферируют, а затем подвергаются атрофии, что приводит к перманентному диабету.
У рыб этот гормон необходим для роста тела в длину [324, 325, 439]. «М
Секреция гормона роста регулируется специфическим высвобождающим фактором гипоталамуса.
Тиреотропный гормон (ТТГ). Этот гормон представляет собой гликопротеид с мол. весом около 30 000, вырабатываемый базофильными клетками; установлена его химическая структура [328]. Основная роль ТТГ — стимуляция функции щитовидной железы во всех ее проявлениях. Гормон повышает количество внутриклеточного коллоида и стимулирует выделение тироксина; он способствует увеличению высоты фолликулярных клеток, поглощению .ими иода и синтезу тиреоидных гормонов. В структуре тиреотропного гормона имеются межвидовые различия [147].
Между гормоном щитовидной железы и ТТГ существуют регуляторные взаимодействия. Первый, блокируя реакцию гипофиза на ТТГ-высвобождающий фактор, подавляет секрецию второго. Хотя эти взаимоотношения служат главным механизмом, регулирующим секрецию тиреотропного гормона, ее подавляют также .адреналин и кортикостероиды. Этим и объясняется снижение активности щитовидной железы при первичных стресс-реакциях, вызванных различными факторами (кроме холода: холодовой стресс оказывает противоположное действие). Таким образом, в обычных условиях стресс стимулирует выделение АКТГ и в тоже время подавляет секрецию тиреотропного гормона. Холодовой стресс составляет исключение — он стимулирует выработку обоих гормонов.
Адренокортикотропные гормоны (АКТГ). Эти гормоны представляют собой полипептиды; они образуются в базофильных клетках. По-видимому, существуют три кортикотропных гормона. Их мол. вес составляет примерно 4500, они содержат до 39 аминокислотных остатков, причем определенная последовательность 24 из них имеет существенное значение. Из этих аминокислот 13 являются общими с меланоцитстимулирующим гормоном (а-МСГ) [209, 204, 265, 379]; и действительно, АКТГ способен вызывать потемнение кожи у серебряного карася [83].
АКТГ стимулирует кору надпочечников, вызывая гипертрофию ее ткани, в особенности пучковой зоны, и усиливая секрецию глюкокортикоидов. На выделение минералокортикоидов он в основном не влияет.
Секрецию АКТГ гипофизом в обычных условиях регулирует гипоталамус при помощи фактора, высвобождающего АКТГ; на выделение этого фактора в свою очередь, вероятно, влияет содержание кортикостероидов в крови. При стрессе секреция АКТГ стимулируется частично нервным путем через гипоталамус, а частично путем возбуждающего воздействия адреналина на центры 28*
436
Глава 21
гипоталамуса и на саму железу. По-видимому, на секрецию АКТГ обычно влияет также средний участок eminentia mediana [159, 160], хотя основной уровень АКТГ поддерживается и без такой регуляции.
Фолликулостимулирующий гормон (ФСГ). Это гликопротеид с мол. весом около 30 000, который, как полагают, вырабатывается бледными базофильными клетками дистальной части аденогипофиза. Его назначение—стимулировать рост фолликулов в яичнике у самок и способствовать сперматогенезу у самцов.
Лютеинизирующий гормон (ЛГ), или гормон, стимулирующий интерстициальные клетки. Это тоже гликопротеид с мол. весом 32 000 [266, 327]; он также, вероятно, образуется в бледных базофилах дистальной части.
Лютеинизирующий гормон стимулирует у самок секрецию яичниками эстрогена и прогестерона, а у самцов — секрецию семенниками андрогенов. Секреция обоих гонадотропных гормонов — ФСГ и ЛГ — регулируется гипоталамусом [338]. В настоящее время установлено, что высвобождающий фактор для этих гормонов вырабатывается в гипоталамусе; его структура выяснена [14]. В регуляции, по-видимому, участвует также задняя часть eminentia mediana. Активность гипоталамуса в свою очередь регулируется содержанием половых гормонов в крови, а также многими другими факторами, например воздействием света и температуры, присутствием особей другого пола и раздражением половых органов.
Пролактин, или лютеотропный гормон (ЛТГ). Этот гормон представляет собой белок, который, как принято считать, вырабатывается красными ацидофильными клетками в дистальной части. Его мол. вес около 25000; обнаружены межвидовые различия, например между пролактинами быка и овцы [162, 263].
У самок млекопитающих пролактин поддерживает существование желтого тела и длительную секрецию прогестерона; однако у разных видов в этом отношении есть некоторые различия. Этот гормон дополняет также действие половых гормонов, обеспечивая развитие молочных желез, так что он наряду с гормоном щитовидной железы и кортикостероидами необходим для нормальной лактации '[298]. Третья возможная роль пролактина — поддержание материнских инстинктов и поведения. Имеются также данные о влиянии пролактина на развитие предстательной железы у самцов [82]; в этом отношении пролактин оказывается синергистом половых гормонов и гонадотропинов.
Выделение пролактина подавляется гипоталамусом, хотя основной уровень его секреции поддерживается, видимо, без регуляции [300]. Во время лактации длительная секреция пролактина зависит от раздражения сосков и возбуждения афферентных путей, идущих к головному мозгу, который в свою очередь стимулирует как активность аденогипофиза, так и секрецию окситоцина
Эндокринные механизмы
437
нейрогипофизом, что приводит к активному выведению молока. Выделение пролактина регулируется главным образом гипоталамическим пролактин-ингибирующим фактором, но возможно, что существует и активирующий фактор.
У других позвоночных пролактин наряду с хорошо известным действием на зобную железу голубя (стимуляция выделения «молочка») усиливает совместно с меланоцитстимулирующим гормоном пигментацию покровов у рыб [326] и стимулирует передвижение тритона (Triturus) к воде в период размножения [176].Он вызывает и другие эффекты [32], например задержку ионов натрия в организме рыб.
Меланоцитстимулирующий гормон (МСГ) [303]. Этот полипеп-тидный гормон секретирует промежуточная часть аденогипофиза. Выделены две разновидности гормона: а-МСГ, состоящий из 13 аминокислотных остатков [257], и ряд различных £-МСГ с цепью из 18—22 аминокислотных остатков [165, 190]. Последовательность из 7 аминокислот (Мет-Глу-Гис-Фен-Арг-Три-Гли), известная как гептапептидное ядро, является общей для мелано-цитстимулирующих и адренокортикотропных гормонов; этим объясняется способность АКТГ вызывать рассеивание меланина.
У некоторых низших позвоночных функция МСГ состоит в том, что этот гормон обусловливает потемнение тела, вызывая рассеивание меланина в меланофорах (гл. 23), сокращение иридофо-ров, усиленный синтез меланина и распад пуринов. Роль МСГ у птиц и млекопитающих менее ясна [303]. Чистый МСГ стимулирует синтез меланина и вызывает потемнение волосяного покрова или кожи у мыши [163] и человека [258]. Помимо своего влияния на пигмент, гормон вызывает и другие эффекты, физиологическое значение которых не вполне ясно. Он оказывает, например, тиреотропное, липолитическое [13], антигонадотропное [69], гипокаль-цемическое [152] и кардиостимулирующее [241] действие. Какую функциональную роль могут играть эти эффекты, неизвестно. Другой возможной функцией МСГ, связанной с тем, что этот гормон способствует адаптации животного к цвету грунта, может быть участие в процессах адаптации вообще. МСГ или АКТГ необходимы для выработки и закрепления определенных условных рефлексов избегания у крыс [105], так что МСГ, по-видимому, облегчает приспособительные реакции. Кроме того, МСГ и АКТГ могут вызывать зевоту и потягивание у собак [133]; возможно, что они играют значительную роль в осуществлении такого рода поведенческих реакций у интактных животных.
Секреция МСГ находится под тормозным контролем гипоталамуса, хотя есть также данные о наличии возбуждающего регуляторного влияния. После пересадки гипофиза у саламандры наблюдается усиленная секреция МСГ [40]. Экстракты гипоталамуса
438
Глава 21
лягушек содержат вещество, подавляющее секрецию МСГ [340], но имеется еще адренэргическое нервное сплетение, которое может быть ответственно за тормозное влияние. Недавно появилось сообщение, что фактор из бычьего гипоталамуса, тормозящий секрецию МСГ, представляет собой пролиллейцилглицинамид [294].
Нейрогипофиз. Этот орган нельзя считать эндокринной железой в том смысле, как другие железы, описанные выше. Он не только образуется как выпячивание дна III мозгового желудочка, но и сохраняет связь с головным мозгом в течение всей жизни. В настоящее время точно установлено, что его гормоны синтезируются в супраоптическом и паравентрикулярном ядрах гипоталамуса, а затем переносятся по нервным волокнам в нейрогипофиз, где и выделяются в капилляры при соответствующем раздражении. Таким образом, это нейросекреторная железа. С помощью электронной микроскопии в клетках нейрогипофиза был выявлен секреторный материал, заключенный в пузырьки. Нейрогипофиз выделяет два гормона—антидиуретический гормон (аргинин- или . лизин-вазопрессин) и окситоцин. Это полипептиды, содержащие 8 аминокислот, 6 из которых общие для обоих гормонов [419—421].
Антидиуретический гормон. Основное действие АДГ у млекопитающих заключается в стимуляции всасывания воды в дистальных почечных канальцах. Обнаружена корреляция между содержанием в нервных клетках АДГ и задержкой воды в организме [449]. АДГ оказывает влияние непосредственно на канальцы, -особенно на петлю Генле, дистальный извитой каналец и собирательные трубки. .Второе, менее существенное действие АДГ направлено на сосудистую систему — он вызывает в основном сокращение артерио-венозных капилляров. Физиологическое значение этого эффекта не выяснено.
Окситоцин. Этот гормон вызывает сокращение гладкой муску-.латуры, особенно в стенке матки и в меньшей степени — мочевого шузыря, желчного пузыря и кишечника.
Окситоцин (питоцин)
^/Глутамин
Изо лейцин \
• | Аспарагин
Тирозин /
Цистин '
Пролин
Лейцин
Глицинамид
Эндокринные механизмы
43S:
Эстрогены усиливают действие окситоцина на мускулатуру матки, а прогестерон противодействует ему. Особую роль окситоцин играет в процессе лактации: он влияет на миоэпителиальные клетки, окружающие альвеолы молочной железы, что приводит к поступлению молока в сосок. Кроме того, он увеличивает проницаемость кожи [358] и мочевого пузыря [359] лягушки, хотя основным гормоном, вызывающим эти эффекты, является вазотоцин.
Двойная регуляция активности нейрогипофиза осуществляется через его ножку: этим путем в железу попадают выделяемые ею> гормоны [67, 315], а также поступают стимулы, приводящие к, переходу этих гормонов в кровь [332]. Способы раздельного регулирования секреции двух гормонов до сих пор неизвестны. Секреция АДГ происходит непрерывно, но ее скорость регулируется осмотическим давлением крови; понижение осмотического давления ослабляет секрецию гормона [311]. Полагают, что рецептором осмотического давления служат клетки самого супраоптического ядра. На секрецию АДГ влияют и многие другие факторы; Повышение секреции наблюдается при стрессе, уменьшении объема циркулирующей крови и действии таких ядов, как морфин и никотин. Увеличение объема циркулирующей крови и охлаждение организма снижают секрецию АДГ.
Окситоцин выделяется во время лактации при раздражении соска. Афферентные нервные импульсы, идущие от молочной железы, возбуждают центр в гипоталамусе. Есть, по-видимому, и другие стимулы, которые через афферентные нервы вызывают секрецию окситоцина: растяжение матки в поздний период беременности, а у некоторых видов также спаривание. Адреналин,, напротив, подавляет секрецию окситоцина.
У млекопитающих имеются два вазопрессина, которые отличаются друг от друга всего лишь одной аминокислотой: у свиньи № бегемота обнаружен лизин-вазопрессин, а у всех других исследо ванных млекопитающих —аргинин-вазопрессин.
Вазопрессин (питрессин) /Глутаминч
Фенилаланин
Тирозин
Аспарагин
Цистин
Пролин
Аргинин.....[Лизин] (у свиньи).
Глицинамид
(у большинства млекопитающих)
440
Глава 21
Действие этих двух вазопрессинов может существенно различаться. Например, аргининовая форма гормона при введении собаке оказывает примерно в 6 раз более сильное антидиуретиче-ское действие, чем лизиновая [360, 415, 416].
У птиц и пойкилотермных позвоночных обнаружены значительные различия в природе и действии нейрогипофизарных пептидов. Например, АДГ понижает кровяное давление у птиц, а окситоцин (вызывает нерестовые реакции у рыбы Fundulus hetero-clitus [440]. У птиц, амфибий и рыб имеется другой гормон — ар-гинин-вазотоцин. Известны еще три гормона: мезотоцин, обнаруженный у примитивных костистых рыб, амфибий и рептилий, изо-тоцин, найденный у костных ганоидов и костистых рыб, и глуми-тоцин — у скатов. Таким образом, число известных нейрогипофизарных пептидов доходит по меньшей мере до семи, что позволяет выдвигать интересные гипотезы относительно их эволюции.
Аргинин-вазотоцин
Аргинин I Глицинамид (у птиц, костистых рыб, амфибий, круглоротых)
АДГ — это, по-видимому, еще один гормон, действие которого связано со стимуляцией аденилатциклазы и последующим образованием циклического АМФ. Изменяя проницаемость изолированного мочевого пузыря жабы для воды, АДГ вместе с тем увеличивает содержание в нем циклического АМФ [185]; с помощью циклического АМФ можно также имитировать влияние АДГ на этот орган [306]. Возможно, что циклический АМФ участвует и в действии АДГ на почки, так как в мозговом веществе почек, где находятся дистальные извитые канальцы, обнаружена АДГ-чувствительная аденилатциклаза [80].
Урогипофиз. Это нейросекреторный эндокринный орган, находящийся в каудальной области спинного мозга костистых рыб [150]. Нейросекреторные клетки спинного мозга (клетки Даль-грена) соединены волокнистым трактом с нейрогемальным органом— урогипофизом. Хотя у костистых рыб этот орган хорошо выражен, у акуловых рыб он отсутствует, что, возможно, отра
Эндокринные механизмы
441
жает их примитивность. По мнению большинства исследователей, урогипофиз участвует в осморегуляции; отмечено влияние его как на перенос натрия, так и на задержку воды в организме. Урогипофиз костистых рыб содержит также вещество, вызывающее сокращение изолированного мочевого пузыря радужной форели [251, 252]; кроме того, в этом органе обнаружены гипер- и гипотензивные агенты.
Гормоны и размножение
Гонадотропины — ФСГ, Л Г и пролактин — встречаются у представителей всех классов позвоночных [164]. В отсутствие гонадотропинов, например после гипофизэктомии, у молодых самок млекопитающих приостанавливается развитие яичников, а у взрослых они уменьшаются в размерах и при этом наступает атрезия уже образовавшихся фолликулов. Избыток ФСГ, напротив, вызывает одновременное созревание большего числа фолликулов. У некоторых млекопитающих, например у кролика и хорька, овуляция происходит только после спаривания или какого-либо другого эффективного стимула. Известно, что такая стимуляция приводит к выделению Л Г, который и вызывает овуляцию [127]. Другим млекопитающим, в том числе и человеку, для овуляции не нужно никакого специального дополнительного стимула. Образование желтого тела на месте лопнувшего фолликула у всех млекопитающих обычно происходит под действием Л Г.
После удаления гипофиза астральные циклы прекращаются. Их удается восстановить путем пересадки гипофиза, но только в том случае, если имплантаты помещают в область серединного возвышения (eminentia mediana) гипоталамуса [301]. Гонадотропины не обладают половой специфичностью. Они регулируют активность как семенников, так и яичников. Удаление гипофиза у самцов приводит к прекращению функции семенников и уменьшению их размеров. У гипофизэктомированных особей активность семенников можно восстановить пересадкой гипофиза или введением его экстрактов. У многих млекопитающих, у которых размножение строго приурочено к определенным временам года, с помощью гонадотропинов можно активировать семенники независимо от сезона. Это было показано на таких животных, как суслик (Citellus) и сурок (Marmota). После сезона спаривания семенники у этих животных обычно возвращаются в брюшную полость и сперматогенез прекращается. Вводя гонадотропный гормон гипофиза или даже хорионический гонадотропин, который в норме имеется только у беременных самок, можно вызвать эти процессы в любое время года, так как последний гормон обладает ЛГ-подобными свойствами. Введением сыворотки жеребых кобыл, а также ФСГ или ЛГ из гипофиза свиньи удалось вызвать увеличение яичников у жабовидной ящерицы (Phrynosoma) [282].
442
Глава 21
С помощью гипофизэктомии показано, что у болотной черепахи функция половых желез и развитие вторичных половых признаков зависят от гипофиза [89].
Если самкам лягушек или жаб вводить мацерированную переднюю долю гипофиза, то у них можно вызвать овуляцию в любое время года, за исключением периода, наступающего сразу после икрометания [351—353]. Препараты гипофизов, взятых от самок, примерно вдвое эффективнее, чем от самцов. При овуляции, которая происходит между 2-м и 4-м днями после введения гипофизарного препарата, наблюдается разрыв фолликулов и выталкивание яиц в результате сокращения гладких мышечных волокон в стенке фолликула. Эти явления можно воспроизвести in vitro ,[349]. Введение экстрактов гипофиза самцам лягушки вызывает у них обнимательный рефлекс. Подобные реакции у амфибий могут быть вызваны также экстрактами гипофиза других амфибий, рыб [448], млекопитающих [458] или даже хорионическим гонадотропином из мочи беременных самок. Опыты с перекрестными инъекциями выявили существенные различия в активном •факторе (ЛГ) разных позвоночных [453].
У некоторых рыб можно вызвать преждевременное икрометание, вводя свежую ткань гипофиза других рыб. У яйцеживородящей рыбы Cnasterodon после введения гипофизарной ткани от Micropagon и Luciopimelodus нерест происходит более чем на две недели раньше обычного. У бразильских рыб Pimelodus и Prochi-lodus, которые обычно мечут икру после сильных дождей, следующих в Бразилии за длительным периодом засухи, икрометание и выделение молок можно было вызвать и во время засухи: реакция наступала спустя 1—3 дня после внутримышечной инъекции ткани гипофиза, взятой у тех же видов. В опытах, проведенных в Висконсине, у форелей Salmo gairdneri и S. fario внутрибрюшинное введение свежей или высушенной в ацетоне ткани гипофиза карпа вызывало созревание икры и молок на 6—7 недель раньше обычного нерестового сезона [194]. ФСГ овцы и сыворотка жеребой кобылы такого влияния не оказывали. У миноги Petromyzon после введения мочи беременной женщины наблюдалось увеличение размеров ооцитов и яичников и преждевременное половое созревание [107]. У рыбы Fundulus гипофизэктомия приводит к атрофическим изменениям в яичниках и семенниках. Если осенью удалить гипофиз, то нормального увеличения гонад весной не происходит. Пересадка гипофизов взрослых Fundulus, производимая в течение месяца с трехдневными интервалами, (вызывает у неполовозрелых особей обоего пола значительную активацию половых желез; при этом у самцов наступают изменения пигментации, характерные для брачного сезона. По-видимому, у Fundulus нерест регулируется непосредственно гипофизом [324].
Андрогены имеются у всех позвоночных. Главная роль их состоит в стимуляции развития и функции придаточных органов
Эндокринные механизмы
443?
мужской половой системы. Частично от них зависят также особенности телосложения самцов, более низкий голос у мужчин, характерное распределение волос, драчливость и многие другие вторичные половые признаки, например брачные мозоли на пальце у самцов лягушек, гребень у петухов и тритонов, брачная окраска и гоноподии у некоторых живородящих рыб. Многие из; этих признаков проявляются главным образом в брачный период.
Разные андрогены отличаются друг от друга по своей общей активности и по относительному влиянию на многочисленные индивидуальные особенности организма; эти различия, кроме того,, могут варьировать от вида к виду, что значительно усложняет-изучение функций андрогенов.
Основным источником андрогенов служат семенники; они обладают также способностью извлекать из циркулирующей крови1 менее активные андрогены и превращать их в более активные вещества. Как показали многочисленные наблюдения, андрогены образуются в интерстициальных клетках семенников. Редукция или исчезновение зачаткового эпителия семенных канальцев под действием рентгеновских лучей или после перевязки семевынося-щего протока, а также при крипторхизме не сопровождаются атрофией интерстициальной ткани семенников, и андрогенная активность при этом полностью сохраняется. У тритона Triton cri-status интерстициальная ткань в семенниках появляется лишь незадолго до наступления брачного периода; при этом определенные участки эпителиальной ткани превращаются в особые железистые тела, и это превращение сопровождается развитием у самцов характерного брачного наряда. Разрушение этой железистой ткани равносильно полной кастрации и предотвращает развитие указанных признаков. У колюшки (Gasterosteus) существует весьма тесная связь между годичным циклом дифференцировки интерстициальных железистых клеток, с одной стороны, и циклом развития вторичных половых признаков и соответствующих форм поведения (окраска тела, выделение слизи для постройки гнезда и брачное поведение) — с другой. Между тем у этого вида не обнаружено никакой связи между последними признаками и спер-матогенной активностью. О том, что сперматогенный эпителий не является существенным источником андрогенов, говорят также опыты с удалением большей части семенников у петухов и исследованием этих органов после их регенерации. Регенерировавшие органы состоят главным образом из семеобразующего эпителия с очень небольшими включениями интерстициальной ткани и соответственно продуцируют очень мало андрогенов.
Эти и многие другие экспериментальные данные позволяют предполагать, что местом образования тестикулярных андрогенов служат интерстициальные клетки.
Андрогены образуются, однако, не только в семенниках. У кастрированных самцов придаточные половые железы можно, под--
444
Глава 21
держивать в состоянии полной функциональной активности, пересадив таким животным ткань яичника. Рост гребней как у петухов, так и у кур стимулируется андрогенами. Яичник курицы обычно вырабатывает андрогены в количестве, достаточном для поддержания определенной степени развития гребня; после удаления яичников гребень атрофируется. Сезонное пожелтение клюва у самцов и самок скворца, так же как и почернение клюва у самцов и самок домового воробья, обусловлено действием андрогенов. Эстрогены в этом отношении неэффективны. У черной кваквы (вид цапель) в брачный период происходят определенные изменения в окраске перьев, одинаковые у обоих полов. Эти и многие другие факты убедительно доказывают, что в яичниках также образуются андрогены.
Еще один источник андрогенов — кора надпочечников. Пересадка ткани этих желез петушкам приводит к преждевременному развитию у них мужских половых признаков и характерному поведению. Кастрация молодых крыс без одновременной полной адреналэктомии не вызывает атрофии придаточных половых структур в течение многих дней. Андрогены обнаруживаются в моче животных и после кастрации. В экстрактах тканей надпочечников найдены вещества андрогенной природы—адреностерон и минералокортикоид дезоксикортикостерон; последний примерно так же активен, как андростерон, а активность первого составляет около 1/5 этой величины.
Нет почти никаких данных о сколько-нибудь значительном накоплении андрогенов в организме. Обычно они быстро разрушаются в печени, и поэтому постоянный уровень их в крови может поддерживаться только при непрерывной секреции. Однако у некоторых рыб из сем. Poeciliidae андрогены, вводившиеся в течение всего лишь нескольких часов, продолжали действовать на протяжении многих дней. Очень высокое содержание андрогенов обнаружено в так называемой гибернационной железе (пигментированной жировой ткани) лесного сурка во время зимней спячки. Андрогены инактивируются главным образом в печени. Андрогенная активность в крови, оттекающей от печени, гораздо ниже, чем в крови, поступающей в этот орган. При этом из андрогенов образуются как совсем неактивные вещества, так и андрогены с несколько более слабым действием, например андростерон, а также обычно и эстрогены. Все эти продукты выводятся через почки.
Была детально изучена связь между андрогенами, развитием характерных мужских органов — гоноподиев—и меньшей величиной тела у самцов рыб из сем. Poeciliidae [410]. После кастрации превращение анального плавника в гоноподий тотчас же прекращается. У кастрированных самцов или нормальных самок под действием андрогенных гормонов развиваются мужские признаки, типичные для данного вида. Показано, что для последовательных
Эндокранные механизмы
445
этапов развития гоноподия требуется постепенное повышение концентрации гормона; чтобы вызвать первый этап, достаточно одной части гормона на 4,2 -1010 частей воды. Обычно гоноподий развивается под влиянием андрогенов, концентрация которых в крови постепенно возрастает в результате активности семенников. Для нормального развития на каждой его стадии нужна некоторая оптимальная концентрация андрогенов; избыточные концентрации подавляют рост и вызывают преждевременную дифференцировку. У жаб обнимательный рефлекс исчезает после кастрации [59].
Эстрогеном называют любое вещество, которое вызывает у взрослых мышей явления, характерные для нормальной течки, в том числе ороговение влагалища. Эстрогены образуются в яичниках у всех позвоночных.
Материнская и эмбриональная части плаценты также богаты эстрогеном, весьма сходным с эстрогенами яичников. Более того, удаление яичников у беременных самок некоторых млекопитающих не приводит к заметным изменениям сроков беременности или к стойкому уменьшению количества эстрогенов, выделяемых с мочой. Расхождение симфиза (изменение, обусловленное в норме совместным действием эстрогенов и прогестерона) и в этих случаях происходит нормально.
Эстрогены образуются также и в семенниках. Действительно, ткань, наиболее богатая эстрогеном, — это семенники жеребца, в которых содержится эстрон. В семенниках источником эстрогенов, так же как и андрогенов, служат, по-видимому, интерстициальные клетки. Эстрогены удалось выделить и из надпочечников. Андрогены служат необходимыми предшественниками для биосинтеза эстрогенов.
В организме эстрогены очень быстро инактивируются, образуя глюкурониды или сульфаты. Как показывает сравнение физиологических эффектов при пересадке ткани яичника в различные участки тела, разрушение эстрогенов происходит главным образом в печени. Содержащая эстрогены кровь, пройдя через печень, оказывается практически лишенной этих гормонов. Разрушение печеночных клеток четыреххлористым углеродом приводит к повышению концентрации эстрогенов в крови и к соответствующим изменениям астрального цикла.
Срезы печени инактивируют эстрадиол in vitro при участии какого-то механизма, чувствительного к цианиду. Прогестерон снижает скорость инактивации эстрогенов и таким образом спо-J собствует поддержанию более высокой их концентрации в кро-ви; при этом значительная часть их может выделяться с мо-i чой. Почки и печень — основные органы, выводящие эстрогены из |организма.
| Разные клетки и ткани сильно различаются по своей чувствительности к эстрогенам, что приводит к градуальному характеру
446
Глава 21
ответов. Кроме того, эти ответы легко обратимы при снижении1 концентрации гормона. Такая обратимость наиболее очевидна в случае вторичных половых признаков и придаточных органов половой системы, которые у многих видов изменяются в течение цикла размножения.
Как и при воздействии андрогенов, специфический порог и характер ответа на гормон определяются внутренними свойствами-реагирующих клеток. При пересадке отдельных тканей в пределах одного организма их реакция остается неизменной. Зависимость реакции от генетической конституции ткани наглядно проявляется в опытах с перекрестными пересадками участков кожи, из которых растут перья (у кур и петухов): характер оперения определяется полом птицы, от которой была взята кожа.
Получено много данных относительно механизма действия; эстрогенов на матку крысы. Мюллер [290] описал последовательность событий, развивающихся после введения эстрогена. В первые 4 ч наблюдаются гиперемия и набухание ткани. В период от 6 до 24 ч происходит накопление РНК, а затем белков. Через 24—72 ч после эстрогенизации отмечается синтез ДНК. В последнее время усилия исследователей сосредоточились на выяснении механизма, благодаря которому эстрогены усиливают синтез РНК, что, по-видимому, является общим звеном в действии многих стероидных гормонов [384]. Хотя не исключено, что в действии* эстрогенов может участвовать циклический АМФ [399], большинство исследователей объясняют влияние этих гормонов присоединением их к цитоплазматическим белкам — рецепторам эстрогенов — и последующим переходом в клеточное ядро. Есть данные в пользу того, что 17р-эстрадиол в цитоплазме клеток матки присоединяется к белку с константой седиментации 9,5S. Затем комплекс гормона с этим белком переходит в ядро, где белок диссоциирует на 53-субъединицы и эстрадиол остается связанным с ними. Полагают, что комплекс эстрадиол — белок запускает здесь-синтез информационной РНК, имеющей отношение к последующим гормональным эффектам [175]. Локализация стероидных гормонов в клетке была также определена методом радиоавтографии производных, меченных тритием [395].
Прогестерон. Этот гормон ответствен главным образом за изменения, связанные с беременностью и родами. Он является также обязательным промежуточным продуктом при биосинтезе стероидов, например в коре надпочечника. Основным источником этого гормона служат желтые тела яичника.
Прогестерон образуется также в надпочечниках. Здесь же вырабатывается дезоксикортикостерон, частично сходный по своему действию с прогестероном. Вводя экстракты надпочечников крольчихам, предварительно обработанным эстрогенами^ можно вызывать типичные изменения в матке, характерные для подготовки к беременности.
Эндокринные механизмы
447
Другой важный естественный источник прогестерона — плацента. У млекопитающих через некоторое время после наступления беременности плацента берет на себя главную роль в выработке прогестерона. Даже после. полного удаления яичников у беременных крыс в их плаценте образуется столько прогестерона, сколько нужно для нормального завершения беременности. У человека этот переход регулирующей функции от желтого тела к плаценте происходит, вероятно, между 79-м и 90-м днями беременности.
Прогестерон не накапливается в организме; он очень быстро исчезает из крови. Введение прогестерона или нормальное образование этого гормона сопровождается появлением в моче соответствующих количеств относительно неактивного спирта прег-нандиола. Судя по имеющимся данным, прогестерон превращается в прегнандиол главным образом в секретирующем эндомет-’рии; это превращение протекает особенно интенсивно в предменструальной (секреторной) фазе овариального цикла и обычно прекращается после удаления матки. Оно сильно замедляется после введения эстрадиола, который, как известно, подавляет развитие предменструальных изменений. По-видимому, предменструальный эндометрий — не единственное место описанного выше превращения гормона; на это указывает выделение прегнандиола самцами и изредка самками после удаления матки. Отметим также, что тельца Станниуса в почках радужной форели способны превращать прогестерон в 11-дезоксикортикостерон [88] и могут, таким образом, играть важную роль в метаболизме стероидов.
Желтые тела, образующиеся из фолликулов как без овуляции (при атрезии), так и после овуляции, заметно варьируют в своем развитии. Наибольшего развития достигает желтое тело при беременности; оно бывает менее развито при лактации и ложной беременности, а наименьших размеров достигают обычные желтые тела, образующиеся при овуляции. Развитие желтых тел зависит от поступления лютеинизирующего гормона (Л Г) из ^гипофиза. Млекопитающие заметно различаются по своей потребности во внешних раздражителях для выработки ЛГ. Показано, что у кроликов адекватным стимулом служит спаривание. У крысы секрецию ЛГ стимулируют сосание и лактация. У таких животных, <ак голуби и кролики, эффективными оказываются даже «психи-1еские» раздражители, например просто присутствие другой лсоби. Последний стимул, по крайней мере для самки голубя, является зрительным: овуляция часто происходит у нее даже при шде собственного отражения в зеркале.
Все исследователи согласны в том, что для возникновения келтого тела необходим Л Г, а для стимуляции его секреторной 1КТИВНОСТИ—ЛГ или ЛТГ, но по вопросу об эндокринных факторах, участвующих в поддержании желтых тел, мнения весьма
448
Глава 21
противоречивы. Половые гормоны способствуют поддержанию желтых тел, вероятно, путем подавления секреции ФСГ гипофизом. Был поставлен ряд опытов с удалением эмбрионов, причем в одних случаях плаценту оставляли на месте, а в других — тоже удаляли; сопоставление результатов ясно показало, что плацента выделяет вещество типа гормона, способствующее сохранению желтого тела. Полученные результаты можно, видимо, объяснить и тем, что половые гормоны плацентарного происхождения подавляют выработку ФСГ. Об участии в этом процессе каких-то дополнительных факторов свидетельствует сохранение желтого тела даже после удаления эмбриона вместе с плацентой, если только матку поддерживают в растянутом состоянии — например, с помощью физиологически инертных парафиновых шариков.
Желтые тела описаны и у пойкилотермных позвоночных, но их наивысшее развитие, по-видимому, связано с живорождением у млекопитающих. По имеющимся данным, они отсутствуют у птиц и яйцекладущих рептилий, но образуются у беременных самок некоторых живородящих змей [7, 49], например у Crotalus и Bothrops, у живородящей ящерицы Zootoca [312] и живородящей бесхвостой амфибии Nectophrynoides [245—248]. Однако роль желтых тел в поддержании беременности четко доказана только у млекопитающих.
Было высказано предположение, что в эволюции желтые тела возникли в связи с процессом «очистки» после овуляции, а прогестерон— это просто один из конечных продуктов [202]. У низших млекопитающих прогестерон обеспечивает задержку яиц в организме до наступления времени их откладки. У сумчатых период беременности точно соответствует фазе желтого тела в эстральном цикле. У крыс и мышей, у которых беременность длится дольше, в сохранении желтых тел участвует лютеотропный гормон (ЛТГ) хориона. У лошадей и приматов, у которых беременность еще продолжительнее, фаза желтого тела сначала удлинилась благодаря действию ЛТГ хориона, а затем соответствующую роль взяла на себя плацента; она вырабатывает не только прогестерон, но и ряд важных дополнительных веществ, например эстрогены и пролактины.
Релаксин. Релаксин — полипептид с мол. весом около 8000 — содержится в желтых телах и может быть выделен из крови беременных крольчих. Спустя 6—12 ч после однократной инъекции релаксин вызывает у морской свинки выраженное расслабление мышц тазового пояса. 'Прогестерон и эстрадиол стимулируют секрецию релаксина у крольчих, если только они не кастрированы и у них не удалена матка. Таким образом, для этой реакции на половые гормоны необходимы яичники и матка. Релаксин может также способствовать расширению шейки матки у беременных женщин перед самыми родами.
Эндокринные механизмы
449
Пролактин (лютеотропный гормон). Выработка молока представляет собой приспособление, развившееся у всех млекопитающих (и у таких птиц, как голубь) для кормления детенышей сразу же после рождения. Не удивительно поэтому, что половые гормоны оказывают значительное влияние на молочные железы. У плода млекопитающего молочные железы увеличиваются под влиянием гормонального комплекса, поступающего с материнской кровью, но после рождения регрессируют и, как правило, остаются в недоразвитом состоянии до наступления половой зрелости, когда их развитие начинают стимулировать собственные гормоны животного. Пролактин играет важную роль в стимуляции образования молока ацинарными клетками молочной железы.
Регуляция репродуктивных циклов и поведения. Гипофиз как источник гонадотропных гормонов служит главным эндокринным органом, который управляет функциями, связанными с размножением. Поэтому мы можем ожидать, что изучение факторов, регулирующих активность этого органа, позволит нам получить очень важные сведения о механизме регуляции половых циклов и приспособительных репродуктивных реакций организма в ответ на изменение окружающей среды. Большое значение имеют в связи с этим «высвобождающие факторы» гипоталамуса.
Гипофиз молодого млекопитающего «амбивалентен» — он может развиваться либо по мужскому, либо по женскому типу в зависимости от того, какая половая железа имеется в организме. У взрослых самок, как правило, гипофиз больше, чем у самцов. Как женские, так и мужские гипофизы обычно вырабатывают относительно много ФСГ и ЛГ. Основное различие между мужским и женским гипофизом — это различие в функциональном цикле. Пересадка взрослой кастрированной самке яичников (но не семенников) тормозит секрецию ФСГ гипофизом; у кастрированного самца секреция ФСГ подавляется только при пересадке семенников. Таким образом, у половозрелых млекопитающих реакция гипофиза обнаруживает половую специфичность, тогда как у новорожденных животных гипофиз в половом отношении индифферентен.
В период роста секреция гонадотропинов у молодых млекопитающих постепенно усиливается вплоть до наступления половой зрелости, когда завершается развитие половых желез; эстрогены или андрогены способствуют окончательной дифференцировке вторичных половых признаков и придаточных структур половой системы. После этого животное становится способным к нормальной репродуктивной деятельности. У позвоночных это обычно циклический процесс, периодичность которого часто бывает связана с годичным солнечным циклом таким образом, чтобы рождение и раннее постнатальное развитие потомства приходилось на наиболее благоприятное время года. Однако у некоторых видов, например у крысы, мыши и человека, периодичность половой 29—2067
450
Глава 21
активности почти или даже вовсе не связана с внешними раздражителями и, по-видимому, всецело обусловлена врожденным ритмом.
Было проведено огромное число исследований, касающихся роли взаимодействующих эндокринных факторов в поддержании нормального репродуктивного цикла у млекопитающих. Эндокринные влияния и взаимодействие важнейших органов, участвующих в циклах половой активности, схематически показаны на рис. 21-3 для самок и на рис. 21-4 для самцов.
У самок гипофиз, выделяя ФСГ, стимулирует развитие фолликулов и интерстициальной ткани яичников, что приводит к усиленной секреции эстрогенов. Возросшая концентрация эстрогенов в крови оказывает такое же влияние, как ФСГ, и способствует дальнейшему развитию яичников. В то же время эстрогены постепенно подавляют секрецию ФСГ и вначале энергично способствуют выделению Л Г гипофизом. Спустя некоторое время эстрогены начинают подавлять и секрецию Л Г. Выделяемый Л Г вызывает овуляцию у многих млекопитающих и у всех стимулирует развитие желтых тел с последующей выработкой ими прогестерона. Хотя ЛГ индуцирует развитие желтых тел, для стимуляции выработки ими прогестерона у некоторых видов нужен еще один гипофизарный гормон — лютеотропин. Полагают, что лютеотропин идентичен пролактину. Прогестерон совместно с эстрогеном еще больше затормаживает секрецию ФСГ гипофизом. Во время беременности прогестерон активирует плаценты развивающихся зародышей, и последние сами начинают выделять большое количество хорионического гонадотропина, очень сходного по своему действию с гипофизарным ЛГ. Гонадотропный гормон хориона, стимулируя выработку гормонов половыми железами, участвует в подавлении секреции обоих гипофизарных гонадотропинов.
Эндокринные взаимодействия, участвующие в мужском половом цикле, не так сложны. Гипофизарный гонадотропин активирует интерстициальную и сперматогенную ткань семенников, и это приводит к повышенному образованию тестостерона (в первой из них). Тестостерон способствует дальнейшему развитию спер-матогенного эпителия и в то же время усиливает влияние ФСГ и ЛГ. Позднее постепенно возрастающие концентрации андрогена в крови подавляют секрецию гипофизарных гонадотропинов. Уменьшение выработки андрогена после подавления активности гипофиза и конце концов приводит к тому, что гипофиз инициирует новый цикл половой активности.
У некоторых животных ежегодный период размножения приурочен к определенному сезону независимо от какого-то конкретного фактора внешней среды. Наличие такого врожденного ритма ясно показано у ряда организмов, у которых брачный период наступает в те же календарные месяцы даже после переселения из северного полушария в южное или наоборот. У других же ви-
Эндокринные механизмы
451
ов после такого переселения время брачного периода довольно быстро изменяется: они приспосабливаются к климатическим условиям новой местности [275]. Легкость подобной адаптации, по-видимому, определяется степенью зависимости полового цикла от факторов среды.
Из всех известных внешних раздражителен, влияющих на репродуктивный цикл, пожалуй, наиболее эффективны свет и фотопериодизм [21, 128, 129, 350, 455]. У самцов некоторых птиц, обитающих в умеренной зоне северного полушария, максимум активности семенников приходится на позднюю весну. Эксперименты на самцах юнко, воробьев и скворцов ясно показали, что постепенным увеличением длительности светового периода (путем добавочного искусственного освещения) можно довести активность семенников до максимума в необычное для этих видов время года. Тщательно проведенные опыты подтвердили, что действующим фактором служит сам свет, а не удлинение периода дневной активности и питания. После окончания одного полового цикла птица неспособна сразу же вновь реагировать на увеличение светового периода [285]. Подобным же образом удлиненный период освещения в течение примерно двух месяцев вызывает у самок хорька течку в зимнее время, когда при нормальном половом цикле они находятся в фазе анэструса. Аналогичные наблюдения были сделаны на мышах Peromyscus. Такие рептилии, как Anolis, Xantu-sia и Lacerta, также реагируют на фотопериод [23]. Среди рыб существенное влияние фотопериодизма отмечено у форелей, ушастых окуней и некоторых гольянов.
Показано, что у полевки Microtus при постепенном укорочении светового периода половые железы уменьшаются в размерах.
Подобные эксперименты указывают, по-видимому, на то, что постепенное изменение длины дня на протяжении годичного цикла представляет собой весьма эффективный раздражитель, определяющий нормальную сезонность размножения у многих позвоночных [454].
В опытах на неполовозрелых утках, у которых гонады обычно реагируют на удлинение светового периода, гипофиз птиц, подвергавшихся воздействию света, обнаруживал большую гонадотропную активность (при пересадке неполовозрелым мышам), чем у контрольных уток. Наиболее эффективным оказался красный и оранжевый свет. Эта реакция может осуществляться без участия глаз: прямое воздействие на гипофиз путем освещения глазницы после удаления глазного яблока также ведет к активации. Если свет ^подводят прямо к гипофизу через кварцевую палочку, то синий свет, обычно оказывающий лишь слабое действие, становится столь же эффективным, как красный или оранжевый. Половые циклы продолжаются у уток и при постоянном освещении, и в темноте, но правильная годовая периодичность исчезает [29]. 29*
452
Глава 21
Аналогичное влияние света на половой цикл, осуществляемое независимо от глаз, обнаружено и у воробьев. Между тем у хорьков перерезка зрительных нервов полностью устраняет действие света; таким образом, посредником в этой реакции у них служит сетчатка глаза.
Необходимым условием нормальной половой функции обычно является сохранение нервной связи гипоталамуса с другими отделами нервной системы. Сохранность нервных связей между гипоталамусом и гипофизом имеет большое значение для определенных сторон регуляции репродуктивного цикла [274], например для овуляции у кролика, но не играет, по-видимому, никакой роли во многих других случаях, когда связь осуществляется, очевидно, гуморальным путем. У белоголовых зонотрихий (воробьиные) в нервных ганглиях срединного возвышения накапливаются секреторные гранулы; при 8-часовом фотопериоде эти гранулы исчезают, а при удлинении фотопериода до 20 ч происходит увеличение семенников [305].
•Показано, что у колюшки важную роль в определении времени развития половых желез и появления вторичных половых признаков играет температура окружающей среды: достаточное повышение температуры быстро вызывает эти изменения. У суслика (Citellus) низкая температура и темнота, обычно сопутствующие зимней спячке, могут активировать половые железы в любое время года. Свет и температура влияют на половой цикл и у рептилий [22].
Большое значение часто имеют и другие факторы. Например, для живущего в экваториальной области ткачика Quelea стимулом к размножению служит период дождей, сопровождающийся появлением богатой зеленой растительности [106].
Эпифиз и размножение. Известно, что эпифиз выделяет мелатонин (М-ацетил-5-метокситриптамин), который оказывает прямое воздействие на меланофоры амфибий, вызывая в них концентрирование меланина [302, 304]. Однако эпифизу приписывают также участие в регуляции размножения. У человека обнаружена связь между преждевременным половым созреванием и наличием непаренхиматозных опухолей эпифиза [226]. Такие опухоли разрушают секреторную паренхиматозную ткань. В связи с этим возникла мысль, что эпифиз может оказывать тормозящее влияние на развитие половых желез. У некоторых животных экстракты эпифиза и мелатонин действительно оказывают сильное тормозное действие на гонады [343]. Такое действие эпифиза усиливается при содержании животного в темноте — как предполагают, вследствие повышения активности фермента, ответственного за синтез меланина,—оксииндол-О-метилтрансферазы [459]. Кроме того, удаление эпифиза ускоряет рост неполовозрелых животных и может привести к кратковременному увеличению репродуктивных органов у взрослых особей. Неясно, однако, вызывает ли
Эндокринныемеханизмы
453
эпифиз эти эффекты непосредственно, поскольку есть данные о тюм, что мелатонин способен влиять на секрецию тропных гормонов гипофиза. Наконец, все еще нельзя утверждать, что у какого-либо вида регуляция размножения светом осуществляется через эпифиз.
Гормоны и координация функций желудочно-кишечного тракта [293]
Интеграция деятельности различных отделов пищеварительной системы при переработке пищи осуществляется у млекопитающих •частично с помощью эндокринных механизмов. Важнейшие гормоны, места их образования и точки воздействия схематически представлены на рис. 21-5.
Слизистая пилорической части желудка в ответ на соответствующий стимул выделяет в кровь гормон гастрин [178]. При поступлении в желудок пищи в пересаженном карманообразном лоскуте желудка, лишенном нервных связей, тоже начинается секреция. В течение долгого времени считали, что гастрин тождествен с гистамином, но теперь установлено, что не содержащие гистамина экстракты слизистой из пилорического отдела желудка стимулируют секрецию сильно кислого сока, обладающего очень слабой пептической активностью и одновременно вызывающего панкреатическую секрецию. Однако есть некоторые основания полагать, что гистамин тоже играет здесь роль гормона '[368]. Путем фракционного осаждения удалось отделить стимулятор желудка гастрин от активатора поджелудочной железы; последний очень напоминает секретин, который был найден в слизистой оболочке двенадцатиперстной кишки. Из слизистой желудка свиньи выделены два чистых пептида, гастрины I и II, которые были затем синтезированы. Они способны стимулировать не только желудок, но и поджелудочную железу.
Слизистая двенадцатиперстной кишки возбуждает секрецию сока поджелудочной железы при посредстве секретина — полипеп-тидного гормона, состоящего из 27 аминокислотных остатков [177]. Прежде считали, что секретин стимулирует главным образом выделение поджелудочной железой воды и бикарбоната, выработка же ферментов контролируется нервной системой; однако никаких веских данных, подтверждающих эту гипотезу, получено не было. Концентрация ферментов <в панкреатическом соке, образующемся в ответ на инъекции секретина, довольно изменчива. Это можно объяснить тем, что в неочищенном экстракте содержится второй гормон. Одна фракция стимулирует главным образом выделение денервированной поджелудочной железой воды и неорганических солей, а другая — секрецию ферментов. Активный фактор этой второй фракции был назван панкреозимином.
Из грубых экстрактов секретина был выделен еще один активный дуоденальный фактор. Это холецистокинин—мощный агент,
454
Глава 21
вызывающий сокращение желчного пузыря. Он оказался тождественным панкреозимину.
И наконец, из слизистой двенадцатиперстной кишки был выделен четвертый фактор — энтерогастрон. Этот фактор подавляет
Рис. 21-5. Схема, показывающая места образования и точки приложения действия важнейших гормонов, координирующих деятельность желудочно-кишечного тракта.
выделение желудком соляной» кислоты и двигательную активность желудка. По-видимому, энтерогастрон оказывает главным: образом антисекреторное действие, а так как в его присутствии желудочный сок, выделяемый под влиянием таких стимуляторов, как гистамин, имеет низкую-кислотность и богат пепсином,, можно предположить, что энтерогастрон подавляет в основном функцию обкладочных клеток желудка, вырабатывающих соляную кислоту.
На основании некоторых опытов выдвигались также гипотезы о существовании других химических координаторов, регулирующих функцию желудочно-кишечного тракта, но они еще не получили достаточного подтверждения.
Окраска и ее сезонные изменения у птиц и млекопитающих
У многих видов птиц самцы и самки имеют разное оперение, и это может быть обусловлено весьма различными механизмами [99, 108]. У домового воробья, например, тип оперения определяется только генетическими факторами, а у фазанов — одновременным действием генов и гормонов. Но, вероятно, у большинства птиц оперение, свойственное одному полу, нейтрально, тогда как особенности другого пола зависят от гормонов, циркулирующих в крови. У обычных домашних кур нейтральным типом, является, по-видимому, оперение самцов, а оперение самок определяется действием эстрогенов. У африканских ткачиков нейтральный тип оперения имеют самки, а самцовый тип является результатом непрямого действия гормонов гипофиза [102, 339, 376, 452]. Среди исследованных до сих пор птиц только у серебристых чаек особенности оперения самцов зависят от действия андрогенов [45].
У большинства «обычных» птиц взрослые особи по окончании периода размножения полностью линяют, сменяя красивый брачный наряд на менее яркое зимнее оперение. Вторая линька^
Эндокринные механизмы
455
обычно затрагивающая не все оперение, происходит весной перед брачным периодом. Эта весенняя линька тем полнее, чем ярче брачный наряд. Время смены оперения зависит от ряда факторов, у некоторых видов, например у африканских ткачиков Руготе-lanox и скворцов, эти изменения, видимо, определяются генетически обусловленным ритмом активности гипофиза [450], хотя этот ритм может отчасти зависеть от длины светлого периода суток [56]. У некоторых других видов, например у диких уток, белоголовых зонотрихий и виргинских куропаток, линьку и смену типа оперения можно вызвать в необычное для них время года, если искусственно увеличивать или уменьшать световой период. Таким образом, сезонные изменения в оперении большинства птиц, подобно перелетам и активности, связанной с размножением, по-видимому, в значительной мере регулируются годичным циклом длины дня.
У некоторых северных птиц и млекопитающих происходит сезонная смена окраски — от коричневой летом к белой зимой. Время этой смены у таких форм, как куропатка Lagopus [207], горностай [35] и заяц-беляк [271], определяется сезонными изменениями длины дня; появление белой окраски можно вызвать у животного и не в обычный сезон, если искусственно создать надлежащие изменения светового периода. Эксперименты на зайцах-беляках [271] указывают на то, что рецепторами при этом служат глаза. Как при естественной, так и при искусственно вызванной смене окраски «физиологически коричневое» состояние организма у беляка связано с высоким содержанием гонадотропинов в крови, а «физиологически белое» — с их пониженной концентрацией. Линька у «физиологически коричневого» животного приводит к образованию коричневой шерсти, а у «физиологически белого» — белой шерсти. Экстракты из цельного гипофиза, содержащие гонадотропные гормоны, вызывают превращение «физиологически белого» животного в «физиологически коричневое», что сопровождается выпадением белых волос. Гипофизэктомия у хорьков прекращает циклическую линьку [34]. К этим изменениям окраски, по-видимому, не причастны никакие другие эндокринные железы, кроме гипофиза. У кастрированных и тиреоидэктомированных зайцев наблюдаются нормальные сезонные изменения цвета шерсти [271].
Еще одним доказательством роли гипофиза в сезонных изменениях окраски служит то, что у белого горностая с удаленным гипофизом после линьки снова вырастают белые волосы, а не коричневые, как у нормальных особей [354]. Инъекция меланоцит-стимулирующего гормона или АКТГ гипофизэктомированным горностаям приводила к росту коричневых волос [355]. Пересадка таким животным гипофиза способствовала линьке и росту коричневых волос — по-видимому, в результате секреции трансплантатом меланоцитстимулирующего гормона.
456
Глава 21
Простагландины [206]
Это группа гормоноподобных веществ, которые первоначально были открыты благодаря их способности понижать артериальное давление и стимулировать сокращение гладкой мускулатуры кишечника и матки. Они химически идентифицированы в семенной жидкости, легких, радужной оболочке, головном мозгу, тимусе и мозговом веществе почки; возможно, что они содержатся и в других тканях. Все они — производные простаноевой кислоты и могут быть разделены на четыре группы: Е, F, А и В. Так как биосинтетическим предшественником их служит одна из незаменимых жирных кислот (простаноевая), возможно, что симптомы недостаточности этих кислот связаны именно с нехваткой простагландинов. Однако это всего лишь догадка. К наиболее важным клиническим свойствам простагландинов относится их способность вызывать аборт. Возможно, что механизм их действия каким-то образом связан с циклическим АМФ. В большинстве случаев (например, в коре надпочечников, щитовидной железе и островках Лангерганса) простагландины имеют тенденцию повышать содержание этого нуклеотида. Однако в некоторых тканях (например, в жировой) они препятствуют повышению концентрации циклического АМФ, которое обычно происходит при гормональной стимуляции.
Простаноевая кислота
Простагландин Ех
Насекомые
Эндокринная система насекомых удивительно сходна по своей функциональной организации с эндокринной системой позвоночных. Над нею тоже господствуют нервные центры, которые отчасти сами являются нейроэндокринными структурами. Группы нейросекреторных клеток мозга, составляющие pars intercerebra-lis, секретируют вещества гормональной природы, выделяемые в
Эндокринные механизмы
457
гемолимфу через парные органы, расположенные позади мозга,— так называемые corpora cardiaca. Такой секреторный комплекс весьма сходен с гипоталамо-нейрогипофизарной системой. Этот комплекс регулирует деятельность эндокринной железы (иногда парной), находящейся в переднегруди, — проторакальной, или .линочной, железы. У большинства насекомых с corpora cardiaca тесно связаны другие железы — так называемые corpora all at а, активность которых тоже отчасти регулируется мозгом. В других отделах нервной системы, особенно в подглоточных ганглиях, также встречаются нейроэндокринные элементы. Таким образом, здесь, как и у позвоночных, нервные и эндокринные органы очень тесно связаны между собой и образуют единую координирующую систему.
Линька, окукливание и метаморфоз у насекомых. Постэмбриональное развитие насекомых включает упорядоченную последовательность стадий, в результате которых личинка превращается во взрослое насекомое, или имаго. Периодический рост, связанный с рядом линек, заканчивается метаморфозом, во время которого особь утрачивает личиночные особенности и приобретает все признаки имаго. У насекомых с неполным превращением, таких, как прямокрылые (например, саранча), тараканы, палочники, полужесткокрылые (например, постельный клоп и клоп-хнщнец Rhod-nius) и стрекозы, переход из личиночного состояния во взрослое происходит постепенно. Развивающиеся молодые особи, которых обычно называют нимфами, после каждой последующей линьки становятся все больше и больше похожими на взрослых. У таких насекомых с неполным превращением последняя линька на стадии нимфы связана, однако, с несколько более значительным изменением, и поэтому именно ее обычно рассматривают как линьку метаморфоза.
У насекомых с полным превращением, таких, как чешуекрылые (бабочки), двукрылые (например, мухи и комары) и жесткокрылые (жуки), личиночные линьки сопровождаются лишь очень небольшим изменением формы и в основном лишь обеспечивают рост. Этих быстро растущих личинок обычно называют безногими личинками (у чешуекрылых — гусеницами). В результате своей последней линьки личинка превращается в куколку. Стадия куколки, длительность которой нередко зависит от внешних факторов (температуры, света) [254, 316], часто включает продолжительный период покоя — диапаузу. Внутри хитинового покрова куколки происходит полная перестройка организма, приводящая к 'образованию взрослой формы. Глубоких качественных различий между двумя описанными типами развития нет; наблюдаемые различия касаются в основном времени, в течение которого осуществляются процессы дифференцировки имаго. У насекомых с неполным превращением эти процессы в значительной своей части растягиваются на весь личиноч-
458
Глава 21
ный период, а у насекомых с полным превращением они происходят в основном на стадии куколки.
Мозговой гормон
Головной мозг
Лроторакальна железа
Нейросекреторные клетки
Гормон проторакальной железы
Ювенильный гормон
Эпидермальные клетки
Corpora cardiaca ^^-Corpora allata
Эпидермальные клетки
Взпослая особь
Рис. 21ч6. Схема эндокринной системы, участвующей в регуляции роста и дифференцировки у насекомых [373].
Число личиночных возрастов в развитии насекомых у разных видов различно, но для каждого вида оно, как правило, постоянно. Для клопа Rhodnius характерны, например, 5 личиночных возрастов, для палочника Carausius — 6, а для мухи Drosophila — 3. В настоящее время твердо установлено, что ход событий в про
Эндокринные механизмы
459
цессе постэмбрионального роста и дифференцировки у насекомых в значительной степени определяется интегрирующим влиянием гормонов, выделяемых эндокринной системой (рис. 21-6) [42, 373, 438].
По-видимому, всем насекомым свойственна единая схема эндокринного механизма, регулирующего рост и дифференцировку [85, 212, 218, 330, 337, 373, 437, 438, 445, 446]: инициирующий линьку стимул, исходящий обычно из внешней среды и различный у разных нидов, вызывает секрецию мозгового гормона, который переносится по аксонам к corpora cardiaca, откуда выделяется в гемолимфу [10, 200, 365, 366, 408]. Недавно было установлено, что мозговой гормон — это, по-видимому, полипептид или белок с небольшим молекулярным весом [161, 214, 462], и это вполне соответствует тому, что нам известно о нейросекреторных веществах у позвоночных. Мозговой гормон активирует проторакальные железы, в которых, как полагают, образуется гормон линьки, называемый также гормоном роста и дифференцировки, или экдизоном. Экдизон—это очень своеобразный растворимый в воде стероид, имеющий следующую структуру [2П]:
ОН
При первоначальной очистке этого гормона было замечено, что ему сопутствует второй компонент, обладающий гормональной активностью; он был назван 0-экдизоном, а главный компонент — •а-экдизоном [62]. р-Экдизон отличается тем, что содержит гидроксильную группу в положении 20; иногда его называют также экдистероном, крустэкдизоном или изоинокостероном [167]. Новейшие исследования показали, что у членистоногих ia-экдизон быстро превращается в р-экдизон и что на самом деле истинным гормоном линьки у насекомых, возможно, является р-экдизон [174]. Интересно отметить, что в отличие от млекопитающих насекомые не могут синтезировать стерины из простых предшественников и поэтому нуждаются в получении стеринов извне. Всеядным насекомым нужен холестерин, а растительноядным — р-си-тостерин [168]. Таким образом, вероятными предшественниками гормона линьки у насекомых служат стерины пищи. В связи с этим следует упомянуть об одном из поразительных открытий последних лет: из растений выделено более 20 веществ (включая «-и Р-экдизоны), обладающих активностью линочного гормона на
460
Глава 21
секомых. Все они —стерины, сходные по своей структуре с экдизоном. Некоторые из них содержатся в тканях растений в гораздо-большей концентрации, чем у насекомых и обладают большей гормональной активностью, чем естественные а- и р-экдизоны. Роль этих стеринов в физиологии растений неизвестна, но было высказано предположение, что они выработались для защиты от насекомых (папоротники и тисы содержат эти «фитоэкдизоны» .в особенно большой концентрации и обладают сравнительно высокой устойчивостью к насекомым).
Экдизон стимулирует процесс линьки, воздействуя на эпидермис, в котором быстро возрастает содержание белка и РНК, увеличивается число митохондрий и элементов эндоплазматической сети [378]. Все эти изменения происходят независимо от того, приводит ли линька к другому личиночному возрасту, к стадии куколки или к выходу имаго. Как показывает исследование гигантских политенных хромосом у некоторых мух, экдизон, по-видимому, может вызывать синтез мРНК в специфических генных локусах [87, 261], и это, возможно, первое звено в цепи событий, ведущих к линьке. Однако, судя по новейшим данным, это непрямой эффект: он связан с изменением проницаемости ядерной оболочки, которое приводит к изменению ионной среды, окружающей хромосомы [243, 262]. Помимо этого экдизон, видимо, стимулирует активность аденилатциклазы в эпидермальной ткани, что говорит о возможной роли циклического АМФ в механизме его действия [9].
Для осуществления линьки у растущих личинок или нимф выделяется еще один гормон, секреция которого также частично регулируется головным мозгом [119, 122, 216, 270, 365]. Этот гормон,, называемый ювенильным, образуется в corpora allata; он подавляет дифференцировку имаго, способствуя формированию личиночных структур. При линьке, ведущей к окукливанию, он выделяется в меньшем количестве, а при следующей — имагинальной— линьке совсем отсутствует. Ювенильный гормон, выделенный из экстрактов взрослого самца бабочки Hyalophora cecropia, оказался производным терпена [348]:
С помощью этого вещества можно воспроизвести все морфогенетические эффекты, вызываемые имплантацией corpora allata, в том числе и развитие яиц (см. ниже). Структура ювенильного гормона Н. cecropia была недавно подтверждена; был также идентифицирован низший гомолог этого вещества, который тоже обладает активностью ювенильного гормона [283]. Поскольку ювенильный гормон препятствует превращению личинки или куколки во
1
Эндокринные механизмы
461
взрослую особь, это вещество можно было бы использовать для борьбы с вредными насекомыми. Получено более 500 аналогов, обладающих активностью ювенильного гормона по отношению к разным видам насекомых [383]. Предполагают, что на молекулярном уровне этот гормон, возможно, действует как антагонист экдизона [90, 261, 318].
В регуляции развития насекомых участвует еще один гормон — гормон диапаузы, выделяемый подглоточным ганглием [154, 192]. Наличие или отсутствие этого гормона у бабочки-самки определяет, наступит ли в развитии отложенных яиц фаза диапаузы. Получены активные экстракты, содержащие этот гормон [192]. Показано, что в гемолимфе только чго выведшихся мух имеется гормон, необходимый для твердения кутикулы у взрослой мухи; он получил название бурсикона [149, 422]. Его химическая структура еще точно не установлена. Вероятно, этот гормон выделяют нейросекреторные клетки центральной нервной системы.
В основе действия эндокринных факторов, регулирующих рост и дифференцировку у насекомых, лежат постепенные изменения в реактивности тканей-мишеней. Эти изменения так же важны для упорядоченной дифференцировки, как и сами гормоны [41—44, 389].
Действие двух гормонов — мозгового и ювенильного — было впервые ясно продемонстрировано в опытах на тропическом клопе Rhodnius prolixus [434—436]. У этого насекомого на каждой личиночной (нимфальной) стадии линьки происходят через определенное число дней после питания 'кровью. Адекватным раздражителем служит растяжение брюшка. После удаления головы эти насекомые могут жить 6—10 месяцев. Как показали опыты с декапитацией, производимой в различные сроки после кор.мления, через несколько дней после него наступает «критический период»: у нимф, декапитированных до этого периода, линька никогда уже не происходила; у тех же, которые подверглись декапитации позднее, линька проходила нормально. Критический период соответствует тому моменту, когда расположенный в голове эндокринный орган выделяет гормон, инициирующий линьку. Это было показано в опытах с телобиозом двух обезглавленных особей (рис. 21-7,А). В условиях такого телобиоза нимфа, например, 4-го возраста, обезглавленная после критического периода, индуцировала линьку у нимфы того же возраста, обезглавленной до критического периода; при этом обе превращались в нимфы 5-го возраста.
Нимфы 5-го возраста после линьки, как правило, превращаются во взрослых особей. Если такую нимфу декапитировали после критического периода и телобиотически соединяли с нимфой 1-го или 2-го возраста, декапитированной до критического периода, то последняя претерпевала линьку, так как в гемолимфе нимфы '5-го возраста содержался необходимый для этого гормональный фак
462
Глава 21
тор. При этом нимфа 1-го или 2-го возраста превращалась сразу во взрослую особь (только меньших размеров), так как у нее еще не выделялся ювенильный гормон. Если нимфу 4-го (т. е. еще не последнего) (возраста декапитировали после критического периода и соединяли с нимфой 5-го возраста, декапитированной до критического периода, то последняя линяла и превращалась в гигантскую нимфу дополнительного, 6-го возраста, вместо того чтобы
Рис, 21-7. Экспериментальный телобиоз и парабиоз у клопа Rhodnius [434, 435].
А. Телобиоз нимф. Б. Телобкоз и парабиоз нимф (4-го и 5-го возрастов) и имаго.
превратиться в имаго. Более того, хотя взрослое насекомое в обычных условиях больше не линяет, у него удавалось вызвать линьку путем тело- и парабиоза с двумя или тремя нимфами 5-го возраста, декапитированными после критического периода. Если же вместо этого использовали нимф 4-го .возраста, (взрослая особь линяла и частично возвращалась к состоянию нимфы (рис 21-7,Б).
Из эксперимента (было очевидно, что в гемолимфе личинки после критического периода содержатся факторы, определяющие линьку, которая может сопровождаться или не сопровождаться метаморфозом. Тщательное изучение результатов, полученных при декапитации нимф 3-го и 4-го возрастов в различные моменты самого критического периода, показало, что при линьке нимф, декапитированных в ранние сроки этого периода, проявляется выраженная тенденция к преждевременному метаморфозу, а у декапитированных позднее такой тенденции нет. Это означает, что во время критического периода у нимф первых четырех возрастов сначала выделяется только гормон линьки, а несколько позже — также и фактор, тормозящий метаморфоз (ювенильный гормон). У клопа Rhodnius этот фактор не образуется только в 5-м возрасте.
Установлено, что источником фактора, подавляющего метаморфоз, т. е. ювенильного гормона, служит corpus allatum; у Rhodnius
Эндокринные механизмы
463
это срединная непарная железа, расположенная в задней части головы. Гормон, вызывающий линьку и дифференцировку, выделяют нейросекреторные элементы, лежащие в pars intercerebralis — заднем срединном участке мозга. Благодаря удлиненной форме головы технически очень легко произвести декапитацию таким образом, чтобы сохранить corpus allatum, удалив мозг. Применяя эту методику в сочетании с телобиозом, а также пересаживая corpora allata и pars intercerebralis нимфам, декапитированным до наступления критического периода, можно по желанию вызывать у Rhodnius либо промежуточную линьку, либо линьку с .метаморфозом.
Роль проторакальных желез и их связь с мозговым гормоном впервые удалось выяснить в экспериментах на чешуекрылых. Со времени первоначальных опытов Копеса [239], изучавшего окукливание и метаморфоз у бабочек, было известно, что в определенный «критический» период передняя часть тела гусеницы выделяет какой-то гормон, необходимый для этих двух процессов. У тутового шелкопряда (Bombyx) ©тот гормон (или какой-то фактор, необходимый для синтеза гормона в другом месте) вырабатывается в проторакальной железе, расположенной в переднегрудном сегменте (протораксе) [153]. Если с помощью перевязки (лигатуры) отделить этот сегмент от всей задней части тела сразу после окукливания, то ©та часть дальше не развивается. Если же лигатуру накладывали через 12—18 ч после окукливания, задняя часть претерпевала нормальный метаморфоз. При пересадке проторакальной железы в заднюю часть куколки, перетянутой за 12—18 ч до критического периода, метаморфоз этой части тела также протекал нормально; в брюшке, отделенном лигатурой, удавалось вызвать метаморфоз путем соединения его гемоцеля с гемоцелем особи, проходящей нормальный метаморфоз, при помощи стеклянной капиллярной трубочки.
У бабочки-сатурнии Hyalophora cecropia после окукливания наступает период диапаузы (покоя), продолжающийся около 5— 6 месяцев. Если в ©то время у куколки удаляли мозг, она навсегда утрачивала способность к метаморфозу, хотя обычно оставалась живой околю года [441]. Если нормальную куколку помещали на полтора месяца в условия низкой температуры (3—5°C), то метаморфоз происходил через месяц с небольшим после того, как ее возвращали в обычные условия. При пар абиотическом соединении куколки, подвергнутой охлаждению, с неохлажденной куколкой обе они одновременно проходили метаморфоз примерно через полтора месяца (рис. 21-8, Л и Б). Имплантаты мозга охлажденной куколки вызывали метаморфоз у куколки, не подвергавшейся охлаждению.
Было показано, что проторакальные железы и мозг влияют на метаморфоз согласованным образом. У Н. cecropia активный фактор мозга образуется внутри каждой его доли, где имеются две
/ /
/—
группы нейросекреторных клеток — медиальная и латеральная. Медиальная группа соответствует клеткам pars intercerebralis полужесткокрылых [443]. Чтобы вызвать метаморфоз у куколки с удаленным мозгом, ей нужно пересадить часть мозга, содержащую обе группы нейросекреторных клеток; это указывает на то, что обе группы клеток совместно участвуют в выработке необходимого вещества. 1
Куколок Н. cecropia разрезали на две части на уровне переднего края 6-го брюшного сегмента и раневые поверхности закрывали пластмассовыми пластинками; полученные таким образом препараты оставались живыми в течение 8 месяцев или дольше. Подвергавшийся охлаждению или лишенный .мозга передний отдел, в который пересаживали охлажденный мозг, претерпевал, нормальный метаморфоз. Задний отдел не подвергался метаморфозу даже тогда, когда в одно брюшко пересаживали несколько трансплантатов охлажденного мозга. Если же такое брюшко присоединяли к переднему отделу, находившемуся в процессе метаморфоза, то и оно подвергалось .метаморфозу. Очевидно, что кроме мозгового гормона нужен еще какой-то фактор. Как выяснилось, источником этого фактора служит проторакальная железа. Это было доказано опытами, в которых вызывали окукливание изолированного брюшка, имплантируя в него охлажденный мозг и проторакальную железу (рис. 21-8, В и Г). Последнюю можно было взять и от охлажденной, и от неохлажденной куколки. Одни проторакальные железы (без мозга) не вызывали метаморфоза. Из этого было ясно, что в нормальных условиях метаморфоз зависит по меньшей мере от двух факторов. Первый из них вырабатывается мозгом и активирует проторакальные железы, которые выделяют второй фактор. Мозговой и ювенильный гормоны не проявляют специфичности не только по отношению к виду, но даже и к отряду насекомых.
У палочника Carausius [323], таракана Leucophaea [120, 363, 365] и кобылки Melanoplus [321]—довольно хорошо изученных форм с неполным превращением — гормон, вызывающий линьку, образуется в мозгу и активирует проторакальные железы. Кроме того, corpora allata у всех этих насекомых вырабатывают ювенильный гормон. У Carausius, имеющего в норме 6 личиночных возрастов, удаление corpora allata в 3-м возрасте приводит к преждевременному метаморфозу, хотя обычно после операции происходят еще две личиночные линьки. Эту задержку объясняют тем, что где-то в организме остается некоторый запас ювенильного гормона. Пересадка corpora allata от нимф Carausius 3-го или 4-го возраста нимфам 6-го возраста может привести к трем или четырем добавочным личиночным линькам, в результате чего получаются гигантские имаго. У Leucophaea, у которой в норме 8 личиночных возрастов, удаление corpora allata в 7-м возрасте до критического периода приводит к линьке с образованием формы,
Эндокринные механизмы 465
напоминающей имаго. Таких особей называют «адультоидами»; они отличаются от взрослых меньшей величиной и более короткими крыльями. Если corpora all ata удалить в 5-м или 6-м возрасте, сначала возникают «преадультоиды», которые превращаются в адультоидов лишь после дополнительной линьки.
Рис. 21-8.
А. Куколка Telea polyphemus в состоянии диапаузы, лишенная мозга (справа) и присоединенная к охлажденной куколке Hyalophora cecropia (слева). Б. Те же животные после метаморфоза. В. Задняя часть куколки в состоянии диапаузы. Г. Метаморфоз части, показанной на фото В, после пересадки в нее охлажденного головного мозга и проторакальных желез от куколки в состоянии диапаузы [441, 442].
У двукрылых — насекомых с полным превращением—гормон, вызывающий окукливание, образуется у личинки в кольцевой железе, расположенной у спинной стороны мозга, между его двумя Долями [182]. Это комплекс, функционально соответствующий corpora allata, corpora cardiaca и проторакальным железам. Роль этой железы легко продемонстрировать, перевязав личинку так, чтобы разделить ее на две части — с головным мозгом и без него [148]. Если эта операция произведена до критического периода, окукливается только часть, содержащая кольцевую железу; если л<е лигатура наложена после наступления этого периода, окукливаются обе части. Пересадка кольцевой железы от личинки последнего возраста личинке первого возраста ведет к преждевре-30—2067
466
Глава 21
менному окукливанию последней. У личинок с лигатурой, наложенной до критического -периода, трансплантаты кольцевой железы вызывают также окукливание той части, которая отделена от собственной кольцевой железы. Гормон этой железы нужен не только для окукливания, но и для метаморфоза [26]. Имагиналь-ные диски, пересаженные в гемоцель взрослой мухи, не подвергаются дифференцировке, если вместе с ними не -пересадить кольцевую железу от личинки позднего возраста [182]. Гормон линьки и ювенильный гормон чешуекрылых действуют и на двукрылых: первый из них вызывает окукливание [26], а второй предотвращает метаморфоз {389].
У чешуекрылых, так же как у полужесткокрылых и прямокрылых, источником ювенильного гормона служат corpora allata. Удаление их у гусениц ранних возрастов ведет к преждевременному окукливанию, а пересадка их от гусениц ранних возрастов гусеницам, готовым к окукливанию, наоборот, заметно задерживает образование куколки [46].
Размножение. Хотя имеется много данных о результатах паразитарной кастрации, хирургической кастрации и пересадки поло-пых желез у насекомых, до сих пор только в отношении одного вида удалось показать, что половые и другие гормоны, циркулирующие в крови, оказывают значительное влияние на дифференцировку вторичных половых признаков. У личинок мужского пола Lampyris noctiluca удаление corpora allata и corpora cardiaca незадолго до 3-й линьки (до половой дифференциации) приводило к феминизации [295]. Полагают, что нейросекреторные клетки у мужских личинок ответственны за развитие апикальной ткани семенника, в которой образуется андрогенный (мужской) гормон [296]. Если эту апикальную ткань пересадить личинке генетически женского пола, из нее разовьется маскулинизированная взрослая особь с измененными первичными и вторичными половыми признаками. Это все, что мы знаем относительно половых гормонов у насекомых.
