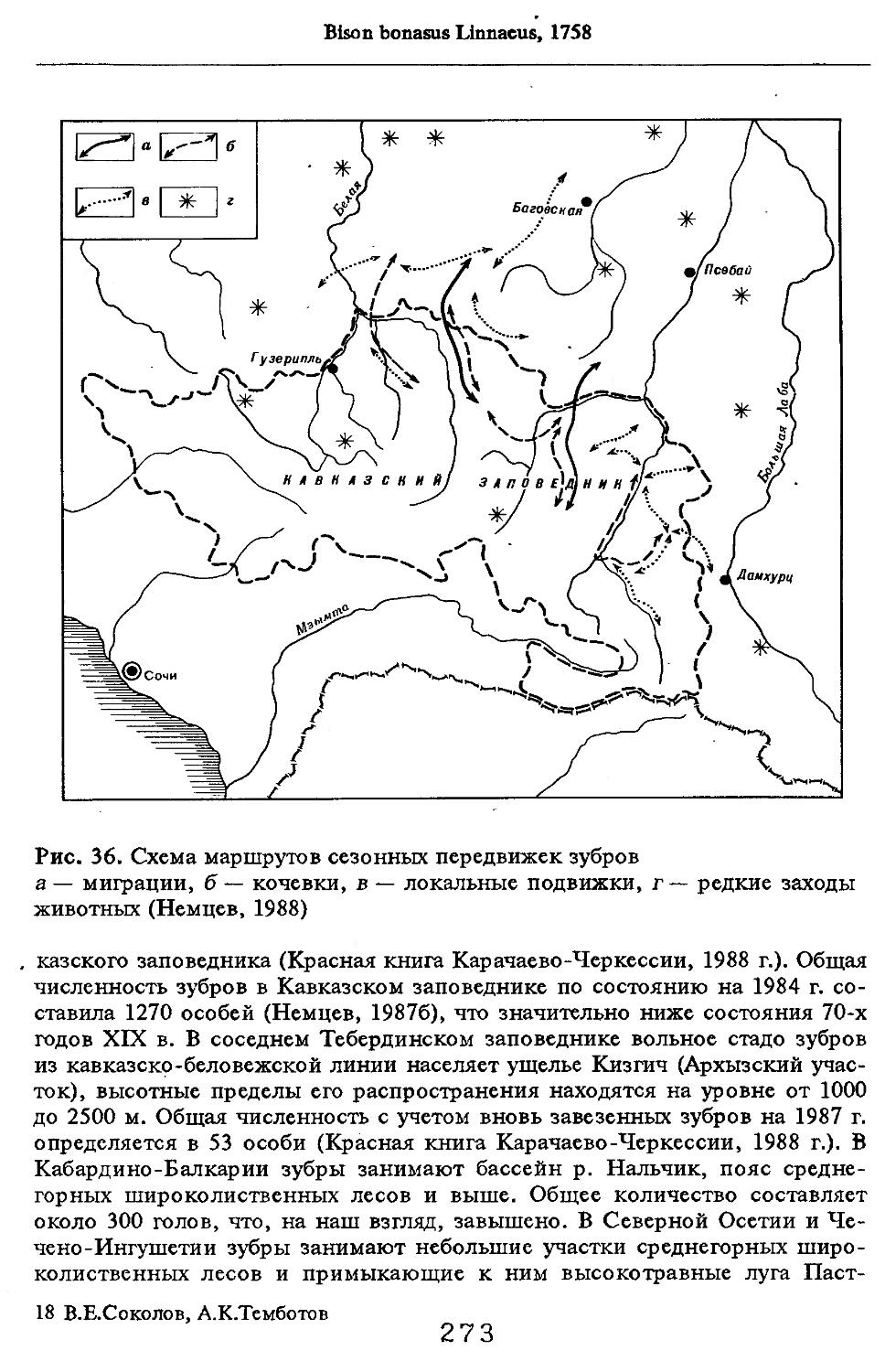
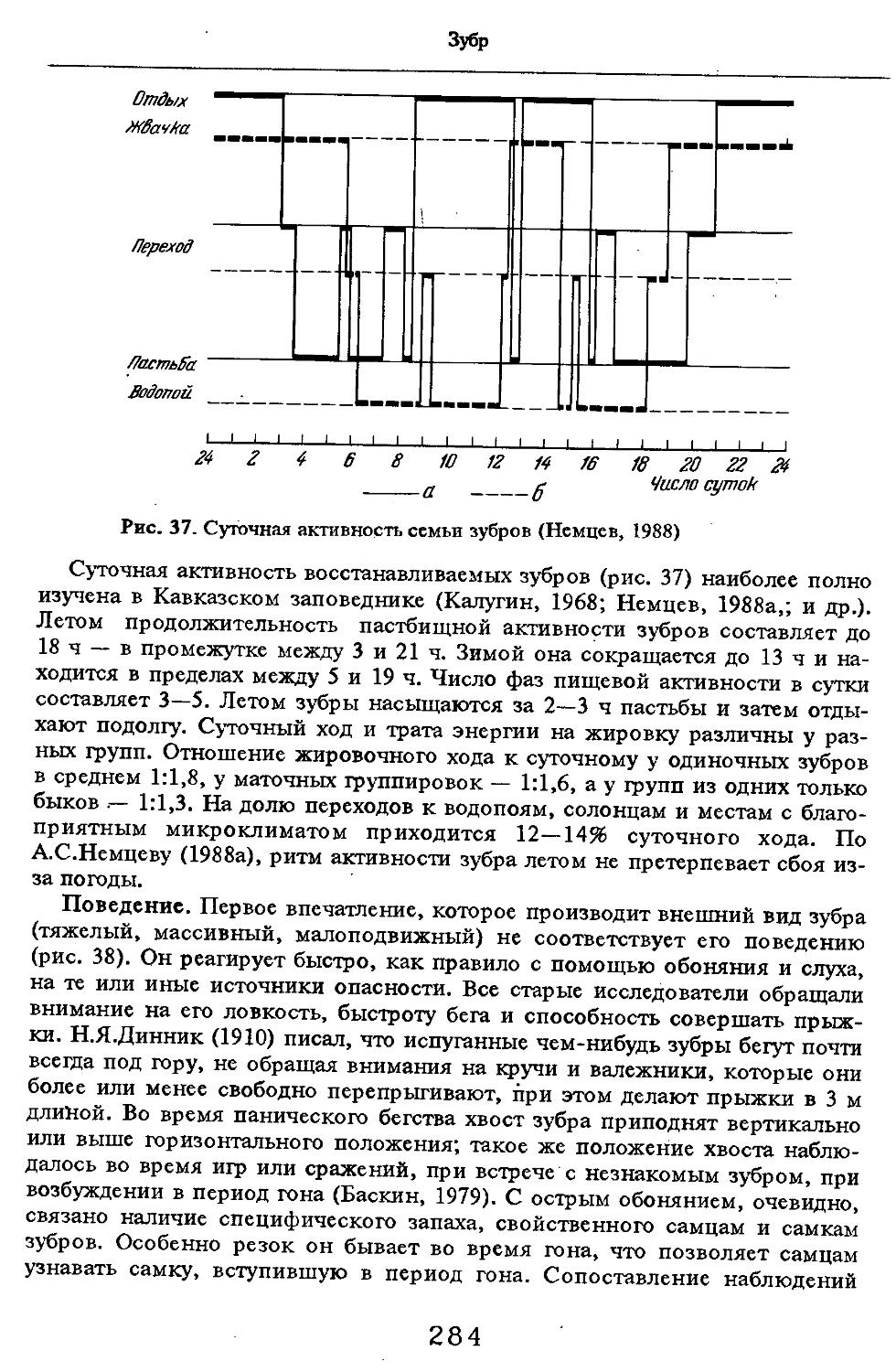
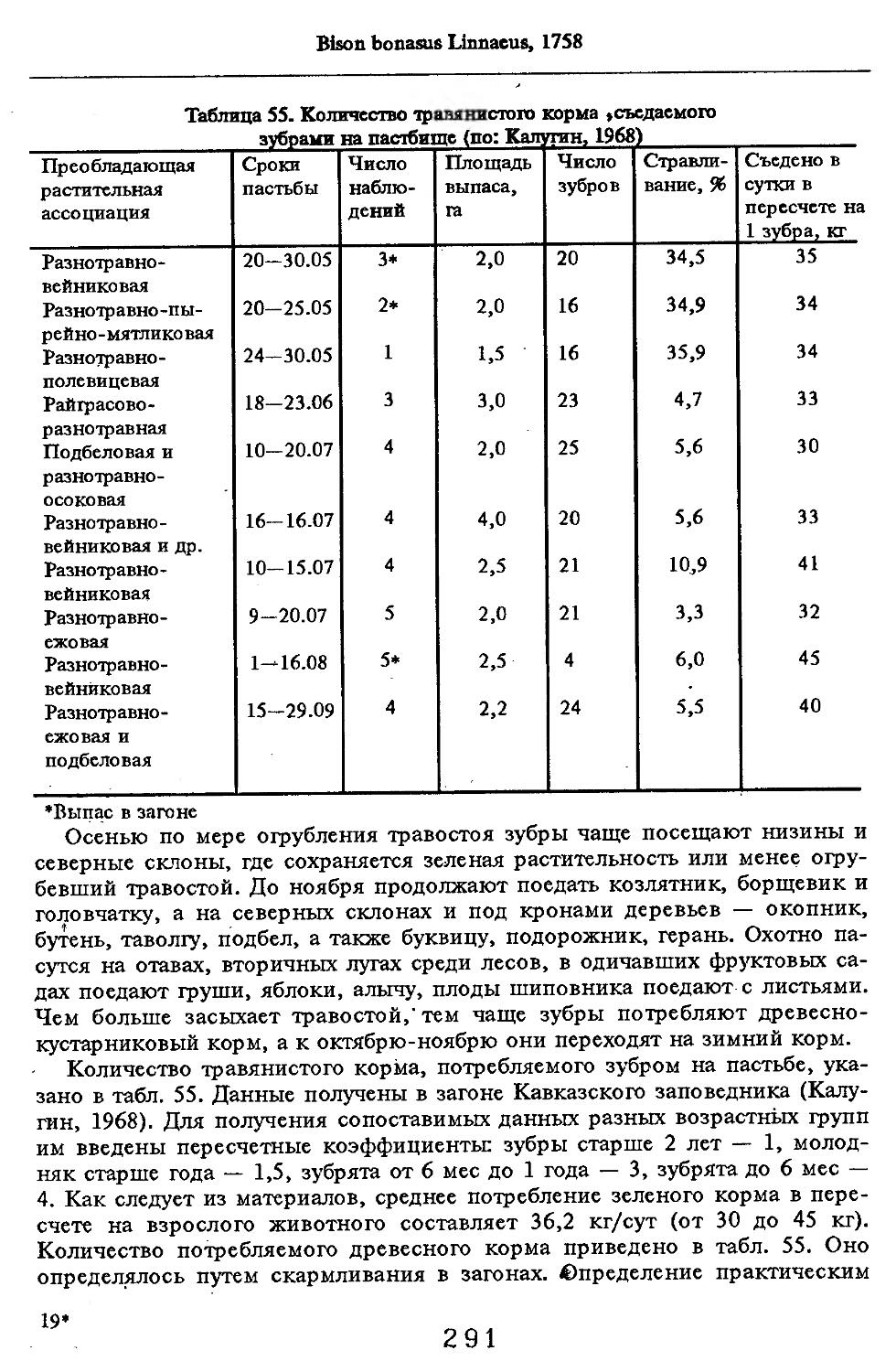
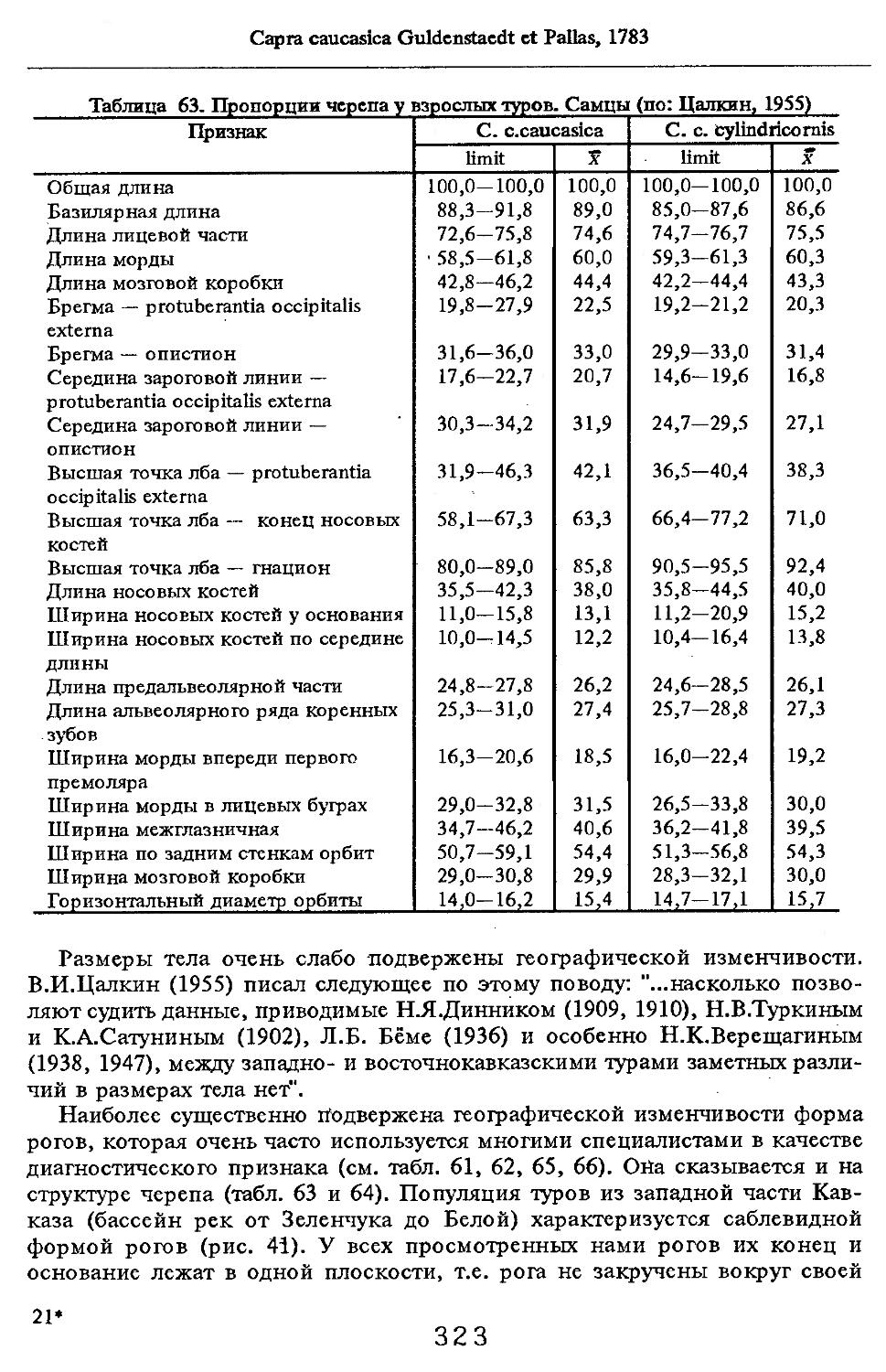
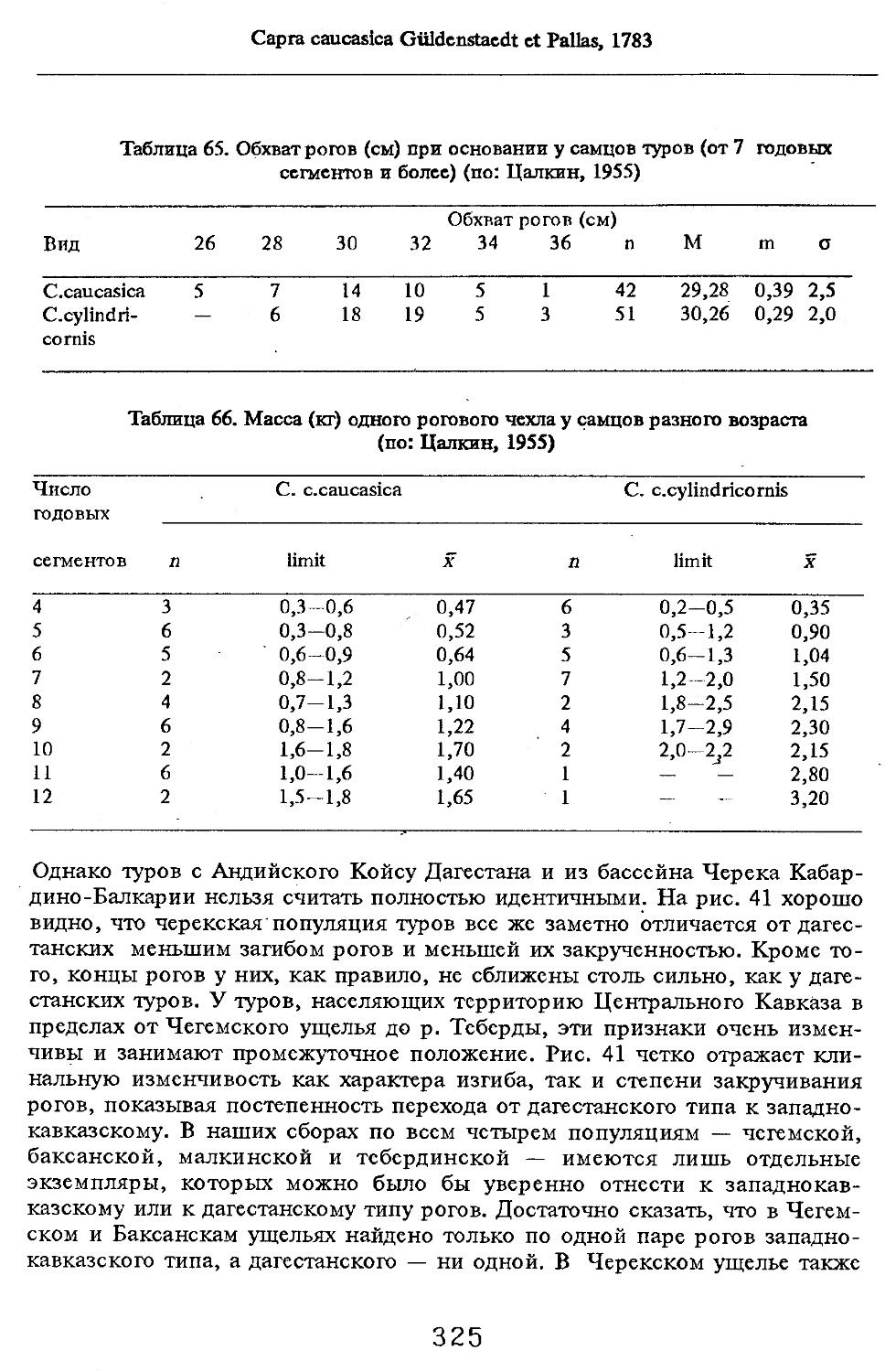
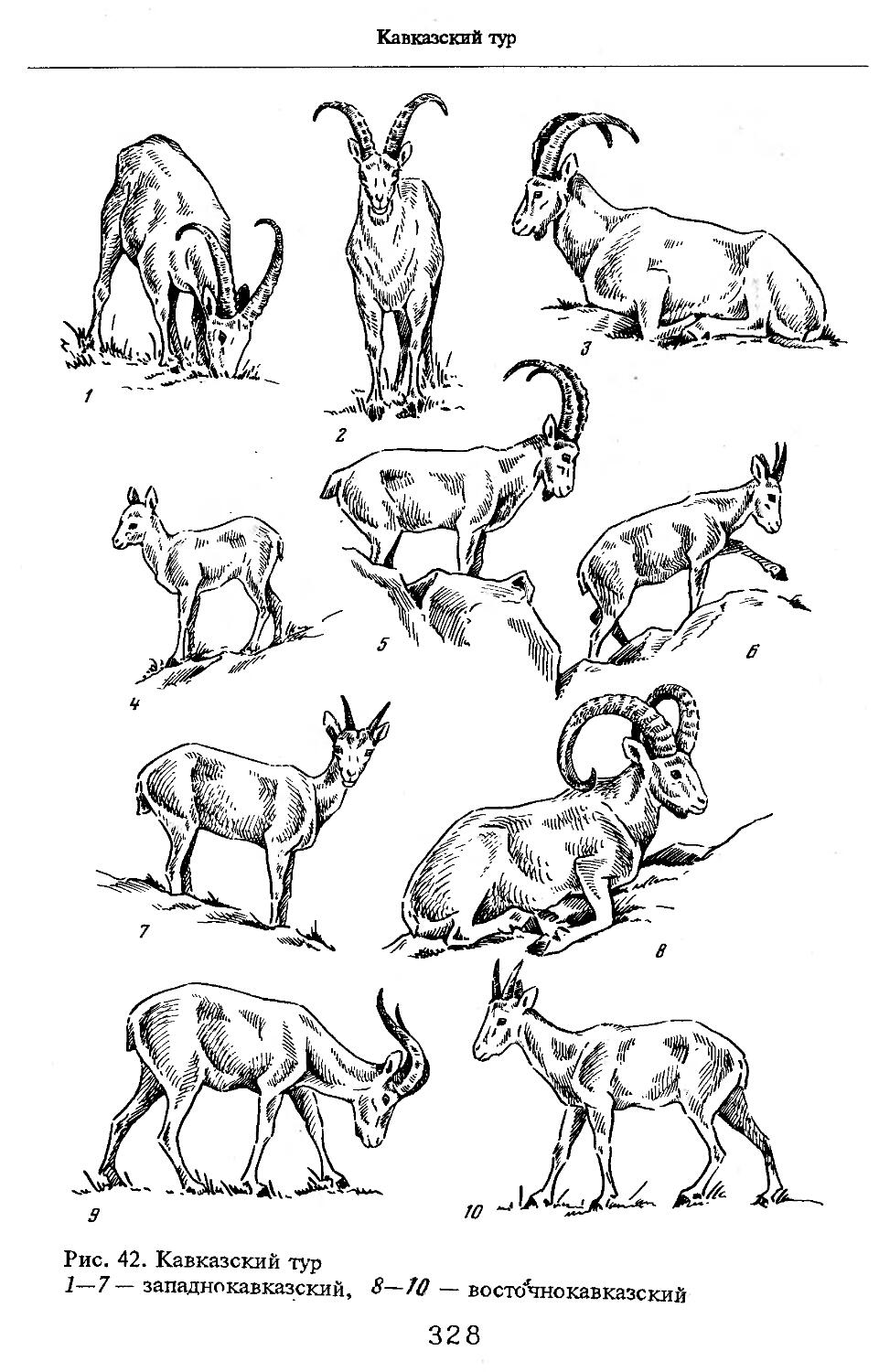
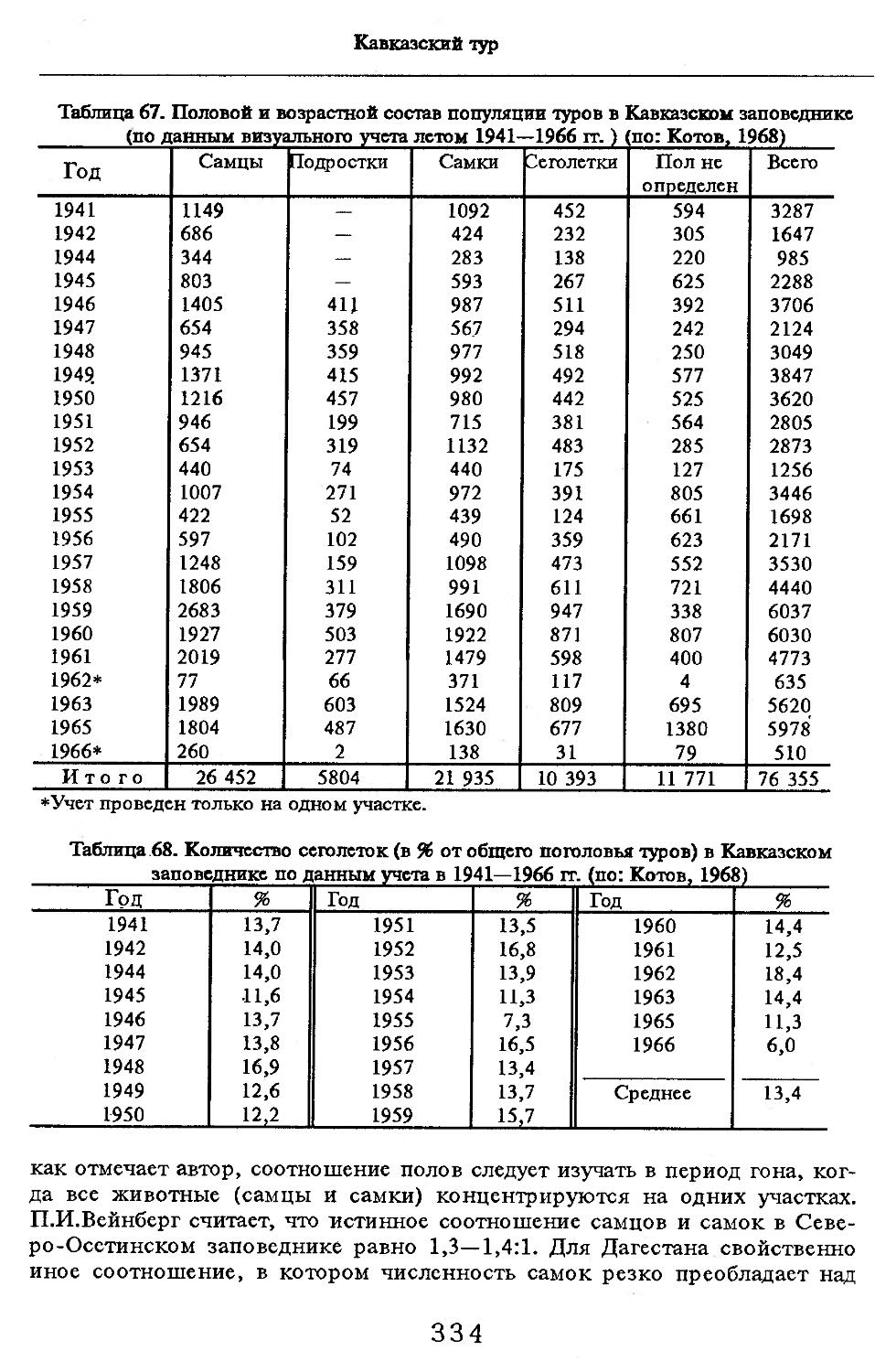
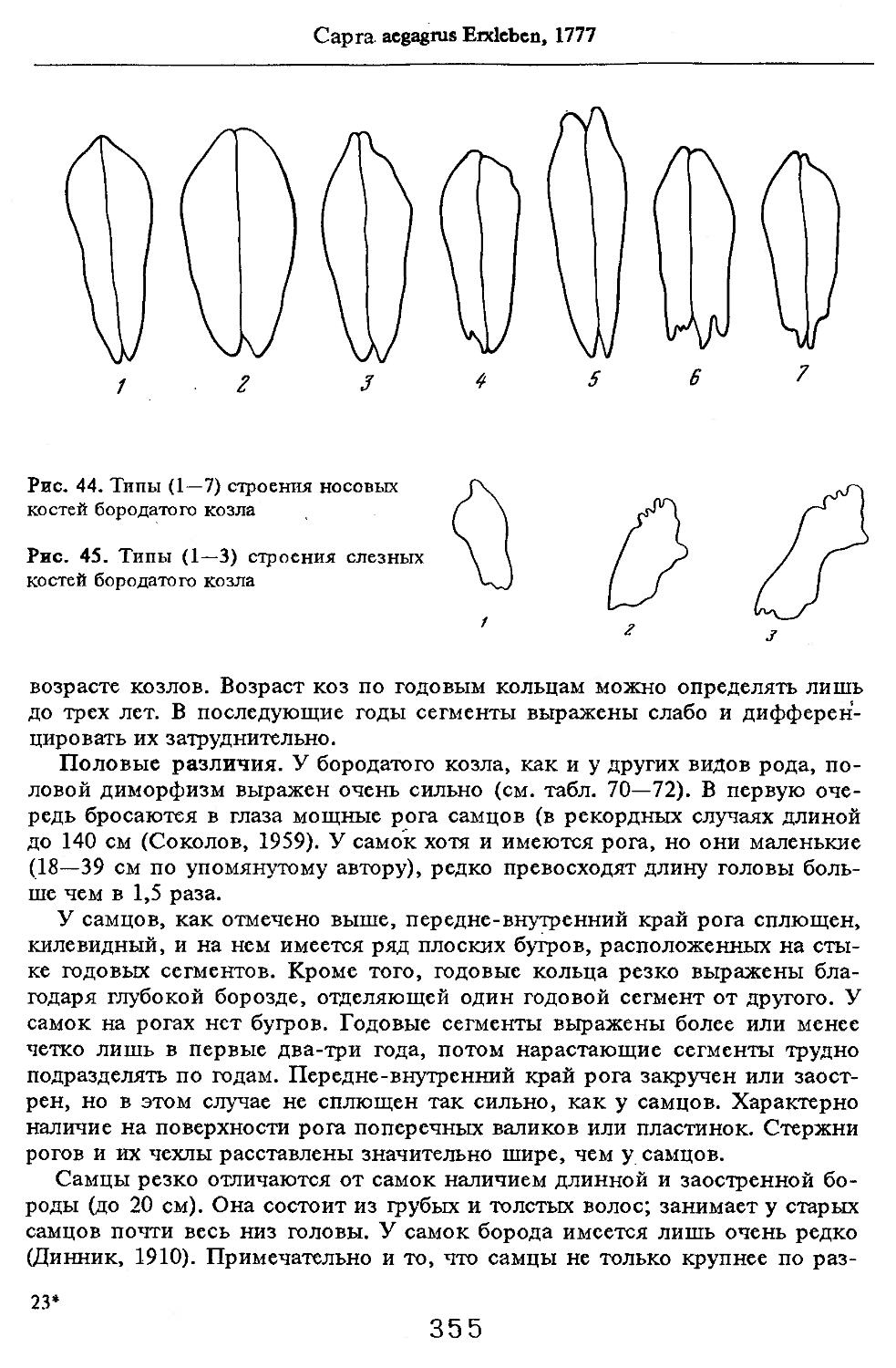
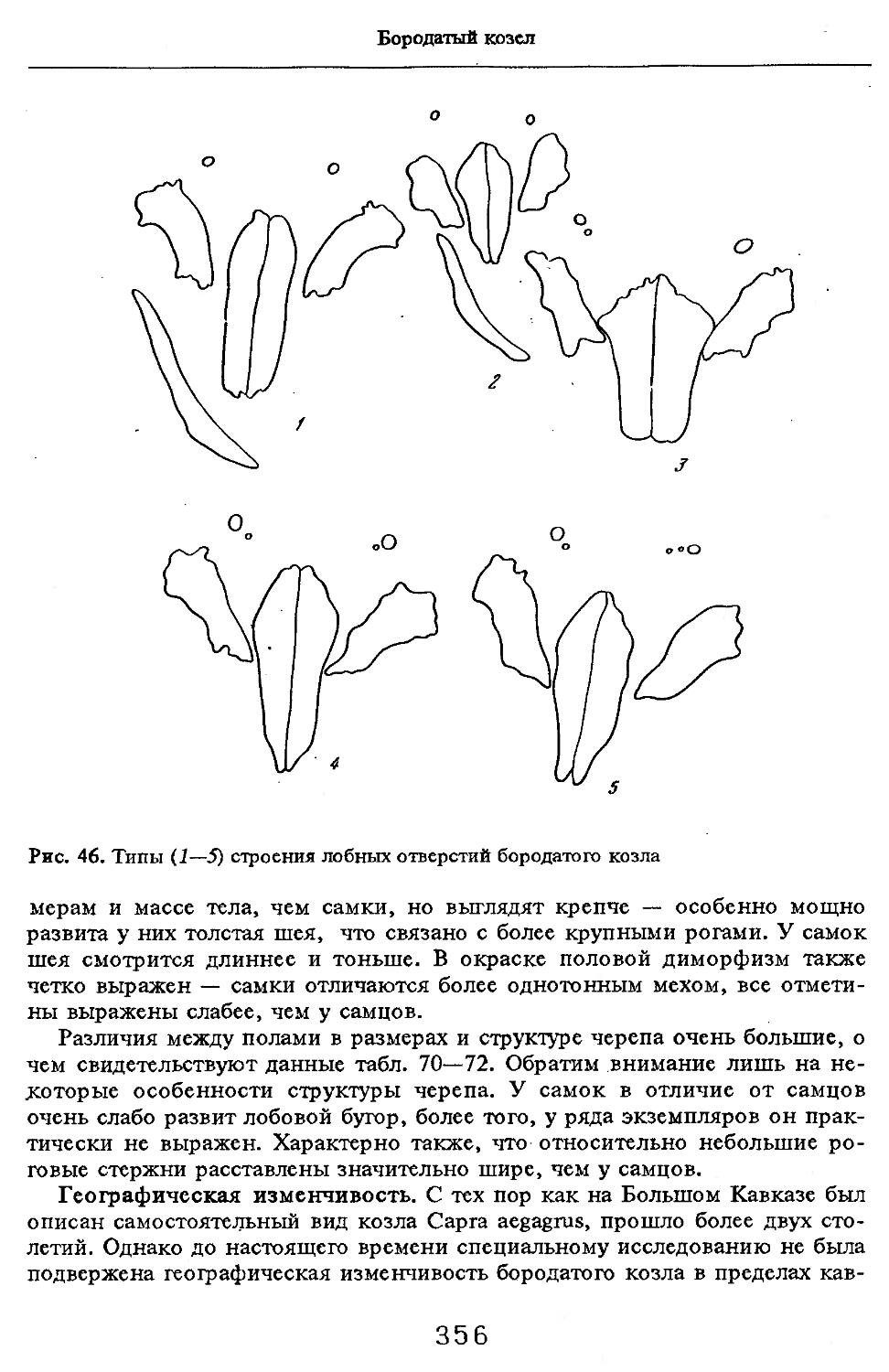
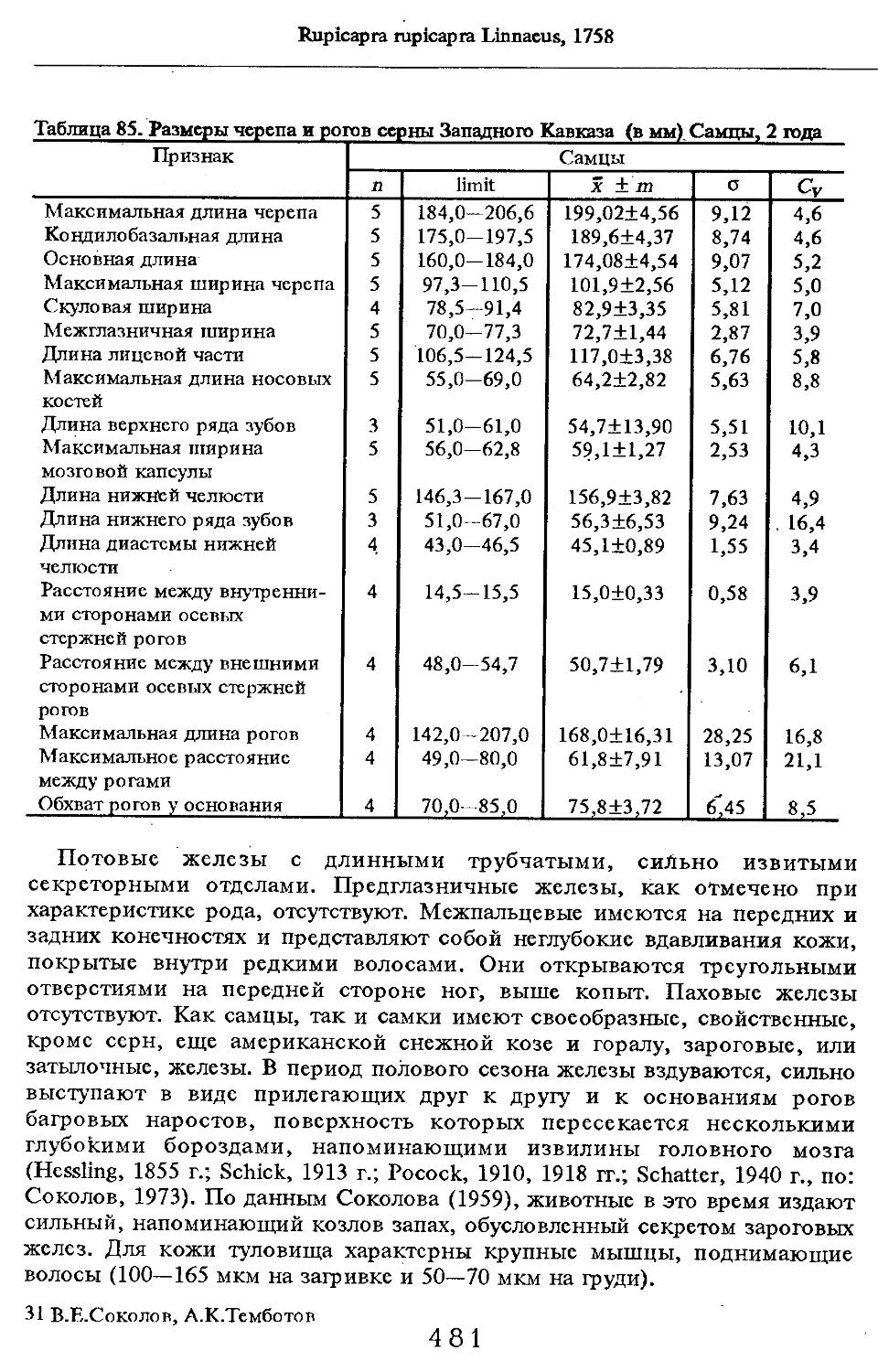
/



























































































































































































































































































































































































































Author: Соколов В.Е. Темботов А.К.
Tags: mammalia млекопитающие копытные
ISBN: 5-02-005450-X
Year: 1993




Text
Позвоночные Кавказа
В.Е.Соколов
А.К.Темботов
Млекопитающие
Копытные
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт эволюционной морфологии и экологии животных им. A JH. Северцова
Позвоночные
Кавказа
В.Е.Соколов
А. К. Темботов
Млекопитающие
Копытные
в
МОСКВА "НАУКА” 1993
УДК 599.33 (479)
Млекопитающие: Копытные // В.Е. Соколов, А.К. Темботов. — М.: Наука, 1993. — 528 с. ISBN5-02-005450-X (Позвоночные Кавказа)
Вторая книга из серии "Позвоночные Кавказа" посвящена копытным и включает описание 12 современных видов, представляющих исключительно важное значение для человека. Для каждого вида обобщены многолетние данные по морфологии, морфо-физиологии, структуре ареала в условиях трехмерного пространства гор, биологии, систематике и закономерностям изменчивости, практическому значению. Значительное место отведено охране уникального генофонда копытных региона.
Табл. 85. Ил. 65. Библиогр. 23 с.
Data based on long-term observations by the authors and on information derived from the literature is presented. Certain specific parameters (morphology, structure of the range, variability and ecology) are considered, both temporally and terms of the three-dimensional space of the mountains, permitting the elucidation of ugulate life features under the specific conditions of the region. Cadastral maps of the ranges for each species are supplemented. The high-altitude limits of species distribution in all the 14 variants of Caucasian zonality are demonstrated in the insets. Information on the practical significance of Caucasian ungulates is summarized and methods for conserving their uniques gene pool and increasing the population of each particular species are proposed.
Ответственный редактор доктор биологических наук Э.В. Ивантер
Рецензенты: академик РАСХН Е.Е. Сыроечковский, доктор биологических наук Л.В. Жирнов
С 1907000000-016.456 — 91- п полугодие
042(02)-93
ISBN5-02-005450-X ® В.Е.Соколов, А.К.Темботов,1993
© Российская академия наук, 1993
ПРЕДИСЛОВИЕ
В жизни людей, населявших территорию Кавказа, значительную роль играли все местные виды копытных млекопитающих. Об этом красноречиво говорят многие факты, в том числе палеонтологические и археологические данные, многочисленные летописи и наскальные рисунки зверей, песни и предания, наконец обширная научная информация, добытая многими поколениями исследователей. Было бы ошибочно полагать, что в век научно-технического прогресса копытные звери потеряли свое значение, что человек может обходиться без них. Наоборот, в самых промышленно развитых странах мира копытные активно вовлекаются в сферу экономической, социальной и духовной жизни. При этом воспроизводство популяций и охрана генофонда поставлены на такую высоту, что сохранение всех видов, в том числе таких гигантов, как зубр и бизон, гарантировано для будущих поколений. Достижение такого уровня природопользования реально только в том случае, если будут познаны закономерности жизни копытных в специфических условиях трехмерного пространства гор и равнин Кавказа.
Между тем многие стороны этой жизни остаются непознанными. Более того, по некоторым формам копытных пробелы в наших знаниях невосполнимы, так как они уничтожены человеком раньше, чем изучены. В этом плане, на наш взгляд, показателен горький опыт с кавказским (аборигенным) зубром. Среди ученого мира спор о существовании зубра на Кавказе был разрешен лишь в 1867 г., когда живой кавказский зубр был доставлен в зоосад Москвы. Последний же кавказский зубр в естественной природе Кавказа был уничтожен в 1927 г. С 1940 г. ведется упорная и очень дорогостоящая работа по восстановлению зубра (уже гибридного!) в регионе, но полученные результаты далеко еще не однозначны. Практический выход всей этой работы, если иметь в виду доход от зубрового хозяйства, мизерный. В Польше, где это дело ведется на должном научном и практическом уровне, за одну голову зубра платят 3 тыс. долл.
Участь кавказского зубра постигла значительно раньше кавказского лося. Тревогу вызывает судьба джейрана, азиатского муфлона, сайгака в Предкавказье и т.д. Не менее остра проблема предотвращения загрязнения генофонда глубоко эндемичного комплекса копытных Кавказа. Во весь рост встал вопрос о сохранении типичных местообитаний для копытных региона. Ведомственный диктат, не выдерживающий даже экономического расчета, грозит непоправимыми последствиями. Достаточно сказать, что акклиматизированный в ряде районов высокогорья Кавказа домашний як "съедает" кавказского тура — уникального эндемика региона, способного давать даже чистой валюты на порядок величин больше, чем яководство в целом.
Нам представляется, что решение теоретических и практических задач во многом сдерживается отсутствием современной монографии по копытным, охватывающей в целом регион. Отдельные работы по тем или иным районам не восполняют пробела, ибо изолированные части не могут обладать
3
Предисловие
свойствами целого — ведь Кавказ с его древней и самобытной фауной сложился как единое целое. Соответственно многие черты морфологических, экологических, биогеографических и других сторон жизни могут быть поняты и учтены лишь при анализе фауны региона как единого целого и при должной оценке местного морфогенеза.
По копытным Кавказа имеется лишь одна монография Н.Я.Динника (1910), которая стала библиографической редкостью и устарела по многим аспектам. Однако значение ее для понимания копытных региона на сегодняшний день трудно переоценить. Большой интерес представляет также монография Н.К.Верещагина (1959), особенно по вопросам истории формирования териофауны Кавказа. Неоценимую роль в изучении копытных сыграли заповедники, из числа которых особенно выделяется Кавказский заповедник. Значительный вклад внесли академические учреждения союзных республик, многие вузы региона, различные научно-практические учреждения и т.д. Немало исследований выполнено такими широко известными териологами страны, как А.Я.Гюльденштедт, П.С.Паллас, А.А.Насимович, В.Г.Гептнер и др.
Особое место в познании териофауны Кавказа, в том числе копытных, занимает коллекционный материал, накопленный многими поколениями специалистов и хранящийся в музеях страны. Среди них особенно велика роль зоомузеев Московского университета и Зоологического института РАН.
Значимость всей этой информации состоит не только в том, что она позволяет подвести итог определенному этапу познания копытных Кавказа, но и обнажает актуальные проблемы и подходы к их решению. Мы надеемся, что это почувствуют читатели данной монографии. Что касается авторов, то мы, как никогда, представляем себе, насколько велика и сложна проблема познания и сохранения уникального генофонда копытных Кавказа для нынешних и будущих поколений.
Выполнение настоящей работы стало реальным благодаря не только нашим многолетним собственным исследованиям, но и работе многих наших коллег, дополнивших трехвековые данные по териофауне Кавказа новыми оригинальными сведениями. Особо хотелось поблагодарить наших постоянных помощников, в том числе Л.М.Арефи, Ф.К.Абидову, Е.А.Барагунову, Р.И.Дзуева, Е.А.Калинину, А.Х.Мамбетова, Т.Х.Срукову, Ф.А. и Э.Ж.Тембо-товых, М.А.Хуштову, О.Ф.Чернову.
Мы выражаем благодарность коллегам, предоставившим первичные данные: В.С.Громову, А.А.Данилкину, В.В.Дурову, А.С.Немцеву, а также Ю.М.Смирину, подготовившему рисунки животных по оригинальным наброскам в природе В.М.Смирина.
В ходе подготовки рукописи к изданию ряд советов был высказан Э.В.Рогачевой, Л.В.Жирновым и Е.Е.Сыроечковским и с благодарностью нами принят.
Artiodactyla Owen, 1848
ОТРЯД ПАРНОКОПЫТНЫЕ ARTIODACTYLA OWEN, 1848
Представители этого отряда характеризуются тем, что III и IV пальцы конечностей, выполняя всю или основную функцию опоры тела, сильно развиты. Остальные пальцы в различной степени редуцированы и располагаются выше средних. Ось конечностей непременно проходит между III и IV пальцами. Все концевые фаланги, независимо от их размеров и расположения, несут копыта.
Внешний облик животных разнообразный, но преобладают виды, адаптированные к быстрому бегу и соответственно длинноногие. Среди горных есть виды, способные к лазанию по скалам. Размеры тела сильно варьируют: высота в холке от 25 см и длина тела от 52 см (карликовые антилопы) до 3 м в холке при общей высоте до 6 м и около 3 м в длине тела (жирафы). Масса тела колеблется от 2—3 до 3200 кг.
Волосяной покров неоднотипный, в большинстве случаев образован остевыми волосами, имеющими хорошо развитую воздухоносную сердцевину. У отдельных видов волосяной покров щетинистый. Есть виды, почти лишенные волос.
Кожные железы хорошо развиты, нередки большие их скопления.
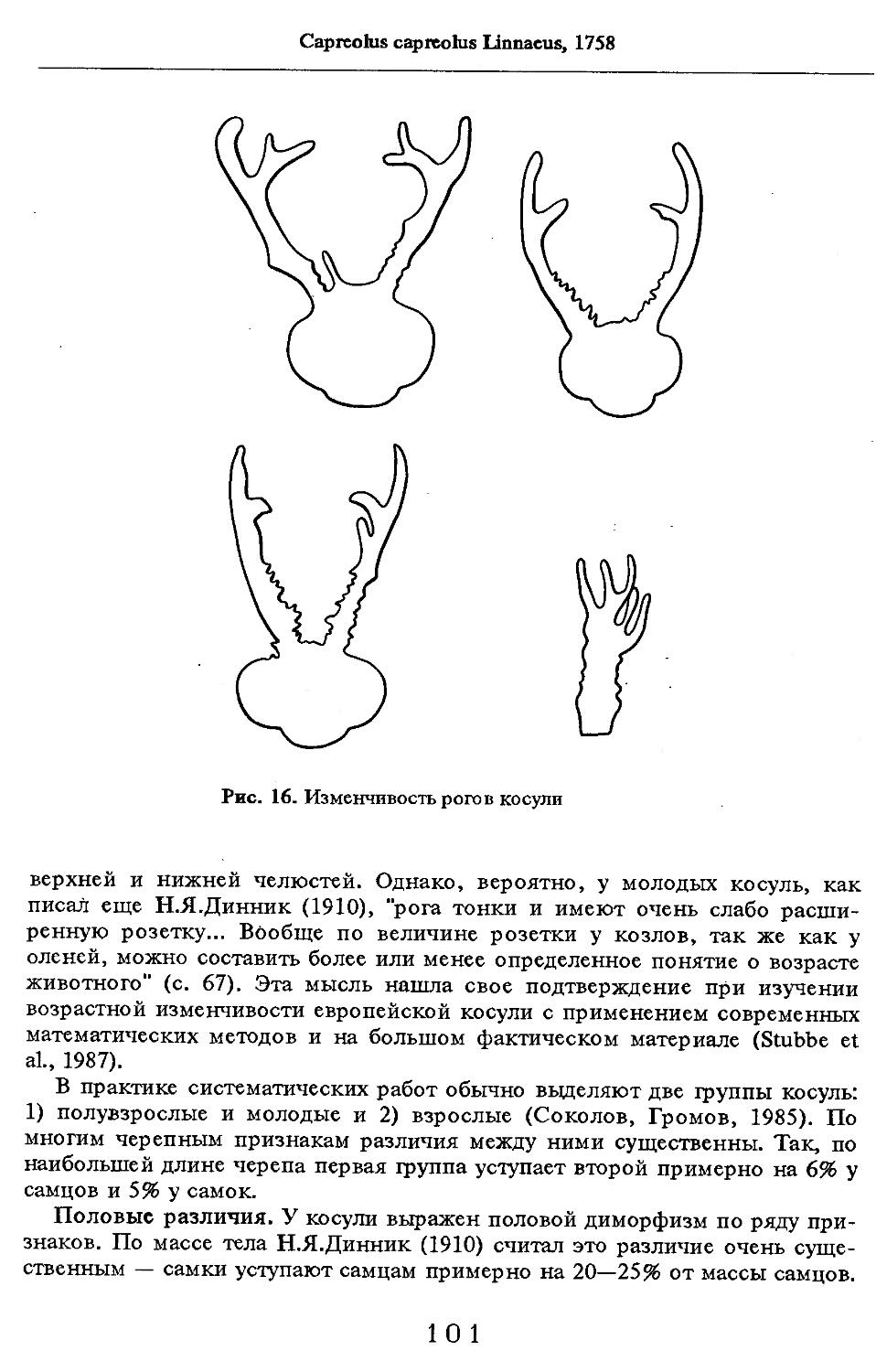
Скелет имеет ряд особенностей. Лицевая часть черепа удлиненная, нередко клиновидная, что связано с увеличением длины челюстей, сужением их передней части. Не менее показательно отсутствие в основаниях крыловидных отростков базисфеноидов can. pterygoideus. Глазница отделена от височной ямы сросшимися отростками лобной и скуловой кости или она остается открытой в той или иной степени. Хорошо развита слезная кость, костный слуховой проход. Хоаны укорочены, редко когда передним краем они заходят дальше М^. Архитектоника нижнечелюстного сочленения с черепом позволяет нижней челюсти совершать движения как боковые, так и переднезадние. На черепе одного или обоих полов многих видов имеются рога постоянные (роговые, на костном стержне) или ежегодно сменяемые (рога костные).
Зубная система подвержена широкой изменчивости. В наиболее полном виде она включает 44 зуба:
3 14 3
I - С - Р — м -.
3 14 3
Аналогичную формулу зубов имеют представители Suidae. У других групп наблюдается редукция резцов верхней челюсти, клыков, предкоренных. В нижней челюсти нередко клыки принимают форму резцов, отделяются от предкоренных диастемой.
В структуре зубов характерно увеличение количества бугорков (с 3 до 4—5, иногда 6), приобретающих округлую форму или вид полулунного гребня.
5
Отряд Парнокопытные
В позвоночном столбе 19—20 пояснично-грудных и 4 крестцовых позвонков. Хвостовой отдел у многих сильно редуцирован, у других может быть хорошо развит (быки). Характерно отсутствие ключицы, третьего вертела на бедре, редуцируются в размере и сокращаются в количестве боковые лучи кисти и стопы, резко усиливаются средние пальцы, удлиняются метапо-дии и фаланги пальцев, образуется цевка (os cannon), что обеспечивает переход от сто похождения к пальце- и фалангохождению. У значительного количества видов локтевая кость недоразвита и срощена с лучевой, так же как малая с большой берцовой.
В связи с питанием травянистой, нередко сухой и 1рубой пищей, а также веточными кормами в зубной и пищеварительной системе происходят существенные изменения. Изменяется ряд коренных и предкоренных зубов, усложняется их жевательная поверхность (гипсодонтизм), желудок обычно сложный с 2—4 камерами, отделенными друг от друга в той или иной степени. Слепая кишка развита, но у отдельных ipynn отсутствует. За исключением семейства оленьих, у парнокопытных имеется желчный пузырь.
Матка двурогая, плацента диффузная или поликотиледонтная. Молочные железы паховые, двух- или четырехдольчатые, редко множественные. Сосков 1—2 пары, иногда до 5—6 пар (свиньи). В году приносят 1—2, реже 4—7 детенышей, иногда количество их превышает 20 (свиньи). Они рождаются обычно хорошо развитыми. В году бывает одна-две линьки.
Копытным свойственна сложная социальная организация. Четко выражены стадность, различные 1руппировки, полигамия, соподчинение особей в группах, территориальность, сезонные и иные перемещения как в горах, так и на равнине; свойственны различные коммуникации, в том числе акустические, химические и др.
Распространение обширное, охватывает Европу, Азию, Африку, Мадагаскар, Северную и Южную Америку, Гренландию, ряд островов Северного Ледовитого океана (Новая Земля, Северные острова, Новосибирские острова и др.). Копытные акклиматизированы и успешно живут в Австралии, Новой Зеландии и многих других районах южного полушария.
Парнокопытные появились в нижнем эоцене Северной Америки и Европы. Свое начало они берут от кондилартр (Condylarthr), тесно связанных с примитивными хищниками. В ходе длительной эволюции парнокопытные приспособились к всевозможным ландшафтным условиям — от тропических до субарктических на равнине, а в горах до субнивального пояса включительно. В итоге образовались весьма разнообразные жизненные формы и таксоны.
Отряд парнокопытных в плане классификации — "трудная" группа, по ряду вопросов единого мнения нет. Это касается как макро-, так и микросистематики. Мы придерживаемся точки зрения, согласно которой в отряде 2 подотряда: нежвачные (Suiformes) и жвачные (Ruminantia). Количество семейств не менее 8: свиные Suidae, пекариевые Tayassuidae, бегемотовые Hippopotamidae, оленьковые Tragulidae, оленьи Cervidae, жирафовые Giraf-fidae, вилороговые Antilocapridae, полорогие Bovidae. Родов 68, видов 171
6
Suidac Gray, 1821
(Соколов, 1979, 1984). Из них в фауне Кавказа представители 3 семейств: свиные, оленьи и полорогие.
Семейство Свиные — Suidae Gray, 1821
Парнокопытные среднего размера с плотным, массивным и несколько сжатым с боков телом, клиновидной головой, которая заканчивается малоподвижным усеченным хоботком с дисковидным терминальным хрящом (пятачок), пронизанным двумя ноздрями. Ноги короткие, четырехпалые, средние пальцы (III и IV) крупные (несущие), а два боковых сдвинуты назад и в обычных условиях не касаются земли, лишь при ходьбе по мягкому грунту животные опираются на них. Ушные раковины хорошо развиты, заостренные, часто с кисточкой на верхнем конце, глаза сравнительно маленькие.
Тело у ряда видов голое, другие виды имеют густой щетинистый покров с подпушью, на верхней стороне шеи и по хребту имеются удлиненные остистые волосы, образующие в холке гриву. Хвост у них относительно длинный, в его дистальной части сильно развиты волосы, образующие кисточку. В
норме окраска тела взрослых животных однотонная, редко пестрая, у молодых полосатая (за исключением бабируссы и домашней свиньи). .Соски брюшные или паховые и брюшные (их от 3 до 6 пар, лишь у бабирусс 1 пара). Специфические кожные железы немногочисленны (Соколов, 1973).
Череп оригинальный — обычно клиновидный, образуется за счет постепенного сужения с боков и снижения верхнего профиля черепа от затылка к носу. Относительно плоские и широкие лобные кости плавно переходят в сужающиеся и поднимающиеся назад теменные кости. Глазница сзади открытая. Слезная кость крупная, обычно с двумя отверстиями для слезного канала. Крыловидная ямка хорошо выражена. Лобные, теменные и верхнезатылочные кости пневматизированы. Костное нёбо длинное, значительно выходит за пределы задней границы зубных рядов. Костные слуховые барабаны увеличены и удлинены сверху вниз. В том же направлении сильно
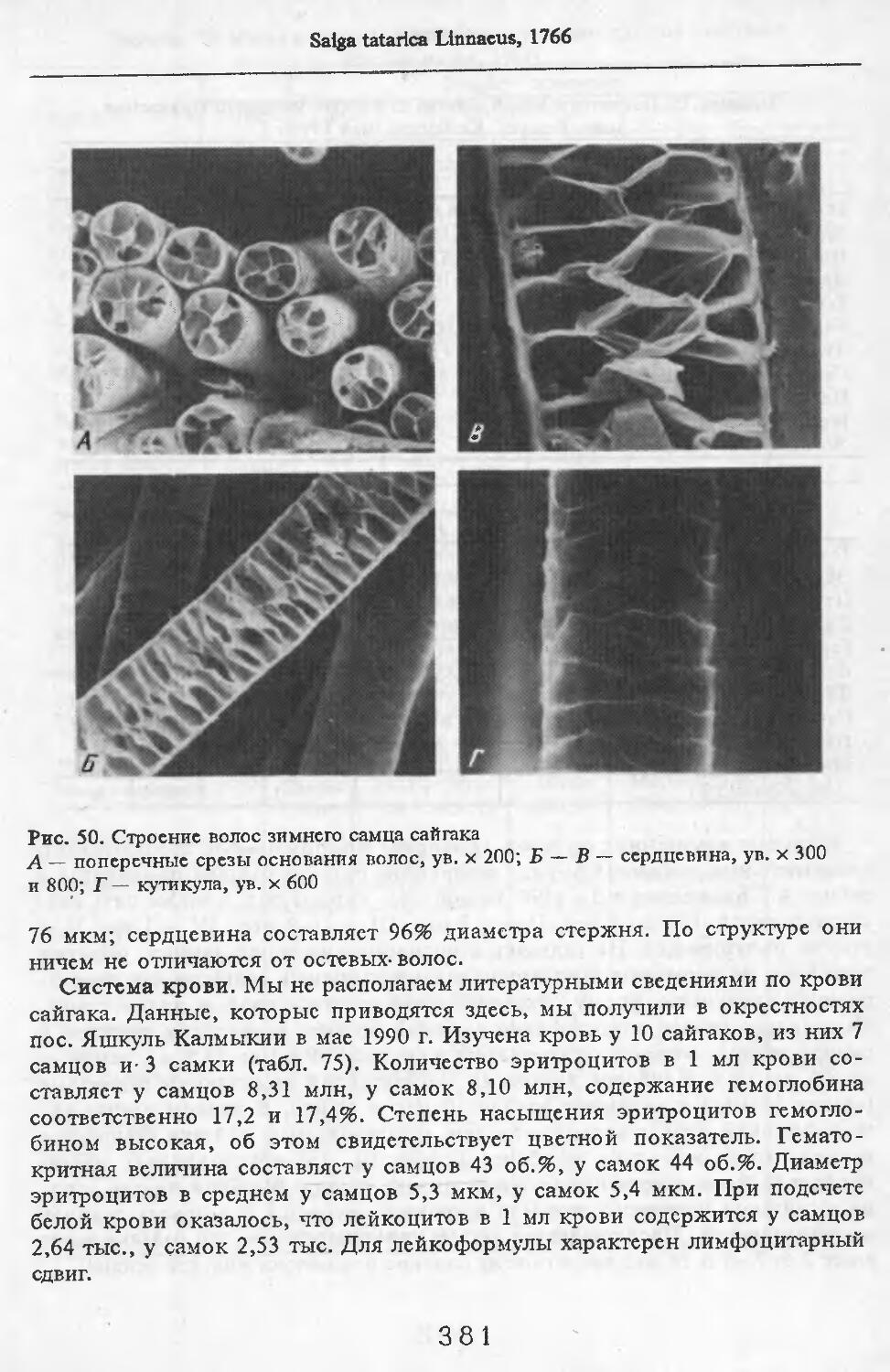
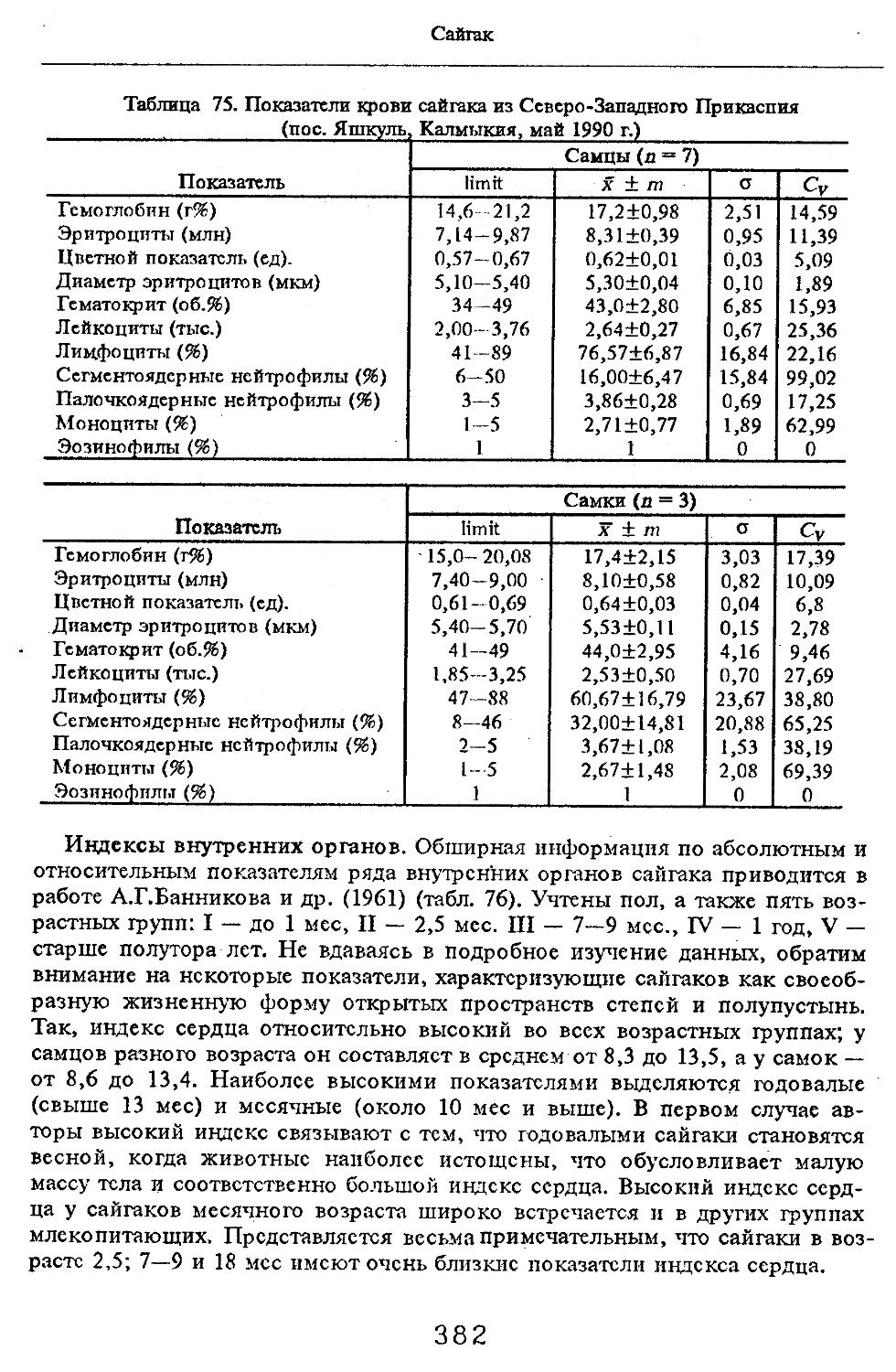
вытянуты сосцевидные отростки.
Зубная формула: л — 3) ] (3 — 4) 3
I-------С - Р --------- М -
(2 — 3) 1 (2 — 4) 3
= 32-44.
Резцы с корнями прилегают плотно друг к другу, расположены почти горизонтально, по размерам составляют убывающий ряд от внутренних к наружным. Верхние клыки крупнее нижних, особенно у самцов. Они растут наружу и назад, а нижние — вверх и назад. Верхние и нижние клыки растут друг против друга, образуя острый угол (у бабируссы клыки растут несколько иначе). Щечные зубы бунодонтные и брахиодонтные. В порядке исключения встречаются гипсодонтные коренные и горизонтальная смена зубов. С возрастом изнашивается эмаль, все зубы исчезают, кроме клыков и последних коренных. В посткраниальном скелете характерно следующее: метапо-
дии на всех конечностях не имеют даже тенденции к слиянию, самостоятельны локтевая и малая берцовая кости. Грудных позвонков 14, поясничных 5—6, крестцовых 4—5.
7
Семейство Свиные
Желудок простой, двухкамерный с небольшим дополнительным кардинальным отделом; слепая кишка простая и короткая. Имеется желчный пузырь. Матка двурогая, плацента диффузная.
Ареал занимает значительную территорию Старого Света — умеренные и южные широты Евразии, Африку с Мадагаскаром. Свиные акклиматизированы в Северной и Южной Америке, Новой Зеландии, Австралии, Тасмании, на Новой Гвинее, на Британских островах.
Приурочены к равнинным и горным лесам, лесостепным и степным ландшафтам, предпочитая влажные, порой заболоченные с тростниковой и древесно-кустарниковой растительностью. Активны преимущественно ночью, держатся труппами до 40—50, иногда до 100 и более особей, однако самцы бородавочника ведут одиночный образ жизни. Звери нередко посещают грязевые ванны. Могут хорошо плавать в воде.
Всеядные, но предпочитают растительные корма, особенно концентрированные (желуди, орешки, плоды, клубни и др.). Полигамны в различной степени, в году дают одно, феже два потомства с 1—14 детенышами в одном помете. Продолжительность беременности 112—175 дней. Лактационный период от 2 до 4 мес. Рост и развитие детенышей интенсивные, половозрело-ста достигают в возрасте 9—15, реже 19 мес. Живут до 20 лет.
Практическое значение разнообразно. Животные служат традиционным объектом охоты, дающим высококалорийное мясо и жир. Вероятно, значительна их биоценотическая роль, но этот аспект изучен недостаточно. Не менее интересна возможность более широкого использования в селекционной работе. Звери могут приносить определенный вред полеводству, садоводству и лесной культуре. Заслуживают внимания и в плане эпизоотологии, ибо они, в частности кабаны, болеют очень многими вирусными, бактериальными и паразитарными заболеваниями (Blancou et al., 1987), возможность передачи которых человеку и домашним свиньям не исключена.
Suidae — это одна из примитивных групп среди нежвачных, представители вымерших подсемейств известны с нижнего олигоцена Европы. Подсемейство Suinae возникло в нижнем плиоцене Евразии. Эволюция шла в сторону увеличения в размерах и превращения брахиодонтных клыков в постоянно растущие, а также усложнения структуры коренных зубов. В настоящее время отдельные виды, проявляя высокую пластичность, легко уживаются с культурой человека.
Семейство Suidae включает 5 родов, из которых на Кавказе обитает один род Sus (кабаны) (Hoffman, 1958; Соколов, 1984).
Род Кабаны — Sus Linnaeus, 1758
Род объединяет всего 4 вида, однотипных по внешнему облику, но разнообразных по величине: от карликовой свиньи (Sus salvanius) с длиной тела 50—65 см, высотой в холке 25—30 см до кабана (Sus scrofa) длиной тела до 205 см и высотой в холке до 120 см.
Голова длинная, до 1/3 длины тела, уши длинные и широкие, глаза маленькие и глубоко сидящие. Передняя часть туловища заметно сплюснута с
8
Sus Linnaeus, 1758
боков, выше и мощнее, чем задняя его часть. Характерно, что средние пальцы способны раздвигаться и при участии боковых пальцев обеспечивается снижение массовой нагрузки на 1 см2 приблизительно в 1,5 раза, т.е. с 500 до 320—390 г (Соколов, 1979).
Кабаны имеют волосяной покров, состоящий из щетины, остевых и пуховых волос. Сильно удлиненные волосы по бокам головы у многих животных образуют "баки", а на шее и хребте "гриву". Линяют один раз в году в весенне-летний период. Окраска тела темно-серая до бурой или черной, иногда со светлой отметиной на морде. Молодые обычно полосатые, наряд взрослых приобретают по достижении четырехмесячного возраста. Соски брюшные и паховые, в количестве 3—6 пар..
Череп длинный, узкий, клиновидный, характерно отсутствие сагиттального гребня. Лобно-теменная область имеет продольно-выпуклую поверхность. Зубная формула полная и этой особенностью род хорошо выделяется из других родов семейства:
3 14 3
I — С - Р — М — = 44.
3 14 3
Даже у молодых могут отсутствовать третья пара резцов и первый верхний предкоренной. При этом последний не имеет молочного предшественника и появляется лишь одновременно с последним коренным. Клыки с незамкнутыми корнями и растут постоянно. У самцов они значительно крупнее, чем у самок. Коренные зубы брахиодонтные.
Ареал охватывает умеренные и южные районы Евразии, Северную Африку, Японию, Тайвань, Хайнань-, Малайский архипелаг до Филиппин, Сулавеси, Молуккские острова, Новую Гвинею и Меланезию (Соколов, 1979). Ландшафтная и биотопическая приуроченность сходна у всех представителей и характеризуется высокой пластичностью. Кабаны занимают пространство от зоны тайги до тропических лесов, явно тяготея к влажным местообитаниям, в том числе густым тростниковым и кустарниковым зарослям по берегам водоемов.
Характеризуются стадным образом жизни, но многие параметры группировок широко меняются в зависимости от внешних и внутренних факторов (сезон года, годовые колебания условий жизни, местообитания, кормовая база, антропогенные факторы, пол, возраст, физиологическое состояние и т.д.). Совершают различные миграции, протяженность которых варьирует в зависимости от конкретных условий. Кабаны всеядны, с широким кругом кормов растительного и животного происхождения. Характерна ограниченная полигамия. В континентальных районах размножение сезонное, а в тропиках (например, бородатая свинья) .сезонности в размножении нет. В помете от 3 до 14 поросят, что зависит от вида, возраста, кормовой базы и т.д. Половозрелость физиологическая наступает на первом году жизни, но самки обычно вступают в размножение на втором году жизни (18—20 мес), а самцы — на четвертом или пятом году. Продолжительность жизни в природе 10—12 лет, в неволе до 20 лет.
Враги — волки, тигры и другие хищные, особенно страдают от них молодые кабаны.
9
Sus Linnaeus, 1758
Хозяйственное значение велико (см. обзор семейства).
Литературные сведения по систематике рода Sus противоречивы; вероятно, наиболее объективно признание четырех видов (Hoffman, 1958; Соколов, 1979, 1984), из которых на Кавказе один вид — Sus scrofa L.
Кабан — Sus scrofa Linnaeus, 1758
Рис. 1
Sus scrofa ferus Gmelin, Гмелин, 1876; Sus europaeus Pallas, Паллас, 1811; Sus scrofa attila Thomas, Адлерберг, 1930.
Коллекционный материал. ЗИН РАН: 43 черепа; МГУ: 40 черепов; КБГУ: 34 черепа, 2 скелета, 50 шкур; КГЗ1 — 140 черепов.
Диагноз. Нежвачные средних и крупных размеров с большой (около 1/3 туловища) клиновидной головой, без перехвата в области предлобья, функционирующими IV и V пальцами; покров из длинных и жестких волос, образующих "гриву"; череп постепенно сужающийся спереди, клыки хорошо развиты, особенно у взрослых самцов.
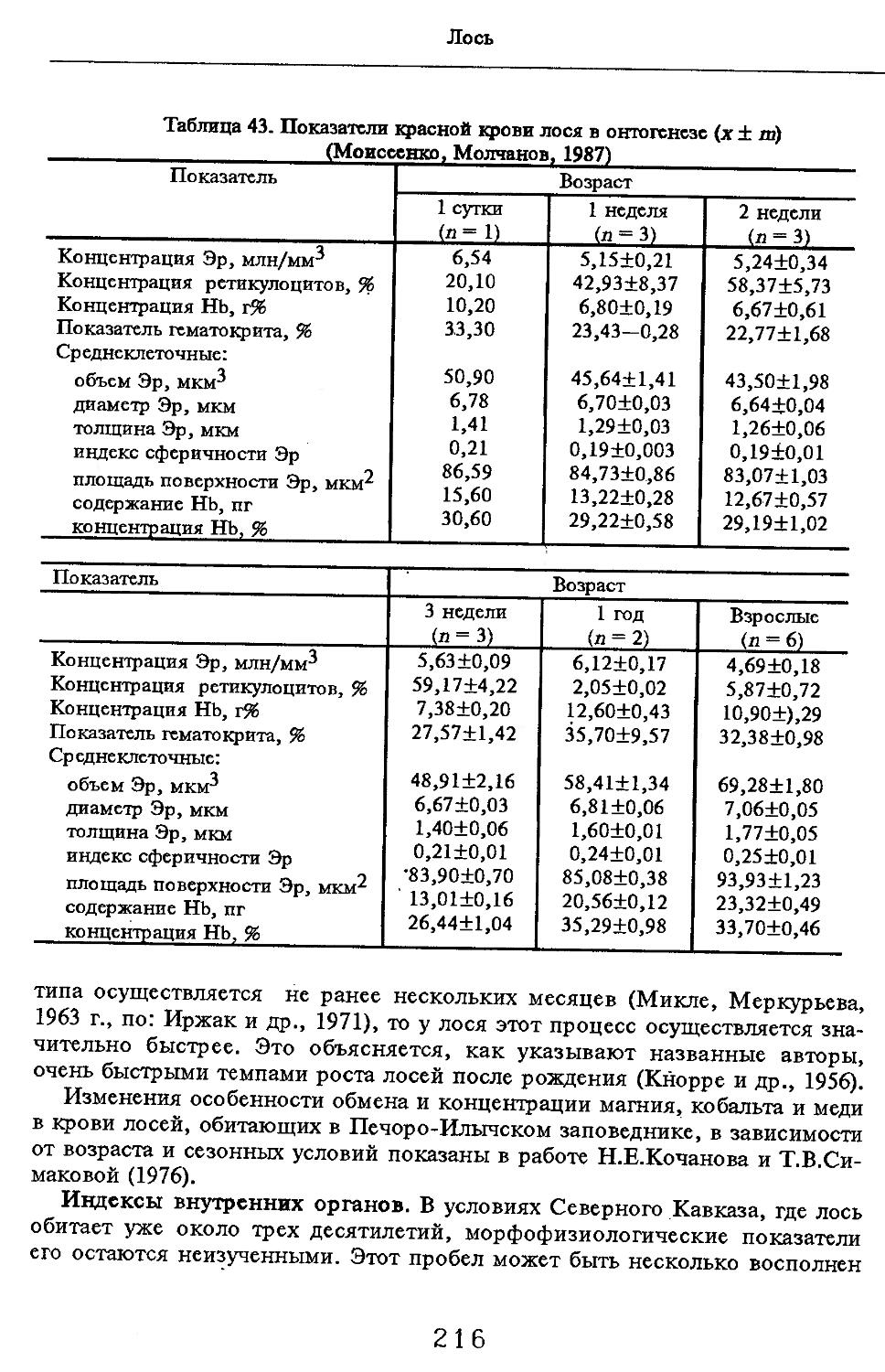
Особенности морфологии и морфофизиологии
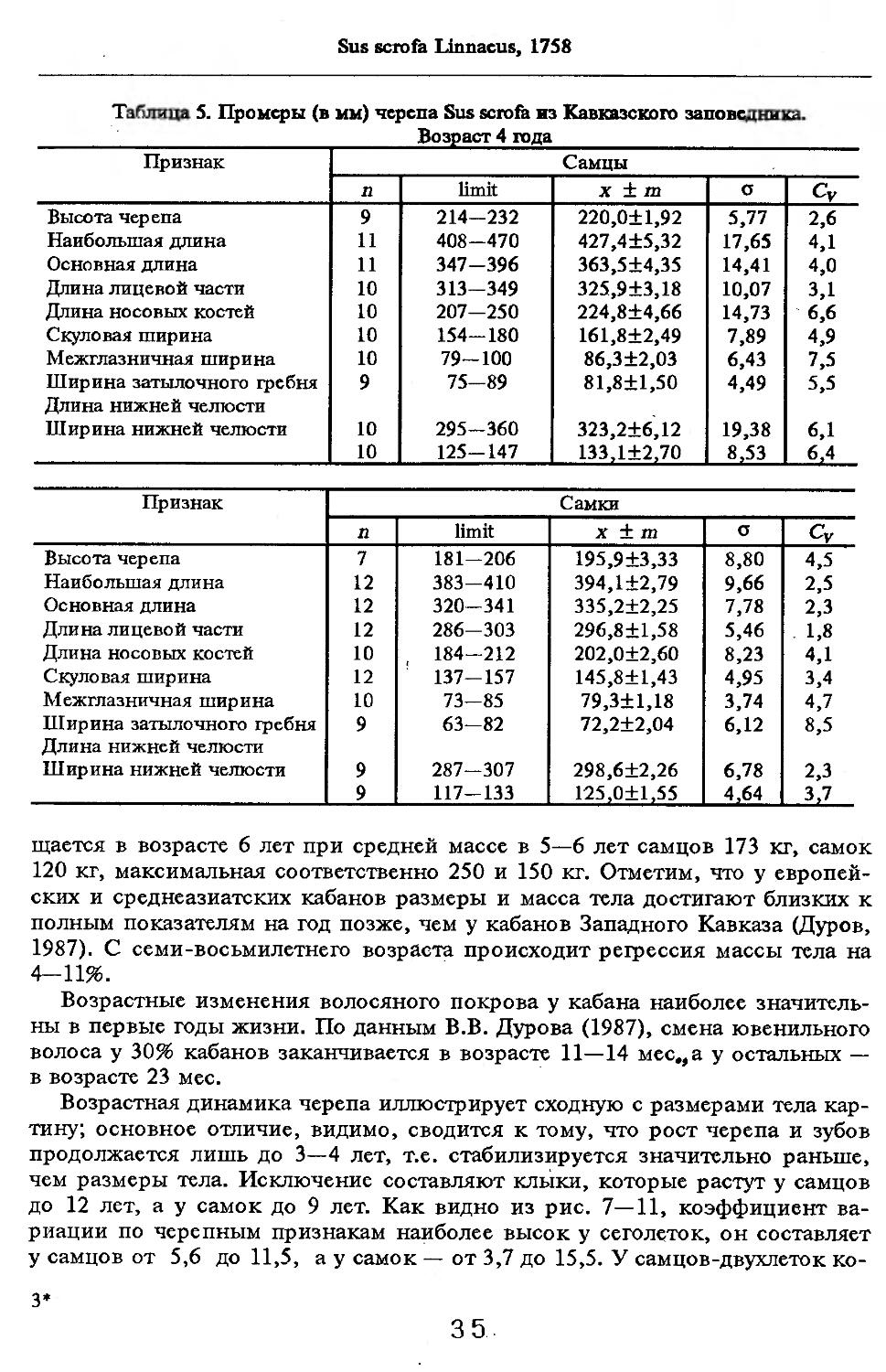
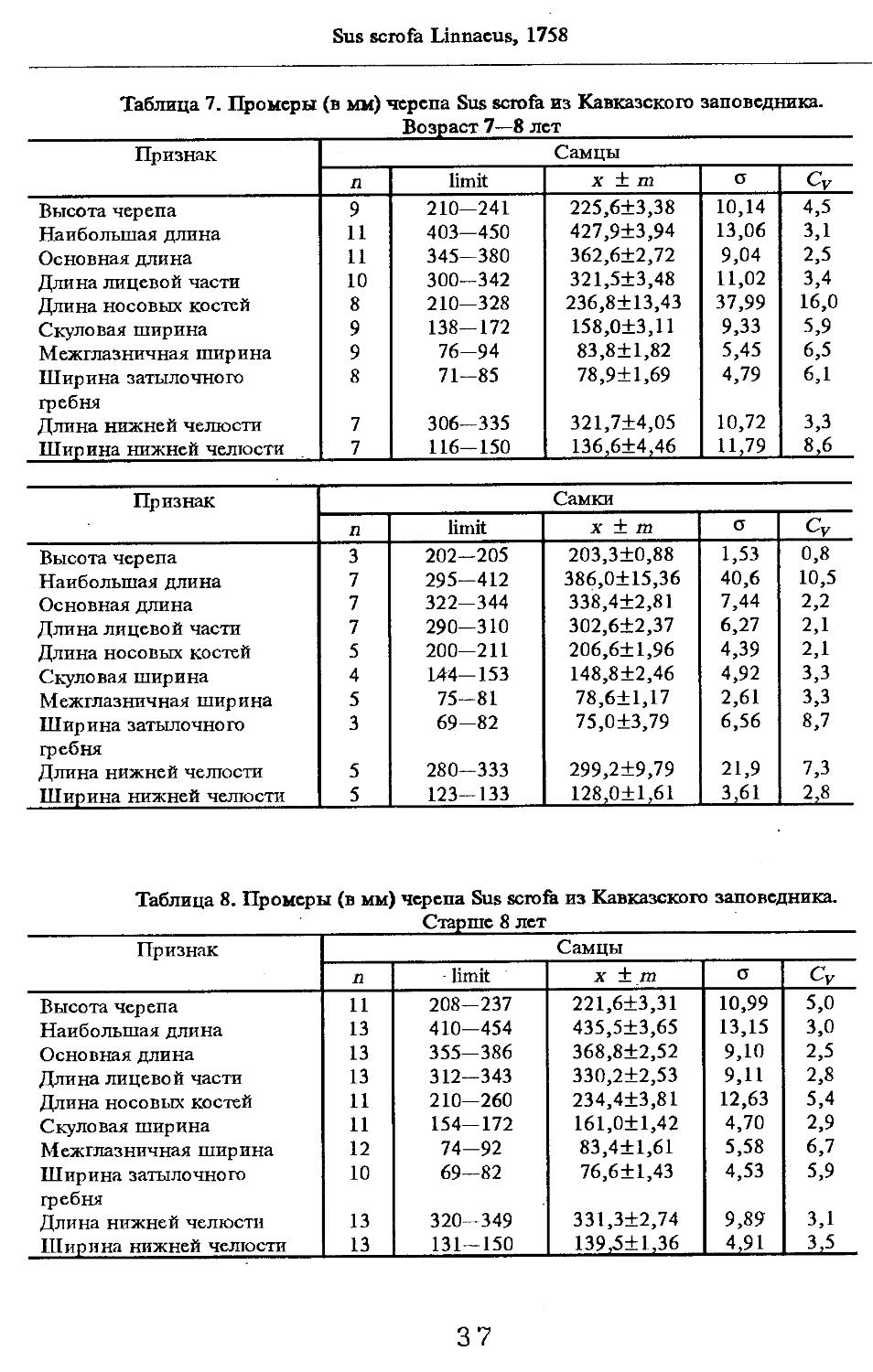
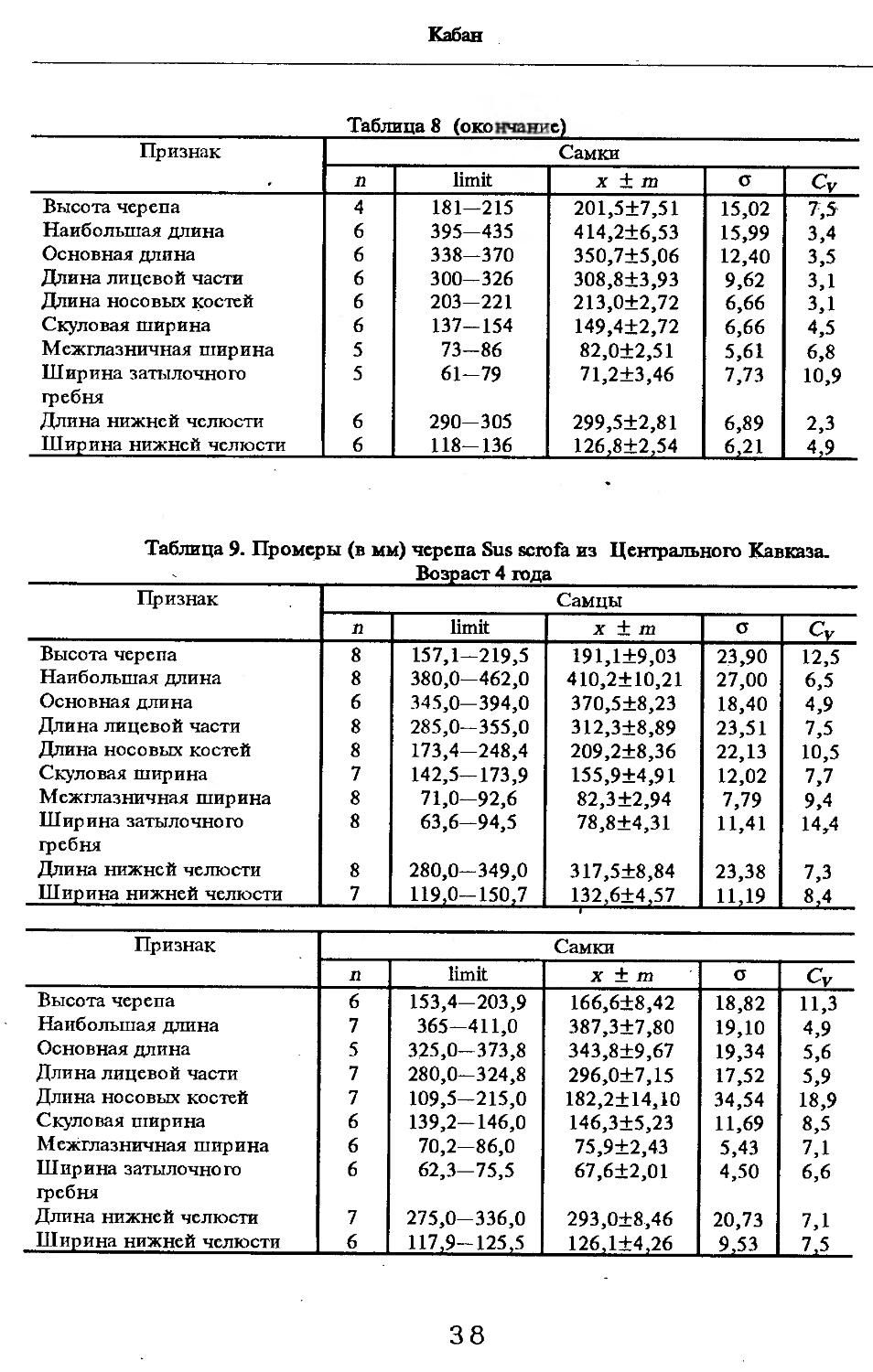
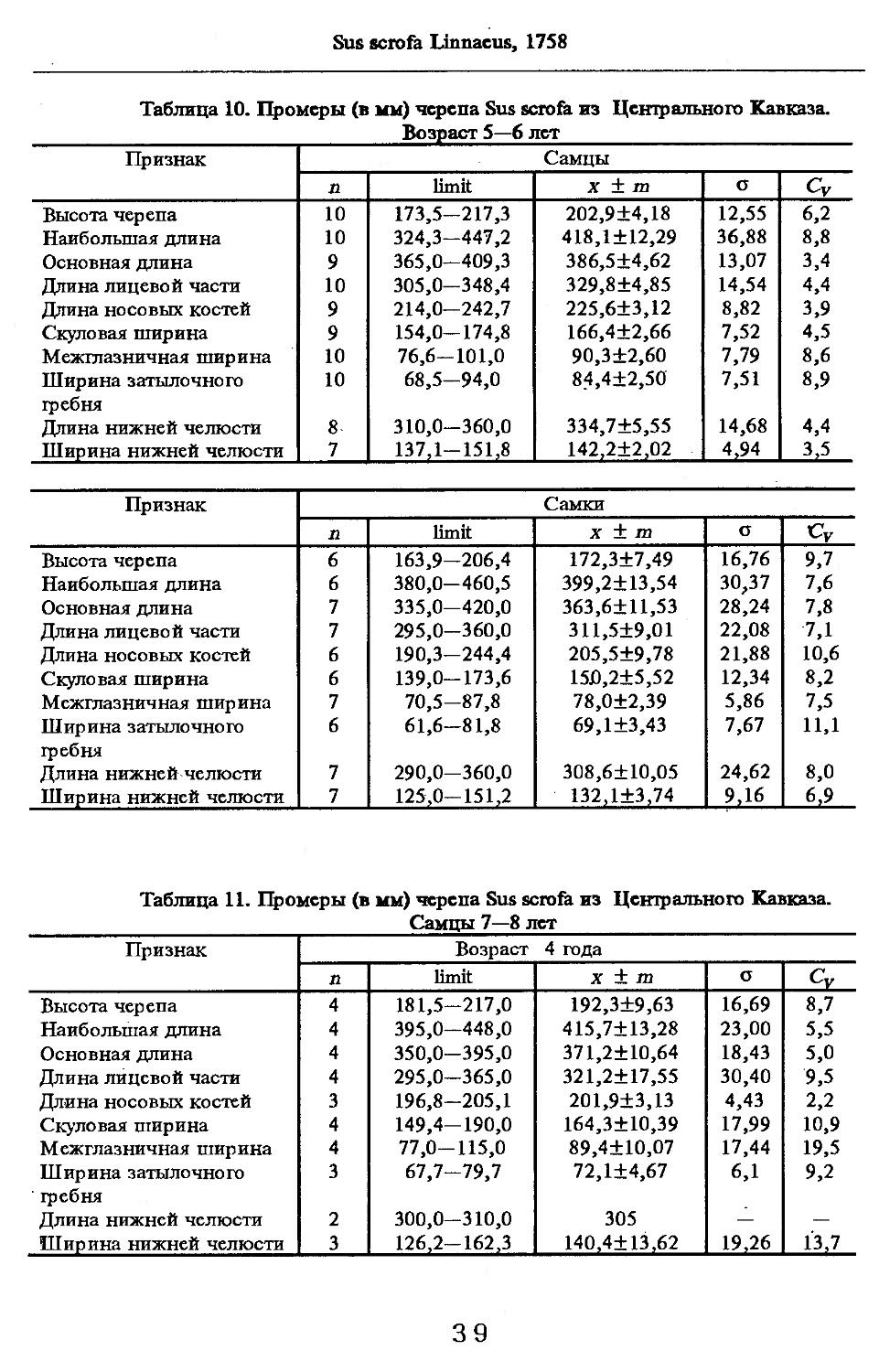
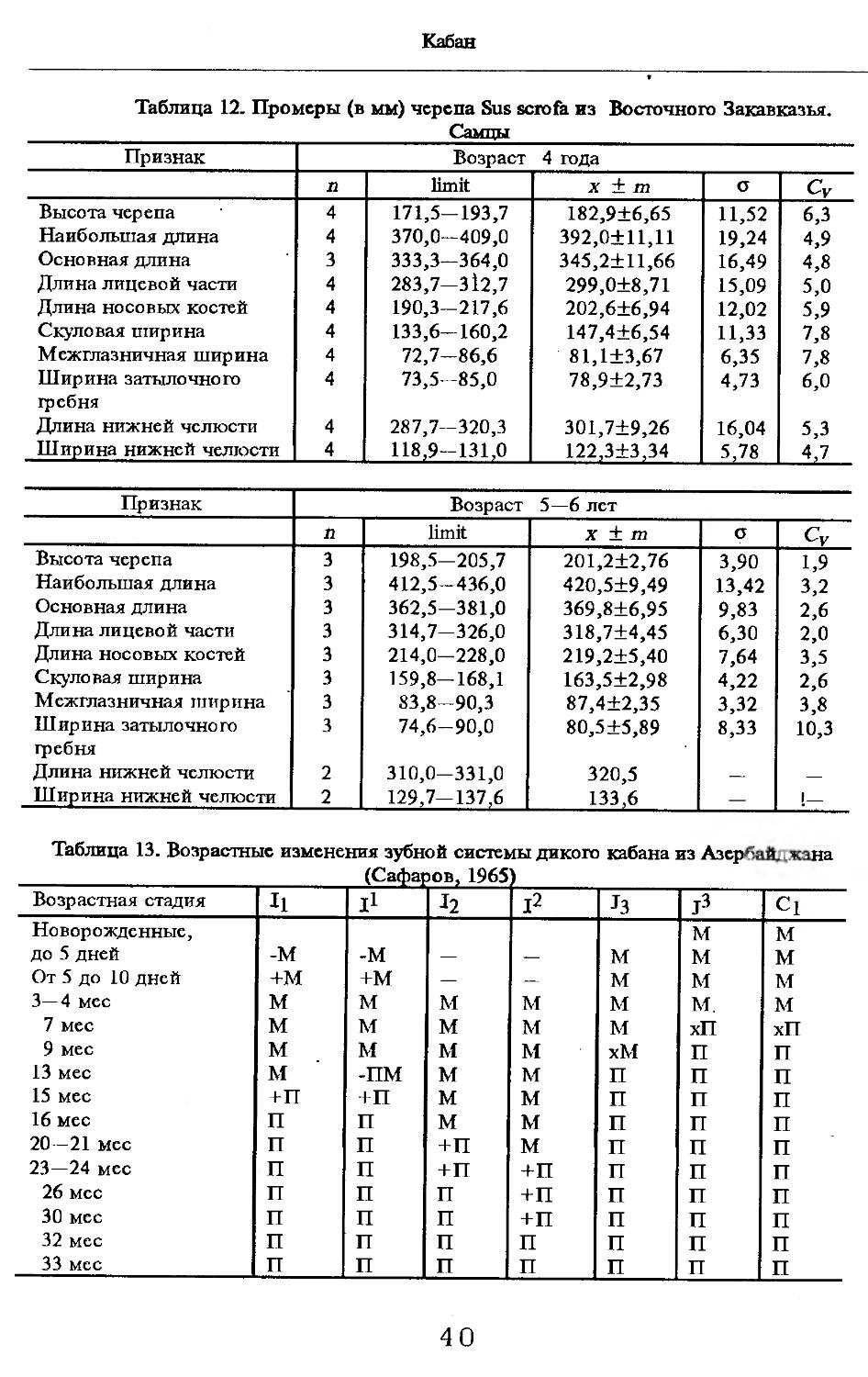
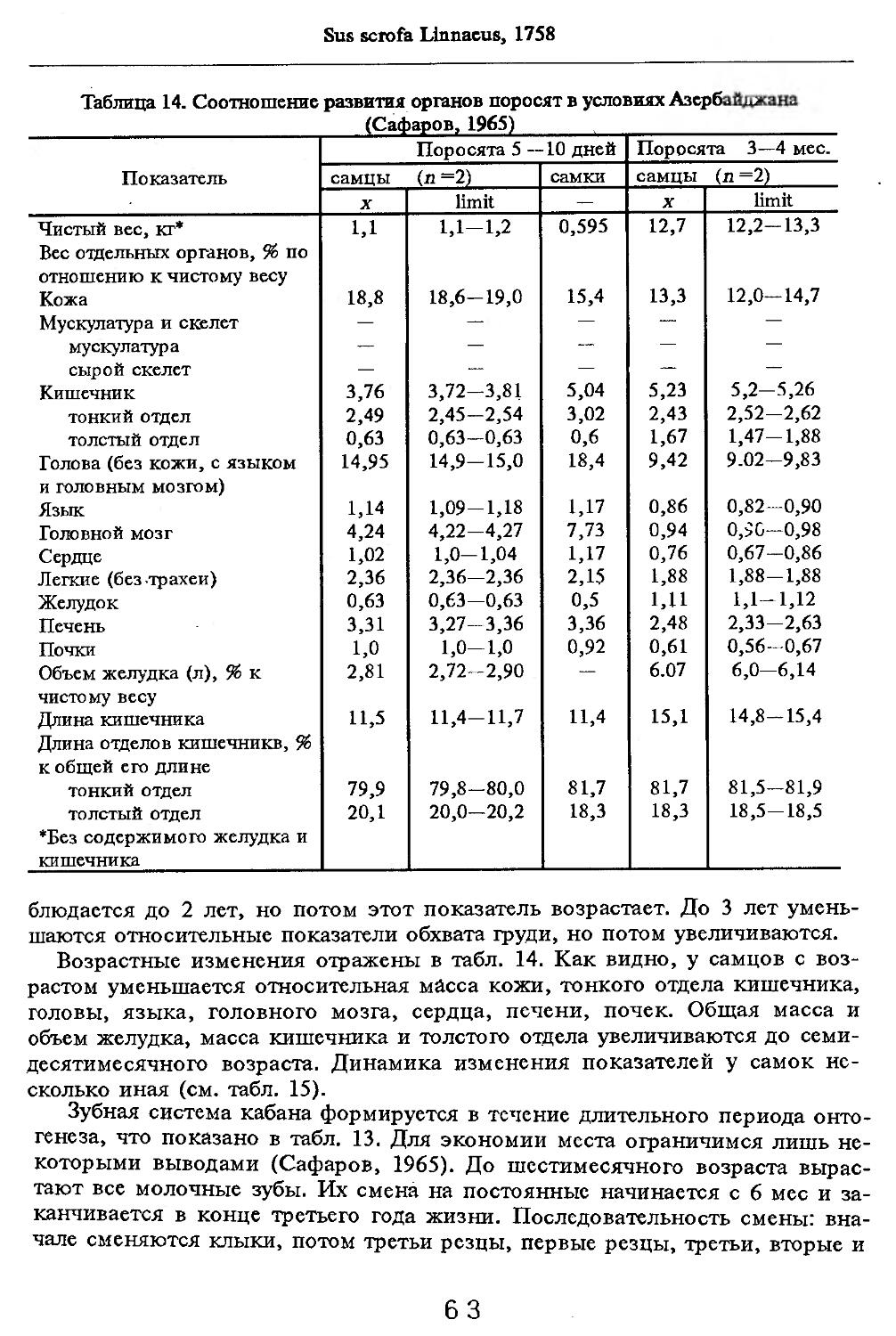
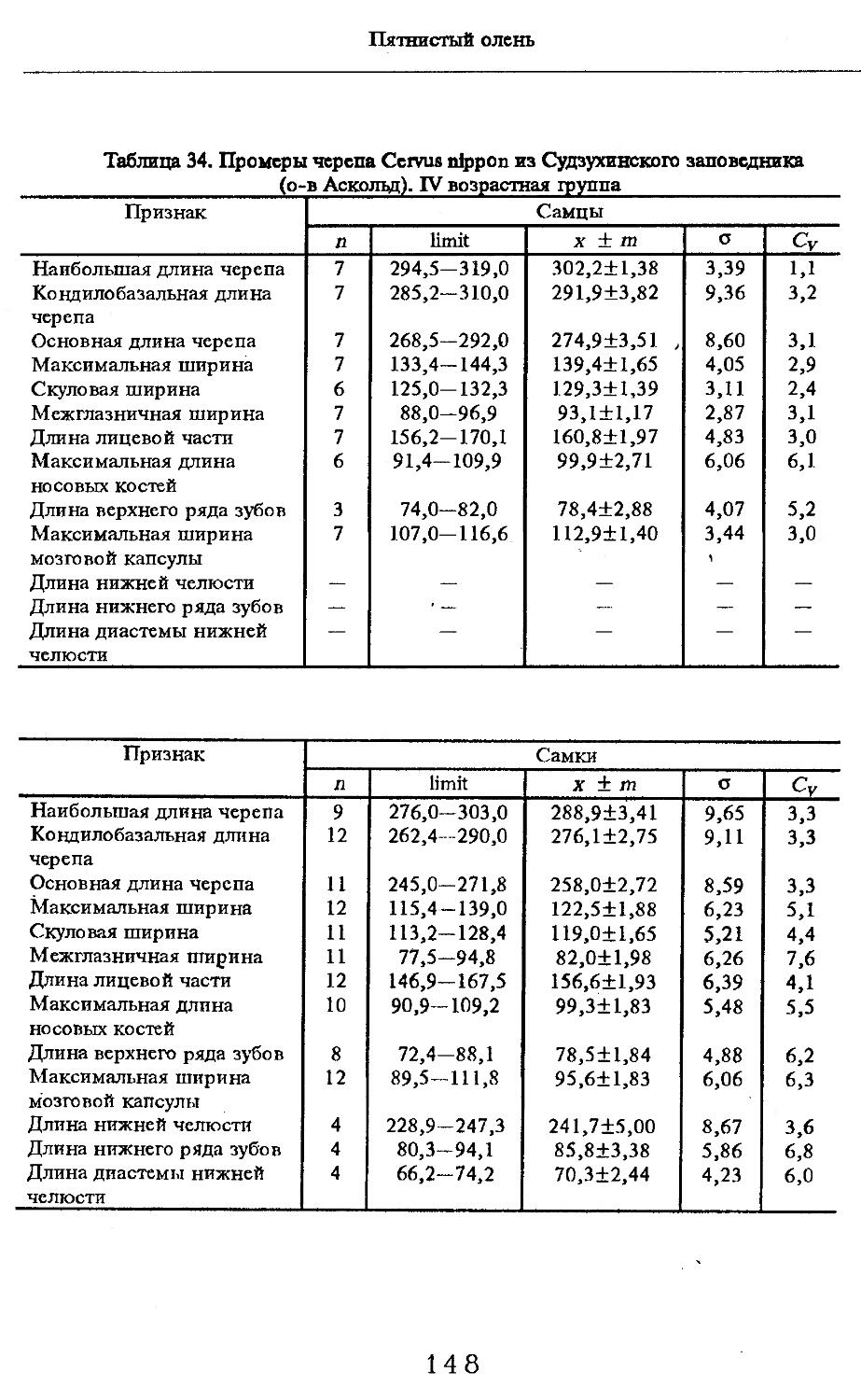
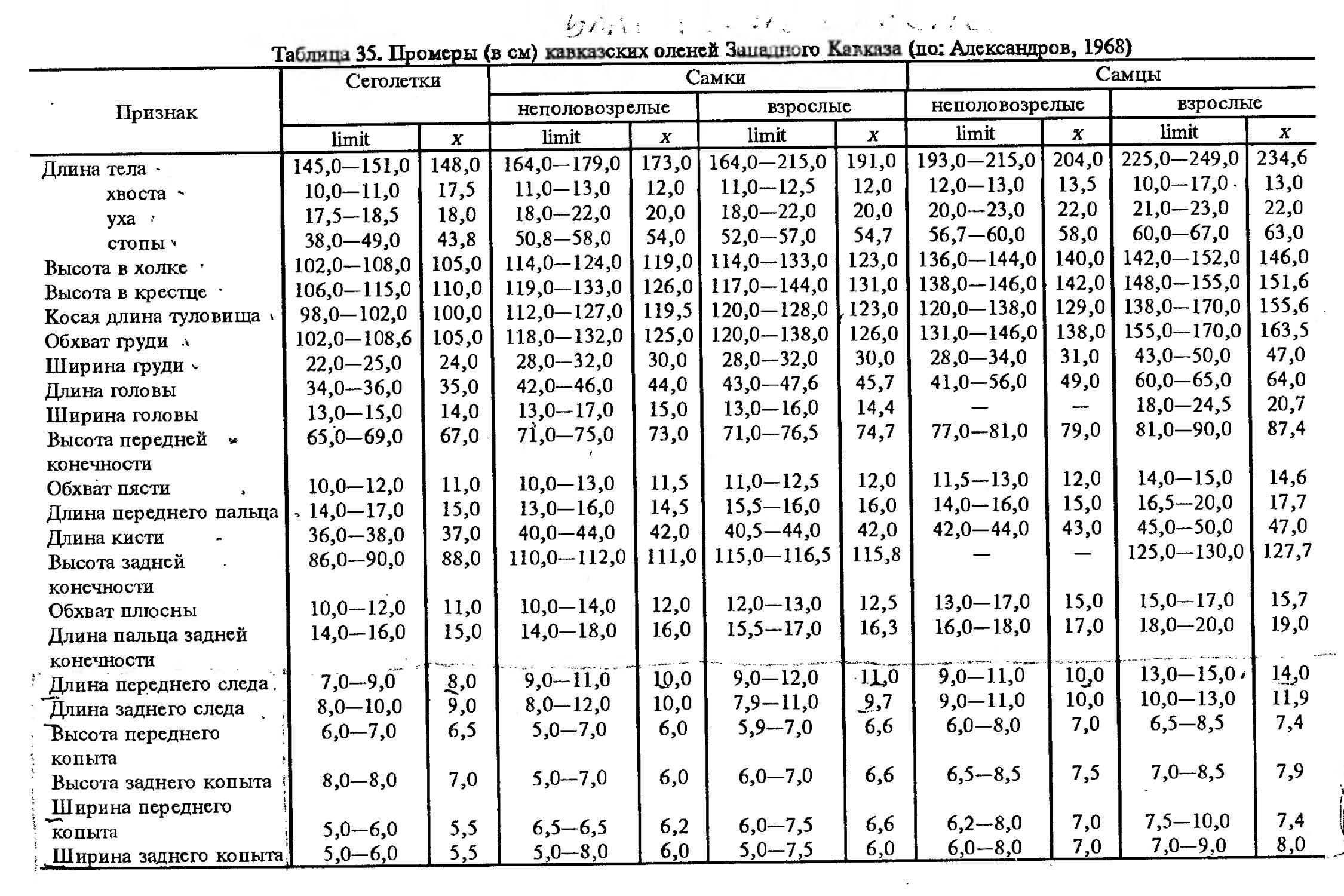
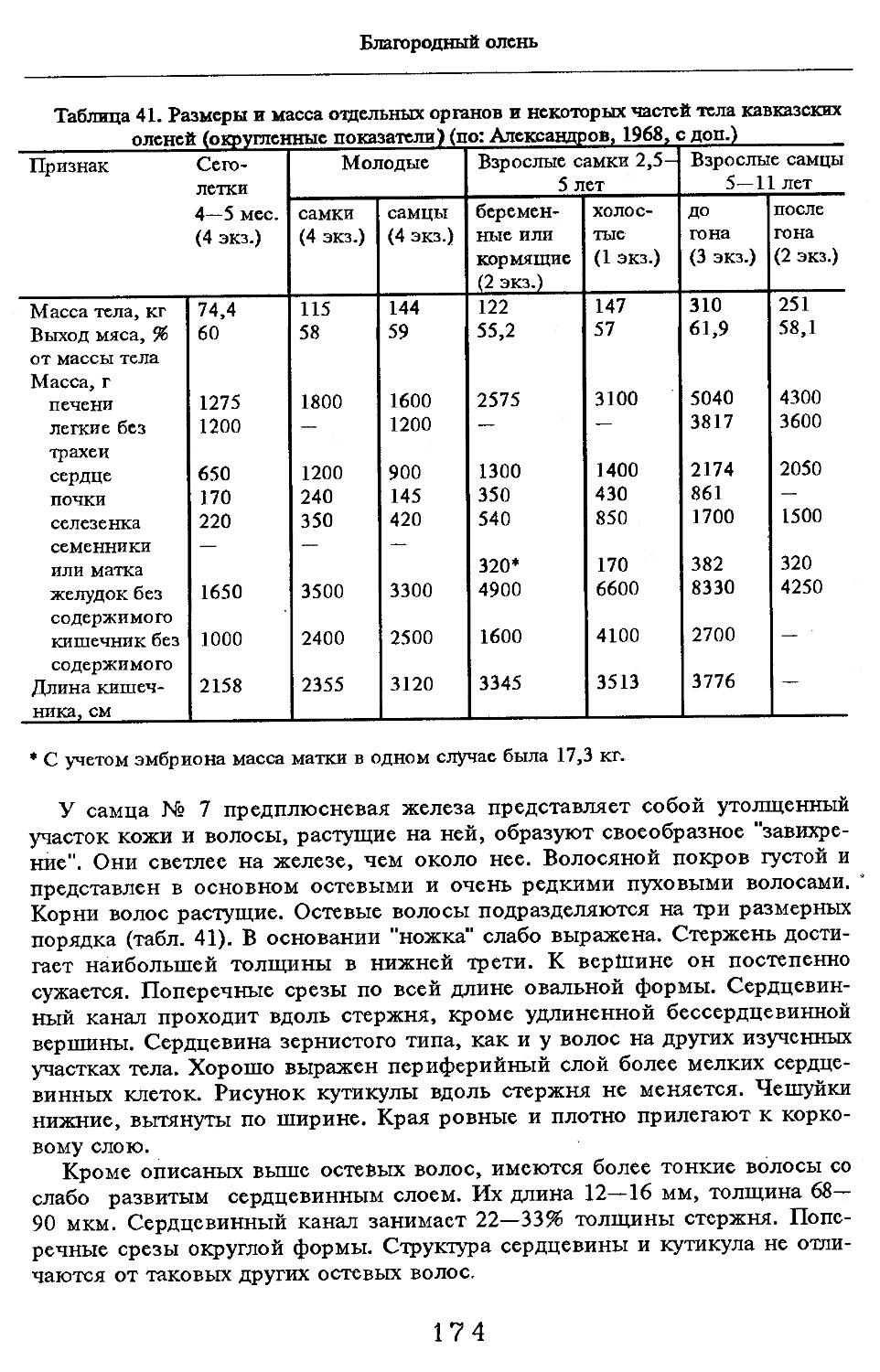
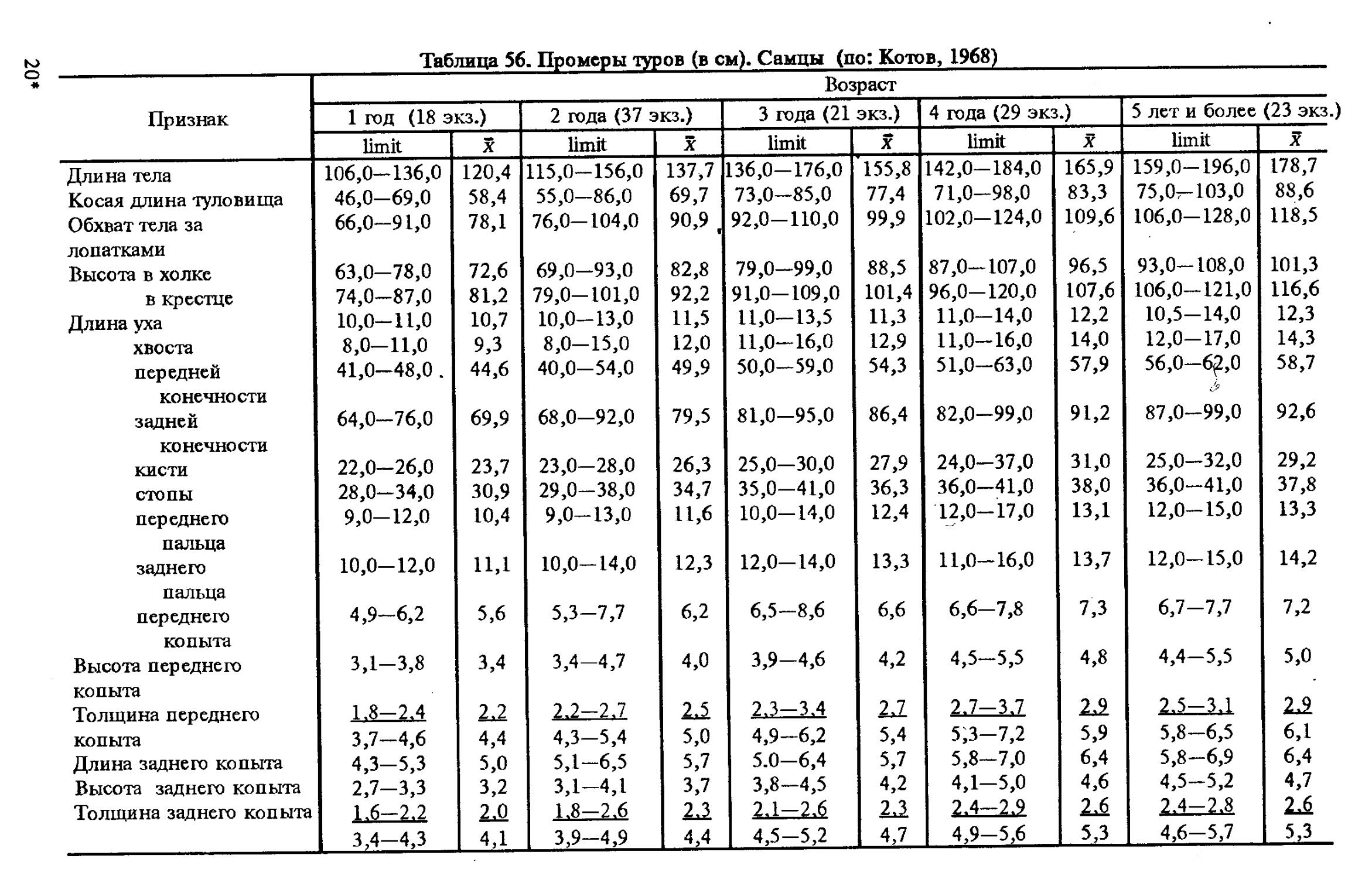
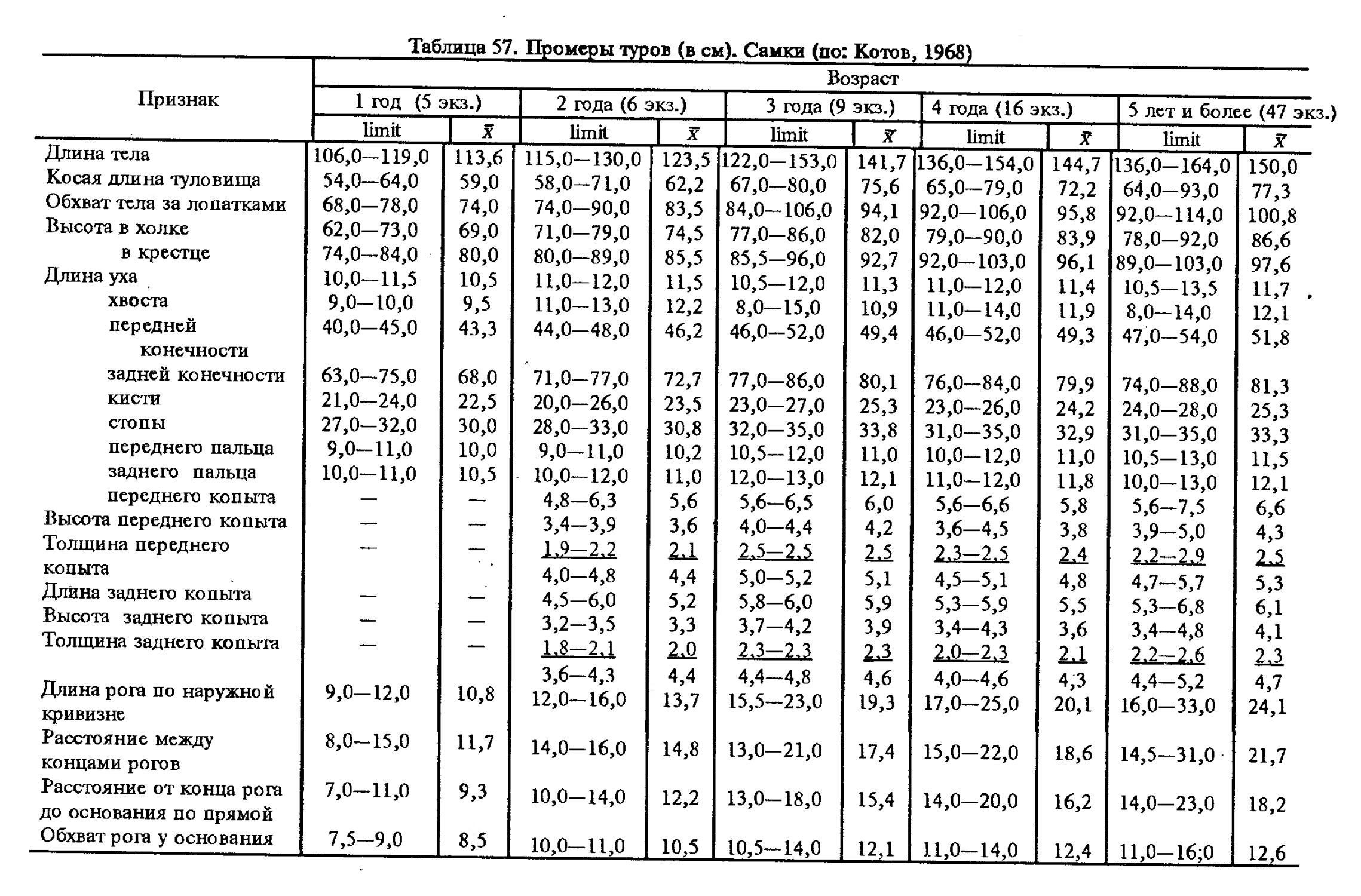
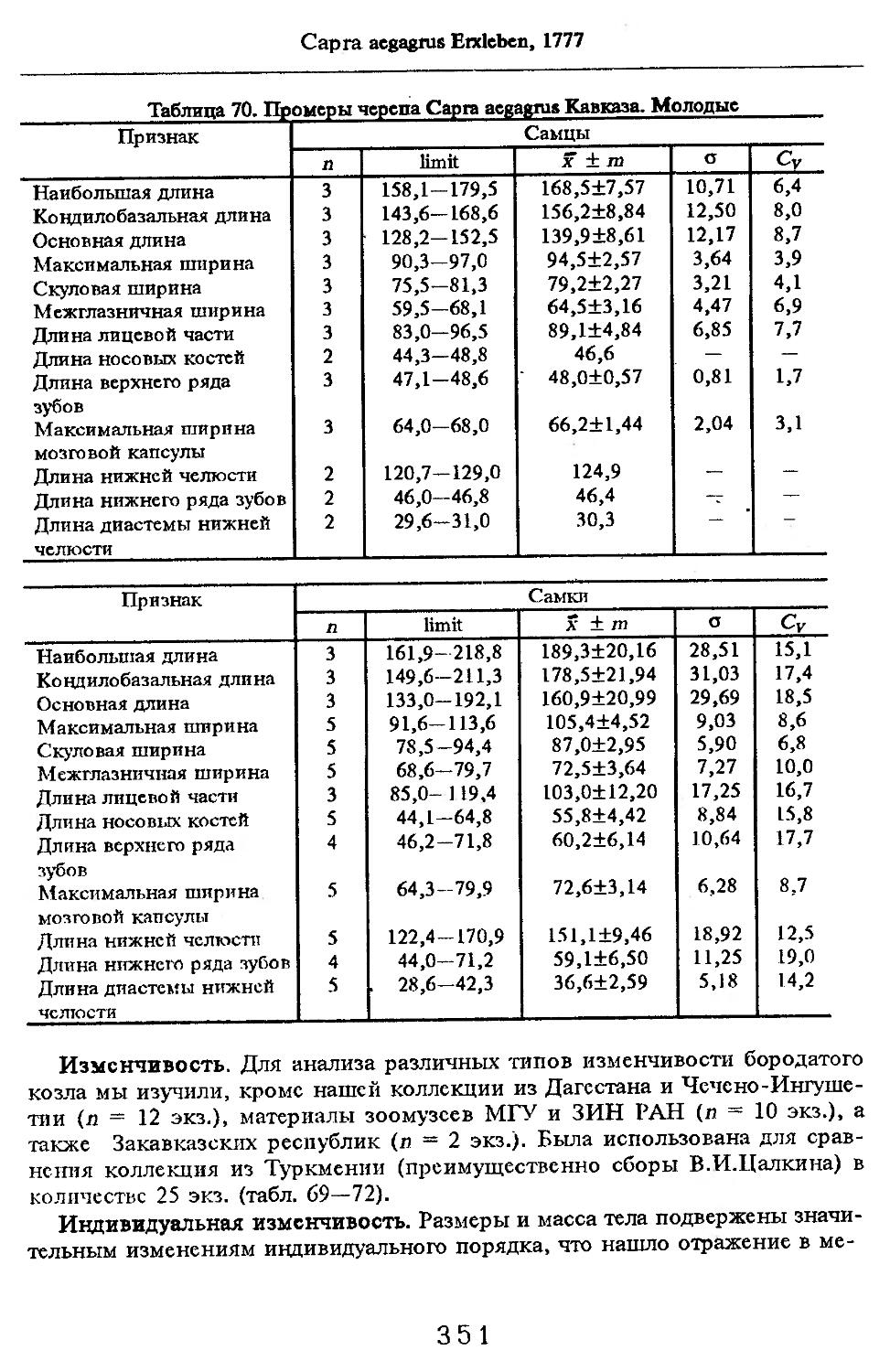
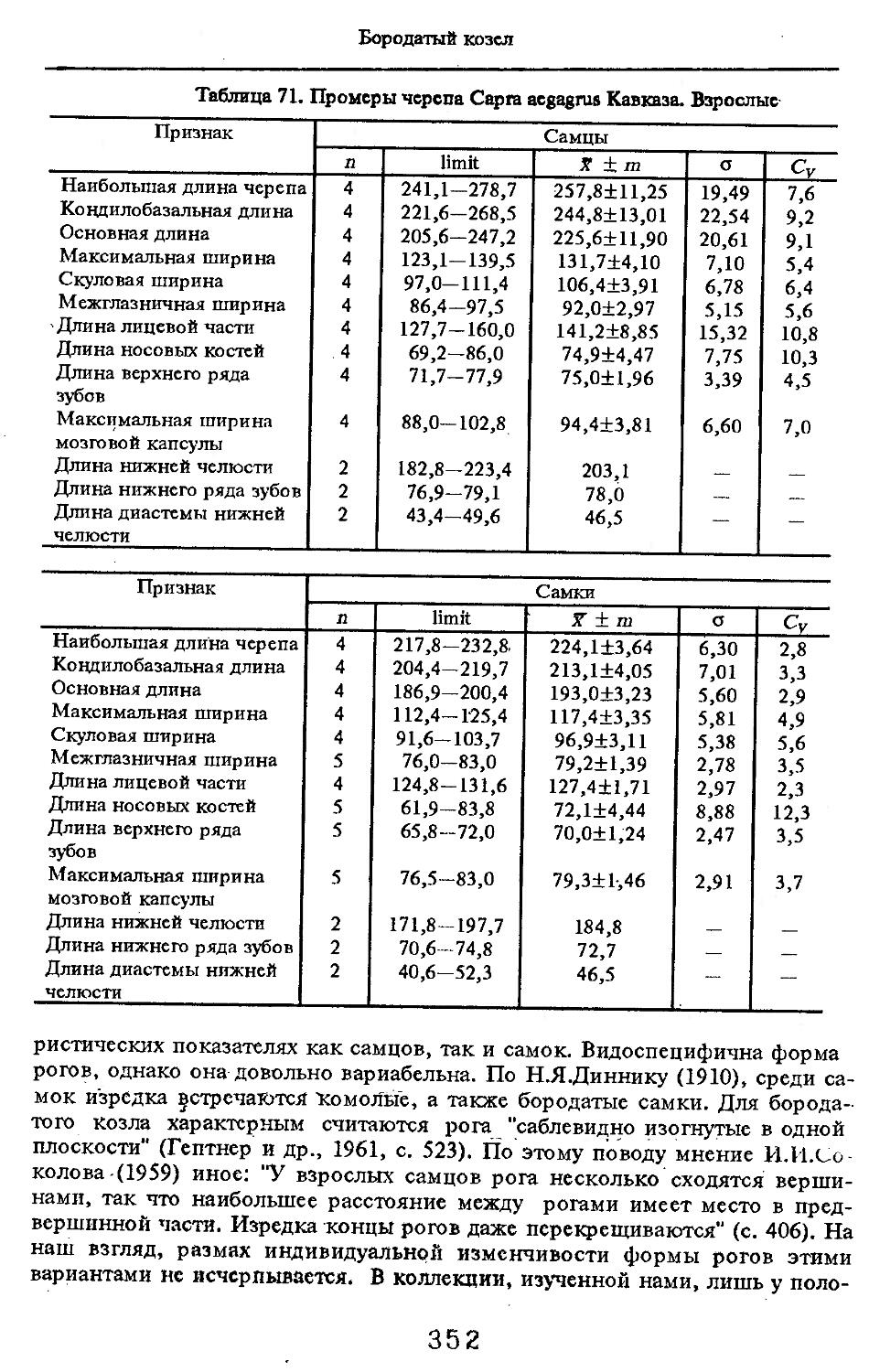
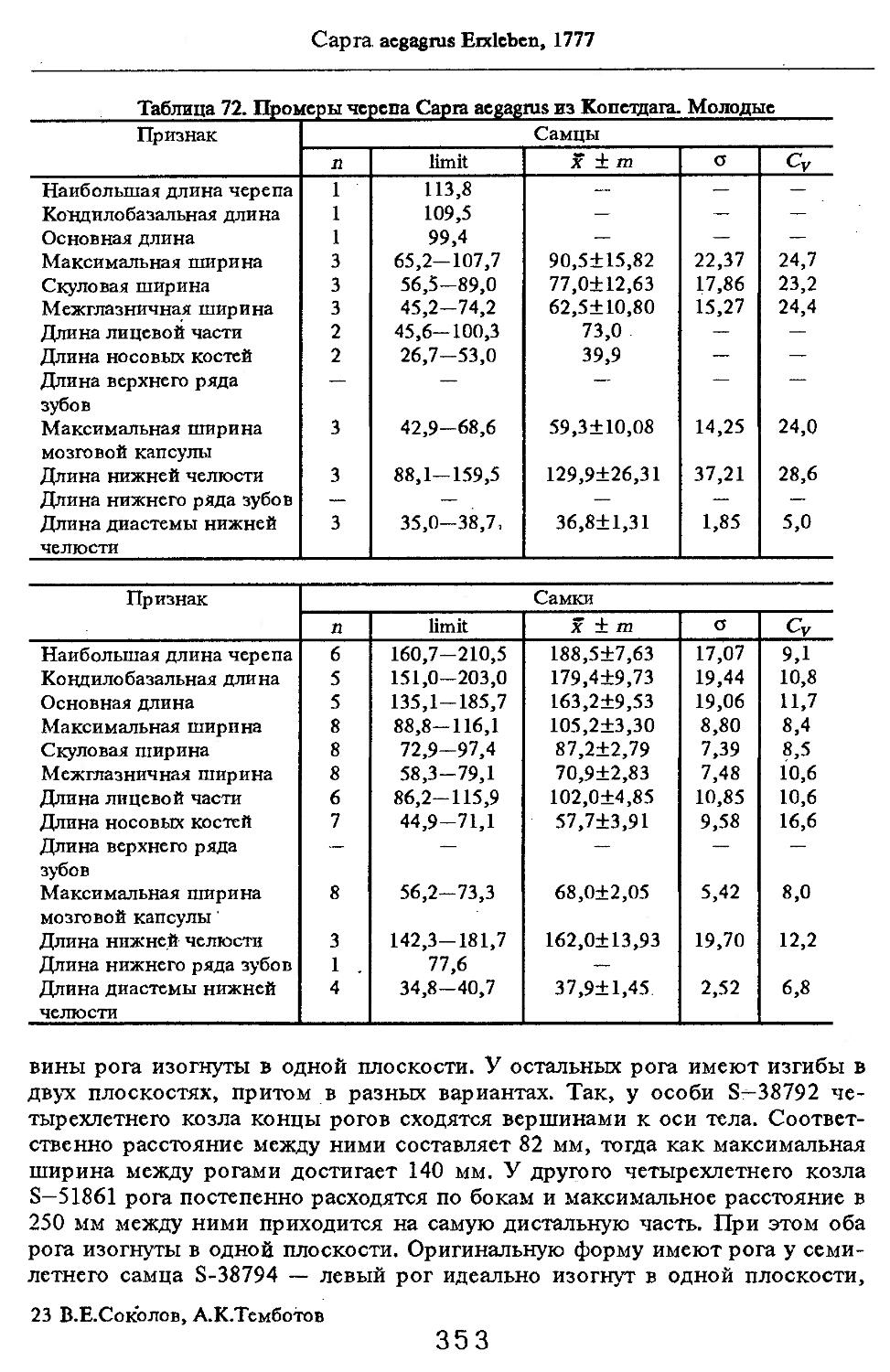
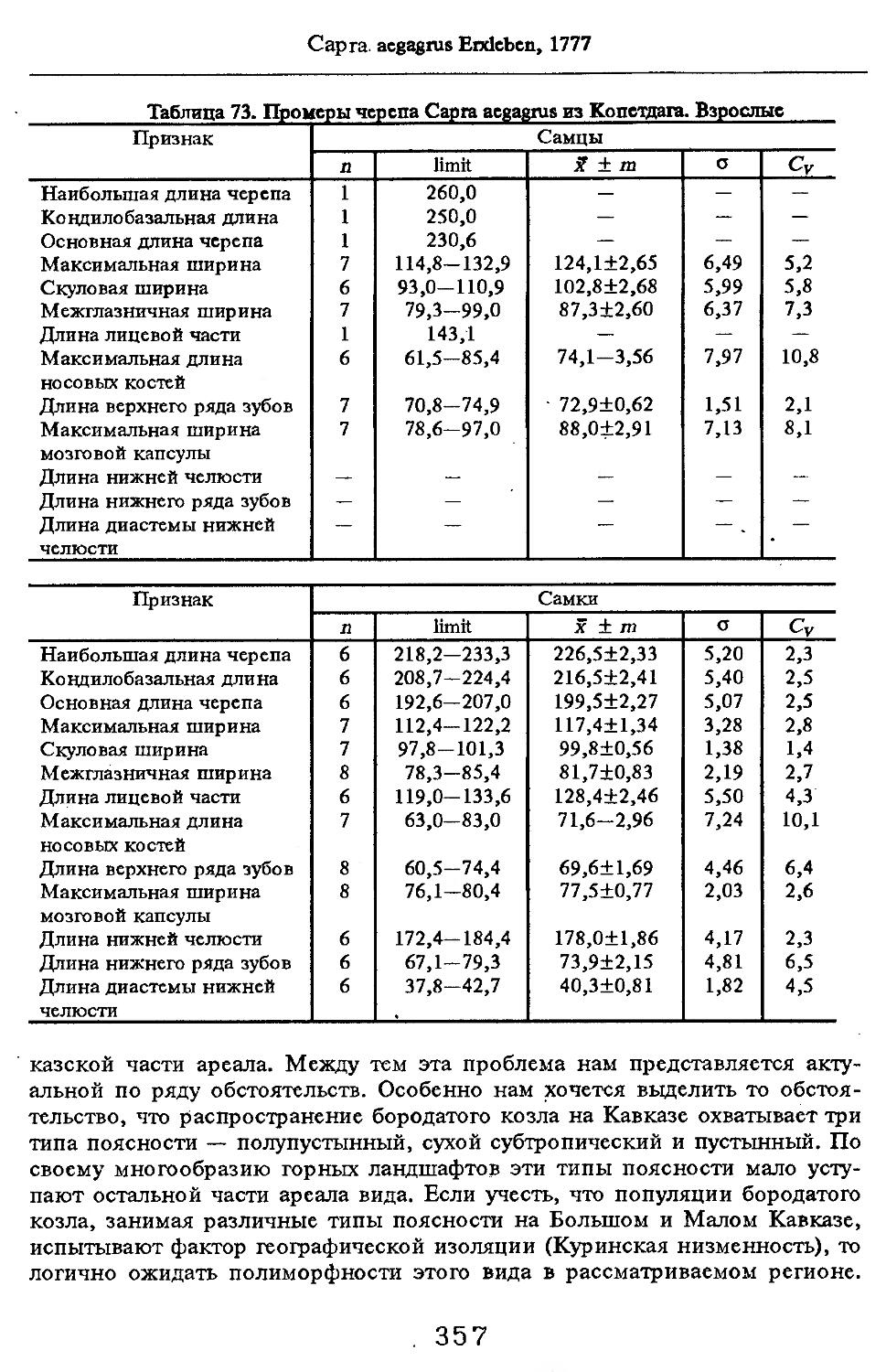
Размеры тела, черепа и отдельных элементов посткраниального скелета. По всем этим параметрам данные сильно варьируют. Наибольшую массу тела самцов (240 кг) зарегистрировал Н.Я.Динник (1910), самок (147,8 кг) — В.В.Дуров и В.Н.Александров (1968). Максимальную длину тела самцов (205 см) и самок (169 см) приводят С.СДонауров и В.П.Теплов (1938). По сведениям этих авторов высота в холке у самцов достигает 120 см, самок — 96 см. Динник (1910) сообщает, что в конце прошлого века в б. Тифлисской губернии был добыт очень крупный кабан, весивший до 320 кг. Наиболее полные данные по размерам черепа приводят С.С.Донауров и В.П.Теплов (1938), в мм: общая длина черепа самцов 467, самок 421, кондилобазальная длина соответственно 414 и 378, скуловая ширина 180 и 158, длина носовых костей 240 и 209, высота черепа с нижней челюстью 258 и 232.
При анализе морфометрических показателей дикого кабана интерес вызывает его отличие от домашней свиньи. В трудах классиков эволюционной теории (Дарвин, Филипченко и др., цит. по: Князев, Тихонов, 1985) приводится ряд особенностей. Так, у культурных рас свиней голова укорачивается и расширяется, что сопровождается изменениями во всем внешнем облике черепа, в том числе снижается его общая масса. Объясняется это значительным снижением функциональной нагрузки на череп в связи с уменьшением подземных кормов у домашней свиньи, столь характерных для диких кабанов. У диких новорожденных поросят масса черепа превышает таковую у домашних в 1,2 раза, а в трехлетием возврасте — в 1,3 раза (Нинов, Хрусталева, 1985). Однако сравнительный анализ черепов диких и домашних и их гибридов, проведенный С.П.Князевым и В.Н.Тихоновым (1985), показал,
1 Кавказский государственный заповедник.
1 о
Sus scrofa Linnaeus, 1758
что краниологические различия между близкими видами и породами более ярко выражены не при сопоставлении непосредственно численных значений размерных показателей черепов, а индексов, вычисленных на их основе, причем не всех, а наиболее характерных (например, отношения высоты черепа к кондилобазальной длине, ширины и длины слезной кости).
Во внешнем облике, как справедливо писал еще Н.Я.Динник (1910), характерно сильное развитие головы и передней части туловища (шеи и труди). По сравнению с ними зад кажется значительно слабее. Различие по высоте в областях холки и крестца превышает 5 % от высоты в холке.
Зубная система. Из трех пар верхних резцов сильнее развита средняя пара, имеющая клиновидную форму и направленная вперед и внутрь. Вторая пара хотя и развита, но значительно меньше первой, вплотную прилегает к ней, тогда как третья пара недоразвита и отделена от них значительным беззубым промежутком. В нижней челюсти первая и вторая пары примерно одинаковы по размерам, долотообразной формы, обе направлены вперед. И здесь третья пара резцов также слабо развита и отделена от других резцов. Клыки сильно развиты, они сидят в челюстных костях отдельно как от резцов, так и от предкоренных.
Обладая постоянным ростом, особенно большие размеры имеют верхние клыки самцов (до 120 мм). У самок длина этих клыков достигает лишь 39 мм. Длина нижних клыков самцов до 109 мм, самок — до 37 мм. Характерно, что задняя грань нижних клыков в отличие от других видов свиней шире наружной и стирается о переднюю поверхность верхних клыков, загнутых в сторону и вверх (Соколов, 1959). Поверхность стирания простирается на верхних и нижних клыках до вершин зубов. Среди коренных меньше всего развиты первые предкоренные (Р1 и Pi), иногда они могут выпадать. Коренные зубы усложняются и увеличиваются в размерах от первых до последних, соответственно самыми крупными и самыми усложненными являются М3 и Мз; они отличаются от других наличием дополнительного ряда бугорков.
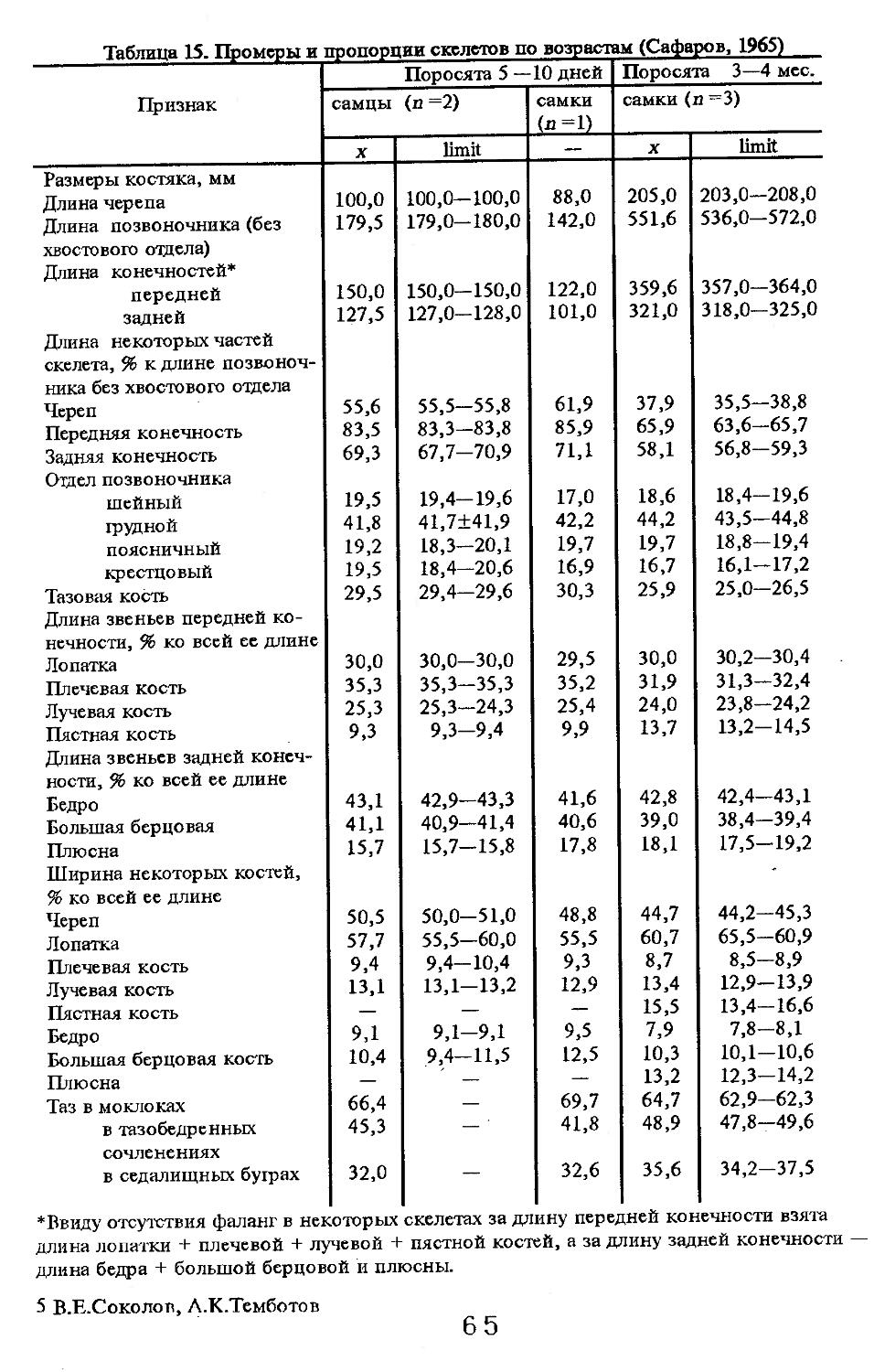
В посткраниальном скелете четко прослеживается ряд адаптивных черт, особенно при его сравнении с домашними Свиньями (Пинов, Хрусталева, 1985). Так, шейный отдел мощно развит, что связано с большим черепом и выполняемой им жизненно важной физической нагрузкой. У дикого кабана относительная масса шейного отдела скелета выше (9,08 %), чем у домашних (8,05 %). Столь значительная разница обнаружена и в относительной массе поясничного отдела (11,3 % у домашних и 7,4 % у диких). Кроме того, поясничных позвонков оказалось у диких (л — 12) 4 (67 %) или 5 (33 %), тогда как у домашних (л = 20) — 6 (20 %) или 7 (80 %). Диким кабанам свойственна большая, относительная масса длинных трубчатых костей, что связано с быстрым передвижением. То же самое наблюдается и при сравнении линейного роста костей скелета. Названные выше авторы считают весьма интересным факт большей плотности длинных трубчатых костей во всех звеньях конечностей у диких'животных. Плотность костей в тазовой конечности больше, чем в грудной, что указывает на ее большую функциональную нагрузку, падающую на тазовые конечности при передвижении живот-
1 1
Кабан
Ростов-на-Дону
Мариуполь
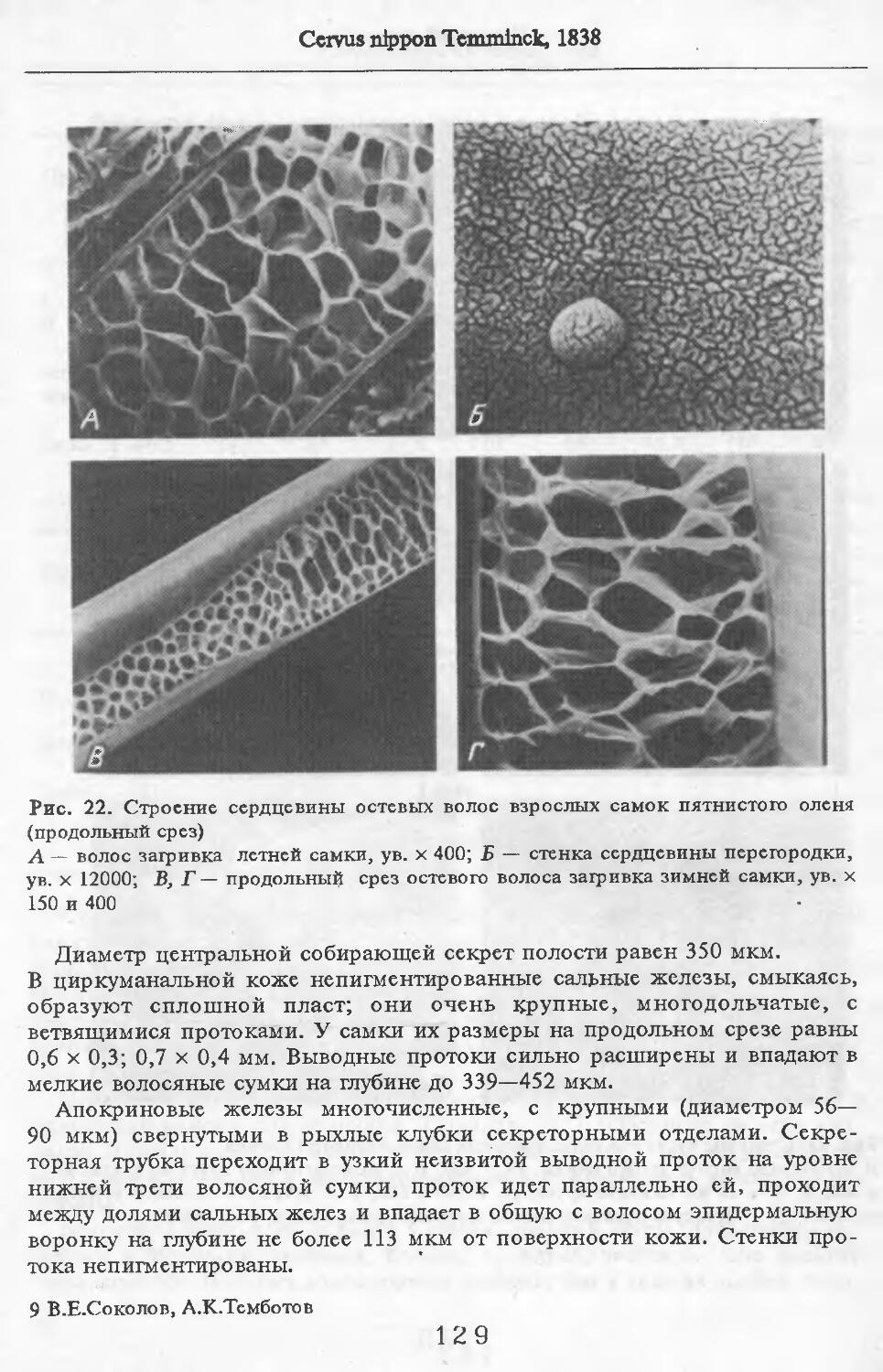
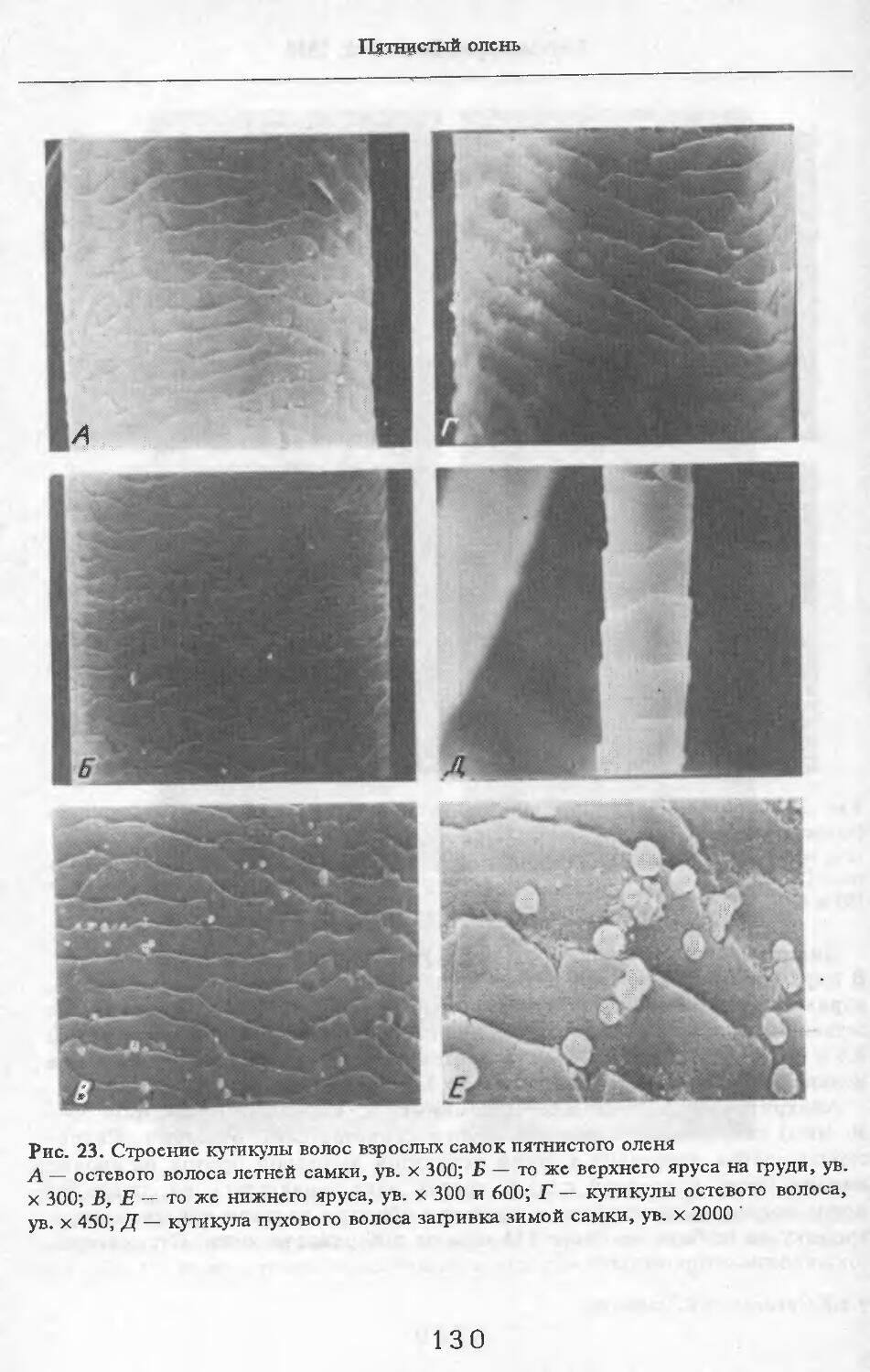
О Заветное
Сарпа
6000 -
6000
4000
3000
2000
1000
о Яшкуль
г- Псоашхо
3255
О
Егорлыксная
О Дивно'е.
