/





























































































































































































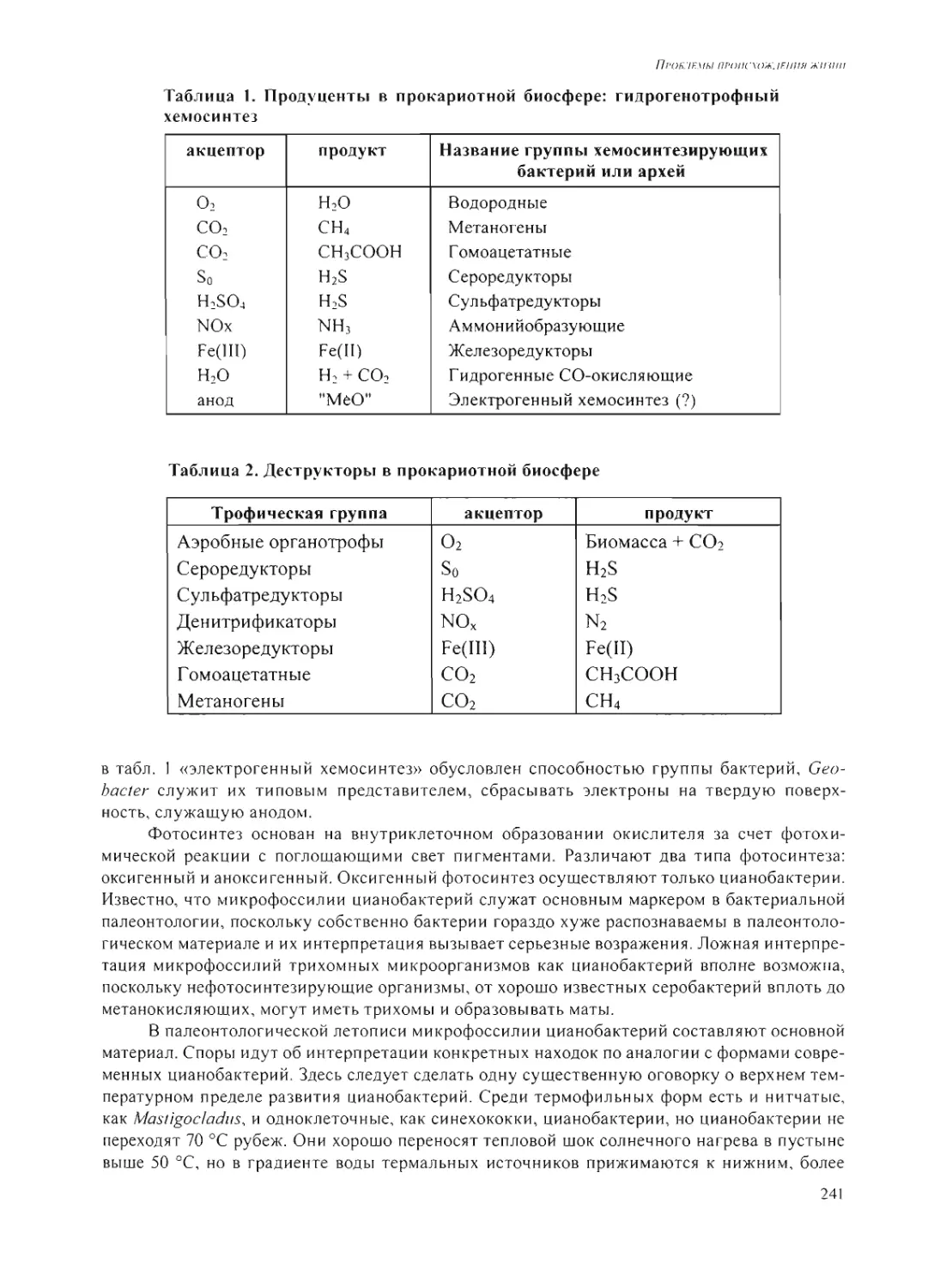

















Author: Григорьев А.И. Добрецов Н.Л.
Tags: астрономия эволюция космохимия
ISBN: 978-5-903825-09-7
Year: 2009




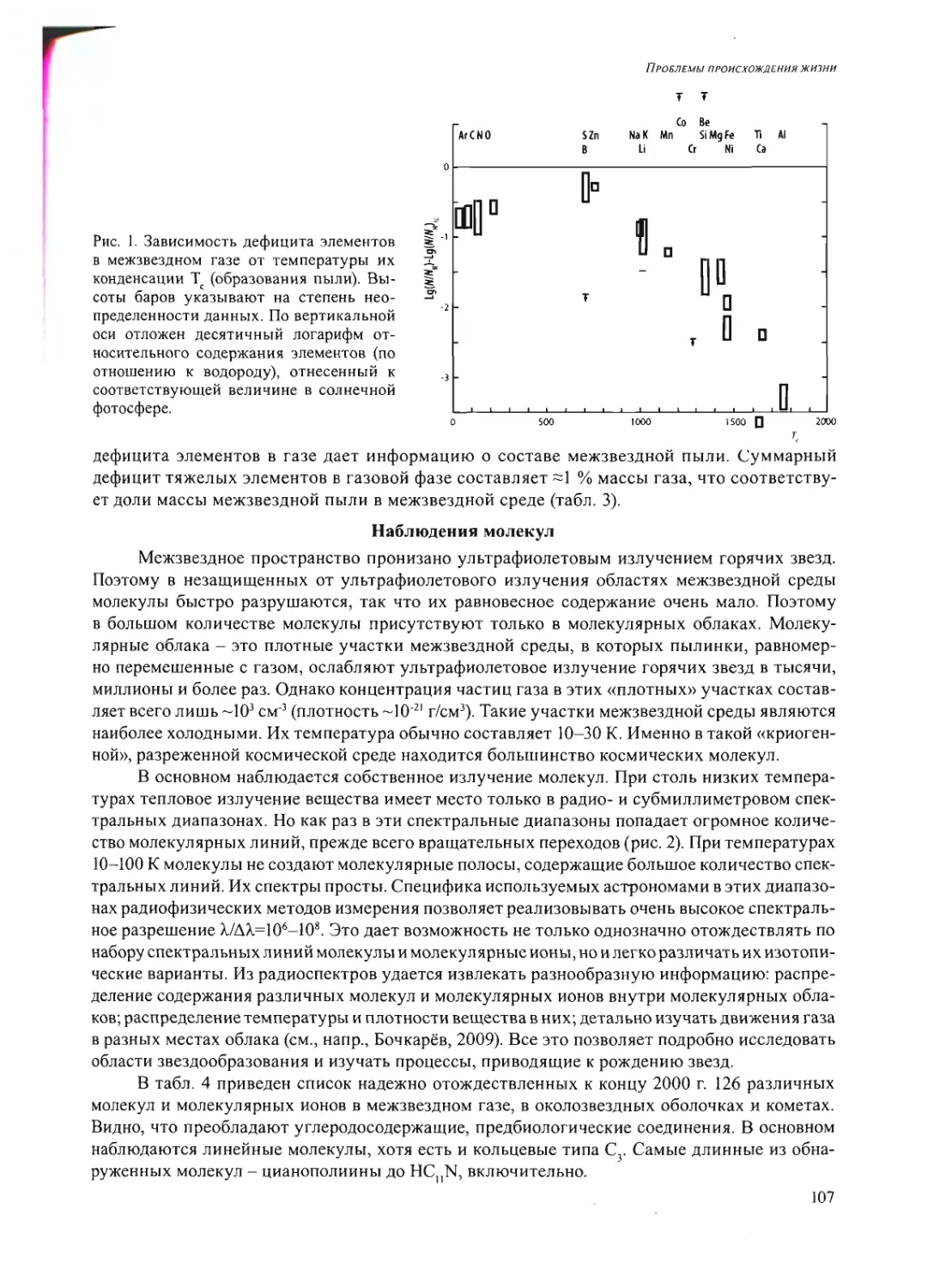
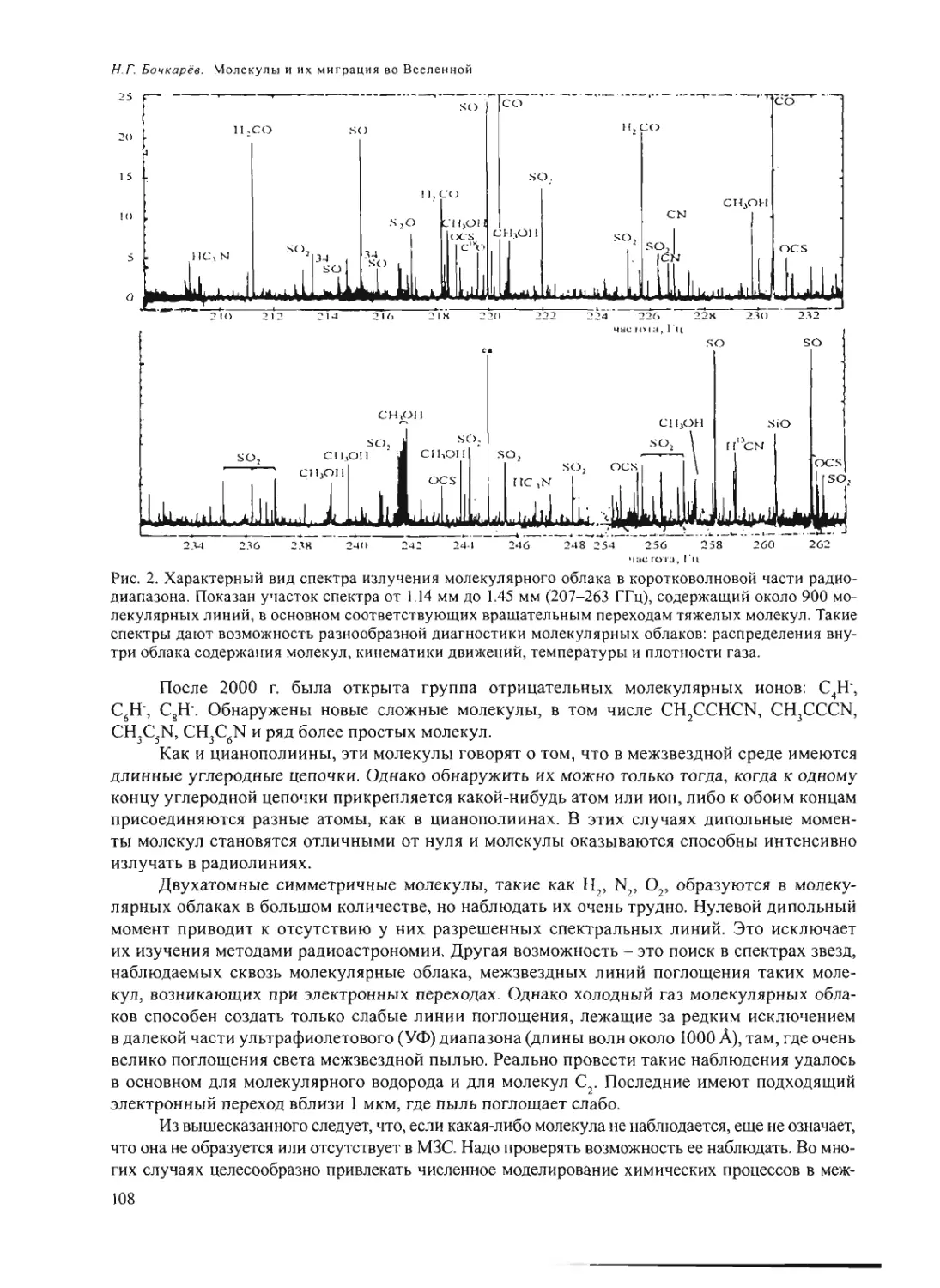
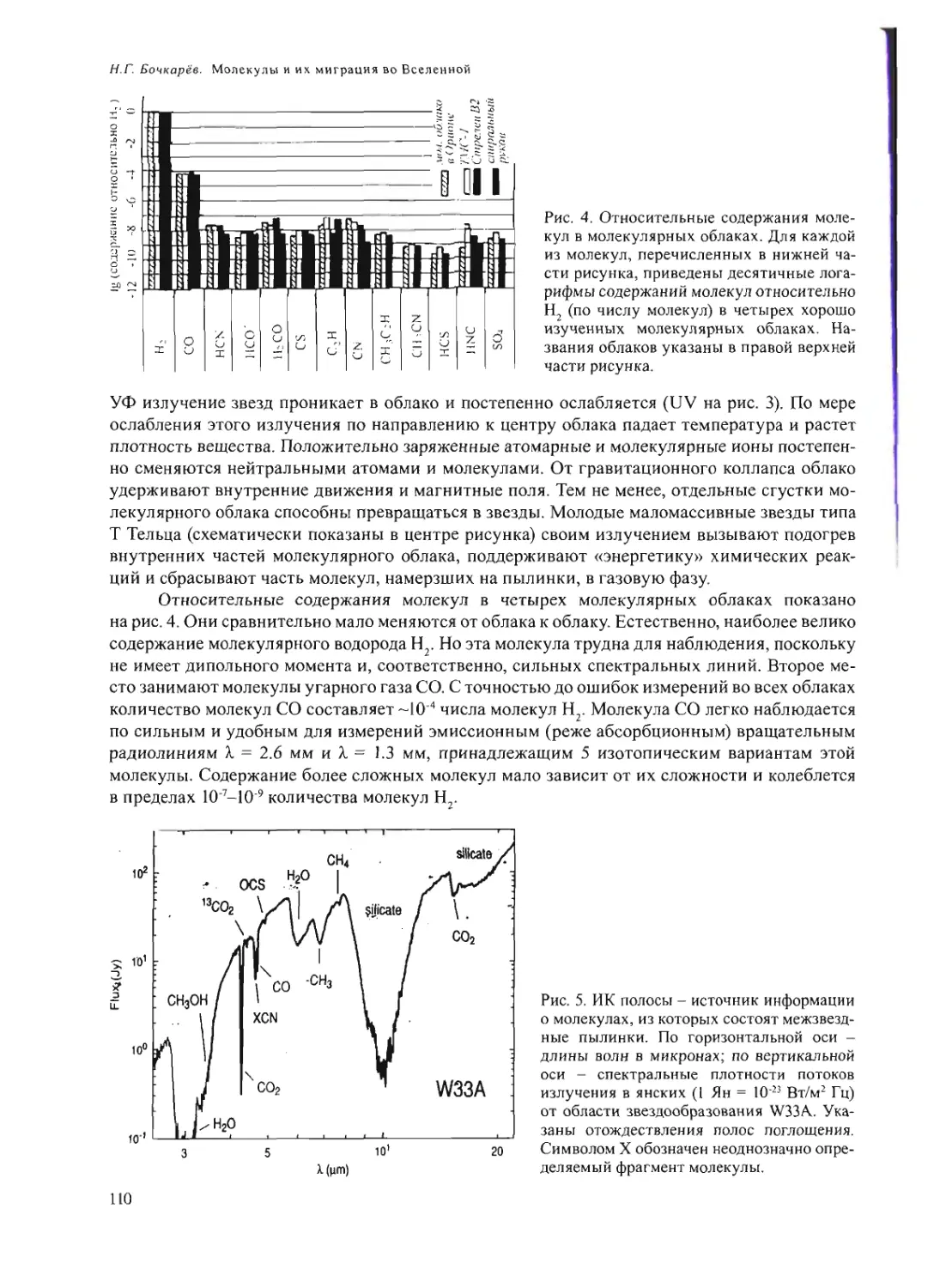
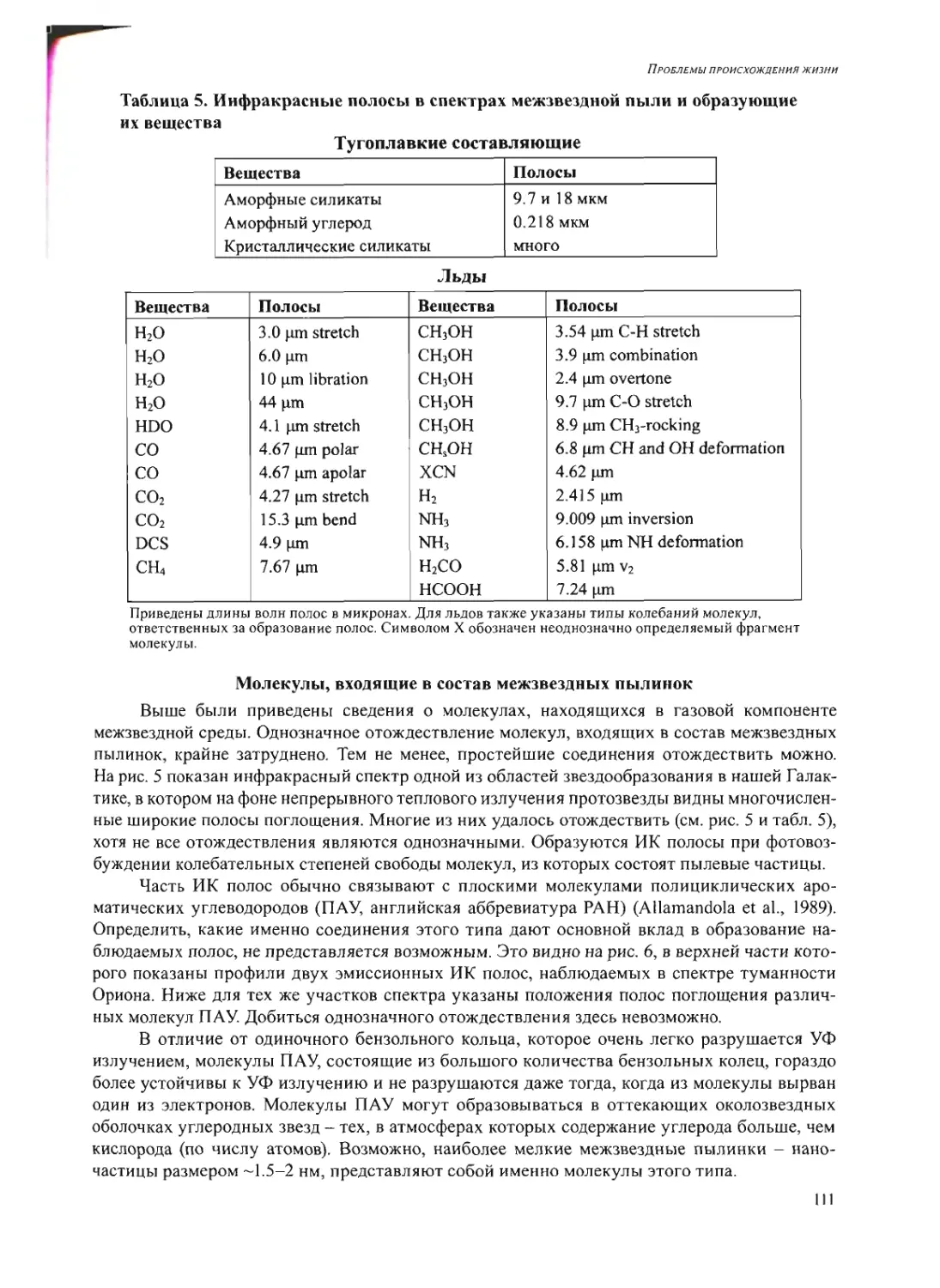
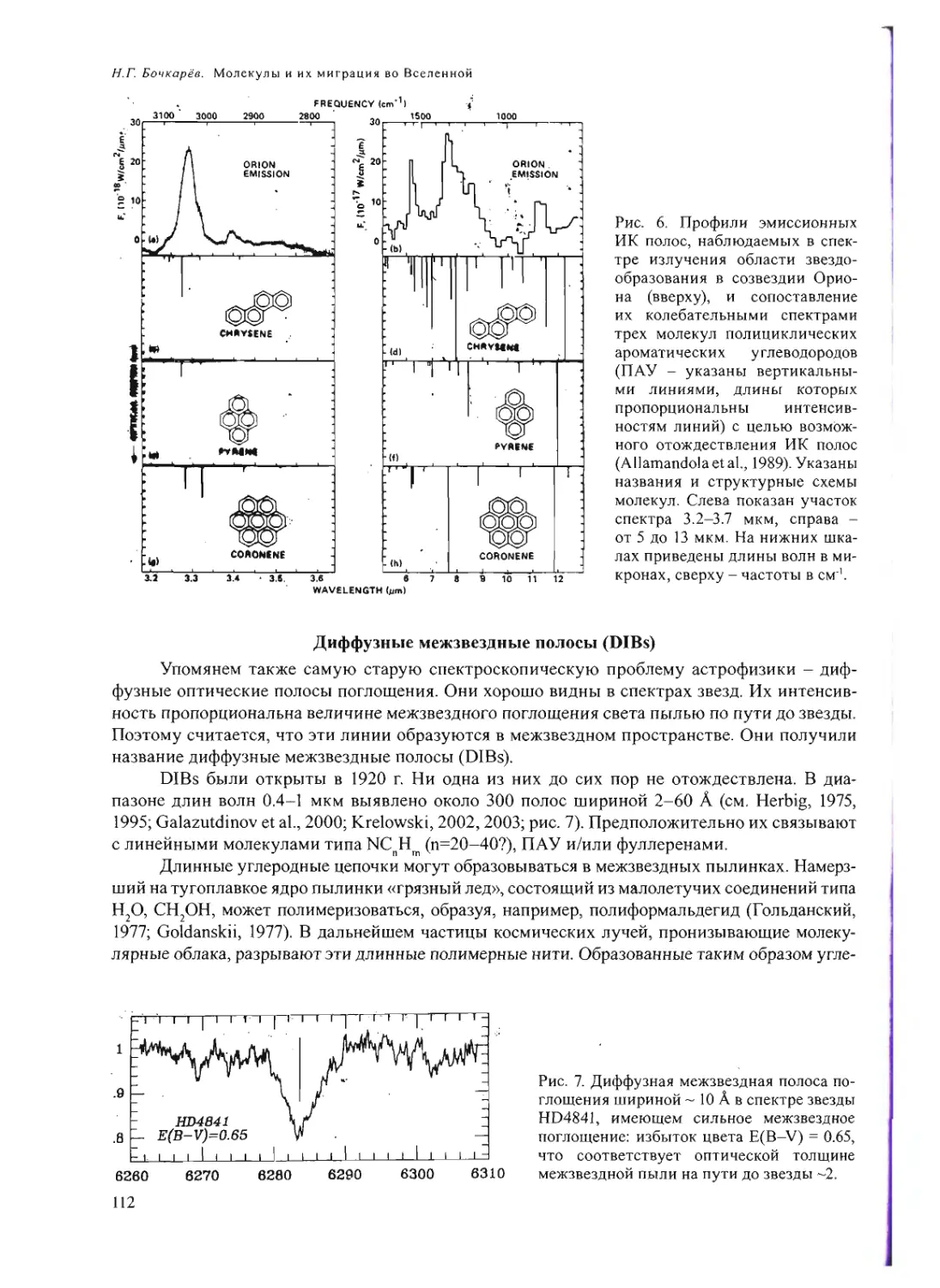
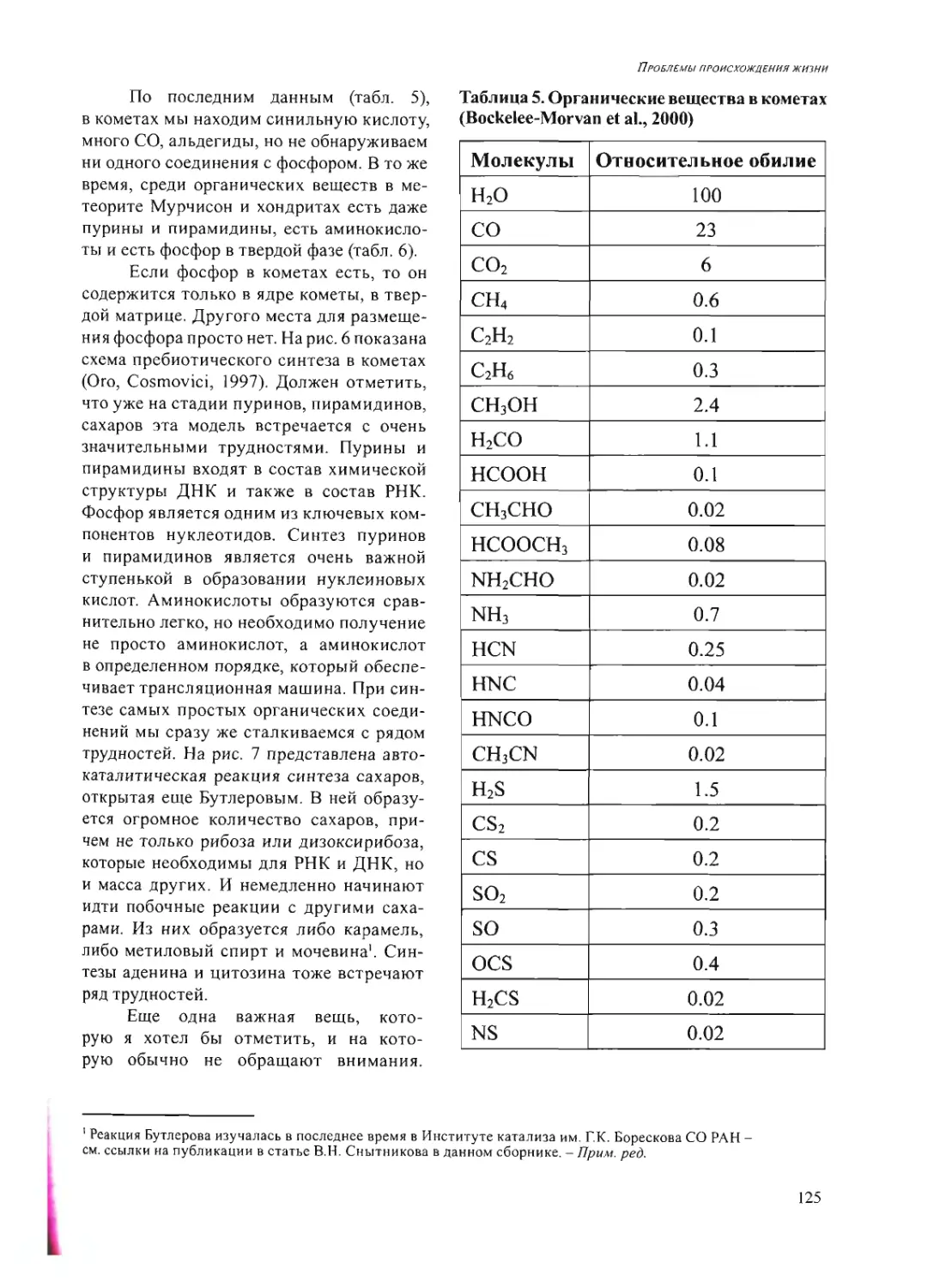
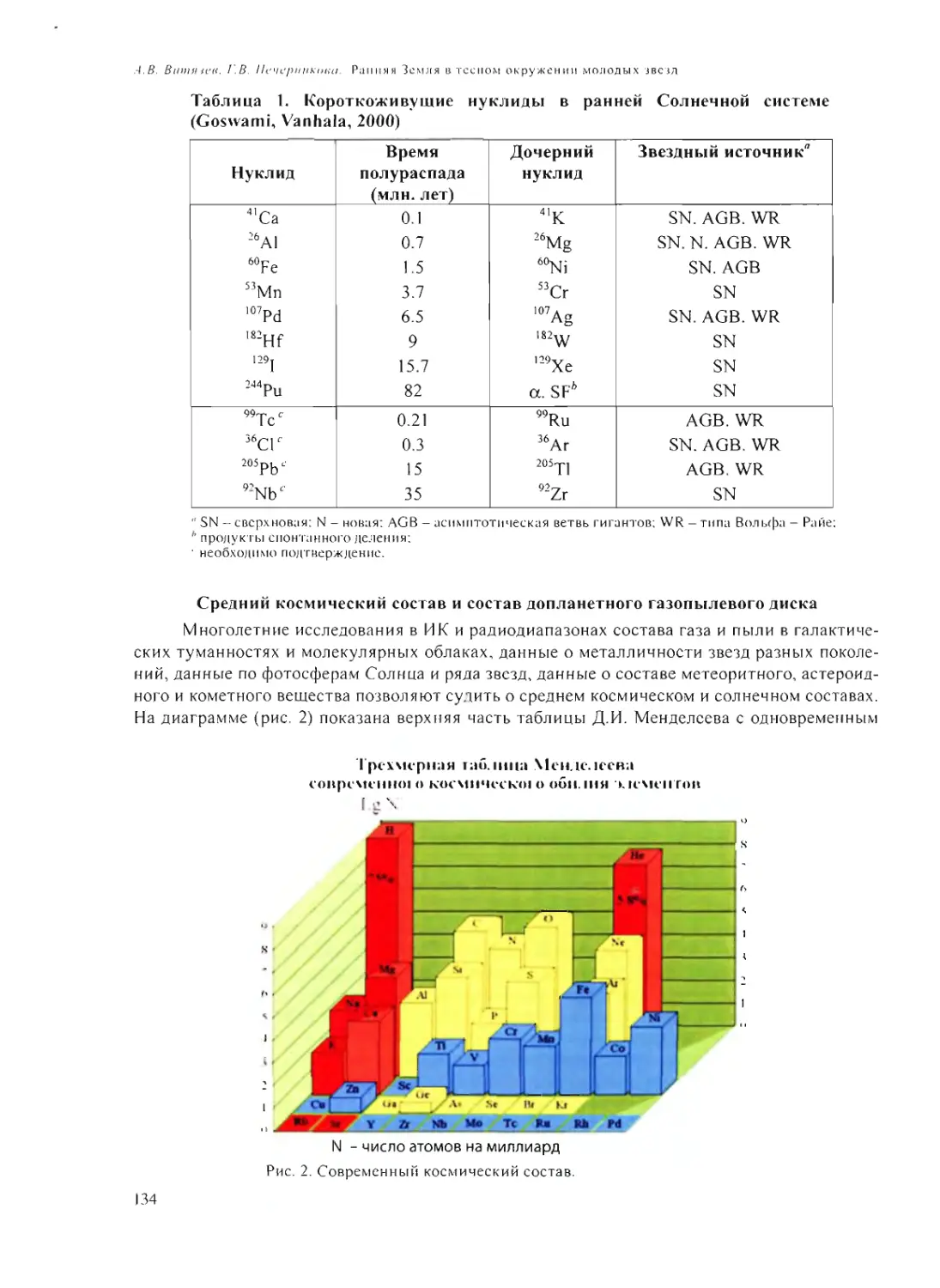
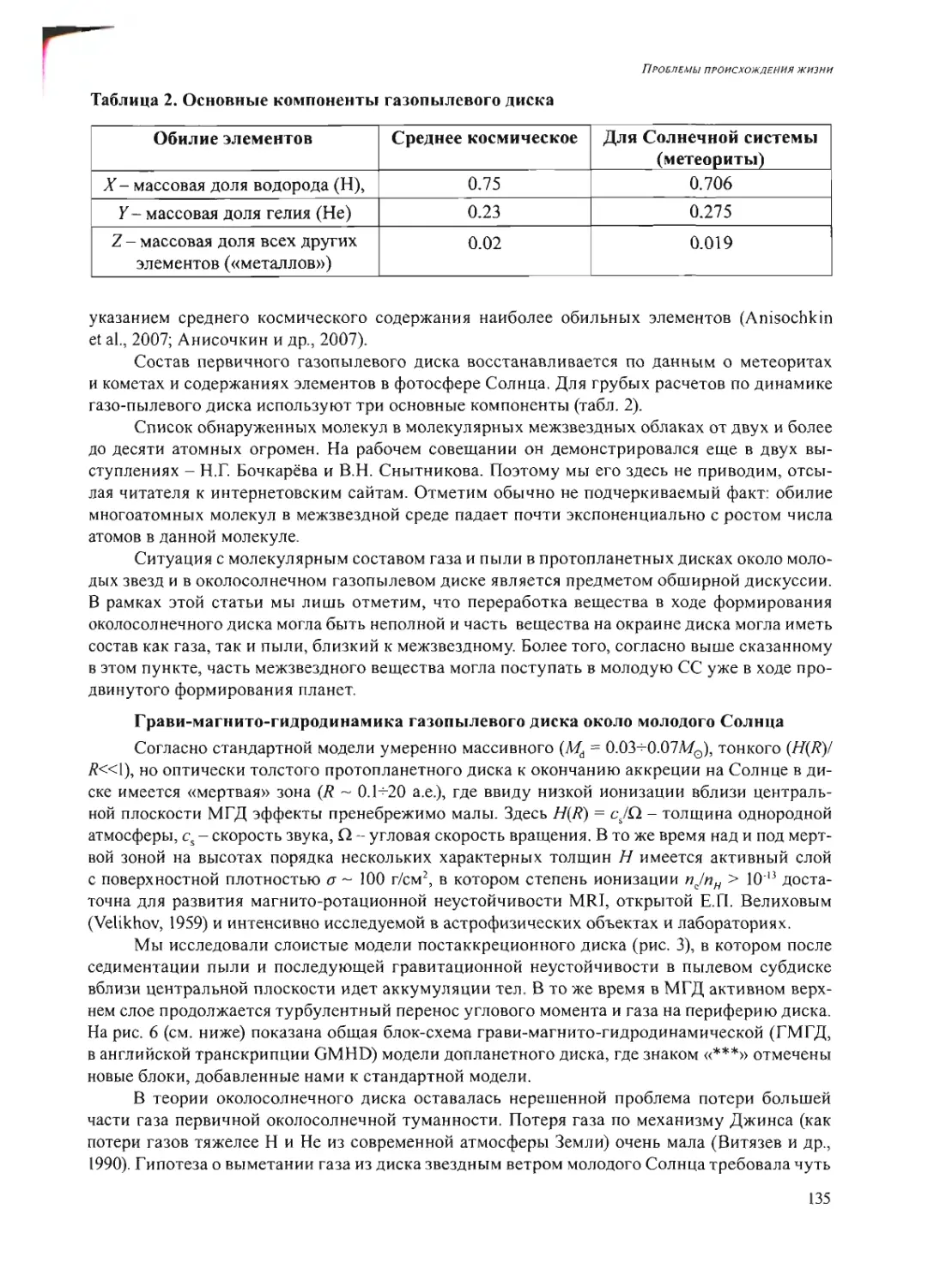
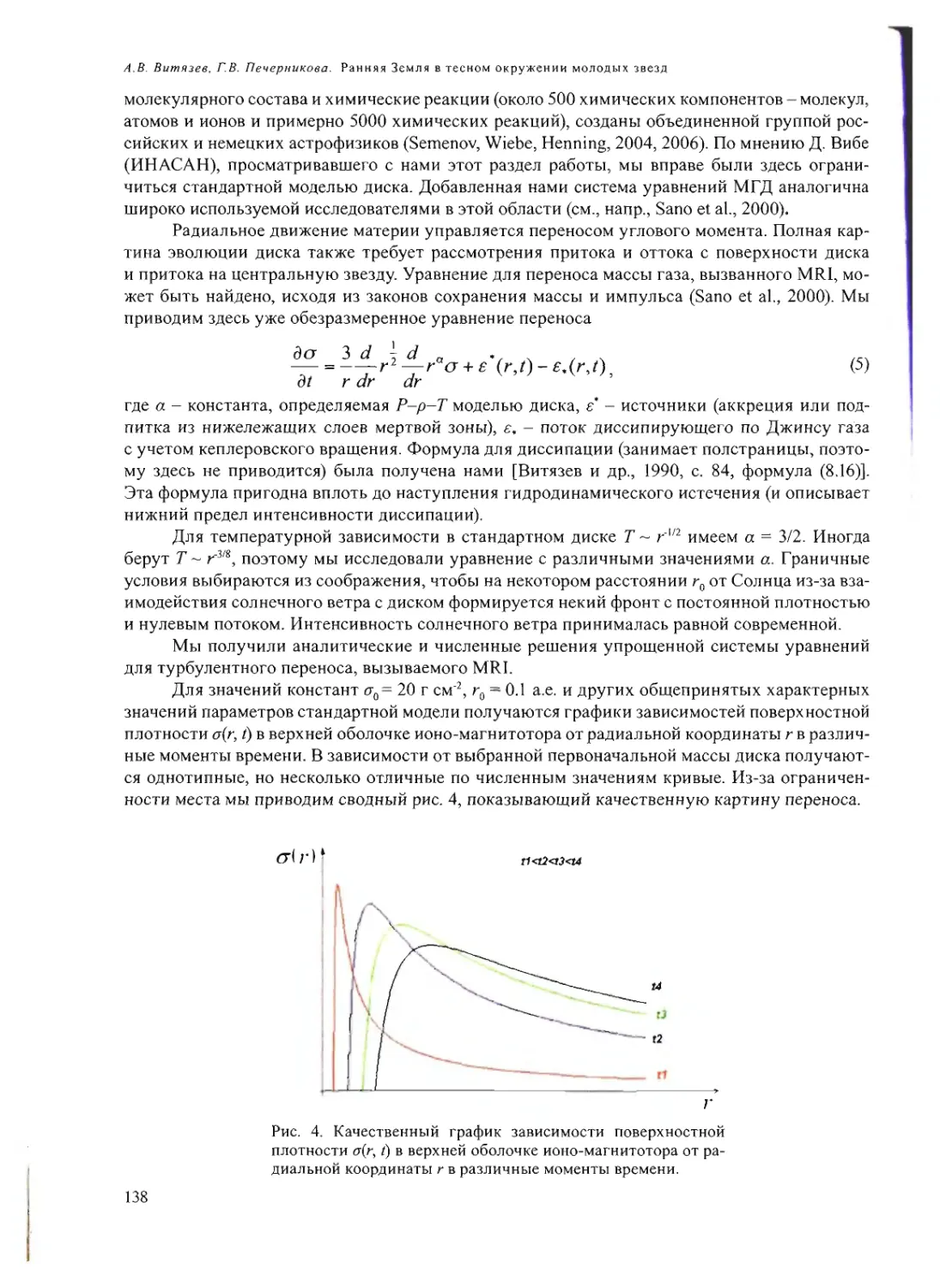
Text
Российская академия наук
Росси Некая академия наук
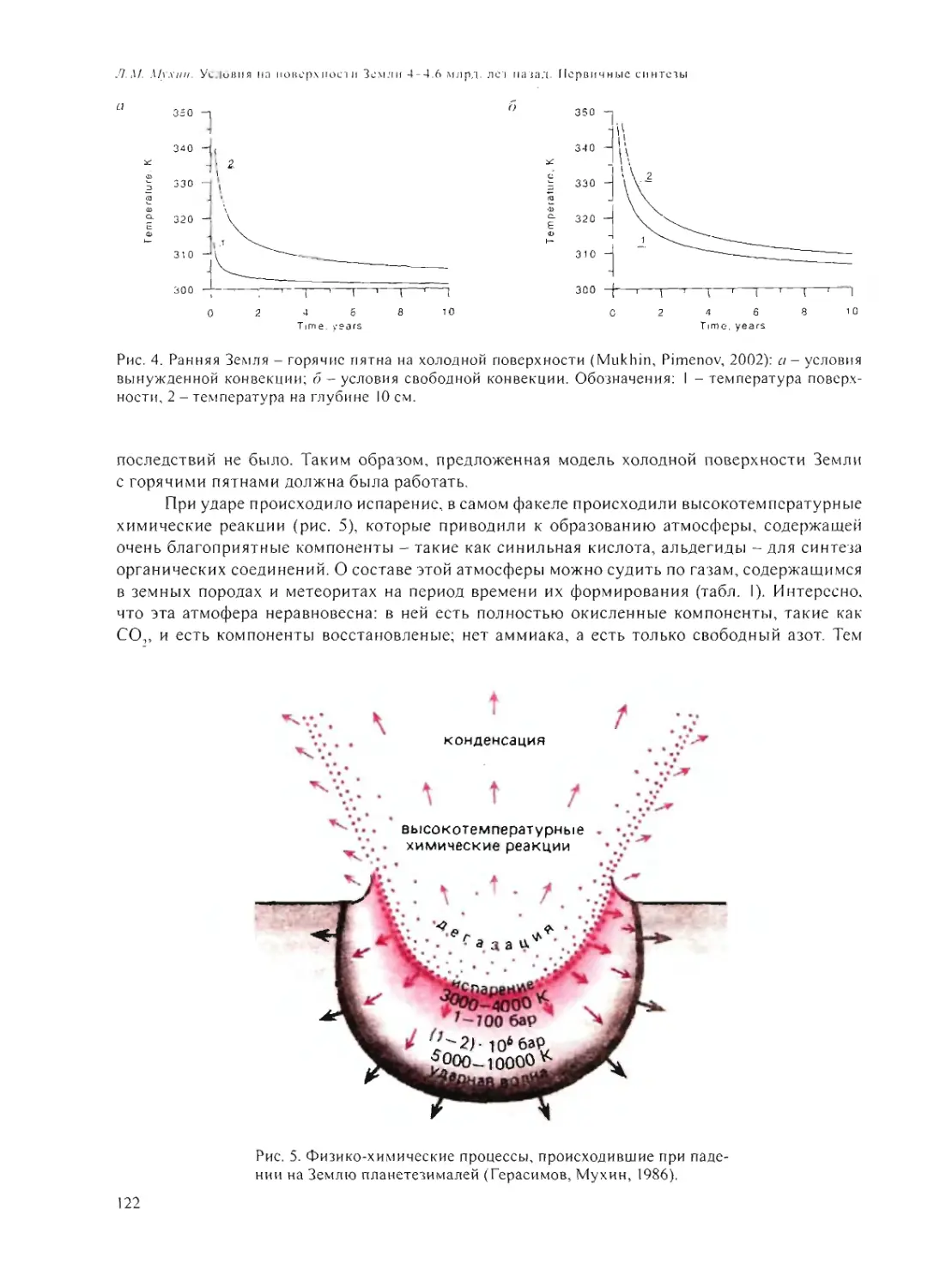
ПРОБЛЕМЫ
ПРОИСХОЖДЕНИЯ
ЖИЗНИ
Москва 2009
ISBN 978-5-903825-09-7
ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ. Российская академия
наук. Сборник научных статей. М.: ПИН РАН, 2009. 258 с. (105 или.,
26 таблиц).
В сборнике представлены статьи по материалам докладов на рабочем
совещании «Проблемы происхождения жизни» (Москва, 22-24 сен-
тября 2008 г.), посвященные различным аспектам изучения вопросов
возникновения и становления органического мира на Земле и во Все-
ленной».
ISBN 978-5-903825-09-7
Редакционная коллегия:
акад. А.И. Григорьев, акад. Н.Л. Добрецов,
акад. Г.А. Заварзин, д.б.н. А.В. Лопатин,
акад. A.IO. Розанов, к.ф.-м.п. В.Н. Снытников,
акад. А.С. Спирин
Рецензенты:
акад. Н.А. Колчанов,
акад. В.Н. Пармон,
акад. С.В. Шестаков
ISBN 978-5-903825-09-7
© Коллектив авторов, 2009
© ПИН РАН, 2009
© А.А. Ермаков (обложка)
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ 7
МОЖНО ЛИ СОБРАТЬ КЛЕТКУ ИЗ ЕЕ КОМПОНЕНТОВ?
А.Б. Четверин 9
ВИРУС - ДО ИЛИ ПОСЛЕ КЛЕТКИ?
В.И. Агол 31
ДРЕВНИЙ МИР РНК
А.С. Спирин 43
ОТ МАРСА К ЕВРОПЕ: ПОИСК БИОСФЕРЫ НА СПУТНИКАХ ПЛАНЕТ-ГИГАНТОВ
Л. В. Ксанфо.малити 60
АСТРОКАТАЛИЗ-АБИОГЕННЫЙ СИНТЕЗ И ХИМИЧЕСКАЯ ЭВОЛЮЦИЯ
НА ДОГЕОЛОГИЧЕСКИХ ЭТАПАХ ФОРМИРОВАНИЯ ЗЕМЛИ
В.И. Снытников 79
МОЛЕКУЛЫ И ИХ МИГРАЦИЯ ВО ВСЕЛЕННОЙ
И. Г. Бочкарёв 102
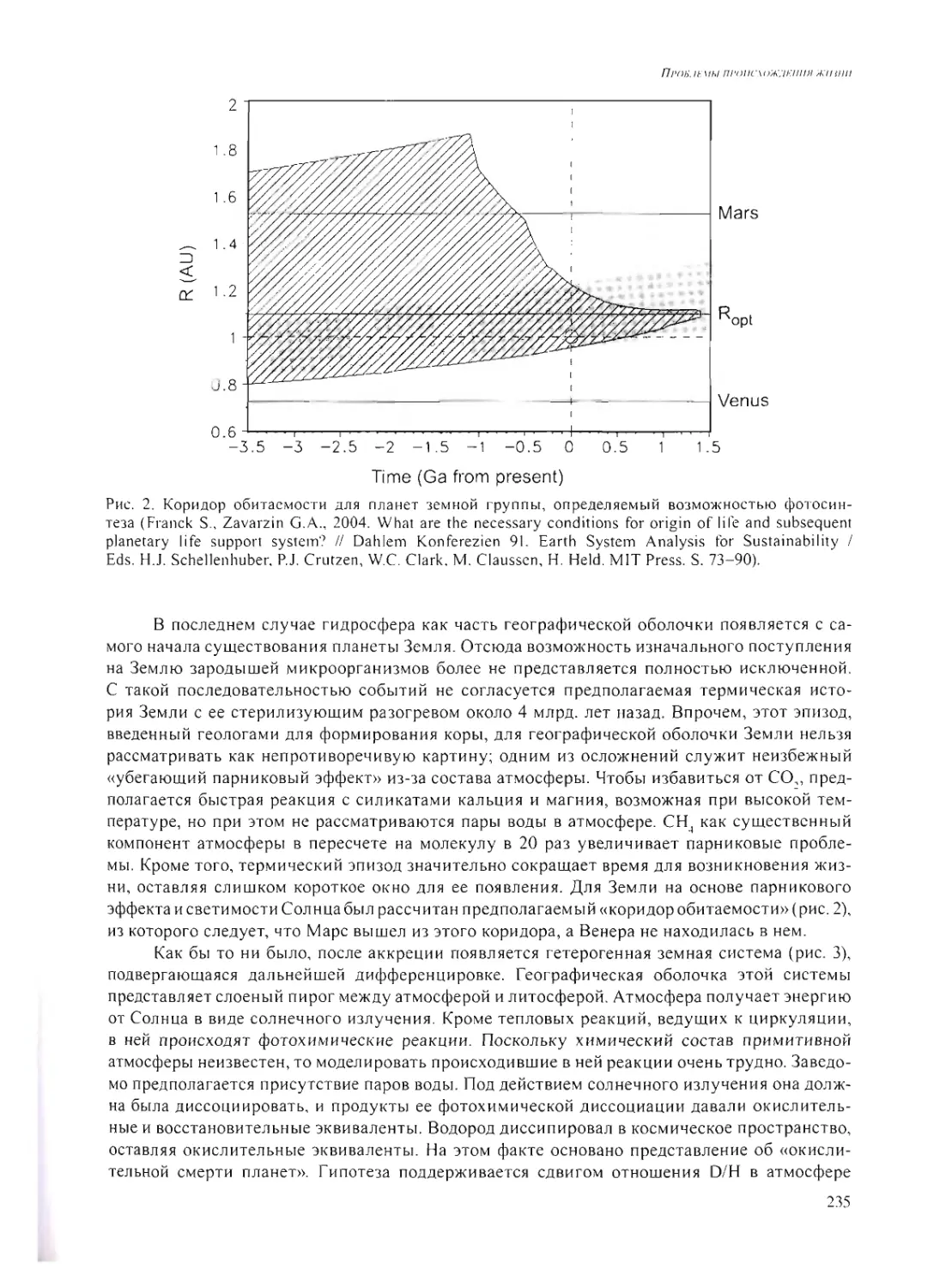
УСЛОВИЯ НА ПОВЕРХНОСТИ ЗЕМЛИ 4-4.6 МЛРД. ЛЕТ НАЗАД.
ПЕРВИЧНЫЕ СИНТЕЗЫ
Л. М. Мухин 120
РАННЯЯ ЗЕМЛЯ В ТЕСНОМ ОКРУЖЕНИИ МОЛОДЫХ ЗВЕЗД
А.В. Витязев, Г. В. Печерникова 131
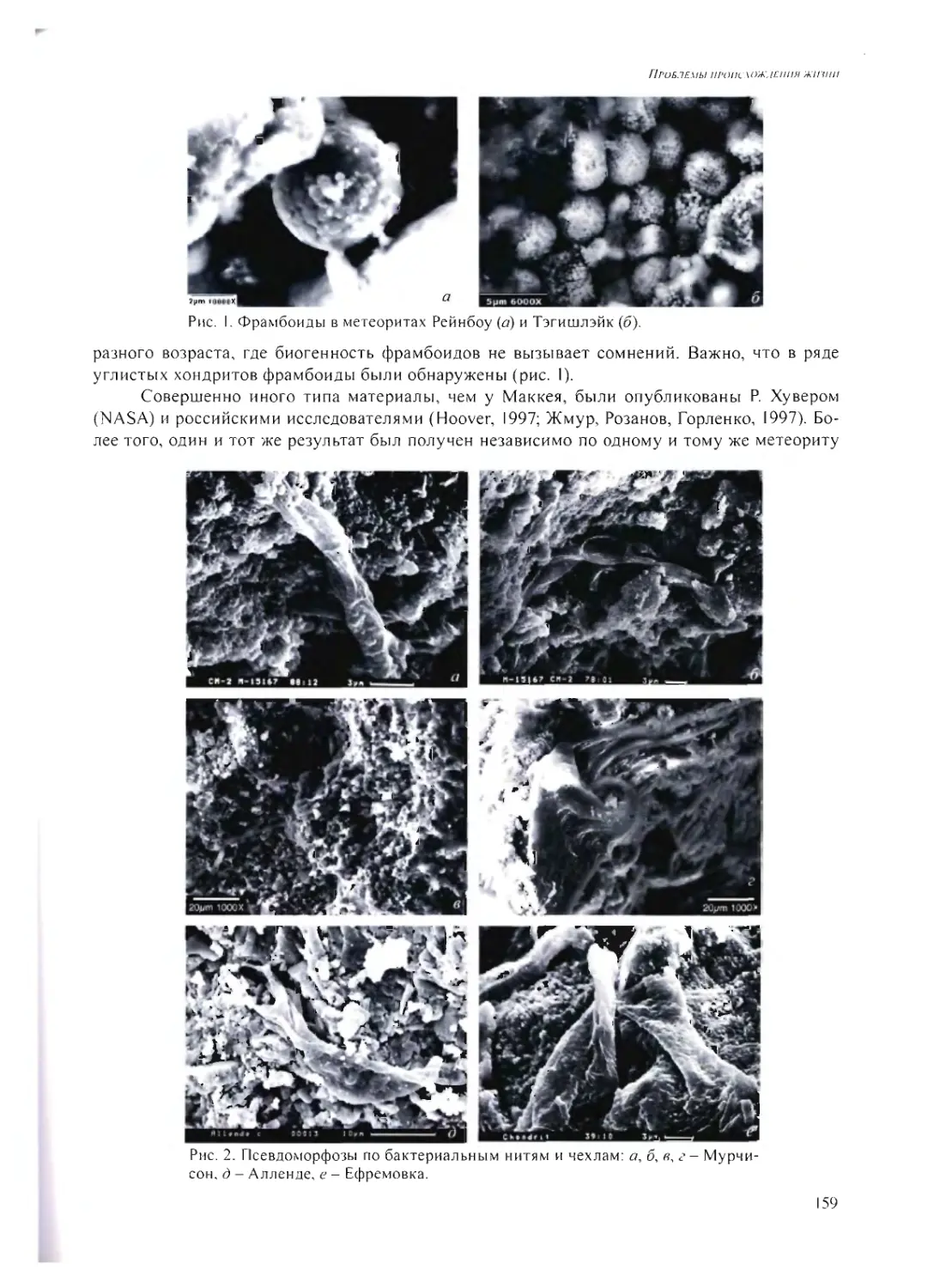
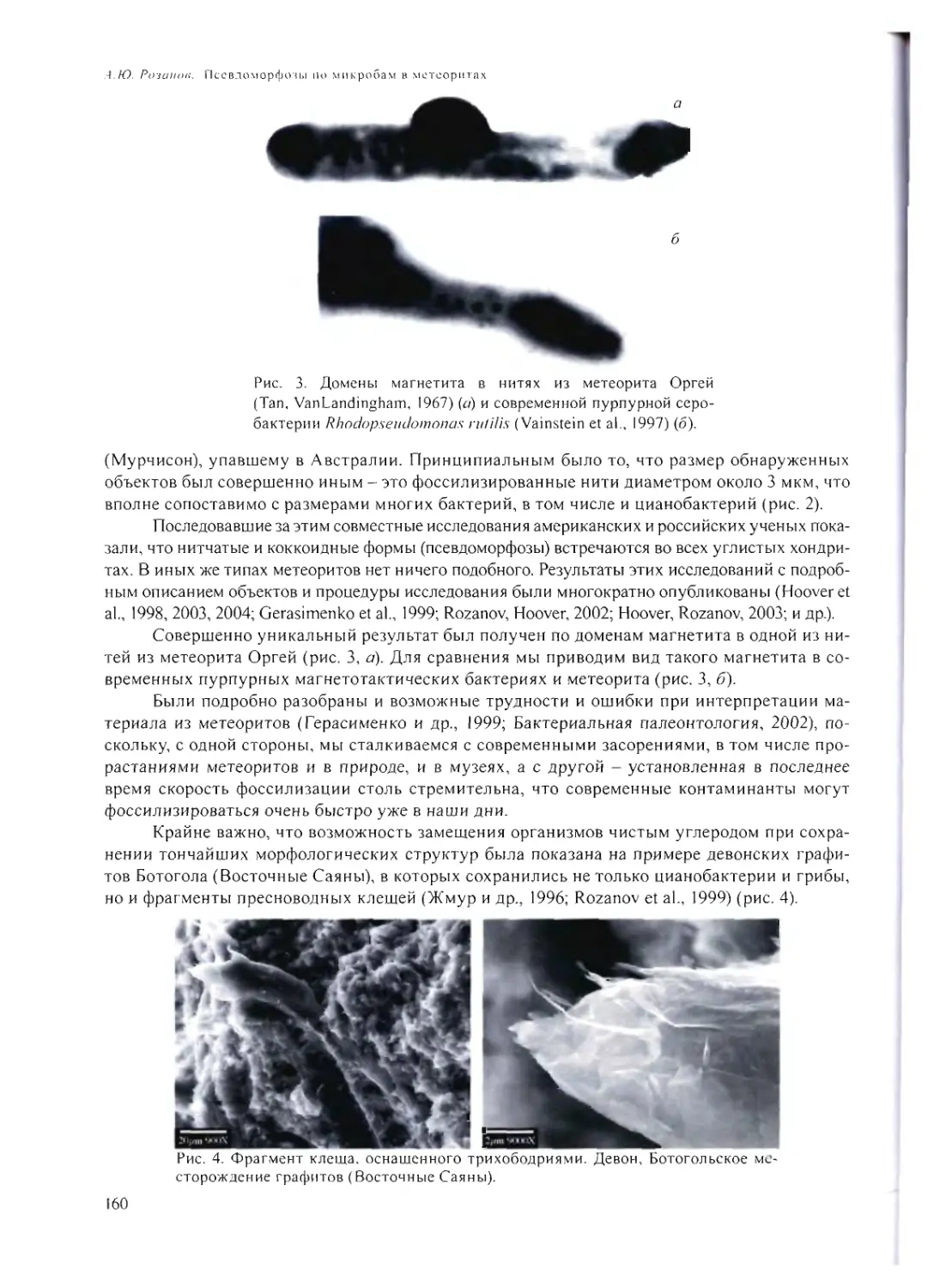
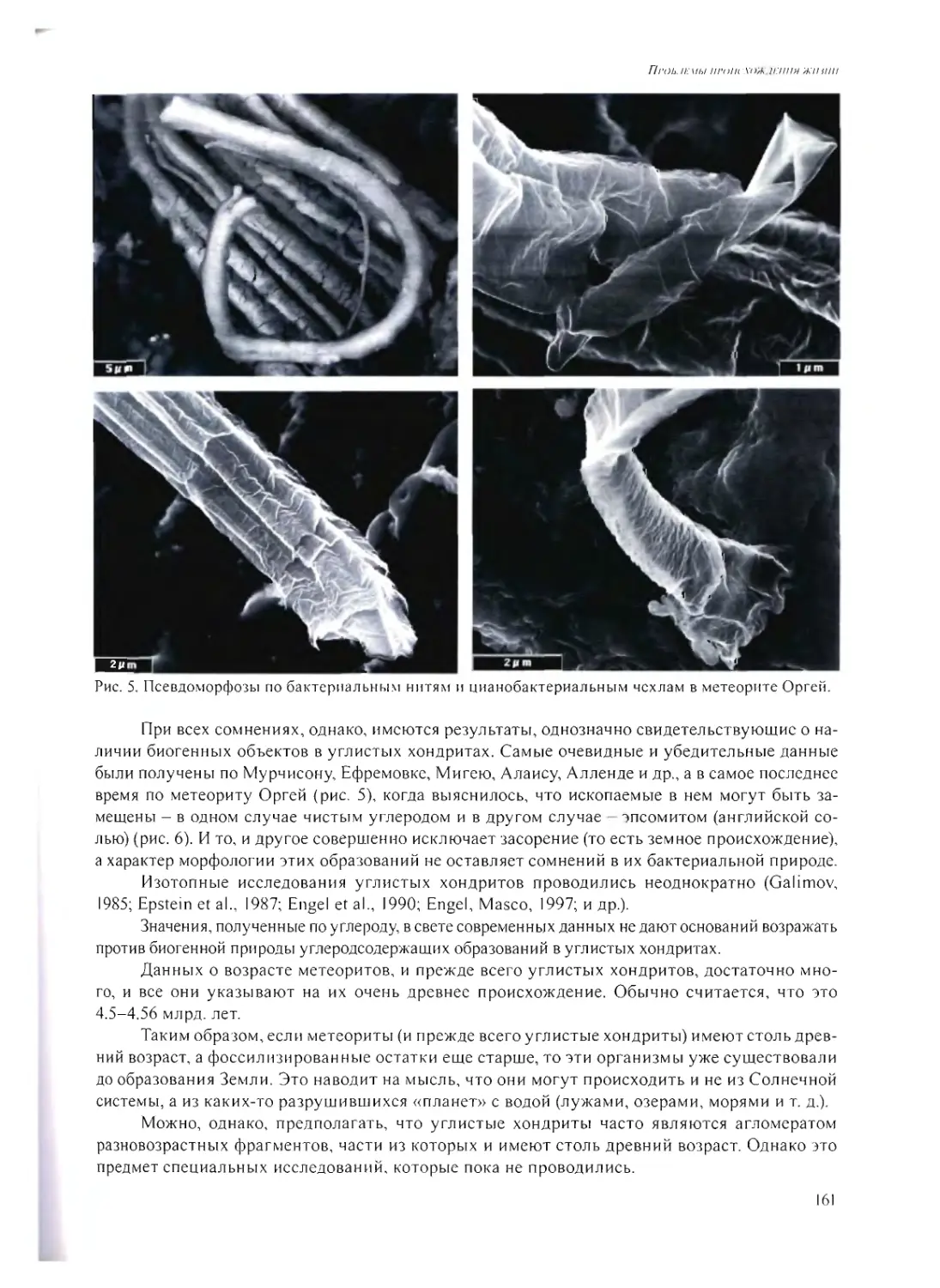
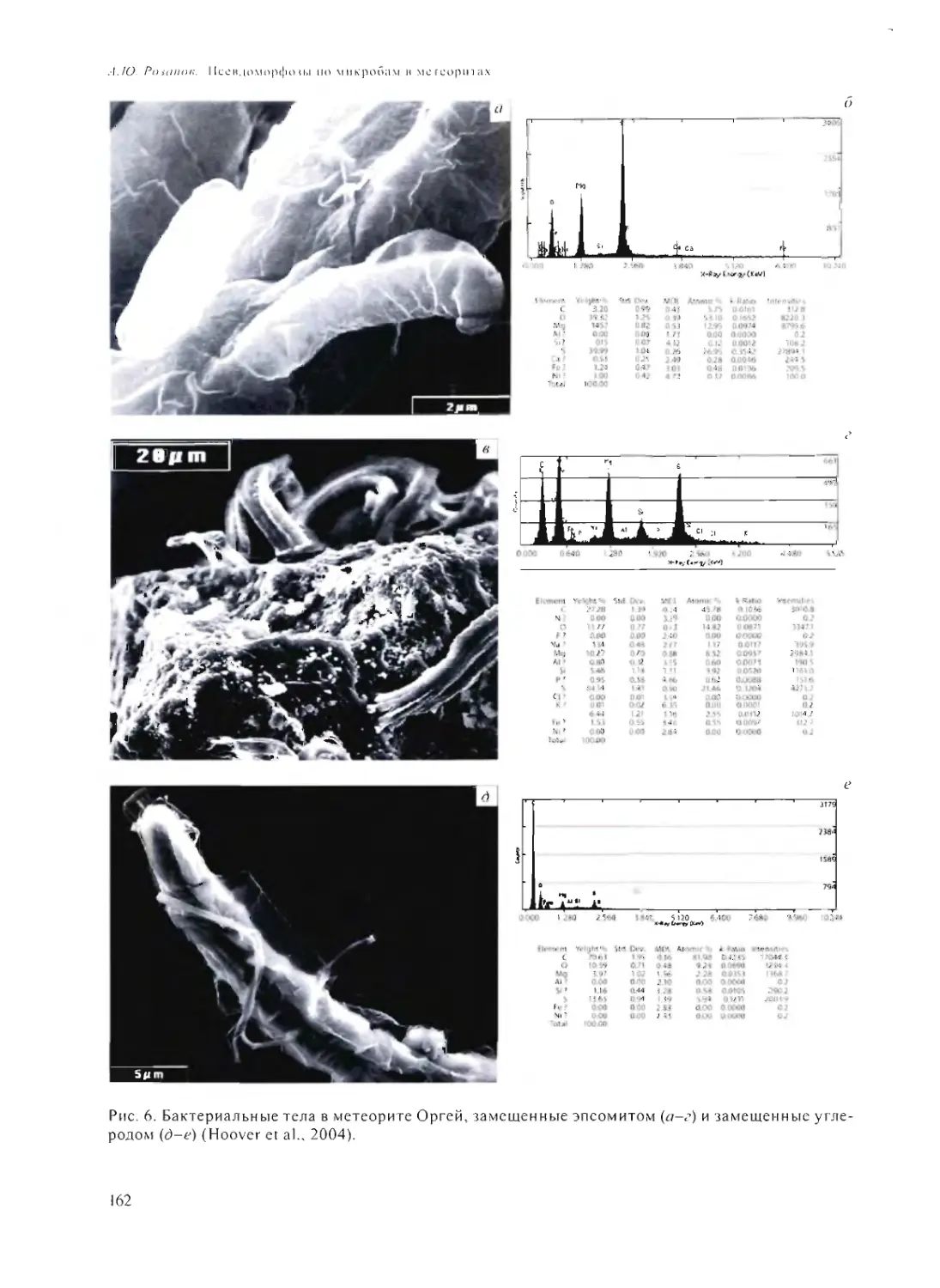
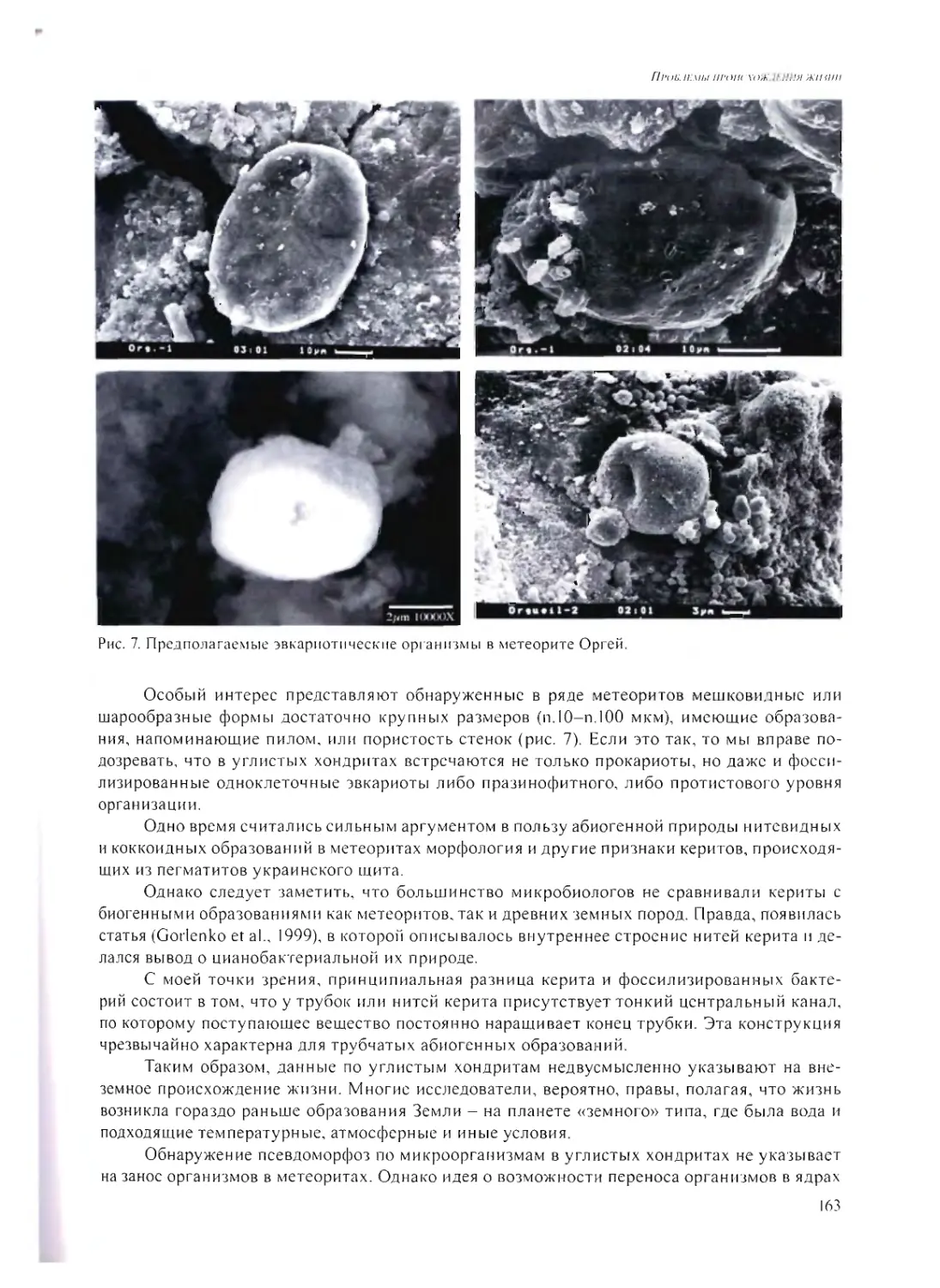
ПСЕВДОМОРФОЗЫ ПО МИКРОБАМ В МЕТЕОРИТАХ
А.Ю. Розанов 158
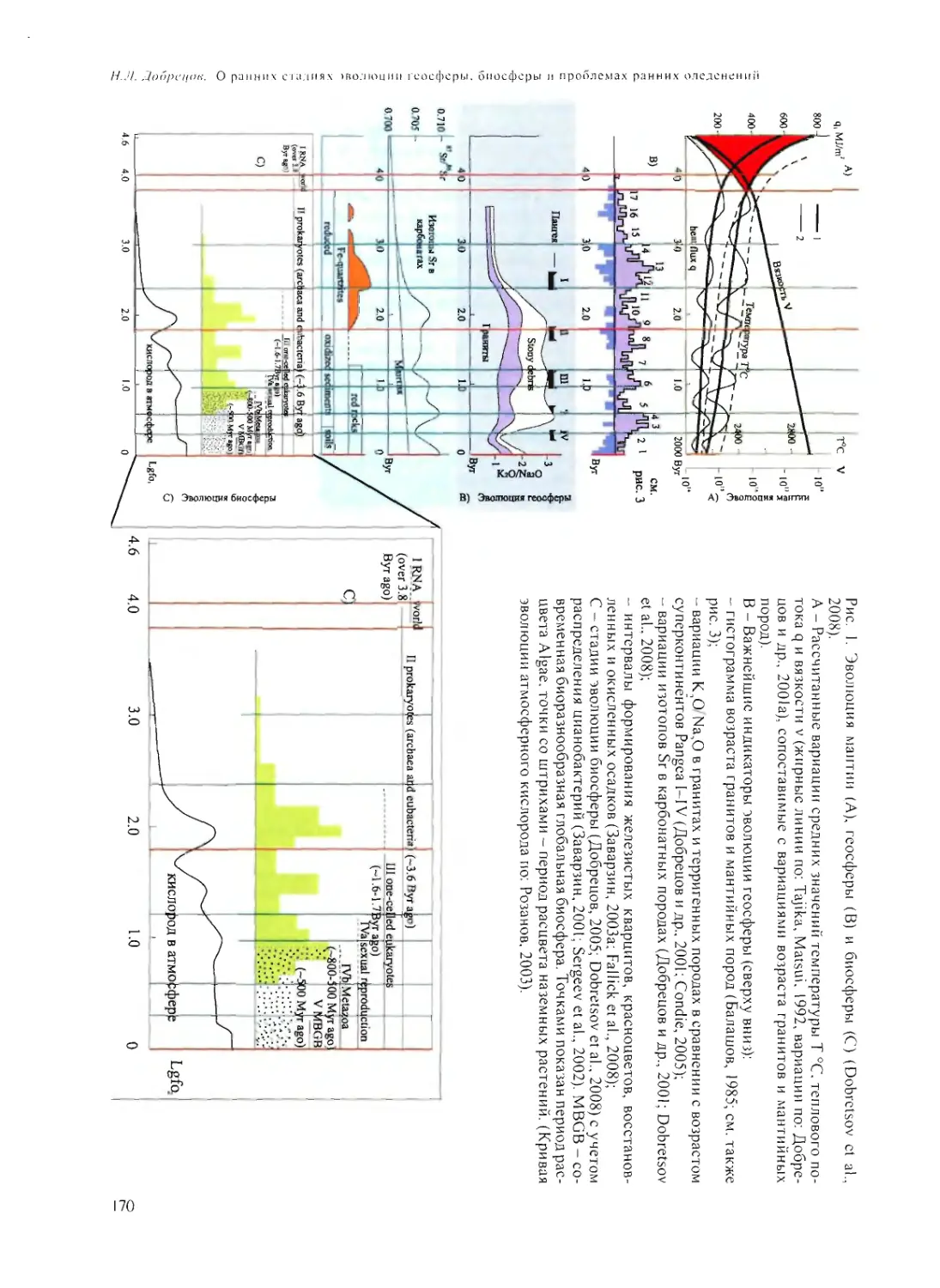
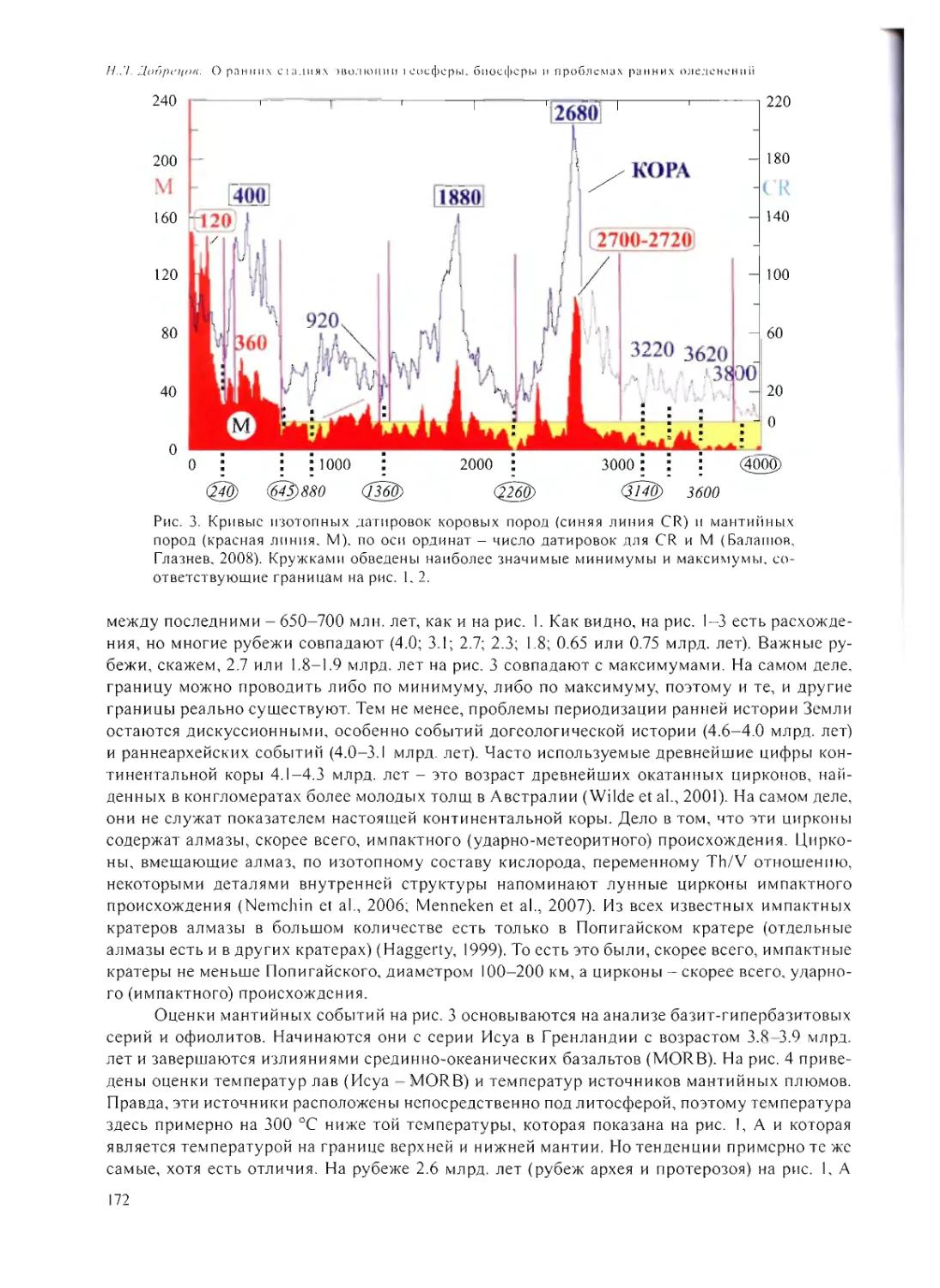
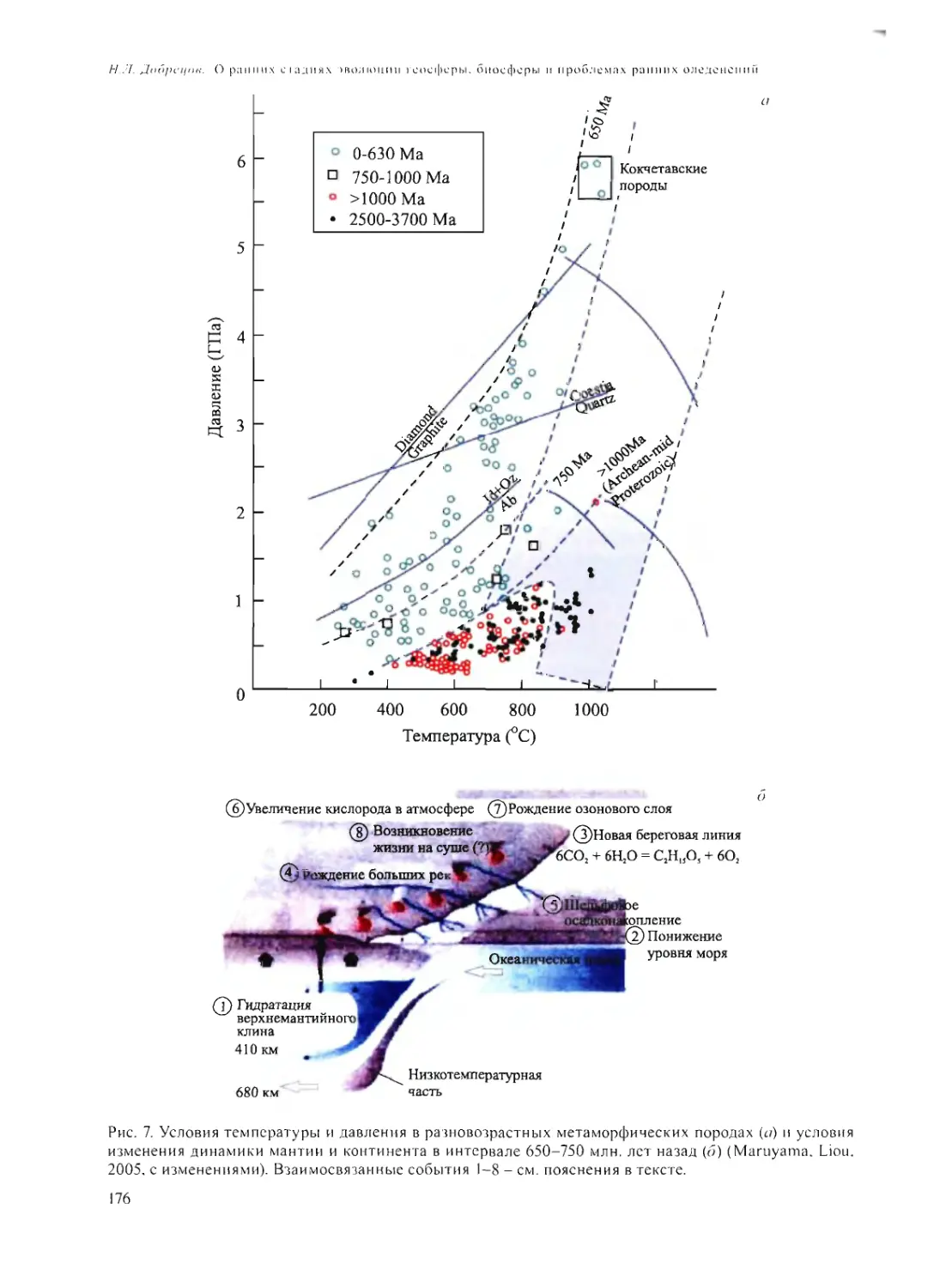
О РАННИХ СТАДИЯХ ЭВОЛЮЦИИ ГЕОСФЕРЫ, БИОСФЕРЫ
И ПРОБЛЕМАХ РАННИХ ОЛЕДЕНЕНИЙ
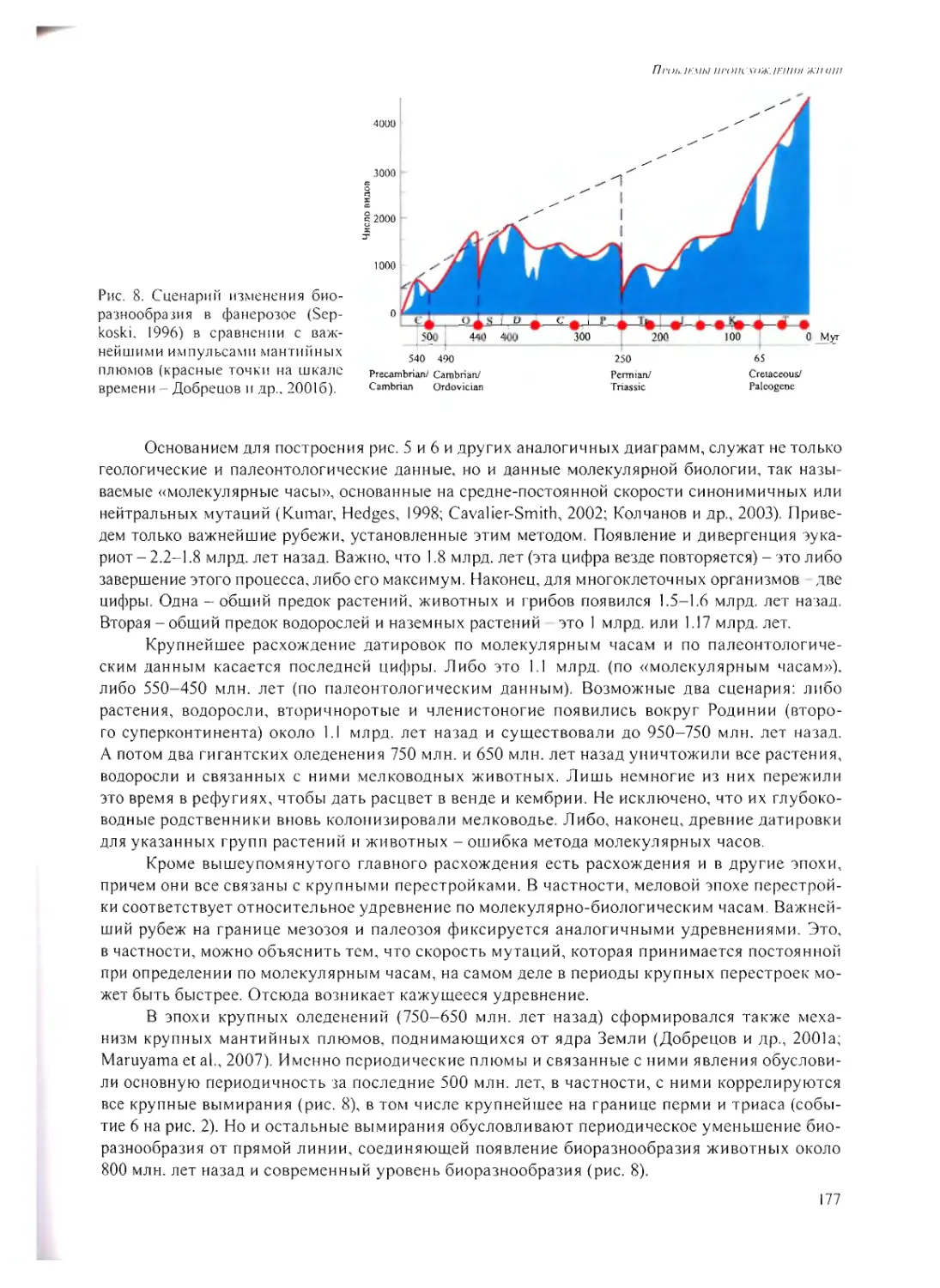
Н.Л. Добрецов 168
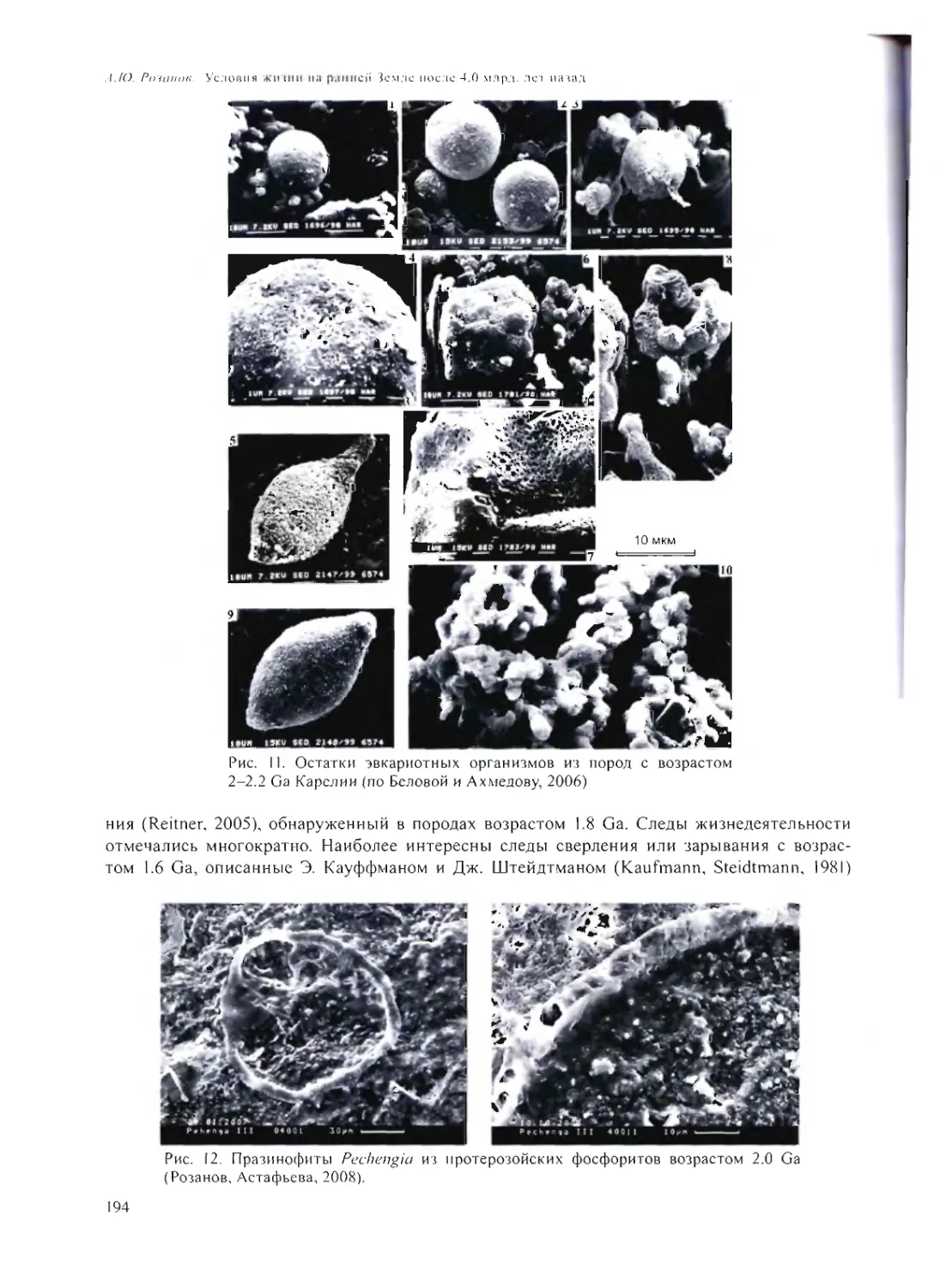
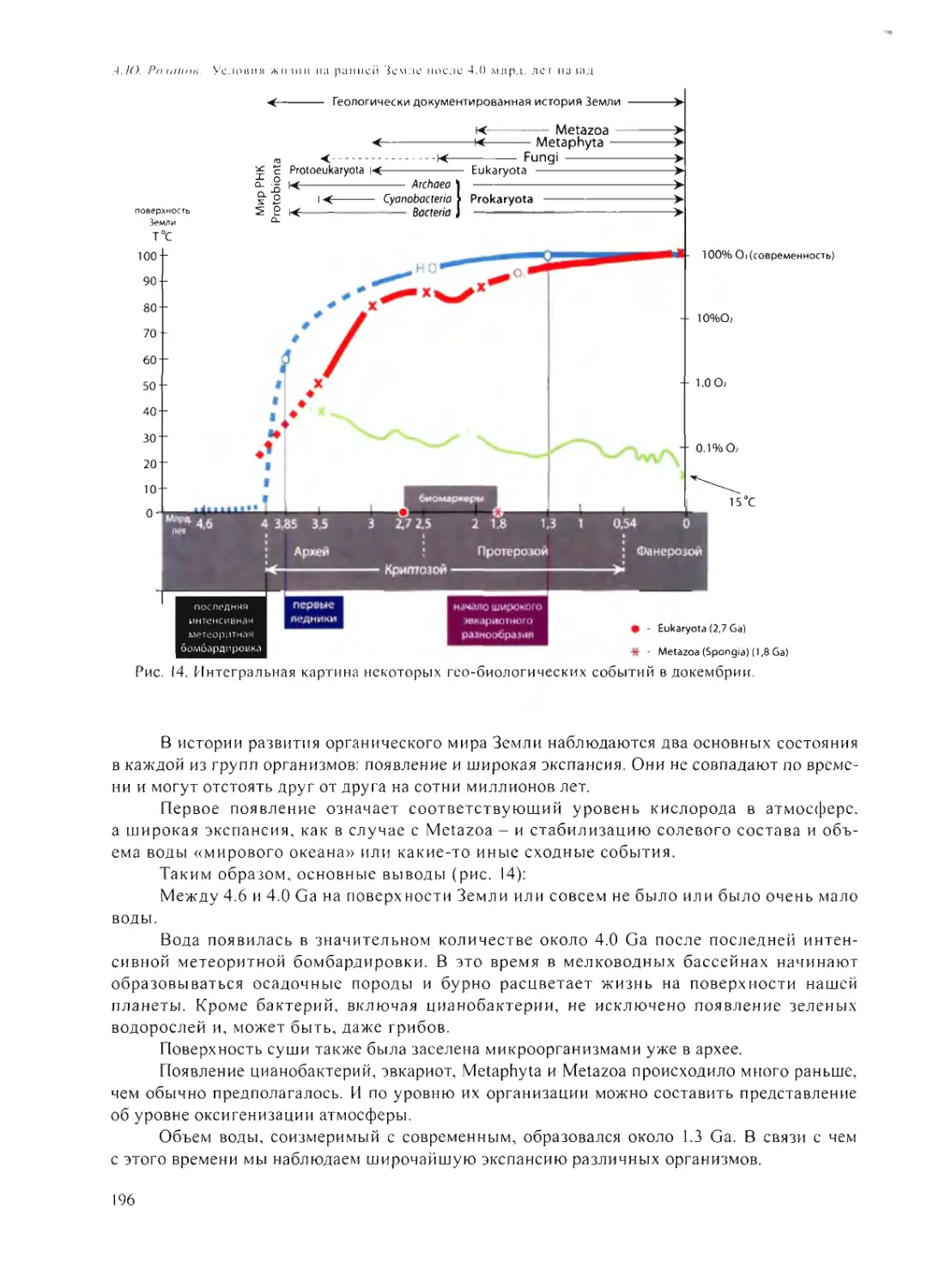
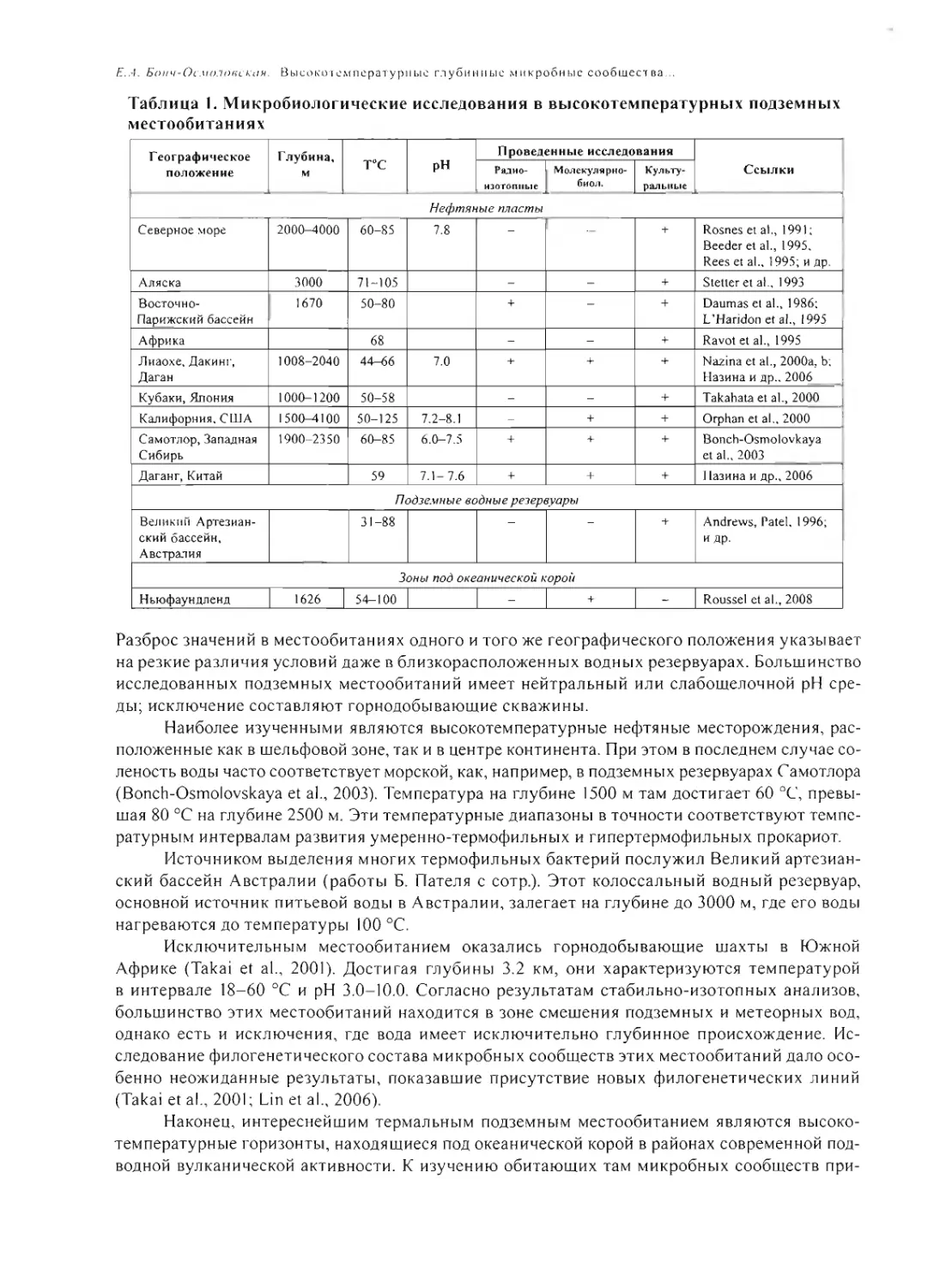
УСЛОВИЯ ЖИЗНИ НА РАННЕЙ ЗЕМЛЕ ПОСЛЕ 4.0 МЛРД. ЛЕТ НАЗАД
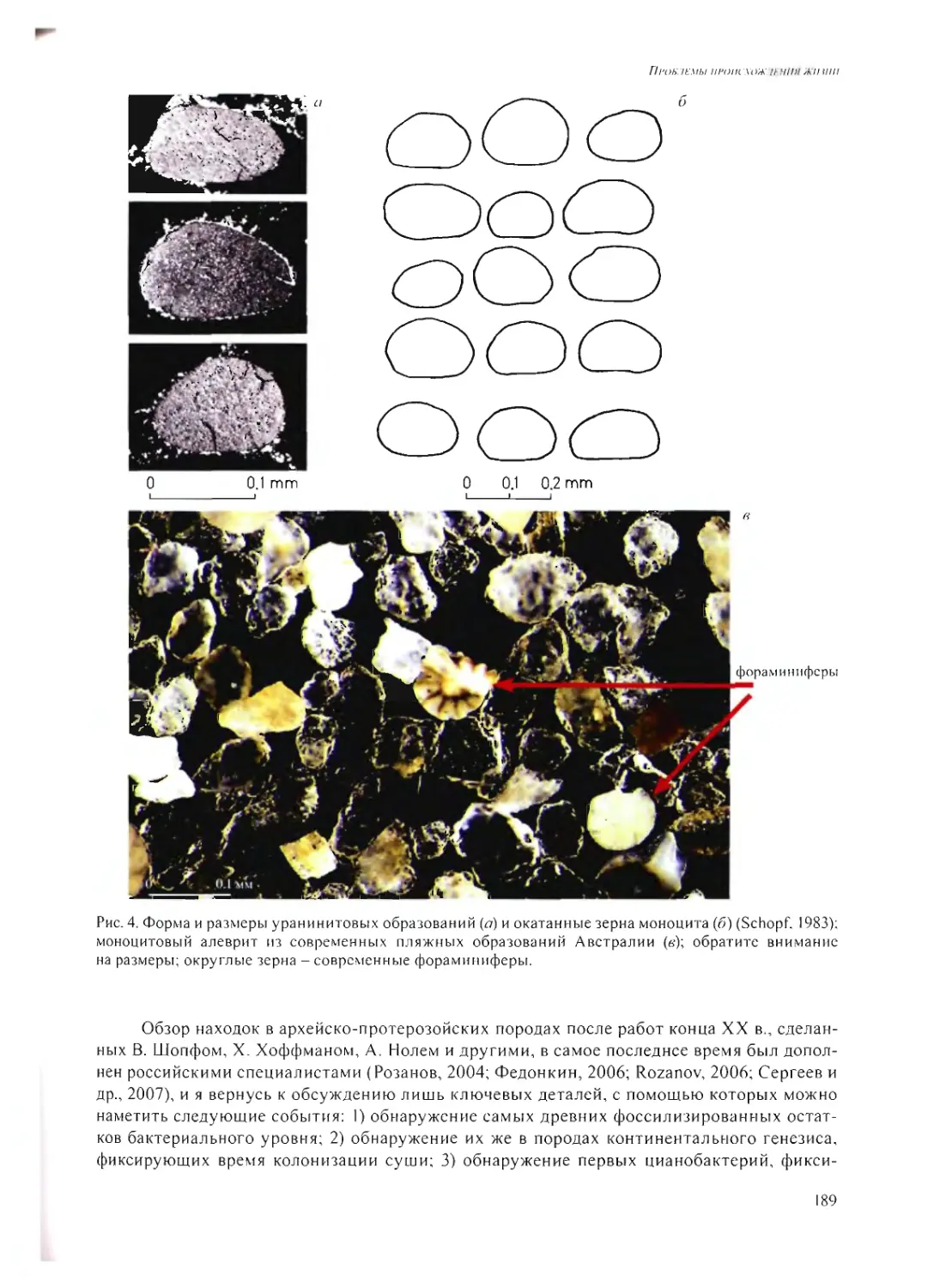
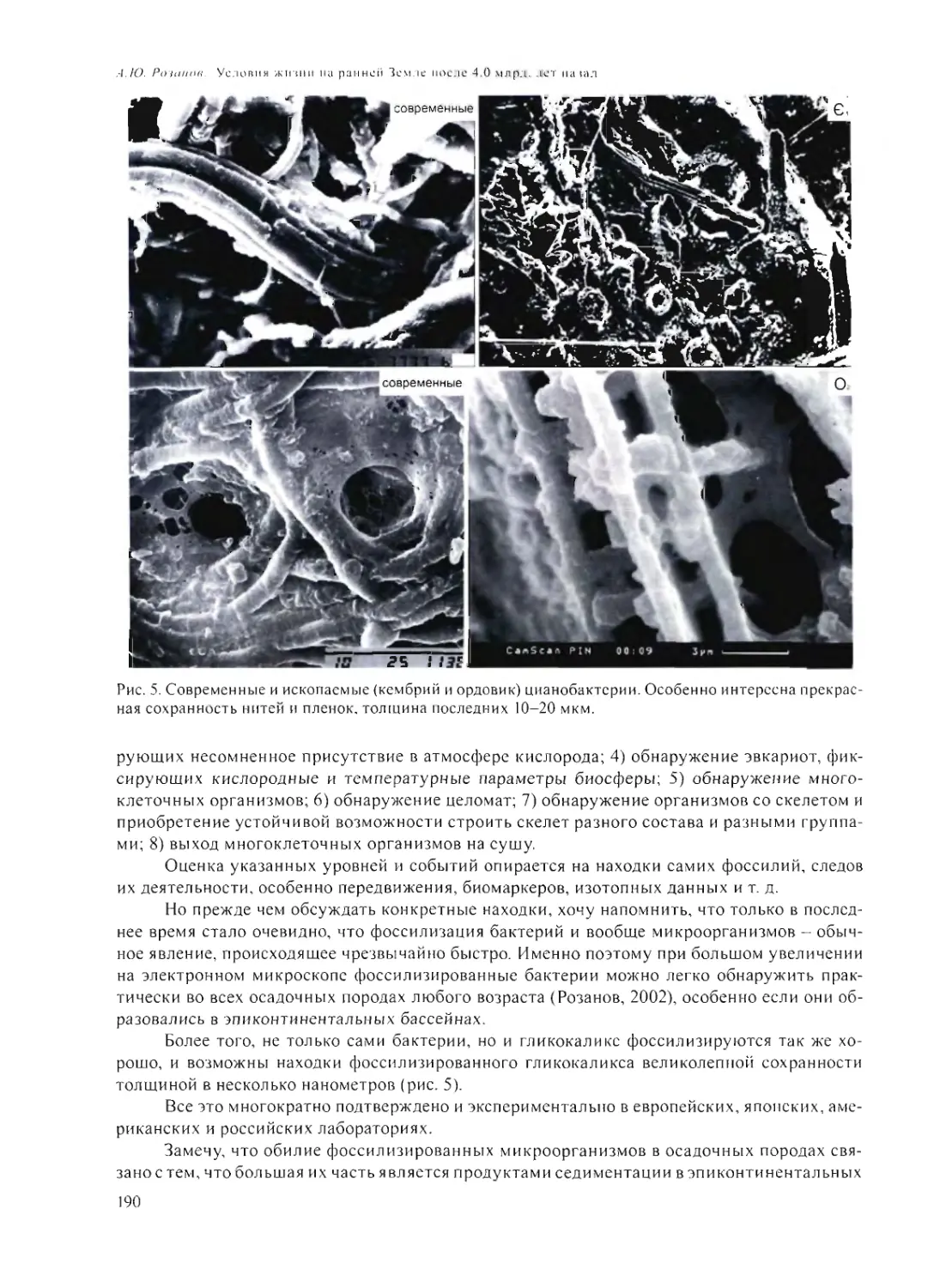
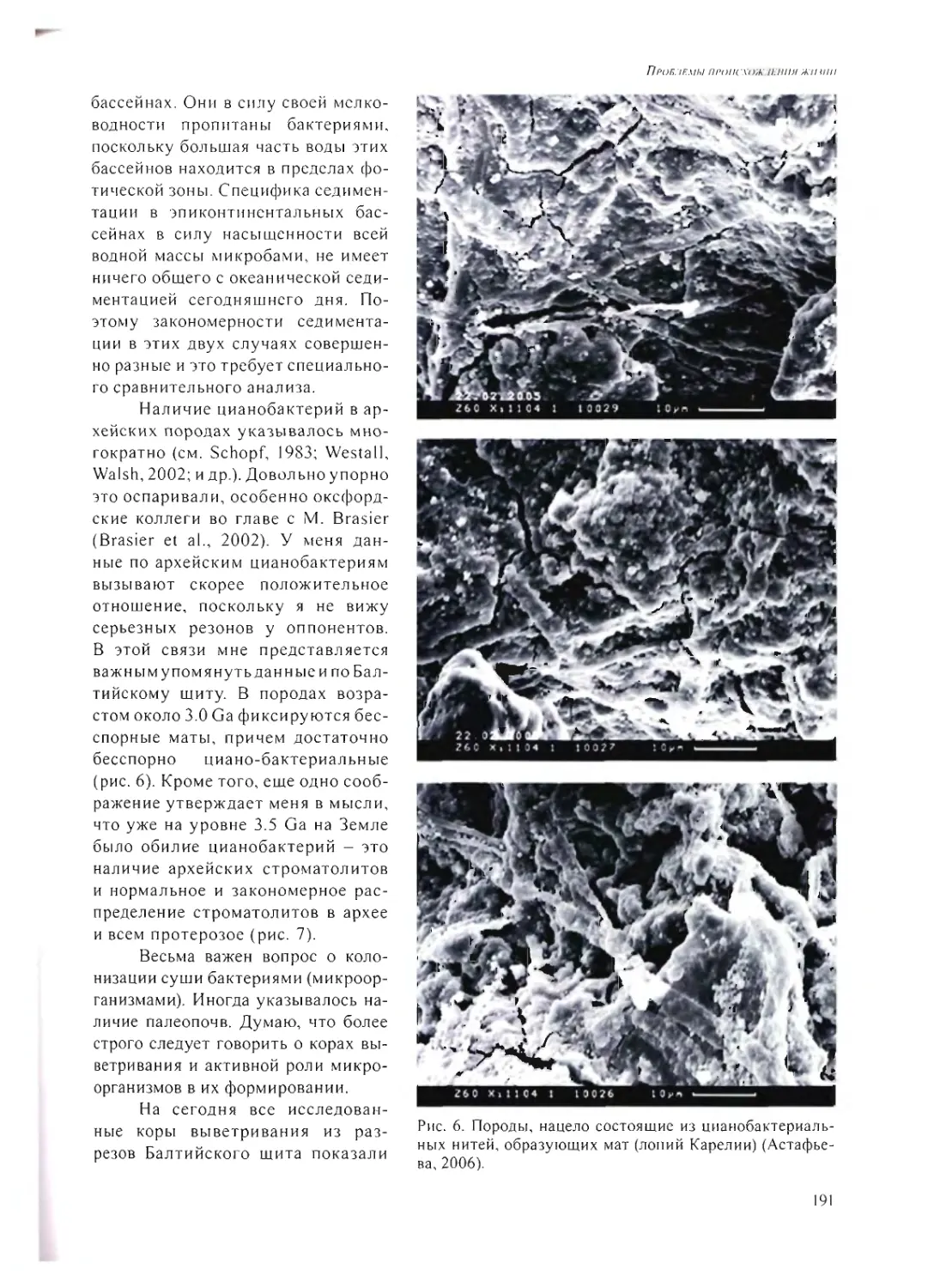
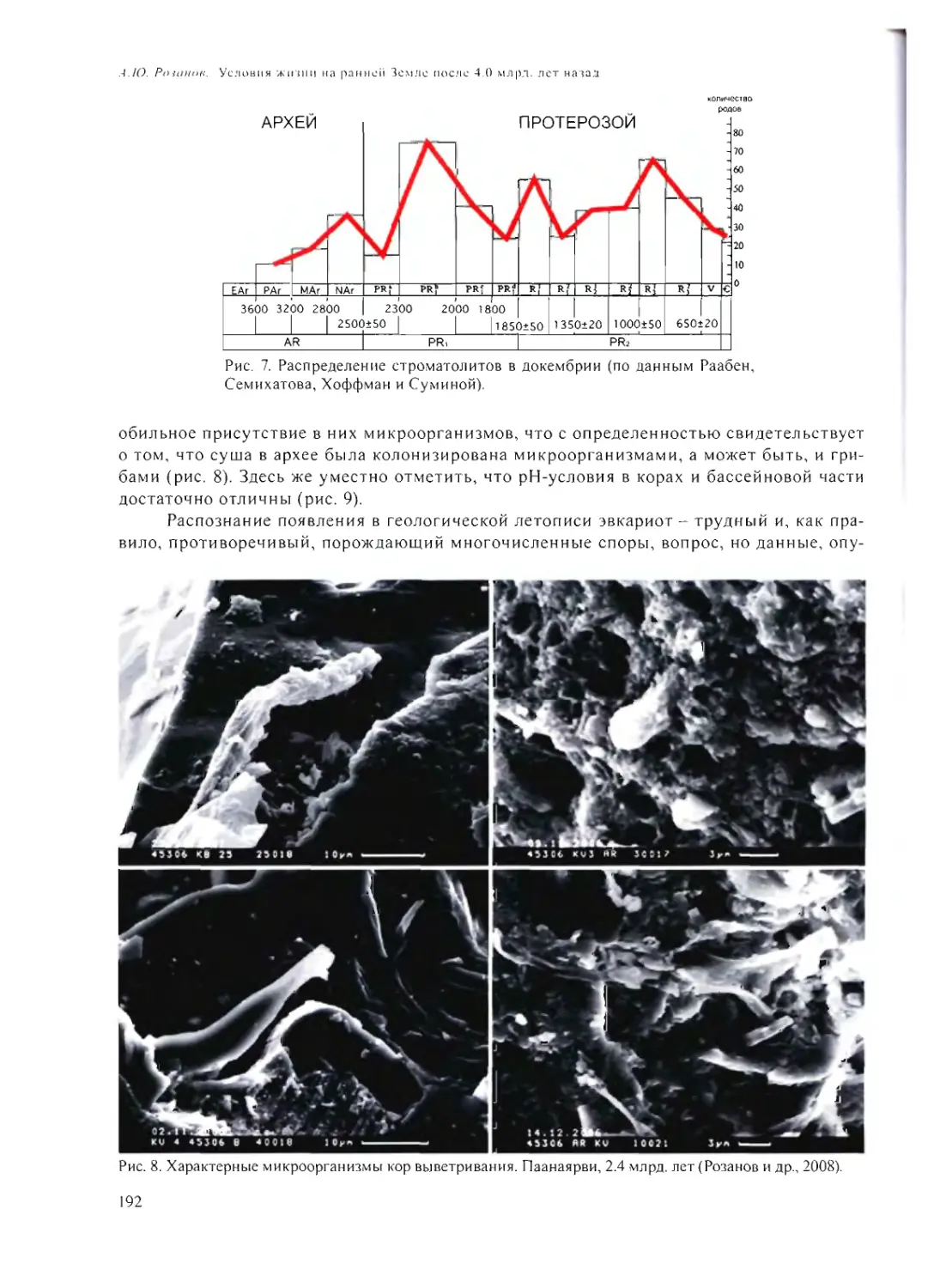
А.Ю. Розанов 185
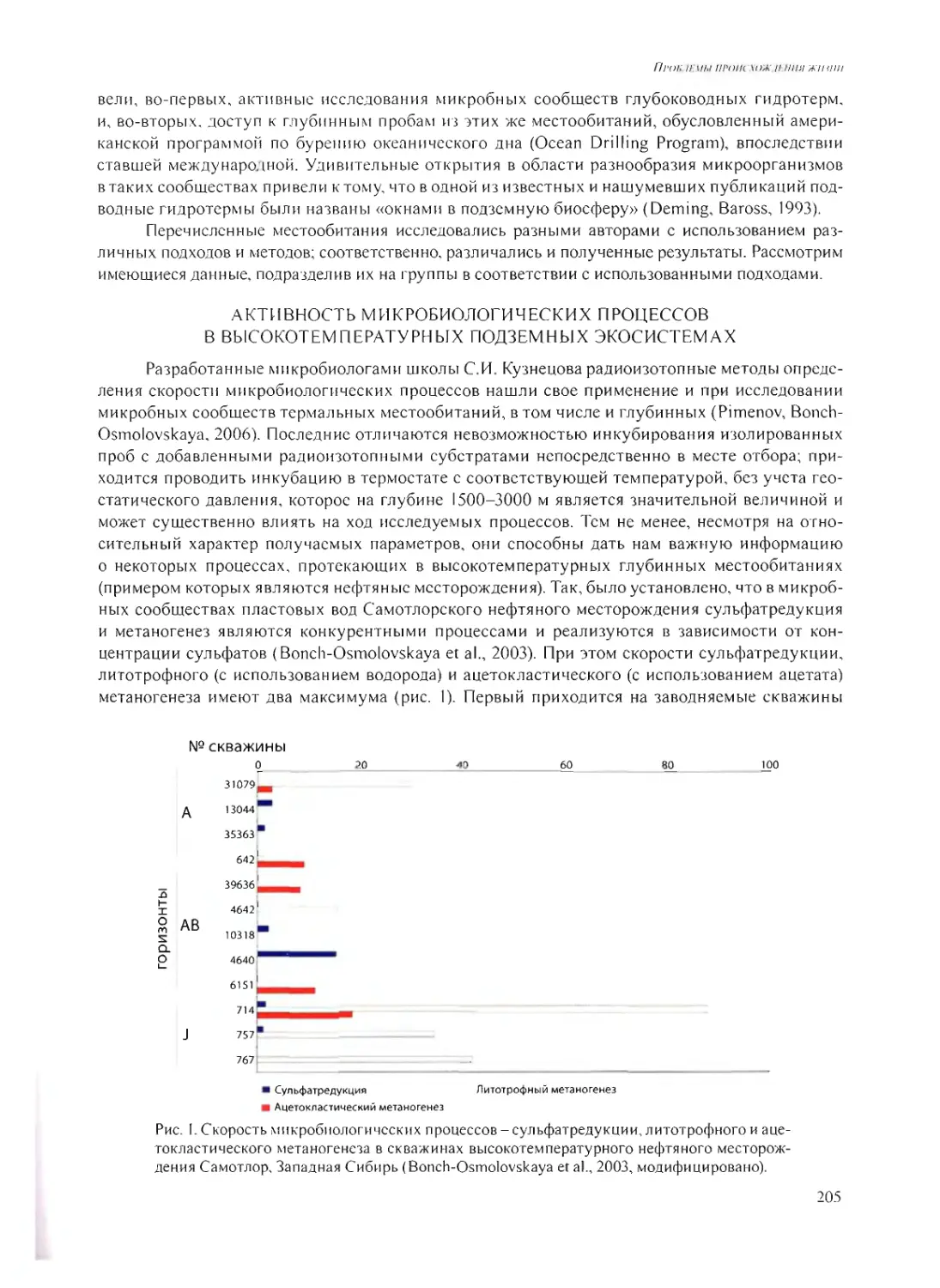
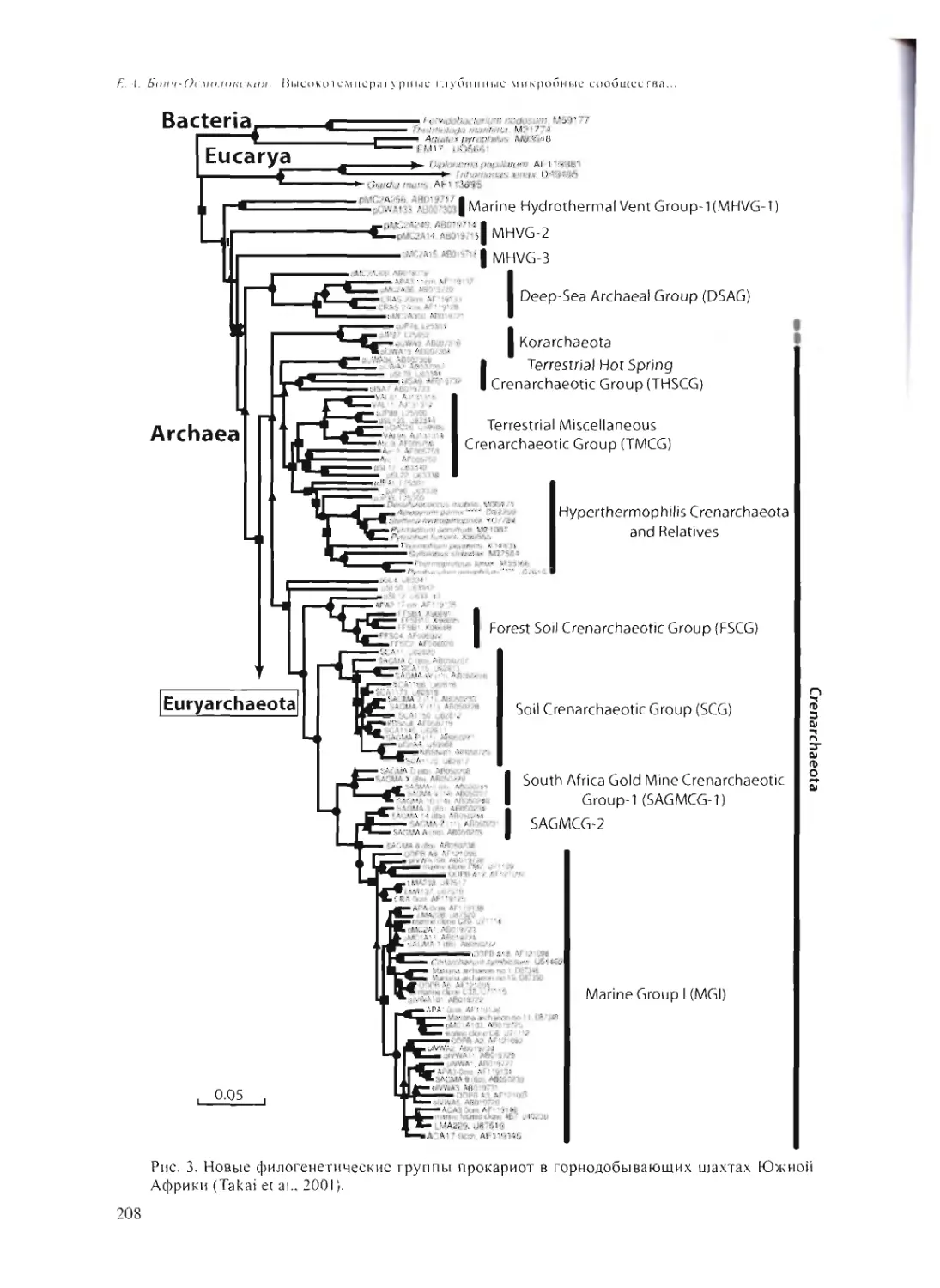
ВЫСОКОТЕМПЕРАТУРНЫЕ ГЛУБИННЫЕ МИКРОБНЫЕ СООБЩЕСТВА -
ВОЗМОЖНЫЙ АНАЛОГ ДРЕВНЕЙШИХ ЭКОСИСТЕМ?