У большинства изученных до -сих пор видов обнаружена гормональная связь между corpora allata и яичниками. Удаление corpora allata на поздних личиночных -стадиях или у молодых взрослых особей приводит к нарушению нормального роста и развития яиц в яичнике. Это установлено в опытах на клопах Rhodnius [435], мухах Calliphora [406], Lucilia [103] и Sarcophaga 103], таракане Leucophaea [364] и многих других насекомых {121]. Пересадка corpora allata особям, у которых они были удалены, восстанавливает у них способность производить нормальные яйца. Однако -подобную связь не удалось обнаружить у палочника Carausius [323] и у непитающихся взрослых бабочек [47]. На возможную роль какого-то мозгового гормона в регуляции размножения указывает наблюдаемая у взрослых особей связь между усиленной нейро-секрецией и -половой активностью [16, 111, 297].
Эндокринные механизмы
467
Гуморальная природа этой связи была ясно продемонстрирована в экспериментах с телобиозом у Rhodnius; эти эксперименты показали, что действующий фактор переносится кровью и что corpora allata оказывают типичное для них действие независимо от своей локализации [435]. Видимо, выделяемый ими фактор влияет на отложение желтка в яйцах, а не на раннее развитие ооцитов. Corpora allata, очевидно, необходимы только в период роста ооцитов и отложения в них желтка в каждом из последовательных репродуктивных циклов. Было показано, что у Melanoplus удаление corpora allata приводит к глубоким изменениям в общем обмене веществ [322]. Это навело некоторых исследователей на мысль, что репродуктивные функции ювенильного гормона носят вторичный характер, первичный же эффект касается основных метаболических процессов. Есть, однако, убедительные данные о том, что у тех насекомых, у которых этот гормон необходим для оогенеза, он стимулирует синтез специфических «женских» белков в жировом теле. Эти белки включаются в желток развивающегося ооцита и нужны для нормального развития яиц [121].
Удаление corpora allata приводит также к замедлению роста и снижению активности некоторых придаточных органов женской половой системы у Calliphora, Melanoplus и Leucophaea-, этот эффект не зависит от присутствия половых желез, так что они не служат здесь посредниками. Имеется, однако, много данных о том, что у насекомых яичники оказывают влияние на corpora allata: удаление яичников приводит к гипертрофии этих органов у Melanoplus, Calliphora ,и Lucilia и, по-видимому, вызывает изменение их функции также и у других насекомых, например у Sarcophaga и Leucophaea.
У самцов удаление corpora allata приводит к менее четким результатам, чем у самок; оно никак не сказывается на образовании семени: оперированные самцы Leucophaea сохраняют способность спариваться с нормальными самками и эффективно оплодотворять их яйца. Однако есть сообщения о том, что удаление corpora allata у Rhodnius и Calliphora приводит к недоразвитию придаточных желез мужской половой системы. Большая часть таких данных касается кол л стернальных желез у тараканов. Коллетериальные железы у Blattella germanica реагируют на ювенильный гормон усиленным синтезом РНК и белков [463]. По-видимому, ювенильный гормон играет важную роль в регуляции накопления глюкозида протокатеховой кислоты, который входит в состав секрета этих придаточных желез [377, 447]. Кастрация самцов Leucophaea и Lucilia не вызывала заметных изменений в corpora allata.
Механизм действия гормонов. Морфологические и биохимические эффекты, обнаруженные после инъекции экдизона или ювенильного гормона различным насекомым, описаны в ряде новейших обзоров [106а, 168а, 460]. Суммируя полученные данные, можно сказать, что экдизон вызывает линьку (аполиз и секрецию 30*
468
Глава 2/
новой кутикулы) и что все биохимические изменения, связанные j с линькой и развитием (например, 'Стимуляция синтеза -белка, РНК и ДНК), обусловлены хотя бы косвенно действием экдизона.. Ювенильный же гормон и множество других веществ, вызывающих -сходные эффекты, препятствуют у многих насекомых переходу во взрослое 'состояние и способствуют морфологическим и биохимическим изменениям, характерным для более ранних -стадий * развития. Ювенильный гормон оказывает свое действие не самостоятельно (за исключением его роли как гонадотропного гормона ;!
у взрослых самок 'многих насекомых), а, по-видимому, ослабляет i
влияние экдизона. Например, экдизон -стимулирует секрецию эпи- | дермальными клетками новой кутикулы, но приобретет ли эта кутикула черты, свойственные личинке, куколке или взрослой особи, зависит от концентрации ювенильного гормона. Однако огромное большинство опубликованных исследований проливает мало света ? на конкретные механизмы действия гормона линьки и ювен-ильно-го гормона. Чтобы все же убедить читателя в том, что некоторые успехи в этом направлении достигнуты, мы рассмотрим ряд новейших работ, которые, возможно, послужат основой для планирования будущих экспериментов, способных дать определенный ответ на вопрос о механизмах действия гормонов.
Для того чтобы вызвать надлежащий эффект, гормон должен попасть туда, где находятся его «мишени» (точки приложения). Это означает, что экдизон должен поступать из проторакальных желез в открытую кровеносную систему, а ювенильный гормон точно так же должен транспортироваться гемолимфой из corpora allata. (Мы будем исходить из предположения, что а-экдизон— ,
это -протормон, синтезируемый проторакальными железами, и что ткани-мишени превращают его в р-экдизон — истинный гормон линьки [225а].) Возможно, что а-экдизон транспортируется, будучи связан со слецифичеокими -белками гемолимфы [117а, 117b], хотя, судя по некоторым данным, этот сравнительно легко растворимый в воде стероид может переноситься гемолимфой в свободном виде [83b]. В отличие от этого ювенильный гормон—классический липид, и он не может транспортироваться в водной среде в свободном состоянии. Действительно, недавние исследования показали, что ювенильный гормон связан со специфическим липопротеидом гемолимфы и, вероятно, переносится из corpora allata к тканям-мишеням именно в виде такого комплекса [433]. Про- £ никает ли этот комплекс в клетку-мишень, или он распадается у ее поверхности, так что в клетку входит только ювенильный гормон, — пока неизвестно.
Согласно имеющимся данным, экдизон попадает во все изученные клетки и ткани [174]; почему же одни клетки — так назы- ,
ваемые «клетки-мишени» — реагируют на него, а другие к нему, *
по-видимому, нечувствительны? Как показали многочисленные работы по эндокринологии млекопитающих, существуют специфиче- -
Эндокринные механизмы
469
ские белки (рецепторы), обладающие способностью связывать определенные гормоны (например, эстрадиол или прогестерон} или их активные метаболиты (5а-дигидротестостерон), и именно эти комплексы рецепторов с гормонами ответственны за действие гормонов на уровне клеточного ядра (см. стр. 446). Благодаря появившейся недавно возможности использовать 3Н-а-экдизон с высокой удельной активностью были выполнены две работы, которые указывают на наличие рецепторов в тканях-мишенях у членистоногих. Эммерих [Н7Ь] показал, что после введения 3Н-а-эк-дизона личинке Drosophila из ее слюнных желез можно выделить два белка (3,6S и 2,OS), с которыми, по-видимому, связан меченый а-экдизон. Можно предполагать, что гормон переходит затем в ядро, где оказывает свое действие как компонент рецепторного комплекса. Когда аналогичный эксперимент был проведен на гепатопанкреасе ракообразного, радиоактивность также была обнаружена в двух белках цитозоля (11,3S и 6,35S) [173а]. Как показало дальнейшее изучение, «меньший» рецепторный белок является субъединицей «большего». Наконец, было установлено, что-радиоактивная метка, связанная с этими рецепторными белками,, принадлежала уже не 3Н-1«-экдизону, а какому-то ранее не описанному метаболиту экдизона, который еще предстоит охарактеризовать. По-видимому, это похоже на то, что происходит у млекопитающих с тестостероном, который превращается в активный метаболит 5а-дигидротестостерон. Приведенные выше данные, хотя они и носят предварительный характер, указывают на возможное наличие в тканях-мишенях членистоногих рецепторов для гормонов. Очень интересно было бы провести аналогичные эксперименты, используя 3Н-р-экдизон с высокой удельной активностью, который пока еще не получен.
Каким образом гормон, воздействуя на клетку-мишень, вызывает те многообразные эффекты, которые можно наблюдать на макроскопическом уровне? Мы этого не знаем, но можно проследить ряд эффектов, проявляющихся в непосредственной близости! от места воздействия. Например, р-экдизон стимулирует синтез: РНК в эпидермальных клетках как in vitro [460а], так и in vivo,, а ювенильный гормон, судя по некоторым данным, подавляет эту-стимуляцию синтеза РНК [90, 318]. Имеются предварительные' сведения о том, что в изолированных ядрах жирового тела синтезируются различные виды РНК в зависимости от гормональных: влияний [90а]. Изящные эксперименты, проведенные на имаги-нальных дисках Drosophila [83а, 152а, 337а], показали, что 0-эк-дизон оказывает быстрое стимулирующее действие на синтез рибосомной РНК (рРНК); эта стимуляция может быть подавлена с помощью ювенильного гормона. Возможно, что стимуляция синтеза рРНК имеет важное значение для переноса новых информационных РНК из ядра в цитоплазму. Эти исследования позволяют предполагать, что р-экдизон и ювенильный гормон действу--
-470
Глава 21
ют на уровне транскрипции. Однако не менее убедительные данные, относящиеся также к молекулярному уровню, говорят, видимо, о том, что по крайней мере ювенильный гормон оказывает ре-тулирующее влияние на события, связанные с трансляцией [213а]. Этот вопрос окончательно еще не разрешен. Возможно, что и гормон линьки, и ювенильный гормон действуют на нескольких уровнях, так как между транскрипцией и трансляцией имеется около 12 потенциальных регуляторных пунктов. Например, как мы только что говорили, получены данные о действии ювенильного гормона на уровне трансляции. В других исследованиях, касающихся эффектов этого гормона, было найдено, что у взрослой самки таракана он может индуцировать синтез de novo специфического белка, необходимого для нормального развития яиц [121а]; он может также индуцировать синтез специфических белков гемолимфы у куколок некоторых чешуекрылых [433а]. Так как индукцию в обоих случаях можно подавить актиномици-,'ном D, ювенильный гормон, очевидно, действует на уровне транскрипции.
Ракообразные1
У ракообразных, так же как у позвоночных и насекомых, эндокринная система сложна и находится в очень тесной функциональной связи с центральной нервной системой [51]. Нейросекреторные клетки в нервных ганглиях вырабатывают гормоны, которые переносятся по аксонам к местам их выделения в кровь. Важными элементами этой системы являются Х-органы, связанные у •большинства ракообразных с ганглиями глазных стебельков, а у остальных — с ганглиями, находящимися в голове. Наиболее изучен Х-орган терминального ганглия и Х-орган чувствительного сосочка [237] (рис. 21-9). Синусную железу глазного стебелька (или головы) обычно считают просто органом, содержащим заполненные гормоном окончания нейросекреторных клеток Х-орга-на [37, 112, 118, 186, 277, 317]. Секреторный материал находится здесь, по-видимому, в пузырьках, ограниченных мембраной [204, 236, 331].
Второй важный эндокринный комплекс включает секреторные нейроны головного мозга и, вероятно, ганглии околопищеводной коннективы с аксонами, проводящими гормоны и оканчивающимися в посткомиссуральных органах (рис. 21-10), где эти гормоны выделяются в кровь {52, 135, 235, 279]. Тому же организационному плану соответствуют клетки брюшной ганглионарной цепочки, выделяющие свой гормон на некотором расстоянии в перикарди-.альные органы у места впадения крупных вен в полость перикарда [5].
1 Раздел написан при участии М. Финтермана.
Рис. 21-9. Схема строения нейроэндокринного комплекса глазного стебелька-Lysmata (Carlisle D. В., Knowles F. G. W., Endocrine Control in Crustaceans,, Cambridge Univ. Press).
1 — Х-орган чувствительного сосочка; 2 — сенсорная пора; 3 — чувствительный нерв; 4 — кон-нектива Х-органа; 5 — терминальный ганглий; 6 — Х-орган терминального ганглия; 7 — тракт, идущий от мозга к Х-органу; 8 — тракт, идущий от Х-органа к синусной железе; 9 — ножка оптической доли; 10 — тракт, идущий от мозга к синусной железе; 11 — внутренний оптический ганглий; 12 — общий тракт к синусной железе; 13 — синусная железа; 14 — наружный оптический ганглий; 15 — ганглионарная пластинка.
Рис. 21-10. Схема строения комплекса нейросекреторно-посткомиссуральыого-органа десятиногого рака Leander (Carlisle D. В., Knowles F. G. W., Endocrine Control in Crustaceans, Cambridge Univ. Press).
1 — пищевод; 2 — комиссура; 3 — посткомиссуральный орган; 4 — сухожилие; 5—мышца?. 5 — эндофрагмальный скелет; 7 — окологлоточная коннектива; 8 — двигательное волокно?.
9 — нейросекреторное волокно.
472 Глава 21
Высказывалось предположение, что нейросекреторные гормоны J ракообразных, подобно гормонам нейрогипофиза у позвоночных, -представляют собой полипептиды. Это согласуется с новейшими ? .данными о том, что такие нейросекреторные вещества, как хроматофоротропные гормоны [131, 231], гормоны, влияющие на пигмент сетчатки [229], гипергликемические гормоны [223], гормон, j тормозящий линьку [341], и гормон, ускоряющий ритм сердца [28, 31], все, по-видимому, относятся к полипептидам. j
Однако не все эндокринные железы ракообразных непосредст- * венно связаны с нейросекрецией. Особняком стоит пара желез в переднем грудном отделе в антеннальном или максиллярном сегменте. Эти железы, так называемые Y-органы [115, 155], не имеют, по-видимому, никакой прямой иннервации. Они регулиру- к ются секретами комплекса желез глазного стебелька. Кроме того, эндокринную функцию выполняют яичники [73]. Существует еще один важный орган — андрогенная железа, морфологически свя- ; занная с семяпроводом [73, 75]. У некоторых равноногих раков •она располагается в непосредственной близости от семенников.
Линька у ракообразных. Десятиногие раки и другие ракообразные в процессе развития обычно претерпевают несколько линек, проходя ряд характерных личиночных стадий; позднее, когда их тело приобретает форму, свойственную взрослому животному, они | продолжают периодически сбрасывать наружный покров, что не--обходимо для роста. J
Линька — процесс очень сложный. Фактически в течение всего | периода роста ракообразное либо готовится к следующей линьке, * либо завершает предыдущую. Линочный цикл можно разделить I .на четыре периода: 1) предлиньку, или проэкдизис — период ак- | дивной подготовки к линьке, которая включает постепенное истон- J •чение кутикулы, запасание в гастролитах или гепатопанкреасе J неорганических составных частей для нового экзоскелета, ускорение регенеративных процессов, отложение гликогена в эпидермальных тканях и много других изменений; 2) линьку, или экди-зис — раскалывание и сбрасывание старой, частично резорбиро- ’ ' ;ванной кутикулы и резкое увеличение размеров тела за счет поглощения воды; 3) послелиньку, или постэкдизис — период быстрого отложения нового хитина и неорганических солей для об- .
разования новой кутикулы с одновременным ростом тканей; 4) межлиньку, или интерэкдизис — период относительного покоя, •когда физиологические процессы, обычно связанные с активной : .линькой, в основном прекратились, но все же происходит накопление резервов в гепатопанкреасе и других местах в ожидании следующей линьки. У некоторых ракообразных, например у Maja и .Carcinus, имеется период терминального роста, переходящий в : непрерывное состояние межлиньки, называемое анэкдизисом.
У ряда видов точно установлена средняя продолжительность ; .отдельных стадий линочного цикла [75, 109, 110, 180, 366, 369]. ।
Эндокринные механизмы
473J
У пресноводного рака Cambarus нет истинной личиночной стадии; вылупляющаяся молодь напоминает взрослых особей в миниатюре. В течение первого года жизни линька происходит каждые 12— 13 дней, вероятно, без сколько-нибудь значительных межлиноч-ных периодов. 'После первого сезона роста обычно совершаются две линьки в год: одна — весной, в конце апреля или в мае, другая — летом, в июле или августе. У этих более зрелых раков пред-линька длится 3—5 недель. В этот период постепенно резорбируется наружный скелет и в передне-боковых стенках кардиального отдела желудка откладываются соли кальция в виде гастролитов. Примерно за неделю до линьки начинает постепенно возрастать потребление кислорода и содержание воды в организме, достигающие максимума во время линьки. В период послелиньки происходят обратные изменения, и на них затрачивается примерно столько же времени, сколько на соответствующие процессы в предлиньке. Затем наступает период межлиньки, продолжительность которого после летней линьки больше, чем после весенней.
Изучение изменений, связанных с линочным циклом у других ракообразных, показало, что кальций, фосфаты, гликоген и липиды запасаются в гепатопанкреасе. Однако запас солей недостаточен для полного затвердения наружного скелета; это делает необходимым быстрое поглощение кальция как с пищей, так и непосредственно из внешней среды в период послелиньки. Гастролиты у таких форм, как речные раки и сухопутный краб Gecarcinus, следует, по-видимому, рассматривать как приспособление к среде, относительно бедной кальцием; у морских ракообразных они, как правило, отсутствуют. Определение дыхательного коэффициента показывает, что если в периоды межлинек он близок к 0,8, то в первые несколько часов после линьки он составляет всего лишь 0,1—0,2; такие низкие величины объясняются фиксацией СО2 во время твердения головогрудного щита; в первые недели после линьки дыхательный коэффициент постепенно возрастает до 0,7— 0,8. Эти данные говорят о том, что сразу после линьки происходит жадное поглощение кальция из окружающей среды и что интенсивность этого поглощения быстро снижается в течение первых нескольких дней.
Известно, что удаление глазного стебелька у Astacus [373], Uca [233], Eriocheir [187], Palaemonetes [50], Cambarus (Orconec-tes) [53] или Cambarus (Procambarus) [388] ведет к более раннему наступлению последующих линек. У молодых особей Cambar us (Procambarus) после такой операции, произведенной в первый год жизни, период между линьками сокращается с 12 до примерно 8 дней (при температуре 20—22°C). Это нельзя объяснить общей оперативной травмой: при других, не менее травматизи-рующих операциях, например после разрушения сетчаток, подобного учащения линек не происходит — наоборот, линька может иногда запаздывать. Причиной наблюдаемого эффекта могло так
474
Глава 21
же быть разрушение важных нервных центров или же удаление эндокринных органов, играющих существенную роль в регуляции линьки. Из этих двух объяснений верным оказалось второе. Если у половозрелого рака удалить оба глазных стебелька, то линька наступит значительно раньше, чем у неоперированных контрольных особей. Если же в брюшко такому раку, лишенному глазных стебельков, пересадить синусную железу от другой особи, то линька наступит позже, чем в контроле [50, 53].
Рис. 2'141. Увеличение размеров гастролита и линька у речных раков [374].
J — после удаления глазных стебельков с включенными в них синусными железами; перелинявшее животное почти тотчас же начинает готовиться к следующей линьке. 1 — первая линька; 2~ вторая линька. II— у раков, лишенных глазных стебельков, которым пересаживали с интервалом в 3—4 дня синусную железу, гастролиты не образуются.
То, что у ракообразных комплекс Х-орган — синусная железа подавляет линьку, подтвердилось при изучении регуляции образования гастролитов (рис. 21-11). Формирование этих конкреций, обычно приуроченное к периоду предлиньки, можно вызвать в другое время удалением обоих глазных стебельков или комплекса их желез [375]. После удаления стебельков у речных раков в период, когда линьки обычно не бывает, например между сентябрем и мартом, меньше чем через 24 ч начинают формироваться гастролиты (при 20°'С); затем они медленно увеличиваются в течение 8—10 дней, после чего быстро вырастают ко времени линьки, наступающей обычно между 15-м и 20-м днями. У особей, выживших после линьки, сразу же начинает формироваться новый комплект гастролитов. Если же особям, у которых были удалены глазные стебельки, пересаживать в брюшко с интервалами в 3— 4 дня синусную железу, то образование гастролитов будет подавлено. Если имплантации синусной железы прекратить, то примерно через неделю снова начнут формироваться гастролиты. Это значит, что пересаженные железы уже не оказывают действия.
В синусных железах ракообразных обнаружены гистологические изменения, коррелирующие с линочным циклом [336]. Непосредственно перед линькой в железах преобладают ацидофильные секреторные гранулы, а после завершения линьки — базофильные.
Эндокринные механизмы
475-
Процесс линьки, вызванный удалением глазных стебельков,, сходен с происходящим у нормальных животных [375]; при этом все связанные с линькой изменения можно предотвратить пересадкой синусных желез, если произвести ее сразу после удаления глазных стебельков. Поэтому кажется вероятным, что все эти изменения находятся под прямым или косвенным влиянием гормона синусной железы, подавляющего линьку. Удаление Y-органов блокирует линьку, а их имплантация восстанавливает способность-к линьке [66, 116, 156]. В Y-органах также наблюдаются циклические гистологические изменения, соответствующие линочному циклу; у крабов, находящихся в фазе анэкдизиса, эти органы сильно уменьшаются в размерах. Взаимоотношения между комплексом Х-орган — синусная железа и Y-органом в процессе регуляции линьки детально изучены. Х-орган терминального ганглия вырабатывает гормон, подавляющий активность Y-органа. Этот-гормон непрерывно вырабатывается в периоды послелиньки и межлиньки. После прекращения его секреции Y-орган снова начинает выделять гормон линьки. Как только произошел полный1 «запуск» предлиньки, тормозящий фактор глазного стебелька утрачивает способность задерживать дальнейший ход событий. Возможно, что гормон линьки нужен не только для инициации предлиньки, но и для успешного завершения ее последующих этапов.
У морского рака Jastis обнаружен гормон, идентичный одному из важнейших гормонов линьки насекомых, — 20-оксиэкдизон ( = 0-экдизон, экдистерон, крустэкдизон) [184]. У голубого краба; (Callinectes') помимо 20-оксиэкдизона найдены еще два стероида — вероятно, инокостерон и макистерон. Инъекция 20-оксиэкдизона и родственных стероидов речному раку Procambarus и крабу Uca ведет к преждевременному наступлению событий, характерных для предлиньки, таких, как аполиз, образование гастролитов, ускорение регенеративного роста конечностей и секреция веществ для построения нового экзоскелета; соответственна период межлиньки сокращается [130, 242, 268, 342]. Окончательное признание экдизонов истинными гормонами линьки у ракообразных будет возможно только после испытания их на особях, лишенных Х-органов.
Регуляция этого «пускового* механизма» у ракообразных весьма сложна и изменчива, так как на наступление линьки влияют самые разнообразные факторы. К ним относятся питание [94, 345], температура, свет и фотопериод [37, 393], наличие паразитов, повреждения и состояние репродуктивной активности. Гормон синусных желез, подавляющий линьку, по-видимому, повинен в неспособности самки Crangon, несущей на себе яйца, линять До тех пор, пока не вылупилась молодь, а также в том, что самки Cambar us (Orconectes), несущие яйца, линяют при ежегодной весенней линьке на несколько недель позже самцов и только после
476
Глава 21
того, как молодь покинула материнские плеоподы. У самок речных раков, несущих яйца, удаление синусных желез точно так же вызывает линьку, как у самцов [375].
Судя по некоторым данным, в регуляции линьки у ракообразных участвуют и другие гормональные механизмы. Сообщалось, например, о факторе, ускоряющем линьку, из Х-органа терминального ганглия и из Х-органа чувствительного сосочка глазного стебелька [63]. Есть основания подозревать, что в регуляции линьки участвует по меньшей мере еще один добавочный фактор, который образуется не в глазных стебельках, а в каком-то другом месте [64, 375]. Тщательное изучение отложения солей кальция в гастролитах показало, что этот процесс подчиняется определенному ритму — он происходит главным образом ночью. В связи с этим интересно, что введение экстракта мозговой ткани или сильное электрическое раздражение перерезанных концов зрительных нервов у животных с удаленными глазными стебельками на неко- ; торое время повышают потребление кислорода. |
Недавние исследования регуляции линьки у личинок коротко- | хвостых раков показали, что регуляторные механизмы у разных I видов могут быть различными. Удаление обоих глазных стебель- 1 ков у личинки краба Rhithropanopeus на стадии гоёа приводило ; к появлению одной или двух добавочных стадий гоёа, но ускорения личиночных линек не наблюдалось [97]. Подобным же образом удаление этих органов на любой стадии развития у Rhithropanopeus не уменьшало длительность стадии мегалопы. То, что у личинок с удаленными глазными стебельками появляются добавочные стадии гоёа, свидетельствует в пользу гипотезы, что какой-то фактор, вырабатываемый в глазных стебельках, может регулировать скорость морфологического развития и может быть частично ответственным за запуск физиологических процессов, завершающихся метаморфозом. Отсутствие ускорения линек в период личиночного развития позволяет предполагать, что комплекс , Х-орган — синусная железа, тормозящий линьку у взрослых осо- ;; •бей, у личинок Rhithropanopeus не функционирует и что линька у них регулируется каким-то другим механизмом. Однако тормозная регуляция линьки становится очевидной на стадии мегалопы у крабов Callinectes [95] и Sesarma [96]: удаление глазных стебельков, произведенное в надлежащее время на этой стадии, приводит к ускорению линьки.
Размножение. Как показали эксперименты, оогенез и особенно -вителлогенез у женских особей креветки Leander (Palaemon) регулируется гормоном, выделяемым синусными железами [313, 314, 401, 461]. По-видимому, этот гормон образуется в Х-органе терминального ганглия [65, 418]. У Leander (Palaemon) конец сезона размножения приходится на позднее лето; при этом яичники резко уменьшаются в размерах, их активность падает и они обычно «остаются в таком состоянии на протяжении осени, зимы и ранней
Эндокринные механизмы
477
весны. Удаление глазных стебельков или одних только синусных желез, произведенное в сентябре или октябре, приводит к очень быстрому увеличению яичников; за 45 дней вес этих органов возрастает примерно в 70 раз (рис. 21-12), и в конце этого периода животное откладывает нормальные яйца. У неоперированных (контрольных) животных яичники за это время почти не увеличиваются. Пересадка синусных желез в брюшко животных с уда-
Рис. 211-112. Увеличение веса яичников у Leander [313].
3 — после ампутации глазного стебелька; II — после удаления синусной железы; III — в Hopgate (контроль); IV—после ампутации глазного стебелька с последующей пересадкой синус-|«ой железы. По оси ординат отложено отношение сырого веса яичников к кубу длины тела. |ленными глазными стебельками подавляет рост яичников — они |становятся даже меньше, чем в контроле. Аналогичная гормональная связь между синусной железой и яичниками, когда синусная железа действует как ингибитор созревания яичников, обнаружена и у краба Uca {55]. По-видимому, так же обстоит дело и у речного рака. Можно предположить, что именно этим путем осуществляется влияние света на репродуктивный цикл [394].
Участие синусной железы в репродуктивной функции проявляется также в следующем факте: у самок речного рака, носящих на своих плеоподах яйца, весенняя линька обычно происходит значительно позже, чем у самцов, — только после того, как молодь переходит к свободному образу жизни. Эта адаптивная реакция, по-видимому, обусловлена действием гормона синусной железы, так как после ее удаления самки, несущие яйца, линяют в одно время с самцами. Фактор, подавляющий линьку, не идентичен фактору, угнетающему функцию яичников [92, 104].
478
Глава 21
Можно привести еще один пример зависимости яичников от эндокринных факторов. У Carcinus половые железы не развиваются в отсутствие Y-органа. Однако после развития гонад этот орган уже не имеет существенного значения для поддержания их функции [11].
Накоплено много данных о влиянии паразитарной кастрации самцов десятиногих раков на такие вторичные половые признаки,, как форма плеопод, хелипед (клешненосных конечностей) и брюшка. При частичной или полной кастрации, к которой обычно приводит заражение паразитами — корнеголовыми раками (паразитические Cirripedia) или бопиридами (паразитические Isopoda),. названные выше части тела обычно не приобретают форму, типичную для самцов; такие особи имеют женский тип строения.
Гипотезы, объясняющие паразитарную кастрацию, можно разбить в основном на две группы: в одних предполагается, что паразит влияет на реализацию генетического определения пола, а согласно другим — он вызывает разрушение семенников или каких-то иных органов с эндокринной функцией. Аргументом в пользу первой группы гипотез служит тот факт, что некоторые десятиногие раки обладают гермафродитными гонадами или такие гонады легко развиваются у них под влиянием паразитов. Возможно,, что у ряда форм, например у Leander (Palaemon), у которых, в результате кастрации вторичные половые признаки не изменяются, генетический механизм определения пола относительно устойчив и мало подвержен влиянию паразитов; у других же, например у Upogebia, этот механизм очень лабилен, и поэтому они легко подвергаются феминизации. Ряд исследователей обратил внимание на то, что у пораженных паразитами самцов часто наблюдается более высокое содержание жира в крови и в печени, характерное обычно для самок. Возникло предположение, что паразит предъявляет к метаболизму хозяина примерно такие же требования, какие в обычных условиях предъявляют развивающиеся в яичнике яйца, и возможно, что при повышении жирового» обмена в организме образуется какое-то вещество, которое влияет одновременно на гонады и на вторичные половые признаки.
Во второй группе гипотез, объясняющих результаты паразитарной кастрации у самцов, предполагается участие какого-то маскулинизирующего гормона. Согласно этим гипотезам, животное под влиянием паразита не феминизируется, а принимает нейтральную форму, которая случайно напоминает больше самку, чем самца. Аргументом в пользу этого предположения служат наблюдения над тремя видами паразитов краба Munida sarsi. Два мелких паразитических ракообразных, Triangulus munidae и Ler-naeodiscus ingolfi, полностью или частично кастрируют краба и вызывают резкие изменения во вторичных мужских половых признаках; значительно более крупный паразит Triangulus boschmai не нарушает функции гонад и не влияет на половые признаки.
Эндокринные механизмы
479"
Эти наблюдения как будто исключают прямое влияние метаболических требований паразита как причину наблюдаемых изменений. Другие исследователи, однако, не смогли обнаружить корреляции между степенью атрофии гонад и степенью подавления
Рис. 21-4'3. Поперечный разрез бокоплава Orchestia, показывающий расположение андрогенных желез [76].
1 — мышцы; 2 — отросток кишки; 3 — гепатопанкреас; 4 — седьмой переопод; 5 — нервная трубка; 6 — половой сосочек; 7 — андрогенная железа; 8 — выносящий проток.
мужских признаков; они высказали предположение, что гормон, о котором идет речь, образуется не в семенниках, а в какой-то другой ткани.
Возможно, что разгадка кроется в наличии андрогенных желез, которые впервые были открыты у Orchestia (рис. 21-13), а позднее найдены почти у всех ракообразных, у которых их искали [74—77]. Удаление этих желез приводит к феминизации, которая обратима: после имплантации желез явления феминизации исче-
480
Глава 21
зают. При пересадке андрогенных желез неполовозрелым самкам наблюдается выраженная маскулинизация с появлением всех соответствующих признаков, включая развитие придаточных органов половой системы. Яичники превращаются в семенники. Даже поведение таких самок напоминает поведение самцов. У ракообразного Lysmata превращение самца в самку совпадает с дегенерацией андрогенных желез [77].
Для половых придатков речного рака характерен сезонный цикл изменений. Эти придатки приходят в функциональное состояние (форма I) во время поздней летней линьки. Это период, когда семенники увеличены и активны. Во время весенней линьки, соответствующей периоду низкой активности половых желез, придатки возвращаются в нефункциональное состояние (форма II). Линька, вызванная экспериментально в зимние месяцы, когда активность семенников также низка, всегда сопровождается образованием формы II. Эти изменения регулируются, по всей вероятности, тем же самым эндокринным комплексом, который регулирует репродуктивную активность у самцов [95].
Паразитарная кастрация самок у ракообразных, как правило, сопровождается разве только незначительными изменениями формы тела и придатков. Однако было отмечено, что при лучевом повреждении яичников выводковые камеры у Asellus и Daphnia не развиваются. У бокоплава Gammarus при действии на яичники паразитического червя Polymorphus minutus или при их облучении происходит утрата способности к развитию типичных маргинальных щетинок оостегитов. Когда сеансы облучения прекращали, способность к развитию маргинальных щетинок восстанавливалась параллельно с восстановлением оогенеза. У самок креветки Leander, кастрированных бопиридами или действием рентгеновских лучей, не развивались ни брюшная инкубаторная камера, ни специальные гуанофоры, связанные с соответствующими брюшными сегментами. Эти и другие наблюдения могут служить убедительными доводами в пользу гипотезы о том, что у этих ракообразных яичники вырабатывают какой-то гормон, способствующий развитию у самок особых изменений, связанных с заботой о потомстве [73]. Судя по некоторым данным, в регуляции нормального полового развития участвуют и другие гормоны, в том числе гормон из глазного стебелька [16, 93, 392]. Очевидно, у обоих полов существуют гормоны нейросекреторного происхождения, тормозящие и стимулирующие деятельность половых желез. У самок эти гормоны действуют непосредственно на яичники, у самцов же они оказывают непрямое влияние, подавляя или активируя функцию андрогенных желез [2, 309].
Перемещение ретинальных пигментов. У многих животных, в том числе позвоночных, ракообразных и насекомых, передвижение глазных пигментов в значительной мере способствует механической адаптации к изменениям интенсивности света. Однако чет
Эндокринные механизмы
481 ”
кие данные о роли гормонов в регулировании этих реакций получены только на ракообразных.
Сложный глаз ракообразного состоит из многих единиц— омматидиев (рис. 21-14). Каждый омматидий обладает тремя функционально различными группами пигмеи-.
Рис. 21-14. Расположение ретинальных пигментов в омматидии глаза креветки Palaemonetes [220].
А — при адаптации к свету; Б — при адаптации к темноте; В — после введения экстракта глазных стебельков креветки, адаптированной к свету, креветке, адаптированной к темноте и содержащейся в темноте. 1 — роговица; 2 — дистальный ретинальный пигмент; 3 — проксимальный ретинальный пигмент; 4 — базальная мембрана; 5 — отражающий пигмент; 6 — раб-дом.
тов. Дистальный ретинальный пигмент — либо оммохром, либо меланин — находится в клетках, окружающих дистально расположенный диоптрический аппарат омматидия. В глазу, адаптированном к темноте, этот пигмент занимает только дистальное положение, а при дневном свете он распространяется проксимально, окутывая весь омматидий до самых элементов ретинулы. Проксимальный пигмент, химически сходный с дистальным, находится в клетках ретинулы; в темноте этот пигмент мигрирует в участок, проксимальный по отношению к ба-31—2067
482
Глава 21
зальной мембране, а на свету он перемещается дистально до встречи с дистальным пигментом. (Поэтому при световой адаптации весь омматидий заключен в поглощающую свет муфту из пигмента. Третий пигмент — отражающий белый — состоит, по-видимому, из пуринов и птеридинов [230]. В темноте он располагается вокруг элементов ретинулы, образуя своего рода tapetum, а на свету переходит в участок, проксимальный по отношению к базальной мембране.
Дистальный пигмент, который обычно называют пигментом радужки, действует подобно радужной оболочке позвоночных — градуально реагирует на свет в широком диапазоне интенсивности [137, 356]. Попытки выяснить, в какой мере пигментные клетки правого и левого глаза одной особи способны к независимой реакции на свет, дали неоднозначные результаты. Более поздние эксперименты привели к выводу, что существует по крайней мере частичная взаимозависимость между двумя глазами: при освещении одного глаза другой — затемненный — тоже в той или иной степени адаптируется к свету. Многочисленные наблюдения показали также, что у многих ракообразных положение одного или нескольких пигментов может претерпевать суточные ритмические изменения, даже если животное днем и ночью находится в одинаковых условиях, особенно в постоянной темноте [139, 426]. У особей Leander (Palaemon), содержавшихся на освещенном черном фоне, отмечалось неравномерное дорсовентральное распределение пигмента — очевидно, в связи с значительно меньшей освещенностью вентральных элементов глаза по сравнению с дорсальными. Эти разнообразные реакции ретинальных пигментов наводят на мысль о том, что регуляция их не так проста и что наряду с прямой реакцией пигментных клеток на освещение в этой регуляции участвуют также эндокринные, а может быть, и нервные факторы.
Комплекс желез глазного стебелька выделяет гормон, влияющий на положение ретинального пигмента [227, 431]; у Palaemo-netes и Cambarellus подобной же функцией обладает подглоточный узел [137]. Этот гормон назвали ретинальным пигментным гормоном, или РПГ. Инъекция экстрактов глазных стебельков, взятых у адаптированных к свету Palaemonetes, особям, адаптированным к темноте и содержащимся в темноте, вызывает у последних миграцию дистального и отражающего пигментов в положение, характерное для адаптации к свету. В глазных стебельках всех исследованных ракообразных обнаружен РПГ в большем или меньшем количестве. О роли этого гормона в перемещениях ретинального пигмента свидетельствует то, что в надглоточных ганглиях Palaemonetes, которых поместили на 14 дней в темноту, содержание РПГ возросло в четыре раза [142]. Имеются, однако, сообщения, что у некоторых грапсоидных крабов суточные изменения в содержании ретинальных пигментов продолжаются даже
Эндокринные механизмы
483
после удаления синусных желез. Гормон из глазных стебельков участвует хотя бы частично в регуляции дистального ретинального пигмента у Leander (Palaemon) и Uca [134, 136].
У речных раков ретинальные пигменты обладают различными порогами реакции на РПГ. На низкие концентрации гормона реагирует только дистальный пигмент, а при более высоких концентрациях начинают перемещаться оба пигмента — дистальный и проксимальный; они мигрируют в положение, характерное для адаптации к свету [228].
Источником РПГ служит комплекс, состоящий из Х-органа терминального ганглия и синусной железы глазного стебелька. Экстракт, приготовленный только из синусной железы, способен вызвать выраженную реакцию со стороны ретинального пигмента.
Подобно хроматофоротропинам глазного стебелька, ретинальный пигмент синусной железы выдерживает кипячение. РПГ, из глазных стебельков десятиногого рака Pandalus — это октадекапептид с мол. весом около 2000, содержащий 12 различных аминокислот [131, 134]. Имеются достаточные основания полагать, что он не идентичен ни одному из главных хроматофоротропинов. Действительно, пигментная система наружных покровов при изменении фона обычно проходит последовательно всю гамму изменений цвета, глаз же отражает постоянную адаптацию к свету, хотя порог реакции ретинальных пигментов на экстракт глазного стебелька значительно выше, чем порог реакции хроматофоров тела. Такого положения, очевидно, не могло бы быть, если бы РПГ был идентичен одному из главных хроматофоротропинов. Поскольку, однако, РПГ до сих пор не удалось отделить от вещества, вызывающего рассеивание пигмента в хроматофорах, было высказано предположение, что адаптацию к свету при помощи дистального ретинального пигмента и рассеивание пигмента хроматофоров вызывает либо одна и та же молекула, либо разные, но очень близкие молекулы [232].
Попытки обнаружить влияние аналогичного фактора на состояние ретинальных пигментов у бабочки Ephestia не привели к успеху. Введение экстрактов из тканей головы особей, адаптированных к свету, бабочкам, адаптированным к темноте или к свету, не вызывало никаких изменений в состоянии пигментов. Экстракты синусных желез ракообразных также не оказывали действия на ретинальные пигменты бабочки.
Существует еще один нейросекреторный гормон, который оказывает адаптирующее к темноте влияние на дистальный пигмент [54, 137]. Показано, что можно вызвать дифференциальную секрецию обоих гормонов с помощью световых стимулов регулируемой длительности и интенсивности [57]. Получены также данные О суточном ритме секреции обоих гормонов [57, 426]. Эти два гормона, влияющие на ретинальный пигмент, удалось разделить Методами электрофореза [140, 141] и гель-фильтрации [138]; оба 31*
484
Глава 21
они вызывают градуальную реакцию в прямом соотношении с введенной дозой. *
Другие явления. Частота сердечных сокращений у ракообразных также находится под влиянием эндокринных факторов. Гормон, образующийся в глазных стебельках, ускоряет работу сердца [430], причем у Palaemonetes его секреция коррелирует с выделением некоторых гормонов, изменяющих окраску тела [374]. Есть
<7
Рис. 21-15. Перикардиальные органы краба Maja и их взаимоотношения с отверстия-ми бранхио,кардиальных вен, открывающимися в перикардиальную полость [5].
1 — нити, на которых подвешены нервные стволы; 2 — ветви, иннервирующие мышцы; 3 — дорсальные кардиальные нервы. Пунктирными линиями показаны нервы от центральных нервных органов.
основания полагать, что активаторы сердца и хроматофоров не идентичны [189, 237]. Более специфический фактор, регулирующий сокращения сердца, вырабатывается элементами нейросекреторной системы, расположенными в перикардиальной полости. Они были названы перикардиальными органами (рис. 21-15). Экстракты из этих органов увеличивают амплитуду сердечных со- кращений и вызывают ряд других эффектов, которые у разных видов различны. Гормон, выделяемый перикардиальными органами — это, по-видимому, свободный полипептид, не связанный с белком-носителем [31]. Удаление глазных стебельков или синусных желез из этих стебельков у Cambarus (Orconectes) и Gecarci-nus быстро приводит к повышению основного обмена, о чем свидетельствует возрастающее потребление О2 [37, 375]. Интенсивность обмена можно уменьшить введением водных экстрактов синусных желез. Инъекция экстракта глазных стебельков краба Uca крабу Callinectes ведет к быстрому повышению содержания сахара в крови примерно с 20 до 80 и более мг% [229]. Максимум достигается в течение одного часа, а затем происходит медленное
Эндокринные механизмы
485
снижение до нормы. Такое действие объясняли присутствием в синусной железе ракообразных диабетогенного фактора, и недавно это было подтверждено в отношении краба Libinia. Этот гормон термолабилен и, по-видимому, представляет собой полипептид [224, 231]; у речного рака Orconectes молекулярный вес его около 10000. Этот гормон в известной мере видоспецифичен: гормон краба оказывает очень слабое действие на речного рака, и наоборот [225]. Есть также данные о том, что у ракообразных имеются диуретический и антидиуретический гормоны, контролирующие водно-солевой баланс [38, 222, 223].
Эндокринные факторы у других животных
У многих других беспозвоночных также наблюдаются годичный и иные ритмы размножения, при которых периоды половой активности чередуются с периодами спада. В большинстве случаев механизмы активации или угнетения половых желез до сих пор неизвестны.
Для многих беспозвоночных, принадлежащих к разным типам, характерен половой диморфизм. Давно уже подозревали, что он по крайней мере отчасти связан с действием гормонов, сравнимых с гормонами половых желез у позвоночных [187]. В большинстве случаев убедительных доказательств этого до сих пор не получено — они имеются только в отношении ракообразных. Тем не менее в литературе описано много фактов, указывающих на возможную роль гормонов в интересующем нас явлении. Сюда относятся: 1) параллельное влияние паразитов на половые железы и половую дифференцировку; 2) параллельные циклические изменения активности половых желез, с одной стороны, и некоторых вторичных половых признаков — с другой; 3) результаты воздействия на гонады или на весь организм рентгеновскими лучами или лучами радия; 4) последствия хирургического удаления половых желез.
Физиологические данные о роли эндокринных механизмов у членистоногих, не относящихся к насекомым или ракообразным, относительно, скудны. У мечехвоста (Limulus) обнаружена корреляция между плотностью нейросекреторных клеток и активностью экстрактов по отношению к хроматофорам [362]. У многоножек [334, 370] и некоторых паукообразных [157, 198] в мозгу имеются нейросекреторные клетки, которые связаны с нейрогемальными структурами, напоминающими такие органы, как corpora cardiaca насекомых и синусные железы ракообразных; были также описаны железистые органы, гистологические изменения которых при линьке указывают на сходство их с проторакальными железами насекомых [117, 217, 253].
У гидры обнаружены нейросекреторные клетки в гипостомаль-ной области. Косвенным указанием на то, что эти клетки являются источником гормона роста, служит тот факт, что экстракты из
486
Глава 21
гипостомальной ткани стимулируют рост и образование новых почек. В гипостомальных экстрактах из половых форм фактора роста гораздо меньше или совсем нет; поэтому было высказана предположение, что в присутствии этого фактора интерстициальные клетки обеспечивают соматический рост, а в его отсутствие-происходит образование половых клеток [60, 61, 256].
Регенерация у планарий, по-видимому, частично регулируется химическими медиаторами, выделяемыми раневой поверхностью. В мозгу также имеются нейросекреторные клетки, и у многоглазой планарии Polycelis мозг необходим для нормальной регенерации глаз [255]. Было высказано предположение, что нейросекреторные факторы, образующиеся в мозгу, стимулируют регенерацию, а также размножение путем деления особи на части [33].
В дорсальных и вентральных ганглиях окологлоточного нервного кольца нематоды Phocanema описаны нейросекреторные клетки, и полученные данные указывают на то, что один из продуктов этих клеток, возможно, имеет отношение к линьке '[101]. У немертин есть очень своеобразная структура, состоящая из нервно-железистых элементов; ее называют церебральным органом, так как она тесно связана с церебральным ганглием, но ее функция еще не выяснена [361].
Из радиальных нервных стволов морской звезды может быть получен экстракт, который, будучи введен в полость луча, стимулирует выведение икры и семени [70, 71, 381]. В обычных условиях соответствующий фактор, по-видимому, выделяется прямо в> окружающую среду, откуда его могут поглощать амбулакральные ножки той же самой и других находящихся поблизости особей. Этот гормон — полипептид с мол. весом около 2600. Полагают, чта у морских звезд какой-то гормон, вырабатываемый яичниками,, индуцирует деления созревания. Есть также данные о том, чта радиальные нервы выделяют гормон, тормозящий выведение половых продуктов, а гормон, стимулирующий этот процесс, эффективен только тогда, когда концентрация тормозного гормона падает.
Известны некоторые факты, касающиеся эндокринной регуляции роста и регенерации у полихет, а также регуляции их размножения [24]. У нереид гормоны, образующиеся в мозгу, регулируют рост ооцитов, а также эпитокию, т. е. развитие половых продуктов только в задней или в передней части тела. Гормональная активность мозга тормозит эти процессы, а их нормальное развитие, по-видимому, зависит от прогрессивного уменьшения концентрации тормозного гормона [86, 113, 195]. Образование спермы у Arenicola также подвергается торможению; мозг стимулирует репродуктивную активность у самок и необходим^ для выведения яиц [210]; необходим он и для нормального сперматогенеза.
Эндокринные механизмы
487
У олигохеты Eisenia для развития половых желез и поддержания клителлума (пояска) требуется присутствие мозга; изменения в нейросекреторных клетках коррелируют со стадиями развития репродуктивной активности [197].
Корреляция между функцией семенников и гистологическими признаками активности нейросекреторных клеток у Hirudo, а также действие экстрактов из мозга пиявок указывают на существование у этих организмов какого-то мозгового гормона, стимулирующего половые железы [183].
Относительно брюхоногих моллюсков есть данные, свидетельствующие о возможном эндокринном контроле оогенеза и овуляции — о регулирующей роли церебральных ганглиев у Lymnaea [221] и ганглиев глазных щупалец у Helix и Arion (у последнего также и церебральных ганглиев) [249, 250, ЗГ9, 382]. Нейроэндокринная регуляция яйцекладки описана у Aplysia [244]. По-видимому, нейросекреторные клетки правого плеврального ганглия вырабатывают гормон, участвующий в регуляции водного обмена ,[259]. Данные об эндокринных функциях у двустворчатых моллюсков довольно скудны. Описана корреляция между репродуктивной активностью и изменениями нейросекреции у Mytilus [269] и Dreissena [8].
У головоногих моллюсков существуют две пары эндокринных желез — бранхиальные и оптические. У осьминога первые из них хорошо развиты и обильно васкуляризованы. Удаление их приводит к последствиям, несколько напоминающим результаты адреналэктомии у млекопитающих; возможна кратковременная частичная компенсация этих последствий путем введения экстрактов, приготовленных из желез [402]. Оптическая железа регулирует развитие репродуктивных функций и, по-видимому, в свою оче-редь^ находится под контролем центральной нервной системы
Заключение
При рассмотрении эндокринных механизмов в сравнительном аспекте бросается в глаза то, что у всех групп животных, у которых выработались эндокринные системы, они выполняют функции одного- и того же общего типа. В таких отдаленных группах, как членистоногие и позвоночные, которые, по мнению большинства зоологов, могли иметь общих предков только на самых ранних этапах эволюции, мы видим сходное распределение интегративных функций между нервной и эндокринной системами и сходную координацию деятельности обеих систем. Как у позвоночных, так и У беспозвоночных рост, дифференцировка, размножение, обмен веществ и функция пигментных клеток в значительной степени регулируются гормонами. Коротко говоря, роль эндокринной системы преобладает в регулировании а) таких функций, для кото
488
Глава 21
рых характерно длительное время реакции (функции, связанные с ростом и дифференцировкой); б) таких процессов, которые требуют длительной непрерывной регуляции (поддержание различных основных проявлений жизнедеятельности и обмена веществ), и в) таких ситуаций, в которых эффекторные органы должны находиться в том или ином состоянии в течение долгого времени (как, например, хроматофоры).
Даже в относительно обширных естественных группах животных обычно нет узкой специфичности гормональных факторов. .У позвоночных, по-видимому, довольно широко распространены одни и те же общие функциональные типы гормонов, но у данного вида они, как правило, вызывают характерный для него тип ответа. Межвидовые различия в тонком химическом строении встречаются главным образом у белковых гормонов. У насекомых такие факторы, как ювенильный гормон, мозговой гормон и энди-зон, взаимозаменяемы не только для разных видов, но и для представителей разных отрядов. То же самое можно сказать о хрома-тофоротропинах, ретинальных пигментных гормонах и гормоне, подавляющем линьку, у различных видов ракообразных.
Отмечаются также некоторые черты сходства между гормонами трех главных групп животных, обладающих эндокринной системой. Corpora cardiaca насекомых вырабатывают вещество, высокоактивное по отношению к хроматофорам ракообразных. Накопилось довольно много данных о действии, которое могут оказывать гормоны беспозвоночных животных на позвоночных и еще больше — о влиянии гормонов позвоночных на беспозвоночных. Многие результаты неясны и противоречивы, и создается впечатление, что они представляют интерес главным образом для фармакологии. По-видимому, хроматофоротропин позвоночных — ме-ланоцитстимулирующий гормон — качественно сходен с аналогичным гормоном ракообразных по своему действию на хроматофоры и по некоторым физико-химическим свойствам, однако есть достаточные основания не считать их идентичными [1, 51].
Как показывают гистологические, а во многих случаях и очень убедительные физиологические данные, у самых разнообразных животных в нервной системе дифференцировались эндокринные элементы — нейросекреторные клетки [158, 188, 366]. Такие клетки описаны, в частности, у гидры, плоских червей, нематод, кольчатых червей, морских звезд, моллюсков, членистоногих и позвоночных. У рыб и земноводных они находятся в преоптическом ядре, а у рептилий и млекопитающих в гомологичных ядрах — супраоптическом и паравентрикулярном. Они входят в состав тесно интегрированного гипоталамо-гипофизарного комплекса, который занимает ключевую позицию в координации деятельности нервной и эндокринной систем.
У насекомых такую связь совершенно аналогичным образом осуществляет комплекс pars intercerebralis—corpora cardiaca.
Эндокринные механизмы 489
— 1
у ракообразных подобную же систему образуют комплексы Х-орган—синусная железа и секреторные клетки мозга—посткомиссуральный орган. Данные, касающиеся пауков, говорят о наличии такой же организации у другой обширной группы животных — паукообразных. В нервной системе мечехвоста (Limulus) обнаружен параллелизм между количественным распределением факторов, способных действовать на хроматофоры ракообразных, и частотой нейросекреторных клеток ['53, 362]. В связи с обилием гистологических и физиологических данных о нейросекреции у столь разнообразных форм весьма вероятно, что наличие нейроэндокринного комплекса окажется общей особенностью всех многоклеточных животных.
У кольчатых червей некоторые крупные ганглиозные клетки дают при окраске хромаффинную реакцию, характерную для тканей, вырабатывающих адреналин, а экстракты этой нервной ткани вызывают при испытании физиологические реакции, характерные для адреналина. У позвоночных мозговое вещество надпочечников образуется в эмбриогенезе из нервной системы.
Коротко говоря, ряд важнейших эндокринных структур произошел, по-видимому, от тканей, обладавших обычной нервной или проводящей функцией и расположенных внутри центральной нервной системы; на каком-то промежуточном этапе этой эволюции клетки начали уже выполнять секреторную функцию, но еще -сохраняли присущую нейронам способность проводить возбуждение, а на конечном этапе специализировались только для эндокринной функции и стали образовывать железистые ткани или органы, обособленные от нервной системы. В тех случаях, где тела •секретирующих клеток по-прежнему лежат в органах нервной системы, используются специальные внутриклеточные средства для переноса выделяемых продуктов за пределы высокоспециализированных «гемато-энцефалических барьеров», к общему сосудистому руслу. Другие эндокринные органы возникли морфологически (но не физиологически) независимо от нервной системы.
Интересно, что у позвоночных гормоны, выделяемые структурами экто- или эктодермального происхождения, представляют собой белки или полипептиды (например, СТГ, ЛГ, АДГ, АКТГ, МСГ, ТТГ) или по крайней мере содержат азот (адреналин); гормоны же, синтезируемые в тканях мезодермального происхождения, относятся к категории стероидов (половые гормоны, гормоны коры надпочечников).
Между гормонами позвоночных и беспозвоночных существуют интересные черты сходства в отношении механизма действия. В действии серотонина на мышечную ткань печеночной двуустки (Fasciola hepatica) посредником служит циклический АМФ. Имеются также данные об участии циклического АМФ в осуществлении некоторых эффектов, вызываемых гормонами насекомых и кроматофоротропинами ракообразных. Кроме того, в действии
490
Глава 21
гормонов насекомых и ракообразных существенную роль играет синтез РНК и белков. Изучая влияние гормонов насекомых на эти процессы, удалось получить очень важные сведения о механизмах действия гормонов вообще.
В литературе можно иногда встретить умозрительные рассуждения на тему о том, какой из интегрирующих механизмов — нервный или эндокринный — появился у животных раньше. Очевидно,, оба механизма, если понимать их в широком смысле, встречаются у всех форм живых организмов, как у одноклеточных, так и у многоклеточных. Процессы возбуждения, проведения и межклеточной химической передачи информации, лежащие в основе-физиологической активности нервной системы, являются общими для всех клеток. Как указывалось в начале этой главы, обоим координирующим механизмам как в филогенезе, так и в онтогенезе должны были предшествовать какие-то организующие и дифференцирующие силы. В процессе развития дифференцировка, обусловленная генами, должна предшествовать деятельности организаторов. Поэтому нет веских оснований считать, что один из координирующих факторов — возбуждение или химическая сигнализация — филогенетически более примитивен. Вероятно, оба типа координирующих механизмов возникли одновременно и развивались параллельно. Появление новых функциональных потребностей в связи с возросшими размерами организма, его многоклеточной структурой и разделением функций между отдельными частями привело к формированию двух высокоспециализированных, но действующих совместно систем — нервной и эндокринной.
СПИСОК ЛИТЕРАТУРЫ
1. Abramowitz A. A., Physiol. Zool. 11: 299—311, 1938. Similarity of crustacean sinus gland hormone to intermedin.
2. Adiyodi K. G., Adiyodi R. G., Biol. Rev. 45: 121—165, 1970. Endocrine control of reproduction in decapod Crustacea.
3. Ahlquist R. P., Amer. J. Physiol. 153: 586—600, 1948. Original elucidation of the adrenergic receptors.
4. Ahlquist R. P., Ann. Rev. Pharmacol. 8: 259—272, 1968. Agents which block adrenergic beta-receptors.
5. Alexandrowicz J. S., J. Marine Biol. Assoc. U.K. 31: 563—580, 1953. Pericardial organs of crustaceans.
6. Allen В. M., Biol. Rev. 13: 1—19, 1938. Hormones and amphibian metamorphosis, review.
7. Amoroso E. C., Ann. Endocrinol. 16: 435—447, 1955. Reptile corpus luteum.
8. Antheunisse L. J., Arch. Neerl. Zool. 16: 237—314, 1963. Neurosecretory activities in mussel Dreissena.
9. Applebaum S. W., Gilbert L. I., Devel. Biol. 27: 165—175, 1972. -Ecdysone stimulation of cyclic AMP.
10. Arvy L., Bounhiol J. B., Gabe M., C. R. Acad. Sci. 236: 627—629, 1953. Neurosecretion by corpora cardiaca; increased brain secretion in adult Bombyx.
11. Arvy L., Echalier G., Gabe M., Ann. Sci. Nat. Zool. Biol. Animale 18: 263— 268, 1956. Gonad regulation by Y-organ in Carcinus.
12. Arvy L., Fontaine M., Gabe M., J. Physiol. 49: 685—697, 1957. Hypothalamic control of ACTH and TSH.
Эндокринные механизмы 491
13. Astwood Е. В., рр. 529—532 in: Handbook of Physiology, Sec. 5, Adipose Tissue, edited by A. E. Renold and G. F. Cahill, Jr. Amer. Physiol. Soc., Washington, D. C., 1965. The pituitary gland and the mobilization of fat.
14. Baba У., Matsuo H., Schally A. V., Biochem. Biophys. Res. Comm. 44: 459— 463, 1971. Structure of the porcine LH- and FSH-releasing hormone.
15. Baggerman B., Arch. Neerl. Zool. 12: 105—318, 1957. Photoperiod and sticklebacks; thyroid increases induction of F. W. selection.
16. Balesdent-Marquet M. L., C. R. Acad. Sci. 236: 1086—1088, 1953. 'Hormone other than ovarian on female sex characters.
17. Barr W. A., pp. 164—238 in: Perspectives in Endocrinology. Hormones in the Lives of Lower Vertebrates, edited by E. J. W. Barrington and C. Barker Jorgenson. Academic Press, New York, 1968. Patterns of ovarian activity.
18. Barrington E. J. W., J. Marine Biol. Assoc. U. K- 36: 1—15, 1957. Protein binding of I in Ciona endostyle.
19. Barrington E. J. W., J. Marine Biol. Assoc. U. K- 37: 117—125, 1958. Amphioxus endostyle cell binding of I.
20. Barrington E. J. W., Hormones and Evolution. English Universities Press, London, 1964, 154 p.
21. Bartholomew G. A., Bull. Mus. Comp. Zool. Harvard 101: 433—476, 1949. Photoperiodism and English sparrow.
22. Bartholomew G. A., Anat. Rec. 106: 49—60, 1950. Light, temperature and reptile sex cycles.
23. Bartholomew G. A., pp. 669—676 in: Photoperiodism and Related Phenomena in Plants and Animals, edited by R. B. Withrow. Amer. Assoc. Adv. Sci., Washington, D. C., 1959. Photoperiodism and reptiles.
24. Baskin D. G., Golding D. W., Biol. Bull. 139: 461—475, 1970. Endocrine control of reproduction in a viviparous polychaete.
25. Beaudoin A. R., Anat. Rec. 125: 247—259, 1956. Differential CNS growth with thyroid.
26. Becker E., Plagge E„ Biol. Zentralbl. 59: 326—341, 1939. Hormonal control of dipteran metamorphosis.
27. Behrens D. K., Staub A., Root M. A., Bromer W. W., Ciba Colloq. Endocrinol. 9: 167, 1956. Glucagon, a polypeptide, crystallized.
28. Belamarich F. A., Terwilliger R. C., Amer. Zool. 6: 101—106, 1966. Cardio-excitor hormone from pericardial organs of Cancer.
29. Benoit J., Assenmacher L., Brard E., C. R. Acad. Sci. 242: 3113—3115, 1956. Cycles in constant light or dark.
SO. Berg O., Gorbman A., Kobayashi H., pp. 302—319 in: Comparative Physiology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Variability in thyroid activity and hormones.
31. Berlind A., Cooke I. M., J. Exp. Biol. 53: 679—686, 1970. Release of a neurosecretory hormone as peptide by electrical stimulation of crab pericardial organs.
32. Bern H. A., Nicoll C. S„ Rec. Prog. Hormone Res. 24: 681—720, 1968. Comparative endocrinology of prolactin.
33. Best J. B., Goodman A. B., Pigon A., Science 164: 565—566, 1969. Brain control of planarian fissioning.
34. Bissonnette T. H., Endocrinology 22: 92—103, 1938. Influence of the hypophysis upon molting in the ferret
35. Bissonnette T. H., Bailey E. E., Ann. N. Y. Acad. Sci. 45: 221—260, 1944. Control of seasonal changes in coat color of the ermine, Mustela.
36. Blaschko H., Hagen P., Welch A. D., J. Physiol. 129: 27—49, 1955. Membranes of vesicles.
37. Bliss D. E., Biol. Bull. 104: 275—296, 1953. X-organ source of sinus gland hormc’nes; hormonal regulation of crustacean metabolism; inhibition of growth and regeneration of crab by light.
38. Bliss D. E., Wang S. M. E., Martinez E. A., Amer. Zool. 6: 197—212, 1966. Water balance in the land crab, Gecarcinus lateralis, during the intermolt cycle.
492
Глава 21
39. Bliss D. E., Welsh J. H., Biol. Bull. 103: 157—169, 1952. X-organ sinus gland in brachyurans.
40. Blount R. F., J. Exp. Zool. 63: 113—141, 1932. Pituitary transplantation and embryonic pigmentation.
41. Bodenstein D., J. Exp. Zool. 123: 189—232, 1953. Hormones in development of Periplaneta.
42. Bodenstein D., J. Exp. Zool. 123: 413—434, 1953. Molting in cockroaches.
43. Bodenstein D., J. Exp. Zool. 124: 105—116, 1953. Molting in cockroaches.
44. Bodenstein D., pp. 197—211 in: Recent Advances in Invertebrate Physiology, edited by В. T. Scheer. University of Oregon Publications, 1957. Importance of development of competence to respond to hormones.
45. Boss W. R., J. Exp. Zool. 94: 181—209, 1943. Hormones and plumage type in gulls.
46. Bounhiol J. J., C. R. Soc. Biol. 126: 1189—1191, 1937. Corpora allata and lepidopteran development.
47. Bounhiol J. J., Z. Zool. Exp. Gen. 81: 54—64, 1939. Hormones in lepidopteran reproduction.
48. Bowers C. Y., Schally A. V., Enzmann F., Boler J., Folkers K., Endocrinology 86: 1143—1153, 1970. Porcine thyrotropin releasing hormone.
49. Bragdon D. E., Lazo-Wasem E. A., Zarrow M. X., Hisaw F. L., Proc. Soc. Exp. Biol. Med. 86: 477—480, 1954. Progesterone in blood of pregnant snakes.
50. Brown E. A., Jr., Anat. Rec. 75 (Suppl): 129, 1939. Sinus gland and molting in PulctcirLOfietes.
51. Brown F. A., Jr., Quart. Rev. Biol. 19: 32—46, 118—143, 1944. Endocrines in crustaceans, review.
52. Brown F. A., Jr., Physiol. Zool. 19: 215—223, 1946. Postcommissural gland.
53. Brown F. A., Jr., Cunningham O., Biol. Bull. 77: 104—114, 1939; 81: 80—95, 1941. Sinus gland hormone and molting in Crustacea; Limulus neuro-secretion.
54. Brown F. A., Jr., Hines M. N., Fingerman M., Biol. Bull. 102: 212—225, 1952. Hormonal control of Palaemonetes retinal pigment.
55. Brown F. A., Jr., Jones G. M., Biol. Bull. 91: 228—232, 1949. Eyestalk-hormo-ne inhibition of ovaries in Uca.
56. Brown F. A., Jr., Rollo M., Auk. 57: 485—498, 1940. Influence of light periods upon plumage type in birds.
57. Brown F. A., Jr., Webb H. M., Sandeen M. I., J. Cell. Comp. Physiol. 41: 123—144, 1953. Control of secretion of two retinal hormones.
58. Brown-Grant K., von Euler C., Harris G. W., Reichlin S., J. Physiol. 126: 1—28, 1954. Thyroid response to cold.
59. Burgos M. H., Rev. Soc. Argent. Biol. 26: 259—371, 1950. Loss of clasp reflex of toads on castration.
60. Burnett A. L., Diehl N. A., J. Exp. Zool. 157: 217—226, 1964. Neurosecretion in Hydra reproduction.
61. Burnett A. L., Diehl N. A., Diehl F., J. Exp. Zool. 157: 227—236, 1964. Neurosecretion in Hydra growth and regeneration.
62. Butenandt A., Karlson P., Z. Naturforsch. 9B: 389—391, 1954. Crystallization of insect molting hormone.
63. Carlisle D. B., Pubbl. Staz. Zool. Napoli 24: 279—285, 1953. Molt accelerating principle of MTGX-organ.
64. Carlisle D. B., Pubbl. Staz. Zool. Napoli 24: 285—292, 1953. Thoracic ganglia accelerate molt in Crustacea.
65. Carlisle D. B., Pubbl. Starz. Zool. Napoli 24: (Suppl.) 79—80, 1954. Intra-axonal transport.
66. Carlisle D. B., J. Marine Biol. Assoc. U. K. 36: 291—307, 1957. Y-organ activity induces molt.
67. Carlisle D. B., pp. 18—19. 2. Internal. Symp. Neurosekr, Springer-Verlag, Berlin, 1958, Intra-axonal transport.
Эндокринные механизмы
493
68. Carpenter М. В., de Roos R., Gen. Comp. Endocrinol. 15: 143—157, 1970. Seasonal changes in androgenic gland of crayfish Orconectes.
69. Cehovic G., C. R. Acad. Sci. 261: 1405—1408, 1965. Antigonadal action of MSH.
70. Chaet A. B., Amer. Zool. 6: 263—271, 1966. Gamete-shedding factors in star-fishes.
71. Chaet A. B., Symp. Zool. Soc. Lond. 20: 13—24, 1967. Gamete-shedding factor of starfish.
72. Chang C. Y., Witschi E., Endocrinology 61: 514—519, 1957. Facilitation of ovulation by cortisone.
73. Charniaux-Cotton H., C. R. Acad. Sci. 239: 780—782, 1954. Ovarian hormone in crustacean; androgenic gland of Crustacea.
74. Charniaux-Cotton H., Ann. Biol. 32: 371—398, 1956. Androgenic gland.
75. Charniaux-Cotton H., Ann. Sci. Nat. Zool. Biol. Animate 19: 411—559, 1957. Molting in Orchestia; androgenic gland of Crustacea.
76. Charniaux-Cotton H., Bull. Soc. Zool. France 83: 314—336, 1958. Crustacean sex differentiation.
77. Charniaux-Cotton H., C. R. Acad. Sci. 246: 2817—2819, 1958. Androgenic gland of crustaceans.
78. Charniaux-Cotton H., Kleinholz L. FL, pp. 135—198 in: The Hormones, Vol. 4, edited by G. Pincus, К. V. Thimann, and E. B. Astwood. Academic Press,. New York, 1964. Hormones in invertebrates other than insects.
79. Chase L. R., Aurbach G. D., Proc. Nat. Acad. Sci. 58: 518—525, 1967. Effect of PTH on cyclic AMP excretion.
80. Chase L. R., Aurbach G. D., Science 159: 545—547, 1968. Renal adenyl cyclases responsive to PTH and ADH.
81. Chase L. R., Fedak S. A., Aurbach G. D., Endocrinology 84: 761—768, 1969. In vitro activation of adenyl cyclase by PTH.
82. Chase M. D., Geschwind I. I., Bern H. A., Proc. Soc. Exp. Biol. Med. 94: 680—683. 1957. Prolactin synergism with sex hormones and gonadotropins. Direct influence of LTH on prostate.
83. Chavin W., J. Exp. Zool. 133: 1—46, 1956. Goldfish melanization by ACTH.
83a. Chihara C. J., Petri W. EE, Fristrom J. W., King D. S., J. Insect. Physiol. 18: 1115—1123, 1972. Ecdysone and juvenile hormone on imaginal discs in vitro.
83b. Chino EL, Gilbert L. L, Siddall J. B., Hafferl W., J. Insect Physiol. 16: 2033— 2040, 1970. Ecdysone transport in lepidopteran hemolymph.
84. Chiu K. W„ Lynn W. G., Leichner I. P„ Biol. Bull. 139: 107—114, 1970. Effect of temperature on thyroid in lizard.
85. Church N. S., Canad. J. Zool. 33: 339—369, 1955. Hormones and insect molt.
86. Clark R. B., Oceanography and Marine Biology, an Annual Review, Vol. 3, edited by H. Barnes, 1965. Endocrine activity in polychaetes.
87. Clever V., Karlson P., Exp. Cell. Res. 20: 623—626, 1960. Ecdysone activation of gene activity.
88. Colombo L., Bern EL A., Pieprzyk J., Gen. Comp. Endocrinol. 16: 74—84,. 1971. Steroid transformations by salmon corpuscle of Stannius.
89. Combescot C„ Bull. Soc. Hist Nat. Afrique Nord 45: 366—377, 1955. Hypo-physectomy and gonad and secondary sex character dependence in reptiles.
90. Congote L. F„ Sekeris С. E„ Karlson P., Exp. Cell. Res. 56: 338—346, 1969. Juvenile hormone and ecdysone on RNA synthesis.
90a. Congote L. F., Sekeris С. E., Karlson P., Z. Naturforsch. 25b: 279—284, 1970. Specific RNA synthesis in isolated fat body nuclei after ecdysone and juvenile hormone stimulation.
91. Copp D. H., pp. 377—398 in: Fish Physiology, Vol. 2, edited by W. S. Hoar and D. J. Randall. Academic Press, New York, 1969. Ultimobranchial glands.
494
Глава 21
92. Comubert G., C. R. Acad. Sci. 238: 952—953, 1954. Distinction between ovary-inhibiting hormone and molting one.
93. Comubert G., Demeusy N., Veillet A., C. R. Acad. Sci. 234: 1405—1407, 1952. Acceleration of sexual development by eyestalk removal.
94. Costolow J. D., Jr., Bookhout C. G., Biol. Bull. 113: 224—232, 1957. Nutrition and molting in barnacles.
95. Costlow J. D., Jr., Gen. Comp. Endocrinol. 3: 120—130, 1963. Eyestalk extirpation and metamorphosis of blue crab megalops.
96. Costlow J. D., Jr., pp. 209—224 in: Some Contemporary Studies in Marine Science, edited by El. Barnes. George Allen and Unwin, London, 1966. Eyestalk extirpation and larval development of the crab Sesarma.
97. Costlow J. D., Jr., Gen. Comp. Endocrinol. 7: 255—274, 1966. Eyestalk extirpation and larval development of mud crab Rhithropanopeus.
98. Courrier R., Cehovic G., C. R. Acad. Sci. 251: 832—834, 1960. Thyrotropic action of MSH.
99. Danforth С. H., Biol. Symp. 9: 67—80, 1942. Hormones and plumage changes in birds.
100. D’Angelo S. A., Traum R. E., Endocrinology 59: 593—596, 1956. Hypothalamic damage disturbs TSH secretion.
101. Davey R. G., Ran S. P., Canad. J. Zool. 46: 893—897, 1968. Neurosecretory function in nematodes.
102. Davis D. E., Science 126: 253, 1957. Effects of castration of starling male singing and aggressiveness.
103. Day M. F„ Biol. Bull. 84: 127—140, 1943. Corpus allatum hormone and dipte-ran reproduction.
104. Demeusy N., Lenel R., C. R. Soc. Biol. 148: 156—158, 1954. Ovary-inhibiting factor and molting hormone.
105. De Wied D., pp. 97—140 in: Frontiers in Neuroendocrinology, edited by W. F. Ganong and L. Martini. Oxford University Press, New York, 1969. Effects of peptid hormones on behavior.
106. Disney H. J., Marshall A. J., Proc. Zool. Soc. Lond. 127: 379—387, 1956. Cycles in equatorial birds.
106a. Doane W. W., in: Developmental Systems: Insects, edited by S. J. Counce. Academic Press, New York, 1972. Review of insect endocrinology.
107. Dodd J. M., Evennett P. J., Goddard C. R., Symp. Zool. Soc. Lond. 1: 77—103, 1960. Reproductive endocrinology of cyclostomes and elasmobranchs.
108. Domm L. V., pp. 227—327 in: Sex and Internal Secretions, edited by W. C. Young. Williams & Wilkins, Baltimore, 1939. Hormones and plumage changes in birds.
109. Drach P„ Ann. Inst. Oceanogr. Paris N. S. 19: 103—391, 1939. Crustacean molt-cycle stages. Brachyura.
110. Drach P„ Bull. Biol. France Belg. 78: 40—62, 1944. Crustacean molt-cycle stages, Natantia.
111. Dupont-Raabe M., Bull. Soc. Zool. France 76: 386—397, 1952. Increased brain secretion in adult phasmids.
112. Durand J. B., Biol. Bull. Ill: 62—76, 1956. X-organ in brachyuran.
113. Durchon M., Montreuil J., Boilly-Marer Y., C. R. Acad. Sci. 257: 1807—1808, 1963. Hormonal activities in polychaetes.
114. Eales J. G., Canad. J. Zool. 41: 811—824, 1963. Seasonal thyroid changes in the salmon.
115. Echalier G., C. R. Acad. Sci. 238: 523—525, 1954. Y-organ and crustacean molt.
116. Echalier G., C. R. Acad. Sci. 242: 2179—2180, 1956. Y-organ and crustacean molt.
117. Eckert M., Acta Soc. Zool. Biochem. 32: 34—38, 1968. Hormonal control of arachnid molt.
117a. Emmerich H., J. Insect Physiol. 16: 725—747, 1970. Binding of ecdysone to hemolymph proteins.