Кропоткин
Тамань![ЛубЖ
58
Туапсе
—32
Сочи
70
-Гагр
78
Владикавказ
Очамчир
Казбеги
Зугдиди
Поти
Кировакан
КХ
0
125
4000 -
131
Нахичевань
3000
127
г. Далидаг 3616
134 !*Гс
г. Караная 2850
г. Меписцкаро 2850
г. Бол-.Г иная д аг 3367
г. Нюмиркей 2477
^Черкесск
Пятигорск
Мартуни 124
ЕЗ
Невинномысск
46
Светлоград
Элиста
КБ
*12
13 1Г18]3
г. Эльбрус 5642
503
Ставрополь
Карачаевск
Благодарный
Буденновск
утаиси
Ахтала
2000
1000 0
А7К
к-з
ц-м
оздок
Ереван
60
1 2
Sus scrofa Linnaeus, 1758
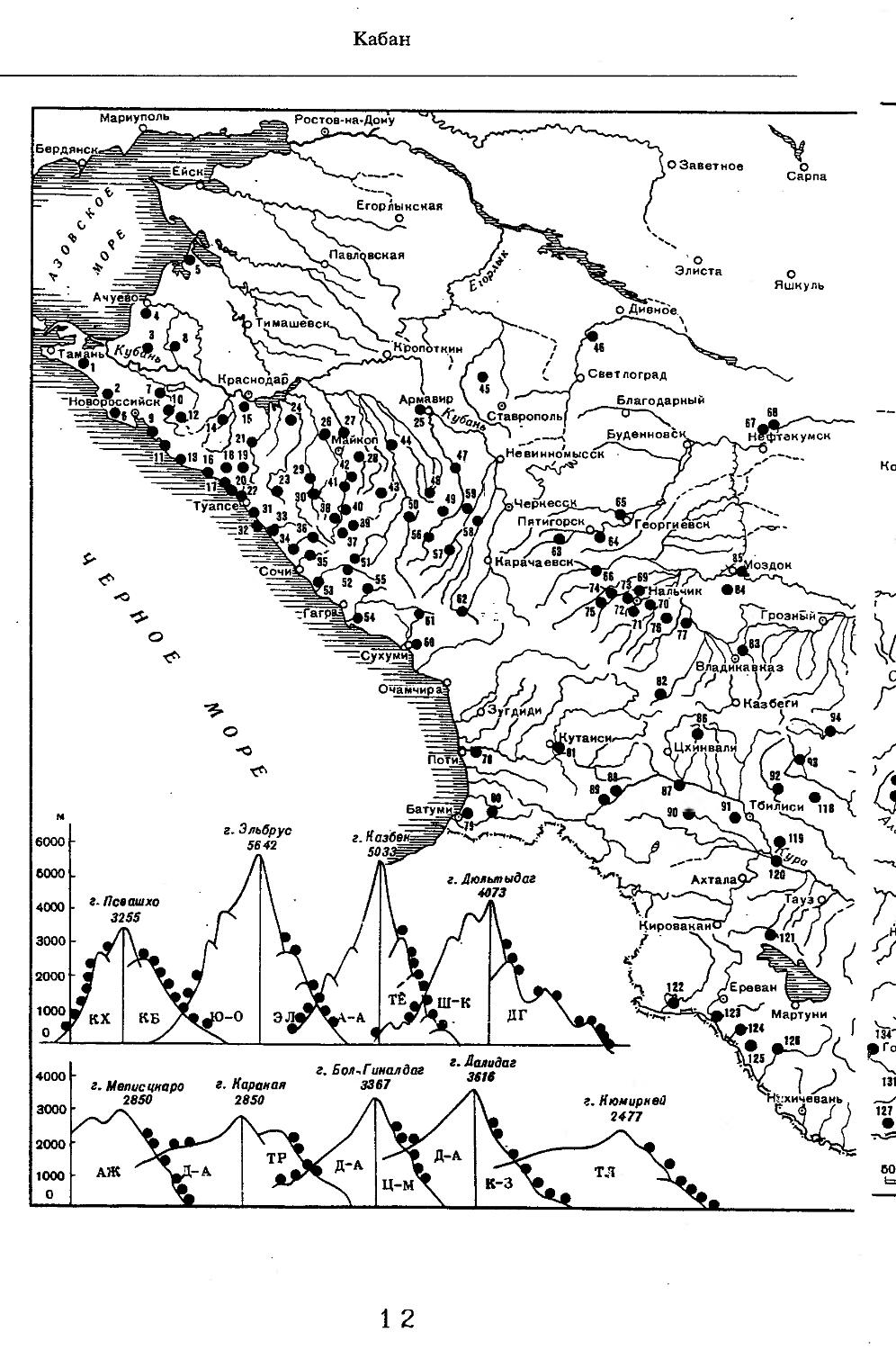
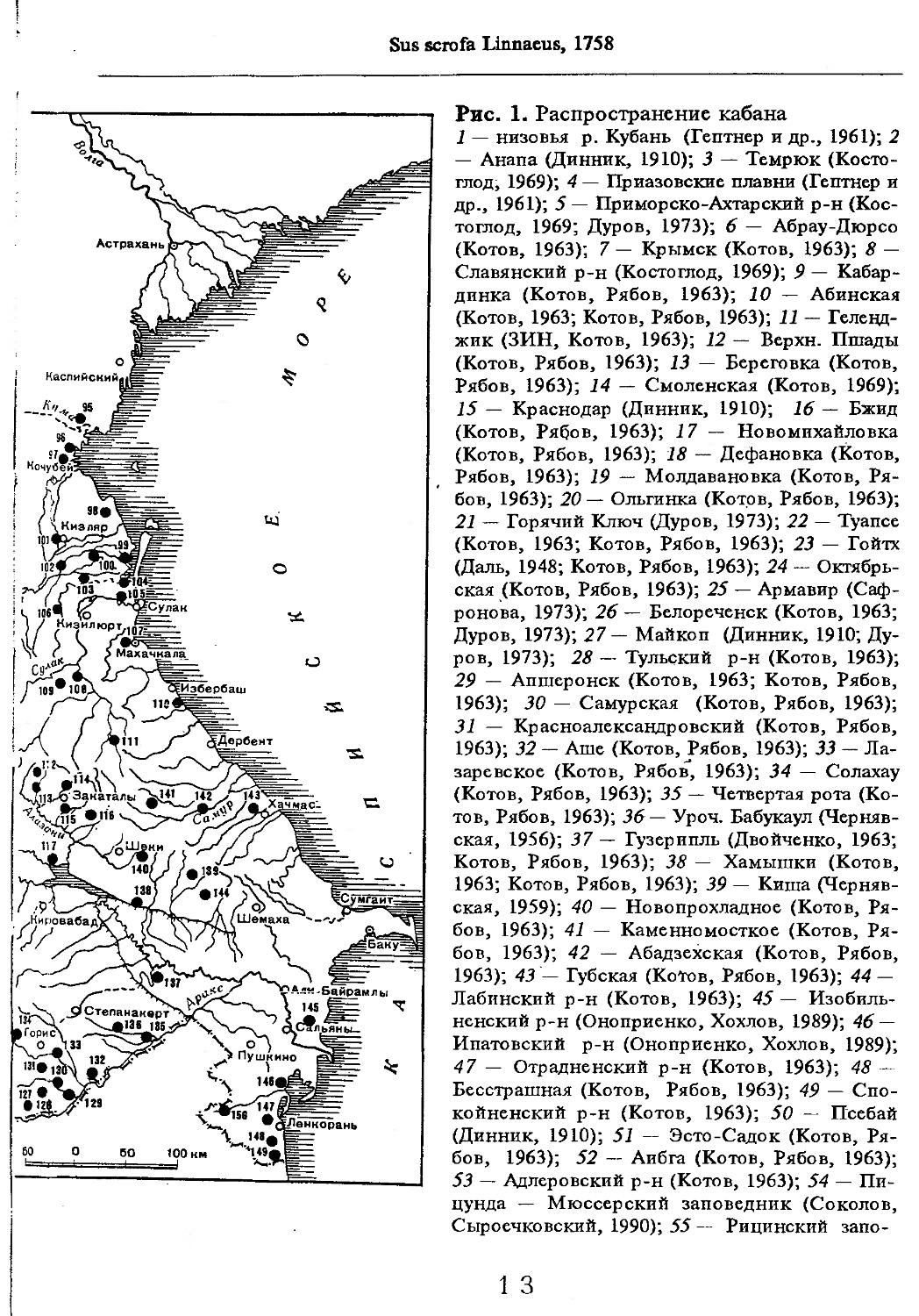
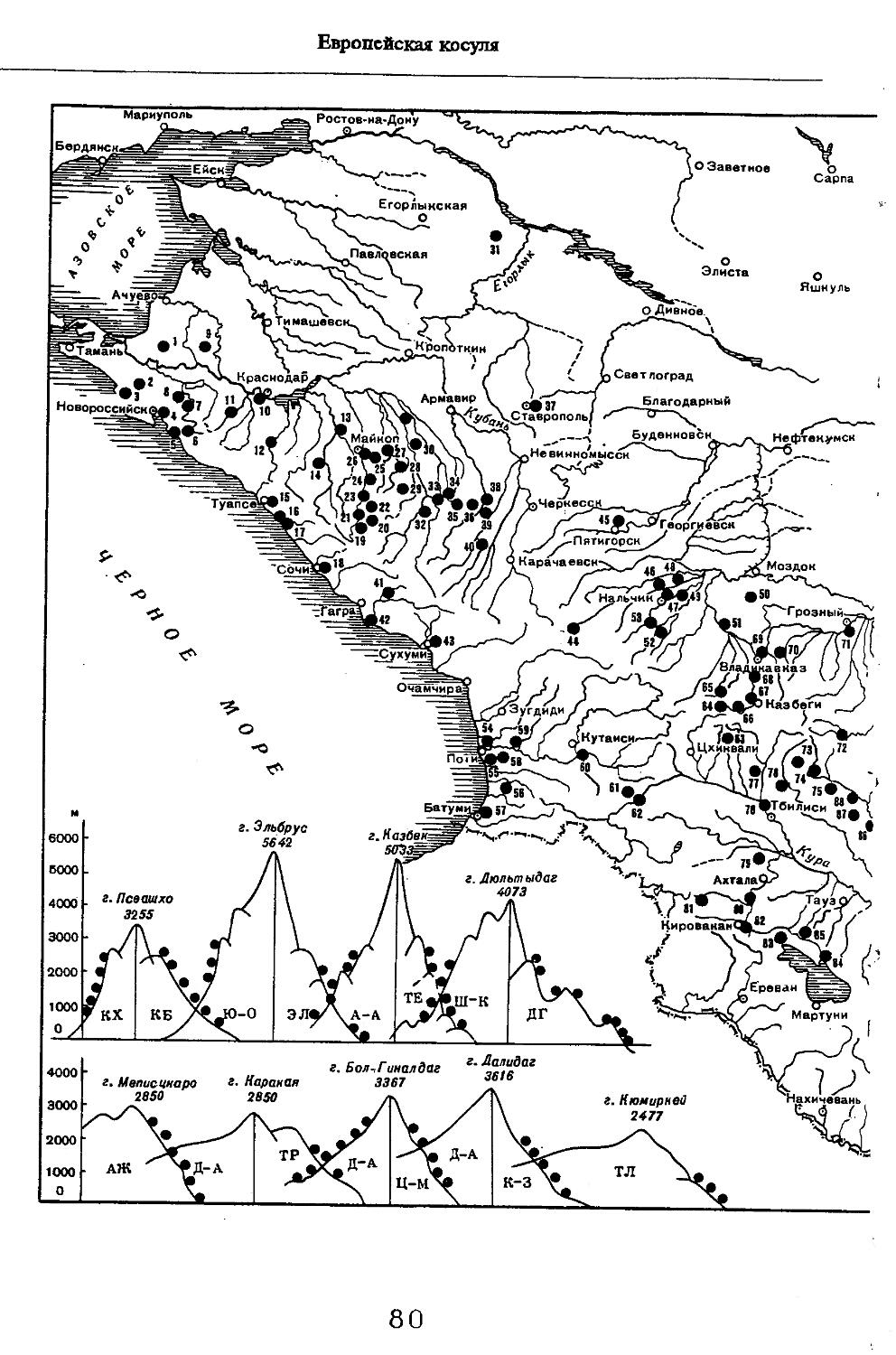
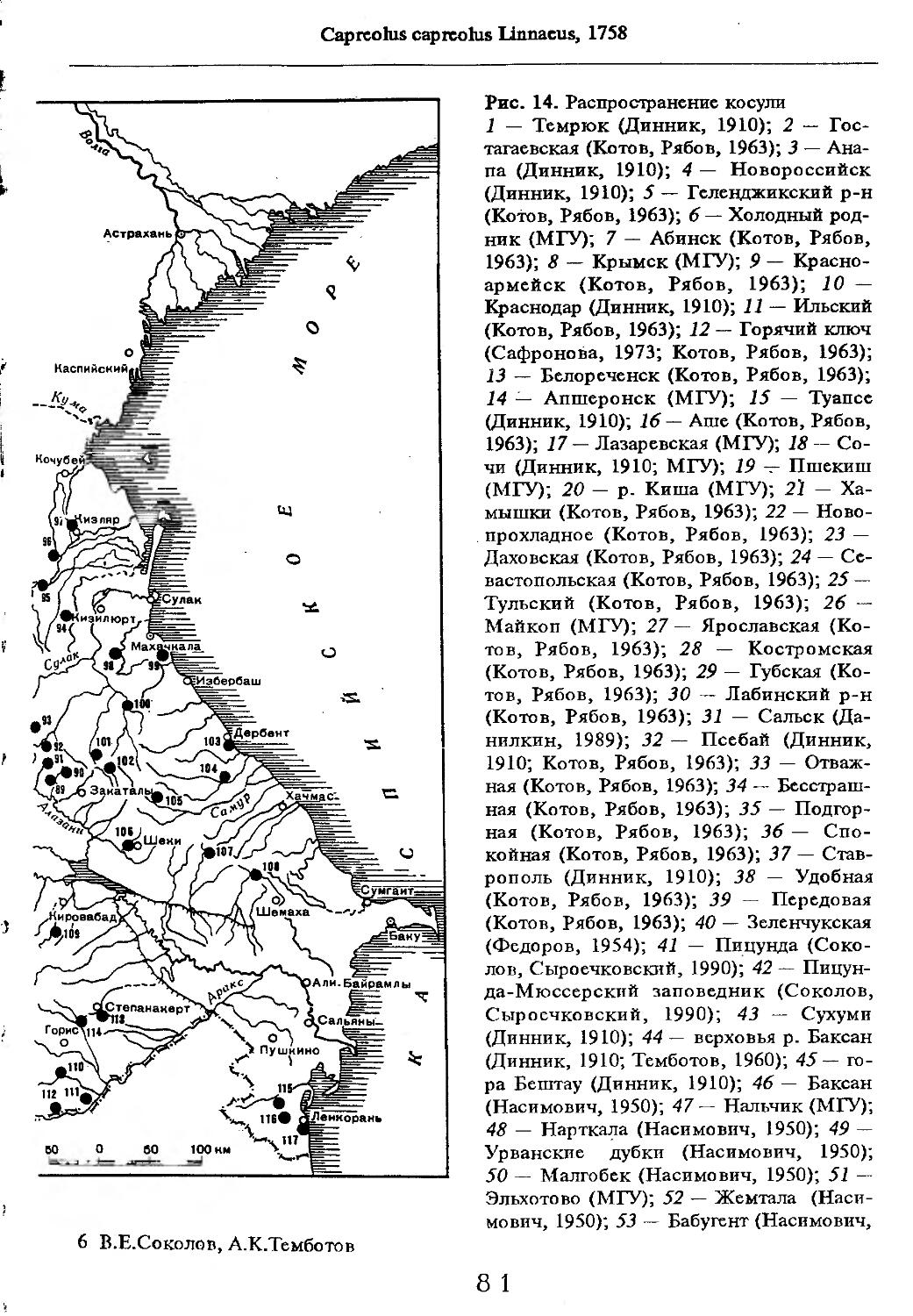
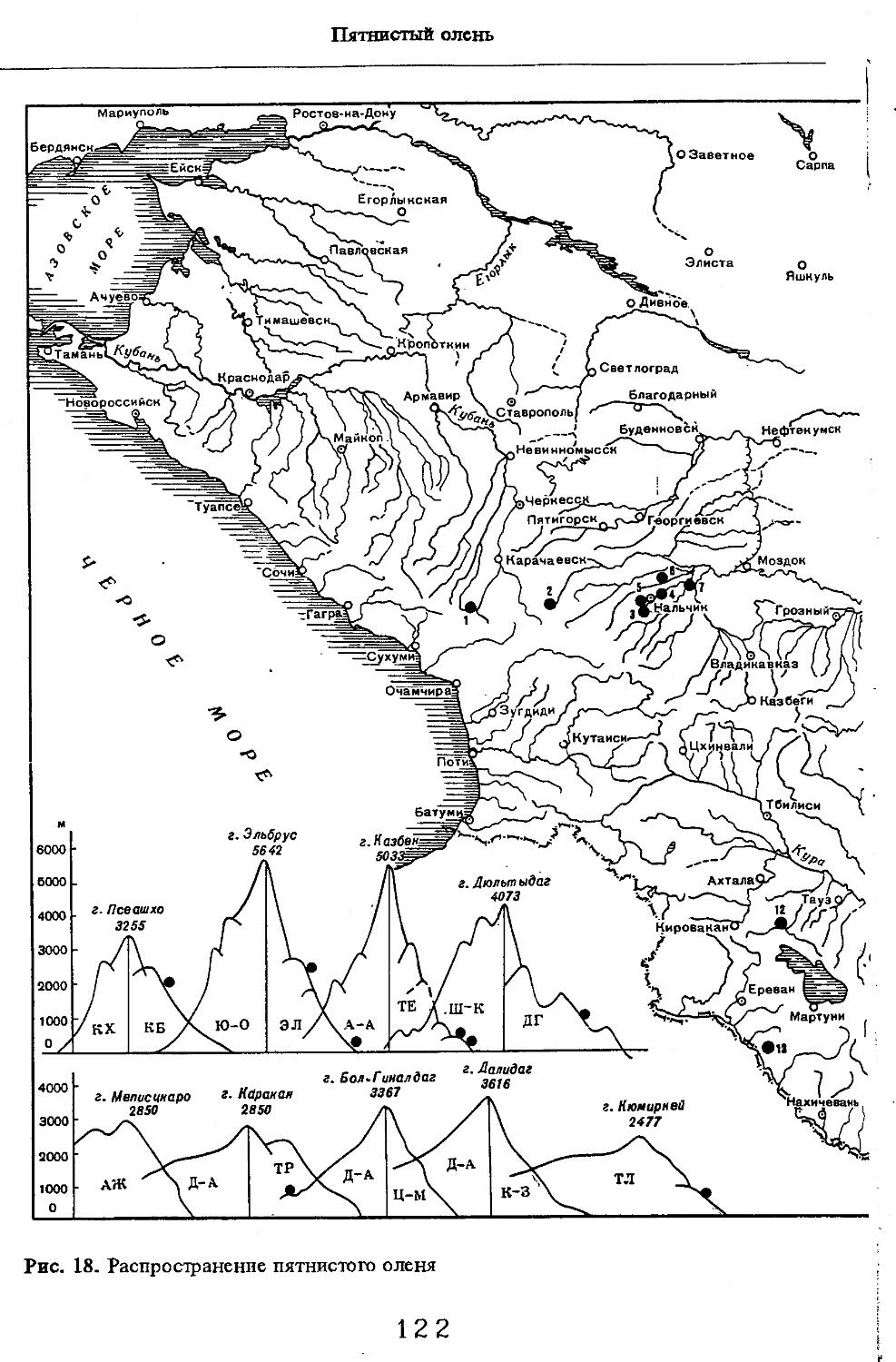
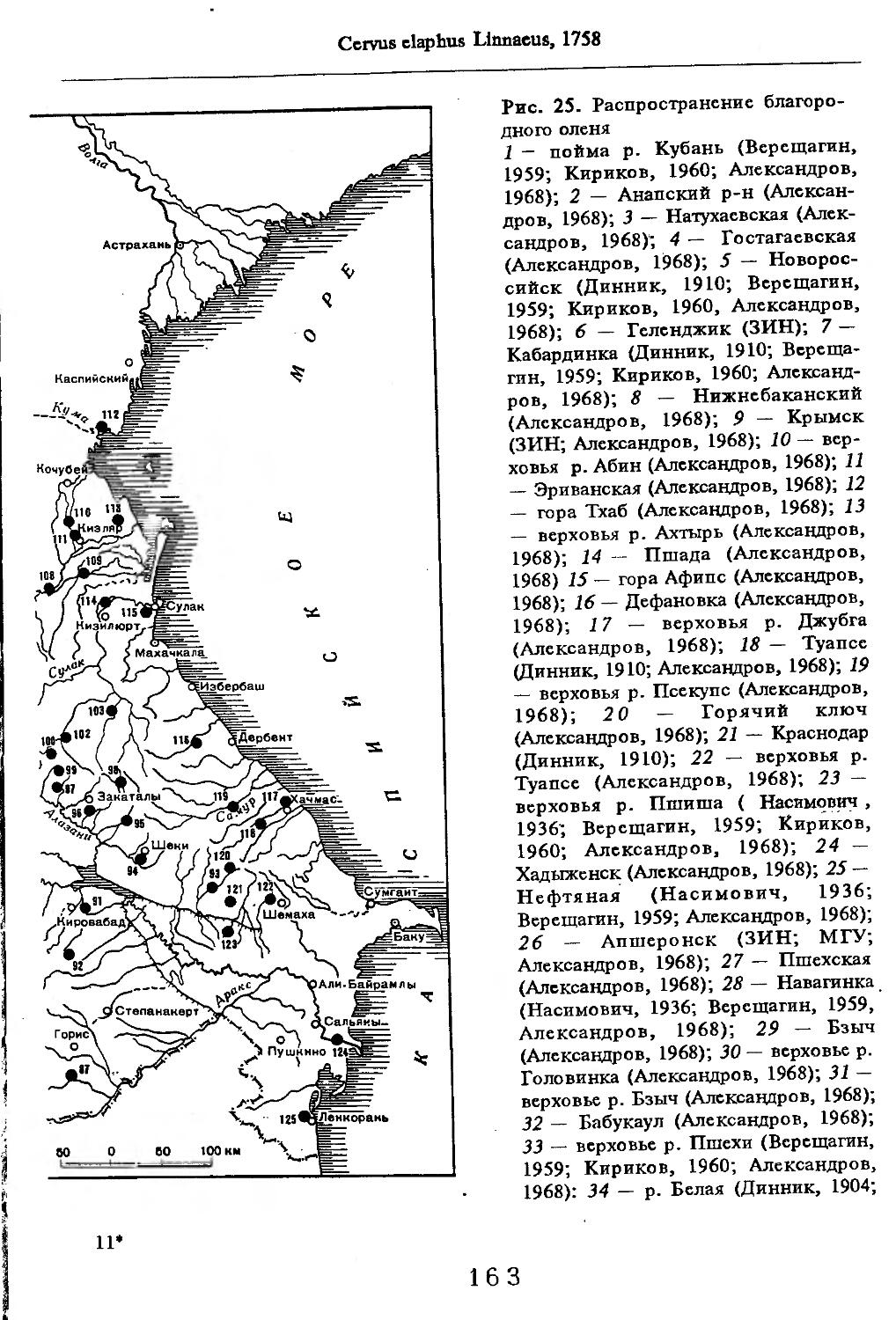
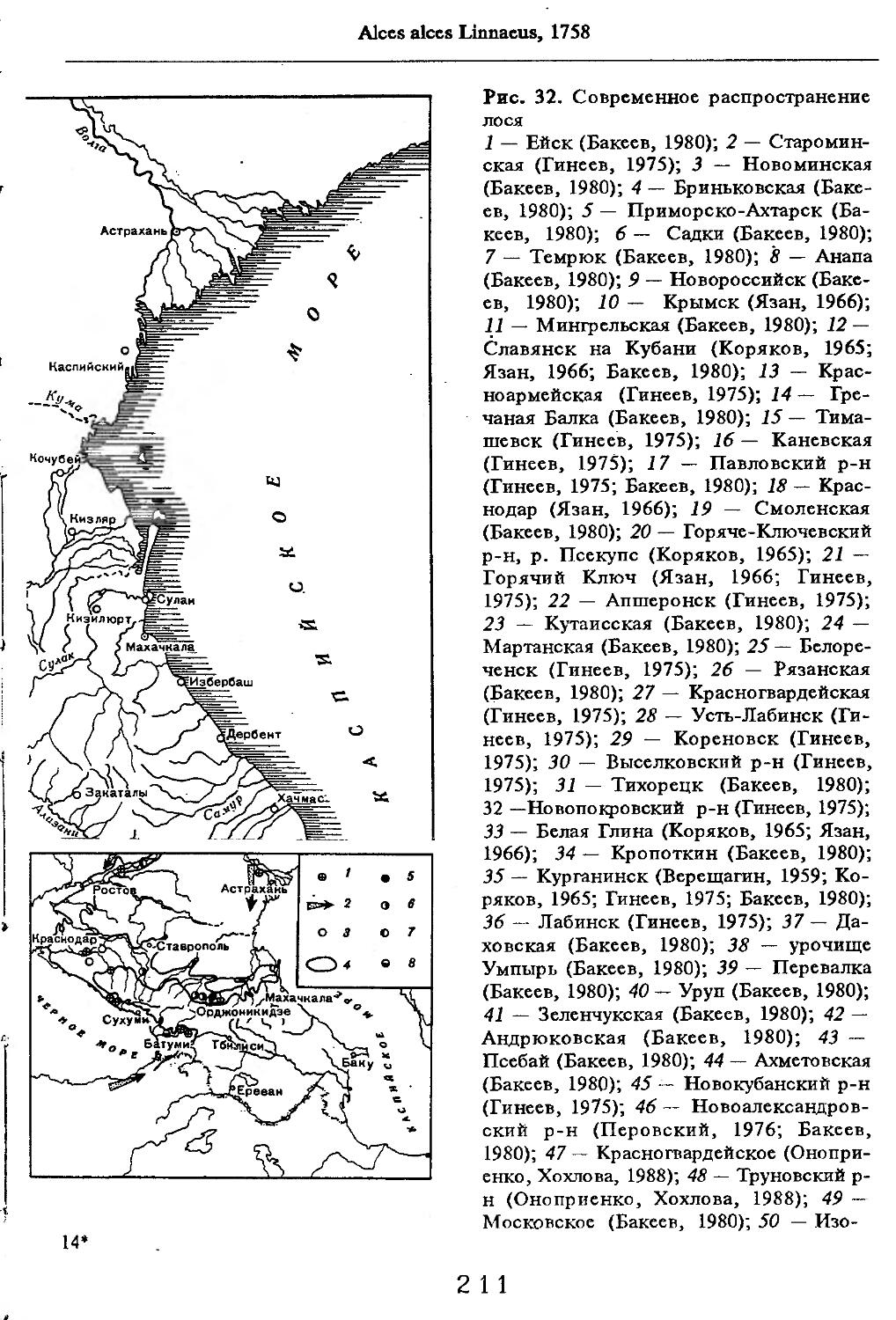
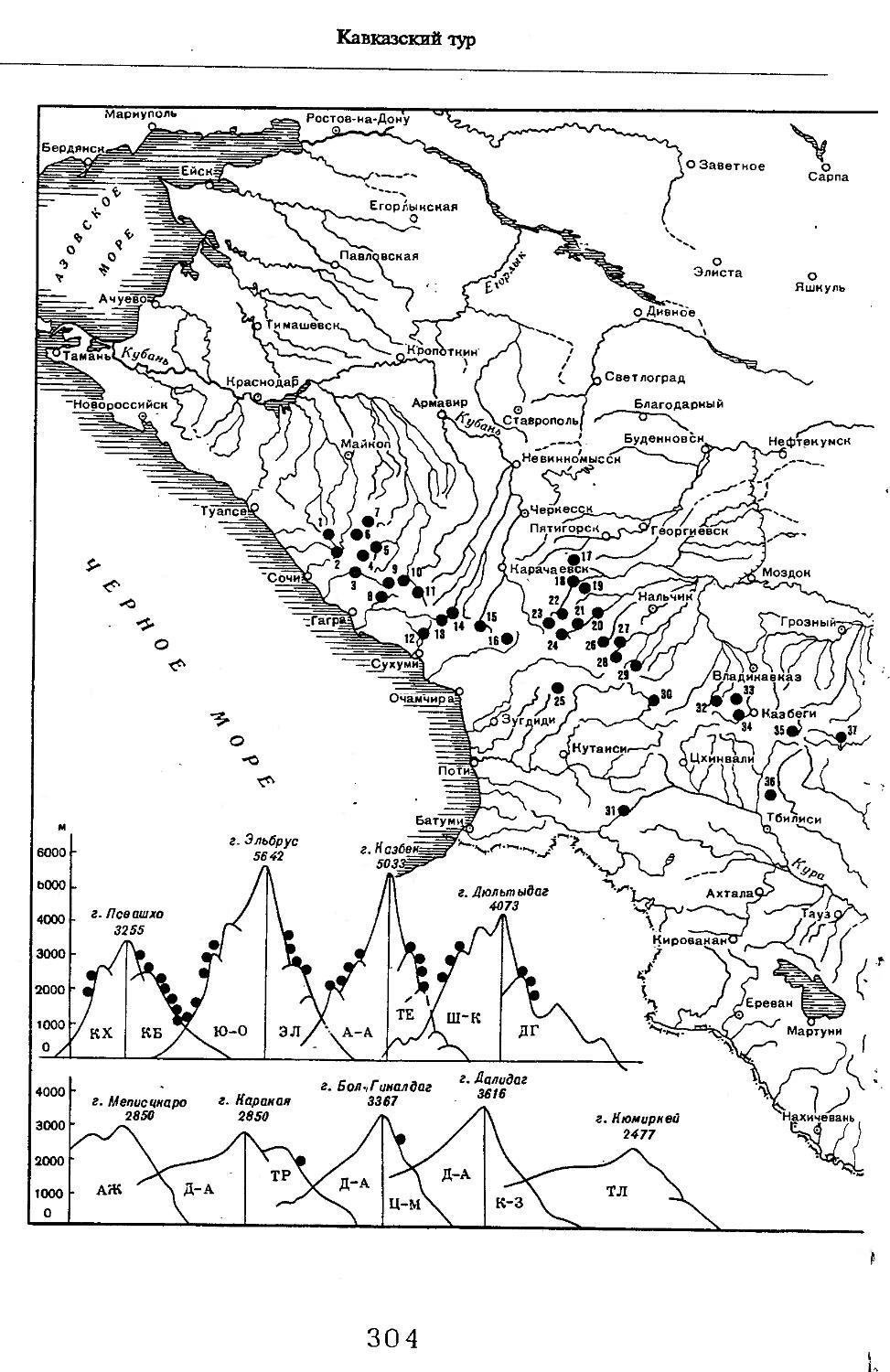
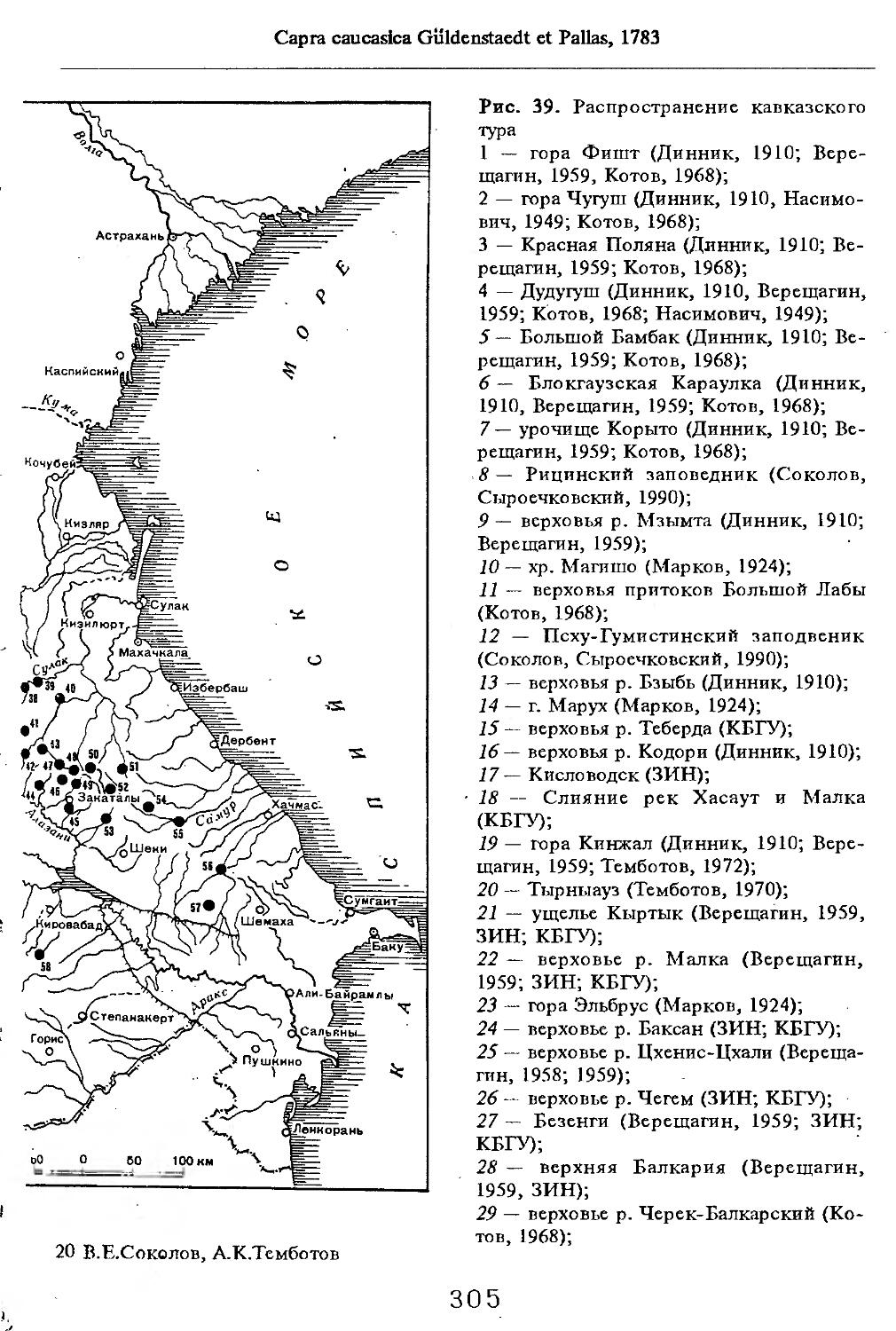
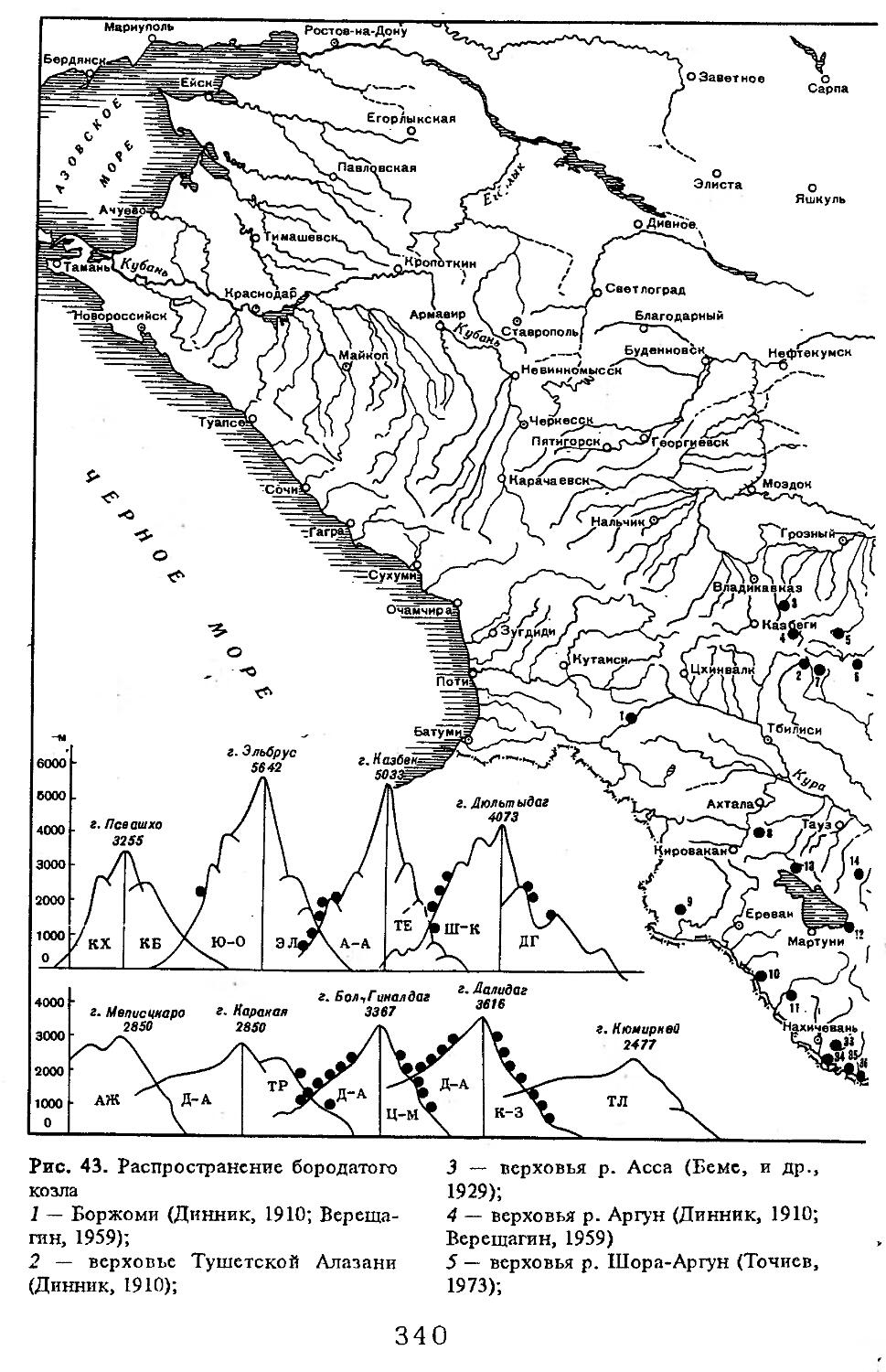
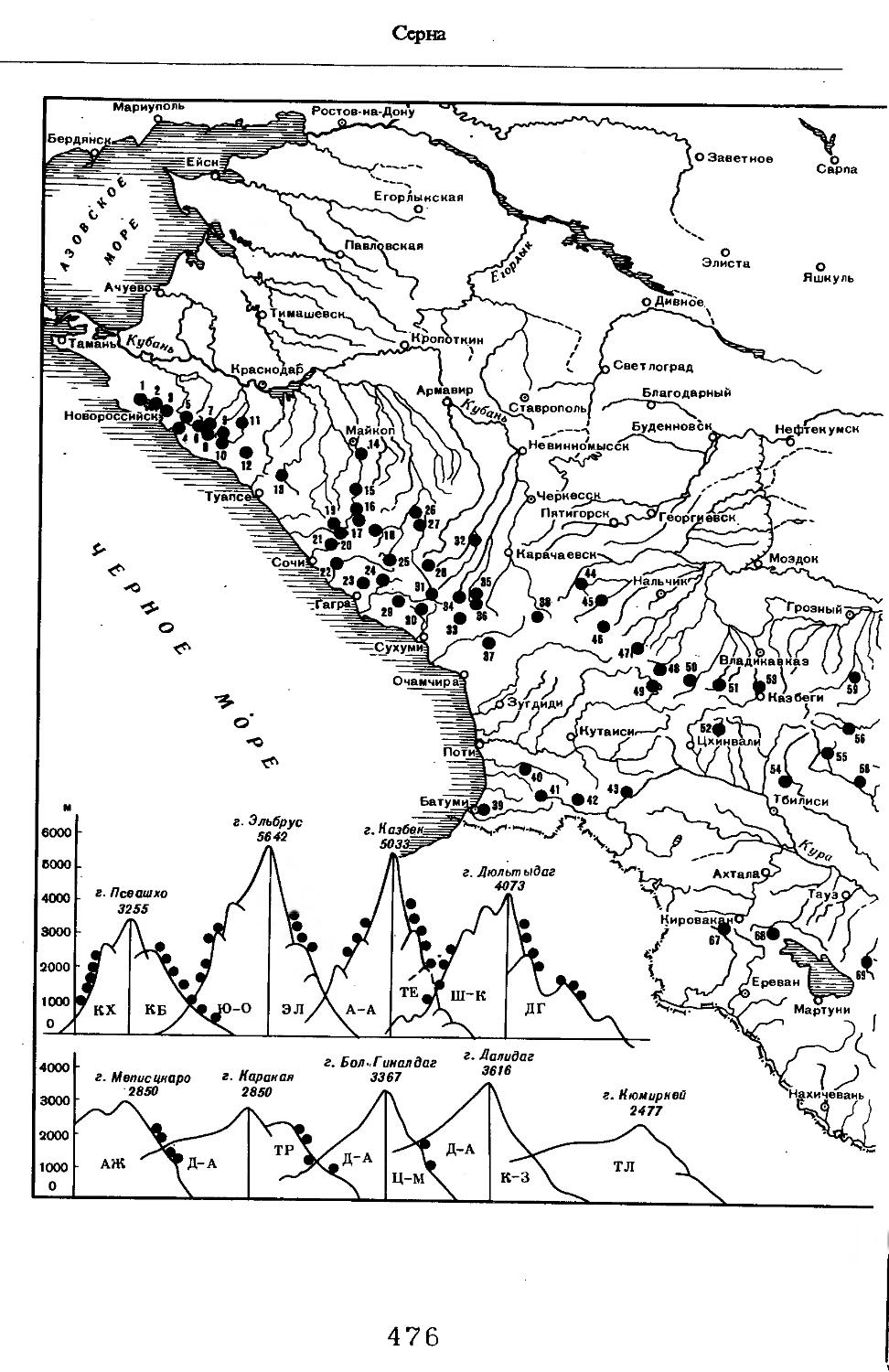
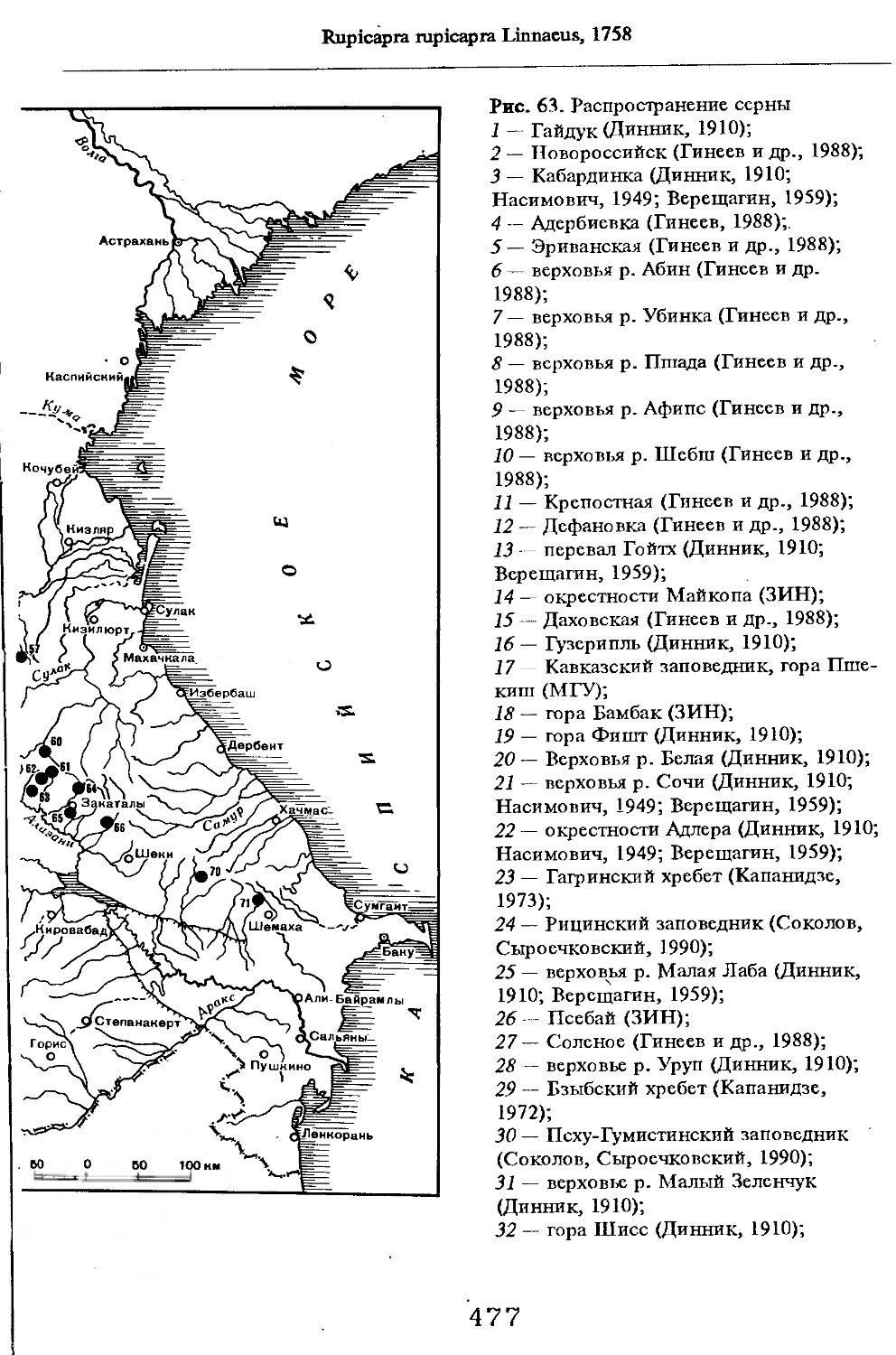
Рис. 1. Распространение кабана
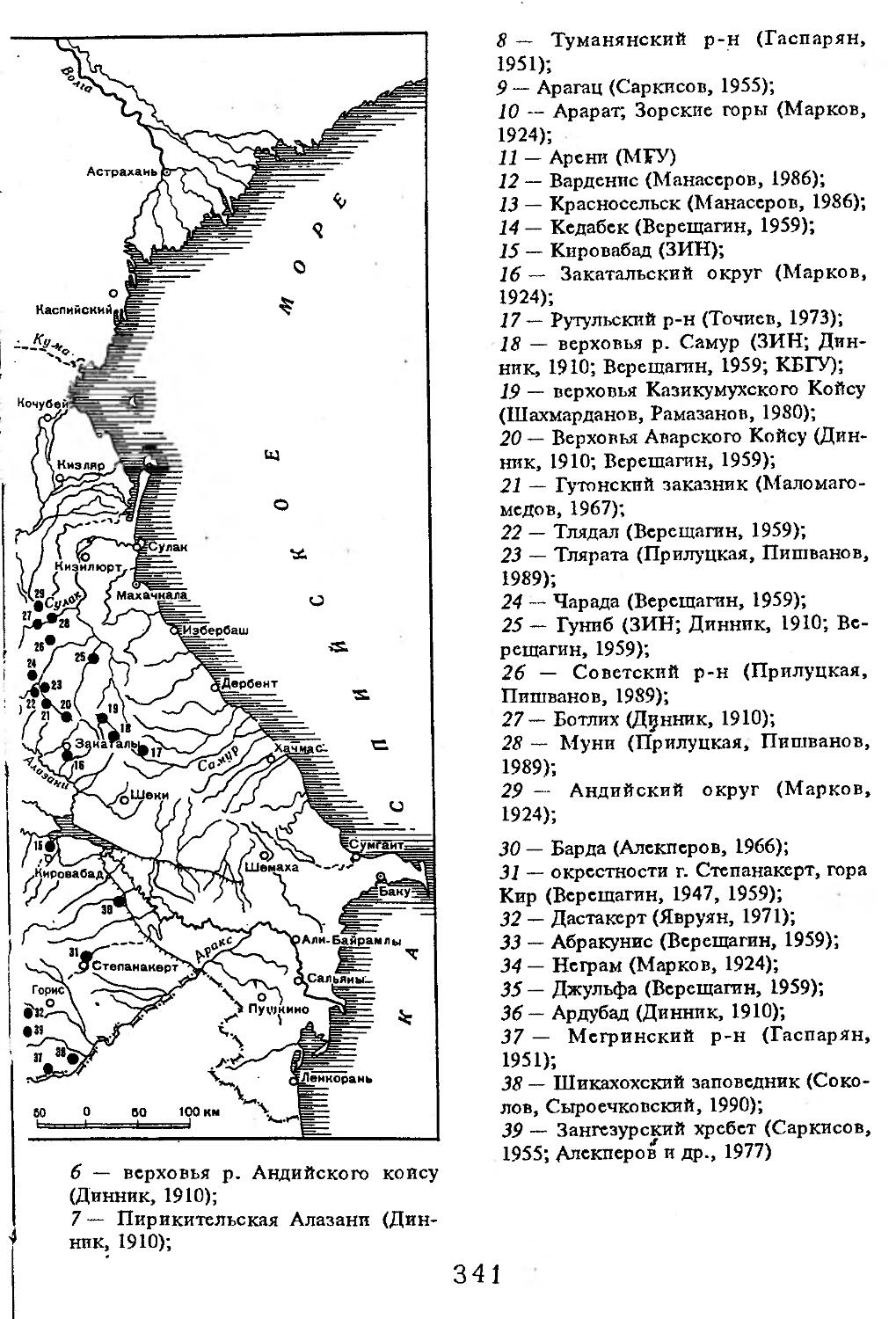
1 — низовья р. Кубань (Гептнер и др., 1961); 2 — Анапа (Динник, 1910); 3 — Темрюк (Косто-глод, 1969); 4 — Приазовские плавни (Гептнер и др., 1961); 5 — Приморско-Ахтарский р-н (Кос-тоглод, 1969; Дуров, 1973); 6 — Абрау-Дюрсо (Котов, 1963); 7 — Крымск (Котов, 1963); 8 — Славянский р-н (Костоглод, 1969); 9— Кабардинка (Котов, Рябов, 1963); 10 — Абинская (Котов, 1963; Котов, Рябов, 1963); 11 — Геленджик (ЗИН, Котов, 1963); 12 — Верхи. Пшады (Котов, Рябов, 1963); 13 — Береговка (Котов, Рябов, 1963); 14 — Смоленская (Котов, 1969); 15 — Краснодар (Динник, 1910); 16 — Бжид (Котов, Рябов, 1963); 17 — Новомихайловка (Котов, Рябов, 1963); 18 — Дефановка (Котов, Рябов, 1963); 19 — Молдавановка (Котов, Рябов, 1963); 20 — Ольгинка (Котов, Рябов, 1963); 21 —- Горячий Ключ (Дуров, 1973); 22 — Туапсе (Котов, 1963; Котов, Рябов, 1963); 23 — Гойтх (Даль, 1948; Котов, Рябов, 1963); 24 •— Октябрьская (Котов, Рябов, 1963); 25 — Армавир (Сафронова, 1973); 26 — Белореченск (Котов, 1963; Дуров, 1973); 27 — Майкоп (Динник, 1910; Дуров, 1973); 28 — Тульский р-н (Котов, 1963); 29 — Апшеронск (Котов, 1963; Котов, Рябов, 1963); 30 — Самурская (Котов, Рябов, 1963); 31 — Красноалександровский (Котов, Рябов, 1963); 32 — Аше (Котов, Рябов, 1963); 33 — Лазаревское (Котов, Рябов, 1963); 34 — Солахау (Котов, Рябов, 1963); 35 — Четвертая рота (Котов, Рябов, 1963); 36— Уроч. Бабукаул (Чернявская, 1956); 37 — Гузерипль (Двойченко, 1963; Котов, Рябов, 1963); 38 — Хамышки (Котов, 1963; Котов, Рябов, 1963); 39 — Киша (Чернявская, 1959); 40 — Новопрохладное (Котов, Рябов, 1963); 41 — Каменномосткое (Котов, Рябов, 1963); 42 — Абадзехская (Котов, Рябов, 1963); 43 — Губская (Котов, Рябов, 1963); 44 — Лабинский р-н (Котов, 1963); 45 — Изобильненский р-н (Оноприенко, Хохлов, 1989); 46 — Ипатовский р-н (Оноприенко, Хохлов, 1989); 47 — Отрадненский р-н (Котов, 1963); 48 — Бесстрашная (Котов, Рябов, 1963); 49 — Спо-койненский р-н (Котов, 1963); 50 — Псебай (Динник, 1910); 51 — Эсто-Садок (Котов, Рябов, 1963); 52 — Аибга (Котов, Рябов, 1963); 53 — Адлеровский р-н (Котов, 1963); 54 — Пицунда — Мюссерский заповедник (Соколов, Сыроечковский, 1990); 55 — Рицинский запо
1 3
Кабан
ведник (Соколов, Сыроечковский, 1990); 56 — Преградная (Котов, Рябов, 1963); 57 — Сторожевая (Котов, Рябов, 1963); 58 — Исправная (Динник, 1910); 59 — Передовая (Динник, 1910); 60 — Сухуми (Чернявский, 1879, Бочарников, 1880); 61 — Псху-Гумистинский заповедник (Соколов, Сыроечковский, 1990); 62 — Тебердинский заповедник (Федоров, 1954; Аренс, 1957; Соколов, Сыроечковский, 1990); 63 — Кисловодск (ЗИН); 64 — Пятигорск (Динник, 1910); 65 — Подгорная (Котов, Рябов, 1963); 66 — Екапцоко (КБГУ); 67 — Нефтекумский р-н (Оноприенко, Хохлов, 1989); 68 — Величаевское (ЗИН); 69 — Нальчик (Динник, 1910; ЗИН; КБГУ); 70 — Черная речка (Темботов, 1960); 71 — Вольный Аул (КБГУ); 72 — Белая речка (КБГУ); 73 — Кенженское лесничество (КБГУ); 74 — Нижний Чегем (Темботов, 1960); 75 — Лашкута (Темботов, 1960); 76 — Ар гуд ан (Темботов, 1960); 77 — Змейская (МГУ); 78 — Поти, (Верещагин, 1959; Соколов, Сыроечковский, 1990); 79 — Батуми (Динник, 1910); 80 — Кинтришский заповедник (Соколов, Сыроечковский, 1990); 81 — Кутаиси (Динник, 1910); 82 — Северо-Осетинский заповедник (Соколов, Сыроечковский, 1990); 83 — Владикавказ (Динник, 1910); 84 — Малгобек (Динник, 1910); 85 — Моздок (Динник, 1910); 86 — Лиаховский заповедник (Соколов, Сыроечковский, 1990); 87 — Горийский р-н (Даль, 1954); 88 — Боржоми (Динник, 1910); 89 — Боржомский заповедник (Соколов, Сыроечковский, 1990); 90 — Малгетии (Соколов, Сыроечковский, 1990); 91 — Тбилиси (Динник, 1910); 92 — Ахметский р-н (Арабули, 1970); 93 — Ахметский заповедник (Соколов, Сыроечковский, 1990); 94 — Турианчайский заповедник (Соколов, Сыроечковский, 1990); 95 — низовье Кумы (Динник, 1910); 96 — Мегринский р-н (Даль, 1954); 97 — Кизлярский участок Дагестанского заповедника (Соколов, Сыроечковский, 1990); 98 — низовье р. Терек (Динник, 1910; Гептнер, Формозов, 1941; Гинеев, 1954); 99 — побережье Аг-раханского залива (Гинеев, 1981); 100 — Манасаул (МГУ); 101 — Кизлярский р-н (Шах-марданов, Спасская, 1975); 102 — Бабаюртовский р-н (Шахмарданов, Спасская, 1975); 103 — плавни р. Аксай (Гинеев, 1971); 104 — плавни р. Акгаш (Гинеев, 1971); 105 — низовье р. Сулак (Динник, 1910; Гептнер, Формозов, 1941; Гинеев, 1954); 106 — Хасавюрт (Гептнер, Формозов, 1941); 107 — Махачкала (Динник, 1910); 108 — Унцукыльский р-н (Шахмарданов, Спасская, 1975); 109 — Хунзахский р-н (Шахмарданов, Спасская, 1975); 110 — Каякент (Динник, 1910); 111 — Магарамкентский р-н (Шахмарданов, Спасская, 1975); 112 — Лагодехский заповедник (Соколов, Сыроечковский, 1990); 113 — Лагодехи (Динник, 1910); 114 — Закатальский заповедник (Соколов, Сыроечковский, 1990); 115 — Закатали (Динник, 1910); 116— Илисунский заповедник (Соколов, Сыроечковский, 1990); 117 — Вашло ване кий заповедник (Соколов, Сыроечковский, 1990); 118 — Телавский р-н (Арабули, 1970); 119 — Гардабанское охотхозяйство (Арабули, 1970); 120 — Караязский заповедник (Соколов, Сыроечковский, 1990); 121 — Дилижанский заповедник (Соколов, Сыроечковский, 1990); 122 — долина р. Араке в Октемберянском р-не (Явруян, 1975); 123 — долина р. Араке в Арташатском р-не (Явруян, 1975); 124 — Арарат (Даль, 1954); 125 — Хосровский заповедник (Соколов, Сыроечковский, 1990); 126 — Арени (Даль, 1948); 127 — Мегринский р-н (Даль, 1954); 128 — Шикахохский заповедник (Соколов, Сыроечковский, 1990); 129 — Басутчайский заповедник (Соколов, Сыроечковский, 1990); 130 — Прирвани (Алекперов, 1966); 131 — Кафанский р-н (Даль, 1954); 132 — Кумлак (Алекперов, 1966); 133 — Кубаталы (Алекперов, 1966); 134 — Сисиан (Даль, 1954); 135 — Кеордмахмудлу (Алекперов, 1966); 136 — Бехманлы (Алекперов, 1966); 137 — Ачкёльский заповедник (Литвинов, 1971; Алекперов, Кулиев, 1987; Соколов, Сыроечковский, 1990); 138 — Турианчайский заповедник (Соколов, Сыроечковский, 1990); 139 — Исмаилинский заповедник (Соколов, Сыроечковский, 1990); 140 — Варташен (Мейр, Схоль, 1954); 141 — Ахтинский р-н (Явруян, 1975); 142— Кусарский р-н (Алекперов, Кулиев, 1987); 143 — Хачмас (Алекперов, Кулиев, 1987); 144 — Исмаиллы (Верещагин, 1959); 145 — Ширванский заповедник (Соколов, Сыроечковский, 1990); 146 — Кызыл-Агачский заповедник (Литвинов, 1981; Соколов, Сыроечковский, 1990); 147 — Ленкорань (Динник, 1910; Верещагин, 1959); 148 — Гирканский заповедник (Соколов, Сыроечковский, 1990); 149 — Астара (Сафаров, 1958); 150 — Ярдамлы (КБГУ)
Sus scrofa Linnaeus, 1758
ного. Это еще раз свидетельствует о большей роли тазовой кости в локомоции.
Окраска меха. Во всех исследованиях, посвященных анализу морфологических показателей кабана, отмечалась высокая внутрипопуляционная изменчивость окраски животных. Обращалось внимание на вариабельность общей окраски тела и отдельных его участков, что зависит от цвета дистальной части щетин. Еще Н.Я.Динник (1910) писал, что щетина имеет черный или почти черный цвет, но рассученные концы ее окрашены в различные цвета (от серо-бурого до почти белого). Эти концы вместе с просвечивающейся основной частью щетин, а также подпушь темно-коричневого, , темно-серого и грязно-белого цвета определяют общую окраску животного и отдельных частей тела. В условиях Западного Кавказа (Дуров, 1987) в зимнем мехе ость у 80 % кабанов черная, на ее вершинах доминируют бурый, темно-бурый или серый цвет; лицевая часть головы имеет бурый, темнобурый или серый цвет, шея — бурый или трязно-коричневый, трива и спина — темно-бурый до черного, бока, и бедра — грязно-бурый, грудь и пах — палево-коричневый; ноги, уши, подбородок и хвост — черный. Более светлые животные по окраске меха сходны с среднеазиатским кабаном, описанным А.А.Слудским (1956).
Около 15 % западнокавказских кабанов (Кавказский заповедник) имеют более светлый цвет (светло-серый цвет ости на боках и шее, черный на ногах, кончике хвоста, ушах).
Окраска меха кабана нами изучена по свежему коллекционному материалу (л = 34 экз.), полученному нами в поясе широколиственных лесов Центрального Кавказа (октябрь-ноябрь 1988 г.). Эта выборка иллюстрирует широкий размах изменчивости как общей окраски животного, так и отдельных участков тела. Все это многообразие окраски можно свести к трем типам: черный, светлый и буроватый. Первый тип характеризуется тем, что щетина на спине и боках черного цвета, лишь кончики волос темцо-коричневые, в итоге животные с такой окраской выглядят очень темными, почти черными. Посветление меха за счет грязновато-белых кончиков волос отмечается в паху, на труди и других участках низа тела. Особи с первым типом окраски в нашей выборке составляют около 10 %. Второй тип резко отличается от первого тем, что щетина с беловатой дистальной частью (около 1/3 длины) покрывает почти всю поверхность тела. Лишь на хребте, ногах, хвосте и некоторых других участках тела и головы (см. описание западнокавказских кабанов) щетина черная. На нижней стороне тела, особенно в области паха, серовато-белый цвет сильно распространен, нередко на всю длину волос. Кабаны со вторым типом окраски в нашей выборке составляют также около 10 %. Третий тип отличается тем, что кончики щетин буроватые с ржавым налетом, на брюхе и особенно в паху окраска волос довольно оригинальна (коричневатая с зеленым налетом). Животные с третьим типом окраски составляют в нашей выборке около 20 %. Остальные экземпляры, в том числе молодняк, составляют промежуточные звенья между тремя описанными типами. В наших сборах не оказалось животных с аномальной окраской.
1 5
Кабан
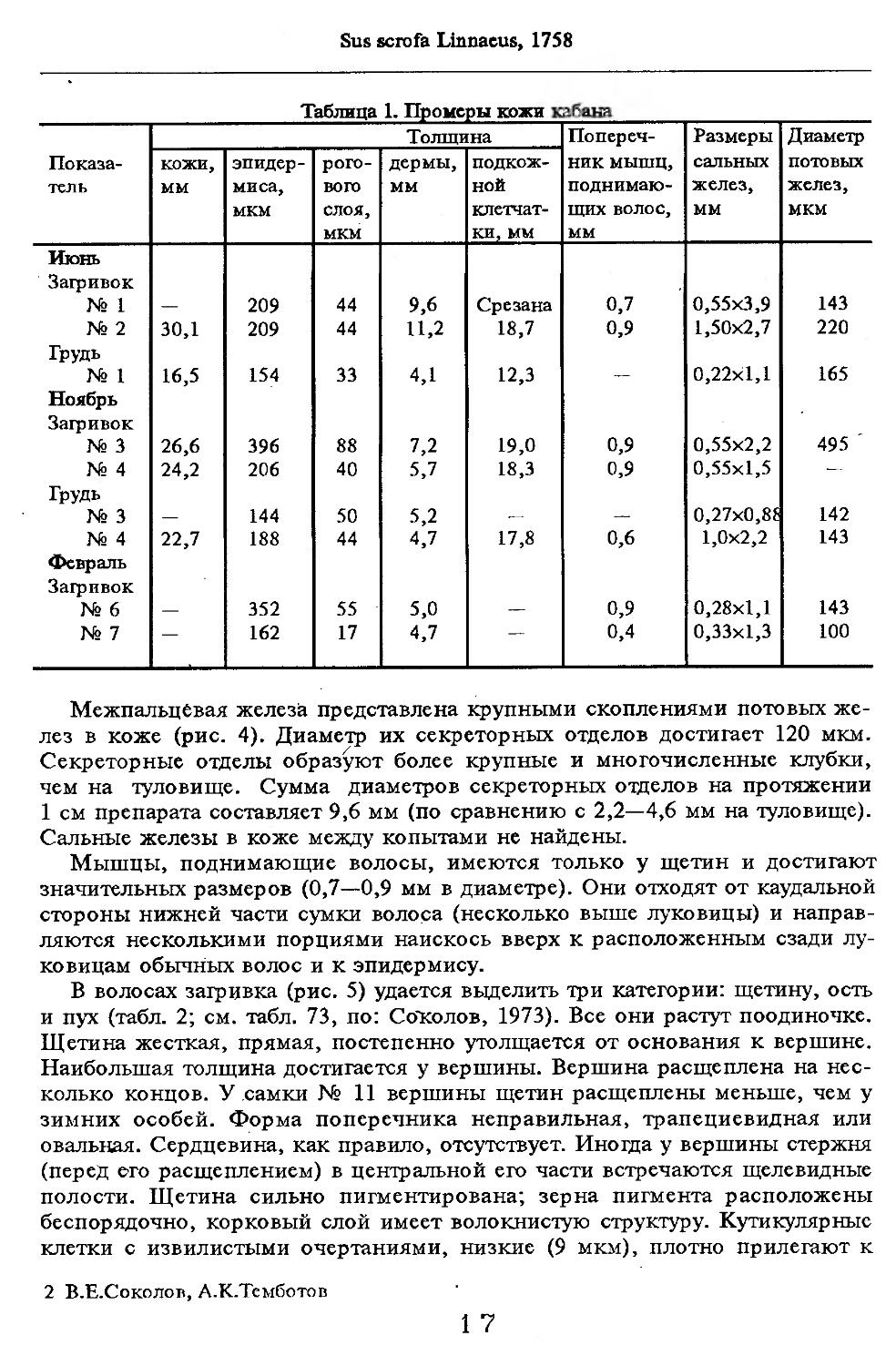
Кожный покров. Изучены два самца, добытые в ноябре в Нальчикском ле-соохотохозяйстве (№ 3 и 4), длиной тела 149 и 170 см и самец и самка (№ 8 и 9), добытые соответственно в ноябре 1989 г. и в июне 1989 г. (№ 10, 11) там же. Для сравнения использованы данные по самцу (№ 1) длиной тела 162 см и самке (№ 2) длиной тела 130 см из Приморско-Ахтарского госохот-хозяйства (июнь), а также по самке (№ 5) длиной тела 160 см и двум сеголеткам (№ 6 и 7) из Калининской области. Результаты сведены в табл. 1 и частично опубликованы ранее (Соколов, 1973).
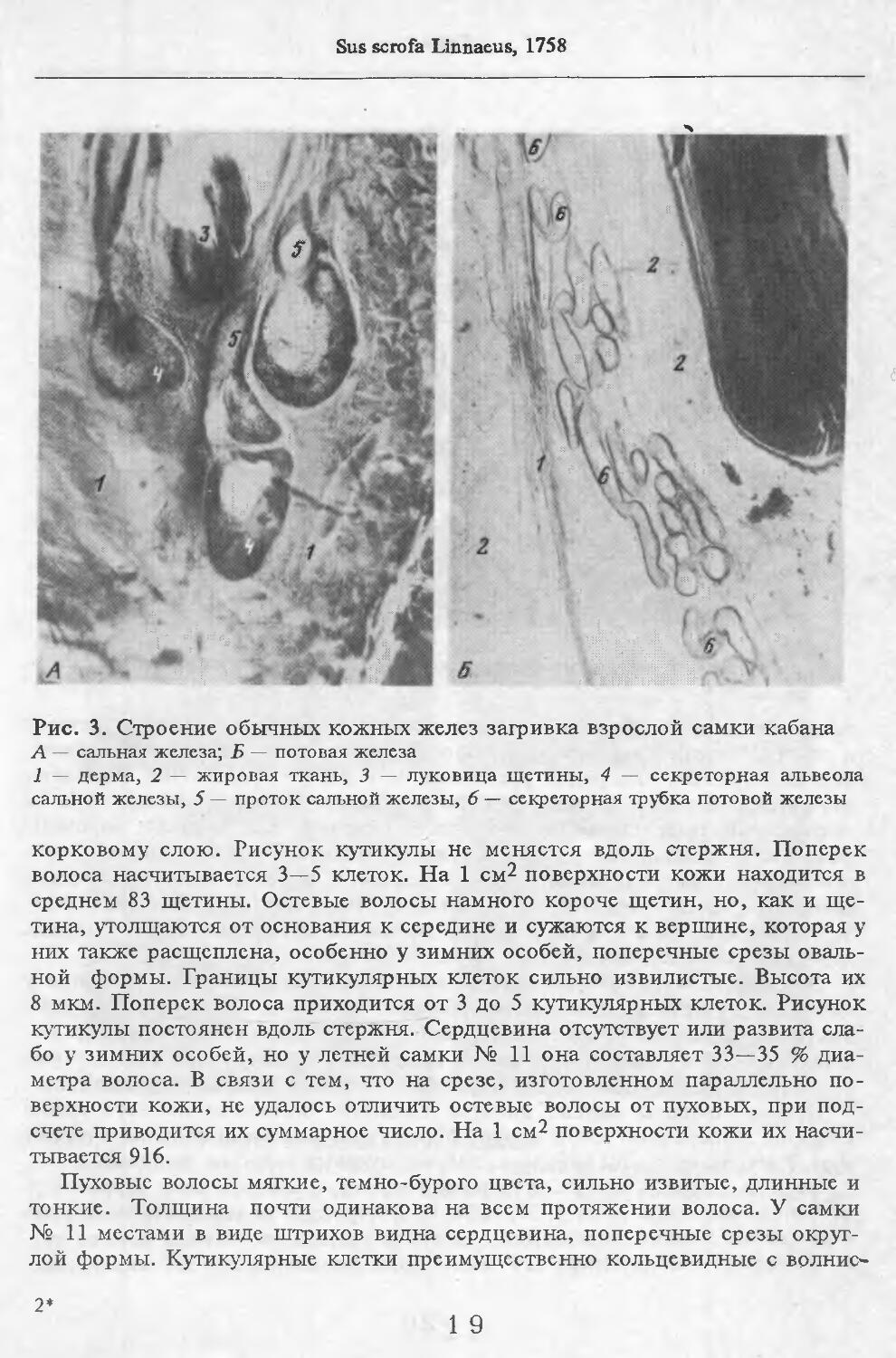
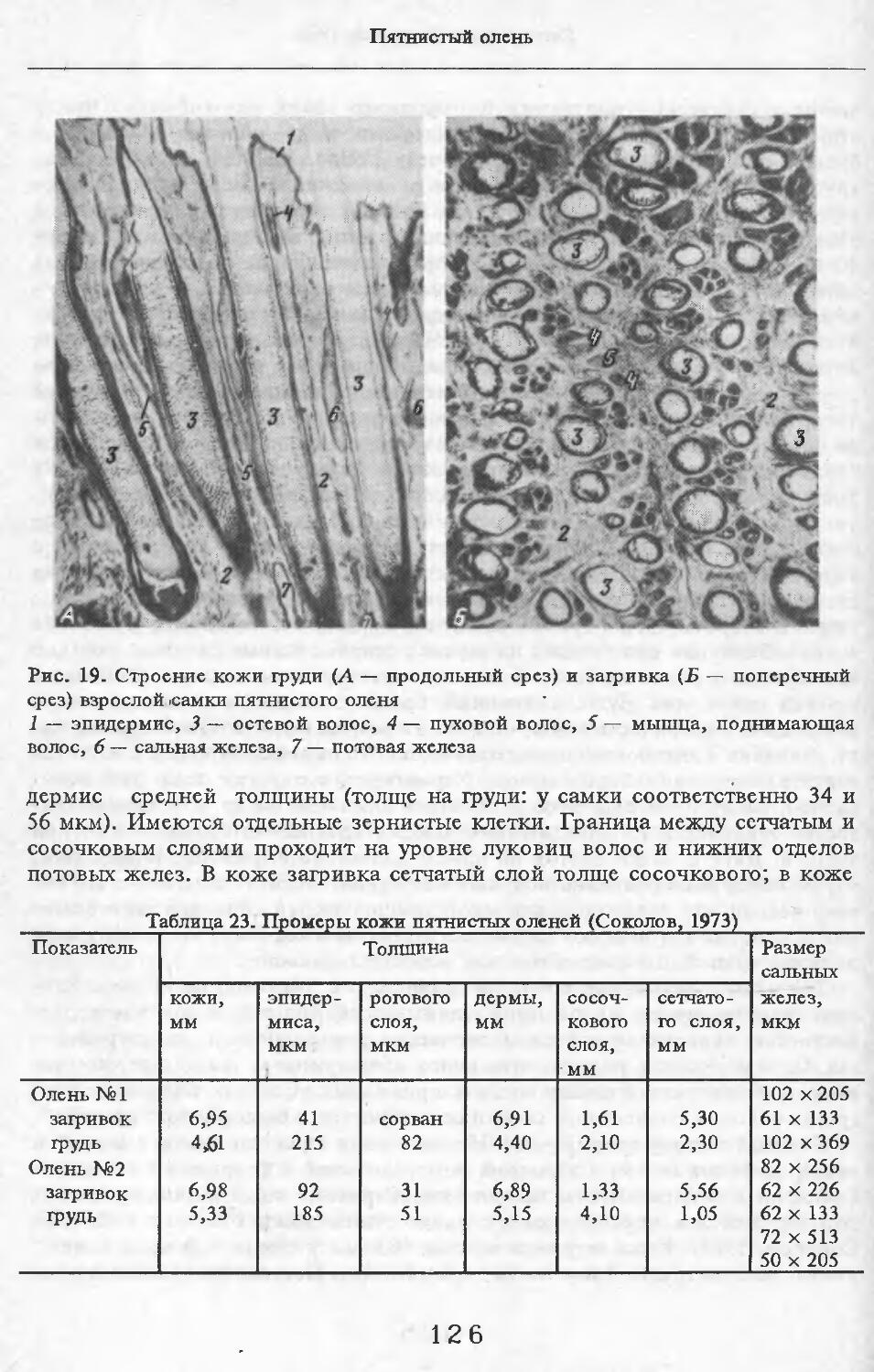
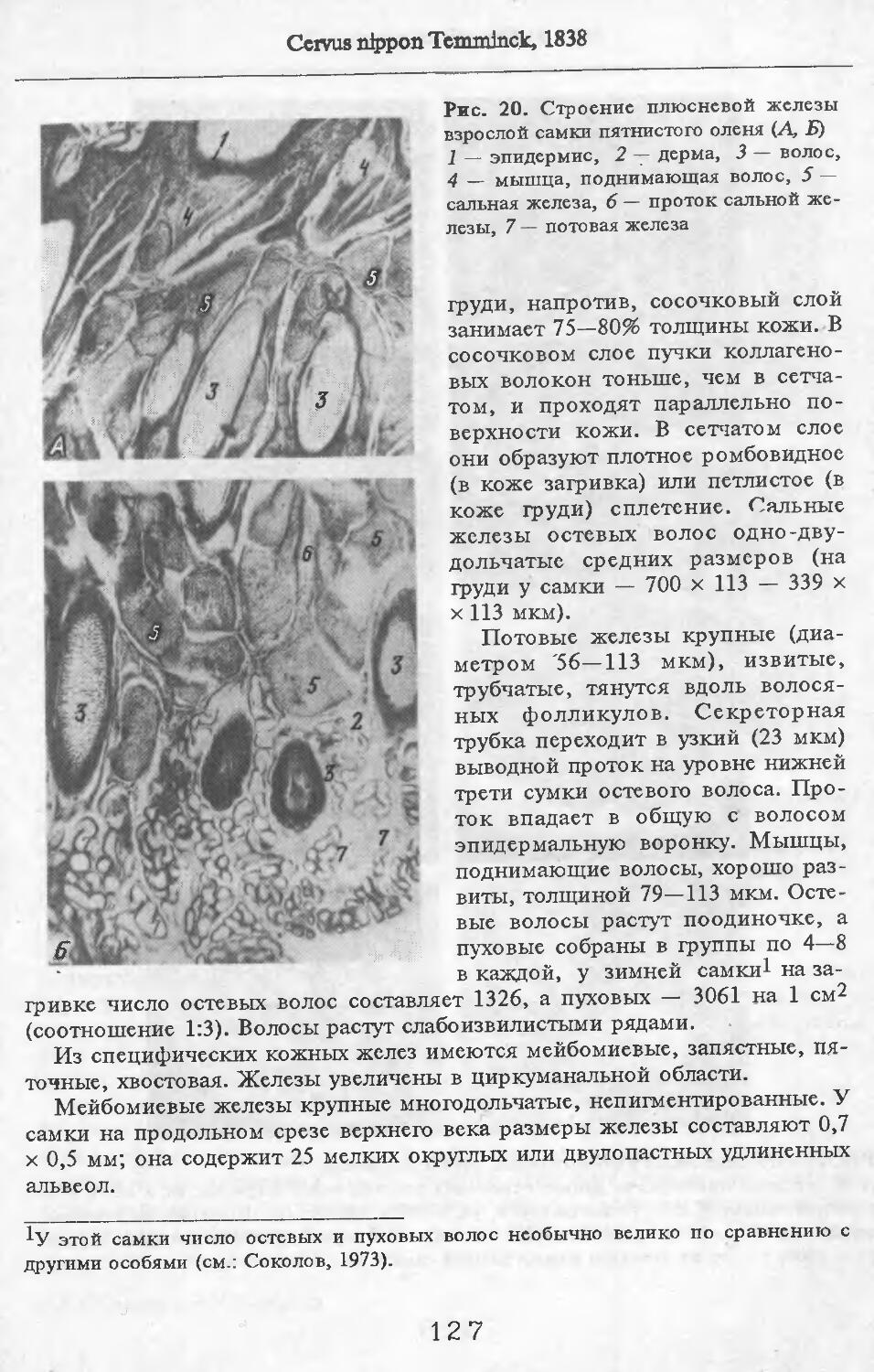
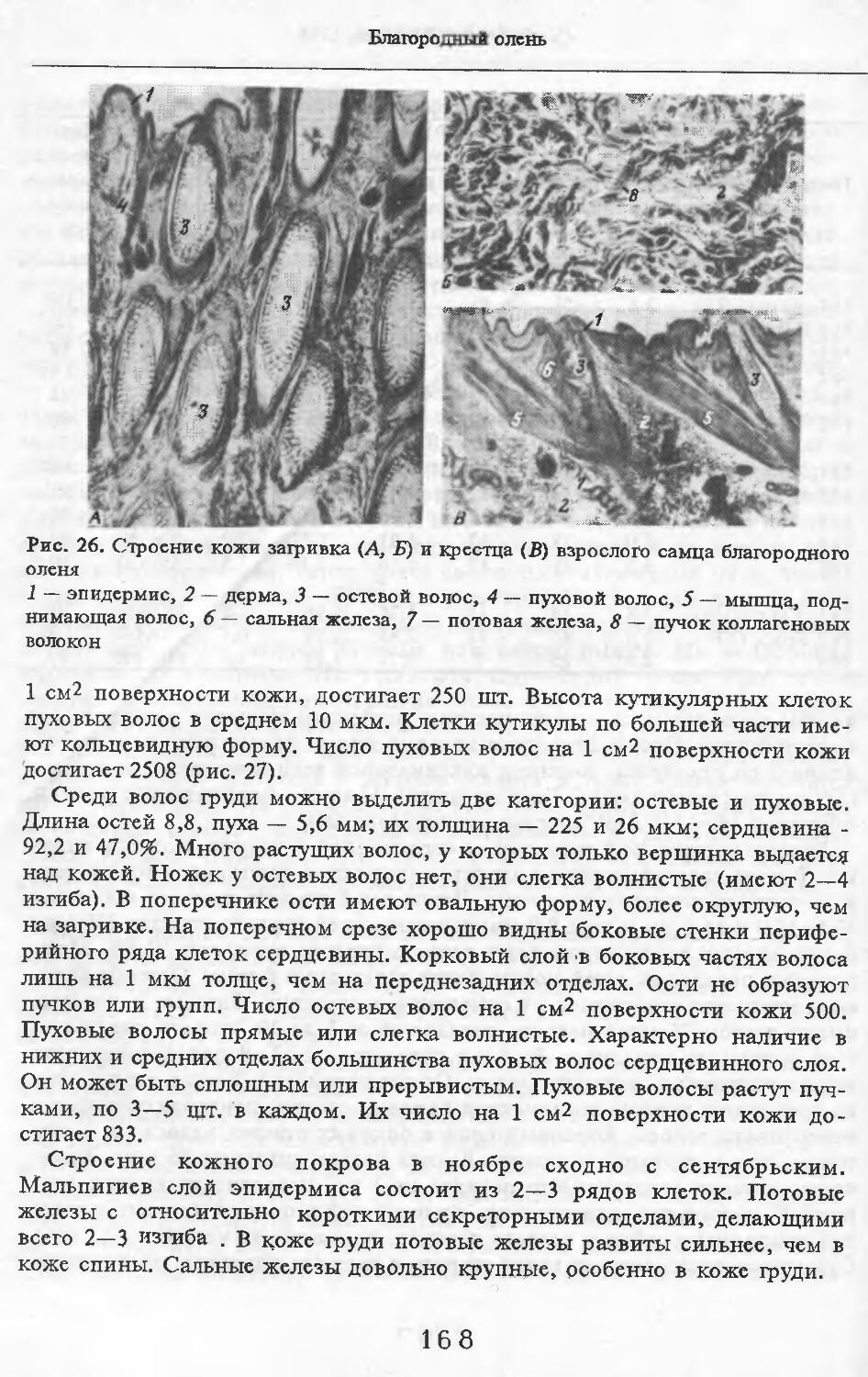
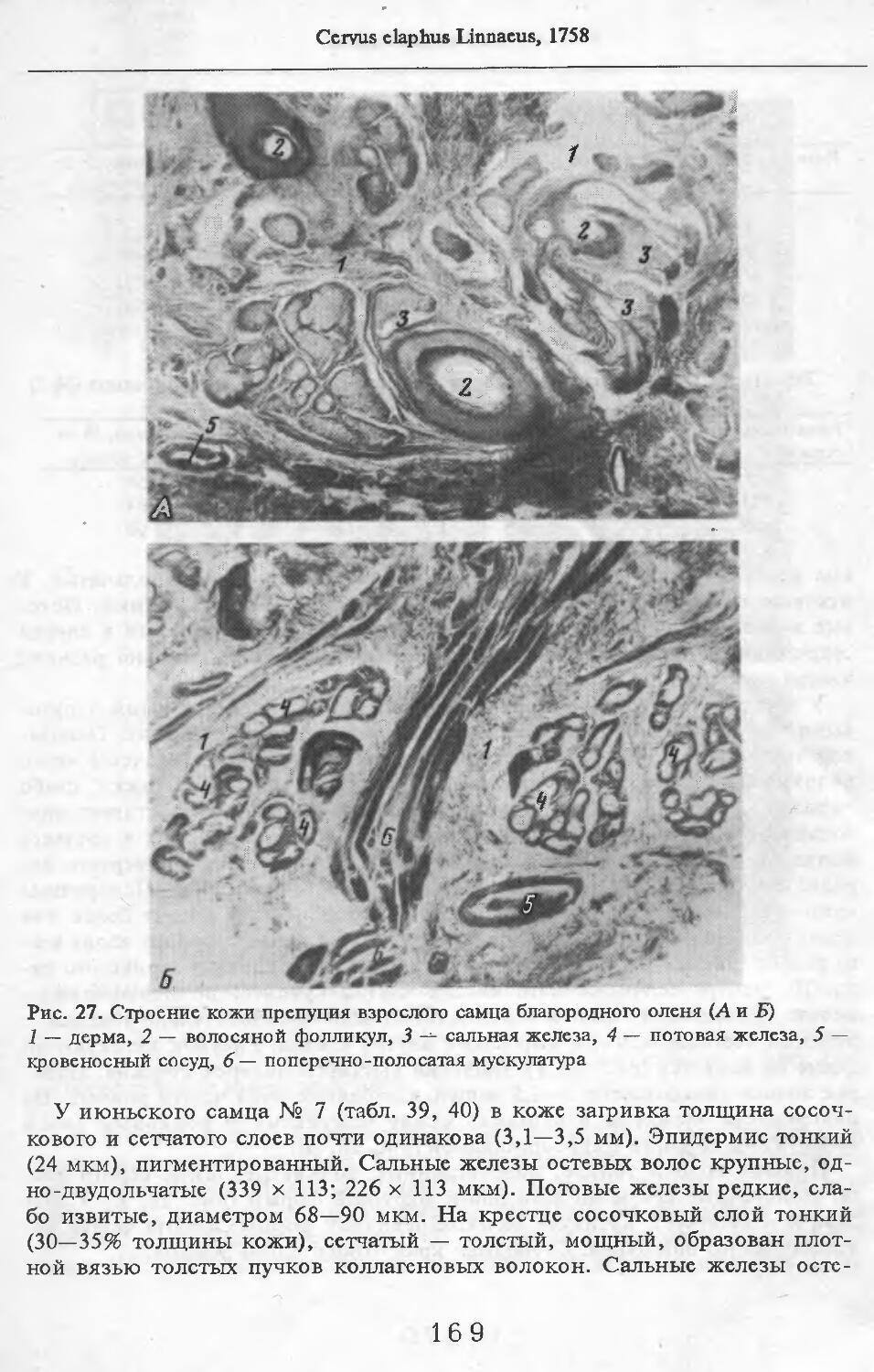
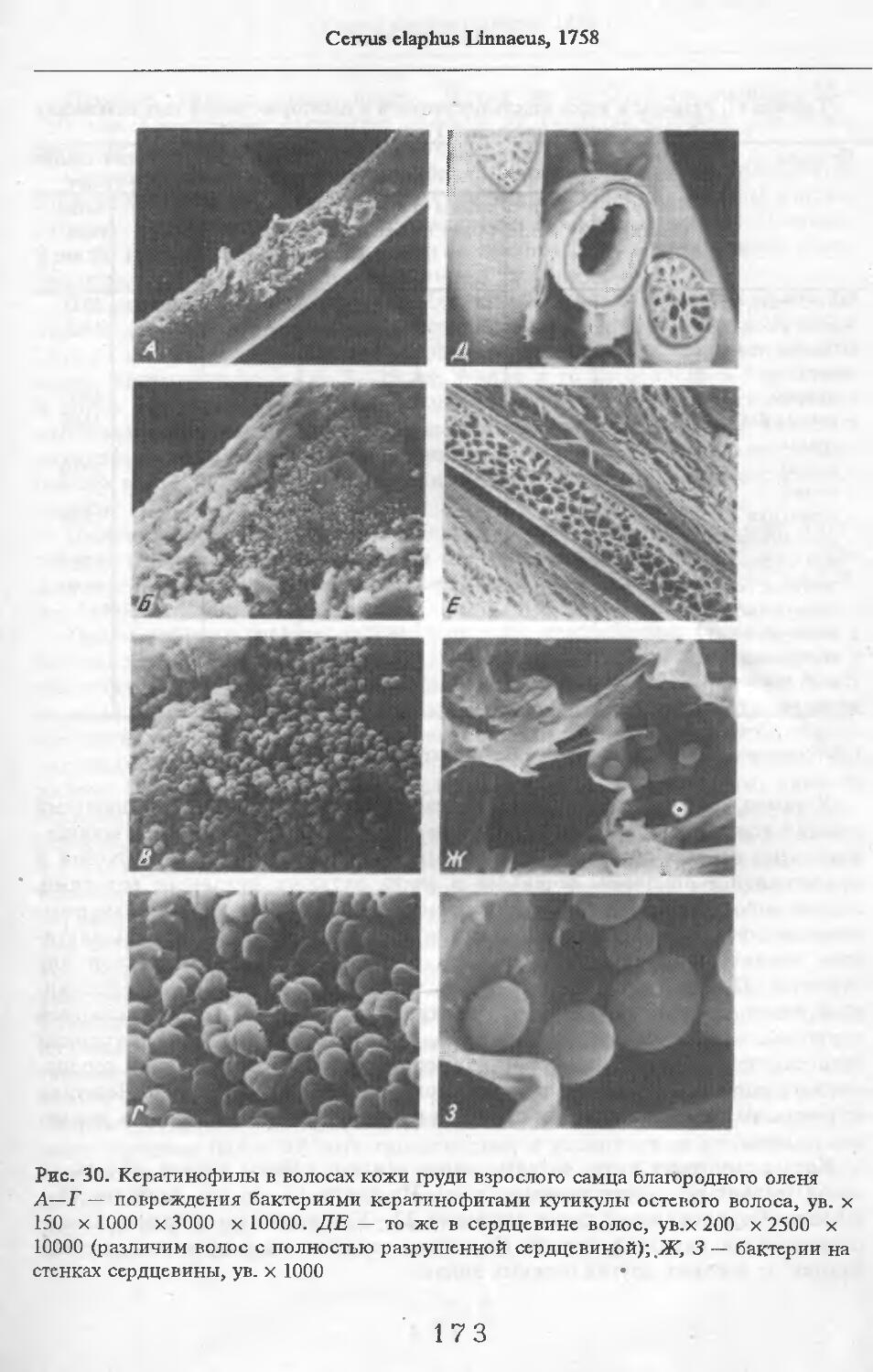
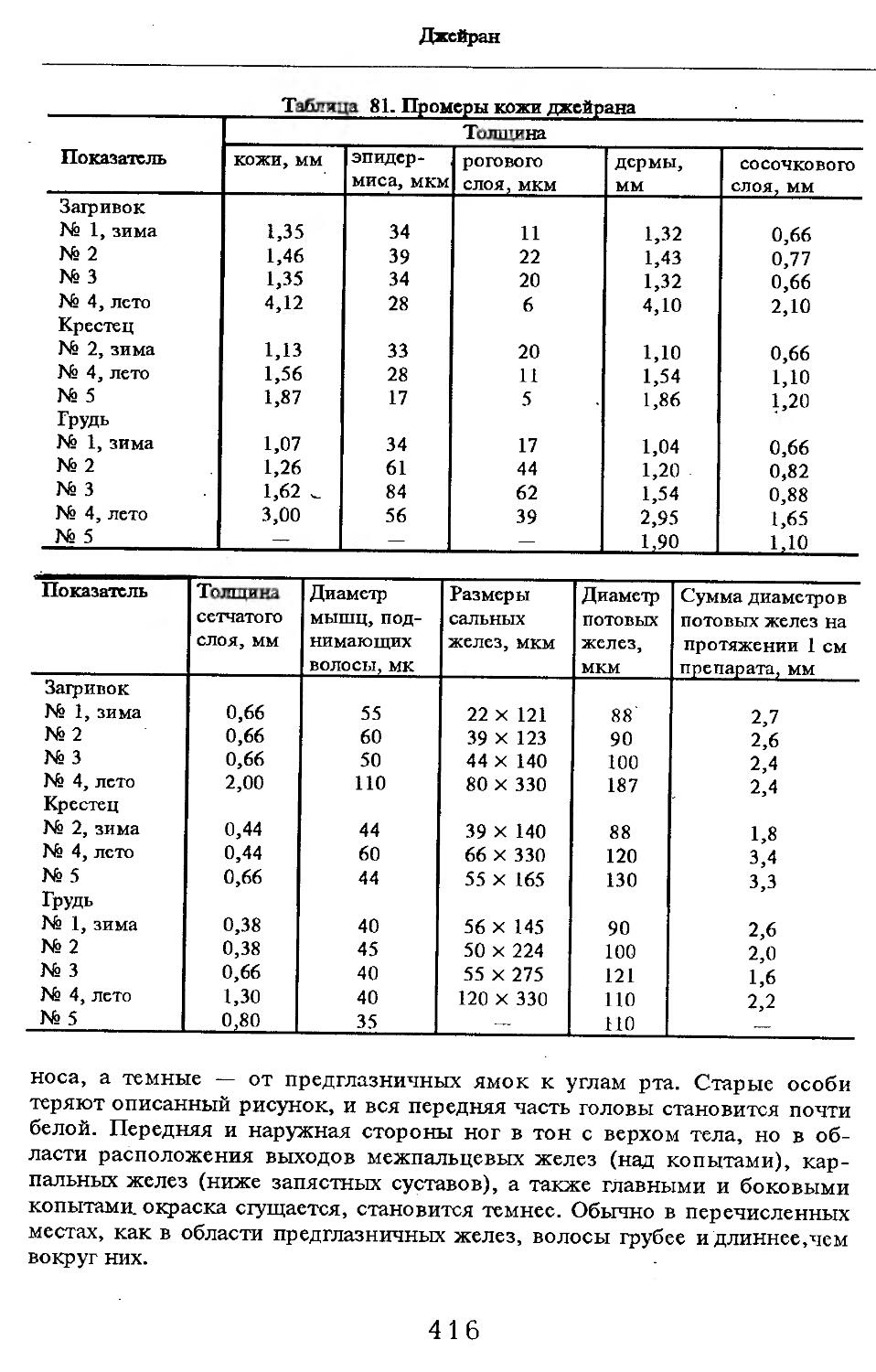
Летом у самцов и самок (№ 1 и 2) эпидермис кожи с небольшими ячейками на внутренней поверхности. Высота их от 88 до ПО мкм. Только местами представлен одно-двухслойный зернистый слой, блестящий слой отсут-> ствует. Базальный слой пигментирован со средней интенсивностью. Роговой слой толстый, многорядный. НиЖняя граница дермы с крупными выступами, вдающимися в подкожную жировую клетчатку. Выступы образуются в районе корней щетин, глубоко опускающихся в кожу. Дерма не подразделяется на слои, так как корни щетин пронизывают всю дерму насквозь. В дерме преобладают пучки коллагеновых волокон, проходящие параллельно поверхности кожи. Их связь довольно рыхлая. В дерме загривка много толстых эластиновых волокон, располагающихся в том же направлении, что и коллагеновые пучки. В дерме труди эластиновые волокна немногочисленны. В подкожной жировой клетчатке очень толстые пучки коллагеновых волокон (иногда целые пласты) проходят параллельно поверхности кожи. Размеры жировых клеток одинаковы на разной глубине клетчатки и достигают максимальных размеров: 80 х 130, ПО х 130 мкм. Сальные железы своеобразны. У каждой щетины с передней и задней стороны находится по огромной многодольчатой железе (рис. 2). Длинный пигментированный проток-железы открывается в волосяную сумку близко к поверхности кожи (примерно в 0,6—0,7 мм от нее). В глубь кожи сальные железы идут на расстояние до 4,2 мм. У обычных волос сальные железы однодольчатые, небольшие, по одной у каждого волоса.
У каждой щетины расположено по потовой железе (рис. 3). Длинные секреторные отделы извиты в клубок, расположенный, как правило, на уровне луковиц щетин. Около клубков желез концентрируются жировые клетки. Диаметр секреторных отделов некоторых желез очень ‘велик.
В литературе приводятся данные о наличии у кабанов предглазничных, карпальных, мейбомиевых, межпальцевых желез, желез на верхней губе над клыками, подбородочной, желез пятачка, половых губ (Соколов, Чернова, 1988). Мы располагаем материалом по карпальной железе кабана № 1 и 11 и межпальцевой — № 3 и 11. Карпальная железа лежит примерно посередине запястья, на его внутренней поверхности. Железа открывается наружу небольшими отверстиями. Помимо крупных клубков типичных потовых желез (диаметр их секреторных отделов достигает 132 мкм), имеются огромные скопления трубчатых желез с более узкими секреторными отделами диаметром до 100 мкм. Они очень сильно извиты и закручены в крупные клубки (см. рис. 3). Трубчатые железы образуют сплошной пласт, разделенный узкими прослойками пучков коллагеновых волокон.
1 6
Sus scrofa Linnaeus, 1758
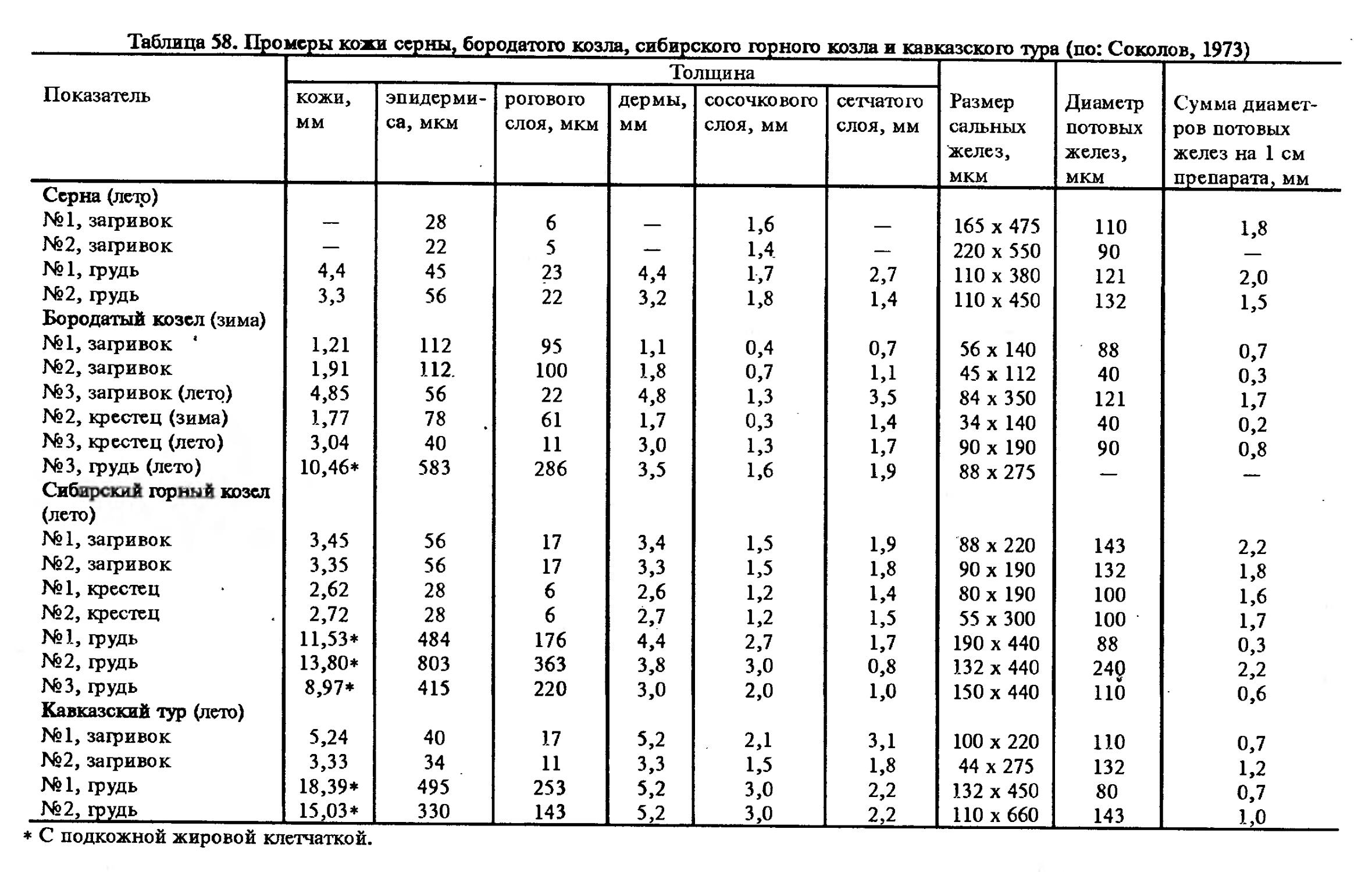
Таблица 1. Промеры кожи клбана
Показатель Толщина Попереч-ник мышц, поднимающих волос, мм Размеры сальных желез, мм Диаметр потовых желез, мкм
КОЖИ, мм эпидермиса, мкм рогового слоя, мкм дермы, мм подкожной клетчат- ки, мм
Июнь
Загривок № 1 — 209 44 9,6 Срезана 0,7 0,55x3,9 143
№ 2 30,1 209 44 11,2 18,7 0,9 1,50x2,7 220
Грудь
№ 1 16,5 154 33 4,1 12,3 — 0,22x1,1 165
Ноябрь Загривок № 3 26,6 396 88 7,2 19,0 0,9 0,55x2,2 495 '
№ 4 24,2 206 40 5,7 18,3 0,9 0,55x1,5 -—
Грудь
№ 3 — 144 50 5,2 .— —. 0,27x0,8? 142
№ 4 22,7 188 44 4,7 17,8 0,6 1,0x2,2 143
Февраль Загривок
№ 6 — 352 55 5,0 — 0,9 0,28x1,1 143
№ 7 — 162 17 4,7 — 0,4 0,33x1,3 100
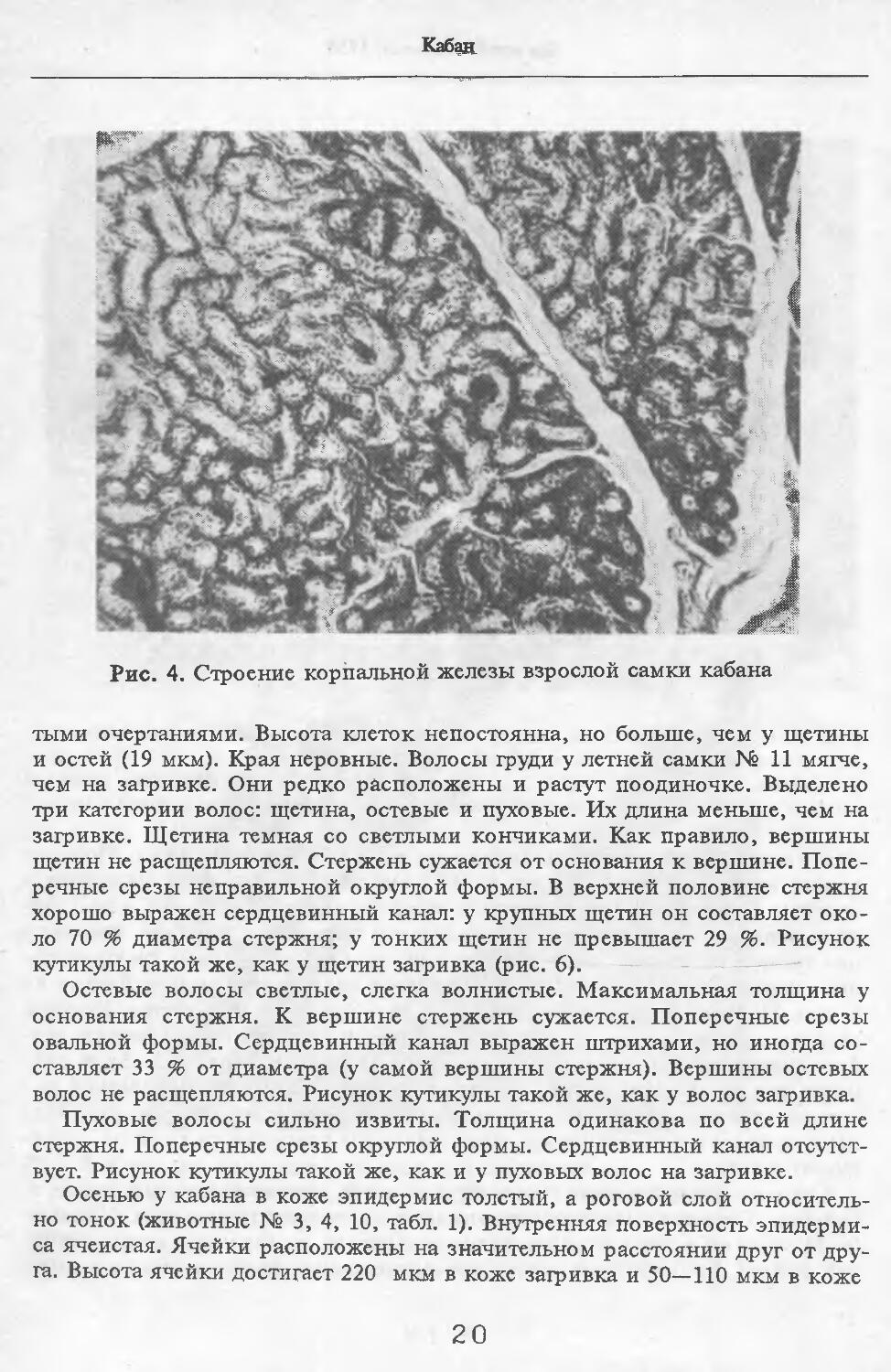
МежпальцСвая железа представлена крупными скоплениями потовых желез в коже (рис. 4). Диаметр их секреторных отделов достигает 120 мкм. Секреторные отделы образуют более крупные и многочисленные клубки, чем на туловище. Сумма диаметров секреторных отделов на протяжении 1 см препарата составляет 9,6 мм (по сравнению с 2,2—4,6 мм на туловище). Сальные железы в коже между копытами не найдены.
Мышцы, поднимающие волосы, имеются только у щетин и достигают значительных размеров (0,7—0,9 мм в диаметре). Они отходят от каудальной стороны нижней части сумки волоса (несколько выше луковицы) и направляются несколькими порциями наискось вверх к расположенным сзади луковицам обычных волос и к эпидермису.
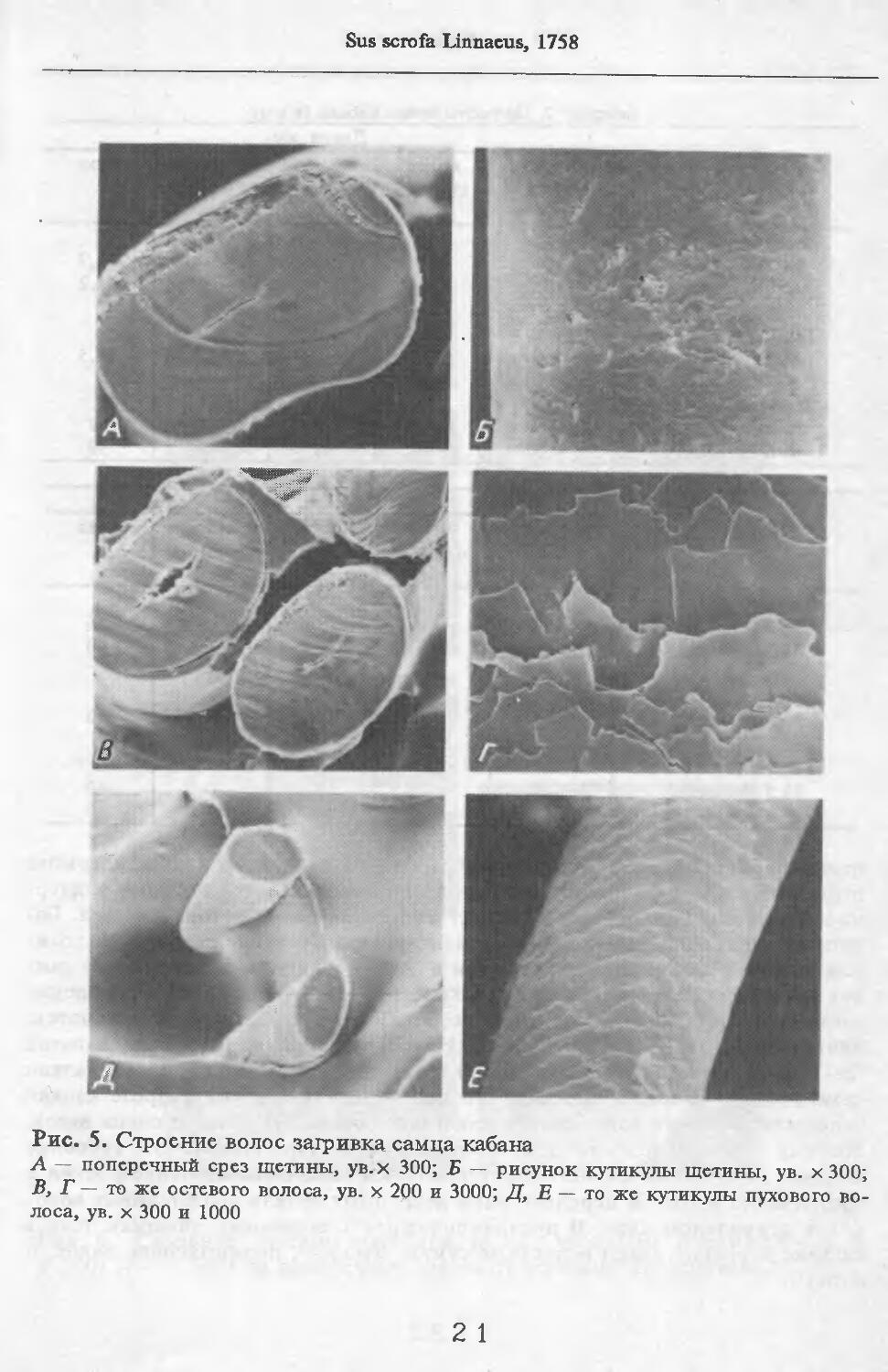
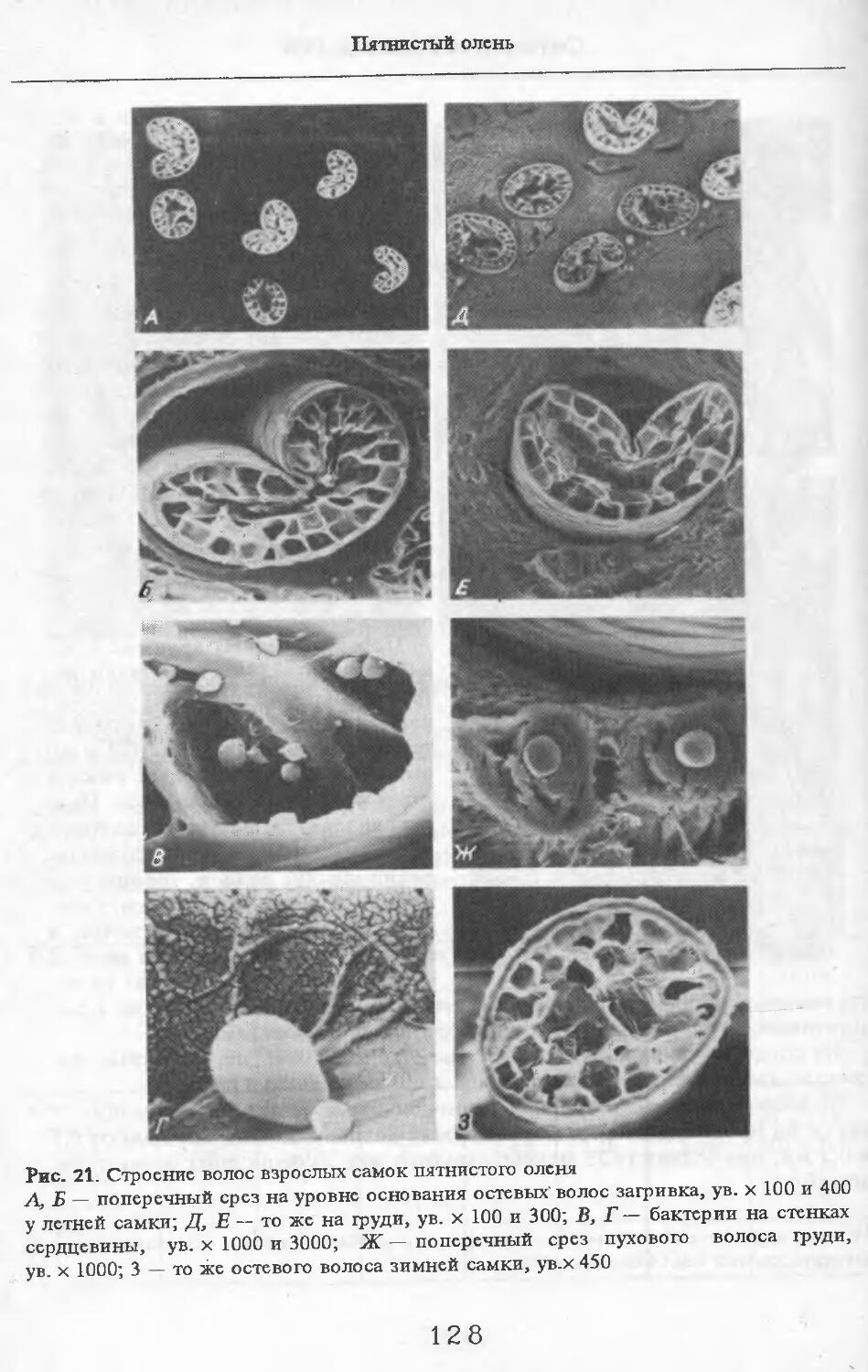
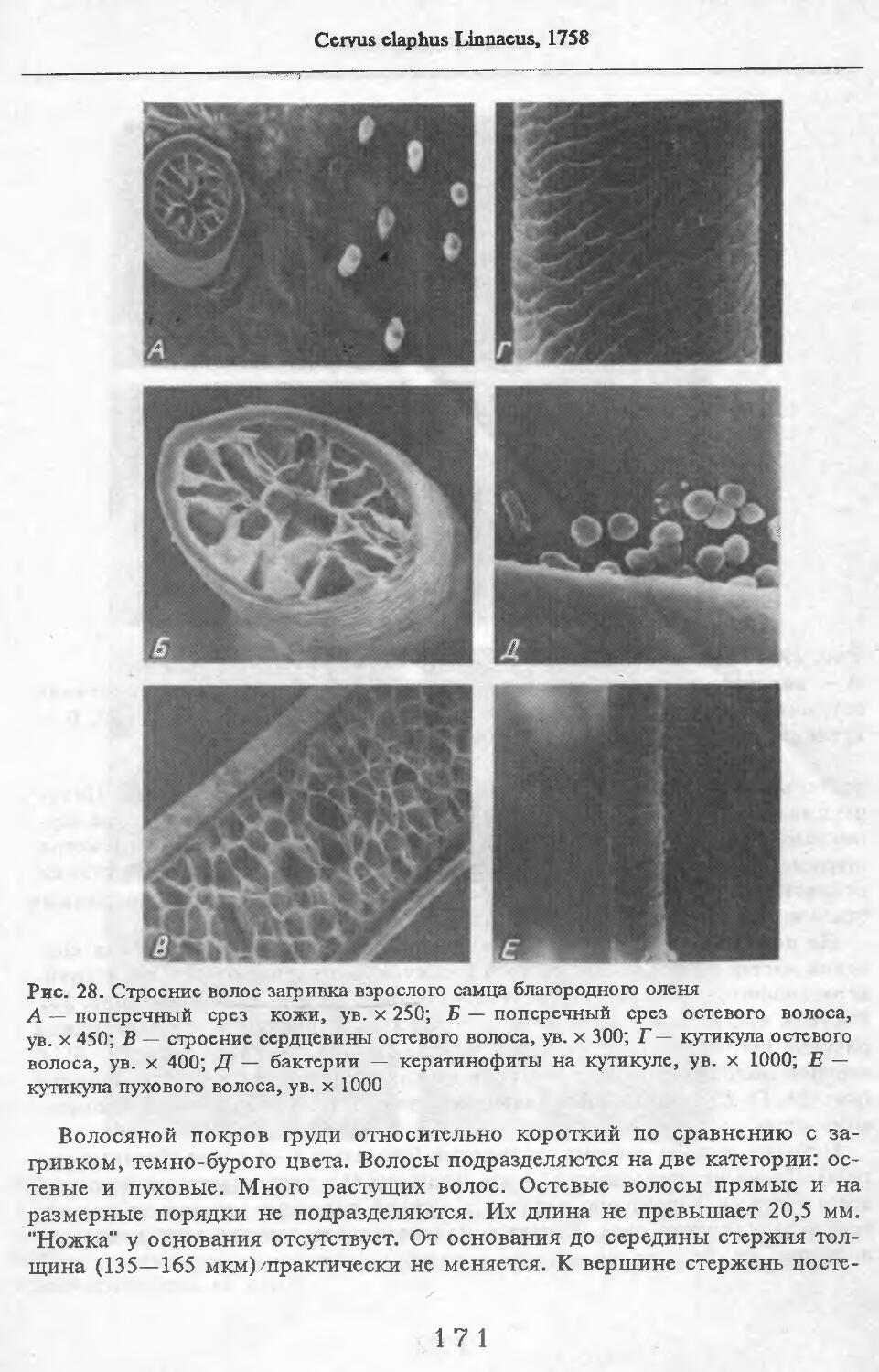
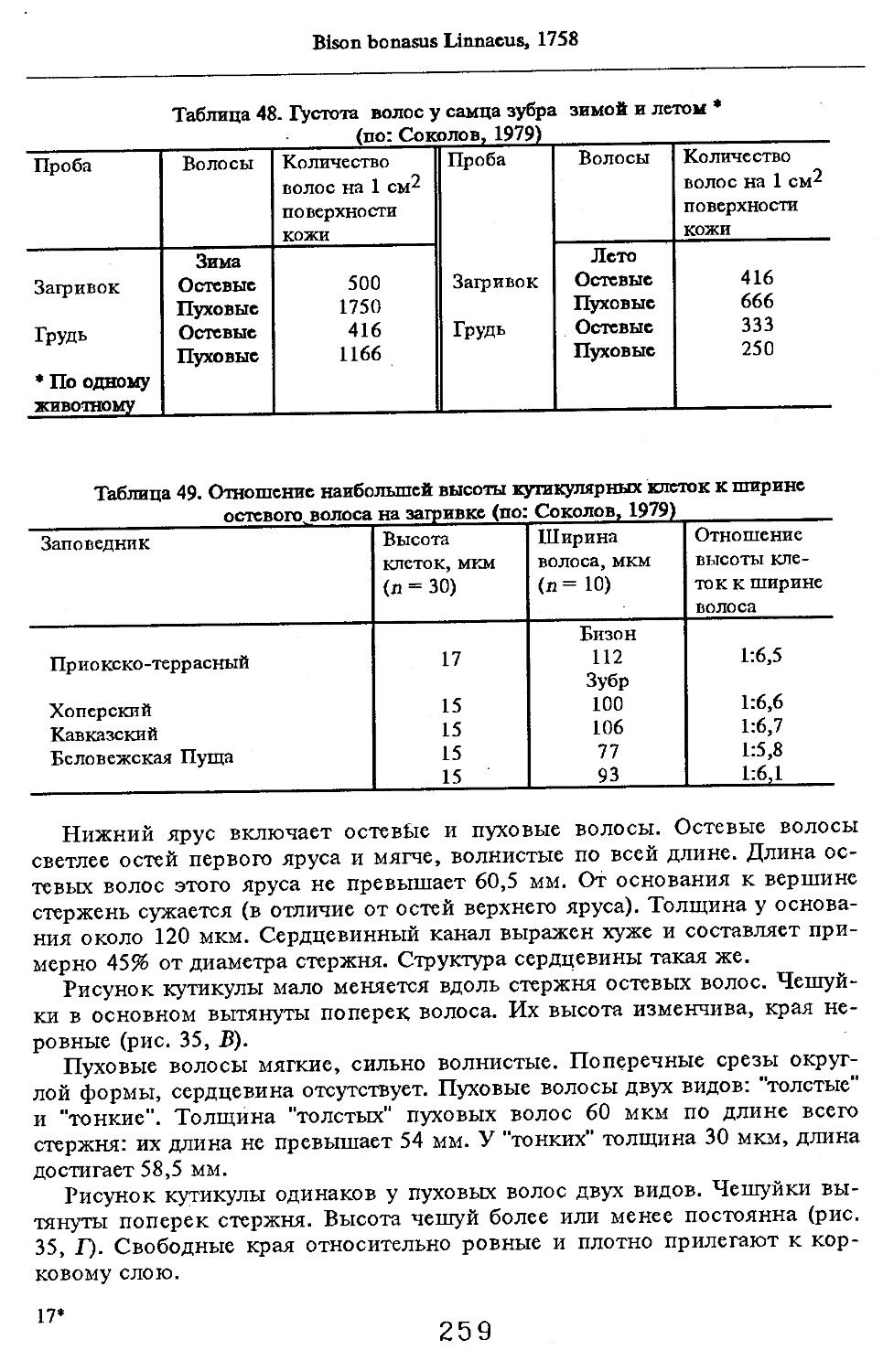
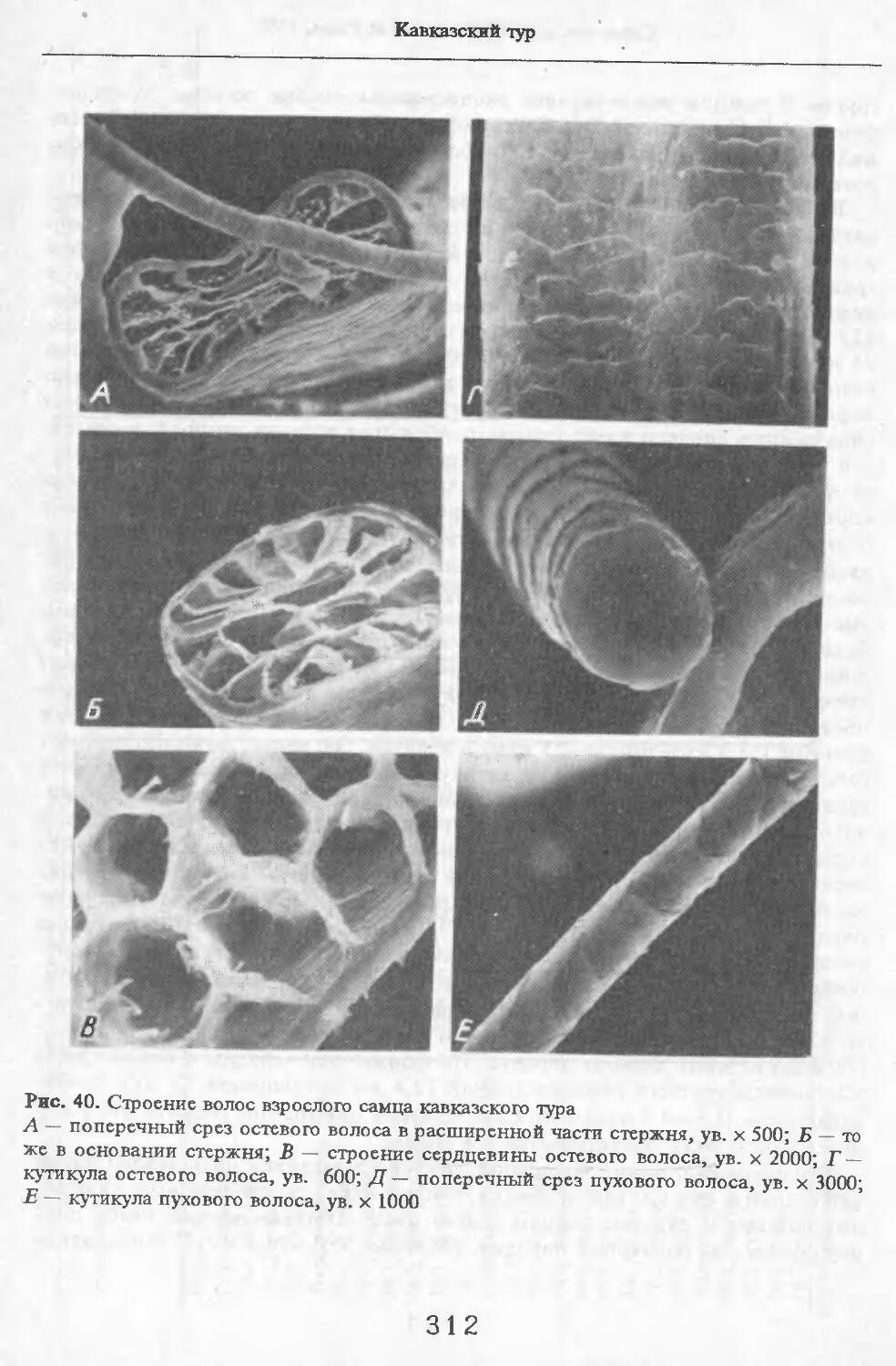
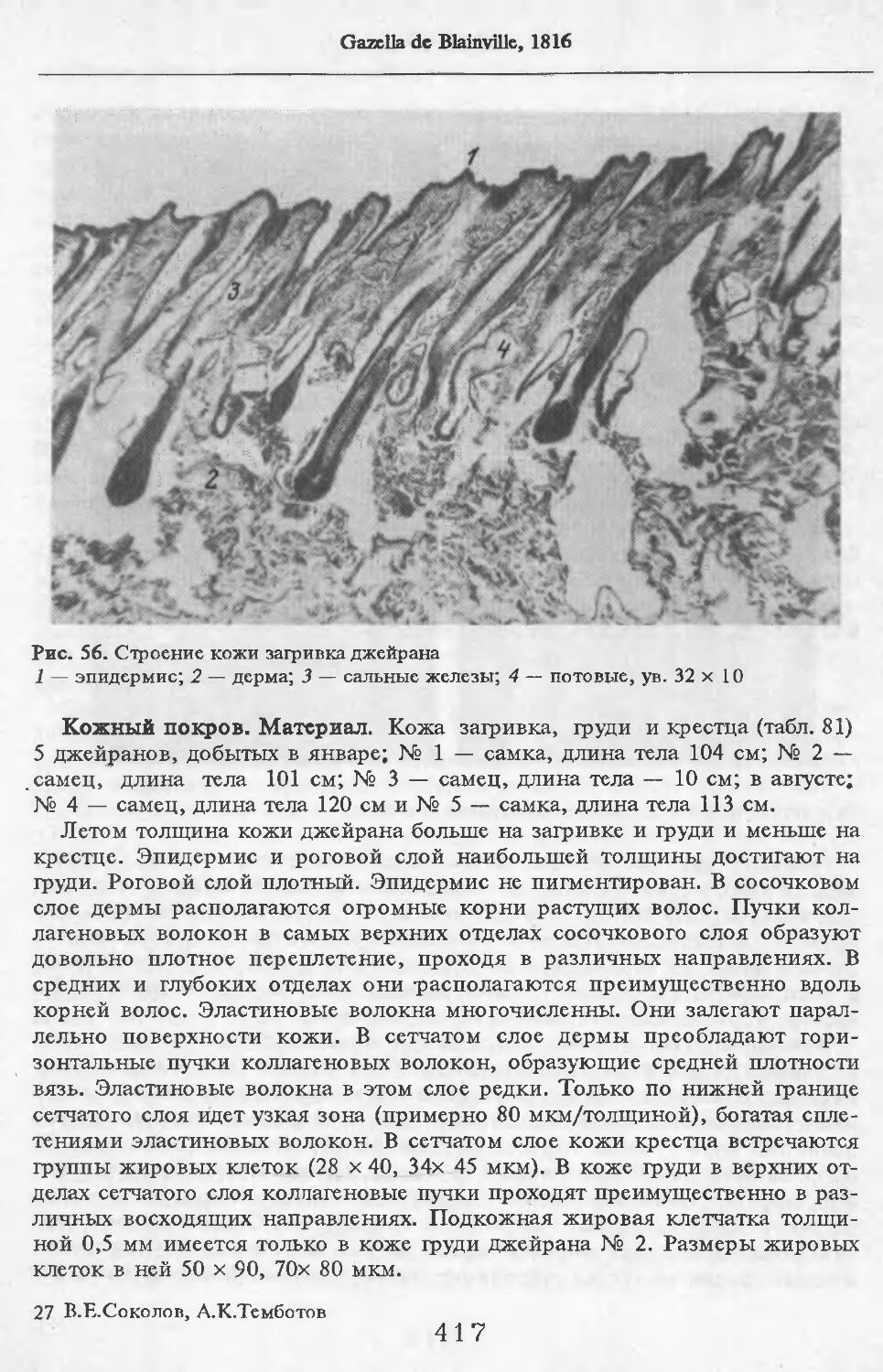
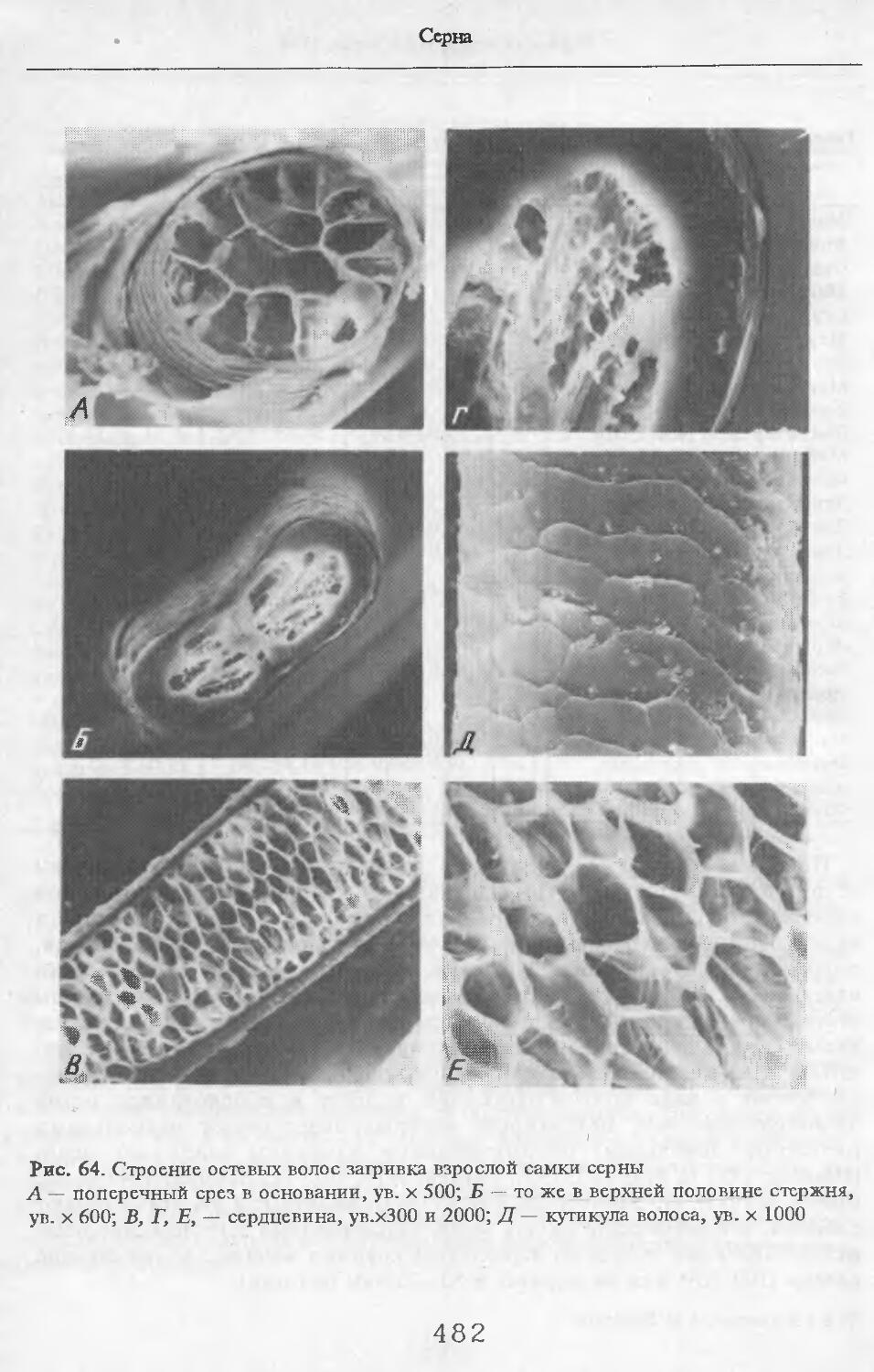
В волосах загривка (рис. 5) удается выделить три категории: щетину, ость и пух (табл. 2; см. табл. 73, по: Соколов, 1973). Все они растут поодиночке. Щетина жесткая, прямая, постепенно утолщается от основания к вершине. Наибольшая толщина достигается у вершины. Вершина расщеплена на несколько концов. У самки № 11 вершины щетин расщеплены меньше, чем у зимних особей. Форма поперечника неправильная, трапециевидная или овальная. Сердцевина, как правило, отсутствует. Иногда у вершины стержня (перед его расщеплением) в центральной его части встречаются щелевидные полости. Щетина сильно пигментирована; зерна пигмента расположены беспорядочно, корковый слой имеет волокнистую структуру. Кутикулярные клетки с извилистыми очертаниями, низкие (9 мкм), плотно прилегают к
2 В.Е.Соколов, А.К.Темботов
1 7
Кабан
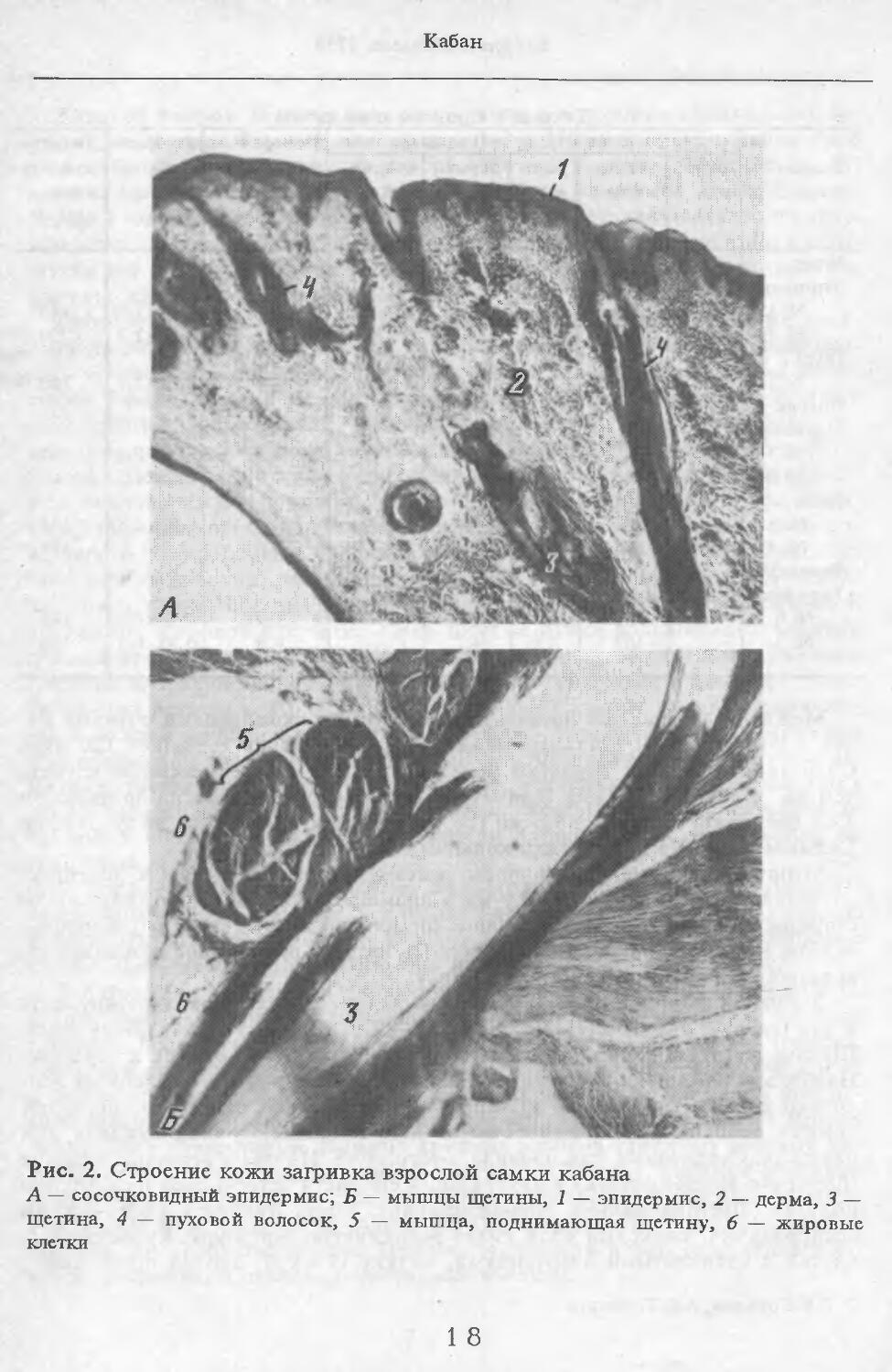
Рис. 2. Строение кожи загривка взрослой самки кабана
А — сосочковидный эпидермис; Б — мышцы щетины, 1 — эпидермис, 2 — дерма, 3 — щетина, 4 — пуховой волосок, 5 — мышца, поднимающая щетину, 6 — жировые клетки
1 8
Sus scrofa Linnaeus, 1758
Рис. 3. Строение обычных кожных желез загривка взрослой самки кабана
А - сальная железа; Б — потовая железа
1 — дерма, 2 — жировая ткань, 3 — луковица щетины, 4 — секреторная альвеола сальной железы, 5 — проток сальной железы, 6 — секреторная трубка потовой железы
корковому слою. Рисунок кутикулы не меняется вдоль стержня. Поперек волоса насчитывается 3—5 клеток. На 1 см2 поверхности кожи находится в среднем 83 щетины. Остевые волосы намного короче щетин, но, как и щетина, утолщаются от основания к середине и сужаются к вершине, которая у них также расщеплена, особенно у зимних особей, поперечные срезы овальной формы. Границы кутикулярных клеток сильно извилистые. Высота их 8 мкм. Поперек волоса приходится от 3 до 5 кутикулярных клеток. Рисунок кутикулы постоянен вдоль стержня. Сердцевина отсутствует или развита слабо у зимних особей, но у летней самки № 11 она составляет 33—35 % диаметра волоса. В связи с тем, что на срезе, изготовленном параллельно поверхности кожи, не удалось отличить остевые волосы от пуховых, при подсчете приводится их суммарное число. На 1 см2 поверхности кожи их насчитывается 916.
Пуховые волосы мягкие, темно-бурого цвета, сильно извитые, длинные и тонкие. Толщина почти одинакова на всем протяжении волоса. У самки №11 местами в виде штрихов видна сердцевина, поперечные срезы округлой формы. Кутикулярные клетки преимущественно кольцевидные с волнис-
2*
1 9
Кабая
Рис. 4. Строение корпальной железы взрослой самки кабана
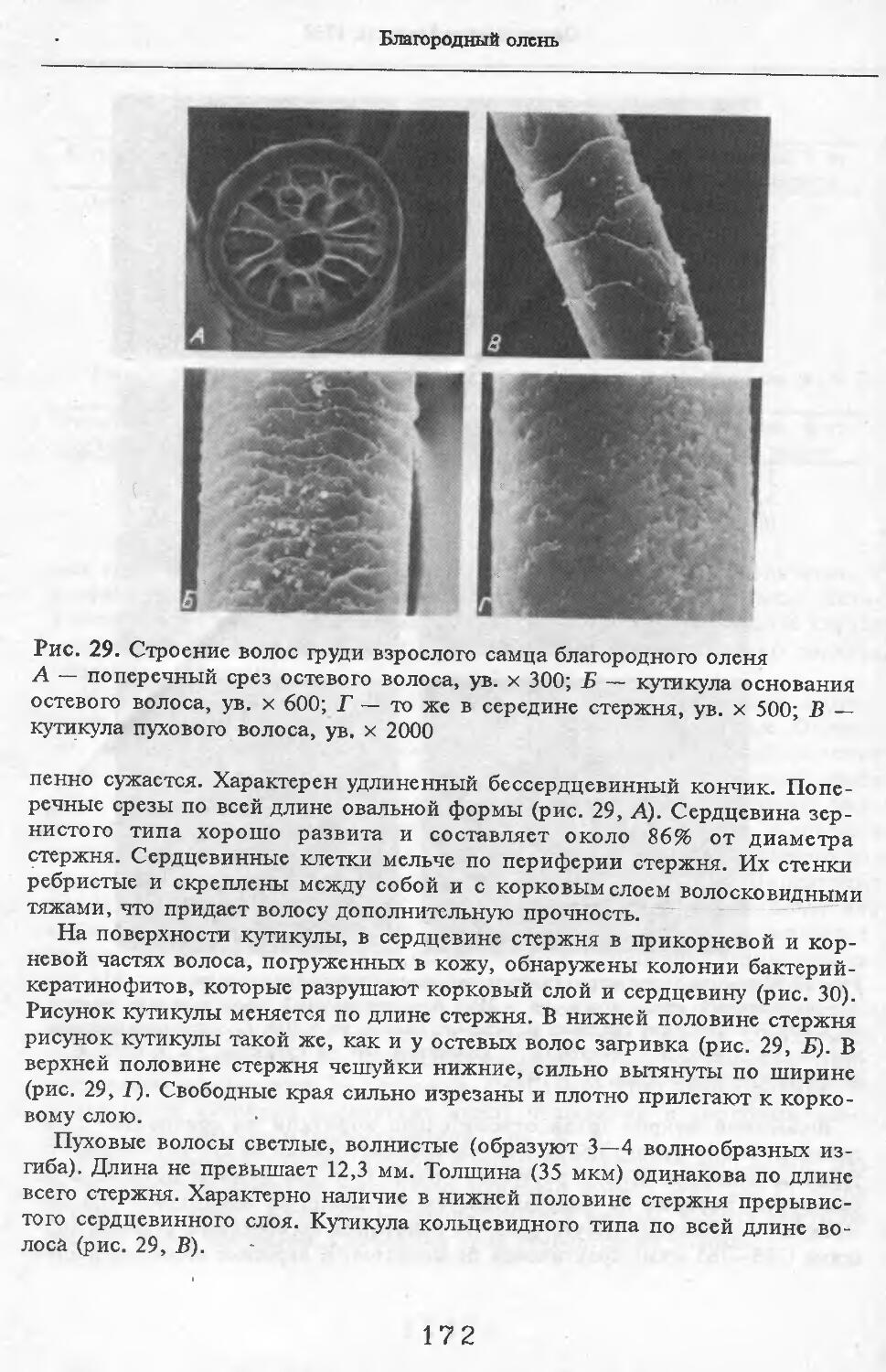
тыми очертаниями. Высота клеток непостоянна, но больше, чем у щетины и остей (19 мкм). Края неровные. Волосы 1руди у летней самки № 11 мягче, чем на загривке. Они редко расположены и растут поодиночке. Выделено три категории волос: щетина, остевые и пуховые. Их длина меньше, чем на завивке. Щетина темная со светлыми кончиками. Как правило, вершины щетин не расщепляются. Стержень сужается от основания к вершине. Поперечные срезы неправильной округлой формы. В верхней половине стх ржня хорошо выражен сердцевинный канал: у крупных щетин он составляет около 70 % диаметра стержня; у тонких щетин не превышает 29 %. Рисунок кутикулы такой же, как у щетин за!ривка (рис. 6).
Остевые волосы светлые, слегка волнистые. Максимальная толщина у основания стержня. К вершине стержень сужается. Поперечные срезы овальной формы. Сердцевинный канал выражен штрихами, но иногда составляет 33 % от диаметра (у самой вершины стержня). Вершины остевых волос не расщепляются. Рисунок кутикулы такой же, как у волос за!ривка.
Пуховые волосы сильно извиты. Толщина одинакова по всей длине стержня. Поперечные срезы округлой формы. Сердцевинный канал отсутствует. Рисунок кутикулы такой же, как и у пуховых волос на за!ривке.
Осенью у кабана в коже эпидермис толстый, а роговой слой относительно тонок (животные № 3, 4, 10, табл. 1). Внутренняя поверхность эпидермиса ячеистая. Ячейки расположены на значительном расстоянии друг от друга. Высота ячейки достигает 220 мкм в коже загривка и 50—110 мкм в коже
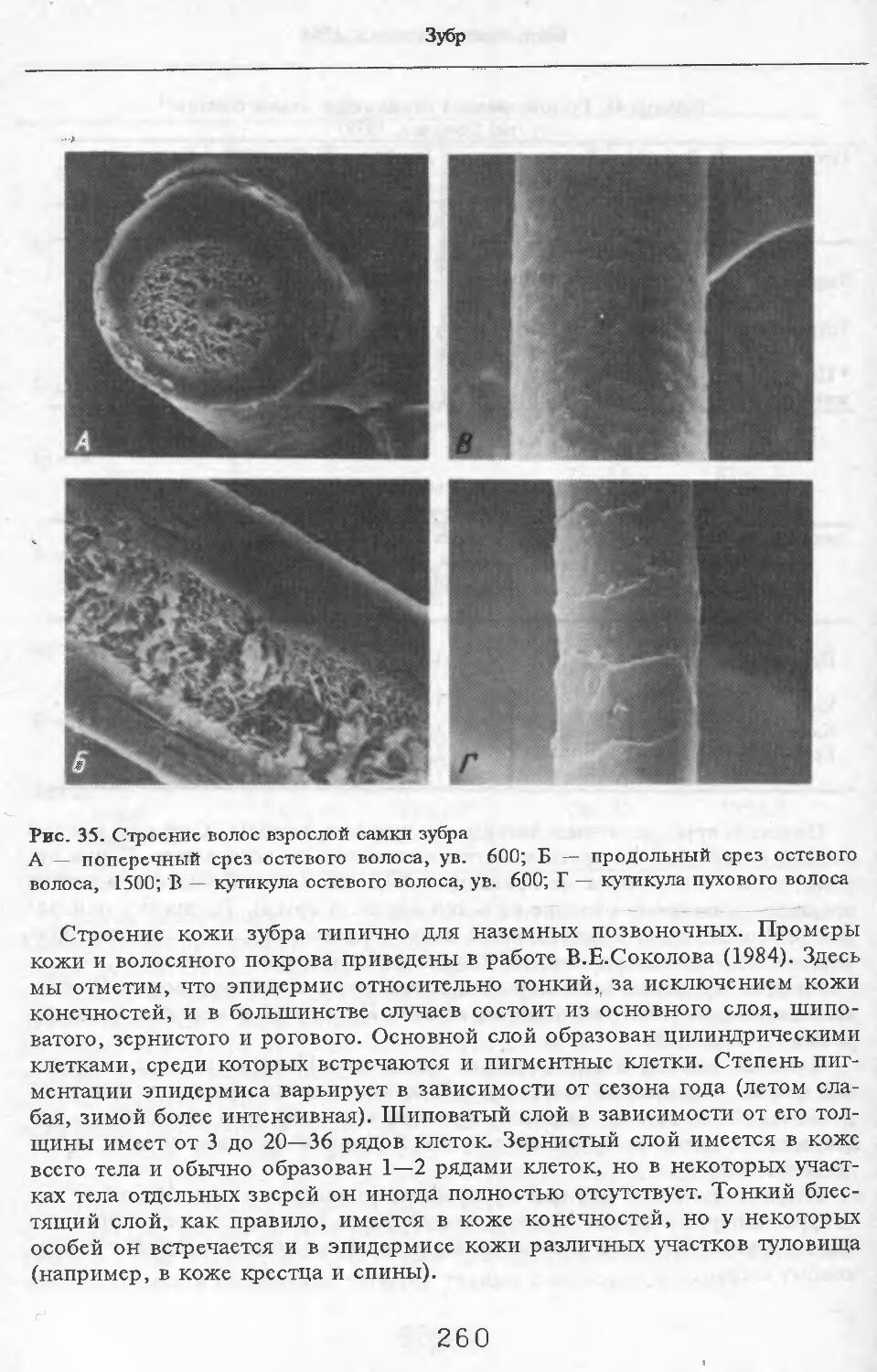
20
Sus scrofa Linnaeus, 1758
Рис. 5. Строение волос загривка самца кабана
А — поперечный срез щетины, ув.х 300; Б — рисунок кутикулы щетины, ув. х 300; В, Г — то же остевого волоса, ув. х 200 и 3000; Д, Е — то же кутикулы пухового волоса, ув. х 300 и 1000
2 1
Кабан
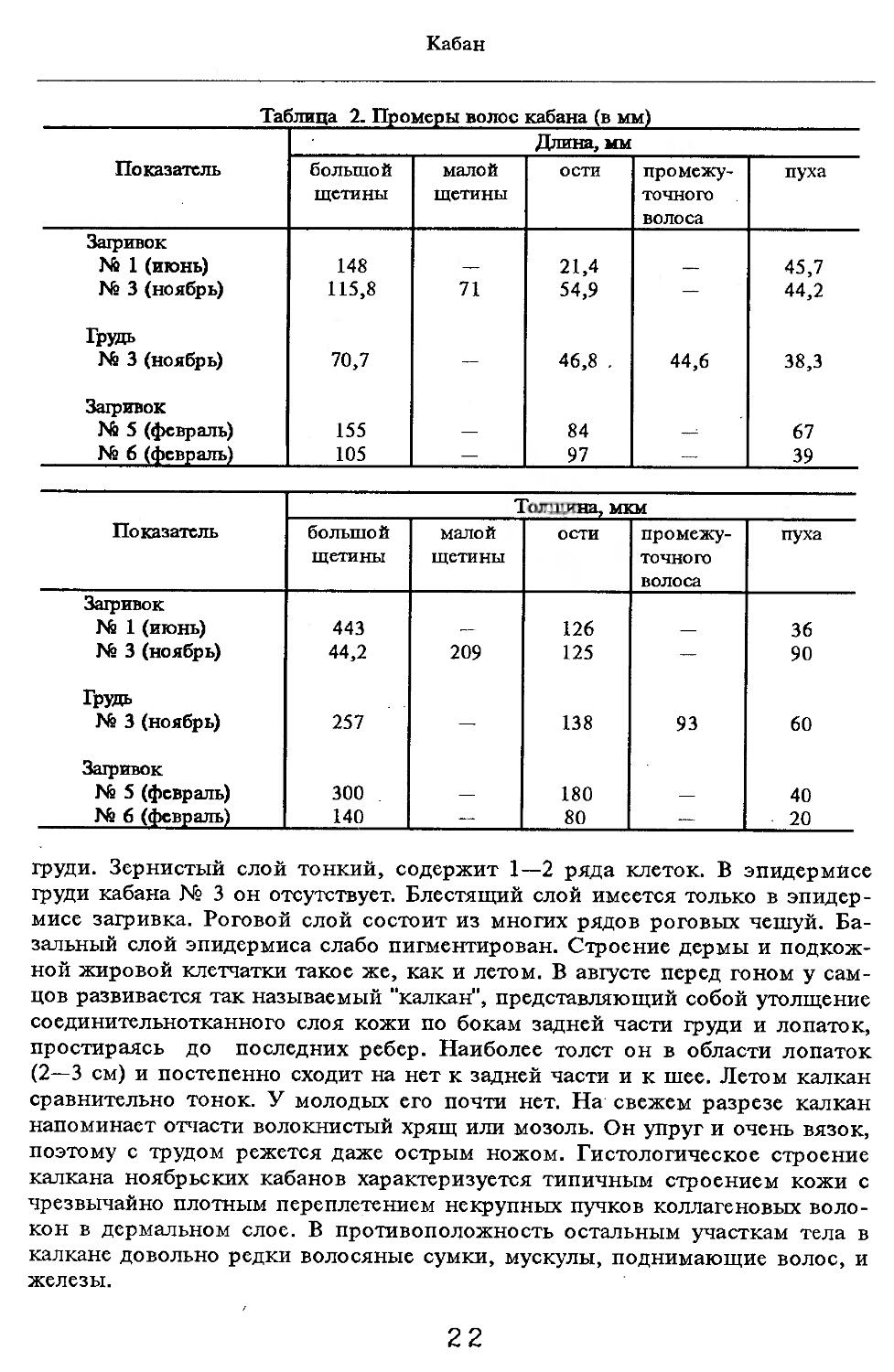
Таблица 2. Промеры волос кабана (в мм)
Показатель Длина, мм
большой щетины малой щетины ости промежуточного волоса пуха
Загривок Na 1 (июнь) Na 3 (ноябрь) 148 115,8 71 21,4 54,9 45,7 44,2
Грудь Ns 3 (ноябрь) 70,7 — 46,8 . 44,6 38,3
Загривок Nil 5 (февраль) Ns 6 (февраль) 155 105 — 84 97 67 39
Показатель Толщина, мкм
большой щетины малой щетины ости промежуточного волоса пуха
Загривок Ns 1 (июнь) 443 — 126 — 36
Ns 3 (ноябрь) 44,2 209 125 — 90
Грудь Ns 3 (ноябрь) 257 — 138 93 60
Загривок Ns 5 (февраль) 300 180 40
Ns 6 (февраль) 140 — 80 — 20
груди. Зернистый слой тонкий, содержит 1—2 ряда клеток. В эпидермисе груди кабана № 3 он отсутствует. Блестящий слой имеется только в эпидермисе загривка. Роговой слой состоит из многих рядов роговых чешуй. Базальный слой эпидермиса слабо пигментирован. Строение дермы и подкожной жировой клетчатки такое же, как и летом. В августе перед гоном у самцов развивается так называемый "калкан", представляющий собой утолщение соединительнотканного слоя кожи по бокам задней части груди и лопаток, простираясь до последних ребер. Наиболее толст он в области лопаток (2—3 см) и постепенно сходит на нет к задней части и к шее. Летом калкан сравнительно тонок. У молодых его почти нет. На свежем разрезе калкан напоминает отчасти волокнистый хрящ или мозоль. Он упруг и очень вязок, поэтому с трудом режется даже острым ножом. Гистологическое строение калкана ноябрьских кабанов характеризуется типичным строением кожи с чрезвычайно плотным переплетением некрупных пучков коллагеновых волокон в дермальном слое. В противоположность остальным участкам тела в калкане довольно редки волосяные сумки, мускулы, поднимающие волос, и железы.
22
Sus scrofa Linnaeus, 1758
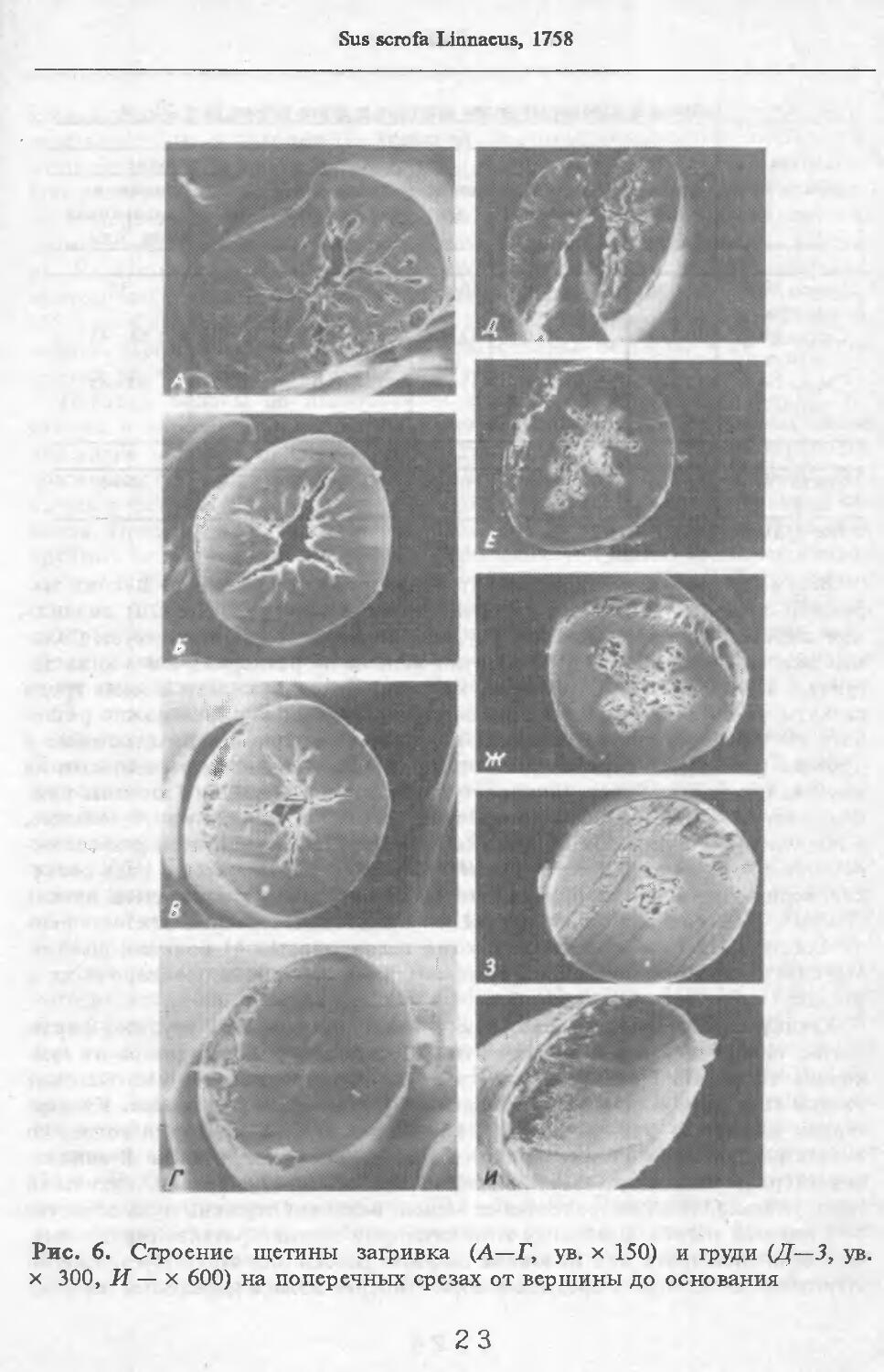
Рис. 6. Строение щетины загривка (А—Г, ув. х 150) и 1руди (Д—3, ув. х 300, И— х 600) на поперечных срезах от вершины до основания
2 3
Кабан
Таблица 3. Промеры* волос загривка и 1руди кабана (л = 10)
Пол, дата добычи Категория волос
щетина остевые пуховые
длина, мм толщина в расширенной гасти, мкм длина, мм толщина в расширенной гасти, мкм длина, мм толщина в расширенной гасти, мкм
Загривок
Самец № 8, 75-98 310-450 36-59 135 35-54 38
ноябрь Самка № 9, 90-130 345-510 52-76 75-120 30-45 34-45
ноябрь Самка № 10, 52-113 315—645 25-46 75-110 12-25 30-42
ИЮНЬ
Грудь
Самка № 11, 31-65 105-225 16-24 75-105 10-14 30-42
ИЮНЬ
*Все промеры сделаны в 1989 г.
Весьма своеобразно оказалось строение кожи уха кабанов. В коже наружной стороны уха имеется довольно толстый слой (1,5—1,8 мм) подкожной жировой клетчатки. В коже внутренней стороны она отсутствует. Сальные железы у кабана № 3 в коже груди меньше по размерам, чем в коже загривка, а у кабана № 2, наоборот, крупнее. Потовые железы в коже груди развиты не так сильно, как в коже загривка. Волосы загривка можно разделить на четыре категории: большие щетины, маленькие щетины, остевые и пуховые (табл. 3). У первых трех категорий волос вершины расщеплены на нитевидные части. Все волосы растут поодиночке. Большие щетины прямые, слегка загнуты назад, в поперечнике округлые (иногда многоугольные, с закругленными углами), сердцевины не имеют. На поперечном разрезе через волос хорошо видны зерна пигмента, концентрирующиеся в виде сосочков, вершинами направленных к центру. Кутикулярные клетки очень низкие (5 мкм). Поперек волоса находится 4—6 клеток. Очертания клеток очень извилистые. На 1 см2 поверхности кожи насчитывается 41 большая щетина. Маленькие щетины по своей форме сходны с большими, но короче их и тоньше.
Кутикулярные клетки выше, чем у волос предыдущей группы, форма клеток такая же, как у больших щетин. При подсчете волос дается их суммарное число. На 1 см2 поверхности кожи их насчитывается 833. Остевые волосы еще короче. Часть из них прямые, часть — слегка извитые. Кутикулярные клетки по форме такие же, как у предыдущей категории волос, но высота их больше. В стержне имеется слаборазвитая сердцевина. В основании стержня она отсутствует. Пуховые волосы можно назвать пуховыми лишь условно, так как их толщина весьма велика. Стержень пуха образует 4—5 пологих изгиба. В волосах этой категории хорошо развита сердцевина, хотя в нижней трети или половине стержня волоса она отсутствует. Высота кутикулярных клеток в среднем 9 мкм. Поперек волоса насчитывается 2—3
2 4
Sus scrofa Linnaeus, 1758
клетки. Волосы на груди разделяются на четыре категории: щетины, ости, промежуточные и пуховые (см. табл. 3). Все они утоньшаются постепенно от основания к вершине и растут поодиночке. Щетина значительно меньших размеров, чем на загривке. Сердцевинный слой в ней отсутствует. Форма поперечника неправильно округлая. На поперечном срезе видно, что пигмент концентрируется в виде сосочков, вершинами направленных к центру. На 1 см2 поверхности кожи растет в среднем 21 щетина. Остевые волосы слабо извитые, в высоту достигают в среднем 46,8 мм, а в толщину 138 мкм. Сердцевина имеется только в верхней половине или трети стержня волоса. Промежуточные волосы извиты сильнее остевых, а по размерам меньше их. Сердцевинный слой развит очень слабо.
Пуховые волосы по извитости не отличаются от промежуточных, но тоньше и короче их. Сердцевина развита очень слабо. У отдельных волос она видна только в виде небольших редких штрихов. На 1 см2 поверхности кожи растет 416 остевых, промежуточных и пуховых волос. У кабанов, добытых в феврале (№ 5—7), строение кожи такое же, как у ноябрьских кабанов. Промеры кожи проведены в табл. 2. В сальных железах встречаются крупные полости (размером до 440 х 880 мкм). Очевидно, это отсекретиро-вавшие дольки желез. Потовые железы развиты очень сильно. Секреторные отделы, хотя и не имеют больших диаметров, сильно извиты в клубочки, которые окружены жировыми клетками. Волосы представлены тремя категориями: щетинами, остевыми и пуховыми (табл. 3). На 1 см2 поверхности кожи загривка 40 щетин у самки и 60 сеголеток, 150 остевых волос у самки и 60 у сеголеток, 1000 пуховых волос у самки и 2160 у сеголеток.
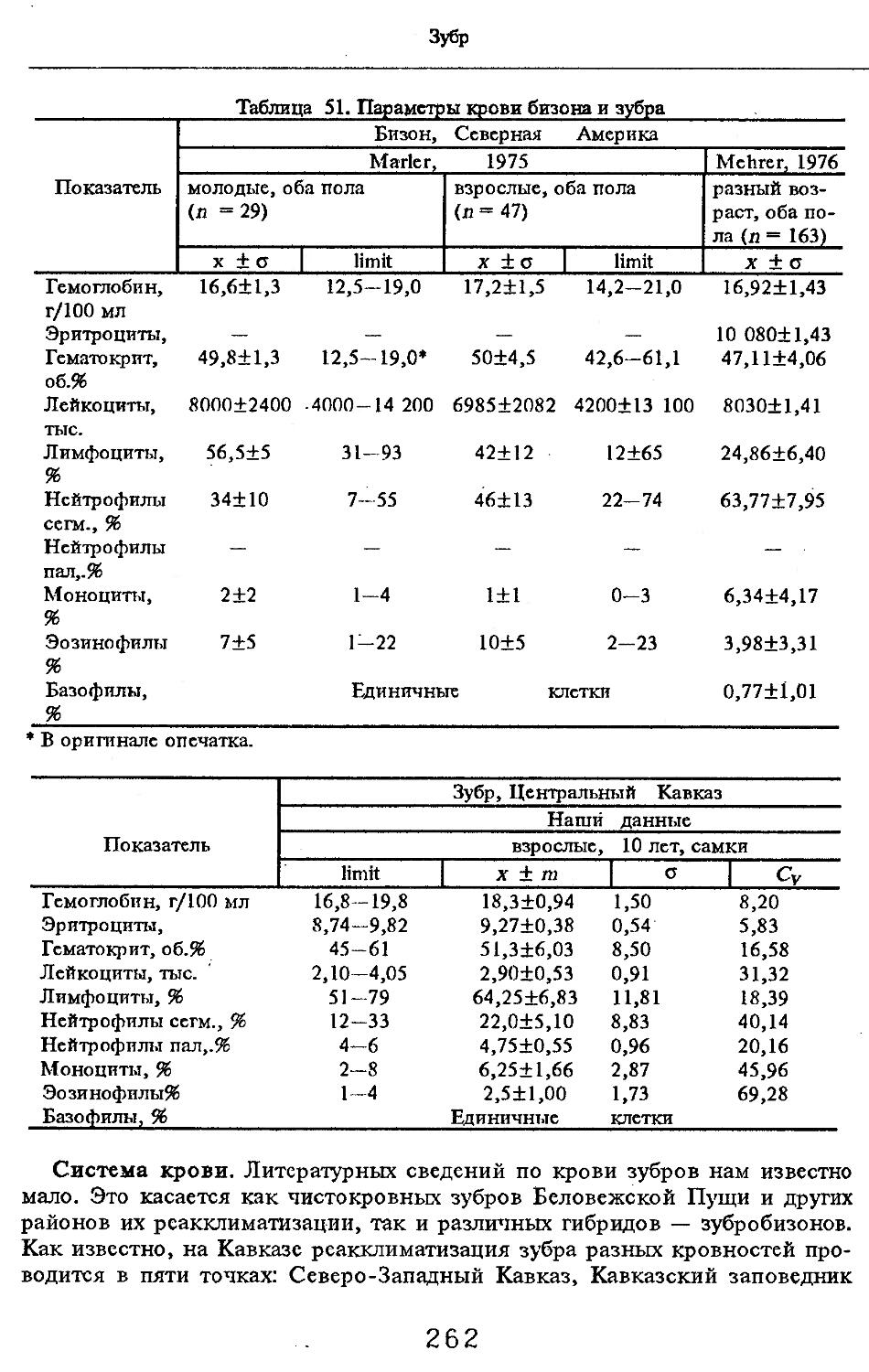
Система крови. Литературные сведения по крови кабана очень малы. Даже в специальной монографии П.А.Коржуева (1964), посвященной анализу качественно-количественных показателей крови позвоночных, нет оригинальных данных; приводятся лишь ссылки на три старых источника по характеристике эритроцитов (Bethe, 1891, Gotze, 1923, Wintrobe, 1933, по: Кор-жуев, 1964). Результаты исследования названных авторов, касающиеся количества эритроцитов, очень близки (6,90 и 6,96 млй в 1 мм3 крови), но заметно расходятся по объему эритроцитов (по Gotze — 56,7 мкм3 ; по Wintrobe — 61,0 мкм3 ).
Изменения гематологических показателей кабана в течение постнатального развития изучены А.Костелеска-Мирха (Kosteleska-Myrcha, 1974). При этом сравнивались кабаны нормальной черной окраски (п = 6 экз.) и светлой окраски (п = 11 экз.). Анализ крови проводился с недельным интервалом, начиная с 4-дневного возраста до 71-дневного возраста у черных кабанов и с 12-дневного до 80-дневного возраста у светлых кабанов. Установлено, что в течение первых 3 мец жизни в крови черных кабанов интенсивно увеличивается количество эритроцитов (3,63—7,71 млн/мм3) и гематокрит (33,0—48,3%), сокращается объем эритроцитов (98,04—50,11 мкм3), увеличивается концентрация гемоглобина (10,1—16,0 %). Показательно, что у светлых кабанов указанные свойства крови первоначально выше, чем у черных кабанов, но в течение второго месяца жизни эти различия нивелируются. Однако у черных кабанов лейкоцитов больше (7,89—18,65 тыс./мм3), чем у
25
Кабан
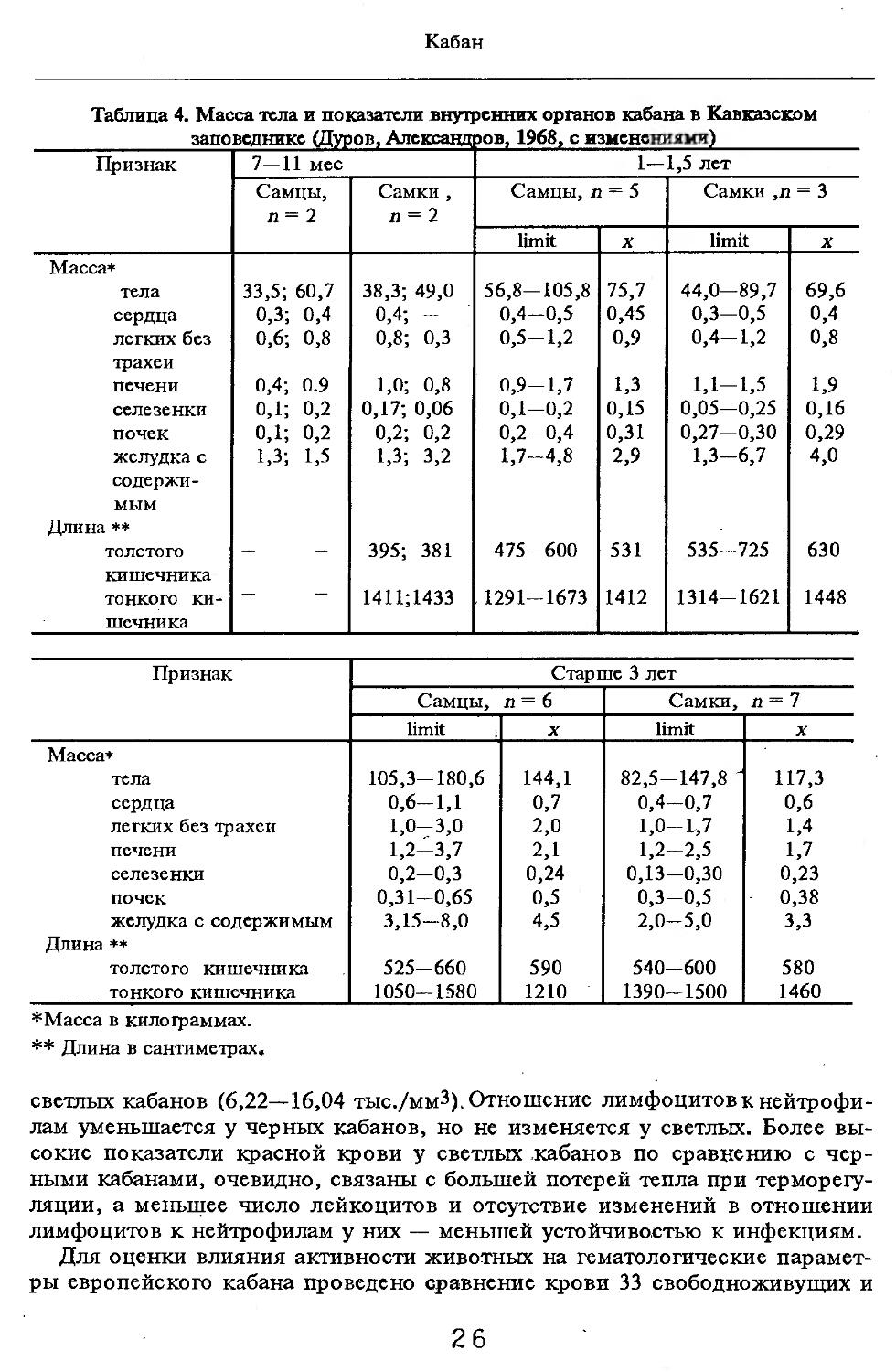
Таблица 4. Масса тела и показатели внутренних органов кабана в Кавказском
заповеднике (Дуров, Александров, 1968, с изменениями)
Признак 7—11 мес 1—1,5 лет
Самцы, п= 2 Самки, п = 2 Самцы, п = 5 Самки ,п = 3
limit X limit X
Масса* тела 33,5; 60,7 38,3; 49,0 56,8-105,8 75,7 44,0—89,7 69,6
сердца 0,3; 0,4 0,4; - 0,4-0,5 0,45 0,3-0,5 0,4
легких без 0,6; 0,8 0,8; 0,3 0,5-1,2 0,9 0,4-1,2 0,8
трахеи печени 0,4; 0.9 1,0; 0,8 0,9-1,7 1,3 1,1-1,5 1,9
селезенки о,1; 0,2 0,17; 0,06 0,1—0,2 0,15 0,05-0,25 0,16
почек 0,1; 0,2 0,2; 0,2 0,2-0,4 0,31 0,27-0,30 0,29
желудка с 1,3; 1,5 1,3; 3,2 1,7-4,8 2,9 1,3-6,7 4,0
содержимым Длина ♦♦ толстого 395; 381 475-600 531 535-725 630
кишечника тонкого ки- 1411;1433 1291-1673 1412 1314-1621 1448
шечника
Признак Старше 3 лет
Самцы, п= 6 Самки, п=7
limit । X limit X
Масса*
тела 105,3-180,6 144,1 82,5-147,8 ' 117,3
сердца 0,6-1,1 0,7 0,4—0,7 0,6
легких без трахеи 1,0-3,0 2,0 1,0-1,7 1,4
печени 1,2-3,7 2,1 1,2-2,5 1,7
селезенки 0,2-0,3 0,24 0,13-0,30 0,23
почек 0,31-0,65 0,5 0,3-0,5 0,38
желудка с содержимым 3,15-8,0 4,5 2,0-5,0 з,з
Длина **
толстого кишечника 525-660 590 540—600 580
тонкого кишечника 1050-1580 1210 1390—1500 1460
*Масса в килотраммах.
** Длина в сантиметрах.
светлых кабанов (6,22—16,04 тыс./мм3). Отношение лимфоцитов к нейтрофилам уменьшается у черных кабанов, но не изменяется у светлых. Более высокие показатели красной крови у светлых кабанов по сравнению с черными кабанами, очевидно, связаны с большей потерей тепла при терморегуляции, а меньшее число лейкоцитов и отсутствие изменений в отношении лимфоцитов к нейтрофилам у них — меньшей устойчивостью к инфекциям.
Для оценки влияния активности животных на гематологические параметры европейского кабана проведено сравнение крови 33 свободноживущих и
26
Sus scrofa Linnaeus, 1758
37 выращенных в загоне (штат Теннесси, США) (Williamson, Pelton, 1976). Обнаружены различия между этими группами (как и возрастные): по количеству эритроцитов, лейкоцитов, тромбоцитов, концентрации гемоглобина, гематокриту, среднеклеточному объему эритроцитов и среднеклеточной концентрации гемоглобина. Дифференциальная картина крови сходна у кабанов всех возрастов в каждой группе, хотя в условиях загона в крови характерен лимфоцитный профиль, а у свободноживущих — нейтрофильный. Половые различия по картине крови оказались незначительными.
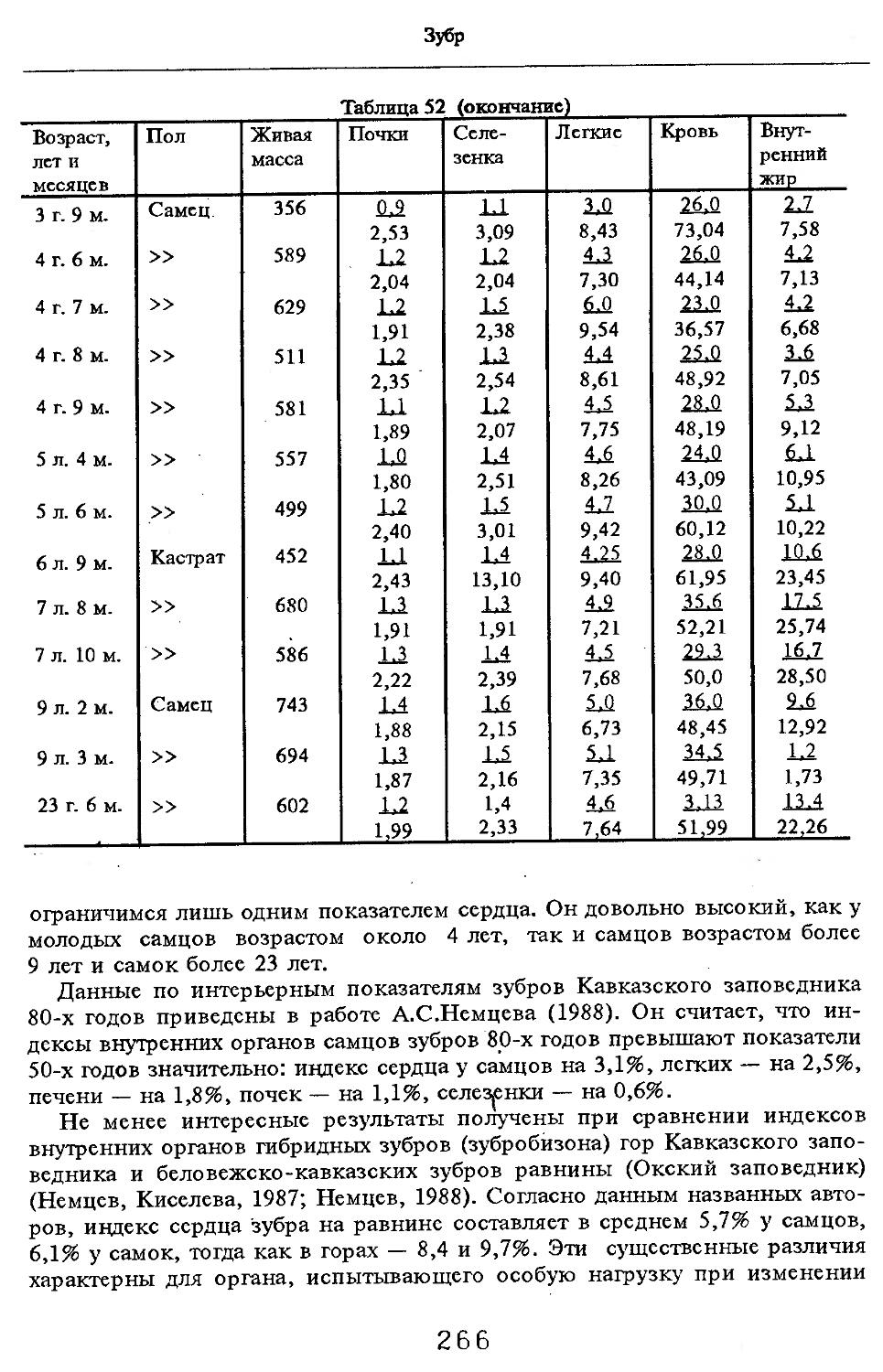
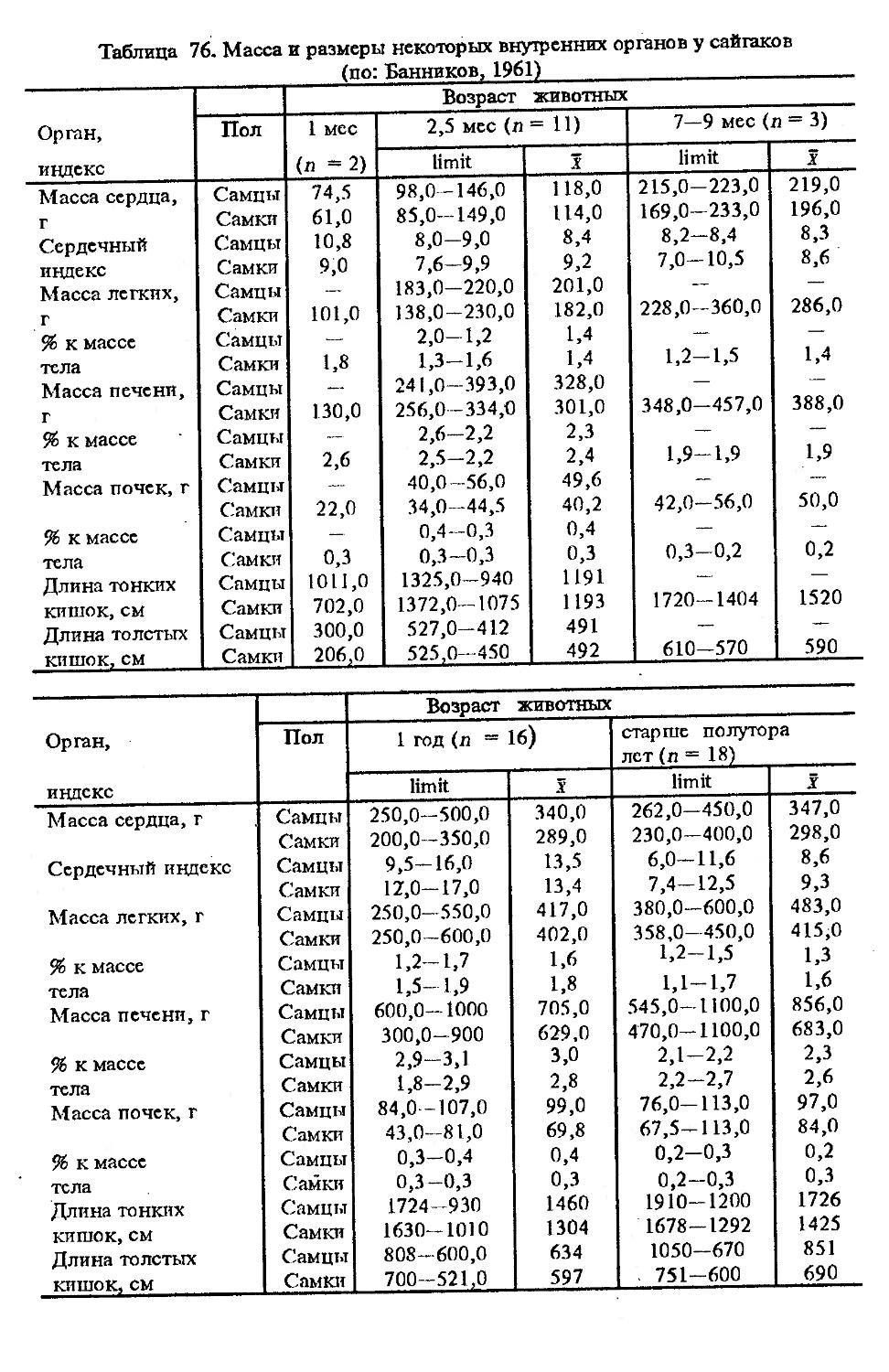
Индексы внутренних органов. В условиях Кавказского заповедника (кубанский вариант) показатели внутренних органов изучены В.В.Дуровым и В.Н.Александровым (1968). Как видно из табл. 4, авторы располагали всего 25 особями, возраст которых варьировал с 7 мес. до ! лет и старше. Объем материала недостаточен для широкого обсуждения, но все же обратить внимание на некоторые стороны целесообразно. Так, хотя абсолютная масса сердца значительно увеличивается за 2—3 года постнатальной жизни, его индекс сокращается почти в 2 раза. По другим изученным внутренним органам закономерно растет с возрастом общая масса, но снижение индексов не столь значительно или вовсе не выражено (почки).
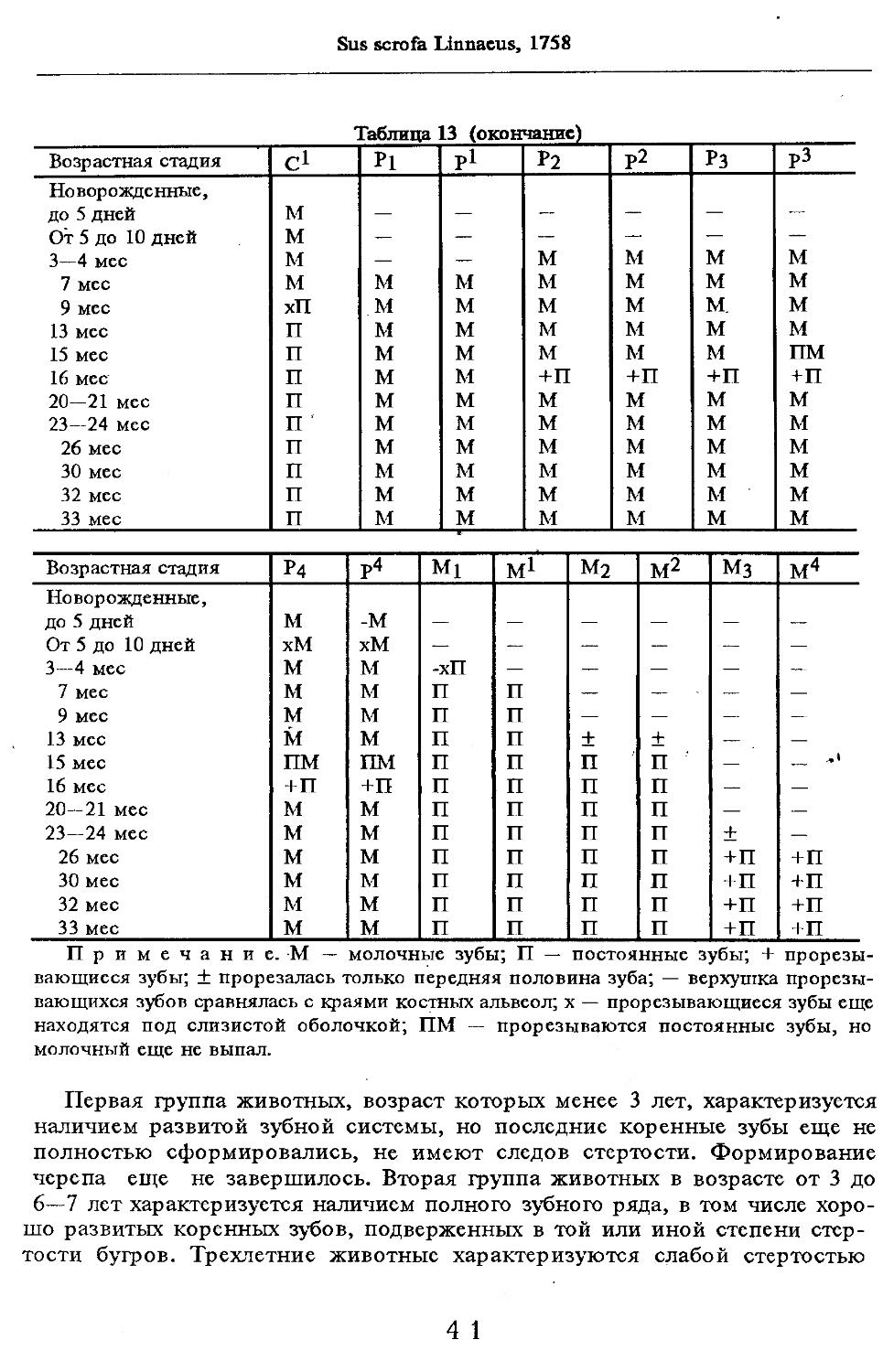
Кариотип. Дикие кабаны и домашние породы свиней цитогенетически изучены хорошо (см. обзор: Трошина^ Тихонов, 1985). Установлено, что диплоидное число хромосом колеблется от 36 до 38, а число плеч постоянно, во всех кариотипах NF = 64.
. Хромосомный полиморфизм обусловлен двумя типами перестроек — центрическим слиянием акроцентриков. В первом случае соединяются 16-я и 17-я пары акроцентриков, а во втором — 15-я и 17-я пары акроцентриков. В кариотипе кабанов можно условно выделить 4 группы: А — крупные субметацентрические (4 пары), Б — акроцентрические (от 4 до 6 пар), В — средние субметацентрические (6 пар) и Г — метацентрические (3 пары). Х-хро-мосома — субметацентрик среднего размера, Y-хромосома — мелкий субме-тацентрик (Тихонов, Трошина, 1974). На наш взгляд, в исследовании названных авторов интерес представляет тот факт, что из 37 изученных кабанов Киргизии 24 животных имели 36 хромосом, а остальные — 37, тогда как в выборке из 10 особей закавказской и европейской популяций 36-хромо-сомных кабанов не было.