Е.А. Бонч-Осмоловская 202
ВОЗНИКНОВЕНИЕ МЕМБРАН
Ю.В. Наточин 215
ПЕРВЫЕ ЭКОСИСТЕМЫ НА ЗЕМЛЕ
ГА. Заварзин 230
ИТОГОВАЯ ДИСКУССИЯ 245
ПРИЛОЖЕНИЕ /. 255
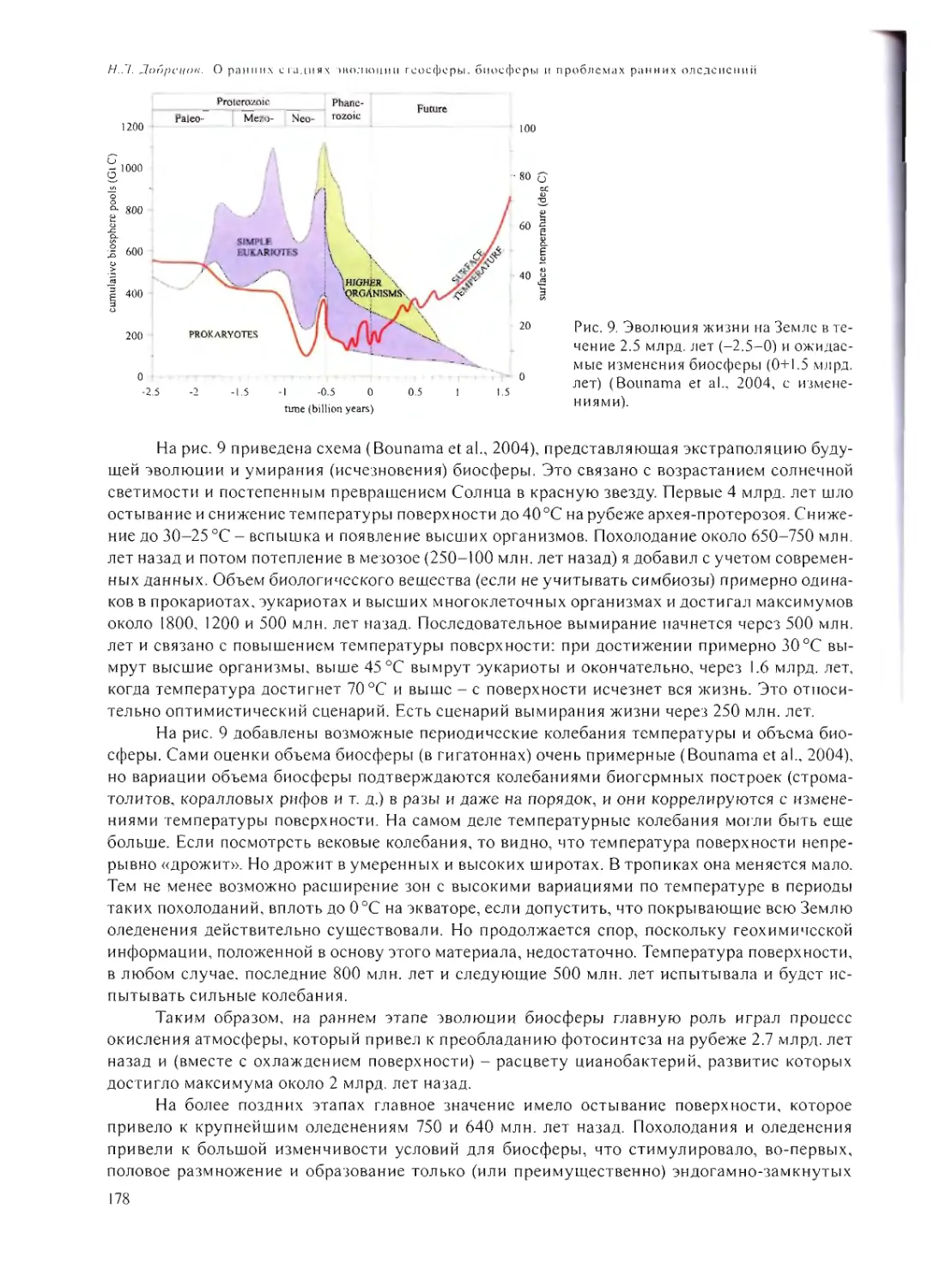
ПРИЛОЖЕНИЕ 2. 257
5
CONTENT
PREFACE 7
CAN A CELL BE ASSEMBLED FROM ITS CONSTITUENTS?
A.B. Chet ver in 9
VIRUS - BEFORE OR AFTER THE CELL?
V.l. Ago! 31
ANCIENT RNA WORLD
A.S. Spirin 43
FROM MARS TO EUROPA: SEARCH FOR A BIOSPHERE ON SATELLITES
OF GIANT PLANETS
L.V. Ksanfomality 60
ASTROCATALYSIS - A BIOGENIC SYNTHESIS AND CHEMICAL EVOLUTION
ON PRE-GEOLOGIC STAGES OF EARTH FORMATION
V.N. Snytnikov 79
MOLECULES IN SPACE AND MIGRATION OF MOLECULES
N.G. Bochkarev 102
CONDITIONS ON THE EARTH 4-4.6 GA. THE PRIMARY SYNTHESES
L.M. Mukhin 120
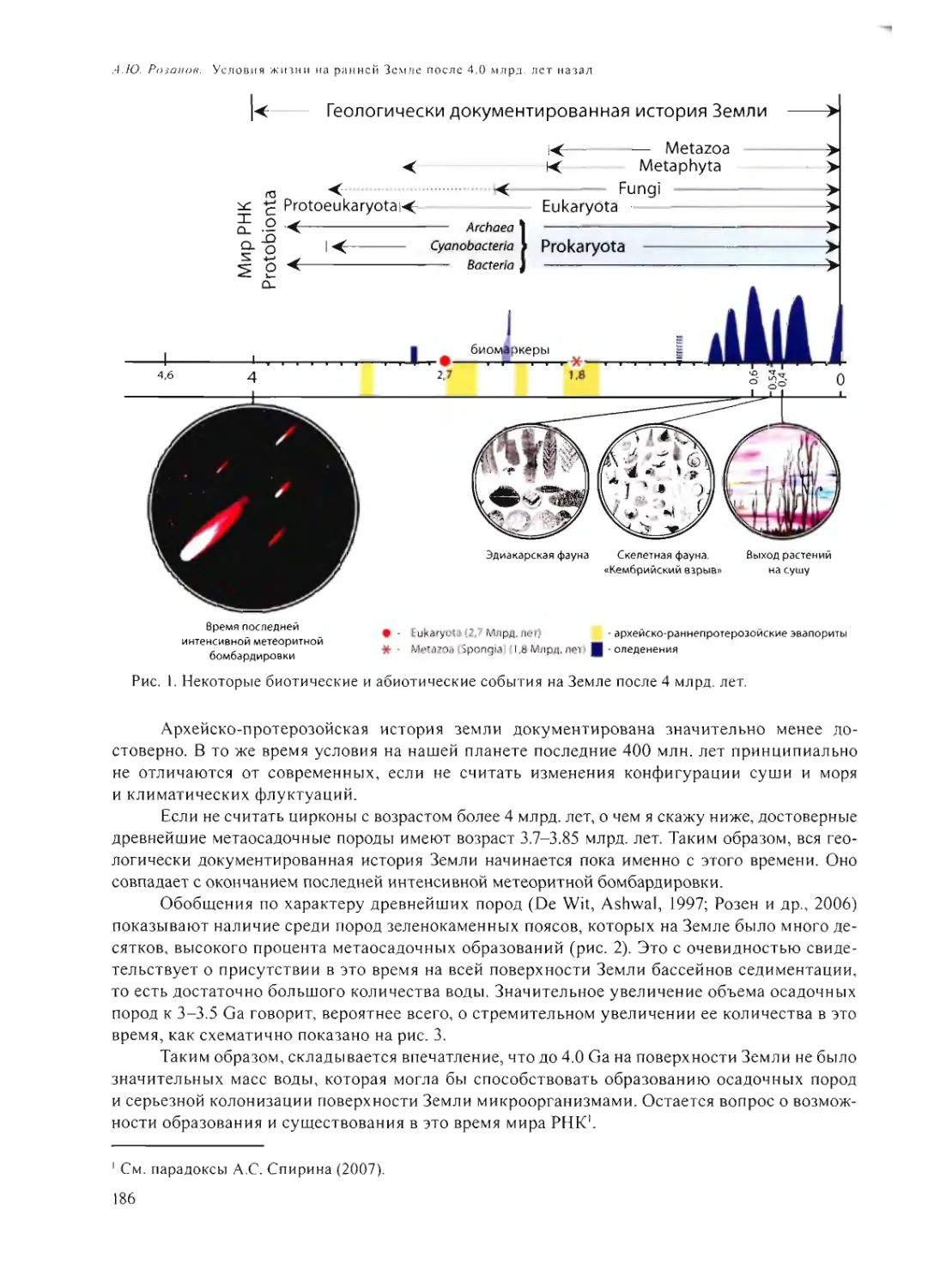
THE EARLY EARTH CRAMPED BY SURROUNDING YOUNG STARS
Vityazev A.V., Pechernikova G.V. 131
PSEUDOMORPHIC MICROBIAL STRUCTURES IN METEORITES
A.Yu. Rozanov 158
ON THE EARLY STAGE OF GEOSPHERE AND BIOSPHERE EVOLUTION,
AND THE PROBLEM OF THE EARLY GLACIATIONS
N.L. Dobretsov 168
LIFE CONDITIONS ON TH E EARLY EARTH AFTER 4.0 GA
A.Yu. Rozanov 185
HIGH-TEMPERATURE, DEEP SUBSURFACE MICROBIAL COMMUNITIES -
ARE THEY THE POSSIBLE ANALOGUES OF ANCIENT ECOSYSTEMS?
E.A. Bonch-Osmolovskaya 202
THE APPEARANCE OF MEMBRANES
Yu.V. Natochin 215
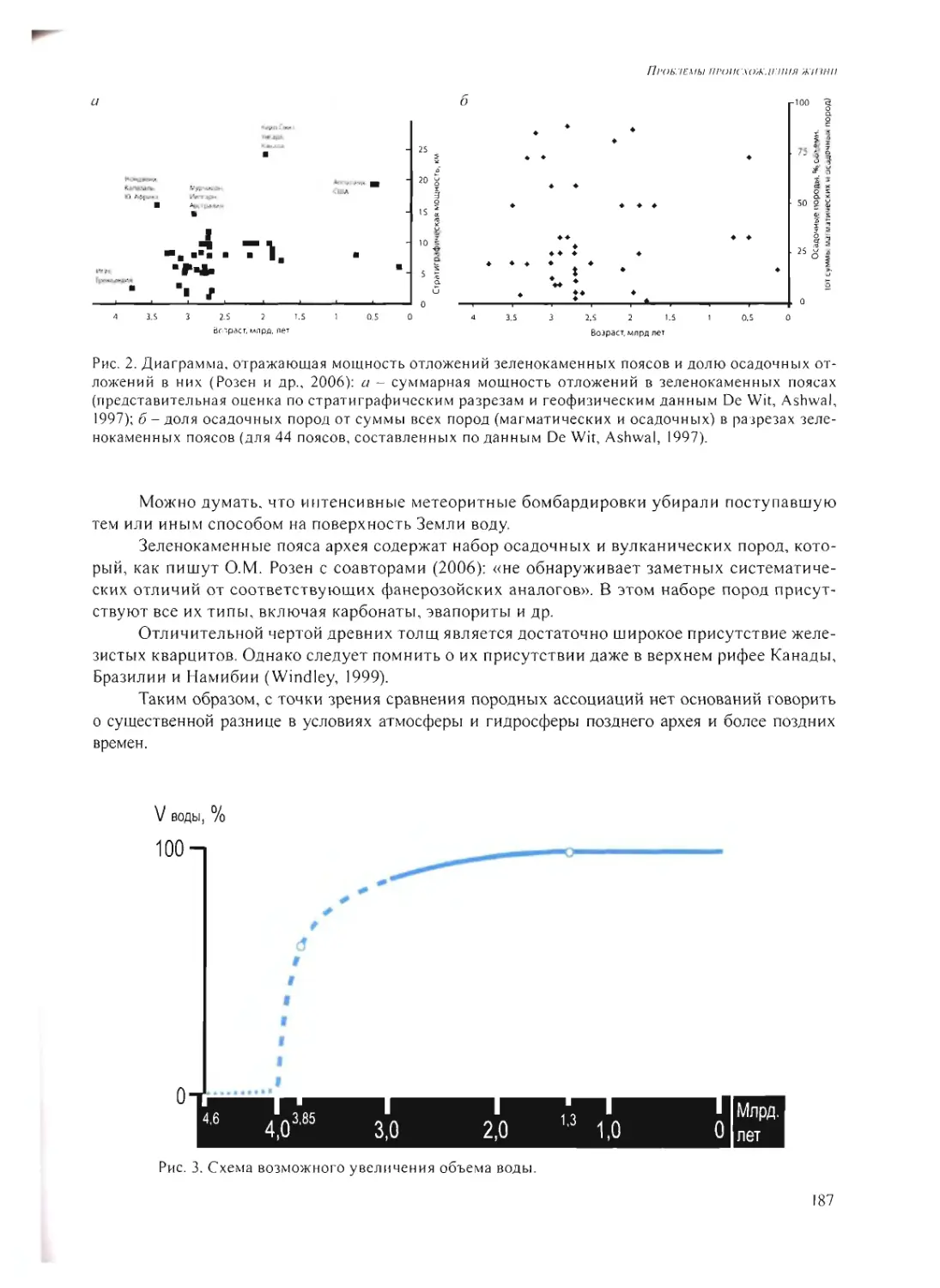
FIRST ECOSYSTEMS ON THE EARTH
G.A. Zavarzin 230
FINAL DISCUSSION 245
APPENDIX 1. 255
APPENDIX 2. 257
6
ПРЕДИСЛОВИЕ
В 2006 г. акад. Н.С. Кардашев и проф. Л.М. Мухин встретились с акад. А.Ю. Розановым
с предложением собрать небольшую группу коллег (коллоквиум) для обсуждения проблем
происхождения жизни. Сформированная рабочая группа включала, кроме упомянутых орга-
низаторов, акад. Н.Л. Добрецова, акад. Г.А. Заварзина, акад. А.С. Спирина, д.б.н. С.В. Рожнова
и д.б.н. А.В. Лопатина. Затем присоединились акад. О.Г. Газенко, акад. А.И. Григорьев, к.ф.-м.н.
В.Н. Снытников. На заседания, которые проводились в ПИН один раз в месяц-полтора, при-
глашались авторитетные специалисты с сообщениями по возникавшим острым проблемам:
чл.-корр. РАН В.Н. Агол, д.ф.-м.н. А.В. Витязев, акад. Ю.В. Наточин, акад. М.Я. Маров и др.
С самого начала возникла идея собрать через некоторое время небольшое совещание
из 30-35 человек.
Участники рабочей группы отдавали себе отчет в том, что совещание должно вклю-
чать не все аспекты исследований, которые так или иначе связаны с происхождением жизни,
а лишь те, где в последнее время появился значимый элемент новизны. Кроме того, было
решено сделать это обсуждение в достаточно узком кругу, предоставив всем участникам воз-
можность затратить максимум необходимого времени на дискуссию.
Мы также отдавали себе отчет в том, что при всех упомянутых выше ограничениях
круг обсуждаемых проблем должен быть достаточно широк и мультидисциплинарен.
Мы договорились, что не станем обсуждать формулировки понятия «жизнь». Мы по-
нимали, что это может увести нас в терминологические дебри. Заседание показало, что каж-
дый из нас связывает начало того, что мы называем «жизнь», с разными этапами физико-
химического развития Вселенной. Однако все это позволяет наиболее продуктивно изу-
чать процесс развития материи и становления жизни и формулировать наиболее сложные
и острые вопросы и проблемы на пути познания этого становления. В этом смысле простая
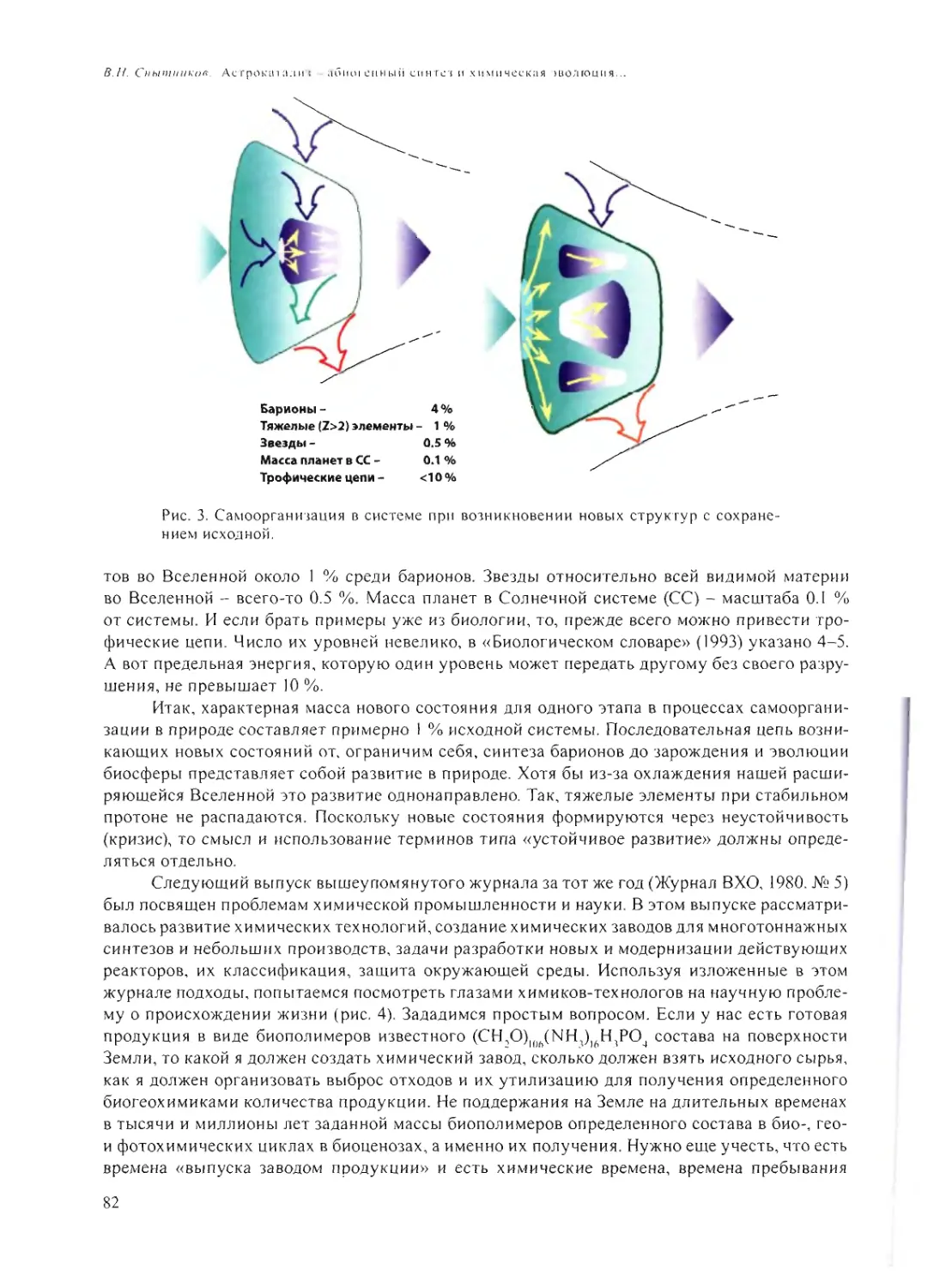
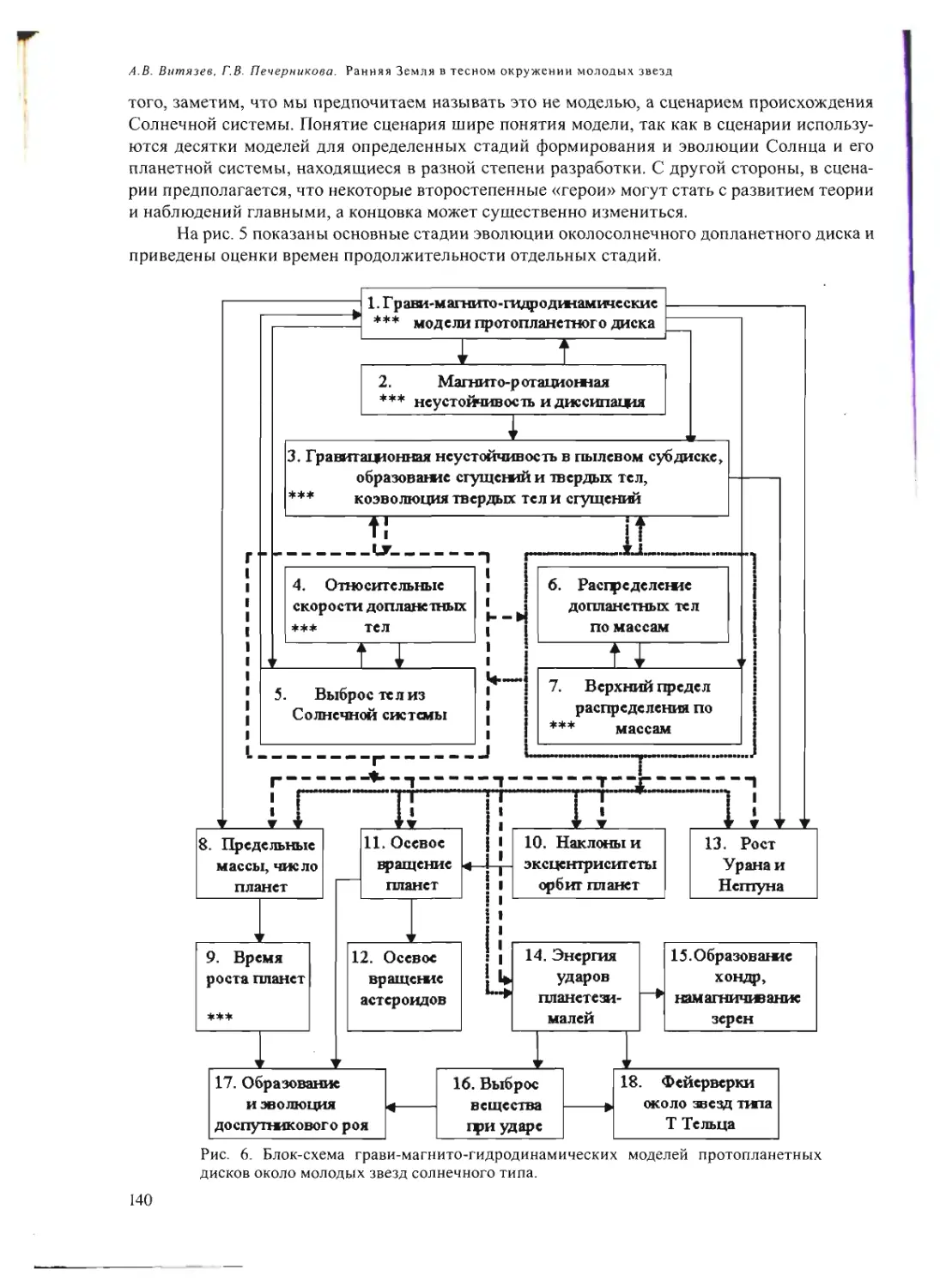
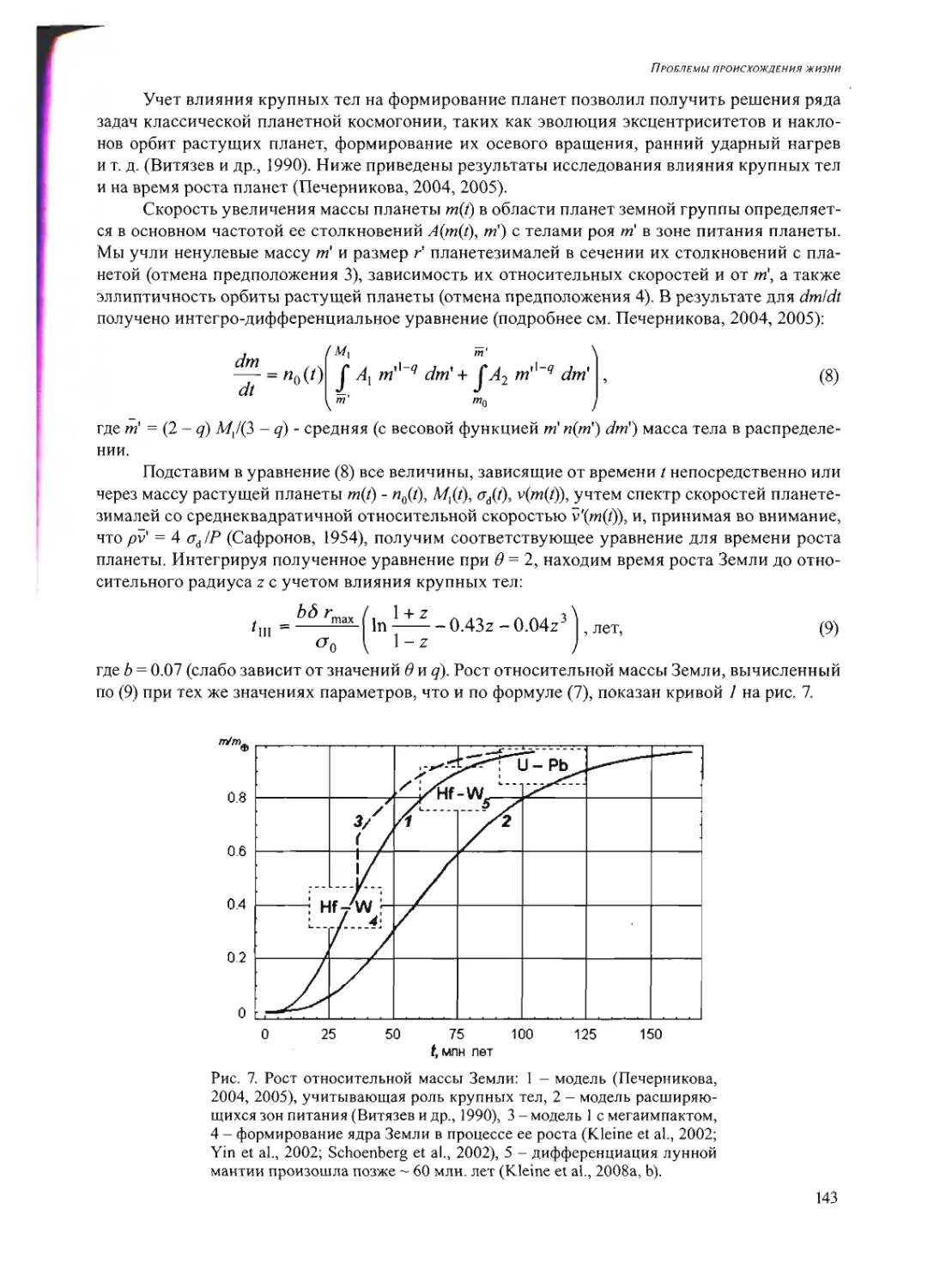
схема (см. рис.) может помочь восприятию сказанного.
Сложность данной проблемы еще и в том, что, в отличие от того, чем занимается на-
ука - повторяющими явлениями, для которых можно построить какие-то закономерности
и выявить отличительные особенности - здесь, по крайней мере, так считается, явление
не повторялось - жизнь появилась один раз. Соответственно, по одной точке построить гра-
фик невозможно, и что-то прогнозировать и предполагать крайне сложно. Такой мудрый чело-
век как Николай Владимирович Тимофеев-Ресовский (он работал у нас в Институте медико-
биологических проблем, проводил ежемесячные семинары, на которые приезжали со всей
Москвы), когда его спрашивали насчет происхождения жизни, говорил: «Вы знаете, я тогда
был еще совсем маленьким, я ничего не помню, поэтому вы спросите лучше у академика
Опарина, он, наверное, знает...». И, несмотря на то, что за последующее время стало кое-что
известно, все равно, к сожалению, это еще очень немного. Но - есть желание узнать больше.
7
Eukarya
Bacteria
Archaea
В результате такое небольшое рабочее совещание было подготовлено. На нем обсуж-
дались следующие блоки проблем: I) возможность сборки клетки из ее компонентов; 2) «ви-
русы - до или после клетки?»; 3) мир РНК и возможность его существования на предземной
стадии; 4) органические вещества во Вселенной и значение астрокатализа в проблеме про-
исхождения жизни; 5) условия на ранней Земле 4.6-4.0 млрд, лет назад; 6) условия жизни
на ранней Земле после 4.0 млрд, лет назад; 7) первичные экосистемы и глубинная биосфера;
8) проблема происхождения мембран.
Обозначенные блоки проблем, естественно, предопределили исключительную муль-
тидисциплинарность совещания, а широкая и неограниченная дискуссия способствовала
значительному сближению специалистов разного профиля и нахождению общего языка. Мы
собрали представителей многих специальностей: здесь были и палеонтологи, и астрофизики,
и «чистые» биологи, и физиологи, и генетики, и молекулярные биологи, физики, химики,
геологи... И всем нам что-то известно лучше, а что-то хуже... Но все хотели бы узнать боль-
ше. Поэтому такая неформальная дискуссия помогла всем вместе как-то продвинуться.
Прошедшее совещание, в котором приняли участие 36 человек (полный список см. при-
ложение), было столь интересным, что созрела идея публикации его материалов. В англий-
ской версии (приложение к «Палеонтологическому журналу») публикуются только доклады,
а в русской версии - и полная дискуссия. Все совещание было записано на видеокамеру и
предполагается изготовить компакт-диски с полной записью докладов и дискуссий. Кро-
ме того, материалы совещания составят основу возможной новой Программы Президиума
РАН.
Сборник посвящен светлой памяти наших дорогих коллег, академика Олега Георгие-
вича Газенко (12.12.1918-17.11.2007) и профессора Льва Михайловича Мухина (24.10.1933-
24.04.2009).
А. И. Григорьев
8
Проблемы нроисхожлкния жиши. М.: ПИН РАН, 2009. С. 9-27
МОЖНО ЛИ СОБРАТЬ КЛЕТКУ ИЗ ЕЕ КОМПОНЕНТОВ?
© 2009 г. А.Б. Четверин
Институт белка РАН
alexch@vega.protres.ru
К настоящему времени составлен примерный список минимального набора
генов, необходимых для функционирования и размножения живой клетки
в максимально благоприятных условиях, разработаны методы полного
химического синтеза минимального генома, а также разработаны бескле-
точные системы, позволяющие осуществлять все биохимические реакции,
составляющих репликацию и экспрессию клеточного генома. Самой се-
рьезной из нерешенных проблем остается обеспечение двух взаимоисклю-
чающих требований: обособления биохимических процессов от окружаю-
щей среды и обмена между клеткой и средой. В этих условиях экспери-
ментальной моделью для сборки клеточных компонентов могут служить
молекулярные колонии (другие названия - наноколонии, полонии), кото-
рые образуются при репликации РНК или ДНК в твердой среде с порами
нанометрового размера. Молекулярные колонии могли также служить до-
клеточной формой компартментализации в мире РНК.
ВВЕДЕНИЕ
Приглашение выступить на рабочем совещании «Проблемы происхождения жизни»
стало для меня неожиданным, потому что я никогда не думал, что то, чем мы занимаемся,
имеет отношение к этой области. Однако в последние годы Александр Сергеевич Спирин
стал цитировать наши результаты в своих работах по происхождению мира РНК (Spirin, 2002;
Спирин, 2005а, б). И тогда я подумал, что то, что мы сделали, действительно имеет к этому
какое-то отношение. В процессе подготовки к докладу мне пришлось забраться в области,
в которых я не являюсь специалистом. Поэтому, если я не совсем точно изложу факты из
пограничных областей, прошу не судить строго. По просьбе редакционной коллегии сборника
во вводной части статьи я привожу краткие определения ключевых понятий и терминов, в
порядке их упоминания.
Ключевые понятия
Экспрессия генома - совокупность биохимических процессов, приводящая к синтезу
функционально активных белков путем декодирования информации, заключенной в нуклео-
тидной последовательности генов. Экспрессия ДНК-генома включает стадии транскрипции
(синтез РНК на ДНК-матрице) и трансляции (синтез белка на РНК-матрице).
9
А.Б. Четверни. Можно ли собрать клетку из ее ком ноненiон0
Репликация - размножение РНК или ДНК путем синтеза копий (реплик) исходной ма-
трицы.
Репликаза - катализатор (белок или РНК), осуществляющий репликацию.
Qfl-репликаза - белок фага Q{3 (бактериального вируса), осуществляющий репликацию
геномной РНК этого фага, а также некоторых других видов РНК.
Рибозим - РНК, способная катализировать ту или иную биохимическую реакцию.
Компартментализация - обособление; здесь: сосредоточение клеточных компонентов
и процессов в некоем объеме (компартменте), изолированном от внешней среды.
Липиды - вещества, содержащие гидрофильную группу («голову») и длинную гидро-
фобную углеводородную часть («хвост»). Формируют мембраны из двух слоев, обращенных
друг к другу гидрофобными, а к водному окружению - гидрофильными поверхностями.
Липосомы - водные пузырьки, окруженные мембраной из двух слоев липидов или ана-
логичных по свойствам веществ.
Нуклеотид - мономер РНК или ДНК, состоящий из одного из четырех видов гетероци-
клических азотистых оснований [А - аденин, G - гуанин, С - цитозин, U - урацил (в РНК) или
Т - тимин (в ДНК)], рибозы (пятну глеродный сахар) и фосфата (остаток фосфорной кислоты).
Полинуклеотид - линейный полимер, состоящий из нуклеотидов, соединенных фосфо-
диэфирными связями, и имеющий сахарофосфатный остов. Различают полирибонуклеоти-
ды (РНК) и полидезоксирибонуклеотиды (ДНК).
Комплементарностъ - дополнительность; здесь - точное структурное соответствие
между двумя полинуклеотидами, способными сформировать двойную спираль посредством
образования изоморфных пар оснований A:U (А:Т в ДНК) или G:C.
Праймер - затравка; здесь - олигонуклеотид, комплементарный участку последова-
тельности РНК- или ДНК-матрицы; добавляя к праймеру по одному нуклеотиду, репликаза
строит из них цепь, комплементарную матрице.
ПЦР (полимеразная цепная реакция) - процесс репликации ДНК с использованием
температуроустойчивой ДНК-зависимой ДНК-полимеразы (ДНК-репликазы), олигонуклео-
тидного праймера и дезоксирибонуклеотидов, включающий периодическое нагревание ре-
акционной среды с целью расплавления двуцепочечной ДНК на составные цепи и последую-
щего охлаждения с целью отжига праймера с матрицей и его удлинения ДНК-репликазой.
Молекулярные колонии (наноколонии) - скопления наномолекул-копий РНК- или ДНК-
матриц, образующиеся вокруг матриц при их репликации в твердой пористой среде, имею-
щей поры нанометрового размера. Как правило, в качестве твердой среды используют гидро-
фильные гели: агарозу или полиакриламид. В зарубежной литературе молекулярные коло-
нии часто называют «полонии» (от «полимеразные колонии»).
Монтмориллонит - глина, слоистый алюмосиликат из группы смектитов. Отличается
большой сорбционной и ионообменной емкостью.
1. МИНИМАЛЬНЫЙ РАЗМЕР КЛЕТОЧНОГО ГЕНОМА
Когда мы говорим о возможности сборки клетки, то, прежде всего, наверное, надо за-
дать вопрос: «А насколько это сложный объект? Какова по сложности может быть самая ма-
ленькая клетка?» Прежде всего, имеется в виду сложность информационная, генетическая.
Можно просто посмотреть, что на этот счет известно.
К)
П РОБЯЕМЫ ПРОИСХОЖДЕНИЯ ж и ши
Так, наименьший геном из свободно живущих организмов имеет фотосинтетическая
родопсин-содержащая бактерия Pelagihacter ubique, составляющая значительную долю био-
массы океана: I 308 759 пар оснований (п.о.) и I 354 генов (Giovannoni et al., 2005).
Однако есть более редуцированные геномы. Размер генома наименьшего культивируе-
мого организма (т. е. способного к размножению в искусственных условиях - in vitro) обли-
гатного паразита Mycoplasma genitalium равен 582 970 п.о.; этот геном состоит из 519 генов
(Fraser et al., 1995).
Наконец, существует рекордсмен по простоте устройства - бактерия Carsonella ruddi'i,
которая является внутриклеточным симбионтом тли. Ее геном имеет размер 159 662 п.о.
и состоит из 211 генов (Nakabachi et al., 2006). Хотя насчет того, можно ли ее рассматривать
в качестве бактерии, есть сомнения: существует дискуссия по этому поводу. Поскольку эту
тварь нельзя культивировать, она может быть рассмотрена как клеточная органелла, кото-
рая берет из клетки не только питательные вещества, но также белки и/или нуклеиновые
кислоты (Tamames et al., 2007). Таким образом, исходя из вышеприведенных фактов, мини-
мальный размер генома клетки должен быть порядка 200-500 тыс. пар оснований и состоять
из 200-500 генов.
Есть и иные подходы для того, чтобы оценить минимально возможный размер клеточ-
ного генома. Существует филогенетический анализ, когда сравнивают геномы разных орга-
низмов и смотрят, что у них есть общего. Считается, что общее - это то, что необходимо, а то,
что отличается, - это то, без чего можно обойтись. Так было определено, что 256 из 519 генов
М. genitalium являются эволюционно консервативными. Вероятно, эти гены представляют
собой минимальный набор, необходимый для функционирования клетки в наиболее бла-
гоприятных условиях: в присутствии полного набора питательных веществ и в отсутствие
какого-либо внешнего стресса (Mushegian et al., 1996).
Еще один подход для определения минимально необходимого набора генов состоит
в том, что случайным образом инактивируют гены бактерии - по одному за раз - и смотрят,
сможет ли такая бактерия жить. Для той же бактерии М. genitalium было показано, что инак-
тивация, путем вставки транспозона, 101 из 482 белок-кодирующих генов не препятствовала
росту клеток в максимально благоприятных условиях. Инактивация же любого из 381 остав-
шихся генов была летальной (Glass et al., 2006). В то же время нельзя сказать, что каждый
из 381 гена действительно необходим, потому что бактерия - это сложная взаимозависимая
система. Удаление какого-то элемента может привести к летальному исходу не потому, что
именно этот элемент необходим, а потому что от него может зависеть функционирование
других элементов.
Наконец, использовали биохимический подход: выясняли, какие биохимические ре-
акции необходимы для жизни клетки и, соответственно, какие для этого нужны ферменты
и структурные элементы. Из такого анализа был сделан вывод, что белки и РНК, осуществляю-
щие минимально необходимые биохимические реакции, кодируются 151 генами (113 тыс. п.о.)
(Forster, Church, 2006, 2007). Это близко к геному С. ruddii (Nakabachi et al., 2006).
2. ПОПЫТКИ СОЗДАНИЯ ИСКУССТВЕННОЙ КЛЕТКИ
В настоящее время вопрос о создании искусственной клетки уже рассматривается
в практической плоскости, а соответствующие проекты щедро финансируются в надежде
на будущую прибыль. Предполагается, что искусственные клетки могут помочь в создании
новых лекарств и технических материалов, а также в решении проблем энергетики (произ-
водство биотоплива) и экологии (утилизация отходов). Появились даже такие понятия как
«синтетическая геномика» и «синтетическая биология».
Основателем синтетической геномики является Крейг Вентер - тот самый, в чьей фир-
ме Celera Genomics расшифровали первый геном человека - геном самого Вентера. А сей-
II
А.Б. Четкерин. Можно ли собрать клетку из се компонентов?
час в Институте Вентера (J. Craig Venter Institute) решают задачу создания искусственного
организма. К настоящему времени сотрудники института трансплантировали ДНК из одной
бактериальной клетки в другую, уничтожив ДНК бактерии-реципиента. Было показано, что
трансплантация всего генома привела к тому, что реципиент приобрел все признаки, опреде-
ляемые геномом донора (Lartigue et al., 2007). А буквально в этом году ими опубликовано
сообщение о химическом синтезе полного генома М. gemtalium - более 500 тыс. п.о. (Gibson et
al., 2008). Пока еще не проверено, действительно ли этот геном является биологически функ-
циональным, но синтетическая часть работы завершена. Это достижение интересно не про-
сто тем, что поставлен рекорд синтеза длинной ДНК, но также и тем, что теперь становится
возможным проектировать искусственные геномы, абсолютно новые, их синтезировать и та-
ким образом лепить организм по своему собственному усмотрению.
Идеологами синтетической биологии являются Энтони Форстер (Anthony С. Forster)
и Джордж Черч (George М. Church). Они определили список из 151 гена для всех белков
и РНК, которые, как они считают, необходимы, чтобы происходила полноценная экспрес-
сия генома. Они также определили группы белков, которые отвечают за разные стадии экс-
прессии генома: от синтеза РНК до посттрансляционной модификации белков. По мнению
авторов, можно взять смесь этих компонентов; добавить туда ДНК, которая будет програм-
мировать синтез новых копий мРНК и, соответственно, белков; загрузить этими компонен-
тами липосомы, и тогда система начнет самовоспроизводиться - репродуцироваться (Forster,
Church, 2006, 2007). В случае успеха это привело бы к сборке живой клетки из ее компонен-
тов. В принципе, на этом можно было бы завершить мое выступление, потому что есть люди,
которые считают: «Да, ответ положительный: собрать клетку из ее компонентов можно».
Однако здесь есть одна проблема, на мой взгляд, существенная. Форстер и Черч забыли
про одну вещь. В свой список они нс включили белки, которые отвечают за синтез липидов,
за построение мембраны. Они не учли того, что в мембране должны быть системы транс-
порта, которые отвечают за связь клетки с окружающей средой. А это проблема серьезная
и до сих пор, как следует из научной литературы, она не решена.
3. НЕОБХОДИМОСТЬ КОМПАРТМЕНТАЛИЗАЦИИ
Действительно ли нужна мембрана? Почему нельзя все компоненты сгрузить в пробир-
ку? Почему это не клетка? В принципе, отдельные этапы экспрессии генома так и осущест-
вляют - в пробирке (/« vitro) происходят и синтез ДНК, и синтез РНК на ДНК, и трансляция
РНК в белок. Будет ли это клетка или нет?
В этой аудитории много специалистов, которые имеют свое представление о том, что
такое клетка и как ее определить. Как мне кажется, главный признак клетки и ее отличие
от просто порции биохимического раствора - это то, что клетка должна быть способной к эво-
люции. Если бы клетка не могла эволюционировать, не могла развиваться и не было бы отбора
в направлении улучшения ее признаков, то никогда бы не возник такой сложный организм как
человек. Конечно, такой вывод можно сделать лишь при допущении, что жизнь произошла
естественным путем. А для того, чтобы шла эволюция, необходима компартментализация -
обособление клетки от окружающей среды. В связи с этим приведу цитаты трех классиков.
Первый из них - это Уолтер Гилберт (Walter Gilbert), который, собственно говоря,
и является автором идеи мира PH К, сформулированной им в 1986 г. в журнале Nature (Gilbert,
1986). Гилберт считает, что компартментализация необходима, чтобы привязать ген к его
продукту. Поскольку отбор происходит, естественно, по свойствам продукта экспрессии
гена, а наследуется собственно ген, то продукт и ген должны всегда быть вместе, иначе эво-
люция невозможна (Gilbert, de Souza, 1999).
Александр Сергеевич Спирин считает, что в отсутствие сегрегации молекул невозможно
избирательное размножение улучшенных вариантов, возникающих путем мутаций или реком-
12
Проблемы происхождения жииш
бинаций, так как одинаково хорошо размножаются все молекулы, содержащиеся в бульоне.
То есть сегрегация нужна, чтобы как-то отличать хорошее от плохого (Спирин, 2005).
Джек Шостак (Jack W. Szostak) приводит такой аргумент. Если возникнет мутантная
репликаза, обладающая улучшенной каталитической активностью и точностью, то она бу-
дет лучше реплицировать другие - худшие - репликазы, но себя реплицировать не сможет.
И даже если она будет реплицирована другой репликазой, то ее дочерние молекулы «разбе-
гутся» и не смогут помочь друг другу (Szostak, 1999).
Можно привести еще много разных аргументов, почему надо отделять, индивидуали-
зировать, обособлять этот компартмент (клетку) от окружающей среды, но думаю, что этого
достаточно.
4. КОМПАРТМЕНТАЛИЗАЦИЯ В ЛИПОСОМАХ
Естественно предложить, что, поскольку существующие клетки окружены мембра-
ной, то давайте сделаем что-нибудь подобное искусственно. На первый взгляд, это сделать
нетрудно. Действительно, оказалось, что из эмульсии липидов довольно легко спонтанно
образуются липосомы, то есть такие пузырьки, которые окружены двухслойной липидной
мембраной. Они способны к самопроизвольному росту путем поглощения новых молекул
или мицелл липидов, а также к делению (после того, как липосома достигает определенного
размера, она начинает самопроизвольно дробиться) (Szostak, Bartel, Luisi, 2001).
Более того, еще в 1995 г. было показано, что если внедрить в липосому РНК
и QP-репликазу (это такой белок, который может реплицировать РНК, то есть синтезировать
РНК на РНК-матрице и, таким образом, размножать РНК очень эффективно), то дробление
липосом будет сопряжено с размножением РНК. Следовательно, новообразующиеся липосо-
мы содержат и репликазу, и РНК (Oberholzer, 1995). Внешне это напоминает деление клеток.
Однако в этом направлении, когда используют липосомы как аналог клеток, есть ряд
существенных проблем, которые до сих пор не решены и не позволяют сделать сборку клет-
ки экспериментальным предметом.
Считается, что происхождение жизни стартовало с мира РНК, где все биохимические
реакции осуществлялись рибозимами (Спирин, 2005). Шостак с соавторами отметили, что
рибозимы имеют оптимальную активность в присутствии высоких концентраций двухва-
лентных катионов. Но в этих условиях липосомы агрегируют, что мешает их росту и деле-
нию (Szostak, Bartel, Luisi, 2001).
Но есть еще более серьезная проблема. Для того чтобы внутри липосомы происходил
синтез ДНК, РНК и белков, необходимо, чтобы внутрь этого компартмента поступали суб-
страты, из которых все синтезируется - нуклеотиды, аминокислоты и другие низкомолеку-
лярные компоненты.
С одной стороны, липидная мембрана изолирует клетку от окружающего простран-
ства, не дает разбегаться высокомолекулярным компонентам, обособляет ее. В этом состоит
ее положительный эффект. Но с другой стороны, мембрана является плохо преодолимым
барьером для низкомолекулярных веществ, которые должны поступать в клетку снаружи.
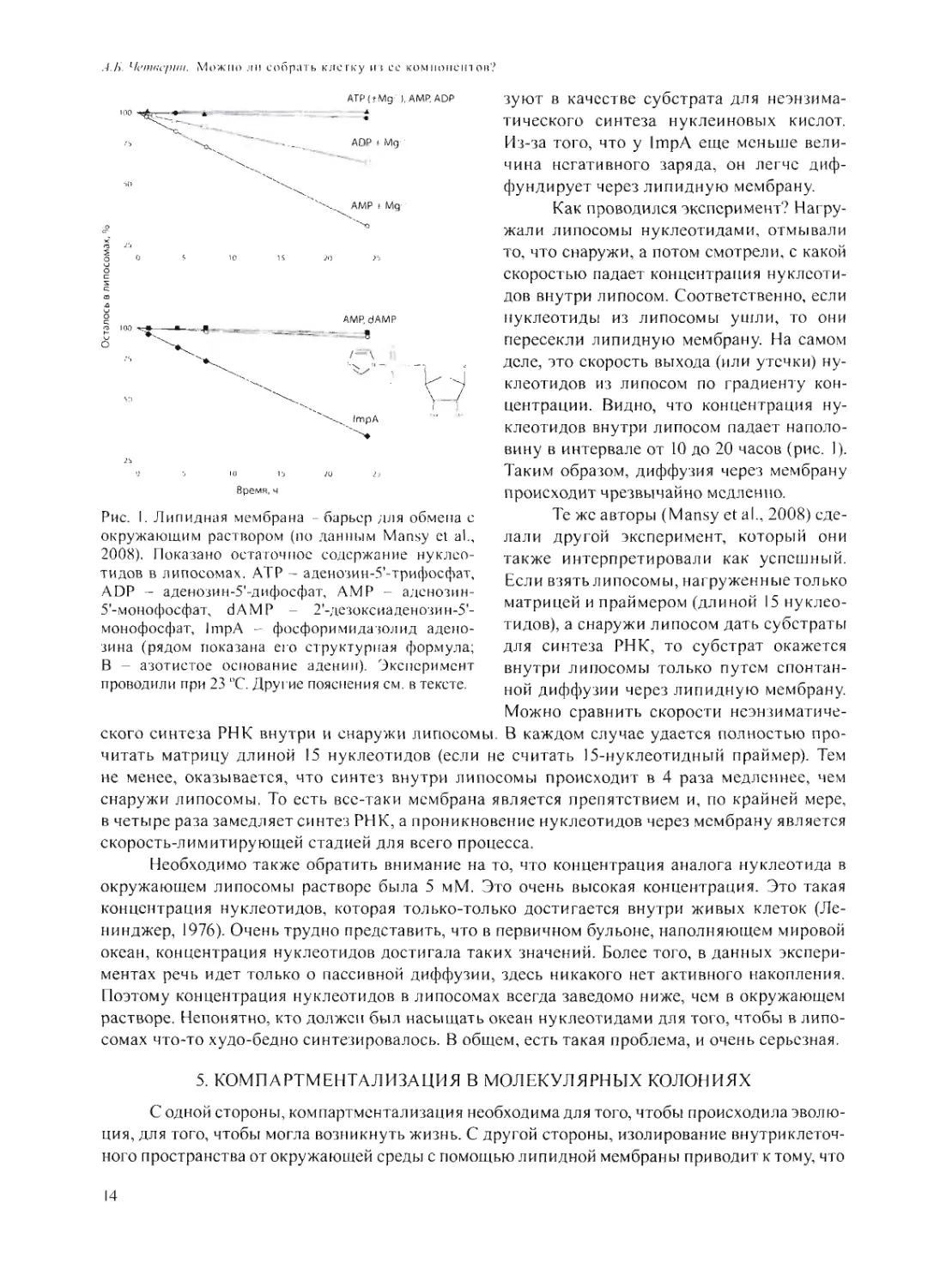
На рис. 1 показаны результаты экспериментов, которые авторы интерпретируют как доказа-
тельство проницаемости мембраны для низкомолекулярных компонентов (Mansy et al., 2008).
Для меня же они являются свидетельством того, что проницаемость мембраны очень низка.
Если посмотреть, как спонтанно диффундируют через липидную мембрану нуклео-
тиды, то оказывается, что АТР (аденозинтрифосфат) совсем не диффундирует. Менее заря-
женные ADP (аденозиндифосфат) и АМР (аденозинмонофосфат) в присутствии ионов Mg2
(который благодаря электростатическим взаимодействиям экранирует негативный заряд на
нуклеотиде) транспортируются несколько лучше. Еще лучше транспортируется ImpA (фос-
форимидазолид аденозина) - активированный аналог нуклеотида, который широко исполь-
13
ATPftMg- ), AMRADP
0 5 К) 1-J 20 2,-
Время, ч
Рис. I. Липидная мембрана - барьер для обмена с
окружающим раствором (по данным Mansy et al.,
2008). Показано остаточное содержание нуклео-
тидов в липосомах. АТР - аденозин-5'-трифосфат,
ADP - аденозин-5'-дифосфат, АМР - аденозин-
5'-монофосфат, dAMP - 2'-дезоксиаденозин-5'-
монофосфат, ImpA - фосфоримидазолид адено-
зина (рядом показана его структурная формула;
В - азотистое основание аденин). Эксперимент
проводили при 23 “С. Другие пояснения см. в тексте.
.-/.Л’, Четкернц. Можно ли собрать клетку из се компонентов?
зуют в качестве субстрата для неэнзима-
тического синтеза нуклеиновых кислот.
Из-за того, что у ImpA еще меньше вели-
чина негативного заряда, он легче диф-
фундирует через липидную мембрану.
Как проводился эксперимент? Нагру-
жали липосомы нуклеотидами, отмывали
то, что снаружи, а потом смотрели, с какой
скоростью падает концентрация нуклеоти-
дов внутри липосом. Соответственно,если
нуклеотиды из липосомы уйти, то они
пересекли липидную мембрану. На самом
деле, это скорость выхода (или утечки) ну-
клеотидов из липосом по градиенту кон-
центрации. Видно, что концентрация ну-
клеотидов внутри липосом падает наполо-
вину в интервале от 10 до 20 часов (рис. I).
Таким образом, диффузия через мембрану
происходит чрезвычайно медленно.
Те же авторы (Mansy et al., 2008) сде-
лали другой эксперимент, который они
также интерпретировали как успешный.
Если взять липосомы, нагруженные только
матрицей и праймером (длиной 15 нуклео-
тидов), а снаружи липосом дать субстраты
для синтеза РНК, то субстрат окажется
внутри липосомы только путем спонтан-
ной диффузии через липидную мембрану.
Можно сравнить скорости нсэнзиматиче-
ского синтеза РНК внутри и снаружи липосомы. В каждом случае удается полностью про-
читать матрицу длиной 15 нуклеотидов (если не считать 15-нуклеотидный праймер). Тем
не менее, оказывается, что синтез внутри липосомы происходит в 4 раза медленнее, чем
снаружи липосомы. То есть все-таки мембрана является препятствием и, по крайней мере,
в четыре раза замедляет синтез РНК, а проникновение нуклеотидов через мембрану является
скорость-лимитирующей стадией для всего процесса.
Необходимо также обратить внимание на то, что концентрация аналога нуклеотида в
окружающем липосомы растворе была 5 мМ. Это очень высокая концентрация. Это такая
концентрация нуклеотидов, которая только-только достигается внутри живых клеток (Ле-
нинджер, 1976). Очень трудно представить, что в первичном бульоне, наполняющем мировой
океан, концентрация нуклеотидов достигала таких значений. Более того, в данных экспери-
ментах речь идет только о пассивной диффузии, здесь никакого нет активного накопления.
Поэтому концентрация нуклеотидов в липосомах всегда заведомо ниже, чем в окружающем
растворе. Непонятно, кто должен был насыщать океан нуклеотидами для того, чтобы в липо-
сомах что-то худо-бедно синтезировалось. В общем, есть такая проблема, и очень серьезная.
5. КОМПАРТМЕНТАЛИЗАЦИЯ В МОЛЕКУЛЯРНЫХ КОЛОНИЯХ
С одной стороны, компартментализация необходима для того, чтобы происходила эволю-
ция, для того, чтобы могла возникнуть жизнь. С другой стороны, изолирование внутриклеточ-
ного пространства от окружающей среды с помощью липидной мембраны приводит к тому, что
14
Проблемы происхождении жити
там никакие клеточные процессы не могут
происходить с нормальными скоростями.
В связи с этим я подумал, а почему
Александр Сергеевич Спирин предложил
мне сделать этот доклад? Может быть,
дело в этом. Нами были изобретены моле-
кулярные колонии (Chetverin, Chetverina,
Munishkin, 1991; Chetverin, Chetverina,
1993, 1997, 2008) или наноколонии (Четве-
рина, Четверин, 2008), так их тоже можно
называть в связи с последними поветрия-
ми. В данном случае действительно умест-
но использовать приставку «нано», потому
что эти колонии состоят из молекул на-
норазмера, то есть ДНК или РНК, и они
(колонии) образуются, если РНК или ДНК
размножают в геле - иммобилизованной
среде, чей полимерный матрикс имеет на-
нометровые поры. Таким образом, размер
молекул примерно сопоставим с размером
пор этого матрикса. Это приводит к тому,
что молекулы оказываются «пойманны-
Рис. 2. Молекулярные колонии (наноколонии)
как способ компартментализации биополимеров
и биохимических реакций в отсутствие мембраны.
Наноколонии (показаны зеленым) формируются,
когда матричные наномолекулы (ДНК или РНК)
размножают в иммобилизованной среде с нано-
метровыми порами - например, в агарозном или
полиакриамидном геле. Каждая колония является
скоплением молекул - копий родительской моле-
кулы, то есть представляет собой молекулярный
клон.
ми» в этом матриксе как мухи в паутине: они «трепыхаются», но никуда не могут деться.
Если молекула прокопировалась (образовалась ее копия), копия остается примерно там же,
где была родительская молекула. Потомство одиночной молекулы образует большое скопле-
ние молекул, которые фактически являются молекулярным клоном. Если посмотреть на эти
скопления, то становится понятным, что они похожи на клетки (рис. 2). Единственное отли-
чие от клеток - это то, что они не окружены липидной мембраной. Тем не менее, здесь имеет
место компартментализация - обособление колоний от окружающей среды.
Наноколонии мы изобрели, работая с Qp-репликазой (Chetverin, Chetverina, Munishkin,
1991), о которой я уже упоминал. Qp-репликаза «засветилась» многими своими сторонами
в проблеме происхождения жизни и эволюции. Qp-репликаза - это РНК-зависимая РНК-
полимераза бактериофага Qp, который паразитирует на клетках Е. coli. Она знаменита тем,
что это самый эффективный фермент, который умеет размножать РНК в пробирке, то есть
вне клетки (Ugarov, Chetverin, 2008). Что она делает? Матрицей у нее является одноцепочеч-
ная РНК, и QP-репликаза, ее считывая, собирает из нуклеотидов комплементарную цепочку
РНК. После окончания цикла считывания, как исходная матрица, так и комплементарный ей
продукт являются матрицами для синтеза своих комплементарных копий. Таким образом,
получается, что из одной матрицы образовалось две матрицы, из двух матриц при следую-
щем копировании - четыре матрицы и так далее. В каждом цикле репликации число ма-
триц удваивается, то есть по экспоненте происходит увеличение числа РНК-матриц (Haruna,
Spiegelman, 1965).
6. «СПОНТАННЫЙ» СИНТЕЗ РНК
Одним необычным, загадочным свойством QP-репликазы был, так называемый, спон-
танный синтез РНК. Оказалось, что если взять очищенную QP-репликазу, смешать ее с чи-
стыми нуклеотидами, не давая никакой матрицы, и проинкубировать в присутствии ионов
Mg2+ при комнатной температуре, а примерно через час подвергнуть эту инкубационную
смесь анализу с помощью электрофореза, то можно увидеть огромное разнообразие больших
15
A.К. Четверни. Можно ли собрать клетку из се компонен тов?
количеств РНК, которые спонтанно синтезировались - формально, без матрицы. Эти РНК
обозначаются RQ (от «Replicable by Qp replicase», то есть реплицируемые QP-репликазой),
а число, которое указано при RQ, отражает длину синтезируемой РИК в нуклеотидах (рис. 3).
Пара сотен нуклеотидов - это примерно середина диапазона RQ РНК, длина которых варьи-
рует от нескольких десятков до тысяч нуклеотидов (Chetverin, Spirin, 1995; Четверин, 1998).
Формально, RQ РНК синтезируются без матрицы. Это дало возможность Манфреду
Айгену (Manfred Eigen) и его сотрудникам утверждать, что они наблюдают в присутствии
Qp-репликазы самозарождение информационных молекул, причем очень быстро - в течение
часа. Из нуклеотидов получаются информационные молекулы, которые способны к эффек-
тивному самовоспроизведению (Sumper, Luce, 1975; Biebricher, Eigen, Luce, 1986).
Естественно возникает вопрос - какова вероятность случайного возникновения такой
молекулы, которая способна реплицироваться QP-репликазой? Оказалось, очень небольшая.
В специальных экспериментах, с помощью так называемой SELEX-процедуры, где генери-
руют случайные последовательности, а потом из них пытаются выбрать реплицируемые
молекулы, оказалось, что реплицируемыми худо-бедно, гораздо хуже, чем спонтанно син-
тезированные РНК, оказываются единичные молекулы из примерно 1012 стартового разноо-
бразия случайных последовательностей длиной 70 нуклеотидов (Brown, Gold, 1995). Таким
образом, вероятность спонтанного возникновения реплицирующихся молекул очень низка.
Тем не менее, в течение часа инкубации с QP-репликазой получается огромное разнообразие
довольно длинных молекул РНК, причем все способны к экспоненциальному размножению
Qp-репликазой.
Откуда берутся и каким образом возникают эти РНК? Мы заподозрили, что дело в за-
грязнении. Однако это надо было продемонстрировать, потому что Айген с сотрудниками
утверждали, что они все сделали очень чисто, что они полностью очистили Qp-репликазу,
и приводили доказательные эксперименты, что их препарат QP-репликазы содержит ме-
нее одной молекулы матрицы. И, тем не менее, они видели синтез РНК (Sumper, Luce, 1975;
Biebricher, Eigen, Luce, 1986).
RQ1 20
RQ223
Рис. 3. Qp-репликаза синтезирует разнообразные
РНК в отсутствие добавленной матрицы («спон-
танно»), QP-репликазу смешивают с ATP, GTP, СТР
и UTP, инкубируют 1 ч при 37 °C и анализируют
с помощью гель-электрофореза. РНК окраши-
вают в геле с помощью бромистого этидия.
Стрелки указывают на мажорные RQ РНК, об-
наруженные в продуктах спонтанного синтеза.
Тогда мы предположили, что мы на-
блюдаем нечто подобное тому, с чем Пастер
имел дело в XIX в. Речь идет о самозарож-
дении жизни в бульоне. До Пастера все го-
ворили: «Бульон является источником жиз-
ни. Ведь если дать постоять прокипяченно-
му бульону, то через неделю обязательно
что-нибудь в нем вырастет». Что сделал
Пастер? Он взял специальную колбу, у ко-
торой верхняя часть была изогнута таким
образом, что ничто более тяжелое, чем воз-
дух, в колбу не могло упасть. После того,
как он прокипятил бульон в такой колбе,
а затем оставил стоять, то в течение меся-
ца там ничего не выросло (Pasteur, 1860).
Таким образом, Пастером было показано,
что бактерии в бульоне самопроизвольно
не возникают, по крайней мере, в процессе
эксперимента, что они попадают в бульон
из воздуха. Там, в воздухе, летают микро-
организмы, которые попадают в бульон
и дают начало росту бактерий в бульоне.
16
Пгоь. n:\thi пгонсхожи-ння жнит
Рис. 4. RQ РНК переносятся по воздуху. В две
чашки Петри заливают агарозу, содержащую
Qp-репликазу и рибонуклеозидтрифосфаты. Одну
чашку оставляют закрытой (слева), а другую от-
крытой (справа). Чашки инкубируют в течение
1 часа при комнатной температуре, затем окраши-
вают агарозу бромистым этидием.
Мы предположили, что у нас про-
исходит что-то подобное. Ведь если бак-
терии могут летать в воздухе, то почему
бы не летать молекулам РНК, которые
гораздо меньше по массе9 И мы сделали
такой эксперимент (Chetverin, Chetverina,
Munishkin, 1991). Решили провести ре-
акцию размножения РНК не в жидкости,
как все до этого делали, а в геле. Исполь-
зовали легкоплавкую агарозу. Эту агарозу
смешали с репликазой и нуклеотидами,
в двух чашках Петри. Одну чашку оста-
вили открытой в течение часа, а другую
инкубировали закрытой. Они стояли ря-
дом на одном и том же столе. После часа
инкубации агарозу покрасили бромистым
этидием. Бромистый этидий связывается
прочно с высокомолекулярными полину-
клеотидами, потому что он - интеркали-
руюший краситель. Он встраивается меж-
ду плоскостями азотистых оснований. Соответственно, одиночные нуклеотиды не способны
связать бромистый этидий, а полинуклеотиды способны. И когда флуоресцирующая краска
оказывается в гидрофобном окружении между плоскостями оснований, то квантовый вы-
ход флуоресценции резко возрастает. Таким образом, в том месте, где синтезировалась РНК,
будет повышенная флуоресценция бромистого этидия. Оказалось, что, действительно, по-
сле инкубации в течение часа в агарозе образуются флуоресцирующие пятна, и этих пятен
гораздо больше в том случае, если чашка была открыта, чем когда она была закрыта. Это,
как нам представляется, является прямым указанием на то, что реплицирующиеся молеку-
лы находятся в лабораторном воздухе и «инфицируют» агарозу с QP-репликазой (рис. 4).
Соответственно, в экспериментах Айгена и сотрудников реплицирующиеся РНК тоже инфи-
цировали реакционные среды, вызывая как будто бы спонтанный синтез РНК, как будто бы
матриценезависимый.
7. ЭВОЛЮЦИЯ РНК В МОЛЕКУЛЯРНЫХ КОЛОНИЯХ
Таким образом, с одной стороны, молекулярные колонии нам помогли найти источник
так называемого спонтанного синтеза РНК, а, с другой стороны, они подвергли сомнению
эксперименты Сола Шпигельмана (Sol Spiegelman), которые были проведены сразу после
того, как Qp-репликаза была обнаружена, и которые до сих пор цитируются как эксперимен-
ты, в которых впервые была показана эволюция молекул в пробирке (Joyce, 2007).
Что было сделано Шпигельманом? Он брал в качестве исходной матрицы геномную
РНК фага Qp. Это длинная одноцепочечная РНК: более 4000 нуклеотидов. Добавлял сколько-
то этой матрицы в пробирку с репликазой и нуклеотидами. Через некоторое время маленькую
порцию переносил в следующую пробирку, потом в следующую, следующую, следующую...
Таким образом, осуществлял последовательные пересевы. И оказалось, что если сделать
20-30 пересевов и проанализировать продукты синтеза, то в продуктах обнаруживается не
высокомолекулярная РНК - та, исходная, 4000 нуклеотидов длиной, - а малые РНК, длиной
около 200-300 нуклеотидов (Chetverin, 1997). Шпигельман и соавторы сделали вывод о том,
что в их экспериментах путем спонтанных делеций происходили изменения РНК в направ-
лении увеличения скорости репликации. И таким образом они имели, буквально, эволюцию
17
.<£>. Четверни. Можно ли собрать клетку ил ее компонентов'?
Нереплицируемые фрагменты RQ РНК
Мембрана
\ с нуклеотидами
реагирующие
5'-фрагмент участки
В З'-фрагмент
Рекомбинация
X
1 [
Реплицируемая РНК
Агароза с Qp-репликазой
Один Оба
из фрагментов фрагмента
Рис. 5. Отбор рекомбинантных молекул РНК по способности реплицироваться.
в пробирке: у них из неэффективно реплицирующейся большой молекулы получились эф-
фективно реплицирующиеся малые молекулы РНК. Однако в свете наших результатов о том,
что так называемый спонтанный синтез вызывается загрязнениями РНК, присутствующими
в воздухе, можно предположить, что в вышеуказанных экспериментах происходила не эво-
люция, а просто вытеснение одних малоэффективно реплицируемых молекул другими более
эффективно реплицируемыми молекулами.
Все-таки, возможно ли создание новой генетической информации на уровне РНК? Мы
попытались ответить на этот вопрос с помощью молекулярных колоний. С этой целью ис-
пользовали два взаимодополняющих фрагмента реплицируемой РНК: 5'-фрагмент (то есть,
5'-концевой фрагмент) и З'-фрагмент. Qp-репликаза может экспоненциально размножать це-
лую молекулу РНК, но не может размножать ее составные фрагменты. Однако если между
фрагментами произойдет реакция (ее называют рекомбинацией) таким образом, что образу-
ется целая молекула, включающая оба фрагмента. - то тогда, возможно, образуется реплици-
руемая молекула (рис. 5). Действительно, так и оказалось. Мы приготовили смесь фрагмен-
тов, посеяли ее на агарозу с репликазой, потом накрыли эту агарозу мембраной, пропитанной
нуклеотидами - субстратами для синтеза РНК. Если только один фрагмент присутствовал
в агарозе, то никаких колоний РНК мы нс видели. Но если в агарозе присутствовали оба
фрагмента, то колонии образовывались. Отсюда мы сделали вывод о том, что, действительно,
между фрагментами происходит реакция таким образом, что из двух молекул получается
одна, то есть образуется новый РНК-геном (Chetverin, 1997).
В вышеописанном эксперименте в реакции, в агарозе, присутствовала QP-репликаза.
Поэтому было непонятно, кто осуществляет рекомбинацию? Сами по себе фрагменты реа-
гируют друг с другом или Qp-репликаза каким-то образом их соединяет? Мы несколько
изменили постановку эксперимента. Оказалось, что если фрагменты окислить: обработать
их перйодатом натрия таким образом, что будут уничтожены З’-гидроксилы, - то тогда Q3-
репликаза не способна промотировать рекомбинацию между такими фрагментами. Мы
инкубировали смесь фрагментов в присутствии ионов Mg2+ без Qp-репликазы, после чего
окисляли реакционную смесь перйодатом, чтобы уничтожить гидроксилы (таким образом,
дальнейшая рекомбинация была невозможна). Затем высевали смесь фрагментов на агарозу,
содержащую Qp-репликазу. Оказалось, что из нереплицирующихся фрагментов образуются
реплицирующиеся молекулы, при этом число таких молекул, которое отражается в числе ко-
лоний РНК, зависело от температуры и времени предынкубации фрагментов до того, как они
были окислены и смешаны с Qp-репликазой (рис. 6). Это прямо говорило о том, что фрагмен-
18
ril'OR.'ll-.Mhl Г1Р()11С\(>ЖЯ1'Н11Яжтни
Температура, °C
Рис. 6. Возникновение реплицируемых молекул РНК в результате спонтанной рекомбинации. Смесь
фрагментов RQ РНК инкубируют в присутствии Mg2’, а затем окисляют перйодатом для уничтоже-
ния З'-гидроксилов. Это подавляет рекомбинацию в присутствии Qp-репликазы, поэтому число коло-
ний РНК становится зависимым от температуры и времени инкубации фрагментов до их контакта с
репликазой. Скорость спонтанной рекомбинации - 101) ч'1 на нуклеотид.
О 16 64
Время инкубации, ч
ты РНК способны спонтанно реагировать в отсутствие всякого белка с образованием более
крупной молекулы (Chetverina et al., 1999).
На самом деле мы наблюдали ничто иное как эволюцию молекул РНК. Происходило
образование нового качества, которое подвергалось отбору и побеждало постольку, посколь-
ку обладало преимуществом - новым признаком: способностью к репликации.
8. РАЗМНОЖЕНИЕ ГЕНОВ В МОЛЕКУЛЯРНЫХ КОЛОНИЯХ
Мы попытались использовать эту систему для того, чтобы размножать не малые RQ
РНК, а целые гены, способные кодировать белки. Казалось бы, ген можно вставить внутрь
реплицируемой молекулы и этот ген будет размножаться с остальными частями реплици-
руемой молекулы. Оказалось, что QP-репликазу нельзя обмануть: она отвергает такие искус-
ственные рекомбинанты и способна размножать полученную РНК, только если вставка не
превышает 50 нуклеотидов. Если больше, то экспоненциальный синтез прекращается: РНК
становится нереплицируемой (Morozov et al., 1993).
Поэтому мы решили обратиться к другой системе экспоненциального размножения по-
линуклеотидов - полимеразной цепной реакции (ПЦР) (Saiki et al., 1995). Существует прин-
ципиальное отличие ПЦР от Qp-репликазы. Qp-репликаза размножает РНК, не используя
никакого праймера или затравки, а для того, чтобы с помощью ПЦР копировать ДНК, надо
дать затравку - олигодезокси рибонуклеотид, который будет отжигаться с З'-концом размно-
жаемой последовательности. Благодаря этому, с помощью ПЦР можно размножить практи-
чески любую ДНК, используя подходящие праймеры.
Еще одним существенным отличием ПЦР от Qp-репликазы является то, что реплика-
ция РНК QP-репликазой происходит при постоянной температуре в диапазоне от 20 до 40 °C,
а для того, чтобы синтезировать ДНК с помощью ПЦР, необходимо периодически нагревать
смесь для расплавления ДНК, потому что матрицей может работать только одноцепочечная
молекула, а продуктом репликации (полимеризации) является двуцепочечная молекула. Для
того, чтобы двуцепочечную молекулу разбить на одноцепочечные, необходимо ее распла-
вить путем нагревания.
Если мы стартуем с двуцепочечной ДНК, то ее надо расплавить, нагрев реакционную
смесь до >90 °C. Если в среде присутствуют олигонуклеотидные праймеры, то при после-
дующем понижении температуры они окажутся отожженными с З'-концевыми участками
одноцепочечных молекул. Далее, если в среде присутствует ДНК-полимераза [термостабиль-
ная, способная работать при достаточно высокой температуре без инактивации (Saiki et al.,
1998)], то она удлинит эти праймеры на их матрицах. Таким образом, стартуя с одной двуце-
19
Четверни. Можно .in собрать k.icikv из ос компонентов?
Гель пропитывают ПЦР-
смесью, содержащей
' генетический материал
ДНК переносят
на нейлоновую мембрану
Осуществляют ПЦР
Мембрану гибридизуют
с меченым зондом
Рис. 7. Выращивание и обнаружение колоний ДНК.
почечной молекулы ДНК, мы получим две такие же молекулы. Повторяя циклы нагревания-
охлаждения, можно в экспоненциальном режиме увеличивать число молекул ДНК.
Чтобы получить с помощью ПЦР колонии ДНК, нельзя использовать агарозу, потому
что агароза при нагревании выше 90 °C обязательно расплавится. Поэтому мы используем
полиакриламидный гель, который полимеризуем в лунках, сделанных в предметном стекле
для микроскопии (Четверина, Четверин, 2008). После формирования геля его промываем
в воде, чтобы убрать все, что не заполимеризовалось и что может потом мешать в ПЦР,
и высушиваем. Непосредственно перед реакцией в лунки наливаем реакционную смесь, ко-
торая содержит ДНК-полимеразу, матрицу, праймеры и нуклеотиды, из которых делается
ДНК. Гель всасывает жидкость, разбухает до исходного объема, и таким образом, убирается
вся свободная жидкость. Важно, чтобы свободной жидкости не было, потому что если есть
свободная жидкость, то молекулы ДНК будут мигрировать и колонии не образуются. Потом
осуществляем ПЦР (несколько десятков циклов нагревания-охлаждения), после чего про-
мокаем гель нейлоновой мембраной. Если ее прогибридизовать с радиоактивным зондом,
который липнет к размноженной ДНК, то там, где оказались размноженные колонии ДНК,
на автографе будут видны темные пятна (рис. 7). Таким образом, можно осуществлять раз-
множение ДНК в виде колоний (Chetverina, 2002).
Оказалось также, что можно наблюдать рост колоний ДНК и в реальном времени
(Samatov, Chetverina, Chetverin, 2006). Для этого реакцию проводят в геле, который заклеен
фольгой с отверстием, через которое можно наблюдать, что в геле происходит, а в гель вво-
дят зонды, которые способны увеличивать флуоресценцию, если образуется ДНК. Напри-
мер, можно использовать так называемые бинарные зонды, которые несут два флуорофора,
один из которых способен поглощать возбуждающий свет и передавать энергию флуорофору
другого зонда, который потом уже излучает свет. В данной ситуации, флуоресценция воз-
можна тогда и только тогда, если эти два флуорофора сближены, а это возможно, только если
образовалась ДНК, с которой эти два зонда могут гибридизоваться рядом друг с другом. Со-
ответственно, сначала (в нулевом цикле) флуоресценция не видна, и только после того, как
произошло более 30 циклов, появляются флуоресцирующие пятна, соответствующие коло-
ниям ДНК (рис. 8).
20
Проблемы пропсхожлкппя жн'нт
О 30 31 32 33 34 35 36 37 38 39 40
Номер цикла ПЦР
Рис. 8. Рост колоний ДНК в присутствии двух гибридизующихся рядом FRET-зопдов
(от «Fluorescence Resonance Energy Transfer»).
Таким способом можно размножать очень большие фрагменты ДНК, такие как ген обе-
лина и ген зеленого флуоресцентного белка длиной примерно 800 и 1500 пар оснований соот-
ветственно. При этом образуется столько колоний, сколько посеяно молекул ДНК (Samatov,
Chetverina, Chetverin, 2005).
9. МОЛЕКУЛЯРНЫЕ КОЛОНИИ КАК АНАЛОГИ ЖИВЫХ КЛЕТОК
Оказалось также, что можно прямо внутри молекулярных колоний, непосредствен-
но в геле, осуществлять экспрессию размноженной ДНК. Чтобы провести транскрипцию,
гель пропитывают транскрипционной смесью, которая содержит ДНК-зависимую РНК-
полимеразу и нуклеотиды, необходимые для синтеза РНК. В каждой колонии синтезируется,
как минимум, 10 РНК-копий на одну молекулу ДНК, то есть колония уже содержит не только
молекулы ДНК, но и молекулы РНК, которые синтезировались с этих молекул ДНК (Samatov,
Chetverina, Chetverin, 2005).
Более того, оказалось, что если пропитать гель компонентами системы трансляции,
то есть рибосомами и прочими компонентами, необходимыми для синтеза белка, то можно
осуществить и трансляцию, то есть в колониях ДНК будет синтезироваться белок. Иными
словами, функционально молекулярные колонии являются аналогами клеток. В молекуляр-
ных колониях может происходить как репликация ДНК, так и полноценная экспрессия гена:
от синтеза РНК до синтеза функционально активного белка (Samatov, Chetverina, Chetverin,
2005; Chetverin, Samatov, Chetverina, 2008).
В отличие от липосом, молекулярные колонии не окружены мембраной. Поэтому
в молекулярную колонию легко диффундируют низкомолекулярные вещества. В то же вре-
мя, полимерный матрикс геля удерживает наномолекулы (крупные молекулы - ДНК, РНК,
белки) на месте и не дает им распространяться по всему реакционному объему. Таким об-
разом, можно достичь компартментализации всех биохимических реакций, которые харак-
теризуют жизнь, в виде молекулярных колоний.
Недавно в одной лаборатории в Китае было продемонстрировано, что колонии ДНК
можно получить с использованием так называемого механизма катящегося кольца (Zhou
et al., 2008). Как и мы, авторы проводили реакцию в полиакриламидном геле, но только вместо
ПЦР использовали другую систему амплификации, способную размножать ДНК в изотер-
мическом режиме и при низкой температуре - подобно размножению РНК QP-репликазой.
21
Чепшсрнн. Можно .ш собран, k.jcikv из ее ком нонен гон?
Это позволяет выращивать колонии ДНК при температуре, совместимой с существованием
обычных (мезофильных) организмов.
Таким образом, молекулярные колонии - «безоболочечные клетки» - можно выращи-
вать в лаборатории, в полиакриламидном геле. Имеет ли это какое-то отношение к проблеме
происхождения жизни? В 1999 г. Шостак первым высказал мысль о том, что молекулярные
колонии, аналогичные гем, что образуются при размножении РНК в агарозе, содержащей
QP-репликазу, могли бы бы ть доклеточной формой компартментализации в мире РНК - во-
обще компартментализации живой материи (Szostak, 1999). В этой ситуации, когда компар-
тменгализация обеспечивается не оболочкой, а относительно малой скоростью диффузии
макромолекул в пористом матриксе по сравнению с низкомолекулярными веществами, как-
ого мы видим на примере полиакриламидного геля, колонии РНК могли бы формироваться
во влажной глине или иных пористых минералах.
10. ЖИЗНЬ ЗАРОДИЛАСЬ В ГЛИНЕ?
Есть разные глины, которые различаются по своим свойствам. Есть каолинит и монт-
мориллонит. Они, с одной стороны, похожи по внешнему виду, но с другой стороны, сильно
различаются по своим свойствам (табл. 1). В частности, удельная поверхность монтморил-
лонита па два порядка больше, чем у каолинита, и емкость по ионному обмену тоже гораздо
больше, примерно на порядок (Ерёмин, 2004). То есть глина это нс просто пористое вещество,
а ионообменник. Монтмориллонит - это слоистый алюмосиликат. Расстояние между слоя-
ми в сухом виде 1-2 нм, а при набухании (пропитывании водой) увеличивается в несколько
раз, что объясняет большую сорбционную емкость монтмориллонита. При этом расстояние
между слоями становится сопоставимым с размерами наномолекул.
Оказалось, что благодаря своим ионообменным и сорбционным свойствам, слоистые
глины могут концентрировать нуклеотиды из окружающего пространства. Если 50 мг гли-
ны (это 20 микролитров, то есть одна пятидесятитысячная часть литра) добавить к одному
литру 15 мкМ АМР, то в пределах суток на этом количестве глины окажется адсорбировано
от 10 % (в присутствии Na ) до 90 % (в присутствии Mg2’) AMP (Ferris, Ertem, Agarwal, 1989).
Иными словами, объем, в котором находился нуклеотид, уменьшился в пятьдесят тысяч
раз. Следовательно, имеет место пятидесятитысячекратное концентрирование нуклеотидов
на глине по сравнению с тем, что было в окружающей среде, то есть локальная концентрация
нуклеотида в глине самопроизвольно в присутствии ионов Mg2 может возрасти до 0.5 М.
Это совсем другие концентрации, чем используются в экспериментах с липосомами [5 мМ
(Mansy et al., 2008)].
Далее оказалось, что слоистые алюмосиликаты могут сорбировать с высоким срод-
ством полинуклеотиды, при этом сорбционная способность (аффинность) полинуклеоти-
да зависит от того, в присутствии какого катиона проводится эксперимент (Franchi, Ferris,
Gallori, 2003). Особенно хорошо алюмосиликаты адсорбируют полинуклеотиды в присут-
ствии кальция и магния. Если концентрация катиона 1 мМ или выше, то одноцепочечные
Таблица 1. Сравнение свойств слоистых алюмосиликатов (по Ferris, Ertem, Agarwal, 1989)
Минерал Плотность, г/см5 Удельная поверхность, м2/г Емкость катионного обмена, мг-экв./г
Каолинит 2.60-2.68 8-20 0.03-0.15
Монтмориллонит 2.35-2.70 600 - 800 0.80-1.50
22
Uрок и:мы прори \ож,и-.пия жпшн
РНК или ДНК сорбируются па тине полностью. Важно, что глины имеют большее сродство
к одноцепочечным молекулам, чем к двуцепочечным (Franchi, Ferris, Gallori, 2003). Это может
иметь отношение к механизму репликации полинуклеотидов, поскольку тем самым стаби-
лизируется одноцепочечное состояние, которое и является реплицируемым. Иными словами,
глины могут выполнять роль SSB-бслков (single strand-binding protein), которые участвуют
в репликации ДНК и препятствуют отжигу комплементарных цепей, связываясь с ними.
Более того, оказалось, что глины не только адсорбируют нуклеотиды и полинукле-
отиды, но и катализируют безматричную полимеризацию нуклеотидов. В присутствии
10-нуклеотидного праймера на глине происходит спонтанное образование РНК длиной
до 40 нуклеотидов и больше, если в качестве субстрата реакции используют активиро-
ванное производное АМР - фосфоримидазолид аденозина. Это при том, что в отсутствие
глины максимально синтезируемая длина - 10 нуклеотидов, причем, в основном, обна-
руживаются только динуклеотиды, потому что скорость полимеризации только в 10 раз
выше скорости спонтанного гидролиза полирибонуклеотидов (Ferris et al., 1996). Если же
активировать нуклеотиды нс имидазолом, а метиладенином, то РНК длиной 40 нуклеоти-
дов образуются на глине в пределах 8 часов (Huang, Ferris, 2006). Это существенно более
высокие скорости, чем при полимеризации нуклеотидов в липосомах (Mansy et al., 2008).
Интересно, что до 80 % продуктов синтеза представлены РНК, в которых нуклеотиды
связаны между собой 3'-5' связями. Это очень важно, так как именно такие связи присут-
ствуют в нормальных природных РНК.
Большинство этих экспериментов было сделано в лаборатории Джеймса Ферриса (James
Р. Ferris), который также показал, что глина придает процессу полимеризации нуклеотидов
определенную хиральную селективность. Так, при использовании рацемической смеси D-
и L-энантиомеров фосфоримидазолида аденозина для синтеза олигонуклеотидов, на монт-
мориллоните образуется соответственно в 1.3, 1.6 и 2.1 раз больше гомохиральных димеров
(D,D и L,L), тримеров (D,D,D и L,L,L) и тетрамеров (D,D,D,D и L,L,L,L), чем статистически
ожидаемая доля (Joshi, Pitsch, Ferris, 2007). Иными словами, при удлинении синтезируемого
полинуклеотида его гомохиральность усиливается.
Наконец, недавно сделаны интересные наблюдения о поведении монтмориллонита
по отношению к простым органическим молекулам. Если метанол нагревать при 300 °C
и 1000 атм, то он разлагается в отсутствие монтмориллонита, но сохраняется в его присут-
ствии. Более того, в этих условиях монтмориллонит катализирует образование из метано-
ла других, более сложных соединений. Отсюда авторы предположили, что в определенную
геологическую эпоху в монтмориллоните могли образовываться предшественники РНК
и других биополимеров (Williams et al., 2005). Также монтмориллонит может защищать ад-
сорбированные рибозимы от разрушения ультрафиолетовым излучением (Biondi et al., 2007).
Возможно, этому есть простое объяснение: то, что глина непрозрачна для ультрафиолета.
Таким образом, существует много свидетельств в пользу того, что монтмориллонит
мог служить чем-то вроде инкубатора для простых и сложных органических молекул, свое-
го рода «первичной маткой» [primordial womb (Williams et al., 2005)] для жизни на Земле.
В этой связи интересно, что монтмориллонит не только повсеместно распространен на Земле,
но и обнаружен в составе метеоритов, а также на Марсе (Poulet et al., 2005).
Все указанные свойства делают монтмориллонит идеальной средой для образования
и роста молекулярных колоний в мире РНК: он гидрофилен, содержит необходимого размера
поры и обладает способностью концентрировать на себе субстраты для синтеза РНК. Более
того, монтмориллонит способен был, в отсутствие клеточной мембраны, решить проблему
совместного наследования разных видов РНК, составляющих смешанную молекулярную ко-
лонию и обладающих взаимодополняющими функциями (Спирин, 2005а, б). Действительно,
колония, выросшая на монтмориллоните, может размножаться путем выветривания (отделе-
23
А.Б. Четверни. Можно .in собран, kjjcjkv hi ее ком попон гон'?
ния частиц) минерала. Множество молекул РНК, адсорбированных на одной оторвавшейся
микрочастице глины, будут перенесены все вместе в другое место и там дадут начало новой
колонии, содержащей все необходимые для ее роста виды РНК.
Теперь вернемся к липосомам. На каком-то этапе эволюции вокруг биомолекул должны
были образоваться липидные мембраны, чтобы дать начало современным клеткам. Оказы-
вается, монтмориллонит мог и здесь сыграть важную роль, потому что он в 100 раз ускоряет
формирование липосом из эмульсии липидов. Более того, частицы монтмориллонита, спо-
собные нести на себе нуклеотиды, РНК и ДНК (см. выше), оказываются включенными внутрь
липосом, то есть окружаются липидной мембраной (Hanczyc, Fujikawa, Szostak, 2003).
Здесь я хочу высказать предположение, что липидная мембрана вокруг реплицирую-
щихся молекул (РНК, ДНК) образовалась на более поздних стадиях эволюции, тогда, когда
стал возможен синтез хотя бы коротких полипептидов, подобных антибактериальным (анти-
микробным) пептидам - цекропину, магаинину и т. д. (Wang Z., Wang G., 2004). Это короткие
(20 -40 аминокислотных остатков) а-спиральные пептиды, способные образовывать поры
в липидных мембранах, экспонируя свои гидрофобные поверхности внутрь мембраны, а ги-
дрофильные - внутрь поры (Hallock, Lee, Ramamoorthy, 2003). Такие поры могли решить про-
блему проницаемости мембраны липосом для низкомолекулярных веществ и, тем самым,
обеспечить потребности синтеза биополимеров внутри липосом.
11. ВЫВОДЫ
1. В настоящее время осуществим полностью химический синтез генома, содержащего
набор всех генов, необходимых для функционирования клетки. Однако отсутствует экспери-
ментальная модель для сборки жизнеспособных клеток de novo.
2. Наноколонии (молекулярные колонии) могли бы стать такой экспериментальной мо-
делью. Они являются функциональными аналогами клеток, позволяющими осуществлять
сборку клеточных компонентов и проверять, какой вариант сборки обеспечивает полноцен-
ную экспрессию генома:
- обеспечивают компартментализацию этих компонентов,
- способны осуществлять различные биохимические реакции, составляющие всю це-
почку экспрессии генов,
- позволяют добавлять и убирать компоненты путем простого пропитывания геля (по-
скольку они лишены оболочки),
- удобны для скрининга (расположены в одной плоскости),
- обеспечивают связь генотипа и фенотипа, что необходимо для естественного отбора,
- способны эволюционировать, то есть образовывать новый генетический материал,
который затем может быть экспрессирован.
3. Наноколонии, растущие в глине, могли быть формой компартментализации на пер-
вых этапах эволюции в мире РНК и в процессе становления белкового мира. Монтморилло-
нит, один из видов глины, способен обеспечивать все условия, необходимые для роста, эво-
люции и размножения колоний РНК, а также для их трансформации в клетки, окруженные
липидной мембраной:
- концентрировать из окружающей водной среды и стабилизировать активированные
рибонуклеотиды,
- катализировать полимеризацию рибонуклеотидов,
- обеспечивать относительную хиральную однородность синтезированных полирибо-
нуклеотидов (РНК),
- иммобилизовать РНК - как матрицы, так и их копии, не давать им уходить в окру-
жающий раствор,
24
Прошпмы црот ,\ож. /кипи >кп <нп
- более прочно связывать однотяжные РНК, чем двутяжные, что стабилизирует спо-
собное к репликации состояние,
- компартментализовать РНК в отсутствие липидных мембран (формировать колонии
РНК),
- удерживать рядом разные виды РНК, которые могли бы создавать смешанную коло-
нию. и обеспечивать их совместное наследование,
- формировать липосомы вокруг колоний РНК.
Я благодарен оргкомитету рабочего совещания «Проблемы происхождения жизни» за
приглашение выступить с этим докладом, а также моей жене и коллеге Е.В. Четвериной за
неоценимую помощь в подготовке рукописи. Работа поддержана грантами РФФИ и програм-
мой Президиума РАН «Молекулярная и клеточная биология».
СПИСОК ЛИТЕРАТУРЫ
Ерёмин Н.П. 2004. 11еметаллические полезные ископаемые. М.: Изд-во МГУ. 459 с.
Ленинджер А. 1976. Биохимия: молекулярные основы структуры и функции клетки. М.: Мир. 960 с.
Спирин А.С. 2005а. Мир РНК н его эволюция // Молекуляр. биология. Т. 39. С. 550-556.
Спирин А.С. 20056. Происхождение, возможные формы существования и размеры первозданных осо-
бей // Палеон гол. журн. № 4. С. 25-32.
Четверни А.Б. 1998. Бактериофаг Qp как объект молекулярной биологии // Успехи биол. химии. Т. 38.
С. 3-75.
Четверииа Е.В., Четверни А.Б. 2008. Наноколонии: обнаружение, клонирование и анализ индивиду-
альных молекул // Успехи биол. химии. Т. 48. С. 3-64.
Biebricher С.К., Eigen М., Luce R. 1986. Template-free RNA synthesis by QP replicase // Nature. V. 321.
P. 89-91.
Biondi E., Branciamore S.. Maurel M.C., Gallon E. 2007. Montmorillonite protection of an U V-irradiated hair-
pin ribozyme: evolution of the RNA world in a mineral environment// BMC Evol. Biol. V. 7. P. S2.
Brown D., Gold L. 1995. Selection and characterization of RNAs replicated by QP replicase // Biochemistry.
V. 34. P. 14775-14782.
Chetverin A.B.. Chetverina HA'. 1997. Method for amplification of nucleic acids in solid media. U.S. Patent
5, 616. 478.
Chetverin A.B., Chetverina HA'. 2008. Molecular colony technique: a new tool for biomedical research and
clinical practice // Prog. Nucleic Acid Res. Mol. Biol. V. 82. P. 219-255.
Chetverin A.B.. Chetverina H.V.. Demidenko A.A.. Ugarov К/. 1997. Nonhomologous RNA recombination in
a cell-free system: Evidence for a transesterification mechanism guided by secondary structure // Cell.
V. 88. P. 503-513.
Chetverin A.B.. Chetverina II.V., Munishkin A.V. 1991. On the nature of spontaneous RNA synthesis by Qp
replicase /' J. Mol. Biol. V. 222. P. 3-9.
Chetverin A.B., Samatov T.R., Chetverina H.V. 2008. Gene cloning and expression in molecular colonies //
Cell-free protein synthesis: methods and protocols / Eds. A.S. Spirin, J.R. Swartz. Weinheim: Wiley-
VCH. P. 191-206.
Chetverin A.B., Spirin AS. 1995. Replicable RNA vectors: prospects for cell-free gene amplification, expres-
sion and cloning // Prog. Nucleic Acid Res. Mol. Biol. V. 51. P. 225-270.
Chetverina HA'., Chetverin A.B. 1993. Cloning of RNA molecules in vitro // Nucleic Acids Res. V. 21. P. 2349-2353.
Chetverina H.V., Demidenko A. A., Ugarov V.I., Chetverin A.B. 1999. Spontaneous rearrangements in RNA
sequences 7 FEBS Lett. V. 450. P. 89-94.
25
.J./i. Четверни. Можно ни собрать kjic i ку из ее компонентов?
Chetverina //J'. Samalov T.R.. Ugarov И/., Chetverin А.В. 2002. Molecular colony diagnostics: detection
and quantitation of viral nucleic acids by in-gel PCR // BioTechniques. V. 33. P. 150-156.
Ferris J. P., F.rtem G.. Agarwal Г.К. 1989. The adsorption of nucleotides and polynucleotides on montmorillo-
nite clay/7 Orig. Life Evol. Biosph. V. 19. P. 153-164.
Ferris J.P.. Hill A.R. Lin R.. Orgel L.E. 1996. Synthesis of long prebiotic oligomers on mineral surfaces /7
Nature. V. 381. P. 59-61.
Forster A.C.. Church G.M. 2006. Towards synthesis of a minimal cell 7/ Mol. Syst. Biol. V. 2. P. 45.
Forster A.C., Church G.M. 2007. Synthetic biology projects in vitro // Genome Res. V. 17. P. 1-6.
Franchi M„ Ferris .J. P.. Gallon E. 2003. Cations as mediators of the adsorption of nucleic acids on clay sur-
faces in prebiotic environments/7 Orig. Life Evol. Biosph. V. 33. P. 1-16.
Fraser С. M„ Gocayne J.D., White O. et al. 1995. The minimal gene complement of Mycoplasma genitalium /.'
Science. V. 270. P. 397-403.
Gibson D.G., Benders G.A., Andrews-PJannkoch C. et al. 2008. Complete chemical synthesis, assembly, and
cloning of a Mycoplasma genitalium genome // Science. V. 319. P. 1215-1220.
Gilbert W. 1986. Origin of life: The RNA world//Nature. V. 319. P. 618.
Gilbert W., de Souza S.J. 1999. Intron and the RNA world // The RNA World. Cold Spring Harbor, New York:
CSHL Press. P. 221-231.
Giovannoni S.J., Tripp H.J.. Givan S. ct al. 2005. Genome streamlining in a cosmopolitan oceanic bacterium //
Science. V. 309. P. 1242-1245.
Glass J. I., Assad-Garcia N.. Alperovich N. et al. 2006. Essential genes of a minimal bacterium // Proc. Natl.
Acad. Sci. USA. V. 103. P. 425-430.
Hallock K.J., Lee D.K.. Ramamoorthy A. 2003. MS1-78, an analogue of the magainin antimicrobial peptides,
disrupts lipid bilayer structure via positive curvature strain /7 Biophys J. V. 84. P. 3052-3060.
Hanczvc M.M., Fujikawa S.M., Szostak J.W. 2003. Experimental models of primitive cellular compartments:
encapsulation, growth, and division // Science. V. 302. P. 618-622.
Haruna Spiegelman S. 1965. Autocatalytic synthesis of a viral RNA in vitro // Science. V. 150. P. 884-886.
Huang W.. Ferris J. P. 2006. One-step, regioselective synthesis of up to 50-mers of RNA oligomers by montmo-
rillonite catalysis // J. Amer. Chem. Soc. V. 128, P. 8914-8919.
Joshi P.C., Pitsch S., Ferris J.P. 2007. Selectivity of montmorillonite catalyzed prebiotic reactions of D, L-
nucleotides // Orig. Life Evol. Biosph. V. 37. P. 3-26.
Jovce G.F. 2007. Forty years of in vitro evolution // Angew. Chem. Int. Ed. Engl. V. 46. P. 6420-6436.
Lartigue C., Glass J.1.. Alperovich N. et al. 2007. Genome transplantation in bacteria: changing one species to
another// Science. V. 317. P. 632-638.
Mansy S.S., Schrttm J.P.. Krishnamurthy M. et al. 2008. Template-directed synthesis of a genetic polymer in
a model protocell /. Nature. V. 454. P. 122-125.
Mills D.R., Peterson R.L.. Spiegelman S. 1967. An extracellular Darwinian experiment with a self-duplicating
nucleic acid molecule // Proc. Natl. Acad. Sci. USA. V. 58. P. 217-224.
Morozov l.Yu., Ugarov III.. Chetverin A.B., Spirin A.S. 1993. Synergism in replication and translation of mes-
senger RNA in a cell-free system // Proc. Natl. Acad. Sci. U.S.A. V. 90. P. 9325-9329.
Mttshegian A.R., Koonin E.V. 1996. A minimal gene set for cellular life derived by comparison of complete
bacterial genomes II Proc. Natl. Acad. Sci. USA. V. 93. P. 10268-10273.
Nakabachi A.. Yamashita A., Toh H. et al. 2006. The 160-kilobase genome of the bacterial endosymbiont
Carsonclla / Science. V. 314. P. 267.
Oberholzer T, Wick R., Lttisi P.L., Biehricher C.K. 1995. Enzymatic RNA replication in self-reproducing
vesicles: an approach to a minimal cell // Biochem. Biophys. Res. Commun. V. 207. P. 250-257.
Pasteur L. I860. Experiences relatives aux generations dites spontanees // C. R. Acad. Sci. V. 50. P. 303-307.
26
Пронлкмы ПРО! К ХОЖДЕНИЯ жн и in
Potilet F.. Bibring J.P.. Mustard J.F et al. 2005. Phyllosilicates on Mars and implications for early martian
climate Nature. V. 438. P. 623 627.
Sciiki R.K.. Gelfand D.H.. Stoffel S. el al. 1988. Primer-directed enzymatic amplification of DNA with a ther-
mostable DNA polymerase // Science. V. 239. P. 487-491.
Saiki R.K.. Scharf S.. Faloona F. et al. 1985. Enzymatic amplification of [3-globin genomic sequences and
restriction site analysis for diagnosis of sickle cell anemia // Science. V. 230. P. 1350-1354.
Samatov T.R.. Chetverina HA'.. Chetverin A.B. 2005. Expressible molecular colonies // Nucleic Acids Res.
V. 33. P. 145.
Samatov T.R., Chetverina HA'.. Chetverin A.B. 2006. Real-time monitoring of DNA colonics growing in a
polyacrylamide gelAnal. Biochem. V. 356. P. 300-302.
Spirin A.S. 2002. Omnipotent RNA // FEBS Lett. V. 530. P. 4-8.
Sumper M.. Luce R. 1975. Evidence for de novo production of self-replicating and environmentally adapted
RNA structures by bacteriophage Q[3 replicase // Proc. Natl. Acad. Sci USA. V. 72. P. 162-166.
Szostak J.W. 1999. Constrains on the sizes of the earliest cells // Size limits of very small microorganisms.
Proc. Workshop, Washington, D.C.: Nation. Acad. Press, P. 120-125.
Szostak J.W.. Bartel D.P.. Lttisi P.L. 2001. Synthesizing life // Nature. V. 409. P. 387-390.
Tamames J.. Gil R.. Latorre A. et al. 2007. The frontier between cell and organelle: genome analysis of Can-
didatus Carsonella ruddii BMC Evol. Biol. V. 7. P. 181.
Ugarov E/„ Chetverin A.B. 2008. Functional circularity of legitimate QP replicase templates // J. Mol. Biol.
V. 379. P. 414-427.
Wang Z.. Wang G. 2004. APD: the antimicrobial peptide database // Nucleic Acids Res. V. 32. P. D590-
D592.
Williams L.B., Canfield B.. loglesonger K.M.. Holloway J.R. 2005. Organic molecules formed in a «primor-
dial womb»//Geology. V. 33. P. 913-916.
Zhou D., Zhang R.. Fang R. et al. 2008. Methylation pattern analysis using high-throughput microarray of
solid-phase hyperbranched rolling circle amplification products // Electrophoresis. V. 29. P. 626-633.
Can a cell be assembled from its constituents?
A.B. Chetverin
At present, there exist provisional list of a minimal set of genes required for the functioning
and multiplication of a living cell under maximally favorable conditions, methods for the complete
chemical synthesis of the minimal genome, as well as cell-free systems that permit all the biochemi-
cal reactions comprising genome replication and expression to be performed. The most serious prob-
lem which remains to be solved is the need to meet two mutually exclusive requirements: isolation
of the biochemical reactions from the environment, and exchange between the environment and the
cell. Under such circumstances, molecular colonies (other names: nanocolonies, polonies), which
form when RNA or DNA is replicated in a solid medium having pores of a nanometer size, can serve
as an experimental model for the assembly of cell constituents. Also, molecular colonies might have
served as a pre-cellular form of compartmentalization in the RNA World.
27
Дискуссия
по пленарному докладу А.Б. Четверина
«МОЖНО ДИ СОБРАТЬ КДЕТКУ ИЗ ЕЕ КОМПОНЕНТОВ?»
Председательствующий А.И. Григорьев
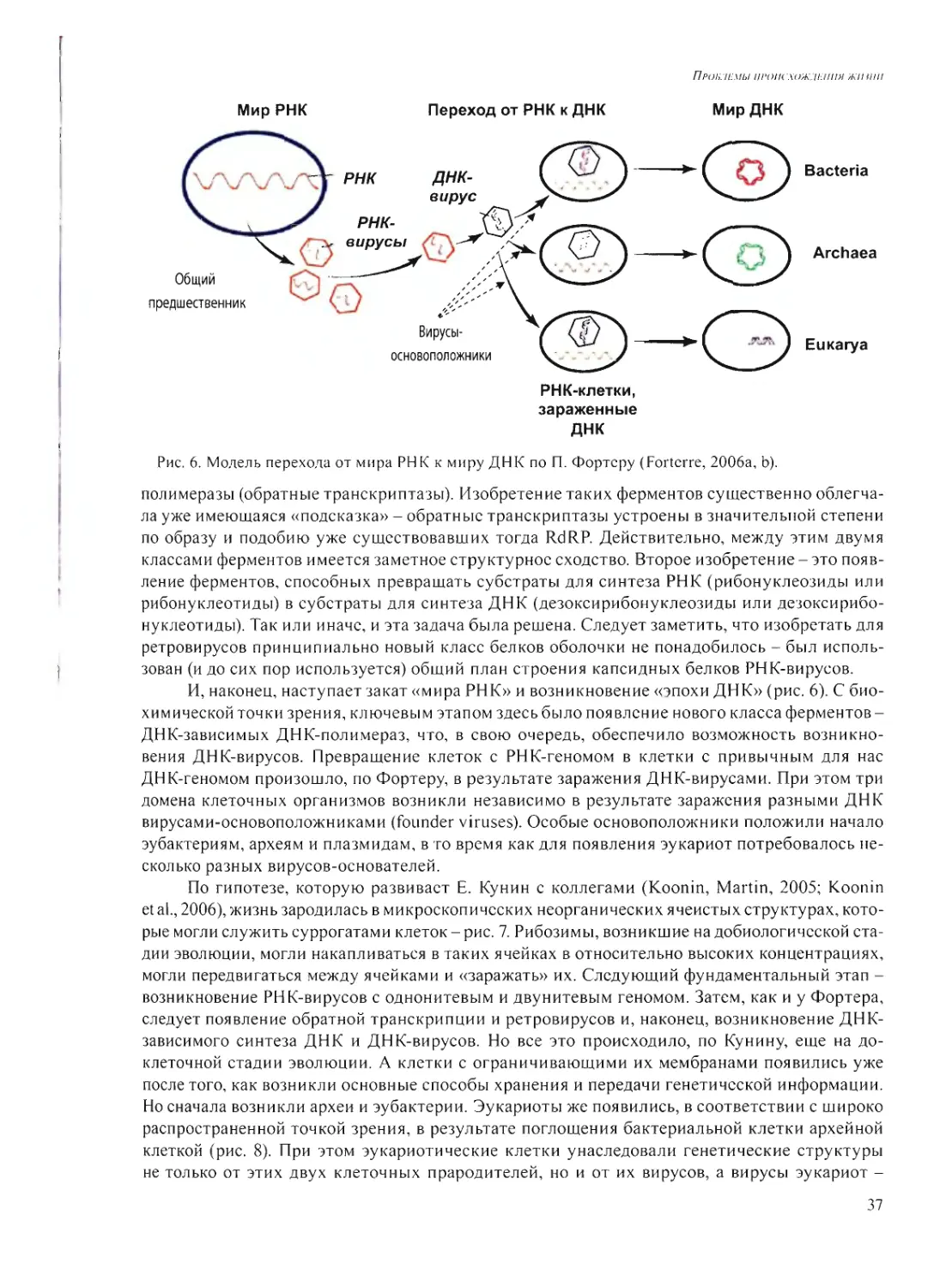
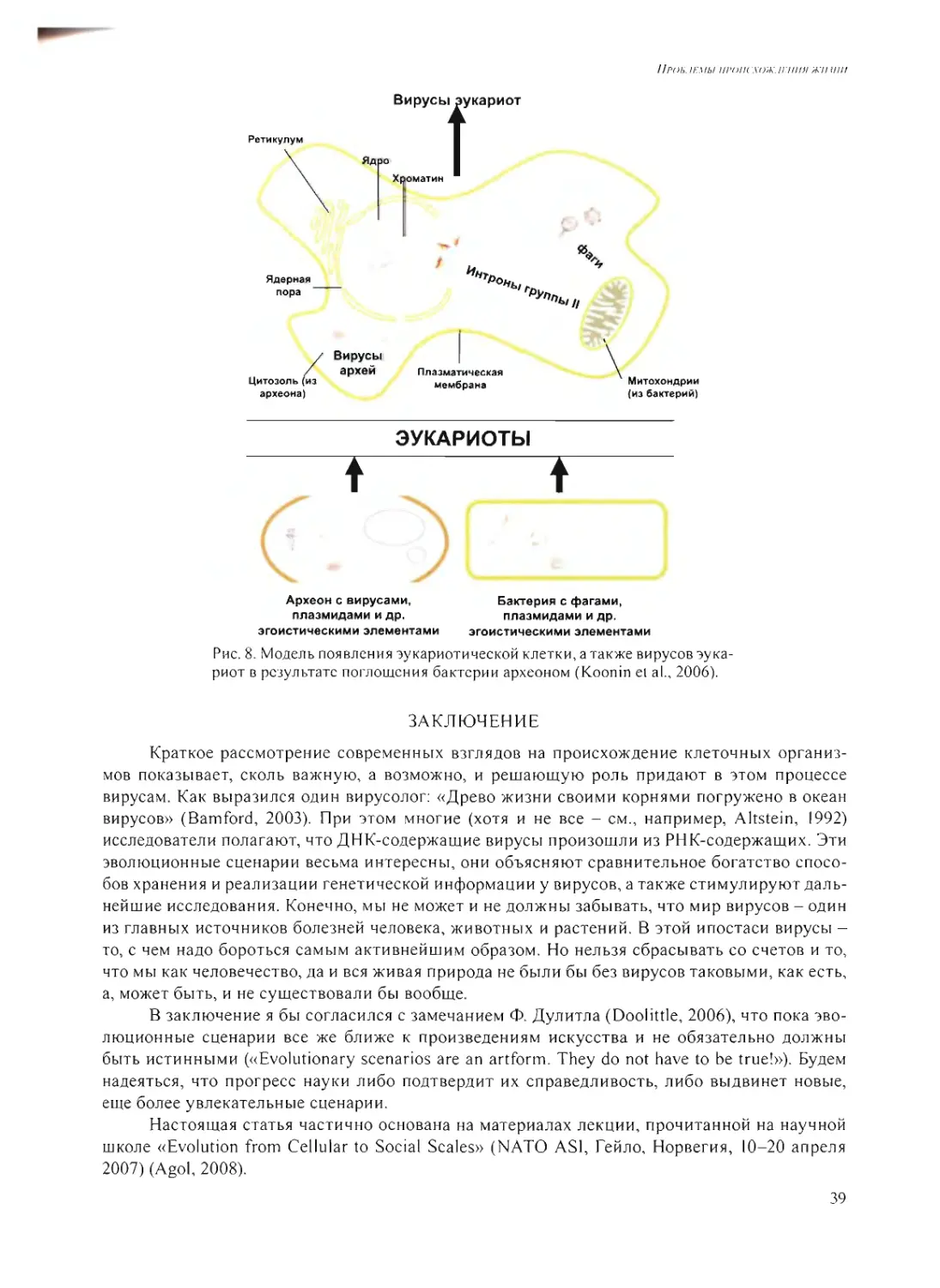
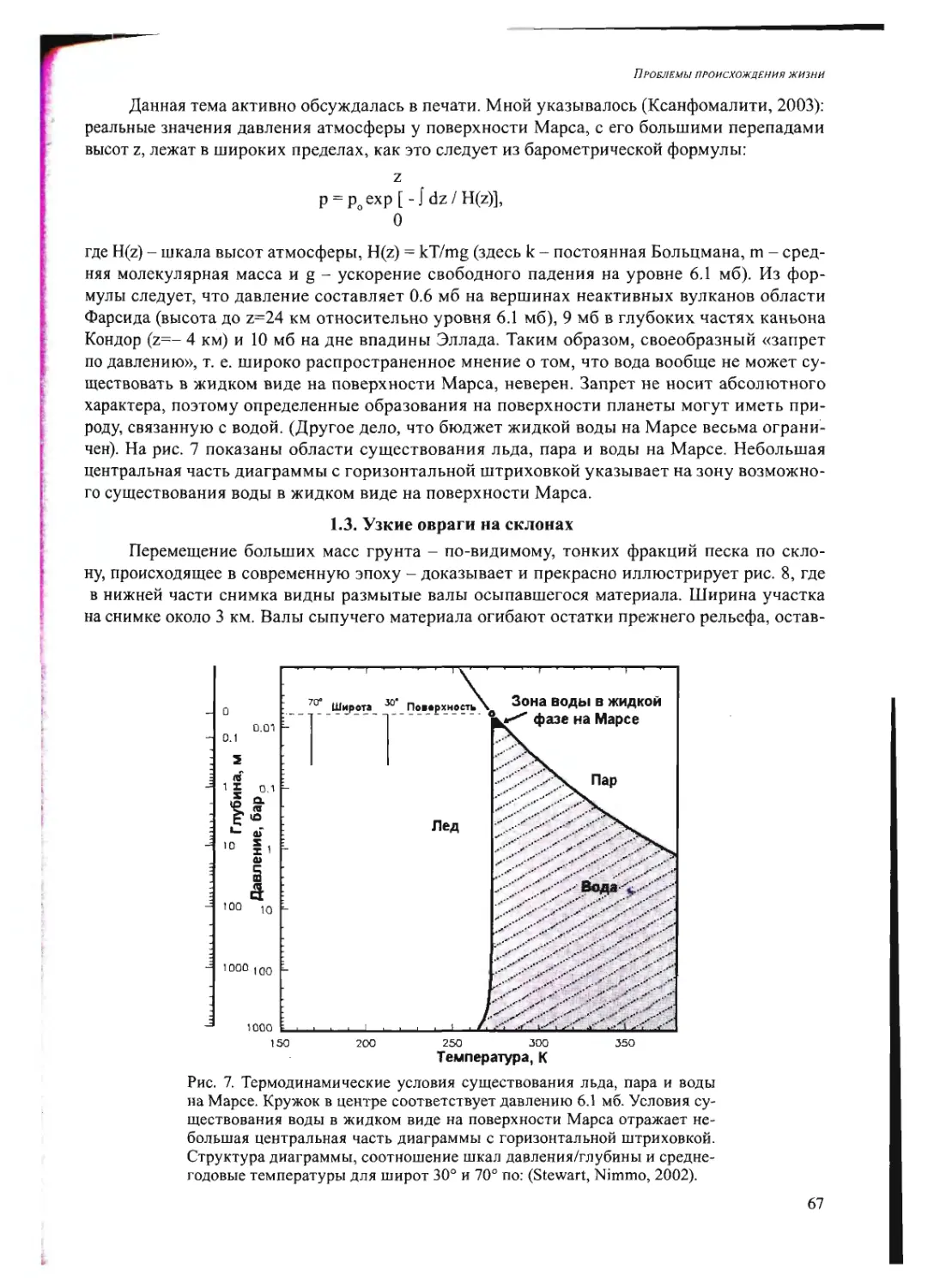
Л.М. Му.хнн: Поясните Ваш тезис о том, что наноколонии, растущие в глине, могли
быть формой компартментализации не только в мире РНК, но и в процессе становления бел-
кового мира?
А.Б.Ч.: Мы показали, что в напоколониях возможно не только размножение РНК или
ДН К, но возможна трансляция, экспрессия нуклеиновых кислот сели гель содержит компо-
ненты, которые катализируют синтез белка на РНК. На каком-то этапе рибозимы приобрели
способность катализировать производство полипептидов - тогда и происходило становление
белкового мира.
В.Н. Спытников: Известно, что алюмосиликаты, в частности, монтмориллонит, явля-
ются хорошими катализаторами. Многие цеолиты используются в промышленности в этом
качестве. И то, что они сорбируют органические соединения - тоже хорошо известный факт.