Эндокринные механизмы
495
117b- Emmerich Н., Z. vergl. Physiol. 68: 385—402, 1970. Binding proteins for ecdysone in Drosophila.
118. Enami M., Biol. Bull. 101: 241—258, 1951. Secretory droplets in X-organ-si-nus gland complex; cyclic changes.
119. Engelmann F., J. Insect Physiol. 1: 257—278, 1957. Ovarian stimulation of brain; inhibition of corpus allatum by brain.
120. Engelmann F„ Z. vergl. Physiol. 41: 456—470, 1959. Prothoracic glands in adult Leucophaea.
121. Engelmann F., The Physiology of Insect Reproduction. Pergamon Press, Oxford, 1970. Review of insect reproduction.
121a. Engelmann F., Arch. Biochem. Biophys. 145: 439—447, 1971. Juvenile hormone induction of female egg protein.
122. Engelmann F., Luscher M., Verhandl. Deutsch. Zool. Ges. Hamburg,, 215— 220, 1956. Nerve inhibition of corpora allata.
123. Epple A., Gen. Comp. Endocrinol. 9: 137—142, 1967. Amphiphil cells in. pancreatic islets of lower vertebrates.
124. Etkin W. N., in: Analysis of Development, edited by В. H. Willier, P. A. Weiss,, and V. Hamburger, W. B. Saunders Co., Philadelphia, 1955. Review of thyroid1 and amphibian development.
125. Etkin w., Gona A. G., J. Exp. Zool. 165: 249—258, 1967. Prolactin-thyroid* antagonism in amphibian development.
126. Etkin W. N., Root R. W., Mofskin В. P., Physiol. Zool. 13: 415—429, 1940. Thyroid and oxygen consumption in fishes.
127. Everett J. W., Endocrinology 58: 786—796, 1956. LH stimulation of ovulation.
128. Farner D. S„ pp. 198—237 in: Recent Studies in Avian Biology, edited by A. Wolfson. University of Illinois Press, 1955. Annual stimulus for bird' migration; independent of gonads or perhaps all hormones.
129. Farner D. S., pp. 717—750 in: Photoperiodism and Related Phenomena in Plants and Animals, edited by R. B. Withrow. Amer. Assoc. Adv. Sci., Washington D. C., 1959. Photoperiodic control of annual gonadal cycles in birds.
130. Faux A., Horn D. H. S., Middleton E. J., Fales H. M., Lowe M. E. Chem. Comm. 175—176, 1969. Moulting hormones of crab during ecdysis.
131. Fernlund P., Biochim. Biophys. Acta 237: 519—529, 1971. Chromactivating hormones of Pandalus borealis; light-adapting hormone.
132. Fernlund P., Josefsson L., Biochim. Biophys. Acta 158: 262—273, 1968. Chromactivating hormones of Pandalus borealis; red-concentrating hormone.
133. Ferrari W., Gessa G. L„ Vargui L., Ann. N. Y. Acad. Sci. 104: 330—343,. 1963. Behavioral effects of ACTH and MSH.
134. Fielder D. R., Rao K. R., Fingerman M., Marine Biol. 9: 219—223, 1971„ Control of distal retinal pigment migration in Uca.
135. Fingerman M., Amer. Zool. 6: 169—179, 1966. Neurosecretory control of pigmentary effectors in crustaceans.
136. Fingerman M., J. Interdiscipl. Cycle Res. 1: 115—121, 1970. Circadian rhythm . of distal retinal pigment migration in Uca.
137. Fingerman M., J. Cell. Comp. Physiol. 50: 357—370, 1957. Evidence for light-and dark-adapting hormones for retinal pigments of crayfish.
138. Fingerman M., Er as now R. A., Fingerman S. W., Physiol. Zool. 44: 119—128, 1971. Retinal pigment light-adapting hormones in Palaemonetes.
139. Fingerman M., Lowe M. E., J. Cell. Comp. Physiol. 50: 371—380, 1957. Daily rhythms of crayfish retinal pigment.
140. Fingerman M., Lowe M. E., Sundararaj B. L, Amer. Midi. Nat. 62: 167—173, 1959. Electrophoretic separation of dark- and light-adapting retinal pigment hormones.
141. Fingerman M., Lowe M. E., Sundararaj B. L, Biol. Bull. 116: 30—36, 1959. Dark- and light-adapting hormones of Palaemonetes retinal pigment.
142. Fingerman M., Mobberly W. C., Jr., Biol. Bull. 118: 393—406, 1960. Hormones controlling the distal retinal pigment of Palaemonetes.
496
Глава 21
143. Foa Р. Р., Galansino G., Pozza G., Rec. Prog. Hormone Res. 13: 473—510, 1957. Properties and role of glucagon.
144. Foglia V. G., Wagner E. M., de Barros M., Marques M., C. R. Soc. Biol. 149: 1660—1661, 1955. Pancreatectomy and hyperglycemia and glucosuria in tortoises.
145. Fontaine M., Mem. Soc. Endocrinol. 5: 69—81, 1956. Thyroxin in osmoregulation in poikilotherms.
146. Fontaine M., Baraduc M. M., Hately J., C. R. Soc. Biol. 147: 214—216, 1953. Thyroid in liver glycogen control in poikilotherms.
147. Fontaine M., Fontaine У. A., J. Physiol. Paris 49: 169—173, 1957. Species differences in TSH.
148. Fraenkel G., Proc. Roy. Soc. Lond В 118: 1—12, 1935. Hormonal control of dipteran development.
149. Fraenkel G., Hsiao C., J. Insect Physiol. 11: 516—556, 1965. Bursicon in flies.
150. Fridberg G., Bern. H. A., Biol. Rev. 43: 175—199, 1968. The urophysis of fishes.
151. Frieden E., Just J. J., pp. 1—52 in: Biochemical Actions of Hormones, Vol. 1, edited by G. Litwack. Academic Press, New York, 1970. Hormonal responses in amphibian metamorphosis.
152. Friesen H., Endocrinology 75: 692—697, 1964. Hypocalcemic effect of MSH.
152a. Fristrom J. W., Raikow R., Petri W., Stewart D., in: Problems in Biology: RNA in Development, edited by E. W. Hanly. University of Utah Press, Salt Lake City, 1969. Ecdysone and juvenile hormone on imaginal discs.
153. Fukuda S„ Ann. Zool. Japan 20: 9—13, 1941. Prothoracic glands and lepi-dopteran metamorphosis.
154. Fukuda S., Proc. Japan. Acad. 27: 272—677, 1951. Diapause hormone from subesophageal ganglion.
155. Gabe M.t C. R. Acad. Sci. 237: 1111—1113, 1953. Y-organ and molt in crustaceans.
156. Gabe M., Ann. Sci. Nat. Zool. Biol. Animale 18: 145—152, 1956. Y-organ and molt in crustaceans.
157. Gabe M., Arch. Anat. Mier. 44: 351—383, 1955. Neurosecretion in arachnids.
158. Gabe M., Neurosecretion, Pergamon Press, New York, 1966.
159. Gamong W. F„ Gold N. I., Name D. M., Fed. Proc. 14: 54, 1955. Absence of corticoids in stress after lesions in median eminence.
160. Ganong W. F., Hume D. M., Endocrinology 55: 474—483, 1954. Basal level of ACTH secretion without hypothalamus.
161. Gersch M., Sturzebecher J., J. Insect Physiol. 14: 87—96, 1968. Protein nature of brain hormone.
162. Geschwind I. I., pp. 421—443 in: Comparative Endocrinology, edited by A. Gorbman, John Wiley & Sons, New York, 1959. Prolactin differences between ox and sheep.
163. Geschwind I. I., Endocrinology 79: 1165—1167, 1966. Action of MSH on mouse hair color.
164. Geschwind I. I., pp. 180—189 in: Progress in Comparative Endocrinology (Gen. Comp. Endocrinol. Suppl. 2), edited by M. R. N. Prasad, 1969. Comparative biochemistry of gonadotropins.
165. Geschwind I. I., Li С. H., Barnafi L., J. Amer. Chem. Soc. 78: 4494—4495, 1956. Structure of |3-MSH. Chemistry of MSH.
166. Gilbert L. I., pp. 67—134 in: The Hormones, Vol. 4, edited by G. Pincus, К. V. Thimann, and E. B. Astwood. Academic Press, New York, 1964. Hormones regulating insect growth.
167. Gilbert L. I., Proc. Third Int. Cong. Endocrinol., pp. 340—346. Chemistry of ecdysones.
168. Gilbert L. I., Adv. Insect Physiol. 4: 69—211, 1967. Lipids and sterols in insects.
I
i
Эндокринные механизмы
497
168а. Gilbert L. I., King D. S., in: Physiology of Insecta, 2nd edition, edited by M. Rockstein. Academic Press, New York, 1973. Review of insect endocrinology-
169. Gorbman A., Proc. Soc. Exp. Biol. Med. 45: 772—773, 1940. Thyrotropic principle in fish pituitary.
170. Gorbman A., Physiol. Rev. 35: 336—346, 1955. Bound I in dermal glands of Hemichorda; bound I with exoskeletal scleroproteins.
171. Gorbman A., pp. 266—282 in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Problems in comparative morphology and physiology of vertebrate thyroid gland.
172. Corbman A., Bern H. A., Textbook of Comparative Endocrinology. John Wiley & Sons, New York, 1962, 468 pp.
173. Gorbman A., Clements M., O’Brien R., J. Exp. Zool. 127: 75—89, 1954. Protein binding of I in stolonic canals, Botryllus.
ПЗз. Gorell T. A., Gilbert L. I., Siddall J. B., Proc. Nat. Acad. Sci. 69: 812—815, 1972. Binding proteins in a crustacean for an ecdysone metabolite.
174. Gorell T. A., Gilbert L. I., Tash J., Insect. Biochem. 2: 94—106, 1972. Uptaka of ecdysone by insect tissues.
175. Gorski J., Toft D., Shyamala G., Smith D., Notides A., Rec. Prog. Hormona Res. 24: 45—72, 1969. Extrogen interaction with uterine receptors.
176. Grant W. C., Grant J. A., Biol. Bull. 114: 1—9, 1958. Prolactin in amphibians; responsibility for water drive in red eft stage of newt Triturus.
177. Grossman M. I., Vitam. Horm. 16: 179—263, 1958. Secretin.
178. Grossman M. I., Gastrin. University of California Press, Berkeley, 1966s 337 pp.
179. Gudernatsch J. F., Arch. Entwick-Mech. 35: 457—483, 1912. Thyroid and amphibian metamorphosis.
180. Guyselman J. B., Biol. Bull. 104: 115—137, 1953. Molting in Uca.
181. Habener J. F., Singer F. R., Deftos L. J., Neer R. M., Potts J. T., Jr., Nature 232: 91—92, 1971. Explanation for the high potency of salmon calcitonin.
182. Hadorn E., Neel J., Arch. Entwick-Mech. 138: 281—304, 1938. Larval ring gland and dipteran development.
183. Hagadorn I. R., Amer. Zool. 6: 251—261, 1966. Neurosecretion in leech reproduction.
184. Hampshire F„ Horn D. H. S., Chem. Comm., 37—38, 1966. Structure of crustecdysone, a crustacean moulting hormone.
185. Handler J. S., Butcher R. W., Sutherland E. W., Orloff J., J. Biol. Chem. 240: 4524—4526, 1965. ADH increases toad bladder cyclic AMP.
186. Hanstrom B., Kungl. svensk. Vetensk. Handl. 16: 1—99, 1937. X-organs and sinus glands.
187. Hanstrom B., Hormones in Invertebrates. Oxford University Press, 1939, 198 p.
188. Hanstrom B., Colston Papers, Butterworths Sci. Publ. London 8: 23—27, 1956. Comparative neurosecretion.
189. Hara J., Ann. Zool. Japan 25: 411—414, 1952. Hormonal regulation of crustacean heart.
190. Harris J. I., Roos P., Nature 178: 90, 1956. Amino acid sequence in pig |3-MSH. Chemistry and structure of MSH.
191. Harris J. I., Sanger F., Naughton M. A., Arch. Biochem. 65: 427—438, 1956. Structure of insulin.
192. Hasegawa K-, Proc. Japan. Acad. 27: 667—671, 1951, Diapause hormone from subesophageal ganglion.
193. Hasegawa K., Nature 179: 1300—1301, 1957. Extract of diapause hormone.
194. Hasler A. D., Meyer R. K., Field H. M„ Endocrinology 25: 978—983, 1939. Hypophyseal gonadotropins in fishes.
195. Hauenschild C., Zool. Anz. Suppl. 27: 111—120, 1964. Neuroendocrine roles in polychaetes.
32—2067
498
Глава 21
196. Henry S., van Dyke H. B., J. Endocrinol. 16: 310—325, 1958. Immunological differences in ICSH.
197. Herlant-Meewis H., in: Advances in Morphogenesis, Vol. 4, edited by M. Abercrombie and J. Brachet, Academic Press, New York, 1964. Neurosecretory roles in oligochaetes.
198. Hertault J., C. R. Acad. Sci. 272: 1981—1983, 1971. Neuroendocrine system of pseudoscorpions.
199. Hickman С. P., Jr., Canad. J. Zool. 37: 997—1060, 1959. Greater thyroid activity in euryhaline fish in more saline medium.
200. Highnam К. C., Quart. J. Mier. Sci. 99: 73—88, 1958. Neurosecretion by corpora cardiaca.
201. Highnam К. C., Hill L., The Comparative Endocrinology of the Invertebrates. American Elsevier Publ. Co., New York, 1969, 270 p.
202. Hisaw F. L., in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Endocrine adaptation of the mammalian» estrous cycle and gestation.
203. Hoar W. S., in: The Physiology of Fishes, edited by M. Brown. Academic Press, New York, 1957. Thyroid and fish development.
204. Hodge M. G., Chapman G. B., J. Biophys. Biochem. Cytol. 4: 571—574, 1958. Vesicles in sinus gland.
205. Hopper A. F., J. Exp. Zool. 119: 105—109, 1952. Low thyroid prolongation of sex immaturity; thyroxin treatment and early sex development — Lebistes.
206. Horton E. W., Prostaglandins. Monographs on Endocrinology, No. 7. Sprin-ger-Verlag, New York, 1972, 197 p. Up-to-date survey of prostaglandins.
207. Host P., Auk 59: 388—403, 1942. Light periods and seasonal plumage changes in the ptarmigan.
208. Houssay B. A., in: Comparative Endocrinology, edited by A. Gorbman. John-Wiley & Sons, New York, 1959. Comparative physiology of the endocrine pancreas.
209. Howard K. S., Shepherd R. G., Eigner E. A., Davies D. S., Bell P. H., J. Amer. Chem. Soc. 77: 3419—1955. Three corticotropins.
210. Howie D. I. D„ Gen. Comp. Endocrinol. 6: 347—361, 1966. Endocrine factors in Arenicola reproduction.
211. Huber R., Hoppe W., Chem. Ber. 98: 2403—2424, 1965. Structure of ecdysone.
212. Ichikawa M., Nishutsutsuji J., Ann. Zool. Japan. 24: 205—211, 1951. Braiff and imaginal development in Lepidoptera.
213. Idler D. R., Truscott B., Steroids 9: 457—478, 1967. Discovery of 1-ct-hydro-corticosterone in ray interrenal.
213a. Ilan J., Ilan J., Patel N., J. Biol. Chem. 245: 1275—1281, 1970. Juvenile hormone action at the translational level.
214. Ishizaki I., Ichikawa M., Biol. Bull. 133: 355—368, 1967. Protein nature of brain hormone.
215. Iwanoff P. P., Mestscherskaja K. A., Zool. Jahrb. Abt. allg. Zool. PhysioL 55: 281—348, 1935. Gonadal hormone in insect reproduction.
216. Johannsen A. S., Nature 181: 198—199, 1958. Inhibition of corpus allatum by brain.
217. Joly R., Bull. Biol. Lance Belg. 100: 379—480, 1966. Molt regulation iff myriapods.
218. Jones В. M., J. Exp. Biol. 33: 174—185, 1956. Brain and prothoracic gland in embryonic molts.
219. Jones I. C., The Adrenal Cortex. Cambridge University Press, 1957. Role of adrenal cortex; annual rhythm.
220. Jones I. C., in: The Neurohypophysis, edited by H. Heller. Butterworth & Co-, London, 1966. Adrenalectomy in winter and summer frogs.
221. loose J., Arch. Neerl. Zool. 16: 1—103, 1964. Neurosecretory activities iff Lymnaea.
222. Kamemoto F. I., Kato K. N., Tucker L. E., Amer. Zool. 6: 213—219, 196& Neurosecretion and salt and water balance in the Annelida and Crustacea-.
Эндокринные механизмы
499
223. Kato К. N>, Kamemoto F. I., Comp. Biochem. Physiol. 28: 665—674, 1969. Neuroendocrines in osmoregulation in grapsid crab Metopograpsus.
224. Keller R., Verh. Deutsch. Ges. Innsbruck, 628—835, 1968. Diabetogenic hormone of Orconectes.
225. Keller R., Z. vergl. Physiol. 63: 137—145, 1969. Species specificity of a crustacean hormone.
225a. King D. S., Gen. Comp. Endocrinol. Suppl. 3: 221—227, 1972. Metabolism of ecdysone.
226. Kitay J. I., Altschule M. D., The Pineal Gland. Harvard University Press, Cambridge, 1954, 280 p.
227. Kleinholz L. H., Biol. Bull. 70: 159—184, 1936. Eyestalk hormone and retinal pigment migration in crustaceans.
228. Kleinholz L. H., Biol. Bull. 75: 510—532, 1938. Hormonal control of crustacean retinal pigment movements.
229. Kleinholz L. H„ Biol. Bull. 99: 454—468, 1950. Hormonal regulation of blood sugar in crustaceans.
230. Kleinholz L. H., Biol. Bull. 109: 362, 1955. Reflecting pigment of crustacean eye.
231. Kleinholz L. H., Amer. Zool. 6: 161—167, 1966. Separation and purification of crustacean eyestalk hormones.
232. Kleinholz L. H„ Gen. Comp. Endocrinol. 15: 578—588, 1970. Separation and purification of crustacean neurosecretory pigmentary-effector hormones.
233. Kleinholz L. H., Bourquin E., Proc. Nat. Acad. Sci. 27: 145—149, 1941. Eyestalks and molting in crabs.
234. Kleinholz L. H., Kimball F„ McGarvey M., Gen. Comp. Endocrinol. 8: 75—81, 1967. Separation of diabetogenic hormone from the crustacean eyestalk.
235. Knowles F. G. W., Proc. Roy. Soc. Lond. В 141: 248—267, 1953. Postcom-m-issural organ.
236. Knowles F. G. W., in: Comparative Endocrinology, edited by A. Gorbman, John Wiley & Sons, New York, 1959. Secretory granules.
237. Knowles F. G. W„ Carlisle D. B„ Biol. Rev. 31: 396—473, 1956. Terms MTGX-, SPX-organ. Crustacean chromatophorotropin polypeptides. Crustacean endocrinology, review.
238. Kollros J. J., pp. 340—350 in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Complex reacting system of amphibian development to thyroid activity.
239. Kopec S„ Biol. Bull. 42: 324—342, 1922. Endocrine control of insect development.
240. Korner A., Proc. Roy. Soc. Lond. В 176: 287—290, 1970. Hormonal control of protein synthesis.
241. Krayer 0., Astwood E. B., Waud D. R., Alper M. H., Proc. Nat. Acad. Sci. 47: 1227—1236, 1961. Effect of ACJH and MSH on heart rate.
242. Krishnakumaran A., Schneiderman H. A., Biol. Bull. 139: 520—538, 1970. Control of molting in mandibulate and chelicerate arthropods by ecdysones.
243. Kroeger H., in Metamorphosis: A Problem in Developmental Biology, edited by W. Etkin and L. I. Gilbert. Appleton-Century-Crofts, New York, 1968. Control of gene activity by ions.
244. Kupferman I., J. Neurophysiol. 33: 877—881, 1970. Neuroendocrine regulation of egg-laying in Aplysia.
245. Lamotte M„ Prurn P., C. R. Soc. Biol. 151: 1187—1191, 1957. Endocrine phy-siology of a viviparous amphibian.
246. Lamotte M., Rey P., C. R. Acad. Sci. 238: 393—395, 1954. Corpora lutea in amphibian.
247. Lamotte M., Rey P., C. R. Soc. Biol. 151: 1191—1194, 1957. Corpora lutea in amphibian.
248. Lamotte M., Rey P., Vilter V., C. R. Soc. Biol. 150: 393—396, 1956. Endocrine physiology of viviparous amphibian.
249. Lane N. J., Quart. J. Mier. Sci. 103: 211—223, 1962. Neurosecretory cells in snail optic tentacles.
32*
500
Глава 21
250. Lane N. J. Quart. J. Mier. Sci. 105: 31—34, 1964. Neurosecretory cells in Helix.
251. Lederis K„ Gen. Comp. Endocrinol. 14: 417—426, 1970. Bioassay of the teleost urophysis.
252. Lederis K, Gen. Comp. Endocrinol. 14: 427—437, 1970. Characterization of the teleost urophysis principle.
253. Legendre R., Ann. Sci. Nat. Zool. 1: 339—474, 1959. Neurosecretory cells in arachnids.
254. Lees A. D., pp. 585—600 in: Photoperiodism and Related Phenomena in Plants and Animals, edited by R. B. Withrow. Amer. Assoc. Adv. Sci., Washington, D. C., 1953. Photoperiodism in insects.
255. Lender T., Klein N., C. R. Acad. Sci. 253: 331—333, 1961. Neurosecretion in Polycelis regeneration.
256. Lentz T. L., Science 150: 633—635, -1965. Neurosecretion and Hydra growth.
257. Lerner A. B., Lee T. H., Amer. Chem. Soc. 77: 1066—1067, 1955. Structure of MSH.
258. Lerner A. B., McGuire J. S., Nature 189: 176—179, 1961. Effect of MSH on human skin color.
259. Lever J., Jansen J., De Vlieger T. A., Proc. K. ned. Akad. Wet. C. 64: 532—542, 1961. Neurosecretion in Lymnaea water regulation.
260. Lewis С. P., ed. 5-Hydroxytryptamine. Pergamon Press, London, 1958. Serotonin (5-hydroxytryptamine) as a neurohumoral agent.
261. Lezzi M., Gilbert L. L, Proc. Nat. Acad. Sci. 64: 498—503, 1969. Gene activation by ecdysone and juvenile hormone.
262. Lezzi M., Gilbert L. L, J. Cell. Sci. 6: 615—628, 1970. Effects of ions on gene activity in isolated chromosomes.
263. Li G. H., in: Symposium on Protein Structure, edited by A. Neuberger. John Wiley & Sons, New York, 1958. Three classes of growth hormone; differences in growth hormones and in sheep and ox prolactin.
264. Li С. H., Persp. Biol. Med. 11: 498—521, 1968. Current concepts of pituitary hormone biochemistry.
265. Li С. H. et al., Nature 176: 687—689, 1955. Three corticotropins.
266. Liao T. H., Pierce J. G., J. Biol. Chem. 245: 3275, 1970. Structure of LH.
267. Lofts B., pp. 239—304 in: Perspectives in Endocrinology, Hormones in the Lives of Lower Vertebrates, edited by E. J. W. Barrington and C. Barker Jorgenson. Academic Press, New York, 1968. Patterns of testicular activity.
268. Lowe M. E„ Horn О. H. S., Galbraith M.. Experientia 24: 518—519, 1968. The role of crustecdysone in molting crayfish.
269. Lubet P., Ann. Sci. Nat. Zool. 18: 175—183, 1956. Neurosecretion and reproduction in Mytilus.
270. Luscher M., Engelmann F., Rev. Suisse Zool. 62: 649—657, 1955. Corpora allata function in Leucophaea.
271. Lyman С. P., Bull. Mus. Comp. Zool. 93: 391—461, 1943. Control of seasonal changes in coat color of the varying hare, Lepus.
272. Lynn W. G., Wachowski H. E., Quart. Rev. Biol. 26: 123—168, 1951. Growth and differentiation effects, thyroid.
273. Maher M. J., Levedahl В. H., J. Exp. Zool. 140: 169—189, 1959. Thyroid effect on lizard oxidative metabolism.
274. Marshall A. J., Mem. Soc. Endocrinol. 4: 75—93, 1955. Hypothalamic control of pituitary in bird.
275. Marshall F. H. A., Biol. Rev. 17: 68—90, 1942. Factors in sex periodicity.
276. Martini L., Ganong W. F., eds. Neuroendocrinology. Academic Press, New York, 1967, 1551 pp.
277. Matsumoto K., Biol. J. Okayama Univ. 4: 103—176, 1958. X-organ in crabs.
278. Matthews L. H., Mem. Soc. Endocrinol. 4: 129—148, 1955. Reptile corpus luteum.
279. Maynard D. M., Biol. Bull. 121: 316—326, 1961. Thoracic neurosecretory structures in Brachyura.
Эндокринные механизмы
501
280. McLennan Н., Synaptic Transmission, 2nd edition. W. В. Saunders Co., Phi ladelphia, 1970, 134 pp.
281. Megusar F., Arch. Entwick-Mech. 33: 462—665, 1912. Eyestalks and molting in Crustacea.
282. Mellish С. H., Meyer R. R., Anat. Rec. 69: 179—189, 1937. Gonadotropic activity in reptiles.
283. Meyer A. S., Hanzmann E., Schneiderman H. A., Gilbert L. I., Boyette M., Arch. Biochem. Biophys. 137: 190—213, 1970. Structure of the two juvenile hormones.
284. Mialhe P„ J. Physiol. Paris 47: 248—250, 1955. Ducks and pancreas.
285. Miller A. H., Condor 56: 13—20, 1954. Refractory period in birds.
286. Miller M. R., Wurster D. H., in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Sensitivity of amphibians to insulin; insensitivity to glucagon; absence of «-cells.
287. Miller M. R., Wurster D. H., Endocrinology 63: 191—200, 1958. Sensitivity of reptiles to glucagon and abundance of «-cells.
288. Molony P. J., Goldsmith L., Canad. J. Biochem. Physiol. 35: 79—92, 1957. Antibodies to administered insulin.
289. Morel F., Aldosterone. J. & A. Churchill, London, 1958.
290. Mueller G. C., Cancer Res. 17: 490—506, 1957. Discussion of steroid hormone mechanism of action.
291. Muhlbach O., Boot L. M., Ann. Endocrinol. 17: 338—343, 1956. Series of pseudopregnancies in mice with extra pituitary subcutaneous-LTH production.
292. Munson P. I., et al., Rec. Progr. Hormone Res. 24: 589—637, 1968. Thyrocalcitonin.
293. Mutt V., Jorpes J. E., Rec. Progr. Hormone Res. 23: 483—503, 1967. Gastrointestinal hormone biochemistry.
294. Nair R. M. G., Rastin A. J., Schally A. V., Biochem. Biophys. Res. Comm. 43: 1376—1381, 1971. MSH release-inhibiting hormone.
295. Naisse J., Arch. Biol. 77: 139—201, 1966. Sex hormone in an insect.
296. Naisse J., Gen. Comp. Endocrinol. 7: 105—110, 1966. Sex hormones in an insect.
297. Nayar R. R., Curr. Sci.*22: 149, 1953. Correlation of insect brain secretion with oviposition.
298. Nelson W. O., Mecanisme Physiologique de la Secretion Lactee. Paris, Publ. de CNRS, 1951. LTH affect mammary tissue.
299. Nickerson M., Ann. N. Y. Acad. Sci. 139: 571—579, 1967. New developments in adrenergic blocking drugs.
300. Nickerson M., Goodman L. S., Fed. Proc. 7: 397—409, 1948. Adrenergic blockage by dibenamine.
301. Nikitovitch-Winer M., Everett J. W., Endocrinology 63: 916—930, 1958. Cycles in rats only when pituitary implants under median eminence.
302. Novales R. R., Ann. N. Y. Acad. Sci. 100: 1035—1047, 1963. Melanophore responses to MSH, melatonin, and epinephrine.
303. Novales R. R., in: Handbook of Physiology, Sec. 7, Endocrinology: The Hypothalamus, edited by K- Knobil. Amer. Physiol. Soc., Washington, D. C., 1971. Physiological actions of MSH.
304. Novales R. R., Novales B. J., Progr. Brain. Res. 10: 507—519, 1965. Melatonin antagonisms on melanophores.
305. Oksche A., Laws D., Rememoto F. L., Farner D. S., Z. Zellforsch. 51: 1—42, 1959. Secretory granules in nerve loops of median eminence of white-crowned sparrows under 8 hours’light.
306. Orloff J., Handler J. A., Biochem. Biophys. Res. Comm. 5: 63—66, 1961. ADH-like effects of cyclic AMP on toad bladder.
307. Oshima R., Gorbman A., Gen. Comp. Endocrinol. 7: 398—409, 1966. Effect _ of T4 on electrical activity in goldfish brain.
308. Oshima R., Gorbman A., Gen. Comp. Endocrin. 7: 482—491, 1966. Further information on thyroxine and goldfish brain.
502
Глава 21
309. Otsu Т., Embryologia 8: 1—20, 1963. Bihormonal control of sexual cycle in Potamon.
310. Page I. H., Physiol. Rev. 38: 277—335, 1958. Serotonin as a neurohumoral agent.
-311. Palay S. L., Anat. Rec. 121: 348, 1955. Vesicles in nerve fibers: the secre-tion.
312. Panigel M., Ann. Sci. Nat. Zool. 18: 569—668, 1956. Reptile corpus luteum; presence in all reptiles, whether oviparous or viviparous.
313. Panouse J., C. R. Acad. Sci. 218: 293—294, 1944. Sinus glands and ovarian growth in crustaceans.
• 314. Panouse J., Ann. Inst. Oceanogr. 23: 65—147, 1946. Sinus glands and ovarian growth in crustaceans.
315. Pardoe A. Y., Weatherall M., J. Physiol. 127: 201—212, 1955. Vasopressin and oxytocin from granules of neurohypophysis.
• 316. Paris О. H., Jr., Jenner С. E., pp. 601—624 in: Photoperiodism and Related Phenomena in Plants and Animals, edited by R. B. Withrow. Amer. Assoc. Adv. Sci., Washington, D. C., 1959. Photoperiodic control of diapause in insects.
• 317. Passano L. M., Physiol. Comp. Oecol. 3: 155—189, 1953. X-organ-sinus gland complex.
• 318. Patel N., Madhaven K„ J. Insect Physiol. 15: 2141—2150, 1969. Juvenile hormone and ecdysone on RNA synthesis.
319. Pelluet D., Lane N. J.. Canad. J. Zool. 39: 789—805, 1961. Neurosecretory roles in slugs.
• 320. Pesetsky I., Kollros J. J., Exp. Cell. Res. 11: 477—482, 1956. Differential CNS growth with thyroid.
321. Pfeiffer I. W., J. Exp. Zool. 82: 439—461, 1939. Corpora allata and reproduction in Orthoptera. Hormonal control of development.
322. Pfeiffer I. W., J. Exp. Zool. 99: 183—233, 1945. Corpus allatum and general metabolism in insects.
323. Pflugfelder O., Z. wiss. Zool. 153: 108—135, 1940. Hormones in Dixippus development and reproduction.
324. Pickford G. E., Bull. Bingham Oceanogr. Coll. 14: 46—68, 1953. Control of spawning in Fundulus; fish response to beef growth hormone.
325. Pickford G. E., Endocrinology 55: 274—287, 1954, Fish growth to fish growth hormone.
-326. Pickford G. E., pp. 404—420 in: Comparative Endocrinology, edited by A. Gorb-man. John Wiley & Sons, New York, 1959. Prolactin cooperation with MSH in fish pigmentation.
327. Pickford G. E., Atz J. W., The Physiology of the Pituitary Gland of Fishes. New York Zoological Society, 1957.
328. Pierce J. G., Endocrinology 89: 1331—1344, 1971. Structure of TSH.
329. Porter G. A., Bogoroch R., Edelman I. S., Proc. Nat. Acad. Sci. 52: 1326— 1333, 1964. Aldosterone action on sodium transport.
330. Possompes B., Arch. Zool. Exp. Gen. 89: 203—364, 1953. Hormones and Cal-liphora metamorphosis.
• 331. Potter D. D., pp. 113—118 in: 2. Int. Symp. Neurosekr., Springer-Verlag, Berlin, 1958. Secretory granules.
332. Potter D. D., Lowenstein W. R., Amer. J. Physiol. 183: 652, 1955. Conduction of impulses by axons of pituitary stalk.
333. Potts J. T., Jr., Aurbach G. D„ pp. 53—67 in: The Parathyroid Glands, edited by P. J. Gaillard, R. V. Talmage, and A. M. Budy. University of Chicago Press, Chicago, 1965. Chemistry of PTH.
334. Prabhu V. R. K„ Z. Zellforseh. 54: 717—733, 1961. Neurosecretory organs of myriapods.
335. Pritchard A. W., Gorbman A., Biol. Bull. 119: 109—119, 1960. Thyroid hormone and dogfish oxygen use.
.336. Pyle R. W., Biol. Bull. 85: 87—102, 1943. Cycles in histological picture in sinus gland and molting in crustaceans.
Эндокринные механизмы бОЭ1
337, Rahm U. Н., Rev. Suisse Zool. 59: 173—237, 1952. Hormones and Sialis development.
337a. Raikow R. B., Fristrom J. W„ J. Insect Physiol. 17: 1599—1614, 1971. Effect of ecdysone on RNA metabolism in imaginal discs.
338- Ralph C. L„ Frapps R. M., Anat. Rec. 130: 360—361, 1958. Gonadal controF via hypothalamus.
339. Ralph C. L., Grinwich D. L., Hall P. F., J. Exp. Zool. 166: 289—294, 1967. Hormones and weaver bird pigmentation.
340. Ralph C. L., Sampath S., Gen. Comp. Endocrinol. 7: 370—374, 1966. Inhibition of MSH release.
341. Rao K. R., Experientia 21: 593—594, 1965. Moult-inhibiting hormone of the-crustacean eyestalk.
342. Rao K. R., Fingerman M., Hays C., Amer. Zool. 11: 644, 1971. a-Ecdysone and 20-hydroxyecdysone on regeneration and molting in Gca.
343. Reiter R. J., Sorrentino S., Jr., Amer. Zool. 10: 247—258, 1970. Pineal gland* and reproduction.
344. Riggs T. R., pp. 157—208 In: Biochemical Actions of Hormones, Vol. 1, edited by G. Litwack. Academic Press, New York, 1970. Hormones and transport" across cell membranes.
345. Roberts J. L., Physiol. Zool. 30: 232—242, 1957. Nutrition and crab molting.
346. Robinson G. A., Butcher R. W., Sutherland E. W., Ann. Rev. Biochem. 37r 149—174, 1968. Cyclic AMP.
347. Robinson G. A., Butcher R. W., Sutherland E. W., Cyclic AMP. Academic Press, New York, 1971, 531 p. Thorough treatment of all aspects.
348. Roller H., Dahm К. H., Sweeley С. C., Trost В. M. Angew. Chem. 79: 190— 191, 1967. Structure of juvenile hormone.
349. Rondell P. A., Wright P. A., Physiol. Zool. 31: 236—243, 1958. Metabolic studies of frog ovulation.
350. Rowan W., Biol. Rev. 13: 374—402, 1938. Light and reproductive cycles.
351. Rugh R., Biol. Bull. 66: 22—29, 1934. Gonadotropic activity of hypophysis in» amphibians.
352. Rugh R„ J. Exp. Zool. 71: 149—162, 1935. Gonadotropic activity of hypophysis in amphibians.
353. Rugh R., J. Exp. Zool. 71: 163—193, 1935. Gonadotropic activity of hypophysis in amphibians.
354. Rust С. C., Gen. Comp. Endocrinol. 5: 222—231, 1965. Hormonal control oF weasel pelage.
355. Rust С. C„ Meyer R. K., Gen. Comp. Endocrinol. 11: 548—551, 1968. Pituitary-grafts and weasel hair color.
356. Sandeen M. I., Brown F. A. Jr., Physiol. Zool. 25: 233, 1952. Retinal pigment response of Palaemonetes to illumination.
357. Sandor T., Gen. Comp. Endocrinol. Suppl. 2: 284—298, 1969. Survey of steroids-throughout the vertebrates.
358. Sawyer W. H., Amer. J. Physiol. 164: 44—48, 1951. Posterior pituitary and* frog skin water permeability.
359. Sawyer W. H., Endocrinology 66: 112—120, 1960. Neurohypophyseal hormones-and water regulation in Rana; neurohypophyseal hormones and frog bladder permeability.
360. Sawyer W. H., Munsick R. A., van Dyke H. B., Circulation 21: 1027—1037, I960. Distribution of posterior lobe hormones.
361. Scharrer B., J. Comp. Neurol. 74: 109—130, 1941. The nemertean cerebraF organ.
362. Scharrer B., Biol. Bull. 18: 96—104, 1941. Neurosecretory cells in Limulus.
363. Scharrer B., Endocrinology 38: 35—45, 1946. Hormonal control of orthopteran* development.
364. Scharrer B., Endocrinology 38: 46—55, 1946. Corpus allatum hormone and? ovarian growth in Orthoptera.
365. Scharrer B., Biol. Bull. 102: 261—272, 1952. Intra-axonal transport in insects; role of corpus cardiacum; nerve inhibition of corpus allatum.
-504
Глава 21
366. Scharrer Е., Scharrer В., Rec. Progr. Hormone Res. 10: 183—240, 1954. Neurosecretion; neuroendocrinology.
367. Scharrer E„ Scharrer B., Neuroendocrinology. Columbia University Press, New York, 1963, 287 p.
368. Schayer R. W., Ivy A. C., Amer. J. Physiol. 189: 369—372, 1957. Histamine as a hormone.
369. Scheer В. T., Scheer M. A. R., Pubbl. Staz. Zool. Napoli 25: 419—426, 1954. Crustacean molt-cycle stages, Natantia.
370. Scheffel H., Zool. Jahrb. 71: 359—370, 1965. Hormone in myriapod molting.
371. Schell-Frederick E., Dumont J. E., pp. 415—463 in: Biochemical Actions of Hormones, Vol. 1, edited by G. Litwack. Academic Press, New York, 1970. Alechanism of action of TSH.
372. Schneiderman H. A., Gilbert L. I., Biol. Bull. 115: 530—535, 1958. Widespread distribution among animals of juvenile hormone activity.
373. Schneiderman H. A., Gilbert L. I., pp. 157—187 in: Cell, Organism and Milieu, edited by D. Rudnick Ronald Press Co., New York, 1959. Substances with juvenile hormone activity among animals.
374. Scudamore H. EL, Trans. Illinois Acad. Sci. 34: 238—240, 1941. Hormonal regulation of crustacean heart rate.
375. Scudamore H. H., Physiol. Zool. 20: 187—208, 1947. Sinus gland and oxygen consumption in crayfishes; hormonal control of gastrolith formation in cray-fishes
376. Segal S. J., Science 126: 1242—1243, 1957. Gonadotropic control of nuptial plumage in African finches.
377. Shaaya E., Bodenstein D., Proc. Nat. Acad. Sci. 59: 1223—1230, 1968. Juvenile hormone on collateral glands.
378. Shappirio D. G., Williams С. M., Proc. Roy. Soc. Lond. В 147: 233—246, 1957. Ecdysone induction of increase in mitochondria and endoplasmic reticulum of epithelial cells.
379. Shepherd R. G. et al., J. Amer. Chem. Soc. 78: 5067—5076, 1956. Three corti> costeroids.
380. Sherwood L. M., Handwerger L. M., McLaurin W. D., banner M., Nature New Biol. 233: 59—61, 1971. Structure of human placental lactogen.
38] . Shirai H., Kanatanui H., Taguchi S., Science 175: 1366—1368, 1972. Action of the starfish gonad-stimulating factor.
382. Simpson L., Bern H. A., Nishioka R. S., Amer. Zool. 6: 123—138, 1966. Neurosecretion in gastropods.
383. Slama K., Ann. Rev. Biochem. 40: 1079—1102, 1971. Review of juvenile hormone analogues.
384. Smellie R. M. S., ed., Biochemical Society Symposium 32, 1971. The Biochemical Society, London, 1971, 178 p. The biochemistry of steroid hormone action.
385. Smith D. C., Matthews S. A., Amer. J. Physiol. 153: 215—221, 1948. Thyroid and oxygen consumption in fishes.
386. Smith D. C. W., Mem. Soc. Endocrinol. 5: 83—98, 1956. Thyroid in poikilotherm osmoregulation.
387. Smith P. E., Anat. Rec. 11: 57—64, 1916. Hypophysis in amphibian metamorphosis.
388. Smith R. L, Biol. Bull. 79: 145—152, 1940. Eyestalks and molting in young crayfishes.
389. Srivastava U. S., Gilbert L. I., Science 161: 61—32, 1968. Juvenile hormone
390. Sroka P., Gilbert L. I., J. Insect Physiol. 17: 2409—2420, 1971. Juvenile hormone and post-emergence egg maturation.
391. Steiner D. F. et al., Rec. Progr. Hormone Res. 25: 207—272, 1969. Proinsulin.
392. Stephens G. C., Biol. Bull. 103: 242—258, 1952. Eyestalk hormone and cement glands.
Эндокринные механизмы
505
393- Stephens G. С., Biol. Bull. 108: 235—241, 1955. Molt induction in crayfish by photoperiod.
394. Stephens G. J., Physiol. Zool. 25: 70—84, 1952. Light influence upon cyclic ovarian activity in crayfishes.
395. Stumpf W. E., Amer. Zool. 11: 725—739, 1971. Autoradiographic localization of estrogen.
396. Sturkie P. D., Avian Physiology. Comstock Publishing Associates, Ithaca, N. Y., 1954. Insulin-resistant reptiles and birds.
397. Sutherland E. W., Rail T. W., J. Amer. Chem. Soc. 79: 3608, 1957. Isolation of cyclic AMP.
398. Swingle W. W., J. Exp. Zool. 36: 397—421, 1922. Thyroid and amphibian metamorphosis.
399. Szego С. M., Davis J. S., Proc. Nat. Acad. Sci. 58: 1711—1718, 1967. Effect of estrogen on uterine cyclic AMP.
400. Takasugi N., J. Fac. Sci. Univ. Tokyo, Sec. IV 7: 605—623, 1956. Rat ovulation by stress.
401. Takewaki K., Yamamoto Y„ Ann. Zool. Japan. 23: 187—190, 1950. Hormonal control of Paratya ovary.
402. Taki I., J. Lac. Fish. Anim. Husb. Hiroshima 5: 345—417, 1964. Branchial gland in cephalopods.
403. Tata J. R., Nature 197: 1167—1168, 1963. Actinomycin inhibits thyroid hormone action.
404. Tata J. R., Progr. Nucleic Acid Res. Mol. Biol. 5: 191—250, 1966. Hormones and nucleic acids.
405. Tata J. R., pp. 89—133 in: Biochemical Actions of Hormones, Vol. 1, edited by G. Litwack. Academic Press, New York, 1970. Regulation of protein synthesis by hormones.
406. Thomsen E., Vidensk. Meddel. Dansk. Naturhist. Foren. 106: 320—405, 1942. Corpus allatum hormone and dipteran reproduction.
407. Thomsen E., J. Exp. Biol. 29: 137—172, 1952. Neurosecretory brain cells and corpus cardiacum in Calliphora.
408. Thomsen E., J. Exp. Biol. 31: 322—330, 1954. Intra-axonal transport in Calliphora.
409. Turner C. D., Bagnara J. T., General Endocrinology, 5th edition. W. E Saunders Co., Philadelphia, 1971, 659 pp.
410. Turner C. L., Physiol. Zool. 15: 263—280, 1942. Gonadal hormones and secondary sex characters of fish.
411. Turtle J. R., Kipnis D. M., Biochem. Biophys. Res. Comm. 28: 797—802, 1967. Adrenergic receptors and cyclic AMP.
412. Vaes G., Nature 219: 939—940, 1968. PHT-like action of cyclic AMF
413. Van der Kloot W. G., Biol. Bull. 109: 276—294, 1955. Controls of diapause in H. cecropia.
414. Van der Kloot W. G., Williams С. M. Behavior 5: 157—174, 1953. Hormonal control of cocoon formation.
415. Van Dyke H. B„ Adamsons K-, Engle S. L., in: The Neurohypophysis, edited by H. Heller. Academic Press, New York, 1957. Differing activities of various vasopressins.
416. van Dyke H. B., Engel S. L., Adamsons K-, Proc. Soc. Exp. Biol, Med. 91: 484—486, 1956. Differing activities of vasopressins.
417. van Tienhoven, A., Reproductive Physiology of Vertebrates. W. B, Saunders Co., Philadelphia, 1968, 498 pp.
418. Veillet A., Cornubert G., Demeusy N., C. R. Soc. Biol. 147: 1264—1265, 1953. MTGX hormone inhibiting ovaries.
419. du Vigneaud V., Science 123: 967—974, 1956. Differences in pig and beef vasopressin.
420. du Vigneaud V. H., Lawler С., Popenoe E. A., J. Amer. Chem. Soc. 75: 4880— 4881, 1953. Octapeptides oxytocin and vasopressin.
421. du Vigneaud V., Lawler H. C., Popenoe E. A., Trippett S., J. Biol. Chem. 205: 949—957, 1953. Octapeptides oxytocin and vasopressin.
Ф06
Глава 21
-422. Vincent J. F. V., J. Insect Physiol. 17: 625—636, 1971. Bursicon in locusts.
423. von Euler U. S., Noradrenaline. Charles C. Thomas, Springfield, Ill, 1956. Noradrenaline at nerve endings; catecholamines.
424. von Hagen F., Zool. Jahrb. Abt. Anat. 61: 467—538, 1936. Thyroid and metamorphosis in fish.
425. Warren M. R., J. Exp. Zool. 83: 127—156, 1940. Thyroxin and oxygen consumption in frogs.
426. Webb H. M., Brown F. A., Jr., J. Cell. Comp. Physiol. 41: 103—122, 1953. Daily rhythm in Palaemonetes retinal pigment.
427. Weber R., Experientia 221: 665—666, 1965. Actinomycin inhibits tail atrophy.
428. Wells M. J., Wells J., J. Exp. Biol. 36: 1—33, 1959. Hormonal control of sexual maturity in Octopus.
429. Wells M. J., Wells J., Nature 222: 293—294, 1969. Pituitary analogue in Octopus.
430. Welsh J, H., Proc. Nat. Acad. Sci. 23: 458—460, 1937. Hormonal influence on crus13.c€З.П hcs.i*t bcs.t
431. Welsh J. H., J. Exp. Zool. 86: 35—49, 1941. Sinus glands and retinal-pigment migration in crayfishes.
432. Welsh J. H., Ann. N. Y. Acad. Sci. 66: 618—630, 1957. Serotonin as a neurohumoral agent.
433. Whitmore E., Gilbert L. I., J. Insect Physiol. 18: 1153—1168, 1972. Lipoprotein transport of juvenile hormone; induction of enzymes.
433a. Whitmore D., Jr., Whitmore E., Gilbert L. I., Proc. Nat. Acad. Sci. 69: 1592— 1595, 1972. Juvenile hormone.
434. Wiggleworth V. B., Quart. J. Mier. Sci. 77: 191—222, 1934. Hormone in insect molt and metamorphosis: Hemiptera.
435. Wigglesworth V. B., Quart. J. Mier. Sci. 79: 91—121, 1936. Hormones in molt and metamorphosis and reproduction: Hemiptera.
436. Wigglesworth V. B., J. Exp. Biol. 17: 201—222, 1940. Hormones in insect molt and metamorphosis: Hemiptera.
437. Wigglesworth V. B., J. Exp. Biol. 29: 561—570, 1952. Hormones and insect molt.
438. Wigglesworth V. B., The Physiology of Insect Metamorphosis. Cambridge University Press, 1954.
439. Wilhelmi A., p. 59 in: The Hypophyseal Growth Hormone, Nature and Actions, edited by R. W. Smith, Jr., О. H. Gaebler, and C. N. H. Long. Blakiston Co., New York, 1955. Crystalline fish growth hormone.
440. Wilhelmi A., Pickford G. E., Sawyer W. H., Endocrinology 57: 243—252, 1955. Spawning in Fundulus by neurohypophysis. Oxytocin and vasopressin as sex controllers; mammalian posterior lobe hormones give spawning in Fundulus.
441. Williams С. M., Biol. Bull. 90: 234—243, 1946. Brain hormones in lepidopte-ran metamorphosis.
442. Williams С. M., Biol. Bull. 93: 89—98, 1947. Roles of brain and prothoracic gland principles in lepidopteran metamorphosis.
443. Williams С. M., Anat. Rec. 99: 671, 1947. Brain and termination of pupal diapause.
444. Williams С. M., Biol. Bull. 94: 60—65, 1948. Prothoracic glands and insect development.
445. Williams С. M., Biol. Bull. 103: 120—138, 1952. Brain and prothoracic gland in insect metamorphosis.
446. Williams С. M., Harvey Leet. 47: 126—155, 1952. Brain and prothoracic gland in larval-pupal molt.
447. Willis J. H., Brunet P. C. J., J. Exp. Biol. 44: 363—378, 1966. Juvenile hormone on colleterial glands.
448. Wills I. A., Riley G. M., Stubbs E. M., Proc. Soc. Exp. Biol. Med. 30: 411—412, 748—786, 1933. Hypophysis and sex behavior in Amphibia.
449. Wingstrand K. G., Arkiv. Zool. Soc. 26: 41—67, 1953. Low neurosecretory material, ADH, and water retention in fetuses and young.
Эндокринные механизмы
507
450. Witschi Е., Wilson Bull. 47: 177—188, 1935. Hypophyseal control of plumage in weaver finches.
451. Witschi E., J. Clin. Endocrinol. 13: 316—329, 1953. Estrogens give adrenal-hyperplasia.
452. Witschi E., Mem. Soc. Endocrinol. 4: 149, 1955. Control of nuptial plumage-in African finches.
453. Witschi E., in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Taxonomic specificity of LH; endocrine basis of reproductive adaptations in birds.
454. Wolfson A., pp. 38—70 in: Comparative Endocrinology, edited by A. Gorbman. John Wiley & Sons, New York, 1959. Photoperiodism and bird physiology.
455. Wolfson A., in: Photoperiodism and Related Phenomena in Plants and Animals, edited by R. B. Withrow. Amer. Assoc. Adv. Sci., Washington, D. C.». 1959. Photoperiodism and annual cycles in birds.
456. Wright P. A., Endocrinology 64: 551—558, 1959. Blood sugar studies in the bullfrog.
457. Wright P. A., Biol. Bull. 119: 351, 1960. Rana ovulation in vitro by steroids.
458. Wright P. A., Hisaw F. L., Endocrinology 39: 247—255, 1946. Ovulation in-frogs with mammalian FSH and LH.
469. Wurtman R. J., Axelrod J., Phillips L. S., Science 142: 1071—1072, 1963a. Light controls melatonin synthesis.
460. Wyatt G. R., in: Biochemical'Actions of Hormones, Vol. 2, edited by G. Lit-wack. Academic Press, New York, 1972. Review of insect endocrinology.
460a. Wyatt S. S„ Wyatt G. R., Gen. Comp. Endocrinol. 16: 369—374, 1971. In vitro stimulation of RNA synthesis in epidermis by ecdysone.
461. Yamamoto Y., Ann. Zool. Japan 28: 92—99, 1955. Ovariectomy influence on sinus gland.
462. Yamazaki M., Kobayashi M., J. Insect Physiol. 15: 1981—1990, 1969. Purification of brain hormone.
463. Zalokar M„ J. Insect Physiol. 14: 1177—1184, 1968. RNA synthesis in colie-ferial glands.
Глава 22
Нематоцисты и их аналоги
Ф. БРАУН
У представителей Cnidaria и некоторых простейших имеются так называемые стрекательные тельца. Эти тельца представляют собой продукт секреции особых клеток. У книдарий стрекательные тельца формируются в книдобластах, или нематоцитах, и называются нематоцистами. К аналогам нематоцистов относятся трихоцисты многих ресничных инфузорий и жгутиконосцев, книдоцисты ряда динофлагеллят, а также полярные капсулы книдоспоридий.
Нематоцисты
Нематоцисты представляют собой очень мелкие внутриклеточные структуры сферической, овальной или веретеновидной формы. Они состоят из капсулы и заключенной в ней полой нити, которая представляет вырост одного конца капсулы, вывернутый наизнанку и закрученный {17, 37, 48]. В нематоцистах обнаружены белки, а также различные моно- и дифенолы, аминокислоты и ферменты [28, 34].. 'Следует отметить, что в состав нематоцистов входят, по-видимому, глутаминовая и аспарагиновая кислоты. Сама капсула, возможно, состоит из какого-то коллагеноподобного вещества [4, 13, 14, 20, 27, 31, 34]. При электронно-микроскопическом исследовании глютинант Corynactis было найдено, что невыстрелившая нить имеет форму сужающейся книзу трубки с глубокими складками, придающими ей вид винта с трехзаходной резьбой. После выброса нить становится гладкой, увеличиваясь в длину почти в 3 раза, тогда как ее диаметр и площадь поверхности остаются почти неизменными. Диаметр основания нити крупных стрепто-лин в 27г раза превышает ее диаметр на вершине (рис. 22-1). Поперечная исчерченность нити имеет период около 190 А, а продольная — около 50 А. Выстреливание нематоциста заключается в быстром выбрасывании нитевидного отростка. У покоящегося нематоциста та его часть, через которую выбрасывается нить, обычно покрыта колпачком, или крышечкой, а шипики нити плотно упакованы, и в момент разряда они направляются вперед, затем поворачиваются назад и удерживают нить в теле жертвы.
Нематоцисты и их аналоги
509
Рис. 22-4. Три нити стр епт олин после выброса [43].
Нити сплющены, так что видны три ряда зубцов вокруг стебля.
Последние широко распростри-
Нематоцисты образуются в интерстициальных клетках в месте своего дефинитивного расположения или же на некотором расстоянии от него. В последнем случае они достигают данного места в результате либо амебоидного движения, либо пассивного переноса, как, например, в процессе нормального роста щупалец гидры [7]. Участок книдобласта, содержащий нематоцист, в конечном счете оказывается на поверхности клетки, и на ее наружном конце образуется щетинковидный отросток — книдоциль. Последний погружен в маленькое кратероподобное углубление, края которого выступают над поверхностью клетки. В периферической части книдобласта дифференцируются щетинковидные опорные палочки, а также фибриллярная сетка, связанная с капсулой нематоциста и простирающаяся от нее проксимально в клетку. Полностью развитый нематоцист Diadumene считается внеклеточным образованием [51, 54].
Нематоцисты Cnidaria делятся на две большие группы: спи-роцисты Zoantharia, представляющие собой тельца, в ряде случаев окрашивающиеся кислыми красителями и снабженные особыми адгезивными нитями, и собственно нематоцисты, окрашивающиеся основными красителями, йены у представителей всего типа кишечнополостных. Нематоцисты отличаются огромным разнообразием по строению их выстреливающих нитей. При этом строение нематоцистов может до некоторой степени служить критерием для уточнения эволюции и систематики кишечнополостных [45, 46]. У вольвент нить образует плотно свернутую спираль и при выстреливании обвивается вокруг щетинок или фибрилл жертвы. У других нематоцистов наконечники обычно открытые и, как полагают, вонзаются в добычу, инъецируя в нее токсичное вещество. Нити таких нематоцистов (например, у глютинант и пенетрант) покрыты шипами, расположенными в 3 спиральных ряда, благодаря которым нити прочно закрепляются в тех тканях, куда они проникли. Способность неко
516
Глава 22
торых нитей прободать ткани настолько велика, что они могут прокалывать даже хитиновую кутикулу мелких организмов.
Нематоцисты некоторых кишечнополостных могут вызывать у человека зуд и другие кожные реакции. Токсичность содержимого нематоцистов некоторых кишечнополостных, таких, как Phy-salia, а также крупных сцифоидных медуз, например Dactilo-metra, настолько велика, что они представляют серьезную опасность для человека. Возникающий при этом ожог иногда даже приводит к смерти. Природа токсинов, связанных с нематоцистами, исследовалась у актиний Metridium [32] и Aiptasia [3], у одного из видов кубомедуз [11] и особенно у Physalia [15, 25, 26]. Активным началом токсина, вероятно, служат пептиды. При испытании токсического эффекта разряда нематоцистов на ракообразных, амфибиях (лягушки, жабы) и млекопитающих показано, что токсин особенно сильно действует на сердечно-сосудистую систему, а также на процессы нервного и мышечного возбуждения.
Книдобласты ведут себя как независимые эффекторы. Нематоцисты выстреливают под действием многих химических веществ, включая сильные кислоты и щелочи [53]. В ответ на обычные раздражения или на строго локализованное электрическое раздражение выстреливают лишь те нематоцисты, которые расположены на участке, непосредственно подвергающемся раздражению [29, 30]. Интенсивное механическое раздражение индифферентными объектами вызывает лишь слабый ответ, тогда как слабого механического раздражения естественной пищей оказывается достаточно, чтобы вызвать сильную реакцию (рис. 22-2). Погружение книдобластов Anemonia в слабый экстракт, приготовленный из обычной для данного вида пищи, который сам по себе не вызывает выстреливания, понижает порог реакции на чисто механические раздражители [29]. Специфическими пищевыми факторами, вызывающими такое повышение чувствительности, являются вещества липоидной природы, адсорбированные на белках. Адсорбционная связь столь прочна, что эти вещества не удается экстрагировать эфиром, и удалить их можно лишь при помощи спирта. По-видимому, те активные химические соединения, к которым чувствительны книдобласты, по своим свойствам напоминают стерины и фосфолипиды и обладают высокой поверхностной активностью. Ввиду того что очень быстрое и полное выстреливание нематоцистов происходит даже в ответ на воздействие сухой пищи, можно предположить, что нормальная реакция носит характер «контактно-химической», причем повышение чувствительности к механическому раздражению под действием химического происходит почти мгновенно.
Чрезвычайно своеобразные особенности возбуждения нематоцистов Anemonia очень точно приспособлены к функциям, выполняемым данными органеллами в естественных условиях. Важное значение того, что нематоцисты выстреливают лишь в тот мо- ,
Нематоцисты и их аналоги
511
мент, когда они вступают в механический контакт с раздражителем, иллюстрирует следующий опыт. Нематоцисты раздражали ваткой, смоченной раствором желчных солей, и ваткой, смоченной экстрактом из обычного пищевого субстрата. В первом случае выстреливание нематоциста вызывается взаимодействием с желчными солями, которые служат очень сильным раздражителем
Рис. 22-2. Выстреливание нематоцистов Anemonia в ответ на различные раздражения .[29].
А. Прикосновение человеческим волосом. Б. Прикосновение чистой стеклянной палочкой (выстреливания не происходит). В. Прикосновение чистой стеклянной палочкой после предварительного повышения чувствительности книдобластов путем помещения их в разбавленную слюну. Г. Прикосновение стеклянной палочкой, смоченной спиртовой вытяжкой из жабр морского гребешка.
ДО ТОГО,
Для книдобласта; поэтому нематоцисты выстреливают как приходят в соприкосновение с раздражителем, и, следовательно, выбрасываемая при этом нить не может прикрепиться к волоконцам ваты, подобно тому как она прикрепляется к организмам, составляющим обычную пищу этого кишечнополостного.
Книдоцили, вероятно, имеют какое-то отношение к процессу возбуждения нематоцистов. Однако они, по-видимому, играют второстепенную роль, поскольку отсутствуют у многих Anthozoa. Можно предположить, что в случае их отсутствия рецепторной областью служит вся поверхность книдобласта [27]. Нематоцисты Можно извлечь из ткани так, чтобы они при этом остались невы-отрелившими, если подвергнуть ткань воздействию ионов К+ или или если орган обработать Mg2+, С1~, Вг_ и анестезирующи-
512
Глава 22
ми веществами в отсутствие этих ионов [33, 51]. Изолированные нематоцисты могут на некоторое время сохранять способность к выстреливанию, которая дольше всего проявляется в одномолярном растворе глицерина, а в изотонических растворах СаС12, NaCl, КС1 и морской воде сохраняется ненадолго [49, 52]. Предполагается, что в этих условиях выстреливание происходит в результате ослабления крышечки [50].
Хотя книдобласты являются независимыми эффекторами, существуют данные о том, что их чувствительность в какой-то мере поддается регуляции. Она может меняться в зависимости от сытости животного [8], времени года [5], совпадения с локомоторной активностью [39] или же, как, например, у комменсала Cal-liactis parasitica, от того, прикреплена ли актиния к хозяину [38, 39]. Дальнейшая судьба выстрелившего нематоциста в теле животного-хозяина регулируется организмом. Предполагается, что в такой регуляции принимают участие окончания нейритов на книдобластах [28].
Механизм выстреливания нематоцистов еще до конца не выяснен. Среди специалистов в этой области наибольшей популярностью пользуются две группы теорий. Они построены на предположении, что выброс нити вызывается повышением давления внутри капсулы. Согласно одной группе теорий, повышение давления при раздражении объясняется поступлением воды в капсулу. При этом допускается, что нематоцист действует как простой осмометр. Показано, что точка замерзания содержимого капсулы— величина порядка —6 °C — становится существенно ниже после выстреливания нити [34]. Считают также, что поступление воды в капсулу связано с гидратацией и набуханием коллоидов внутри нематоциста в ответ на изменение pH, что влечет за собой набухание содержащегося в ней коллоидного материала. Согласно теориям другой группы, повышение давления внутри капсулы вызывается сокращением либо самой капсулы, либо связанной с ней фибриллярной сетки. Возможно также, что среди многочисленных типов нематоцистов различных видов кишечнополостных можно найти механизмы, соответствующие всем этим теориям. Нематоцисты Metridium, по-видимому, выстреливают вследствие поступления воды; выстреливание нематоцистов Physalia, очевидно, связано с сокращением фибрилл нематобласта.
Функционирующие нематоцисты были обнаружены также у плоского червя Microstotnum, у некоторых поликлад и у голожа-берника Aeolis [21—23]. Однако, как удалось установить, нематоцисты попали в организм этих животных от кишечнополостных, которые служат им пищей. Интересно отметить, что, «присваивая» нематоцисты кишечнополостных, эти черви и моллюски проявляют избирательность и используют нематоцисты только определенных типов. Так, Microstotnum переваривает вольвенты и использует пенетранты (крупные стрекательные капсулы). Голожаберни-
Нематоцисты, и их аналоги 513
ки, поедающие гидрополипов Pennaria, используют только высокоэффективный тип нематоцистов, так называемый микро-базальный, и не берут другие типы. В организме новых обладателей число присвоенных нематоцистов может быть весьма значительным, и они, по-видимому, служат прекрасным оборонительным оружием. Это явление связано, очевидно, с потерей реактивности нематоцистов по отношению к новому хозяину. Такая же потеря реактивности характерна для рака-отшельника и актинии, находящихся в симбиотических отношениях [2, 38].
Аналоги нематоцистов
В эктоплазме многих видов инфузорий содержатся особые выстреливающие тельца — трихоцисты [1, 34]. Трихоцисты имеют веретеновидную форму и расположены наклонно или под прямым углом к поверхности тела. При этом они либо равномерно распределены по всей поверхности'тела животного (например, у Paramecium), либо ограничены определенными участками (на хоботке у Dileptus). У Actinobolina трихоцисты располагаются на концах длинных щупалец и во время кормления выставляются наружу. Книдоцисты панцирного жгутиконосца Polykrikos лежат глубоко в цитоплазме.
При электронно-микроскопических исследованиях выяснен ряд деталей происхождения, развития и структуры аналогов нематоцистов, а также регенеративной способности организмов при воспроизводстве последних [6, 9, 10, 24, 36, 55, 56]. У Frontonia трихоцисты закладываются, по-видимому, в макронуклеусе или поблизости от него, а затем перемещаются к своему дефинитивному местоположению в эктоплазме, причем в процессе этого перемещения их организация усложняется. Сильное ультрафиолетовое облучение приостанавливает формирование трихоцистов [12]. Подобно нематоцистам, трихоцисты, как правило, имеют форму капсулы, содержащей прикрепленную к ней одним концом трубку; как и у нематоцистов, нить при (выстреливании вырывается наружу, подобно тому как выворачиваются пальцы перчатки [34]. У Paramecium покоящиеся трихоцисты состоят из овального тельца (длиной около 2—3 мкм и диаметром 2/з мкм) и покрытого колпачком наконечника несколько меньшего диаметра. При выстреливании трихоцисты наконечник с колпачком отделяется от тела; при этом стержень, соединяющий наконечник с телом, вытягивается и разрывается [6].
Выстреливание трихоциста можно легко вызвать при помощи ряда химических веществ (кислот или оснований), механических воздействий (давление) или электрических раздражений (разряд конденсатора, индукционный удар). Экстракт герани, по-видимо-Му, является высокоэффективным химическим агентом, вызывающим выстреливание трихоциста у Paramecium, что, возможно,,
33—2067
514
Глава 22
происходит благодаря содержанию в нем танниновой кислоты [42]. С увеличением силы электрического раздражения увеличивается число выстреливших трихоцистов [41]. Весь цикл выстреливания происходит очень быстро, занимая лишь несколько миллисекунд. Выстреливший трихоцист имеет обычно игловидную форму и в 10 или более раз длиннее покоящегося. У Paramecium
Рис. 22-3. Электронная микрофотография выстреливших трихоцистов Paramecium (напыление хромом) ['19].
длина выстрелившего трихоциста может достигать 40 мкм. Трихоцисты Paramecium обладают двойным лучепреломлением, что свидетельствует об ориентации вытянутых субмикроскопических частиц вдоль длинной оси органеллы. При электронно-микроскопическом изучении выпрямленных нитей выстреливших трихоцистов Paramecium была обнаружена характерная поперечная исчерчен-ность с периодом 600—650 А (рис. 22-3) и отсутствие пограничной мембраны. Судя по имеющимся данным, стержень состоит из вытянутых поперечноисчерченных белковых фибрилл, обнаруживающих периодическую структуру, отчасти напоминающую коллагеновые волокна. Общий характер исчерченности органеллы, по-видимому, отражает правильное взаимное расположение этих фибрилл. На конце стержня располагается относительно плотное шиповидное тельце, которое, по-видимому, имеет белковую природу или состоит из элементов с большим атомным весом [18, 19, 35].
Нет почти никаких данных, позволяющих судить о механизме выстреливания трихоцистов, а также о том, в какой степени трихоцисты могут активироваться другими раздражителями, кроме прямого действия внешнего раздражителя. Трихоцисты, по-видимому, связаны с системой нитей, которая, по мнению некоторых исследователей, обладает проводимостью, свойственной нервным сетям. Механизм выстреливания, возможно, связан с гидратацией
Нематоцисты и их аналоги
515
каких-то белков внутри трихоцистов или же с быстрым током воды за счет осмоса, возникающего в ответ на раздражение этих органелл.
Роль трихоцистов у простейших еще полностью не установлена. Многие авторы считают, что они обладают защитной функцией. Это кажется сомнительным в отношении таких видов, как Paramecium, поскольку хищные инфузории Didinium с готовностью заглатывают их даже после выстреливания многими из трихоцистов. Более вероятно, что у таких видов трихоцисты функционируют в качестве органов прикрепления [19]. Если жертва соприкасается с участками тела инфузорий Dileptus или Actinobolina, покрытыми трихоцистами, то она мгновенно парализуется; поэтому можно предполагать, что выстрелившие трихоцисты несут какое-то токсичное вещество. Трихоцисты у панцирных жгутиконосцев Polykrikos и Nematodinium очень сходны по своему строению с нематоцистами кишечнополостных, и их обычно называют книдоцистами. У этих видов можно обнаружить трихоцисты, находящиеся на разных стадиях дифференцировки [16]. Трихоцисты Noctiluca, по-видимому, имеют митохондриальное происхождение [44].
Другой тип органелл, очень сходных с нематоцистами кишечнополостных, — это полярные капсулы споровиков, принадлежащих к отрядам миксоспоридий и актиномиксидий. В пищеварительном тракте организма хозяина под действием пищеварительных ферментов эти капсулы, подобно нематоцистам, выбрасывают заключенные в них спирально закрученные полые нити. Такие нити служат для временного прикрепления паразита к тканям хозяина. У большинства микроспоридий вся спора, по-видимому, ведет себя как полярная капсула, причем содержимое целой споры выбрасывается через выворачивающуюся трубку.
СПИСОК ЛИТЕРАТУРЫ
1. Anderson Е., J. Protozool. 9: 380—395, 1962. Chilomonas trichocysts.
2. Berner L., Bull. Soc. Zool. France 78: 221—226, 1953. Nematocyst response in commensal relationship.
3. Blanquet R., Comp. Biochem. Physiol. 25: 893—902, 1968. Toxity of sea anemone nematocyst.
4. Boisseau J. P., Bull. Soc. Zool. France 77: 151—169, 1952. Chemical contents of nematocysts.
5. Bouchet С., C. R. Acad. Sci. (Paris) 252: 327—328, 1961. Seasonal variation in cnidoblast responsiveness.
o. Bouck G. B., Sweeny В. M., Protoplasma 61: 205—223, 1966. Fine structure of dinoflagellate trichocysts.
7. Burnett A. L„ Lentz T., Ann. Soc. Zool. Malacol. Belg. 90: 281—293, 1960. Nematocyst migration in Hydra.
8. Burnett A. L., Lentz T., Warren N., Ann. Soc. Zool. Malacol. Belg. 90: 247—267;
I960. Nutrition and nematocyst discharge.
9. Doroshewski M., Golinska K., Acta Protozool. 4: 343—350, 1967. Trichocyst regeneration in Dileptus.
33*
516
Глава 22
10. Dragesco J., Auderset G., Baumann M., Protistologica 1: 81—90, 1965. Structure and genesis of trichocysts of Dileptus.
11. Endean R. C., Duchemin, McColm D., Fraser E. H., Toxicon 6: 179—204, 1969. Nematocysts in a cubomedusan.
12. Haller G. de, Heggeler B. ten, Protistologica 5: 115—120, 1969. Inhibition of trichocyst formation by ultraviolet light.
13. Hamon M., Nature 176: 357, 1955. Chemical nature of nematocysts.
14. Hamon M., Bull. Soc. d’Hist. Nat. 1’Afrique du Nord 46: 169—179, 1955. Chemistry of nematocysts.
15. Hastings S. G., Larsen J. B., Lane С. E., Proc. Soc. Exp. Biol. Med. 125: 41—45, 1967. Effects of Physalia nematocyst toxin on dog cardiovascular system.
16. Hovasse R., Arch. Zool. Gen. 102: 189—198, 1963. Trichocysts of Polykrikos.
17. Hyman L. H., The Invertebrates. McGraw-Hill Book Co., New York, 1940. Trichocysts and nematocysts.
18. Jakus M. A., Exp. Zool. 100: 457—485, 1945. Trichocysts; structure and properties.
19. Jakus M. A., Hall С. E., Biol. Bull. 91: 141—144, 1946. Electron micrograhs of trichocysts.
20. Johnson F. B., Lenhoff H. M., J. Histochem. Cytochem. 6: 394, 1958. Nature of Hydra nematocyst capsule.
21. Earling T. G., Acta Zool. Fennica 116: 3—28, 1966. Nematocysts in turbella-rians.
22. Hepner W. A., J. Morphol. 73: 297—311, 1943. Nematocysts in nudibranchs.
23. Hepner W. A., Gregory W. C., Porter R. J., Zool. Anz. 121: 114—124, 1938. Nematocysts of Microstomum.
24. Hudo R. R., Daniels E. W., J. Protozool. 10: 112—120, 1963. Fine structure of microsporidian spore.
25. Larsen J. B., Lane С. E., Toxicon 4: 199, 1966. Effect of nematocyst toxin on land crabs, Cardisoma, and myogenic rat heart.
26. Larsen J. B., Lane С. E., Toxicon 8: 21—23, 1970. Action of Physalia toxin on frog nerve and muscle.
27. Lenhoff H. M., Hline E. S., Hurley R., Biochem. Biophys. Acta 26: 204—205, 1957. Chemistry of Hydra nematocyst.
28. Lentz T. L., wood J. G„ J. Histochem. Cytochem. 12: 37, 1964. Neurite endings on cnidoblasts in hydra and sea anemones.
29. Pantin C. F. A., J. Exp. Biol. 19: 294—310, 1942. Control of nematocyst discharge.
30. Parker G. H., Van Alstyne M. A., J. Exp. Zool. 63: 329—344, 1932. Control and mechanism of nematocyst discharge.
31. Phillips J. H., Nature 178: 932, 1956. Chemical nature of Metridium nematocysts.
32. Phillips J. H„ Abbott D. P., Biol. Bull. 113: 296—301, 1957. Toxin of Metridium nematocyst.
33. Picken L. E. R., Quart. J. Mier. Sci., 94, 203—227, 1953. Corinactis nematocysts.
34. Picken L. E. R., Skaer R. J., Symp. Zool. Soc. Lond. 16: 19—50, 1966. Review of researches on nematocysts.
35. Potts B., Biochem. Biophys. Acta 16: 464—470, 1955. Electron microscopic study of trichocysts.
36. Puytorac P. de, Acta Protozool. 2: 147—152, 1964. Ultrastructure of ciliate trichocyst.
37. Robson E. A., Quart. J. Mier. Sci., 94: 229—235, 1953. Electron microscopic study of nematocysts.
38. Ross D. M., Proc. Zool. Soc. Lond. 134: 43—57, 1960. Nematocyst response in commensal relationships.
39. Ross D. M., Sutton L., J. Exp. Biol. 41: 751—757, 1964. Locomotion and nematocyst discharge.
40. Saunders J. T., Proc. Cambridge Phil. Soc. (Biol. Sci.) 1: 249—269, 1925. Function of trichocysts of Paramecium.
Нематоцисты и ах аналоги
517
41 Schmitt F. О., Hall С. Е., Jakus М. A., Biol. Symp. 10: 261—276, 1943. Trichocysts.
42. Schuster F. L„ Prazak B., Ehret C. F., J. Protozool. 14: 483—485, 1967. Tricho-cyst discharge in Paramecium.
43. Skaer R. H., Picken L. E. R., Phil. Trans. Roy Soc. Lond. В 250: 131—164, 1965. Structure and discharge of nematocyst of Corynactis.