Все 38-хромосомные кариотипы кабанов европейских и азиатских популяций одинаковы по характеру дифференциальной окраски..При этом полностью соответствуют кариотипу домашних свиней (Орлов, Булатова, 1983).
Структура и динамика ареала
Видовой ареал занимает обширную территорию — Европу до 60—55° с.ш., Азию до 55—50’ с.ш., а на юг простирается, до Индии и Шри-Ланки включительно, занимает значительную часть островной области от Малакки до Соломоновых островов; включает также Северо-Восточную и Северо-Западную Африку (Гептнер и др., 1961).
27
Кабан
Распространение на Кавказе. Кабан был и остается одним из наиболее обычных и широко ар сальных промысловых животных местной териофауны, но структура ареала в исторический период подвергалась очень большим изменениям. Временные ее сдвиги при этом носили глубокие следы регионального порядка. Раньше и быстрее всего, по Н.К.Верещагину (1959), сокращение численности и распространения кабана происходило на Армянском нагорье, где из-за сухости климата обезлесение горных склонов происходило быстро, а отсутствие религиозного запрета на мясо кабанов не oipa-ничивало их отстрел. В сухих нагорьях Ирана и Турции, где мусульманское население не.употребляет кабанов в пищу, численность этих животных остается и по сей день значительно выше, чем в Армении. По этой же причине больше кабанов оставалось в Азербайджане, Дагестане, Кабардино-Балкарии и ряде других районов. Об обилии кабанов на Кавказе еще в XVIII и XIX вв. свидетельствуют многочисленные записки путешественников, кавказских краеведов и натуралистов. По всем данным сохранению кабанов во многих районах Грузии с христианским населением способствовало обилие лесов и труднодоступных заболоченных мест.
Наиболее значительное сокращение численности и распространения кабана началось со второй половины прошлого века после завоевания Кавказа, сопровождавшегося варварским уничтожением лесов, а также интенсивным заселением равнин и предгорья русскими. Тем не менее еще в начале XX в. область распространения кабана, по данным Н.Я.Динника (1910), оставалась очень обширной. По его данным, кабан встречался всюду, где только сохранились леса. На Северном Кавказе нижнюю границу ареала он проводил по линии населенных пунктов; Анапа — южнее Краснодара — Пшехская — севернее Майкопа — Каладинская — Передовая — Исправная — Усть-Джегутинская — леса Бекешевской — Нальчик — Моздок — Малго-бек — Владикавказ — Махачкала — побережье Каспия до Апшеронского полуострова. Кроме того, кабан встречался в низовьях Кумы, Терека, Сула-ка. Лет 40 тому назад обитал в лесах Ставропольской возвышенности, в районе Пятигорья. На лесистом Черноморском побережье кабаны встречались всюду вплоть до Батуми. Обычным был в б.Кутаисской и Тифлисской губерниях. Н.Я.Динник располагал скудными данными по остальной территории Закавказья, но отмечал изобилие кабана в Лагодехи, Закаталье, в долинах Куры, Алазани, в заболоченных и заросших камышами озерах Муган-ской степи и в камышах по р. Араке, почти во всех лесах б. Елизаветполь-ской губернии. Далее, в б. Карской области они были нередки недалеко от Сарыкамыша (около с. Арсеняк). Около Караклиса и Дилижана кабан уже не встречался, но изредка попадался вблизи перевала от Дилижана к Севану. Очень много кабанов сохранилось к началу текущего столетия в Ленкоранской низменности и в лесах Талыша (Верещагин, 1959).
Распространение кабана в первой половине XX в. Сокращение ареала, охватывающего в конце прошлого века всю территорию Кавказа, достигло своего предела в 30-х годах. Эти годы можно принять за время максимального сокращения кабана не только на Кавказе, но и во всей стране (Гептнер и др., 1961). Сокращение распространения сопровождалось падением плотнос
28
Sus scrofa Linnaeus, 1758
ти населения и появлением многократных разрывов, порою отдельные популяции разделяло обширное пространство. В то время в кавказской части ареала кабанов не стало в лесах северо-западного Карабаха и на северном макросклоне Малого Кавказа вплоть до Грузии. Лишь на небольшом участке долины р. Акера-Чай и лесистого северного склона Зангезурского хребта сохранились кабаны, поселение которых поддерживалось иммигрантами из северного Ирана (Верещагин, 1959). Значительное сокращение ареала кабана в Закавказье имело место в конце 40-х годов, что было вызвано подготовкой ложа для Мингечаурского водохранилица по р. Куре (от Иори и Алазани до Мингечура), а затем закрытием плотины и соответственно иссушением долины Куры ниже г. Евлах.
Сокращение распространения кабана в результате антропогенного воздействия в Закавказье в первой половине XX в. затронуло также Талыш, горные леса которого изобиловали этим зверем в прошлом (Динник, 1910). Несколько благополучнее было с кабаном в Западном Закавказье — лесистая и заболоченная местность Колхидской низменности и горные леса Большого и Малого Кавказа сохраняли более благоприятные защитные и кормовые условия, чем сухие субтропики Восточного Закавказья. По данным И.Д.Чхи-квишвили (1939), в Абхазии кабан "до сих пор весьма обыкновенен, а местами даже многочислен" (с. 19). Однако, К.К.Флеров (1927), обследовавший долину р. Чороха в этом районе в 1924 п, отмечал очень низкую численность кабана. Судя по литературным данным, на Северном Кавказе наблюдалась аналогичная динамика ареала. В западной части при всем сокращении его численности и распространения кабана оставалось больше, чем в центральном и восточном районах. Бесспорно, положительную роль сыграл Кавказский заповедник, где имеются благоприятные условия для кабана и в связи с высотными перемещениями поддерживалась численность за его пределами, в том числе в предгорьях и низовьях р. Кубань. Подробности размещения в этой части Западного Кавказа не освещены в литературе, но, по данным В.Г.Гептнера и др. (1961), в середине 50-х годов нижняя граница кабана спускалась до низовьев р. Кубань и выходила на приазовские плавни.
В пределах эльбрусского варианта (бассейн Р- Малка и верховья Кубани) в первой половине XX в. кабан был уничтожен во многих местах. Даже в Те-бердинском заповеднике до 1947 г. этот вид не втречался (Аренс, 1957). Лучшие условия были в терском варианте, но и здесь обширные пространства лесостепного и степного поясов кабаны, видимо, не заселяли. Лишь изредка попадались в приречных лесах, особенно низовьях рек с камышовыми зарослями (р. Терек и др.). Основное поголовье водилось в леенрм поясе (Туров, 1926; Формозов, 1926; Нанисв, 1956? Тсмботов, 1960, 1972).
К Дагестану в рассматриваемый период относятся работы В.Г.Гептнера и А.Н.Формозова (1941), проводивших исследования в 1924—1926 гг. на равнине и в горах. В то время кабан здесь встречался широко и был одним из обычных видов. Однако обширное пространство Внутреннего Дагестана не было им заселено, лишь в верховьях рек с древесно-кустарниковой растительностью кабан изредка встречался- При этом, по мнению названных авторов, поселения в высокогорьях были изолированы от основного, приуроченного к
29
Кабан
лесистому склону передовых хребтов и низовьям рек, начиная от Самура на юге, до Кумы на севере. Во многих местах, особенно где встречались непроходимые приречные леса, камышовые и кустарниковые заросли, кабан был многочислен. К излюбленным местам охоты на кабанов относились не только приморские угодья и низовья рек (Терек, Сулак, Прорва и др.), но и окрестности Хасавюрта. Даже в лесу г. Тарки-Тау, близ г. Махачкалы, встречался кабан. В 30—40-х годах кабан был уничтожен во многих местах равнин и предгорий Дагестана, что привело к разрыву его поселений.
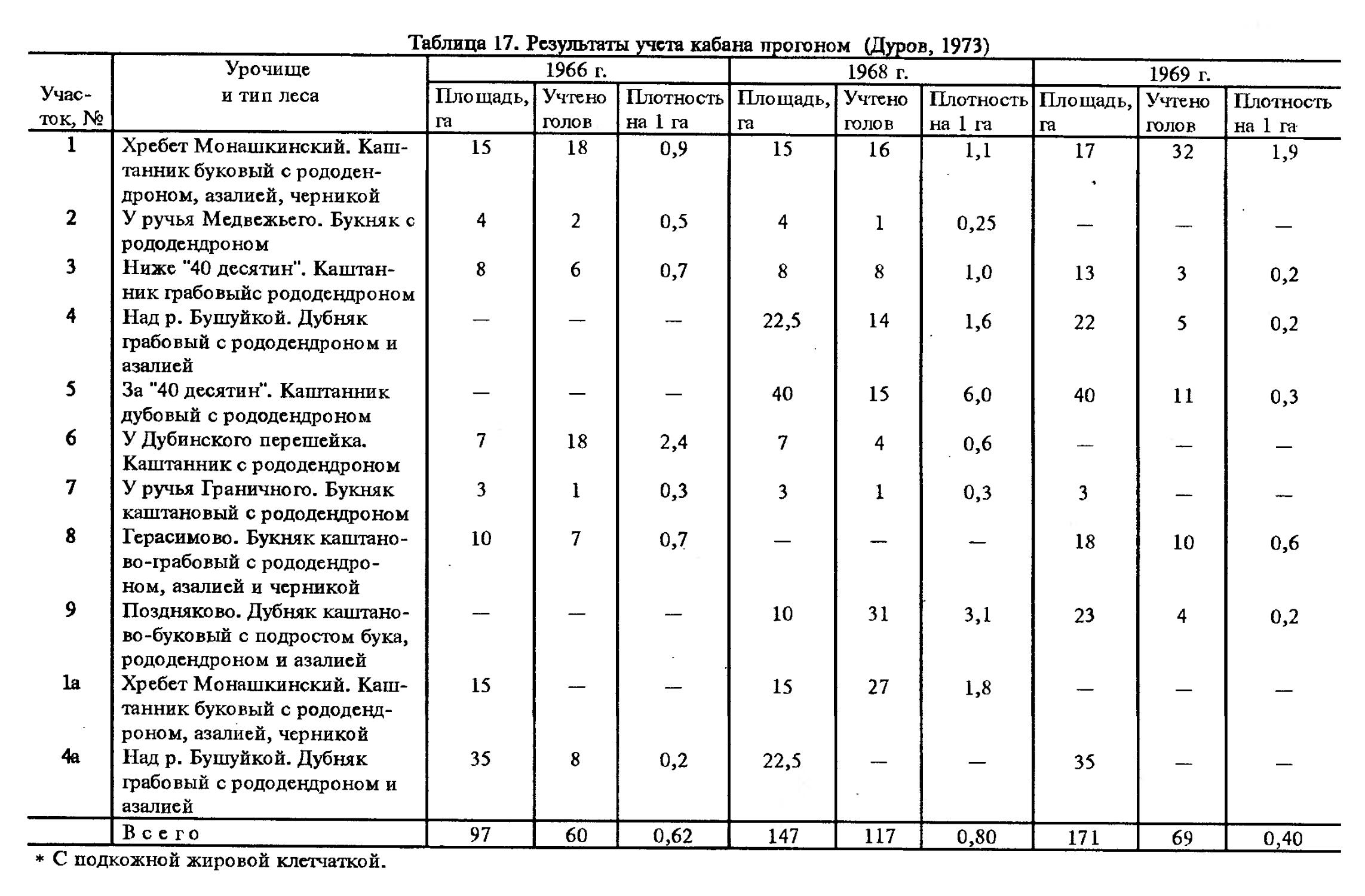
Современное распространение (вторая половина XX в.). На большей части ареала, в особенности в европейских регионах страны, Казахстане и на юге Западной Сибири отмечалось увеличение площади распространения и возрастание численности кабана (Гептнер и др., 1961). Сходная тенденция в динамике ареала имеет место на Кавказе, однако в различных районах она реализуется по-разному, нередко даже наблюдается обратное явление — сокращение распространения и снижение численности. По мнению В.В.Дурова (1973), на Северо-Западном Кавказе северная граница распространения кабана сместилась на юг, в предгорья, в среднем на 50 км по сравнению с тем, что было в середине 50-х годов согласно В.Г.Гептнеру и др. (1961). Теперь кабан не встречается в пойме р. Кубань и на Таманском полуострове, граница его проходит южнее г. Майкопа, станиц Пшехской, Бжедуховской, Смоленской, г. Крымска, которые по сообщению В.А.Котова являются крайними северными точками распространения кабана в 60-х годах. За последние два десятилетия вновь отмечено расширение распространения кабана в Краснодарском крае, что вызвало появление его в некоторых районах равнины (Дуров, 1987). Южная граница на широте г. Анапы, огибая предгорные леса, тянется вдоль береговой линии Черного моря вплоть до Абхазии. В приазовских плавнях поселение кабана занимает берег Азовского моря от лимана Курчанского до Ахтарского. Внутри очерченной территории в ряде мест кабан уничтожен или вытеснен, что связано со строительством дорог, лесоразработкой, появлением новых населенных пунктов.
В соседнем Ставропольском крае, судя по данным С.М.Федорова (1954), Л.Е.Аренса (1957) и др., кабан восстановил былое распространение. Так, начиная с 1947 г. он стал регулярно попадаться в Тебердинском заповеднике, расселился по всем долинам, вплоть до Водораздельного хребта. По долинам рек Хадюка, Аксаута и Кубани обитает много кабанов. В настоящее время кабан в Тебердинском заповеднике — обычный вид; численность его составляет 6,2 особи (17.01. 1986) — 8,7 особей (19.11. 1987) на 1000 га. Численность на всей территории заповедника составляет 143—200 особей (Летопись Тебердинского заповедника). В предгорьях и на равнине Ставропольского края кабан заселяет почти всю территорию, но, за исключением лесных поясов, распространение его пятнистое. По речным долинам, лесонасаждениям и камышовым зарослям кабан спускается с лесистых гор и пред-горьев до Каспийского побережья. По данным Ставропольского краевого управления за 1988 и 1989 гг., из 34 районов края кабан зарегистрирован в 28 районах, в том числе Красногвардейском, Изобильненском, Ипатовском, Нефтекумском и др. При этом в ряде лесных массивов его численность
30
Sus scrofa Linnaeus, 1758
превышает допустимые размеры (Оноприенко, Хохлов, 1989). Сходные данные нами получены в бассейне р. Малка Кабардино-Балкарии (эльбрусский вариант).
В бассейне Терека (терский вариант) кабан имеет широкое горизонтальное и высотное размещение. Оптимум ареала находится в поясе горных лесов, содержащих плодоносящие деревья и кустарники (бук, дуб, фруктарни-ки и др.), вторичные луга и небольшие обрабатываемые поля. За последние годы заметную положительную роль сыграло совершенствование работы лесоохотничьих хозяйств, улучшение охраны и воспроизводства кабана. В итоге этот вид проник в горы до 3000—3500 м над ур. моря, а по интразональ-ным ландшафтам спустился в Восточное Предкавказье до Каспийского побережья (Темботов, 1982; Точиев, 1988).
По данным Л.И.Прилуцкой и Ю.В.Пишванова (1988), кабан в Дагестане широко распространен начиная с низменности до высокогорья. Однако во многих местах поселения спорадичные, особенно во Внутреннем Дагестане и сухих полупустынях Прикаспия. Более оптимальные условия сохранились в лесах Передовых хребтов, в низинных лесах и камышовых зарослях морского побережья. В частности, А.М.Гинеев (1971) зарегистрировал кабанов в плавнях Терека, Аксая, Акташа, Сулака, а также изредка среди бурунов Тер-ско-Кумской низменности.
Современному распространению кабана в Кубо-Хачмасской части Азербайджана посвящена работа Х.М.Алекперова и С.М.Кулиева (1987), выполненная в 1981—1983 гг. По их данным, в этом регионе кабан обитает в низменных лесах вдоль побережья Каспия (от Хачмаса до Дагестана), в предгорных лесах и кустарниках Кусарского и Кубинского районов, а также в Ку-сарском заказнике с прилегающими территориями. Обитаемая площадь в регионе составляет 20—30 тыс. га с численностью кабана в среднем за 3 года 673 особи.
По закавказской части ареала кабана современных данных мало. В западной Грузии он встречается редко, но занимает все высотные пояса от Колхидской низменности (Арабули, 1973) до субальпийского пояса. В восточной Грузии живет в пойменных лесах Куры, Алазани и Иори, но численность его в этих местах сильно сократилась. Кабан сохранился в Гардабанском и Чиа-урском охотхозяйствах, а также в Вошлованском заповеднике с прилегающей территорией. В связи с интенсификацией сельского хозяйства кабаны оттеснены в горные леса Большого Кавказа, где они освоили, кроме упомянутой территории заповедника, также Телавский и Ахметский районы (Арабули, 1970). По Дж.Ратани (1989), кабаны обитают на плоскогорье Месхет-Джава-хета. Из Азербайджана и Армении данные фрагментарны. Основная область обитания этого вида здесь приурочена к горным лесам Большого Кавказа, оттуда он проникает в субальпийский пояс, особенно в летний период. Что касается нижней границы, то пока трудно ее провести однозначно. На Малом Кавказе распространение кабана, видимо, остается спорадичным, но более обширным, чем это имело место в 30-х годах.
По данным С.К.Даля (1954), это животное населяет леса Кафанского, Мегринского, Горийского и Сисианского районов; далее на запад в неболь-
3 1
Кабан
шом количестве встречается на Айоцдзорском хребте (окрестности Амагу, Джула, Канлыгеля) и в редколесьях южного склона Гегамского хребта (окрестности Гелджгина, Манкука, Хосрова в Вединском районе). Судя по учетным данным Э.Г.Явруяна (1975), в ряде этих мест кабан остается редким. Известны случаи захода вплоть до лесных участков на хребте Цахкуняц (Ах-тинский район). В 50-х годах кабаны были обычны в пойме Аракса от Арта-шатского до Октемберянского района. Чаще регистрировался в окрестностях Зангибасара, в долине р. Вохча и на южном склоне Метринского хребта. Отдельные поселения кабана известны в низовьях Куры. Так, в Кызыл-Агач-ском заповеднике кабан занимает более 20 тыс. га с плотностью около 35 голов на 1000 га (Литвинов, 1981).
Современные тенденции антропогенных изменений ареала. Кабан — экологически пластичное животное, относительно легко приспосабливается к измененным ландшафтам, в том числе к сельскохозяйственным угодьям. При этом общая тенденция антропогенной трансформации горных и равнинных экосистем Кавказа, в особенности мезофитизация и облесение сухих мест, благоприятна для кабана. Иными словами, на Кавказе кабан имеет определенные экологические предпосылки для расширения области распространения. Однако особенности его поведения, в том числе роющая деятельность, делают его несовместимым с агроэкосистемами. Поэтому. необходимость контролировать как его распространение, так и численность совершенно очевидна.
Учитывая изложенное, будущая структура ареала кабана на Кавказе нам представляется весьма мозаичной и обширной: сравнительно небольшие участки с высокой плотностью населения будут чередоваться с обширными территориями, лишенными кабанов. Заселенными могут (и должны) быть горные леса, луга, различные древесно-кустарниковые насаждения, а также долины крупных рек, болотистые места, камышовые заросли и т.д. Существование кабанов недопустимо рядом с сельскохозяйственными полями, занимающими большие площади в регионе. Эта тенденция четко выражена в настоящее время, особенно благодаря организации целого ряда заповедников, охотхозяйств, строительству водоемов с последующим развитием мощных камышовых зарослей, наличию приморских и заболоченных угодий, плавней и т.д.
Систематика и изменчивость
Систематическое положение. О положении кабана в системе рода Sus нет противоречивых мнений среди систематиков; его принадлежность к Sus scrofa можно считать общепринятой. Очевидно также, что этот вид объединяет все формы кабана, населяющие всю территорию нашей страны. Мнение Ю.А.Филипченко (1933 г., цит. по: Соколов, 1959) о видовой самостоятельности восточных форм, названных Sus orientalis, лишено основания при отсутствии не только морфологических критериев (Соколов, 1959), но и иммунологических (Тихонов и др., 1985) и цитогенетических (Трошина, Тихонов, 1985). Что касается внутривидовой систематики, то по сей день нет удовлетворительного ее решения. Описано большое число форм (до 26—28), а
32
Sus scrofa Linnaeus, 1758
количество имен достигает 50 (Гептнер и др., 1961). Со времени Г.П.Адлер-берга (1933 г., по: Бобринский и др., 1944) географическая изменчивость вида не подвергалась ревизии. Между тем, по И.И.Соколову (1959), внутривидовая систематика кабана настоятельно требует ревизии, так как некоторые формы не заслуживают выделения их в качестве подвидов. В частности, по его мнению, очень сомнительна необходимость разделения популяций кабанов Кавказа, низовьев Волги и Средней Азии. На наш взгляд, такому суждению не соответствуют данные иммуногенетических и цитогенетических исследований, проведенных на значительном фактическом материале (Тихонов и др., 1985; Трошина, Тихонов, 1985). Более того, эти данные свидетельствуют о целесообразности признания самостоятельными подвидами не только кабанов Средней Азии и Восточной Европы, но и Северного Кавказа и Закавказья. Об этом будет сказано подробнее ниже.
Изменчивость. Для анализа различных аспектов изменчивости кабана мы изучили 170 экз. с северного и южного макросклонов поясов темнохвойных и широколиственных лесов Кавказского заповедника (кубанский вариант), пояса широколиственных лесов окрестностей г. Нальчика (терский вариант) и ряда других точек Северного Кавказа и Закавказья. Наряду с серийными выборками изучены также отдельные экземпляры, происходящие из различных районов Кавказа. Как видно, материал отражает широкий спектр ландшафтных условий и позволяет" оценить размах изменчивости кабана в масштабе региона.
Индивидуальная изменчивость. Окраска волосяного покрова подвержена широкой индивидуальной изменчивости. На это обращал внимание еще Н.Я.Динник (1910), приводя в качестве примера зверей, добытых в Дагестане. Один кабан выделялся очень светлой окраской, особенно задняя часть, другой — темно-бурой окраской; зверь, добытый вблизи Каякента, был черным и только на голове щетина была заметно светлее, серо-бурого цвета. Наконец, в окрестностях Губденя попался один зверь, имевший окраску вовсе не темно-бурого цвета, а светлую, рыжеватую, серую или светло-серо-бурую. Голова была особенно светлой, грязно-рыжеватого цвета.
Как известно, в природе изредка встречаются кабаны с пятнистой окраской тела. Показано (Andrzcjewski, 1974), что такая окраска определяется генетически, проявляется у гомозиготных по фактору пятнистости особей, тогда как гетерозиготы окрашены нормально. По мнению автора, в изученной им популяции (Кампиноский национальный парк близ Варшавы), видимо, 20% осо-бей гетерозиготны по фактору пятнистости.
В коллекционном материале, изученном нами, не было особей с пятнистой окраской. Не располагаем мы также аналогичными литературными данными, если не считать гибридов диких кабанов и домашних свиней, обладающих рядом признаков (поля светлой ости на шкуре — пятнистость, светлые копыта и пятак, висячие уши, короткая зимняя ость и др.) (Дуров, 1987). В предгорьях Краснодарского края такие гибриды составляют 4%. Что касается соотношения светлых и темных морфотипов, то в выборке из 34 особей, происходящих из окрестностей Нальчика, они составили по 10%.
3 В.Е.Соколов, А.К.Темботов
3 3
Кабан
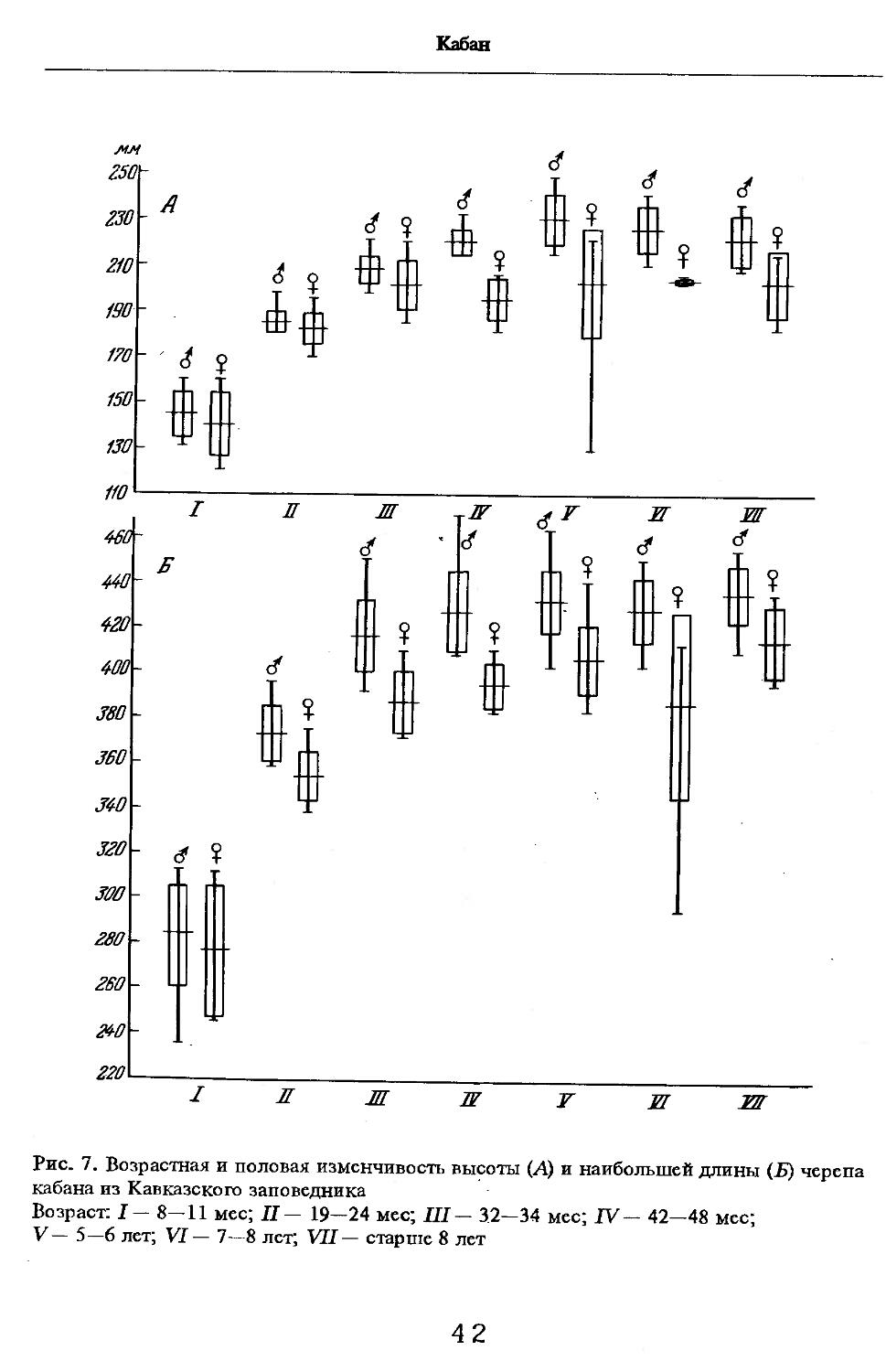
Значительной индивидуальной изменчивости подвержены масса и размеры тела, что зависит от многих факторов, в том числе от состояния кормовой базы, условий роста и развития как в эмбриональный, так и постэмбриональный периоды и т.д. Сеголетки имеют наибольший вес в годы урожая основных кормов (бук, дуб и др.). В неурожайные годы масса тела может уменьшиться на 47%, а размеры — на 4—5% (Дуров, 1987). Кроме того, условия жизни сеголеток накладывают глубокий отпечаток на рост и развитие в последующие периоды, определяя тем самым высокую индивидуальную изменчивость животных. В кавказском заповеднике около 5% в разных популяциях составляют особи больших размеров и массы (длина тела самцов до 198—205 см, масса — до 250 кг; самок соответственно до 180 см и 150 кг). Эти животные характеризуются также светлой, пепельно-серой однотонной остью, короткими ушами, малой вислозадостью, большой вогнутостью профиля черепа, большой длиной тела.
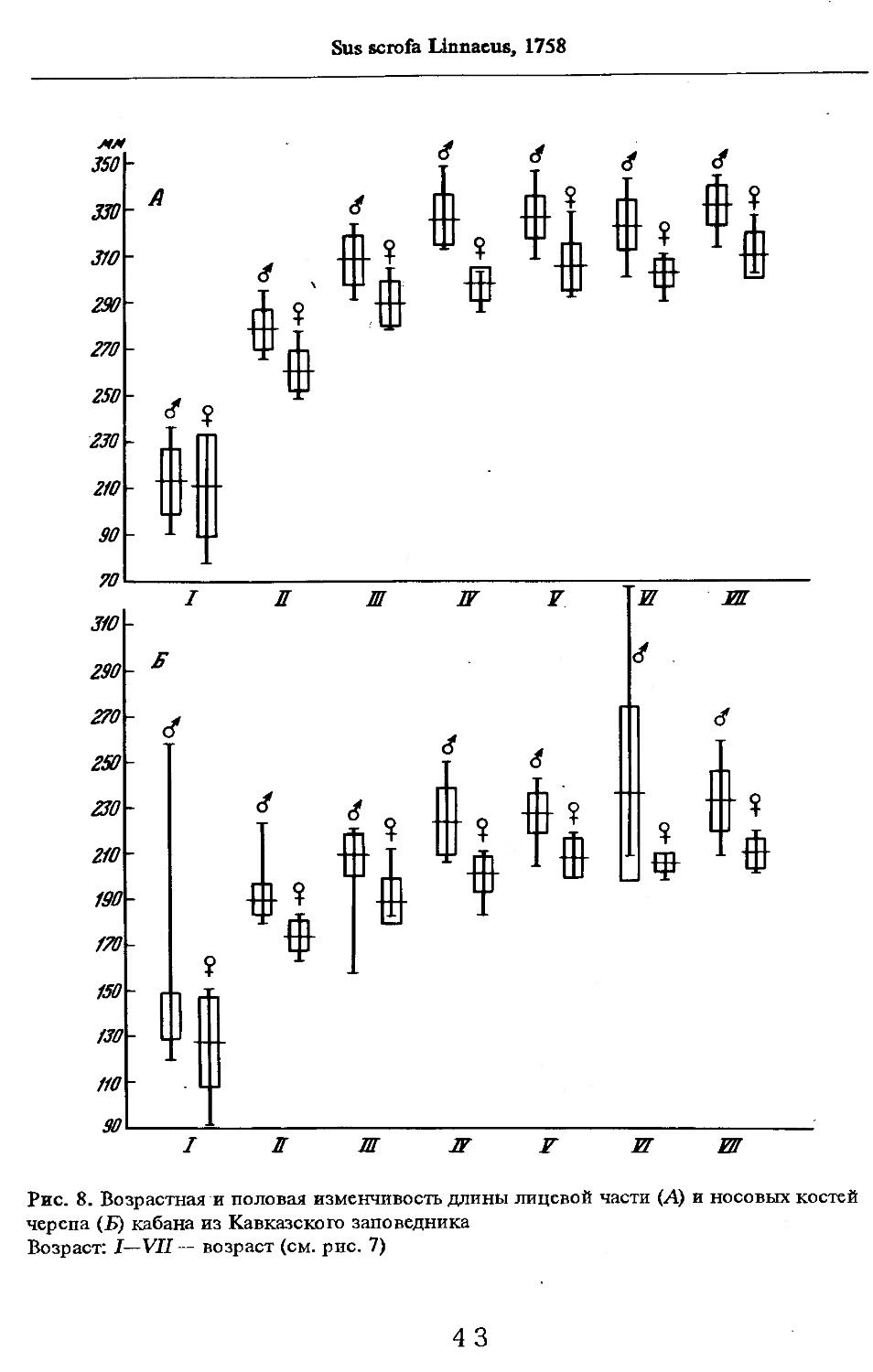
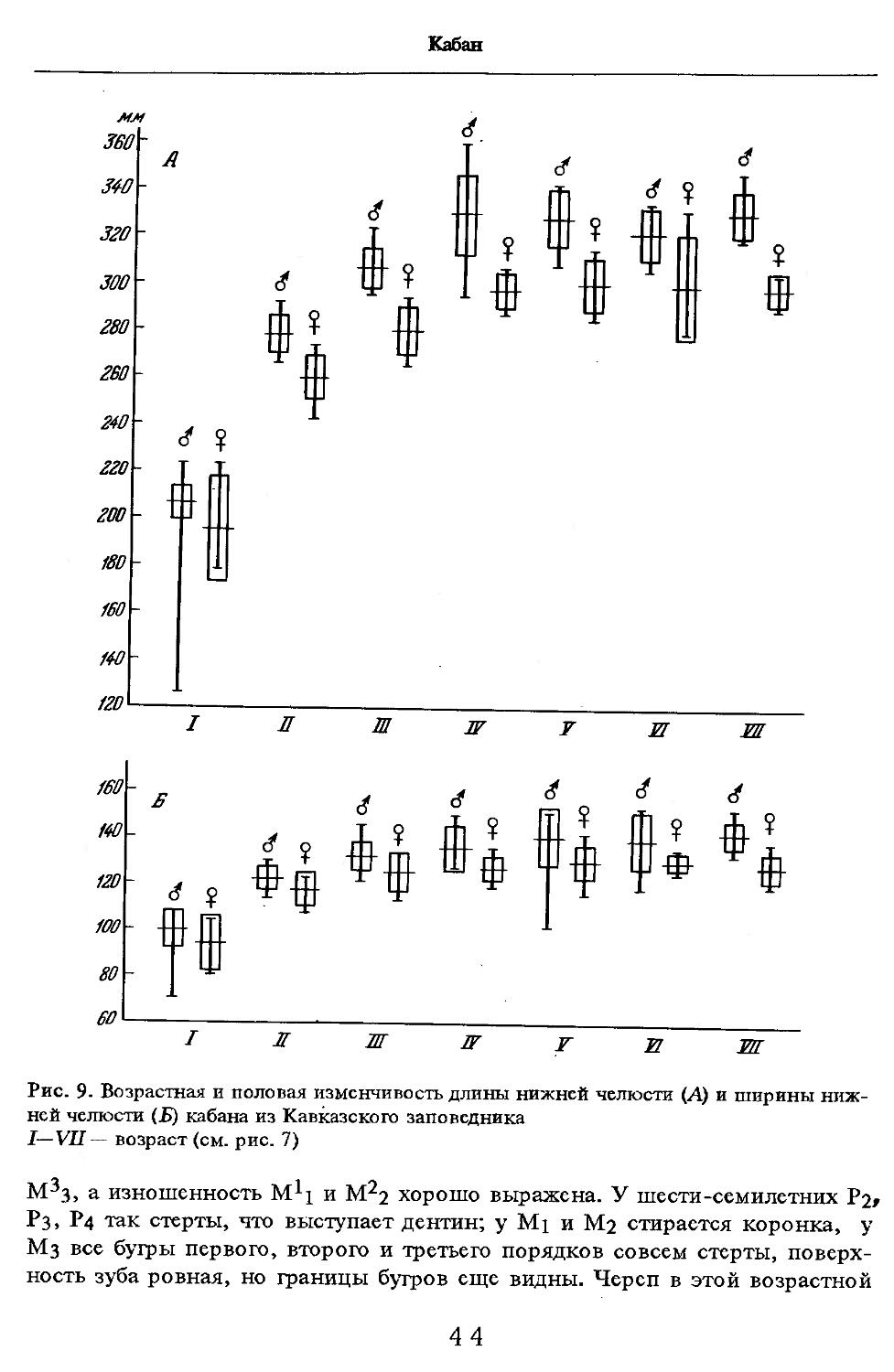
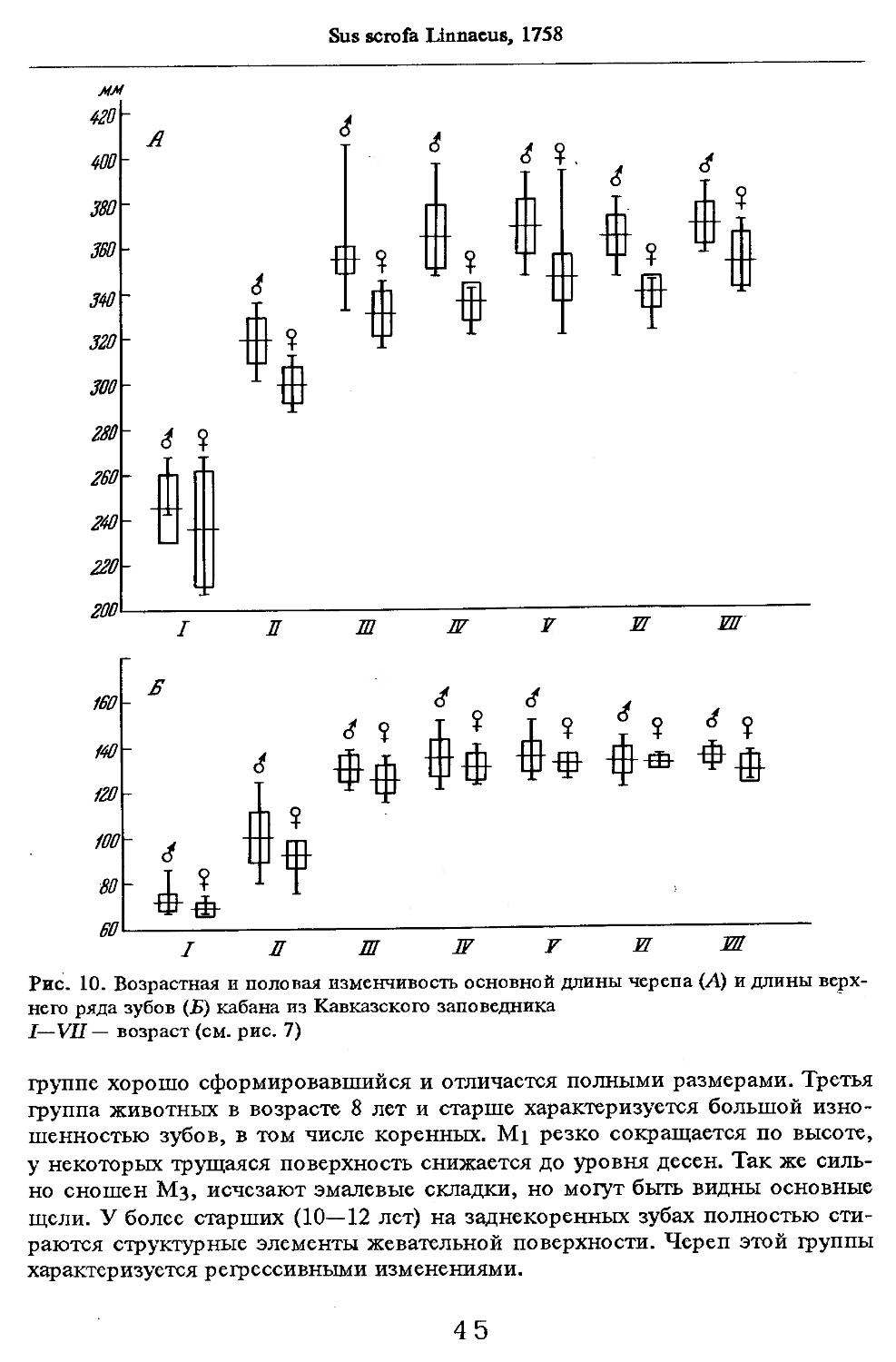
Об индивидуальной изменчивости структуры черепа можно судить по коэффициенту вариации, приведенному в табл. 5—12.
Так, в нашей коллекции есть экземпляр, левая верхняя челюсть которого имеет 5 предкоренных зубов, а правая — 4. Индивидуальная изменчивость фенов у кабанов Кавказа не изучена, но, судя по нашему коллекционному материалу (п — 17), она довольно высока. Даже у двух произвольно отобранных черепов различными оказались верхнечелюстные отверстия — у одного их два и они средних размеров, а у другого одно более крупное отверстие.
Что касается зубной системы, то и здесь индивидуальная изменчивость не менее значительна; особенно вариабельны клыки взрослых кабанов. Н.Я.Динник (1910) приводит описание двух черепов из Псебая (Кавказский заповедник), резко выделявшихся среди других формой и размерами клыков. Эти клыки были так сильно развиты, что загнулись кольцом, достигли верхней поверхности носовых костей, проросли сквозь них, а на одном черепе проникли в носовую полость и были хорошо видны через носовое отверстие.
Возрастная изменчивость. Наиболее полные данные по возрастной изменчивости кабана получены в условиях Западного Кавказа. Они освещены в диссертационной работе В.В.Дурова (1987) и в табл. 5—13. Аналогичная работа по кабану в Украинских Карпатах выполнена Н.С.Гунчаком (1978). В обеих выборках, происходящих из сходных по природным условиям (типу поясности) горных районов, определение возраста и соответственно анализ возрастной изменчивости проводились по одной и той же методике (Козло, Никитенко, 1967; Козло, 1975). Это дает возможность провести анализ возрастной изменчивости в сравнительном плане.
Возрастная динамика массы и размеров тела кабана характеризуется очень высокой интенсивностью в первый год жизни. Если новорожденные имеют массу всего 0,6—1,15 кг (Сафаров, 1965), то у сеголеток она в среднем составляет 45,4 кг (Дуров, Александров, 1968). У двухлеток по сравнению с сеголетками масса тела удваивается. У самок размеры и масса, близкие к предельному (9’6%), достигаются в 3 года, а у самцов в 4 года. Рост тела прекра-
34
Sus scrofa Linnaeus, 1758
Таблица 5. Промеры (в мм) черепа Sus scrofa из Кавказского заповедника.
______________________________Возраст 4 года_________________________
Признак Самцы
п limit х ± т G Су
Высота черепа 9 214-232 220,0±1,92 5,77 2,6
Наибольшая длина 11 408—470 427,4±5,32 17,65 4,1
Основная длина 11 347-396 363,5±4,35 14,41 4,0
Длина лицевой части 10 313-349 325,9±3,18 10,07 3,1
Длина носовых костей 10 207-250 224,8±4,66 14,73 6,6
Скуловая ширина 10 154-180 161,8±2,49 7,89 4,9
Межглазничная ширина 10 79-100 86,3±2,03 6,43 7,5
Ширина затылочного гребня 9 75-89 81,8±1,50 4,49 5,5
Длина нижней челюсти
Ширина нижней челюсти 10 295-360 323,2±6,12 19,38 6,1
10 125—147 133,1±2,70 8,53 6,4
Признак Самки
п limit х +т О Су
Высота черепа 7 181-206 195,9 ±3,33 8,80 4,5
Наибольшая длина 12 383-410 394,1±2,79 9,66 2,5
Основная длина 12 320-341 335,2±2,25 7,78 2,3
Длина лицевой части 12 286-303 296,8±1,58 5,46 1,8
Длина носовых костей 10 184-212 202,0±2,60 8,23 4,1
Скуловая ширина 12 137-157 145,8+1,43 4,95 3,4
Межглазничная ширина 10 73-85 79,3±1,18 3,74 4,7
Ширина затылочного гребня 9 63-82 72,2±2,04 6,12 8,5
Длина нижней челюсти
Ширина нижней челюсти 9 287—307 298,6±2,26 6,78 2,3
9 117-133 125,0±1,55 4,64 3,7
щается в возрасте 6 лет при средней массе в 5—6 лет самцов 173 кг, самок 120 кг, максимальная соответственно 250 и 150 кг. Отметим, что у европейских и среднеазиатских кабанов размеры и масса тела достигают близких к полным показателям на год позже, чем у кабанов Западного Кавказа (Дуров, 1987). С семи-восьмилетнего возраста происходит рецессия массы тела на 4—11%.
Возрастные изменения волосяного покрова у кабана наиболее значительны в первые годы жизни. По данным В.В. Дурова (1987), смена ювенильного волоса у 30% кабанов заканчивается в возрасте 11—14 мес., а у остальных — в возрасте 23 мес.
Возрастная динамика черепа иллюстрирует сходную с размерами тела картину; основное отличие, видимо, сводится к тому, что рост черепа и зубов продолжается лишь до 3—4 лет, т.е. стабилизируется значительно раньше, чем размеры тела. Исключение составляют клыки, которые растут у самцов до 12 лет, а у самок до 9 лет. Как видно из рис. 7—11, коэффициент вариации по черепным признакам наиболее высок у сеголеток, он составляет у самцов от 5,6 до 11,5, а у самок — от 3,7 до 15,5. У самцов-двухлеток ко-
3*
3 5
Кабан
Таблица 6. Промеры (в мм) черепа Sus scrofa из Кавказского заповедника.
__________________________Возраст 5—6 лет___________________________
Признак Самцы
п limit х ± т О Су
Высота черепа 16 215-249 229,5±2,82 11,30 4,9
Наибольшая длина 17 403-463 433,9±3,34 13,75 3,2
Основная длина 17 346-392 367,7±2,83 11,65 3,2
Длина лицевой части 16 308-346 325,7±2,34 9,36 2,9
Длина носовых костей 16 206-244 229,2±2,30 9,19 4,0
Скуловая ширина 17 154-178 165,5±1,70 7,00 4,2
Межглазничная ширина 15 83—100 89,5±1,39 5,40 6,0
Ширина затылочного 15 72-104 86,1±2,38 9,21 10,7
1ребня
Длина нижней челюсти 9 308-343 329,4±3,98 11,93 3,6
Ширина нижней челюсти 10 100-148 137,5±4,42 13,98 10,2
Признак Самки
п limit х ± т О Су
Высота черепа 12 128-221 202,7±7,29 25,26 12,5
Наибольшая длина 14 383—440 406,2+3,91 14,61 3,6
Основная длина 14 320-393 345,6+4,58 17,12 5,0
Длина лицевой части 13 292—328 305,2+2,66 9,60 3,1
Длина носовых костей 12 188—220 208,9+2,60 9,01 4,3
Скуловая ширина 14 147-163 153,8±1,13 4,21 2,7
Межглазничная ширина 12 74-86 81,1±0,99 3,42 4,2
Ширина затылочного 12 70-86 76,3±1,51 5,25 6,9
гребня
Длина нижней челюсти 11 285-315 300,3±3,03 10,06 3,4
Ширина нижней челюсти 11 114-139 127,2±2,14 7Д1. 5,6
эффициент вариации показателей черепа остается высоким, а у самок заметно выравнивается. Судя по нашим данным, аналогичное выравнивание коэффициента вариации у самцов наблюдается в трехлетием возрасте. Более старшие возрастные труппы характеризуются относительно низким коэффициентом вариации. Сходная динамика врзрастной изменчивости морфологических признаков выявлена у кабанов, обитающих в горах Украинских Карпат (Гунчак, 1978) и Беловежской Пуще (Козло, 1975).
Возрастные изменения зубов довольно сложные, о чем свидетельствуют результаты обработки нами коллекционного материала и литературные данные (Козло, Никитенко, 1967; Козло, 1975). По мнению названных авторов, анализ состояния зубов позволяет с достаточно высокой точностью различать 10 возрастных групп. О них более подробно будет сказано в разделе, посвященном росту и развитию. Здесь мы остановимся на характеристике трех возрастных групп, специфику которых необходимо учитывать при решении систематико-таксономических и микроэволюционных проблем; I — молодые (растущие), II — взрослые (стабилизировавшиеся) и III — старые (регрессирующие).
36
Sus scrofa Linnaeus, 1758
Таблица 7. Промеры (в мм) черепа Sus scrofa из Кавказского заповедника.
____________________________Возраст 7 8 лет_________________________
Признак Самцы
п limit х ± т О Су
Высота черепа 9 210-241 225,6±3,38 10,14 4,5
Наибольшая длина 11 403-450 427,9±3,94 13,06 3,1
Основная длина 11 345-380 362,6±2,72 9,04 2,5
Длина лицевой части 10 300-342 321,5±3,48 11,02 3,4
Длина носовых костей 8 210-328 236,8±13,43 37,99 16,0
Скуловая ширина 9 138-172 158,0±3,11 9,33 5,9
Межглазничная ширина 9 76-94 83,8 + 1,82 5,45 6,5
Ширина затылочного 8 71-85 78,9+1,69 4,79 6,1
гребня
Длина нижней челюсти 7 306-335 321,7±4,05 10,72 3,3
Ширина нижней челюсти 7 116-150 136,6±4,46 11,79 8,6
Признак Самки
п limit х ± т О Су
Высота черепа 3 202—205 203,3±0,88 1,53 0,8
Наибольшая длина 7 295-412 386,0±15,36 40,6 10,5
Основная длина 7 322-344 338,4±2,81 7,44 2,2
Длина лицевой части 7 290-310 302,6±2,37 6,27 2,1
Длина носовых костей 5 200-211 206,6±1,96 4,39 2Д
Скуловая ширина 4 144—153 148,8±2,46 4,92 з,з
Межглазничная ширина 5 75-81 78,6±1,17 2,61 3,3
Ширина затылочного 3 69-82 75,0+3,79 6,56 8,7
гребня
Длина нижней челюсти 5 280—333 299,2+9,79 21,9 7,3
Ширина нижней челюсти 5 123-133 128,0±1,61 3,61 2,8
Таблица 8. Промеры (в мм) черепа Sus scrofa из Кавказского заповедника.
___________________________Старше 8 лет___________________________
Признак Самцы
п limit х ± т О Су
Высота черепа 11 208-237 221,6+3,31 10,99 5,0
Наибольшая длина 13 410-454 435,5+3,65 13,15 3,0
Основная длина 13 355-386 368,8+2,52 9,10 2,5
Длина лицевой части 13 312-343 330,2+2,53 9,11 2,8
Длина носовых костей 11 210-260 234,4±3,81 12,63 5,4
Скуловая ширина 11 154-172 161,0±1,42 4,70 2,9
Межглазничная ширина 12 74-92 83,4±1,61 5,58 6,7
Ширина затылочного 10 69-82 76,6±1,43 4,53 5,9
гребня
Длина нижней челюсти 13 320-349 331,3+2,74 9,89 3,1
Ширина нижней челюсти 13 131-150 139,5+1,36 4,91
37
Кабан
Таблица 8 (окончание)
Признак Самки
п limit х ± т О Су
Высота черепа 4 181-215 201,5±7,51 15,02 7,5
Наибольшая длина 6 395-435 414,2±6,53 15,99 3,4
Основная длина 6 338-370 350,7±5,06 12,40 3,5
Длина лицевой части 6 300-326 308,8±3,93 9,62 3,1
Длина носовых костей 6 203-221 213,0+2,72 6,66 3,1
Скуловая ширина 6 137-154 149,4±2,72 6,66 4,5
Межглазничная ширина 5 73-86 82,0±2,51 5,61 6,8
Ширина затылочного 5 61-79 71,2±3,46 7,73 10,9
гребня
Длина нижней челюсти 6 290-305 299,5±2,81 6,89 2,3
Ширина нижней челюсти 6 118-136 126,8±2,54 6,21
Таблица 9. Промеры (в мм) черепа Sus scrofa из Центрального Кавказа.
-_____________________Возраст 4 года_________________________
Признак Самцы
п limit х ± т О Су
Высота черепа 8 157,1-219,5 191,1+9,03 23,90 12,5
Наибольшая длина 8 380,0-462,0 410,2+10,21 27,00 6,5
Основная длина 6 345,0—394,0 370,5+8,23 18,40 4,9
Длина лицевой части 8 285,0-355,0 312,3±8,89 23,51 7,5
Длина носовых костей 8 173,4-248,4 209,2+8,36 22,13 10,5
Скуловая ширина 7 142,5-173,9 155,9+4,91 12,02 7,7
Межглазничная ширина 8 71,0-92,6 82,3+2,94 7,79 9,4
Ширина затылочного 8 63,6-94,5 78,8+4,31 11,41 14,4
гребня
Длина нижней челюсти 8 280,0—349,0 317,5+8,84 23,38 7,3
Ширина нижней челюсти 7 119,0-150,7 132,6±4,57 11,19
Признак Самки
п limit х + т о Су
Высота черепа 6 153,4-203,9 166,6±8,42 18,82 11,3
Наибольшая длина 7 365-411,0 387,3±7,80 19,10 4,9
Основная длина 5 325,0—373,8 343,8±9,67 19,34 5,6
Длина лицевой части 7 280,0—324,8 296,0±7,15 17,52 5,9
Длина носовых костей 7 109,5-215,0 182,2±14,10 34,54 18,9
Скуловая ширина 6 139,2-146,0 146,3±5,23 11,69 8,5
Межглазничная ширина 6 70,2-86,0 75,9±2,43 5,43 7,1
Ширина затылочного 6 62,3-75,5 67,6±2,01 4,50 6,6
гребня
Длина нижней челюсти 7 275,0-336,0 293,0±8,46 20,73 7,1
Ширина нижней челюсти 6 117,9-125,5 126,1±4,26 9,53 _L5_
38
Sus scrofa Linnaeus, 1758
Таблица 10. Промеры (в мм) черепа Sus scrofa из Центрального Кавказа.
_________________________Возраст 5—6 лет________________________
Признак Самцы
Л limit х ± т о Су
Высота черепа 10 173,5-217,3 202,9±4,18 12,55 6,2
Наибольшая длина 10 324,3—447,2 418,1+12,29 36,88 8,8
Основная длина 9 365,0-409,3 386,5±4,62 13,07 3,4
Длина лицевой части 10 305,0-348,4 329,8+4,85 14,54 4,4
Длина носовых костей 9 214,0-242,7 225,6±3,12 8,82 3,9
Скуловая ширина 9 154,0-174,8 166,4±2,66 7,52 4,5
Межглазничная ширина 10 76,6-101,0 90,3±2,60 7,79 8,6
Ширина затылочного 10 68,5-94,0 84,4±2,50 7,51 8,9
гребня
Длина нижней челюсти 8 310,0-360,0 334,7+5,55 14,68 4,4
Ширина нижней челюсти 7 137,1-151,8 142,2±2,02 4,94 3,5
Признак Самки
Л limit х ± т о Су
Высота черепа 6 163,9-206,4 172,3+7,49 16,76 9,7
Наибольшая длина 6 380,0-460,5 399,2±13,54 30,37 7,6
Основная длина 7 335,0-420,0 363,6±11,53 28,24 7,8
Длина лицевой части 7 295,0—360,0 311,5±9,01 22,08 7,1
Длина носовых костей 6 190,3-244,4 205,5+9,78 21,88 10,6
Скуловая ширина 6 139,0-173,6 15П,2±5,52 12,34 8,2
Межглазничная ширина 7 70,5-87,8 78,0+2,39 5,86 7,5
Ширина затылочного 6 61,6-81,8 69,1±3,43 7,67 11,1
гребня
Длина нижней челюсти 7 290,0-360,0 308,6±10,05 24,62 8,0
Ширина нижней челюсти 7 125,0-151,2 132,1+3,74 9,16 6,9
Таблица 11. Промеры (в мм) черепа Sus scrofa из Центрального Кавказа.
________________________Самцы 7—8 лет________________________
Признак Возраст 4 года
Л limit X ± пз о Су
Высота черепа 4 181,5-217,0 192,3±9,63 16,69 8,7
Наибольшая длина 4 395,0—448,0 415,7±13,28 23,00 5,5
Основная длина 4 350,0-395,0 371,2+10,64 18,43 5,0
Длина лицевой части 4 295,0-365,0 321,2+17,55 30,40 9,5
Длина носовых костей 3 196,8-205,1 201,9±3,13 4,43 2,2
Скуловая ширина 4 149,4-190,0 164,3+10,39 17,99 10,9
Межглазничная ширина 4 77,0-115,0 89,4±10,07 17,44 19,5
Ширина затылочного 3 67,7-79,7 72,1±4,67 6,1 9,2
' гребня
Длина нижней челюсти 2 300,0-310,0 305 — —
Ширина нижней челюсти 3 126,2-162,3 140,4±13,62 19,26 13,7
39
Кабан
Таблица 12. Промеры (в мм) черепа Sus scrofa из Восточного Закавказья. ________________________________Самцы___________________________________
Признак Возраст 4 года
п limit х ± т о Су
Высота черепа 4 171,5-193,7 182,9±6,65 11,52 6,3
Наибольшая длина 4 370,0-409,0 392,0±11,11 19,24 4,9
Основная длина 3 333,3-364,0 345,2±11,66 16,49 4,8
Длина лицевой части 4 283,7-312,7 299,0±8,71 15,09 5,0
Длина носовых костей 4 190,3-217,6 202,6±6,94 12,02 5,9
Скуловая ширина 4 133,6-160,2 147,4±6,54 11,33 7,8
Межглазничная ширина 4 72,7-86,6 81,1±3,67 6,35 7,8
Ширина затылочного 4 73,5-85,0 78,9+2,73 4,73 6,0
гребня
Длина нижней челюсти 4 287,7—320,3 301,7±9,26 16,04 5,3
Ширина нижней челюсти 4 118,9-131,0 122,3+3,34 5,78
Признак Возраст 5—6 лет
л limit х ± т с Су
Высота черепа 3 198,5-205,7 201,2+2,76 3,90 1,9
Наибольшая длина 3 412,5-436,0 420,5+9,49 13,42 3,2
Основная длина 3 362,5-381,0 369,8+6,95 9,83 2,6
Длина лицевой части 3 314,7-326,0 318,7+4,45 6,30 2,0
Длина носовых костей 3 214,0—228,0 219,2+5,40 7,64 3,5
Скуловая ширина 3 159,8—168,1 163,5±2,98 4,22 2,6
Межглазничная ширина 3 83,8-90,3 87,4±2,35 3,32 3,8
Ширина затылочного 3 74,6-90,0 80,5+5,89 8,33 10,3
гребня
Длина нижней челюсти 2 310,0-331,0 320,5 — —
Ширина нижней челюсти 2 129,7-137,6 133,6 — !—
Таблица 13. Возрастные изменения зубной системы дикого кабана из Азербайджана ___________________________ (Сафаров, 19651____________________________
Возрастная стадия 11 I1 12 12 1з I3 С1
Новорожденные, м м
до 5 дней -М -М — — м м м
От 5 до 10 дней +м +м — — м м м
3—4 мсс м м м м м м. м
7 мес м м м м м хП хП
9 мес м м м м хМ п п
13 мес м -пм м м п п п
15 мес +п +п м м п п п
16 мес п п м м п п п
20—21 мсс п п +п м п п п
23—24 мес п п +п +п п п п
26 мес п п п +п п п п
30 мес п п п +п п п п
32 мсс п п п п п п п
33 мсс п п п п п п п
40
Sus scrofa Linnaeus, 1758
Таблица 13 (окончание)
Возрастная стадия С1 Р1 в1 Р2 Р2 Рз Р3
Новорожденные,
до 5 дней м — — — — — —
От 5 до 10 дней м -— — — — — —
3—4 мес м — — м м м м
7 мес м м м м м м м
9 мес хП м м м м м. м
13 мес п м м м м м м
15 мес п м м м м м ПМ
16 мес п м м +п +п +п +п
20—21 мес п м м м м м м
23—24 мсс П ' м м м м м м
26 мес п м м м м м м
30 мес п м м м м м м
32 мес п м м м м м м
33 мес п м м м м м м
Возрастная стадия Р4 Р4 Ml м1 М2 м2 Мз м4
Новорожденные,
до 5 дней М -М — — — — — —
От 5 до 10 дней хМ хМ .— — — — — —
3—4 мес М м -хП — — — — —
7 мес М м П п — — — —
9 мес м м П п — — — —
13 мес м м П п + + .— —
15 мес ПМ ПМ п п п п —
16 мес +п +п п п п п — —
20—21 мес м м п п п п — —
23—24 мес м м п п п п + —
26 мес м м п п п п +п +п
30 мес м м п п п п +п +п
32 мес м м п п п п +п +п
33 мес м м п п п п +п +п
Примечание. M — молочные зубы; П — постоянные зубы; + прорезывающиеся зубы; ± прорезалась только передняя половина зуба; — верхушка прорезывающихся зубов сравнялась с краями костных альвеол; х — прорезывающиеся зубы еще находятся под слизистой оболочкой; ПМ — прорезываются постоянные зубы, но молочный еще не выпал.
Первая труппа животных, возраст которых менее 3 лет, характеризуется наличием развитой зубной системы, но последние коренные зубы еще не полностью сформировались, не имеют следов стертости. Формирование черепа еще не завершилось. Вторая труппа животных в возрасте от 3 до 6—7 лет характеризуется наличием полного зубного ряда, в том числе хорошо развитых коренных зубов, подверженных в той или иной степени стертости бутров. Трехлетние животные характеризуются слабой стертостью
4 1
Кабан
Рис. 7. Возрастная и половая изменчивость высоты (А) и наибольшей длины (Б) черепа кабана из Кавказского заповедника
Возраст: 1- 8-11 мес; II- 19-24 мес; III- 32-34 мес; IV — 42—48 мес;
V— 5—6 лет; VI— 7—8 лет; VII— старше 8 лет
42
Sus scrofa Linnaeus, 1758
230 -
210-
90 -
Рис. 8. Возрастная и половая изменчивость длины лицевой части (А) и носовых костей черепа (Б) кабана из Кавказского заповедника
Возраст: I—VII— возраст (см. рис. 7)
4 3
Кабан
Рис. 9. Возрастная и половая изменчивость длины нижней челюсти (А) и ширины нижней челюсти (Б) кабана из Кавказского заповедника
I—VII— возраст (см. рис. 7)
М3з, а изношенность М1! и М22 хорошо выражена. У шести-семилетних Р2, Р3) Р4 так стерты, что выступает дентин; у Mj и М2 стирается коронка, у М3 все бугры первого, второго и третьего порядков совсем стерты, поверхность зуба ровная, но границы бугров еще видны. Череп в этой возрастной
44
Sus scrofa Linnaeus, 1758
Рис. 10. Возрастная и половая изменчивость основной длины черепа (А) и длины верхнего ряда зубов (Б) кабана из Кавказского заповедника
I— VII — возраст (см. рис. 7)
группе хорошо сформировавшийся и отличается полными размерами. Третья группа животных в возрасте 8 лет и старше характеризуется большой изношенностью зубов, в том числе коренных. Mi резко сокращается по высоте, у некоторых трущаяся поверхность снижается до уровня десен. Так же сильно сношен Мз, исчезают эмалевые складки, но могут быть видны основные щели. У более старших (10—12 лет) на заднекоренных зубах полностью стираются структурные элементы жевательной поверхности. Череп этой группы характеризуется регрессивными изменениями.
4 5
Кабан
Рис. 11. Возрастная и половая изменчивость межглазничной ширины (А), скуловой ширины (Б), длины нижнего ряда зубов (А) и ширины затылочного гребня (Г) кабана из Кавказского заповедника
I— VII— возраст (см. рис. 7)
4 6
Sus scrofa Linnaeus, 1758
Половые различия. Во всех работах, посвященных морфометрическим параметрам кабана Кавказа, анализ данных проводился с учетом пола. Таковы исследования Н.Я.Динника (1910), С.С.Донаурова и В.П.Теплова (1938), М.А.Сафарова (1965), В.В.Дурова и В.Н.Александрова (1968), В.В.Дурова (1987) и др. Анализ литературных данных и нашего материала позволяет очертить целый круг существенных различий между самцами и самками кабанов. Хотя различий по массе и размерам тела в труппе сеголеток между полами практически нет, в последующем вследствие расхождения в интенсивности роста и развития начинает все больше проявляться половой диморфизм. В Карпатах, по Н.С.Гунчаку (1978), различие по массе между полами четко проявляется начиная с трехлетнего возраста. Как отмечено выше, в Кавказском заповеднике самки достигают показателей размеров, близких к предельному, на один год раньше, чем самцы. Из внешних признаков обращает на себя внимание то, что самки обычно стройнее и менее массивные, чем самцы, особенно в области труди. Весьма характерен для половозрелых самцов так называемый "калкан", представляющий утолщение кожи необычной прочности и служащий для защиты во время жестоких драк в период гона, когда он достигает максимальной толщины.
Половые различия в структуре черепа не выражены в течение первых двух лет, они четко проявляются в трехлетием возрасте и сохраняются до конца жизни. Рис. 7—11 четко иллюстрируют, насколько линейные показатели черепа самцов превышают таковые самок. По форме черепа самки характеризуются более ровным профилем верха, тогда как у самцов он обычно вогнутый. Что касается полового диморфизма зубной системы, то, на наш взгляд, особый интерес представляет возможность определения пола на основе параметров клыков. Хотя в литературе давно известно, что клыки у самцов больше, чем у самок, наиболее удобным показателем половой принадлежности оказалась форма поперечного среза верхнего клыка и соответственно форма костного гнезда в челюсти. У самцов они имеют трапециевидную форму, тогда как у самок — форму треугольника (Mayer, Brisbin, 1988). Не менее интересен, по мнению этих авторов, тот факт, что у самцов есть, а у самок нет эмали на корнях верхних и нижних клыков. На клыках самок в отличие от самцов хорошо выражена граница между верхним слоем эмали и нижним (корневым) цементом. Методика определения пола по форме поперечного среза клыков пригодна и для молодых животных в возрасте от 30 до 51 недель.
Изменчивость в связи с антропогенным прессом. Еще Н.Я.Динник (1910) писал, что в последнее время в тех местах, где кабанов преследуют, редко можно встретить свинью тяжелее 5—7 пудов и кабана тяжелее 8 пудов. Это подтвердили С.С.Донауров и В.П.Теплов (1938). Сравнение средних размеров кабанов, добытых в Кавказском заповеднике в 30-х годах, с таковыми, приведенными Н.Я.Динником (1910), не выявило каких-либо существенных различий. Сравнение этих данных с материалом из окрестностей г. Владикавказа, где охота довольно интенсивна, показало иные результаты. В Осетии средние размеры кабанов оказались существенно ниже, чем в условиях заповедника (в Осетии средняя масса тела 63 кг, максимум 144 кг, а в Кавказском
47
Кабан
заповеднике соответственно 70 и 178 кг) (Донауров, Теплов, 1938). Как видно, стихийный характер промысла кабана, имеющий место в ряде районов Кавказа, может иметь нежелательный селекционный эффект. В этой связи интерес представляет хорологическая изменчивость кабана на Кавказе.
Изменчивость кабана во времени (четвертичный период) на Кавказе рассмотрена Н.К.Верещагиным (1959). Согласно его данным, с верхнего плиоцена до современности проходит однонаправленное изменение — уменьшение размеров, о чем можно судить по сокращению ширины нижней челюсти позади клыков с 73 до 46 мм.
Географическая изменчивость. Для анализа изменчивости кабана в трехмерном пространстве гор Кавказа мало литературных данных, нет соответствующего коллекционного материала. Поэтому приходится ограничиваться лишь отдельными ее параметрами. Для определения особенностей морфометрии западнокавказских кабанов сравним их с популяциями Украинских Карпат. Такое сравнение представляет определенный интерес по ряду обстоятельств: при значительном географическом удалении друг от друга (около 1200 км) и территориальной изоляции они занимают сходные ландшафтные условия (тип поясности) и, по мнению многих териосистематиков, относятся к одному подвиду S. s. attila (Адлерберг, 1930; Соколов, 1959; и др.). Кроме того, из обоих районов имеется хороший сравнительный материал, нашедший обобщение в ряде научных работ (по Кавказу: Донауров и Теплов, 1938; Дуров и Александров, 1968; Дуров, 1987; по Карпатам: Гунчак, 1978).
По данным В.В.Дурова (1987), кавказский кабан по цвету волос темнее европейского, уссурийского и среднеазиатского. Но в кавказских и карпатских популяциях широко представлены обе цветовые вариации покрова: черновато-бурая с серым или рыжеватым оттенком и черная с сероватым или темно-бурым оттенком. Реальность существования этих вариаций признавали Н.Я.Динник (1910), П.Г.Козло (1975) и др. Что касается процентного соотношения их в разных районах, то сравнительных данных нет. Кроме указанных двух цветовых вариаций, в Европе регулярно встречается третья — пестрая (черно-белая или бело-черная). Кабаны с такой окраской нередки в Польше и ГДР, у нас они зарегистрированы в Беловежской Пуще (Козло, 1975; And-rzejewski, 1974; Andrzejewski et al., 1975; и др.). Пестроокрашенных кабанов мы не встречали на Кавказе, аналогичных сообщений в литературе также не находили. Если учесть, что пестрая окраска волосяного покрова отмечена в различных возрастных группах и, как указано выше, имеет генетическую природу, то ее отсутствие или низкая ее концентрация в популяциях Кавказа — явление не случайное. На наш взгляд, заслуживает внимания более подробное изучение распространения пестроокрашенных кабанов в пределах ареала вида, учитывая роль гибридизации с домашними свиньями.
Сравнительный анализ массы тела показал, что кавказские кабаны заметно уступают карпатским. По С.С.Донаурову и В.П.Теплову (1938), масса тела кабана в среднем на Западном Кавказе 166 кг у самцов и 68 кг у самок. В.В.Дуров (1987) в тех же условиях получил более высокие показатели: 173 и 120 кг. В Карпатах взрослые самцы имеют в среднем 202 кг, самки 133 кг (Гунчак, 1978). Кабаны из популяции Беловежской Пущи значительно усту
48
Sus scrofa Linnaeus, 1758
пают предыдущим — самцы весят 135 кг, самки 99 кг (Козло, 1975). Близкие результаты получены и по другим меристическим данным, в том числе по черепным. Так, кондилобазальная длина взрослых самцов Западного Кавказа 388 мм, самок — 347 мм, а в Карпатах — 393 и 361 мм соответственно. Эти показатели в Беловежской Пуще — 389 и 349 мм. Как видно, в сходных ландшафтных условиях Западного Кавказа и Карпат формируется очень близкий фенотип, а в иных (Беловежская Пуща, хотя и территориально близкая к Карпатам) кабаны существенно отличаются от обеих популяций по массе тела.
Географическую изменчивость кабана в пределах Кавказа мало кто обсуждал; мы располагаем оригинальным материалом, чтобы проследить динамику показателей массы и размеров тела с запада на восток в связи с очень большим градиентом ландшафтных условий Кавказа. Мнение М.А.Са-фарова (1965) о том, что в Азербайджане встречаются относительно крупные кабаны, видимо, нуждается в подтверждении новым материалом. Если по его данным масса взрослых самцов составляет 142 кг, а самок — в среднем 98 кг, то на Западном Кавказе она достигает соответственно 173 и 120 кг.
Самые крупные черепа Также встречаются у зверей из популяции Западного Кавказа (см. табл. 5—12). На Центральном Кавказе и Восточном Закавказье череп кабана заметно меньше.
Заслуживают внимания данные В.В. Дурова (1987) о том, что между кабанами южного и северного макросклонов Западного Кавказа существуют морфологические различия. У первых, названных "черноморской популяцией", шерсть более темная, на 5% меньше вислозадость, на 6% больше размеры затылочных гребня и кости, чем в "кубанской популяции". Автор объясняет такое расхождение адаптацией первых к густым колхидским лесам, более крутым склонам и плотной почве, что обусловливает более сильное развитие шейного отдела. Кроме того, он полагает, что на границе названных "популяций" сложилась высокогорная "субпопуляция", в которой размеры тела и черепа кабанов крупнее обычных на 7%, что может быть связано с проявлением гетерозиса или адаптации к суровым условиям высокогорья. С этой мыслью созвучно мнение Н.Я.Динника (1910) о том, что высотная поясность накладывает отпечаток на морфологию и другие параметры кабана, следствием чего явилось образование "двух пород кабана": одни звери меньшего роста, с относительно коротким туловищем, большой головой и грудью, длинными ногами; особи другой формы имеют большой рост, относительно длинное туловище, менее развитые голову и грудь. Первые имеют более темную окраску, а вторые — светлую. Различна их ландшафтная приуроченность: первые живут среди высоких и крутых гор, а вторые — в местах более низких и ровных. Наконец, они различаются и по нраву; считается, что высокогорные кабаны более агрессивны, чем равнинно-предгорные. Однако высотная дифференциация, о которой пишут названные авторы, плохо согласуется с миграциями, столь характерными для кабанов Кавказа.
Для познания закономерностей географической изменчивости и микроэво-люционного процесса рассматриваемого вида в целом и кавказских популяций в частности наиболее продуктивными оказались иммуногенетические ис-
4 В.Е.Соколов, А.К.Темботов
4 9
Кэбам
следования (Тихонов и др., 1985). Заслуживает внимания то, что, несмотря на внешнюю морфологическую близость (во всяком случае, пока невозможна морфологическая дифференциация) северокавказских и закавказских кабанов, они существенно отличаются иммуногенетически. Полиаллельная система Е северокавказских популяций более родственна с восточноевропейскими, чем с закавказскими. Кроме того, у северокавказских кабанов обнаружена большая концентрация аллеля aegil, отмечавшегося до настоящего времени лишь у популяции уссурийского кабана. Из 4 аллелей системы Е, обнаруженных у дикого кабана изученных 'популяций, самую высокую концентрацию образуют аллели Ee<Jghkmn и Ecdgjmn Частота первого аллеля у восточноевропейских кабанов составляет 0,71, северокавказских — 0,56, закавказских — 0,53, а второго аллеля у восточноевропейских — 0,24, северо-кавказских — 0,33, закавказских — 0,47. Аллель bdkm имеется у центральноевропейских (0,30) и восточноевропейских (0,05), но не выявлена у всех кавказских, среднеазиатских и уссурийских кабанов.
С помощью системы L обнаружена иммуногенетическая дифференциация на уровне подвидов, а также в пределах подвида, например между восточноевропейским и центральноевропейским, а также северокавказским и закавказским кабанами. Более того, ленкоранская и Щекинская (северо-западная граница Азербайджана с Грузией) существенно (статистически достоверно) различаются по встречаемости антигенов Ld и Lh, концентрации аллеля Lagim и частоте генотипа Ladhi/bcgi (соответственно 44,83 и 14,82%). В целом закавказская популяция кабана существенно отличается от северокавказской. Достоверно различаются также кубанская и черноморская популяции.
По диаллельной системе G мономорфизм свойствен только центральноевропейским кабанам, а всем остальным подвидам, в том числе кавказским популяциям, свойствен полиморфизм. При этом концентрация его в географическом плане носит клинальный характер — антиген Gb отсутствует в центральноевропейской популяции, а в восточноевропейской составляет около 5%; в закавказской и северокавказской популяциях — 84—93%, среднеазиатской — 98%, а дальневосточной — 100%. Противоположную закономерность имеет концентрация аллеля Ga.
Диаллельная система D четко дифференцирует также северокавказских и закавказских кабанов.
Изложенные данные убедительно свидетельствуют о том, что кабан на Кавказе подвержен значительной географической и популяционной изменчивости, при которой различия генетического порядка между северокавказскими и закавказскими популяциями значительны. Однако слабая изученность географической изменчивости морфологических признаков не позволяет подвергнуть ревизии внутривидовую систематику Sus scrofa и внести уточнение в положение в ней кавказских популяций. До выполнения такой работы мы, вслед за предыдущими териологами, относим все популяции кабанов Кавказа к подвиду S. s. attila Thomas.
50
Sus scrofa Linnaeus, 1758
Экология
Ландшафтная приуроченность. Биотопы. На Кавказе кабан занимает очень широкий круг равнинных и горных ландшафтов. Однако, судя по многочисленным литературным данным и нашим наблюдениям, наиболее оптимальными в настоящее время являются предгорные и низкогорные широколиственные и смешанные леса с обилием диких плодоносящих деревьев и кустарников, обеспечивающих оптимальные кормовые и защитные условия. Набор таких лесов с геоботанических позиций весьма разнообразен, но для анализа экологии кабана их можно свести к следующим: чистые и смешанные насаждения дуба, бука, каштана, грецкого ореха, лещины, диких груш и яблонь и др. В различных районах Кавказа преобладает тот или иной тип широколиственных лесов, но, как правило, они привлекают не только высококалорийной пищей и хорошими убежищами, но и наличием различных водоемов (рек, родниковых источников, озер, заболоченных мест), значительным периодом бесснежного или малоснежного времени, разнообразием лугов и высокотравья, небольших сельхозугодий и т.д. Не менее благопри-' ятными были и в ряде районов остаются речные долины равнин как Северного Кавказа, так и Закавказья, отличающиеся буйным ростом дубняков, фруктарников, околоводной и болотной растительности, непроходимых терновников, облепихи, лещины, переплетенных всевозможными лианами. В условиях субтропиков незаменимые защитные условия для кабана создают по сей день заросли держи-дерева, понтийской ежевики и др. К благо приятным ландшафтам, очевидно, относятся сохранившиеся поныне лиманные заросли Приазовья и Прикаспия, где кабан остается обычным видом. Такие места занимают, по данным В.Е.Костоглода (1969), в Приазовье часть территории Приморско-Ахтарского, Темрюковского и Славянского районов, общей площадью 90 тыс. га. По Каспийскому побережью поселения кабана сохранились в плавнях Аграханского залива, низовьях Терека, Сулака, Самура, в Ку-ба-Хачмасском районе, в Кызыл-Агачском заповеднике и ряде других мест Куринской низменности (Гинеев, 1971; Литвинов, 1981; Алекперов, Кулиев, 1987; и др.). В круг благоприятных ландшафтов для кабана, видимо, следует отнести субтропические лесные низменности Закавказья — Колхидскую, Ленкоранскую и Алазано-Авторанскую. В сильно измененных лесах этих мест доминируют длиннолистный дуб, граб, ильм, черная ольха с обилием колючих лиан. Лесные поляны с папоротником орляком, понтийской ежевикой, держи-деревом и другими насаждениями и болотистые места и топи служат надежной защитой для кабана. Во всех перечисленных ландшафтах кабаны могут жить, поддерживая высокую плотность населения в течение всего года. Достаточно сказать, что в урожайные годы кабаны кормятся плодами дуба, бука, каштана с октября по июнь (Дуров, 1973; наши данные).
Наряду с указанными ландшафтами кабаны заселяют темнохвойные леса; особенно буково-пихтовые, субальпийские и альпийские луга, иногда "забираются на скалы, поднимающиеся на 9 тысяч футов, заходя таким образом на места, обитаемые сернами и турами" (Динник, 1910, с. 35). Однако продолжительность и время пребывания кабанов в разных биотопах существен
4*
5 1
Кабан
но меняются, что определяется характером снежного покрова, кормовой базы и т.д. Как высоко пластичное животное кабан успешно осваивает сельско-хозяйственные угодья, проникая по долинам рек, лесистым балкам и лесонасаждениям глубоко в степную зону и полупустыню Восточного Предкавказья и Кура-Араксинскую низменность. Нередко кабан заходит на кормохранилища (силосные ямы, стога сена и соломы и др.), в отдельные животноводческие и иные постройки, места свалок пищевых отходов и т.д. Словом, кабан становится элементом антропогенного ландшафта, обнаруживая склонность к синантропизации.
Во всех перечисленных ландшафтах имеются жизненно важные и соответственно особо предпочитаемые биотопы. По этому поводу Динник (1910) писал, что хотя кабаны, попадаются и на местах более или менее открытых, их любимым местопребыванием служат самые непроходимые лесные чащи, камыши или высокий как лес густой бурьян. Зимой в лесах зверь чаще всего забирается в непроходимые заросли вечнозеленого самшита или кавказской пальмы, в заросли кавказских рододендронов, сохраняющих немало зеленой листвы, в густой орешник, дубняк с зеленью или мелкий густой ельник. Особенно кабаны любят скрываться в непроходимой чаще колючих и вьющихся кустарников, в том числе держи-дерева, ежевики, ожины, ломоноса и т.д. Летом часто забираются в терновые заросли или густой грушняк. В таких местах кабаны держатся днем, а ночью бродят всюду. Следы их деятельности, в том числе многочисленные порой, часто встречаются по опушкам леса и вблизи них, на открытых больших полянах, на субальпийских лугах и криволесьях, на различных посевах, особенно кукурузы и картофеля. Ландшафтное и биотопическое размещение кабана в условиях Кавказского заповедника отражено в статье В.В.Дурова (1973).
Миграции. Характерной чертой кабана являются сезонные перемещения и кочевки, вызванные изменениями кормовой базы и характером распределения снежного покрова. Конкретные аспекты этого явления в условиях Кавказа освещены в ряде работ (Динник, 1910; Логинов, 1936; Насимович, 1936, 1939, 1955; Донауров, Теплов, 1938; Чернявская, 1956; Котов, Рябов, 1963; Дуров, 1973, 1987; и др.).
Большая часть этой информации относится к Западному Кавказу, преимущественно к Кавказскому заповеднику. В условиях этого заповедника летом кабаны распространены широко, по всем горным поясам, включая альпийский до 2500 м над ур. моря. Однако большая часть поголовья сосредоточена на субальпийских лугах, а в лесных поясах остается небольшое количество. Так, по учетным данным Дурова (1973), в июне, июле и первой половине августа (1968—1970 гг.) в лесном поясе северного макросклона Главного Кавказского хребта на 100 км маршрута встречаемость кабана составила всего 1—2 особи, а в субальпийском поясе — до 95 особей; в среднем за май-август — 70 голов. В лесных поясах южного макросклона за этот же период в среднем на 100 км маршрута встречаемость составила всего 7,3 головы, т.е. почти в 10 раз меньше, чем на субальпийских лугах. Зимой основная масса животных держится в лесных поясах, особенно в тех местах, где имеется обилие плодов бука, дуба и фруктарников. В снежный период года мало
52
Sus scrofa Linnaeus, 1758
кабанов остается в горах выше 1500 м, основное поголовье держится ниже 1200 м. Соответственно диапазон высотных перемещений кабана в обычные годы при урожае орехоносов и фруктарников составляет около 1000—1500 м по вертикали.
Перемещение кабанов с субальпийских лугов в лесные пояса на северном макросклоне обычно происходит во второй половине сентября. На южном макросклоне появление кабанов в предгорных и горных фруктарниках Черноморского побережья, как правило, наблюдается на месяц раньше (во второй половине августа). При неурожае или малом урожае орехоносов и фруктарников животные спускаются еще ниже, в предгорье, дубравы, а при отсутствии желудей в этих местах они вынуждены совершать более продолжительные ми1рации. Большие переходы, вплоть до предгорий, совершаются в многоснежные зимы, как это было неоднократно с 1948 по 1954 г. В отдэе-стностях Гузерипля в январе 1950 г. температура достигала -34,8”, а снеговой покров доходил до 106 см. Животные, фактически лишенные корма, мигрировали далеко, даже заходили в населенные пункты предгорий и нередко погибали в большом количестве. Аналогичное наблюдалось и в Ставропольском крае (Двойченко, 1955; Котов, Рябов, 1963).
Обратное перемещение из мест зимовок на субальпийские луга наблюдается в мае-июне. Животные перемещаются постепенно вверх, поедая вытаивающие прошлогодние орешки бука и зеленую растительность по мере их появления. Они задерживаются в пределах лесных поясов 2—3 недели и лишь в конце июня широко заселяют субальпийские луга. Мигрируют обычно по определенным маршрутам, но при вмешательстве человека, особенно браконьерстве, кабаны меняют маршрут и диапазон миграции.
На Центральном Кавказе и в Закавказье сезонные митрации кабана, по данным А.А.Насимовича (1955), в существенных чертах мало чем отличаются от сезонных перемещений, наблюдаемых на Западном Кавказе. Так, в бассейне Терека такие перемещения имеют протяженность до 60 км. Аналогичные данные получены нами по Кабардино-Балкарии. В Закатальском и Лагодехском заповедниках кабаны держатся летом до 2000—2200 м над ур. моря, при хорошем урожае орехоносов и обычном снеговом покрове зимой находятся в лесном поясе довольно высоко. В снежные зимы (1950 г., февраль) кабаны вынуждены были спуститься в долину Алазани. Подобное явление отмечалось также в Западном Закавказье. В снежную зиму 1879/80 г. в результате переселения с гор в Колхидскую низменность количество кабанов сильно возросло в окрестностях Сухуми и Батуми (Чернявский, 1879 г.; Бочарников, 1880 г., цит. по: Насимович, 1955). Сезонные кочевки в прошлом наблюдались также в Талышских горах (Бекашенко, 1902 г., по: Насимович, 1955). С этих гор при выпадении там больших снегов кабаны в массе спускаются в низменность, достигая приморских тростников. После зимовки в этих местах кабаны весной вновь возвращаются в горы. Протяженность митрации составляет не менее 30 км.
Как видно, высотные митрации кабана на Кавказе — явление обычное, обусловленное в первую очередь экологическими факторами (пища, снеговой покров). Однако каких-либо существенных перемещений кабаны, живу
53
Кабан
щие в тростниках Приазовья, Прикаспия и дельтах рек, впадающих в моря, не совершают. Они ведут оседлый образ жизни, особенно в наши дни в связи с их изоляцией сельхозугодьями от горных популяций.
Логова, купальни. В условиях Кавказского заповедника для строительства логовищ избираются места чаще всего (40 из 70 найденных логовищ) у стволов пихт, которые обеспечивают надежную защиту от снега и дождя (Донауров, Теплов, 1938). Нередко можно встретить логово в густых зарослях рододендрона, папоротника, в чащах, в камышах, высокотравье, самшите, в. буреломе и других укромных местах. По Диннику (1910), свиньи, особенно каба-ны-одинцы, очень любят забираться на скалы и вообще на каменистые кручи, если по ним растет лес или кустарник. Такие лежки он очень часто находил зимой. Лежки нередко располагались на узких карнизах, над и под которыми громоздились отвесные скалы.|Устройство лежек изменчиво, наиболее сложное строение имеет выводковая, при которой в углубление, в земле натаскана трава, листья, хвоя или ветки, и внутри этой кучи образуется ‘Тнездо", иногда прикрытое сверху ветками с листьями. Длина лежки около 2 м, ширина 1 м, высота 0,5 м. Аналогичные лежки строят также одиночные самцы и супоросные самки. В теплую и сухую погоду групповые лежки очень простые — в рыхлой и голой земле делаются углубления, в них кабаны отдыхают и спят. В сырую или снежную погоду они сгребают в кучу рылом и конечностями листья, ветки, бурьян, хвою и в них ложатся] Такие лежки зимой чаще попадаются на хорошо прогреваемых склонах. Наши и литературные данные говорят о том, что|одну и ту же лежку кабаны используют подряд несколько раз очень редко^ Обычно они их строят ежедневно, однако при сильных морозах и глубоком снеге кабаны не могут часто менять свои лежки, j
Групповые лежки делают семейные объединения, но старые кабаны вместе со свиньями никогда не ложатся. Лежки могут быть расположены в кучах и скирдах сена, соломы и всевозможных отходов во время уборки зерновых. I
Купальнями для кабанов служат грязные ямы, илистые источники, лужи на лесных дорогах, заболоченные берега водоемов и т.п. В чистых водоемах они практически не купаются. Потребность в приеме грязевых ванн большая, особенно в теплый период года и во время гона. Однако и зимой взрослые кабаны купаются. Обычно они принимают ванну вечером, редко днем. Излюбленные ванны кабаны посещают регулярно. После купания они чешутся о стволы деревьев, и вблизи ванн можно обнаружить постоянные чесалки — деревья, лишенные значительной части коры на высоте роста кабана. Ванны и чесалки нередко соединены тропами кабанов.
Суточная активность. Поведение (рис. 12). На образ жизни кабана глубокий отпечаток накладывают погодно-климатические и антропогенные факторы. В теплый период года и при отсутствии преследования со стороны человека кабаны бродят не только ночью, но и в течение дня. Подобное явление отмечалось как в прошлом, так и в настоящее время. В субальпийском поясе и по верхней границе лесов, где редко бывает человек, мы не раз регистрировали пасущиеся утром и вечером небольшие группы кабанов. Вероятно, в полуденные часы они уходят в укромные места на свои лежки и
54
Sus scrofa Linnaeus, 1758
Рис. 12. Кабан
отдыхают там. Как справедливо отмечает Н.Я.Динник (1910), в туманную и дождливую погоду кабаны могут бродить по пастбищу целый день. В тех местах, где воздействие антропогенного фактора сильно выражено, кабаны покидают свои лежки лишь после наступления темноты, иногда даже полуночи, а с рассветом вновь возвращаются в свои убежища, малодоступные для человека (густые заросли кустарников, глухие места ущелий и балок с вы-сокотравьем, лесами, камышовые крепи лиманов и т.д.). Зимой короткое время дня ори проводят в этих же убежищах. В горах они любят забираться на скалы, каменистые кручи с древесно-кустарниковой растительностью и отдыхать там. По данным Н.Я.Динника (1910), почти всю длинную зимнюю ночь кабаны и свиньи проводят на ногах, ходят быстро. Экспериментальные исследования показали коррелятивную зависимость двигательной активности кабана от температуры окружающей среды: понижение температуры приводит к повышению двигательной активности (Blasetti et al., 1988).
55
Кабан
Кабан — очень чуткое животное, с особенно хорошо развитыми обонянием и слухом. Опытные охотники знают, что запах человека, идущего по ветру, он чует на расстоянии сотни метров. Однако в ненастную погоду иногда удается подойти к нему почти вплотную (Котов, Рябов, 1963). С помощью тонкого чутья самец узнает и находит самку, готовую к спариванию. Не только у взрослых, но и у малышей хорошо развито обоняние — благодаря ему они узнают своих матерей.
У кабана плохо развито зрение; не замечая неподвижно стоящего человека, он может близко подойти к нему.
Кабаны очень заботятся о потомстве, начиная с гнездового периода до перехода молодняка к самостоятельной жизни. У них наблюдается высшая форма проявления инстинкта материнства — новорожденные могут кормиться молоком другой матери, оставшиеся сиротами поросята могут быть приняты лактирующей самкой.
В отличие от других копытных у кабанов наблюдается сложная социальная организация. Структурированность популяции очень высокая, выражены все ранги — от одиночек до семейных групп и групповых объединений. Это определяется физиологическим состоянием, половой принадлежностью, возрастом, кровным родством, погодой, состоянием кормовой базы. Четко выражена территориальность, широко меняющаяся в зависимости от тех же факторов. О них подробнее будет сказано в соответствующих разделах.
Линька. Судя по литературным данным (Динник, 1910; Козло, 1975), сезонная смена волос кабана проходит в разных местах европейской части б. СССР в близкое время. Ее начало приходится на последнюю декаду марта или на начало апреля. В конце мая или начале июня животные фактически оголяются и становятся уязвимыми для кровососущих насекомых. Они спасаются от докучливых насекомых в густых зарослях кустарников, папоротника, в грязевых ваннах, под кучами хвороста и т.д. Линька задерживается у ослабевших животных — подранков, проболевших и др. Обычно в конце июня кабаны уже покрываются редкой щетиной длиной всего около 2 см, светлой, буровато-серой окраски, многие волосы остаются светлыми и придают животному светлый цвет. Грубые волосы растут быстро, уже в начале августа они имеют длину около 4 см, а в сентябре уже достаточно длинные, мало отличимые от зимнего наряда. Длина остистых волос в октябре из области холки у животных с Центрального Кавказа в Нашей коллекции составляет 11—13,7 см. К этому времени вырастают пуховые волосы. В условиях Беловежской Пущи новый волосяной покров достигает полного развития в октябре-ноябре (Козло, 1975).
Последовательность смены волос отображена на рис. 13 (Козло, 1975). Как правило, замена старых волос начинается одновременно ца морде, за ушами, у основания хвоста, в области скакательных суставов, затем охватывает грудь, брюхо, конечности, а в последнюю очередь бока, огузок, шею.
Питание. Кабан — всеядное животное. Круг его пищевых объектов включает надземные и подземные части растения, плоды орехоносных деревьев и фруктарников, листья, побеги и даже кору деревьев и кустарников, всевозможные сельскохозяйственные культуры, особенно корнеплодные и куку
56
Sus scrofa Linnaeus, 1758
рузу; почвообитающих беспозвоночных, в частности личинок насекомых, дождевых червей, различных позвоночных, в том числе норных мелких млекопитающих. Кабан не брезгует падалью, пищевыми отбросами, имеет место каннибализм.
Анализ литературных данных и наших наблюдений свидетельствует о том, что основу питания кабана на Кавказе, как и в других частях ареала, составляют растительные корма. Список видов травянистых растений, включающий на Западном Кавказе до 37 видов, приводят С.С.Донауров и В.П.Теплов (1938). По В.В.Дурову (1987), этот список включает 197 видов растений и 16 видов животных кормов. Из поедаемых растений чаще встречаются 40 видов, из которых предпочитаются иваш-чай, орляк крымский, мытник Нордманна, лабазник шестилепестный, ежа сборная, бубенчик, порезник, борщевик понтийский,
тмин кавказский, мятлик длиннолистный, тростник обыкновенный. В Беловежской Пуще установлено, что кабан поедает около 100 видов растений, в том числе 73
Рис. 13. Последовательность линьки у кабана (штриховкой показаны облинявшие участки тела) (Козло, 1975)
травянистых и кустарниковых растений,
13 видов деревьев и кустарников, 10 видов культурных растений. Среди растений, поедаемых кабаном, кроме ценных кормовых, имеются ядовитые и лекарственные растения, малокалорийные и почти балластные, т.е. набоо
растений характеризует кабана как зверя очень пластичного в отношении
пищи.
Растительные корма поедаются в течение года, особенно их роль велика в тех многочисленных районах Кавказа, где сильных морозов нет и часто встречаются растения, вегетирующие, в течение всего года.
Список видов кормов животного происхождения, судя по всем данным, не меньше, чем растительных. Два-три десятка видов животных, приводимых в литературе, можно удвоить, утроить. Трудно назвать животных, доступных кабану, которые бы им не поедались. К числу поедаемых относятся многие почвообитающие животные (от личинок насекомых и дождевых червей до всевозможных норных грызунов), птицы, гнездящиеся на земле, пресмыкающиеся, земноводные и др. В столь разнообразных природных условиях, какие имеются на Кавказе, мы еще плохо представляем, насколько существенную роль могут играть "нетрадиционные" корма в жизни кабана. Об этом свидетельствуют многие литературные данные, обзор которых приводят О.С.Русаков и Е.К.Тимофеева (1984). Однако это не исключает, что списки видов растений и животных, наиболее употребляемых кабаном в разных
57
Кабан
частях ареала, в целом сходные и включают ограниченное количество видов. При этом соотношение растительных и животных кормов не меняется принципиально — животные корма поедаются кабаном регулярно и часто во всех частях ареала, но в небольших количествах, в пределах 2—9% всей их пищи (Козло, 1975; Henry, Couley, 1972; Schaffer, 1979, по: Русаков, Тимофеева, 1984).
О суточной норме пищи кабана в условиях Кавказа данных мало. Видимо, она меняется широко не только в связи с возрастом и физиологическим состоянием организма (самцы во время гона фактически не питаются), но зависит и от пола животного, сезона года, ландшафтных условий обитания, наличия и характера пищи и т.д. При хорошем урожае бука один кабан в среднем потребляет около 3 кг буковых орешков в день (Дуров, 1987). С.С.Донауров и В.П.Теплов (1938) отмечают, что осенью во время жировок кабан съедает за сутки около 1000 желудей. П.Генов (1981) полагает, что в среднем взрослый самец в сутки съедает 4,3 кг пищи, а самка — 3,0 кг. В Беловежской Пуще, по Т.Б.Саблиной (1955), вес содержимого желудка кабанов колеблется от 0,1 до 7 кг, в Кавказском заповеднике, по В.В.Дурову и В.Н.Александрову (1968), у самцов — 3,15—8,0 кг (х = 4,5), самок — 2,0— 5,0 кг (х = 3,3). С.С.Донауров и В.П.Теплов (1938) считают, что для наполнения желудков взрослых кабанов требуется 2—3 кг пищи.
Кормовая база кабана на Кавказе, на наш взгляд, хорошая на большей части ареала, и животные изымают незначительную часть природных кормов. По расчетным данным В.В.Дурова (1987), в годы урожая кабан питается буковыми орешками до 240 дней, и на одну голову требуется до 720 кг этих плодов. Если допустить, что плотность кабанов достигает 40—45 особей на 1000 га, то они могут использовать 3,8% урожая плодов орехоносов (при урожае 780—1500 кг на 1 га и сохранении орешков до апреля). Дикие фруктовые деревья дают урожай от 600 до 2500 т на 1000 га, и кабаны съедают в среднем 10%. Словом, в урожайные годы на Кавказе достаточно богата кормовая база и кабаны нагуливают жир очень быстро; в конце осени они имеют уже толстый жировой слой.
Зимний период на Кавказе обычно теплый. Если и бывают сильные морозы и мощный снеговой покров, то они осложняют нахождение и добычу корма кабанам обычно на непродолжительное время. Как правило, кабаны в достаточном количестве находят в течение зимы желуди, буковые орешки, всевозможные корневища, особенно папоротников и ДР- При этом на кормежках кабаны могут использовать объекты питания на 66—87% (злаки на 36—55%), наиболее полно — иван-чай, орляк крымский, мытник, лабазник, ежевику. В очень холодные зимы на лесных полянах южных склонов корневища злаков съедаются на '60—80%. На субальпийских и альпийских лугах корни поедаемых растений используются на 35—92% (Дуров, 1987).
Питание кабана подвержено широкой изменчивости по сезонам года, что объясняется соответствующими изменениями во всех районах Кавказа, в том числе субтропических. В весенне-летний период в питании кабана доминируют зеленые части растения, что зарегистрировано почти во всех частях ареала. Хотя зеленые растения поедаются круглый год, наибольшее значение
58
Sus scrofa Linnaeus, 1758
они имеют в конце весны и летом, в течение 4—5 мес. Наряду с этим кабан поедает много корневищ, клубней, луковиц, а также сохранившиеся плоды бука, дуба, каштана, ореха и др. Ведущую роль в питании кабана вегетативные части растений играют на равнине, особенно в условиях камышовых и тростниковых зарослей Приазовья и Прикаспия, а в горных лесах их заменяют плоды орехоносных деревьев.
Как на равнине, так и в горах весной кабан добывает животный корм, особенно почвообитающих животных. Нередко можно встретить разрытую кабаном землю в лесах, по опушкам, на полянах, где кабан добывал себе пищу. На таких копанках мы часто находили подстилку гнезд грызунов и их остатки. По мере созревания фруктов в питании кабана заметную роль начинают играть плоды — сначала созревшая дикая черешня, затем алыча, потом яблоки, труши, кизил и др. По возможности звери отбирают зрелые плоды. В поисках таких плодов кабаны могут совершать значительные переходы. В конце сентября — октябре созревают желуди и становятся основным кормом. Когда созревают буковые орешки, они приобретают ведущую роль в питании кабана в горных лесах всего Кавказа.
Осенью и зимой кабаны охотно посещают сельскохозяйственные поля, особенно кукурузные. Эта культура относится к их излюбленным кормам. По Н.Я.Диннику (1910), они с жадностью набрасываются на гречиху, просо, охотно пожирают пшеницу, ячмень, но картофель кабаны не любят и нс едят. В настоящее время от кабанов сильно страдают и картофельные плантации. За рубежом ставятся опыты по выявлению менее привлекательных для них сортов'с целью сохранить урожай. Во всяком случае, в предгорных и горных районах Кабардино-Балкарии от кабанов приходится спасать посевы картофеля не менее, чем посевы кукурузы. В течение зимы и ранней весны при урожае орехоносов в пищевом рационе кабана ведущая роль принадлежит плодам бука, дуба, каштана, грецкого ореха, лещины, а также корням и вегетирующим частям растений. Почвообитающие беспозвоночные зимой из-за низкой температуры почвы мигрируют глубже и бывают мало доступны кабанам. Во многих районах Кавказа'грызуны становятся их жертвами в течение всего года.
При отсутствии концентрированных кормов в виде плодов орехоносов кабаны вынуждены удовлетворяться второстепенными кормами — корневищами различных трав и кустарников{ ветками и зелеными листьями, грибами, омелой и др. К вынужденным кормам относятся сухйе трава и листья, мхи, всевозможные отходы полевых культур и т.д. Если такой неурожай осложняется многоснежьем, то кабаны оказываются в критических условиях, что вызывает гибель большого количества животных, в первую очередь молодняка. Спасти их можно лишь искусственной подкормкой.
Воспроизводство.’и. структура популяции
Размножение. Сведения о воспроизводстве популяции в условиях Кавказа содержатся в ряде региональных^. работ, но наиболее полные у Н.Я.Динника (1910), ЕЛ.Маркова (1932), В;В.Логинова (1936), С.С.Донаурова и В.ПЛепло-
59
Кабан
ва (1938), В.В.Дурова (1987). Исследованиями охвачены как Северный Кавказ, так и Закавказье, но большая часть работ посвящена Западному Кавказу, преимущественно популяции Кавказского заповедника.
В соответствии с названной литературой и нашими полевыми наблюдениями гон кабана растянут и занимает 2—3 мес (ноябрь-январь). Вероятно, это явление видоспецифичное, о чем свидетельствует информация из различных зон Европы (Русаков, Тимофеева, 1984), из Беловежской Пущи (Козло, 1975), черноземных районов РФ (Сухорослов, 1972); согласно им продолжительность гона в названных районах определяется, как правило, также 2—3 мес. Все имеющиеся данные говорят и о том, что календарные сроки могут быть смещены в ту или другую сторону в зависимости от внешних и внутренних факторов (упитанность, состояние кормовой базы, климатических условий и т.д.). При оптимальном состоянии организма и среды сроки гона резко сокращаются. В обычных условиях гон на Северном Кавказе наступает раньше, в первых числах ноября, что связано с более ранним похолоданием, а в Закавказье, где дольше держится теплая погода, заметно позже — с конца ноября по конец декабря, захватывая часть января (Марков, 1932). Н.Я.Динник (1910) сообщает, что течка у самок кабана в Боржоми наблюдалась в январе, но это сообщение, вероятно, относится не к началу, а больше к концу гона. Более того, в пределах одного только Кавказского заповедника сроки начала размножения могут сильно расходиться на его различных участках — в одном и том же 1925 г. гон кабана начался на северном участке 5 ноября, а на восточном — 5 декабря (Донауров, Теплов, 1938).
{Начало размножения знаменуется многими биологическими явлениями, наблюдаемыми в различных популяциях и разными исследователями. Например, самцы становятся очень возбужденными, активно разыскивают и отвоевывают в жестокой борьбе самок, метят территорию, занятую ими (делают засечки на стволах деревьев, зачесы с оставлением секрета кожных желез и др.), силой изгоняют более слабых самцов из стада и с территории группы, на протяжении всего периода гона не принимают пищу, преследуя самок и побуждая их к спариванию, издают хриплые протяжные и низкие звуки. Драки между самцами настолько жестоки, что могут кончаться смертельным исходом для одного из соперников, tea время гона самцы сильно худеют и теряют до 20% массы тела. Как адаптация к схваткам у секачей сильно развивается калкан из очень прочной соединительной ткани в области шеи и груди толщиной 2—3 см. Из физиологических сдвигов характерна гиперфункция кожных желез, запах секрета которых человек может воспринимать на значительном расстоянии. Высокая активность самцов сопровождается выделением пота, слюны изо рта, частым питьем воды, купанием в холодных лужах. При этом кабаны становятся менее осторожными. Происходит увеличение семенников и соответствующих желез в 1,5—2 раза по сравнению с периодом покоя (Козло, 1975).
В жизни самок в этот период происходит также немало изменений: растут группы, включающие обычно близкородственные семьи, из стада изгоняются сеголетки, которые жили с ними до начала гона (и возвращаются в свои
60
Sus scrofa Linnaeus, 1758
семьи после завершения гона), самки уединяются в более глухие и защищенные места, куда самцы привлекаются специфическим запахом. Между самцами происходят схватки, и победитель формирует гарем. Он находится при гареме до оплодотворения всех самок, готовых к спариванию (обычно в течение 2—3 недель), после этого он покидает гарем и переходит к другим труппам самок. Физиологическая готовность самок к спариванию длится всего 2 дня в году, и такую самку секач узнает по запаху, выделяемому ею. Яловость самок кабана — явление частое, иногда охватывающее, видимо, до 50% самок (Донауров, Теплов, 1938).
Беременность длится около 4 мес (Динник, 1910; Донауров, Теплов, 1938; Дуров, 1987), по другим авторам — колеблется от 112 до 140 дней, притом у молодых она короче, чем у старых (Соколов, 1979; Kenneth, 1943, по: Русаков, Тимофеев, 1984).
Перед опоросом одна, иногда две или три самки вместе устраивают гнездо, которое "своей формой напоминает гнездо птиц" (Динник, 1910, с. 53), и в нем рождаются детеныши. Появление поросят на Западном Кавказе отмечалось с 10—15 марта до конца мая в зависимости, как и сроки спаривания, от комплекса внутренних и внешних факторов.
Плодовитость. Все ее параметры подвержены широкой изменчивости. Так, в конце XIX и начале XX в., когда в популяциях встречались нередко вполне взрослые кабаны, количество эмбрионов на одну самку, по Н.Я.Диннику (1910), колебалось от 5 до 10, в среднем 7,6 (л = 8). В 30-е годы текущего столетия в связи с массовым браконьерством произошло омоложение популяции, количество эмбрионов на самку резко снизилось, составляя от 3 до 7, в среднем 4,4 на самку (Марков, 1932; Логинов, 1936; Донауров, Теплов, 1938). В настоящее время в условиях Кавказского заповедника самки кабана приносят от 1 до 11 поросят, обычно 4,9—5,9 (в среднем 5,1) (Дуров, 1987). Эти данные, как видно, согласуются с мнением о том, что у кабана четко выражена возрастная дифференциация плодовитости. На эту особенность обращали внимание почти все вышеупомянутые авторы. При этом, по мнению В.В.Дурова (1987), плодовитость самок увеличивается до возраста 8 лет, а П.Г.Козло (1975) считает оптимальным возрастом размножения кабана от 4—5 до 9—10 лет. На число эмбрионов у беременной самки большое влияние оказывает упитанность, и соответственно оно бывает разным в урожайные и неурожайные годы ведущих кормовых растений, особенно орехоно-сов, а также в зависимости от характера зимы и доступности кормов, состояния популяции (период роста или спада численности) и многих других факторов.
Плодовитость кабана широко меняется также в географическом плане, в том числе в широтно-долготном направлениях. Обзор соответствующей литературы содержится у О.С.Русакова и Е.К.Тимофеевой (1984).
Максимальное количество детенышей в помете, как правило, не превышает 10, что соответствует количеству сосков у самки кабана. Как редкий случай, находили беременную самку с 14 нормально развитыми эмбрионами (Русаков, Тимофеева, 1984).
6 1
Кабан
Рост и развитие. В условиях Кавказа эмбриональный период онтогенеза нс изучен, но по другим регионам страны (в том числе в европейской части) опубликован целый ряд работ (Курносов, Петрищев, 1977; Петрищев, 1980; Петрищев, Бородин, 1985; и др.). Выявлено, что в зародышевом периоде, продолжающемся 22 сут, масса зародыша увеличивается в 555 раз (с массы зиготы в 0,00045 до 0,21 г); в следующем — предплодном — периоде (23—38-е сутки) она увеличивается в 26 раз, в третьем — плодном — периоде (39— 114-е сутки) — в 146 раз. Как видно, увеличение массы эмбриона особенно интенсивно в первом и третьем периодах. Неравномерный характер носит также линейный рост эмбриона и пропорций тела (индексов тела), что обусловлено различиями в темпе увеличения отдельных частей тела. По сравнению с домашними свиньями этапы интенсивного роста эмбриона дикого кабана сдвигаются на более ранние сроки.
Постнатальный период роста и развития кабана в условиях Азербайджана изучен М.А.Сафаровым (1965). Его данные (табл. 14), как и данные других авторов, свидетельствуют о том, что интенсивность роста молодняка хотя и ниже, чем в плодном периоде, на порядок величин, но поросята растут довольно быстро. Тем не менее в отличие от других копытных они могут следовать за матерью лишь через неделю после рождения. Вероятно, слишком малые размеры новорожденных (1,5% от массы тела матери), несмотря на хорошо сформировавшийся вид, выступают основной причиной некоторой задержки активного передвижения. Быстрый рост поросят обеспечивается высококачественным молоком свиньи, содержащим большое количество белка (5,0—7,6%), лактозы (3,6—5,6%), фосфора, кальция, различных витаминов и кислот (Грачев,.Галанцев, 1973). При этом кормление поросят молоком продолжается в течение 3—4 мес. Хотя поросята в первые дни жизни теряют в массе, но уже к трех-четырехмесячному возрасту их масса возрастает с 0,6—1,15 кг у новорожденных до 14 кг, т.е. увеличение массы происходит более чем в 17 раз. К восьми-девятимесячному возрасту масса тела составляет 45—50 кг, а на втором году жизни отдельные хорошо упитанные самцы могут достигать 70 кг, самки — 60 кг. Вполне взрослые имеют массу до 142 кг у самцов и до 98 кг у самок. Увеличение размеров тела идет менее интенсивно, чем массы. Длина тела и высота в холке к десятимесячному возрасту увеличиваются в 3 раза, длина головы — в 2,5 раза, обхват труди — в 3,5 раза. В последующем до взрослого состояния темп роста замедленный, соответствующие показатели увеличиваются всего в 1,5 раза (табл. 15). По другим районам ареала, в том числе по Северо-Западу б. СССР, получены аналогичные данные — увеличение линейных размеров и массы тела происходит наиболее интенсивно до 2 лет, а затем с каждым годом снижается (Русаков, Тимофеева, 1984).
Возрастная динамика пропорций тела иная. Так, отношение длины тела к высоте в холке со дня рожздения до десятимесячного возраста увеличивается, после этого возраста рост длины тела отстает от роста высоты в холке. У взрослых самок продолжается рост относительной длины тела. Относительная длина головы самцов уменьшается с возрастом, у самок то же самое на-
62
Sus scrofa Linnaeus, 1758
Таблица 14. Соотношение развития органов поросят в условиях Азербайджана _______________________ (Сафаров, 1965)_________, ________________
Показатель Поросята 5—10 дней Поросята 3—4 мес.
самцы (п =2) самки самцы (п=2)
X limit — X limit
Чистый вес, кг* Вес отдельных органов, % по отношению к чистому весу 1,1 1,1—1,2 0,595 12,7 12,2—13,3
Кожа 18,8 18,6-19,0 15,4 13,3 12,0—14,7
Мускулатура и скелет — — — — —
мускулатура — — — — —
сырой скелет — — — — —
Кишечник 3,76 3,72-3,81 5,04 5,23 5,2-5,26
тонкий отдел 2,49 2,45-2,54 3,02 2,43 2,52-2,62
толстый отдел 0,63 0,63-0,63 0,6 1,67 1,47-1,88
Голова (без кожи, с языком и головным мозгом) 14,95 14,9—15,0 18,4 9,42 9.02-9,83
Язык 1,14 1,09-1,18 1Д7 0,86 0,82-0,90
Головной мозг 4,24 4,22-4,27 7,73 0,94 0,90—0,98
Сердце 1,02 1,0-1,04 1,17 0,76 0,67-0,86
Легкие (без трахеи) 2,36 2,36—2,36 2,15 1,88 1,88-1,88
Желудок 0,63 0,63—0,63 0,5 1,11 1,1-1,12
Печень 3,31 3,27-3,36 3,36 2,48 2,33-2,63
Почки 1,0 1,0-1,0 0,92 0,61 0,56-0,67
Объем желудка (л), % к чистому весу 2,81 2,72-2,90 — 6.07 6,0—6,14
Длина кишечника Длина отделов кишечникв, % к общей его длине 11,5 11,4-11,7 11,4 15,1 14,8-15,4
тонкий отдел 79,9 79,8—80,0 81,7 81,7 81,5-81,9
толстый отдел *Без содержимого желудка и кишечника 20,1 20,0—20,2 18,3 18,3 18,5-18,5
блюдастся до 2 лет, но потом этот показатель возрастает. До 3 лет уменьшаются относительные показатели обхвата груди, но потом увеличиваются.
Возрастные изменения отражены в табл. 14. Как видно, у самцов с возрастом уменьшается относительная масса кожи, тонкого отдела кишечника, головы, языка, головного мозга, сердца, печени, почек. Общая масса и объем желудка, масса кишечника и толстого отдела увеличиваются до семи-десятимесячного возраста. Динамика изменения показателей у самок несколько иная (см. табл. 15).
Зубная система кабана формируется в течение длительного периода онтогенеза, что показано в табл. 13. Для экономии места ограничимся лишь некоторыми выводами (Сафаров, 1965). До шестимесячного возраста вырастают все молочные зубы. Их смена на постоянные начинается с 6 мес и заканчивается в конце третьего года жизни. Последовательность смены: вначале сменяются клыки, потом третьи резцы, первые резцы, третьи, вторые и
6 3
Кабан
Таблица 14 (окончание)
Поросята 3—4 мес Поросята 7—10 мес
Показатель самки (п =3) самцы (л =12) самки(л =4)
х limit X limit X limit
Чистый вес, кг* Вес отдельных органов, % по отношению к чистому 12,2 12,0-12,4 39,6 31,9-53,5 33,6 28,6-39,5
весу Кожа 15,4 13,3-17,0 9,84 6,84-14,0 10,6 9,76-11,2
Мускулатура и скелет 63,2 61,2-65,0 58,1 46,4—65,5 56,2 52,4—60,7
мускулатура — — 49,9 49,7-53,2 — —
сырой скелет — — 14,7 12,3-17,2 — —
Кишечник 6,49 6,25-6,69 4,13 3,01-5,17 4,76 4,1-5:24
тонкий отдел 3,07 2,91-3,38 1,4 0,93-1,75 1,58 1,26-1,74
толстый отдел 2,07 2,01-2,18 1,67 1,18—2,01 1,94 1,77-2,16
Голова (без кожи, с языком и головным мозгом) 9,75 9,67-9,83 7,66 4,75-9,77 9,37 6,7-11,8
Язык 0,84 0,8-0,87 0,33 0,25-0,37 0,33 0,32-0,34
Головной мозг 0,94 0,92—0,97 0,35 0,31-0,38 0,37 0,34-0,4
Сердце 0,82 0,79-0,87 0,61 0,48—0,65 0,63 0,51-0,73
Легкие (без трахеи) 2,28 1,83-2,62 2,04 1,45-2,35 2,34 2,09—2,46
Желудок 1,32 1,25-1,42 1,06 0,78—1,44 1,23 1,06-1,38
Печень 2,8 2,58-3,08 1,74 1,33-2,6 2,39 1,46—3,14
Почки 0,66 0,61—0,72 0,39 0,31-0,62 0,43 0,35—0,62
Объем желудка (л), % к 6,4 6,09-6,66 3,92 2,8-5,1 4,87 4,43-5,38
чистому весу Длина кишечника Длина отделов кишечники, % к общей его длине 14,2 13,9-14,5 13,7 11,5-15,5 14,3 13,7-14,6
тонкий отдел 82,1 81,8-82,6 76,2 71,0-81,3 76,2 75,8-76,6
толстый отдел 17,9 17,4-18,2 23,7 18,7—29,0 27,3 23,4—24,2
*Без содержимого желудка
и кишечника
четвертые предкоренные и в последнюю очередь вторые резцы. Первые
предкоренные (Р11) не имеют молочных предшественников и соответст-
венно не меняются. Смена зубов на нижней челюсти происходит быстрее на 2—3 мес, чем на верхней. Коренные зубы начинают прорезываться в пяти
шестимесячном возрасте и заканчивают свое развитие не раньше трехлетнего
возраста.
Рост и развитие черепа происходит своеобразно, существенно отличаясь от возрастной динамики размеров и массы тела. Данные по Кавказскому заповеднику, Кабардино-Балкарии и Азербайджану (см. рис. 7—11) свидетельствуют о том, что интенсивный рост черепа происходит лишь до 3—4 лет, а затем практически прекращается, имеют место даже регрессивные изменения в связи со старостью. Вероятно, на севере ареала рост черепа происходит дольше — до 5 лет на Северо-Западе б. СССР (Русаков, Тимофеева, 1984). Исключением следует считать массу черепа и размеры клыков, которые увеличиваются до старости.
64
Таблица 15. Промеры и пропорции скелетов по возрастам (Сафаров, 1965)
Признак Поросята 5—10 дней Поросята 3—4 мес.
самцы (и =2) самки (г=1) самки (п =3)
X limit — X limit
Размеры костяка, мм Длина черепа 100,0 100,0—100,0 88,0 205,0 203,0-208,0
Длина позвоночника (без 179,5 179,0-180,0 142,0 551,6 536,0—572,0
хвостового отдела) Длина конечностей* передней 150,0 150,0-150,0 122,0 359,6 357,0-364,0
задней 127,5 127,0-128,0 101,0 321,0 318,0-325,0
Длина некоторых частей скелета, % к длине позвоночника без хвостового отдела Череп 55,6 55,5-55,8 61,9 37,9 35,5—38,8
Передняя конечность 83,5 83,3—83,8 85,9 65,9 63,6-65,7
Задняя конечность 69,3 67,7-70,9 71,1 58,1 56,8-59,3
Отдел позвоночника шейный 19,5 19,4-19,6 17,0 18,6 18,4-19,6
трудной 41,8 41,7±41,9 42,2 44,2 43,5—44,8
поясничный 19,2 18,3-20,1 19,7 19,7 18,8-19,4
крестцовый 19,5 18,4-20,6 16,9 16,7 16,1-17,2
Тазовая кость 29,5 29,4-29,6 30,3 25,9 25,0-26,5
Длина звеньев передней конечности, % ко всей ее длине Лопатка 30,0 30,0-30,0 29,5 30,0 30,2—30,4
Плечевая кость 35,3 35,3—35,3 35,2 31,9 31,3-32,4
Лучевая кость 25,3 25,3-24,3 25,4 24,0 23,8—24,2
Пястная кость 9,3 9,3-9,4 9,9 13,7 13,2-14,5
Длина звеньев задней конечности, % ко всей ее длине Бедро 43,1 42,9-43,3 41,6 42,8 42,4-43,1
Большая берцовая 41,1 40,9-41,4 40,6 39,0 38,4-39,4
Плюсна 15,7 15,7-15,8 17,8 18,1 17,5-19,2
Ширина некоторых костей, % гл всей ее длине Череп 50,5 50,0-51,0 48,8 44,7 44,2—45,3
Лопатка 57,7 55,5—60,0 55,5 60,7 65,5-60,9
Плечевая кость 9,4 9,4—10,4 9,3 8,7 8,5-8,9
Лучевая кость 13,1 13,1-13,2 12,9 13,4 12,9—13,9
Пястная кость — — — 15,5 13,4-16,6
Бедро 9,1 9,1-9,1 9,5 7,9 7,8-8,1
Большая берцовая кость 10,4 9,4-11,5 12,5 10,3 10,1-10,6
Плюсна — — — 13,2 12,3-14,2
Таз в моклоках 66,4 — 69,7 64,7 62,9-62,3
в тазобедренных 45,3 — • 41,8 48,9 47,8—49,6
сочленениях в седалищных бухрах 32,0 — 32,6 35,6 34,2-37,5
*Ввиду отсутствия фаланг в некоторых скелетах за длину передней конечности взята длина лопатки + плечевой + лучевой + пястной костей, а за длину задней конечности — длина бедра + большой берцовой и плюсны.
5 В.Е.Соколов, А.К.Темботов
65
Кабан
Таблица 15 (окончание)
Поросята 7—10 мес. Подсвинки
Признак самцы (п =2) самцы (о =2) самки
(П=1)
X limit X limit —
Размеры костяка, мм Длина черепа 285,0 — 338,5 330,0-347,0 344,0
Длина позвоночника (без хвостового отдела) Длина конечностей* 788,0 757,0-820,0 881,5 868,0-895,0 -—
передней 540,5 498,0-583,0 600,0 576,0-624,0 589,0
задней 479,0 447,0-511,0 528,0 511,0-545,0 515,0
Длина некоторых частей скелета, % к длине позвоночника без хвостового отдела Череп 37,6 38,3 38,0-38,7
Передняя конечность 68,4 65,7-71,1 68,0 66,3-69,7 —
Задняя конечность Отдел позвоночника 60,6 59,0-62,3 59,8 58,8-60,9 —
шейный 19,9 19,2—20,7 19,1 19,0-19,3 —
грудной 43,7 43,6-43,9 43,1 43,0-43,2 —
поясничный 19,9 19,8—20,1 19,6 19,0-20,2 —
крестцовый 16,3 15,3-17,4 18,2 17,3-19,0 —
Тазовая кость 27,3 26,1-28,6 27,3 26,8—27,9 —
Длина звеньев передней конечности, % ко всей ее длине Лопатка 30,1 30,0—30,3 30,3 30,3—30,4 30,4
Плечевая кость 30,6 30,5—30,7 30,5 30,4—30,6 30,6
Лучевая кость 24,6 24,3—24,9 25,1 25,0-25,2 25,1
Пястная кость 14,6 14,6—14,7 14,0 14,0-14,1 13,9
Длина звеньев задней конечности, % ко всей ее длине Бедро 42,6 42,3-43,0 42,6 42,2—43,1 42,9
Большая берцовая 39,2 39,1-39,3 39,3 39,1-39,5 39,6
Плюсна 18,1 17,6—18,6 18,0 17,8-18,3 17,5
Ширина некоторых костей, % ко всей ее длине Череп 39,7 38,7 37,2-40,3 38,7
Лопатка 62,9 59,6-66,3 58,0 56,6-59,4 62,6
Плечевая кость 9,0 8,9-8,1 9,3 8,9-9,7 8,9
Лучевая кость 13,1 12,7—13,6 12,9 12,8-13,1 12,1
Пястная кость 15,9 15,0-16,5 14,2 12,5-16,0 15,8
Бедро 8,2 8,2-8,2 8,7 8,3—9,1 8,6
Большая берцовая кость 9,8 9,4—10,3 9,6 9,3-10,0 9,8
Плюсна 14,9 14,4—15,5 14,2 13,0-15,4 15,5
Таз в моклоках 70,4 70,2-70,7 70,2 68,4-72,1 69,5
в тазобедренных 48,5 47,6—49,5 48,0 47,6—48,5 50,8
сочленениях
в седалищных буграх 43,5 42,4-22,6 41,8 41,2—42,4 44,5
66
Sus scrofa Linnaeus, 1758
Таблица 16. Абсолютные промеры отдельных частей посткраниального скелета ______________________по возрастам (Сафаров, 1965)_____________________
Наименование отделов и костей Новорожденны! Поросята 3—4 мсс. Поросята 10 мес. Взрослые
X limit X limit — X limit
Отделы позвоночника шейный 35,0 35-35 104,0 102-105 170 207,5 200-215
грудной 75,0 75—75 244,6 240-249 360 457,5 450—465
поясничный 34,5 33-36 105,6 101—110 165 206,2 200-215
крестцовый 35,0 33-37 92,0 90-94 125 195,0 190-200
Длина звеньев передней конечности лопатка 45,0 45-45 109,0 108-110 175 247,2 236—248
плечевая 53,0 53-53 115,0 112-118 178 215,5 214-218
лучевая 38,0 38—38 86,3 85-88 145 185,5 183-188
пястная 14,0 14-14 49,3 48-52 85 92,2 90-96
Длина звеньев задней конечности бедренная 55,0 55-55 137,6 135-140 220 263,2 258-267
большая бер- 52,5 52-53 125,3 122-128 201 242,2 241-244
цовая плюсна 20,0 20-20 58,0 56-61 90 100,7 96-106
таз 53,0 53-53 143,0 142—144 235 292,5 287-297
Темп роста отдельных краниологических признаков существенно различается. Наиболее интенсивно из линейных показателей увеличивается длина носовых, лобных и челюстных костей. В основном за их счет происходит увеличение длины черепа. Наиболее низким темпом роста вьщеляется межглазничная ширина; по сравнению с длиной черепа медленно увеличиваются его высота и ширина. Адаптивный смысл быстрого роста носовой части головы легко понять, если учесть, что поросята буквально через неделю начинают выходить из логова и пытаются скусывать зеленые растения. В возрасте 2 мес они уже ведут активный поиск пищи в почве. Половые различия в темпе роста размеров и массы черепа небольшие и не .по всем параметрам заметны.
Посткраниальный скелет растет быстрее, чем череп. Это относится и к конечностям, но они отстают в темпе роста от позвоночника (табл. 16). Словом, линейный рост отдельных элементов скелета в постнатальный период проходит различно — те части, которые у новорожденных достигли высокого уровня развития, растут медленнее, чем не достигшие такого уровня (Сафаров, 1965).
Исследования В.В.Дурова (1987) показали, что в горах Кавказского заповедника на рост и развитие кабанов сильно влияют колебания урожайности плодов. В неурожайные годы кабаны истощаются, замедляется рост и развитие молодняка. В годы урожая масса и размеры сеголеток в течение зимы
5*
67
Кабан
увеличиваются на 4—15%, в годы неурожая масса уменьшается на 47%, а размеры на 4—5%. Условия зимовки в первый год жизни накладывают глубокий отпечаток на дальнейший рост особей более старшего возраста.
Структура популяции. Высокий уровень структурированности популяций кабана регистрировался во всех районах ареала, где проводились исследования биологии этих животных. Общепризнано, что по степени организации популяции кабан выделяется среди других копытных Палеарктики, и это служит видоспецифичной особенностью. Структура популяции кабана на Кавказе обсуждалась в ряде работ (Динник, 1910; Донауров, Теплов, 1938; Дуров, 1987; и др.).
На основании наблюдений в горах Кавказского заповедника В.В.Дуров (1987) выделяет семь, по его мнению устойчивых, социальных единиц: 1) самки взрослые с поросятами; 2) самцы взрослые; 3) самки взрослые с поросятами и подсвинками; 4) самки взрослые с подсвинками; 5) самки взрослые без молодняка; 6) подсвинки; 7) поросята. Семейная ipynna образуется из самцов и гуртов (самки с поросятами), включая молодняк, на период гона. Справедливо широко распространенное мнение о том, что особи из второй и третьей единиц составляют фундамент всех 1руппировок кабана. На долю самок с поросятами приходится 49% структурных единиц, взрослых самцов — 20%, самок с поросятами и подсвинками — 7,5, самок с подсвинками — 7, самок взрослых без молодняка — 5, подсвинок — 5, поросят — 2,5, взрослых самок и самцов (в том числе гуртов) — 4%. Это соотношение может меняться в пределах от 12 до 17% в зависимости от ряда биологических факторов — размножения, гибели животных разных возрастов, территориальных перемещений, изгнания двухлеток из семьи и т.д. За пределами заповедной территории очень большое влияние на структуру популяции оказывает охотничий пресс, в том числе спортивная охота или мясозаготовка (Русаков, Тимофеева, 1984). Динамика структуры популяции кабана в зависимости от конкретных условий высотно-поясной структуры горных ландшафтов Кавказа остается неразработанной.
Одним из критериев оценки благополучия популяции является показатель стадности, отражающий относительную плотность населения в различных частях ареала, он может использоваться для выявления ориентировочной численности кабана на данной территории (Донауров, Теплов, 1938; Козло, 1975; и др.). В свое время Н.Я.Динник (1910) писал, что обычно кабаны живут небольшими табунами, которые состоят иногда из 3—4 неделимых групп, а иногда из 10—20. Раньше, отмечал он, когда зверей беспокоили меньше, они собирались в 1руппы по 40 особей и более, но теперь подобное отмечается редко. Анализ данных по стадности кабана в 20—30-х годах, проведенный С.С.Донауровым и В.П.Тепловым (1938) в Кавказском заповеднике, свидетельствует о том, что показатель стадности (среднее число животных, приходящееся на одну встречу для данного месяца) наиболее высок в ноябре и декабре, составляя 6,6 и 10,2. В этот период формируется "гарем", включающий несколько самок. Стадность высока также в марте (8,0), что объясняется объединением на участках с обильным урожаем орехоносов яловых свиней и изгнанных беременными самками подсвинок. Относительно высо
68
Sus scrofa Linnaeus, 1758
кая стадность зарегистрирована и в сентябре (7,2), когда происходит концентрация животных на небольших участках с дикорастущими фруктарниками. В остальное время показатель стадности колебался от 3,0 до 5,2; очевидно, они формируются из отдельных семей.
В 40-х годах (Котов, Рябов, 1963) в Кавказском заповеднике возросла стадность кабана, достигнув в ряде мест показателя, имевшего место во времена Н.Я.Динника. Положительно сказался, судя по всему, заповедный режим.
Территориальная структура популяции, как следует из данных В.В.Дурова (1987), в Кавказском заповеднике выражена четко и отражает многие стороны экологии кабана — состояние кормовой базы, физиологическое состояние организма, социальное положение и т.д. Так, самцы от 4 лет и старше оседлы, имеют участки обитания в 0,4—1,0 тыс. га, которые охраняются, метятся засечками на молодых деревцах, мочой и пометом. Обычно эти участки располагаются по верхней полосе горнолесных поясов и включают различные типы лесов и макросклонов, Логова здесь располагаются в верхних полосах крутых склонов, как правило, в снежный период, под кронами шатровых пихт или сосен южных склонов. Логова используются в течение 1—3 недель, потом строятся новые. Зимой индивидуальный участок сокращается на 40—60е,’^а в многоснежные зимы даже на 80—90%. Более того, в конце особенно голодных зим 3—4 истощенных секача мотут поселиться на индивидуальном участке одного местного самца. При гибели самца его индивидуальный участок в течение года занимают другие.
Самки с молодняком менее привязаны к территории, составляя номадную часть популяции, участки их меньше участков самцов (от 0,1 до 0,7 тыс. га, что зависит от кормовой базы). Гурты обычно держатся ниже поселения самцов, логовом им служат заросли кустарников, подроста хвойного леса. Представители других структурных группировок (неполовозрелые самцы, самки-подсвинки) занимают периферию участков гуртов, реже участков самцов.
Половая и возрастная структура. Оба параметра популяции очень вариабельны, что зависит от целого комплекса внутренних и внешних факторов среды (см. обзор: Русаков, Тимофеева, 1984). Высокой изменчивостью можно, видимо, объяснить различия в данных за 20—30-е годы (Донауров, Теп-лов, 1938) и 80-е годы (Дуров, 1987), а также наличием региональных особенностей в популяции Западного Кавказа по сравнению с другими регионами ареала. Так, в 20—30-х годах среди эмбрионов и новорожденных явно преобладали самки (69%) над самцами (31%) (л = 16). Среди взрослых животных (л = 1101), отстрелянных не избирательно, соотношение полов было близко 1:1 (самки — 52%, самцы — 48%). В 80-х годах выявлены несколько иные соотношения: в эмбрионный период самки составляли 56%, самцы — 44% (л = 109); в постнатальный период, наоборот, самок было 48%, а самцов — 52% (л = 486). В целом по всей стране среди новорожденных поросят самки составляют от 60 до 70% (Гептнер и др., 1961). Динамика соотношения полов является, очевидно, популяционным механизмом авторегуляции численности популяции — при снижении доли самцов складывается ре
6 9
Кабан
альная предпосылка к увеличению численности, а при возрастании — наоборот (Лобанов и др., 1972; Шварц, 1969; и др.).
Возрастной состав в Кавказском заповеднике изучен В.ВДуровым (1987). В соответствии с его данными в годы урожая плодов в январе-марте 24% популяции составляют самцы, 28 — самки, 15 — подсвинки, 33% — сеголетки. При неурожае это соотношение выглядит иначе: соответственно 40% — самцы, 25 — самки, 15 — подсвинки, 25% — сеголетки. Как видно, число взрослых самцов увеличивается. В малоснежные зимы при неурожаях плодов доля самцов возрастает до 53%, а самок и сеголеток уменьшается до 47%. К началу опороса (апрель) и в годы неурожая самцы составляют 25%, самки — 35%, молодняк — 40%. Такое соотношение возрастов, по мнению автора, наиболее оптимально, ибо за ним следует максимальное увеличение репродукции популяции. Как видно, формирование оптимальной возрастной и половой структуры популяции кабана происходит в депрессивные годы, обусловленные кормовой базой.
Враги, паразиты, конкуренты, смертность. Главный враг кабана в условиях Кавказа — волк. Об этом писал еще Н.Я.Динник (1910). По данным В.П.Теплова (1938), в Кавказском заповеднике остатки кабана в экскрементах волка составляют 37,6%, а рыси — всего 4,4% (Котов, 1958). В.В.Дуров (1987) считает, что на долю волка приходится не менее 16% погибших особей кабанов. В эту цифру не включены поросята, съеденные волками полностью, и те крупные свиньи, остатки которых невозможно обнаружить в крепях. Пресс волка особенно увеличился в связи с прекращением борьбы с ним. В период с 1966 по 1976 г. волки и медведи ежегодно изымали около 1000 поросят, 250 годовалых и 200 взрослых кабанов (7—10 особей на 1 хищника). В последние годы из-за возобновления борьбы с волком отход сократился на 40%. Доля кабана в пищевом рационе хищника составляет 26%; зимой она увеличивается до 37%. На долю медведей приходится 7,2% погибших кабанов, а на Черноморском побережье, где они заменяют волков, этот показатель выше в 2,4 раза. Не меньше кабанов уничтожается волком в тростниковых зарослях устьев рек и морских берегов. Так, в Кызыл-Агач-ском заповеднике Азербайджана одна волчья семья из 6—7 животных удовлетворяет свою потребность в корме за год на 66,7% за счет "продукции" группы кабанов численностью не менее 80—90 особей (Литвинов, 1981). Влияние волка особенно велико в зимние многоснежные периоды, когда истощенные животные становятся добычей хищников. Легко они справляются с больными животными и поросятами.
За последние годы, очевидно, возросла роль одичавших собак, особенно в равнинно-предгорных районах Кавказа. Эта проблема заслуживает специального изучения, так как количество бродячих собак растет, а меры регулирования их численности недостаточно изучены.
Из инфекционных болезней дикие кабаны подвержены заболеванию чумой свиней, которая передается от домашних свиней, вступающих в контакт с дикими. Эпизоотия чумы, например, зарегистрирована в Кавказском заповеднике в 1935—1936, 1946—1949, 1971—1975, 1980—1984 гг. За последние две эпизоотии на территории Краснодарского края от чумы погибло около
70
Sus scrofa Linnaeus, 1758
2 тыс. кабанов, что составило 12% от общего поголовья (Дуров, 1987). Давно известно отрицательное влияние ящура, передаваемого от домашних свиней к диким (Динник, 1910).
В условиях Кавказа паразитофауна кабана включает, как минимум, 28 видов, из которых 19 видов зарегистрированы в Кавказском заповеднике. Степень поражения и роль отдельных видов паразитов различны в разных ландшафтных районах. Так, в Кавказском заповеднике трихинеллезом поражено 3,6% кабанов, а в азовских плавнях — до 25%. Из эктопаразитов чаще всего встречаются вши Hematopinus suis L., что отмечено также и в Беловежской Пуще (Донауров, Теплов, 1938; Козло, 1975; Дуров, 1987; и др.). Немало страдают кабаны от кровососов (комаров, слепней и др.), особенно в период линьки, спасение от них кабаны находят лишь в грязевых ваннах и густых зарослях растений.
Имеет место гибель кабанов от различных природных и антропогенных факторов, в том числе от пожаров в тростниковых зарослях.
Список конкурентов кабана на Кавказе немалый: это медведь, олень, косуля, многие виды грызунов (белка, соня-полчок и др.), но при хорошем урожае орехоносов и фруктарников эта пищевая конкуренция не острая.
Смертность кабана, как и других многоплодных животных, высокая, особенно в первый год жизни. Между тем у кабана инстинкт материнства и забота о потомстве выражены сильнее, чем у кого-либо из копытных. Поражает то, с какой самоотверженностью самка защищает своих поросят, как легко она принимает осиротевших поросят и берет на себя всю заботу о них наравне со своими, насколько тонки и биологически целесообразны взаимоотношения особей всех возрастов и т.д. Тем не менее в Кавказском заповеднике, например, в период лактации (с мая по август), по данным В.В.Дурова (1987), гибнет 12 % поросят, с сентября по ноябрь — 26 %, с декабря по март, когда обычно лежит снег, отход составляет 24 %. Кроме того, около 9 % выводков гибнет полностью. Эти средние показатели варьируют, естественно, в зависимости от многих факторов. Так, при оптимальной кормовой базе гибнет на 11—15 % меньше, чем в годы неурожая. Не менее значительна роль эпизоотии — минимальное количество поросят в выводке (2,7 особи) зарегистрировано в 1932 г. после эпизоотии чумы, а наибольшее (5,5 особи) приходилось на I960 г., которому предшествовали 5 лет урожаев орехоносов.
Высока смертность кабана в многоснежные годы, особенно молодняка, самок и по разным причинам ослабевших животных. Аналогичных фактов в литературе довольно много.
Численность и ее динамика. Для кабана как многоплодного животного характерна периодическая динамика численности и плотности населения во времени и пространстве. Благодаря существованию заповедного режима хро-нологические изменения наиболее полно изучены на Западном Кавказе, что нашло отражение в ряде публикаций (Динник, 1910; Котов, Рябов, 1963; Дуров, 1973, 1987; и др.) (см. табл. 11). Анализ всего имеющегося материала позволяет заключить, что в-зависимости от периодичности плодоношения орехоносов и фруктарников, погодных условий, пресса хищников и эпизо-
7 1
Таблица 17. Результаты учета кабана прогоном (Дуров, 1973)
Участок, № Урочище и тип леса 1966 г. 1968 г. 1969 г.
Площадь, га Учтено голов Плотность на 1 га Площадь, га Учтено голов Плотность на 1 га Площадь, га Учтено голов Плотность на 1 га
1 Хребет Монашкинский. Каш-танник буковый с рододендроном, азалией, черникой 15 18 0,9 15 16 1,1 17 32 1,9
2 У ручья Медвежьего. Букняк с рододевдроно м 4 2 0,5 4 1 0,25 — — —
3 Ниже "40 десятин". Каштан-ник грабовыйс рододендроном 8 6 0,7 8 8 1,0 13 3 0,2
4 Над р. Бушуйкой. Дубняк грабовый с рододендроном и азалией — — — 22,5 14 1,6 22 5 0,2
5 За "40 десятин". Каштанник дубовый с рододендроном — — — 40 15 6,0 40 11 0,3
6 У Дубинского перешейка. Каштанник с рододендроном 7 18 2,4 7 4 0,6 — — —
7 У ручья Граничного. Букняк каштановый с рододендроном 3 1 0,3 3 1 0,3 3 — —
8 Герасимово. Букняк каштаново-грабовый с рододендроном, азалией и черникой 10 7 0,7 — — — 18 10 0,6
9 Поздняково. Дубняк каштаново-буковый с подростом бука, рододендроном и азалией — — — 10 31 3,1 23 4 0,2
1а Хребет Монашкинский. Каштанник буковый с рододендроном, азалией, черникой 15 — — 15 27 1,8 — — —
4а Над р. Бушуйкой. Дубняк грабовый с рододендроном и азалией 35 8 0,2 22,5 — — 35 — —
Всего 97 60 0,62 147 117 0,80 171 69 0,40
* С подкожной жировой клетчаткой.
Sus scrofa Linnaeus, 1758
отий численность кабана в Кавказском заповеднике менялась в 3,5-кратном, а плотность населения — 5-кратном размерах (табл. 17). Наиболее высокая численность в этом заповеднике (2500 голов) последовала за десятилетним периодом отсутствия суровых зим и снятием пресса хищников (1955— —1965 гг.). Наиболее низкая численность приходится на голодные годы, при повышении пресса хищников и эпизоотии чумы (1972—1973 гг.). В эти годы численность кабанов упала на 40 % и восстановление проходило медленно (тянулось до 1984 г. по: Дуров, 1987).
Если судить по материалам В.А.Кузякина (1989), то 1981 и 1982 гг. характеризовались повышенной численностью кабана в Краснодарском крае, составляя соответственно 7,7 и 7,6 тыс. голов. В пределах заповедника существование такого количества кабанов невозможно; очевидно, имел место повсеместный подъем численности кабанов в крае. В последующие годы (1983—1985 гг.) численность кабана в крае постепенно снижалась до уровня 5,3 тыс.
Географический аспект динамики численности кабана — трудный и слабо изученный вопрос экологии кабана Кавказа. Как общая закономерность, на наш взгляд, заслуживают внимания сдвиги в пространственной организации видового населения и соответствующие изменения в его численности и плотности, происходящие с северо-запада на юго-восток (т.е. влияние структуры поясности на количественные показатели населения вида). На данном этапе познания этого явления, видимо, можно констатировать асинхронность динамики численности в разных секторальных отрезках горных систем региона. Судя по 1рафику, составленному на основе учетных данных по Северному Кавказу, с 1981 по 1985 г. в Краснодарском крае наблюдалось постепенное уменьшение численности кабана (с 7,7 до 5,3 тыс.), в Ставропольском крае она была стабильной, на уровне около 4 тыс. в течение первых 4 лет, а в 1985 г. резко снизилась до 1 тыс. Сходная динамика отмечалась и в Северной Осетии. Пик численности кабана в Кабардино-Балкарии приходится на 1981 г., а в Чечено-Ингушетии — на 1984 г. Причины столь резких расхождений в динамике численности в указанных районах мы склонны объяснять асинхронными изменениями кормовой базы, снежности зимой и другими абиотическими факторами. Однако этот вопрос нуждается в специальном изучении, ибо имеющихся данных все еще мало.
Практическое значение. Во всех союзных и автономных республиках, а также в обоих краях Кавказа кабан занимает как по численности, так и значению в народном хозяйстве ведущее место среди копытных. Значение кабана как объекта спортивной охоты нельзя недооценивать. Кабан привлекает внимание охотников в первую очередь из-за мяса и сала, выход которых превышает 50 %. Наибольший выход, до 56 %, имеют двух-трехлетние животные (Козло, 1975). Кроме мяса и сала кабан дает ряд второстепенной продукции: кожу для кожевенной промышленности, волосы для изготовления щеток или для набивки седел и др. Клыки крупных секачей ценятся как охотничьи трофеи. Если учесть, что в урожайные годы рекомендуется изымать 25 % общего числа кабанов, а в неурожайные — до 50 % (Дуров, 1973), то нетрудно себе представить объем продукции, получаемой от каба
73
Семейство Оленьи
на в регионе в целом и в отдельных районах. На наш взгляд, этот объем может быть значительно увеличен, ибо имеются благоприятные условия на обширной территории. Для этого нужно лучше использовать опыт других районов (Прибалтика, Западная Европа).
Семейство Оленьи — Cervidae Gray, 1821
Жвачные копытные различных размеров, обычно легкого и изящного телосложения, с ветвистыми рогами (за исключением водяных оленей) у самцов, самки имеют рога лишь у северного оленя. Конечности длинные, изящные и крепкие, приспособлены к большой физической нагрузке; у большинства видов четко выражен половой диморфизм по ряду признаков.
Масса тела колеблется почти десятикратно (от 7 кг у пуду до 600—800 кг у лосей), размеры тела от крупного зайца до быков лосей с длиной тела более 3 м, высотой в холке до 235 см.
Рога в отличие от полорогих костные (мезодермальные), образуются из выростов лобных костей (пеньков постоянных), проходят стадии соединительнотканную, а затем хрящевую и костную. Растущие рога (панты) имеют кожный покров с короткими волосами и пронизаны кровеносными сосудами и нервами. После завершения окостенения рогов кожный покров отмирает и сбрасывается, оставляя на их поверхности следы кровеносных сосудов. Рост рогов осуществляется на верхушках, окостенение идет от основания к дистальной части. Отделение старых рогов от лобных костей происходит у основания, обозначенного бугристым пояском.
Рога, за исключением родов Elaphodus, Mazama и Pudu, ветвистые, но вначале появляются в виде шпилек, а затем с возрастом усложняются за счет появления боковых и венечных отростков. Количество их у разных видов колеблется от 2 до 20 и более. Однако прямой зависимости между ко-» личеством отростков (сложностью строения) и возрастом животного нет. Кроме того, на размере рогов и количестве отростков сказываются многие экологические факторы, в том числе условия питания, но видоспецифич-ность структуры рогов все же сохраняется.
Окраска волосяного покрова взрослых обычно однотипная, темная, лишь у отдельных видов наблюдаются светлые пятна. У примитивных видов в молодом возрасте пятнистая окраска — явление обычное. Многим видам свойственно околохвостовос светлое "зеркало". Волосы имеют мощно развитый сердцевинный воздухоносный слой, они короткие, плотно прилегают друг к другу’. Характерно наличие хорошо развитых специфических кожных желез, образующих скопления предглазничные, межпальцевыс, на внутренней стороне плюсны и скакательного сустава под хвостом, иногда на лбу. Мускусную железу имеют кабарги, у оленей их нет.
В черепе пневматизация костей слабая, имеется только в верхней челюсти, в слезной и переднем отделе лобной кости. В структуре его, по
7 4
Corvidae Gray, 1821
И.И.Соколову (1959), характерно наличие двойного отверстия носослезного канала, которое находится вблизи или у самого края глазницы. На внешней поверхности имеются предглазничные ямки. Сагиттальный гребень не развит. Этмоидальные отверстия большие. Орбиты больших размеров и направлены в стороны, тогда как слуховой барабан обычно небольшой, всегда полый. Зубная формула:
О 0—1 3 3
I— С ---- Р - М - = 32-34.
3 13 3
Клыки обычно слабо развиты и не выделяются среди других, пред-коренных всегда 3, коренные зубы у подавляющей части видов брахио-донтные, приспособленные к питанию мягкой травянистой растительностью, листвой и побегами кустарников и деревьев.
В посткраниальном скелете специфические изменения происходят в структуре конечностей. Так, характерно увеличение длины передних ног, и лишь у примитивных видов сохраняется значительное преобладание длины задних конечностей над передними. Показательно и то, что во II и V пальцах полностью редуцированы метатарзальные кости, от соответствующих метакарпальных остаются только их верхние и нижние части (Гептнер и Др., 1961).
Из анатомических особенностей заслуживает внимания дальнейшая дифференциация отделов желудка. Плацента характеризуется небольшим количеством котилодонов. Нет желчного пузыря.
Ареал оленей обширный, занимает Евразию на север до Арктики, а на юг — до тропиков включительно; олени распространены также в Северной Африке, Северной и Южной Америке, акклиматизированы в Австралии и Новой Зеландии. Места обитания связаны с лесостепными, лесными и отчасти тундровыми ландшафтами равнин и соответствующих поясов гор.
Животные активны круглогодично, нередко круглосуточно или в разное время дня и ночи, имеются виды типично ночные. Многим видам свойственны суточные и сезонные миграции, иногда охватывающие значительные расстояния.
Основу питания оленей, как правило, составляют различные мезофильные и отчасти гигрофильные растения: разнотравья, нежные побеги, кора и листва деревьев и кустарников и др. Словом, эволюция проходила в сторону замены всеядности на питание мягкой растительной пищей.
Самки, как правило, моноэстральные, но могут быть полиэстральными. Период гона обычно приходится на осень-зиму. Беременность длится примерно от 5 до 10 мес.
Размножение отличается от представителей предыдущего семейства (свиные) относительно низкой плодовитостью (от 1 до 4—7 детенышей), выживаемость потомства более высокая. Наряду’ с другими адаптивными особенностями существенную роль играет то, что детеныши появляются на свет хорошо развитыми, способными успешно следовать за матерью в день рождения.
75
Семейство Оленьи
Практическое значение представителей семейства разнообразно и значительно. Они служат объектами промысловой и спортивной охоты, дающими ценное и вкусное мясо, кожу, сырье для фармацевтической промышленности (панты и др.), декоративный материал в качестве трофея и т.д. Практикуется полувольное содержание, проводятся опыты по доместикации. Не менее важно эстетическое значение как природного объекта, обладающего красотой и изяществом. Будучи мало прихотливыми, нередко они представлены в зоопарках, а также в культурных ландшафтах вблизи населенных пунктов.
Эволюция оленей уходит своими корнями в нижний олигоцен и, видимо, связана с трагулидами (Tragulidae). Примитивные олени были мелкими, безрогими, с четырехпалыми конечностями. Прогрессивные изменения происходили в сторону увеличения размеров, появления рогов (преимущественно в качестве брачного наряда), совершенствования адаптации конечностей к быстрому бегу, к питанию мягкой растительной пищей и т.д. В настоящее время олени хотя немногочисленная, но прогрессирующая группа. Семейство включает до 12 родов, из которых на Кавказе встречаются три: косули Capreolus, настоящие олени Cervus и лоси Alecs.
Род Косули — Capreolus Gray, 1821
Олени мелких размеров: длина тела 100—150 см, хвоста 1—2 см, высота в холке 65—100 см, масса тела 20—59 кг. Телосложение стройное, голова небольшая, уши длинные, глаза большие. На конце морды в области ноздрей большое голос поле. Рудиментарный хвост скрыт в меху и не виден снаружи. Ноги высокие и тонкие, передние короче задних. Копыта на средних пальцах узкие, небольшие, края заостренные. Боковые пальцы очень слабо развиты и не достают при ходьбе землю. Рога небольшие, превышают длину’ головы не более чем в 1,5—2 раза, их стволы прямые, обычно трех-ветвистыс. Ствол рога в поперечном сечении округлый, его поверхность от основания до переднего отростка покрыта многочисленными бугорками и удлиненными выступами.
Волосяной покров, как и у других оленей, жесткий. Окраска его одноцветная, летом рыхлая на верхней стороне, светлеющая на брюхе, зимой серая. Хорошо выраженное околохвостовое "зеркало" обычно белого цвета с желтоватым налетом. Оно не заходит выше корня хвоста. Новорожденные телята имеют пятнистую окраску, хорошо развитые предплюсневые железы, межпальцевые железы лучше развиты на задних конечностях, чем на передних. Предглазничные железы небольшого размера. У косуль 2 пары сосков.
Череп относительно небольшой, лицевой его отдел несколько укороченный. Носовые кости нормальной длины. Нет отверстий на переднем крае межчелюстных костей, очень слабо выражена ямка предглазничной железы на слезной кости. Этмоидальные отверстия большого размера.
76
Caprcohis Gray, 1821
Зубная формула:
0 0 3 3 „
I— С — Р - М - = 32.
3 13 3
Естественный ареал рода нс выходил за пределы Евразии ни в прошлом, ни в текущий период, т.е. составляет группу палеоэндемиков и автохтонов Старого Света. Современное распространение охватывает обширную полосу от Пиренейского полуострова, Англии и Скандинавии на Западе до Дальнего Востока и Корейского полуострова на востоке. Северная граница ареала в Западной Европе достигает Полярного круга, а в Восточной Европе и Сибири опускается до уровня 58—60’ с.ш. Лишь в области Уральских гор и Забайкалья вновь расширяется меридионально. Южная храница идет до Средиземноморья, северных районов Ирана, Ирака, Афганистана, огибая пустыни Средней Азии, вновь спускается до горных систем Тянь-Шаня, Монголии, северо-востока Китая и Корейского полуострова. Как видно, восстановленный ареал рода приурочен к умеренному поясу Евразии, тяготеет к зонам смешанных и широколиственных лесов, лесостепья. Антропогенные изменения современного распространения очень большие; хотя косули легко уживаются в ахроэкосистемах, во многих регионах их не стало, но в других они остаются обычными, наиболее многочисленными из всех копытных.
В природных и антропогенных условиях предпочтение отдается мозаичным экосистемам (разреженные леса, чередующиеся с лугами и вырубками, островные леса и лесополосы, сельскохозяйственные поля, сады, пойменные луга и леса и т.д.). При этом косули тесно привязаны к своим индивидуальным участкам. Сезонные миграции, наблюдаемые в ряде равнинных и горных областей, связаны с изменениями условий жизни. Летом косули встречаются поодиночке, обычно утром и вечером, осенью они образуют смешанные стада до 20—30 голов, а зимой распадаются на небольшие группы (2—4 особи), круглосуточно активны. Питаются различными травами, листьями и побегами деревьев и кустарников. Пользуются природными и искусственными солонцами. Гон наблюдается в июле — октябре, иногда в ноябре, что зависит от климатических условий. Характерна ограниченная полигамия. Беременность длится около 9 мес., но может варьировать от 6 до 10 мес. Имеет место латентная стадия в развитии оплодотворенной яйцеклетки. Рождение телят приходится на май—июнь. Рождается 2, реже 3 теленка. Продолжительность лактации 2—3 мес. Самки живут до 11—12, самцы — до 16 лет.
Косули — ценный объект охоты, и в ряде районов страны и за рубежом их добывают в больших количествах. Во многих регионах нашей страны при условии обеспечения некоторых охотоведческих и охранных мероприятий их численность могла бы быть значительно увеличена.
По современным данным в роде два вида: европейская косуля С. сар-rcolus L. и сибирская косуля'С. pygargus Pall. В фауне Кавказа один вид.
77
Европейская косуля
Европейская Косуля — Capreohis caprcolus Linnaeus, 1758 Рис. 14
С. caprea Grey, Сатунин, 1909; Динник, 1910; С. vulgaris Pall. var. cauca-sica, Динник, 1910.
Коллекционный материал: ЗИН РАН — 28 черепов, 1 рог; МГУ — 27 черепов, 3 рога; КБГУ — 2 черепа.
Диагноз. Размеры относительно небольшие. Масса тела взрослых 20— 34 кг, длина тела 100—150 см, наибольшая длина черепа в среднем 202,5 мм, индекс ширины черепа 48,7 %, на переносье светлое пятно, лоб темнее тела, волосы на метатарзальных железах более темные, чем на задних конечностях; 2 п = 70, нет микрохромосом.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. По размерным показателям косули региона весьма неоднородны. В различных популяциях Северного Кавказа и Закавказья многие предыдущие и современные зоологи отмечали как довольно крупных, близких по размерам к сибирским популяциям, соответственно названных "сибирской" или "кавказской" косулей, так.и заметно меньших, иногда уступающих мелким формам европейской косули, получивших название "европейской косули" (Динник, 1910). Различия между ними существенные, в природе их можно различать свободно. По данным названного автора, если первые имеют массу тела около 30 кг, а длину тела до 150 см, то вторые соответственно 20 кг и 105 см. Они различаются также по окраске. В зимнее время крупные косули обычно имеют серую окраску, а мелкие — рыжеватую. Как отмечено выше, такие симпатрические морфотипы образуют и хищные млекопитающие Кавказа (медведи, ласки). Однако задача определения видовых пределов европейской косули в регионе сильно осложнена в результате акклиматизации сибирской косули (Capreohis pygargus) в Краснодарском и Ставропольском краях. В коллекциях Зоологических музеев МГУ (Москва) и ЗИН РАН (Санкт-Петербург) содержатся несколько черепов из этих мест, сходных по многим признакам с черепами сибирской косули (Соколов, Громов, 1985, 1988).
Если с учетом такого положения посмотреть на соотношения размеров косули из разных частей ареала вида, то легко заметить, что по всем морфологическим признакам в пределы вида С. capreohis укладываются и северокавказские, и закавказские популяции. Так, по данным В.Е.Соколова и В.С.Громова (1985, 1988), наибольшая длина черепа взрослых самцов Центральной Европы составляет 192,7, Кавказа — 202,9, Прибалтики — 205,1 мм. Аналогичные соотношения наблюдаются по многим другим черепным признакам, в том числе по наибольшей ширине черепа (89,6:93,0:94,8 мм), межглазничной ширине (51,3:54,8:54,9 мм), наибольшей длине носовых костей (57,0:61,8:62,1 мм), длине диастемы нижней челюсти (40,7:43,2:44,4 мм) и т.д. Как видно, по многим черепным параметрам косули Кавказа занимают промежуточное положение между центральноевропейскими и прибалтийскими
78
Capreolus capreolus Linnaeus, 1758
популяциями. Иная последовательность наблюдается при сравнении длины верхнего ряда зубов (56,1:57,0:56,3 мм), длины нижней челюсти (152,0:161,5:160,9 мм) и длины нижнего ряда зубов (64,3:65,8:64,5 мм). По этим 3 из 13 изученных параметров черепа кавказские косули превышают остальных. Однако не обнаружены "статистически достоверные различия между косулями Кавказа и Прибалтики ни в размерах, ни в пропорциях черепа" (Соколов, Громов, 1985, с. 741). Такое заключение подтверждается также результатами сравнительного анализа размеров черепа взрослых самок Кавказа, Центральной Европы и Прибалтики.
Анализ размерного соотношения различных популяций европейской и сибирской косуль, проведенный В.Е.Соколовым и В.С.Громовым (1985, 1988), позволил определить видовые особенности по габитуальным показателям. Оказалось, что по массе и размерам тела отличие европейских косуль, в том числе кавказских популяций, от сибирских очень велико. В среднем масса тела косули на Кавказе (л = 15) составляет 25,3 кг, длина тела 116,0 см; в Прибалтике соответственно 28,8 кг и 119,6 см (п = 74 и 112); в Центральной Европе длина тела (л = 547) в среднем 112,0 см. В 9 различных популяциях сибирской косули, обитающих на равнине и в горах от Урала до Приморского края, масса тела (л = 65) варьирует в среднем от 34,8 до 49,4 кг, длина тела (л = 69) — от 129,0 до 144,7 см. По всем промерам черепа самцы европейской косули, в том числе кавказской, заметно уступают аналогичным самцам сибирской косули (р < 0,001). Достоверны также различия по пяти индексам черепа: наибольшей ширины черепа (0,49; р < 0,001; кроме косуль Тянь-Шаня), скуловой ширины (0,46; р < 0,001; кроме косуль Тянь-Шаня), относительной длины верхнего ряда зубов (0,30; р < 0,01; кроме косуль Урала и Восточной Сибири), ширины лицевой части (0,90; р < 0,001; кроме косуль Тянь-Шаня) и ширины мозговой капсулы (0,32; р < 0,05). Сходные результаты получены и по самкам. Как видно, европейские косули отличаются от сибирской более мелким, но относительно широким черепом, сравнительно коротким верхним рядом коренных зубов, широкой лицевой частью и широкой мозговой капсулой. Положение кавказской косули среди других популяций европейской и сибирской косуль по трем основным признакам (наибольшая длина и ширина и относительная ширина черепа) наглядно показывает рис. 15 (Соколов, Громов, 1985, с. 743).
Зубная- система. По имеющимся у нас данным, между европейской и сибирской косулями нет качественных различий по структуре зубной системы. Из количественных особенностей европейской косули обращено внимание на относительно короткий верхний ряд коренных зубов (Соколов, Громов, 1985). По описанию зубной системы И.И.Соколовым (1959) у косули, как правило, отсутствуют верхние клыки. Иногда они появляются у молодых, но держатся они недолго. Резцы асимметричны; средняя пара значительно шире боковых, наружные углы оттянуты в сторону. Зубы второй пары относительно узкие, а третья пара и клыки тонкие и изогнутые наружу. Коренные зубы брахиодонтные; коронки этих зубов широкие у основания и сужены к вершине зуба. В качестве характерных особенностей
79
Европейская косуля
80
Caprcolus capreohis Linnaeus, 1758
6 В.Е.Соколов, А.К.Темботов
Рис. 14. Распространение косули
1 — Темрюк (Динник, 1910); 2 — Гос-тагаевская (Котов, Рябов, 1963); 3 — Анапа (Динник, 1910); 4 — Новороссийск (Динник, 1910); 5 — Геленджикский р-н (Котов, Рябов, 1963); 6 — Холодный родник (МГУ); 7 — Абинск (Котов, Рябов, 1963); 8 — Крымск (МГУ); 9 — Красноармейск (Котов, Рябов, 1963); 10 — Краснодар (Динник, 1910); 11 — Ильский (Котов, Рябов, 1963); 12 — Горячий ключ (Сафронова, 1973; Котов, Рябов, 1963); 13 — Белореченск (Котов, Рябов, 1963); 14 — Апшеронск (МГУ); 15 — Туапсе (Динник, 1910); 16 — Аше (Котов, Рябов, 1963); 17 — Лазаревская (МГУ); 18 — Сочи (Динник, 1910; МГУ); 19 — Пшекиш (МГУ); 20 - р. Киша (МГУ); 2'1 - Ха-мышки (Котов, Рябов, 1963); 22 — Новопрохладное (Котов, Рябов, 1963); 23 — Даховская (Котов, Рябов, 1963); 24 — Севастопольская (Котов, Рябов, 1963); 25 — Тульский (Котов, Рябов, 1963); 26 — Майкоп (МГУ); 27 — Ярославская (Котов, Рябов, 1963); 28 — Костромская (Котов, Рябов, 1963); 29 — Губская (Котов, Рябов, 1963); 30 — Лабинский р-н (Котов, Рябов, 1963); 31 — Сальск (Данилкин, 1989); 32 — Псебай (Динник, 1910; Котов, Рябов, 1963); 33 — Отважная (Котов, Рябов, 1963); 34 —- Бесстрашная (Котов, Рябов, 1963); 35 — Подгорная (Котов, Рябов, 1963); 36 — Спокойная (Котов, Рябов, 1963); 37 — Ставрополь (Динник, 1910); 38 — Удобная (Котов, Рябов, 1963); 39 — Передовая (Котов, Рябов, 1963); 40 — Зеленчукская (Федоров, 1954); 41 — Пицунда (Соколов, Сыроечковский, 1990); 42 —- Пицун-да-Мюссерскпй заповедник (Соколов, Сыроечковский, 1990); 43 — Сухуми (Динник, 1910); 44 — верховья р. Баксан (Динник, 1910; Темботов, I960); 45 - гора Бештау (Динник, 1910); 46 — Баксан (Насимович, 1950); 47 — Нальчик (МГУ); 48 — Нарткала (Насимович, 1950); 49 — Урванские дубки (Насимович, 1950); 50 — Малгобек (Насимович, 1950); 51 — Эльхотово (МГУ); 52 — Жемтала (Насимович, 1950); 53 — Бабугент (Насимович,
8 1
Европейская косуля
1950); 54 — Чаладиди (Насимович, 1950); 55 — Сатаплийско-Колхидский заповедник (Соколов, Сыроечковский, 1990); 56 — Махарадзе (МГУ); 57 — Батуми (Динник, 1910); 58 — среднее течение р. Печоры (Насимович, 1950); 59 — Цхакая (МГУ); 60 — Аджаметский заповедник (Соколов, Сыроечковский, 1990); 61 — Боржомский заповедник (Соколов, Сыроечковский, 1990); 62 — Боржоми (ЗМ Грузии); 63 — Лиахский заповедник (Соколов, Сыроечковский, 1990); 64 — верховья р. Терек (Динник, 1910); 65 — Север о-Осетинский заповедник (Соколов, Сыроечковский, 1990); 66 — Каз-бегский заповедник (Соколов, Сыроечковский, 1990); 67 — Казбегский р-н (Арабули, 1966); 68 — Терское (ЗМ МГУ); 69 — Владикавказ (МГУ, Динник, 1910); 70 — Сунженское (МГУ); 71 — Грозный (Динник, 1910); 72 — Ахметский заповедник (Соколов, Сыроечковский, 1990); 73 — Бацарский заповедник (Соколов, Сыроечковский, 1990); 74 — северо-восточная часть Кахетского хр. (Соколов, Сыроечковский, 1990); 75 — Телави (ЗМ Грузии); 76 — Ахметский р-н (ЗМ Грузии); 77 — Душети (ЗМ Грузии); 78 — Сагурамо-Марнамджварский заповедник (Соколов, Сыроечковский, 1990); 79 — Шулавери (Марков, 1924); 80 — Алавердский р-н (Даль, 1954); 81 — Сте-панаван (Даль, 1954); 82 — Кироваканский р-н (МГУ); 83 — Дилижанский заповедник (Соколов, Сыроечковский, 1990); 84 — Красносельск (Даль, 1954); 85 — Иджеванский р-н (Даль, 1954); 86 — Сигнахи (ЗМ Грузии); 87 — Гурджаани (ЗМ Грузии); 88 — Кварели (ЗМ Грузии); 89 — Лагодехи (ЗМ Грузии); 90 — Закатальский заповедник (Соколов, Сыроечковский, 1990); 91 — Лагодехский заповедник (Соколов, Сыроечковский, 1990); 92 — верховья Аварского койсу (Динник, 1911); 93 — верховья Андийского койсу (Динник, 1911); 94 — Хасавюрт (Динник, 1910); 95 — Шелковская (Гептнер, Формозов, 1941); 96- Старогладковская (Гептнер, Формозов, 1941); 97 — Кизляр (Динник, 1910); 98 — Бунакок (Гептнер, Формозов, 1941); 99 — Каспийский р-н (ЗМ Грузии); 100— Гуниб (Динник, 1910); 101 — верховья р. Каракойсу (Динник, 1910); 102 — верховья р. Самур (Динник, 1911); 103 — Дербент (Радде, 1899); 104 — Белиджи (Гептнер, Формозов, 1941); 105— Рутул (Гептнер, Формозов, 1941); 106 — Нухинский р-н (Лавров, 1929; Дементьев, 1933); 107 — Исмаиллинский заповедник (Соколов, Сыроечковский, 1990); 108 — Пиркулинский заповедник (Соколов, Сыроечковский, 1990); 109 — Гейгёльский заповедник (Соколов, Сыроечковский, 1990); 111 — Басутчайский заповедник (Соколов, Сыроечковский, 1990); ПО — Кафан (МГУ); 112 — Мегринский р-н (Даль, 1954); 113 — Шуша (ЗИН); 114 — Лачин (Алекперов, 1966); 115 — Гардуш (Алекперов, 1966); 116 — ущелье Виляшчай (Верещагин, 1959); 117 — ущелье Вазарчая (Верещагин, 1959); 118 — Шахгач (ЗИН)
названный автор приводит такие: "Между передней и задней половиной заднекоренных зубов, на внутренней стороне у верхних и на наружной у нижних, у основания коронок расположены зачаточные бугорки; на верхних зубах они могут отсутствовать. Передне-внутренняя долька последнего нижнего премоляра отделена от передне-наружной почти до основания коронки; обе задние дольки соединены друг с другом и с передне-наружной уже у вершины зуба. Второй нижний премоляр (Ру) вместо передне-внутренней дольки обычно имеет складки на середине и впереди главной (передне-наружной) доли зуба. На переднем нижнем премоляре (Рз) имеются только слабо развитые задние и передняя складки" (Соколов, 1959, с. 151).
Морфологическое своеобразие европейской косули, в том числе кавказской, хорошо прослеживается в размерах и структуре рогов. При сход-
82
Capreolus capreolus Linnaeus, 1758
Рис. 15. Положение кавказской косули среди других популяций европейской и сибирской косули
Самцы и самки косули по трем признакам: наибольшая длина черепа (1), наибольшая ширина черепа (II) и индекс наибольшей ширины черепа (III): 1 — Центральная Европа, 2 — Прибалтика, 3 — Кавказ, 4 — Северо-Восточная Украина, 5 — Урал, 6 — Северо-Восточный Казахстан, 7 — Тянь-Шань, 8 — Северо-Западная Монголия, 9 — Алтай, 10 — Восточная Сибирь, 11 — Северо-Восточная Монголия, 12 — Приамурье, 13 — Приморский край (Соколов, Громов, 1985)
стве характера ветвления и количества отростков рога кавказской косули, как и вида в целом, значительно уступают рогам сибирской по размерам. Еще Н.Я.Динник (1910) писал, что на Кавказе им не встречены рога длиной более 33 см, тогда как в Сибири рога козлов в 35—40 см не составляют редкости. Показательно, что длина стержней рогов в Центральной Европе (л = 397) составляет в среднем всего 16,9 см. в Прибалтике (л == 485) — 20,1 см. Раскид рогов составляет соответственно 8,1 и 10,1 см. В 10 различных популяциях сибирской косули средняя длина стержней варьирует в пределах от 25,3 (Приморский край) до 34,0 см (Тянь-Шань). Раскид рогов
6*
83
Европейская косуля
в тех же популяциях составляет 13,8 и 30,4 см. Эти различия сохраняются и при сравнении индексов длины рогов (отношение длины стержней к наибольшей длине черепа) и индекса раскида рогов (отношение длины раскида к длине стержней рогов).
Окраска тела подвержена изменчивости, но тем не менее ее видоспе-цифичность, очевидно, может служить одним из ведущих диагностических признаков (Громов, 1986). Еще Н.Я.Динник (1910) обращал внимание на ряд особенностей, свойственных косуле Кавказа. Так, если у сибирской косули голова и туловище одного цвета, то у косули Кавказа они окрашены различно. Летом большая часть головы покрыта черно-бурым волосом, кончики которого рыжевато-серого цвета, в результате формируется желтобурый или рыжевато-бурый тон. На лбу окраска приобретает более темный цвет. Спина и бока туловища желтовато-рыжие или золотисто-рыжие. В пахах и за локтями волосы почти белые. Зимой цвет гораздо бурее или серее. На голове растет более или менее однообразная по окраске серо-бурая шерсть. Ноги зимой и летом покрыты светло-рыжей шерстью. В это время четко выражено белое пятно на задней стороне бедер. В ряде последующих работ в качестве видовой особенности европейской косули указывали на светлое, нередко почти белое носовое пятно на переносье, а также на коричневый, бурый и черновато-бурый цвета волос, покрывающих метатарзальные железы. В результате образуется контрастирующая окраска на задних конечностях, тогда как у сибирской косули волосы на метатарзальных железах и задних конечностях обычно одного цвета (Громов, 1986).