С точки зрения работоспособности катализаторов проблема заключается в том, что стоит им
только что-то адсорбировать - и после этого крайне сложно происходит десорбция (все поры
закрываются). Как же в Вашем случае будет происходить десорбция из пор этих длинных
и сложных молекул?
А.Б.Ч.: Я предполагаю, что десорбировать полинуклеотиды там и не надо. Монону-
клеотиды сорбируются обратимо. Полинуклеотиды сорбируются практически необратимо.
Но это и не нужно, так как глина может разбиваться на мелкие фрагменты, кусочки, эти
частички глины могут переноситься на другое место - вместе с теми РНК, которые были на
них сорбированы. На новом месте, прилипая к еще «незаселенному» куску глины, они могут
дать место новой колонии РНК.
В.Н. Спытников: Так ведь активный центр на алюмосиликатах уже закрыт.
А.Б.Ч.: Правильно. Но колония может расти по принципу грибницы - своей перифери-
ей, там, где активные центры еще нс забиты.
М.Я. Маров: Вы говорили о бактериальном геноме. Геном - это каталог генов. Вы упо-
мянули, что можно налить компоненты в пробирку и собрать геном. Возникает вопрос -
все-таки любой каталог организован - эта организация, она определяется некой внутренней
упорядоченностью, которая возникает в геноме?
А.Б.Ч.: Мы берем пробирку, в которой есть все компоненты для синтеза РНК, ДНК
и белка, кидаем туда сколько-то молекул матрицы, начинается синтез ДНК, на ней - РНК,
а потом синтез белка. Для этого процесса никакой особой организации, кроме последователь-
ности полинуклеотидов, не надо. Идет синтез белка. Александр Сергеевич Спирин показал,
что если все это поместить в реактор и добавлять туда новые компоненты и убирать продукты
жизнедеятельности, то теоретически этот процесс может происходить бесконечно долго.
М.Я. Маров: Для клетки понятно - есть некая матрица, которая это все выстраивает.
А вот для генома это все-таки что - система сама знает, в какой последовательности полину-
клеотиды выстраивать?
А.Б.Ч.: Она не знает. Если есть матрица, то вопрос, откуда взялась матрица? Об этом
лучше расскажет Александр Сергеевич Спирин. Относительно того, имеет ли значение вза-
имное расположение генов внутри хромосомы - это существенно для высокоорганизован-
ных организмов, скажем, для эукариот. В случае бактерий этот эффект минимален. Для про-
стейшей клетки эти проблемы не имеют значения - должны быть гены, последовательность
28
Проблемы нронсхожления жизни
полинуклеотидов, которые обеспечивают синтез полипептида, который способен собираться
во что-то и давать функционирующий белок.
М.Я. Маров: Это в какой-то мерс отражает концепцию, сейчас все более и более при-
нимаемую в качестве некой парадигмы, что в нелинейных открытых системах происходит
некая упорядоченность из первичной хаотической структуры. Отражается ли это в данной
ситуации на таком макромолекулярном уровне?
А.Б.Ч.: Думаю, что да.
В.Н. Снытников: У химиков есть понятие «выхода». Каков выход собранных клеток
из компонентов?
А.Б.Ч.: Для биологии выход гораздо менее важен, чем для химии, потому что в био-
логии, если что-то родилось в количестве одной миллиардной или одной триллионной доли
от всей массы, но при этом это что-то получило способность эффективно размножаться, этот
успех будет закреплен.
С.В. Рожнов: Можно ли создать из необходимых компонентов нс просто клетку, а такую
клетку, из которой в дальнейшем могло бы возникнуть все разнообразие органического мира?
А.Б.Ч.: Не знаю. Это совершенно особая тема.
С.В. Шестаков: Из Вашего доклада следует, что и без Qp-репликазы возможно само-
произвольное образование полинуклеотидов, и очень длинных. Эти полинуклеотиды уже
могут быть матрицей для репликации этих же молекул. Возникает вопрос, на каком этапе мы
должны думать о возможности упорядоченной структуры полинуклеотидов для создания
предпосылок генетического кода? Ведь синтез этих молекул у Вас - неупорядоченный, это
просто химическая реакция.
А.Б.Ч.: Не знаю. Это все очень сложные вопросы. Рассматривая вопрос, вынесенный
в название доклада, я предполагаю, что все компоненты, из которых планируется создать
клетку - в наличии, имеется их некий склад. Вопрос: можно ли собрать клетку из ее компо-
нентов? Ответ: если есть готовые компоненты - да, можно. Можно ли создать клетку есте-
ственным путем, путем эволюции, неконтролируемой экспериментатором - я не знаю.
С.В. Шестаков: Химическим способом можно создать геном, пусть и очень маленький.
А.Б.Ч.: Да, но мы сами пока еще нс можем придумать последовательность ДНК, кото-
рая была бы жизнеспособна. Чтобы запустить реакцию, нам необходимо поместить эту ДНК
в «сосуд», который уже содержит все необходимые белки, чтобы запустить воспроизведе-
ние, размножение и экспрессию этой ДНК. Если мы дали этот первичный толчок белками,
то после этого будут синтезироваться новые белки. Но первоначальные белки-то мы берем
из живой клетки.
С.В. Шестаков: Если у Вас образовались многочисленные разнообразные полинуклео-
тиды, они могут взаимодействовать между собой, удлиняется полинуклеотидная цепочка.
К чему это увеличение размера полинуклсотидной цепочки приведет?
А.Б.Ч.: Если в результате такого складывания образуется репликаза, которая потом
станет копировать, появится механизм для дальнейшего усложнения и усовершенствования.
Достаточно запустить реакцию.
А.Б. Рубин: Если все получившееся в результате эксперимента поместить в липосому,
то это все будет работать как клетка. Но в живой клетке есть гетерогенность, есть последо-
вательность процессов.
А.Б.Ч.: Главным образом это относится к клеткам совершенным, эукариотическим,
у которых есть цитоскелет, органеллы, ядро отделено от цитоплазмы и т.д. В бактериальной
клетке самая сложная структура - это мембрана, пожалуй. Проблема структурной организа-
29
ции мембраны нс решена. Поэтому я предлагаю в качестве временной меры для эксперимен-
тов использовать наноколонии.
Л.В. Ксанфома.-тти: Сорбционные свойства монтмориллонита обладают температур-
ной избирательностью?
А.Б.Ч.: Здесь я не специалист.
В.Н. Спытников: Да, обладают.
В.Н. Агол: Шостак показал в вышедшей неделю назад статье, что при определенном
составе липосомы термостабильны, при высокой температуре резко повышается проницае-
мость нуклеотидов, и создается возможность осуществлять полимеразную цепную реакцию
в липосомах.
А.Б.Ч.: То, что в липосомах можно делать ПЦР, было показано несколько лет назад.
Но я не думаю, что были такие большие скачки температуры.
Л.М. Мгхин: Какое значение имеет тот факт, что на глинах нельзя строить бесконечные
цепочки (там ведь порядка 40 нуклеотидов)?
А.Б.Ч.: Это безматрично. Да, можно спонтанно синтезировать 40 нуклеотидов, а потом
путем рекомбинаций сделать больше.
30
П роняемы происхождения жиши. M.: ПИН РАН. 2009. С. 31-40
ВИРУС-ДО ИЛИ ПОСЛЕ КЛЕТКИ?
©2009 г. В.И. А гол
Институт полиомиелита и вирусных энцефалитов им. М.Г1. Чумакова РАМН
и Московский государственный университет им. М.В. Ломоносова
agol(2r. belozcrsky.msu.ru
Кратко рассмотрена природа вирусов и фундаментальное отличие механиз-
мов их репродукции от клеточного деления. Способы хранения и выраже-
ния генетической информации у вирусов значительно богаче, чем у клеток.
По современным представлениям, «мир PH К» возник раньше «мира ДН К».
В свете этих представлений рассмотрены гипотезы о происхождении виру-
сов, - как та. по которой сначала появились клетки с РНК-геномом, а затем
уже вирусы, так и та, которая постулирует первичное возникновение РНК-
содержащих вирусов. И в том, и другом случае предполагается, что ДНК-
геномы впервые появились у вирусов и что вирусы сыграли решающую
роль в возникновении архей,эубактерий и эукариот.
ВВЕДЕНИЕ
Вирусы - это генетические элементы, существующие в двух формах. Во-первых, пас-
сивная внеклеточная форма - это молекулы ДНК или РНК, обычно заключенные в бслок-
содсржащую оболочку. Можно сказать, что вирусы - просто некий класс весьма сложных
химических соединений. Например, вирус полиомиелита может быть представлен такой
формулой - C,,2652H4,)2,ssN,)x;45O|,||y(P7S()|S,U0 (Wimmer et al., 1993). Другая форма существова-
ния вирусов - активная, внутриклеточная и обычно способная к выражению генетической
информации. В такой форме вирусы - живые объекты, обладающие наследственностью
и способные к эволюции.
Вирусы - наиболее многочисленный класс биологических объектов. По некоторым
оценкам, на нашей планете существует около 10” вирусных частиц (Breitbart, Rohwer, 2005).
Полагают, что в одних лишь вирусах, обитающих в океанах и морях, содержится —2х 10* тонн
углерода, что примерно эквивалентно содержанию этого элемента в 7*107 голубых китов. Со-
ответственно, если эти вирусы выстроить в цепочку, то ее длина составит ~107 световых лет
(—100 диаметров нашей Галактики) (Suttle, 2005, 2007).
Вирусы не только многочисленны, но и чрезвычайно разнообразны. В килограмме мор-
ского осадка может находиться порядка миллиона различных вирусных генотипов (Breitbart,
Rohwer, 2005). Вирусы могут иметь палочковидную или нитевидную форму, обладать куби-
31
В.И. Аго.т. Вирус до или после клетки'*
Рис. I. Разнообразие форм и размеров вирусов. Масштаб (если не указано иначе) - 100 нм.
ческой или спиральной симметрией, представлять собой плеоморфные образования и т. д.
Некоторое представление о разнообразии форм и размеров вирусов дает рис. 1. Наиболее
крупный из известны,х вирусов - мимивирус - вполне сопоставим по размерам с мельчайши-
ми микроорганизмами, такими, например, как микоплазма.
Вирусный геном может быть представлен как однонитевыми, так и двунитсвыми моле-
кулами РНК или ДНК, причем эти молекулы могут быть линейными или кольцевыми (рис. 2).
Как известно, клеточные организмы имеют только двунитевые ДНК-геномы. Количество ну-
клеотидов в геномах известных РНК-вирусов варьирует от немногим более полутора тысяч
до -32000, а количество генов - от 1 до полутора десятков. В ДНК-геномах вариации более
значительны - если минимальное число нуклеотидов в них не сильно отличается от такового
у РНК-вирусов, то известный сегодня максимум это ~1,2*106 пар нуклеотидов, а число ге-
однонитевая
двунитевая
кольцевая
линейная
линейная
/
*
Рис. 2. Схематическое изображение структуры и формы вирусных геномов.
32
Проблемы происхождения жиши
Клеточное деление
(не обязательно симметричное
и бинарное)
Рис. 3. Различия между механизмами деления клеток и размножения вирусов.
Дизъюнктивная репродукция вирусов
нов колеблется от 2 до 1200. Размер ДНК и число генов у самых крупных вирусов в несколько
раз превышает эти параметры у мельчайших бактерий (например, Buchnera aphidicola), архей
{Nanoarchaeon equitans') и эукариот (Guillardia theta).
Если вирусы столь разнообразны, то что же их объединяет и отличает от всех других
(клеточных) биологических организмов? Таких фундаментальных признаков два. Первый -
это особенности метаболизма: вирусы неспособны синтезировать белок из-за отсутствия
трансляционного аппарата, а также аккумулировать химическую энергию в виде АТР
и подобных соединений из-за отсутствия дыхания, гликолиза или фотосинтеза (в отли-
чие от вирусов даже мельчайшие паразитические микроорганизмы умеют синтезировать
белки и производить АТР). Поэтому вирусы - облигатные паразиты, т. е. их размножение
может происходить только в клетке-хозяине. Облигатный паразитизм входил во многие
определения вирусов. Сейчас необходимо сделать некоторые уточнения. Так, показана
принципиальная возможность репродукция вирусов в бесклеточных экстрактах (Molla
et al., 1991). Тем не менее, абсолютная зависимость вирусов от клеточных компонентов
и клеточных источников энергии остается непреложным фактом. Показана также возмож-
ность лабораторного химического синтеза жизнеспособного вирусного генома (Cello et al.,
2002), но размножаться этот геном может опять-таки только либо в живых клетках, либо
в бесклеточных экстрактах. Таким образом, фундаментальное различие метаболизма виру-
сов и клеточных организмов сохраняет силу.
Второй фундаментальный признак - сам способ размножения. Размножение вирусов
основано на так называемом дизъюнктивном механизме, при котором вирус-специфические
белки и нуклеиновые кислоты накапливаются в виде отдельных пулов, из которых в конеч-
ном счете они черпаются для сборки дочерних вирусных частиц или их главного компо-
нента - нуклеопротеидной сердцевины (рис. 3). В противоположность этому, размножение
клеток происходит путем деления (чаще всего, но не обязательно - бинарного). На каких-
то этапах своей жизненной истории геном некоторых вирусов может включаться в состав
клеточной хромосомы. В этом случае вирусный геном существует как интегральный ком-
понент этой хромосомы и подвергается удвоению при клеточном делении. Однако, после-
дующее образование вирусного потомства (т. е. собственно репродукция вируса) всегда
происходит по каноническому дизъюнктивному механизму.
33
/?.//. J.'O.L Вирус ДО ИЛИ после kjicikii?
СТРАТЕГИИ ХРАНЕНИЯ И ЭКСПРЕССИИ ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ ВИРУСОВ
Уже на сравнительно ранней стадии изучения молекулярной биологии вирусов ста-
ло ясно, что эти биологические объекты использую! непредвиденно разнообразные способы
хранения и выражения генетической информации. Как уже упоминалось, вирусные геномы -
в отличие от клеточных - могут быть представлены различными формами нуклеиновых кис-
лот. Удобную и сейчас общепринятую - классификацию вирусов, принимающую во вни-
мание это фундаментальное разнообразие, предложил Д. Балтимор в 1971 г. (Baltimore, 1971)
(рис. 4). Ключевую позицию в этой классификации занимает вирусная информационная или
матричная РНК (мРНК), которой Балтимор приписал положительную (+) полярность. Пози-
тивными стал!! называть и тс цепи геномных нуклеиновых кислот (РНК и ДНК), у которых
полярность соответствует полярности мРНК. Цепочки же РНК или ДНК, которые компле-
ментарны вирусной мРНК, имеют, соответственно, негативную (-) полярность. В итоге, ви-
русы были разбиты на 6 групп или классов — вирусы с позитивным РНК-геномом, вирусы
с негативным РНК-геномом, вирусы с двунитевым PHК-геномом, вирусы с двунитевым ДНК-
геномом, вирусы с однонитевым ДНК-геномом (в этом случае полярность цепей не имеет
принципиального значения) и ретровирусы, использующие механизм обратной транскрип-
ции (т. е. синтез ДНК на матрице РНК).
Вскоре после публикации классификации Балтимора мы задумались над тем, исчер-
пывают ли включенные в нее вирусы все возможные способы хранения и выражения гене-
тической информации. На основе достаточно простых молекулярно-биологических постула-
тов - прежде всего, принципе комплсментарности при синтезе нуклеиновых кислот - была
предложена теоретическая система, которая предусматривала возможность существования
и некоторых тогда неизвестных классов вирусов (Агол, 1974; Agol, 1974) (рис. 5). Наиболее
важными «новичками» были вирусы с ДНК-геномом, цикл репродукции которых должен
был включать обратную транскрипцию. Такие предсказанные вирусы действительно вскоре
были обнаружены - сейчас их называют ретроидные вирусы или парарстровирусы. К ним
относятся такой важный патоген, как вирус гепатита В, а также ряд растительных вирусов,
например, вирус мозаики цветной капусты. Вирусы с позитивным РНК-геномом и двуните-
Комплементарный синтез
Вирусные геномы.
Вирусы с
позитивным
РНК-геномом
( (-)РНК ')
С(-)рнк
(±)РНК
Вирусы с
двунитевым
РНК-геномом
Вирусы с
негативным
РНК-геномом
(+)РНК
Ретровирусы
Вирусы с
двунитевым
ДНК-геномом
мРНК = (+)РНК
(-)ДНКД+)ДНК
Вирусы с
однонитевым
ДНК-геномом
Рис. 4. Классификация вирусов, предложенная Д. Балтимором (Baltimore, 1971, модифицировано).
34
И роняемы iipoiii хождения ЖИ Hill
Вирусные геномы । Интермедиаты
с однонитевым
ДНК-геномом
,НК вфмРНК
Вирусы с двунитевым
РНК-геномом
мРНК <^м!(±)РНК (Т)РНК
мРНК ^1(±)ДНК| (±)ДНК
мРНК
Вирусы с негативным
РНК-геномом
Г(-)рнк1 (+)рнк
мРНК <»!(±)ДНК онДНК
Ретровирусы
Рис. 5. Система вирусов (Агол, 1974, модифицировано). Генетические системы вирусов, изображен-
ных на голубом фоне, были предсказаны, а впоследствии обнаружены.
вым ДНК-геномом были формально разбиты на два класса каждый - в зависимости от того,
являются ли промежуточные (образующиеся при репликации) формы геномной нуклеино-
вой кислоты однонитевыми или двунитевыми. Все эти классы сейчас «заселены».
Таким образом, у вирусов - в отличие от клеточных организмов - реализуются все теоре-
тические возможные способы хранения и выражения генетической информации. Это важнейшее
обстоятельство, несомненно, имеет прямое отношение к проблеме происхождения вирусов.
«РОДСТВЕННИКИ» ВИРУСОВ
У вирусов есть многочисленные и достаточно близкие родственники это «эгоистиче-
ские» генетические элементы (как ДНК, так и РНК), которые, подобно вирусам, полностью
зависят от клеточных механизмов синтеза белков и генерации энергии. К ним относят вирои-
ды (инфекционные, но не способные кодировать белки, относительно низкомолекулярные
кольцевые РНК), плазмиды (разнообразные нехромосомные ДНК- и РНК-элементы), транс-
позоны (мобильные генетические элементы, перемещающиеся между различными участка-
ми генома), ретротраспозоны (мобильные элементы ДНК, перемещающиеся при участии ме-
ханизма обратной транскрипции) и интроны (участки РНК, которые выщепляются из «зре-
лых» молекул РНК в процессе сплайсинга, а также соответствующие им участки ДНК).
Эти элементы могут быть интегрированы в клеточные ДНК или РНК или существовать
в виде самостоятельных (до известной степени) молекул. Некоторые из них способны размно-
жаться в клетке и мигрировать между клетками. И хотя в отличие от вирусов у них нет белок-
содержащих оболочек и стабильных неклеточных форм, с вирусами их объединяет фунда-
ментальная черта - они используют те же механизмы репликации и выражения генетической
информации, которые были рассмотрены в предыдущем разделе. Поэтому есть все основания
объединить эти элементы с вирусами в один общий домен биологических объектов. Это поло-
жение также весьма существенно при рассмотрении происхождения вирусов.
35
В.Н. Аго‘1. Вирус - до пли после клетки?
ПРОБЛЕМА ПРОИСХОЖДЕНИЯ ВИРУСОВ
Эволюционные взаимоотношения между вирусами и клеточными организмами - один
из ключевых вопросов проблемы происхождения жизни. Нередко эволюцию трех главных
живых доменов или «царств» - эубактерий, архей и эукариот - изображают в виде единого
древа (хотя полного консенсуса по этому вопросу нет). Каково место вирусов на этом древе?
И есть ли, вообще, такое место? Более того, правомерно ли представлять эволюцию вирусов
в виде единой ветви какого-то древа или даже просто отдельного древа?
Проблема происхождения вирусов, естественно, привлекала к себе внимание с момен-
та их открытия Д.И. Ивановским в конце XIX в. Обсуждение этого интригующего вопроса
в течение первых — 100 лет вынуждено носило чисто спекулятивный характер (см., например,
Luria, Darnell, 1967; Agol, 1976). Тем не менее, было высказано несколько интересных и в то
время привлекательных гипотез. Схематически их можно разбить на три группы: вирусы -
это «сбежавшие» из клетки «сумасшедшие» гены; это клетки, дегенерировавшие до предела
паразитизма; и это доклсточныс генетические элементы. В то время последняя точка зрения
была наименее популярна, но, как мы увидим, оказалась наиболее близкой нашим тепереш-
ним представлениям.
СОВРЕМЕННЫЕ ВЗГЛЯДЫ НА ПРОИСХОЖДЕНИЕ ВИРУСОВ
Наиболее распространенные современные гипотезы происхождения вирусов основа-
ны на двух крупных достижениях молекулярной биологии: во-первых, открытии рибозимов
(молекул РНК, обладающих ферментативной активностью) и формулировании теории «мира
РНК», постулирующей, что РНК была «изобретена» раньше ДНК и белков, и, во-вторых,
успехах геномики (расшифровки нуклеотидной последовательности огромного числа кле-
точных и вирусных геномов). Общим для большинства современных гипотез является пред-
ставление о об очень значительном, если не решающем, вкладе вирусов в генетическую ин-
формацию клеточных организмов.
В качестве примера таких взглядов кратко рассмотрим две в чем-то перекрывающиеся,
но, все же, существенно различные гипотезы происхождения вирусов и их роли в эволюции
клеточных организмов. Одна из гипотез принадлежит П. Фортеру (Forterre, 2006а, Ь). Фран-
цузский исследователь постулирует, что последний общий предшественник всего живого
(Last Universal Common Ancestor или LUCA) представлял собой клетку с РНК-геномом, в
которой «жили» РНК-содержащие вирусы (рис. 6). Важно, что для репликации клеточного и
вирусного геномов в этом случае требуется один и тот же класс ферментов - РНК-зависимые
РНК-полимеразы (RNA-dependent RNA polymerase; RdRP). Предполагается, что современ-
ные РНК-вирусы унаследовали этот ключевой фермент с тех древнейших времен. Действи-
тельно, RdRP большинства известных вирусов с позитивным однонитевым или двунитевым
РНК-геномом проявляют столь существенное сходство, что можно говорить об их общем
происхождении. Для возникновения РНК-вирусов потребовалось изобрести капсид, т. е. бел-
ковую оболочку. Опять-таки, белки оболочки многих вирусов с однонитевым и двунитевым
РНК-геномом обладают принципиально сходной структурной организацией, несмотря на то,
что аминокислотные последовательности этих белков могут иметь между собой мало обще-
го (см. также Bamford, 2003). Отсюда выводится предположение, что эти капсидные белки
имеют общего прародителя.
Следующим фундаментальным эволюционным событием, по гипотезе Фортера, было
возникновение ДНК (рис. 6). Постулируется, что первые ДНК появились, в результате син-
теза на матрице РНК, т.е. в результате обратной транскрипции, и появились они у вирусов,
которые по современной терминологии называют ретровирусами (рис. 4, 5). Возникновение
ретровирусов сопряжено с двумя принципиальными новшествами. Во-первых, потребова-
лись ферменты, умеющие осуществлять обратную транскрипцию, т.е. РНК-зависимые ДНК-
36
Проблемы происхождения жпшп
Мир РНК
Переход от РНК к ДНК
Мир ДНК
Bacteria
Archaea
Еикагуа
РНК-клетки,
зараженные
ДНК
Рис. 6. Модель перехода от мира РНК к миру ДНК по П. Фортсру (Fortcrre, 2006а, Ь).
полимеразы (обратные транскриптазы). Изобретение таких ферментов существенно облегча-
ла уже имеющаяся «подсказка» - обратные транскриптазы устроены в значительной степени
по образу и подобию уже существовавших тогда RdRP. Действительно, между этим двумя
классами ферментов имеется заметное структурное сходство. Второе изобретение - это появ-
ление ферментов, способных превращать субстраты для синтеза РНК (рибонуклеозиды или
рибонуклеотиды) в субстраты для синтеза ДНК (дезоксирибонуклеозиды или дезоксирибо-
нуклеотиды). Так или иначе, и эта задача была решена. Следует заметить, что изобретать для
ретровирусов принципиально новый класс белков оболочки не понадобилось - был исполь-
зован (и до сих пор используется) общий план строения капсидных белков РНК-вирусов.
И, наконец, наступает закат «мира РНК» и возникновение «эпохи ДНК» (рис. 6). С био-
химической точки зрения, ключевым этапом здесь было появление нового класса ферментов-
ДНК-зависимых ДНК-полимераз, что, в свою очередь, обеспечило возможность возникно-
вения ДНК-вирусов. Превращение клеток с РНК-геномом в клетки с привычным для нас
ДНК-геномом произошло, по Фортеру, в результате заражения ДНК-вирусами. При этом три
домена клеточных организмов возникли независимо в результате заражения разными ДНК
вирусами-основоположниками (founder viruses). Особые основоположники положили начало
эубактериям, археям и плазмидам, в то время как для появления эукариот потребовалось не-
сколько разных вирусов-основателей.
По гипотезе, которую развивает Е. Кунин с коллегами (Koonin, Martin, 2005; Koonin
et al., 2006), жизнь зародилась в микроскопических неорганических ячеистых структурах, кото-
рые могли служить суррогатами клеток-рис. 7. Рибозимы, возникшие на добиологичсской ста-
дии эволюции, могли накапливаться в таких ячейках в относительно высоких концентрациях,
могли передвигаться между ячейками и «заражать» их. Следующий фундаментальный этап -
возникновение РНК-вирусов с однонитевым и двунитевым геномом. Затем, как и у Фортера,
следует появление обратной транскрипции и ретровирусов и, наконец, возникновение ДНК-
зависимого синтеза ДНК и ДНК-вирусов. Но все это происходило, по Кунину, еще на до-
клеточной стадии эволюции. А клетки с ограничивающими их мембранами появились уже
после того, как возникли основные способы хранения и передачи генетической информации.
Но сначала возникли археи и эубактерии. Эукариоты же появились, в соответствии с широко
распространенной точкой зрения, в результате поглощения бактериальной клетки архейной
клеткой (рис. 8). При этом эукариотические клетки унаследовали генетические структуры
не только от этих двух клеточных прародителей, но и от их вирусов, а вирусы эукариот -
37
В.И. Агоп. Вирус до или после клетки'?
Появление клеток
с ДНК-геномами
Доклеточные
«изобретения»
Изобретение ДНК-
зависимого синтеза ДНК
Изобретение ДНК и РНК-
зависимого синтеза ДНК
Изобретение белков
Phi Л \Л/лг1г4
RNA World
Изобретение РНК
Вирусы Бактериофаги
архей
RNA-DNA Retro
World
ДНК-вирусы,
плазмиды
Ретровирусы,
ретроидные
вирусы и
элементы
Вирусы с
(+)РНК и (±)РНК
Эгоистичные
рибозимы
Рис. 7. Модель перехода от доклеточного мира РНК к археям и бактериям по Е. Кунину (Koonin et al.,
2006).
от фагов и вирусов архей. Этот вывод подкрепляется сравнительным анализом нуклео-
тидных последовательностей и белковых структур таких ключевых ферментов, как RdRP,
обратные транскриптазы, некоторые ферментов синтеза ДНК, а также капсидных белков.
Таким образом, по этому сценарию, клетки произошли позже вирусов и на их основе. Что
же касается самих вирусов, то их разные группы могут не иметь общего предшественника (т. е.
не являются монофилетичными), но, в то же время, в каком-то смысле, все они - родственники.
Общим для этих двух гипотез эволюционной истории является представление о том,
что современные вирусы унаследовали от древнейшего мира многие молекулярные механиз-
мы, которые не сохранились у клеточных организмов. Именно поэтому способы транскрип-
ции и репликации у вирусов существенно более разнообразны по сравнению с таковыми
у клеточных организмов (рис. 5). Однако возникновение клеток не означало прекращения
обмена генетической информацией между миром вирусов и миром клеточных организмов.
Такой обмен, по-видимому, продолжался на протяжении всей истории биосферы. В частно-
сти, можно полагать, что некоторые вирусы приобрели у клеток вспомогательные ферменты
синтеза РНК и ДНК, а также, что очень важно, протеазы. Пример относительно более свежих
вирусных заимствований у клеток - белки, взаимодействующие с компонентами врожденно-
го и приобретенного иммунитета, особенно характерные для крупных вирусов.
Не останавливался и поток генов от вирусов к клеткам. Здесь следует упомянуть
об эндогенных вирусах - остатках генов ретровирусов, интегрировавших в клеточные хро-
мосомы. По некоторым оценкам, более половины генома млекопитающих имеет вирусное
происхождение, если учитывать как самые древнейшие, так и более новые приобретения.
38
Проб. !кмы пропс хож. и ная жи n/if
Вирусы эукариот
Ядро
Хроматин
Ядерная
пора
Цитозоль (из
археона)
Вирусы
архей
Плазматическая
мембрана
Нк,Ъуп
Митохондрии
(из бактерий)
ЭУКАРИОТЫ
Археон с вирусами, Бактерия с фагами,
плазмидами и др. плазмидами и др.
эгоистическими элементами эгоистическими элементами
Рис. 8. Модель появления эукариотической клетки, а также вирусов эука-
риот в результате поглощения бактерии археоном (Koonin el al., 2006).
ЗАКЛЮЧЕНИЕ
Краткое рассмотрение современных взглядов на происхождение клеточных организ-
мов показывает, сколь важную, а возможно, и решающую роль придают в этом процессе
вирусам. Как выразился один вирусолог: «Древо жизни своими корнями погружено в океан
вирусов» (Bamford, 2003). При этом многие (хотя и не все - см., например, Altstein, 1992)
исследователи полагают, что ДНК-содержащие вирусы произошли из РНК-содержащих. Эти
эволюционные сценарии весьма интересны, они объясняют сравнительное богатство спосо-
бов хранения и реализации генетической информации у вирусов, а также стимулируют даль-
нейшие исследования. Конечно, мы не может и не должны забывать, что мир вирусов - один
из главных источников болезней человека, животных и растений. В этой ипостаси вирусы -
то, с чем надо бороться самым активнейшим образом. Но нельзя сбрасывать со счетов и то,
что мы как человечество, да и вся живая природа не были бы без вирусов таковыми, как есть,
а, может быть, и не существовали бы вообще.
В заключение я бы согласился с замечанием Ф. Дулитла (Doolittle, 2006), что пока эво-
люционные сценарии все же ближе к произведениям искусства и не обязательно должны
быть истинными («Evolutionary scenarios are an artform. They do not have to be true!»). Будем
надеяться, что прогресс науки либо подтвердит их справедливость, либо выдвинет новые,
еще более увлекательные сценарии.
Настоящая статья частично основана на материалах лекции, прочитанной на научной
школе «Evolution from Cellular to Social Scales» (NATO AS1, Гейло, Норвегия, 10-20 апреля
2007) (Agol, 2008).
39
В.П. Вирус ло пли после клетки?
СПИСОК ЛИТЕРАТУРЫ
Ачл/ В.И. 1974. О системе вирусов // Усн. соврем, биол. Т. 77. С. 9-29.
Ago/ )’./. 1974. Towards the system of viruses.' Biosystcm. V. 6. P. 113 132.
Ago/ IL 1976. An aspect of the origin and evolution of viruses//Origins of Life. V. 7. P. 119-132.
Jgo/ 1.1. 2008. The origin and evolution ofviruses // Evolution from Cellular to Social Scales / Eds. A.T. Skjel-
torp, AV Belushkin. NATO Science for Peace and Security. Ser. B: Physics and Biophysics. Springer.
P. 91 8.
A/tstein A.D. 1992. The protocellular concept of the origin of viruses // Semin. Virol. V. 3. P. 409-417.
Baltimore D. 1971. Expression of animal virus genomes // Bacteriol. Rev. V. 3. P. 235-241.
Bamford D.H. 2003. Do viruses form lineages across different domains of life? // Res. Microbiol. V. 154.
P. 231 -236.
Breitbart M.. Rohwer F. 2005. I lore a virus, there a virus, everywhere the same virus? // Trends Microbiol.
V. 13. P. 278-284.
Cello J.. Pau! .4.1'.. Wimmer E. 2002. Chemical synthesis of poliovirus cDNA: generation of infectious virus
in the absence of natural template ’ Science. V. 297. P. 1016-1018.
Doolittle W.F. 2006. Reviewer’s report I. In Koonin et al., 2006 // Biology Direct. V. I. P. 29.
Forterre P. 2006a. Three RNA cells for ribosomal lineages and three DNA viruses to replicate their genomes:
a hypothesis for the origin of cellular domain // Proc. Natl. Acad. Sci. USA. V. 103. P. 3669- 3674.
Forterre P. 2006b. The origin ofviruses and their possible roles in major evolutionary transitions // Virus
Res. V. 117. P. 5 16.
Koonin E.V., Martin W. 2005. On the origin of genomes and cells within inorganic compartments " Trends
Genet. V. 21. P. 647-654.
Koonin E.V., Senkevich T.G.. Dolja V.V. 2006. The ancient virus world and evolution of cells // Biology Direct.
V. I. P. 29.
Luria S.E., Darnell .J. E. 1967. General virology. 2nd cd. New York: John Wiley and Sons. 512 p.
Motia A., Paid A. K, Wimmer E. 1991. Cell-free, denovo synthesis of poliovirus//Science. V. 254. P. 1647-1651.
Suttle C.A. 2005. Viruses in the sea//Nature. V. 437. P. 356-361.
Suttle C.A. 2007. Marine viruses - major players in the global ecosystem // Nature Rev. Microbiol. V. 5.
P. 801-812.
Wimmer E.. Helfen C.U., Cao X. 1993. Genetics of poliovirus // Annu. Rev. Genet. V. 27. P. 353-436.
Virus - before or after the cell?
V.l. Agol
The nature of viruses and fundamental distinction of the mechanism of their reproduction
from the cellular division are briefly considered. The ways of storage and expression of viral
genetic information are much more diverse compared to those of cells. According to current views,
the «RNA World» predated the «DNA World». In this perspective, hypotheses on viral origin
according to which RNA viruses had emerged after or before the cells with RNA-genomes are
considered. In any case, it is proposed that the DNA-genomes had originated in viruses and that
viruses played a decisive part in the appearance of Archea, Eubacteria, and Eukarya.
40
Проблемы происхождения ж и ши
Дискуссия
но пленарному докладу В.И. Агола
«ВИ РУС-ДО ИЛИ ПОСЛЕ КЛЕТКИ?»
Председательствующий Л. М. Мухин
А.В. Тутуков: Почему так много видов вирусов существует одновременно, что обеспе-
чивает их многообразие и многообразие их экологичсских ячеек?
В.И.А.: Я придерживаюсь той точки зрения, что эволюционные сценарии - это вид ис-
кусства, не обязательно все должно быть правдой. Полагаю, что первично разнообразие ви-
русов ничем нс сдерживалось, ничем не ограничивалось, они имели все возможные способы
репликации и транскрипции. Зачем они рекомбинировали, приобретали новые гены, либо
в результате дупликации, либо от других вирусов, либо из клетки... Так что, по-моему, во-
прос заключается в том, сеть ли ограничения этому разнообразию. Вирусы удивительно
устойчивы. Можно в их геноме очень консервативные последовательности заменять совер-
шенно случайными - мы это делаем - и отбирать вирусы, которые будут жизнеспособны.
Возможностей для изменений вирусного генома очень много.
А.В. Тутуков: В астрономии многочисленные типы звезд оказалось возможным располо-
жить в эволюционной последова тельности и стало ясно, что многие типы - это одна и та же звез-
да, наблюдаемая в разные фазы развития. Нельзя ли что-то подобное применить к вирусам?
В.И.А.: Таксономия и классификация вирусов оперирует семействами и надсемей-
ствами, внутри которых можно построить дерево, основанное на монофилетических связях.
Но поскольку возможен горизонтальный перенос генетической информации, построить об-
щее дерево не удастся. Я бы гак сказал: дерево построить нельзя, а кустарник - пожалуйста.
А.В. Тутуков: Неужели Чарльз Дарвин так плох, что в докладе ни разу не прозвучал
термин «естественный отбор»?
В.И.А.: Когда мы говорим об эволюции конкретных вирусов - например, вируса полио-
миелита - естественный отбор имеет большое значение.
Л.М. Мухин: И есть критерии отбора?
В.И.А.: Должна выжить популяция вируса. При этом популяция бывает генетически
очень гетерогенной. И иногда для воспроизводства популяции нужно, чтобы между ее от-
дельными представителями непременно существовало взаимодействие. То есть эволюция
происходит на уровне популяции, а не ее частей или членов.
Л.В. Ксанфомалити: Можно ли дать какие-то временные оценки, сколько каждая ста-
дия занимала, исходя из вероятности того или иного процесса?
В.И.А.: Я точно не берусь. Есть люди, которые этим занимаются. Мы разбираем невос-
производимые события. Скорость эволюции конкретных вирусов оценить можно.
А.С. Спирин: Как Вы думаете, если бы в космосе носились вирусы, бактерий бы
не было, и мы исходили бы из внеземного происхождения жизни, возможно ли попада-
ние на Землю такой комбинации вирусов, из которой в принципе могла бы возникнуть
живая клетка, жизнь?
В.И.А.: Действительно, вирусы существенно более устойчивы физически, можно пред-
положить, что они могут переноситься в космическом пространстве (хотя таких данных,
естественно, нет). И если на Земле были подходящие условия, то я не вижу большой разницы,
были ли они созданы на Земле или прилетели.
А.Д. Алытитейн: Как Вы представляете происхождение ДНК-содержащих вирусов
из РНК-содержащих? Должны возникнуть минимум три фермента: тот, который будет
41
делать дезоксирибонуклеотиды, тот, который будет делать код ДНК на матрице РНК, и тот,
который будет считывать ДНК с РНК.
В.И.А.: Проше всего с обратной транскриптазой, она очень похожа на PH К-зависимую
РНК-полимеразу. Что касается ферментов, которые синтезируют ДНК на матрице РНК, они,
в общем, тоже не из другого теста. В общем, инженерных трудностей здесь нет, есть труд-
ности скорее идеологические - зачем это все появлялось, какое это давало преимущество?
И эти ферменты не обязательно должны появиться одновременно. Что-то может появиться
и не использоваться или использоваться для чего-то другого.
М.Я. Маров: Вирус можно представить как некую субстанцию, паразитирующую
на клетке. Можно ли говорить о том, что природа создала вирус как своего рода механизм
естественного отбора для более сложных организмов?
В.И.А.: Я бы так ответил - создано не для этого, но природой используется и для этого.
С.В. Шестаков: Когда Вы говорите о происхождении и эволюции вирусов, то Вы,
конечно, имеете в виду очень ранние этапы эволюции биосферы. Но, говоря об эволюции
современных вирусов, вероятно, целесообразнее этот процесс называть не эволюцией, а из-
менчивостью.
В.И.А.: Это вопрос скорее семантический. Конечно, в этом случае речь идет о микро-
эволюции. Но в то же время, я хочу отмстить, что и в наше время вирусы подвергаются эво-
люции -тот же вирус иммунодефицита человека, вирус гриппа. Когда вирус получает нового
хозяина, он эволюционирует, это в некотором смысле макроэволюционное событие. То есть
пока, например, вирус полиомиелита остается вирусом полиомиелита, это микроэволюция,
дрейф и так далее, но когда один вирус становится другим, происходит качественный скачок -
это уже эволюция.
Л.М. Мухин: Необходима начальная информация о начальном геноме, иначе все это -
искусство. Откуда ее взять?
В.И.А.: Это центральный вопрос - где начинается смысл. Если взять случайную после-
довательность, когда появится жизнеспособный геном? Число сценариев ограничено, число
возможностей бесконечно.
А.Б. Четверин: В экспериментах стартовали со случайных последовательностей дли-
ной в несколько десятков нуклеотидов. Надо стартовать с 1012, чтобы что-то получилось.
В.И.А.: Но там уже был отбор, смысл, внесенный экспериментатором.
42
Нроелемы происхождении жити. М.: ПИН РАН. 2009. С. 43-55
ДРЕВНИЙ МИР РНК
©2009 г. А.С. Спирин
Институт белки РАН
н Московский государственный университет им. М.В. Ломоносова
spirin@vega.protres.ru
Основываясь на современных представлениях о многофункциональности
РНК, автор рассматривает ряд новых механизмов, участие которых в про-
исхождении и эволюции древнего мира РНК могли играть критическую
роль. Так, реакция спонтанной трансэстерификации полирибонуклеоти-
дов, открытая А.Б. Четвериным с сотрудниками, могла приводить к удли-
нению первичных коротких олигорибонуклсотидов и к генерации вариан-
тов последовательностей для последующего естественного отбора случай-
но возникающих функционально активных молекул. Далее, образование
смешанных молекулярных колоний РНК на влажных твердых средах типа
глин могла обеспечить компартментализацию ансамблей функционально
активных РНК в отсутствие оболочек и мембран, что было необходимо
для дальнейшей эволюции формирующегося мира РНК. Систематическое
экспоненциальное обогащение популяции РНК функционально лучшими
молекулами за счет попеременного растворения колоний при затоплении
и образования новых колоний при подсушивании первобытных водоемов
(«первобытный естественный SELEX») могло быть главным двигателем
эволюционного процесса в мире РНК.
ВВЕДЕНИЕ
Главной проблемой в происхождении жизни является вопрос о возникновении ап-
парата наследственности, а, следовательно, о возникновении «вещества наследственно-
сти» - нуклеиновых кислот. Обе нуклеиновые кислоты — рибонуклеиновая кислота (РНК)
и дезоксирибонуклеиновая кислота (ДНК) - химически представляют собой линейные (не-
разветвленные) полимеры, построенные из четырех сортов нуклеозидных остатков (соеди-
нений пятиуглеродного сахара и азотистого пуринового или пиримидинового основания),
соединенных между собой фосфатными группами. Фосфатные группы соединяют угле-
водные части нуклеозидов в цепь, так что цепь нуклеиновой кислоты может быть описана
как пснтозо-фосфатный полимер с азотистыми основаниями в качестве боковых групп.
Основное различие химических структур РНК и ДНК состоит в том, что углеводным ком-
понентом РНК является рибоза, а в ДНК - дезоксирибоза, дегидроксилированное производ-
ное рибозы.
43
.4.С. Спирин. Древний мир РНК
Воспроизведение /
генов ДНК
(редупликация) I
Копирование генов
(транскрипция)
V
РНК
Кодирование структуры белков
(трансляция)
БЕЛОК
Рис. 1. Центральная догма молекулярной биологии.
Однако пространственная структура этих двух близкородственных полимеров разли-
чается принципиально: ДНК - это жесткая двойная спираль, образованная за счет парного
взаимодействия бок о бок двух полидезоксирибонуклеотидных цепей, а РНК - одноцепочеч-
ная гибкая молекула. ДНК представляет собой гены, и ее двуспиральпая пространственная
структура необходима для точного копирования своей собственной первичной структуры
(специфической последовательности четырех сортов мономеров цепи) в процессе компле-
ментарной редупликации (Watson, Crick, 1953). Долгое время считалось, что функция РНК
состоит лишь в копировании (транскрипции) одной из цепей ДНК для того, чтобы служить
генетической матрицей для синтеза белков. Представление об этом потоке генетической
информации от ДНК через РНК к белкам и его необратимости было сформулировано как
«центральная догма молекулярной биологии» (рис. 1). Впоследствии, однако, утверждение
о необратимости потока информации от ДНК к РНК было подвергнуто ревизии: было откры-
то, что в ряде случаев - в первую очередь при вирусных инфекциях - наблюдается явление
«обратной транскрипции», когда на матрице РНК синтезируется комплементарная цепь ДНК
(Baltimore, 1970; Temin, Mizutani, 1970). При этом признание строгой необратимости потока
информации от нуклеиновых кислот к белкам продолжает быть основой всей современной
генетики и молекулярной биологии.
МНОГОФУНКЦИОНАЛЬНОСТЬ РНК
Итак, вплоть до недавнего времени за нуклеиновыми кислотами, и в том числе за РНК,
признавались лишь генетические функции (рис. 2). Главная генетическая функция - функция
комплементарной репликации (т. с. репликации через образование комплементарной поли-
нуклеотидной цепи) при участии катализирующего полимеризацию фермента - безусловная
прерогатива ДНК, но оказалось, что она присуща и РНК. Выше было упомянуто об открытии
способности РНК служить матрицей для синтеза ДНК в процессе обратной транскрипции.
Еще раньше была продемонстрирована функция воспроизведения своей структуры - репли-
кация РНК на матрице РНК - прежде всего, на примере репликативного цикла ряда РНК-
содержащих вирусов (Baltimore et al., 1966; Spiegelman, Haruna, 1966). Такую репликацию
РНК, независимую от ДНК и запрещенную в большинстве случаев в циклах нормальных
клеток современных организмов, можно рассматривать как реликт, сохранившийся в совре-
менном мире благодаря вирусам как реликтовым генетическим агентам, выводящим репли-
44
Проблемы происхождения жизни
\ /
\ /
РНК
Репликативная функция:
копирование генов,
обратная транскрипция,
саморепликация
Кодирующая функция:
программирование белкового синтеза
Белки
Рис. 2. Генетические функции РНК.
Рис. 3. Конформации и конформацион-
ные переходы высокополимерной РНК
в зависимости от ионной силы и темпера-
туры. Внизу справа - конформация ком-
пактной глобулы (Spirin, 1960).
кацию и трансляцию из под контроля клеточной ДНК. В настоящее время выясняется, что
реликтовую репликацию РНК можно обнаружить и в нормальных клетках: многие живот-
ные и растения несут гены, кодирующие РНК-зависимые РНК-полимеразы или их гомологи,
а недавно открытые регуляторные микроРНК и интерферирующие РНК являются, возмож-
но, их субстратом для комплементарной репликации.
Другая генетическая функция - кодирование белков - не требует особых комментари-
ев. РНК (мРНК) кодирует белки не только «наравне» с ДНК, но именно она непосредственно
участвует в процессе декодировки, служа матрицей для синтеза белков. Таким образом, обе
генетические функции ДНК - репликативная и кодирующая - оказываются присущи как
ДНК, так и РНК (рис. 2).
Если кодирующие функции так или иначе свя-
заны в основном с полинуклеотидами как линей-
ными полимерами, то функции белков в основном
зависят от их трехмерной структуры, т. е. специфи-
чески свернутой, по большей части компактной (гло-
булярной) конформации. Поэтому принципиально
важным было установление того факта, что высоко-
полимерные полинуклеотидные цепи РНК, как и по-
липептидные цепи белков, способны самосворачи-
ваться в компактные структуры (Спирин и др., 1959;
Spirin, 1960; Kisselev et al., 1961; Богданова и др., 1962)
(рис. 3) и что их сворачивание приводит к специфи-
ческим конформациям молекул РНК (Vasiliev et al.,
1978; Vasiliev, Zalite, 1980). С другой стороны, в раз-
витии современных представлений о функциях РНК
решающим было открытие некодирующих РНК.
Оказалось, что кодирующие РНК составляют лишь
малую долю клеточных РНК, а основная часть РНК
представлена некодирующими РНК, куда относятся
в первую очередь рибосомные РНК (Belozersky, Spirin,
1958; Brenner et al., 1961; Spiegelman, 1961). Именно
45
.1.С. Спирин. Древний мир РНК
Рис. 4. Кодирующая РНК (мРНК) и два основ-
ных типа некодирующих РНК (рибосомные
РНК и тРНК).
для высокополимерных некодирующих РНК
двух рибосомных субъединиц было показано,
что их специфическое компактное самосвора-
чивание задает специфическую форму каждой
из субъединиц и, в конечном счете, определяет
конформацию рибосомы (Vasiliev et al., 1986).
Таким образом, так же как и белки, РНК спо-
собны образовывать специфические третичные
структуры, т.е. обладают структурной и фор-
мообразующей функцией.
Способность РНК к формированию ком-
пактных трехмерных структур, как и в слу-
чае белков, дает основу для специфического
взаимодействия с другими молекулами - как
макромолекулами, так и малыми лигандами.
Другими словами, для молекул РНК, сверну-
тых в специфическую глобулу и тем самым соз-
дающих на своей поверхности уникальный пространственный узор, приходится допустить
возможность функции молекулярного узнавания, как и у белков. Пожалуй, первыми извест-
ными «узнающими» РНК можно считать тРНК, выполняющие адапторную роль в биосин-
тезе белка (рис. 4). Эти среднего размера компактно свернутые молекулы РНК поочередно
и очень избирательно взаимодействует с рядом макромолекулярных структур в клетке: сна-
чала с аминоацил-тРНК-синтетазой, связанной с аминоациладенилатом как активированной
формой аминокислоты, затем, уже неся на себе ковалентно присоединенный аминоацильный
остаток, с фактором элонгации EF1, вместе с которым она поступает в рибосому. Хотя на этом
пути несомненно реализуются функции специфического узнавания молекулами тРНК других
макромолекул, долгое время все же молчаливо принималось, что основную роль здесь играет
узнавание тРНК со стороны белков - ферментов, факторов трансляции и рибосомных бел-
ков. В 1980-х гг. английским ученым Э. Кандлиффом (Cundliffe, 1986) было впервые заявлено
о способности структурированных участков рибосомной РНК специфически узнавать малые
лиганды ненуклеиновой и небелковой природы. Он представил экспериментальные данные
в пользу избирательного взаимодействия (связывания) именно участков свернутой рибосом-
ной РНК, а не рибосомных белков, с рядом антибиотиков рибосомного действия - тиостреп-
тоном, эритромицином, аминогликозидами (стрептомицином, канамицином, неомицином).
Через 10 лет были представлены прямые структурные данные о специфическом связывании
аминогликозидных антибиотиков районом малой (16S) рибосомной РНК (Fourmy et al., 1996).
Окончательное признание самых широких возможностей у РНК узнавать другие молекулы
и весьма специфично взаимодействовать с ними пришло благодаря аптамерам - небольшим
по размерам синтетическим РНК, получаемым путем отбора из многих вариантов нуклеотид-
ных последовательностей с помощью процедур так называемой «бесклеточной эволюции»,
«эволюции в пробирке» (Ellington, Szostak, 1990; Tuerk, Gold, 1990). Оказалось, что можно
отобрать и размножить РНК, обладающие способностью избирательно связывать практиче-
ски любой вид молекул, начиная от низкомолекулярных органических соединений и кончая
различными индивидуальными пептидами и белками (см. обзоры Gold et al., 1995; Puglisi,
Williamson, 1999). Другими словами, РНК, как и белки, действительно в полной мере могут
обладать функцией специфического молекулярного узнавания.
На способности РНК к специфическому молекулярному узнаванию базируется и ката-
литическая функция РНК. Однако, на протяжении всей предшествующей истории биохимии
утверждалось, что биохимический катал из-«прерогатива» исключительно белков-ферментов.
46
Проблемы происхождения жиши
Мультифункциональность РНК
(1) Кодирующая функция:
программирование белкового синтеза линейными последовательностями
полирибонуклеотидов.
(2) Репликативная функция:
репликация генетического материала через комплементарные
последовательности полинуклеотидов.
(3) Структурообразующая функция (формирование трехмерных структур):
самосворачивание линейных полирибонуклеотидов в уникальные компактные
конформации.
(4) Функция специфического узнавания лигандов:
специфические пространственные взаимодействия с другими
макромолекулами и малыми лигандами.
(5) Каталитическая функция:
специфический катализ химических реакций рибозимами.
Рис. 5. Основные типы функциональной активности молекул РНК в живой природе.
Поэтому и все теории происхождения жизни вынуждены были исходить из первичности белков
как макромолекул, абсолютно необходимых для возникновения биохимического метаболиз-
ма. Открытие каталитической функции РНК в американских лабораториях Т. Чека и С. Алт-
мана (Kruger et al., 1982; Guerrier-Takada et al., 1983) перевернуло все прежние представления
об исключительной роли белков не только в возникновении жизни, но и в понимании самого
явления жизни. По аналогии с белками-ферментами - энзимами - каталитические РНК были
названы рибозимами. По-видимому, почти все рибозимы, естественно существующие в живой
природе в клетках современных организмов, так или иначе участвуют в процессах, связан-
ных с превращениями полинуклеотидных цепей самих РНК. Однако оказалось возможным
создавать и искусственные рибозимы с более широким спектром катализируемых реакций
(Cech, Golden, 1999). Кроме того, как выясняется из всей совокупности данных по структуре
рибосом и особенностей катализируемой рибосомой реакции образования пептидных связей
в процессе биосинтеза белка, каталитический центр этой реакции (пептидил-трансферазный
центр рибосомы) формируется определенным доменом большой рибосомной РНК, без уча-
стия рибосомных белков, т. е. имеет рибозимную природу (Nissen et al., 2000).
Итак, именно после открытия каталитической функции РНК поменялась парадигма,
и взоры биохимиков и молекулярных биологов обратились к РНК как самому «самодостаточ-
ному» биополимеру. В самом деле, молекулы РНК оказались способны делать все то, что де-
лают белки - складываться в специфические структуры и определять формообразование био-
логических частиц, с большой точностью узнавать другие макромолекулы и малые лиганды
и взаимодействовать с ними, наконец, осуществлять катализ ковалентных превращений узна-
ваемых молекул (рис. 5). Таким образом, РНК является уникальным биополимером, которому
свойственны как функции ДНК, так и функции белков. Конечно, белки делают все это более
эффективно и разносторонне, чем РНК. Но зато белки в принципе «не умеют» самовоспроиз-
водиться - не существует никаких собственных белковых механизмов для воспроизведения
их структуры, кроме как через РНК. В то же время РНК содержит все необходимые структур-
ные предпосылки для точного воспроизведения ее собственной структуры (см. выше).
47
А.С. Сннрин. Древний мир РНК
ГИПОТЕЗА О ДРЕВНЕМ МИРЕ РНК
Открытие каталитически активных РНК - рибозимов - в 1982-1983 гг. сыграло решаю-
щую роль в формулировании и развитии гипотезы о том, что РНК как самодостаточный по-
лимер могла бы и воспроизводиться, и функционировать в глубокой древности на Земле или
других космических объектах до появления клеточных форм жизни. Можно предположить,
что молекулы РНК могли бы обходиться не только без ДНК как генетического вещества,
но и без белков для осуществления катализа важных синтетических и метаболических реак-
ций. Идея древнего безбелкового мира РНК как возможного предшественника современной
жизни на Земле была окончательно сформулирована в 1986 г. (Gilbert, 1986) и быстро при-
обрела многочисленных сторонников. В настоящее время гипотеза о том, что жизнь начи-
налась с молекул РНК и их ансамблей, является почти общепринятой (см. сборники «Мир
РНК» - «The RNA World» - под редакцией Gesteland et al., 1993, 1999, 2006). Таким образом,
термин «мир РНК» широко используется теперь для обозначения древней, пребиотической
ситуации на Земле, имевшей место около 4 млрд, лет назад, когда самореплицирующиеся
молекулы РНК или их ансамбли могли существовать и эволюционировать без белков.
Таким образом, согласно существующим представлениям, в древнем мире РНК не
было ни белков, ни ДНК, а лишь ансамбли различных молекул РНК, выполняющих разные
вышеперечисленные функции. Однако вопрос о возникновении такого мира на Земле - один
из самых трудных в теориях происхождения жизни. Можно предполагать, что первичные
олигорибонуклеотиды возникали из абиогенно образующихся монорибонуклеотидов или
их активированных производных путем полимеризации на поверхностях глин и глинопо-
добных минералов. Возможна была также и предшествующая химическая эволюция ну-
клеотидоподобных и олигонуклеотидоподобных соединений. В любом случае, появление
олигорибонуклеотидов должно было быть отправной точкой появления мира РНК. Однако,
для дальнейшего развития было необходимо, чтобы абиогенный синтез олигорибонуклео-
тидов, основанный на редких случайных событиях, был дополнен постоянным механизмом,
который бы мог генерировать варианты этих олигомеров и удлинять их (при сильной тенден-
ции к их спонтанной химической и физической деструкции). Элонгация коротких олигори-
бонуклеотидов в полирибонуклеотиды представляется абсолютно необходимым условием
для образования компактно свернутых структур со свойствами специфического узнавания
лигандов и каталитическими активностями, а генерация вариантов в популяции абиогенных
олиго- и полирибонуклеотидов требуется для того, чтобы дать шансы для случайного воз-
никновения нужных функциональных, в том числе каталитических, активностей.
В течение долгого времени не было предложено какого-либо удовлетворительного ре-
шения этой проблемы. Около 15 лет назад А.Б. Четвериным и Е.В. Четвериной с сотрудни-
ками (Chetverin et al., 1991; Chetverina, Chetverin, 1993) был разработан метод молекулярного
клонирования РНК: из единичных молекул РНК, помещенных на поверхность геля, содержа-
щего катализатор репликации (в данном случае вирусную РНК-зависимую РНК-полимеразу)
и рибонуклеозид-трифосфаты, оказалось возможным выращивать колонии молекул РНК,
идентичных исходной молекуле (рис. 6). Позднее метод был применен для регистрации еди-
ничных событий, происходящих с популяцией РНК в растворе, и была впервые эксперимен-
тально показана способность молекул РНК к спонтанной перестройке их нуклеотидных по-
следовательностей в отсутствие каких-либо ферментов и рибозимов (Chetverina et al., 1999).
Открытая спонтанная реакция характеризовалась следующими особенностями. Во-первых,
цепи РНК в растворе при температурах от 5 до 37 °C время от времени обменивались частя-
ми своих последовательностей; обмен мог осуществляться как между разными молекулами
(транс-перестройки), так и внутри одной и той же молекулы (цис-перестройки). Во-вторых,
эти перестройки были неспецифичны по отношению к последовательности и могли происхо-
дить в любом месте цепей. В-третьих, в отличие от рибозимных и ферментативных реакций,
48
rii'db.и.мы прош \4)x,ii:iin>i жи<нн
а также реакций самокатализируемого сплайсинга,
З'-гидроксилы не участвовали в этой спонтанной ре-
акции, а части РНК реагировали друг с другом вну-
тренними районами. Реакция оказалась зависима от
присутствия Mg2 . Скорость спонтанных перестроек
была невысока - одно событие в час на миллиард ну-
клеотидов; это означает, что 0.002-0.02 % цепей РНК
с длиной 800-8000 нуклеотидных остатков спон-
танно перестраиваются в популяции РНК в течение
24 часов. Реакция не требует никаких других компо-
нентов, кроме самой РНК и Mg2', и, таким образом,
может рассматриваться как присущее РНК химиче-
ское свойство и должна происходить в водной среде
повсюду, как в живой, так и в неживой природе.
Это открытие, очевидно, имеет прямое от-
ношение к проблеме возникновения древнего мира
РНК. Прежде всего, спонтанные перестройки ну-
клеотидных последовательностей могли быть
основным эффективным механизмом непрерывной
генерации вариантов олиго- и полирибонуклеоти-
дов до эры рибозимов. Более того, элонгация оли-
Рис. 6. Колонии молекул РНК, выросшие
на влажной поверхности агарозы или по-
лиакриламидного геля в присутствии
рибонуклеозид-трифосфатов и катализа-
тора комплементарной репликации РНК
(РНК-зависимой РНК-полимеразы фагафР)
(Chetverin et al., 1991; Chetverina, Chetverin.
1993).
горибонуклеотидов в полирибонуклеотиды путем спонтанной трансэстерификации, проис-
ходящей в этой реакции, представляется гораздо более реальной возможностью в то время,
по сравнению с последовательным ростом цепи, требующим специального механизма, по-
стоянной защиты от деградации удлиняемого полинуклеотида и постоянного притока энер-
гии. Именно этим путем и могли появляться длинные полирибонуклеотиды на заре возник-
новения мира РНК.
Появление достаточно длинных полирибонуклеотидов и генерация вариантов
за счет спонтанных цис- и транс-перестроек должна была привести к случайному появ-
лению рибозимов, и критичным должно было быть появление в популяции РНК рибози-
ма, катализирующего процесс комплементарной репликации РНК. Это - принципиальное
условие для того, чтобы размножить - амплифицировать - единичные молекулы случайно
возникших в популяции вариантов и сохранить их для эволюции. Другими словами, по-
явление механизмов РНК-катализируемой репликации РНК должно рассматриваться как
первое и необходимое условие для начала эволюции мира РНК. В ряде экспериментов была
экспериментально показана возможность создания рибозимов, осуществляющих лиги-
рование олигонуклеотидов на комплементарной матрице (Dotidna, Szostak, 1989; Doudna
et al., 1991, 1993) или полимеризующих короткие олигонуклеотиды или мононуклеоти-
ды путем удлинения олигонуклеотидной затравки на комплементарной матрице (Bartel,
Szostak, 1993; Ekland, Bartel, 1996; Johnston et al., 2001). С появлением рибозимов, катали-
зирующих полимеризацию РНК на матрице РНК - хотя бы одной молекулы на популяцию
молекул РНК в каком-то небольшом водоеме - мир РНК обретал свою сущность как само-
сохраняющаяся и развивающаяся материя на древней Земле.
КОМПАРТМЕНТАЛ ИЗАЦИЯ КАК УСЛОВИЕ БИОЛОГИЧЕСКОЙ ЭВОЛЮЦИИ
Возникновение и существование мира РНК на Земле естественно требовало жидкой
водной среды с нейтральной pH и растворенными солями одновалентных металлов (в пер-
вую очередь К' и Na") и Mg2'. Скорее всего это были мелкие водоемы и лужи («Дарвиновские
пруды»), где могли концентрироваться абиогенно возникающие органические вещества; оке-
49
.1.С. Спирин. Дренпий мир PI I К
анскис просторы вовсе не годились для этого. (Впрочем, как полагает большинство геологов
и палеонтологов, в то время океаны на Земле, по-видимому, еще и не существовали). При-
сутствие РНК-рспликазной активности в водной среде PH К-содержащей лужи или пруда да-
вало в результате эффект амплификации всех олиго- и полирибонуклеотидов этого водоема,
т.е. рост общей популяции молекул РНК. Однако, на этом этапе еще не могло быть никакого
отбора «лучших» и, стало быть, никакой биологической эволюции.
Дело в том, что в гаком случае эффективный РНК-рсплицирующий рибозим, присут-
ствующий в луже, одинаково хорошо должен был амплифицировать как редкие молекулы
РНК, обладающие какими-либо полезными для популяции свойствами (например, адсорб-
ция из среды или катализ синтеза нужных веществ), так и основную массу неактивных, бал-
ластных молекул РНК. Чтобы естественный отбор начал работать, необходима была какая-то
форма компартментализации, обособления отдельных ансамблей РНК, в которых рибозимы
и их продукты удерживались бы вместе. Только тогда естественный отбор мог отличить те
РНК, чей продукт лучше, и те ансамбли, чьи РНК функционально лучше дополняют друг
друга. Лучшие обособленные ансамбли РНК - первозданные особи - должны расти быстрее
других, перерастать других, тем самым обеспечивая отбор лучших.
По-видимому, впервые идея «обособления от окружающей среды» как предпосыл-
ки для того, чтобы воспроизводящаяся система макромолекул могла эволюционировать,
была выдвинута А.И. Опариным (1924). В качестве наиболее примитивного способа пер-
вичной компартментализации, который мог бы осуществляться в присутствии абиогенно
возникающих полипептидов и белков, он предложил коацервацию - образование капелек
концентрированного золя в разбавленной среде с растворенными макромолекулами. Это
было логично в рамках гипотезы Опарина о начале жизни на основе абиогенного синтеза
белка (хотя в конце концов ученый мир пришел к пониманию того, что на основе белков
без нуклеиновых кислот невозможно создать самовоспроизводящуюся систему). Однако,
согласно гипотезе о древнем мире РНК, этот мир был «однополимерным», и по причине
отсутствия полипептидов, полисахаридов и других полимеров, способных к коацервации,
да еще и в достаточно больших концентрациях, компартментализания ансамблей РНК
в виде коацерватных капель кажется маловероятной. Тем не менее, для того чтобы каждый
ансамбль PH К мог существовать как система, наследовать приобретенные признаки, полез-
ные для всей системы, и эволюционировать, его РНК-реплицирующий рибозим, лиганд-
связывющие РНК, нуклсотид-синтсзирующис рибозимы и продукты синтезов должны
быть, очевидно, как-то связаны, объединены в пространстве. Поэтому в большинстве
теорий происхождения жизни возникновение каких-то примитивных ограничивающих
мембран или хотя бы поверхностей раздела фаз постулируется необходимым условием на-
чала эволюции, в том числе эволюции ансамблей РНК. И в этом состоит одна из основных
трудностей таких теорий.
Возможна, однако, и очевидная альтернатива обособлению за счет мембран или по-
верхностям раздела фаз. Как уже указывалось выше, А.Б. Четвериным с сотрудниками была
экспериментально показана способность молекул РНК формировать молекулярные колонии
на гелях или других влажных твердых средах, если на этих средах им предоставлены усло-
вия для репликации (рис. 6). Смешанные колонии РНК на твердых или полутвердых по-
верхностях, без каких-то ни было мембран, и могли быть первыми эволюционирующими
бесклеточными ансамблями, где одни молекулы выполняли генетические функции (репли-
кацию молекул РНК всего ансамбля), а другие формировали структуры, необходимые для
успешного существования (например, такие, которые адсорбировали нужные вещества из
окружающей среды) или были рибозимами, ответственными за синтез и подготовку субстра-
тов для синтеза РНК (Спирин, 2003). Такая бесклеточная ситуация создавала условия для
очень быстрой эволюции: колонии РНК не были отгорожены от внешней среды и могли легко
50
Проняемы нроисхож.ишпя ж и ши
обмениваться своими молекулами - своим генетическим материалом. Легкое распростране-
ние молекул РНК через среду, в том числе атмосферную, также было продемонстрировано
в прямых экспериментах (Chetverin cl al., 1991).
Эта альтернатива представляется вероятной потому, что образование колоний РНК
легко себе представить как естественное следствие подсыхания лужи, населенной молекула-
ми РНК: на влажной поверхности глины, в тех местах, где оказывался РНК-реплицирующий
рибозим, молекулы PH К, осевшие на поверхность, должны были амплифицироваться и обра-
зовывать колонии при условии, что необходимые органические вещества (предшественни-
ки пуринов, пиримидинов, рибозы и т. д.) и высокоэнергетические фосфаты присутствовали
иатой же поверхности. Таким путем могли образовываться смешанные колонии РНК с раз-
личными функциональными активностями. Такой ансамбль молекул РНК в виде растущей
смешанной колонии мог быть успешным в своем существовании и росте, если он включал
в себя (1) лигапд-связывающис PH К для избирательной адсорбции и аккумуляции необходи-
мых веществ из окружающей среды, (2) набор рибозимов, катализирующих метаболические
реакции для синтеза нуклеотидов и их активированных (фосфорилированных) производных,
и (3) рибозим, катализирующий комплсмсн гарную репликацию всех РНК колонии.
Наиболее серьезным следствием компартментализации РНК в форме смешанных ко-
лоний было появление механизма естественного отбора: колонии, содержащие РНК, более
активные и более подходящие друг другу (функционально дополняющие друг друга) мог-
ли расти быстрее, становиться больше и гем самым «перерастать» другие колонии, вытес-
нять их. Таким образом, образование компартментализованпых ансамблей функционально
дополняющих друг друга РНК в качестве особей, способных расти и конкурировать друг
с другом, представляется вероятным без окружающих их мембран или оболочек другого
типа, и даже без четкой границы раздела.
ЭВОЛЮЦИЯ ДРЕВНЕГО МИРА РНК ПУТЕМ «СИСТЕМАТИЧЕСКОГО
ЭКСПОНЕНЦИАЛ ЬНОГО ОБОГАЩЕНИЯ» (SELEX)
В 1990 г. одновременно тремя группами ученых в США (Tuerk, Gold, 1990; Ellington,
Szostak, 1990; Robertson, Joyce, 1990) была опубликована методология бесклеточной селекции
или бесклеточной эволюции в пробирке молекул некодирующих РНК, позволившая быстро
получать в лаборатории нужные синтетические функционально активные РНК, специфически
взаимодействующие с определенными лигандами (аптамеры) или катализирующие опреде-
ленные химические реакции (искусственные рибозимы), а также совершенствовать функции
таких РНК. Оказалось, что эта методология может указать тот путь, которым, возможно, шла
естественная эволюция древнего мира РНК. Суть методологии «эволюции в пробирке» состо-
ит в многократном чередовании стадий селекции и амплификации РНК («Systematic Evolution
of Ligands by Exponential enrichment», SELEX). После того как в лаборатории А.Б. Четверина
была экспериментально показана возможность существования, роста и амплификации моле-
кул РНК в форме колоний на влажных твердых средах, можно было предположить, что по-
добное чередование процессов селекции и амплификации могло иметь место на первобытной
Земле (Спирин, 2005). Действительно, важным свойством молекулярных колоний РНК явля-
ется то, что они не имеют никакой ограничивающей их оболочки, удерживающей молекулы
внутри компартмента: молекулы РНК удерживаются вместе только благодаря их локализации
на твердой поверхности в отсутствие перемешивания и конвекции, типичных для жидкой
среды. Это - временное состояние, и любое затопление должно приводить к растворению
колоний и перемешиванию всех РНК в общей луже.
Исходя из вышесказанного, можно предложить следующий сценарий «естественного
первобытного SELEX’a». (1) Когда функционально различные молекулы РНК, включая РНК-
реплицирующий рибозим, оказываются в одной луже, вся популяция РНК количественно
51
.4.С. Спирин. Древний мир РНК
АБИОГЕННЫЕ РИБОНУКЛЕОТИДЫ
ОЛИГОРИБОНУКЛЕОТИДЫ, ТРАНСЭСТЕРИФИКАЦИЯ
Ф
ПОЛИРИБОНУКЛЕОТИДЫ, РЕКОМБИНАЦИИ
Ф
УЗНАВАНИЕ ЛИГАНДОВ, КАТАЛИТИЧЕСКИЕ АКТИВНОСТИ
ДРЕВНИЙ
МИР РНК:
Эволюция
в циклах
репликации -
селекции
(древний
природный
SELEX)
КАТАЛИТИЧЕСКАЯ РЕПЛИКАЦИЯ РНК: <
КОММУНЫ МОЛЕКУЛ РНК В “ДАРВИНОВСКИХ” ЛУЖАХ
ПОДСУШИВАНИЕ
ЗАТОПЛЕНИЕ
ОТБОР (СЕЛЕКЦИЯ) ПОДХОДЯЩИХ ДРУГ ДРУГУ РНК:
РОСТ СМЕШАННЫХ КОЛОНИИ РНК НА ВЛАЖНЫХ ТВЕРДЫХ ПОВЕРХНОСТЯХ,
ВОЗНИКНОВЕНИЕ ДОКЛЕТОЧНЫХ АНСАМБЛЕЙ
"УНИВЕРСАЛЬНЫЙ
ПРЕДШЕСТВЕННИК"
ВОЗНИКНОВЕНИЕ
СОВРЕМЕННОЙ
ЖИЗНИ
ПОЯВЛЕНИЕ ПРОНИЦАЕМЫХ МЕМБРАННЫХ ОБОЛОЧЕК,
f РИБОЦИТЫ
I ВОЗНИКНОВЕНИЕ АППАРАТА СИНТЕЗА БЕПКОВ.
у УНИВЕРСАЛЬНЫЙ ГЕНЕТИЧЕСКИЙ КОД.
* ВОЗНИКНОВЕНИЕ ДНК И РНК
ПРОГЕНОТЫ
Г
КЛЕТКИ, ОРГАНИЗМЫ
Рис. 7. Сценарий возникновения и эволюции мира РНК (Спирин, 2005).
увеличивается, а благодаря реакциям спонтанной трансэстерификации (реакция Четвери-
на) и ошибкам репликации увеличивается и разнообразие молекул. (2) Когда лужа подсы-
хает, молекулы РНК оказываются на влажной поверхности глины или другого минераль-
ного субстрата, и при наличии в данном месте молекулы РНК-реплицирующего рибозима
и нескольких других молекул РНК, обеспечивающих связывание нужных веществ и ката-
лиз нужных реакций, образуется и растет смешанная колония РНК; наиболее успешные
колонии (т. е. колонии с наиболее активными и лучше всего дополняющими друг друга
молекулами РНК) растут быстрее других. (3) Последующее затопление подсушенного во-
доема или его части растворяет колонии, и в общем водоеме опять начинается общая ам-
плификация, но уже в популяции, обогащенной «хорошими», т.е. активными и функцио-
нально дополняющими друг друга молекулами. Таким путем чередующиеся затопления
и подсушивания РНК-содержащих водоемов (луж) обеспечивают систематическое обога-
щение популяции РНК функционально лучшими молекулами («систематическая эволюция
путем экспоненциального обогащения»). Именно таким путем и мог развиваться и эволю-
ционировать древний мир РНК. Две ипостаси этого мира РНК сосуществовали на Земле
и переходили друг в друга: коммунальные сообщества размножающихся и разнообразя-
щихся индивидуальных молекул РНК, растворенных в водной среде луж, где реализовал-
ся принцип «от каждого по способностям, каждому по потребностям», и конкурирующих
особей в виде смешанных колоний РНК, временно существующих и растущих на влажных
поверхностях. Общая гипотетическая картина существования и эволюции древнего мира
РНК дана на рис. 7.
Таким образом, циклы амплификации-селекции вышеописанного типа могли быть
основной формой существования эволюционирующего древнего мира РНК на первобытной
Земле. Эволюционный процесс должен был быть исключительно быстрым благодаря трем
52
Проблемы проис'хож.ц-.ния жити
обстоятельствам. Во-первых, непрерывные спонтанные рекомбинации и перестройки моле-
кул РНК, а также низкая точность примитивных механизмов репликации, обеспечивали ши-
рочайшее поле вариантов для отбора. Во-вторых, свободный латеральный перенос и обмен
молекулами РНК между колониями через воду и атмосферу делал любые полезные иннова-
ции достоянием всех и позволял колониям быстро совершенствоваться в течение короткого
времени их существования. В-третьих, экспоненциальное обогащение всей популяции «луч-
шими» молекулами РНК в циклах амплификации-селекции создавало мощный эволюцион-
ный двигатель для всего коммунального мира РНК в целом.
В свое время К. Вуз (Woesc, 1998) выдвинул концепцию «Универсального Предше-
ственника» живых существ на Земле. Он предположил, что предшественники современных
организмов - «прогеноты» - представляли собой примитивные особи, лишенные полно-
ценной внешней мембраны или любого другого барьера, который бы препятствовал сво-
бодному обмену генетического материала между ними. Они характеризовались высоким
уровнем мутаций (ошибок репликации) из-за примитивности и несовершенства механиз-
мов репликации генетического материала и интенсивным латеральным обменом генами
и продуктами, когда любые продукты и инновации одних становились достоянием всех.
Тем самым они существовали как коммуна, и этот коммунальный мир отличался очень
высокой скоростью эволюции. Именно этот коммунальный мир «прогенотов», без строго
очерченных индивидуальностей, Вуз предложил считать универсальным предшественни-
ком всех живых организмов на Земле. В результате эволюционного распада этого единого
коммунального мира обособились три главные линии клеточных живых существ - бакте-
рии (эубактерии), археи (архсбактерии) и эуакарии (эукариоты), - обладающие одинаковым
генетическим кодом - кодом своего универсального предшественника. В настоящее время,
в свете всех данных и представлений о мире РНК, представляется возможным передать
роль Универсального Предшественника коммунальному сообществу колоний-ансамблей
РНК, временно существующих на твердых или гелеобразных поверхностях первобытной
Земли, не ограниченных физически никакими мембранами и фазовыми разделами и пото-
му свободно обменивающихся как генетическим материалом, так и продуктами катализи-
руемых реакций, а затем растворяющихся в общей коммуне и заново собирающихся в но-
вых комбинациях молекул. Как уже указывалось выше, такая коммуна должна была очень
быстро эволюционировать. Во всяком случае, если мир РНК возник и эволюционировал
в клеточные формы жизни на Земле, то весь путь эволюции до индивидуальных организ-
мов с клеточной структурой, ДНК и современным аппаратом белкового синтеза должен
был быть пройден менее чем за полмиллиарда лет, - во временной промежуток между воз-
никновением планеты Земля 4.6 млрд, лет назад и появлением на Земле первых клеточных
организмов современного типа 3.8-3.9 млрд, лет назад.
Конечно, вполне вероятной альтернативой этому сценарию может быть космическое
происхождение жизни на Земле: примитивные клеточные формы жизни - скорее всего бак-
терии и/или археи - были занесены на Землю из ближнего или далекого космоса, возможно,
как раз в период интенсивной метеоритной бомбардировки между 4.6 и 4 млрд, лет назад,
а в дальнейшем на Земле происходила лишь их эволюция в высокоразвитые многоклеточные
организмы. В последнее время именно эта точка зрения приобретает все больше сторонни-
ков, так как само возникновение и существование безбелкового мира РНК на первобытной
Земле сталкивается со многими, нс разрешимыми пока, проблемами (Спирин, 2007). Тем
не менее, это не снимает принципиального вопроса о возникновении клеточной формы жиз-
ни из мира РНК, а лишь переносит его на другие космические объекты.
53
J.С. Cmipini. Древний мир РНК
СПИСОК ЛИТЕРАТУ РЫ
Богданова Е.С.. Гаврилова Л.П., Дворкин ГА., Киселев Н.А., Спирин А.С. 1962. Изучение макро-
молекулярной структуры высокополимерной (рибосомальной) рибонуклеиновой кислоты
из Escherichia coli 7 Биохимия. Т. 27. С. 387-402.
Опарин А.11. 1924. Происхождение жизни. М.: Московский рабочий. 71 с.
Спирин А.С. 2003. Рибонуклеиновые кислоты как центральное звено живой материи // Вести. РАН.
Т. 73. С. 117-127.
Спирин А.С. 2005. Мир РНК и его эволюция // Мол. Биология. Т. 39. С. 550-556.
Спирин А.С. 2007. Где. когда и в каких условиях мог возникнуть и эволюционировать мир РНК?
Палеонтол. журн. № 5. С. 11-19.
Спирин А.С, Гаврилова Л.П., Бреслер С.Е., Мосевицкий М.И. 1959. Изучение макромолекулярной
структуры инфекционной рибонуклеиновой кислоты из вируса табачной мозаики // Биохимия.
Т. 24. С. 938-947.
Baltimore D. 1970. RNA-dependent DNA polymerase in virions of RNA tumour viruses // Nature. V. 226.
P. 1209-1211.
Baltimore D., Girard M., Darnell J.E. 1966. Aspects of the synthesis of poliovirus RNA and the formation
of virus particles // Virology. V. 29. P. 179-189.
Bartel D.P.. Szostak J.W. 1993. Isolation of new ribozymes from a large pool of random sequences // Science.
V. 261. P. 1411-1418.
Belozersky A.N., Spirin A.S. 1958. A correlation between the compositions of deoxyribonucleic and ribonu-
cleic acids // Nature. V. 182. P. 11 l-l 12.
Brenner S., Jacob F. Meselson M. 1961. An unstable intermediate carrying information from genes to ribo-
somes for protein synthesis // Nature. V. 190. P. 576-581.
Cech T.R., Golden B.L. 1999. Building a catalytic active site using only RNA // The RNA World. 2 cd. / Eds.
R.F. Gesteland, T.R. Cech, J.E Atkins. N.Y.: Cold Spring Harbor Lab. Press. P. 321-347.
Chetverin A.B., Chetverina H., Munishkin A.V. 1991. On the nature of spontaneous RNA synthesis by
QP replicase 7 J. Mol. Biol. V. 222. P. 3-9.
Chetverina H.E, Chetverin A.B. 1993. Cloning of RNA molecules in vitro // Nucleic Acids Res. V. 21.
P. 2349-2353.
Chetverina H.V., Demidenko A.A., Ugarov El., Chetverin A.B. 1999. Spontaneous rearrangements in RNA
sequences // FEBS Lett. V. 450. P. 89-94.
Cundlijfe E. 1986. Involvement of specific portions of ribosomal RNA in defined ribosomal functions: a study
utilizing antibiotics // Structure, function, and genetics of ribosomes / Eds. B. Hardesty, G. Kramer.
New York: Springer-Verlag. P. 586-604.
Doudna J.A., Couture S„ Szostak J.W. 1991. A multisubunit ribozyme that is a catalyst of and template
for complementary strand RNA synthesis// Science. V. 251. P. 1605-1608.
Doudna J.A., Szostak J.W. 1989. RNA-catalysed synthesis of complementary-strand RNA //Nature. V. 339. P. 519-522.
Doudna J.A., Usman N., Szostak J.W. 1993. Ribosome-catalyzed primer extension by trinucleotides: a model
for the RNA-catalyzed replication of RNA // Biochemistry. V. 32. P. 111-2115.
EklandE.H., Bartel D.P. 1996. RNA-catalysed RNA polymerization using nucleoside triphosphates//Nature.
V. 382. P. 373-376.
Ellington A., Szostak J. 1990. In vitro selection of RNA molecules that bind specific ligands // Nature. V. 346.
P. 818-822.
Fourmy D„ Recht M.I., Blanchard S.C., Puglisi J.D. 1996. Structure of the A site of E. coli I6S rRNA com-
plexed with an aminoglycoside antibiotic // Science. V. 274. P. 1364-1371.
Gesteland R.F, Atkins J.F. (eds.) 1993. The RNA World. I ed. N.Y.: Cold Spring Harbor Lab. Press. 630 p.
Gesteland R.F., Cech T.R., Atkins J. F. (eds.) 1999. The RNA World. 2 ed. N.Y.: Cold Spring Harbor Lab. Press. 709 p.
Gesteland R.F., Cech T.R., Atkins J.F. (eds.) 2006. The RNA World. 3 ed. N.Y.: Cold Spring Harbor Lab.
Press. 768 p.
54
Прок. //:.\//>/ игоисх'ож.н-.пия жпти
Gilbert Ж 1986. Origin of life: the RNA world Nature. V. 319. P. 618.
Gold L., Poliskv R.. Uhlenheck O.. Yarns M. 1995. Diversity of oligonucleotide functions // Ann. Rev. Bio-
chem. V. 64 .P. 763 797.
Gnerrier-Takada C.. Gardiner K., March T. 1983. The RNA moiety of ribonuclease P is the catalytic subunit
of the enzyme Cell. V. 35. P. 849-857.
Johnston H’A’.. Unran P.J.. Lawrence M.S. et al. 2001. RNA-catalyzcd RNA polymerization: accurate and
general RNA-templated primer extension Science. V. 292. P. 1319 1325.
Kisse/ev N.,4.. Gavrilova L.P.. Spirin A.S. 1961. On configurations of high-polymer ribonucleic acid macro-
molecules as revealed by electron microscopy // J. Mol. Biol. V. 3. P. 778-783.
Kruger K.. Grabowski P.J.. Zang A.J. et al. 1982. Self-splicing RNA: autoexcision and autocyclization of the
ribosomal RNA intervening sequence of Tetrahymena // Cell. V. 31. P. 147-157.
Nissen P„ Hansen J.. Ban N. et al. 2000. The structural basis of ribosome activity in peptide bond synthesis ii
Science. V. 289. P. 920 930.
Pttglisi J.D.. Williamson J. R. 1999. RNA interaction with small ligands and peptides // The RNA World. 2 ed.
Eds. R.F. Gesteland, T.R. Cech, J.F. Atkins. N.Y.: Cold Spring Harbor Lab. Press. P. 403-425.
Robertson D.L., Jovce G.F. 1990. Selection in vitro of an RNA enzyme that specifically cleaves single-stran-
ded DNA ' Nature. V. 344. P. 467-468.
Spiegehnan S. 1961. The relation of informational RNA to DNA // Cold Spring Harbor Symp. Quant. Biol.
V. 6. P. 75-90.
Spiegehnan S.. Haruna 1. 1966. A rational for an analysis of RNA replication // Proc. Natl. Acad. Sci. USA.
V. 55. P. 1539-1554.
Spirin A.S. 1960. On macromolecular structure of native high-polymer ribonucleic acid in solution . J. Mol.
Biol. V. 2. P. 436-446.
Temin H.M.. Mizutani S. 1970. RNA-dcpendent DNA polymerase in virions of Rous sarcoma virus // Nature.
V. 226. P. 1211-1213.
Tuerk C„ Gold L. 1990. Systematic evolution of ligands by exponential enrichment// Science. V. 249. P. 505-510.
Pasiliev V.D.. Selivanova O.M., Koteliansky V.E. 1978. Specific self-packing of the ribosomal I6S RNA //
FEBS Lett. V. 95. P. 273v276.
Vasiliev V.D., Serdyuk LN., Gudkov A.T. Spirin A.S. 1986. Self-organization of ribosomal RNA //Structure,
function, and genetics of ribosomes / Eds. B. Hardesty, G. Kramer. N.Y.: Springer. P. 128-142.
Fasiliev V.D., ZaHte O.M. 1980. Specific compact self-packing of the ribosomal 23S RNA // FEBS Lett. V. 12. P. 101-104.
Watson J. D., Crick F.H.C. 1953. Genetical implications of the structure of deoxyribose nucleic acid // Nature.
V. 171. P. 964-967.
Woese C.R. 1998. The universal ancestor// Proc. Natl. Acad. Sci. USA. V. 95. P. 6854-6859.
Ancient RNA World
A.S. Spirin
The recently discovered capability of RNA to rearrange their sequences in solution by spon-
taneous cis- and trans-transesterification reactions may help to solve the problems of polyribonucle-
otide elongation and variant generation in the ancient, pre-enzyme and pre-ribozyme era. The earlier
demonstrated formation and growth of RNA colonies on gels and other moist solid surfaces shows
the possibility of compartmentalization of molecular ensembles without an enfolding membrane.
This can solve the problems of concentration/dilution of substrates and products in an aqueous me-
dium on the prebiotic Earth, retention of the «gene» and the «trait» together, and thus identification
and selection of the best «genes» (RNAs) for the evolution of RNA ensembles. The application of the
principle of «systematic evolution by exponential enrichment» (SELEX) to the idea of periodically
dissolving and reforming RNA colonies (as alternating amplification and selection stages, respec-
tively) can explain very fast progressive evolution of the ancient RNA world.
55
Дискуссия
по пленарному докладу А.С. Спирина
«ДРЕВНИЙ МИР РНК»
Председательствующий А.Ю. Розанов
АД. Алыпштейн: Как Вы решаете проблему хиральности?
J.C.C.: Не решаю. Обратите внимание, я не рассматриваю вопрос абиогенного синтеза
мономеров, а хиральность - это проблема того времени. Хиральность относится к мономе-
рам, к полимерам она имеет отношение уже с точки зрения закрученности спирали вправо
или влево.
Н.Л. Добрецов: Гипотеза «пересыхающих луж» - эти условия могли перемежаться
с другими, где все бурлило и кипело.
А.С.С.: Важно, чтобы в каждой «луже» происходили циклы.
Н.Л. Добрецов: Может рассматриваться подземное происхождение жизни или на дне
океана (черные курильщики).
А.С.С.: Мне больше всего нравятся промежуточные варианты. В любом случае, по-
верхность имеет одно колоссальное преимущество - разнос материала. Инновации в одной
«луже» быстро становятся достоянием другой «лужи». Молекулы при высыхании еще
и в атмосфере переносятся. Но в принципе, возможны некие подземные инкубаторы, с более
стабильной температурой, отсутствием смены дня и ночи.
Г.А. Заварзин: У меня экологические замечания. Я боюсь, что Александр Сергеевич
не делает различий между лужей и прудом. Пруд образуется за счет латерального переноса
влаги, а лужа имеет дождевое происхождение, в результате в луже всегда создаются ультра-
пресные омбротрофные условия, где соответствующих ионных условий быть не может. Аль-
тернативой для Вас является почвенный раствор, который как раз составляет переменную
влажность и высыхание. Слово «почвы» я использую в смысле «soil».
А.С.С.: Дело в том, что для того, чтобы происходил SELEX, ионные условия не столь
уж критичны. По отношению к ионным условиям РНК довольно нетребовательна, была бы
матрица, это ведь не микроорганизмы. Да, pH должен быть 7, плюс-минус 0.2-0.3. Ионы же.
особенно анионы, не так важны, а общая ионная сила может быть от 0.01 до 0.5.
Г.А. Заварзин: Когда Вы говорите, что нужна высокая скорость размножения рибозимов,
я в этом не уверен и думаю, что, может быть, выживание важнее скорости размножения.
А.С.С.: Я сказал не о скорости размножения, я сказал о скорости эволюции. А скорость
эволюции зависит от количества циклов.
М.Я. Маров: Мое замечание касается известного геохимического и связанного с ним во-
дного парадокса. Расцвет мира РНК происходит сразу после завершения максимальной фазы
бомбардировки Земли и других планет земной группы. Уже в протопланетном диске, в горя-
чей зоне формирования этих планет при температуре 800-1000 К, с неизбежностью возника-
ет сильное фракционирование летучих и обеднение летучими, включая вероятный дефицит
воды. Соответственно, возникает парадокс происхождения земной атмосферы и гидросферы.
В Институте Келдыша нами была развита концепция возможного притока значительного
объема летучих за счет миграции ледяных тел типа комет из внешних областей Солнеч-
ной системы, прежде всего, из пояса Койпера. Компьютерное моделирование показало, что
количество воды, которое может быть приобретено за счет этого механизма, соизмеримо
с объемом земной гидросферы, или даже немного превышает его. Практически те же оцен-
ки получаются для Венеры и Марса. Поэтому мне очень симпатично Ваше предположение
о том, что должен быть какой-то внешний источник, который мог компенсировать дефицит
56
Проблемы происхождения жизни
летучих. Получается вполне естественное объяснение - существовал некий привнесенный
извне водный резервуар, который был составляющим элементом гетерогенной аккреции
Земли на завершающей стадии аккумуляции и, возможно, ее предбиологической эволюции.
С.В. Рожнов: Все-гаки, мир РНК был миром живых существ или миром, обогащенным
особым типом химических соединений?
А.С.С.: А Вы можете дать мне определение жизни?
А.Ю. Розанов (председательствующий): Нет, этим сейчас не нужно заниматься...
А.С.С.: Но задан вопрос, ответ на который зависит от этого определения.
С.В. Рожнов: Можно подойти с другой точки зрения. Когда появилась эукариотная
биосфера, она нс заместила собой прокариотную, а наложилась на нее. А прокариотная био-
сфера еще раньше наложилась на мир РНК?
А.С.С.: Наложилась полностью. Мир РНК живет в нас, он стал внутриклеточным. По-
давляющая часть разновидностей РНК - некодирующие, с неизвестными нам функциями.
Теперь мир РНК действительно в тепличной обстановке существует в каждой нашей клетке.
В.Н. Снытников: Вы сказали, что для того, чтобы провести синтезы, не нужно много
времени, все это проходит в течение суток. И все-таки Вы затребовали 100 миллионов лет
на эти синтезы. Как соотносятся скорость этих реакций и эти 100 миллионов лет?
А.С.С.: Для того, чтобы перейти от колонии РНК к клетке, нужно очень много чего еще
сделать, проэволюционировать. РНК копируется быстро - из одной молекулы за один час
создается 1012 копий. Но нужно сформировать структуры, мембрану - нужна клетка. Четыре
миллиарда лет назад мы встречаем уже бактериальные клетки.
А.Ю. Розанов: А какая разница - 100 лет или 100 миллионов лет?
А.С.С.: 100 лет недостаточно.
И.Л. Добрецов: Чтобы создать мембрану, нужны миллионы лет.
А.Ю. Розанов: Но какой-нибудь критерий измерения времени можно предложить?
Н.Л. Добрецов: Можно только сказать, что многократно больше, чем первичный синтез
и отбор.
А.Б. Макалкин: Увеличение амплитуды колебаний температуры замедлит процесс син-
теза? Почему интервал от 20 до 40 °C, а не от 0 до 50?
А.С.С.: При 50 °C сильно (на порядок) увеличивается скорость деградации полирибону-
клеотидов. При низких температурах реакции сильно замедлятся. При 20 °C замораживается
плавление спиралей РНК.
Л.В. Ксанфомалити: Следует ли думать что-нибудь о другой, неуглеродной форме
жизни?
А.С.С.: Раньше этот вопрос много обсуждался, в основном по поводу кремния. Но пока
мы ничего об этом не знаем, как можно это обсуждать. Есть факт - углеродная жизнь. Если
найдется другая форма, будем спекулировать.
А.Д. Альтштейн: С моей точки зрения, мир РНК никогда не существовал, это идея,
ведущая в тупик. Самый главный момент в происхождении жизни - это не образование
различных олигонуклеотидов и полинуклеотидов, самое главное, чтобы образовался ген -
кодирующий, самовоспроизводящийся и способный эволюционировать. Такой ген — это
один полинуклеотид с одной полимеразой, которая должна быть процессивной. Сейчас
показано экспериментами по РНКзимам, что действительно возникают полимеразы, но все
они дистрибутивные, это значит, что для того, чтобы они осуществили синтезы, их должно
быть сразу много, а это утопия. Одна молекула процессивной полимеразы, если она возник-
57
ла с кодирующей сс матрицей, будет эту матрицу размножать, и такая система - один ген,
одна полимераза - это и будет первое живое существо, способное к самовоспроизведению
и эволюции. Так вот, по тем моделям, которые предлагает гипотеза мира РНК, возникно-
вение такой системы невозможно. Короткие (до ста нуклеотидов) молекулы РНК не могут
быть процессивными полимеразами. Нет ни одного примера процессивной полимеразы на
основе чистой РНК. Рибосома, которая является фактически процессивной полимеразой
(она полимеризует аминокислоты), состоит из РНК и белков. Не исключено, что большие
молекулы РНК (сотни нуклеотидов) могут свернуться так, что будут двигаться по матрице,
синтезируя комплементарную нить. Но это значит, что сразу должен образоваться длин-
ный ген и длинная полимераза. А здесь вступает в действие другой запрет - хиральность.
До ста нуклеотидов еще можно как-то проскочить, какие-то редкие молекулы могут об-
разоваться, можно найти какой-нибудь механизм. Но при образовании молекул из сотен
или тысяч нуклеотидов невозможно преодолеть правило хиральности (стереохимическую
ингибицию синтеза полинуклеотида). Таким образом, этот первый организм, состоящий из
гена и полимеразы, не мог возникнуть на основе модели мира РНК. Мы имеем здесь ситуа-
цию, похожую на ту, которая была с теорией Опарина. Она была очень важной, на ее основе
возник очень большой экспериментальный материал, она принесла много знаний, но она не
могла в принципе объяснить происхождение жизни, так как не учитывала гены. Гипотеза
мира РНК также дала очень много, особенно в изучении рибозимов, она учитывает необхо-
димость объяснить возникновение генов, но нс может объяснить происхождение первого
гена, а значит - первого живого организма. Сообщество полинуклеотидов, необходимое
для возникновения первого живого организма по гипотезе мира РНК - это совершенно
неправдоподобная идея. Без процессивных полимераз оно возникнуть не могло. Следует
также отметить, что гипотеза мира РНК не дает правдоподобного объяснения переходу от
РНКовых организмов к ДНК-РНК-белковым клеткам. Не объясняется возникновение ге-
нетического кода и трансляции даже на уровне правдоподобной гипотезы. Поэтому гипо-
теза мира РНК не может приблизить нас к пониманию механизма происхождения жизни.
Я думаю, что рибозимы возникли на ранних этапах эволюции, но уже после возникновения
протоклеток. Первичными же ферментами были белки, а первые организмы имели нуклео-
протеидную природу.
А.С. Спирин: Думаю, что Вы не совсем правы, так как до сих пор все, кто работает
с рибозимами, пытались получить лишь каталитическую активность. Однако, когда мы име-
ем дело с рибосомой или с полимеразой, надо по-другому ставить эксперименты. Надо, что-
бы полимераза была полимеразой, то есть обладала не только каталитической активностью,
но и активностью конвейерной молекулярной машины, как, впрочем, и в случае с рибосомой.
Действительно ли нужны длинные молекулы - не знаю, никто этого не проверял. Для рибо-
зимов минимальные размеры находятся в пределах ста нуклеотидов. Сейчас исследователи
приходят к тому, что рибосома - это машина конвейерного типа, и именно поэтому она долж-
на быть по крайней мере двух- трехкомпонентной. При этом вся структура и все функции ри-
босомы основываются на ее молекулах высокополимерной РНК, а рибосомные белки играют
лишь сугубо подсобную роль. Думаю, что если начать ставить направленные эксперименты
по созданию или поиску молекулярных машин на основе РНК, то надо делать двух- или
трехкомпонентные рибозимы, а какой там минимальный размер, я не знаю, но не думаю, что
РНК-машина должна обязательно состоять из длинных полимеров. Стонуклеотидный поли-
нуклеотид уже способен складываться в третичную структуру, образовывать аптамеры, про-
являть каталитическую активность. Почему же машинной функции у него не может быть?
Молекулярная машина - это довольно простая вещь, главное, чтобы она могла существовать
в двух альтернативных конформационных состояниях. Одно конформационное состояние
требует лиганда, скажем, это закрытое, сомкнутое состояние; когда лиганд разрушается или
58
ПРОБЛЕМЫ ПГ-()И( ХОЖДЕНИЯ ЖИ <1Ш
изменяется химически, возникает второе - открытое, разомкнутое состояние. Что такое по-
требление ГТФ или АТФ в молекулярных машинах? Это связывание лиганда, но так как он
ГТФ или АТФ, он легко распадается с выделением большого количества свободной энергии,
возникает новое состояние. И вот идет такая конформационная пульсация, обеспечивающая
конвейерную работу макромолекулы. Кстати, существуют такие минимальные машины
в виде трехдоменпых молекул белка (например, фактор элонгации Ти). То же самое может
быть достигнуто в случае РНК. Я не думаю, что это должна быть длинная молекула. Надо
попробовать сделать машину, а объединение молекулярной машины с каталитической актив-
ностью - это и сеть уже подобие процессивной полимеразы или рибосомы. Давайте делать.
Пока не было попыток сделать - это не значит, что сделать невозможно. Можно нарисовать
гипотетическую схему, как перейти от мира РНК к синтезу белков. Я делал доклад в Москов-
ском университете - «Происхождение рибосом»... Там я основывался на идее о самосборке
относительно коротких РНК, с тем чтобы создать из молекул конвейерную (процессивную)
молекулярную машину, одновременно обладающую и каталитической активностью. Разные
блоки как субъединицы - тоже вероятный путь усложнения структуры на пути возникно-
вения жизни, это еще один способ компартментализации и агрегации, которая дает новое,
эволюционно выигрышное качество.
А.Д. Алътштейи: Но как быть с хиральностью?
А.С. Спирин: Хиральность создается мономерами. Их синтез происходит на более ран-
ней стадии.
А.Д. Альтштейн: Но если у вас несколько сотен нуклеотидов, как они получились
одной хиральности из рацемической смеси, это невозможно!
А.С. Спирин: Они уже в растворе были одной хиральности. Абиогенный синтез уже
произошел. Есть данные, что на некоторых глинах 80 процентов нуклеотидов образуются
одной хиральности, то ли левых, то ли правых. Это меня устраивает. Проблема хиральности
решается до появления мира РНК. Я очень благодарен Альтштейну, потому что он обострил
идею молекулярной машины, до сих пор все говорили только о каталитических активностях.
Да, для того чтобы перейти в настоящее живое существо, в мир РНК, необходимо еще соз-
дать молекулярную машину. А о них почти не говорили.
59
Пгон. чкмы нрошлож.н-нпя жпяш. M.: НИН РАН. 2009. С. 60-76
ОТ МАРСА К ЕВРОПЕ: ПОИСК БИОСФЕРЫ НА СПУТНИКАХ
ПЛАНЕТ-ГИГАНТОВ
© 2009 г. Л.В. Ксанфомалити
Институт космических исследований РАН
ksanf@iki.rssi.ru
Почти 50 лет длятся исследования планеты Марс с помощью космических
аппаратов. К главным задачам исследований относится поиск признаков
возможного существования на планете жизни. В предлагаемой статье рас-
сматриваются некоторые результаты экспедиций к Марсу и научные задачи
будущих исследований Европы, одного из четырех галилеевых спутников
Юпитера, возможная обитаемость которой также обсуждается в новых
проектах. Физические условия на Марсе и Европе сравниваются с условия-
ми на Земле.
1. МАРС
Среди кандидатов на обитаемость планет земной группы на первом месте всегда оста-
вался Марс, благодаря некоторому сходству физических условий и относительной близости
планеты к Земле. Марс - удобный объект для астрономических наблюдений, при которых
легко различаются крупные детали его поверхности. Так, существование полярных шапок
Марса отмечал еще В. Гершель в XVI11 в. В конце XIX в. П. Ловелл сообщил о своем от-
крытии «каналов» на Марсе и вероятном существовании обитателей планеты, что вызвало
небывалый интерес общественности. В 1897 г. в русском переводе вышла книга французско-
го популяризатора науки К. Фламмариона «Живописная астрономия». В главе, посвящен-
ной планете Марс, говорилось: «Человеческий мир Марса, вероятно, значительно опередил
нас во всем и достиг большого совершенства... они построили города и научились всяким
искусствам». Работы Ловелла немало способствовали огромному интересу публики к Марсу.
Вскоре было показано, что результаты Ловелла связаны с оптической иллюзией. Но, несмо-
тря на это, гипотеза обитаемости Марса сохранилась надолго.
ЕЕ Результаты экспериментов на Марсе
В последней трети XX в. и в первом десятилетии XXI в. к Марсу были отправлены
многочисленные экспедиции автоматов, главной задачей которых был поиск признаков жиз-
ни. Сегодня, спустя 44 года после первых снимков Марса, сделанных с космического аппа-
рата, единственным полученным ответом остается «признаков жизни не обнаружено», хотя
оптимисты продолжают предлагать все новые места и новые методы поисков. Сами методы
60
ПрОЪ.И \/А/ 11Р()1К ХЧ)'Ж.1ЫШЯ жииш
становятся более изощренными и вместо поиска жизни обращаются к поиску следов жизни,
возможно, давно исчезнувшей. Отметим наиболее существенные результаты проведенных
экспериментов.
Два аппарата «Викинг» («Viking», США), опустившиеся на поверхность Марса
в 1976 г., доставили туда великолепные биохимические лаборатории, ориентированные
на поиск признаков метаболизма микроорганизмов, подобных земным. Несмотря на то, что
жизнь амино-нуклеино-кислотного (земного) типа в экспериментах на «Викингах» на Марсе
найти не удалось (Klein et al., 1992), полученные результаты стали основой для последую-
щих миссий. Эксперименты в биолабораториях аппаратов «Викинг», направленные на поиск
микроорганизмов на Марсе, должны были ответить на три вопроса:
1. Есть ли процессы усвоения углекислого газа на свету (фотосинтез);
2. Обнаруживается ли метаболизм микроорганизмов (обмен веществ);
3. Происходит ли газообмен с атмосферой.
Вероятность обнаружения микроорганизмов, аналогичных земным, оценивалась
в 40 %. Результаты были неоднозначными и, скорее всего, отражали сложный химизм грунта
Марса, активируемого солнечной ультрафиолетовой радиацией. Только эксперимент с мече-
ным газообменом дал результаты, которые можно было бы считать частично положительны-
ми. При первом обогащении грунта водой и питательным веществами с меченым углеродом
ЦС было зарегистрировано выделение меченого углекислого газа |4СО.,. Но при новом экс-
перименте эффект не повторился. Более однозначными и решающими оказались результаты
пиролитического эксперимента, где проба грунта постепенно разогревалась до высокой тем-
пературы, а отходившие газовые продукты анализировались масс-спектрометром и газовым
хроматографом. Любая известная форма жизни (или ее следы) при пиролизе выделяет орга-
нические летучие вещества. Исследовались образцы, взятые с глубины грунта от 4 до 6 см.
Чувствительность приборов к органическим составляющим достигала 10 '°. Для сравнения
с экспериментом, авторы указывали, что при анализе 0.1 г антарктического (земного) грунта
обнаруживалось более 20 органических соединений.
Рис. I. Поверхность Марса в районе долины Арес.
6)
Л.В. l\cLin<l>().\tLi.'iii»ni. От Марса к Европе: поиск биосферы на спутниках планет-гигантов
Рис. 2. В миссии «Mars Exploration Rover» самоходные аппараты искали следы древних водоемов
на поверхности Марса. Кратер Бигль на пути аппарата «Opportunity».
Однако никаких органических соединений в эксперименте обнаружено не было. В це-
лом, по итогам трех экспериментов по метаболизму и эксперимента по пиролизу результаты
поиска микроорганизмов были признаны отрицательными.
Позже в литературе высказывалась мысль, что этот отрицательный результат нельзя
относить ко всей планете, что он может быть локальным. Г. Левин, один из авторов экспери-
ментов на «Викингах», полагал, что его результаты все же говорят в пользу жизни на Мар-
се (Kein et al., 1976). Однако эти результаты широко обсуждались специалистами, которые
считают, что суперокислители, содержащиеся в грунте Марса, могли дать такой эффект без
метаболизма каких-либо микроорганизмов.
Давление углекислотной атмосферы Марса у поверхности примерно в 150 раз меньше
земного. Такая атмосфера и отсутствие магнитного поля не в состоянии защитить поверх-
ность планеты от радиации. Несильное магнитное поле Марса исчезло около 4 млрд, лет
назад. Расчеты показывают, что радиация проникает в грунт и стерилизует его на большую
глубину, вплоть до нескольких метров.
С другой стороны, если бы на Марсе жизнь однажды все же возникла, уничтожить ее
было бы очень непросто. Жизнь не только приспосабливается к окружающей среде, но и при-
спосабливает ее к себе. Поэтому многие высказывают мнение, что возникшая когда-то жизнь
на Марсе могла бы исчезнуть лишь под действием каких-то совершенно катастрофических об-
стоятельств. Поэтому, если бы она сейчас существовала, ее было бы трудно не обнаружить.
Дальнейшие эксперименты, проведенные в последние десятилетия, были не столь пря-
молинейными и касались, главным образом, физико-химических свойств грунта. В 1997 г.
в районе долины Арес совершил посадку аппарат «Pathfinder», включавший подвижный мо-
62
Проблемы нропсхожлепия ж и inn
Рис. 3. Мелкие глобулы (конкреции) образовались в соленой воде древнего водоема
на Марсе. Район посадки аппарата «Opportunity».
дуль, который мог передвигаться по поверхности, удаляясь примерно на 10 м от посадочного
аппарата. Выбор места посадки был связан с тем, что поиск следов жизни целесообразно ве-
сти в районах, где присутствуют следы потоков воды. Долина Арес была широкой рекой или
водоемом около 2 млрд, лет назад. Современный вид поверхности в месте посадки показан
на рис. 1. Песок, пыль и камни разных размеров повторяют вид поверхности в районах посад-
ки «Викингов», но крупномасштабные детали указывают на следы мощных потоков и даже
острова, существовавшие в ту эпоху. Аппарат исследовал химический состав камней. Ника-
ких выводов о существовании древних форм жизни сделано не было (Lakdawalla, 2008).
После аппаратов «Викинг» прямых поисков жизни больше не проводилось. Миссия
НАСА в 2004 г. «Mars Exploration Rovers» была посвящена другой проблеме: существовали
ли обширные водоемы или оксаны на поверхности Марс в далеком прошлом. Многие спе-
циалисты считали, что сами очертания рельефа ясно указывают на древние берега таких
океанов, но другие предлагали иные объяснения, как ветровую эрозию или что-либо другое.
В ходе миссии были обнаружены овраги со следами недавних потоков (Malin, Edgett, 2000).
Два самоходных аппарата (ровера), названные «Opportunity» и «Spirit», опустились на по-
верхность Марса и начали длительную работу в 2004 г., путешествуя на значительные рас-
стояния и передавая изображения поверхности (рис. 2) и сведения о проведенных исследова-
ниях (Kerr, 2006). Роверы проработали уже 5 лет; они представили, в частности, достаточно
убедительные доказательства того, что в ранние эпохи своей истории на Марсе действитель-
но существовали обширные водоемы. Аппараты нашли минерал гематит, который формиру-
ется в присутствии воды. Поверхность в районе работы «Opportunity» включает множество
мелких круглых зерен (глобул) размерами менее сантиметра (рис. 3), которые напоминают
конкреции на дне земных океанов и однозначно указывают на некогда существовавший здесь
большой водоем или даже океан. При необходимости аппараты могли специальным инстру-
ментом срезать участок камня (круг на рис. 3) и провести его химический анализ.
63
Л.В. Ксиифами.тпт. От Марса к Европе: поиск биосферы на спутниках плане!-гигантов
Подробное исследование свойств грунта показало, что вода в древних водоемах содер-
жала много растворенных солей. Было высказано предположение, что, как было установлено
ровером «Opportunity», высокие концентрации растворенных веществ во влажной среде ран-
него Марса, вероятно, исключали возникновение и эволюцию любых микроорганизмов. Как
известно, высокая концентрация растворенных солей может использоваться как консервант.
Аппарат «Phoenix» (США, 2008 г.) проводил исследования в полярном районе Марса.
Исследовался реголит на небольшой глубине, в частности, исследовались типы и концентра-
ция антиоксидантов. Километровые слои льда воды и пыли образуют северную полярную
шапку Марса (рис. 4), на границе которой опустился аппарат. В отличие от южной полярной
шапки, примесей льда СО, здесь практически нет. При посадке аппарата струя газа из тор-
мозного двигателя сдула тонкий слой пыли. Под ним находилась значительная масса льда.
В числе экспериментов было микроскопирование, но подробности исследований не сообща-
лись. Было установлено содержание в грунте перхлоратов, которые, наряду с высокой соле-
ностью влаги в грунте, также могут быть серьезной помехой для возникновения и развития
жизни (Johnson, 2008).
Считается, что возникновение земной жизни связано с обширными водоемами. Ор-
битальный аппарат «Mars Express» Европейского космического агентства вышел на орбиту
спутника Марса в 2003 г. Спутник оснащен камерами высокого разрешения, что позволяет
видеть на поверхности образования 10-метровых размеров. В частности, камеры даже заре-
гистрировали положение на Марсе аппаратов, доставленных туда в прошлые годы.
Аппаратом «Mars Express» проводилось минералогическое картирование значи-
тельной части планеты (Fletcher, 2008), и, при существенном разнообразии минерально-
го состава, карбонаты (соли угольной кислоты), широко распространенные на Земле, все
же найдены не были. Это важный результат, поскольку на нашей планете именно в их за-
Рис. 4. Северная полярная шапка Марса образована толстыми слоями льда воды и пыли. Аппа-
рат «Phoenix» исследовал физико-химические свойства грунта в полярном районе.
64
Проблемы пропс хож jehidi жи шн
Рис. 5. Аппарат «Mat s Express» передал изображение обширной равнины, которая могла быть ледяной
поверхностью древнего океана. Пыль, покрывающая всю поверхность планеты, придает ей типич-
ный красноватый цвет.
лежах сосредоточено основное количество углерода. Более того, аппарат «Mars Express»
не подтверждает больших запасов углекислого газа (например, в конденсированном виде),
достаточных для существенных изменений массы атмосферы планеты, и соответственно,
преобразования климата планеты благодаря парниковому эффекту. Этот результат остается
в противоречии с постоянно упоминаемой в литературе гипотезе о теплой эпохе раннего
Марса, когда возникновение жизни, как предполагается, было возможно.
Астрогеология подразделяет историю Марса на три эпохи: нойскую, гесперийскую
и амазонскую, продолжающуюся в наши дни. Нойская соответствует насыщенному рельефу
(пик образования 3.8-3.9 млрд, лет назад, тяжелая метеоритная бомбардировка, 700 млн. лет
после образования Марса и вообще всей Солнечной системы). Гесперийская эпоха харак-
теризуется невысокой плотностью метеоритной бомбардировки. Ее завершение очень осто-
рожно можно датировать 2-3 млрд, лет назад. Амазонская эпоха - это низкая плотность ме-
теоритных кратеров, разительные отличия южного и северного полушарий и формирование
современного климата Марса (Head et al., 1999).
На одном из снимков аппарата «Mars Express» видна обширная гладкая равнина с не-
многочисленными метеоритными кратерами (рис. 5), что указывает на сравнительно позднее
ее появление. Верхний окрашенный слой - это, конечно, пыль. Но под ней просматриваются
плоские блоки протяженностью в десятки и даже в сотню километров. Из сравнения с ви-
дом ледовых полей Арктики авторы сделали вывод, что это - поверхность замерзшего моря
или небольшого океана, возникших, все же, в эпоху более мягкого климата Марса, вероятно,
65
J/.В. Кссшфомилити. Oi Марса к Епропс: поиск биосферы на спутниках планет-гигантов
Рис. 6. Странные отверстия площадью в тысячи
квадратны,х метров обнаружены на древни,х вул-
канических склонах Марса в районе Фарсида.
в начале амазонской эпохи. Если эти результа-
ты подтвердятся, а условия для возникновения
жизни на Марсе действительно существовали,
то именно такие водоемы могли быть обитае-
мыми, хотя бы на уровне микроорганизмов.
Обнаруженное аппаратом «Mars
Express» ничтожное количество метана
в атмосфере Марса (около 10 ppb) могло бы
свидетельствовать о наличии жизни на пла-
нете. Дело в том, что метан в атмосфере не-
прерывно разрушается за счет фотодиссо-
циации. Поэтому его запасы в марсианской
атмосфере должны непрерывно пополняться,
либо в результате жизнедеятельности микро-
организмов, либо в процессах геологической
активности, например, в реакциях серпен-
тинизации. Для поддержания концентрации
10 ppb в атмосфере на Марсе должен присут-
ствовать источник мощностью около Зх|0к г
метана в год. Таким источником могла бы быть тектоническая деятельность, остаточный
вулканизм,геотермальная активность.
Как известно, большая масса земных микроорганизмов обитает в глубоких слоях грун-
та. Высказывалось предположение, что такой же может быть среда обитания жизни на Мар-
се. В этой связи большой интерес вызвало открытие в 2006-2007 гг. странных образований
на склонах древних вулканических конусов в районе Фарсида. Район Фарсида, характеризу-
ющийся огромной массой вулканических выбросов, возник, по-видимому, в амазонскую эпо-
ху. Высота вулканических конусов превышает 20 км. На их склонах обнаружены загадочные
отверстия (рис. 6) диаметром несколько сотен метров, неустановленной глубины. Тепловое
излучение из отверстий (исходящее, по-видимому, от их дна) соответствует усредненному
суточному излучению окружающей поверхности. Появившиеся в околонаучной литературе
сообщения о «теплой среде» на дне полостей и их возможной связи с обитаемостью Марса
были вскоре опровергнуты. Происхождение полостей остается неизвестным. Не исключено,
что это результат вытаивания или испарения грунтовых льдов.
1. 2. Вода в жидкой фазе на поверхности Марса
После первых измерений с аппаратов «Viking» возникла и стала быстро развивать-
ся идея о том, что водные запасы Марса сконцентрированы в подпочвенной мерзлоте
(и полярных шапках) планеты (Soderblom, Wenner, 1978; Carr, 1979, 1981, 1996; Squires
et al., 1992; Clifford, 1993). Часто утверждается, что жидкая вода, необходимая для жизни
и метаболизма, не может существовать на поверхности Марса и никогда там не появля-
ется (Hecht, Vasavada, 2006). Присутствие жидкой воды на поверхности многие авторы
считают исключенным не только из-за низких средних температур на Марсе (Т —220 К),
но и по причинам, определяемым термодинамическими свойствами системы [лед-вода-
водяной пар]: при давлении 6.1 мб и ниже, вода кипит при любой температуре, допускаю-
щей ее жидкое состояние. Разумеется, парциальное давление водяного пара (р =n кТ),
из-за малого отношения смеси, составляет ничтожную долю давления атмосферы Мар-
са, состоящей почти полностью из углекислого газа. Сама величина 6.1 мб, принятая
для «средней» поверхности планеты, соответствует тройной точке диаграммы состояния
воды [лед-вода-водяной пар] при 0.01 °C, и была выбрана как удобная абстракция для сред-
него давления у референтной эквипотенциальной поверхности.