44. Soyer M.-O, Protistologica 5: 327, 334, 1969. Trichocysts of Noctiluca (dinoflagellate).
45. weill R-> Trav. Station Zool. Wimereux 10, 11, I, II, 1934. Nematocyst classification.
46. Werner B., Helg. wiss. Meeresunters. 12: 1—39, 1965. Cnidarian nematocysts, and systematics and evolution.
47. Wood J. G., Lentz T. L., Nature 201: 88—90, 1964. Amines in sea anemone and hydra nematocysts.
48. Yanagita T. M„ J. Fac. Sci. Tokyo Univ. 6: 97—108, 1943. Nematocyst discharge.
49. Yanagita T. M., Nat Sci. Report Ochanomizu Univ. 2: 117—123, 1951. Physiology of isolated nematocysts.
50. Yanagita T. M., Japan J. Zool. 12: 363—375, 1959. Responses of isolated nematocysts.
51. Yanagita T. M., J. Exp. Biol. 36: 478—494, 1959. Extrusion of nematocysts.
52. Yanagita T. M., J. Fac. Sci. Tokyo Univ. 8: 381—400, 1959. Response of isolated nematocysts.
53. Yanagita T. M., Wada T., Nat. Sci. Report Ochanomizu Univ. 4: 112—118, 1953. Nematocyst discharge by acids and bases.
54. Yanagita T. M., Wada T., Cytologia 24: 81—97, 1959. Physiology of acontia and nematocysts.
55. Yusa A., J. Protozool. 10: 253—262, 1963. Fine structure of regenerating Paramecium trichocysts.
56. Yusa A., J. Protozool. 12: 51—60, 1965. Fine structure of Frontonia trichocysts.
w
Глава 23
Хроматофоры и изменение окраски
Ф. А. БРАУН МЛАДШИЙ'
i
< s
Введение
Способность к изменению окраски в результате перемещения пигментов в определенных клетках покровных тканей или в некоторых органах широко распространена среди животных. Из позвоночных этой способностью обладают многие виды круглоротых, рыб, амфибий и рептилий, из беспозвоночных — многие высшие ракообразные, головоногие моллюски, пиявки, а также некоторые виды насекомых, иглокожих и полихет. Изменение окраски наблюдалось у одной из эвглен (простейшие). Еще в IV в. до н. э. Аристотель описал эффектные изменения окраски от черной до зеленой у хамелеона и быстрые изменения окраски у осьминога. Несколько позже Плиний описал это явление для рыб, наблюдая изменение окраски тела у подыхающей кефали. Среди амфибий способность к изменению окраски впервые была обнаружена у лягушки, а среди ракообразных — у креветки Hippolyte [179].
Уже давно установлено, что сравнительно быстрые изменения окраски у головоногих моллюсков связаны в основном с деятельностью специальных органов, расположенных в коже, которые были названы хромофорами. Позднее было показано, что изменения окраски у хамелеона [202], лягушки [11] и ракообразных [261] вызваны перемещением пигментов в специальных органах, которые получили название хроматофоров.
Брюкке [53] исследовал физиологические механизмы изменения окраски у хамелеона, Пуше [242] —у ракообразных и рыб; Гембл и Кибл [159, 160] —у ракообразных. По мнению всех этих авторов, активность хроматофоров либо регулируется нервной системой, либо возникает непосредственно в ответ на внешние воздействия. Предположение об участии в изменении окраски гормонов впервые возникло после того, как было обнаружено, что при инъекции лягушке адреналина цвет ее кожи бледнеет [188]. Тщательные обзоры ранних работ по хроматофорам сделаны ван Рин-берком [257] и Фуксом [118]. Позднее появились общие обзоры Хогбена [141], Паркера [214, 230] и Фингермана [89, 90, 92],
1 При участии М. Фжнгермаяа-.
Хроматофоры и изменение окраски
519
также обзоры по отдельным группам: по позвоночным [240, 3121, насекомым [262] и ракообразным [35, 58, 172]. Пигменты животных описаны и в ряде других обзоров [109, 128].
Строение и способ действия хроматофоров
Хроматофоры представляют собой специальные пигментные клетки, расположенные в коже, а нередко и в некоторых более глубоких тканях животного. Хроматофоры обладают способностью вызывать перераспределение содержащегося в них пигмента, что сопровождается изменением общей окраски животного. Если пигмент сконцентрирован в маленький шарик (зернистое состояние), то он почти не влияет на общую окраску, если же он распределен ио большой поверхности (сетчатое состояние), то окраска всего животного принимает его оттенок.
Концентрирование и диспергирование пигмента называют физиологическим изменением окраски. Хроматофоры могут также влиять на окраску животного в результате того, что пигмент в них образуется и накапливается или же разрушается и исчезает. Этот механизм называют морфологическим изменением окраски.
Физиологические изменения окраски. Существуют хроматофоры двух основных типов. Хроматофор одного из них, обнаруженный у головоногих моллюсков, представляет собой сложный орган, который состоит из клетки, содержащей пигмент, и связанного с ней большого числа радиально расположенных гладких мышечных волокон (рис. 23-1). Однако для большинства животных, способных к изменению окраски, характерны хроматофоры другого, более общего вида. Такой хроматофор состоит из одиночной клетки или небольшого синцития, обычно сильно разветвленных, в которых изменения в распределении пигмента происходят за счет внутриклеточных перемещений.
Хроматофор головоногих моллюсков представляет собой одноядерную клетку, наполненную пигментом, который находится в очень эластичном внутриклеточном мешке. Эластичные свойства хроматофора обусловлены этим мешком, а не клеточной мембраной. Когда мешок сокращается, мембрана становится складчатой [61]. От центральной клетки, располагаясь в плоскости кожи, отходит в радиальных направлениях от 6 до 20 или больше одноядерных гладких мышечных волокон. Все волокна хроматофора обычно сокращаются одновременно, растягивая маленькую сферическую пигментированную клетку в диск, диаметр которого в 15—20 раз больше, чем у исходной клетки. После расслабления радиальных волокон центральная пигментированная клетка вновь принимает сферическую форму за счет эластичности мешка. По имеющимся данным, к каждому мышечному волокну подходит одно нервное волокно, разветвленное окончание которого распределяется по всей поверхности мышцы [140]. Моторные концевые
520
Глава 23
пластинки отсутствуют, и поэтому кураре не блокирует передачу с двигательного нерва. Природа вещества, участвующего в передаче сигнала по нервному волокну, не определена [106]. Скорость сокращения мышечных волокон в ответ на электрическое раздражение близка к скорости сокращения волокон поперечнополосатой мышцы (гл. 16).
О химической природе пигментов головоногих моллюсков почти ничего не известно. Оказывается, пигменты их хроматофоров
Рис. 23-1. Хроматофор головоногого моллюска [24]. А. Пигмент сконцентрирован. Б. Пигмент диспергирован.
представляют собой оммохромы [269]. У осьминога Octopus обнаружены хроматофоры двух типов: содержащие красновато-коричневый и желтый пигменты. У кальмара Loligo содержатся хроматофоры трех типов: с коричневым, красным и желтым пигментом. Под хроматофорами располагается неподвижный слой пигмента, отражающего свет. Благодаря этим хроматофорам головоногие моллюски способны изменять окраску быстрее, чем другие животные.
Хроматофоры других животных состоят либо из одной клетки (например, у большинства позвоночных), либо из группы близко расположенных клеток или синцития (например, у ракообразных). Раньше считали, что хроматофоры представляют собой амебоидные клетки: при втягивании псевдоподий масса пигмента собирается в маленький шарик, а при усиленном выпускании псевдоподий пигмент диспергируется. Теперь общепризнано, что хроматофор имеет постоянную древовидную форму, а гранулы пигмента либо сконцентрированы в центре хроматофора, образуя зер-нистую массу, либо рассеяны с разной плотностью по всем сложным ветвям хроматофора (рис. 23-2) [194], создавая видимую окраску животного. Мэтьюз и др. сообщили, что ими были обнаружены хроматофоры, в отростках которых пигмент находите^
Хроматофоры и изменение окраски
521
в зернистом состоянии. Это сообщение подтверждается данными электронной микроскопии [21]. Выявлено также удивительное единообразие состояния пигмента в разных частях клетки, вплоть до самых мелких ответвлений хроматофоров [27, 237, 288].
Надо сказать, что у различных групп животных, у разных видов и даже у отдельных особей хроматофоры различаются по
Рис. 23-2. Серия фотографий одиночного белого хроматофора Palaemonetes, позволяющих проследить концентрирование пигмента при перенесении животного на черный фон и возврат пигмента в прежнее диспергированное состояние при перенесении животного обратно на белый фон [26].
форме, типу пигмента и реакциям на раздражение. Известны монохроматические, дихроматические и полихроматические хроматофоры — в зависимости от того, содержат ли они один, два или более пигментов. Хроматофоры ракообразных относятся, как правило, к типу дихроматических или полихроматических, при этом каждый пигмент обычно диспергируется в свои собственные отростки, а когда оба пигмента концентрируются в центре хроматофоров, каждый из них сохраняет известную обособленность. При реакциях этих хроматофоров на изменение окраски фона содержащиеся в одном и том же хроматофоре пигменты могут Действовать в значительной степени независимо друг от друга [29, 176].
У некоторых насекомых функции хроматофоров выполняют клетки эпидермиса. При потемнении кожи коричнево-черный пигмент, содержащийся в этих клетках в виде мелких скоплений,
522
Глава 23
лежащих под слоем равномерно распределенных желтого и зеленого пигментов, перемещается ближе к поверхности и переходит в диспергированное состояние (рис. 23-3). Красный пигмент при этом диспергирует из небольших сферических скоплений, образуя сплошной слой. По-видимому, силы, которые участвуют в концентрировании и диспергировании пигментов в хроматофорах, ничем
Рис. 23-3. Схемы срезов эпидермиса Carausiiis, адаптированного к светлому (Л) и темному (Ь) фону [123].
Самые крупные точки (3) — зерна коричнево-черного пигмента, средние (2) — зерна красно* го, а самые мелкие (/) — желто-зеленого пигмента.
не отличаются от сил, действующих в клетке при изменении ее формы.
У Euglena rubra в ответ на повышение интенсивности света красный пигмент (гематохром) перемещается из более глубоких участков клетки, где он лежит под хлоропластами, ближе к поверхности и одновременно переходит в диспергированное состояние, что и приводит к изменению окраски от зеленой к красной [154].
Хроматофоры ракообразных содержат несколько различных пигментов. У разных видов, а иногда и у отдельных особей одного вида, но живших в разных условиях, встречаются различные наборы пигментов. Представители ракообразных, по-видимому, всегда содержат желтый и красный пигменты. К ним относятся различные каротиноиды, которые часто связаны с белками [183]. Кроме того, в хроматофорах ракообразных почти всегда присутст-
Хроматофоры и изменение окраски
523
ует белый отражающий пигмент. Большинство длиннохвостых десятиногих раков содержат прозрачный синий пигмент, представляющий собой комплекс каротиноида с белком [309]. Под действием тепла или спирта, воздействующих на участок поверхности животного, содержащего синий пигмент, последний быстро превращается из водорастворимого синего пигмента в жирорастворимый красный. В хроматофорах некоторых ракообразных имеется черный или коричневато-черный пигмент. Такого рода пигмент, обнаруженный у Crago (Crangon) и у равноногих, например у Ligia, по-видимому, представляет собой оммохром, а у некоторых крабов— меланин [128].
Хроматофоры позвоночных сходны с хроматофорами ракообразных. Однако в отличие от последних они обычно состоят из одной клетки и, как правило, содержат один пигмент; в большинстве случаев это меланин, а все примеры изменения окраски в этой группе связаны с активностью меланофоров. У многих позвоночных имеются также отражающие белые хроматофоры, называемые лейкофорами, иридолейкофорами или антаугофорами [116]. Часто встречаются хроматофоры, содержащие жирорастворимый красный пигмент (эритрофоры) или желтый пигмент (ксантофоры); их называют также липофорами. Эти пигменты относятся к каротиноидам [108]. Помимо упомянутых основных типов пигментных клеток, иногда встречаются переливчатые синевато-зеленые тела — иридофоры, окраска и изменение цвета которых зависят от особенностей их структуры и определяются формой, расположением и перемещениями тонких пластинчатых кристаллов.
Нередко в создании окраски у позвоночных, так же как и у головоногих моллюсков, помимо собственно хроматофоров, участвует неподвижный слой беловатого или желтоватого пигмента. Этот слой либо целиком определяет окраску животного (в тех случаях, когда активные хроматофоры практически не участвуют в создании окраски), либо участвует в этом совместно с хроматофорами. У таких видов, как ящерица Anolis, центральные тела меланофоров расположены под этим инертным слоем пигмента, а при потемнении животного в ответ на соответствующее раздражение меланин перемещается в отростки меланофоров, оказываясь ближе к поверхности тела, чем этот светлый слой, и таким образом маскирует его [165] (рис. 23-4).
Скорость физиологического изменения окраски ограничивается скоростью механической реакции эффекторного органа и управляющего им механизма, причем для различных животных эти скорости неодинаковы. Кальмар и рыба-белка Holocentrus способны изменению окраски за доли секунды, а рыбам es требуется для этого несколько минут. Про
цесс изменения окраски занимает от одного до нескольких часов У многих ракообразных, насекомых, а также у сомика Ameiurus;
максимальному Puridulus и Lebis
624
Глава 23
камбалам, угрю Anguilla, пластиножаберным и амфибиям для него требуется несколько дней.
При измерении физиологических изменений окраски пытались использовать ряд методов; критический обзор этих методов сделал Паркер [229]. Однако ни один из них не дает возможности полностью дифференцировать те изменения окраски животного»
Рис. 23-4. Чешуйки ящерицы Anolis даны в поперечном разрезе, с тем чтобы можно было видеть состояние хроматофоров [165].
Л и Б. Состояние хроматофоров, при котором образуется коричневатая окраска. В и Г. Со* стояние хроматофоров, при котором образуется зеленая окраска.
в которых участвуют морфологические факторы, от чисто физиологических. Поэтому измерения стараются проводить в течение достаточно коротких периодов времени, чтобы исключить возможность существенного влияния морфологических изменений на получаемые результаты.
В одной группе методов за показатель степени дисперсности темных пигментов принимается просто общее изменение цвета животного. К таким методам относятся визуальные, при которых окраска животного просто описывается как темная, светлая или имеющая промежуточный оттенок или же субъективно оценивается [33] по четырех- или пятибалльной шкале.
Неизбежную субъективность таких оценок удается до некоторой степени устранить с помощью фотометрирования, определяя
Хроматофоры и изменение окраски Ь2о
либо долю падающего света, отражаемую единичным участком поверхности кожи [207, 316], либо долю света, пропускаемого изолированными чешуйками рыб [283].
Другая группа методов основана на оценке изменений самих хроматофоров. Один из этих методов, использованный для исследования хроматофоров рыб и ракообразных, состоит в измерении изменений диаметра отдельных хроматофоров [27, 30, 289]. В другом довольно широко применяемом методе различают следующие
Рис. 23-5. Разные состояния меланофоров в зависимости от степени дисперсности пигмента [1192].
А — зернистое; Б — зернисто-звездчатое; В — звездчатое; Г *— сетчато-звездчатое; Д — сетчатое.
состояния хроматофоров [192, 280] (рис. 23-5): 1) зернистое, 2) зернисто-звездчатое, 3) звездчатое, 4) сетчато-звездчатое, 5) сетчатое. Достоинство такой системы состоит в том что при известном навыке, для того чтобы получить числовые значения, которые можно использовать в различных сравнительных исследованиях, достаточно быстрого осмотра животного.
Описанные методы иногда дополняются микрофотографиями [217] или быстрой (часто тепловой) фиксацией животного для получения временных или постоянных препаратов кожи [30, 317].
Морфологические изменения окраски. Морфологические изменения окраски представляют собой изменения количества пигмента, содержащегося в организме животного или в его кожном покрове. При нормальных приспособительных изменениях окраски животных физиологические и морфологические изменения происходят одновременно. Морфологические изменения могут привести как к увеличению содержания пигмента в каждой пигментной Клетке, так и к увеличению числа активных хроматофоров на единицу поверхности кожи [28, 209]. Количественные исследования таких изменений проводились соответственно методами двух ти-Пов: 1) определением изменений числа активных хроматофоров На ^единицу площади поверхности тела и 2) определением изменений общего содержания пигмента в организме животных с помощью экстракции и последующего колориметрирования пигмеи-
526
Глава 23
та в экстрактах (рис. 23-6). Хороший обзор по морфологическим изменениям окраски у позвоночных сделал Самнер [296]; природа морфологических изменений окраски у беспозвоночных, по-видимому, такая же.
Создается впечатление, что между физиологическими и морфологическими изменениями окраски существует тесная функциональная связь, которую отмечали многие исследователи [15, 160].
нис. 23-6. Изменение концентрации красного пигмента у Palaemonetes в зависимости от фона [28].
А. Уменьшение концентрации на белом фоне. Б. Увеличение концентрации пигмента на черном фоне.
Когда пигмент в течение длительного времени находится в хро-матофоре в концентрированном состоянии, то при этом, по-видимому, его количество обычно уменьшается; и наоборот, при диспергированном состоянии пигмента его образование, по-видимому, увеличивается. Эта зависимость известна как закон Бабака [15]. Поэтому могут существовать две возможности: 1) образование или разрушение пигмента зависят от степени его диспергирования, 2) регуляция физиологических и морфологических изменений окраски происходит параллельно при помощи общих регуляторных механизмов. Вторая возможность представляется более вероятной, особенно потому, что такие механизмы могли бы обеспечивать даже возникновение новых хроматофоров.
Xроматофоры и изменение окраски
Ь27
Изменения окраски у животных
На хроматофорную систему животных оказывает влияние целый ряд различных факторов.
Температура. Температура сильно влияет на окраску многих животных. Низкая температура обычно вызывает потемнение окраски у Hyla [71] и у Phrynosoma [224]. Повышение температуры приводит к концентрированию темного пигмента и вследствие этого — к посветлению окраски [224]. Среди беспозвоночных такого единообразия не наблюдается: Callinectes и Palaemonetes при повышении температуры светлеют [81]; креветка Macrobrachium и равноногое Idothea темнеют как при повышении, так и при понижении температуры [83, 198]; в отличие от них Hippolyte и ~ Uca в обоих случаях обычно бледнеют [43]; палочник Carausius имеет черную окраску при 15 °C и становится зеленым при 25 °C.
Влажность. Изменение влажности сказывается на окраске Carausius: при высокой влажности он темнеет [123]. У Rana во влажной среде наблюдается потемнение окраски, в сухой, наоборот, — посветление [256].
Тактильное раздражение. Складывается впечатление, что тактильное раздражение в общем мало влияет на хроматофоры. Ранее полагали, что изменение окраски у древесной лягушки в значительной степени определяется текстурой поверхности, на которой она находится: считалось, что лягушка становится темной на неровной поверхности и зеленой — на гладкой. Теперь общепризнано, что это не так. Отмечалось, что тактильное раздражение присасывательных дисков некоторых головоногих моллюсков рефлекторно влияет на хроматофоры [291]. По данным Шлипера [266], ракообразное Hyperia galba, паразитирующее на медузах, бледнеет, когда прикреплено к своему хозяину, и становится темным, если плавает свободно. Считают, что прикрепление к любой поверхности, черной или белой, вызывает у этого животного реакцию побледнения, которая, несомненно, является приспособительной, поскольку обычный хозяин его совсем прозрачный.
Общее возбуждение. У ряда животных общее возбуждение также, видимо, влияет на хроматофоры. Для кальмара или каракатицы, возбужденных присутствием хищника, например большого краба, характерна необычайная игра цветов. Изменение окраски часто производит впечатление волн, пробегающих быстро и плавно по поверхности тела животного. Эти переливы цветов, по-види-мому, часто оказывают непосредственное влияние на поведение Животных в брачный период.
Рептилии и некоторые рыбы при возбуждении также изменяют окраску. У жабовидной ящерицы Phrynosoma при сильном .возбуждении окраска бледнеет. Лягушка Xenopus при возбуждении темнеет; введение ей адреналина вызывает такую же реакцию [56, 150, 161]. Когда ящерица Anolis вступает в поединок с сопер
528
Глава 23
ником или когда ее внезапно раздражают, она приобретает характерную пятнистую окраску [165].
Свет. Из всех факторов внешней среды наибольшее влияние на состояние хроматофорной системы животных оказывает свет. На преобладающее большинство животных свет воздействует главным образом через глаза, центральную нервную систему и различные эфферентные пути — нервные, гуморальные или и те и другие
Рис. 23-7. Зависимость среднего диаметра меланофора гольяна Ericymba от логарифма интенсивности падающего света (Д) и от отношения интенсивности падающего света к интенсивности отраженного света, попадающего в глаз животного |(5) [30].
одновременно. Значение глаз для реакции хроматофоров четко выявляется при ослеплении животного: при этом изменение окраски немедленно прекращается или же резко изменяет свой характер. Изменения окраски, регулируемые посредством зрения, известны под названием вторичных реакций хроматофоров в отличие от первичных реакций, которые возникают при непосредственном действии света на хроматофоры или же при рефлекторном влиянии на рецепторы, расположенные вне глаз.
У большинства взрослых животных преобладают вторичные реакции. В широком диапазоне интенсивностей света эти изменения определяются отношением количества света, падающего сверху непосредственно на глаз, к количеству света, отраженного от фона, на котором находится данный организм. На освещенном черном фоне, когда это отношение велико, животное темнеет, а на освещенном белом фоне, где это отношение мало, — бледнеет неза
Хроматофоры и изменение окраски 529
висимо от общей освещенности. Отношение интенсивности падающего света к интенсивности отраженного света, попадающего в глаз животного, часто хорошо коррелирует со степенью диспергирования черного пигмента [30, 300] (рис. 23-7, Б) или количеством меланина, образующегося в меланофорах [296]. Более того, любая из этих реакций существенно меняется с изменением интенсивности света в широком диапазоне общей освещенности.
Приспособительные морфологические изменения окраски в ответ на изменение цвета фона отмечались для хроматофоров ракообразных [28], а также для меланофоров [297], липофоров [3] и иридолейкофоров [298] различных видов рыб. Например, если камбалу ослепляли или помещали в черные садки и освещали снизу, то на ее брюшной поверхности, обычно лишенной пигмента, появлялись активные мел анофоры [212].
Во многих случаях можно создать видимость черного фона, прикрыв животным нижние половины глаз; такие опыты были проведены с палочником Carausius [13, 244], креветками Palaemonetes [132] и Leander (Palaemon) [133] и с костистыми рыбами [57].
Механизм влияния света на вторичные цветовые реакции у животных, очевидно, специально приспособлен к тому, чтобы обеспечить животное необходимой защитной или маскирующей окраской применительно к фону. Можно предполагать, что эта функция лучше развита у глубоководных животных, а также у тех, которые большую часть времени проводят в неподвижности, прикрепившись к каким-либо субстратам, но не у животных, ведущих подвижный образ жизни.
Приспособление к цвету фона во многих случаях не сводится просто к посветлению или потемнению кожи на светлом и темном фоне соответственно. Много лет назад была описана адаптация десятиногого рака Hippolyte ко многим цветам и оттенкам фона [203]. Позднее отмечалось, что Crago (Crangon) может приспосабливать свою окраску не только к черному и белому, но также к красному и желтому фону [176]. У Palaemonetes была обнаружена способность в течение нескольких дней изменять свою окраску, приспосабливаясь к черному, белому, красному, желтому, синему или зеленому фону [27]. Крабы P.ortunus ordwayi [4] и Planes [139] способны приспосабливаться к черному, белому, красному и желтому фону. Головоногие моллюски Sepia и Octopus, имеющие, помимо иридофоров, черные, оранжевые и желтые хроматофоры, обнаруживают поразительные цветовые реакции на окраску фона [181].
Среди позвоночных наиболее удивительные примеры приспособительной окраски можно наблюдать у некоторых видов камбал. У Paralichthys albiguttus, например, были описаны изменения окраски на красном, зеленом, желтом и синем фоне [184, 191]. Более того, эта рыба способна подражать довольно сложному ри-34—2067
530 Глава 23
сунку фона, что значительно усиливает покровительственную функцию окраски. Многие другие костистые рыбы и амфибии могут приобретать желтый оттенок в ответ на появление желтоватых тонов в окраске фона.
Все эти адаптации к цвету фона связаны с соответствующими дифференцированными перемещениями различных пигментов внутри хроматофоров, причем такая физиологическая активность усиливается дополнительными морфологическими изменениями окраски и наличием неподвижного слоя пигментов в покровных тканях. У некоторых животных обнаружена способность изменять распределение или количество имеющихся у них различных пигментов более или менее независимо друг от друга, что свидетельствует о сложности механизмов, регулирующих активность хроматофоров. Отсутствие пигментов соответствующих цветов ограничивает возможности вида в отношении диапазона различных фонов, к которым они могут приспособиться. Например, у Crago (Crangon) отсутствует синий пигмент, что не позволяет ему приспособиться ни к синему, ни к зеленому фону. У Palaemonetes имеются красный, желтый и голубой пигменты, поэтому он может путем соответствующего перераспределения этих пигментов принять почти любую окраску, в том числе и черную. .В то же время некоторые виды, имеющие пигмент данного цвета, все же не могут приспособиться к фону такого же цвета. Примером может служить палочник Carausius, который, видимо, не способен принимать красноватый цвет, хотя его кутикула содержит красный пигмент.
Для того чтобы животное могло адаптироваться к цвету фона, его органы зрения должны быть способны различать не только падающий и отраженный свет, но и свет разных длин волн, причем именно той частью сетчатки, которая возбуждается отраженным светом.
Первичные реакции хроматофоров у большинства половозрелых высших животных подчинены вторичным реакциям. Это становится особенно очевидным в экспериментах с ослеплением животного. Многие животные и после ослепления сохраняют способность к значительному диспергированию и концентрированию своих пигментов на свету и в темноте соответственно, однако обычно при этом утрачивается реакция на цвет фона. Примеры хроматофорных реакций у ослепленных животных описаны для всех основных групп, обладающих способностью изменять окраску. Поразительное исключение из указанного правила описано для ряда прямокрылых: оказывается, они сохраняют способность к приспособительной реакции на цвет фона [74, 75].
Окраска, по-видимому, не зависит от характера пищи. Считается, что клетки эпидермиса действуют в качестве независимых эффекторов [162]. Степень влияния первичных реакций по сравнению со вторичными изменяется от вида к виду или даже в пре
Хроматофоры и изменение окраски 531
делах одного вида (применительно к различным пигментам). Хроматофоры личинок Crangon обладают, вероятно, только первичными реакциями на освещение [235], в то время как хроматофоры крабов Ocypode [247] и Rhithrop апо peas [187] проявляют как первичные, так и вторичные реакции на свет. Черные и красные хроматофоры краба Ocypode имеют первичные и вторичные реакции на свет, а белые — только первичные [248]. Более того, способность краба Ocypode к изменению окраски варьирует в зависимости от возраста особи; она меньше у более зрелых особей по сравнению с молодыми. Долгосрочная адаптация раков и Ocypode к черному или белому фону приводит к значительному уменьшению их способности изменять окраску при смене фона [94, 97, 249].
Наиболее общий механизм первичной реакции связан с неким непосредственным воздействием света на хроматофоры. В этом случае хроматофор действует как независимый эффектор. Первичные реакции такого типа наблюдались: 1) у хроматофоров позвоночных после полной денервации пигментных клеток в результате перерезки и последующего перерождения нерва, 2) в локальных реакциях на свет у видов, хроматофоры которых в норме лишены иннервации, 3) у молодых особей, хроматофоры которых еще не находятся под контролем нервной системы.
Изменение окраски в ответ на световое раздражение можно рефлекторно вызвать и в отсутствие глаз. На примере Phoxinus было показано, что в этой реакции роль приемника играет средний мозг [263]. У личинок миноги Lampetra рецепторным органом в реакции хроматофоров служит шишковидное тело, тогда как у взрослой особи преобладающую роль в этой реакции играют глаза [321].
У большинства взрослых животных, способных изменять окраску, последняя в широком диапазоне освещенности больше зависит от цвета фона, чем от общей освещенности. При полном отсутствии света обычно наблюдается посветление животного, но часто не до такой степени, как на освещенном белом фоне. Некоторые виды, например Crago (Crangon) или Xenopus, в тени приобретают промежуточную окраску, частично диспергируя имеющиеся у них темные пигменты. Следует отметить, что у Crangon нет меланина; его темным пигментом является оммохром. Оказалось, что свет не влияет на окраску гольяна Ericymba на черном фоне, пока освещенность не превышает 19 лк; при меньших значениях освещенности средний диаметр меланофоров линейно зависит от значений логарифма падающего света вплоть до освещенности 0,0058 лк, которая действует на окраску животного так же, как и полная темнота [30] (рис. 23-7, Л).
Суточные ритмы. Постоянный суточный ритм активности представляет собой еще один важный фактор, который влияет на Функцию хроматофоров >[293]. У многих видов животных сохра
34*
532
Глава 23
няются специфические суточные ритмы изменения окраски, даже если содержать животных в среде с постоянной температурой и освещенностью, и особенно в постоянной темноте. Такие ритмы могут сохраняться в течение довольно долгого времени. У Idothea, содержащейся в полной темноте, ритм сохранялся по меньшей мере в течение 8—9 нед [198]. У Uca влияние этого ритма, выражающегося в побледнении окраски ночью и потемнении днем [9, 47], настолько сильное, что изменения фона или освещенности вызывают лишь незначительные изменения окраски животного»
Рис. 23-8. Различные фазы окраски краба Uca pugnax (темная днем и бледная в ночное время).
(рис. 23-8) [43]. Сообщалось о некоторых интересных особенностях ритмического механизма, управляющего хроматофорами Uca.
Сроки потемнения и посветления окраски в суточных циклах, которые сохраняются при содержании животных в постоянных лабораторных условиях, можно сдвинуть за несколько дней путем соответствующего изменения светового и температурного режима [48, 292, 313]. В результате такого сдвига фазы ритма потемнение и посветление окраски будут наступать в любое заданное время суток, причем этот новый суточный ритм будет сохраняться до тех пор, пока он вновь не будет перестроен. Более того, можно вызвать даже более тонкие сдвиги в характере суточных изменений окраски и их диапазоне, которые затем сохранятся при постоянных лабораторных условиях [40, 45]. Механизм, задающий этот постоянный суточный ритм, не зависит от температуры в диапазоне, равном по меньшей мере 20 °C [47], и его точность не нарушается даже в необычных условиях среды (например, когда крабов перевозили самолетом на 50° западнее [49], в результате
Хроматофоры и изменение окраски 533*
чего длительность суток увеличивалась для них примерно до 27,5 ч).
Установление того факта, что при некоторых условиях вновь приобретенный ритм изменения окраски (сдвинутый по фазе ш> отношению к нормальному) постепенно возвращается назад — к ритму с нормальными сроками появления таких изменений, — привело к предположению о существовании по меньшей мере двух, задающих ритм центров [48]. Чтобы добиться постоянного сдвига первоначального ритма, необходимо перестроить оба центра.
Получены данные, свидетельствующие о том, что сдвиг фаз-изменения окраски под действием света и температуры зависит' от суточного ритма чувствительности крабов [38, 292, 313] к этим факторам: крабы реагировали на увеличение освещенности или повышение температуры только во время ночной фазы своего нормального ритма. Было высказано мнение, что это и есть механизм, при помощи которого крабы настраивают свой ритм изменений окраски в соответствии с сменой дня и ночи в природе [36].
На постоянный суточный ритм изменения окраски накладывается другой ритм — приливный, период которого равен 12,4 ч [39,. 79, 80, 86]; в результате модулирующего действия этого ритма? примерно два раза в месяц возникают флуктуации в суточных циклах изменения окраски. Приливные ритмы изменения окраски,, период которых связан с фазами луны, также, по-видимому, сохраняются в течение неопределенно долгого времени в тех постоянных условиях, которые поддерживаются в лаборатории во время экспериментов, и обнаруживают те же необычные свойства, которые отмечались выше для суточных ритмов [50]. Известно, что» приливные ритмы изменения окраски согласованы с местным временем отливов, когда вследствие отступления воды обнажаются? укрытия, в которых обитают крабы [80, 85, 99].
По имеющимся данным, ритмы изменения цвета, существующие у других животных, видимо, таковы же, как и у крабов. Постоянные ритмы изменений окраски обнаружены не только у многих других ракообразных, но и у позвоночных, например у миног Lampetra [321], гольяна Phoxinus, личинок саламандры, лягушек,, ящерицы Phrynosoma [253], хамелеона [323], а также у палочника Carausius и у морского ежа Diadema [200].
В тех случаях, когда изменения, связанные с суточными ритмами, маскируются вторичными реакциями на освещенный фон,, скрытое влияние ритма проявляется в повышенной скорости тех. реакций, которые соответствуют фазе ритма в данный момент, ю в замедленной скорости реакций, направленных в противоположную сторону. Так, многие животные легче приспосабливаются к освещенному белому фону во время ночной фазы и к освещенному черному фону днем или проявляют в период ночной и дневною фаз различную восприимчивость к инъекции гормонов, вызывающих изменение окраски.
534
Глава 23
Функциональная организация системы хроматофоров
Кольчатые черви. Окраска некоторых видов кольчатых червей бледнеет в темноте и темнеет при освещении. Это отмечалось для полихеты Nereis (Platynereis) dumerilii [136] и пиявок Piscicola geometra [152, 153], Protoclepsis tessellata [294], Hemiclepsis mar-ginata [152, 153], Glossiphonia complanata [152, 153, 314] и Placobdella parasitica и P. rugosa [287]. Ни у одного из этих видов не обнаружено какой-либо реакции на цвет фона.
Хоботная пиявка Placobdella parasitica (распространенный вид, паразитирующий на черепахах центральных областей США) имеет пигментные клетки трех типов, которые влияют на пятнистую коричнево-белую окраску ее тела. Количество белых пятен у разных особей весьма различно; у одних животных белая почти вся спина, у других имеется лишь несколько белых боковых сосочков и короткая белая черточка посредине спины на передней части тела. Клетки, содержащие зерна бледно-желтого пигмента, располагаются вдоль характерной продольной полосы посредине спины, а также на многочисленных светлых сосочках по бокам тела, где они образуют скопления на каждом сегменте. Физиологических реакций у этих клеток не обнаружено. По всем более темным участкам тела распределены сравнительно крупные хроматофоры, содержащие темный зеленоватый пигмент, который принимает участие в физиологическом изменении окраски. Пигмент третьего типа, красновато-коричневый, содержится в хроматофорах, которые много меньше зеленоватых и расположены ближе к поверхности. Этот пигмент также принимает участие в изменении окраски. Зеленый пигмент в отличие от красновато-коричневого растворим в спирте. У европейской пиявки Protoclepsis tessellata имеется три пигмента — коричневый, зеленый и желтый, а средиземноморская полихета Nereis (Platynereis) содержит иридо-форы и кирпично-красный и коричневато-фиолетовый хроматофоры. Имеется циркадный ритм миграции этих пигментов. Они диспергированы гораздо больше днем, чем ночью. Этот ритм, по-видимому, имеет эндогенную составляющую [104, 255].
Окраска пиявки Placobdella parasitica, так же как и у других аннелид, способных к физиологическому изменению окраски, бледнеет в темноте и темнеет на свету. Процесс этот длится около 1 я и зависит в основном от зеленоватого пигмента, который концентрируется в темноте и диспергируется на свету.
Некоторые данные указывают на то, что функция зеленых хроматофоров регулируется нервной системой. Обезглавливание, а также любое повреждение нервной системы приводит к потемнению содержащихся в темноте животных (которые в норме имеют светлую окраску). Это потемнение сохраняется на протяжении многих часов. Если у пиявки, находящейся в темноте и имеющей равномерную светлую окраску, раздражать электрическим
Хроматофоры и изменение окраски 535
током переднюю или заднюю половину тела, то вся поверхность тела животного темнеет. Если же этот опыт повторить с особью, у которой нервная цепочка посередине перерезана, то потемнеет только та часть тела, которая подвергалась раздражению, а по другую сторону от места перерезки тело останется светлым. О непосредственном действии света на хроматофоры известно мало.
Цва глаза, расположенные на переднем конце тела, играют, по-видимому, активную роль в реакциях животного, однако способность к этим реакциям зависит не только от них. У обезглавленных животных также обнаруживаются типичные изменения окраски, хотя и более замедленные. Если животных перенести из темноты на свет и сразу же накрыть одну половину тела непрозрачной бумагой, то накрытая половина в значительной степени сохранит окраску, характерную для адаптации к темноте, тогда как ненакрытая полностью адаптируется к свету. Опыт даст особенно наглядные результаты, если накрыть не переднюю, а заднюю половину тела. Такого рода опыты указывают на участие в цветовых реакциях фоторецепторов, размещенных по всему телу и действующих через сегментарные рефлексы.
Хроматофоры Placobdella parasitica, содержащие красновато-коричневый пигмент, реагируют на раздражение независимо от зеленых, и какой-либо определенной реакции на цвет фона или интенсивность света обнаружить у них не удается. Состояние красновато-коричневого пигмента и в норме не остается постоянным. Когда этот пигмент диспергирован, интенсивным раздражением животного обычно удается вызвать его концентрирование. У животных с перерезанной нервной цепочкой реакция на электрическое раздражение (точно так же как и в случае зеленых хроматофоров) не распространяется дальше места перерезки. Это позволяет думать, что функционирование красновато-коричневых хроматофоров также регулируется нервной системой, но что возбуждение вызывает в них концентрирование пигмента, а не диспергирование (как в зеленых хроматофорах).
Иглокожие. Окраска морских ежей Arbacia pustulosa, Centros-tephanus longispinus, Diadema antillarum и Diadema setosum светлеет при перенесении животных со света в темноту [166, 199, 200, 307, 319]. Морской еж Arbacia pustulosa имеет на свету (как на белом, так и на черном фоне) черноватую окраску, а при перенесении в темноту становится коричневым. Centrostephanus longispinus при помещении в темноту через 1—2 ч изменяет окраску с темно-пурпуровой на серую. Под микроскопом в отдельных амбулакральных ножках морского ежа видно большое число красновато-коричневых хроматофоров; у животных, адаптированных к темноте, пигмент в этих клетках сконцентрирован, а у животных, адаптированных к свету, он находится в диспергированном состоянии. Изменения окраски у этих морских ежей происходят вследст-вие перемещения пигментов внутри определенных хроматофоров,
536
Глава 23
но при этом также имеют место и морфологические изменения. Хроматофоры амбулакральных ножек, помещенных на столик микроскопа, реагируют на освещение и затемнение так же, как хроматофоры целого животного. Это позволило предположить, что хроматофоры реагируют на свет либо непосредственно, либо путем локальных рефлексов, подобных тем, которые участвуют в движениях амбулакральных ножек. Опыты с освещением отдельных точек на поверхности тела животного доказали, что хромато-<форы реагируют на свет непосредственно.
Однако у американского вида рода Arbacia (Arbacia punctula-4а) не удалось обнаружить изменений окраски, подобных описан-дым выше [215].
Головоногие моллюски. Многие головоногие моллюски вследствие особого устройства хроматофоров способны удивительно -быстро изменять свою окраску. Такие изменения могут быть вызваны раздражителями различных типов, среди которых свет — один из наиболее эффективных.
Функция хроматофоров регулируется, главным образом нервной системой. Перерезка нерва, иннервирующего определенный участок тела, приводит к немедленному прекращению всех цветовых реакций в этом участке. Давно известно, что при перерезании коннективы мантии [ПО] хроматофоры соответствующей половины тела утрачивают способность к реакции и вся эта половина обесцвечивается. Поскольку соединения между мантийным ганглием и хроматофорами при этом не нарушаются, ясно, что ганглий -сам по себе принимает лишь незначительное участие в регулировании реакций и центр регулирования располагается в высших отделах нервной системы. В церебральном ганглии имеется, по-видимому, центр торможения хроматофорной системы [239, 270]; после его разрушения или инактивации наступает тоническое расширение хроматофоров. Функция центра торможения состоит, как полагают, в управлении цветовым центром [271], расположенным в центральном ганглии, который в свою очередь действует через двигательные центры, обнаруженные в подглоточном ганглии. Каждый из двигательных центров управляет хроматофорами соответствующей половины тела.
Глаза — главные органы чувств, влияющие на нервные центры. Удаление обоих глаз не приводит к исчезновению цветовых .реакций, но реакции, сохраняющиеся после этого, уже не имеют •приспособительного характера. Если удален только один глаз, то реакции хроматофоров соответствующей части тела оказываются “ослабленными.
Другим важным источником влияния на центры, регулирующие окраску животного, служат присоски на конечностях. Если их полностью удалить, то хроматофоры в значительной мере утрачивают тонус, что приводит к посветлению окраски [291]. Однако удаление обоих глаз и присосок не ведет к окончательному подав
Хроматофоры и изменение окраски
537
лению реакций хроматофоров. После такой операции сильные раздражители все еще вызывают изменения окраски, вероятно, вследствие раздражения органов типа тактильных или органов-равновесия.
Хроматофоры, расположенные на самой нижней части тела (в тех случаях, когда животное соприкасается с субстратом), всегда сокращены сильнее, чем хроматофоры остальной части тела [271]. Можно было ожидать, что это связано с особенностями освещения, но такое объяснение неверно, поскольку освещение снизу не снимает этого эффекта. Скорее всего это локализованное сокращение хроматофоров представляет собой реакцию на изменение положения тела, вызванную рефлекторным возбуждением тактильных рецепторов через центральную нервную систему.
Известно, что в крови головоногих моллюсков присутствуют такие вещества, как тирамин и бетаин. Первое из них, подобно адреналину, вызывает повышение тонуса двигательных центров, что приводит к потемнению окраски [272]. В отличие от него бетаин, как и пилокарпин или ацетилхолин, понижает тонус хроматофоров посредством возбуждения центра торможения. При переливании крови от животных с более темной окраской (Eledone или Octopus macropus) животным с более светлой окраской (О. vulgaris) последние темнеют [273]. Сходные результаты можно получить в опытах с парабиозом. По имеющимся данным, у животных с более темной окраской концентрация тирамина в крови ъыше. Удаление задней слюнной железы, которая, как известно, служит важным источником тирамина в крови этих животных, приводит к побледнению окраски и полной утрате тонуса хроматофорами [273]. Потемнение можно вновь вызвать инъекцией растворов тирамина. Отсюда следует, что тирамин (а, вероятно, также и бетаин) действует как гуморальный фактор, который через нервные центры изменяет общий тонус хроматофоров.. Более того, опыты с денервированными хроматофорами [274, 275] показывают, что тирамин и бетаин, действуя на сами хроматофоры, усиливают их тонус. Следовательно, они способны влиять на хроматофоры как опосредованно, так и непосредственно. Однако эти результаты, полученные Серени, подверг критике Таки, который обнаружил, что удаление задней слюнной железы не приводит к изменению окраски у головоногих моллюсков, с которыми он работал [302]. Общая концепция гормонального регулирования изменения окраски у этих животных должна быть подкреплена, таким образом, в дальнейших исследованиях.
Относительно медленное гуморальное влияние дополняется более быстро действующим нервным механизмом, при посредстве которого происходят быстрые изменения окраски. В результате определенных наблюдений была высказана гипотеза, что у хроматофоров имеется двойная иннервация. Если изолировать часть покровной ткани Loligo вместе с содержащимися в ней хромате-
538
Глава 23
форами и оставить ее на некоторое время, то хроматофоры вначале сократятся, а затем несколько расширятся и останутся в этом состоянии. Сокращение хроматофоров можно вызвать электрическим раздражением. В этом случае электрическое раздражение вызывает, видимо, эффект торможения и снижает тонус мышц хроматофоров [24]. Если же раздражать нерв хроматофора или непосредственно сам хроматофор одиночными импульсами тока, то это приведет лишь к одиночным подергиваниям мышцы хроматофора. Ряд быстро следующих друг за другом электрических раздражений вызывает тетанус и расширение хроматофоров. На основании этого был сделан вывод, что мышцы хроматофоров Lo-ligo имеют двойную иннервацию (возбуждающие и тормозные нервы). Однако торможение, возможно, обусловлено торможением Введенского, а не функционированием ингибиторного аксона. Более того, электрофизиологические исследования не подтвердили наличия тормозной иннервации. За все поведение мышечных клеток хроматофоров можно считать ответственным действие моторных нейронов i[105, 106].
Насекомые. Многие насекомые способны изменять окраску в ответ на внешние раздражения. Чаще всего это происходит вследствие морфологических изменений. К числу факторов, от которых зависят такие изменения, относятся температура, влажность, освещенность и общая активность насекомого. Лишь немногие виды насекомых способны к сравнительно быстрым физиологическим изменениям окраски.
Интенсивность окраски куколок многих бабочек зависит от того, при какой температуре они выращиваются — высокой или низкой; чем ниже температура, тем темнее окраска. У Vanessa влияние температуры осуществляется через головной отдел; это доказывается тем, что если разные части тела содержать при разной температуре, то общая окраска куколки будет определяться температурой головного отдела [124]. Как показали опыты с гусеницами на стадии, предшествующей окукливанию, окраска обычно определяется гормонами. Точно установлено участие экдизона, который выделяют проторакальные железы; кроме того, вероятно, действует также и ювенильный гормон [55, 137]. Сходное влияние температуры на степень развития темных пигментов наблюдалось у осы Habrobracon {457, 268] и клопа Perillus [168].
У Pieris brassicae окраска определяется меланином кутикулы, белым пигментом клеток эпидермиса и зеленым пигментом более глубоких тканей. На окраску куколок Р. brassicae влияет также фон. На черном или красном фоне они имеют серовато-белую окраску, а на зеленом или оранжевом становятся светло-зелеными (так как в этих условиях подавляется образование черного и белого пигментов) [67]. Освещение куколок светом разных длин волн оказывает на них такое же действие, как и разный цвет фона. В цветовой реакции существенную роль играют глаза или не
Хроматофоры и изменение окраски 539
которые другие структуры головы; после удаления глаз или обезглавливания способность к реакции утрачивается и животные на свету ведут себя так же, как в темноте [25].
У перелетной саранчи Locusta migratoria способность приспосабливать окраску тела к цвету фона до известной степени ограничена [77]. В результате изменения количества желтоватого и черного пигментов саранча может приобретать желтовато-белую, коричневую или черную окраску. Количество образующегося черного пигмента зависит, по-видимому, от соотношения падающего на глаз и отраженного глазом света. Количество желтоватого пигмента определяется, как можно думать, спектральным составом падающего света; пигмент образуется быстрее, когда в падающем свете преобладают длинные волны (550—660 нм), и медленнее,, когда преобладают короткие (450—500 нм). В период объединения в стаи насекомые приобретают более темную окраску, что, вероятно, связано с более интенсивной метаболической активностью. При возвращении к одиночному образу жизни саранча снова светлеет. Посветление можно предотвратить, поддерживая насекомое в состоянии постоянного возбуждения [77].
У сравнительно немногих насекомых изменения окраски можно-объяснить перераспределением пигментов в пигментных клетках или хроматофорах. Известны быстрые физиологические изменения окраски воздушных мешков Corethra (Chaoborus), в которых имеются специальные пигментные клетки [64, 190]. На черном фоне пигмент диспергируется и пигментные клетки равномерно распределяются по воздушным мешкам. На белом фоне пигмент концентрируется, и создается впечатление, что клетки собираются к одной стороне мешков. В этих реакциях принимают участие глаза и мозг насекомого; диспергирование темного пигмента осуществляется под действием гормонального фактора, присутствующего» в мозге и в меньшем количестве в прилежащих телах [64, 265].
Разносторонние исследования цветовых реакций были проведены на Carausius morosus. Гиподермальные клетки у этого вида содержат четыре пигмента: коричневый (меланин), оранжевокрасный и желтый (липохромы) и зеленый [123, 265]. Коричневый и оранжево-красный пигменты активно концентрируются и диспергируются в клетках под действием внешнего раздражения. Зеленый и желтый пигменты такой способности не обнаруживают. Поэтому находимые в природе зеленые разновидности в отличие °т коричневых не способны к физиологическим изменениям окраски. Особи с коричневой окраской обычно бывают темными ночью и более светлыми днем вследствие соответственно диспергирования и концентрирования коричневого и оранжево-красного пигментов. Эти изменения иногда не зависят от непосредственного влияния света, так как типичные суточные циклы сохраняются и У насекомых, постоянно содержащихся в темноте [1123, 265]. Осве-Щая насекомое ночью и оставляя его в темноте днем, можно обра-
540
Глава 23
тить этот ритм; тогда в постоянной темноте будет сохраняться новый ритм. $
Известно, что повышенная влажность также вызывает потемнение тела. Используя этот факт, Гирсберг [123] остроумным способом доказал, что данный раздражитель действует на хромато-4>оры косвенным путем, через афферентные нервы, мозг, какой-то эндокринный орган и, наконец, гормональный агент, циркулирующий в крови. Если заднюю половину тела светлоокрашенной особи
Рис. 23-9. Опыт с влажной камерой, показывающий, что повышенная влажность вызывает потемнение тела у Carausius [123].
Потемнение наступает в ответ на раздражение брюшной области. Оно начинается с головного отдела и распространяется по телу только до лигатуры, наложенной на переднюю часть груди.
Carausius поместить в небольшую влажную камеру, то через 30— <50 мин все тело насекомого приобретет темную окраску (рис. 23-9); сначала потемнеет передняя часть, которая находится вне камеры, в сухом воздухе, а затем потемнение постепенно распространится на все тело. После извлечения из камеры через 1—2 ч насекомое снова светлеет. Если повторить опыт, предварительно наложив тугую лигатуру на переднюю часть груди, то потемнение распространится только до места перевязки. В таком же опыте на светлоокрашенном животном, у которого аккуратно перерезана брюшная нервная цепочка между подглоточным и первым грудным ганглиями или между под- и надглоточным ганглиями, потемнения не наступает вообще; если же перевернуть животное и поместить во влажную камеру не заднюю часть его тела, а голову, то сначала потемнеет голова, а затем потемнение распространится по всему телу, как у неоперированного насекомого. Нервные пути у насекомого с лигатурой все еще обладают способностью проводить в переднюю часть тела нервные импульсы, возникающие в нервных окончаниях брюшного участка, который раздражается повышенной влажностью, но выделяемый в передней части гормон не может диффундировать за лигатуру. После перерезки нервной цепочки нервные импульсы уже не могут поступать в мозг и стимулировать выделение гормона. Однако образование гормона можно вызвать раздражением нервных окончаний, лежащих кпереди от места перерезки, и тогда активное вещество будет переходить в заднюю часть тела.
Результаты пересадки участков кожи от одного насекомого к ? другому представляют новое доказательство того, что физиологи-
Хроматофоры и изменение окраски___________544
веские изменения цвета у Carausius контролируются главным образом гормональным фактором [151]. Через 2—3 дня пересаженные ткани изменяют свою окраску точно так же, как и ткани насекомого, к которому они пересажены. Вряд ли можно предполагать, что за столь короткий период трансплантированная ткань могла приобрести иннервацию от организма-хозяина, тем более «что и в нормальной эпидермальной ткани не удается обнаружить признаков иннервации.
Глаза Carausius играют существенную роль в реакциях на свет £13]. Если перерезать зрительные нервы или закрыть поверхность глаза, то реакция нэ свет исчезнет.
При постоянной влажности и освещенности Carausius реагирует на цвет фона: насекомое темнеет на черном и красном фоне и светлеет на белом и желтоватом. Реакция на фон определяется соотношением интенсивности света, падающего на дорсальную и вентральную части глаз. Закрывание нижней половины глаза приводит к такому же потемнению тела, как и помещение на черный фон.
Мозг, по-видимому, является главным источником гормона, регулирующего изменения окраски. После удаления мозга животное становится бледно-серым. Введение мозгового экстракта вызывает потемнение окраски насекомых [65, 66]. Активный гормон, названный С-веществом, удалось выделить электрофоретическим методом [173].
У Carausius наблюдаются также и морфологические изменения окраски в ответ на изменение освещения, цвета фона и влажности; регулируются они, по-видимому, тем же механизмом, что и физиологические изменения. Для морфологических изменений требуется, однако, значительно более длительное раздражение [13, 124]. Высказывалось предположение, что вещества, обычно влияющие на физиологические изменения, перестраивают общий обмен веществ в организме насекомого, а уж это в свою очередь приводит к дальнейшим изменениям окраски в результате образования или разрушения пигмента.
Суточные изменения окраски у некоторых богомолов [117] тесно связаны с суточными движениями пигментов сетчатки в латеральных участках сложного глаза.
Ракообразные. У ракообразных встречаются чрезвычайно любопытные примеры цветовых реакций. Хроматофоры большинства представителей этой группы содержат белый, красный, желтый, а часто также черный, коричневый и синий пигменты. В результате соответствующей перегруппировки отдельных пигментов в хро-матофорах ракообразные способны довольно точно воспроизводить цвет фона, т. е. того субстрата, на котором они в данный момент находятся.
Хотя первые исследователи полагали, что функция хроматофоров регулируется нервной системой, никому еще не удавалось убе-
542
Глава 23
дительно продемонстрировать наличие нервных окончаний в хро-матофорах или доказать непосредственное влияние перерезки нер- , вов на реакцию хроматофоров у неповрежденного животного. Кол- I лер [175, 176] впервые отчетливо показал на Crago vulgaris | (Crangon crangon), что вещество, активно регулирующее реакцию I хроматофоров, содержится в крови. Он обнаружил, что если от г
животного, находящегося на черном фоне и имеющего темную (
окраску, перелить кровь животному, адаптировавшемуся к белому | фону, то и это животное потемнеет. Однако переливанием крови J от светлых особей к темным вызвать посветление не удалось. Не ч удалось, следовательно, доказать и существование вещества, вы- | зывающего посветление. Коллер наблюдал также, что перелива-ние крови от животного, приспособившегося к желтому фону, вы- J зывает пожелтение у животного, находящегося на белом фоне. J
Перкинс [237] установил, что денервация какого-либо участка поверхности тела Palaemonetes ни в какой мере не влияет на ре- | акции этого участка при помещении животного на черный или 1
белый фон; однако при прекращении притока крови любой учас- |
ток тела немедленно утрачивает способность к цветовым реакци- | ям. После восстановления притока крови способность к изменению | цвета сразу восстанавливается и данный участок приобретает | окраску, свойственную в этот момент телу. Эти результаты были I истолкованы как указание на то, что вещества, регулирующие диспергирование и концентрирование пигмента, переносятся к от- j дельным хроматофорам кровью. Опыты с введением экстрактов различных органов показали, что глазной стебелек у Palaemone- | tes содержит вещество, вызывающее концентрирование красного пигмента и диспергирование белого, т. е. в конечном счете побледнение окраски животного [238]. В результате удаления глазного стебелька наступает устойчивое потемнение окраски животного. Эти наблюдения были вскоре подтверждены Коллером [177] на Crago (Crangon), Leander (Palaemon) и Processa. Опыты с пере- j крестной инъекцией свидетельствуют о том, что данный гормо- а нальный фактор не обладает ни видовой, ни родовой специфичностью. После этих первых работ многим исследователям удалось i показать, что источником вещества, влияющего на состояние хроматофоров, служит либо глазной стебелек, либо (у некоторых видов) передняя часть груди.
Гормональная функция глазного стебелька особенно хорошо изучена у десятиногих раков. В зависимости от роли, которую играет у них глазной стебелек, десятиногих раков можно разделить на три группы (рис. 23-10). ‘В группу I входят роды Palaemonetes, Penaeus, Hippolyte, Leander (Palaemon), Orconectes и Cam-barellus. Хроматофорная система представителей этой группы со- 1
держит обычно красный, желтый, синий и белый пигменты. Труп- <
па II включает только один род Crago (Crangon). Эти животные [ имеют весьма сложную систему пигментации (не менее восьми
Хроматофоры и изменение окраски
543
типов хроматофоров с различной реакцией), что позволяет им изменять не только оттенок или общую интенсивность окраски, но также в определенной степени и цветовой рисунок. Хроматофоры у животных этой группы содержат черный, коричневый, красный, желтый и белый пигменты. Группа III объединяет большинство изученных крабов. В нее входят Eriocheir, Hemigrapsus, СаШпес-tes, Еса и Sesarma [10, 100]. Наиболее хорошо изучен манящий
Группа I Группа II
(Palaemonetes) (Сгадо)
Начальная Конечная реакция стадия
Гриппа 111 (Uca)
Рис. 23-flO. Схема, иллюстрирующая окраску ракообразных и состояние преобладающих хроматофоров в каждой из трех различно реагирующих групп при уда-: лении глазных стебельков (Д) и при. последующем введении экстракта из глазных стебельков или синусной железы (Б) [35].
---->_цельный экстракт (водорастворимый материал);--—> фракция, нерастворимая в спирте; - - - - —> фракция, растворимая в спирте.
краб Uca, хроматофорная система которого содержит обычно черный, красный, желтый и белый пигменты.
После удаления глазного стебелька у представителя любой из этих трех групп наблюдается характерная для данной группы реакция. У животных группы I красный и желтый пигменты диспергируются. Вследствие этого животные быстро темнеют и уже через 1—2 ч становятся совершенно темными (хотя обычно и не настолько темными, как при нормальной реакции на черный фон). В таком состоянии они могут оставаться сколь угодно долго. Белый пигмент обычно на какое-то время концентрируется, а затем Может вновь изменить свое состояние.
У Crago (Crangon) (группа II) изменения, возникающие после Удаления глазного стебелька, носят, как правило, более сложный Характер. Сначала наступает временное потемнение тельсона и брюшных отростков с одновременным посветлением остальной
544
Глава 23
части тела. Такая окраска сохраняется в течение 30—60 мин. Затем следует полное посветление тельсона и брюшных отростков и потемнение всего остального тела, которое приобретает при этом промежуточную и пятнистую окраску. Белый пигмент сначала концентрируется, а затем переходит в промежуточное состояние.
Краб Uca (группа III) после удаления глазных стебельков довольно быстро светлеет; большая часть его черных хроматофоров становится пятнистой, а белые обычно приобретают сетчатый вид. Животное остается в этом состоянии без существенных изменений неопределенно долгое время. Мы видим, таким образом, что удаление глазных стебельков может приводить у ракообразных к трем разным результатам: потемнению тела, некой промежуточной окраске и посветлению. В любом из этих состояний животные практически утрачивают способность реагировать на цвет фона или освещение.
После инъекции экстракта глазных стебельков животным группы I эти последние быстро светлеют [84, 87, 102, 264]. У животных группы II при этом светлеет все тело, включая тельсон и брюшные отростки. У представителей группы III, напротив, наблюдается потемнение тела [23, 72, 192]. Такое резкое различие у животных трех групп объясняется главным образом различной реакцией соответствующих хроматофорных систем на гормоны глазного стебелька; опыты показали, что экстракты глазных стебельков, полученные от животных какой-нибудь одной группы, вызывают у представителей любой другой группы реакцию, характерную для них при введении гомологичного экстракта. Так, например, экстракты глазных стебельков животного группы I вызывают потемнение у животного группы III (у которого глазные стебельки удалены).
Хроматофоротропные гормоны глазных стебельков обычно выделяются синусной железой (рис. 21-9), что убедительно подтвердил Ханстрем [132]. Эта железа была описана еще до работ Хан-стрема [131, 278] и в то время получила название «кровяной железы», ибо вначале считали, что она гомологична кроветворной железе глазных стебельков Crago (Crangon), которую ошибочно рассматривали как источник активного вещества [178]. Синусная железа обнаружена практически у всех ракообразных, у которых ее когда-либо пытались искать. Ханстрем испытывал влияние экстракта глазных стебельков в многочисленных опытах на Uca, Palaemonetes и Penaeus, у которых их собственные глазные стебельки были удалены.
Он отметил, что экстракты не обнаруживали хроматофоротропной активности, если их получали от тех видов, у которых синусные железы расположены в головном отделе близко к мозгу, — Gebia и Hippa (Emerita). У некоторых других видов для получения экстрактов брали срезы глазных стебельков на разных уровнях. Экстракты оказались при этом активными во всех тех слу-
Хроматофоры иизменение окраски
545
чем у животных
чаях, когда срезы захватывали синусную железу. Остальные части стебелька были относительно неактивными. Используя различия в анатомическом строении глазных стебельков, Ханстрем подобрал ряд вариантов таким образом, что каждый из остальных органов стебелька (помимо синусной железы) в каком-то случае находился в неактивном или почти неактивном участке стебелька. Гистологические признаки секреторной активности имелись только в одном органе, так называемом Х-органе. Однако срезы, содержавшие этот орган, но не содержавшие синусной железы, не обнаруживали хроматофоротропной активности, а удаление Х-органа из глазного стебелька не вызывало снижения активности. На основании всего этого был сделан вывод о том, что синусная железа в глазном стебельке — единственный источник гормона, регулирующего распределение черного и красного пигментов у изученных животных.
Заключения Ханстрема подтвердил Браун [32], которому удалось выделить синусную железу и приготовить из нее экстракты. Эти экстракты обладали почти такой же активностью, как и глазной стебелек в целом, хотя их объем составлял лишь около 1 % всей ткани стебелька. При введении экстракта интенсивность реакции оказывалась всего лишь на 20% ниже, с нормальным глазным стебельком. Это было проверено как на красных хроматофорах Palaemonetes, так и на черных хроматофо-рах Ucci. Таким образом, синусная железа в глазном стебельке служит основным источником гормонов для этих двух весьма различных хроматофоров. Пересадка синусной железы в вентральный брюшной синус Palaemonetes приводит к побледнению окраски животного. Это побледнение сохраняется в 100 раз дольше, чем при инъекции экстракта в.количестве, эквивалентном одной железе.
Общепризнано, что синусные железы не являются местом синтеза хроматофоротропных гормонов. Скорее можно думать, что эти гормоны образуются в телах нейросекреторных клеток, входящих в состав комплекса, именуемого Х-органом (см. гл. 21). Синусная железа, образованная расширенными окончаниями этих нейросекреторных клеток, представляет собой только место накопления и освобождения активных гормонов. Такая функциональная связь между Х-органами и синусными железами, первоначально установленная на основе изучения гормонального регулирования линьки, анализа морфологических связей и наблюдений над фактическим прохождением гранул секрета в синусную железу по системе Х-орган—аксоны, была экспериментально подтверждена тем фактом, что удаление Х-органа из креветки Leander (Palaemon) подавляет все цветовые реакции (черный пигмент животного остается при этом в диспергированном состоянии) [234].
Комплекс, состоящий из Х-органа и синусной железы, вырабатывает более одного хроматофоротропного гормона. Впервые ^5—2067
546
Глава 23
это было обнаружено при изучении влияния, которое оказывали экстракты из глазных стебельков ракообразных семи родов на концентрирование красного пигмента у Palaemonetes и диспергирование черного у Uca [44]. Отношение эффективности действия экстрактов на хроматофоры двух типов (черные Uca и красные Palaemonetes) было обозначено через U/P.
Величины U/P, найденные для стебельков и синусных желез из разных источников, неодинаковы. Так, например, эта величина для синусной железы Crago (Crangon) относительно высока, а для синусной железы Palaemonetes, напротив, низка. Для Uca получено промежуточное значение. Величины U./P у семи изученных видов не обнаруживают какой-либо корреляции с размерами животных или с относительной эффективностью экстрактов (при испытании их на хроматофорах Uca). Для объяснения этих данных предполагают, что синусные железы отличаются друг от друга по относительному содержанию двух веществ: 1) гормона, вы-зывающего преимущественно потемнение у Uca (Uca darkening hormone, UDH), и 2) гормона, вызывающего преимущественно посветление у Palaemonetes (Palaemonetes lightening hormone. PLH). Эта гипотеза получила подтверждение, когда было уста новлено, что из синусных желез животных каждого вида можно получить две активные фракции: одну — растворимую в спирте, и другую — нерастворимую в нем. Первая фракция характеризуется очень низким значением U/P (что указывает на преобладание PLH), а вторая —* высоким (соответствует большему содержанию UDH). Надлежащим . образом комбинируя эти фракции, можно получить то значение U/P, которое характерно для железы данного вида. Эти результаты показывают, что все виды, представляющие группы I, II и III, содержат два хроматофоротропных гормона глазного стебелька, которые присутствуют в разных соотношениях.
Синусные железы у изученных ракообразных из группы III не обладают активностью особого типа, присущей железам видов, входящих в группы I и II. Экстракты из глазных стебельков и си нусных желез этих последних видов, если их ввести Crago (Cram gon), вызывают у него уже через 3—4 мин посветление тельсона и уроподов [37, 52]. Синусные железы видов группы III этой способностью не обладают, хотя их глазные стебельки способны вызывать посветление самого тела Grago, точно так же как и глаз ные стебельки двух других групп. Вещество, имеющееся у живот ных групп I и II, но отсутствующее у представителей группы III названо гормоном, вызывающим посветление «хвоста» Crago (Crago «tail»-lightening hormone, CTLH). Хотя в синусных желе зах ракообразных встречается не менее трех хроматофоротропных гормонов, полученные данные можно объяснить одним из следующих допущений: 1) виды животных, принадлежащие к группам I и II, имеют три активных вещества, а представители группы III —
Хроматофоры и изменение окраски___________547
только два; 2) все виды содержат только два вещества, но физиологические свойства одного из этих веществ у групп I и II, с одной стороны, и у группы III — с другой, различны.
Глазные стебельки — не единственное место образования хроматофоротропных гормонов у ракообразных. Хорошо известно, что невозбужденные особи Crago, у которых глазные стебельки удалены, довольно часто обнаруживают случайные изменения окраски. Раздражая перерезанные концы зрительных нервов электрическим током или каким-либо другим способом, можно вызвать у этих животных цветовые реакции в результате образования циркулирующих в крови веществ. При этом у ракообразных группы I окраска бледнеет. Однако, в то время как обычно под влиянием гормонов синусной железы белый пигмент диспергируется, в этом случае он, наоборот, концентрируется. Потемнение у представителей группы III в аналогичном опыте также связано с концентрированием белого и диспергированием черного пигмента в отличие от того, что происходит у тех же животных под влиянием гормонов синусной железы. Реакции Crago (группа II), по-видимому, более сложны, характер ответа этого животного на раздражение электрическим током концов перерезанного зрительного нерва позволяет предположить, что у ракообразных существует еще один дополнительный источник хроматофоротропных гормонов. При слабом раздражении темнеет все тело животного, при более интенсивном темнеют только тельсон и уроподы, а остальная часть тела, наоборот, светлеет. Очевидно, какие-то хромато-форотропные гормоны, вырабатываемые вне глазных стебельков, могут в одних случаях действовать как антагонисты веществ, вырабатываемых синусной железой, а в других — как их синергисты.
Инъекция экстрактов из органов центральной нервной системы некоторых видов группы I вызывает посветление у животных с удаленными глазными стебельками и концентрирование белого пигмента (рис. 23-11) [26, 51, 84, 88, 149, 169]. Экстракты из органов нервной системы Uca также стимулируют диспергирование красного и концентрирование белого пигмента. Экстракты, полученные из Crago (группа II), вызывают потемнение тельсона и Уроподов, посветление остальной части тела и концентрирование белого пигмента. Следовательно, инъекция экстрактов из органов Центральной нервной системы животным с удаленными глазными стебельками приводит во всех трех группах к тем же результатам, к каким приводит сильное раздражение стебельков глаз у нормальных животных.
Активность, обусловливающая потемнение тельсона и уропо-Дов, а также посветление тела у Crago, сосредоточена главным образом в органе головного отдела, так называемом посткомиссуральном органе (гл. 21) [33, 170]. Однако и остальная часть нервной системы тоже обладает известной активностью, ответственной за посветление тела. Этот факт, а также то обстоятельство, что 35*
1
548
Глава 23
растворимая в спирте фракция тритоцеребральных комиссур вызывает сильное посветление тела и не вызывает потемнения хвоста, тогда как нерастворимая в спирте фракция вызывает сильное потемнение и тела, и хвоста, указывает на присутствие в нервной системе Crago двух хроматофоротропных гормонов (рис. 23-12). Один из них вызывает посветление тела и не вызывает посветле-ния хвоста у Crago; поэтому он был назван гормоном, вызывающим посветление тела Crago (Crago body-lightening hormone,
.„Группа I , Группа II Группа III
(Palaemonetes) (Crago) (Uca)
Начальная Конечная
Рис. 23-11. Схема, иллюстрирующая окраску ракообразных и состояние преобладающих хроматофоров в каждой из трех различно реагирующих групп при удалении глазных стебельков (А) и при последующем введении экстрактов из органов нервной системы (Б) [35].
•---> цельный экстракт (водорастворимый материал);--—> фракция, нерастворимая
в спирте;----—> фракция, растворимая в спирте.
CBLH); другой гормон в присутствии CBLH приводит к потемнению только хвоста, но в отсутствие CBLH вызывает потемнение всего тела; он получил название гормона, вызывающего потемнение Crago (Crago darkening hormone, CDH). Резкое различие в реакциях, наблюдаемых у этих видов при сильном и слабом раздражении концов перерезанного зрительного нерва, есть, по-видимому, следствие освобождения обоих веществ при сильном раздражении и только одного (CDH) —при слабом.
Сравнительное изучение влияния на Crago экстрактов из нервной системы различных видов группы I позволяет заключить, что все эти виды содержат и CBLH и CDH, хотя ни у одного из них локализация данных веществ не совпадает с их локализацией у Crago [42]. Обычно эти гормоны распределены по всей нервной системе (Palaemonetes, Cambarus, Homarus) или сосредоточе-
Хроматофоры и изменение окраски
549
Нь1 в определенных ее участках, отличных от тритоцеребральных комиссур (например, в задней части грудной цепочки у Pagurus и других Anomura). В нервной системе некоторых видов, особенно в некоторых участках нервной системы определенных видов, активность CDH сильно превышает активность CBLH. Экстракты из этих участков вызывают потемнение у Crago (у некоторых глазные стебельки удалены) столь же эффективно, как и спирторас-
Рис. 23-12. Внешний вид трех особей Crago (первоначально имевших почти одинаковую окраску) после введения им трех различных растворов [41].
А. Морская вода (контроль, изменений не наблюдалось). Б. Цельный экстракт (водорастворимая фракция тритоцеребральных комиссур). В. Нерастворимая в спирте фракция.
творимая фракция тритоцеребральных комиссур Crago (например, брюшной цепочки омара и рака).
Активность CDH брюшной цепочки омара (рис. 23-13), по-видимому, локализуется главным образом в оболочке, а не в ганглиозных клетках или нервных волокнах. Представители группы I в отличие от представителей группы III, очевидно, не содержат CDH ни в центральной нервной системе, ни в какой-либо иной части тела. Однако наличие вещества, вызывающего потемнение У Crangon, можно обнаружить, если подвергнуть глазные стебельки гель-фильтрации, при которой это вещество отделяется от естественного красителя [96]. В глазных стебельках Crangon CDH был обнаружен с помощью этой же техники [279]. Активность CBLH обнаружена в нервной системе всех изученных ракообразных.
Установлено, что у животных из рода Uca (группа III) имеется в нервной системе не менее двух хроматофоротропных гормонов [260]. Один из них вызывает диспергирование черного пигмента, а Другой — концентрирование белого. На это указывает сравнение относительного влияния экстрактов из мозга, околопищеводных коннектив и грудной цепочки животных, у которых были удалены
550
Глава 23
глазные стебельки. Экстракты из мозга и грудной цепочки вызы-
вают сильное диспергирование черного и одновременно белого. пигментов (или вообще не влияют на последний, если он уже находится в диспергированном состоянии). Экстракты же из коннек-
тив обусловливают слабое диспергирование черного пигмента и концентрирование белого. Эти различные результаты нельзя воспроизвести простым подбором-концентраций экстрактов. Имеющиеся данные явно указывают-на присутствие двух хроматофоротропных гормонов. Существуют следующие две возможности: 1) экстракты всех трех тканей вызывают диспергирование черного и концентрирование белого пигмента, но мозг и грудная цепочка содержат еще какое-то вещество антагонистического действия, которое вызывает диспергирование белого пигмента; 2) все экстракты вызывают диспергирование и белого,, и черного пигментов, но только-
коннективы содержат вещества,
Рис. 23ч13. Доказательство двойного (регулирования функции красных хро-матофоров у Palaemonetes [51].
Реакция красных хроматофоров на введение экстрактов посткомиссуральиых органов и брюшной нервной цепочки. У животных удаляли глазные стебельки и вводили им экстракт посткомиссуральных органов, а затем (через 30 мин) одной половине животных (I) вводили экстракт брюшной нервной цепочки, а другой (//) — морскую воду (эти животные служили контролем).