Кожный покров. Изучено 2 взрослых самца (длина тела самца № 1 — 96 см, № 2 — 121 см), добытых в октябре 1960 г. в Нальчикском лесоохотничьем хозяйстве. Этот материал положен в основу как настоящего обзора, так и одной из предыдущих работ (Соколов, 1973). Промеры кожи приведены в табл. 18.
В эпидермисе мальпигиев слой состоит из 2—3 рядов клеток. Зернистых и блестящих слоев нет. Роговой слой тонкий и плотный. Эпидермис не пигментирован. Волосы, растущие с удлиненными и расширенными корнями волос. Сосочковый слой дермы вследствие этого утолщен и составляет более 50 % толщины дермы. Эластиновые- войокна многочисленны в верхней половине сосочкового слоя, где располагаются горизонтально, почти отсутствуют в сетчатом слое, за исключением зоны на границе с подкожной мускулатурой.
Небольшие сальные железы находятся по одной с каудальной стороны у каждого волоса. На груди сальные железы (120 х 440 мкм) более крупные, чем на спине (55 х 380; 10 х 440 мкм).
Хорошо развитые потовые железы сильно извиты и располагаются вдоль волосяных влагалищ, вплоть до луковиц волос. На спине (диаметр секреторных отделов 66—80 мкм) потовые железы крупнее, чем на труди (диаметр 55 мкм). Мсжпальцсвыс специфические железы задних конечностей имеют вид глубокого мешочка с узким выводным отверстием, а железы передних конечностей — полого вдавления.
84
Caprcolus caprcolus Linnaeus, 1758
Таблица 18. Промеры кожи европейской косули из Нальчикского лесоохотничьего хозяйства Кабардино-Балкарии (Соколов, 1973)
Показа- Толщина
тель ______________________________
КОЖИ, мм эпидермиса, мкм рогового слоя,мкм дермы, мм СОСОЧКОВОГО слоя, мм сетчатого слоя, мм
Загривок
№ 1 4,30 28 6 4,27 2,17 2,10
№ 2 2,45 17 4 2,43 1,78 0,65
Крестец
№ 1 3,70 17 3 3,68 2,08 1,60
№ 2 2,70 17 5 2,68 1,98 0,70
Грудь
№ 1 2,50 17 5 2,48 1,48 1,00
Изучена межпальцевая железа передней конечности косули № 2. Трубчатые железы очень сильно развиты и опускаются в дерму, значительно глубже волосяных луковиц. Диаметр секреторных отделов не крупнее, чем в коже туловища, но эти части желез немного длиннее и очень сильно извиты. Сумма диаметров трубчатых желез, подсчитанных на одном уровне на протяжении 1 см препарата, у межкопытной железы составляет 9,8 мм, тогда как на загривке косули № 2 — 1,1, на крестце — 1,2 и на груди 0,6 мм. Выводные протоки трубчатых желез открываются в волосяные воронки. Сальные железы достигают также очень крупных размеров — 0,13 х 0,9 мм. Сальные железы многодольчатые. Эпидермис в области межкопытных желез довольно толст и состоит из 3—4-рядного мальпигиевого слоя, однорядного зернистого, тонкого блестящего и многослойного рогового. На внутренней поверхности эпидермиса имеются небольшие впадины и выступы. Эти данные в общем совпадают с ранее опубликованным материалом (Solger, 1978, Schaffer, 1940, Arvy, Bommclear, 1960, по: Соколов, 1973). У косули, кроме того, имеются циркуманальные железы (Рососк, 1923, по: Соколов, 1973; Соколов, Чернова, 1988) и плюсневые железы (Соколов, Данилкин, 1981). Последние представляют собой небольшие участки утолщенной кожи на внутренней стороне плюсны, покрытые более длинными волосами, склеенными в основании. На поверхности кожи выделяется черноватый, нс обладающий запахом (для человека) секрет. На голове, шее, груди, в предглазничной. области развит "кожный щит" — поле утолщенной кожи с увеличенными железами (Соколов, Данилкин, 1981). Предглазничные железы рудиментарные, не функционирующие.
Мышцы, поднимающие волосы, хорошо развиты, толщиной 80—165 мкм. Волосы загривка косуль можно подразделять на две категории: остевые двух порядков и пуховые. Длина остей первого порядка 23,7 мм, остей второго порядка — 16,1 мм, пуха — 16,0 мм; толщина соответственно 286, 143 и 15 мкм; сердцевина у остей первого порядка занимает 95,7 %. Имеются также отдельные редкие длинные (65 мм), почти прямые волосы, выступающие из общего покрова (Флеров, 1928), которые нами не обнаружены. Ножек у волос нет. Остевые волосы групп и пучков не образуют. Число остей пер
85
Европейская косуля
вого порядка на 1 см2 поверхности кожи составляет 583., В поперечнике они имеют почти округлую форму. Хорошо выражен периферический слой клеток сердцевины, примыкающей к корковому слою., Кутикулярные клетки остей первого порядка некольцевидные, высотой 17 мкм. Пуховые волосы в численном отношении преобладают над другими группами. На 1 см2 поверхности кожи их растет 1000 штук. Располагаются пуховые волосы группами по 2—3 штуки. Пучков не образуют. Кутикулярные клетки высотой 10 мкм по большей части кольцевидные. Волосы труди подразделяются на две категории — остевые и пуховые. Длина остей 25,3, пуза — 14,7 мм; толщина — 179 и 14 мкм, сердцевина у остей занимает 95,2 %. "Ножки" отсутствуют. В поперечнике ости округлые, периферический слой клеток сердцевины выражен слабо. Пуховые волосы прямые. Они образуют слабо-выраженные пучки по 4—8 волос в каждом. Пучки пуховых волос располагаются преимущественно позади или по бокам остевых. На 1 см2 поверхности кожи насчитывается 583 остевых волоса и 1749 пуховых.
Система крови. Показатели периферической крови косули изучены Н.Н.Булатовой (1962). У взрослых самок (п = 3) в среднем количество эритроцитов оказалось 10,25 млн, гемоглобина — 14,8 г%, гематокрита — 35,3 %, цветной показатель — 0,44 ед., диаметр эритроцитов — 5,2 мкм, средний объем эритроцитов — 30,7 мкм3. У самцов (п = 2) ряд параметров крови имел более высокие значения: эритроцитов — 13,65 млн, гемоглобина — 17,6 г%, гемато!дэита — 42 %, средний объем эритроцитов — 34,4 мкм3. Цветной показатель заметно ниже (0,39 ед.), почти равными оказались диаметры эритроцитов обоих полов. Выявленные расхождения картины крови самцов и самок считать следствием проявления полового диморфизма мало оснований из-за недостатков фактического материала. Однако приведенные выше гематологические данные однозначны в том, что кислородная емкость крови косули довольно высокая, значительно превосходит многих диких и домашних животных. Словом, кислородная емкость крови вполне соответствует тому активному образу жизни, который ведет косуля.
Индексы внутренних органов. Литературных сведений по этому вопросу мало. Наиболее интересные данные получены по массе надпочечников косули в условиях Баварии (Bertram, 1974). Анализ 238 особей из 6 провинций дифференцированно по полам и возрастам (6 групп) позволил установить определенную связь между массой надпочечников и окружающей средой (плотность популяции, взаимоотношения особей внутри се и т.д.), а также между массой надпочечников и возрастом животных. Выявлена зависимость и от состояния здоровья косули (инфекция и др.).
Кариотип. Хромосомные исследования косули обоих видов проведены во многих частях ареала, охвачен широкий спектр ландшафтных условий (Соколов и др., 1978; Марков и др., 1985; Zemahle, 1980; и др.). На основании анализа этих многочисленных данных установлена видоспсцифичность кариотипа. У европейских косуль в кариотипе, кроме больших хромосом, имеются добавочные -хромосомы в количестве от 1 до 12. Все аутосомы обоих видов — акроцентрики, образующие по величине постепенно убывающий ряд от первой до последней пары.
86
Capreolus capreolus Linnaeus, 1758
Цитогенетические исследования европейской и сибирской косуль и их гибридов выявили, что наследование микрохромосом является неправильным: при скрещивании животных с различным количеством микрохромосом число их у гибридного потомства получается ниже теоретически ожидаемого (Zemahle, 1980, по: Соколов, Громов, 1988). Соответственно количество микрохромосом не может быть стабильным не только у подвидов, но и в отдельных популяциях косули.
Структура и динамика ареала
Географическое распространение европейской косули, "облик которой всеми исследователями трактуется как лесостепной" (Соколов, Данилкин, 1981, с. 18), связано с южной окраиной лесной зоны и лесостепью, частью со степной зоной и горными лесами (Гептнер и др., 1961). Соответственно первичный, восстановленный (т.е. максимальный в исторический период) ареал охватывал почти всю Европу и Малую Азию на восток до Волги и Северного Ирана. Граница этого ареала на западе простирается до Англии включительно, на север — до Средней Скандинавии, Финского залива, устья Камы и верховья Печоры; на юг — до побережья Средиземного моря, Сицилии (на других островах отсутствовал), северной части Сирии и Ирана, Палестины на юг до Генисаретского (Тивериадского) озера и Хайфы (гора Кармель) и север Ирана, в том числе г. Эльбурс (Lehmann, 1988).
В период максимальной депрессии ареала вида, которая приходится на конец XIX и начало XX в. (до 20-х годов), распространение европейской косули, так же как и сибирской, резко сократилось, распалось на отдельные части, образовав изолированные популяции: западную, крымскую, кавказскую и др. (Гептнер и др., 1961).
Распространение на Кавказе. Характеризуется очень сложной динамикой как в историческое время, так и в наши дни. Хотя с XVII в. повсеместно выражено сокращение ареала и плотности населения косули, тем не менее в этом процессе можно выделить три этапа: 1) до 70-х годов XIX в.; 2) конец ХЕХ и до 30-х годов XX в.; 3) с 40-х годов до наших дней. Первый этап характеризуется тем, что хотя антропогенный пресс значительно возрос и происходило сокращение ареала и численности косули, но она сохранялась на большей части территории, особенно в горах, и оставалась многочисленной. На втором этапе настолько возрос этот пресс, что происходило резкое сокращение как распространения, так и численности. Эти показатели достигли минимальных значений в 20-х годах. Третий, современный этап знаменует положительные сдвиги по восстановлению былого распространения косули на Кавказе. Было бы ошибочно полагать, что динамика ареала во времени не зависела от конкретных ландшафтных условий, от характера высотно-поясной структуры.
В период максимального распространения косули встречались не только в горных и предгорных лесах, но проникали далеко на равнину. Такие особенно благоприятные условия были на Западном Предкавказье, благодаря наличию в то время приречных лесов, кустарниковых зарослей и других
87
Европейская косуля
благоприятных мест обитания косули. Сходные условия были также в низовьях Дона, что способствовало слиянию кавказской и европейской популяций. Кстати, костные остатки косули обнаружены при раскопках крепости Саркел (нижнее течение Дона), датированной X—Х1И вв.н.э. (Верещагин, 1959; Соколов, 1959; Гептнср и др., 1961). В конце XIX и начале XX в. косули встречались на Западе до Темрюка, в окрестностях Анапы, Туапсе, где по Н.Я.Диннику (1910) они заходили даже в сады. В районе эльбрусского варианта, несмотря на менее благоприятные условия жизни, в конце XIX в. косуля водилась в лесах Ставропольской возвышенности, гор Бештау и Железной (Динник, 1910); по долине р. Малка проникала высоко в горы, занимая леса глубоких балок и склонов гор Харабуз и Мушт. Там была добыта старая самка (Россиков, 1887). На обширных плато из-за отсутствия лесов косули не встречались, их не было по этой причине и в предгорьях.
В области терского варианта (бассейн Терека) в прошлые века косуль было очень много. По К.Н.Россикову (1887), настоящими мсстамиобитания косули являются лесистые Черные горы. По Н.Я.Диннику (1910), много косуль было в окрестностях Нальчика, Владикавказа, Грозного, Хасавюрта, в верховьях Баксана, Терека и других горных рек. Что касается нижней границы ареала, то она спускалась по приречным лесам вплоть до берега Каспия.
В Дагестане, отличающемся своеобразным поясным спектром, рассматриваемый вид был широко распространен еще во времена Н.Я.Динника. По его данным (Динник, 1911, 1910), косуля занимала все высотные пояса, начиная от низовых лесов приморской полосы до верховьев Самура и четырех Койсу. Он приводит следующие места обитания: Хасавюрт, Кизляр, леса Койтаго-Табасаранского округа, Кюринский и Аварский округа, окрестности аулов Зиберхаль и Будты Андийского округа, глухие ущелья Гу-нибского округа, склоны гор Богасского и Главного хребтов. В конце 90-х годов прошлого века косуль добывали в березовых лесах близ Верхнего Гуниба.
Закавказская часть ареала косули занимала не менее обширную территорию, чем на Северном Кавказе. Она простиралась от Черноморского побережья (Новороссийск, Сочи, Сухуми, Батуми) до Талыша включительно. Высотные пределы ареала в горах Закавказья, судя по всем имеющимся данным, были так же значительны, как и на Северном Кавказе.
В период наибольшей депрессии численности и распространения косули, наблюдавшейся в начале XX в. не только на Кавказе, но и в других районах б. СССР, кавказская часть ареала претерпела большие изменения. Во многих местах как равнины, так и гор косули были выбиты. В самом начале текущего столетия она была уничтожена в лесах Ставропольской возвышенности, Пятигорья, на Кабардинской равнине, по долине р. Сунжа, на всем протяжении Восточного Предкавказья, за исключением приречных лесов и кустарниковых зарослей долин Терека и Сулака. В.Г.Гептнер и А.Н.Формозов (1941), обследовавшие Дагестан в 1924—1926 гг., писали, что по долине Терека косули попадаются по лесным угодьям и местами
88
Capreolus capreolus Linnaeus, 1758
обыкновенны. Так, их еще довольно много у станиц Старогладковской и Шелковской. Если не считать таких отдельных поселений по долинам крупных рек, то нижняя граница ареала в летний период была смещена вверх до 1500 м над ур. моря (Верещагин, 1959).
Изменения, происходившие в горах Северного Кавказа в начале XX в., трудно оценить, но, вероятно, они сводились к резкому усилению пятнистости ареала. Наиболее полные данные, относящиеся к этому периоду, имеются по горному Дагестану благодаря исследованиям В.Г.Гептнера и А.Н.Формозова (1941). По их данным, еще в 1924 г. две—три пары косуль держались в березовой роще близ Верхнего Гуниба, часто попадались в Ка-рахском лесу на р. Бецор, притоке Кара-Койсу; в верховьях Аварского койсу в лесах над аулом Корда в 1924 г. были еще довольно многочисленны. На Богосском хребте, где косуль было много во времена Н.Я.Динника, они не были обнаружены, тогда как в лесах окрестностей сел Рутул и Лучек эти животные попадались постоянно. Хотя Н.Я.Динник не находил косуль в районе Буйнакска, они встречались здесь в период работы В.Г.Гептнера и А.Н.Формозова.
В Закавказье наиболее благоприятные условия сохранились в то время на Западе (колхидский и аджарский варианты), однако и здесь их было больше в средней трети лесных поясов. Ниже из-за густой сети населенных пунктов косуль было мало, выше они также были малочисленны, но здесь больше сказывался отрицательно глубокий снеговой покров. Сходный характер размещения косули отмечался и в соседних вариантах (юго-осетинский и триа-летский). В Восточном Закавказье, как отмечает Н.К.Верещагин (1959), косули населяют преимущественно нижнюю треть пояса горных лесов с зарослями кизила, боярки, грабинника. В буково-грабовых лесах они попадаются близ полян с гигантскими папоротниками и ежевикой. Верхнюю опушку горных лесов и прилежащие к ним субальпийские луга заселяли редко. Значительно чаще и в течение всего года косули регистрировались в дубово-лапинных лесах Алазино-Авторанской низменности, притом зимой больше, чем летом. Что касается Кура-Араксинской низменности, в том числе тугаев, тростниковых зарослей и других оазисов, то косуль в этих местах не находили. О широком распространении и высокой плотности населения косули в Геокчаевском, Нухинском и Закатальском районах писали еще П.ПЛавров (1929) и Г.П.Дементьев (1933). По их данным, в 20-е годы нашего века очень мало косуль было в б. Курдистанском уезде и в Нагорном Карабахе. В последующие 30—40-е годы распространение и численность косули в горных лесах Малого Кавказа, видимо, стало заметно увеличиваться, но тем не менее она оставалась в депрессии, и в связи с этим охота на нее была запрещена (Даль, 1954). На обширной территории Джа-вахетско-Армянского нагорья практически косуль не было. В Талыше они встречались редко в поясе горных лесов, притом лишь по некоторым осветленным ущельям. Как видно из изложенного, в конце XIX и первой половине XX в. ареал косули значительно сократился, в виде нссплошной ленты он тянулся по низкогорщям-срсднегорным лесам вдоль всего горного Кавказа, а нижняя граница ареала сместилась вверх в предгорья. На равнине
89
Европейская косуля
и в низовьях косули отмечались только в отдельных районах, а в высокогорьях — обычно в летний период.
Современный этап динамики структуры ареала косули на Кавказе, как и во многих других районах страны, характеризуется четко выраженной тенденцией увеличения распространения и численности. Положительно сказываются рсакклиматизациоиныс мероприятия, улучшение охраны, в.том числе расширения площадей, охваченных заповедниками, национальными парками, госзаказниками. Однако в целом ряде мест процесс восстановления косули проходит не так успешно, как в других районах Европы.
Переходя к обзору материалов на основе высотно-поясной структуры горных экосистем, отметим, что по комплексу исторических и ландшафтных факторов довольно оптимальные условия для обитания косуль имеются в кубанском варианте. Распространение и численность косуль в этом варианте в пределах Краснодарского края по состоянию на 1956—1958 гг. анализированы В.А. Котовым и Л.С. Рябовым (1963). Ниже приводятся их данные.
В отмеченные годы северная граница распространения на Западном Кавказе проходила по линии, соединяющей следующие населенные пункты: Передовая (севернее от нее) — Подгорная — Бесстрашная (севернее от нее) — Губская — Костромская — Мохошевская — Кужорская (южнее от нее) — Белореченск (севернее от него) — Северская (южнее от нее) — Абинская — Гостагаевская. Южная граница простиралась до самого берега Черного моря, а на запад — до окрестностей Анапы (восточные районы). В пределах очерченной территории косули нет в высокогорной части Главного Кавказского хребта и его северных отрогов. В частности, этого животного нет в верховьях рек Большая и Малая Лаба с притоком Уруштен, Белая с притоком Киша.
Что касается состояния численности в то время, то сведения были противоречивы: одни считали, что косуль стало очень мало (Слепов, 1956), другие, наоборот, — много (Навозова, 1955). Достаточно однозначные данные получены В.А. Котовым и Л.С. Рябовым (1963). Оказалось, что в Отрадненском и Спокойненскрм районах косули малочисленны. Они отмечались лишь в южной лесной части, но имели Место заходы в степную часть по балкам, поросшим терновником и боярышником. Аналогичные заходы регистрировались до станицы Удобной, но заметно реже, чем до 1950 г. Чаще всего они имели место в верховьях рек Малый и Большой Те-гинь. В большом количестве косули обитали в южной части Лабинского района. В 1956 г. южнее станицы Отважной за один день можно было поднять до десятка косуль. В окрестностях станицы Ахметовской численность была ниже. Если не считать высокогорной части, то косули были обычны в Псебайском, Ярославском, Тульском и Апшеронском районах. Наиболее высокая численность в перечисленных районах зарегистрирована в окрестностях Псебай, Авдрюки, Бугунжа, Шедок, Севастопольская, Даховская, Ново-Прохладное, Хамышки и др.
Косули широко встречаются по всей территории Белореченского, Горяче-Ключевского, Северского и Абинского районов, за исключением севср-
90
Capreohis capreohis Linnaeus, 1758
ной части. В егерском участке Горяче-Ключевского района на 1000 га приходится до 9 голов. В Северском и Абинском районах численность косуль резко сократилась после зимы 1953/54 г. Редки в Крымском районе, а в Анапском отмечалось некоторое увеличение численности косуль. На южном макросклоне Главного Кавказского хребта чаще встречаются в Семигор ье, Сукко, а также в Геленджикском районе. Снижена численность в большинстве районов Краснодарского края, расположенных по Черноморскому побережью. На правом берегу Кубани, в Красноармейском районе (Красный лес) косули встречались до 1954 г., но в многоснежном 1953/54 г. их почти всех истребили. В 1958 г. для восстановления поголовья сюда была завезена партия сибирских косуль.
"В целом в предгорьях и горных районах Краснодарского края численность косуль за последние годы значительно сократилась. Особенно тяжелой для них оказалась чрезвычайно многоснежная и продолжительная зима 1953/54 г. В феврале 1954 г. в окрестностях г. Майкопа высота снежнего Петрова достигала 80—100 см. Такая же картина наблюдалась и в других-районах края. В эту зиму огромное количество беспомощных животных погибло от собак и браконьеров. Численность популяции в результате истребления косуль была настолько подорвана, что, несмотря на запрет охоты, она до сих пор восстанавливается крайне медленно" (Котов, Рябов, 1963, с. 143-144).
В 60—70-х годах, судя по литературным данным (Дуров, 1980; Темботов, 1982), восстановление численности косули на Западном Кавказе проходило плохо. Более того, в пределах Краснодарского края за эти годы нижняя граница ареала отодвинулась в горы на северном макросклоне на 20—50 км, а по Черноморскому побережью — на 2—5 км: Даже при заповедном режиме продолжалось резкое сокращение распространения косули. Так, если с 1930 по 1971 г. она занимала высоты от 800 до 2300 м над ур. моря и 15 участков Кавказского заповедника площадью'60 тыс. га, то за 1972—1975 гг. количество таких участков сократилось до 9 (20000—25000 га), а в 1976—1977 гг. до 5 (1000 га) (Дуров, 1980). Организация Краснодарского лесоохотничьего хозяйства в 1958 г., видимо, мало положительного внесла в расширение ареала и восстановление численности косули в.предгорно-равнинной части. Завоз сибирской косули из Сибири и Казахстана в 1958—1961 гг. и выпуск их на Краснодарском участке (правый берег Кубани, Красноармейский участок) отодвинул северо-западную границу косули вниз и позволил ей вновь заселить степи Западного Предкавказья. Наличие обширных лесопольных пространств могло иметь существенный эффект, однако, судя по данным В.А.Кузякина (1989), хотя рост поголовья косули в Краснодарском крас и наметился, но идет медленно (увеличение с 4,3 тыс. в 1981 г. до 5,0 тыс. в 1985 г.).
В соседнем Ставропольском крае и Тебсрдинском заповеднике положение с косулей было еще хуже. С.М.Федоров (1954) отмечал, что косули в крае обитают лишь в горных буковых лесах Черкессии. Косули, водившиеся в лесах окрестностей Ставрополя, хищнически уничтожены. Что касается горных лесов, то в 1949/50 г. на территории Тебердинского
9 1
Европейская косуля
заповедника обитало всего 10 особей, но в 1952 г. поголовье возросло до 15 особей (Аренс, 1957). Они встречались в Джемагатской и Мухинской, в области слияния рек Чоначхира и Аманауз. Маловероятным представляется утверждение Л.ЕАрснса (1957) о том, что "в долине реки Аксаута и некоторых его притоков косуль много” (с. 119). В дальнейшем, очевидно, косуль в Тсбердинском заповеднике не стало. А.М.Тарасов (1973), проработавший ряд лет в этом заповеднике, в списке копытных заповедника по состоянию на 1972 г. косули вообще не упоминает. Вне заповедника появление косули в Ставропольском крае, вероятно, обусловлено реакклиматизацией (Емельянов, Рыбалов, 1968), однако процесс восстановления косули проходил плохо. В 1971—1973 гг. численность ее была ниже уровня оптимального в 5 раз (Ильинский, Ладова, 1976). В 1980-х годах, очевидно, наметился рост численности косули в Ставропольском крае, о чем свидетельствуют данные В.А.Кузякина (1989): если в 1981 г. косуль в крае было 1,6 тыс. особей, то в 1982 г. — 2,0, в 1983 г. — 2,7, в 1984 г. — 3,5, в 1985 г. — 4,0 тыс. Как видно, в крае за 5 лет численность косули удвоилась.
В бассейне р. Малка, где представлен эльбрусский вариант поясности, условия жизни для косули ограниченны, ибо обширные плато Вольских пастбищ и прилегающих районов безлесны, интенсивно используются летом для выпаса многочисленных отар в сопровождении домашних собак. Косули могут удержаться лишь в глубоких ущельях, где сохранились небольшие леса. Однако и здесь для них не совсем благоприятные условия из-за ограниченности лесопокрытой площади и глубокого снегового покрова зимой. Тем не менее опыт работы Ставропольского госохотхозяйства (организовано в 1968 г., расположено в бассейне Кумы, занимает предгорья) свидетельствует о том, что искусственное лесоразведение, необходимое в здешних условиях, может способствовать восстановлению косули не только в районе Кавказских Минеральных Вод, но и на значительной территории Вольского района Кабардино-Балкарии.
Благоприятны условия жизни для косуль в бассейне Терека (терский вариант поясности), где по сей день сохранились хорошие леса из бука, храба, дуба и других широколиственных пород и различные кустарниковые заросли. Этот пояс широколиственных лесов тянется по всему Меловому хребту и северному макросклону Скалистого хребта на протяжении Кабардино-Балкарии, Северной Осетии и Чечено-Ингушетии. В этих лесах в 40-х годах косуля была обычной. По А.А. Насимовичу (1950), в "Урванских дубках" (12 км от Нальчика) в январе 1949 г. за один день на небольшой площади леса были подняты собаками в общей сложности 6—7 косуль. Также много косуль зарегистрировано им несколько выше — в окрестностях Бабугента и Жемталы. В настоящее время положение дел с косулями в Кабардино-Балкарии нс столь благополучно, как в 40-х годах. Более того, несмотря на организацию Нальчикского лссоохотничьего хозяйства (1957 г., центральная часть Северного Кавказа), Кабардино-Балкарского заповедника (1976 г.) и несколько позже Национального парка в Приэлбрусье и ряда заказников, резко сократилось распространение и численность косули в Кабардино-Балкарии: с 1,8 тыс. в 1981 г. до 0,6 тыс. голов в 1985 г. Как видно,
92
Capreolus capreolus Linnaeus, 1758
сокращение трехкратное. Лучше обстоит дело в соседних. республиках: в Северной Осетии численность се за эти годы осталась почти неизменной (на уровне 1,2—1,6 тыс.), а в Чечено-Ингушетии зарегистрирован значительный ее рост (с 1,6 до 2,9 тыс.) (Кузякин, 1989). В настоящее время нижняя граница ареала в терском варианте проходит по линии Баксан — Нарткала — Малгобск — Грозный. Верхняя простирается до субальпийских лугов (летний период).
Сведения о распространении косули по Закавказью в 40—50-х годах обобщены Н.К.Верещагиным (1959). В соответствии с этими данными, косули широко населяли Западное Закавказье (колхидский и аджарский варианты) начиная от Колхидской низменности до верхней границы лесов и субальпийских лугов. Однако глубокий снег вытеснял их из высокогорья. Например, в ущельях Верхней Сванетии, в частности по Ингуру, их фактически нет. Распространение косуль в Колхидской низменности и по Черноморскому побережью нс сплошное, во многих местах они вовсе отсутствуют, что связано с густой сетью населенных пунктов. В 40—50-х годах немного косуль обитало в колхидских ольховых топях в окрестности Чаладиди и в среднем течении Пичори. Потийские охотники добывали в этих местах до 50 косуль ежегодно. По данным В.А.Котова и Л.С.Рябова (1963), для большинства районов Черноморского побережья характерно уменьшение численности косуль. Так, в окрестностях с. Аше Лазаревского района до 195Q г. косули встречались в 2 раза чаще, чем в 1957 г. У Чертова Моста того же района до 1941 г. 2—3 охотника могли добыть до 5—6 косуль, а в 1957 г. встречались лишь отдельные следы. Тем не менее на карте распространения косули на южном макросклоне Главного Кавказского хребта, приводимой этими авторами, нижняя граница проходит по берегу моря, что соответствует положению ее в грузинской части ареала. При всем этом оптимум ареала косули в Западном Закавказье 50-х годов находился в среднегорном лесном поясе. Такое положение, судя по всем имеющимся данным, сохранилось и в последующие годы на всем протяжении Западного Закавказья.
В Восточном Закавказье современное распространение косули приурочено к горнолссным поясам Большого и Малого Кавказа, лишь в Алазано-Авторанской низменности спускается до равнины. Однако ее нет в настоящее время в Кура-Араксинской низменности. По южному склону Главного (Водораздельного) хребта в 50-е годы косули были обычны в нижней трети лесного пояса, где нередки заросли кизила, боярышника и грабинника. Верхняя граница простиралась летом до 2400 м над ур. моря (Верещагин, 1959). На Малом Кавказе косули регистрировались всюду, где имелись значительные лиственные леса. По данным названного автора, по ущельям рек Дебеда и Акстафа они живут летом в субальпийском высокотравье, вблизи уцелевших островных лесов, занимая высоты 1800—2000 м над ур. моря. В Карабахе косуль в то время было много в сохранившихся буковых лесах бассейна Тсртер-Чая. С.К.Даль (1954) приводит более обширную территорию Армении, занятую косулей в 50-х годах, чем Н.К.Версщагин. Она включает леса Степанованского, Алавердского, Кировоканского, Иджеванского, Шамшадинского и Красносельского районов, особенно много косуль, как
9 3
Европейская косуля
полагает цитируемый автор, в Кафанском и Мегринском районах, т.е. косули обитают по Малому Кавказу до самой его юго-восточной части (до 1раницы с Ираном). Высотные пределы распространения составляют от 700 до 2200 м над ур. моря. На южных склонах Армянского нагорья в рассматриваемый период косули не было. В Талыше косули, очевидно, занимают весь горнолесной пояс, но подробности их распространения в этих местах не изучены. Ареал европейской косули простирается на восток за пределы государственной границы б. СССР, в частности, она недавно зарегистрирована в лесах Эльбруса (Lehmann, 1988).
В последующие годы наиболее полные сведения по распространению косули в Закавказье получены А.Б.Арабули (1966). Сбор данных им проводился в Восточной Грузии с 1959 по 1965 г. В соответствии с этими данными ареал косули занимает южный склон Главного Кавказского хребта от Южной Осетии до Азербайджана в пределах от 500 до 2200 над ур. моря. Ряд авторов проводили верхнюю 1раницу до 2300—2500 м (Енукидзе, 1953; и др.). Оптимум ареала находится в среднем лесном поясе (900—1000 м). Как отмечено выше, косули в предыдущие десятилетия обитали в пойменных лесах и на горных пастбищах. Однако в 60-х годах в этих местах косули не встречались. А.Б.Арабули вообще не встречал косуль юго-восточнее Тбилиси — в Цителцкарском и Марнеульском районах, а также в Казбегском районе и в лесах Тушетии. На Триалетском и Циви-Гамборском хребтах косули не проникают выше 1700—1800 м над ур. моря из-за отсутствия лесов, хотя рельеф и почвы для них подходящи (Арабули, 1966).
Современные тенденции антропогенных изменений ареала. Как следует из приведенного выше обзора, современный этап динамики ареала косули на Кавказе характеризуется определенной стабилизацией. Этому способствуют не только наличие благоприятных условий жизни в регионе, но и практические мероприятия по восстановлению численности косули (реакклиматизация в Краснодарском и Ставропольском краях и др.). На наш взгляд, реально дальнейшее расширение ареала и заметное увеличение численности косули почти повсеместно. Этому, видимо, будут благоприятствовать как перспективные планы народного хозяйства региона, так и особенности экологии косули (Темботов, 1972).
Европейскую косулю, как известно, считают типичным представителем лесостепных ландшафтов. Действительно, на Кавказе она отдает предпочтение комплексным экосистемам, где островные широколиственные леса чередуются с открытыми луговыми или степными полянами. Соответственно в горных лесах Кавказа, приобретающих лесолуговой облик, на всем их протяжении косуля может найти самые благоприятные условия (малоснежные зимы, широкий набор пищевых объектов и др.). Вероятно, столь же благоприятными окажутся предгорья и равнины, где с каждым годом увеличиваются лесные полосы, расширяется посадка лесов по долинам рек. Словом, обширные пространства Предкавказья и Кура-Араксинской низменности приобретают лесопольный характер, что в других частях ареала европейской косули обеспечивает процветание полевого экотипа этого вида (опыты разведения косули в Чехословакии и других европейских государ
94
Capreolus capreolus Linnaeus, 1758
ствах). Что касается увеличения численности и распространения косули в горах, то предпосылки для этого в различных вариантах различны. Видимо, в горах колхидского и аджарского вариантов они ограничены из-за большого снегового покрова зимой, тогда как в восточнозакавказских этот фактор не имеет такого значения. То же самое можно сказать и в отношении восточно-северокавказского типа поясности. Бесспорно, что косули здесь могут в летний период подниматься по склонам до 2500 м и выше; мы их находили на этой высоте в районе верховья Баксана (Темботов, 1960). Близкие данные получены и в условиях высокогорья Карабаха, Зангезура и других гор восточного Кавказа (Даль, 1954; Аалекперов, 1966; и др.). Для нас вне сомнения то, что косули могут быть расселены широко в различных районах переднеазиатского типа поясности, в особенности джавахето-армянского варианта.
При восстановлении косули путем реакклиматизации, на наш взгляд, необходим строгий подход к подбору популяции-"донора" с таким расчетом, чтобы завозить в горные районы лесной экотип, а в предгорья и равнины с "лесопольным" ландшафтом — полевой экотип. На наш взгляд, малая эффективность реакклиматизационных мероприятий на Северном Кавказе, особенно в Краснодарском и Ставропольском краях, обусловлена завозом сибирской косули. На современном этапе разработки систематики рода Capreolus недопустимо искусственное расширение зоны межвидовой гибридизации.
Систематика и изменчивость
Систематическое положение. По поводу систематики рода Capreolus и положения в нем кавказских популяций существуют противоречивые мнения. Известный знаток териофауны Кавказа К.А.Сатунин (1901, 1906) считал, что на Кавказе встречаются два вида косули: Capreolus pygargus Pall, и С. vulgaris Fitz. К первому виду он относил популяции Северного Кавказа, а ко второму — закавказские. Однако, говорил он, форма и некоторые особенности рогов северокавказской косули отличают ее от типичных сибирских представителей этого вида, хотя все же она ближе к ней, чем к европейскому виду. Нет нужды доказывать, насколько близко стоял к истине КА.Сатунин, особенно в оценке промежуточного положения северокавказских популяций между двумя названными видами. В последующих исследованиях, включая современные, это положение подтверждалось новыми данными, расхождение касалось в основном таксономического ранга — одни признавали самостоятельность этих видов, другие низводили их в ранг подвидов. Так, в своей монографии Н.Я.Динник (1910) приводит два вида косули: Capreolus pygargus Pall, ("кавказская дикая коза") и Capreolus саргеа Gray ("обыкновенная дикая коза"). По его словам "Capreolus саргеа Gray, без сомнения, представляет особый вид, сильно отличающийся от кавказской дикой козы гораздо меньшим ростом, несколько иной окраской, маленькими рогами и иным географическим распространением". По его мнению, ареал этого вида не ограничен Закавказьем, а простирается на Север
95
Европейская косуля
ный Кавказ, в том числе Ставропольскую возвышенность, т.е. распространение обоих видов симпатрическое. Эти суждения Н.Я.Динника, на наш взгляд, заслуживают внимания, ибо решение проблемы "вид или подвид" в отношении косули Кавказа непосредственно упирается в необходимость выявления территориального соотношения этих форм. Поэтому географический критерий вида всегда присутствовал во всех работах, посвященных косулям Кавказа.
Реальность существования европейской (С. capreolus L.) и сибирской (С. pygargus Pall.) косуль как самостоятельных видов признавал К.К.Флеров (1928), но в последующем появился целый ряд работ, низводивших их до подвидов одного вида Capra capreolus L. (Бобринский и др., 1944, 1965; Пастернак, 1955; Соколов, 1959; Громов и др., 1963; Гептнер и др., 1961). Признавая значительную амплитуду морфологической изменчивости косули по В.Г.Гептнеру и др. (1961), в пределах б. СССР выделяются две группы подвидов этого вида: европейская и сибирская. К первой ipynne отнесены косули Закавказья, а ко второй — популяции, населяющие северные склоны Главного Кавказского хребта от Новороссийска и Сочи на западе до Каспийского побережья Дагестана на востоке (от низовья Терека до Дербента); их выделили в отдельный подвид С.с. caucasicus Dinnik.
В последние два десятилетия интерес к систематике рода Capreolus резко возрос, что нашло отражение в многочисленных работах, осветивших многие параметры видов, в том числе морфологические, биохимические, иммунологические, гибридологические, гео1рафические и кариологические.
Морфологическое своеобразие европейских и сибирских косуль, по В.Е.Соколову и В.С.Громову (1988), вне сомнения. Европейская косуля заметно уступает сибирской в массе, размере тела, черепа и рогов, отличается деталями окраски меха и по целому ряду других признаков. При этом различия у самцов выражены в большей степени, чем у самок. Показательно, что, как отмечали предыдущие исследователи (Флеров, 1952; Гептнер и др., 1961; Громов, 1986), многие морфологические признаки являются альтернативными и можно рекомендовать их в качестве систематических (особенно окраска меха). По другим, особенно по краниологическим признакам и размерам рогов, между этими видами обнаружен хиатус (см. обзор: Соколов, Громов, 1988).
Очевидно, достаточно высок систематико-таксономический вес кариотипических параметров, в особенности наличие микрохромосом у сибирской и отсутствие их у европейской косули. Вероятно, заслуживает внимание и то, что число микрохромосом сибирской косули, варьирующее, как отмечено выше, от 1 до 12, не может быть надежным маркером не только подвидов, но и отдельных популяций (Соколов, Громов, 1988).
В кариологическом плане косули Кавказа мало изучены, что существенно осложняет оценку других морфологических признаков северокавказских и закавказских популяций. Это в первую очередь касается равниннопредгорных, ибо они, по В.Е.Соколову и В.С.Громову (1985), являются смешанно-гибридной популяцией европейских и сибирских косуль. На это указывали и предыдущие исследователи. В частности, В.Г.Гептнер с соав
96
Caprcolus capreohis Linnaeus, 1758
торами (1961) отмечали, что закавказская (С.с. caprcolus) и северокавказская (С.с. caucasica) встречаются вместе на западе и востоке Кавказских хребтов, имеют место заходы отдельных особей в ареалы друг друга, что ведет к широкой гибридизации. "Весьма возможно, что описываемая раса (т.е. С.с. caucasica) вообще имеет в значительной мере гибридный характер и образовалась в результате столкновения двух потоков заселения Кавказа косулей — севера сибирской, в Закавказье из Малой Азии — европейской" (с. 202). При этом он обращал внимание на относительно крупные размеры тела и черепа отдельных особей косули с Северного Кавказа, что усиливает зону налегания меристических показателей обоих видов. С таким суждением согласуются кариологические данные В.Е.Соколова и А.А.Данилкина (1981) — из семи изученных косуль Кавказа у одной оказались две микрохромосомы, характерные для сибирской косули. Такое же положение обнаружено и по левобережью Днепра (Днепропетровская обл.), где обитают крупные косули, сходные с северокавказскими. Из девяти изученных там косуль семь имели одну—две микрохромосомы, что дает им основание считать этих зверей гибридными. Это, по их мнению, первый случай обнаружения гибридов европейской и сибирской косуль в природе.
Гибридная зона европейских и сибирских косуль искусственно расширяется, видимо, в связи с завозом и акклиматизацией последних из Казахстана и Сибири. Это, очевидно, значительно осложнит и без того непростую диагностику косуль на Кавказе на основе меристических показателей и кариотипов.
В плане оценки реальности существования двух видов косуль в б. СССР однозначными оказались результаты гибридологического анализа (Штуббе, Брухгольц, 1979; Соколов, Громов, 1985). Показательно, что репродуктивная изоляция между европейской и сибирской косулями обусловливается различными причинами. Так, нормальные роды не получаются у значительной части самок европейской косули при их гибридизации с самцами сибирской косули. Наблюдается стерильность части гибридных самцов первого поколения. Происходит также нарушение нормального контакта между половыми партнерами в гибридных парах, поэтому успешное покрытие самок европейской косули происходило только в 50—60% случаев. Самки сибирской косули не спаривались с самцами европейской косули. Выявлены другие этологические особенности, определяющие их репродуктивную изоляцию. Не менее интересными оказались исследования особенностей звуковой сигнализации европейской и сибирской косуль (Соколов и др., 1987). Так, у европейских косуль не обнаружено аналогов скуления самцов и сдвоенных писков самок, характерных для сибирских косуль в период гона. Европейские косули издают звуки более высокие по частоте.
Географический аспект критериев вида также выражен хорошо; оба вида имеют собственные ареалы, лишь в отдельных районах (на Северном Кавказе) он "смазан".
На основании изложенного можно считать доказанным, что европейским и сибирским косулям присущи все атрибуты для признания их в качестве самостоятельных видов: Caprcolus caprcolus и С. pygargus. Существование
7 В.Е.Соколов, А.К.Тсмботов
97
Европейская косуля
обоих видов косули на Кавказе отрицать нельзя, тем более после завоза и реакклиматизации сибирских косуль в пределах ареала кавказских популяций европейской косули. Из-за недостаточности данных пока невозможно очертить ареалы этих видов на Кавказе.
Что касается изученности внутривидовой изменчивости и систематики косули на Кавказе, то единого мнения по этому вопросу в литературе нет. Н.Я.Динник (1910) на основании сравнительного анализа популяций косули Северного Кавказа и Сибири пришел к выводу о существовании особой формы сибирской косули на Кавказе и назвал ее Capreolus pygargus Pall, var. caucasica. Эта форма затем была возведена в ранг подвида С.с. саи-casicus Dinnik и отнесена в "сибирскую группу" подвидов (Гептнер и др., 1961). Закавказские косули, по мнению этого автора, относятся к номинальной форме — С.с. capreolus Lin., к "европейской группе" подвидов. Реальность существования двух подвидов косули на Кавказе признавал известный систематик И.И.Соколов (1959), однако северокавказских не выделял в самостоятельный таксон, а относил к С.с. pygargus. Мнения как В.Г.Гептнера, так и И.И.Соколова разделяют многие отечественные и зарубежные систематики.
К принципиально иному мнению пришли В.Е.Соколов и В.С.Громов (1985, 1988), проводившие исследования морфологической изменчивости обоих видов косуль с применением современных методов статистической обработки систематических признаков. Суть этого суждения состоит в том, что европейские косули, распространенные как в Западной Европе, так и в европейской части б. СССР и на Кавказе, сходны по строению черепа и рогов, по массе и окраске тела. Популяционные различия так невелики, что нет основания для их разделения на подвиды. Это касается и степени различия между закавказскими и северокавказскими косулями — статистический анализ черепных признаков показал отсутствие между ними достоверных различий как в размерах, так и в пропорциях. При таком положении необходимо, по мнению этих авторов, учитывать существование гибридной популяции (или группы популяций) косули в Краснодарском и Ставропольском краях, как и в юго-восточных областях Украины.
Как видно, по систематике косули Кавказа выполнена значительная работа, однако еще остаются нерешенные проблемы. Так, на наш взгляд, мало еще однозначных данных по характеру распространения сибирской косули на Кавказе, о возможном ее влиянии путем частичной гибридизации на фенотип местных популяций европейской косули. Действительно ли экземпляры косули из Эльбурса, описанные ЕЛехманомДЬеЬтапп, 1988), относятся к сибирской косуле и подтверждают тем самым реальность существования в Закавказье и на Северном Кавказе Capreolus pygargus caucasica Din., как это следует из работы Н.Я.Динника (1910) и процитированной выше работы. Разрешить эти вопросы, видимо, возможно лишь на основе новых данных, полученных не только с использованием морфологических, но и цитогенетических и биохимических методов исследования.
Изменчивость. Различные аспекты изменчивости косули на Кавказе обсуждались рядом териологов, в том числе К.А.Сатуниным (1906), Н.Я.Дин-
98
Capreolus capreolus Linnaeus, 1758
ником (1910), K.K. Флеровым (1928, 1952), Ф.А.Пастернаком (1955), И.И.Со-коловым (1959), В.Г.Гептнером и др. (1961), А.Б.Арабули (1966), В.Е.Со-коловым и В.С.Громовым (1985, 1988) и др. Список литературы можно заметно увеличить за счет работ, в которых данная проблема хотя и не обсуждалась специально, но в них содержится ценный фактический материал по изменчивости косули на Кавказе.
Полевые наблюдения и сборы коллекции, хранящиеся в различных музеях страны, проведены с охватом широкого спектра ландшафтных условий: лесостепи Ставропольской возвышенности, пояса широколиственных лесов восточно-северокавказского типа поясности (Кабардино-Балкария, Северная Осетия, Чечено-Ингушетия и Дагестан), темнохвойных и широколиственных лесов бассейна Кубани, влажных субтропиков Западного Закавказья, сухих субтропиков Восточного Закавказья, влажных субтропиков восточного побережья Каспийского моря. Размах изменчивости морфологических параметров вида может быть оценен на имеющемся материале с Кавказа, однако для статистической обработки его применительно к высотно-поясной структуре ландшафтов он недостаточен.
Индивидуальная изменчивость. Литературные и оригинальные данные, которыми мы располагаем, свидетельствуют о том, что в условиях Кавказа косуля подвержена значительной индивидуальной изменчивости. Особенно велика, на наш взгляд, эта изменчивость по массе тела. Среди любой популяции в пределах одной возрастной группы встречаются как мелкие, так и крупные особи. Размах варьирования массы тела может достигать двукратной величины. На Западном Кавказе Н.Я.Диннику (1910) попадались косули вполне взрослые с трехветвистыми рогами, масса которых колебалась в пределах от 16 до 32 кг. По его мнению, кавказские косули этого района могут иметь массу до 40 кг. Генетическая природа такого разнообразия не изучена, но фенотипическая обусловленность массы тела давно находится в поле зрения териологов и охотников. Еще Н.Я.Динник (1910) отмечал, что на массу тела, рост тела и развитие рогов положительно влияют избыток пищи и относительное спокойствие, обеспечиваемое режимом охраны природы Западного Кавказа. Кроме того, существенное влияние на фенотип оказывают условия местообитания — косули в горных местообитаниях, по его наблюдениям, бывают обычно меньше тех, которые живут на равнине или в предгорьях Кавказа. На обширных охотничьих трофеях в Чехословакии показаны факторы, определяющие широчайший размах индивидуальной изменчивости экстерьерных параметров европейской косули: качество и количество пищи, минеральное питание, факторы постнатального развития, плотность популяции, паразиты, различные повреждения и травмы и т.д. (Necas, 1975). По мнению Н.Я.Динника (1910), косули, живущие в привольных лесистых местах Западного Кавказа, 1д>упнее и имеют более сильно развитые рога, чем косули Восточного Кавказа, отличающегося более бедной растительностью. Многочисленный материал Ф.А.Пастернака (1955) по европейской и сибирской косулям убедительно свидетельствует о том же: в благоприятных экологических условиях встречаются крупные косули, а в неблагоприятных — мелкие.
7*
99
Европейская косуля
Индивидуальной изменчивости сильно подвержены не только размеры рогов, но и их структурные параметры. Мерой этой изменчивости может служить коэффициент изменчивости, изученный Ф.А.Пастернаком (1955) в масштабе всего ареала европейской и сибирской косуль. Так, общая длина рога имеет коэффициент вариации в Закавказье 11,2, а в Беловежском заповеднике 9,7, что, видимо, объясняется различием в степени разнообразия условия жизни сравниваемых популяций. Не менее вариабельны относительные величины отростков. Они у первого, второго и третьего отростков северокавказских косуль составляют соответственно (по отношению к общей длине рога) от 15,2 до 31%, от 28,1 до 42,4% и от 10 до 27,1%. Эти индексы меньше колеблются в Закавказье. Не менее изменчивы степень развития и количество бороздок и перлов на стержнях рогов, а также количество боковых отростков. По Н.Я.Диннику (1910), располагавшему значительным количеством полевых данных, на Кавказе самыми типичными рогами являются те, которые имеют по 3 отростка на каждом роге. Восьми-или десятиконечные рога, образующиеся в результате раздвоения одного или двух верхних концов рога, встречаются редко. Динник располагал рогами, у которых на правом роге, кроме пяти нормально расположенных отростков, был еще длинный шестой, выросший из главного стержня в двух дюймах от розетки. Могут быть различные вариации, например может отсутствовать первый отросток при наличии второго и третьего или рог разделяется на несколько ветвей (5—10, даже более 20) у самой розетки, образуя как бы кустик (рис. 16). Количество таких ветвей может быть больше, и они могут иметь разнообразную форму (Necas, 1975). В нашей коллекции имеется оригинальный экземпляр (S-11952), у которого оба больших рога несходны друг с другом и резко отличаются от типичных. Левый рог длиной в 38 см не имеет никаких отростков, правый — длиной 33 см — имеет один боковой отросток длиной около 9 см. Он отходит от стержня в средней части. Все эти изменения рогов обусловлены мутационно или фенотипически (недостаток пищи, минеральных веществ, повреждение семенников и др.).
Индивидуальная изменчивость окраски тела значительна. Это относится к общей окраске, а также отдельным участкам. Известно, что мелкие косули, встречающиеся почти во всех популяциях Кавказа, характеризуются зимой желтовато-серой окраской, а летом рыжей, тогда как крупные косули заметно темнее. Крайние формы изменения окраски, как альбинизм и меланизм, видимо, не встречаются на Кавказе или очень редки.
Возрастная измечивость. Этот вопрос рассматривается нами более подробно в разделе "Рост и развитие". Здесь обратим внимание на те особенности возрастной изменчивости, которые должны быть приняты во внимание при решении систематико-таксономических вопросов. По массе и размерам .тела косуля в условиях вольера достигает максимального уровня в 3,5 года (Соколов, Громов, 1988). Что касается размеров и структуры рогов, то И.И.Соколов (1956) решительно отвергает возможность их применения для оценки возрастных изменений косули. По его мнению, для этой цели наиболее приемлемы сроки появления, смены и стирания коренных зубов
100
Capreolus capreolus Linnaeus, 1758
Рис. 16. Изменчивость рогов косули
верхней и нижней челюстей. Однако, вероятно, у молодых косуль, как писал еще Н.Я.Динник (1910), "рога тонки и имеют очень слабо расширенную розетку... Вообще по величине розетки у козлов, так же как у оленей, можно составить более или менее определенное понятие о возрасте животного" (с. 67). Эта мысль нашла свое подтверждение при изучении возрастной изменчивости европейской косули с применением современных математических методов и на большом фактическом материале (Stubbe et al., 1987).
В практике систематических работ обычно выделяют две труппы косуль: 1) полувзрослые и молодые и 2) взрослые (Соколов, Громов, 1985). По многим черепным признакам различия между ними существенны. Так, по наибольшей длине черепа первая труппа уступает второй примерно на 6% у самцов и 5% у самок.
Половые различия. У косули выражен половой диморфизм по ряду признаков. По массе тела Н.Я.Динник (1910) считал это различие очень существенным — самки уступают самцам примерно на 20—25% от массы самцов.
1 0 1
Европейская косуля
Очевидно, такие результаты можно получить лишь при сравнении минимального показателя самок с максимальными показателями самцов. По современным литературным источникам (см. обзор: Соколов, Громов, 1981), самцы тяжелее самок в среднем лишь на 2—3 кг. Так, в Чехословакии масса тела самцов (п — 50) в среднем 24,0 кг, а самок (л = 17) — 22,6 кг, в Болгарии (л = 52 и 73) соответственно 26,3 и 23,9 кг; в Латвии (л = 74 и 49) — 28,8 и 26,7 кг. В условиях Восточной Грузии самцы (л = 15) весят в среднем 25,3 кг, а самки — 24,3 кг. Сходные данные получены по размерам тела, в том числе по его длине. В той же Чехословакии самцы (л = 50) имеют в среднем длину тела 115,6 см, а самки — 114,4 см, в ГДР (л = 547 и 379) соответственно 112,0 и 111,0 см, в Болгарии (л = 43 и 55) — 117,1 и 115,0 см, в Латвии (л = 112 и 176) — 119,6 и 118,9 см. В Восточной Грузии, по данным А.Б.Арабули (1966), самцы (л = 15) оказались несколько меньше (116 см), чем самки (л = 16) (117,6 см). Возможно, что на Кавказе половой диморфизм у косули по размерам тела выражен менее резко, чем в европейской части ареала. Фрагментарные данные, которые содержатся в литературе, не противоречат такому суждению (Котов, Рябов, 1963; и др.).
У европейской косули не выявлены половые различия ни в зимнем, ни в летнем мехе (Гептнер и др., 1961), а также в расцветке волос (Громов, 1986).
Для самцов обоих видов косули характерны рога и соотстствующис костные выросты на лобных костях. У самок в норме отсутствуют рога, но на их месте нередко появляются бугорки на лобных костях. Изредка встречаются рогатые самки.
Половые различия хорошо выражены и по размерам черепа — у самцов череп крупнее, нежели череп самки. Из 13 параметров черепа косуль Кавказа, изученных нами и В.Е.Соколовым и В.С.Громовым (1985)^ самцы заметно превосходят самок по 10 параметрам (табл. 19). Такого превосходства нет по длине лицевой части, верхнего и нижнего рядов зубов и диастемы. При сравнении пропорции черепов самок и самцов количество признаков, по которым различаются разные полы, резко сокращается. Тем не менее данные однозначно свидетельствуют о том, что череп самцов относительно широк, но укорочен.
В конфигурации черепа выявлен целый ряд признаков, иллюстрирующих половой диморфизм (Пастернак, 1955).
Географическая изменчивость. Как отмечено выше, в териологической литературе по Кавказу давно обсуждаются различные аспекты систематики, в том числе внутривидовой, имеющей прямое отношение к проблеме географической изменчивости. Однако до настоящего времени нет однозначных данных ни по одному вопросу, относящемуся к закономерностям этой изменчивости в условиях трехмерного пространства гор Кавказа. Главная причина, обусловившая такое положение, состоит в недостатке фактического материала. Во времена К.А.Сатунина и Н.Я.Динника, судя по публикациям этого периода и музейным материалам, коллекций по косулям Кавказа было очень мало. В последующем их количество возросло совсем немного. Так, Ф.А.Пастернак (1955), выполнивший большую работу по обоим видам и с охватом всей территории б.СССР, располагал всего 4 экз.
102
Таблица 19. Промеры (в мм) черепа Capreolus capreolus из Предкавказья. Взрослые
Признак Самцы
п limit X ± т О Су
Наибольшая длина 9 202,0—230,0 212,8±2,96 8,38 3,9
Кондилобазальная длина 9 190,0—206,0 198,1±1,92 5,44 2,7
Основная длина 9 178,0—203,0 185,9±2,62 7,42 4,0
Максимальная ширина 12 92,0-105,0 97,2±1,14 3,79 3,9
Скуловая ширина 13 81,0-95,0 89,1±1,09 3,78 4,2
Межглазничная ширина 14 52,0—63,0 58,5±0,94 3,40 5,8
Длина лицевой части 9 103,0-122,0 110,6±1,92 5,43 4,9
Длина носовых костей 12 62,0—83,0 71,2±1,94 6,42 9,0
Длина верхнего ряда 13 55,0—73,0 59,9±1,61 5,56 9,3
зубов Максимальная ширина мозговой капсулы 14 52,0-69,0 62,3±1,14 4,12 6,6
Длина нижней челюсти 11 163,0-192,0 171,2±2,68 8,49 5,0
Длина нижнего ряда зубов 11 62,0—84,0 67,6±2,18 6,89 10,2
Длина диастемы нижней 11 44,0—55,0 47,9±0,95 3,0 6,3
челюсти
Признак Самки
п limit X ± т а Су
Наибольшая длина 7 192,0-228,7 207,5±4,99 12,23 5,9
Кондилобазальная длина 6 177,8-200,0 191,5±3,65 8,17 4,3
Основная длина 7 167,4—196,7 181,4±3,78 9,25 5,1
Максимальная ширина 14 81,5—96,9 88,7±1,18 4,27 4,8
Скуловая ширина 14 81,5-90,8 86,6±0,84 3,02 3,5
Межглазничная ширина 14 46,9-74 53,1±1,85 6,69 12,6
Длина лицевой части 7 95,8-115,9 107,0±2,75 6,75 6,3
Длина носовых костей 9 53,4—74,7 64,7±2,66 7,52 11,6
Длина верхнего ряда 14 51,0-69,0 60,1±1,27 4,60 7,6
зубов
Максимальная ширина 14 52,2-64,9 58,5±1,21 4,36 7,4
мозговой капсулы
Длина нижней челюсти 14 151,2-181 163,2±2,53 9,11 5,6
Длина нижнего ряда зубов 14 55,0—82,0 66,7±1,63 5,88 8,8
Длина диастемы нижней 14 38,9—51,0 44,7±1,14 4,11 9,2
челюсти
Таблица 20. Промеры (в мм) черепа Capreolus capreolus из Предкавказья. Молодые
Признак Самцы
п limit X ± т а Су
Наибольшая длина 4 189,5-206,2 196,8±4,83 8,37 4,2
Кондилобазальная длина 4 175,4-194,2 184,6±5,13 8,89 4,8
Основная длина 4 162,8-182,6 172,6±5,30 9,19 5,3
Максимальная ширина 6 75,0—98,3 87,1±4,51 10,09 11,6
Скуловая ширина 6 76,2—91,0 84,6±2,29 5,12 6,0
Межглазничная ширина 7 43,7-57,5 50,9±2,28 5,58 11,0
Длина лицевой части 4 92,9—104,7 99,1±3,12 5,41 5,4
Длина носовых костей 5 44,0—67,0 54,9±4,36 8,73 15,9
Длина верхнего ряда зубов 7 49,3-65,4 54,7±2,42 5,94 10,08
Максимальная ширина мозговой капсулы 7 45,8-63,5 55,4+2,84 6,95 12,5
Длина нижней челюсти 7 132,5—164,4 150,9±5,59 13,7 9,1
Длина нижнего ряда зубов 7 49,3-67,3 57,9+2,91 7,13 12,3
Длина диастемы нижней челюсти 7 34,1-46,5 40,7±1,95 4,78 11,7
Таблица 20 (окончание)
Признак Самки
п limit Я ± т О Су
Наибольшая длина 1 — — —
Ковдилобазальная длина 1 — — —
Основная длина 1 — — —
Максимальная ширина 4 65,0—86,0 76,4±5,67 9,83 12,9
Скуловая ширина 4 66,5—84,0 75,8±4,91 8,51 11,2
Межглазничная ширина 4 38,2—51,0 44,9±3,50 6,07 13,5
Длина лицевой части 1 — — — —
Длина носовых костей 3 31,7-58,0 41,8 ±8,19 14,19 33,9
Длина верхнего ряда зубов 4 40,8—61,0 50,2±б,24 10,81 21,5
Максимальная ширина мозговой капсулы 4 45,5-58,0 52,4±3,79 6,57 12,5
Длина нижней челюсти 3 108,0-155,0 126,0±13,49 23,36 18,5
Длина нижнего ряда зубов 3 40,8-72,0 51,5±10,25 17,76 34,5
Длина диастемы нижней челюсти 2 26,0—39,0 32,5±6,50 9,19 28,3
Таблица 21. Промеры (в мм) черепа Caprcolus caprcolus из Закавказья. Взрослые
Признак Самцы
п limit Я ± т О сг
Наибольшая длина 19 183,0—206,0 199,8±1,23 5,22 2,6
Ковдилобазальная длина 19 175,0—193,0 188,1±0,95 4,03 2,1
Основная длина 19 163,0-180,0 175,9±0,87 3,68 2,1
Максимальная ширина 23 84,0—98,0 90,9 ±0,70 3,29 3,6
Скуловая ширина 21 77,0—91,0 85,6±0,79 3,55 4,1
Межглазничная ширина 24 47,0-63,0 53,5±0,68 3,28 6,1
Длина лицевой части 21 91,0-107,0 102,9±0,81 3,64 3,5
Длина носовых костей 24 46,4—70,0 58,2±1,11 5,32 9,1
Длина верхнего ряда зубов 24 52,0-62,9 57,7±0,58 2,77 4,8
Максимальная ширина 23 55,0-63,0 60,5 ±0,48 2,25 3,7
мозговой капсулы
Длина нижней челюсти 12 148,0-167,0 158,2±1,67 5,55 3,5
Длина нижнего ряда зубов 14 57,0-72,0 65,1±0,96 3,47 5,3
Длина диастемы нижней 14 36,5—46,0 42,5 ±0,89 3,20 7,5
челюсти
Признак Самки
п limit X ± т а сг
Наибольшая длина 14 187,0—204,0 197,4±1,29 4,64 2,4
Ковдилобазальная длина 14 176,0-193,0 186,3±1,23 4,43 2,4
Основная длина 14 162,0-180,0 173,7±1,29 4,64 2,7
Максимальная ширина 18 74,0—89,0 86,0±0,87 3,57 4,2
Скуловая ширина 18 73,5-87,0 83,2±0,81 3,35 4,0
Межглазничная ширина 19 45,5-52,0 49,6±0,40 1,70 3,4
Длина лицевой части 14 93,0—106,0 101,1±1,12 4,03 4,0
Длина носовых костей 18 50,0—65,0 57,9±0,90 3,72 6,4
Длина верхнего ряда зубов 19 50,1-61,0 58,0±0,70 2,96 5,1
Максимальная ширина мозговой капсулы 19 51,0—63,0 58,2±0,69 2,93 5,0
Длина нижней челюсти 5 130,5-159,0 151,0±5,89 11,78 7,8
Длина нижнего ряда зубов 5 52,4-68,0 62,6±3,08 6,15 9,8
Длина диастемы нижней челюсти 5 , 31,4-42,0 38,8±2,13 4,25 11,0
Capreolus capreolus Linnaeus, 1758
Таблица 22. Промеры (в мм) черепа Capreolus capreolus из Закавказья. Молодые
Признак Самцы
п limit X + т о CF
Наибольшая длина 7 156,0-204,0 183,5±6,38 15,62 8,5
Кондилобазальная длина 7 148,0—192,0 173,3±6,08 14,89 8,6
Основная длина 7 136,0-178,0 161,3+5,92 14,50 9,0
Максимальная ширина 11 57,0-91,0 81,9±3,12 9,88 12,1
Скуловая ширина 10 55,0-91,0 78,9+3,29 9,86 12,5
Межглазничная ширина 12 40,0—54,0 48,8+1,08 3,57 7,3
Длина лицевой части 7 54,0-103,0 88,2+6,63 16,25 18,4
Длина носовых костей 8 37,0-58,0 49,4±2,60 6,87 13,9
Длина верхнего ряда зубов 11 47,0—63,0 56,1±1,45 4,59 8,2
Максимальная ширина мозговой капсулы 12 51,0-62,0 57,0±0,90 2,98 5,2
Длина нижней челюсти 8 122,0-160,0 145,3+4,49 11,89 8,2
Длина нижнего ряда зубов 7 52,0-71,0 63,2+2,84 6,95 11,0
Длина диастемы нижней челюсти 8 32,0-41,0 38,3+1,02 2,71 7,1
Признак Самки
п limit х + т О Су
Наибольшая длина 5 129,0-185,0 163,2+11,95 23,90 14,6
Кондилобазальная длина . 4 122,0-174,0 156,5+14,02 24,28 15,5
Основная длина 4 112,0-163,0 146,0+13,75 23,82 16,3
Максимальная ширина 7 60,0-85,0 74,0+4,54 11,13 15,0
Скуловая ширина 7 61,0-83,0 72,6±4,22 10,34 14,2
Межглазничная ширина 8 34,0-71,0 48,1±3,95 10,45 21,7
Длина лицевой части 4 54,0-92,0 79,2+10,21 17,69 22,3
Длина носовых костей 6 30,0-61,0 48,5+4,65 10,41 21,5
Длина верхнего ряда зубов 7 31,0,0-60,0 52,6+4,03 9,88 18,8
Максимальная ширина мозговой капсулы 6 52,0-59,0 56,8±1,11 2,48 4,4
Длина нижней челюсти 4 100,0-154,0 132,2±13,37 23,16 17,5
Длина нижнего ряда зубов 4 28,0-68,0 56,0±10,84 18,78 33,5
Длина диастемы нижней челюсти 4 25,0-40,0 35,0±3,97 6,88 19,6
с Северного Кавказа и 11 экз. с Закавказья. В.А.Котов и Л.С.Рябов (1963), проработавшие многие годы в Кавказском заповеднике, приводят промеры всего 3 самцов и 1 самки косуль из Краснодарского края. Несколько больше фактического материала, видимо, имел А.Б.Арабули (1966), выполнивший диссертационную работу по косуле Восточной Грузии (47 экз.). Морфометрический анализ черепа разных популяций косули В.Е.Соколов и В.С.Громов (1985) провели по материалу, содержащему 17 экз. с Северного Кавказа и 13 экз. с Закавказья. Мы имели возможность изучить 57 экз., в том числе 21 с Северного Кавказа и 36 из Закавказья (табл. 19—22).
105
Европейская косуля
Несмотря на ограниченность коллекционного материала и неоднозначность литературных данных, анализ их позволяет усматривать определенную зависимость географической изменчивости косули на Кавказе от высотно-поясной структуры горных ландшафтов. Еще Н.Я.Динник (1910) писал о том, что на Большом Кавказе с- запада на восток происходит уменьшение размеров косули. Многие исследователи от К.А.Сатунина (1906) до многих териологов наших дней (Пастернак, 1955; Соколов, 1959; Гептнер и др., 1961; и др.) считают, что косули в Закавказье заметно меньше, чем на Северном Кавказе. Так, по данным Ф.А.Пастернака (1955), в среднем общая длина черепа закавказских косуль составляет 200,3 мм, а на Северном Кавказе — 225 мм. Кроме габитуальных параметров черепа, этот автор приводит ряд структурных признаков его, подверженных географической изменчивости, и соответственно обосновывает правоту своих предшественников. Как отмечено выше, совершенно к противоположному выводу пришли В.Е.Соколов и В.С.Громов (1985), располагавшие большей выборкой коллекционного материала и применившие современные методы статистической обработки этого материала. Вывод их сформулирован однозначно — нет достоверных различий между косулями Северного Кавказа и Закавказья ни в размерах, ни в пропорциях черепа. Исключение составляют несколько черепов из Краснодарского края, которые по ряду признаков имеют сходство с сибирской косулей. Более того, по мнению этих авторов (Соколов, Громов, 1988), кавказские и европейские косули различаются недостаточно, чтобы их разделить на подвиды. Мы считаем такое заключение бесспорным. Данные, сведенные в табл. 19—22, показывают существенные расхождения по целому ряду краниологических параметров. Бесспорно, что косули Северного Кавказа заметно крупнее своих сородичей Закавказья. Однако, чтобы поставить точки над i, на наш взгляд, необходимы специальные исследования с применением различных методов анализа параметров вида.
До получения новых данных, следуя мнению В.Е.Соколова и В.С.Громова (1988), европейских косуль Кавказа мы относим к номинальному подвиду.
Экология
Ландшафтная приуроченность. Биотопы. Морфофункциональные и экологические особенности европейской косули характеризуют ее как типичный вид лесостепной зоны. По мнению М.КЛимофеевой (1986), косуля — исконный обитатель лесостепных дубрав, в которых она находит наиболее благоприятные условия существования в течение круглого года. Все имеющиеся данные говорят о том, что именно в этой зоне находится оптимум ареала. На север, в лесные зоны она проникает в основном по разреженным лесам и лесным полянам, а на юг, в степную зону — по лесистым долинам рек и балок, а также по искусственным лесонасаждениям. В горах территориальное ее размещение также связано с ландшафтами лесолугового и лесостепного характера. Н.Я.Динник (1910) писал, что чем больше в
106
Caprcolus capreohis Linnaeus, 1758
лесу полян, тем больше они нравятся косулям. Такого же мнения придерживается А.А.Насимович (1939). Все имеющиеся палеонтологические данные, которыми мы располагаем, свидетельствуют, что именно в этих условиях европейская косуля сформировалась как вид, что соответствует костным остаткам, прослеженным со среднего плиоцена, расположенным в пределах восстановленного ареала. При всей экологической пластичности косули она избегает тундру, обширные болота, сплошные таежные массивы, обширные степные, полупустынно-пустынные пространства, лишенные древесно-кустарниковых массивов, а также альпийский, субнивальный и нивальный пояса высокогорья.
При оценке ландшафтной приуроченности европейской косули необходимо учитывать влияние антропогенного фактора. Ф.А.Пастернак (1955) считает, что настоящие лесные и тем более горные биотопы выступают для косули лишь рефугиями, избранными в основном под влиянием оттеснения из равнин деятельностью человека. На наш взгляд, это не относится к лесистым предгорьям и среднегорьям Кавказа. Более того, по мнению Н.Я.Динника (1910), косули любят гористые места, пересекающиеся впадинами и балками, по которым протекают речушки и ручьи с чистой и свежей водой, особенно там, где имеются поляны. При этом они заселяют как хвойные леса, так и лиственные. Словом, современная ландшафтная приуроченность косули сильно трансформирована под влиянием человека, и без оговорки судить об исконных и предпочитаемых ландшафтах нельзя.
Выбор местообитания у косули широко меняется, что зависит от совокупного действия многих абиотических и биотических факторов (климат, погода, рельеф, кормовые и защитные условия, плотность населения популяции, деятельность человека и т.д.). Тем не менее связь косули с древесно-кустарниковой и лесной растительностью четко выражена и сохраняется в течение круглого года.
Лесные угодья предпочтительны тогда, когда насаждения многопородные, многоярусные, разреженные и чередуются лугами, степями и сельскохозяйственными полями. Особенно привлекательны для косули дубовые леса, когда они сильно разрежены и под деревьями растет чистая невысокая трава, состоящая из различных злаков, осоки и т.д. Такие биотопы, расположенные на Западном Кавказе в предгорьях от 300 до 1000 м над ур. моря, благоприятны для косули не только летом, но и зимой (Насимович, 1955). К основным биотопам косули относятся смешанные широколиственные леса, которым отдается большее предпочтение, нежели высокоствольным хвойным лесам. Мало предпочтительны высокоствольные леса верхней полосы лесного пояса. По Н.К.Верещагину (1959), основная масса косули в бесснежный или малоснежный период держится в Закатальском заповеднике в буково-трабовых лесах с полянами и осинником в пределах 700—1800 м над ур. моря. Глубинные участки ущелий с буковыми лесами (1200—1600 м) заселяются значительно реже. Еще в прошлом веке к аналогичному заключению пришел К.Н.Россиков (1887), обследовавший долину р.-Малка от г. Прохладного до подножия горы Эльбрус. В субальпийских лесах Харабуз и Мушт, расположенных в глубоких
107
Европейская косуля
ущельях, он добыл старую самку, но в предгорьях (окрестности с. Малка), где нет лесов, косуль не встречал. Они обитают в низовьях Баксана. Настоящими местами обитания косуль Россиков считал смешанные широколиственные леса "Черных гор" бассейна Терека. Это мнение было подтверждено позднее Н.Я.Динником (1910), А.А.Насимовичем (1950), нами (Темботов, 1960, 1970, 1982)и др.
К основным летним биотопам косуль многие териологи относят разреженные высокоствольные букняки, хорошо продуваемые летом (соответственно здесь меньше насекомых-кровососов), где фон субстрата и окраска косули сходны (косуля мало заметна врагам) и имеется хороший обзор, что позволяет косуле своевременно заметить приближение врага. По мнению С.К.Даля (1948), распространение косули на Памбакском хребте Армении связано именно с буковыми лесами, как старыми высокоствольными, так и молодыми с подростом. Зимой, видимо, буковые леса менее благоприятны для косули из-за глубокого снегового покрова, но на Кавказе, особенно на Центральном и Восточном, такой покров держится обычно недолго. В условиях Азербайджана основными местообитаниями косули считаются кизилоорешниковые ассоциации низкогорья, особенно зимой. В летний период косуля может заселять, по мнению Н.Я.Динника (1910), все типы лесов, включая субальпийские, что нами отмечалось в наши дни неоднократно и притом в различных районах Кавказа. Экологическая пластичность косули настолько велика, что она может жить в ольховых топях Колхидской низменности (Верещагин, 1959), но, видимо, это обусловлено антропогенным прессом. Очевидно, положительное отношение косули к островным лесам степей и лесостепных предгорий, о чем красноречиво говорят результаты успешной реакклиматизации косули в Ставрополье (Емельянов, Рыбалов, 1967).
Наши данные и многочисленные литературные источники однозначны в том, что косули предпочитают лесосеки, поросшие молодым лесом, но сплошную непролазную чащу, которая так нравится кабанам, косули избегают (Динник, 1910). Охотно заселяются косулями различные кустарниковые заросли ивняка, боярышника, шиповника и др. По балкам с такими зарослями косуля спускается в предгорную степь бассейна Уруп (Котов, Рябов, 1963). Там, где ее не преследуют, косуля успешно обитает в лесных насаждениях равнины и в пойменных лесах, в том числе по рекам Терек, Самур и др. В условиях Восточной Грузии (сухой субтропический тип поясности), по А.Б.Арабули (1966), местообитания косули представлены следующими пятью типами лесов: 1) равнинные и пойменные леса (200—500 м над уровнем моря): 2) дубово-трабовые леса (500—1000 м); 3) буковые леса (1000—2100 м); 4) субальпийские леса (1800—2200 м); 5) смешанные леса (700—2000 м). Редка в полно возрастных дубово-вязовых лесах, отсутствует в шибляке (сухо), избегает чисто хвойные леса, в зарослях вечнозеленых кустарников (типа понтийской ежевики) обычно не встречается. Отношение косули к одним и тем же насаждениям может сильно меняться. Так, радиотелеметрическое наблюдение, проведенное в Баварии, показало, что при сухой погоде дневки косуль располагаются в
108
Capreolus capreolus Linnaeus, 1758
молодом подросте или на участках возобновления в зрелом лесу, при влажной — в зрелом лесу в течение всего дня. В августе-сентябре косули встречаются в различных типах леса, но ночью — только в старолесье (K6'nig, 1987).
К местообитаниям косули относятся открытые участки с весьма разнообразной травянистой растительностью и полевыми культурами. Излюбленными являются лесные поляны с сочной растительностью. По Н.Я.Дин-нику (1910), некоторые сильно заросшие бурьяном поляны косули особенно любят и здесь их можно найти чаще, чем на других полянах. Летом они часто ложатся в густых высоких бурьянах, нередко встречающихся на Кавказе по лесным опушкам и полянам. Предпочитаемы также заросли папоротника орляка, но обычно они ложатся по окраинам полян, а не в середине обширных сплошных зарослей. Любят заросли папоротника, перемешанные с мелким дубняком или полосами рододендровых кустарников. В начале текущего столетия много косуль было среди степи, заросшей высоким бурьяном (окрестности Змейской, Северная Осетия). К летним биотопам относятся различные травянистые экосистемы субальпийского пояса, в том числе субальпийское высокотравье. Здесь в бесснежный период косуля находит достаточно корма и хорошие защитные условия. Предпочтение к высокотравью усиливается с запада на восток, что, очевидно, обусловлено ростом сухости горных ландшафтов в том же направлении. Вероятно, прав А.А.Насимович (1939) в том, что косуля на открытых альпийских лугах бывает только проходом. Зимой высокогорья, в том числе субальпийское высокотравье, становятся мало пригодными для жизни косули, что вызывает местные кочевки.
Местообитания косули на Кавказе мало связаны с агроэкосистемами, как это имеет место в Западной Европе, в Прибалтике. В этих местах, кроме лесной расы,, широко встречаются косули, тесно связанные с полевым ландшафтом и характеризующиеся как морфологическими, так и экологический особенностями (Падайга, 1965, Prossinagg, 1988; и др.). По нашему мнению, на Кавказе имеются необходимые предпосылки для заселения косулей многих равнинно-предгорных районов с полеводством и садоводством (Краснодарский и Ставропольский края, Дагестан и др.).
Миграции. При всей привязанности косуль к территории они вынуждены, кроме биотопических перемещений, совершать более или менее дальние сезонные перемещения. В условиях Западного Кавказа (кубанский вариант) это явление наиболее полно изучено А.А.Насимовичем (1955). По его данным, летом косули имеют широкие пределы высотного распространения, достигая альпийского пояса, хотя основная масса косуль сосредоточена в широколиственном поясе гор, т.е. не выше 1200—1400 м над ур. моря. На зиму косули откочевывают ниже по склону гор и большей частью живут в дубовых и смешанных лесах, богатых лиственным подлеском и подростом, обычно не выше 1000 м. Держатся на участках, где снеговой покров, как правило, не выше 30 см. Перемещение сверху вниз происходит в течение октября и ноября. Начало его совпадает обычно с первым значительным снегопадом и заканчивается, когда на высоте 1500 м
109
Европейская косуля
устанавливается постоянный снежный покров. При малоснежных зимах косули могут оставаться высоко в горах. Обратная кочевка выше в горы происходит в апреле-мае. Протяженность высотных перемещений в одну сторону колеблется от 5 до 30 км, иногда до 40—50 км.
В связи с климатическими условиями особо значительные высотные миграции косули происходят в Западном Закавказье (влажносубтропический тип поясности). Такие большие кочевки, обусловленные режимом снежного покрова в прошлом, наблюдали нередко в Абхазии (Чернявский, 1879; Юхнович, 1887) и Мегрелии (Радде, 1899; Насимович, 1955). После выпадения большого снега косули появлялись иногда даже в ближайших окрестностях Сухуми. На Центральном и Восточном Кавказе, где значительно меньше осадков, нижняя граница многоснежья располагается довольно высоко.Соответственно высотные перемещения косули в этих районах незначительны. Зимой в Кабардино-Балкарии еще К.Н.Россиков (1887) наблюдал перемещение косуль с Черных гор в низовья Баксана, а А.А.На-симович (1955) — в Черек-Балкарском ущелье сверху вниз по ущелью. В суровые и многоснежные зимы местные кочевки косули наблюдались И в Предкавказье. Так, Н.Я.Динник (1910) описывает перемещение косуль из лесов Ставропольской возвышенности в долину Кубани. Что касается характера сезонных миграций косуль в Дагестане, то сведениями о них мы не располагаем.
В Восточной Грузии, преимущественно в пределах юго-осетинского варианта, по данным А.Б.Арабули (1966), более или менее значительные высотные кочевки косули совершают, когда выпадает большой снег и наступают внезапные морозы в горах. В обычные зимы косули перемещаются на юго-восточные слабо заснеженные скалы, где благодаря солнечной радиации и ветрам снега мало и имеется корм в виде травянистой и древесной растительности. Результаты учета следов косули, проведенного Н.К.Верещагиным (1959) в Закатальском заповеднике в разные сезоны года, показывают значение снежного покрова в горах Восточного Кавказа:
Дата учета Южные отроги хребтов. Буково-грабовый лес с полянами и осинником, 700-1800 м Глубинные участки ущелий с буковым лесом, 1200-1600 м Верхняя опушка леса и субальпийские луга, 2000—2400 м
22-26,VIII1935 г. 21-26,XI 1935 г. 8 2 1
(снег 15—30 см) 1.1 1936 г. (снег 16 3 —
30—50 см и более) 7 1 —
Как известно, половые и возрастные особенности ландшафтной и био-топической приуроченности, как и в равной степени их сезонные проявления, у косуль значительны. Однако мы не располагаем аналогичными данными по Кавказу.
1 1 0
Capreolus capreolus Linnaeus, 1758
Убежища. Лежки. Как отмечено выше, косуля спасается от жестких климатических и иных условий жизни, сменяя различные биотопы, в том числе разнообразные типы лесов, лугов, лесных полян или субальпики, бурьяны, кустарники, иногда камышовые заросли в предгорьях и на равнине. Географический аспект смены предпочитаемых убежищ нуждается, на наш взгляд, в самостоятельном изучении и с учетом высотно-поясной структуры горных ландшафтов Кавказа. В подтверждение можно сослаться на отдельные примеры. Так, на Западном Кавказе, где влажность климата очень высока, косуля как летом, так и зимой любит ложиться в редких дубовых или дубово-грабовых лесах (Динник, 1910; Насимович, 1939). В Армении, отличающейся сухим климатом, в летнее время лежки косули чаще встречаются в разреженных буковых лесах, тогда как в дубовых — заметно реже (Даль, 1948).
По Н.Я.Диннику (1910), косули зимой часто ложатся на солнечных пригревах и могут находиться там продолжительное время. Прежде чем ложиться, они разгребают снег до самой земли, даже при толщине снегового покрова не менее 70 см. В такой глубокой лежке над снегом видна только голова косули. Нередко также разгребает опавшие листья. Характерно, что косуля ложится аккуратно, нс разметываясь, а подобрав под себя ноги, на груди и животе. На бок не ложится, кроме как будучи сильно пораненной.
Сезонная и суточная активность. Поведение. Анализ обширной литературы и результатов собственных наблюдений В вольере и полевых условиях свидетельствует о том, что экология и поведение европейской и сибирской косуль очень сходны (Соколов, Данилкин, 1981). Еще в начале прошлого века, когда не вызывала сомнения реальность существования обоих видов на Кавказе, известный полевой исследователь и заядлый охотник Н.Я.Дин-ник (1910) не обнаружил каких-либо существенных отличий в образе жизни между этими видами, кроме как различия в ареалах и, как отмечено выше, по морфологии. Зимой и летом для косули характерен полифазный тип активности (2—11, в среднем 5—8 раз в сутки) (Соколов, Данилкин, 1981). В условиях Кавказа пищевая активность косули начинается перед рассветом, а к 7—8 ч желудки наполняются непереваренной пищей. В полдень, особенно в летние жаркие дни, она малоактивна, отсиживается в тени или в высоких травостоях, где меньше кровососущих насекомых. Активность вновь повышается вечером, в 17—18 ч. Ночью активность резко снижается, косули обычно находятся в лежках. Летом продолжительность кормежки редко длится больше 1 ч, но зимой увеличивается в 1,4 раза. В пасмурные дни или в безопасных местах дневная активность возрастает, но там, где ее преследуют, она вынуждена перейти на пастьбу утром и вечером.
На вид косуля — миловидное (рис. 17), доверчивое и миролюбивое животное, но это справедливо по отношению к самкам и молодым животным. Самцы с появлением рогов и достижением половозрелости становятся агрессивными, злыми, неуживчивыми. В это время они опасны даже для взрослого человека, не говоря о детях. Немало литературных данных (см. обзор: Соколов, Данилкин, 1981), свидетельствующих о том, что между
1 1 1
Европейская косуля
Рис. 17. Европейская косуля
1 1 2
Caproleus caproleus Linnaeus, 1758
уровнем агрессивности самцов и размерами семенников и содержанием в них тестостерона существует прямая коррелятивная связь. Соответственно агрессивность самцов возрастает летом, а зимой падает до минимума. Агрессивное поведение, связанное с размножением, у самок проявляется по отношению к прошлогодним телятам за три недели до отела. В это время мать, отгоняя от себя молодняк, вынуждает его мигрировать.
Косулям свойственны и другие элементы агонистического поведения, как демонстрация "силы", угрозы, ложные нападения, борьба и поведение после боя. Известно маркировочное поведение в летний период: ольфакторная и ольфакторно-оптическая метки; возможно, ту же роль играют звуковые сигналы. Ориентировочно-исследовательское и оборонительное поведение включает ряд элементов: ориентировочные позы, лай, "топтание", сигнальные прыжки, уточнение опасности обонянием, бегство и затаивание. Этой проблеме посвящена обширная литература как у нас в стране, так и за рубежом (см. обзор: Соколов, Данилкин, 1981).
Линька. По этому вопросу существуют различные взгляды. Так, В.Г.Гептнер с соавторами (1961) и некоторые другие авторы признают у косули одну весеннюю линьку. Н.Я.Динник (1910), И.И.Соколов (1959) и др. считают, что косуля меняет волосяной покров дважды в году — весной и осенью. Суть разногласий терминологического порядка — если под линькой понимать только замену старых волос на новые (как весной), то у косули одна линька. Но в случае, когда за линьку признается качественное изменение волосяного покрова за счет отрастания летних волос и обновление его новыми (как это происходит осенью), то у косули две линьки. Мы придерживаемся второго мнения.
Фенология линьки косули на Кавказе зависит от многих факторов, в том числе от возраста, пола, упитанности, здоровья, а также от ландшафтных условий — высоты местности, климата и т.д. Так, в теплых районах Кавказа, по Н.Я.Диннику (1910), линька начинается в марте, возможно даже в конце февраля. В горах — на месяц с лишним позже, чем на низменности (Радде, 1899). В середине апреля на Западном Кавказе косули несут много еще старых волос, но в мае они совершенно "красные", т.е. имеют летний мех. Осенняя линька начинается в августе и заканчивается в октябре. Осенняя линька продолжительнее, так как короткие летние рыжие волосы отрастают медленно, превращаясь в редкую ость зимнего меха (длиной 5—7 см). Между этими волосами прорастают более короткие, толстые и сильно волнистые зимние волосы, соответствующие пуху других млекопитающих.
Питание. Состав пищи европейской косули очень разнообразен. В условиях лесостепи Тульской области, где питание косули изучалось более 20 лет (Тимофеева, 1^86), установлено около 140 видов растений, поедаемых ею. Среди них 39 видов деревьев и кустарников, 95 травянистых растений, несколько видов лишайников, мхов и грибов. В Восточной Грузии, по А.Б.Арабули (1966), ассортимент поедаемых косулей растений включает не менее 120 видов, в том числе 40 видов древесно-кустарниковых, 16 злаковых, 61 разнотравья и 3 вида грибов. В Армении питание косуль изучал
8 В.Е.Соколов, А.К.Темботов
1 1 3
Европейская косуля
С.КДаль (1948). Он приводит список кормов, включающий всего 12 видов растений. Интересные сведения по Кавказу содержатся в монографии Н.Я.Динника (1910), хотя он приводит мало конкретных видов. Все имеющиеся данные свидетельствуют о том, что состав предпочитаемых видов кормовых растений меняется в зависимости от сезона года, ландшафтных и биотопических условий и т.д. В начале весны косуля охотно поедает раннецветущие растения, с появлением листвы и молодых побегов деревьев и кустарников значительную долю составляют веточный корм и листья. Однако по мере огрубления побегов и листьев заметно увеличивается- доля разнотравья. По Н.Я.Диннику (1910), они очень любят отаву и вообще молодую траву. Осенью видное место в пищевом рационе составляют грибы, ягоды, буковые орехи, желуди, дикие яблоки и груши и т.д. Зимой велико значение зимне-зеленых растений, как ежевика, омела, зимний хвощ, плющ, горная овсяница. Однако основную массу составляет ветошь, которую косуля может копытить из-под снега толщиной до 10—15 см. Из-под снега косуля достает также желуди, более привлекательные плоды и листья, зелень и другие растения.
Суточное потребление корма неустойчиво, что зависит от запаса его в природе, сезона года, его перевариваемости. Весной и летом, когда имеется обильная зеленая растительность, косуля потребляет за 1 день 3—4 кг фитомассы. При переходе на веточный корм косуля съедает за сутки от 1,5 до 2,7 кг, телята — 0,6—0,9 кг. Зимой, когда косуля питается преимущественно пожухлыми растениями, суточная норма снижается до 2 кг, что связано с уменьшением перевариваемости пищи. Дело в том, что, чем меньше питательность и усвояемость кормов и соответственно меньше запас энергии, поступающей в организм, тем меньше корма поедается. Так, весной перевариваемость фитомассы возрастает до 73,5% против 10% зимой (Тимофеева, 1986).
В пищевом поведении косули характерна избирательность в поедании растений — степень поедания отдельных видов нередко определяется не обилием их в природе, а предпочитаемостью их перед другими. К таким излюбленным кормам относятся кизил, жасмин, клен, бересклет, дуб, ива, ежевика, черника и др. Из разнотравья — клевер, люцерна, крестовик, борщевик, осока, злаки и др. Набор предпочитаемых растений .может сильно меняться в разных частях ареала. Ухудшение кормовой базы приводит к сдвигам в составе поедаемых растений. По В.Г.Гептнеру и др. (1961), питание сосной означает напряженное положение с зимними кормами. Тоже самое можно сказать и о ели — косуля поедает ее в тех случаях, когда плотность популяции чрезмерно высока и кормов не хватает.
Косуля для кормления иногда посещает сельскохозяйственные угодья, находящиеся вблизи лесов. Судя по литературным данным и нашим наблюдениям, она редко удаляется на большое расстояние от леса. Только однажды (осенью 1987 г.) мы зарегистрировали косулю в окрестностях с. Чегем-П (Кабардино-Балкария), примерно в 10 км от предгорных лесов. Однако нам не известен случай, чтобы где-либо на Кавказе косуля закрепилась сколько-нибудь долго на сельскохозяйственных полях, что указывало
1 1 4
Capreolus capreolus Linnaeus, 1758
бы на возможность естественного формирования полевого экотипа, столь характерного для многих районов Западной Европы и Прибалтики.
На полях косуля поедает широкий круг полевых и садовых культур, различные породы деревьев и кустарников лесополос, овощные культуры. В связи с этим, на наш взгляд, заслуживают внимания данные польских экологов о том, что полевой экотип европейской косули не только не приносит ущерба посевам, а даже способствует повышению урожайности и увеличению биомассы растений при условии, если плотность косуль не превышает 10 особей на 100 га (Пеловский, 1976, Kaluzinski, 1976, по: Соколов, Данилкин, 1981). Такое положительное воздействие обусловлено особенностями пищевого поведения косули — она постоянно перемещается по участку, срывая с растения только 1—2 листочка или обкусывая соцветие, после чего растение кустится и масса его увеличивается в 1,5—2 раза. С такого растения без вреда для него косуля может брать пищу еще несколько раз. Что касается кормовых достоинств полевых культур, то опасность для косули могут представлять отдельные сорта. Так, исследования, проведенные в Австрии и ФРГ, показали, что в районах интенсивного возделывания рапса участились случаи гибели косуль, так как при систематическом его поедании он вызывает у косуль гемолитическую анемию. В крови остается только 50% здоровых эритроцитов. У погибших косуль отмечалось пенистое брожение в рубце, заполненном зеленой массой рапса. Заболевание чаще наблюдается зимой. При нормальном питании больные косули выздоравливают через 4 недели. Очевидно, рапс опасен только для косуль (Оп-derscheka et al., 1987а, б; Onderscheka, 1988).
Воспроизводство и структура популяции
Размножение. По Н.Я.Диннику (1910), на Кубани гон косули начинается с самых первых чисел августа или в конце июля. Продолжается он около месяца или немного более, заканчивается в начале сентября. В Восточной Грузии косули спариваются с мая по сентябрь, в основном в июне-августе (Арабули, 1966). По С.К.Далю (1954), в Армении период гона косули длится от конца июля до середины сентября. Во время гона самец довольно aipec-сивен, может ударить рогами самку, старается удержать ее на своей территории. Это длительное ухаживание, а затем погоня обычно завершаются бегом самки и за ней самца вокруг дерева или кустарников, где происходит спаривание изможденных партнеров. Многократный пробег по кругу оставляет сильно выбитую тропу. Самке удается иногда уйти от самца и спрятаться, но он находит ее по следам с помощью обоняния. Один самец может оплодотворить несколько самок. Во время гона самец сильно худеет, теряет осторожность, издает характерные "гонные" звуки; в качестве оборонительной реакции самки издают несколько свистящих звуков (Соколов, Данилкин, 1981). Между самцами нередки драки, часто оканчивающиеся травмами, даже смертью. После окончания гона к самке возвращаются прибылые, восстанавливается семейная труппировка.
8*
1 1 5
Европейская косуля
В начале беременности оплодотворенное яйцо находится в диапаузе, развивается очень медленно, в начале зимы эмбрион простым глазом не заметен, лишь в декабре — середине января его можно увидеть. Этому периоду соответствует начало интенсивного развития плода, сопровождающееся повышением активности косули (соответствует ложному гону куньих). В окрестностях пос. Гузерипль Кавказского заповедника в конце февраля 1954 г. масса двух изученных эмбрионов составила 125 и 160 г, а 28 апреля 1953 г. — 679 и 721 г. Перед рождением детеныши весят около 1 кг (Котов, Рябов, 1963). Массовый отел происходит в мае или в начале июня. Соответственно продолжительность беременности достигает 10 мес; она варьирует за счет длительности латентного периода.
Для отела самки выбирают лучшие по кормовым и защитным условиям угодья — опушки и лесные поляны с высоким травостоем. В густом травостое самка рожает, прячет детенышей и кормит их, периодически возвращаясь с пастьбы. Косулята первые дни неподвижно лежат на земле. Через неделю они могут следовать за матерью.
Сперматогенез у косули в Восточном Закавказье начинается, по А.Б.Ара-були (1966), в апреле и усиленно продолжается до конца августа. В сентябре он замедляется, в октябре в канальцах придатка мало сперматозоидов, в ноябре-декабре они вовсе отсутствуют. Половой активности соответствует размер семенников — в июле масса его около 32 г, размеры 55 х37 мм, а в декабре соответственно 6,8 г, 35 х 20 мм.
Плодовитость. Самка ежегодно приносит от одного до трех косулят. Чаще бывает два, очень редко три. У молодых обычно рождается один детеныш.
Рост и развитие. В постнатальном развитии европейской косули в условиях вольера В.С.Громов (1988) выявил два периода роста. Первый — от рождения до 6—7 мес. — характеризуется быстрым темпом нарастания массы и размеров тела (в 13—15 раз увеличивается масса, в 2,7—2,9 раза — размеры тела по сравнению с показателями при рождении). Второй период составляет от 6—7 мес. до 3—3,5 лет и отличается замедлением роста и сезонными колебаниями массы и размеров тела. За это время масса тела увеличивается всего на 50%. Размеры тела растут на 15—20%. При достатке полноценных кормов косули в возрасте 3,5 лет практически достигают максимальной массы тела. В природе самки косуль приступают к размножению на 2-м году жизни, первое потомство приносят соответственно на 3-м году жизни (Ellenberg, 1978; Stubbe, Passarge, 1979; Соколов, Данилкин, 1981, по: Громов, 1988). Половозрелость самцов наступает также на 2-м году жизни (к 14—15 мес.), но возможна и раньше; в полугодовалом возрасте наблюдались кратковременные первые стадии сперматогенеза (Цаплюк, 1977; Short, Mann, 1965, 1966 гг., по: Соколов, Данилкин, 1981).
Возрастные изменения окраски у косули значительны. Молодые рождаются пятнистыми, но к месячному возрасту пятнистость обычно исчезает (Соколов, 1959). В возрасте 4 мес. молодые окрашены, как взрослые (Гепт-нер и др., 1961). В.С.Громов (1986) считает, что к 2—2,5 мес. окраска становится однотонной, как у взрослых животных.
1 1 6
Capreolus capreolus Linnaeus, 1758
Изменения структуры рогов и зубной системы в связи с возрастом изучены И.И.Соколовым (1956) на основе анализа значительного коллекционного материала. Этот материал позволил решительно опровергнуть существовавшее мнение о зависимости развития рогов, в том числе количества отростков, от возраста животного. Достаточно сказать, что из 17 черепов самцов, возраст которых по состоянию зубов соответствовал 12—18 мес., рога с простыми спичками имеются у 5 экз.; с двумя концами на каждом роге — у 2 экз.; у трех черепов один рог имел два, а второй — три конца; у 6 самцов были вполне развитые и с тремя концами рога. Один самец имел рога в виде длинных спичек, покрытые шерстью" и с зачатками всех трех отростков. Как видно, рога у косули достигают полного развития в возрасте 11—18 мес. и обладают характерным для вида числом ветвей — по три на каждом роге. Однако такое состояние достигается разными животными в разном возрасте.
Развитие массы тела и рогов и длина их стержней достигают максимальных величин в возрасте 7 лет (Necas, 1975).
В отличие от рогов зубная система в постнатальный период жизни претерпевает закономерные изменения и на|основе анализа сроков появления, смены и стирания коренных зубов верхней и нижней челюстей с большой достоверностью можно определить возраст косули. Для целей определения возраста косули не могут служить резцы, так как в их стирании нс существует какой-либо закономерности. И.И.Соколов (1956) показал, что в течение 17 мес. в зубной системе происходят определенные изменения ежемесячно, что позволило ему выделить 17 возрастных стадий. Основные положения возрастных изменений зубной системы таковы.
1. Новорожденные имеют все молочные зубы (резцы и предкоренные).
2. Молочные резцы сменяются на постоянные, начиная с внутренней пары (11), в возрасте от 5 до 11—12 мес. При этом наружная пара резцов (13) и клыки (С) сменяются в последнюю очередь и почти одновременно.
3. Молочные предкоренные на постоянные сменяются в возрасте 13—14 мес. почти одновременно в обеих челюстях.
4. Прорезывание коренных через слизистую оболочку ротовой полости начинается с трехмесячного возраста, причем вначале передних (М 1 и Mi), а потом задних (М3 и Мз). В нижней челюсти оно заканчивается на 3—4 недели раньше, чем в верхней.
5. Полного развития зубная система достигает в возрасте 15 мес. жизни животного.
Стирание зубов и соответствующие возрастные изменения в их структуре, по Ф.А.Пастернаку (1955), у косули наступают поздно. В возрасте 1,5 лет предкоренные зубы не имеют следов стертости. По достижении 3 лет эти следы появляются на гребнях первого Р и первого М. В четырехлетием возрасте стерты гребни первого и второго Р и первого М. К 5—6 годам полностью стираются гребни всех Р и частично гребни второго М. В 7 лет полностью стираются гребни второго М, в 8—9 лет — третьего
11 7
Европейская косуля
М. У 10—12-летних жующая поверхность зубов совершенно гладкая. Известна продолжительность жизни косули до 18 лет.
Возрастные изменения структуры черепа значительны (Пастернак, 1955), но в условиях Кавказа не изучены.
Структура популяции. В литературе по этому вопросу данных практически нет ни по одному району Кавказа. Лишь А.Б.Арабули (1966) приводит некоторые сведения по возрастному и половому составу косули в условиях Восточной Грузии. Среди отстрелянных 47 косуль соотношение полов было 1:1. Взрослых оказалось 69,8%, а молодых — 30,2%. В группе взрослых самцов и самок было почти одинаковое количество, а среди молодых животных самцов было немного больше (56,3%), чем самок (43,7%). Иные результаты получены по материалам количественного учета (159 данных). На долю взрослых приходилось 81,1%, молодых — 18,9%. Среди взрослых (п = 68) преобладали самки (52,7%). Автор приходит к выводу, что соотношение полов косули на обследованной территории неблагополучное, поскольку оно должно быть по крайней мере 1 <С:2 j . Неблагополучен, по его мнению, также возрастной состав популяций косули в Восточной Грузии.
Враги, паразиты, смертность, конкуренты. У косули много врагов, особенно у козлят. Из всех хищников наиболее опасны волк и рысь, которые уничтожают косуль начиная с первых дней жизни. Сильно страдают косули от волков в снежные зимы. Они становятся жертвой шакалов, численность которых резко возросла на Северном Кавказе. В желудке медведей находили остатки косуль. Косулят пожирают лисицы, дикие кошки, енотовидные собаки. Н.Я.Динник (1910) полагает, что куница также, видимо, нападает на косулят. Роль хищных птиц, очевидно, мала. Что касается одичавших собак, которых становится все больше, то они, вероятно, наносят существенныйущерб поголовью косули.
Косули подвержены целому ряду заболеваний, в том числе ящуру, сибирской язве, чуме рогатого скота, туберкулезу и др. В Восточной Грузии А.Б.Арабули (1966) отметил следующие заболевания: глазное (Leucoma), челюстных костей (Osteomielitis), ороговение сосков щечной слизистой оболочки (Hiperceratosis) и злокачественная опухоль семенников (Seminoma).
По данным Д.П.Рухлядева (1964), у косуль найдено 113 видов паразитических червей. Из них на Кавказе зарегистрировано 39, в том числе 9 видов на северо-западе Кавказа, по 18 видов в Азербайджане и Армении. В Восточной Грузии А.Б.Арабули (1966) выявил 15 видов, причем из 40 вскрытых косуль 21 оказалась зараженной гельминтами. Наиболее часты Congylo-ncma pulchrum и Dictyocaulus eckerti.
Многочисленными бывают эктопаразиты. В частности, большинство добытых в Восточной Грузии косуль были поражены массой эктопаразитов (Sipoptera cervi и Ixodes ricinus) (Арабули, 1966). Повсеместно на Кавказе косули страдают от слепней, комаров и мошек.
Из абиотических факторов очень велика отрицательная роль снегового покрова, затрудняющего передвижения косуль. Известны случаи, когда из-
1 1 8
Caprcolus caprcolus Linnaeus, 1758
за суровости зимы и обилия волков погибало 49—80% приплода (Саблина, 1955 г., по: Гептнер и др.,-1961). Для популяции косули Западного Кавказа очень тяжелой оказалась чрезвычайно многоснежная и продолжительная зима 1953/54 г. Снежный покров достигал местами 80—100 см (февраль, окрестности Майкопа). В эту зиму большое количество беспомощных животных погибло от браконьеров и собак (Котов, Рябов, 1963).
Очевидно, наиболее значительную конкуренцию косулям составляют благородный и пятнистый олени.
Численность и ее динамика. Данные, которыми мы располагали, свидетельствуют о том, что на Кавказе повсеместно косуля относительно редка, что в первую очередь объясняется массовым браконьерством. Достаточно сказать, что за редким исключением в предгорных и среднегорных районах Кавказа сохранились благоприятные условия для косули — разреженные леса, вторичные луга, лесосеки, чередующиеся небольшими полями сельскохозяйственных культур. И тем не менее численность косуль в этих местах низка, а в ряде районов она отсутствует совсем.
Возможно, в будущем на косуле отрицательно будет сказываться химизация сельского хозяйства и промышленное загрязнение среды.
Практическое значение. Косуля относится к ценным и перспективным видам охотничье-промысловых животных Кавказа. Каждый зверь дает чистого мяса в среднем 15 кг, что составляет около 60% живой массы. Косуля бывает наиболее упитанной летом до гона или в конце осени — начале зимы. В это время, по словам Н.Я.Динника (1910), отдельные самцы дают чистого мяса (не считая головы и ног) до 2 пудов, бывало даже 2,5 пуда. Жира можно собрать более 2 кг. По вкусовым качествам мясо косули уступает ряду других копытных, но бесспорно относится к высококачественным продуктам.
Шкуры косули идут на различные нужды, в том числе из них делают шубы, спальные мешки, коврики, потники для седел и т.д. Не менее важна косуля как охотничий трофей, ее рога могут быть красивым украшением.
Отрицательную роль косуля играет в полеводстве, лесном хозяйстве, в том числе лесонасаждениях, что ощущается лишь при очень высокой численности животных.
Многолетнее слежение за развитием отдельных экземпляров подроста бука, граба, сосны и ели показало, что многократное скусывание косулей основных и боковых побегов не снижает интенсивности роста и качества древесины (Meidel, 1987). В разделе "Питание" было отмечено, что по данным польских экологов косули в условиях культурного ландшафта стимулируют прирост фитомассы ряда полевых культур.
Бонитировка угодий для косули на Кавказе не проведена, но наши полевые наблюдения и отдельные литературные данные (Арабули, 1966) говорят о том, что количество косуль на Кавказе можно увеличить в 5—10 раз.
1 1 9
Род Олени
Род Олени — Cervus Linnaeus, 1758
Копытные различных размеров, высота в холке 60—165 см, длина тела 105—265 см. Самки несколько меньше самцов. Телосложение легкое, стройное, сравнительно длинные и крепкие конечности, передние несколько короче задних. Конечности четырехпалые, хотя боковые пальцы недоразвиты, но имеют скелет фаланг. Шея умеренной длины, обычно с удлиненными волосами, голова с заостренной лицевой частью, длина ее превосходит ширину более чем в 2 раза. На конце морды имеется обширный безволосый участок. Уши заостренные, длинные и прямостоящие. Рога с одним или двумя надглазничными отростками, размеры и степень их ветвистости сильно варьируют. Рога обычно в поперечном сечении округлые (у лани на конце уплощенные), поверхность стволов шероховатая, но выростов, как у косули, нет. Хвост хорошо развит, выступает из мехового покрова, всегда виден, но не достигает скакательных суставов. Окраска взрослого животного круглый год одноцветная или пятнистая (только в летнее время). Как правило, хорошо развито белое "зеркало" на задней поверхности ляжек. Окраска молодых в первые месяцы жизни пятнистая. Линяют 2 раза в году. Самцы сбрасывают рога обычно весной, а к осени рога вновь полностью вырастают и происходит чистка рогов от кожи. Характерны многочисленные специфические кожные железы: предглазнич-ные, межпальцевые, запястные и пяточные (могут отсутствовать), хвостовые (имеются у немногих видов).
В черепе лицевая часть развита умеренно, максимальная его ширина составляет более 40% длины. Лобные кости крупные, а теменные уменьшены. Межчелюстные кости по своей длине обычно не превышают длину ряда верхнекоренных зубов, они всегда соприкасаются с носовыми костями, которые не укорочены. Слезные кости с глубокими ямками для предглаз-ничных желез. Этмоидальные отверстия крупные. Костные слуховые барабаны средних размеров.
Верхний клык обычно имеется у самцов, но слабо развит. Исключение составляет подрод Dama. У самок верхний клык наблюдается редко.
Зубная формула:
Диплоидный набор палеарктических видов, как и для большинства оленей, характеризуется большим количеством: 64—68 при постоянном числе плеч, равном 68. Y-хромосома варьирует по морфологии, а Х-хро-мосома у всех видов акроцентрическая (Орлов, Булатова, 1983).
Ареал рода обширный, состоит из двух разобщенных частей, которые расположены в Старом и Новом Свете. В Евразии северная граница приближается или местами (Западная Европа и Восточная Сибирь) пересекает 60° с.ш., а на юг простирается до Западной Африки, Малой Азии, Ирана, Индии, Тайваня, Филиппин, Японских островов. В Новом Свете северная
120
Cervus Linnaeus, 1758
граница достигает также 60’ с.ш., а южная — до 35’ с.ш. (до Мексики). В ряде районов, в том числе Европы, олени были уничтожены. В настоящее время ведется большая работа по их реакклиматизации. Акклиматизированы в новых местах вплоть до Южной Америки и Новой Зеландии.
Олени населяют леса различных широтных зон на равнине и высотных поясов в горах. Живут небольшими группами (до 10 голов), взрослые самцы большую часть года проводят отдельно от самок. Активны утром и вечером. Характерны горизонтальные и высотные миграции. Питаются летом разнообразной растительной пищей, а зимой преимущественно древесно-веточными кормами. Регулярно посещают естественные и искусственные солонцы. Полигамы; самцы в жестокой драке отбивают несколько самок. Спаривание, сопровождаемое ревом самцов, происходит обычно осенью. Беременность длится около 8 мес., телята рождаются весной или в начале лета. Самки приносят по одному теленку, достаточно хорошо развитому и вскоре после рождения способному следовать за матерью. Поло во зрелость наступает в двухлетнем возрасте, но самцы вступают в размножение значительно позже, после достижения полного физического развития.
Главный враг оленей — волк, но на них нападают и другие хищники, в том числе рысь, медведь и шакал.
Олени как объект спортивной и промысловой охоты высоко ценятся, в ряде районов страны и за рубежом относятся к числу ведущих охотничьих животных. Высоко ценятся неокостеневшие рога оленей — панты, используемые для изготовления лечебного препарата — пантокрина. Для этой цели широко практикуется полувольное содержание пятнистых оленей и марала.
Систематика рода нуждается в дальнейшей разработке, так как нет единого мнения в отношении объема рода, реальность существования отдельных видов все еще спорна. В отечественных сводках приводятся от 12 до’16 видов (Флеров, 1952; Соколов, 1959;.Гептнер и др., 1961; Соколов, 1979; и др.). В фауне Кавказа 2 вида, один из них аборигенный, а второй — акклиматизированный.
Пятнистый олень — Cervus nippon Temmlnck, 1838
Рис. 18
С. dybowskii Taczanovski, 1864 (цит. по: Гептнер и др., 1961).
Коллекционный материал. МГУ: 131 черепов; КБГУ: 57 черепов, 12 полных скелетов, 5 шкур, 30 проб волос с 30 животных.
Диагноз. Олени средних размеров, высота в холке в среднем около 120 см, длина хвоста соответствует длине уха. Летний мех животных всех возрастов пятнистый, зеркало небольшое и не распространяется выше корня хвоста.
1 2 1
Пятнистый олень
Рис. 18. Распространение пятнистого оленя
122
Cervus nippon Temmlnck, 1838
1 — Теберда, долина Азчек (Иня-кова, 1957);
2 — Вольские пастбища (данные авторов);
3 — Белореченское отделение зверосовхоза "Майский" (данные авторов; КБГУ);
4 — Нальчикское лесоохотхозяй-ство (данные авторов);
5 — Кенженское лесничество (КБГУ);
6 — Баксанский гос. заповедник (данные авторов);
7 — зверосовхоз "Майский" (данные авторов);
8 — пос. Дылым, Казбегский лесхоз (Шахмарданов, Рамазанов, 1980);
9 — Алмахское лесохозяйство (Шахмарданов, Рамазанов, 1980);
10 — Алтыагач (данные авторов);
11 — Гирканский заповедник (Соколов, Сыроечковский, 1990);
12 — Дилижанский заповедник (Соколов, Сыроечковский, 1990);
13 — Хосровский заповедник (Гамбарян; 1959; Гамбарян и др., 1963; Соколов, Сыроечковский, 1990)
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Аборигенная популяция пятнистого оленя в Южном Приморье (Лазовский заповедник) характеризуется более крупными размерами, чем было принято считать в монографиях, опубликованных до 60-х годов нашего века. Так, по данным В.Е.Присяж-нюка (1981), масса тела оленя этой популяции составляет в среднем у самцов (л = 5) 134,2 кг, у самок (л = 6) 91 кг.
12 3
Пятнистый олень
В.Г.Гептнер и др. (1961) приводят соответственно 117,4 кг у самцов (п = 5) и 73,1 кг у самок (п = 12). Длина тела аборигенов 188,7 см у самцов, (п = 8) и 171,1 см у самок (л = 10), а по В.Е.Присяжнюку — 173 см у самцов (л = 5) и 162 см у самок (л = 12). Для определения видовых параметров экстерьера пятнистого оленя, на наш взгляд, определенный интерес представляют показатели интродуцированных животных в различных районах страны, в том числе в горных лесах Кабардино-Балкарии. Наши данные свидетельствуют о том, что масса и размеры тела при условии паркового содержания с обильным кормлением достигают тех показателей, которые характерны для популяции Южного Приморья. Большие колебания массы тела, имеющие место в разных хозяйствах, определяются условиями содержания, а не генотипической изменчивостью вида.
Размеры черепа и посткраниальный скелет изучены недостаточно. По И.И.Соколову (1959), череп пятнистого оленя имеет длину от 265 до 335 мм. В условиях Центрального Кавказа, где пятнистый олень находит благоприятные условия жизни, наибольшая длина черепа составляет у самок III возрастной группы в среднем 290,8 мм.
Приведенные выше данные свидетельствуют о том, что даже самые крупные формы пятнистых оленей, в том числе парковые, не достигают размеров кавказских благородных оленей. По размерам тела и черепа разрыв между этими двумя видами в условиях Кавказа (при одинаковых ландшафтах) очень большой. В частности, по длине тела он составляет в среднем 50 см, а по длине черепа — около 20 см. Как видно, тезис "длина черепа больше или меньше 335 мм" (Соколов, 1959; Громов и др., 1963; и др.) вполне применим для дифференциации настоящих оленей Кавказа. Но если брать для сравнения весь спектр внутривидовых форм С. elaphus, то пятнистый олень окажется сходным с некоторыми мелкими расами этого вида (Гептнер и др., 1961). Что касается различия экстерьера пятнистых оленей и лани (Cervus dama), то оно незначительно, зона трансгрессии по массе и размерам тела широкая.
Рога у пятнистого оленя по сравнению с благородным оленем, особенно кавказским подвидом, уплощенные. У типичных рогов 4 конца: надглазничный, средний и 2 концевых. Иногда бывает 5 отростков в связи с появлением третьего концевого отростка. Тогда образуется примитивная крона рогов. Ствол рогов растет вертикально и в сечении округлый. У благородного оленя много концов (не менее 5), имеются 2 надглазничных, верхушка у хорошо развитого рога имеет не менее 2 отростков. В целом они образуют красивую крону и очень отличаются от рогов пятнистого оленя. Не менее существенны различия структуры рогов пятнистого оленя и лани (Соколов, 1959; Громов и др., 1963; и др.).
Окраска меха в условиях горных лесов Кабардино-Балкарии остается типичной. В стаде из Ц5 тыс. голов и в коллекционном материале, полученном нами в названных условиях, нам не удалось выявить каких-либо существенных сдвигов, обусловленных местными факторами. Летний мех (июль-август) характеризуется видоспецифичной яркой пятнистостью, которая ни в коем случае не соизмерима с той пятнистостью, которая отме
124
Ccrvus nippon Tcmminck, 1838
чается у кавказской популяции благородного оленя или гибридов между этими видами. Общий фон верхней стороны тела — бледно-красноватобурый, сиена жженая по А.С.Бондарцеву (1954). На этом фоне имеются крупные чисто-белые пятна. Форма и расположение белых пятен сильно варьируют. Исключением являются те, которые образуют два параллельных ряда по спине и шее. Особенно изменчивы пятна, образующие нижние ряды на боках; они могут сливаться и образовывать довольно длинные белые пятна. Между спинными белыми рядами пятен довольно часто шерсть темного цвета. У отдельных особей эта окраска четко выражена, образует черную полосу по спине и верху шеи. Черные волосы у многих животных интенсивно развиты и в области хвоста. У одних они образуют полукольцо шириной до 6 см у корня хвоста, постепенно сужающегося вниз по задней части бедер. У других животных вместо полукольца образуются две полосы по бокам, а в области корня хвоста нет черных волос. Между этими двумя типами окраски зада можно найти целый ряд переходов. Многие животные таких черных рисунков не имеют вовсе. Окраска хвоста меняется широко, что связано с вариацией соотношения черных, белых и бледно-красноватобурых волос на верхней стороне хвоста. Окраска шеи не контрастирует с окраской тела; она лишь немного светлее, не столь резко выражены белые пятна, они сводятся на нет по мере приближения к голове. От боков к нижней стороне шеи окраска значительно светлее, тогда как на голове темнее. Особенно она темнеет на верхней стороне ушных раковин, нередко приобретая черную окраску, особенно заметную по их краям. Отдельные участки горла, низ груди, подмышки, брюхо, нижняя часть хвоста, как и внутренняя поверхность ушей, обычно имеют светлую, почти белую окраску. Окраска конечностей мало отличается от окраски туловища, если не считать отсутствия белых пятен. Характеризуя окраску тела пятнистых оленей, следует, на наш взгляд, обратить внимание на то, что каждый волос в отдельности имеет один цвет: бледно-красновато-бурый, белый или черный. Двух и более цветов на одном волосе не выражено. Кроме того, мы не обнаружили заметного изменения тональности от основания к кончику волоса, что характерно для многих видов зверей. У кавказского благородного оленя мы нередко наблюдали пятнистую окраску, но пятна у него не такие ярко-белые и крупные, как у пятнистого оленя.
Как видно, пятнистый олень по сравнению с благородным оленем Кавказа характеризуется рядом примитивных особенностей, в том числе пятнистостью окраски во взрослом состоянии и упрощенной структурой рогов. Однако близкое родство этих видов общепринято, что подтверждается также гибридизацией между ними в природных условиях Кавказа и ряда других районов совместного обитания пятнистого и благородного оленей.
Кожный покров (рис. 19—23). Изучили кожу взрослого самца и взрослой самкц, добытых летом, и взрослой самки, добытой в феврале в Кабардино-Балкарии в Белореченском лесничестве. Строение кожи и специфических кожных желез в целом сходно с ранее описанными особями (табл. 23; Соколов, 1973). Кожа загривка толстая (6,4 мм у самца, 6,0 мм у самки), толще, чем на труди (4,8 у самца, 4,6 у самки). Нссегментированный эпи-
125
Пятнистый олень
Рис. 19- Строение кожи труди (А — продольный срез) и загривка (Б — поперечный срез) взрослой самки пятнистого оленя
1 — эпидермис, 3 — остевой волос, 4 — пуховой волос, 5 — мышца, поднимающая волос, 6 — сальная железа, 7 — потовая железа
дермис средней толщины (толще на груди: у самки соответственно 34 и 56 мкм). Имеются отдельные зернистые клетки. Граница между сетчатым и сосочковым слоями проходит на уровне луковиц волос и нижних отделов потовых желез. В коже загривка сетчатый слой толще сосочкового; в коже
Таблица 23. Промеры кожи пятнистых оленей (Соколов, 1973)
Показатель Толщина Размер сальных желез, мкм
КОЖИ, мм эпидермиса, мкм рогового слоя, мкм дермы, мм сосочкового слоя, мм сетчато -го слоя, мм
Олень № 1 102 х205
загривок 6,95 41 сорван 6,91 1,61 5,30 61 х 133
грудь Олень №2 4,61 215 82 4,40 2,10 2,30 102 х369 82 х 256
загривок 6,98 92 31 6,89 1,33 5,56 62 х 226
ГРУДЬ 5,33 185 51 5,15 4,10 1,05 62 х 133 72 х 513 50 х 205
126
Ccivus nippon Tcmminck, 1838
Рис. 20. Строение плюсневой железы взрослой самки пятнистого оленя (А, Б) 1 — эпидермис, 2 — дерма, 3 — волос, 4 — мышца, поднимающая волос, 5 — сальная железа, 6 — проток сальной железы, 7 — потовая железа
груди, напротив, сосочковый слой занимает 75—80% толщины кожи. В сосочковом слое пучки коллагеновых волокон тоньше, чем в сетчатом, и проходят параллельно поверхности кожи. В сетчатом слое они образуют плотное ромбовидное (в коже загривка) или петлистое (в коже груди) сплетение. Сальные железы остевых волос одно-двудольчатые средних размеров (на груди у самки — 700 х 113 — 339 х х 113 мкм).
Потовые железы крупные (диаметром '56—113 мкм), извитые, трубчатые, тянутся вдоль волосяных фолликулов. Секреторная трубка переходит в узкий (23 мкм) выводной проток на уровне нижней
трети сумки остевого волоса. Проток впадает в общую с волосом эпидермальную воронку. Мышцы, поднимающие волосы, хорошо развиты, толщиной 79—113 мкм. Остевые волосы растут поодиночке, а пуховые собраны в группы по 4—8 в каждой, у зимней самки! на за-
гривке число остевых волос составляет 1326, а пуховых — 3061 на 1 см2 (соотношение 1:3). Волосы растут слабоизвилистыми рядами.
Из специфических кожных желез имеются мейбомиевые, запястные, пяточные, хвостовая. Железы увеличены в циркуманальной области.
Мейбомиевые железы крупные многодольчатые, непигментированные. У самки на продольном срезе верхнего века размеры железы составляют 0,7 х 0,5 мм; она содержит 25 мелких округлых или двулопастных удлиненных
альвеол.
!у этой самки число остевых и пуховых волос необычно велико по сравнению с
другими особями (см.: Соколов, 1973).
127
Пятнистый олень
Рис. 21. Строение волос взрослых самок пятнистого оленя
А, Б — поперечный срез на уровне основания остевых' волос загривка, ув. х 100 и 400 у летней самки; Д, Е — то же на груди, ув. х 100 и 300; В, Г — бактерии на стенках сердцевины, ув. х 1000 и 3000; Ж — поперечный срез пухового волоса груди, ув. х 1000; 3 — то же остевого волоса зимней самки, ув.х 450
128
Ccrvus nlppon Tcmmlnck, 1838
Рис. 22. Строение сердцевины остевых волос взрослых самок пятнистого оленя (продольный срез)
А — волос загривка летней самки, ув. х 400; Б — стенка сердцевины перегородки, ув. х 12000; В, Г — продольный срез остевого волоса загривка зимней самки, ув. х 150 и 400
Диаметр центральной собирающей секрет полости равен 350 мкм.
В цир кум анальной коже непигментированные сальные железы, смыкаясь, образуют сплошной пласт; они очень крупные, многодольчатые, с ветвящимися протоками. У самки их размеры на продольном срезе равны 0,6 х 0,3; 0,7 х 0,4 мм. Выводные протоки сильно расширены и впадают в мелкие волосяные сумки на глубине до 339—452 мкм.
Апокриновые железы многочисленные, с крупными (диаметром 56— 90 мкм) свернутыми в рыхлые клубки секреторными отделами. Секреторная трубка переходит в узкий неизвитой выводной проток на уровне нижней трети волосяной сумки, проток идет параллельно ей, проходит между долями сальных желез и впадает в общую с волосом эпидермальную воронку на глубине не более ИЗ мкм от поверхности кожи. Стенки протока непигментированы.
9 В.Е.Соколов, А.К.Темботов
129
Пятнистый олень
Рис. 23. Строение кутикулы волос взрослых самок пятнистого оленя
А — остевого волоса летней самки, ув. х 300; Б — то же верхнего яруса на груди, ув. х 300; Б, Е — то же нижнего яруса, ув. х 300 и 600; Г — кутикулы остевого волоса, ув. х 450; Д — кутикула пухового волоса загривка зимой самки, ув. х 2000
130
Cervus nippon Tcmminck, 1838
Таблица 24. Промеры волосяного покрова пятнистого оленя (остевые волосы)
Проба Размерный порядок остевых волос
I II III
длина, мм толщина, мкм сердце-вина, % длина, мм толщина, мкм сердце-вина, % длина, мм толщина, мкм сердце-вина, %
I II Загривок Грудь Зима 92,5 225 88 82,6 165 90 47,3 150 90 65,0 195 85 58,9 240 87 50,2 217 89 Лето 44,2 180 75 37,9 210 78 29,2 225 79 I Ярус 56,5 142 93 51,0 170 88 41,2 170 88 II ярус 22,2 285 89 19,0 255 88 17,1 230 87
Проба Размерный порядок остевых волос
IV V
длина, мм толщина, мкм сердце-вина, % длина, мм толщина, мкм сердце-вина, %
I II Загривок Грудь Зима 37,5 120 89 28,3 90 89 37,7 189 88 30,5 157 87 Лето 17,3 180 79 13,5 90 80 I ярус 34,9 135 89 25,7 120 88 II ярус 16,0 210 86 11,3 200 86
Сходно с циркуманальными железами развиты железы половых губ. У самки некоторые из сальных желез замещаются или побужены в лимфоидную ткань, островки которой многочисленны и в сосочковом слое. По-видимому, здесь идет воспалительный процесс. В отличие от циркум-анальной кожи кожа половых губ содержит крупные волосяные сумки, снабженные мощными мышцами, а утолщенный эпидермис имеет мелкие ячеи на внутренней поверхности.
Строение запястной пяточной и межпальцевых желез вполне сходно с описанным ранее у других особей (Соколов, 1973). Обращает на себя внимание мощное развитие мышц, поднимающих волосы. Они могут состоять из нескольких фасций и чашевидно окружать нижнюю часть волосяной луковицы.
Волосяной покров на загривке у самки, добытой летом, представлен остевыми и пуховыми волосами. Волосы, вероятно, растущие. Они темного цвета, прямые. Пуховых волос гораздо меньше, чем у зимних особей. Осте-
9*
1 3 1
Пятнистый олень
вые волосы условно можно подразделить на пять размерных порядков (табл. 24). Длина первого размерного порядка не превышает 45 мм. От основания к вершине стержень сужается.
Волосяной покров на труди представлен двумя ярусами. Первый ярус — более светлый, длинные остевые волосы, которые подразделяются на пять размерных порядков (вероятно, остатки зимнего меха) длина которых колеблется 25,7 до 56,5 мм. Волосы второго, более низкого яруса (длина 11,3 до 22,2 мм) темнее и однотонно пигментированы по всей длине. Различимы также пять размерных порядков остей. Волосы второго яруса значительно толще (210—285 мкм) остей первого (120—170 мкм).
В остальном структура волосяного покрова у всех изученных проб сходна между собой. Поперечные срезы у остевых волос в основании округлой или сжато-бобовидной формы (рис. 21,а,б,д,с,з), в верхней половине стержня овальной формы. Сердцевинный канал очень хорошо развит по длине всего стержня (рис. 22). Сердцевина зернистого типа. Сердцевинные клетки плотно прилегают друг к другу в виде сот. По периферии они мельче по размеру, чем в центре сердцевинного канала. Размеры сердцевинных клеток колеблются от 8 х8 до 40 х24 мкм (ширина высота) независимо от сезона. Стенки клеток относительно тонкие, при больших увеличениях различимо их ячеистое строение (рис. 22,6). На них располагаются многочисленные бактерии-кератинофиты (рис. 21,г). Рисунок кутикулы мало меняется по длине стержня. Чешуйки вытянуты по ширине стержня. Высота чешуй более или менее постоянна. В нижней половине стержня свободные края относительно ровные, в верхней — с небольшими выемками и зазубринами и плотно прилегают к корковому слою (рис. 23).
Категория пуховых волос хорошо развита в зимних пробах. Длина их достигает 22 мм. Толщина одинакова по всей длине стержня (примерно 45 мкм). Волосы серого цвета, сильно извиты. Поперечные срезы округлой формы (рис. 21,ж). Сердцевинный канал чаще всего отсутствует или'фрагментарен. Кутикула кольцевидного типа. По строению пуховые волосы в летних пробах ничем не отличаются от зимних. Они лишь более редкие и короткие.
Плюсневая железа взрослого летнего самца выглядит как утолщение кожи, покрытое длинными рыжеватого цвета волосами со слипшимися от секрета корнями. Верхушки волос не покрыты секретом и не стерты. Волосяной покров представлен лишь одной категорией остевых волос, которые можно разделить на размерные порядки. В каждом размерном порядке выделяются "толстые" и "тонкие" волосы, которые схожи по длине, но отличны по форме, толщине стержня и степени развития сердцевинного слоя. У тонких волос стержень слегка расширяется от основания к середине, к вершине он вновь сужается. У толстых волос в прикорневой зоне выражена "ножка", далее в нижней трети стержень имеет наибольшую толщину, после чего сужается к вершине (табл. 25).
132
Ccrvus nippon Tcmminck, 1838
Таблица 25. Промеры остевых волос плюсневой железы взрослого летнего самца (выборочные данные)
Длина, мм Тонкие Толстые
толщина , сердцевина, толщина, сердцевина,
мкм % к толщине мкм % к толщине
стержня стержня
40,8 120 50 250 69
31,5 135 48 270 67
29,1 105 43 165 73
26,8 135 51 170 72
21,5 105 47 140 75
17,3 150 40 300 80
и Т.Д.
По структуре сердцевина зернистого типа и ничем не отличается от сердцевины остевых волос у описанных выше проб. Можно лишь отметить, что выступающая (покрывающая железу) часть стержня у длинных волос в поперечнике имеет овальную форму и сердцевинный канал выражен хуже, чем в нижней половине стержня. Свободные края чешуй кутикулы на этом участке волоса неровные, высота чешуй изменчива. У коротких волос по всей длине и в нижней части у длинных чешуйки вытянуты по ширине стержня. Высота чешуй постоянна, края ровные и плотно прилегают к корковому слою.
Длина волос рядом с железой не превышает 22 мм. Волосы прямые. От основания к вершине стержень сужается. Толщина у основания волос сильно колеблется от 75 до 210 мкм. Сердцевина зернистого типа и развита по длине всего стержня. В нижней половине стержня она составляет около 70% от диаметра стержня, в верхней — 48%. Имеются также мелкие волоски, длина которых не превышает 5 мм, толщина 30—60 мкм в расширенной незначительно части в середине стержня. Сердцевина хорошо развита (58% от диаметра) по длине всего стержня, кроме прикорневой зоны, где она отсутствует. Рисунок кутикулы в коротких волосах практически не меняется по длине. Высота чешуй постоянна, края ровные. В более длинных волосах в верхней половине стержня наблюдаются неровности свободных краев чешуй кутикулы.
Система крови. Анализ крови пятнистых оленей мы проводили в Белореченском оленарии Кабардино-Балкарии в августе и декабре 1989 г. Всего исследовано 97 оленей в возрасте 4—6 лет. Кровь брали у 40 самцов из рогов при срезе, а у 15 самцов и 42 самок — из уха. Летом у самцов содержание гемоглобина составляет (табл. 26) в среднем 14,5 %, количество эритроцитов — 10,36 млн, гематокрит — 45,7 об.%. Эритроциты некруп-
133
Пятнистый олень
Таблица 26. Показатели гфови пятнистого оленя в условиях Центрального Кавказа _________________(окрестности Нальчика, 900 и над ур. моря)______________
Самцы, кровь из уха 7.08. 89 г.
Показатель п limit х ± т о Су
Гемоглобин (г%) 15 12,6-16,2 14,5±0,34 1,26 8,72
Эритроциты (млн) 14 8,20-12,10 10,36+0,27 0,97 9,34
Цветной показатель (ед.) 14 0,36-0,50 0,42+0,07 0,04 9,17
Диаметр эритроцитов (мкм) 13 4,0-4,8 4,3±0,07 0,26 5,94
Гематокрит (об.%) 15 40-54 45,7±1,10 4,13 9,04
Лейкоциты (тыс.) 15 0,95-7,60 3,73+0,55 2,05 54,97
Лимфоциты (%) 15 25-63 38,47+3,14 11,76 30,56
Сегментоядерные нейтрофилы (%) 15 30—66 54,13+3,16 11,81 21,81
Палочкоядерные нейтрофилы (%) 15 1-4 1,93±0,34 1,28 66,32
Моноциты (%) 15 1-4 2,40±0,24 0,91 37,93
Эозинофилы (%) 15 1-6 3,47±0,44 1,64 47,41
Базофилы (%) 15 0-2 0,53±0,17 0,64 120,00
Показатель Самцы, кровь из рога 2.08. 89 г.
п limit х + т о Су
Гемоглобин (г%) 40 13,2-19,0 15,6+0,26 1,60 10,25
Эритроциты (млн) 40 10,16±14,68 12,55±0,21 1,29 10,28
Цветной показатель (ед.) 38 0,31-0,47 0,38±0,01 0,04 11,00
Диаметр эритроцитов (мкм) 32 3,6-4,6 4,1±0,04 0,25 6,11
Гематокрит (об.%) 37 35-57 ‘41,80+0,94 5,66 13,55
Лейкоциты (тыс.) 32 3,05-7,50 4,93+0,21 1,16 23,62
Лимфоциты (%) 38 24-61 39,61+1,45 8,79 22,19
Сегментоядерные нейтрофилы (%) 38 30-68 53,32+1,58 9,63 18,05
Палочкоядерные нейтрофилы (%) 38 1-6 2,68+0,22 1,32 49,15
Моноциты (%) 38 1-6 3,16+0,23 1,42 44,94
Эозинофилы (%) 38 1-3 1,37+0,10 0,59 43,00
Базофилы (%) 38 0-1 0,11+0,05 0,31 282,74
Показатель Самки, кровь из уха 19.12. 89 г.
п limit х ± т о Су
Гемоглобин (г%) 42 7,40-14,20 11,73±0,27 1,74 14,84
Эритроциты (млн) 42 6,24-10,95 8,96+0,21 1,31 14,62
Цветной показатель (ед.) 42 0,25-0,52 0,40+0,01 0,06 15,00
Диаметр эритроцитов (мкм) 26 3,6-4,9 4,28-0,06 0,31 7,24
Гематокрит (об.%) 42 18-46 33,09+1,03 6,60 19,93
Лейкоциты (тыс.) 40 1,75-10,00 3,29±0,23 1,47 44,68
Лимфоциты (%) 40 30-85 58,80±2,71 16,92 28,78
Сегментоядерные нейтрофилы (%) 40 10-64 34,37+2,69 16,77 48,79
Палочкоядерные нейтрофилы (%) 40 1-7 2,55-0,28 1,72 67,45
Моноциты (%) 40 0-4 1,45±0,16 1,01 69,66
Эозинофилы (%) 40 1-8 2,68+0,31 1,95 72,76
Базофилы (%) 40 0—2 0,20±0,08 0,52 260,00
134
Cervus nippon Tcmminck, 1838
ные, 4,3 мкм. Сравнение картины крови из уха и их рогов показывает значительные расхождения по ряду параметров. Так, содержание гемоглобина и количество эритроцитов достоверно выше в крови, взятой из рогов, но эритроциты мельче, о чем свидетельствует низкий гематокрит и меньшая насыщенность гемоглобином.
Показательно, что содержание гемоглобина, количество эритроцитов и гематокрит у самок в зимний сезон ниже, чем у самцов летом, что, видимо, связано с сезонными изменениями этих показателей.
При детальном изучении морфологии эритроцитов у пятнистых оленей нами обнаружен факт серповидно-клеточности эритроцитов. Около 50—75% эритроцитов крови у пятнистых оленей имеет серповидную форму. Впервые факт серповидно-клеточности и других аномальных эритроцитов у оленей установлен в 1840 г. (Gulliver, по: Kitchen et al., 1964).
В 1949 г. Полинг (Pauling) и его сотрудники показали, что у больных серповидно-клеточной анемией (всегда гомозиготных по дефектному гену) имеется гемоглобин S и нет гемоглобина А. В то же время у носителей признака серповидно-клеточности (гетерозиготны по гену серповидно-клеточности) оба гемоглобина содержатся примерно в равных количествах.
Белая iqpoBb. Количество лейкоцитов у пятнистых оленей (табл. 26) составляет в среднем из уха 3,73 тыс. у самцов, а из рогов 4,93 тыс., что достоверно выше. При действительном изучении разновидностей лейкоцитов установлено, что сегментоядерные нейтрофилы у самцов составляют в среднем 53,32% из рогов и 54,13% с уха и значительно преобладают над лимфоцитами в летний период. А у самок в зимнее время преобладают лимфоциты над сегментоядерными нейтрофилами. Все это свидетельствует о сезонных преобразованиях в качественном составе крови.
Индексы внутренних органов. Литературные сведения по пятнистым оленям немногочисленные и неполные, но тем не менее представляют определенный интерес для оценки специфики вида и адаптивных его возможностей. На скалистом острове Аскольд (Японское море) 5 самцов и 7 самок пятнистого оленя в возрасте от 1,5 до 10 лет изучены В.Е.Присяж-нюком (1973) (табл. 27, 28). Индексы внутренних органов определялись по отношению к чистой массе тела животного. Результаты показали, что по абсолютной массе сердца самцы превосходят самок, а по индексу, наоборот, уступают (у самцов 6,56 %о, у самок 7,63 %о). То же самое обнаружено по индексу головного мозга. Индексы легких у самцов и самок сходны и незначительно изменяются по сезонам года. Почти сходными также были индексы селезенки обоих полов в группе взрослых животных, но они оказались меньше, чем у двухлетних оленей. Индекс печени летом больше, чем в начале зимы, что, по мнению автора, связано с подготовкой самцов к спариванию, а у самок — с лактацией. Очевидно, резерв быстромобильных питательных веществ в столь напряженный период жизни, как лактация и жестокая борьба самцов за самок, адаптивно оправдан. Сходные результаты получены также по индексам почек, что логично, если учесть тесную трофическую взаимосвязь между печенью и почками. Индекс надпочечников летом меньше, чем зимой. У самок индекс поджелудочной же-
135
Таблица 27. Абсолютная масса внутренних органов пятнистых оленей с о-ва Асколвд (по: Присяжнюк, 1973)
Дата отстрела Возраст, лет Масса
живая, кг убойная, % к живой массе сердца, г легких, г печени,г
Самцы
22.07 2 79,0 59,0 480 1090 1251
2.08 6 115,8 60,9 644 1095 2120
25.11 1,5 56,5 52,7 300 480 749
27.11 2,5 77,0 52,0 486 873 1020
3.12 2,5 78,5 54,0 395 1228 1180
— 2,2-2,5* 78,2 55,0 453 1063 (1100)**
Самки
26.07 2 62,3 65,6 495 989 1395
29.07 10 75,0 58,0 464 685 1394
22.11 1,5 65,0 52,8 365 463 900
22.11 6,5 81,5 57,4 540 862 1150
28.11 7,5 70,0 54,8 422 708 880
29.11 4,5 72,5 51,9 491 625 960
30.11 8,5 94,5 54,5 547 790 1048
— 4,5-10* 78,6 55,3 493 732 (1009)**
Дата отстрела Возраст, лет Масса
селезенки, г почек, г мозга, г желудка с содержимым, кг кишечника с содержимым, кг
Самцы
22.07 2 258 179 218 12,4 7,1
2.08 6 275 391 . 253 15,8 7,3
25.11 1,5 151 100 200 7,9 4,4
27.11 2,5 209 161 196 12,2 5,9
3.12 2,5 208 138 209 9,9 5,7
__ 2,2-2,5* (208) (149) 207 (11,0) (5,8)
Самки
26.07 2 251 204 215 8,1 з,з
29.07 10 170 211 191 14,9 4,7
22.11 1,5 197 108 204 7,1 3,6
22.11 6,5 287*** 190 200 10,7 4,7
28.11 7,5 207 136 202 9,5 5,1
29.11 4,5 144 142 199 9,3 5,1
30.11 8,5 228 207 234 15,6 6,0
— 4,5-10* (193) (169) 205 (И>2) (5,0)
*3десь и в табл. 28 приведены средние показатели для животных указанного возраста.
**3десь и в табл. 28 в скобках приведены средние величины показателией, которые подвержены значительной сезонной изменчивости; учтены показатели 2 самцов и 4 самок, добытых в ноябре и декабре.
***3десь и в табл. 28: гипертрофия селезенки вызвана ранением оленя. При вычислении средней величины показателя масса селезенки данного оленя не учитывалась.
Cervus nippon Tcmminck, 1838
Таблица 28. Относительная масса внутренних органов пятнистых оленей
с о-ва Асколвд (по: Присяжнюк, 1973)
Дата отстрела Возраст, лет Чистая масса животного ,кг Масса, % к чистой массе животного
головного мозга сердца легких селе- зенки печени поджелудочной железы
Самцы
22.07 2 63,6 3,43 7,54 17,34 4,05 19,67 1.01
2.08 6 98,1 2,57 6.56 11,16 2,80 21,61 0,77
25.11 1,5 46,1 4,56 6,52 10,74 3,27 16,25 0,89
27.11 2,5 61,9 3,16 7,85 14,11 3,37 16,47 0,89
3.12 2,5 65,2 3,21 6,06 18,83 3,19 18,59 0,63
— 2,2-2,5* 63,6 3,27 7Д5 16,76 (3,28)** (0.76)
Самки
26.07 2 53,7 3,99 9,21 18,41 4,67 25,97 1,07
29-07 10 59,6 3,20 7,78 11,49 2,85 23,39 1,05
22.11 1,5 56,4 3,61 6,49 8,20 3,49 15,95 0,65
22.11 6,5 69,2 2,89 7,80 12,45 4,14 16,61 0,62
28.11 7,5 58,1 3,47 7,27 12,19 3,56 15,14 —
29.11 4,5 60,4 3,29 8,12 10,41 2,38 15,89 0,72
30.11 8,5 76,1 3,07 7,18 0,38 2,99 13,77 0,64
— 4,5-10* 63,7 3,18 7,63 11,38 2,96 (15,35) (0,66)
Дата отстрела Возраст, лет Чистая масса животного, кг Масса, % к чистой массе животного
почек щитовидной железы надпочечников гипофиза эпи- физа хвостовой железы
Самцы
22.07 2 63,6 2,81 0,070 0,049 0,0100 0,0011 —
2.08 6 98,1 3,99 0,051 0,043 0,0130 0,0012 0,51
25.11 1,5 46,1 2,17 0,100 0,066 0,0110 0,0009 0,98
27.11 2,5 61,9 2,60 0,086 0,048 0,0098 0,0014 0,55
3.12 2,5 65,2 2,11 0,065 0,050 0,095 0,0008 0,64
— 2,2-2,5* 63,6 (2,30) 0,073 0,049 0,0098 0,0011 0,59
Самки
26.07 2 53,7 3,81 0,055 0,045 0,0140 0,0024 1,04
29.07 10 59,6 3,54 0,059 0,044 0,0130 0,0008 1,18
22.11 1,5 56,4 1,90 0,069 0,033 0,0092 0,0028 0,92
22.11 6,5 69,2 2,74 6,059 0,075 0,0100 0,0022 0,94
28.11 7,5 58,1 2,35 0,062 0,063 0,0130 0,0017 1,11
29.11 4,5 60,4 2,36 0,077 0,050 0,0099 0,0015 1,90
30.11 8,5 76,1 2,71 0,073 0,052 0,0100 0,0013 0,72
— 4,5-10* 63,7 (154) 0,066 (0,060) (0,0110) (0,0019) 1,17
137
Пятнистый олень
Таблица 28 (окончание)
Дата отстрела Возраст, лет Чистая масса животно го, кг Масса, % к чистой массе животного
тимуса внутр и полостного жира околопочечного жира околохво-стового жира языка
Самцы
22.07 2 63,6 — — 3,46 — —
2.08 6 98,1 — — 2,24 20,38 —_
25.11 1,5 46,1 0,28 60,73 7,74 5,75 3,94
27.11 2,5 61,9 0 31,21 1,87 0 3,10
3.12 2,5 65,2 0,088 47,25 8,75 7,20 3,07
— 2,2—2,5* 63,6 — 39,23 — — 3,08
Самки
26.07 2 53,7 0,85 — 2,74 — —.
29.07 10 59,6 0 — 3,45 — —
22.11 1,5 56,4 0,64 50,88 5,24 8,98 2,50
22.11 6,5 69,2 0 51,52 12,57 6,02 2,89
28.11 7,5 58,1 0 26,79 7,18 4,13 2,99
29.11 4,5 60,4 0,05 55,39 9,00 10,59 2,98
30.11 8,5 76,1 0 66,62 12,61 19,84 2,82
— 4,5-10* 63,7 — (50,08) (10,34) (10,14) (2,90)
лезы летом выше, чем зимой (соответственно 1,05 и 0,66%), что зависит, очевидно, от сезонной смены кормов. Индекс хвостовой железы почти в 2 раза выше у самок, чем у самцов. Хорошо развит тимус у молодых.
Кариотип. Для пятнистых оленей, как и других видов оленей Палеарктики, характерно высокое диплоидное число, равное 68, при постоянном количестве плеч. Кроме того, у этого вида обнаружен хромосомный полиморфизм, вызванный перестройками типа центрического соединения. Соответственно диплоидное число равно 64—68. Х-хромосома, как у всех видов, акроцентрическая, Y-хромосома варьирует (Орлов, Булатова, 1983).
Структура и динамика ареала
Видовой ареал занимает обширную территорию Восточной Азии от Уссурийского края на севере до Южного Китая и Северного Вьетнама на юге, на восток до Японских островов и Тайваня включительно. В пределы Советского Союза заходит лишь северо-восточная окраина ареала, охватывая очень небольшую территорию на самом юге Дальнего Востока (Гептнер и др., 1961). За последнее столетие распространение и численность аборигенного пятнистого оленя настолько были сокращены в связи с хищническим его уничтожением, что вид был занесен в Красную книгу СССР. В настоящее время пятнистый олень как вид исключен из Красной книги, однако его аборигенная популяция остается в списках исчезающих животных.
Мероприятия по искусственному расширению ареала пятнистых оленей в нашей стране проводятся давно. В дореволюционное время пятнистых
138
Cervus nippon Tcmminck, 1838
оленей завозили в Асканию-Нова дважды. В 1926 г. группу оленей завезли в Забайкалье из Приморья (Дорогостайский, 1930). В последующие годы оленей поселили в Ильменском заповеднике (юго-западнее Челябинска), Бузулукском (близ Бузулука), в Куйбышевском заповеднике в Жигулях, Мордовском (к юго-западу от Арзамаса, близ Темникова), Окском заповеднике (по р. Пра, к востоку от Рязани), Хоперском заповеднике (на р. Хопер, близ Борисоглебска) и др. (Гептнср и др., 1961). Искусственное расселение продолжается по сей день, охватывая различные ландшафтные районы от Карельского перешейка до южных 1раниц страны. Результаты акклиматизации в разных местах различны: в одних он не закрепился (например, Забайкалье), а в других "прочно вошел в местную фауну" (Черкасская область) (Евтушсвский, 1977, с. 7).
На Кавказе мероприятия по акклиматизации пятнистого оленя ведутся более 50 лет. Первый выпуск проводился в 1938 г. в Тебердинском заповеднике. Судя по данным Л.Е.Аренса (1957), через год количество оленей увеличилось с 59 до 76 особей, в 1940 г. их стало 96, в 1941 г. — 128—133, а в 1942 г. — 147—150 оленей. В последующем численность пятнистых оленей резко упала, что, видимо, связано с браконьерством в годы войны. Уже в 1948 г. в заповеднике насчитывалось всего 7—8 оленей. С 1949 г. наблюдалось некоторое увеличение поголовья оленей, которое достигло в 1952 г. 48 особей и вновь начало снижаться с 1953 г. По А.П.Иняковой (1957), в том году было учтено 36 голов, в 1954 г. — 21, а в 1955 г. была зарегистрирована лишь одна точка рева в долине Азгека. В последующие годы встречи с пятнистым оленем в пределах Тебердинского заповедника не регистрировались. По всем данным, олени разбрелись и были уничтожены браконьерами. В частности, в 50-х годах мы встретили пятнистого оленя на Зольских пастбищах, примерно в 50 км от Тебердинского заповедника.
Вторично в Ставропольский край пятнистый олень завезен в 70-х годах, но на этот раз в целях паркового разведения в совхозе Эльбурчанский Карачаево-Черкесии. С этой же целью олени получены в Майском зверосовхозе Кабардино-Балкарии.
В Краснодарском крае акклиматизация пятнистых оленей начата в 1979 г. Выпуск оленей общей численностью в 135 голов проводился четырьмя партиями. Звери прижились, в начале 80-х годов зарегистрировано около 150 оленей. По данным В.А.Кузякина (1989), поголовье пятнистых оленей в Краснодарском крае возросло в 1985 г. до 600 особей.
Завоз и акклиматизация оленей в Кабардино-Балкарии начаты в 1966 г., когда в пределах Нальчикского лесоохотничьего хозяйства организовали оленеводческое отделение Майского зверосовхоза. Выпуск производился трижды, общим числом 101 особь. В 1975 г. партию в 50 оленей выпустили из оленария на волю в леса Нальчикского лссоохотничьего хозяйства. Кроме них, из оленария неоднократно убегали олени, что поддерживало поголовье дикарей. В целях расселения пятнистых оленей в других лесных угодьях Кабардино-Балкарии осуществлялся выпуск, в том числе 40 животных,
139
Пятнистый олень
в Баксанском госзаказнике. Словом, наряду с парковым содержанием оленей в Кабардино-Балкарии практикуется вольное их разведение. Результаты работы по разведению оленей в условиях вольеры нам известны, о них будет подробнее сказано ниже. Что касается пятнистых оленей на воле, в лесах Кабардино-Балкарии имеющиеся данные неоднозначны. Они не погибли. В 1980 г. их было до 120, а в 1985 г., по данным В.А.Кузякина (1989), поголовье достигло почти 600 особей.
Белореченское отделение Майского зверосовхоза поставляло пятнистых оленей и за пределы Кабардино-Балкарии. Большая партия в 343 оленя из этого отделения была доставлена в Дагестан в 1970 г. (в Казбековский мех-лесхоз, пос. Дылым). По климатическим и ландшафтным условиям районы Дылым и Белая Речка очень сходны, поэтому получение положительных результатов реально. Другую партию оленей из Кабардино-Балкарии завезли в Алмахское хозяйство Дагестана, где организована оленеферма. Вокруг обеих оленеферм обитало на воле небольшое число пятнистых оленей, которым удалось сбежать из вольер. Как видно, на Северном Кавказе пятнистый олень в настоящее время занимает ряд районов. Все они приурочены к поясу горных лесов, где животные содержатся как в условиях вольер, так и на воле.
Пятнистого оленя для акклиматизации завозили также и в Закавказье. Так, в 1952 г. 13 голов оленей из Уссурийского края были получены Управлением охотхозяйства Азербайджана. Их выпустили в огороженный загон, расположенный в Фундаганском лесном массиве, недалеко от с. Алтыагач. Более крупную партию (77 оленей) выпустили в Талыше в пределах Гир-канского заповедника. Здесь акклиматизация прошла удачнее, чем в районе первого выпуска. Олени хорошо освоились, держатся преимущественно северо-западных участков заповедника. В основных местах обитания на 1000 га приходится 0,17 оленя. Общее поголовье к началу 80-х годов достигло 200 пятнистых оленей.
В Армении акклиматизация пятнистых оленей начата в 1954 г. (Гамбарян, 1959; Гамбарян и др., 1963; Унанян, 1975; и др.). Тогда завезли 18 голов и выпустили в лесу Хосровского заповедника, численность их постепенно росла. Второй раз, в 1958 г., был осуществлен дополнительный выпуск 19 оленей в тех же хосровских лесах. Хотя поголовье колебалось в широких масштабах, ежегодно отмечался приплод, и, по мнению некоторых местных териологов, этот факт подтверждает реальность акклиматизации пятнистых оленей в сухих районах юго-востока Кавказа. Для освоения северных лесных массивов Армении в 1969—1970 гг. большую партию в 200 оленей выпустили в Куйбышевском лесничестве Дилижанского заповедника, в Арзакандское и Азизбековское охотхозяйства. В 1972 г. группу пятнистых оленей выпустили также в Азизбековском охотхозяйстве Армении. Всего в этой республике к началу 80-х годов насчитывалось более 500 пятнистых оленей (Богачев, 1989).
Насколько нам известно, пятнистого оленя выпускали лишь в лесах Восточного Закавказья, на территорию Грузии этот вид не завозили.
140
Cervus nippon Tcmminck, 1838
Итак, на Кавказе создана целая сеть акклиматизационных точек по вселению пятнистого оленя, достаточная для обеспечения успешного расселения этого вида во все подходящие районы.
Современные тенденции антропогенных изменений ареала. Коренной ареал пятнистого оленя, как отмечено выше, резко сократился под влиянием человека. Что касается рукотворного ареала, складывающегося с помощью человека в наши дни, то по площади он превосходит во много раз территорию, занятую этим видом в нашей стране. Результаты акклиматизации в различных районах б. СССР и за рубежом свидетельствуют о том, что пятнистый олень — довольно пластичный вид и может освоить значительную площадь на Кавказе. Однако, на наш взгляд, многое зависит от уровня постановки работы по охране и воспроизводству охотничье-про-мысловых животных, особенно от того, как успешно изживается браконьерство, насколько разумно регулируется численность хищных, особенно волков, насколько оперативно и эффективно человек оказывает помощь оленям в отдельные критические периоды года (подкормка в многоснежные зимы и т.д.). Что касается абиотических факторов среды, то, на наш взгляд, в пределах предгорных и среднегорных широколиственных лесов они не будут лимитирующими. Вопрос в другом — насколько целесообразно вообще вселение в местную, здесь весьма богатую и разнообразную фауну Кавказа пятнистого оленя, когда имеется кавказский благородный олень. Очевидно, не следует засорять генофонд уникальной фауны Кавказа, ведь пятнистый и благородный олени в природе скрещиваются.
Систематика и изменчивость
Систематическое положение. Почти все специалисты сходятся на том, что пятнистый олень С. nippon представляет собой наиболее примитивный вид подрода Cervus. Он четко обособлен от других видов и характеризуется рядом специфических признаков. По В.Г. Гептнеру и др. (1961), внешне он похож на индийских пятнистых оленей, составляющих другой подрод (Axis), однако отличается от них как остеологическими признаками, так и строением рогов.
Среди современных териологов страны нет также разногласий и в отношении внутривидовой систематики. Общепринято выделение 4 подвидов, из которых на территории б. СССР обитает C.n. hortularum Swinhoe, 1864. Однако это не означает, что уже нет проблем в познании закономерностей изменчивости, в анализе систематико-таксономического веса тех или иных особенностей морфологических и других признаков. На наш взгляд, интерес к этой проблеме возрастает в связи с акклиматизацией пятнистого оленя в разных физико-географических условиях, приведшей к появлению многочисленных изолятов. Более того, превращение этого вида в полудо-машнее животное, парковое его содержание и селекционная работа, которая ведется с ним, очевидно, приведут к интенсивной дивергенции.
1 4 1
Пятнистый олень
Изменчивость. Изучение различных аспектов изменчивости пятнистого оленя в условиях Кавказа нам представляется довольно интересным в теоретическом и практическом отношениях. Дело в том, что этот вид приживается в различных ландшафтных районах Кавказа и слежение за адаптивными сдвигами в организме животного и его экологией позволит глубже понять норму реакции вида, совершенствовать зоотехническую и селекционную работу.
Наиболее полные данные по рассматриваемому вопросу мы получили в Белореченском оленарии (под Нальчиком). В литературе появились сообщения, относящиеся к этому хозяйству (Дормидонтов, 1977). Для оценки наших и литературных данных считаем целесообразным коротко описать ландшафтные условия содержания пятнистых оленей в данном районе.
Олснарий площадью в 3,5 тыс.га находится на склонах Лесистого хребта в интервале высот 600—1100 м над уровнем моря, т.е. геоморфологическая амплитуда составляет 500 м. Преобладают склоны северо-западных и юго-восточных экспозиций. По характеру рельефа эту территорию, как и все урочище Нарт-Бора, можно разделить на две части: пологую с уклоном до 6° на северо-запад в центральной и юго-восточной частях и изрезанную балками с крутизной склонов 10—15° в северо-западной части. Имеются р. Нарт-Бор, родники. Почвы горные серые лесные и горные дерново-карбонатные с фрагментами горных намытых и горных лугово- болотных почв. Свыше 80% площади оленария занято широколиственным буково-кленово-ясеневым и буково-карагачево-липовым лесом. Травянистая растительность пастбищ представлена мезофильными и сухими разнотравнозлаковыми, разнотравными лугами, вейниково-люцерновыми, разнотравно-узко-листномятликовыми лугами. По мнению некоторых териологов, весной и в первой половине лета погода в здешних местах сходна с погодой в Приморье. Что касается зимы, особенно мощности и продолжительности снежного покрова, то, как правило, этот фактор не выступает в качестве лимитирующего для оленей, в том числе пятнистых.
Индивидуальная изменчивость. Пятнистые олени в условиях Белореченского оленария характеризуются широкой индивидуальной изменчивостью. Это в равной степени относится к массе и размерам тела, черепа и окраске меха. Вероятно, в первую очередь сказывается отсутствие жесткого естественного и искусственного отбора. Известны наблюдения, согласно которым минимальная масса взрослых пятнистых оленей составляет всего 57,6 кг, в другом случае зверь пал при массе 62,1 кг. Возможно, что в тяжелых кормовых условиях самцы способны терять до 35—45% обычной зимней массы, но если снижение зимней массы тела приближается к 50% первоначального значения, то они гибнут (Присяжнюк, 1981).
Индивидуальная изменчивость рогов одновозрастных животных высока в Белореченском оленарии. В табл. 29—34 отражены колебания размеров черепа в разрезе пола и возраста. Большая индивидуальная вариация нами зарегистрирована в окраске меха. При всей видоспецифичности пятнистой окраски тела широко колеблются размеры белых пятен, особенно по бокам тела, изменчиво их расположение. Особенно лабильна степень раз-
142
Cervus nippon Temminck, 1838
Таблица 29. Промеры черепа Cervus nippon из Белореченского лесничества _____________________(Нальчик). П возрастная группа_________________
Признак Самцы
п limit х ± т о Су
Наибольшая длина черепа 5 272,2-308,0 295,3+8,32 18,30 6,2
Кондилобазальная длина 5 260,0-305,0 284,4±8,14 16,27 5,7
черепа Основная длина черепа 4 240,4-288,0 265,7+11,34 19,64 7,4
Максимальная ширина 5 112,4—138,3 128,8+4,94 9,87 7,7
Скуловая ширина 5 114,3-130,3 123,2±3,21 6,42 5,2
Межглазничная ширина 5 71,2-96,0 85,3+4,59 9,17 10,8
Длина лицевой части 5 144,6-172,3 159,2+5,11 10,21 6,4
Максимальная длина носовых костей 3 98,0-108,0 100,6+4,62 6,54 6,5
Длина верхнего ряда 5 78,8-89,9 84,7+2,18 4,35 5,1
зубов Максимальная ширина 4 92,8-115,3 105,4+5,38 9,32 8,8
мозговой капсулы Длина нижней челюсти 5 213,4-255,0 235,4±7,51 15,01 6,4
Длина нижнего ряда зубов Длина диастемы нижней 5 77,5-97,0 90,0+3,69 7,38 8,2
челюсти 5 57,5-65,4 62,9±1,59 3,17 5,0
Признак Самки
п limit х + т G Су
Наибольшая длина черепа 23 268,4-296,0 281,7+1,65 7,76 2,8
Кондилобазальная длина черепа 23 256,8—283,3 271,0+1,59 7,44 2,7
Основная длина черепа 23 240,7-263,5 252,6+1,43 6,70 2,7
Максимальная ширина 23 108,0-125,0 117,0+1,10 5Д4 4,4
Скуловая ширина 22 109,7-122,9 115,6+0,98 4,49 3,9
Межглазничная ширина 23 71,0-83,5 76,5+0,68 3,25 4,2
Длина лицевой части 23 145,2-163,0 153,0+1,15 5,51 3,6
Максимальная длина носовых костей 21 76,4-108,3 96,1+1,55 6,95 7,2
Длина верхнего ряда зубов 23 76,7-90,9 84,9±0,76 3,66 4,3
Максимальная ширина мозговой капсулы 22. 87,6-100,5 92,3±0,74 3,40 3,7
Длина нижней челюсти 20 209,0-235,8 224,4±1,74 7,58 3,4
Длина нижнего ряда зубов 23 84,4—99,3 90,6+0,83 3,97 4,4
Длина диастемы нижней челюсти 23 54,3—68,8 61,9+0,69 3,31 5,3
1 4 3
Пятнистый олень
Таблица 30. Промеры черепа Cervus nippon из Белореченского лесничества
(Нальчик). Самки
Признак III возрастная группа
п limit х + т а Су
Наибольшая длина черепа 22 277,9—324,0 290,8±2,07 9,70 3,34
Ковдилобазальная длина черепа 22 263,3-313,0 278,9+1,99 9,36 3,36
Основная длина черепа 21 236,4—293,5 259,2±2,28 10,46 4,03
Максимальная ширина 23 111,9-126,4 120,8±0,69 3,30 2,73
Скуловая ширина 22 113,3-124,1 119,2±0,65 3,06 2,56
Межглазничная ширина 23 73,6—88,0 79,8+0,75 3,61 4,52
Длина лицевой части 22 147,0-181,6 157,3±1,49 6,98 4,43
Максимальная длина носовых костей 21 92,5—110,8 99,6±0,95 4,36 4,38
Длина верхнего ряда зубов 23 79,0-92,3 83,0±0,73 3,50 4,20
Максимальная ширина мозговой капсулы 22 89,7+98,6 94,4±0,53 2,48 2,62
Длина нижней челюсти 22 219,9+258,8 231,9±1,61 7,57 3,20
Длина нижнего ряда зубов 23 84,2-100,4 9О,О±О,79 3,81 1,20
Длина диастемы нижней челюсти 23 61,4-74,0 65,9±0,62 2,98 4,50
Признак I возрастная группа
п limit х + т а Су
Наибольшая длина черепа 4 192,5-262,2 229,3±21,14 36,62 16,0
Кондилобазальная длина черепа 4 94,9-253,8 196,5±43,73 75,74 38,5
Основная длина черепа 4 166,2-235,6 202,4±19,91 34,49 17,0
Максимальная ширина 5 87,5-112,4 99,6+5,74 11,47 11,5
Скуловая ширина 4 91,0-113,4 99,8±6,13 10,61 10,6
Межглазничная ширина 5 53,0-69,4 61,7±3,57 7,13 11,6
Длина лицевой части 4 95,5-140,7 119,1±13,83 23,95 20,1
Максимальная длина носовых костей 5 55,3-88,2 71,3±7,44 14,88 20,9
Длина верхнего ряда зубов 5 54,4-77,2 66,9±5,31 10,61 15,9
Максимальная ширина мозговой капсулы 5 66,5-82,4 75,8±3',42 6,83 9,0
Длина нижней челюсти 5 149,0-208,4 182,2±13,46 26,91 14,8
Длина нижнего ряда зубов 5 53,3-74,3 70,0±7,89 15,77 22,5
Длина диастемы нижней челюсти 5 39,5-51,7 47,2±2,88 5,76 12,2
144
Cervus nippon Tcmminck, 1838
Таблица 31. Промеры черепа Cervus nippon из Судзухинского заповедника __________________(о-в Аскольд). I возрастная группа_______________
Признак Самцы
п limit х ± т О Су
Наибольшая длина черепа 7 148,3-264,3 212,0+22,96 56,24 26,5
Кондилобазальная длина черепа 6 138,6-254,4 195,2+25,42 56,83 29,1
Основная длина черепа 6 128,0-230,6 177,8+22,32 49,91 28,1
Максимальная ширина 9 64,4-113,1 93,0+7,09 20,05 21,6
Скуловая ширина 7 73,9-112,5 100,9+5,72 14,02 13,9
Межглазничная ширина 9 42,0-71,5 58,7+4,49 12,71 21,7
Длина лицевой части 7 56,8-143,3 105,1+15,42 37,78 35,9
Максимальная длина носовых костей 4 31,7-85,0 58,0+16,58 28,72 49,5
Длина верхнего ряда зубов 9 42,3-90,2 63,0+5,91 16,73 26,6
Максимальная ширина мозговой капсулы 7 50,3-88,0 73,1+6,78 16,60 22,7
Длина нижней челюсти 4 122,5-218,1 182,9+24,06 41,68 22,8
Длина нижнего ряда зубов 4 39,2—90,4 61,0+14,39 24,92 40,8
Длина диастемы нижней челюсти 5 19,1-63,9 43,0+9,48 18,95 44,1
Признак Самки
п limit х ± т О Су
Наибольшая длина черепа 17 259,3-277,9 239,4+6,46 25,82 10,8
Кондилобазальная длина черепа 15 208,0-265,7 231,1+4,47 16,73 7,2
Основная длина черепа 16 136,7-248,2 208,6+6,49 25,13 12,0
Максимальная ширина 19 67,8—113,6 99,3+2,19 9,30 9,4
Скуловая ширина 19 69,5-119,3 100,6+2,26 9,60 9,5
Межглазничная ширина 19 43,3-72,4 62,3+1,41 5,99 9,6
Длина лицевой части 17 72,6-150,8 123,6+4,36 17,45 14,1
Максимальная длина носовых костей 15 67,5-88,4 78,2+2,25 8,42 10,8
Длина верхнего ряда зубов 18 56,2—81,2 69,0+2,03 8,39 12,2
Максимальная ширина мозговой капсулы 19 57,0-92,0 78,8+1,78 7,55 9,6
Длина нижней челюсти 13 123,4-208,8 184,9+6.34 21,97 П>9
Длина нижнего ряда зубов 11 54,3-83,1 66,8+2,91 9,19 13,8
Длина диастемы нижней челюсти 13 30,5-60,0 52,1+2,26 7,83 15,0
10 В.Е.Соколов, А.К.Темботов
145
Пятнистый олень
Таблица 32. Промеры черепа Cervus nippon из Судзухинского заповедника _________________(о-в Асколвд). П возрастная группа________________
Признак Самцы
п limit х ± т О Су
Наибольшая длина черепа 7 263,8-301,0 283,3±5,03 12,31 4,3
Кондилобазальная длина черепа 8 252,2-287,7 271,3±4,21 11,14 4,1
Основная длина черепа 8 235,8—267,6 252,1±3,84 10,15 4,0
Максимальная ширина 9 111,6-130,5 121,6+2,03 5,75 4,7
Скуловая ширина 10 112,0-125,5 117,4±1,54 4,61 3,9
Межглазничная ширина 10 71,1-90,2 79,8±2,11 6,32 7,9
Длина лицевой части 8 134,9-160,6 149,5±3,15 8,34 5,6
Максимальная длина носовых костей 9 84,8-104,0 92,4±2,31 6,53 7,1
Длина верхнего ряда зубов 9 82,2-90,8 86,8±0,94 2,66 3,1
Максимальная ширина мозговой капсулы 10 90,8-107,8 97,4±2,23 6,69 6,9
Длина нижней челюсти 5 213,8-235,0 223,9±3,96 7,91 3,5
Длина нижнего ряда зубов 5 90,3-97,8 92,4±1,44 2,87 3,1
Длина диастемы нижней челюсти 5 56,5—62,5 59,5+1,21 2,42 4,1
Признак Самки
п limit х ± т О Су
Наибольшая длина черепа 15 266,2—308,0 282,2+3,06 11,46 4Д
Кондилобазальная длина черепа 17 252,6-292,9 268,0+2,47 9,89 3,7
Основная длина черепа 17 234,6-273,4 248,9±2,28 9,12 3,7
Максимальная ширина 18 108,8-121,8 113,1±0,78 3,22 2,8
Скуловая ширина 17 107,5-120,0 112,3±0,83 3,31 2,9
Межглазничная ширина 18 67,5-79,0 71,4±0,68 2,79 3,9
Длина лицевой части 17 114,9-176,8 148,9+3,72 14,87 10,0
Максимальная длина носовых костей 17 80,9-110,7 97,4+2,18 8,73 9,0
Длина верхнего ряда зубов 18 82,2-92,7 86,3±0,73 2,99 3,5
Максимальная ширина мозговой капсулы 17 81,4-92,4 8&,1±0,81 3,23 3.7
Длина нижней челюсти 8 212,2-233,9 221,1±2,68 7,09 3,2
Длина нижнего ряда зубов 7 88,8-97,9 93,2± 1,40 3,43 3,7
Длина диастемы нижней челюсти 8 53,6-67,6 59,9±1,79 4,74 7,9
146
Cervus nippon Temminck, 1838
Таблица 33. Промеры черепа Cervus nippon из Судзухинсксго заповедника _________________(о-в Аскольд). Ш возрастная группа________________
Признак Самцы
п limit х ± m О Су
Наибольшая длина черепа 8 254,6-315,0 298,6+6,61 18,69 6,26
Ковдилобазальная длина черепа 10 243,7-301,0 287,6+5,15 16,28 5,66
Основная длина черепа 10 '224,0—283,1 268,6+5,25 16,61 6,18
Максимальная ширина 10 104,2-143,4 132,3±3,61 11,40 8,62
Скуловая ширина 10 104,1-134,2 123,2±2,84 8,99 7,29
Межглазничная ширина 10 63,0-94,7 86,8±3,01 9,57 11,02
Длина лицевой части 10 133,4-170,4 159,5±3,22 10,19 6,39
Максимальная длина носовых костей 10 80,8-112,2 100,9±2,90 8,71 8,63
Длина верхнего ряда зубов 10 74,2-90,8 83,9±1,98 6,25 7,45
Максимальная ширина мозговой капсулы 10 81,3-118,3 107,4±3,16 9,99 9,31
Длина нижней челюсти 6 198,4-242,9 232,0±6,92 16,95 7,31
Длина нижнего ряда зубов 6 74,1-98,6 90,7±3,53 8,64 9,52
Длина диастемы нижней челюсти 6 61,8—69,6 65,4±1,26 3,08 4,71
Признак Самки
и limit х ± т о Су
Наибольшая длина черепа 40 272,8-310,0 289,6±1,43 9,05 3,1
Ковдилобазальная длина черепа 41 254,-294,1 276,7±1,36 8.76 3,2
Основная длина черепа 41 241,3-274,5 258,1±1,25 8,02 3,1
Максимальная ширина 44 112,0-160,0 121,2+1,11 7,35 6,1
Скуловая ширина 42 111,2-123,4 117,6+0,48 3,10 2,6
Межглазничная ширина 44 70,1-91,4 79,4±0,58 3,86 4,9
Длина лицевой части 42 144,1-173,3 157,7±1,14 7,39 4,7
Максимальная длина носовых костей 41 85,0-109,0 98,8±0,94 6,01 6,1
Длина верхнего ряда зубов 37 72,6-93,0 83,2+0,71 4,29 5,1
Максимальная ширина мозговой капсулы 42 85,3-108,1 93,5±0,61 3,96 4,2
Длина нижней челюсти 26 216,5±245,1 232,7± 1,69 8,60 3,7
Длина нижнего ряда зубов 25 81,6-99,8 91,2+0,85 4,24 4,6
Длина диастемы нижней челюсти 26 56,1-78,0 64,8±0,98 5,02 7,7
10*
147
Пятнистый олень
Таблица 34. Промеры черепа Cervus nippon из Судзухинского заповедника _________________(о-в Асколвд). IV возрастная группа_______________
Признак Самцы
п limit X ± ш о Су
Наибольшая длина черепа 7 294,5—319,0 302,2±1,38 3,39 1,1
Кондилобазальная длина 7 285,2—310,0 291,9+3,82 9,36 3,2
черепа Основная длина черепа 7 268,5-292,0 274,9+3,51 , 8,60 3,1
Максимальная ширина 7 133,4-144,3 139,4+1,65 4,05 2,9
Скуловая ширина 6 125,0-132,3 129,3+1,39 3,11 2,4
Межглазничная ширина 7 88,0-96,9 93,1+1,17 2,87 3,1
Длина лицевой части 7 156,2-170,1 160,8+1,97 4,83 3,0
Максимальная длина носовых костей 6 91,4-109,9 99,9+2,71 6,06 6,1
Длина верхнего ряда зубов 3 74,0—82,0 78,4+2,88 4,07 5,2
Максимальная ширина мозговой капсулы 7 107,0—116,6 112,9+1,40 3,44 3,0
Длина нижней челюсти — — — — —
Длина нижнего ряда зубов — ' — — — —
Длина диастемы нижней — — — — —
челюсти
Признак Самки
п limit х + т О сг
Наибольшая длина черепа 9 276,0-303,0 288,9+3,41 9,65 3,3
Кондилобазальная длина черепа 12 262,4-290,0 276,1+2,75 9Д1 з,з
Основная длина черепа 11 245,0-271,8 258,0+2,72 8,59 3,3
Максимальная ширина 12 115,4-139,0 122,5+1,88 6,23 5,1
Скуловая ширина 11 113,2-128,4 119,0+1,65 5,21 4,4
Межглазничная ширина 11 77,5-94,8 82,0+1,98 6,26 7,6
Длина лицевой части 12 146,9-167,5 156,6+1,93 6,39 4,1
Максимальная длина носовых костей 10 90,9-109,2 99,3+1,83 5,48 5,5
Длина верхнего ряда зубов 8 72,4-88,1 78,5+1,84 4,88 6,2
Максимальная ширина мозговой капсулы 12 89,5-111,8 95,6+1,83 6,06 6,3
Длина нижней челюсти 4 228,9-247,3 241,7+5,00 8,67 3,6
Длина нижнего ряда зубов 4 80,3-94,1 85,8+3,38 5,86 6,8
Длина диастемы нижней челюсти 4 66,2—74,2 70,3+2,44 4,23 6,0
148
Таблиц* 35. Промеры (в см) кавказских оленей Зццвддиго Кавказа (до: Александров, 1968)
Признак Сеголетки Самки Самцы
неполовозрелые взрослые неполовозрелые взрослые
limit X limit X limit X limit X limit X
Длина тела - 145,0-151,0 148,0 164,0-179,0 173,0 164,0-215,0 191,0 193,0-215,0 204,0 225,0-249,0 234,6
хвоста * 10,0-11,0 17,5 11,0-13,0 12,0 11,0-12,5 12,0 12,0-13,0 13,5 10,0-17,0- 13,0
уха ? 17,5-18,5 18,0 18,0-22,0 20,0 18,0—22,0 20,0 20,0-23,0 22,0 21,0-23,0 22,0
стопы * 38,0-49,0 43,8 50,8-58,0 54,0 52,0-57,0 54,7 56,7-60,0 58,0 60,0-67,0 63,0
Высота в холке ' 102,0-108,0 105,0 114,0-124,0 119,0 114,0-133,0 123,0 136,0-144,0 140,0 142,0-152,0 146,0
Высота в крестце • 106,0-115,0 110,0 119,0-133,0 126,0 117,0-144,0 131,0 138,0-146,0 142,0 148,0-155,0 151,6
Косая длина туловища > 98,0-102,0 100,0 112,0-127,0 119,5 120,0-128,0 123,0 120,0-138,0 129,0 138,0-170,0 155,6
Обхват груди л 102,0-108,6 105,0 118,0—132,0 125,0 120,0-138,0 126,0 131,0-146,0 138,0 155,0-170,0 163,5
Ширина груди >- 22,0-25,0 24,0 28,0-32,0 30,0 28,0-32,0 30,0 28,0-34,0 31,0 43,0-50,0 47,0
Длина головы 34,0-36,0 35,0 42,0-46,0 44,0 43,0-47,6 45,7 41,0-56,0 49,0 60,0-65,0 64,0
Ширина головы 13,0-15,0 14,0 13,0-17,0 15,0 13,0-16,0 14,4 — — 18,0-24,5 20,7
Высота передней « 65,0-69,0 67,0 li, 0-75,0 73,0 71,0-76,5 74,7 77,0-81,0 79,0 81,0-90,0 87,4
конечности Обхват пясти 10,0-12,0 11,0 10,0-13,0 11,5 11,0-12,5 12,0 11,5-13,0 12,0 14,0-15,0 14,6
Длина переднего пальца , 14,0-17,0 15,0 13,0-16,0 14,5 15,5—16,0 16,0 14,0-16,0 15,0 16,5-20,0 17,7
Длина кисти 36,0-38,0 37,0 40,0-44,0 42,0 40,5-44,0 42,0 42,0—44,0 43,0 45,0-50,0 47,0
Высота задней 86,0-90,0 88,0 110,0-112,0 111,0 115,0-116,5 115,8 — — 125,0-130,0 127,7
конечности Обхват плюсны 10,0-12,0 11,0 10,0-14,0 12,0 12,0-13,0 12,5 13,0-17,0 15,0 15,0-17,0 15,7
Длина пальца задней 14,0-16,0 15,0 14,0-18,0 16,0 15,5-17,0 16,3 16,0-18,0 17,0 18,0-20,0 19,0
конечности
: Длина переднего следа. 7,0-9,0 .8,0 9,0-11,0 10,0 9,0—12,0 J 1,0 9,0-11,0 10,0 13,0-15,0 > 14,0
Длина заднего следа 8,0-10,0 9,0 8,0-12,0 10,0 7,9-11,0 9,7 9,0-11,0 10,0 10,0-13,0 11,9
^Высота переднего j 6,0-7,0 6,5 5,0-7,0 6,0 5,9-7,0 6,6 6,0-8,0 7,0 6,5-8,5 7,4
копыта ? Высота заднего копыта 1 8,0-8,0 7,0 5,0-7,0 6,0 6,0-7,0 6,6 6,5-8,5 7,5 7,0-8,5 7,9
j Ш ир и на п ер ед него - копыта 5,0-6,0 5,5 6,5-6,5 6,2 6,0-7,5 6,6 6,2-8,0 7,0 7,5-10,0 7,4
i. Ширина заднего копыта 5,0-6,0 5,5 5,0-8,0 6,0 5,0-7,5 6,0 6,0-8,0 7,0 7,0-9,0 8,0
Пятнистый олень
вития черной полосы по спине — от интенсивно развитой до слабо выраженной или полного ее отсутствия. Как отмечено выше, очень большой изменчивости подвержен также черный рисунок вокруг зеркала — от хорошо развитого, имеющего вид полумесяца, до фрагментов его или полного исчезновения рисунка. По нашим наблюдениям, олени одной 1руппы очень несходны по окраске горла и нижней стороны шеи. У одних она бывает однотонной, значительно светлее окраска верхней стороны, у других хорошо заметны одно или два серовато- белых поля диаметром до 10 см.