66
Проблемы происхождения жизни
Данная тема активно обсуждалась в печати. Мной указывалось (Ксанфомалити, 2003):
реальные значения давления атмосферы у поверхности Марса, с его большими перепадами
высот z, лежат в широких пределах, как это следует из барометрической формулы:
z
р = роехр [ -Jdz/H(z)],
0
где H(z) - шкала высот атмосферы, H(z) = kT/mg (здесь к - постоянная Больцмана, m - сред-
няя молекулярная масса и g - ускорение свободного падения на уровне 6.1 мб). Из фор-
мулы следует, что давление составляет 0.6 мб на вершинах неактивных вулканов области
Фарсида (высота до z=24 км относительно уровня 6.1 мб), 9 мб в глубоких частях каньона
Кондор (z=- 4 км) и 10 мб на дне впадины Эллада. Таким образом, своеобразный «запрет
по давлению», т. е. широко распространенное мнение о том, что вода вообще не может су-
ществовать в жидком виде на поверхности Марса, неверен. Запрет не носит абсолютного
характера, поэтому определенные образования на поверхности планеты могут иметь при-
роду, связанную с водой. (Другое дело, что бюджет жидкой воды на Марсе весьма ограни-
чен). На рис. 7 показаны области существования льда, пара и воды на Марсе. Небольшая
центральная часть диаграммы с горизонтальной штриховкой указывает на зону возможно-
го существования воды в жидком виде на поверхности Марса.
1. 3. Узкие овраги на склонах
Перемещение больших масс грунта - по-видимому, тонких фракций песка по скло-
ну, происходящее в современную эпоху - доказывает и прекрасно иллюстрирует рис. 8, где
в нижней части снимка видны размытые валы осыпавшегося материала. Ширина участка
на снимке около 3 км. Валы сыпучего материала огибают остатки прежнего рельефа, остав-
Температура, К
Рис. 7. Термодинамические условия существования льда, пара и воды
на Марсе. Кружок в центре соответствует давлению 6.1 мб. Условия су-
ществования воды в жидком виде на поверхности Марса отражает не-
большая центральная часть диаграммы с горизонтальной штриховкой.
Структура диаграммы, соотношение шкал давления/глубины и средне-
годовые температуры для широт 30° и 70° по: (Stewart, Nimmo, 2002).
67
Л.В. Ксанфомегштн. От Марса к Европе: иоиск биосферы на спут никах планет-гигантов
Рис. 8. Осыпи грунта и нитевидные овраги (показаны стрел-
ками) на склоне кратера на Марсе (424°S, 158.2°W). Овраги
похожи на следы земных горных рек, но в отличие от земных
оврагов, они не расширяются, а сужаются вниз по склону
(MGS МОС Release No. МОС2-320, NASA/JPL/MSSS).
ляя обнаженный склон. Такие же осыпи можно видеть и в других районах Марса; они извест-
ны со времен миссии «Viking» (1976 г.).
Но наряду с осыпями сыпучего материала, на рис. 8 можно видеть не отмечавшиеся
прежде (из-за недостаточного разрешения) образования. Это тонкие нитевидные километро-
вые овраги или борозды, спускающиеся по склону (показаны стрелками на рис. 8). Их шири-
на в узкой части составляет всего десятки и единицы метров. Овраги очень похожи на про-
моины земных горных рек или ручьев, но в отличие от земных оврагов, они не расширяются,
а сужаются вниз по склону. Поэтому они не могли возникнуть под действием камнепада
или крупномасштабного селя. Тем более они не могли образоваться под действием пылевых
оползней, которые засыпают все овраги. Зато именно жидкие потоки (воды или какой-то
другой жидкости) легко могли бы образовать такие промоины. В работах (Malin, Carr, 1999;
Malin, Edgett, 2000) указывается именно на источники грунтовой воды как среды, форми-
рующей овраги, промоины и другие образования такого рода на Марсе. По их данным, узкие
овраги или протоки достаточно часто встречаются в полосе широт 30°N-70°S. Они мор-
фологически подобны склоновым руслам земных рек и не перекрываются более поздними
образованиями (такими как песчаные дюны). Ширина (и, вероятно, глубина) оврагов близка
к 10-20 м, а протяженность составляет от нескольких сотен метров до километров (Malin,
Edgett, 2000). Авторы еще одной работы на ту же тему (Palermo et al., 2001) обработали сотни
снимков камеры МОС и также получили доказательства современных следов грунтовых вод.
Их источники, согласно обеим работам (Malin, Edgett, 2000; Palermo et al., 2001), находятся на
крутых склонах долин и кратеров, на глубине 150-500 м под уровнем окружающей поверх-
ности. Но по данным второй работы (Palermo et al., 2001) широта районов, где в основном
сосредоточены источники, лежит в пределах от 30°S до 30°N.
68
/1роб:н:мы происходи пня aihiiii
Рис. 9. Склон кратера с протоками на Марсе (39°S, 166°W). В правой нижней
части снимка бассейн шириной около 600 м (MGS МОС Release No. МОС2-
320, NASA/JPL/MSSS).
На рис. 9 представлен другой район с подобными оврагами. Их ширина от единиц
метров до 10-20 м. Они также не расширяются, а сужаются вниз по склону. На первый
взгляд, это кажется парадоксальным, если овраг образован потоком. Но можно предложить
простое объяснение: если грунтовая вода действительно образовала ключ и поток вышел
на склон и устремился вниз, то в условиях Марса масштаб развивающейся промоины будет
зависеть, прежде всего, от температуры поверхности и температуры потока.
Описываемые Э. Палермо и др. (Palermo et al., 2001) объекты обнаружены, в основ-
ном, в полосе широт ± 20-30°. Если температура поверхностного слоя составляет типичные
для экваториальной зоны Марса 240-260 К или ниже, поток, спускаясь по склону, должен
постепенно впитываться в сухой морозный грунт и замерзать. Образуется ложе канала
из промерзшего грунта, по которому поток устремляется дальше, впитываясь, наращивая
промерзшее ложе и охлаждаясь. Поэтому, в отличие от земных склоновых рек, потоки на
Марсе сужаются, спускаясь по склону. При переходе воды с температурой 0 °C в фазу льда
выделяется почти 80 ккал/кг. Теплоемкость грунта близка к 0.2, поэтому промерзшее ложе
потока может получиться достаточно толстым, если ключ существует достаточно долго.
Как ведет себя грунт Марса при увлажнении и сколько при этом поглощается тепла, точно
не известно, но баланс отдаваемого тепла должен включать текущие теплопотери в об-
разующемся ледяном ложе канала и более медленные процессы излучения и отдачи тепла
в атмосферу. Температура истекающей воды также неизвестна, но она не может быть высо-
кой, и, как показано выше, энтальпия воды не играет существенной роли.
Часто утверждается, что жидкая вода на поверхности Марса немедленно испаряет-
ся. Это недоразумение. Роль испарения пренебрежимо мала и ее нетрудно оценить. Пусть
давление р в данном районе составляет 8 мб, а температура кипения Т(р) = 4 °C, соглас-
но рис. 7. При температуре TL потока, например, 10 °C, истекающая вода должна кипеть,
с уменьшением ее энтальпии. Какая-то доля потока Ц будет потеряна при испарении
с понижением его температуры до Т.(р) = 4 °C (или до 0 °C при давлении 6.1 мб) и с пре-
кращением кипения. Поскольку теплота парообразования составляет qv = 539 ккал/кг,
а теплоемкость с = 1 ккал/(кг °C),
N =[TL-T(p)] caq / qv.
69
.4.В. Ксанфол1и:1итн. От Марса к Ь.нронс: поиск биосферы на спутниках плане г-ги гаптов
Рис. 10. Протяженность следа потока
на склоне Марса (см. рис. 9) достигает
6 км. Для земных грунтов потемнение
соответствует увлажнению. Предпо-
ложительно, темный след относится
к более позднему источнику (MGS
МОС M0807686b. NASA/JPL/MSSS).
Роль испарения в этом примере сведется к тому, что
потерянная часть Nv составит 1.1 %. Таким образом,
сколько-нибудь значительная часть истекающей воды
испариться не может.
Рис. 9 представляет склон кратера, богатого
склоновыми протоками. В нижней части снимка на-
ходится изрезанной формы бассейн. Внешняя гра-
ница бассейна выделяется светлой окантовкой. Ве-
роятно, это ледяная кромка. Поверхность бассейна,
по сравнению с примыкающей поверхностью, глад-
кая; возможно, это лед. В верхней (по снимку) части
бассейна видны два-три следа, соответствующие
многократному понижению уровня поверхности.
Сток воды через края бассейна образовал второй,
внешний контур (в нижней части снимка). Два таких
же, но меньших по размерам контура можно видеть
в левой части снимка. Источников жидкой фазы,
пополняющих бассейн, на снимке видно несколько.
Вероятно, главный источник находится справа над
бассейном. Это вытянутое образование с шестью на-
правлеными вниз отростками. По-видимому, вода
стекает вдоль отростков. Более мелкие структуры
того же вида видны слева над бассейном и, вероятно,
связаны с наиболее широким протоком вдоль склона.
Форма промоин на рис. 9, соответствующая крутому
склону, указывает, что поток должен нести с собой
значительное количество грунта. Горизонтальная ось
снимка около 1500 м. Размер бассейна около 600 м, а
площадь около 0.3 км2.
Никакие песчаные запруды на Марсе нс смог-
ли бы удержать большие массы воды, даже с учетом
втрое более низкой силы тяжести на планете. Но если
грунт очень холодный, поступающая вода, впитыва-
ясь в морозный грунт, могла бы быстро создать чаши
из льда и промерзшего грунта, обладающие опреде-
ленной прочностью. По существу, это тот же предло-
женный выше механизм, который объясняет сужение
протоков вдоль склона.
Возраст образований на рис. 9 не может быть
большим. Вполне вероятно, что комплекс источни-
ков и бассейн действуют в наши дни. На это указы-
вает чистая (без отложений пыли), насколько можно
судить по снимку, кромка бассейна, примыкающий
к нему второй контур и четкие нитевидные протоки на склонах. В ряде случаев наблюда-
ются следы потоков, уходящие на несколько километров, а также повторно появляющиеся
свежие следы, пересекающие прежние истечения (рис. 10). Подобных образований найде-
но много (Ксанфомалити. 2003). Жидкая вода локально появляется во многих точках пла-
неты, спорадически или даже периодически, хотя в целом бюджет жидкой воды на Марсе
весьма ограничен.
70
Проблемы происхождения жизни
1. 4. Поиск следов жизни на Марсе: заключение
Итак, длительные поиски обитаемости Марса положительных результатов пока не
дали. На какой-то стадии исследований утверждалось, что важным шагом было бы обна-
ружение на Марсе воды в жидкой фазе. Но и после этого задача осталась столь же далекой
от разрешения, а никаких следов жизни найти не удается.
Это следует считать удивительным, так как в последние десятилетия было доказано,
что планеты постоянно обмениваются осколочным материалом (Melosh, 1984), образующим-
ся при ударах крупных метеоритов. Результаты поиска следов жизни в марсианских SNC
метеоритах широко известны (McKay et al., 1996). Материал с поверхности Земли в эпоху
интенсивной метеоритной бомбардировки также выбрасывался в космос в виде вторичных
метеоритов и нес в себе элементы земной биосферы. Поэтому метеориты, подобные марси-
анским SNC, попадали не только с Марса на Землю, но и с Земли на Марс. Если учесть, что
присутствие микроорганизмов на земных образцах неизбежно, а на Марсе следы их не на-
ходят, уместен вопрос: не обладает ли Земля каким-то исключительным свойством, которое
нас окружает, но которого мы не замечаем?
2. ЕВРОПА
Четыре планеты-гиганта Солнечной системы окружены многочисленными спутника-
ми. Число зарегистрированных спутников превышает 100, но если рассматривать относи-
тельно крупные тела, диаметром более 1000 км, то их всего 14. Все они обладают внешними
ледяными оболочками разной протяженности. Равновесные температуры на их орбитах на-
столько низки, а запасы тепла так ограничены, что их глубокие слои должны были перейти
в твердую фазу в пределах менее 1 млрд, лет их существования. Однако имеются убеди-
тельные свидетельства того, что под ледяной оболочкой у многих из них сохраняется оке-
ан в жидкой форме. Необходимым для этого источником тепла является рассеяние энергии
в приливных взаимодействиях.
2.1. Физические характеристики Европы
Второй галилеев спутник Юпитера Европа по диаметру вдвое меньше Марса, а по мас-
се менее 1 % массы Земли (масса Марса 11 % массы Земли). Основные физические данные
Европы: средняя плотность 2.97 гхсм’3, радиус тела 1569 км, масса 4.87х1025 г, ускорение сво-
бодного падения 132 см/с2, температура поверхности 69-100 К, альбедо поверхности 0.7-0.9,
имеются следы атмосферы с плотностью <10-9 г-см3. Период обращения вокруг Юпитера
3.551 сут., орбитальный радиус 670 900 км. Спутник находится в поясе наиболее интенсив-
ной радиации. Вращение синхронное (Европа всегда повернута к Юпитеру одной стороной).
Спутники Ио, Европа и Ганимед находятся в орбитальном резонансе - их орбитальные пе-
риоды соотносятся как 1:2:4.
Европа относится к ледяным спутникам. Предполагается, что под ледяной оболоч-
кой, «корой», толщиной 10-30 км находится глобальный океан глубиной не менее 100 км.
На его дне залегают горные породы, а в центре, возможно, находится небольшое металличе-
ское ядро (Ксанфомалити, 1997). У Европы имеется магнитное поле необычной структуры.
Поверхность Европы покрыта сетью пересекающихся линий, представляющих собой
разломы и трещины ледяной оболочки и застывшую жидкую среду, выходившую сквозь
них на поверхность. Система трещин и ледяных полей (рис. 11) очень напоминает структу-
ру ледяных полей Северного Ледовитого океана. Поверхность Европы относительно ровная,
с небольшими холмами и длинными грядами и долинами. Хотя предполагается, что поверх-
ность Европы претерпевает постоянные изменения, но сравнение космических снимков не
выявило каких-либо изменений. По оценкам, возраст поверхности не превышает 30 млн. лет.
Таким образом, Европа обладает высокой геологической активностью.
71
Л.В. Ксииф<>мс1.1И1}Н1. О г Марса к Кироне: поиск биосферы на спутниках планет-гигантов
Рис. 11. Поверхность Европы покрыта бесчисленными разломами и трещи-
нами в ледяной оболочке, возникающими под действием приливных сил.
Присутствие кислорода в следах атмосферы объясняется фотодиссоциацией медленно
испаряющегося льда и быстрой диссипацией водорода.
2.2. Подледный океан Европы
Говорить о возможности существования каких-то форм жизни можно только в под-
ледном океане Европы. Ледяная кора спутника, по разным гипотезам, может быть толщиной
минимум 1 км, максимум - до дна. Несмотря на популярность всех трех моделей строения
Европы (относительно тонкая, 10-30 км, ледяная оболочка; частично промерзший вглубь
океан; и полностью промерзшая среда; рис. 12), модель с относительно тонкой ледяной обо-
лочкой представляется наиболее реалистичной. Она объясняет и фрагментированные ледя-
ные поля, и трещины, и рассеяние мощного притока энергии, возникающей в приливных
взаимодействиях системы спутников Европа-Ио и Европа-Юпитер. Если глубина океана
50-60 км, давление на дне должно быть 600 кг/см:.
Сильным аргументом в пользу существования океана говорит и необычное маг-
нитное поле Европы. Напряженность поля Европы имеет переменный характер, причем
магнитные полюса расположены вблизи экватора спутника и постоянно перемещают-
ся, а изменения напряженности и ориентации магнитного поля синхронизированы с про-
хождением Европы через магнитное поле Юпитера (Kivelson, 1995). Это можно объяс-
нить лишь наличием токопроводящей жидкости под поверхностью спутника: его движе-
ние в сильном магнитном поле Юпитера вызывает электрические токи в соленом океане
Европы, что и создает необычное магнитное поле спутника. Если бы поле было связано
с ядром спутника, оно было бы стабильным и имело бы меньшую напряженность.
Существование подповерхностного океана подтверждается также структурой крупномас-
штабных трещин глубиной около I км, имеющих глобальную протяженность. Было показано
(Schenk et al., 2008), что трещины глубиной от 300 м до 1.5 км возникли при перемещении (поворо-
те) полюсов спутника примерно на 80°, причем ледяная оболочка двигалась независимо от ядра.
Достаточно высокая температура в глубоких слоях Европы объясняется рассеянием
приливной энергии в орбитальной соизмеримости со спутником Ио, и в меньшей степени -
приливным воздействием Юпитера. Приливное рассеяние-мощный источник энергии; основ-
ная ее часть рассеивается в Ио, но и на Европу приходится ее существенная часть. Полный пе-
риод Т повторения приливных эффектов определяется известным соотношением 1/Т(|о) - 1/Т
и составляет, благодаря соизмеримостям, почти точно орбитальный период Европы. Кроме при-
ливных эффектов определенный вклад энергии создает процесс серпентинизации, химическое
72
связывание воды с горными породами в верхнем
слое ядра спутника. Серпентинизация увеличива-
ет объем горных пород и выделяет тепло. Наконец,
какое-то рассеяние энергии происходит и в упоми-
навшихся выше магнитны,х взаимодействиях. Во
всяком случае, Европа располагает достаточными
источниками энергии, чтобы поддерживать хотя
бы часть своих запасов воды в жидком состоянии,
подобно тому, как это происходит на других ледя-
ных спутниках (McKay, Matson, 2008).
По составу воды ее выходы на поверхность
из трещин и разломов неоднородны. По данным
спектроскопии, окрашенные участки поверхно-
сти предположительно включают примеси серной
кислоты, а также других сернистых и железистых
веществ. Спектральный анализ линейных обра-
зований и пятен на поверхности показал наличие
в них солей, в частности, сульфата магния, кото-
рый содержится и в земных океанах. Обнаружены
также следы перекиси водорода и сильных кислот.
Таким образом, массы воды, проникающие на по-
верхность из разломов, неоднородны по составу.
Не исключено, что подледный океан не имеет гло-
бальной протяженности.
Рис. 12. Существующие модели строения
Европы.
2.3. Возможная обитаемость подледного океана Европы
В подледном океане Европы, куда не попадает солнечный свет, где невозможен фото-
синтез и отсутствует кислород, может существовать микрофлора. На Земле можно найти
примеры таких экзотических экосистем. Роль i идротермальных глубоководных источников
в возникновении жизни можно отнести и к Европе, где также могут присутствовать глубоко-
водные гидротермальные источники. В гипотезе о происхождении жизни в районах глубо-
ководных термальных источников жизнь на Земле возникла, когда химически обогащенные
жидкости какими-то механизмами апвеллинга стали подниматься с нижних морских эта-
жей. Химическую энергию могли доставлять реакции восстанавливающих газов, например,
водорода или сульфида водорода, которые выделялись из придонных источников, в контакте
с подходящим окислителем, например, углекислым газом.
Гипотетический океан Европы должен близко напоминать условия жизни в глубинах
земных океанов, где населенность различными видами живых существ очень высока. В зем-
ных океанах их концентрация наблюдается у гидротермальных придонных океанических ис-
точников, от которых поднимается обогащенная растворенными минеральными солями горя-
чая вода. В такой среде обитает не только микрофлора, но и достаточно крупные животные.
На Земле найдены три экосистемы, на аналогах которых могла бы базироваться жизнь
на спутнике Юпитера Европе. Две системы основаны на метаногенезисе и принадлежат
к древней группе бактерий Archaea, которые живут в бескислородных условиях. В глубоко
залегающих вулканических породах в районе р. Колумбия (район водопадов Айдахо-Фолс)
существуют две такие экосистемы, которые получают энергию от химических реакций в раз-
личных горных породах. Третья экосистема существует за счет энергии, возникающей при
радиоактивном распаде горных пород. Она была найдена глубоко под поверхностью в шахтах
Южной Африки, на глубине 3 км под поверхностью Земли в зоне разлома (Lin et al., 2006). Ми-
кроорганизмы здесь живут без света, без кислорода и без исходных органических веществ.
73
Л.В. Ксанфомалити. От Марса к Европе: поиск биосферы на спутниках планет-гигантов
Перечисленными примерами, конечно, не исчерпываются возможности существова-
ния форм жизни на Европе. Теорий возникновения жизни на Земле много, но ни одна не
объясняет, как воспроизвести зарождение земной жизни в лаборатории и насколько уместно
распространять их на Европу, хотя теория «первичного бульона» и теория гидротермальных
придонных океанических источников наиболее близки к тому, что нам известно о Европе.
Начиная с античности, у исследователей нет идеи, более популярной, чем поиск жизни
в других мирах. Когда развитие технологии позволило осуществить полеты к другим плане-
там, поиск жизни (или следов жизни) на них стал рабочим разделом национальных научных
программ космических исследований. В планах космических агентств уже появляются про-
екты исследований Европы, рассчитанные на следующее десятилетие XXI в. Исследователям
предстоит преодолеть серьезные технические трудности, такие как необходимость проник-
нуть сквозь многокилометровую толщу льда, при температуре 80 К обладающего прочно-
стью гранита, выполнить исследования и прислать на Землю их результаты. Многие задачи
миссии к Европе сегодня кажутся практически невыполнимыми. Но работа уже ведется.
Европейское и Российское космические агентства наметили экспедицию из несколь-
ких космических аппаратов к Юпитеру и Европе. Европейский проект намечает выведение
на орбиты Юпитера и Европы двух космических аппаратов, а российский проект предпо-
лагает включение в экспедицию еще одного, спускаемого аппарата для посадки в одном
из разломов на поверхности Европы. После посадки аппарат начнет поиск следов жизни
в колонке полуметрового слоя льда. Проект получил название «Лаплас». Предполагается, что
он войдет в программу Европейского космического агентства на период с 2015 по 2025 гг.
3. ФАКТОРЫ, ОГРАНИЧИВАЮЩИЕ ВОЗНИКНОВЕНИЕ, СУЩЕСТВОВАНИЕ
И ЭВОЛЮЦИЮ ЖИЗНИ НА ПЛАНЕТАХ, ПОДОБНЫХ ЗЕМЛЕ
В марте 2009 г. на орбиту спутника Земли выведен аппарат «Kepler» (США), предназна-
ченный для поиска экзопланет, обладающих комфортными для возникновения жизни условия-
ми. «Комфортность» физических условий понимается применительно именно к нашей, амино-
нуклеино-кислотной форме жизни. В качестве вероятной среды обитания приняты гипотети-
ческие планеты, аналогичные группе Земли. Планеты-гиганты (газо-жидкой природы), если
не принимать всерьез некоторые фантастические гипотезы, требованиям земной биосферы не
удовлетворяют. Однако таким требованиям могут отвечать спутники планет-гигантов.
В литературе обычно приводятся следующие ограничения: с одной стороны, суще-
ствование жидкой воды, с другой - конденсация углекислого газа (что в подавляющем
большинстве случаев не позволяет развиться парниковому эффекту). С учетом послед-
него, такая зона охватывает интервал планетных орбит от уровня «саморазгоняющегося»
парникового эффекта (для Солнечной системы это случай Венеры), до, примерно, орби-
ты Марса. Оба ограничения, по существу, температурные. Коагуляция белков происходит
при температурах >65 °C; следовательно в комфортных зонах температурный интервал
должен быть 273<Т<340 К.
Можно напомнить, что в моих работах (Ксанфомалити, 1995, 2008; Ksanfomality, 2002)
были сформулированы наиболее очевидные факторы, ограничивающие возникновение, су-
ществование и эволюцию жизни на планетах земного типа. Масса планеты или спутника
накладывает дополнительные ограничения при переходе к многоклеточным формам. Это
ограничения энергетического характера, зависящие от уровня гравитации. Энергетически
выгодны небольшие размеры организмов, так как их энерговооруженность обратно пропор-
циональна примерно квадрату характерных линейных размеров. С другой стороны, доста-
точно сложные животные не могут быть маленькими. Но слишком крупные виды вынуж-
дены существовать в эволюционном тупике, образуемом положительной обратной связью
энергетика-масса-пища. Ограничения очевидны, и о них природа ясно говорит зоологией:
животных, больших, чем синие киты и гигантские рептилии, на Земле не было.
74
Проблемы происхождения жизни
Уровень гравитации планеты должен быть умеренным, а период вращения не слишком
длительным; слишком долгая ночь приведет к глубокому падению температуры окружаю-
щей среды. По тем же причинам эксцентриситет орбиты должен быть умеренным, а наклон
экватора к плоскости орбиты не слишком большим.
Эволюция жизни земного типа требует, чтобы атмосфера планеты обладала способно-
стью отфильтровывать внешнюю жесткую радиацию. Вместе с тем, атмосфера должна про-
пускать к поверхности фотоны с энергией Е = 1-3 эВ, а плотность лучистой энергии должна
быть достаточной, например, для поддержания фотосинтеза. Для поддержания жизни земного
типа, планета, наряду с притоком лучистой энергии, должна обладать наличием других (хи-
мических) источников энергии, например, окислительной среды и окисляемых материалов.
Если исходить из гипотезы возникновения жизни в водоемах, необходимы открытые
водные пространства и наличие континентов или крупных островов. В атмосфере должен
присутствовать водяной пар.
Для развития жизни земного типа планета должна обладать достаточно близким и массив-
ным спутником, который вызывает приливы и побуждает морские организмы осваивать сушу.
Планета должна обладать вулканизмом и/или тектоникой плит. В районах активного
вулканизма значительно увеличивается концентрация в водных растворах веществ, необхо-
димых для существования жизни.
История планеты должна включать эпохи интенсивного выпадения крупных метеори-
тов или другие космические катастрофы, которые стимулируют обновление видов и появле-
ние среди них наиболее приспособленных.
ЗАКЛЮЧЕНИЕ
Планета Земля представляет собой уникальное сочетание физических свойств, необ-
ходимых для возникновения, существования и эволюции жизни. Возможно ли подобное со-
четание на планетных системах других звезд - станет ясным не раньше, чем планеты земной
группы будут там реально обнаружены. Перечисленные выше требования образуют стран-
ный многомерный лабиринт, причем лишь узкие интервалы многих параметров и их сочета-
ния могут обеспечить условия, необходимые для возникновения амино-нуклеино-кислотной
формы жизни и ее эволюции в разумные формы. Анализ условий на других телах Солнечной
системы показывает, что Земля действительно обладает свойствами, не повторяющимися ни
на одной из других планет земной группы.
СПИСОК ЛИТЕРАТУРЫ
Ксанфомалити Л.В. 1995. Характеристики планет, ограничивающие возможное возникновение
и развитие на них жизни И Астроном, вести. Т. 29. № 5. С. 399-404.
Ксанфомалити Л.В. 1997. Парад планет. М.: Наука. Физматлит. 256 с.
Ксанфомалити Л.В. 2003. Потоки и бассейны на Марсе // Астроном, вести. Т. 37. № 5. С. 435-452.
Ксанфомалити Л.В. 2008. Факторы, ограничивающие возникновение, эволюцию и существование
жизни на планетах, подобных Земле И Тр. научн. конф. Иркутского научного центра СО
РАН «Развитие жизни в процессе абиотических изменений на Земле» (18-20 марта 2008 г.,
пос. Листвянка). С. 212-224.
Carr М.Н. 1979. Formation of Martian flood features by the release of water from confined aquifers // J. Geo-
phys. Res. V. 84. P. 2995-3007.
Carr M.H. 1996. Water on Mars. New York: Oxford Univ. Press. 229 p.
Carr M.H., Clow G.D. 1981. Martian channels and valleys: their characteristics, distribution and age // Icarus.
V. 48. P. 91-117.
75
Л.В. Ксамфомалити. От Марса к Европе: поиск биосферы на спутниках планет-гигантов
Carr М.Н, Schaber G.G. 1977. Martian permafrost features // J. Geophys. Res. V. 82. P. 4039-4054.
Clifford S.M. 1993. A model for the hydrologic and climatic behavior of water on Mars H J. Geophys. Res.
V. 98. P. 10973-11016.
Fletcher K. 2008. ESA’s report to the 37 COSPAR meeting. SP-1312. Montreal, Canada, July 2008.
P. 59-66, 155-157.
Head J.W., Hiesinger H, Ivanov M.A. et al. 1999. Possible ancient oceans on Mars: evidence from Mars
Orbiter Laser Altimeter data // Science. V. 286. P. 2134-2137.
Hecht M.H., Vasavada A.R. 2006. Transient liquid water near an artificial heat source on Mars // Mars. V. 2.
P. 83-96.
Johnson J. 2008 // Los Angeles Times, http://www.latimes.com/news/printedition/asection/la-sci-phoenix6-
2008aug06,0,4986721 .story.
KerrR. 2006. Mars orbiter’s swan song: the Red Planet is A-Changin’ // Science. V. 314. P. 1528-1529.
Kivelson M.G. 1995. The physics of planetary magnetospheres // Introduction to space physics / Eds. M.G. Ki-
velson, C.T. Russel. Cambridge: Cambridge Univ. Press. 588 p.
Klein H.P., Horowitz N.H, Bieman K. 1992. The search for extant life on Mars // Mars / Eds. H.H. Kieffer
et al. Tucson: Univ. Ariz. Press. P. 1221-1233.
Klein HR, Levin G.V. 1976. The Viking biological investigation: preliminary results // Science. V. 194. P. 99-105.
Ksanfomality L. V. 2002. Extrasolar planets as a case for life beyond the Earth // Proc. SPIE. P. 4939.
Lakdawalla E. 2008. Phoenix sol 30 update: alkaline soil, not very salty, «nothing extreme» about it! // The
Planetary Society weblog. Planetary Society, http://www.planetary.org/blog/article/00001526/. Re-
trieved on 2008-06-26.
Lin L.-H, Wang P.-L., Rumble D. et al. 2006. Long-term sustainbility of a high-energy, low-diversity crustal
biome // Science. V. 314. P. 479-482.
Malin M.C., Carr M.H. 1999. Ground-water formation of Martian valleys //Nature. V. 397. P. 589-591.
Malin M.C., Edgett K.S. 2000. Evidence for recent groundwater seepage and surface runoff on Mars // Sci-
ence. V. 288. P. 2330-2335.
McKay D.S., Gibson E.K., Keprta T.K.L. et al. 1996. Search for past life on Mars: possible relic biogenic acti-
vity in Martian meteorite ALH84001 // Science. V. 273. P. 924-930.
McKay C., Matson D. 2008. A perspective on Life on Enceladus: a world of possibilities, http://saturn.jpl.nasa.
gov/mission/introduction.
Melosh H.J. 1984. Impact ejection, spallation, and the origin of meteorites // Icarus. V. 99. P. 28-41.
PalermoE., EnglandJ., MooreH. 2001. Martian waterstainsordustslides?//http://home.attbi.com/~palermo63/
MarsAnomalies/MarsSociety _Seeps_Paper.pdf.
Soderblom L.A., Wenner D.B. 1978. Possible fossil water liquid-ice interfaces in the Martian crust // Icarus.
V. 34. P. 622-637.
Squires S.W., Clifford S.M., Kuzmin R.O. et al. 1992. Ice in the Martian regolith // Mars / Eds. H.H. Kieffer
et al. Tucson: Univ. Ariz. Press. P. 523-554.
Stewart S.T., Nimmo F. 2002. Surface runoff features on Mars: testing the carbon dioxide formation hypo-
thesis // J. Geophys. Res. V. 107. № E9. P. 5069.
From Mars to Europa: search for a biosphere on satellites of giant planets
L.V. Ksanfomality
Research of the planet Mars by means of spacecrafts lasts for almost 5 decades. A possible
existence of life concerns to the main tasks of the researches. In the paper some results of missions
to Mars and scientific goals of the future investigations of Europa, one of four galilean satellites
of Jupiter, are considered; their possible habitability is considered in new projects. Physical condi-
tions on Mars and Europa are compared to the Earth’s conditions.
76
Проблемы происхождения жизни
Дискуссия
по пленарному докладу Л.В. Ксанфомалити
«ОТ МАРСА К ЕВРОПЕ: ПОИСК БИОСФЕРЫ НА СПУТНИКАХ ПЛАНЕТ-ГИГАНТОВ
В СИСТЕМЕ СОЛНЦА И ДРУГИХ ЗВЕЗД»
Председательствующий А.Ю. Розанов
М.Я. Маров: Какова вероятность сочетания природных свойств у других планетных
систем, делающих их потенциально пригодными для существования жизни?
Л.В.К.: Чудовищно низкая. Но есть пример нашей собственной планеты.
А.Ю. Розанов: В 1995 году не было известно ни одной такой системы, а сегодня извест-
но уже триста.
М.Я. Маров: Это так, но эти открытия, как и модельные оценки, тоже говорят об очень
низкой вероятности повторения конфигурации Солнечной системы.
А.В. Тутуков: Вы уверенно исключили Марс из числа возможных носителей жизни.
Почему, ведь Вы показали, что там есть вода?
Л.В.К.: Однажды возникшую жизнь очень тяжело уничтожить. Если бы на Марсе она
была, мы бы ее уже увидели.
А.Ю. Розанов: Есть множество свидетельств присутствия остатков организмов в мар-
сианских метеоритах. И если на поверхности Марса слишком плохо, жизнь может быть
в глубинах.
В.Н. Снытников: Почему все-таки считается, что метеориты из Антарктиды, о которых
Вы говорили, происходят именно с Марса, а не из пояса астероидов?
Л.В.К.: На основании изотопов благородных газов.
А.С. Спирин: Вы сказали, что есть бактерии, берущие энергию от радиоактивного рас-
пада. Я не очень это понимаю.
Л.В.К.: Есть соответствующая ссылка.
ГА. Заварзин: Радиоактивный распад служит источником водорода.
А.С. Спирин: Это другое дело. Второй вопрос в отношении Марса. Вот Вы говорите,
что если бы жизнь на Марсе была, она бы проявилась. Что Вы называете проявлением - нуж-
но, чтобы марсиане ходили?
Л.В.К.: Но какие-то видимые следы должны быть. Ведь эволюция идет путем приспо-
собления.
А.С. Спирин: Они и приспособились, предположим, ушли под поверхность.
М.Я. Маров: Я согласен, что большое количество архебактерий могло сохраниться на
этой планете в более глубоких слоях, этого мы исключить не можем. Кстати, с разбившимися
космическими аппаратами (а таких было немало) на Марс могли попасть земные микробы,
и есть две гипотезы, что с ними стало: они либо распространились в условиях сильно дина-
мичной атмосферы планеты, либо не смогли выжить.
ГА. Заварзин: Леонид Васильевич, Ваш доклад, по-моему, ставит крест на всяких иде-
ях о так называемом «parent body» - исходном космическом теле, с которого на Землю что-то
было занесено, и крест на всех панспермиях. Вы говорите, что в обозримых окрестностях
космоса нет тела, сходного с Землей. Следовательно, нет тела, с которого на Землю могла бы
быть перенесена жизнь.
А.Ю. Розанов: Оно нам неизвестно, но, может быть, когда-нибудь оно было.
ГА. Заварзин: Для «когда-нибудь» у нас есть определенный временной интервал -
77
менее четырех миллиардов лет назад, так как нам надо вместить это событие во времена
до появления жизни на Земле. И если для этого периода у нас нет исходного космического
тела, то вся проблема закрывается.
М.Я. Маров: Речь может идти, например, о том, что изначально «колыбелью жизни»
был Марс. А затем, в результате его столкновения с крупным телом и достаточно большой
эффективности транспорта вещества в Солнечной системе, жизнь могла быть перенесена
на Землю. Исключить такой сценарий мы не можем.
Г.А. Заварзин: Мне-то как раз это предположение очень нравится, я с этим согласен, но,
по-моему, Леонид Васильевич показал, что этого не было...
78
Проблемы происхождения жизни. М.: ПИН РАН, 2009. С. 79-9
АСТРОКАТАЛИЗ - АБИОГЕННЫЙ СИНТЕЗ И ХИМИЧЕСКАЯ ЭВОЛЮЦИЯ
НА ДОГЕОЛОГИЧЕСКИХ ЭТАПАХ ФОРМИРОВАНИЯ ЗЕМЛИ
© 2009 г. В.Н. Снытников
Институт катализа им. ГК. Борескова СО РАН
snyt@catalysis.nsk.su
Биосфера на Земле появилась в результате самоорганизации вещества при
возникновении Солнечной системы. С использованием методов изучения
самоорганизации и на основе имеющихся естественнонаучных данных вы-
яснено, что основными этапами химической эволюции в становлении био-
сферы являются «холодная предыстория жизни» в плотных молекулярных
облаках, «допланетный хемоценоз», «мир РНК» в околосолнечном диске
и первичные биоценозы из протоклеток (жизнь) на допланетных телах. Мас-
са первичной биосферы на поверхности ранней Земли оценена в 2.4* 1019 кг
по углероду. Деструкция первичной земной биомассы и биогеохимические
циклы привели за 2.5 млрд, лет к «планете бактерий» с 2.0*1015 кг биоты
в протерозое с кислородной атмосферой. Установлены основные параме-
тры - давление, температура, состояние каталитической твердой фазы -
на этих этапах возникновения жизни. Показано, что абиогенный синтез
пребиотического вещества осуществлялся в Солнечной системе в гранди-
озных масштабах с участием практически каждого атома всех элементов
в наночастицах - катализаторах. Отбор среди каталитически активных
наночастиц происходил по способности синтезировать высокомолекуляр-
ные соединения в допланетном диске. Допланетная химическая эволюция
шла с участием автокатализаторов, начиная уже с таких простых веществ
как этилен и гликолевый альдегид. Первичный синтез автокатализаторов
осуществлялся через внешнее высокоэнергетическое воздействие, к приме-
ру, посредством ультрафиолетового излучения.
Проблема происхождения жизни - это крупнейшая проблема, которая нам досталась
в наследство от XX в. В первую очередь мне было интересно ответить на вопрос, что изме-
нилось в исследованиях по этой проблеме и в чем было максимальное продвижение за по-
следние 30 лет. Почему взят отрезок времени именно в 30 лет?
Дело в том, что в 1980 г. два вышедших подряд выпуска журнала Всесоюзного хи-
мического общества им. Д.И. Менделеева (Журнал ВХО, 1980. № 3-4) были полностью по-
священы происхождению и эволюции жизни. В № 3 напечатана одна из последних статей
А.И. Опарина. В этих же номерах был затронут практически тот же круг вопросов, который
обсуждался и на совещании «Проблемы происхождения жизни» в 2008 г. Кроме того, два
автора тех выпусков - Л.М. Мухин и Н.С. Кардашев - участвовали в наших заседаниях.
79
B.H. CiibDHiii/Koe. Астрокатализ - абиогенный синтез и химическая пюлюция...
Примеры:
1 .Земля и ее биосфера
---► “Система” ----►
L То
т,>т0
AS<0
Космос
* Т-2.7К
Физика процессов
A t /i .0
,/> d!
ц=‘Ю ^^' =11' ' - '
<// At C Tt.
TAS=O
2."Курильщик"
I =WA-
VS,. •- о
:()
I i-SA’
Земля - замкнутая
система, биосфера
на ее поверхности
и на глубине -
открытая система.
Рис. I. Примеры структурообразующих систем с биоценозами. S - экспортируемая энтропия,
8 - производство энтропии в системе в неравновесных процессах.
Первое, что обращает на себя внимание - интенсивное изучение дальнего и ближнего
космоса. Это уже нашло свое отражение в выступлениях на нашем совещании. Второе. На се-
годня весьма сильно развиты информационные подходы, что ранее проходило под термином
«кибернетика». Третье. Стало редким обсуждение проблем термодинамики и второго закона
возрастания энтропии для изолированных систем. Почему? По моему мнению, интерес пере-
местился в область нелинейных неравновесных процессов в открытых системах (Эбелинг
и др., 2001). Кроме того, найдено несколько полезных и важных примеров, на которых на ка-
чественном уровне можно понять эти проблемы. Остановимся здесь чуть подробнее.
Рассмотрим либо замкнутую систему, либо открытую систему, в которую попада-
ет поток энергии. Ровно тот же поток энергии уходит из этой системы. Но приходит энергия
с большей температурой, чем та, с которой энергия уходит. В этом случае система экспорти-
рует энтропию. Примеры таких систем известны достаточно хорошо (рис. 1). Прежде всего,
это наша Земля, на которую падает излучение от Солнца с температурой 6000 К. В свою оче-
редь Земля излучает энергию в космос, который имеет температуру 2.7 К. Излучение Земли
в космос определяется в основном атмосферой. Меньшую часть излучает поверхность Земли.
Взяв для оценки значение 300 К температуры поверхности, мы получаем величину энтропии
dSc/dt=-6* 10’4 Вт/К, которую Земля непрерывно экспортирует в космос в единицу времени. Еще
один пример это литосфера Земли. Через нее высокотемпературная энергия из ядра Земли
выходит на охлаждаемую атмосферой поверхность Земли. Приведем соответствующую оцен-
ку для экспорта энтропии dS /dt=-IO" Вт/К для литосферы. И последний пример - обычные
вулканы и подводные «черные курильщики», для которых эта величина не столь значительна.
Тем самым, повторю давно сделанный вывод из термодинамики, что Земля как физиче-
ское тело - это замкнутая система, теряющая энтропию на временах своего существования.
Биосфера на ее поверхности и на глубине это открытые системы. Во всех указанных при-
мерах в системах возможна самоорганизация (развитие, усложнение) в границах, которые
определяются балансом между экспортом энтропии и ее ростом внутри в неравновесных
процессах. Интересный и тут же возникающий вопрос. Связанные сегодня между собой био-
ценозы в наземных, глубинных подземных и в подводных вулканических системах имеют
одного родоначальника или же они возникли и развивались автономно друг от друга?
80
Проблемы происхождения жимп!
Рис. 2. Самоорганизация в системе при возникновении новой структуры с разрушением исходной.
Следует отметить еще одно направление исследований, в котором имеется значительное
продвижение. Это проблемы самоорганизации в приложениях к различным объектам, начиная
космологическими явлениями и вплоть до событий социальных и исторических. Происхожде-
ние и эволюция биосферы - одна из таких проблем. Задачи в этой области являются принципи-
ально нестационарными, динамическими. Часто исследования ведутся на качественном, описа-
тельном уровне, хотя основы для количественных моделей уже заложены (Эбелинг и др., 2001).
С общефизической точки зрения подход к решению проблемы самоорганизации, как
выше было уже отмечено, состоит в рассмотрении открытой системы. Открытая система
в квазистационарном состоянии подстраивается под приход и уход энергии или соответ-
ствующей массы. Насколько полно выполняется баланс между приходом и уходом зависит
от конкретного объекта. Примером может служить биосфера с неполной замкнутостью цик-
ла по углероду. Если приход энергии компенсирует ее потери, то и система может оказать-
ся стабильной по своему внутреннему состоянию даже при значительном экспорте энтро-
пии. При изменении внешних условий или внутренних параметров прихода-ухода энергии
система двигается к состоянию, в котором начинают развиваться неустойчивости (рис. 2).
Неустойчивое состояние означает, что все обычно небольшие и затухающие флуктуации,
присутствующие в этой системе, начинают резко усиливаться и достигают макроуровня.
Из этих макросостояний остаются только те, которые соответствуют новым условиям, внеш-
ним и внутренним. Происходит отбор и, конечно, он естественный.
Исходная система может перестроиться вся в целом с потерей исходного состояния,
как схематично показано на рис. 2. Но перестройка может пройти и так, что внутри исходной
системы может возникнуть какое-то новое состояние или новый уровень (рис. 3). В этом слу-
чае считается, что в исходной системе произошло усложнение. В литературе иногда встреча-
ется термин «упорядочивание» (Климентович, 1990; Галимов, 2008) в смысле возникновения
коллективного согласованного движения в противоположность хаотическому. Приведу при-
меры такого сорта усложнений.
Барионы имеют массу в 4 % от всей гравитирующей материи. Тяжелые элементы среди
всех барионов с большей, чем у водорода и гелия, атомарной массой. Таких тяжелых элемен-
81
В. IЧ. Си ыш пи кок. Ас г рок in ал и < а 6 и 01 сними синтез и химическая твоя го ц и я...
Барионы- 4%
Тяжелые (Z>2) элементы - 1 %
Звезды - 0.5 %
Масса планет в СС - 0.1 %
Трофические цепи - <10%
Рис. 3. Самоорганизация в системе при возникновении новых структур с сохране-
нием исходной.
тов во Вселенной около I % среди барионов. Звезды относительно всей видимой материи
во Вселенной - всего-то 0.5 %. Масса планет в Солнечной системе (СС) - масштаба 0.1 %
от системы. И если брать примеры уже из биологии, то, прежде всего можно привести тро-
фические цепи. Число их уровней невелико, в «Биологическом словаре» (1993) указано 4-5.
А вот предельная энергия, которую один уровень может передать другому без своего разру-
шения, не превышает 10 %.
Итак, характерная масса нового состояния для одного этапа в процессах самооргани-
зации в природе составляет примерно 1 % исходной системы. Последовательная цепь возни-
кающих новых состояний от, ограничим себя, синтеза барионов до зарождения и эволюции
биосферы представляет собой развитие в природе. Хотя бы из-за охлаждения нашей расши-
ряющейся Вселенной это развитие однонаправлено. Так, тяжелые элементы при стабильном
протоне не распадаются. Поскольку новые состояния формируются через неустойчивость
(кризис), то смысл и использование терминов типа «устойчивое развитие» должны опреде-
ляться отдельно.
Следующий выпуск вышеупомянутого журнала за тот же год (Журнал ВХО, 1980. № 5)
был посвящен проблемам химической промышленности и науки. В этом выпуске рассматри-
валось развитие химических технологий, создание химических заводов для многотоннажных
синтезов и небольших производств, задачи разработки новых и модернизации действующих
реакторов, их классификация, защита окружающей среды. Используя изложенные в этом
журнале подходы, попытаемся посмотреть глазами химиков-технологов на научную пробле-
му о происхождении жизни (рис. 4). Зададимся простым вопросом. Если у нас есть готовая
продукция в виде биополимеров известного (CH,O)|0h(NH,)|6H1PO4 состава на поверхности
Земли, то какой я должен создать химический завод, сколько должен взять исходного сырья,
как я должен организовать выброс отходов и их утилизацию для получения определенного
биогеохимиками количества продукции. Не поддержания на Земле на длительных временах
в тысячи и миллионы лет заданной массы биополимеров определенного состава в био-, гео-
и фотохимических циклах в биоценозах, а именно их получения. Нужно еще учесть, что есть
времена «выпуска заводом продукции» и есть химические времена, времена пребывания
82
Прои. ii'Aib! прот \ож п-ния жтни
1. Сколько требуется сырья?
2. Как подводится и снимается энергия?
Рис. 4. Происхождение жизни как первичное производство биополимеров.
реагентов в реакторах и передвижения реагентов из реактора в реактор. Эти времена синтезов
органических соединений составляют сутки, максимум, что можно себе представить - годы,
десятки, но не тысячи лет. За тысячи и более лет скорее происходит деструкция, разложение
органических соединений. Далее, один из важнейших вопросов - куда мы должны девать
отходы? Не менее важный вопрос с точки зрения реакторов и химических технологий - как
подводится, используется и отнимается энергия в комплексе эндо- и экзотермических реак-
ций? Такая постановка вопроса не рассматривалась ранее, не исключение и упомянутые вы-
пуски журнала ВХО им. Д.И. Менделеева, посвященные происхождению и эволюции жизни.
На мой взгляд, это перспективный методологический подход к изучению проблемы абиоген-
ного синтеза пребиотического вещества, поскольку происхождение жизни выходит за рамки
последних 3.9 млрд, лет, которые геологически документированы для Земли. Кроме того, хи-
мические технологии требуют как экспериментальную демонстрацию своих процессов по-
следовательно на лабораторном, пилотном и опытно-промышленном уровнях, так и проведе-
ния количественных инженерных расчетов. Все это возвращает далекие от реальности идеи
и фантазии на прокрустово ложе базовых законов сохранения в балансовых соотношениях.
Итак, «проблема химического завода» - рис. 5. На «планете бактерий» (Заварзин, 2008)
углерода биологического органического вещества несколько единиц на 10'* г. Если я добавлю
сюда воду, азот, то получаю число примерно I019 г состава ((CH,O)|0ft(NH3)|6H3PO4)n. Л.М. Му-
хин (1980) утверждал, что выходы в синтезах нуклеотидов, сахаров и других важных компо-
нентов составляют значения в 0.01 % по массе. Обычно выходы полинуклеотидов при пере-
ходе к «миру РНК» также указываются в 0.01 %. Мной не найдены литературные сведения
о возможных выходах продукции при переходе от «мира РНК» к простейшим клеткам и
далее к «планете бактерий». И я принял (но здесь требуется уточнение, стоит вопрос) то же
самое значение 0.01 % по массе, что соответствует двум стадиям самоорганизации в этом пе-
реходе. А теперь собираем все порядки. Итого на трех основных этапах выход составил I О12,
двенадцать порядков! Для того чтобы на поверхности Земли получить 10|у г бактерий, в ка-
честве сырья на входе «химического завода» нужно иметь 10” г углерод-азот-кислородного
сырья. А если по космической распространенности элементов я добавлю еще два порядка
по массе водорода и гелия, то необходимое общее количество сырья на входе «завода» 10”’ г,
что всего в два раза меньше массы современного Солнца. Число астрономическое.
S3
В.Н. Снытников. Астрокатализ - абиогенный синтез и химическая эволюция...
Нуклеотиды
Сахара
10
Исходное вещество
1031г (10“г)
Рис. 5. Проблема «химического завода».
Органическое вещество
биосферы до 1019г
В учебниках по химическим технологиям написано, что нужно делать, чтобы умень-
шить это число, количество исходного сырья. Для этого мы должны кардинально увеличить
выходы особенно на первых стадиях, решив как минимум две проблемы. Во-первых, найти
наиболее эффективные синтезы с катализаторами. И, во-вторых, создать для них экономичные
реакторы, которые еще должны быть согласованы по массо- и теплообмену по всей цепочке
синтезов. Мы должны согласовать абсолютно всю цепочку синтезов от сырья из простейших
соединений до конечной продукции клетки и «планеты бактерий». Если у нас возникнет за-
тор в одном хотя бы месте, то это будет означать, что вся цепочка, весь «завод» не работает.
В этом случае на временах в миллиарды лет заведомо происходит как деструкция наработан-
ных полупродуктов, так и разрушение самого «завода». Что еще мы можем сделать с точки
зрения химических технологий? Мы можем сменить исходное сырье и вместо инертных, труд-
но активируемых молекул - СН4, СО,, N, - использовать реакционноспособные соединения.
Тогда их нужно найти. И зададим еще один, возможно, самый главный, или ключевой, вопрос.
А может, и нет ничего страшного в этом числе Ю” г, и в происхождении жизни действительно
участвовало вещество, по массе сравнимое с Солнцем? Положительный ответ переводит про-
блему происхождения жизни из геохимии в астрохимию и астрофизику.
Современный космохимический ответ на вопрос об исходных высоко реакционноспо-
собных соединениях на первый взгляд очень прост. Даже вся Земля, а не только ее атмос-
фера и поверхность с привнесенным метеоритным дождем, не имела и не имеет указанной
массы исходного вещества для первичного абиогенного синтеза органических соединений. А
вот молекулярные облака и околозвездные диски сразу предоставляют необходимое сырье.
В его состав помимо метана, воды, диоксида углерода входят многие активные и в обычных
условиях неустойчивые соединения, которые идентифицированы астрофизиками и астрохи-
миками. Среди этих соединений можно отметить цианводород, формальдегид, аммиак, мо-
нооксид углерода, формамид и другие молекулы и многие радикалы. Поэтому здесь не надо
придумывать никаких молний, вулканов и тому подобных плазмохимических, электродуго-
вых, фото- и прочих реакторов с их экстремальными условиями, к примеру, расплавленных
лав или суперкритических сред для активации инертных соединений. А при этом - с разру-
шением сложных молекул и стерилизацией среды. Это примерно так же, как нас заставили
84
Пгт, // 1/6/ Ill’OiJi ,///;/ Ж! КИИ
Рис. 6. Распад облака при развитии неустойчивости Джинса: бесстолкновительная динамика, иллю-
стрирующая формирование звезд; компьютерное моделирование: а и б-два момента времени.
бы производить этилен из метана, вместо того, чтобы воспользоваться другим источником
сырья - этаном в составе добываемого природного газа. Дегидрированием этана получают
в мире основное количество этилена в относительно мягких условиях. Точно так же и мы
должны воспользоваться этим подходом. Однако надо помнить, что водород с гелием, угле-
род, азот и кислород - самые распространенные элементы во Вселенной. И сложность здесь
в том, что на поверхности или вообще на Земле по этим элементам имеется огромная не-
хватка. Поэтому нам необходимо привлечь дополнительные данные по образованию ман-
тии, ядра, коры на ранних этапах (Адушкин и др., 2008), чтобы ответить на вопрос, когда
и в каких процессах Земля потеряла эти элементы. Или же потеря произошла на этапе форми-
рования протопланеты и Солнечной системы? И очень важно, что в наших целях мы должны
в первую очередь иметь дело с соединениями и потоками массы именно из этих пяти элемен-
тов. Таким образом, исходное сырье для абиогенного синтеза пребиотических соединений
надо находить в плотных молекулярных облаках.
Эти облака представляют собой место зарождения звезд вместе со своими околозвезд-
ными дисками. В околозвездных дисках формируются планеты. Околозвездные протопла-
нетные диски интенсивно изучаются астрофизическими методами. В настоящее время обна-
ружено много звезд, имеющих планеты. Звезды в молекулярных облаках формируются при
развитии гравитационной неустойчивости. Она возникает, если температура и плотность
облака зададут такой длину Джинса, которая станет меньше характерного размера моле-
кулярного облака. Облако с массой в десятки тысяч масс Солнца начинает распадаться на
отдельные сгустки числом несколько тысяч и более, которые становятся зародышами звезд.
Рис. 6 иллюстрирует этот процесс нашим компьютерным пространственно трехмерным мо-
делированием (Кукшева, Снытников, 2007). К этим результатам надо относиться только как
к иллюстрации, так как расчеты проведены для физически бесстолкновительной модели,
в то время как распад молекулярного облака на начальных этапах происходит в газодина-
мических условиях. И только на более поздних временах сгустки переходят к бесстолкно-
вительной динамике. Тем не менее, расчеты дают представление о возникновении сгустков,
их взаимодействии с поглощением друг друга и даже дисковых структур. В правом нижнем
углу рис. 6 имеется место, где возникла вращающаяся структура или же двойная система.
Итак, молекулярные облака распадаются на отдельные сгустки. Гравитационный кол-
лапс ведет к звездообразованию в сгустках. Результатом коллапса становится протозвезда
вместе с вращающейся структурой вокруг нее, которая и получила название околозвездно-
го аккреционного диска. Диск, начиная с определенного момента времени оседания пыли
к экваториальной плоскости, становиться достаточно тонким, а звезда типа нашего Солнца
примерно за один миллион лет набирает основную свою массу. При этом за вторую половину
85
В.Н. С и ы in и и кои. Ас тро к а I а .1 и i - aonoi ен и ы ii синтез
Гравитационный
коллапс
аввада-кокон,
10*-10* лет
Дегазация Земли
дифференциация,
преобразования земной коры
500 млн. лет
Рои первичных тел,
бомбардировка,
дегазация тел.
Солнечная система
60 млн. лет
Геологические
образцы с возрастом
3.9 млрд, лет
Рис. 7. Основные этапы смены внешних условий для абиогенного синтеза органиче-
ских соединений на Земле.
этого времени звезда набирает ориентировочно половину своей массы. Отсюда у нас есть осно-
вание считать, что через протопланетный диск прошла эта масса и что природа в этот времен-
ной интервал пользовалась солнечными значениями масс и их соответствующими потоками.
В дисках могут сформироваться планеты. В современной Солнечной системе помимо
планет обнаружено много тел естественного происхождения. Их число в последнее время
с развитием наблюдательных методов только возрастает. И сейчас, спустя 4.56 млрд, лет от
своего зарождения, в Солнечной системе множество астероидов движется между орбитами
Марса и Юпитера. Как же возникла такая система как Солнечная?
Основные этапы ее возникновения приведены на рис. 7 с точки зрения смены внешних
условий для абиогенного синтеза органических соединений (Снытников, 20076). Как уже
выше было сказано, на первом этапе в молекулярном облаке происходит гравитационный
коллапс. В отдельном сгустке он идет за времена 104-105 лет, в результате чего формируется
протозвезда-кокон с массой менее 0.) массы Солнца. Протозвезда продолжает набирать массу
и по достижению 0.1 массы Солнца в ядре протозвезды зажигаются термоядерные реакции.
Вокруг протозвезды формируется тонкий аккреционный диск, через который на протозвез-
ду примерно I млн. (максимум - 10 млн.) лет продолжает поступать вещество. Возрастаю-
щее излучение от звезды и солнечный ветер сдувает окружающее вещество молекулярного
облака и водород с гелием из протопланетного диска. К этому времени в диске сформиро-
вались зародыши будущих планет, планетезимали, и возник рой первичных тел Солнечной
системы. Из роя первичных тел и зародышей планет по последним оценкам (Маров и др.,
2008) за времена масштаба 60 млн. лет в столкновительных процессах аккумуляции тел на
протопланеты произошло превращение последних в планеты. На следующем этапе проис-
ходили дифференциация вещества Земли и преобразование земной коры. В это же время
продолжалась дегазация с затуханием своей интенсивности. Образцы пород, с которых на-
чинается геологически документированная летопись на Земле, имеют возраст 3.8-3.9 млрд,
лет. Мало кто сейчас сомневается в том, что к этому времени на поверхности Земли уже
существовала биосфера из микроорганизмов. Поэтому, если жизнь зародилась в Солнечной
системе, то для этого наиболее подходят этап протопланетного аккреционного диска или
этап роя первичных тел и формирования планет.
Вернемся к околосолнечному диску. По современным данным (Маров и др., 2008) сред-
нее по углу квазистационарное давление водорода и гелия на земном радиусе от протозвезды
86
Проблемы происхождения жизни
составляло до 10'4 атмосфер и выше, уменьшаясь к орбитам современных Юпитера и Сатурна
до 10б атмосфер. Температура газа падала от примерно 1000 К на астрономической единице
к 100 К на удалении орбиты Сатурна. Околосолнечный диск представлял собой термодина-
мически открытую систему, которая поглощала энергию протозвезды, а на ранних этапах
и аккреционной оболочки. Эта энергия переизлучалась в остаточное окружение молекуляр-
ного облака и космос. На фоне этого потока энергии движение газопылевой массы на прото-
солнце осуществлялось в турбулентном режиме с возможностью самоорганизации при пере-
ходе от одного режима к другому, более высокоорганизованному (Климонтович, 1990).
Что происходило с пылью, которая двигалась вместе с газом при своей массе пример-
но в 1 % от газа? Заметим, что пылинки имеют сложную слоистую структуру. В среднем
если диаметр пыли составляет сотню нанометров, то внутреннее ядро из тугоплавких окси-
дов кремния, магния, железа составляет несколько нанометров. Далее идут слои из веществ
в порядке их летучести. Летучие органические соединения, вода, метан находятся на поверх-
ности. Под поверхностью находятся указанные выше сложные органические соединения, при-
чем их больше обнаруженных количеств в молекулярных облаках. В молекулярных облаках
регистрируют соединения в газовой фазе, в то время как львиная доля органических соеди-
нений сконденсирована на пылинках и составляют эти пылинки. Еще надо отметить, что при
вращении газа с пылью вокруг протозвезды значения центрифужного параметра достигает
огромных по земным меркам значений в 1000-2000 для пыли относительно газа. (В земных
условиях этих значений пока не достигли). Поэтому при поступлении газа и пыли с пери-
ферии околозвездного диска пыль начинала накапливаться в диске. При своем движении из
холодных внешних областей диска во внутренние разогретые области пылинки теряли свою
шубу из органических соединений. Ядра пылинок очищались. Но на ядрах могли происходить
и гетерогенные реакции синтеза органических соединений из СО, водорода, соединений азо-
та, если такие наночастицы обладали каталитической активностью. Состав наночастиц можно
восстановить, исходя из распространенности элементов во Вселенной. Он приведен в наших
предыдущих работах (напр., Снытников, 2007а) вместе с составом газа. Фактически это со-
ставы классических катализаторов для синтеза Фишера-Тропша получения сложных углево-
дородов из СО и водорода и синтеза аммиака из азота и водорода. Надо подчеркнуть, что все
окружающее нас вещество, вся Земля и тяжелые элементы Солнца находились в виде этих на-
ночастиц. При этом в наночастицах из тысяч атомов в качестве активного центра работал или
был оттестирован на работоспособность не просто какой-либо элемент, а каждый атом даже
самого редкого элемента!! Это поддерживает утверждение о необходимости для биоценоза
всех элементов (Меклер, 1980). К примеру, отдельные атомы вольфрама, молибдена, ванадия
и так далее, влияют на свойства наночастицы. На важность этих элементов для становле-
ния биосферы вновь было обращено внимание (Dobretsov et al., 2008; Федонкин, 2008). Отбор
в протопланетном диске по активности катализаторов был осуществлен на этом этапе в совер-
шенно невообразимых для химических технологий масштабах. Но для конечного продукта,
биоценозов, каталитическая суперактивность наночастиц разрушительна.
Конечно, нужно было экспериментально подтвердить это предположение о высокой ка-
талитической активности наночастиц состава природной распространенности элементов. Для
этого в Институте катализа СО РАН была построена установка (Snytnikov et al., 2007), на кото-
рой методом лазерного испарения были приготовлены наночастицы из природных минералов
и метеоритов, в частности, из метеорита Царев. Образцы наноматериалов были отданы в две
независимые лаборатории Института катализа СО РАН для определения их активности в реак-
ции синтеза углеводородов из СО и водорода. Все приготовленные наноматериалы оказались
активны в этой реакции (Хасин, Снытников, 2005). А один из образцов показал активность,
не уступающую активности промышленного катализатора. Далее, все это означает, что в ука-
занных условиях протопланетного диска наночастицы синтезировали большое количество
87
В.Н. Снытников. Астрокатализ - абиогенный синтез и химическая эволюция...
различной органики. И все неорганические частицы с диаметром в сантиметры, на которых
обычно заканчивается укрупнение неорганических материалов (на больших масштабах неор-
ганические материалы в условиях диска плохо соединяются между собой), реально были по-
гружены в органическую связку, матрицу или пропитаны органическими соединениями, кле-
ем. Фактически, это были кусочки пластилина, которые в столкновениях между собой могли
легко слипаться. Тогда характерный масштаб, на котором заканчивается обычный рост частиц,
составляет не сантиметры, а метры. Тем самым на этой уже относительно поздней стадии дис-
ка с температурой порядка 300-500 К на земном радиусе мы имеем газ и набор метровых в диа-
метре тел, двигающихся слабостолкновительно вокруг протозвезды. Слабостолкновительно
означает, что тела могут столкнуться между собой на временах, сравнимых со временем одно-
го оборота вокруг протозвезды. Такие столкновения происходят с космическими скоростями,
большими скорости звука в твердом теле, и они приводят к дроблению тел.
Отметим, что органической компонентой и гетерогенными каталитическими реак-
циями в околозвездных дисках многие исследователи обычно пренебрегают, уделяя основ-
ное внимание газовой компоненте (Маров и др., 2008). Это справедливо в расчетах тем-
пературы и давления газовой компоненты по диску. Но в эволюции пылевой компоненты
безусловно важную роль сыграли, в частности, реакции полимеризации на наночастицах
с появлением комплекса разных полимеров. В качестве примера можно указать полиэти-
лен, мономер которого этилен синтезируется из СО и Н2 наряду с другими углеводородами
и молекулами воды.
Далее мы задались следующим вопросом, и в этом мы также были оригинальны по
сравнению с исследователями московской и других школ (Маров и др., 2008). Существует
ли физический механизм, который за предельно быстрые времена порядка нескольких лет
приводит к формированию зародышей планет из первичных тел метрового размера? По-
чему год или несколько лет? Ответ прост, А.С. Спирин в своем выступлении на нашем со-
вещании потребовал для синтезов «мира РНК» часы, возможно, несколько суток. Но диск
функционирует миллион лет, и нужно связать астрофизические и химические времена.
Отсюда, имеются ли в протопланетном диске существенно более быстрые процессы, чем
процесс, определяющий этот миллион лет существования диска? Ответ на этот вопрос
оказался «да». Но для ответа пришлось рассматривать гравитационную неустойчивость
в двухфазной среде из газа и первичных тел, в которой из последних формируются зароды-
ши планет. Данный вид неустойчивости отличается от неустойчивости Тоомре, которую
анализировали М.Я. Маров и др. (2008). Мы рассмотрели длину Джинса в такой среде и по-
казали, что при эффективной энергии, характеризующей разброс тел по скоростям, близ-
кой к температуре газа, длина Джинса может уменьшиться на много порядков (Снытников,
20076). Возникает условие гравитационного собирания многих первичных тел в зародыши
планет. Этот процесс коллективный и бесстолкновительный. Определенный и изученный
аналог такого процесса можно найти в физике ускорителей элементарных частиц. Он из-
вестен как охлаждение легкими электронами тяжелых протонов. В изучаемом процессе
при трении тел о газ разброс тел по скоростям снижается, и в какой-то момент времени
возникают условия для гравитационного коллективного коллапса тел в газе.
Развитие такого явления в околозвездном протопланетном диске на предельно корот-
ких по астрофизическим меркам временах очень сложно наблюдать, зато можно рассчитать,
промоделировать методами вычислительной физики (Снытников и др., 2004). Но для этого
нужно научиться решать сложную систему уравнений математической физики. Эта система
включает в себя уравнение Больцмана для функции распределения в шестимерном фазовом
пространстве с самосогласованном гравитационным полем. Для гравитационного поля име-
ется уравнение Пуассона, а для газовой компоненты - газодинамические уравнения. Система
для численного решения очень сложна и требует ресурсов суперкомпьютеров. При числен-
88
Прок, //.л/л/ npiHK хожл-нпя >кп ши
Рис. 8. Логарифм поверхностной плотности пылевой (слева) и газовой (справа) компоненты про-
топланетного диска без учета химической реакции.
ном моделировании мы сталкиваемся со сложными математическими проблемами, главная
из которых состоит в необходимости решать не просто нестационарную задачу, а задачу на
самоорганизацию, физически неустойчивую в линейном приближении. Для решения этой
проблемы были разработаны новые методы и подходы, более подробно можно ознакомиться
с ними в наших публикациях (Snytnikov, Kuksheva, 2007).
Результаты ряда вычислительных экспериментов можно увидеть на рис. 8. В резуль-
тате развития гравитационной неустойчивости в околозвездном диске формируются сгуст-
ки, причем за времена всего несколько десятков оборотов - лет. Сгущения могут двигаться
и в обратную сторону по отношению к общему направлению движения. Тем самым эти
сгустки представляют собой волны плотности, нелинейные солитоноподобные структуры.
Но в таких структурах коагуляция может протекать на существенно более коротких време-
нах по сравнению со средними условиями в диске. Расчеты выполнены по разным програм-
мам с разными алгоритмами решения уравнений, что входит в условие получения достовер-
ных результатов вычислительных экспериментов. На рис. 9 представлены кадры из фильмов
с логарифмом плотности по частицам и газу в случае, когда на температуру газа оказывает
влияние экзотермическая реакция образования молекулы водорода из его атомов на твер-
Рис. 9. Логарифм поверхностной плотности пылевой (слева) и газовой (справа) компоненты про-
топланетного диска с учетом химической реакции.
89
В.Н. Сны/нннков. Ac'i рокаra.iкз aonoi спиый синтез и химическая эволюция...
у-координата
Рис. 10. Распределение логарифма плотности частиц по экваториальной плоскости диска - фоновый
график. Слева вверху - аналогичная величина с увеличением для нижнего сгустка. Справа вверху -
распределение логарифма плотности газа в том же самом сгустке плотности. Внизу - изменение мас-
сы твердой фазы на единицу площади (поверхностная плотность) для нижнего сгустка.
дых частицах. В варианте без учета этой реакции сгустки сколлапсированного вещества
появляются в диске, а при дополнительном тепловыделении такие сгустки отсутствуют.
Это показывает, что на ранних этапах при большом содержании атомарного водорода или
при высоких температурах структуры могут не возникать. Параметры среды в солитоно-
подобных волнах сильно отличаются от средних значений в диске. Так, на рис. 10 приведен
график с распределением плотности по твердой фазе в сгустке. Эта плотность в максимуме
на 4-6 порядков выше фонового значения. Более точно я сказать не могу, так как в самом
сгустке эту амплитуду мы считаем с низкой точностью. В самих сгустках давление по
газу также во столько же раз отличается от фона. Если принять удаление сгустка от про-
тозвезды за астрономическую единицу с абсолютным значением давления 10’4 атмосферы,
то в сгустке мы получили давление до ста атмосфер.
Попробуем взглянуть на сгусток с точки зрения химических технологий с их реактора-
ми и охарактеризуем его с использованием классификации химических реакторов (Слинько,
1980). Это реактор с кипящей твердой фазой в газе. Частицы-катализаторы поступают из
диска в реактор. Они некоторое время, порядка суток, ведут реакции в сгустках, а затем сно-
ва выходят в зону с низким давлением и высокой радиацией от протозвезды, где освобожда-
ются от летучих соединений. Происходит регенерация поверхности. Известно, что реакторы
90
Проблемы происхождения жизни
с кипящим катализатором относятся к одному из самых эффективных реакторов химических
технологий. Правда, они редко встречаются на реальном производстве по причине своей до-
роговизны, сложности в управлении, истирании катализаторов и внутренних поверхностей.
Но природа на эти обстоятельства могла не обращать своего внимания.
Самая главная особенность этого реактора - это возможность быстрого переноса ве-
щества внутри реактора путем обмена телами с наработанным веществом на каталитически
активных поверхностях, а также и сложным органическим веществом при десорбции в газ
и вновь адсорбции на тела из газовой фазы. Важны и процессы слипания - разрушения тел
при их столкновениях между собой. Вот именно этот реактор, место и время я хотел бы
предложить для «мира РНК». При быстром перемещении первичных тел внутри реактора
должно неизбежно происходить их смачивание и высушивание. Смачивание происходит
в зонах, где начинает конденсироваться вода. При развитой внутренней поверхности рыхлых
и пористых первичных тел вода проникает по порам внутрь тел. При уходе первичных тел из
областей конденсации воды происходит испарение воды в условиях атмосферы из водорода
и гелия. При надлежащем подходе такой процесс может быть воспроизведен эксперимен-
тально и методами математического моделирования. В то же время внутри сгустка темпера-
тура меняется незначительно даже при протекании экзотермических реакций, характерных
для превращений сложных органических соединений, к примеру, синтеза Фишера-Тропша,
по причине замечательно высокой теплопроводности водорода и гелия, из которых состоит
в основном окружающий газ. Высокая теплопроводность должна выровнять любые темпе-
ратурные неоднородности и снизить колебания температур, несмотря на окружающую раз-
реженную среду вокруг реактора. Эффект здесь может быть примерно такой, как в холодном
плотном молекулярном облаке, которое окружено горячим корональным газом. Таким об-
разом, это весьма сложный реактор для синтеза химических соединений, реактор высокого
внутреннего давления и кипящего катализатора. Про эти реакторы известно, что высокие
давления повышают эффективность синтезов целевых продуктов органического синтеза, как
это и делается в химической промышленности. Далее встает вопрос об эффективности хи-
мических процессов на начальных стадиях абиогенного синтеза органических соединений.
Первые стадии наиболее важны с точки зрения общего выхода конечных предбиологических
соединений и общей массы биосферы.
В созданном реакторе могут происходить самые разнообразные явления. При про-
текании химических реакций вместе с диффузией и теплопроводностью возникают про-
цессы самоорганизации (Пригожин, 1985), бифуркации, периодические и хаотические
химические реакции. Другая идея - возникновение отбора среди автокатализаторов, что
было предложено В.Н. Пармоном (2002) независимо от более ранней работы (Эбелинг
и др., 2001). В цитированном выпуске журнала ВХО напечатана статья об отборе среди ав-
токаталитических систем (Руденко, 1980). В рамках этих идей в ПК СО РАН исследовалась
автокаталитическая реакция Бутлерова и реакция дегидрирования легких алканов, в част-
ности, этана в газовой фазе. Последняя реакция была выбрана из-за высокого содержания
алканов на поверхности Титана, спутника Сатурна.
В реакции Бутлерова интерес представляет возможность селективного синтеза рибозы.
У нас в институте изучили механизм этой реакции. Мы выяснили, что в отличие от усто-
явшихся представлений об автокаталитичности этой реакции ее механизм является цепным
с вырожденным разветвлением цепей (Симонов и др., 20076) без какого-либо отбора среди
автокатализаторов. В реакции образуются до 57 групп продуктов. И наш вывод по этой ре-
акции состоит в том, что избирательный синтез рибозы невозможен. Другая идея состояла
в использовании катализатора, который бы включал в себя фосфор. Оказалось, что из низших
моносахаридов в присутствии катализаторов могут образовываться жизненно важные угле-
воды с суммарным выходом до 59 %. Причем основные компоненты, которые ведут реакцию,
91
В.Н. Спытников. Астрокатализ - абиогенный синтез и химическая эволюция...
это появляющиеся из-за распада углеводов гликолевый и глицериновый альдегиды. Мы также
показали, что в исходно чистом формальдегиде никаких синтезов не происходит. В реагентах
со следами сахаров синтезы идут. В чистых, без затравок - нет (Симонов и др., 2007а).
Далее мы задались вопросом, можем ли мы получить эти затравки при протекании фо-
тохимических реакций. Для этого мы исходный раствор формальдегида подвергли излуче-
нию ArF-ультрафиолетового лазера с длиной волны излучения 193 нм. Это примерно 6.3 эВ.
В экспериментах было получено, что реагент, конечно, начинает разрушаться. Но необходи-
мая затравка из гликолевого и глицеринового альдегидов получается, а при некоторых усло-
виях и с весьма высокими выходами 4-5 % (Pestunova et al., 2005). Здесь ничего удивитель-
ного нет. Из большого числа работ по высокоэнергетическому воздействию на простейшие
реагенты, в том числе электронными или ионными пучками, в разных условиях, скажем,
и в ледяной матрице, эти соединения были получены. Мы также попытались в одном реакто-
ре объединить фотоинициацию реакций и каталитический синтез сахаров из формальдеги-
да. Это у нас получилось с выходами С5-углеводов до 40 %. В целом синтез идет из формаль-
дегида (Pestunova et al., 2008).
Аналогичный подход мы попытались осуществить и для синтеза углеводородов. Воз-
можен ли в простой цепной реакции селективный синтез этилена при дегидрировании этана?
Можно ли повысить выход пропилена? Проблема состояла в необходимости осуществить реак-
цию без стенок, которые играют роль катализаторов и на которых могут гибнуть или появлять-
ся радикалы. Мы собрали у себя установку, в которой излучением СО2-лазера нагревали реа-
генты из газов, а далее анализировали состав получаемых смесей. Нагрев осуществлялся при
поглощении этиленом энергии лазерного излучения. Неожиданно для себя мы выяснили, что
классическая цепная реакция дегидрирования этана является автокаталитической по этилену
(Снытников и др., 2009). Начальное количество этилена для проявления автокаталитичности
можно оценить в 4-5 % при атмосферном давлении. Этот этилен может быть наработан путем
известной стадии распада этана на два метильных радикала. Но далее более предпочтитель-
ной становиться реакция появления метильного и пропильного радикалов при столкновении
молекул этана с этиленом. При этом повлиять на селективность выхода пропилена или ацети-
лена в 2 % при высоких степенях конверсии этана весьма проблематично, поскольку появле-
ние указанного канала реакции перекрывает другие реакции. Ацетилен - энергонасыщенная
молекула, открывающая путь к появлению цианацетилена и широкой гаммы органических
соединений (Трофимов и др., 2005) в околозвездной среде. Следовательно, нужен катализатор,
классическое предназначение которого состоит в изменении путей синтеза соединений.
Таким образом, существуют пути эффективного синтеза сложных органических соеди-
нений, в частности, сахаров, из простейших соединений при включении катализаторов или
автокатализаторов. При этом стадии автокаталитического синтеза должен предшествовать
(или как-то сочетаться) этап разрушительного грубого воздействия, в которых активизиру-
ются все возможные пути синтезов и появляются, не взирая на издержки, качественно новые
соединения в минимальных количествах. Но из этих количеств на относительно больших
временах в автокаталитических реакциях нарабатываются уже основная масса этих веществ
с формированием, так сказать, хемоценоза. По-видимому, это некий общий принцип услож-
нения и самоорганизации химической системы.
Еще один вопрос, который часто задается А.Ю. Розановым и Н.Л. Добрецовым, о тем-
пературе на поверхности примитивной Земли. Этот вопрос, если на него посмотреть шире, о
температуре примитивных тел при их динамике и слипании в зародыши планет. Для ответа
на этот вопрос в рамках нашей гипотезы об астрокатализе мы должны включить в наши
математические модели неупругие столкновения тел. Эти столкновения особенно важны на
последних десятках млн. лет из 60 млн. лет существования протопланетного диска. Переход
к модели Сафонова-Шмидта роя тел с крупными зародышами планет меняет используемую
92
Проблемы происхождения жизни
физическую модель. Учет столкновений со слипанием математически осуществляется запи-
сью интеграла столкновений в правую часть уравнения Больцмана для функции распреде-
ления тел по координатам, скоростям и теперь еще и по массам. Температура нагрева тел пря-
мо связана с энергией, которая может выделиться при неупругом столкновении двух тел. Эта
энергия рассчитывается достаточно точно из закона сохранения энергии и при отнесении на
массу нового объединенного тела является тепловым эффектом. Теплопроводность на границе
тела-газ и теплопотери излучением не учитываются. В расчете взято 1 млн. частиц, а ядро
столкновений (коагуляции) в интеграле увеличено по абсолютной величине, чтобы не считать
эти 60 млн. оборотов. Это можно сделать, так как ядро, связанное с характерным временем,
входит в интеграл линейно. Что получилось в результате? Тела при своей динамике сохраняют
круговые траектории. Картина динамики тел мало отличается от многих других работ в этой
области. Интерес представляют следующие графики рис. 11. Эти графики характеризуют ди-
намику нагрева пылевой компоненты протопланетного диска за счет неупругих столкновений.
По оси ординат отложена энергия, запасенная в теле, в зависимости от массы тела для трех мо-
ментов времени. На график выводилась только представительная выборка, характеризующая
функцию распределения частиц по массам. На ранней стадии большое количество мелких тел
поглотило много энергии, и тела были нагреты до высоких температур. Они могли плавиться
в результате удара. На более поздней стадии появились тела со значительно большей массой,
но они не обладают высокой температурой. Массивные тела не накапливают энергии. Но на
этих временах продолжает оставаться много укрупнившихся тел, которые несут высокую
энергию. На больших временах (рис. 11, третий график) количество относительно небольших
нагретых тел сокращается, в основном остались холодные тела. Среди крупных массивных
тел нет нагретых. При этом при моделировании всегда остается небольшое количество нагре-
тых некрупных тел. Вывод из этих данных такой. Крупные тела не проходили стадию своего
общего разогрева за счет бомбардировки. Но многие метеориты могли расплавляться. Исполь-
зование таких метеоритов как определителей температур на начальных временах для планет и
других крупных тел может привести к ошибочным выводам. Должен подчеркнуть, что вывод
основан на использовании в расчетах лишь закона сохранения энергии. И, скорее всего, этот
вывод сложно изменить. Таким образом, поверхность Земли как и других планет при фор-
мировании не была расплавленной. На ней отсутствовали высокие температуры от аккреции
других первичных тел, несмотря на то, что некоторые метеоритные данные могут продемон-
стрировать высокие значения температуры с плавлением материала. Поэтому на поверхности
Земли с начала ее формирования существовали условия для развития биосферы.
Итак - наша основная гипотеза астрокатализа, идея которой возникла довольно давно,
в начале 1990-х гг. Первое сообщение о ней на конференции было сделано в 1996 г. (Snytnikov
et al., 1996), а основательные публикации вышли в 2007 г. (Снытников, 2007а, б). Гипотеза
состоит в том, что абиогенный каталитический синтез органических соединений проходил
непосредственно при формировании первичных тел и протопланет при развитии гравита-
ционной коллективной неустойчивости с одновременным объединением многих малых тел.
Или, кратко, катализ сформировал планеты. И основные выводы по нашей работе. Если
по гипотезе Опарина-Холдейна синтез органических соединений произошел на поверхности
Земли, то сейчас стало ясно, что химическая предбиологическая эволюция прошла в допла-
нетной околозвездной среде в первый миллион лет. Эта та цифра, которую хотел услышать
от меня А.Ю. Розанов. Зародыши планет формировались в местах каталитического синтеза
органических соединений благодаря гравитационным неустойчивостям.
В последовательности этапов возникновения жизни неорганическое вещество с актив-
ными каталитическими центрами увеличилось в своих размерах от наночастиц до тел в ты-
сячи километров. Но при этом каталитические неорганические материалы последовательно
уступали свое место катализаторам из органических соединений, вплоть до белков и фер-
93
В Н. Снытников. Астрокатализ - абиогенный синтез и химическая эволюция...
Т=10 оборотов
масса частицы
Рис. 11. Тепловая энергия укрупняющихся тел в зависимости от их массы для трех моментов времени.
Увеличенное ядро коагуляции в интеграле столкновений.
ментов. Неорганическое вещество меняло свою роль и влияние. Место и роль конкретных
элементов, формировавших активные каталитические центры, в возникновении жизни, по-
видимому, могут быть определены в настоящее время по их воздействию на внутриклеточ-
ные реакции в прокариотах и по способности ряда микроорганизмов избирательно концен-
трировать многие элементы.
Здесь мне необходимо отметить высказанное ранее предположение (Войткевич, 1979,
с. 125), что «жизнь на Земле существует столько, сколько существует и сама Земля». Идея,
что «химическая эволюция на предпланетной стадии в Космосе и протопланетном обла-
ке заходит так далеко, что жизнь возникает практически сразу после сформирования зем-
ноподобной планеты», была отмечена как «наиболее радикальная точка зрения» (Цицин,
94
Проблемы происхож.п-.ния жиши
Время, лет О
ПротоСолнце
с
Взрыэ Протозвезда-
Сверхновой кокон
4.56 млрд.лет
назад
106
аккреционным диском
ю5
6-107 6.6-1O0
Солнце Планеты Начало
геологической
летописи Земли,
3.9 млрд, лет назад
Рис. I2. Происхождение жизни как процесс самоорганизации.
1980). Она почти 30 лет оставалась вне основного внимания исследователей, по-видимому,
из-за недостаточного объема естественнонаучных данных, в частности, астрофизических
и космохимических, по процессам в «протопланетных облаках».
Итак, происхождение жизни может быть представлено следующими основными эта-
пами (рис. I2). Плотное молекулярное облако с его туннельными реакциями «холодной
предыстории жизни» (Гольданский, I997) предоставляет исходные вещества для образова-
ния звезд вместе с их протопланетными дисками. Это первый, физический этап. На втором
этапе в околозвездных дисках из высокоактивных веществ синтезировались органические
соединения с участием всевозможных по своим свойствам наночастиц. Подчеркну еще раз,
что именно в наночастицах состава космической распространенности элементов было пред-
ставлено все наше земное вещество, вещество планет и тяжелые элементы Солнца. Синтез
проходил с преимущественным сохранением в диске высокомолекулярных соединений. Но
только если они формировали крупные первичные тела. Обратно, отбор происходил и среди
катализаторов по способности синтезировать именно высокомолекулярные соединения. Все
низкомолекулярное и легкое вещество потоком водорода и гелия уносилось на протосолн-
це в термоядерную топку, источник энергии для «химического завода». На этом этапе, по-
видимому, появились нуклеотиды, сахара, липиды, белки, тяжелые углеводороды, азотистые
основания. Здесь было представлено все, что могло быть синтезировано абиогенным путем.
На этом промежутке времени был сформирован первичный «допланетный хемоценоз». Его
состав менялся во времени и в зависимости от расстояния от протозвезды. Большое влияние
на состав оказывали температуры плавления, испарения и тому подобные свойства соеди-
нений. Общую эффективность синтезов на стадии допланетного хемоценоза по отношению
к выходу сложных соединений из первичной органики, скорее всего, следует оставить
в 0.01 %. Первичная масса вместе с водородом и гелием - масса Солнца. Характерные време-
на существования этих синтезов - сотни тысяч лет. По-видимому, кометы могут нести нам
информацию об этапе «допланетного хемоценоза» на своих расстояниях от Солнца.
Следующий этап - формирование из первичных крупных тел зародышей планет. Это
происходит при высоких давлениях в сгустках, в этих обнаруженных нами реакторах с кипя-
щими метровыми зернами катализатора. На этом этапе среди всех возникающих реакторов
есть такие, в которых происходит переход от «допланетного хемоценоза» к «миру РНК».
95
В.Н. Спытников. Астрокатализ - абиогенный синтез и химическая эволюция...
«Мир РНК» может появиться в умеренных по температурам условиях и, следовательно,
на вполне определенных удалениях от протозвезды. Умеренные температуры - это диапазон
от замерзания воды до ее испарения. И в этих условиях периодические увлажнение и высы-
хание поверхности тел становятся просто неизбежными. Отбор среди реакторов по темпе-
ратурным условиям, по активности катализаторов для дальнейшего усложнения органиче-
ских молекул (по способности их полимеризовать), образовать километровые планетезимали
и зародыши планет, а вместе с тем и перейти к «миру РНК», также можно оценить в 0.01 %
по массе вещества. Этот этап самый быстрый, масштаба нескольких лет для одного реактора
уединенной волны, иными словами, самогравитирующего сгустка вещества. Помимо сгуст-
ков, вещества в околозвездном диске еще много, не на одну планету. Вне сгустков, реакторов
уединенной волны, «допланетный хемоценоз» эволюционирует в сторону преимущественно
деструкции органических соединений, возможное свидетельство о которой и, возможно, об ис-
ходном составе первичного вещества находится в углистых хондритах. Методически, с точки
зрения разделения труда, на этом этапе мы передаем свои данные по абиогенному синтезу ис-
следователям «мира РНК». В этом месте у нас возникает совпадение с данными А.С. Спирина
о необходимости интенсивного обмена тел с периодически высыхающей поверхностью. Воз-
можные свидетельства этого этапа, кроме углистых хондритов, могут находиться в наименее
плотных астероидах и спутниках планет (Маров и др., 2008; Адушкин и др., 2008).
Переход от «мира РНК» к сообществу простейших одноклеточных организмов нами
не рассматривается. Но я возьму на себя смелость высказать, с использованием звучавшей
в зале на совещании терминологии, «экстремистское» или, как говорили 30 лет назад, «ра-
дикальное» предположение. Оно состоит в том, что в какой-то момент времени из-за своей
конденсации жидкая вода стала постоянно присутствовать на поверхности некоторых пла-
нетезималей и протопланет. И именно жидкая вода стала той причиной, по которой из всего
богатства «мира РНК» оказались отобраны те соединения, которые в своей совокупности,
сформировав оболочку - мембрану, смогли противостоять такому крайне разрушительно-
му реагенту как вода. Все остальное богатство «мира РНК» погибло. А вот что осталось,
то получило толчок к дальнейшему формированию сообществ одноклеточных организмов
(биоценозов), способных использовать воду и выживать в новых катастрофических и других
изменениях среды обитания с капельно-жидкой водой. Все отходы оказались отправлены
в космос, Юпитер и Сатурн как «в третьи и развивающиеся страны», и в топку на Солнце.
И если прав А.Ю. Розанов (Герасименко и др., 1999) относительно биоморфных микрострук-
тур в метеоритах с возрастом 4.56 млрд, лет, то первичный биоценоз возник и существовал
именно на допланетной стадии. Этот биоценоз перешел к «планете бактерий», естественно,
с большими потерями органического вещества, большая часть которого подверглась де-
струкции, а что-то оказалось погребено в земных толщах.
Переход от первичного биоценоза к «планете бактерий» прошел в следующие два этапа,
которые идентифицируются по биогеохимическим циклам в географической оболочке Земли
(Заварзин, 2001). На первом этапе основная первичная биомасса подверглась деструкции:
(CH2O),06(NH3)16H3PO4 -Э 53 СН4 + 53 СО2 + 8 N2 + 24 Н2 + [Н3РО4] .
Легкие компоненты - Н2, Не, СН4 - Земля утратила. На сегодня существуют доводы
полагать, что атмосферный азот является продуктом разложения первичной биомассы. Тогда
массовое содержание азота в атмосфере к концу этого этапа стало близким к современному
значению 4*1018 кг. Отсюда первичная земная биомасса может быть оценена в 2.4х1019 кг по
углероду. Выделившееся 1.2*1019 кг СО2 и абиогенные СО2 связались в древнем «содовом оке-
ане» с отложением свыше 6*1019 кг карбонатов (Заварзин, 2006). Значит, масса абиогенного
СО2 в древнейшей атмосфере примерно 4.8*1019 кг по углероду, а давление в ней оценивается
по порядку величины в 10 атмосфер. На этом же этапе осадочные породы закрыли древней-
шие породы с возрастом в 4.56 млрд, лет, возможно, с погружением последних вместе с оса-
96
Проблемы происхождения жизни
дочными породами в мантию, метаморфизацией и вновь выносом на поверхность. Отметим,
что на Земле современное количество воды 1.3><1021 кг. Не меньшее количество содержалось
и в палеоокеане. При отсутствии воды на Земле объемное отношение СО2 и N2 в атмосфере
было бы свыше 20, как на Венере и Марсе.
Атмосферы Венеры и Марса имеют главными компонентами 95 % СО2 и 3-5 % N2
по объемному содержанию. Предполагая, что содержание азота в атмосфере этих планет
связано, как и для Земли, с разложением первичной биомассы с отношением элементов
С : N = 6, получим, что свыше 60 % СО2 в их атмосферах является абиогенным. Другие ком-
поненты - Н,, Не, СН4, Н2О - потеряны этими планетами. Вода в виде льда на прото-Марсе
при его низкой температуре не обеспечила сток СО, в карбонаты. На прото-Венере связыва-
ния СО2 не произошло из-за ее высокой температуры и воды в виде пара. Отсутствие воды
в жидком виде не запустило на этих планетах важнейший геохимический процесс - углекис-
лотное выщелачивание, связывание углекислоты в карбонаты и появление осадочных пород
и, вместе с тем, питания микроорганизмов, что привело к исчезновению органического веще-
ства на поверхности Марса и Венеры.
На последнем из рассматриваемых этапов становления «планеты бактерий» примерно
2 млрд, лет назад произошло формирование окислительной атмосферы (Розанов, 2008, с. 49).
Первичная продукция образовывалась в реакциях фотосинтеза
СО2 + Н2О -» [СН2О] + О2,
которая не была сбалансирована с деструкцией в биогеохимическом цикле. Это привело
к накоплению избыточной мортмассы, которая в настоящее время представлена в основном
керогеном в 1.5*1О18 кг - рассеянным углеродом осадочных пород. Масса керогена пример-
но соответствует накопившемуся кислороду атмосферы в 1><1O’S кг. Массу самой биосферы
на этом этапе определить очень сложно. Поэтому оценим совместную эффективность двух
последних этапов самоорганизации. Тогда для этапа деструкции первичной биомассы с вы-
ходом на фотосинтезирующий биоценоз и переходом к «планете бактерий» с ее современной
массой 2х1015 кг получается эффективность 2х]015 кг / 2.4хЮ19 кг ~ 0.01 %, что совпадает
с обычной эффективностью двух этапов самоорганизации. Итак, на рубеже примерно 2 млрд,
лет масса биосферы достигла современного значения, и появилась «планета бактерий».
Работа выполнена по программе Президиума РАН № 18-2 по направлению академика
В.Н. Пармона, которому автор приносит искреннюю благодарность за поддержку и обсужде-
ния исследований по астрокатализу. Автор выражает признательность сотрудникам иссле-
довательского коллектива СО РАН, в частности, Института катализа им. Г.К. Борескова, где
в основном была выполнена эта работа. Многие идеи обсуждались на встречах у академика
А.Ю. Розанова в дискуссиях с академиками Г.А. Заварзиным, А.С. Спириным, А.И. Григо-
рьевым, Н.С. Кардашевым, Н.А. Колчановым, С.В. Шестаковым, которым автор признателен
за критику и полезные замечания.
СПИСОК ЛИТЕРАТУРЫ
Адушкин В.В., Витязев А.В., Печерникова ГВ. 2008. В развитие теории происхождения и ранней эво-
люции Земли // Проблемы зарождения и эволюции биосферы / Ред. Э.М. Галимов. М.: Либро-
ком. С. 275-296.
Биологический энциклопедический словарь. 2003 / Ред. М.С. Гиляров. М.: Большая Российская энци-
клопедия. 864 с.
Войткевич ГВ. 1979. Химическая эволюция солнечной системы. М.: Наука. 240 с.
Галимов Э.М. 2008. Концепция устойчивого упорядочения и АТФ-зависимый механизм происхожде-
ния жизни // Проблемы зарождения и эволюции биосферы / Ред. Э.М. Галимов. М.: Либроком.
С. 23-32.
97
В.Н. Снытников. Астрокатализ - абиогенный синтез и химическая эволюция...
Герасименко Л. М., Жегалло Е.А., Жмур С.И., Розанов А.Ю., Хувер Р. 1999. Бактериальная палеонтоло-
гия и исследования углистых хондритов // Палеонтол. журн. № 4. С. 103-125.
Гольданский В.И. 1997. Нетрадиционные механизмы внеземного образования органических соедине-
ний // Изв. АН. Сер. хим. № 3. С. 409-417.
Журн. ВХО. 1980. Т. 25. № 3-4.
Журн. ВХО. 1980. Т. 25. № 5.
Заварзин Г.А. 2001. Становление биосферы // Вести. РАН. Т. 71. № И. С. 988-1001.
Заварзин Г.А. 2006. Алкалофильное микробное сообщество как аналог наземной биоты протерозоя И Эволю-
ция биосферы и разнообразия. К 70-летию А.Ю. Розанова. М.: Т-во научных изданий КМК. С. 97-119.
Заварзин Г.А. 2008. Планета бактерий // Вести. РАН. Т. 78. № 4. С. 328-336.
Климентович Ю.Л. 1990. Турбулентное движение и структура хаоса. М.: Наука. 320 с.
Кукшева Э.А., Снытников В.Н. 2007. Параллельный алгоритм и программа Key7D для решения неста-
ционарных трехмерных задач гравитационной физики // Вычислительные технологии. Т. 12.
№ I. С. 35-44.
Маров М.Я., Колесниченко А.В., Макалкин А.Б. и др. 2008. От протосолнечного облака к планетной си-
стеме: модель эволюции газопылевого диска И Проблемы зарождения и эволюции биосферы /
Ред. Э.М. Галимов. М.: Либроком. С. 223-273.
Меклер Л.Б. 1980. О происхождении живых клеток: эволюция биологически значимых молекул - пе-
реход химической эволюции в биологическую. Новый подход к проблеме И Журн. ВХО. Т. 25.
№ 4. С. 460-473.
Мухин Л.М. 1980. Химическая эволюция И Журн. ВХО. Т. 25. № 4. С. 412-417.
Пармон В.Н. 2002. Пребиотическая фаза зарождения жизни // Вестн. РАН. Т. 72. № 11. С. 976-983.
Пригожин И. 1985. От существующего к возникающему. М.: Наука. 328 с.
Розанов А.Ю. 1996. Цианобактерии и, возможно, низшие грибы в метеоритах // Соросовский образо-
вательный журнал. № 11. С. 61-65.
Розанов А.Ю. 2008. Происхождение и эволюция биосферы: важнейшие результаты 2004-2007 гг. /
Отв. ред. Н.Л. Добрецов. Новосибирск: Изд-во СО РАН. С. 49.
Руденко А.И. 1980. Эволюционная химия и естественноисторический подход к проблеме происхожде-
ния жизни // Журн. ВХО. Т. 25. № 4. С. 390-404.
Симонов А.Н, Матвиенко Л.Г, Пестунова О.П. и др. 2007а. Селективный синтез эритрулозы и 3-пен-
тулозы из формальдегида и дигидроксиацетона, катализируемый фосфатами в нейтральной
водной среде // Кинетика и катализ. Т. 48. № 4. С. 550-555.
Симонов А.Н, Пестунова О.П., Матвиенко Л.Г, Пармон В.Н. 20076. Природа возникновения автоката-
лиза в реакции Бутлерова И Кинетика и катализ. Т. 48. № 2. С. 261-270.
Слинько М.Г 1980. Химические процессы и реакторы // Журн. ВХО. Т. 25. № 5. С. 531-536.
Снытников В.Н. 2007а. Абиогенный синтез пребиотического вещества для биосферы Земли как ста-
дия самоорганизации на астрофизической и палеонтологической шкале времени // Палеонтол.
журн. № 5. С. 1-8.
Снытников В.Н. 20076. Абиогенный допланетный синтез пребиотического вещества // Вестн. РАН.
Т. 77. № 3. С. 218-226.
Снытников В.Н, Вшивков В.А., Кукгиева Э.А. и др. 2004. Трехмерное численное моделирование нестацио-
нарной гравитирующей системы многих тел с газом // Письма в астрономический журнал. № 2.
С. 146-160.
Снытников В.Н, Мищенко Т.И., Снытников Вл.Н. и др. 2009. Автокаталитическое газофазное деги-
дрирование этана в «бесстеночном» реакторе И Кинетика и катализ (в печати).
Трофимов Б.А., Малькина А.Г, Шемякина О. А. и др. 2005. Цианацетилен и его производные // Журн.
орган, химии. Т. 41. № 6. С. 903-909.
98
Проблемы происхождения жизни
Федонкин М.А. 2008. Роль водорода и металлов в становлении и эволюции метаболических систем //
Проблемы зарождения и эволюции биосферы / Ред. Э.М. Галимов. М.: Либроком. С. 417-438.
Хасин А.А., Снытников В.Н. 2005. Особенности состава продуктов синтеза Фишера-Тропша на мате-
риале метеорита Царёв // Тр. Междунар. семинара «Происхождение и эволюция биосферы».
26-29 июня 2005 г. Новосибирск: Институт катализа СО РАН. С. 158-159.
Цицин Ф.А. 1980. Космические факторы возникновения и развития жизни и разума во Вселенной //
Журн. ВХО. Т. 25. № 4. С. 435-439.
Эбелинг В., Энгель А., Файстель Р. 2001. Физика процессов эволюции. М.: УРСС. 326 с.
Dobretsov N.L., Kolchanov N. A., Suslov V.V. 2008. On important stages of geosphere and biosphere evolution//
Biosphere origin and evolution / Eds. N. Dobretsov, N. Kolchanov, A. Rozanov, G. Zavarzin. Springer.
P. 3-23.
Pestunova O.P., Simonov A.S., Snytnikov V.N. et al. 2005. Putative mechanism of the sugar formation on
prebiotic earth initiated by UV-radiation // Advances Space Res. V. 36. № 2. P. 214-219.
Pestunova O.P., Simonov A.N., Snytnikov V.N., Parmon V.N. 2008. Prebiotic carbohydrates and their deri-
vates // Biosphere origin and evolution / Eds. N. Dobretsov, N. Kolchanov, A. Rozanov, G. Zavarzin.
Springer. P. 103-118.
Snytnikov V.N., Kuksheva E.A. 2007. Studing instability of 3D collisionless systems on stochastic traejecto-
ries // Collective phenomena in macroscopic systems / Eds. G. Bertin, R. Pozzolli. World Sci. Publ. Co
Inc. P. 280-285.
Snytnikov V.N., Snytnikov Vl.N, Dubov D.A. et al. 2007. Production of nanomaterials by vaporizing ceramic
targets irradiated by a moderate-power continuous-wave CO2 laser // J. Applied Mechanics and Techni-
cal Physics. V. 48. № 2. P. 292-302.
Snytnikov V.N., Vshivkov V.A., Parmon V.N. 1996. Mathematical modeling of non-stationary physico-chemical
processes in natural protoplanetary catalytic reactor// 13 Int. Con. on Chemical Reactors. Novosibirsk,
June 18-21, 1996. Abstracts. Pt II. P. 226-227.
Astrocatalysis - abiogenic synthesis and chemical evolution
on pre-geologic stages of Earth formation
V.N. Snytnikov
The Earth atmosphere was given rise to by the matter self-organization after the formation
of the Solar System. Studies of self-organization on the basis of the available natural science data
allow main stages of the chemical evolution of the biosphere to be revealed; these are cold prehis-
tory of life in dense molecular clouds, pre-planetary chemocenosis, RNA world in the circumsolar
disk, and primary biocenoses of protocells (life) on protoplanetary bodies. The estimated mass of
the primary biosphere on the early Earth surface was 2.41019 kg expressed as carbon. In 2.5 billion
years, destruction of the primary Earth biomass led to the bacteria planet with 2.01015 kg of biota in
Proterozoic with oxygen atmosphere. Main parameters (pressure, temperature, state of the catalytic
solid phase), which were characteristic of these stages of life origin, are established. The tremendous
scale of abiogenic synthesis of prebiotic matter is shown to involve practically each atom of all the
elements of nanoparticles-catalysts. Selection of catalytically active nanoparticles was based on their
capability of synthesizing high-molecular compounds in the protoplanetary disk. Autocatalysts took
part in the protoplanetary chemical evolution beginning from such simple compounds as ethylene
and glycol aldehyde. Primary synthesis of autocatalysts was initiated by external high-energy im-
pact, for example under the action of ultraviolet radiation.
99
Дискуссия
по пленарному докладу В.Н. Снытникова
«АСТРОКАТАЛИЗ - АБИОГЕННЫЙ СИНТЕЗ И ХИМИЧЕСКАЯ ЭВОЛЮЦИЯ
НА ДОГЕОЛОГИЧЕСКИХ ЭТАПАХ ФОРМИРОВАНИЯ ЗЕМЛИ»
Председательствующий А.Ю. Розанов
В.А. Царёв: Мне кажется очень привлекательной идея космического происхождения
жизни. Вы в качестве исходного объекта принимаете газово-пылевое облако, в котором есть
гранулы и прочее.
В.Н.С.: Исходным считается протопланетный аккреционный диск на стадии примерно
от полумиллиона до миллиона лет, когда протозвезда еще набирает массу.
В.А. Царёв: Мое замечание сводится к следующему. Надо начинать с облака, оно об-
ладает гравитационной устойчивостью, чтобы оно превратилось в диск, его надо встряхнуть.
Такая встряска достигается при взрыве сверхновой в относительной близости. Мне кажется,
стоит привлечь эту стадию в Вашу модель, потому что Вы одновременно получите еще одну
вещь - при взрыве сверхновой испускается большое число нейтронов, нейтроны распадают-
ся, возникает поток поляризованных электронов, которые облучают протосолнечное облако,
действуя вместе с ударной волной, тем самым можно получить хиральную асимметрию пер-
вичной органики.
В.Н.С.: Вот что здесь мне не совсем нравится. Характерные времена в наших моде-
лях связаны с временами протекания химических реакций, это очень короткие времена (от
нескольких часов!) по сравнению с теми интервалами, которыми оперируют большинство
астрофизиков. Это времена нескольких десятков оборотов. За это время проходится стадия
от размеров частиц порядка сантиметра до тел величиной несколько километров и более.
Времена у нас очень маленькие.
В.А. Царёв: Но радиационный фактор действует практически мгновенно, электроны
летят со скоростью света.
В.Н.С.: Да, но времени-то прошло уже почти миллион лет с момента взрыва сверхно-
вой, который дал толчок для формирования Солнечной системы.
В.А. Царёв: Тогда есть вторая возможность - это облако, в котором уже есть органика,
и оно готово распасться на глыбы, оно еще облучается взрывом другой сверхновой - они же
происходят время от времени в Галактике. Иначе не решается вопрос с хиральностью.
В.Н.С.: Может быть...
Н.Л. Добрецов: Вашу модель нужно верифицировать. Верификация всех таких моделей
проводится на материале метеоритов. Пытались ли Вы как-то это сделать? По метеоритам
получается, что в центре облака был чистый газ, по периферии - только пыль, в пограничной
области - смесь пыли и газа. Ваша модель должна соответствовать данным по метеоритам.
Ничего другого, кроме метеоритов, у нас нет.
В.Н.С.: То, о чем Вы говорите - это одна из более поздних стадий существования прото-
планетного диска. Стадия, которую мы рассматриваем - примерно от первого полумиллиона
лет до одного миллиона лет. В это время есть еще и газ, и твердое вещество, они вместе, газ
является носителем, который несет твердое вещество в центр. Это все - еще до метеоритов.
Н.Л. Добрецов: Бомбардировка космическими телами древней Земли продолжалась
около 600 миллионов лет, Луна образовалась от удара гигантского метеорита, часть Земли
была уничтожена, там ничего первичного не осталось. Вот когда бомбардировка кончилась
четыре миллиарда лет назад, тогда и жизнь зародилась. Затем, Вы не объясняете возникно-
100
Проблемы происхождения жизни
вение полинуклеотидов, а более примитивная органика не так существенна. Поэтому Ваша
модель мне кажется настолько упрощенной, что ее трудно куда-либо применить.
В.Н.С.: Я все время подчеркиваю, что ее можно применить только там, где она приме-
нима - на стадии, когда происходит накопление органического вещества.
Н.Л. Добрецов: Но как ее можно верифицировать, где найти остатки того, что в это
время образовалось? Вы говорите, метеориты не подходят. Между тем, 99.9 процентов метео-
ритов имеет один возраст - 4.56 миллиарда лет.
В.Н.С.: Правильно, он более поздний. Что касается верификации этой модели. Есть
огромное количество астрофизических данных - исследование облаков, зарождающихся
протозвезд с их дисками, на основе которых можно верифицировать эти модели. Главное -
все синтезы произошли в первый миллион лет.
Н.Л. Добрецов: Но почему удлинились нуклеотидные цепочки? Кирпичики - добио-
логический синтез - могли возникнуть, а синтез длинных цепочек стал возможен лишь на
более поздней стадии, когда появилась вода.
В.Н.С.: Мое утверждение таково - все синтезы произошли быстро.
А.С. Спирин: Без воды не могло быть РНК и ее структур. Никаких молекул нуклеино-
вых кислот без воды нельзя сделать. Мир РНК в Вашей модели оказался до воды.
В.Н.С.: Я не утверждал, что не было воды до мира РНК. Я сказал, что переход от мира
РНК к планете бактерий произошел не тогда, когда появилось немного воды (сухо/чуть влаж-
но), а тогда, когда ее стало огромное количество.
А.С. Спирин: Для мира РНК нужно столько же воды, сколько и для бактерий, а то и
больше.
В.Н.С.: Можно полагать, что вода была.
А.С. Спирин: Есть еще один момент. Когда Вы рисуете свой “химический завод”, это
никакого отношения к периоду мира РНК не имеет, потому что там выходы - не одна сотая
процента, а девяносто процентов, начиная с появления рибозимов и так далее. Отходов таких
нет. Когда начинается биологический синтез, выход повышается до девяноста процентов.
В.Н.С.: Поэтому я поставил там вопросы. Я обязательно внесу коррективы.
А.Б. Макалкин: Соотношение углерода в кометах к таковому даже в самых богатых
углистых хондритах - пять-десять к одному, то есть метеориты действительно недополучили
углерод. Если бы роль органики была так велика, это было бы очень важно для космогони-
ческих моделей образования планет, так как там существует проблема быстрого перехода от
сантиметровых размеров частиц к размерам планетезималей. В этом переходном интервале
размеров уже не работает механизм физического слипания и еще не работает самогравитация
планетезималей. Добавление значительного количества органики было бы большим подспо-
рьем для теории образования планет в преодолении этого барьера из-за повышенной адгези-
онной способности ряда природных органических соединений. Но имеются сомнения в связи
с относительно малым содержанием органики в метеоритах по сравнению с кометами.
А.Ю. Розанов (председательствующий): С углистыми хондритами не все так просто.
А.Б. Макалкин: Да, летучая органика в метеоритах могла быть потеряна на более позд-
них стадиях, во времена ударов крупных тел.
В.Н.С.: Это совсем другие времена и совершенно другие процессы.
А.В. Витязев: Какая часть газа собралась в звезды? Должно быть менее одного про-
цента.
В.Н.С.: Согласен.
101
Проблемы происхождения жизни. М.: ПИН РАН, 2009. С. 102-119
МОЛЕКУЛЫ И ИХ МИГРАЦИЯ ВО ВСЕЛЕННОЙ
© 2009 г. Н.Г. Бочкарёв
Государственный астрономический институт им. П.К. Штернберга МГУ
boch@sai.msu.ru
В статье кратко описан набор молекул, наблюдаемых в различных астроно-
мических объектах (исключая планеты Солнечной системы и их спутники),
главным образом, в межзвездной среде. Рассмотрены различные механиз-
мы пространственной миграции молекул с переносом молекулярных соеди-
нений от одной планетной (протопланетной) системы к другой. Показано,
что за космологическое время перенос молекул возможен на расстояния до
100 млн. световых лет. Кратко обсуждена гипотеза Ф. Хойла и Ч. Викрама-
синга о биологической природе некоторых межзвездных пылинок.
ВВЕДЕНИЕ
Астрономы обычно рассматривают научные проблемы с самых широких позиций.
Проблема происхождения жизни - не исключение. Оказалось, что многие особенности стро-
ения Вселенной и физических законов, которые в ней реализуются, могут быть объяснены
так называемым «антропным принципом», согласно которому мы видим Вселенную именно
такой, какая она есть, потому что будь она другой, ее, возможно, некому было бы видеть.
И это не просто слова, а достаточно подробные исследования того, каковы должны быть
физические законы, чтобы смогли сформироваться и быть устойчивыми атомы, молекулы,
звезды и другие структуры во Вселенной, необходимые для появления и развития жизни
[см., напр., книгу И.Л. Розенталя (1984)]. Для этого, например, нужно, чтобы пространство
было трехмерным, а время одномерным. Жизнь на углеродной основе была бы невозмож-
на, если бы энергия резонанса ядерной реакции образования углерода путем слияния трех
альфа-частиц (3*4Не—>|2С) отличалась от существующего значения всего на миллиардные
доли своей величины. В этом случае в природе практически не было бы углерода.
В междисциплинарной кооперации с самых широких позиций обсуждаются воз-
можности существования иных форм жизни, вопросы о том, на каких астрономических
объектах могут быть условия, подходящие для жизни вообще и для образования высших
форм жизни. Рассматриваются возможные пути эволюции внеземных цивилизаций, их по-
иска (SETI - Search for Extraterrestrial Intelligence) и связи с ними (CETI - Communication
with Extraterrestrial Intelligence), см. монографии И.С. Шкловского (1987), Д. Голдсмита
и Т. Оуэна (1983), Л.М. Гиндилиса (2004). Для обсуждения этого круга вопросов в Между-
народном астрономическом союзе, объединяющем около половины всех профессиональных
102
астрономов мира, была создана комиссия № 51 по биоастрономии. В мире работают несколь-
ко научных учреждений этого профиля. Однако в целом эта тематика пока составляет очень
малую часть астрономических исследований.
Совещание «Проблемы происхождения жизни» рассматривало более узкую проблему -
происхождение земной жизни. Успехи палеонтологии показывают, что одноклеточные организ-
мы появились на Земле около 4 млрд, лет назад, по-видимому, в течение всего ~100 млн. лет, что
может оказаться недостаточным для возникновения столь сложных образований. Это привело
к возрождению интереса к гипотезе панспермии - занесения на Землю жизни из космоса. Хотя
эта гипотеза не решает проблемы происхождения жизни, она облегчает ее, увеличивая количе-
ство потенциальных мест зарождения жизни во много раз. Из каких мест и как возможен пере-
нос жизни - это вопрос в основном к астрономам. В настоящее время еще невозможно делать
надежные оценки и приходится ограничиваться в основном качественными суждениями.
Оргкомитет совещания просил рассказать о молекулах в космосе и возможных путях
миграции сложных молекул во Вселенной. Ниже эта проблема освещена максимально широ-
ко, хотя, возможно, не все излагаемые вопросы будут немедленно востребованы участниками
совещания. Мы не будем касаться образования и миграции молекул в пределах Солнечной
системы, в том числе во время ее формирования из протопланетного диска, так как эти вопро-
сы освещены в других докладах.
1. МОЛЕКУЛЫ В КОСМОСЕ
Космические объекты, в которых обнаружены или могут существовать молекулы
В табл. 1 приведен набор космических объектов и типы среды, в которых изучаются
молекулы. Знаком «+» указано, что молекулы изучаются (наблюдаются или хотя бы пред-
сказаны теоретически); «(+)» - можно ожидать молекулы. Здесь границы фаз - это преиму-
щественно поверхности пылевых частиц, а также границы между твердой (или жидкой) по-
верхностью небесного тела и его атмосферой. Твердое тело - тело планеты или иного косми-
ческого объекта, включая кору нейтронных звезд, а также пылевые частицы.
Жидкая фаза известна лишь на Земле и нескольких спутниках планет. Марс также
хранит следы жидкой фазы, но там она, по-видимому, исчезла, по крайней мере, 2-3 млрд,
лет назад. Не исключена возможность того, что жидкая вода могла или даже может сейчас
присутствовать во внутренних частях наиболее крупных кометных ядер (Цицин, Чепурова,
2003), а также ледяных астероидов, найденных во внешнем поясе астероидов - поясе Койпе-
ра. Предполагается, источником нагрева служит распад радиоактивных изотопов.
За пределами Земли, ее атмосферы, Луны, а также метеоритов пока не удалось ото-
ждествить молекулы, содержащие более 13 атомов, хотя имеются косвенные признаки су-
ществования более сложных соединений, содержащих примерно от 20 до 60, а возможно,
и больше атомов.
Таблица 1. Объекты и типы среды, в которых изучаются молекулы (Бочкарёв, 2006)
Типы среды Планеты н их спутники Кометы; межпланет- ная среда Солнце холоди. звезды (G,K,M...BD) Нейтрон- ные звезды Околозв. обол.: М, Of,WR,N,SN МЗС Межгалак- тическая среда AGN OSO Ранняя Вселен- ная
Плазма 4- 4- 4- — (+) 4- (+) (+) 4-
Нейтр. газ + 4- ± — 4- + 4- 4- 4-
Граница фаз + (+) — (+) (+) + (+) (-) —
Жидкость 4- - — — - — — — —
Твердое тело 4- 4- - (+) 4- 4- (+) 4- (-)
BD - коричневые карлики; МЗС - межзвездная среда; AGN, QSO - активные ядра галактик, квазары;
WR - звезды Вольфа-Райе; N - новые звезды; SN - сверхновые звезды; Of- горячие звезды высокой светимости
с сильным истечением вещества (звездным ветром); G, К, М - спектральные классы звезд.
103
Н.Г Бочкарёв. Молекулы и их миграция во Вселенной
Где можно ожидать присутствие сложных молекул?
Ниже перечислены виды объектов, на которых обнаружено или может ожидаться при-
сутствие сложных молекул (в том числе углеродосодержащих).
• Планеты и их спутники, имеющие атмосферу;
• Ядра комет и ледяные астероиды;
• Астероиды и метеоритные тела типа углистых хондритов;
• Протозвездные, околозвездные и протопланетные диски;
• Околозвездные оболочки вокруг холодных звезд с интенсивным истечением
вещества;
• Газовая и конденсированная (пылевая) компоненты молекулярных облаков;
• Межзвездные пылинки;
• Атмосферы наиболее холодных звезд:
- остывших до Т = 300-500 К белых карликов;
- холодных нейтронных звезд (?).