под влиянием которых концентрируется белый пигмент. Экспериментально показано, что реализуется первая возможность [252]. Гормон, вызывающий концентрирование черного пигмента, найден у Uca. Следовательно, распределение черного-пигмента контролируется двумя антагонистическими гормонами.
Клетки, содержащие красный пигмент, также контролируются
двумя веществами, диспергирующими и концентрирующими пигмент [34]. Вещество, диспергирующее красный и белый пигменты в глазных стебельках Uca, было отделено от вещества, концен-
трирующего пигмент, но пока не ясно, может ли то или другое диспергировать (или концентрировать) более чем один из указанных пигментов [95]. Гормоны, концентрирующие красные и белые пигменты в глазных стебельках Crangon, вероятно, отличаются друг от друга [279].
Многочисленные опыты с перекрестной инъекцией у Crago, Uca и Palaemonetes, в которых изучалось распределение в нервных
Хроматофоры и изменение окраски 551
системах различных видов животных гормонов CDH и CBLH, также гормонов, вызывающих диспергирование черного (Uca dispersing hormone, UDH) и концентрирование белого пигмента v Uca (Uca white-concentrating hormone, UWCH), довольно ясно показали, что CDH и UDH не могут быть одним и тем же веществом. То же относится, по-видимому, к CBLH и UWCH. Правда, последнее еще не доказано окончательно. Быть может, CBLH и yWCH все-таки идентичны, так как существует известный качественный параллелизм в распределении этих веществ у трех изученных видов. Однако некоторые количественные различия в активности заставляют все же сомневаться в их идентичности.
До тех пор пока мы не научимся полностью отделять эти активные вещества друг от друга, определение индивидуальной роди каждого из них будет наталкиваться на очень большие трудности. В ранних попытках разделить хроматофоротропные вещества ракообразных использовалась методика электрофореза на бумаге [88, 93, 173, 174]. С помощью этой методики было установлено, что гормоны из мозга и околопищеводных коннектив рака Cambarellus, диспергирующие и концентрирующие красный пигмент, имеют при pH в интервале от 7,4 до 7,5 противоположные заряды, — и + соответственно [88, 93]. Позднее были предприняты попытки выделить и очистить хроматофоротропины у ракообразных, используя гель-фильтрацию, ионообменную хроматографию и диск-электрофорез [78, 167, 250].
Гормон, изолированный из глазных стебельков Pandalus borealis, концентрирующий красный пигмент, представляет собой пептид, состоящий из восьми аминокислот: Асп, Глу, Гли, Лей, Фен, Про, Сер и Три [78]. Гормон, который вызывает диспергирование пигмента в меланофорах Uca, также, по-видимому, пептид, поскольку он инактивируется протеолитическими ферментами [19, 236]. Он инактивируется также окисляющими веществами, такими, как перекись водорода и надмуравьиная кислота, причем активность окисленного гормона может быть полностью восстановлена обработкой его тиогликолятом [251].
При длительной адаптации раков к определенному фону накапливается, по-видимому, тот гормон (потемнения или посветле-кия), который в данных условиях не используется. При изменении фона количество пигмент-диспергирующих и пигмент-концентри-РУющих гормонов в крови меняется. Например, у рака, находящегося на^ черном фоне, в крови больше гормона, диспергирующего 9yaggjII] пигмент> чем У рака, находящегося на белом фоне [94, Таким образом, в настоящее время мы знаем только, что первая система десятиногих раков содержит по меньшей мере три или четыре хроматофоротропных гормона. Их роль в общих хромати-ских реакциях целостного организма для многих видов еще не Женена. Представляется правдоподобным, однако, что за слож
552
Глава 23
ное управление пигментными системами ракообразных ответственны хроматофоротропные гормоны нервной системы, а также гормоны синусных желез.
Характер изменений меланофоров у равноногих Ligia oceanica при переходе от света к темноте, а также при смене фона позволяет предположить, что эти пигментные клетки находятся под двойным гормональным контролем, т. е. что один гормональный фактор вызывает их потемнение (вещество В), а другой — посветление (вещество W) [285]. Хроматофорные системы равноногих очень разнообразны по своей способности реагировать на изменения фона или освещенности. Установлено, что введение экстрактов из тканей головы животного вызывает концентрирование темного пигмента у многих видов Sphaeroma, Idothea и Ligia [59, 205, 211], у некоторых других видов Ligia оно приводит к диспергированию темного пигмента [83]. Есть данные о том, что у Tracheli-pus имеются гормоны обоих типов [196]. Таким образом, опыты с введением экстрактов из тканей головы подтверждают предположение о том, что темные хроматофоры у равноногих регулируются, как и у многих других ракообразных, парой антагонистических гормонов. Так как после удаления органа Белончи (тесно связанного с оптическими долями и синусными железами у Sphaeroma и, вероятно, гомологичного Х-органу десятиногих раков) пигмент остается в полностью диспергированном состоянии, было высказано предположение, что именно этот орган служит источником вещества, вызывающего концентрирование пигмента [240].
У представителя ротоногих Squilla [101, 171] экстракты из синусных желез посткомиссуральных органов и органов центральной нервной системы вызывают диспергирование темного пигмента. Коричневато-черный хроматофорный пигмент Squilla, который имеет свойства оммохрома, концентрируется максимально в глазных стебельках особей.
Эксперименты, поставленные с целью выяснить механизм действия хроматофоротропных гормонов ракообразных на молекулярном уровне, показали, что гормоны, концентрирующие красный пигмент Palaemonetes, вызывают гиперполяризацию трансмембранного потенциала эритрофора по мере концентрации пигмента [111]. Первичное действие этого гормона, по-видимому, состоит в стимуляции натриевого насоса, который вызывает замену ионов натрия внутри хроматофора на ионы калия снаружи. Это приводит к высокому отношению К+: Na+ и в известной степени вызывает запуск механизма концентрирования красного пигмента [91] -
Позвоночные. Хроматофорные системы всех позвоночных (у которых такие системы имеются) в основных чертах сходны. Представляется целесообразным рассмотреть эволюцию хроматофорных систем внутри каждой систематической группы позвоночных в отдельности.
Xроматофоры и изменение окраски
553
Амфибии. На ранних стадиях развития у амфибий можно обнаружить период, когда у них имеются только первичные реакции на свет: животные темнеют на свету и светлеют в темноте. Впервые такие реакции описал Бабак [14] для очень молодых аксолотлей. Позднее их наблюдали у очень молодых особей Rana pi-piens [148] и Ambystoma [185]. Глаза не принимают участия в этих реакциях [311]. Было высказано предположение, что в данный период развития глаза еще не функционируют. У других амфибий (к ним относятся Bombinator и Hyla [14], а также Хепо-pus [311]), вероятно, имеют место вторичные реакции, обусловленные раздражением органов зрения сразу после вылупления. Первичные ответы при этом остаются, но вторичные реакции доминируют.
При исследовании физиологии изменения окраски у амфибий какой-либо непосредственной нервной регуляции меланофоров кожи обнаружить не удалось Давно известно, что бледная окраска у гипофизэктомированных головастиков сохраняется неопределенно долгое время [286]. Этот факт служит веским доводом в пользу того, что в гипофизе у этих животных вырабатывается гормон, регулирующий дисперсию меланина. Хогбен и Уинтон [146, 147] тщательно изучили связь гипофиза с изменениями цвета у Rana. Оказалось, что после гипофизэктомии животные становились бледными и затем утрачивали способность изменять цвет. Удаление передней доли гипофиза не сопровождалось сколько-нибудь заметным изменением реакции на цвет фона. Вытяжки из задней доли очень активно вызывали потемнение у бледноокра-шенных лягушек. Попытки вызывать специфические реакции хроматофоров раздражением или, наоборот, перерезкой нерва оказались безуспешными. Все это позволило Хогбену и Уинтону сделать вывод, что изменения окраски у Rana регулируются одним гормоном, образующимся в задней доле гипофиза, а концентрация этого гормона в крови в свою очередь регулируется раздражителями, поступающими из внешней среды через глаз. Это заключение было подтверждено в опытах по переливанию крови от темных особей Rana светлым [233].
Позднее очень подробные исследования, объектом которых послужила Xenopus [144, 145], также не обнаружили прямой иннервации меланофоров. У Xenopus, как и у Rana, гипофизэктомия сопровождается посветлением окраски, а инъекция экстрактов из задней доли гипофиза приводит к ее потемнению. Детальное кри-ическое изучение характеристик изменения меланофоров при перенесении Xenopus с белого фона на черный и обратно, с черного Т?На в темноту и обратно и с белого фона в темноту и обратно Ни^С 23-14) показало, что наблюдаемые явления нельзя объяс-Га ь> исходя из существования лишь одного вещества. Предпола-НазТ’ ЧТ°“ ГИП0Физ вырабатывает два гормона, один из которых, ванный веществом В, управляет дисперсией меланина (мела-
554
Глава 23
нофорстимулирующий гормон, melanophore-stimnlating hormone, MSH), а другой, названный веществом W, влияет на концентрирование меланина (меланин-концентрирующий гормон, melanineconcentrating hormone, МСН). Все полученные данные можно объяснить усилением секреции вещества W при стимуляции периферических элементов сетчатки и усилением секреции вещества В при стимуляции центральных элементов сетчатки (рис. 23-15),
Рис. 2344. Изменения состояния меланофоров у Xenopus при перенесении животного с белого на черный фон и обратно-, а также с черного фона в темноту [145],
Более того, из полученных данных следовало дополнительное । предположение, что вещество W выделяется в кровь п исчезает ( из нее медленнее, чем вещество В. Эти два вещества обладают ' антагонистическим действием. Согласно данной гипотезе, веще- ' ство В всегда секретируется при освещении, но при слабом осве- • щении и в темноте секреция ослабляется. Механизм реакции на белый и черный освещенный фон включает изменение количества вещества W, следовательно, при этом изменяется отношение B/W.
Очень медленные реакции, наблюдаемые при перенесении животного с освещенного белого фона в темноту и обратно, можно, ' по-видимому, объяснить соответственно уменьшением или увеличением содержания обоих этих веществ-антагонистов одновремен-| но. При переносе из темноты на освещенный черный фон мелано- | форы проходят через такую фазу, в которой пигмент диспергирован в большей степени, чем обычно на таком фоне. Этот факт можно объяснить более быстрой секрецией вещества В по сравнению с веществом V/. Затем по мере увеличения количества вещества W до нормы состояние меланина приближается к характерному конечному состоянию дисперсии. Точно так же при перенесении с освещенного черного фона в темноту меланофоры проход^ , через фазу большего концентрирования пигмента, чем в конечг,,А' состоянии, очевидно, благодаря более быстрому уменьшению держания вещества В по сравнению с веществом W.
Xроматофоры и изменение окраски
555
Прямое доказательство существования вещества W у Xenopus было получено из наблюдений над реакциями животных на внешние раздражения после различных операций на гипофизе [145]. При удалении передней доли гипофиза животное отвечает на раздражение точно так же, как нормальное животное. Когда же удаляют переднюю долю и область воронки, пигмент в меланофорах максимально диспергирован и реакции
Рис. 23-15. Схемы, иллюстрирующие две гипотезы гормональной регуляции мелаиофоров у Xenopus [145].
Одна гипотеза предполагает существование механизма с одним гормоном (А), другая — с двумя (Б).
на фон отсутствуют. При удалении задней доли у животных наблюдается максимально возможное побледнение окраски. Источник гормонов, вызывающих потемнение, несомненно, расположен в промежуточной доле гипофиза [12, 145]. Удаление всего гипофиза приводит пигмент в слегка диспергированное состояние и сопровождается утратой способности реагировать на раздражение. Все эти факты подтверждают гипотезу о существовании двух активных веществ, причем область воронки, видимо, ответственна за производство вещества W, а задняя или промежуточная доля— за производство вещества В. В пользу существования двух активных факторов можно привести данные, полученные в опытах с введением вещества В гипофизэктоми-рованным особям Xenopus, а также нормальным животным и животным, у которых удалена только задняя доля (рис. 23-16). Как и следовало ожидать, в двух последних случаях для вызывания заданного
ответа требуется большая доза, чем у животных с полной гипо-физэктомией, у которых меньшая доза вызывает такое же действие. Последнее наблюдение наиболее логично объясняется тем, что антагонист веществу MSH отсутствует.
Однако существование гормона гипофиза, вызывающего по-^-Ветление у Xenopus, было поставлено под сомнение, поскольку не исключено, что Хогбен и Слоум не полностью удалили промежуточную долю гипофиза. Когда это было сделано, меланин действительно полностью концентрировался, а не оставался на некоем ромежуточном, субмаксимальном уровне, как об этом сообщили
556
Глава 23
эти авторы [155]. Выделение MSH, вероятно, регулируется в Хе-nopus возбуждением и торможением нервной системы [156].
МСН был выделен путем фракционирования вытяжек, содержащих MSH, этиловым спиртом [70, 73]; полагают, что источником МСН служит гипоталамус.
MSH вызывает потемнение у Rana pipiens, R. clamitans и Ну-la, а адреналин приводит, напротив, к посветлению их окраски
Рис. 23-16. Реакция меланофоров Xenopus, помещенных на белый фон, на инъекцию одинаковых доз экстрактов гипофиза [145].
I — реакция гипофизэктомированных животных; II — реакция нормальных особей; III — реакция животных, у которых удалена только задняя доля гипофиза.
[204, 316]. Эти реакции имеют место также в изолированной коже лягушки in vitro [316]. Ионы натрия необходимы для MSH-pe- ‘ акции [207], тогда как иодацетат, подавляющий гликолиз, тормозит ее [316]. Тироксин вызывает потемнение окраски у Xenopus, что объясняется, по-видимому, непрямой стимуляцией секреции MSH [60]. АКТГ индуцирует дисперсию меланина у Hyla [70]. Некоторые данные указывают на то, что кожа лягушки сама содержит меланоцит-концентрирующий фактор [277]. Головастики Xenopus при помещении в темноту светлеют. Если же удалить у них шишковидное тело, то эта реакция не происходит. Это доказывает, что посветление окраски обусловлено мелатонином — гормоном, выделяемым этой железой в амфибиях [16, 17].
Аденозин 3': 5'-монофосфат (циклический АМФ) имитирует действие MSH на коже Rana [22]. По-видимому, обычно он служит посредником при действии MSH. Известно, например, что MSH стимулирует синтез циклического АМФ в коже лягушки [2].
Рыбы. Вероятно, лучше всего хроматофорная система и изменения окраски изучены у рыб. У них имеются хроматофоры нескольких типов. Наиболее обычны и заметны меланофоры. Шир0' ко распространены также ксантофоры, содержащие желтый пигмент, эритрофоры, содержащие красный пигмент, и иридофоры, содержащие белый пигмент. Кроме того, у многих рыб имеются
Хроматофоры и изменение окраски 557
хроматофоры, содержащие небольшие скопления мерцающих плоских кристаллов, которые придают им синевато-зеленую окраску, эти хроматофоры получили название иридосом.
Наблюдаемые у рыб реакции на свет и темноту, а также на белый и черный цвет фона обусловлены в основном активностью меланофоров. У рыб, так же как и у амфибий, на ранних стадиях развития наблюдается замена первичных цветовых реакций вторичными. Такое чередование в механизме реакций обнаруживается у молодых особей Регса и Salmo [68, 206], Macropodus [303] и Hoplias [197]. В то же время у многих других рыб стадия первичных реакций, по-видимому, отсутствует, у них имеют место сразу вторичные реакции на свет. К числу таких рыб относятся Fundulus [126, 318], Lebistes, Xiphophorus, Gambusia [303], Scyllium [311] и Mustelus [219]. У многих видов рыб, у которых в норме имеют место вторичные реакции на свет, после ослепления происходит возвращение к первичным реакциям [303].
Вторичные цветовые реакции у рыб полностью зависят от глаз. В этих реакциях участвуют нервные пути, идущие в центральную нервную систему, а оттуда или к эндокринным железам, действующим на хроматофоры посредством выделяющихся в кровь гормонов, или через эфферентные нервные пути непосредственно к хроматофорам, где нервными окончаниями выделяются химические медиаторы. Гуморальный и нервный механизмы во многих случаях могут действовать совместно, причем регулирование обоих механизмов отличается большим разнообразием. Паркер [228], ведущий исследователь изменений окраски у животных, делит рыб на три группы по степени прямой иннервации меланофоров. Двунейронные хроматофоры имеют двойную иннервацию с волокнами для диспергирования и концентрирования пигментов. Мононейронные хроматофоры имеют одиночную иннервацию, активность которой приводит всегда к концентрированию пигмента. Вторичные реакции анейронных хроматофоров, не имеющих иннервации, стимулируются исключительно химическими веществами, присутствующими в крови и лимфе.
Для огромного большинства исследованных к настоящему времени костистых рыб характерны, по-видимому, двунейронные меланофоры. 'С помощью электрического раздражения определенных точек или путей центральной нервной системы или же периферических нервов легко продемонстрировать характер действия нервов одного из этих типов, а именно тех, которые управляют концентрированием пигмента. Площадь поверхности кожи, окраска которой бледнеет при таком раздражении, соответствует величине участка кожи, иннервированного этими нервными элементами. Более того, после денервации какого-либо участка кожи раздражение нервных путей до точки денервации не будет сопровождаться посветлением окраски в этой области, хотя в прилегающих Участках кожи этот процесс может происходить. Таким образом,
558
Глава 23
побледнение окраски, сопровождающее раздражение нервов у жи« водных, обусловлено локализованными реакциями в области нервных окончаний, а не действием легкодиффундирующих веществ, переносимых с кровью. Как полагают [225], действие волокон, стимулирующих концентрацию пигмента, осуществляется посредством адреналиноподобного вещества, поступающего из нервных окончаний.
Существование волокон второго типа, стимулирующих диспергирование пигмента, было выявлено в опытах на меланофорах
Рис. 23-17. Возникновение темной полосы на хвостовом плавнике адаптированной к белому фону особи Fundulus после перерезки радиальных нервов (Л) и вторил-ное потемнение исчезнувшей полосы после повторного разреза, сделанного дистальнее первого (Б) [216].
хвостового плавника у многих костистых рыб. Если у костистой рыбы, например у Fundulus, помещенной на освещенный белый фон, перерезать группу радиальных нервов хвостового плавника, то иннервируемая этими волокнами полоса хвоста быстро темне-ет, а затем, по истечении нескольких дней, вновь бледнеет [216] (рис. 23-17). Если теперь дистально и на некотором расстоянш от первого разреза вновь перерезать эти волокна, то побледневшая полоса опять пройдет этот период потемнения. Такое поведс ние позволяет высказать предположение, что наблюдаемая реак ция меланофора является результатом повторного раздраженш диспергирующих пигмент нервных волокон, которые были рассе чены и возбуждены при первой операции. Наблюдаемое явление нельзя объяснить тем, что были рассечены волокна, стимулирую щие концентрирование пигмента, как это можно было предполо жить, судя по реакции на первый разрез. Если понижением темпе ратуры блокировать нервное проведение между местом поперечно го разреза и меланофорами, то реакция исчезает [221]. Подобное потемнение прежде бледной полосы при повторной перерезке нервов исследователи наблюдали у многих костистых рыб (Holoceni rus [222], Parasilurus [193], Pterophyllum [305], Ameiurus [226] и Gobius [114]). Активацию меланин-диспергирующих волокон хвоста у сомика также можно вызвать электрическим раздражением [232].
Целый ряд других данных как морфологического, так и физиологического характера послужил дальнейшим подтверждением
Хроматофоры и изменение окраски 559
гипотезы о двойной иннервации меланофоров костистых рыб. Бал-ловиц [18] еще в 1893 г. ясно показал, что меланофоры окуня несут нервные окончания более чем от одного волокна. Таким образом, имеется анатомическое обоснование для заключения, к которому впоследствии пришли на основании результатов физиологических экспериментов. В электронно-микроскопических исследованиях также обнаружили пресинаптические окончания меланофоров Lebistes двух типов: одни содержали пузырьки диаметром 500 А, а другие — диаметром 1000 А [120]. Кроме того, был проведен анализ реакций хроматофоров на границах денервированных хвостовых полос Fundulus [201] и на участках вблизи регенерирующей границы нервных волокон во время реиннервации денервированных полос [6], когда животное темнеет на черном фоне и светлеет на белом. Эти наблюдения позволили предположить, что многие меланофоры, расположенные на регенерирующих границах, имеют нервные волокна только одного типа, а не обоих сразу, как это имеет место в норме. В некоторых из этих меланофоров происходит быстрое концентрирование пигмента и очень медленное диспергирование его, в других — наоборот. Результаты изучения действия различных лекарственных препаратов на хроматофоры Phoxinus [125] и Fundulus [282] также подтверждают предположение о двойной иннервации меланофоров. Циклический АМФ оказывает меланин-диспергирующее действие в Fundulus [208]. Нервные передатчики (пигмент-диспергирую-щие и пигмент-концентрирующие), возможно, действуют, управляя уровнем циклического АМФ в этих меланофорах.
По-видимому, диспергирующие волокна действуют на мслано-форы посредством выделения ацетилхолина, который, как известно, вызывает диспергирование меланина у рыб при условии, что его разрушение предотвращается инъекцией эзерина. В самом деле, как показал анализ содержания ацетилхолина в коже у адаптированных к черному особей Ameiurus [225] и Ophiocephalus. это вещество присутствует в концентрации около 0,078 мкг на 1 г кожи. Это примерно соответствует той концентрации ацетилхолина, которая при инъекции его рыбе, обработанной эзерином, еще не токсична, но уже вызывает хорошо выраженную реакцию со стороны меланофоров. Однако некоторые исследователи утверждают, что ацетилхолин или вообще не вызывает. [2], или в лучшем случае вызывает слабую меланпн-дпспергирующую реакцию у костистых рыб [135].
Мононейронные меланофоры обнаружены у акул Mustelus и Squalus. Если сделать поперечный разрез на конце грудного плавника акуйы, имеющей промежуточную окраску кожи, то дистальнее места разреза появится светлая полоса. После того как такие полосы потемнеют, можно вызвать их повторное посветление, сделав новый разрез [220]; они вновь появляются также при электрическом раздражении кожных нервов [218]. Эти полосы
560
ifлава 23
повторяют распределение перерезанных нервов и не обязательно соответствуют расположению кровеносных сосудов. Более того, появление светлых полос можно вызвать поперечным разрезом на плавнике, к которому прекращен доступ крови. Все эти факты указывают на нервную регуляцию меланофоров, функция которой состоит в стимуляции концентрирования пигмента. Пока нет каких-либо данных, говорящих о том, что у этих животных имеются диспергирующие пигмент нервные волокна.
Паркер [227] постулировал, что пигмент-концентрирующие волокна влияют на меланофоры, выделяя химический медиатор, который он назвал селахином. Это вещество можно выделить путем экстракции кожи животного (обладающего нормально бледной окраской) эфиром или оливковым маслом, но не водой. Введение такого экстракта адаптированным к темному животным вызывает временное посветление окраски, очень медленно распространяющееся от места инъекции.
У других исследованных акуловых рыб, Raja [223], Scyllium [320], и у миноги Lampetra [321] непосредственной иннервации меланофоров не обнаружено; поэтому считается, что они обладают анейронными меланофорами.
У рыб вещества, циркулирующие в крови, обычно действуют совместно с нервной системой на вторичные реакции меланофоров, возникающие в ответ на световые раздражения. У более примитивных позвоночных, таких, как круглоротые и пластиножаберные, у которых имеются лишь анейронные хроматофоры, реакции обусловлены только действием гормонов. Среди этих гормонов важная роль принадлежит пигмент-диспергирующему фактору, выделяемому гипофизом; это вещество В Хогбена, или MSH (те-lanine-stimulating hormone) [322]. Оно выделяется задней долей гипофиза. Согласно наблюдениям Лундстрема и Барда [189], окраска гипофизэктомированных особей Mustelus бледнеет и остается такой впоследствии. Потемнение можно вызвать вновь инъекцией экстракта из задней доли гипофиза. Позже такой же эффект вещества задней доли гипофиза был обнаружен и у других пластиножаберных — Raja и Scyllium [142], а также у миноги Lampetra [321]. Введение шишковидного комплекса в Geotrea вызывает локальное побледнение, так же как и внутрибрюшинная инъекция мелатонина [69].
Имеются данные, говорящие о том, что у Scyllium и Raja второй нейрогуморальный фактор гипофиза действует как пигмент-концентрирующее вещество [142]. Данные о роли этого вещества, вызывающего побледнение окраски животного, были получены при изучении характеристик реакций меланофоров на изменения цвета фона и интенсивности света, а также при исследований влияния гипофизэктомии на состояние пигментов и характер их реакций в ответ на инъекцию экстракта задней доли. Общие методы получения экспериментальных данных и их обработки в этих
Хроматофоры и изменение окраски 561 опытах были такими же, как и при изучении регулирования меланофоров амфибий. Согласно бигуморальной концепции Хогбена и его коллег, состояние меланофоров у этих рыб определяется относительным содержанием MSH и МСН (меланин-концентриру-юший гормон, melanine-concentrating hormone) в крови в каждый данный момент, а оно в свою очередь зависит от того, что при раздражении сетчатки глаза происходит неодинаковое возбуждение ретинальными импульсами различных структур в дорсовен-тральном направлении; отсюда и различная скорость секреции этих двух веществ.
Гормон гипофиза, принимающий участие в диспергировании меланина, по-видимому, также участвует в большей или меньшей степени в нормальных изменениях цвета большинства костистых рыб. У угря Anguilla наблюдаются замедленные изменения цвета, для завершения которых требуется несколько дней [206]. У Anguilla, несмотря на наличие диспергирующих и концентрирующих нервных волокон, цветовые изменения, по-видимому, определяются в основном гормонами, причем полагают, что действие MSH дополняется действием МСН [310]. О том, что прямая иннервация играет определенную роль в изменениях цвета у угря, можно судить по реакции на цвет фона после гипофизэктомии.
У сомика Ameiurus MSH имеет несколько меньшее значение для нормальных изменений цвета. Гипофизэктомированные особи Ameiurus продолжают реагировать на белый или черный фон, но при этом на черном фоне достигается только промежуточная степень потемнения [5]. Вместе с тем инъекция экстракта задней доли приводит к тому, что животные полностью чернеют. Отсюда можно сделать вывод, что в нормальных реакциях на черную окраску фона переносимый с кровью MSH дополняет действие диспергирующих нервных волокон. Об активности МСН у этой рыбы данных пока не имеется. Нанностома Nannostomus beckfor-di anomalus имеет различные дневные и ночные метки. Мелатонин вызывает ночную окраску, концентрируя пигмент в одних меланофорах и диспергируя в других [254].
Многочисленные исследования, проведенные на карпозубой рыбе Fundulus, показали, что у нее преобладает нервный механизм управления меланофорами. Изменения окраски происходят очень быстро: через 1—2 мин достигается почти максимальное изменение цвета. У особей, подвергнутых гипофизэктомии, эти реакции сохраняются [195]. Более того, инъекция экстрактов задней Доли гипофиза особям с бледной окраской, помещенным на белый фон, не сопровождается значительным потемнением животных. Поскольку эти экстракты вызывают потемнение в денервированных участках кожи, приходится заключить, что обнаруженный в гипофизе этих животных MSH в норме оказывает некоторое воздействие на их меланофоры [8, 164]. Однако его влияние, вероят-н°, проявляется только в экстремальных условиях достаточно дли-36—2067
562
Глава 23
тельной адаптации к темному фону. У Macropodus, как показало изучение влияния гипофиза после нервной стимуляции меланофоров, MSH не принимал сколько-нибудь заметного участия в описываемых реакциях [308].
Все изученные к настоящему времени костистые рыбы, по-видимому, образуют определенную последовательность с точки зрения соотношения нервной и гуморальной регуляции хроматофоров у них. Была выдвинута не лишенная оснований гипотеза, согласно которой гуморальная регуляция является филогенетически
Рис. 23-18. Схема регулирующего механизма меланофоров у угря Anguilla [228]. ДС — дорсальная часть сетчатки; ВС — вентральная часть сетчатки; ЦНС — центральная нервная система; Адр. — адренэргические волокна; Хол. — холинэргические волокна: К. и Л. — кровь и лимфа; ДМ — диспергированный меланин, КМ — концентрированный меланин.
более старой формой регуляции по сравнению с нервной, появившейся в более поздний период эволюционного развития этих рыб [311]. Прямая нервная регуляция, обеспечивающая более быструю реакцию на цвет фона, стала возможной благодаря тому, что более быстрыми стали изменения самих меланофоров. Типичный механизм нервной регуляции меланофоров костистых рыб в схематическом виде представлен на рис. 23-18.
На основании анализа характеристик реакции меланофоров рыб на действие некоторых газообразных отравляющих веществ было выдвинуто предположение о наличии двух по-разному реагирующих типов меланофоров; у сомиков имеется один из них, а чешуйчатые рыбы содержат другой [245]. Ионы натрия, по-видимому, чрезвычайно эффективно вызывают диспергирование меланина у Fundulus [119]; их роль в данном случае сходна с той, которую они выполняют при действии MSH в лягушках.
Значительно меньше, чем о меланофорах, известно о регуляции эритрофоров, ксантофоров, иридофоров и иридосом. Эритрофоры рыбы-белки Holocentrus представляют собой быстро реагирующие
Хроматофоры и изменение окраски 563
эффекторные органы [222], с помощью которых рыба может изменить цвет от красного до белого примерно за 5 с и через 20 с вновь принять прежнюю окраску. Эти изменения вызываются при изменении цвета фона с белого на черный и наоборот. -Перерезка нервных стволов у рыбы, помещенной на белый фон, приводит к диспергированию красного пигмента. Через короткий отрезок времени красные области опять обесцвечиваются, но при повторном разрезе, сделанном дистальнее первого, вновь возникает потемнение, что говорит о наличии нервных волокон, управляющих диспергированием пигмента. Существование концентрирующих пигмент нервных волокон можно выявить путем электрического раздражения продолговатого мозга, которое приводит к быстрому концентрированию пигмента всех иннервированных эритрофоров. Экспериментально денервированные клетки не обнаруживают такой реакции. Адреналин вызывает концентрирование красного пигмента. Экстракты из гипофиза других рыб-белок при введении их нормально адаптированным к светлому или темному фону особям не оказывают никакого действия. Отсюда, видимо, следует, что эритрофоры Holocentrus в норме регулируются исключительно с помощью нервного механизма [284]. Напротив, эритрофоры Phoxinus находятся под контролем вещества, выделяемого гипофизом [125, 134].
Ксантофоры Fundulus имеют двойную иннервацию, включающую концентрирующие и диспергирующие нервные волокна [ИЗ]. В концентрировании пигмента, которое происходит, если эту рыбу трогать руками, принимает участие также гормональный фактор — по-впдимому, адреналин. Эта последняя реакция протекает одинаково быстро как в случае денервированных, так и в случае иннервированных ксантофоров. В то же время внутрибрюшинная трансплантация гипофиза особям, подвергнутым гипофиз - ^омии, вызывает диспергирование пигмента в денервированных не а сферах независимо от окраски фона. Такой имплантат преп тствует также нормальному концентрированию пигмента в иннервированных клетках, наступающему при помещении животного на иний пли белый фон [115]. Итак, можно, по-видимому, считать, что в норме на состояние хроматофоров у этой рыбы оказывают влияние несколько факторов. Меланофоры и ксантофоры у Fundulus реагируют на цвет фона независимо друг от друга [7]. Гипофиз-эктомия не оказывает действия на меланофоры Gillichthys, но приводит к концентрации пигмента в его ксантофорах. Пролактин стиммулирует диспергирование желтого пигмента ксантофоров [258]. Выделение пролактина обычно регулируется ингибиторным фактором, образующимся в мозге. Выделение MSH в Ictalurus, по-видимому, также контролируется определенными ингибиторами [163].
Многие рыбы, в том числе Fundulus, имеют слой белых отражающих хроматофоров, называемых гуанофорами или иридолей-36*
л
564 Глава 23
кофорами [112]; эти хроматофоры, видимо, принимают участие в физиологических изменениях цвета при приспособительных реакциях данной рыбы на фон. Они продолжают реагировать на цвет фона после гипофизэктомии, а также после введения большой дозы эрготамина (с целью снять реакцию сопутствующих меланофоров). Адреналин вызывает диспергирование пигмента, содержащегося в этих хроматофорах [210]. На основании экспериментов с Bathygobius, включающих гипофизэктомию и перерезку нервов, был сделан вывод, что активность иридолейкофоров подчиняется в значительной степени нервным регуляторным механизмам, хотя гормональное влияние также имеет место [116].
Иридосомы играют лишь пассивную роль в приспособительных цветовых реакциях, становясь более или менее заметными вследствие деятельности хроматофоров других типов. Обычно эти тела зеленого или синего цвета. Они весьма чувствительны к определенным раздражителям внешней среды, обратимо изменяя свой цвет в сторону красной части спектра при возбуждении и в обратную сторону при снятии возбуждения. Эта реакция прямая, в ней не принимают участия координирующие механизмы животного — ни нервные, ни гуморальные [107].
Рептилии. У рептилий, как и у рыб, система регулирования меланофоров характеризуется различными степенями сочетания гормональных и нервных влияний.
Клейнхольц [165] провел интенсивное изучение реакции меланофоров у Anolis. Окраска этих ящериц изменяется от ярко-зеленой до темно-коричневой, причем обычно первая окраска появляется у животных, помещенных в освещенный белый контейнер, а вторая — в освещенный черный. При перемещении с белого фона на черный реакция полностью заканчивается в течение 5—10 мин. Изменение цвета в обратную сторону обычно требует от 20 до 30 мин. Эта реакция на изменение окраски фона исчезает при двустороннем ослеплении животного. Однако ослепленные животные все еще способны изменять цвет (они темнеют на свету и светлеют в темноте), что обусловлено первичными реакциями.
После классических работ Брюке [53], исследовавшего изменения окраски у хамелеона, для которого он показал существование регулирования хроматофоров симпатической нервной системой, в течение многих лет продолжали считать, что хроматофоры рептилий в основном регулируются именно таким образом. В результате изучения Phrynosoma [253] возникли серьезные основания предполагать, что нервная система — не единственный путь регулирования цветовых изменений у рептилий. Было обнаружено, что окраска ящерицы Hemidactylus бледнеет после гипофизэктомии и что Anolis в результате такой же операции приобретает неисчезающую ярко-зеленую окраску [165] (рис. 23-19). Такие животные перестают темнеть при перенесении на черный фон или на яркий свет. Однако потемнение у них легко вызвать путем
Хроматофоры и изменение окраски
565
введения экстрактов, полученных либо из целого гипофиза рыб, либо из промежуточной доли гипофиза лягушек или рептилий. У гремучей змеи диспергирование пигмента меланофоров наступает в результате действия какого-то фактора гипофизарного происхождения [246].
Рис. 23-119. Цветовые реакции у Anolis [165].
Нормально окрашенная темная особь (А) и гипофизэктомированиая светлоокрашенная особь (Б).
Рассечение спинного мозга в нескольких местах или перерезка седалищного нерва не оказывают никакого влияния на нормальные цветовые реакции у Anolis. Попытки гистологическими методами обнаружить нервные окончания на меланофорах постоянно оканчивались неудачей. Оказалось далее, что пересаженные участки кожи очень скоро начинают приобретать способность к изменениям окраски, синхронным с изменениями ее у реципиента. При прекращении снабжения кровью любого участка кожи примерно через 15 мин наступает его побледнение.
Роль гипофиза и надпочечников у этих животных можно выявить путем электрического раздражения; при этом один электрод
566
Глава 23
помещают в рот животного, а другой — в клоаку. Если подвергнутых такому раздражению животных, у которых произошло побледнение окраски, поместить в темноту, то они приобретают постоянную темно-коричневую окраску, причем денервированные области реагируют точно так же, как иннервированные. Если таким же образом раздражать животных, подвергнутых гипофиз-эктомии, то они приобретают характерную пятнистую окраску, причем сами по себе гипофизэктомия и адреналэктомия не сопровождаются подобной реакцией. Более того, инъекция адреналина или экстракта собственных надпочечников животного в растворе Рингера вызывает появление характерной пятнистости. Животные после адреналэктомии светлеют на белом фоне. На основании всех этих, а также некоторых других наблюдений можно заключить, что у Anolis диспергирование меланина обусловлено действием MSH, образующегося в промежуточной доле гипофиза; постепенное исчезновение MSH из крови приводит к посветлению окраски. Быстрое обесцвечивание, возникающее обычно после электрического раздражения или возбуждения, по-видимому, является результатом вазоконстрикторного эффекта, а в некоторых случаях обусловлено действием адреналина. В условиях стресса стимуляция альфа-адренэргических рецепторов катехоламинами, приводит к возбуждению побледнения, за которым следует возбуждение потемнения вследствие стимуляции р-адренэргических рецепторов [130]. Диспергирование меланина в Anolis вызывается также циклическим АМФ [127]. Для действия MSH в Anolis необходимо присутствие кальция [306].
Влияние света на состояние меланофоров у интактных особей Anolis, видимо, незначительно, и действие его проявляется в основном через глаз.
Получены данные, свидетельствующие о том, что у представителя игуанид Phrynosoma на реакции меланофоров оказывает влияние как гуморальная, так и нервная регуляция. Ранние работы Редфилда [253], изучавшего этот вид, впоследствии были подтверждены и значительно развиты Паркером [224]. Обычная окраска Phrynosoma — серая, с характерными черными пятнами неправильной формы. Эти пятна не изменяют своего цвета, тогда как лежащие между ними области при соответствующем раздражении изменяют цвет от темно-серого до бледного серовато-белого. Эти изменения обусловлены главным образом активностью меланофоров. Потемнение окраски животного наступает примерно через 15 мин, посветление же занимает почти в два раза больше времени.
Установлено, что Phrynosoma темнеет на освещенном черном фоне, а также при низкой температуре и при очень сильном освещении. На освещенном белом фоне, при высокой температуре и в темноте Phrynosoma светлеет.
Меланофоры обычно иннервируются пигмент-концентрирующи-
Хроматофоры и изменение окраски,567’
ми нервными волокнами. Раздражение седалищного нерва вызывает посветление окраски соответствующей задней ноги. Электрическое раздражение неба или клоаки приводит к быстрообратимому посветлению всего тела животного. Если затем денервировать какой-либо участок — ногу или боковую часть — и вновь провести такую же стимуляцию, то все тело, за исключением денервированных областей, посветлеет, хотя меланофоры денервированных участков и не утрачивают способности концентрировать пигмент при введении адреналина. Пигмент-диспергирующие волокна у этого животного обнаружены не были.
После гипофизэктомии окраска Phrynosoma, так же как и у Anolis, бледнеет и остается такой неопределенно долгое время. Инъекция экстрактов гипофиза Phrynosoma вызывает сильное потемнение окраски у бледноокрашенных особей гипофизэктомиро-ванных животных. Более того, введение дефибринированной крови, взятой у особи с темной окраской, в ногу бледноокрашенного животного приводит к потемнению последнего.' Все эти факты убедительно доказывают участие MSH гипофизарного происхождения в реакции, ведущей к потемнению окраски животного. О действии пигмент-концентрирующего гормона можно также судить на том основании, что адреналин и экстракты надпочечников Phrynosoma вызывают сильное обесцвечивание темноокрашенных особей. Присутствие аналогично действующего вещества в крови адаптированных к свету животных проявляется в том, что их де-фибринированная кровь при инъекции в ногу темноокрашенной особи вызывает посветление последней. Уже давно было известно, что если животных перевернуть на спину, насильно удерживая их в этом положении, или каким-либо другим способом вызвать у них энергичное сопротивление, то окраска животных быстро светлеет. Сравнивая влияние денервации и прекращения доступа крови на появление подобного посветления, можно показать, что в этом процессе принимает участие переносимое с кровью вещество. Все эти наблюдения позволяют с очевидностью заключить, что побледнение окраски может быть вызвано факторами двух типов — нервных и гуморальных, — каждый из которых сам по себе способен вызвать такую реакцию.
Диспергирующий гормон гипофиза и концентрирующий гормон надпочечников действуют непосредственно на меланофоры; каждый из этих гормонов полностью проявляет свою активность после перерезки и последующей дегенерации нервных волокон.
Влияние температуры, света и темноты на меланофоры Phrynosoma также прямое; при этом меланофоры реагируют как независимые эффекторы. Если действовать раздражителями на специфические области полностью денервированных участков тела, можно вызвать локальные реакции. Поскольку реакция не исчезает даже после отмирания иннервирующих волокон, можно считать, что аксон-рефлексы не принимают в ней участия.
568
Глава 23
Координация активности меланофоров у хамелеонов Chamae-leo или Lophosaura в отличие от игуан происходит, по-видимому, исключительно за счет нервной регуляции [ 143]. «Перерезка нерва приводит к потемнению обычно иннервированной области. Эти нервы относятся к так называемой вегетативной нервной системе. С помощью электрического раздражения нервов легко вызвать концентрирование пигмента в меланофорах. Изменения, наблюдаемые после перерезки нервов, были объяснены исчезновением тонических импульсов, достигающих пигментных клеток [259, 323]. До сих пор не получено веских доказательств в пользу того, что пигмент активно диспергируется вторичной системой нервных волокон, как это, видимо, имеет место у многих костистых рыб.
О возможной роли гормонов в изменении цвета у хамелеонов почти ничего не известно, однако тот факт, что при действии на это животное попеременно света и темноты [53] у него появляется соответствующий рисунок, говорит против сколько-нибудь существенного значения гуморальных факторов.
В меланофорах интактных животных на свету происходит диспергирование, а в темноте — концентрирование пигмента. В темных участках, возникших в результате произведенной денервации, такие реакции отсутствуют. Зуннд и Эйр [323] использовали эти данные для доказательства того, что реакции меланофоров носят чисто рефлекторный характер. Паркер [224], однако, указал на возможность противоположного объяснения, которое состоит в том, что пигментные клетки у этих форм, возможно, реагируют непосредственно, когда их состояние не определяется каким-либо доминирующим механизмом.
Итак, у рептилий, как и у рыб, видимо, имеется целый ряд механизмов координации активности хроматофоров — от систем с гормональной регуляцией, которую, по-видимому, следует считать примитивной формой регуляции, к системам с совместным гормональным и нервным управлением и далее, вплоть до систем с преобладанием нервной регуляции.
Функциональное значение хроматофоров и изменения окраски
Поскольку реакции хроматофоров представляют собой главным образом реакции на цвет или на освещенность фона, естественно было предположить, что изменения цвета вносят значительный вклад в маскирующую окраску животного, облегчающую защиту или нападение, и таким образом увеличивают его шансы на выживание. К числу немногих примеров экспериментального подтверждения того, что изменение цвета действительно увеличивает вероятность выживания животных (кстати, эту точку зрения часто оспаривают), можно отнести данные Самнера [295], который показал, что рыбы, у которых было достаточно времени для адаптации и изменения окраски, реже становились добычей хищных
Хроматофоры и изменение окраски
569
птиц, чем неадаптированные животные. Другой пример представлен личинками насекомых Acrida и Oedipoda, которые меньше истребляются птицами, когда они находятся на фоне, соответствующем их приспособительной окраске [76]. Далее было четко показано, что рыбы, адаптированные к черному, чаще стремятся
Рис. 23-20. Выбор черного фона (%) для каждого из 8 видов подопытных рыб, адаптированных к белому (/) и черному (//) фону [46].
1 — Notropis blenniuss; 2 — Corregonus nigripinnis; 3 — Ericymba buccata; 4 — ’Notropls whypplei; 5 — Campostoma anomalum; 6 — Lepomis humilis- 7 — Pimephales noiatus; 8 — рыба* луна.
выбрать черный цвет фона, когда им предоставляется возможность выбора между белым и черным, чем особи, адаптированные к белому фону [46, 191] (рис. 23-20), причем у Ericymba быстрота изменения выбора при изменении фона приблизительно соответствует быстроте изменения окраски кожи (рис. 23-21).
У рыб с более быстрыми и выраженными изменениями окраски выбор, видимо, в большей степени определяется адаптацией к цвету фона, чем у животных с менее выраженными изменениями цвета. Рак Cambarus (Orconectes) также, видимо, способен адаптироваться и выбирать соответствующий фон [31].
Хроматофоры, по-видимому, несут также функцию защиты от вредного действия яркого света. У ряда животных, например у не
-570
Глава 23
которых пиявок и морских ежей, диспергирование содержащегося в хроматофорах пигмента происходит только на ярком свету и не связано с защитной адаптацией к определенному фону. Черный и белый пигменты краба Uca обычно диспергированы на свету в дневное время и концентрированы в темноте ночью частично благодаря присущему этим животным ритму, частично как реакция
Бремя после изменения фона, мин
'Pirc. 23-21. Скорость изменения выбора черного фона (%) при перенесении Ericymba с черного на белый фон и наоборот [46].
на общую освещенность. Меланофоры Uca наиболее чувствительны к части спектра, соседней с ультрафиолетовой (300—400 нм). Эта часть спектра более чем в 200 раз эффективнее для вызывания прямого диспергирования меланина, чем обычный свет [62]. В этой связи небезынтересно отметить, что хроматофоры таких ракообразных, как Latreutes fucorum, Leander tenuicornis и Hip-polyte acuminata, обитающих на саргассовых водорослях, в условиях яркого света обильно насыщены отражающим белым пигментом. При ярком освещении этот пигмент сильно диспергирован, так что образуется непрерывный слой, очень эффективно рассеивающий свет. Поскольку природа первичных цветовых реакций животных такова, что на свету происходит диспергирование пигмента, а в темноте — его концентрирование, представляется возможным предположить, что светозащитная функция хроматофоров является примитивной, а их функция в создании защитной окраски животных появилась позже, в процессе эволюционного
Хроматофоры и изменение окраски 571’
развития хроматофорных систем. Образование меланина в коже человека при воздействии яркого солнечного света представляет собой в сущности явление такого же характера.
Хроматофорам приписывается также функция терморегуляции [20, 213]. Известно, что ящерица Phrynosoma, обитательница пустынь, имеет светлую окраску ночью и в полдень и темную — рано утром и к вечеру [180]. Подобные наблюдения позволили заключить, что у этого животного хроматофоры принимают участие в терморегуляции. Ящерица, очевидно, способна к адаптивной регуляции поглощения и излучения тепла в различное время дня благодаря активности своих хроматофоров. В поддержку этой точки зрения можно привести данные наблюдений большого числа исследователей по изменению цвета у рептилий; было отмечено, что при повышении температуры тела примерно до 40 °C происходит концентрирование меланина, а при понижении температуры до 5 °C — его диспергирование [224]. У краба Uca при повышении, температуры тела примерно от 25 до 30 °C также наблюдается тенденция к концентрированию черного пигмента [43]. Таким образом, очевидно, регулируется в нужном направлении количество света и тепла, поглощаемых черным пигментом. В поддержку такого вывода имеются экспериментальные свидетельства. Через 5 мин после экспозиции при дневном освещении темной особи Uca, меланин которой перед этим был максимально диспергирован, температура ее тела оказалась на 2 градуса выше, чем у особей с бледной окраской, имеющих максимально концентрированный меланин [315]. Избирательное отражение света от кожи лягушки и от дорсальной поверхности головогруди Uca, когда лучше отражаются более длинные волны спектра света, а также корреляция изменения отражения и изменений окраски подтверждают вывод о терморегуляторной роли хроматофоров [63, 315].
Еще одна функция изменений окраски у животных, как полагают, связана с размножением; у некоторых животных, например у Anolis [129], такие изменения наступают в брачный период. Во время спаривания у Anolis происходит поразительное изменение цвета от зеленого до коричневого. Подобные явления наблюдаются также в период размножения у некоторых костистых рыб и головоногих моллюсков, однако значение этих изменений окраски в брачный период не известно [231]. Особые брачные морфологические изменения окраски иногда связывают со временем выведения потомства.
Заключение
Изменения окраски, обусловленные перемещением пигментов^ а также увеличением или уменьшением их количества в особых органах, называемых хроматофорами, обнаружены у многих рептилий, амфибий, костистых и пластиножаберных рыб, круглоро
572
Глава 23
тых, ракообразных, насекомых, головоногих моллюсков, аннелид и иглокожих. Эти изменения окраски в большинстве случаев представляют собой характерное адаптивное приспособление животного к цвету фона посредством функциональной связи между органами зрения и хроматофорами, хотя на изменения окраски могут влиять также общее освещение, температура, влажность, кожные раздражения, эндогенные ритмы и многие другие факторы.
Рефлекторные пути, берущие свое начало в сетчатке глаза, которые ответственны за регулирование изменений окраски у всех ракообразных, насекомых, амфибий и многих видов рыб и рептилий, включают секрецию гормонов. Как циркулирующие в крови гормоны, так и непосредственная иннервация играют важную роль у большинства других видов; исключением, по-видимому, являются хамелеон (рептилии), Fundulus и Macropodus (костистые рыбы), а возможно, еще и пиявки, у которых рефлекторная регуляция имеет преобладающее или исключительное значение. Реакции хроматофорной системы на свет имеют место у большинства животных и в отсутствие глаз — либо как результат прямой реакции хроматофоров, действующих как независимые эффекторы, либо рефлекторно благодаря функционированию других световых рецепторов, например шишковидного комплекса. Последний обусловливает только реакции на интенсивность освещения, так что животное темнеет при повышенной освещенности и светлеет при низкой, но не может обеспечить реакции на цвет фона.
Итак, хроматофорной системе приписывают несколько функций: во-первых, защитная и агрессивная окраска, во-вторых, терморегуляция, в-третьих, защита от интенсивного освещения и, наконец, в-четвертых, брачная расцветка.
СПИСОК ЛИТЕРАТУРЫ
1. Abbott F. S., Can. J. Zool. 46: 1149—1161, 1968. Drugs and biogenic substances on Fundulus melanophores.
2. Abe K-, Butcher R. W., Nicholson W. E., Baird С. E., Liddle R. A., Liddle G. W., Endocrinology, 84: 362—368, 1969. Cyclic AMP as mediator for frog melanocytes.
3. Abramowitz A. A., Proc. Nat. Acad. Sci., 21: 132—137, 1935. Morphological color change, Fundulus xanthophores.
4. Abramowitz A. A., Proc. Nat. Acad. Sci., 21: 677—691, 1935. Adaptation to colored backgrounds, Crustacea.
5. Abramowitz A. A., Biol. Bull. 71: 259—281, 1936. Hypophysis and color change in Ameiurus.
6. Abramowitz A. A., Proc. Nat. Acad. Sci., 22: 233—238, 1936. Dual innervation of teleost melanophores.
7. Abramowitz A. A., Amer. Nat., 70: 372—378, 1936. Independence of action of melanophores xanthophores, Fundulus.
8. Abramowitz A. A., Biol. Bull., 73: 134—142, 1937. Role of hypophysis in Fundulus color change.
9. Abramowitz A. A., Abramowitz R. K., Biol. Bull., 74: 278—296, 1938. Diurnal rhythm and chromatophore state, Crustacea.
10. Aoto T., Annot. Zool. Japan, 34: 193—196, 1961. Melanophore regulation in Sesarma.
Xроматофоры и изменение окраски
573
II. Ascherson, Arch. Anat. Physiol, wiss. Med., 7: 15—23, 1840. Color change, frog.
12. Atwell W. J., Holley E., J. Exp. Zool., 73: 23—42, 1936. Pituitary in amphibian color change.
13. Atzler M., Z. vergl. Physiol., 13: 505—533, 1930. Relation of eyes to Carausius color change.
14. Babak E.. Arch. ges. Physiol., 131: 78—118, 1910. Color change in young Amphibia.
15- Babak E., Arch. ges. Physiol., 149: 462—470, 1913. Relationship of morphological to physiological color change, vertebrates.
16. Bagnara I. T., Science, 132: 1481—1483, 1960. Pineal in amphibian color change.
17. Bagnara J. T., Gen. Comp. Endocrinol., 3: 86—100, 1963. Pineal in amphibian color change.
18. Ballowitz E., Z. wiss. Zool., 56: 673—706, 1893. Innervation of melanophores of teleosts.
19. Bartel С. K., Rao K. R., Fingerman M„ Comp. Biochem. Physiol., 38A: 17—36r 1971. Melanin-dispersing factors in Uca.
20. Bauer V., Z. allg. PhlysioL, 16: 191—212, 1914. Melanophores in heat regulation.
21. Bikie D., Tilney L. G., Porter K. R., Protoplasma, 61; 322—345, 1966, Microtubules in fish melanophores.
22. Bitensky M. W., Burstein S. R., Nature, 208: 1282—1284, 1965. Cyclic AMP and frog melanophores.
23. Bowman T. E„ Biol. Bull., 96: 238—245, 1949. Chromatophore hormones in Hemigrapsus.
24. Bozler E., Z. vergl. Physiol., 8: 371—390, 1929. Cephalopod chromatophores and their innervation.
25. Brecher L., Arch. Mikr. Anat., 102: 501—548, 1924. Color change in insect pupae.
26. Brown F. A., Jr., Proc. Nat. Acad. Sci., 19: 327—329, 1933. Chromatophorotro-pins from the crustacean central nervous system.
27. Brown F. A., Jr., J. Morphol., 57: 317—333, 1935. Adaptation to colored backgrounds. Crustacea. Behavior of chromatophores in color change in Palaemonetes.
28. Brown F. A., Jr., Biol. Bull., 57: 365—380, 1934. Morphological color change: Palaemonetes.
29. Brown E. A., Jr., J. Exp. Zool., 71: 1—15, 1935. Functional independence of pigments, crustaceans.
30. Brown F. A., Jr., Biol. Bull., 70: 8—15, 1936. Total illumination and chromatophore state, fishes; ratio of incident to reflected light and color change; measurement of chromatophore activity.
31. Brown F. A., Jr., Ecology, 20: 507—516, 1939. Adaptive background selection by crayfishes.
32. Brown F. A.. Jr., Physiol. Zool. 13: 343—355, 1940. Crustacean sinus gland and color change.
33. Brown F. A.., Jr., Physiol. Zool., 19: 215—233, 1946. Central nervous sources of crustacean chromatophorotropins: measurement of chromatophore activity.
34. Brown F. A., Jr., Biol. Bull., 98: 218—226, 1950. Responses of Uca red chromatophores.
35. Brown F. A., Jr., Action of Hormones in Plants and Invertebrates. New York, Academic Press, 1952. Review of crustacean color change.
36. Brown F. A., Jr., Science, 130: 1535—1544, 1959. Timing of biological rhythms.
37. Brown F. A., Jr., Ederstrom H. E., J. Exp. Zool., 85: 53—69, 1940. Chromatophorotropins from the crustacean central nervous system.
38. Brown F. A., Jr., Fingerman M., Hines M. N„ Biol. Bull., 106: 308—317, 1954. Shifting phases of color change rhythm.
39. Brown F. A., Jr., Fingerman M., Sandeen M. I., Webb H. M., J. Exp. Zool., 123: 29—60, 1953. Tidal and monthly rhythms of color change in Uca.
574
Глава 23
40. Brown F. A., Jr., Hines M. N., Physiol. Zool., 25: 56—70, 1952. Persistent modifications of daily color change cycles.
41. Brown F. A., Jr., Klotz I. M., Proc. Soc. Exp. Biol. Med., 64: 310—313, 1947. Chemical separation of chromatophorotropins from Crustacean central nervous, system.
42. Brown F. A., Jr., Saigh L. M., Biol. Bull., 91: 170—180, ,1946. Central nervous sources of chromatophorotropins, Crustacea.
43. Brown F. A., Jr., Sandeen M. J., Physiol. Zool., 21: 361—371, 1948. Uca colo* response to light and temperature: light intensity and Uca color change.
44. Brown F. A., Jr., Scudamore H. H., J. Cell. Comp. Physiol., 15: 103—119, 1940., Two chromatophorotropins from crustacean sinus glands.
45. Brown F. A., Jr., Stephens G. C., Biol. Bull., 101: 71—83, 1951. Persistent modification of daily color-change cycles by photoperiod.
46. Brown F. A., Jr., Thompson D. H., Copeia, 3: 172—181, 1937. Adaptive backgro-and their innervation.
47. Brown F. A., Jr., Webb H. M., Physiol. Zool., 21: 371—381, 1948. Diurnal rhythm in Uca color change.
48. Brown F. A., Jr., Webb И. M., Physiol. Zool., 22: 136—148, 1949. Shifting by light of phases of color-change rhythm.
49. Brown F. A., Jr., Webb H. M., Bennett M F., Proc. Nat. Acad. Sci., 41: 93— 100, 1955. Color change rhythms during change of longitude.
50. Brown F. A., Jr., Webb H. M., Bennett M F., Sandeen M. I., Physiol. Zool. 27: 345—349, 1954. Temperature independence, tidal rhythm of color change.
51. Brown F. A., Jr., Webb H. M., Sandeen M. I., J. Exp. Zool. 120: 391—420, 1952. Dual control of Palaemonetes red chromatophores.
52. Brown F. A., Jr., Wulff V. J., J. Cell. Comp. Physiol. 18: 339—353, 1941. Chromatophorotropins from the crustacean central nervous system.
53. Brilcke E., Denkschr. Acad. Wiss. Wien. 4: 179—210, 1852. Color changes in the Chameleon.
54. Buchholz R., Arch. Anat. Physiol, wiss. Med., pp. 71—81, 1863. Color changes, fishes.
55. Buckmann D., J. Insect. Physiol. 3: 159—189, 1959. Control of prepupal color change in Cerura caterpillars.
56. Buergers A. C. J., Boschman T., Van der Kam er J. C., Acta Endocrinol. 14: 72—82, 1953. Xenopus darkening by adrenin and in excitement.
57. Butcher E. O., J. Exp. Zool. 79: 275—297, 1938. Relation of portions of eyes to color changes, fishes.
58. Carlisle D. B„ Knowles F. G. W., Endocrine Control in Crustaceans. Cambridge University Press, 1959, 120 p.
59. Carstam S. P., Suneson S., Kungl. Fysiogr. Sallskap. Lund. Forhandl. 19: 1—5, 1949. Isopod chromatophorotropins.
60. Chang C. Y., Science 126: 121—122, 1957. Thyroxine and Xenopus melanopho-res.
61. Cloney R. A., Florey E., Z. Zellforsch. 89: 250—280, 1968. Ultrastructure of cephalopod chromatophore organs.
62. Coohill T. P., Bartell С. K, Fingerman M., Physiol. Zool. 43: 232—239, 1970. Ultraviolet and visible light effects directly on Uca rnelanophores.
63. Deanin G. G., Steggerda F. R., Proc. Soc. Exp. Biol. Med. 67: 101—104, 1948. Light reflection from frog skin.
64. Dupont-Raabe M., Arch. Zool. Exp. Gen. 86: 32—39, 1949. Regulation of color change in Corethra.
65. Dupont-Raabe M„ C. R. Acad. Sci. 228: 130—132, 1949. Chromatophorotropins
66. Dupont-Raabe M„ Ann. Biol. 32: 247—282, 1956. Mechanism of color change in
67. Darken B„ Arch. Mikr. Anat. 99: 222—389, 1923. Color change in insect pU'
68. ^Duspiva F„ S.-B. Acad. Wiss. Wien Math.-Nat. KI. 140: 553—596, 1931. Color change in young fishes.
Хроматофоры и изменение окраски
575
69. Eddy J. М. Р., Strahan R., Gen. Comp. Endocrinol. 11: 528—534, 1968. Pineal influences upon chromatophores of lampreys.
70. Edgren R. A., Proc. Soc. Exp. Biol. Med. 85: 229—230, 1954. Hyla darkening by ACTH.
71. Edgren R. A., Proc. Soc. Exp. Biol. Med. 87: 20—23, 1954. Hyla color change and temperature.
72. Enami M., Biol. Bull. 100: 28—43, 1951. Neurosecretion and color change in Eriocheir.
73. Enami M., Science 121: 36—37, 1955. A melanocyte-concentrating factor.
74. Ergene S., Z. vergl. Physiol. 32: 530—551, 1950; 34: 69—74, 159—165, 1952;
35: 36—41, 1953; 37: 221—229, 1955; 38: 311—316, 1956. Adaptation to colored backgrounds in Orthoptera.
75. Ergene S., Zool. Anzeiger. 153: 110—113, 1954. Color change in adult Oedaleus without molting.
76. Ergene S., Mitt. Zool. Mus. Berlin 29: 127—133, 1953. Selective value of background color adaptation.
77. Faure J. C., Bull. Ent. Res. 23: 293—405, 1932. Color change in locusts.
78. Fernlund P., Josefsson L., Biochim. Biophys. Acta 158: 262—273, 1968. Chro-mactivating hormones of Pandalus borealis.
79. Fingerman M, Biol. Bull. 109: 255—264, 1955. Tidal rhythms of color change in Callinectes.
80. Fingerman M., Biol. Bull. 110: 274—290, 1956. Tidal rhythms of color change in Uca; phasing of color-change tidal rhythm by tides.
81. Fingerman J. Exp. Zool. 133: 87—106, 1956. Response of Callinectes melanophores to background, light and temperature.
82. Fingerman M., Science 123: 585—586, 1956. Uca pigment-concentrating factor.
83. Fingerman M., Tulane Stud. Zool. 3: 139—148, 1956. Hormones and Ligia chromatophores.
84. Fingerman M., Tulane Stud. Zool. 5: 137—148, 1957. Dual control of Cambarel-lus red chromatophores; background responses in Cambarellus.
85. Fingerman M., Biol. Bull. 112: 7—20, 1957. Plasing of tidal rhythm of color change in nature.
86. Fingerman M., Amer. Nat. 91: 167—178, 1957. Review of crustacean tidal rhythms of color change.
87. Fingerman M„ Amer. Midi. Nat. 60: 71—83, 1958. Chromatophore system of Orconectes (Faxonella).
88. Fingerman M., Tulane Stud. Zool. 7: 21—30, 1959. Electrophoretic separation of crayfish chromatophorotropins.
89. Fingerman M., Internal. Rev. Cytol. 8: 175—210, 1959. Review: physiology of chromatophores.
90. Fingerman M., The Control of Chromatophores. New York, Pergamon Press, 1963, 184 p.
91. Fingerman M., Amer. Zool. 9: 443—452, 1969. Cellular aspects of the control of crustacean color changes.
92. Fingerman 714,, Ann. Rev. Physiol. 32: 345—372, 1970. Comparative physiology: chromatophores.
93. Fingerman 714., Aoto T., J. Exp. Zool. 138: 25—50, 1958. Electrophoretic separation of color-change hormones of crayfish.
94. Fingerman M., Aoto T., Physiol. Zool. 31: 193—208, 1958. Crayfish hormones and long-term color adaptation.
95. Fingerman 714., Bartell C. K., Krasnow R. A., Biol. Bull. 140: 376—388, 1971. Chromatophorotropins from Limulus and Uca.
96. Fingerman 714., Fingerman S. W., Amer. Zool. 1971 A Crrmgon-darkening substance in brachyurans.
97. Fingerman M., Lowe M. E., Physiol. Zool. 30: 216—231, 1957. Crayfish colorchange ability facilitated by use, sluggish with disuse.
'98. Fingerman M., Lowe M. E., Tulane Stud. Zool. 5: 149—171, 1957. Rate of hormone secretion following background change in Cambarellus.
576
Глава 23
99. Fingerman М., Lowe М. Е., Mobberly W. С., Limnol. Oceanog. 3: 271—282, 1958. Phasing of tidal rhythm of color change. *
100. Fingerman M„ Nagabhushanam R„ Philpott L., Biol. Bull. 120: 337—347, 1961. Physiology of Sesarma melanophores.
101. Fingerman M., Rao K. R., Physiol. Zool. 42: 138—147, 1969. Physiology of brown-black chromatophores of Squilla.
102. Fingerman M„ Sandeen M. I., Lowe M. E„ Physiol. Zool. 32: 128—149, 1959. Color-change hormones of Palaemonetes.
103. Fingerman M., Tinkle D. W., Biol. Bull. 110: 144—152, 1956. Response of white chromatophores of Palaemonetes to light, temperature and background.
104. Fischer A., Z. Zellforsch. 65: 290—312, 1965. Chromatophores and color-change in Platynereis.
105. Florey E., Comp. Biochem. Phisiol. 18: 305—324, 1966. Nervous control of cephalopod chromatophores.
106. Florey E„ Kriebel M. E., Z. vergl. Physiol. 65: 98—130, 1969. Electrical and mechanical responses of Loligo chromatophore muscle.
107. Foster K. W., Proc. Nat. Acad. Sci. 19: 535—540, 1933. Control of iridosome changes in Fundulus.
108. Fox D. L., Ann. Rev. Biochem. 16: 443—470, 1947. Chemical nature of pigments.
109. Fox D. L., Animal Biochromes and Structural Colors. Cambridge University Press, 1953.
110. Fredericq L., Arch. Zool. Exper. Gen. 7: 535—583, 1878. Control of color change, Cephalopods.
111. Freeman A. R., Connell P. M., Fingerman M., Comp. Biochem. Physiol. 26: 1015—1029, 1968. Electrophysiological study of Palaemonetes red chromatophores.
112. Fries E. F. B., Proc. Nat. Acad. Sci. 28: 396—401, 1942. Control of Fundulus leucophores.
113. Fries E. F. B., Biol. Bull. 82: 261—272, 1942. Control of Fundulus xanthopho-res.
114. Fries E. F. B., Biol. Bull. 82: 273—283, 1942. Dual innervation of teleost melanophores.
115. Fries E. F. B., Physiol. Zool. 16: 199—212, 1943. Control of Fundulus xantho-phores.
116. Fries E. F. B., J. Morphol. 103: 203—254, 1958. Physiology of Fundulus reflecting chromatophores.
117. Friza F., Z. vergl. Physiol. 8: 289—336, 1928. Diurnal color changes in man-tids.
118. Fuchs R. F., Winterstein, Handb. vergl. Physiol. 4, pt. 1: 1189—1656, 1914. Color change, review.
119. Fugh R., J. Fac. Sci., Tokyo, Sec. IV, 8: 371—380, 1959. Melanin dispersion in Fundulus by sodium ions.
120. Fujii R., Novales R. R., Amer. Zool. 9: 453—463, 1969. Cellular aspects of control of fish color changes.
121. Gabritschevsky E., J. Exp. Zool. 47: 251—267, 1927. Color changes in spider,
122. Gamble F. W., Keeble F. W„ Quart. J. Mier. Sci. 43: 589—698, 1900. Color changes, crustaseans; influence of temperature.
123- Giersberg FL, 7. vergl. Physiol. 7: 657—695, 1928. Color change in Carausius; influence of humidity.
124. Giersberg H., Z. vergl. Physiol. 9: 523—552, 1929. Temperature and morphological color changes, insects.
125. Giersberg H„ Z. vergl. Physiol. 13: 258—279, 1930. Control of erythrophores of Phoxinus; influence of drugs.
126. Gilson A. S., Jr., J. Exp. Zool. 45: 415—455, 1926. Color changes in young
127. Goldman J. M., Hadley M. E., Brit. J. Pharmacol. 39: 160—166, 1970. Mela-nophore stimulating hormone and cyclic AMP in melanophore responses.
Хроматофоры и изменение окраски 577
l28. Goodwin Т. W., рр. 101—140 in: The Physiology of the Crustacea, Vol. 1, edited by T. H. Waterman. New York, Academic Press, 1960.
129. Hadley С. E., Bull. Mus. Comp. Zool. 69: 108—114, 1929. Color changes during mating of lizards.
130. Hadley M. E., Gildman J. M., Amer. Zool. 9: 489—504, 1969. Physiological color changes in reptiles.
131. Hanstrdm B., Zool. Jahrb. Abt. Ontog. Tiere. 56: 387—520, 1933. The crustacean sinus gland.
132. Hanstrdm B., Kungl. Svenska Vetenskap. Handl. 16: 1—99, 1937. Relation of portions of eyes to color changes, Crustacea. Crustacean sinus gland and color change.
133. Hanstrdm B„ Kungl. Fysiogr. Sallsk. Handl. N. F. 49: 1—10, 1938. Relation of portions of eyes to color changes, Crustacea.
134. Healey E. G., J. Exp. Biol. 31: 473—490, 1954. Color change control in Pho-xinus.
135. Healey F. G., Ross D. M., Comp. Biochem. Physiol. 19: 545—580, 1966. Drug effects on the background color response of Phoxinus.
136. Hempelmann F., Z. wiss. Zool. 152: 353—383, 1939. Color change, po-lychaetes.
137. Hidaka T., Annot. Zool. Japan 29: 69—74, 1956. Control of pupal color in Lepidoptera.
138. Hill A. V., Parkinson J. L., Solandt D. Y., J. Exp. Biol. 12: 397—399, 1935. Measurement of chromatophore activity.
139. Hitchcock H. B., Biol. Bull. 80: 26—30, 1941. Adaptation to colored backgrounds, Crustacea.
140. Hofmann F. G., Arch. Mikr. Anat. 70: 361—413, 1907. Innervation, cephalopod chromatophore.
141. Hogben L. T., The Pigmentary Effector System. Edinburgh, Oliver and Boyd, 1924, 152 p.
142. Hogben L. T., Proc. Roy. Soc. Lond. В 120: 142—158, 1936. Hypophysis in elasmobranch melanophore control.
143. Hogben L. T., Mirvish L., Brit. J. Exp. Biol. 5: 295—308, 1928. Control of color changes in chameleons.
144. Hogben L., Slome D., Proc. Roy. Soc. Lond. В 108: 10—53, 1931. Mechanism of chromatophore control, Xenopus.
145. Hogben L., Slome D„ Proc. Roy. Soc. Lond. В 120: 158—173, 1936. Mechanism of chromatophore control, Xenopus.
146. Hogben L., Winton F. R., Proc. Roy. Soc. Lond. В 94: 151—162, 1922. Hypophysis and amphibian color change.
147. Hogben L„ Winton F. R., Proc. Roy. Soc. Lond. В 95: 15—30, 1923. Hypophysis and amphibian color change.
148. Hooker D., Amer. J. Anat. 16: 237—250, 1914. Color change in young Rana.
149. Hosoi T., J. Fac. Sci. Imp. Univ. Tokyo 3: 265—270, 1934. Chromatophorotropins from the crustacean central nervous system.
150. Hudson B., Bentley G. A., Lancet 1: 775, 1955. Adrenin darkening in Xenopus.
151. Janda V., Zool. Anz. 115: 177—185, 1936. Color change in skin transplants in Disippus.
152. Janzen R., Z. Morph. Okol. Tiere 24: 327—341, 1932. Color change, leeches.
153. Janzen R., Zool. Anz. 101: 35—40, 1932. Color changes, leeches; Hemiclepsis and Glossiphonia.
154. Johnson L. P., Jahn T., Physiol. Zool. 15: 89—94, 1942. Color changes, Euglena.
155. Jorgenson С. B., Gen. Comp. Endocrinol. 2: 610, 1962. Hypophysectomy influences on melanophores of Xenopus.
156. Jorgenson С. B., Larsen L. O., Nature 186: 641—642, 1960. Control of colour change in amphibians.
157. Kaestner H., Arch. Entw.-Mech. Org. 124: 1—16, 1931. Temperature and morphological color change, insects.
37—2067
578
Глава 23
158. Kalmus FL, Z. vergl. Physiol. 25: 494—508, 1938. Diurnal rhythm in Carausius color change.
159. Keeble F. IT., Gamble F. W., Phil. Trans. Roy. Soc. Lond. В 196: 295—388, 1904. Relationship of morphological to physiological color change, crustaceans.
160. Feeble F. W., Gamble E. W„ Phil. Trans. Roy. Soc. Lond. В 198: 1—16, 1905. Color changes, crustaceans.
161. Ketterer B., Remilton E., J. Endocrinol. 11: 7—18, 1954. Xenopus darkening by adrenin and in excitement.
162. Key K. H. L., Day M. R., Australian J. Zool. 2: 309—363, 1954. Primary color change in grasshopper, Kosciuscola.
163. Khokhar R., Experientia 27: 340—341, 1971. Inhibitory control of pars intermedia function in the teleost Ictalurus.
164. Kleinholz L. H., Biol. Bui. 69: 379—390, 1935. Role of hypophysis in Fundulus color changes.
165. Kleinholz L. H., J. Exp. Biol. 15: 474—499, 1938. Control of color changes in Anolis.
166. Kleinholz L. EL, PubbL Staz. Zool. Napoli 17: 53—57, 1938. Color change, echinoderm.
167. Kleinholz L. EL, Gen. Comp. Endocrinol. 14: 578—588, 1970. Isolation of crustacean chromatophorotropins.
168. Knight H. H., Ann. Ent. Soc. Amer. 17: 258—272, 1924. Temperature and morphological color change, insects.
169. Knowles F. G. W., Physiol. Comp. Oecol. 2: 284—296, 1952. Color changes after sinus gland removal.
170. Knowles F. G. W., Proc. Roy. Soc. Lond. В 141: 248—267, 1953. Postcommis-sural organs and color change.
171. Knowles F. G. W., Pubbl. Staz. Zool. Napoli 24 (suppl.): 74—78, 1954. Color change control in Squilla.
172. Knowles F. G. W., Carlisle D. B., Biol. Rev. 31: 396—473, 1956. Review of crustacean color change.
173. Knowles F. G. W., Carlisle D. B., Dupont-Raabe M., J. Marine Biol. Assoc. U.K. 34: 611—635, 1955. Chemical properties of Carausius chromatophorotro-pin.