Белореченские пятнистые олени остаются весьма разнородными по этологии. Среди них четко выделяются различные группы по агрессивности, осторожности и враждебности к человеку. Они заметны как во время пастьбы и подкормки, так и во время съема пантов. В тех хозяйствах, где усиленно ведется искусственный отбор, произошли существенные этологические изменения содержащихся в парках оленей (Галкин, 1987).
Возрастная изменчивость. Для изучения внутривидовой изменчивости Cervus nippon нами исследованы две выборки: Белореченское лесничество (Кабардино-Балкария) и Судзухинский заподведник (пос. Аскольд). Выбор точек не случаен. Особи Белореченского лесничества представляют прямых потомков пятнистых оленей Дальнего Востока, которые были завезены в 1968 г. для вольерного содержания. С тех пор пополнения популяции из Кабардино-Балкарии другими особями не было, ее особи на протяжении всего времени были полностью изолированы. В связи с этим вполне понятен интерес к сравнению этих двух выборок.
При изучении изменчивости Cervus nippon мы выделили четыре возрастные группы:
Г"~1 — молодняк, у которого идет смена зубов;
II — молодые особи, у которых завершилась смена зубов и еще не появилась стертость, а если и появилась, то незначительная на предкоренных;
III — взрослые особи, у которых коренные и предкоренные имеют ту или иную степень стертости;
IV — старые особи, у которых очень сильно стерты все зубы, их поверхность сглажена в разной степени.
Цифровой материал по Cervus nippon представлен в табл. 30—35. Как видно, характер возрастных изменений у самцов и самок из Судзухинского заповедника сходен. Увеличение размеров черепа происходит вплоть до Ш возрастной группы, за исключением длины верхнего и нижнего ряда зубов, которые достигают максимальных размеров у особей II возрастной труппы. После этого, как видно из табл. 30—35, происходит закономерное уменьшение средних значений этих признаков, что объясняется стиранием с возрастом коронок первых резцов, и последнего моляра. Увеличение других 10 признаков черепа значительно, что видно при сравнении трупп особей I и II, II и III. Различия достоверны при всех вариантах анализа. Дальнейший рост размеров черепа прекращается как у самцов, так и самок, что видно при сравнении особей III и IV возрастных трупп.
150
Cervus nippon Temminck, 1838
Аналогичный характер возрастной изменчивости нами выявлен у самок из Белореченского лесничества (данных по самцам нет).
Половая изменчивость. Половая изменчивость краниометрических признаков нами изучена у особей из Судзухинского заповедника всех четырех возрастных групп, а из Белореченского лесничества — во II возрастной группе.
Самки первой возрастной группы из Судзухинского заповедника незначительно крупнее самцов почти по всем изученным признакам, за исключением скуловой ширины (см. табл. 31). В старших возрастных группах различия увеличиваются (см. табл. 32—34). Так, во II возрастной группе самцы значительно крупнее по следующим признакам: максимальной ширине черепа, скуловой и межглазничной ширине, а также ширине мозговой капсулы. В III возрастной группе различия аналогичны, однако они близки к достоверным и по кондилобазальной, и по основной длине. Различия по этим признакам и наибольшей длине усугубляются в самой старшей возрастной группе.
В условиях Белореченского лесничества у нас была возможность сравнения самцов и самок II возрастной труппы. Ситуация оказалась аналогичной: самцы крупнее самок, причем различия достоверны по максимальной ширине черепа и скуловой ширине. Близко к достоверному различие по межглазничной ширине.
Таким образом, череп самцов с возрастом становится более крупным, чем у самок. Причем различия касаются признаков ширины черепа (максимальная ширина черепа, скуловая и межглазничная ширина), а также длины и ширины мозговой капсулы.
Географическая изменчивость. В условиях Кавказа эта изменчивость не изучена. Нет еще соответствующего коллекционного материала. Из сказанного ранее вполне понятен интерес к сравнению особей из Белореченского лесничества и Судзухинского заповедника. Как видно из табл. 30—35, самцы (II возрастная труппа) из Белореченского лесничества несколько крупнее таковых из Судзухинского заповедника, однако эти различия несущественны. Различия между самками также несущественны, за исключением таковых по трем признакам (максимальная ширина черепа, скуловая и межглазничная ширина) во II возрастной труппе. Самки из Белореченского лесничества по этим признакам достоверно крупнее, чем у сородичей из Судзухинского заповедника.
Подвиды. Как отмечено выше, пятнистые олени, акклиматизированные на Кавказе, происходят из Приморского края и относятся к одному подвиду — С. п. hortularum Swinhoe.
Экология
Ландшафтная приуроченность. Биотопы. В пределах коренного ареала пятнистый олень приурочен к дубово- и кедрово-широколиственным лесам Приморья. По Р.В. Дормидонтову (1977), эти животные держатся в горных местностях с сильно пересеченным рельефом, богатой древесно-кустар
1 5 1
Пятнистый олень
никовой и луговой растительностью, а также речками и ручейками. Летом предпочитают луговые поляны среди лесов. При выпадении большого снега совершают перемещения с гор к приморью.
В местах акклиматизации пятнистые олени сохранили свою привязанность к горным широколиственным лесам и лесным полянам, однако в различных частях Кавказа они занимают различные типы лесов и полян. В Талыше они обитают во влажных субтропических лесах из железного дерева, бука, грецкого ореха, многочисленных лиан. В Хосровском заповеднике Армении пятнистые олени держатся на западных и северо-западных склонах гор на высоте 1700—1800 м над уровнем моря, где представлено мелкорослое редколесье южного типа с преобладанием дубово-грабовых насаждений, перемежающееся рощами можжевельника и лугами (Гамбарян, 1959; Айрумян, 1962). В Дилижанском заповеднике (Армения) и в Нальчикском лесоохотничьем хозяйстве (Карбардино-Балкария) пятнистые олени живут на воле в буково-храбовых лесах среднегорья, имеющих луговые поляны с обильной травянистой растительностью. Как видно, пятнистые олени адаптируются к широкому спектру ландшафтов Кавказа, но при этом сохраняют тяготение к умеренно-влажным биотопам. В сухом редколесье Хос-ровского заповедника в летнее время пятнистые олени держатся в более низинных и влажных местах.
Миграции. В условиях Приморского края пятнистые олени совершают различные перемещения. Они вызываются обычно сезонными изменениями условий жизни (см. обзор: Гептнер и др., 1961). Так, в связи со снего- . падами, что нередки в феврале-марте, пятнистые олени вынуждены спускаться с гор ближе к морю, где меньше снега. В это время большая часть популяции держится в прибрежной полосе шириной всего около 3 км. После таяния снегов они рассредоточиваются и в мае переходят на летние местообитания. На характер территориального размещения существенное влияние оказывают кровососы. При массовом появлении гнуса и клещей олени выходят к морским побережьям, на открытые и хорошо проветриваемые места.
В условиях Кавказа пятнистые олени, по нашим наблюдениям, так же как на родине, совершают кочевки. Даже на небольшой территории Белореченского оленария в летнее время они держатся преимущественно на северо-западных и юго-восточных склонах, где растут мощные леса, чередующиеся с луговыми полянами. При этом пятнистые олени населяют весь интервал высот, охваченный О1радой оленария (до 1100 м над уровнем моря), широко передвигаются в поисках более-кормных мест, благоприятных условий отдыха, прячутся в подлесках и высокотравье и т.д. Зимой в связи с появлением снегового покрова пятнистые олени перемещаются на южный, хорошо прогреваемый макросклон, занимают в основном безлесный участок, расположенный недалеко от кормушки.
В Хосровском заповеднике Армении, где бывают суровые и многоснежные зимы, зарегистрированы вынужденные сезонные кочевки пятнистых оленей (Гамбарян, 1959; Айрумян, 1962). В летне-осенний период пятнистые олени отмечались в основном на территории Хосровского леса,
152
Cervus nippon Tcmminck, 1838
преимущественно в верхнем течении одноименной реки в пределах 1700— 1800 м над уровнем моря. В этих местах почвенный покров более влажный, дольше вегетирует травянистая растительность, что и привлекает пятнистых оленей. Теплой и малоснежной зимой пятнистые олени регистрировались в пределах Хосровского леса, но в очень суровую зиму 1955/56 г., когда слой снега достигал 50—110 см, пятнистые олени вынуждены были спуститься по долине одноименной реки до с. Карабахлар, где нет лесов. Весной они снова вернулись в Хасровский лес.
Суточная активность. Поведение (рис. 24). На равнине в условиях вольного содержания суточная активность пятнистых оленей ритмична в течение всего теплого периода года. В погожие дни пик активности приходится на утренние и вечерние часы. Зимой такой ритмичности не наблюдается. В дождливые дни и во время снегопадов они малоактивны.
Данные по Северному Кавказу, накопившиеся у нас и в литературе (Дормидонтов, 1977) , показывают, что суточная активность пятнистых оленей в новых, условиях жизни остается сложной, зависимой от сезона года, погоды и других внешних и внутренних факторов. Довольно четко суточный режим соблюдается летом при хорошей погоде: утром пятнистые олени пасутся 3—4 ч, потом отлеживаются в кустарниках, лесных массивах и в высокотравье, а вечером вновь возобновляют пастьбу. Пастьба отмечалась также в первой половине ночи. В дождливую погоду они малоактивны, в основном пасутся в промежутках между дождями. Зимой режим суточной активности нарушается, увеличивается дневная активность.
В условиях вольерного содержания Белореченского оленария, по нашим наблюдениям, суточная ритмика сильно нарушена. Хотя животных подкармливают утром и вечером, дневная активность не только зимой, но и летом хорошо выражена. Даже в жаркую погоду июля 1989 г. (до +36°) мы наблюдали пасущихся оленей в поддень.
Пятнистые олени, акклиматизированные на Кавказе, остаются очень чуткими и пугливыми. При хорошем зрении и обонянии слух у них тонкий. Уши, большие, как локаторы, постояно начеку и в движении. При малейшем шорохе олени готовы сорваться с места. Среди пасущихся оленей мы всегда отмечали особо чутких животных, выполнявших роль сторожа. Осторожность и пугливость пятнистые олени сохранили и в оленарии, хотя они родились там и постоянно общаются с людьми при кормлении и уходе за ними. Однажды в загоне Белореченского оленария пятнистый олень был настолько напуган, что стрелой бросился на деревянную стену и головой насквозь пробил доску толщиной в 40 мм. При всей пугливости,. по словам оленеводов, олени могут нападать на людей, защищаясь в критических условиях.
Линька. В условиях Северного Кавказа пятнистый олень, как и на родине, линяет дважды в году — весной и осенью. Раньше линяют здоровые упитанные животные, а линька слабых задерживается.
Питание. Количество видов растений, поедаемых пятнистым оленем в заповедниках европейской части СССР и Зауралье, превышает 380. Из них
153
Пятнистый олень
Рис. 24. Пятнистый олень
154
Cervus nippon Temminck, 1838
лишь восьмую часть составляют те, которые используются ими в пищу в Приморском крае. При этом есть виды, предпочитаемые в одних районах и избегаемые в других. Существенные расхождения выявлены и по соотношению различных групп кормов. В Приморском крае, в частности, роль древесно-кустарниковой растительности в летнем рационе значительно выше, чем в названных заповедниках; основу питания в это время составляет преимущественно высокотравье (Ильина, 1956; Гептнер и др., 1961). Как видно, пятнистый олень в выборе пищи довольно пластичен, что немаловажно иметь в виду при акклиматизации в различных регионах.
Питание пятнистых оленей на Кавказе изучалось в ряде районов: в Тебердинском заповеднике (Курапова, Степанов, 1940; Князев, 1946), в Белореченском оленарии (Дормидонтов, 1977; наши данные), в Азербайджане (Алекперов и др., 1982).
Установлено, что в Тебердинском заповеднике олени охотно поедают желуди и плоды бука (животные достают их даже зимой из-под снега), ветки лещины, дуба, осины, ивы, ильма и особенно бересклета и других кустарников. Из травянистых растений предпочтительно относятся к борщевику, таволге, манжетке, лапчатке и др.
О высокой пластичности пятнистого оленя в питании свидетельствуют наблюдения Р.В.Дормидонтова (1977), проведенные в Белореченском оленарии Кабардино-Балкарии. На территории оленария зарегистрировано 219 видов растений, из которых пятнистые олени поедали 167. При этом 78 видов учтены как поедаемые впервые. С наибольшим удовольствием поедаются листья и побеги бука, граба, ежевики, козьей ивы, липы, калины, мушмулы и сливы. Из травянистых растений предпочитаются бухарник шерстистый, вика, девясил высокий, ежа, козлятник, недотрога, овсяница, клевер, свербиг, тимофеевка, язвенник. О роли тех или иных групп растений можно судить по ряду показателей. Так, несмотря на обильное кормление сеном и комбикормами и наличие лугов с хорошим травостоем, подрост всех типов лесных формаций уничтожен на всей территории, вследствие чего исключено естественное возобновление лесов. Показательны также результаты наблюдений в первый год акклиматизации. В начал'е марта 1968 г., когда пятнистых оленей впервые выпустили в буковых лесах Белореченского лесничества, значительная часть территории еще находилась под снегом, и лишь по юЖным склонам бугров появились проталины. В этих условиях животные получали достаточный корм (сено и комбикорма), но тем не менее пятнистые олени потребляли регулярно буковые орешки, прошлогодние побеги букового подроста, кустарники разных видов. В конце марта снег сошел почти полностью, в низинах зазеленело разнотравье и пятнистые олени стали больше пастись на полянах. На полянах пятнистые олени паслись в течение всего теплого периода года, пока не выпал снег.
В Азербайджане в составе кормов пятнистого оленя отмечено 100 видов растений, которые относятся к 61 роду и 14 семействам.
155
Пятнистый олень
Воспроизводство и структура популяции
Размножение. В Тебердинском заповеднике большинство пятнистых оленей участвовало в размножении с 5—10 октября до середины ноября (Курапова, Степанов, 1940). Рев отдельных рогачей регистрировали значительно раньше. На родине пятнистые олени вступают в гон с 29 сентября по 11 октября. Гон заканчивается к 5—8 ноября. Известен случай, когда одиночный бык приступил к реву 10 сентября (Бромлей, 1956).
Парковое содержание в новых местах внесло большие изменения в биологию размножения пятнистых оленей. В частности, сроки гона и соответственно время рождения телят сильно расширились. В Белореченском оле-нарии Кабардино-Балкарии, по данным местных оленеводов, рев рогачей и спаривание с оленухами продолжаются до декабря. В Азербайджане, по данным Х.М.Алекперова и др. (1982), гон происходит с сентября по конец октября. В Хосровском заповеднике Армении на высоте 1600—1800 м, в густой части леса, рев пятнистых оленей отмечался в начале октября (1958—1960 гг.). Рев более регулярен в период с 21 октября по 6 ноября. Он был слышен в основном только по утрам, продолжительность его 10—12 с, с перерывами в 5—10 мин. По вечерам и ночью рев отмечался редко (Айрумян, 1962).
Как на воле, так и в оленариях формирование гаремов (3—4 самки) сопровождается драками между самцами (в том числе со спиленными рогами). Победивший самец ревностно охраняет свой гарем, мало ест, сильно худеет. Максимальная масса семенников во время гона составляет 80,32 ± 6,8 г. После размножения этот показатель снижается в среднем до 24,5 ± 2,5 г (Стекленев, 1978). Самки во время гона фактически не теряют в массе.
Продолжительность беременности самок составляет обычно 7,5 мес, основная масса телится в мае-июне. Отдельные телята появляются даже поздней осенью. В Азербайджане отелы наблюдались с мая по середину июля (Алекперов и др., 1982). На сроки размножения, как показали наши наблюдения в оленарии, существенно влияют внешние условия, питание и др. Для отела оленухи выбирают укромные места — труднопроходимые лесонасаждения, высокотравье по опушкам леса, каменистые места, поросшие кустами, и т.п.
Плодовитость. Самка пятнистого оленя в естественных условиях, как правило, приносит одного теленка, двойни бывают исключительно редко. Однако в условиях паркового содержания возможно увеличение процента двоен. В зоопарке Аскания-Нова он составляет не более 1—2%, здесь же зарегистрировано рождение тройни (Стекленев, 1978). Плодовитость популяций сильно колеблется в зависимости от количества яловых оленух, при этом в оленариях яловость бывает значительно выше, чем в природе. В отдельных хозяйствах она составляет 30 и даже 70% (Гептнер и др., 1961).
Рост и развитие. Телята пятнистых оленей рождаются крупными (телята-самцы до 7,3 кг, самки до 6,2 кг), но в первые дни беспомощны, плохо держатся на ногах. Тем не менее через день могут следовать за матерью, а через 10—20 дней начинают самостоятельно пастись. Молодые остаются с
156
Cervus nippon Tcmminck, 1838
матерью до весны следующего года и дольше. В Приморье к 9-12 мес жизни самцы достигают массы тела 49—59 кг, самки — 48—55 кг, а через 16 мес соответственно 70—75 и 65—70 кг. Интенсивный рост продолжается в течение 2 лет, затем прирост массы тела замедляется, особенно у самок. Молодые быстро набирают массу в конце лета и осенью, а зимой рост почти прекращается. Масса тела самцов максимальна в 7—10 лет, самок — к 4—6 годам. На 10-м месяце жизни у самцов пятнистых оленей на голове появляются "дудки", на месте которых в апреле вырастают первые неветвя-щиеся рожки, потом через год появляются ветвящиеся рога. Максимальное развитие рогов зарегистрировано к 10—12 годам жизни. Сброс рогов в Приморье происходит в апреле или мае. В конце мая-июне появляются панты (см. обзор: Гептнер и др., 1961).
Примерно в те же сроки или с задержкой на месяц этот процесс происходит на Кавказе (Курапова, Степанов, 1940). По нашим данным, в Белореченском оленарии сброс основания спиленных рогов, появление и созревание пантов, соответственно срезка их продолжаются до августа включительно, т.е. процесс образования рогов у разных особей сильно растянут.
Смена ювенильной окраски на взрослую происходит осенью, после линьки, но у разных животных сроки линьки бывают весьма различные.
Формирование зубной системы происходит долго — лишь в возрасте 1—1,5 лет происходит смена молочных зубов на постоянные, а в 2 года сменяются коренные.
Максимальная продолжительность жизни 18—21 год, но в природных условиях не более 11—14 лет (см. обзор: Гептнер и др., 1961).
Структура популяции. В Азербайджане на начало 80-х годов насчитывалось 200 пятнистых оленей (Гирканский заповедник). Структура популяции выглядела так: 29,4% самцов, 50,0% самок, 20,6% молодняка. В Хос-ровском заповеднике Армении учет пятнистых оленей с помощью вертолета проведен в апреле 1961 г. ВсегоУчтено 7 групп общим числом 58 голов (вероятно, их было 62—65), из них 16 голов составляли самцы всех возрастов, кроме прибылых 1960 г. рождения, остальные были самки и прибылые.
В Хосровском заповеднике Армении пресс со стороны волков привел к некоторой стабилизации структуры популяции. С 1968 г. число пятнистых оленей в заповеднике колеблется в пределах 80—120 голов. Структура хос-ровской популяции включает 20% взрослых самцов, 40% взрослых самок и 40% молодняка (Богачев, 1989).
В Белореченском оленарии, по данным оленеводов, на 1 сентября 1989 г. содержится 1106 пятнистых оленей. Из них 390 (43,1%) рогачей, 39 (4,3%) перворожек (до 2,5 лет), 502 (45,4%) оленухи, 50 (4,5%) оленушек (до 2,5 лет) и 125 (11,3%) телят (до 1,5 лет).
Враги, паразиты, конкуренты, смертность. Наиболее опасным врагом пятнистых оленей на Кавказе является волк, особенно в многоснежные зимы. Нападения волка на пятнистых оленей отмечались неоднократно в Хосровском заповеднике. В этом же заповеднике волосы пятнистого оленя обнаружены в помете леопарда (окрестности с. Анд). В экскрементах же рыси
157
Пятнистый олень
волос оленей обнаружено не было (Гамбарян, 1959; Айрумян, 1962). При парковом содержании не меньшую опасность для пятнистых оленей представляют бродячие собаки. На молодых пятнистых оленей могут нападать медведь, шакал, лесная кошка, лисица.
Гельминтофауна пятнистых оленей включает немного видов, но зараженность их печеночной двуусткой высока на Центральном Кавказе. В условиях питомников выявлены 4 вида плоских глистов, 3 вида ленточных и 29 круглых (Абрамов, 1954 г., по: Гептнер и др., 1961). Пятнистые олени сильно страдают от клещей как на воле, так и в вольерах. Роль кровососущих насекомых в регионе не оценена, но, бесспорно, значительна, так как в ряде мест акклиматизации многочисленны комары, мошки, носоглоточные оводы, которые при парковом содержании животных, по данным разных авторов, могут нанести им большой урон (на воле пятнистый олень свободен от носоглоточных оводов). В ряде районов пятнистые олени страдают от различных болезней, в том числе от ящура, сибирской язвы, туберкулеза и т.д. В Тебердинском заповеднике зарегистрирован случай гибели пятнистых оленей от геморрагической септицемии (Ильина, 1956).
Конкурентами пятнистого оленя на Кавказе могут выступать благородный олень (Кавказский, Тебердинский и другие заповедники), косуля (лишь в отдельных местах), кабан (в ряде районов Кавказа).
Смертность. Смертность пятнистых оленей может быть очень высокой от разных причин. Так, первый завоз и выпуск оленей в Хосровский заповедник оказался неудачным из-за волков, постоянно уничтожающих оленей (Айрумян, 1962). Хищники и браконьеры уничтожили пятнистых оленей в Тебердинском заповеднике (Темботов, 1972).
Численность и ее динамика. На Кавказе практикуется разведение пятнистых оленей на воле и в оленариях. Вольное содержание проводится на охраняемых территориях — заповедниках, заказниках-, охотхозяйствах. Парковое разведение ведется в зоопарках, оленариях, зверосовхозах. В первом случае благополучие пятнистых оленей зависит больше от природных факторов и животные более уязвимы, чем во втором, когда человек берет на себя ряд функций (подкармливает, защищает от хищников и т.д.).
Соответственно динамика численности пятнистого оленя в природе и оленариях существенно различается. Так, в Краснодарском крае акклиматизированные в 1979 г. 135 пятнистых оленей увеличили свою численность к началу 80-х годов лишь до 150 голов. В последующем наблюдалась тенденция снижения численности до 60 в 1985 г. За 1981—1985 гг. в Кабардино-Балкарии поголовье пятнистых оленей вне Белореченского оленария сократилось в 20 раз (со 160 до 8 голов) (Кузякин, 1989). Вероятнее всего численность пятнистых оленей на воле, в лесах Нальчикского лесоохотничьего хозяйства, поддерживается за счет тех животных, которым удается сбежать из оленария. По данным этого же автора, в Дагестане за рассматриваемый период лишь в 1983 г. зарегистрировано 30 свободноживущих пятнистых оленей, а в остальные годы встреч не было. Очевидно, пятнистый олень прижился В Гирканском заповеднике, где отмечается рост поголовья и достигнута плотность населения 0,17 оленей на 1000 га основных
158
Cervus nippon Temminck, 1838
местообитаний (Алекперов и др., 1982). Что касается Армении, то поголовье пятнистых оленей, как отмечено выше, стабилизировалось в 80-х годах на низком уровне численности.
Динамику численности оленей в условиях загона лучше всего проиллюстрировать на примере Белореченского оленария (под Нальчиком). За период его существования с 1966 по 1989 г. он превратился в большое хозяйство с числом пятнистых оленей до 1300—2000 голов на территории загона площадью до 3,5 тыс. га. В 60-х годах имел место массовый падеж оленей. Причинами послужили отсутствие должного ухода, плохое кормление, перегрузка пастбищ и их деградация. В естественных условиях главными причинами снижения численности пятнистых оленей являлись хищники, в основном волк, глубокий снежный покров, затрудявший передвижения животных, наконец, необузданное браконьерство. Остальные факторы, в том числе инфекции, не выступали, как правило, лимитирующими численность оленей.
Практическое значение. В настоящее время основная масса пятнистых оленей на Кавказе сосредоточена в парковых хозяйствах, в том числе зверосовхозах. В природе их мало. Так, в 1977 г. по Северному Кавказу числилось 1500 пятнистых оленей, разводимых в полувольном состоянии. На воле обитало всего около 100 оленей. К 80-м годам количество пятнистых оленей возросло в одном только Белореченском оленарии Кабардино-Балкарии до 2000 голов, тогда как в природе — до 120 (Богачев, 1989). Летом 1989 г. в этом хозяйстве обитало 1300 пятнистых оленей. Основная цель содержания пятнистых оленей в этом, как и в других аналогичных хозяйствах, состоит в том, чтобы получать панты — молодые неокостеневшие рога. Панты были главной причиной истребления пятнистых оленей на их родине, точно так же как и причиной акклиматизации во многих районах страны.
Из пантов получают лекарственный препарат пантокрин — тонизирующее средство, высоко ценимое в китайской медицине. В одном только Белореченском оленарии получают за год 190 кг первосортных пантов, 26 кг второго сорта и 3,6 кг третьего сорта. К первосортным относятся молодые панты, имеющие надглазничный отросток и начинающее образовываться утолщение на конце главного, растущего ствола ("вторые шишки"), но когда образование среднего отростка еще не началось, т.е. панты имеют возраст около 48—55 дней с момента сбрасывания старых рогов (Соколов, 1959).
Следует отметить положительную зависимость между возрастом пантового оленя и пантовой продуктивностью до семилетнего возраста, которая затем становится отрицательной (Zhou, Wu, 1988). Названные авторы установили также положительную связь между массой тела и пантовой продуктивностью.
Мясо пятнистых оленей характеризуется высокими вкусовыми качествами, и как дополнительную продукцию пантового хозяйства его нельзя сбрасывать со счетов.
159
Благородный олень
При оценке практического значения пятнистых оленей на Кавказе, на наш взгляд, особо следует учесть, что оленеводство с загонной системой содержания животных все больше вступает в острое противоречие с интересами охраны горных лугов и лесов. Дело в том, что селекционная работа, направленная на повышение пантовой продукции, базируется на усилении инстинкта стадности и соответственно плотности животных на единицу площади (Галкин, 1987). Это неминуемо ускоряет процесс разрушения экосистем, что недопустимо в столь густонаселенном и малолесном регионе, как Кавказ. На наш взгляд, пятнистый олень не может быть в дальнейшем рекомендован ни для паркового, ни для вольного содержания на Кавказе.
Благородный олень — Cervus elaphus Linnaeus, 1758
Рис. 25
Cervus elaphus Linnaeus, Паллас, 1811; Gray, 1850, 1872; Blasius, 1857, Fitzinger, 1873-1874; Brooke, 1873; Radde, 1886; Lydekker, 1898, 1915; Сатунин, 1903; Beninde, 1937; Cervus maral Ogilby, 1840; Sclater, 1872; Gray, 1872; Ce-верцов, 1873; Ward, 1878; Radde, 1886; Satunin, 1896; Carvus wallichi Cuveir, Gray, 1850; Cervus wapiti Barton, Северцов, 1873; Cervus caspicus Brook, 1874, P.Z.S.L.; Cervus caucasica Winaus, Winaus, 1914.
Коллекционный материал. ЗИН РАН: 21 череп, 52 рога, 2 шкуры; МГУ: 13 черепов, 10 рогов, 5 шкур. •
Диагноз. Олень относительно крупного размера, высота в холке более 120 см, телосложение крепкое, шея толстая, голова длинная, рога крупные, простые, "кавказского типа”, летний мех слабо пятнистый. Зеркало крупное, занимает круп выше основания хвоста, хвост короче длины уха.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Масса тела самцов 260—342 кг, самок 133,3—160 кг, длина Дела соответственно 225—249 см и 164—215 см, высота в холке 142—152 см и 114—133 см (Александров, 1968).
Из этих и приведенных в табл. 35 и 36 данных следует, что на Кавказе благородный олень имеет крупные размеры и массу, значительно превосходящие таковые не только пятнистого оленя, но и своих европейских сородичей. Более того, по мнению Н.Я.Динника (1910), кавказские олени по размерам не уступают иногда американским вапити и алтайскому маралу. Однако В.Г.Гептнер и В.И.Цалкин (1947, с. 53) не разделяют эту точку зрения, считая, что хотя кавказские олени "несомненно превосходят по размерам среднеевропейских оленей, но в то же время заметно уступают маралам, занимая как бы промежуточное положение между двумя последними формами". При оценке приведенных выше меристических данных необходимо учесть, что они относятся к западнокавказским популяциям, отличающимся более крупными размерами, чем восточнокавказские.
160
Cervus elaphus Linnaeus, 1758
Таблица 36. Масса тела кавказского оленя (в кг) из района Кавказского заповедника
(по: Александров, 1968)
Масса
Возраст Число эк- самцов самок
земпляров
limit limit X
До 1 сут 6 7,0-11,1 9,1 7,2—9,7 8,7
1 мес 15 18,4-29,5 24,8 21,5-23,1 22,2
4—6 мес 4 74,7-75,6 75,2 71,0—74,4 73,7
1,5—2,5 лет 7 120,0-162,0 144,0 103,8—128,5 115,0
2,5—5 лет 5 275,6-286,2 296,8 133,3-160,0 147,0
5—11 лет 6 260,0-342,0 310,0 — —
Череп кавказских оленей обладает рядом особенностей, выявленных наиболее полно В.Г.Гептнером и В.И.Цалкиным (1947) при анализе различных индексов его (в процентах по отношению к кондилобазальной длине). Так, скуловая ширина у кавказских оленей составляет 40,5%, тогда как во всех других изученных популяциях не менее 43%. Межглазничная ширина наиболее узка у кавказских и китайских оленей — 34,5 и 34,0% соответственно, а у других — 36,0% и более. Так же четко кавказские олени выделяются наибольшей узостью черепа, измеренной по задним стенкам орбит (40,5% у кавказских, 42,5% и более у других). Наоборот, длина рыла максимальна у кавказских (61,5%), однако по рассматриваемому показателю все изученные формы, за исключением бухарских оленей (57,5%), близки друг к другу. Характерна также ширина затылочных мыщелков — у кавказских она равна всего 49,0%, а в других популяциях не менее 54,0%. Словом, у кавказских оленей череп относительно вытянутый с несколько удлиненным рылом и суженный, что характерно как для самцов, так и для самок. Однако по мнению названных авторов структура черепа в общих чертах у различных гео-1рафических форм настоящих оленей весьма сходна. "Отличия в этих признаках меньше, чем в общих размерах, окраске и прежде всего в строении рогов ... Существенно, черепа рас, объединяемых некоторыми авторами в видовые комплексы, нередко отличаются друг от друга сильнее, нежели от рас другого вида" (Гептнер, Цалкин, 1947, с. 64).
Рога у благородного оленя в отличие от пятнистого сложные, количество отростков не менее 5, обычно их 8—9, максимально до 66 концов (Шилин-гер, 1930 г., по: Соколов, 1959). При минимальном количестве отростков (глазной, ледовый, средний и два концевых) присутствие ледового, по Гентнеру и др. (1961), является характерной чертой вида. В остальном структура рогов весьма изменчива, подвержена широкой индивидуальной и географической изменчивости (табл. 37). Тем не менее Гептнер и Цалкин (1947) считают возможным свести все многообразие рогов благородного оленя к трем типам: 1) среднеевропейский; 2) хангуловый (центральноазиатский), 3) мараловый. Кавказские благородные олени ближе всего к первому типу, но существенно отличаются "особой личной изменчивостью: одни особи имеют рога с кроной, другие без нее. Эта комбинация европейского типа и маралового типа в личной изменчивости" (с. 17). (Подробнее см. раздел "Изменчивость").
Н В.Е.Соколов, А.К.Темботов 16 1
Благородный олень
Мариуполь
Ростов-на-Дону
Бердянск
О Заветное
О
О Яшкуль
г. Эльбрус
5642
о Сарпа
О Элиста
Егорлыкская
О Дивное.
Благодарный
-Гагр
—Сухуми
Зугдиди
• 12
6000 -
503
6000 -
хтала
4000 -
3000
2000
1000
Ю-0
КБ
4000
Нахичевань
3000
^Черкесск
Пятигорск
г. Далидаг
3616
г. Каракал 2850
г. Кюмиркей 2477
г. Мелис цкаро 2850
г. БолчГиналдаг
3367
Ереван
Мацони
винно'мысск
3?
-
Сочи
г. Псеашха
3255
кх
30
Очамчир
Ставрополь
Свет лоград
Буденновск
2000
1000 0
К-3
ц-м
А
Нефтекумск
Моздок
Грозы
162
Cervus elaphus Linnaeus, 1758
Рис. 25. Распространение благородного оленя
1 — пойма р. Кубань (Верещагин, 1959; Кириков, I960; Александров, 1968); 2 — Анапский р-н (Александров, 1968); 3 — Натухаевская (Александров, 1968); 4 — Гостагаевская (Александров, 1968); 5 — Новороссийск (Динник, 1910; Верещагин, 1959; Кириков, I960, Александров, 1968); 6 — Геленджик (ЗИН); 7 — Кабардинка (Динник, 1910; Верещагин, 1959; Кириков, 1960; Александров, 1968); 8 — Нижнебаканский (Александров, 1968); 9 — Крымск (ЗИН; Александров, 1968); 10 — верховья р. Абин (Александров, 1968); 11 — Эриванская (Александров, 1968); 12 — гора Тхаб (Александров, 1968); 13 — верховья р. Ахтырь (Александров, 1968); 14 — Пшада (Александров, 1968) 15 — гора Афипс (Александров, 1968); 16 — Дефановка (Александров, 1968); 17 — верховья р. Джубга (Александров, 1968); 18 — Туапсе (Динник, 1910; Александров, 1968); 19 — верховья р. Псекупс (Александров, 1968); 2 0 — Горячий ключ
(Александров, 1968); 21 — Краснодар (Динник, 1910); 22 — верховья р. Туапсе (Александров, 1968); 23 — верховья р. Пшиша ( Насимович, 1936; Верещагин, 1959; Кириков, 1960; Александров, 1968); 24 — Хадыженск (Александров, 1968); 25 — Нефтяная (Насимович, 1936; Верещагин, 1959; Александров, 1968); 26 — Апшеронск (ЗИН; МГУ; Александров, 1968); 27 — Пшехская (Александров, 1968); 28 — Навагинка (Насимович, 1936; Верещагин, 1959, Александров, 1968); 29 — Бзыч (Александров, 1968); 30 — верховье р. Головинка (Александров, 1968); 31 — верховье р. Бзыч (Александров, 1968); 32 — Бабукаул (Александров, 1968); 33 — верховье р. Пшехи (Верещагин, 1959; Кириков, 1960; Александров, 1968): 34 — р. Белая (Динник, 1904;
11*
163
Благородный олень
Александров, 1968); 35 — Новопрохладное(Александров, 1968); 36 — Даховская (Насимович, 1936; Верещагин, 1959; Александров, 1968); 37— Майкоп (ЗИН, Динник, 1910; Верещагин, 1959; Кириков, 1960; Александров, 1968); 38 — Тульский р-н (Александров, 1968); 39 — Лабинский р-н (Александров, 1968); 40 — Ходзь (Александров, 1968); 41 — Красная поляна (Александров, 1968); 42 — г. Абаго (МГУ; Александров, 1968); 43 — Ри-цинский заповедник (Соколов, Сыроечковский, 1990); 44 — верховья р. Большая Лаба (Александров, 1968); 45 — Псху-Гумистинский заповедник (Соколов, Сыроечковский, 1990); 46 — Архызское лесничество (Маков, 1938); 47 — Теберда (Инякова, 1957; Сафронова, 1973; Тарасов, 1977); 48 — верховье р. Уруп (Александров, 1968); 49 — Нижняя Ермолаевка (Александров, 1968); 50 — Уруп (Александров, 1968); 51 — Усть-Джегутинск (Динник, 1910; Темботов, 1972); 52 — Претрадная (Верещагин, 1959; Александров, 1968); 54 — Курджиново (Александров, 1968); 53 — Соленое (Насимович, 1936; Верещагин, 1959; Александров, 1968); 55 — Иваново (Александров, 1968); 56 — Пссбай (Александров, 1968); 57 — Передовая (Динник, 1910; Верещагин, 1959; Кириков, 1960); 58 — низовье р. Кодори (Верещагин, 1959); 59 — среднее течение р. Ингури (Верещагин, 1959); 60 — верховье р. Кодори (Соколов, Темботов, 1989); 61 — верховье р. Малки (Темботов, 1972); 62 — Кисловодск (Динник, 1910); 63 — низовье р. Ингури (Верещагин, 1959); 64 — дельта р. Риони-(Соколов, Темботов, 1989); 65 — Батуми (Динник, 1910; Верещагин, 1959); 66 — Боржомский заповедник (Марков, 1938); 67 — Боржоми (Марков, 1924); 68 — верховье р. Риони (Соколов, Темботов, 1989); 69 — Стыр-Дигора (Наниев, 1956); 70 — Нарт (Наниев, 1956); 71 — верховье р. Ингури; 72 — Нальчикское лесоохотхозяйство (Темботов, 1972); 73 — Нальчик (Динник, 1910; Верещагин, 1959); 74 — Алагир (Динник, 1910); 75 — Цей (Наниев, 1956); 76 — Владикавказ (Динник, 1910); 77 — Балта (Наниев, 1956); 78 — Пасанаури (Динник, 1910; Верещагин, 1959); 79 — Ли-ахвекий заповедник (Соколов, Сыроечковский, 1990); 80— Ананури (Динник, 1910; Верещагин, 1959); 81 — Сагурамо-Мариамджварский заповедник (Соколов, Сыроечковский, 1990); 82 — Тбилиси (Динник, 1910; Верещагин, 1959); 83 — Алгетский заповедник (Соколов, Сыроечковский, 1990); 84 — Шулавери (Марков, 1924); 85 — Караязский заповедник (Соколов, Сыроечковский, 1990); 86 — Каракилиса (Марков, 1924); 87 — Кафан (Даль, 1950): 88 — Азизбеков (Межлумян, Гукасян, 1986); 89 — Дилижан (Айру-мян, Гаспарян, 1977); 90 — оз. Севан (Межлумян, Гукасян, 1986); 91 — Кировабад (Верещагин, 1959); 92 — Гейгёльский заповедник (Соколов, Сыроечковский, 1990); 93 — Калацджак (Верещагин, 1959); 94 — Нуха (Марков, 1924); 95 — Илисунский заповедник (Соколов, Сыроечковский, 1990); 96 — Закаталы (Марков, 1924); 97 — Лагодехи (Марков, 1924); 98 — Закатальский заповедник (Марков, 1938; Соколов, Сыроечковский, 1990);/99 — Лагодехский заповедник (Марков, 1938; Соколов, Сыроечковский, 1990); 100 — Тлядал (Динник, 1910); 101 — Ахметский заповедник, Туметский участок (Соколов, Сыроечковский, 1990); 102 — Тляратинский район (Прилуцкая, Пишванов, 1988); 103 — Гуниб (Динник, 1910); 104 — Щелковский район (Точиев, 1973); 105 — Грозный (Марков, 1924); 106 — р. Сунжа вверх до г. Грозного (Верещагин, 1959; Точиев, 1973); 107 — Гудермес (Верещагин, 1959; Точиев, 1973); 108 — Гребенская (Верещагин, 1959; Точиев, 1973); 109 — Бабаюртовский р-н (Прилуцкая, Пишванов, 1988); 110 — Тарумов-ка (Прилуцкая, Пишванов, 1988); Ill — Кизлярский р-н (Прилуцкая, Пишванов, 1988); 112 — низовье р. Кумы (Верещагин, 1959; Кириков, 1960; Александров, 1968); 113 — низовье р. Терек (Верещагин, 1959; Кириков, 1960; Александров, 1968); 114 — Кизилюртовский р-н (Прилуцкая, Пишванов, 1988); 115 — низовье р. Сулак (Верещагин, 1959; Кириков, 1960; Александров, 1968); 116 — Маджалис (Динник, 1910); 117— Хач-масские низовые леса (Верещагин, 1959); 118 — Кубинский р-н (Верещагин, 1959); 119 — Кусары (Верещагин, 1959); 120 — Исмаиллинский заповедник (Соколов, Сыроечковский, 1990); 121 — Исмаиллы (Верещагин, 1959); 122 — Пиркулинский заповедник
164
Cervus elaphus Llnnaeus>1758
Таблица 37. Размеры (в см) рогов кавказского оленя Западного Кавказа ______(196 рогов, в основном непарных) (по: Александров, 1968)___
г— ? 1 Признак Число с , , иростков на рогах
Общая длина рога 77,5 96,9 105,1 104,7 103,0 106,1 111,0
Длина первого отростка 24,0 31,3 34,4 35,3 41,0 36,7 39,1
Длина второго отростка 23,4 27,0 30,7 32,1 34,5 35,6 34,4
Расстояние между основанием первого и второго 24,2 25,2 25,4 26,3 26,5 23,5 24,5
отростков Обхват основания рога 12,5 20,4 21Д 24,0 24,0 22,5 23,2
Обхват ствола над первым отростком 10,2 13,5 16,7 19,9 17,0 16.6 18,1
Масса одного рога, кг — 2,4 2,9 3,9 4,0 4,2 5,1
Окраска тела благородного оленя имеет некоторые видоспецифические особенности. В частности, характерно отсутствие пятнистости во все периоды жизни, кроме ювенильного. Лишь у некоторых географических форм в летнем мехе отдельных особей могут быть пятна. Пятнистость чаще всего проявляется у кавказского оленя, реже у бухарского, у других рас не отмечена или очень редко встречается (Гептнер и др., 1961). Видовым признаком считается сильное развитие зеркала, которое распространяется на верх тазовой области вплоть до корня хвоста включительно. Вероятно, не менее характерно большое разнообразие общей окраски тела и отдельных частей, которое нередко использовали в качестве диагностического признака (форма, размер и окраска зеркала, окраска морды, губ, подбородка и т.д.). Многочисленные исследования, в особенности Гептнера и Цадкина (1947), убедительно свидетельствуют о том, что характерные черты вида и внутривидовых форм настоящих оленей слагаются из ограниченной труппы комбинирующихся признаков. При этом окраска тела четко подвержена географической изменчивости, подчиняется правилу Глогера: посветление окраски идет в общем направлении с запада на восток и отчасти с севера на юг; интенсивно окрашенные ("красные") особи свойственны окраинноматериковым районам (английский олень и изюбр), а светлые — внутрима-териковым (центральноазиатские).
В этом ряду кавказские благородные олени располагаются ближе к среднеевропейским, однако обладают своеобразной комбинацией признаков. Анализу специфики окраски оленей Кавказа посвящен целый ряд исследований отечественных и зарубежных териологов, но особой тщательностью выделяется работа Н.Я.Динника (1910). Его описание практически невозможно подвергнуть ревизии, так как старый материал несет глубокий след времени хранения в коллекции, а новые сборы малочисленны и фрагментарны. Поэтому ниже приводится обзор окраски тела кавказских оленей преимущественно на основании описания Н.Я.Динника с некоторыми дополнениями.
При всем сходстве со среднеевропейскими оленями кавказские олени заметно светлее, что обусловлено заменой красноватой окраски на рыжую,
165
Благородный олень
рыжеватую; кроме того, плечи и бедра темно-бурые, иногда даже черноватые. Для кавказских оленей обычное явление, когда у самцов и самок различных возрастов хорошо выражена пятнистость окраски, степень развития которой меняется очень широко — от полного отсутствия до четко обозначенных рядов на спине. Мы имели возможность изучить такого пятнистого быка, происходящего из среднегорных лесов Кабардино-Балкарии. Животное находилось в отличном состоянии как по упитанности, так и по размерам тела. Желто-рыжеватые пятна четко выделялись из общего коричневого фона, образуя на спине ряды. Количество рядов, судя по литературным данным и нашим наблюдениям, меняется широко — от полного отсутствия до четырех рядов. Столь же широко изменчива окраска зеркала — она может быть белой, но с рыжеватым оттенком или чисто-белой, а также неоднородной: над корнем хвоста рыжеватой или желтоватой, переходящей внизу на бедрах в белую. Вдоль позвоночного столба на шее и спине тянется темно-бурая или даже черная полоса, интенсивность которой снижается от головы к хвосту’. Иногда этой полосы нет или она слабо выражена. Окраска верхней стороны шеи сходна с туловищем или немного светлее; на боках рыжевато-серая или пепельно-серая, а на нижней стороне светло-серо-бурая. Горло имеет светло-рыжевато-серый цвет, иногда белый. Голова значительно серее, чем туловище. У некоторых животных на темени имеется черная полоса. По бокам головы окраска волос светлеет. Вокруг глаз и рта волосы белесые или светло-рыжие. На подбородке шерсть также беловатая. По углам рта черно-бурые пятна. Уши летом снаружи покрыты редкими черными волосами, а изнутри — длинными волнистыми волосами белого цвета. Ресницы и редкие волосы вокруг глаз длинные и черного цвета. Короткий хвост голый снизу, а сверху покрыт рыжей шерстью. Низ тела в области живота и груди рыжевато-белого цвета. Ноги с передней стороны бурые или темно-бурые, а с боков и сзади значительно светлее. У самок различие в окраске ног и туловища выражено слабо. Копыта черно-рогового цвета. Сезонный, возрастной, индивидуальный аспекты окраски тела значительны, и о них будет сказано ниже.
Кожный покров. Изучена кожа загривка, крестца и груди (табл. 38) оленей, добытых на Кавказе (в сентябре № 1 — взрослый самец и в июне № 7 — взрослый самец; для сравнения 15—16 ноября в Крымском госохот-хозяйстве самки: № 2 — длина тела 179 см, № 3 — 169 см, № 4 — 198 см, № 5 — 206 см, № 6 — 193 см).
Роговой слой эпидермиса кожи у самца оленя № 1 в сентябре плотный. Эпидермис слабо пигментирован. Сосочковый слой дермы толще сетчатого. В сосочковом слое пучки коллагеновых волокон образуют между собой плотное сплетение. Эластиновые волокна не очень многочисленны и залегают параллельно поверхности кожи. Только на самой границе с подкожной мускулатурой имеются пучки эластиновых волокон, проходящих параллельно поверхности кожи. Сальные железы, как правило, очень малы, у многих из-за волос не видны совсем (лишь некоторые из них достигают крупных размеров). Наиболее крупные и сильно извитые потовые
166
Cervus claphus Linnaeus, 1758
Таблица 38. Промеры кожи благородного оленя (по: Соколов, 1973)
Показатели Толщина Размеры сальных желез, мкм Диаметр потовых желез, мкм
КОЖИ эпидермиса, мкм рогового слоя, мкм дермы, мм СОСОЧКОВОГО слоя, мм сетчатого слоя, мм
№1, Кавказ (IX) 3,5 28 11 Загривок 3,47 2,27 1,2 66x190 120
№2 Крым (XI) 3,6 45 28 3,55 1,85 1,7 120x550 143
№3 3,3 34 17 3,26 1,86 1,4 80x130 80
№4 5,7 Сор ван 5,7 3,5 2,2 60x660 110
№5 :5,0 39 17 4,96 2,36 2,6 44x190 44
№6 3,9 50 34 3,85 1,75 2,1 66x165 80
№1, Кавказ (IX) 3,8 39 17 Крестец 3,76 2,66 1,1 55x440 187
№2 Крым (XI) 4,7 45 22 4,65 2,85 1,8 66x220 165
№3 4,3 34 17 4,26 2,36 1,9 50x440 90
№4 5,0 28 12 4,97 2,77 2,2 50x330 80
№5 5,2 39 17 5,16 1,86 з,з 55x330 80
№1, Кавказ (IX) 3,8 39 11 Грудь 3,76 1,96 1.8 130x495 100
№2 Крым (XI) 2,5 45 22 2,45 1,75 0,7 120x440 120
№3 2^3 25 7 2,27 _LJ_ 110x440 90
железы располагаются в коже крестца, несколько меньших размеров — в коже загривка. Сумма диаметров потовых желез, подсчитанных на протяжении 1 см препарата, достигает максимальной величины также на крестце (3,6 мм по сравнению с 2,7 мм на труди). Мышцы, поднимающие волосы, толщиной 160—220 мкм, хорошо развиты (рис. 26).
Волосы у оленя № 1 растущие, с сердцевиной в корнях. Ножек у волос нет. В коже загривка можно выделить волосы двух категорий: остевые пер-вого-третьего порядков и пуховые. Длина остей первого-третьего порядков: 49,6; 27,0 и 16,9 мм и пуха 8,0 мм; толщина остей первого порядка 258 мкм и пуха 15 мкм; сердцевина в остях первого порядка составляет 91,8%. Между всеми порядками остей можно найти переходные формы. Остевые волосы постепенно утончаются от основания к вершине. Остевые и пуховые имеют изгибы. У остей первого порядка их от 8 до 10, второго порядка — 4—6, у третьего порядка — 3—4 и у пуховых — 2—3. В поперечнике ости имеют вытянутую овальную форму. От периферийных клеток сердцевины на препаратах хорошо сохраняются боковые стенки, перпендикулярные к поверхности волоса. Корковый слой в боковых отделах волоса на 3 мкм толще, чем в передних и задних. Высота клеток кутикулы 18 мкм. Количество остевых волос первого порядка на 1 см2 поверхности кожи достигает 333. Очертания кутикулярных клеток остей второго и третьего порядков напоминают остевые первого порядка, но высота их меньше — 15 мкм. Суммарное число остевых волос второго и третьего порядков, растущих на
167
Благородный олень
Рис. 26. Строение кожи загривка (А, Б) и крестца (В) взрослого самца благородного оленя
1 — эпидермис, 2 — дерма, 3 — остевой волос, 4 — пуховой волос, 5 — мышца, поднимающая волос, 6 — сальная железа, 7 — потовая железа, 8 — пучок коллагеновых волокон
1 см2 поверхности кожи, достигает 250 шт. Высота кутикулярных клеток пуховых волос в среднем 10 мкм. Клетки кутикулы по большей части имеют кольцевидную форму. Число пуховых волос на 1 см2 поверхности кожи достигает 2508 (рис. 27).
Среди волос груди можно выделить две категории: остевые и пуховые. Длина остей 8,8, пуха — 5,6 мм; их толщина — 225 и 26 мкм; сердцевина -92,2 и 47,0%. Много растущих волос, у которых только вершинка выдается над кожей. Ножек у остевых волос нет, они слегка волнистые (имеют 2—4 изгиба). В поперечнике ости имеют овальную форму, более округлую, чем на загривке. На поперечном срезе хорошо видны боковые стенки периферийного ряда клеток сердцевины. Корковый слой в боковых частях волоса лишь на 1 мкм толще, чем на переднезадних отделах. Ости не образуют пучков или трупп. Число остевых волос на 1 см2 поверхности кожи 500. Пуховые волосы прямые или слегка волнистые. Характерно наличие в нижних и средних отделах большинства пуховых волос сердцевинного слоя. Он может быть сплошным или прерывистым. Пуховые волосы растут пучками, по 3—5 щт. в каждом. Их число на 1 см2 поверхности кожи достигает 833.
Строение кожного покрова в ноябре сходно с сентябрьским. Мальпигиев слой эпидермиса состоит из 2—3 рядов клеток. Потовые железы с относительно короткими секреторными отделами, делающими всего 2—3 изгиба . В коже труди потовые железы развиты сильнее, чем в коже спины. Сальные железы довольно крупные, особенно в коже груди.
168
Cervus elaphus Linnaeus, 1758
Рис. 27. Строение кожи препуция взрослого самца благородного оленя (Ли Б)
1 — дерма, 2 — волосяной фолликул, 3 — сальная железа, 4 — потовая железа, 5 — гровеносный сосуд, 6— поперечно-полосатая мускулатура
У июньского самца № 7 (табл. 39, 40) в коже загривка толщина сосочкового и сетчатого слоев почти одинакова (3,1—3,5 мм). Эпидермис тонкий (24 мкм), пигментированный. Сальные железы остевых волос крупные, одно-двудольчатые (339 х 113; 226 х 113 мкм). Потовые железы редкие, слабо извитые, диаметром 68—90 мкм. На крестце сосочковый слой тонкий (30—35% толщины кожи), сетчатый — толстый, мощный, образован плотной вязью толстых пучков коллагеновых волокон. Сальные железы осте-
169
Благородный олень
Таблица 39. Промеры волос загривка взрослого самца (№ 7)
благородного оленя (л == 10)
Категория волос Длина, мм Толщина стержня, мкм Сердцевина, % от толщины волоса
Ость порядка
I 47,2±1,9 260 87
II 40,3±0,7 240 87
III 35,5±1,2 225 87
IV 29,3±1,5 180 84
Пуховые 10,0±22,0 15-30 —
Таблица 40. Промеры остевых волос предплюсневой железы взрослого самца (№ 7) благородного оленя (л = 10)
Размерный порядок Длина, мм Толщина стержня, мкм Сердцевина, % от толщины волоса
I 27,8+1,8 180 58
II 22,3+1,5 188 64
III 17,2+1,6 150 60
вых волос круглые (226 х 226; 113 х 113 мкм), трех-четырехдольчатые. У пуховых волос они также хорошо различны, 56 х 36; 56 х 45 мкм. Потовые железы многочисленные, как и у самца № 1, со свернутыми в клубки секреторными отделами. Мышцы, поднимающие волосы, сильно развиты, иногда двухвершинные.
У летнего самца № 7 волосяной покров представлен остевыми и пуховыми волосами. Волосы относительно жесткие, слабо волнистые. Основание темно-бурое. Кончики остевых волос рыжие. Остевые волосы четко подразделяются на четыре размерных порядка (табл. 39). "Ножка" слабо выражена. У остей первого размерного порядка стержень достигает наибольшей толщины в начале рыжего участка. У остей второго и третьего порядков толщина мало меняется по длине стержня. У остей четвертого порядка стержень постепенно сужается от основания к вершине. Поперечные срезы овальной формы (рис. 28, А, Б). Корковый слой развит более или менее равномерно со всех сторон. Сердцевинный канал проходит вдоль всего волоса (рис. 28, В). В "ножке" он отсутствует. Сердцевина зернистого типа. По центру сердцевинного канала клетки крупнее, по периферии — мельче. Поверхность стенок сердцевинных клеток относительно гладкая, с редкими волосковидными выростами внутрь клетки. Рисунок кутикулы по длине не меняется (рис. 28, Г). Чешуйки вытянуты поперек стержня. Поперек волоса укладывается 2—2,5 чешуи. Свободные края чешуи ровные. На поверхности кутикулы в складках между чешуйками и корковым слоем обнаружены колонии микроорганизмов (рис. 28, Д).
Пуховые волосы тонкие, малочисленные, волнистые, темно-серого цвета. Поперечные срезы по всей длине округлой формы (рис. 28, Е). Сердцевина отсутствует. Кутикула полукольцевидной формы. Высота чешуй более или менее постоянна. Свободные края относительно ровные.
170
Cervus elaphus Linnaeus, 1758
Рис. 28. Строение волос загривка взрослого самца благородного оленя
А — поперечный срез кожи, ув. х 250; Б — поперечный срез остевого волоса, ув. х 450; В — строение сердцевины остевого волоса, ув. х 300; Г— кутикула остевого волоса, ув. х 400; Д — бактерии — кератинофиты на кутикуле, ув. х 1000; Е — кутикула пухового волоса, у в. х 1000
Волосяной покров груди относительно короткий по сравнению с загривком, темно-бурого цвета. Волосы подразделяются на две категории: остевые и пуховые. Много растущих волос. Остевые волосы прямые и на размерные порядки не подразделяются. Их длина не превышает 20,5 мм. "Ножка" у основания отсутствует. От основания до середины стержня толщина (135—165 мкм)/практически не меняется. К вершине стержень посте-
1 7 1
Благородный олень
Рис. 29. Строение волос труди взрослого самца благородного оленя
А — поперечный срез остевого волоса, ув. х 300; Б — кутикула основания остевого волоса, ув. х 600; Г — то же в середине стержня, ув. х 500; В — кутикула пухового волоса, ув. х 2000
пенно сужается. Характерен удлиненный бессердцевинный кончик. Поперечные срезы по всей длине овальной формы (рис. 29, А). Сердцевина зернистого типа хорошо развита и составляет около 86% от диаметра стержня. Сердцевинные клетки мельче по периферии стержня. Их стенки ребристые и скреплены между собой и с корковым слоем волосковидными тяжами, что придает волосу дополнительную прочность.
На поверхности кутикулы, в сердцевине стержня в прикорневой и корневой частях волоса, погруженных в кожу, обнаружены колонии бактерий-кератинофитов, которые разрушают корковый слой и сердцевину (рис. 30). Рисунок кутикулы меняется по длине стержня. В нижней половине стержня рисунок кутикулы такой же, как и у остевых волос загривка (рис. 29, Б). В верхней половине стержня чешуйки нижние, сильно вытянуты по ширине (рис. 29, Г). Свободные края сильно изрезаны и плотно прилегают к корковому слою.
Пуховые волосы светлые, волнистые (образуют 3—4 волнообразных изгиба). Длина не превышает 12,3 мм. Толщина (35 мкм) одинакова по длине всего стержня. Характерно наличие в нижней половине стержня прерывистого сердцевинного слоя. Кутикула кольцевидного типа по всей длине волоса (рис. 29, В).
172
Cervus elaphus Linnaeus, 1758
Рис. 30. Кератинофилы в волосах кожи труди взрослого самца благородного оленя А—Г — повреждения бактериями кер ати но фитами кутикулы остевого волоса, ув. х 150 х 1000 х 3000 х 10000. ДЕ — то же в сердцевине волос, ув.х 200 х 2500 х 10000 (различим волос с полностью разрушенной сердцевиной); .Ж, 3 — бактерии на стенках сердцевины, ув. х 1000
173
Благородный олень
Таблица 41. Размеры и масса отдельных органов и некоторых частей тела кавказских
оленей (округленные показатели) (по: Александров, 1968, с доп.)
Признак Сего- Молодые Взрослые самки 2,5- Взрослые самцы
летки 5 лет 5— 11 лет
4—5 мес. самки самцы беремен- ХОЛОС- До после
(4 экз.) (4 экз.) (4 экз.) ные или тыс гона гона
кормящие (2 экз.) (1 экз.) (3 экз.) (2 экз.)
Масса тела, кг 74,4 115 144 122 147 310 251
Выход мяса, % 60 58 59 55,2 57 61,9 58,1
от массы тела
Масса, г
печени 1275 1800 1600 2575 3100 5040 4300
легкие без 1200 — 1200 — — 3817 3600
трахеи сердце 650 1200 900 1300 1400 2174 2050
почки 170 240 145 350 430 861 —
селезенка 220 350 420 540 850 1700 1500
семенники — — —
или матка 320* 170 382 320
желудок без 1650 3500 3300 4900 6600 8330 4250
содержимого кишечник без 1000 2400 2500 1600 4100 2700 —
содержимого Длина кишеч- 2158 2355 3120 3345 3513 3776 —
ника, см
* С учетом эмбриона масса матки в одном случае была 17,3 кг.
У самца № 7 предплюсневая железа представляет собой утолщенный участок кожи и волосы, растущие на ней, образуют своеобразное "завихрение". Они светлее на железе, чем около нее. Волосяной покров густой и представлен в основном остевыми и очень редкими пуховыми волосами. Корни волос растущие. Остевые волосы подразделяются на три размерных порядка (табл. 41). В основании "ножка" слабо выражена. Стержень достигает наибольшей толщины в нижней трети. К вершине он постепенно сужается. Поперечные срезы по всей длине овальной формы. Сердцевинный канал проходит вдоль стержня, кроме удлиненной бессердцевинной вершины. Сердцевина зернистого типа, как и у волос на других изученных участках тела. Хорошо выражен периферийный слой более мелких сердцевинных клеток. Рисунок кутикулы вдоль стержня не меняется. Чешуйки нижние, вытянуты по ширине. Края ровные и плотно прилегают к корковому слою.
Кроме описаных выше остевых волос, имеются более тонкие волосы со слабо развитым сердцевинным слоем. Их длина 12—16 мм, толщина 68— 90 мкм. Сердцевинный канал занимает 22—33% толщины стержня. Поперечные срезы округлой формы. Структура сердцевины и кутикула не отличаются от таковых других остевых волос.
174
Cervus claphus Linnaeus, 1758
Пуховые волосы очень редки. Длина их 14—21,3 мм, толщина 38— 180 мкм. Сердцевина составляет 26—92% от диаметра стержня. По структуре они не отличаются от волос, покрывающих железу.
Строение кожного покрова в ноябре сходно с сентябрьским. Мальпигиев слой эпидермиса состоит из 2—3 рядов клеток. Потовые железы с относительно короткими секреторными отделами, делающими всего 2—3 изгиба. В коже труди потовые железы развиты сильнее, чем в коже спины. Сальные железы довольно крупные,особенно в коже труди.
Специфические кожные железы благородного оленя многочисленны. На нижней стороне хвоста оленя располагается подхвостовая железа (Schaffer, 1940 г., по: Соколов, 1973). Она образована удлиненными долями сальных желез, открывающимися по 5—10 шт. в одно и то же волосяное влагалище. В длину эти железы достигают 0,8 мм и залегают в нишах, образованных мышцами, сокращение которых выдавливает секрет наружу. Под волосами находится слой апокриновых трубчатых желез. Они имеют секреторные отделы в 75—140 мкм в поперечнике, сильно закрученные в поперечные клубки.
Циркумкаудальный желе'зистый орган оленей был известен еще Аристотелю. Орган образован крупными скоплениями трубчатых желез, имеющих в поперечнике в среднем 80 мкм (Rapp, 1839; Zietschmann, 1903; Schaffer, 1940).
Предплюсневая железа до сих пор была подробно описана только у белохвостого оленя (Hershkovitz, 1958; Quay, 1959) и более поверхностно у некоторых других видов оленей (Zierschmann, 1903; Pocock, 1923). У изученной нами предплюсневой железы оленей (№ 2, 3), расположенной на внешней стороне верхней части предплюсны, кожа немного утолщена и растущие волосы образуют своеобразное "завихрение". Сальные й потовые железы сильно увеличены в размерах: сальные до 0,44 х 1,1 мм, диаметр секреторных отделов потовых желез — до 200 мкм. Сумма диаметров секреторных отделов потовых желез на протяжении 1 см препарата достигает 6,6 мм по сравнению с 0,6—1,9 мм на туловище. Железы открываются в волосяную сумку. Хорошо виден постепенный переход от обычных желез, окружающих железистое поле, к крупным железам внутри поля. Укрупненные сальные железы образуют скопление на меньшей площади, чем потовые. Клубки потовых желез находятся на уровне волосяных луковиц. Строение этой железы у благородного оленя оказалось идентичным таковой белохвостого оленя (Hershkovitz, 1958; Quay, 1959). Кожных выпячиваний, или мешков, между пальцами у благородного оленя нет (у некоторых других видов оленей они имеются). В коже между пальцами передней ноги оленей № 3 и 4 находятся огромные скопления сальных и трубчатых желез. Сальные железы очень крупные (0,6 х 1 мм), много дольчатые, некоторые особо крупные (0,9 х 0,9 мм), гроздевидные с несколькими протоками, открывающимися в одном месте в волосяную сумку. Секреторные отделы трубчатых желез сильно извиты в клубки. Размеры клубков до 0,7 х 0,9 мм. Диаметр секреторных отделов достигает НО мкм. Протоки открываются в волосяные влагалища.
175
Благородный олень
Строение межпальцевой железы благородного оленя сходно со строением межпальцевой железы других видов оленей (Schafler, 1940). Межпальцевая железа задней ноги оленей № 3 и 4 представляет собой такие же крупные скопления альвеолярных и трубчатых желез, как и на передних конечностях, но сальные железы здесь имеют несколько меньшие размеры (0,44 х 0,80 мм), тогда как потовые образуют основную железистую ткань. Диаметр их секреторных отделов достигает 120 мкм. Клубки трубчатых желез достигают размеров 0,8 х 1,0 мм. Сумма диаметров на 1 см препарата составляет 8 мм. Предглазничная железа у оленей № 3 и 4 представлена кожистым впячиванием в ямку слезной кости черепа. Кожистый мешочек имеет размеры до 3,8 см в длину, 1,27 см в ширину и 1,9 см в глубину и покрыт изнутри редкими короткими волосами, луковицы которых находятся примерно на 1,1 мм от поверхности кожи. Многочисленные альвеолярные сальные железы образуют настоящие грозди и в глубину почти достигают волосяных луковиц. В одно место сумки волоса открываются 2—3 сальные железы (или 2—3 лопасти одной железы). Размеры желез достигают 0,9 х 1,1 мм. Трубчатые железы небольшие, хотя расположены они довольно часто. У большинства из них диаметры секреторных отделов невелики, но у некоторых увеличены до 110 мкм. У самца № 7 в коже препуция сальные и потовые железы также увеличины. Первые шести-се-мидольчатые размером до 350 х 350 мкм — имеют крупные, диаметром 79—113 мкм, вводные протоки, их окружают круглые волосяные фолликулы на глубине 700—900 мкм от поверхности кожи. Вторые — с широкими (диаметром 56—90 мкм), закрученными в плотные клубки секреторными отделами. Размеры клубков на поперечных следах составляют 0,9 х 0,5; 0,5 х 0,5 мм к корковому слою. Корковый слой в боковых отделах волоса на 3 мкм толще, чем на передних и задних. Среднее число остевых волос на 1 см2 поверхности кожи достигает 583. Ости располагаются по поверхности кожи нечетко выраженными рядами. Пуховые волосы сильно извиты. Число пуховых волос на 1 см2 поверхности кожи 1166. Пуховые волосы растут пучками по 2—3 волоса в каждом. Система крови остается нами не изученной.
Индексы внутренних органов. Специальных исследований по индексам внутренних органов благородных оленей не проводилось. В литературе имеются данные по абсолютным показателям внутренних органов 20 особей из Кавказского заповедника (Александров, 1968) и 3 особей из Азербайджана (Адиев, 1965). При учете возраста, пола и физиологического состояния выборки становятся мизерными (см. табл. 41) и по ним невозможно составить однозначные суждения. Тем не менее, на наш взгляд, эти данные указывают на определенные закономерности. Судя по данным табл. 41, абсолютные показатели внутренних органов с возрастом увеличиваются, что характерно и для других видов зверей. Индексы этих органов изменяются в обратном направлении, что также является широко распространенной закономерностью. Показательно, что абсолютные показатели изученных внутренних органов значительно выше у холостых самок, чем у беременных и кормящих самок, так же как у взрослых самцов до гона по сравнению с теми же
176
Cervus elaphus Linnaeus, 1758
взрослыми самцами после гона. При сравнении индексов внутренних органов получаются иные результаты, что можно объяснить различием интенсивности изменения массы тела и массы органов.
Кариотип. Для благородных оленей, как и для других представителей палеарктических видов Cervidae, характерно большое количество хромосом — в диплоидном наборе 68 хромосом. Кариологический анализ значительного количества благородных оленей (л=22) из трех популяций Западной Германии (Herzog, 1987) показал, что количество хромосом этого вида устойчиво и не подвержено популяционной изменчивости. За исключением первой пары аутосом и Х-хромосомы, все аутосомы представлены тело-центриками (соотношение длины плеч меньше, чем 1:4). Первая пара аутосом и Y-хромосома, самая мелкая в наборе, являются метацентриками. Во всех хромосомах, за исключением первой пары, в перицентромерных участках имеются хорошо выраженные С-гетерохроматиновые блоки разной величины. Х-хромосома содержит также терминальные сегменты С-гетеро-хроматины. Парицентромерная область первой пары аутосом слабо окрашивается С-методом. Названный автор провел также анализ всех хромосом западногерманских благородных оленей G-методом, что позволило идентифицировать каждую хромосому. Как и предыдущие кариологические данные (Орлов, Булатова, 1983), данные по западногерманским оленям свидетельствуют о геотрафической изменчивости Y-хромосом.
Структура и динамика ареала
Видовой ареал (восстановленный) охватывает лесные, лесостепные, степные и отчасти пустынные ландшафты равнин и гор Старого Света в пределах от 25—30 до 55—60° с.ш., а также в Северной Америке от 35 до 50—60° с.ш. (Гептнер и др., 1961). Современный ареал вида по сравнению с исходным резко сокращен — в широтном и высотном направлениях он сильно сужен и занимает лишь отдельные регионы зоны широколиственных и смешанных лесов и лесных горных поясов Южной и Западной Европы, южных районов Скандинавского полуострова, западных районов Белоруссии, Крыма, Кавказа, Северной Африки, Азии (Малой, Средней, Центральной), Южной и Восточной Сибири, Дальнего Востока; в Северной Америке — умеренной зоны.
Распространение на Кавказе. Все имеющиеся данные говорят о том, что благородный олень населял обширную территорию перешейка. Несмотря на интенсивную охоту, проводившуюся человеком различными способами, этот вид оставался наиболее распространенным и обычным охотничье-про-мысловым до последних десятилетий XIX в. (Шильдер, 1895, 1897; Динник, 1904, 1909; и др.). Состояние ареала в начале текущего столетия наиболее подробно и на профессиональном уровне описано Н.Я.Динником (1910). По его данным, на Северном Кавказе ареал благородного оленя выглядел следующим образом. В кубанском варианте олень встречался всюду, где были значительные леса (это преимущественно горные леса). Оттуда он
12 В.Е.Соколов, А.К.Тсмботов
177
Благородный олень
заходил высоко в горы, практически до вечных снегов, летом до 3000 м, а зимой до 1600- 1700 м над ур. моря. Нижняя граница его распространения проходила по линии Горячий Ключ — местность севернее Майкопа — Передовая — Усть-Джегутинск. Кроме того, благородный олень обитал на равнине в так называемом Красном лесу, расположенном в районе г. Краснодара по правому берегу Кубани. По Черноморскому побережью Западного Кавказа олени спускались почти до уровня моря и встречались в окрестностях г. Туапсе. В центральной части Северного Кавказа (эльбрусский и терский варианты) они встречались спорадично, но около Нальчика, Алагира, Владикавказа и Воздвиженской олени не были редкими. Очевидно, нижняя граница ареала в то время здесь проходила по линии Кисловодск — Нальчик — Грозный. К тому времени благородные олени были уничтожены на Ставропольской возвышенности, но по долине Терека они проникали до его устья и встречались по Каспийскому побережью, где в то время леса не были редкостью. Судя по описанию Н.Я.Динника, в Дагестане олени встречались не во всех районах, а преимущественно в западных, в частности в Андийском и Гунибском районах. По его словам, "часто встречаются олени в Кайтаго-Табасаранском округе в очень лесистой местности, недалеко от Маджалиса. Они держатся также и на горах Дешлагира, но в очень ограниченном количестве" (Динник, 1910, с. 97). В высокогорье Дагестана, которое лежит вблизи Главного Кавказского хребта и прорезается четырьмя ущельями р. Койсу, олени попадались спорадично. Их вовсе не было по Казикумухскому и Кара-Койсу из-за отсутствия здесь лесов, но по Аварскому Койсу и его притокам, отличавшимся самой высокой лесистостью, их было довольно много. Олени тогда регистрировались около Анцуга в лиственных и хвойных лесах, вблизи Тлядаля в буковых лесах и т.д.
Закавказская часть ареала в начале текущего столетия, очевидно, была также обширной, как и на Северном Кавказе, однако она обладала существенными региональными особенностями. Так, в Западном Закавказье (колхидский и отчасти аджарский варианты) олени практически не жили в верховьях рек Риони, Ингури, Цхенис-Цкали и других местах, где обычно выпадает много снега и снежный покров держится долго. Это специфическая черта природы обоих вариантов, накладывающая глубокий отпечаток на всю биоту (Соколов, Темботов, 1989).
Что касается среднегорья, предгорья и Колхидской низменности, то благородный олень встречался там широко. Однако, видимо, были и незаселенные места (например, дельта Риона); редко встречались они вблизи Батуми, но в 23 км от Батуми к Атвину попадались гораздо чаще. Неравномерное размещение оленей в Западном Закавказье, особенно в нижних поясах гор и низменности, было обусловлено антропогенными факторами.
Юго-восточнее Сурамского хребта, в б. Тифлисской губернии (триалет-ский и юго-осетинский варианты), олени были распространены значительно шире, чем в Западном Закавказье. Они населяли, хотя и в ограниченном количестве, все большие леса, в том числе окрестности г. Тбилиси, долины рек Арагви (станция Ананура, Пасанаури), Алазани, Куры и т.д.
178
Cervus elaphus Linnaeus, 1758
Оленей было много вблизи Боржоми (в бывшем великокняжеском имении).
Сведения Н.Я.Динника по Азербайджану весьма ограниченны. Он отмечал, что олени встречаются там в очень немногих местах. Н.К.Верещагин (1959) приводит более полные данные, согласно которым оленей не стало в Хачмасских низовых лесах в начале нашего столетия, а выше, в лесах Кубинского района, они перевелись в 80-х годах XIX в. В тугайных лесах и камышах среднего и нижнего течения р. Куры они встречались еще в конце прошлого столетия. Есть достоверные данные, что олени жили по окраинам камышей Шильянского озера. В Карабахе, по ущельям Дзегамчая и Шамхора, олени исчезли позже — в 20-х годах текущего столетия. В конце XIX в. они были обычны в окрестностях с. Чайкенд (южнее Кировобада). Озеро Маралгель (близ оз. Гейгель) получило свое название, видимо, из-за обилия в свое время оленей в этих местах.
Что касается Армении, то и здесь олени были широко распространены в конце прошлого столетия; так, в Зангезуре в 90-х годах оленей и косуль было так много, что некоторые охотники убивали до 150—200 голов за год (Мелик-Шахназаров, 1898 г., по: Верещагин, 1959). Новые данные, полученные С.К.Межлумяном и Р.Х.Гукасяном (1986), также свидетельствуют о том, что благородный олень был широко распространен в лесах северной, восточной и южной Армении еще в конце прошлого века. Численность их была особенно высока на восточных склонах Гегамского хребта, в Азиз-бековском, Ехегнадзорском районах, в окрестностях (оз. Парзлич и в Ка-фанском районе. Видимо, особенно много оленей было в районе оз. Севан, где они играли важную роль в жизни еще древних людей. На высоте 2000—3200 м очень часто встречаются наскальные изображения кавказского благородного оленя.
По данным С.К.Даля (1950), последний олень на горе Аде-Теле был убит в 1860—1865 гг. В северной части бассейна оз. Севан, на склонах Севанского хребта, эти животные просуществовали дольше, чем на его южной и юго-западной сторонах, на склонах Гегамского (Аглаганского) хребта. По сведениям того же автора (Даль, 1953), в районе г. Кафана олень был убит еще в 1912 г. Примерно в начале XX в. оленей не стало в Зангезуре и во всей Армении, хотя там и могли иметь место случайные их заходы из Грузии и Азербайджана (Даль, 1954). В Талыше, по Н.К.Верещагину (1959), последний олень убит в 1912 г.
Как видно из изложенного, в Восточном Закавказье оленей стало очень мало еще в конце XIX в. или в самом начале XX в. В этих местах олень к тому времени сохранился лишь в отдельных, обычно охраняемых участках лесных поясов восточнозакавказского типа (Боржоми, Закаталь, Лагодехи).
В первой четверти XX в. ареал и численность оленей продолжали интенсивно сокращаться. В 1930—1935 гг., по наблюдениям А.А.Насимовича (1936), на равнине и в предгорьях Западного Кавказа оленей уже не было. Нижняя граница их распространения отступила значительно южнее линии Псебай — Даховская — Самурская — Новагинская. На западе эта линия доходила лишь до р. Пшиш. Основное поголовье было сосредоточено в Кав
12*
179
Благородный олень
казском заповеднике. Восточнее этого заповедника олени еще встречались в 40-х годах текущего столетия в окрестностях Преградной и Микоян-Шахора (Верещагин, 1959). В центральной части Северного Кавказа они были уничтожены повсеместно уже в начале 30-х годов. В Северной Осетии последнего оленя убили в 1928 г. (Бёме, 1936; Наниев, 1956). К этому времени, очевидно, олени сохранялись в Дагестане лишь в небольшом количестве в низовьях Терека и Сулака (Гептнер, Формозов, 1941).
В Западном Закавказье, по данным Н.К.Верещагина (1959), сокращение распространения и численности благородного оленя отмечалось в начале I мировой войны. При интенсивной рубке леса в Колхидской низменности и окружающих ее гор олени были уничтожены или оттеснены из низовьев рек Риони, Кодори и Ингури в среднегорья. Больше всего оленей в пределах колхидского и аджарского вариантов сохранилось в то время в лесах среднего течения Ингури. Они встречались в 30-х годах по восточным отрогам Аджаро-Имертинского хребта, населяя буковые леса Боржомского заповедника. Летом они держались на Ахалкалакском и Джавахетском нагорьях. В Восточном Закавказье олени встречались в Караязском лесу и на южном макросклоне Большого Кавказа в пределах от верховья Алазани до р. Катехичай. Много оленей было в Закатальском заповеднике (в 40-х годах было 1000 животных). Немало оленей зарегистрировано в тот период и восточнее,, в Куткашенеком и Исмаиллинском районах, где высотное их распространение охватывало территории от низменности до гор субальпики. Фактическое местонахождение относится к районам селений Чухур-Кабала, Исмаилла, Каланджан, Рушан-Кенд и др.
Вторая половина XX в. характеризуется положительными сдвигами как в распространении, так и в численности благородного оленя в ряде регионов Кавказа. Заметное расширение ареала оленя по сравнению с предвоенными годами на Северо-Западном Кавказе наметилось к середине 50-х годов. По данным В.Н.Алексацдрова (1968), олени стали регистрироваться к югу и северу от прежних траниц, притом вдали от заповедников Краснодарского и Ставропольского краев. Площадь ареала их в этой части Кавказа составила около 2 млн га. Западный предел распространения к тому времени находился на стыке Анапского, Новороссийского и Крымского районов. Отдельные животные периодически заходили до окрестностей станиц Гостагаевская и Натухаевская. В окрестностях станицы Шапсугская и хутора Николаевский (Крымский район) небольшое количество оленей держалось постоянно. В 1962—1963 гг. кратковременные заходы отдельных животных отмечались близ г. Афонки (Новороссийский район). Распространение в Абинском районе занимало верховья рек Абин, Ахтырь и Хабль, окрестности станицы Крепостная. К юго-востоку ареал расширялся, достигая в 1962 г. территории Горяче-Ключевского района (Кутаис, Горячий Ключ, Имер-тинская). На южном склоне Главного Кавказского хребта ареал переходил в смежные угодья Геленджикского и Туапсинского районов. Места нахождения: склоны гор Тхаб и Афипс, верховья рек Пшада, Вулон и Джубга. В Апшеронском районе больше всего оленей было в верховьях рек Кур-джипса и Пшехи, а также их притоков — Тогота, Тогопса, Тугупса, Цице и
180
Cervus elaphus Linnaeus, 1758
Серебрянка. Изолированное поселение благородного оленя в настоящее время имеется по правому берегу Кубани в так называемом Красном лесу (Краснодарское лесоохотничье хозяйство). Здесь было выпущено 90 особей; реакклиматизация проходит успешно, возникла необходимость жесткого регламентирования численности из-за ограниченной площади лесного массива в степной зоне (Темботов, 1972).
Южная граница ареала в Краснодарском крае проходила в 60-х годах севернее с. Пшада, Урочища Черный аул, Гирляково и др. Ежегодно, хотя и редко, отмечались в верхнем и среднем течении рек Аше и Псезуапсе, но более часто по долине р. Головинка. Совсем были р'едки в верховьях рек Бзыча и Мзымта. В отдельные многоснежные зимы (например 1962/63 и 1963/64 гг.) олени изредка спускаются до пос. Красная поляна. На рубеже Краснодарского и Ставропольского краев и Абхазии оленей в 60-х годах не отмечали. К северу от границ Кавказского заповедника олени заходят до линии населенных пунктов Даховская — Ново-Прохладное — Батовская — Псебай. Олени постоянно обитают в верховьях рек Ходзь, Бугунжа, по среднему течению Малой Лабы, а также притоков Закана, Дамхурца и др. Встречаются также по Зеленчуку и Урупу. Севернее населенных пунктов Нижняя Ермоловка и Хасаут-Греческий оленей в 60-х годах обычно не регистрировали. В настоящее время благородные олени в Карачаево-Черкесии имеют тенденцию к сокращению численности и соответственно распространения. Они встречаются в наши дни лишь в горных лесах верховья Большой Лабы и ее притоков, а также по Урупу, Большому Зеленчуку и Теберде. В Тебердинском заповеднике олени обитают в основном по долине р. Кизгыч. Там насчитывается всего 50—60 голов-(Красная книга Карачаево-Черкесии, 1988).
К юго-востоку от Тебердинского заповедника благородные олени встречаются изредка (Темботов, 1972). В верховьях р. Малка мы регистрировали следы и остатки оленей в 50-х и в последующие годы. Видимо, олени периодически заходят в эти места из Тебердинского заповедника, но становятся легкой добычей браконьеров. В пределах Кабардино-Балкарии благородные олени живут постоянно в лесистом поясе, особенно часто встречаются в Нальчикском лесоохотничьем хозяйстве. Реакклиматизированы также в Северной Осетии и Чечено-Ингушетии, где занимают отдельные районы торных лесов. Благородные олени и здесь прижились успешно, и дальнейшее их существование зависит от уровня охраны от волков в снежные годы и от браконьеров. В Дагестане благородный олень в наши дни встречается в низменности (Нагайский, Тарумовский, Кизлярский, Баба-юртовский и Кизилюртовский районы) и в высокогорье (Тляратинский и Цунтинский районы) (Прилуцкая, Пишванов, 1988). Поселение благородных оленей в торах поддерживается за счет их миграции на лето из Лаго-дехското и Закатальского заповедников. С выпадением снегов олени возвращаются обратно в заповедники. Такие перемещения благородных оленей, видимо, закономерны, о чем свидетельствуют и данные Н.К.Вере-щагина (1959).
1 8 1
Благородный олень
О современном распространении благородного оленя в Закавказье однозначных сведений мало. Это касается в первую очередь западных районов (колхидского и аджарского вариантов). Очевидно, положительное влияние Кавказского заповедника на поселения благородных оленей южного макросклона Большого Кавказа незначительно и немного выходит за пределы Краснодарского края. По данным Госкомприроды Грузии в 1988 г. четыре оленя было зарегистрировано в Рицинском заповеднике. Судя по данным А.Б.Арабули (1975), Т.Бараташвили (1980), положение благородного оленя в Боржомском заповеднике в наши дни остается неудовлетворительным. Снижение темпа воспроизводства, начатое в 40-х годах, продолжалось до 80-х годов включительно. Отрицательно сказывается комплекс неблагоприятных факторов, в том числе близкородственное скрещивание, выпас скота, рубка леса, неконтролируемая охота, пресс волка и др. Благородные олени в этом районе занимают горные пояса с 800 до 2000 м над ур. моря. Ранней весной и поздней осенью олени держатся в альпийском поясе, зимой редко поднимаются выше 1800 м над ур. моря. Высотное размещение оленей зависит от продолжительности пребывания домашнего скота на горных пастбищах, а зимой — от уровня снегового покрова. Благородный олень в наши дни испытывает значительный антропогенный пресс и в других районах Грузии. Г.В.Мамукелашвили (1987) пишет: "Это ценное животное всего 20—30 лет назад часто встречалось в предгорных и пойменных лесах Лагодехского района. Однако из-за бессистемной рубки леса они полностью уничтожены и сохранились только в заповеднике и охотничьих хозяйствах". Сокращение численности копытных, в том числе оленей, зарегистрировано также в Сагурамском заповеднике Грузии (близ Мцхета), что связано, по мнению З.Сихарулидзе (1987), не с кормовой базой, а с миграцией и браконьерством. Вне заповедников, видимо, нигде в Грузии олень не сохранился (Мамукелашвили, 1989). По данным Госкомприроды ГССР, во всей республике по состоянию на 1988 г. благородных оленей было учтено 1755 голов.
В пределах Азербайджана в рассматриваемый период олени сохранились в Нуха-Закатальской части южного склона Большого Кавказа, к западу от верхнего течения р. Гирдиманчай до течения Мазымчай (Алиев, 1965). Охарактеризовать распространение более подробно и с охватом последних лет мы не можем из-за недостатка материалов.
Современные тенденции антропогенных изменений ареала. Кавказского благородного оленя мы относим к числу перспективных охотничье-промысловых животных Кавказа. Такое суждение, на наш взгляд, справедливо с учетом как особенностей его биологии, так и современных ландшафтных условий региона. Наши наблюдения и многочисленные литературные данные, в том числе Н.Я.Динника (1910), А.А.Насимовича (1936, 1955), В.Н.Алсксандрова (1968) и др., свидетельствуют о том, что местные олени характеризуются высокой пластичностью. В прошлые века, когда охотничий пресс был невелик, эти олени процветали почти во всех регионах Кавказа: на равнине, от степей и полупустынь Предкавказья и Куринской низменности до влажных субтропиков Колхиды и Ленкорани; в горах, от предгорьев с весьма различными ландшафтами до высокогорья всех типов
182
Cervus elaphus Linnaeus, 1758
поясности. Словом, ландшафтный спектр нормального существования кавказского оленя, на наш взгляд, включает все многообразие антропогенных экосистем. Следовательно, многократно раздробленную и сильно разобщенную топографию современного ареала благородного оленя на Кавказе следует считать результатом прямого преследования этих животных человеком или косвенного воздействия на них антропогенного фактора. При снятии этих факторов на Кавказе олень станет вновь одним из массовых видов копытных. Кроме того, в силу многообразия ландшафтного покрова Кавказа и высокой подвижности кавказских благородных оленей последние могут легко выбрать для себя оптимальные места обитания, в частности в зависимости от высоты снежного покрова, экспозиции склона гор, типа растительности (Chang, Xiao, 1988).
Систематика и изменчивость
Систематическое положение. Настоящие олени (Cervus) — в систематикотаксономическом плане довольно сложная группа, что обусловлено, высокой изменчивостью при обширном ареале. Что касается положения популяции благородного оленя Кавказа, то проблема заметно усугубляется. Сказывается влияние самобытной истории формирования природы Кавказа, положение его на стыке Европы и Азии, пестрота ландшафтов и многое другое. Уже на заре становления систематики рассматриваемой группы встал вопрос о месте кавказских оленей в роде Cervus. П.С.Палласу (Pallas, 1811, по: Гептнер, Цалкин, 1947) было известно о широкой изменчивости благородных оленей России, но он считал возможным объединить их в один вид Cervus elaphus. Однако в 1940 г. Оджильби (Ogilby) описал оленей Северного Ирана в качестве самостоятельного вида Cervus maral. Из особенностей этого "нового вида" он приводил более крупные размеры, превосходящие размеры западноевропейского благородного оленя. В связи с этой работой возник вопрос: к какому виду относятся кавказские олени — к европейскому С. elaphus или североиранскому С. maral? Этот вопрос многие систематики разрешили просто — оба вида объединили в один под названием Cervus elaphus, сохранив за каждым ранг подвида и соответствующие названия (см. обзор: Гептнер, Цалкин, 1947). Однако были и те, которые выделяли кавказских и североиранских благородных оленей в один самостоятельный вид Cervus maral (Schlater, 1872 г.; Fitzinger, 1873, 1874 гг., Gray, 1972 г.). Принципиально иную позицию занимал Г.И. Радде (Radde, 1886 г.), согласно которому на Кавказе представлены оба вида — С. elaphus и С. maral.
Мнение о существовании двух видов оленей на Кавказе разделял К.А.Са-тунин (1896 г.). Однако после появления монографии Р.Лидеккера (Lydek-ker, 1887 г.), согласно которой на Кавказе С. maral низводится в ранг подвида С. elaphus, К.А.Сатунин (1903) изменил свою точку зрения и кавказских благородных оленей относил к С. elaphus, выделяя их в качестве самостоятельного подвида (С. elaphus maral). Такая точка зрения не всех систематиков удовлетворила. Некоторые териологи (Winaus, 1914 г., по:
183
Благородный олень
Гептнер, Цалкин, 1947) выделяют кавказских оленей в самостоятельный вид с названием Cervus caucasicus.
В нашей отечественной литературе утвердилось мнение, согласно которому кавказские и европейские благородные олени относятся к одному виду Cervus elaphus (Динник, 1910; Бобринский, 1932; Бобринский и др., 1944; Гептнер, Цалкин, 1947; Соколов, 1959; Гептнер и др., 1961; Павлинов, Россолимо, 1987 и др.).
Изменчивость. Для анализа различных аспектов изменчивости благородного оленя в условиях трехмерного пространства гор Кавказа очень мало фактического материала. Ни по одной популяции мы не имеем такой выборки, которая с учетом пола и возраста соответствовала бы требованиям биологической статистики. Более или менее удовлетворительные данные, обобщенные В.Н.Александровым (1968), получены лишь по Западному Кавказу. Эти материалы положены в основу анализа различных сторон изменчивости с дополнением своих и литературных данных.
Индивидуальная изменчивость. Размах индивидуальной изменчивости кавказского благородного оленя велик. Масса тела самцов и самок даже в оптимальных условиях Кавказского заповедника сильно варьирует (см. табл. 41). Четко сказывается влияние физиологического состояния организма, кормовой базы и других факторов среды. Достаточно сказать, что у самцов 5—11 лет различие в массе тела до и после гона может составлять в среднем около 50 кг, а межу кормящими и холостыми самками — до 20 кг в среднем. По данным В.Н.Александрова (1968) и нашим, за время гона самцы теряют в среднем до 19—20% от первичной массы. По В.Г.Гептнеру и В.И.Цалкину (1947), в период рева быки Западной Европы теряют до четверти своей массы. При этом изменяются не только жировые запасы, но и масса большинства внутренних органов. Так, у 5—11-летних быков масса печени до гона составляет в среднем 5040 г, а после гона — 4300 г. В существенных пределах меняются размеры тела как самцов, так и самок.
Наряду с фенотипической изменчивостью у благородного оленя выявлена широкая генотипическая изменчивость белков. Так, в условиях Шотландии у этого вида оленя определено 8 полиморфных локусов, изменчивость которых прямо связана с ювенильной смертностью. В частности, гетерозиготы по Idh=2 отличаются лучшей выживаемостью, чем оба типа гомозигот, не различающихся в этом отношении. Такая же связь выявлена и по локусу Tfr (Pemberton et al., 1988).
Формы и размеры рогов, как свидетельствуют наши и литературные данные, сильно вариабельны, что зависит от многих внешних и внутренних факторов. Во времена Н.Я.Динника, когда кавказский благородный олень процветал, масса рогов обычно составляла 7—8 кг, отдельные экземпляры достигали более 14 кг, имея, при этом до 18 отростков на каждом роге. В наши дни, по данным В.Н.Александрова (1968), на Западном Кавказе, где сохранились наиболее оптимальные условия жизни для благородного оленя, чаще всего попадаются олени с рогами с 6—7 отростками, редко с 9—11 (см. табл. 37). Вероятно, сказывается ряд экзогенных факторов. Еще Н.Я.Динник (1910) указывал, что, чем сильнее и крупнее олень, тем лучше
184
Cervus elaphus Linnaeus, 1758
развиты его рога. У слабых и больных животных или питающихся плохо рога бывают тонкие и вообще невелики. По его наблюдениям в боржом-ском парке, обнесенном проволочной отрадой, из-за перенаселенности и недостатка зеленого корма олени отличаются малым ростом по сравнению с теми, которые живут в соседних лесах.
В литературе имеются данные, показывающие влияние зимнего питания на развитие рогов у благородного оленя — плохая упитанность самцов зимой может оказывать влияние на массу пантов через более поздние сроки сбрасывания рогов (Muir, Sykes, 1988). Влияние пищевого фактора прослеживается и в ряду поколений. «По-видимому, "севанская популяция" оленей, пережившая четвертичные похолодания в наиболее благоприятных условиях, и дала особо крупные и разнообразные формы>> (Межлумян, Гукасян, 1986, с. 119).
Анализ индивидуальной изменчивости окраски кавказского благородного оленя представляет определенный интерес, так как она широко используется в качестве одного из важных признаков для видовой и подвидовой систематики настоящих оленей (Гептнер, Цалкин, 1947). Характерной особенностью окраски кавказского благородного оленя, видимо, следует считать относительно высокую индивидуальную изменчивость. В частности, наши и многочисленные литературные данные свидетельствуют о том, что среди кавказских взрослых оленей чаще встречается пятнистость, чем среди других популяций. Однако степень развития этой пятнистости меняется широко. Вероятно, она не зависит от упитанности животного, как это полагает В.Н.Александров (1968), который считает, что пятнистость сильнее выражена у голодающих оленей. В нашем материале имеется рогач из Нальчикского лесоохотничьего хозяйства, крупного роста и хорошо упитанный, с четко выраженной пятнистостью. Неустойчива окраска зеркала и его окаймления. Зеркало может быть чисто-белым или беловатым со слабым рыжеватым налетом. То же самое наблюдается по интенсивности развития черного окаймления — от темно-бурого до черного цвета. Нередко окаймление на крупе может вообще отсутствовать и окраска зеркала переходит в окраску спины. При желании перечень аналогичных деталей окраски тела благородного оленя, подверженных индивидуальной изменчивости, можно продолжить.
Возрастная изменчивость. По кавказскому благородному оленю мы не располагаем однозначными данными, позволяющими определить возраст, при котором морфологические признаки вида и внутривидовых форм наиболее полно выражены. Судя по данным В.Н.Александрова (1968), в условиях Западного Кавказа размеры и масса тела благородного оленя увеличиваются до 5 лет включительно. Максимальные показатели приходятся на период старше 5 до И лет. По В.Г.Гептнеру и др. (1961), полного развития маралы достигают несколько раньше — к 4—5 годам. Очень старые животные претерпевают регрессивные изменения. Наши и литературные данные свидетельствуют о том, что аналогичная динамика свойственна возрастной изменчивости размеров и структуры рогов. Так, к 15—16 жс*.фор-мируются рога в виде простой шпильки длиной 30—50 см (изредка конец
18 5 i :
Благородный олень
раздваивается). На третьем году жизни рога имеют по 3—4 отростка (надглазничный и средний); в возрасте до 6—7 лет. Полного развития рога достигают в 7—8 лет; в последующие годы, до 11 лет, идет рост массы главного ствола, но количество отростков не увеличивается. С 11—12 лет рога претерпевают регрессивные старческие изменения — уменьшается масса рогов, сокращается количество отростков. Раньше других исчезает второй надглазничный отросток.
С учетом стадии роста и развития рогов еще в начале текущего века были предложены методы определения возраста благородного оленя (Туркин, Сатунин, 1902), но общепризнанными эти методы не стали. И.И.Со-колов (1959) считал их мало пригодными для определения возраста благородных оленей. Недавно предложен новый, более точный метод определения возраста благородных оленей (Lockow, 1987). Метод основан на анализе взаимосвязи показателей длины рогов, величины их обхвата у основания и возраста.
По И.И.Соколову (1959), с определенной точностью возраст благородного оленя можно установить по зубам, в особенности по резцам. Как известно, олененок рождается со всеми молочными резцами. Примерно в годовалом возрасте первая пара молочных резцов сменяется на постоянные, в двухгодовалом — сменяется вторая пара, в трехгодовалом — третья пара, а в четырехгодовалом — клыки нижней челюсти. В возрасте 6 лет начинают стираться режущие края резцов. В 7 лет обнажается дентин на первой паре, в 8 лет — на второй паре, в 9 лет — на третьей и в 10 лет — на клыках. О дальнейших возрастных изменениях судят по степени стертости зубов. К возрасту 20—23 лет зубы стачиваются до верхушек корней.
Возрастные изменения окраски тела значительны. Как известно, оленята имеют пятнистую окраску. Она может сохраняться и у взрослых. По В.Н.Александрову (1968), пятнистость чаще сохраняется у молодых самок, задерживающихся в развитии. Однако мы располагаем фактическим материалом, свидетельствующим, что пятнистость окраски спины может сохраняться у нормально развитых животных и притом у вполне взрослых рогачей. Нередко также молодые отличаются от взрослых темной окраской спины и бедер.
Половые различия. Половой диморфизм у благородного оленя выражен резко и затрагивает многие морфологические показатели вида. По массе тела различие между полами одного возраста очень большое, особенно оно разительно в старших возрастных группах. В возрасте 1,5—2,5 лет масса самки составляет около 80% от массы самца того же возраста, но в возрастной группе 5—11 лет масса тела самки может быть вдвое меньше массы тела самца. Соответственно в предбрачный период самец дает до 200 кг, а самка до 100 кг высококачественного мяса (Александров, 1968). Значительные различия выявлены и по размерам тела. Длина тела и высота в холке, четко характеризующие общие размеры тела благородного оленя, у самок на 15—20% меньше, чем у самцов соответствующего возраста. Еще больше различия по показателям объема грудной клетки и
186
Cervus elaphus Linnaeus, 1758
ширины груди — самки по этим частям тела меньше самцов на 30%. На большие половые различия кавказского оленя, как и других его сородичей, обращали внимание и предыдущие исследователи, но при этом отмечали, например Н.Я.Динник (1910, с. 103), что иногда попадаются самки относительно крупного размера: "...нередко это можно заметить, смотря даже издали на табунок ланей: среди десятка их иногда одна или две оказываются сильно отличающимися своим большим ростом от всех прочих".
Выявлен также ряд других экстерьерных признаков, отличающих самок от самцов. По В.Г.Гептнеру и В.И.Цалкину (1947), в летнем мехе у самок i7' коричневый оттенок выступает заметно сильнее, чем у самцов. Лучше А развита также темная полоса на шее и спине у самок, чем у самцов. В зимнее время на шее самцов развивается грива. Кроме того, общая окраска зимнего меха самцов более контрастная, чем у самок.
Половой диморфизм у благородного оленя, очевидно, не исчерпывается только перечисленными особенностями. При наличии достаточного материала и организации специальных исследований можно значительно расширить объем признаков и особенностей, характерных для самцов и самок. В качестве подтверждения можно сослаться на половые различия благородного оленя покомпонентам сыворотки крови (Kent et al., 1980), тазовых костей северного оленя (Tyler, 1987) и т.п.
Географическая изменчивость. Анализ этой изменчивости на материале до 40-х годов провели В.Г.Гептнср и В.И.Цалкин (1947). На наш взгляд, бесспорно доказано, что кавказский благородный олень по многим морфологическим признакам (масса и размеры тела, структура и величина рогов, окраска тела и др.) занимает промежуточное положение между среднеевропейским и алтайским подвидами благородного оленя.
Что касается географической изменчивости кавказского благородного оленя в пределах кавказской части ареала, то по этому поводу пока трудно высказать однозначное суждение. Нет еще достаточной выборки фактического материала, хотя бы для сравнительного анализа двух-трех удаленных друг от друга популяций. Поэтому до настоящего времени остается в силе замечание Н.К.Верещагина (1959, с. 438) о том, что <<геотрафическая изменчивость современного оленя С. elaphus maral в пределах Кавказа не прослежена в достаточной мере и, в сущности, "затушевана" индивидуальной и возрастной изменчивостью».
Для подхода к решению данной проблемы, на наш взгляд, интерес представляют наблюдения, проведенные териологами на Кавказе. Н.Я.Динник (1910) по этому вопросу высказывался достаточно определенно. Он считал, что самые крупные олени встречаются на Кавказе в Кубанской области, именно в ее майкопской части. Объяснял он это тем, что здесь находятся самые благоприятные условия жизни для оленей — богатая и разнообразная кормовая база, не сухой мезоклимат, хорошие защитные условия, мало их тревожит человек. Поэтому в кубанских лесах встречались быки ростом с крупную лошадь. Он полагал, что и рога у них тоже развиваются гораздо сильнее, чем у оленей прочих местностей Кавказа. Судя по его данным, олени южных частей Закавказья и соседних с ними районов Турции и
187
Благородный олень
Ирана заметно мельче, они отличаются более однообразным рыжим цветом шерсти в осеннее время. "На всем теле они имеют шерсть однообразного ржаво-бурого цвета; такого же цвета шерсть покрывает и нижнюю часть передних и задних ног, где у оленей Северного Кавказа она всегда бывает бурой" (Динник, 1910, с. 107).
Точку зрения Н.Я.Динника (1910) в отношении как характера географической изменчивости, так и оценки экологических факторов этой изменчивости полностью разделяет Н.К.Верещагин (1959). Что касается работы Ф.Ф.Алиева (1965), то он не вносит ничего нового в рассматриваемую проблему.
Разделяя неудовлетворенность коллег состоянием внутривидовой систематики Cervus elaphus в целом, а также степенью изученности географической изменчивости этого вида на Кавказе, мы считаем целесообразным до выполнения специального исследования объединять, как и другие систематики, в один подвид С.е. maral все популяции благородных оленей Кавказа.
Экология
Ландшафтная приуроченность. Биотопы. Благородный олень как вид экологически высоко пластичный способен жить в весьма различных ландшафтах равнин и горных областей. Он свободно проникает в пустыню (бухарский олень), обитает в степях (асканийский олень), широко заселяет лесостепь и зону широколиственных и смешанных лесов (среднеевропейский олень), заходит в таежную зону (изюбр), успешно осваивает все высотные пояса горных областей от равнинных степей и влажных субтропиков низменностей до альпийского высокогорья (кавказский олень). При всей эвризональности рассматриваемого вида, очевидно, ландшафтный оптимум благородного оленя приурочен к широколиственным и смешанным лесам и лесостепям равнин и соответствующих поясов горных областей.
На Кавказе былое распространение и ландшафтный спектр С. elaphus maral были весьма обширные. Ископаемые остатки этого оленя прослеживаются с нижнего плейстоцена почти по всей территории Кавказа, начиная с Ленкоранской и Колхидской низменностей до степей Западного Предкавказья (Верещагин, 1959). Словом, кавказский олень формировался при очень широком ландшафтном разнообразии равнин и гор. Еще в начале века, несмотря на большую антропогенную нагрузку, кавказский олень населял весьма разнообразные ландшафты Кавказа. Н.Я.Динник (1910) по этому поводу писал, что область высотного распространения оленей на Кавказе была велика; там, где сплошные леса доходили до самого берега моря, олени спускались очень низко. Например, в Дагестане олени живут по сей день в низовьях Терека, по берегу Каспия. На Западном Кавказе олени часто держались на высоте 150—200 м над уровнем моря, но их было гораздо больше на высоте 1000—1500 м над уровнем моря. Чтобы избавиться от комаров, слепней и других насекомых, в летнее время они поднимались высоко в горы и держались среди скал, осыпей, недалеко от
188
Cervus elaphus Linnaeus, 1758
вечных снегов. "В конце июня 1899 г. в верховьях ручья, впадающего в Малую Лабу, я видел за полдня штук 30 ланей и нескольких оленей на совершенно открытых и безлесных местах на высоте от 2400 до 2700 м над уровнем моря" (Динник, 1910, с. 99). Более всего олени любят, по его мнению, большие леса, особенно если в них находятся поляны, покрытые хорошей травянистой растительностью.
Хотя в наши дни ареал кавказского оленя сильно сократился и трансформировался, а характер территориального размещения искажен мощным антропогенным прессом, еще хорощо прослеживается широкий размах био-топической приуроченности. Наши.наблюдсния и обстоятельные исследования А.А.Насимовича (1936, 1939, 1950), В.Н.Александрова (1968) свидетельствуют о том, что на Западном Кавказе местообитания оленя в теплый период года остаются разнообразными. Однако предпочтение в это время они отдают высокогорным местообитаниям (субальпийские и альпийские луга, субальпийское криволесье и парковые леса, чередующиеся высокотравными лугами). Кормовая база в этих условиях богатая и разнообразная, достаточная для поддержания высокого уровня численности копытных, в том числе оленя. Иная ситуация складывается зимой, особенно в многоснежные годы, в связи с сокращением на 60—70% благоприятных местообитаний (зимовок). На Западном Кавказе такие места зимовки сосредоточены в поясе лесов. В малоснежные годы зимовки оленей могут быть на ветренных склонах альпийского пояса, где оленям доступна ветошь, составляющая значительную долю в их питании. Благодаря именно неравномерному распределению снегового покрова в высокогорьях возможно существование многих копытных.
Территориальная организация населения кавказского оленя в других заповедниках Кавказа, расположенных в горах, имеет много общего с тем, что наблюдается в Кавказском заповеднике, но очень своеобразна биотопи-ческая приуроченность благородного оленя в северной части Дагестана. Здесь олени обитают в плавнях Терека, Аксая и Акташа круглый год.
Миграции. На Кавказе, где в течение года резко меняются погодные и климатические факторы, благополучие оленя, как и ряда других копытных, зависит от его подвижности, смены местообитания. В горах с очень пестрыми ландшафтами не требуется дальних перемещений, чтобы попасть из мест с низким бонитетом в более благоприятные местообитания. Достаточно переместиться всего на несколько сот метров или в крайнем случае на 1 км вниз или вверх, чтобы преодолеть любой из лимитирующих факторов (в том числе кормовые и метеорологические).
Сезонные кочевки оленя наиболее полно изучены в Кавказском заповеднике (Насимович, 1936, 1939, 1955; Чернявская, 1956; Александров, 1968). В летний период олени встречаются во всех поясах гор, но чаще всего держатся по верхней опушке темнохвойных лесов, в субальпийском и альпийском поясах, в пределах от 1500 до 2500 над уровнем моря. Зимой они концентрируются преимущественно в поясе широколиственных и темнохвойных лесов, притом самки и молодняк на уровне 700—1300 м, а взрослые самцы несколько выше — от 1000 до 1700 м. На этом размещении,
189
Благородный олень
очевидно, сказывается высота снежного покрова, составляющая обычно 30—40 см в первых местообитаниях и 50—60 см и больше во вторых местообитаниях. В малоснежных поясах (например, предгорные) благородные олени распределены почти равномерно по всей территории, а в многоснежных они выбирают теневые склоны, где снег остается рыхлым и подснежный корм доступен животным. В тех районах высокогорья, где нет больших снегопадов, часть благородных оленей может зимовать в субальпийском и альпийском поясах, занимая бесснежные участки. Зимовка самок и молодняка зарегистрирована на высоте 1700—2200 м над уровнем моря (хр. Пшекиш, бассейн Киши). При выпадении снега в этих местах олени спускаются вниз, в пояс лесов. Высоко в горах, в полосе верхней траницы лесов (полоса парковых лесов), где снеговой покров может достигать 70—100 см, могут зимовать лишь взрослые самцы.
Хронология, а также интенсивность сезонных перемещений определяются погодными условиями. Обычно перемещение оленей в места зимовок нередко совпадает с разгаром гона, что приходится на конец сентября или начало октября, а иногда продолжается в начале декабря или даже дольше. При ранних похолоданиях и больших снегопадах олени спускаются с высокогорья раньше и в более сжатые сроки. Однако они нередко мигрируют в два-три этапа по мере увеличения снегового покрова. В первую очередь мигрируют самки и молодняк, в последнюю — взрослые самцы. Обратная миграция оленей в высокогорья начинается в период сильного таяния снега в лесном поясе, что приходится обычно на апрель. При этом самцы, яловые самки и прошлогодний молодняк митрируют вверх раньше. Отелившиеся самки остаются в лесном поясе еще 2—4 нед., чтобы окрепли оленята. Где-то к середине июня они успевают подняться в субальпийский и альпийский пояса и присоединиться к стадам оленей, поднявшихся в высокогорья раньше. В лесном поясе остаются лишь немногие олени. Общая протяженность высотных миграций оленей на Западном Кавказе составляет в разных частях горных хребтов от 5—10 до 40 км, преодолевая высоту по вертикали 1000—1500 м.
В других частях гор Кавказа сезонные митрации дают аналогичную картину. Еще Г.И.Радде (1899), отмечал, что в Сванетии и Абхазии в связи с глубокоснежьем олени зимой покидают горы и перемещаются в прибрежную часть гор. В Центральном Закавказье (Боржомский заповедник) олени занимают летом высоты от 800 до 2000 м над уровнем моря. Ранней весной и поздней осенью держатся в альпийском поясе, ибо в это время домашний скот находится внизу. Зимой олени редко поднимаются до 1800 м над уровнем моря (Бараташвили, 1980). В Лагодехском заповеднике летом олени занимают верхний пояс лесов, где ночью пасутся в субальпийском редколесье и на полянах, а днем отдыхают в глубоких ущельях, в буковых лесах с папоротниковым покровом и в субальпийском высокотравье, выбирая западные и северные экспозиции гор (Мамукелашвили, 1987). Из Лагодех-ского заповедника, по данным ряда зоологов, олени летом переходят Главный хребет и занимают высокогорные луга Дагестана. К зиме олени вновь возвращаются в Грузию (Гептнер, Формозов, 1941; Прилуцкая, Пишванов,'
190
Cervus elaphus Linnaeus, 1758
1988). Осенью олени занимают верхнюю полосу горных лесов до завершения рева (примерно до середины октября), после чего спускаются ниже в среднюю полосу горных лесов. Сначала спускаются самки с молодняком, а потом самцы.
На незаповедной территории (в горах Кабардино-Балкарии) олени вытеснены с большей части горных пастбищ, заметно сокращен круг заселенных ими биотопов. Основное поголовье оленей почти круглый год находятся в поясе лесов.
Убежища. Лежки. В жизни кавказского оленя очень важное значение имеют места отдыха, лежки как взрослых, так и молодняка в разные периоды года. На заповедных территориях, где животных тревожат меньше, чем на других территориях, защитные условия местообитания играют подчиненную роль по сравнению с кормовой базой. Поэтому в летний период самки и прошлогодний молодняк на дневку располагаются там же, где они пасутся. Нередко, спасаясь от кровососов, они выбирают хорошо продуваемые ветром места или ложатся на снежники. Самки с оленятами устраивают свои лежки в более защищенных местах, таких как высокотравье, заросли рододендронов, субальпийские березняки. Совсем молодые оленята прячутся в папоротниках и на лесных полянах.
Взрослые самцы во время роста рогов прячутся в зарослях березы, клена и в других субальпийских лесах, имеющих густой подлесок. Одни и те же лежки используются продолжительное время, что легко заметить по вы-битости травостоя. Такие лежки хорошо скрыты, и человеку трудно подойти к ним незамеченным. Известны случаи, когда для отдыха олени уходили в пихтарники или в субальпийское высокотравье.
В тех местах, где животных тревожат часто, места пастьбы и отдыха бывают сильно разобщены. Для лежки в таких случаях выбираются глубокие и труднодоступные ущелья. Как отмечал еще Н.Я.Динник (1910), днем они почти никогда нс показываются на открытых местах и выходят пастись на поляны и лесные опушки только с наступлением темноты. На таких открытых участках они ложатся только ночью. Самки менее осторожны, чем самцы; нередко их можно видеть недалеко от дорог и людных мест.
Сезонная и суточная активность. Поведение (рис. 31). Кавказский олень, как и представители других популяций, ведет активный образ жизни в течение года и круглосуточно. Однако его активность меняется в широких пределах в зависимости от многих внешних и внутренних факторов. В жаркую летнюю пору олени наиболее активны в утренние и вечерние часы, ночью пастьба периодически возобновляется, в пасмурную погоду они могут пастись и днем. Дневная активность нередка в высокогорьях Кавказского заповедника, где погода прохладна и малолюдно.
Суточная активность зимой, особенно при недостатке корма, заметно возрастает. Пасущихся оленей мы встречали в разное время суток, в том числе и в дневные часы. При всем этом олени сохраняют осторожность и ведут скрытый образ жизни, особенно в тех местах, где их тревожит человек. Рогачи теряют скрытность и осторожность во время рева. Когда возбуждение быка достигает своего апогея, пишет Н.Я.Динник (1910), он
1 9 1
Благородный олень
Рис. 31. Благородный олень
192
Cervus elaphus Linnaeus, 1758
ревет очень часто и в глухих местах всю ночь, утром и вечером настолько увлеченно, что охотник, подходя к нему, может сделать более полусотни шагов, прежде чем олень заметит его. В поискав самок ревущий самец делает переходы, значительно превышающие индивидуальные участки вне периода размножения. Однако поведение оленя сильно меняется в зависимости от конкретной обстановки, в частности от того, насколько его тревожит человек, какова плотность населения самок и т.д. По В.Н.Александ-рову (1968), в местах со значительной концентрацией самцов рев проходит более энергично. В районах с низкой численностью рев регистрировался значительно реже, при этом в ненастную погоду’ он может совсем прекратиться. Увеличивается и доля совсем не ревущих самцов, составляя более 20% (7% при высокой численности). В ненастную погоду рев может совсем прекратиться.
Судя по литературным данным и нашим полевым наблюдениям, у оленя в равной степени хорошо развиты органы зрения, обоняния и слуха, поэтому его застать врасплох или подойти к нему близко практически невозможно. Н.Я.Динник (12.10) отмечал, что олень относится к самым осторожным, бдительным и чутким животным. По ветру он может почуять человека более чем за 300—400 м, малейший шум или шорох заставляет лежащего оленя подняться на ноги.
Линька. Сроки сезонной смены волос у кавказского оленя, как и других форм, сильно растянуты, что зависит от погоды, пола, возраста, упитанности и т.д. В условиях Кавказского заповедника весенняя линька в мягкие зимы начинается в первой декаде февраля, в марте и апреле она становится массовой. Обычно первыми приступают к линьке взрослые самцы, они же раньше ее и заканчивают. В 1960, 1961 и 1963 гг. большинство самцов к маю имело летний наряд. Линька сильно задерживается у беременных и кормящих самок, у молодых, а также исхудавших животных. У таких животных зимний мех сохраняется до конца мая и начала июня. Линька у них полностью заканчивается лишь в июле. Формирование зимнего меха начинается в августе или начале сентября, постепенно летняя окраска приобретает рыжевато-серый цвет, к ноябрю олени становятся серо-бурыми (Александров, 1968).
Последовательность смены волос, очевидно, видоспецифична; зимний мех сменяется вначале на голове или конечностях, потом на бедрах, брюхе, спине и, наконец, на зеркале, крестце и шее (Цалкин, 1944; Гептнер и др., 1961).
Питание. Сведения о качественном и количественном составе- кормов кавказского оленя содержатся в ряде работ. Они охватывают различные точки Кавказа, но наиболее полные данные получены в Кавказском заповеднике (Александров, 1968) и Лагодехском заповеднике (Мамукелашвили, 1987). Эти данные интересны не только своей полнотой, но и тем, что они относятся к различным типам поясности: степной тип, к которому принадлежит Кавказский заповедник, и сухой субтропический (полупустынный) тип поясности — Лагодехский заповедник. Как известно, первый заповедник находится на Западном Кавказе, а второй — на Восточном. Сравнительный
13 В.Е.Соколов, А.К.Тсмботов
193
Благородный олень
анализ данных названных авторов с привлечением доступной нам литературы позволит, на наш взгляд, приблизиться к оценке адаптивных возможностей вида в целом.
Видовой состав растений, поедаемых кавказским оленем в обеих точках, очень обширный — на Западном Кавказе зарегистрировано 260 видов растений, а на Восточном — 261 вид растений.
При сопоставлении списков растений, поедаемых оленями в Кавказском и Лагодехском заповедниках, нетрудно заметить, что по количеству видов в обоих районах доминируют травянистые растения: в Кавказском заповеднике их 182 вида (70% от общего количества видов), а в Лагодехском — 87 видов (54%). На долю древесно-кустарниковой растительности приходится соответственно 51 вид (20%) в первом районе и 31 вид (19%) во втором районе. Остальные группы составляют 25 видов (10%) и 43 вида (27%) соответственно.
Сезонный аспект состава кормов в обоих районах Кавказа сходный. Весной с появлением зеленой растительности на хорошо прогреваемых участках лесного пояса олени начинают питаться ею интенсивно. По В.Н.Александрову (1968), олени в это время настолько жадно потребляют свежую зелень, что нередко они утрачивают присущую им осторожность и пасутся в течение всего светлого периода дня. На лесных полянах животные поедают мятлик, типчак, осоку, коротконожку, вейник, а в лесу — пролеску, черемшу, особенно листья злаков. Наряду с травянистой растительностью олени объедают распускающиеся почки и побеги клена, бересклета, ясеня, бука, рябины и других деревьев. Однако доля их в питании заметно ниже, чем зимой. Лишь поздней весной травянистая пища уступает древесно-кустарниковой, включающей не только побеги, но и кору ильма и пихты. Кора также поедается при тяжелых условиях зимовки. По данным Г.В.Мамукелашвили (1987), в условиях Лагодехского заповедника весной на первом месте стоят молодые побеги древесных пород: граба, липы, козьей ивы и бузины черной. Из травянистых растений весной активно поедают стебли, листья и цветы кавказского бутеня, городского гравилата, глухой крапивы, безлистой чины, различные клевера, большой полевицы, лесной коротконожки, пестрого костра, кустарниковидного вейника, люцерны, мятликов и др.
Летом, когда от снегов освобождаются высокогорные пояса и появляется свежий травостой, олени перемещаются вверх и пасутся на субальпийских и альпийских лугах. В Кавказском заповеднике в июне и июле в питании доминирует зеленая травянистая растительность, среди которой преобладают злаки: костер, ежа, тимофеевка, лисохвот, просяник, овсяница кавказская и др. Охотно олени поедают различные бобовые, разнотравье, листья и побеги деревьев и кустарников, в том числе рябины, березы, шиповника, кавказской черники, смородины, кавказского рододендрона и др. Состав кормов самок с оленятами, задерживающихся в лесном поясе, несколько отличается от кормов тех оленей, которые раньше перемещаются в высокогорье. Показательно, в частности, что отелившиеся самки первое время, по В.Н.Александрову (1968), охотно поедают папоротники, что отмечалось
1 9 4
Cervus elaphus Linnaeus, 1758
неоднократно. Более того, такое предпочтение папоротников отмечалось и в тех местах, где изобилуют излюбленные растения оленей.
В летнем корме оленя Лагодехского заповедника доминируют также травянистые растения, среди которых особо предпочитаемыми являются луговая овсяница, глухая крапива, хмелевидная люцерна, разные клевера, лекарственная валериана, лесной и луговой мятлики, бородавник и др. Папоротники в списке излюбленных или предпочитаемых кормов отелившихся оленей, приводимом (Г.В.Мамукелашвили (1987), отсутствуют.
Наблюдения в Кавказском заповеднике свидетельствуют о том, что в горах к концу лета злаки теряют свою привлекательность для оленей из-за огрубления, тогда как бобовые и разнотравье становятся ведущими. Осенью многие травянистые растения высыхают и мало привлекают оленей. Предпочитаемыми оказываются крупностебельные растения, листья которых еще сохраняют свежесть. Среди них могут оказаться такие виды, которые раньше не привлекали внимание оленей (чемерица, аконит и др.). Они хорошо едят листья бутеня, головчатки, васильков, купавки, астранции, володушки, цицербиты, бодяка, кипрея и др. На сенокосах и лесных полянах оленей привлекает отава злаков. По сравнению с летним периодом резко возрастает роль веточного корма, в том числе листьев и побегов ивы, клена, осины, ясеня и других в поясе лесов, а в субальпийском — побегов кизильника, березы, барбариса, шиповника, обыкновенной черники и брусники. После созревания фруктарников в питании оленей большое значение приобретают концентрированные корма. Хотя больших миграций оленей в места с обилием урожая дикоплодных культур не зарегистрировано (Чернявская, 1956), они охотно поедают плоды груши, менее охотно яблони, алычи и др. Особое место в питании оленей занимают буковые орешки, желуди, каштан. Н.Я.Динник (1910) отмечал, что буковых орешков олень поедает много не только осенью, но и зимой.
На питание зимними кормами в Кавказском заповеднике олени переходят в ноябре-декабре, при этом доля веточного корма резко возрастает. Состав его сильно варьирует в зависимости от конкретных условий — типа местообитаний, характера растительности, глубины снежного покрова и др. В малоснежные годы и в малоснежных районах Кавказа (Лагодехский заповедник) и зимой в питании оленей значительную долю составляет травянистая растительность, в особенности осока. Охотно поедают олени такие виды растений, которые сохраняют зимой зеленые листья. Из древесных и кустарниковых растений они предпочитают побеги дуба, бука, лещины, граба, осины, березы. В поясе темнохвойных лесов сокращается видовой состав кормов, который включает ожину, овсяницу, падуб, плющ, побеги хвойных, лишайники, мхи и др. В малоснежную зиму олени выходят в пояса субальпийских и альпийских лугов, где кормятся пожухлой растительностью. Такие перемещения оленей наблюдали сотрудники Кавказского заповедника (Насимович, 1939, 1955; Александров, 1968). Олени здесь кормятся в январе, феврале, марте. Кроме высохшей травы, олени поедают такие кустарники и кустарнички, как рододендрон, брусника, • черника, можжевельник. Хорошо поедают лишайники. В Лагодехском заповеднике,
13*
19 5
Благородный олень
для которого характерна теплая и малоснежная зима, в поясах лесов олени имеют возможность питаться круглый год зеленой растительностью, в том числе плющом, несколькими видами мхов и осоки, ежевикой (около семи видов), овсяницей, глухой крапивой, луковичным мятликом и др. при всем этом и здесь зимой основным кормом оленей остается древесно-кустарниковая растительность, особенно побеги клена, граба, ясеня, черной бузины (Мамукелашвили, 1987).
Суточная потребность кавказского оленя в кормах в природных условиях изучена плохо. По В.Н.Александрову (1968), летом она не превышает 35—40 кг зеленой массы. В зимний период при благоприятных условиях олень поедает около 6 кг корма в сутки, считая в воздушно-сухой массе. Потребность в кормах меняется не только по сезонам, но и по полам и возрастам. На единицу массы тела самки и молодняк потребляют больше, чем самцы. Взрослые самцы перед началом гона питаются интенсивно, а во время гона потребляют очень мало корма.
Как видно из изложенного, кавказский олень употребляет в пищу значительное количество видов древесно-кустарниковой и травянистой растительности. Бесспорно, он не ограничивается 260—261 видами, приводимыми В.Н.Александровым (1968) и Г.В.Мамукелашвили (1987). Если суммировать виды, указанные этими и другими авторами (Динник, 1904, 1910; Насимович, 1939 и др.), то список растений, поедаемых кавказским оленем, превысит 300 видов. Если к этому добавить, что на большей части территории Кавказа зимы малоснежные, а преобладающие ландшафты комплексные с явным доминированием широколиственных лесов и луговых полян, то можно уверенно сказать, что кормовая база для кавказского оленя разнообразная и устойчивая. Это позволит значительно увеличить поголовье оленей при условии надлежащей охраны и воспроизводства популяций.
В питании кавказского оленя важное значение имеют минеральные вещества, ибо нехватка их ощущается повсеместно. Этот аспект питания освещен в целом ряде работ, в том числе А.А.Насимовича (1938), В.НАлек-сандрова (1968) и др. На Кавказе олени используют различные источники минеральных веществ: сухие солонцы в субальпийском и альпийском поясах и мокрые в лесных поясах. Олени посещают минеральные источники в темные часы суток, так как на этих местах их подстерегает немало врагов. Но там, где обеспечена безопасность животных например, в Кавказском заповеднике, на солонцах), оленей можно видеть и днем. Солонцы, посещаемые различными копытными, легко обнаружить: к ним протоптаны тропинки длиной в несколько километров (длина троп зависит от ландшафтных условий), минерализованные места выбиты, лишены растительности, углублены иногда до полуметра.
Воспроизводство и структура популяции
Размножение. Общепризнано, что наиболее ярким внешним проявлением наступления половой активности оленя является подача голоса половозрелыми самцами (так называемый рев). Большой знаток копытных Кавказа
196
Cervus elaphus Linnaeus, 1758
Н.Я.Динник (1910), приводя очень подробную характеристику этого явления, писал, что самым интересным периодом в жизни оленя является время его рева, когда поведение оленя меняется коренным образом. Олень, по своей сути, очень осторожный и скрытный, в период рева выдает себя очень громким голосом. В разгаре рева олень настолько теряет инстинкт самосохранения, что к нему можно приблизиться. "В это время, — пишет Н.Я.Динник (1910, с. 125), — в глухих местах олень ревет почти целые сутки и делается как бы сумасшедший. Он утрачивает свою обычную осторожность и иногда не обращает даже внимание на сделанный по нему выстрел".
Характер рева зависит от многих факторов. Наиболее интенсивно он проходит в середине гона, а в начале и конце значительно слабее. Плохая погода, видимо, угнетает оленя, и соответственно в такое время он мало ревет или вовсе не подает голоса, тогда как в хорошую погоду его активность резко повышается. В это время, если его человек не тревожит, он ревет в глухих местах почти круглосуточно, лишь в поддень умолкает на 3—4 ч (Динник, 1910). По наблюдениям А.А.Насимовича (1936), В.Н.Алек-сандрова (1968) и других, в период рева олени на Западном Кавказе придерживаются верхней полосы темнохвойного и субальпийского лесов, выходя для рева на лесные опушки и луга. На день они спускаются ниже, выби-fj рая для дневки более укромные и затемненные места. __
Самец во время рева занимает определенные участки — "точки", диаметром всего несколько метров. Таких точков у одного самца несколько, они находятся близко друг от друга, иногда в 0,5 км и посещаются так часто, что на них выбивается трава и оголяется земля. Олень оставляет их в случае привлечения к себе самок, когда следует за ними, или во второй половине рева при отсутствии самок на данной территории. Во время рева олень выбивает не только землю на точках, но часто бьет рогами вблизи растущ^,деревь^ и кустарникам, сбивая с них кору и ветки. Агрессивность проявляется и по отношению друг к другу, без самок стычки самцов незначительны, но в борьбе за гарем они могут быть смертельными. В Кавказском заповеднике количество погибших во время драк самцов составляет около 3% от общего количества погибших (Александров, 1968). Между тем, по сведениям Н.Я.Динника (1910), кавказский олень менее агрессивен, чем европейский или сибирский подвиды. За весь период гона самцы питаются очень мало и очень сильно теряют в массе, мясо таких быков имеет неприятный вкус. Самки во время гона не прекращают интенсивно питаться и соответственно сохраняют хорошую упитанность, что биологически оправдано, если учесть предстоящую длительную беременность, роды и выкармливание оленят молоком в течение нескольких месяцев. Самцы и самки активно посещают солонцы, а после гона забрасывают их до весны.
Продолжительность периода гона составляет до 2—2,5 мес. По А.А.На-симовичу (1936), массовый рев оленей в Кавказском заповеднике происходит с десятых чисел сентября по десятые числа октября, максимум его приходится на период между 20 сентября и и 5 октября. Однако отдельные животные могут значительно раньше начать и кончить соответственно
197
Благородный олень
раньше, а другие — немного позже, вплоть до ноября-декабря. На сроки гона существенное влияние оказывают местные условия, в результате чего они могут быть сдвинуты в ту или другую сторону почти на 2—3 нед. Так, в Кавказском заповеднике в заметном количестве олени начинали реветь с 15 сентября в бассейне Лабы, где местность выше, а климат континентальнее; в бассейне Белой рев такой интенсивности регистрировался позже на несколько дней. На южном склоне Главного хребта, в районе бассейна Голо-винки, рев отмечался на 10 дней позже, чем в бассейне Лабы (Насимович, 1941). В Восточном Закавказье (Лагодехский и Закатальский заповедники), где климат значительно мягче, чем на Западном Кавказе, массовый рев обычно приходится на конец сентября или первую декаду октября (Гептнер и др., 1961).
Во втором периоде к самцам присоединяются самки. По Н.Я.Диннику (1910), с одним самцом бывает часто 5—6 самок, а иногда до десятка, а по А.А.Насимовичу (1936) их обычно 3—4 в октябре, 1—2 в сентябре. Сходные результаты получены В.НАлександровым (1968): во второй декаде сентября на каждого самца в среднем приходится 1,6 самки, в третьей декаде этого же месяца — 2,4, а в первой половине октября — 3,4 самки. В 1954—1966 гг. самое крупное стадо самок, зарегистрированное в Кавказском заповеднике с одним самцом, включало 16 голов. По его данным, в те же годы в конце брачного периода меньше 25% общего количества зарегистрированных самцов не имели самок. Такое положение, как полагает автор, свидетельствует о нормальном соотношении половых групп оленя в Кавказском заповеднике.
Самки находятся с самцами в течение октября, а иногда и ноября, а потом отделяются. Очевидно, те самки, которые раньше покрываются самцами, соответственно и раньше покидают гарем. Покрытие самок в Кавказском заповеднике происходит во второй период рева, иногда позже, когда олени кончают реветь. Неоднократно было зарегистрировано преследование самцами самок или нераспавшиеся гаремы до конца ноября (Насимович, 1936).
Точных данных о проценте участия самцов в размножении получить порою невозможно, хотя бы потому, что при гареме наряду с его обладателем нередко бывает молодой самец, который может покрыть самку тогда, когда взрослые соперники дерутся между собой или в начале, а также в конце рева (Насимович, 1936).
Продолжительность беременности благородного оленя различными исследователями оценена по-разному. По А.А.Насимовичу (1936), она составляет 240—250 дней, по К.К.Флерову (1952) — 245—269, по В.И.Цалкину (1944) — 238 дней, по И.И.Соколову (1959) — 247—248 дней, по В.Г.Гепт-неру и др. (1961) — 238—245 дней. Как видно, это различие значительное, достигающее 31 дня. Пока трудно сказать, что лежит в его основе — неточность наблюдений или влияние на продолжительность беременности состояния организма оленя или погодно-климатических факторов.
В Кавказском заповеднике отел происходит с 20-х чисел мая до 10-х чисел июня, иногда сроки рождения оленят могут быть сдвинуты в
198
Cervus elaphus Linnaeus, 1758
зависимости, например, от состояния прошлой осени и рева оленей. Соответственно отел в различных условиях Кавказа может проходить в различные сроки (Насимович, 1936). Многочисленные наблюдения, проведенные позже (Александров, 1968) в том же заповеднике, свидетельствуют о том, что массовое появление молодняка отмечается с 15 мая по 10 июня, иногда оленята появляются в начале мая, а наиболее поздний срок появления зафиксирован 18 июля 1963 г. Для Грузии приводятся слишком растянутые сроки рождения оленят — с мая по июль (Курашвили др., 1981).
Перед отелом самка отделяется от стада и держится в местах с лучшими защитными условиями; телята рождаются в высоких зарослях разнотравья, в густой лесной чаще и других аналогичных биотопах, где беспомощный олененок может быть спасен от многочисленных врагов.
Плодовитость. Самка благородного оленя приносит за год, как правило, одного теленка (Динник, 1910; Насимович, 1936), очень редко бывает двойня. В.Н.Александров (1968) в Кавказском заповеднике трижды наблюдал самок, у которых было по два теленка-сосунка, что составляет менее 1% общего числа самок с приплодом, встреченных им в те годы. Этот исследователь считает, что в природе двойни на самом деле встречаются несколько чаще, чем это предполагается. В сводке В.Г.Гептнера и др. (1961) приводятся данные о том, что в отдельных районах ареала доля самок с двойнями достигает 2%.
О плодовитости кавказского оленя можно судить по доле яловых самок в популяции, которая может быть значительной (Насимович, 1936). По подсчетам В.Н.Александрова (1968), проведенным в Кавказском заповеднике, на 100 половозрелых самок в 1963 г. приходилось 62 олененка, а в 1964 г. — 60. Очевидно, что число самок, участвовавших в размножении и давших приплод, было значительно больше; какая-то часть молодняка могла оказаться неучтенной, а другая могла погибнуть. В целом у благородного оленя процент яловости может меняться широко: в европейской части СССР от 25% (Крымский заповедник) до 44% (Воронежский заповедник), а у западноевропейских оленей обычные размеры яловости составляют 20—25% (см. обзор: Гептнер и др., 1961).
Рост и развитие. Оленята рождаются беспомощными, с массой тела от 7 до 11 кг, вначале они не могут держаться на ногах, но по истечении всего нескольких часов после рождения они уже встают и начинают сосать молоко. Но за матерью олененок способен передвигаться через неделю. До этого периода самка вынуждена оставлять его и пастись в радиусе 100—200 м, возвращаясь для кормежки 4—6 раз в день (Гептнер и др., 1961). Позднее, по истечении недели, оленята довольствуются двух-трехразовым кормлением. При искусственном кормлении коровьим молоком (Александров, 1968) оленята в возрасте 8—15 дней с массой тела 13—18 кг потребляют 2,5—3 л молока в сутки, а 16—30-дневные с массой тела 17—25 кг — от 3,5—4 до 4,5 кг молока. Растут они быстро, при таком режиме искусственного питания среднесуточный привес в течение первого месяца жизни достигает 450—500 г, живая масса тела увеличивается на 200—300%. Судя по данным названного автора, в естественных условиях оленята растут еще
199
Благородный олень
быстрее. Они в 2—3-недельном возрасте начинают поедать травянистую растительность, а в месячном пасутся со взрослыми, посещают естественные и искусственные солонцы, периодически 1рызут землю. Однако оленята сосут молоко долго, о чем говорят многие фактические данные. В ноябре и декабре в желудке молодых обнаружено свернувшееся молоко; самка, убитая уже в январе, оказалась кормящей. Продолжительный период выкармливания молоком, видимо, способствует тому, что сеголетки в сентябре-октябре мало отличаются от годовиков по размерам и массе тела (масса тела сеголеток осенью 65—70, иногда 80 кг). В последующем рост и развитие молодняка продолжается интенсивно, как видно из табл. 41, в 1,5—2,5 года они имеют массу тела, превышающую половину массы 5—11-летних. Увеличение массы тела почти прекращается к 6—7 годам.
Половозрелости самки достигают на 2-м году жизни. Искусственно выращенные, но потом выпущенные в природу приступали к размножению в 1,5 года. От самцов такого же возраста получали нормальное потомство. Однако полного физиологического развития самки достигают в 3 года, а самцы в 4—5 лет. Большинство самцов приступают к реву на 3-м году жизни, но они еще не в состоянии конкурировать со взрослыми самцами.
Формирование рогов начинается с конца первого года жизни, когда на лобных костях вырастают пеньки, не сменяемые в течение всей жизни оленя. На этих пеньках в возрасте 12—14 мес появляются рога, как правило не имеющие ветвей "спички”. Соответственно таких молодых самцов называют спичаками или шилками. Первая смена рогов у молодых самцов происходит весной, значительно позже (почти на 1,5 мес), чем у старых рогачей. После первой смены "спичек” появляются рога с 3—4 отростками, а в последующие годы до старческого возраста с каждым годом увеличивается масса, размерные показатели и количество отросткод на каждом роге.
На Кавказе, как правило, олень сбрасывает старые рога в конце зимы, по преимуществу в определенных участках лесного пояса. По Н.Я.Диннику (1910), в таких местах бывшей Кубанской охоты один человек мог найти до 10 рогов, тогда как в других местах рога вовсе не попадались. После сбрасывания рогов в первые же дни начинают расти новые рога, вначале очень мягкие, чувствительные, обильно снабженные кровью, покрытые тонкой кожей с нежными волосами. Рога растут интенсивно, по Н.Я.Диннику (1910), через 2—3 нед после сбрасывания старых рогов новые достигают около 30 см в длину, а в середине мая увеличиваются до 70 см. В конце мая нижняя часть рога начинает уже окостеневать, но’ даже в первой половине июня концы рогов остаются мягкими. Кожа,на рогах почти всех оленей сохраняется до середины июня, а во второй половине июля они начинают обдирать ее. Первые дни после удаления кожи рога белые, но позднее они делаются более или менее темными. Концы рогов со временем оттачиваются о стволы и ветки, превращаясь в острые, гладкие и светлые, как слоновая кость. Наблюдения, проведенные в Московском зоопарке, свидетельствуют, что продолжительность периода роста рогов сходна у разных форм благородного оленя, т.е. видоспецифична: процесс формиро
200
Cervus elaphus Linnaeus, 1758
вания рогов занимает 102—119 дней, после этого в течение 30—40 дней идет их окостенение, очистка рогов от кожи осуществляется за 1—3 нед. Рога окостеневают за 3—4 нед до гона. Известны случаи, когда в загонах олень в течение жизни давал 23 пары рогов. Судя по тем же данным, зарегистрирована максимальная продолжительность жизни благородного оленя до 30 лет (Гептнер и др., 1961).
Индивидуальное развитие черепа и зубной системы проходит сложно, поэтапно, что позволяет использовать морфологические их изменения для определения возраста животного.
Постнатальное развитие волосяного покрова и его окраски у кавказского оленя сходно с другими формами благородного оленя — ювенильная окраска, характеризующаяся пятнистостью, сменяется на первый зимний мех в конце лета. Характерно, что пятнистость в той или иной степени может сохраняться, как показали наши данные, у вполне взрослых самцов и самок.
Структура популяции. Наиболее полно изучена в Кавказском заповеднике А.А.Насимовичем (1936), В.Н.Александровым (1968). Однако, и эти результаты должны быть восприняты с рядом оговорок, ибо встречаемость разных полов и возрастов широко меняется в зависимости от сезона, погоды, местности, физиологического состояния животных, методов учета и т.д. Тем не менее в данных названных авторов четко прослеживается заметная изменчивость, половой и возрастной структуры популяции по годам и периодам. В 40-х годах текущего столетия в ipynne взрослых соотношение полов было 1:1 или близко к нему. Такое же соотношение полов было у оленят возрастом до 1 мес, отловленных для искусственного выращивания. Начиная с 1954 г. встречаемость самок стала стабильно более высокой, нередко составляя 60% и даже больше. В 60-х годах это соотношение самцов и самок стало близко к 1:2. В Лагодехском заповеднике, по устному сообщению Я.К. Бадридзе, в 1979 г. соотношение полов благородного оленя было 1:4, но в 1988 г. оно стало 3:4 (сказалось сокращение численности волков, элиминировавших самцов, ослабевших после гона).
В отношении возрастной структуры данные относятся к двум периодам: с 1924 по 1932 г. (Насимович, 1936) и с 1941 по 1966 г. (Александров, 1968). В первом периоде было зарегистрировано 700 самок и молодняка оленей, при этом в трехлетие 1924—1926 гг., с июня по август включительно молодняк составлял 27%, в 1927—1929 гг. тоже с июня по август — 24% и в 1930—1932 гг. — 20%. В среднем за 9 лет — 24%. В осенний период (сентябрь-октябрь) доля молодняка составила в 1924—1926 гг. 20%, в 1927— 1929 гг. — 24%, в 1930—1932 гг. — 16%. В среднем за 9 лет — 20%, т.е. ниже на 4%. В период с 1924 по 1932 г., как следует из приведенных данных, на каждого олененка приходилось 4—5 самок. Во втором периоде, с 1941 по 1966 г., такое соотношение молодняка и самок зарегистрировано 13 раз, соотношение одного молодого и более 5 самок отмечено всего 5 раз, а соотношение, когда на одного молодого приходилось менее 4 самок, — 4 раза (по трем годам данные отсутствуют).
201
Благородный олень
Данные по Кавказскому заповеднику позволяют судить о степени агрегации кавказских оленей по годам и сезонам года. По Н.Я.Диннику (1910), в конце XIX и начале XX в. на Западном Кавказе стадо самок в 8—10 голов было обычным явлением. На Умпыре, впадающем в Малую Лабу, он однажды встретил самок с оленятами в 26 голов, на Большом Бамбаке одних самок 35—36 голов, а в верховьях Уруштена видел стадо в 70—75 голов. Больших стад самцов не находил, хотя однажды зимой встретил 13 голов вместе, а егери зарегистрировали стадо самцов в 40—50 голов. В 20-х годах текущего столетия, когда Кавказский заповедник был только что организован, стадо оленей в 4—5 голов было редкостью. Но в 1933—1935 гг., т.е. через десять лет существования заповедника, стали попадаться более крупные табуны (от 4 до 13 голов). Но в этот период численность оленей была ниже, чем в начале века, однако в последующие годы, в том числе в 60-х годах, численность оленей стала выше, чем в начале века. Это сказалось и на численности оленей в стадах. Так, на альпийских склонах г. Армовки нередко стали отмечать группы оленей по 40—50 голов. Однажды было встречено стадо оленей в несколько сот голов. Такие крупные группы встречаются все же редко, обычно в стаде бывает не более 5—7 оленей.
Стадность выше всего зимой и весной, когда сужается круг благоприятных биотопов. Ближе к лету, в период 'появления молодняка, стадность сокращается, так как самки с молодняком ходят обособленно или небольшими 1руппами. Крупные стада в это время — явление редкое. Летом самцы живут группами, нередко по 8—12, а в отдельных случаях до 20 и более голов в стаде. В августе-октябре в связи с наступлением гона эти группы распадаются. Низкий показатель стадности и во время гона (сентябрь-октябрь). К ноябрю-декабрю стадность вновь увеличивается.
Смешанные стада из самцов и самок обычно попадаются во время гона и непродолжительное время после его окончания. В ноябре эти смешанные стада распадаются, олени одного пола образуют обособленные группы (Александров, 1968).
Группировки благородных оленей Лагодехского заповедника, где резко возросла в настоящее время доля самцов, включают во время гона около 3, иногда до 6—7 конкурирующих быков и всего около 4 самок (устное сообщение Я.К. Бадридзе).
Враги, паразиты, конкуренты, смертность. Наиболее опасны для оленей волки. В Кавказском заповеднике, где волков было много в 30-х годах, от них погибало более 60% молодняка (Теплов, 1938). Несмотря на значительное сокращение численности волка в этом заповеднике, в 60-х годах на его долю приходилось более 30% числа всех известных случаев гибели оленей (Александров, 1968). Столь же ощутимый вред причиняет волк кавказскому оленю и в других районах Кавказа (Арабули, 1975). В настоящее время гибель оленей от волка резко сократилась в связи с уничтожением хищника.
От волков олени больше всего страдают зимой, особенно в многоснежье и весной, когда копытные наиболее истощены. В летне-осеннее время упитанные животные и при наличии разнообразных защитных условий
202
Cervus elaphus Linnaeus, 1758
погибают значительно меньше. Из 30 зарезанных волками в Кавказском заповеднике самки и второгодки составили 56%, взрослые самцы — 24% и сеголетки — 20%. Высокая доля взрослых самцов вызвана тем, что во время гона они сильно истощаются и становятся легкой добычей волков (Александров, 1968). Очень низкая численность волков, по Я.К. Бадридзе (устное сообщение), имеет в Лагодехском заповеднике отрицательные последствия — отелившиеся самки перестали поедать послед и организм молодняка недополучает гормоны, необходимые для нормального развития.
К числу врагов кавказского оленя относится рысь, в питании которой это копытное составляет заметную часть: остатки оленя встречены в 8,3—8,8% случаев (Котов, 1958). В 1957 г. случаях гибели оленя, зарегистрированных в Кавказском заповеднике в 1938—1966 гг., на долю рыси приходится 6 случаев (3,8%). Значительно чаще убивают оленей медведи, которые за тот же период уничтожили 16 (10,2%) оленей (Александров, 1968). Конечно, все эти данные значительно занижены, так как найти в горных лесах и скалах (куда загоняют волки оленей) остатки оленей нелегко, особенно тогда, когда жертвой становится молодняк. От них порой ничего не остается или же небольшие остатки от них легко растаскивают другие хищные звери и птицы. Известен случай нападения хищных птиц на оленят (Насимович, 1936), но бесспорно, что они не представляют для оленей серьезной опасности.
Паразитофауна кавказского оленя довольно разнообразна. В летнее время олени сильно страдают от кровососущих насекомых, в особенности комаров, слепней, оводов и др. Спасаясь от них, олени взбираются высоко в горы, вплоть до ледников, где они проводят день. Для лежки они выбирают хорошо обдуваемые возвышенности или высокий травостой, заросли кустарников, где меньше кровососов. На оленях паразитируют иксодовые клещи. Количество гельминтов, обнаруженных у кавказского оленя, составляет 14 видов, из которых наиболее патогенным является Onchocerca bovis, поражающий суставы и подкожную мускулатуру. Dictyocaulus относится также к широкораспространенным и патогенным гельминтам, вызывающим легочные заболевания (Рухлядев, 1964).
Кавказский олень подвержен ряду заболеваний, в том числе ящуру, сибирской язве, некробациллезу, туберкулезу некротическому, параплазмозу и др. В литературе известны тр-и случая эпизоотии среди оленей Западного Кавказа.
Конкурентами оленя по кормовым ресурсам выступают различные копытные, такие, как кабан, косуля, тур, серна, местами зубр, а иногда и другие животные, питающиеся одинаковыми с кавказским оленем древеснокустарниковыми и травянистыми растениями. Однако, на наш взгляд, ни одно из этих животных на Кавказе не служит для численности оленей лимитирующим фактором из-за богатства и разнообразия растительного покрова, с одной стороны, и малочисленности копытных — с другой. Исключение составляют домашние животные, когда из-за перевыпаса горных лугов и лесов на обширной территории природные экосистемы деградируют и теряют свою продуктивность.
203
Благородный олень
Говоря о смертности кавказского оленя в результате перечисленных выше факторов, следует добавить, что определенную роль играют также травмы в период гона, различные абиотические факторы, такие, как высокий снеговой покров, паводки и др.
Численность и ее динамика. В конце XIX в. благородных оленей было много в бывшей Кубанской охоте и прилегающих районах. Об этом писали как зоологи, так и охотники (Шильдер, 1895, 1897; Динник, 1909; и др.). Олени были многочисленны или обычны не только на охраняемых территориях (Кубанская охота), но и в тех районах, где такого режима не было, например в лесах под Нальчиком (Темботов, 1960). В лесах Западного Кавказа, по словам Н.Я.Динника, за день ему приходилось видеть 2—3 десятка оленей, а в лесах Центрального Кавказа олени так часто встречались, что во время рева в близлежащих селах спать было трудно из-за производимого ими шума. В период революции территория нынешнего Кавказского заповедника была пристанищем всевозможных банд и численность оленя там катастрофически упала.
По А.А.Насимовичу (1936), в 1924 г., т.е. в год объявления этой территории Кавказским заповедником, оленей было так мало, что в течение целого дня можно было и не встретить ни одного следа оленя. В ряде точек заповедника он был вообще истреблен. Например, в окрестностях пос. Хамышки восстановление рева было зафиксировано лишь в 1934 г. За десять лет существования заповедника поголовье оленей заметно возросло, достигнув в 1934 г., как минимум, около 900 голов, плотность населения на 1000 га составляла 2,6—3,0 особей. Через 5 лет эти показатели увеличились более чем в 2 раза (в 1939 г. оленей было 2280 голов, с плотностью 7,6 особей на 1000 га). Во время войны и в последующие годы численность оленей сократилась, но в 1952 г. плотность населения резко возросла (до 11,5 голов на 1000 га). На численности оленей сказалось браконьерство и рост численности волков, а повышение плотности вызвано сокращением площади заповедника более чем на 60%, при котором на небольшой территории скопилось много оленей (Александров, 1968). С 1958 г. наметился быстрый рост как количества оленей, так и плотности населения (с 1360 особей в 1955 г. до 8000 в 1966 г., с плотностью 30 голов на 1000 га). Причем в ряде частей заповедника была достигнута предельная плотность. Положительно сказались увеличение территории заповедника до 262,5 тыс. га, усиление охраны и борьба с волками.
Приведенные данные, на наш взгляд, однозначно иллюстрируют причины и следствия положения дел с благородными оленями на всем Кавказе.
Практическое значение. У народов Кавказа олень издавна славится как объект охоты. Приемы высокопоставленных людей, как свидетельствуют исторические данные и различные летописи, обычно сопровождались охотой и в перечне трофеев видное место занимал кавказский олень. Цари и их приближенные за день охоты добывали до 100—150 оленей. Высоко ценятся оленье мясо и рога. По Н.Я.Диннику (1910), обычно масса туши более или менее крупного оленя без кожи и внутренностей составляет
204
Alecs Gray, 1821
220—250 кг. На Западном Кавказе в настоящее время убойная масса во время наибольшей упитанности оленя определяется в 60—62%, соответственно взрослый самец может дать до 200, а самка до 100 кг высококачественного мяса (Александров, 1968).
В дореволюционное время окостеневшие рога шли на экспорт в Западную Европу в качестве трофейных украшений. Для этих целей на Кавказ съезжались скупщики из многих мест, в том числе из Парижа. По данным Н.Я.Динника (1910), за хорошие рога платили от 20 до 30 рублей, иногда и больше, что ускорило уничтожение оленя в ряде районов. Известны факты, что в восточной части бывшей Кубанской охоты в год собирали до 300 пудов оленьих рогов (Александров, 1968).
Оленьи шкуры не.имели такого спроса, как рога, но всегда использовались в домашнем хозяйстве кавказцев. Что касается пантов, то нет данных о сколько-нибудь широком их использовании для этих целей. Между тем есть немало оснований полагать, что фармацевтические качества пантов кавказского оленя не уступают таковым марала и заслуживают внимания.
Значительна роль оленя в лесном хозяйстве. Известно, что при высокой численности он серьезно препятствует лесовозобновлению, влияет на породный состав, выбивает травостой и т.д. Однако такой опасности в настоящее время на Кавказе практически нет, лишь в отдельных местах заповедников может иметь место высокая плотность населения оленей (особенно зимой) и соответственно возможно их отрицательное воздействие на растительность.
Род Лоси — Alces Gray, 1821
Самые крупные олени из всего семейства Cervidae. Длина тела до 300 см, высота в холке более 170 см, масса тела может превышать полтонны. Массивное, неуклюжее и относительно короткое тело опирается на длинные ноги. Голова мощная, вытянутая, с хорошо развитым лицевым отделом. Глаза большие. Морда на переднем конце расширена благодаря сильно увеличенной носовой области. Верхняя губа большая, нависает над нижней. Уши большие, длинные, широкие и заостренные. Хвост маленький, но четко виден. У многих особей на горле имеется кожный вырост, развитый сильнее у самцов, чем у самок. Хорошо развитые предглазничные межпальцевые железы имеются лишь на задних конечностях, тарзальные и метатарзальные железы небольшие и расположены высоко на внутренней стороне ног.
Копыта узкие и раздвигающиеся, приспособленные к мягкому грунту, боковые пальцы хорошо развитые, посажены заметно низко и служат дополнительной опорой при передвижении по болоту. Волосы на шее и холке длиннее, чем на остальной части тела. Отростки на лобных костях отходят горизонтально, что специфично для лосей. Лопатообразные рога
205
Род Лоси
расположены также горизонтально, лишь кончики отростков загибаются вверх. Самцы рогатые, самки комолые.
Окраска тела молодых и взрослых не пятнистая, брюхо и ноги значительно светлее боков и спины, нет околохвостового зеркала. Половой диморфизм в окраске меха отсутствует.
Череп крупный, сильно вытянутый, однако носовые кости короткие (почти на 1/3 короче длины верхнего ряда зубов). Наоборот, межчелюстные кости очень длинные, но не соприкасаются с носовыми. Носовое отверстие очень большое (длина его больше длины верхнего ряда зубов). Задняя часть сошника низкая и не разделяет отверстия хоан. Слуховые барабаны малы и не выступают над засуставным отростком. Сильно вытянута нижняя челюсть.
Зубная формула:
1-С-Р-М- = 32.
3 13 3
Характерно отсутствие в норме верхних клыков, увеличение мощности коренных зубов (они широкие, низко посажены, боковые стенки сближены так, что площадь их жевательной поверхности меньше, чем основания зубов).
Лоси — типичные представители таежной зоны Европы, Азии и Северной Америки. Северная граница восстановленного ареала далеко выходит как в Старом, так и Новом Свете на Полярный круг. Это особенно выражено в азиатской части ареала, где лось проникает на север почти до низовьев сибирских рек (Оби, Лены, Енисея и др.).
За последнее столетие ареал рода претерпел большие изменения под антропогенным воздействием. В американской части ареала распространение лося интенсивно сокращалось и продолжает сокращаться, но в СССР в 50-х годах были приняты серьезные практические шаги, приведшие к восстановлению былого ареала как за счет северной, так и южной границ. По соответствующим экологическим руслам лось проник в лесостепную и степную зоны, местами закрепился там.
Местообитания лосей связаны с лесными и кустарниковыми насаждениями, но в отдельные периоды года лоси могут "отрываться" от лесной зоны и далеко проникать в тундру, достигая берегов северных морей, а на юге — в лесостепную зону, осваивая островные и приречные леса. Более того, в наши дни лось заселяет зоны степей и полупустынь через берега различных водоемов с гитрофильным высокотравьем и древесно-кустарниковой растительностью. Наряду с равнинными биотопами лоси занимают горные леса, отдавая предпочтение местности со спокойным рельефом — пологим склонам хребтов, платообразным вершинам и широким межгорным долинам. Местами в горы проникают высоко — в Алтае и Саяне летом они поднимаются до 2200—2400 м над уровнем моря. Зимние местообитания не всегда совпадают с летними, сезонная смена местообитаний нередко сопровождается значительными переходами — к зиме из тундры в лесотундру, где достаточно веточного корма; с западного макросклона Урала (Прсдуралье) на восточный (Зауралье), отличающийся относи
206
Alecs Gray, 1821
тельным малоснежьем; в горах с более высоких и снежных спускаются в широкие горные долины (Восточный Алтай). Протяженность таких сезонных переходов составляет нередко сотни километров. Зимние местообитания определяются наличием и доступностью древесно-кустарниковой растительности, а также благоприятным снежным покровом (лоси избегают мест со снегом глубиной более 70—80 см) (Гептнер и др., 1961).
Суточная активность меняется широко. Она зависит от состояния организма самого животного и окружающей его экологической обстановки. Так, в летнюю жару и в пору обилия кровососов лоси ведут сумеречный и ночной образ жизни — днем отлеживаются в кустарниках, в высоком травостое, в более прохладных и укромных местах, нередко в сырых и заболоченных старицах. Зимой активны и днем, притом в течение суток они ложатся несколько раз и несколько раз кормятся.
В период гона ритмичность суточной активности нарушается и лосей можно встретить в любое время дня и ночи.
Питание довольно разнообразное. В условиях СССР лоси поедают деревья, кустарники и травянистые растения, относящиеся к 175 родам и 250 видам. Очевидно, число видов растений, в действительности поедаемых лосем, значительно больше и будет расти по мере изучения его биологии в различных частях ареала. Набор кормов, особенности пищевого поведения и соответствующее территориальное распределение широко меняются по сезонам года. Так, зимой резко возрастает доля веточного корма, а летом — травянистой растительности, в том числе водной и околоводной. Лоси посещают солонцы, преодолевая нередко большие расстояния.
Размножение лосей имеет много общего с представителями рода Cervus. С некоторой оговоркой лосей можно отнести к полигамам — хотя они не образуют гаремы, но в период гона один самец покрывает несколько самок (Кнорре, 1953). Между самцами бывают драки из-за самок, нередко они кончаются смертельным исходом. Возбужденный самец теряет осторожность, становится агрессивным, мало ест, нарушается его суточная активность. Период гона длится около 1,5—2 мес, но основная масса спаривается в течение 10—20 дней. Беременность длится до 240 дней. Сроки размножения меняются географически и в зависимости от погодных условий. Плодовитость также изменчива, но обычно доля двоен в одном помете значительна (от 10—25% у американского лося до 42—92% в разных частях ареала европейско-азиатского лося). Тройни бывают очень редко.
Рождение телят в южных частях ареалав б-СССР происходит с конца апреля по конец мая, но возможны отклонения. Отел в местах, где лосей тревожат, происходит в укромных, мало посещаемых местах, таких, как густые заросли кустарников, на болотах, по берегам рек и т.д. Рост и развитие телят проходят интенсивно — с третьего дня после рождения они ходят свободно, а через 5 дней человеку их трудно догнать; в двухнедельном возрасте хорошо плавают. Лактация продолжается 3,5—4 мес, иногда дольше. Зеленый корм начинает потреблять через две недели после рождения. Масса тела новорожденного от 6 до 16 кг, за 6 мес она увеличивается почти в 10 раз, достигая в среднем 120—130 кг, иногда до 160—206 кг. Полного
207
Лось
физического развития лоси достигают на четвертом году жизни; увеличение массы тела у самцов прослежено до десятилетнего возраста. Расцвет сил бывает в возрасте 6—12 лет. Продолжительность жизни до 20 лет, возможно даже превышает 25 лет (Гептнер и др., 1961).
Лоси становятся способными к спариванию на вторую осень жизни, но основная масса самок вовлекается в размножение на 3-м году, а самцов — на 3—4 году жизни.
Лоси имеют важное практическое значение, главным образом как объект промысловой охоты. Особо ценится высококачественное мясо, выход которого составляет более 60% от общей массы тела. С одного лося получают до 30 кг внутреннего и подкожного жира. Шкура толстая и прочная, служит сырьем для различных целей. Рога представляют интерес как охотничьи трофеи. Целый ряд особенностей лосей привлекает внимание селекционеров к возможностям их одомашнивания (высокий темп роста и развития, плодовитость, высокая жирность молока, дешевые корма и т.д.).
Лоси как естественные компоненты лесных экосистем заслуживают специального изучения. При высокой численности они отрицательно влияют на древесную растительность, особенно на лесные питомники (состав пород, лесовозобновление, продуктивность т.д;). Полеводству обычно не вредят.
Род Alces включает лишь один современный вид Alces alces L., населяющий Европу, Азию и Северную Америку. Ряд зарубежных авторов рассматривает как самостоятельные виды американских и европейских лосей. В пределах СССР нижнеплейстоценовые лоси составляли, очевидно, отдельный вид — Alces latifrons Dawk.
Лось — Alces alces Linnaeus, 1758
Рис. 32
A.a. europaeus, Burnett, 1830;
A. savinus, Fischer, 1834;
A.a. caucasicus, Верещагин, 1955.
Коллекционный материал. ЗИН РАН: 7 черепов; МГУ: 5 черепов.
Диагноз. В роде один современный вид.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Экстерьерные данные по вымершим лосям Кавказа отсутствуют, но сохранившиеся в пещерах Осетии позднеплейстоценовые черепа относятся к западноевропейской группе вида A. alces. К этой же группе принадлежат, очевидно, современные лоси Предкавказья, являющиеся выходцами из Европы (Коряков, 1965; и др.). Из-за отсутствия описания экстерьера местных лосей мы ниже приводим данные по европейской форме, позаимствовав их у К.К.Флерова (1952): длина тела обычно 250—270 см, высота в загривке 175—185 см, масса тела 320—400 кг (иногда до 480 кг). Рога крупные,
208
Alecs alces Linnaeus, 1758
нередко с широкими лопатами, их размах до 135 см, а масса без черепа иногда более 20 кг. Самки немного уступают самцам, особенно по массивности головы и толщине шеи.
Телосложение очень специфичное и резко выделяется среди современных видов семейства Олени по многим параметрам. Из-за мощного развития носовых хрящей, верхних губ и больших ноздрей голова значительно сужена в средней части. Однако такое внешнее строение головы не соответствует структуре черепа, которая в общих чертах остается сходной с черепами других видов настоящих оленей. Предглазничные железы сравнительно с размерами головы небольшие. Шея короткая, но сильно развитая, на ее верхней стороне волосы удлинены, а под горлом часто имеется "серьга" — кожный вырост, покрытый длинными волосами. Наибольшего развития серьга достигает у 3—4-летних лосей. Ряд длинных волос на шее простирается на холку, образуя вместе большой горб. Спина прямая, зад покатый, хвост короткий (около 8 см). Хорошо развиты боковые пальцы, при ходьбе они достигают земли.