Белые карлики и нейтронные звезды являются заключительными этапами ядерной эволю-
ции звезд. Они рождаются горячими, но не имеют источников нагрева и со временем остывают,
становясь трудно обнаружимыми. Поэтому компактные звезды этих типов, достаточно холодные
для образования на них сложных молекул, пока не найдены, хотя их должно быть много.
Космологическая эволюция и космическая распространенность
химических элементов
Вопрос о том, какие молекулы могут образовываться, определяется не только физически-
ми условиями и химическими свойствами веществ, но и тем, какое их количество содержится
в космическом веществе. Содержание химических элементов в космосе определяется эволюци-
ей окружающего нас мира - Вселенной, и несет в себе информацию об этой эволюции.
Вселенная образовалась около 15 млрд, лет назад в результате Большого взрыва. Ядра
атомов водорода и гелия возникли в первые минуты расширения Вселенной. Сотни тысяч
лет спустя вещество остыло до температуры, когда атомы Н и Не стали нейтральными.
Углеродом и следующими за ним в периодической системе химическими элементами (ча-
сто называемыми в отличие от Н и Не тяжелыми элементами или металлами) космическое
вещество начало обогащаться на миллиард лет позже, когда во Вселенной сформировались
первые звезды. Они были массивными, короткоживущими (<10 млн. лет) и до наших дней
не сохранились. В конце своей эволюции они выбросили в окружающий газ продукты проис-
ходившего внутри них термоядерного горения - углерод и более тяжелые элементы.
Около 12-13 млрд, лет назад, когда содержание тяжелых элементов достигло уровня
~10'4-10'5 от современного, образовались самые старые из наблюдаемых сейчас звезд и со-
держащие их галактики (включая нашу Галактику). По мере завершения своей эволюции
эти звезды продолжали обогащать межзвездный газ внутри галактик тяжелыми элементами.
Чем меньше масса звезды, тем медленнее она эволюционирует и тем дольше живет.
Последующие поколения звезд формировались из межзвездного вещества с все боль-
шим содержанием тяжелых элементов по отношению к водороду и гелию. Солнце родилось
около 5 млрд, лет назад из межзвездного вещества, 2 % массы которого составляли химиче-
ские элементы тяжелее гелия. Сейчас межзвездное вещество, сосредоточенное в основном
в диске нашей Галактики (наблюдаемом как Млечный путь), содержит около 4 % тяжелых
элементов, т. е. вдвое больше, чем при образовании Солнца.
В межзвездной среде Галактики наблюдаются сравнительно небольшие вариации содер-
жания углерода и более тяжелых элементов, по-видимому, не превышающие примерно 3-4 раз.
Основная доля объема Галактики и других галактик, как и большинство астрономических объ-
ектов в галактиках (кроме старых звезд), а также вне галактик, имеют более или менее универ-
сальное содержание элементов, характерное для атмосферы Солнца. Относительное содержа-
104
Проблемы происхождения жизни
ние наиболее распространенных элементов (по числу атомов, а не в долях массы) представлено
в табл. 2. Содержание элементов тяжелее железа мало, < 10 s по числу атомов.
Из старых звезд к настоящему времени не успели завершить эволюцию лишь звезды
с массами менее ~0.8 массы Солнца. Элементный состав их атмосфер не изменился, т. е. со-
держание углерода, кислорода и других элементов в атмосферах старых звезд очень мало.
Исследование экзосолнечных планет указывает, что для образования планет необхо-
димо достаточно высокое содержание тяжелых элементов. У звезд, содержащих примерно
втрое меньше тяжелых элементов, чем на Солнце, пока планеты не обнаружены. Заметим,
что в атмосферах небольшой части звезд, более горячих, чем Солнце, имеются сильные от-
клонения содержания отдельных элементов от средней космической распространенности,
прежде всего, избытки редких элементов - бария, ртути, лантанидов. Хотя эти избытки мо-
гут достигать 5 и даже иногда 6 порядков величины, они, по-видимому, не представляют
интереса для темы данного обзора. После исчерпания ядерного топлива звезды сбрасывают
значительную часть своей массы либо плавно, создавая околозвездные оболочки и так назы-
ваемые планетарные туманности (название историческое, к планетам отношения не имеет),
либо при взрыве звезды, наблюдаемой как явление сверхновой звезды. Центральная часть за-
канчивающей свою эволюцию звезды превращается в очень плотный объект: белый карлик,
нейтронную звезду или черную дыру. Сброшенное вещество содержит продукты ядерной
переработки, т. е. обогащено тяжелыми элементами, и пополняет ими межзвездную среду
в галактиках. Обогащение элементами группы железа и более тяжелыми происходит практи-
чески только при взрывах сверхновых.
Звезды около 90 % времени своей эволюции проводят на стадии главной последователь-
ности, когда в их недрах идут ядерные реакции горения водорода и образования гелия. После
исчерпания в ядре звезды водородного топлива ядро сжимается, а внешние части звезды рас-
ширяются. Диаметр звезды возрастает во много раз, а температура поверхности падает - звез-
да становится красным гигантом или сверхгигантом с температурой поверхности Т < 4000 К.
Этот этап занимает около 10 % всего времени активной жизни звезды - пока в ее недрах про-
исходят экзотермические ядерные реакции (ядерная эволюция). В ядрах красных гигантов
и сверхгигантов гелий превращается в углерод и более тяжелые элементы (иногда вплоть
до железа). Преимущественно образуются четно-четные изотопы (см. примечание к табл. 2).
Тем временем внешние части звезды медленно (со скоростью -10-50 км/с) разлетают-
ся, создавая охлаждающуюся при расширении околозвездную оболочку - идеальное место
для интенсивного образования как сложных молекул (в том числе, вероятно, фуллеренов С60
и др.), а также формирования наночастиц - тугоплавких зерен размером примерно до 100 нм.
Таблица 2. «Нормальное» содержание элементов в космическом веществе по числу
атомов относительно водорода, соответствующее составу атмосферы Солнца
Атомный номер Элемент Распростра- ненность Атомный номер Элемент Распростра- ненность
1 Н 1 12 Mg 2.6 XI0’5
2 Не 0.1 14 Si 3.3 x 10'5
6 С 3.3 XI О’4 16 S 1.6ХЮ'5
7 N 0.9 XI О’4 18 Ar 0.6 xi 0’5
8 О 6.6 Х10'4 20 Ca 0.2 xi 0’5
10 Ne 0.8 XI О’4 26 Fe 4.0 Xl 0’5
В природе преобладают четно-четные изотопы четных элементов: |2С, |60,20Ne, 24Mg, 28Si, 32S, ..., '6Fe ...
105
Н.Г. Бочкарёв. Молекулы и их миграция во Вселенной
Расширение околозвездных оболочек приводит к выносу молекул и пылинок в окружающую
межзвездную среду.
При догорании гелия ядро одиночной звезды (не являющейся компонентом тесной
двойной звездной системы) с массой меньше ~10 масс Солнца сжимается, превращаясь в
горячий белый карлик. Часть образованных в ядре тяжелых элементов выносится расши-
ряющейся оболочкой в межзвездную среду.
Эволюция более массивных звезд и некоторых маломассивных двойных звездных си-
стем завершается мощным взрывом и образованием нейтронных звезд и черных дыр. Взрыв,
наблюдаемый как явление сверхновой звезды, сопровождается образованием всех элементов
тяжелее железа вплоть до трансурановых и выносом их ударной волной наружу в межзвезд-
ное пространство.
Из обогащенного тяжелыми элементами межзвездного газа рождается новое поколе-
ние звезд. Такой круговорот вещества в галактиках приводит к постепенному обогащению
Вселенной элементами тяжелее гелия.
Поскольку звезды образуются не поодиночке, а группами (скоплениями, ассоциациями,
комплексами) в так называемых очагах звездообразования, наиболее массивные из родившихся
совместно звезд успевают взорваться как сверхновые и «запачкать продуктами своей жизнедея-
тельности» протопланетные диски близлежащих звезд солнечной массы и менее массивных. Ве-
роятно, такому воздействию подверглась на ранних этапах своего развития Солнечная система.
Обсуждавшаяся на совещании интенсивная астероидная бомбардировка «молодой» Земли
могла произойти в результате прохождения одной из звезд вблизи Солнечной системы. Вероятно,
это была одна из звезд, родившихся одновременно с Солнцем в том же очаге звездообразования.
Основные компоненты межзвездной среды (МЗС)
Подавляющая часть объема Галактики и других галактик занята межзвездной средой -
материей, заполняющей пространство между звездами внутри галактик. Именно из меж-
звездной среды рождаются звезды и окружающие их протопланетные диски, а затем планет-
ные системы. Причем на ранних стадиях образования звезд и протопланетных дисков моле-
кулярный состав вещества сохраняется таким, каким он был в наиболее холодных участках
межзвездной среды, из которых образуются протозвезды.
Основные компоненты межзвездной среды приведены в табл. 3. Заметим, что все эти
компоненты существуют не сами по себе, а любая их пара взаимосвязана, причем в боль-
шинстве случаев взаимодействие сильное. Поскольку в межзвездной среде осуществляется
примерное равенство плотности энергии движений газа (кинетической энергии), магнитно-
го поля и заряженных частиц (космических лучей), межзвездная среда представляет собой
очень сложно структурированный, динамичный объект (см., напр., Бочкарев, 1985).
Таблица 3. Основные компоненты межзвездной среды
1. Газ (атомы, молекулы, ионы, электроны) ~ 99% массы
2. Пыль ~ 1% массы
3. Магнитные поля ~ 0% массы
4. Космические лучи ~ 0% массы
5. Электромагнитное излучение ~ 0% массы
Дефицит тяжелых элементов в межзвездном газе
Содержание элементов в газовой фазе межзвездной среды заметно отличается
от указанной в табл. 2 средней распространенности: наблюдается дефицит тяжелых эле-
ментов, достигающий для Fe, Ni, Ti, Са, Ai трех порядков величины. На рис. 1 видно,
что величина дефицита зависит от температуры Т его конденсации, т. е. перехода из га-
зообразной в твердую фазу с образованием межзвездных пылинок. Поэтому величина
106
Проблемы происхождения жизни
т т
Рис. 1. Зависимость дефицита элементов
в межзвездном газе от температуры их
конденсации Т (образования пыли). Вы-
соты баров указывают на степень нео-
пределенности данных. По вертикальной
оси отложен десятичный логарифм от-
носительного содержания элементов (по
отношению к водороду), отнесенный к
соответствующей величине в солнечной
фотосфере.
Со Be
ArCNO SZn Na К Мп Si MgFe Ti Al
В Li Cr Ni Ca
0 500 1000 1500 0 2000
дефицита элементов в газе дает информацию о составе межзвездной пыли. Суммарный
дефицит тяжелых элементов в газовой фазе составляет ~1 % массы газа, что соответству-
ет доли массы межзвездной пыли в межзвездной среде (табл. 3).
Наблюдения молекул
Межзвездное пространство пронизано ультрафиолетовым излучением горячих звезд.
Поэтому в незащищенных от ультрафиолетового излучения областях межзвездной среды
молекулы быстро разрушаются, так что их равновесное содержание очень мало. Поэтому
в большом количестве молекулы присутствуют только в молекулярных облаках. Молеку-
лярные облака - это плотные участки межзвездной среды, в которых пылинки, равномер-
но перемешенные с газом, ослабляют ультрафиолетовое излучение горячих звезд в тысячи,
миллионы и более раз. Однако концентрация частиц газа в этих «плотных» участках состав-
ляет всего лишь ~103 см-3 (плотность ~10'21 г/см3). Такие участки межзвездной среды являются
наиболее холодными. Их температура обычно составляет 10-30 К. Именно в такой «криоген-
ной», разреженной космической среде находится большинство космических молекул.
В основном наблюдается собственное излучение молекул. При столь низких темпера-
турах тепловое излучение вещества имеет место только в радио- и субмиллиметровом спек-
тральных диапазонах. Но как раз в эти спектральные диапазоны попадает огромное количе-
ство молекулярных линий, прежде всего вращательных переходов (рис. 2). При температурах
10-100 К молекулы не создают молекулярные полосы, содержащие большое количество спек-
тральных линий. Их спектры просты. Специфика используемых астрономами в этих диапазо-
нах радиофизических методов измерения позволяет реализовывать очень высокое спектраль-
ное разрешение А./ДХ=106-108. Это дает возможность не только однозначно отождествлять по
набору спектральных линий молекулы и молекулярные ионы, но и легко различать их изотопи-
ческие варианты. Из радиоспектров удается извлекать разнообразную информацию: распре-
деление содержания различных молекул и молекулярных ионов внутри молекулярных обла-
ков; распределение температуры и плотности вещества в них; детально изучать движения газа
в разных местах облака (см., напр., Бочкарёв, 2009). Все это позволяет подробно исследовать
области звездообразования и изучать процессы, приводящие к рождению звезд.
В табл. 4 приведен список надежно отождествленных к концу 2000 г. 126 различных
молекул и молекулярных ионов в межзвездном газе, в околозвездных оболочках и кометах.
Видно, что преобладают углеродосодержащие, предбиологические соединения. В основном
наблюдаются линейные молекулы, хотя есть и кольцевые типа Сг Самые длинные из обна-
руженных молекул - цианополиины до HCHN, включительно.
107
Н.Г Бочкарёв. Молекулы и их миграция во Вселенной
час гота, Гц
Рис. 2. Характерный вид спектра излучения молекулярного облака в коротковолновой части радио-
диапазона. Показан участок спектра от 1.14 мм до 1.45 мм (207-263 ГГц), содержащий около 900 мо-
лекулярных линий, в основном соответствующих вращательным переходам тяжелых молекул. Такие
спектры дают возможность разнообразной диагностики молекулярных облаков: распределения вну-
три облака содержания молекул, кинематики движений, температуры и плотности газа.
После 2000 г. была открыта группа отрицательных молекулярных ионов: С4Н,
С6Н_, С8Н’. Обнаружены новые сложные молекулы, в том числе CH2CCHCN, CH3CCCN,
CH3C5N, CH3C6N и ряд более простых молекул.
Как и цианополиины, эти молекулы говорят о том, что в межзвездной среде имеются
длинные углеродные цепочки. Однако обнаружить их можно только тогда, когда к одному
концу углеродной цепочки прикрепляется какой-нибудь атом или ион, либо к обоим концам
присоединяются разные атомы, как в цианополиинах. В этих случаях дипольные момен-
ты молекул становятся отличными от нуля и молекулы оказываются способны интенсивно
излучать в радиолиниях.
Двухатомные симметричные молекулы, такие как Н2, N , О2, образуются в молеку-
лярных облаках в большом количестве, но наблюдать их очень трудно. Нулевой дипольный
момент приводит к отсутствию у них разрешенных спектральных линий. Это исключает
их изучения методами радиоастрономии. Другая возможность - это поиск в спектрах звезд,
наблюдаемых сквозь молекулярные облака, межзвездных линий поглощения таких моле-
кул, возникающих при электронных переходах. Однако холодный газ молекулярных обла-
ков способен создать только слабые линии поглощения, лежащие за редким исключением
в далекой части ультрафиолетового (УФ) диапазона (длины волн около 1000 А), там, где очень
велико поглощения света межзвездной пылью. Реально провести такие наблюдения удалось
в основном для молекулярного водорода и для молекул С2. Последние имеют подходящий
электронный переход вблизи 1 мкм, где пыль поглощает слабо.
Из вышесказанного следует, что, если какая-либо молекула не наблюдается, еще не означает,
что она не образуется или отсутствует в МЗС. Надо проверять возможность ее наблюдать. Во мно-
гих случаях целесообразно привлекать численное моделирование химических процессов в меж-
108
Проблемы происхождения жи:
Таблица 4. Список 126 молекул, обнаруженных к ноябрю 2000 г.
в газовой фазе в межзвездном, околозвездном и кометном
веществе
2 A1F А1С1 C2 CH CH+ CN
CN+ со CO+ CP cs CSi
Н2 НС1 HF KC1 LiH n2+
NH NO NS NaCl OH OH+
PN S2 SH SO SO+ SiN
SiO SiS
3 С3 C2H C2O C2S CH2 co2
HCN HCO HCO+ HCS+ Нз + HOC+
Н2О H2S HNC HNO MgCN MgNC
nh2 c-SiC2 n2h+ n2o NaCN ocs SO2
4 с-С3Н 1-C3H C3N C3O C3S c2H2
СН3 HCCN HCNH+ HNCO HNCS HOCO+
Н2СО H2CN H2CS H3O+ NH3 SiC3
5 С5 C4H C4Si 1-C3H2 c-C3H2 CH2CN
сн4 HC3N hc2nc HCOOH H2CHN H2COH+
Н2С2О h2ncn HNC3 SiH4
6 С5Н C2H4 CH3CN CH3NC CH3OH CH3SH
НС4Н HC3NH+ HC3HO hconh2 1-H2C4 CSN
7 с6н с-СДО ch2chcn CH3C2H hc5n HCOCHs nh2ch3
8 СН2ОНСНО Н2с6 CH3COOH CH3C3N HCOOCH3 C7H hc6h
9 СН3С4Н CH3CH2CN (СНз)2О CH3CH2OH hc7n c8H
10 (СНз)2СО
11 hc9n
13 HC„N
В левой колонке указано число атомов в молекуле. Буквой «с» отмечены
циклические молекулы, буквой «I» - линейные.
звездной среде. При таком моделировании также возникает немало проблем. Обсуждение вопросов,
связанных с межзвездной химией, можно найти в работах автора (Бочкарёв, 2009) и других иссле-
дователей (Каплан, Пикельнер, 1979; Далгарно, 1979, 1986; Tielens, 2005; Lemaire, Combes, 2007).
Многие молекулы обнаружены в сотнях и тысячах различных объектов, хотя большин-
ство других удалось пока наблюдать лишь в
облаках. Чувствительность современных
радиоастрономических измерений позво-
ляет изучать молекулы не только в нашей
Галактике, но и в других галактиках. В на-
стоящее время в других галактиках найдено
около 50 различных молекул. Наша Галак-
тика не уникальна по количеству межзвезд-
ного вещества в молекулярной форме. Есть
галактики, у которых доля массы, приходя-
щаяся на молекулы, как заметно больше, так
и значительно меньше, чем в Галактике.
На рис. 3 приведена схема строе-
нескольких наиболее изученных молекулярных
г~юо к;
ч-100/см]
сн’
сн’
; I 5 20 К:
N |Г Р- Г
СН'| -
N NH, :
N'?NN NH’ С,II _
CH"
ОПГ
CHjCOOHN2 НС, i N с,’н.
СО COCOCOCOCQ А
С С С С С (М
С'С С*С’ НС£Х
нсо* -
щ <cscs
s s cs
s s
S' S ’
H, S" s
сн .... .
I'c.n CH
.CH4
<cs ,0 о о о о о о
* b.o.o.o,o.o,
нс
Н,СО' . .
Н,0
Н.СО
ния молекулярного облака. Показано, что
Рис. 3. Схема строения молекулярного облака.
N
S
109
Н.Г. Бочкарёв. Молекулы и их миграция во Вселенной
Рис. 4. Относительные содержания моле-
кул в молекулярных облаках. Для каждой
из молекул, перечисленных в нижней ча-
сти рисунка, приведены десятичные лога-
рифмы содержаний молекул относительно
Н2 (по числу молекул) в четырех хорошо
изученных молекулярных облаках. На-
звания облаков указаны в правой верхней
части рисунка.
УФ излучение звезд проникает в облако и постепенно ослабляется (UV на рис. 3). По мере
ослабления этого излучения по направлению к центру облака падает температура и растет
плотность вещества. Положительно заряженные атомарные и молекулярные ионы постепен-
но сменяются нейтральными атомами и молекулами. От гравитационного коллапса облако
удерживают внутренние движения и магнитные поля. Тем не менее, отдельные сгустки мо-
лекулярного облака способны превращаться в звезды. Молодые маломассивные звезды типа
Т Тельца (схематически показаны в центре рисунка) своим излучением вызывают подогрев
внутренних частей молекулярного облака, поддерживают «энергетику» химических реак-
ций и сбрасывают часть молекул, намерзших на пылинки, в газовую фазу.
Относительные содержания молекул в четырех молекулярных облаках показано
на рис. 4. Они сравнительно мало меняются от облака к облаку. Естественно, наиболее велико
содержание молекулярного водорода Н2. Но эта молекула трудна для наблюдения, поскольку
не имеет дипольного момента и, соответственно, сильных спектральных линий. Второе ме-
сто занимают молекулы угарного газа СО. С точностью до ошибок измерений во всех облаках
количество молекул СО составляет ~10-4 числа молекул Н2. Молекула СО легко наблюдается
по сильным и удобным для измерений эмиссионным (реже абсорбционным) вращательным
радиолиниям А, = 2.6 мм и 1= 1.3 мм, принадлежащим 5 изотопическим вариантам этой
молекулы. Содержание более сложных молекул мало зависит от их сложности и колеблется
в пределах 10-7—10 9 количества молекул Н2.
Рис. 5. ИК полосы - источник информации
о молекулах, из которых состоят межзвезд-
ные пылинки. По горизонтальной оси -
длины волн в микронах; по вертикальной
оси - спектральные плотности потоков
излучения в янских (1 Ян = Ю’23 Вт/м2 Гц)
от области звездообразования W33A. Ука-
заны отождествления полос поглощения.
Символом X обозначен неоднозначно опре-
деляемый фрагмент молекулы.
ПО
Проблемы происхождения жизни
Таблица 5. Инфракрасные полосы в спектрах межзвездной пыли и образующие
их вещества
Тугоплавкие составляющие
Вещества Полосы
Аморфные силикаты Аморфный углерод Кристаллические силикаты 9.7 и 18 мкм 0.218 мкм много
Льды
Вещества Полосы Вещества Полосы
Н2О 3.0 pm stretch CH3OH 3.54 pm C-H stretch
Н2О 6.0 pm СНзОН 3.9 jim combination
Н2О 10 jim libration CH3OH 2.4 jim overtone
Н2О 44 pm CH3OH 9.7 pm C-O stretch
HDO 4.1 |im stretch CH3OH 8.9 pm CH3-rocking
СО 4.67 pm polar CHSOH 6.8 pm CH and OH deformation
СО 4.67 pm apolar XCN 4.62 jim
со2 4.27 pm stretch H2 2.415 pm
со2 15.3 pm bend NH3 9.009 jim inversion
DCS 4.9 pm NH3 6.158 pm NH deformation
сн4 7.67 pm H2CO 5.81 pm v2
HCOOH 7.24 pm
Приведены длины волн полос в микронах. Для льдов также указаны типы колебаний молекул,
ответственных за образование полос. Символом X обозначен неоднозначно определяемый фрагмент
молекулы.
Молекулы, входящие в состав межзвездных пылинок
Выше были приведены сведения о молекулах, находящихся в газовой компоненте
межзвездной среды. Однозначное отождествление молекул, входящих в состав межзвездных
пылинок, крайне затруднено. Тем не менее, простейшие соединения отождествить можно.
На рис. 5 показан инфракрасный спектр одной из областей звездообразования в нашей Галак-
тике, в котором на фоне непрерывного теплового излучения протозвезды видны многочислен-
ные широкие полосы поглощения. Многие из них удалось отождествить (см. рис. 5 и табл. 5),
хотя не все отождествления являются однозначными. Образуются ИК полосы при фотовоз-
буждении колебательных степеней свободы молекул, из которых состоят пылевые частицы.
Часть ИК полос обычно связывают с плоскими молекулами полициклических аро-
матических углеводородов (ПАУ, английская аббревиатура РАН) (Allamandola et al., 1989).
Определить, какие именно соединения этого типа дают основной вклад в образование на-
блюдаемых полос, не представляется возможным. Это видно на рис. 6, в верхней части кото-
рого показаны профили двух эмиссионных ИК полос, наблюдаемых в спектре туманности
Ориона. Ниже для тех же участков спектра указаны положения полос поглощения различ-
ных молекул ПАУ. Добиться однозначного отождествления здесь невозможно.
В отличие от одиночного бензольного кольца, которое очень легко разрушается УФ
излучением, молекулы ПАУ, состоящие из большого количества бензольных колец, гораздо
более устойчивы к УФ излучению и не разрушаются даже тогда, когда из молекулы вырван
один из электронов. Молекулы ПАУ могут образовываться в оттекающих околозвездных
оболочках углеродных звезд - тех, в атмосферах которых содержание углерода больше, чем
кислорода (по числу атомов). Возможно, наиболее мелкие межзвездные пылинки - нано-
частицы размером —1.5-2 нм, представляют собой именно молекулы этого типа.
111
Н.Г Бочкарёв. Молекулы и их миграция во Вселенной
Рис. 6. Профили эмиссионных
ИК полос, наблюдаемых в спек-
тре излучения области звездо-
образования в созвездии Орио-
на (вверху), и сопоставление
их колебательными спектрами
трех молекул полициклических
ароматических углеводородов
(ПАУ - указаны вертикальны-
ми линиями, длины которых
пропорциональны интенсив-
ностям линий) с целью возмож-
ного отождествления ИК полос
(Allamandolaetal., 1989). Указаны
названия и структурные схемы
молекул. Слева показан участок
спектра 3.2-3.7 мкм, справа -
от 5 до 13 мкм. На нижних шка-
лах приведены длины волн в ми-
кронах, сверху - частоты в см1.
Диффузные межзвездные полосы (DIBs)
Упомянем также самую старую спектроскопическую проблему астрофизики - диф-
фузные оптические полосы поглощения. Они хорошо видны в спектрах звезд. Их интенсив-
ность пропорциональна величине межзвездного поглощения света пылью по пути до звезды.
Поэтому считается, что эти линии образуются в межзвездном пространстве. Они получили
название диффузные межзвездные полосы (DIBs).
DIBs были открыты в 1920 г. Ни одна из них до сих пор не отождествлена. В диа-
пазоне длин волн 0.4-1 мкм выявлено около 300 полос шириной 2-60 А (см. Herbig, 1975,
1995; Galazutdinov et al., 2000; Krelowski, 2002, 2003; рис. 7). Предположительно их связывают
с линейными молекулами типа NCnHm (п=20-40?), ПАУ и/или фуллеренами.
Длинные углеродные цепочки могут образовываться в межзвездных пылинках. Намерз-
ший на тугоплавкое ядро пылинки «грязный лед», состоящий из малолетучих соединений типа
Н2О, СН2ОН, может полимеризоваться, образуя, например, полиформальдегид (Гольданский,
1977; Goldanskii, 1977). В дальнейшем частицы космических лучей, пронизывающие молеку-
лярные облака, разрывают эти длинные полимерные нити. Образованные таким образом угле-
Рис. 7. Диффузная межзвездная полоса по-
глощения шириной ~ 10 А в спектре звезды
HD4841, имеющем сильное межзвездное
поглощение: избыток цвета Е(В-У) = 0.65,
что соответствует оптической толщине
межзвездной пыли на пути до звезды ~2.
Проблемы происхождения жизни
родные цепочки попадают в газовую компоненту молекулярного облака. Как отмечалось выше,
ПАУ могут формироваться в холодных оттекающих оболочках звезд красных гигантов. Пред-
полагается, что фуллерены также могут вырастать в расширяющихся оболочках этих звезд.
2. МИГРАЦИЯ МОЛЕКУЛ И ПЫЛИ ВО ВСЕЛЕННОЙ
Как могут перемещаться сложные молекулы в космическом пространстве
Молекулы, кроме наиболее устойчивых типа ПАУ, быстро (за 10-10 000 лет) разруша-
ются УФ излучением звезд (Бочкарёв, 2009). Нужны «транспортные средства», защищающие
их от разрушения. Какие они? Что может служить защитным экраном?
Молекулы, находящиеся в межзвездном газе (т. е. не входящие в состав пылинок), мо-
гут мигрировать только вместе с молекулярным облаком, медленно приближаясь к центру
Галактики. Там они надежно экранированы содержащейся в облаке пылью от губительно-
го УФ излучения и в процессе звездообразования переходят из межзвездного пространства
в околозвездные газопылевые диски.
Молекулы могут перемещаться внутри межзвездных пылинок, хотя в этом случае они
не так хорошо защищены от УФ излучения. Миграция пылинок рассмотрена ниже.
Другими «транспортными средствами» для миграции молекул в космосе могут служить
более крупные объекты: микрометеориты (межпланетные пылинки размером 10-100 мкм), ме-
теороиды (метеорные и метеоритные тела величиной от 0.1 мм до 10 м), астероиды (> 10 м),
планеты, а также ядра комет. Ядра комет приходят из очень далекой периферии Солнечной си-
стемы - из пояса Оорта. Внутри планетной системы траектория движения ядра кометы испыты-
вает возмущающее влияние гравитационных полей планет-гигантов (прежде всего, Юпитера).
Если из-за этих возмущений тело потеряло часть своей энергии, его орбита становится эллип-
тической и тело захватывается планетной системой. Возможна и обратная ситуация, когда тело
приобретает энергию и навсегда покидает Солнечную систему, т. е. теряет связь с Солнцем.
Поскольку кометные ядра - это многокилометровые глыбы «грязного льда», который
может содержать внутри себя мелкие тугоплавкие тела типа метеороидов, после испарения
кометного ядра остаются пылинки, метеорные и метеоритные тела. Под действием грави-
тационных возмущений со стороны планет Солнечной системы они постепенно заполняют
всю орбиту, по которой двигалась комета. Такие образования называются метеорными роя-
ми. Вследствие гравитационных возмущений часть вещества метеорного роя также может
покинуть Солнечную систему. Эти процессы приводят к возникновению кометных и метеор-
ных тел, свободно блуждающих в межзвездном пространстве. Находясь внутри блуждаю-
щих тел, сложные молекулы могут вместе с ними попадать в окрестности других звезд, и
наоборот, фрагменты внешних частей протопланетных дисков других звезд могут залетать
в Солнечную систему. Тем самым осуществляется обмен веществом между протопланет-
ными дисками (между планетными системами).
Заведомо «чужеродными» могут быть метеориты, возраст которых больше возраста
Солнечной системы. К сожалению, автору не удалось найти в литературе ни одного такого
примера. Другим критерием внесолнечного происхождения метеорных тел является высокая
скорость их движения (превышающая скорость убегания из Солнечной системы). Радиолока-
ция метеорных следов показывает, что такие частицы существуют (Hajdukova, 2008).
Галактики - это гравитационно-связанные системы, состоящие из большого числа
звезд. Например, наша Галактика - типичная крупная галактика (см. рис. 8) - насчитывает
более 150 млрд, звезд. Некоторые из этих звезд при сравнительно теснь1х сближениях друг
с другом совершают гравитационные маневры, в результате которых могут покинуть пределы
Галактики вместе со своими планетными системами. Через время ~1 млрд, лет они могут до-
стичь другой галактики и войти в ее состав. Последнее время появилось довольно много наблю-
дательных данных о звездах-бегунах, чьи скорости больше скорости убегания из Галактики.
113
Н.Г Бочкарск. Молску.1ы и их миграция во Вселенной
Рис. 8. Спиральные галактики, похожие на нашу Галактику: а - вид «плашмя»; б - вид почти «с ребра».
Это приводит к «испарению» из Галактики небольшой части ее звездного населения. Наблюда-
ются и звезды-бегуны, влетающие из межгалактического пространства внутрь Галактики. Оче-
видно, что вместе с планетными системами звезд-бегунов межгалактические перелеты могут
совершать и сложные молекулярные соединения, находящиеся в этих планетных системах.
В последнее время стало ясно, что за время жизни галактик (~10 млрд, лет), они неод-
нократно сталкиваются. При этом крупная галактика способна полностью поглощать более
мелкие. Это наиболее эффективный способ переноса вещества из одной галактики в другую.
Наиболее интенсивно он происходит в скоплениях галактик (рис. 9, и). При взаимодействии
галактик примерно одинаковых масс (рис. 9, б) возникает не только обмен звездами и межз-
вездным веществом сталкивающихся галактик, но и выброс значительной части звездного
населения в межгалактическое пространство. В процессе столкновения и слияния галак-
тик перемещаются из одной галактики в другую и находящиеся внутри галактик сложные
органические соединения.
Пекулярные скорости как отдельных галактик в целом, так и выбрасываемых из них
фрагментов, содержащих звезды, не превышает -3000 км/с, т. е. ~1 % скорости света. Поэтому
за время существования галактик (—10—13 млрд, лет) обмен органическим веществом мог прои-
зойти на расстояниях, не превышающих —100 млн. световых лет, что составляет примерно 1 %
радиуса наблюдаемой части Вселенной. Такой объем содержит примерно 1/106 часть всех га-
лактик, т. е. -10" IО' галактик. По-видимому, названные цифры можно рассматривать как верх-
ний предел расстояний, в пределах которых может происходить панспермия, т. е. перенос «за-
родышей жизни» с одной планеты на другую. Высокая эффективность может осуществляться
лишь в пределах одного скопления галактик, т.е., между 100-10000 галактик, охватывающих
Ю1”- 1014 планетных систем. Наша Галактика не является членом скопления, поэтому частич-
ный обмен вещества мог охватить объем, не превышающий объем Местной Группы галактик.
Рис. 9. Галактики: а- скопление галактик; б- взаимодействующие галактики; взаимодействие часто
кончается слиянием.
114
Проь.ц- '.fh! llPOIICMKK.t ПИЯ ЖИИИ1
В научной литературе иногда обсуждается вопрос о возможности ускорения пылевых
частиц до релятивистских, т.е. близких к скорости света, скоростей (см., напр., Dasgupta,
1980). Такие частицы могли бы переносить содержащиеся в них молекулы на еще большие
расстояния. Однако если процесс ускорения может происходить плавно, то механизмов плав-
ного торможения таких гипотетических частиц не найдено. Они погибают, врезаясь в какое-
нибудь препятствие. В этом случае вероятность сохранения возможно имеющихся в этих
пылинках молекул равна нулю.
Перенос на большие расстояния возможен, если справедлива гипотеза существова-
ния так называемых «кротовых нор», интенсивно развиваемая в последние годы академиком
Н.С. Кардашевым, чл.-корр. РАН И.Д. Новиковым и А.А. Шацким (см., напр., Новиков, Карда-
шев, Шацкий, 2007). Речь идет о возможности существования топологических особенностей
пространства-времени, допускающих прямую связь сколь угодно отдаленных участков Все-
ленной (или даже других вселенных) друг с другом. Такие образования, если они существуют,
должны быть подобны черным дырам, но с некоторыми особенностями, которые пока далеки
от возможности экспериментального изучения. Не выяснены до конца и многие теоретические
вопросы, связанные с возможностью существования и функционирования «кротовых нор».
Межзвездная пыль, ее образование и эволюция
Строение пылинок непосредственно из наблюдений практически не определяется.
Из сведений о распределении размеров, составе, происхождении и эволюции пылинок сле-
дует, что они состоят из мелких аморфных или кристаллических ядер силикатов, графита
и, возможно, окислов металлов (а на рис. 10) и намерзших на них оболочек грязного льда (б) -
смеси молекул воды с многочисленными примесями.
Характерный размер ядрышек - 0.01-0.05 мкм. Плотность вещества в них составляет
несколько г/см'. Вокруг имеется толстая (~0.1 мкм) рыхлая оболочка из льда и намерзших
газов с низкой плотность (' I г/см'') и пленка высокомолекулярных соединений.
Конспективно эволюция космических пылинок может быть описана следующим
образом:
- образование тугоплавких ядер и мельчайших пылинок в виде молекул полицикли-
ческих ароматических углеводородов (ПАУ) происходит главным образом в оттекающих
околозвездных оболочках старых звезд;
- далее идет нарастание оболочки «грязного льда» - в молекулярных облаках (десятки
млн. лет);
Рис. 10. Строение межзвездной пылинки: а -
тугоплавкое ядрышко размером в сотые доли
мкм; б - оболочка грязного льда; с - пленка
из высокомолекулярных соединений.
Рис. 11. Межпланетная пылинка - результат
слипания (коагуляции) межзвездных пылинок.
Длина масштабной линейки справа внизу соот-
ветствует 1 мкм.
115
Н.Г Бочкарёв. Молекулы и их миграция во Вселенной
- разрушение ледяных оболочек происходит каждые —100 млн. лет при попадании
пылинок в области звездообразования;
- ядра пылинок разрушаются в ударных волнах от взрывов сверхновых звезд или при
попадании пылинки внутрь рождающейся звезды; среднее время жизни ядер >10 млрд, лет
(порядка возраста Вселенной!);
- в протопланетных дисках рост пылинок имеет место за счет коагуляции при медленных
столкновениях (рис. 11); пылинки могут дорастать до размера метеороидов, ядер комет, планете-
земалей и т. д.
При количественном анализе перечисленных стадий эволюции пылинок необходимо
принимать во внимание широкий набор процессов, прежде всего тех, которые представлены
в табл. 6, а в некоторых случаях и ряд других.
Таблица 6. Основные процессы на поверхности и внутри ледяных оболочек пылинок
1) Адсорбция (физическая и химическая).
2) Миграция по поверхности (Hollenbach, Salpeter, 1970, 1971).
3) Тепловая десорбция.
4) Образование молекул - основной канал образования Н, и некоторых других молекул в
современной межзвездной (М3) среде.
5) Хемодесорбция - выброс Н2 и некоторых других молекул в М3 газ.
6) Полимеризация (Гольданский, 1975, 1977; Goldanskii, 1977; Hoyle, Wickramasinghe, 1977) -
образование длинных углеродных цепочек С .
7) Химические взрывы - разрушение ледяных оболочек пылинок, выброс молекул в М3 газ.
8) Десорбция космическими лучами, УФ и рентгеновским излучением - выброс фрагментов
Сп-цепочек в М3 газ.
9) Разрушение при столкновениях - образование молекул в плазме микровзрыва, возникающего
при столкновении пылинок с частицами космических лучей или между собой (Managadze, 2007;
Brinckerhoff et al., 2005).
10) Налипание электронов при столкновениях их с пылинками.
11) Фотоэффект, т. е. выброс электронов из тела пылинки при поглощении УФ-фотона.
Гипотеза Хойла и Викрамасинга
Ф. Хойл и Ч. Викрамасинг (Hoyle, Wickramasinghe, 1979,1980) около 30 лет назад выска-
зали предположение, что часть межзвездных пылинок может быть одноклеточными организ-
мами: бактериями или фитопланктоном. В работе 1979 г. они основывались на соответствии
положения и формы широкой спектральной особенности (покрывающей диапазон длин волн
X от 4000 до 5000 А) зависимости от л. коэффициента поглощения света межзвездной пылью
абсорбционным свойствам пигментов фитопланктона. В статье 1980 г. они показали, что по-
лоса поглощения пыли в области 3 мкм, которая обычно связывается со льдом в оболочках
пылинок (см. рис. 5 и табл. 5), обусловлена поглощением света бактериями, находящимися
в межзвездной среде, поскольку это дает лучшее совпадение с наблюдаемыми профилями
этой полосы. Согласно их работе, до 60 % пылинок могут состоять из бактерий.
Эта гипотеза возродила интерес к возможности панспермии. Но аргументы авторов
нельзя считать убедительными ввиду неоднозначности интерпретации широких спектраль-
ных деталей (это видно на примере рис. 6). Поэтому гипотеза была встречена большинством
специалистов скептически. Однако она не противоречит наблюдениям (по крайней мере, если
доля бактерий среди пылинок не так велика). К тому же выяснилось, что одноклеточные ор-
ганизмы очень устойчивы к существующему в космическом пространстве ионизующему из-
лучению. Поэтому исключать из рассмотрения гипотезу Хойла и Викрамасинга не следует.
Внутри молекулярных облаков такие органические «пылинки» (подобно тугоплавким
ядрам обычных межзвездных пылинок) могут обрастать ледяными оболочками. Эти оболочки
могут играть роль дополнительного экрана, защищающего органическое ядро таких пылинок
116
Проблемы происхождения жизни
Рис. 12. Траектории частиц при оттоке газа
от плоскости спиральной галактики в межга-
лактическуюсреду («галактический ветер») (а)
и траектории движения газа в «галактических
фонтанах» (б), когда газ не имеет достаточной
энергии, чтобы покинуть галактику. Горизон-
тальная черта - галактическая плоскость,
ЦГ - галактический центр.
от излучений, способных повредить находящийся внутри замороженный организм. Органи-
ческие ядра могут пережить эпохи испарения оболочек (нагрев до 100-200 К в областях звез-
добразования), но, в отличие от тугоплавких ядер пылинок, не способны пережить нагрев до
1000-2000 К. Однако нагрев пылевых частиц до таких температур происходит очень редко.
Движение пылинок
Тела, способные переносить молекулы, такие, как крупные пылинки (> 1 мкм), метео-
роиды и небольшие ядра комет пока не удается наблюдать. Поэтому пока нет надежной ин-
формации об их количестве и движении. В отличие от них межзвездные пылинки размером
0.01-1 мкм) давно и интенсивно изучаются. Пылинки движутся в основном вместе с межз-
вездным газом. Из-за неоднородности распределения в Галактике наиболее ярких горячих
звезд пылинки под действием светового давления могут прибретать скорость до 2-10 км/с
относительно газа (Tarafdar, Wickramasinghe, 1976), а в некоторых случаях ускоряться (так
называемым бетатронным механизмом) до -30-100 км/с.
В галактиках с бурным звездообразованием пылинки выносятся вместе с газом в меж-
галактическое пространство (рис. 12). Если они попадают внутрь плотного скопления галак-
тик, то разрушаются горячим межгалактическим газом скопления, если нет - могут через
~1 млрд, лет попасть в другую галактику.
ЗАКЛЮЧЕНИЕ
Молекулы существуют в космическом пространстве в широком наборе астрономиче-
ских объектов. К настоящему времени в газовых средах отождествлено около 150 различных
молекул примерно в 250 изотопических вариантах. Самые крупные из найденных молекул
содержат 13 атомов. Основная часть молекул содержит атомы углерода, эти молекулы яв-
ляются простейшими органическими соединениями. Молекулы обнаружены в атмосферах
холодных звезд, в атмосферах планет и их спутников, но наибольшее их количество и раз-
нообразие найдено в молекулярных облаках межзвездной среды и в оттекающих оболочках
звезд, находящихся на поздних стадиях эволюции. Имеются косвенные указания на суще-
ствование в межзвездной среде более крупных молекул, содержащих 20-60 атомов, в виде
длинных углеродных цепочек, полициклических ароматических углеводородов и, возможно,
фуллеренов, но надежных отождествлений таких молекул нет.
Молекулы могут перемещаться в пространстве на значительные расстояния, если они вхо-
дят в состав пылинок или более крупных тел. Наиболее радикальной является гипотеза Хойла и
Викрамасинга, согласно которой некоторые пылинки могут представлять собой одноклеточные
организмы. Эта гипотеза не подтверждена, но и не опровергнута ни наблюдениями, ни теорети-
ческими расчетами.
Они состоят из тугоплавких ядер и намерзших на них ледяных оболочек. Время жизни
ядер пылинок сравнимо с временем жизни галактик, т. е. они постепенно накапливаются
117
1
Н.Г Бочкарёв. Молекулы и их миграция во Вселенной
в течение всего времени эволюции Вселенной. Пылинки могут перемещаться внутри галак-
тик из одной области звездообразования в другую, а при достаточной мощности процес-
сов звездообразования в галактике - выноситься галактическим ветром в межгалактическое
пространство. Однако если пылинки попадают в горячий газ скопления галактик, они там
быстро разрушаются. Если же они выносятся из галактики, не принадлежащей крупному
скоплению галактик, то через время ~1 млрд, лет они могут попасть в другую галактику.
Другими «транспортными средствами» для молекул, защищающими их от губитель-
ного действия ультрафиолетового излучения, могут служить метеорные, метеоритные тела
и ядра комет, астероиды и планеты, некоторые из которых должны блуждать в пространстве
между звездами и время от времени проникать в планетные системы. Блуждающими могут
быть также отдельные звезды и их группы, выброшенные из галактик в процессе взаимодей-
ствия галактик между собой. В этом случае звезды переносятся из одной галактики в дру-
гую вместе с окружающими их планетными системами и могут переносить за космологиче-
ское время (~1010 лет) сколь угодно сложные молекулы и другие объекты как биологической,
так и иной природы на расстояния вплоть до десятков миллионов световых лет. Наиболее
эффективно этот процесс происходит при слиянии (мержинге) галактик.
СПИСОК ЛИТЕРАТУРЫ
Бочкарёв Н.Г. 1985. Магнитные поля в космосе. М.: Наука. 208 с.
Бочкарёв Н.Г. 2006. Энциклопедия низкотемпературной плазмы. Сер. Б. Справочные приложения, базы
и банки данных. Т. V-1. Диагностика низкотемпературной плазмы. Ч. I. М.: Янус-К. С. 239-323.
Бочкарёв Н.Г. 2009. Основы физики межзвездной среды. 2 изд. М.: УРСС. 352 с.
Гиндилис Л.М. 2004. SETI: Поиск Внеземного Разума. М.: Физматлит. 647 с.
Гольданский В.И. 1975. Явление квантового низкотемпературного предела скорости химических ре-
акций // Усп. химии. Т. 44. № 12. С. 2121-2149.
Гольданский В.И. {911. Механизм образования полиформальдегида в межзвездном пространстве И
ДАН. Т. 235. С. 1053-1055.
Голдсмит Д., Оуэн Т. 1983. Поиски жизни во Вселенной. М.: Мир. 488 с.
Далгарно А. 1979. Химия межзвездной среды // На переднем крае астрофизики / Ред. Ю. Эвретт. М.:
Мир. С. 371-404.
Далгарно А. 1986. Роль электрон-ионных и протон-ионных столкновений в астрофизике // Физика ион-
ионных и электрон-ионных столкновений / Ред. Ф. Бруайр, Дж. Мак-Гоуэн. М.: Мир. С. 11-45.
Каплан С.А., Пикельнер С.Б. 1979. Физика межзвездной среды. М.: Наука. 531 с.
Новиков И.Д., Кардашев Н.С., Шацкий А.А. 2007. Многокомпонентная Вселенная и астрофизика кро-
товых нор // Усп. физ. наук. Т. 177. № 9. С. 1017-1023.
Розенталь И.Л. 1984. Элементарные частицы и структура Вселенной, М.: Наука. 112 с.
Цицин Ф.А., Чепурова В.М. 2003. О структуре кометных ядер // Кометы, астероиды, метеоры, метеориты,
астроблемы, кратеры / Ред. К.И. Чурюмов. Тр. 2 междунар. конф. КАММАК 02. Винница. С. 26-30.
Шкловский ИС. 1987. Вселенная, жизнь, разум. М.: Наука. 6 изд. (перераб.). 319 с.
Allamandola L.J., Tielens A.G.G.M., Barker J.M. 1989. Interstellar polycyclic aromatic hydrocarbons -
the infrared emission bands, the excitation/emission mechanism, and the astrophysical implications //
Astrophys. J. Suppl. Ser. V. 71. P. 733-775.
Brinckerhoff W.B., Managadze G.G., ChumikovA.E., ManagadzeN.G. 2005. Processing and synthesis of pre-
biotic chemicals in hypervelocity impacts // Lunar and Planetary Science. V. 36. P. 1377-1378.
118
Проблемы происхождения жизни
Dasgupta А.К. 1980. Relativistic and non-relativistic dust grains // Astrophys. and Space Sci. V. 67 P. 247-254.
Galazutdinov G.A., Musaev F.A., Krelowski J., Walker G.A.H. 2000. Narrow diffuse interstellar bands.
A survey with precise wavelengths // Publ. Astron. Soc. Pacific. V. 112. P. 648-690.
Goldanskii V.l. 1977. Mechanism for formaldehyde polymer formation in interstellar space // Nature. V. 268.
P. 612-613.
Hajdukova M. 2008. The occurrence of interstellar meteoroids in the vicinity of the Earth // Organic matter in
space. I AU Symp. V. 251. P. 347-348.
Herbig G.H. 1975. The diffuse interstellar bands. IV - The region 4400-4850 A // Astrophys. J. V. 196.
P. 129-160.
Herbig G.H. 1995. The diffuse interstellar bands // Ann. Rev. Astron. Astrophys. V. 33. P.19-74.
Hottenbach D., Salpeter E.E. 1970. Surface adsorption of light gas atoms // J. Chem. Phys. V. 53. P. 79-86.
Hottenbach D., Salpeter E.E. 1971. Surface recombination of hydrogen molecules // Astrophys. J. V. 163.
P. 155-164.
Hoyle F., Wickramasinghe C. 1977. Polysaccharides and the infrared spectra of galactic sources // Nature.
V. 268. P. 610-612.
Hoyle F„ Wickramasinghe C. 1979. On the nature of interstellar grains // Astrophys. Space Sci. V. 66. P. 77-90.
Hoyle F, Wickramasinghe C. 1980. The identification of the 3ц spectral feature in galactic infrared sources //
Astrophys. Space Sci. V. 68. P. 499-503.
Krelowski J. 2002. Organic molecules and the diffuse interstellar bands // Adv. Space Res. V. 30. P. 1395-1407.
Krelowski J. 2003. How to identify diffuse band carriers? // Solid state astrochemistry / Eds. V. Pirronello, J.
Krelowski, G. Manico. NATO Sci. Ser. И. V. 120. Dordrecht: Kluwer. P. 147-174.
Lemaire J.L., Combes F. (eds.) 2007. Molecules in Space and Laboratory. Paris. 134 p.
Managadze G. 2007. A new unicersal mechanism of organic compaunds synthesis during prebiotic evolution //
Planetary and Space Science. V. 55. P. 134-140.
Tarafdar S.P., Wickramasinghe N.C. 1976. The role of Lyman-alpha photons in the interstellar medium //
Astrophys. Space Sci. V. 39. P. 19-30.
Tielens A.G.G.M. 2005. The physics and chemistry of the interstellar medium. Cambridge Univ. Press. 495 p.
Molecules in Space and migration of molecules
N.G. Bochkarev
Molecules observed in different astronomical objects (with the exception of the Solar Sys-
tem planets and their satellites) are briefly described, giving special attention to molecules in the
interstellar medium. Different ways of spatial migration of molecules and their transferring from
one planetary (proto-planetary) system to another are considered. The possibility of transferring
of interstellar molecules over distances up to 100 million light years in cosmological time is stated.
F. Hoyle and C. Wickramasinghe hypothesis on biological nature of some interstellar dust grains
is briefly discussed.
119
Проблемы происхождения жизни. М.: ПИН РАН. 2009. С. 120-128
УСЛОВИЯ НА ПОВЕРХНОСТИ ЗЕМЛИ 4-4.6 МЛРД. ЛЕТ НАЗАД.
ПЕРВИЧНЫЕ СИНТЕЗЫ*
© 2009 г. Л.М. Мухин
Институт космических исследований РАН
4-4.6 млрд, лет назад поверхность Земли могла быть в основном относи-
тельно холодной, с отдельными горячими пятнами, обусловленными им-
пактными событиями. Такая сильно гетерогенная по температуре система
могла обеспечить возможность процессов первичных синтезов органиче-
ских соединений.
Мы говорим о том, что трудно дать определение Жизни. Может быть, этого не нужно
делать, но стоит отметить, чем отличается живое от неживого. Живое отличается от неживого
наличием совершенно уникальной машины, которая состоит из двух частей (компонентов) -
транскрипции и трансляции. Такой механизм есть в любой живой системе. В связи с этим
мне очень понравилась диаграмма, которую я нашел в одной из американских энциклопедий.
Это такой пазл, состоящий из отдельных квадратиков, где есть практически все составляю-
щие проблемы происхождения жизни на Земле (рис. 1). Здесь есть проблема возраста жиз-
ни на Земле, есть источники энергии, необходимые для синтеза органических соединений
и т. д. Эта схема ставит перед исследователями, занимающимися проблемами происхождения
жизни, некоторые задачи. Я думаю, что одной из первых важнейших задач является возраст
Земли, который сейчас можно определить достаточно точно, благодаря высокотемператур-
ным включениям в метеоритах. Это самое древнее, что мы сейчас имеем. Совсем недавно,
порядка 10 лет тому назад, появились данные по цирконам. Это самые древние минералы
на Земле, один из них имеет возраст 4.4 млрд, лет, более молодые цирконы - 3.8 млрд, лет
и менее. Их возраст определялся соотношением изотопов свинца (207РЬ/206РЬ) в этих образо-
ваниях (рис. 2). Интересно, что именно на уровне 3.8-3.9 млрд, лет назад закончилась так
называемая последняя тяжелая бомбардировка, которая происходила до этого времени. Это
накладывает некоторые ограничения на проблему происхождения жизни. По изотопии кис-
лорода в этих цирконах был сделан очень важный вывод, что уже более 4 млрд, лет назад
на Земле была вода. В сочетании с анализом следовых элементов это достаточно убедитель-
ное доказательство того, что на Земле в те времена была прагидросфера.
* Профессор Лев Михайлович Мухин - один из активнейших организаторов рабочего совещания «Проблемы
происхождения жизни» - скончался 24 апреля 2009 г. Учитывая намерение Льва Михайловича подготовить
статью по материалам его доклада, редколлегия сборника взяла на себя смелость опубликовать выступление
Л.М. Мухина по его видеозаписи, адаптировав содержание к письменной речи и иллюстрировав материалами
презентации. Ответственность за возможные недостатки полностью лежит на редакторах сборника.
120
Проь.н мы прояс.мькл нпя жпши
Origin of Life on Earth
<5 C-ef/oi t/o/i of
tion of
W at^r
Thermal
Ponds
Carbonaceous
Hydrothermal
Phosphorus P,
Me teOrrti
Infalls
Pfedic
Probability of
►
igh tnuiy
1
Laborator-
Models
4
Hyper th er ma ph he
Li fe Forms
Carbon C,
Hydrogen H,
Lipjds
A < on ih t s
Synthesis
on Earth
Рис. I. Проблематика происхождения жизни на Земле. Объяснения в тексте.
Во время формирования Земли происходили высокоскоростные удары (рис. 3), и поэто-
му очень многие ученые считали, что поверхность Земли была горячей и даже расплавлен-
ной. Но это не совсем так. Если принять во внимание то обстоятельство, что тепло сбрасы-
валось не с помощью простой, регулярной свободной конвекции, а с помощью вынужденной
конвекции, то оказывается, что на достаточно холодной поверхности Земли были горячие
пятна (рис. 4). И такая сильно гетерогенная по температуре система обеспечивала возмож-
ность для процесса синтеза органических соединений. При ударах тел размером порядка
500 км и более выкипали океаны, сильно менялся климат Земли и ничего живого не оста-
валось. Но если ударные тела были размером порядка 10-20 км, то таких катастрофических
!"'PI>/;i“'l'bagelM;i)
Рис. 2. Возраст древнейших цирконов
(Cavosic et al., 2004).
Рис. 3. Формирование Земли (Герасимов. Мухин. 1986).
121
,7.Д/. Мухин. Ус ЮВ11Я на поверх пост и Земли 4-4.6 млрд, лет назад. Первичные синтезы
Рис. 4. Ранняя Земля - горячие пятна на холодной поверхности (Mukhin, Pimenov, 2002): а - условия
вынужденной конвекции; о - условия свободной конвекции. Обозначения: I - температура поверх-
ности, 2 - температура на глубине 10 см.
последствий не было. Таким образом, предложенная модель холодной поверхности Земли
с горячими пятнами должна была работать.
При ударе происходило испарение, в самом факеле происходили высокотемпературные
химические реакции (рис. 5), которые приводили к образованию атмосферы, содержащей
очень благоприятные компоненты - такие как синильная кислота, альдегиды - для синтеза
органических соединений. О составе этой атмосферы можно судить по газам, содержащимся
в земных породах и метеоритах на период времени их формирования (табл. I). Интересно,
что эта атмофера неравновесна: в ней есть полностью окисленные компоненты, такие как
СО,, и есть компоненты восстановленью; нет аммиака, а есть только свободный азот. Тем
Рис. 5. Физико-химические процессы, происходившие при паде-
нии на Землю планетезималей (Герасимов, Мухин, 1986).
122
Проблемы происхождения жизни
Таблица 1. Химический состав реликтовых газов (Mukhin et al., 1989)
Initial concentration (wt%) Mass loss (mg) Gases (IO'6 g)
Sample С s H2 n2 CO CO2 HC+ SO, H2S HCN CHjCHO CO/ COt CO+CO,/ НС
Delection limit 0.2 0.1 0.1 0.02 0.02 0.1 0.08 0.06 0.02
Не атмосфера
Augite ND ND 10.8 <0.2 0.4 3.3 2.7 0.7 <0.1 <0.08 <0.06 0.02 1.2 8.6
Базальт 0.04* <0.01* 20.1 <0.2 0.5 3.5 5.9 0.6 <0.1 <0.08 <0.06 -0.02 0.6 16
Перидотиты 0.03* 0.03* 10.2 2.6 0.1 25.4 21.4 1.3 ND ND ND ND 1.2 36
Г аббро 0,05* 3.04* 14.0 19.6 0.9 36.3 28.3 2.6 240 1.8 0.7 0.2 1.3 25
L5 («Царев») ND 1.92" 7.3 6.2 1.9 36.3 19.8 2.5 17.7 ND 0.2 -0.08 1.8 22
СЭО (Канзас) 0.61” 2.06” 10.5 18.6 1.9 236 94.3 12.2 <0.1 ND ND -0.04 2.5 27
Н2О атмосфера
Augite 9.1 ND ND 25.6 9.0 8.2 <0.1 <0.08 <0.06 <0.02 2.8 4.2
L5 («Царев») 7.1 ND ND 16.9 10.8 7.0 <0.1 162 ND -0.1 1.6 4.0
СЭО (Канзас) 9.7 ND ND 266 46.4 25.6 <0.1 149 0.4 -0.2 5.8 12
ND - not determined
Ccrtiified by IGEM Acad. Sci. USSR
+ -1IC, the sum of hydrocarbons.
не менее, эти данные приводят к выводу о том, что 3.8-3.9 млрд, лет назад не было никакой
восстановительной атмосферы. Наблюдаются следы кислорода, который должен был образо-
вываться во время высокотемпературных процессов и очень быстро реагировать с другими
компонентами взрыва. Это важно потому, что в присутствии кислорода не проходят никакие
абиогенные синтезы органических соединений.
Органические вещества образуются не только в Солнечной системе, они образуются во
всем цикле формирования планет из протопланетного облака. Вообще, на самом деле космос
просто «набит» органикой. В 2006 г. было известно 146 межзвездных молекул. Среди них
очень много органических соединений, но нет фосфора1. Таким образом, органические моле-
кулы из космоса дают не очень много для модели происхождния жизни.
Накоплению предшественников органических соединений на поверхности ранней Зем-
ли придавалось очень большое значение. Основным источником добиологической органики
считались кометы (Chyba, Sagan, 1997) (табл. 2). Но здесь вновь возникает проблема фосфора.
Таблица 2. Источники добиологического органического
вещества на ранней Земле (Chyba, Sagan, 1997)
Земные источники (kgyr-’)
УФ фотолиз 3 х 108
Электрические разряды 3 х 107
Ударные воздействия 4х 102
Гидротермальные источники 1 х 108
Внеземные источники (kgyf1)
IPDs 2х 108
Кометы 1 х 10"
Всего 10"
1 В настоящее время в молекулярных облаках обнаружены такие соединения как СР, PN, РО - см. статью
Н.Г. Бочкарёва в данном сборнике и Итоговую дискуссию, с. 252 -253. - Прим. ред.
123
Л.М. Мухин. Условия на поверхности Земли 4-4.6 млрд, лет назад. Первичные синтезы
Таблица 3. Обилие химических элементов во Все-
ленной (Chang, 2007)
Элемент Долей на миллион
Водород 739 000
Гелий 240 000
Кислород 10 700
Углерод 4 600
Неон 1 340
Железо 1 090
Азот 950
Кремний 650
Магний 580
Сера 440
Фосфор 14
Все прочие 650
Распространенность этого элемента во Вселенной на два порядка меньше, чем распростра-
ненность углерода (табл. 3). В то же время в живых системах фосфора больше, чем серы, т. е.
он занимает очень значительное место. К примеру, в организме человека отношение по массе
углерода к фосфору составляет 15 (табл. 4). Вопрос - почему и как живые организмы нака-
пливали фосфор, и как он оказался в достаточном количестве для этого накопления?
При анализе полученного вещества межзвездных пылинок фосфора в них пока
не обнаружено.
Таблица 4. Химический состав тела человека (Chang, 2007)
Элемент Процент от массы
Кислород 65
Углерод 18
Водород 10
Азот 3
Кальций 1.5
Фосфор 1.2
Калий 0.2
Сера 0.2
Хлор 0.2
Натрий 0.1
Магний 0.05
Железо, кобальт, медь, цинк, йод Селен, фтор <0.05 каждый <0.01 каждый
124
Проблемы происхождения жтни
По последним данным (табл. 5),
в кометах мы находим синильную кислоту,
много СО, альдегиды, но не обнаруживаем
ни одного соединения с фосфором. В то же
время, среди органических веществ в ме-
теорите Мурчисон и хондритах есть даже
пурины и пирамидины, есть аминокисло-
ты и есть фосфор в твердой фазе (табл. 6).
Если фосфор в кометах есть, то он
содержится только в ядре кометы, в твер-
дой матрице. Другого места для размеще-
ния фосфора просто нет. На рис. 6 показана
схема пребиотического синтеза в кометах
(Oro, Cosmovici, 1997). Должен отметить,
что уже на стадии пуринов, пирамидинов,
сахаров эта модель встречается с очень
значительными трудностями. Пурины и
пирамидины входят в состав химической
структуры ДНК и также в состав РНК.
Фосфор является одним из ключевых ком-
понентов нуклеотидов. Синтез пуринов
и пирамидинов является очень важной
ступенькой в образовании нуклеиновых
кислот. Аминокислоты образуются срав-
нительно легко, но необходимо получение
не просто аминокислот, а аминокислот
в определенном порядке, который обеспе-
чивает трансляционная машина. При син-
тезе самых простых органических соеди-
нений мы сразу же сталкиваемся с рядом
трудностей. На рис. 7 представлена авто-
каталитическая реакция синтеза сахаров,
открытая еще Бутлеровым. В ней образу-
ется огромное количество сахаров, при-
чем не только рибоза или дизоксирибоза,
которые необходимы для РНК и ДНК, но
и масса других. И немедленно начинают
идти побочные реакции с другими саха-
рами. Из них образуется либо карамель,
либо метиловый спирт и мочевина1. Син-
тезы аденина и цитозина тоже встречают
ряд трудностей.
Еще одна важная вещь, кото-
рую я хотел бы отметить, и на кото-
рую обычно не обращают внимания. 1
Таблица 5. Органические вещества в кометах
(Bockelee-Morvan et al., 2000)
Молекулы Относительное обилие
Н2О 100
СО 23
со2 6
сн4 0.6
С2н2 0.1
с2н6 0.3
СН3ОН 2.4
Н2СО 1.1
нсоон 0.1
СНзСНО 0.02
НСООСНз 0.08
nh2cho 0.02
NH3 0.7
HCN 0.25
HNC 0.04
HNCO 0.1
CH3CN 0.02
H2S 1.5
cs2 0.2
cs 0.2
so2 0.2
so 0.3
ocs 0.4
H2CS 0.02
NS 0.02
1 Реакция Бутлерова изучалась в последнее время в Институте катализа им. Г.К. Борескова СО РАН -
см. ссылки на публикации в статье В.Н. Снытникова в данном сборнике. - Прим. ред.
125
Л.М. Мухин. Условия на поверхности Земли 4-4.6 млрд, лет назад. Первичные синтезы
Таблица 6. Органические вещества из метеорита Мур-
чисон и нескольких хондритов (Ehrenfreund et al., 2002)
Классы органических веществ Концентрация в миллионных долях (мг/кг)
Аминокислоты
- соединения типа CCs в метеорите Мигеи 17-60
- соединения типа CCs в метеорите Ивуна ~5
Алифатические гидрокарбонаты >35
Ароматические гидрокарбонаты 3319
Фуллерены >100
Карбоксиловые кислоты >300
Гидрокарбоксиловые кислоты 15
Пурины и пиримидины 1.3
Спирты 11
Сульфоновые кислоты 68
Фосфоновые кислоты 2
Если нам нужно синтезировать РНК или ДНК, то мы должны иметь кооператив ну-
клеотидов, то есть они должны быть примерно в равных пропорциях и синтезировать-
ся по одинаковой схеме. Даже при синтезе аденина, который из этой группы нуклео-
тидов синтезируется легче всего, возникает ряд существенных сложностей. В частно-
сти, синтез аденина требует концентраций цианистого водорода по меньшей мере 0.01 М,
что совершенно нереально в условиях примитивной Земли. Кроме того, аденин подвержен
гидролизу (время полужизни для деаминации при температуре 37 °C, pH 7 - около 80 лет).
Следовательно, нельзя ожидать накопления аденина в условиях любого «пребиотическо-
Cometary
Precursors
H,O + NH
Monomers
HCN
HCN
H;O + NH:
HC,N
H2O + Urea
HCHO
H, + Nickel - Iron Grains . . .
CO --------———Fatty Acid
Fischer-Tropsch Processes
Amino acids
Purines
Pyrimidines
Ribose (-5P)
Protocell
+ Choline or Serine
(liposomes)
Рис. 6. Схема пребиотического синтеза в кометах (Oro, Cosmovici, 1997).
126
Проблемы происхождения жизни
СН2ОН-СНО
СН2ОН«СНО
сн2он-снон-снон- сно СН2ОН**снон-сно
А
V V
сн2он-снон-с-сн2он <СН2° СН2ОН-С-СН2ОН
II II
о о
Рис. 7. Схема автокаталической реакции синтеза сахаров.
го супа». К тому же, взаимодействие аденина и урацила слабо и неспецифично, и поэтому
никогда нельзя ожидать его функционирования в любой специфической схеме узнавания
при хаотических условиях «пребиотического супа».
Представляется очень важным вывод Л. Оргейла (Orgel, 2004). Он говорит, что неиз-
бежное заключение исследования синтеза нуклеотидов состоит в том, что нет убедительных,
общих пребиотических синтезов каждого из нуклеотидов. Крайне ограниченное количество
реакций дает высокий выход продуктов; а те, что дают, обычно производят сложные смеси
продуктов. Я уже продемонстрировал это на примере синтеза сахаров. Здесь необходимо за-
метить, что вопрос хиральности является исключительно сложным для проблемы синтеза,
потому что как только мы имеем рацемическую смесь сахаров, сразу терминируется рост
следующего звена. Не менее важным мне кажется заключение Дж. Джойса (Joyce, 1989), ко-
торый считал, что наиболее разумным предположением является то, что жизнь не началась
с РНК, а переход к миру РНК и происхождение жизни в целом полны неопределенностей
и отсутствием экспериментальных данных.
Проблема хиральности до сих пор не решена, и все смеси, которые образуются в экс-
периментах по предбиологическому синтезу, являются рацемическими смесями. И можно
сказать, что это обстоятельство стоит на пути решения проблемы происхождения мира РНК,
потому что еще до его появления необходимо получить нерацемические смеси. Еще одной
проблемой являются упомянутые побочные реакции, которые сильно мешают выделению
основного продукта.
В заключение приведу некоторые соображения насчет уровня содержания кислорода
в примитивной атмосфере - она на начало геологической летописи не была восстановитель-
ного характера, и уже 3.5 млрд, лет назад содержала значительно количество кислорода. Это
дает возможность поставить здесь барьер на временной шкале органических синтезов.
Закончить я бы хотел одним любопытным фактом. Была жуткая вспышка энтузиазма в
1950-х гг. на первой конференции по происхождению жизни. После этого 90 % работ по этой
проблематике поддерживали эволюционную идею. Однако трудности этого вопроса оказа-
лись столь велики, что в 2006 г. число публикаций, посвященных искусственному зарожде-
нию жизни на Земле (т. е., связанному с идеей творения), оказалось близко к 80 %. Таково
мировоззренческое значение проблем происхождения жизни...
127
Л.М. Мухин. Условия на поверхности Земли 4-4.6 млрд, лет назад. Первичные синтезы
СПИСОК ЛИТЕРАТУРЫ
Герасимов М.В., Мухин Л.М. 1986. Как и когда появились атмосфера и океан? // Природа. № 8.
С. 22-32.
Bockelee-Morvan D., Lis D.C.. Wink J.E. et al. 2000. New molecules found in comet C/1995 01 (Hale-Bopp).
Investigating the link between cometary and interstellar material // Astron, and Astrophys. V. 353.
P. 1101-1114.
Cavosie A.J., Wilde S.A., Liu D. et al. 2007. Internal zoning and U-Th-Pb chemistry of Jack Hills detrital
zircons: a mineral record of early Archean to Mesoproterozoic (4348-1576 Ma) magmatism // Precam-
brian Res. V. 135. № 4. P. 251-279.
Chang R. 2007. Chemistry. 9 ed. New York: McGraw-Hill. 1061 p.
Chyba C.F., Sagan C. 1997. Comets as a source of prebiotic organic molecules for the early Earth / Eds. P.J. Tho-
mas, C.P. McKay: New York: Springer-Verlag. P. 147-174.
Ehrenfreund P.. Irvine W.M., Becker L. et al. 2002. Astrophysical and astrochemical insights into the origin
of life//Repts. Prog. Physics. V. 65. P. 1427-1487.
Joyce G.F. 1989. RNA evolution and the origin of life H Nature. V. 338. P. 217-224.
Mukhin L.M., Gerasimov M., Safonova E. 1989. Origin of precursors of organic molecules during evaporation
of meteorites and mafic terrestrial rock // Nature. V. 340. P. 46-48.
Mukhin L.M., Pimenov K.Y. 2002. Impact hot spots on the cold surface of the early Earth // Planetary
and Space Science. V. 50. P. 41-43.
Orgel L.E. 2004. Prebiotic chemistry and the origin of the RNA world /7 Crit. Rev. Biochem. Mol. Biol. V. 39.
P. 99-123.
Oro J., Cosmovici C.B. 1997. Comets and life on the primitive Earth // Astronomical and biochemical origins
and the search for life in the Universe. Proc. 5 Intern. Conf. Bioastronomy. Capri. 1996. P. 97-120.
Conditions on the Earth 4-4.6 GA. The primary syntheses
L.M. Mukhin
During the period of 4-4.6 GA the Earth surface could be relatively cold but with isolated hot
spots caused by impacts. Such heterogeneous system regarding the temperate could provide a pos-
sibility of primary syntheses of organic compounds. '•
128
Проблемы происхождения жизни
Дискуссия
по пленарному докладу Л.М. Мухина
«УСЛОВИЯ НА ПОВЕРХНОСТИ ЗЕМЛИ 4.6-4.0 МЛРД. ЛЕТ НАЗАД:
ПЕРВИЧНЫЕ СИНТЕЗЫ»
Председательствующий А.С. Спирин
М.Я. Маров: Откуда следует, что океан был уже четыре с половиной миллиарда лет
назад? Согласно известным мне геологическим данным и моделям, он появился где-то 3.8,
может быть, 3.9 млрд, лет назад. Действительно, в этой зоне аккреционного диска изначально
была высокая температура (около тысячи градусов, как показывают наши термодинамиче-
ские расчеты), и, следовательно, происходило обеднение летучими. К этому же времени от-
носится, вероятно, появление первых гидросиликатов. Другой вопрос - почему у Вас только
отдельные очаги на первичной Земле горячие? Вы полагаете, что изначально Земля была
холодной и горячие области обусловливались только очень крупными импактами?
Л.М.М.: Заключение о том, что гидросфера возникла достаточно рано, сделано на осно-
ве анализа изотопов кислорода в цирконах. Дельта 18О оказалась между 7 и 9 (а не 5, как в
магме), сделан вывод, что это были какие-то гранитные расплавы, которые получались от
взаимодействия магмы с водой. Нет другого решения этой проблемы.
М.Я. Маров: Но циркон - не летучий элемент, и такая связка могла быть, скажем, в
межпланетном пылевом веществе, так же, как и в корунде. Поэтому, с моей точки зрения, это
не является доказательством.
Л.М.М.: На мой взгляд, с цирконами все в порядке, они не могли быть привнесены, они
очень древние. В сочетании с возрастом, данные по изотопии кислорода и другие данные
коррелируют.
М.Я. Маров: Теперь о проблеме фосфора. На Землю летучие могли быть привнесены не
только кометами, но и астероидами, в частности, достаточно большой фракцией астероидов,
содержащих гидратированные минералы. Поэтому фосфор может быть найден.
Л.М.М.: Согласен. Но проблема существует.
А.Б. Рубин: Поясните по поводу содержания кислорода и окисления железа.
Л.М.М.: Соотношение двухвалентного и трехвалентного железа показывает, что был
молекулярный кислород.
М.Я. Маров: Возможно, изначально было много FeO, который, переходя в ядро, обе-
спечивал приток тяжелого элемента для роста ядра. Кислород высвобождался и параллельно
связывался в Fe2Or
Л.М.М.: Он очень быстро связывался.
А.В. Витязев: В кометах отношение D/Н очень не подходит для земного океана. В ко-
метах плохо с фосфором. Слабая надежда на неисследованные кометы семейства Юпитера. В
общем, идея, что большая часть воды была принесена кометами, сейчас выглядит слабее.
Л.М.М.: Да, абсолютно правильно.
А.В. Тутуков: Я специально занимался температурой Земли на стадии ее формирова-
ния. На этапе 300 миллионов лет средняя поверхностная температура была бы только 100
градусов Кельвина. Для того, чтобы повысить температуру до 1 000 градусов, нужно, чтобы
время аккреции сократилось до 30 миллионов лет. Если Земля образовывалась из пыли, ее
температура была низкой, но от ударов крупных тел локально повышалась и до одной тыся-
чи и до нескольких тысяч градусов.
Л.М.М.: Я согласен с Вами. Критическим параметром здесь является время аккреции.
129
А.Ю. Розанов: Куда исчезли геологические породы древнее четырех миллиардов лет?
Л.М.М.: Есть цирконы, им 4.4 миллиарда лет.
А.Ю. Розанов: Это несколько десятков зерен. Где сами породы?
Л.М.М.: У меня нет ответа.
Н.Г Бочкарёв: Как Вы относитесь к работам Георгия Георгиевича Манагадзе по им-
пактному образованию сложных соединений, в том числе воды - происходит атомизация
ударного объекта, потом заново образование молекул. Им было показано, что все океаны
могли образоваться импактным образом.
Л.М.М.: Георгий Георгиевич - блистательный инженер, но то, что он пишет в своих
статьях, по меньшей мере, сомнительно...
М.Я. Маров: Отношение D/H в кометах действительно вдвое выше, чем в земных океа-
нах, но это результаты относятся только к долгопериодическим кометам, а что касается ко-
роткопериодических, в частности, семейств Юпитера, Нептуна, об этом мы пока ничего не
знаем.
Л.М.М.: Да, это существенно.
130
Проблемы происхождения жизни. М.: ПИН РАН, 2009. С. 131-155
РАННЯЯ ЗЕМЛЯ В ТЕСНОМ ОКРУЖЕНИИ МОЛОДЫХ ЗВЕЗД
© 2009 г. А.В. Витязев, Г.В. Исчерникова
Институт динамики геосфер РАН
avit@idg.chph.ras.ru
Данные о физико-химических условиях на ранней Земле важны для спе-
циалистов, работающих по проблеме зарождения биосферы. Обсуждается
последняя модификация теории формирования Земли и планет. Рассма-
триваются возможные последствия формирования Солнца в области звез-
дообразования в тесном окружении соседних молодых звезд. Приводится
обобщение классической задачи о темпе роста Земли и планет с новыми
оценками, позволяющими согласовать результаты как по долгоживущим
(уран-свинцовым), так и короткоживущим (гафний-вольфрам) космохро-
нометрам. Обсуждается модель ранней эволюции Земли, учитывающая
как новые динамические оценки, так и последние геохимические данные
(древнейшие австралийские цирконы, реликтовая плеяда ксенона). Указа-
ны проблемы теории ранней эволюции Земли, по которым пока не удается
получить удовлетворительного решения.
ВВЕДЕНИЕ
Продвижение в решении проблемы зарождения земной биосферы так или иначе зави-
сит от ограничений, следующих из моделей образования планет. Идеи межзвездной или меж-
планетной панспермии или самозарождения биоты на Земле требуют оценок астро-физико-
химической обстановки на нашей планете в первые полмиллиарда лет. Модели ранней Земли
основываются на нашем понимании процессов, приведших к формированию Земли и планет.
Предлагавшиеся модели «огненно-жидкой» и конкурентной «холодной» первичной Земли
базировались на тех или иных космогонических представлениях, появлявшихся после зна-
менитой гипотезы Канта-Лапласа.
В 1970-1980 гг. благодаря совместным усилиям российской (московской), японской,
американской и европейской школ был построен так называемый «стандартный сценарий
происхождения Солнечной системы» (описание см. Hayashi et al., 1985; Витязев и др., 1990;
более поздние обзоры, включая Wikipedia). В статье эта модель представлена схематически,
как часть новой более общей модели (см. ниже рис. 6).
Первый раздел посвящен обобщенному подходу к исследованию происхождения око-
лозвездных протопланетных систем в гигантских молекулярных облаках. Здесь мы обсуж-
даем разные возможности образования Солнечной системы (СС), формирующейся в тесном
131
A.B. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
окружении других молодых звезд (Pechernikova, Vityazev, 2007). Здесь же объясняются новые
понятия - плазменные оболочки газопылевых дисков около молодых звезд - «ионоторы» и
«магнитоторы». Поскольку к системе уравнений грави-гидродинамики добавляется система
уравнений Максвелла для ионизованной среды, модификация стандартной модели названа
нами «грави-магнито-гидродинамической моделью». В этом разделе, по сути, предлагается
принципиальное решение старой (весьма острой) проблемы о потере первичным газопыле-
вым диском более 90 % его первоначальной массы.
Во втором разделе дается краткое описание «стандартного сценария формирования
планетной системы Солнца». Приводятся новые оценки по уточненной формуле для скоро-
сти роста Земли. Полученная короткая шкала роста твердых планет на ранней стадии по-
зволяет интерпретировать показания Hf-W-геохронометра как указание на плавление и диф-
ференциацию в родительских планетезималях в первые десятки миллионов лет жизни СС.
С другой стороны, подтверждена оценка общей длительности завершающего этапа роста
Земли - 100 млн. лет.
В третьем разделе приводится описание агрегированной модели ранней Земли. Мы
продолжаем придерживаться полученного нами ранее вывода о том, что Земля не была ни-
когда ни «огненно-жидкой», ни «холодной». Нагреваемая ударами падающих тел, массы
крупнейших из которых могут на заключительных стадиях составлять до 1 % от массы пла-
неты, растущая планета временами имеет гигантские очаги расплавов. В этих гигантских
подземных, но все же локальных, морях магмы должна происходить дифференциация. При
определенных условиях, описываемых критерием «теплового взрыва», энергии гравитаци-
онной дифференциации становится достаточно, чтобы локальная дифференциация перешла
в глобальную. Обсуждаются процессы формирования примитивных атмосферы и гидро-
сферы. Здесь же приводится сводная схема блоков, составляющих референц-модель ранней
эволюции Земли (см. ниже рис. 8).
В Заключении приводятся остающиеся нерешенными основные проблемы моделей
ранней Земли. Имея в виду общую направленность сборника, мы сочли необходимым огра-
ничиться минимумом формул, отсылая читателя к соответствующим публикациям.
1. ФОРМИРОВАНИЕ ПЛАНЕТНЫХ СИСТЕМ В ТЕСНОМ ЗВЕЗДНОМ ОКРУЖЕНИИ
Астрофизические и космохимические аспекты
Одним из важнейших открытий за последние менее чем полстолетия является обнару-
жение множества планетных систем около других звезд в Галактике. К настоящему времени
найдено более 300 экзопланет около -100 молодых звезд.
Более того, обнаружены и исследуются тысячи газопылевых, вероятно, протопланет-
ных, дисков около молодых звезд. Таким образом, спустя два с половиной столетия после
знаменитых работ Канта-Лапласа, идея о происхождении планет из околосолнечного газо-
пылевого диска обрела экспериментальное подтверждение. Да, у нас нет возможности свои-
ми глазами увидеть раннюю Солнечную систему. Но мы можем наблюдать рождение планет-
ных систем в современной Галактике около звезд солнечного класса. Можно сказать, что мы
не знаем, каким был этот дуб в молодости, но мы теперь видим деревья в нашей дубраве на
разной стадии их жизни.
В последние два-три года мы пытаемся сделать следующий шаг: восстановить астро-
физическую обстановку вблизи формирующейся Солнечной системы. Понимание того, что
звезды не формируются поодиночке, а возникают группами в огромных туманностях типа
знаменитой туманности Ориона, было достигнуто более полувека назад. Однако послед-
ствия такого совместного рождения планетных систем для нашей Солнечной системы по
сути остаются не исследованными. Здесь имеется несколько важных подпроблем. Первая -
как долго наша СС находилась в туманности и, вероятно, затем в звездном скоплении типа
132
Проелемы upoik xoajehhh жниш
Проплайд (схема)
Протопланетный диск
Рис. I. Схема протопланетного диска (проплайда) (с/) и артвидение протопланетного диска, обдувае-
мого звездным ветром близкого голубого гиганта (б).
Плеяд или Гиад (без газопылевого окружения)9 Какие космохимические и динамические
следствия тесного звездного окружения имели место для молодой СС?
В связи с первым вопросом заметим, что характерные времена жизни туманностей, в
которых рождаются звезды с планетными системами (и без них), порядка десятков миллио-
нов лет. Голубые гиганты своим звездным ветром рассеивают газо-пылевую составляющую,
и на месте туманности остаются звездные скопления. Звездные скопления распадаются го-
раздо медленнее, и известны NGC 752, М 67, Гиады, Ясли, существующие от первых до мно-
гих сотен миллионов лет.
В связи со вторым вопросом следует обратиться к характеристикам астрофизической
обстановки у наблюдаемых протопланетных дисков в молодых туманностях. На рис. I по-
казана схема типичного протопланетного диска около молодой звезды солнечного класса,
который испытывает сильнейшее влияние звездного ветра, рентгеновского и УФ излучения
от близкого голубого гиганта. Говоря коротко, в молодую СС с уже сформировавшимися
крупными телами и протопланетами в первые десятки миллионов лет после образования
Солнца может по-прежнему поступать свежий материал от окружающего звездного населе-
ния. По массе в сравнении с массой протопланетного диска это может быть и незначительная
доля. То есть на химический состав молодой СС влияние привноса вещества может быть
пренебрежимо малым.
Если инжектируются свежие радионуклиды короткоживущих элементов (см. табл. I),
по дочерним которых мы хронометрируем ранние стадии эволюции СС, это может быть су-
щественно! Кроме того, если в молодой СС уже есть сформировавшиеся планеты и другие
меньшие тела (астероиды и кометные ядра), помимо потока солнечного ветра и солнечного
излучения может быть важным звездный ветер и космические лучи из ближайших окрест-
ностей, действующие на приповерхностные слои.
Напомним, что космические лучи - это массивные частицы, протоны, ядра гелия и
т. д., фотоны (гамма, рентген, ультрафиолет и ИК). Общая плотность энергии ~1 эВ/см\ Для
сравнения - плотность светового излучения звезд ~ 0.3 эВ/см3. Во время формирования СС в
туманности плотность излучения могла быть на два-три порядка выше.
Итак, мы считаем важным обратить внимание на возможность перенормировки космо-
и гео- хронометров ранних стадий эволюции СС.
133
.-I.В. Внтяи'в. Г.В. Псчирпикоян. Ранняя Земля в тесном окружении молодых звезд
Таблица 1. Короткоживущие нуклиды в ранней Солнечной системе
(Goswami, Vanhala, 2000)
Нуклид Время полураспада (млн. лет) Дочерний нуклид Звездный источник"
41Са 0.1 4,К SN. AGB. WR
26А1 0.7 26Mg SN. N. AGB. WR
60Fe 1.5 60Ni SN. AGB
53Мп 3.7 53Cr SN
l07Pd 6.5 l07Ag SN. AGB. WR
'82Hf 9 l82W SN
129j 15.7 ,29Xe SN
244Pu 82 a. SF6 SN
адТсс 0.21 99Ru AGB. WR
36C1£ 0.3 36Ar SN. AGB. WR
205pbr 15 AGB. WR
92Nbc 35 92Zr SN
" SN - сверхновая: N - новая: AGB - асимптотическая ветвь гигантов; WR - типа Вольфа - Райе;
продукты спонтанного деления;
' необходимо подтверждение.
Средний космический состав и состав допланетного газопылевого диска
Многолетние исследования в ИК и радиодиапазонах состава газа и пыли в галактиче-
ских туманностях и молекулярных облаках, данные о металличности звезд разных поколе-
ний, данные по фотосферам Солнца и ряда звезд, данные о составе метеоритного, астероид-
ного и кометного вещества позволяют судить о среднем космическом и солнечном составах.
На диаграмме (рис. 2) показана верхняя часть таблицы Д.И. Менделеева с одновременным
Трехмерная таблица Менделеева
современно! о космически! о обилия >. icmciitoh
Рис. 2. Современный космический состав.
134
Проблемы происхождения жизни
Таблица 2. Основные компоненты газопылевого диска
Обилие элементов Среднее космическое Для Солнечной системы (метеориты)
X - массовая доля водорода (Н), 0.75 0.706
Y - массовая доля гелия (Не) 0.23 0.275
Z - массовая доля всех других элементов («металлов») 0.02 0.019
указанием среднего космического содержания наиболее обильных элементов (Anisochkin
et al., 2007; Анисочкин и др., 2007).
Состав первичного газопылевого диска восстанавливается по данным о метеоритах
и кометах и содержаниях элементов в фотосфере Солнца. Для грубых расчетов по динамике
газо-пылевого диска используют три основные компоненты (табл. 2).
Список обнаруженных молекул в молекулярных межзвездных облаках от двух и более
до десяти атомных огромен. На рабочем совещании он демонстрировался еще в двух вы-
ступлениях - Н.Г. Бочкарёва и В.Н. Снытникова. Поэтому мы его здесь не приводим, отсы-
лая читателя к интернетовским сайтам. Отметим обычно не подчеркиваемый факт: обилие
многоатомных молекул в межзвездной среде падает почти экспоненциально с ростом числа
атомов в данной молекуле.
Ситуация с молекулярным составом газа и пыли в протопланетных дисках около моло-
дых звезд и в околосолнечном газопылевом диске является предметом обширной дискуссии.
В рамках этой статьи мы лишь отметим, что переработка вещества в ходе формирования
околосолнечного диска могла быть неполной и часть вещества на окраине диска могла иметь
состав как газа, так и пыли, близкий к межзвездному. Более того, согласно выше сказанному
в этом пункте, часть межзвездного вещества могла поступать в молодую СС уже в ходе про-
двинутого формирования планет.
Грави-магнито-гидродинамика газопылевого диска около молодого Солнца
Согласно стандартной модели умеренно массивного (Д, = 0.03-^0.07A/Q), тонкого
/?«1), но оптически толстого протопланетного диска к окончанию аккреции на Солнце в ди-
ске имеется «мертвая» зона (7? ~ 0.1 ^20 а.е.), где ввиду низкой ионизации вблизи централь-
ной плоскости МГД эффекты пренебрежимо малы. Здесь H(R) = с XI - толщина однородной
атмосферы, cs - скорость звука, Q - угловая скорость вращения. В то же время над и под мерт-
вой зоной на высотах порядка нескольких характерных толщин Н имеется активный слой
с поверхностной плотностью а ~ 100 г/см2, в котором степень ионизации nJnH > 10 13 доста-
точна для развития магнито-ротационной неустойчивости MRI, открытой Е.П. Велиховым
(Velikhov, 1959) и интенсивно исследуемой в астрофизических объектах и лабораториях.
Мы исследовали слоистые модели постаккреционного диска (рис. 3), в котором после
седиментации пыли и последующей гравитационной неустойчивости в пылевом субдиске
вблизи центральной плоскости идет аккумуляции тел. В то же время в МГД активном верх-
нем слое продолжается турбулентный перенос углового момента и газа на периферию диска.
На рис. 6 (см. ниже) показана общая блок-схема грави-магнито-гидродинамической (ГМГД,
в английской транскрипции GMHD) модели допланетного диска, где знаком «***» отмечены
новые блоки, добавленные нами к стандартной модели.
В теории околосолнечного диска оставалась нерешенной проблема потери большей
части газа первичной околосолнечной туманности. Потеря газа по механизму Джинса (как
потери газов тяжелее Н и Не из современной атмосферы Земли) очень мала (Витязев и др.,
1990). Гипотеза о выметании газа из диска звездным ветром молодого Солнца требовала чуть
135
А.В. Витязев, ГВ. Печерникова. Ранняя Земля в тесном окружении молодых звезд
Звездный
(солнечн>й)
ветер
Рис. 3. Схема сечения молодого Солнца и околосолнечного протопла-
нетного газопылевого диска.
ли не стократной по сравнению с околосолнечным диском массы, т. е. еще одной солнечной
массы, что противоречит физике Солнца.
Еще одной загадочной и необъясненной проблемой является факт наличия намагни-
ченности метеоритных зерен. Лабораторные исследования естественной остаточной намаг-
ниченности (Natural Remanent Magnetization - NRM) многих фрагментов сотен метеорит-
ных образцов указывают на существование в допланетном диске магнитных полей 1-500
тТ. Неупорядоченное расположение намагниченных фрагментов в метеоритных образцах
свидетельствует о намагничивании, произошедшем до их объединения в единые агрегаты.
Было предложено несколько механизмов генерации магнитных полей в весьма различаю-
щихся моделях газопылевых дисков (сжатое межзвездное в молодом Солнце, генерируемое
в недрах раннего Солнца, самогенерация на аккреционной стадии и т. д.). Но они не смогли
обойти проблему «мертвой зоны».
Вынос газа на периферию протопланетного диска
Ниже мы рассмотрим возможность диссипации всего газа из диска благодаря магни-
торотационной неустойчивости (Vityazev et al., 2006; Зимина, Витязев, 2007). Этот вариант
соответствует случаю, когда формирующаяся СС находится вдали от голубых гигантов ком-
плекса звездообразования и процесс фотоиспарения несущественен. Сразу скажем, что в тер-
минах известной модели a-дисков Шакуры-Сюняева, значение ass в случае MRI (для тонкой
внешней оболочки с crg~100 г/см2) оказывается -0.01-0.1.
Впервые MRI было упомянуто в неастрофизическом контексте Е.П. Велиховым в 1959 г.
при рассмотрении стабильности потока Куэтта идеальной гидромагнитной жидкости. Его ре-
зультат был позже обобщен Чандрасекаром в 1960 г. 30 лет спустя эти ранние результаты были
полностью оценены, когда было установлено (Balbus, Hawley, 1991), что слабые магнитные
поля могут существенно изменить характер устойчивости аккреционных дисков.
Вг ~ 2
Неустойчивость требует наличия слабого (подтеплового, то есть -< с ) магнит-
ного поля. За таковое можно взять солнечное. MRI также требует, чтобы угловая скорость
3Q2 --
убывала наружу (-----< 0), что всегда выполняется в кеплеровских дисках (Q ос г 2 ).
3 In г
136
Проблемы происхождения жизни
В ряде работ (см., напр., Gammie, 1996) показано, что МГД турбулентность в диске,
порождаемая MRI, эффективна, если магнитное число Рейнольдса
2
V
Rew= —al, (1)
где 7 - коэффициент магнитной вязкости, va - альвеновская скорость. Rew может быть пере-
£
2 • 102 Т 2
писано, как функция электронной плотности хе и температуры Т. Так как , то
Хе
v 2 сН
Re w = — >= —
Qr] fir)
ДОУ хе У Н \ 1
/3 Д10’13 0.1 а.е. j &
(2)
где /? ~10-100 - плазменный параметр, Н - характерная высота атмосферы.
Для удовлетворения этого неравенства доля электронов должна быть больше хе= 10‘13
на г = 1 а.е. Согласно (Sano et al., 2000) для п = 10'5 см‘3 MRI требует степень ионизации
в 30 000 раз выше, чем стандартное значение £ся « LO 'V. В области образования молодых
звезд может быть на два порядка больше. Все равно не хватает двух порядков.
Для простой оценки степени ионизации можно использовать формулу пн £ = neni а,
пе ~ И-, где пн - концентрация нейтрального водорода, пе - концентрация электронов, nj - кон-
центрация ионов, а - параметр рекомбинации, который может быть найден по формуле
, л . 0.7
/1о4ал
«„(?)= 2.5-10
(3)
1015 см-3, пе = 106 см-3 , —► хе = 10“9;
10'3 см-3, пе = 105 см 3 , —► хе = 10-8 ;
1015 см-3, пе = 10s см-3 , —> хе = 10“10;
1013 см-3, пе = 104 см-3 ,—>*, = 10“9;
Т
\ /
Для Т ~ 300 К имеем а 10 " см3с '. Отметим, что мы не учитываем рекомбинацию
на пыли. Считается, что она быстро опускается к центральной плоскости (а = аА/).
Найдем степень ионизации: хг = пе/пн для одной и двух характерных высот атмосферы
диска:
сн = Ю14 С1, для пн =
для пн =
= 10“'6 с"1, для пн =
для пн =
Таким образом, видно, что если и взять на два-три порядка меньше, т. е. подняться
на одну-две характерные высоты в область низких плотностей, то требуемая степень иони-
зации достигается.
Оценим величину магнитного поля. Зная степень ионизации для температурного рас-
пределения Т ~ r'v\ согласно Сано и др. (Sano et al., 2000), легко находится оценка величины
магнитного поля в гауссах:
' г
1 а.е.
2G.
(4)
Кратко об основных уравнениях для переноса массы газа, вызванного MRI. В осно-
ву рассмотрения нами взята стандартная осесимметричная модель (д/д<р = 0 в среднем)
P(r, z,i)-p (г, z, /) - T(r, z, f) (давления, плотности, температуры) распределений в гравитирую-
щем газопылевом диске, находящемся в квазикеплеровом вращении около молодого Солн-
ца (описанная, напр., Hayashi et al., 1985; Витязев и др., 1990). Самые последние модели дис-
ка, основанные на наблюдениях в инфракрасном и милллиметровом диапазонах длин волн
дисков у звезд типа Т Тельца, учитывающие вертикальные градиенты температуры, ионно-
137
А.В. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
молекулярного состава и химические реакции (около 500 химических компонентов - молекул,
атомов и ионов и примерно 5000 химических реакций), созданы объединенной группой рос-
сийских и немецких астрофизиков (Semenov, Wiebe, Henning, 2004, 2006). По мнению Д. Вибе
(ИНАСАН), просматривавшего с нами этот раздел работы, мы вправе были здесь ограни-
читься стандартной моделью диска. Добавленная нами система уравнений МГД аналогична
широко используемой исследователями в этой области (см., напр., Sano et al., 2000).
Радиальное движение материи управляется переносом углового момента. Полная кар-
тина эволюции диска также требует рассмотрения притока и оттока с поверхности диска
и притока на центральную звезду. Уравнение для переноса массы газа, вызванного MRI, мо-
жет быть найдено, исходя из законов сохранения массы и импульса (Sano et al., 2000). Мы
приводим здесь уже обезразмеренное уравнение переноса
= ——г2 — гасг + е*(г,/) - (5)
dt г dr dr
где а - константа, определяемая Р-р-Т моделью диска, е - источники (аккреция или под-
питка из нижележащих слоев мертвой зоны), е, - поток диссипирующего по Джинсу газа
с учетом кеплеровского вращения. Формула для диссипации (занимает полстраницы, поэто-
му здесь не приводится) была получена нами [Витязев и др., 1990, с. 84, формула (8.16)].
Эта формула пригодна вплоть до наступления гидродинамического истечения (и описывает
нижний предел интенсивности диссипации).
Для температурной зависимости в стандартном диске Т ~ г'12 имеем а = 3/2. Иногда
берут Т~ гу&, поэтому мы исследовали уравнение с различными значениями о.. Граничные
условия выбираются из соображения, чтобы на некотором расстоянии г0 от Солнца из-за вза-
имодействия солнечного ветра с диском формируется некий фронт с постоянной плотностью
и нулевым потоком. Интенсивность солнечного ветра принималась равной современной.
Мы получили аналитические и численные решения упрощенной системы уравнений
для турбулентного переноса, вызываемого MRI.
Для значений констант аа = 20 г см'2, г0 = 0.1 а.е. и других общепринятых характерных
значений параметров стандартной модели получаются графики зависимостей поверхностной
плотности <т(г, /) в верхней оболочке ионо-магнитотора от радиальной координаты г в различ-
ные моменты времени. В зависимости от выбранной первоначальной массы диска получают-
ся однотипные, но несколько отличные по численным значениям кривые. Из-за ограничен-
ности места мы приводим сводный рис. 4, показывающий качественную картину переноса.
Рис. 4. Качественный график зависимости поверхностной
плотности <т(г, /) в верхней оболочке ионо-магнитотора от ра-
диальной координаты г в различные моменты времени.
138
Проблемы nrout .\<>,k.;miu>i жиши
Оценим характерное время г турбулентного переноса: г ~-/?О;М ~ 0.1г Ю’1'- 10' лет. Так
как оболочка ионо-магнитотора подпитывается из центральных, на порядок более плотных,
областей, то общее время диссипации газа из диска составляет порядка нескольких млн лет
(эта оценка сделана для .г. - 10"'). Итак:
I) Умеренно массивные протопланетные околозвездные диски имеют внешние оболоч-
ки с достаточной для возбуждения MRI степенью ионизации. Нами введены термины ионо-
магнитоторы для характеристики подобных образований (ср. ионосферы и магнитосферы
планет и звезд). Отличие магнитоторов протопланетных околозвездных дисков в том, что
магнитное поле в них не обязательно генерируется центральным телом.
2) Показано, что величина поля и параметры, характеризующие турбулентный перенос
вследствие MRI, могут быть существенными для перестройки структуры диска за приемле-
мое с астрофизической точки зрения время - 1-10 млн. лет).
3) Ввиду эффективного переноса газа истечение газа с периферии диска может про-
исходить за характерные времена порядка миллионов лет с формированием распределения
остающейся массы по закону г'22. Характерное время диссипации газа из диска может со-
ставлять ~ 1 — 10 млн. лет, что согласуется с астрофизическими данными времени суще-
ствования околозвездных газопылевых дисков. Дано возможное решение задачи о потере
порядка 90 % массы из околосолнечного протопланетного диска.
4) Оценки возможной величины магнитного поля около первых единиц Гаусса, что
важно в связи с проблемой источника намагниченности метеоритных зерен.
2. СТАНДАРТНЫЙ СЦЕНАРИЙ ФОРМИРОВАНИЯ СОЛНЕЧНОЙ СИСТЕМЫ
В 1970-1980 гг. была построена так называемая «стандартная модель происхождения
Солнечной системы» (СМПСС). Учитывая характер сборника, мы даем лишь сжатое описа-
ние модели, опуская почти всю математическую часть, уравнения, формулы и т. д. Кроме
Рис. 5. Стандартная модель эволюции газопылевого допланетного диска около молодого Солнца. Сле-
ва сверху вниз (1-4) - опускание пыли к центральной плоскости и образование пылевого субдиска,
уплощение пылевого субдиска, гравитационная неустойчивость в нем и его распад на пылевые сгу-
щения, сжатие пылевых сгущений и образование роя плотных тел астероидных размеров. Справа
(5-8) - объединение планетезималей в планеты: появление крупных тел, ударный нагрев, сильный
метаморфизм, плавление и испарение вещества, реконденсация, диссипация газа из СС и аккреция
его на Юпитер и Сатурн, образование облака Оорта и пояса Койпера, завершение процесса формиро-
вания планет и спутниковых систем (Левин, 1964; Витязев и др., 1990).
139
А.В. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
того, заметим, что мы предпочитаем называть это не моделью, а сценарием происхождения
Солнечной системы. Понятие сценария шире понятия модели, так как в сценарии использу-
ются десятки моделей для определенных стадий формирования и эволюции Солнца и его
планетной системы, находящиеся в разной степени разработки. С другой стороны, в сцена-
рии предполагается, что некоторые второстепенные «герои» могут стать с развитием теории
и наблюдений главными, а концовка может существенно измениться.
На рис. 5 показаны основные стадии эволюции околосолнечного допланетного диска и
приведены оценки времен продолжительности отдельных стадий.
Рис. 6. Блок-схема грави-магнито-гидродинамических моделей протопланетных
дисков около молодых звезд солнечного типа.
140
Проблемы происхождения жизни
Длительность стадий - это исходно динамические оценки, подтвержденные позже кос-
мохронометрией:
1,2----- 100 000 лет
3, 4---1 000 000 лет
5,6---10 000 000 лет
7, 8 - ~ 100 000 000 лет.
На рис. 6 показаны ключевые моменты строящейся теории. Каждый блок в схеме - это,
по сути, отдельное направление теоретических и/или экспериментальных исследований для
объяснения наблюдаемых данных. Обычно это десятки и сотни работ различных междуна-
родных исследовательских коллективов. На схеме показаны также связи между отдельными
направлениями и зависимость решения одних задач и проблем от других. Блоки 7-11 в свое
время являлись проблемами классической космогонии. Их принципиальное решение было
получено в рамках упомянутой «стандартной модели» (Витязев и др., 1990). Надо сказать,
что некоторые современные компьютерные анимации пока не могут дать ответы на клас-
сические вопросы о том, почему планетная система такая, какой мы ее видим. Отметим,
что по блокам 3-7, 13-17 дискуссия продолжается, ведутся компьютерные расчеты по все
усложняющимся моделям. Блоки 1 и 2 представляют последнюю модификацию сценария,
который обсуждается ниже. Блок 18 - это наше предложение астрофизикам искать вспыш-
ки, вызванные макроимпактами в формирующихся планетных системах около современных
молодых звезд.
Обобщение классической задачи о темпе роста Земли и планет
Характерные времена формирования планет - один из фундаментальных вопросов
планетной космогонии. Время роста Земли - важнейший параметр и для наук о Земле. Оцен-
ки возраста Солнечной системы (4.6 млрд, лет) и общей продолжительности формирования
Земли (около 100 млн. лет) подтверждены изотопными данными. Однако скорость роста Зем-
ли на ранних стадиях, которая является одним из определяющих моментов ее дальнейшей
эволюции, не соответствовала показаниям Hf-W геохронометра. (Надо сказать, что дискус-
сия по интерпретации этого геохронометра продолжается (см., напр., Kleien et al., 2008а, b).
Первая оценка скорости роста массы Земли в результате объединения «отдельных
небольших тел и более мелких частиц» была предпринята О.Ю. Шмидтом в 1945 г. Исходя
из этой оценки, он получил продолжительность формирования Земли I ~ 7-109 лет, что пре-
вышает возраст Солнечной системы. В.С. Сафронов (1954) при выводе скорости увеличе-
ния массы планеты учел барометрическое распределение плотности в допланетном диске
и гравитационное сечение столкновения планеты, что более чем на порядок увеличило
скорость роста. Соответственно, полученное им в 1954 г. время формирования Земли со-
ставило t ~ 108 лет, что впоследствии было подтверждено изотопными данными. Его фор-
мула скорости роста Земли
dm 4л(1 + 20) ( т \ 2
—-------1----------------г (О
dt Р у т® J
Здесь т и г - масса и радиус растущей планеты, Р - кеплеровский период обраще-
ния, <т0 - начальная поверхностная плотность твердого вещества в зоне питания планеты,
(1 - т/т .) - фактор уменьшения поверхностной плотности вещества в результате его вычер-
пывания растущей планетой, т:. - современная масса планеты (Земли), 0 - параметр Саф-
ронова: средняя относительная скорость планетезималей в зоне питания растущей планеты
равна v'2=Gm/0 г. В предположении постоянства 0 и плотности растущей планеты S после ин-
тегрирования (6) в (Сафронов, 1954, 1969) было получено время Z, роста планеты до величины
относительного радиуса z = r/r-,:. При 0 = 3, <70 = 10 г/см2 и <5 = 5.52 г/см3 время роста Земли до
т = 0.97т#. (z = 0.99) получилось равным 108 лет.
141
A.B. Витязев, ГВ. Печерникова. Ранняя Земля в тесном окружении молодых звезд
Формула (6) была получена при следующих предположениях:
1) зона питания планеты замкнута (нет обмена веществом);
2) полное слипание (объединение масс падающих тел и растущей планеты);
3) массы т' падающих на планету тел малы (т' « т},
4) орбита растущей планеты - круговая (ее относительная скорость v(m) = 0), для
остальных тел относительная скорость определяется формулой v'2 = Gm/Or,
5) поверхностная плотность твердого вещества в зоне питания планеты ^уменьшает-
ся (в результате его вычерпывания растущей планетой т) как ad = cr0 (1 -
Но откуда растущая планета «могла знать», до какой массы ей расти?
Нами (Витязев и др., 1978) была разработана самосогласованная модель расширяющих-
ся и перекрывающихся зон питания и рассмотрен рост крупнейших тел. Полуширина зоны
питания D/?(Z) тела с массой ди(/) определяется средним эксцентриситетом ё орбит основной
массы тел на расстоянии R от Солнца D7?(/) = ё(/) R ~ v'(t) R!VK, где VK - кеплеровская круговая
скорость. Так как v'(/) 0(2 r(t), то ё(/) r(t) и D7?(Z) ос r(t). Следовательно, ширина кольцевой
зоны питания 2D/? и масса находящегося в ней вещества Q увеличиваются пропорционально
r(z), в то время как w(Z) °с r’(Z). На ранней стадии роста w(Z)/g(Z)«l, но со временем все боль-
шая часть вещества сосредотачивается в m(t), отношение m(f)/Q(f) растет, и поверхностная
плотность твердого вещества в зоне питания планеты уменьшается от своего начального
значения <зй как <rd(Z) = <т0 [1 - m(f)/Q(t)]. Рост планеты прекращается, когда отношение m(t)/Q(f)
достигает единицы, т. е. т(1) —> ттт, где wmax определяется (в первом приближении по ё(/))
параметрами допланетного диска, и m(t)IQ(f) = (m(t)/ тта$п. Для зоны Земли /итах = т • .
Без 5-го предположения ВС. Сафронова, т. е. когда современная (максимальная) масса
планеты определяется параметрами самого допланетного диска, получено время роста пла-
неты до относительного радиуса z (в годах):
ln 1 + Z
2(1 + 20)сго 1 - Z ’
(7)
Рост массы Земли, вычисленный по формуле (7) при в = 2, а0 = 10 г/см2 и средней плот-
ности <5 = 4.5 г/см3, показан кривой 2 на рис. 7.
Распределение по массам и относительным скоростям в рое допланетных тел явля-
ется важнейшей характеристикой, определяющей динамику формирования планетной си-
стемы (Сафронов, 1969; Витязев и др., 1990). Распределение по массам допланетных тел
обычно принимается в виде простой степенной зависимости n(m\ t) dm' = n0(jt) (т')4 dm',
(1 < q < 2), где w0(Z) - коэффициент нормировки. Нормируем n(m', t) на плотность конденси-
рованного вещества в зоне питания рл и при тй« М, и т0- верхний и нижний пределы
распределения) получаем w0(Z) = (2 - q) М} 9-2 рл. Таким образом, распределение по массам
определяется не только видом функции п(т'), но и значением верхнего предела распре-
деления А/,. Ранее в работах по теории аккумуляции принималось т' « т (см. выше -
3-е предположение Сафронова). Нами (Печерникова, Витязев, 1979) в рамках модели ра-
стущих и объединяющихся зон питания получена зависимость массы наибольшего после
растущей планеты тела ди, или, согласно В.С. Сафонову и Е.В. Звягиной (Safronov, Zvjagina,
1969), практически совпадающей с т{ величины верхнего предела распределения А/, от m(t)
в виде M^i)/m(t) ~ m^tymtt) ~ 1 - m(t)IQ(t). Переходим от отношения m(f)/Q(t) к переменной z
и получаем A/,(z) (1 - z1) m(t).
Можно видеть, что на ранних стадиях аккумуляции планет, когда z«l, крупнейшие
тела зоны питания сравнимы по массе с растущей планетой (вопреки предположению 3).
Лишь когда m(t) начинает составлять заметную долю от Q(t) и становится существенным
уменьшение поверхностной плотности вещества из-за вычерпывания, растущая планета на-
чинает опережать по массе остальные тела в своей зоне.
142
Проблемы происхождения жизни
Учет влияния крупных тел на формирование планет позволил получить решения ряда
задач классической планетной космогонии, таких как эволюция эксцентриситетов и накло-
нов орбит растущих планет, формирование их осевого вращения, ранний ударный нагрев
и т. д. (Витязев и др., 1990). Ниже приведены результаты исследования влияния крупных тел
и на время роста планет (Печерникова, 2004, 2005).
Скорость увеличения массы планеты w(/) в области планет земной группы определяет-
ся в основном частотой ее столкновений A(m(t), т') с телами роя т' в зоне питания планеты.
Мы учли ненулевые массу т' и размер г' планетезималей в сечении их столкновений с пла-
нетой (отмена предположения 3), зависимость их относительных скоростей и от т', а также
эллиптичность орбиты растущей планеты (отмена предположения 4). В результате для dm/dt
получено интегро-дифференциальное уравнение (подробнее см. Печерникова, 2004, 2005):
, /М\ т'
—— = п0 (/) J* m''~q dm' + JA2 m''~q dm'
m0 t
(8)
где rri = (2 - q) - q) - средняя (с весовой функцией m' n(m') dm') масса тела в распределе-
нии.
Подставим в уравнение (8) все величины, зависящие от времени / непосредственно или
через массу растущей планеты m(t) - n0(t), crd(7), v(w(/)), учтем спектр скоростей планете-
зималей со среднеквадратичной относительной скоростью v'(w(0)> и, принимая во внимание,
что pv' = 4 cjd/P (Сафронов, 1954), получим соответствующее уравнение для времени роста
планеты. Интегрируя полученное уравнение при 6 = 2, находим время роста Земли до отно-
сительного радиуса z с учетом влияния крупных тел:
/ш = Ь6 (In - 0.43z - 0.04z3 , лет, (9)
сг0 i-z )
где b = 0.07 (слабо зависит от значений в и q). Рост относительной массы Земли, вычисленный
по (9) при тех же значениях параметров, что и по формуле (7), показан кривой 1 на рис. 7.
t, млн лет
Рис. 7. Рост относительной массы Земли: 1 - модель (Печерникова,
2004, 2005), учитывающая роль крупных тел, 2 - модель расширяю-
щихся зон питания (Витязев и др., 1990), 3 - модель 1 с мегаимпактом,
4 - формирование ядра Земли в процессе ее роста (Kleine et al., 2002;
Yin et al., 2002; Schoenberg et al., 2002), 5 - дифференциация лунной
мантии произошла позже ~ 60 млн. лет (Kleine et al., 2008а, b).
143
A.B. Витязев. Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
На рис. 7 можно видеть, что в предложенной нами модели учет роли крупных тел су-
щественно ускорил набор массы на ранней и основной стадиях роста планеты: массы Марса
(~ 0.1 т ) Земля достигает примерно за 17 млн. лет. К 50 млн. лет ее масса оказывается вдвое
большей, чем в ранних моделях. Однако время роста до 0.97 т(т. е. до г = 0.99 г ,) осталось
прежним ~ 108 лет.
Если на ранней стадии происходила не твердотельная аккумуляция, а планетезимали
сначала росли в виде постепенно сжимающихся газо-пылевых сгущений вплоть до масс по-
рядка двух-трех лунных, то время роста Земли до /и(/) на последующих стадиях сокращается
примерно на 10 млн. лет.
Новая модель дает времена роста масс тел на ранних стадиях, согласующиеся с изотоп-
ными данными образования тел с массами Весты и Марса. Решение проблемы трехкратно-
го расхождения космохимической и динамической шкал формирования Земли предложено
нами (Vityazev et al., 2003; Витязев, Печерникова, 2003). Образование тысячекилометровых
тел уже в первые миллионы лет и их разогрев короткоживущим 26А1 (время полураспада
0.72 млн. лет) и ударами до температуры плавления их недр позволяют интерпретировать по-
казания Hf-W-геохронометра как указание на плавление и дифференциацию в родительских
планетезималях и отсутствие дальнейшей существенной регомогенизации этого материала
в ходе последующей аккумуляции планет земной группы.
3. РАННЯЯ ЗЕМЛЯ. ПРОЦЕССЫ НА ЗАКЛЮЧИТЕЛЬНЫХ СТАДИЯХ
ФОРМИРОВАНИЯ ЗЕМЛИ И ЕЕ РАННЯЯ ЭВОЛЮЦИЯ
Проблеме состояния «первичной» Земли и ее эволюции в первые 500 млн. лет посвя-
щены тысячи публикаций. Отсутствие образцов пород указанных возрастов делает пробле-
матичным выбор рабочей модели из десятка гипотез, предложенных хотя бы в последнее
десятилетие. Мы занимаемся этой проблемой с конца 1980-х гг., когда нам стало ясно, что
Земля испытала существенный нагрев, частичное (неоднократное) плавление существенной
части ее вещества уже в ходе роста планеты. На рис. 8 дана общая схема строящейся тео-
рии, где каждый отдельный блок, по сути, представляет собой отдельную проблему, каждой
из которых посвящены десятки, а по изотопии и сотни исследовательских работ. Глав-
ной все же пока остается проблема дифференциации Земли на два основных резервуара -
ядро и мантию. В современной Земле они составляют 99.5 % по массе вещества и не менее
по энергетике. Мы не приводим известных профессионалам в этой области систем урав-
нений. В определенной мере они были выписаны в работах В.П. Мясникова и его школы
более тридцати лет назад (см., напр., Мясников, Фадеев, 1980). Проблемы описания возник-
новения примитивных геосфер лежат не в трудности создания численных 2D или 3D моде-
лей динамики ранней Земли. Проблема состоит в согласовании многочисленных указаний
(и при этом все же недостаточных) на общий характер начального состояния Земли со сторо-
ны астрофизики, планетологии, геохимии и геологии.
Проблема крупнейших тел: мегаимпакт или макроимпакты?
Земля - дифференцированная планета. Если о современном состоянии геосфер мы
знаем относительно много, то о времени их образования - пока удручающе мало. В первую
очередь важно определить, когда произошло расслоение (дифференциация) планеты на два
основных резервуара - ядро и мантию. Основатель московской школы планетной космогонии
академик О.Ю. Шмидт надеялся, что после создания работающей модели формирования Зем-
ли будут получены «начальные условия» для разработки моделей ее эволюции. В 1960-е гг.
В.С. Сафроновым (Сафронов, 1969), столь много внесшим в создание стандартной модели,
были получены оценки распределения температуры в формирующейся Земле. По его оценке,
к моменту завершения роста Земли ее температура была всюду ниже температуры плавле-
144
Проблемы происхождения жизни
Рис. 8. Блок-схема модели Земли в катархее.
ния с температурным завалом (800 К) в центре, максимумом (1500 К) на глубинах порядка
500 км и спадом к поверхностной температуре порядка 0 °C.
В наших ранних работах (1970-е гг.) с С.В. Маевой была построена компьютерная мо-
дель дифференциации Земли, в которой после разогрева долгоживущими U-Th-40K спустя
несколько сотен млн. лет происходит дальнейший нагрев, достижение температуры плав-
ления и начало дифференциации на железо и силикаты. Надо сказать, что в те годы не было
изотопных данных, позволяющих проверить эти модели. Существовали геохимические ги-
потезы (А.П. Виноградов, М.Н. Clark, К.К. Turekian, L.A. Grossman), о первичном возник-
новении железных ядер, на которые потом выпадает мантийный материал. Они были со-
вершенно неудовлетворительны с точки зрения динамики формирования планет. Однако
следует сказать, что гипотеза гетерогенной аккреции имела свои аргументы. Не исключено,
что некоторые ее моменты надо будет учесть в дальнейших исследованиях. Отметим, что
модель позднего формирования земного ядра вступала в противоречие с открытием ядра у
Марса с его древней поверхностью и верой всех планетологов, что Меркурий тоже прошел
раннюю дифференциацию.