174. Knowles F. G. W„ Carlisle D. B., Dupont-Raabe M., C. R. Acad. Sci. 242: 825, 1956. Chemical properties of crustacean chromatophorotropin.
175. Koller G., Verh. deutsch. zool. Gesell. 30: 128—132, 1925. Mechanism of chro-matophore control, Crustacea.
176. Koller G., Z. vergl. Physiol. 5: 191—246, 1927. Functional independence of pigments, crustaceans; color adaptations; mechanism of chromatophore control.
177. Koller G., Z. vergl. Physiol. 8: 601—612, 1928. Mechanism of chromatophore control. Crustacea.
178. Koller G„ Z. vergl. Physiol. 12: 631—667, 1930. Hormones in crustacean color change.
179. Kroyer H., Kong. Dansk. Videnskap. Selskabet 9: 209—361, 1842. Color changes, Hippolyte.
180. Kruger P., Kern H., Arch. ges. Physiol. 202: 119—138, 1924. Melanophores and heat regulation.
181. Kuhn A., Z. vergl. Physiol. 32: 572—598, 1950. Background adaptation in Sepia and Octopus.
.182. Kuhn A., Heberdey R. F„ Zool. Anz. 231: suppl. 4, 1929. Adaptation to colored backgrounds, Cephalopoda.
183. Kuhn A., Lederer E., Ber. deutsch. chem. Ges. 66: 488—495, 1953. Chemical nature pigments, crustaceans.
184. Kuntz A., Bull. U. S. Bur. Fish. 35: 1—29, 1916. Adaptation to colored backgrounds, fishes.
185. Laurens H., J. Exp. Zool. 16: 195—210, 1914. Color change in young Ambly stoma.
Хроматофоры и изменение окраски 579
186. Laurens Н., J. Exp. Zool. 18: 577—638, 1915. Color change in young Ambly-stoma.
187. Lawinski L., Pautsch F., Acta Biol. Med. Soc. Gedan. 9: 5—14, 1965. Chromatophores and their behaviour in the crab, Rhithropanopeus.
188. Lieben S., Centralbl. Physiol. 20: 108—117, 1906. Adrenaline and color change, frog-
189. Lundstrom H. M., Bard P., Biol. Bull. 62: 1—9, 1932. Hypophysis in elasmobranch melanophore control.
190. Martini E., Achundow L, Zool. Anz. 81: 25—44, 1929. Color changes in Co-rethra.
191. Mast S. O., Bull. U. S. Bur. Fish. 34: 173—238, 1916. Adaptation to colored backgrounds, fishes; adaptive background selection.
192. Matsumoto K., Biol. J. Okayama Univ. 1: 234—248, 1954. Neurosecretino and color change in crustaceans; color-change hormones of Eriocheir.
193. Matsushita K., Sci. Rys. Imp. Univ. Sendai, 4, Biol. 13: 171—200, 1938. Dual innervation of teleost melanophores.
194. Matthews S. A., J. Exp. Zool. 58: 471—486, 1931. Mechanism of pigment migration in chromatophores, fish.
195. Matthews S. A., Biol. Bull. 64: 315—320, 1933. Color changes in hypophysecto-mized Fundulus.
196. McWhinnie M. A., Sweeney H. M., Biol. Bull. 108: 160—174, 1955. Color change in Trachelipus.
197. Mendes E. G., Bol. Gac. Filos. Gien. Letr. Univers. Sao Paulo, 15. Zool. 6: 285—299, 1942. Color change in young fishes.
198. Menke F., Arch. ges. Physiol. 140: 37—91, 1911. Diurnal rhythm and chromatophore state, Crustacea; influence of temperature on chromatophores.
199. Millott N., Nature 170: 325—326, 1952. Color change daily rhythm in Diadema.
200. Millott N., Experientia 9: 98—99, 1953. Color changes in Diadema in response to illumination.
201. Mills S. M., J. Exp. Zool. 64: 231—244, 1932. Dual innervation of teleost melanophores.
202. Milne-Edwards H., Ann. Sci. Nat. Sec. 2, Zool. 1: 46—54, 1834. Color change, Chameleon.
203. Minkiewicz R., Bull. Acad. Sci. Cracovie, 918—929 (November), 1908. Adaptation to colored backgrounds, Crustacea.
204. Mussbichler A., Umrath K„ Z. vergl. Physiol. 32: 311—318, 1950. Effect of MSH and adrenin on Hyla melanophores.
205. Nagano T., Sci. Rep. Tohoku Univ. ser. 4, 18: 167—175, 1949. Color changes in-isopods.
206. Neill R. M., J. Exp. Biol. 17: 74—94, 1949. Color change in young fishes; color change in Anguilla.
207. Novales R. R., Physiol. Zool. 32: 15—28, 1959. MSH action on isolated frog skin.
208. Novales R. R„ Fujii R., J. Cell. Physiol. 75: 133—136, 1970. Melanin-dispersing effect of cyclic AMP in Fundulus.
209. Odiorne J. M„ Proc. Nat. Acad. Sci. 19: 329—332, 1933. Morphological color change, fishes.
210. Odiorne J. M., Proc. Nat. Acad. Sci. 19: 750—754, 1933. Adrenaline and teleost leucophores.
211. Okay S., C. R. Ann. Arch. Soc. Turqu. Sci. Phys. Nat. 12: 101, 1946. Color change hormones in isopods.
212. Osborn С. M., Proc. Nat. Acad. Sci. 26: 155—161, 1940. Induced pigmentation on ventral surface of flounder.
213. Parker G. FL, J. Exp. Zool. 3: 401—414, 1906. Melanophores and heat regulation.
214. Parker G. H., Biol. Rev. 5: 59—90, 1930. Chromatophores review.
215. Parker G. EL, Proc. Nat. Acad. Sci. 17: 594—596, 1931. Color change, chino-; derm. ; •
37*
580
Глава 23
216. Parker G. H., Proc. Nat. Acad. Sci. 20: 306—310, 1934. Dual innervation of Fundulus melanophores.
217. Parker G. H., Proc. Amer. Philosoph. Soc. 75: 1—10, 1935. Measurement of chromatophore activity.
218. Parker G. H., Biol. Bull. 68: 1—3, 1935. Innervation of Mustelus melanophores.
219. Parker G. H., Biol. Bull. 70: 1—7, 1936. Color change in young elasmobranchs.
220. Parker G. H., Biol. Bull. 71: 255—258, 1936. Single innervation of Mustelus melanophores.
221. Parker G. H„ Cold Spring Harbor Symp. Quant. Biol. 4: 358—370, 1936. Coldblock of chromatophore nerves.
222. Parker G. H., Proc. Nat. Acad. Sci. 23: 206—211, 1937. Control of erythrophores of Holocentrus; dual innervation of melanophores.
223. Parker G. H., Proc. Amer. Philosoph. Soc. 77: 223—247, 1937. Control of elasmobranch melanophores.
224. Parker G. H., J. Exp. Biol. 15: 48—73, 1938. Control of color changes in Phrynosoma; influence of temperature.
225. Parker G. H., Proc. Amer. Philosoph. Soc. 83: 379—409, 1940. AcCh and fish color change.
226. Parker G. H., Proc. Amer. Philosoph. Soc. 85: 18—24, 1941. Dual innervation of teleost melanophores.
227. Parker G. FL, J. Exp. Zool. 89: 451—473, 1942. Chemical mediator in Mustelus melanophore control.
228. Parker G. H., Quart. Rev. Biol. 18: 205—227, 1943. Vertebrate color changes, review.
229. Parker G. H., Biol. Bull. 84: 273—284, 1943. Methods of measurement of color changes.
230. Parker G. H., Animal Colour Changes and Their Neurohumors. New York, Cambridge University Press, 1948, 377 p.
231. Parker G. H., Brower H. P., Biol. Bull. 68: 4—6, 1935. Seasonal development of nuptial secondary sex coloration, Fundulus.
232. Parker G. FL, Rosenblueth A., Proc. Nat. Acad. Sci. 27: 198—204, 1941. Electrical stimulation of melanophore nerves.
233. Parker G. FL, Scatterty L. E„ J. Cell. Comp. Physiol. 9: 297—314, 1937. Hormonal control Rana color change.
234. Pasteur C„ C. R. Acad. Sci. 246: 320—322, 1958. Effect of X-organ extirpation on Leander (Palaetnon) color change.
235. Pautsch F., Bull. Internat. Acad. Polon. Sci. Classe Sci. Math. Nat. ser. В 7: 511—523, 1951. Primary response in Crangon zoea.
236. Perez-Gonzalez M. D., Biol. Bull. 113: 426—441, 1957. Crustacean chromacti-vator destroyed by chymotrypsin.
237. Perkins E. B., J. Exp. Zool. 50: 71—105, 1928. Pigment migration in chroma-tophores, crustacean. Mechanism of chromatophore control.
238. Perkins E. B., Snook T., J. Exp. Zool. 61: 115—128, 1938. Mechanism of chromatophore control, Crustacea.
239. Phisalix C., Arch. Physiol. Norm. Path., ser. 5, 6: 92—100, 1894. Control of color change, cephalopods.
240. Pickford G. E., Atz J. W., The Physiology of the Pituitary Gland of Fishes. New York Zoological Society, 1957.
241. Pigeault N., C. R. Acad. Sci. 246: 487—489, 1958. Organ of Bellonci and isopod color change.
242. Pouchet G., J. Anat. Physiol. 12: 1—90, 113—165, 1876. Color changes, crustaceans, fishes.
243. Poulton E. B., The Colors of Animals. New York, D. Appleton and Co., 1890, 360 p.
244. Priebatsch I., Z. vergl. Physiol. 19: 453—485, 1933. Relations of portions of eyes to color changes, insects.
245. Prosser C. L., von Limbach B., Bennett G. W., Physiol. Zool. 20: 349—354, 1947. Reactions of fish chromatophore to war gases.
Хроматофоры и изменение окраски 581
246. Rahn Н., Biol. Bull. 80: 228—237, 1941. Hypophysis and color change: in rattlesnake.
247. Rao K. R., Experientia 23: 231, 1967. Crustacean larval chromatophore responses to light and endocrines.
248. Rao K. R., Zool. Jahrb. Physiol. 74: 247—291, 1968. Chromatophorotropin and color change variations during the life of the crab, Ocypode.
249. Rao K. R., Gen. Comp. Endocrinol. 12: 547—585, 1969. Influence of prolonged adaptation to a background upon responses og crab chromatophores.
250. Rao K. R., Fielder D. R., Fingerman M„ Amer. Zool. 10: 495, 1970. Melanindispersing substances from the eyestalks of Uca.
251. Rao K. R., Fielder D. R., Fingerman M., Amer. Zool. 10: 495—496, 1970. Effects of oxidation and reduction on melanin-dispersing material from Uca eyestalks.
252. Rao K. R., Fingerman M., Bartell С. K., Biol. Bull. 133: 606—617, 1967. Physiology of the white chromatophores in Uca.
253. Redfield A. C., J. Exp. Zool. 26: 275—333, 1918. Control of color change in Phrynosoma; diurnal rhythm of color change.
254. Reed B. L., Life Sci., Part II: 961—973, 1968. Melatonin in circadian color changes in pencil fish.
255. Rbseler 1., Z. vergl. Physiol. 70: 144—174, 1970. Die Rhythmik der Chroma-tophoren des Polychaeten Platynereis dumerilii. [Chromatophore rhythms in Platynereis.]
256. Rowlands A., J. Exp. Biol. 29: 127—136, 1952. Humidity and Rana color change.
257. Van Rynberk G„ Ergebn. Physiol. 5: 347—571, 1906. Color change, review.
258. Sage M., J. Exp. Zool. 173: 121—128, 1970. Prolactin role in color change in the teleost, Gillichthys.
259. Sand A., Biol. Rev. 10: 361—382, 1935. Control of color changes in chameleons.
260. Sandeen M. I., Physiol. Zool. 23: 337—352, 1950. Chromatophorotropins of Uca.
261. Sars G., Histoire naturelie des Crustaces d’eau douce de Norvege. Christiania, 1867, 145 p. Color changes, Crustaceans.
262. Scharrer B., Action of Hormones in Plants and Invertebrates. New York, Academic Press, 1952. Review of insect color change.
263. Scharrer E., Z. vergl. Physiol. 7: 1—38, 1928. Pineal body and color changes, fishes.
264. Scheer В. T., Scheer M. A. A., Pubbl. Staz. Zool. Napoli 25: 397—418, 1954. Color changes in Leander (Palaemon).
265. Schliep W., Zool. Jahrb. Physiol. 30: 45—132, 1910. Color change in Carausius; duirnal rhythm in Carausius color change.
266. Schlieper C., Z. vergl. Physiol. 3: 547—557, 1926. Color change in Hyperia.
267. Schlottke E., Z. vergl. Physiol. 3: 692—736, 1926. Temperature and morphological color change, insects.
268. Schlottke E., Z. vergl. Physiol. 20: 370—379, 1934. Temperature and morpholo-Sical color change, insects.
chwinck I., Naturwissenschaften 40: 365, 1953. Uber den Nachweis eines Redox-Pigmentes (Ommochrom) in der Haut von Sepia officinalis. [Pigments in Sepia skin.]
270. Sereni E., Boll. Soc. Ital. Biol. Sper. 2: 377—381, 1927. Control color change, cephalopods.
271. Sereni E., Z. vergl. Physiol. 8: 488—600, 1928. Inhibitory color-change center, cephalopods. Postural chromatophore reflexes, cephalopods.
272. Sereni E„ Boll. Soc. Ital. Biol. Sper. 3: 707—711, 1928. Betaine and tyramine in cephalopod color change.
273. Sereni E., Boll. Soc. Ital. Biol. Sper. 4: 749—753, 1929. Salivary gland (tyramine) and cephalopod color change.
274. Sereni E., Z. vergl. Physiol. 12: 329—503, 1930. Color change mechanism, cephalopods.
582 Глава 23
275. Sereni Е., Biol. Bull. 59: 247—268, 1930. Direct action of betaine and tyramine on cephalopod chromatophores.
276. von Siebold К. T. E., Die Susswasserfische von Mittleuropa. Leipzig, 1863*..
431 p. Color change, fishes.
277. Sieglitz _G., Z. vergl. Physiol. 33: 99—124, 1951. Melanin-concentrating factor in frog skin.
278. Sjogren S., Zool. Jahrb. Abt. Anat. Ontog. Tiere 58: 145—170, 1934. The crustacean sinus gland.
279. Skorkowski E. F„ Marine Biol. 8: 220—223, 1971. Chromatophorotropic hormones from the eyestalk of Crangon.
280. Slome D., Hogben L. T„ South African J. Sci. 25: 329—335, 1928. Measurement, of chromatophore activity.
281. Smith D. C., Biol. Bull. 58: 193—202, 1930. Temperature and color change-, (Z rust а.сез
282. Smith D. C., J. Exp. Zool. 58: 423—453, 1931. Autonomic drugs and teleost color change.
283. Smith D. C., J. Cell. Comp. Physiol. 8: 83—87, 1936. Measurement of chroma--tophore activity.
284. Smith D. C., Smith M. T., Biol. Bull. 67: 45—58, 1934. Control of erythropores of Holocentrus.
285. Smith H. G., Proc. Roy. Soc. Lond. В 125: 250—263, 1938. Dual control of isopod melanophores.
286. Smith P. E., Anat. Rec. 11: 57—64, 1916. Hypophysis and amphibian color change.
287. Smith R. I., Physiol. Zool. 15: 410—417, 1942. Mechanism of color change, leeches.
288. Spaeth R. A., Anat. Anz. 44: 520—524, 1913. Pigment migration in chroma--' tophore, fish.
289. Spaeth R. A., Amer. J. Physiol. 41: 597—602, 1916. Measurement of chromatophore activity.
290. Steggerda F. R., Soderwall A. L., J. Cell. Comp. Physiol. 13: 31—37, 1939. Dual humoral control of melanophores, Rana pipiens.
291. Steinach E., Pfliigers Arch. Physiol. 87: 1—37, 1901. Tactile stimuli and color change, cephalopods.
292. Stephens G. C., Physiol. Zool. 30: 55—59, 1957. Phase shifting of color change rhythm by temperature cycles; sensitivity rhythm.
293. Stephens G. C„ Amer. Nat. 91: 135—152, 1957. Review of crustacean daily rhythms of color change.
294. Steschegolew G. G., Rev. Zool. Russe 7: 149—166, 1927. Color change, leeches.
295. Sumner F. B., Amer. Nat. 69: 245—266, 1935. Protective value of hish color change.
296. Sumner F. B., Biol. Rev. 15: 351—375, 1940. Morphological color change in fishes and amphibians, review.
297. Sumner F. B., Biol. Bull. 84: 195—205, 1943. Morphological color changes* Girella, Fundulus.
298. Sumner F. B., Proc. Nat. Acad. Sci. 30: 285—294, 1944. Morphological color change, guanine.
299. Sumner F. B., Doudoroff P„ Biol. Bull. 84: 187—194, 1943. Assay of melanin in fishes.
300. Sumner F. B., Keys A. B., Physiol. Zool. 2: 495—504, 1929. Ratio of incident to reflected light and color change.
301. Suneson S., Kungl. Fysiogr. Sallsk. Handl. N. F. 58: 5, 1947. Color change system of Idothea.
302. Taki L, J. Fac. Fish. Anim. Husb. Hiroshima Univ. 5: 345—417, 1964. Physiology of branchial gland in Cephalopoda.
303. Tomita G., J. Shanghai Sci. Inst. IV 2: 237—264, 1936. Color change in young fishes; color change in blinded teleosts.
Хроматофоры и изменение окраски
583
.304. Tomita G., J. Shanghai Sci. Inst. IV 4: 1—8, 1938. Dual innervation of teleost melanophores.
305. Tomita G., J. Shanghai Sci. Inst. IV 5: 151 —178, 1940. Dual innervation of teleost melanophores.
306. Vesely D. L., Hadley M. E., Science 173: 923—925, 1971. Calcium requirement for melanophore-stimulating hormone action on melanophores.
307. von Uexkull J., Z. Biol. 34: 319—339, 1896. Color change, echinoderm.
308. Umrath K., Watcher H., Z. vergl. Physiol. 33: 129—141, 1951. Control of Macropodus melanophores.
309. Verne J., Arch. Morph. Gen. Exp., 1923, 168 p. Chemical nature of pigments, crustaceans
310. Waring H., Proc. Roy. Soc. Lond. В 128: 343—353, 1940. Dual hormonal control of Anguilla melanophores.
311. Waring H., Biol. Rev. 17: 120—150, 1942. Vertebrate color changes, review.
312. Waring H., Landgrebe F. W., in: The Hormones, edited by G. Pincus and К. V. Thimann, New York. Academic Press, 1950. Vertebrate color change, review.
313. Webb H. M., Physiol. Zool. 23: 316—336, 1950. Shifting by light, phases of color change rhythm.
314. Wells G. P., Nature 129: 686—687, 1932. Color change, leeches.
315- Wilkens J. L., Fingerman M., Biol. Bull. 128: 133—141, 1965. Temperature relationships and Uca body coloration.
316. Wright P. A., Physiol. Zool. 28: 204—218, 1955. Isolated Rana melanophore response to MSH and adrenin.
317. Wykes U., J. Exp. Biol. 14: 79—86, 1937. Measurement of chromatophore activity.
318. Wyman L. C., J. Exp. Zool. 40: 161—180, 1924. Color change in young fishes.
319. Yoshida M., J. Exp. Biol. 33: 119—123, 1956. Color change in Diadema, a direct chromatophore response.
320. Young J. Z., Quart. J. Mier. Sci. 75: 571—624, 1933. Control of elasmobranch melanophores.
321. Young J. Z„ J. Exp. Biol. 12: 254—270, 1935. Pineal body in color change, Lampetra; control of Lampetra melanophores.
322. Zondek B„ Krohn H„ Klin. Wchnschr. 11: 405—408, 1932. Intermedin in melanophore responses.
323. Zoond Z., Eyre J., Phil. Trans. Roy. Soc. Lond. В 223: 27—55, 1934. Control of color change in chameleons; diurnal rhythm of color changes.
Глава 24
Биолюминесценция
Ф. БРАУН
Способность к свечению очень широко распространена среди бактерий, грибов, водорослей и животных [38, 43—45], и, по-видимому, в ходе эволюции возникала неоднократно. Среди животных светящиеся виды описаны почти для всех основных типов — от простейших до хордовых. Не обнаружены светящиеся виды лишь для Mesozoa, Entoprocta, Phoronidea, Echiuroidea, Sipunculoidea, Brachiopoda, Onychophora, Linguatulda и Tardigrada. Способность к люминесценции у отдельных групп животных или в пределах одной группы носит случайный характер и не подчиняется какой бы то ни было закономерности: она может наблюдаться у одних видов данного рода и отсутствовать у других. Практически все известные светящиеся организмы — морские или сухопутные. Фосфоресцирующие морские виды можно обнаружить среди глубоководных, прибрежных и планктонных организмов. Единственными описанными в литературе светящимися пресноводными животными являются жук-светляк и улитка, а среди пещерных животных — личинка одного из двукрылых.
Люминесценция у животных является результатом хемилюминесцентных реакций, в ходе которых происходит окисление субстрата. Такие реакции обычно связаны с присутствием в цитоплазме люминесцирующей ткани особых гранул (возможно, органелл). У многих организмов вещества, участвующие в фотогенных реакциях, выводятся в окружающую среду, где эти реакции и осуществляются. Такой способ генерирования света называется внеклеточной люминесценцией в отличие от внутриклеточной, при которой испускание света происходит внутри клетки. У животных с внеклеточной люминесценцией светогенерирующие органы представляют собой одноклеточные или многоклеточные железы, которые выделяют секрет наружу. Иногда в светящихся железах имеются два типа секреторных клеток, содержащих фермент и субстрат (например, у рачка Cypridina); в процессе генерации света участвуют оба типа клеток.
У высших животных с внутриклеточной люминесценцией прослеживается общая тенденция к развитию специализированных
Биолюминесценция
585
(фотогенных органов. У простейших люминесцирующие гранулы рассеяны по всей цитоплазме, а у высших животных фотогенные клетки или органы сосредоточены в определенных местах. У таких высших животных, как головоногие, ракообразные, насекомые и рыбы, светящиеся клетки составляют только часть органа, который помимо этих клеток имеет поглощающие и отражающие свет слои (светофильтры), преломляющие тельца и нервные окончания. Такие органы отдаленно напоминают фоторецепторы.
Далеко не всегда бывает легко установить, обладает ли светящееся животное способностью самостоятельно генерировать свет, поскольку люминесценция часто возникает в результате присутствия в этом организме или на его поверхности люминесцирующих бактерий. Одним из критериев, позволяющих отличать системы, содержащие бактериальные симбионты, от систем, способных к истинной самолюминесценции, является то, что у первых свечение происходит непрерывно, тогда как у вторых оно прерывистое и возникает лишь в ответ на внешнее раздражение. Из этого правила имеется, однако, немало исключений. Многие истинно фотогенные клетки животных дают непрерывное свечение. С другой стороны, люминесценция бактериального происхождения может производить впечатление прерывистой, как, например, у рыбы Pho-toblepharon-, в этом случае симбиотические светящиеся бактерии находятся в специальном светогенерирующем органе, обладающем подвижным фильтром, способным «выключать» свет.
Распространенность биолюминесценции среди животных. Среди простейших имеется много люминесцирующих морских видов, например морские радиолярии и динофлагелляты. Из последних наиболее известны ночесветка (Noctilucd) и Gonyaulax [53]. Генерирующие свет гранулы рассеяны у них по всей цитоплазме, но особенно многочисленны по периферии клетки. Noctiluca, как правило, вспыхивает в ответ на механическое или электрическое раздражение, которое вызывает медленный, градуальный, подобный генераторному электрический потенциал, генерирующий в свою очередь потенциалы действия, распространяющиеся по закону «все или ничего». Сильная электрическая стимуляция вызывает с очень । коротким латентным периодом синхронную вспышку всей поверхности клетки. Возникающая в обычных условиях более продолжительная вспышка клетки обусловлена, по-видимому, временем I проведения по ней возбуждающего потенциала действия с после-I дующим вовлечением микросом, содержащих люминесцирующие вещества [32].
У Gonyaulax, Pyrodinium и Pyrocystis биолюминесценция возникает как спонтанно, так и при возбуждении; ее относительная , интенсивность меняется в зависимости от времени суток [4].
Среди кишечнополостных также известно много светящихся нидов — гидроидные полипы [74—76], медузы [23], сифонофоры, морские перья, актинии [84, 85, 89]. Люминесценция у них возни
586
Глава 24
кает только в ответ на раздражение. Естественным стимулом является, вероятно, механическое раздражение. Первой отвечает непосредственно раздражаемая область, а затем от нее распространяется волна свечения. Электрические изменения, связанные с люминесценцией, были описаны у Obelia [73]. У медузы Pelagia расстояние, на которое распространяется волна люминесценции, прямо пропорционально силе раздражения. Свечение у некоторых кишечнополостных имеет, по-видимому, отчасти внеклеточную природу, поскольку с поверхности тела животных можно легко удалить светящуюся слизь. У морских перьев, актиний и гидромедуз оно может быть также и внутриклеточным: у гидромедуз светогенерирующие клетки расположены по краю кольцевого канала.
К люминесценции способны, по-видимому, все гребневики (Cte-nophora) [18, 47]. У них генерирующие свет клетки расположены вдоль меридиональных гастроваскулярных каналов вблизи половых клеток. Светятся гребневики в ответ на раздражение, причем на свету эта реакция подавляется [58]. Если после пребывания на дневном свету животных перевести в темноту, способность к свечению за короткое время полностью восстанавливается. Из= гребневика Mnemiopsis был выделен фотопротеид, который в отличие от других фотопротеидов обладает светочувствительностью — свет его инактивирует [50].
Известна только одна светящаяся немертина — Emplectonema kandai. Фотогенные клетки, внутриклеточно продуцирующие свет, распространены у этого червя по всему телу. Местное тактильное раздражение вызывает локальные ответы; при растягивании животного возникает генерализованная реакция.
Среди кольчатых червей люминесценция наблюдается только-у наземных олигохет и морских полихет. Многие дождевые черви, например Eisenia submontana, при раздражении выбрасывают-светящуюся слизь, которая может выходить из ротового или анального отверстия или через дорсальные поры. У полихеты Chaetopterus [78—80, 82] проявляется поразительная способность к люминесценции: при малейшем раздражении начинает светиться большая часть ее поверхности (рис. 24-1). Фотогенные клетки расположены в гиподерме вместе со слизистыми клетками, причем и те и другие выделяют свой секрет наружу (рис. 24-2, В). Свечение регулируется нервной системой; раздражение головной части тела червя вызывает распространение световой волны по направлению к хвостовой части. У непрозрачного пелагического-кольчеца Tomopteris фотогенные органы представляют собой особые видоизмененные нефридиальные воронки. У светящихся кольчатых червей свет генерируется в перекрывающихся дорсальных чешуйках определенных клеток [81, 83]. Раздражение любой части их тела вызывает волну света, которая проходит вверх и вниз.-от точки возбуждения, причем назад волна свечения распростра
Б иолюминесценция
587
няется легче, чем вперед, указывая тем самым на участие в этом процессе частично поляризованной нервной системы.
Довольно много светящихся видов среди членистоногих. Это в основном ракообразные [26] и насекомые, иногда многоножки [25] и паукообразные. Важные сведения об основных химических процессах, протекающих в биолюминесцирующих системах, были
получены именно при изучении жука-светляка. Светогенерирующий орган у Cypridina представляет -собой большую железу, расположенную вблизи ротового отверстия. Известно два вида секреторных клеток — одни продуцируют субстрат (люциферин), а другие — фермент (люциферазу), и то и другое в виде маленьких гранул. При мышечном сокращении эти гранулы выбрасываются наружу, т. е. люминесценция в этом случае относится к внеклеточному типу.
Способностью к биолюминесценции обладают также светящиеся веслоногие раки. У них источником выбрасываемых в морскую воду активных веществ являются маленькие группки зеленоватых секреторных клеток, расположенных в различных частях тела.
светящегося рачка Cypridina и
Рис. 24-.1, Морская полихета Chaetop-terus i[44].
А. Вид сверху. Б. Люминесценция особи в темноте.
Многие эвфаузииды и креветки обладают достаточно высокоразвитыми органами [3] с отражательным слоем и линзой, морфологически связанными с генерирующими свет клетками (рис. 24-2,Л). Эти органы разбросаны по всей поверхности тела. Деятельность многочисленных люминесцентных органов регулируется, по-видимому, нервной системой, поскольку они могут возбуждаться в определенной последовательности в направлении от головной части тела к хвостовой, как, например, у Sergestes. У глубоководной креветки Acanthephyra purpurea, помимо типичных фотофоров (светящихся органов) возле рта, имеются железы, из которых наружу с силой выбрасываются светящиеся вещества; это позволяет креветке, окружив себя светящимся облачком, скрываться от хищников.
Среди насекомых способностью к люминесценции обладают представители лишь нескольких отрядов. Описано несколько видов многохвосток Collembola, которые светятся непрерывно, хотя интенсивность люминесценции изменяется в зависимости от степе-
588
Глава 24
Рис. 24-2. 1
А. Срез через фотогенный орган глубоководной креветки Sergestes. Б. Срез через фотогенный орган головоногого моллюска Rondeletia. В. Срез через люминесцирующую область эпидермиса полихеты Chaetopterus. 1 — линзы; 2 — полоска соединительной ткани; 3 — гиподерма: 4 — пигментный слой; 5 — эпителий линз; 6 — светящиеся клетки; 7—базальная мембрана; 8 — отражающий слой; 9 — сумка; 10 — разрядившиеся фотогенные клетки; 11 — кутикула; 12 — слизистые клетки.
ни возбуждения особи. Обнаружена способность к люминесценции у личинки грибного комара Ceratoplanus, а также у личинки и взрослой особи грибного комара Bolitophila. Свечение, по .всей вероятности, возникает у них в мальпигиевых канальцах.
Самые известные и поразительные примеры свечения организ
Б иолюминесценция
589 "
мов можно найти среди Coleoptera, особенно среди светляков Lampyridae и щелкунов Elateridae. У этих насекомых фотогенный орган, как правило, расположен вентрально, в задней области брюшка. Этот орган состоит из двух клеточных слоев — брюшного слоя фотогенерирующих клеток и дорсального, или внутреннего, слоя «отражающих» клеток. Отражающие клетки содержат какие-то мелкие частицы, очевидно соли мочевой кислоты. Этот слой, по-видимому, играет для фотогенных клеток роль рассеивающего отражателя, одновременно являясь дорсальным щитком.: Фотогенный орган пронизан множеством трахеальных трубочек, которые оканчиваются в многочисленных трахейных концевых клетках. Фотогенные клетки не имеют нервных окончаний. Детальное строение этих органов подробно описано в ряде работ [10, 42, 102].
Свечение светляка регулируется нервной системой. Замечательной особенностью деятельности этих органов, свидетельствующей о существовании нервной регуляции, является синхронное вспыхивание некоторых тропических светляков [7]. Было установлено, что у самцов таиландской разновидности Pteroptyx malaccae вспышки отличаются исключительной ритмичностью: 560 ±6 мс при 28°C, и происходят синхронно с точностью до ±20 мс [И]. Поскольку минимальное время, которое проходит от момента восприятия светового сигнала глазом до передачи его на световой орган, составляет 60—80 мс, следует ожидать, что внутренние индивидуальные сигналы, регулирующие каждую массовую вспышку, должны возникать в ответ на завершение предыдущего цикла.
У южноамериканского щелкуна Pyrophorus имеется пара лю-минесцирующих органов зеленоватгоо цвета, расположенных дорсолатерально на переднегруди, и оранжево-желтый орган на брюшной поверхности первого брюшного сегмента. У южноамериканского «железнодорожного червя» Phrixothrix голова светится красноватым светом, а по всему телу сегментарно расположены зеленоватые люминесцирующие пятнышки. Североамериканская разновидность родственного ему Phengodes (рис. 24-3) по своим фотогенным органам напоминает южноамериканскую с той лишь разницей, что у нее отсутствует красный головной орган, а свечение непрерывное, в то время как «железнодорожный червь» светится только в ответ на возбуждение.
Некоторые виды многоножек выделяют светящуюся слизь в основном таким же образом, как и светящиеся дождевые черви. Был описан также светящийся морской паук.
Моллюск-камнеточец Pholas имеет ряд фотогенных желез, выделяющих секрет в сифон [87]. У некоторых голожаберных моллюсков по всему телу рассеяны вспыхивающие при соответствующей стимуляции клетки. Среди моллюсков эта способность наиболее развита у головоногих. У многих из них, например у кальмара Loligo, светящиеся органы сообщаются с внешней средой, в ко-
590
Глава 24
торой живут и размножаются светящиеся бактерии (т. е., по всей видимости, сам по себе кальмар не люминесцирует). У большин-
ства других свет генерируется клетками самого животного {рис. 24-2). У одного из кальмаров (Heteroteuthis} есть непарный светящийся орган, который открывается в полость мантии, куда через сифон при раздражении выбрасывается светящееся облачко.
Рис. 24-3. Личинка самки жука Phen-godes [44].
А. Вид сверху. Б. Вид сбоку. В. Светящееся животное в темноте.
клеточные фотогенные органы, минесценция возникает в ответ У хордовых способностью
У некоторых видов кальмаров, например у светящегося кальмара Watsenia, имеется сложная система фотогенных органов внутриклеточного типа, разбросанных по всему телу. В частности, у Watsenia на конце каждой конечности имеется по три светящихся органа. Эти органы содержат мелкие палочкообразные структуры размером примерно 2—4 мкм, которые раньше ошибочно считали бактериями [88].
Еще более высокая степень дифференциации наблюдается у таких глубоководных форм, как Lycoteuthis diadema. У них раз-
личные люминесцирующие орга-
мы испускают свет до четырех разных цветов.
У иглокожих все светящиеся виды относятся к офиурам. У некоторых из них имеются однорассеянные по всему телу, и лю-на любое раздражение.
к свечению обладают лишь пер-
вичнохордовые и рыбы. Некоторые виды Balanoglossus вырабатывают светящуюся слизь. Среди оболочников наиболее хорошо известна люминесценция колониальных видов Pyrosoma; эти орга-
низмы люминесцируют в ответ на раздражение, причем волна фотогенной активности распространяется по всей колонии. В связи с тем что в такой колонии активность может быть вызвана вспышкой света, свечение одной из особей, очевидно, и служит
стимулом для естественного распространения активности в колонии от одной люминесцирующей особи к соседним.
Множество светящихся видов обнаружено среди акуловых и особенно костистых рыб (главным образом среди глубоководных). У некоторых из них, например у Photoblepharon, Anomalops, Physiculus, Leiognathus и Monocentris, свет генерируется симбиотическими бактериями, находящимися в особых органах, локализован-
Биолюминесценция
591
<9
ных в нескольких специфических участках. У Malacocephalus светящиеся бактерии выделяются при возбуждении из центральных мешочков на вентральную поверхность тела. Astronesthes и StomP as снабжены хорошо развитыми головными световыми органами с линзами и пигментированными «глазными бокалами». У некоторых рыб свечение в ответ на раздражение развивается медленно. У Porichthys латентный период при стимуляции симпатического нерва на уровне спинного мозга равен 8—10 с [37]. Многократное электрическое раздражение области, непосредственно прилегающей к фотофорам, вызывает их свечение с минимальной латентностью в 1,1 с. В качестве медиаторов в этом процессе выступают, по-видимому, адреналиноподобные вещества, поскольку, во-первых, при локальной инъекции норадреналина или амфитамина (агента, вызывающего выделение адреналина) латентный период не превышает 5 с, а во-вторых, в нейро-фотоцитарном слое фотофоров гистохимическими методами обнаружены катехоламины [15],
Физические аспекты биолюминесценции. Испускание света обычно сопровождается выделением тепла, однако при биолюминесценции этого не происходит — она представляет собой холодное свечение. С физической точки зрения различают два типа све-тоиспускания: тепловое излучение, при котором испускание света зависит от температуры и возникает в результате теплового возбуждения электронов, и люминесценцию, не связанную с выделением тепла, при которой происходит специфическое возбуждение электронов. При биолюминесценции энергия, необходимая для такого возбуждения, вырабатывается в ходе химической реакции.
При тепловом излучении (огонь или электрическая лампочка) цвет испускаемого света зависит от температуры — чем выше температура, тем больше энергия испускаемых фотонов. В отличие от теплового излучения при люминесценции (люминофор телевизионного экрана, флуоресцентные лампы) цвет определяется природой молекулы, которая, специфически поглотив энергию, отдает ее в процессе электронного перехода, испуская фотон. Таким образом, энергия электронного перехода определяет цвет. При биолюминесценции различные организмы испускают свет самых разнообразных оттенков, от голубого до красного (рис. 24-4 и табл. 24-1).
У некоторых организмов, например у североамериканского «железнодорожного червя» Phrixothrix, различные органы одного и того же животного имеют разную окраску. Эти различия в окраске, возможно, обусловлены присутствием цветных мембран, выполняющих роль фильтров, однако обнаружить их до сих пор не удалось. Не исключено, что эти различия можно отнести и к собственно биолюминесцентной реакции, в частности к различиям в специфичности люциферазы [90, 91].
В связи с особой ролью светоиспускания определенный цвет биолюминесценции, очевидно, имеет в каждом конкретном случае
592
Глава 24
приспособительное значение. Голубой свет, как правило, испускается морскими видами и по спектральному составу близок свету, проникающему на глубину 30 и более метров, а также согласуется с чувствительностью органа зрения многих морских рыб [19, 27, 28, 107]. Хотя интенсивность свечения, испускаемого фотогенными органами, относительно мала, тем не менее темноадаптированным глазом оно легко воспринимается. Интенсивность биолюминесценции удобнее всего выражать числом фотонов (квантов), прямо пропорциональным числу возбужденных молекул (см. ни-
Рис. 24-4. Спектры биолюминесценции ряда организмов in vivo.
I — Gonyaulax polyedra; II — Photobacterium fischeri; III — Obelia geniculata; IV — Otnphalia flavida; V — Photuris penns у Iv ante a; VI — Photinus scintillans. Обратите внимание на различия в Лтах и ширине кривых, а также на существенные различия в ^тах люминесценции двух видов светляков.
же). Одиночный изолированный фотогенный орган рыбы-мичмана Porichthys испускает около 109 квант-с-1; длительность свечения составляет почти 1 с; всего, таким образом, за одну .вспышку испускается 109 квантов [2]. Рыба-мичман снабжена сотнями специфических органов, напоминающих пуговицы (отсюда и ее название), расположенных вдоль вентральной и латеральной поверхностей тела. Фотогенный орган жука-светляка гораздо ярче — за одну вспышку он испускает 1014—1015 фотонов; длительность вспышки около 1—2 с, в то время как один вид динофлагеллят (Gonyaulax) испускает всего 108—1010 фотонов за время свечения такой же длительности. Штаммы бактерий, обладающие ярким свечением, могут испускать до 104 квантов в 1 с; за время 104 с (т. е. около трех часов) для клетки размером всего в несколько микрон это весьма значительная величина.
Эффективность биолюминесценции может быть выражена несколькими способами. Энергетический коэффициент рассчитывается как процент теоретически рассчитанной энергии химической
•Биолюминесценция
593 "
Таблица 24-1
Длины воли максимумов люминесценции некоторых светящихся организмов. Во всех случаях, за исключением кишечнополостных, окраска свечения in vitro практически такая же, как и у живых организмов.
У кишечнополостных, кроме того, in vivo имеется еще одно люминесцирующее вещество
Животные К, нм
in vivo in vitro
Ракообразные (Cypridina) Динофлагелляты (Gonyaulax) Бактерии (Photobacterium, Achromobaite) Кишечнополостные (Aequorea, Obelia, Renilla) Грибы (Omphalia) Светляки (Photinus и другие виды) Phrixothrix 470 480-490 508 525 550—580 (красное свечение) ~600 465 470 490 460—485 520 550—580
реакции, переходящей в свет. Величины, приводимые для таких коэффициентов, являются расчетными и обычно довольно высоки— между 10 и 50% [91]. Более целесообразно экспериментально измерять квантовый выход хемилюминесценции — число фотонов, приходящихся на каждую молекулу окисленного субстрата. У жука-светляка эта величина составляет около 0,86, т. е. почти все молекулы люциферина, вступающие в реакцию, приводят к испусканию фотонов. В других биолюминесцентных системах также были отмечены высокие значения квантовых выходов — 0,27 у медузы Aequorea и 0,3 у бактерий. И наконец, эффективность люминесценции может быть описана на основе чувствительности детектора, т. е. на основе того, в какой степени спектр испускания света совпадает со спектральной чувствительностью воспринимающего элемента. Это обычно называется эффективностью свечения. Глаз со спектральной чувствительностью, характерной для глаза человека, воспринимает свечение жука-светляка Photuris с эффективностью около 92%. Для голубого свечения рачка Cypridina эффективность свечения в этом случае значительно ниже — около 20%. Следует, однако, всегда иметь в виду, что при оценке эффективности свечения необходимо учитывать экологические факторы, и, хотя мы до сих пор не располагаем всей совокупностью сравнительно-физиологических данных по этому вопросу, можно думать, что спектр биолюминесцентного свечения окажется весьма близким спектральной чувствительности органов зрения тех организмов, с которыми светящиеся виды взаимодействуют.
Химические основы генерации света. Биолюминесценция представляет собой хемилюминесценцию, катализируемую фермента
38—2067
594
Глава 24
ми. В ходе хемилюминесцентной реакции выделяется энергия, которая не теряется в виде тепла и не сопряжена с какими-либо реакциями синтеза, а идет на специфическое возбуждение молекул, способных выделять энергию в виде фотонов.
Энергия фотона Е зависит от цвета, т. е. частоты (длины волны) света и определяется по формуле E = hx, где h — постоянная Планка, a v — частота. Для биолюминесценции, при которой частота генерируемого света лежит в области, соответствующей длинам волн от 450 до 600 нм (рис. 24-4), эта энергия составляет 65—45 ккал/моль. Это довольно большая энергия; например, считающаяся высокой энергия гидролиза АТФ равна примерно 7 ккал/моль, т. е. почти на порядок меньше. Знание биохимических и ферментативных механизмов этих реакций имеет очень большое значение для понимания физиологических процессов биолюминесценции, особенно в связи с энергетикой этих процессов.
Уже в середине прошлого столетия французский химик Дюбуа [31] установил, что способность химического вещества к люминесценции сохраняется и in vitro, если его быстро заморозить или высушить. В опытах со светящимся моллюском Pholas Дюбуа показал, что люминесценция возникает в ходе реакции окисления молекулярным кислородом субстрата, для которого он придумал название люциферин (светоносный); реакция катализируется ферментом — люциферазой. <В приведенном ниже уравнении химической реакции этот субстрат обозначен буквой L; звездочка в правой части уравнения означает, что он находится в электронно-возбужденном состоянии; затем происходит переход в основное состояние и испускается фотон:
Люцифераза (Е)
Ь + О2-----------Е —L*,
Е—L* -----> E+L-|-/zv.
Время жизни такого возбужденного состояния чрезвычайно мало — от 10~9 до 10-8 с. Если считать, что возбуждение возникает в ходе ферментативной реакции, то разумно предположить, что свечение происходит до того, как L* отщепляется от фермента. Таким образом, правильнее считать, что в возбужденном состоянии находится фермент-субстратный комплекс Е—L*.
Термины «люциферин» и «люцифераза» употребляются теперь как общие обозначения субстрата и фермента, участвующих в любой . биолюминесцентной реакции. Несмотря на то что разные классы или группы организмов содержат различные специфические виды молекул, общий механизм реакции весьма сходен и поэтому может быть представлен вышеприведенным общим уравнением. У нескольких организмов биохимические механизмы реакции изучены весьма детально. В 1966 году удалось установить структуру люциферина у рачка Cypridina, выбрасывающего и фер-
Биолюминесценция
595 “
мент, и субстрат в морскую воду [61—64]. Активный центр соединения (рис. 24-5) содержит триптамин, аргинин и L-изолейцин. Судя по данным хроматографии на бумаге, удельной адсорбции и временным параметрам люминесценции синтетический люциферин
Рис. 24-5. Последовательность реакций светогенерации у Cypridina.
оказался весьма сходным с природным. Была очищена и частично охарактеризована и люцифераза Cypridina. Ее мол. вес равен примерно 50 000, и она не содержит ни атомов металла, ни простети-ческой группы. Известно [103], что в ходе биолюминесцентной реакции поглощается одна молекула кислорода и выделяется одна молекула СО2 на каждую молекулу вступившего в реакцию люциферина; квантовый выход хемилюминесценции равен примерно 0,3 [59]. При этом предполагается, что реакция с кислородом происходит так, как это показано на рис. 24-5, и что через несколько промежуточных стадий образуются СО2 и возбужденный продукт. Этот продукт неустойчив и расщепляется с образованием а-метилмасляной кислоты и этиолюциферина. Поскольку прозе*
Глава 24 .........
Люциферин
СР-Р—О—Р—О—Р—о—сн
1 Аденин N
Рибоза
но он
АТФ
Люцифераза
' *
о"
о—р—о—рибоза- аденин
I
о
+ СО, И АМф
о
о
I о
Желтое свечение
,(М) 560нм
Рис. 24-6. Реакция взаимодействия люциферина светляка с АТФ и последующее окисление промежуточного продукта.
дукты в организме не восстанавливаются, вопроса об их повторном использовании или обмене не возникает.
У жука-светляка реакция протекает внутриклеточно и регулируется нервной системой. Если световой орган светляка погрузить в воду, то свечение растворимой фракции будет продолжаться в течение нескольких минут. Свечение можно восстановить добавлением АТФ. В настоящее время известно, что жук-светляк содержит люциферин (рис. 24-6), который активируется в ходе реакции с АТФ, образуя люцифериладенилат; подобные реакции происходят между аминокислотами и АТФ и представляют собой первую стадию синтеза белка. Как и у Cypridina, на последующих этапах реакции на одну окисленную молекулу люциферина погло
Биолюминесценция
597
щается одна молекула кислорода и образуется одна молекула СОз- В этом случае квантовый выход хемилюминесценции значительно выше — 0,89. Все приведенные на рис. 24-6 промежуточные стадии были предложены на основании опытов с применением изотопов [29].
Люцифераза жука-светляка была получена в кристаллическом виде в 1956 г. [20, 36], и ее химическая структура сейчас изучена достаточно хорошо. Подобно люциферазе Cypridina, ее активность никак не связана с присутствием металла или простетической группы. Мол. вес фермента равен примерно 100 000; молекула состоит из двух (возможно, различных) субъединиц и имеет один активный центр. Как уже упоминалось, большое внимание было уделено изучению различий в спектрах люминесценции жуков-светляков разных видов; вероятно, эти спектры в значительной степени зависят от структуры фермента или его конформации или от обоих факторов одновременно [68, 69]. Предположение о том, что различия в спектрах обусловлены люциферином, было отвергнуто как опытами, в которых использовался синтетический люциферин, так и тем, что, хотя люциферины из различных жуков-светляков одинаковы, спектры люминесценции при этом могут быть разными. В чем именно в данном случае состоит различие между люциферазами, пока неясно.
Молекулярный механизм контроля вспышки жука-светляка, длящейся обычно около 0,5 с, остается невыясненным. Одним из необходимых требований для «темнового» состояния является разделение реагирующих веществ — компартментализация или какой-либо другой способ предотвращения вступления их в реакцию друг с другом в темноте. Фотогенные клетки, несомненно, содержат все химические компоненты, необходимые для свечения, и для решения указанной проблемы крайне интересно и важно было бы располагать сведениями об их внутриклеточном распределении в интактной клетке.
Биолюминесценция у бактерий представляет собой третий, явно непохожий на предыдущие как с точки зрения химических процессов, так и клеточной организации и регуляции случай биолюминесценции. Это внутриклеточное, непрерывное свечение; в основе его лежит окисление восстановленного кофактора флавина —• рибофлавин-5'-фосфата, известного также как флавинмононуклеотид (ФМН). Эта система, будучи непосредственно связанной с кофакторами, принимающими участие в окислении субстратов, обеспечивает передачу электронов прямо на молекулу кислорода; энергия при этом расходуется на генерирование света, а не на синтез АТФ, как это имеет место при окислительном фосфорилировании.
Более детально механизм этой реакции не изучен, однако полагают, что она аналогична реакциям для смешанных оксидаз,, так как для светоиспускания требуется также альдегид с длинной
5598
Глава 24
углеродной цепью (более 6 атомов углерода) (см. ниже). Как и ранее, электронно-возбужденное состояние обозначается Е—ФМН* (комплекс фермент—флавин), и в этом случае для такого обозначения существует дополнительное основание, поскольку ФМН флуоресцирует зеленым светом с 7.юах 530 нм, в то время как спектр люминесценции бактерий сдвинут в синюю область (490 нм). Такой сдвиг может быть обусловлен взаимодействием с белком или промежуточными фермент-связанными формами.
Флавинредуктаза
НАД • Н+ (или НАДФ • Н) + ФМН + Н+---------------* НАД + ФМН •
, Люцифераза
ФМН.Н2 + ИСНО + О2--------------* Е—ФМН* 4-RCOOH +Н2О
(восст. флавин- (альдегид) (рибофлавин-5-
'• мононуклеотид) фосфат)
Чистая бактериальная люцифераза была получена в кристаллическом виде и подробно охарактеризована. Она представляет собой простой белок (мол. вес 79 000), состоящий из двух неидентичных субъединиц: ю (мол. вес 42 000) и р (мол. вес 37000) [34, 52, 56]. Было показано, что активный центр фермента локализован на а-субъединице; роль р-субъединиц до сих пор не выяснена; возможно, они важны для солюбилизации системы [70, 71].
Одним из наиболее интересных физиологических явлений, связанных с люминесцентной системой бактерий, является то, что синтез люциферазы протекает, по-видимому, в достаточной степени независимо от синтеза других белков. Так, например, в только что инокулированной культуре в течение первых нескольких часов люцифераза не синтезируется; синтез начинается при достижении определенной плотности клеток и протекает со скоростью, в несколько раз превышающей скорость их размножения, а затем внезапно прекращается [77]. Эта квазинезависимая регуляция активности люциферазного гена может быть отнесена на счет симбиотической роли бактерий: в этом случае их собственное свечение развивается внутри специализированного фотогенного органа хозяина [49].
Второе, еще более резкое ограничение активности гена люциферазы наблюдается у спонтанно темновых мутантов, у которых активность гена полностью подавлена и синтеза люциферазы не происходит. Однако ген при этом не теряется и не претерпевает необратимых изменений, поскольку легко могут быть получены ревертанты [60]. Это — второй вид регуляции, очень важный для организма, который способен существовать в одном из двух состояний — симбиотическом, когда свечение является необходимым, и в свободной форме, когда люминесценция отсутствует.
Весьма детально были изучены также биохимические процессы, лежащие в основе люминесценции у некоторых видов кишечнополостных. Из кораллов Renilla Кормье с сотр. [20, 21] выделили
I
Биолюминесценция 599
субстрат — люциферин — и ферменты, показав, что процесс распадается на две следующие стадии:
Са2+
ДФА + Щ,—х---------> LH2 + ФАФС
Фермент 1
LH„+ Оо ------* Свет.
На первой стадии активация люциферина (ЬН2) заключается в удалении одной группы (х), идентифицированной как сульфат; эта реакция стимулируется кофактором 3',5'-дифосфоаденозином (ДФА), при этом образуется активированный люциферин и фос-фоаденозилфосфорилсульфат (ФАФС). Активированный люциферин по структуре весьма сходен с люциферином Cypridina, и при его окислении, химический механизм которого также напоминает механизм окисления люциферина Cypridina, наблюдается свечение.
Среди других кишечнополостных необычный тип белка был выделен сначала из медузы Aequorea и сидячей медузы Halistaurea [93], а затем и из других видов [51, 75]. Этот белок начинает светиться при простом смешении с ионами кальция:
Са2+ Фотобелок---> Свет.
Процесс in vitro протекает очень быстро [52] и представляет собой вспышку длительностью около 1 с при 20 °C с высоким квантовым выходом. За исключением белка и кальция, никаких других веществ, в том числе и кислорода, для этой реакции не требуется. Хастингс и Морин [51] постулировали единую схему биолюминесценции у кишечнополостных, в которой фотобелок является относительно стабильным промежуточным продуктом цепи реакций, сходных с теми, которые были описаны для Renilla. На последнем этапе превращений фотобелок взаимодействует с кислородом; для активации заключительной внутримолекулярной реакции, в ходе которой выделяется энергия, необходим кальций. Этот процесс представляет особый интерес с физиологической точки зрения, поскольку кальций используется и в других системах, например для регуляции некоторых специфических реакций, в частности мышечного сокращения. Весьма знаменателен тот факт, что здесь удается выделить «активируемый» кальцием биохимический промежуточный продукт. Интересно, что в большинстве случаев [76] биолюминесцентная система кишечнополостных локализована в клеточных гранулах или органеллах, где, по-видимому, синтезируются и хранятся эти промежуточные вещества; мембраны этих гранул или органелл, очевидно, участвуют в процессе свечения.
Опыты с кишечнополостными привели к еще -более удивительному наблюдению, а именно к обнаружению процесса переноса энергии [76]. У тех светящихся организмов, для которых удалось
€00
Глава 24
получить спектральные данные, спектр биолюминесценции животного организма такой же, как и при изолированных биохимических реакциях, протекающих согласно уравнениям 1 и 2.
(1) А + В---* C* + D
(2) С* --> C + ftv (Ар голубой)
(3) С*4~Е---> Е*4-С
(4) Е*---> E+/1V (Х2, зеленый)
У некоторых видов люминесцирующих кишечнополостных свет, '.испускаемый живым организмом, зеленый, в то время как in vitro свечение изолированной системы синее (табл. 24-1). Это указывает на то, что энергия от молекулы возбужденного продукта (С*) передается другому веществу (Е), которое затем люминесцирует при переходе из возбужденного в основное состояние. Далее было показано, что участвующие в этом процессе компоненты локализованы внутри каких-то субклеточных частиц, которые можно выделить, причем активные компоненты люминесцирующей in vivo •системы останутся интактными.
Еще одной системой, включающей в себя частицы или органеллы, является биолюминесцентная система динофлагеллят, одноклеточных морских организмов, отвечающих на стимуляцию короткой вспышкой длительностью 0,1 с. Хастингс и др. (см. [20, 55]) обнаружили, что крупная, быстро седиментирующая субклеточная частица (коэффициент седиментации ~15 ООО S) при простом снижении pH от 8 до 6,7 может испускать вспышку света, очень напоминающую вспышку in vivo. Предполагают, что эта •частица существует в аналогичной форме in vivo и что она ответственна за люминесценцию клетки, обусловленную каким-то процессом на уровне мембраны, который на непродолжительное время вызывает увеличение проницаемости ее для протонов [30]. Как люциферин, так и люцифераза могут быть выделены из над-юсадочной жидкости и, по-видимому, являются компонентами активного биолюминесцентного комплекса [33]. «Разряженный» комплекс после инкубации его со свежим субстратом (люциферином) при pH 8,0 с последующим понижением pH может вызвать -повторную вспышку.
Биохимия процессов, протекающих в других светящихся системах, изучена пока весьма поверхностно.
Было показано, что биолюминесценция грибов включает по крайней мере две стадии: 1) восстановление дегидро- или оксилюциферина при участии НАД-Н (НАДФ-Н) с растворимым ферментом и 2) окисление восстановленного продукта кислородом с помощью связанного с мембранами фермента с генерированием света [1].
Пресноводное блюдечко Latia из Новой Зеландии является «единственным известным светящимся животным, весь жизненный
Биолюминесценция
60>
цикл которого протекает в пресной воде. Его люциферин был выделен и очищен; удалось установить его эмпирическую формулу — С15Н24О2. Мол. вес люциферазы Latia равен 173000; система требует также присутствия кофактора, названного пурпурным белком (мол. вес 39 000). Квантовый выход (т. е. число фотонов, испускаемых на 1 реагирующую молекулу) при 8 °C для люциферазы? равен 0,63, а для люциферина — 0,0068 [95, 98, 99].
Люминесцентная система кольчатого червя Chaetopterus содержит фотобелок, который удалось получить в кристаллическом виде (мол. вес 184 000). Для свечения, как оказалось, необходимо-также присутствие двух кофакторов (один из которых подобен нуклеопротеиду, а другой — липиду), перекиси (Н2О2 или органической перекиси), кислорода и двухвалентного железа [94, 97].
Люминесцентная система эвфаузииды Meganyctiphanes состоит из двух компонентов, один из которых содержит фотобелок и какое-то до сих пор не установленное светящееся вещество [96,. 100].
Среди изученных рыб у видов Acropoma, Cleidopus, Gazzcg Leiognathus, Paratrachychthys и Siphamia, а также, возможно, у Photoblepharon и Anomalops свечение определяется присутствием симбиотических бактерий [40]. В светящихся органах Apogon,. Parapriacanthus и Pempheris бактерии не обнаружены. Перекрестные реакции систем «люциферин — люцифераза» у Apogon и Parapriacanthus с системой Cypridina указывают, что, возможно,, образование люциферина рыб обусловлено питанием их рачками-Cypridina [39]. Как следует из различий в кинетике реакций и иммунологических тестов, люциферазы Apogon и Cypridina неодинаковы даже при поверхностном рассмотрении, что указывает нагих эндогенное происхождение в каждой группе [106].
Сравнительное изучение строения светящихся органов различных животных показало, что у Pholas и Latia, эвфаузиидов и некоторых видов рыб имеется по два типа железистых клеток. У других организмов (например, у полихет) имеется только один1 тип клеток. Согласно данным электронной микроскопии, разделение реагирующих веществ, участвующих в биолюминесцентных реакциях, может осуществляться с помощью стенок микротрубочек [3].
Регуляция биолюминесценции. У животных свечение, как правило, происходит не непрерывно, а возникает время от времени в ответ на внешнее раздражение [9, 86]. Это положение настолько общее, что в случаях непрерывного свечения животного неизбежно возникает мысль о присутствии внутри или на поверхности тела животного симбиотических, патогенных или случайных светящихся бактерий; исключение составляет, по-видимому, лишь Phen-godes.
Существует три основных типа регуляции свечения у животных. Два первых относятся к экстраклеточной люминесценции,
1602
Глава 24
В тех случаях, когда светящиеся вещества при раздражении выделяются из фотогенного мешочка в окружающую морскую воду (например, у головоногого Heteroteuthis, рачка Cypridina и креветки Acanthephyra'), регуляция является опосредованной и осуще-ставляется при помощи типичных нервно-мышечных механизмов. ’ У животных (светящиеся морские полихеты Chaetopterus, многоножки, некоторые кишечнополостные и моллюск Pholas), которые при определенных условиях выделяют светящуюся слизь на поверхность тела, регуляция секреторного процесса осуществляется прямым нервным, а в некоторых случаях — эндокринным возбуждением. Регуляция третьего типа имеет место у тех многочисленных животных, у которых люминесценция является внутриклеточной, z например у таких хорошо известных видов, как ночесветка, медузы, гребневики, насекомые и некоторые виды рыб.
В этом случае имеется некий механизм, посредством которого возбуждение клетки приводит к свечению уже имеющихся в ней генерирующих свет фотогенных веществ. Это достигается с помощью: а) быстрого выделения небольших количеств одного из необходимых фотогенных веществ, вступающих в реакцию внутри клетки; б) кратковременного изменения внутриклеточной среды, благоприятствующего люминесцентным реакциям, например изменение содержания кислорода, воды или ионов водорода; в) изменения направления путей химических превращений в сторону, благоприятную для протекания хемилюминесцентных реакций.
В настоящее время мы можем лишь строить догадки о том, каким образом животное в ответ на раздражение дает очень короткие и интенсивные вспышки света, которые по скорости нарастания и спада интенсивности могут соперничать с лучшими лампами накаливания. У многоклеточных организмов вспышки обычно имеют рефлекторный характер и возникают в ответ на раздражение тактильных рецепторов или фоторецепторов. В этих рефлекторных процессах могут участвовать как нервная система, так и гормональные механизмы. У кишечнополостного Renilla свечение после локального раздражения распространяется по всему организму со скоростью, характерной для проведения возбуждения в нервной сети. Чрезмерное раздражение может приводить к такой степени возбуждения, при которой люминесценция продолжается достаточно долго. У гребневика Mnemiopsis тактильные рецепторы, раздражение которых рефлекторно вызывает свечение, расположены вдоль нервных проводящих путей под рядами ресничных гребешков. В реагирующем нейроэффекторном механизме обнаружено явление облегчения, сходное с тем, которое наблюдается в нервно-мышечных синапсах.
Регуляция люминесценции изучалась главным образом на жуках-светляках [13, 15, 16, 17]. У этих насекомых наряду с другими раздражителями ответную вспышку возбуждают зрительные сигналы. Фотогенный орган иннервирован (рис. 24-7 и 24-8),
Биолюминесценция
603",
однако нервные волокна доходят лишь до трахеальных окончаний внутри его. Иннервируется, очевидно, какой-то механизм, локализованный в трахеальных концевых клетках [41, 42, 102]. Эта система адренэргична [101].
Для объяснения процесса регуляции вспышек у светляка были выдвинуты две основные гипотезы. Согласно одной из них, вспыш-
Рис. 24-7. Трехмерная модель фотогенного органа светляка (из John В. Buck,. «Unit Activity in Firefly Lanterns», in Frank H. Johnson and Yata Haneda, Bioluminescence in Progress, Princeton University Press, p. 462, 1966).
Видны цилиндры с нервными волокнами и расположенные вокруг каждого цилиндра в виде розеток 12 фотоцитов; 1 — вентральная поверхность «фонарика» светляка; 2 — цилиндр; 3— дифференцированная зона; 4 — дорсальный слой; 5 — фотоцит; 6 — концевая клетка трахеи; 7 — трахеола; 8 — трахея; 9 — нерв.
ка определяется быстрым поступлением кислорода в фотогенные клетки, находящиеся в состоянии аноксии. Считается, что это достигается либо путем прямой нервной регуляции поступления кислорода в трахеальные концевые клетки, либо увеличением количества метаболитов в фотогенной ткани при раздражении с последующим осмотическим выведением воды из концевых частей трахеальных трубочек, благодаря чему кислород доставляется прямо к железистым клеткам. Присутствие кислорода делает воз-
€04
Глава 24
можным окисление продуктов обмена, вновь уменьшая осмотическое давление в клетке и обеспечивая новое наполнение водой трубочек, а также повторное выведение кислорода. Существование такого механизма регуляции содержания кислорода подтвержда-
Рис. 24-8. Схематическое изображение тангенциального среза через фотогенный орган Photuris (гистол101гичеокое строение) [42].
Указаны средние размеры соответствующих структур. В цилиндрах содержатся трахеи и концевые органы; от цилиндров радиусами расходятся фотофоры, соединяющие цилиндры между собой. 1 — нерв; 2 — нервное окончание; 3 — концевая клетка трахеи; 4 — трахея; ДЗ — дифференцированная зона.
стся еще и тем, что самое яркое свечение люминесцирующей железы исходит из областей, расположенных в непосредственной близости к концам трахеальных трубочек. Однако система клапанов концевой клетки не играет здесь существенной роли, поскольку некоторые насекомые, особенно личинки светляков, хотя бы в небольшой степени генерирующие прерывистое свечение (хо-
Биолюминесценция
605
тя и не истинное вспыхивание), не имеют развитых трахеальных концевых клеток. И разумеется, быстрое вспыхивание может возникать у многих организмов, лишенных трахеальной системы.
Вторая гипотеза постулирует прямое нервное возбуждение ге-Л нерирующих свет клеток специальными фотогенными нервами (рис. 24-9). Такое возбуждение охватывает и регуляцию люминесценции у животных, не имеющих трахеальной системы.
У рыб Porichthys и Echiosto-та нормальные рефлекторные пути регуляции вспыхивания в системе фотогенных органов включают симпатическую иннервацию. При введении адреналина также наблюдается медленная фотореакция.
В результате целого ряда наблюдений удалось установить, что яркий свет подавляет свечение животных, однако о механизмах такого подавления в боль
Рис. 24-9. Спонтанные и вызванные электрической стимуляцией вспышки фотогенного органа Photuris pennsyl-vanica и соответствующие потенциалы действия, зарегистрированные на вентральной поверхности светящегося органа [16].
А. Спонтанная вспышка. Б. Электрическое раздражение мозга (артефакт раздражения — слева в начале записи) с последующим нервным,разрядом и вспышкой.
шинстве случаев почти ничего не известно; мы не знаем, осуществляется ли оно непосредственным влиянием на систему фотогенных реакций или же подавляются механизмы, связанные с рефлекторным возбуждением. Данные, полученные, в частности, на гребневиках, указывают, что причиной подавления может быть непосредственное разрушение фотогенного материала внутри светогенерирующих клеток. Экстракты, выделенные из особей, подвергавшихся облучению солнечным светом, практически не лю-
минесцировали. Кроме того, после пребывания на ярком свету •свечение таких животных, как гребневики и Renilla, возобновлялось лишь через несколько минут после перенесения их в темноту.
Уже неоднократно сообщалось о четкой суточной ритмичности изменения способности к свечению у ряда животных — светляка Photinus [5], морского желудя Ptychodera [22], динофлагеллята Gonyaulax [54, 101] и, вероятно, медузы Pelagia [12, 55]. Было
606
Глава 24
показано, что такой ритм сохраняется в течение нескольких дней непрерывного пребывания их в темноте.
Люминесцентная реакция на раздражение у гребневиков и гидромедуз быстро ослабевает и восстанавливается точно таким же образом, как и в случае других типичных сенсорно-нейроэффектор-ных механизмов.
Функциональная роль биолюминесценции. Роль люминесценции в жизни многих светящихся организмов полностью еще не выяснена. Тем не менее можно предположить, что эта функция имеет определенное значение для выживания и появилась в процессе отбора, поскольку у большинства способных к свечению высших животных развились для этой цели специальные клетки и органы. Возможно также, что свечение является частью общего механизма, с помощью которого организм отвечает на раздражение. Поразительным примером, иллюстрирующим это явление, может служить агрессивная мимикрия у самок некоторых видов Photuris к некоторым видам Photinus. Привлеченные таким образом самцы пожираются [65, 67].
Люминесценции у животных обычно приписывают три основные функции. Одной из них является привлечение пищи [65].
Вторая — защитная; в этом случае люминесценция служит для предостережения, отпугивания или отвлечения хищников, помогая светящемуся животному скрыться, как это имеет место у выпускающей светящееся облако глубоководной морской креветки Acanthephyra и кальмара Heteroteuthis. Существование такой функции подтверждает характер нервной регуляции свечения у кольчатого червя Acholoe. Если перерезать этого червя пополам, имитируя нападение хищника, то яркое свечение наблюдается только в его задней части, в то время как свечение передней гаснет. Это явление обусловлено, по-видимому, тем, что таким способом более жизнеспособной передней части обеспечивается возможность скрыться от хищника. Аутотомированные чешуйки этого червя также ритмично вспыхивают. Яркое свечение, наблюдаемое у некоторых рыб после инъекций адреналина, также подтверждает представление о защитной роли люминесценции. Высказывались также интересные предположения о существовании у люминесценции несколько видоизмененной защитной функции: возможно, стимуляция свечения у одной особи группы служит предупреждением для других особей и позволяет им избежать опасности. Интересная роль биолюминесценции была описана на примере рыбы-пони. Небесно-голубая люминесценция брюшка этой рыбы помогает ей маскироваться от более глубоководных хищников [48].
Лучше всего обосновано представление о сигнальной роли люминесценции для встречи самцов и самок в брачный период. Особенно хорошо выражена эта способность у светящегося червя
Б иолюминесценция
607
Odontosyllis с Бермудских островов [35]. Самки, у которых наблюдается четкий лунный ритм, в период спаривания появляются на морской поверхности, где и плавают небольшими кругами, испуская яркое свечение. Самцы плывут из глубины прямо к светящимся самкам и присоединяются к их брачному «танцу», после чего и самцы, и самки одновременно выделяют в воду гаметы. Если за короткое время ни один самец не присоединится к светящейся самке, ее свечение постепенно угасает, но после небольшого перерыва фотогенные органы самки снова активизируются и люминесцирующий «танец» повторяется. Отмечалось, что самцы, направляющиеся прямо к светящейся самке, но не успевшие приблизиться к ней до окончания свечения, прекращали свое направленное движение и разбредались до тех пор, пока свечение самки не возобновлялось.
Другие яркие примеры использования свечения в качестве сигнала для привлечения особей в брачный период были описаны у различных видов светляков [6]. У многих видов самки, иногда даже бескрылые, сидят в траве, в то время как самцы летают вокруг них, подлетая к тем самкам, которые подают сигналы в ответ на их вспышки. Разные виды светляков различаются по характеру свечения, т. е. по частоте, общему числу вспышек, цвету, интенсивности и длительности каждой вспышки. Самка отвечает только на вспышки самца своего вида и вполне определенным для данного вида образом. Например, сигналом у самцов Photinus greeni служат две раздельные вспышки длительностью 1,1 и 1,7 с при 27 °C. Частота возникновения таких парных вспышек или длительность каждой вспышки особой роли при этом не играют. Ответная реакция самки возникает через 0,8—1,1 с после второй вспышки самца [12]. Самец продолжает сигнализировать самке до тех пор, пока оба не встретятся. Привлекающий световой ответ самки можно имитировать с помощью вспышки света, соответствующей типичным ответам самки данного вида.
Было описано много других примеров роли биолюминесценции в брачном поведении животных, однако большинство из них требует более убедительных доказательств или более тщательного экспериментального изучения.
Какое функциональное значение может иметь биолюминесценция у бактерий и грибов, представить трудно. Следует, вероятно, считать, что биолюминесценция возникла как побочный процесс, связанный с окислительными механизмами клетки, и сохранилась у многих животных, в частности у низших, несмотря на то что этот признак для выживания значения не имел [46]. Впоследствии это явление специфическим образом стало использоваться у высших животных и оказалось тем полезным свойством, которое приобрело право на существование.
608
Глава 24
у СПИСОК ЛИТЕРАТУРЫ
1. Airth R. L., Foerster G. Е., Behrens Р. Q., pp. 203—223 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Fungal bioluminescent system.
2. Baguet F., Case J., Biol. Bull. 140: 15—27, 1971. Porichthys photophore excitation.
3. Bassot J.-M., pp. 557—610 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Fine structure of luminous organs.
4. Biggley W. H., Swift E., Buchanan R. J., Seliger H. H., J. Gen. Physiol. 54: 96—122, 1969. Bioluminescence in marine dinoflagellate.
5. Buck J. B., Physiol. Zool. 10: 45—58, 1937. Diurnal rhythm in Photinus luminescent mecanism.
6. Buck J. B., Physiol. Zool. 10: 412—419, 1937. Luminescence in firefly mating.
7. Buck J. B., Quart. Rev. Biol. 13: 301—314, 1938. Sychronous flashing of fireflies.
8. Buck J. B., Ann. N. Y. Acad. Sci. 49: 397—482, 1947. Luminescence in fireflies.
9. Buck J. B., pp. 323—332 in: The Luminescence of Biological Systems, edited by F. H. Johnson. Amer. Assoc. Adv. Sci., Washington D. C., 1955. Control of bioluminescence.
10. Buck J. B., pp. 459—474 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. The firefly lantern.
11. Buck J. B., Buck E., Science 159: 1319—1327, 1968. Mechanism of rhythmic synchronous firefly flashing.
12. Buck J. B., Buck E., Biol. Bull. 142: 195—205, 1972. Bioluminescent signals in courtship of Photuris greeni.
13. Buck J. B., Case J., Biol. Bull. 121: 234—256, 1961. Control of firefly flashing.
14. Buck J. B., Case J., Hansen F. E., Jr., Biol. Bull. 125: 251—269, 1963. Control of firefly flashing.
15. Case J., Baguet F., Burns J., Amer. Zool. 10: 504, 1970. Photophore control in Porichthys.
16. Case J., Buck J. B„ Biol. Bull. 125: 234—250, 1963. Electrical activity in firefly flash.
17. Case J., Trinkle M. S„ Biol. Bull. 135: 476—485, 1968. Light inhibition of Photuris flashing.
18. Chang J. J., J. Cell. Comp. Physiol. 44: 365—394, 1954. Luminescence of Mne-miopsis.
19- Clarke W. D., Nature 198: 1244—1246, 1963. Functional significance of bioluminescence.’
20. Cormier M. J., Hori K., Kreiss P., pp. 349—362 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Light production in sea pansy, Renilla.
21. Cormier M. J., Karkhanes Y. D., Hori K., Biochem. Biophys. Res. Comm. 38: 962—964, 1970. Bioluminescent system of Renilla.