Череп большой, сильно вытянутый в длину, особенно в ростральной области. Общая длина черепа в пределах вида колеблется от 540 до 620 мм, скуловая ширина — от 200 до 240 мм (Флеров, 1952). Размеры черепа вымершего кавказского лося сравнительно небольшие. По данным Н.К.Вере-щагина (1959), если общая длина черепа самцов в Ленинградской области составляет около 555 мм, то на Северном Кавказе — 538 мм, длина носовых костей соответственно 103 и 104 мм, длина заглазничной области 160 и 180 мм, длина зубного ряда 137 и 150 мм и т.д. (табл. 42). В структуре черепа, по мнению К.К.Флерова (1952), имеется ряд видоспецифичных особенностей. Так, межчелюстные кости длинные, почти в 1,5 раза длиннее зубного ряда верхней челюсти. Сильно вытянут передний отдел верхней челюсти. Носовые кости, наоборот, очень короткие, более чем вдвое короче носового отверстия. В отличие от других оленей у лося передний отдел черепа резко скошен сверху вниз. Слезные кости сравнительно узкие и длинные, мала и неглубока слезная ямка, также относительно малы и узки слуховые капсулы. Нижняя челюсть характеризуется удлинением в передней части. Зубная формула в пределах рода устойчива, но видоспецифичность строения отдельных зубов значительна (Флеров, 1952). По данным этого автора, значительны также особенности скелета, выработанные в связи с жизнью в лесах и болотах. В частности, боковые пальцы сравнительно велики, кости конечности сильно удлинены, мала величина мягкой пятки в сравнении с общей длиной копыта, что связано с хождением в условиях мягкого грунта. К тому же все четыре пальца могут широко раздвигаться; связки, соединяющие их, очень прочные.
Рога лося, при всей их изменчивости, сохраняют свою оригинальность. Так, пеньки и основания рогов по отношению к лобным костям расположены горизонтально, а не вертикально, как у настоящих оленей. В типичном случае рог недалеко от основания сильно расширяется, приобретая лопатовидную форму (тип рога "лопата"). От "лопаты" отходят от 8 до 18 пальцевидных отростков, направленных вперед, в стороны и вверх, а не
14 В.Е.Соколов, А.К.Темботов
209
Лось
Мариуполь
Ростов-на-Дону
Бердянс
О Заветное
Сарпа
Элиста
Яшкуль
174
Егорлынская
• 33
Новороссийск^
Сочи
'Гагр
Владикавказ
Очамчира
Казбеги
Зугдиди
Поти
6000
503
6000
Ахтала
4000
3000
2000
1000
4000 •
Нахичевань
3000
г. Далидаг 3616
г. Кюмиркей 2477
г. Меписцкаро 2850
г. БолчГиналдаг
3367
г. Каракал 2850
60 пЧеркесок
Пятигорск
ополь
Б2 Буденновск 70
г. Эльбрус
5642
ивно'е.
Снетлоград
Благодарный
*$1
Карачаевск
2000
1000 0
АЖ
к-з
Ц-М
Моздок
Грозный
Ереван
Мартуни
210
Alces alces Linnaeus, 1758
14*
Рис. 32. Современное распространение лося
1 — Ейск (Бакеев, 1980); 2 — Староминская (Гинеев, 1975); 3 — Новоминская (Бакеев, 1980); 4 — Бриньковская (Бакеев, 1980); 5 — Приморско-Ахтарск (Бакеев, 1980); 6 — Садки (Бакеев, 1980); 7 — Темрюк (Бакеев, 1980); 8 — Анапа (Бакеев, 1980); 9 — Новороссийск (Бакеев, 1980); 10 — Крымск (Язан, 1966); 11 — Мингрельская (Бакеев, 1980); 12 — Славянск на Кубани (Коряков, 1965; Язан, 1966; Бакеев, 1980); 13 — Красноармейская (Гинеев, 1975); 14 — Гречаная Балка (Бакеев, 1980); 15 — Тима-шевск (Гинеев, 1975); 16 — Каневская (Гинеев, 1975); 17 — Павловский р-н (Гинеев, 1975; Бакеев, 1980); 18 — Краснодар (Язан, 1966); 19 — Смоленская (Бакеев, 1980); 20 — Горяче-Ключевский р-н, р. Псекупс (Коряков, 1965); 21 — Горячий Ключ (Язан, 1966; Гинеев, 1975); 22 — Апшеронск (Гинеев, 1975); 23 — Кутаисская (Бакеев, 1980); 24 — Мартанская (Бакеев, 1980); 25 — Белореченск (Гинеев, 1975); 26 — Рязанская (Бакеев, 1980); 27 — Красногвардейская (Гинеев, 1975); 28 — Усть-Лабинск (Гинеев, 1975); 29 — Кореновск (Гинеев, 1975); 30 — Выселковский р-н (Гинеев, 1975); 31 — Тихорецк (Бакеев, 1980); 32 —Новопокровский р-н (Гинеев, 1975); 33 — Белая Глина (Коряков, 1965; Язан, 1966); 34 — Кропоткин (Бакеев, 1980); 35 — Курганинск (Верещагин, 1959; Коряков, 1965; Гинеев, 1975; Бакеев, 1980); 36 — Лабинск (Гинеев, 1975); 37 — Да-ховская (Бакеев, 1980); 38 — урочище Умпырь (Бакеев, 1980); 39 — Перевалка (Бакеев, 1980); 40 — Уруп (Бакеев, 1980); 41 — Зеленчукская (Бакеев, 1980); 42 — Ацдрюковская (Бакеев, 1980); 43 — Псебай (Бакеев, 1980); 44 — Ахметовская (Бакеев, 1980); 45 — Новокубанский р-н (Гинеев, 1975); 46 — Новоалексацдров-ский р-н (Перовский, 1976; Бакеев, 1980); 47 — Красногвардейское (Оноприенко, Хохлова, 1988); 48 — Труновский р-н (Оноприенко, Хохлова, 1988); 49 — Московское (Бакеев, 1980); 50 — Изо
2 1 1
Лось
бильный (Бакеев, 1980); 51 — оз. Сенгилеевское (Бакеев, 1980); 52 — Убеженская (Бакеев, 1980); 53 — Николаевская (Бакеев, 1980); 54 — Успенский р-н (Бакеев, 1980); 55 — Ставрополь (Язан, 1966; Перовский, 1976; Бакеев, 1980); 56 — Бешпагир (Бакеев, 1980); 57 — Кочубеевское, Темнолесская (Бакеев, 1980); 58 — Хабаз (Оноприенко, Хохлова, 1988); 59 — Курсавка (Оноприенко, Хохлова, 1988); 60 — Крым-гиреевское (Бакеев, 1980); 61 — Султановское (Бакеев, 1980); 62 — Круглолесское (Бакеев, 1980); 63 — Ипатовский р-н р. Калаус (Перовский, 1976); 64 — Гофицкое (Бакеев, 1980); 65 — Ипатово (Оноприенко, Хохлова, 1988); 66 — оз. Маныч-Гудило (Бакеев, 1980); 67 — р. Маныч (Язан, 1966); 68 — Апанасенковский р-н (Оноприенко, Хохлова, 1988); 69 — Арзгир (Бакеев, 1980); 70 — Томузловское (Бакеев, 1980: Оноприенко, Хохлова, 1988); 71 — Зеленокумск (Бакеев, 1980); 72 — Георгиевск (Перовский, 1976; Оноприенко, Хохлова, 1988); 73 — Буденовок (Бакеев, 1980); 74 — Надзорное (Оноприенко, Хохлова, 1988).
Карта-врезка. Распространение лося в прошлом (по: Верещагин, 1959)
1—2 — Alces aff. alces L.: 1 — верхний плейстоцен, 2 — возможные пути вселения; 3 — Famanalces sp., нижний плейстоцен-верхний плиоцен; 4—8 — кавказский лось (Alces alces caucasicus): 4 — вероятный ареал в историческую эпоху, 5 — литературные данные начала XIX в., 6 — XV—XIX вв. н.э., 7 — VIII—XIII вв. н.э., 8 — IV—I вв. до н.э.
Таблица 42. Промеры черепа лося (в мм) (по: Верещагин, 1949)
Самцы Самки
Признак Ставро- Осетия, Ленинград- Осетия, Ленинград-
ПОЛЬ, п=3 ская обл., п=4 ская обл.,
п= =1 п=4 п=3
Общая длина — 538 555 — 539
Acrocr.-Bregma 121 109 118 110 111
Br.-Nasion 138 112 137 129 135
N.-Prostion 326 323 2 305
Длина носовых 111 103 104 98 89
костей
Длина заглазничной 190 160 180 154 163
части
Длина зубного 145 137 150 138 142
ряда
Ширина височная 102 94 103 93 94
Ширина слуховая 157 144 142 131 133
Наибольшая ширина
носовых костей
сзади 85 79 71 73 65
спереди 70 60 59 54 56
Наибольшая ширина 220 203 214 206 194
лба
Наименьшая. 160 144 146 147 137
межорбитная
ширина
Ширина верхнече- 148 138 143 140 131
люстная
в области
2 1 2
Alces alces Linnaeus, 1758
назад, как у лани (Соколов, 1959). Нередко встречаются лоси с "оленьим" типом рога, у которых не выражены лопаты. По В.Г.Гептнеру с соавторами (1961), наряду с указанными двумя крайними вариантами структуры рогов выделяется промежуточный тип, представляющий тот же тип "лопаты", но отличающийся от первого типа степенью развития "лопаты” и другими деталями. Как отмечают эти авторы, выделение различных форм лопатовидных рогов осложняется переходными формами.
Окраска тела темная, черно-бурая или черная, особенно по бокам тела и шеи. По хребту проходит черная полоса. Окраска головы сходна с окраской туловища. Лишь на конце морды волосы бывают значительно светлее (рыжеватого или серого тона). Нижняя половина конечностей и живот, как и верхняя губа, светло-серого, почти белого цвета. Характерная для многих видов оленей пятнистость у лосей отсутствует в .течение всего онтогенеза. Нет также прианального зеркала.
Кожный покров. Исследовали кожу и специфические кожные железы взрослого самца, добытого в июне. Строение кожи загривка в-целом сходно с таковым у ранее описанных лосей, добытых в Подмосковье (Соколов, 1973) и в Мантуровском районе Костромской области (Соколов, Чернова, 1988).
Эпидермис загривка средней толщины (34—45 мкм) с хорошо выраженным роговым слоем, сильно пигментирован, содержит отдельные зернистые клетки. Дерма плотная, с многочисленными эластиновыми волокнами на границе с подкожной мускулатурой. Сосочковый слой занимает 45% толщины кожи. Подкожная жировая клетчатка имеется; размеры жировых клеток составляют 34 х 34; 45 х 56 мкм. Сальные железы остевых волос крупные (226 х875; 339 х900 мкм), многодольчатые, могут располагаться в два яруса с каждой стороны волоса. У пуховых желез имеются одно-двудольчатые мелкие (34 х 34; 34 х 56 мкм) сальные железки.
Потовые железы многочисленные, с крупными (диаметром 79—113 мкм) сильно извитыми секреторными отделами.
Волосяной покров представлен остевыми и пуховыми волосами. Остевые волосы толстые, с тонким корковым слоем, с хорошо развитой сердцевиной. Длина остевых волос достигает 68,2 мм, толщина 261 мкм. Длина пуховых волос 2,5 см; они волнообразно извиты или скручены. Форма поперечника и остевых и бесссрдцевинных пуховых волос округлая.
Из специфических кожных желез у лосей имеются мейбомиевые, пред-глазничная, предплюсневая, межпальцевые (Соколов, 1973; Соколов, Чернова, 1988). Железы также увеличены на губах, в циркуманальной коже и на дорсальной и .вентральной поверхностях хвоста. Мы исследовали мейбомиевые и предплюсневую железу.
Мейбомиевые железы хорошо развиты, крупные: размеры отдельной многодольчатой железы на продольном срезе верхнего века достигают 2,8 х 1,2 мм. Число крупных (226 х 226; 339 х226 мкм) округлых одно- или двух-трехдольчатых альвеол составляет 50—60 в каждой железе. Диаметр центрального протока, собирающего секрет от альвеол.невелик — 158 мкм.
213
Лось
Предплюсневая железа располагается на внутренней стороне предплюсны и обозначена "завихрением" длинных волос. Толщина кожи здесь составляет 6,4 мм. Эпидермис толстый, содержит зернистый и блестящий слои. Сосочковый слой занимает 73% толщины кожи и содержит, помимо крупных волосяных луковиц, мощные (диаметром 498—560 мкм) мышцы. Размеры крупных многодольчатых сальных желез могут достигать 1 х0,3; 1,2 х0,4 мм.
Система крови. Изучению различных параметров крови лося посвящен ряд работ отечественных и зарубежных авторов. Однако сведения, содержащиеся в них, нередко противоречивы и несопоставимы. В сводке А.А.Куд-рявцева и Л.А.Кудрявцевой (1974) приведены следующие показатели: количество эритроцитов 6,5—8,5 млн (х = 7,5), гемоглобина 14,0—18,0 г% (х = 16,0), цветной показатель 1,1—1,35 ед. (х — 1,25), размер эритроцитов 5,8—6,6 мкм (х = 6,2), объем эритроцитов 39,0 мкм3, лейкоцитов 7,5—9,5 тыс. (х= 8,5). Лейкоцитарная формула: базофилы 0,0—0,5 (х = 0,0), эозинофилы 5,0—9,0 (х= 7,0), нейтрофилы палочкоядерные 2,0—6,0 (х= 4,0), нейтрофилы сегментоядерные 48,0—58,0 (54,0), лимфоциты 28,0—42,0 (х = 32,0), моноциты 1,0—5,0% (х = 3,0). Сходные данные цитирует в своей монографии П.А.Коржуев (1964): количество эритроцитов 7,70 млн (Ponder et al., 1928), диаметр эритроцитов в (мкм): 5,8 (Wormley, 1888), 6,2 (Ponder, 1934), 6,5 (Gulliver, 1875). Результаты исследования Е.П.Кнорре и Е.К.Кнорре (1959) по крови лося оказались иными: количество эритроцитов составило в среднем (л = 2) 5,27 млн лейкоцитов 4,0 тыс., гемоглобина 8,2 г% (по Сали, 48%). Пока трудно объяснить, чем обусловлены столь большие различия — малой выборкой, биотическими или абиотическими факторами (местные условия, сезон года, возраст, состояние организма). Между тем немало данных по лосю свидетельствуют о лабильности показателей крови в зависимости от указанных выше факторов. Данные Е.П.Кнорре и Е.К.Кнорре (1959) показывают, что при сравнительно небольших различиях по полу возрастные изменения очень большие. Так, трехмесячные лосята (2 самца и 2 самки) в августе имели эритроцитов 6,9—9,5 млн (Я = 7,7), лейкоцитов 8,0—9,3 тыс. (х = 8,6), гемоглобина 10,0—12,6 г% (х = 11,5), по Сали, 59—74% (х = 68). У годовалых животных (2 самца и 2 самки) показатели крови близки к таковым трехлетних: эритроцитов 5,2—5,5 млн (х = 5,4), лейкоцитов 4,4—4,8 тыс. (х = 4,6), гемоглобина 9,9—10,5 г% (х = 10,2), по Сали, 58—62% (х = 60). Существенные возрастные изменения обнаружены в лейкоцитарной формуле. В частности, среднее число нейтрофилов в трехмесячном возрасте составило 33%, в годовалом — 41, в трехлетием — 37%. Миэлоциты не обнаружены, юные встречены лишь в трехмесячном возрасте. Наибольшее количество лимфоцитов зарегистрировано в трехмесячном возрасте (х = 61), в годовалом — 53, в трехгодовалом — 56. Моноциты не обнаруживают определенной возрастной изменчивости, тогда как количество базофилов заметно увеличивается с возрастом. По мнению названных авторов, большие сдвиги вызывает характер пищи. У трехмесячного лосенка-самца, выкормленного молоком матери, количество эритроцитов оказалось значительно выше, чем у лосенка того же возраста и пола, но питавшегося коровьим молоком. В первом случае эритроцитов было 9,5 млн,
214
Alces alces Linnaeus, 1758
гемоглобина 12,6 r% (74% по Сали), а во втором — соответственно 6,9 млн и 10,0 г% (59% по Сали).
По ряду показателей крови иными оказались результаты исследования. Л.И.Иржака и В.В.Гладилова (1981), проведенные в мае-июне 1979 г. в том же Печорр-Илычском заповеднике. Так, на протяжении первого месяца жизни у лосят увеличивается количество эритроцитов, хотя гематокрит и число ретикулоцитов остаются постоянными. Однако взрослые животные характеризуются высоким гематокритом и содержанием гемоглобина при меньшем числе ретикулоцитов по сравнению с лосятами первого месяца жизни. Авторы считают, что в целом с возрастом увеличивается количество эритроцитов и содержание гемоглобина, что сопровождается повышением кислородной емкости крови.
На очень большом фактическом материале показано (Franzmann, Zere-sche, 1978), что на Аляске показатели крови существенно меняются, особенно в связи с условиями среды и состоянием организма (пол, возраст, сезоны года, разные места и разные популяции). Четкой зависимости от перечисленных факторов не выявлено лишь по содержанию в 100 мл азота мочевины (12,5 мг), мочевой кислоты (0,41 мг), холестерола (85 мг), билирубина (0,46 мг), глутамин-оксалат-трансаминазы (173 мг). Пол (только в период гона) и возраст накладывают отпечаток на все показатели крови, кроме содержания фосфора, глюкозы, а- и Р-глобулинов. Лактация существенно снижает содержание фосфора. Уровень возбуждения животного, сопровождающегося повышением ректальной температуры, значительно сказывается на содержании глюкозы, молочной дегидрогеназы, альбумина и глобулина. Благоприятные условия жизни вызывают увеличение кальция, фосфора, глюкозы, общего белка, альбумина, Р-глобулина, гемоглобина и гематокрита. В таких условиях гематокрит составляет 50%, гемоглобин — 18,6 г, кальций, — 10,4 мг, фосфор — 5,2 мг, общий белок — 7,5 г, альбумин — 4,5 г, Р-глобулин — 0,7 г, глюкоза — 140 г (в 100 мл).
Закономерная динамика состава красной, крови и некоторых морфометрических и расчетных показателей эритроцитов у лосей в неонатальный период выявлена Н.А.Моисеенко и Н.Н.Мочаловым (1987) (табл. 43). Ими показана также гетерогенность гемоглобина лося и ее возрастные .изменения, что было обсуждено еще раньше Л.И.Иржаком и Э.В.Качмарчиком (1971). Данные Н.А.Моисеенко и Н.Н.Мочалова существенно отличаются от таковых у Е.П.Кнорре и Е.К.Кнорре (1959). Согласно данным табл. 43, хотя обе работы выполнены в одном и том же Печоро-Илычском заповеднике, показатели крови по ряду позиций расходятся, особенно у годовалых лосей. Вероятно, сказались сезон года и физиологическое состояние животных. Результаты работы по электрофорезу на агаре показали, что подвижность гемоглобина различна у 2,5-месячных лосят, новорожденных в возрасте 1—7 сут и взрослых животных. Наиболее подвижны гемоглобины плодов, наименее — взрослых лосей. В период новорожденное™ наблюдается возврат к более ранним типам гемоглобина. Деления на фракции не отмечалось. При этом, на наш взгляд, примечательно, что если у крупного рогатого скота сближение тина гемоглобина у телят с гемоглобином взрослого
215
Лось
Таблица 43. Показатели красной крови лося в онтогенезе (х ± т) _______________(Моисеенко, Молчанов, 1987)_______________
Показатель —— Возраст
1 сутки <Л= 1) 1 неделя (л = 3) 2 недели (л = 3)
Концентрация Эр, млн/мм^ 6,54 5,15±0,21 5,24±0,34
Концентрация ретикулоцитов, % 20,10 42,93±8,37 58,37±5,73
Концентрация НЬ, г% 10,20 6,80±0,19 6,67+0,61
Показатель гематокрита, % 33,30 23,43-0,28 22,77±1,68
Среднеклеточные: объем Эр, мкм^ 50,90 45,64±1,41 43,50±1,98
диаметр Эр, мкм 6,78 6,70±0,03 6,64+0,04
толщина Эр, мкм 1,41 1,29±0,03 1,26±0,06
индекс сферичности Эр 0,21 0,19+0,003 0,19+0,01
86,59 84,73+0,86 83,07+1,03
площадь поверхности Эр, mkmz 15,60 13,22+0,28 12,67+0,57
содержание НЬ, иг 30,60 29,22±0,58 29,19+1,02
концентрация НЬ, %
Показатель Возраст
3 недели (л = 3) 1 год (л = 2) Взрослые (л = 6)
Концентрация Эр, млн/мм^ 5,63 ±0,09 6,12±0,17 4,69 ±0,18
Концентрация ретикулоцитов, % 59,17±4,22 2,05±0,02 5,87±0,72
Концентрация НЬ, г% 7,38±0,20 12,60±0,43 10,90±),29
Показатель гематокрита, % Ср едне клеточные: 27,57±1,42 35,70±9,57 32,38±0,98
объем Эр, мкм^ 48,91±2,16 58,41±1,34 69,28±1,80
диаметр Эр, мкм 6,67±0,03 6,81+0,06 7,06±0,05
толщина Эр, мкм 1,40±0,06 1,60±0,01 1,77±0,05
индекс сферичности Эр 0,21±0,01 •83,90+0,70 0,24±0,01 85,08+0,38 0,25±0,01 93,93+1,23
площадь поверхности Эр, mkmz 13,01+0,16 20,56+0,12 23,32+0,49
содержание НЬ, пг концентрация НЬ, % 26,44±1,04 35,29±0,98 33,7О±О,46
типа осуществляется не ранее нескольких месяцев (Микле, Меркурьева, 1963 г., по: Иржак и др., 1971), то у лося этот процесс осуществляется значительно быстрее. Это объясняется, как указывают названные авторы, очень быстрыми темпами роста лосей после рождения (Кнорре и др., 1956).
Изменения особенности обмена и концентрации магния, кобальта и меди в крови лосей, обитающих в Печоро-Илычском заповеднике, в зависимости от возраста и сезонных условий показаны в работе Н.Е.Кочанова и Т.В.Симаковой (1976).
Индексы внутренних органов. В условиях Северного Кавказа, где лось обитает уже около трех десятилетий, морфофизиологические показатели его остаются неизученными. Этот пробел может быть несколько восполнен
216
Alces alces Linnaeus, 1758
данными по южным районам Восточной Европы, особенно из тех, откуда лось проник в Предкавказье. Такие данные привлекательны не только тем, что эти лоси близки генетически, но и тем, что они существуют в довольно сходных ландшафтных условиях, и со временем станет возможным рассмотрение морфофизиологии лося в спектре широтной зональности восточной Европы и Северного Кавказа.
В связи с изложенным наш выбор остановился на данных А.П.Варнакова (1980), проводившего изучение морфологических параметров лося (л =131 особь) из двух популяций: южной и северной. Фактический материал позволил автору провести сравнительный анализ в четырех возрастных ipynnax: сеголетки, 2,5 года, 3,5 года и 6,5—7,5 лет. В различных половых и возрастных ipynnax показатели внутренних органов лосей южной популяции заметно ниже, чем у представителей северной популяции. При этом максимальная относительная масса органов характерна для телят обеих популяций, что является широко распространенной возрастной особенностью для млекопитающих. Однако относительная масса печени телят-самок южной популяции в 1,5 раза ниже, чем северной. С возрастом это различие несколько уменьшается. Индекс сердца довольно стабильный — во всех возрастных ipynnax он примерно одинаковый. Что касается тенденции возрастной динамики индексов печени, почек, легких, то она сходна в обеих популяциях и характеризуется их снижением к 2,5 годам, а затем постепенно увеличением к 6,5—7,5 годам. Популяционное различие выражено в индексе сердца — у средневозрастных самцов южной популяции он выше, чем у животных северной. Кроме того, индекс легких у самцов с возрастом снижается, но у животных северных районов ареала это снижение несколько интенсивнее, чем в южных районах.
Кариотип лося, по материалам из Скандинавии, включает 68 хромосом. В аутосомах лишь одна двуплечая хромосома, тогда как все остальные акроцентрические. Х-хромосома субметацентрическая, У-хромосома акроцентрическая. Американские лоси характеризуются диплоидным числом 70, что объясняется увеличением аутосом за счет одной пары акроцентриков (Орлов , Булатова, 1983).
Структура и динамика ареала
Ареал вида очень обширный, занимает леса умеренного пояса Евразии и Северной Америки; он соответствует ареалу рода Alces.
Распространение на территории б. СССР также обширное, большая часть видового ареала находится здесь, восстановленный ареал занимает почти всю страну, кроме полупустынь, пустынь и гор Средней Азии (Гептнер и др., 1961).
Динамическое состояние ареала лося в историческое время — уникальный "природный эксперимент", не сводимый лишь к одному антропогенному фактору. Как справедливо отмечено В.Г.Гептнером, проблема увеличения и сокращения ареала, пульсация его границ, формы расселения и пути освоения новых ландшафтных условий четко проявляются на истории
217
Лось
ареала лося. Научных работ по этой проблеме много, достаточно ознакомиться с материалами трех всесоюзных совещаний по копытным, проведенных в Москве (1975, 1979, 1989 гг.), чтобы представить, насколько обширен поток информации по данному виду. Не вдаваясь в обобщения новых сведений о распространении и состоянии популяций, отметим, что лось как вид процветает в масштабе всей страны. Возможности по расширению ареала и повышению численности лосей, особенно на южных перифериях, неисчерпаемы.
Распространение на Кавказе. Местные популяции лося на Кавказе, судя по всем имеющимся данным, просуществовали до начала XIX в. Обзор этого материала приводится в различных сводках (Верещагин, 1959; Гептнер и др., 1961; и др.). В соответствии с ними восстановленный ареал лося на Кавказе охватывал ряд районов Северного Кавказа и Западного Закавказья (рис. 32, врезка). Видимо, ареал широкой полосой простирался от низовьев Дона по всей Прикубанской низменности, переходя дальше в предгорья и среднегорья Западного Кавказа. Есть все основания полагать, что лось занимал Ставропольскую возвышенность, предгорно-среднегорные пояса Центрального Кавказа вплоть до Андийского хребта, а также прилегающие равнины с лесостепными ландшафтами. Более того, лоси, видимо, проникали по долине Терека до Каспийского побережья и по зарослям тростников Западного Прикаспия до низовьев Волги. О возможном существовании территориальных связей кавказских лосей в прошлом с сородичами Восточной Европы через побережья Черного и Азовского морей на западе и побережье Каспийского моря на востоке указывают современные пути митрации лосей из Русской равнины в Предкавказье. Такому суждению соответствуют не только исторические данные по лосю, но и былое распространение лесной и кустарниковой растительности в Предкавказье.
Восстановленный ареал лося в Западном Закавказье простирался по всему Черноморскому побережью, охватывая, вероятно, и соответствующие ландшафты турецкого побережья. Насколько были широки высотные пределы распространения в колхидском и аджарском вариантах поясности, трудно судить. Что касается низменности, то палеозоологические данные указывают на обитание лосей по всей ее территории вплоть до окрестностей г. Кутаиси. Видимо, они встречались в предгорьях и среднегорьях, опоясывающих всю Колхидскую низменность.
Описанный ареал относится к периоду XV—XVII вв. и началу XVIII в. Интенсивное истребление лося на Кавказе происходило в XVII и XVHI вв., однако, судя по данным Н.И.Бурчака-Абрамовича и Л.А.Ширвашидзе (1967), лоси встречались до начала XX в. в ряде мест Западного Кавказа (район г. Гатра, р. Афипс). Когда был уничтожен последний аборигенный лось Кавказа, трудно сказать.
Вторичное заселение Кавказа лосем происходило, вероятно, в самом начале второй половины XX в. Местные жители Западного Кавказа сообщали о том, что в лесах Черноморского побережья Краснодарского края (севернее г. Сочи) лоси попадались еще в 1950-е годы, а выше в горах — в 1960-е годы (Бурчак-Абрамович, Ширвашидзе, 1967). В те же годы зарегистриро
218
Alces alces Linnaeus, 1758
вано стремительное продвижение лося на юг. В 1982—1983 гг. он прочно вошел в состав фауны лесных участков между Доном и Северским Донцом. Одна взрослая самка, по данным Б.Корякова (1965), забрела в район оз. Ма-ныч-Гудило в 1960 г. В 1962 г. лось был обнаружен в горных лесах Курга-нинского района Краснодарского края.
К этому времени относится сообщение охоткорреспондента о встрече двух самок и одного самца в лесах Горяче-Ключевского района. Годом позже лоси зарегистрированы в ряде точек Западного Предкавказья (окрестности с. Белая Глина, ст. Брюховецкая, г. Славянск).
Освоение лосями новых районов Северного Кавказа происходило не только с низовьев р. Дона, но и на всем протяжении южной границы ареала вида от Азовского до Каспийского морей. По данным Ю.П.Язана (1966), лоси заходят в Краснодарский край из Ростовской области и Ставропольского края, в Ставрополье — из Ростовской области и Калмыкии, в Калмыкию — из Ростовской, Астраханской и Волгоградской областей, в Астраханскую область — в основном из Волгоградской области. Эта "генеалогия кавказской популяции" представляет интерес и в плане экологии, в частности для оценки степени пригодности нынешних условий жизни лося на Кавказе.
Наиболее интенсивно продвижение лосей на юг, по всем имеющимся данным, происходило вдоль восточного побережья Азовского и Черного морей, где нередки камышовые заросли, древесно-кустарниковая растительность, в том числе лесополосы, необрабатываемые земли. Заходы лосей в Приазовские степи становятся обычным явлением еще в 1968 г., а летом 1971 г. их следы зарегистрированы в плавнях лимана Кущеватого и в лесополосе того же района. В следующем году здесь были отмечены 3 лося (Бакеев, 1980). По свидетельству того же автора, в конце 1975 г. недалеко от р. Челбас лосей было свыше 10 особей, в 1976 г. в Челбасском лесничестве наблюдались 6 особей. В 1977 г. их здесь не стало. Отдавая явное предпочтение побережью моря, с 1972 по 1977 г. лоси появлялись на берегу моря почти ежегодно (близ хутора Садки Приморско-Ахтарского района). Аналогичное явление регистрировалось в ряде других точек (ст. Ольгинская, хутор Гречная Балка, ст. Бриньковская на берегу Бейсугского лимана и ДР-)-
На Черноморское побережье лоси проникли, очевидно, из Крымского и Анапского районов. Об этом свидетельствует заход лосей в Крымский район еще в 1965 г. (Язан, 1966), а через 8 лет — в окрестности поселков Горное и Греческое., в 18 км от Новороссийска (Бакеев, 1980). Освоение степных пространств Краснодарского края, судя по имеющимся данным, начиналось также из поселений на Азовском побережье. Миграционными путями служили долины степных речек, впадающих в Азовское море. По А.М.Гинееву (1975), расселяясь по р. Ея вверх, лоси заселили Новопокровский и Новокубанский районы. По его данным, лоси зарегистрированы в окрестностях станиц и городов: Староминская, Каневская, Тимашевск, Красноармейская, Кореновск, Усть-Лабинск, Красногвардейская, Белореченск, Горячий Ключ и далее Апшеронск, Курганинск, Лабинск. Наиболее
219
Лось
крупные группы лосей (5—14 особей) отмечались в 70-х годах в шести районах. К этому времени относится их интенсивное продвижение по всему левобережью Кубани. В нижнем течении р. Пшиш одиночного лося видели еще в 1965 г. В начале 1976 г. около 6 лосей жили в Кубанском, Тверском и Апшеронском лесничествах.
По данным Ю.Н.Бакеева (1980), горные леса Западного Кавказа лоси заселяли крайне медленно. В окрестностях ст. Рязанская Белореченского района лосей встречали с 1972 г., в Даховском заказнике (верхнее течение Белой) в 1977 г. обитали два лося. Сообщалось также, что в Белореченском районе основное поголовье лосей сосредоточено в Кубанском, Комсомольском и Фадеевском лесничествах. Менее полные сведения имеются по восточным районам Закубанья. В литературе имеются сведения о встрече трех лосей в окрестностях станицы Ахметовской по Большой Лабе в 1971 г. (Гинеев, 1973) и близ района пос. Псебай (на горе Куцая) в 1976 г. Здесь в 1977 г. был убит самец, а лосенок позднее погиб во время миграции по Малой Лабе, в окрестностях пос. Перевалка (Бакеев, 1980).
В Ставропольском крае впервые лось отмечен в 1965 г. под самым Ставрополем. Весной того же года на левом берегу Маныча, близ с. Дивное, был обнаружен труп двухлетнего лося. Возможно, что эти две точки указывают миграционные пути лося на восток по Кума-Манычской депрессии и на юг по р. Егорлык, о чем свидетельствует, по Ю.Н.Бакееву (1980), появление лосей на одних и тех же местах, составляющих единый маршрут. Видимо, первый путь реализован раньше, чем второй. Севернее Ставрополя, где расположены Изобильненский, Красногвардейский и Ново-Александровский районы, лоси стали регистрироваться с 1968 г. Тогда хорошо упитанный зверь был встречен в степи между селами Московское и Донское (пойма Ташлы, правый приток Егорлыка). В 1971 г. лося видели возле оз. Сенгелеевского, в окрестности Ставрополя. Особенно заметными стали передвижения лосей в период с 1967 по 1972 г. (Перовский, 1976), появление их в Кочубеевском районе (Надзорненское хозяйство, что на правом берегу Кубани) отмечено в 1976 г. Упомянутый выше автор полагает, что с кочевками лосей по системе Маныч-Егорлык связано заселение приречных лесов Кубани к югу от Ставрополя и освоение ими горных районов Карачаево-Черкесии. Сначала были заселены леса Невинномысского района, потом Зеленчукский район, где в 1975 г. встречен трехлетний самец в окрестностях пос. Уруп. В последующем звери могли попасть в Кавказский и Тебердинский заповедники.
Заселение Ставропольской возвышенности происходило и с востока по р. Калаус: появившись в районе с. Дивного в 1965 г., лосиха была зарегистрирована в Ипатовском районе в 1972 г., а затем близ с. Октябрьского (левый берег Калауса) и в Петровском районе, в 50 км от Ставрополя. Известна зимовка лося в сезон 1975/76 г. в сосняке с. Бешпагир Грачевского района. Поднимаясь вверх по Калаусу, лоси могли заселить водоразделы Калауса и Кумы. В 1975 г. их видели близ с. Гофицкое, на р. Буйвол, а годом раньше трех зверей в окрестностях Круглолесского Александровского района. В 1972 г. в лесах Кумы животные почти одновременно появились в
220
Alces alces Linnaeus, 1758
Георгиевском и Советском районах, где они стали размножаться (Перовский, 1976; Бакеев, 1980).
Вероятно, из лесов р. Кумы лоси проникали неоднократно в. лесостепное предгорье Горячеводска Ставропольского края и Зольский район Кабардино-Балкарии. Один зверь был убит браконьерами в окрестностях с. Залукокоаже в 1975 г. В настоящее время в Кабардино-Балкарии лосей нет. Что касается Ставропольского края, последние литературные данные (Оноприенко, Хохлова, 1988) свидетельствуют о том, что в ряде районов края лоси стали постоянными жителями и размножаются успешно. Наиболее крупное поселение имеется на землях плодосовхоза "Лесная дача" колхоза им. Ленина, где в 1987 г. было учтено 28 лосей. Относительно стабильна численность лосей в пойме р. Кумы в Советском и Георгиевском районах, где имеются приречные леса. Единичные встречи в 1986—1988 гг. были в Курсавском, Хабезском, Предгорном, Красногвардейском, Апана-сенковском, Турклинском и других районах.
Положение лосей в Восточном Предкавказье плохо изучено. Известно, что лоси по Манычу продвинулись далеко на восток, в 1972 г. встречены в окрестностях с. Арзгир, они появлялись также в бассейне Терека Северной Осетии в 1976 г. (Бакеев, 1980). Ряд исследователей (Язан, 1965; Гинеев, 1975) указывали на продвижение лосей по пойме Волги до низовья. Дальнейший путь по западному побережью Каспия, видимо, осложняется обширными полупустынными и степными просторами, слабым развитием лесонасаждений и т.д.
Современные тенденции антропогенных изменений ареала. Данные, которыми мы располагаем, свидетельствуют о том, что на Северном Кавказе, как и во многих районах страны, сохраняется положительная тенденция увеличения численности и площади ареала лося. Есть основания относить их к труппе перспективных видов копытных Кавказа. При надлежащей охране лоси могут стать объектом спортивной и промысловой охоты в таких районах, как горы и равнины Краснодарского и Ставропольского краев, Кабардино-Балкарии, Северной Осетии, Чечено-Ингушетии, Грузии и ряда районов Азербайджана, Армении и Дагестана. Нам представляется, что общая тенденция антропогенных изменений ландшафтов, сопровождающаяся расширением лесопосадок на равнине и рубкой леса в горах и соответственно увеличением ландшафтов с лесостепным обликом, положительно скажется на условиях жизни лосей на Кавказе. Как известно, такие изменения зоны лесов Евразии способствовали увеличению численности и ареала лося (Гептнер и др., 1961).
Систематика и изменчивость
Систематическое положение. Систематика рода Alces дискутировалась долго. Противоречивыми были мнения как в отношении объема рода, так и количества внутривидовых форм. В старых работах, включая публикации начала текущего века, признавалось несколько видов лосей. К.К.Флеров (1934) все формы лосей свел в две группы, придав каждой ранг самостоя
221
Лось
тельных видов: 1) европейский лось Alces alces L. и 2) американский лось Alces americanus Clinton. Впоследствии (Флеров, 1952) он свел обе группы в один вид Alces alces L. В последние десятилетия мнение о монотипичности рода стало общепризнанным (Бобринский и др., 1944, 1965; Соколов, 1959; Гептнер и др., 1961; Соколов, 1979, 1984; Corbet, 1978; Corbet, Sill, 1980; Но-nacki et al., 1982). Значительная работа проведена также по упорядочению внутривидовой систематики. В результате количество подвидов сократилось в 3—4 раза, однако реальность существования некоторых из них вызывает до сих пор разногласия. Так, К.К.Флеров (1952) признает 3 подвида: обыкновенный А.а. alces, американский А.а. americanus и кавказский А.а. саи-casicus. И.И.Соколов (1959) выделяет, кроме первых двух подвидов, еще уссурийского лося А.а. bedfordi. По В.Г.Гептнеру и др. (1961), на территории б. СССР реально существуют 4 подвида: европейский А.а. alces (син. обыкновенный); восточносибирский, или якутский, А.а. pfizenmajeri; кавказский А.а. caucasicus и уссурийский А.а. cameloides. Кроме этих подвидов, он допускает существование 2—3 подвидов, в том числе двух американских: А.а. gigas и А.а. americanus. Существует и противоположное мнение, в частности Корбет (Corbet, 1978) сводит все внутривидовые формы лося Евразии и Америки лишь к 3 подвидам: А.а. alces, А.а. americanus и А.а. caucasicus.
Изложенное выше свидетельствует о том, что внутривидовая изменчивость лося нуждается в дополнительных исследованиях, ибо существующая информация не отражает еще всего спектра изменчивости. В этом плане, на наш взгляд, определенный интерес представляют адаптивные изменения лося в связи с освоением новых ландшафтных условий.
Изменчивость. Данные по изменчивости лосей, как аборигенных, так и молодых вселенцев Кавказа, малочисленны. Как известно, Н.К.Верещагин (1948, 1949, 1959) располагал 8 черепами кавказского лося, обнаруженными им в осетинских святилищах по р. Урух (левый приток Терека). Время накопления этого материала, по мнению автора, относится к XV—XIX столетиям. Ландшафтные условия расположения пещер и в соответствии с этим, очевидно, местообитания вымерших лосей относятся к поясу широколиственных лесов Центрального Кавказа (терский вариант). Иными словами, мы имеем дело, вероятно, с лосями, обитавшими в среднегорных условиях. Автор располагал также одним экземпляром лося, череп которого экспонировался в Ставропольском музее. Происхождение его неизвестно.
По современным лосям, обитающим в наши дни на Северном Кавказе, у нас нет фактического материала. Кроме отдельных фрагментов, отсутствуют также литературные данные.
Несмотря на малую выборку и дефекты черепов, данные, приведенные в табл. 42, позволяют рассмотреть некоторые аспекты изменчивости кавказского лося Аа. caucasicus.
Индивидуальную и возрастную изменчивость мы лишены возможности рассмотреть из-за отсутствия данных и фактического материала.
Половые различия, судя по табл. 42, прослеживаются четко по всем параметрам. Как следует из этих данных, по двум признакам из 14 провести
222
Alces alces Linnaeus, 1758
сравнения нельзя (общая длина черепа и N-Prostion). По остальным 12 промерам самцы заметно превосходят самок. К ним относятся длина носовых костей, длина заглазничной части, височная ширина, скуловая ширина, наибольшая ширина носовых костей сзади и спереди. По остальным параметрам (наибольшая ширина лба, наименьшая межорбитальная ширина, ширина верхнечелюстная в области М1 и др.), наоборот, самки превосходят самцов. Говорить о степени достоверности различия не приходится из-за недостатка материала.
Географическая изменчивость. В пределах Северного Кавказа оценить гео-1рафическую изменчивость кавказского лося нет возможности, хотя единственный экземпляр лося, сохранившийся в Ставропольском музее, стоит особняком по сравнению с коллекцией из Северной Осетии. Как указывал Н.К.Верещагин (1949), рога этого экспоната, по возрасту старого, невелики, но основания их очень толстые, выше венчика имеются своеобразные зачатки отростков, направленных вниз. Подобных отростков он не обнаружил у 136 пар рогов современных лосей Евразии и Америки.
В табл. 42 представлены данные по географической изменчивости и особенности структуры черепа кавказского лося. Как видно, кавказские лоси по сравнению с лосями Ленинтрадской области меньше по ряду параметров. Это хорошо прослеживается как в ipynne самок, так и самцов. Так, у самцов на Кавказе общая длина черепа в среднем 538 мм, а в Ленинградской области — 555 мм. Словом, череп A. alces caucasicus, по Н.К.Верещагину (1959), обладал ярко выраженными европейскими признаками и принадлежал сравнительно мелким зверям. Однако он обладал рядом особенностей, в том числе был относительно широк, особенно в предглазничной и лобной частях, но в области носовой части сужен сильнее, чем у лосей севера.
Характер изменчивости современной популяции лосей Северного Кавказа, сформировавшейся в последние десятилетия из выходцев Южной Европы, остается неизученным. В условиях сильной подвижности местных лосей постоянный приток извне, вероятно, осложняет формирование определенного фенотипа, характерного для Северокавказского региона. Возможности такой изменчивости нельзя исключать, если учесть высокую пластичность вида. К тому же имеющиеся отдельные полевые наблюдения соответствуют такому суждению. Так, среди учтенных лосей Северного Кавказа (примерно 90—100 особей) встречались животные с лопатовидными и оленевидными рогами в соотношении примерно 9:1 (Гинеев, 1975). Как известно, степень развития рогов и количественное соотношение разных типов рогов меняется не только внутри популяции, но и географически (Флеров, 1952; Соколов, 1959; Гептнер и др., 1961; Раков, 1966; Приедитис, 1979; и др.).
С учетом истории формирования ареала лося в прошлом и в настоящее время и морфологических особенностей вымерших и ныне живущих животных мы считаем необходимым выделять на Кавказе два подвида: 1) кавказский лось A. alces caucasicus и 2) европейский лось A. alces alces.
223
Лось
Подвиды
1. А.а. caucasicus Verestschagin, 1955. Тип из ущелья р. Урух, левый приток Терека, пояс горных лесов Центрального Кавказа. Мелкая форма. Длина черепа самцов в среднем 538 мм, длина зубного ряда в среднем у самцов 137 мм, самок — 138 мм. Верхние отростки межчелюстных костей на концах расширены, изредка доходят до носовых костей. В ростральной области лицевая часть сужена сильнее, чем у европейского лося. Лопата рогов развита весьма слабо. Неизвестными остаются размеры, окраска тела не описана.
Распространение — горные леса Северного Кавказа и Западного Закавказья, прилегающие к ним предгорья и лесистые ландшафты низменности и долин рек (Сулак, Терек, Кубань, Риони и др.) вплоть до их впадения в моря.
2. A. alces alces Linnaeus, 1758. Тип из Швеции. Средних и крупных (но не самых крупных) размеров, с хорошо развитыми рогами лопатовидного типа.
Длина черепа самцов 550—580 мм. Небные отростки межчелюстных костей впереди сошника узкие, образуют углубление желобообразной формы. Верхние отростки межчелюстных костей не сужены кверху, а лопатообразно расширяются, заканчиваясь тупо (Флеров, 1952; Гептнер и др., 1961).
Распространение на Кавказе — Предкавказье, горные леса Мелового и отчасти Скалистого хребтов. Продолжается расширение ареала.
Экология
Ландшафтная приуроченность. Биотопы. Можно считать общепризнанным, что лось является типичным представителем лесной зоны Евразии и Северной Америки. Выход за ее пределы и освоение смежных широтных зон как на севере, так и на юге осуществляется по соответствующим лесным ландшафтам. Лось обладает рядом адаптивных особенностей, обеспечивающих, как отмечает К.К.Флеров (1952), возможность передвигаться по местам, заваленным буреломом, по кочкарникам с высоким травостоем, переходить вброд достаточно глубокие водоемы, ходить по рыхлому и глубокому снегу, мало доступному основному врагу лосей — волку. Не менее важны приспособления к добыванию, перетиранию и перевариванию древесно-веточного корма, различной мезофильной, околоводной и болотной растительности.
Что касается характера биотопического размещения, то оно охватывает широкий круг разнообразных экосистем. Набор их существенно меняется в зависимости от сезона года, состояния кормовой базы, защитных условий и Т.д.
В оптимуме ареала, каковым является Ленинградская область, большое значение в жизни лося, по данным Е.К.Тимофеевой (1974), играют зарастающие вырубки. В этой группе биотопов лоси держатся почти круглый год. По возрасту, характеру зарастания и составу пород они настолько разнообразны, что обеспечивают лосей излюбленными кормами и защитой. В
224
Alecs alecs Linnaeus, 1758
снежный период роль вырубок несколько снижается. Зимой лоси выбирают кормные местообитания с благоприятными микроклиматическими условиями, защищенные от хищников. Таковы густые подросты старше 10 лет, разновозрастные сосняки, особенно в многоснежные зимы, спелые ельники и т.Д. Анализ материалов тропления (3,5 тыс. суточного хода), полученных в 1966—1980 гг. на значительной территории Восточной Европы (Червонный, 1989), показал, что в большинстве округов независимо от их географического положения ведущую роль в жизни лося выполняют вырубки, молодые сосняки и ивняково-пойменно-болотный комплекс. На их долю приходится около половины всех угодий, населяемых лосем зимой. В условиях Литвы, где исследования проводились в 1976—1978 гг. в 9 лесах-стационарах, зимой лоси концентрируются в чистых сосняках и постоянно посещают сосняки I—II класса возраста (степень предпочитаемо ста 1,5—5), низинные болота (1,8), ельники I класса возраста (1,7), места рубки лиственных (1,7) и сосны (1,6), верховые болота (1,4). По значимости первое место принадлежит чистым соснякам II класса возраста (34,7%), за ним следуют такие же сосняки, но IV—XII классов возраста (12,6%), сосняки с примесью лиственных Ш—XII классов возраста (11,4%) и верховые болота (Белейшие и др., 1980). В бассейне среднего течения Оби (Белов, Юдин, 1980) наиболее высокая численность лосей зимой отмечена в долине реки с елово-пихтово-кедровыми, сосновыми, осиново-березовыми лесами и сосново-сфагновыми болотами. Второе место занимают заболоченные поверхности междуречий с растительностью из кедровых и березово-осиново-сосновых лесов и осоково-сфагновыми и грядово-мочажными болотами. Почти все учтенные лоси находились в местах с разреженным древостоем или на открытых заболоченных землях.
Весной лоси предпочитают сухие холмистые вырубки с возобновляющейся березой. Здесь раньше, чем где-либо, начинает вегетацию травянистая и древесно-кустарниковая растительность. В этих условиях лоси могут задержаться очень долго. Летом заселяют почти все типы вырубок, в том числе двух-трехлетние заросли осины, березы, иван-чая и др. По значимости для лося в бесснежный период второе место занимают открытые моховые и травянистые болота заболоченные кустарниковые заросли среди леса, сырые луга, чередующиеся с обширными зарослями кустарников. Считается, что привязанность лося к сырым и топким местам, к берегам водоемов является характерной чертой его экологии, что накладывает отпечаток на биотопическое размещение лося в бесснежный период. К благоприятным местообитаниям относятся смешанные и лиственные леса, где лоси держатся почти круглый год. В пору, когда глубокий снег и частые насты затрудняют передвижение лосей на открытых биотопах (.лесные луга, вырубки и др.), они переходят в лиственные леса.
В зимних биотопах лося, по В.В.Червонному (1989), в условиях южной тайги на долю лиственных пород с преобладанием осины приходится половина их. Роль названных пород еще больше возрастает в более южных широтных зонах. В экологических нишах лося в широколиственных лесах доля лиственных пород по отношению к хвойным возрастает почти в
15 В.Е.Соколов, А.К.Темботов
225
Лось
2 раза, а в лесостепной и степной зонах — в 3—4 раза. В жизни лося в последних двух зонах в 2—4 раза возрастает значение облесенных пойм. Упомянутый автор считает, что внутренняя неоднородность экологической ниши лося, как правило, определяется обилием используемых видом ресурсов, хотя к ряду пород, в том числе сосне, осине и др., по данным Е.К.Ти-мофеевой, разным ивам, а в особенности к определенным ландшафтам лось проявляет избирательность.
Посещение лосями сельскохозяйственных угодий (поля зерновых, овощных и других культур, огороды и др.) в Ленинградской области имеет место, но, видимо, носит случайный характер (Тимофеева, 1974). По данным этого же автора, глухих ельников с плохо развитым подлеском лоси обычно избегают. Как отмечено в ряде других литературных источников, лось избегает также скалистые, сильно пересеченные горные местности, отдавая предпочтение пологому, относительно спокойному рельефу.
На Северном Кавказе ландшафтное и биотопическое размещение лося отражает высокую пластичность вида. По Ю.Н.Бакееву (1980), обобщившему литературные и собственные данные по расселению лося в регионе, лоси чаще всего отмечаются по речным долинам, в том числе степных рек, таких, как Ея, Челбас, Егор лык, Калаус и др. Нередки случаи нахождения их на берегу лиманов, на различных участках побережья Азовского моря с тростниками, в плавнях рек, по берегам оз. Маныч и др. Лоси охотно поселяются в лесонасаждениях, в том числе лесополосах. В Ставропольском крае (Центральное Предкавказье) первые лоси отмечены весной 1973 г. в пойменном лесу Кубани и, судя по данным Л.Г.Оноприенко и З.Х.Хохловой (1988), приречные леса служат местообитаниями лосей постоянно и круглогодично. Такое же значение имеют лесные массивы на водоразделах (Томуз-ловский лес Александровского района и др.), но более благоприятными для обитания этого вида в пределах края авторы считают горные лесные массивы. Интерес представляет сообщение названных авторов о том, что самый крупный очаг пребывания лосей расположен на землях плодосовхоза "Лесная дача" и колхоза им. Ленина (на границе Труновского и Ипатовского районов), где они поселились в лесополосах и фруктовых садах. В отличие от Ленинградской области заселение лосями сельхозугодий на Северном Кавказе, очевидно, явление не случайное, более того, лесопольный облик благодаря хорошо развитым лесополосам и садам здесь привлекает лосей. Лосей отмечали на посевах колосовых, кукурузы и др.
Сведений о сезонной смене местообитаний, как и о различиях в территориальном размещении разных полов и возрастных групп, по Северокавказскому региону нет. Между тем аналогичных данных по другим районам, частично отмеченных выше, немало в литературе. Как характерная черта экологии вида отмечается, что в бесснежный период лоси чаще рассеяны, а зимой в связи с сокращением площади благоприятных условий, чаще всего из-за глубокого снегового покрова, они концентрируются на отдельных участках, образуя стада. На юге, в том числе на Северном Кавказе, этот фактор, видимо, играет второстепенную роль, ибо здесь снеговой покров выше
226
Alces alces Linnaeus, 1758
50 см — явление редкое и непродолжительное как на равнине и в предгорьях, так и в среднегорьях всего региона.
Миграции. Характерной чертой экологии лосей считается их способность совершать территориальные перемещения, вызванные сезонными изменениями условий жизни, в первую очередь снежным покровом. Однако способность лосей питаться надснежными частями древесно-кустарниковой растительности и древесными лишайниками, а также возможность преодолевания глубокого снежного покрова обеспечивают оседлую жизнь на значительной территории ареала (большая часть центральных районов европейской части страны и Восточной Сибири). Обобщение многочисленных литературных и собственых наблюдений позволило А.А.Насимовичу (1955) выявить условия, при которых лоси совершают зимние миграции. Так, лоси в большинстве случаев зимой остаются там, где держались летом, совершая только небольшие перемещения из одних стаций в другие в тех областях, где максимальная высота снежнего покрова в среднем не превышает 50 см. К таким местным кочевкам можно отнести перемещения не только на равнине, но и в горах, где резко сказывается климатический фактор. В частности, в Жигулевском заповеднике массовые миграции лосей с равнинного плато южной части территории в горные районы наблюдаются при снежном покрове толщиной более 40 см (Белянин, 1980).
В районах, где максимальная толщина снегового покрова составляет в среднем 70 см и выше и продолжителен многоснежный период, лоси концентрируются на сравнительно небольшой (по сравнению с летом) территории с меньшим слоем снега или совершают миграции в другие районы.
По мнению А.А.Насимовича (1955), наиболее значительны миграции при двух условиях:
1) в глубокоснежных областях севера, где лоси обитают на равнине, откуда мигрируют на зиму в менее снежные районы, преодолевая иногда до 300 км (например, верховья Печоры, низовья Оби и др.);
2) в отдельных горных областях с высоким снежным покровом, где отдельные хребты выполняют роль климатораздела, что позволяет лосям откочевать в районы снежной тени, преодолевая до 100—150 км (Урал, Баргу-зинский хребет).
Переход в места зимовки обычно растянут до двух и более месяцев (в разных местах с октября, даже сентября или ноября до декабря или даже января). Начало массовых перемещений совпадает с первыми большими снегопадами, когда в летних местообитаниях снег достигает толщины 25— 45 см. Нередко небольшая часть популяции остается зимовать в летних местообитаниях или покидает их лишь при достижении толщины снегового покрова 60—70 см.
Весенняя перекочевка в места летнего обитания проходит в сжатые сроки и совпадает с периодом интенсивного таяния снега.
Изложенные выше особенности экологии лося позволяют составить прогноз о возможном размахе высотного размещения лося на Кавказе, о том, что в площадь потенциального ареала этого вида можно включать значительную часть гор региона.
15*
227
Лось
Убежища, лежки. На различные экстремальные факторы лось реагирует не только этологически, но также путем изменения местопребывания, используя различные биотопические условия. Так, в период массового лета слепней большую часть дня они лежат на окраинах болот, по берегам различных водоемов, на различных сырых и хорошо продуваемых местах (Тимофеева, 1974). Когда дуют зимой сильные ветры, лоси забираются в густые насаждения хвойных молодняков (Гептнер и др., 1961). От хищников и холодов спасаются в еловых лесах, где меньше ветра и рыхлый снег.
Лежки, как правило, устраиваются там, где лось находился на жировке. Лоси предпочитают более открытые и возвышенные участки. По наблюдениям в Ленинградской области (Тимофеева, 1974), основанным на анализе 799 лежек, лежка расположена так, чтобы обеспечивался хороший обзор. Особенно это справедливо зимой, но летом, вероятно, лось больше пользуется слухом, чем зрением. По данным упомянутого автора, в снежный период лежки, располагающиеся на холмах и других возвышенностях, составляли 31%, на 1ранице леса и открытого пространства — 10%, в понижениях и западинах — всего 3%. В бесснежный период размещение лежек по перечисленным местам составило соответственно 9, 2 и 1%.
Взрослые лоси в группе ложатся вместе, но на расстоянии не ближе 2— 20 м друг от друга. Головы располагают независимо от направления ветра. Лишь вспугнутые или потревоженные звери ложатся под ветер, к источнику запаха потенциального врага. Маленькие лоси ложатся близко к матери, даже прижавшись к ней. В зависимости от погодных условий, мощности снегового покрова, а также пола и возраста животного количество лежек в сутки может достигать 15 (Тимофеева, 1974). Продолжительность нахождения в лежке меняется. Ночью зверь находится в лежке 75—80% времени, а днем лишь 35—40%. При сильном морозе лось, погрузившись в глубокий снег до головы, может лежать несколько часов (Кнорре, 1953).
Сезонная и суточная активность. Поведение (рис. 33). Сезонный аспект активности хорошо выражен у лося. Он четко отражает реакцию на динамику ряда абиотических и биотических факторов среды. С этими изменениями сопряжена и суточная активность. В целом лось характеризуется как малоподвижное животное, особенно это проявляется в суровые многоснежные зимы. При морозе 40—50° лоси лежат в снегу многими часами, снижая тем самым расход тепла. Зимой лось резко сокращает протяженность суточного хода, который составляет всего 0,9—3,5 км, в среднем 1,3 км. Сходные данные приводят многие авторы (см. обзор: Тимофеева, 1974). Соответственно сокращается площадь, занимаемая животными. Лосиха с лосенком в течение двух суток держалась на участке всего 1,3 га. В районе Новосибирского Академгородка табунок лосей держался на площади 0,7— 0,8 км2 в течение 3,5 мес. Температура опускалась днем лишь до -15°', а ночью до -30° (Телегин и др., 1980). На офаниченной площади лоси могут держаться в конце зимы — начале лета, когда на открытых участках образуется наст, затрудняющий передвижения животного.
228
Alecs alces Linnaeus, 1758
Рис. 33. Лоси
229
Лось
Суточная активность зимой, как правило, имеет два пика — с рассвета до 10 ч и вечером с 15—16 ч, однако при сильных морозах она может сократиться до 1—2 ч, но при кочевке лоси бодрствуют до 14 ч (Язан, 1961).
В теплый бесснежный период, как свидетельствуют данные многих авторов, суточная активность лосей полифазная — жировка чередуется с отдыхом в течение целых суток, но, видимо, четко выражено повышение ее в ранние утренние и вечерние часы. Летом, когда температура значительно повышается, как и во время лёта кровососущих, резко сокращается дневная активность. На кормежку лоси выходят с наступлением сумерек и снижением температуры, а в лежку возвращаются не позже 6—7 ч утра (Гептнер и др., 1961).
Многие авторы обращают внимание на большие сдвиги в суточной активности, вызванные состоянием гона. Так, половозрелые самцы в это время деятельны круглые сутки, однако пик в зорях сохраняется. Особенно проявляются сдвиги в сокращении времени жировок и отдыха. На кормежку возбужденный бык теряет всего 2% активного времени (Тимофеева, 1974). Во время гона значительно повышается также подвижность половозрелых самок исвязянныхс ними сеголеток.
В условиях загона (65 га) бюджет сезонной активности лося изучен канадскими специалистами в 1982—1984 гг. (Renccker, Hudson, 1989). Установлено, что в годовом цикле время, затраченное на питание, составляет в среднем 10 ч в день, а на жвачку — зимой 12,6 ч, летом (в мае) 2,8 ч. Продолжительность жвачки зависела от содержания клетчатки в кормах. В зимнее, осеннее и весеннее время лоси, как правило, кормились на рассвете и вечерней заре. На это тратилось в среднем 107 мин. Выявлена полифаз-носуь цикла жировок и отдыха с 4—7 циклами в сутки.
Поведение лося многие исследователи характеризуют как меллите лыше.. Обычно он передвигается широкими шагами, обращается в беглищь вынужденно. Пасется обычно стоя, реже на ходу. В отдельных местообитаниях лоси могут стоять долго, объедая сочную травянистую растительность или привлекательный кустарник. Более активны на кормежке лосихи с лосятами. По отношению к людям лоси не проявляют атрессивности, хотя могут быть совершенно неожиданные выпады. Особенно опасны раненые лоси. От волков взрослые лоси защищаются активно, передними ногами могут убить хищника.
Брачное поведение довольно сложное, самцы становятся агрессивными, нередко теряют свойственную им осторожность, близко подпускают человека, но впоследствии могут нападать на него.|Наиболее показательной осо-(бенностью брачного поведения является изготовление "тонных ямок", появление которых связывают с началом гона. Образуются они в связи с тем, что самки перед течкой издают специфический запах, который возбуждает ' самца. В тех местах, где лосиха оставила мочу со специфическим запахом, бык копает землю и сам потом оставляет свою мочу. Запах мочи такого f возбужденного быка неприятный, сохраняется днями и чувствуется на рас-кстоянии до 20 м.'По Р.Ж.Байдавлетову (1988), поведение самцов в период гона включает ряд элементов: подход или подбегание к лосихе, назо-
230
Alces alecs Linnaeus, 1758
генитальный и назо-назальный контакты, тыкание в тело мордой, хождение друг за другом, как правило по кругу, бодание, уринацию флеммование и садку.
У лосей хорошо прослежено оборонительное поведение, которое включает соответствующую позу, испуг, поиск, уточнение источника раздражения, подачу голоса, затаивание, бегство.
В наиболее ответственные периоды жизни лося звуковыми сигналами являются: писки и стоны новорожденных лосят, звуки взрослой лосихи во время кормления лосят, а также в период гона, сигналы взрослых самцов, направленные на поддержание связи с самками-партнерами и самцами-соперниками. Общее число функционально-типологических вариантов звуковых реакций лося, вероятно, не превышает 10 (Богомолова и др., 1984).
Ограниченность места не позволяет рассматривать и другие аспекты поведения лося.
Линька. У лося в году бывает лишь одна смена волос, начало которой приурочено к весне и которая соответственно именуется весенней. Однако линька проходит так медленно, что охватывает даже лето. В ряде мест ареала вида, в том числе в Якутии (Егоров, 1965), первые признаки смены волос зарегистрированы в апреле. Наиболее интенсивно линька проходит, по В.Г.Гептнеру и др. (1961), в мае и июне, притом остатки изношенного зимнего шерстного покрова сохраняются еще в июле. В августе встречаются лоси с полным летним мехом, который отличается от зимнего короткой длиной, отсутствием Подшерстка, темной, почти черной окраской. К зиме шерсть отрастает, появляется подшерсток. Сроки линьки сильно варьируют не только географически, но и в зависимости от пола, возраста, состояния организма. Как правило, у взрослых самцов и яловых самок линька проходит интенсивнее, чем у беременных самок и молодых.
Линька обычно начинается на голове и ногах, потом переходит на бока, заканчиваясь на спине и задней части тела.
Питание. Имеется обширная литература, посвященная особенностям качественного и количественного состава кормов лосей в различных частях обширного ареала. К настоящему времени известно, что лоси потребляют в пищу представителей 149 родов покрытосеменных растений, из них травянистых ПО родов, 39 деревьев и кустарников. Они поедают также представителей 5—6 родов голосеменных (сосновые, можжевельник, тис и др.), более 5 родов папоротникообразных, 3—4 родов лишайников, 11 родов трибов, водорослей, иногда и мхи. Всего набирается не менее 175 родов растений, включающих не менее 250 видов (Данилов, 1958). По А.А.Калецкому (1968), по всей территории б. СССР в рационе лося представлено не менее 355 видов растений. Думается, что этим числом не ограничивается состав кормов лося, более того, оно значительно увеличивается за счет местных видов растений, поедаемых лосем в новых районах. В пользу такого суждения говорят многие литературные источники (см. обзор: Гептнер и др., 1961; Калецкий, 1968; Тимофеева, 1974; и др.). Как видно, качественный состав кормов лося весьма разнообразный и свидетельствует о высокой пластичности вида. Однако круг поедаемых видов растений в одном районе
231
Лось
невелик. Так, в Ленинградской области список кормов лося включает всего 30 видов деревьев и кустарников, 65 видов травянистых и кустарниковых растений, принадлежащих 79 родам и 37 семействам (Тимофеева, 1974), в Западном Алтае — 34 вида деревьев и кустарников, 68 видов травянистых, кустарниковых растений и несколько видов водорослей, мхов и грибов (Байдавлетов, 1988). В летнем питании лося в Волгоградской области определены всего 12 видов деревьев и кустарников, злаки, повойник, кукуруза (Варнаков, 1980). Нетрудно понять причину столь резкого сокращения набора кормов в условиях степи и лесостепи — это урбанизация ландшафтов и обеднение флоры и растительности.
При всем размахе географической изменчивости состава кормов лосей четко выражена предпочитаемость этих зверей к определенным группам растений/ В их питании особенно велика роль представителей семейств ивовых, кипрейных, розоцветных, березовых, сосновых, аронниковых, горечавковых (Тимофеева, 1974). Соотношение тех или иных кормов в разные сезоны меняется очень широко и довольно однотипно в разных районах страны. Весенний период характеризуется разнообразием рациона, определяемым значительным увеличением доли травянистой растительности в сочетании с древесно-кустарниковой. Из трав лоси охотно поедают злаки, осоку, ландыш, сныть, разнотравье, некоторые хвощи, всходы озимых и т.д. С появлением листвы на деревьях лоси потребляют в значительном количестве разные виды ив, березу, осину и др. Летом в связи с огрублением вегетативных частей и снижением кормовых достоинств постепенно сокращается доля травянистой растительности, из рациона выпадают ранневесенние травы. Увеличивается значение листвы деревьев и кустарников, а также пнрофильной и прибрежной растительности (иван-чай, болотные травы и др.). Осенью еще больше обедняется набор кормов, ибо теряют пищевые качества и болотные растения, однако в ряде районов (особенно в более южных) возобновляется осенняя вегетация травянистой растительности. Лесные лужайки и прибрежная растительность становятся привлекательными для лосей. Тем не менее резко возрастает доля мягко породных, особенно ивы, сохраняющихся зелеными дольше других. По данным Е.К.Тимофеевой (1974), листья и побеги этих кустарников в Ленинградской области составляют около 64% всех поедов. В это время отмечаются поеды безлистных побегов осины и коры лиственных пород, позднее к ним прибавляются побеги сосны, можжевельника и др. Роль их в зимнем рационе лосей значительна, ибо зимой питательность сосны и можжевельника не уступает таковой ивы. Однозначные данные по этому вопросу содержатся в ряде литературных источников (Корякин, 1961; Томмэ, 1964). Так, сосновая хвоя содержит протеина 5,5—7,7%, жира 5,3—12,2%, клетчатки 13,6—33,2%. Значение имеет также то, что хвойные вегетируют в течение года, сохраняют высокое содержание витаминов: в 1 кг хвои содержится до 50 мг каротина. Содержание питательных веществ в хвое можжевельника также высокое: 6,7—7,3% протеина, 6,2—9,0% жира, 29,0—33,7% клетчатки. В различных частях растения содержится до 8% дубильных веществ, значительное количество эфирных масел, особенно в плодах.
232
Alces alces Linnaeus, 1758
Сравнительный анализ питания лосей в разных частях ареала (Тимофеева, 1974) показал, что соотношение древесно-кустарниковых пород и травянистых растений в летней пище лосей существенно меняется в зависимости от конкретных ландшафтных условий. В Карелии, например, веточный корм составляет 60%, а травянистая растительность — 40%, в Ленинградской области соответственно 48,9 и 51,1%, в Печоро-Илычском заповеднике — 81,5 и 15,5%, в Дарвиновском — 79,6 и 20,4%.
В отношении суточного рациона лося существуют различные мнения, но в отечественной литературе большинство авторов считает, что взрослый лось в день съедает 30—40 кг сочных растений, двухгодовалые звери — около 15 кг, а сеголетки — 2,4 кг. Этот рацион к осени снижается у взрослых до 20 кг, у двухгодовалых до 12 кг, у сеголетков, наоборот, увеличивается до 7,7 кг. Зимой при переходе на веточный корм взрослые животные съедают в сутки в среднем около 13 кг, двухгодовалые — 11,2 кг, сеголетки — 7,7 кг (Кнорре, 1959; Калецкий, 1967; и др.).
Для анализа возможной кормовой базы лося в новых условиях Северного Кавказа, особенно Предкавказья, интерес представляют данные А.П.Варна-кова (1980) по питанию лося в степях и лесостепи Волгоградской области. На основании разбора содержимого желудков и определения остатков растений установлено, что основу питания лосей в названной области составляют различные ивы — 91,7% встреч. На втором месте стоит клен татарский (63,9%), а потом осина и дуб. Автор обращает внимание на то, что, помимо прочих видов, в пробах встречался и краснотал (до 53,3%), известный как не поедаемый лосем в других районах. Относительно часто встречались травы, в частности злаки (22,2%). Охотно поедаемый в других районах бересклет бородавчатый зарегистрирован лишь в 5,5% встреч. На долю сосны приходилось всего 8,3%, что объясняется отсутствием в районе исследования посадок сосны.
Основу питания телят составляют ивы (100%), а встречаемость вяза в 4 раза больше, чем у взрослых (15,4 и 4,4%). Яблоня и бересклет бородавчатый отмечаются почти в 1,8 раза больше у телят, чем у взрослых (соответственно 7,7 и 4,4%). У телят не обнаружены такие виды, как тополь, терн, шиповник, липа, которые составляют у взрослых второстепенные или случайные корма.
Минеральное питание. Лоси, как и многие другие копытные, посещают места, где отмечается повышенное содержание солей, но в разных частях ареала они пользуются ими в различной степени. Более того, обзор литературы по этому вопросу, проведенный в монографии В.Г.Гептнера и др. (1961), показывает, что на значительной территории ареала лоси вообще не пользуются естественными солонцами. Таковы европейская территория России, Западная Сибирь, Алтай, Саяны. Редко пользуются'Ими лоси в Иркутской области, а в Якутии, Приамурье и на Сихоте-Алине лоси активно посещают солонцы в течение бесснежного периода. Сходные данные имеются по Восточной Сибири и Приморскому краю. Установлено также, что в Западном Алтае лоси активно посещают природные солонцы, содержащие много растворимых веществ, в том числе такие элементы, как натрий,
233
Лось
кальций, магний (Байдавлетов, 1984, 1988). Посещение солонцов может начинаться ранней весной. Так, в 1983 г. самка с теленком и молодая лосиха выходили на солонец 8-12 апреля, когда там еще лежал снег 50—80 см толщиной. Активное солонцевание почти всего населения лосей Западного Алтая наблюдается со второй половины мая до конца июня. В сентябре-октябре, особенно в связи с выпадением снега, животные солонцуют очень редко.
Воспроизводство и структура популяции
Размножение. Литературных сведений по размножению вымершего кавказского лося нет. Отсутствуют такие данные и по современным европейским лосям, обитающим на Северном Кавказе. В связи с таким положением мы ниже приводим краткий обзор основных особенностей размножения вида, проецируя их на конкретные ландшафтные условия жизни популяции. Представляется, что такая информация будет полезна при изучении экологии лося в условиях кавказского региона.
"""Начало половой активности самцов легко установить по обломанным и ободранным деревцам и кустарникам, а также по своеобразным голосам — “стонам", издаваемым возбужденными быками. Аналогичные следы на деревцах появляются и в связи с чисткой рогов от омертвевшей шкуры. Для этого лось обтирает свои рога о ветвистые деревья и кустарники на высоте до 1,5—2,0 м от земли. Особенно показательно появление гонных ям, указывающее на то, что в охоту вступила также и лосиха. Все это происходит в местах, где они обычно обитают в конце лета — по окраинам болот, зарастающим вырубкам и гарям, на лугах и т.д. Сроки появления первых признаков начала гона неодинаковы в разных районах ареала. Так, в Ленинградской области поврежденные деревья и кустарники отмечаются в конце июля — начале августа, а гонные ямы — в конце августа — начале сентября (Тимофеева, 1974); на Западном Алтае соответственно в конце августа — начале сентября и'в первой декаде сентября (Байдавлетов, 1988). На течение гона оказывает влияние целый ряд факторов, в том числе погодные условия, состояние популяции и др. Гон продолжается 1,5—2 мес., но у большинства лосей заканчивается через 10—20 дней. Однако единицы стонущих и дерущихся быков встречали в Ленинградской области до ноября включительно. Имеющиеся наблюдения свидетельствуют о том, что самцы и самки различных возрастов включаются в размножение в разное время — первыми начинают спаривание взрослые животные, а потом лишь созревшие молодые.
Долгое время считали, что лоси относятся к типичным моногамам, ибо во время гона чаще видели одного самца с одной самкой. Реже встречались иные соотношения: самка с двумя, тремя и даже с шестью самцами или один самец с несколькими самками (см. обзор: Гептнер и др., 1961). Последующие исследования показали что лось является типичным полигамом: после покрытия самки, самец бросает ее и начинает преследовать другую
234
Alces alces Linnaeus, 1758
самку, вступившую в охоту позже. Самец не несет никакой заботы о потомстве.
Схватки между самцами за право овладения самкой не столь часты, как среди других видов оленей, но бывают жестокие, кончающиеся гибелью одного из дерущихся.
Продолжительность беременности лося в Печоро-Илычском заповеднике составляет 225—237 дней (Кнорре, 1953). По данным М.В.Кожухова (1989), во всех случаях датированного покрытия продолжительность беременности лосих была в пределах от 215 до 230 дней. Роды у лосих происходят в соответствии со сроками покрытия и продолжительностью беременности в разное время — в мае или начале июня. Рождение в более поздние сроки — явление, видимо, редкое.
Перед родами лосиха ведет одиночный образ жизни, она отгоняет от себя своих прошлогодних телят, которые вынуждены существовать самостоятельно. Непосредственно перед родами лосиха уединяется, и наблюдение в природе за родами практически невозможно (Altman, 1963, String-ham, 1974, по: Богомолову, Курочкин, 1984).
Плодовитость. Анализ обширного фактического материала, проведенного М.В.Кожуховым (1983), показал, что плодовитость лосих в значительной степени зависит от их возраста. Репродуктивная способность лосих сохраняется до глубокой старости — до 15—17 лет. При этом части лосих свойственно одноплодие, другим же с 1—2-го отела и до 14—15-летнего возраста — двуплодие. Факты многоплодия (3 и 4 эмбриона у одной лосихи) приводятся различными авторами и зарегистрированы в ряде районов видового ареала, но преимущественно в средней полосе и западных районах страны, отличающихся более мягкими и малоснежными зимами. Многоплодие крайне редко встречается в экстремальных условиях Средней и северной тайги. Исследования, проведенные в Дарвинском заповеднике (Филонов, Калецкая, 1984), показали, что такие параметры размножения, как число сеголеток на 100 самок с приплодом и доля самок от всех взрослых зверей, характеризуются повышенной стабильностью. Высокой изменчивостью отличаются такие сопряженные показатели Плодовитости, как двойни у самок и доля сеголеток в популяции, что свидетельствует об их высокой зависимости от условий жизни.
Нет сомнений в зависимости плодовитости лося от сложного сочетания ряда факторов, при этом конкретные взаимодействия их в разных популяциях или в одной и той же, но в разные годы меняются очень широко. Из абиотических факторов нам хотелось особо обратить внимание на состояние зимы, а из внутрипопуляционных — на численность популяции (корреляция положительна). Из этологических особенностей лосих важно учесть, что они легко принимают осиротевших лосят, проявляя равную заботу как о своих, так и о приемышах.
Рост и развитие. Многочисленные литературные данные свидетельствуют о значительной роли физических показателей новорожденных в постнатальной жизни лося. Более того, как показали новые данные М.В.Кожухова (1989), важным фактором выживания лосят является их масса тела при
235
Лось
рождении. По данным Е.П.Кнорре и Е.К.Кнорре (1956), в условиях Печо-ро-Илычского заповедника эта масса колеблется в пределах от 6 до 16 кг; Последующие исследования М.В. Кожухова (1989) свидетельствуют, что в природе того же заповедника отдельным лосихам свойственно крупноплодно. Лосята с массой до 19 кг неоднократно рождались у одной и той же лосихи. Среди двоен были такие, которые имели суммарную массу от 30 до 34 кг, а два бычка-близнеца — 16,5 и 19 кг.
Значительные колебания в массе тела новорожденных в пределах одной популяции, как свидетельствуют данные названных авторов, зависят, кроме индивидуальных особенностей родителей, также от условий зимовки, состояния здоровья лосихи, ее возраста, упитанности и др. При этом лосихи достигают физического и физиологического расцвета к 5—6-летнему возрасту и сохраняют его до 12—13 лет. В этом возрастном промежутке наблюдается максимальная масса тела и упитанность к периоду гона, наиболее высокая молочная продуктивность и соответственно рождение более полноценных и жизнеспособных лосят. Регулярность размножения, как считает М.В.Кожухов (1989), и высокий показатель плодовитости у лосей сохраняются до 15—17 лет. По Ю.П.Язану (J972), наибольшей плодовитостью обладают лосихи в возрасте 4—9 лет.
Рост и развитие лося наиболее полно изучены в условиях Печоро-Илычского заповедника Е.П.Кнорре и Е.К.Кнорре (1956). Согласно их наблюдениям, новорожденные лосята растут очень быстро, особенно в течение первого полугодия. Достаточно сказать, что за это время масса тела новорожденного увеличивается в 10 раз, т.е. лосята по достижении 6 мес, имеют массу тела до 160 кг и как исключение даже до 200 кг. Прямое и косвенное влияние на интенсивность роста молодняка оказывают не только метеорологические условия, но и кормовая база. Вся многолетняя практика, по мнению Е.П.Кнорре (1939), доказывает, что лучше растут и развиваются те лосята, которые раньше и в большем количестве поедают зеленую растительность.
С наступлением осени и по мере увядания зеленой растительности замедляется рост массы тела, а с наступлением зимы совсем прекращается. Она остается почти неизменной в течение зимне-весеннего периода, поэтому массы тела шестимесячного и годовалого лося почти одинаковы. Зимняя депрессия жизнедеятельности лося отражается не только на массе тела, но и на его размерах. При этом наиболее резко она проявляется на севере ареала, где сильнее выражены неблагоприятные зимние условия (высота, глубина снежного покрова, наличие наста и т.д.). В этих условиях количество поедаемого корма снижается по сравнению с летом в 4—5 раз, падают пульс и дыхание, а также температура тела. Е.П.Кнорре и др. (1956) считают, что зимнюю остановку роста массы и размеров тела нельзя сводить лишь к одному факту истощения организма, в ней проявляется и сезонная адаптация лося, позволяющая ему существовать в суровых условиях зимы. На второе лето жизни интенсивное увеличение массы тела продолжается, и при сочетании благоприятных факторов среды прирост массы тела за одно лето может достигать 150 кг и более. Соответственно к 1,5 годам жизни
236
Alecs alces Linnaeus, 1758
лось может иметь массу в 350 кг. К числу благоприятных условий для роста молодняка относится дождливое и прохладное лето. Наименее благоприятно жаркое и сухое лето. На третьем году жизни увеличение массы тела замедляется и ее абсолютный прирост никогда не достигает уровня первого и второго года жизни. Рост массы тела завершается на четвертом году жизни, однако сохраняет сезонную динамику с максимумом осенью и минимумом весной. Амплитуда этой сезонной динамики составляет до 80 кг и больше. Наиболее резко она выражена у быков, лишь за время гона они теряют до 17% от своей максимальной массы, тогда как лосихи теряют за этот осенний период всего лишь не более 5% от своей массы.
Лоси достигают половозрелости осенью второго года жизни, но это не означает, что они в этом возрасте приступают к размножению. Как правило, самки отгуливаются только на третью осень, а самцы в покрытии самок до своего полного физического развития (до 4 лет) участвуют случайно или нерегулярно.
Продолжительность жизни лося, возможно, превышает 25 лет, как об этом писал еще А.Черкасов (1867; по: Гептнеру и др., 1961).
Ювенильный наряд лосенка не пятнистый, как у ряда видов других оленей, но тем не менее он резко отличается от взрослого. Лосята имеют рыжую окраску’ шерсти, а не серовато-бурую, к тому же ноги у них не светлее, чем туловище. Смена ювенильной окраски на взрослую происходит к осени — в августе, сентябре и, возможно, позже — в зависимости от ряда внешних и внутренних факторов (Гептнер и др., 1961).
Рога у самцов закладываются в виде бугорков в 4—5-месячном возрасте, а вырастают в конце первого — начале второго года жизни. Они, как правило, не ветвистые, в виде шпильки, лишь изредка вильчатые. Вторые рога у лосей развиваются на третьем году жизни, имеют вильчатое строение. Хорошо развитые рога с лопатой наблюдаются лишь у животных не ранее пятилетнего возраста. По А.А.Приедитису (1979), в Латвии наилучшие рога свойственны быкам в возрасте 7—11 лет, когда лоси достигают наибольшей массы. Им выявлена прямая корреляция между массой тела и размерами рогов. Она наиболее резко выражена у взрослых особей, ибо рога с большой массой могут вырасти у животных, у которых завершено развитие скелета (Bubenik, 1959, по: Приедитис, 1979). Здесь уместно отметить, что к трехмесячному возрасту плода череп в основном становится костным (Петров, 1964). Что касается окостенения остального скелета, то, по данным того же автора, появление очагов окостенения у лося проходит в том же порядке, что и у крупного рогатого скота. Однако у лося очаги окостенения появляются раньше, чем у крупного рогатого скота.
Возрастные изменения при всей видоспецифичности не идентичны в различных частях ареала. Как известно, лоси рождаются с хорошо развитыми молочными зубами,а также прорезывающимися предкоренными. Формирование постоянных резцов в условиях Печоро-Илычского заповедника завершается в полтора года (Кнорре, 1949). У американского лося первый коренной прорезывается на 10—14-й неделе, при этом нижнекоренной несколько раньше верхнекореиного. К 4—6 мес. эти зубы функционируют
237
Лось
полностью. В возрасте 6—7 мес появляется второй коренной зуб; достигнув 13—16 мес, лоси, как правило, теряют все молочные зубы, а в 16—19 мес завершается формирование коренных зубов (Питерсон, 1955, по: Гептнер и др., 1961).
После формирования зубной системы наиболее значительные изменения в структуру зубов вносит процесс изнашивания, что отражается в степени их стертости. На этом основана методика определения возраста лосей; использование размеров зубов мало перспективно. Однако характер и степень изношенности зубов неодинаковы в различных частях ареала, и поэтому необходимо провести модификацию существующих методик применительно к условиям Северного Кавказа. Для этого, очевидно, приемлема методика, предложенная В.Д.Херувимовым и И.И.Орловым (1967). По мнению названных авторов, таблицы определения возраста лосей по зубам нижней челюсти "тамбовской популяции" можно использовать во всех южных областях ареала лося в пределах европейской территории б. СССР.
По определению возраста лосей наиболее обнадеживающие результаты получены на основании анализа поперечных срезов корней зубов лося. Исследование Р. Балейшиса (1970) крашенных гематоксилином Эрлиха и эозином ^поперечных срезов резцов и клыков (п = 118) подтвердило мнение Серджента и Пимлота (Sergeant, Pimlott, 1959, по: Белейшие, 1970) о том, что’слои цемента в 1-й паре резцов являются годовыми. Число слоев совпадает с возрастом животного по годам. Во 2-й и 3-й парах резцов и в клыках первый годовой слой цемента формируется к 2 годам. При определении возраста лосей, считает автор, по срезам корней этих зубов к имеющемуся количеству годовых слоев нужно прибавить 1 год.
Структура популяции. Литературные данные по этому вопросу весьма противоречивы, что обусловлено методическими трудностями получения однозначных данных. Так, определение половой структуры популяции на основе статистики заготовки может быть искажено в ту или другую сторону. При лицензионной охоте, как известно, отстреливаются самые крупные животные. Установка может предусматривать преимущественный отстрел определенных возрастов и полов, что исказит результаты расчета (соотношение полов в заготовке не будет соответствовать пропорциям, реально существующим в природе). Расчет половой и возрастной структуры по регистрации встреч также имеет свои издержки. Сказывается динамика активности животных в зависимости от их физиологического состояния: повышенная активность самцов во время гона, самок и телят после отела. Известно также, что молодые лосихи чаще приносят телочек, в возрасте 4—5 лет — оба пола в равном количестве, а в старом возрасте — преимущественно бычков (Шубин, Язан, 1959; Юргенсон, 1968). Изменения половой и возрастной структуры популяции в связи с кочевками, сезонными миграциями, погодно-климатическими условиями также велики (см. раздел "Миграции"). Чтобы проиллюстрировать, насколько сильно варьируют показатели структуры популяции лося и разнообразны факторы, обусловливающие их, ограничимся 2—3 примерами.
238
Alces alces Linnaeus, 1758
По данным Е.К.Тимофсевой (1974), в популяции лосей Ленинградской области неизменно преобладают самцы независимо от метода учета пола: по встречам в природе (п = 570), по данным отстрела (л = 13 950), по исследованию эмбрионов (л = 60). Сходные результаты получены во Владимирской области (Сысоев, 1968): среди взрослых (л = 630) самцы составляли 51,5%, самки — 48,5%, а среди эмбрионов (л = 51) соответственно 58,5% и 41,5%. В условиях Печоро-Илычского заповедника, по материалам Ю.П.Язана (1961), у стельных лосих доля бычков колеблется по годам в пределах от 43,6 до 56,5%. Бычки преобладают обычно после суровой зимы, когда ослабевшие лоси нс успевают накопить жира, столь необходимого в период гона. В среднем за ряд лет соотношение полов эмбрионов близко к 1:1. Обзор обширной литературы, иллюстрирующей преобладание в одних районах самцов, а в других самок, приводится в ряде сводок (Гептнер и др., 1961; Тимофеева, 1974; и др.).
С учетом различных факторов Е.К.Тимофеева (1974) приходит к выводу, что возрастная структура популяции лосей Ленинградской областицмеет следующий состав (в%): телята до года — 20; 1,5 года — 10; 2,5—3,5 года — 25; 4,5—5,5 лет — 19; 6,5—7,5 лет — 16; 8,5—9,5 лет — 6; 10 лет и старше — 4. Сравнение этой структуры с литературными данными по другим районам страны позволило обнаружить близость ее с тем, что наблюдалось в районе Верхней Печоры, Финляндии, на Кольском полуострове, в Карелии, Эстонии. В более южных районах страны (Окский заповедник, Тамбовская и Витебская области, Татария, Украина и др.) явно преобладает молодняк. Такое омоложение популяции обусловлено высокой плодовитостью стада и лучшей выживаемостью молодняка в течение зимы.
При оценке возрастной структуры популяции лося, на наш взгляд, особое внимание заслуживает анализ динамики численности популяции. Данные О.С.Русакова (1975) убедительно свидетельствуют о том, что на разных фазах динамики численности возрастная структура меняется значительно. Так, в период роста и пика численности доля молодняка была наибольшей, а в фазе резкого снижения — наименьшей. В противоположном направлении менялась доля взрослых.
Данных о возрастной и половой структуре лося на Северном Кавказе очень мало. По мнению А.М.Гинеева (1975), в предгорьях региона в начале 70-х годов обитало около 90—100 лосей. Из них взрослые самки составляли 45,7%, взрослые самцы — 37,1%, а молодняк — 17,2%. Эти данные соответствуют, очевидно, этапу становления новой популяции, росту численности за счет молодняка, родившегося на месте, снижению притока мигрантов и увеличению доли местных животных. Такое суждение согласуется также с полевыми наблюдениями, согласно которым телята, родившиеся на месте, встречаются на обширной территории региона (Бакеев, 1980; Оноприенко, Хохлова, 1988; Гинеев, 1989).
Враги, паразиты, конкуренты, смертность. В пределах ареала лося в Евразии наиболее опасными хищниками считаются волк, бурый медведь и местами в тайге росомаха. Оценке роли каждого из этих хищников и доли, изымаемой ими в популяции лося, посвящена обширная литература. Здесь
239
Лось
мы обратим внимание лишь на отдельные стороны отношения хищник-жертва, которые могут иметь место на Кавказе.
При нынешнем уровне численности волков и медведей, видимо, они будут играть лишь санитарную и селективную роль. Волки для лося опасны зимой, когда появляется наст, который ставит волка в более выгодное положение, чем лося, — нередко наст выдерживает весовую нагрузку волка, а лось проваливается и становится жертвой хищников. Деятельность волков (стаи в 8—10 особей) при наличии более доступных жертв не снижает численности за счет прямого уничтожения, однако в условиях Башкирского заповедника, например, способствует оттеснению лосей в более глубокоснежные места и на смежную территорию. Словом, волк оказывает негативное влияние на территориальную организацию популяции (Гордиюк, 1980). В теплое время года, очевидно, больше всего от волков страдают телята и годовалые, а в сезон охоты — подранки. Бурый медведь нападает на лося в основном весной и осенью, особенно когда он в многоснежную раннюю весну пробуждается и вынужден бродить повсеместно в поисках пищи. Напав на след лося, он может преследовать жертву много километров (Гептнер и др., 1961). Из других хищных зверей, видимо, некоторый вред могут приносить шакалы, численность которых высока во многих районах Кавказа. В период отела возможны нападения шакалов на лосят. В литературе также содержатся данные о нападении рыси на лосят, но если учесть, что этих хищников нет на равнине и предгорьях, а в горах их стало мало, то можно не принимать в расчет возможный вред, причиняемый этим хищником лосю на Северном Кавказе.
О складывающейся ситуации для лося на Северном Кавказе очень мало данных, но некоторое представление дают наблюдения Л.Г.Оноприенко и З.И.Хохловой (1988). Из 10 известных случаев гибели лосей 6 были подстрелены браконьерами, 1 погиб под поездом, 3 — по неизвестным причинам (предполагается, что они отравлены ядохимикатами).
Для благополучия лося большое значение имеют различные экто- и эндопаразиты, в первую очередь гельминты. Число паразитических червей, обнаруженных в организме лося Палеарктики, превышает 38 видов. Хотя количество видов гельминтов в отдельно взятом районе невелико, интенсивность инвазии и поголовье зараженных лосей бывают очень высокими. Например, в Сосновском лесоохотхозяйстве Ленинградской области лоси заражены гельминтами поголовно. У 20 изученных животных выявлено 12 видов (3 вида трематод, 3 вида цестод и 6 видов нематод). При этом интенсивность инвазии лишь только одного паразита Ostertagia antipini достигла 3040—19 520 экз., в среднем на одного лося 14 488 паразитов. Обнаружен гемопаразит Bunostomum trigonocephalus, вызывающий анемию и прогрессирующее истощение. Лоси очень сильно страдают от носоглоточного овода (Gephenomyia ulrichi), слепней (Tabanus tarandinus, Т. arpadi, Т. tropicus, Т. lapponi), мух-кровососок (Lepoptena cervi). На лосях паразитируют также клещи (Dermacentor pictus и др.). Имеются основания относить лося к прокормителям иксодовых клещей (Ixodes persulcatus и др.), тем самым он поддерживает природный очаг клещевого энцефалита. Более того, лось
240
Alces alces Linnaeus, 1758
играет основную роль в распространении этой инфекции. Этому вопросу посвящена обширная литература, обзор которой приводится в монографиях В.Г.Гептнера и др. (1961), Е.К.Тимофеевой (1974) и др.
Конкурентов у лося, очевидно, много. Об этом свидетельствуют не только многочисленные литературные данные, но и характер биотопического распространения, кормовые ресурсы, отношение к домашним животным и т.д. Однако наиболее острые конкурентные отношения складываются с оленями, особенно тогда, когда численность последних высока. Имеются наблюдения, что при повышении численности других копытных, в том числе оленей, происходило сокращение численности лосей в Беловежской Пуще (Карпов, 1903; Врублевский, 1912, 1927; и др.). Такая ситуация может сложиться на Кавказе, на наш взгляд, на сравнительно небольшой территории среднегорных лесов Западного и отчасти Центрального Кавказа, особенно в заповедниках и прилегающих районах. Наиболее острые прямые и косвенные конкурентные отношения могут возникнуть с домашними животными. Дело в том, что на всем Кавказе на равнине и в горных лесах выпасают скот, местами травостой выбит как на полянах, так и под пологом, особенно угрожают вездесущие козы. Это наблюдается и на водоразделах, и по долинам рек, и по берегам водоемов. Соответственно будут осложнения не только в связи с истощением кормовой базы, но и ухудшением эпизоотологической обстановки. Аналогичное положение выявлено, например, в Белоруссии (Шималов, 1980). Здесь среди диких и домашних животных наиболее распространенными болезнями являются болезни паразитарной этиологии. Наибольший ущерб популяции лося наносят парафасциолоцеоз, экстенсивность инвазии которого составляет в отдельные годы свыше 60%. Из печени больных животных в среднем выделяли 3—4 тыс., максимум 23 тыс. экз. паразитов.
Говоря о смертности лося и роли хищников и паразитов в его жизни, необходимо учесть, что в итоге не эти факторы являются решающими. В период массового размножения вида элиминация части популяции не подрывает популяцию, а, наоборот, стимулирует воспроизводство ее. Так, по данным В.В.Лавровского (1989), в Окском заповеднике из 58 лосей, добываемых волком в среднем за год, 45% особей были явно ослабленными и неосторожными. При оценке роли прямого влияния человека то же самое можно бы сказать и в отношении охоты, если бы она была строго контролируемой и планируемой. Человек, вооруженный нарезным оружием и другими техническими средствами, представляет большую опасность для любого вида животных.
Немаловажную опасность для лося в условиях Кавказа представляют ядохимикаты, применение и хранение которых в регионе нуждаются в квалифицированном исполнении.
Отрицательная роль абиотических факторов в жизни лося многогранна, значение отдельных факторов нами рассмотрено в предыдущих разделах.
Численность и ее динамика. Однозначных данных по численности лося на Северном Кавказе все еще мало, нередко они противоречивы. Как правило,
16 В.Е.Соколов, А.К.Тсмботов
24 1
Лось
результаты учета Госохотинспекции оказываются несколько завышенными, чем данные, опубликованные в печати.
Все материалы по численности лося на Северном Кавказе относятся к двум краям — Краснодарскому и Ставропольскому. По мнению А.М.Гине-ева (1975), в самом начале 70-х годов в регионе общая численность ориентировочно определялась в 90—100 особей, из них в Краснодарском крае 60—70 лосей. Остальные 30 животных, видимо, обитали в Ставропольском крае. По данным Л.Г.Оноприенко и З.И.Хохловой (1988), в 1986—1988 гг. численность лосей в Ставропольском крае составляла около 70—80 особей. В Краснодарском крае, по данным В.А.Кузякина («1989), имело место увеличение численности лося до 150 особей в 1981 г. Такой же была численность и в 1982 г., но в 1983 г. она упала до 100 особей и на этом уровне держалась до 1985 г. включительно. Согласно учетным данным Госохотинспекции, в Краснодарском крае в 1984 г. поголовье лосей оценивалось в ПО особей, в 1985 г. — 132 особи, в 1986 г. оно достигло 175 особей и это количество сохранилось в 1987 и 1988 гг.
Площадь, пригодная для обитания лося в Краснодарском крае, определена в 150 тыс. га, плотность населения лося оценивалась в 1984 и 1988 гг. соответственно в 0,9 и 0,8 особей/1000 га, а в 1986 и 1987 гг. — 1,2 осо-бей/1000 га.
В Ставропольском крае, по данным Госохотинспекции, для жизни лося пригодна территория площадью в 71 тыс. га. Плотность населения, выявленная по результатам послепромысловых учетов, составляла в 1989 г. от 0,8 особей/1000 га (Малокарачаевский район) до 2,0 особей/1000 га в Александровском районе.
Для оценки состояния численности лося на Северном Кавказе и перспектив ее изменений в будущем интерес представляют данные В.Е.Размах-нина и Д.Н.Сабурова (1980). Согласно этим данным, в подтаежных, лесостепных и степных регионах Центра, Поволжья, Северо-Западного и Центрально-Черноземного экономических районов повышение численности лося отмечалось в начале 60-х и 70-х годов, а период спада — в середине-конце 60-х годов. Как отмечают авторы, при достаточно высоком уровне среднемноголетней плотности лося 3,5—5,0 особей/1000 га за этот период минимум составил 1,5—2 особи/1000 га, а максимум — до 7—12 особей/1000 га.
Здесь уместно добавить, что спектр ландшафтных условий перечисленных выше экономических районов Европы и Северного Кавказа, на наш взгляд, соизмерим и соответственно возможны соизмеримые плотности населения лося.
Практическое значение. Лось издавна известен как важнейший объект промысловой и спортивной охоты во всей лесной зоне Евразии. Об этом свидетельствуют многочисленные данные археологии, архивных летописей и литературы различных периодов.
Из продукции лося особо высоко ценится мясо, выход которого составляет от 60—62,5 до 64—65%, возможно до 70% живой массы. Жир, который можно собрать с одного лося, достигает 30 кг (Гептнер и др., 1961; Тимофеева, 1974). Максимальная упитанность быков наблюдается осенью перед го
242
Bovidae Gray, 1821
ном, а во время гона они теряют до 17% от исходной массы и к тому же меняются вкусовые качества мяса. Самки дольше сохраняют упитанность (в гоне теряют в 3—4 раза меньше, чем быки) и соответствующие показатели мяса вплоть до середины зимы. Яловые самки еще более устойчивы к зимним условиям и сохраняют запас энергии дольше.
Кожа лося высокого качества, толстая и прочная, издавна используется для изготовления различных вещей. Очень высоко ценятся рога в качестве охотничьего трофея.
Проводится определенная работа по одомашниванию лося, обладающего рядом положительных качеств: высокая плодовитость, быстрый рост молодняка, хорошие вкусовые качества мяса и молока, перспективность как вьючного животного таежной зоны, способность питаться дрсвесно-кустарниковой растительностью и т.д. Однако пока эта работа не вышла за пределы экспериментов,
Имеется обширная литература, посвященная отрицательной роли лося в лесонасаждениях. Там, где происходит чрезмерное увеличение численности, вред, наносимый лосем лесопосадкам, бывает очень значительным.‘Задача состоит в том, чтобы разумно управлять численностью лося, сводят к минимуму причиняемый им вред.
В условиях Северного Кавказа имеются сообщения о вреде, наносимом лосем садоводству, полеводству и лесопосадкам. Более того, обитание лосей в условиях равнинно-предгорной части малоперспективно (Оноприенко, Хохлова, 1988). На наш взгляд, вопрос требует более тщательного анализа.
Как отмечено выше, лось представляет определенную опасность в эпидемиологическом и эпизоотологическом плане, но в регионе этот аспект значения лося не изучен.
Семейство полорогие — Bovidae Gray, 1821
Жвачные копытные, разнообразные по массе и размерам тела, жизненным формам и ландшафтной приуроченности. Это самое обширное семейство парнокопытных. Размах изменчивости экстерьерных показателей достигает 1—2 порядков величин. Так, карликовая антилопа Neotragus pyg-maeus имеет массу всего 2—3 кг и высоту’ в холке около 25 см, тогда как зубр Bison bonasus весит более 1000 кг, имеет высоту’ в плечах до 200 см. Телосложение от изящного и легкого до массивного и тяжелого. Обычно конечности высокие, рога не ветвистые и не сменяемые, покрыты роговыми чехлами эпидермального происхождения; они растут не с верхушки, как у оленей, а от основания, и при этом рост постоянный, но с определенной периодичностью. Формы, размеры и характер закрученности рогов сильно изменчивы, но они никогда не образуют отростков. На их поверхности образуются бугры, продольные ребра, кольца, складки, отражающие динамику роста и развития животных. Рога обычно имеются у обоих полов, но у самок они меньше по размерам.
16
243
Семейство Полорогие
Конечности как правило четырехпалые, боковые (II и V) пальцы слабо развиты, иногда совсем редуцированы, несут небольшие койыта. Передние ноги длиннее задних и лишь у наиболее примитивных видов семейства наблюдается преобладание длины задних над передними конечностями. По И.И.Соколову (1959), характерно также сращение (слияние) в запястье головчатой кости с малой многоугольной и в заплюсне — кубовидной с ладьевидной и второй клиновидной с третьей.
Окраска тела весьма разнообразна, варьирует от белой до черной, но чаще она без цветовых узоров. Нередко встречаются виды, обладающие характерной светлой расцветкой прианальной области тела ("зеркало"). Половые и возрастные различия окраски у полорогих выражены в меньшей степени, чем у оленей.
Волосяной покров подвержен широкой изменчивности, что соответствует величине спектра ландшафтных условий жизни. У одних видов, например у овцебыка, волосы очень длинные и с обилием подшерстка, а у других, как у ряда видов газели, довольно короткие и негустые.
Кожный покров хорошо развит. У ряда видов(зубров и бизонов) кожа очень толстая; толщина ее в области затривка составляет около 0,3% длины тела (Соколов, 1973). Характерно наличие многих специфических кожных желез: пр ед глазничных, межроговых, паховых, межпальцевых, хвостовых и т.д. Они играют исключительную роль как средство химической коммуникации, нередко видоспецифичны, что находит отражение в степени развития желез, особенностях расположения на теле, характере запаха и т.д. Очень сильно бывает развита хвостовая железа (например, у кавказских туров). Словом, морфологические признаки, связанные с кожными железами, представляют значительный интерес как в плане систематики, так и экологии полорогих. У рассматриваемых копытных 1—2 пары сосков.
Череп обладает рядом структурных особенностей, что обусловлено переходом полорогих от пассивной защиты к активной. По И.И.Соколову (1959), рога как орудия защиты и нападения увеличиваются в размере и прочности, ими обладают не только самцы, но обычно и самки. Несущие их лобные кости усиливаются, теменные кости несколько смещаются назад, преобразуя зароговую часть, происходит увеличение излома оси черепа, а также пневматизация лобных костей и стержней рогов, тесное соединение или полное слияние костей черепа, особенно зароговой части, и т.д. Характерно, что в слезной кости сильно развита лицевая часть с ямкой для пр ед глазничной железы и без нее. Обычно имеется только одно отверстие слезного канала. Этмоидальные отверстия отсутствуют или развиты слабо. Черепные кости сильно пневматизированы. Межчелюстная кость обычно сравнительно невелика, верхнечелюстная очень крупная.
Зубная формула:
0 0 2-3 3
I—С — Р ------ М - = 28-32.
3 -1 2-3 3
Характерно отсутствие верхних резцов и клыков, нижние резцы долотообразные, на относительно длинных шейках и корнях, в альвеолах сидят несколько свободно и подвижно. Слабо развит или совсем отсутствует вто
244
Bovidae Gray, 1821
рой предкоренной на верхней и нижней челюстях. Верхний клык отсутствует, и его появление рассматривается как аномальное явление. Коренные зубы гипселодонтные и тетраселадонтные (четырехлунчатые). Характерно, что обычно коронки зубов выше, чем у оленьих; они не растут постоянно, ибо корни их замкнутые.
В числе характерных анатомических признаков прогрессивных трупп полорогих укажем на сложный желудок, четко дифференцированный на 4 отдела: рубец, сетку, книжку и сычуг. Как правило, наличие желчного пузыря, поликотиледонной плаценты, двухдольчатой или четырехдольчатой молочной железы. У penis многих видов имеется нитевидный отросток.
Распространение полорогих очень обширное. Восстановленный ареал занимает всю Африку, за исключением Мадагаскара, Европу (кроме Британских островов) до юга Скандинавского полуострова, Финского залива, Верхней Волги, Самарской Луки, а также юг Урала и Западной Сибири, большую часть Средней и Восточной Сибири и Дальний Восток. На остальной части Азии полорогие заселяют всю южную часть материка, включая многие прилегающие острова. В Новом Свете ареал занимает большую часть Северной Америки на юг до Калифорнии, Флориды и Северной Мексики, Арктический архипелаг, северное и восточное побережья Гренландии.
Современное распространение полорогих значительно сокращено в связи с антропогенным прессом, изменением природных экосистем, неумеренным промыслом и т.д.
Местообитания весьма разнообразны, включают очень широкий круг биотопов от субарктики до влажных тропиков. Полорогие живут в тундре, различных лесах, в степях, полупустыне и пустыне различных материков. В горы проникают выше всех других млекопитающих, достигая высоты 5500 м над ур. моря. Однако наибольшее разнообразие видов и форм семейства связано с открытыми ландшафтами.
В экологическом и эволюционном отношениях самым характерным является окончательный переход на питание растительной пищей, нередко более грубой и жесткой, чем у оленей. Характерно, что полорогие большей частью собирают корм с поверхности земли вместе с твердыми частицами почвы, песка и пыли, что ускоряет износ зубов. Такое положение обусловило появление гипсодонтии (Соколов, 1959). С характером пищи связано, по мнению названного автора, увеличение размеров заднекоренных зубов, на которые падает больше нагрузки, увеличение прочности зубов, усиление мышц, связанных с актом перетирания пищи.
Полорогие большей частью полигамы, однако имеются и моногамы. Самцы ряда видов во время размножения образуют гаремы самок. Сезонность размножения полорогих в тропиках обычно не выражена, тогда как на севере это явление обычное. Беременность длится 4—11 мео, в одном помете бывает от одного до 4—5 детенышей. Рождаются они обычно хорошо сформировавшимися, способными через короткое время подняться на ноги и следовать за матерью.
Практическое значение велико и разнообразно. Они составляют неотъемлемый компонент многих экосистем, способны давать значительную
245
Род Зубры
животную продукцию при минимальных издержках для охраны природы. На Кавказе к таковым можно отнести туров, сайгаков и др. Многие виды служат объектом спортивной охоты и в ряде районов мира широко используются с этой целью. Трудно переоценить роль полорогих для человека как предков домашних животных и в селекционной работе по их дальнейшему породному совершенствованию. Эта селекционная работа ведется и по сей день. Велико эстетическое значение полорогих. Дикие полорогие известны как источники инфекционных болезней человека и домашних животных, могут иметь отрицательное значение в полеводстве и других областях сельского хозяйства. Но эти и другие стороны вреда полорогих несравнимы с разнообразным положительным их значением.
Эволюция полорогих находится в стадии расцвета, о чем свидетельствует богатство, разнообразие и широчайшее распространение видов данного семейства. В этом плане равных ему нет среди других семейств отряда парнокопытных и тем более непарнокопытных. Корни этого семейства ведут к нижнеолигоценовым оленькам (Tragulidae), промежуточное звено между ними и настоящими полорогими неизвестно, но в среднем миоцене Европы уже обитали примитивные антилопы.
Родиной полорогих следует считать Евразийский континент, на стыке которого с Африкой лежал первичный центр расселения полорогих (Соколов, 1959).
Систематика полорогих недостаточно разработана, положение целого ряда групп не обосновано фактическим материалом. По В.Е.Соколову (1979), в семействе 43 рода, объединяющих до 109 видов, а по J.H.Honacki и др. (1982), соответственно 45 родов и 122 вида. В современной фауне Кавказа содержатся 6 родов: зубры Bison, горные козлы Capra, газели Gazella, бараны Ovis, серны Rapicapra, сайгаки Saiga. Они объединяют 7 видов, или 50% всех парнокопытных региона (названия таксонов по: Соколов, 1984).
Род Зубры — Bison Н. Smith, 1827
Род включает самых крупных животных из семейства полорогих. Телосложение тяжелое, длина тела достигает 220—308 см, длина хвоста 50—110 см, высота в холке 140—200 см. Масса тела 430—1000 кг. Самки меньше самцов. Характерно мощное развитие передней части тела, а задняя менее развита. Холка высокая, горбовидная, спина поката к хвосту. Относительно короткая и массивная голова с широким лбом посажена низко. Рога имеются у самцов и самок, но у последних меньше; они относительно небольшие, но очень прочные, широкие у основания, круглые в поперечном сечении, поверхность гладкая, серо-бурого или буро-черного цвета. Копыта крупные, по цвету сходны с рогами.
В волосяном покрове характерна его неоднородность на поверхности тела — на передней части тела он длинный и косматый, а на задней — короткий и плотно прилегающий. Окраска его зимой от серо-бурой до рыжебурой, буро-коричневой и темно-бурой, а летом становится заметно
246
Bison H. Smith, 1827
светлее. Смена зимнего волосяного покрова, длинного и густого, на летний, короткий и жесткий (без подпушки), происходит весной. Осенняя линька не выражена у зубра, но свойственна бизону. На морде и на конечностях отсутствуют специфические кожные железы.
В структуре черепа характерно сокращение длины и увеличение ширины мозгового отдела, что обусловлено развитием роговых выростов на лобных костях и усилением нагрузки на мышцы головы и шеи. По К.К.Флерову (1979), наибольшая ширина лба в области орбит составляет обычно у самцов не менее 65% и около 60% у самок по отношению к основной длине черепа. Кости черепа и стержни рогов сильно пневматизированы, края орбит заметно выдаются в стороны, особенно у старых особей. Слезные кости без ямок для предглазничных желез. Нет этмоидальных отверстий. Межчелюстные кости не достигают носовых костей.
Зубная формула:
1 ° с 2 Р 2 м — = 32.
3 13 3
Молочных зубов 20; врс резцы, клыки и предкоренные имеют молочных предшественников, лишь коренные не имеют таковых. Характерно, что верхние коренные гипссДодонтные с дополнительной призмой на внутренней стороне.
В посткраниальном скелете показательно резкое увеличение остистых отростков трудных и седьмого шейного позвонков, связанных с мощными шейными мышцами.
Диплоидный набор зубра и бизона включает 60 хромосом.
Восстановленный ареал рода Bison состоит из двух частей, одна из которых находится в Северной Америке, а другая — в Европе. В обеих частях животные уничтожались беспощадно, находились на грани полного уничтожения. Сейчас предприняты широкомасштабные мероприятия по их сохранению.
Экология зубров и бизонов в условиях естественной и не ограниченной искусственными преградами природы фактически осталась неизученной. В природе их уничтожили раньше, чем изучили. Многочисленные литературные источники, которыми в настоящее время мы располагаем, получены в загонах, заповедниках и других изолированных клочках земли. Естественно, в таких условиях такие гиганты, как зубр и бизон, способные к быстрому перемещению и обладающие очень тонким чутьем, не могут не реагировать на стесненное их положение. Вероятно, они "чувствуют", что находятся в руках человека и соответственно несут на себе печать условий, создаваемых человеком. В частности, столь характерное явление, как миграции бизонов в американских прериях, которые отмечались до прихода европейцев в Америку, в настоящее время уже не существуют. Сокращение типов обитаемых биотопов не могло не сказаться на качественном и количественном составе пищи, оказались также сдвинутыми сроки размножения, изменены структура популяции, поведение и т.д. Спору нет, что многие биологические особенности сохранились, но не зная того, что было, трудно говорить о том, что осталось. Тем не менее анализ разнообразных
247
Зубр
данных, проведенный группой специалистов (Зубр 1979), позволил восстановить многие черты морфологии и экологии в условиях природы как зубра, так и бизона. Эта монография освобождает нас от необходимости обзора особенностей экологии р. Bison. Здесь лишь отметим, что зубры являются уникальным памятником природы, их судьба полностью находится в руках человека и, следовательно, велика наша ответственность за их сохранение.
По современным данным в роде два вида: бизон (Bison bison L.) и зубр (В. bonasus L.) В фауне Кавказа представлен только второй вид.
Зубр — Bison bonasus Linnaeus, 1758
Рис. 34
Bison europaeus, Виноградов; Bos bonasus caucasicus, Туркин, Сатунин, 1902; Сатунин, 1903; Bison bonasus var. caucasica Grave, 1906, Bison kauka-sicus, Hilzheimer, 1909; Bison caucasicus Hilzheimer, 1909; Bos (Bison) caucasicus, Lydekker, 1913.
Коллекционный материал. ЗИН РАН: 8 черепов, 2 шкуры; МГУ: 4 черепа; КГЗ: 102 черепа, 83 скелета; КБГУ: 5 черепов, 4 шкуры, 1 скелет.
Диагноз. Bison с уплощенным телом, относительно легкой головой (наибольшая длина черепа быков 490—580 см), линия спины за горбом прямая, рога резко изогнуты, основная длина черепа быков 449—489 см, расстояние от нижнего края орбиты до края альвеолы М3 значительно меньше длины коренных и предкоренных, рельеф лобной кости с буграми и впадинами, в М3 передне-внутренняя складка резко не отделена.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Провести подробный анализ видовых параметров Bison bonasus на Кавказе пока нереально. Местный подвид Bison bonasus caucasicus уничтожен в 20-х годах XX в., т.е. еще до того как его изучили. Восполнить этот пробел в наши дни уже невозможно. Между тем имеющиеся фрагментарные данные говорят о том, что аборигенная форма зубра характеризовалась целым рядом морфологических и биологических особенностей (Сатунин, 1903; Динник, 1910; Флеров, 1979, и др.).
С 40-х годов нашего века ведется акклиматизация межвидового гибрида (Bison bonasus и Bison bison) в Кавказском заповеднике (Западный Кавказ) и Нальчикском лесоохотничьем хозяйстве (Центральный Кавказ). Несколько позже начата акклиматизация другого гибрида, но уже внутривидового (В. bonasus bonasus и В. bonasus caucasicus) в Тебердинском заповеднике и в двух заказниках Северной Осетии и Чечено-Ингушетии.
В настоящее время в горных условиях Кавказа наиболее полные сведения получены в Кавказском заповеднике, где обитает зубробизон (“горный зубр"). По другим районам данных очень мало. Мы свели данные по размерам тела и черепа горных зубров в табл. 44—45. Что касается характе
248
Bison bonasus Linnaeus, 1758
ристик и видовых параметров Bison bonasus, то она приводится ниже по природным формам, преимущественно беловежского зубра Bison bonasus bonasus и вымершего аборигенного кавказского зубра B.b. caucasicus. Лишь для восстановления пробелов (кариотип, картина крови и некоторые другие параметры) в обзор включены данные по гибридным зубрам. Характеристика последних приводится в разделе "Изменчивость".
Масса тела В. bonasus, по И.И. Соколову (1959), составляет у самцов 430—1000 кг, длина тела 234—350 см, высота в холке 158—195 см. Самки несколько меньше самцов — 370—400 кг. Длина тела взрослых самок около 240—290 см, высота в холке до 160 см. Более высокие показатели экстерьера приводят различные авторы, но, видимо, они или преувеличены, или относятся к гибридам. Что касается аборигенного кавказского зубра, то данных по нему очень мало. Весь имеющийся фактический материал говорит о том, что он был относительно легкого телосложения и более мелким, чем беловежский зубр. Масса тела была до 700 кг, изредка до 800 кг, в среднем 480 кг, а высота в холке около 160 см. Высота в крестце у В. bonasus меньше высоты в холке на 20—25 см у самцов, на 12—15 см у самок.
Размеры черепа зубра варьируют. У беловежского зубра, по К.К.Флерову (1979), общая длина черепа 500—580 мм, ширина лба наименьшая (позади орбит) — 220—285 мм. Череп кавказского зубра мельче: общая длина 490—560 мм, ширина лба наименьшая (позади орбит) 220—250 мм.
По размерам черепа оба подвида В. bonasus заметно уступают бизону В. bison, у которого наибольшая длина черепа достигает 630 мм. Однако между обоими видами имеется значительная зона трансгрессии. Более четкие различия между названными видами и в равной степени между подвидами одного вида обнаруживаются при сравнении структуры черепов (Флеров, 1979). По данным названного автора, лицевая часть короче у зубров, чем у бизонов, а лоб более узкий, менее массивный и менее выпуклый. Поверхность его неровная, имеет впадины и бугры, однако степень выраженности этого признака у зубра подвержена большим возрастным и половым изменениям. В роде Bison объем лобных полостей минимален в черепе В. bonasus, но среди подвидов этого вида наименьшими лобными полостями выделяется B.b. caucasicus, имеющий самую легкую голову. Характерно также, что лицевая часть черепа у зубра ниже, чем у бизона — расстояние от нижнего края глазницы до края лунки М3 намного меньше, чем общая длина всех коренных. У бизона это расстояние больше длины коренных зубов, так как лицевая часть высокая. Носовая полость минимальна у B.b. caucasicus (по данным названного автора), расстояние от нижнего края орбиты до края альвеолы М3 намного меньше длины всех коренных зубов (приблизительно на 1/3). Такая особенность связана, очевидно, с более теплым климатом Кавказа и меньшими перемещениями, чем у беловежского зубра, не говоря уже о бизоне. В связи с низкой носовой полостью у B.b. caucasicus верхние носовые отростки intermaxillaria достигают носовых костей. Из черепных особенностей зубра укажем еще на размеры хоаны — у В. bonasus она ниже и уже (ширина ее у нёба равна длине М2), чем у В. bison
249
Зубр
250
Bison bonasus’ Linnaeus, 1758
Рис. 34. Современное распространение зубра
1 — Майкоп (ЗИН, Гинеев, 1988);
2 — Даховская (Гинеев и др., 1988);
3 — Гузерипльское лесохозяйство (Гинеев и др., 1988);
4 — урочище Блокгаузская караулка (Калугин, 1968);
5 — верховье р. Белая (Марков, 1924);
6 — окрестности Кишинского зубропарка (ЗИН);
7 — Псебайский гос. заказник (Гинеев и др., 1988);
8 — г. Бамбак (Калугин, 1968);
9 — ущелье Малой Лабы (Сатунин, 1914);
10 — истоки р. Мзымта (Сатунин, 1914);
11 — истоки р. Бзыбь (Сатунин, 1914);
12 — г. Марух (Марков, 1924);
13 — г. Пшиш (Марков, 1924);
14 — Архызское лесничество (Тарасов, 1973);
15 — верховье р. Уруп (Динник, 1910);
16 — Зеленчукская (Радде, 1899);
17 — верховье р. Кумы (Pallas, 1891);
18 — верховье р. Кодори (Башкиров, 1939);
19 — подножье г. Эльбрус (Сатунин, 1903);
20 — Белореченское лесничество (КБГУ);
21 — Нальчикское лесоохотхозяйство (КБГУ);
22 — Ахсарисар (Наниев, 1956);
23 — Урухское ущелье (ЗИН);
24 — заказчик "Цейский" (Липкович, 1989);
25 — Северо-Осетинский заповедник (Соколов, Сыроечковский, 1990);
26 — Чечене-Ингушский заказник (Ли-пович, 1989);
27 — верховье р. Ардон (ЗИН, Темботов, 1972).
Карта-врезка. Распространение зубра в прошлом (по: Верещагин, 1959)
1—2 — первобытный зубр; 1 — верхний плейстоцен, 2 — средний плейстоцен, 3 — таманский зубр, нижний плейстоцен и верхний плиоцен; 4—8 — кавказский зубр: 4 — начало XX в., 5 — пункт гибели последних зубров, 6 — коллекционные экземпляры XIV—XVIII вв., 7 — литературные сведения XIV—XIX вв., 8 — ранний голоцен
251
Зубр
Таблица 44. Убойная и живая масса тела зубров Кавказского заповедника __________________в 1952 — 1965 гг. (Калугин, 1968)________________
Возраст, Число Живая масса Убойная масса
лет зубров КГ кг | %
Самцы
До 1 года 1 129 67 51,9
2-3 2 321-347 161-171 49,2-50,4
3-4 2 356-706 222-366 51,8-62,9
4-5 7 511-691 255-418 50,0-63,7
5-6 5 466-688 237-356 50,8-52,8
6-7 10 421-708 224-356 50,0-55,9
7-8 4 542 - 276 50,9
8-9 1 671 346 56,0
9-10 2 694-743 338-394 48,6-53,0
10-11 1 1150 601 52,2
Кастраты
6-7 1 451 272 58,8
7-8 2 629-679 370-389 54,4—64,’8
10-11 3 606-694 331-403 53,8-57,9
11-12 1 781 414 53,0
Самки
23-24 1 602 310 5С6
(ширина се у нёба больше длины М2). Уместно отметить, что по этому признаку самцы и самки не различаются.
При оценке краниологических различий между зубром и бизоном, на наш взгляд, заслуживает внимания то, что они прослеживаются на ранних стадиях постэмбрионального развития. По К.К.Флерову (1979), они сводятся к следующему: в возрасте 1—3 дней череп В. bison крупнее, чем у В. bison.
Из черепных признаков зубра, отличающих его от бизона, заслуживает внимания относительно узкая слезная кость, которая заостренным зубцом вклинивается между верхнечелюстной и носовой костями (Соколов, 1959).
Существенные видовые различия выявлены в структуре нижней челюсти. У В. bonasus ветви нижней челюсти имеют высоту’, равную общей длине первого и второго коренного этих челюстей. Кроме того, они относительно толстые и вздутые. У В. bison ветви нижней челюсти значительно выше — высота их на уровне М3 много больше, чем общая длина первого и второго коренных зубов нижней челюсти; они не вздутые и относительно плоские.
Структура резцов зубра и бизона существенно различна. В частности, по К.К.Флерову (1979), резцы у зубра долотообразны и очень мало расширены. Второй и третий резцы и клык имеют на лингвальной стороне зубов воротничок. У бизона эти воротнички выражены лишь на клыках, резцы расширены и отогнуты наружу. Строение коренных сходное, однако М3 у В. bonasus более упрощенный, чем у бизона (передне-внутренняя складка его резко отделена).
252
Bison bonasus Linnaeus, 1758
Таблица 45. Промеры тела (в м) взрослых зубров Кавказского заповедника, отстрелянных или павших в 1975 — 1984 гг. (данные А.С.Немцева)
Промер Самцы Самки
limit (л = 43) х — $х limit (л = 32) х ±
Длина тела 2,60-2,96 2,79±0,31 1,92-2,63 2,25±О,О26
хвоста 0,38-0,64 0,51+0,029 0,47-0,73 0,60±0,022
головы 0,49-0,67 0,51±0,016 0,42-0,59 0,50±0,019
уха 0,16-0,17 0,16+0,023 0,14-0,16 0,16±0,029
СПИНЫ 0,52-0.76 О,64±О,О23 0,66—0,82 0,71±0,021
поясницы 0,49-0,82 0,67±0,034 0,37-0,65 0,56±0,026
зада 0,49-0,63 0,56±0,014 0,39-0,57 0,48±0,018
косая туловища 1,53-1,93 1,73±0,040 1,50-1,82 1,65±0,032
передней ноги 1,60-1,72 1,66±0,011 1,29-1,69 1,48+0,048
КИСТИ 0,28-0,56 0,42±0,024 0,30-0,46 0,38+0,012
ступни 0,61-0,71 0,66±0,009 0,61-0,73 0,67±0,010
задней ноги 1,60-1,87 1,76+0,020 1,33-1,78 1,55+0,034
Высота в холке 1^67—1,80 1,75±0,013 1,54-1,65 1,60±0,008
плеча 1,12-1,24 1,18±0,010 0,98-1,20 1,00+0,025
спины 1,33-1,69 1,51±0,031 1,34-1,59 1,46±0,027
поясницы 1,28-1,48 1,38±0,017 1,24-1,37 1,30±0,020
в крестце 1,28-1,52 1,40+0,025 1,26-1,45 1,35+0,026
седалищных 1,15-1,31 1,23±0,013 1,16-1,24 1,20+0,007
бугров
Обхват туловища 2,19-2,69 2,48±0,043 1,90-2,13 2,04±0,012
ПЯСТИ 0,22-0,28 0,25-0,005 0,21-0,27 0,24+0,005
плюсны 0,18-0,26 0,22±0,006 0,18-0,25 0,22±0,004
Ширина головы 0,34-0,41 О,39±О,ОО5 0,25—0,36 '0,30+0,009
в плечах 0,44-0,62 0,53±0,016 0,38-0,53 0,45±0,018
поясницы 0,48-0,70 0,59±0,022 0,47-0,65 0,57±0,016
таза 0,37-0,47 0,42±0,009 0,39-0,58 0,48±0,014
зада 0,24-0,36 0,30+0,010 0,31-0,50 0,40+0,022
Видовые особенности, а также специфичность В. b. caucasicus отчетливо выражены в структуре роговых стержней и чехлов. В отличие от бизона роговые стержни зубра относительно длинные, но слабо направлены назад, концы их не доходят до заднего края черепа, не отогнуты вниз ниже лобной поверхности. B.b. caucasicus отличается от B.b. bonasus тем, что роговые стержни опущены до уровня лба меньше, но отогнуты назад сильнее. По данным К.К.Флерова (1979), у самцов кавказского зубра верхняя сторона основания стержней находится на уровне лобной поверхности или немного ниже (менее 5 мм у самцов, не более 9 мм у самок). Для В. bonasus характерен сильный изгиб рогов, особенно резко выраженный у B.b. caucasicus. У самок кавказского зубра, у которых эта особенность выражена еще сильнее, рога изгибаются резко наружу, потом вверх, внутрь и назад; в результате концы сходятся к середине черепа, иногда достигая средней линии лба.
253
Зубр
На сравнительно большую длину рогов и значительную изогнутость рогов B.b. caucasicus издавна обращали внимание многие зоологи (Сатунин, 1903; Туркин, Сатунин, 1902; Динник, 1910, и др.)- Н.Я.Динник писал, что рога одного старого быка,измеренного им, имели окружность основания 33 и 35 см, а более молодого — 31 см. Длина этих же рогов по наружной поверхности колебалась от 33 до 42 см.
Из внешних признаков часто упоминается строение копыт. У зубра передние копыта почти прямые, концы их не сходятся и не налегают друг на друга. У бизона они имеют серпообразный изгиб и налегают друг на друга — концы наружных пальцев налегают на концы внутренних. Длина передних копыт взрослого быка беловежского зубра 90—105 мм, ширина одного пальца около 40 мм. Длина боковых копыт 40—55 мм. У B.b. caucasicus копыта короче и выше, чем у B.b. bonasus. Длина передних копыт у самцов составляет 70—85 мм, боковых пальцев 25—35 мм. Длина задних копыт 75—90 мм, боковых — 25—35 мм (Флеров, 1979).
Посткраниальный скелет видов и отдельных форм р. Bison изучен многими морфологами и освещен в литературе достаточно полно (см. обзор: Решетов, Суханов, 1979). Здесь мы обратили внимание лишь на отдельные особенности. В шейном отделе характерно сильное развитие остистых отростков шестого и особенно седьмого позвонков, однако у зубра, в том числе B.b. caucasicus, они в 2 раза короче, чем у бизона. Грудной отдел, включающий 14 позвонков, содержит ряд позвонков с очень удлиненными остистыми отростками. Среди них выдается по своей длине у B.b. bonasus и B.b. caucasicus второй и третий, а у бизона таковым оказался первый грудной позвонок. Длина остистых отростков грудных позвонков B.b. bonasus у самцов, по данным названных авторов, в среднем такова (мм): I позвонок — 37,5, II - 39,4, III - 38,3, IV - 35,9, V - 34,1. У B.b. caucasicus соответственно 33,9; 37,5; 36,6; 34,8; 32,5 см. У B.b. bison 52,1; 50,6; 47,8; 43,8; 39,9 см. Поясничный отдел включает пять позвонков, сходных у всех современных видов и форм р. Bison. Крестец также состоит из сросшихся пяти позвонков, абсолютная и относительная длина его минимальны у B.b. bonasus. В грудной клетке как зубра, так и бизона по 14 ребер, отличие зубра от бизона сводится к тому, что у второго ребра большая кривизна, чем у первого. Соответственно у зубра грудная клетка более плоская, чем у бизона.
Что касается поясов конечностей, то обращается внимание на некоторое усиление плечевого пояса зубра, его облегчение, особенно у кавказского зубра. Характерно, что лопаточная ось у зубра изогнута значительно меньше, чем у бизона. Тазовые кости зубра длиннее, но уже и легче, чем у бизона, в частности крыло подвздошной кости шире у второго вида, чем у первого. В отношении конечностей видовые различия в соотношении длины передних и задних ног не выявлены — как и у многих копытных задние конечности длинее передних. Однако отношение длины конечностей к длине туловищного отдела позвоночника у разных видов различно. Наиболее высокий показатель по этому индексу свойствен зубру (он более "длинноног"). Характерна также меньшая массивность костей конечностей зубра,
254
Bison bonasus Linnaeus, 1758
чем у бизона. Среди внутривидовых форм зубра более массивным костяком пояса передних конечностей выделяется кавказский зубр. Кроме того, кости конечностей этого подвида относительно короткие, что соответствует жизни в условиях гор.
Из костей конечностей видов р. Bison, очевидно, определенный систематико-таксономический вес может иметь таранная кость. Данные А.В.Стар-кина (1988) показывают, что бугорок внутреннего гребня таранной кости хорошо выражен у бизона, но отсутствует у зубра. Этот бугорок имеется у целого ряда вымерших и современных копытных открытых ландшафтов (сайгак, тур, северный олень и др.).
Внешний облик видов р. Bison существенно варьирует. У зубра в отличие от бизона голова менее массивная, горб умеренной высоты, грудная клетка, хотя и массивная, но более плоская, профиль спины слабо изогнут, зад относительно хорошо развит и не так опущен, как у бизона. Экстерьер кавказского зубра аналогичен беловежскому зубру, но более легкий. Для зубра характерен относительно длинный хвост, достигающий скакательного сустава (у бизона не достигает). Длина его 50—60 см, а с кистью длинных волос на конце — 80—100 см (Соколов, 1959).
Волосяной покров на теле очень неравномерный. Наиболее длинные волосы имеются на темени и между рогами, образуя челку (у бизона челка больше). У кавказского зубра челка короче, чем у беловежского, ее волосы извитые. На щеках волосы короче, но сильно удлинены на нижней стороне головы, образуют бороду, переходящую в подвес нижней части шеи и труди. У самцов длина волос и подвеса к старости уменьшается, а у коров, наоборот, увеличивается. Старые быки бывают лишены бороды и подвеса. У кавказского зубра борода не длиннее 250 мм, подвес около 150 мм. Волосы на плечах и лопатках короче, чем на гриве, но длиннее, чем на задней части тела. Относительно длинные волосы имеются вдоль хребта, образуя гребень. У беловежских зубров этот гребень сплошной, у кавказского несколько прерывается над лопатками. Характерно, что волосяной покров туловища кавказского зубра более извитый, а на брюхе коров нет длинных волос, что характерно для беловежского зубра. Существенно различаются оба подвида характером волосяного покрова хвоста — у беловежского зубра, особенно зимой, хвост от самого корня покрыт длинными волосами ("лошадиный хвост"), а у кавказского проксимальная часть хвоста имеет всегда, как у быков, так и у коров, короткие и курчавые волосы. Внутренняя сторона проксимальной части длиной 2/3 хвоста у кавказского почти голая. У беловежского эта голая полоса короче и составляет 1/3 длины хвоста.
Окраска зубра довольно однотонная, обычно буро-коричневая. При этом задняя часть тела светлее передней. Заметно выделяются насыщенностью темной окраски бока, низ головы, борода и подвес. Аналогичная темно-коричневая окраска имеется на конечностях, а также по средней линии шеи и спины. Более светлая окраска имеется на холке и плечах. Волосы подшерстка светлее ости, они имеют желтовато-бурый цвет. Окраска волос у самцов и самок почти одинаковая.
255
Зубр
Таблица 46. Длина волос зубра (в мм) (по: Соколов, 1979)
Категории ВОЛОС
Проба Направляющие Остевые I порядка
limit | х ± т limit | х + т
Самка, июль 1974 г.
Загривок 46-67 67* 36-40 39±0,7
Крестец 49-65 56±2,7 40-49 44+1,5
Бок тела 26-33 29+1,0 26-30 28±0,6
Брюхо 25-47 36±3,4 19-27 23±0,9
Самец, январь 1975 г.
Загривок 61-71 67±3 63—86 79+3,0
Крестец 69—82 72±2,6 63-84 78+4,1
Бок тела 38-48 41±1,6 28-37 32+0,7
Брюхо 52—76 76* 53-57 56±1,1
оборв
Бизон самец, Приокско-террасный заповедник »
сентябрь 1974 г.
Загривок 59-102 102* 58-85 85*
Категории волос
Проба Остевые II порядка Пух
limit | X ± т limit | х + т
Зубр самка, июль 1974 г.
Загривок 24—28 25±0,6 23-29 26+0,8
Крестец 36-41 39+0,9 24-33 27±1,1
Бок тела 12-13 12,35+2,4 10-12 11+0,3
Брюхо — — 17-33 29+0,6
Зубр самец, январь 1975 г.
Загривок 68-80 74+2,2 39-43 40±1,1
Крестец 36-38 37±0,7 31-41 35+1,9
Бок тела 30-36 33±0,8 30-34 33+0,5
Брюхо 48-56 51±1,3 37-52 42±2,2
Бизон самец, Приокско-террасный заповедник,
сентябрь 1974 г.
Загривок — — 62-75 67+2,1
* Максимальная
длина
Кожный покров. Материал различных участков кожного покрова 4 самцов зубробизона в возрасте 2—4 лет, улучшенных зубров, с кровностью 58/64 зубра + 6/64 бизона, добытых в феврале в Кавказском заповеднике и взрослой самки, добытой в Кабардино-Балкарии в Нальчикском лесохозяй-стве в феврале 1990 г. Для сравнения использованы 3 самца тех же возрастов, VII—VIII поколения поглотительного скрещивания на зубра со следами крови бизона, добытых в августе в Хоперском заповеднике. По морфологии волосы самцов зубра подразделяются на три категории: направляющие, остевые и пуховые.
256
Bison bonasus Linnaeus, 1758
Таблица 47. Толщина волос зубра и сердцевины в наиболее широкой части стержня волоса (в мкм) (по: Соколов, 1979)
Категориия ВОЛОС
Проба Направляющие остевые I порядка
limit | М ± т limit | М + т
Зубр самка, июль 1974 г.
Загривок
ВОЛОС 95-123 116±8,1 67-112 97±6,6
сердцевина 50-67 63±3,9 34-84 52+2,9
Крестец
ВОЛОС 129-140 132+4,5 129-140 131±3,7
сердцевина 78-95 59+2,3 84-112 197±4,3
Бок тела
ВОЛОС 95-112 205±3,0 61-78 68+1,9
сердцевина 56—73 66+1,4 16-50 41±4,5
Брюхо
ВОЛОС 166-118 110+1,7 84-101 90+2,6
сердцевина 45-67 58±2,7 45-62 52+2,1
Зубр самец, январь 1975 г.
Загривок
ВОЛОС 129-140 127±8,1 90-112 93±3,4
сердцевина 56-84 69+4,0 34-67 46+5,0
Крестец
ВОЛОС 129-140 131±3,1 67-112 88+6,3
сердцевина 62-78 74±2,6 17-67 33+8,9
Бок тела
ВОЛОС 84-129 108+6,0 73-101 85±4,4
сердцевина 28-72 50+6,1 17-39 27+2,2
Брюхо
ВОЛОС 112-157 12б±8,2 90-106 98±2,9
сердцевина 50-106 70-9,9 17-39 23+2,3
Направляющие волосы длинные, толстые, слегка изогнутые, на всем протяжении имеют примерно одинаковую толщину, утончаясь к вершине и основанию. Поперечник направляющих волос имеет округлую форму, кутикулярный рисунок некольцевидной формы. В отличие от пуховых направляющие волосы имеют сердцевину.
Остевые волосы слабо извитые, статистически достоверно разделяются на два размерных порядка. Длина остевых первого порядка и направляющих волос примерно одинакова. Остевые второго порядка значительно короче (табл. 46). Верхняя часть стержня остевого волоса расширена в небольшую гранну и в поперечнике овальна. Кутикула остевых волос некольцевидной формы. В области гранны наружный край кутикулы имеет мелкие зазубрины, а в прикорневой зоне ровный.
Пуховые волосы наиболее многочисленные (табл. 47—49), сильно извитые, тонкие и короткие. Форма поперечника сходна с таковой направляющих волос. Сердцевина пуховых волос прерывистая или отсутствует. Рисунок кутикулярных.чешуй не кольцевидной формы. Волосы растут поодиночке, не образуя пучков.
17 В.Е.Соколов, А.К.Темботов
257
Bison bonasus Linnaeus, 1758
Таблица 47 (окончание)
Проба Категории волос
остевые II порядка пуховые
limit I х ± т limit | х ± т
Зубр самка, июль 1974 г.
Загривок
ВОЛОС 67-78 74±2,3 50-62 58±1,6
сердцевина 22-39 27±2,5 22-28 26±1,1
Крестец
волос 84-112 92±4,0 44,8-50,4 45±0,7
сердцевина 45-78 57±5,5 16,8-22,4 17±0,7
Бок тела
ВОЛОС 67-101 80±4,7 22-45 34±3,8
сердцевина 45-62 52±4,5 И 22±0
Брюхо
ВОЛОС — — 78-84 82+1,3
сердцевина — — 50-62 52±6,0
Зубр самец, январь 1975 г.
Загривок
ВОЛОС 56—62 59±1,2 34-50 38±3,0
сердцевина 17-28 18,7±19 Участки прерывистой
сердцевины
Крестец
волос 39 — 56 47±4,9 39-50 46+1,9
сердцевина 11 11±0 6-11 10±1,1
Бок тела
ВОЛОС 62-84 69+3,1 39-50 45+1,3
сердцевина 17-28 19+1,3 6-11 11±0
Брюхо
волос 62-73 66±1,7 39-45 43±2,8
сердцевина 11 Участки прерывистой сердцевины
У самки волосяной покров представлен остевыми и пуховыми волосами. Категория направляющих волос в данных пробах нс обнаружена. Мех высокий, густой. Общая окраска темно-бурая. Волосы расположены в два яруса. Самый верхний ярус составляют длинные темные волосы, однотонно пигментированные по всей длине. Они относительно жесткие, волнистые. Их длина колеблется от 85 до 120 мм. По длине их условно можно разделить на пять размерных порядков. По толщине они более или менее одинаковы. От основания к вершине стержень слегка расширяется. Толщина у основания колеблется в пределах от 105 до 120 мкм, в верхней половине стержня около 135 мкм. У некоторых остевых волос вершины расщеплены (секутся). Поперечные срезы по всему стержню округлой формы. Сердцевинный канал хорошо выражен вдоль всего стержня и составляет 67% от его диаметра. Корковый слой развит неравномерно вокруг стержня. Сердцевинный канал сдвинут вбок от центра (рис. 35, Б). Сердцевина имеет сетчатое строение и образована губчатой тканью с мелкими ячеями.
258
Bison bonasus Linnaeus, 1758
Таблица 48. Густота волос у самца зубра зимой и летом * _________________(по: Соколов, 1979) ________г——
Проба Волосы Количество волос на 1 см2 поверхности кожи Проба Волосы Количество волос на 1 см^ поверхности кожи
Загривок Зима Остевые 500 Загривок Лето Остевые 416
Грудь Пуховые Остевые 1750 416 Грудь Пуховые Остевые 666 333
♦ По одному животному Пуховые 1166 Пуховые 250
Таблица 49. Отношение наибольшей высоты кутикулярных клеток к ширине ______________остевого^волоса на загривке (по: Соколов, 1979) ___________
Заповедник Высота клеток, мкм (п = 30) — у t Ширина волоса, мкм (п= 10) Отношение высоты клеток к ширине волоса
Бизон
Пр ио кско -терр асный 17 112 1:6,5
Зубр
Хоперский 15 100 1:6,6
Кавказский 15 106 1:6,7
Беловежская Пуща 15 77 1:5,8
15 93 1:6,1
Нижний ярус включает остевЬте и пуховые волосы. Остевые волосы светлее остей первого яруса и мягче, волнистые по всей длине. Длина остевых волос этого яруса не превышает 60,5 мм. От основания к вершине стержень сужается (в отличие от остей верхнего яруса). Толщина у основания около 120 мкм. Сердцевинный канал выражен хуже и составляет примерно 45% от диаметра стержня. Структура сердцевины такая же.
Рисунок кутикулы мало меняется вдоль стержня остевых волос. Чешуйки в основном вытянуты поперек волоса. Их высота изменчива, края неровные (рис. 35, В).
Пуховые волосы мягкие, сильно волнистые. Поперечные срезы округлой формы, сердцевина отсутствует. Пуховые волосы двух видов: "толстые" и "тонкие". Толщина "толстых" пуховых волос 60 мкм по длине всего стержня: их длина не превышает 54 мм. У "тонких" толщина 30 мкм, длина достигает 58,5 мм.
Рисунок кутикулы одинаков у пуховых волос двух видов. Чешуйки вытянуты поперек стержня. Высота чешуй более или менее постоянна (рис. 35, Г). Свободные края относительно ровные и плотно прилегают к корковому слою.
17*
259
Зубр
Рис. 35. Строение волос взрослой самки зубра
А — поперечный срез остевого волоса, ув. 600; Б — продольный срез остевого волоса, 1500; В — кутикула остевого волоса, ув. 600; Г — кутикула пухового волоса
Строение кожи зубра типично для наземных позвоночных. Промеры кожи и волосяного покрова приведены в работе В.Е.Соколова (1984). Здесь мы отметим, что эпидермис относительно тонкий,, за исключением кожи конечностей, и в большинстве случаев состоит из основного слоя, шиповатого, зернистого и рогового. Основной слой образован цилиндрическими клетками, среди которых встречаются и пигментные клетки. Степень пигментации эпидермиса варьирует в зависимости от сезона года (летом слабая, зимой более интенсивная). Шиповатый слой в зависимости от его толщины имеет от 3 до 20—36 рядов клеток. Зернистый слой имеется в коже всего тела и обычно образован 1—2 рядами клеток, но в некоторых участках тела отдельных зверей он иногда полностью отсутствует. Тонкий блестящий слой, как правило, имеется в коже конечностей, но у некоторых особей он встречается и в эпидермисе кожи различных участков туловища (например, в коже крестца и спины).
260
Bison bonasus Linnaeus, 1758
Таблица 50. Сумма диаметров (средняя, в мм) потовых желез, измеренных на уровне волосяных луковиц на протяжении 1 см препарата у самцов зубров
____________________ (по: Соколов, 1979)_______________________________
Проба Зима (п-4) Лето (П=3) Проба Зима (л =4) Лето (л=3)
Загривок 1,0 2,17. Брюхо под крестцом 3,37 5,62
Середина спины 0,99 1,60 Локоть 1,91 2,18*
Крестец -1,23 1,94 Запястье 1,82** 2,08*
Бок под загривком Середина бока тела 1,43 0,41* 2,23 2,46* У копыта передней ноги 0,39** —-
Бок тела под крестцом 1,00 2,05 Колено 1,15** 2,24
Грудь 1,58 2,21 Пятка 1,25** 2,01
Середина брюха 2,12 2,13 У копыта задней ноги 0,39** —
♦Среднее по 2 экз. **Среднее по 3 экз.
Дерма образована плотной вязью пучков коллагеновых волокон и эластиновыми волокнами. Эластиновые волокна в коже туловища более многочисленны, чем в коже конечностей. Пучки коллагеновых волокон с глубиной утолщаются, а на границе с подкожной мускулатурой вновь становятся тоньше. Жировые клетки в дерме, как правило, отсутствуют. Граница между сосочковым и сетчатым слоями проходит на уровне луковиц волос или нижних отделов потовых желез.
В сосочковом слое относительно тонкие пучки коллагеновых волокон проходят в основном параллельно поверхности кожи. В этом же направлении располагаются и тонкие эластиновые волокна. В коже труди и копыт имеются различной высоты дермальные сосочки. В верхних и средних частях сетчатого слоя пучки коллагеновых волокон проходят в самых разнообразных направлениях, но в глубоких слоях вновь занимают горизонтальное положение. Здесь встречаются толстые эластиновые волокна, которые нередко образуют сплетения и даже рыхлые пучки. Иногда в глубоких отделах сетчатого слоя попадаются небольшие труппы жировых клеток, достигающих размеров 56 х 84, 56 х 78 мкм. Жировые клетки в подкожной клетчатке, как правило, встречаются редко.
Сальные железы обычно состоят йз нескольких лопастей. Располагаются они позади волос, и их протоки открываются в волосяной фолликул. Сальные железы в коже туловища крупнее, чем в коже конечностей. Летом размеры сальных желез в некоторых участках тела сильно увеличиваются (например, кожа труди и крестца).
Потовые железы имеют более или менее извитые секреторные отделы и прямые выводные протоки, открывающиеся в волосяные воронки позади волос, недалеко от поверхности кожи или прямо на поверхности кожи рядом с волосяной воронкой. Диаметр секреторных отделов потовых желез примерно одинаков по всему телу. Более многочисленные и сильно извитые потовые железы встречаются в коже брюха. Летом потовые железы в основном больших размеров, активно секретирующие (табл. 50).
26 1
Зубр
Таблица 51. Параметры крови бизона и зубра
Бизон, Северная Америка
Marler, 1975 Mehrer, 1976
Показатель молодые, оба пола взрослые, оба пола разный воз-
(л = 29) (л = 47) раст, оба пола (л = 163)
X ± о limit X ± О | limit X ± о
Гемоглобин, г/100 мл 16,6±1,3 12,5-19,0 17,2±1,5 14,2-21,0 16,92±1,43
Эритроциты, — — — — 10 080±1,43
Гематокрит, об.% 49,8+1,3 12,5-19,0* 50±4,5 42,6—61,1 47,11±4,06
Лейкоциты, 8000+2400 4000—14 200 6985+2082 4200±13 100 8030±1,41
ТЫС.
Лимфоциты, % 56,5+5 31—93 42+12 12+65 24,86+6,40
Нейтрофилы сегм., % 34±10 7—55 46±13 22-74 63,77±7,95
Нейтрофилы пал,.% — — — — — .
Моноциты, % 2 ±2 1-4 1±1 0-3 6,34±4,17
Эозинофилы % 7+5 1-22 10+5 2-23 3,98±3,31
Базофилы, Единичные клетки 0,77±1,01
%
♦ В оригинале опечатка.
Показатель Зубр, Центральный Кавказ
Наши данные
взрослые, 10 лет, самки
limit х ± т О Су
Гемоглобин, г/100 мл 16,8—19,8 18,3+0,94 1,50 8,20
Эритроциты, 8,74-9,82 9,27±0,38 0,54 5,83
Гематокрит, об.% 45-61 51,3±6,03 8,50 16,58
Лейкоциты, тыс. 2,10-4,05 2,90±0,53 0,91 31,32
Лимфоциты, % 51-79 64,25±6,83 11,81 18,39
Нейтрофилы сегм., % 12-33 22,0+5,10 8,83 40,14
Нейтрофилы пал,.% 4-6 4,75±0,55 0,96 20,16
Моноциты, % 2-8 6,25+1,66 2,87 45,96
Эозинофилы% 1-4 2,5±1,00 1,73 69,28
Базофилы, % Единичные клетки
Система крови. Литературных сведений по крови зубров нам известно мало. Это касается как чистокровных зубров Беловежской Пущи и других районов их рсакклиматизации, так и различных гибридов — зубробизонов. Как известно, на Кавказе реакклиматизация зубра разных кровностей проводится в пяти точках: Северо-Западный Кавказ, Кавказский заповедник
262
Bison bonasus Linnaeus, 1758
(основатель популяции — зубробизон), в верховьях р. Архыз, Тебердинский заповедник (основатель — чистокровный зубр кавказско-беловежской линии), Нальчикское лесоохотничье хозяйство, Кабардино-Балкария (основатель зубробизон с примесью крови кавказско-беловежской линии), Цейс-кий заказник, Северная Осетия (основатель из кавказско-беловежской линии), Ассиновский заказник, Чечено-Ингушетия (основатель из кавказско-беловежской линии). Из всех этих точек литературные сведения по крови мы имеем лишь по Кавказскому заповеднику (Камьянов, 1969). Недавно мы провели изучение крови зубра из Кабардино-Балкарии (Нальчикское лесоохотничье хозяйство). Таким образом, гематологические сведения имеются по зубрам Западного и Центрального Кавказа, где обитают гибриды с различной кровностью. К сожалению, мы не располагаем данными по крови чистокровного зубра ни с Кавказа, ни с равнин Европы. Лучше обстоит дело с гематологической изученностью американского бизона Bison bison. По этому виду выполнен ряд работ (Marler, 1975; Mehrer, 1976; Haines et al., 1977). Как сравнительный материал эти данные представляют определенный интерес.
Литературные и наши оригинальные данные по гематологии реакклима-тизируемых зубров на Кавказе сведены в табл. 51. Для их оценки необходимо сделать некоторые пояснения. Мы проводили сбор материала в поясе среднегорных широколиственных лесов Нальчикского лесоохотничьего хозяйства в феврале 1990 г. Высота местности здесь около 1000 м над ур. моря. Зубры находились в дикой природе и добывали себе корм сами, т.е. животные несли полную нагрузку, в том числе физическую, в связи с жизнью в горах и притом в снежную зиму. В горах Кабардино-Балкарской АССР зубры живут с 1959 г., однако и в последующие годы сюда были дополнительные завозы. Тем не менее необходимо брать в расчет существование популяции этих зубров в течение трех десятилетий в горах. Кстати, зубры первой партии (основатели) были взяты из Кавказского заповедника, где они прошли начальные этапы адаптации к горам. Именно в это время у них были проведены исследования крови Б.Т.Камьяновым (1969). Однако зубры содержались тогда в загоне, им давали подкормку и их существование более напоминало жизнь домашних животных, чем “дикарей".
К сожалению, литературные и наши данные по анализу крови восстановленных на Кавказе зубров еще недостаточны, чтобы сделать обобщающие выводы. Однако предварительные замечания представляют интерес. Судя по материалам табл. 51, на Центральном Кавказе (Нальчикское лесо-охотничье хозяйство) кислородная емкость крови зубров значительно выше, чем на Западном Кавказе (Кавказский заповедник). Причину, видимо, можно усматривать в различии продолжительности адаптации к горным условиям и физической нагрузки на животных. Влияние сезона года в данном случае, по-видимому, не могло сказаться, так как пробы брались зимой. То же самое можно сказать и в отношении роли пола животного. У американского бизона среди одновозрастных животных половые различия в картине крови не были выявлены (Marler, 1975). Было отмечено несколько достоверных различий (Р < 0,5) между взрослыми (от 2 лет и старше) и
263
Зубр
молодыми (от 9 мес до 2 лет). Возрастные изменения у бизона обнаружены и другими исследователями. Так, по данным H.Haines et al. (1977), у взрослых бизонов (2—3-летних) кислородная емкость равна 22,2 ± 0,5 мл О2/ЮО мл крови, концентрация НЪ — 17,1 ±0,5 г%, гематокрит — 45,6 ± 1,0%. У телят (10—40 нед соответственно 17 мл Оо/ЮО мл крови, 13,6 ± 0,6 г% и 41,1%. Число эритроцитов у взрослых 3,6 хЮ6 мм3, их среднеклеточный объем 52 мкм3. Среднеклеточный НЪ в эритроцитах у взрослых 19,1 ± 0,8, а у телят 15,9 ± 0,6 мг/клетка.
Статистически достоверные различия в крови 163 американских бизонов выявлены только в возрастных группах. В целом как у самцов, так и самок кислородная емкость крови бизонов высока: концентрация НЪ составляет 16,92 ± 1,43 мн/мм3, гематокрит — 47,11 ± 4,06%, число эритроцитов — 10,08 ± 1,43 мн/мм3, число лейкоцитов — 8,03 ± 1,41 тыс/мм. Лейкоцитарная формула: нейтрофилы — 63,77 ± 7,95%, лимфоциты — 24,86 ± 6,40, моноциты — 6,34 ± 4,17, эозинофилы — 3,98 ± 3,31, базофилы — 0,77 ± 1,01%.
Достоверно более высокая кислородная емкость крови бизона по сравнению с домашними полорогими того же района Северной Америки объясняется способностью бизонов к быстрому бегу (Marler, 1975). Что касается показателей крови зубров Центрального Кавказа, то и здесь, очевидно, сказывается суммарный эффект преадаптации и повышения функции красной крови. Некоторые минимальные данные у одной особи (самка) нашей выборки, видимо, были связаны с патологией. Она была тощей, но причин этому не было установлено. Известно, что гемоглобин, объем клеточных элементов, количество эритроцитов значительно ниже у фасциолезных буйволов, чем у контрольных (Chaudhri et al., 1988).
Индексы внутренних органов. Интерьерные показатели зубров в горах Кавказа более или менее полно могут быть охарактеризованы лишь на примере зубров Кавказского заповедника. Хотя здесь основателями популяции являются гибриды, но изучение, формирования особенностей морфо-физиологии этих животных в горах представляет определенный интерес. Тем более что получены данные, отражающие различные этапы натурализации восстанавливаемых зубров.
Первые данные, обобщенные С.Г.Калугиным (1968) и Б.Т.Камьяновым (1969), относятся к периоду 1952—1968 гг. Интерьерные особенности в 80-х годах по сравнению с 50-ми годами анализированы А.С.Немцевым (1988а). При оценке данных 50-х годов необходимо учитывать, что все животные были бракованными, и трудно судить, насколько по ним можно охарактеризовать оставшихся в то время в живых и их потомков. Не менее важно учесть, что отстрел зубров производился в разное время и животные были различной упитанности.
Данные С.Г.Калугина, приведенные в табл. 52, относятся к периоду с октября по декабрь и началу января, т.е. после завершения гона. Взрослые самцы были наименее упитанные, а масса молодых самцов и самок мало чем отличается от осенних. Как известно, зубры имеют максимальные по-
264
Bison bonasus Linnaeus, 1758
Таблица 52. Показатели массы тела (живая масса) и органов (в кг) зубробизона Кавказского заповедника в 1952—1962 гг. (по: Калугин, 1968) ____________
Возраст, лет и месяцев Пол Живая масса Голова Кожа Печень Сердце Язык
3 г. 9 м. Самец 356 17,7* 49,72 6U 168,54 10,96 2.05 5,76 14 2,81
4 г. 6 м. >> 589 39,90 52 98,47 6,7 11,38 4,92 2,21
4 г. 7 м. >> 629 24,6 39,11 68. 108,11 6,0 9,54 4,61 1,91
4 г. 8 м. >> 511 23.0 45,01 41 84,15 6,8 13,31 6,07 2,35
4 г. 9 м. » 581 24,4 42,00 51 98,11 10,67 5,51 2,24
5 л. 4 м. >> 557 42,19 52 105,92 6.3 11,31 3.0 5,39 2,15
5 л. 6 м. » 499- 24,2 48,50 60 120,24 6,5 13,03 6,61 2,40
6 л. 9 м. Кастрат 452 20,4 45,13 54 110,29 4.7 10,40 3.0 6,64 2,88
7 л. 8 м. >> 680 25,0 36,76 75 110,29 7,5 11,03 3,7 5,44 1,4 2,06
7 л. 10 м. >> 586 22.0 37,54 56 85,32 6.8 11,60 5,29 Ы6 1,98
9 л. 2 м. Самец 743 25,3 34,05 71 98,25 2*2 12,38 4,0 5,3& 14 1,88
9 л. 3 м. >> 694 24,8 35,73 71 102,315 24 12,97 3,8 5,48 1,87
23 г. 6 м. >> 602 21.6 35,88 2 98,01 8,1 13,46 5,48 1,99
*Над чертой — в килограммах, под чертой — в процентах.
казатели массы тела в июле-августе. С такими оговорками обратим внимание на ряд показателей. Так, убойная масса самцов разных возрастов в процентах от живой массы колеблется от 48 до 68%. Она не ниже соответствующих показателй крупного рогатого скота. Максимальная убойная масса выявлена у самцов 3- и 4-летнего возраста. С 7—8-летнего возраста у самцов убойная масса несколько снижается, что объясняется, как это было сказано выше, значительной потерей массы тела в связи с гоном. Кастраты, изученные автором, имели большую убойную массу (53—64%). Живая и убойная масса животных одного и того же пола и возраста значительно варьировали, что вызвано их индивидуальными особенностями.
Абсолютные и относительные показатели внутренних органов зубров Кавказского заповедника периода 1952—1968 гг. приведены в табл. 52. Учитывая, что для каких-либо обобщающих суждений выборка очень мала,-
265
Зубр
Таблица 52 (окончание)
Возраст, лет и месяцев Пол Живая масса Почки Селезенка Легкие Кровь Внутренний жир
3 г. 9 м. Самец 356 О 1 д 3,0 26,0 22
2,53 3,09 8,43 73,04 7,58
4 г. 6 м. » 589 12 12 4Д 26,0 К2
2,04 2,04 7,30 44,14 7,13
4 г. 7 м. >> 629 1.2 и. 6.0 23.0 4,2
1,91 2,38 9,54 36,57 6,68
4 г. 8 м. » 511 12 1.3 4Л 25,0 3,6
2,35 2,54 8,61 48,92 7,05
4 г. 9 м. » 581 11 1,2 4,5 28,0 5,3
1,89 2,07 7,75 48,19 9,12
5 л. 4 м. >> 557 1,0 1Л 4.6 24,0 о 1
1,80 2,51 8,26 43,09 10,95
5 л. 6 м. » 499 ы и. 4,7 30,0 5Д
2,40 3,01 9,42 60,12 10,22
6 л. 9 м. Кастрат 452 Ы 1Д 4.25 28,0 10,6
2,43 13,10 9,40 61,95 23,45
7 л. 8 м. >> 680 1,з 1 д 4,9 35.6 17,5
1,91 1,91 7,21 52,21 25,74
7 л. 10 м. >> 586 1,3 1,4 4,5 29,3 16,7
2,22 2,39 7,68 50,0 28,50
9 л. 2 м. Самец 743 1Д Кб 5,0 36,0 9,6
1,88 2,15 6,73 48,45 12,92
9 л. 3 м. >> 694 1Д 1Л 5Д 34,5 1.2
1,87 2,16 7,35 49,71 1,73
23 г. 6 м. >> 602 К2 1,4 4,6 3,13 13,4
1,99 2,33 7,64 51,99 22,26
ограничимся лишь одним показателем сердца. Он довольно высокий, как у молодых самцов возрастом около 4 лет, так и самцов возрастом более 9 лет и самок более 23 лет.
Данные по интерьерным показателям зубров Кавказского заповедника 80-х годов приведены в работе А.С.Немцева (1988). Он считает, что индексы внутренних органов самцов зубров 80-х годов превышают показатели 50-х годов значительно: индекс сердца у самцов на 3,1%, легких — на 2,5%, печени — на 1,8%, почек — на 1,1%, селезенки — на 0,6%.
Не менее интересные результаты получены при сравнении индексов внутренних органов гибридных зубров (зубробизона) гор Кавказского заповедника и беловежско-кавказских зубров равнины (Окский заповедник) (Немцев, Киселева, 1987; Немцев, 1988). Согласно данным названных авторов, индекс сердца зубра на равнине составляет в среднем 5,7% у самцов, 6,1% у самок, тогда как в горах — 8,4 и 9,7%. Эти существенные различия характерны для органа, испытывающего особую нагрузку при изменении
266
Bison bonasus Linnaeus, 1758
парциального давления кислорода. На показателях не мог не сказаться тот факт, что беловежско-кавказские зубры, обитающие на равнине, крупнее, чем восстанавливаемые в горах Западного Кавказа. С другой стороны, полувековое обитание в горах, несомненно, могло привести к гипертрофии сердца, что столь характерно для горных популяций широко распространенных видов. С таким заключением согласуются данные, полученные еще в 50-х и 60-х годах С.Г.Калугиным (1968). Как отмечено выше, ивдекс сердца всех изученных зубров не достигал 7%. Все эти данные пока следует рассматривать как предварительные.
Сравнительное изучение биохимического статуса зубров Кавказского заповедника и Беловежской Пущи в зимний период проведено С.С.Елфимо-вой и др. (1989). В качестве интегральных показателей уровня метаболизма углеводов оценивалось содержание глюкозы в крови, активность глюкозо-6-фосфатазы, щелочной фосфатазы и содержание гликогена в печени и мышцах. Оказалось, что у кавказских и беловежских зубров содержание глюкозы в крови почти одинаково. Изменение ее содержания отмечено в связи с возрастом и полом (более высокое у молодых и самцов, а у самок ниже в среднем на 33%). В печени беловежских зубров концентрация гликогена составила 2,4±0,3 мг/г, в мышцах — 1,7±0,2 мг/г ткани. У зубров Кавказского заповедника содержание гликогена в печени оказалось в 3 раза выше и соответственно активность глюкозо-6-фосфатазы и щелочной фосфатазы ниже, чем у беловежского. Иными словами, энергетические запасы кавказского зубра, обитающего в горах, в 3 раза выше, чем у зубра равнины с относительно благополучными условиями жизни. Как известно, в зоологической литературе немало данных, свидетельствующих о том, что в экстремальных условиях (например, в Субарктике; Шварц и др., 1969) содержание гликогена увеличено и соответственно возрастает индекс печени. Словом, относительно высокий индекс печени зубров 80-х годов, адаптированных в течение 50 лет в горах Западного Кавказа, имеет физиолого-биохимическую основу. Мы разделяем точку зрения авторов в том, что эти данные нужно рассматривать также в качестве предварительных и они нуждаются в дальнейшем подтверждении.
Кариотип. Согласно первоописанию (Mclander, 1959) хромосомный набор зубра, подтвержденный последующими исследованиями (Шумов, 1970— 1980 гг., по: Шумов, 1980; Орлов, Чудиновская, 1979; и др.) и нашими данными, включает 60 хромосом. Столько же хромосом у бизона и крупного рогатого скота. Х-хромосома всех названных животных сходна и легко идентифицируется, так как она единственная двуплечая хромосома (субме-тацентрик), имеющая крупный размер (почти равна первой паре аутосом). У-хромосома зубра также субметацентрик, но очень мелкая, почти в 3 раза меньше Х-хромосомы. У-хромосома крупного рогатого скота также мелкая субметацентрическая хромосома, а у бизона — в отличие от них мелкий акроцентрик. Однако существуют данные о том, что У-хромосома бизона может сильно варьировать, быть и акроцентриком и субметацентриком (Wurster, Benirschke, 1968 г., по: Орлов, Чудиновская, 1979). Все аутосомы как зубра, так и бизона акроцентрики, по размерам образующие непре
267
Зубр
рывный и слабоубываюхций длинный ряд. Соответственно количество плеч хромосом обоих видов одинаково и равно 62. У домашнего крупного рогатого скота также 2п = 62.
Кариотипы современных зубров Кавказа независимо от местообитания и основателя (В. bonasus х В. bison и B.b. bonasus х B.b. caucasicus) имеют один и тот же кариотип (2п = 60, NF = 62) и ничем не отличаются от такового описанных выше беловежского зубра и бизона. К сожалению, мы не располагаем дифференциально окрашенными хромосомами местных зубров. Между тем такая информация могла бы быть полезной в решении вопросов систематики и эволюции зубров Кавказа. Так, по данным В.Н.Орлова и Г.А.Чудиновской (1979), по характеру поперечной исчерченности можно идентифицировать с первой до двадцатой пары эдэомосом. Остальные в силу очень мелких размеров практически не поддаются идентификации. Анализ дифференциально окрашенных хромосом позволил установить, что все первые 20 хромосом, за исключением 2, 10, 12 и 14-й пар, по G-полосам сходны у зубра, бизона и крупного рогатого скота, что свидетельствует об их гомологии. С другой стороны, авторы не смогли констатировать с уверенностью видовых различий по G-полосам.
Для решения систематико-таксономических задач Северной Осетии обнадеживающие результаты получены при изучении районов ядрышковых организаторов и гетерохроматина у трех видов Bovidae, в том числе бизона (Mayer et al., 1987).
Структура и динамика ареала
Ареал вида в историческое время охватывал обширную территорию Западной, Центральной и Восточной Европы. По В.Г.Гептнеру и др. (1961), зубр как обитатель разреженных лесов, лесостепей и отчасти степей с приречными и водораздельными лесами проникал на север до Балтийского моря, примерно до уровня Рижского залива. Оттуда северная граница проходила южнее Москвы, далее на восток захватывала область Курска и Воронежа. Возможно, что зубр обитал по левому берегу Дона и правобережью Волги, в районе Волгограда. Южная граница простиралась до Азовского и Черного морей. От этого основного ареала было оторвано поселение зубра на Кавказе. Н.К.Всрсщагин (1959), В.Г.Гептнср и др. (1961) считают, что в середине прошлого века кавказская популяция зубра имела связь с их сородичами Восточной Европы по долине Дона. В.И.Громов (1935), наоборот, полагает, что еще в конце ледникового периода произошел разрыв единого ареала зубров, населявших Европу и Кавказ.
Распространение на Кавказе. Зубр относится к местным, аборигенным животным региона. Костные остатки древних зубров плейстоценового возраста (Bison priscus) и современных голоценового возраста обнаружены в ряде мест Предкавказья, Большого и Малого Кавказа и Закавказья, вплоть до оз. Севан. Вероятно, они проникали на юг до Талыша и Эльбурса. Мы разделяем точку зрения К.К.Флерова (1979) о том, что Bison bonasus caucasicus, как и древние зубры, занимали в прошлом весь Кавказ, в том числе
268
Bison bonasus Linnaeus, 1758
Закавказье, а также хребты Северного Ирана. В доисторическое время ареал его сократился очень резко. О былом распространении зубра в тот период данных мало, но те фрагменты, которые имеются, согласуются с мнением о широком его распространении в прошлом. Так, летописец Рашид эд-Дин оставил описание охоты на зубров в Талыше, организованной Абага-ханом в 1275—1276 гг. и Газан-Ханом в 1301—1302 гг. Возможно, зубры в этих местах сохранялись даже до XVIII—XIX вв. (Menetries, 1832).
Существование зубра в Западном Закавказье до прошлого века признают Н.В.Туркин и К.А.Сатунин (1902). Согласно им, еще 200 лет до нашего времени Архангело Ламбрети упоминал о "диком буйволе" из Мингрелии. По мнению Г.И.Радце (1899), в Абхазии зубры встречались до XIX в., чему решительно возражал Н.Я.Динник (1910). Вероятно, уже в конце XIX и начале XX в. зубров не стало по всему Черноморскому побережью и в горах Западного Закавказья. Н.Я.Динник (1910) писал, что мнение ряда зоологов о том, что зубры встречаются в Абхазии и спускаются почти до берегов Черного моря, совершенно необосновано. В верховьях Сочи, Шахе, Мзымты или Кодора "я следов их никогда не видел, и все охотники и полесовщики этих мест говорили мне в один голос, что зубры здесь не водятся вовсе" (с. 148). Однако надо полагать, что в Закавказье зубр дольше всего продержался именно в Абхазии, о чем говорят данные самого Н.Я.Динника. Периодическое появление зубра в этих местах из Кубанской области отмечал сам Н.Я.Динник. Необходимо учесть и то, что наиболее мощные леса и благоприятные условия жизни и защиты для зубра по сей день сохранились на южном и северном макросклонах Западного Кавказа.
Что касается истории зубра на Северном Кавказе, где это животное просуществовало до 20-х годов XX в., то она во многом поучительна. Такое гигантское и уникальное животное, как зубр, тысячелетиями существовало в горах древнего Кавказа, на него устраивали ханские охоты и его убивали на протяжении многих столетий, но фактический материал в виде одной шкуры попал в руки специалиста только в 1835 г. По этому единственному экземпляру академик Бэр в 1836 г. составил описание кавказского зубра. Однако это сообщение не убедило специалистов в существовании на Кавказе зубра. Еще в 60-х годах XIX в. бытовало мнение, что на Кавказе обитает не зубр, а индийский бык, -возможно гяур (проф. Усов). Спор разрешился только в 1867 г., когда живой кавказский зубр был доставлен в зоосад Москвы. В самом Кавказском регионе первый достоверный музейный материал попал благодаря Г.И.Радце в музей Грузии только в 1869 г. О зубре и связанных с ним противоречивых толках было известно научной общественности России и Западной Европы давно. В частности, Жан де Люк, описывая в 1625 г. черкесов и их страну, простиравшуюся в то время от Темрюка до Дербента, отмечал, что черкесы вместо стаканов пользовались рогами диких буйволов (Жан де Люк, 1878 г., по: Кириков, 1979). Очевидно, имелся в виду зубр. Совершенно определенно о зубре на Кавказе было сказано в указе кабинета русского царя, датированном 31 декабря 1739 г. Астраханский обер-комендайт обязывался прислать в Москву, в Измайловский зверинец, живых зверей. В дополнение к этому приказу читаем: "Еще из
269
Зубр
вестно нам, что в Кабардс есть дикие быки и кдосы, которые по-тамошнему называются домбаи, того ради имеете вы всячески стараться, не жалея на то употребить несколько из казны нашей денег, чтобы тамошные князья одного рода бычков и телок молодых по 5 или 10 велели ловить... от-йравлять с прочими зверьми в Москву, и о том же имеете вы от себя писать и к Шамхалу Горскому, также в Кизлярскую крепость к коменданту и Эльмурзе Черкасскому, чтобы в ловле и присылке оных бычков и телок старание приложили" (Полное собрание законов Российской империи с 1649 года. Т. 10, с. 7994).