145
A.B. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
В конце 1970-х гг. нами (Печерникова, Витязев, 1979, 1980) были пересмотрены резуль-
таты работ (Зиглина, 1976; Зиглина, Сафронов, 1976), посвященных оценке эксцентриситетов
и наклонов орбит растущих планет. Был сделан вывод о присутствии в зонах роста планет
крупных (тысячекилометровых) тел. Стало ясно, что верхний предел на размеры тел (100 км
по В.С. Сафронову), падающих на Землю, занижен на порядок (на три порядка по массе).
С этого момента стало ясно, что оценки температуры первичной Земли В.С. Сафронова за-
нижены в два-три раза. Следует отметить, что в известных монографиях (Сорохтин, Уша-
ков, 1991, 2002) эволюция Земли до сих пор рассматривается с начальными условиями по
В.С. Сафронову. Теперь можно с уверенностью говорить, что эти взгляды не соответствуют
современным данным. Так в 1995-1997 гг. несколькими группами на основе анализа изотоп-
ных систем Sm-Nd в SNC метеоритах показано, что дифференциация Марса - образование
ядра и выделение коры - произошла в течение первых 100 млн. лет. Указанные выше данные
по Hf-W нескольких исследовательских групп подтвердили наш вывод о ранней дифферен-
циации планетезималей и планет.
Мы продолжаем придерживаться полученного нами ранее вывода (Витязев и др., 1990;
Витязев, Печерникова, 1996) о том, что Земля не была никогда ни «огненно-жидкой», ни
«холодной». Нагреваемая ударами падающих тел со спектром масс n(ni) ос т'4, где q < 2,
а верхний предел («крупнейшее» тело после планеты), меняясь со временем, может на заклю-
чительных стадиях составлять до 0.01 от массы планеты, растущая планета временами имеет
гигантские очаги расплавов - высокотемпературные пятна на общем фоне субсолидусных
температур. В этих подземных гигантских, но все же локальных, морях магмы должна про-
исходить дифференциация. При определенных условиях, описываемых критерием «тепло-
вого взрыва» энергии гравитационной дифференциации становится достаточно, чтобы эта
дифференциация перешла в глобальную.
При имеющемся согласии по стандартной модели образования планет между западной
(в основном американской) и российской школами планетной космогонии есть существен-
ное расхождение по одному весьма важному вопросу (пп. 5, 6, 17 на рис. 6). Это - величина
верхнего предела в распределении масс падающих на Землю тел. Согласно гипотезе мега-
импакта образование системы Земля-Луна произошло вследствие столкновения первичной
Земли с планетой (Тейя) с массой порядка массы Марса (0.1 А/ф). Согласно нашей модели ма-
кроимпактов (Витязев, Печерникова, 1996; Vityazev, Pechernikova, 1998) массы крупнейших
падающих тел на поздних стадиях роста Земли не превышали массы Луны. Следствия для
этих двух подходов различаются кардинально. В отличие от нашей модели, первичная Зем-
ля после мегаимпакта представляет собой сплошной магматический океан. Формирование
Луны по американской модели было вызвано последствием этого единичного удара и прои-
зошло в результате аккреции выброшенного вещества менее, чем за несколько тысяч лет. По
нашей модели формирование Луны происходило в течение миллионов лет за счет вещества,
выброшенного с растущей Земли при макроимпактах и тел протопланетного диска (Витязев,
Печерникова, 1996). Следует сказать, что эта модель, как и другие модели, недостаточно раз-
работана. В геохимическом плане она имеет те же недостатки, что и модель мегаимпакта.
В динамическом плане не показана ее осуществимость: могут ли небольшие выбросы быть
удержаны на орбите? Но этот же вопрос относится и к модели мегаимпакта.
Представляется, что выбор в пользу тех или иных моделей должен будет сделан
на основании соответствия динамических параметров и космохимических данных на-
блюдаемым значениям.
В статье А.А. Боярчука и др. (1998) были приведены оценки максимально возможно-
го размера тела, падающего на Землю, исходя из имеющегося аналитического решения для
классической задачи двух тел. Сделан вывод, что удар тела размером с Марс ведет в случае
Земли к эксцентриситету ее орбиты, на порядок превышающему современный. Иными сло-
146
Проблемы происхождения жизни
вами, модель мегаимпакта с массой Тейи порядка массы Марса, по мнению указанных авто-
ров, не проходит. Мы рассмотрели обобщение классической задачи столкновения двух тел
в поле центрального тела с учетом потери части энергии и вещества при столкновении.
Диссипативные столкновения протопланетных тел - к синтезу небесной механики
и теории кратерообразования
При высокоскоростных (5-40 км/с и более) столкновениях тел происходит изменение
их внутренней энергии Ем, кинетической энергии Е и орбитального L и осевого Ls момен-
тов. В теории кратерообразования пренебрегают изменением орбитальной энергии мишени,
в то время как в задачах небесной механики не учитывают диссипативные потери энергии
и вещества. Используя физически интересную область малости ряда параметров, мы (Lazarev,
Vityazev, 2005,2006) получили аналитические выражения, связывающие коэффициенты дис-
сипативных потерь и динамические параметры тел после столкновения.
Постановка задачи. Пусть в сфере Хилла (г<7?и) центрального тела массы Мс (Солн-
ца MQ или планеты Мр) происходит столкновение двух тел с массами М} и Мг (Мг / «1).
В неинерциальной системе с центром в М} скорость тела перед столкновением V, где Г7ИК«1,
Ек = (GMJR)'11, Е7Ее»1, Уе = (2СЛ/|/г|)|/2 - скорость убегания с М}.
При столкновении часть вещества М+ тела М2 присоединяется к Мх (М+< М2,
часть вещества М4 из кратера покидает сферу Хилла образовавшегося тела
с массой (Л/, + Л/+) со скоростью Часть е, кинетической энергии удара му1/! идет на дро-
бление (si t ), выброс 2), плавление (ei 3) и другие диссипативные потери. При отсутствии
диссипативных потерь коэффициент биллиардности ц = 1, с ростом диссипативных потерь
ц = 1 — es —> 0. Обозначим е, 2 = MjM,, et, = £+ = VJV.
Решение задачи. В отличие от классической задачи двух тел небесной механики,
в уравнении для момента импульса появляется второй член справа: Му = (Л/, + AL+)V2 - М4У4.
И, например, если до столкновения тела и М2 двигались в одной плоскости, и тело М, име-
ло круговую орбиту, то для приращения эксцентриситета тела М} после столкновения мы,
используя известные интегралы задачи двух тел, получаем:
Де = ji/!(e, 2 - et,„ ej (1 - £,>2) + О(р, е, 2, е4 „ е+...). (10)
При ц = 1, е4|, £+ = 0 имеем классический случай. Для более сложных начальных усло-
вий (нецентрального удара и т.д.) формулы отягощены тригонометрическими функциями,
но вполне обозримы (Lazarev, Vityazev, 2005).
Из нашей оценки следует, что столкновения первичной Земли с телами размером
с Луну, но не Марса, не противоречат наблюдаемым эксцентриситету и наклону орбиты си-
стемы Земля-Луна. Получено решение ограниченной задачи двух тел небесной механики,
учитывающее изменение внутренней энергии тел при их столкновении. Путем интегрирова-
ния по функциям распределения тел по скоростям и массам теперь можно получать оценки
интегральных потерь в системах взаимодействующих тел.
Спектр масс и скоростей падающих тел, состав тел и его вариации
В наших расчетах по стандартному сценарию формирования Земли получены спектр масс
и скоростей тел, формировавших планету, и оценен темп роста планеты. Достаточно наглядное
представление о массах тел в зоне растущей планеты и частоты их падений дает рис. 9.
Осредненный состав тел в зоне планет земной группы, включая пояс астероидов, со-
гласно современной парадигме близок к составу хондритов. В то же время давно показано,
что ни из одного отдельного класса углистых (CI, CM, CV, СО), обыкновенных (Н, L, LL),
энстатитовых (ЕН, EL) и других хондритов и ахондритов Землю построить нельзя. Можно
подобрать смесь из известных классов метеоритов, представленных в земных коллекциях,
удовлетворяющую одновременно изотопным, геохимическим и геофизическим критериям
147
А.В. Витязев. Г.В. Печерникова.
Ранняя Земля в тесном окружении молодых звезд
Масса растущей Земли
Й 0.99 0.999 0.9999 0.99999
200 250 300 350 400 450 500
Время t (млн. лет)
Рис. 9. Уменьшение со временем частоты столкновений ра-
стущей Земли с планетезималями в интервале диаметров:
1 — 0.5 1 км, 2 — 1 2 км, 3 - 2 4 км, 4-4^8 км, 5 - 8 -Н6 км,
6 - 16 32 км, 7 - 32 64 км, 8 - 64 + 128 км, 9 - 128 256 км.
(Витязев, Печерникова, 1989; Печерникова, Витязев, 1989), но нет гарантии, что существенная
доля вещества была привнесена планетезималями в среднем хондритового, но в остальном
(содержание железа, степень окисленности и т.д.) весьма вариабельного состава, которые либо
полностью выпали на планеты, либо еще не обнаружены, например, в поясе астероидов.
Сводка данных по массовым расчетам ударов падающих тел
Диапазон размеров тел, падавших на растущую планету, огромен - от субмикронных
размеров для межпланетной и межзвездной пыли до сотен км в поперечнике для крупнейших
тел, т. е. составлял 13 порядков. Физика ударных процессов для падающих тел в диапазоне
1-10 км изучалась в последние годы многими исследователями в связи с проблемой формиро-
вания кратеров на поверхностях Земли и других планет. Для заключительных стадий расту-
щей Земли необходимо важны оценки последствий падений достаточно крупных - 10-100 км -
тел, так как именно они определяли в основном характер процессов на этой стадии. Понятно,
что процесс носит вероятностный характер, средняя частота падений показана на рис. 9.
На основании наших оценок присутствия крупных тел в зонах формирования планет было
предложено провести в ИДГ РАН рассчеты последствий падений тел диаметром от 1 км до со-
тен км, оценить размеры формирующихся кратеров, количество испаренного и расплавленного
вещества ударника, океанической воды, коры и мантии, выброшенного испарившегося и выпав-
шего сконденсированного на поверхность планеты вещества (Печерникова, Витязев, 2005).
В ИДГ РАН ведутся массовые расчеты процессов, сопровождающих падения тел, в до-
статочно широких диапазонах масс и размеров тел, скоростей падения, углов вхождения в
атмосферу, состава тел. Расчеты ведутся для различных вариаций свойств мишени, толщины
и состава коры, присутствия океана той или иной глубины, различных вариантов теплового
профиля и реологических свойств верхней оболочки. Для ранних стадий эволюции Земли су-
щественна относительно высокая частота падений, т. е. кумулятивные эффекты множествен-
ных ударов. Поэтому нами была поставлена цель - получение параметризованных формул для
интегральных оценок количества ударных расплавов, доли испаренного вещества и вещества,
выбрасываемого на высотные орбиты, эрозии атмосферы и т.п. при падении множества тел.
148
Проблемы происхождения жизни
Рис. 10. Отношение расплавленной массы
мантии (кружки) и коры (квадраты) к массе
ударника при ударах астероидов со скоростью
20 км/с (тонкие линии) и комет со скоростью
50 км/с (толстые линии) (Светцов, 2007).
Рис. 11. Отношение испаренной массы мантии
(кружки) и коры (квадраты) к массе ударника
при ударах астероидов со скоростью 20 км/с
(тонкие линии) и комет со скоростью 50 км/с
(толстые линии) (Светцов, 2007).
Согласно численным расчетам (Shuvalov, Dypvik, 2004; Svetsov, 2006; Светцов, 2007,
2008), испаренная масса верхней оболочки в единичном ударе составляет 0.07—0.1 от массы
ударника для 50-км тел (для астероидных тел, имеющих скорости около 15-20 км/с). Испа-
ренное породообразующее вещество быстро (сутки) остывает, конденсируется и выпадает на
планету. Очень малая доля его (порядка первых процентов) может уходить в межпланетное
пространство. Отношение массы расплавленной мантии к массе ударника для тех же тел со-
ставляет от 10 для 50-км тел до 100 для 400-км. Результаты этих расчетов подтверждают вы-
вод, полученный нами ранее (Витязев и др., 1990; Печерникова, Витязев, 2005). При падении
меньших тел объемы расплавов быстро уменьшаются с уменьшением размера ударника, и
выпадение части вещества в виде пыли и мелких обломков на растущую планету не приво-
дит к существенному нагреву и плавлению. На основной стадии роста планеты, когда были
достигнуты скорости падения больше 3-5 км/с и в то же время доля крупных тел по массе
составляла более половины, верхняя оболочка планеты в местах ударов крупных тел пре-
терпевала неоднократное плавление.
Сводка по энергетическим источникам и процессам теплопереноса
Сводка по энергетическим источникам и процессам теплопереноса дана в табл. 3.
Краткое пояснение к табл. 3. Энергия короткоживущих изотопов выделяется в первые мил-
лионы лет и существенна для нагрева крупнейших планетезималей и центральных областей
протопланет. Гравитационная энергия является самым мощным источником на стадии роста
планет, однако лишь ~10 % остается захороненной в недрах. В то же время энергия гравита-
ционной дифференциации в ходе возникновения двух крупнейших резервуаров - ядра и ман-
тии - почти полностью идет на нагрев недр. Во второй части таблицы приведены значения
эффективного числа Нуссельта, указаны крайние значения, достигаемые в разное время, для
соответствующих геосфер. Из табл. 3 видно, что на стадии роста Земли действовали удар-
ное перемешивание и адвективные течения, сопровождавшие уход тяжелой фракции в ядро
и на порядки превышавшие по мощности современные геодинамические течения.
Наша сводка по энергобалансу ранней Земли, приведенная в табл. 3 (Печерникова, Ви-
тязев, 2005; Адушкин, Витязев, 2007), по существу, есть «визитная карточка» Земли в первые
полмиллиарда лет ее истории.
149
А.В. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
Таблица 3. Энергобаланс Земли в первые 500 млн. лет
ИСТОЧНИКИ Эрг
1 Короткоживущие радиоактивные Al26, Fe60 1037
2 Долгоживущие радиоактивные U, Th, К40 4x1037
3 Энергия ударов падающих тел 2x1039
4 Энергия упругого сжатия 1037
5 Энергия дифференциации 1.5x1038
6 Энергия приливной диссипации 1037
7 Энтальпия падающих планетезималей 1037
8 Энергия химических реакций *и фазовых переходов* 1037
9 Солнечная энергия ИК - УФ + солнечный ветер 2.5х1041
*в зависимости от эндо- или экзотермичности знак « - » или « + »
ПРОЦЕССЫ ТЕПЛО-МАССОПЕРЕНОСА Nu
1 Ударное перемешивание 102-105
2 Адвективные течения 1О2-1О5
3 Конвективный теплоперенос 10
4 Кондуктивный теплоперенос, Диффузионный массоперенос I 1
5 Флюидный 7
6 Приливное пенеплирование 10
Ударный выброс на гелио- и геоцентрические орбиты 1-10% 44
Дифференциация на ядро, мантию и примитивную кору
Оценка суммарного выделения энергии при расслоении первично однородной пла-
неты на два основных физических и химических резервуара Земли была сделана впервые
в 1949 г. выдающимся российским геофизиком в Институте физики Земли Евгением Никола-
евичем Люстихом еще при О.Ю. Шмидте. Позже делались неоднократные попытки уточнить
эту оценку и у нас и за рубежом, но цифра 1,5><1038 эрг, полученная Е.Н. Люстихом, остается
предпочтительной. Следующие вопросы можно сформулировать так: каково распределение
этой энергии по радиусу планеты и темп выделения этой энергии?
Итак, важно указать время начала дифференциации планеты на силикатную и же-
лезную оболочки и ответить на основной вопрос - когда сформировалась основная масса
земного ядра? К концу 1960-х гг. была популярна модель известного американского физика
У. Ельзассера, исходившего из модели первично «холодной» Земли, разогревающейся в по-
следующий 1-1.5 млрд, лет за счет радиоактивных урана, тория и калия с достижением тем-
пературы плавления на глубинах порядка 500 км (со ссылкой на упомянутую кривую началь-
ной температуры Земли, полученную В.С. Сафроновым в 1959 г.). Схемы позднего образова-
ния ядра неоднократно обсуждались в упоминавшихся работах О.Г. Сорохтина с коллегами
из Института океанологии РАН. В наших работах 1980-х гг. был совершен переход к «те-
плой модели первичной Земли» с ранним образованием ядра (Витязев и др., 1990). Нами еще
в 1972 г. было показано, что если в планете образуется слой расплава с мощностью поряд-
ка 300 км, то энергии, выделяющейся при дифференциации этого слоя, уже достаточно
для поддержания условий плавления вышележащих и нижележащих слоев и перехода
150
Проблемы происхождения жизни
к глобальной дифференциации (Витязев, 1974, 2004). Самоподдерживающаяся гравитаци-
онная дифференциация наступает при числе Gd > GdKpHT= 3.22, где Gd = 'Drgch1vaEIAlRT^ *.
Для Земли условие выполняется при массе планеты тр = 0.02-0.04 т д. Было ясно, что усло-
вия появления такого слоя достигаются, когда масса Земли становится больше массы со-
временной Луны, но темп развития дифференциации и времени образования земного ядра
строго оценить не удается. Дело в том, что в соответствующих уравнениях гидродинамики,
описывающих отделение «железной» компоненты от «силикатной», присутствует важный
параметр - коэффициент вязкости, который даже для современной конвектирующей мантии
оценивается (для разных слоев) с точностью до порядка величины. Время дифференциации
в первом приближении обратно пропорционально значению этого коэффициента. Поэтому
мы можем получить сто миллионов лет и несколько миллиардов для динамической оценки
скорости роста земного ядра. Было показано (Azbel et aL, 1993), что ранний рост ядра согла-
суется с данными по уран-свинцовой изотопной системе. Но лишь с появлением нанотехно-
логии по измерениям в системе Hf-W в рамках одностадийной модели стало ясно, что земное
ядро формировалось практически одновременно с ростом планеты, а именно по Hf-W систе-
ме - началась в первые 30-50 млн. лет, одновременно с основной стадией роста планеты (Yin
et al., 2002; Kleine et al., 2002; Schoenberg et al., 2002).
Дифференциация примитивной мантии и образование примитивной коры
В эксперименте, проведенном в ИДГ РАН по СВЧ нагреву, плавлению и дифференциа-
ции образцов метеорита Царев (L6) (Zetzer, Vityazev, 1996), был получен результат, свидетель-
ствующий о том, что при дифференциации примитивного земного вещества на «железную»
и «силикатную» компоненты, в силикатной части происходит вторичная дифференциация
с выделением малой доли весьма «кислого» (обогащенного SiO2) материала - вероятно, ана-
лога примитивной коры. В ряде недавних работ геохимиков предлагаются варианты образо-
вания (по данным Sm-Nd - системы в первые 100-300 млн. лет) так называемых EDR - Early
Depleted Reservoir и EER - Early Enriched Reservoir - примитивную кору, ныне, по-видимому,
в D” ** (Boyet, Carlson, 2005); древняя первичная кора, обогащенная Th, U, К (9 Twt = 9 тера-
ватт = 9* 1012 Вт), Rb в D” (Tolstikhin, Hofmann, 2005).
В ближайшие годы следует ожидать решения проблемы оценки массы и состава прими-
тивной земной коры. На сегодня оценка, что примитивная кора могла составлять 10 % массы
современной континентальной, представляется разумной, но требующей перепроверки.
Первичные атмосфера и гидросфера
Состоянию первичных атмосферы и гидросферы, столь интересующих специалистов
по происхождению биосферы, в последние полстолетия посвящены тысячи работ. Поддер-
живаемое фактически большинством астрофизиков мнение о меньшей (на 20-25 %) светимо-
сти молодого (в первые сотни млн лет) Солнца привело к обсуждению вероятных составов
первичной атмосферы, вызывающих парниковый эффект. Это известные работы Карла Сага-
на, его ученика Чайбы и др. Так как этим вопросам, вероятно, будет посвящена статья в этом
сборнике нашего классика профессора Л.М. Мухина, мы опускаем обсуждение этой пробле-
мы. Считаем важным отметить следующий выдающийся результат работ международного
коллектива по австралийским цирконам - гидросфера и атмосфера Земли существовали уже
в первые 150 млн. лет.
* Dr - разность плотностей матрицы и тяжелых включений, g - сила тяжести, с - доля объема, занимаемая
включениями, h - толщина слоя, v0 - начальная скорость эффективной дифференциации, Е - энергия активации,
/ - коэффициент обычной теплопроводности, R - универсальная газовая постоянная, 7) - температура начала
эффективной дифференциации.
** Область D” - погранслой между нижней мантией и жидким ядром, толщиной порядка 200 км.
151
A.B. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
Согласно интерпретации данных по изотопии кислорода (|6О/17О/18О) в цирконах Jack
Hills (Peck et al., 2001), 4.2, 4.3 и 4.4 млрд, лет назад на Земле была вода в жидкой фазе. Объемы
ее пока оценить не удается (ручейки, озера в кратерах, мелкие моря?). Согласно работам фран-
цузской школы (1960-1980 гг.) и известного специалиста по изотопии ксенона Р. Озимы, ксенон
в атмосфере Земли имеет возраст порядка 4.0 и более млрд. лет. Наша интерпретация содержаний
изотопов инертных газов, в частности, изотопной плеяды Хе (Pechernikova et al., 2003; Vityazev
et al., 2005) указывает на то, что примитивная атмосфера сформировалась в ходе заключи-
тельных стадий образования Земли. Оценка ее массы и состава пока представляет проблему.
Однако в сочетании с данными по австралийским цирконам можно уверенно говорить о том,
что давление на поверхности ранней Земли было выше необходимого для существования при-
митивной гидросферы (тройная точка для Н2О), т. е. порядка и выше современного для Марса.
На стадии аккумуляции Земли, когда поток падающих тел был достаточно велик, па-
раметры ранней атмосферы в значительной степени определялись ударной дегазацией и эро-
зией. В.В. Светцовым (2007) путем численного моделирования ударов астероидов и комет
с размерами от 100 м до 10 км были оценены потери атмосферы на Земле с ее современной
атмосферой и Марсе с более плотной атмосферой из углекислого газа, которая могла быть
на ранних стадиях его эволюции. Численное моделирование вертикальных ударов прово-
дилось путем решения двумерных уравнений газовой динамики в цилиндрических коорди-
натах, начиная со стадии входа тела в атмосферу. Получены приближенные аппроксимирую-
щие выражения потерь массы атмосферы и ударника в зависимости от их параметров и силы
тяжести на планете. Характерные величины уносимой массы атмосферы при ударах астерои-
дов (километровых и более размеров) оказываются порядка 0.01-0.1 от массы ударника, а при
ударах долгопериодических комет - порядка 0.1-1 от массы кометы.
Согласно расчетам, параметры ранней атмосферы после окончания роста планеты
существенно зависят от того периода аккумуляции, когда масса планеты составляет более
99 % от ее конечной величины. Более ранний период забывается, а в более поздний интен-
сивность притока массы недостаточна для существенного изменения массы образовавшей-
ся до этого атмосферы.
Отметим, что результаты вычислений зависят от таких точно не известных входных па-
раметров, как содержание атмофильных элементов в падающих телах, распределение по массам
и скоростям падающих тел, размеры наибольших ударников. При вариации входных данных
в допустимых пределах установившиеся к концу аккумуляции (99.9 % массы планеты) атмосфе-
ры имеют давление более 1 бар. Более совершенные модели должны учитывать взаимодействие
выделяющихся при ударах газов с атмосферой и породами верхнего слоя планеты, физико-
химическое состояние ранних атмосфер, сток газов в водные бассейны и учет глубинной дега-
зации примитивной части вещества, полученного планетой на ранних стадиях аккумуляции.
ЗАКЛЮЧЕНИЕ
1. Обнаружение протопланетных дисков, экзопланет и планетных систем около мо-
лодых звезд и их исследования позволяют надеяться на уточнение условий на поверхности
ранней Земли и других планет.
Луна хранит следы «поздней бомбардировки» (Late bombardment) 3.7-3.9 млрд, лет
назад. Чем она была вызвана? Наше предположение - это выброс Солнечной системы
из звездного скопления. Это предположение должно быть либо опровергнуто, либо поддер-
жано динамическими расчетами и другими данными.
2. Новые технологии обнаружения остатков короткоживущих радиоактивных элементов
(2бА1, 60Fe, 182Hf и т.д.) позволяют хронометрировать отдельные стадии формирования и ранней
эволюции планет с точностью до 1 млн. лет. Данные по австралийским цирконам и по плеяде изо-
топов Хе свидетельствуют о наличии атмосферы и гидросферы, начиная с 4.4 млрд, лет назад.
152
Проблемы происхождения жизни
Но масса первичной гидросферы, состав и масса первичной атмосферы пока не уста-
новлены. Вероятно, проблему формирования атмосфер надо решать одновременно для Марса,
Земли и Венеры. В частности, необходимо решить проблему выхода Венеры на парниковый
режим. И понять, почему Земля избежала этого исхода. Это задача ближайшего десятилетия.
В работе использованы результаты исследований сотрудников ИДГ РАН, выполнен-
ных по проекту «Процессы на завершающей стадии аккумуляции Земли» (руководитель
акад. В.В. Адушкин) в рамках программы Президиума РАН «Проблемы зарождения биосфе-
ры Земли и ее эволюции». Авторы благодарны проф. Л.М. Мухину и акад. РАН А.Ю. Розано-
ву за приглашение к участию в рабочем совещании «Проблемы происхождения жизни».
СПИСОК ЛИТЕРАТУРЫ
Адушкин В.В., Витязев А.В. 2007. Происхождение и эволюция Земли: современный взгляд // Вести.
РАН. Т. 77. № 5. С. 396-402.
Анисочкин В.М., Печерникова Г.В., Витязев А.В. 2007. Эволюция химического состава Вселенной (4D
таблица Менделеева) // Физические поля и динамика взаимодействующих геосфер. Сб. научн.
тр. ИДГ РАН / Отв. ред. С.Б. Турунтаев. М.: ГЕОС. С. 317-320.
Боярчук А.А., Рускол Е.Л., Сафронов В.С,, Фридман А.М. 1998. Происхождение Луны: спутниковый рой
или мега-импакт? И ДАН. Т. 361. № 4. С. 481-484.
Витязев А.В. 1974. Неизотермическая неустойчивость гравитационной дифференциации. Препринт
КФАН. ИФЗ АН СССР. 24 с.
Витязев А.В. 1983. Модели образования и ранней эволюции планет земной группы //Геохимия радио-
генных изотопов на ранних стадиях эволюции Земли. М.: Наука. С. 42-60.
Витязев А.В. 2004. Тепловые взрывы в ранней Земле // Физика горения и взрыва. Т. 40. № 6. С. 121-125.
Витязев А.В., Печерникова ГВ. 1989. Оценки состава допланетных тел, формировавших Землю, по их
плотностям и изотопному составу кислорода // 13 Всесоюзн. симпоз. «Изотопы в гидросфере».
М.: Ин-т водных проблем АН СССР. С. 93-94.
Витязев А.В., Печерникова ГВ. 1996. Ранняя дифференциация Земли и проблема лунного состава //
Физика Земли. № 6. С. 3-16.
Витязев А.В., Печерникова ГВ. 2003. Происхождение геосфер: новые результаты и остающиеся про-
блемы // Геофизические процессы в нижних и верхних оболочках Земли. Сб. научн. тр. ИДГ
РАН / Отв. ред. Ю.И. Зецер. М.: ИДГ РАН. Кн. 2. С. 13-25.
Витязев А.В., Печерникова Г.В., Сафронов В.С. 1978. Предельные массы, расстояния и времена акку-
муляции планет земной группы И Астроном, журн. Т. 55. С. 107-112.
Витязев А.В., Печерникова ГВ.. Сафронов В.С. 1990. Планеты земной группы: происхождение и ран-
няя эволюция. М.: Наука. 296 с.
Зиглина И.Н. 1976. Влияние сближений тел роя с растущей планетой на эксцентриситет ее орбиты //
Астроном, журн. Т. 53. Вып. 6. С. 1288-1294.
Зиглина И.Н., Сафронов В.С. 1976. Осреднение эксцентриситетов орбит тел, объединяющихся в пла-
нету // Астроном, журн. Т. 53. Вып. 2. С. 429-435.
Зимина Г.И., Витязев А.В. 2007. Магнито-гидродинамика околозвездных газопылевых дисков // Тр.
X конф, молодых ученых «Современные проблемы в астрофизике и физике космической плаз-
мы». БШФФ-2007. Иркутск: Изд-во ИСЗФ СО РАН. С. 212-214.
Левин Б.Ю. 1964. Происхождение Земли и планет. М.: Наука. 116 с.
Мясников В.П., Фадеев В.Е. 1980. Гидродинамические модели эволюции планет земной группы. М.:
ВИНИТИ. 207 с.
Печерникова ГВ. 2004. Крупные тела сокращают шкалу времени роста Земли // Динамика взаимодей-
ствующих геосфер. Сб. научн. тр. ИДГ РАН / Отв. ред. Ю.И. Зецер. М.: ИДГ РАН. С. 217-223.
153
A.B. Витязев, Г.В. Печерникова. Ранняя Земля в тесном окружении молодых звезд
Печерникова ГВ. 2005. Время роста Земли // Докл. АН. Т. 401. № 3. С. 391-394.
Печерникова ГВ., Витязев А.В. 1979. Массы крупнейших тел и дисперсия скоростей при аккумуляции
планет // Письма в «Астроном, журн.». Т. 5. С. 54-59.
ПечерниковаГВ.. Витязев А.В. 1989. 8О|7-8О18-р ограничения на состав планетезималей, формирующих
Землю //12 Всес. симпоз. по стабильным изотопам в геохимии. М.: ГЕОХИ АН СССР. С. 91-92.
Печерникова Г.В., Витязев А . В. 2005. Импакты и эволюция ранней Земли // Катастрофические воздей-
ствия космических тел / Ред. В.В. Адушкин, И.В. Немчинов. М.: Академкнига. С. 251-265.
Сафронов В.С. 1954. О росте планет в протопланетном облаке // Астроном, журн. Т. 31. С. 499-510.
Сафронов В.С. 1969. Эволюция допланетного облака и образование Земли и планет. М.: Наука. 244 с.
Светцов В.В. 2007. Потери и пополнение атмосферы за счет ударов космических тел на Земле и Марсе //
Астроном, вести. Т. 40. № 1. С. 33-46.
Светцов В.В. 2008. О возможности происхождения океана в результате поставки воды кометами на
Землю // Локальные и глобальные проявления воздействий на геосферы. Сб. научн. тр. ИДГ
РАН / Отв. ред. С.Б. Турунтаев. М.: ГЕОС. С. 162-169.
Сорохтин О.Г, Ушаков С.А. 1991. Глобальная эволюция Земли. М.: Изд-во МГУ. 446 с.
Сорохтин О.Г, Ушаков С.А. 2002. Развитие Земли. Учебник / Ред. В.А. Садовничий. М.: Изд-во МГУ. 560 с.
Anisochkin V.M., Vityazev A.V., Pechernikova G.V. 2007. Evolution of abundance of chemical elements in the
Universe - 4D Mendeleev’s table // Origin and evolution of biosphere and photonics of nucleic acids.
Abstr. Intern. Symp. Terskol. P. 39.
Azbel I.Ya., Tolstikhin I.N., Kramers J.D., Pechernikova G.V., Vityazev A.V. 1993. Core formation during
homogeneous Earth accretion: a dynamic approach // Geochim. Cosmochim. Acta. V. 57. № 12.
P. 2889-2898.
Balbus S.A., Hawley J.F. 1991. A powerful local shear instability in weakly magnetized disks. I - Linear
analysis // Astrophys. J. V. 376. P. 214-233.
Boyet M„ Carlson R.W. 2005. 142Nd Evidence for early (>4.53 Ga) global differentiation of the silicate Earth И
Science. V. 309. № 5734. P. 576-581.
Gammie C.F. 1996. Layered accretion in T Tauri disks // Astrophys. J. V. 457. P. 355-362.
Hayashi C, Nakazawa K„ Nakagava Y. 1985. Formation of the solar system // Protostars and Planets II / Eds.
D.C. Black, M.S. Matthews. Tucson: Univ. Arizona Press. P. 1155-1176.
Kleine T, Miinker C., Mezger K., Palme H. 2002. Rapid accretion and early core formation on asteroids and
the terrestrial planets from Hf-W chronometry // Nature. V. 418. P. 952-955.
Kleine T, Touboul M„ Bourdon B., Palme H, Wieeler R. 2008a. Hafnium-Tungsten chronometry of lunar differentia-
tion // Goldschmidt Conference Abstr. A480.
Kleine T, Touboul M., Burkhardt C, Bourdon B. 2008b. Dating the first —100 Ma of the solar system: from the
formation of CAIs to the origin of the Moon // Goldschmidt Conference Abstracts. A480.
Lazarev M.P., Vityazev A.V. 2005. Dissipative collisions of asteroid-sized bodies // Vernadsky-Brown Micro-
symp. 42. Abstr. CD.
Lazarev M.P., Vityazev A.V. 2006. Dissipate collisions of asteroid-sized bodies // Geophys. Res. Abstracts,
EGU. Vienna, Austria. V. 8. 07128.
Pechernikova G. V, Vityazev A. V. 2007. Origin of the Solar system in compact area of star formation of Orion nebula
type // Origin and evolution of biosphere and photonics of nucleic acids. Abstr. Intern. Symp. Terskol. P. 21.
Pechernikova G.V, Vityazev A.V, Bashkirov A.G. 2003. Early Earth and Хе-missing problem solution //
Vernadsky-Brown Microsymp. 38. Abstr. P. 34.
Peck W.H., Valley J.W., Wilde S.A., Graham CM. 2001. Oxygen isotope and rare elements in 3.3 to 4.4 Ga
zircons: ion microprobe evidence for high 818O continental crust and oceans in the Early Archean //
Geochim. Cosmochim. Acta. V. 65. № 22. P. 4215-4229.
Safronov V.S., Zvjagina E.V 1969. Relative sizes of largest bodies during the accumulation of planets//Icarus.
V. 10. P. 109-118.
154
Проблемы происхождения жизни
Sano Т., Miyama S.M., Umebayashi Т, Nakano N. 2000. Magnetorotation instability in protoplanetary disks. II.
Ionization state and unstable regions // Astrophys. J. V. 543. P. 486-501.
Schoenberg R., Kamber B.S., Collerson K.D., Eugster O. 2002. New W-isotope evidence for rapid terrestrial
accretion and very early core formation // Geochem. Cosmochem. Acta. V. 66. № 17. P. 3151-3160.
Semenov D., Wiebe D., Henning Th. 2004. Reduction of chemical networks. II. Analyses of the fractional
ionization in protoplanetary discs // Astron, and Astrophys. V. 417. P. 93.
Semenov D., Wiebe D., Henning Th. 2006. Gas-Phase CO in protoplanetary discs: a challenge for turbulent
mixing // Astrophys. J. V. 647. P. L57.
Shuvalov V., Dypvik H. 2004. Ejecta formation and crater development of the Mjolnir impact // Meteoritics and
planetary science. V. 39. № 3. P. 467-479.
Svetsov V.V. 2006. Numerical modelling of large asteroidal impacts on the Earth // Int. J. Impact Eng. V. 29.
P. 671-682.
Tolstikhin I., Hofmann A.W. 2005. Early crust on top of the Earth’s core // Physics of the Earth and Planetary
Interiors. V. 148. P. 109-130.
Velikhov E.P. 1959. Stability of an ideally conducting liquid flowing between cylinders rotating in a magnetic
field // J. Exptl. Theoret. Phys. V. 36. P. 1398-1404.
Vityazev A.V, Bashkirov A.G., Pechernikova G.V. 2006. Gravity-MHD of gas-dust disk near young Sun: dis-
sipate stage // 37 LPSC. Sess. 118. № 1860.
Vityazev A.V., Pechernikova G.V. 1998. Macroimpacts and crust and atmosphere forming in early terrestrial
history // Planetary Systems: the long view / Eds. L.M. Celnikier, J. Van Tran Thanh. France: Edit.
Frontieres. P. 95-97.
Vityazev A.V., Pechernikova G. V, Bashkirov A.G. 2003. Accretion and differentiation of terrestrial protoplan-
etary bodies and Hf-W chronometry // LPSC. Sess. 85. № 1656.
Vityazev A. V, Pechernikova G.V., Bashkirov A.G. 2005. «Missing Xenon» problem and climate of the early
Earth//36 LPSC. Sess. 118. №1719.
Vityazev A.V., Pechernikova G.V., Bashkirov A.G. 2006. Dissipate stage of protoplanetary disk. Gravity-MHD
effects // EGU General Assembly 2006, Vienna, Austria, 02-07 April 2006, ID-NR: EGU06-A-06661.
Tin Q.Z., Jakobsen S.B., Yamashita K. et al. 2002. A short timescale for terrestrial planet formation from
Hf-W chronometry of meteorites // Nature. V. 418. P. 949-952.
Zetzer J.I., Vityazev A.V. 1996. Experiment «Tsarev» and differentiation of chondritic bodies // Earth, Moon,
and Planets. V. 72. № 1-3. P. 474-480.
The Early Earth cramped by surrounding young stars
A.V. Vityazev, G.V. Pechernikova
Early Earth in close environment of young stars. The physics-chemical conditions on the early
Earth are very important for the specialists in the area of problems of origin of the biospheres. The
last modification of the theory of the Earth and planets formation is being researches. The effects of
the Sun formation in the region of star formation in close surrounding of neighboring new stars are
being discussed. The results of an investigation of ion- and magnetic-torus of protoplanetary discs
near young stars are presented. A generalization of classical problem of the growth of the Earth and
the planets allows us to derive estimations of the timing of massive bodies that are consistent with
both long-lived (U-Pb) and short-living (Hf-W) isotopic data. Furthermore the early Earth evolu-
tion model taking into account the new calculations results and the last geochemical data (ancient
Australian zircons, relict xenon pleiad) is being discussed. In the conclusion variety of early Earth’s
problems are being discussed, which has not yet been able to obtain a satisfactory solution.
155
Дискуссия
по пленарному докладу A.B. Витязева, Г.В. Печерниковой
«РАННЯЯ ЗЕМЛЯ В ТЕСНОМ ОКРУЖЕНИИ МОЛОДЫХ ЗВЕЗД»
Председательствующий А.С. Спирин
М.Я. Маров: Число Нуссельта, для сравнения с обычной теплопроводностью, считалась
для какой глубины?
А.В.В.: Естественно, это порядковая оценка в среднем на всю мантию, включая
нижнюю.
М.Я. Маров: Второй вопрос более серьезный. Вы солидаризируетесь со Львом
Михайловичем Мухиным в отношении раннего океана. Вы говорите, что океан и гидросфера
существовали в первые 150 миллионов лет.
А.В.В.: Присоединяюсь ко Льву Михайловичу, Уайльду, Пеку и так далее. На основании
цирконов и ксенона.
М.Я. Маров: Тогда возникает такой вопрос. До выхода Солнца на Главную
последовательность его светимость была существенно меньше (~ 0.75 от современной) и,
следовательно, равновесная температура ранней Земли бы ла ниже ~ 240 К, отвечающей полной
светимости. Чтобы появилась жидкая вода, необходимо повысить начальную температуру
примерно на 30°, что можно обеспечить за счет обычно привлекаемого парникового
эффекта, но сомнительно, что такой механизм мог компенсировать еще большую разницу в
температурах и обеспечить существование океана на этой стадии. А обращение к еще более
раннему этапу эволюции Солнца, включающего высокую светимость на цикле Хаяши,
нереалистично, поскольку еще не аккумулировались планетные тела в диске.
А.В.В.: Нужно просить астрофизиков рассказать нам о раннем Солнце. Дополнительный
источник тепла - падающие планетезимали, но это, конечно, только «горячие пятна».
Л.М. Мухин: Цифры 3.8-3.9 млрд, лет относительно бомбардировки означают время ее
окончания?
А.В.В.: Тяжелый вопрос. Половина исследователей считают, что в это время был ее
пик, другая половина полагает, что она к этому времени заканчивается. Я полагаю, что не
заканчивается. Надо честно сказать, в архее мы имеем семь-восемь слоев, дающих указания
на мегаимпакты. В остальном мы судим об этом косвенно, по Луне.
Л.М. Мухин: Относительно температуры поверхности и первичных океанах. Атмосфера
образовывалась не только за счет импактов, была же и обычная дегазация, поэтому вполне
мог работать парниковый эффект.
А.В.В.: Согласен. Хочу добавить про импакты. Пока планета не превысила радиуса
нескольких лунных, скорости ударов были меньше пяти километров в секунду-это вызывало
просто дробление и нагрев на первую сотню градусов. А вот когда планета уже размером с
Марс, тогда, в нулевом приближении, упавшее тело плавит свой объем. Будет дегазация.
М.Я. Маров: Но дело в том, что будет мало удерживаться легколетучих атмофильных
элементов. Вот в чем проблема.
В.Н. Снытников: Среди источников тепла Вы указали распад радиоактивных элементов.
Хорошо известно то, что даже на поверхности Земли существовал ядерный реактор - реактор
Окло (в Габоне). Кроме того, примерно 7-10 лет назад командой Титова из Новосибирска был
проделан ряд работ, которые показали возможность функционирования ядерного реактора
на границе ядра и мантии. Не могли бы Вы это прокомментировать?
156
Проблемы происхождения жизни
А.В.В.: Речь идет о том, возможны ли были на планетах или, может быть, даже на
планетезималях, процессы самопроизвольной концентрации минералов, богатых ураном,
торием и калием? И тогда были бы локальные очаги... Ну, гипотезы выдвигать можно...
Подождем геонейтрино, они нам точно все это покажут...
А.В. Тутуков: Отмечается периодичность развития живого на Земле - периоды расцвета,
угасания. Что могло бы быть внешней причиной этой периодичности, в том числе массовых
вымираний?
А.В.В.: Мы сейчас закончили такую работу, где проанализировали магматическую -
коровую и мантийную - активность, массовые вымирания и импактные кратеры. Некоторые
вымирания совпадают с ними, другие - нет. Моя сегодняшняя оценка - массовые вымирания
на 20 процентов обусловлены экзогенными факторами (в основном импактами, про
сверхновые и гамма-всплески пока не хотелось бы говорить) и на 80 процентов - геологической
активностью Земли.
Б.М. Шустов: Мой вопрос о возможных земных метеоритах на Луне.
А.В.В.: Земные метеориты следовало бы поискать на Луне (по изотопии), такая
вероятность есть, хотя и небольшая.
С.В. Рожнов: Каков минимальный размер падающего на Землю космического тела,
способного полностью стерилизовать Землю, уничтожить все живое?
А.В.В.: Когда есть океан, достаточно тела порядка 100-150 километров для испарения
его верхних слоев, чтобы уничтожить весь океан - тела 400-500 километров. В гадее такие
тела падали на Землю.
157
Проблемы происхождения жизни. М.: ПИН РАН, 2009. С. 158-165
ПСЕВДОМОРФОЗЫ ПО МИКРОБАМ В МЕТЕОРИТАХ
© 2009 г. А.Ю. Розанов
Палеонтологический институт им. А.А. Борисяка РАН
aroza@paleo.ru
Данные по углистым хондритам, имеющиеся сегодня, показывают присут-
ствие в них псевдоморфоз по микробам. Из возраста метеоритов следует,
что жизнь возникла до образования Земли.
Как показывает практика последних 10-15 лет, результаты биологических исследований
метеоритов оказались чрезвычайно важными, хотя часто и достаточно противоречивыми.
Строго говоря, уже в 1975 г. после выхода капитальной сводки Б. Надя (Nagy, 1975)
можно было бы утверждать, что в углистых хондритах содержатся фоссилизированные
остатки организмов как минимум бактериального уровня организации.
Однако научная общественность, особенно американские астрономы, встретили рабо-
ты Надя и его коллег в штыки. Эта отрицательная реакция докатилась до Европы и СССР,
и занятие углистыми хондритами как источником биологической информации стало счи-
таться практически неприличным.
Новая волна обсуждений этих вопросов возникла в связи с публикациями Д. Маккея с
коллегами (McKay et al., 1996), работами Р. Хувера (Hoover, 1997) и российских исследовате-
лей (Жмур, Розанов, Горленко, 1997; Zhmur, Rozanov, Gorlenko, 1997).
В первой из них (McKay et al., 1996) описывались продолговатые вытянутые черве-
образные сегментированные ископаемые из марсианского метеорита ALH 84001. Однако
сразу многие исследователи обратили внимание на слишком малые размеры объекта (250 нм
в длину), что поставило под сомнение биогенную природу этих образований.
Мне удалось в лаборатории у Дэвида Маккея в Хьюстоне видеть весь материал из мар-
сианских образцов, и я могу лишь выразить сожаление, что лучшие и безусловно биогенной
природы образцы так и не были опубликованы.
Д. Маккей и его коллеги одновременно исследовали кристаллографические осо-
бенности найденного в этих образцах магнетита, показав его специфику (McKay et al.,
2000). В это же время были опубликованы статьи (Astafieva, Hoover, Rozanov, 2004;
Астафьева, Розанов, Хувер, 2005) по кристаллографическому характеру магнетита из
разных образований, где показано, что биогенный магнетит фрамбоидов значительно
отличается от абиогенного.
Я не стал бы настаивать на безусловности таких заключений, но они должны быть при-
няты во внимание, поскольку эта специфика была неоднократно отмечена в земных породах
158
Проблемы hi-oiкложого/и>1 жичии
Рис. I. Фрамбоиды в метеоритах Рейнбоу (<з) и Тэгишлэйк (б).
разного возраста, где биогенность фрамбоидов не вызывает сомнений. Важно, что в ряде
углистых хондритов фрамбоиды были обнаружены (рис. 1).
Совершенно иного типа материалы, чем у Маккея, были опубликованы Р. Хувером
(NASA) и российскими исследователями (Hoover, 1997; Жмур, Розанов, Горленко, 1997). Бо-
лее того, один и тот же результат был получен независимо по одному и тому же метеориту
Рис. 2. Псевдоморфозы по бактериальным нитям и чехлам: о, б, «, г - Мурчи-
сон. д - Алленде, е - Ефремовка.
159
А.Ю. Ричиипн. Псевдоморфозы но микробам в метеоритах
Рис. 3. Домены магнетита в нитях из метеорита Оргей
(Tan, VanLandingham, 1967) (ci) и современной пурпурной серо-
бактерии Rhodopxeudomonas rulilis (Vainstein et al., 1997) (6).
(Мурчисон), упавшему в Австралии. Принципиальным было то, что размер обнаруженных
объектов был совершенно иным - это фоссилизированные нити диаметром около 3 мкм, что
вполне сопоставимо с размерами многих бактерий, в том числе и цианобактерий (рис. 2).
Последовавшие за этим совместные исследования американских и российских ученых пока-
зали, что нитчатые и коккоидные формы (псевдоморфозы) встречаются во всех углистых хондри-
тах. В иных же типах метеоритов нет ничего подобного. Результаты этих исследований с подроб-
ным описанием объектов и процедуры исследования были многократно опубликованы (Hoover et
al., 1998, 2003, 2004; Gerasimenko et al., 1999; Rozanov, Hoover, 2002; Hoover, Rozanov, 2003; и др.).
Совершенно уникальный результат был получен по доменам магнетита в одной из ни-
тей из метеорита Оргей (рис. 3, а). Для сравнения мы приводим вид такого магнетита в со-
временных пурпурных магнетотактических бактериях и метеорита (рис. 3, б).
Были подробно разобраны и возможные трудности и ошибки при интерпретации ма-
териала из метеоритов (Герасименко и др., 1999; Бактериальная палеонтология, 2002), по-
скольку, с одной стороны, мы сталкиваемся с современными засорениями, в том числе про-
растаниями метеоритов и в природе, и в музеях, а с другой - установленная в последнее
время скорость фоссилизации столь стремительна, что современные контаминанты могут
фоссилизироваться очень быстро уже в наши дни.
Крайне важно, что возможность замещения организмов чистым углеродом при сохра-
нении тончайших морфологических структур была показана на примере девонских графи-
тов Ботогола (Восточные Саяны), в которых сохранились не только цианобактерии и грибы,
но и фрагменты пресноводных клешей (Жмур и др., 1996; Rozanov et al., 1999) (рис. 4).
Рис. 4. Фрагмент клеща, оснащенного трихободриями. Девон, Ботогольское ме-
сторождение графитов (Восточные Саяны).
160
Пгоь. немы и Pol к \i»k.пенни ж и ши
Рис. 5. Псевдоморфозы по бактериальным нитям и цианобактериальным чехлам в метеорите Оргей.
При всех сомнениях, однако, имеются результаты, однозначно свидетельствующие о на-
личии биогенных объектов в углистых хондритах. Самые очевидные и убедительные данные
были получены по Мурчисону, Ефремовке, Мигею, Алаису, Алленде и др., а в самое последнее
время по метеориту Оргей (рис. 5), когда выяснилось, что ископаемые в нем могут быть за-
мещены - в одном случае чистым углеродом и в другом случае эпсомитом (английской со-
лью) (рис. 6). И то, и другое совершенно исключает засорение (то есть земное происхождение),
а характер морфологии этих образований не оставляет сомнений в их бактериальной природе.
Изотопные исследования углистых хондритов проводились неоднократно (Galimov,
1985; Epstein et al., 1987; Engel et al., 1990; Engel, Masco, 1997; и др.).
Значения, полученные по углероду, в свете современных данных не дают оснований возражать
против биогенной природы углеродсодержащих образований в углистых хондритах.
Данных о возрасте метеоритов, и прежде всего углистых хондритов, достаточно мно-
го, и все они указывают на их очень древнее происхождение. Обычно считается, что это
4.5-4.56 млрд. лет.
Таким образом, если метеориты (и прежде всего углистые хондриты) имеют столь древ-
ний возраст, а фоссилизированные остатки еще старше, то эти организмы уже существовали
до образования Земли. Это наводит на мысль, что они могут происходить и не из Солнечной
системы, а из каких-то разрушившихся «планет» с водой (лужами, озерами, морями и т. д.).
Можно, однако, предполагать, что углистые хондриты часто являются агломератом
разновозрастных фрагментов, части из которых и имеют столь древний возраст. Однако это
предмет специальных исследований, которые пока не проводились.
161
AJO. Роиток. Псевдоморфозы по микробам в метеоритах
Рис. 6. Бактериальные тела в метеорите Оргей, замещенные эпсомитом (а-г) и замещенные угле-
родом (д-е) (Hoover et al.. 2004).
162
Проб. п:мы Прош \<>ж .1 пая жия/н
Рис. 7. Предполагаемые эвкариотические организмы в метеорите Оргей.
Особый интерес представляют обнаруженные в ряде метеоритов мешковидные или
шарообразные формы достаточно крупных размеров (п. 10-п.100 мкм), имеющие образова-
ния, напоминающие пилом, или пористость стенок (рис. 7). Если это так, то мы вправе по-
дозревать, что в углистых хондритах встречаются не только прокариоты, но даже и фосси-
лизированные одноклеточные эвкариоты либо празинофитного, либо протистового уровня
организации.
Одно время считались сильным аргументом в пользу абиогенной природы нитевидных
и коккоидных образований в метеоритах морфология и другие признаки керитов, происходя-
щих из пегматитов украинского шита.
Однако следует заметить, что большинство микробиологов не сравнивали кериты с
биогенными образованиями как метеоритов, так и древних земных пород. Правда, появилась
статья (Gorlenko et al., 1999), в которой описывалось внутреннее строение нитей керита и де-
лался вывод о цианобактериальной их природе.
С моей точки зрения, принципиальная разница керита и фоссилизированных бакте-
рий состоит в том, что у трубок или нитей керита присутствует тонкий центральный канал,
по которому поступающее вещество постоянно наращивает конец трубки. Эта конструкция
чрезвычайно характерна для трубчатых абиогенных образований.
Таким образом, данные по углистым хондритам недвусмысленно указывают на вне-
земное происхождение жизни. Многие исследователи, вероятно, правы, полагая, что жизнь
возникла гораздо раньше образования Земли - на планете «земного» типа, где была вода и
подходящие температурные, атмосферные и иные условия.
Обнаружение псевдоморфоз по микроорганизмам в углистых хондритах не указывает
на занос организмов в метеоритах. Однако идея о возможности переноса организмов в ядрах
163
А.Ю. Розанов. Псевдоморфозы по микробам в метеоритах
ледяных комет представляется весьма правдоподобной. Исследования организмов в антар-
ктических льдах и вечной мерзлоте Сибири показали, что в анабиозе многие организмы мо-
гут находиться миллионы лет и, следовательно, могут путешествовать в космосе длительное
время и на большие расстояния.
Таким образом, мы сегодня ясно можем говорить только о времени появления жизни
на Земле, а не о ее происхождении (зарождении). Во всяком случае, это справедливо для по-
следних 4.0 млрд, лет, то есть документированной геологической истории.
Работа выполнена по Программе Президиума РАН «Происхождение биосферы и эво-
люция гео-биологических систем» (подпрограмма II), гранту РФФИ № 08-04-00484 и науч-
ной школе НШ-4207.2008.5.
Я хотел бы поблагодарить Р. Хувера, М.М. Астафьеву, Е.А. Жегалло, С.И. Жмура
и Д. Маккея за плодотворные беседы и предоставленные материалы.
СПИСОК ЛИТЕРАТУРЫ
Астафьева М.М., Розанов А.Ю., Хувер Р. 2005. Фрамбоиды: их структура и происхождение// Палеон-
тол. журн. № 5. С. 1-7.
Бактериальная палеонтология. 2002 / Ред. А.Ю. Розанов. М.: ПИН РАН. 188 с.
Герасименко Л. М., Жегалло Е.А., Жмур С.И., Розанов А.Ю„ Хувер Р. 1999. Бактериальная палеонтоло-
гия и исследования углистых хондритов // Палеонтол. журн. № 4. С. 103-125.
Жмур С И., Розанов А.Ю., Горленко В.М. 1997. Литифицированные остатки микроорганизмов в угли-
стых хондритах И Геохимия. № 1. С. 66-68.
Жмур С.И, Розанов А.Ю., Лобзова Р.В., Жегалло Е.А. 1996. Об источнике углерода графитовых руд
Ботогольского щелочного массива (Восточный Саян) // Докл. АН. Т. 348. С. 360-362.
Astafieva М.М., Hoover R., Rozanov A. Yu. 2004. Framboidal structures in Earth rocks and in astromaterials //
Proc. SPIE. V. 5163. P. 36-47.
Engel M.H., Masco S.A. 1997. Isotopic evidence for extraterrestrial non-racemic amino acids in the Mur-
chison meteorite H Nature. V. 389. P. 265-268.
Engel M.H., Masco S.A., Silfer J.A. 1990. Carbon isotope composition of individual amino acids in the Mur-
chison meteorite H Nature. V. 348. P. 47-49.
Epstein S., Krishnamurthy R.V., Cronin J.R. et al. 1987. Unusual stable isotope ratios in amino acid and car-
boxylic acid extracts from the Murchison meteorite // Nature. V. 326. P. 477-479.
Galimov E.M. 1985. The biological fractionation of isotopes. New York: Acad. Press. 261 p.
Gerasimenko L.M., Hoover R.B., Rozanov A.Yu. et al. 1999. Bacterial paleontology and studies of carbona-
ceous chondrites // Paleontol. J. № 4. P. 103-125.
Gorlenko V.M., Zhmur S.I., Osipov G.A. et al. 1999. Microbial nature of fibrous kerite of Volyn // Proc. SPIE.
V. 3755. P. 83-95.
Hoover R.B. 1997. Meteorites, microfossils and exobiology // Proc. SPIE. V. 3111. P. 115-136.
Hoover R.B., Jerman G., Rozanov A.Yu., Davies P.C.W. 2003. Biomarkers and microfossils in the Murchison,
Tagish Lake and Rainbow memteorites // Proc. SPIE. V. 4859. P. 5-31.
Hoover R. В., Jerman G., Rozanov A.Yu., Sipiera P.P. 2004. Indigenous microfossils in carbonaceous meteorites//
Proc. SPIE. V. 5555. P. 1-17.
Hoover R.B., Rozanov A.Yu. 2003. Microfossils, biominerals and chemical biomarkers in meteorites // Proc.
SPIE. V. 4939. P. 10-27.
Hoover R.B., Rozanov A.Yu., Zhmur S.I., Gorlenko V.M. 1998. Further evidence of microfossils in carbona-
ceous chondrites // Proc. SPIE. V. 3441. P. 203-216.
McKay D.S., Gibson E.K., Thomas-Keprta J. et al. 1996. Search for past life on Mars: possible relic biogenic
activity in Martian meteorite ALH 84001 // Science. V. 273. P. 926-930.
164
Проблемы происхожиения жизни
McKay D.S., Steele A., Allen С. et al. 2000. Mars immunoassay life detection instrument (MILDI) // Lunar and
Planet. Inst. Contrib. № 1062. Pt 2. P. 219-220.
Rozanov A.Yu., Hoover R.B. 2002. Bacterial paleontology and astrobiology H Proc. SPIE. V. 4495. P. 283-
294.
Rozanov A. Yu., Zhegallo E.A., Hoover R.B. 1999. Microbiota of the Botogol graphites // Proc. SPIE. V. 3755.
P. 38-47.
Tan W.C., VanLandingham S.L. 1967. Electron microscopy of biological-like structures in the Orgueil car-
bonaceous meteorite // Geophys. J. Roy. Astron. Soc. V. 12. P. 237.
Vainshtein M., Suzina N., Sorokin V. 1997. A new type of magnet-sensitive inclusions in cells of photosyn-
thetic purple bacteria // System. Appl. Micribiol. V. 20. P. 182-186.
Zhmur S.I, Rozanov A.Yu., Gorlenko V.M. 1997. Lithified remnants of microorganisms in carbonaceous chon-
drites // Geochem. Intern. V. 35. P. 58-60.
Pseudomorphic microbial structures in meteorites
A.Yu. Rozanov
The modern data on carbonaceous chondrites prove the presence of pseudomorphic microbial
structures in this type of meteorites. The age of the meteorites suggests that life has originated be-
fore the formation of the Earth.
165
Дискуссия
по пленарному докладу А.Ю. Розанова
«ПСЕВДОМОРФОЗЫ ПО МИКРОБАМ В МЕТЕОРИТАХ»
Председательствующий ГА. Заварзин
М.А. Федонкин: Изучение биоморфных структур в метеоритах имеет достаточно дли-
тельную историю. Почему все-таки, по Вашему мнению, все эти факты не стали фактами
науки, почему они не были вовлечены в активное обсуждение? В чем корень недоверия?
А.Ю.Р.: Я бы сказал так: в определенных кругах есть сильное беспокойство, что день-
ги NASA пойдут не в ту сторону. И там были предприняты жесточайшие меры по этому
поводу.
А.С. Спирин: Но все-таки хоть какая-то критика в литературе была?
А.Ю.Р.: Нет! По Маккею было много, а у нас - ничего, будто этого и нет. А нами на эту
тему были опубликованы два десятка статей, это попало в две монографии, всё на англий-
ском языке...
М.А. Федонкин: Я подумал, что, может быть, дело в том, что протокол неочевиден.
Когда 10 лет назад австралийцы опубликовали статью о биомаркерах из пород возрастом
2.7 миллиарда лет, рукопись была около сотни страниц по протоколу самого эксперимента-
как бурили, как химически маркировали буровой раствор, как чистили вакуумные камеры
и так далее. Были описаны все меры предосторожности в отношении контаминации. И я ду-
маю, что в Вашем случае отсутствие критики может быть связано с тем, что некоторые вещи
невозможно проверить, протокол неочевиден.
А.Ю.Р.: Я просто дам Вам эти статьи, протокол абсолютно очевиден, подробно описа-
но, как все делалось.
М.А. Федонкин: Не все, видимо. Ну, допустим, берете Вы метеоритное вещество, ведь
оно при ударе сплавляется, переплавляется, разбрасывается...
А.Ю.Р.: Ничего подобного. Метеорит небольших размеров внутри даже не разогревает-
ся. История падения и изучения метеоритов многократно описаны.
Г.А. Заварзин: Могу добавить относительно Мурчисона и Оргея. Материалы по Оргею
были опубликованы в 1864 г. По эллипсу рассеяния кусков метеорита была определена об-
ласть, откуда он прилетел. Он действительно растворяется в воде. Гувер все время говорит
про эпсомит, но это не всегда так, по элементарному анализу там получается и магния тио-
сульфат... Как Мурчисон упал в овчарню, тоже известно... Но здесь другой вопрос. Если на
Земле на камнях с возрастом, скажем, два с половиной миллиарда лет, как в Прионежье, разо-
вьется какой-то микроб, то это не значит, что возраст микроба два с половиной миллиарда
лет. Гувер утверждает, что биоморфные структуры сингенетичны самому метеориту... Это
еще вопрос, они могли образоваться позднее, на древнем материале.
М.Я. Маров: Насколько четко отождествлено, что эти углеродные образования являют-
ся действительно ископаемыми организмами?
А.Ю.Р.: Морфологически это обосновывается совершенно надежно.
М.Я. Маров: Как известно, открытие становится открытием тогда, когда оно независи-
мо подтверждено в нескольких лабораториях мира. Есть классический пример - холодный
термоядерный синтез, который был закрыт. Как в этом случае? Это принципиальный вопрос,
который может существенно повлиять на наши представления об источниках микроорга-
низмов, поэтому, с моей точки зрения, на это должны были бы наброситься все сопричаст-
ные. ..
166
Проблемы происхождения жизни
А.Ю.Р.: Это было получено в трех местах - в Хьюстоне, в Хансвилле и здесь у нас в
Палеонтологическом институте, на одном и том же материале (Мурчисон). Гувер потому и
оказался здесь. Реакция была такой - когда Маккей опубликовал свою статью, президент
США Клинтон добился выделения пятнадцати миллиардов долларов на эту программу, а
мое выступление в Президиуме РАН привело к тому, что мне дали шесть должностных еди-
ниц для Межинститутской лаборатории, без обеспечения зарплатой...
Н.Г. Бочкарёв: Радиоуглеродный анализ этих углеродных образований делался?
А.Ю.Р.: Да, получено очень много данных.
С.В. Шестаков: Алексей Юрьевич, Ваши утверждения, что в углистых хондритах
были найдены следы, возможно относящиеся к эукариотам, основаны исключительно на
морфологическом анализе?
А.Ю.Р.: Да, но здесь этого вполне достаточно.
С.В. Шестаков: Мне кажется, что чистой морфологии все-таки недостаточно, ведь
есть очень сложные прокариоты...
А.С. Спирин: Иногда парадигма сменяется через два десятилетия. Единицы работают,
а потом все ахают...
М.Я. Маров: Особенно в условиях, когда консерватизм мешает объективному вос-
приятию новых фактов.
А.С. Спирин: Вокруг грантов сплачиваются единомышленники, чему-то новому про-
биться крайне трудно. Вопрос о происхождении жизни - - одна из самых консервативных и
инерционных областей науки.
В.Н. Снытников: Были ли другие возражения по статье Маккея, кроме размеров объ-
ектов?
А.Ю.Р.: А это очень серьезные возражения.
А.С. Спирин: Там получалось, что размеры меньше рибосомы, а значит, это неживое.
М.А. Федонкин: Были не только морфологические возражения, но и геохимические. В
частности, в статье Галимова.
А.Ю.Р.: Это все весьма своеобразные возражения, я бы не стал их обсуждать.
А.С. Спирин: Но все-таки я готов поддержать Сергея Васильевича Шестакова, что
одной морфологии мало, нужно что-то еще.
А.Г. Пономаренко: Тогда палеонтологии вообще бы не существовало.
А.Б. Макалкин: Обратите внимание, что углистые хондриты - это единственный тип
метеоритов, в котором содержится химически связанная вода. Наиболее распространена
модель, что эти гидросиликаты образовались в среде с жидкой водой. Соответственно,
предполагается, что родительские тела углистых хондритов имели среду с жидкой водой,
что делает их условия благоприятными для образования жизни.
А.Ю.Р.: Не обязательно, чтобы они там образовывались, достаточно, чтобы они там
просто жили.
167
Проблемы происхождения жизни. М.: ПИН РАН. 2009. С. 168-181
О РАННИХ СТАДИЯХ ЭВОЛЮЦИИ ГЕОСФЕРЫ, БИОСФЕРЫ
И ПРОБЛЕМАХ РАННИХ ОЛЕДЕНЕНИЙ
© 2009 г. Н.Л. Добрецов
Объединенный институт геологии, геофизики и минералогии СО РАН
dobr@uiggm.nsc.ru
Ранние стадии эволюции геосферы и биосферы определяются тремя взаимо-
связанными причинами: 1) непрерывное остывание поверхности и недр (ман-
тии) Земли (снижение средней температуры мантии в 1.5-2 раза, поверхност-
ной в 3-4 раза, среднего теплового потока на порядок, вязкости на 3 порядка);
2) непрерывное ступенчатое окисление поверхности, наиболее существенное
в интервале 3.8-1.8 млрд, лет назад; 3) периодические и сопряженные колеба-
ния условий в геосфере и биосфере разного масштаба и природы. Важнейши-
ми рубежами этой эволюции являются рубежи около 4 млрд, лет назад (рож-
дение мощной, но неоднородной земной коры, начало жизни), около 3 млрд,
лет назад (появление сильного магнитного поля, усиление фотосинтетиче-
ской активности), около 1.8-1.9 млрд, лет назад (появление окисленной ат-
мосферы, первого суперконтинента, возможно первых суперплюмов от ядра),
около 0.75 млрд, лет назад (ускорение субдукции, «обводнение» верхней ман-
тии, поднятие континентов с обширной сушей, шельфами, большими реками,
первые крупные оледенения). Значение и корреляция событий самой ранней
истории (до и около 4 млрд, лет назад), событий около 750 млн. лет назад -
предмет острой дискуссии. В позднем архее и раннем протерозое (до 1.8 млрд,
лет назад) в биосфере преобладали цианобактерии, динамика и максимумы
развития которых фиксируются по широкому развитию строматолитовых
построек в карбонатных породах (вначале преимущественно доломитовых).
На рубеже 700-750 млн. лет назад появились сильные и частые оледенения
как отражение остывания Земли. Возможно, максимальным было оледенение
около 640 млн. лет назад, для которого дискутируется модель замерзшей Зем-
ли (snowball). С оледенениями также связан вопрос о появлении и эволюции
скелетов в животном мире. Эти события коррелируются и объясняются круп-
ными эндогенными изменениями. Одно из важнейших событий в эндогенной
истории - старт около 750 млн. лет назад периодического проявления мантий-
ных струй (суперплюмов), объясняющих дальнейшую периодизацию эволю-
ции биосферы. В заключение приведена экстраполяция будущей эволюции и
последовательного исчезновения сегментов биосферы по мере превращения
Солнца в красную звезду и разогрева поверхности Земли.
В первой части статьи я остановлюсь на эволюции Земли и геосферы и важнейших со-
бытиях этой эволюции.
Начнем с диаграммы (рис. 1), отражающей три основных тенденции в эволюции гео-
сферы и, соответственно, взаимосвязанной эволюции биосферы (Dobretsov et al., 2008). Пер-
168
Проблемы происхождения жизни
вая - это непрерывное остывание недр (мантии) и поверхности Земли (Tajika, Matsui, 1992;
Добрецов, 2003). Остывание мантии показано на этой диаграмме в виде снижения средней
температуры (в 1.5-2 раза с 3 000 °C по 2 100 °C), среднего теплового потока (почти на по-
рядок) и повышения вязкости (на 3 порядка). Вторая тенденция - это непрерывное окисление
поверхности Земли, приводящее, в частности, к осаждению железистых кварцитов (т. е. маг-
нетита), в интервале 3.2-1.8 млрд, лет, а затем, после 1.8 млрд, лет к появлению красноцветов
и гематита (по Заварзину, 2003а). Третья тенденция - периодическое колебание всех параме-
тров, включая температуру недр и поверхности, а также взаимосвязанных параметров (До-
брецов и др., 2001а). На рис. 1 выделены зелеными и красными линиями главные интервалы
в этой периодичности. Природа этой периодичности и границы тех периодов, которые можно
наметить - это один из предметов дискуссии. Красное поле в интервале 4.6-3.8 млрд, лет
иллюстрирует вывод, что начиная с 3.8 млрд, лет эволюция недр Земли была единой, не-
смотря на возможность разных начальных условий (Tajika, Matsui, 1992). Что касается взаи-
мосвязанной эволюции биосферы и геосферы, то я об этом еще буду говорить не один раз.
Но отражение взаимосвязанного ступенчатого развития - это ступенчатое окисление, по-
казанное внизу на рис. 1. Возможные минимумы и максимумы окислительного потенциала
допускает, в частности, А.Ю. Розанов (2003). Вероятно, намеченные границы надо будет про-
двигать в сторону некоторого удревнения (Rozanov, 2006).
Для сравнения приведем совершенно независимую оценку эволюции Земли (рис. 2),
полученную под руководством японского исследователя Ш. Маруямы (Maruyama, Liou, 2005;
Maruyama et al., 2007). Им выделены семь важнейших событий в эволюции биосферы и гео-
сферы. Первое - рождение Земли (4.55 млрд, лет), второе (около 4 млрд, лет) - начало жизни,
рождение континентальной коры, инициирование плейт-тектоники. Насчет континенталь-
ной коры и плейт-тектоники я сомневаюсь, а то, что рождение жизни произошло около это-
го рубежа - сейчас уже стало привычным допущением (Schidlowski, 1988; Zavarzin, 2008)
и совпадает с нашей границей (рис. 1). Третье событие (около 2.7 или 3.0 млрд, лет) - появ-
ление сильного «магнитного динамо» (слабое магнитное поле было и до этого), связанное
с появлением твердого внутреннего ядра, а также усиление фотосинтетической активности.
Фотосинтез появился раньше, но в полную меру заработал около 2.7 млрд, лет назад. Причем
японские авторы связывают это в том числе с резким усилением магнитного поля. Четвертое
событие (около 1.9 млрд, лет) - появление первого суперконтинента, который был связан
с карельской фазой складчатости и, возможно, в связи с появлением постперовскитовых фаз
на границе жидкого ядра и нижней мантии и появлением обогащенного слоя, который по-
том стал рождать плюмы и суперплюмы. Это объяснение тоже пока неоднозначное, но то,
что граница номер 4 - это время появления суперконтинента и целого ряда других магма-
тических и тектонических событий, - с этим можно согласиться. Можно двигать ее ближе
к 1.8 млрд, лет, но это уже в пределах точности измерений на этот период.
Пятый рубеж около 0.75 млрд, лет - очень важный рубеж, о котором я еще буду го-
ворить. Существенно ускорилась субдукция и вода в виде водных минералов снова стала
поступать в мантию, до этого мантия успела «высохнуть». В связи с усилением субдук-
ции и разуплотнения обводненной мантии произошло поднятие континентов и появились
обширная суша и шельф, большие реки и т. д. (см. ниже рис. 7). Одна из крупнейших пе-
рестроек в термальном режиме и, соответственно, условий на поверхности около 750 млн.
лет назад прошла в литературе незамеченной. И по мнению Ш. Маруямы и др. (Maruyama
et al., 2007), здесь еще огромное поле работы. Шестое событие на палеозойско-мезозойской
границе (250 млн. лет назад) - массовое вымирание, связанное с действием суперплюма,
породившего сибирские траппы, а также и эмейшаньские траппы в Китае и Вьетнаме и, на-
верное, еще много других крупных базальтовых излияний в древних океанах, реликты кото-
рых не сохранились. Седьмое событие (последние миллионы лет) - это появление человека
169
Н.Л. Лооре ион. О ранних стадиях >ro.iiouhii геосферы. биосферы и проблемах ранних оледенений
Рис. I. Эволюция мантии (А), геосферы (В) и биосферы (С) (Dobretsov et al.,
2008).
А - Рассчитанные вариации средних значений температуры Т °C. теплового по-
тока q и вязкости v (жирные линии по: Tajiks, Matsui, 1992, вариации по: Добре-
цов и др., 2001а), сопоставимые с вариациями возраста гранитов и мантийных
пород).
В - Важнейшие индикаторы эволюции геосферы (сверху вниз):
- гистограмма возраста гранитов и мантийных пород (Балашов, 1985; см. также
рис. 3);
- вариации К,О Na,О в гранитах и терригенных породах в сравнении с возрастом
суперконтине’нтов Pangea I—IV (Добрецов и др., 2001; Condie, 2005);
- вариации изотопов Sr в карбонатных породах (Добрецов и др., 2001; Dobretsov
et al., 2008);
- интервалы формирования железистых кварцитов, красноцветов, восстанов-
ленных и окисленных осадков (Заварзин, 2003а; Fallick et aL, 2008);
С - стадии эволюции биосферы (Добрецов, 2005; Dobretsov et al., 2008) с учетом
распределения цианобактерий (Заварзин, 2001; Sergeev et al., 2002). MBGB - со-
временная биоразнообразная глобальная биосфера. Точками показан период рас-
цвета Algae, точки со штрихами - период расцвета наземных растений. (Кривая
эволюции атмосферного кислорода по: Розанов, 2003).
Lgfq
4.6 4.0 3.0 2.0 1.0
Проьярмы црош xo. - . жтни
Phanerozoic
Hadean
Archean
Proterozoic
OXS 0.25 0065
4.5S Ga
0.55
У/С P/T
КЛ
P M
0. '
0.75
1.9
First supel tontinent
о
0.75
Plate te
rtonics
Return-fl' 'v uf svdw<it«ri
- Density-stratified
Dyna mo started
Magma
ocean
Double-layered
convection
Magma
ocean
Whole
convection
Magma I
ocean ]
Plume
tectonics
Birth of
solid inner core?
Earl;
31 noosphere
ЯГ
>, C02)
Ocean
H,O-CO
□ntinental crust
Heraatite precip I atior
Rapid increase of salinity
H,0
-mantle
Biggest 7 events
Рис. 2. Важнейшие события в истории Земли (Maruyama et al., 2007, с уточнениями): 1. Рождение
Земли: 2. Рождение жизни, континентальной коры, возможное начало тектоники плит; 3. Рождение
внутреннего ядра, усиление магнитного поля и выделения фотосинтетического кислорода в атмосфе-
ре: 4. Рождение первого достоверного суперконтинента, появление эукариот; 5. Начало возвращения
морской воды в верхнюю мантию и результирующее появление обширных континентов выше уровня
моря; 6. Pz/Mz массовое вымирание; 7. Появление человека и вымирания в голоцене.
и его взаимодействие с биосферой, следствием которого могли стать массовые вымирания
в голоцене. Пунктиром на рис. 2 показаны еще промежуточные рубежи, о которых говорится
в тексте. И тут тоже получились интервалы по 600-700 млн. лет, как на рис. 1.
Для сравнения с рис. 1 и 2 приведены диаграмма Ю. А. Балашова и В.А. Глазнсва (2008)
с максимумами и минимумами изотопных возрастов (рис. 3). На рис. 1 она показана в упро-
щенном виде, в верхней части сектора В (эволюция геосферы).
Верхняя синяя кривая-это число возрастных определений гранитоидов и других пород
континентальной коры, красная - мантийных пород. Кружками отмечены те рубежи, которые
близки к отмеченным выше. Минимумы: 240 млн. лет, 645 млн. лет, 1.36 млрд, лет, 2.26 млрд,
лет, 3.14 млрд, лет, 4 млрд. лет. Интервалы между древнейшими минимумами ООО млн. лет,
171
Рис. 3. Кривые изотопных датировок коровых пород (синяя линия СР) и мантийных
пород (красная линия. М). по оси ординат - число датировок для CR и М (Балашов,
Глазнев. 2008). Кружками обведены наиболее значимые минимумы и максимумы, со-
ответствующие границам на рис. 1. 2.
между последними - 650-700 млн. лет, как и на рис. I. Как видно, на рис. 1-3 есть расхожде-
ния, но многие рубежи совпадают (4.0; 3.1; 2.7; 2.3; 1.8; 0.65 или 0.75 млрд. лет). Важные ру-
бежи, скажем, 2.7 или 1.8-1.9 млрд, лет на рис. 3 совпадают с максимумами. На самом деле,
границу можно проводить либо по минимуму, либо по максимуму, поэтому и те, и другие
границы реально существуют. Тем не менее, проблемы периодизации ранней истории Земли
остаются дискуссионными, особенно событий догеологической истории (4.6-4.0 млрд, лет)
и раннеархейских событий (4.0-3.1 млрд. лет). Часто используемые древнейшие цифры кон-
тинентальной коры 4.1-4.3 млрд, лет - это возраст древнейших окатанных цирконов, най-
денных в конгломератах более молодых толщ в Австралии (Wilde et al., 2001). На самом деле,
они не служат показателем настоящей континентальной коры. Дело в том, что эти цирконы
содержат алмазы, скорее всего, импактного (ударно-метеоритного) происхождения. Цирко-
ны, вмещающие алмаз, по изотопному составу кислорода, переменному Th/V отношению,
некоторыми деталями внутренней структуры напоминают лунные цирконы импактного
происхождения (Nemchin et al., 2006; Menneken et al., 2007). Из всех известных импактных
кратеров алмазы в большом количестве есть только в Попигайском кратере (отдельные
алмазы есть и в других кратерах) (Haggerty, 1999). То есть это были, скорее всего, импактные
кратеры не меньше Попигайского, диаметром 100-200 км, а цирконы — скорее всего, ударно-
го (импактного) происхождения.
Оценки мантийных событий на рис. 3 основываются на анализе базит-гипербазитовых
серий и офиолитов. Начинаются они с серии Исуа в Гренландии с возрастом 3.8 3.9 млрд,
лет и завершаются излияниями срединно-океанических базальтов (MORB). На рис. 4 приве-
дены оценки температур лав (Исуа - MORB) и температур источников мантийных плюмов.
Правда, эти источники расположены непосредственно под литосферой, поэтому температура
здесь примерно на 300 °C ниже той температуры, которая показана на рис. 1, А и которая
является температурой на границе верхней и нижней мантии. Но тенденции примерно те же
самые, хотя есть отличия. На рубеже 2.6 млрд, лет (рубеж архея и протерозоя) на рис. I, А
172
Прок.n:\ihi ni'oiu \ож.и:пия жпши
Рис. 4. Эволюция потенциаль-
ной мантийной температуры
(plume sours) и температуры
верхнемантийных плюмовых
расплавов в сравнении с со-
держанием в них FeO. Серии
Isua (Исуа. Гренландия): NP
- район Северный блок, Пил-
бара; Австралия, CI - Кли-
вервилл, Пилбара; НМ. KI
- протерозойские комплексы
в Австралии: MORB - совре-
менные базальты срединно-
океанических хребтов (Komija.
2007). Справа показана магне-
зиальиость пород (Mg=85-90
%). Соответствующие содер-
жания FeO в мантии 10 и 8 %.
более интенсивно снижалась температура, а потом она перестала снижаться. А на рис. 4 тем-
пература подкорковых очагов варьировала на одном уровне, около 1500 °C, а потом, после
2500 млн. лет, систематически стала снижаться до 1300 °C. Поведение FeO в мантии и темпе-
ратуры лав менялись синхронно. До рубежа архея-протерозоя концентрация железа в мантии
варьировала незначительно, а после этого непрерывно снижалась. Магнезиальность (Mg на
рис. 4 - отношение Mg к сумме Mg+Fe) росла от 85 до 90 %). Этот вывод несколько неожидан-
ный. Можно ожидать, что железо (особенно в верхней мантии) в результате его истощения и
за счет появления разнообразных дифференцированных серий, уходит из мантии, что объ-
яснено ниже изменением конвекции в мантии.
Теперь перейдем к эволюции биосферы и ее корреляции с эволюцией геосферы.
На рис. 1, С показаны главные рубежи и корреляция, а на рис. 5 - сценарий этой эволюции
[У/ ДАРВИНОВСКАЯ ЭВОЛЮЦИЯ
’ (- 800-500 млн. лет назад)
МНОГОКЛЕТОЧНЫЕ ОРГАНИЗМЫ
I
Половой процесс + эндогамно замкнутые виды
ДНК- -РНК—- белок + мембрана КЛЕТКА
3 РНК + белок
I
2 РНК-олигонуклеотиды РНК-полинуклеотиды
I
1 Синтез РНК на монтмориллоните (или панспермия?)
МИР РНК
(~ 4.1 млрд, лет назад)
Рис. 5. Сценарий главных стадий зарождения и эволюции жизни (Добрецов, 2005; Dobretsov et al., 2008).
173
Н.Д. Добрецов. О ранних и кцнях irc.iioiiiiii i сосфсры. биосферы и проблемах ранних оледенении
Стратиграф,
шкала
1850
С Cambrian - взрыв биоразнообразия
550
650
850
1000
1150
1350
-1650
2000 «'
2500 )
3000 3 g
Возраст, q Ordovician - биосферная эволюция
млн. лет
5 S
< ф
Земная кора и гидросфера
(округлые цирконы как доказательство)
3 5 Вут gпоявление фотосинтеза
* д предполагаемое)
2 7 зуг . появление \
7 w фотосинтеза =
(документировано)
2.0 Вут_______________
Железистые
кварциты
2.4 Вут
4200 <
Циклы
0.16
0J4
Климати-
ческие
^Ъезскелетные
ногоклеточные
Metazoa
Половое
размножение
Важнейшие оледенения
палеошироты
90°
Число континентов
Рис. 6. Появление и развитие фотосинтеза и рост биоразнообразия в сравнении с важ-
нейшими оледенениями в докембрии и изменением числа континентов (от I - Пангеи,
до 6). Максимум железистых кварцитов (Заварзин, 2001, 20036), рост биоразнообразия
эукариот (Fedonkin, 2003).
Huron
Griquatuan
PR]
Witwaters rand
(Добрецов, 2005; Dobretsov et al., 2008). Выделены четыре стадии эволюции биосферы мир
РНК, прокариотический мир, эукариоты, многоклеточные организмы с «дарвиновской эво-
люцией». Многие границы здесь надо, вероятно, удревнить, например, появление мира РНК
(на рис. I, С и 5 - 3.8 млрд, лет) - теперь, скорее, 4.0 или 4.I млрд, лет (Розанов, 2003, 2006;
Ferris, 2005). На этой первой, доклеточной стадии (т. е. в мире РНК) уже появились белки, так
как клеточные протомембраны, в частности, их осмотические помпы, строились с участием
белков. Синтез АТФ в клетках обеспечивается белковыми помпами - древнейшей натрие-
вой и более поздней протонной. Для этих сложных белковых конструкций нужно время на
их создание и эволюцию (Chetverin, 1999; Колчанов и др., 2003; см. доклад А.С. Спирина).
Поэтому рубеж около 4 млрд, лет - важнейший, поскольку знаменует исчезновение докле-
точной (Bernal, 1967) и появление белково-клеточной формы жизни. Возможно, удревнения
заслуживают другие границы, как считает А.Ю. Розанов (2003; Rozanov, 2006). Например,
прокариоты появились не 3.6, а 3.8 3.9 млрд, лет назад, эукариоты - не I.7 млрд, лет назад,
а раньше 1.8 млрд, лет назад. Один из вопросов, который далее будет обсуждаться, - где про-
водить границу появления животных - I млрд, или 750 млн., или 650 млн. лет назад.
Следующий вопрос - была ли эволюция бактерий, как и когда возникли археи
и эубактерии (Добрецов и др., 20016; Заварзин, 20036; Шестаков, 2003; Zavarzin, 2008).
I74
rii’ob. iEMbi hi*oiic.\oa.ieiiii>i /ки ат
В позднем архее и раннем протерозое (до 1.8 млрд, лет), безусловно, преобладали циано-
бактерии. Но они существуют и сейчас без заметных морфологических и биохимических
изменений (Sergeev et al., 2002). Максимальное развитие цианобактерий было около 2 млрд,
лет назад (рис. I, С), судя по широкому развитию в это время строматолитовых построек
в карбонатных (преимущественно доломитовых) породах (окаменевшие цианобактериальные
маты). Сначала преобладали анаэробные, а потом с 2.5 2.7 млрд, лет аэробные бактерии (гра-
ница архея-протерозоя), но зафиксированный массовый оксифотосинтез начался 2.7 млрд, лет
назад (рис. 2, 6). Появился он, возможно, уже 3.5 млрд, лет назад (рис. 6). Железистые фор-
мации, образование магнетитовых осадков в океане - отражение этого процесса. На рис. I, В
я повторил оценки Г.А. Заварзина (2003а, б; Zavarzin, 2008).
Теперь обратимся к изменению климата и самым ярким его проявлениям - оледене-
ниям. Как видно из рис. 6, самое раннее зафиксированное оледенение - на границе архея-
протерозоя. Следует обратить внимание, что в это время произошла смена общемантий-
ной конвекции на двухслойную (Добрсцов и др., 2001а). Поэтому изменился режим обмена
вещества. В частности, это объясняет, почему содержание железа в мантии до этого было
постоянным (рис. 4) - вся мантия перемешивалась. Как только верхнемантийная конвекция
обособилась, верхняя мантия стала истощаться, терять железо за счет дифференцированных
серий, включая граниты (в т. ч. железистые граниты, характерные для раннепротерозойских
серий). Одновременно двухслойная конвекция привела к остыванию поверхности и построе-
нию новой климатической системы. Но все же в раннем и среднем протерозое оледенения
были редкими. Лишь на рубеже 700-750 млн. лет назад появились очень сильные и частые
оледенения. Возможно, максимальным было ранневендское оледенение около 640 млн. лет
назад (Добрецов, Чумаков, 2003). Для них как раз и применяют определение «snowball» -
замерзшая Земля, похожая на снежный шар (Hoffman, Schrag, 2002; Maruyama, Liou, 2005).
Первые гипотезы об этом родились из чисто геохимических данных, а далее главным доказа-
тельством того, что ледниковые отложения были вблизи экваториальных областей, служили
уточненные палеомагнитные данные (но здесь еще много дискуссионного).
Диаграмма, приведенная на рис. 7 одно из объяснений важности рубежа 750 млн. лет
назад. На ней обобщены данные по породам из зон субдукции, извлеченным с разных глубин
в разное время (Maruyama, Liou, 2005). Красные точки это древние породы (древнее I млрд,
лет), им соответствуют очень высокотемпературная граница и малые глубины. Давление здесь
не превышает 10 кбар. Белые квадраты - это комплексы с возрастом от 1000 до 750 млн. лет.
Зеленые точки включают более молодые породы (менее 750 млн. лет), извлеченные из глубин
более 200 км, то есть соответствующих давлению 60-50 кбар. Они возникли в палеозое и
мезозое, в частности, кокчетавские алмазеодержащие породы с возрастом 530-540 млн. лет
(Dobretsov, Shatsky, 2004; и др.). Японские авторы (Maruyama, Liou, 2005; Maruyama etal., 2007)
сдвигают эту границу - то 540, то 750-800 млн. лет. В любом случае, с 750 млн. лет назад на-
чалась интенсивная и ускоренная (в разы) субдукция, обусловившая и быстрое извлечение по-
род с больших глубин. И вместе с субдуцирующим материалом в мантию снова стала посту-
пать вода. До этого высокотемпературными расплавами из мантии вся вода была извлечена.
Усиление субдукции дало вспышку островодужного магматизма, массовое поступление СО,
в атмосферу и ее окисление. И главное - разуплотнение обводненной мантии приподняло все
континенты и островные дуги. Они стали более существенно выступать над уровнем океана,
расширился шельф, появились крупные реки, крупные дельты и т. д. По данным Ш. Маруямы
(Maruyama et al., 2007), все это произошло в интервале 650-750 млн. лет назад (рис. 7).
С оледенениями связан вопрос об эволюции скелетов. Впервые хитиновые (белковые)
скелеты появились на границе протерозоя. Минеральные скелеты - фосфатные, карбонат-
ные, силикатные, появились лишь через 1.5-2 млрд. лет. Почему? Это вопрос нерешенный,
возможно, это результат частых неблагоприятных обстановок, связанных с оледенениями.
175
H.'l. Joopcifuti. О ранних с i алиях эволюции геосферы. биосферы и проблемах ранних оледенений
а
6
° 0-630 Ма
□ 750-1000 Ма
° >1000 Ма
• 2500-3700 Ма
2
О
200 400 600 800 1000
Температура (°C)
@Увеличение кислорода в атмосфере (7)Рождение озонового слоя
(8) Возникновение
(3)Новая береговая линия
+ 6НгО = С2Н„О5 + 6Ог
Понижение
уровня моря
(7) Гидратация
680 км
клина
410км
Низкотемпературная
часть
Рис. 7. Условия температуры и давления в разновозрастных метаморфических породах (а) и условия
изменения динамики мантии и континента в интервале 650-750 млн. лет назад (б) (Maruyama. Liou.
2005, с изменениями). Взаимосвязанные события 1-8 - см. пояснения в тексте.
176
rjl'Dh. lEMhl НРОНСХОЖ.И'ИНЯ ЖП <Jill
Рис. 8. Сценарий изменения био-
разнообразия в фанерозое (Sep-
koski. 1996) в сравнении с важ-
нейшими импульсами мантийных
плюмов (красные точки на шкале
времени - Добрецов и др.. 20016).
Precambrian/ Cambrian/
Cambrian Ordovician
Permian/
Triassic
Cretaceous/
Paleogene
Основанием для построения рис. 5 и 6 и других аналогичных диаграмм, служат не только
геологические и палеонтологические данные, но и данные молекулярной биологии, так назы-
ваемые «молекулярные часы», основанные на средне-постоянной скорости синонимичных или
нейтральных мутаций (Kumar, Hedges, 1998; Cavalier-Smith, 2002; Колчанов и др., 2003). Приве-
дем только важнейшие рубежи, установленные этим методом. Появление и дивергенция эука-
риот-2.2-1.8 млрд, лет назад. Важно, что 1.8 млрд, лет (эта цифра везде повторяется) - это либо
завершение этого процесса, либо его максимум. Наконец, для многоклеточных организмов две
цифры. Одна - общий предок растений, животных и грибов появился 1.5-1.6 млрд, лет назад.
Вторая-общий предок водорослей и наземных растений это 1 млрд, или 1.17 млрд. лет.
Крупнейшее расхождение датировок по молекулярным часам и по палеонтологиче-
ским данным касается последней цифры. Либо это 1.1 млрд, (по «молекулярным часам»),
либо 550-450 млн. лет (по палеонтологическим данным). Возможные два сценария: либо
растения, водоросли, вторичноротые и членистоногие появились вокруг Родинии (второ-
го суперконтинента) около 1.1 млрд, лет назад и существовали до 950-750 млн. лет назад.
А потом два гигантских оледенения 750 млн. и 650 млн. лет назад уничтожили все растения,
водоросли и связанных с ними мелководных животных. Лишь немногие из них пережили
это время в рефугиях, чтобы дать расцвет в венде и кембрии. Не исключено, что их глубоко-
водные родственники вновь колонизировали мелководье. Либо, наконец, древние датировки
для указанных групп растений и животных - ошибка метода молекулярных часов.
Кроме вышеупомянутого главного расхождения есть расхождения и в другие эпохи,
причем они все связаны с крупными перестройками. В частности, меловой эпохе перестрой-
ки соответствует относительное удревнение по молекулярно-биологическим часам. Важней-
ший рубеж на границе мезозоя и палеозоя фиксируется аналогичными удревнениями. Это,
в частности, можно объяснить тем, что скорость мутаций, которая принимается постоянной
при определении по молекулярным часам, на самом деле в периоды крупных перестроек мо-
жет быть быстрее. Отсюда возникает кажущееся удревнение.
В эпохи крупных оледенений (750-650 млн. лет назад) сформировался также меха-
низм крупных мантийных плюмов, поднимающихся от ядра Земли (Добрецов и др., 2001а;
Maruyama et al., 2007). Именно периодические плюмы и связанные с ними явления обуслови-
ли основную периодичность за последние 500 млн. лет, в частности, с ними коррелируются
все крупные вымирания (рис. 8), в том числе крупнейшее на границе перми и триаса (собы-
тие 6 на рис. 2). Но и остальные вымирания обусловливают периодическое уменьшение био-
разнообразия от прямой линии, соединяющей появление биоразнообразия животных около
800 млн. лет назад и современный уровень биоразнообразия (рис. 8).
177
Н.. /. 'loo рецок.
О ранних сгадиях
эволюции геосферы.биосферы и
проблемах ранних оледенении
tune (billion years)
Рис. 9. Эволюция жизни на Земле в те-
чение 2.5 млрд, лет (-2.5-0) и ожидае-
мые изменения биосферы (0+1.5 млрд,
лет) (Bounama et al., 2004, с измене-
ниями).
На рис. 9 приведена схема (Bounama et al., 2004), представляющая экстраполяцию буду-
щей эволюции и умирания (исчезновения) биосферы. Это связано с возрастанием солнечной
светимости и постепенным превращением Солнца в красную звезду. Первые 4 млрд, лет шло
остывание и снижение температуры поверхности до 40°C на рубеже архея-протерозоя. Сниже-
ние до 30-25 °C - вспышка и появление высших организмов. Похолодание около 650-750 млн.
лет назад и потом потепление в мезозое (250-100 млн. лет назад) я добавил с учетом современ-
ных данных. Объем биологического вещества (если не учитывать симбиозы) примерно одина-
ков в прокариотах, эукариотах и высших многоклеточных организмах и достигал максимумов
около 1800, 1200 и 500 млн. лет назад. Последовательное вымирание начнется через 500 млн.
лет и связано с повышением температуры поверхности: при достижении примерно 30°C вы-
мрут высшие организмы, выше 45 °C вымрут эукариоты и окончательно, через 1.6 млрд, лет,
когда температура достигнет 70 °C и выше - с поверхности исчезнет вся жизнь. Это относи-
тельно оптимистический сценарий. Есть сценарий вымирания жизни через 250 млн. лет.
На рис. 9 добавлены возможные периодические колебания температуры и объема био-
сферы. Сами оценки объема биосферы (в гигатоннах) очень примерные (Bounama et al., 2004),
но вариации объема биосферы подтверждаются колебаниями биогермных построек (строма-
толитов, коралловых рифов и т. д.) в разы и даже на порядок, и они коррелируются с измене-
ниями температуры поверхности. На самом деле температурные колебания могли быть еще
больше. Если посмотреть вековые колебания, то видно, что температура поверхности непре-
рывно «дрожит». Но дрожит в умеренных и высоких широтах. В тропиках она меняется мало.
Тем не менее возможно расширение зон с высокими вариациями по температуре в периоды
таких похолоданий, вплоть до 0°С на экваторе, если допустить, что покрывающие всю Землю
оледенения действительно существовали. Но продолжается спор, поскольку геохимической
информации, положенной в основу этого материала, недостаточно. Температура поверхности,
в любом случае, последние 800 млн. лет и следующие 500 млн. лет испытывала и будет ис-
пытывать сильные колебания.
Таким образом, на раннем этапе эволюции биосферы главную роль играл процесс
окисления атмосферы, который привел к преобладанию фотосинтеза на рубеже 2.7 млрд, лет
назад и (вместе с охлаждением поверхности) - расцвету цианобактерий, развитие которых
достигло максимума около 2 млрд, лет назад.
На более поздних этапах главное значение имело остывание поверхности, которое
привело к крупнейшим оледенениям 750 и 640 млн. лет назад. Похолодания и оледенения
привели к большой изменчивости условий для биосферы, что стимулировало, во-первых,
половое размножение и образование только (или преимущественно) эндогамно-замкнутых
178
ПРОБЛЕМЫ ПРО! к ХОЖ.^.ЦИЯ ЖИ <1111
видов (или к рубежу 1.5 млрд, лет назад, или 1-1.2, или 0.75 млрд, лет назад), и, во-вторых, -
развитию разных форм минерального скелета, начиная с 550 млн. лет назад.
После формирования главных крупны,х таксонов в кембрии-ордовике (550-450 млн.
лет назад), основное значение для видообразования и усложнения видов и их приспособляе-
мости к внешним условиям явились периодические изменения климата и внешней среды,
иллюстрируемые рис. 8, обусловленные, в том числе (или преимущественно?) периодически-
ми мантийными крупными струями (суперплюмами). Примеры изменения биоразнообразия,
обусловленные изменениями климата, хорошо изучены на Байкале (Грачев, 2002; Добрецов,
2003). Один из наиболее ярких примеров - крупнейшее вымирание на границе перми и триа-
са (рис. 8), вызванное Сибирским и Эмейшанским суперплюмами. Механизм этого вымира-
ния (таяние ледников и кислотные дожди, вызванные выбросом огромного количества серы)
обсуждался неоднократно (Добрецов, Чумаков, 2001; Добрецов, 2003; Isozaki, 2007).
В заключение вернемся к основным проблемам зарождения и эволюции биосферы. Где
появилась жизнь - на Земле или в космосе? Есть доказательства, что правдоподобно внезем-
ное происхождение жизни (панспермия), вытекающее из изучения марсианских метеоритов
и космохимических моделей. Более вероятная альтернатива - зарождение жизни в гидротер-
мальных струях типа современных черных или белых курильщиков.
Второе важнейшее событие - появление и развитие фотосинтеза. Бактерии изменя-
ли мир, т. к. за счет фотосинтеза они способствовали окислению поверхности. Окисление
на ранних этапах шло и без бактерий за счет диссипации водорода в космос. Водород един-
ственный элемент, который Земля теряла всегда, а особенно на ранних стадиях, когда она
была горячая. Зная примерную диссипацию сегодня и учитывая более высокую температу-
ру в прошлом, можно рассчитывать диссипацию в архее и посчитать количество водорода.
У меня получилось, что в архее, на рубеже 4 млрд, лет назад, атмосфера была водородно-
метановая. Не было никакого свободного кислорода.
Ну, а третье важнейшее событие это появление половой репродукции и натураль-
ной селекции. Это важнейший рубеж, который привел к появлению многоклеточных
организмов, а затем к тому естественному отбору, который нам известен как дарвиновский
отбор. Обычно проводится рубеж около 1.2 млрд, лет назад. Но есть и другие данные, что это
рубеж может быть 1.5 млрд, лет назад, а по мнению академика А.Ю. Розанова - еще раньше.
Это один из важнейших вопросов, которые надо уточнить. Дело в том, что в ископаемых
отпечатках мы не можем достоверно отличить колониальные организмы от многоклеточных.
Поэтому здесь важнее уточнить изотопные смешения. Как только появились специализирован-
ные многоклеточные организмы, появились специфические белки. Вероятно, прав академик
М.А. Федонкин (Fedonkin, 2003), что в это же время произошла и смена активных центров в
белках. Сегодня это железо, никель, частично медь и цинк. А в прошлом должны были быть
трехвалентные катионы - вольфрам, молибден, ванадий. Мы должны искать изотопные доказа-
тельства и найти существенное смешение именно изотопов молибдена, ванадия, вольфрама. Се-
годня это уже возможно инструментально. Есть черные сланцы (продукты деятельности сначала
бактерий, а потом и более сложных организмов), в них и надо искать этот рубеж. Наряду с тем,
чтобы развивать бактериальную палеонтологию и другие методы и с их помощью реконструи-
ровать последовательность появления специфических белков, специфических тканей, нейронов,
нервной системы и, наконец - сознания. Таковы возможные темы будущих исследований.
В заключение хочу поблагодарить академика А.Ю. Розанова за приглашение принять
участие в совещании и многолетнее сотрудничество в программе «Происхождение и эволю-
ция биосферы», а также моих коллег и неоднократных соавторов - академика Н.А. Колчано-
ва и В.В. Суслова - за сотрудничество и плодотворные дискуссии, в том числе способство-
вавшие улучшению данной статьи.
179
Н.Л. Доирсцок. О ранних cia.iHMx эволюции геосферы, биосферы и проблемах ранних оледенении
СПИСОК ЛИТЕРАТУРЫ
Балашов Ю.А 1985. Изотопно-геохимическая эволюция мантии и коры Земли. М.: Наука. 222 с.
Балашов Ю.А.. Глашев В.A. 2008. Геохимические и изотопные особенности наиболее ранних горо-
образования и дифференциации мантии Земли // ТИНЕТА. № 4. С. 22-27.
Грачев М.А. 2002. О современном состоянии экосистемы озера Байкал. Новосибирск: СО РАН. 156 с.
. {обрецов Н.Л. 2003. Корреляция биологических и геологических событий в истории Земли и возмож-
ные механизмы биологической эволюции Палеонтол. журн. № 6. С. 4-15.
Добрецов Н.Л. 2005. О ранних стадиях зарождения и эволюции жизни Вести. Всерос. об-ва генетич.
селекц. Т. 9. № 1. С. 43-54.
Добрецов Н.Л.. Кирдяшкин А.Г.. Кирдяшкин А. А. 2001а. Глубинная геодинамика. Новосибирск: Изд. ГЕО. 409 с.
Добрецов Н.Л.. Чумаков Н.М. 2001. Глобальные периодичности в эволюции литосферы и биосферы И
Глобальные изменения природной среды / Род. Добрецов Н.Л., Коваленко В.Н. Новосибирск:
Изд. ГЕО. С. 11-27.
Добрецов Н.Л.. Шестаков С.В.. Шумный В.К.. Каныгин А.В. 20016. Проблемы происхождения и эволю-
ции жизни Вести. Всерос. об-ва гснст. селекц. № 17. С. 2-6.
Заварзин Г.А. 2001. Образование биосферы // Вести. РАН. Т. 71. С. 988-1001.
Заварзин Г.А. 2003а. Лекции по природоведческой микробиологии. М.: Наука. 348 с.
Заварзин ГА. 20036. Становление системы биологических циклов // Палеонтол. журн. № 6. С. 16-24.
Колчанов Н.А.. Суслов В.В.. Шумный В.К. 2003. Молекулярная эволюция генетических систем // Пале-
онтол. журн. № 6. С. 617-629.
Розанов А.Ю. 2003. Ископаемые бактерии, седимснтогенез и ранние стадии эволюции биосферы
Палеонтол. журн. № 6. С. 41-49.
Шестаков С.В. 2003. О ранних этапах биологической эволюции с позиции геномики ' Палеонтол.
журн. № 6. С. 50-57.
Bernal J.D. 1967. The origin of Life. L.: Weidenfeld and Nicolson. 345 p.
Bounama C. Bloh IK von. Franck S. 2004. Das Endo des Raumschiffs Erde Spektriim der Wissenschaft.
Oktober2004. P. 100-107.
Cavalier-Smith T. 2002. Origins of the machinery of recombination and sex < Heredity. V. 88. P. 125-141.
Chetverin A.B. 1999. The puzzle of RNA recombination • FEBS Lett. № 460. P. 1-5.
Condie K.C. 2005. Earth as an evolving planetary system. L.: Elsevier Acad. Press. 447 p.
Dobretsov ML. Kolchanov N.A.. Suslov I'.l'. 2008. On important stages of geosphere and biosphere evolution // Bio-
sphere origin and evolution I Eds. N. Dobretsov, N. Kolchanov, A. Rozanov, G. Zavarzin. Springer. P. 3-24.
Dobretsov N.L., Shatsky F.S. 2004. Exhumation of high-pressure rocks of rhe Kokchctov massif: facts and
models // Lithos. V. 78. P. 307-318.
Fallick A.E.. Melezhik Г.А.. Simonson B.M. 2008. The ancient anoxic biosphere was not as we know it "Biosphere
origin and evolution / Eds. N. Dobretsov. N, Kolchanov, A. Rozanov, G. Zavarzin. Springer. P. 169-188.
Fedonkin M.A. 2003. The origin of the Metazoa in the light of the Proterozoic fossil record II Paleontol. Res.
V. 7. P. 9-41.
Ferris J.P. 2005. Mineral catalysis and synthesis: montmorillonite catalyzed formation of RNA // Elements.
V. I. P. 145-149.
Haggerty S.E. 1999. A diamond triology: superplumes, supcrcintinents and supernovae// Science. V. 285. P. 851-860.
Hoffman P.F.. Schrag D P. 2002. The snowball Earth hypothesis: testing the limits of global change ' Terra
nova. V. 14. P. 129-155.
Isozaki Y. 2007. Plume winter scenario for biosphere catastrophe: the Permo-Triassic boundary case ' Super-
plumes: beyond plate tectonics Eds. D.A. Yuen et al. Springer. P. 409-439.
Komiva T. 2007. Material circulation through time: chemical differentiation within the mantle and secular variation
of temperature and composition of mantle // Superplume / Eds. D.A. Yuen et al. Springer. P. 187-234.
180
ПрОЬ. HMtid ,Н‘1)И1М)Ж ."ИНН ЖИ Ц.(Л
Kumar S.. Hedges S.B. 1998. A molecular timescale for Vertcbrata evolution // Nature. V. 392. P. 917-920.
Maruyama Sb.. Lion J.G. 2005. From snowball to Phancrozoic Earth . Intern. Geol. Rev. V. 47. P. 775-791.
Maruyama Sir. Yuen DM.. Windlev B.F. 2007. Dynamics of plumes and superplumes through time Super-
plume Eds. D.A. Yuen et al. Springer. P. 441-502.
Menneken M.. Hemehin .4..4.. Geisler Th. et al. 2007. Hadean diamonds in zircons from Jack Hills. W. Aus-
tralia // Nature. V. 448. P. 917-921.
Nemchin A.A.. Pidgeon R.T. Whitehouse M.J. 2006. Re-evaluation of the origin and evolution of ' 4,2 GA
zircons from the Jack Hills metasedimentary rocks Earth Planet. Sci. Lett. V. 244. P. 218-233.
Rozanov A. Yu. 2006. Precambrian geobiology Paleontol. J. V. 40. Suppl. 4. P. S434-S443.
Schidlowski M.,4. 1988. 3,8 billion year old record of life from carbon in sedimentary rocks. Nature. V. 333. P. 313-318.
Sepkovski J..J. 1996. Pattern of Phanerozoic extinction: a perspective from global data bases//Global events
and event stratigraphy / Ed. O.H. Wallister. Berlin: Springer. P. 35-51.
Sergeev V.N., Gerasimenko L.M., Zavarzin G..4. 2002. Proterozoic history and present state of Cyanobacteria //
Microbiology. V. 71. № 6. P. 725-740.
Tajika E.. Matsui V. 1992. Evolution of terrestrial proto CO, atmosphere coupled with thermal history
of Earth Earth Planet. Sci. Lett. V. 113. P. 251-266.
Wilde S.A., Valiev J.W., Peek W.H.. Graham C.M. 2001. Evidence from detrital zircons for the existance
of continental crust and ocean on the Earth 4,4 Gyr ago Nature. V. 409. P. 175-178.
Zavarzin G.A. 2008. Microbial biosphere // Biosphere origin and evolution / Eds. N. Dobrctsov, N. Kolchanov,
A. Rozanov, G. Zavarzin. Springer. P. 25-44.
On the early stage of geosphere and biosphere evolution,
and the problem of the early glaciations
N.L. Dobretsov
The earliest stages of Geosphere and Biosphere evolution are controlled by three interrelated
factors: 1) the ongoing cooling-down of the Earth surface and mantle (decrease of the mean mantle
temperature in 1.5-2 times, of the mean surface temperature in 3-4 times, of the mean heat flow in one
order of magnitude, of the viscosity in three orders of magnitude); 2) the ongoing step-by-step oxida-
tion of the surface, more intensive during the period of 3.8-1.8 Ga; 3) periodic and conjugate fluctua-
tions of geosphere and biosphere conditions of various scale and nature. The most important moments
in this evolution were the episodes about 4 Ga (origin of vigorous but heterogeneous Earth crust, be-
ginning of life), about 3 Ga (origin of strong magnetic field, increase of photosynthetic activity), about
1.8-1.9 Ga (origin of oxidized atmosphere, the first super continent, possibly the first superplums of the
core), about 0.75 Ga (speeding-up of subduction, «irrigation» of the upper mantle, upraise of continents
with waste land, shelfs, large rivers and first glaciations). The significance and correlations of the earli-
est events (before and near 4 Ga) and the events about 0.75 Ga are strongly debatable. During the late
Archaean and Early Proterozoic (up to 1.8 Ga) the biosphere was dominated by cyanobacteria, which
dynamics and maximums of distribution were marked by strong development of stromatolite buildups
in carbonate rocks (mostly dolomitic at the beginning). Since the period of 700-750 Ma the strong and
frequent glaciations started on the Earth reflecting its cooling-down. Possibly the strongest glaciation
took place about 640 Ma, this period is argued as a «Snowball Earth». The glaciations are sometimes
connected with the problem of the animals’ skeleton origin and its evolution. All these events correlate
to each other and can be explained by the major endogen changes. One of the most important phenom-
ena in the geological history is a periodic activation of mantle flows (superplums) started 750 Ma. This
phenomenon explains the further periodization of biosphere evolution. The extrapolation of the future
evolution and consecutive collapse of biosphere segments due to the transformation of the Sun into a
red star and heating of the Earth surface is given in the conclusion.
181
Дискуссия
по пленарному докладу Н.Д. Добрецова
«О РАННИХ СТАДИЯХ ЭВОЛЮЦИИ БИОСФЕРЫ
И ПРОБЛЕМАХ РАННИХ ОЛЕДЕНЕНИЙ»
Председательствующий Г.А. Заварзин
М.Я. Маров: Прежде всего, позвольте сделать один короткий комментарий. Вы упре-
кнули физиков и математиков, что не сосчитаны модели, связанные с реконструкцией дисси-
пативного механизма. На самом деле, все это не так просто. Дело в том, что Вы, по-видимому,
говорите о джинсовской диссипации, чисто тепловой - ее сосчитать легко. Но на начальных
этапах эволюции мог существовать другой специфический род диссипации, когда при до-
вольно высоких температура,х быстро улетает водород и в потоке он увлекает за собой дру-
гие, более тяжелые элементы. Без такого механизма не удается, например, объяснить потерю
древнего океана на Венере. Его предложил и обосновал известный американский планетолог
Дональд Хантен (Donald М. Hunten, University of Arizona, Lunar and Planetary Laboratory).
Но я хочу сказать, что моделировать этот гидродинамический механизм диссипации доволь-
но трудно - и сложно, таким образом, реконструировать историю потери летучих.
Н.Л.Д.: Я действительно посчитал только джинсовскую диссипацию - и то оказалось,
что к рубежу 4.2 млрд, лет назад резко преобладали водород и метан. А если еще добавить -
значит, водорода было еще больше. Для меня важна минимальная оценка.
М.Я. Маров: Хорошо, у меня есть другие вопросы. Вы говорите о сильно возросшей роли
механизма субдукции на рубеже около 750 млн. лет назад. Как известно, различные модели
по-разному датируют начало глобальной тектоники. Как Вы связываете возрастание роли
субдукции с механизмом развития глобальной тектоники, на каком рубеже это происходило?
Н.Л.Д.: По вопросу, когда началась субдукция, когда началась тектоника плит, есть це-
лый ряд независимых данных. Первое прямое доказательство существования океанической
коры - это офиолиты. Океаническая кора, включая дайковые комплексы, пиллоу-лавы, осад-
ки океанического типа, известна начиная с двух миллиардов лет или немного древнее. Вто-
рое - субдукция. Она должна быть равна или больше скорости спрединга. Сегодня скорость
спрединга достигает 16 см в год, в зонах субдукции поглощение до 20 см в год. Обратное дей-
ствие при завершении субдукции - извлекаются глубинные породы, в которых содержатся
такие индикаторы высоких давлений, как алмаз, коэсит, специфические гранаты и так далее.
Так вот, только после 750 млн. лет назад появились эти признаки - породы, извлеченные
с глубин в 150-200 км, что косвенно говорит о скорости субдукции, близкой современной.
Наиболее показательный такой пример с алмазосодержащими породами - это кокчетавские
породы с возрастом около 540 млн. лет. Есть объекты в Мали (самые древние, 750 млн. лет,
там нет алмазов, только коэсит), потом в Китае, Норвегии и другие.
М.Я. Маров: Соответственно, тогда же заработал активный карбонатно-силикатный цикл?
Н.Л.Д.: Конечно! Весь этот конвейер активно заработал.
М.Я. Маров: А когда Вы говорите о начале глобальной тектоники два миллиарда лет
назад, это как раз то, что отвечает перестройке конвекции от сплошной к раздельной в ниж-
ней и верхней мантии9
Н.Л.Д.: В принципе, такая постановка вопроса правомочна. Но один из ведущих пе-
трологов и геофизиков - Ш. Маруяма - немного иначе трактует, ближе к моей точке зрения.
Субдукция, если она нс очень интенсивная, складирует весь материал в зоне С, между глу-
бинами 400 и 650 км, и там накапливается связанная вода. И объем воды в слое С больше,
чем объем воды в Мировом океане! Сплошная субдукция, до границ ядра, сейчас невозмож-
182
Пронлг.мы нроисхож-П-шия жшнп
на, потому что слишком вязкая нижняя мантия. Но тяжелый материал проникает и тянет
за собой воду и легкоплавкие материалы. Соответственно, вода совершенно меняет условия
магматизма в нижней мантии, а опустившийся материал скапливается в слое D2 на границе
нижней мантии и ядра и рождает суперплюм. Начиная с рубежа 750 млн. лет назад пошли
суперплюмы, они заменили общемантийную конвекцию.
М.Я. Маров: Каков вклад антропогенного фактора в температурном тренде?
Н.Л.Д.: Я думаю, что антропогенный фактор больше работает как триггерный меха-
низм, сегодня, по моим оценкам, он составляет менее 20 %. Но он чрезвычайно важен, так как
работает, накладываясь на другие факторы, а там всюду нелинейный эффект.
Ю.В. Наточшк Что Вы имели в виду, когда говорили об осмотическом насосе?
Н.Л.Д.: Я имел в виду белковые помпы, в том числе накачивающие в клетку калий,
который в большом количестве появился на поверхности Земли после 1.8 млрд, лет назад.
Поэтому и животные появились, когда появился избыток калия, это одна из причин.
С.В. Шестаков: Николай Леонтьевич, когда Вы говорили о фотосинтезе, вы наверняка
имели в виду оксигенный фотосинтез, связанный с деятельностью цианобактерий, продуци-
рующих кислород. Но ведь есть очень много форм фотосинтеза, не связанных с выделением
кислорода - аноксигенный фотосинтез и не связанная с фотосинтезом фотореакция протео-
родопсина, например. С точки зрения биолога, цианобактерии - это вершина биохимической
эволюции. В Ваших схемах речь всегда идет об оксигенном фотосинтезе?
Н.Л.Д.: Цианобактериальные колонки в архее неоднородны, внутри были оксигенные
прослои, снаружи - аноксигенные. Я хотел бы подчеркнуть, что в докембрии накапливались
огромные мощности строматолитов, если пересчитать на единицу времени, масса биосферы
могла быть выше современной.
С.В. Шестаков: Вы говорили об оледенениях. Насколько глубоко они воздействовали?
Н.Л.Д.: Оледенение глубоко не проникает, внутреннее тепло Земли не меняется. Иногда
отрицательная температура распространяется на несколько сотен метров. Контакт пород и льда
в среднем от -5 до 0 °C. Но первое, что прекращается - выветривание и связывание СО, в карбо-
наты. Поэтому начинает работать обратная связь - СО,, выбрасываемый вулканами, накаплива-
ется в атмосфере и лед начинает таять. По нашим расчетам, такая ситуация может существовать
не более пяти миллионов лет. Но Маруяма предположил, что, поскольку не было субдукции,
то и СО почти не выделялся, темпы выделения СО, вулканами были на два порядка ниже -
поэтому у него получилось 500 миллионов лет. Тут слишком большая неопределенность.