22. Crozier W. J., Anat. Rec. 20: 186—187, 1920. Diurnal rhythm in Ptychodera luminescence.
23. Davenport D., Nicol J. A. C., Proc. Roy. Soc. Lond. В 144: 399—411, 1955. Hydromedusoid luminescence.
24. Davenport D., Nicol J. A. C., Proc. Roy. Soc. Lond. В 144: 480—496, 1955. Luminescence of sea pens.
25. Davenport D., Wootton О. M., Cushing J. E.t Biol. Bull. 102: 100—110, 1952. Luminous millipede.
26. Dennell R„ J. Linn. Soc. 42: 393—406, 1955. Luminescence in Crustacea, Decapod a.
27. Denton E., Phil. Trans. Roy. Soc. Lond. В 258: 285—313, 1970. Nature of reflectors in bioluminescence.
28. Denton E., Sci. Amer. 224: 64—72, 1971. Biological significance of color of luminescence.
Биолюминесценция 609
29. De Luca M., Dempsey M. E., Biochem. Biophys. Res. Comm. 40: 117, 1970. Firefly bioluminescent reaction.
30. DeSa R., Hastings J. W., J. Gen. Physiol. 51: 105—122, 1968. Luminescent system in Gonyaulax.
31. Dubois R., C. R. Soc. Biol. 39: 564—566, 1887. Luciferin and luciferase in Py-phorus and Pholas.
32. Eckert R., pp. 269—300 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Electrophysiology of lumi-прясрпср in N ortilnca
33. Fogel M„ Hastings J. W., Arch. Biochem. Biophys. 142: 310—321, 1971. Gonyaulax luminescent system.
34. Friedland J. M., Hastings J. W., Proc. Nat. Acad. Sci. 58: 2336, 1967. Bacterial 1 ucif er ase
35. Galloway T. W., Welch P. S., Trans. Amer. Mier. Soc. 30: 13—39, 1911. Luminescence of Odontosyllis during mating.
36. Green A. A., McElroy W. D., Biochem. Biophys. Acta 20: 170—176, 1956. Crystal luciferase.
37. Greene C. W., Greene H. H., Amer. J. Physiol. 70: 500—507, 1924. Luminescence in Porichthys; action of adrenaline.
38. Haneda Y., pp. 335—385 in: The Luminescence of Biological Systems, edited by F. H. Johnson. Amer. Assoc. Adv. Sci., Washington D. C., 1955. Luminous organisms of Far East.
39. Haneda Y., Johnson F. H., Shimomura O., pp. 533—545 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Origin of luciferin in fishes.
40. Haneda Y., Tsugi F. I., Science 173: 143—145, 1971. Light Production in the luminous fishes Photoblepharon and Anomalops.
41. Hanson F. E., J. Insect Physiol. 8: 105—111, 1962. Control of firefly flashing.
42. Hanson F. E., Miller J., Reynolds G. T., Biol. Bull. 137: 447—464, 1969. Subunit coordination in fireflies.
43. Harvey E. N„ The Nature of Animal Light. J. B. Lippincott, Philadelphia, 1920, 182 pp.
44. Harvey E. N., Living Light. Princeton University Press, 1940, 328 pp.
45. Harvey E. N., Bioluminescence. Academic Press, New York, 1952, 649 pp.
46. Harvey E. N., Fed. Proc. 12: 597—606, 1953. Evolution of bioluminescence.
47. Harvey E. N., Chang J. J., Science 119: 581, 1954. Luminous response of Mnemiopsis.
48. Hastings J. W., Science 173: 1016—1017, 1971. Bioluminescence as camouflage in pony fish.
49. Hastings J. W„ Mitchell G., Biol. Bull. 141: 261—268, 1971. Photogenic organ for symbiotic bacteria.
50. Hastings J. W., Morin J. G„ Biol. Bull. 135: 422, 1968. Calcium activated bioluminescent proteins of Mnemiopsis and Obelia.
51. Hastings J. W., Morin J. G., Biochem. Biophys. Res. Comm. 37: 493—498, 1969. Renilla bioluminescence.
52. Hastings J. W„ Mitchell G„ Mattingly P. H„ Blinks J. R., Van Leeuwen M„ Nature 222: 1047—1050, 1969. Aequorin and calcium response.
53. Hastings J. W., Sweeney В. M., J. Cell. Comp. Physiol. 49: 209—226, 1957. Diurnal rhythms in light reactants.
54. Hastings J. W„ Sweeney В. M„ Biol. Bull. 115: 440—458, 1958. Diurnal rhythm of Gonyaulax light.
55. Hastings I. W., Vergin M., DeSa R„ pp. 301—329 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Particulate bioluminescent system in Gonyaulax'. «scintillon».
56. Hastings J. W., Weber K., Friedland J., Eberhard A., Mitchel G. W., Gunsalus A., Biochemistry 8: 4681, 1969.
^7. Heymans C., Moore A. R., J. Gen. Physiol. 6: 273—280, 1923. Diurnal rhythm in Pelagia luminescene.
39—2067
610
Глава 24
58. Heymans С., Moore А. R., J. Gen. Physiol. 7: 345—348, 1925. Inhibition of cte-nophore luminescence by light.
59. Johnson F. H., Shimomura O., Saiga Y., Gershman L., Reynolds G. T., Waters J. R., J. Cell. Comp. Physiol. 60: 85—104, 1962. Quantum efficiency in Cypridina.
60. Reynan A., Veeder C„ Hastings J. W„ Biol. Bull. 125: 382, 1963. Studies on the survival of dark and bright mutants of luminescent bacteria in seawater.
61. Rishi Y., Goto T., Hirata Y., Shimomura O., Johnson F. H., pp. 83—113 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Cypridina luciferin.
62. Rishi Y., Goto T., Hirata Y., Shimomura 0., Johnson F. H., Tetrahedron Letters 29: 3427, 1966. Cypridina luciferin.
63. Rishi Y., Goto T., Inoue S., Sugiura S., Rishimoto H., Tetrahedron Letters 29: 3445, 1966. Cypridina luciferin.
64. Rishi Y., Goto T„ Eguchi S., Hirata Y., Watanabe E., Aoyarna T., Tetrahedron Letters 29: 3434, 1966. Cypridina luciferin.
65. Lloyd J. E., Science 149: 653—654, 1965. Aggressive mimicry in Photuris.
66. Lloyd J. E., Misc. Publ. University of Michigan, No. 1930, 1966, 93 p. Studies on the flash communication system in Photinus fireflies.
67. Lloyd J. E., Ann. Rev. Entomol. 16: 97—122, 1971. Bioluminescent communication in insects.
68. McElroy W. D., pp. 463—505 in: The Physiology of Insecta, Vol. 1, edited by M. Rockstein. Academic Press, New York, 1964. General review.
69. McElroy W. D., De Luca M., Travis J., Science 157: 150, 1967. Conformational changes in bioluminescent enzymes.
70. Meighen E. A., Nicoli M. Z., Hastings J. W., Biochemistry 10: 4062—4068, 1971. Bacterial luciferase.
71. Meighen E. A., Nicoli M. Z., Hastings J. W., Biochemistry 10: 4069—4073, 1971. Chemically modified bacterial luciferase.
72. Moore A. R., J. Gen. Physiol. 9: 375—381, 1926. Diurnal rhythms in Pelagia luminescence.
73. Morin J. G„ Cooke I. M., J. Exp. Biol. 54: 707—721, 1971. Electrical activity in Obelia bioluminescence.
74. Morin J. G., Cooke I. M., J. Exp. Biol. 54: 723—735, 1971. Bioluminescent system of Obelia.
75. Morin J. G., Hastings J. W., J. Cell. Physiol. 77: 305—311, 1971. Coelenterate and ctenophore photoprotein and Ca++.
76. Morin J. G., Hastings J. W., J. Cell. Physiol. 77: 313—338, 1971. Coelenterate and ctenophore photoprotein and calcium.
77. Nealson R„ Markowitz A., J. Bacteriol. 104: 300, 1970. Bacterial luciferase synthesis.
78. Nicol J. A. C., J. Marine Biol. Assoc. U. K. 30: 417—431, 1952. Chaetopterus light glands.
79. Nicol J. A. C„ J. Marine Biol. Assoc. U. K. 30: 433—452, 1952. Nerve control of light production in Chaetopterus.
80. Nicol J. A. C., J. Marine Biol. Assoc. U. K. 31: 113—144, 1952. Nerve control of light production in Chaetopterus.
81. Nicol J. A. C., J. Marine Biol. Assoc. U. K- 32: 65—84, 1953. Luminescence in polynoid worms.
82. Nicol J. A. C., J. Abarine Biol. Assoc. U. K. 33: 173—175, 1954. Chaetopterus luminescence.
83. Nicol J. A. C., J. Marine Biol. Assoc. U. K. 33: 225—255, 1954. Light control in polynoid worms.
84. Nicol J. A. C„ J. Exp. Biol. 32: 299—320, 1955. Luninescence in Renilla.
85. Nicol J. A. C., J. Exp. Biol. 32: 619—635, 1955. Nerve control of luminescence in Renilla.
86. Nicol J. A. C„ pp. 299—319 in: The Luminescence of Biological Systems, edited by F. H. Johnson. Amer. Assoc. Adv. Sci., Washington, D. C., 1955. Control of luminescence.
Биолюминесценция 6П
87. Nicol I. А. С., J. Marine Biol. Assoc. U. К. 37: 43—47, 1958. Character of Pho-las light.
88. Okada Y. K., pp. 611—625 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Fine structure of Wat-senia luminous systems.
89. Parker G. H„ J. Exp. Zool. 31: 475—513, 1920. Spread of luminescence in Renilla.
90. Seliger H. H., McElroy W. D., Proc. Nat. Acad. Sci. 52: 75—81, 1964. Enzyme structure and conformation in bioluminescence.
91. Seliger H. H., McElroy W. D., J. Marine Res. 26: 244—255, 1968. Diurnal rhythms in bioluminescence.
92. Seliger H. H., Morton R. A., pp. 253—314 in: Photophysiology, Vol. 4, edited by A. C. Giese. Academic Press, New York, 1968. Specificity of luciferase; efficiency of photosynthesis.
93. Shitnotnura 0., Johnson F. FI., Saiga Y., Science 140: 1339—1340, 1963. Luminescence in Aequorea.
94. Shimomura 0., Johnson F. EE, pp. 495—522 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Luminous system of Chaetopterus.
95. Shimomura 0., Johnson F. H., Haneda Y., pp. 391—404 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Isolation of Latia luciferin.
96. Shimomura O., Hohnson F. H., Piochemistry 6: 2293—2306, 1967. Bioluminescent system of euphausid.
97. Shimomura O., Johnson F. FI., Science 159: 1239—1240, 1968. Isolation of Chaetopterus photoprotein.
98. Shimomura O., Johnson F. H., Biochemistry 7: 2574—2580, 1968. Luciferase and protein cofactor in Latia.
99. Shimomura O., Johnson F. H., Biochemistry 7: 1734—1738, 1968. Luciferin of Latia.
100. Shimomura O., Johnson F. H., Proc. Nat. Acad. Sci. 59: 475—477, 1968. Photoprotein system in euphausid.
101. Smalley K. N., Comp. Biochem. Physiol. 16: 467—477, 1965. Adrenergic transmission in Photinus light organ.
102. Smith D. S., J. Cell Biol. 16: 323—359, 1963. Fine structure of firefly luminescent organ.
103. Stone H., Biochem. Biophys. Res. Comm. 31: 386, 1968. Cypridina luminescent reaction.
104. Sweeney В. M., Hastings J. W., J. Cell. Comp. Physiol. 49: 115—128, 1957. Diurnal rhythm in Gonyaulax light.
105. Swift E., Taylor W. R., J. Physiol. 3: 77—81, 1967. Luminescence in Pyro-cystis.
106. Tsuji F. L, Haneda Y., pp. 137—149 in: Bioluminescence in Progress, edited by F. H. Johnson and Y. Haneda. Princeton University Press, 1966. Luciferases of Cypridina and Apogon.
107. Tyler J. E., Bull. Scripps Inst. Oceanogr. 7: 363—411, 1960. Color in bioluminescence and depth in ocean.
39*
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
, I*' •
Абдоминальный рецептор II 357 Автотрофные организмы I 242 Агломерулярная почка I 117—118 Адаптация, аллометрическая I 398 — к высоте I 412—413 — к зимней спячке I 453—454 — к механическому раздражению
II 354—355
— к нырянию I 408—412
— к пищевому рациону I 310—319 — к солености I 52—56, 126, 580 — к температуре см. Температурная адаптация
— темновая II 543—546
Аденилатциклаза I 472, 473, 514, 516;
II 330
— в сетчатке II 517
— и действие гормонов III 412, 418, 421, 423, 440
Аденогипофиз III 431, 433—438 Аденозинмонофосфат см. АМФ Аденозинтрифосфат см. АТФ Адреналин II 327, 424, 425; III 357, 361, 381
— влияние на фосфорилирование I 513
-----на жабры I 194
— и мышечная дрожь II 177—178 — секреция III 424—426
Адренокортикотропный гормон (АКТГ) I 155; II 143; III429, 430, 435—437
Адренэргические нервы III 263, 264, 357, 359, 361
— рецепторы II 328—329; III 259, 261, 357, 424
-----в сердце III 383
Азот, выделение I 536—591
— круговорот I 311—312
Акклиматизация температурная II 154—159
Акклимация II 107—117
— к гипоксии I 403
— к температуре I 451, 470; II 151—154
— осмотическая II 54
— ферментов II 117—125
Аккомодация возбуждения II 298
Аксолотль, объем яиц I 41
Аксонема III 327, 328
Аксоны, гигантские II 273, 299—300
— двойное лучепреломление II 298
— синапсы на них II 310
Аксоподии III 306, 314
АКТГ см. Адренокортикотропный гормон
Активный транспорт I 32—34, 63—
64, 131, 177—199, 202—217, 229—
231, 547; II 274, 388
Актин III 165, 180—182, 244, 245
— в ресничках III 328
се-Актин III 184
Актинии III 118, 254, 332—333
Актиномицин D III 417
Актомиозин III 176, 180, 182, 313
Акулы I 112
— коллаген I 222
— кровообращение III 359, 360
— терморецепторы II 133
— эластин III 360
Аланин I 66, 111, 184, 488, 538
Аллигатор, аммиак I 558
— кровообращение III 371
— мочеотделение I 132
— нервная система II 355; III 87,91
— обмен I 577
Аллостерические ферменты I 509, 589—591
Альгиназа I 315
Альдолаза I 501—503, 521
Альдостерон I 150, 155—156; III429
— структура III 428
Альфа-волны III 68, 69
— десинхронизация III 131
Алюминий I 177, 221
Амакриновые клетки II 547, 554— 555
Амбулакральные ножки III 366, 535, 536
Амеба I 180
— рецепция II 352, 433
— фототаксис II 478, 480
Амебоидное движение III 303—317
-----теории III 310—313
Амилаза в слюне I 287, 300
— в поджелудочной железе I 297
Амилоза I 297
Амилорид I 206
п-Аминогиппуровая кислота I 129, 145, 200
Предметный указатель
613
Аминокислоты, абиотический синтез I 243
в аксонах II 273
— в гемоглобине II 19—21
— в культуральных средах I 250— 251
— вкус II 443
— всасывание I 320, 330—332
— гомеостаз I 317
— и синтез белков I 484
— как нервные медиаторы II 315— 336
— катаболизм I 431, 477, 536—544, 560
— осмотическая концентрация I 66, 183—185
— перенос I 331—332
— потребность в них I 248, 253— 260, 571
— реабсорбция I 109
— токсичность I 254
— у насекомых I 95
— фильтрация в почках I 145
Аминопептидазы I 306
Аминоптерин I 272
Аммиак, выделение и детоксикация I 570—575
Аммониотелия I 536, 537, 558
Аммония ионы, выделение I 536, 545—549, 553, 555, 557—561, 570— 571
— обмен на натрий I 205 -----при пищеварении 1311
Ампулы Лоренцини II 133; III 292, 293, 295, 296
Ампулярные сердца III 372—378 АМФ (см. также Циклический АМФ) — влияние на ферменты I 507, 520, 521
Амфибии, зрение III 89—95, 255
— кровообращение I 588; III 359, 370, 383, 418, 419
— легкие I 368—369
— лимфатические сердца III 372, 385
— метаболизм I 126, 454—455, 577, 580, 581; II 45; III 419
— метаморфоз III 418, 419
— нервная система III 62, 74—75, 89—96, 107—112, 262, 361
— осморегуляция I 125—131
— рецепторы II 391, 448—451, 454— 456, 485, 491
— условные реакции III 119
— хроматофоры III 553—556
— эндокринная система I 153—154, 388, 491; III 418, 419, 423, 427,440 Анаболизм I 430—432
— и цикл Кребса I 483—484
Анадромные рыбы I 111
Анальные папиллы I 96
Анаэробиоз у рыб I 451—452
— углеводный обмен I 432—443
Анаэробный катаболизм I 432
Ангиотензин I 155; III 358, 429
Андрогенная железа III 472, 480
Андрогены III 426, 432—433, 443—
444
Андростендион III 432
Анемотакгис II 367
Аннелиды (кольчатые черви), гемоглобин II 6, 52—53
— дыхание I 354
— жгутики III 324
— жидкости тела III 346, 364
— люминесценция III 586
— мышцы III 165
— нервная система II 300, 333; III
12, 13, 40—43, 118
— обмен веществ I 200—201, 551;
II 572; III 201
— сердца III 400
— условные реакции III 118
— хитин I 316
— хроматофоры III 534—535
Аноксия, толерантность у черепах I 454—457
Ансерин I 262
Антаугофоры III 523
Антеннальная железа I 81, 197
Антенны, механорецепторы II 365
Антидиуретический гормон I 130, 149, 151, 153; III 438—440
-----и тироксин III 417
-----у ракообразных III 485
Антидромное проведение II 309
Антиплектическое движение ресничек III 335
Антифризы I 450; II 88, 93
Анэкдизис III 472
Апатит I 222
Апокринные железы II 160
Аппозиционный глаз II 519
Арагонит I 222
Арахидоновая кислота I 263, 474
Аргиназа I 219, 221, 567, 572, 591
Аргинин I 66, 184
— биосинтез I 554, 562, 563, 566, 567, 590
— метаболизм I 571—572, 577, 580, 591
Аргинин-вазопрессин I 131, 138, 153—
154; III 439, 440
Аргининкиназа III 200
Аргининфосфат III 200, 201
Аррениуса кривые I 471; II 111
Артемия (Artemia) I 57
— осморегуляция I 73—75, 215
614
Предметный указатель
Артерио-венозные анастомозы (шунты) III 358
Аскорбиновая кислота I 273—274
Аспарагиновая кислота III 508
Аспартат в цикле мочевины I 484, 553
Асфиксия III 363
Асцидии I 221—222
— как осмоконформеры I 44
— мышцы III 173
Атропин I 325
Аттрактанты II 435—438
АТФ I 349
— в ресничках III 328, 342
— в синапсах II 315, 318, 336; III 261
— в электрическом органе III 290
— взаимодействие с АМФ I 511,519 — и гликолиз I 456—457, 519—520 — и мышечное сокращение III 176, 179, 183, 197, 199
— и транспорт натрия I 203
— при реакции с люциферином III 594, 596, 597
АТФ/АДФ, отношение I 481
АТФаза (см. также Магний-АТФаза; Натриево-калиевая АТФаза) I 208, 213—216, 229, 290
— влияние свинца I 222
-----температуры II 99
— в мышцах III 176, 180, 185, 217— 222
Ацетат и липогенез I 492—494
Ацетаттиокиназа I 493
N-ацетиласпартат (в нейронах) II 273
N-ацетилглутаминовая кислота I 590
N-ацетилглюкозамин I 448, 450
6-Ацетилглюкозаминидаза I 317
Ацетил-КоА I 430, 431, 452, 461, 466—469; II 319, 320
Ацетилхолин I 371; II 312, 318—325, 327
— в электрическом органе III 290
— влияние на выработку условных рефлексов III 133
-----на мембраны II 291
-----на нервно-мышечные соединения III 34, 35, 203, 241, 259
-----на носовые железы I 139
-----на реснички III 338
----- на рецепторы I 371
-----на сердце III 382, 383, 389
-----на сосуды I 194; III 357, 361
Ацетилхолинэстераза II 315, 319—325
Ацетоксициклогексимид III 132
Ацидоз I 561—562
Ацилкарнитины I 461, 462
Ацил-КоА I 237, 430, 431, 461, 467, 492
Ацинарные клетки I 289
Ашоффа правило II 214
Бабака закон III 526
Бабочки, летательные мышцы I 470-
III 237
— метаболизм, биохимия I 263, 300
— метаморфоз III 457
— рецепторы II 434
Базальная мембрана I 145; II 518
Базальное тельце III 327, 328
Базальные ядра III 62
Баклан I 138
Бактерии II 98—99
— барофильные II 368
— изоферменты I 504—505
— кишечные I 313, 314
— люминесценция III 584, 585, 597, 598
— метаболизм I 394
— симбионты I 310—311, 314; III 585
— фототаксис II 477—478
— хеморецепция II 432
Барабанная лестница II 386
— перепонка II 385, 391
— струна II 458
Барометрическое давление I 351
Барорецепторы II 364; III 356, 357, 361
Барракуда I 416
Белки, абиотический синтез I 243— 244
— биосинтез I 484; II 118, 216; III 132, 412, 417
— в ресничках III 327, 328, 341
— переваривание I 301—306
— сократительные III 176—184, 313
Беллончи орган III 552
М-белок III 184
Беспозвоночные, азотистый обмен I 542—544, 551—552, 570—571
— гемоглобин II 6—7, 21, 52—57
— гормоны и метаболизм I 491—492
— ионная регуляция I 186—192
— карбогидразы I 299—301
— кровообращение III 364—369
— мышцы III 170—176, 200, 201, 250—251, 264—266
— осморегуляция I 88—111
— привыкание III 128----130
— рецепторы равновесия II 408— 411
— сложные глаза II 518—537
— фоторецепторы II 481, 508, 513
— хеморецепция II 432—447
— эстеразы I 308
[Iредметный указатель
615
Бесхвостые амфибии, глаза II 544
маутнеровские клетки II 300
— метаболизм I 125—131, 408, 581
— нервная система III 262
условные реакции III 119
Бесчелюстные, осморегуляция I 111—
112
Бетаин, влияние на хроматофоры
III 537
— как заместитель холина I 183,184
Биологические ритмы II 210—254
(см. также Суточные ритмы) Биолюминесценция III 584—606 — амеб III 312
Биотин I 266, 268—269
Биполярные клетки II 547, 553
Блуждающий нерв I 325; III 383, 384
Бобр, потеря воды I 140, 149
Богомолы, ходьба III 51
Бокаловидные клетки I 290
Боковая линия II 373—376, 392, 407;
III 293
Бокоплавы I 66—70, 81—82, 385; III
479, 480
Бомбикол II 435, 463
Бонелли дин III 201
Бор I 221
Бора эффект I 415; II 11, 31—32, 36, 42—46
Брадикардия при гипоксии I 359
— при погружении в воду I 408—
412, III 362
— у рыб и черепах III 363
Бром I 218
Брюхоногие моллюски, дыхание I
367, 408
—• — метаболизм I 551, 570
-----нервная система III 29—31, 129
-----объем гемоцеля III 352
— — осморегуляция I 47
-----сердце III 385
— — тепловая кома II 102—103
-----ферменты I 315, 317, 571
-----фильтры ресничные III 326
Брюшные ганглии I 372, 373
Бунгаротоксин III 205
Бурая жировая ткань I 472—474;
II 173—174
Бурундук, зрительная система III 99
Буферная емкость крови II 68—69
Вазомоторная регуляция III 357
Вазомоторные адаптации II 158—159
Вазопрессин I 146—151, 153—155;
III 439—440
— влияние на сосуды III 357
Вазотоцин I 127; III 439
Вакуолярное пищеварение I 285—286
Валиномицин I 547
Ванадий I 221
Ванадоциты I 221
Веберовы косточки (веберов аппарат) II 385, 395
Верблюд, гемоглобин II 49
— испарение воды I 140—143
Веретена мышечные II 360—363
Вестибулярный лабиринт II 411—417
— нерв II 412, 415
Вибрационная чувствительность II 3gg____зуд
Витамин А I 274—275; II 488, 489, 493
— В6 I 539
— В12 I 267—268
— С I 273—274
— D I 224, 275—276; III 411
— Е I 276—277, 414
— К I 276
Витамины I 219, 222, 249
— водорастворимые I 265—274
— жирорастворимые I 274—277
— фильтрация в почке I 145
Вкусовые нервы II 457; III 103
— рецепторы (см. также Сенсиллы;
Хеморецепторы)
-----для воды I 150; II 456—457, 462
Влажность I 101
— влияние на окраску III 527
— гигрорецепция I 102
Внеклеточное пищеварение I 286— 292
— пространство I 181, 349
Внутриклеточное пищеварение I 285—286
Вовлечения реакция III 69
Вода, вкусовые рецепторы для нее
I 150; II 456—457, 462
— и гормоны III 428, 438
— обмен в организме I 140—144;
III 349
— растворимость газов в ней I 350
— связанная I 28, 41
— термические свойства II 85—86
— течение в жабрах I 362, 365
— тяжелая II 216
Водородные ионы (pH), влияние аммиака I 545—546
-------на дыхательную функцию крови II 62
------- на реснички III 342
-----и гликолиз I 442
----- и секреция газа I 453
----- и температура II 125
616
Предметный указатель
— связи I 28—29; II 461
-----при замерзании воды II 88
Водоросли, симбиоз I 318
— фототаксис II 478
Водянистая влага II 537
Возбудительный постсинаптический потенциал (ВПСП) II 285, 287— 290; III 202, 204, 217, 228— 222, 264
--------в электрическом органе III 289
Воздух, содержание кислорода I 349—350
Воздушные мешки I 369
Волокна гигантские см. Гигантские аксоны
— миелиновые II 300
— моховидные III 82
— параллельные III 82
— сократительные III 164
— фузимоторные III 360, 362
Ворвань I 471
Ворсинки I 289—290
ВПСП см. Возбудительный пост-синаптический потенциал
Временная константа клетки II 269—270; III 742
Всасывание (продуктов переваривания) I 328—334
Вставочные нейроны II 526; III 31—36, 73
Выделение см. Экскреция
Выделительные органы беспозвоночных I 191
Вызванные потенциалы III 66—71, 128, 131
Газель I 141; II 160
Газовая железа I 415, 417
Газовый пузырь I 368
Газы, растворимость I 350
— секреция I 414—417
Галактоза, перенос I 328—329
Галогены I 218
Гамма-аминомасляная кислота (ГАМК) II 333—335, 359; III 20, 87, 232—233, 236, 247, 397
Гаммариды I 67—69, 81
Ганглиозные клетки II 547, 548, 555—557; III 89
Гармин II 326
Гастриксин I 302
Гастрин I 325; III 442, 453
Гастролиты I 224; III 472, 473 S
Гебста тельца II 355
Геккон, почки I 133
Гексаметоний II 321
Гексокиназа I 498—501
Гельминты I 88—89, 437—439
Гем II 5, 11, 30
— структура II 8—9
Гематин I 261—262
Геминовые пигменты I 219
Гемоглобины I 219, 253, 261, 408
— влияние на биение ресничек III 341
— насыщение I 415—417, 485; II 5—21, 27—35, 42—59, 72—75
— структура II 8—10
— у бобовых растений II 7
Гемолимфа II 302
— и осмотическая регуляция I 95, 96, 98, 99, 101, 103, 105, 108, 444
— циркуляция III 347
Гемоцель III 347
Гемоцианин I 219; II 7—8, 16, 22—
26, 59—64
Гемоэритрин II 6, 7, 16, 26—27, 64— 65
Гендерсона — Г ассельбаха уравнение II 65
Генераторные потенциалы II 282— 284
Ген-продюсер I 587
Гены II 494
— гемоглобина II 18, 19, 49
— регуляция активности I 585, 587
Геотаксис II 408—409
Геофизические факторы II 223—227, 238, 243—245
Гепатопанкреас I 192, 218, 219, 288, 300—301
— и линька III 472, 473
Гетеротермные организмы II 84—85, 141
Гетеротрофные организмы I 242
Гиалиновый слой III 308
Гиалуроновая кислота в кутикуле I 449
Гибернационная железа III 444
Гигантские аксоны II 299—300; III
12—29, 57
Гигрорецепторы I 102; II 439
Гидра I 89—90, 323; II 268, 433; III
25, 118
Гидравлические механизмы III 366
Гидроидные полипы III 23
Гидроксоний (ион) I 28
Гидролазы I 317
Гидростатический скелет III 366—
369
Гидростатическое давление I 35,415, 418; III 366
Гиперкальцемия III 421
Гиперосмотическая регуляция I 43,
52—56, 78—83, 90, 112, 125, 151
Гиперосмотический раствор I 36
Гиперпирексия II 162
Предметный указатель
617
Гиперполяризация II 262, 352, 503, 512
Гипоксия I 359, 365, 403
— влияние на гемоглобин II 57—59
— у рыб III 362, 363
Гипоосмотическая регуляция I 70— 78, 116, 125
Гипоосмотический раствор I 36
Гипоталамус, гормоны I 153
— и кровяное давление III 362
— и терморегуляция II 142—143,
177—178
— и эмоции III 104
Гипофиз III 441, 449—452, 553
— гормоны I 151—155, 157
Гиппокамп III 70, 71, 104—105
Гирудин III 201
Гистамин III 353, 356, 357
Глаза позвоночных II 537—541
— сложные II 518—524
Глазные пятна II 480
— стебельки III 49, 473—475, 482—
484, 544—546
Глиальная оболочка П 969
Гликоген I 287, 296, 444—450,
обо—507, 512, 013
— в мышцах III 199
— в фоторецепторах II 484
Гликогенолиз III 424
Гликогенсинтетаза и гликогенфосфо-рилаза I 436, 512—518
Гликокаликс I 290, 291
Гликолиз I 404, 432—457, 488, 520— 522
Гликопротеидные антифризы II 93— 94
Гликопротеиды I 449
Глиоксилатный цикл I 247, 483
Глицеральдегид-З-фосфат I 447
Глииеральдегид-З-фосфатдегидроге-
наза I 448, 502
Глицериды I 332—334
Глицерин I 184, 444, 450; II 90
а-Глицерофосфатиый цикл I 447— 448
Глицин I 66, 184; II 90, 334, 432
Глия II 303
Глобин II 5
— структура II 8—9
Гломерулярная фильтрация I 144—
146
Глотание III 74
Глумитоцин I 153; III 440
Глутамат I 184, 477; II 432
— влияние на нейроны и мышцы II
335; III 232, 236, 397
Глутамат дегидрогеназа I 477, 539—
540, 542—543
Глутамин I 66
Глутаминовая кислота I 66
-----в нематоцистах III 508
----- как синаптический передатчик
II 335
Глутаминсинтетаза I 546, 556—558, 568, 574
Глутатион, у гидры I 323; II 433
Глюкагон I 326, 490, 516, 517; III 423 424
Глюкоза I 117, 287, 490; II 90
— реабсорбция I 63, 82, 92, 130, 145
а-Глюкозидаза I 300
Глюкозо-6-фосфат I 445, 518; III423
Глюкокортикоиды I 487—490; III 426—428
Глюконеогенез I 432—457, 488, 522, 539, 560—561
6-Глюкуронидаза I 315
Годичные ритмы II 235
Головной мозг (см. также Центральная нервная система)
-----позвоночных I 456, 557, 563;
II 328; III 61—71
-----синтез аргинина I 405, 563
-----функции при охлаждении II 146
Головоногие моллюски I 44
-----жаберные сердца III 372
-----зрение II 528, 532—534
-----кровообращение III 348, 365
-----люминесценция III 589
-----нервная система II 300, 323;
III 12, 36—40
-----осморегуляция I 44, 183, 191
-----хроматофоры III 250, 519, 520, 536—538
-----эндокринные железы III 487
Голотурии I 44, 356; III 251, 348
— гидростатический скелет III 366
— сердце III 372
Голубь, метаболизм в полете I 407
Гольджи клетки III 85—86
Гольдмана уравнение I 32; II 272
Гольян III 76
Гомарин I 51
Гомойотермия, развитие II 165
Гомойотермные животные II 84, 85, 141—174
Гонадотропины III 441—442
Горизонтальные клетки II 547, 554
Гормон роста III 434—435
Гормоны III 411—415
— и лактация I 494; III 436—437, 449
— и метаболизм I 155, 388, 487— 494; III 484—485
— и окраска III 541—568
— и осморегуляция I 82, 97, 124, 151—157; III 485
618
Предметный указатель
— и размножение III 441—453, 466— 467, 476—480
— и термогенез II 143
Гребешок (моллюск) III 251
Гребневики I 44; II 409; III 325, 586
Грудной ганглий III 237
Гуанидиновые основания III 201
Гуанин, выделение (гуанотелия) I 537, 570, 575
Гуаниннуклеотиды I 539
Губки I 89—90
— миоциты III 164, 252—254, 324
Губоногие, трахеи I 372
Давление, в плавательном пузыре I 4(5; II 369
— в системе кровообращения I 144; III 353—363
— влияние на поведение II 367—369
— гидростатическое I 415, 418; II 368
— интерстициальное III 358
— критическое I 401
— осмотическое I 29—30, 33
— рецепторы II 364; III 356—357
Дальгрена клетки III 440
Двойственности зрения теория II 541—544
Двоякодышащие рыбы I 368, 375, 580; II 70
----- гемоглобин II 45
-----летняя спячка I 453—454, 581
Двукрылые, зрение II, 524
— метаморфоз III 457
— полет I 522; III 236, 237
— синаптические связи II 526—527
Двупарноногие I 99, 372
Двустворчатые моллюски I 191; III 29—31, 129, 352
Дегидратазы I 541
Дегидратация I 99, 140—141; 1189; III 357
Дезоксикортикостерон I 155; III 428, 447
Дезоксирибонуклеаза I 316
Дезоксирибонуклеиновая кислота см.
ДНК
Дезоксихолевая кислота I 307
Дейтерированная вода I 41; II 216
Декаметоний III 205
Декарбоксилирование I 452
Декси оплектическое движение III
335
Дельфины I 141, 470
— ныряние III 362
Денервационная гиперсенситивностъ III 114
Деполяризация II 262, 282—284, 352, 510, 512
Десмонемы II 263
Детоксикация I 562—584
Децибел, определение II 384
Джонстоновы органы II 370, 378
Диамокс I 416
Диапауза I 551; III 457—458
Диапаузы гормон III 461
Дибенамин II 289
Дигиталис I 203
Диизопропилфторфосфат (ДФФ) Н 319
Дииодтирозин I 218
Динеин III 328, 341
Динофлагелляты III 508, 585, 600
Диоксиацетонфосфат (ДОАФ) I 447—448
Дисфасилитация III 10
Диурез I 146—148, 154, 222
Дифосфоглицерат II 31
Дифосфопиридиннуклеотид (ДПН)
I 270
Диффузионные легкие I 367
Диффузия I 31, 34, 145—146, 153— 154, 559
— коэффициент I 31, 352
— облегченная II 42
— обменная I 213
ДНК I 587; II 216, 368
— и биологические ритмы II 247, 248
Дождевой червь I 327, 404
----гемоглобин II 53
----детоксикация азота I 570
— — кровообращение III 368
----мышцы III 173, 248—250
----нервная система II 300, 305,
330, 333; III 13, 40, 118
----осморегуляция I 91—94
----рецепторы II 351, 433—434, 480
------ тигмотаксис II 367
----ферменты I 301, 316, 543
----фототаксис III 118
Доннана равновесие I 33
Дофамин II 325—328
Дрожь II 177, 178
Дрозофила I 99, 101, 102; II 437;
III 458
— действие Ог I 413
— и геомагнитное поле III 297
— пищевые потребности I 259—263, 269, 272
— потребление Ог I 382, 389
— циркадный ритм вылупления II 480
Дыхальца I 99, 101, 372—374; II 138, 139
— иннервация III 235
Дыхание I 349—421, 478
— и терморегуляция II 163—164
Предметный указатель
619
— роль крови II 5—75
Дыхательный центр III 78
Дэйла правило II 322
Евстахиевы трубы II 385
Ехидна III 101
Жаба, гипофиз III 442
— легкие I 388 !
— мочевой пузырь I 208, 457 '' 1
— тектум III 81
Жаберные сердца I 200
Жаберный «насос» I 360
Жабры I 62, 64, 65, 354—367, 486, 559
— функция АТФазы I 456
Жаворонок I 137
Жвачные, метаболизм I 270, 492— 494
— пищеварение I 247, 310—315
Жгутики III 324—329
— хеморецепторов II 433
Жгутиковые I 242; III 324, 327, 508
Железо I 177, 219, 261; II 8
Железопорфирины II 5, 7, 9 (см.
также Гемоглобины)
Желтое тело III 447, 448
Желудок, мускулатура III 258
Желчные соли I 307—308
Жидкости тела, обмен I 31
----объем III 349—351
----отсеки III 348, 349, 352
---- состав I 185—192
Жираф, артериальное давление III 356
Жирные кислоты, всасывание I 158
----и обмен глюкозы I 452, 489—
490
----незаменимые I 262—263
----образование у жвачных I 311
---- окисление I 460—462, 477
----роль в терморегуляции II
121—122, 143, 153
----синтез I 462—468, 492—494
Жировой обмен I 457—474
Жиры см. Липиды
Жужжальца II 418
Задняя кишка у насекомых I 101, 102, 105—110
Закрытопузырные рыбы I 368, 414;
II 369
Замораживание, устойчивость II 88—95
Заяц, гормоны и окраска меха III 455
— миоглобин II 44
Звездчатые клетки III 86
Звуковая рецепция II 376—418
— сигнализация II 396—398
Зеин I 253, 271
Зимнеспящие животные II 153; III 381
Зимняя спячка I 195; II 85, 134, 171, 180—187
Зимогеновые гранулы I 289, 294
Златоглазка, тимпанальный орган II 378
Змеи, мышцы III 168
— органы чувств II 372, 397, 546
— осморегуляция I 131, 133—135
— сердце III 371
— терморегуляция II 133, 139
Зоб I 287—288, 300
Зрачок II 537; III 544, 546
Зрение, молекулярная основа II 516—518
— цветное II 497, 531
— центральные процессы III 89—99 — электрофизиология II 503—518 Зрительные пигменты II 487—503 -----перемещение II 520, 544
Иглокожие I 44, 45, 265; III 251, 265
— амбулакральные ножки III 366, 535—536
— кровообращение II 6; III 346
— нервная система II 323; III 28— 29, 118
— ферменты I 304, 308
Избегания реакции III 18, 22, 48,57
Изоосмотический раствор I 36
Изотонический раствор I 36
Изотоническое сокращение III 192— 194
Изотопы, кинетика I 34—35
Изотоцин I 153; III 440
Изоферменты I 496—508, 539
— и акклимация II 119—120
Иммунофлуоресцентный анализ II
21
Импульсная реакция нейрона II 376
Ингибитор Казаля I 303
— Куница I 303
Ингибиторы активного транспорта I 34
— потенциалов II 279—282
Индолилалкиламины II 325
Индукция ферментов I 318
Инозинмонофосфат I 556, 563, 564
Инозитгексафосфат II 35
Инокостерон III 475
Инсулин I 487—490, 499; III 422,423
Интерренальная ткань III 426
Интрафузальные волокна II 362 Инулин, выделение I 62, 81—82, 91— 93, 117, 133, 197, 199
620
П редметный указатель
— использование в физиологических исследованиях I 181; III 349
— переваривание насекомыми I 300
— роль в осморегуляции I 130, 131, 133, 300
Инфузории (см. также Парамеция)
I 247, 248
— замерзание II 89
— трихоцисты III 508
Иод I 218; III 416, 418
Иодацетат I 404
Иодид III 416, 418
Ионная регуляция I 178—185, 192— 196, 203—217
Ионные насосы I 203—204
Ионный состав жидкостей тела I-187—190
-----клеток I 179—182, 229
-----мочи I 198—199
Ионофоры I 547, 548
Ионы (см. также Активный транспорт)
— влияние на гемоглобин II 33—35
— и вкусовые рецепторы II 363, 460, 461
— концентрация в жидкостях тела I 187—190
Испарение воды I 125—126, 131— 132; II 160, 161
Калиевая проводимость II 296; III 204, 236, 381
Калиевый насос I 103; II 386
Калий II 262
— активный транспорт I 108—ПО
— выделение I 138—139, 198—200; III 429
— и перенос аммиака I 547
— реабсорбция I 201
— у членистоногих I 106, 197; II302
Каллионимус I 362, 364
Кальмар I 183; III 523, 527
— гемоцианин II 25, 60
— желудок III 264
— зрение I 275; II 487, 515; III 250
— мышцы III 173
— нервная система I 41; II 273,285—
287, 300; III 15—16, 129
Кальциевая проводимость II 290
Кальциевые скелеты I 223
Кальций, активный транспорт I 215
— в аксоне II 273, 279, 287
— в ресничках III 328
— в сердечной мышце III 381, 382
— в тканях и плазме I 179—180
— влияние на гормоны I 47, 55,193, 197, 216—218; III 421
-----на мышцы III 177, 182—187, 218
----- на постетаническую потенциа-цию III 127
-----на фосфорилазу I 518, 520
— роль в фоторецепции II 511
— у улитки I 91
— у Branchiostoma III 205
Кальцит I 223
Кальцитонин III 421
Кальциферол I 275
Кальция карбонат в мышцах I 222___224
Камбала I 116, 117
Каракатица (Sepia) I 417; II 330;
III 37, 527
Карбамоилфосфатсинтетаза I 552— 556, 565—566, 577, 581
Карбоангидраза I 205; II 66, 67, 71, 72
Карбогидразы позвоночных I 297— 299
Карбоксипептидаза I 305—306; II 11
Карнитин I 262, 469
— и транспорт жирных кислот I 461
— у жуков I 469
Карнитинацетилтрансфераза I 461
Карнитинпальмитилтрансфераза I 461, 462
Каротидный синус III 356
6-Каротин I 274
Каротиноиды III 523
Карп, дыхание I 363, 365, 451, 452
Катаболизм I 430 (см. также Аминокислоты)
— гормонов III 416
— и цикл лимонной кислоты I 477
Катадромные рыбы I 111
Каталаза I 541
Катализ, регуляция I 509—525
Катепсины I 301
Катехоламины, в сердце III 385
— влияние на обмен глюкозы I 490
----------триглицеридов I 458,472
— как нервные медиаторы II 325— 331; III 357
— подавление нервного возбуждения III 261
Катехол-о-метилтрансфераза II 326
Кванты II 475; III 594
«Кванты» медиатора II 285, 319;
III 203
Кенгуровая крыса I 143, 144
Кератин, в скелетах I 222
— переваривание I 317
а-Кетоглутарат I 431, 452, 538—540
Киноцилин II 373, 409
Кислород I 349—421 (см. также
Гипоксия; Оксигенация)
— влияние на сердце III 363
редметный указатель
621
— и терморегуляция II 127—128, 144—145, 152—154
— потребление, влияние гормонов I 388; III 416, 419, 484
—-------температуры II 111
— секреция I 453
Кислородная емкость II 35—37
Кислотно-щелочной баланс I 42, 559;
II 68, 573
Кистеперые рыбы I 111, 192, 576, 577
Китообразные, водный обмен I 150, 153
— жировой обмен I 471
— ныряние I 408; III 362
— слуховой орган II 399
— терморегуляция II 168
— устойчивость к аноксии I 454
— эхолокация II 401, 407
Кишечник I 288
— мускулатура III 257—258
Кишечнодышащие, гигантские аксоны II 300
Кишечнополостные I 246—247; III 346
— жгутики III 324
— люминесценция III 585—586, 600
— нервные функции II 299; III 24— 28, 118
— осморегуляция I 44, 89—90
— рецепторы II 409, 433
— сократительные клетки III 252— 254
— химотрипсин I 304
Кишечные бактерии I 313
— паразиты I 317—318, 320
Клейдониевые яйца I 583
Клест I 135 . v • .
Клетки, контакты III 381 1
— форма I 43
Клетки-зерна III 82
Клиренс клубочковый I 129, 130
Клубочковая зона III 426
Книдобласт III 508—509
Книдоциль III 509
Кобаламин I 267—268
Кобальт I 220
Кожное дыхание I 354, 355
Кожные жабры I 356
Колбочки II 481—487
Колибри I 382, 406; II 171—173
Коллаген I 222; II 89, 99, 100
Коллагеназа I 304
Коллетериальная железа III 467
Коллигативные свойства I 29—30
Коловратки III 324, 346
— осморегуляция I 90
Колоколовидная сенсилла II 351
Колорадский жук II 437—438
Колхицин III 314, 315
Кольцевая железа III 465—466
Кольцевое проведение III 27, 243 > Кольчатые черви см. АннелиД' Кома температурная II 102-- 133 Комары I 95—98, 102 — люминесценция III 588 — рецепция II 370, 436, 439
Константа длины клетки II 269— 270; III 212
— седиментации II 14—17
Контраст (зрение) II 533
Конхиолин I 222
Концентрирующая способность почек I 147—151
Кооперативность I 519
Копрофагия I 310, 314
Копытные I 471
— потеря воды I 140—141
— реакция на температуру II 161— 163
Кора большого мозга III 65—67, 79, 97, 101—103
------- роль в обучении III 125 -------стимуляция и рост III 133—
134
— надпочечников II 143; III 417, 426—430, 444
Коралловые полипы I 223
-----нервная сеть III 23 Корзинчатые клетки III 86
Корненожки I 85
Кортиев орган II 386
Кортизол I 155; III 427
Кортизон, структура III 427 Кортикостероиды III 430
Кортикостерон I 155; III 427
Кортикотропин (АКТГ) I 154; II 143; III 429—430
Костистые рыбы, ацетилхолин III 361
-----гормоны III 419, 420, 426, 440
-----желчные соли I 309
-----магний I 192
-----обмен аргинина I 577
-----объем крови III 352
-----осморегуляция I 114—125
Костные ганоиды 1 111
Кофермент А I 270, 333, 462 Кохлеарное ядро III 100
Кошачья акула, мышцы III 217 -----нервная система III 76, 87 ----- ректальная железа I 114
Кошка, локомоция III 205
— мигательная перепонка III 355
— мышцы III 114
— нервная система III 70, 71, 73, 85, 97, 105, 112
— рефлексы II 135
— рецепторы II 134, 176, 356. 363, 364, 372, 456
622
Предметный указатель^
Крабы I 69—72, 197, 199; III 529, 531—533
— биологические ритмы II 229—230
— гемоцианин II 62, 64
— глазные стебельки I 156, 388;
I II 484
— глутаматдегидрогеназа I 543
— жабры I 357, 359
— линька III 475, 476
— миохордальные органы II 364
— рецепторы II 371, 410
— хроматофоры III 523, 542—543
Крахмал I 295—297
Креатинин I 117
Креатинкиназа III 199
Креатинфосфат I 520; III 199—201
Кребса цикл см. Цикл лимонной кислоты
Креветки I 72—78; II 301
— эндокринная система III 473, 476, 478, 481—484
Кристаллический конус II 518
— стебелек I 288, 300—301
— тракт II 519
Кристы полукружного канала II 412, 413
— митохондрий I 485
Критическая температура II 416
Кровообращение III 346—401
Кровь, дыхательные функции II 5—74
— объем III 349—353
— содержание аммиака I 550—551
-----сахара I 444; III 423, 484 Кровяное давление III 353—363 Крокодилы (см. также Аллигатор)
I 131—134, III 371
Кролик, гемоглобин II 48
— пищеварение I 314
— потребность в аргинине I 562
— рецепторы II 453, 457—459
— тропонин III 184
Круглоротые (бесчелюстные) I 111, 289; III 359, 360, 385
— гормоны I 152, 153; III 427
— нервная система III 65, 76, 77, 81
Круглые черви см. Нематоды
Крыса, акклимация II 152, 153
— водный баланс I 142, 146—147
— дыхание I 371
— метаболизм I 252, 262—263, 268, '270—271, 324, 331
— мышцы III 217
— недостаточность рибофлавина I 269—270
— потребность в аминокислотах
I 252, 270, 331
----- в металлах I 220
Ксантиноксидаза I 220, 556, 590 [
Ксантофоры III 523
Ксерофиты I 37
Ксилан I 310
Ксиланаза I 315
Ксилоза I 329
Культуральные среды I 224—228, 249—252
Куриный эмбрион, развитие нервных функций III 108
-----сердце III 378
-----снабжение кислородом II 49,
51
Куры, гемоглобин II 20, 49—50
— мышцы III 219
— почка I 137—138
— хеморецепторы I 371
Кутикула I 100—101, 448—450
Лактаза I 295, 298, 300
Лактат, у черепах I 455
Лактатдегидрогеназа (ЛДГ) I 432, 438—439, 453; II 120; III 216
— изоферменты I 503—504
Лактация I 492, 494, 495; III 436, 439
Лактоза I 494—496
Лангергансовы островки III 422
Ланцетник (Branchiostoma) I 218
— мышцы III 171, 176, 205, 214, 246
— сердце III 372
Латеральное коленчатое тело III96
Легкие I 367—371; II 263
Лед, плотность I 28
Лейдига клетки III 432
Лейкофиты I 246
Лейкофоры III 523
Лейкоциты, движение III 305, 306
— при зимней спячке II 184
Лейцин 1 111, 184
Ленивцы I 313; III 205
Ленточные черви, всасывание пищи I 317
— — гигантские волокна II 300
Леоплектическое движение III 335
Летательные мышцы III 236—240
Летучие мыши, зимняя спячка II 183
----крикотиреоидная мышца III
206
----- оцепенение II 170—171
----- переваривание пектина I 315
-----устойчивость к аммиаку I 548—549
-----фонорецепция II 401—408
Лецитин I 308; II 124
Леша — Нихана синдром I 564
Лигнин I 310
7редметный указатель
623 -
Лизерговой кислоты диэтиламид II 333
Лизин-вазопрессин I 153; III 439
Лизосомы I 286, 301, 589
Лизофосфатидазы I 460
Лимбическая система III 65, 104
Лимфатические пути III 348
— сердца III 348, 372
Линолевая кислота I 263
Линочная железа III 457
Линька у зайца III 455
— у птиц III 454—455
— у членистоногих III 456—466, 472—476
Липаза I 300
Липиды, биосинтез I 492—494
•— в плавательном пузыре I 416
— влияние температуры II 119—120
— и акклимация II 121—124, 153
— обмен I 457—474; II 154
ос-Липоевая кислота I 273
Липофоры III 523
Лировидные органы II 371
Литоральная зона, обитатели I 69—
70, 118, 441, 451; II 228
Лобоподии III 306
Локомоция I 406—408; III 79—81
Лошадь, пищеварение I 314
Лунные ритмы II 227—235, 242
Люминесценция III 584—606
Люмиродопсин II 490
Лютеинизирующий гормон (ЛГ) III
431—433, 436, 441, 447, 449
Лютеотропный гормон (ЛТГ) III
436, 449
Люцифераза III 587, 591, 594—599
Люциферин III 587, 594
Лягушка, влияние адреналина II 282
— выработка рефлексов III НО—112
— гипофиз III 442
— дыхание I 354—355
— кожа I 42
— кровообращение, сердце III 359, 360, 378, 384
— мышцы I 41; II 362, 363; III 196, 199, 204, 206, 211, 212
— нервная система II 396; III 70, 75, 85, 89—91, 93, 94
— рецепторы II 345, 363, 417, 490, 491, 494; III 91
Лягушка-бык см. Rana catesbeiana
Магний I 177, 180, 199, 215, 221; II 5, 25
— в мышцах III 183
— в ресничках III 328
Магний-АТФаза (Mg-АТФаза) I 216, 221; III 183
Магнитные поля II 242—243, 245
Магния карбонат I 223
Макистерон III 475
Малат I 439, 493
Малонил-КоА I 463, 466, 467
^Мальпигиевы сосуды I 96—97, 102— 104, 109
Мальтаза I 298—299
Марганец I 221
Матка, мускулатура III 254—256
Маутнеровские нейроны II 310; III 19—23, 77, 78
Медиаторы II 312—336; III 133
— высвобождение III 286
— у ракообразных III 232—233 Медузы I 44, 90, 190, 216; III 510 Медь I 219; II 7, 23
Межжелудочковая перегородка НЕ 371
Межклеточные соединения II 263— 269
Межклиночный период III 472
Мезотрофы I 242
Мезофиты I 37
Мезоциты I 153; III 440
Меланин III 523, 554
Меланин-концентрирующий гормон (МСН) III 554—556
Меланофорстимулирующий гормон (MSH) III 554—556
Меланофоры III 523
Меланоцитстимулирующий гормон (МСГ) III 437, 438, 554
Мелатонин III 452
Мембрана (ы) (см. также Рецепторы; Центральная нервная система)
— влияние аммиака I 547—548
— возбудимые II 134—137, 261—339
— и амебоидное движение III 316, 317
— лецитин II 124
— плазматическая II 261
— полунепроницаемые I 29 ‘
— постсинаптическая III 203, 204
— физическое состояние I 471 ’’
Мембранные константы II 269
— потенциалы II 272—294 Меромиозин III 179—180 Местный потенциал II 274—277 Метаболизм (см. также Кислород, потребление) I 377—382, 408
— и гормоны I 388, 491; III 484— 485
— суточные изменения II 225—226 — эволюция I 565—584
— Qio II 105—106
Метаболическая вода I 142 Метаболические пути -----аргинина — мочевины 571—573
624
Предметный указатель
----------детоксикации аммиака I 552 -----использования отходов I 564 — точки разветвления I 510—511 Металлы I 177, 219—222
-Метаморфоз, роль тироксина I 588 — V насекомых III 457—466 .Метародопсин II 490
Метатрофы I 242
Метионин I 259, 260
Метахолин I 139
Метахронизм III 333—337 а-Метилмасляная кислота III 596 Мех как термоизолятор II 181 Механорецепторы, типы II 350—352 Мечехвост (Limulus), глаз II 514, 528, 532—534
— дыхание I 360
— мышцы III 169, 232
— нейроэндокринные клетки III 485
— 5-окситриптамин II 333
— пигменты крови II 8, 24, 53, 62, 64
— проприоцептивные нейроны II 364
— сердце III 371
Мигательная перепонка III 355
Мидия (Mytilus), ацетилхолин III 389
— дыхание I 356
— и ионы I 183
— и соленость I 47—50, 387
— парамиозин III 174
— реснички I 322, III 330
— термоустойчивость II 90, 101
— трансаминазы I 544
Миелиновая оболочка II 269, 300— 301
Микроворсинки I 289, 290, 297
Микросомы III 218
Микроспектрофотометрия II 497
Микротрубочки III 314—315
Микрофиламенты III 314—315
Микрофонные потенциалы II 373, 3gg____391
Миксины I 111, 184; III 76, 360
Миксомиозин III 313
Миндалины (amygdala) III 104—105
Минералокортикоиды III 426, 428
Миниатюрные потенциалы II 286;
III 203
Миноги I 111—112; III 76, 214
Минутный объем III 374—376
Миоглобин II 6—9, 14, 17, 42—43, 54; III 215
Миозин III 165, 166, 174, 177—180, 219, 244, 245
— АТФаза III 217—219 > ;
Миоид II 544
Миоинозит I 183 I.?,
Миофибриллы III 165, 168, 244
Митохондриальные органы II 364
Митохондрии, биохимия I 447, 460 461, 475, 478, 479, 482, 540, 547— 548, 552, 554; II 326
— в различных клетках II 312, 484'
III 167, 170, 202, 420
— гигантские I 486
— окисление жирных кислот I 461— 462
— проницаемость I 486
— роль в терморегуляции II 124 153, 154
— структура I 484—485
— у насекомых III 460
Митральные клетки (нейроны) II 458—459
Михаэлиса — Ментен уравнение II 460
Многощетинковые черви см. Поли-хеты
Модуляторы ферментов I 509—510, 519—520
— влияние температуры II 124—125
Мозговое вещество надпочечника
III 357, 423—426
Мозжечок III 65, 81—89
— клетки Пуркжнье III 69—70, 79
— у электрических рыб III 300
— эволюция III 66
— электрическая активность III 71
Молибден I 220
Моллюски, анаэробиоз I 441
— гемоцианин II 7—8, 24, 62
— двустворчатые I 47, 90, 315; III 251
— жабры I 356
— жидкости тела III 346, 347, 352
— замерзание II 90
— кровеносная система III 364, 387
— миоглобин II 54
— мышцы III 183, 186, 217, 240—246
— нервная система II 291—294, 302—303, 333; III 29—40
— осморегуляция I 47—50, 90—91
— раковина I 223; II 71
— сердце III 371, 387—390, 396, 397
— статоцисты II 409
— углеводный обмен I 440—443
Молоточек II 385
Молочная кислота, влияние на сосуды III 358
Молочные железы (и гормоны) III 432, 439
Моноаминоксидаза II 326
Моноамины II 325
Моржи I 150
Морская вода I 38, 84—89, 186
— свинья I 410, 411
Предметный указатель
625
Морские ежи, мышцы III 251 -----нервная система III 28—29 -----обмен I 393, 397
-----осморегуляция I 41, 42, 44
-----пищеварение I 315
----- потребление кислорода I 356
— звезды I 44—45, 356; II 368; III 486
-----амбулакральные ножки III 366
----- нервная система III 28
----- обучение III 118
-----реснички III 326
Морской слон III 362
Мотонейроны II 360; III 41, 72, 73, 128
— у аплизии III 33
Моторная концевая пластинка III 201—204
Мочевая кислота I 537, 555, 569,570
Мочевина (см. также Цикл мочевины) I 185, 561—563
— выделение I 537, 571—572
— роль в осморегуляции I 129, 130, 580
— у амфибий I 126, 130
— у жвачных I 311
— у пластиножаберных рыб I 112— ИЗ
Мочевой пузырь, мускулатура III 255, 263
-----реабсорбция I 121, 129—131
Мочеотделение, у крокодила I 132 — у ракообразных I 63, 81—82
Мочеточник, мускулатура III 257
Муравьи, рецепторы II 371
— феромоны II 436
Мускариновый рецептор II 321; III 259
Мускульный желудок I 288
Муха мясная, инициация полета I 522
----- использование сахара I 247, 249
-----потребление кислорода I 389
-----хеморецёпция II 439, 461, 462
Мухи I 247, 249
— рецепторы II 365, 522—523, 526
Мучной червь, пищевые потребности I 253
Мшанки, кровообращение III 346
Мышечное сокращение, биохимия
III 199—201
-----и возбуждение III 187—188
-----изометрическое III 165, 192, 194
----- изотоническое III 165, 192
-----независимость от кислорода III 197
----- теории III 176—177, 184
-----тоническое и фазическое III 165
Мышечные веретена II 360—363
— рецепторы II 356—363
Мышца-замыкатель III 240—246
Мышцы III 164—270
— гладкие III 359
— дрожь II 144, 158
— митохондрии I 485—486
— потенциалы покоя II 272—274
— стимуляция при деформации II 351
— термолабильность II 99
— фазное сокращение III 241
Мышь I 140, 144, 147
— гемоглобин II 43
— кровообращение III 349
— питание I 248
Мюллеровские клетки III 77
Навигация, у животных I 238—240 НАД-Н I 432, 447—448, 476—478,539 НАДФ-Н I 493; II 488 Надглоточный ганглий III 57—61 Надпочечники, барорецепторы III 356
— кора III 417, 426—432, 444
— мозговое вещество II 327; III357, 423—426
Напряжение (мышцы) III 167, 187— 191, 193—196
Насекомые, антенны II 365, 370, 371 — гемоглобин II 55
— жабры I 359
— жидкости тела III 352
— защитные вещества II 436
— липидный обмен I 468—470
— люминесценция III 587—588 — мышцы III 170, 190, 233—240 — нервная система II 301—302,
324—325, 334; III 55—56, 59—61, 124, 234—236
— обмен веществ I 264—265, 267, 269, 468—470, 562; III 458
— осморегуляция I 95—ПО
— полет, летательные мышцы 1469;
II 417—418; III 51—52, 236—240 — размножение III 466—467 — рецепторы II 132, 356, 365—366, 370
-----фонорецепция II 376—383
-----фоторецепция II 519, 521— 527, 534—537
-----хеморецепция II 434—448, 453 — сердце III 372, 398—400
— таксисы II 367
— температурная адаптация II 137— 139
40—2067
626
Предметный указателъ\
— терморегуляция II 90, 103, 127— 129
— трахеи I 372
— ферменты I 300, 301, 308, 542, 574
— хроматофоры III 521, 538—541
— эндокринная система III 456— 470
Насосы ионные см. Натриево-калие-вый насос; Натриевый насос
Натриевая проводимость II 296
-----в двигательной концевой пластинке III 204
— — в мышцах III 227, 382
— — в электрических органах III 291
Натриево-калиевая АТФаза (Na+K+-
АТФаза) I 124, 139, 211, 456—457;
II 116—147, 185; III 290
Натриево-калиевый насос I 37, 203—205
Натриевый насос I 229; II 272—273, 283, 297, 359; III 11, 381
Натрий I 177—231
— в гемолимфе II 302
— в сетчатке II 504, 506, 509, 512
— выделение I 116, 118—120, 138, 217; III 429
— и перенос сахаров I 328—329
— поглощение I 81, 82, 91, 97, 112, 114, 131, 134
— при лактазной недостаточности I 299
— реабсорбция I 131, 133, 137, 150, 201
— транспорт I 107, 118—119, 146, 215, 486, 545, 547; II 302
Нейрогипофиз I 152—153; III 438— 440
Нейромасты II 373, 374
Нейрон — отметчик спайков II 378, 379
Нейроны (см. также Гигантские нервные волокна)
— вставочные III 61, 378
— действие температуры II 135
— монополярные II 524, 531
— пирамидные III 70,79
— потенциал покоя II 272
— проприоцептивные II 364
— пуринэргические II 355, III 261
— Реншоу II 287, 322
— связи между ними II 304—308;
III 109—110
— сетчатки II 547
— центробежные II 526; III 61
Нематоды (круглые черви) I 89, 308, 318
— гемоглобин II 56
— кровообращение III 346
Нематоцисты III 508—515
Немертины, гемоглобин II 7
— кровообращение III 348
— нейроны II 300
— реснички III 324
Непарнокопытные, пищеварение I 314
Нервная сеть III 23—29, 118
— система, развитие III 107—115
— ткань I 551
Нервы, миелиновые II 269, 300—301
— проведение II 298—301
— пуринэргические II 335; III 261
— ракообразных I 551
— стимуляция при деформации II 351
— фотогенные III 605
Нереиды (см. также Nereis spp.) Ill 250
Нернста уравнение I 32—33, 215
Нефридии I 92—94, 197, 200
Нефрон I 129, 145 ।
Ниацин I 270—271
Никель I 221
Никотинадениндинуклеотид см.