Нам представляется достоверным сообщение академика Ловича, обнаруженное потом П.Палласом (Pallas, 1831. Т. 1, по: Кириков, 1979), о том, что в 1776 г. в верховьях р. Кумы, недалеко от горы Бештау, был убит огромный зубр. В том же сообщении приведены сведения АЛ.Гюльдснштедта о нахождении им в пещерах долины р. Урух черепа зубра. Позднее этот уникальный материал изучали специалисты, в том числе В.И.Наниев (1956), который пришел к убеждению, что в пределах Северной Осетии зубры жили до конца XVIII — начала XIX в. С таким заключением согласуются и данные Н.Я.Динника (1899, 1910). Уместно отметить, что среди черепов из пещеры Дигоризед оказался один череп, помеченный "1833 г.“. Н.Я.Динник указывает также, что его поиски следов зубра или очевидцев в разных местах бассейна Уруха и прилегающих районов не увенчались успехом. Лишь старики рассказывали, что их отцы и деды охотились на зубров. Таким образом, надпись на черепе зубра "1833 г.", очевидно, указывает, что в 30-х годах XIX в. добыча зубра на Центральном Кавказе была редкостью, событием для охотников. Вероятно, такое же положение наблюдалось в то время и в бассейнах рек Малка и Теберда. В этом районе, выделенном нами в отдельный эльбрусский вариант поясности, порой совершенно недоступные леса были в глубоких ущельях, а платообразные вершины хребтов представляли уникальные луга — пастбища. В отличие от В.Г.Гептнсра и др. (1961), учитывая былую растительность и орографию района Пятигорье-Эльбрус, мы считаем заслуживающим внимания сообщение академика Ловича о добыче зубра в верховьях Кумы в 1776 г. Думается, что прав и Н.Я.Динник (1910), говоря, что название ущелья Домбай-Ольген, расположенного в верховьях Тебсрды, свидетельствует также о существовании в прежние времена по соседству с Эльбрусом зубров-домбаев. Однако зубры в этом районе погибли, видимо, раньше, чем в бассейне Терека, не говоря уже о бассейне Кубани. Дело в том, что лесистые ущелья и балки бассейна рек Малка-Теберда имели островной характер, они разобщены платообразными вершинами Мелового и Скалистого хребтов. Они издавна использовались горцами в качестве летних пастбищ, и пастухи здесь постоянно охотились на диких зверей.
Изложенные выше и другие многочисленные факты говорят о том, что к середине XIX в. зубр на большей части территории Кавказа был уничтожен, сохранившись лишь на северо-западном Кавказе. Судьба кавказского зубра в этом районе Кавказа прослежена лучше и отражена в ряде охотничьих записок и специальных исследований. Характеризуя вторую полови
270
Bison bonasus Linnaeus, 1758
ну XIX в., Н.Я.Динник писал в 1910 г., что в течение последних 30—40 лет область распространения зубров сужается чуть не с каждым годом; в 60-х годах зубры встречались в верховьях Большого Зеленчука, в долине Иркы-за, а в 40-х — в верховьях Урупа. В 1884 г., путешествуя по Большой Лабе, Динник находил тропы зубров в вековых лесах Загдана, расположенного вблизи истоков названной реки. Генерал Кеслер сообщал, что в бассейне Лабы, в горных лесах, зубры бродили стадами (Миддендорф, 1869).
Сходные данные приводит А.Ф.Виноградов (1870). Согласно им в 60-х годах XIX в. зубры встречались в лесах верховьев рек Малого и Большого Зеленчука, Урупа и Лабы. Количество их составляло около 2000 особей. По имеющимся данным, через три десятилетия это поголовье сократилось в 4 раза, что было связано не только с браконьерством, но и промышленной заготовкой леса. Знатоки фауны Кавказа — Н.Я.Динник, К.А.Сатунин и др., отмечали, что зубры не выносят деятельности человека в связи с лесоразработкой "и поэтому, если начинают близ занимаемого ими участка рубить лес, они сейчас же покидают его и переселяются в другое спокойное место" (Туркин, Сатунин, 1902)ЛОрганизация в 90-х годах Кубанской великокняжеской охоты на территории в 522 тыс. десятин нарушила спокойствие зубров. Они были загнаны в глухие, малодоступные горные леса и, судя по всем данным, находились в угнетенном состоянии.
Зубра нередко характеризуют спокойным, любопытным, даже флегматичным, но очень умным животным. Несмотря на эти его черты, в 90-х годах прошлого века увидеть зубра в лесах Кубанской охоты стало крайне сложным делом. Судя по очеркам В.Шильдера (1895), в кубанских охотах князья бывали задействованными до 86 человек (1894 г.), но встречи с зубром были крайне редки. По его словам, известный английский^спортсмен и охотник Д.СЛитльдел три месяца в 1888 г. безрезультатно гонялся за зубром. Цели своей он достиг лишь в 1891 г. благодаря "знаменитому лезгину Лабазану, полуразбойнику, браконьеру и знатоку этих мест" (с. 9). Бык и корова, добытые им, были отправлены в Британский музей.
Литературные данные, в том числе Д.Филатова (1912), свидетельствуют о том, что к концу первого десятилетия XX в. зубры встречались только между верховьями рек Белая и Лаба в количестве всего нескольких сот голов. Более точное определение поголовья зубров, по мнению названного автора, было невозможно. Данные его показывают, что к тому времени зубры были оттеснены в пояс темнохвойных лесов, где они лучше скрывались от людей благодаря соответствию окраски тела и фона субстрата. В неподвижном состоянии, несмотря на отромные размеры, зубра увидеть в таких лесах было трудно. По данным Н.Я.Динника (1910), область высотного размещения кавказского зубра простиралась примерно от 1000 до 2300 м над ур. моря.
Период тражданской войны и интервенции, а также процветание браконьерской охоты предопределили судьбу кавказского зубра. Организация в 1924 г. Кавказского заповедника на месте бывшей Кубанской охоты не смогла уже спасти его — слишком подорвана была численность зубра в горах Кавказа, впрочем как и в Беловежской Пуще, не говоря уже о
271
Зубр
Карпатах. Хронология гибели последних зубров на Кавказе трактуется по-разному, но, очевидно, к истине близки данные И.Башкирова (1939): сведения о добыче зубра в 50 км западнее г. Майкопа (окрестности ст. Линейной) отмечены в 1921 г., близ г. Гефо — в 1923 г., на Алоусе и Мастакане, верховья Кодора, — в 1925—1926 гг. По мнению других териологов, последний зубр был убит в 1927 г. (Соколов, 1959; Заблоцкий, Заблоцкая, 1987). Во всяком случае, с 1928 г. никаких достоверных фактов о том, что в естественной природе сохранился кавказский зубр, в печати не появлялось. В 1923—1945 гг. во всем мире живых зубров Bison bonasus оставалось 54 головы известного происхождения. Они содержались в частных руках и зоопарках Европы. Среди них чудом оказался один самец кавказского зубра В. bonasus caucasicus. Он был молодым бычком отловлен на Западном Кавказе в 1908 г. и содержался в Штеллингене (Германия).
Современные тенденции антропогенных изменений ареала. Новый этап в истории зубра на Кавказе начат в 1940 г., когда из Аскании-Нова в Кавказский заповедник был завезен межвидовой гибрид зубра и бизона (В. bonasus X В. bison). К этому времени в СССР не было чистокровных зубров. Ныне живущие в Кавказском заповеднике зубры являются потомками этих гибридов (зубробизонов). Из Кавказского заповедника зубробизонов завезли в Кабардино-Балкарию (Нальчикское лесоохотничье хозяйство) в 1959 г. Позднее в этом хозяйстве дополнительно были выпущены зубры, но уже другой линии — кавказско-беловежской (В. bonasus bonasus х В. bonasus caucasicus). В итоге в Нальчикском лесоохотничьем хозяйстве создана сложная гибридная популяция зубров. В других районах Северного Кавказа реакклиматизация зубров проводилась за счет чистокровных кавказско-беловежских зубров (В. bonasus caucasicus х В. bonasus). Местами выпуска являются Архызский участок Тебердинского заповедника (1968), Цейский заказник Северной Осетии (1964) и Ассиновский заказник Чечено-Ингуше'Тии, в верховьях .Фортанги, близ границы с Северной Осетией (1969). В этих трех точках проводились дополнительные выпуски, но каждый раз завозились только чистокровные зубры из кавказско-беловежской линии.
Во всех районах выпуска зубры существуют по сей день, но натурализация их проходит неодинаково. Наиболее значительное поселение сформировалось на северо-западе Кавказа с основным поголовьем в пределах Кавказского заповедника, где основателями популяции являются гибриды зубробизонов. По последним данным (Немцев,. 1988а), зубры в этом районе занимают около 140 тыс. га заповедной и незаповедной территории. Эта территория находится в верховьях рек Белая и Лаба (рис. 36). Высотные пределы распространения на заповедной территории составляют 620—2900 м над ур. моря, вне заповедной — от 470 до 2300 м над ур. моря. Как видно, современное распространение восстанавливаемого зубра на Северо-Западном Кавказе близко к состоянию ареала аборигенного зубра в конце XIX'в., но все же меньше его, так как северо-западная и юго-восточная ipa-нйцы ареала более сужены. Однако имеет место тенденция к расширению ареала. Так, на левобережье Большой Лабы часто заходят зубры из Кав-
272
Bison bonasus Linnaeus, 1758
Рис. 36. Схема маршрутов сезонных передвижек зубров
а — миграции, б — кочевки, в — локальные подвижки, г — редкие заходы животных (Немцев, 1988)
казского заповедника (Красная книга Карачаево-Черкессии, 1988 г.). Общая численность зубров в Кавказском заповеднике по состоянию на 1984 г. составила 1270 особей (Немцев, 19876), что значительно ниже состояния 70-х годов XIX в. В соседнем Тебердинском заповеднике вольное стадо зубров из кавказскр-беловежской линии населяет ущелье Кизгич (Архызский участок), высотные пределы его распространения находятся на уровне от 1000 до 2500 м. Общая численность с учетом вновь завезенных зубров на 1987 г. определяется в 53 особи (Красная книга Карачаево-Черкессии, 1988 г.). В Кабардино-Балкарии зубры занимают бассейн р. Нальчик, пояс среднегорных широколиственных лесов и выше. Общее количество составляет около 300 голов, что, на наш взгляд, завышено. В Северной Осетии и Чечено-Ингушетии зубры занимают небольшие участки среднегорных широколиственных лесов и примыкающие к ним высокотравные луга Паст-
18 В.Е.Соколов, А.К.Темботов
273
Зубр
бищного хребта. Подробности их размещения не изучены, численность их в Северной Осетии — 212 особей, а в Чечено-Ингушетии — 29 особей (Лип-кович, 1988).
Будущее современных зубров на Кавказе специалисты представляют по-разному. Одни считают, что необходимо уничтожить всех гибридных зубров (потомков зубробизона), заменив их чистокровными животными кавказско-беловежской линии. Это мнение много лет отстаивает М.А. Заблоцкий, и он имеет своих последователей. На наш взгляд, оно заслуживает внимания, если учесть, что чистокровные зубры рано или поздно вступят в контакт с гибридными и тогда все население зубров со временем окажется гибридным. Эта угроза особенно велика на Центральном Кавказе, где бок о бок живут чистокровные зубры в Северной Осетии и гибриды в Кабардино-Балкарии. Исходя из этого, мы считаем, что в первую очередь эту угрозу необходимо снять, отстрелив гибридов Нальчикского лесоохотничьего хозяйства и заменив их чистокровными зубрами кавказско-беловежской линии.
Противоположное мнение, исходящее в первую очередь от сотрудников Кавказского заповедника (Калугин, 1967, 1968, 1975; Немцев, 1983, 19876, 1988а; и др.), сводится к тому, что гибридные зубры весьма жизнеспособны, "подогнаны" к условиям гор, хорошо изучены и более перспективны для существования на Кавказе. Наоборот, чистокровные зубры Центрального Кавказа, как и Тебердинского заповедника, судя по состоянию численности и размещению, не имеют перспектив либо из-за отсутствия естественных предпосылок для акклиматизации, либо из-за ошибок, допущенных при разведении. По мнению А.С.Немцева (19876), оценка перспектив увеличения численности и расширения ареала чистокровных зубров на Кавказе затруднена отсутствием планомерного изучения. С последним доводом трудно не согласиться. Однако мы не можем разделять мнение, что горные леса Северного Кавказа и ряда лесных районов Закавказья непригодны или малоперспективны для обитания чистокровных зубров кавказско-беловежской линии. Биологические особенности зубра (Raczynski, 1980) и современные ландшафтные условия Кавказа служат благоприятными предпосылками для восстановления зубра на Кавказе. Главные препятствия к достижению такой цели мы видим в отсутствии хозяйского подхода к природным ресурсам как единому целому, процветании браконьерства и других субъективных факторах. Вопрос о правомочности существования гибридов зубробизонов на Кавказе требует дальнейшего обсуждения.
Систематика и изменчивость
Систематическое положение. Таксономический статус современных зубров Европы не вызывает сомнения, при всей общности морфологии, морфофизиологии , цитогенетики и происхождения от одного общего предка целесообразность возведения в ранг самостоятельных видов зубра (Bison bonasus) и бизона (Bison bison) достаточно обоснована.
274
Bison bonasus Linnaeus, 1758
Что касается внутривидовой систематики и микроэволюционных процессов, связанных с акклиматизацией и реаклиматизацией, а также гибридизацией в новых ландшафтных условиях, то мы далеки от однозначных выводов. В этом плане заслуживает обсуждения положение внутривидовых таксонов Bison bonasus (в том числе В. bonasus caucasicus), ревизия систематико-таксономического веса тех или иных признаков. Вынужденно, сознательно или бессознательно, проводится серия природных экспериментов для их решения. С учетом такого подхода мы ниже приводим обзор суждений систематиков о реальности подвида В. bonasus caucasicus и оценки систематических его признаков.
Как отмеченов выше, первый экземпляр кавказского зубра попал в руки академика К.М.Бэра, составившего первое описание этого животного, отмечая его отличие от беловежского (литовского) зубра. На заседании Академии наук 21 декабря 1863 г. он доложил о своих выводах (по: Туркин, Сату-нин, 1902):
1) у кавказского экземпляра рога тоньше и короче, расстояние между рогами и ширина лба меньше, что связано с полом животного (шкура коровы);
2) цвет шерсти кавказской шкуры светлее и содержит много серых волос;
3) волосы на передних частях тела короче и курчавы только на лбу и отчасти на затылке, что обусловлено влиянием сезона (линьки) и возрастом животного;
4) копыта и ложные копыта гораздо' короче у кавказского зубра, что легко объясняется жизнью в горах;
5) концы рогов немного изгибаются назад;
6) вдоль спины идет весьма ясная темная полоса, чего нет у литовских зубров.
По мнению К.М.Бэра, отмеченные им морфологические особенности недостаточны для выделения нового подвида, однако его оппоненты, приводя еще дополнительные (ширина лба по отношению к длине черепа) признаки, находили, что сумма внешних и черепных особенностей кавказского зубра достаточно значительна и она скорее указывает на отличие его от номинального подвида, чем на их ‘сходство. Эту мысль наиболее полно изложили Н.В.Туркин и К.А.Сатунин (1902), что позднее легло в основу описания нового подвида зубра — Bison bonasus caucasicus Satunin (Сатунин, 1903).
Н.Я.Динник (1910), хорошо знавший копытных Кавказа, считал, что кавказский зубр отличается от беловежского многими особёностями: "...он несколько меньше ростом, имеет более тонкие ноги, не такие большие и притом сравнительно узкие копыта, не так сильно развитую триву и вообще волосяной покров на передней половине туловища и на голове, немного более тонкие рога и более светлый цвет шерсти ... шерсть, покрывающая заднюю половину его туловища, а также бедра и голени, закручивается в красивые мелкие завитки" (с. 144).
18'
275
Зубр
Особенности кавказского зубра, отмеченные Бэром, Туркиным, Сату-ниным и Динником, долго переходили из монографии в монографию, нередко даже в усеченном варианте (особенно черепные признаки) или несколько расширенно (размеры тела), без детального анализа краниологических показателей (Соколов, 1959; Гептнер и др., 1961; и др.). В моногра-фии "Зубр" (1979) этот пробел значительно восполнен К.К. Флеровым. Как отмечено нами в обзоре черепа и посткраниального скелета Bison bonasus, им выявлен целый ряд особенностей, свойственный кавказскому зубру. В характеристике подвида В. bonasus caucasicus акцентируются некоторые из них, в том числе:
— лоб, как правило, плоский, очень мало выпуклый, всегда присутствуют два вздутия при основании роговых стержней;
— опускание роговых стержней до уровня лба меньшее, чем у Bison bonasus bonasus;
— предглазничная яма выражена ясно, верхний край слезной кости в виде валика и поставлен вертикально. Мыщелки часто с рудиментами переднего загиба;
— носовая полость относительно мала, соответственно расстояние от нижнего края орбиты до края альвеолы много меньше длины ряда предкоренных и коренных зубов верхней челюсти;
— на слабо выражены бугор и валик на maxillare (tuber maxillare);
— нёбные отростки intermaxillaria по дорзальной поверхности вогнуты продольно, в общем кость длиннее, чем у B.b. bonasus, ее тупой конец подходит к nasalia ближе, чем длина М^, иногда даже соприкасается;
— рога у коров сильно загнуты внутрь, концы их близко сходятся, у быков они изогнуты крутой спиралью.
Что касается внешних признаков, то они мало отличаются от беловежских, но экстерьер более легкий у кавказского зубра.
Суммируя изложенное, можно заключить, что морфологическое своеобразие кавказского зубра довольно значительно, возведение в ранг подвида В. bison caucasicus обосновано. Однако, на наш взгляд, немалый интерес в теории и практике представляет анализ систематико-таксономического веса перечисленных и иных признаков вида Bison bonasus в целом и отдельных внутривидовых форм его. В частности, очевидно, закономерен вопрос, связанный с характером наследования отдельных признаков в различных районах Кавказа с варьирующими ландшафтными условиями, а также сходством или различием основателей популяции. Решение этого вопроса требует значительного материала, которого в настоящее время нет. Но мы считаем, что каждый экземпляр зубра Кавказа должен анализироваться под этим углом зрения.
Изменчивость. По В. bonasus caucasicus наиболее полные данные содержатся в работах К.К.Флерова (1932, 1965а, 1979) и В.И.Наниева (1956). Что касается изменчивости современных зубров Кавказа, то в Кавказском заповеднике она изучена рядом специалистов (Камьянов, Нусов, 1967; Камья-нов, 1969; Калугин, 1968; Немцев, 1982, 1985, 1988а; Немцев, Киселева, 1987).
276
Bison bonasus Linnaeus, 1758
Наряду с литературными данными мы имели возможность изучить 29 черепов из пещер Северной Осетии: 27 экз. из пещеры Дигоризад и 2 экз. из Кувандона у с. Лезгор, а также 4 экз. с Северо-Западного Кавказа. По современным зубрам, потомкам зубробизоновой линии, значительный коллекционный материал изучен в Кавказском заповеднике (102 черепа и элементы скелета от 83 особей). Из Нальчикского лесоохотничьего хозяйства, где обитают гибриды от двух линий (зубробизона и беловежско-кавказского зубра), нами получены 4 черепа и 1 полный скелет. Анализ всего материала позволяет нам составить некоторые суждения об изменчивости зубров разного происхождения и в различных районах.
Индивидуальная изменчивость. По массе тела кавказского зубра у нас нет данных. Неплохой материал имеется по черепам этого зубра, особенно по взрослым животным из пещер Северной Осетии (12 самцов и 12 самок). У самцов коэффициент вариации нами проанализирован по 29 параметрам черепа. Самой высокой вариабельностью отличается соотношение верхней поверхности лба и стержней — захождение стержня ниже поверхности лба (Cv—60, 76). Очень высока изменчивость наименьшей ширины височной ямы (Cv—40), а также длины анатомической лицевой оси, горизонтального поперечника стержня, длины темени. Более стабильны показатели длины анатомической мозговой оси черепа, наибольшая ширина затылка (на гребне) и др. Сходные результаты по индивидуальной изменчивости черепа В. bison caucasicus получены и по самкам. Однако по отдельным параметрам прослеживаются некоторые особенности. Так, длина анатомической лицевой оси более вариабельна у самок, чем самцов.
Индивидуальная изменчивость краниологических признаков у Bison bonasus caucasicus лесного пояса Северной Осетии7изученная на 98 черепах и 8 роговых чехлах из горной Дигории, бассейна р. Урух (пещеры и святилища Дигоризад, близ с. Задалеск, кувйндоны сел. Верхнего и Нижнего Лезгора), подробно рассмотрена В.И.Наниевым (1956). Обнаружен широкий размах показателей структуры роговых стержней, лобной поверхности продольной лобной впадины, впадины в области брегмы, боковых ямок лобной поверхности; продольной возвышенности, лежащей между лобной впадиной и надглазничной бороздой, надглазничной борозды, верхней поверхности орбиты, срединной лобной поверхности, теменной и затылочной области, орбиты, слезной и межчелюстной костей.
Возрастная изменчивость. Для зубров, как и для других видов Bovinae, характерны очень большие возрастные изменения в морфологии. Их обзор выходит за рамки нашей работы. Интересующимся этим вопросом рекомендуем монографию "Зубр" (1979), здесь же обратим внимание на те стороны возрастных изменений, которые должны быть учтены при решении систематике-таксономических и микроэволюционных задач.
В экстерьере зубра, как и многих других видов полорогих, наиболее значимые возрастные изменения происходят в структуре роговых чехлов. По К.К.Флерову (1979), у молодых примерно до 2,5 лет роговые чехлы матовочерного цвета, исчерченные продольными трещинами. Потом они стано
277
Зубр
вятся гладкими и черного цвета. Ниже этой гладкой верхушки начинают появляться годовые кольца (валики и борозды). У старых экземпляров эти кольца покрывают около половины или большую часть поверхности рогов. Один раз в жизни (на 10—11-м году) имеет место замена ювенильного сегмента, появившегося первым. У самцов остатки этого ювенильного сегмента опоясывают в виде хомутика конец постоянного рога. У самки они совсем сбрасываются, не оставляя следов. У старых зубров, особенно самцов, концы рогов притуплены, становятся короче, чем у более молодых. С учетом количества годовых колец и степени притупления вершин постоянных рогов можно определить возраст зубров.
Для определения возраста используют также закономерности формирования зубной системы: к двум годам сменяется Ij, в 2,5 года — I2, в 3 года — I3, в 4 года — Сц моляризацией Р4 завершается становление зубной системы, дальнейшие изменения отражаются, как и у других полорогих, в усилении стертости зубов и жевательных поверхностей.
Из многочисленных возрастных изменений структуры черепа и посткраниального скелета важно учесть сдвиги в пропорциях отдельных элементов черепа. Так, с возрастом сокращается зароговая часть черепа, а лицевая удлиняется. Молодые самцы и самки 3—4 лет отличаются от более старых почти полным отсутствием трубкообразных орбит. У самцов сращение костей черепа заканчивается к 8—9 годам, а у самок — после окончания половой жизни и прекращения лактации.
Половые различия. Наличие рогов у самцов и самок, сходства окраски и ряда других параметров экстерьера затушевывает половой диморфизм. Однако различие в деталях очень велико. Так, самки заметно меньше самцов, голова легче, лицевой отдел ее длиннее; рога тоньше и сильнее изогнуты вовнутрь. У быков с возрастом борода и особенно подвес сильно редуцируются, а у самок, наоборот, они увеличиваются. Половые различия в размерах и пропорциях черепа значительны. Обратим внимание еще раз на то, что самки долго сохраняют ювенильные черты. По данным К.К.Флерова (1979), у Bison bonasus caucasicus отношение ширины лба к основной длине черепа у самцов составляет 66,5—70,3, а у самок — 56,8—63,8. Индекс наименьшей ширины затылка соответственно 51—53,5 и 42,5—49. Существенный половой диморфизм выражен в структуре роговых стержней — у самцов они мощнее (длиннее и толще), уплощены у основания, а у самок, как и молодых, они круглые в сечении.
Географическая изменчивость. Наиболее полно она изучена К.К.Флеро-вым (1979). Что касается аборигенного кавказского зубра, населявшего лесные пояса гор, то он четко характеризовался как горное животное, тонко "подогнанное" к горным условиям. Эта особенность, как отмечено выше, наложила отпечаток на весь экстерьер, на краниологические и посткраниологические параметры (см. обзор вида, приведенный выше). Наши и литературные данные (Наниев, 1956) свидетельствуют о том, что зубры на Северном Кавказе имеют относительно малую изменчивость. Во всяком случае черепа из пещер Северной Осетии (Центральный Кавказ) фактически
278
Bison bonasus Linnaeus, 1758
не отличаются от таковых сородичей Западного Кавказа. По этому поводу В.И.Наниев писал, что мнение некоторых современных исследователей фауны Кавказа о том, что зубры Северной Осетии по отношению к зубрам Западного Кавказа составляют более мелкую расу, результатами изучения 98 черепов и 8 роговых чехлов не подтверждается. Наоборот, зубры, встречавшиеся в Северной Осетии, ничем не отличаются от типичного Bison bonasus caucasicus Sat., который еще недавно (1927) встречался на Западном Кавказе. Этому свидетельствуют результаты детального краниологического изучения материала, согласно которому черепа зубров Северной Осетии по своим абсолютным размерам и морфологическим особенностям ничем не отличаются от черепов кавказских зубров с Кубани.
Для оценки географической изменчивости и географического формообразования интерес представляют данные по формированию популяций восстанавливаемого зубра в различных районах Северного Кавказа. Как отмечено выше, в нашем распоряжении наиболее значительная выборка имеется по Кавказскому заповеднику, где обитают потомки зубробизонов (В. bonasus х В. bison). Для удобства в дальнейшем этих зубров мы будем называть "горные зубры", акцентируя на том, что становление новой формы происходит в горах бассейна Кубани. Называть их "горными зубрами" нам представляется правомочным.
Специальные исследования сотрудников Кавказского заповедника, проведенные в разные этапы становления популяции (Камьянов, Нусов, 1967; Камьянов, 1969; Немцев, 1982, 1985, 1988а; Немцев, Киселева, 1987, и др.), а также наши данные позволили составить определенное суждение о специфике горного зубра. Как видно из табл. 45, 46, длина тела горного зубра на период 1975—1984 гг. составила в среднем 279 см, что меньше беловежского на 6 см, не говоря уже о бизоне. По А.С.Немцеву (1988а); горный зубр по основным линейным размерам тела уступает беловежским и беловежско-кавказским зубрам на 10—30%. Что касается отношения горного зубра к аборигенному (вымершему) кавказскому зубру, то траншрессия промеров и пропорций тела между ними увеличилась до 76,1 и 79,4% соответственно. За три с половиной десятилетия у горных зубров появились черты аборигенных зубров: относительная длина передних конечностей по сравнению с задними увеличилась, уменьшилась ширина головы, а массивность пясти, объем груди и степень скрученности рогов возросли.
В структуре черепа также выявлены существенные изменения. Расчеты А.С.Немцева показали, что по абсолютным размерам черепа евклидова дистанция между горными и аборигенными зубрами сократилась с 1958 г. в 2 раза. Это расстояние на 10% меньше такового между горным и беловежским зубрами. При этом внутрипопуляционная изменчивость горного зубра остается высокой, почти в 2—3 раза больше, чем у чистокровных зубров.
Учитывая систематике-таксономический вес абсолютной и относительной ширины лба, отметим, что горные зубры более узколобые, чем чистокровные беловежские зубры и бизоны. Однако, по А.С.Немцеву (1988 г.), они значительно превосходят по этому параметру аборигенных кавказских зубров. Аналогичное положение выявлено при анализе других параметров
279
Зубр
черепа (ширина темени, длина носовых костей и др.)- Заслуживает внимание то, что ряд признаков черепа аборигенного кавказского зубра, такие, как рудиментарный загиб кондилюсов, желобок на нёбных отростках межчелюстных костей, соприкосновение носовых и межчелюстных костей и др., обнаружены у большей части коллекции горных зубров. Пока трудно сказать, что эти признаки действительно отражают микроэволюционные сдвиги в популяции горных зубров или указывают на низкий систематикотаксономический вес их. Во всяком случае, примечательно заключение А.С.Немцева (1988а) о том, что примерно 40 лет назад по индексам черепа нельзя было отличить горных зубров от беловежско-кавказских, но в настоящее время они достоверно на 5%-ном уровне значимости различаются по 4 из 10 изученных краниологических пропорций. Однако этого, видимо, недостаточно для таксономического обособления горных зубров.
На данном этапе на Кавказе мы выделяем один подвид B.b. caucasicus Sat., уничтоженный человеком в 1926 или 1927 г. Гибридам разных кров-ностей, обитающих в настоящее время на Кавказе, видимо, преждевременно придавать систематическое название.
Экология
Ландшафтная приуроченность. Биотопы. Большая часть специалистов, занимавшихся изучением зубров, едина в суждении о том, что аборигенный кавказский зубр (Bison bonasus caucasicus) формировался как естественный компонент горно-лесных и горно-луговых ландшафтов Кавказа. К сожалению, имеется мало литературных данных по анализу территориального размещения аборигенного зубра в горах Кавказа. Наиболее полные сведения содержатся у Н.Я.Динника (1910). Они основаны преимущественно на сообщениях охотников или служителей Кубанской охоты. Сам Н.Я.Динник застал кавказского зубра за 2—3 десятилетия до его уничтожения, а Д.Филатов (1910, 1912)-— еще позже.
В 80-х годах XIX в. Н.Я.Динник (1884—1910 гг.) проводил полевые наблюдения за аборигенным зубром на ссверо-Западном Кавказе, собрал информацию от разных лиц, участвовавших в отлове зубренка для зоопарка Москвы. По его данным, кавказские зубры занимали в разные времена года довольно широкий крут ландшафтов и биотопов от среднегорных широколиственных лесов до субальпийского пояса. Высотные пределы распространения составляли от 900 до 2100 м над ур. моря. Нередко следы зубров находили в альпийском поясе, даже на снежниках, например на фирновом поле Санчарского перевала, что в верховьях Большой Лабы (Россиков, 1890). В прежние годы, предшествующие наблюдениям Н.Я.Динника, зубры, несомненно, спускались и в предгорье (см. обзор ареала).
Зубрам, как и многим другим копытным Кавказа, была свойственна сезонная смена местообитаний". По рассказам охотников и моим личным наблюдениям, — писал Динник (1910), — летом они очень любят держаться на границе лесного и субальпийского поясов, где в течение всего лета зеленая травянистая растительность, высокотравные луга чередуются парковы
280
Bison bonasus Linnaeus, 1758
ми лесами из горного клена, нередки заросли березы, рододендрона. Здесь всегда меньше кровососов, потому что дует ветер и прохладнее. Из этих мест утром и вечером зубры выходили пастись на альпийские луга, а в дождливую погоду спускались вниз, в ельники и пихтарники и отлеживались под густой кроной хвойных деревьев. Под таким кровом земля оставалась сухой, покрытой слоем опавшей хвои. Такие места, судя по следам зубров (пометы, следы копыт, лежки), посещались часто и многими животными. Именно в таких условиях зубры находили лучшую защиту от людей, ибо фон субстрата и окраска животных точно соответствовали друг ДРУГУ-"
Очевидно, такая гармония да плюс еще некоторые черты этологии помогали зубрам дольше сохраниться в поясе темнохвойных лесов. Вероятно, поэтому Д.Филатов (1910, 1912), обследовавший состояние кавказского зубра на Северо-Западном Кавказе в 1909—1911 гг., пришел к выводу, что в летнее время распространение этих животных отраничено поясом темнохвойных лесов. По его мнению, зубры в то время кормились в основном на лесных полянах с высоким травостоем, на склонах балок, поросших борщевиком и белокопытником. Эти места, по Д.Филатову, привлекательны летом для зубров тем, что и здесь также прохладно, тень, сравнительно мало кровососущих насекомых, меньше тревожат их люди. На высокогорные луга субальпийского и альпийского поясов зубры могли появляться лишь до прихода домашних животных на летние пастбища и после их ухода оттуда. Однако сам Д.Филатов не находил зубров выше пояса темнохвойных лесов. Как видно, в конце XIX и начале XX в. ландшафтное и биотопичес-кое размещение кавказского зубра было отраничено как снизу (лесозаготовки), так и сверху (скотоводство), что повлекло за собой резкое сокращение не только численности, но и круга обитаемых биотопов.
Переходя к обзору ландшафтного и биотопического размещения современных горных зубров (потомки зубробизоновой линии) на Северо-Западном Кавказе, следует отметить, что натурализация их в горных условиях проходит успешно. Постепенно с переводом их на вольное содержание эти зубры экологически замещают аборигенных кавказских зубров, и в настоящее время круг населяемых ими биотопов довольно широк. На наш взгляд, он соизмерим с тем, что было у аборигенного кавказского зубра в 80-х годах XIX в. В летнее время горный зубр, по А.С.Немцеву (1988а), занимает (в порядке убывания степени предпочитаемости) следующие биотопы: папоротниково-разнотравный пихтарник, березняк разнотравный, овсяницевый пихтарник, папоротнике во-разнотравный буко-пихтарник и др. На летнее размещение горного зубра влияние оказывают, кроме пищевого фактора, такие, как защитные условия, наличие источников воды, благоприятные микроклиматические условия и т.д. В зимнее время горные зубры предпочитают биотопы с обилием древесно-веточных и травянистых кормов. Таковы злаковые осинники с коэффициентом предпочитаемости 33,7%, букняки овсяницевые (21,5%) и разнотравно-ожиновые (13,4%), а также овсяницевый буко-пихтарник (12,6%).
281
Зубр
Ландшафтное и биотопическое размещение горных зубров на Центральном Кавказе сходно с тем, что имеет место на Северо-Западном Кавказе. Наши данные и литературные источники (Рашкевич, 1973; Варзиев, Курятников, 1980; Темботов, 1980; Темботов, Шхашамишев, 1984; Литвинова, 1980; Брагин, 1987; Липкович, 1988) свидетельствуют, что современное размещение зубров приурочено к разным типам широколиственных лесов и вторичных лугов с высоким травостоем. Среднегорья с такими лесами, чередующиеся лугами, зубры занимают наиболее продолжительное время. Летом нередко они поднимаются выше до траницы лесов и горных лугов, но расширение летних выпасов домашнего скота отраничивает круг населяемых зубром биотопов, подрывает основу нормального его существования.
Миграции. Для аборигенного кавказского зубра была характерна высокая подвижность, но наиболее значительные перемещения наблюдались по сезонам года. В прежние времена, когда человек преследовал зубров меньше, а домашнего скота на летних пастбищах паслось не так много, регулярно регистрировались высотные миграции зубров. Вслед за отступающими снегами и появлением зеленой растительности зубры поднимались выше в горы до верхней траницы лесных поясов, субальпийского криволесья и парковых лесов, чередующихся субальпийским высокотравьем. Н.Я.Динник (1910) и ряд других естествоиспытателей, работавших в XVIII и XIX вв. на Кавказе, регистрировали следы пребывания зубра в альпийском и субнивальном поясах (Россиков, 1890; и др.). С выпадением снегов в высокогорьях зубры спускались ниже, вплоть до предгорьев. Наряду с высотными миграциями отмечались переходы с северного макросклона Западного Кавказа на южный, в область верховий рек Сочи, Бзыбь и др. Н.Я.Динник отмечал, что зубры в поисках соленых источников в сухое время года отправлялись за много верст. Обилие зубрового помета и сильно истоптанная земля свидетельствовали, как часто такие места посещали зубры. Подобные тропы и солонцы Н.Я.Динник находил часто на горе Абаго.
Миграции и значительные кочевки формируются на Кавказе и у восстанавливаемых горных зубров. На формирование их, по А.С.Немцеву (1988а), влияние оказали многоснежные зимы. Широкие передвижки зубров в предгорье осенью отмечаются раз в 8—10 лет, причем в них вовлекается около половины всех зубров Кавказского заповедника. Протяженность миграционного пути составляет около 30 км, и ориентированы они с юго-востока на северо-запад. Перепад высоты местности при этом иногда достигает, по А.С.Немцеву (1988а), около 2000 м, а по С.Г.Калугину (1968) — до 2600 м. Сигналом для начала миграции обычно служат высокий снеговой покров (около 30—40 см глубиной) и уменьшение доступности кормов., Названные авторы считают, что в малоснежные зимы у зубров отсутствуют постоянные участки обитания, они последовательно сменяют одни угодья другими, близлежащими, без дальних переходов, и этот тип миграционной активности наиболее характерен для современных горных зубров. При этом протяженность миграции за сезон обычно не превышает 20—30 км при среднесуточной скорости перемещения в 1 км. Обратный путь зубров из мест зимовки в высокие пояса гор вслед за отступающими снегами также не
282
Bison bonasus Linnaeus, 1758
снегами также не носит характер дальних переходов. В долине Малой Лабы сезонные перемещения горного зубра сведены до минимума, протяженность их не превышает сотни метров. Фактически зубры ведут оседлый образ жизни, что обусловлено наличием высоких хребтов вокруг долины, недоступных им. Если учесть, что высотные миграции, охватывающие ряд поясов (от предгорьев до альпийского и даже нивального), свойственны многим горным животным, в том числе уничтоженному аборигенному кавказскому зубру, то ограниченность сезонных перемещений современного горного зубра нам представляется показателем несовершенства его адаптивных механизмов.
Лежки. Н.Я.Динник (1910) дает описание мест отдыха аборигенного кавказского зубра. По его данным, в ненастную погоду зубры спускались в ельники и под "шатром" могучих елей и пихт находили надежный кров от самого сильного дождя и отдыхали на сухой земле, осыпанной высохшей хвоей. Под такими деревьями, отмечал Н.Я.Динник, часто встречались по одной или по две огромных лежки, а в долине Шиши под небольшой труппой пихт им было найдено восемь таких лежек. Нередко лежки зубров располагались на рыхлой земле у корней вывороченных бурей пихт, буковых и других деревьев. Судя по состоянию земли на лежках и вокруг них, автор с уверенностью полагал, что одни и тс же лежки зубры посещали изо дня в день. Иногда несколько зубров выбирали для отдыха одно место, и в этом случае трава выбивалась полностью, а земля превращалась в пыль. Старые зубры выбивали землю копытами, валялись там, а вокруг них бегали, играли молодые.
Что касается лежек современных горных зубров, то они располагаются не столь избирательно: отдыхающих животных находят не только под кронами хвойных деревьев, но и по опушкам лесов и на полянах.
Сезонная и суточная активность. По аборигенному кавказскому зубру конкретных данных мало. Лишь отдельные фрагменты имеются в трудах Н.Я.Динника (1897, 1910), Д.Филатова (1910), в различных записках и воспоминаниях служителей Кубанской охоты. Максимальная активность зубров наблюдалась в период гона, когда взрослые самцы в поисках самок совершали переходы общей протяженностью около 6 км в сутки.- Значительные переходы зубры совершали и тогда, когда они испытывали опасность со стороны человека. В литературе имеются данные о переходе зубра с северного макросклона Главного Кавказского хребта на южный. В спокойной обстановке зубры могли подолгу находиться в районе одной лесной поляны.
Суточная активность кавказского зубра зависела от состояния погоды и ряда других факторов (кровососущих насекомых и др.). Обычно летом зубры паслись утром и вечером, нередко и ночью. Спад активности днем, видимо, был обычным явлением. В летнюю жару зубры скрывались под кронами деревьев на лежках или стояли здесь же, пережевывая жвачку. На вечернюю пастьбу или водопой зубры уходили за 3—4 ч до захода солнца.
283
Зубр
Переход
Отдых
Мачка
Пастьда
Водопой
Рис. 37. Суточная активность семьи зубров (Немцев, 1988)
24 Z 4 6 8 10 12 14 18 18 20 22 21
а _________Число суток
Суточная активность восстанавливаемых зубров (рис. 37) наиболее полно изучена в Кавказском заповеднике (Калугин, 1968; Немцев, 1988а,; и др.). Летом продолжительность пастбищной активности зубров составляет до 18 ч — в промежутке между 3 и 21 ч. Зимой она сокращается до 13 ч и находится в пределах между 5 и 19 ч. Число фаз пищевой активности в сутки составляет 3—5. Летом зубры насыщаются за 2—3 ч пастьбы и затем отдыхают подолгу. Суточный ход и трата энергии на жировку различны у разных групп. Отношение жировочного хода к суточному у одиночных зубров в среднем Г.1,8, у маточных группировок — 1:1,6, а у групп из одних только быков .— 1:1,3. На долю переходов к водопоям, солонцам и местам с благоприятным микроклиматом приходится 12—14% суточного хода. По А.С.Немцеву (1988а), ритм активности зубра летом не претерпевает сбоя из-за погоды.
Поведение. Первое впечатление, которое производит внешний вид зубра (тяжелый, массивный, малоподвижный) не соответствует его поведению (рис. 38). Он реагирует быстро, как правило с помощью обоняния и слуха, на те или иные источники опасности. Все старые исследователи обращали внимание на его ловкость, быстроту бега и способность совершать прыжки. Н.Я.Динник (1910) писал, что испуганные чем-нибудь зубры бегут почти всегда под гору, не обращая внимания на кручи и валежники, которые они более или менее свободно перепрыгивают, при этом делают прыжки в 3 м длиной. Во время панического бегства хвост зубра приподнят вертикально или выше горизонтального положения; такое же положение хвоста наблюдалось во время игр или сражений, при встрече с незнакомым зубром, при возбуждении в период гона (Баскин, 1979). С острым обонянием, очевидно, связано наличие специфического запаха, свойственного самцам и самкам зубров. Особенно резок он бывает во время гона, что позволяет самцам узнавать самку, вступившую в период гона. Сопоставление наблюдений
284
Bison bonasus Linnaeus, 1758
старых авторов (Виноградов, 1872; Динник, 1910; Филатов, 1910, 1912) и новых данных (Калугин, 1968; Немцев, 1988а; Перерва, 1989а) дают основание полагать, что зубру свойствен сложный комплекс поведения, особенно связанный с размножением. Отдельные фрагменты его нашли отражение в различных публикациях. В период гона самцы маркируют территорию, для чего рогами сдирают кору деревьев, обтираясь о стволы оставляют здесь свой запах. Они становятся агрессивными, отгоняют соперников, самок и молодняк от вступившей в период гона самки. Последние также становятся возбужденными, агрессивными, отгоняют от себя других самок и телят. Самец неотступно следует за своей избранницей, подолгу простаивает рядом с самкой, периодически обнюхивает ее, нередко "флемует", бодает ее, нередко вспрыгивает на нее. Ответ самки вначале агрессивный, но потом она сама начинает бодать самца, закидывает голову на спину' самцу, трется о него, После спаривания бык неподвижно стоит и тяжело дышит, иногда ложится, а самка чаще ложится.
Из сложного комплекса формирования поведения телят и в связи с этим поведения матери обратим внимание лишь на некоторые. Нахождение кавказской зубрицы с телятами в первые дни их жизни отдельно от других, видимо, связано с необходимостью для телят запоминать запах, внешний облик и голос матери. Зубрица защищает теленка от хищника, проявляет попытки агрессии (но редко реализует их) к человеку, приближающемуся к ее теленку. При безвыходном положении, оставив теленка, убегает. Сами зубрята не боятся человека, не убегают от него. Более того, как отмечают многие исследователи, зубрятам свойственно любопытство’. У них четко выражена склонность к играм, что отмечено как у кавказского, так и беловежского зубра.
У аборигенного кавказского зубра зарегистрирована целая система оборонительного поведения. Очевидно, многие ее стороны усилились в последние десятилетия существования местного зубра на Кавказе, что явилось следствием преследования его со стороны человека. Думается, с таким суждением согласуются данные старых авторов, хотя на первый взгляд в них содержатся противоречивые мнения (Динник, 1899, 1910; Сатунин, 1898; Филатов, 1910, 1912 и др.). Самое большое возбуждение у зубра вызывал человек — почуяв его, зубр быстро поднимал голову, чтобы определить по запаху, с какой стороны приближается опасность, и затем “сломя голову" бежал в противоположную сторону. .Уходил быстро и далеко, не давая возможности охотнику приблизиться. При интенсивном преследовании кавказские зубры оставляли район своего обитания и переходили в другой район, притом перемещались они цепочкой на десятки километров. Об этом можно было судить по следам. Бывало и так, что стадо разбивалось на ряд мелких групп и они разбегались в разные стороны. Д.Филатов (1912) писал, что в годы массового преследования зубры из среднегорья Кубанской охоты переходили горные хребты на юг, на запад, вниз, в предгорья. Этим, очевидно , можно объяснить имевшее место появление зубра на Черноморском побережье, близ г. Сухуми, а в Предкавказье — недалеко от горы Бештау. Стрессовое состояние зубра или неопределенность реакции
285
Зубр
7
Рис. 38. Зубры беловежский (1-7) и чистокровный кавказский (8, 9)
286
Bison bonasus Linnaeus, 1758
на источник опасности находило отражение в различных действиях: при уходе от опасности зубры периодически останавливались, прислушивались, регулярно испражнялись. Пытаясь узнать, насколько реальна опасность, отдельные особи выходили из группы, чтобы лучше почуять обстановку. Если преследования не было, зубры постепенно возвращались к прежнему месту, при этом издавали специфичный звук, сходный с хрюканьем. Рев издавали лишь раненые звери. Что касается нападения на человека, даже будучи ранеными, зубры совершали это лишь изредка, и притом взрослые самцы. Судя по данным С.Г.Калугина (1968), у гибридных зубров в период привыкания к персоналу нападение на людей случалось нередко. С переводом их на вольное содержание и ростом пугливости зубров эта агрессивность снижалась и они реже нападали на человека, предпочитая уходить от него. Активность оборонительного поведения ослабляется у зубриц с зубренком.
Интерес представляет сообщение Д.Филатова (1912) о том, что оборонительная реакция аборигенного кавказского зубра была вариабельна по сезонам года. Весной зубры были менее пугливы, летом по мере усиления браконьерства становились очень осторожными, осенью в связи с размножением инстинкт к самосохранению вновь падал.
Учитывая, что многие ритуальные элементы поведения зубров видоспецифичны и соответственно могут быть экстраполированы на разные подвиды, в том числе уничтоженного кавказского зубра, обратим внимание на разнообразие форм угрозы (Перерва, 1989а): у!рожающий выпад (head-on-threat), кивки головой (nod-threat) и предупреждение (preliminaries). Первые две формы относятся к простой демонстрации, а третья представляет сложный комплекс, используемый зубрами в редуцированной форме. Некоторые формы поведения, связанные с экологией популяций, будут затронуты в соответствующих разделах.
Линька. По данным старых авторов, в том числе Н.Я.Дин-ника (1910), смена волос у аборигенного кавказского зубра происходила лишь один раз в году и приходилась на начало лета. Еще в середине мая встречались животные в зимнем мехе — с длинными и густыми волосами, особенно ос? тями на гриве, челке, бороде л т.д. Их выпадение становилось заметным в первой декаде июня, когда темп замены волос
287
Зубр
Таблица 53. Сроки линьки зубров различного пола и возраста (Калугин, 1968)
Пол и возраст Число наблюдений Начало линьки Конец линьки Продолжительность линьки, мес.
Взрослые самцы 29 II декада февраля Май 3-3,5
Взрослые самки 93 III декада марта Июль 4-5
Молодняк старше года 108 II и III декады марта Июнь—июль 3-4-
сильно ускорялся. Однако и в конце июня встречались стада, в которых самки, казалось, были еще в зимнем меху. 22 июня были встречены зубры в зимних волосах. Более того, во второй половине июня иногда замечались быки, полностью не перелинявшие, у которых наряду с редкими короткими и новыми волосами были старые длинные волосы. Отрастание нового волосяного покрова особенно интенсивно проходило в сентябре.
Топография смены волос у кавказского аборигенного зубра была, видимо, сходна с таковой беловежского: выпадение волос начиналось с шеи, головы и ног и затем переходило на грудь, бока, спину, в том числе. В последнюю очередь линяла задняя часть тела. Здоровые и упитанные животные линяли раньше, равномернее и без образования плешин. У ослабленных и больных линька задерживалась.
Сведения по линьке восстанавливаемых зубров наиболее полно содержатся в работе С.Г.Калугина (1968). По его данным, зимний мех у зубров Кавказского заповедника очень хорошо развит. Он длинный, густой и настолько теплый, что животные ложатся на снег и лежат по нескольку часов, не считаясь с низкой температурой. Лишь во время больших снегопадов они иногда уходят под кроны елей и пихт. Зубр линяет раз в году, смена волос происходит в более растянутые сроки; в зависимости от пола и возраста они колеблются от II декады февраля до III декады июля (табл. 53). Не линяют сеголетки.
Последовательность смены волос сходна с таковой у кавказского зубра — раньше линяют шея, передняя часть туловища и бока. Несколько позже волосы сменяются на спине, задней части туловища и брюхе. В последнюю очередь линяют маклаки и хвост. В отличие от кавказского зубра волосы у горного зубра могут выпадать целыми пучками, образуя плешины на теле, что указывает, возможно, на примесь крови бизона (у бизонов зимние волосы выпадают клочками).
В Кавказском заповеднике полный летний наряд горные зубры приобретают перед гоном, в августе. Подшерсток начинает расти с сентября.
Питание. В старых источниках по кавказскому зубру мало данных по питанию, но при всей неполноте они представляют исключительный интерес. Впервые Н.Я.Динник (1899, 1910) обратил внимание на роль папоротников в рационе кавказского зубра. Он писал, что присутствие зубров в данной
288
Bison bonasus Linnaeus, 1758
местности узнается по огромным следам, тропам, лежкам, экскрементам, а также по объеденным ими папоротникам, которых не потребляет ни домашний скот, ни водящиеся здесь дикие звери. Несмотря на специфический запах папоротников, зубры потребляют их с удовольствием даже в летнее время, когда не ощущается недостатка в пище. По этой причине в тех местах, где обитают зубры, часто можно видеть поляны с более или менее сильно обгрызанными папоротниками. По данным Н.Я.Динника, зубры охотно едят кору некоторых деревьев, особенно рябины и ильма. Особенно возрастает роль коры в питании зубра зимой, когда нет травы, а также весной, когда начинается сокодвижение и кора легко отделяется и обладает лучшими кормовыми свойствами. Он наблюдал, что и летом зубры часто обдирают кору с деревьев, хотя вокруг имеется много свежей травы. Кроме коры рябины и ильма, зубр ест кору пихты. Диннику приходилось часто видеть столетние пихты, у которых толстая кора была съедена зубрами до самой древесины. О том, что это зубровые поеды, легко можно было судить по широким резцовым следам. "Конечно, — писал он, — летом на полянах и горных лугах зубры едят самые разнообразные виды травянистой растительности". По Диннику (1910), Филатову (1910, 1912), в теплый период года зубры питаются преимущественно такими видами растений, как белокопытник, калужница, недотрога, овсяница, костер, а также листья ежевики, борщевика и др. Общий набор видов растений, поедаемых кавказским зубром, достигал 40. Однако авторы указывали, что этим количеством не ограничивается разнообразие растительной пищи кавказского зубра, но отсутствие специальных исследований не позволяло составить полную характеристику питания кавказского зубра.
Что касается питания современных зубров Кавказа, то систематические исследования их питания проводились в Кавказском заповеднике (см. обзор: Калугин,. 1968). По другим районам современного распространения зубров сведений меньше, но они представляют также интерес в плане географической изменчивости состава кормов. В 60-х годах в Кавказском заповеднике количество растений, поедаемых горным зубром, составило 325 видов. Как отмечает С.Г.Калугин (1968), это количество может увеличиться до 400 видов по мере расширения круга обитаемых ландшафтов и биотопов. Данные указанного автора свидетельствуют о том, что горные зубры проявляют избирательность не только к видам растений, но и- к вегетативным частям. Из древесных кормов наиболее ценными для них являются листья, содержащие много протеина (12—16%), сырого жира (3—5%), безазотистых экстрактивных (47—54%) и минеральных веществ (СаО — 0,9—1,6%) и относительно мало клетчатки (12—17%). Листья значительно питательнее, чем, например, трава мелких пастбищ. Высокой питательностью характеризуются кустарниковые корма. Зубры отдают предпочтение бересклету, иве и осине, содержащим более высокий процент протеина и кальция.
Состав кормов существенно меняется не только по сезонам года, но и по местообитаниям. Зимой в желудке зубров больше всего обнаружено зеленых подснежных растений, таких, как ежевика, овсяница. Второе место
19 В.Е.Соколов, А.К.Тсмботов
289
Зубр
Таблица 54. Потребление зубрами древесного корма (по: Калугин, 1968)
Порода Часть дерева Время наблюдений Число зубров Съедено в пересчете на 1 взрослого зубра в сутки, кг
Осина (3)* Концы ветвей с листьями Май 1953 (5)»» 2 2,4
Осина (3) Кора Апрель 1953 (5) 2 0,9
Ива (2) Концы ветвей с листьями Июнь 1953 (2) 3 2,2
Ива (2) Кора Июнь 1953 (2) 3 1,1
Бересклет (3) Концы ветвей с листьями Апрель 1954 (3) 1 2,7
Бересклет (3) Кора Апрель 1954 (3) 1 1,4
Пихта (2) Кора Май 1954 (2) 4 0,5
Ильм (4) Листья с ветками Май 1954 (4) 6 3,2
Ильм (4) Кора Май 1954 (24 6 1,0
* Число закладок.
**Число дней.
занимают высохшая трава, кора деревьев и кустарников. Лишь в начале зимы и в небольшом количестве поедают сухие листья и сохранившиеся желуди (последние едят хорошо, но их мало в местах обитания зубра Кавказского заповедника). В отличие от оленей зубр не ест орешки бука. Сравнение состава кормов Кишинского и Умпырского районов показало, что в первом основными зимними кормами, являются ежевика, осина и ильм, а во втором — овсяница, ветошь различных трав на южных склонах, граб и осина. Весенний набор кормов в том же Кавказском заповеднике включает вначале (март первая половина апреля) вейник, ежу, пырей, мятлик, пролеску, типчак, подмаренник, воронец, ясколку и др. В наибольшем количестве поедают черемшу, нередко встречающуюся на лесных полянах и в редколесьях. Значительное место ранней весной в питании занимает, как видно по данным табл. 54, древесно-веточный корм, в том числе кора лиственных деревьев и пихты. Во втором периоде весны усиливается роль тех трав, вегетация которых идет интенсивно, в том числе листьев злаков, осоки, щавеля, а также ядовитых для домашнего скота воронца и черемицы. С мая в пищевом рационе зубра усиливается роль зеленых листьев. Летом в питании зубров, находящихся на верхней границе лесных поясов, значительно возрастает доля травянистой растительности по отношению к древесно-кустарниковой. В это время зубры поедают борщевик, лигустикум крылатый, ежу сборную, райграс французский высокий, мятлик и вейники, козлятник восточный и чину луговую. Когда зубры выходят на высоту 2300—2400 м над ур. моря, они поедают овсяницу, щучки, тимофеевку альпийскую и др. Роль древесно-кустарниковой растительности летом заметно падает до ранга второстепенности.
290
Bison bonasus Linnaeus, 1758
Таблица 55. Количество травянистого корма ^съедаемого
зубрами на пастбище (по: Калугин, 1968)
Пр ео бладающая растительная ассоциация Сроки пастьбы Число наблюдений Площадь выпаса, га Число зубров Стравливание, % Съедено в сутки в пересчете на 1 зубра, кг
Разнотравно -вейниковая 20-30.05 3* 2,0 20 34,5 35
Разнотравно-пырейно - мятлико вая 20-25.05 2* 2,0 16 34,9 34
Разнотравно -полевицевая 24-30.05 1 1,5 16 35,9 34
Райграсово-разнотравная 18-23.06 3 з,о 23 4,7 33
Подбеловая и разнотравно -осоковая 10-20.07 4 2,0 25 5,6 30
Разнотравно-вейниковая и др. 16-16.07 4 4,0 20 5,6 33
Разнотравно-вейниковая 10-15.07 4 2,5 21 10,9 41
Разнотравно -ежовая 9-20.07 5 2,0 21 3,3 32
Разнотравно-вейниковая 1-16.08 5* 2,5 4 6,0 45
Разнотравно-ежовая и подбеловая 15-29.09 4 2,2 24 5,5 40
♦Выпас в загоне
Осенью по мере огрубления травостоя зубры чаще посещают низины и северные склоны, где сохраняется зеленая растительность или менее огрубевший травостой. До ноября продолжают поедать козлятник, борщевик и головчатку, а на северных склонах и под кронами деревьев — окопник, бутень, таволгу, подбел, а также буквицу, подорожник, герань. Охотно пасутся на отавах, вторичных лугах среди лесов, в одичавших фруктовых садах поедают груши, яблоки, алычу, плоды шиповника поедают с листьями. Чем больше засыхает травостой,'тем чаще зубры потребляют древеснокустарниковый корм, а к октябрю-ноябрю они переходят на зимний корм.
Количество травянистого корма, потребляемого зубром на пастьбе, указано в табл. 55. Данные получены в загоне Кавказского заповедника (Калугин, 1968). Для получения сопоставимых данных разных возрастных трупп им введены пересчетные коэффициенты: зубры старше 2 лет — 1, молодняк старше года — 1,5, зубрята от 6 мес до 1 года — 3, зубрята до 6 мес — 4. Как следует из материалов, среднее потребление зеленого корма в пересчете на взрослого животного составляет 36,2 кг/сут (от 30 до 45 кг). Количество потребляемого древесного корма приведено в табл. 55. Оно определялось путем скармливания в загонах. Определение практическим
19*
291
Зубр
путем выявило, что в пастбищный период (с мая по октябрь) без подкормки один зубр может прокормиться на площади 3,5 га, где на 2 га поляны приходится 1,5 га леса.
Питание зубра кавказско-беловежской линии в Цейском заказнике изучено Н.А.Литвиновой (1980). Работа выполнена в среднегорье Центрального Кавказа, где представлены пырейно-разнотравные луговые степи с участками лиственного леса, широколиственные леса с подлеском из лещины, серой ольхи, буково-трабовые леса, субальпийские и альпийские луга и горные степи. Учет поедаемых растений велся на пробных площадках и маршрутах, где отмечалось число повреждений, их степень и характер.
На территории заказника в 33 тыс. га, занятой зубрами, зарегистрировано поедание ими в летний период 47 видов травянистых растений и 22 видов деревьев и кустарников. Зимний рацион включает 21 вид деревьев и кустарников. Отсутствие трав в составе зимних кормов автор статьи объясняет значительным снежным покровом, имевшим место во время полевых работ, который, видимо, исключал возможность поедания их зубрами.
В списке летних кормов основное значение, по мнению автора, имело разнотравье (69% повреждений). Среди трех видов бобовых поедался один — козлятник восточный, процент повреждений которого составлял 94,5. Значительно реже поедали папоротники (30,6%) и злаки (22,7% повреждений). Среди разнотравья к предпочитаемым (основным) кормам относилось семейство сложноцветных, а среди этого семейства — телекия (95,7% повреждений). К таким кормам принадлежали также представители губоцветных, некоторые виды колокольчиковых и бальзаминовых. Из кустарников в летних кормах основными являлись ежевика, черная и травянистая бузина, ива козья. Доля повреждений их соответственно: 84,9; 100, 99,2; 98,3%. Зимой на первое место выходили бузина черная (91,2%) и бересклет (99,1%). Лещина, которая в летнем питании отмечалась изредка, в зимнем питании была представлена довольно часто (82,0%). Из деревьев предпочтение отдавалось летом липе, клену, ильму, ipa6y. При этом у липы и клена поедались листья, у осины в основном кора, а у остальных — побеги с листьями. Зимой ведущим являлась липа (92,9% повреждений), потом ильм (88,2%), траб (83,9%), верба (87,0), осина (77,8%). У липы зубры поедали только кору, а у всех других — как побеги, так и кору. Как и в Кавказском заповеднике, зубры в Цейском заказнике повреждают побеги дикой яблони и трецкого ореха, а у алычи, кроме побегов, еще и кору.
В Нальчикском лесоохотничьем хозяйстве нами изучено в феврале 1990 г. содержимое желудков 4 взрослых самок. Масса желудка с содержимым доходила до 100 кг, доминировали остатки деревьев и кустарников, составившие по массе около 92,5%. Среди них были остатки дуба, граба, бука, ежевики. Остатков папоротника было около 7,5%. Из травянистой растительности, на долю которой приходилось около 1%, попадались определенно листья осоки, листья клена.
Обзор кормов зубра, приведенный выше, на наш взгляд, свидетельствует о том, что зубр, сохраняя основные видовые черты питания и соответствующего поведения, располагает на Кавказе хорошей кормовой базой.
292
Bison bonasus Linnaeus, 1758
В жизни зубров, как вымерших аборигенных, так и восстанавливаемых современных, большое значение имеет минеральное питание. В растительной пище, используемой зубрами, таких веществ недостаточно. Соответственно возникает необходимость в естественных и искусственных солонцах.
В старых работах по аборигенному кавказскому зубру заметное место отводилось описанию солонцевания. Н.Я.Динник (1910) писал, что зубры очень любят соленую воду и в сухое время года отправляются к соленым источникам за много верст. К некоторым солонцам идут тропы со всех сторон, напоминая торные дороги. Судя по сведениям, которыми располагал Н.Я.Динник, зубры не пьют соленую воду, а лижут или едят соленую землю, выедая большие ямы. По данным С.Г.Калугина (1968), полученным в Кавказском заповеднике в 1953—1955 гг., суточная потребность в соли в разные сезоны года у зубра — от 15 до 53 г. Обычно повышенная потребность в солях отмечается весной (с марта по начало июня). С июня эта потребность падает, но в августе вновь повышается и с некоторыми снижениями держится высоко в сентябре-ноябре. Сезонный характер минерального питания отмечался как у вымерших, так и у восстанавливаемых зубров. Увеличение или снижение потребности в соли связано с количеством потребляемого зеленого корма, бедного минеральными солями, а также состоянием организма (развитие плода, лактация и т.д.). Однако соль нужна зубрам в течение всего года и соответственно они активно разыскивают природные и искусственные солонцы.
Для утоления жажды зубры посещают речки и ручьи. В жаркое время дня, по данным С.Г.Калугина (1968), они пьют воду не менее 2 раз в течение суток, а в пасмурную и дождливую погоду водопои посещают раз в 2—3 дня. Во всех случаях зубры потребляют лишь чистую воду и не пьют из луж. При обилии влаги зубры утоляют жажду росой на траве, а зимой — за счет снега. По данным названного автора зубру необходимо воды около 50 л/сут.
Воспроизводство и структура популяции
Размножение. Ряд ценных наблюдений по размножению аборигенного кавказского зубра содержится в старых работах (Винотрадов, 1872; Динник, 1910; Филатов, 1910, 1912). Согласно данным Н.Я.Динника, время течки приходилось на август и, вероятно, на первую половину сентября. Об этом свидетельствовали, по мнению названного автора, многие факты. Так, отстрелянные в это время самцы были худые, кишечник и желудок почти пустые; наблюдение за живыми быками показало, что они сильно возбуждены (копытили землю, обтирались о деревья, били их рогами и т.д., однако без рева). По А.Ф.Винотрадову (1870), гон у аборигенных кавказских зубров длился всего около трех недель (начавшись в августе). У современных (восстановленных) зубров Кавказского заповедника продолжительность течки и яра сильно растянуты, что обусловлено, по С.Г.Калу-гину (1968), подкормкой и оптимизированными условиями. Случаи течки в
293
Зубр
период с 1946 по 1954 г. отмечались с июля по январь включительно. Перевод на вольное содержание в связи с натурализацией сопровождался сокращением продолжительности гона. Течка у одной коровы продолжается всего 1—2 дня, зависимость ее от возраста животного не установлена (Калугин, 1968), однако могут быть колебания (Jaczewski, 1958 г., по: Баскин, 1979). В охоту, как правило, старые самки приходят раньше, чем молодые. Беременность зубриц в условиях загона продолжается в среднем 9 мес. В Кавказском заповеднике она варьирует от 261 до 283 дней. О сроках появления телят аборигенного кавказского зубра некоторые данные сообщал Н.Я.Динник (1910). Так, 10 мая (по новому стилю) охотники, занимавшиеся отловом молодых зубрят, на небольшой территории видели трех коров, причем каждая имела по маленькому зубренку. Двумя днями раньше видели одного только что родившегося теленка, а 17 мая зарегистрировали достаточно окрепшего зубренка, способного догонять уходящую корову. 21— 22 мая егери также видели несколько маленьких зубрят. Автор заключает, что в конце XIX в. зубрята появлялись на свет в пределах одного месяца (обычно в мае).
У восстанавливаемых горных зубров сроки появления новорожденных по мере усиления натурализации сокращаются. По А.С.Немцеву (1988а), в 1956—1959 гг. зубрята появлялись в районе 12 июня. Стандартное отклонение дат рождения составляло 30±3 дня. К 1985 г. средняя дата сдвинулась на 21 мая и заметно сократилось отклонение от нее (23 ± 1 день).
Плодовитость. В помете, как правило, один теленок, двойни встречались крайне редко. Н.Я.Динник (1910) утверждал, что никто из охотников на Кавказе не видел у зубрицы более одного теленка. При этом, отмечал он, имеющиеся наблюдения свидетельствуют о том, что у многих коров телята рождаются не каждый год. Соответственно летом попадается много коров без зубрят и далеко не часто приходится видеть след коровы вместе со следом теленка. Такого же мнения был и Д.Филатов (1910). Очевидно, отел раз в 2—3 года был обычным явлением на Кавказе, хотя при хорошем содержании зубры дают потомство ежегодно (Заблоцкий, 1957). В потомстве обычно бывает одинаковое количество самцов и самок, но в зависимости от состояния кормовой базы преобладают особи одного или другого пола (Калугин, 1968).
Способность к размножению самки сохраняют до 20, а при благоприятных условиях до 25 лет. В природе первый отел зубриц наблюдается на четвертом или пятом году жизни, но при содержании с подкормкой и на хороших пастбищах в Кавказском заповеднике отел их отмечался и в трехлетием возрасте. Интересные расчеты о влиянии натурализации зубров на интенсивность размножения приводит А.С.Немцев (1988а). Так, в среднем за период с 1960 по 1980 г. рождаемость зубров составила 31%, но в последние годы сократилась до 20% и репродуктивный потенциал популяции снизился до 3,3%. Отел на 6-м году жизни стал нормой. Нормой стал и двух-трехлетний перерыв между деторождениями.
Рост и развитие. Теленок рождается хорошо сформировавшимся и через час после рождения уже становится на ноги, а через полтора часа может
294
Bison bonasus Linnaeus, 1758
следовать за матерью. Вероятно, в природных условиях телята в первые дни оставались на месте, а самка держалась вблизи. В недельном возрасте или раньше теленок с матерью присоединяется к стаду. Коровы, отелившиеся рано, прекращают кормить молоком телят пятимесячного возраста, но при позднем отеле — до года и больше. Траву теленок начинает есть в возрасте 19—22 дней. Рога появляются с первой осени жизни (Гептнер и др., 1961).
В Кавказском заповеднике, по С.Г.Калугину (1968), новорожденные имеют массу 19—24 кг. Первотелки обычно приносят относительно мелких телят. Бычки, как правило, тяжелее телочек. Развитие их (особенно в первые месяцы жизни) очень интенсивное.
Масса тела (л = 14 экз.) к полугоду достигает у бычков 110—130 кг, у телочек — 90—110 кг.
Развитие теленка сопровождается изменениями в телосложении. Новорожденный по сравнению со взрослым более высоко но гий, зад несколько приподнят, туловище короткое, узкое, и неглубокое, голова короткая и широкая. К 5 или 6 годйм жизни эти различия нивелируются у самцов, а у самок — к 4—5 годам, т.е. ко времени окончания физиологического развития.
Структура популяции. Сведения по различным параметрам популяции аборигенного кавказского зубра содержатся в ряде старых работ (Диннйк, 1910; Филатов, 1912 и др.). В отношении стадности Н.Я.Динник отмечал, что зубры не любят одиночества, даже старые самцы редко бродят в одиночку. Чаще всего зубры держались группами в 6—7 голов. Такая ipynna, по его данным, состояла обычно из молодых самцов (2—3 лет), телят и старых коров. Сходные данные приводит Д.Филатов (1912). Наряду с мелкими труппировками встречались более крупные. Так, в начале текущего столетия под Слесарней видели стадо из 18 голов, среди которых были два старых быка, около десятка коров и почти столько же телят в возрасте от одного до 2 лет. В 1900 г. зарегистрировано стадо в 14 голов, состоящее из старых коров, телят и сравнительно взрослых самцов. Нестарые самцы также живут нередко группами в 6—8 голов. Больших стад зубры на Кавказе не образуют (Динник, 1910). Однако, по данным этого автора, в лесах по Алаусу и Уруштену зарегистрировано вместе 20 зубров, которые за редким исключением были старые быки. В том же Уруштене, но в другом месте, встречено около 40 голов, а в долине Абаго и Малчепы — 50 зубров. К сожалению, автор не детализировал состав и характер их группировки.
О сезонной изменчивости структуры популяции аборигенного кавказского зубра данных почти нет. Н.Я.Динник в начале мая встретил ipynny из трех коров с маленькими телятами.
По структуре популяции восстанавливаемого зубра Кавказа имеется ряд публикаций, но наиболее корректные результаты получены по Кавказскому заповеднику (Калугин, 1968; Немцев, 1980а, б, 1987а, 1988). Социальная организация аборигенного и восстанавливаемого зубров довольно сходна. Так, самцы с трехлетнего возраста и старые самки держатся поодиночке. На их долю приходится около 13,2% всего населения (по Немцеву, этот пока
295
Зубр
затель был один и тот же в 70-х и 80-х годах). Семьи представляют в среднем 6,0% населения, в их состав входят самка с 1—3 молодыми. Маточные компании из 4—12 голов (самки с потомством) составляют 28,2% всех особей популяции и являются наиболее устойчивыми труппами. Самцы образуют труппы из 2—10 знакомых зверей, непрочно связанных между собой социально и территориально. Смешаные стада, объединяющие знакомых зубров из ряда семей и компаний в количестве 13—35 особей, составляют 35,8% от общего поголовья. Скопления, отличающиеся очень непрочными связями, объединяющие до 35—120 животных, образуются на митрационных путях, в местах зимовок и яра. Как указывал НЛ.Динник (1910), такие скопления во время кочевок отмечались и у аборигенных кавказских зубров. В иерархии соподчинения особей в популяции наиболее жесткие стычки (до смерти одной из сторон) отмечались и в прошлом, и в настоящем, как правило, лишь во время гона. Доминантам в стаде является самец, но вожаками-лидерами в труппах — старые и опытные самки.
В работах А.С.Немцева (1988а, б), где обобщены результаты многолетней работы коллектива Кавказского заповедника, подвергнута анализу половая и возрастная структура горного зубра во времени и пространстве. Так, соотношение полов длительное время находилось в пользу самок, но колебалось от 0,54 ± 0,064 в 1960 г. до 0,55 ± 0,053 в 1965 г. и 0,57 ± 0,044 в 1980 г. до 0,50 ± 0,049 в 1985 г. Минимальное количество самцов за пределами Кавказского заповедника там, где имеются неосвоенные кормовые ресурсы. Оно составляет всего 12,6% от поголовья в целом. Наоборот, доля быков наиболее высока (39,8%) в долине Малой Лабы, где хуже всего с кормовой базой в зимнее время. Соотношение полов новорожденных хотя и варьирует по годам, но в среднем составляет 1:1. Так, в условиях вольерного содержания (1941—1964 гг.) из 556 зубрят на долю самцов приходилось 50,3%, а самок — 49,7%. На соотношение полов новорожденных, как известно, влияет целый ряд факторов. От быков 5—6-летнего возраста рождается больше телочек, тогда как от более молодых или более старых — бычки. Сказывается также возраст самок: от молодых зубриц рождается больше телочек. У коров старых и среднего возраста рождается примерно одинаковое количество бычков и телочек.
Возрастная структура горного зубра характеризуется тем, что в 1985 г. население зубров заповедника включало 11,5% сеголеток, 9,2% годовалых телят, 393% самок старше 2 лет и 40% самцов. Однако наибольшее количество сеголеток (25%) зарегистрировано в долине Малой Лабы, где выше всего смертность новорожденных зубрят,-по сравнению с другими районами .заповедника. С такой оценкой согласуется очень низкий процент годовалых телят — всего 7,5%. Адаптивный смысл высокой рождаемости, восполняющей убыль новорожденных в связи с худшими условиями жизни, очевиден. Данные названного автора показывают, что в хороших условиях жизни, какими является долина р. Киша, соотношение долей новорожденных и годовалых телят обратное таковому в долине Малой Лабы.
Враги, паразиты, смертность, конкуренты. У взрослых и здоровых зубров врагов, видимо, не было в прошлом и нет в настоящее время. Волк опасен
296
Bison bonasus Linnaeus, 1758
для молодняка, однако co стороны коров и других взрослых обеспечивается активная защита их от хищников. За время вольного разведения зубров в Кавказском заповеднике обнаружено лишь два случая гибели сеголеток от волков (Калугин, 1968). Что касается хищнической роли медведя, рыси или других хищных, то по ним достоверных данных нет. Видимо, на Кавказе хищники не были и не могут быть лимитирующими или регулирующим численность зубра факторами.
Все имеющиеся данные свидетельствуют о том, что на Кавказе в отношении зубров велика отрицательная роль инфекций, особенно общих с домашними животными. Во многих старых источниках (см. обзор: Калугин, 1968) приводятся факты гибели или заболевания зубров ящуром, сибирской язвой, пастереллезом, эмфизематозным карбункулом, плевропневмонией, туберкулезом, бруцеллезом, столбняком и бешенством. Из них чаще всего зубры гибнут от ящура и пастереллеза. Так, на Западном Кавказе зубры гибли от эпизоотии ящура, имевшей место среди домашнего рогатого скота в 90-х годах XIX в. и в начале XX в. (1907 и 1919 гг.) (Динник, 1909; Башкиров, 1939). Зубрам свойственны такие протозойные заболевания, как пи-раплазмоз и трипанозоллез.
Паразитических червей у зубров и их гибридов, по Д.П.Рухлядеву (1967), 43 вида. Из них условно-специфическими могут быть названы только три вида. Общих с домашним крупным рогатым скотом 27 видов, общих с овцами и козами — 26, с оленями — 17, косулями — 21, муфлонами — 16, сернами — 15, турами — 11, кабанами — 6, волками и собаками — 2 вида. Особенностями гельминтофауны, по мнению названного автора, является отсутствие у них эхинококков, тизаниезий, стилезий, авителлин, однообразие видов легочных нематод, а также меньшее количество видов гельминтов, чем у других представителей жвачных.
Гельминтофауна зубров и их гибридов формировалась под влиянием мероприятий по акклиматизации, реакклиматизации, доместикации и гибридизации. Соответственно видовой состав гельминтов получился очень пестрым и сборным.
В Кавказском заповеднике, по Д.П.Рухлядеву (1964), у зубров выявлены следующие гельминты: Fasciola hepatica L., F. gigantica Cob., Paramphi-stomum spec. Ruch., цестоды Multiceps multiceps L., нематоды Neoascoris vitulorum Trav., Ostertagia spec. Ruch., Cooperia oncophora R., Nematodiris spec. Ruch., Dictuocaulus vivparus‘R., D. spec. Ruch., Trichocephalus ovis Ab., Thelazia rhadesi. Д.П.Козлов и Н.С.Назарова (1979), вскрывшие двух зубров в Кавказском заповеднике, обнаружили 8 видов гельминтов: F. gigantica, Liorchis scotilae, Moniezia benedeni, Bunostomum phlebotomum, Oftcrtagia ostertagia, Cooperia memasteri, Haemonchus contortus, H. placei.
С.Г.Калугин (1968), располагавший значительным фактическим материалом, считает, что случаи высокой инвазированности зубров в Кавказском заповеднике редки. Кроме того, у всех 60 вскрытых в разное время года зубров не обнаружены личинки кожного и носоглоточного оводов, что, по мнению названного автора, объясняется особенностями строения кожи зубра, ее толщиной и прочностью. Из эктопаразитов обнаружены
297
Зубр
иксодовые и некоторые другие клещи. Зарегистрировано нападение на зубров слепней, мошек, мокрецов, мух-жигалок и др.
Конкурентами зубра в питании могут быть олени, косули, туры, серны, кабаны, однако по видовому составу поедаемых растений ближе всего стоят зубры и олени (Насимович, 1939; Александров, 1958). По данным названных авторов у этих двух видов число общих поедаемых растений достигает 150 видов. Кроме того, оба вида сходны в сезонном питании (Калугин, 1968). Дальнейшее изучение конкурентных отношений зубра с другими копытными Северо-Западного Кавказа показало (Немцев, 1988а), что из всех видов растений, поедаемых копытными, в рацион кубанского зубра входит 90,8%, оленя — 94,6%, косули — 91,3%, кабана — 48,7% видов. Лишь 5,1% видов травянистых растений свойственны только зубрам, тогда как общие с оленями составляют около 90%. Однако полевые наблюдения, проведенные рядом сотрудников Кавказского заповедника, свидетельствуют о том, что ни один из перечисленных видов не вытесняет другого, ибо механизмы из популяционного самоотраничения превалируют над межвидовым антагонизмом (Немцев, 1988а). Что касается отношений зубра и домашних животных, то весьма отрицательно сказываются не только эпизоотологические последствия их контакта, но и деградация растительности, вызванная чрезмерной нагрузкой последних на горные луга. В Северной Осетии и Чечено-Ингушетии стада зубров поставлены в трудные условия в связи с повышенной пастбищной нагрузкой домашнего скота на горные луга (Брагин, 1987; Лип-кович, 1989).
Смертность. По данным С.Г.Калугина (1968), в Кавказском заповеднике отход в стаде в результате инфекций и инвазии составил 10,7% всех случаев гибели зубров. Из инфекций отмечены паратиф сеголеток и пастереллез сеголеток и взрослых.
Гибель зубров в прошлом имела место в многоснежные зимы из-за истощения (Динник, 1909, 1910; Филатов, 1910 и др.). Имеет место гибель молодняка при переходе рек, от ушибов и ранений от быков и коров во время гона и яра, затаптывания зубрят бегущими в испуге зубрами. В целом гибель сеголеток в Кавказском заповеднике по разным причинам достигает 10%, а с вычетом гибели от инфекций эта цифра составит 7% (Калугин, 1968). Что касается гибели взрослых от стихийных бедствий, то их число незначительно.
Численность и ее динамика. Более или менее достоверная информация о численности кавказского аборигенного зубра относится к последним десятилетиям его существования на Западном Кавказе (конец XIX и начало XX в.). Как отмечено выше, в середине прошлого столетия распространение этого зубра на Кавказе было сужено так, что его юго-восточная граница не выходила из бассейна Теберды. Вероятно, зубры населяли верховья рек Шахэ, Сочи, Мзымты, Псоу, Бзыби, Кодора и Ингура. Словом, местный кавказский зубр занимал лишь Северо-Западный Кавказ и поголовье его определялось в пределах 2000 особей (Винотрадов, 1870). В 90-х годах XIX в., по данным Н.Я.Динника (1910), распространение зубров было резко сокращено, сохранившись в основном между верховьями рек Белой и Лабы.
298
Bison bonasus Linnaeus, 1758
Численность определялась приблизительно в 500 голов. Организация в то время Кубанской охоты несколько затормозила катастрофическое падение численности зубра, но приостановить его уничтожение не удалось. Во всяком случае зубры оставались вытесненными в пределы пояса темнохвойных лесов, не являющегося оптимальным горным ландшафтом для них, и численность их в 10-х годах XX в., очевидно, не превышала нескольких сот. Социальные потрясения в связи с революцией и гражданской войной обусловили падение численности зубров с катастрофической скоростью в 20-х годах нашего века. Учреждение Кавказского заповедника в 1924 г. не смогло противостоять этому падению и полному уничтожению подвида в дикой природе. В 40-х годах чистокровный кавказский зубр оставался в единственном числе и, как отмечено выше, дал начало кавказско-беловежской линии.
Главные причины гибели кавказского аборигенного зубра — браконьерство с использованием нарезного оружия и лесоразработка, сузившая его распространение до пояса темнохвойных лесов. Велико было, очевидно, количество скота, выпасаемого на горных лугах.
Динамика численности горного зубра характеризуется очень быстрым ростом в первые десятилетия: с 1940 по 1965 г. поголовье этого зубра в Кавказском заповеднике выросло с 5 до 449 особей (Калугин, 1968). Наиболее интенсивный рост наблюдался при подкормочно-выпасном режиме (1946— 1953 гг.). В этих условиях за 8 лет ежегодный прирост в среднем определялся в 25,9% (от 14,6 до 50%), и в итоге стадо зубров увеличилось в 5 раз. При загонно-выпасной системе (1940—1945 гг.) оно возросло за 6 лет в. 2,4 раза, средний ежегодный прирост составил 15,2%. При вольном содержании (с 1954 по 1965 г.) за 12 лет средний ежегодный прирост несколько возрос (17,0%), а стадо возросло в 6,7 раз. В последние 3 года прирост составил 10,3—14,1%, что, по С.Г.Калугину (1968), является нормальным темпом роста численности зубра в природных условиях Западного Кавказа. Падение численности происходило в многоснежные зимы 1967—1968 гг., в результате чего поголовье зубров в Кавказском заповеднике сократилось на 24%. По А.С.Немцеву (19886), с этой зимы началось стабильно-флуктуационное колебание численности зубров. При этом интенсивность их прироста с тех пор, по его данным, колебалась в отдельные годы от +12% до —21%. По расчетам Немцева, экспоненциальная скорость роста после выпуска из загона достигла 0,22, а удвоение численности наблюдалось за 3 года. В последующие годы скорость увеличения поголовья медленно снижалась и в 1976—1985 гг. в среднем составила 0,061 на заповедной территории, а за ее пределами — 0,138. Общая численность зубра на Северо-Западном Кавказе составляет около 1300 голов. Она по математическим расчетам стремится к 1600 ± 150 (Немцев, 19886). Из абиотических условий лимитирующим фактором выступают условия зимовки, которые сказываются на благополучии зубров через кормовую базу.
Что касается численности зубров в других районах Северного Кавказа, то данных пока мало и они не всегда однозначны. Зубры благополучно прижились в Цейском заказнике Северной Осетии, где уже обитает стадо в
299
Зубр
230 голов. Однако территория, занимаемая ими, мизерна, соответственно концентрация их возросла до 14 зубров на 1000 га. Там наметилось отрицательное воздействие зубров на растительность, что приведет к падению численности самих животных. Это положение усугубляется организацией на лугах Пастбищного хребта круглогодичного выпаса лошадей (Липкович, 1989).
Зубры, завезенные в Нальчикское лесоохотничье хозяйство в 1’959 г., также прижились и в настоящее время их поголовье достигло 300 особей. Рост их численности сдерживается в основном низким уровнем охраны и ведения охотничьего хозяйства.
В Тебердинском заповеднике и госзаказнике Чечено-Ингушетии численность зубров очень низка (менее 50 особей в каждом).
Подводя итоги по современному состоянию численности зубров на Северном Кавказе, следует отметить, что за пределами Кавказского заповедника значительная территория лесного пояса и субальпики остается не освоенной зубрами и главным препятствием в этом освоении является человек.
Практическое значение. Теоретическое и практическое значение зубра, замечательного животного гор Кавказа, трудно переоценить. Роль кавказского аборигенного зубра как природного компонента горно-лесных и горно-луговых экосистем осталась совершенно не изученной. Сейчас лишь можно полагать, что поскольку это животное формировалось в составе местных лесов и лугов много тысячелетий, то оно необходимо было для этих лесов и лугов. Восстановленный горный зубр в условиях Кавказского заповедника при относительно высокой его численности не выступает в роли решающего фактора дигрессивных сукцессий ни в одном типе лесных ценозов. Более того, на вырубках зубр способствует возобновлению пихты (Голгофская и др., 1987; Александров, 1958). Не менее важно, что зубр не вступает в такое конфликтное взаимоотношение с аборигенными копытными Кавказа, которое исключало бы существование какого-либо вида. Соответственно благоприятная экологическая ниша зубра на значительной территории Кавказа может быть заполнена этим видом без существенного ущерба для местной фауны, флоры и природных экосистем. Значит, при освоении пастбищного пространства зубром численность его можно значительно увеличить. Конкретную цифру допустимого количества зубров без специальной бонитировки угодий каждого ландшафтного района Кавказа трудно назвать. Невозможно соответственно определить предполагаемый экономический эффект в рублях от зубрового хозяйства. Уместно сказать, что там, где практикуется валютный отстрел зубров, каждая лицензия стоит 3 тыс. долл. (Duda, 1987). Если учесть это, то без ущерба для популяции горного зубра в одних только районах, прилегающих к Кавказскому заповеднику, можно изымать 23—28 зубров в год (Немцев, 1988) и получать 69—84 тыс. долл, ежегодно. При такой постановке вопроса необходимо решить проблему гибридных зубров, дать однозначную оценку престижности лицензионной охоты на потомство зубробизонов, определить, насколько оправданно предложение ряда ученых (Заблоцкий, Заблоцкая, 1987 и др.) о
300
Capra Linnaeus, 1758
замене их чистокровными зубрами кавказско-беловежской линии. Чем быстрее будут решены эти теоретические вопросы, тем быстрее можно реализовать и практический вопрос об организации спецохотхозяйств по разведению зубров на Кавказе.
Еще Н.Я.Динник (1910) отмечал, что мясо кавказских и беловежских зубров, хотя и крупноволокнистое, но не уступает мясу крупного рогатого скота. На наш взгляд, хорошо приготовленное блюдо из мяса зубра вкуснее, чем из говядины.
При оценке зубра как представителя дикой фауны особого внимания заслуживает его значение в селекционно-племенной работе, а также в эстетическом воспитании людей.
Род Горные козлы — Capra Linnaeus, 1758
Полорогие мелких и средних размеров. Масса тела 35—150 кг, длина тела 100—170 см, длина хвоста 10—20 см, высота в холке 65—115 см. По массе и размерам тела самки меньше самцов. Характерно тяжелое телосложение с относительно короткими и массивными конечностями, короткой и сильной шеей, короткой, нормальной, без вздутий и хобота, головой. Уши заостренные и короткие. Хвост короткий, но больше длины уха, кожа на нижней стороне голая. Рога имеются у самцов и самок, но у последних они развиты слабее. Форма рогов изменчива — от саблевидной до спирально изогнутой. В поперечном сечении они имеют форму круга, треугольника, четырехугольника, линзы и капли. Поверхность рогов негладкая, годовые кольца хорошо выражены, могут быть большие бугры. Волосяной покров на теле не лохматый, примерно равномерной длины, лишь на нижней стороне под челюстями могут быть удлиненные волосы, образуя "бороду", а также под шеей и на груди длинные волосы сильно выдаются в виде подвеса. Верхняя губа покрыта волосами, голыми остаются лишь края ноздрей и срединная полоска на передней части. В коротком и густом меховом покрове много пуховых волос, отрастающих на зиму. Смена волос происходит обычно один раз, в весенне-летнее время. Окраска чаще всего одноцветная, коричневая или серая, на зиму темнеет, а летом становится светлее. Окраска конечностей на фронтальной стороне изредка бывает неоднородной благодаря появлению светлых и темных рисунков.
Кости черепа сильно пневматизированы. Лицевая часть его развита нормально, но зароговая удлинена и изогнута в лобно-теменной области. Этмоидальное отверстие, как правило, отсутствует, но если оно имеется, то развито слабо. Длинные отростки межчелюстных костей вклиниваются между носовыми и верхнечелюстными костями или примыкают к ним. Слезные кости не имеют ямок предглазничных желез.
Зубная формула:
I — С — Р — М- =32.
3 13 3
301
Род Горные козлы
Зубы гипсодонтные, что особенно сильно проявляется в структуре резцов (они в виде долота). На наружной поверхности верхних предкоренных не выражены задние и средние вертикальные ребра (Гептнер и др., 1961).
По И.И.Соколову (1959), для козлов в отличие от представителей рода баранов характерна укороченная, но широкая метаподия. Ширина верхнего края пястной кости не менее 20% от наибольшей ее длины. Длина пястной кости не более 73% длины лучевой кости. Длина плюсневой кости больше 60—61% длины берцовой.
В размерах тела, скелета, рогов и окраске четко выражен половой диморфизм. Окраска тела самок обычно скромнее, чем у самцов.
Из анатомических особенностей заслуживают внимания хорошо развитые органы обоняния, слуха и зрения. Имеются два функционирующих соска, иногда еще два недоразвитых. У взрослых самцов имеются сильно развитые подхвостовые железы, выделяющие во время половой активности сильно пахнущие секреты. У самок их нет. Как у самок, так и у самцов нет межпальцевых, паховых и предглазничных кожных желез.
У всех видов рода козлов, у которых изучена кариология, диплоидное число хромосом равно 60. Число плеч аутосом 58. Х-хромосома — акро-центрик, У-хромосома нестабильна. По структуре изученности хромосом кариотип рода Capra можно считать примитивным, ибо он сходен с предковой формой, от которой произошли представители Caprinae (Орлов, Булатова, 1983).
Ареал рода Capra занимает горы Южной Европы, Северо-Западной Африки, Передней, Средней и Центральной Азии и Северо-Западной Индии. Этот обширный ареал разбит на ряд изолированных массивов, что объясняется разобщенностью гор и ландшафтной приуроченностью козлов к горам. Второе название р. Capra — каменные козлы (Гептнер и др., 1961). Антропогенные изменения ландшафтов и охотничий пресс усилили дизъюнктивность ареала рода.
Местообитания козлов связаны со скалистыми участками гор на различной высоте до 5000 м над ур. моря. Как морфология, так и вся биология козлов тонко ''подогнаны" к жизни в скалистых местах. Хозяйственная деятельность человека значительно сузила круг биотопов, занятых различными видами рода. В одних случаях они оттеснены в высокогорья, труднодоступные для человека и домашних животных, в других — в глубокие ущелья, а в третьих — в удаленные от поселения человека места.
Суточная активность изменчива по сезонам года: летом они активны -преимущественно рано утром и вечером допоздна, а зимой активность многофазная, включая дневные часы.
Питаются козлы в летнее время различными травянистыми растениями, в том числе злаками, осоками (несколько видов), сложноцветными и др., а также побегами деревьев и кустарников. В зимнем рационе доминируют высохшая трава, ветви, хвоя, кора, почки кустарников и деревьев. Соотношение тех или иных кормов изменяется по сезонам года, в разных частях ареала и в зависимости от видовой принадлежности.
302
Capra caucasica Gilldcnstacdt ct Pallas, 1783
Размножение и связаное с ним поведение (драки между самцами и т.д.) наблюдаются в октябре-январе. Беременность продолжается 21—25 нед. Деторождение приходится на весну (март-май). Козленок рождается хорошо развитым, на второй день жизни способен следовать за матерью, очень проворный. Козлята начинают потреблять траву с месячного возраста, но лактация продолжается около полугода. Половой зрелости обычно достигают в 1,5 года. Живут козлы до 12—18 лет, изредка до 20 лет. Врагами козлов являются хищные звери (волк, рысь, в отдельных местах леопард и снежный барс).
Практическое значение как объекта спортивной и промысловой охоты значительно. В прошлом это значение было несравненно большим, чем в наши дни, однако некоторые виды остаются перспективными и при разумном их использовании могут давать значительный экономический эффект.
Систематика рода Capra нуждается в дальнейшем изучении. В ряде моно-трафий выделяются 8 видов козлов (Гептнер и др., 1961; Соколов, 1973, 1984; Corbet, Still, 1980, и др.). И.И.Соколов (1959) признает существование до 10 видов. По мнению одного из авторов настоящей сводки (Темботов, 1970), туры Кавказа составляют один вид и- соответственно количество видов рода козлов должно быть сокращено на один вид. Из козлов на Кавказе представлены два вида: кавказский тур Capra caucasica Guld. et Pall, и бородатый козел Capra aegagrus Erxl.
Кавказский тур — Capra caucasica Guldenstaedt et Pallas, 1783 Рис. 39
Capra caucasica Guld., Guldenstaedt, 1779: Верещагин, 1938, 1947; Ellerman, Morrison-Scott, 1951; Соколов, 1959, Темботов, 1970, 1974; Capra cylindri-comis Blyth, Blyth, 1841; Динник, 1910; Лидекер, 1931; Цалкин, 1955; Верещагин, 1959; Гептнер и др., 1961; Schaller, 1977; Aegoceros pallasii Rouil., Rouiller, 1841; Capra severtzovi Menz., Мензбир, 1887; Лидекер, 1913; Capra raddei Mats., Sitzungsber, 1901; Capra dinniki Sat., Сатунин, 1905; Capra ibex L., Ellerman, Morrison-Scott, 1951; Haltenorth, 1963; Corbet, 1978; Corbet, Still, 1980.
Коллекционный материал. ЗИН РАН: 68 черепов с роговыми чехлами; МГУ: 17 черепов с роговыми чехлами; КБГУ: 88 черепов с роговыми чехлами, 56 отдельных роговых чехлов, 1 скелет.
Диагноз. Крупных размеров, рога у самцов массивные, от саблевидной до гетеронимной формы. В поперечном сечении рог округлый, округло-треугольный. На спине имеется темный ремень различной степени интенсивности. Борода обычно короткая, если удлинена, то лишь зимой.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Все формы кавказского тура довольно крупные. Масса тела до 155 кг, длина тела до 196 см, высота в холке до 108 см. Самки значительно меньше
303
Кавказский тур
Мариуполь
Ростов-на-Дону
О Заветное
Сарпа
Яшкуль
2000 •
1000 -
Егорлыксная
О Дивное
Благодарный
Пятигорск
Сочи
Нальчик
Очамчира
Зугдиди
36
6000
503
ЬООО
Ахтала
4000
3000
КХ
4000
Нахичевань
3000
г. Далидаг 3616
г. Кюмиркей
2477
г. Мелис чкаро
2850
Не винно'мыссн
Элиста
Ставрополь
Карачаевск
Светлоград
Буденновск
Нефтекумск
Моздок
Тбилиси
г- Эльбрус
5642
г. fl се аш ха 3255
Ю-0
2000
1000 о
г. БоЛчГ инал&аг
3367
г. Каракал
2850
Ц-М
Ереван
Мартуни
304
Capra caucasica Guldenstaedt et Pallas, 1783
20 В.Е.Соколов, А.К.Темботов
Рис. 39. Распространение кавказского тура
1 — гора Фишт (Динник, 1910; Верещагин, 1959, Котов, 1968);
2 — гора Чугуш (Динник, 1910, Насимович, 1949; Котов, 1968);
3 — Красная Поляна (Динник, 1910; Верещагин, 1959; Котов, 1968);
4 — Дудугуш (Динник, 1910, Верещагин, 1959; Котов, 1968; Насимович, 1949);
5 — Большой Бамбак (Динник, 1910; Верещагин, 1959; Котов, 1968);
6 — Блокгаузская Караулка (Динник, 1910, Верещагин, 1959; Котов, 1968);
7 — урочище Корыто (Динник, 1910; Верещагин, 1959; Котов, 1968);
8 — Рицинский заповедник (Соколов, Сыроечковский, 1990);
9 — верховья р. Мзымта (Динник, 1910;
Верещагин, 1959);
10 —- хр. Магишо (Марков, 1924);
11 — верховья притоков Большой Лабы (Котов, 1968);
12 — Псху-Гумистинский заподвеник (Соколов, Сыроечковский, 1990);
13 — верховья р. Бзыбь (Динник, 1910);
14 — г. Марух (Марков, 1924);
15 — верховья р. Теберда (КБГУ);
16 — верховья р. Кодори (Динник, 1910);
17 — Кисловодск (ЗИН);
18 — Слияние рек Хасаут и Малка (КБГУ);
19 — гора Кинжал (Динник, 1910; Верещагин, 1959; Темботов, 1972);
20 — Тырныауз (Темботов, 1970);
21 — ущелье Кыртык (Верещагин, 1959, ЗИН; КБГУ);
22 — верховье р. Малка (Верещагин, 1959; ЗИН; КБГУ);
23 — гора Эльбрус (Марков, 1924);
24 — верховье р. Баксан (ЗИН; КБГУ);
25 — верховье р. Цхенис-Цхали (Верещагин, 1958; 1959);
26 — верховье р. Чегем (ЗИН; КБГУ);
27 — Безенги (Верещагин, 1959; ЗИН;
КБГУ);
28 — верхняя Балкария (Верещагин, 1959, ЗИН);
29 — верховье р. Черек-Балкарский (Котов, 1968);
305
Кавказский тур
30 — верховье р. Риони (Верещагин, 1959);
31 — Боржомский заповедник (Соколов, Сыроечковский, 1990);
32 — Северо-Осетинский заповедник (Соколов, Сыроечковский, 1990);
33 — гора Казбек (Марков, 1924);
34 - Казбегский заповедник (Соколов, Сыроечковский, 1990);
35 — верховья р. Аргун (ЗИН);
36 — Ахметский р-н (Кулиев, 1989);
37 - Ахметский заповедник (Соколов, Сыроечковский, 1990);
38 — Агвали (КБГУ);
39 — Ботлихский р-н (Абдурахманов, 1971);
40 — Советский р-н (Абдурахманов, 1971);
41 — верховья Авдийского Койсу (Абдурахманов, 1971);
42 — верховья Аварского Койсу (Абдурахманов, 1971);
43 — Тляратинский р-н (Рамазанов, Шахмарданов, 1980);
44 — Лагодехи (ЗИН);
45 — Закатальский округ (Марков, 1924; Верещагин, 1959);
46 — Лагодехский заповедник (Марков, 1924; Соколов, Сыроечковский, 1990);
47 — Гутонский заказник (Маломагомедов, 1967);
48 — Камилух (ЗИН);
49 — Закатальский заповедник (Верещагин, 1938, 1959; Соколов, Сыроечковский, 1990; ЗИН);
50 — верховья Каракойсу (Абдурахманов, 1971);
51 — верховья Казикумухского Койсу (Абдурахманов, 1971);
52 — верховья р. Самур (Верещагин, 1959; Абдурахманов, 1971);
53 — Илисунский заповедник (Соколов, Сыроечковский, 1990);
54 — Рутульский р-н (Шахмарданов, Рамазанов, 1980);
55 — Ахтинский р-н (Шахмарданов, Рамазанов, 1980);
56 — гора Бабаданг (Разевич, 1904);
57 — Исмаиллинский р-н (Верещагин, 1959; Кулиев, 1989);
58 — Гейгельский заповедник (Соколов, Сыроечковский, 1990)
самцов (см. раздел "Изменчивость"). Основная длина черепа самцов 228— 287 мм, самок — 205—234 мм. Размеры и форма рогов подвержены очень большой изменчивости и соответственно будут рассмотрены ниже (табл. 56, 57).
Размеры отдельных элементов посткраниального скелета мы приводим по одному экземпляру пятилетнего тура-самца, отловленного в ущелье Безенги Кабардино-Балкарии (Центральный Кавказ). Длина тазовых костей 255 мм, минимальная ширина тазовых костей 64 мм, максимальная ширина тазовых костей 112 мм, длина бедра 254 мм, длина плеча 295 мм, длина лопатки (без хряща) 208 мм, ширина лопатки 136 мм.
Окраска волосяного покрова. Летняя окраска спины взрослых особей варьирует от рыжевато-серой до рыжевато-бурой. Нижняя сторона тела, внутренняя сторона конечностей, нижняя сторона хвоста имеют более светлую окраску: грязно-белую или желтовато-серую. Подшерсток и основание остевого волоса спины серого цвета. Подшерсток вдоль хребта имеет темно-серую окраску, а остевой волос на концах — бурую, в результате чего создается впечатление, что вдоль хребта тянется слегка заметная темная
306
ю Таблица 56. Промеры туров (в см). Самцы (по: Котов, 1968)
* Возраст
Признак 1 год (18 экз.) 2 года (37 экз.) 3 года (21 экз.) 4 года (29 экз.) 5 лет и более (23 экз.)
limit X limit X limit X limit X limit X
Длина тела 106,0-136,0 120,4 115,0-156,0 137,7 136,0-176,0 155,8 142,0-184,0 165,9 159,0-196,0 178,7
Косая длина туловища 46,0—69,0 58,4 55,0-86,0 69,7 73,0-85,0 77,4 71,0-98,0 83,3 75,0—103,0 88,6
Обхват тела за 66,0-91,0 78,1 76,0-104,0 90,9 < 92,0-110,0 99,9 102,0-124,0 109,6 106,0-128,0 118,5
лопатками
Высота в холке 63,0-78,0 72,6 69,0-93,0 82,8 79,0-99,0 88,5 87,0-107,0 96,5 93,0-108,0 101,3
в крестце 74,0-87,0 81,2 79,0-101,0 92,2 91,0-109,0 101,4 96,0-120,0 107,6 106,0-121,0 116,6
Длина уха 10,0-11,0 10,7 10,0-13,0 11,5 11,0-13,5 11,3 11,0-14,0 12,2 10,5-14,0 12,3
хвоста 8,0-11,0 9,3 8,0-15,0 12,0 11,0-16,0 12,9 11,0-16,0 14,0 12,0-17,0 14,3
передней 41,0-48,0 . 44,6 40,0-54,0 49,9 50,0-59,0 54,3 51,0-63,0 57,9 56,0-6g,0 58,7
конечности &
задней 64,0-76,0 69,9 68,0-92,0 79,5 81,0-95,0 86,4 82,0-99,0 91,2 87,0-99,0 92,6
конечности
кисти 22,0-26,0 23,7 23,0-28,0 26,3 25,0-30,0 27,9 24,0-37,0 31,0 25,0-32,0 29,2
стопы 28,0-34,0 30,9 29,0-38,0 34,7 35,0-41,0 36,3 36,0-41,0 38,0 36,0-41,0 37,8
переднего 9,0-12,0 10,4 9,0-13,0 11,6 10,0-14,0 12,4 12,0—17,0 13,1 12,0-15,0 13,3
пальца
заднего 10,0-12,0 Н,1 10,0-14,0 12,3 12,0-14,0 13,3 11,0-16,0 13,7 12,0-15,0 14,2
пальца
переднего 4,9-6,2 5,6 5,3-7,7 6,2 6,5-8,6 6,6 6,6-7,8 7,3 6,7-7,7 7,2
копыта
Высота переднего 3,1-3,8 3,4 3,4-4,7 4,0 3,9-4,6 4,2 4,5-5,5 4,8 4,4-5,5 5,0
копыта
Толщина переднего 1,8-2,4 12 2.2—2.7 2,5 2,3-3,4 2,7 2.7-3,7 2.9 15=3.1 2.9
копыта 3,7-4,6 4,4 4,3-5,4 5,0 4,9-6,2 5,4 5,3-7,2 5,9 5,8-6,5 6,1
Длина заднего копыта 4,3-5,3 5,0 5,1-6,5 5,7 5.0-6,4 5,7 5,8-7,0 6,4 5,8-6,9 6,4
Высота заднего копыта 2,7-3,3 3,2 3,1-4,1 3,7 3,8-4,5 4,2 4,1-5,0 4,6 4,5-5,2 4,7
Толщина заднего копыта 1,6-2,2 2,0 1,8-2,6 2,3 2,1-2.6 2.3 14—22 16 2,4-2,8 2,6
3,4-4,3 4,1 3,9-4,9 4,4 4,5-5,2 4,7 4,9-5,6 5,3 4,6-5,7 5,3
Таблица 57. Промеры туров (в см). Самки (по: Котов, 1968)
Возраст
Признак 1 год (5 экз.) 2 года (6 экз.) 3 года (9 экз.) 4 года (16 экз.) 5 лет и более (47 экз.
limit 1 х limit 1 * limit 1 * limit 1 * limit I *
Длина тела 106,0-119,0 113,6 115,0-130,0 123,5 122,0-153,0 141,7 136,0-154,0 144,7 136,0-164,0 150,0
Косая длина туловища 54,0-64,0 59,0 58,0-71,0 62,2 67,0-80,0 75,6 65,0-79,0 72,2 64,0—93,0 77,3
Обхват тела за лопатками 68,0-78,0 74,0 74,0-90,0 83,5 84,0-106,0 94,1 92,0-106,0 95,8 92,0-114,0 100,8
Высота в холке 62,0-73,0 69,0 71,0-79,0 74,5 77,0-86,0 82,0 79,0-90,0 83,9 78,0-92,0 86,6
в крестце 74,0-84,0 80,0 80,0-89,0 85,5 85,5-96,0 92,7 92,0-103,0 96,1 89,0-103,0 97,6
Длина уха 10,0-11,5 10,5 11,0-12,0 11,5 10,5-12,0 п,з 11,0-12,0 11,4 10,5-13,5 11,7 .
хвоста 9,0-10,0 9,5 11,0-13,0 12,2 8,0-15,0 10,9 11,0-14,0 11,9 8,0-14,0 12,1
передней 40,0-45,0 43,3 44,0-48,0 46,2 46,0-52,0 49,4 46,0-52,0 49,3 47,0-54,0 51,8
конечности задней конечности 63,0-75,0 68,0 71,0—77,0 72,7 77,0-86,0 80,1 76,0-84,0 79,9 74,0-88,0 81,3
кисти 21,0-24,0 22,5 20,0-26,0 23,5 23,0-27,0 25,3 23,0-26,0 24,2 24,0-28,0 25,3
стопы 27,0-32,0 30,0 28,0-33,0 30,8 32,0-35,0 33,8 31,0-35,0 32,9 31,0-35,0 33,3
переднего пальца 9,0-11,0 10,0 9,0-11,0 10,2 10,5-12,0 11,0 10,0-12,0 11,0 10,5-13,0 11,5
заднего пальца 10,0-11,0 10,5 10,0-12,0 11,0 12,0-13,0 12,1 11,0-12,0 11,8 10,0-13,0 12,1
переднего копыта — — 4,8-6,3 5,6 5,6-6,5 6,0 5,6-6,6 5,8 5,6-7,5 6,6
Высота переднего копыта — — 3,4-3,9 3,6 4,0-4,4 4,2 3,6-4,5 3,8 3,9-5,0 4,3
Толщина переднего — — 1,9—2,2 2.1 2,5—2,5 2.5 2.3-2.5 2Л 2,2—2,9 2,5
копыта 4,0-4,8 4,4 5,0-5,2 5,1 4,5-5,1 4,8 4,7-5,7 5,3
Длина заднего копыта — — 4,5-6,0 5,2 5,8-6,0 5,9 5,3-5,9 5,5 5,3-6,8 6,1
Высота заднего копыта — — 3,2-3,5 з,з 3,7-4,2 3,9 3,4-4,3 3,6 3,4-4,8 4,1
Толщина заднего копыта — — 1,8-2,1 2,0 2,3—2.3 2.3 2,0—2,3 2,1 2,2—2,6 2,3
Длина рога по наружной 9,0-12,0 10,8 3,6-4,3 12,0-16,0 4,4 13,7 4,4-4,8 15,5-23,0 4,6 19,3 4,0-4,6 17,0-25,0 4,3 20,1 4,4-5,2 16,0-33,0 4,7 24,1
кривизне Расстояние между 8,0-15,0 11,7 14,0-16,0 14,8 13,0-21,0 17,4 15,0-22,0 18,6 14,5—31,0 21,7
концами рогов Расстояние от конца рога 7,0-11,0 9,3 10,0-14,0 12,2 13,0-18,0 15,4 14,0-20,0 16,2 14,0-23,0 18,2
до основания по прямой Обхват рога у основания 7,5-9,0 8,5 10,0-11,0 10,5 10,5-14,0 12,1 11,0-14,0 12,4 11,0-16;0 12,6
Capra caucasipa Guldenstaedt et Pallas, 1783
полоса. Окраска головы на лобной части более темная — бурая или буровато-рыжая. Конец морды несколько светлее. Хвост покрыт жесткими длинными волосами темно-бурого цвета, который постепенно переходит к туловищу в окраску тела.
Зимний волосяной покров несколько светлее. При этом окраска его варьирует от серовато-бурой до грязновато-белой.
Структура черепа, по И.И.Соколову (1959), характеризуется тем, что расхождение роговых отростков более широкое, чем у других видов козлов. К характерным признакам автор относит также сильный излом профиля черепа в области стыка между лобной и зароговой частями. Лоб широкий — межглазничная ширина, как правило, больше расстояния от переднего края хоан до нижнего края затылочного отверстия. В.Г.Гептнер и др. (1961) полагают, что череп кавказского тура, будучи сходным с черепом сибирского козла, имеет специфические особенности: наиболее высокая точка лба расположена вблизи переднего грая основания рогов, лоб в поперечном направлении плоский, а в продольном вогнутый, зароговая часть укороченная и на ее верхней стороне нет вогнутостей. Костные стержни массивны, но укорочены.
Зубная система малооригинальна. Предкоренные зубы, по И.И.Соколову (1959), у кавказского тура сильно редуцированы, длина рядов этих зубов на верхней челюсти взрослых животных составляет не более одной трети длины всего ряда коренных зубов.
Кожный покров (табл. 58). Изучены два взрослых самца, добытых 26 июня, и взрослый самец, добытый в ноябре 1988 г. в Кабардино-Балкарии в Безенгийском ущелье. Для сравнения приводятся данные сибирского горного козла Capra sibirica.
Толщина кожи на загривке изученных зверей довольно различна. Более> сходны у них толщина эпидермиса и рогового слоя. Мальпигиев слой состоит из 2—3 рядов клеток. Местами в виде прерывистых штрихов виден зернистый слой. Блестящий слой отсутствует. Роговой слой плотный. Эпидермис пигментирован со средней интенсивностью. Хорошо видны пигментные клетки. Толщина кожи на груди значительная. Эпидермис и роговой слой толстые. Эпидермис на внутренней поверхности имеет ячеи высотой до 220 мкм. Зернистый слой есть лишь местами. Плотный роговой слой очень сходен по внешнему виду с блестящим. Базальный слой эпидермиса пигментирован со средней интенсивностью. Растущие волосы обусловили сильное развитие сосочкового слоя. Пучки коллагеновых волокон в нем проходят в разных направлениях. В сетчатом слое дермы загривка преобладают восходящие пучки коллагеновых волокон. В сетчатом слое дермы груди коллагеновые пучки образуют сложное и плотное переплетение, проходя под различными утлами к поверхности кожи. В глубоких отделах сетчатого слоя располагается много групп жировых клеток. Подкожная жировая клетчатка достигает на груди значительной толщины (9,5—12,7 мм). Сальные железы, крупные в коже груди и более мелкие в коже загривка, расположены по одной позади остевых волос. Потовые железы мешковидные, нсизвитые в коже загривка и трубковидныс, слегка извитые в коже
309
Таблица 58. Промеры кожи серны, бородатого козла, сибирского горного козла и кавказского тура (по: Соколов, 1973)
Показатель Толщина Размер сальных желез, мкм Диаметр потовых желез, мкм Сумма диаметров потовых желез на 1 см препарата, мм
КОЖИ, мм эпидермиса, мкм рогового слоя, мкм дермы, мм сосочкового слоя, мм сетчатого слоя, мм
Серна (летр) №1, загривок — 28 6 — 1,6 — 165 х 475 110 1,8
№2, загривок — 22 5 — 1,4 — 220 х 550 90 —
№1, грудь 4,4 45 23 4,4 1,7 2,7 110 х 380 121 2,0
№2, грудь 3,3 56 22 3,2 1,8 1,4 110 х 450 132 1,5
Бородатый козел (зима) №1, загривок ‘ 1,21 112 95 1,1 0,4 0,7 56 х 140 88 0,7
№2, загривок 1,91 112 100 1,8 0,7 1,1 45 х 112 40 0,3
№3, загривок (летр) 4,85 56 22 4,8 1,3 3,5 84 х 350 121 1,7
№2, крестец (зима) 1,77 78 61 1,7 о,3 1,4 34 х 140 40 0,2
№3, крестец (лето) 3,04 40 11 з.о 1,3 1,7 90 х 190 90 0,8
№3, грудь (лето) 10,46* 583 286 3,5 1,6 1,9 88 х 275 — —
Сибирский горный козел (лето) №1, загривок 3,45 56 17 3,4 1,5 1,9 88 х 220 143 2,2
№2, загривок 3,35 56 17 з,з 1,5 1,8 90 х 190 132 1,8
№1, крестец 2,62 28 6 2,6 1,2 1,4 80 х 190 100 1,6
№2, крестец 2,72 28 6 2,7 1,2 1,5 55 х 300 100 1,7
№1, грудь 11,53* 484 176 4,4 2,7 1,7 190 х 440 88 0,3
№2, грудь 13,80* 803 363 3,8 з,о 0,8 132 х 440 240 2,2
№3, грудь 8,97* 415 220 3,0 2,0 1,0 150 х 440 116 0,6
Кавказский тур (лето) №1, загривок 5,24 40 17 5,2 2,1 3,1 100 х 220 110 0,7
Ь1з2, загривок 3,33 34 11 з,з 1,5 1,8 44 х 275 132 1,2
№1, грудь 18,39* 495 253 5,2 3,0 2,2 132 х 450 80 0,7
№2, грудь 15,03* 330 143 5,2 3,0 2,2 110 х 660 143 1,0
* С подкожной жировой клетчаткой.
Capra caucasica Guldenstaedt et Pallas, 1783
груди. У самцов около хвоста расположены особые железы, усиленно функционирующие в период спаривания и обусловливающие специфический запах козлов (Соколов, 1959). Толщина мыщц, поднимающих волосы, достигает 50—132 мкм.
Волосы загривка летних туров подразделяются на две категории: остевые двух порядков и пух. Кроме того, из кожи торчит довольно много вершин растущих волос. Все волосы ножек не имеют. У остей первого порядка гранны нет, они слегка извилистые в направлении спереди назад и немного уплощены в этом же направлении. Их длина 45,1 мм, толщина 132 мкм, сердцевина 84,8%. Клетки кутикулы имеют в высоту в среднем 25 мкм. Поперек волоса помещается от 2 до 3—4 клеток. По-видимому, это оставшиеся зимние волосы. Остевые волосы второго порядка значительно короче предыдущих (длина 20,0 мм). Это летние волосы. Они имеют уплощенную спереди назад гранну, наибольшая ширина которой достигает 159 мкм. Гранна находится в верхней трети волоса. В поперечнике она имеет форму овала с впячиванием на задней стороне. В боковых отделах корковый слой вдвое толще, чем в передне-заднем (12 мкм по сравнению с 6 мкм). Высота клеток кутикулы почти не уступает предыдущим волосам и равна 24 мкм. Поперек волоса располагается 2—3 клетки. На 1 см2 поверхности кожи насчитывается в среднем 588 остей первых двух порядков. Остевые волосы третьего порядка очень малы (длина 1,3 мм, толщина 32 мкм). Гранна у них хорошо развита. Сердцевина отсутствует в основании и вершине волоса и имеется только в гранне. Волосы этого порядка растут пучками (точнее, группками), по 2—3 в каждом пучке. На 1 см2 поверхности кожи их насчитывается 1750. Пуховые волосы очень редки. Они длинные (55,8 мм), тонкие (23 мкм) и извитые. Очевидно, они представляют собой остатки зимнего пуха. Их кутикулярные клетки в высоту достигают в среднем 21 мкм, что почти равно толщине всего волоса. Клетки, как правило, кольцевидной формы. Волосы груди принадлежат к одной категории остевых волос, которая делится на три порядка. Между ними можно найти переходные формы. Гранны и ножек они не имеют. Растут они поодиночке, располагаясь слабо выраженными рядами. Остевые волосы первого порядка длиной 22,9 мм, толщиной 132 мкм, с сердцевиной 73,9%, слегка расширены в средней части, не извитые, немного загнутые назад. У большинства вершины обломаны. Форма их поперечника неправильно округлая. На 1 см2 поверхности кожи насчитывается 583 волоса первого и второго порядка. Остевые волосы второго порядка короче (16,8 мм) и тоньше (78 мкм) остевых первого порядка. По форме они сходны с ними. Остевые волосы третьего порядка длиной 12,4 мм и толщиной 78 мкм слабоизвилистые (1,5—2,5 извилин). Сердцевина в них хорошо развита. На 1 см2 поверхности кожи насчитывается 416 волос.
У ноябрьского самца волосяной покров на загривке представлен двумя категориями: остевыми и пуховыми. Общий окрас светло-бежевый. Основания остевых и пуховые волосы серого цвета. Остевые волосы слабо подразделяются на размерные порядки. Их длина 55,1—97,3 мм. У самых длин-
3 1 1
Кавказский тур
Рис. 40. Строение волос взрослого самца кавказского тура
А — поперечный срез остевого волоса в расширенной части стержня, ув. х 500; Б — то же в оснований стержня; В — строение сердцевины остевого волоса, ув. х 2000; Г — кутикула остевого волоса, ув. 600; Д — поперечный срез пухового волоса, ув. х 3000;
Е — кутикула пухового волоса, ув. х 1000
312
Capra caucasica Gflldcnstaedt ct Pallas, 1783
ных волос (90—97 мм) стержень от основания к вершине сужается. Толщина у основания около 180 мкм. У коротких (55—89 мм) стержень слегка расширяется от основания к верхней своей половине, где толщина колеблется от 120 до 170 мкм. К вершине он сужается. Ножка у всех волос отсутствует. Поперечные срезы у основания неправильной округлой формы (рис. 40, б). В верхней половине стержня, вдоль него, с дорсальной и вентральной сторон проходят желобки. С дорсальной стороны желобок выражен лучше, чем с вентральной, поэтому поперечные срезы в этой части стержня имеют бобовидную форму (рис. 40, а).
Сердцевинный канал хорошо выражен вдоль всего стержня и составляет 76—80% его диаметра. Сердцевина зернистого типа, крупноячеистая (рис. 40, в). Сердцевинные клетки неправильной формы. Сердцевинные перегородки тонкие. Их поверхность слабо-бугристая. Перегородки укреплены волосковидными отростками.
Рисунок кутикулы вдоль стержня практически не меняется (рис. 40, г). Высота чешуй более или менее постоянна. Свободные края слегка волнисты с небольшими выемками и плотно прилегают к корковому слою.
Категория пуховых волос хорошо выражена. Их длина достигает 50 мм. Толщина одинакова вдоль всего стержня (20 мкм). Они сильно извитые. Поперечные срезы округлой формы (рис. 40, д). Сердцевина отсутствует. Рисунок кутикулы вдоль стержня не меняется. Она кольцевидного типа. Чешуйки высокие (21 мкм). Свободные края относительно ровные (рис. 40, е). Стержень слегка расширяется от основания к верхней своей половине, где толщина колеблется от 120 до 170 мкм. К вершине он вновь сужается. "Ножка" у всех волос отсутствует. Поперечные срезы у основания неправильной округлой формы (рис. 40, в). В верхней половине стержня вдоль него с дорсальной и вентральной сторон проходят желобки. С дорсальной стороны желобок выражен лучше, чем с вентральной, поэтому поперечные срезы в этой части стержня имеют бобовидную форму (рис. 40,а).
Система крови. Сведений по кавказскому туру очень мало. По одной самке из Дагестана приводятся данные Н.Н.Булатовой (1962), согласно которым кислородная емкость крови кавказского тура очень высокая, вполне соизмерима с таковой автохтонных видов высокогорья, как лама и уриал. Количество эритроцитов составляет 14,34 млн в 1 мм^, содержание гемоглобина 12,4 г%, гематокрит 30,0 об.%, цветной показатель 0,26 ед., диаметр эритроцитов 3,54 мкм, средний объем эритроцитов 20,9 мкм’. Кавказский тур сильно уступает по количеству эритроцитов винторогому козлу (25,77 млн) и туру (21,92 млн), по гемоглобину уступает первому (17,1 г%) и почти одинаков со вторым (12,7 г%).
Индексы внутренних органов. Морфофизиологические показатели кавказского тура специально нс изучены, но очень ценные данные по абсолютным показателям внутренних органов содержатся в работе В.А.Котова
313
Кавказский тур
Таблица 59. Масса отдельных органов и размеры некоторых из них кавказских туров Западного Кавказа. Самцы (по: Котов и др., 1968)
Признак Возраст
1,5—2 мес (1 экз.) 3 года (1 экз.) 5 лет, истощенный после зимовки (1 экз.) от 6 до 18 лет (15 экз.)
Общая масса тела 9,2 64,6 Масса (кг) 81,2 123,5-155,0 (142,8)
Шкура 0,8 4,5 7,6 8,8-12,4 (11,6)
Убойный вес туши 5,6 35,8 45,2 71,7-94,1 (83,6)
Выход мяса (%) 60,2 55 55,6 52,8-63,9 (58,7)
Внутренний жир Нет — Нет 1,3-7,1 (4,8)
Голова с языком 0,55 3,2 7,5 9,5-15,4 (11,4)
Язык 0,1 — — 0,2-0,3 (0,2)
Сердце 0,1 — 0,5 0,6-1,1 (0,7)
Легкие без трахеи 0,2 — 1,8 1,5-2,6 (1,9)
Печень 0,25 — 1,9 1,1-2,4 (1,7)
Селезенка 0,09 — 0,1 0,3-0,4 (0,3)
Почки 0,1 — 0,3 0,3-0,4 (0,3)
Семенники — — 0,1 0,1-0,3 (0,2)
Все отделы желудка с 0,6 10,5 — 11,5-19,5 (15,6)
содержимым Содержимое всех 0,4 — — 7,1-17,0 (12,6)
отделов желудка Тонкий отдел 0,5 — — 1,0-1,6 (1,4)
кишечника с содержимым Толстый отдел 0,3 1,4-2,8 (2,3)
кишечника с содержимым Тонкий отдел 1400 — Длина (см) 2220-2300 (2260)
кишечника Толстый отдел 400 — 1200-1300 (1250)
кишечника Длина семенника 1,6 — — 7,0-7,2 (7,1)
Ширина семенника . - ,0.9 — 4,6-4,6 (4,6)
314
Capra caucasica Gtlldenstaedt ct Pallas, 1783
Таблица 60. Масса отдельных внутренних органов и размеры кавказских туров Западного Кавказа. Самки (по: Котов,1968)
Признак Возраст
сеголетки (1 экз) 3 года (1 экз) взрослые (9 экз)
Общая масса 14,1 Масса (кг) 35,0 58,0-70,7 (65,5)
Шкура 1,2 — 4,6-5,2 (4,9)
Убойный вес туши 6,8 20,0 31,6-39,0 (35,1)
Выход мяса (%) 48,2 57,0 48,7-56,5 (53,1)
Внутренний жир 0,1 — 1,0-1,2 (1,1)
Голова с языком 0,7 — 2,5-2,6 (2,6)
Язык — — 0,1-0,2 (0,15)
Сердце 0,1 — 0,3-0,45 (0,4)
Легкие, без трахей 0,3 — 1,1-1,9 (1,4)
Печень 0,4 — 0,8-1,3 (1,0)
Селезенка 0,1 — 0,1-0,3 (0,2)
Почки 0,1 — 0,2-0,2 (0,2)
Матка — — 0,1-0,2 (0,1)
Все отделы желудка 2,4 — 8,5-14,5 (Ю,1)
Тонкий отдел — Длина (см) 2300—2600 (2500)
кишечника Толстый отдел — — 1050-1150 (1100)
кишечника
(1968). Как видно из данных табл. 59, абсолютная масса сердца 6—18-летних самцов составляет от 600 до 1100 г, в среднем 700 г. Если учесть, что средняя масса тела 142,8 кг, то индекс сердца составляет 4,90%. Этот показатель интересно сравнить с данными по пятилетнему самцу, истощенному после зимовки: масса сердца 500 г при массе тела 81,2 кг, соответственно индекс сердца 6,16%. Хотя выборка состоит всего из одной особи, видно, что потеря в массе тела приводит к увеличению индекса сердца. Сходную динамику иллюстрируют индексы легких, печени, почек. По селезенке получены противоположные результаты: как по абсолютной, так и относительной массе потеря резерва питательных веществ сопровождается уменьшением селезенки. Очевидно, это связано с анемичным состоянием истощенного организма тура.
Индексы внутренних органов самок, судя по данным табл. 60, имеют существенные особенности. Показательно, что индекс сердца очень высокий (6,10%), т.е. сходен больше с индексом сердца истощенного самца, чем с таковым нормально упитанного. По другим индексам внутренних органов столь разительных половых различий, видимо, нет.
315
Кавказский тур
Несмотря на ряд изъянов в расчетах по индексу сердца и другим внутренним органам, неполноту количества интерьерных признаков, можно заключить, что морфофизология кавказского тура четко укладывается в рамки показателей автохтонных горных видов млекопитающих.
Кариотип кавказского тура сходен с таковыми других видов коз.
Структура и динамика ареала
Туры — эндемичные животные Большого Кавказа, распространение их охраничсно Главным, Боковым и Скалистым хребтами. О прежнем обитании этих козлов на Малом Кавказе нет никаких данных, в том числе археологических и палеонтологических (Верещагин, 1959).
В пределах Северного Кавказа, как в прошлом, так и в наши дни, расположена большая часть ареала этого вида и вместе с тем находится основное поголовье кавказских туров.
Западная 1раница современного ареала туров проходит по линии гор Абаго, Атамажи и Чугуш (Котов, 1968). В середине XIX в., по мнению Н.Я.Динника (1910), они, вероятно, обитали и западнее этих мест — на горах Фишт и Оштен. На востоке их ареал выходит за пределы Дагестана и простирается до истоков р. Пирсагат Азербайджана, что к северу от горы Шемахи (Верещагин, 1938).
Нижнюю храницу распространения кавказского тура в настоящее время трудно очертить: имеющиеся сведения недостаточны и порой противоречивы. На Западном Кавказе его ареал очень узкий и занимает в основном Главный и Боковой хребты: на первом хребте он сплошной, а на втором носит "пятнистый характер". Северным пределом его распространения считают горы Белый Камень, Дзювя и Ачешбок (Насимович, 1939; Котов, 1968). Известны отдельные заходы туров в низовья р. Киши, западнее горы Дудугуш, а также на горы Хатавитця, на правом берегу Малой Лабы (Насимович, 1939). По долине р. Аксаута, правого притока Малого Зеленчука, туры не спускаются ниже с. Горного (Аренс, 1957). В районе Эльбруса ареал кавказского тура расширяется и охватывает Главный, Боковой и Скалистый хребты. Утверждение Н.Я.Динника (1910) о том, что на горах Кинжал и Бермамыт в его время туров не было, нс совсем верно (Темботов, 1970). По глубокому ущелью р. Малки, мало посещаемому людьми и сельскохозяйственными животными, туры прослежены нами значительно ниже горы Кинжал, вплоть до впадения р. Хасаут в Малку. Из этих мест в нашей коллекции имеется 5 экз., из которых в 1960 г. добыто два. Не исключено, что туры во времена Н.Я.Динника заходили туда.
В Баксанском ущелье туры обитают на Главном и Боковом хребтах, спускаясь вниз до г. Тырныауза. На Скалистом хребте их, очевидно, уже нет, но в Черекском ущелье они заселяют его, начиная от высоких вершин Ак-Кая, Кара-Кая, Соух и Царицы и до верхней границы леса (Айунц, 1964). В пределах Северной Осетии распространение туров, видимо, охра-нйчсно Главным и Боковым хребтами, а на Скалистый хребет они нс заходят (Нанисв, 1956; Залиханов, 1967). Судя по нашим наблюдениям и
316
Capra caucasica Gfildenstaedt ct Pallas, 1783
рассказам местных жителей, в Чечено-Ингушетии ареал тура сильно сужен и ограничен осевыми хребтами. В Дагестане, по данным М.Г.Абдурахманова (1973), распространение тура простирается узкой полосой от горы Дик-лоемта с ее отрогами до горы Базардюзю. Западными пунктами их распространения являются горы Заинкорт, Диклосмта и Шавиклдс, а на юго-востоке — на несколько километров восточнее горы Базардюзи (Абдурахманов, 1973).
Распространение кавказского тура в Закавказье оставалось до 30-х годов XX в. очень слабо изученным. Показательно мнение Н.Я.Динника (1910), обобщившего наряду с личными данными все имеющиеся к началу текущего столетия литературные и иные сведения. Он, как и его коллега Разе-вич (1904), полагал, что юго-восточная граница ареала на Большом Кавказе простирается до горы Бабадаг. Возможно, туры проникали еще восточнее — у горы Гюмишлю, т.е. до 48° ЗГ в.д. (Верещагин, 1938), но лишь зимой, когда нет домашнего скота. Н.Я.Динник указывал на обитание туров в верховьях Гсокчая, на горах против Закаталы и Лагодсхи. В отношении сообщения Г.И.Радде о том, что туры водятся и в горах к северу от Шемахи, Н.Я.Динник допускал подобное распространение туров как явление случайное, так как горы здесь для них низкие. В отношении Грузии Н.Я.Динник считал, что во всех высоких горах Сванстии и соседних с ней горах обитает Capra caucasica, а по словам Г.И.Радде, они встречаются и в верховьях Цхенис-Цкали. Охотники сообщали ему, что туры встречаются часто также в верховьях Риона, а раньше попадались и в стороне от него, например на горе Шоде. На южном макросклоне Западного Кавказа Capra severtzovi встречается в верховьях Мзымты, Бзыби и "даже Кодора, истоки которого отделяются от Кубанской области более высокими горами" (Динник, 1910, с. 186).
Чтобы оценить изложенную информацию и состояние изученности структуры ареала кавказского тура в Закавказье до первых десятилетий текущего века, достаточно, на наш взгляд, привести мнения специалистов того времени о наличии или отсутствии тура на Малом Кавказе. Так, Н.Я.Динник (1910) писал, что этот вопрос по сей день не решен, но он, как и К.А.Сатунин, полагал, что туры живут только на Большом Кавказе. Однако "очень мало известны .горы юго-западного угла Кавказа, т.е. горы окрестностей Батума и вообще Чорохского края ... поэтому нельзя сказать с уверенностью, что там туров нет..." (с. 176). К.О. Кесслер (1877), обследовавший Закавказье в 1875 г., указывал на существование кавказского тура в б. Елизаветпольской губернии и вообще на Малом Кавказе.
Положительные сдвиги в познании структуры ареала кавказского тура внес Н.К.Верещагин (1938), проводивший кратковременные полевые исследования в горах Азербайджана с 1935 до 1937 г. Ему удалось установить, что летнее распространение кавказского тура на юго-восток простирается до вершины Айвазель, в 3 км к востоку от горы Гюмишлю (высота ее 2700 м абс. высоты). Здесь туры живут на северном склоне с альпийскими лугами. Начиная с Гюмишлю и далее на северо-запад они занимают и южные скло
317
Кавказский тур
ны, особенно часто в снежное время; тогда они заселяют островные леса верховья рек Пирсагат и Гердыманчай на уровне 1800 м. В этих местах в июне 1937 г. зарегистрированы вблизи вершины Кара-Ятаг в двух ущельях южного направления 4 самки с 6 молодыми и отдельно 3 самца (4—7 лет) в течение всего двух полудневных экскурсий. Судя по количеству следов и экскрементов, автор полагал, что здесь бывало много туров. На северо-восточном склоне, находящемся в пределах Азербайджана, траница' летнего обитания проходит на уровне 2000—2200 м, занимая верховья рек Гиль-чинчай, Конахксндчай, Карачай, Кубихчай, через вершины Уюг (2567 м), Ах-Задун (2820 м), Курукли (2563 м) и на северных склонах горы Шахдаг, в среднем течении Кусарчая. На южном склоне хребта тур непрерывно встречается в летнее время в лесной зоне и вблизи перевалов, от вершины Айвазель на запад до Мазымчая в Азербайджане и дальше в Грузии, "где распространение его достаточно точно нам пока не известно" (Верещагин 1938, с. 8).
По мнению Н.К.Версщагина, особенно много туров было в ущельях притоков рек Гсокчай, Вандамчай, Демирапаранчай, Чалминчай, Дашгил-чай, Шинчай, Курмухчай и к западу от Закатал-Катехчая, Белоканчая и Мазымчая. В то время во всех этих местах были малотрансформированные человеком буково-трабовые леса. На наш взгляд, особый интерес представляет карта распространения кавказского тура в юго-восточной части Кавказа, составленная им и приведенная в названной выше работе (Верещагин, 1938).
О современном распространении кавказского тура в Закавказье сведений очень мало. В Азербайджане, по С.М. Кулиеву (1989), он занимает горы начиная от Исмаиллинского и Куткашснского районов на юге до границы с Грузией на западе. На юго-востоке туры зарегистрированы, в частности, на склонах гор Бабадаг (3629 м над ур. моря), Гюмишлю (2700 м), Базардюзи (4466 м) и Шахдаг (4243 м). В Грузии, по данным Главного управления заповедников и охотничьего дела Грузии, по состоянию на 1988 г., туры встречаются в пяти горных районах: Лагодехском, Казбекском, Гумистанском, Ах-мстском и Рицинском. Подробности их размещения ни в Азербайджане, ни в Грузии нам не известны. По сообщениям местных специалистов, положение с турами благополучно лишь в пределах заповедников. За их пределами туры встречаются спорадично, занимая малодоступные места альпийского, субальпийского и лесного поясов.
Современные тенденции антропогенных изменений ареала. Все имеющиеся данные говорят о том, что кавказский тур — горный, автохтонно-эндемичный вид Большого Кавказа — характеризуется высокой жизнеспособностью, обладает широкой адаптивной зоной. При рачительном отношении к этому виду вполне реально сочетание развитого скотоводства и сохранения тура. Необходимо восстановить былое распространение кавказского тура почти на всей территории высокогорий Большого Кавказа. Об этом красноречиво говорят опыты ряда хозяйств, в том числе Нальчикского лесоохотничьего хозяйства Кабардино-Балкарии.
318
Capra caucasica GiUdcnstaedt ct Pallas, 1783
Систематика и изменчивость
Систематическое положение. Кавказские туры привлекают к себе внимание исследователей вот уже 200 лет, именно с тех пор, когда академику И.А.Гюльденштедту стало известно о существовании этих животных в горах Кавказа. За этот продолжительный период накоплен большой фактический материал, отражающий различные стороны жизни этих замечательных животных. Количество выполненных по этому виду научных работ включает свыше сотни названий, среди которых немало работ, принадлежащих зарубежным ученым. Много также работ, посвященных внутривидовой изменчивости и систематике кавказских туров. Тем не менее единого мнения по систематике туров нет.
Современное представление о географической изменчивости вида, систематическом ранге отдельных форм и их распространении можно свести к трем хруппам:
1) все формы кавказских туров относятся к двум эндемичным видам Кавказа — западнокавказскому Capra caucasica и восточнокавказскому Capra cylindricornis. Первый из них населяет Западный Кавказ, второй — Восточный Кавказ. В пределах Центрального Кавказа, примерно от Эльбруса до Казбека, происходит налегание ареалов обоих видов и частичная гибридизация между ними (Цалкин, 1955; Верещагин, 1959; Гептнер и др., 1961; Абдурахманов, 1973; Вейнберг, 1984; и др.);
2) кавказские туры образуют две географические формы, граница распространения между которыми проходит в области верховьев Ингури и Кубани. Западной форме придается ранг подвида и его относят к виду Capra ibex, объединяющему туров Западного Кавказа с альпийскими и*сибирскими козлами. Форма, населяющая Центральный и Восточный Кавказ, составляет один монотипичный и эндемичный для Кавказа вид — Capra caucasica (Schwarz, 1935; Ellerman, Morrison-Scott, 1951);
3) все популяции кавказских туров относятся к одному эндемичному для Большого Кавказа виду с 3—4 подвидами; западнокавказским С. caucasica severtzovi, населяющим Западный Кавказ; центральнокавказским С. caucasica caucasica, занимающим Центральный Кавказ от Эльбруса до горы Шха-ра, и дагестанским С. caucasica cylindricornis, распространение которого охватывает Восточный Кавказ к юго-востоку от Шхары. Четвертый подвид — тур Динника (С. caucasica dinniki) — весьма сомнителен (Верещагин, 1938, 1947; Соколов, 1959; и др.). Н.Я.Динник (1910) считает все четыре подвида самостоятельными видами.
Многолетние полевые исследования и значительный коллекционный материал убеждают нас в том, что ключ к решению сложных вопросов систематики кавказских туров и к выявлению закономерностей их географической изменчивости лежит во всестороннем изучении популяций туров Центрального Кавказа. Именно в этой части Кавказа находится стык ареалов различных ранее описанных форм. Результаты этих исследований позволили прийти к выводу, что на Кавказе распространен один эндемичный вид — Capra caucasica (Темботов, 1970).
319
Кавказский тур
Изменчивость. Вопросы изменчивости кавказских туров интересовали многих зоологов. Так, хорошо освещена возрастная и половая изменчивость туров с учетом географии. По западнокавказскому туру это работы В.И.Цалкина (1955), В.А.Котова (1966), по восточнокавказскому — работы Н.К.Верещагина (1938), П.И.Вейнберга (1984). Однако центрально кавказский тур при этом выпадал из поля зрения, что, видимо, обусловлено его происхождением. Как уже ранее было отмечено, многие зоологи считали этого тура гибридной формой, в результате чего к нему обращено не столь значительное внимание. К тому же не было достаточного коллекционного материала по этой форме.
Возрастная изменчивость. По мнению многих авторов, средняя продолжительность жизни кавказского тура составляет 14—15 лет. Однако встречаются особи, которые живут более 20 лет. Так, в 1908 г. кавказскому туру, убитому Демидовым Сан Донато, было 22 года. При этом возраст определялся по годичным кольцам роговых чехлов, которые хорошо выражены. Этот способ, по мнению Н.К.Верещагина (1938), можно использовать при определении продолжительности жизни тура и срока его воспроизводительной способности. Однако эта методика не пригодна для самок. В этой же работе автор дает методику определения возраста туров и по стертости зубов: как по резцам, так и по коренным зубам в возрасте от 10 дней до 13 лет. Она приводится ниже. Смена и стирание резцов идет следующим образом:
10 дней — прорезаются 2—4 резца;
1 год — появляются все 8 резцов;
2 года — имеется 1-я пара резцов постоянных и 2—4-я пары стачивающихся молочных резцов;
3 года — имеются 1-е две пары постоянных резцов, начинает сменяться 3-я пара резцов, имеется 4-я пара молочных резцов;
4 года — 1-е две пары постоянных резцов значительно стерты, 4-я пара сменяется;
5 лет — начинает стираться 3-я пара резцов, прорезалась 4-я пара постоянных резцов;
6 лет — все резцы постоянные, 4-я пара стерта незначительно;
12 лет — все резцы сильно стерты, при этом форма стертой поверхности овальная.
Стирание коренных и предкоренных и их смена идет следующим образом:
10 дней — 1-й пррдкоренной прорезается из челюсти, 2-й и 3-й — из десен на обеих челюстях;
• 1 год — 1-е два предкоренных значительно стерты, 3-й начинает стираться, на верхней челюсти прорезался 1-й моляр, 2-й прорезается; на нижней челюсти 1-й коренной прорезался из десен, а 2-й — из челюсти;
2 года — начинают сменяться 1-е два предкоренных и 3-й предкоренной на нижней челюсти; 1-й коренной сильно стерт на обеих челюстях; 2-й на верхней челюсти начинает стираться в передней части, а на нижней челюсти стираются обе части 2-го коренного;
320
Capra caucasica Guldenstaedt et Pallas, 1783
3 года — все подкоренные верхней челюсти сменились, на нижней появляется 1-й предкоренной, 2-й и 3-й сильно стерты. На верхней челюсти равномерно стерты 1-й и 2-й коренные, 3-й появляется; 3-й коренной нижней челюсти начинает стираться;
4 года — три верхних предкоренных еще имеет лунки, все коренные равномерно стертые, 1-й предкоренной нижней челюсти нс стерт; 2-е два предкоренных несколько стерты, все три коренных стираются;
5 лет — все три верхних предкоренных значительно стерты, а коренные имеют лунки; 1-й предкоренной нс стирается, 3-й коренной стерт слабо;
б лет — 1-й предкоренной нижней челюсти начинает стираться, все остальные зубы находятся на разной степени стертости;
12 лет — стертость всех зубов как верхней, так и нижней челюстей настолько сильная, что отсутствуют лунки всех предкоренных и первых коренных верхней и нижней челюстей.
Скорость роста самцов и самок туров различна. По данным В.А.Котова (1968), П.И.Всйнберга (1984), самки прекращают расти уже к трехлетнему возрасту. Самцы же растут до 5 лет. П.И.Вейнбсрг (1984) утверждает, что самцы дагестанских туров растут в течение всей жизни.
Существенно изменяются с возрастом рога самцов туров. При этом изменяются не только количественные характеристики рогов, но и их форма. В табл. 61 и 62 приводятся наши данные по размерам рогов разных возрастов у туров Кавказа. Изменение формы рогов находится в прямой зависимости от мест обитания. Так, у западнокавказских особей рога с возрастом увеличиваются в размерах, тогда как форма практически не изменяется и остается саблевидной. У особей из восточной части Кавказа идет увеличение длины рогов, а также существенно меняется с возрастом их форма — появляется, а затем хвсличивается степень закручснности вокруг своей оси, рога не лежат в одной плоскости. В отличие от восточнокавказских туров у центральнокавказских степень закручснности с возрастом увеличивается не столь существенно.
Таблица 61. Общая длина рогов (см) у самцов разного возраста
Число годовых сегментов Западный Кавказ Центральный Кавказ Восточный Кавказ
п limit X п limit X п limit X
3 3 26—33 29,0 2 35-42 38,5 8 37-50 41,9
4 — — — 7 37-57 45,0 3 33-51 42,3
5 3 40-52 47,0 7 42-57 50,4 2 45-61 53,0
6 2 40-56 48,0 6 42-64 56,2 6 61-69 64,8
7 3 60-82 68,0 5 57-70 63,8 3 58-73 66,0
8 3 67-69 67,7 2 57-69 63,0 — — -г-
9 — — — 6 57-75 67,0 4 70-86 78,3
10 11 5 68-78 71,6 7 66-89 72,9 3 78-85 80,7
12-13 1 95 — 6 72-91 77,8 3 77-88 82,7
21 В.Е.Соколов, А.К.Темботов
32 1
Кавказский тур
Таблица 62. Обхват рогов (см) при основании у самцов разного возраста
Число годовых сегментов Западный Кавказ Центральный Кавказ Восточный Кавказ
п limit Я п limit X п limit X
3 3 18-20 19,0 2 21-23 22,0 8 20,5-26 22,9
4 — — .— 7 23,5-27 25,1 3 22-29 25,7
5 3 23,5-25 24,2 9 23-29 26,4 1 26 —
6 2 21,5-30 25,8 5 26-28 27,0 6 25,5-29,5 27,8
7 3 27-31 29,7 6 25-31,0 28,0 3 26-30 28,0
8 3 27-28 27,7 2 26-29,5 27,8 — — —
9 — — — 6 24,5-31 28,5 4 25-31,5 29,4
10-11 6 27,5-33 30,1 8 26-30 28,8 3 26-32 28,3
12-13 1 30 — 5 27-32 29,0 3 27-31 29.0
Как отмечалось ранее, у самцов увеличивается и длина бороды. Борода появляется у них в возрасте 2,5 лет (Вейнберг, 1984) и с возрастом приближается к концу челюсти.
Половые различия. Этому вопросу также посвящен ряд работ (Цалкин, 1955; Абдурахманов, 1973; Вейнберг, 1984). Однако они носят большей частью региональный характер. Нами проведена попытка объединения всех этих данных и собственных.
Половой диморфизм выражен значительно, как и у многих других представителей копытных.
По нашим данным, размеры тела самок значительно уступают таковым самцов. Так, одновозрастные самки почти вдвое уступают самцам по массе тела. 6—9-летние самки весят в среднем 56,4 кг, тогда как 6—11-летние самцы имеют среднюю массу тела 99,8 кг. Об этом же свидетельствуют данные Н.К.Верещагина (1938) по дагестанскому туру и В.И.Цалкина (1955) по западнокавказскому туру (см. табл. 59 и 60).
Как и у многих представителей копытных, различия касаются размеров рогов. У самок они очень маленькие и не превышают в длину 33 см, тогда как у самцов их длина зависит от возраста: чем старше животное, тем длиннее рога.
Третьим существенным моментом в половом диморфизме является наличие у самцов бороды, которая появляется в 2,5 года.
Интересно отметить и тот факт, что половые различия касаются не только экстерьерных признаков, но и характера возрастной изменчивости. Это относится к скорости роста животных разного пола. Как уже отмечено выше, самки значительно раньше достигают предельных размеров (уже к трехлетнему возрасту), тогда как самцы растут всю жизнь.
Географическая изменчивость. По данным В.И.Цалкина (1955) и П.И.Вейнберга (1984), окраска меха кавказского тура подвержена географической изменчивости, что выражено в разном ходе смены окраски меха в связи с сезоном года. Так, у дагестанских туров, по мнению авторов, зимняя окраска меха более темная, чем летняя. Турам с западного Кавказа присуща зимняя светлая окраска и более темная летняя окраска меха.
322
Capra caucasica Guldenstaedt et Pallas, 1783
Таблица 63. Пропорции черепа у взрослых туров. Самцы (по: Цалкин, 1955)
Признак С. c.caucasica С. с. cylindricornis
limit X limit X
Общая длина 100,0-100,0 100,0 100,0-100,0 100,0
Базилярная длина 88,3-91,8 89,0 85,0-87,6 86,6
Длина лицевой части 72,6-75,8 74,6 74,7-76,7 75,5
Длина морды • 58,5-61,8 60,0 59,3-61,3 60,3
Длина мозговой коробки 42,8-46,2 44,4 42,2-44,4 43,3
Брегма — protuberantia occipitalis externa 19,8-27,9 22,5 19,2-21,2 20,3
Брегма — опистион 31,6-36,0 33,0 29,9-33,0 31,4
Середина зароговой линии — protuberantia occipitalis externa 17,6—22,7 20,7 14,6—19,6 16,8
Середина зароговой линии — опистион 30,3-34,2 31,9 24,7-29,5 27,1
Высшая точка лба — protuberantia occipitalis externa 31,9-46,3 42,1 36,5—40,4 38,3
Высшая точка лба — конец носовых костей 58,1-67,3 63,3 66,4-77,2 71,0
Высшая точка лба — гнацион 80,0-89,0 85,8 90,5-95,5 92,4
Длина носовых костей 35,5-42,3 38,0 35,8-44,5 40,0
Ширина носовых костей у основания 11,0-15,8 13,1 11,2-20,9 15,2
Ширина носовых костей по середине длины 10,0—14,5 12,2 10,4-16,4 13,8
Длина предальвеолярной части 24,8-27,8 26,2 24,6-28,5 26,1
Длина альвеолярного ряда коренных зубов 25,3-31,0 27,4 25,7-28,8 27,3
Ширина морды впереди первого премоляра 16,3-20,6 18,5 16,0-22,4 19,2
Ширина морды в лицевых буграх 29,0-32,8 31,5 26,5-33,8 30,0
Ширина межглазничная 34,7-46,2 40,6 36,2-41,8 39,5
Ширина по задним стенкам орбит 50,7-59,1 54,4 51,3-56,8 54,3
Ширина мозговой коробки 29,0-30,8 29,9 28,3-32,1 30,0
Горизонтальный диаметр орбиты 14,0-16,2 15,4 14,7-17,1 15,7
Размеры тела очень слабо подвержены географической изменчивости. В.И.Цалкин (1955) писал следующее по этому поводу: "...насколько позволяют судить данные, приводимые Н.Я.Динником (1909, 1910), Н.В.Туркиным и К.А.Сатуниным (1902), Л.Б. Бёме (1936) и особенно Н.К.Верещагиным (1938, 1947), между западно- и восточнокавказскими турами заметных различий в размерах тела нет".
Наиболее существенно подвержена географической изменчивости форма рогов, которая очень часто используется многими специалистами в качестве диагностического признака (см. табл. 61, 62, 65, 66). Она сказывается и на структуре черепа (табл. 63 и 64). Популяция туров из западной части Кавказа (бассейн рек от Зеленчука до Белой) характеризуется саблевидной формой рогов (рис. 41). У всех просмотренных нами рогов их конец и основание лежат в одной плоскости, т.е. рога не закручены вокруг своей
21*
323
Кавказский тур
Таблица 64. Пропорции черепа у взрослых туров. Самки (по: Цалкин, 1955)
Признак С. c.caucasica С. с. cylindricorms
limit X limit f
Общая длина 100,0-100,0 100,0 100,0-100,0 100,0
Базилярная длина 86,1-88,3 87,4 85,7-87,2 86,5
Длина лицевой части 77,0—78,8 78,0 76,8-80,0 78,7
Длина морды 60,3-63,3 61,2 59,2-63,3 61,1
Длина мозговой коробки 41,9-44,2 42,9 40,2-43,1 42,1
Брегма — protuberantia occipitalis externa 24,0-27,5 25,6 20,7-25,7 23,9
Брегма — опистион 32,5-35,2 33,8 29,0-32,3 31,8
Середина зароговой линии — protuberantia occipitalis externa 27,8-31,2 28,5 23,6-29,2 26,7
Середина зароговой линии — 35,4-38,0 36,6 32,0-35,6 33,6
ОПИСТИОН
Высшая точка лба — protuberantia occipitalis externa 37,6-40,0 39,0 33,8—39,2 37,0
Высшая точка лба — конец носовых костей 60,0-65,0 63,0 60,8-69,2 64,4
Высшая точка лба — гнацион 82,7-86,8 84,6 84,1-88,5 86,3
Длина носовых костей 38,3-43,0 41,1 39,3-44,0 41,0
Ширина носовых костей у основания 9,7-13,6 П,7 10,1-12,9 12,1
Ширина носовых костей по середине 8,9-11,2 10,4 9,1-11,1 10,2
ДЛИНЫ Длина предальвеолярной части 26,2-28,2 27,0 25,2-28,7 26,6
Длина альвеолярного ряда коренных 27,8-31,2 30,0 28,6-32,8 30,6
зубов Ширина морды впереди первого 16,0-17,3 16,8 14,9—18,4 1'8,1
моляра Ширина морды в лицевых буграх 29,6-31,4 30,6 28,7-33,1 30,1
Ширина межглазничная 36,9-43,3 39,1 35,7-40,7 37,8
Ширина по задним стенкам орбит 52,7-57,4 54,5 52,5-56,0 54,1
Ширина мозговой коробки 28,3-30,6 29,3 27,1—30,8 29,5
Горизонтальный диаметр орбиты 16,4-17,9 16,8 16,5-18,8 J_7?6_
оси. Кроме того, по мере удаления от основания рога постепенно расходятся, в результате чего наибольшее расстояние между рогами приходится на концевую часть. Рога дагестанских туров резко отличаются от описанного типа рогов западнокавказских туров. Все экземпляры дагестанских туров в наших сборах имеют чашеобразную форму рогов: они направлены сначала вверх, а потом сильно сближаются. Кроме того, для всех дагестанских туров специфично закручивание рогов вокруг своей оси примерно на 180°. Этот признак мы наблюдали у большого количества экземпляров разных возрастов, происходящих из различных районов. Самым северо-западным районом распространения туров с таким типом рогов, очевидно, следует считать Черекское ущелье, где нами отмечены туры с рогами в основном дагестанского типа.
324
Capra caucasica Giildenstaedt ct Pallas, 1783
Таблица 65. Обхват рогов (см) при основании у самцов туров (от 7 годовых сегментов и более) (по: Цалкин, 1955)
Обхват рогов (см)
Вид 26 28 30 32 34 36 n М то
C.caucasica 5 7 14 10 5 1 42 29,28 0,39 2,5
C.cylindri- - 6 18 19 5 3 51 30,26 0,29 2,0
cornis
Таблица 66. Масса (кг) одного рогового чехла у самцов разного возраста (по: Цалкин, 1955)
Число годовых сегментов С. c.caucasica С. c.cylindricomis
п limit X п limit X
4 3 0,30,6 0,47 6 0,2-0,5 0,35
5 6 0,3-0,8 0,52 3 0,5-1,2 0,90
6 5 0,6-0,9 0,64 5 0,6-1,3 1,04
7 2 0,8-1,2 1,00 7 1,2-2,0 1,50
8 4 0,7-1,3 1,10 2 1,8-2,5 2,15
9 6 0,8-1,6 1,22 4 1,7-2,9 2,30
10 2 1,6-1,8 1,70 2 2,0—2^2 2,15
11 6 1,0-1,6 1,40 1 — — 2,80
12 2 1,5-1,8 1,65 1 — 3,20
Однако туров с Андийского Койсу Дагестана и из бассейна Черека Кабардино-Балкарии нельзя считать полностью идентичными. На рис. 41 хорошо видно, что черекская популяция туров все же заметно отличается от дагестанских меньшим загибом рогов и меньшей их закрученностью. Кроме того, концы рогов у них, как правило, не сближены столь сильно, как у дагестанских туров. У туров, населяющих территорию Центрального Кавказа в пределах от Чегемского ущелья до р. Теберды, эти признаки очень изменчивы и занимают промежуточное положение. Рис. 41 четко отражает кли-нальную изменчивость как характера изгиба, так и степени закручивания рогов, показывая постепенность перехода от дагестанского типа к западнокавказскому. В наших сборах по всем четырем популяциям — чегемской, баксанской, малкинской и тебердинской — имеются лишь отдельные экземпляры, которых можно было бы уверенно отнести к западнокавказскому или к дагестанскому типу рогов. Достаточно сказать, что в Чегемском и Баксанскам ущельях найдено только по одной паре рогов западнокавказского типа, а дагестанского — ни одной. В Черекском ущелье также
325
Кавказский тур
Рис. 41. Географическая изменчивость рогов кавказского тура (Темботов, 1970) 1 — верховья Андийского койсу; 2 — Черекское ущелье; 3 — Чегемское ущелье;
4 — Баксанское ущелье; 5 — верховье Теберды; 6 — бассейны Зеленчука и Лабы
326
Capra caucasica Guldcnstacdt ct Pallas, 1783
найдена одна пара рогов с типично западнокавказским характером загиба и такой же формой закручивания. Все остальные, которыми мы располагаем, имеют дагестанский тип строения.
Структура поверхности роговых чехлов также подвержена значительной географической изменчивости. Популяция туров из западных районов Кавказа, как правило, характеризуется наличием валиков на передней поверхности чехлов; у дагестанских за редким исключением они не выражены. Бе-зенгийские туры по этому признаку не отличимы от дагестанских. Популяция из Чегема, Баксана, Малки, Теберды, наоборот, отличается пестрой картиной строения передней поверхности валиков. В отличие от западнокавказских туров, с которыми они сближаются по этому признаку, поперечные валики не узкие, а имеют более широкие основания, в результате чего они кажутся более массивными, чем у западнокавказских (рис. 42).
Такой признак, как строение передней поверхности роговых стрежней, также признававшийся систематиками в качестве диагностического, подвержен географической изменчивости. В частности, дагестанские туры характеризуются наличием глубоких бороздок на передней поверхности роговых стержней, тогда как у западнокавказских их нет, поверхность стержней у них гладкая. Наш материал подтверждает то же самое. Кроме того, хорошо выраженные борозды на стержне рогов у туров из восточной половины Кавказа прослеживаются на запад до Черека включительно. Что касается туров, обитающих на территории от Чегема до Теберды, то у них можно встретить все варианты архитектуры стержня рогов — от чисто гладкой поверхности стержня до хорошо выраженной ребристой. При этом экземпляры с гладкой поверхностью стержней составляют от 20 до 40%.
Таким образом, морфологические показатели кавказских туров подвержены широкой геотрафической изменчивости. Наиболее четкую закономерность в характере изменчивости имеет форма рогов, в особенности степень их закручснности.
Исходя из общих закономерностей изменчивости кавказского тура в условиях высотно-поясной структуры горных ландшафтов Кавказа, мы выделяем три подвида: 1) Capra caucasica caucasica Guldcnstacdt et Pallas; 2) C.c.severtzovi Menzbier; 3) C.c.cylindricomis Blyth.
Подвиды
1. Capra caucasica caucasica Guldcnstacdt et Pallas — центральнокавказский тур. Рога у самцов отходят вверх и в сторону, потом загибаются назад, постепенно расходятся в стороны так, чтобы, описав небольшую дугу, вновь сблизиться концами. В целом рог образует полудугу, конец которой приподнят от плоскости, где находится основание и средняя часть рога. Расстояние между концами рогов меньше, чем в средней части, составляя около 49—53 см у старых козлов.
Распространение — Центральный Кавказ, в верховьях рек Малка-Черек на северном и Ингури на южном макросклонах.
327
Кавказский тур
Рис. 42. Кавказский тур
7 западнокавказский, 8—JQ — востбчнокавказский
328
Capra caucasica Giildenstaedt et Pallas, 1783
2. C. c. severtzovi Menzbier — западнокавказский тур. Рога относительно короткие и толстые, саблевидные, и все изгибы лежат почти в одной плоскости. Расстояние между концами больше, чем в средней части, превышает 60 см у старых животных.
Распространение — Западный Кавказ, от верховьев рек Тебсрда на северном и Кодори на южном макросклонах до западных траниц ареала вида.
3. С. с. cylmdricornic Blyth. — восточнокавказский тур. Рога чашеобразные, сильно закругленные — сначала направлены вверх, наружу, потом назад более или менее горизонтально, загибаются внутрь и снова вверх (загиб гетеронимный).
Распространение — Восточный Кавказ, от верховьев Уруха на северном и Риони на южном макросклонах до юго-восточной границы ареала.
Экология
Ландшафтная приуроченность. Биотопы. Все наши полевые наблюдения и большинство литературных данных свидетельствуют о том, что кавказский тур как самостоятельный вид сформировался в условиях высокогорья Кавказа. С таким суждением четко согласуются также его мофрологичскис, морфофизиологические, в том числе гематологические, показатели, рассмотренные нами выше.
Благоприятные ландшафтные условия складываются при наличии не только продуктивных горных лугов и лесных насаждений в качестве пастбищ, но и высоких, малодоступных для человека и хищных зверей скал (для спасения от врагов), различных солонцов и участков с повышенным содержанием минеральных веществ (для минерального питания), источников воды, как горные речки, ручейки и водоемы (водопои), всевозможных навесов, карнизов, пещер и др. (для отдыха, ягнения и т.д.). Все эти важные для туров компоненты горных ландшафтов наиболее часто представлены в субальпийском поясе. Соответственно туры при отсутствии антропогенного пресса чаще встречаются в этом поясе гор, чем в других поясах. Так, по данным И.Ф.Попковой (1967), в Закатальском заповеднике из нескольких тысяч туров, учтенных летом 1953 г., на долю субальпийских и альпийских лугов приходилось 77,6% встреч, а 22,4% — в лесном поясе. Сходные данные получены в Кавказском и Тебердинском заповедниках (Эквтимишвили, 1953; Котов, 1968; нашишанные).
Высотные пределы распространения туров и соответственно спектр населяемых турами горных ландшафтов очень широкие — от 800 до 4000 м и выше, где представлены пояса широколиственных и темнохвойных лесов, субальпийский, альпийский, субнивальный и нивальный пояса.
За пределами заповедников территориальная организация видового населения сильно изменена деятельностью человека. Так, в верховье Малки часть туров оттеснена в высокогорье северного макросклона Эльбруса, где животные держатся на рубеже альпийского и субнивального поясов. Другая часть занимает глубокое, скальное ущелье с обилием высокотравья и широколиственных лесов (Темботов, 1969, 1970). В горном Дагестане, по
329
Кавказский тур
данным В.Г.Гептнера и А.Н.Цалкина (1971), распространение туров аналогичное.
Миграции. У кавказских туров наблюдаются сезонные и суточные перемещения, обусловленные состоянием кормовой базы, ее доступностью, наличием снегового покрова, факторами беспокойства человеком, кровососущими насекомыми и т.д.
Летом, с появлением зеленой растительности, туры поднимаются- на субальпийские луга и выше. Зимой, после выпадения снежного покрова, животные спускаются в леса. Интересно отметить, что в тех районах, где лесной пояс нс развит, он практически и не играет роли в жизни туров. Но там, где имеются лесные участки, туры встречаются в них даже летом, о чем свидетельствуют данные П.И.Вейнберга по Севере-Осетинскому заповеднику.
В зависимости от варианта поясности несколько отклоняется от общей схемы описанный выше ход сезонных миграций. По данным В.А.Котова (1968), в зимнее время туры на Западном Кавказе делятся как бы на две экологические группы: одна, представленная преимущественно самками с молодняком, зимует в лесной части ареала, а другая, представленная большей частью взрослыми самцами, остается в субальпике. С наступлением весны обе группы встречаются на верхней границе леса, где ранней весной появляется травянистая растительность. Затем объединенная группа туров поднимается вверх вслед за появлением растительности в высокогорье. Таким образом, большая часть туров в июне-июле встречается в альпийском или субнивальном поясах. С сентября начинается разделение туров на две группы. При этом, как отмечает В.А.Котов (1968), сезонные миграции на большие расстояния для западнокавказского тура не отмечаются. Они ограничиваются несколькими километрами, а иногда и сотнями метров.
По нашим данным (Темботов, 1984), распространение туров в верховьях Малки (Центральный Кавказ) носит несколько иной характер, обусловленный выпасом домашнего скота в горах. Они здесь в течение всего пастбищного сезона вынуждены находиться в глубоких ущельях, а зимой, после отгона домашних животных, выходят на плато и тут пасутся.
Сезонные миграции туров в Северо-Осетинском заповеднике имеют свои особенности (Вейнберг, 1984). Так, осенью начиная с октября наблюдается постепенное перемещение туров в лесной пояс. Сначала спускаются самки с молодняком, а затем через месяц — взрослые самцы. При этом большая часть туров проводит зиму в лесу. В феврале — начале марта животные покидают лесной пояс и переходят на луга, где к этому времени стаивает снег. С началом вегетации снова перемещаются в лес, а затем с появлением зеленой растительности поднимаются на субальпийские луга.
Наряду с сезонными миграциями кавказскому туру свойственны и суточные перемещения, которые определяются такими факторами, как кормовая база, наличие убежищ от хищников и воздействие паразитов. Суточные митрации совершаются и зимой, и летом. Однако летом они более выражены, чем зимой, так как преодолеваются большие расстояния и высоты местности.
330
Capra caucasica Giildcnstacdt ct Pallas, 1783
По данным В.А.Котова (1968), западнокавказские туры в условиях Кавказского заповедника активны в светлое время суток: в утренние часы с 3 до 9 ч и в послеобеденное время с 15 ч до наступления темноты. В самые жаркие часы дня и ночью туры находятся на лежках. Как отмечает автор, суточная активность самок и самцов одинакова, однако обнаружено различие в активности животных на склонах с различной экспозицией. Переходы туров Западного Кавказа колеблются от нескольких метров до нескольких километров в зависимости от рельефа.
По данным П.И.Вейнберга (1984), суточная активность дагестанских туров отличается от таковой западнокавказских. Как отмечает автор, туры Север о-Осетинского заповедника активны в сумеречное и ночное время. К 17 ч животные спускаются на пастбища и пасутся всю ночь, а ранним утром (к 4—5 ч) возвращаются на лежку. Цикличность суточных миграций хорошо выражена у взрослых самцов. Самки и молодые самцы пасутся и в дневное время. Взрослые самцы совершают переходы протяженностью в среднем до 1500 м, тогда как у самок и молодых самцов переходы не столь большие (300—500 м в среднем).
- Активность туров в течение дня подвержена сезонной изменчивости. Летом животные пасутся в сумерках и ночью, хотя у самок отмечена пастьба и в течение дня. Зимой же туры пасутся преимущественно днем.
Интересно заметить тот факт, что направление переходов туров на северном и южном макросклонах Большого Кавказа различно. Как уже отмечалось выше, для туров северного макросклона характерно то, что пастбища находятся ниже, чем места лежек. У животных южного макросклона наблюдались переходы другого направления. Пастьба проходит на больших высотах. В зимнее время туры не совершают суточных переходов.
Линька. Туры линяют 2 раза в год. Весенняя линька у западнокавказских туров, по данным В.А.Котова (1968), начинается с конца, апреля. Заканчивается она у большинства животных в середине июля, хотя встречаются такие особи, у которых летний мех появляется уже к началу июля., У восточнокавказских туров весенняя линька начинается и кончается несколько раньше — с первых чисел марта до первой половины июля. При этом самки дагестанских туров заканчивают линьку значительно раньше, уже к концу июля (Вейнберг, 1984).
Шерсть сменяется начиная с подшерстка, который клочьями счесывается с тела. Затем сменяется остевой волос. Линька западнокавказских туров начинается с головы (летний мех появляется на щеках и за ушами), затем волосы сменяются на лопатках, боках, бедрах, на нижних частях ног. Позже всего линяют спина, круп и лоб (Котов, 1968). Летний мех представлен коротким жестким остевым волосом. Летняя окраска туров Западного Кавказа, как отмечалось ранее, более темная, чем зимняя (Цалкин, 1955; Котов, 1968). По данным Н.К.Верещагина (1938) и П.И.Вейнберга (1984), у туров Восточного Кавказа обнаружена иная смена окраски. Летний мех у них более светлый в сравнении с зимним.
Вторая линька, т.е. осенняя, начинается у дагестанских туров с середины сентября (у нас имеются данные по линьке туров с Восточного Кавказа).
33 1
Кавказский тур
Зимний мех густой, с хорошо выраженным подшерстком, остевой волос зимой по длине превышает летний в 2—2,5 раза и составляет 55—60 мм. Пух также очень длинный и достигает примерно той же длины, что и остевой волос, но в силу своей большой извитости имеет высоту около 25 мм (Верещагин, 1938).
Питание. Питание туров разнообразное и резко меняется в разные сезоны года. В теплый период года животные поедают преимущественно травянистые растения, при этом незначительную долю в рационе составляет веточный корм. В зимний период доля веточного корма резко возрастает, а во второй половине зимы он становится основным. Период, в течение ко-торго туры потребляют веточный корм, равен примерно 180 дням (Котов, 1968). Список поедаемых турами Западного Кавказа растений включает в себя 195 видов. К наиболее предпочитаемым травянистым растениям относятся злаки: костер пестрый, лисохвост, луговик извилистый, овсяницы, тимофеевка; помимо перечисленных злаков туры предпочитают манжетки, бодяк и некоторые другие. Злаки, по данным некоторых авторов, составляют 80—90% в рационе туров. В рационе дагестанского тура также основную долю составляют злаки. Однако, по данным М.Г.Абдурахманова (1977), список поедаемых растений более значителен (256 видов) в сравнении с таковым западнокавказского тура. При этом наиболее предпочитаемыми из травянистых растений являются: овсяницы горная и овечья, тимофеевки альпийская и луговая, лисохвост пушистоцветковый, душистый колосок, вейник, горошек альпийский, камнеломки, купавки, тмины.
Интересно отметить тот факт, что предпочитаемый рацион зависит не столько от избирательности туров, сколько от обилия и флористического состава пастбищ, о чем свидетельствуют результаты исследований многих авторов (Котов, 1968; Абдурахманов, 1973, 1977; Вейнберг, 1984).
Грибы, лишайники и различные кустарники входят в рацион не только западнокавказских туров, но и восточнокавказских (Залиханов, 1967; Вейнберг, 1984).
В питании туров большое значение имеет наличие естественных солонцов в связи с нехваткой натриевых солей в растительном корме. Туры охотно посещают искусственные солонцы, которые представляют собой разбросанные комья соли на пастбищах.
Воспроизводство и стуруктура популяции
Размножение. Половая зрелость самок наступает к 2,5 годам (Верещагин, Т938). В.А.Котов (1968) сообщает о том, что самки кубанских туров вступают в размножение в возрасте старше 3 лет. Однако отмечены случаи более раннего полового созревания — в 1,5—2 года (Джанашвили, 1948). Самцы также могут достигать по'ловозрелости к 2,5 годам, хотя, как утверждает Н.К.Верещагин (1938), расцвет половой функции у них наступает к 6 годам.
Гон туров на Западном и Восточном Кавказе, по данным В.А.Котова (1968) и М.Г.Абдурахманова (1973), в Кавказском заповеднике и Дагестане длится со второй половины ноября до конца декабря. В холодные зимы он
332
Capra caucasica Gtildcnstaedt ct Pallas, 1783
может растягиваться и до конца января (Абдурахманов, 1973). В Северо-Осетинском заповеднике гон туров отмечен несколько позже — в декабре и длится до начала января (Вейнберг, 1984).