С.В. Шестаков: Но, тем не менее, во время сильных оледенений деятельность растений
невозможна, цианобактерий - тоже...
Н.Л.Д.: Даже в сплошном леднике должны были оставаться оазисы - рефугии (напри-
мер, около вулканов). Поэтому жизнь в отдельных оазисах могла сохраняться, в том числе
и высокоорганизованная...
Г.А. Заварзин: Для гипотезы «Snowball Earth» хорошей моделью является современная
Арктика, окраины Антарктиды, где развивается достаточно мощная подледная жизнь... Там
много бактерий, не все фотосинтезирующие, в зависимости от толщины слоя... Так что во-
обще с гипотезой «Snowball Earth» нужно обращаться аккуратно... Для меня непонятно, по-
чему синхронно существовали строматолиты и образовывались доломиты?
Н.Л.Д.: Расцвет доломитов был предопределен изменением состава Мирового океана.
Нужно посчитать баланс, почему накапливался магний, каков был уровень карбонатной ком-
пенсации. Нужно смотреть весь баланс.
М.А. Федонкин: Это может быть связано с физиологией?
183
ГА. Заварзин: Доломит в основном не биогенный. Первичный доломит удается полу-
чить с алкалофильными сульфатредукторами, но это другая ситуация.
Н.Л.Д.: Часть доломитов органогенная, они присутствуют в строматолитах.
Н.Г Бочкарёв: Какова роль импактных событий в поздней эволюции Земли? Могло ли
вращение мантии объясняться ударом крупного тела? Могло ли появление азиатских трап-
пов объясняться импактами?
Н.Л.Д.: Конвекция мантии обусловлена тепловой, гравитационной неустойчивостью,
тут метеоритов не требуется. Активная бомбардировка метеоритами до рубежа 4.2 млрд, лет
подтверждается содержанием алмазов в цирконах, свидетельствующим об импактах. Потом
массовых бомбардировок не было. Отдельные падения были, скажем, Попигайский кратер -
скорее всего, это след удара кометы, так как в этом гигантском кратере метеоритного вещества
не найдено. Трапповые поля не вызывались импактами, они объясняются моделью плюма.
В.Н. Спытников: Николай Леонтьевич, не кажется ли Вам, что у нас пока нет объектив-
ных данных для того, чтобы определить время существования мира РНК? Он был заведомо
раньше 4.56 млрд, лет назад, и, скорее всего, не на Земле?
Н.Л.Д.: Не знаю. Бомбардировки исключают наличие биосферы, могли быть только от-
дельные очаги жизни. Мне кажется, что биосфера возникла после массовой бомбардировки.
Время для мира РНК остается между окончанием бомбардировок и первым появлением бак-
териальных матов. Этот интервал -4.2-3.9 млрд, лет назад.
М.А. Федонкин: Относительно того, что водород и метан, вероятно, составляли основу
древней атмосферы. Выясняется, что было около десятка источников водорода на ранней Зем-
ле. Второе. Сейчас как будто появляется все больше данных в пользу того, что металлическое
(железно-никелевое) ядро на 95 % сформировалось очень рано, чуть ли не в первые сто милли-
онов лет после аккреции. Если так, то магнитное поле тоже существовало с этого времени?
Н.Л.Д.: Дополнительные источники водорода несущественны. Первичная атмосфера
была захвачена еще из солнечной небулы, остальные источники не имели большого значе-
ния. Сильное магнитное поле появилось только тогда, когда появилось твердое внутреннее
ядро, слабое поле было и до этого. Есть расчеты, что во внутреннем ядре 20 % углерода,
значит, в растворенном ядре в основном должен быть не водород, а метан. Тогда этим будет
определяться состав мантии.
М.А. Федонкин: Один объем никеля «вдыхает» 17 объемов водорода (в твердом виде).
Но углерод в межатомное пространство металла так не войдет, как он мог попасть в ядро?
Н.Л.Д.: В алмазных наковальнях сейчас достигаются давления, близкие к давлению
на внешней границе жидкого ядра. Растворимость водорода и метана в жидком железе ока-
залась сопоставимой.
ГА. Заварзин: Мы ушли в такие глубины... И все-таки, жизнь - это поверхностное
явление, ее развитие идет в пределах географической оболочки...
Н.Л.Д.: Поверхностное и приповерхностное. Все-таки можно заключить, что более ком-
фортные условия для первичной жизни были не прямо на поверхности, где масса всяких воз-
мущающих факторов, а в каких-то пещерах, прослоях, кавернах на небольшой глубине от по-
верхности, где нет жесткого ультрафиолетового излучения, резких колебаний температуры,
где создается некий термостат. В таких условиях и могли возникнуть первые очаги жизни.
184
Hi’in, n мы 11РО11СХЧЖ.1И1111Я жимп. М.: НИИ РАН. 21X19. С. 1X5-198
УСЛОВИЯ ЖИЗНИ НА РАННЕЙ ЗЕМЛЕ ПОСЛЕ 4.0 МЛРД. ЛЕТ НАЗАД
© 2009 г. А.Ю. Розанов
Палеонтологический институт им. А.А. Борисяка РАН
aroza@paleo.ru
Уровень организации ископаемых организмов, находимых в докембрии —
наиболее достоверный индикатор состояния биосферы и показатель ее па-
раметров, таких как состав атмосферы, средние температуры поверхности
Земли и т. д. Предполагается, что появление цианобактерий, одноклеточных
и многоклеточных эвкариот и целомат наблюдается в геологической исто-
рии Земли значительно раньше, чем считалось прежде.
При обсуждении проблем происхождения жизни огромное значение имеют наши зна-
ния и представления об условиях на ранней Земле. В этой самой древней истории, вероятно,
принципиальный рубеж пролегает около 4 млрд, лет назад (Ga) - рубеж геологически до-
кументированной и недокументированной истории Земли. Об истории Земли и возможных
условиях на ее поверхности до 4 Ga сказано в этом сборнике в статьях Л.М. Мухина, А.В. Ви-
тязева и Г.В. Печерниковой, частично Л.В. Ксанфомалити.
Об условиях на Земле после 4 Ga существует огромная литература. Модели эволю-
ции Земли после 4 Ga многочисленны, частично это показано Н.А. Добрецовым (2003, 2005;
Dobretsov et al., 2006).
Основные построения, особенно для начального этапа от 4 Ga до рифея (1.6 Ga),
в основном базируются на геохимических, петрологических и тектоно-геофизических дан-
ных. В меньшей степени учитываются общегеологические, в том числе литологические, точ-
нее, седиментологические данные.
Несколько огрубляя, к настоящему времени у основной массы исследователей сложилось
впечатление о том, что история атмосферы состоит из двух этапов: этап восстановительной ат-
мосферы (бескислородной) и этап кислородной атмосферы, который начинается около 2.6 Ga.
Кроме того, существует устойчивое убеждение в достаточно высоких температурах на Земле
(100 и более градусов Цельсия). Это совершенно невероятное представление, особенно, если
говорить о средней температуре поверхности Земли (напоминаем, что сейчас она 15 °C).
Верхний предел рассмотрения интересующего нас интервала зафиксирован известны-
ми хорошо описанными событиями, вошедшими во все учебники: экспансией бесскелетной
эдиакарской фауны (0.6 Ga), обнаруженной на всех континентах, появлением кембрийской
скелетной фауны, когда многие организмы приобрели возможность строить разнообразный
по составу скелет (так называемый «кембрийский взрыв») примерно 530-540 млн. лет назад
и появлением около 400 млн. лет назад растений на суше (рис. 1).
185
А.Ю. Розанов. Условия жизни на ранней Земле после 4.0 млрд, лет назад
Геологически документированная история Земли
i<—- — Metazoa >
< < Metaphyta >
.... F11 n n i ч.
время последней ,
к . • - L-ukaryotj Млрд, лет» - археиско-раннепротерозоиские эвапориты
интенсивном метеоритном , .
бомбардировки *' Ме^гп.,. >ре>пЛ1Э 1.8 Млрд, ле, | - оледенения
Рис. 1. Некоторые биотические и абиотические события на Земле после 4 млрд. лет.
Архейско-протерозойская история земли документирована значительно менее до-
стоверно. В то же время условия на нашей планете последние 400 млн. лет принципиально
не отличаются от современных, если не считать изменения конфигурации суши и моря
и климатических флуктуаций.
Если не считать цирконы с возрастом более 4 млрд, лет, о чем я скажу ниже, достоверные
древнейшие метаосадочные породы имеют возраст 3.7-3.85 млрд. лет. Таким образом, вся гео-
логически документированная история Земли начинается пока именно с этого времени. Оно
совпадает с окончанием последней интенсивной метеоритной бомбардировки.
Обобщения по характеру древнейших пород (De Wit, Ashwal, 1997; Розен и др., 2006)
показывают наличие среди пород зеленокаменных поясов, которых на Земле было много де-
сятков, высокого процента метаосадочных образований (рис. 2). Это с очевидностью свиде-
тельствует о присутствии в это время на всей поверхности Земли бассейнов седиментации,
то есть достаточно большого количества воды. Значительное увеличение объема осадочных
пород к 3-3.5 Ga говорит, вероятнее всего, о стремительном увеличении ее количества в это
время, как схематично показано на рис. 3.
Таким образом, складывается впечатление, что до 4.0 Ga на поверхности Земли не было
значительных масс воды, которая могла бы способствовать образованию осадочных пород
и серьезной колонизации поверхности Земли микроорганизмами. Остается вопрос о возмож-
ности образования и существования в это время мира РНК1.
1 См. парадоксы А.С. Спирина (2007).
186
Проблемы пропс хождения жизни
Рис. 2. Диаграмма, отражающая мощность отложений зеленокаменных поясов и долю осадочных от-
ложений в них (Розен и др., 2006): а - суммарная мощность отложений в зеленокаменных поясах
(представительная оценка по стратиграфическим разрезам и геофизическим данным De Wit, Ashwal,
1997); б-доля осадочных пород от суммы всех пород (магматических и осадочных) в разрезах зеле-
нокаменных поясов (для 44 поясов, составленных по данным De Wit, Ashwal, 1997).
Можно думать, что интенсивные метеоритные бомбардировки убирали поступавшую
тем или иным способом на поверхность Земли воду.
Зеленокаменные пояса архея содержат набор осадочных и вулканических пород, кото-
рый, как пишут О.М. Розен с соавторами (2006): «не обнаруживает заметных систематиче-
ских отличий от соответствующих фанерозойских аналогов». В этом наборе пород присут-
ствуют все их типы, включая карбонаты, эвапориты и др.
Отличительной чертой древних толщ является достаточно широкое присутствие желе-
зистых кварцитов. Однако следует помнить о их присутствии даже в верхнем рифее Канады,
Бразилии и Намибии (Windley, 1999).
Таким образом, с точки зрения сравнения породных ассоциаций нет оснований говорить
о существенной разнице в условиях атмосферы и гидросферы позднего архея и более поздних
времен.
V воды, %
Рис. 3. Схема возможного увеличения объема воды.
187
А.Ю. Рошнон. Условия жизни на ранней Земле после 4.0 млрд, лег назад
Представления о существенной разнице состава атмосферы базируются на ряде
фактов, достоверность которых вызывает сомнения.
Присутствие воды, точнее, ее поступление на поверхность Земли в интервале вре-
мени 4.6 4.0 Ga неоднократно обсуждалось. Мне представляются весьма вероятными
представления М.А. Марова (Marov, Ipatov, 2001; Маров, Ипатов, 2005) о поступлении
на Землю воды с кометами в количестве, соизмеримом с объемом современного океана.
Однако единовременно такого количества воды, вероятно, никогда до 4 Ga, а скорее,
до 1.3 Ga', не было. И максимум, на что можно рассчитывать, это вода в небольших
лужах-озерцах и трещинах под поверхностью. Вопрос о том, куда девается такая боль-
шая масса воды, следует обсуждать отдельно.
Несколько слов о самых древних цирконах, возраст которых определяется
до 4.2 Ga - даже 4.4 Ga.
Изотопические исследования кислорода не могут свидетельствовать об обломочном ха-
рактере зерен циркона и его окатанности в воде. Остроугольная форма и размеры (до 300 мк)
изображенных зерен тем более не допускают такого толкования. Гораздо более вероятным
является вулканическое (пепловое) или эоловое их происхождение.
Теперь о восстановительной атмосфере архея и об уранинитах. Описанные и изо-
браженные образования уранинита интерпретируются как окатанные обломки и срав-
ниваются с окатанными моноцитами (рис. 4, а, б; Schopf, 1983). Изображения зерен ура-
нинита, имеющих размер менее 0.15 мм, говорят о любом ином характере их образо-
вания, но не окатанности, поскольку их размер меньше размера окатывания обломков.
Для сравнения привожу фотографии современного пляжного осадка из того же района,
что и окатанные моноциты. Все зерна остроугольные, а округлые образования - это
фораминиферы (рис. 4, <?). Зерна уранинита в этом случае могут быть нераскристаллизо-
ванными аморфными коллоидными образованими или даже скорее фоссилизированны-
ми бактериальными сгустками. Необходимо проведение дополнительных исследований
на электронном микроскопе при больших увеличениях.
Оценка различных признаков восстановительной атмосферы весьма неоднозначна.
К примеру очень показателен график, приведенный в книге под редакцией В. Шопфа (Schopf,
1983, с. 284, рис. 11, 12), в котором сведены известные на то время точки с геологическими
и геохимическими данными - показателями, как полагали авторы, восстановительных или
окислительных обстановок. Противоречивость приведенных материалов привела к тому, что
одни и те же районы (местонахождения) по разным признакам оказываются в разных полях.
Если же из этого рисунка убрать ураниниты, то подтверждения восстановительных условий
не останется вовсе.
Традиционные представления о восстановительной атмосфере отражаются, естествен-
но, и на интерпретации уровня организованности находимых в архее и нижнем протерозое
ископаемых остатков организмов. Они же порождают фантастические (Brasier et al., 2002)
обсуждения характера ископаемых остатков возрастом предположительно 3.5 Ga из архея
Австралии, описанные в свое время В. Шопфом и другими исследователями.
Мы полагаем, что для оценки параметров биосферы, таких как содержание кислоро-
да, температура поверхности, крайне важно исходить, прежде всего, из уровня организации
организмов, найденных в архейско-протерозойских толщах.
Так, например, наличие цианобактерий предопределяет принципиальное присутствие
атмосферного кислорода, а присутствие, например, Metazoa и тем более целомат говорит
о наличии кислорода в количествах, соизмеримых с современными.
'М.А. Жарков (2005), вероятно, справедливо полагает на основании распределения эвапоритов во времени, что
солевой режим оксана и соответственно объем воды мирового оксана начал стабилизироваться около 1.3 Ga.
188
Проблемы hpoik муж. /л чин жи ши
О 0.1 mm 0 0.1 0.2 mm
Рис. 4. Форма и размеры уранинитовых образований (а) и окатанные зерна моноцита (б) (Schopf. 1983);
моноцитовый алеврит из современных пляжных образований Австралии (в); обратите внимание
на размеры; округлые зерна - современные фораминиферы.
Обзор находок в архейско-протерозойских породах после работ конца XX в., сделан-
ных В. Шопфом, X. Хоффманом, А. Нолем и другими, в самое последнее время был допол-
нен российскими специалистами (Розанов, 2004; Федонкин, 2006; Rozanov, 2006; Сергеев и
др., 2007), и я вернусь к обсуждению лишь ключевых деталей, с помощью которых можно
наметить следующие события: I) обнаружение самых древних фоссилизированных остат-
ков бактериального уровня; 2) обнаружение их же в породах континентального генезиса,
фиксирующих время колонизации суши; 3) обнаружение первых цианобактерий, фикси-
189
.1.10. Розанов. Условия жизни на ранней Земле после 4.0 млрд, лют па *ал
Рис. 5. Современные и ископаемые (кембрий и ордовик) цианобактерии. Особенно интересна прекрас-
ная сохранность нитей и пленок, толщина последних 10-20 мкм.
руюших несомненное присутствие в атмосфере кислорода; 4) обнаружение эвкариот, фик-
сирующих кислородные и температурные параметры биосферы; 5) обнаружение много-
клеточных организмов; 6) обнаружение целомат; 7) обнаружение организмов со скелетом и
приобретение устойчивой возможности строить скелет разного состава и разными группа-
ми; 8) выход многоклеточных организмов на сушу.
Оценка указанных уровней и событий опирается на находки самих фоссилий, следов
их деятельности, особенно передвижения, биомаркеров, изотопных данных и т. д.
Но прежде чем обсуждать конкретные находки, хочу напомнить, что только в послед-
нее время стало очевидно, что фоссилизация бактерий и вообще микроорганизмов - обыч-
ное явление, происходящее чрезвычайно быстро. Именно поэтому при большом увеличении
на электронном микроскопе фоссилизированные бактерии можно легко обнаружить прак-
тически во всех осадочных породах любого возраста (Розанов, 2002), особенно если они об-
разовались в эпиконтинентальных бассейнах.
Более того, не только сами бактерии, но и гликокаликс фоссилизируются так же хо-
рошо, и возможны находки фоссилизированного гликокаликса великолепной сохранности
толщиной в несколько нанометров (рис. 5).
Все это многократно подтверждено и экспериментально в европейских, японских, аме-
риканских и российских лабораториях.
Замечу, что обилие фоссилизированных микроорганизмов в осадочных породах свя-
зано с тем, что большая их часть является продуктами седиментации в эпиконтинентальных
190
бассейнах. Они в силу своей мслко-
водности пропитаны бактериями,
поскольку большая часть воды этих
бассейнов находится в пределах фо-
тической зоны. Специфика седимен-
тации в эпиконтинентальных бас-
сейнах в силу насыщенности всей
водной массы микробами, не имеет
ничего общего с океанической седи-
ментацией сегодняшнего дня. По-
этому закономерности седимента-
ции в этих двух случаях совершен-
но разные и это требует специально-
го сравнительного анализа.
Наличие цианобактерий в ар-
хейских породах указывалось мно-
гократно (см. Schopf, 1983; Westall,
Walsh, 2002; и др.). Довольно упорно
это оспаривали, особенно оксфорд-
ские коллеги во главе с М. Brasier
(Brasier et al., 2002). У меня дан-
ные по архейским цианобактериям
вызывают скорее положительное
отношение, поскольку я не вижу
серьезных резонов у оппонентов.
В этой связи мне представляется
важнымупомянутьданныеи по Бал-
тийскому шиту. В породах возра-
стом около 3.0 Ga фиксируются бес-
спорные маты, причем достаточно
бесспорно циано-бактериальные
(рис. 6). Кроме того, еще одно сооб-
ражение утверждает меня в мысли,
что уже на уровне 3.5 Ga на Земле
было обилие цианобактерий - это
наличие архейских строматолитов
и нормальное и закономерное рас-
пределение строматолитов в архее
и всем протерозое (рис. 7).
Весьма важен вопрос о коло-
низации суши бактериями (микроор-
ганизмами). Иногда указывалось на-
личие палеопочв. Думаю, что более
строго следует говорить о корах вы-
ветривания и активной роли микро-
организмов в их формировании.
На сегодня все исследован-
ные коры выветривания из раз-
резов Балтийского шита показали
П POF.'lF.Mhl ПРОНСХОЖ ШИПЯ Ж!пни
Рис. 6. Породы, нацело состоящие из цианобактериаль-
ных нитей, образующих мат (лопий Карелии) (Астафье-
ва, 2006).
191
А.Ю. Розанов. Условия жизни на ранней Земле после 4.0 млрд, лет назад
Рис. 7. Распределение строматолитов в докембрии (по данным Раабен,
Семихатова, Хоффман и Суминой).
обильное присутствие в них микроорганизмов, что с определенностью свидетельствует
о том, что суша в архее была колонизирована микроорганизмами, а может быть, и гри-
бами (рис. 8). Здесь же уместно отметить, что pH-условия в корах и бассейновой части
достаточно отличны (рис. 9).
Распознание появления в геологической летописи эвкариот - трудный и, как пра-
вило, противоречивый, порождающий многочисленные споры, вопрос, но данные, опу-
Рис. 8. Характерные микроорганизмы кор выветривания. Паанаярви, 2.4 млрд, лет (Розанов и др., 2008).
192
ПI'i >}i. \/А/ А7>;л‘ . ,'///>/ ЖН Hill
Рис. 9. Схема соотношения кор выветривания и бассейновых отложений, в том числе карбонатов, гео-
морфологических особенностей и pH среды.
бликованные Б.В. Тимофеевым (1982) вызывают, как нам представляется, незаслуженное
недоверие. Я уже писал о переизучении этого материала как в коллекциях Института
геологии и геохронологии докембрия РАН, так и повторно собранных из тех же ме-
стонахождений в конкретных разрезах. Сферические формы диаметром до 100-150 мк
и трубки диаметром до 30-40 мкм правдоподобно считать остатками эвкариотных ор-
ганизмов (рис. 10).
Бесспорные эвкаритические организмы были описаны М.Ю. Беловой и А.М. Ахме-
довым (Belova, Akhmedov, 2006) из отложений возрастом 2-2.2 Ga (рис. II) и из фосфо-
ритов возраста 2 Ga печенегской серии района Печенги (рис. 12) Последняя находка до-
статочно уверенно определяется как празинофиты, и названа нами Pechengia (Розанов,
Астафьева, 2008).
В исследовании Беловой и Ахмедова описаны своеобразные органические остатки,
часть из которых может оказаться кандидами грибов или протистами.
обратить внимание на присутствие Leiosphaeridia со
По непонятным причинам иногда род
Leiosphaeridia объявлялся сборным (напр.,
Сергеев и др., 2007), объединяющим оболоч-
ки эвкариотного происхождения и оболочки
колоний прокариот. Однако, замечу, что объ-
единение в Leiosphaeridia гладких форм и со
складками смятия не может быть признано до-
статочно корректным. Leiosphaeridia - оболоч-
ки со складками смятия, и это в силу специфи-
ки материала оболочки исключает отнесение
их к прокариотным образованиям. Поэтому
только при наличии складок смятия позво-
лительно употребление названия Leiosphaeri-
dia. При их отсутствии определение не может
считаться достоверным.
Появление в палеонтологической летопи-
си многоклеточных эвкариотных организмов и
тем более целомат определяет высокий, поч-
ти равный современному, уровень насыщения
атмосферы кислородом. Фоссилизированные
Metazoa могут быть определены по их остат-
кам, по следам их жизнедеятельности, особенно
следам движения и ископаемым биомаркерам.
Из известны,х на сегодня данных по биомарке-
рам отметим биомаркер губкового происхожде-
Хочу особенно
складками смятия.
Рис. 10. Фоссилии излопия (Тимофеев, 1982;
переизучено в 2006 г. в ПИН).
193
А.Ю. PoituioK. Условия жизни ни paibiicii Земле после 4.0 млрд. леч назад
Рис. II. Остатки эвкариотных организмов из порол с возрастом
2-2.2 Ga Карелии (по Беловой и Ахмедову, 2006)
ния (Reitner, 2005), обнаруженный в породах возрастом 1.8 Ga. Следы жизнедеятельности
отмечались многократно. Наиболее интересны следы сверления или зарывания с возрас-
том 1.6 Ga, описанные Э. Кауффманом и Дж. Штейдтманом (Kaufmann, Steidtmann, 1981)
Рис. 12. Празинофиты Pechengia из протерозойских фосфоритов возрастом 2.0 Ga
(Розанов, Астафьева, 2008).
194
Hroh. a. \ib/ прошлое ...ним жишн
Рис. 13. Udokanicr. и - форма Udokanicr. б- пришлифовка продольного сечения трубок.
из Северной Америки, и следы ползания, собранные Б.Б. Шишкиным из пород с возрастом
1.4 Ga северо-запада Сибирской платформы (Rozanov, 2006).
Наиболее интересной является находка Udokania. С момента, когда Udokania была
описана А.М. Лейтесом, прошло много лет. Первоначально Р.Ф. Беккером, В.В. Меннером
и Б.С. Соколовым она была признана реальным ископаемым целентератного или поли-
хетного уровня организации. Затем Л.И. Салопом Udokania была объявлена кристаллами
скаполита (Салоп, 1982). Переизучение (Саютина, Вильмова, 1990) показало несомненную
их биологическую и, более того, метазойную природу. Однако это серьезно расходилось
с моделями эволюции биосферы, принятыми большинством западных ученых, а затем и рос-
сийских (Сергеев и др., 2007), поскольку возраст удокании около 2 Ga.
Парадокс ситуации состоял в том, что данные о Udokania упорно игнорировались
самыми известными специалистами по докембрию. Болес того, никто не дал себе труда
пересмотреть многочисленные материалы, хранящиеся в том числе и в ПИН. В начале
третьего тысячелетия четко обозначилось, что те специалисты, которые изучали и пере-
изучали конкретный каменный материал, не сомневались в метазойной природе удо-
каний. Те же, кто каменного материала нс смотрел, занимали молчаливую, но четкую
абиогенную позицию.
Я неоднократно обращал внимание моих коллег, что простое замалчивание проблемы
с Udokania не может далее продолжаться. И наконец лсд тронулся. Московские коллеги (Сер-
геев и др., 2007, с. 32) написали: «Мнения о природе Udokania и ассоциированных структу-
рах расходятся. Одни исследователи считают их остатками многоклеточных животных эдиа-
карского облика, другие трактуют как абиогенные формы, а третьи относят к биогенным
образованиям, не уточняя их биологической природы (Саютина, Вильмова, 1990; Розанов,
2003,2004; Синица и др., 2003, и ссылки в этих работах). Комплексный анализ этих дисковид-
ных (выделено мной - A.Pi) образований, проведенный А.А. Терлеевым, А.А. Постниковым,
Б.Б. Кочневым, К.Е. Наговициным, Д.В. Гражданкиным и А.М. Станевичем (2006), показал,
что они являются отпечатками сложно организованных колоний одноклеточных микро-
организмов - бактерий, грибов, эукариотных водорослей». Читатель, посмотрев на рис. 13,
на котором приведено фото Udokania, по последней фразе о «дисковидных образованиях»
и «животных эдиакарского облика», приведенной в цитируемой работе, легко представит
себе странность всей этой ситуации.
Новосибирские коллеги, на которых ссылаются москвичи, действительно изучали
каменный материал, но собранный ими, и также не переизучили типовые коллекции.
Таким образом. Metazoa на уровне 2 Ga это факт, с которым невозможно не считаться.
И это, кстати, хорошо согласуется с данными по биомаркерам (см. рис. I и 14).
195
I. /(). Pn шипи. Ус. i о is и я ж inn n 11 a ранней Jcm.ic nnc.ic 4.0 млрд, лег па вад
Геологически документированная история Земли
Prokaryota
100 -
90-
80-
70 -
60-
50-
40-
30-
20 -
10-
0 4
1 0%О;
1.0 01
0.1 % О,
поверхность
Земли
100% Or (современность)
• - Eukaryote (2,7 Ga)
бмомармсры
2.7 2,5
2 1,8
Фанерозой
Археи
бомбардировка
первые
ледники
начало широкого
)вм ар потного
разнообразия
последняя
интенсивная
Протерозой
Криптозой---------------
К ----------Metazoa
<--------------К---------Metaphyta
<-----------------к------------ Fungi --------
Protoeukaryota К------------ Eukaryota
К-----------------Archaea 1 -----------
* - Metazoa (Spongia) (1,8 Ga)
Рис. 14. Интегральная картина некоторых гео-биологических событий в докембрии.
В истории развития органического мира Земли наблюдаются два основных состояния
в каждой из групп организмов: появление и широкая экспансия. Они не совпадают по време-
ни и могут отстоять друг от друга на сотни миллионов лет.
Первое появление означает соответствующий уровень кислорода в атмосфере,
а широкая экспансия, как в случае с Metazoa - и стабилизацию солевого состава и объ-
ема воды «мирового океана» или какие-то иные сходные события.
Таким образом, основные выводы (рис. 14):
Между 4.6 и 4.0 Ga на поверхности Земли или совсем не было или было очень мало
воды.
Вода появилась в значительном количестве около 4.0 Ga после последней интен-
сивной метеоритной бомбардировки. В это время в мелководных бассейнах начинают
образовываться осадочные породы и бурно расцветает жизнь на поверхности нашей
планеты. Кроме бактерий, включая цианобактерии, не исключено появление зеленых
водорослей и, может быть, даже грибов.
Поверхность суши также была заселена микроорганизмами уже в архее.
Появление цианобактерий, эвкариот, Metaphyta и Metazoa происходило много раньше,
чем обычно предполагалось. И по уровню их организации можно составить представление
об уровне оксигенизации атмосферы.
Объем воды, соизмеримый с современным, образовался около 1.3 Ga. В связи с чем
с этого времени мы наблюдаем широчайшую экспансию различных организмов.
196
Прог>:п:мы пгонсхож иная ж пит
Средние поверхностные температуры не могли отличаться от современных более, чем
на 15-25 ’С.
Мир РНК, если таковой существовал, мог быть только до 4.0 Ga или, может быть,
до образования Земли.
Вероятность возникновения жизни именно на Земле крайне мала.
Работа выполнена по Программе президиума РАН «Происхождение биосферы
и эволюция гео-биологических систем» (подпрограмма II), гранту РФФИ № 08-04-00484
и научной школе Н Ш-4207.2008.5.
СПИСОК Л ИТЕРАТУ РЫ
Астафьева М.М. 2006. Археи Карелии и бактериальная палеонтология .. Эволюция биосферы и био-
разнообразия. К 70-летию А.Ю. Розанова. М.: Т-во научных изданий КМ К. С. 120-128.
Добрецов Н.Л. 2003. Корреляция биологических и геологических событий в истории Земли и возмож-
ные механизмы биологической эволюции Палеонтол. журн. № 6. С. 4-15.
Добрецов Н.Л. 2005. О ранних стадиях зарождения и эволюции жизни // Вести. Всерос. об-ва генетич.
селекц. Т. 9. № I. С. 43-54.
Жарков М.А. 2005. Эволюция эвапоритов в докембрии в связи с преобразованиями биосферы и хими-
ческого состава мирового океана. Статья 1. Эвапориты архея и раннего протерозоя ..' Страти-
графия. Геол, корреляция. Т. 12. № 2. С. 19-29.
Маров М.Я.. Ипатов С.И. 2005. Миграция пылевых частиц и доставка летучих на планеты земной
группы // Астроном, вести. Т. 39. № 5. С. 374-380.
Розанов А.Ю. (ред.) 2002. Бактериальная палеонтология. М.: ПИН РАН. С. 188.
Розанов А.Ю. 2003. Ископаемые бактерии, седиментогенез и ранние стадии эволюции биосферы
Палеонтол. журн. № 6. С. 41-49.
Розанов А.Ю. 2004. Бактериальная палеонтология, седиментогенез и ранние стадии эволюции био-
сферы 'Современные проблемы геологии. М.: Наука. С. 448-462.
Розанов А.Ю., Астафьева М.М. 2008. Празинофиты (зеленые водоросли) из нижнего протерозоя Коль-
ского полуострова // Палеонтол. журн. № 4. С. 90-93.
Розанов А.Ю.. Астафьева М.М., Вревский А.Б. и др. 2008. Микрофоссилии раннедокембрийских конти-
нентальных кор выветривания Фенноскандинавского щита //Отечеств, геол. № 3. С. 83-90.
Розен О.М.. Аббясов А.А.. Аксаментова Н.В. и др. 2006. Седиментация в раннем докембрии: типы
осадков, метаморфизованные осадочные бассейны, эволюция терригенных отложений. М.: На-
учный мир. 398 с. (Тр. Геол, ин-та. Т. 569).
Салоп Л.И. 1982. Геологическое развитие Земли в докембрии. Л.: Наука. С. 1-128.
Саютина Т.А.. Вильмова Е.С. 1990. Проблематичные ископаемые Udokania Leites из протерозойских
отложений Забайкалья Палеонтол. журн. № 3. С. 100-105.
Сергеев В.Н, Семихатов М.А., Федонкин М.А. и др. 2007. Основные этапы развития докембрийского
органического мира: сообщение I. Архсй и ранний протерозой Стратиграфия. Геол, корреля-
ция. Т. 15. № 2. С. 25-46.
Синица С.М.. Вильмова Е.С.. Туранова Т.К. 2003. Протерозойские экосистемы удоканского комплек-
са Удокан: геология, рудогеиез, условия освоения. Новосибирск: Наука. 128 с.
Спирин А.С. 2007. Когда, где и в каких условиях мог возникнуть и эволюционировать мир РНК?
Палеонтол. журн. № 5. С. 11-19.
Терлеев А.А., Постников А. А.. Кочнев Б.Б. и др. 2006. Раннепротерозойская биота из удоканской серии
западной части Алданского щита (Россия) ' Эволюция биосферы и биоразнообразия. К 70-ле-
тию А.Ю. Розанова. М.: Т-во науч, изданий КМК. С. 271-281.
197
А.Ю. Pt>j(iito«. Условия жизни на ранней Земле после 4.0 млрд, лет назад
Тимофеев Б.В. 1982. Микрофитофоссилии раннего докембрия. Л.: Наука. 128 с.
Федонкин М.А. 2006. Две летописи жизни: опыт сопоставления (палеобиология и геномика о ран-
них этапах эволюции биосферы) Проблемы геологии и минералогии. Сыктывкар: Геопринт.
С. 331-350.
Belova M.Yu.. Akhmedov А.М. 2006. Petsamomyces - new genus of organic walled microfossils in carbona-
ceous deposits of Early Proterozoic of Kola peninsula •'/ Paleontol. J. № 5. P.
Brasier M.D., Green O.K., Jephcoat A.P. et al. 2002. Questioning the evidence for Earth’s oldest fossils •
Nature. V. 416. P. 76-81.
Dobretsov N.L., Kolchanov N.F.. Suslov V.V. 2006. On the early stages of the evolution of the geosphere and
biosphere // Paleontol. J. V. 40. Suppl. 4. P. S407-S424.
Hoffman H.J. 2000. Archaean stromatolites as micribial archives, microbial sediments. B.: Springer.
P. 315-327.
Kauffman E G., Steidtmann JR. 1981. Are these the oldest metazoan trace fossils? 7 J. Paleontol. V. 55. № 5.
P. 923-947.
Marov M. Ya., Ipatov S.I. 2001. Volatiles inventory and formation of planetary atmospheres F Collisions in the
Solar System I Eds. M.Ya. Marov, H. Rickman. Kluwer Acad. Publ. 223 p.
Reitner J. 2005. Precambrian-Cambrian sponge critical interval - geobiological insights in old animals // Acta
Micropaleontol. Sin. № 22. Suppl. P. 160-161.
Rozanov A.Yu. 2006. Precambrian geobiology // Paleontol. J. V. 40. Suppl. 4. P. S434-S443.
Schopf J.W. (ed.). 1983. Earth’s earliest biosphere; its origin and evolution. Princeton Univ. Press. 544 p.
Westall F„ Walsh M.M. 2002. Early Archean fossil bacteria // Bacterial paleontology. Moscow: PIN RAS.
P. 84-90.
Wind/ey B.F. 1999. The evolving continents. Chichester: John Wiley and Sons. 526 p.
Wit M.J. de, Ashwal L.D. 1997. Greenstone belts. Oxford: Claredon Press. 809 p.
Life conditions on the Early Earth after 4.0 Ga
A.Yu. Rozanov
The organization level of fossil organisms from the Precambrian is the most reliable guide
for the biosphere condition, an indicator of its various parameters like the atmosphere composition,
mean temperature of the planet surface, etc. The origin of cyanobacteria, protists, eukaryotes and
coelomates is observed much earlier in the geological history than it was believed recently.
198
Проблемы нроислои, и ппп липни
Дискуссия
по пленарному докладу А.Ю. Розанова
«УСЛОВИЯ ЖИЗНИ НА РАННЕЙ ЗЕМЛЕ ПОСЛЕ 4.0 МЛРД. ЛЕТ НАЗАД»
Председательствующий Г.А. Заварзин
М.А. Федонкин: Вы показали архейские остатки возрастом три миллиарда лет. Этолопий,
метаморфическая порода, там пластические деформации, растяжение и так далее. А трубочки
эти выглядят как новенькие. Этот момент настораживает. В древних толщах архея в Норвегии
и Швеции в гранитах обнаружили зерна плагиоклаза, пронизанные туннелями - несколько
дециметров этих туннелей в одном зерне. Это делают современные литотрофные бактерии.
А.Ю.Р.: У них очень маленький диаметр, нанометровый. Те же, которые я называл -
по меньшей мере, на порядок больше, два-три микрона и чуть больше, в диаметре.
М.А. Федонкин: Но есть всякие игольчатые, волосовидные минералы и так далее. Вот да-
ется такая картинка с трубочками, тут бы и дать к ней анализ, что-то еще, кроме морфологии.
А.Ю.Р.: Анализы на микрозонде делаются всегда. Если Вы предложите, что сделать
еще, я сделаю, но тут пока нечего предложить.
М.А. Федонкин: В таких случаях, когда «экстраординарные заявления требуют
экстраординарных аргументов» - по выражению Карла Сагана - все-таки приходится рас-
кошеливаться и на время, и на средства, в таких случаях начинают мерить все, что угод-
но, биомаркеры, отличия от матрицы, используют рамановский микроскоп и так далее. Это
слишком сильное заявление (эукариоты в очень древних породах или марсианских метеори-
тах), и одной морфологии здесь недостаточно.
А.Ю.Р.: Но что еще нужно? Есть двойная стенка, оболочка с порами, этого вполне
достаточно.
М.А. Федонкин: Например, наши коллеги, Весталл и другие, при исследовании в пер-
вую очередь органостенных, но и минерализованных тоже, архейских объектов пытаются
искать реликты органических соединений. Или Энди Ноэл - он берет одну микрофоссилию
и сканирует ее лучом, лазерным или электронным, и делает карту концентрации равных эле-
ментов. Этих карт может быть несколько по разным элементам. Потом они накладываются,
и вдруг выявляются внутренние субклеточные структуры, идет картирование одной клетки
по разным элементам. Тогда их аргументы обретают новую дополнительную силу.
А.Ю.Р.: Такие «карты» мы с Ричардом Хувером делали неоднократно, и это опублико-
вано. У меня только один вопрос - Вы ставите под сомнение печенгию?
М.А. Федонкин: Не знаю, честно говоря.
А.Ю.Р.: Если ставите под сомнение именно Pechengia, то наш разговор непродуктивен.
Ю.В. Наточин: Можно комментарий? Элементы не помогут, потому что все уже вымыто.
М.А. Федонкин: Некоторые вещи минерализуются, остаются металлоорганические
соединения. Они, конечно, искажены, потеряли первичную структуру, но именно по этим
обрывкам подчас восстанавливают биохимические компоненты.
М.Я. Маров: Говорит ли все это об эволюционном тренде в развитии биосферы,
об архейской древности сложноорганизованных форм? Каков главный вывод?
А.Ю.Р.: Главный вывод состоит в том, что появление сложноорганизованных форм
произошло значительно раньше, чем мы считали. И это означает, что количество кислорода
199
в атмосфере Земли стало достаточно большим значительно раньше, чем мы предполагали (рань-
ше, чем 2.5 миллиарда лет назад). И это говорит о том, что средние температуры поверхности не
могли быть такими чудовищными, как это обычно представляется. Следствий еще очень много.
М.Я. Маров: Это говорит еще и о том, что Земля потрясающе организованная при-
родная система, в ней задействованы такие фантастически глубокие механизмы обратных
связей, которые поддерживают вполне определенную стабильность. Это очень интересно
и это может быть одним из главных выводов.
ГА. Заварзин: С этим можно сразу согласиться. Работают столь мощные обратные свя-
зи, что система не выходит из коридора.
А.Ю.Р.: Да, и за четыре миллиарда лет никаким событиям не удалось прервать разви-
тия биосферы. А сбой у нас в мышлении бывает, когда мы пытаемся рассуждать, что будет
с биосферой, применительно к нашим - человеческим - целям и интересам.
С.В. Шестаков: Алексей Юрьевич, можно согласиться с Вашими доводами относи-
тельно палеомикробиологии. Но с моей точки зрения, «крамолой» в Ваших представлениях
является уверенность в том, что к трем с половиной миллиардов лет назад можно отнести
появление эукариот. Дело в том, что даже у современных микроорганизмов мы можем найти
примеры очень сложных клеточных организаций, которые могут давать картины, схожие
с теми, которые Вы трактуете как эукариоты.
А.Ю.Р.: Три с половиной миллиарда я не называл в отношении эукариот, но 2.9-3.0 да.
С.В. Шестаков: Около двух - да, я согласен.
А.Ю.Р.: Два - это бесспорно, против печенгии спорить бессмысленно. Относительно
трех миллиардов свидетельствуют тимофеевские данные, которые мы перепроверили.
С.В. Шестаков: Хорошо, дело даже не в датировках. Просто нужна уверенность в том,
что описываемые вами формы могут быть отнесены к эукариотам. Для этого нужно хорошо
представлять организацию не только современных, но и вымерших прокариот и эукариот,
а эти знания еще очень не полны.
А.Ю.Р.: Сергей Васильевич, если Вы сможете мне привести пример прокариота, ко-
торый имеет двойную стенку, пронизанную порами, да еще имеет размер в 130 микронов -
я Вам буду благодарен. Я такого не знаю.
С.В. Шестаков: Я берусь показать...
В.Н. Спытников: Алексей Юрьевич, если мы попытаемся фоссилизировать современ-
ные аналоги изображенных Вами организмов, за какое время они дадут приблизительно
такой же элементный состав?
А.Ю.Р.: Фоссилизировать что-нибудь живое можно почти мгновенно, любой состав
можно получить в течение нескольких часов. Многочисленные эксперименты были проде-
ланы Людмилой Михайловной Герасименко и Галиной Тихоновной Ушатинской, а ранее -
Крыловым, Заварзиным и другими.
В.Н. Снытников: И этот образец не будет отличаться от древнего?
А.Ю.Р.: Будет морфологически отличаться, потому что он современный.
В.Н. Снытников: Я имею в виду химический состав. В химии существует понятие
«встречного синтеза», когда нужно подтвердить какое-то вещество, это вещество синтезиру-
ется и сравнивается с получаемым. Нельзя ли применить аналогичный прием здесь?
А.Ю.Р.: Если я возьму современный празинофит, его фоссилизирую, доведу до такого
200
Прог //. ил/ пронсхож.н.ния жишп
состояния, чтобы получился такой же спектр, что Вы мне скажете - «Ну и что?» И больше
ничего. Это не будет доказательством чего-либо.
М.А. Федонкин: Алексей Юрьевич, а это массовый материал?
А.Ю.Р.: Да, массовый. Вы видели снимки на томографе.
Г.А. Заварзин: Что я хотел бы сказать. Пункт первый. Видим ли мы то, что видим на
экране? Видим. Второй пункт - как мы эти изображения интерпретируем? Мы можем гово-
рить, что это ветвистые минеральные образования по кавернам, по трещинам. Но уйти от
самого факта существования такого объекта мы не можем - это не подделка, не имитация.
Мы можем говорить, что это образовалось на породе из лопия позже, что это контаминант,
который так же фоссилизировался - но так можно сказать о единичном образце, о массовом
материале так сказать гораздо труднее (однородное загрязнение более сомнительно). В этом
случае будет смещена интерпретация по времени. Следующий аргумент относится к требо-
ванию получить какой-то дополнительный материал к одной показанной картинке. Он может
дать что-то дополнительное. Но есть очень старое правило логики, восходящее к Платону:
для того, чтобы потерять истину, достаточно говорить на эту тему слишком долго и много.
Это одно из первых правил софистики. Поэтому дополнительный материал может сработать,
а может и увести в сторону. Конечно, эти дополнительные материалы могут быть ценны, важ-
ны, их, естественно, надо получать, но не нужно забывать, что это так называемые сопутству-
ющие доказательства, а не первичные. Теперь еще относительно изображений. Я морфолог
тоже, и я, вообще говоря, верю тому, что я вижу, хотя догматические предпосылки у нас очень
разные. Далее история с кислородом. Сергеев из Геологического института по своим образцам
не согласится с цианобактериями древнее 2.5 или 2.7 миллиардов лет. Кислород в стопроцент-
ной концентрации появляется с первого момента существования цианобактерий. И кислоро-
да возле цианобактериальных сообществ будет много. И придется объяснять, как анаэробы
справятся с этим стопроцентным окислительным стрессом. Сколько времени понадобилось,
чтобы из пузырька кислорода произвелось воздействие на всю атмосферу? Это не короткий
период. Поэтому отсутствие кислорода в атмосфере не является ограничением для времени
появления цианобактерий и для появления довольно крупных аэробных организмов, которые
могли бы жить в непосредственной близости от скопления цианобактерий (но только днем,
так как ночью они будут находиться в среде сероводорода). Поместить сравнительно крупные
существа доли миллиметра, до миллиметра - в цианобактериальный мат, по-моему, можно.
Мат отрывается от грунта, слегка всплывает, а под ним пузыри кислорода - образуется такая
прослойка - несколько миллиметров, даже до сантиметра, в котором можно дышать и жить.
Но это не целоматный уровень, конечно. Так что этот пункт не так критичен. Поверить всем
тем интерпретациям, которые дает Алексей Юрьевич, я смогу только после очень большой
паузы, после долгого «переваривания» и осознания. С ходу поверить не получится. Еще об
эукариотах. Игорь Николаевич Крылов в свое время дал мне шлифы, на которых была видна
отличная эукариотная структура. Все получалось. Потом оказалось, что это оболочки чехлов
цианобактерий. Так что над этими картинками я немножко подумаю... И относительно ми-
нимальных размеров, которые могут иметь прокариоты и цианобактерии, в частности. Есть
такой организм - носток Рябушинского, колония цианобактерий размером с мелкую сливу,
которая перекатывается в пресных озерах (я наблюдал ее в вулканических озерах на Камчат-
ке), если ее раздавить, фоссилизировать, такая колония может имитировать очень многое. Так
что природа предоставляет нам очень много возможностей ложных интерпретаций. Но я по-
вторяю - отрицать то, что мы видим, нельзя, надо объяснять.
201
Пнж :> мы пгоисмл:;ымя ,KHMnt. Л/,: ПИН РАН. 2009. С. 202-212
ВЫСОКОТЕМПЕРАТУРНЫЕ ГЛУБИННЫЕ МИКРОБНЫЕ СООБЩЕСТВА -
ВОЗМОЖНЫЙ АНАЛОГ ДРЕВНЕЙШИХ ЭКОСИСТЕМ?
© 2009 г. Е.А. Бонч-Осмоловская
Институт микробиологии им. С.И. Виноградского РАН
lbo@mail.ru
Данные микробиологических, молекулярно-биологических и радиоизотоп-
ных исследований указывают на существование в глубинных слоях под-
земной биосферы активных и многокомпонентных микробных сообществ.
В настоящем обзоре рассматривается лишь определенная группа таких
сообществ - те, которые развиваются при высоких (60 °C и выше) темпе-
ратурах. Наиболее исследованными высокотемпературными подземными
экосистемами являются нефтяные месторождения, подземные водные ре-
зервуары (например, Великий артезианский бассейн Австралии), глубин-
ные горнодобывающие шахты (Южная Африка), высокотемпературные
горизонты, находящиеся под океанической корой в районах современной
подводной вулканической активности. Эти микробные сообщества значи-
тельно отличаются друг от друга по составу, исходному энергетическому
субстрату, ведущим микробиологическим процессам. Однако для того,
чтобы их можно было рассматривать в качестве аналогов первичных эко-
систем Земли, необходимо доказать их энергетическую независимость
от современной биосферы.
ВВЕДЕНИЕ
Исследования последнего десятилетия подтвердили сенсационные заявления, сделан-
ные в начале декады: в глубинных слоях земной оболочки, ранее считавшихся стерильными,
присутствуют активные и многокомпонентные микробные сообщества (Parkes et al., 1994,
2000; Whitman et al., 1998; Wellsbury et al., 2002; Teske, 2005). Однако остается неясным, как
в этих экосистемах функционируют циклы основных биогенных элементов, и что в них яв-
ляется первичным источником энергии. Если бы удалось доказать, что такие сообщества
используют энергетические субстраты, окислители и источник углерода нефотосинтетиче-
ского происхождения, их можно было бы рассматривать как аналог первичных экосистем
Земли, развивавшихся вне поверхности Земли, жизнь на которой в отсутствие атмосферы
была невозможна из-за жесткого ультрафиолетового излучения.
В настоящем кратком обзоре мы коснемся лишь определенной группы таких микроб-
ных сообществ - тех, которые развиваются при высоких температурах (60 °C и выше). Ми-
кробные сообщества современных вулканических местообитаний часто рассматриваются
как экосистемы, базирующиеся на хемо-, а не фотосинтезе (Amend, Shock, 2001; Spear et al.,
202
Проблемы происхождения лиши
2005). Действительно, в них обитает большое число уже описанных и исследованных в ла-
бораторных культурах анаэробных литоавтотрофных термофильных прокариот, подходя-
щих на роль первичных продуцентов в таких экосистемах. Это многочисленные водород-
используюшие термофильные и гипертермофильные прокариоты: метаногены, сульфат-,
тиосульфат- и сероредукторы, нитрат- и железоредукторы (Слободкин и др., 1999). Другая
группа возможных первичных продуцентов в термофильных хемосинтезирующих сообще-
ствах представлена анаэробными гидрогеногенными карбоксидотрофами, использующи-
ми моноксид углерода как единственный источник углерода и энергии и окисляющими его
до СО, с одновременным образованием водорода из воды (Sokolova et aL, 2009). Активность
анаэробных литоавтотрофов в гидротермальных местообитаниях различного типа под-
тверждается прямыми радиоизотопными экспериментами (Бонч-Осмоловская и др., 1999;
Pimenov, Bonch-Osmolovskaya, 2006; Слепова и др., 2007).
Тем не менее, гидротермальные местообитания, в том числе и высокотемпературные,
находятся под значительным влиянием (в той или иной форме) современной биосферы. Го-
рячие источники кальдеры Узон на Камчатке достаточно хорошо изучены геохимиками
и микробиологами различных специальностей: установлена высокая скорость идущей в них
микробной ассимиляции неорганического углерода и присутствие разнообразных термо-
фильных и гипертермофильных литоавтотрофных прокариот (Бонч-Осмоловская и др., 1999,
2004; Бонч-Осмоловская, 2004). Однако глубинный флюид магматического происхождения
составляет лишь до 15 % воды горячих источников Узона; остальная часть замещается ме-
теорными водами, несущими растворенное органическое вещество с окружающих ненагре-
ваемых участков почвы, покрытых высшей растительностью (Zavarzin et al., 1989). В мелко-
водных морских гидротермах, располагающихся на литорали, энергетическим субстратом
термофильных микробных сообществ является, конечно же, аллохтонное органическое ве-
щество, в изобилии туда поступающее (Намсараев и др., 1994). И даже в глубинных морских
гидротермах, где идет активнейший хемосинтез, основанный на использовании энергии
серных соединений свободноживущими и симбионтными хемолитоавтотрофами, связь с со-
временной биосферой происходит за счет участия кислорода, поступающего в сообщества
с холодной океанической водой (Jannasch, Mottl, 1985; Karl, 1995). Поэтому все чаще при по-
иске автономных микробных сообществ выдвигаются гипотезы о их существовании в под-
земной биосфере, изолированной от наземного мира, в котором и энергетический субстрат
(органическое вещество), и окислитель (кислород) создаются в результате преобразования
энергии солнечного света при фотосинтезе.
Высокие температуры в глубинных местообитаниях могут быть обусловлены как
близостью к магматическому ядру, так и гидротермальной активностью. В первом случае
на глубине 1500-3000 м температура повышается до 60-100 °C, что создает идеальные (есте-
ственно, при наличии доноров и акцепторов электронов) условия для развития термофиль-
ных микробных сообществ. Высокотемпературные глубинные местообитания вулканиче-
ского происхождения присутствуют, по всей видимости, во многих, если не во всех, районах
современной вулканической активности и могут располагаться на относительно небольшой
глубине. Однако и те, и другие становятся доступными только в процессе бурения, которое
крайне редко диктуется исключительно научными целями, что, естественно, резко ограни-
чивает имеющуюся в настоящее время информацию об этих системах.
ОСНОВНЫЕ ТИПЫ ВЫСОКОТЕМПЕРАТУРНЫХ ПОДЗЕМНЫХ ЭКОСИСТЕМ
Характеристики высокотемпературных подземных экосистем, исследованных микро-
биологами в разные годы и разными методами даны в табл. 1. Из нее видно, что подземные
местообитания значительно различаются по основным физико-химическим характеристи-
кам (pH, температура, соленость); при этом температура не прямо коррелирует с глубиной.
203
Е.А. Бонч-Осмолокская. Высокотемпературные глубинные микробные сообщества...
Таблица 1. Микробиологические исследования в высокотемпературных подземных
местообитаниях
Географическое положение Глубина, м Т«с рн Проведенные исследования Ссылки
Радио- изотопные Молекулярно- биол. Культу- ральные
1 Нефтяные пласты
Северное море 2000-4000 60-85 7.8 - — + Rosnes et al., 1991; Beeder et al., 1995, Rees et al., 1995; и др.
Аляска 3000 71-105 - - + Stetter et al., 1993
Восточно- Парижский бассейн 1670 50-80 + - + Daumas et al., 1986; L’Haridon et al., 1995
Африка 68 - - + Ravot et al., 1995
Лиаохе, Дакинг, Даган 1008-2040 44-66 7.0 + + Nazina et al., 2000a, b; Назина и др.. 2006
Кубаки, Япония 1000-1200 50-58 - - + Takahata et al., 2000
Калифорния. США 1500-4100 50-125 7.2-8.1 - + 4- Orphan et al.. 2000
Самотлор, Западная Сибирь 1900-2350 60-85 6.0-7.5 + + + Bonch-Osmolovkaya et al.. 2003
Даганг, Китай 59 7.1-7.6 + + + 11азина и др., 2006
Подземные водные резервуары
Великий Артезиан- ский бассейн, Австралия 31-88 - - Andrews, Patel. 1996; и др.
Зоны под океанической корой
Ньюфаундленд 1626 54-100 - - Roussel et al., 2008
Разброс значений в местообитаниях одного и того же географического положения указывает
на резкие различия условий даже в близкорасположенных водных резервуарах. Большинство
исследованных подземных местообитаний имеет нейтральный или слабощелочной pH сре-
ды; исключение составляют горнодобывающие скважины.
Наиболее изученными являются высокотемпературные нефтяные месторождения, рас-
положенные как в шельфовой зоне, так и в центре континента. При этом в последнем случае со-
леность воды часто соответствует морской, как, например, в подземных резервуарах Самотлора
(Bonch-Osmolovskaya et al., 2003). Температура на глубине 1500 м там достигает 60 °C, превы-
шая 80 °C на глубине 2500 м. Эти температурные диапазоны в точности соответствуют темпе-
ратурным интервалам развития умеренно-термофильных и гипертермофильных прокариот.
Источником выделения многих термофильных бактерий послужил Великий артезиан-
ский бассейн Австралии (работы Б. Пателя с сотр.). Этот колоссальный водный резервуар,
основной источник питьевой воды в Австралии, залегает на глубине до 3000 м, где его воды
нагреваются до температуры 100 °C.
Исключительным местообитанием оказались горнодобывающие шахты в Южной
Африке (Takai et al., 2001). Достигая глубины 3.2 км, они характеризуются температурой
в интервале 18-60 °C и pH 3.0-10.0. Согласно результатам стабильно-изотопных анализов,
большинство этих местообитаний находится в зоне смешения подземных и метеорных вод,
однако есть и исключения, где вода имеет исключительно глубинное происхождение. Ис-
следование филогенетического состава микробных сообществ этих местообитаний дало осо-
бенно неожиданные результаты, показавшие присутствие новых филогенетических линий
(Takai et al., 2001; Lin et al., 2006).
Наконец, интереснейшим термальным подземным местообитанием являются высоко-
температурные горизонты, находящиеся под океанической корой в районах современной под-
водной вулканической активности. К изучению обитающих там микробных сообществ при-
Прок u:\ihi про//схож и пня ж/н/ш
вели, во-первых, активные исследования микробных сообществ глубоководных гидротерм,
и, во-вторых, доступ к глубинным пробам из этих же местообитаний, обусловленный амери-
канской программой по бурению океанического дна (Ocean Drilling Program), впоследствии
ставшей международной. Удивительные открытия в области разнообразия микроорганизмов
в таких сообществах привели к тому, что в одной из известных и нашумевших публикаций под-
водные гидротермы были названы «окнами в подземную биосферу» (Deming, Baross, 1993).
Перечисленные местообитания исследовались разными авторами с использованием раз-
личных подходов и методов; соответственно, различались и полученные результаты. Рассмотрим
имеющиеся данные, подразделив их на группы в соответствии с использованными подходами.
АКТИВНОСТЬ МИКРОБИОЛОГИЧЕСКИХ ПРОЦЕССОВ
В ВЫСОКОТЕМПЕРАТУРНЫХ ПОДЗЕМНЫХ ЭКОСИСТЕМАХ
Разработанные микробиологами школы С.И. Кузнецова радиоизотопные методы опреде-
ления скорости микробиологических процессов нашли свое применение и при исследовании
микробных сообществ термальных местообитаний, в том числе и глубинных (Pimenov, Bonch-
Osmolovskaya, 2006). Последние отличаются невозможностью инкубирования изолированных
проб с добавленными радиоизотопными субстратами непосредственно в месте отбора; при-
ходится проводить инкубацию в термостате с соответствующей температурой, без учета гео-
статического давления, которое на глубине 1500-3000 м является значительной величиной и
может существенно влиять на ход исследуемых процессов. Тем не менее, несмотря на отно-
сительный характер получаемых параметров, они способны дать нам важную информацию
о некоторых процессах, протекающих в высокотемпературных глубинных местообитаниях
(примером которых являются нефтяные месторождения). Так, было установлено, что в микроб-
ных сообществах пластовых вод Самотлорского нефтяного месторождения сульфатредукция
и метаногенез являются конкурентными процессами и реализуются в зависимости от кон-
центрации сульфатов (Bonch-Osmolovskaya et al., 2003). При этом скорости сульфатредукции,
литотрофного (с использованием водорода) и ацетокластического (с использованием ацетата)
метаногенеза имеют два максимума (рис. 1). Первый приходится на заводняемые скважины
Сульфатредукция Литотрофный метаногенез
Ацетокластический метаногенез
Рис. I. Скорость микробиологических процессов - сульфатредукции, литотрофного и аце-
токластического метаногенеза в скважинах высокотемпературного нефтяного месторож-
дения Самотлор, Западная Сибирь (Bonch-Osmolovskaya et al., 2003, модифицировано).
205
F.A. Бонч-Ослк>:1О1икия. Высоко гем нсратурн ыс иублнныс микробные сообщества...
Рис. 2. Влияние сульфата на скорость сульфатредукции (а) и молекулярного водорода на скорость
литотрофного метаногенеза (г5) в скважинах высокотемпературного нефтяного месторождения Са-
мотлор, Западная Сибирь.
поверхностных горизонтов, второй (основной) - на скважины глубинных юрских горизонтов,
откуда нефть изливается под естественным давлением. Можно с значительной долей уверен-
ности предположить, что именно в этих скважинах при температуре 84 °C процессы сульфат-
редукции и метаногенеза осуществляет аборигенная гипертермофильная микрофлора.
Исследование влияния на скорость процессов добавления дополнительного количества
сульфатов или введения водорода в газовую фазу инкубационных флаконов показало, что при-
сутствие сульфатов стимулирует сульфатредукцию, в то время как добавление водорода нс
влияет на скорость метаногенеза (рис. 2). Первое явление может быть объяснено тем, что
сульфатредукторы при лимитации сульфатами могут участвовать в других процессах, напри-
мер, синтрофном окислении органического вещества путем передачи электронов метаноге-
нам. При добавлении дополнительного количества сульфатов эта микрофлора переключалась
на их восстановление с использованием тех же доноров. Стимуляцию литотрофного мета-
ногенеза и сульфатредукции водородом можно было бы ожидать, считая что он компенси-
рует потери присутствующего в пластовых водах растворенного водорода за счет дегазации
при поднятии проб на поверхность и снижении давления. Однако, так как этот эффект нс на-
блюдался, приходится предположить, что литотрофный метаногенез является не результатом
использования водорода глубинного происхождения, а терминальной реакцией анаэробного
разложения органического вещества, и численность присутствующих в пробе метаногенов
соответствует количеству образующегося при брожении органических субстратов водорода.
Таким образом, и сульфатредукция, и литотрофный метаногенез в микробных сообществах
высокотемпературных нефтяных месторождений являются не первичными процессами, да-
ющими начало микробному сообществу, существующему за счет хемосинтеза, а, наоборот,
завершают анаэробную деструкцию органического вещества, видимо, захороненного в этих
горизонтах в более древние геологические периоды.
ФИЛОГЕНЕТИЧЕСКИЙ АНАЛИЗ ПОДЗЕМНЫХ ТЕРМОФИЛЬНЫХ
МИКРОБНЫХ СООБЩЕСТВ
Молекулярно-биологические методы, получившие развитие в последние десятилетия,
дали возможность экспрессной оценки разнообразия микроорганизмов в природных место-
обитаниях без этапа культивирования. Кратко суть этих методов заключается в следующем:
с помощью полимеразной цепной реакции гены 16S рибосомальной РНК (общепризнанный
филогенетический маркер) всех микроорганизмов, присутствующих в природном образце,
сначала выделяются, затем тем или иным способом разделяются и по их последовательности
определяется филогенетическое положение присутствующих в пробе организмов. В случае
наземных и морских гидротерм применение этих методов обнаружило присутствие в этих со-
206
ПРОИ-Н~Л1Ы Г1Р()1!С.\'ОЖ,1) НИЯ Ж НИИ!
обществах большого количества новых организмов, неизвестных в лабораторных культурах.
Оказалось, что подземные термальные местообитания по филогенетическому составу микро-
организмов сильно различаются.
Микробные сообщества высокотемпературных нефтяных месторождений состоят в
основном из представителей уже известных родов. Применение микрочипа с нанесенными
на нем люминисцентными зондами, специфичными к I6S рРНК 17 родов или групп родов
термофильных архей и бактерий, позволило обнаружить в пластовых водах юрского гори-
зонта (глубина 2384 м, температура 84 °C) представителей 12 из них (Bonch-Osmolovskaya
et al., 2003). Это гипертермофильные археи рода Thermococcus, представители относящихся
к глубоким филогенетическим ветвям бактерий порядков Thermotogales и Aquificales (также
гипертермофилы), литотрофные метаногены родов Methanothermobacter, сульфатредуцирую-
щие бактерии рода Desulj'olomaculum, анаэробные органотрофы рода Thermoanaerobacter и
некоторые другие (рис. 2). Близкие результаты были получены путем анализа библиотек кло-
нов генов 16S рРНК из высокотемпературных нефтяных месторождений Калифорнии (Orphan
et al., 2000) и Китая (Назина и др., 2006).
Совершенно иные результаты дал филогенетический анализ термофильных микробных
сообществ, развивающихся в горнодобывающих шахтах в Южной Африке (Takai et al., 2001).
Путем анализа библиотек клонов генов I6S рРНК были обнаружены присутствующие там
археи типа Crenarchaeota, представляющие новые филогенетические линии (рис. 3). При этом
наиболее разнообразными были микробные сообщества, развивающиеся в шахтах, не овод-
няемых поверхностными водами метеорного происхождения. Все это дало основания пред-
положить, что в исследуемых глубинных экосистемах развивается аборигенное сообщество,
состоящее из микроорганизмов с неизвестное пока физиологией, возможно, литоавтотрофов.
В сходном местообитании близкого географического расположения (также Южная Афри-
ка, горнодобывающая шахта глубиной 2.8 км) была обнаружена еще одна микробная экосистема,
также уникальная - биопленка, состоящая практически из одного микроорганизма - «Candidates
Desulforudis audaxiviator» (Lin et al., 2006). Роста этой бактерии в лабораторных условиях пока
получить не удалось, но уже получена полная последовательность ее генома, из которой следует,
что «Desulforudis audaxiviator» является умеренно-термофильной литоавтотрофной сульфатре-
дуцируюшей бактерией, использующей молекулярный водород как энергетический субстрат и
углекислоту как источник углерода, причем пути фиксации углекислоты сходны с архейными.
РАЗНООБРАЗИЕ И ЧИСЛЕННОСТЬ КУЛЬТИВИРУЕМЫХ ОРГАНИЗМОВ,
ВЫДЕЛЕННЫХ ИЗ ВЫСОКОТЕМПЕРАТУРНЫХ ПОДЗЕМНЫХ ЭКОСИСТЕМ
Параллельно с применением «прямых» методов исследования микробиологических
процессов и филогенетического состава микроорганизмов в высокотемпературных глубин-
ных экосистема,х велась и работа по изучению их традиционными культуральными методами.
Ряд гипертермофильных архей и бактерий, широко известных по микробным сообществам
морских гидротерм, был выделен из проб в высокотемпературных нефтяных месторождени-
ях шельфовой зоны: на Аляске (Stetter et al., 2003), в Северном море (Nielsen, Torsvik, 1996;
Stetter et al., 2003), в Калифорнии (Orphan et al., 2000) и в Японии (Takahata et al., 2000). Среди
них были органотрофные археи родов Thermococcus и Pyrococcus, метанобразующие и суль-
фатредуцируюшие археи (роды Methanothermococcus и Archaeoglobus}, органотрофные бак-
терии порядка Thermotogales). Еще более удивительным было выделение этих же организмов,
типичных обитателей морских гидротерм, из пластовых вод с соленостью уровня морской, но
расположенных в центре континента: нефтяного резервуара Парижского бассейна (L’Haridon
et al., 1995), месторождения Самотлор в Западной Сибири (Bonch-Osmolovskaya et al., 2003).
Эти изоляты либо принадлежали к уже известным видам, либо представляли новые виды.
Так, из проб юрских горизонтов Самотлорского месторождения был выделен новый вид ги-
207
. Высоко i см пера ( у рн ые r.i убин н ые микробные сообщества...
Bacteria,---с=— ;
ч f >‘/г. '>>>.. .%Лу: <:ИВ
Eucarya ................ Д|,
* * “ Г. ч. I)
< : У ... ' | Marine Hydrothermal Vent Group-I(MHVG-I)
---------------- \| MHVG-2
------------------Л1«*И- 'JmHVG-3
Deep-Sea Archaeal Group (DSAG)
| Korarchaeota
I Terrestrial Hot Spring
Crenarchaeotic Group (THSCG)
Terrestrial Miscellaneous
Crenarchaeotic Group (TMCG)
Hyperthermophilis Crenarchaeota
and Relatives
Рис. 3. Новые филогенетические группы прокариот в горнодобывающих шахтах Южной
Африки (Takai et al.. 2001).
Crenarchaeota
208
Рис.4. Клетки Thermococcus sibiricns - анаэ-
робной органогрофной гипертермофильной
археи, выделенной из нагнетательной скважи-
ны Самотлорского нефтяного месторождения.
Западная Сибирь. Масштаб 0.5 мкм.
Пг<>Г,.1КМЫ ПП)1И 11.411>1 Ж1К11И
пертермофильных архей Thermococcus sibiricns (Miroshnichenko et al., 2001). Этот организм
с клетками кокковидной формы (рис. 4) растет при температуре от 40 до 95 °C, сбраживая
пептиды в присутствии элементной серы, которая восстанавливается в сероводород. Можно,
однако, предположить, что, как и у остальных представителей рода Thermococcus, восстанов-
ление серы у этого организма играет важную роль лишь при лабораторном культивировании
в закрытых сосудах, так как позволяет избежать ингибирующего действия водорода - основ-
ного восстановленного продукта в отсутствие серы (Бонч-Осмоловская, Мирошниченко,
1994). Другие новые органотрофные термофильные прокариоты, выделенные из пластовых
вод Самотлорского нефтяного месторождения, относились к порядку Thermotogales и пред-
ставляли новые виды в родах Thermosipho и Petrotoga (L’Haridon et al., 2002a, b).
Сходные микробные сообщества были обнаружены под океанической корой в районе
глубинной гидротермальной активности (Deming, Baross, 1993; Takai, Horikoshi, 1999). Куль-
туральные методы показали доминирование Thermococcales - органотрофных гипертермо-
фильных архей (Takai et al., 2004). Однако при применении молекулярно-биологических ме-
тодов к этому же сообществу выявилось значительное преобладание метаногенов порядка
Methanococcales. Об активности последних свидетельствовал и легкий изотопный состав
присутствующего метана, говорящий о его биологическом происхождении.
Численность органотрофных термофильных прокариот в высокотемпературных нефтя-
ных месторождениях оказалась неожиданно высокой - 104-106 клеток в мл. Это подтверждает
сделанное на основании радиоизотопных экспериментов предположение о том, что микроб-
ные сообщества нефтяных месторождений используют энергию захороненного органическо-
го вещества. Однако недавние данные, полученные Н.М. Шестаковой и Т.Н. Назиной (Ше-
стакова, 2007), указывают на совершенно особую экологическую функцию, которую могут
играть в природных сообществах даже представители хорошо изученных таксонов. Авторам
этой работы удалось показать, что представитель органотрофного рода Thermoanaerohacier
в присутствии водород-использующего метаногена способен анаэробно окислять ацетат. Та-
ким образом, было найден агент, или, вернее, агенты ацетокластического метаногенеза, заре-
гистрированного в высокотемпературных нефтяных месторождениях. Полученные данные
указывают также на то, как далеки от полноты наши познания о физиологии даже легко куль-
тивируемых в лабораторных условиях микроорганизмов; экстраполяция их на природные
системы может не приблизить нас к пониманию истинного положения вещей.
Другим способом раскрытия экологической роли микроорганизмов, в том числе некульти-
вируемых, является метагеномный анализ микробных сообществ и выявление функциональных
генов, которые могут указать нам на метаболические пути, реализуемые в сообществе. В случае
«Desulforudis audaxiviator» такой анализ явился, по существу, определением полной геномной по-
следовательности единственного организма, растущего, фактически, в виде чистой культуры. Не-
сомненно, подобные работы будут проводиться и с другими, болеее сложными экосистемами.
209
Е..-i. Бонч-Осмоловския. Высокотемпературные глубинные микробные сообщества...
ЗАКЛЮЧЕНИЕ
В заключении мы собираемся вернуться к вопросу, поставленному в заглавии этого
обзора: могут ли микробные сообщества высокотемпературных подземных экосистем рас-
сматриваться как аналоги первичных биоценозов Земли? Как мы уже увидели, эти сообще-
ства очень сильно различаются, в первую очередь по исходному энергетическому субстрату.
Микробные сообщества нефтяных месторождений, по видимости, являются органотрофны-
ми и используют органическое вещество если не самой нефти, то более доступное, захоро-
ненное вместе с окружающей породой. Это органическое вещество, скорее всего, являлось
продуктом фотосинтеза. Таким образом, подземные сообщества, ассоциированные с нефтя-
ными месторождениями, не зависят от современного фотосинтеза, но зависят от продуктов
фотосинтеза, происходившего в предшествующие геологические периоды.
Что же может являться энергетическим субстратом подземных анаэробных микроб-
ных сообществ? Конечно же, в качестве основного такого субстрата рассматривается водород
(Nielsen, Takai, 2005). Известно, что он может образовываться из базальта при реакции с во-
дой и поддерживать рост водород-используюгцих прокариот, например метаногенов (Stevens,
McKinley, 1995). Другим источником водорода является радиолиз воды - этот процесс
предлагается в качестве основного для уникальной экосистемы, образуемой «Desulforudis
audaxiviator». В последнем случае остается непонятным, почему вместе с «Desulforudis
audaxiviator» не развивается полноценное микробное сообщество, использующее органи-
ческое вещество первичного продуцента. Возможно, какие-то ограничения накладываются
особыми физико-химическими условиями, исключающими развитие всех прочих организ-
мов, кроме «Desulforudis audaxivialor».
Однако в случае высокотемпературных микробных сообществ основным источником
водорода все-таки остаются ювенильные газы вулканического происхождения. Как уже было
сказано, водород часто рассматривается как возможный источник энергии для анаэробных
хемосинтетиков в гидротермах различного типа. Однако приток аллохтонного органического
вещества в этих системах не позволяет считать их независимыми от современной биосферы.
Возможно, наиболее близким к искомым гипотетическим сообществам будут микробные эко-
системы гидротермальных местообитаний под океанической корой, где высокая численность
литоавтотрофных метаногенов по сравнению с органотрофами указывает на возможность хе-
мосинтетического происхождения первичного органического вещества в этой экосистеме.
СП ИСОК Л ИТЕРАТУ РЫ
Бонч-Ос.моловская Е.А. 2004. Изучение термофильных микроорганизмов в Институте микробиоло-
гии РАН 7 Микробиология. Т. 73. С. 644-658.
Бонч-Ос.моловская Е.А.. Мирошниченко М.Л. 1994. Влияние молекулярного водорода и элементной
серы на метаболизм экстремально-термофильных архебактерий рода Thermococcus//Микробио-
логия. Т. 63. С. 777-782.
Бонч-Ос.моловская Е.А., Мирошниченко М.Л., Слободкин А.И. и др. 1999. Биоразнообразие анаэробных
литотрофных прокариот в наземных гидротермах Камчатки . Микробиология. Т. 68. С. 398-406.
Бонч-Ос.моловская Е.А., Мирошниченко М.Л., Соколова Т.Г, Слободкин А.И. 2004. Термофильные ми-
кробные сообщества: новые физиологические группы, новые местообитания ' Тр. Ин-та микро-
биол. Вып. 12. С. 29-40.
Назина Т.Н., Шестакова Н.М., Григорьян А.А. и др. 2006. Филогенетическое разнообразие и актив-
ность анаэробных микроорганизмов высокотемпературных горизонтов нефтяного месторожде-
ния Даган (КНР) // Микробиология. Т. 75. С. 1-12.
На.мсараев Б.Б.. Бонч-Ос.моловская Е.А.. Мирошниченко М.Л. и др. 1994. Микробиологические процес-
сы круговорота углерода в мелководных гидротермах Юго-Западной окраины Тихого океана
Микробиология. Т. 63. С. 100-111.
210
Проб. п-чы нронсхогк.и.нпя жптп
Сленова ТВ., Русинов И.И., Соколова Т.Г. и др. 2007. Радиоиозотопное измерение интенсивности транс-
формации СО анаэробными термофильными прокариотами “ Микробиология. Т. 76. С. 594-601.
Слободкин А.И.. Заварзина Д. Г. Соколова Т.Г. Бонч-Осмоловская Е.А. 1999. Диссимиляционное вос-
становление неорганических акцепторов электронов термофильными анаэробными прокариота-
ми И Микробиология. Т. 68. С. 600-622.
Шестакова Н.М. 2007. Филогенетическое разнообразие и активность микроорганизмов высокотемпе-
ратурных нефтяных пластов. Диес. канд. биол. н. М. 24 с.
Amend J.P., Shock E.L. 2001. Energetics of overall metabolic reactions of thermophilic and hyperthermophi-
lic Archaea and Bacteria " FEMS Microbiol. Revs. V. 25. P. 175-243.
Andrews K.T, Pate! B.K.C. 1996. Fervidobacterium gondwanense sp. nov., a new thermophilic anaerobic bac-
terium isolated from nonvolcanically heated geothermal waters of the Great Artesian Basin of Australia //
Int. J. Syst. Bacteriol. V. 46. P. 265-269.
Boeder.J., Torsvik T. Lien T 1995. Thermodesuljorhabdits norvegieus gen. nov., sp. nov., a novel thermophilic
sulfate-reducing bacterium from oil field water Arch. Microbiol. V. 164. P. 331 336.
Bonch-Osmolovskaya E.A.. Miroshnichenko M.L.. Lebedinsky A. V. et al. 2003. Radioisotopic, culture-based,
and oligonucleotide microchip analyses of thermophilic microbial communities in a continental high-
temperature petroleum reservoir , Appl. Environ. Microbiol. V. 69. P. 6143-6151.
Daumas S.. Goeneche O., Bianchi A. 1986. Activites metaboliques des bacteries des eaux geothermales
du Dogger et du trias du Bassin Parisien // C. R. Acad. Sci. Paris. Ser. 3. V. 301. P. 295-297.
Deming J. W., Baross J. A. 1993. Deep-sea smokers: windows to a subsurface biosphere? ,, Geochim. Cosmo-
chim. Acta. V. 57. P. 3219-3230.
Jannasch H.W., Mottl M.J. 1985. Geomicrobiology of deep-sea hydrothermal vents // Science. V. 229. P. 717-725.
Karl D.M. 1995. The microbiology of deep-sea hydrothermal vents. Boka Raton: CRC Press. 299 p.
Lin L.-H., Wamg P.-Z., Rumble D. etal. 2006. Long-term sustainability ofa high-energy, low-diversity crustal
biome .. Science. V. 314. P. 479.
L'Haridon S., Miroshnichenko M L., Hippe H. et al. 2002a. Thermosipho geolei sp. nov., a thermophilic bacte-
rium isolated from a continental petroleum reservoir in Western Siberia Int. J. System. Evol. Microbiol.
V. 52. P. 1327- 1334.
L’Haridon S., Miroshnichenko M.L., Hippe H. et al. 2002b. Petrotoga olearia sp. nov. and Petrotoga sibirica
sp. nov., two thermophilic bacteria isolated from a continental petroleum reservoir in Western Siberia //
Int. J. Syst. Evol. Microbiol. V. 52. P. 1715-1722.
L'Haridon S., Reysenbach A.-L.. Glenat P.D. et al. 1995. Hot subterranean biosphere in a continental oil res-
ervoir ' Nature. V. 337. P. 223-224.
Miroshnichenko M.L.. Hippe H., Stackebrandt E. et al. 2001. Isolation of hyperthermophilic Archaea in West-
ern Siberia high temperature oil reservoir and characterization of Thermococcus sibiricns sp. nov. //
Extremophiles. V. 5. P. 85-91.
Nazina T.N., Xue Y-F, Wang X-Y. et al. 2000a. Microorganisms of the high-temperature Liaohe oil field
of China and their potential for MEOR // Res. Environm. Biotechnol. V. 3. P. 149-160.
Nazina T.N.. Xue Y-F., Wang X-Y. et al. 2000b. Diversity and activity of microorganisms in the Daqing oil field of
China and their potential for biotechnological applications // Res. Environm. Biotechnol. V. 3. P. 161-172.
Nielsen K., Takai K. 2005. Hydrogen-driven subsurface lithoautotrophic microbial ecosystems (SliMEs):
do they exist and why should we care? Trends in Microbiol. V. 13. P. 405-410.
Nilsen R.K., Torsvik T. 1996. Methanococcus thermolithotrophicus isolated from North Sea oil field reservoir
water//Appl. Environm. Microbiol. V. 62. P. 728-731.
Orphan V.J., Taylor L.T., Hajenbradl D.. Delong E.F. 2000. Culture-dependent and culture-independent char-
acterization of microbial assemblages associated with high-temperature petroleum reservoirs .. Appl.
Environm. Microbiol. V. 66. P. 700-711.
Parkes R.J., Cragg B.A.. Bale S.J. et al. 1994. Deep bacterial biosphere in Pacific Ocean sediments // Nature.
V. 371. P. 410-413.
211
Е.А. Бонч-Осмо.ювския. Высокотемпературные глубинные микробные сообщества...
Parkes R.J.. Cragg В.A.. Wellsbury Р. 2000. Recent studies of bacterial populations and processe in subsea-
floor sediments: a review Hydrogeology J. V. 8. P. 11-28.
Pimenov N.A.. Bonch-Osmolovskaya E.A. 2006. In situ activity studies in thermal environments Methods
in microbiology i Eds. F.A. Rainey, A. Oren. Elsevier. Acad. Press. P. 29-54.
Ravot G., Magot M.. Fardeau M.-L. et al. 1995. Thermotoga elfii sp. nov., a novel thermophilic bacterium from
an African oil-producing well ' Int. J. Syst. Bacteriol. V. 45. P. 308-314.
Rees G.N., Grassia G.S.. Sheehy A.J. et al. 1995. Desulfacinum infernttm gen. nov., sp. nov., a thermophilic
sulfate-reducing bacterium from a petroleum reservoir ' Int. J. Syst. Bacteriol. V. 45. P. 85-89.
Rosnes J.T., Torsvik T. Lien T. 1994. Sporeforming sulfate-reducing bacteria isolated from North Sea oil field
waters Appl. Environm. Microbiol. 1994. V. 57. P. 2302-2307.
Roussel E.G., Cambon-Bonavita M.-A.. Querellou J. et al. 2008. Extending the subsea-floor biosphere .. Sci-
ence. V. 320. P. 5879-5882.
Sokolova T. Henstra A-М.. SiptnaJ. et al. 2009. Diversity and ecophysiological features of thermophilic car-
boxydotrophic anaerobes “ FEMS Microbiol. Ecol. V. 68. P. 131-141.
Spear J. R., Walker J.J.. McCollom T.M., Pace N.R. 2005. Hydrogen and bioenergetics in the Yellowstone
geothermal ecosystem Proc. Natl. Acad. Sci. USA. V. 102. P. 2555-2560.
Stetter K.O.. Huber R., Bloechl E. et al. 1993. Hyperthermophilic archaea are thriving in deep North Sea and
Alaskan oil reservoirs // Nature. V. 365. P. 743-745.
Stevens T.O.. McKinley J.P. 1995. Lithoautotrophic microbial ecosystems in deep basalt aquifers // Science.
V. 270. P. 450-455.
Takahata V. Nishijima M„ Hoaki T, Maruyama T. 2000. Distribution and physiological characteristics of hyper-
thermophiles in the Kubiki oil reservoir in Niigata, Japan // Appl. Environm. Microbiol. V. 66. P. 73-79.
Takai K., Gamo T. Tsunogai U. et al. 2004. Geochemical and microbiological evidence for a hydrogen-based
subsurface lithiautotrophic microbial ecosystem (HyperSLiME) beneath an active deep-sea hydrother-
mal fluid Extremophiles. V. 8. P. 268-282.
Takai K.. Horikoshi K. 1999. Genetic diversity of Archaea in deep-sea hydrothermal vent environments /
Genetics. V. 152. P. 1285-1297.
Takai K., Moser D.P.. DeF/aun M. et al. 2001. Archaeal diversity in waters from deep South African gold
mine H Appl. Environm. Microbiol. V. 67. P. 5754-5760.
Teske A. 2005. The deep subsurface biosphere is alive and well . Trends in Microbiol. V. 13. P. 402-404.
Wellsbury P.. Mather Parkes R.J. 2002. Geomicrobiology of deep, low organic carbon sediments in the
Woodlark Basin, Pacific Ocean FEMS Microbiol. Ecol. V. 42. P. 59-70.
Whitman W.B., Coleman D.C., Wiebe J. W. 1998. Prokaryotes: the unseen majority U Proc. Natl. Acad. Sci.
USA. V. 95. P. 6578-6583.
Zavarzin G.A., Bonch-Osmolovskaya E.A., Svetlichnyi V.A. 1989. Extreme thermophiles in hydrotherms /7
Recent advances in microbial ecology / Ed. T. Hattori. Japan Sci. Soc. Press. P. 33-37.
High-temperature, deep subsurface microbial communities - are they the possible analogues
of ancient ecosystems?
E.A. Bonch-Osmolovskaya
Microbiological, molecular and radioisotopic data indicate the presence of active and multi-
component microbial communities in the deep layers of subsurface biosphere. In this review only
some of such communities are considered - those that are active at high (60 °C and higher) tem-
peratures. Those most studied of them are the oil wells, subsurface water reservoirs (e.g. the Great
Arthesian Basin in Australia), deep mines (South Africa), high temperature horizons located beneath
the oceanic crust in the regions of submarine volcanic activity. These microbial communities differ
significantly in their composition, energy source and dominating microbiological processes.
However, if proposed as the analogues of primary Earth ecosystems, they should to be proved
to be autonomous of modern Earth biosphere.
212
Проблемы нроисяожлення лиши
Дискуссия
по пленарному докладу Е.А. Бонч-Осмоловской
«ГЛУБИННАЯ БИОСФЕРА»
Председательствующий Л. М. Мухин
В.Н. Снытников: Мне показалось, что Вы в своем докладе ни разу не указали конкрет-
ные значения давления. Правильно ли я понимаю, что если глубина три километра, то это не
менее 600 атмосфер9 А если вода, то я могу посчитать давление по столбу воды?
Е.А.Б.-О.: Да, это все так. Поэтому, извлекая части этих сообществ, мы грубейшим обра-
зом нарушаем существующие там условия. Видимо, мы отсекаем основную популяцию микро-
бов, так же как, исследуя воду, отсекаем всех тех, кто прочно привязан к грунту. Мы должны все
время отдавать себе отчет о том, что мы изучаем пока только какую-то «верхушку айсберга».
Ю.В. Наточин: Как измеряли возраст грунтовых вод?
Е.А.Б.-О.: Точно не знаю, думаю, что по стабильному изотопному составу.
Л.М. Мухин: Скажите, пожалуйста, какое отношение глубинная биосфера имеет к про-
блеме происхождения жизни?
Е.А.Б.-О.: Я бы считала, что как аналогия. Пытаясь исследовать потоки вещества и
энергии в живых системах, мы обращаемся к таким изолированным сообществам как к со-
временным моделям.
Л.М. Мухин: А каково происхождение этих ценозов? Это следы наружной биосферы
или они возникли совершенно независимым путем на этих глубинах?
Е.А.Б.-О.: Эти микробные сообщества образовались из поверхностных. Их очень ши-
рокий оптимум температуры и pH - это какой-то древний фенотип, оставшийся еще со вре-
мен их пребывания на поверхности и в силу их очень медленной эволюции сохранившийся -
почему так интересно было бы посмотреть их полный геном и сравнить его с таковым гидро-
термальных микробов.
М.Я. Маров: Елизавета Александровна, у меня два связанных вопроса. Первый: если
предельная температура, как Вы показали, 122 градуса, то можно ожидать наличие биосфе-
ры до глубины шесть километров, поскольку рост температуры в коре примерно 20 градусов
на километр. Но, если я правильно понимаю, с такой глубины никакой экскавации не было?
Е.А.Б.-О.: Есть Кольская скважина, но интересующих нас данных по ней нет.
М.Я. Маров: Конечно, очень интересно, сколь глубоко простирается эта граница, и ме-
няются ли там условия, свойственные такой закрытой системе. Мой второй вопрос следую-
щий. Я с удивлением услышал о том, что глубинная биосфера может составлять около 80
процентов биомассы Земли. Известный американский ученый Том Голд в статье, опублико-
ванной в 1992 году в журнале «Science», утверждал, что глубинная биосфера по массе может
значительно превышать то, что мы имеем на поверхности. Приведенные Вами цифры - это
что, изменившиеся воззрения?
Е.А.Б.-О.: Я думаю, что это связано с новыми методами и возможностями, например,
сейчас доказано, что эти клетки активно функционируют.
М.Я. Маров: То есть раньше это было спекулятивно, а сейчас в значительной мере ме-
тодично?
Е.А.Б.-О.: Да.
М.А. Федонкин: Я бы хотел узнать о водородном обмене, его роли. Насколько он широ-
ко распространен? Не является ли, скажем, деятельность органотрофов борьбой за водород,
выбиранием водорода из органики?
213
Е.А.Б.-О.: С водородом проблемы нет, есть много его источников, проблема в окислите-
ле, потому что если нет кислорода, то нет и почти всего остального. С серой тоже не совсем
понятно. Это считается древним видом дыхания. Но если элементная сера напрямую возго-
няется из глубин, она достигает поверхности в конформации, недоступной микробам, в виде
кристаллов. Микробы с удовольствием поедают ту серу, которая образуется из сероводорода
в его реакции, опять же, с кислородом. Так что сера тоже под вопросом. И главный акцептор
электронов — это СО,, которого много. Здесь есть две возможности - во-первых, образование
метана, во-вторых, образование ацетата. И образование ацетата до сих пор остается в тени.
М.А. Федонкин: Радиолиз может вызывать разложение воды с выходом водорода.
Е.А.Б.-О.: Водород образуется из базальтов, в реакции с водой.
М.А. Федонкин: Да, источников очень много. А вот, скажем, цикл водорода и окиси
углерода, СО?
Е.А.Б.-О.: СО - это тоже совершенно отдельная статья. СО это обязательный компо-
нент гидротермальных газов, и огромное количество организмов может использовать СО, и
ничего при этом другого им не нужно, только СО и вода. А водород образуется в результате
этой реакции. Я не совсем поняла Вас про органотрофов, почему они борются за водород, у
них есть такой прекрасный органический субстрат. Вообще, побывав на этом совещании, я
стала думать, что, если действительно возможен абиогенный синтез органических веществ,
то тогда может быть, что первыми организмами были органотрофы, потому что их внутрен-
нее устройство гораздо проще, им не нужны мембраны, не нужны всякие сложности... Суб-
стратное фосфорилирование - и все... Кстати, на Камчатке довольно много ацетата в гидро-
термах, и при высоких температурах в черных курильщиках тоже идет абиогенный синтез
ацетата. Правда, ацетат не используется без акцептора, а какая-то более сложная органика
вряд ли может абиогенно синтезироваться... В общем, тут есть над чем подумать, в том числе
и с точки зрения возможностей первичных метаболизмов...
М.Я. Маров: Откуда СО на глубине?
Е.А.Б.-О.: СО идет как ювенильный газ из магмы, а кроме того, образуется в результате
биологических процессов. С СО тоже нет проблем.
Л.М. Мухин: Ювенильность любой составляющей надо доказывать. Потому что масса
компонентов в вулканических газах, гидротермах возникает за счет разложения осадочных
пород.
Е.А.Б.-О.: Абсолютно согласна.
А.Ю. Розанов: Елизавета Александровна, как Вы относитесь к публикациям Оборина и
компании и их схеме подземной биосферы?
Е.А.Б.-О.: К сожалению, я плохо их знаю.
А.Ю. Розанов: Вы постоянно говорите, что это у вас закрытые системы. Но если Вы
связываете их с нефтяными или артезианскими бассейнами, то это не закрытые системы.
Е.А.Б.-О.: Как только мы доберемся в закрытую систему, она перестанет быть закры-
той.. .
М.Я. Маров: Она закрытая в том смысле, что прямо не выходит на поверхность, а на-
ходится в каких-то определенных резервуарах или «карманах»...
А.Ю. Розанов: Если это бассейны, то где-то они выходят на поверхность...
Е.А.Б.-О.: Открытые или закрытые системы - для циклов.
214
Прор.ц.мы 11р()ис.\ож:и-:ння жизни. М.: ПИН РАН. 2009. С. 215-228
ВОЗНИКНОВЕНИЕ МЕМБРАН
© 2009 г. Ю.В. Наточин
Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН
natochin@iephb.ru
Рассмотрены вопросы о физико-химических факторах среды, в которой
возникла жизнь, о появлении протоклеток, их оболочки, а затем плазмати-
ческой мембраны. Обоснована гипотеза о появлении первых клеток в водо-
емах. где доминировали соли калия и магния, поскольку они необходимы
для синтеза белков, а без этого нс построить мембрану клетки. В последую-
щем произошла адаптация этих клеток к внешней водной среде, подобной
океанической, в которой превалировали соли натрия. Этот этап эволюции
требовал функционирования плазматической мембраны, способной обе-
спечить ионную асимметрию цитоплазмы клеток по отношению к внеш-
ней среде. На этом этапе развития жизни у предшественников животных
в плазматической мембране стали действовать механизмы удаления ионов
натрия и накопления ионов калия. Проблема развития многоклеточных
была решена разным способом у животных и растений: у животных сфор-
мировалась система жидкостей внутренней среды, которые обеспечивали
стабильные физико-химические условия с внешней поверхности плазмати-
ческой мембраны. Ионы натрия явились стимулом образования полярной
клетки, где в плазматической мембране с одной стороны клетки сосредо-
точены натриевые каналы, а в другой - натриевые насосы, что обеспечило
развитие функций всасывания, выделения, дыхания. Формирование жид-
костей внутренней среды стало предпосылкой становления гомеостаза,
условий биологического прогресса мира животных.
ВВЕДЕНИЕ
Возникновение плазматической мембраны можно считать одним из ключевых событий
в происхождении и развитии жизни на Земле. Очевидно, что живому существу необходимы
для существования молекулы органических веществ, генетический код или его прообраз,
синтез полимеров биологически значимых веществ, но столь же ясно, что они должны быть
собраны вместе, отделены, изолированы от окружающего мира в единое целое для создания
некоей первичной целостной структурированной системы, иначе говоря, требуется появление
оболочки, мембраны. Такое событие, безусловно, столь же важно для развития живых су-
ществ, как и наличие остальных ключевых факторов, входящих в понятие живого.
Следующее важнейшее условие возникновения жизни, которое не только обычно не
обсуждается, его часто и не упоминают, касается неорганических компонентов среды, где
215
Ю.В. Наточин. Возникновение мембран
появилась жизнь. Необходим ответ на вопрос о физико-химических условиях водной среды,
где происходило формирование первых живых существ. Обсуждение и хотя бы принципиаль-
ное решение этого вопроса имеет безусловное значение для понимания природы зарождения
жизни и ее дальнейших шагов. Задача этого доклада состоит в том, чтобы обсудить вопрос
о возможном электролитном составе первичной среды зарождения жизни, способах изоляции
появившихся протоклеток от внешней среды и дальнейшего их развития. Иными словами -
ключевое значение в этой проблеме имеет ответ на вопросы о физико-химических параметрах
среды и возникновении плазматической мембраны клетки. Следует сказать, что в литерату-
ре имеется огромное количество работ, касающихся строения, химического состава, функ-
ционального значения плазматических мембран. Если до средины XX в. обсуждался вопрос
о сорбционных свойствах протоплазмы как альтернативе мембран, то с того времени, когда
начались исследования с применением методов электронной микроскопии, стало возможным
представить строение мембран. Биохимические и биофизические подходы дали факты, касаю-
щиеся функциональной организации мембран. В этом докладе речь пойдет о вероятных усло-
виях и факторах, определивших возникновение мембран на заре биологической эволюции.
Очевидно, что ряд физико-химических параметров среды, в которой появились первые
живые существ, не только оказал определяющее влияние на их становление, но на их основе
формировались основные явления жизни, базовые функции исходных, доклеточных форм
жизни. Нет сомнения в том, что водная среда была и фоном, и важнейшим компонентом
первичной жизни, самой сценой для химических процессов организации жизни. Основная,
можно сказать, единственная концепция в литературе о среде появления жизни, сформу-
лированная в результате анализа данных, накопленных в XX в., состоит в формуле: жизнь
возникла в море. «Столь же несомненно, что первые живые существа появились не в пре-
сной воде, но в растворе солей Na, К, Са и Mg. Иначе нельзя объяснить тот факт, что клетки
всех животных от самых простых до самых сложных, какова бы ни была среда их обитания,
содержат в себе эти ионы и погибают, когда они отсутствуют. В настоящее время никто
не сомневается в том, что жизнь возникла в воде океана палеозойской эры, содержащей
определенные соотношения одновалентных и двухвалентных катионов, которые с полным
основанием могут быть названы биологическими» (Гинецинский, 1963, с. 9). «Жизнь за-
родилась в море. Как показывают данные геохимии, ионный состав морской воды не пре-
терпел существенных изменений с раннекембрийского периода, хотя ее общая соленость,
по-видимому, несколько возросла» (Проссер, 1977, с. 177). «Жизнь возникла в море. Хи-
мический состав морской воды определен составом земной коры, физическими и химиче-
скими свойствами ее компонент, а, следовательно, химическим составом планеты. Высокая
вероятность возникновения жизни именно в море подчеркивалась почти всеми. Мне также
кажется это почти бесспорным» (Шноль, 1981, с. 95). Цитирование подобных высказываний
можно было бы продолжить, но их суть ясна.
Верно ли это утверждение? Имеется ряд, как нам кажется, логически бесспорных аргу-
ментов, которые дают основания, чтобы признать неправомерность предположения о возникно-
вении жизни в море, подобном современному, море, где доминировали ионы натрия. Аргументы,
которые будут приведены ниже, не позволяют принять эту точку зрения и основаны на доводах,
главный из которых в том, что плазматическая мембрана не могла первично появиться в океа-
нической воде, где превалируют ионы натрия, а без такой мембраны нет одного из обязательных
условий для последующей адаптации клеток к натриевой океанической среде, к жизни в пре-
сной воде и на суше (Наточин, 2005, 2006). При возникновении «первой» клетки (протоклетки)
ее содержимое надо было отделить от «неклетки», в цитоплазме протоклетки необходимо было
иметь критическую концентрацию нуклеотидов, аминокислот, ионов, всего того, что необходи-
мо для синтеза белков, метаболизма. При обсуждении функций протоклетки следует учитывать
не только наличие органических веществ, но и неорганические ее компоненты.
216
Проблемы пропс хожн-лпя жисни
В 1960-е гг. стало ясно, что ключевые реакции, связанные с синтезом белка, требуют
присутствия таких неорганических ионов как магний и калий. В среде для синтеза белка in
vivo и in vitro необходимы 5-20 мМ Mg-' и 100 мМ К.*, ионы Na+ «являются антагонистами,
ингибируя работу белок-синтезируюшей системы» (Спирин, Гаврилова, 1971, с. 152). Реак-
ция транспептидации на рибосоме нуждается в присутствии в среде ионов Mg2+ и К? (или
NH4‘) (Maden, Monro, 1968; Спирин, Гаврилова, 1971). Сказанное выше дает нам возможность
заключить, что при условии сохранения базовых молекулярных механизмов синтеза белка
в ходе эволюции жизни на Земле принципиально неизменными, плазматическая мембрана
с включенными в нее молекулами белка не могла возникнуть в среде, где превалировали
ионы натрия. Из известных данных, из сказанного выше ясно, что соли натрия доминируют
в водах современного океана (Салоп, 1982; Холленд, 1989). Без плазматической мембраны
не могла возникнуть и клетка в ее современном понимании, она была бы незащищенной,
ее компоненты были бы разрозненными, она не могла бы адаптироваться к средам с раз-
личным ионным составом, разной осмоляльностью, разной концентрацией неорганических
веществ. Следовательно, необходимо предложить иную схему последовательности событий,
происходивших на начальных этапах эволюции жизни, чем появление жизни в среде, подоб-
ной современной океанической. Речь идет прежде всего о возникновение жизни в морской
воде, если ее состав был подобен современному океану.
Суть нашей гипотезы состоит в том, что средой возникновения первых форм жизни
могли быть водоемы с доминированием магния и калия в качестве катионов, поскольку
в этих условиях могли протекать процессы синтеза белка, а затем могли сформироваться
белковые компоненты клетки, включая первичную плазматическую мембрану протоклетки.
Аргументами в пользу этой гипотезы могут быть данные об элетролитном составе клеток
и внеклеточных жидкостей различных групп современных животных, анализ функций суще-
ствующих различных типов мембран. Очевидно, что наше заключение может быть основано
только на косвенных данных, прямых сведений о событиях, которые были около 4 млрд, лет
назад, нет. Дополнительными аргументами могли бы быть модельные системы, математиче-
ское моделирование этих процессов.
СРЕДА ВОЗНИКНОВЕНИЯ ПРОТОКЛЕТОК
Можно принять, что солевой состав среды в период зарождения первых форм жизни
должен был представлять собой водные растворы, в которых концентрация ионов благопри-
ятствовала биохимическим процессам, необходимым для реализации базовых жизненных
функций, в частности, синтеза белков. Конечно, в настоящее время нет прямых данных для
суждения о физико-химических условиях среды в период зарождения жизни, но некоторые
заключения об электролитном составе этой среды могут быть сделаны. Консерватизм при-
роды в отношении базовых принципов построения живых систем должен был проявиться
в качественном сходстве фундаментальных условий реализации жизненных функций в ряду
последующих поколений от первых ее форм до ныне живущих особей.
Анализ геологического прошлого планеты был необходим для понимания физико-
химических условий среды, где сформировалась жизнь, но данные палеогеохимии в наше
время недостаточны, чтобы оценить ионный состав среды в эпоху возникновения живых
существ. Для этого необходима информация о событиях, которые были около 4 млрд, лет
назад. Поэтому попытаемся использовать иной подход, чем наши предшественники, имея
в виду в последующих рассуждениях и анализе фактов использовать физиологический под-
ход для ответа на поставленный вопрос.
Без разностороннего физиологического анализа становления жизни на Земле вряд ли
может быть осуществлено глубокое проникновение в механизмы ее развития. Первые живые
организмы, возникшие около 4 млрд, лет назад, можно обозначить как протоклетки (Нато-
217
Ю.В. Нипючин. Возникновение мембран
Таблица I. Возможное усложнение функций плазматической мембраны в ходе эволюции
Этапы эволюции Сохранение органических молекул Селективность к ионам металлов Стабилизация рн Регулируемая водная проницаемость
Начальный + - - -
Промежуточный + + + -
Современный + + + +
чин, 2005), чтобы не соотносить их с конкретными видами современных или ископаемых ор-
ганизмов. Этот термин использован нами, чтобы охарактеризовать стадию развития живых
существ, еще не имевших плазматической мембраны, но уже обладавших оболочкой, огра-
ничивающей переход в окружающую среду новообразованных органических молекул этого
организма и обеспечивающих их сохранение в клетке. Есть все основания полагать, что в про-
токлетке под оболочкой концентрация одновалентных ионов и осмотическое давление не отли-
чались от окружающей среды. Любые отличия этих параметров от внешней среды требовали
бы наличия специальных мембранных макромолекул типа ионных насосов, ионных каналов,
водных каналов. Но их возникновение требует синтеза этих специальных белков в клетке, что
трудно предполагать на начальном этапе возникновения жизни, их не могло быть до появления
протоклетки, их синтез требовал создания соответствующих биохимических машин.
Анализируя имеющиеся данные, можно признать, что возрастающее значение плазма-
тических мембран, усложнявшихся в ходе эволюции, привело к выполнению части или всех
следующих ее функций:
- сохранение макромолекул внутри клетки,
-создание и поддержание оптимума концентрации ионов,
- электрогенез,
- встраивание рецепторов для регуляции функций клетки,
- осмотическая зашита при изменении осмоляльности среды,
- стабилизация pH.
В настоящее время имеются в клетках разные типы мембран, на начальном этапе раз-
вития это могли быть и менее специализированные образования (оболочки), в числе пред-
ставленных в настоящее время могут быть названы следующие:
- плазматическая,
- митохондриальная,
- щелевая,
- ядерная.
Селективность мембран связана с их способностью различать молекулы по размеру, за-
ряду, форме. Наличие в плазматических мембранах специальных белков - каналов для воды
(аквапорины), ионов (натриевые, калиевые и др.), котранспортеров, ионных насосов - обеспечи-
вает возможность поддержания внутри клеток строго определенного состава неорганических и
органических веществ. Постепенно в ходе эволюции происходило увеличение числа функций,
выполняемых плазматическими мембранами у клеток различной специализации (табл. I).
ОСМОЛЯЛЬНОСТЬ И КОНЦЕНТРАЦИЯ ИОНОВ В КЛЕТКАХ
И ВНЕКЛЕТОЧНОЙ ЖИДКОСТИ
О концентрации ионов в воде морей, озер, заливов, рек, в клетках и во внеклеточной
жидкости у современных животных можно судить по данным прямых измерений (табл. 2),
которые говорят об исключительно широком диапазоне отличий состава сред, к которым
адаптированы представители разных групп беспозвоночных и позвоночных животных.
Сопоставление концентрации тех же неорганических элементов в гемолимфе, сыворотке
218
Проблемы происхождения жияш
Таблица 2. Концентрация ионов в некоторых морях, реках и озерах, ммоль/л
Водоем Na К Са Mg
Баренцево море 465 1 1.5 11.6 53.5
Белое море (Чупинский залив) 225 6 5.85 34
Океан (усредненные данные) 468 9.9 10.2 53.5
оз. Балхаш (восточный) 42 2 3.35 8
оз. Байкал 0.18 0.025 0.1 0.12
р. Волга 1.1 0.08 0.7 0.4
Таблица 3. Концентрация и доля ионов натрия и калия в осмоляльности гемолимфы
и сыворотки крови некоторых животных и человека
Объект исследования Осмоляльность, мосм/кг Na, ммоль/л K, ммоль/л
Человек 287±1 142±0.4 4.5±0.04
Собака 297±2 148±2 4.1±0.2
Крыса (Вистар) 301±3 149±1 4.9±0.2
Голубь Columbia Hvia 295±5 147±1.7 2.4±0.3
Черепаха Tesludo horsfieldi 343±6 135±2.5 5.5±0.3
Лягушка Rana temporaria 222±2 108±3 2.6±0.2
Горбуша Oncorhynchus gorbitscha 342±2 154±2 2.5±0.48
Минога Lamperta fhtvialUis 235±2 120±3 3.2±0.3
Миксина Myxine glutinosa 1152 549 11.1
Мидия Mytilu edulis (соленость 26 %o) 704±7 331±11 7.5±0.3
Жемчужница Margaritana margantij'era 30.2±l.l 14.1±0.4 0.39±0.02
Перловица обыкновенная Unto pectorium 51.4±1.6 14.4±0.6 0.47±0.06
Перловица клиновидная Unio tumidu\ 49.3±3.2 12.9±0.9 0.41±0.04
Живородка E'iviparus 104±3.6 31.8±1 1.65±0.1
Таблица 4. Концентрация некоторых катионов в клетках
Объект исследования Na, ммоль/л К, ммоль/л
Клетка мышцы млекопитающих 12 155
Крыса, волокно m. gastrocnemius 16.2±1.2 163±3.7
Neclitfiis, клетка проксимального канальца нефрона [1] 42.2±2.2 58.7±2.3
Amphiuma, клетка дистального канальца нефрона [1] 21.3±2.8 53.8±3.8
Примечание. Данные [I] по Khuri, 1979.
крови (табл. 3), клетках (табл. 4) и тканях, содержащих как клетки, так и внеклеточную жид-
кость (табл. 5), дает основание для суждения о тех ограничениях, которые вносит жизнь по
отношению к среде обитания. В этих рассуждениях не учитывается и гетерогенность содер-
жания ионов в различных компартментах клетки.
В морской воде доминируют ионы натрия, высока концентрация ионов магния, зна-
чительно ниже концентрация ионов калия и кальция (табл. 2). В пресной воде концентрация
натрия в десятки раз ниже, чем в океанической воде, ниже и концентрация других ионов.
219
Ю.В. Наточим. Возникновение мембран
Таблица 5. Содержание катионов (мкмоль/г сухого вещества) в тканях животных
Объект исследования Na К Mg
Крыса, почка, кора 73.9±5.2 80.9±1.9 9.2±0.25
Наружное мозговое вещество 103±3 81,4±2.1 9.5±О.З
Внутреннее мозговое вещество 191±16 65.2±1.5 6.2±0.25
Семенник 64.8±3.1 65.6±2.6 7.25±0.1
Печень 31.8±1.6 88.2±2.0 11.6±0.8
Пчела Apis mellifera, глаз 23,9±6.6 101±2.0 24.5±7.5
Треска Gadus morphua, мышца 26.3±2.8 129±3.9 12.1±0.46
Береговая улитка Liltorina Hltorea, мышца 126±8 94±6 39±4
Асцидия Goniocarpa ruslica, стенка тела 183±7 85.6±4.6 23.6±1.1
Русский осетр Acipenser gii/densladli. мышца 25.3±1.1 91.3±3.6 10.6±0.5
Таракан Periplanela americana, крыловая мышца 21.7Д4.8 85.6±2.6 —
Пилильщик Rhadinoceraea micas, мышца 11.0±1.6 72.7±2.8 —
В сыворотке крови представителей всех классов позвоночных доминируют ионы
натрия, концентрация ионов калия в 25-35 раз ниже (табл. 3).
Если сопоставить концентрацию катионов в гемолимфе, сыворотке крови и тканях са-
мых разнообразных организмов, среди которых моллюски, насекомые, рыбы пресноводные
и морские, млекопитающие, то ясно, что в клетках эукариот, как хорошо известно, преоб-
ладают ионы калия. В тканях доля натрия ниже, а калия выше, чем в гемолимфе или в сы-
воротке крови, что обусловлено их накоплением в клетках. У простейших выявляется та
же особенность: в клетке Amoeba proteus (мкмоль/мг сухого вещества) К - 0.376, Na - 0.026,
Са - 0.063, Mg - 0.217 (цит. по Проссер, 1977). В этих клетках в 14.5 раза выше содержание
ионов калия, чем натрия. В клетках мышц млекопитающих (табл. 4) K/Na соотношение состав-
ляет 12.9. Следовательно, у совершенно разных по уровню развития и специализации клеток
животных выявляется сходство этого существенного физико-химического параметра. Это по-
зволяет полагать со значительной долей вероятности, что в клетках удерживается доминиро-
вание данного катиона (калия), что имеет принципиальное значение для физико-химических
особенностей внутриклеточной среды с момента их формирования, то есть миллиарды лет.
КОНЦЕНТРАЦИЯ ИОНОВ В СЫВОРОТКЕ КРОВИ НЕКОТОРЫХ ЖИВОТНЫХ
И ЧЕЛОВЕКА
Сказанное выше свидетельствует о том, что становление клеток было связано с воз-
никновением мембраны, с необходимостью определенной концентрации различных ионов по
обеим ее сторонам, наконец, наличием в мембране специальных механизмов, молекуляр-
ных устройств, обеспечивающих поддержание оптимальных физико-химических условий
внутриклеточного содержимого. У животных и человека формируется специализирован-
ная система, благодаря которой достигается все более высокая степень постоянства соста-
ва околоклеточной жидкости. Если на начальных этапах эволюции среда обитания была по
физико-химическим характеристикам подобна внеклеточной среде, то при формировании
многоклеточных животных происходит становление системы жидкостей внутренней среды,
внеклеточных жидкостей (Smith, 1943):
- околоклеточная жидкость,
- кровь, гемолимфа,
- ликвор,
- перилимфа,
- эндолимфа.
220
ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ ж и :пи
Система стабилизации объема и состава внеклеточной жидкости сыграла в эволюции
животных важную роль. Приведенные выше факты свидетельствуют об исключительном
значении именно в эволюции животных и человека выбора ионов натрия в виде контр-
катиона для внутриклеточного катиона калия. В то же время у исходных форм, которые
служили началом развития мира растений, натрий не был использован для осуществления
этих базисных функций. У позвоночных, особенно у млекопитающих и человека, наряду
со стабилизацией физико-химических параметров плазмы крови, внеклеточной жидкости,
дополнительно стабилизирована среда функционирования центральной нервной системы,
что обеспечивается специальной жидкостью, находящейся за гемато-энцефалическим барье-
ром, имеется и ряд других физиологических механизмов, например гемато-офтальмический
барьер, во внутреннем ухе образуются эндолимфа и перилимфа.
В отношении доминирующего катиона во внеклеточной жидкости имеется лишь не-
сколько исключений. К ним относится гемолимфа ряда видов насекомых (Natochin, Parnova,
1989), в которой выше концентрация ионов калия, чем ионов натрия. Однако и в этих случаях
концентрация ионов калия в клетках мышц насекомых с калиевой гемолимфой тоже выше,
чем в гемолимфе. В клетке этих организмов, как и у насекомых с «натриевой» гемолимфой,
K/Na отношение было выше, чем в гемолимфе. Так, в мышце пилильщика это отношение
равно 2.7. Чтобы обеспечить столь высокий градиент ионов калия у насекомых с «калиевой»
гемолимфой были использованы нестандартные механизмы адаптации. У этих насекомых
повышена осмоляльность гемолимфы за счет накопления в ней никомолекулярных органи-
ческих веществ (аминокислот, моносахаров), что создает возможность для поддержания бо-
лее высокого уровня калия в клетках.
ГОМЕОСТАЗ
Со времен К. Бернара центральная идея физиологии состоит в признании положения,
что постоянство внутренней среды служит основой свободной жизни (Харди, 1986). Про-
токлетка могла существовать в стабилизированных физико-химических условиях внешней
среды, касающихся осмоляльности, концентрации ионов калия, магния, pH. Эти параме-
тры практически не должны были, а вернее не могли отличаться от внутриклеточного со-
держимого, поскольку это было основным условием реализации функций клетки. По мере
эволюции Земли живые особи стремились распространиться по все более разнообразным
ареалам, средам с разной соленостью, осуществили выход на сушу. Среди этапных со-
бытий в становления жизни было возникновение протоклетки и ее оболочки, появление
плазматической мембраны, формирование клетки эукариот, затем появление многоклеточ-
ных животных. Физиологическая эволюция сопровождалась появлением системы внекле-
точных жидкостей, которые создали предпосылки для стабилизации физико-химических
параметров среды около плазматических мембран. Это достижение эволюции многокле-
точных животных следует особенно подчеркнуть, так как оно определило саму возмож-
ность последующей стабилизации условий во внутренней среде для животных. Чем выше
на эволюционной лестнице находится организм, тем более развита система стабилизации
физико-химических параметров внутренней среды особей, а тем самым среды около плаз-
матической мембраны у всех клеток организма. Проведенные нами исследования позво-
лили выявить наиболее жестко стабилизированные физико-химические параметры сыво-
ротки крови позвоночных, достигших наиболее высокого уровня совершенства у человека
(Natochin, Chernigovskaya, 1997). К этим показателям относится осмоляльность, концентра-
ция ионов натрия и ионизированного кальция. Объяснение очевидно - от осмоляльности
крови, концентрации в ней ионов натрия зависит объем каждой клетки, биоэлектрические
процессы на плазматической мембране. Ионы кальция определяют регуляцию многих про-
цессов в клетке.
221
Ю.В. Ниточин. Возникновение мембран
Особое значение в сохранении и поддержании постоянства состава внутренней среды
принадлежит выделительным органам. К их числу относятся не только почки, но и солевые
железы, жабры, клетки которых способны к секреции ионов во внешнюю среду, удалению
углекислоты и ряда иных веществ. Ниже дан перечень этих органов у представителей разных
классов позвоночных:
- круглоротые (почки, жабры),
- рыбы (жабры, почки, мочевой пузырь, ректальная железа),
- амфибии (почки, кожа, мочевой пузырь, легкие),
- рептилии (почки, солевые железы, легкие),
- птицы (почки, солевые железы, легкие),
- млекопитающие (почки, потовые железы, легкие).
В большинстве случаев функция выделительных органов основана на двухэтапном
принципе - в просвет канальца происходит ультрафильтрация плазмы крови или гемолимфы,
а затем ионы Na‘ и многие другие вещества при участии ]Ча+-зависимых механизмов возвра-
щаются в нужном количестве в жидкости внутренней среды. Двухэтапная схема опреснения
действует и в целом организме - животные пьют морскую воду, всасывают в кишечнике ионы
и воду, а соли натрия секретируют солевыми железами, что приводит к опреснению крови.
В XIX в. и особенно в первой половине XX в. внимание физиологов занимали про-
блемы происхождения жизни в связи с выяснением развития основных физиологических
систем. После того, как было сформулировано понятие о физико-химических параметрах
внутренней среды по отношению к среде внешней, понятие о гомеостазе, можно представить
ряд этапов эволюции живого в следующей последовательности:
- протобионт,
- протоклетка,
- плазматическая мембрана,
- клетка эукариот,
-дифференцировка клеток,
- организм животного, внутренняя среда.
Особого внимания заслуживает вопрос о способах стабилизации околоклеточной сре-
ды. Поддержание Na/K градиента требует создания системы жидкостей внутренней среды,
регуляции баланса натрия, осмоляльности внеклеточной жидкости. В этом участвует не-
сколько регуляторных систем:
- нервная система,
- эндокринная система,
- аутакоиды.