НАД-Н
Никотиновая кислота I 270—271
Никотиновые рецепторы II 321; III 259
Ниобий I 221
Новая кора III 62
-----эволюция III 81
Новокаин см. Прокаин
Норадреналин II 178, 315, 326, 327;
III 259, 261, 424
— действие на бурый жир I 173
-----на сосуды III 357
— и термогенез II 143
Носовая железа I 134, 138
Ночные бабочки II 376—381; III 53, 55, 235
Нуклеиновые кислоты, абиотический синтез I 243—244
— переваривание I 311, 316
Нуклеотиды, синтез I 563—564
Ныряние I 408—412, 454—455; III 362—363
— и гемоглобин II 43—44
Обезвоживание см. Дегидратация,
Испарение воды
Обезьяны, гемоглобин II 20
— осморегуляция I 146
— терморегуляция II 160
Обкладочные клетки I 294—295, 325
Облегчение III 25, 227, 228
— в электрическом органе III 291
— гетеросинаптическое III 10, 34, 127—130
Предметный указатель
627 “
- — нервно-мышечной передачи III 195, 223
— - синаптическое III 9—10
Облегченная диффузия I 34; II 42
Обмен веществ (см. также Метабо-
лизм)
— - интенсивность I 389—399
— основной I 376, 453; III 484
— стандартный I 377—382, 385, 386
Обменная диффузия I 34
Оболочники, сердце III 372, 390—392
Обонятельная система II 458—459, 462; III 103
Обучение III 116—117, 120—126,
130—132
Овальное окно II 385
Овцы, водный обмен I 140, 142, 143
— гемоглобин II 20, 43, 57, 58
— ионный состав крови I 178
— терморегуляция II 160
Однопроходные I 292; II 169
Озон II 477
Окислительное фосфорилирование I 478, 491, 547
-----разобщение I 472—474
Окись углерода (угарный газ) II 8, 47
Окраска тела, изменения III 519—533 Оксалоацетат I 430, 452, 467, 484, , 492, 544
Оксигенация гемоглобина II 8, 27— 35
Оксидазы аминокислот I 541—542 Окситоцин I 151, 153; III 438—440 5-окситриптамин (серотонин) II 178, 331—333; III 106, 241, 243—245, 261
— влияние на мышечное сокращение III 241
-----на сосуды III 357 5-окситриптофан II 331—333 Оксифильные клетки III 420 Окукливание, роль гормонов III 457 Олива нижняя III 84
Олигосахариды I 296, 297
Олигохеты I 91, 300, 354—356
— гемоглобин II 6, 52
— кровообращение III 348
— люминесценция III 586
Олово I 221
Омар (Homarus), азотистый обмен
I 543, 551
— влияние температуры II 101, 113, 134
— водно-солевой обмен I 51, 63, 82
— дыхание I 359
— ионы I 184, 191, 192, 197
— нервная система I 41; II 301, 334;
III 11
— рецепторы II 364
— сердце III 393—394 Омматидии II 487, 518, 528 . Оммохромы III 520 Онихофоры I 572—573
Оогенез, гормональная регуляция III 476
Оперон I 587
Опоссум, терморегуляция II 169
Опсин II 490, 491, 494—500
Оптическая (зрительная) доля II 524—526
-----у насекомых III 61
Оптические патроны II 525, 526
Оптический диск II 537
Оптокинетическая реакция II 535—
536; III 49
Х-органы III 470, 545
Y-органы III 472
Орнитинтранскарбамилаза I 552, 566
Ортодромное проведение II 309
Осел I 141, 143
Осмоконформеры I 39, 44—52, 63, 98
Осмоль I 29
Осмометр I 40
Осморегуляция I 39, 45—154
— и гормоны I 151—157; III 420, 428, 429, 440
— роль аминокислот I 66 Осморецепторы I 149—151 Осмос I 33
Осмотическая концентрация I 29— 30, 37—39, 124, 127, 128, 132—133, 137, 140, 146
Осмотический гомеостаз I 40—41
— градиент I 146
— стресс I 39
Осмотически неактивный объем клеток I 40—41
Осмотическое давление I 29
Осфрадий II 434
Осьминог (Octopus), ацетилхолин II 323
— гемоцианин II 60
— глаз III 120
— катехоламины II 330
— кровь, объем, давление III 352, 364
— мозг III 38, 120—123
— сердце III 388
— статоцисты II 409
— тирамин III 537
— хроматофоры III 529, 537
Отвыкание III 127—128
«Откашливание» у ракообразных и рыб I 359, 360
40*
628
Предметный указатель
Открытопузырные рыбы I 368, 414; II 369
Оцеллярный нерв II 531
Оцепенение I 406; II 169—173, 180—181
— и температурная акклимация II 113—114
Палочки II 481—487 >
Пальмитил-КоА I 468
Память III 123—125, 132, 133
Панкреатические клетки I 294
— липазы I 458
Панкреатический сок, pH I 294—295
Панкреозимин I 325; III 411, 453
Пантотеновая кислота I 271
Панцирная щука I 114
Паразиты I 317, 404, 413, 437—440
— гемоглобин II 56—57
— осморегуляция I 86—89
Параллельные волокна III 82
Парамеция (Paramecium) I 248
— гемоглобин II И, 57
— замораживание II 89
— осморегуляция I 85—88
— поведенческие реакции II 367, 408; III 117
— реотаксис II 367
— реснички III 330, 335
— трихоцисты III 513—514
Парамиозин III 174, 244, 245
Параподии I 355
Парасимпатические нервы III 259
Парафлагеллярное тельце II 481
Парафолликулярные клетки III 421
Паращитовидные железы III 420— 421
Пастера эффект I 453
Паукообразные III 366
— выделение гуанина I 570, 575
— дыхание I 367, 372
— механорецепторы II 371
— осморегуляция I 51
— сердце III 368—400
Пачини тельца II 352—354
Пейсмекер III 378, 381, 382, 395
Пектин I 310, 315
Пектиназа I 315
Пеликан, носовая железа I 138
Пеллагра I 271
Пентобарбитурат I 206
Пентозный цикл I 433, 435—436
Пентозофосфатный шунт I 451
Пепсин I 302
Пепсиноген I 302
Пептидазы I 304—306
Переаминирование I 538—540
Передний мозг III 65, 104
Перекрестная иннервация III 219
Переохлаждение I 29, 124, 450, 451;
II 88—95
Перепончатокрылые III 119, 236
Перикард, фильтрация в его полость
I 91, 200
Перикардиальные органы III 484
— сумки I 71
Перилимфа II 386
Периректальные (мальпигиевы) со-
суды I 109
Перитрофпческая оболочка (мем-
брана) I 291, 449
Пероксисомы I 541
Петля Генле I 144—145
Печень, детоксикация аммиака I 552—558
— разрушение гормонов III 444
— синтез аргинина и пуринов I 562—564
Пигменты крови II 5—75
Пикротоксин II 334; III 232
Пилорические придатки I 294
Пиноцитозные пузырьки III 174
Пирамидные нейроны (в гиппокампе) III 70
Пиридоксальфосфат I 539
Пиридоксин I 271
Пиримидины I 261, 484
Пируват I 430—434, 436, 438, 439, 444
Пируваткарбоксилаза (у жвачных) I 434, 492
Пируваткиназа I 434, 438, 440, 496— 498
Питание I 241—279
Пищеварение I 285—336
— эволюция I 288
Пищеварительные соки I 325
— ферменты I 292—309
Пищеварительный тракт I 286
---мускулатура III 257—259, 262
Пищевод III 259
Пиявки I 324; II 368; III 487
— кровообращение III 348
— мышцы III 201, 250
— нервная система II 303; III 40, 41
— осморегуляция I 91—94
— сердце III 372
— фоторецепторы II 509
Плавательный пузырь I 368, 414— 418, 453
— и слух II 385, 394—395
Плавучесть I 414 *
Плазма крови, ионный состав ' I 179—180
---объем III 349
Плазмазоль III 306
Плазмалемма III 308
Предметный указатель
629- ~
Плазматическая мембрана II 261, 484
Пламенные клетки I 197
Планарии III 117, 486
— реотаксис II 367
Планка постоянная II 475; III 594
Пластиножаберные I 112—114, 124— 125; III 264, 325, 524
— гормоны I 152, 153
— образование мочевины I 577
— рецепторы II 133
— электрические органы III 293, 295
Пластичность (нервной системы) III 115
Пластрон I 374, 408
Плацента, гормоны III 432, 445—447
Плеченогие, раковина I 223
Плоские черви, гемоглобин II 7
Пневмостом I 367
Поведенческие реакции, влияние пролактина III 436—437
-----и метаболизм I 66—67, 101— 102, 367, 405—408
-----и передний мозг III 104—106
-----и рецепторы II 365—371, 375—
376, 392—408, 417—418, 438—441, 444, 449—457, 531
-----нервные механизмы III 5, 59
----- развитие III 106—107
-----связанные с размножением
III 449—452
-----стойкие изменения III 115— 117
-----у рыб с электрическим органом III 297—300
Погонофоры I 320
Поджелудочная железа I 292; III 422—423, 453
-----ферменты I 293—294
Подглоточный ганглий III 457, 461, 536
Подоциты I 145
Пойкилотермия II 84
Пойкилотермные организмы I 385;
II 85, 105, 132—133, 137—141
Полет у насекомых II 367; III 51— 52, 236—240
Полигон толерантности II 98
Полисахариды I 288, 295—296
Полихеты (многощетинковые черви)
I 191, 223, 224; III 265
— анаэробиоз I 355
— гемоглобиж II 6, 7, 54
— геотаксис II 408
— жабры I 354
— кровообращение III 348
— люминесценция III 586
— мышцы III 201
— нервная система II 300; III 13г 15, 40—42
— осморегуляция I 51—56
— реотаксис II 367
— эстеразы I 308
Полосы Сорэ II 13
Полужесткокрылые, развитие III457
Полунасыщение гемоглобина II 29, 38—43
— ферментов I 34
Полупроницаемые мембраны I 29
Порфирины II 5
Порфиропсин II 489, 491—494
Поры (в мембранах) I 43, 145
Послелинька (у ракообразных) III 472
Посткомиссуральные органы III 470-Постоянная времени см. Временная константа
Постсинаптическая мембрана III 203, 204
Постсинаптический потенциал (ПСП)
II 285—294; III 227, 231, 289
Посттетаническая потенциация III 126, 127
Потенциал(ы), в рецепторах II 352—353, 356, 361, 367
— покоя II 272—274; III 381
S-потенциалы II 554
Потовые железы II 160—161
Почки I 113
— агломерулярные I 117—118, 121 — амфибий I 129—131
— гигантские митохондрии I 486
— и обмен веществ I 562, 563
— кур I 137
— млекопитающих I 144—151, 456
— ракообразных см. Антеннальная железа
— синтез аргинина I 562
— эволюция I 196—197
— эласмобранхий I 113—114
Прегнандиол III 447
Прелюмиродопсин II 490
Препотенциалы действия II 274—284
Пресмыкающиеся см. Рептилии Привыкание III 116, 127—130 Приливы II 227—228
Приматы, вкусовая рецепция III457 — зрительная система II 498; III96 — менструальный цикл II 233 — мозжечок III 88
— слух II 399
Пробит II 96
Прогестерон I 495; III 426, 431, 446—448
Происхождение жизни I 242
Прокаин I 205
Прокариоты I 567, 586
630
Предметный указатель
Пролактин I 154, 155; III 419, 431, 436—438, 447, 449
Пролин I 66, 111, 184
Промежуточный мозг III 62
Проницаемость I 32—33, 1—44,59— 61, 83—84, 89—90, 123—124, 140, 153, 216—217
Пропионил-КоА I 430
Пропионовая кислота (у жвачных)
I 311; II 445
Простагландины I 474; III 456
Простейшие, движение III 164, 305, 306, 333
— действие кислорода I 413
— люминесценция III 584, 585
— осморегуляция I 84—88
— питание и метаболизм I 261, 389, 571
— реотаксис II 367
— условные реакции III 117
— фагоцитоз I 321
— хеморецепция II 432
— циркуляция жидкостей III 346
Протеазы I 300—304, 317
Противоточные системы I 145, 354, 365, 417, 453; II 51, 130, 158
Протоплазма, движение III 305, 307, 308, 346
Проторакальные железы III 457,463
Прямокрылые, полет III 237
Псевдоподии, типы III 306
Псевдоцель III 347
Птицы II 577
— акклимация к холоду II 155
— гиперпноэ II 68
— гомойотермия II 166
— дыхание I 369—371
— зрение II 485, 499, 500, 544, 548; III 95
— метаболизм I 135—139, 195, 275, 405, 541, 554—557, 562, 577
— мышцы II 363; III 206, 212
— нервная система III 62, 65, 81, 88, 95, 100, 125
— ориентация II 238—240, 244—245
— половые признаки III 443, 444
— реакция на погружение в воду I 409
— сезонные изменения окраски III 455
- сердце III 375, 378, 383, 385
— слух II 391, 412; III 100
— терморегуляция II 163—164
— условные реакции III 120
— хеморецепция II 456; III 95
Пуриннуклеотидный цикл I 544
Пуриновый путь I 563, 565, 567-—570, ’573—575
Пуринотелия I 570, 575
Пурины I 261, 484, 554, 573—575
Пуринэргические нервы II 336; III
261
Пуркинье волокна (в сердце) III 380. 381
— клетки II 289, 322, 334; III 7, 69, 82, 85, 87
— сдвиг II 541
Пуромицин III 132
Пути биосинтеза см. Метаболические пути
Пучковая зона III 426
Пчелы I 247; II 383, 417
— зрение II 521—522, 530, 531
— летательные мышцы I 469
— обучение III 119
— рецепторы II 370, 436; III 297
— сердце III 398
— терморегуляция II 129—130
— ферменты I 300
Рабдом II 487, 518, 528
Равновесие, рецепторы II 408—418
Радиолярии III 306, 585
Размножение, месячные ритмы II 233
— роль гормонов III 441—453, 466, 476
— членистоногих III 466—467, 476— 485
Рак речной, гормоны III 483, 485
-----дыхание I 358, 359
-----кишечник III 264
-----метаболизм I 551; II НО
-----мышцы III 169—170, 225, 226 -----нервная система II 266, 300, 305, 309; III 16—18, 46, 129, 323, 324
------ осморегуляция I 78—82
-----половые придатки III 480
-----сенсорные процессы II 364— 365
-----терморецепторы II 132
Раковины, состав I 223
Ракообразные, гемоглобин II 7, 55, 59
— гемоцианин II 62
— дыхание I 356—359
— кровообращение, сердце III 8, 352, 364, 365, 392—398
— люминесценция III 584, 586
— мышцы III 115, 169, 222—233, 264
— нервная система II 300, 334—336, 534—536; III 12, 16—18, 45—51, ПЗ. 223—232
— осморегуляция I 51, 56—84
[Iредметный указатель
ба»
— рецепторы II 351, 356—360, 370, 371\ 410—411, 434, 501, 510
— ферменты I 299, 300, 315—317, 543, 551
— эндокринная система I 388; III 470—485, 541—552
Ранний рецепторный потенциал II 514—516
Распределительные системы III
6—9
Расслабление (мышцы) III 205,
217—218, 240—246
Растворимости коэффициент I 350
Растения, биологические ритмы II 236—237
— гемоглобин II 7, 13
— геотропизм II 408
— замерзание II 89
— кремний I 222
— осмотическое давление I 37
— паразиты II 433
— фотопериодические реакции II 477, 478, 480
— эволюция II 438
— экдизоны III 460
ахит I 275
Реабсорбция I 129—134, 196, 197, 199
— влияние ртути I 222
Регуляция см. Осмо-, Терморегуляция и отдельные регулируемые процессы
— роль изоферментов I 496—525
Резерпин II 326, 332
Рейснеровская мембрана II 386
Ректальные железы I 114, 212, 213
Ректальный транспорт воды I 102—
ПО
Релаксин III 448
Ренин I 155; III 357, 358
Реннин I 302, 319
Рентгеноструктурный анализ (гемоглобина) II 17
Реншоу клетки II 287, 322; III 7,72
Реобаза II 274
Реотаксис II 367
Рептилии (пресмыкающиеся), гликолиз I 454—457
— гормоны III 423, 427
— желтые тела III 448
— кровообращение, сердце III 359, 362, 371, 372, 383
— легкие I 368, 371
— нервная система III 62, 104,
262, 263
— осморегуляция I 131—135
— рецепторы II 363, 389, 391, 485
— терморегуляция II 128, 139—141
Реснички II 262; III 324—342
— и питание I 321
— структура III 327—329
— у коловраток I 90
— хемочувствительность II 433
Ретикулоподии III 306
Ретиналь II 488—490
Ретинальный пигментный гормон III 482, 483
Ретиномоторные процессы II 544
Ретинула II 487, 518, 527
Рефлексы сегментарные III 40
— сердечно-сосудистые III 356
— у беспозвоночных III 30—31, 48—51, 55—57
Рефлектометрия глазного дна II 49В
Рецепторы (см. также Абдоминальный рецептор, Баро-, Термо-, Фоно-, Фото-, Хеморецепторы)
— гормонов III 469
— гравитации II 365
— давления II 357
— кислорода III 363
— насекомых I 102
— прикосновения II 363
— равновесия II 408—418
— растяжения II 356—360
Рибазолфосфат I 267
Рибонуклеаза I 316
Рибонуклеиновая кислота см. РНК
Рибофлавин I 266, 269—270
Рибофлавин-5'-фосфат (и люминесценция) III 597
Ритмы размножения III 485
РНК I 287, 587, 589
— и гормоны III 417, 419, 460, 469
— и обучение III 132—134
— и синтез гемоглобина II 58
— синтез II 216, 248; II 412, 417
— транспортные II 588
РНКаза в поджелудочной железе I 306
РНК-активатор I 587
Роговица II 518, 537
Родопсин II 487—488, 490—494
Ротовой «насос» I 360, 368
Ртуть I 221—222
Рубец I 310—311
Рукокрылые см. Летучие мыши
Рута эффект I 416, 417; II 32—33, 45—46
Рыбы (см. также Закрытопузырные рыбы; Открытопузырные рыбы; Плавательный пузырь)
— ацетилхолинэстераза II 323
— боковая линия II 373—376
— веберов аппарат II 385
— гемоглобин II 6, 44—47
— головной мозг III 62, 65, 81, 82, 87
— дыхание I 360—367
632
Предметный указатель
— звуковые рецепторы II 392—395
— кровообращение III 359, 361
— лабиринт II 412—415
— липидный обмен I 470
— люминесценция III 590—592
— маутнеровские клетки II 300; III 19—23
— мышцы III 168, 180, 214, 255
— осморегуляция I 111—125
— поведенческие реакции II 127— 128, 367, 368, 407
— проприоцепторы II 363
— сенсорные пути III 89—95
— сердце III 359, 370, 373, 377, 378, 384—386
— температурная акклимация I 470;
II 104, 114—117, 125—126
— терморецепторы II 132—133
— углеводный обмен I 450—454
— фоторецепторы II 485, 491, 544
— хеморецепторы II 448—449, 454
— хроматофоры III 556—564
— электрические II 308
— эндокринная система I 152; III 416, 419—420, 440—443
Сакса орган III 299
Саранча (Locusta) II 382—383; III 539
— жирные кислоты I 469
— зрение II 530
— кишечник III 264
— лактаза I 300
— мышцы III 234—236
— нервная система III 52, 56, 61, 234—236
— обмен воды I 100, 107
— рецепторы I 102; II 367, 377,445, 536; III 53, 237
Сарколемма III 165—167
Саркомеры III 166—168
Саркоплазматический ретикулум III
166, 167, 186, 221
Сахара, вкус 247; II 461
— всасывание I 328—330
Сахараза I 299—300
Сахаразо-изомальтазный комплекс
I 298
Сахароза I 295
Сверчок, нервная система III 18, 59
— рецепторы II 132, 377
— стрекотание II 380; III 65, 60
Свет II 474, 475, 477
— адаптация к нему II 543, 544; III 480—481
— влияние на окраску II 487—490;
III 528—531
-----на ритмы II 217—218
— и проводимость мембран II 516 — поляризованный II 503, 528, 530 Светляки, свечение III 55, 587, 589, 592—597
Свинец I 221—222
Свиньи, пищеварение I 314, 316
Секретин I 325; III 453
Селен I 221—222
Сельдь, гемоглобины II 18
— реакция на звук II 395
Семенник, гормоны III 432—433, 443—445
Сенсиллы I 102; II 351, 365, 370, 440—445
Сенсорный потенциал II 352, 354
Септированные синапсы II 263
Сердечно-сосудистая система III 353—401
Сердечные гликозиды I 204
Сердца, ампулярные III 372—378
— добавочные III 360, 372
— жаберные III 372
— трубкообразные III 371—372
Сердце III 369—401, 484
— барорецепторы в нем III 356
Серебро I 221
Серебряный карась, влияние температуры II 96, 127, 136
-----гемоглобин II 47
-----дыхание I 382—384, 402, 403, 451, 452
-----зрение II 499; III 124
-----нервная система II 456; III
19, 20, 23, 76, 81, 88, 91, 104, 128
----- перенос сахаров I 329
-----фосфолипиды II 121
Серин, дезаминирование I 541
Серингидратаза I 503
Сериновые протеазы I 302—304
Сернобык I 141; II 160
Серотонин см. 5-окситроптамин
Серповидноклеточная анемия II 11, 21
Сетчатка II 481—487, 537—539,
547—557
Симбиоз I 223, 246—247, 270
Симбиотические организмы I 276, 310, 311, 314, 316, 318
Симплектическое движение III 335 Синапсы III 126—130 — в нервной сети III 24 — в сетчатке II 548—551
— в фоторецепторах беспозвоночных II 524, 527
— в электрорецепторах III 294
— ленточные II 315, 550
— поляризация III 6—9 — структура II 312—315 — тормозные II 297 • . ,
П редметный указатель
633
— химические II 263, 284—287,
311—336
— электрические II 309
Синаптические ленты II 315, 550
— потенциалы II 285—290
— пузырьки III 202
Синаптосомы II 312
Синерезис III 311
Синтетаза жирных кислот I 462, 466
Синусная железа III 470, 477, 483, 545, 546
Сипункулиды (Golfingia) I 44—46, 190, 354
— ацетилхолин II 323
— кровообращение III 346
— мышцы III 174, 201
Система переноса электронов I 477, 478, 511
Сифонофоры I 417
Скаты I 153; II 133
Скафогнатиты I 356, 359
Скелет (химический состав) I 223— 224, 253
Склера II 537
Склеропротеины I 218
Скорпион, кислородная емкость крови II 64
— мышцы III 173, 231
— рецепторы II 364
Скотопический глаз II 520
Слизистая кишечника I 294—295
Слизь I 290, 321
Сложный глаз II 518—524
Слуховое ядро III 100
Слуховой нейрон II 391
— проход II 384—385 г
Слуховые косточки II 385
— пути II 399; III 99—101
Слюна I 287
Слюнные железы, мембраны II 267
Смертность, кривые II 95—97
Сократимые белки III 176—184
— волокна III 164
— системы III 310
Сократительные вакуоли I 85—87, 89, 197
Солевые железы I 139, 212, 456, 486 Соленость I 38, 66, 68, 69, 123,388;
II 113
— влияние на амеб III 306
Солнечники III 306
Солоноватая вода, адаптация к чей
I 38—39, 52—56
Соляная кислота I 292
Сом, электрорецепторы III 293, 296
Соматосенсорные пути III 101—103
Соматотропный гормон III 434—435
Сон III 106
Сорбит I 450
Сосальщики I 317—318; III 173
Сосудистая оболочка II 537
Сосудистое сопротивление III 353, 355
С осу до двигательные центры III 78
Спайки II 277, 298
— в механорецепторах II 352, 354, 356, 357, 361, 365, 371
— в мышцах III 187, 205, 206, 217, 227, 231, 236, 243, 248, 258, 262
— в обонятельном эпителии II 455
— в сердце III 378, 381, 393
— в слуховых нейронах II 379, 389, 511
— в фоторецепторах II 509
— в электрических органах III 290
Спектр поглощения гемоглобинов II 11—13
----- пигментов фоторецепторов II 480
Спинальный шок III 73
Спинной мозг III 65, 71—78
-----мотонейроны III 113
-----терморецепторная функция II 176—180
Спинномозговая жидкость I 185
Спироцисты III 509
Спонгин I 218, 222
Спячка (см. также Зимняя спячка)
— летняя у двоякодышащих рыб I 453, 581
Средний мозг III 62
Средняя кишка I 288, 289, 300
Ставермана коэффициент I 43
Стайлса — Кроуфорда эффект II 540 Стандартный обмен II 105, 117, 127 -----у разных животных I 377—382 Статоцисты II 409—411, 535 Стеногалинность I 39, 44
Стереоцилии II 373
Стерины, у насекомых I 263—265;
III 459
Стероиды, структура III 426, 430— 431
Стигма (глазное пятно) II 481
Стоматогастральный ганглий III 49
Страус I 139; II 164
Стрекозы I 372; III 236, 457
Стрекотание II 380—382
Стремечко II 385
Стремечковая мышца II 385
Стрихнин II 334
Стронций I 224
Сукцинат I 430, 433, 438—440
Сукцинатоксидаза I 439
Сульфат I 190—191, 197
Сумм анионный потенциал II 389— 390
€34
Предметный указатель
Суммация синаптическая III 9—10
Сумчатые I 313, 394; II 169; III 101—102
Суперпозиционный глаз II 519
Суслик (atelias') II 155, 157, 556
— зимняя спячка II 181, 184, 185
— стимуляция гонад III 441, 452
Суточные ритмы II 212—213
— — биолюминесценции III 405— 406
---- изменения окраски III 531— 533
Сухожильные органы II 360—362
Сухопутные крабы I 69—70, 357,359
Сцинк I 131
Тактильные окончания II 354—356
Таламус III 69, 94
Тараканы, водный баланс I 99, 101, 104, 107—108
— дыхание I 372
— метаболизм I 255, 388
— мышцы III 170, 234, 235
— нервная система III 18—19, 43, 59, 109, 124, 236
— пищевые потребности I 253, 264
— половой аттрактант II 436
— рецепторы II 132, 434, 365—368, 514, 531
— сердце III 399
— температура тела II 137
— ферменты I 300, 315
— ходьба III 51
— эндокринные органы III 466, 467
Тарантулы III 45
Таурин I 45
Тегментум III 62
Тектум III 62
Телобиоз III 461, 467 ,
Телэнцефалон III 65
Темновая адаптация II 542—546
Температура II 84—191
— влияние на гемоглобин II 46, 62
-----на окраску III 527, 538
----на потребление кислорода I 385
----на размножение III 452
---- на реснички III 342
----на ритмы II 226—227
— — на сенсорный потенциал II 354
----щитовидную железу III 417— 418
Температурная адаптация I 385— 386, 451; II 95—132
— акклимация I 445, 507—508; II 107—126, 159—165
Температурный коэффициент (Q
II 105—107, 109
— — для активности ресничек III 342
Тепловой шок II 162—163
Теплоизоляция II 155—157, 160
Теплоотдача II 85, 160—161
Теплопроводность II 148, 149, 151
Теплопродукция I 472; II 144; III 197—198
Теплота испарения I 28, 85, 160—161
Термиты I 315—316; II 129
Термогенез II 143, 145, 173—174
Термонейтральная зона II 146—148 Терморегуляция II 134, 137—141,146 — модели II 178—180
— нервные механизмы II 175—180
Терморецепторы II 22—26, 132—133, 135, 175—180
Тестостерон III 432—433
Тетраэтиламмоний II 279, 296
Тетродотоксин II 279, 296, 352; III 204
Тиамин I 265
Тигмотаксис II 367
Тимпанальные мышцы III 236—240
Тимпанальный орган II 377, 378
Тиоктовая кислота I 273
Тиоцианат III 349
Тир а мин III 537
Тиреоглобулин III 416
Тиреокальцитонин I 156
Тиреотропный гормон. (ТТГ) III 417—418, 435
Тирозин II 325, 326 г
Тироксин I 218, 388; III 416, 417
— влияние на метаболизм I 491
— — на холодоустойчивость II 126
— и метаморфоз I 588
Тиронины III 416
Титан I 221
Ткачик I 137
Тли, питание I 317
а-Токоферол 276—277
Толерантность к температуре II 96 Толстая кишка, мускулатура III259 Тонические мышцы III 165
Тонкотелая обезьяна I 313
Торможение, в глазу II 513, 514, 532—534
— пре- и постсинаптическое III 10, 225
— рецепторов растяжения III 357
— реципрокное перекрестное III 21
Тормозные постсинаптические потенциалы (ТПСП) II 285
Торпидность см. Оцепенение
Трансаминирование I 478
Трансдезаминирование I 540—544
Трансмуральная стимуляция III 261
Транспортные РНК II 588 ,
Предметный указатель
635
Трахеи I 372—374
Трахейные жабры I 359
Трегалаза I 298, 301, 446
Трегалоза I 444—445, 491
Трегалозофосфатсинтетаза I 446
Треонин I 541
Триглицериды I 307, 333, 458—459, 469
Триметиламиноксид I 112, 113, 183, 184
Триозофосфатдегидрогеназа I 433, 502—503, 521
Триозофосфаты I 430, 501—502
Трипсин I 302—304, 590; III 179,180
Трипсиноген I 302
Тритированная (меченая) вода I 41— 42, 131, 141
Трифосфопиридиннуклеотид I 270
Трихогенная клетка II 365
Трихоцисты III 508, 513
Тропомиозин III 174, 182—184, 244
Тропонин III 182—184
d-Тубокурарин II 321
Тубулин III 327
Тунцы, чудесная сеть II 130—131
Турацин I 220
Турбеллярии, реснички III 324, 332
Тургор I 33, 37
Тюлень I 141—142, 147, 411; II 168
— ныряние II 44; III 362
Уабаин, влияние на потенциал действия II 283, 505, 506
-----на транспорт ионов I 64, 65, 124, 130, 139, 203—207, 210, 212, 215
Углеводный баланс I 248—249
Углеводы, метаболизм у насекомых I 443—450
— переваривание I 295—296
Углекислота (двуокись углерода), в воздухе 350—351
— в крови II 65, 68
— в мышце III 235
— и гемоглобин II 31
— и обмен веществ I 246, 371, 372, 411, 570
— при аноксии I 452
Углерод, потребность в нем I 246— 249
Угорь I 116, 117; III 75
— выделение солей I 115, 194
— газовая железа I 416
— дыхание I 354, 457
— кровообращение II 6, III 361
— лимфатические сердца III 368
Ударный объем III 375—376
УДФ-галактоза I 494—495
Укачивания синдром II 417
Улитка (уха) II 385—388, 392
Улитки (см. также Брюхоногие моллюски)
— гемоглобин II 54
— гемоцианины II 25, 60
— геотаксис II 408
— мышцы III 251
— рецепторы II 390, 392
Ультимобранхиальные железы III 421—422
Уреогенез I 129
Уреогенные животные I 560
Уреотелия I 536, 537
Уридиндифосфатацетилглюкозамин (УДФАГ) I 507
Уридиндифосфатглюкоза (УДФГ)
I 445—446, 507
Уриказа I 568
Урикотелия I 536, 537, 581—582
Урогипофиз I 155; III 440—441
Уроподы, иннервация III 47
Ускорения рецепторы II 409
Условный рефлекс (см. также Обучение) III 116—120, 125, 126,
130_132
Утка I 138—139; II 355
— ныряние I 409; III 362
— рецепторы II 363
— стимуляция гонад III 451
Утомление III 13, 15, 206
Ушная раковина II 384
Фагоцитоз I 322
Фазные мышцы III 165
Фактор роста нервной системы III
114
Факультативные анаэробы I 432, 440
Фарадея число I 31
Фасеточные глаза II 522, 523
Ферменты, акклимация I 507, 588—
591; II 114—125, 153
— денатурация при замс>раживании
II 89
— и клеточные структуры I 524—525
— и осморегуляция I 124
— индукция I 318
— обновление I 589
— пищеварительные I 286, 292—309
— регуляция I 487, 509, 588—591
Феромоны II 436, 463
Физостигмин см. Эзерин
Фика закон I 31
Фиксация напряжения II 278, 509
Фитопланктон I 321
Фитохром II 480
Фитоэкдизон III 460
Флавинадениндинуклеотид (ФАД)
I 269
*636
Предметный указатель
Флавинмононуклеотид I 269; III 597____598
Флоридзин I 63, 82, 117, 130
Фовеа см. Центральная ямка
Фолиевая кислота I 272—273
Фолликулостимулирующий гормон
(ФСГ) III 431, 433, 436
— влияние эстрогенов III 430
— и размножение III 441
Фонорецепция II 350, 376—408
Фораминиферы I 223; III 306
Форель (см. также Salmo), блуждающие нервы III 263 '
— жабры I 360
— кровообращение III 360, 385
Форониды II 7; III 348
Фосфагены III 199—201
- Фосфатидазы I 460
Фосфатидилхолины I 459, 460
Фосфатидилэтаноламины I 459—460
Фосфатидная кислота I 459
Фосфоаргинин I 573
Фосфоенолпируват (ФЕП) I 437— 443
Фосфоенолпируваткарбоксикиназа I 434, 438—442, 488, 490, 492
Фосфолипиды I 42, 416, 459—460
Фосфор, круговорот I 312
Фосфорилирование I 221, 472—474, 512, 514
— окислительное I 478, 491, 547
Фосфорная кислота, в кишечнике на-
секомых I 294
Фосфофруктокиназа I 434, 519—524
Фотогенный орган III 592, 601—604
к Фотон II 475, 477
Фотопериодизм I 387; II 236; III 451
- -Фотопериодические реакции II 475, 477
Фотопические глаза II 519—521
Фотопротеин III 599
Фоторецепторы III 535
— пигменты II 487—503
— рабдомерные II 481, 487, 508—512
— ресничные II 480
— структура II 482—486
— электрофизиология II 503—618
Фоторецепция II 474—559
Фотосинтез I 242, 245—246, 248; II 475, 477
Фототаксис II 477—478, 480
Фототропизм I 242; II 477
Фруктоза I 295, 329, 497, 498
Р-Фруктозидазная активность I 300
Фруктозодифосфат (ФДФ) I 447, 501, 502, 505—507
Фруктозодифосфатаза (ФДФаза) I 492, 505, 507, 522—524
Фтор I 218
Фузимоторные волокна II 360, 362
Фумарат I 433, 439
Хамелеон, изменения окраски III533 Хеморецепторы I 371; II 432—474 — беспозвоночных II 432—433 — позвоночных II 448—460
— электрические реакции II 441— 447, 454—460
Хеморецепция, клеточные механизмы I 569—575
Хемотаксис II 433
Хемотрофные организмы I 242
Хиломикроны I 333
Химическая эволюция I 243—244
Химотрипсин I 302, 304, 590
Химотрипсиноген I 303, 590
Хинин II 461
Хиноны II 436
Хирономиды I 95—96
— акклимация II 109
— биологические ритмы II 233
— гемоглобин II 17, 53, 55—56
— действие кислорода I 413
— мембраны II 267
— полет III 237
Хитин I 100, 222, 316—317, 444,448— 450, 505—507
Хитиназа I 317, 450
Хитобиаза I 317, 450 сс
Хитон II 223
— гемоцианин II 62
— объем гемолимфы III 352
Хлор I 218
Хлорид (ионы С1_), выведение I 116, 215
— и деполяризация II 282
— и минералокортикоиды III 428
— и осморегуляция I 41, 92—93, 95, 131
— поглощение I 52—55, 93, 111— 116, 209
— реабсорбция I 81
— у насекомых I 96
— у ракообразных I 63, 64, 197,211
Хлоридные клетки I 121, 122
Хлорная проводимость II 296, 297, 359
Хлорокруорин II 6, 7, 21—23, 59
«Хлоролаб» II 499
Хлорофилл II 5, 480
Хлорофлагелляты I 246—247
Ходьба III 73
— у амфибий III 75
— у членистоногих III 45, 51
Холдена эффект II 32
Холевая кислота I 307, 309
П редметный указатель
637
Холестерин I 42, 263—265, 308—309; III 459
— в плавательном пузыре I 416
Холецистокинин I 325; III 411, 453— 454
Холин I 260—261; II 318, 319; III 381
Холинацетилаза II 323
Холинэргические рецепторы II 320, 321; III 356, 359
Хомячок (хомяк), замораживание
II 94
— зимняя спячка II 181—184
— терморегуляция II 134—135, 153, 181
Хордотональная сенсилла II 351
Хордотональный орган II 351, 370
Хроматофоротропные гормоны III 483, 546, 548—551
Хроматофоры III 518—538
— мышечные волокна в них III 250
Хромаффинные клетки II 289; III 385
Хромофор II 488
Хронон II 247-—248
Хрусталик II 537—541
Хрящевые рыбы, осморегуляция I 111—114, 124
Цветное зрение, пигменты II 497, 531
Цельноголовые I 577
Целлюлоза, в скелетах I 222 — переваривание I 310, 314—316 Центральная нервная система I 185;
III 5—140
-------зрительные функции II 524—526
—------и гормоны III 414—415
------- и рецепторы II 392, 480
—------обонятельные функции II
458
—------ракообразных III 547
------- толерантность к аноксии I 455______________________________457
— ямка II 537—539, 542
Церулоплазмин I 219
Цианид I 53
Цианкобаламин I 268
«Цианолаб» II 499
Цианопсин II 499
Цикл аргинин — мочевина I 554, 562—563, 565, 571—572, 575, 582 -------г- ферменты у разных животных I 577—580
— лимонной кислоты (Кребса; трикарбоновых кислот) I 272, 416, 431, 439, 467, 474—484, 554
— мочевины I 552—554, 561—563, 571—572
Цикл мочевой кислоты I 552, 554— 556, 569
Циклический АМФ (см. также Аде-нилатциклаза) I 513, 514; II 330; III 412, 421, 423, 556, 559
----- и зрение II 517
----- регуляция активности ферментов I 516—518, 520, 589
Цилиарные образования II 480—487
Цинк I 220
5а-ципринолсульфат I 309
Циркадные ритмы II 213—223, 230, 480
Циркуляция жидкостей в организме .111’346—402
Цирры III 329
Цистерны, в мышечных волокнах III 167
— в синапсах II 312
Цитохромы I 478; II 5
Цитрат I 467, 480—482, 493, 519
Цитратсинтаза I 439
Цитруллин I 554, 563, 567
Чайки I 138—139
Часы биологические II 212, 245—252
— навигационные II 238—240
Человек, биологические ритмы II 233 234
— вкус II 456, 459, 461
— водный обмен I 141, 143, 144; III 349
— высотная адаптация I 412—413
— гемоглобин II 13, 14, 17, 19—21, 57, 58
— гормоны III 441, 447
— дыхание I 371
— желчные соли I 307
— зрение (и глаз) II 417, 537—539, 544, 546
— кровообращение (и сердце) III 349, 382
— мозг III 65—69, 105
— ныряние I 410, 411; II 43—44
— пищеварение I 287, 289
— пищевые потребности I 220, 248, 252, 253, 259, 263, 268, 271, 272
— потребление кислорода I 382
— слух II 385
— спинномозговая жидкость I 185
— терморегуляция II 143, 159—163
— токсичность кислорода I 414
— укачивание II 417
— энергетический обмен I 470
Черви, кровяное давление III 368, 369
Черепахи I 131—135
— выделение азота I 582
— гемоглобин II 30
638
Предметный указатель
Черепахи гипоталамус III 362
— дыхание I 369
— зрительные пигменты II 500
— кровообращение III 359, 362, 363
— обоняние I 369
— сердце III 371
— устойчивость к аноксии I 369,404, 409, 454—457
Чешуекрылые см. Бабочки
Членистоногие, гемоцианин II 7, 24
— жидкости тела III 346
— локомоция III 45, 231
— люминесценция III 587
— нервная система II 325; III 43— 61, 124, 222—223
— рецепторы II 351, 365, 369, 518— 537, 558
— тропонин II 183
Чудесная сеть II 130—131, 158
— в плавательном пузыре I 415— 417, 453
Шиншилла I 149
Щелевое соединение мембран II 263—269
Щеточная каемка I 290, 301
Щитовидная железа III 415—420
— влияние на обмен веществ I 388
— и термогенез II 143
— у Fundulus I 155
Эвригалинность I 39, 67, 125
Эзерин (физостигмин) II 319, 322;
III 203 252
Экдизон ’ill 459—461, 468, 469, 475, 538
Экзопептидазы I 305—306
Эккринные железы II 160
Экскреция аммиака, внепочечная I 559
— воды I 60—61
— ионов I 196—201
— солей I 113—114, 134
— у насекомых I 102—ПО
Экстрафузальная иннервация II 363
Эксцентрические клетки II 518
Эктоплазма III 306, 311, 312
Эктотермия II 85
Эластаза I 303
Эластин I 303; III 355, 359
Эластолипопротеиназа I 303
Эластомуказа I 303
Электрический насос I 204; II 297
— орган III 287—302
— гликолиз I 457, 520, 521
— синтез ацетилхолина II 319
Электрогенез, типы II 296—297
Электрокардиограмма (ЭКГ) Щ 378—381
Электролиты, фильтрация в ночке I 144, 145
Электр о локация III 299
Электромагнитные поля и биологические ритмы II 223, 243—245
Электромагнитный спектр II 474
Электронная микроскопия мембран
II 263—269
-----порфиринов II 22—24
Электроольфактограмма (ЭОГ) И 454—455
Электроретинограмма II 515
Электрорецепторы III 292—297
Электротоническая связь II 262— .269. 305—311, 528—531; III 299
Электроциты III 289—292
Электрофорез (гемоглобинов) II 17—19
Электроэнцефалограмма (ЭЭГ) III 66—68
Элементы химические I 177, 218, 222
Эллипсоид (в палочках) II 484
Эмульгаторы I 308—309
Эндокохлеарный потенциал II 388
Эндокринные органы III 485—487
-----насекомых III 457—467
-----позвоночных III 415—441
-----ракообразных III 470—485
Эндолимфа II 388
Эндопаразиты I 317—318, 320; II 56—57
Эндопептидазы I 301
Эндоплазма III 306, 310
Эндотермия II 85
Энергия активации II 86—88
Энтерогастрон III 454
Энтерокиназа I 304—305
Энтерокринин I 326
Эпителиальные клетки целомического мешка I 81
Эпителий, пищеварительный I 290— 292
— транспорт ионов I 204—216
Эпифиз III 452
Эритрокруорин II 11
«Эритролаб» II 499
Эритропоэтин II 57—58
Эритрофоры III 523
Эритроциты I 40, 42, 43, 178—181, 203, 456, 564; II 6—74
— аномальные II 21
— ацетилхолинэстераза II 322
— влияние кислорода II 57—59
— замораживание II 92
— строение II 6
Эстеразы I 308
Эстрадиол III 430 : ,•
Предметный указатель
639
Эстрогены III 426, 430—432, 445—446
Эстрон III 430
Этиолюциферин III 596
Эукариоты I 587—588
Эупноэ I 369
Эхиуриды II 6; III 368
Эхолокация II 398, 401—408
Ювенильный гормон III 460, 468— 470, 538
Юкстагломерулярные клетки I 155; III 357
Юссинга уравнение I 32
«Яблочный» фермент I 493
Яд змеиный I 318
Ядерный магнитный резонанс II 89 Ядро (и амебоидное движение) III 315—317
Яичник, секреция гормонов III 430— 432, 445
Яйца, осморегуляция I 83—84, 583
Ямкоголовые змеи, терморецепторы II 133
Ящерицы I 131—135, 369, 407, 582; III 205, 371
— потребление кислорода I 388
— рецепторы II 361, 363, 379
— сердце II 112
— центральная нервная система III 104
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ И ТЕРМИНОВ
Термины
Amygdala III 104
Area centralis II 546
— tensor tympani II 385
— tenuiSsimus III 195
Bulbus arteriosus III 354
Conus arteriosus III 372
Corpora allata III 457, 458, 462—468
— cardiaca III 457—459, 465
— pedunculata III 457
Corpus cerebelli III 82
Dishabituation III 34
Eminentia mediana III 415, 441
Felderstructur III 167, 171, 215, 266
Fibrillenstructur III 167
Filia olfactoria II 452
Ganglium opticum (осьминога)’ III 37
Gap junctions II 263
Granulosa III 431
Nervus opticus (осьминога) III 37
— palatinus (осьминога) III 37
— rectus capitis (осьминога) III 37
— sympaticus III 37
Nexus II 263
Nuclei emboliformes III 82
— fastigiae III 82
Nucleus caudatus II 328
— globosus III 82 <
Pars distalis III 415, 434
— intercerebralis III 456, 463
— intermedia III 414, 434 *
— nervosa III 415
— tuberalis III 434
Putamen II 328
Rebound contraction III 261, 263
Habituation III 34
Lobus pedunculatus III 37
Macula occludens II 263
Medulla terminalis III 57
Musculi (mm.) abductores mandibu-lae I 363
— levatores hyomandibulae I 363, 364
Musculus (m.) adductor mandibulae I 364
-----operculi I 363, 364
— biceps brachii III 200
— dilator operculi I 363, 364
— expansor secundariorum II 329
— gastrocnemius III 216
— gracilis III 206
— hyohyoideus I 363, 364
— iliofubularis II 361
— protractor hyoideus I 363, 364
— soleus III 216
— sternohyoideus I 363, 364
Sacculus II 412, 420
Scala media II 386
— tympani II 386
— vestibuli II 386
Sinus venosus III 372
Stria vascularis II 386
Subfornix II 313
Taenia coli III 194, 195, 210, 213
Tapetum lucidum II 546
Theca lutea III 430
Tight junctions II 263
Torus semicircularis II 397, III 100
Tractus opticus (осьминога) III 37
Utriculus II 412, 415, 417, 420
Vas deferens II 329
Vulvuli cerebelli III 82
2оёа III 476
Zona occludens II 263, 264
641
Указатель латинских названий и терминов
Названия организмов
Acanthepyra III 587, 606
Acartia tonsa I 54
Acerina I 115; II 373
Achaearanea II 371
Achatina I 200 '
Acheta I 99 • -
Acholoe III 606 -1
Acillus II 129
Acipenser III 384 1 ‘ 4 •
Acmaea I 47
Acrida III 569
Acris II 114 A < •'
Acropora III 26, 601
Actinobolina III 513, 515 •
Actinoloba III 325, 332
Aedes I 95, 105 . '
Aedes aegypti I 96, 97, 206, 259, 261;
II 436
Aedes argenteus I 96
Aedes detritus I 96, 97
Aeolis III 512
Aequorea III 593, 599
Agrotis I 255; III 237
Aiptasia III 510 !
Alectrion III 337
Allogromia III 312
Allolobophora I 92 ' -
Alma I 356
— emini II 52 '
Amblyrhynchus I 135
Ambystoma I 389; III 107, 553
— opacum I 375
— tigrinum I 205 ’
Ameiurus II 393—395, 415; III 103, 107,
523, 558, 559, 561
Amoeba I 84; 478, 480
— dubia III 306
— Umax III 306
— mira I 86 ;
- r- proteus I 88, 180; III 306, 309, 313—
317 .
— verrucosa I 87; III 306, 309
Atnphibolurus I 134; II 140
Amphileptus I 87
Amphioxus I 286
Amphiprion II 433
Amphiuma II 44; III 371
Amplelorin II 436
Anagasta II 90
Anas I 139 1 .
Anax III 398 •
Ancylostoma I 89 i
Anemonia III 25, 510, 511
Anguilla I 121, 123, 214, 217, 367; И ‘
449; III 103, 524, 561, 562
— japonica II 13, 14 Л
— vulgaris I 115; II 449 •
Anisodoris II 284
Anodonta I 90, 91, 183; II 302, 332;
433; III 30, 364, 366, 388
Anolis III 451, 523, 527, 564—567, 571
— carolinensis III 416
A nomalops III 590, 601
Anopheles I 98; II 378; III 402
— bellator I 102
— homunculus I 102
Antheraea II 447
Anthonomus I 263, 274
Aphrodite III 13
Apis II 486, 527
Aplocheilus II 376
Aplodontia I 140
Aplysia I 47, 218; II 132, 135, 272, 281, 284, 286, 291—293, 297, 313, 324, 330, 333„ 409, 434; III 29, 129, 130, 387, 388, 487
— depilnas I 317
Apogon Ill 601
Aquipecten I 51
Arbacia I 40, 389; III 265, 312, 536
— pustulosa III 535
Archidoris I 218
Arctonoe II 433
Arenicola I 51, 286, 355; II 6, 11, 13, 17, 21, 30, 37, 53, 72; III 13, 265, 367, 368, 372, 400, 486
— ecaudata III 42
— marina I 51; III 42 -
Argyrotaenia I 255, 263
Ariolimax III 388
Arion I 367; III 264
Armadillidium II 137
Artemia I 38, 73—76, 214, 215, 279, 320, 387, 396, 397; II 6, 55, 59, 73; III 402
Ascaris I 89, 308, 413, 437—439, 441;
II, 7, 11, 13, 56, 57, 73; III 174, 247
— lumbricoides I 437
Aschelminthes II 246
Ascidia I 44
Asellus III 373, 480
Astacus I 60, 63, 78, 359; III 392, 473
Astasia I 248
Asterias I 44, 45, 190, 320, 387
— rubens I 45
Astroconger II 375
Astronesthes III 591
Astroscopus III 289, 290, 297—299
Aurelia I 223; III 24, 326
Bacterium megaterium II 121
Baiomys II 172 -
41—2067
642
4
Указатель латинских названий и терминов
Balaenoptera physalus I 410
Balanoglossus III 590
Balanus I 320, 359; II 90
— balanoides I 51; II 101, 110
— nubilus I 295
Bathygobius II 449; III 563
Bdelloura I 193, 320
Belostoma III 399
Bipalium kewense I 570, 571
Birgus I 70
Blaberus III 235
Blarina II 401
Blastophagus II 437
Blatta I 101; II 137, 138
Blattella I 253, 255, 264, 269; II 103
— germanica III 467
Blennius I 118
Blepharisma III 329
Bolitophila III 588
Bombinator III 553
Botnbyx I 258; II 435, 436, 444, 463;
III 463
— mori I 450
Bothrops III 448
Bracon II 90
— cephi I 450
Branchiostoma II 6, 281; III 171, 174, 176, 205, 214
Brioboa praetiosa I 101
Buccinium II 434; III 30 :.
Bufo I 132; II 101; III 349
— boreas I 127, 128 •“
— bufo I 123, 128, 129; III 422
.— cognatus I 126
— marginus I 123, 126, 131, 155; 206; III 75
— punctatus I 126
— v allice ps II 13
.— viridis I 127—129
Busycon II 8, 24, 60—62; III 251
Caenorhabditis I 264, 414
Caiman I 131, 369
Calamactis HI 25
Calanus I 322
Calliactis III 24, 27
— parasitica II 299; III 512
Callianassa II 24
— californiensis I 58
Callinectes I 59, 64, 65, 72, 357, 387;
II 114, 134; III 475, 476, 484, 527, 543
Callionymus 1 403
Calliphora I 102, 103, 107, 255, 294, 300, 327, 387; II 88. 110, 324, 370, 378, 464, 508, 531; III 466, 467
Calypte II 171
Calytocephalella I 205 , .
Cambarellus III 482, 542, 551
Cambarus III 473, 475, 484, 548, 569
Cancer I 71, 309; II 24, 63, 434; III 228
— antennarius I 51
— magister II 62
Caprimulgus II 172
Carabus I 99
Carassius I 205 (см. также Серебряный карась)
Carausius I 103, 123; II 114; III 43, 51, 234, 464, 466, 522, 527—533, 540, 541
— morosus II 302; III 539
Carcharhinus I 113
Carcinides II 434
Carcinus I 58, 60—66, 81, 93, 124, 191, 193, 199, 299, 357, 359, 387, 404; II 335, 364; III 365, 397, 472
— maenas I 299, 543, 551
Cardisoma I 69, 70, 72, 211; II 62
— guanhumi I 551
Cardita II 54
Cardium I 356
Caretta I 132
Caudina I 193; II 6
Celerio II 376; III 54, 352
Centrostephanus longispinus III 535
Cephas II 90
Ceratoplanus III 588
Chaenocephalus II 47
Chaetopterus I 320, 321; III 13, 586— 588, 601, 602
Chamaeleo III 568
Chaoborus III 539
Chaous carolinensis III 306, 311
Chelonia III 363
Chelydra I 306 ’
Chilomonas 1 246, 248, 267, 270, 271
Chilonycteris II 403
Chimaera I 327
Chironomus II 11, 55, 56, 59, 109, 265, 267; III 237
— salinarius I 96
— thummi I 96; II 17, 56
Chiton III 388
Chlamydomonas I 246, 261
Chlorohydra viridissima I 89, 90
Chrysopa II 378
Chrysops I 300
Ciona I 221; II 324; III 391, 392
Citellus II 155, 181; III 441
— tereticaudus II 155
Clarias I 375
Clinocardium III 388
Clupea II 393, 395
— harengus II 18
Clymenella I 286, 408
Cnasterodon III 442
Указатель латинских названий и терминов
643> ”
Coenobit а I 71
Coleonyx II 396
Collembola I 95
Collocalia II 389
Collotheca II 433
Colpoda I 270, 271; III 330,335,
Columba I 407
Compus niveus II 128
Conolophus I 135
Convoluta I 286; II 228
— roscoffcensis I 318
; Corcyra I 263
I Cordylophora III 24
! Corethra III 539
Corixa I 100, 360
; — dentipes I 95
; Corophium II 368
Corynactis III 508
Cottus III 214
— bubalis I 119, 120
— morio I 119, 120
— scorpius I 118, 120
Coturnix II 143
Crago cm. Crangon
Crangon I 57, 72, 219; III 475, 523, 529 -531, 542—544, 546—550
Craspedacusta I 90
Crassostrea I 47, 51, 223, 322, 387; III
180, 248
Crepidula I 321
Crithidia I 250, 255, 261, 263
Crotalus I 577; III 488
Crocodylus I 133
Cryptochiton I 330; II 8, 62
Cryptomphallus II 291, 293
Cryptotympana III 398
Ctenolepisma I 315
Ctenosaura I 134, 577, 582
Cucumaria I 356; II 6, 34
Culex I 95
— pipiens I 96
Cyanea III 24
Cycloporus I 286
Cypridina III 584, 587, 593, 597, 599» 601, 602
Cyprinodon II 102 ’
Cystophora I 410
Cyzicus II 17
Dactylometra III 510
Danaus II 128
Daphnia I 322; II 6, 98; HI 373, 397,
480
Dardanus III 27
Dasypus II 169
Dendrostomum I 45; П
Dermestes I 262, 26-4 .
Diadema I 387; II 478; III 29, 533, 535
Diadumene II 114; III 509
Dictyota II 233
Didelphis II 169
Didinium III 515
Difflugia III 309
Dileptus III 513, 515 /
Diodor a II 62 4
Diphyllobothrium I 404
Dipodomys I 141, 143; III 349 '
Dipsosaurus I 134; II 140
Discophrya I 88
Dobsonia II 170
Dolabella III 388, 389
Doris I 47
Doroceras II 114
Dreissena III 487
Dreissensia I 387
Drosophila I 95, 99, 101, 102, 260-^ 263, 269, 272, 389, 413; II 437, 480; III 297, 458, 469
— melanogaster I 259, 262, 269, 542
— persimUis I 102
— pseudoobscura I 102
Dugesia I 193; III 296
Dysdercus II 438
Dytiscus I 327, 374; II 524; III 398
Echinococcus I 439
Echinus III 251
Eigenmannia III 291, 294, 296, 297»
300
Eisenia I 92; III 107, 487
— submontana HI 586
Electrophorus I 375, 457, 520; II 320;
III 287, 290, 297, 299
Eledone I 183; III 389, 537
Emplectonema kandai III 586
Ensis III 30, 366
Ephestia I 99, 263; III 483
Ephydatia I 321
Ephydra cinerea I 98 ’
— riparia I 98
Epilachna II 437
Eptatretus II 30
Eptesicus I 354; II 404
— fuscus I 548; II 170
Ericymba III 528, 531, 569, 570
Erinaceus II 182; III 101
Eriocheir I 43, 57, 63, 65, 66, 193, 194_
196, 387; III 473, 543
• — sinensis I 82
Eriphia 1 387
Escherichia I 394
— coli I 466, 471, 504, 586; II 99, 124»
432, 462
Esox II 415
41*
644.
Estigmene I 274 « , •
Etroplus I 396 i.
Eucioraris I 397 : ; ;:.--
Euglena I 525; III 427 »
- — gracilis I 267 1 : I.
- — rubra III 522 t i.
Eumeces I 387 n.j
Eumops II 171
Eunice II 238
Eupagurus III 231
Euplotes I 248; II 262; III 329, 337
Eurypelma III 398
Euscorpius II 365
Fasciola I 404
• — hepatica I 286; III 489
Eorcips III 237
Formica rufa II 129; III 119
Front onia III 513
Fundulus I 83, 111, 117, 118, 121, 122, 155, 156, 195, 213, 214, 216, 415; II 94, 267, 374; III 168, 385, 419, 442, 523, 557—559, 561—563, 572
— heteroclitus I 120, 121, 154; II 120;
III 440
— kansae I 121 ; •. .
Fungia I 320
Fusitriton II 62
Galleria I 263; II 103; III 398, 399 — mellonella I 575 •
Gambusia II 557
Gammanus I 65, 67, 68, 219, 384; III
480
— duebeni I 57, 67, 68, 72, 193, 387
- — lacustris Г 67
— locusta I 67, 68, 81
— oceanicus I 385; II 112
— pulex I 67, 68, 81, 193 н
— tigrinus I 68
— zaddachi I 68
Gasteropelecus II 309, 310; III 77, 78
Gasterosteus I 154; 420, 443
Gastrimargus II 137 .
Gastrophilus II 7, 13, 16, 47, 71
Gecarcinus I 70—72, 199; II 64; Ш
473, 484
Gehydra I 131
Gillichthys I 374, III 563
Girella II 126
Glaucoma' I 272
Glis II 187
Glossina II 139 ,
Glossiphonia complanata III 534 . \
Glossoscolex II 53; III 364, 400
Glycera II II, 17, 54; III 13, 266,, 368
Gnathocerus I 262
Указатель латинских названий и терминов -—-----------------nWHWiin i . nf.il ...
Gnathophausia I 402
Gobiesox II 45 "
Gobio II 449
Gobius I 213; III 558
Golfingia I 45, 46, 193; II 7, 26, 64;
III 174, 190, 243, 246, 251—253,
369
Gonyaulax II 216; III 585, 592, 605
Gopherus I 132, 133; II 30
Grus grus I 289
Gryllus II 378
Gryphaea III 389
Gunda I 46
Gymnarchus III 291, 297
Gymnotus III 291, 293, 295, 296
Habrobracon I 413; III 538
H alicarcinus australis I 59 '
Haliotis I 200, 411; III 388
Halistaurea III 599
Halmiscus I 77
Harmothoe II 356; III 15, 42
Hartmannella III 314
Heliothis I 274; II 437
— zea I 258
Heliozoa I 221
Helix I 219, 221, 265, 287, 300, 367;
II 23, 62, 70, 75, 281, 291, 293, 331, 333, 335; III 29, 30, 36, 190, 192, 251, 388—390, 487
— aspersa I 571; II 25
— pomatia II 22, 25, 60
Hemiclepsis marginata III 534
Hemidactylus III 564
Hemigrapsus I 64, 70, 71, 199, 385, 387, 398; II 113; III 543
— nudus I 59, 61, 63
— oregonensis I 59; II 101 “
Hemisqilla III 222, 223, 227
Heterocephalus I 394, 405
Heterodontus III 385
Heterometrus II 64
Heteroteuthis III 590, 602, 606
Heteroxenia III 26
Hippocampus III 168
Hippolyte III 518, 527, 529, 542, 570
Hirudo I 94, 200, 324; II 6, 511; III 201, 487
Holocentrus II 393; III 214, 523, 558, 562, 563
Holothuria I 356
Homarus I 51, 63, 191, 192, 359; II 72, , 113; III 392, 395, 548
— vulgaris I 543, 551
Hoplias III 557
Hyalophora I 373; II 129; 111'400 — cecropia-I <211; 373; 382, 444, ’449, 450; III 399, 460, 463—465 ik >
Указатель латинских названий и терминов
645
Hyas I 51, 191
Hydra I 90, 158, 323; II 268
.— viridis I 89
Hydrobia I 47
Hydrobius I 100
Hydrolagus I 153, 185
Hyla III 527, 533, 556
Hylemya I 251
— antique I 259
Hymenoleptis I 89, 251, 255, 271, 334
Hyperia galba III 527
Hypopomus III 291, 300
Ichthyomyzon II 30
Ictalurus II 132, 374, 449, 454, III 563
Idothea I 76; III 527, 532
Idus II 109
Iguana I 131, 384, 385; II 139, 140
Ips cetnbrae I 247
Ixodes I 101
Jasus I 66; III 475
Kakabekia umbellata I 545
Lacerta I 132, 582; III 451
Lacistrophychus I 247
Laeonereis culvert I 52
Lagodon II 47
Lagopus II 164; III 455
Lampetra I 155, 201; II 30; III 531,
533, 560
— fluviatilis I 112; II 18
— planeri I 112; II 45
Lampyris noctiluca III 466
Larus I 139
Latia III 600, 601
Laticauda I 134
Latimeria 1 111, 192, 577
Leander III 471, 476—478, 480, 482,
483, 529, 542, 545, 570
Lebistes II 102; III 523, 557, 559
Leiognathus III 590, 601
Leishmania I 262, 267, 274
Lepidobactracus I 126 > ‘
Lepidochelys I 135
Lepidosiren I 375, 580
Lepomis II 111
Leptodactylus I 208
Lernaeodiscus ingolfi III 478
Leuciscus II 133
Leucophaea III 464, 466, 467
Libinia I 59, 64, 387; II 72, 112; III
484
Ligia I 77, 219; II 137; III 392
— oceanica I 76; III 552
Limax I 367; II 126, 333; III 388
Limnodrilus I 355
Limnophilus I 97, 98, 100
I i m tirin'li I 994
Limulus I 51, 359, 404; II 8, 22, 24, 62, 64, 70, 72, 73, 284, 333, 364, 514, 518^ 527, 528, 532; III 169, 232, 371, 392— 394, 396—398, 484, 489
Lingula II 7, 64
Liomys I 140, 141
Lithobius II 132
Littorina I 47, 320; II 90, 98, 102, 110
— neritoides II 98
Locusta I 99, 100, 102, 107; II 367, 370, 378, 445, 446; III 235, 237
— migratoria II 132; III 539
Loligo I 183; II 24, 60, 61, 70; III 108*
180, 250, 520, 537, 589
Lophius 1 117, 214
Lophortyx II 164
Lophosaura III 568
Loxostege I 263
Lucilia II 138, 445, 446; III 466, 467
— cuprina I 551
Luciopimelodus III 442
Lumbricus см. Дождевой червь
— terrestis I 543, 570, 572
Lygosoma I 397
Lymnaea I 91, 215; II 54; III 389, 390,.
487
Lysmata III 471, 480
Macaca I 146
Macrobrachium III 527
Macroderma II 170
Macropipus I 60, 61
Macropodus III 557, 562, 572
Macropus II 169
Maemalon I 332
Magelona II 6
Magicicada II 382
Maja I 51, 59, 65, 191, 219, 387; II 70;
III 472, 483
Malaclemys centrata I 132, 134
Malacocephalus III 591
Malacosoma II 90, 440
Malapterurus II 308; III 287, 291, 298
Manduca II 129
Marinogammarus finmarchicus I 68
Marmosa II 172
Marmota III 441
Marphysa II 13
Megaderma II 403
Meimura III 238
Melanophila II 132
Melanoplus II 111; III 398, 464, 46T
646
Указатель латинских названий и терминов
fMpniппр I 72
Mercenaria I 356, 387, 397; II 54, 332;
III 265, 388, 389
Mesidotea I 76
Mesocricetus II 181
Metachirus II 169
Metapenaeus I 387
Metridium II 114; III 332, 333, 510, 512
— marginatum III 325
Miamiensis I 87 j
Microciona I 320 ’ I
.Microdipodops II 172 1
Micropagon III 442 |
Microstomum III 512 ;г
Microtus I 405; II 149; III 451 •?
-— montanus I 394
Mirounga III 362
Mnemiopsis III 586, 602
Modiolus I 47, 48, 51, 387; II 99; III
389 /
— modiolus I 544 /
Moina II 25 I
Molgula I 44, 221, 320, 574 /
— manhattensis I 570 ' /
Monas III 331 |
Monema flavescens I 450 Д
Monocentris III 590 / \
Mormyrus I 288 /
Mugil I 288
Munida sarsi III 478 /
Mus II 457
Musca I 264, 294, 387; II 103, 325, 486
Mustelus III 557, 559, 560
Муа I 49; III 342, 367, 388.
Myotis I 548; II 170, 171, 187, 401, 403—405
Mytilus см. Мидия /
— edulis I 387 /
Myxicola II 299; III 13 '
Myxine I 111, 184, 201, II 46
Л
/
Nannostomus beckfordi anomalus Д1 561 /
Nanomia III 24 /
Nassarius II 434; III 325 /
Matrix I 133; II 29, 139 /
Mavanax II 307; Ш 36 ' I
Neanthes III 13 /
— lighti I 52 y'
— virens I 52 /
Mecrophorus II 464
Mectophrynoides III 448 /
Necturus I 93, 129, 130,/185, 209; II
44, 303, 374, 503, 550, 552; III 418
Megaprion I 113
Nemachilus III 103 . i
Nematodinium III 515
Nematodirus II 13
Neoceratodus I 375, 580, 581; II 45
Neoconocephalus II 129; III 235
Neotoma I 313
Nephrops I 184, 191
Nephthys II 54, 65, 368; III 43, 329, 335
Nereis I 52—56, 124; II 30; III 15, 42, .58, 118, 368
— diversicolor I 52—56, 191, 216, 218;
II 53
— dumerilii III 534
— limnicola I 52—54
— pelagica I 51, 52
— succinea I 52, 54
— vexillosa I 52
Nerita II 102
Nictimene albiventer I 406
Nippostrongylus I 89; II 7, 56
Noctiluca III 515, 585
Notonecta I 360, 374; II 370, 527
Obelia III 253, 586 !
Octopus см. Осьминог
— dofleini I 551
— macropus III 537
— vulgaris III 537
Ocypode I 65, 71, 199, 359, 387; II
114, 371; III 531
Odontosyllis III 607
Oedipoda III 569
Onchidiutn I 47
Oncopeltus I 251, 389; II 101
Oncorhynchus I 116, 214, 397
— kisutch I 115, 194
Oniscus asellus I 543, 570
Onychotnys toridus I 292
Opalina II 262; III 335—337
Ophiocephalus III 87, 214, 559
Opsanus I 117, 217; II 115, 118
— tau I 117, 201
Orchestia III 479
Orconectes I 78; II 368; III 473, 475,
484, 485, 542, 569
Ornithodorus I 324
Orocharis III 235
Oryzaephilus I 255, 277
Ostrea I 183, 220, 322, 356; III 332,
342
Otala I 571, 574
Pachygrapsus I 57, 61, 64, 66, 70, 71, 191, 193, 194, 197, 199, 387, 396; II 109; III 227, 232
— crassipes I 70
Pacificastacus I 78, 79
Указатель латинских названий и терминов
647
Pagurus I 51; III 27, 549
Palaemon I 73 (см. также Leander)
Palaemonetes I 57, 60, 65, 387; III 473, 481, 482, 484, 521, 526—530, 542— 546, 548, 550, 552
— varians I 72, 73
Palatnis I 135
Palinurus I 51
— vulgaris I 543
Pandalus I 359; III 483
Panulirus III 223, 392
Papilio II 437 ? '
Paragordius III 176, 246 -*
Paralichthys albiguttus III 529
Paralithodes I 51; III 229
Paramecium см. Парамеция
— aurelia I 250, 255; II II
— multimicronucleatum I 261
Parapriacanthus III 601
Parasilurus III 558
Patagona II 173
Patella I 219
Pecten I 220, 356; II 434, 479, 481, 513;
III 173, 180, 192, 217, 240, 248, 341
Pectinophora II 435
Pediculus II 138
Pelagia III 586, 605
Pelmatohydra oligatis I 89
Pelomyxa I 38, 394
— carolinensis I 85, 88, 180
— palustris III 306
Penaeus I 66; III 542
— setiferus I 295; III 544
Penicillium notatum I 545
Pennaria III 513
Peranema III 339
Perea I 115; III 557
Perillus III 538
Perinereis cultrifera I 52, 53
Periophthalamus I 408
Peripat us III 45
Periplaneta I 99, 101, 102, 107, 108, 315, 372; II 103, 132, 137, 324, 365, 435, 531; HI 19, 234, 235, 399
— americana I 449, 575
Perisoreus II 155
Perognathus I 141; II 162, 172
Peromyscus II 155, 457; III 384, 451
— crinitus I 144, 394, 405
— maniculatus II 155
Petrochirus II 371
Petromyson I 111; 13, 30, 46, 310, 449;
III 76, 128, 442
— marinus II 14, 18, 45
Phacoides II 54
Phalaenoptilus I 405; II 164, 171, 172
Phengodes III 589, 590, 601
Pheretima I 92; II 98
— posthuma I 92
Phialidium III 24
Phoca I 141
— vitulina I 150, 410
Phocanema III 486
Pholas III 589, 601, 602
Pholis I 124
— gunnelus I 119, 120
Phormia I 102, 255, 258, 262, 263, 267,. 324, 373, 389; II 365, 440, 442; III 218
Photinus III 605, 606, 607
Photoblepharon III 585, 590, 601
Phoxinus II 136, 369, 395, 449; III
531, 533, 559, 563
Phrixothrix III 589, 591
Phrynosoma I 133; II 139; III 441, 527, 533, 564, 566, 567, 571
Phyllomedusa sauvagii I 582
Physa III 337
Physalia III 510, 512
Physarum polycephalum III 309, 310, 313, 316, 317
Phy set er macrocephalus I 410
Physiculus III 590
Pieris II 437, 528; III 538
Pimelodus III 442
Pis cicola geometra III 534
Pista III 13
Pituphis I 133
Pizonyx I 142
Placobdella parasitica III 534, 535
Planes III 529
Planocera III 29
Planorbis I 367; II 7, 54, 59
Plasmodium I 404
Platichthys I 121, 123, 210, 213, 216, 217
Platynereis II 233
Platy pleura III 237
Platypus II 169
Pleurobrachia III 325, 326
Pleuronectes I 123
Podarke II 433
Podophthalmus II 535; III 223
Polistotrema I 111; II 34
Polycelis III 486
Polukrikos III 513, 515
Polytnorphus minutus III 480
Polypterus I 153
Polythoa III 26 \
Polytoma caeca I 246
Poneroplax III 264
Porcellio I 219
— scaber I 570
Porichthys II 395; III 591, 592,.605 \
Porites III 26
Porthetria II 435
Portunus I 51, 60, 191
— ordwayi III 529 \ \ \ \ \
648 .'Указатель латинских названий и.термин^
Potamilla II 7, 59
Potamon I 63, 71, 387
Potamotrygon I 113, 580
Procambarus I 82; II 478, 486, 536;
III 473, 475
Procerodes ulvae I 46, 47
Process a III 542
Prochilodus III 442
Prodenia II 378, 435
Prostoma II 57
Protoclepsis tessellata III 534
Protopar се II 437, 441
Protopterus I 153, 154, 368, 580, 584
Psammomys I 148 1
Pseudacris sequita I 125
Pseudemys 1 131—133, 409; II 13, 141,
500, 503
Pseudopleuronectes I 214; II 94
Pseudotrichonympha III 228
Pterophyllum III 558
Pteroptyx malaccae III 589 /
Pterostichus II 90
Ptychodera III 605 r. /
Pyrocystis III 585 ( ,/ i?
Pyrodinium III 585 7.
Pyromelanox III 455 (
Pyrophorus III 589, 590 . /'\
Pyrosoma III 590 /
Quelea III 452
Raja I 153; II 392, 415; III 295, 560
— clavata II 412, 416
— eglanteria I 113
— erinacea I 113
Rana I 123, 132, 133, 389; III 107, 363, 527, 553
— cancrivora I 127—129, 584
— catesbeiana (catasbiana) I 130, 329, 562, 585; II 18, 28; III 349
— clamitans I 125; III 418, 556
— esculenta I 123, 195; II 16
— pipiens I 125; II 396, 450; III 376, 419, 553, 556
— septentrionalis I 125 у ь
Rangia I 49
Rapana III 387
Renilla III 25, 598, 602, 605
Rhinolophus II 402—404
Rhithropanopeus I 59, 60, 124; III 476, 531
Rhodnius I 102, 105, 156, 375, 389, 449;
II 127, 132; III 457, 458, 461—463, 466, 467
— prolixus III 461 . s
Rousettus II 407 . . i ... ,. .
Sabellaria III 330 i:
Saccobranchus I 375
Salmo I 83, 201, 217; II 103, 118; 1И
420, 557
— clarki I 398
— far io III 442
— gairdneri I 116, 205, 361, 365; II 46, 112; III 363, 442
— salar II 454
Salmonella I 394
Salvelinus I 403; II 110
— fontinalis I 401; II 45, 98, 118
Sarcophaga III 352, 398, 466, 467
Sauromalus I 131, 134, 354; II 112, 140
Scaphiopus hammondi I 125
Sceloporus I 134
— occidentalis III 417
Schistocephalus II 110
Schistocerca I 102, 105; II 128, 137, 235, 382, 527; III 56, 191
— gregaria I 449; II 138
Schistosoma mansoni I 318
Scorpaenichthys II 45, 52
Scyliorhinus I 113; III 107, 295
— canicula III 422
Scyllium III 384, 557, 560
Sebastodes II 29, 45
Selasphorus I 383
Sepia I 183, 264, 417; II 72, 330; III 39, 529
Sergestes III 587, 588
Ser pula II 59
Serranus I 117, 121, 123, 210, 213, 217
Sesarma I 69, 70, 72; II 231, 232; III 476, 543
Sialis I 83, 97
Sicyases III 363 . •
Sigur a distincta I 95 r ’ -
— lugubris I 95
Silpha obscura II 128
Siphonosoma ingers II 65
Sipunculus II 7, 26, 64; III 201, 369
Sorex II 149
Sphaeroma III 532
Sphenodon I 528; II 140
Sphenomorphus I 131
Sphinx I 373; II 527, 536, 537; III 399
Spirographis II 23, 30
Spirorbis II 7, 23
Spirostomum I 85
Spisula II 330, 514; III 30, 251, 265
Spongilla I 89, 321
Squalus I 114, 195, 201, 214; II 46, 51;
III 559
— suckleyi III 416
Squilla I 359; II 356; III 392, 393, 395—397, 552
Указатель латинских названий и терминов
*49
Steatornis I 398 ?
Stenella III 66 ' ... .
Stenostomum III 332, 337
Stentor III 164
Sternarchella III 297
Sternanchus III 291 I
Sternopygus III 291
Sternothaerus I 408
Stichopus I 332
Stomias III 591
Stomoxys I 294
Stomphia III 26, 27
Streptococcus cremoris II 118
Strigomonas I 267, 274
Strombus I 200
Strongylocentrotus I 182
Strophocheilus I 183
— oblong us I 571
Stychopus I 330, 332
Styela I 221
Stylonichia I 248
Symbranchus I 375
Sy near is I 73
Synsepalum II 456
Toxotes III 103
Trachelipus III 552
Trachysaurus I 132; II 397
Trematomus I 124; II 47, 93, 118, 1201
Triangulus boschmai III 478
— munidae III 478
Triatoma I 262
Tribolium I 101, 251
Trichinella I 404
Trichomonas I 413
— columbae I 264
Tricosurus III 81
Trionyx I 131, 132
Triops II 55
Triton cristatus III 443
Tritonia III 29, 33, 36
Triturus I 154; II 16, 18; III 437
Tropidurus I 133
Trypanosoma I 262, 267, 274; III 33>
— cruzi I 404; II 323
Tubifex I 355, 413; II 6, 52
Tubipora III 26
Tubularia III 25
Tursiops I 141; II 401
— gilli I 470
Ту to II 398
Tachyglossus III 101
Tadarida brasiliensis I 5.48
Tapes II 332
Tautoga II 45
Tealia III 25
Telphusa fluviatilis I 78
Tenebrio I 100, 101, 110, 253, 263, 267, 269, 327, 389; II 103, 138, 225, 235, 296, 330
— confusum I 102, 263
— molitor I 109, 263
Terebellidae I 355
Teredo I 315; III 366
Testudo I 132
Tetrahymena I 180, 220, 221, 247, 248, 250, 253, 261, 263, 264, 269—273, 278, 401; II 7, 89, 323
__ J 255
— Thamnophis II 51, 105, 139; III, 168, 212
— elegans I 582
Themiste I 46
Therapon II 395
Thermobia I 101
Thyone II 16; HI 251, 265, 366
Tilapia I 120, 217; III 124
Tiliqua II 136, 137, 361, 363
Tinea II 415; III 263, 384
— tinea I 367
Tomopteris III 326, 586
Torpedo II 314, 320; III 287, 289, 290, 299
Uca I 57, 65, 70—72, 197, 304, 387; II 112, 114, 220, 228—232, 235, 371,. 408; III 473, 477, 483, 527, 532, 543— 546, 549—551, 570, 571
Umbra III 420
Unio III 240
Unionicola II 433
Upogebia I 58; III 478
Uranoscopus I 121, 213, 217
Urechis II 6, 37, 70; III 369
Urodacus III 398
Uromastrix I 132
Uronychia III 337
Uta I 131
Vamp у rum II 403
Vanessa III 538
Var anus I 134, 195
Varuna I 69
Venus III 240
Vespa orientalis I 304
Viviparus I 200
Vorticella III 164
Watsenia III 580
Xantusia III 451
650 Указатель латинских названий и терминов
Xenopsylla I 101, 323; II 138 Xiphophorus I 154; II 126; III 557 <<
Xenopus I 42, 123, 131, 369, 560, 584, Xyleborus I 264
585; II 268, 375; III 91, 107, 110,
112, 527, 531, 553—556 -
— laevis I, 127, 551, 559 Zapus II 155
Xiphister I 117—119 Zootoca III 448
ОГЛАВЛЕНИЕ
Глава 15. Центральная нервная система. Л. Проссер. (Перевод
Н. Ю. Алексеенко.)............................. 5
Методы исследования............................................. 5
Общие свойства нервной интеграции........................ 6
Системы гигантских волокон........................... 12
Нервные сети.............................................23
Функции ганглиев у моллюсков.............................29
Физиология ганглионарных нервных систем лестничного типа 40
Основные черты морфологии нервной системы позвоночных 61
Вызванные потенциалы и ритмическая, электрическая активность головного мозга...............................66
Двигательные системы позвоночных..............................71
Мозжечок.................................................61
Центральные сенсорные пути у позвоночных.................89
Центры эмоций в переднем мозге, участвующие в самосохранении и воспроизведении............................104
Сон...........................................................106
Развитие нервных функций................................106
Стойкие изменения функции центральной нервной системы 115
Заключение..............................................136
Список литературы........................................140
Глава 16. Мышцы. Л. Проссер. (Перевод И. А. Ломова.) . . 164
Гистологические типы мышц...............................165
Сократительные белки и теории сокращения................176
Механические свойства, «активное состояние» и теплопродук- 188
ция.......................................................
Нервная активация быстрых скелетных мышц позвоночных 201
Скорость сокращения мышц................................205
Множественная иннервация мышц у членистоногих, особенно
у ракообразных......................................222
Неосциллирующие мышцы насекомых.........................233
Резонирующие летательные тимпанальные мышцы насекомых 236
Запирательные мышцы и задержанное расслабление . . . 240
Мышцы с диагональной исчерченностью.....................247
"Соматические мышцы с неиспорченными короткими волокнами 250
Гладкие мышцы позвоночных...............................254
Висцеральные мышцы беспозвоночных.......................264
Заключение..............................................266
Список литературы ............................................. 270
Глава 17. Электрические органы и электрорецепторы. Л. Прос-
сер. (Перевод Ю. Г. Каминского и И. А. Ломова.) 287
Оглавле&¥&
Генерация электрических разрядов..........................287
Электрорецепция ......................................... 292
Поведенческие реакции и нервная регуляция.................297
Помехозащита..............................................300
Заключение................................................301
Список литературы.........................................302
Глава 18. Амебоидное движение. Дж. Андерсон. (Перевод Ю. Г. Каминского и И. А. Ломова.).........................305
Типы амебоидных клеток и псевдоподий.....................305
Общая картина амебоидного движения.......................308
Теории амебоидного движения..............................310
Сократительные белки.....................................313
фибриллы, микрофиламенты и микротрубочки.................314
Роль ядра................................................315
Роль мембраны............................................316
Список литературы........................................318
• Глава 19. Реснички. Ф. Браун. (Перевод М. 3. Ароновой.) . . 324
Строение ресничек ...................................... 327
Характеристика движения ресничек.........................329
Регуляция и координация активности ресничек..............333
Теории движения ресничек.................................338
Энергия движения ресничек................................341
Влияние факторов окружающей среды на активность ресничек 342
Список литературы........................................343
Глава 20. Циркуляция жидкостей в организме. Л. Проссер. (Перевод Д. Л. Длигач.)...................................346
Типы циркуляторных систем.................................346
Жидкостные отсеки.........................................348
Давление и ток жидкости в циркуляторных системах позвоночных ...............................................353
Периферическая циркуляция у беспозвоночных................364
Типы сердец...............................................369
Ритм сердечной деятельности и минутный объем .... 373
Системные сердца позвоночных..............................378
Сердца моллюсков.........................................387
Сердца оболочников ..................................... 390
Сердца ракообразных и Limulus . 392
Сердца пауков, скорпионов и насекомых....................398
Сердца аннелид ......................................... 400
Заключение................................................401
Список литературы.........................................402
Глава 21. Эндокринные механизмы. Р. Новалес, Л. Гилберт, Ф. Браун. (Перевод Л. Г. Лейбсона и Э. М. Плисецкой.) ............................411
Позвоночные.............................................415*
Насекомые................................................456
Ракообразные........................................... 470
Эндокринные факторы у других животных....................485
Заключение............................................. 487
Список литературы . . . . ...... . . . .... . 490*
Оглавление 663
Глава 22. Нематоцисты и их аналоги. Ф. Браун. (Перевод М. 3. Ароновой.).........................................508
Нематоцисты..............................................508
Аналоги нематоцистов ................................... 513
Список литературы .......................................515
Глава 23. Хроматофоры и изменение окраски. Ф. А. Браун младший. (Перевод И. С. Гольдина.) .... 518
Введение.............................................518
Строение и способ действия хроматофоров..............519
Изменения окраски у животных.........................527
Функциональная организация системы хроматофоров . . . 534
Функциональное значение хроматофоров и изменения окраски 568
Заключение...........................................571
Список литературы ...................................... 572
Глава 24. Биолюминесценция. Ф. Браун. (Перевод Д. Н. Островского.) ..............................................584
Список литературы ...................................... 608
Предметный указатель.................................612
Указатель латинских названий и терминов..............64G
УВАЖАЕМЫЙ ЧИТАТЕЛЬ!
Ваши замечания о содержании книги, ее оформлении,, качестве перевода и другие просим присылать по адресу: 129820, Москва, И-110, ГСП, 1-й Рижский пер., д. 2». издательство «Мир»
СРАВНИТЕЛЬНАЯ ФИЗИОЛОГИЯ животных -7
(Том 3)
Редактор Е. А. Яновская Художник В. Медников ‘ Технический редактор Н. Д. Толстякова Корректор Н. В. Спичкина
Сдано в набор 24.08.77.
Подписано к печ. 21.12.77.
Бумага кн.-журн.: 60Х90'/1б=20,50 бум. л.
41 печ. л. Уч.-изд. л. 51,66. Изд. № 4/8826 Зак. 2067. Цена 3 р. 80 к.
ИЗДАТЕЛЬСТВО «МИР» Москва, 1-й Рижский пер., 2
Московская типография № И Союзполиграфпрома при Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли.
Москва, 113105, Нагатинская ул., д. 1.
Опечатки
Том Страница Строка Напечатано Следует читать
I 30 10 св. де Врие Де Фриз
I 220 10 св. Ostopus Octopus
I 315 22 св. иглокожих эхиурид
II II 335 360 10/1I св. подпись к рис. 12—5 7 св. моуральная m. ibiofubularis моноуральная т. iliofibularis
II 369 6 св. Закрытопузырные Открытопузырные
III 217 7 св. гликолитически гликолитических
HI 372 14 св. сокращающейся сокращающийся