Чем выше стоит организм в эволюционном ряду, тем более совершенна система ре-
гуляции, тем меньше отклонения от стандартного значения. Сравнение концентрации на-
трия в сыворотке крови у представителей различных классов позвоночных показало, что
вариабельность этого параметра у миног, лягушек, кур составляет от 1.9 до 4.8 %, у человека
она наименьшая - 0.6 %. Дело не в отличиях абсолютного значения концентрации ионов,
эти величины имеют видовую принадлежность, а речь о величинах отклонений, они зависят
от эффективности деятельности систем регуляции:
Lampetra fluviatilis CV : 4.8 %;
Rana temporana PNa:IOl, CV : 1.9 %;
Gallus domesticus P. :149, CV : 3.4 %;
Homo sapiens Pv' :I41, Na ’ CV : 0.6 %.
Следует объяснить значение этих измерений, расчетов, исследований применительно
к теме доклада. Возникновение мембраны, в частности, обеспечило стабилизацию концен-
трации ионов, физико-химических условий для всей совокупности процессов внутриклеточ-
ного метаболизма. Создание системы жидкостей внутренней среды, стабилизации их соста-
ва легли в основу прогресса царства животных. Ниже будут перечислены некоторые пара-
222
Проб y мы hpoik хождения жпиш
метры, поддержание которых на постоянном уровне обеспечивает эффективность условий
в околоклеточной среде, в среде окружения мембраны каждой клетки:
- стабилизация осмоляльности,
- волюморегуляция,
- стабилизация ионного состава,
- стабилизация pH,
- регуляция артериального давления,
- регуляция эритропоэза.
Одним из кажущихся простым, но не решенным остается вопрос о том, почему у боль-
шинства животных концентрация ионов калия в клетках и ионов натрия во внеклеточной
жидкости находится в пределах 0.1-0.5 М. Это объясняли в ряде работ тем, что древний
океан имел меньшую соленость, чем современный, и кровь позвоночных сохранила те же
значения концентрации натрия и ряда других ионов, ту же соленость (Гинецинский, 1963).
Недавно было показано, что для клеток существенно и вещество, вызывающее гиперосмо-
ляльность. Если повышение осмоляльности обусловлено NaCl выше критического значения,
то отмечается повреждение молекул ДНК, в той же концентрации мочевина не оказывает
этого эффекта (Kiiltz, Chakravarty, 2001). Из этого следует, что существует зона концентра-
ций натрия в околоклеточной среде, в которой возможно осуществление физиологических
функций без дополнительных адаптаций. У многоклеточных существ появились на одном
из этапов эволюции жидкостные фазы тела, возникли физиологические системы стабилиза-
ции объема и состава этих жидкостей.
Развитие исходных форм протоклеток с калиевой цитоплазмой в связи с появлением
натриевых водоемов только тогда имело перспективы адаптации к среде в новых условиях,
если они обретали плазматическую мембрану. Эта структура с наличием в ней натриевых
насосов и натриевых каналов создавала предпосылки для приспособления к новым ареалам,
включая пресные, солоноватые воды и океан. Выживание клеток в «натриевой» внешней сре-
де, объединение однотипных клеток привело к появлению колониальных форм, стало но-
вым этапом развития. Естественно, что у таких организмов следующим шагом могла быть
морфо-функциональная дифференцировка клеток, она давала преимущества этим организ-
мам в поиске пищи, выживании.
ДОЛЯ ИОНОВ НАТРИЯ В ОСМОЛЯЛЬНОСТИ ВНЕКЛЕТОЧНОЙ ЖИДКОСТИ
СЫВОРОТКИ КРОВИ У ЖИВОТНЫХ И ЧЕЛОВЕКА
Натрий служит основным катионом сыворотки крови, внеклеточной среды у большин-
ства животных, он доминирует в океане, морской воде. Принято, что ионы натрия с сопрово-
ждающими анионами определяют осмоляльность жидкостей внутренней среды у многокле-
точных животных. В то же время известно, что у некоторых организмов в сыворотке крови
высока концентрация мочевины (эласмобранхии), аминокислот и сахаров (высшие насекомые).
Проведенные нами исследования показали, что концентрация ионов натрия в гемолимфе, плаз-
ме крови может отличаться во много раз - от десятков ммоль/л до нескольких сотен ммоль/л.
Чтобы дать адекватный ответ на вопрос о доле ионов натрия в осмоляльности, была рассчитана
доля ионов натрия в осмоляльности сыворотки крови, гемолимфы, морской воды. В усреднен-
ной морской воде концентрация натрия равна 470.2 ммоль/л, по данным, приводимым Л. Прос-
сером (1977), осмотический эквивалент этой морской воды составляет 0.949 ммоль/л. Тем са-
мым доля ионов натрия в суммарной концентрации осмотически активных веществ составляет
49.5 %. Аналогичные расчеты доли концентрации ионов натрия в осмоляльности сыворотки
крови требуют учета высокомолекулярных веществ, занимающих часть объема жидкости.
Поэтому в гемолимфе и сыворотке крови корректно сопоставлять осмоляльность, а не осмо-
лярность, что нами и было выполнено (расчет велся в ммоль/кг Н,О, а не ммоль/л).
223
К).В. Наточин. Возникновение мембран
Полученные результаты показали, что у млекопитающих (человека, собаки, крысы)
доля натрия в осмоляльности сыворотки крови составляет 49 %, у представителей других
классов позвоночных - птиц, рептилий, амфибий, рыб - она близка 47 % (колеблется в преде-
лах от 44.6 % до 48.8 %). В то же время осмоляльность сыворотки крови этих животных и че-
ловека различается в несколько раз - от 1152 до 222 мосм/кг Н,О. Не менее поучительно, что
у беспозвоночных - у морских двустворчатых моллюсков и у пресноводных двустворчатых
моллюсков, имеющих самую низкую осмоляльность из всех изученных нами организмов
(жемчужница) - доля натрия в осмоляльности составляет тоже около 47 %. У жемчужниц
осмоляльность гемолимфы в 37 раз ниже осмоляльности гемолимфы баренцевоморских ми-
дий и воды в этом море. Это сходство доли натрия в осмоляльности у столь разных организ-
мов заслуживает специального анализа.
ФИЗИКО-ХИМИЧЕСКИЕ ОСОБЕННОСТИ ИОНОВ НАТРИЯ И КАЛИЯ
Натрий и калий имеют одинаковый заряд, несколько отличаются радиусы гидратиро-
ванного и дегидратированных ионов (табл. 6), но нет точно сформулированного представле-
ния, какой из параметров обусловил то, что калий стал доминирующим внутриклеточным
катионом, а натрий - внеклеточным, что именно определило выбор природы.
Таблица 6. Некоторые характеристики катионов
Параметры Na+ К+ Са2+ Mg2+
Атомная масса 22.99 39.1 40.08 24.3
Радиус (А) 0.95 1.33 0.99 2.75
Подвижность 5.19 7.62 3.08 2.75
Заряд 1 1 2 2
Диаметр гидратированного иона (А) 5.6 3.8 9.6 10.8
Диаметр дегидратированного иона (А) 0.98 1.33 1.06 1.81
Логически следует, что качественные характеристики катионов определили востре-
бованность этих катионов у исходных форм протоклеток, с которых начиналась жизнь на
Земле. Протоклетки, вероятно, были окружены оболочкой, отличие от современных клеток
состояло, по-видимому, в том, что внутри и вне клеток были одинаковые физико-химические
условия, оптимальные для первых проявлений жизни. Объясняется это тем, что еще не было
физиологических систем для создания ионной асимметрии. Это были первые формы жизни,
но еще не могло быть мембранных механизмов для поддержания ионной асимметрии, отли-
чий ионного состава клеток по отношению к внеклеточной среде. Клетки могли жить только
в этой среде. Для формирования таких молекулярных машин, которые служили бы для под-
держания ионной асимметрии, необходимо было возникновение, синтез мембранных макро-
молекул типа ионных насосов, ионных каналов, а затем и водных каналов, аквапоринов:
ионные каналы мембран:
Na+,
К’,
Сад
СР ;
котранспортеры:
Na - глюкоза - котранспортер,
Na'-аминокислота - котранспортер,
224
Проблемы происхоукления 'лопни
Na’. К 2СГ - котранспортер,
Na , СГ - котранспортер;
аквапорины:
- хрусталик,
-проксимальный каналец,
- собирательная трубка (люминальная мембрана),
- собирательная трубка (базолатеральные мембраны),
- собирательная трубка (базолатеральные мембраны), легкие,
- экзокринные железы,
- почка,
- сперма,
- поджелудочная железа.
РОЛЬ ПЛАЗМАТИЧЕСКОЙ МЕМБРАНЫ У МНОГОКЛЕТОЧНЫХ РАСТЕНИЙ
И ЖИВОТНЫХ
Одно из предположений может состоять в том, что предки, общие для растений и жи-
вотных, обитали в «калиевой» среде, а затем приспособились к иным по соотношению ионов
средам. В этой связи следует проанализировать три факта, на которые не обращали внима-
ния: 1) осмоляльность околоклеточной жидкости и внутриклеточной жидкости; 2) содержа-
ние калия и натрия в организме растений и животных; 3) внеклеточная жидкость, жидкости
внутренней среды как самостоятельные физиологические проблемы.
Проведенные нами расчеты на основании имеющихся данных свидетельствуют о том,
что содержание калия у растений в сотни раз превышает общее количество натрия в организ-
ме человека, у которого доля калия несколько ниже, чем натрия (табл. 7, 8).
Таблица 7. Содержание некоторых элементов (мкмоль/г
сухого вещества) у растений (Epstein, 1977)
С 40 000
к 250
Са 125
Mg 80
Na 0.4
Zn 0.3
Таблица 8. Содержание некоторых элементов (мкмоль/г
сухого вещества) у человека (Москалев, 1985)
с 31 147
к 84.3
Са 585
Mg 18.6
Na 103.1
Zn 0.082
Сопоставление приведенных выше данных позволяет придти к заключению, что име-
ются огромные отличия в соотношении этих катионов. У растений К/Na+ отношение со-
ставляет 625, у человека оно равно 0.82. Очевидно, что такие отличия обусловлены работой
ионных насосов плазматических мембран.
225
Ю.В. Ниточин. Возникновение мембран
Как и у животных, в плазмалемме растений найдены калиевые, кальциевые каналы, ани-
онные каналы, среди них имеются каналы потенциал-зависимые, активируемые растяжением,
светом и рядом иных факторов (K_rol, Trebacz, 2000). Как и в мембранах клеток животных, про-
ницаемость плазматической и вакуолярной мембран для воды регулируется при участии аква-
поринов (Kjellbom et al., 1999). В организме сосудистых растений имеются внутриклеточные,
до- и постфлоэмный, до- и постксилемный межклеточные системы транспорта жидкости.
ДИФФРЕНЦИРОВКА ПЛАЗМАТИЧЕСКИХ МЕМБРАН
Физиологическим стимулом дифференцировки плазматических мембран клеток так-
же могло быть влияние ионов натрия. Наличие в мембране натриевых каналов и натрие-
вых насосов, вероятно, было исходным пунктом для дифференцировки клеток. У исходных
клеток эти макромолекулы случайно распределены в плазматической мембране, но возмож-
ны случаи, когда будет наблюдаться скопление в одной их части ионных каналов, а в дру-
гой - ионных насосов. Это случайное перераспределение каналов и насосов может лежать
в основе появления полярной клетки, асимметричной клетки, у которой в мембране на одной
стороне будут преимущественно находиться ионные каналы, а на другой - натриевые насо-
сы, Na, К-АТФаза. Этот процесс предопределил появление клеток эпителия. У современных
организмов такая особенность присуща клеткам кожи амфибий, клеткам ряда отделов пи-
щеварительного тракта, выделительных и ряду других органов. Но в многоклеточных орга-
низмах имеются и симметричные клетки, например, эритроциты, в них в этом отношении
плазматическая мембрана однородна. Следовательно, связанные с транспортом ионов натрия
макромолекулы плазматической мембраны могли стать и источником формирования новой
морфо-функциональной организации клеток, клеточной дифференцировки, появления кле-
ток эпителия. Возникновение разных по дифференцировке клеток выражено у многоклеточ-
ных, например, кишечнополостных.
Роль ионов натрия нашла выражение не только как стимул для возникновения вместо
оболочки плазматической мембраны, отграничивающей жидкости с разным ионным соста-
вом, но появление в мембране натриевых насосов и каналов могло стать исходным пунктом
клеточной дифференцировки, а затем органогенеза. По нашему предположению, во всех этих
случаях ключевую роль играли Na-зависимые физиологические процессы. К ним относится
появление элетрогенеза мембран, формирование в плазматической мембране Na-зависимых
котранспортеров, обеспечивающих симпорт в клетку глюкозы, аминокислот, некоторых не-
органических ионов. Эти молекулярные механизмы обеспечивали новые возможности эво-
люции клетки и ее участие в построении многоклеточного организма. Детальнее рассмотрим
физиологическое значение этих Na-зависимых функций макромолекул.
\а',К'-АТФ-аза плазматической мембраны обеспечивала воссоздание калиевой ци-
топлазмы клетки в натриевой внешней среде. Возникновение электрического потенциала
на плазматической мембране этой клетки позволило в последующем реализовать ряд функ-
ций и прежде всего возможность возникновения нервной клетки, быстрой передачи информа-
ции, объединение системы управления клетками в организме как в целостном образовании.
Возможно, что синтез в такой клетке физиологически активных веществ в конечном сче-
те привел к появлению нейросекреции, возникла система гуморального контроля функций
в многоклеточном организме, эндокринная система в сочетании с нервной системой.
№’,К'-АТФ-аза, обеспечивая удаление ионов Na из клетки, создавала предпосылки для
формирования Na'-зависимого механизма поступления в клетку органических и неорганиче-
ских веществ. Ключевым звеном в системе натрий-зависимого котранспорта становится ион
Na*. При участии этого механизма формируются специализированные эпителии, сыгравшие
ведущую роль в органогенезе при формировании сорбирующих эпителиев покровов, всасыва-
ющего эпителия пищеварительного тракта, эпителиев экскреторных органов, солевых желез.
226
Проблемы нроисхож и.нпя лиши
ЗАКЛЮЧЕНИЕ
В новейших работах, касающихся эволюции живого (Diamond, 1993; Gould, 1993; Га-
лимов, 2001; Розанов. 2003; Kutschera, Niklas, 2004; Чайковский, 2006), оставался вне рассмо-
трения вопрос о неорганических факторах среды возникновения жизни. Представленные
в докладе материалы позволяют придти к выводу, что одним из высоко значимых событий
в эволюции живого, по-видимому, было появление протоклеток с калиевой цитоплазмой и по-
следующая их адаптация к внешней водной среде, в которой доминировали ионы натрия. Этот
этап был связан с таким важнейшим событием как возникновение плазматической мембраны,
способной обеспечивать ионную асимметрию клетки по отношению к внешней среде. Этот
этап эволюции предшественников животных был связан с появлением в мембране механизма
удаления ионов натрия и накопления ионов калия, что стало предпосылкой сохранения калие-
вой цитоплазмы с противоионом натрия во внешней среде. Проблемы многоклеточности была
решена разным способом у животных и растений. У животных длительный дальнейший пери-
од эволюции, вероятно, происходил в морской среде, наличие ионов натрия во внешней среде
при наличии калия внутри послужило исходным условием электрогенеза, возникновения по-
лярной, асимметричной клетки, что дало стимул для дифференцировки тканей, возникнове-
ния эпителия. Электрогенез этих клеток служил безусловной физиологической предпосылкой
появления нервной клетки, возбудимой клетки, становления нервной системы, а потому были
обеспечены исключительные преимущества у таких существ в координации функций, реак-
ции животных на сиюминутные изменения во внешней среде. Наличие нервных клеток, ней-
росекреции, аутакоидов было основой формирования эндокринных регуляторов. Натрий как
стимул образования полярной клетки, где в плазматической мембране одной стороны клет-
ки могли быть сосредоточены натриевые каналы, а в другой - натриевые насосы, обеспечил
основу развития таких функций как всасывание, пищеварение, выделение, дыхание. Форми-
рование системы жидкостей внутренней среды с доминированием в них ионов натрия стало
предпосылкой становления системы гомеостаза, физико-химического постоянства жидкостей
внутренней среды - обязательного условия для развития высших функций нервной системы.
Сказанное выше позволяет предположить, что выбор натрия как противоиона для калия дал
толчок, стимул биологического прогресса мира животных.
Работа поддержана фондом Ведущих научных школ (грант НШ № 4414-2008.4) и про-
граммой № 18 Президиума РАН «Происхождение и эволюция биосферы».
СПИСОК ЛИТЕРАТУРЫ
Галимов Э.М. 2001. Феномен жизни: между равновесием и нелинейностью. Происхождение и принци-
пы эволюции. М.: Едиториал УРСС. 256 с.
Гинецинский А.Г. 1963. Физиологические механизмы водно-солевого равновесия. М.-Л.: Изд. АН СССР. 428 с.
Москалев Ю.И. 1985. Минеральный обмен. М.: Медицина. 288 с.
Наточим К).В. 2005. Роль ионов натрия как стимула в эволюции клеток и многоклеточных животных //
Палеонтол. журн. № 4. С. 19-24.
Наточим Ю.В. 2006. Физико-химические детерминанты физиологической эволюции: от протоклстки
к человеку Рос. физиол. журн. им. И.М. Сеченова. Т. 92. № 1. С. 57-71.
ПроссерЛ. \9Т1. Неорганические ионы Сравнительная физиология животных. Т. 1. М.: Мир. С. 177-240.
Розанов А.Ю. 2003. Бактериальная палеонтология, седиментогенез и ранние стадии эволюции био-
сферы Палеонтол. журн. № 6. С. 41-49.
Салоп Л.И. 1982. История Земли в докембрии. Л.: Недра. 343 с.
Спирин А.С., Гаврилова Л. П. 1971. Рибосома. М.: Наука. 256 с.
227
К).В. Нипычин. Bothпкповснис мембран
Харди Р. 1986. Гомеостаз. М.: Мир. 81 с.
Хопленд X. 1989. Химическая эволюция океанов и атмосферы. М.: Мир. 552 с.
Чайковский Ю.В. 2006. Наука о развитии жизни. Опыт теории эволюции. М.: Т-во научных изданий КМК. 712 с.
Шноль С.Э. 1979. Физико-химические факторы биологической эволюции. IM.: Наука. 263 с.
Diamond.]. 1993. Evolutionary physiology , The logic of Life / Eds. C.A.R. Boyd, D. Noble. Oxford: Oxford
Univ. Press. P. 89-111.
Epstein E. 1977. Mineral nutrition of plants: principles and perspectives. NY: Wiley. 342 p.
Gould S. J. 1993. Evolution of organisms // The logic of Life / Eds. C.A.R. Boyd, D. Noble. Oxford: Oxford
Univ. Press. P. 15-42.
Khuri R.N. Electrochemistry of the nephron 1 Transport organs / Ed. G. Giebisch. Berlin: Springer. 1979. P. 47-95.
Kjellbom P.. Larsson C.. Johansson /. et al. 1999. Aquaporins and water homeostasis in plants Trends Plant
Sci. V. 4. № 8. P. 308-314.
Krol E.. Trehac: K. 2000. Ways of ion channel gaiting in plant cells // Ann. Botany. V. 86. P. 449-469.
Kitltz D., Chakravarty D. 2001. Hyperosmolality in the form of elevated NaCI but not urea causes DNA da-
mage in murine kidney cells // Proc. Natl. Acad. Sci. USA. V. 98. № 4. P. 1999-2004.
Kutschera U.. Niklas K.J. 2004. The modern theory of biological evolution: an expanded synthesis// Natur-
wissenschaften. V. 91. № 6. P. 255-276.
Maden B.R.. Monro R.E. 1968. Ribosome-catalyzed peptidyl transfer. Effects of cations and pH value ' Eur.
J. Biohem. V. 6. P. 309-316.
Natochin Yu.V., Chernigovskaya T.V. 1997. Evolutionary physiology: history, principles ' Comp. Biochem.
Physiol. V. 118A. P. 63-79.
Natochin Yu.V. Parnova R.G. 1987. Osmolality and electrolyte concentration of hemolymph and the problem of
ion and volume regulation of cells in higher insects// Comp. Biochem. Physiol. V. 88A. № 3. P. 563-570.
Smith H. 1943. The evolution of the kidney ' H. Smith Lectures on the kidney. Kansas: Univ. Kansas. P. 1-24.
The appearance of membranes
Yu.V. Natochin
The problem of physico-chemical factors of the environment in which life has been origi-
nated, the appearance of protocells and their envelops and subsequently of the plasma membrane
is considered. The hypothesis has been substantiated about the appearance of the first cells in the
water reservoirs in which potassium and magnesium salts are predominant, as they are necessary
for protein synthesis, while without this the cell membrane cannot be built. Later on, there occurred
adaptation of these cells to the external water environment resembling the sea water, ocean, in which
sodium salts prevailed. This stage of evolution needed function of the plasma membrane able to
provide ion asymmetry of the cytoplasm with respect to environment. At this stage of the life devel-
opment, in the plasma membrane of progenitors of animals, mechanisms of sodium ion removal and
of potassium ion accumulation began to act. The problem of development of multicellular organisms
was solved by different way in animals and in plants: in animals, there was formed a system of the
internal medium fluids that provided stable physico-chemical conditions from the external surface of
the plasma membrane. Sodium ions have become a stimulus of formation of the asymmetric, polar
cell, in whose plasma membrane on one side of the cell the sodium channels are located, whereas on
the other side - the sodium pumps; this provided development of functions of absorption, excretion,
and respiration. Formation of the internal medium fluids has become a prerequisite for establishment
of homeostasis, conditions of biological progress of the animal kingdom.
228
Прон, ir\!hi updip кож. w имя ж и <ни
Дискуссия
по пленарному докладу Ю.В. Наточина
«ВОЗНИКНОВЕНИЕ МЕМБРАН»
Председательствующий Л.М. Мухин
Л.М. Мухин: Что такое «калиевая среда»'?
Ю.В.Н.: Водоем, в котором концентрация калия (КД такая же, какая нужна для синтеза
белка, для реализации биохимических процессов, лежащих в основе жизненных явлений.
Л.М. Мухин: А где ее взять?
Ю.В.Н.: В небольших богатых солями калия водоемах, где породы могли отдавать сор-
бированные соли калия. Там могла зародиться жизнь. Там должна была быть выше концен-
трация ионов калия и магния (Mg2 ), но мала концентрация ионов натрия (Na ). Нужно пре-
валирование калия и магния над ионами других металлов.
А.В. Тутуков: Имеет ли клетка «часы»? Определяют ли они время жизни клеток?
Ю.В.Н.-. Эта проблема сейчас активно разрабатывается. «Часы» есть, они связаны с
циклами, с суточной активностью.
А.Б. Макалкин: Как растения выживают там, где очень много натрия, в солончаках, в
океане и так далее?
Ю.В.Н.: Есть множество вторичных адаптаций, в отличие от животных у растений нет
богатой натрием внеклеточной жидкости, их клетки поглощают калий.
А.С. Спирин: Любое количество калия может содержать почвенный раствор. Это интер-
стициальная вода. И еще. Для чего и почему возник биосинтез белка? Белки первично были
необходимы для создания мембраны! Важно, чтобы они были трансмембранны, могли встра-
иваться в мембрану и пронизывать ее. Для этого природой был создан специальный пептидо-
проводящий канал - белок, состоящий из десяти трансмембранных альфа-спиралей, а между
ними, как внутри трубки отверстие с пробкой. Но самое удивительное, что этот белок иден-
тичен во всех структурных деталях у архей, у эубактерий и у всех эукарий (эукариот). Это
самая древняя белковая система! Еще один момент, относящийся к тому, что жизнь началась
с мира РНК. Калий, благодаря своему ионному радиусу, меньше всего нарушает структуру
воды, он встраивается в ее структуру, а вода встраивается в структуру РНК. И ион калия
(но не натрия!) встраивается в структуру РНК. Натрий же ее дестабилизирует, разрушает.
Пространственная структура РНК совместима с калием, но не с натрием. Калий-натриевая
селективность-антиселективность построена на различиях в ионном радиусе. Магний тоже
необходим для пространственного структурирования РНК. Это связано не только с ионным
радиусом, но еще с координационным числом магния как двухвалентного катиона.
Ю.В.Н.: И это очень хорошо дополняет то, что я говорил.
С.В. Рожнов: Не могли бы Вы сказать несколько слов о кальциевом балансе? Не была
ли кембрийская скелетная революция следствием какого-то смещения этого баланса?
Ю.В.Н.: Кальций в организмах существует в разных формах: ионной и связанной (с
белком, с другой органикой). Концентрация ионов кальция в клетках на четыре порядка
меньше, чем в околоклеточной среде. Он оказался крайне важным регулятором разных кле-
точных функций. Наши исследования показали, что концентрация ионизированного каль-
ция в плазме крови относится к числу самых точно поддерживаемых констант в живом орга-
низме. Системы регуляции обеспечивают перенос избытка кальция в скелет либо удаление
из организма.
229
Пн>ь.., I//; ...имжии..!. ,M.: Ill/li РАН. 2009. С. 2.10-243
ПЕРВЫЕ ЭКОСИСТЕМЫ НА ЗЕМЛЕ
© 2009 г. Г.А. Заварзин
Институт .микробиологии и м. С.И. Виноградского РАИ
zavarzin@inmi.host.ru
Предположения о происхождении жизни на Земле должны исходить
из представления, что обитаемость предшествует обитанию. Требуется со-
ставить представление о возможной жизнеобеспечивающей экосистеме,
прежде чем строить предположения о первом организме. Экосистема обя-
зательно должна включать первичных продуцентов-автотрофов и гидро-
генотрофия представляется адекватным типом физиологии для примитив-
ных экосистем. Рассматривая жизнь на Земле, следует различать возникно-
вение организмов in siiu и появление организмов извне и с космическими
телами в рамках концепции жизни как универсального феномена Вселен-
ной. В случае транспорта жизни с космическими телами нет ограничения
на перенос сообщества, а не единичной клетки. В случае транспорта со-
общества с «грязным льдом» проблема состоит в соответствии сообщества
и его экосистемы на родительском теле с условиями на примитивной Земле,
а не в функциональной дивергенции от первичного предка. Последующие
события находятся в рамках наблюдаемой палеонтологически эволюции
и могут быть описаны как биогеохимическая сукцессия без дополнитель-
ных спекуляций.
Рассматриваемая проблема происхождения жизни относится к виртуальной области
гипотетических построений, которые должны быть непротиворечивыми и соответствовать
фактическим данным, полученным из актуалистического материала. Гипотетические пред-
ставления о происхождении жизни развивают представители самых разных дисциплин, ча-
сто ограничивающие свое внимание какой-либо узкой проблемой, например, абиогенным
происхождением нуклеотидов. В таких построениях авторы исходят из некоторых презумп-
ций, редко четко сформулированных, но чаще находящихся в области невысказанных по-
ложений, или, хуже, игнорируемых для простоты картины. Поэтому внимание должно быть
обращено не на детали, иногда очень впечатляющие своей изысканностью, а на исходные
посылки.
Относительно предыстории биосферы есть серия упрощенных представлений, как бы
очевидных и само собой разумеющихся, которые, находясь за рамками собственно исследо-
вания, ограничивают поле логических построений, и, соответственно, поисков. Будучи не-
явными и вместе с тем базовыми, они оказывают определяющее влияние на поле возможных
гипотез. Многие из этих догм у неспециалиста восходят к усвоенным чуть ли не в школе
230
Пробиемы iipi)1!cxo}k;if.hii}i жншн
представлениям. Такие упрошенные представления о развитии биосферы можно упорядо-
чить по логической альтернативной схеме. Первая по порядку альтернатива касается рамок
системы:
I. Земля - замкнутая по материальному балансу система, открытая в космос только
по энергии, получаемой от Солнца;
11. Земля - открытая в космос по материальному балансу система.
Альтернатива 1 относится ко всей наблюдаемой геологической истории Земли
в 3.8 млрд. лет. Замкнутость системы ограничивается потоком космической пыли в верхние
слои атмосферы, оцениваемой в 1012 г/год. Этот порог можно принять при оценке матери-
альных потоков в биосфере. Для каждого элемента в каждый момент существования соблю-
дается материальный баланс. Для биосферы есть, однако, существенное ограничение. Био-
сфера находится внутри географической оболочки Земли. Эту оболочку уже нельзя считать
замкнутой, поскольку происходит обмен с глубинными слоями, обусловленный тектоникой,
и потоки между географической оболочкой и корой необходимо принимать во внимание
для значительных промежутков времени, условно говоря - миллионолетних. Говоря о ба-
лансе, следует учитывать, что на всем протяжении своей истории Земля открыта в космос
по энергетическому балансу.
Альтернатива II определенно относится к периоду аккреции и догеологической исто-
рии Земли в первый миллиард лет ее существования. В этот период Земля открыта по мате-
риальному балансу и формируется из материала космического происхождения. Поскольку
биосфера находится в пределах географической оболочки, то начало жизни на Земле следует
отнести к периоду формирования этой оболочки.
Каждая из альтернатив достаточна для определенных условий и оказывается недоста-
точной для других. Вместе с тем, существуют и нарушения альтернатив, и их комбинации,
требующие компромиссов в подходах.
В рамках альтернативы I развивалось все представление о биосфере Земли и, в осо-
бенности, о содержащемся в биосфере биоразнообразии. Такое представление достаточно
для описания настоящего в рамках согласованной, «целесообразной» системы. Слово «целе-
сообразная» вызывает множество споров из-за разного его толкования. В действительности
речь идет о наблюдаемой в природе взаимно согласованной системе явлений. В XV11I в. для
этого эмпирического обобщения употребляли слово Ubereinstimmung. Оно точнее отражает
наблюдаемые факты, чем даже Zweckmapigkeit, отражающее причинную зависимость. За-
метим, что в английском языке не оказалось соответствующего слова. Взаимная согласован-
ность включает понятие естественного отбора - исключение несогласованного.
Для расширенного, включающего историю и прошлое, понимания системы необходимо
включение времени и понятия развития («эволюции»). Развитие обусловлено внутренними за-
кономерностями системы; развитие системы может быть упорядочено последовательностью
возникновения от простого к сложному. Традиционно предполагается, что для биоразно-
образия механизмом возникновения служит дивергенция единичного объекта, позволяющая
построить универсальное корневое дерево - порядок. Механизм ветвления обусловлен мута-
цией и отбором и рассматривается в традиционных взглядах на эволюцию живых существ.
Для геосферы принимается несколько иное представление о развитии на основании вну-
тренних сил с эволюцией Земли от первоначального состояния под действием двух факторов:
А. Актуалистической интерпретации изменений географической оболочки под дей-
ствием постоянно действующих факторов с ведущим процессом «выветривание-седименто-
генез». Этот процесс обусловлен циркуляцией подвижных сред - атмосферы и гидросферы -
с основным вкладом тепловой энергии Солнца. Такая интерпретация достаточна для отно-
сительно малого геологического времени и безусловно достаточна для настоящего в смысле
«Глобальных изменений природной среды и климата».
231
/'.А. Заварзин. Первые жосис гемы на Земле
К постоянно действующим геологическим факторам в другой временной шкале сле-
дует отнести дрейф континентов, приводящий к геологическому рециклу и метаморфизму
пород, но этот периодический процесс имеет движущей силой внутренние причины, обу-
словленные энергией Земли. В особенности это относится к состоянию дна океана.
Б. Однонаправленная динамика глубоких слоев Земли от первоначально нагретого
(или временно разогревшегося) к остывающему состоянию с характерными тектоническими
циклами - волнами. По отношению к биосфере и вмещающей ее географической оболочке
тектоника воспринимается как внешний возмущающий процесс. Она находится в иной вре-
менной шкале. Эти представления необходимы для понимания эволюции биосферы, но на-
сколько они согласуются со временем происхождения жизни?
Представление о земной системе как замкнутой по материальному балансу очевидно
достаточно и необходимо для настоящего, но оно так же необходимо и для описания каждого
состояния прошлого. Более того, установление материального баланса служит критерием
достаточности подхода. Характерное время составляет интервалы в десятки миллионов лет
в зависимости от доступной детализации.
Достаточность сложившихся согласующихся взглядов на саморазвитие замкнутой си-
стемы Земли не вызывает сомнений, ни для геологической составляющей, ни для биологиче-
ской. Согласование взглядов достигается ограничением рассматриваемого периода времени,
вне которого находятся внешние к существующей системе факторы. Справедливость такого
подхода основывается событиями фанерозоя, и понимание его постепенно распространяется
на все более ранние периоды протерозоя. Ранние периоды развития прокариотной биосферы
остаются в области неопределенности, но описываются именно в условиях замкнутого мате-
риального баланса.
Вторая альтернатива (И) основана на представлении о Земле как открытой в кос-
мос по материальному балансу системе. В материальный баланс не входит постоянное
поступление в верхние слои атмосферы космического материала порядка I012 г/год; здесь
рассматриваются величины гораздо большего масштаба и, следовательно, необходимо
иметь количественную границу. Но проблема состоит не только в количестве, но и в не-
равномерности поступления. Крупные импакты даже меньшего количества материала
рассматриваются как нерегулярные катастрофические события в рамках альтернативы 1,
допустимой для периода ранее 3.8 млрд, лет назад. Напротив, предшествующий период
аккреции Земли и последующая история импактов рассматривает Землю как открытую
в космос по материальному балансу систему. Эти события находятся в рамках астрофизики
и астрономической шкалы времени, хотя собственно аккреция занимает очень короткий
промежуток. Описывается этот период в рамках астрофизики и ее, преимущественно рас-
четных, представлений. Из рассматриваемых здесь альтернативных гипотез существен-
ны две: 1) гипотеза гомогенной «пылевой» аккреции и 2) гипотеза гетерогенной аккреции
относительно крупных тел со значительным вкладом импактных событий. Эти взгляды
не следует противопоставлять как несовместимые.
Начало жизни на Земле может быть представлено в виде двух альтернатив:
I) Появление первых организмов - бактерий
или
2) Появление первых экосистем, включающих организмы и среду их обитания.
В такой постановке вопроса ложным является союз «или». По умолчанию обычно под-
разумевают появление первых организмов. Но они не могут существовать вне среды своего
обитания, с которой они находятся в состоянии обмена веществом и энергией. Поэтому с са-
мого начала следует рассматривать экосистему, а не организм в отдельности, как это обычно
пытаются делать, отвлекая от сути проблемы детализирующими вопросами о составе орга-
низма и происхождении его частей-компонентов. Отсюда следует другой вопрос:
232
Проб, и мы происхож ц.ния жншп
Что предшествует: условия обитания или же биота? На него нужно дать однозначный
ответ:
обитаемость предшествует обитанию.
Условия обитания представлены географической оболочкой планеты.
Географическая оболочка вмещает биосферу.
Географическая оболочка есть порождение геосферы.
Начало жизни связано с бесконечными малопродуктивными спорами об определении
жизни. Очевидно, что жизнь существует, когда есть первые организмы. Можно ли назвать
жизнью то, что было до того, относится к области неопределенности. Если есть организмы,
то есть и жизнь. Но попытка ограничить проблему такой постановкой явно несостоятельна,
хотя и излюблена экспериментаторами. Организм в принципе способен существовать только
в рамках экосистемы. Эти два понятия связаны между собой в единство и одно немыслимо
без другого. Организм и экосистема связаны между собой обменом, без которого нет жиз-
недеятельности. Экосистема появляется с появлением жизнеспособного организма. Абстра-
гирование от экосистемы полностью лишает смысла все построения о природе первичных
живых существ. Их можно сразу же откладывать в сторону как недостаточные.
Происхождение жизни имеет два аспекта. Для начала жизни на Земле можно постро-
ить дилемму, описывающую наблюдаемые или предполагаемые события: возникновение
или появление жизни? Оба варианта дают в свою очередь ряд возможностей, которые следует
обсуждать.
1. Возникновение жизни.
1.1. Возникновение жизни на Земле.
1.2. Возникновение жизни на ином космическом теле - «parent body» - молчаливо пред-
полагающее всеобщность жизни во Вселенной.
1.2.1. Возникновение жизни в пределах Солнечной системы.
1.2.2. Возникновение жизни вне Солнечной системы.
2. Появление жизни на Земле.
2.1. Панспермия - перенос жизни из космического пространства на Землю.
2.1.1. Средство транспорта - космическая пыль, метеориты, кометы.
2.1.2. Время транспорта на астрономические расстояния.
2.1.3. Способ существования во время транспорта.
2.2. Совместимость условий на ином космическом теле («parent body») с условиями
на примитивной Земле.
Появление жизни на Земле есть эмпирический факт, для которого установлено при-
близительное время этого события не позже 3.5 млрд, лет назад. Очевидно, что появление
жизни не рассматривает возникновения жизни вообще. Более того, возникновение жизни на
ином космическом теле, возможно, при иных условиях, с необходимостью требует переноса
на Землю и описания его возможностей. В течение XX столетия строились разные умо-
зрительные гипотезы возникновения жизни на Земле, иллюстрируемые лабораторными мо-
делями. В настоящее время эти гипотезы встречают две трудности. Во-первых, краткость
времени для возникновения жизни от периода аккреции до появления первых экосистем
с микроорганизмами. Период, в который можно поместить происхождение жизни, сокраща-
ется от 500 млн. лет до 200 млн. лет по разным оценкам. Достаточно ли это время для по-
явления первых организмов? Во-вторых, представление о существовании внеземной жизни
сейчас приобретает силу молчаливого консенсуса, и не только для оправдания дорогостоя-
щих поисков жизни вне Земли с помощью космических аппаратов. Оно получает поддержку
в находках бактериоморфных микрофоссилий в определенной группе метеоритов. Отрицать
их существование невозможно, это факт, но интерпретации могут быть разными. Сходство
микрофоссилий в метеоритах с заведомо биогенными микрофоссилиями, хорошо изучен-
233
Г.А. Закир:шн. Первые экосистемы на Земле
ними в бактериальной палеонтологии, очень велико. Абиогенное происхождение бактерио-
морфных минеральных образований требует экспериментального обоснования, а не общих
слов и субъективного нежелания признать их биогенную природу.
Биологами географическая оболочка Земли как открытая в космос не рассматривается.
«Пришельцы из космоса» для эволюции макроорганизмов не относятся к числу рассматри-
ваемых возможностей. Это справедливо и для прокариотной биосферы вплоть до ее воз-
никновения. Биогеохимическая сукцессия представляется достаточным обоснованием для
объяснения крупномасштабных явлений в биосфере и изучается на фактическом материале.
Привнес извне микроорганизмов другого рода, чем существующие, например, архей в бакте-
риальную биосферу, не относится к числу всерьез рассматриваемых. Исключение составляет
гипотеза панспермии, касающаяся прокариот, прежде всего на стадии их появления. Она ста-
новится альтернативой земной форме биопоэза - происхождения организмов саморазвитием
химической системы.
Основанием для нового всплеска интереса к космическим источникам служат:
- установление присутствия на космических телах органических веществ, согласую-
щихся до известной степени с биохимией современных организмов;
- установление наличия морфологически распознаваемых микрофоссилий или мине-
ральных псевдоморфоз по телам микроорганизмов на космических телах - метеоритах;
- выявление разнообразия космических тел с особым вниманием к ледяным телам.
Предположение о вечности жизни во Вселенной допускает возможность ее появления
на Земле уже на стадии аккреции. По времени такое предположение проходит, поскольку
жизнь регистрируется к концу метеоритной бомбардировки. Для аккреции, которая проис-
ходит очень быстро, с набором массы в течение около 100 млн. лет, есть разные варианты
(рис. 1), которые можно интерпретировать и как гипотетические стадии от пылевого обла-
ка до планетезималей, в число которых с все большей убежденностью включают кометы
как крупные глыбы «грязного льда». Из представленных схем в нашем рассмотрении наи-
больший интерес имеет последняя. Основным источником летучих на планетах-гигантах
и их регулярных спутниках предполагаются ледяные планетезимали, образовавшиеся
в околосолнечном протопланетном диске в зоне питания этих планет и содержавшие в своем
составе конденсаты и клатраты различных газов. Эта гипотеза противопоставляется тра-
диционной картине тепловой дегазации, которая имеет место на более поздних стадиях
и наблюдается как факт. «Грязный лед», а затем не испарившаяся гидросфера предоставляют
возможное место изначального поступления микробиоты.
Аккреция
Компоненты Земли после аккреции
Космическая
пыль
Газовый
материал
Желе шый
материал
Ледяной
материал
Каменный
материал
Рис. 1. Варианты (или стадии) аккреции, ведущие к образованию гидросферы как условию развития
биосферы. Гидросфера образуется либо при ударной дегазации, либо при привносе крупных ледяных
тел. Условия для развития биосферы отсутствуют (а). Заключительная стадия аккреции с форми-
рованием в результате дифференциации атмосферы, гидросферы, поверхностных слоев литосферы,
входящих в географическую оболочку планеты как вместилища биосферы (б).
234
rii'oii.iEMhi гп'()!1С\ож:1К1111я жтпн
Time (Ga from present)
Рис. 2. Коридор обитаемости для планет земной группы, определяемый возможностью фотосин-
теза (Franck S., Zavarzin G.A., 2004. What are the necessary conditions for origin of life and subsequent
planetary life support system? // Dahlem Konferezien 91. Earth System Analysis for Sustainability /
Eds. H.J. Schellenhuber. P.J. Crutzen, W.C. Clark. M. Claussen, H. Held. MIT Press. S. 73-90).
В последнем случае гидросфера как часть географической оболочки появляется с са-
мого начала существования планеты Земля. Отсюда возможность изначального поступления
на Землю зародышей микроорганизмов более не представляется полностью исключенной.
С такой последовательностью событий не согласуется предполагаемая термическая исто-
рия Земли с ее стерилизующим разогревом около 4 млрд, лет назад. Впрочем, этот эпизод,
введенный геологами для формирования коры, для географической оболочки Земли нельзя
рассматривать как непротиворечивую картину; одним из осложнений служит неизбежный
«убегающий парниковый эффект» из-за состава атмосферы. Чтобы избавиться от СО,, пред-
полагается быстрая реакция с силикатами кальция и магния, возможная при высокой тем-
пературе, но при этом не рассматриваются пары воды в атмосфере. СН4 как существенный
компонент атмосферы в пересчете на молекулу в 20 раз увеличивает парниковые пробле-
мы. Кроме того, термический эпизод значительно сокращает время для возникновения жиз-
ни, оставляя слишком короткое окно для ее появления. Для Земли на основе парникового
эффекта и светимости Солнца был рассчитан предполагаемый «коридор обитаемости» (рис. 2),
из которого следует, что Марс вышел из этого коридора, а Венера не находилась в нем.
Как бы то ни было, после аккреции появляется гетерогенная земная система (рис. 3),
подвергающаяся дальнейшей дифференцировке. Географическая оболочка этой системы
представляет слоеный пирог между атмосферой и литосферой. Атмосфера получает энергию
от Солнца в виде солнечного излучения. Кроме тепловых реакций, ведущих к циркуляции,
в ней происходят фотохимические реакции. Поскольку химический состав примитивной
атмосферы неизвестен, то моделировать происходившие в ней реакции очень трудно. Заведо-
мо предполагается присутствие паров воды. Под действием солнечного излучения она долж-
на была диссоциировать, и продукты ее фотохимической диссоциации давали окислитель-
ные и восстановительные эквиваленты. Водород диссипировал в космическое пространство,
оставляя окислительные эквиваленты. На этом факте основано представление об «окисли-
тельной смерти планет». Гипотеза поддерживается сдвигом отношения D/Н в атмосфере
2.35
Г.А. hieupnin. Первые жосиосмы на Земле
атмосфера
Географическая
оболочка
диссипация Н и
окислительная
Фотолиз воды смерть атмосферы
Н2О -> н + он . фотохимические
Биота окислители 1 ОН + Р
восстановители
Термолиз воды
FeMgSiO4 + Н2О = 1/3Fe2O4 + 1/ЗН2 +
1/3 MgSi2O, + 1/3SiO2
Серпентинизация оливина
геосфера
Рис. 3. Вода в географической оболочке планеты как место разви-
тия биосферы. Влияние абиогенных факторов на состояние гео-
графической оболочки и ее трансформация биотой.
Венеры в пользу дейтерия. Для биолога существенно то обстоятельство, что атмосфера мог-
ла служить источником вымываемых из нее окислительных эквивалентов, продуктов ста-
билизации ОН. Биологи гипнотизированы идеей об отсутствии защитного экрана от уль-
трафиолета при отсутствии О. в примитивной бескислородной атмосфере. Проблема эта
в значительной степени надуманная, поскольку ультрафиолет может поглощаться разными
веществами, в особенности Fe(lll), о высокой концентрации которого на примитивной Земле
есть прямые свидетельства в виде железорудных формаций первых осадочных отложений.
Таким образом, обитатели гидросферы от УФ были защищены, и его прямое воздействие на
микроорганизмы как обитателей микрониш не является критическим фактором. Мощность
фотохимического «окислительного дождя» может быть недостаточной для количественного
обоснования предполагаемых процессов, но не может быть исключена из рассмотрения.
В литосфере происходит термическое разложение воды при контакте с нагретыми гор-
ными породами. Окислы остаются на месте, а водород выходит при дегазации в область гео-
графической оболочки. В результате биота оказывается в слое между источником Н, внизу
и возможным источником окислителей сверху. Для обмена микроорганизмов это оптималь-
ная ситуация, когда они располагаются в геохимическом барьере и могут использовать энер-
гию окислительно-восстановительных реакций.
Коль скоро принято представление о экосистеме как исходной для рассмотрения,
то необходимо рассматривать и существующее в ней сообщество. Сообщество основано
на трофической пирамиде, в основании которой лежат продуценты - автотрофные организ-
мы. Пищевая пирамида изображается в виде треугольника. Каждая ступень в трофической
пирамиде обусловливает снижение биомассы на каждом последующем уровне примерно
на I0 % - это очень условная величина, но она дает представление о необходимой ширине
основания. Итак, чтобы получить представление о сообществе, следует построить пирамиду
с указанием первичных продуцентов и источников энергии для них. Поскольку первичные
продуценты - автотрофы, то проблемы источников углерода не возникает, они поступают
из геосферных резервуаров. Проблему составляют источники энергии. Первичные проду-
центы для сообщества могут быть представлены немногими массовыми видами. Следующая
группа проблем возникает в связи с понятием биоразнообразия. О нем существует вполне
устоявшееся и обоснованное фактами представление о возрастающем разнообразии по мере
продвижения к высшим уровням и усложнения во времени.
236
ПРОЬ.П МЫ ПРОШ XOA.-ILHHM Ж1ПНН
Филогенетическая
система
Функциональная
система
Универсальный
общий предок
Трофическая пирамида
Автономное
сообщество
Рис. 4. Филогенетическое и функциональное развитие биоразнообразия. Мо-
нофилетическое и полифилетичсское построения. При монофилетическом
построении биоразнообразие возникает в результате дивергенции потомков
общего предка соответственно расширяющимся возможностям экологиче-
ских ниш. При полифилетическом построении изначальным является само-
поддерживающееся сообщество и существенную роль в развитии биоразно-
образия имеют комбинаторные генетические события.
В результате возрастание биоразнообразия можно отразить в виде треугольника, опи-
рающегося на вершину. Филогенетики изображают это представление в виде ветвящего-
ся дерева, исходящего из общего корня - универсального общего предка, как изображено
на рис. 4. Экологи возражают - первичное сообщество должно включать функционально раз-
личные организмы, иначе оно не может существовать на Земле сколько-нибудь длительно
из-за исчерпания субстратов и накопления продуктов жизнедеятельности. Если предпола-
гать панспермию, то нет никаких оснований, кроме предвзятой идеи, исходить из заноса
единичного организма, а не некоего минимального сообщества.
Для экосистемы существует ряд вариантов потоков вещества (рис. 5). Простейший случай
представляет однонаправленный проток вещества от источника к стоку. Результатом оказыва-
ется исчерпание резервуара источника и наполнение резервуара стока. Пополнение резервуара
Рис. 5. Варианты эволюции экосистем: а- однонаправленная с накоплением продукта и исчерпанием
резервуара; б- циклическая замкнутая по материальному балансу; «-сукцессия, не полностью зам-
кнутая по материальному балансу с изменением условий среды обитания.
237
ГА. Закирзин. Первые экосистемы на Земле
источника происходит за счет внешних абиогенных процессов, например, дегазации. Примером
таких локальных условий могут служить газогидротермы в областях вулканизма и населяющие
их микробные сообщества с хемосинтезом как источником первичной продукции С .
Противоположностью проточной экосистеме служит замкнутый цикл вещества,
в котором продукты разлагаются («минерализуются») деструкторами и вновь используют-
ся в обмене сообщества за счет внешней энергии. Этот вариант относится к сообществам
на основе фотосинтеза. Сообщество с замкнутым циклом вещества автономно и может су-
ществовать неограниченно долго в замкнутом пространстве при поступлении энергии извне.
Если судить по циклу органического углерода, то биосфера по большому счету представля-
ет именно такую автономную экосистему, где продукция уравновешивается деструкцией.
Однако могут быть и малые автономные сообщества - аквариум в запаянном стеклянном
сосуде может представлять такой устойчивый микрокосм в течение многих лет.
В действительности полная замкнутость цикла не реализуется и происходит нако-
пление продуктов как в прямых реакциях для захороненного органического углерода, так
и в биотически опосредованных реакциях накопления минеральных веществ, наиболее на-
глядно - карбонатов как биогенных осадочных пород. В результате реализуется биогеохими-
ческая сукцессия - основная движущая сила эволюции биосферы как целостной системы.
Ни «мир РНК», ни мир вирусов не выдерживают критерия автономности, обуслов-
ленного материальным и энергетическим балансом в экосистеме. Можно представить себе
«мир РНК» заключенным внутрь клеточной оболочки как важную, или важнейшую, часть
организма, в котором ДНК играет роль депо, а белки - роль основных катализаторов необхо-
димых реакций, связанных прежде всего с мембраной. Но самая попытка подменить целое
его частью заведомо обречена. Тем более не приходится рассматривать мир вирусов в роли
продуцентов. В связи с этим хотелось бы сделать оговорку. Громадный объем материала
о биологии микробной клетки поступает из области медицины. Медицинскую микробиоло-
гию, имеющую дело с паразитами, трудно представить себе как единую систему, в то время
как хозяин, включая его иммунную систему, представляется целостной организацией. Меди-
цинская микробиология представляет набор частных случаев, каждый из которых обуслов-
лен вмешательством паразита в какой-либо узел целостной системы. Поэтому она служит
плохой отправной точкой для исследования первичной биосферы, как бы ни были упрощены
рассматриваемые ею представители мира прокариот.
Самоподдержание биосферы основано на биогеохимических циклах, в первую оче-
редь биогенных элементов С, N, Р и сопряженных с ними циклов других элементов - серы,
железа, кальция. Биогеохимические циклы катализируются микроорганизмами, практиче-
ски без участия других форм жизни, обмен которых однообразен. Циклы не могут осу-
ществляться одним организмом, а требуют кооперативного взаимодействия функционально
различных групп организмов. Поэтому сообщество с самого начала следует рассматривать
как кооперативную систему.
Ведущим циклом биосферы является цикл органического углерода. Вследствие посто-
янства состава биомассы с ним сопряжены жестким количественным отношением циклы
других биогенных элементов. Приоритеты микробной биогеохимии определяются масшта-
бами, то есть количеством вещества, вовлеченного в процесс. Нижней границей служит поток
10’- г/год (космическая пыль), значимы потоки 1015 г/год (~1017 г/год для Сорг), динамические
резервуары >10'* г. Крупномасштабные изменения в резервуарах ведут к биогеохимической
сукцессии, составляющей основу эволюции биосферы. Таким образом, эволюция биосферы
не сводится к эволюции биоты, вложенной в эволюцию геосферы, точнее - географической
оболочки. Представление о «физиологии Земли» с созданием оптимальных для биоты усло-
вий основано на преувеличении значения обратных связей в биосфере по сравнению с пря-
мыми. Эволюция геосферы, даже в узком смысле географической оболочки, зависит от вну-
238
Проблемы иропсхожлелшя жтни
тренних причин теплового режима Земли и от внешних астрономических причин, прежде
всего от Солнца, от тепловой машины циркуляции атмосферы и океана. Биота обусловливает
трансформацию веществ, первоисточником которых служит геосфера.
Виртуальная эволюция биоты строится на реконструкции филогении по сходству
гена(ов). Сейчас она для низших организмов связана исключительно с операционной систе-
мой идентификации по малой рибосомальной РНК. В рассматриваемом функциональном
аспекте возможности существования в экосистеме разработанная филогенетическая клас-
сификация дает лишь некоторые корреляции, к которым приходится относиться с большой
осторожностью. Так, утверждение, что геохимически важный процесс нитрификации осу-
ществляется протеобактериями, оказывается поколебленным открытием нитрифицирующих
архей. Если это крайний случай отсутствия корреляции функции и, соответственно, спосо-
ба существования, то точно такая же ситуация вырисовывается в большинстве экосистем
для большинства их обитателей. Геном представляется мозаичным и функциональные гены
слабо коррелируют с генами внутреннего хозяйства. Здесь причина в исходной посылке:
для филогенетических построений были избраны гены, в минимальной степени зависящие
от внешних условий, не адаптивные. Но оборотной стороной такого выбора оказывается, что
существенные для существования свойства не отражаются филогенетической классифика-
цией. Она превосходна для целей идентификации, но мало полезна для выяснения возмож-
ности существования носителя данного гена в природных условиях. Функциональные роли
в экосистемах жестко определены системными требованиями, в то время как состав испол-
нителей может меняться. Отсюда следует парадоксальное заключение, что филогенетиче-
скую классификацию нельзя считать единственно «естественной». Если считать естествен-
ными свойства, обеспечивающие существование в природе, то к ним следует отнести в пер-
вую очередь обеспечивающую дискретность особи форму, обусловливающие энергодающий
обмен реакции, способ самовоспроизведения. В результате, по крайней мере, в мире прока-
риот, понятия филогении и эволюции резко расходятся. Словесная подмена одного аспекта
другим ведет к глубокому взаимонепониманию. Эволюция связана со способом существо-
вания организмов в меняющейся экосистеме. Она регистрируется палеонтологическими
в широком смысле - данными о временной последовательности событий.
Современная виртуальная эволюция микробов строится на реконструкции филогении
по сходству гена(ов) без учета экосистемных требований. Виртуальная эволюция микроб-
ной биосферы в прошлом строилась на сравнительном материале и представлении о после-
довательности обмена микробов от простого к сложному, от брожения к фотоавтотрофии.
Для трофической пирамиды в основании должна лежать автотрофия.
Реальная эволюция ранней биосферы начала выясняться с появлением бактериальной
палеонтологии. Если формы микроорганизмов могут быть идентифицированы на основании
микрофоссилий - и в этом отношении накоплен обширный материал, особенно для циано-
бактерий, - то их обмен остается предметом косвенных доказательств, наиболее убедитель-
ными воспринимаются соотношения изотопов в продуктах.
Деятельность микробиоты оценивается по производимым ею химическим изменени-
ям. Геохимическая деятельность палеомикробиоты фиксируется в сингенетичном минера-
лообразовании. Наибольшее внимание привлекли минералы сульфидов как явно связанные
с деятельностью специфической группы сульфидогенов с сероводородом как продуктом
энергетического обмена. Но вообще следует признать, что микробная минералогия еще
не получила должного развития - она оказалась слишком междисциплинарной. Микробио-
логи пытались ограничиться гомогенными химическими реакциями, по возможности игно-
рируя твердую фазу. Поскольку микробы живут в воде, такое представление было достаточ-
ным. Но при переходе к биопленкам как важнейшей форме существования микробного мира
игнорирование твердой фазы оказалось явно необоснованным.
239
Г. Л. Заварит. Первые экосистемы на Земле
Химическая деятельность микробиоты включает как прямой катализ реакций,
так и опосредованное изменение среды, в которой минеральные соединения принимают со-
стояние, соответствующее термодинамическим полям устойчивости соединений. На это
накладываются закономерности кристаллизации, в значительной степени зависящие от со-
стояния среды, для которого немаловажное значение имеют бактериальные слизи - необхо-
димый компонент биопленок. В общем микробное минералообразование следует последо-
вательности: растворенные соединения коллоиды —> аморфные осадки —>• «эфемерные»
(незрелые) минералы —> кристаллизация в устойчивое состояние. Формирование условий,
соответствующих полям устойчивости минеральных соединений, не обязательно связано
с прямым катализом реакции микроорганизмами.
Поскольку экосистема зависит от первичных продуцентов, то необходимо рассмотреть
два варианта первичной продукции органического вещества: хемосинтез и фотосинтез.
Хемосинтез осуществляется за счет окислительно-восстановительной реакции, кото-
рая дает энергию для ассимиляции СО,. Существовало убеждение, что хемосинтез обладает
слишком малой эффективностью по продукции биомассы. Это представление было разру-
шено открытием глубоководных «оазисов», где первоисточником жизни служит хемосинтез,
основанный на окислении сероводорода. При этом, однако, обычно остается за рамками вни-
мания тот факт, что окислитель поступает с конвекционным потоком воды океана с высоким
содержанием растворенного О, фотосинтеза. Следовательно, хемосинтез на дне океана ока-
зывается вторичным по отношению к фотосинтетическим оксигенным продуцентам. Точно
такая же ситуация имеет место по отношению к СН4 окисляющим метанотрофам как пер-
вичным продуцентам - для окисления и ассимиляции СН4 они нуждаются в О,. Анаэробное
окисление СН4 консорциумами микроорганизмов пока не принимается как существенный
источник биомассы, хотя на выходах газа на дне моря образуются мощные биопленки мета-
нотрофных сообществ.
Наибольший интерес представляют Н, окисляющие гидрогенотрофы, для которых сер-
пентинизация изверженных пород служит постоянным источником водорода, по-видимому,
на всем протяжении геологической истории. Это тот случай, когда биота в экосистеме мо-
жет существовать на протоке и замыкание трофической цепи в цикл не представляется не-
обходимым. Окисление Н, не ведет к накоплению токсических продуктов. Самоотравление
продуктами отсутствует. Но поступление Н, при реакции с перегретыми породами предпо-
лагает развитие термофилов при высокой температуре. Поэтому гидрогенотрофы с абиоген-
ным источником водорода вынуждены идти в глубинную биосферу, прижимаясь к ее ниж-
ней границе, определяемой зоной конденсации паров воды. Гипертермофилы рассмотрены
в отдельном докладе.
Отсюда при рассмотрении первичных микробных сообществ следует особое внима-
ние уделить энергетике гидрогенотрофов, основанной на ключевом ферменте - гидрогеназе.
Гидрогеназы, осуществляющие обратимую реакцию Н, <-> 2Н’ + 2е_, представляют энзима-
тическую основу для энергетики и хемосинтеза, и аноксигенного фотосинтеза. Отсюда мы
приходим к важному выводу:
гидрогенотрофия как вероятный исходный тип метаболизма первичных продуцентов.
Но гидрогеназа представляет только половинку электрохимической ячейки, поскольку
для ее работы необходим окислитель. Для глубинной биосферы набор возможных окислите-
лей очень ограничен.
Из перечисленных в табл. I и 2 особое значение по возможным масштабам процесса
следует уделить метаногенам, сульфидогенам, железоредукторам. Из них последние стали
объектом особенно интенсивных исследований в настоящее время. Значимость их для рас-
сматриваемой проблемы возрастает в связи с тем, что в древнейших осадочных породах
на Земле формации Исуа представлены окислы железа. Обозначенный знаком вопроса
240
Проблемы происхождения жииш
Таблица 1. Продуценты в прокариотной биосфере: гидрогенотрофный
хемосинтез
акцептор продукт Название группы хемосинтезирующих бактерий или архей
о2 Н2О Водородные
со2 СН4 Метаногены
со2 СН3СООН Гомоацетатные
So H2S Сероредукторы
H2SO4 H2S Сульфатредукторы
NOx NH3 Аммонийобразующие
Fe(III) Fe(II) Железоредукторы
H2O н2 + co2 Гидрогенные СО-окисляющие
анод "MeO" Электрогенный хемосинтез (?)
Таблица 2. Деструкторы в прокариотной биосфере
Трофическая группа акцептор продукт
Аэробные органотрофы о2 Биомасса + CO2
Сероредукторы So H2S
Сульфатредукторы H2SO4 H2S
Денитрификаторы NOX n2
Железоредукторы Fe(IIl) Fe(II)
Гомоацетатные CO2 CH3COOH
Метаногены CO2 CH4
в табл. 1 «электрогенный хемосинтез» обусловлен способностью группы бактерий, Geo-
bacter служит их типовым представителем, сбрасывать электроны на твердую поверх-
ность, служащую анодом.
Фотосинтез основан на внутриклеточном образовании окислителя за счет фотохи-
мической реакции с поглощающими свет пигментами. Различают два типа фотосинтеза:
оксигенный и аноксигенный. Оксигенный фотосинтез осуществляют только цианобактерии.
Известно, что микрофоссилии цианобактерий служат основным маркером в бактериальной
палеонтологии, поскольку собственно бактерии гораздо хуже распознаваемы в палеонтоло-
гическом материале и их интерпретация вызывает серьезные возражения. Ложная интерпре-
тация микрофоссилий трихомных микроорганизмов как цианобактерий вполне возможна,
поскольку нефотосинтезирующие организмы, от хорошо известных серобактерий вплоть до
метанокисляющих, могут иметь трихомы и образовывать маты.
В палеонтологической летописи микрофоссилии цианобактерий составляют основной
материал. Споры идут об интерпретации конкретных находок по аналогии с формами совре-
менных цианобактерий. Здесь следует сделать одну существенную оговорку о верхнем тем-
пературном пределе развития цианобактерий. Среди термофильных форм есть и нитчатые,
как Mastigocladus, и одноклеточные, как синехококки, цианобактерии, но цианобактерии не
переходят 70 °C рубеж. Они хорошо переносят тепловой шок солнечного нагрева в пустыне
выше 50 °C, но в градиенте воды термальных источников прижимаются к нижним, более
241
Г.А. Заварзин. Первые экосистемы на Земле
холодным слоям. В общем это понятно, поскольку фотосинтезирующие организмы развива-
ются на освещенной субаэральной поверхности.
Интерпретация микрофоссилий в метеоритах как цианобактерий предоставляет
возможность рассматривать появление жизни на Земле вместе с «кусками грязного льда»
на очень ранней стадии. Если стартовой точкой оказывается циано-бактериальное сообще-
ство, прибывшее неизвестно откуда, неизвестно как, то для интерпретации последующих
событий на Земле вполне достаточно представление о биогеохимической сукцессии. Имею-
щиеся фактические данные укладываются в такую схему, не оставляя места для виртуаль-
ного творчества. Речь может идти о масштабах и скоростях изменений, их приуроченности
к определенным биотопам, постепенного развития разнообразия биоты.
Для бактерий и минеральных форм имеется общее «пространство геометрических воз-
можностей», которое частично перекрывается. В области перекрытия интерпретация неких
форм как микрофоссилий может быть ложной. Не все, что выглядит как бактерия, являет-
ся таковым. Ситуация усугубляется тем, что минерал, например опал, заполняет полость,
оставленную разложившимся микроорганизмом, создавая слепок (псевдоморфозу), и именно
он служит наблюдаемым объектом. Химический анализ в этом случае приводит к ложным
заключениям: например, нет ни углерода, ни фосфора, значит, это не остаток микроорга-
низма. Химики, с их принципиально неморфологическим мышлением, с задачей опознания
не справляются и начинают искать химические маркеры. Морфологи, напротив, сдержано
относятся к химическим маркерам, отчетливо сознавая, как могут мигрировать растворы
в пористой среде, например от нефтематеринской породы к коллектору. Реальная эволюция
ранней биосферы прослеживается на основе бактериальной палеонтологии. Вместе с тем не
все, где обнаруживаются достоверные бактерии, создано ими. В итоге: бактериальная пале-
онтология представляет специальную область знаний, где необходимы профессиональные
навыки и крайне опасно поспешное суждение дилетантов.
В течение длительного времени существовало противопоставление как первичных
продуцентов хемосинтетиков и фотосинтетиков. Между последними находятся организ-
мы, использующие промежуточную возможность: аноксигенные фотосинтетики, анаэробно
окисляющие восстановленные соединения: сероводород, водород, органические вещества.
В последнее время к ним прибавились окисляющие железо и, даже, соединения мышьяка.
Примечательное исключение пока составляет метан. Окисление Н, следует считать наибо-
лее интересной функцией по причинам, уже обсуждавшимся для водородного хемосинтеза.
Такие организмы по своему обмену могут служить исходными для разнообразных типов ме-
таболизма. Их анаэробиоз делает их сообщества независимыми от оксигенного цианобакте-
риального фотосинтеза. Издавна существует предположение, что серные пурпурные бакте-
рии могли создать сульфатный океан в бескислородных условиях. Такое же предположение
существует для образования окислов железа в архее.
Наконец, в последнее время установлена многочисленность водных аэробных органо-
трофных бактерий, содержащих хлорофиллы и составляющих 10-50 % бактериального на-
селения океана. Функционально хлорофилл-содержащей пигментной системы недостаточно,
чтобы обеспечить рост организмов. Хлорофилл образуется ими и в темноте. Пока что функ-
цию хлорофилла описывают как «квази-фотосинтез», подобный тому, который имеет место
у галофильных архей дополнительный источник энергии.
Подведем итоги:
I. Существует принципиальная разница в постановке вопроса о происхождении жизни
между а) возникновением жизни и б) появлением жизни на Земле. Только последнее является
экспериментальным фактом.
2. Виртуальная задача о происхождении жизни должна основываться на описании
автономной экосистемы, в которой находится организм, поскольку без обмена невозможна
242
Проблемы hpoik хожаепня жииш
жизнедеятельность. Происхождение организмов следует заменить вопросом о происхожде-
нии экосистемы.
3. Обмен организма в экосистеме зависит от энергодающей мембраны, отделяющей его
от среды. Возможность существования безмембранного организма есть предмет больших
сомнений.
4. Автономные экосистемы должны включать первичных продуцентов - автотрофов.
5. Из первичных продуцентов возможны два типа обмена: хемосинтез и фотосинтез.
6. Универсальным биоэнергетическим механизмом представляется окисление водоро-
да - гидрогенотрофия. Основой его служит гидрогеназа.
7. Представление о возможности появления жизни на Земле в результате панспермии,
если его принять, не имеет ограничений в отношении возможности переноса сообщества,
а не одиночной клетки, как обычно подразумевается.
8. Сообщество как стартовая точка появления жизни на Земле дает возможность описа-
ния последующих наблюдаемых событий как биогеохимической сукцессии.
First ecosystems on the Earth
G.A. Zavarzin
Origin of life is a field of conventional presumptions which in least should not be contradictory
or incompatible with the larger system. Speculation on the origin of life on the Earth should take in
account that habitability precedes habitation. Outline of possible life supporting ecosystem is needed
previously to the speculations about first organisms. Ecosystems as well as biosphere are located
within geographic envelope - geosphere. Ecosystem should ultimately include prime producers and
hydrogenotrophy seems to be the type of physiology most appropriate for the primitive ecosystems.
Considering life on Earth one should distinguish between possibilities on the origin of organisms
in situ or appearance of organisms from and with cosmic bodies in frames of the concept of life in
the Universe. Concept of appearance leaves origin in the mist of unknown. In the second case there
is no limitation in transportation of community rather than a single cell. In the case of community
transportation with «dirty ice» the problem is the conformity of community and its ecosystem on
the parent body with environment on primitive Earth rather than in functional divergence from the
first ancestor. The subsequent events are observed in paleontological record and might be described
as biogeochemical succession without any additional suppositions.
243
Дискуссия
по пленарному докладу Г.А. Заварзина
«ПЕРВЫЕ ЭКОСИСТЕМЫ НА ЗЕМЛЕ»
Председательствующий М.Я. Маров
Л.М. Мухин: Автономное сообщество должно быть неким образом связано с
универсальным общим предком. Доказательством этому я назову универсальность
генетического кода.
Г.А.З.: Это образец глубоко укоренившегося мышления. Если мы имеем транспорт, и
появление жизни связано с транспортом, то у нас нет никакого запрета на получение некоего
сообщества или отдельного организма. Поэтому я помещаю в самое начало сообщество,
которое очень напоминает глинистые маты в метеорите Оргей.
А.С. Спирин: Вновь по поводу универсального общего предка. Вопрос может быть в
терминологии. Что такое этот предок? Это одна особь?
Г.А.З.: Это так в понятиях монофилии.
А.С. Спирин: Карл Вуз использует термин «универсальный предшественник», имея в
виду не особь, а сообщество. Отсюда он выводит универсальный генетический код.
Г.А.З.: Как только он делает этот шаг к сообществу (а Вы в своих построениях вынуждены
делать такой же шаг на выходе из мира РНК), сразу же нет монофилии.
А.С. Спирин: Не нужна нам монофилия - из болота может расти своими корнями сразу
много разных деревьев.
М.Я. Маров (председательствующий): Поэтому скорее надо говорить об автономных
сообществах, а не о сообществе.
Г.А.З.: Да.
А.С. Спирин: Универсальный предшественник - это автономное сообщество,
универсальный предок все запутывает.
Г.А.З.: Позволю себе отметить, что, начиная с эукариот, все живое есть продукт
очень большой деградации. Дело в том, что смысл жизни, смысл существования особей -
в точном самовоспроизведении. Это достигается у бактерий. Появление организмов с
половым процессом представляет собой нарушение основного принципа жизни - принципа
самовоспроизводства. Вместо того, чтобы воспроизвести себя, вы вынуждены подбирать
что-то другое и получать нечто третье, которое на вас совсем не похоже и выглядит по
особенному.
М.Я. Маров: Были достаточно масштабные американские проекты «Биосфера-1» и
«Биосфера-2», но там, возможно, из-за неаккуратно проведенного эксперимента, накопились
контаминанты, несовместимые с автономным существованием системы. Мой вопрос
следующий. Ваши системы поддерживают или отвергают возможность их существования
как полностью замкнутых?
Г.А.З.: Мои системы неавтономны в энергетическом плане. В их основе фотосинтез. Но
в них еще и плохо работала деструкция, образовывалось много полисахаридов.
М.Я. Маров: Еще один принципиальный вопрос. Говоря о неустойчивости систем, что
Вы имели в виду - то, что они действительно неустойчивые в силу их внутренней природы
или то, что они несамоподдерживающиеся?
Г.А.З.: Каждая система должна перейти в биогеохимическую сукцессию. В этом смысле
я говорил о неустойчивости.
244
Проблемы происхождения жичнп
ИТОГОВ А Я ДИСКУССИЯ
ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ
Председательствующий А.Ю. Розанов
А.Ю. Розанов: Дорогие коллеги! Мы хотим создать программу, посвященную пробле-
мам происхождения жизни. Три дня наших заседаний показали, что у нас есть шанс сделать
нечто разумное и двигаться вперед. Некоторые направления обозначились достаточно четко,
некоторые пока еще вырисовываются. Для того, чтобы дело двигалось, оно должно быть опу-
бликовано - обличено в бумажные или электронные формы. Мое изначальное предложение -
сделать книжку, в которой будут напечатаны и доклады, и дискуссия. Георгий Александро-
вич Заварзин предлагает сделать это все на диске, что тоже возможно. Но самое главное, это
нужно сделать быстро и не растягивать на много лет.
СВ. Шестаков. Тезисы о проблемах биологической эволюции
Глубокоуважаемые коллеги! Позвольте изложить три общих соображения, которые
касаются темы нашего собрания, а именно - возникла ли жизнь на Земле или она привнесе-
на извне, а если на Земле, то когда и каким образом эволюционировала. Продолжая обсуж-
дение доклада Георгия Александровича Заварзина, хочу уточнить ряд понятий, которыми
мы пользуемся здесь в аудитории, весьма разнообразной по научным интересам. Во-первых,
следует подчеркнуть, что биологическая эволюция - это эволюция организмов и их сооб-
ществ на уровне популяции. Другой круг вопросов касается «добиологической эволюции»,
абиогенной эволюции макромолекул, способных реплицироваться, собираться в различные
комплексы и даже обеспечивать сопряженные метаболические реакции. Однако при от-
сутствии мембранных структур и автономности такие конгломераты являются открытыми
системами. Это прогеноты, «предклетки», которые представляют собой то, что называется
«коммунальным хозяйством», где еще нет фиксированных геномов. Прогеноты нестабильны
и находятся в постоянном обмене генетическим материалом в результате горизонтальных
переносов. Из этого единого генного пула происходит сборка дифференцированных геномов
и формирование первичных клеток, которые подвергаются действию селективных факторов
энергетической и экофизиологической выгоды.
Я упомянул об этом для того, чтобы перейти к рассмотрению трех тезисов, первый из
которых совершенно ясен. Он заключается в том, что даже если в какой-то форме жизнь поя-
вилась на Земле извне, неважно, из Солнечной системы, от звезд Галактики и т. д., все равно
остается открытым вопрос, как же она где-то возникла? И тогда нужно четко определить, по-
чему бы это не могло произойти и на Земле? Мое впечатление от большинства прослушанных
докладов таково - на определенных этапах формирования Земли на самом деле были все не-
обходимые условия для инициации добиологической эволюции и последующего зарождения
жизни на нашей планете. Лев Михайлович Мухин сказал такую фразу, что весь «космос забит
органикой». Стало быть, многие органические соединения вполне могли быть привнесены на
Землю и/или синтезироваться при определенных температурных, геохимических и прочих
условиях. Это относится не только к нуклеотидам и аминокислотам, но и к сложным поли-
мерам с вершиной в виде мира РНК и предпосылок для появления первичного генетического
кода. Таким образом, на мой взгляд, нет достаточно весомых логических оснований для того,
чтобы отдавать предпочтение гипотезе панспермии о привнесении жизни извне.
Тезис второй. Современные представления о вирусах, неспособных размножаться вне
организма хозяина, говорят скорее в пользу того, что вирусы возникли позже появления клеток
(это традиционная точка зрения), или, по крайней мере, вместе с ними. «Эволюция» вирусов -
это история коэволюции с хозяином. Вместе с тем, несомненно, вирусы играют огромную
роль в горизонтальном переносе генетической информации и эволюции биосферы. Исходя из
245
I l'i оговая дискуссия
большого сходства вирусных геномов с мобильными элементами, можно полагать, что вирусы
представляют собой геномные сегменты, «сбежавшие» из генома прокариот или эукариот и за-
хватившие с собой часть генов, необходимых вирусам для собственного воспроизведения. От-
давая должное красоте гипотезы первичности доклеточного возникновения вирусов (Koonin Е.
et al., 2006. Biol. Direct. V. 1: 29), изложенной В.И. Аголом, еще нельзя найти достаточно убеди-
тельных аргументов, позволяющих полагать, что основные домены, царства живого - археи,
эубактерии и эукариоты - возникли из разных типов провирусов, так же как и рассматривать
вирусы в качестве источников появления жизни на Земле по сценарию панспермии.
Третий тезис касается проблемы происхождения первичной клетки. Широкое распро-
странение получил постулат о том, что клетка возникла единожды и затем через «узкое гор-
лышко» отбора и началась биологическая эволюция. Вместестем допусти май другая гипотеза,
согласно которой в разных частях планеты при разных локальных условиях (геохимических,
физико-химических, климатических и т. д.) первичные протоклетки могли возникать незави-
симо и неоднократно (Шестаков С.В., 2003. Палеонтол. журн. № 6: 50). Эти предшественники
клеток конвергировали по единому принципу, подчиняясь правилам соотношения поверхно-
сти и объема, формирования мембран и т. п. В этом смысле можно предположить, что археи
и бактерии, сходные по морфологии, но различные по аппарату репликации, транскрипции и
трансляции, могли действительно возникнуть независимо друг от друга. Хорошую поддержку
получила схема происхождения эукариот в результате соединения и взаимодействия геномов
и метаболических сетей архей и эубактерии определенного типа. Из сказанного следует уже
получивший широкое признание вывод о том, что не было единственного универсального од-
нокорневого предшественника. После десяти последних лет бурных дискуссий образовалось
два лагеря, к одному из которых (доминирующему) относятся тс, кто традиционно считает
вертикальное наследование основой эволюции и придерживается иерархической филогении.
Сторонники второго лагеря придают первостепенное значение множественным горизонталь-
ным переносам, т. е. реализации сценария сетчатой эволюции, прежде всего для прокариот и
низших эукариот (Doolittle W.F., Bapteste Е., 2007. PNAS. V. 104: 2043). Таким образом, сегодня
происходит серьезная ревизия теории биологической эволюции. Главная задача заключается
в том, чтобы определить конкретное соотношение вклада и темпов вертикальной, горизон-
тальной и редукционной эволюции для различных таксономических групп. За счет горизон-
тальных переносов идет эволюция организмов с высоким рекомбинационным потенциалом,
тогда как у микробов с низким уровнем рекомбинации преобладает вертикальная эволюция
(мутации, бифуркация, отбор). При горизонтальном переносе происходят интенсивные гене-
тические обмены и геномные перестройки, приводящие к образованию мозаичных «химер-
ных» геномов, при филогенетическом анализе которых затрудняется расшифровка путей ви-
дообразования (Шестаков С.В., 2007. Экол. генетика. Т. 5, № 2: 12).
В нашем совещании участвуют представители многих наук - астрофизики, химики,
биологи, геологи, что очень познавательно и стимулирует к междисциплинарному взаимо-
действию. Вместе с тем, дискуссия показала ограниченность наших знаний вообще и недо-
статочную информированность каждого из нас в смежных научных областях. Поэтому пред-
почтение тем или иным гипотезам, которые мы обсуждаем, скорее отражает нашу веру или
желание верить в какую-то идею, соединяющую разные ветви естествознания и, конечно,
не имеющую ничего общего с догматами креационизма, которые, вообще-то, имеют чисто
антропогенное происхождение, в отличие от тех законов, которые реально действуют в при-
роде. Оставляя в стороне тему божественного креационизма, я хочу сказать несколько слов
о другом креационизме. Ведь можно рассматривать не только вопросы происхождения жиз-
ни на Земле или поиски жизни на других планетах, но и обсуждать перспективы экопоэза как
задачи искусственного создания условий для жизни. Этот термин был предложен известным
генетиком Робертом Хейнсом, статья которого так и называется: «Экопоэз - поиграем в Бога
246
Ilf4)b.iiМЫ НРОНСХОЖ t, ‘П!Я ЖННШ
на Марсе» (Haynes R.H., 1989. J. Biol. Sci. in Space. V. 3, № I: 101). По отношению к Марсу
земляне являются инопланетянами, так почему бы нам на определенном этапе развития тех-
нологий не поставить вопрос об использовании Марса как полигона для экспериментально-
го изучения проблемы происхождения жизни. Речь идет не столько о внедрении сложных
экосистем или поселении человека на Марсе. Я имею в виду то, что можно было бы назвать
созидательным креационизмом, призванным обеспечить условия для зарождения жизни на
другой планете. Мои слова являются данью уважения к сторонникам теории панспермии,
но только к схеме с метеоритами, кометами и другими потенциальными переносчиками я
бы добавил и то, о чем говорят сюжеты некоторых научно-фантастических рассказов, то
есть о том, что, может быть, на Землю жизнь пришла в результате творчества кого-то (может
быть, с других планет), кто ставил эксперименты, направленные на создание условий для
естественного зарождения жизни на Земле. Вот таким странным эссе хочу завершить свое
полушутливое изложение гипотезы «направленной панспермии».
Л.М. Мухин: Сергей Васильевич, в последней части Вашего выступления были серьез-
ные предложения по поводу Марса. Есть работа Криса Маккея (McKay С.Р., 1982. Terraforming
Mars. J. Brit. Interplanet. Soc. V. 35: 427), где используется термин «terraforming» для процесса
создания условий для жизни на других планетах. Что Вы тут скажете?
С.В. Шестаков: Кристофер Маккей и Роберт Хейнс были партнерами и вместе про-
двигали разработку конкретных программ освоения. Да, такие теоретические программы
разрабатываются в надежде на будущее. И честно говоря, эта часть моего выступления была
предназначена коллегам из Института медико-биологических проблем. Кроме того, у меня
сложилось впечатление, что многие участники нашего собрания мало знают об этой области
космического естествознания. Поэтому я счел необходимым об этом упомянуть.
А.Ю. Розанов: Одно маленькое замечание по поводу панспермии. Есть две крайние по-
зиции: панспермия - это «жизнь есть всегда и везде», по Вернадскому, и панспермия - это
транспорт. Обратите внимание, что эта проблема должна обсуждаться особым образом.
В.Н. Снытников: Уважаемые коллеги, я бы хотел обратить Ваше внимание вот на какое
обстоятельство. Казалось бы совершенно элементарный с современной точки зрения экс-
перимент Миллера-Ури, проведенный в 1950-х гг., до сих пор упоминается в публикаци-
ях, хотя внешне его результаты довольно-таки тривиальны, что в неравновесных условиях
да еще под воздействием плазмо-химических реакций могут формироваться более сложные
органические молекулы. Вот какое у меня замечание или даже предложение. В последнее
время российская программа исследования космоса, как ближнего, так и дальнего, находится
в достаточно плачевном состоянии по сравнению с европейской или американской програм-
мами. Поэтому материала не хватает. Например, метеоритный материал с предполагаемыми
псевдоморфозами можно было бы получить непосредственно в космосе. Академии следова-
ло бы занять более активную позицию в этом вопросе. Второе предложение - может быть,
уже настала пора перейти к обсуждению экспериментов, непосредственно связанных с про-
блемой возникновения жизни. При всем том, что я выслушал, мне кажется, что несколько
реальных экспериментов, которые можно было бы сейчас проводить, уже можно наметить.
И если правильно сформулировать программу такого сорта экспериментальной деятельно-
сти в этом направлении, то можно было бы рассчитывать на получение в дальнейшем более
сильных результатов. Я предлагаю обсуждать именно программу проведения различного
типа экспериментов, правда, это не должно выливаться в проблемы органического синтеза,
чем часто грешат зарубежные работы. Спасибо за внимание!
М.Я. Маров: Прежде всего, я хочу выразить признательность организаторам за пригла-
шение принять участие в данном рабочем совещании и с удовлетворением особенно отметить
то, что оно проводилось в «неформальном формате». В результате докладчики имели возмож-
ность достаточно подробно излагать проблемы, обозначенные в названиях докладов, и была
247
Итоговая дискуссия
хорошая возможность их подробно обсуждать. В свою очередь, обсуждения обнажили многие
ключевые проблемы, которые заслуживают дальнейшего внимательного изучения. И, конечно,
важно, чтобы космохимические и биологические направления исследований получили даль-
нейшее развитие и были в числе будущих программ фундаментальных исследований, поддер-
живаемых грантами Российской академии наук. Я очень надеюсь, что, подобно тому, как мы
это делали три предыдущих года, эти исследования продолжатся в рамках новых программ
или подпрограмм. У участников, безусловно, есть очень серьезный задел и есть вполне обозри-
мые перспективы развития данных направлений. Это первое, о чем мне хотелось сказать.
Второе. Я услышал здесь много нового от биологов, что важно для расширения соб-
ственного мировоззрения и понимания стоящих перед нами задач. Но это также важно с точки
зрения выявления некоторых общих закономерностей, проявляющихся в различных областях
знаний, что имеет вполне определенный философский смысл. С позиций механика и физи-
ка обсуждавшаяся проблематика имеет самое непосредственное отношение к стохастической
динамике открытых нелинейных диссипативных систем. Это фундаментальное направление,
восходящее к пионерским работам Анри Пуанкаре и получившее развитие в трудах Ильи При-
гожина, бурно развивается в современном мире. В 2006 г. с академиком Алексеем Максимо-
вичем Фридманом мы выпустили книжку, посвященную астрофизическим дискам, в которой
большое внимание уделено стохастическим процессам. В 2009 г. выходит моя книга, напи-
санная совместно с моим учеником и близким коллегой Александром Владимировичем Ко-
лесниченко, посвященная динамике турбулентных газов в космических и природных средах
и процессам возникновения упорядоченности в хаотических системах. В ней, в частности,
показано, что внутри сложных природных комплексов, открытых взаимодействию с окру-
жающей средой, заложены процессы самоорганизации, и развит математический аппарат для
моделирования таких сред. Замечательно, что в хаотической, изначально незапрограммиро-
ванной диссипативной системе, обменивающейся веществом, импульсом и энергией с окру-
жающей средой, выстраиваются вполне определенные островки упорядоченности. Так, в тур-
булентных средах, при определенных значениях ключевых параметров, вы можете наблюдать,
как в самой системе появляются упорядоченные структуры. Может показаться парадоксаль-
ным, но в вихрях, периодически возникающих в турбулентной среде, молекулы, которые про-
извольным образом ведут себя в ламинарном течении, оказываются более упорядоченными.
Другие примеры возникновения порядка из хаоса -- коллективные взаимодействия в кольцах
планет, атмосферная динамика, образование галактик и галактических кластеров и многое
другое. Другими словами, самоорганизация заложена в самой системе. На совещании я полу-
чил дополнительные подтверждения существованию такой парадигмы и для биологических
систем. В частности, мое внимание привлек доклад Александра Борисовича Четверина, в ко-
тором обсуждалась возможность «собрать клетку». С моей точки зрения, последовательность
выстраивания полинуклеотидов и полипептидов при построении генома, равно как и спо-
собность рибозима катализировать производство полипептидов, есть не что иное, как основа
первичного упорядочения при становлении белкового мира. Говоря физическим языком, это
баланс энтропии внутри сложной системы: наряду с ее ростом, связанным с хаотичностью,
одновременно происходит приток в систему отрицательной энтропии (негэнтропии) за счет
внутренней упорядоченности. Здесь можно усмотреть прямую (хотя, вероятно, и достаточно
грубую) аналогию с упомянутыми мною турбулентными течениями. Однако в биологических
системах все несравненно сложнее - самоорганизация происходит в огромных сообществах
супрамолекул, объединенных многочисленными функциональными связями.
Такой подход близок моему пониманию самой проблемы происхождения жизни, кото-
рое следует рассматривать как непрерывный процесс упорядочения при росте уровня само-
организации. Одновременно можно думать, что в изначально хаотической среде на предбио-
логической стадии эволюции система находилась в квазиустойчивом состоянии. Накопление
248
Пт>оИ п мы прот \ож. Иная жп <ни
неустойчивости в подобной системе за счет внешних воздействий обычно приводит к перехо-
ду ее в новое состояние путем бифуркации, и таких последовательных бифуркаций, вообще
говоря, может быть много. Понятие устойчивости также лежит в основе стохастической ди-
намики. С моей точки зрения, появление жизни на Земле можно уподобить бифуркации, воз-
никшей за счет внешнего источника - привноса некой новой субстанции, как возмущающего
агента, в изначальную среду. Вот почему я рассматриваю столь важной роль комет в эволю-
ции Земли, с которыми, помимо летучих, могли быть (и несомненно были) занесены другие
разнообразные соединения. Мне представляется, что такой фактор мог оказать решающее
воздействие на предбиологическую среду и ее состояние устойчивости, принципиально из-
менившееся с зарождением жизни. Именно с этих позиций я воспринимаю те основные вы-
воды, которые были сделаны на совещании. Спасибо.
Л.М. Гиндилис: Прежде всего, я хотел бы отметить, что необходимость в таком со-
вещании явно назрела. В СССР проводились совещания и конференции по SETI, и на них
обсуждалась проблема происхождения жизни. Особенно плодотворным в этом плане было
обсуждение на советско-американской конференции 1971 г. в Бюракане, где присутствовали
Карл Саган, Френсис Крик, а из наших специалистов по происхождению жизни - Лев Ми-
хайлович Мухин. Но специальных совещаний по происхождению жизни, насколько я помню,
не было. А за рубежом такие совещания довольно регулярно проводились, даже издается
журнал «Origins of Life». Так что это совещание действительно очень нужное. Информации
на нем было много. Это хорошо. Но коэффициент понимания, во всяком случае, у меня, весь-
ма мал. С одной стороны, это неизбежно на подобных междисциплинарных совещаниях;
с другой - всем нам надо стремиться, по возможности, говорить на языке междисциплинар-
ного общения, чтобы облегчить жизнь друг другу.
Меня немного удивило высказывание Николая Леонтьевича Добрецова, который ска-
зал, что, мол, туманности это хорошо, но нас интересует происхождение жизни на Земле.
Полагаю, он сам так не думает, это скорее было чисто эмоциональное высказывание. Я глу-
боко убежден, что мы никогда не поймем, как произошла жизнь на Земле, если не будем при-
нимать во внимание космические факторы. Поэтому организаторы конференции безусловно
правы, что не ограничились геологической точкой зрения и пригласили также астрофизиков.
В конце концов, Земля произошла из космического газопылевого облака, и не учитывать
это невозможно. Здесь был очень содержательный обзор Николая Геннадиевича Бочкарёва
по межзвездной среде, но я также обратил внимание, что астрофизические факторы, так или
иначе, затрагивались во многих докладах.
Я хотел бы также высказать некоторые соображения по поводу панспермии. Когда
эти идеи высказывались еще до С. Аррениуса, в XIX в. (Г. Гельмгольц, У. Томсон и др.),
все представлялось ясным. Считалось, что Вселенная, бесконечная в пространстве, су-
ществует вечно. В этой вечно существующей, стабильной Вселенной всегда существова-
ла жизнь на тех или иных телах. При разрушении этих тел семена жизни переносились
на другие планеты. Но сейчас мы знаем, что Вселенная возникла из сингулярного со-
стояния, и условия в ранней Вселенной, где не было ни молекул, ни атомов, полностью
исключает существование жизни, во всяком случае, в известной нам форме. Значит, она
должна была все-таки возникнуть в определенный момент времени, если не на Земле,
то где-то во Вселенной. В связи с этим я хотел бы обратить внимание на очень интересную
работу Александра Дмитриевича Панова из Института ядерной физики МГУ (жаль, что
его не было среди участников совещания). Эволюция планетарной системы проходит че-
рез последовательность различных этапов (или фаз) с фазовыми переходами между ними.
Как убедительно показал Панов, последовательность фазовых переходов в эволюции пла-
нетарной системы с момента возникновения жизни и далее (на биологической и социаль-
ной стадиях) обладает свойством масштабной инвариантности, т.е. продолжительность
249
последовательных фаз образует убывающую геометрическую прогрессию. Если экстра-
полировать эту закономерность назад (во времени), то для предбиологической эволюции
требуется 5-7 млрд, лет, что существенно превышает время от формирования Земли как
планеты до появления на ней жизни. Отсюда Панов выдвигает предположение, что пред-
биологическая эволюция началась на каких-то первопланетах в Галактике, а затем про-
дукты предбиологической эволюции были перенесены на другие планеты, в том числе
на Землю, в процессе панспермии. Он рассчитал модель, из которой следует, что за два
галактических оборота (400 млн. лет) продукты предбиологической эволюции распро-
странились на всю Галактику. Ввиду того, что это время существенно меньше процесса
естественной предбиологической эволюции (5-7 млрд, лет), Панов пришел к выводу, что
предбиологическая химическая эволюция на отдельных планетах не могла протекать не-
зависимо. Это приводит к тому, что жизнь возникает практически одновременно на всех
планетах, где созрели подходящие условия для существования жизни (ранее такую мысль
высказывал Всеволод Сергеевич Троицкий, но из чисто умозрительных соображений).
При этом жизнь возникает на одной молекулярной основе, с единым генетическим кодом
и с одной хиральностью. Думается, что это очень существенный вывод. Возможно, идеи
Панова в чем-то спорные, но их необходимо обсуждать.
Сергей Васильевич Шестаков говорил об «экопоэзе» и о возможности того, что
жизнь могла быть занесена на Землю в результате творчества неких разумных существ с
других планет. Я хотел бы заметить, что такая гипотеза (гипотеза о направленной панспер-
мии) была высказана Ф. Криком и Л. Оргейлом в 1973 г. (Crick Е, Orgel L., 1973. Directed
Panspermia. Icarus. V. 19, № 3: 341). А еще раньше подобные мысли высказывали Дж. Холдейн
и К.Э. Циолковский. В отличие от своих предшественников, которые исходили из общих
умозрительных соображений. Крик и Оргейл пытались обосновать гипотезу направленной
панспермии с биологических позиций. Одним из важнейших аргументов в пользу этой гипо-
тезы они считают универсальность генетического кода.
Лев Михайлович Мухин в своем докладе упомянул о проблеме Дизайнера или Кон-
структора Вселенной. Мне показалось, что эта мысль была воспринята с большим скепти-
цизмом и даже отторжением. Хочу сказать, что в среде космологов она воспринимается бо-
лее спокойно. Возможно, на такое восприятие повлияла проблематика антропного принципа.
Известно высказывание крупнейшего современного физика и космолога Джона Уилера: «Не
замешан ли человек в проектировании Вселенной более радикальным образом, чем мы это
себе представляем?» (Уилер Дж., 1978. Выступление в дискуссии. Космология: теория и на-
блюдения. М.: 368). Еще раньше Фред Хойл, известный английский астрофизик, утверждал:
«Здравая интерпретация фактов дает возможность предположить, что в физике, а также в
химии и биологии экспериментировал «сверхинтеллект», и что в природе нет слепых сил,
заслуживающих доверия». В том же духе высказался недавно Мартин Рис. По его мнению,
идеи, связанные с Мультиверсом, приводят к нс вполне обычному заключению, что «мы яв-
ляемся порождениями некоторой высшей или сверхъестественной силы» (Рис М. Внутри ма-
трицы. http://www.astronet.ru/db/msg/1190835). Эти примеры можно продолжить. Более того,
учитывая, что в момент возникновения Вселенная имела ничтожные размеры, некоторые
физики и космологи начинают задумываться над тем - нельзя ли создать Вселенную в лабо-
ратории? Но, если наши физики пытаются сделать это на бумаге, то другие более развитые
существа могли сделать это на практике. Поэтому под Конструктором Вселенной совсем не
обязательно (и даже не следует) подразумевать некую Внеприродную Сущность. Это может
быть коллективный разум высокоразвитых существ, прошедших эволюцию в предыдущих
циклах Вселенной или в других вселенных.
В связи с докладом Алексея Юрьевича Розанова у нас возникла острая дискуссия
о морфологии: в какой степени можно доверять выводам, полученным на основе чисто мор-
250
Проблемы пронсхожлгиня жншн
фологических методов и не следует ли добавить к ним какие-то другие методы? Я думаю,
что другие методы (если они есть) всегда полезны. Но в какой мере можно базироваться
на морфологии? Вспоминается, что в 1950-х гг. два наших выдающихся астронома Гри-
горий Абрамович Шайн в Крыму и Василий Григорьевич Фесенков в Алма-Ате успешно
занимались изучением морфологии газовых туманностей, и ими были получены важные
выводы, в частности, о характере магнитного поля в туманностях. А Борис Александро-
вич Воронцов-Вельяминов с помощью микроскопа исследовал карты Паломарского атласа
и открыл многие ранее неизвестные типы галактик. Сам я морфологией никогда не за-
нимался, я занимался спектрофотометрией. Но, в конце концов, что такое спектрограм-
ма? Это же тоже изображение, и опытные спектроскописты по виду спектра, без всякой
фотометрии, могут сделать заключение о природе исследуемого объекта, например, о спек-
тральном классе звезды. Я думаю, что на самом деле, морфологические методы использу-
ются гораздо чаще, чем мы привыкли думать.
В заключение я хотел бы поддержать предложение Валерия Николаевича Снытникова
о получении метеоритного материала непосредственно в космосе. Это можно делать с помо-
щью ловушек метеорной пыли, устанавливаемых на космических аппаратах. Но метеорную
пыль можно (и нужно) собирать также на поверхности Земли. Думаю, в ней можно будет
найти много интересного.
А.В. Витязев: Это одно из немногих совещаний, где я получил очень много нового
и в очень приятной обстановке. Но если я «съел» очень много, то я совершенно это не «пере-
варил». Хорошо, что будут диски, еще лучше, если будут публикации. Некоторые утвержде-
ния, с которыми я внутренне не согласен, я бы хотел увидеть в напечатанном виде. Но мне
кажется, что совещание удалось.
М.А. Федонкин: Алексей Юрьевич Розанов сказал, что мы не должны заниматься опре-
делением, что такое жизнь. Но есть такая простая вещь: в основе жизни лежит универсаль-
ный процесс, который позволяет нам говорить на одном языке - электрон переходит от до-
нора к акцептору, это универсальный язык физики, который лежит и в основе обсуждаемых
нами проблем. И если следовать предложению Валерия Николаевича Снытникова по поводу
экспериментов, то, конечно, нам больше всего здесь не хватает физиков. Все реакции в сво-
ей основе - электрохимические, и сейчас бактерии встраиваются в электрохимические гра-
диенты в среде. Нам нужно подумать, как эти процессы возникали, как жизнь удерживает
энергию и отдает (в отличие от неживых тел). То есть исследовать больше саму жизнь, чем,
скажем, морфологию, включая морфологию молекул. Это мое главное пожелание.
А.Б. Макалкин: Я хотел бы сказать о некоторых физических вопросах, относящихся
к ранним стадиям эволюции Солнечной системы. Во-первых, где в ранней Солнечной систе-
ме могли реализоваться физические условия, необходимые для самых ранних стадий прсд-
биологической эволюции? Об этом говорил Александр Сергеевич Спирин. Органические
соединения сейчас определяют из астрономических наблюдений не только в газовой фазе
межзвездной среды и молекулярных облаков, но и в пылевых частицах таких облаков и око-
лозвездных дисков, а также изучая вещество тел Солнечной системы. Значительная часть
органических соединений - полициклические ароматические и алифатические углеводоро-
ды. Итак, органических соединений в исходном веществе, из которого образовались первые
допланетные тела, было много. В докладе Александра Сергеевича прозвучали следующие
требования к обозначенным условиям. Первое - поддержание температуры около 300 К.
Второе - наличие жидкой воды. Третье - желательное наличие монтмориллонита, одного
из водосодержащих силикатов. По имеющимся данным и основанным на них моделям, все
эти условия реализовывались в родительских телах углистых хондритов. Вероятно, они так-
же выполнялись в планетезималях размерами от ста километров, во внешней части Сол-
нечной системы, в области образования планет-гигантов и, возможно, в койперовом поясе.
251
Итоговая дискуссия
В 2003 г. мы опубликовали статью по поводу происхождения гидросиликатов, обнаружен-
ных на некоторых телах пояса Койпера по наблюдениям их спектров отражения. Были оце-
нены времена эволюции таких тел. Гидросиликаты обнаружены также на поверхности мно-
гих тел в поясе астероидов, причем не только на астероидах С-типа, похожих по веществу
на углистые хондриты, но и на некоторых расслоенных астероидах (магматических типов),
которые значительно отличаются по спектру и веществу от углистых хондритов. Есть загад-
ка в том, откуда на этих астероидах появились гидросиликаты. Гипотеза такая - они были
принесены при соударениях с другими телами, поступившими из внешней части пояса асте-
роидов или даже из области образования Юпитера. О чем свидетельствует наличие гидроси-
ликатов? Большинство исследователей считают, что для их образования нужна жидкая вода,
а в протопланетном облаке из-за низкого давления нет жидкой фазы, там происходит су-
блимация твердого вещества в газообразное. Но жидкая вода могла быть в планетезималях
с размерами от нескольких десятков километров. Как достигались необходимые темпера-
туры? Основным источником нагрева планетезималей сейчас считаются короткоживущие
радиоактивные изотопы, прежде всего - алюминий-26, потому что его больше других. Пери-
од полураспада алюминия-26 - около 700 тыс. лет. Поэтому, если планетезимали с радиусом
от ста километров (а для зоны родительских тел углистых хондритов - от нескольких десят-
ков километров) смогли образоваться за время не более двух миллионов лет, пока хватает
тепла, то океан внутри них мог просуществовать до замерзания в течение нескольких мил-
лионов лет. Возникает вопрос - если там что-то такое могло начать развиваться, то этого
периода - в несколько миллионов лет - достаточно или нет?
А.Ю. Розанов: Вполне.
А.С. Спирин: Мало.
А.Ю. Розанов: Смотря с чего начинать.
А.С. Спирин: Смотря чем кончить.
А.Ю. Розанов: Не выше, чем протистом.
А.С. Спирин: Микробом. А можно вопрос? Наверное, когда я перечислял необходимые
условия, забыл о фосфоре. Его в космической органике и в целом ряде космических тел нет.
Надо искать фосфор. Без фосфора ничего нельзя сделать, потому что не только нуклеиновые
кислоты, не только репродукция, но и вся энергетика живой системы базируется на фосфоре.
А.Б. Макалкин: Может быть, найдут фосфоросодержащие добиологические органиче-
ские соединения...
А.С. Спирин: Хотя бы просто фосфор найти.
В.Н. Снытников: Соединение углерод-фосфор (СР) спектрально установлено в меж-
звездной среде. Кроме того, в углистых хондритах есть фосфор.
А.Ю. Розанов: Хорошо, это мы перепроверим.
А.С. Спирин: А в земной коре где есть фосфор?
А.Ю. Розанов: Везде. И его много.
А.В. Витязев: 0.08 % в земной коре.
А.С. Спирин: Значит, Земля не может быть в этом уникальна?
А.Ю. Розанов: Конечно.
А.Б. Макалкин: Он и в космосе не самый редкий элемент...
А.С. Спирин: Тогда почему в пылевых облаках он не обнаружен?
А.Б. Макалкин: А это уже вопрос к методике. Там пока не всё умеют находить.
ГА. Заварзин: Но совершенно точно сформулирован вопрос - выдайте фосфор.
А.Б. Макалкин: В молекулярных облаках уже обнаружена молекула PN, а в межзвезд-
252
Проблемы происхождения жизни
ной среде и молекула РО. Из наблюдений линии поглощения ионизованного фосфора счита-
ют, что межзвездная среда не обеднена этим элементом.
Я бы хотел еще несколько слов сказать по поводу условий для появления жизни на ран-
ней Земле. Было высказано две позиции. С одной стороны, что температура на ранней Земле
уже через 100 миллионов лет после ее образования была близка к современной. С другой сто-
роны - была мощная бомбардировка, которая длилась еще несколько сотен миллионов лет,
причем не факт, что она все время шла по убывающей, возможно, что около четырех милли-
ардов лет назад, незадолго до окончания, был всплеск ее активности. По моему мнению, не-
смотря на то, что температурные условия, возможно, были в среднем довольно стабильными
и близкими к современным, бомбардировка могла привести к задержке появления жизни на
Земле, так как периодически вызывала масштабные катастрофические события.
М.Я. Маров: В связи с этой проблемой важно соотношение между привносом вещества
падающими телами и обусловленными ими катастрофическими событиями.
В.Н. Снытников: У меня вопрос. Споры микробов после тяжелой космической бомбар-
дировки могут остаться?
Г.А. Заварзин: Споры в основном приспособлены против высыхания, а не против на-
гревания. Но это неважно. Микробы выживут и так.
А.Ю. Розанов: Для корректности я бы уточнил теперь тему нашего заседания как «Не-
которые вопросы происхождения жизни и появления ее на Земле».
253
11 то г о в а я дискус сия
Заключительное слово
Высокочтимые коллеги, какая-то часть из Вас оказалась чрезвычайно устойчивой к
потоку информации и этот самый поток информации выдержала, оставшись в зале. На что я
бы хотел обратить ваше внимание. Очень удачно, что совещание было немногочисленным,
это дало необходимую свободу общения. Поэтому, если предполагается создание следующей
программы, то перед этим можно было бы провести семинар, немного с другим составом,
учитывая те лакуны знаний, которые тут у нас оказались ясны. Это организационное пред-
ложение. Далее следует психологическое наблюдение. Программа Президиума РАН № 18
оказалась весьма продуктивной по производству членов Академии. Можно сделать вывод,
что люди, участвовавшие в этой программе, принадлежат к людям, действительно извне ува-
жаемым. Второй психологический вывод - совещание было весьма междисциплинарным, и
каждый пришел со своей психологической догмой (не парадигмой, потому что парадигма -
это противопоставление догме), при этом психологическое совмещение - понять, что гово-
рит другой - конечно, очень затруднено. Мгновенное взаимопонимание на таких собраниях
невозможно. Но мы здесь, в общем-то, смогли получить представление о способах мышления
смежников - от бесформенного мышления химика (для которого формы не существует) до
сугубо оформленного мышления морфологов. И есть еще третья категория это люди, кото-
рые верят в то, что формула представляет объективную реальность, что написанная формула
уже есть реальность, независимо от параметров, которые в нее вставляют. Сухой остаток:
необходимость проведения следующего семинара и доброе пожелание пытаться понимать
тех, кого мы не понимаем.
ГА. Заварзин
254
Прок. if.mki происхождения жиши
ПРИЛОЖЕНИЕ 1
Рабочее совещание «Проблемы происхождения жизни»,
22-24 сентября 2008 г.
Рабочее совещание «Проблемы происхождения жизни» состоялось 22-24 сентября
2008 г. в Палеонтологическом институте им. А.А. Борисяка РАН (Москва).
22.09.2008 г. совещание открыл вице-президент РАН акад. А.И. Григорьев. Про-
грамма этого дня заседаний включала пленарные доклады чл.-корр. РАН А.Б. Четвери-
ма («Можно ли собрать клетку из ее компонентов?»), чл.-корр. РАН В.И. Агола («Вирус -
до или после клетки9»), акад. А.С. Спирина («Древний мир РНК»), д.ф.-м.н. Л.В. Ксанфомалити
(«От Марса к Европе: поиск биосферы на спутниках планет-гигантов в системе Солнца
и других звезд») и к.ф.-м.н. В.Н. Снытникова («Астрокатализ - абиогенный синтез и хими-
ческая эволюция на догеологических этапах формирования Земли»), В дискуссии с кратким
дополнительным докладом («Молекулы во Вселенной» и «Миграция молекул и пыли во Все-
ленной») выступил д.ф.-м.н. Н.Г. Бочкарёв. В обсуждении приняли активное участие акад.
М.Я. Маров, акад. С.В. Шестаков, акад. А.В. Спирин, акад. Н.Л. Добрецов, акад. Г.А. Завар-
зин, акад. А.Ю. Розанов, чл.-корр. А.Б. Рубин, чл.-корр. В.И. Агол, чл.-корр. А.Б. Четверин,
проф. Л.М. Мухин, к.ф.-м.н. В.Н. Снытников, д.б.н. С.В. Рожнов, д.ф.-м.н. Л.В. Ксанфомали-
ти, проф. А.В. Тутуков, проф. А.Д. Альтштейн, д.ф.-м.н. Н.Г. Бочкарёв, к.ф.-м.н. А.Б. Макал-
кин, д.ф.-м.н. В.А. Царёв, д.ф.-м.н. А.В. Витязев.
23.09.2008 г. состоялись пленарные доклады проф. Л.М. Мухина («Условия на по-
верхности Земли 4.6-4.0 млрд, лет назад: первичные синтезы»), д.ф.-м.н. А.В. Витязева
и д.ф.-м.н. ГВ. Печерниковой («Ранняя Земля в тесном окружении молодых звезд»), акад.
Н.Л. Добрецова («О ранних стадиях эволюции биосферы и проблемах ранних оледене-
ний») и акад. А.Ю. Розанова («Псевдоморфозы по микробам в метеоритах» и «Условия жиз-
ни на ранней Земле после 4.0 млрд, лет назад»), В дискуссии активно участвовали акад.
М.Я. Маров, акад. А.Ю. Розанов, акад. С.В. Шестаков, акад. А.В. Спирин, акад. М.А. Федон-
кин, акад. Г.А. Заварзин, акад. Ю.В. Наточин, чл.-корр. А.Б. Рубин, чл.-корр. С.В. Шустов,
д.ф.-м.н. А.В. Витязев, проф. А.В. Тутуков, д.ф.-м.н. Н.Г. Бочкарёв, проф. Л.М. Мухин,
к.ф.-м.н. В.Н. Снытников, д.б.н. С.В. Рожнов.
Заседания 24.09.2008 г. включали пленарные доклады д.б.н. Е.А. Бонч-Осмоловской
(«Глубинная биосфера»), акад. Ю.В. Наточина («Возникновение мембран») и акад. Г.А. За-
варзина («Первые экосистемы на Земле»). В обсуждении приняли участие акад. Ю.В. На-
точин, акад. М.Я. Маров, акад. М.А. Федонкин, акад. А.Ю. Розанов, акад. С.В. Шестаков,
акад. А.В. Спирин, проф. Л.М. Мухин, к.ф.-м.н. В.Н. Снытников, проф. А.В. Тутуков,
к.ф.-м.н. А.Б. Макалкин, д.б.н. С.В. Рожнов. В итоговой дискуссии выступили акад. А.Ю. Ро-
занов, акад. С.В. Шестаков, проф. Л.М. Мухин, к.ф.-м.н. В.Н. Снытников, акад. М.Я. Маров,
к.ф.-м.н. Л.М. Гиндилис, д.ф.-м.н. А.В. Витязев, акад. М.А. Федонкин, к.ф.-м.н. А.Б. Макал-
кин, акад. Г.А. Заварзин.
Оргкомитет рабочего совещания: акад. О.Г Газенко, акад. А.И. Григорьев,
акад. Н.Л. Добрецов, акад. Г.А. Заварзин, акад. Н.С. Кардашев, д.б.н. А.В. Лопатин,
проф. Л.М. Мухин, д.б.н. С.В. Рожнов, акад. А.Ю. Розанов, к.ф.-м.н. В.Н. Снытников,
акад. А.С. Спирин.
255
Проб. './л/ и рот \ож : /но/ ;к и ai/i
Список участников совещания
«Проблемы происхождения жизни»
- Агол В.И., чл.-корр. РАН, Институт физико-химической биологии им. А.Н. Белозерского МГУ;
- Альтштейн А.Д., проф., д.б.н., Институт биологии гена РАН;
- Бонч-Осмоловская Е.А., д.б.н., Институт микробиологии им. С.Н. Виноградского РАН;
- Бочкарёв Н.Г., д.ф.-м.н., Государственный астрономический институт им. П.К. Штернберга МГУ;
- Быстрицкий А.А., к.б.н.. Институт биологии гена РАН;
- Витязев А.В., д.ф.-м.н., Институт динамики геосфер РАН;
- Герасименко Л.М., д.б.н., Институт микробиологии им. С.Н. Виноградского РАН;
- Гиндилис Л.М., к.ф.-м.н., Государственный астрономический институт им. П.К. Штернберга МГУ;
- Григорьев А.И., академик, вице-президент РАН;
- Добрецов Н.Л., академик, Объединенный институт геологии, геофизики и минералогии СО РАН;
- Заварзин Г.А., академик, Институт микробиологии им. С.Н. Виноградского РАН;
- Кардашев Н.С., академик, Физический институт РАН;
- Корицкая Е.А., к.ф.-м.н., Институт астрономии РАН;
- Ксанфомалити Л.В., д.ф.-м.н., Институт космических исследований РАН;
- Лопатин А.В., д.б.н., Палеонтологический институт им. А.А. БорисякаРАН;
- Макалкин А.Б., к.ф.-м.н., Институт физики Земли им. О.Ю. Шмидта РАН;
- Маров М.Я., академик, Институт геохимии и аналитической химии им. В.И. Вернадского РАН;
- Мухин Л.М., проф., д.ф.-м.н., Институт космических исследований РАН;
- Наточин Ю.В., академик, Институт эволюционной физиологии и биохимии им. РЕМ. Сеченова РАН;
- Оганесян Г.А., д.б.н., Институт эволюционной физиологии и биохимии им. И.М. Сеченова РАН;
- Павлов Д.С., академик, Институт проблем экологии и эволюции им. А.Н. Северцова РАН;
- Печерникова Г.В., д.ф.-м.н., Институт динамики геосфер РАН;
- Пономаренко А.Г., д.б.н., Палеонтологический институт им. А.А. Борисяка РАН;
- Рожнов С.В., д.б.н., Палеонтологический институт им. А.А. Борисяка РАН;
- Розанов А.Ю., академик, Палеонтологический институт им. А.А. Борисяка РАН;
Рубин А.Б., чл.-корр. РАН, Биологический факультет МГУ;
- Снытников В.Н., к.ф.-м.н., Институт катализа им. ГК. Борескова СО РАН;
- Спирин А.С., академик, Институт белка РАН;
- Тутуков А.В., проф., д.ф.-м.н., Институт астрономии РАН;
- Ушатинская Г.Т., д.б.н., Палеонтологический институт им. А.А. Борисяка РАН;
- Федонкин М.А., академик. Институт геохимии и аналитической химии им. В.И. Вернадского РАН;
- Царёв В. А., д.ф.-м.н., Физический институт РАН;
- Четверин А.Б., чл.-корр. РАН, Институт белка РАН;
- Четверина Е.В., д.б.н., Институт белка РАН;
- Шестаков С.В., академик, Институт обшей генетики им. Н.И. Вавилова РАН;
- Шустов Б.М., чл.-корр. РАН, Институт астрономии РАН.
256
Проблемы происхождения жия-ш
ПРИЛОЖЕНИЕ 2
ПРОГРАММА ЗАСЕДАНИЙ
рабочего совещания
«ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ»
22 сентября 2008 г.
9.30: Регистрация
10.00: Вступительное слово
А.И. Григорьев
10.15: Можно ли собрать клетку из ее компонентов?
А.Б. Четверни
11.15: Вирус - до или после клетки?
В. И. Агол
12.15: Древний мир PH К
А.С. Спирин
13.15: Перерыв
14.30: От Марса к Европе: возможная биосфера на спутниках планет-гигантов в системе
Солнца и других звезд
Л. В. Ксанфомалити
15.30: Астрокатализ - абиогенный синтез и химическая эволюция на догеологических
этапах формирования Земли
В.Н. Снытников
16.30: Дискуссия
23 сентября 2008 г.
10.30: Условия на поверхности Земли 4.6-4.0 млрд, лет назад: первичные синтезы
Л.М. Мухин
11.30: Ранняя Земля в тесном окружении молодых звезд
А.В. Витязев. ГВ. Печерникова
12.30: Псевдоморфозы по микробам в метеоритах
А.Ю. Розанов
13.30: Перерыв
14.30: О ранних этапах эволюции биосферы и проблемах ранних оледенений
Н.Л. Добрецов
15.30: Условия жизни на ранней Земле после 4.0 млрд, лет назад
А.Ю. Розанов
16.30: Дискуссия
24 сентября 2008 г.
10.30: Глубинная биосфера
Е.А. Бонч-Осмоловская
11.30: Возникновение мембран
Ю.В. Наточин
12.30: Перерыв
13.30: Первичные экосистемы на Земле
Г.А. Заварзин
14.30: Итоговая дискуссия и заключение
Регламент
пленарный доклад - 40 минут;
на вопросы и обсуждение дополнительно 20 минут, выступление в рамках дискуссии - 10 минут.
257
Утверждено к печати Ученым советом Палеонтологического института им. А.А. Борисяка РАН
НАУЧНОЕ ИЗДАНИЕ
ПРОБЛЕМЫ ПРОИСХОЖДЕНИЯ ЖИЗНИ
ПИН РАН, 2009 г.
Ответственные редакторы:
А.Ю. Розанов, А.В. Лопатин, В.Н. Снытников
Компьютерная верстка: М.К. Емельянова
Подписано в печать 2Х октября 2009 г. Формат бОхХХ/Х. Гарнитура «Таймс».
Печать офсетная. Бумага мелованная.
30 усл. п.л.. 16 уч.-изд. л.
Тираж 500 зкз.
Учреждение Российской академии наук
Палеонтологический институт им. А.А. Борисяка РАН (ПИН РАН)
Москва, Профсоюзная, 123
Издано при финансовой поддержке проф. X. Раусинга
Published by financial support of Prof. H. Rausing
Отпечатано в типографии
«Лакшсри Принт»
Москва, ул. Архитектора Власова, д. 21
ISBN 978-5-903825-09-7
9
785903
825097