Беременность самок, по А.П.Иняковой (1957), длится 150—160 дней, а по Н.К.Верещагину (1938) — 160—165 дней. Самки перед родами отдаляются от стада, при этом они пасутся на верхней границе леса или в скалах субальпийского пояса. Рождение турят на Западном Кавказе приходится на начало мая — середину июня (Котов, 1968); как отмечает автор, самая ранняя самка с сеголетком им была встречена 3 мая. Для Тебердинского заповедника были отмечены случаи очень позднего появления турят — в начале августа (Эквтимишвили, 1954). Рождение турят на Восточном Кавказе, по данным Н.К.Верещагина (1938), наблюдается с 20-х чисел мая по 20 июня.
Самки приносят по одному туренку. Хотя, как отмечает ряд авторов (Верещагин, 1938; Котов, 1968; и др.), имеет место и рождение двоен.
Многие самки с турятами все лето ведут одиночный образ жизни, избегая взрослых особей (Вейнберг, 1984).
Что касается яловости самок, В.А.Котов (1968) пишет, что "в период рождения молодняка в 1962—1966 гг. было поймано 185 половозрелых самок в возрасте 4 лет и старше, из них яловых было в среднем 5,3%, по отдельным годам 3,2—6,9%”. Большинство самок приносят приплод ежегодно. Причина такого интенсивного размножения, по мнению автора, заключается в том, что, во-первых, для популяций кубанских туров свойственна высокая смертность на ранних этапах жизни и, во-вторых, самки поздно достигают половозрелости.
Рост и развитие. Рост и развитие самцов и самок кавказских туров первые три года идут очень интенсивно. Самки к трем годам прекращают расти, тогда как самцы, как было отмечено ранее, по данным некоторых авторов (Верещагин, 1938; Вейнберг, 1984), продолжают постепенно расти всю жизнь.
По данным Н.К.Верещагина (1938), вес новорожденных турят достигает 5—6 кг. 10-дневный туренок весил 8 кг (Верещагин, 1938). Новорожденные турята могут следовать за матерью сразу после рождения; новорожденных турят можно поймать лишь в течение первых часов. Данные по возрастным изменениям морфометрических признаков описываются в разделе "Возрастная изменчивость".
Структура популяции. Половозрастная структура кавказских туров, достаточно характеризующая состояние популяций, сильно меняется с продвижением с северо-запада на юго-восток. Так, по данным В.А.Котова (1968), соотношение взрослых самцов и самок по визуальным подсчетам в период с 1941 по 1966 г. в Кавказском заповеднике сильно колебалось. Самцы обычно преобладали по численности, при этом соотношение менялось от 1:1 до 1,82:1. И лишь в 1948, 1952 и 1955 гг. самок было больше. Соотношение самцов и самок в эти годы было 0,97:1, 0,58:1 и 0,96:1 соответственно.
В субальпийском поясе Северо-Осетинского заповедника в летний период, по данным П.И.Всйнберга (1984), самцы преобладают по численности над самками: в 1977 г. — 1,9:1; в 1978 г. — 1,6:1; в 1979 г. — 1,7:1. Однако,
333
Кавказский тур
Таблица 67. Половой и возрастной состав популяции туров в Кавказском заповеднике
(по данным визуального учета летом 1941—1966 гг.) (по: Котов, 1968)
Год Самцы Подростки Самки Сеголетки — ? - Пол не определен Всего
1941 1149 — 1092 452 594 3287
1942 686 — 424 232 305 1647
1944 344 — 283 138 220 985
1945 803 — 593 267 625 2288
1946 1405 411 987 511 392 3706
1947 654 358 567 294 242 2124
1948 945 359 977 518 250 3049
1949. 1371 415 992 492 577 3847
1950 1216 457 980 442 525 3620
1951 946 199 715 381 564 2805
1952 654 319 1132 483 285 2873
1953 440 74 440 175 127 1256
1954 1007 271 972 391 805 3446
1955 422 52 439 124 661 1698
1956 597 102 490 359 623 2171
1957 1248 159 1098 473 552 3530
1958 1806 311 991 611 721 4440
1959 2683 379 1690 947 338 6037
1960 1927 503 1922 871 807 6030
1961 2019 277 1479 598 400 4773
1962* 77 66 371 117 4 635
1963 1989 603 1524 809 695 5620
1965 1804 487 1630 677 1380 5978
1966* 260 2 138 31 79 510
Итого 1 26 452 5804 21 935 10 393 11 771 76 355
♦Учет проведен только на одном участке.
Таблица ,68. Количество сеголеток (в % от общего поголовья туров) в Кавказском ________заповеднике по данным учета в 1941—1966 гг. (по: Котов, 1968)____
Год % . — ".X. Год % Л — э Год %
1941 13,7 1951 13,5 1960 14,4
1942 14,0 1952 16,8 1961 12,5
1944 14,0 1953 13,9 1962 18,4
1945 11,6 1954 11,3 1963 14,4
1946 13,7 1955 7,3 1965 11,3
1947 13,8 1956 16,5 1966 6,0
1948 16,9 1957 13,4
1949 12,6 1958 13,7 Среднее 13,4
1950 12,2 1959 15,7
как отмечает автор, соотношение полов следует изучать в период гона, когда все животные (самцы и самки) концентрируются на одних участках. П.И.Вейнберг считает, что истинное соотношение самцов и самок в Северо-Осетинском заповеднике равно 1,3—1,4:1. Для Дагестана свойственно иное соотношение, в котором численность самок резко преобладает над
334
Capra caucasica Giildenstaedt et Pallas, 1783
численностью самцов (Абдурахманов, 1973). Автор приводит соотношение полов для трех урочищ: Гвериб (0,5:1), Дарциб (0,7:1), Дюльтыдаг (0,3:1). Такое различное числовое соотношение взрослых самцов и самок связано, видимо, с тем, что на неохраняемых территориях Центрального и Восточного Кавказа браконьерами производится избирательный отстрел взрослых самцов (Абдурахманов, 1973).
Численное соотношение разных возрастных групп на территории Кавказского заповедника приводится в табл. 66—68 по В.А.Котову (1968). Процентное соотношение сеголеток за 24 года (1941—1965 гг.) в среднем за год составило 13,4%. При этом оно колебалось в пределах от 6 до 18,4%. Однако, как отмечает автор, эта цифра в действительности несколько выше в результате того, что в летний период самки с козлятами распространены в лесном и субальпийском поясах, а подсчет производился в летний период и без учета первых. Соотношение возрастных rpyrfn для Северо-Осетинского заповедника приводится в работе П.Н. Вейнберга (1984). Как следует из его данных, наибольшее количество сеголеток приходится на осень. Соотношение взрослых самок и сеголеток в этот период наименьшее — 1,55:1. Оно заметно увеличивается зимой (1,73:1) и летом (1,91:1) из-за гибели сеголеток. Соотношение взрослых самок и подростков носит иной характер. Минимальное соотношение приходится на летний период (2,98:1), максимальное — на зиму (6,86:1), т.е. численность подростков больше всего летом.
Интересна структура распределения туров внутри ареала. П.И.Вейнберг (1984), исходя из состава стада, выделяет для Северо-Осетинского заповедника следующие группы: 1) группа самок, в которой, помимо сеголеток, могут быть и молодые самцы; 2) группа молодых самцов; 3) группа взрослых самцов, среди которых имеется хотя бы один взрослый; 4) смешанная группа, в состав которой обязательно входят одна или несколько самок и хотя бы один взрослый самец.
Встречаемость разных групп меняется по сезонам года, что обусловлено репродуктивным циклом животных. Так, труппа взрослых самцов в период гона объединяется с труппой самок. К труппе самок в период гона присоединяется и труппа молодых самцов.
Враги, паразиты, смертность, конкуренты. К числу естественных врагов кавказских туров относятся представители хищных млекопитающих — леопард, медведь, рысь, волк для взрослых особей и хищных птиц — беркут, ягнятник для турят.
Результаты исследований копытных Кавказского заповедника К.В.Жу-равлевой и Ю.О.Раушснбах (1939) показали, что туры больше других копытных животных поражены гельминтами. Так, процент зараженных особей был наивысшим весной (98,4%) и наименьшим зимой (65,3%). В табл. 69 приводится перечень видов гельминтов, найденных в тех или иных органах туров. Упомянутые авторы отмечают, что случаев смерти из-за инвазии гельминтами не обнаружено. По данным П.И.Вейнбсрга (1984), у восточнокавказских туров обнаружены 32 вида эндопаразитов.
335
Кавказский тур
Таблица 69. Виды гельминтов, обнаруженные у кубанского тура (по данным Д-П-Рухлядева, Т.Э.Родоная и З.С.Эквтимишвили) (Котов, 1968)
V ~ * ? — Вид XXXJ z V — , Л Локализация
Dicrocoelicum lanceatum Stives et Hassal, 1896 Moniezia benedeni Moniez, 1879 Coenurus cerebralis Leske, 1780 Cysticercus tenuicollis Pallas, 1879 Chabertia ovina Fabricius, 1788 Ostertagia circumcineta Stadclmann, 1984 O. trifurcata Ransom, 1907 O. dahurica Orloff, Belova et Gnedina, 1931 Ostertagia occidentalis Ransom, 1907 Muellerius capillaris Mueller, 1889 Neostrongylus linearis Marotel, 1913 Trichocephalus ovis Abildgaard, 1795 Ostertagia ostertagia Stiles, 1892 Marschallagia marschalli Ransom, 1906 Nematodirus spathiger Raill, 1896 N. davtiani Grigorian, 1949 Protostrongylus habmaieri Schulz, Orloff et Hutai, 1934 Skrjabinema ovis Sktjobin, 1915 Печень Тонкий кишечник Головной мозг Серозные покровы брюшной полости Толстый кишечник Сычуг Мелкие бронхи То же Слепая кишка Сычуг Легкие Толстая кишка
Н.К.Верещагин (1938) утверждает, что при тщательном изучении 15 экз. туров Закатальского заповедника им не было обнаружено ни одного случая инвазии гельминтами.
Значительный вред, по данным В.А.Котова (1968) и П.И.Вейнберга (1984), турам приносят личинки носоглоточных оводов. Паразитирование этих личинок может привести к летальному исходу. Личинки оводов могут проникать в головной мозг тура.
Естественными причинами гибели туров могут служить несколько факторов: хищники, паразиты, различные заболевания (в том числе заболевания домашнего скота), а также снежные лавины. Наши данные, как и В.А.Котова (1968), М.Г.Абдурахманова (1973), П.И.Вейнберга (1984), свидетельствуют, что самым существенным фактором, определяющим значительные изменения численности, являются снежные лавины. Под ними ежегодно на всей территории Кавказского заповедника погибает около 300 туров (Котов, 1968). В Кабардино-Балкарии под лавинами гибнет около 370 животных (Залиханов, 1967). Нередко под лавинами большей частью погибают взрослые самцы, ибо весной, в лавиноопасный период, они держатся в субальпийском поясе и первыми осваивают выскогорье, а самки и молодые козы в это время обитают преимущественно в лесном поясе, где лавинная опасность практически отсутствует.
Гибель туров от хищников в разных районах Кавказа различна. Например, роль волка значительна в Дагестане. По данным М.Г.Абдурахманова (1973), здесь 20% туров от общего отхода погибает от волков. Рысь, по мнению автора, также может существенно регулировать численность туров.
336
Capra caucasica Giildcnstacdt ct Pallas, 1783
Медведь, для которого высотные пределы распространения значительны (он поднимается до 3000 м над ур. моря), также может быть опасным для туров (Вейнберг, 1979). Аналогичная картина наблюдается и в других районах Кавказа.
Рассматривая вопрос о заболеваниях туров, следует отметить, что он плохо изучен. Однако известны случаи заболевания туров чумой, чесоткой и другими болезнями домашнего скота (крупного и мелкого рогатого скота) (Верещагин, 1938; Насимович, 1949а).
По данным В.А.Котова (1968), в условиях Кавказского заповедника естественными конкурентами являются серна и в меньшей степени олень. Автор отмечает, что из 195 видов растений, которые поедаются турами, 55 встречаются в питании серны. В восточной части ареала конкурентом кавказского тура является безоаровый козел.
В значительной степени конкурентом кавказского тура на протяжении всего ареала служит домашний скот, как крупный, так и мелкий. Домашний скот (колхозный и частный) выпасается на естественных местах обитания туров, тем самым ограничивая его кормовую базу и вытесняя его в недоступные ущелья.
Численность. Численность туров наиболее высока в заповедниках, находящихся в пределах кубанского варианта поясности. В Кавказском заповеднике к концу 60-х годов численность туров составляла более 15 тыс. голов. Плотность их населения в альпийском поясе определялась от 50,6 до 414,1 экз. на 1000 га. В Кабардино-Балкарии на 1975 г. насчитывалось 5500 голов (Равкин, 1975), в Северной Осетии и Чечено-Ингушетии на 1972 г. обитало 3800 и 2000 туров соответственно (Вейнберг, 1984), в Азербайджане 8 тыс. туров (Кулиев, 1981), в Грузии 12 тыс. (Капанадзе, 197-8). В 1988 г. в Грузии учтено всего около 2 тыс. голов.
Как отмечают многие авторы, численность тура значительно выше на территориях заповедников, а в сопредельных районах намного ниже.
Низкая численность туров в незаповедных районах определяется двумя основными причинами. Первая — браконьерство, вторая — использование пастбищ, местообитания туров под выпас домашнего скота. Причем часто скот выпасается нерационально, неравномерно, с перегрузками на отдельных пастбищах. По мнению П.И.Вейнберга (1984), к этим двум факторам следует добавить промышленное освоение горных туров, туризм и альпинизм, оказываю 1цие отрицательное воздействие на численность туров.
Практическое значение. Во все времена и во всех районах Большого Кавказа кавказские туры служили объектом охоты, доставлявшим не только очень вкусное мясо и жир, кожу и роговые чехлы для украшения и различных поделок, особенно в качестве кубка для вина, но истинное наслаждение от большой физической нагрузки во время охоты. Однако в настоящее время значительное количество туров сохранилось только в пределах заповедников, где охота, естественно, запрещена. За их пределами на большей части территории туры стали редкими, и охотятся на них в основном браконьеры. Между тем имеются реальные предпосылки для широкого восстановления распространения этого вида в высокогорьях Большого
22 В.Е.Соколов, А.К.Темботов
337
Capra aegagrus Erxlcbcn, 1777
Кавказа и доведения плотности до уровня промысловой. Для этого нужно снять или значительно смягчить влияние ряда лимитирующих факторов.
1. Искоренить браконьерство, имеющее место почти во всех районах ареала.
2. Необходимо провести с учетом специфики каждого района экономические расчеты для определения наиболее рационального пути использования высокогорных лугов, исключить узковедомственный подход к ресурсам природы.
3. С учетом характера географической изменчивости и популяционной структуры вида создать сеть резерватов, обеспечивающих сохранение генофонда в многообразии, что необходимо, на наш взгляд, для поддержания высокой жизнеспособности кавказских туров.
Мы убеждены в том, что если подходить по-хозяйски и комплексно к природным ресурсам высокогорья Кавказа, то кавказские туры могут стать объектом спортивной охоты, особенно в случае организации иностранного туризма.
Бородатый козел Capra aegagrus Erxleben, 1777 Рис. 43
Antilope gazella, Gmelin, 1788; Capra caucasica, Gray, 1843.
Коллекционный материал. ЗИН РАН: 9 черепов, 2 рога;МГУ: 8 черепов; КБГУ: 6 черепов.
Диагноз. В роде самый мелкий, масса тела менее 55 кг; саблевидные рога плоские, на передней поверхности крупные и острые бугры. Длинная и густая борода и ремень на спине черного цвета.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Самый мелкий из козлов. Масса тела самцов 35—38 кг, самок 26—30 кг. Длина тела до 150 см, высота в холке 80—90 см. В отдельных сводках приводятся более высокие цифры. Так, по мнению ряда авторов, длина тела козла может достигать 155 см, а масса — 55 кг (Даль, 1951; Соколов, 1959; Гептнер и др., 1961; Гаспарян, 1975; и др.). По данным названных авторов, основная длина черепа у самцов достигает 255 мм, у самок — 185—222 мм. Как видно, бородатый козел по размерам значительно меньше, чем кавказский тур. Голова сравнительно небольшая, носовой отдел прямой, лоб выпуклый, особенно у самцов. Уши небольшие, длиной 10—11 см. Весьма характерно, что у бородатого козла зад выше, чем передняя часть тела. Длина хвоста с волосами всего 17—20 см, без волос — 12—13 см. Экстерьерные особенности дополняет относительно короткая шея, что более сильно выражено у самцов, чем у самок.
Сравнительный анализ скелета кавказского тура, бородатых и домашних коз, проведенный Н.М.Ахмедовым и Л.М.Саркисовой (1987), подтвердил видоспецифичность ряда параметров экстерьера. Так, по отношению к длине позвоночника шейный отдел 1—7-летних животных составляет у туров
338
Capra aegagrus Enclcben, 1777
от 24,48—29,10%, у бородатых козлов 23,47—27,77%, у домашних коз 23,DO-25,0%. Относительная длина трудного отдела в годовалом возрасте у туров 37,75%, бородатых козлов 35,82%, домашних коз 33,27%. Аналогичная картина сохраняется и в других возрастных группах. Высокий индекс четко выражен у диких животных в годовалом возрасте, а у домашних на втором или третьем году жизни. Относительная длина поясничного отдела всех возрастов у туров 22,44—22,55%, бородатых козлов 23,80—24,82%, домашних коз 25,19—27,19%. Особенно показательно, на наш взгляд, соотношение длины скелета передних и задних конечностей. Разница между ними (задние больше передних) наибольшая у бородатого козла и составляет у различных возрастов от 12,03 до 14,28%, тогда как у туров — от 10,76 до 12,00%. У домашних коз в годовалом возрасте она составляет 12,80%, но к 7 годам снижается до 8,00%.
Количество данных, иллюстрирующих структурные изменения посткраниального скелета бородатого козла в связи с жизнью в горах, можно значительно увеличить. Справедливо мнение,, что быстрый темп формирования скелета, в том числе конечностей и грудной клетки, является адаптивной предпосылкой для активной жизни бородатого козла с первых же дней постнатального периода. Этой особенностью обладает такой же типично горный вид — кавказский тур, а также многие другие виды, вовлекающиеся в активную жизнь в очень молодом возрасте (Соколов, Темботов, 1989).
В экстерьере бородатого козла весьма примечательны рога. Они резко отличаются от рогов тура тем, что роговые чехлы плоские, передняя часть заостренная, изогнута серпообразно, на внешнем (переднем) крае имеется редкий ряд крупных бугров, тоже заостренных спереди. Задняя сторона рогов закругленная, без бугров и резких профилей. Характер изгиба специфический — у основания рога почти прямые, слабо наклонены назад и по бокам, в средней части сохраняется пологий наклон и раскид рогов, лишь в конце они круто закручиваются и заметно сходятся внутрь. За исключением дистальной части, весь рог лежит в одной плоскости. Хорошо выражены годовые кольца, кроме них, на всем протяжении рогов, за исключением верхушки (ювенильной части), расположены поперечные рисунки. Они придают своеобразный вид поверхности рогов. Цвет рогового вещества от серовато-бурого до почти черного. Длина рога по внешнему изгибу составляет до 100 см, иногда до 110 см (Динник, 1910), по другим данным — до 140 см (Соколов, 1959). В наиболее широкой части ширина рога достигает 10,7 см.
Что касается рогов самок, то в их структуре совсем мало оригинального. Они намного меньше, чем у самцов. Так, у 6-летней самки (S-5060) рога более чем в 2,5 раза меньше, чем у 3-летнего самца (S-5O55). На поверхности нет бугров, но четко выражены годовые кольца, а между ними имеется различное количество колец. Характерный признак — заостренный передний край выражен, но менее четко, чем у самцов. Рожки с внутренней стороны почти плоские, а снаружи выпуклые. Общая структура рогов самок и самцов сходная, но концевые части у самок не столь круто загнуты, как у самцов.
22*
339
Рис. 43. Распространение бородатого козла
1 — Боржоми (Динник, 1910; Верещагин, 1959);
2 — верховье Тушетской Алазани (Динник, 1910);
3 — верховья р. Леса (Беме, и др. 1929);
4 — верховья р. Аргун (Динник, 1910: Верещагин, 1959)
5 — верховья р. Шора-Аргун (Точиев 1973);
340
6 — верховья р. Авдийского коису (Динник, 1910);
7 — Пирикительская Алазанп (Динник, 1910);
8 — Туманянский р-н (Гаспарян, 1951);
9 — Арагац (Саркисов, 1955);
10 — Арарат; Зорские горы (Марков, 1924);
11 — Арени (МГУ)
12 — Варденис (Манассров, 1986);
13 — Красносельск (Манассров, 1986);
14 — Кедабск (Верещагин, 1959);
15 - Кировабад (ЗИН);
16 — Закатальский округ (Марков, 1924);
17 — Рутульский р-н (Точиев, 1973);
18 — верховья р. Самур (ЗИН; Динник, 1910; Верещагин, 1959; КБГУ);
19 — верховья Казикумухского Койсу (Шахмарданов, Рамазанов, 1980);
20 — Верховья Аварского Койсу (Динник, 1910; Верещагин, 1959);
21 — Гутонский заказник (Маломаго-мсдов, 1967);
22 — Тлядал (Верещагин, 1959);
23 — Тлярата (Прилуцкая, Пишванов, 1989);
24 — Чарада (Верещагин, 1959);
25 — Гуниб (ЗИН; Динник, 1910; Верещагин, 1959);
26 — Советский р-н (Прилуцкая, Пишванов, 1989);
27 — Ботлих (Дрнник, 1910);
28 — Муни (Прилуцкая, Пишванов, 1989);
29 — Андийский округ (Марков, 1924);
30 — Барда (Алекперов, 1966);
31 — окрестности г. Степанакерт, гора Кир (Верещагин, 1947, 1959);
32 — Дастакерт (Явруян, 1971);
33 — Абракунис (Верещагин, 1959);
34 — Ншрам (Марков, 1924);
35 — Джульфа (Верещагин, 1959);
36 — Ардубад (Динник, 1910);
37 — Мегринский р-н (Гаспарян, 1951);
38 — Шикахохский заповедник (Соколов, Сыроечковский, 1990);
39 — Зангезурский хребет (Саркисов, 1955; Алекперов и др., 1977)
341
Бородатый козкл
Череп бородатого козла, по В.Г.Гептнеру и др. (1961), довольно стройный, вытянутой формы, с небольшими глазницами, роговые стержни сжаты с боков, передний их край заостренный. По И.И. Соколову (1959), характерно наличие вздутий в виде бугров на лбу, перед основаниями роговых стрежней. Соответственно средняя часть лба находится выше краев глазниц. У самок лоб почти плоский. Ширина лба меньше расстояния от переднего края хоан до нижнего края затылочного отверстия. Зароговая’часть черепа относительно длинная, так как стержни рогов не сдвинуты назад — передний край стержней и задние края глазниц лежат на одной линии или перекрываются. Названный автор считает также характерными относительно короткие и широкие носовые кости. Максимальная их ширина составляет 32—49% собственной длины. Передний край носовых костей заходит за вертикальную линию, проведенную дорзо-вентрально перед Р1.
В зубной системе бородатого козла по сравнению с другими видами пре-молярии менее редуцированы. Длина их в верхней челюсти по отношению к длине всего ряда зубов составляет 33—37%.
Окраска меха. Изучена по коллекционному материалу зоомузеев МГУ и ЗИН РАН. Полученные данные согласуются с утвердившимся в литературе мнением о том, что окраска тела бородатого козла обладает, как и форма рогов, высоким систематико-таксономическим весом. На наш взгляд, очень показательны отдельные детали окраски, в том числе наличие черного ремня на спине, черная или черноватая окраска бороды, низа головы, передней стороны труди, "хомута", на кончиках ушей, на хвосте, передней стороне ног, боков на транице с брюхом. Остальная часть тела иного цвета. Летом на боках и спице окраска красноватая, рыжевато-бурая, светлеющая по бокам шеи. Верх головы и губы буро-коричневые, лишь на краях губ окраска белая. Окраска белая, или беловато-серая, или белая с желтоватым оттенком на животе, труди, в паху, на внутренней стороне ног, ягодиц. Характерно наличие белого кольца над коленной мозолью.
Окраска меха подвержена различным видам изменчивости, о чем будет сказано в соовстствующем разделе.
Кожный покров. Описание по 2 самцам (№ 1 и 2) и одной самке (№ 3) приводятся по В.Е.Соколову (1973).
Мальпигиев слой зимой состоит преимущественно из двух рядов клеток. Зернистый и блестящий слои отсутствуют. Роговой слой очень толстый, рыхлый. Эпидермис нс пигментирован. В сосочковом слое дермы пучки коллагеновых волокон, образуя плотную вязь, проходят в разных направлениях. Эластиновые волокна довольно многочисленны, горизонтальные. В сетчатом слое преобладают восходящие коллагеновые пучки, слабо переплетенные. Эластиновые волокна в небольшом количестве имеются только в глубоких отделах слоя.
Сальные железы невелики, но с крупными выводными протоками, шаровидно вздутые перед впадением в волосяное влагалище. У пуховых волос видны только выводные протоки. Потовые железы небольших размеров, их секреторные отделы узкие, слабоизвитые. Особенно слабо развиты потовые железы в коже загривка козла № 2, у которого секреторные отделы имеют
342
Capra aegagrus Erxlebcn, 1777
уплощенные нефункционирующие железистые клетки. В коже крестца потовые железы находятся в редуцированном состоянии.
Мышцы, поднимающие волосы, имеют в поперечнике 33—55 мкм. Волосяной покров бородатого козла состоит из упругих слабоволнистых, сравнительно высоких волос и тонкого извитого подшерстка (Соколов, 1959). Длина остевых волос в зимнем мехе составляет 5—6 см. В летнем мехе подшерсток отсутствует, остевые волосы упругие, плотно прилегают к коже. Волосы затривка козлов четко подразделяются на две категории: остевые и пуховые.
Остевые волосы длинные (35,4 мм), тонкие (109 мкм), с небольшой гран-ной, прямые или слегка извитые. Форма поперечника волос — уплощенно-овальная. Сердцевина занимает 84,3%. В боковых частях волоса корковый слой на 5 мкм толще, чем спереди и сзади. Высота кутикулярных клеток 27 мкм. Очертания кутикулярных клеток извилистые. Клетки сильно вытянуты поперек волоса. Поперек волоса располагается 1—3 клетки. На 1 см2 поверхности кожи растет 670 остевых волос.
Пух очень длинный (38,7 мм) и тонкий (11 мкм). Он сильно извилист и закручен в спираль или даже клубок. В связи с этим, хотя пуховые волосы и длиннее остей, они не доходят до их вершины. В пухе сердцевина отсутствует. Высота клеток кутикулы 11 мкм. Кутикулярные клетки кольцевидной формы. Пуховые волосы растут пучками по 5—12 волос в каждом. Пучки располагаются каудальнее остевых волос. На 1 см^ поверхности кожи насчитывается 4166 пуховых волос. Выделяются слабовыраженные группы: 2—3 остевых волоса и 2—3 пучка пуховых волос.
Летом кожа значительно толще, чем зимой. Эпидермис в коже загривка тоньше, чем зимой (особенно роговой слой). Однако мальпигиев слой немного толще и состоит из 3—4 рядов клеток. Зернистый и блестящий слои отсутствуют. Роговой слой плотный. Эпидермис интенсивно пигментирован. В коже груди эпидермис и особенно роговой слой очень толстые. Эпидермис имеет выступы (высотой до 220 мкм), вдающиеся в дерму. Имеется зернистый слой, образованный 4—5 рядами клеток, и толстый блестящий слой. Мощный роговой слой плотный. В ряде мест базальный слой эпидермиса слабо пигментирован. Сосочковый слой дермы пронизан широкими корнями волос. Пучки коллагеновых волокон и эластиновые волокна в коже затривка такие же, как зимой. В сетчатом слое коллагеновые пучки переплетаются под различными углами к поверхности кожи в плотную и сложную вязь. Эластиновые волокна образуют сплетение до 1,2 мм толщины на границе с подкожной мускулатурой. В коже груди сосочковый слой почти равен по толщине сетчатому. Пучки коллагеновых волокон в сосочковом и в верхних и средних частях сетчатого слоя проходят в различных восходящих направлениях, а в глубоких отделах сетчатого — преимущественно горизонтально. Эластиновые волокна в сосочковом слое немногочисленны, а в сетчатом — очень редки. В глубоких отделах сетчатого слоя встречаются группы жировых клеток.
343
Бородатый козел
Для груди характерна толстая подкожная жировая клетчатка, занимающая 63,4% толщины кожи. Сальные железы более крупные, чем зимой (особенно в коже груди). У каждой ости встречаются две железы, расположенные в два яруса. Их выводные протоки отстоят друг от друга на расстоянии 250—300 мкм. Потовые железы увеличены, извиты сильнее, чем зимой.
Среди волос загривка можно выделить две категории: остевые и пуховые. У остевых волос ножки нет, гранна развита слабо, их длина 11,6 мм, толщина 143 мкм, сердцевина 85%, форма поперечника овальная. Сердцевина отсутствует только в кончике и основании. Пуховые волосы имеют веретеновидную форму с довольно значительным расширением в середине. Длина их незначительна (1,5 мм), толщина 55 мкм. У них длинные ножки и кончик, в которых сердцевинного слоя нет. В средней расширенной части сердцевина развита хорошо (85%). Пуховые волосы растут пучками по 2—11 штук в каждом. Пучки располагаются каудальнее остевых волос. Остевые и пуховые волосы образуют слабовыраженные группы: 2—3 ости и 2—3 пучка пуха. На 1 см2 поверхности кожи насчитывается в среднем 666 остевых и 2833 пуховых.
Волосы груди подразделяются на две категории: остевые и пуховые. Волосы ножек не имеют. Ости длиной 12,0 мм, толщиной 127 мкм, прямые, с небольшой гранной и обломанными вершинами. Форма поперечника остевых волос овальная, почти округлая. Сердцевинный слой развит относительно слабо (62,4%). В боковых отделах волоса корковый слой толще, чем в передне-задних, примерно на 10 мкм. Остевые растут поодиночке в количестве 666 на 1 см2 поверхности кожи. Длина пуха 7,0 мм, толщина 32 мкм. Во всех пуховых волосах имеется сердцевинный слой. У отдельных пуховых волос сердцевина развита слабо и представлена в виде штрихов. Пуховые волосы растут преимущественно пучками по 2—3 в каждом пучке. На 1 см2 поверхности кожи их насчитывается 666 штук. Остевые и пуховые волосы расположены слабовыраженными рядами.
В связи с тем что Шаффер (Schaffer, 1940 г., по: Соколов, 1973) указывал на наличие у козлов железистых образований в области затылка, у козы, добытой летом, была исследована кожа с этого места головы. Как потовые, так и сальные железы имеют здесь нормальные размеры, не отличающиеся от желез туловища.
Система крови. Мы не располагаем данными по крови бородатых козлов, но для понимания механизмов их адаптации к экстремальным условиям жизни приводим сведения по картине крови других видов рода Capra. Так, Н.Ш.Булатова (1962), изучившая особенности крови высокогорных животных, обращает внимание на очень широкий диапазон изменчивости показателей дыхательной функции крови коз. Винторогий козел Capra falconeri обладает огромным количеством эритроцитов — до 25,77 млн в 1 мм3 (изучен самец). У другого вида, горного сибирского козерога Capra sibirica в 1 мм3 крови содержится у самцов 15,20 млн, у самок 12,10 млн. Показатели домашней козы (Capra hircus) занимают промежуточное положение между
344
Capra aegagrus Erxleben, 1777
названными выше двумя видами. По ряду источников (Malassez, 1877 г., Наует, 1879 г., Gotze, 1923 г., по: Коржуев, 1964; Булатова, 1953),- количество эритроцитов у С. hircus составляет от 18,00 до 19,00 млн/мм3. Высокое количество эритроцитов в крови домашней козы, видимо, показывает не только на ее высокую активность, но и родственную близость с винторогим козлом. В широких пределах у коз меняется объем эритроцитов, что говорит о вариабельности гемодинамических свойств крови. По Н.Ш.Була-товой (1953), объем одного эритроцита у винторогого козла составляет всего 14,5 мкм3, тогда как у козерога — 24,6 мкм3. У козы Oreamnos ameri-canus эритроциты оказались по размерам средними по сравнению с указанными видами (20,3 мкм3). Содержание гемоглобина наиболее высокое у винторогого козла (17,1 г%), высока также кислородная емкость крови (22,9 об.%). У козерога эти параметры ниже — 15,2 г% гемоглобина и 22,9 об.% кислородной емкости. Оба показателя очень низкие у домашней козы (9,9 г% и 13,3 об.%). О количестве крови диких коз сведений у нас нет, а у домашней козы (п = 30 экз.) оно составляет в среднем 7-,0 мм3 на 100 г массы тела (Courtice, 1943 г., по: Коржуев, 1963).
Индексы внутренних органов. Изучены у 7 бородатых козлов, добытых в Лагодехском заповеднике (Кулиев, 1981). Для сравнения использованы данные по 3 особям кавказского тура, происходящим из тех же мест названного заповедника. Результаты этой работы для широкого обсуждения недостаточны, но все же видно, что уровень адаптации к условиям высокогорья Восточного Кавказа совершенный и согласуется с параметрами крови, свойственными козам. С данными морфофизиологии перекликаются результаты изучения относительной массы общей мускулатуры у бородатых козлов по сравнению с домашними козами (Ахмедов, Керимова, 1987). У первых она составляет 27,6% в годовалом возрасте, в трехлетием — 44,6% и к семилетнему — 50,8%. У вторых эти показатели соответственно таковы: 23,5; 26,1; 31,1%. Обращает внимание степень развитости относительной массы мускулатуры передней конечности бородатого козла — ее показатель в годовалом возрасте 3,7%, в трехлетием — 6,2%, в семилетием — 6,9%. У домашних коз соответственно 3,3; 3,9 и 4,4%.
Мощная мускулатура передних конечностей бородатого козла соответствует показателям индексов внутренних органов и уровню адаптации к горам.
Кариотип. У бородатого козла из Нахичеванской АССР хромосомный набор изучен у 4 самцов и 3 самок (Кулиев, Мамедов, 1974). Количество хромосом 60, все они акроцентрики. Х-хромосома — крупный акроцентрик, У-хромосома не идентифицируется достоверно. Показательно, что у одного из самцов обнаружена транслокация робертсоновского типа.
Структура и динамика ареала
Видовой ареал охватывает территорию на западе от острова Крит в Средиземноморье до западного Синда на востоке. Северная граница простирается до Восточного Кавказа и Южной Туркмении, а южная — до гор Ан-
345
Бородатый козел
тиливана, Сирийской пустыни, Ирана (Гептнер и др., 1961). Естественный ареал вида сильно изменен человеком как в результате истребления в ряде мест, так и акклиматизационных мероприятий.
В СССР встречается на Кавказе и в Туркмении.
Распространение на Кавказе ограничено восточной частью кавказских хребтов, центральной и восточной частями Малого Кавказа. Переходя к более подробному обзору распространения с учетом его динамики во времени, следует отметить, что пресс охоты очень давно оказывал большое влияние на структуру ареала, однако фрагментарность данных не позволяет с достаточной полнотой проследить эти сдвиги во времени.
На Северном Кавказе распространение бородатого козла идет на запад до верховьев Аргун. На эту точку траницы ареала указывал еще Н.Я.Динник (1910), а затем Н.К.Верещагин (1959). В коллекции КБГУ имеется один экземпляр из этого пункта. Западнее названного пункта, в том числе на Крестовом перевале и Казбеке, никаких следов обитания бородатого козла не обнаружено (Темботов, 1972). Останков этого вида не найдено и в осетинских пещерах — бывших святилищах (Верещагин, Наниев, 1949). Судя по литературным данным, бородатый козел был широко распространен в Дагестане, по верховьям Андийского, Аварского и Казикумухского Койсу, а также р. Самура. В начале XX в. он спускался до Ботлиха и Гуниба. В 60-е годы бородатого козла во Внутреннем Дагестане не стало, но по осевым хребтам он продолжал встречаться до Самура. По З.А.П1ахмарданову и Х.М.Рамазанову (1980), в 70-х годах распространение бородатого козла установлено лишь в трех районах: Ботлихском, Гунибском и Рутульском. В настоящее время, как свидетельствуют материалы Л.И.Прилуцкой и Ю.В.Пишванова (1989), дизъюнктивность ареала в Дагестане еще больше усилилась, значительно сократилась площадь распространения. Он полностью исчез в Лакском районе. В Ботлихском районе, на правобережье Андийского Койсу, от селений Муни, Орта-Коло до с. Цолода, где расположены урочища Радул-Даг, Копи-Даг, сохранилось небольшое поселение в 40—60 особей. Сведения, упомянутые выше (Шахмарданов, Рамазанов, 1980), об обитании бородатого козла в Гунибском районе не подтверждаются, возможно его и не было здесь. По всем имеющимся данным, животное сохранилось в верховьях рек Аварского и Андийского Койсу и только в Аварском, Цумадинском, Цунтинском, Тляратинском, Советском и Ботлихском районах. К фактическим местам нахождения относятся: урочище Нарсен-Дох, ручей Букизатола, Раздах, близ с. Верхний Хваршини, хребет Мильчич, ручьи Хецрах, Хемангер, окрестности сел Хваини, Инхо-квари, Хварши. Более крупное поселение находится на Богосском хребте в Путинском районе. Они зарегистрированы по р. Сабакунис-Хеви, урочище Окотех-Керу, на хребте Кабза, по лесным урочищам р. Китлярта, район селений Шаури, Хамаитли, вблизи горы Хорокича (Мокок), по лесным массивам р. Кидеро, в окрестностях с. Бежта, верховьях рек Нахкостльтляр, Симбирис-Хеви и Куцурако. В Советском районе отмечен в верховьях Аварского Койсу и по ее притокам Ратлуб-Ор, Темир-Ор, по хребту Хе-цериб, в урочищах Тликатль, Цономеэр, Фарси, Муццариб и в районе горы
346
Capra aegagrus Erxlcbcn, 1777
Обода. В Тляратинском районе он встречен по урочищам, прилегающим к р. Джурмут.
В Чечено-Ингушетии Н.Я.Динник (1910) встречал этого козла неоднократно в "Терской области, именно в верховьях Аргуна (горная Чечня). В ущелье р. Кий, также принадлежащей бассейну Аргуна, безоаровые козлы до сих пор водятся в изобилии... к западу от Военно-Грузинской дороги они не встречаются вовсе" (с. 164). По А.М.Батхиеву (1980, 1989), в Чечено-Ингушетии еще в 30-х годах бородатый козел занимал обширное пространство от Ассы вдоль Скалистого хребта к верховьям рек Чанты-Аргун и Шаро-Аргун. В горной Ингушетии в то время бородатый козел добывался на горе Герчог (одна из вершин Скалистого хребта). В настоящее время распространение бородатого козла в Чечено-Ингушетии значительно сократилось; сохранился он лишь в верховьях рек Чанты-Аргун, где сосредоточены два поселения в ущельях Кериго и Маисты-Хи. Небольшое количество бородатых козлов держится изолированно друг от друга на горе Байсты-Хи. По Т.Ю.Точиеву (1973), козлы водятся в ущельях Цеси и Ху-ландой, находящихся в верховьях р. Шора-Аргун. Что касается Скалистого хребта и верховья Ассы, то этих животных там не стало.
В закавказской части ареала бородатый козел имеет сходную с Северным Кавказом картину пространственной организации видового населения: отсутствует в западнозакавказском типе поясности, а в восточнозакавказском обитает спорадически. Еще Н.Я.Динник (1910) отмечал, что он встречал бородатого козла в Тушетии и Хевсурии. "На горах, — писал он, — окаймляющих Тушстскую и Пирикотельскую Алазань и принадлежащих как Главному Кавказскому хребту, так и Пирикительскому, безоаровые козлы встречаются довольно часто, в особенности в тех местах, где на этих горах растет лес" (с. 165). О бородатых козлах, обитающих в Тушетии и Хевсурии, упоминает также Н.КВерещагин (1959). При этом восточная граница, по его данным, проходит по верховью Гердыманчай, против с. Ис-маиллы. Занимают бородатые козлы верхнюю часть лесного пояса. В настоящее время в Грузии бородатый козел сохранился лишь в самой восточной части республики, на границе с Азербайджаном. По А.Б.Арабули (1989), бородатый козел в Грузии обитает на небольшой территории (около 11 тыс. га) Большого Кавказа, в ущелье Пирикительской Алазани (северные склоны) и на Малом Кавказе (Аренанский хребет).
Большая часть кавказского ареала бородатого козла находится на Малом Кавказе. Кроме гор близ Боржоми, Н.Я.Динник (1910) относил к области распространения этого вида Карскую область, где "козлы встречаются в довольно значительном количестве на горах, находящихся недалеко от Са-рыкамыша, Кагызмана... Еще чаще они попадаются в Эриванской губернии, причем довольно много живет их на горах недалеко от Ордубада. Встречаются они в южной части Елисаветпольской губернии" (с. 165). Он не находил их в Ленкорани и Талышских горах, что совпадало с данными К.А.Сатунина. Что касается западной части Малого Кавказа, то Н.Я.Динник не имел однозначных данных. Возможно, писал он, бородатые козлы попадаются в горах бассейна Чороха. Если учесть дружбу его с К.А.Сатуниным,
347
Бородатый козел
известным териологом Кавказа, то можно с уверенностью сказать, что эта точка зрения и неоднозначность мнения были общие для обоих специалистов.
О распространении бородатого козла в Закавказье более уверенное суждение высказывал Н.К.Верещагин (1947, 1959), занимавшийся успешно в 30—40-х годах изучением териофауны Кавказа. По его мнению, рассматриваемый вид обитал в то время в пихтовых лесах верховья р. Чорох, спорадично — на Аджаро-Имеретинском хребте и на западных отрогах Триалет-ского хребта, вблизи Боржоми, а далее на востоке козел занимал горы Армянского нагорья. На Шахдагском хребте обычен в верховьях Шамхора, вблизи вершины Кашкар (3379 м), на горе Кяпаз (над оз. Гейгель) и под вершиной Муровдаг (3740 м). Небольшое число животных обитало в 1940 г. и в осыпях среди буковых лесов восточной оконечности Шахдагского хребта (юго-восточный склон горы Кечалдаг). Восточная граница ареала бородатого козла в этой части Кавказа проходила по горе Кире (2743 м), над Степанакертом.
На Армянском нагорье бородатые козлы в 50-х годах текущего столетия обитали в Скалистых горах на западе от р. Азат до хребтов Зангезурского, Мегринского и Баргушатского. Северным пределом распространения считав ются южные склоны Гегамского и Варденисского хребтов (Даль, 1951). В 70-х годах прошлого столетия эти козлы встречались в окрестностях Еле-новки на Севане. С этого места имеется фактический материал в виде черепа. В 1950 г. больше всего бородатые козлы встречались на Урцком и Айоцдзорском хребтах. При этом на Урцком хребте в 1950 г. зарегистрированы "очаги" распространения: 1) Кармир-Кар; 2) поселение на северном склоне указанного хребта в области Каньска; 3) ущелье Джайнамдара; 4) урочища Хачалаг, Сарай-Булаг, Маралбахан, Демаглы. Козы занимали на Урцком хребте 33,5 км^, где их насчитывалось 124 козы. По мнению К.М.Гаспаряна (1975), "очаговость" распространения не жесткая, бородатые козлы могут встречаться более или менее равномерно. Антропогенный фактор поддерживает спорадичность поселения, однако фактами это мнение не подтверждено.
В Нахичевани, по Н.К.Верещагину (1959), распространение рассматриваемого вида спорадично. В 1947 г. отдельные группы, изолированные друг от друга, зарегистрированы на полупустынных вершинах Аливджадаг, Иль-янлыдаг, вблизи селений Абракунис, Боянур, Ханчая. По Зангезурскому хребту и южным его отрогам животные отмечены к востоку до р. Аракса. Особенно много их было в горах между Джульфой и Ордубадом, на высоте 1000—1200 м. Они занимали юго-восточный склон Мегринского хребта, вплоть до Охчичая. Небольшое стадо обитало вблизи Акулиса и Ордубада.
Сведения о высотных пределах распространения бородатого козла в Армянской ССР приводит С.К.Даль (1954), согласно которому вид занижает пояса гор от 550 до 3200 м. Так, в 1940 г. следы этих козлов отмечены на Памбакском хребте, в скалах урочища Кер-Оглы, на высоте 2410 м, а в
348
Capra acgagrus Erxleben, 1777
1943 г. на склоне г. Арагац найден труп козы 4—5 лет, разорванный волками.
Ряд фактических данных о распространении бородатого козла приводит Х.М.Алекперов (1966). В частности, на Карабахском хребте бородатые козлы обитали в 60-х годах в ряде пунктов: урочище Гырхкыз, на скалистых вершинах Делидага (2196 м), Муровдага (3419 м), Кечалдага (2232 м), немного особей на вершине Кашкара (3378 м), на горе Кяпаз у оз. Гейгелы. Они отмечены также на горных массивах хребта Капыджик, Ордубадский район. Указание Н.К.Верещагина (1959) об обитании бородатых козлов на горе Кире (2143 м) относится к периоду до 30-х годов. Здесь последний бородатый козел убит в 1944 г. Вторичное заселение ими этих земель осложняется отгонным животноводством; тем не менее в Нахичеванской АССР в 70-х годах отмечались положительные сдвиги в распространении и численности бородатых козлов (Кулиев, 1974). Их поголовье в то время составляло до 1500—1800 особей, однако занимали они исключительно субальпийский и альпийский пояса.
Новыми данными по бородатому козлу Малого Кавказа, относящимися к нашим дням, мы не располагаем.
Современные тенденции антропогенных изменений ареала. Имеющиеся данные говорят о том, что во многих районах Кавказа, где не так давно еще обитал бородатый козел, его не стало в последние десятилетия. По данным А.М.Батхиева (1989), в Чечено-Ингушетии он сохранился в верховьях Чанты-Аргуна, а в Дагестане, по А.Я.Закариеву (1982), Л.И.Прилуцкой и Ю.В.Пишванову (1989), также оттеснен к труднодоступным местам, и основное поголовье сохранилось в верховьях Аварского Койсу, в пределах Сунженского района. Аналогичная картина наблюдается и в Грузии. Там бородатый козел встречается также в высокогорьях Пирикотельского хребта, на его северном макросклоне, и, видимо, поголовье поддерживается за счет мигрантов из Чечено-Ингушетии. На Малом Кавказе, в пределах Грузии, очевидно, этого вида уже нет (Курашвили и др., 1981). Данные А.Б.Ара-були (1989) по этому району нуждаются в уточнении. Вероятно, был прав Н.К.Верещагин (1959), который определял время уничтожения бородатого козла в районе Боржоми 20-ми годами текущего столетия. Симптоматично, что в данных Главного управления заповедниками и охотничьего дела Грузии по видовому составу и численности копытных республики на конец 1988 г. не оказалось сведений по. бородатому козлу. Сходная тенденция современных изменений распространения этого вида отмечается и в районах восточной части Малого Кавказа. Лишь С.М.Кулиев (1974) сообщает, что в Нахичеванской АССР в связи с усилением охраны распространение и численность бородатого козла стали расти, но тем не менее и здесь этот вид остается оттесненным в субальпийский и альпийский пояса, преимущественно в малодоступные места высокогорья.
Как видно, распространение бородатого козла на Кавказе интенсивно сокращается. Он стал редким видом териофауны региона. Если не будут приняты срочные меры по его защите, уникальный вид будет уничтожен.
349
Бородатый козел
Систематика и изменчивость
Бородатый козел относится к числу немногих видов млекопитающих, систематика которых относительно меньше дискутируется, чем по другим труппам видов. Реальность существования самостоятельного вида Capra Algagrus Егх. в настоящее время, насколько нам известно, не вызывает сомнения ни у одного систематика (Туркин, Сатунин, 1902; Динник, 1910; Бобринский и др., 1944, 1965; Соколов, 1959; Гептнер и др., 1961; Соколов, 1973, 1984; Corbet, 1978; Corbet, Still, 1980; Honacri et al., 1982; и др.). Одним из неопровержимых доказательств его видовой самостоятельности, на наш взгляд, является отсутствие скрещивания с теми видами рода, с которыми бородатый козел обитает совместно. На Кавказе, например, мы имеем налегание ареалов бородатого козла и кавказского тура на всем протяжении гор Восточного Кавказа. Без оговорок их можно считать симпатрическими видами в кавказской части ареала. Аллопатричными они становятся на Центральном и Западном Кавказе (область обитания кавказского тура) и на Малом Кавказе (нет туров). Расхождение (не полное) имеется в биото-пической приуроченности, однако оно больше связано с влиянием антропогенных факторов, чем с биологическими особенностями этих видов. В.Г.Гептнер и др. (1961) справедливо считают, что деятельность человека объясняет тот факт, что кавказский тур в настоящее время больше придерживается высокогорья (верхняя часть субальпийского пояса и выше до нивального), а бородатый козел чаще встречается в горных лесах, в том числе субальпийского высокогорья. Наши и современные литературные данные свидетельствуют о том, что в настоящее время местообитания обоих видов все больше сближаются, так как человек оттесняет их в высокогорья. Только там они могут спастись от браконьеров. В прошлом, когда охотничьего пресса было меньше, оба вида спускались до пояса широколиственных лесов. Словом, непреодолимых биотопических разобщений между ними не было в прошлом, нет и сейчас. Однако в природе ни мы, ни наши предшественники не регистрировали гибридов между бородатым козлом и кавказским туром. Указание Е.В.Пфиценмайра (1915) о встрече такого гибрида в Закавказье, на которое ссылаются систематики (Соколов, 1959; Гептнер и др., 1961; и др.), остается не подтвержденным. Мы не отрицаем возможности гибридизации между разными видами коз, в том числе бородатого козла с турами или с домашними козами. Однако подобные явления в природе, видимо, очень редки и не могут никак привести к появлению смешанных популяций. Среди млекопитающих Кавказа мы выявили виды, например среди Pitymus, которые дают помеси, но сохраняют все видовые параметры, и существование таких близкородственных видов не вызывает сомнения среди систематиков.
Много сложнее обстоит дело с родом Capra, по которому среди ученых нет единого мнения о взаимоотношениях между отдельными видами. Тем не менее реальность С. aegagrus признается всеми систематиками, в том числе теми, кто сводит количество видов рода до минимума (Haltentort, 1963).
350
Capra aegagrus Erxleben, 1777
Таблица 70. Промеры черепа Capra aegagrus Кавказа. Молодые
Признак Самцы
п limit х ± т а Су
Наибольшая длина 3 158,1-179,5 168,5±7,57 10,71 6,4
Ковдилобазальная длина 3 143,6—168,6 156,2±8,84 12,50 8,0
Основная длина 3 128,2-152,5 139,9 ±8,61 12,17 8,7
Максимальная ширина 3 90,3-97,0 94,5±2,57 3,64 3,9
Скуловая ширина 3 75,5-81,3 79,2+2,27 3,21 4,1
Межглазничная ширина 3 59,5-68,1 64,5±3,16 4,47 6,9
Длина лицевой части 3 83,0-96,5 89,1±4,84 6,85 7,7
Длина носовых костей 2 44,3-48,8 46,6 — —
Длина верхнего ряда зубов 3 47,1-48,6 ‘ 48,0+0,57 0,81 1,7
Максимальная ширина мозговой капсулы 3 64,0—68,0 66,2±1,44 2,04 3,1
Длина нижней челюсти 2 120,7-129,0 124,9 .— —
Длина нижнего ряда зубов 2 46,0—46,8 46,4 — —
Длина диастемы нижней 2 29,6-31,0 30,3 — —
челюсти
Признак Самки
п limit х ± т а Су
Наибольшая длина 3 161,9-218,8 189,3±2О,16 28,51 15,1
Ковдилобазальная длина 3 149,6-211,3 178,5+21,94 31,03 17,4
Основная длина 3 133,0-192,1 160,9+20,99 29,69 18,5
Максимальная ширина 5 91,6-113,6 105,4+4,52 9,03 8,6
Скуловая ширина 5 78,5-94,4 87,0+2,95 5,90 6,8
Межглазничная ширина 5 68,6-79,7 72,5±3,64 7,27 10,0
Длина лицевой части 3 85,0- 119,4 103,0±12,20 17,25 16,7
Длина носовых костей 5 44,1-64,8 55,8+4,42 8,84 15,8
Длина верхнего ряда 4 46,2-71,8 60,2+6,14 10,64 17,7
зубов Максимальная ширина мозговой капсулы 5 64,3-79,9 72,6+3,14 6,28 8,7
Длина нижней челюсти 5 122,4-170,9 151,1 ±9,46 18,92 12,5
Длина нижнего ряда зубов 4 44,0-71,2 59,1+6,50 11,25 19,0
Длина диастемы нижней 5 . 28,6-42,3 36,6+2,59 5,18 14,2
челюсти
Изменчивость. Для анализа различных типов изменчивости бородатого козла мы изучили, кроме нашей коллекции из Дагестана и Чечено-Ингушетии (п — 12 экз.), материалы зоомузсев МГУ и ЗИН РАН (л = 10 экз.), а также Закавказских республик (л = 2 экз.). Была использована для сравнения коллекция из Туркмении (преимущественно сборы В. И. Палки на) в количестве 25 экз. (табл. 69—72).
Индивидуальная изменчивость. Размеры и масса тела подвержены значительным изменениям индивидуального порядка, что нашло отражение в ме-
35 1
Бородатый козел
Таблица 71. Промеры черепа Capra aegagrus Кавказа. Взрослые
Признак Самцы
п limit Я +т G сг
Наибольшая длина черепа 4 241,1-278,7 257,8+11,25 19,49 7,6
Кондилобазальная длина 4 221,6-268,5 244,8±13,01 22,54 9,2
Основная длина 4 205,6-247,2 225,6+11,90 20,61 9,1
Максимальная ширина 4 123,1-139,5 131,7+4,10 7,10 5,4
Скуловая ширина 4 97,0-111,4 106,4±3,91 6,78 6,4
Межглазничная ширина 4 86,4-97,5 92,0±2,97 5,15 5,6
Длина лицевой части 4 127,7-160,0 141,2±8,85 15,32 10,8
Длина носовых костей 4 69,2-86,0 74,9+4,47 7,75 10,3
Длина верхнего ряда 4 71,7-77,9 75,0+1,96 3,39 4,5
зубов Максимальная ширина мозговой капсулы 4 88,0—102,8 94,4±3,81 6,60 7,0
Длина нижней челюсти 2 182,8-223,4 203,1 — —
Длина нижнего ряда зубов 2 76,9-79,1 78,0 — —
Длина диастемы нижней 2 43,4-49,6 46,5 — —
челюсти
Признак Самки
п limit У ± m G Q
Наибольшая длина черепа 4 217,8-232,8. 224,1±3,64 6,30 2,8
Кондилобазальная длина 4 204,4-219,7 213,1±4,05 7,01 3,3
Основная длина 4 186,9-200,4 193,0±3,23 5,60 2,9
Максимальная ширина 4 112,4-125,4 117,4±3,35 5,81 4,9
Скуловая ширина 4 91,6-103,7 96,9±3,11 5,38 5,6
Межглазничная ширина 5 76,0-83,0 79,2+1,39 2,78 3,5
Длина лицевой части 4 124,8-131,6 127,4+1,71 2,97 2,3
Длина носовых костей 5 61,9-83,8 72,1+4,44 8,88 12,3
Длина верхнего ряда 5 65,8-72,0 70,0+1,24 2,47 3,5
зубов
Максимальная ширина 5 76,5-83,0 79,3±1,46 2,91 3,7
мозговой капсулы
Длина нижней челюсти 2 171,8-197,7 184,8 — —
Длина нижнего ряда зубов 2 70,6-74,8 72,7 — —
Длина диастемы нижней 2 40,6-52,3 46,5 — —
челюсти
ристических показателях как самцов, так и самок. Видоспецифична форма рогов, однако она довольно вариабельна. По Н.Я.Диннику (1910), среди самок изредка встречаются 'комолые, а также бородатые самки. Для бородатого козла характерным считаются рога "саблевидно изогнутые в одной плоскости" (Гептнер и др., 1961, с. 523). По этому поводу мнение И.И.Со-колова (1959) иное: "У взрослых самцов рога несколько сходятся вершинами, так что наибольшее расстояние между рогами имеет место в предвершинной части. Изредка концы рогов даже перекрещиваются" (с. 406). На наш взгляд, размах индивидуальной изменчивости формы рогов этими вариантами нс исчерпывается. В коллекции, изученной нами, лишь у поло-
352
Capra aegagrus Erxlcben, 1777
Таблица 72. Промеры черепа Capra aegagrus из Копетдага. Молодые
Признак Самцы
п limit х ± т G Су
Наибольшая длина черепа 1 113,8 — — —-
Кондилобазальная длина 1 109,5 — — —
Основная длина 1 99,4 — — —
Максимальная ширина 3 65,2-107,7 90,5±15,82 22,37 24,7
Скуловая ширина 3 56,5-89,0 77,0+12,63 17,86 23,2
Межглазничная ширина 3 45,2-74,2 62,5+10,80 15,27 24,4
Длина лицевой части 2 45,6-100,3 73,0 — —
Длина носовых костей 2 26,7-53,0 39,9 — —
Длина верхнего ряда зубов — — — -— —
Максимальная ширина мозговой капсулы 3 42,9-68,6 59,3+10,08 14,25 24,0
Длина нижней челюсти 3 88,1—159,5 129,9+26,31 37,21 28,6
Длина нижнего ряда зубов — — — — —
Длина диастемы нижней челюсти 3 35,0-38,7, 36,8±1,31 1,85 5,0
Признак Самки
п limit х + т о Су
Наибольшая длина черепа 6 160,7-210,5 188,5+7,63 17,07 9,1
Кондилобазальная длина 5 151,0-203,0 179,4+9,73 19,44 10,8
Основная длина 5 135,1-185,7 163,2+9,53 19,06 11,7
Максимальная ширина 8 88,8—116,1 105,2+3,30 8,80 8,4
Скуловая ширина 8 72,9-97,4 87,2+2,79 7,39 8,5
Межглазничная ширина 8 58,3-79,1 70,9+2,83 7,48 10,6
Длина лицевой части 6 86,2-115,9 102,0+4,85 10,85 10,6
Длина носовых костей 7 44,9-71,1 57,7+3,91 9,58 16,6
Длина верхнего ряда — — —- — —
зубов Максимальная ширина 8 56,2-73,3 68,0+2,05 5,42 8,0
мозговой капсулы' Длина нижней челюсти 3 142,3-181,7 162,0+13,93 19,70 12,2
Длина нижнего ряда зубов 1 . 77,6 —
Длина диастемы нижней 4 34,8-40,7 37,9+1,45 2,52 6,8
челюсти
вины рога изогнуты в одной плоскости. У остальных рога имеют изгибы в двух плоскостях, притом в разных вариантах. Так, у особи S—38792 четырехлетнего козла концы рогов сходятся вершинами к оси тела. Соответственно расстояние между ними составляет 82 мм, тогда как максимальная ширина между рогами достигает 140 мм. У другого четырехлетнего козла S—51861 рога постепенно расходятся по бокам и максимальное расстояние в 250 мм между ними приходится на самую дистальную часть. При этом оба рога изогнуты в одной плоскости. Оригинальную форму имеют рога у семилетнего самца S-38794 — левый рог идеально изогнут в одной плоскости,
23 В.Е.Соколов, А.К.Темботов
353
Бородатый козел
тогда как правый рог со второго сегмента изогнут внутрь. Если его положить внешним боком на стол, конец рога приподнимается на 40 мм.
Как справедливо отмечает И.И.Соколов (1959), у бородатого козла, особенно самцов, основания рогов сильно сближены, но и этот признак довольно вариабельный. Расстояние между килями рогов у самого основания колеблется у самцов от 9,2 до 49,6 мм. При этом значительна индивидуальная изменчивость в пределах одной возрастной группы и одного пола.
Очень высокая индивидуальная изменчивость свойственна длине годового прироста рогов у разных особей независимо от пола и возраста. Так, в нашей выборке прирост рогового чехла на третьем году жизни у разных особей самцов составил от 117 до 185 мм, на четвертом году — от 93 до 170 мм, на пятом году — от 90 до 131 мм и т.д.
Изменчивы также количество и особенно соотношение бугров на киле рога. Обычно более массивными бывают те бугры, которые появляются на 3—6 годах; до и после они слабо обозначены или вовсе отсутствуют. Вариабельна также толщина рогов, которой придается большой систематике-таксономический вес (Цалкин, 1950). У туркменских козлов обхват рогов взрослых самцов у основания рога составляет от 18 до 23 см. Диапазон изменчивости этого параметра у кавказских козлов меньше (22,4—25,0 см). О степени вариабельности толщины рога лучше судить при сравнении одновозрастных животных. Так, у двух козлов из Туркмении возрастом по 4 года толщина рога оказалась 74 х 44 и 73,3 х 46,1 мм. У двух девятилетних козлов Восточного Кавказа соответственно 86 х 49 и 83,2 х 48,7 мм.
Определенным диапазоном варьирования характеризуется окраска меха. Как справедливо указывал И.И.Соколов (1959), у некоторых взрослых самцов темный хомут почти не выражен. У отдельных животных отсутствуют также темные полосы по бокам тела. У некоторых особей имеются темные полосы, идущие от глаз к углам рта. Среди темных волос спинного ремня у некоторых особей появляются белые волосы.
В структуре черепа многие показатели подвержены широкой индивидуальной изменчивости. Так, весьма характерный для бородатого козла лобный бугор у самцов одного и того же возраста развит не одинаково. Экземпляр S-51861 (четырехлетний самец) отличается мощным лобным бугром. У другого четырехлетнего самца S-38792 лобный бугор развит слабо.
Индивидуальную изменчивость носовых и слезных костей можно свести к семи типам (рис. 44, 45). Из них чаще всего встречаются по носовым костям 1-й и 6-й типы, а по слезным — 3-й. Для иллюстрации степени индивидуальной изменчивости нами проанализированы также неметрические признаки (количество лобных и глазничных отверстий). Многообразие различных сочетаний количества и форм отверстий мы сводим к 5 типам (рис. 46). Наиболее часто представлены 1-й и 3-й типы.
Возрастная изменчивость. Специальных исследований не проводилось. Данные, которыми мы располагаем (табл. 70—73), говорят о том, что эта изменчивость сходна с таковой кавказского тура. Очень ярко возрастные изменения отражены на рогах самцов. Наличие годовых сегментов, разделенных хорошо заметным профилем, позволяет однозначно судить о воз-
354
Capra aegagrus Erxleben, 1777
Рис. 44. Типы (1—7) строения носовых костей бородатого козла
Рис. 45. Типы (1—3) строения слезных костей бородатого козла
возрасте козлов. Возраст коз по годовым кольцам можно определять лишь до трех лет. В последующие годы сегменты выражены слабо и дифференцировать их затруднительно.
Половые различия. У бородатого козла, как и у других видов рода, половой диморфизм выражен очень сильно (см. табл. 70—72). В первую очередь бросаются в глаза мощные рога самцов (в рекордных случаях длиной до 140 см (Соколов, 1959). У самок хотя и имеются рога, но они маленькие (18—39 см по упомянутому автору), редко превосходят длину головы больше чем в 1,5 раза.
У самцов, как отмечено выше, передне-внутренний край рога сплющен, килевидный, и на нем имеется ряд плоских бугров, расположенных на стыке годовых сегментов. Кроме того, годовые кольца резко выражены благодаря глубокой борозде, отделяющей один годовой сегмент от другого. У самок на рогах нет бутров. Годовые сегменты выражены более или менее четко лишь в первые два-три года, потом нарастающие сегменты трудно подразделять по годам. Передне-внутренний край рога закручен или заострен, но в этом случае не сплющен так сильно, как у самцов. Характерно наличие на поверхности рога поперечных валиков или пластинок. Стержни рогов и их чехлы расставлены значительно шире, чем у самцов.
Самцы резко отличаются от самок наличием длинной и заостренной бороды (до 20 см). Она состоит из грубых и толстых волос; занимает у старых самцов почти весь низ головы. У самок борода имеется лишь очень редко (Динник, 1910). Примечательно и то, что самцы не только крупнее по раз-
23*
355
Бородатый козел
Рис. 46. Типы (1—5) строения лобных отверстий бородатого козла
мерам и массе тела, чем самки, но выглядят крепче — особенно мощно развита у них толстая шея, что связано с более крупными рогами. У самок шея смотрится длиннее и тоньше. В окраске половой диморфизм также четко выражен — самки отличаются более однотонным мехом, все отметины выражены слабее, чем у самцов.
Различия между полами в размерах и структуре черепа очень большие, о чем свидетельствуют данные табл. 70—72. Обратим внимание лишь на некоторые особенности структуры черепа. У самок в отличие от самцов очень слабо развит лобовой бугор, более того, у ряда экземпляров он практически не выражен. Характерно также, что относительно небольшие роговые стержни расставлены значительно шире, чем у самцов.
Географическая изменчивость. С тех пор как на Большом Кавказе был описан самостоятельный вид козла Capra aegagrus, прошло более двух столетий. Однако до настоящего времени специальному исследованию не была подвержена географическая изменчивость бородатого козла в пределах кав-
356
Capra, aegagrus Erxleben, 1777
Таблица 73. Промеры черепа Capra aegagrus из Копстдага. Взрослые
Признак Самцы
п limit S ± т G Cv
Наибольшая длина черепа 1 260,0 — — —
Кондилобазальная длина 1 250,0 — — —
Основная длина черепа 1 230,6 — — —
Максимальная ширина 7 114,8-132,9 124,1+2,65 6,49 5,2
Скуловая ширина 6 93,0-110,9 102,8+2,68 5,99 5,8
Межглазничная ширина 7 79,3—99,0 87,3+2,60 6,37 7,3
Длина лицевой части 1 143,1 — — —
Максимальная длина 6 61,5-85,4 74,1-3,56 7,97 10,8
носовых костей
Длина верхнего ряда зубов 7 70,8-74,9 72,9+0,62 1,51 2,1
Максимальная ширина 7 78,6-97,0 88,0±2,91 7,13 8,1
мозговой капсулы
Длина нижней челюсти — — — — —
Длина нижнего ряда зубов — — — — —
Длина диастемы нижней — — —- — —
челюсти
Признак Самки
п limit X + го о Cv
Наибольшая длина черепа 6 218,2—233,3 226,5+2,33 5,20 2,3
Кондилобазальная длина 6 208,7-224,4 216,5+2,41 5,40 2,5
Основная длина черепа 6 192,6-207,0 199,5±2,27 5,07 2,5
Максимальная ширина 7 112,4-122,2 117,4+1,34 3,28 2,8
Скуловая ширина 7 97,8-101,3 99,8+0,56 1,38 1,4
Межглазничная ширина 8 78,3-85,4 81,7+0,83 2,19 2,7
Длина лицевой части 6 119,0-133,6 128,4+2,46 5,50 4,3
Максимальная длина 7 63,0-83,0 71,6-2,96 7,24 10,1
носовых костей
Длина верхнего ряда зубов 8 60,5-74,4 69,6+1,69 4,46 6,4
Максимальная ширина 8 76,1-80,4 77,5±0,77 2,03 2,6
мозговой капсулы
Длина нижней челюсти 6 172,4-184,4 178,0+1,86 4,17 2,3
Длина нижнего ряда зубов 6 67,1-79,3 73,9+2,15 4,81 6,5
Длина диастемы нижней 6 37,8-42,7 40,3+0,81 1,82 4,5
челюсти
казской части ареала. Между тем эта проблема нам представляется актуальной по ряду обстоятельств. Особенно нам хочется выделить то обстоятельство, что распространение бородатого козла на Кавказе охватывает три типа поясности — полупустынный, сухой субтропический и пустынный. По своему многообразию горных ландшафтов эти типы поясности мало уступают остальной части ареала вида. Если учесть, что популяции бородатого козла, занимая различные типы поясности на Большом и Малом Кавказе, испытывают фактор географической изоляции (Куринская низменность), то логично ожидать полиморфности этого вида в рассматриваемом регионе.
357
Бородатый козел
Правомочность такого суждения вытекает из противоречивых данных разных авторов. Так, К.К.Флеров (1932) считал, что бородатый козел Кавказа и Туркмении неоднородны, различаются по размерам тела и черепа. При этом более мелкие туркменские козлы должны быть отнесены к индийской расе (С. aegagrus blythi). Что касается положения козлов Малого Кавказа, то этот вопрос остался вне поля зрения. В последующем этот пробел затронут С.К.Далем (1945). Он показал, что козлы из Сарайбулакского хребта по размерам черепа ближе к туркменским козлам, нежели к кавказским. Исследования В.И.Цалкина (1950) показали, что туркменские бородатые козлы обладают признаками, достаточными для выделения их в самостоятельный подвид С. aegagrus turcmenicus. Наиболее существенные различия между этим и номинальным подвидом обнаруживаются, по его мнению, при сравнении толщины рогов. Он писал: "У самцов с Кавказа рога весьма мощны и обхват их при основании у взрослых зверей достигает 25 см (л = 45 экз., limit 20-25 см, х = 22,43 см)... Ни один Из исследованных нами 45 экз. не имели обхвата рогов менее 20 см. Что же касается козлов из Копетдага, то у них большая часть особей имеет обхват рогов менее 20 см. Такая величина обхвата рогов, ниже минимальных размеров, констатированных у самцов с Кавказа, наблюдается примерно у 55% туркменских экземпляров. Максимальные размеры у последних не превышают 23 см (limit 18—23 см; х - — 19,80, т — 0,22, о = 1,34). В среднем различия рогов составляют 2,63 см, a Midff составляет 8,76. Таким образом, бородатые козлы из Туркмении отличаются от кавказских значительно более тонкими рогами. Еще более тонкие рога свойственны бородатым козлам Синда и Белуджистана..." (с. 324).
Что касается размеров черепа, то В.И.Цалкин утверждал, что между кавказским и туркменским козлами нет никакого различия: и пределы варьирования и средние арифметические общей длины черепа у них вполне сходные. Аналогичные результаты им получены также по самкам.
Столь категоричное утверждение В.И.Цалкина малообоснованно.
Анализ литературных данных и коллекционного материала по бородатому козлу последних лет свидетельствует о том, что положение закавказских бородатых козлов не столь однозначно,- как это представлялось В.И.Цалкину (1951). По размерам черепа и рогов животные из Малого Кавказа ближе к туркменским, чем к козлам Большого Кавказа. Однако мы считаем преждевременным выделять на Кавказе два подвида. Нужен дополнительный материал, до его получения и соответствующей ревизии мы вслед за другими систематиками на Кавказе выделяем один номинальный подвид.
Экология
Ландшафтная приуроченность. Биотопы. Бородатый козел обладает, как отмечено выше, целым рядом морфологических и физиологических особенностей, позволяющих успешно жить на разных гипсометрических уровнях. Для него высота местности и горный рельеф не выступают в роли
35 8
Capra aegagrus Erxleben, 1777
физической преграды. Более того, как справедливо отмечают К.А.Айрумян и К.М.Гаспарян (1977), бородатый козел тонко приспособлен к скальным условиям жизни, с чрезвычайной легкостью передвигается по камням, совершая большие прыжки в любом направлении, перемещается по совсем узким карнизам и над крутыми обрывами. У одного из авторов данной монографии на даче содержался козленок с 2—3-недельного возраста. Повзрослев, в 4—5 мес, он поражал людей своей ловкостью в движениях: будучи на улице, чтобы выпросить лакомства в виде хлеба или зерна, часто прыгал на высокий (около 1,6 м) и очень узкий (всего 5—6 см) подоконник, ухитрялся какое-то время задерживаться на нем, показывая себя человеку’, ухаживавшему за ним. При этом он ни разу не задел стекла окон и не разбил их. Для него ворота высотой более 2 м не представляли никакой преграды — он легко перемахивал через них. Однако названные выше авторы, возможно, правы в том, что бородатый козел не приспособлен к длительному скоростному бегу на равнине и становится вдали от скал легкой добычей пастушьих собак.
Современное и былое распространение, а также видовые параметры характеризуют бородатого козла как широко распространенный вид пустынного и отчасти полупустынного типов поясности. При наличии скального рельефа, по нашим и литературным данным, эти козлы могут жить на весьма разных высотах — на Кавказе от 500 до 3900 м (Гаспарян, 1975).
Современная приуроченность к горным ландшафтам Кавказа определяется во многом отношением человека к этому животному. Известно, что в местах, где бородатого козла мало тревожат, явное предпочтение он отдает среднегорным участкам, где скалистые безлесные места чередуются более пологими, занятыми древесно-кустарниковой растительностью.
Еще в начале века Н.Я.Динник (1910) писал, что в Дагестане, где бородатый козел подвергается сильному преследЬванию, его можно встретить часто вблизи ледников и вечных снегов. Между тем он, обращая внимание на биотопическое разобщение туров и бородатого козла, отмечал, что бородатые козлы держатся в горах ниже туров, любят не голые утесы, а скалы, чередующиеся местами, покрытыми лесом и кустарниковыми зарослями. Часто они встречаются в таких местах, где туры вовсе не живут. Более того, по его данным, недалеко от Сарыкамыша, Ордубада и в Закаспийском крае они живут в местах совершенно безлесных. Сходные данные приводят и териологи, занимавшиеся изучением бородатого козла в Дагестане в последующие годы (Гептнер, Формозов, 1941; Верещагин, 1959; Шахмарданов, Рамазанов, 1980; Закариев, 1982; Прилуцкая, Пишванов, 1989; и др.).
В Чечено-Ингушетии, где представлен более мезофильный поясной спектр (терский вариант поясности), сведения по биотопическому размещению бородатого козла наиболее полны у А.М.Батхиева (1989). По его мнению, что соответствует и нашим наблюдениям, этот вид приурочен к труднодоступным скалистым местам, покрытым участками с зарослями кустарников. При этой обычно козел выбирает продольные скальные гребни, ориентированные от вершины хребтов вниз к реке. Высота таких
359
Бородатый козел
гребней, по данным названного автора, составляет, как правило, от 20—30 до 100—150 м. Между такими гребнями располагаются травянистые склоны; каменистая и крутая нижняя часть гребней используется для убежищ и ночевок, тогда как верхняя половина служит местом кормежки. В биотопах с крутыми изрезанными склонами А.М.Батхиев зарегистрировал наибольшее количество учтенных животных (38%). В кустарниках, в том числе •можжевельниковых, 21—23%, а на среднекрутых и сглаженных склонах отмечено 16%. На безлесных массивах встречено всего 2%.
В условиях пустынного типа поясности (джавахетско-армянский вариант) бородатый козел в прошлом был фоновым и полиландшафтным видом. Об этом свидетельствуют многочисленные костные остатки в пищевых отбросах и наскальные рисунки, обнаруженные на разных уровнях местности, в том числе в бассейне оз. Севан, и датируемые началом 3-го тысячелетия до н.э. (Даль, 1949; Верещагин, 1959; Гаспарян, Межлумян, 1969; и др.). В настоящее время,’ когда ландшафтное и биотопическос размещение копытных сильно трансформировано, оказался резко суженным круг экосистем, населяемым бородатым козлом. Более того, выбор местообитания чаще обусловлен защитным фактором, нежели кормовыми и иными.
По данным С.К.Даля (1948), на Айоцдзорском хребте летом эти козлы живут в скалах с кустарниками и осыпями, в зоне фриганоидной растительности, реже в горных степях и на горных лугах или прямо около участков ее на высоте 1250—2600 м. Этот же автор (Даль, 1944) считает, что на Са-райбулагском (Урцском) хребте представлены наиболее типичные местообитания бородатого козла. К такому заключению пришел также К.М.Гас-парян (1975). Мы не раз побывали в этом районе и на основе названных выше источников и личных полевых наблюдений приводим краткое описание Урцского хребта и соседних горных массивов. На наш взгляд, для данного района в целом характерен типичный поясной спектр пустынной зоны, где пустынные ландшафты простираются в горы до 1200—1350 м на северном макросклоне и до 1500—1600 м на южном макросклоне. Выше этого пояса находится пояс аридного редколесья, занимающий полосу от 1200—1300 до 1600—1700 м над ур. моря, а над ним следует пояс можжевеловых и дубовых лесов в пределах от 1500—1600 до 2400—2500 м. Субальпийский и альпийский пояса занимают сравнительно небольшую территорию Зангезурского хребта и др. Из орографических особенностей обратим внимание на холмистое предгорье, которое переходит в Араратскую равнину. В среднегорье значительно возрастает изрезанность рельефа, появляются скальные гряды и отдельные скалы, чередующиеся глубокими ущельями. С повышением высоты местности увеличиваются скалистость, пересеченность и каменные нагромождения. Нередки пещеры, карнизы, впадины и террасы. Почвенный и растительный покров с высотой приобретает мозаичный характер, отдельные участки на выровненных участках горных хребтов и их отрогов невелики по площади.
Район используется человеком давно в качестве пастбищ до самых верхних границ горных лугов; обрабатываемые земли имеются до высоты 2150 м, некоторые виды дикой растительности местное население упо
360
Capra aegagrus Erxleben, 1777
требляет для еды. Словом, антропогенная нагрузка на горные ландшафты, в том числе местообитания бородатого козла, велика вплоть до гребней хребтов. При таких условиях нормальное существование копытных возможно лишь при бережном отношении к ним со стороны человека и преимущественно на землях, непригодных для сельского хозяйства.
Миграции. В литературе практически отсутствуют сведения о территориальных перемещениях бородатого козла в естественных, нетрансформиро-ванных человеком природных условиях Кавказа. Однако, разделяя мнение К.М.Гаспаряна (1975), можно допустить, что в прошлом, когда был относительно сплошной ареал, занимавший обширную территорию Малого Кавказа, в том числе многоснежные высокогорные районы, бородатый козел совершал осенние сезонные миграции: высотные — с высоких многоснежных вершин в менее снежные низкие места; переход с северных на южные макросклоны с более теплым и меньшим снежным покровом; горизонтальные перемещения из северных районов ареала в более южные с пересечением р. Аракса. Весной направление миграций было обратным. В настоящее время, когда бородатый козел уничтожен на большей части ареала, сезонные миграции сохранились лишь на северной части Зангезурского и Айоцдзор-ского хребтов. Высокогорья этих хребтов, населяемые бородатыми козлами летом, зимой покрываются высоким снегом, вынуждая копытных спуститься вниз. По наблюдениям К.М.Гаспаряна (1975), сезонные миграции бородатых козлов в северной части горы Капутджухе выражаются в откочевке зимой на пониженные участки, а также на южные, юго-восточные и юго-западные макросклоны. В особо суровые зимы аналогичные сезонные миграции они совершают и в Гарнинской части Хосровского заповедника. В таких ситуациях часть поголовья бородатого козла выходит за пределы заповедника, оказываясь в зоне более населенной человеком и менее охраняемой. Другая часть, переместившись на южный макросклон, остается в пределах заповедника, хотя там местами довольно значителен снеговой покров. То, что в питании бородатого козла значительную долю составляют древесно-кустарниковые корма, а их в природе достаточное количество, делает, по мнению названного автора, этот вид менее зависимым от снегового покрова. На большей части современного ареала бородатого козла климатические условия вполне благоприятны для оседлой жизни.
Убежи1ца. Лежки. Как отмечено выше, обычными местами обитания бородатого козла можно считать скалы, различные каменистые биотопы. В скалах козлы легче уходят от опасности, чем на ровных местах. Каждый раз, будучи напутанными, они покидают такие ровные места и уходят в скалы, где они "теряются" благодаря соответствию окраски тела животного цвету камней. Нередко они затаиваются в камнях, прячутся в многочисленных пещерах, нишах и под навесами. Увидеть таких козлов нелегко, что заметно осложняет, например, проведение их авиаучета (Явруян и др., 1971).
Лежки бородатого козла располагаются, как правило, также в скалах. С.К.Даль (1948) обнаружил много лежек по небольшим террасам и выступающим утесам водораздела рек Ахта и Джул (Айоцдзорский хребет). В верховьях Ванкдараси того же района по узким коридорам, образовавшимся
36 1
Бородатый козел
между вертикально расположенными пластами каменных пород, также было много лежек козлов. У краев этих коридоров нередко можно видеть длинные ‘'потоки” глинисто-щебнистого грунта, выбитого копытами козлов. Очевидно, животных привлекают в такие коридоры благоприятные для них микроклиматические условия. В жаркие дни здесь прохладно, менее тревожно.
Сезонная и суточная активность. Изменения ритма жизни бородатого козла по сезонам года с учетом местных условий жизни' на Кавказе нс изучены. Логично, однако, допустить, что они должны быть значительны нс только в связи с периодикой физиологии животных, но и благодаря большим колебаниям погоды по сезонам года.
Суточная деятельность бородатого козла летом в нормальных и естественных условиях при отсутствии фактора беспокойства характеризуется четко выраженной двухфазной активностью (Даль, 1944, 195Г, Гаспарян, 1975; и др.). Жаркие часы дня, по данным названных авторов, бородатые козлы проводят в тени, под навесами скал или в пещерах, где больше влаги и прохладнее. Зимой, когда ухудшаются кормовые условия, активные животные регистрировались днем и ночью. Во время гона отмечаются нарушения суточного ритма у половозрелых животных, особенно самцов. Аналогичное явление имеет место и после гона — истощенные самцы восстанавливают потери энергетических ресурсов за счет увеличения продолжительности пд4тьбы. Старым самцам, обладающим большим "жизненным" опытом, свойственна сумеречная пастьба в течение года.
Поведение (рис. 47). Н.Я.Динник (1910) характеризует бородатого козла ловким, как туры, горным животным. Подобно турам они легко взбираются на отвесные скалы, смело спускаются в страшные пропасти, могут пробираться по самым опасным тропинкам и делать самые смелые прыжки. Они имеют прекрасное обоняние, тонкий слух, зоркие глаза и высокую чуткость. Однако, по мнению этого автора, бородатый козел в осторожности заметно уступает турам, может подходить близко к жилью; если они живут в более или менее лесистых местах, то и подкрадываться к ним значительно легче, чем к турам.
К.М.Гаспарян (1975), проводивший в наши дни специальные наблюдения за жизнью и поведением бородатого козла в Армении, считает его довольно флегматичным и малоподвижным в естественной и спокойной обстановке. На пастьбе перемещается медленно, делает короткие переходы с кормового участка на другой, чередует пастьбу отдыхом и играми. В мае 1965 г. за один день наблюдений суточные перемещения козла не выходили за пределы 300 м по горизонтали и 75—100 м по вертикали. При расположении мест кормежки далеко от скал (места убежищ) горизонтальные перемещения сокращаются, а вертикальные увеличиваются до 150—200 м и более. В этих случаях бородатые козлы спускаются и поднимаются по каменистым склонам ловко, проворно, в каждом их движении чувствуется уверенность, решительность. Указанный автор отмечает, что пасущиеся бородатые козлы перемещаются беспорядочно, во всех направлениях, сохраняя при этом чуткость. Периодически они прекращают пастьбу и замирают в самых
362
Capra aegagrus Erxleben, 1777
Рис. 47. Бородатый козел
363
Бородатый козел
разнообразных позах, обнюхивая воздух и прислушиваясь. Напуганное животное очень часто прерывает кормежку, чтобы увидеть или почуять опасность, резко поднимает голову, может в любой позе стоять до получаса, сосредоточив все внимание на источнике беспокойства. При обнаружении опасности животные в рассыпную или труппками непременно уходят в скалы, пользуясь нередко для отхода одними и теми же тропками. К.М.Гаспарян пишет, что на этой особенности основана охота на бородатого козла с нагоном. На постоянных тропах нередко устраивают засады одиночные охотники.
Во время отдыха стада отдельные особи остаются начеку, выполняя роль сторожа.
Поведение половозрелых самцов резко меняется во время гона. А.М.Бат-хисв (1989) отмечает, что в Чечено-Ингушетии внешняя картина гона видо-специфична. Самцы активно разыскивают самок, перемещаясь по всей поверхности хребта. Они становятся очень возбужденными, теряют всякую осторожность, могут подойти к спокойно стоящему человеку на расстояние до 10 шагов, не замечая его. Нередко по 3—4 самца ходят вместе.
Суточная активность бородатого козла в настоящее время сильно трансформирована под влиянием человека на большей части ареала. Особенно велика в этом роль горного животноводства. Как справедливо отмечает К.М.Гаспарян (1975), бородатый козел мало тревожится из-за появления домашнего скота, в том числе баранов, наиболее распространенных в горах Кавказа. Домашние овцы менее агрессивны, чем бородатый козел. В условиях вольера он в состоянии владеть всеми атрибутами доминанта среди нескольких домашних овсц. Упомянутый нами выше приученный к домашним условиям козленок свободно загонял в угол вольера 5—6 овец и один насыщался привлекательным для него кормом. Столь же решительным он был и по отношению к домашней собаке средних размеров. Однако, как отмечено выше, на ровных местах бородатые козлы могут стать жертвой сторожевых собак, которые обычно сопровождают отары овец на горных пастбищах. Опаснее всего вооруженные пастухи и необузданное браконьерство, что вынуждает бородатых козлов изменить привычный нормальный суточный ритм. В условиях Армении, по К.М.Гаспаряну (1975), во время выпаса стад встретить пасущихся бородатых козлов с раннего утра до вечера практически невозможно. В таких случаях они пасутся очень рано, на рассвете, обычно не выходя из скал, на небольших клочках земли, разбросанных среди камней. Когда появляются отары овец, бородатые козлы уходят на лежки и лишь в 18—19 ч возобновляют пастьбу, в том числе на задерненных землях под скалами. В это время года, как указывает упомянутый выше автор, бородатые козлы пасутся нередко и ночью.
Серьезным фактором беспокойства для бородатого козла служит сбор местным населением съедобных трав. "Начиная с апреля и до июля сборщики трав с раннего утра до 16—17 ч бродят по каменистым кручам южных склонов Урцского хребта, собирая эозимию, синеголовник, резак... непрерывно перекликаясь. В период сбора трав нам лишь несколько раз
364
Capra aegagrus Erxleben, 1777
удалось зарегистрировать козлов, спокойно пасущихся или отдыхающих" (Гаспарян, 1975, с. 98).
Не менее отрицательно сказываются на нормальном ритме жизни бородатых козлов интенсивные геологические изыскания, широко распространяющийся туризм, а также увеличение площади обрабатываемых земель и т.д. Перечень факторов, нарушающих исторически сложившуюся суточную активность, можно продолжить. В каждом конкретном районе набор их как и сила влияния отдельных факторов широко варьируют, что еще более усугубляется внутригрупповыми особенностями бородатых козлов.
Линька. Фенология и закономерности сезонной смены волос бородатого козла в условиях Кавказа изучены слабо. Имеющиеся литературные сведения относятся в основном к условиям одного джавахетско-армянского варианта пустынного типа. Отдельные наблюдения принадлежат С.К.Далю (1944, 1949) и более развернутые исследования — К.М.Гаспаряну (1975). Мы разделяем точку зрения последнего автора о том, что весенняя линька бородатого козла в Армении довольно растянута; животные на разных стадиях линьки встречаются с апреля по июнь. Коз в зимней шерсти, встреченных С.К.Далем (1944) до 19 мая 1939 г., не следует рассматривать как характерное для вида позднее начало линьки. Наблюдений о последовательности смены зимних волос на летние на теле животных мало. Что касается поведения бородатых козлов, способствующего ускоренному избавлению от старых волос, то ряд фактов приводит К.М.Гаспарян. В частности, на протяжении мая-июня бородатые козлы часто обтираются об камни, крутые склоны, трутся об кору и ветки. Нередко они вычесывают старые волосы ногами, рогами, выщипывают зубами, катаются в песчаной почве, переворачиваются на спину и т.д. Во всех этих действиях легко просматриваются корни (истоки) поведения домашних коз, подтверждая еще раз их генетическое родство с бородатыми козлами. Появление зимнего покрова проходит менее заметно. Не выявлены какие-либо специфические черты поведения или факты, говорящие о выпадении волос и реакции на это животных. Вероятно., происходит постепенное появление подпушки и длинных остистых волос.
Как видно, следуя нашему пониманию линьки (см. линьку косули), у бородатого козла наблюдается две линьки в году — весенняя и осенняя. Что касается фенологии этих явлений, ее динамики в ландшафтно-географическом плане и в зависимости от организма бородатого козла, конкретных данных мало, если не считать общих закономерностей, свойственных многим копытным.
Питание. В ряде работ содержатся сведения по питанию бородатого козла, полученные в различных частях ареала, относящихся к разным типам поясности: к полупустынному (терский вариант, Чечено-Ингушетия) и к пустынному (джавахстско-армянский вариант, Армения). Данные по первому типу содержатся в работах Т.Ю.Точисва (1968, 1973), Т.Ю.Точиева, А.М.Батхиева (1980), А.М.Батхиева (1989); по второму — С.К.Даля (1944, 1948, 1949, 1951), А.А.Саркисова (1950), К.М.Гаспаряна (1964). Сведения по Большому Кавказу, к сожалению, фрагментарны и недостаточны для
365
Бородатый козел
сравнения и выявления региональных особенностей питания бородатого козла. В связи с этим ниже приводится характеристика кормовых растений по второму району, полученная на Урцском хребте. Упитывая, что кормовой режим бородатого козла, как и других копытных, а также качественный и количественный состав зависят от конкретных условий, в том числе фенологии растительного покрова, очевидно, целесообразно дополнить характеристику Урцского хребта.
На северном макросклоне названного хребта доминирует горно-степная растительность, а на южных — нагорно-ксерофитная, в том числе фригано-идного типа, содержащая такие характерные виды, как миндаль, астрагалы, акантолимон, эфедра, прангос, чабрец, чистец и др. В предгорьях южного склона продуктивность пастбищ низка, не высоки также защитные условия. Поэтому здесь бородатые козлы бывают редко. На северных предгорьях они встречаются чаще, но посещают их неохотно из-за выпаса домашних животных. На верхней полосе хребта защитные условия благоприятны, но очень скудна растительность, ибо здесь доминируют скалы. Основные пастбища расположены в средней полосе хребта.
В связи с очень резкими сезонными изменениями растительного покрова во всех поясах Урцского хребта кормовой режим бородатого козла меняется широко в пространственно-временном аспекте. Пастбища наиболее привлекательны и продуктивны для бородатого козла с марта по июнь. Раньше всего зелень, преимущественно побеги злаков, появляется на южных склонах, и соответственно козы перемещаются на эти склоны и питаются злаками. Через некоторое время разнообразие растительного корма увеличивается за счет беллевалии, маков, василистника, пролески, гусиного и гадючьего луков, птицемлечника, спаржи и др. В это время вплоть до конца мая бородатые козлы выкапывают и скусывают у самого корня эозимию. Во второй половине рассматриваемого периода они хорошо поедают гладиолус, катран, хохлатку, марену, желтушник, ромашку, смолевку, терескен, козлобородник, козолец и марь. Среди деревьев и кустарников они предпочитают шиповник, вишню, миндаль. Редко или плохо поедают эфедру, прангос, чистец, коровяк и др.
Очень большие изменения в кормовом режиме бородатого козла вносит сухой период года в связи с высыханием травянистой растительности с последующим се исчезновением. Этот период занимает длительное время — с июля по октябрь, и в кормовом режиме бородатого козла ведущее значение приобретают деревья и кустарники, а из травянистых растений они поедают преимущественно те, которые остаются зелеными. По данным К.М.Гаспаряна (1964), в течение всего периода козлы лучше поедают миндаль, вишню, каркас, шиповник, жостер, марену, маки, синеголовник, аллохрузу, микролофус, василек, молокан, свинчатник, аспарагус и др.
Наиболее критическим периодом жизни бородатого козла в отношении питания является период с ноября по февраль, что объясняется отсутствием свежей растительности и ненастьем (нередки холодные дожди и снегопады). Животные вынуждены довольствоваться пожухлой растительностью и веточным кормом. В этом время зарегистрировано поедание таких видов, как
366
Capra aegagrus Erxlebcn, 1777
маки, крупина, синеголовник, васильки, ахиллея, аллохруза, терескен, эфедра, кохия и др. Видимо, круг поедаемых растений зимой определяется в основном степенью их доступности. Бородатые козлы поедают до 160 видов растений, относящихся к 119 родам и 35 семействам.
В горной Чечено-Ингушетии Т.Ю.Точиев (1968) зарегистрировал поедание бородатым козлом 102 видов растений, среди которых больше всего было злаков (27 видов), бобовых (12 видов) и др. Это количество видов в последующем было увеличено еще на 50 (Точисв, Батхиев, 1980). Анализ этого "голого" списка затруднителен, периодизация кормового режима, позаимствованная у К.М.Гаспаряна, на наш взгляд, мало приемлема в условиях терского варианта хотя бы потому, что фенология растительности горных поясов терского и джавахетско-армянского вариантов, населяемых бородатым козлом, существенно различна.
Как известно, с режимом питания копытных связаны солонцевание и водопой. По бородатому козлу таких данных мало, хотя С.К.Даль (1951) сообщал, что в апреле илиае зарегистрировал посещение ими солонцов, но не было замечено поедания снега или водопоя. По словам охотников, эти козлы посещают родники Урцского хребта (Гаспарян, 1975).
Воспроизводство и структура популяции
Размножение. Еще Н.Я.Динник (1910) писал, что спаривание у бородатых козлов происходит поздней осенью — в ноябре или, может быть, в начале декабря. Гон сопровождается драками половозрелых самцов между собой. Козлята появляются в основном в конце мая и в начале июня. Он встречал в начале июля очень маленьких козлят как в природе, так и среди отловленных охотниками для выращивания в домашних условиях. Эти наблюдения относятся к Чечено-Ингушетии. По наблюдениям Т.Ю.Точисва (1973), проведенным в том же районе, спаривание козлов происходит в декабре: начинается в первых числах и заканчивается в конце этого месяца. В ущелье Кериго (Чечено-Ингушетия), по данным А.М. Батхиева (1989), гон бородатого козла начинался с 10—13 декабря 1978 г. и достигал разгара в конце того же месяца. Спад наступал лишь в первых числах января.
На южном макросклонс Центрального Кавказа (Горная Тушстия), по наблюдениям З.С.Эквтимишвили (1954), сроки размножения бородатого козла широко меняются в зависимости от погодных условий: начало от середины ноября (1949) до 13 декабря (1951), а конец от 4 января (1951) до 18 января (1952 г.). В обычные годы начало гона падает на декабрь, однако, видимо, бывает и очень рано (Джанашвили, 1947).
В условиях Армении, по К.М.Гаспаряну (1975), гон бородатого козла начинается во второй декаде декабря и сопровождается активными действиями половозрелых самцов: интенсивным поиском самок, преследованием их, драками между самцами и формированием гарема. О жестокости схваток половозрелых самцов можно судить по рогам — их переднее ребро редко остается не поврежденным, выбитыми оказываются бугры. На это обращали внимание и другие авторы.
367
Бородатый козел
По мере угасания интенсивности спаривания самцы начинают отделяться от самок, однако некоторое время они остаются вместе. Общая продолжительность существования совместных (смешанных) стад составляет около одного месяца, иногда до двух (Эквтимишвили, 1954; Гаспарян, 1975). Однако и позже можно видеть смешанные группы, когда на протяжении ряда дней среди самок встречаются отдельные самцы или, наоборот, самки среди самцов. Как правило, отделившиеся беременные самки, особенно в последние дни беременности, ведут уединенную жизнь, встречаются по одиночке или небольшими труппками (2—3 особи, редко больше). Окот- происходит в укромных и труднодоступных местах. По Н.Я.Диннику (1910), на Большом Кавказе в Чечне козлята появляются в основном в конце мая и в июне, а в более южных районах, отличающихся несколько более теплым климатом, — в апреле. Эти данные совпадают с наблюдениями З.С.Эквтимишвили (1954), согласно которым в зависимости от сроков спаривания окот бородатых коз в Тушетии происходит с середины апреля до конца июня. В Армении, как следует из данных К.М.Гаспаряна (1975), козлята появляются начиная со второй половины мая — наиболее ранняя встреча следов молодняка зарегистрирована 24 мая. Беременные козы отмечались вплоть до середины июня. А.А.Саркисов (1944) в той же Армении(Урцский хребет) в 1930 г. отловил 33 козленка в возрасте 3—6 дней в конце апреля, т.е. рожденных в середине апреля. Как видно, фенология- воспроизводства популяции достаточно лабильна и правильно оценена Н.Я.Динником (1910). Мнение его о том, что продолжительность беременности составляет около 5 мес, очевидно, также справедливо.
Плодовитость. Количество козлят в одном помете один, два и очень редко три (Динник, 1910; Гаспарян, 1975). Тройни редко наблюдаются и у домашней козы. В природе не так редко встречаются козы с тремя козлятами, но нужно иметь в виду, что среди них могут иметься и приемыши, потерявшие матерей.
Судя по всем имеющимся данным, в период окота нередко встречаются половозрелые самки без козлят. Допуская многофакторность этого явления, видимо, прав К.М.Гаспарян (1975), связывающий его с высоким процентом яловости. На Урцскомхребте, по его мнению, соотношение беременных и родивших коз к взрослым яловым примерно равно одному к двум или даже к трем. Автор связывает высокий процент яловости с антропогенным прессом, в том числе с отгонным животноводством, барконьерством, изменениями в возрастной структуре популяции и др.
Рост и развитие. Козленок рождается полностью сформировавшимся, но в первые часы беспомощен, и мать проявляет к нему повышенное внимание — держится вблизи, часто кормит. Через 2—3 дня после рождения козленок свободно следует за матерью и поймать его довольно сложно. Через неделю он ходит по самым кручам. Молодняк остается при матери более года. Самцы становятся половозрелыми на втором году жизни, но, видимо, в воспроизводство вовлекаются значительно позже из-за старших и более сильных конкурентов (не ранее 3—4 лет). Самки приносят потомство в возрасте 3 лет.
368
Capra aegagrus Erxleben, 1777
Структура популяции. Наиболее полные данные имеются поУрцскому хребту Армении, где проводился улет бородатых козлов в разные годы. В 1939 г. из 52 встреченных С.К.Далем (1944) бородатых козлов 29, или 60,4%, было самцов и 19, или 39,6%, самок. Кроме них, отмечено 4 годовалых животных, пол которых было определить затруднительно. В мае 1950 г. он насчитал там же 354 бородатых козлов, среди которых самцы составляли 50,5%, самки — 49,5% (Даль, 1951). По данным К.М.Гаспаряна (1975), на Урцскомхрсбте в 60-х годах количество самцов колебалось от 61 до 72% от общего числа зарегистрированных бородатых козлов, не считая молодняка текущего года и в отдельных случаях приплода прошлого года, при условии, если он достаточно четко отличался.
В Чечено-Ингушетии, по А.М.Батхиеву (1989), среди всех встреченных в верховьях Аргуна бородатых козлов, на долю самцов приходится 25%, самок — 65%; сеголеток — 10%. Как видно, общая картина соотношения полов на Северном Кавказе и в Армении сходна. То же самое можно сказать и в отношении возрастной структуры — наУрцском хребте в мае 1939 г. (Даль, 1944) сеголетки составляли 7,7% от общего количества учтенных коз (52 головы). В мае 1950 г. там же популяция включала 36,5% молодых в возрасте 1—2 лет, самцов и самок старше 2 лет — 63,5% (Даль, 1951).
Размер стада служит хорошим показателем благополучия популяции. В первые десятилетия нашего века на Большом Кавказе и в Закавказье встречались стада бородатых козлов в 30 голов и более (Динник, 1906, 1910; Даль, 1944; и др.). В настоящее время такие стада сохранились только на заповедной территории. Так, по К.А.Айрумяну и К.М.Гаспаряну (1977), в Хосровс отмечены стада до 35 особей, тогда как на остальной территории Армении в среднем размер стада не превышает 10 особей. В Чечено-Ингушетии (Батхиев, 1989) эти козлы встречаются, как правило, небольшими группами — от одной до пяти особей, около 76% от общего количества встреч. Стада по пять голов встречаются редко (около 24%).
Враги, паразиты, конкуренты, смертность. В.Г.Гсптнср и др. (1961) относят к врагам бородатого козла рысь, волка, леопарда, некоторых хищных птиц для молодняка. По А.М.Батхиеву (1989), самыми опасными врагами этих коз в Чечено-Ингушетии являются волк и рысь. Он указывает также на вред, который может быть нанесен пернатыми хищниками молодняку. Однако автор не приводит фактического материала для подтверждения своего мнения о роли хищников в жизни бородатого козла. То же самое относится к заявлению о том, что в основном естественный отход зависит от случайной гибели животных от селей, лавин, неудачных падений и травматизма.
В Армении К.М.Гаспарян (1975) считает единственным хищником, способным нанести ущерб поголовью бородатых козлов, леопарда. Если иметь в виду, что этого хищника практически не стало на Кавказе, то и этот лимитирующий фактор отпадает. Что касается волка, то, по мнению названного автора, его роль ничтожна (при просмотре фекалий волков остатки бородатого козла не обнаружены). Автор справедливо отмечает, что данный факт нс отрицает возможности гибели козлов от волков. Специальных исследований по воздействию волков на бородатых козлов не
24 В.Е.Соколов, А.К.Темботов
369
Бородатый козел
проводилось. Не известны также случаи гибели бородатых козлов от стихийных бедствий (сели, лавины и др.). Конечно, Урцкий хребет, где работали зоологи Армении, и Центральный Кавказ — это разные по рассматриваемому фактору районы. Самым важным конкурентом бородатого козла в наши дни являются домашние животные, а врагом — нередко человек.
Паразитические черви бородатого козла Армении изучены Г.А.Гри-горяном (1949). Материалом послужили 26 животных, отстрелянных в 1944—1948 гг. в Микоянском и Вединском районах.
При полном гельминтологическом вскрытии по Скрябину и анализе легких, сердца, всего желудочно-кишечного тракта, печени, селезенки, желчного пузыря, почки, головного мозга в 8 случаях, поджелудочной железы'и мочевого пузыря в 12 случаях у бородатого козла Армении выявлены следующие виды:
Экстенсивность инвазии,%
Cystocaulus nigrescens (Jcrke, 1911) Schulz, Orloff et Kuttas, 1933 7,6
Protostrongylus davtiani (Ssavina, 1940) Davtian, 1949 92,3
P. muraschkinzewi (Davtian, 1940) 34,5
P. kochi Schulz, Orloff ct Kuttas, 1933 11,5
Mullerius capillaris (Muller, 1889) Davtian, 1949 3,8
Trichostrongylus colubriformis (Giles, 1892) 7,6
T. vitrinus Looss, 1905 15,4
T. probolurus (Railliet, 1896) Kalantarjan, 1928 7,6
Ostertagia (Grosspiculagia) occidentalis (Ransom, 1907) Orloff, 1933 42,3
O. circumcincta (Stadelmann, 1894) 26,9
Marshallagia marshalli (Ransom, 1907) Orloff, 1933 69,2
M. orientalis (Bhalcrao, 1932) Travassos, 1937 34,5
Nematodirus oiratianus Rajewskaja, 1929 22,9
N. spathiger (Railliet, 1896) Rail!, et Henry, 1909 42,3
Nematodirus abhormalis May, 1920 30,7
N. filicollis (Rud., 1802) Ransom, 1907 23,0
Chabertia ovina (Fabricius, 1788) Railliet et Henry, 1909 19,2
Trichocephalus ovis (Abildgaard, 1795) Skrj. et Schulz, 1934 19,2
Skrjabincma ovis (Skijabin, 1915) Skij. et Schulz, 1934 30,7
Thysaniezia ovilla (Rivolta, 1878) Skrjabin, 1926 3,8
Nematodirus davtiani nov. sp. 61,5
Ostertagia (Grosspiculagia) aegagri nov. sp. 3,8
На Большом Кавказе гельминты 4 бородатых козлов изучены А.Я.За-кариевым (1982), показавшим значительную зараженность животных червями из классов цестод и нематод. В пищеварительной и дыхательной системах обнаружены 4 вида гельминтов: Monierzia expansa, Chabcrtina ovina, Nematodirus junctispicularis, Trichostrongylus vitrinus.
Бедность видового состава гельминтов автор объясняет отсутствием возможных промежуточных хозяев.
Численность и ее динамика. По этому вопросу НЛ.Динник (1910) писал, что в ущелье Кий, принадлежащем бассейну р. Аргун, бородатые козлы "до
370
Capra aegagrus Erxleben, 1977
сих пор водятся в изобилии". Хотя этих козлов практически уже не было в безлесных верховьях Кази-Кумухского Койсу, Кара-Койсу, но в Аварском Койсу, где растет в верховьях много лесов, сохранилось в то время "много дичи, включая и безоаровых козлов... Часто попадаются... на Богосском хребте и в верховьях Андийского Койсу (с. 164). Но в горах, окаймляющих Тушинскую и Пирикительскую Алазань и принадлежащих как Главному Кавказскому хребту, так и Перикительскому, безоаровыс козлы встречаются довольно часто, в особенности в тех местах, где на горах растет лес... Еще чаще они попадаются в Эриванской губернии, причем довольно много живет их на горах недалеко от Ордубада" (с. 165). Эта информация, дополненная сведениями о стадности (см. выше), свидетельствует о том, что бородатый козел был фоновым видом горных ландшафтов полупустынного и пустынного типов поясности Кавказа. В настоящее время, как было отмечено в разделе "Ареал", он исчез во многих местах, сохранился на отдельных изолированных участках, перешел из группы фоновых к редким, исчезающим видам. Для такой оценки современного состояния популяций имеются серьезные основания, в том числе учетные данные, полученные в разных частях ареала. Сознавая неоднородность и неравноценность этих данных, а также сопутствовавших их получению методических изъянов, в том числе указаных Э.Г.Явруяном и др. (1971), ниже приводим некоторые показатели.
В Чечено-Ингушетии (терский вариант поясности), по учетам А.М.Батхиева (1980), проведенным в декабре, январе и феврале 1978/79 г., общее количество бородатых козлов определялось в 350—360 голов. По его мнению и данным Т.Ю.Точиева (1973), лет 15 назад в республике этих козлов было 450—500 голов. В Дагестане численность бородатого козла выше: по Т.Д.Хехневой (1972), она составляет около 700 особей, а по данным Л.И.Прилуцкой и Ю.В.Пишванова (1989) — около 1000 особей. Весьма желательно подтвердить наличие роста численности бородатого козла в Дагестане. В Грузии численность этого вида по А.Б.Арабули (1989) составляет около 300 особей на площади 11 тыс. га. По данным К.А.Айрумяна и К.М.Гаспаряна (1977), в Армении ориентировочно 400—700 бородатых козлов. Больше всего бородатых козлов в Азербайджане, где наиболее многочислен вид в Нахичевани. По мнению Х.М.Алскперова и др. (1973), здесь численность рассматриваемого вида определяется в 1800—2000 особей, но она сокращается интенсивно, что связано, по мнению авторов, с освоением высокогорных пастбищ для домашнего скота.
Суммируя изложенные выше данные, а также учитывая современные тенденции роста животноводческого освоения гор Восточного Кавказа и Закавказья и состояния охраны природы, трудно рассчитывать на положительные сдвиги в численности бородатого козла на Кавказе. Это возможно только при реализации специальных мер пресечения браконьерства, упорядочения отношения животноводства и охраны животного мира.
Практическое значение. В прошлом бородатый козел, как отмечено выше, был многочислен в ряде районов и служил ценным объектом охо4ы. Мясо высокого вкусового качества, употреблялось местным населением как
24*
371
Сайгак
деликатесная еда; шкура использовалась в хозяйстве, а рога в качестве охотничьих трофеев и украшения. В настоящее время в связи с резким сокращением численности, охота на бородатого козла запрещена. Однако, на наш взгляд, на Кавказе имеются реальные условия для значительного восстановления распространения и численности бородатого козла, а также его роли как объекта спортивной охоты. При оценке значения этого вида коз следует брать в расчет, что он является родоначальным видом домашних коз и может быть использован и в будущем для селекционных целей.
Род Сайгаки — Saiga Gray, 1843
Размеры тела небольшие, примерно с домашнюю козу, но туловище удлиненное, ноги тонкие и относительно короткие, шея длинная и заканчивается непропорционально большой головой. Примечательно наличие сильно горбатой и вздутой лицевой части, что обусловлено резким расширением носовой полости. Передняя часть морды сужена, свисает вниз в виде хоботка, соответственно округленные ноздри сближены и направлены вниз. Уши короткие с округленными дистальными краями. Хвост короткий. Средние копыта крупные, боковые — небольшие. Самцы с рогами, расположенными на голове вертикально, по длине примерно равные длине головы, неправильной лигообразной формы, желтовато-белого цвета. На поверхности рогов имеются поперечные кольца в количестве до 18—20 шт. Самки безрогие. Летний мех низкий и относительно редкий. Цвет желтовато-рыжий на спине, а к брюху постепенно светлеет. По хребту цвет меха темнеет. Околохвостовое зеркало не выражено. Зимой мех длиннее и гуще, его окраска становится светлой, глинисто-серой. В году линяет два раза — весной и осенью. Из специфических кожных желез имеются небольшие пред глазничные, паховые, запястные и межпальцевые. Сосков две пары.
Своеобразна структура черепа, особенно лицевой части: носовые кости
очень короткие, они направлены вперед и несколько наверх, непропорционально черепу наружные носовые отверстия большие, в их образовании принимают участие слезные кости, формируя передне-боковые края. Такое
положение слезных костей среди полорогих отмечается только у сайгаков. Ямка для предглазничной железы на слезной кости выражена слабо. Межчелюстные кости развиты слабо, имеют оригинальную форму — они не
достигают носовых костей, изогнуты дугообразно сверху вниз, несколько
расширены в дистальной части. Верхнечелюстные кости также не сопри-
касаются с носовыми костями. Глазницы имеют трубкообразные края.
Отсутствуют этмоидальные отверстия.
Зубная формула:
J о с о р 3(3) 3 1 2(3)
3 М - = 30(32).
372
Saiga tatarica Linnaeus, 1766
В зубной системе характерно, что последний нижний предкоренной имеет открытую переднюю и замкнутую заднюю долинки (Соколов, 1959).
Ареал занимает Восточную Европу, Среднюю и Центральную Азию. Типичный представитель открытых степей и полупустынь, соответственно вся организация и биология сайгаков несут глубокие следы адаптивного характера. В характеристике рода Saiga уместно указать его положение среди Caprinac и становление таксона в историческом аспекте.
Сайгаки составляют довольно изолированную группу, не имеющую близкого родства ни с одним другим родом полорогих. По В.Г.Гептнеру и др. (1961), они наиболее близки к роду Pantholops, представленному в современной фауне одним видом, живущим на высокогорных равнинах Тибета (от 4500 до 4700 м над ур. моря). Однако различия между ними очень большие, что во многом, видимо, связано с особенностями ландшафтных условий . Род Saiga, очевидно, занимает промежуточное положение между Caprinac и Antilopinae и, возможно, ближе к последнему, что дает повод ряду специалистов относить сайгаков к антилопам. В этой связи интересен тот факт, что по структуре кожных желез сайгаки близки к роду Ovis. Что касается истории становления рода Saiga, то палеозоологические данные отрывочные, относятся к плейстоцену и голоцену и нс позволяют достаточно однозначно составить суждение по этому вопросу. Как отмечает Н.К.Всре-щагин (1959), устойчивость морфологических признаков сайгаков на протяжении со среднего плейстоцена до наших дней свидетельствует о том, что корни рода уходят в плиоцен, возможно даже в миоцен. Остается загадкой отсутствие близкородственных форм в плиоценовых отложениях всей Палеарктики. Говоря о плейстоценовых и голоценовых остатках, следует отметить, что они обнаружены на огромной территории ,— от Англии на Западе до Аляски на востоке, на север до Новосибирских островов, на юг до Восточного Закавказья. Появление сайгаков на Кавказе датируется средним плейстоценом (Верещагин, 1959).
В роде Saiga один вид — Saiga tatarica L.
Сайгак — Saiga tatarica Linnaeus, 1766 Рис. 48
Capra tatarica Lin, Диннсй, 1766; Antilope Saiga Rail, Паллас, 1766; Antilo-pc Scythica Pall., Паллас, 1767; Capra Sayga Lin., Forster, 1768; Cernas Colus., Okcn, 1816.
Коллекционный материал, КБГУ: 10 черепов, 9 полных скелетов и 9 шкур.
Диагноз. Полорогие размером с домашнюю козу, самцы с лирообразными рогами, нс превышающими размеров длины головы. Самки комолые. Большая и горбатая лицевая часть заканчивается хоботком. Слезные кости увеличены и участвуют в образовании переднего края носового отверстия.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. Экстерьерные показатели сайгаков Западного Прикаспия по состоянию на
373
Сайгак
Рис. 48. Распространение сайгака
374
Saiga tatarica Linnaeus, 1766
1 — оз. Сенгелевское (Динник, 1910; Верещагин, 1959); 2 — окрестности Ставрополя (Россиков, 1887; Динник, 1910; Верещагин, 1959; Банников и др., 1961); 3 — Старомарьевка (Динник, 1910); 4 — Тахта (Динник, 1910, Верещагин, 1959); 5 — Бурукшун (Динник, 1910); 6 — о-в Ма-ныч-Гудило, западный берег (Банников и др., 1961); 7 — о-в Маныч-Гудило, восточный берег (Банников и др., 1961); 8 — Светлоград (Динник, 1910); 9 — Рагули (Динник, 1910; Верещагин, 1959); 10 — Арзгир (Динник, 1910; Верещагин, 1959); 11 — окрестности Пятигорска (Банников и др., 1961); 12 — Низовье р. Малки (Россиков, 1887; Верещагин, 1959; Темботов, I960; Банников и др., 1961); 13 — окрестности Грозного (Банников и др. 1961); 14 — устье р. Сулак (Верещагин, 1959; Банников и др., 1961); 15 — Бабаюртов-ский р-н (Прилуцкая, Пишванов, 1988); 16 — Кизляр (Верещагин, 1959); 17 — А1раханский полуостров (Банников и др., 1961); 18 — о-в Чечень (Банников и др., 1961); 19 — Терекли-Мектеб (Гептнер, Формозов, 1941); 20 — Кочубеевский р-н (Спасская, Абдулмананов, 1980); 21 — о-в Тюлений (Банников и др., 1961); 22 — Ачикулак (Верещагин, 1947); 23 — Нефтекумск (Верещагин, 1959); 24 — среднее течение р. Кумы (Кузнецов, 1949); 25 — устье Кумы (Сатунин, 1912; Адольф, 1950, 1952); 26 — район Черных земель (Верещагин, 1959); 27 — окрестности Яшкуль (КБГУ)
1978 г. приводятся в работе В.Е. Присяжнюка и А.В. Проняева (1988). Согласно этим данным, живой вес взрослых самцов составляет 31—58 кг (х = 42,7), длина тела 123—144 см (х = 136,5), высота в холке 75—83 см ()? = 80,2), длина хвоста 9—14 см (х = 11,1), высота уха 7—9 см (х = 7,8). Взрослые самки имеют живую массу 22—31 кг (х = 27,2), длину тела 116—128 см (х= 121,7), высоту в
375
Сайгак
Таблица 74. Промеры черепов взрослых сайгаков калмыцкой популяции
(по: Проняев, 1983)
Признак Самцы (п = 23)
limit X ± о
Длина черепа наибольшая 239,3-258,1 247,99±5,84
Кондилобазальная длина черепа 225,6-250,1 236,81±6,93
Основная длина черепа 212,7-235,7 223,06+6,37
Длина лицевой части черепа 137,4-217,0 205,91±6,62
" мозговой части черепа 89,1-97,7 94,33+2,15
" лобноносовых костей 97,4-112,5 105,10+4,27
носового отверстия 139,2-152,2 145,63±4,05
" • прсдальвсолярная 60,6—72,5 63,90±2,60
Ширина носового отверстия 30,6-38,7 34,90+2,04
Межглазничная ширина 82,2—97,8 90,50+3,86
Ширина черепа в области орбит 122,9-136,0 129,80±3,20
Ширина мозговой коробки 66,4-73,4 69,26±2,09
Мастоидная ширина черепа 76,3—87,0 80,66+2,49
Ширина морды над Р^ 27,3-36,1 30,79±2,27
в лицевых буграх 72,6-82,4 77,89+3,00
Вертикальный диаметр глазницы 39,3-44,0 41,92±1,0б
Горизонтальный диаметр глазницы 40,5-45,0 42,66±1,15
Ширина твердого неба у Р- 28,1-41,2 32,53±3,17
М3 35,7-48,6 29,4-35,7 44,37±2,82 32,16+1,87
Высота слухового пузыря 63,2-73,7 68,01+2,80
Длина верхнего ряда зубов 16,3-23,6 20,71+1,68
ряда предкоренных зубов 43,8-50,5 46,83 + 1,99
ряда коренных зубов 36,4-41,4 38,92+1,55
тела затылочной кости Ширина тела затылочной кости 22,2-29,7 25,81±2,99
холке 67—77 см (х = 73,4), длину хвоста 7—19 см (х = 10,0), высоту уха 6,5—8 см (х = 7,1). Размеры тела сайгаков, приводимые Н.Я.Динником (1910), укладываются в пределы, названные выше. Так, длина тела составляет до 132 см, высота в холке равна 80 см, длина хвоста около 9 см, длина уха 10,5 см. Ковдилобазальная длина черепа, по А.Г.Банникову и др. (1961), у взрослых самцов (h = 30) равна 222,3—250 мм (х = 240,2), самок (й = 30) — 205,4—228,6 мм (х = 217,8). Данные Проняева (1983) несколько ниже (табл. 74).
В структуре черепа характерно своеобразное строение лицевого отдела: резко сокращена длина носовых косей, отсутствуют швы между ними и лобными костями, удлинена слезная кость и участие их в образовании носового отверстия, короткие межчелюстные кости, не соприкасающиеся с носовыми, отсутствие этмоидального отверстия, трубкообразныс глазницы, суженная с боков ростральная часть черепа, на конце согнутая немного вниз, особенно проксимальная часть межчелюстных костей, отсутствие пазух в лобных костях.
376
Saiga tatarica Linnaeus, 1766
Таблица 74 (окончание)
Признак Самки (п = 50)
limit У ± О
Длина черепа наибольшая Кондилобазальная длина черепа Основная длина черепа Длина лицевой части черепа ” мозговой части черепа " лобноносовых костей " носового отверстия " предальвеолярная Ширина носового отверстия Межглазничная ширина Ширина черепа в области орбит Ширина мозговой коробки Мастоидная ширина черепа Ширина морды над " в лицевых буграх Вертикальный диаметр глазницы Горизонтальный диаметр глазницы Ширина твердого неба у Р? М3 Высота слухового пузыря Длина верхнего ряда зубов " ряда предкоренных зубов " ряда коренных зубов “ тела затылочной кости Ширина тела затылочной кости 204,8-233,6 195,4-226,7 184,4-219,9 164,6-192,1 76,5-84,2 84,0-96,6 115,2-137,2 52,1-74,4 25,0-36,4 67,8-80,5 104,0-116,6 59,2-66,6 66,1-75,9 20,9-29,8 61,8—73,4 36,0-40,1 36,4-41,0 21,9-32,2 33,7-43,2 25,7-34,0 60,3-71,8 14,0-35,1 34,9-50,8 28,9-35,7 18,0-24,5 221,50±6,45 212,92+6,64 200,59±6,77 179,28±5,77 80,53±2,12 88,95±2,80 125,53±5,02 58,79±3,74 30,74±2,11 74,11+2,97 111,05±2,68 62,88±1,88 69,68±2,44 24,92±2,21 67,15±2,65 38,22±1,12 39,07±1,15 27,21±2,16 37,72±2,07 29,84±2,12 67,2.9 ±3,11 20,81±2,03 45,88±2,66 31,84±1,35 21,44±1,42
Зубная система. По И.И.Соколову (1959), А.Г.Банникову и др. (1961), отмечен ряд особенностей: заднекоренные зубы гипсодонтные, верхние — с узкими стилями на наружной поверхности. Прсдкоренные зубы редуцированы и соответственно в нижней челюсти у номинальной формы только два предкоренных. Последний предкоренной зуб имеет открытую переднюю долинку. Резцы гипсодонтные, длинные, но не узкие; граница между корнем и коронкой выражена слабо.
, Зубная формула:
1-С-Р-М- = 30.
3 12 3
Строение рогов довольно своеобразное. Согласно данным АТ.Банникова и др. (1961), у вполне сформировавшихся самцов (в возрасте 19 мес и старше) общая длина рогов составляет 28—38 см, чаще 32 см. У самок они лишь намечаются в виде костных бугорков высотой до 5 мм и остаются под кожей. В типичном виде рога самцов неправильно лировидные, располагающиеся почти целиком над глазницами, слегка наклонены назад, от основания до середины они расходятся в стороны, затем постепенно сближаются, перед вершиной вытягиваясь параллельно друг другу или вновь ра
377
Сайгак
зойдясь в стороны. Кончики рогов нередко загнуты внутрь. Характерно отсутствие полного соответствия формы одного рога с другим. Обычно они имеют ряд различий по деталям. Цвет рогов видоспецифичсн, обычно полупрозрачный, желтовато-белый, светло-восковой. Лишь самые кончики бывают темные. На поверхности рогов от самого основания и до верхней трети имеются резко выдающиеся и волнообразно изогнутые кольцевые валики в количестве до 20—22 шт., отстоящие друг от друга на 1—2 см. Последние дистальные 3—4 валика не всегда образуют полный круг из-за слабого развития. Верхняя часть рогов, лишенная кольцевых валиков, обычно гладкая и более прозрачная, чем остальная часть. Поперечное сечение рогов округлое, продольный диаметр несколько превосходит поперечный.
Для посткраниального скелета характерно, что длина пястной кости больше длины плечевой кости. Передние конечности массивнее задних. Угол зацепной части рогового копыта на передней конечности равен 67°, на задней — 55°, что объясняется одновременным выбрасыванием передней и задней ног одной стороны (иноходь).
Как следствие адаптации к быстрому и прямолинейному бегу на равнине тазобедренный сустав приобрел четко выраженную блоковидную форму. Он функционирует в основном по оси сгибания и разгибания. Соответственно большее развитие имеют флексоры и экстензоры сустава.
Окраска тела в летнем наряде желтовато-рыжая на верхней стороне, белая или почти белая на брюхе и груди. Более интенсивно окрашенная полоса проходит по спине и шее до затылка. Такое же сгущение отмечается на крестце и лопатках, они могут быть коричневого цвета. Передняя часть головы трязно-белая, лоб и темя темно-рыжие, иногда почти коричневые. Вокруг глаз окраска светлее — светловато-рыжая; щеки песочного или рыжеватого цвета. На губах и вокруг ноздрей расположены длинные вибриссы черного цвета. Окраска хвоста буровато-желтая, мало выделяется на общем фоне верха тела. На задней части бедер отсутствует светлое зеркало. Ноги спереди рыжевато-коричневые, их внутренняя сторона светлее. На суставах, особенно передних ног, имеются коричневые пятна. Темно-коричневые волосы имеются также по наружному краю ушей. Детали окраски тела сайгака определяются степенью пигментации верхней половины волос. Что касается нижней части, то она в основном серого цвета.
Зимний мех сайгаков очень светлый, глинисто-серый. В отличие от летних волос, имеющих серое основание, у зимних волос нижняя половина чисто-белого цвета. Эта окраска волос выше постепенно переходит в светло-рыжеватые кончики. Окрашенные кончики на фоне белого придают общей окраске спины глинистый отлив. По А.Г.Банникову и др. (1961) и нашим материалам, на боках мех светлеет, особенно на нижней стороне тела. Темное пятно размером около 7 х 40 см имеется в области крестца. Волосы, образующие его, почти в 2 раза длиннее светлых волос остальной части спины. Аналогичные темные и удлиненные волосы имеются также на щеках под глазами (так называемые бакенбарды). Остальная часть щеки как и бока шеи серые с буроватым оттенком.
378
Saiga tatarica Linnaeus, 1766
Рис. 49. Строение остевых волос взрослого летнего самца сайгака
А — поперечный срез в середине стержня, ув. х 6006; Б — то же на вершине, ув. х 600; В — сердцевина, ув. х 1000; Г — микроорганизмы на стенке сердцевинных клеток, ув. к 2000; Д — кутикула вентральной стороны стержня, ув. х 600; £- то же с дорсальной стороны, ув. х 300
379
Сайгак
Кожный покров. Изучили волосяной покров загривка взрослых самцов, добытых соответственно в мае 1980 г. и в ноябре 1978 г. в Калмыкии. У самца, добытого в мае, происходит смена зимнего меха на летний. В структуре волосяного покрова у самца, добытого в мае, четко различимы два яруса волос. Верхний ярус — остатки зимнего меха. Нижний ярус (летний мех) представлен одной категорией остевых волос трех размерных порядков. Длина волос этих размерных порядков: 13,8; 11,6 и 9,3 мм. Стержни слабоволнисты. Характерен тонкий бессердцевинный кончик вершины волоса. В прикорневой зоне хорошо выражена "ножка". У остей первых двух размерных порядков вверх от "ножки" стержень слегка расширяется и наибольшей толщины (соответственно 150 и 135 мкм) достигает у вершины. У остей третьего размерного порядка стержень максимально утолщается (125 мкм) сразу после "ножки". К вершине он постепенно сужается. В верхней половине стержня, с дорсальной стороны проходит желобок. Поперечные срезы основания волоса имеют овальную форму. В середине — овальную форму со слегка вогнутой одной стороной (рис. 49, А). У самой вершины — бобовидную форму с глубокой выемкой (рис. 49, Б). Сердце-вина хорошо развита по всему стержню и составляет 90% его диаметра. Корковый слой равномерно развит со всех сторон стержня. Сердцевина крупноячеистая. Обычно высота и ширина клеток равны. Стенки сердцевинных клеток гладкие. Вертикальные или радиальные межклеточные перегородки часто бывают укреплены небольшими волосковидными выростами, вдающимися в полость клетки (рис. 49, Б). На поверхности стенок видны колонии микроорганизмов (рис. 49, Г). Рисунок кутикулы вдоль стержня и с вентральной и дорсальной сторон одинаков. Чешуйки вытянуты поперек стержня. Их высота постоянна. Свободные края равны и плотно прилегают к корковому слою (рис. 49, Д).
Самец, добытый в ноябре, имеет уже перелинявший зимний мех. Он густой, серо-бежевого цвета, представлен также одной категорией остевых волос, но четырех размерных порядков. Зимний мех (размерные порядки: 42,3; 46,8; 32,1 и 25,2 мм) более чем в 3 раза длиннее летнего. Толщина (105 мкм) у всех размерных порядков одинакова и слабо меняется вдоль стержня. Поперечные срезы основания круглой формы (рис. 50, А). Желобок, проходящий с дорсальной стороны стержня, мельче, чем у летних остевых волос. Поперечные срезы в верхней половине стержня имеют бобовидную форму со слабо выраженной выемкой на дорсальной стороне. Сердцевинный канал очень хорошо развит по длине всего стержня и со-.ставляет около 95% его диаметра. В прикорневой зоне (область "ножки") и у самой вершины сердцевина также отсутствует. Сердцевинные клерки в отличие от таковых у летнего самца сплющены сверху вниз (рис. 50, В). Стенки клеток имеют гладкую поверхность без волосковидных выростов. Рисунок кутикулы вдоль стержня нс меняется. Высота чешуй изменчива, края ровные (рис. 50, Г).
Кроме основной категории остевых волос, в зимнем мехе также встречаются единичные волоски, длина которых не превышает 22 мм, толщина
380
Saiga tatarica Linnaeus, 1766
Рис. 50. Строение волос зимнего самца сайгака
А — поперечные срезы основания волос, ув. х 200; Б — В — сердцевина, ув. х 300 и 800; Г — кутикула, ув. х 600
76 мкм; сердцевина составляет 96% диаметра стержня. По структуре они ничем не отличаются от остевых- волос.
Система крови. Мы не располагаем литературными сведениями по крови сайгака. Данные, которые приводятся здесь, мы получили в окрестностях пос. Ящкуль Калмыкии в мае 1990 г. Изучена кровь у 10 сайгаков, из них 7 самцов и- 3 самки (табл. 75). Количество эритроцитов в 1 мл крови составляет у самцов 8,31 млн, у самок 8,10 млн, содержание гемоглобина соответственно 17,2 и 17,4%. Степень насыщения эритроцитов гемоглобином высокая, об этом свидетельствует цветной показатель. Гематокритная величина составляет у самцов 43 об.%, у самок 44 об.%. Диаметр эритроцитов в среднем у самцов 5,3 мкм, у самок 5,4 мкм. При подсчете белой крови оказалось, что лейкоцитов в 1 мл крови содержится у самцов 2,64 тыс., у самок 2,53 тыс. Для лейкоформулы характерен лимфоцитарный сдвиг.
38 1
Сайгак
Таблица 75. Показатели крови сайгака из Северо-Западного Прикаспия .______________(пос. Яшкуль, Калмыкия, май 1990 г.)______________
Показатель Самцы (д = 7)
limit х ± т с Су
Гемоглобин (г%) 14,6 21,2 17,2+0,98 2,51 14,59
Эритроциты (млн) 7,14—9,87 8,31 ±0,39 0,95 11,39
Цветной показатель (ед). 0,57-0,67 0,62+0,01 0,03 5,09
Диаметр эритроцитов (мкм) 5,10-5,40 5,30+0,04 0,10 1,89
Гематокрит (об.%) 34-49 43,0+2,80 6,85 15,93
Лейкоциты (тыс.) 2,00—3,76 2,64±О,27 0,67 25,36
Лимфоциты (%) 41-89 76,57+6,87 16,84 22,16
Сегментоядерные нейтрофилы (%) 6-50 16,00±6,47 15,84 99,02
Палочкоядерныс нейтрофилы (%) 3-5 3,86+0,28 0,69 17,25
Моноциты (%) 1-5 2,71±0,77 1,89 62,99
Эозинофилы (%) 1 1 0 0
Показатель Самки (п = 3)
limit х ± т G Су
Гемоглобин (г%) 15,0- 20,08 17,4±2,15 3,03 17,39
Эритроциты (млн) 7,40-9,00 8,10+0,58 0,82 10,09
Цветной показатель (ед). 0,61 — 0,69 0,64+0,03 0,04 6,8
Диаметр эритроцитов (мкм) 5,40—5,70 5,53+0,11 0,15 2,78
Гематокрит (об.%) 41-49 44,0+2,95 4,16 9,46
Лейкоциты (тыс.) 1,85-3,25 2,53±0,50 0,70 27,69
Лимфоциты (%) 47-88 60,67 + 16,79 23,67 38,80
Сегментоядерные нейтрофилы (%) 8-46 32,00±14,81 20,88 65,25
Палочкоядерные нейтрофилы (%) 2-5 3,67±1,08 1,53 38,19
Моноциты (%) 1-5 2,67+1,48 2,08 69,39
Эозинофилы (%) 1 1 0 0
Индексы внутренних органов. Обширная информация по абсолютным и относительным показателям ряда внутренних органов сайгака приводится в работе А.Г.Банникова и др. (1961) (табл. 76). Учтены пол, а также пять возрастных групп: I — до 1 мес, II — 2,5 мес. III — 7—9 мес., IV — 1 год, V — старше полутора лет. Не вдаваясь в подробное изучение данных, обратим внимание на некоторые показатели, характеризующие сайгаков как своеобразную жизненную форму открытых пространств степей и полупустынь. Так, индекс сердца относительно высокий во всех возрастных группах; у самцов разного возраста он составляет в среднем от 8,3 до 13,5, а у самок — от 8,6 до 13,4. Наиболее высокими показателями выделяются годовалые (свыше 13 мес) и месячные (около 10 мес и выше). В первом случае авторы высокий индекс связывают с тем, что годовалыми сайгаки становятся весной, когда животные наиболее истощены, что обусловливает малую массу тела и соответственно большой индекс сердца. Высокий индекс сердца у сайгаков месячного возраста широко встречается и в других группах млекопитающих. Представляется весьма примечательным, что сайгаки в возрасте 2,5; 7—9 и 18 мес имеют очень близкие показатели индекса сердца.
382
Таблица 76. Масса и размеры некоторых внутренних органов у сайгаков
_________________ (по: Банников, 1961)__________________________
Возраст ЖИВОТНЫХ
Орган, Пол 1 мес 2,5 мес (п = 11) 7—9 мес (п = 3)
индекс (л =2) limit X limit X
Масса сердца, Самцы 74,5 98,0-146,0 118,0 215,0-223,0 219,0
Г Самки 61,0 85,0-149,0 114,0 169,0-233,0 196,0
Сердечный Самцы 10,8 8,0-9,0 8,4 8,2-8,4 8,3
индекс Самки 9,0 7,6-9,9 9,2 7,0-10,5 8,6
Масса легких, Самцы — 183,0-220,0 201,0 — —
Г Самки 101,0 138,0-230,0 182,0 228,0-360,0 286,0
% к массе Самцы — 2,0-1,2 1,4 — —
тела Самки 1,8 1,3-1,6 1,4 1,2-1,5 1,4
Масса печени, Самцы — 241,0-393,0 328,0 — —
Г Самки 130,0 256,0 -334,0 301,0 348,0-457,0 388,0
% к массе Самцы — 2,6-2,2 2,3 — —
тела Самки 2,6 2,5-2,2 2,4 1,9-1,9 1,9
Масса почек, г Самцы — 40,0-56,0 49,6 — —
Самки 22,0 34,0-44,5 40,2 42,0-56,0 50,0
% к массе Самцы — 0,4-0,3 0,4 — —
тела Самки 0,3 0,3-0,3 0,3 0,3-0,2 0,2
Длина тонких Самцы 1011,0 1325,0-940 1191 — —
КИШОК, см Самки 702,0 1372,0-1075 1193 1720-1404 1520
Длина толстых Самцы 300,0 527,0-412 491 — —
кишок, см Самки 206,0 525,0-450 492 610-570 590
Возраст животных
Орган, Пол 1 год (п = 16) старше полутора лет (п = 18)
индекс limit 5 limit X
Масса сердца, г Самцы 250,0-500,0 340,0 262,0-450,0 347,0
Самки 200,0—350,0 289,0 230,0-400,0 298,0
Сердечный индекс Самцы 9,5-16,0 13,5 6,0-11,6 8,6
Самки 12,0-17,0 13,4 7,4-12,5 9,3
Масса легких, г Самцы 250,0-550,0 417,0 380,0-600,0 483,0
Самки 250,0—600,0 402,0 358,0-450,0 415,0
% к массе Самцы 1,2-1,7 1,6 1,2-1,5 1,3
тела Самки 1,5-1,9 1,8 1,1-1,7 1,6
Масса печени, г Самцы 600,0-1000 705,0 545,0-1100,0 856,0
Самки 300,0-900 629,0 470,0-1100,0 683,0
% к массе Самцы 2,9—3,1 з,о 2,1-2,2 2,3
тела Самки 1,8-2,9 2,8 2,2-2,7 2,6
Масса почек, г Самцы 84,0—107,0 99,0 76,0-113,0 97,0
Самки 43,0-81,0 69,8 67,5-113,0 84,0
% к массе Самцы 0,3-0,4 0,4 0,2-0,3 0,2
тела Самки 0,3 - 0,3 0,3 0,2-0,3 0,3
Длина тонких Самцы 1724-930 1460 1910-1200 1726
кишок, см Самки 1630-1010 1304 1678-1292 1425
Длина толстых Самцы 808-600,0 634 1050-670 851
кишок, см Самки 700-521,0 597 751-600 690
Сайгак
Сердце сайгаков, как и сердце антилоп (Muri, 1870 г., по: Банников и др., 1961), конусовидное, вершина его заостренная.
Для легких характерна резко выраженная дольчатость с большим числом дополнительных борозд и вырезок. Масса легких составляет 1,3—1,6% от массы тела взрослых и 1,4—2% от массы тела молодых животных. Легким сайгаков свойствен ряд анатомических особенностей, направленных на приспособление бронхов и трахеи для обеспечения интенсивного дыхания при быстром беге.
Масса печени и почек значительна, что вполне соответствует интенсивному обмену веществ. Как и показатели сердца, индексы печени и почек мало подвержены возрастным изменениям.
Кариотип. По данным В.Н.Орлова и Н.Ш.Булатова (1983), двойной набор хромосом включает 60 элементов. Все аутосомы и Х-хромосома акроцентрические. У-хромосома — маленький метацентрик. Хромосомный набор, судя по морфологии, довольно упрощенный.
Структура и динамика ареала
Ареал вида в прошлое историческое время (восстановленный ареал) занимал обширную территорию степей и полупустынь Евразии от пред-горьев Карпат на западе до Монголии и Западного Китая на востоке. Анализу гсохрафичсского распространения сайгаков в историческое время посвящен целый ряд научных работ. Обзор этой литературы содержится в монографии А.Г.Банникова и др. (1961). Западная граница ареала в XVII в., очевидно, достигала Южного Буга и Прута (примерно 25° в.д.). Северо-западный угол распространения находился на пересечении 25° в.д. и 50° с.ш. Северная граница проходила южнее Киева, вблизи Усмана, Тамбова, Куйбышева, Уфы до Оренбурга, Кустаная, выходя на Барабинские степи и далее по предгорьям Алтая и через Зайсанскую котловину уходила в Китай. Южная траница простиралась до Черного и Азовского морей, предгорьев Кавказа, нижнего течения Амударьи, среднего течения Сырдарьи, предгорьев Каратау и Алатау и уходила в Китай. В то время изолированная популяция встречалась в Монголии.
В период максимальной депрессии численности (20-е годы XX в.) сайгак сохранился лишь в отдельных районах — в глухих местах Калмыцкой и Волго-Уральской степи, на Устюрте, в Приаральских Каракумах, в Прибалхашье и котловине Зайсана, а также в двух районах Монголии.
Распространение на Кавказе. Восстановленный ареал за исторический период на Кавказе охватывал все Предкавказье и значительную часть Кура-Араксинской низменности. В западном Предкавказье сайгак встречался на юге до Кубани. Очевидно, на левобережье его не было из-за лесистости предгорьев, но что касается среднего Предкавказья, то реальные предпосылки для более широкого распространения здесь были. Отсутствие лесных поясов в эльбрусском варианте не только в наши дни, но и в прошлом позволяло сайгаку проникать далеко в предгорья. Об этом косвенно свидетельствует широкое проникновение их в предгорья до Минераловодских
384
Saiga tatarica Linnaeus, 1766
лакколитов в 50-х годах нашего столетия. В восточной части Предкавказья сайгак вплотную подходил к восточному макросклону Терского хребта, а далее на юг, видимо, продвигался по Каспийскому побережью, с тем чтобы занять равнинные пространства Восточного Закавказья. Провести сколько-нибудь подробное описание восстановленного ареала сайгака этой территории невозможно из-за отсутствия данных.
Резкое сокращение сайгаков в Европе после XVII в. охватило и кавказскую часть ареала, однако это происходило здесь медленнее, чем на остальной территории. Так, во времена К.Н.Россикова (1887) сайгаки еще водились в низовьях Малки (окрестности г. Прохладного).По Н.Я.Диннику (1910), в конце XIX в. сайгаки проникали на юг до Ставрополя, их отстреливали недалеко от оз. Сенгелеевское и с. Старо-Марьевское. Они были обычны в окрестностях Арзгира, Рагулея, Туркменской Ставки. Однако сайгаки исчезли в то время вблизи Большой Кугульты, Джалги, Бурукшуна, Тахты. В 20-х годах нашего века сайгаки сохранились лишь местами на Черных Землях (между низовьями Волги и Кумы). Что касается-более южных районов, В том числе Терско-Кумской полупустыни, то там сайгаки исчезли в 30-х годах (Верещагин, 1959).
Мероприятия по восстановлению сайгаков, начатые в 20—30-х годах, приостановили уничтожение вида, более того, способствовали восстановлению былого распространения и численности в ряде районов ареала, в том числе в Калмыкии и прилегающих районах. В 40-х годах северная граница простиралась до 48* с.ш., западная — до Ергеней (44° в.д.), юго-западная — до оз. Маныч, а южная — до р. Кумы. В 50—60-х годах продолжалось дальнейшее расселение сайгаков с охватом Сальских и Карано-гайских степей, Терско-Кумской полупустыни, в том числе-Аграханско го полуострова, островов Чечень и Тюлений. Судя по карте А.Г.Банникова и Л.ВЖирнова (Банников и др., 1961), граница области постоянного обитания сайгаков в то время проходила на западе вблизи пересечения 48° с.ш. и 44° в.д., восточного берега оз. Маныч-Гудило; на юге по линии южнее Арзгир, окрестности Бажигана и южнее Черного рынка. Граница забегов в суровые зимы и в период летних засух на западе доходила до Цимлянского моря, западнее Зимовников и западных берегов оз. Маныч-Гудило; на юге — до окрестностей Ставрополя, Пятигорска, южнее Грозного и низовья Сулака. В середине 50-х годов площадь обитания сайгаков на правом берету Волги определялась примерно в 100 тыс; км2 (Максймук и др., 1987), а к 1958 г. — 100—110 тыс. км2 (Максймук и др., 1989). В ее состав входила почти вся территория Калмыкии и пограничных районов Астраханской и Ростовской областей, Ставропольского края и Дагестана.
С конца 50-х и в 60-е годы на всем Северо-Западном Прикаспии происходило интенсивное освоение человеком мест обитания сайгаков, что привело к сокращению территории, занятой сайгаком, в 1968 г. до 35 тыс. км2, а численность за 10 лет уменьшилась с 800 тыс. до 85 тыс. особей. Это было, связано с распашкой в западных и центральных районах около 40—50% пастбищных угодий, а также с увеличением поголовья домашних
25 В.Е.Соколов, А.К.Тсмботов
385
Сайгак
животных до 5 млн голов. Прекращение промысла в 1969—1970 и 1972 г. и сокращение поголовья домашних животных до 3,5 млн несколько улучшили положение сайгаков, соответственно их ареал и численность стали расти. К 1977—1978 гг. площадь, занятая сайгаками, возросла до 56 тыс. км2, а численность — до 600—700 тыс. особей (Максимук и др., 1989). По данным Л.И.Прилуцкой и Ю.В.Пишванова (1988), в 70-х годах на Бажиганских песках Ногайского района размножалось до 100 особей. В соответствии с учетными данными за 1977—1979 гг. на территории Северо-Западного Дагестана обитало около 8 тыс. сайгаков (Газалиев, 1980). В суровые зимы они мшрировали в Ногайский и Тарумовский районы, а при особо неблагоприятных условиях — в Кизлярский и Бабаюртовский районы, достигая по ледовому покрову островов Тюлений и Чечень.
Однако начиная с середины 70-х годов экологическая обстановка стала ухудшаться из-за мелиоративных работ, расширения пахотных земель, увеличения поголовья скота до 4,5 млн, что привело, по мнению названных авторов к снижению численности сайгаков в 2,5 раза. К 1981 г. площадь обитания сайгаков составляла всего 30—50 тыс. км2, а численность — около 400 тыс. особей. В 1981—1982 гг. в связи с дальнейшим развитием орошения и обводнения и прогрессирующим опустыниванием ареал сайгака еще сократился и составил около 20»23 тыс. км2, а численность достигла 160—200 тыс. особей. На указанной территории наряду с сайгаками выпасаются 1,9 млн голов овец. Снижение промысловой нагрузки и полный запрет промысла в 1984, 1986 гг. не вывели сайгаков из состояния депрессии.
По состоянию на 1990 г. сайгаки распространены лишь в самой восточной части Калмыкии (устное сообщение работников Калмыцкого госпромхоза). Что касается сайгаков, акклиматизированных в 1950 г. на о-ве Булла, а затем на о-ве Глиняной Каспийского моря (Верещагин, 1959; Алиев, 1961), то сведениями о современном их состоянии мы не располагаем.
Современные тенденции антропогенных изменений ареала. Как известно, в настоящее время резко возросла интенсивность хозяйственной деятельности человека на всем протяжении Прикаспийской низменности. Прямое и косвенное влияние этой деятельности на сайгаков настолько велико, что "угрожает полным уничтожением калмыцкой популяции этого вида" (Дежкин, 1986, с. 144). Наши полевые наблюдения и литературные данные, в том числе Л.В.Жирнова (1977, 1982 и др.), А.В.Максимука и др. (1987), Е.Я.Зуева (1986) и др., свидетельствуют о том, что депрессия численности и резкое сокращение ареала сайгаков обусловлены многими факторами. Однако ведущими являются антропогенные: распашка целинных и залежных земель, рост поголовья скота Калмыкии (с 1,9 млн в 1961 г. до 3 млн в 1978 г.), увеличение объема строительных и монтажных работ по обводнению засушливых районов, плохая организация учета численности и завышенная добыча, строительство оросительных каналов без должного учета экологии сайгаков, промысел сайгаков несовершенными методами, процветание браконьерства, вытеснение животных из характерных мест обитания в
386
Saiga tatarica Linnaeus, 1766
низкопродуктивные угодья, нередко в слабо закрепленные пески, и т.д. Снять или смягчить эти факторы — дело сложное и трудное, но необходимое, хотя бы на охраниченной территории Восточной Калмыкии, где сохранились необрабатываемые земли. И в этих условиях следует вынести решение, которое предусматривало бы комплексное использование пастбищных угодий Калмыкии и прилежащих районов сухих степей и полупустынь в интересах как домашнего животноводства, так и промыслового сайгаководства. Как справедливо отмечено в ряде работ, такое сочетание экономически выгоднее, чем использование первичной продукции пастбищных угодий Калмыкии в одном только животноводстве.
Систематика и изменчивость
Систематическое положение. Сайгаки составляют морфологически четко очерченный таксон, однако их положение в системе Bovidae в зоологической литературе трактуется по-разному. Расхождение взглядов имеет глубокие корни. Исторический анализ становления современных взглядов на макросистематику, проведенный в работе А.Г.Банникова и др. (1961), достаточно подробный, поэтому нет нужды повторять его. Здесь лишь отметим, что сайгаков одни ученые относят к роду Сарга, другие — к Antilope, третьи — к Gazella, большинство систематиков выделяют их в самостоятельный род Saiga.
Противоречивы мнения и о положении сайгаков в системе выше рода. Некоторые специалисты выделяют сайгаков в самостоятельное семейство, другие рассматривают их в ранге подсемейства, занимающего промежуточное положение между антилопами и .баранами, третьи относят их к подсемейству антилоп, большинство же вслед за Г.Симпсоном (Simpson, 1945) к подсемейству баранов Caprinae (Соколов, 1959; Гептнер и др., 1961; Corbet, 1978; Honack et al., 1982; и др.). Сайгаков относят к Caprinae в связи с тем, что у них резцы гипсодонтные, глазницы трубкообразные, basiocli-pitale плоские и короткие. Однако, по мнению А.Г.Банникова и др. (1961), сайгаки обладают также рядом особенностей, отличающих их как от Caprinae, так и от антилоп. Среди них имеются примитивные и специализированные черты (строение рогов, отсутствие пазух в лобных костях, сильное развитие зароговой части черепа и др.). Все это, по мнению названных авторов, свидетельствует о самостоятельном развитии рода Saiga как минимум с миоцена и служит достаточным основанием для придания сайгакам статуса самостоятельного подсемейства Saiginae с одним родом и одним видом.
Что касается внутривидовой систематики, то здесь мнения специалистов совпадают; вид Saiga tatarica включает два современных подвида (S.t. tatarica и S.t. mongolica), хорошо различающиеся морфологически и занимающие обособленные ареалы.
Морфологическое своеобразие монгольских сайгаков побудило А.Г.Бан-никова (1946) выделить их вначале как самостоятельный вид, но впоследствии он низвел его в ранг подвида (Банников, 1951). Новые кариологические
25
387
Сайгак
данные не противоречат такому суждению. Мы считаем достаточно обоснованным рассматривать род Saiga монотипическим, относящимся к подсемейству Caprinae, а вид S. tatarica с двумя подвидами: S.t. tatarica и S.t. mongolica. В пределах Северного Кавказа обитает номинальный подвид.
Изменчивость. Повышение численности и открытие промысла сайгаков с 50-х годов дали возможность многим специалистам провести разносторонний анализ изменчивости вида. Можно считать, что сайгак в этом плане уже изучен неплохо, опубликован ряд интересных работ, позволяющих выявить особенности популяций сайгаков Северо-Западного Прикаспия. Среди литературных источников наиболее значительный и разнообразный материал содержится в публикациях А.В. Проняева и соавторов (Проняев, 1984, 1985а, б, 1986; Проняев, Аксенов, 1987, 1988; Проняев, Рожков, 1988; Присяжнюк и др., 1986; Присяжнюк, Проняев, 1988). За основу нашего обзора изменчивости сайгаков были положены эти и некоторые наши данные. Сбор коллекционного материала мы проводили в Калмыкии в 1990 г. В целях повышения "чистоты" данных, используемых ниже для характеристики экстерьера сайгака, мы выбрали те, которые получены в период их максимальной численности (Калмыкия, Калмыцкий госпромхоз, октябрь 1978 г., п = ПО экз. и Актюбинская область, Иргизский госпромхоз, октябрь 1975 г.). При анализе других данных в тексте приводится ссылка на автора.
Индивидуальная изменчивость. В группе взрослых сайгаков масса тела самцов варьирует от 31 до 58 кг, самок — от 22 до 31 кг, коэффициент вариации у самок несколько ниже. Менее вариабельны размеры тела, особенно высота в холке, длина передней и задней конечностей, обхват труди и др. Сильно варьируют длина хвоста, высота уха, обхват шеи, длина и ширина копыт на всех конечностях, обхват пясти и др. При оценке других данных по индивидуальной изменчивости экстерьера сайгаков следует учитывать способность животных разных экологических поколений (рожденных в разные годы) адаптироваться к быстро меняющимся условиям жизни (Проняев, Аксенов, 1987). Согласно данным этих авторов, особенно динамична группа особей, включающая крупных животных (М+). Они, видимо, гетерозиготны и способны приспосабливаться значительно лучше, чем особи группы средних (М°) и мелких (М‘). Изменение в III сезоне по сравнению с II, например, величины труппы М° происходит в 1,66 раза, а труппы М+ в 9 раз.
Очень высокой индивидуальной изменчивостью характеризуются рога. В коллекциях, просмотренных нами, было больше сильно различающихся рогов, чем сходных. Это касается как формы, так и массивности рогов. Так, в нашей коллекции экземпляр S-2960 отличается относительно короткими, но массивными рогами (индекс толщины, т.е. отношение длины обхвата чехла у основания к длине по внешнему изгибу составляет 0,64). У экземпляра S-12260 этот показатель намного ниже — всего 38. Высока также вариабельность формы рогов. У первого экземпляра выражен лишь один изгиб, приходящийся к средней части рога. В результате оба рога
388
Saiga tatarica Linnaeus, 1766
лежат почти в одной плоскости, изгиб направлен по внешним сторонам. Рога второго экземпляра: в верхней трети рога сближаются, потом постепенно расходятся, а верхушки закручиваются внутрь и вперед. В итоге рога находятся в 2—3 плоскостях, максимальная ширина между рогами находится ниже середины, а минимальная, если не считать основания, — между кончиками рогов. Высокая изменчивость отмечается и по ряду других морфологических показателей рогов (цвет кончиков, количество кольцевых валиков и др.).
Отмечая высокую изменчивость рогов в последние годы, уместно обратить внимание, что частота редких аллелей за период с 1979 по 1986 г. резко сократилась (Проняев, РоЖков, 1988). По данным авторов, аллели, встречавшиеся раньше в калмыцкой популяции с частотой менее 0,07, не выявлены через 7 лет после первого исследования. Имеются три аллеля Tf-системы и один аллель Amy-2-системы. Популяция за это время обновилась полностью 3 раза. Предполагается, что частоты редких аллелей снижались в связи с действием стабилизирующего отбора, роль которого усиливается в годы депрессии численности. Следствием такого отбора, как известно, является сокращение фено- и генотипического разнообразия.
Возрастная изменчивость. Сайгак как вид характеризуется высокой скоростью роста и развития. Массу тела взрослых молодые животные (как самцы, так и самки) достигают в 19-месячном возрасте. В последующие годы жизни изменения массы тела носят сезонный, а не возрастной характер. Размеры тела взрослых животных достигаются также в 19-месячном возрасте. С возрастом происходят значительные изменения в пропорциях тела. Новорожденные имеют большую величину индекса длинноногости и самую низкую величину индексов формата и широкотелости (Банников и др., 1961). Индекс длинноногости падает с возрастом, а формата и широкотелости, наоборот, увеличивается. Уже к семимесячному возрасту молодняк по пропорции тела сходен с взрослыми.
В отличие от массы и размеров тела окраска животного мало подвержена возрастным изменениям. После двухнедельного возраста сайгачата сходны с взрослыми по общей окраске меха. Возрастные изменения сводятся лишь к усилению неоднородности окраски верха тела.
Что касается рогов, То полного роста они достигают в 19 мес. В это время они имеют уже соответствующие взрослым длину, толщину и количество колец на роговых чехлах.
Размеры черепа подвержены очень большим возрастным изменениям. Достаточно сказать, что череп с переменной скоростью растет до трехлетнего возраста. В этом возрасте достигается наибольшее значение массы черепа: у самцов она превышает массу черепа новорожденного в 22,5 раза, а у самок — в 4,5 раза! Возрастная изменчивость пропорции черепа проходит довольно сложно, ибо отдельные его показатели формируются с различной скоростью. Так, размеры черепа шестимесячных сайгаков значительно уступают полуторалетним, но по длине ряда предкоренных зубов, наоборот, превышают.” Четко прослеживается удлинение лицевого отдела, а
389
Сайгак
мозговая часть уменьшается. В целом характерные для взрослых пропорции черепа формируются в 19-месячном возрасте.
Возрастные изменения зубной системы подробно описаны в монотрафии А.Г.Банникова и др. (1961), в ней приводится определитель возраста сайгака-по зубам. Согласно этой таблице можно условно выделить ряд возрастных классов, характеризующихся определенными признаками.
1. Новорожденные — все 20 зубов молочные, среди них
2. Полуторагодовалые — молочные предкоренные заменены на постоянные, появляются все коренные.
3. Двухлетние — закончена замена молочных резцов и клыков на постоянные.
4. Трехлетние — требни М, совсем стираются или остаются неболь-шие следы на задней доле. Щели сохранены на обеих долях. Внутренняя поверхность зацепов совсем плоская, хорошо отшлифованная. Гребни и щели отсутствуют между полулуниями Mj и на передней доле М^.
5. Четырехлетние — на жевательной поверхности М щель небольшая, но хорошо заметная. Щели отсутствуют на Mj, а также на передней доле М2 и задней доле М3. Изнутри зацепы отполированы, а также верхняя половина средних и верхняя треть окрайков.
6. Пяти-шестилетние — следы требней отсутствуют на М. Отшлифованы все резцы.
7. Семи-восьмилетние — коронка на М совсем стерта; начато стирание корней зуба, сильно сношены требни на М2 и М^, почти исчезли на них щели. Нет щелей также на М] и М2, а на М3 щель имеется лишь на средней доле М3.
8. Девяти-десятилетние — М^ стерт до десен, иногда он разрушен. Полость исчезла на передней доле М2. На задней доле этого зуба полость или исчезла совсем или сохранилась, но потеряла подковообразную форму.
Возрастные классы после трех лет определяются по степени стертости зубов приблизительно.
Половые различия. У сайгаков, как и у других стадных животных, резко выражен половой диморфизм. Это явление затрагивает все морфологические и другие показатели вида. По массе тела самцы и самки различаются достоверно во всех возрастных группах. По А.Г.Банникову и др. (1961), новорожденные самцы тяжелее самок на 200 г. Этот разрыв с возрастом увеличивается. За весь период постнатального развития масса самцов возрастает в 11—13 раз, а самок всего в 8—10 раз. По данным тех же авторов, новорожденные самцы в среднем на 4 см длиннее самок; высота в холке превышает на 1,7 см, а в маклаках на 1,2 см. Несколько больше и обхват труди. В возрасте 4 лет и старше самцы длиннее самок на 14,6 см, высота в холке на 8,5 см, а в маклаках на 7,5 см. Половой диморфизм четко проявляется и в динамике возрастных изменений экстерьера.
390
Saiga tatarica Linnaeus, 1766
Так, в течение первого месяца размеры тела самцов увеличиваются гораздо быстрее, чем соответствующие параметры самок. На втором и третьем месяцах самки растут интенсивнее, чем самцы. После зимней депрессии рост самок возобновляется на 2,5—3 мес позже, чем у самцов (связано с беременностью и лактацией).
Половые различия в массе и структуре черепа очень велики, что в первую очередь связано с наличием рогов у самцов. В целом череп самок до глубокой старости носит признаки инфантильности. Особенности черепа самцов начинают формироваться с двухмесячного возраста, когда на лобных костях появляются бугорки, которые в последующем вырастут в костные стержни рогов. С появлением рогов у самцов поднимаются области лобных костей, составляющих основу стержней. Соответственно округлая поверхность лба превращается в выпуклую, тогда как у самок сохраняется ювенильный облик. По обеим сторонам фронтального шва формируются впадины, а сам шов становится выпуклым. Выпуклым становится также венечный шов. В связи с появлением рогов и усилением шейной мускулатуры зароговая часть черепа становится шероховатой и угловатой. Аналогичные изменения наблюдаются, как правило, у всех копытных, обладающих рогами.
Статистическая обработка 224 черепов из калмыцкой популяции позволила А.В.Проняеву (1983) показать динамику становления полового диморфизма по структуре черепа в Северо-Западном Прикаспии. В труппе сайгачат из 25 изученных признаков достоверное раличие при Р < 0,001 выявлено по 17 признакам, при Р < 0,01 — по трем признакам, при Р < 0,05 — по одному признаку. Различие отсутствует лишь по четырем признакам (ширина мозговой коробки, ширина твердого неба у М3, длина верхнего ряда предкоренных зубов и длина ряда коренных зубов). В группе полуторалетних различия достоверны по всем изученным признакам (при Р < 0,001 — 24 признака, при Р < 0,01 — один признак). В труппе взрослых высокодостоверными (Р < 0,001) остаются различия по 22 изученным признакам, тогда как по трем признакам (длина зубного ряда и его частей) различия нивелируются.
На наш взгляд, общебиологический интерес представляют половые различия в динамике относительной массы черепа самок по сезонам года (Банников и др., 1961). Тогда как у самцов к 3 годам масса черепа увеличивается в 20 раз по сравнению с новорожденными, а потом остается неизменной до конца жизни, у самок она возрастает лишь до 1,5 лет. Затем относительная масса черепа колеблется по сезонам. Весной она снижается до минимума, восстанавливаясь к осени, началу периода размножения. Авторы полагают, что значительная часть костного вещества черепа матери идет на формирование скелета эмбриона.
Географическая изменчивость. Все имеющиеся данные говорят о том, что на всем протяжении Западного Прикаспия (правобережье Волги) обитает одна панмиксическая популяция. Об этом свидетельствуют многие особенности жизни сайгаков, в том числе высокая динамичность пространственной организации видового населения, большие сезонные и годовые
39 1
Сайгак
изменения степени агрегации, широкий обмен особями между группи-ровками в результате регулярных и непериодических миграций, вызванных природными стихиями, и др. (Банников и др., 1961; Баскин, 1978; Жирнов, 1982; Максимук и др., 1989; и др.). Популяцию Западного Прикаспия, види7 мо, правомочно называть "калмыцкой", ибо в пределах Калмыкии сохранилось основное поголовье сайгаков в наши дни. Судя по картам Л.В.Жир-нова (Банников и др., 1961), в годы пика численности на этой территории и прилегающих районах, по-видимому, обитала большая часть сайгаков.
Как следует из изложенного выше, географический аспект изменчивости сайгака на Северном Кавказе может быть анализирован в настоящее время лишь при сравнении с другими популяциями, в том числе сайгаками Казахстана. Тем более значительный фактический материал, вполне сравнимый с таковыми Западного Прикаспия, уже имеется в литературе (Проняев, 1983; 1985; Проняев, Рожков, 1988; Присяжнюк, Проняев, 1988).
Экстерьер сайгаков калмыцкой популяции независимо от пола и возраста отличается более высоким индексом относительной длины передней и задней конечностей, длинно но гостью, меньшим значением индекса массивности, быстроаллюрностыо, сбитостью. Что касается абсолютных показателей экстерьера, то по значительному числу количественных признаков всех половых и возрастных групп сайгаки калмыцкой популяции уступают казахстанским сайгакам.
Популяционные различия по черепу, судя по данным А.Г.Банникова и др. (1961), А.В. Проняева (1983), также четко прослеживаются. В группе молодых самцы калмыцкой популяции меньше своих казахстанских сверстников по 17 из 25 промеров. По остальным промерам различий между популяциями не обнаружено. Самки молодых сравниваемых популяций достоверно различаются по 19 промерам. При этом особи калмыцкой популяции меньше, чем казахские. По шести промерам различий между популяциями не обнаружено.
В полуторалетнем возрасте уменьшается число признаков, по которым различаются популяции; черепа самцов казахской популяции превосходят черепа калмыцких сайгаков лишь по длине мозговой части и длине тела затылочной кости. Однако самцы калмыцкой популяции превосходили своих сверстников казахской популяции по ширине носового отверстия, ширине морды в области Р2 и лицевых бугров. По всем остальным промерам черепа различия не выявлены. Самки казахской популяции превооходили также калмыцких по кандилобазальной и наибольшей длине черепа, длине лобно-носовых костей, ширине мозговой части черепа, межглазничной ширине, а также по ширине в области орбит, мастоидной ширине и некоторых других. У калмыцких сайгаков полугодовалого возраста ширина морды над вторым предкоренным зубом значительно больше, чем у казахских.
В группе взрослых межпопуляционные различия выявлены по ряду промеров. Самцы калмыцких сайгаков имеют более длинную лицевую часть черепа, ширины носового отверстия, ширину морды в лицевых буграх, обоих диаметров глазницы, большую ширину твердого неба на уровне Р2. Они уступают достоверно самцам казахской популяции по мастоидной ширине
392
Saiga tatarica Linnaeus, 1766
черепа и по всем трем промерам зубного ряда. По самкам калмыцкие популяции уступают казахским самцам по ширине и длине мозговой коробки, длине лобно-носовых костей; по ширине черепа, твердого неба у М^; у них также значимо ниже показатели длины тела затылочной кости и ряда коренных зубов.
На основании указанного выше можно заключить, что с возрастом снижается количество промеров черепа, по которым сравниваемые популяции различаются. В строении морды калмыцких животных проявляются элементы инфантилизма. Наивысшие раличия между популяциями выявлены в труппах шестимесячных сайгачат. Однако все эти различия популяционного уровня, возможно связаны с новыми условиями, сложившимися в последнее время. Они недостаточны для таксономического обозначения, и соответственно справедливо мнение систематиков, объединяющих "все популяции Прикаспия, Казахстана и Средней Азии в один номинальный подввд.
Экология
Ландшафтная приуроченность. Биотипы. Морфологические и экологические особенности характеризуют сайгаков как типичных представителей равнинных степей и полупустынь. Многие специалисты обыкновенно указывают, что сайгаки избегают не только горы, но и места с пересеченным рельефом, в том числе холмистые, заросшие, песчаные барханы. Такие места могут быть заселены лишь вынужденно, в связи с холодной и вет-ренной погодой. Отсутствие в горах сайгаков не связано с несовершенством адаптивных механизмов к гипоксии, наоборот, кислородная емкость крови у этого вида, по нашим данным, высокая. Лимитирующим фактором, как справедливо отмечают А.Г.Банников и др. (1961), следует считать рельеф — там, где имеется ровная местность, сайгаки могут жить независимо от высоты местности: в Прикаспийской полупустыне они живут у моря, в условиях Казахстана — до 600 м, а в Монголии поднимаются до 900—1600 м над ур. моря.
Ландшафтные условия жизни сайгаков на протяжении обширного ареала характеризуются определенным набором жизненных форм растений, однако составляющие виды и видовые комплексы широко меняются. Фоновыми видами всюду выступают такие дерновинные злаки, как типчак, ковыль, житняк, а также полукустарнички — полыни, прутняк, ромашник и др. В Прикаспии, как и в восточных районах ареала, много мест занимают эфемеры и эфемероиды, особенно обильно растет живородящий мятлик, обычные тюльпаны, местами господствуют луки и. солянки разных видов. Оценивая запасы растительной массы биотопов сайгака, А.Г.Банников и др. (1961) называли цифры от 2 до 7 ц/га. Однако в настоящее время в связи с пастбищной детрадацией урожайность этих биотопов резко упала. По данным А.В.Максимука и др. (1987), в Яшкульском и Черноземельском районах, где сохранилась большая часть сайгаков, площадь сбитых пастбищ составляет 64—83% при средней урожайности естественных угодий всего 1,2—1,4 ц/га.
393
Сайгак
Характер биотопического размещения сайгаков широко меняется в течение года в зависимости от фенологических явлений в природе и биологии вида (Бакеев, Формозов, 1955; Даль и др., 1956, 1958; Жирнов 1959, 1982; Банников и др., 1961; Фадеев, Иванов, 1988; и др.). Видовой особенностью сайгаков, как и многих других стадных животных, является кочевой образ жизни. Лишь при таком использовании сухих степей и полупустынь возможно существование многотысячных стад сайгаков, равномерное освоение ими растительных ресурсов, не подрывающее естественное их возобновление. В связи с этим биотопическое размещение сайгаков и их динамику по сезонам года можно понять только при учете закономерностей митрации и местных кочевок.
Миграции. Среди сезонных миграций сайгаков наибольшей регулярностью, массовостью и направленностью отличаются весенние перемещения, связанные с деторождением. Еще с прошлого века регистрировались массовые перемещения сайгаков в северном направлении, где расположены "родильные дома". АТ.Банников и др. (1961) отмечали, что основные пути кочующих стад сайгаков в Центральной Калмыкии проходят по идеальным равнинам на отрезке от Халхуты до Яшкуля (близ Утты). Более восточные районы сайгаки избегают из-за бугристости рельефа и развеянных песков. Передвижение основной массы сайгаков приходится на апрель. С.К.Даль и др. (1958) в полупустыне северо-восточной части Ставрополя отмечали передвигающиеся стада сайгаков в центральные районы ареала в 20-х числах апреля 1955 г. Через 2—3 дня передвижения сайгаков приобрели массовый характер. Сайгаки шли в места размножения круглые сутки. А.Г.Банников и др. (1961), проводившие наблюдения в Калмыкии в 1957—1959 гг., первые перемещения стад сайгаков зарегистрировали в конце марта — начале апреля. По их мнению, массовые передвижения бывают в первой декаде апреля, а к середине месяца основная масса сайгаков уже находится в Сарпинской низменности, где расположены наиболее устойчивые "родильные дома". Сайгаки перемещались в основном рано утром и в первой половине дня. Во второй половине дня животные отдыхали, паслись. В последние 2—3 сут сайгаки двигались ночью, особенно в предрассветные часы. Скорость перемещения сайгаков меняется в разные года и в зависимости от погодных, кормовых условий, состояния животных. Пасущиеся стада передвигаются со скоростью 3—6 км/ч, вблизи проезжих дорог — около 20 км/ч. Зарегистрирована скорость перемещения в 40—50 км/ч (Банников и др., 1961).
При медленном движении сайгаки идут небольшими 1руппами, а при стремительном — большими скоплениями. В 1958 г., по данным названных авторов, скопление сайгаков содержало около 60—100 тыс. голов. В 1957— 1959 гг. сайгаки перемещались как небольшими труппами, так и скоплениями по нескольку тысяч голов. Весенние митрации завершаются выбором удобного места ягнения, каким является Сарпинская низменность. Это место отличается тем, что поверхность земли ровная, легко обозримая, по ней легко перемещаться не только взрослым животным, но и молодняку, имеются богатые злаковые пастбища, а также водопои.
394
Saiga tatarica Linnaeus, 1766
"Родильные дома" сайгаки покидают через 2—3 нед. По данным АТ.Бан-никова и др. (1961), начало массовой кочевки самок с молодняком на Сар-пинской низменности зарегистрировано 10—15 мая. В 1957 и 1958 гг. животные откочевывали на юг и юго-запад, а в 1959 г. на запад. Двигались медленно, со скоростью 5—10 км/ч, нередко большими скоплениями (150—200 тыс. особей; 1957 г., Центральная Калмыкия). Преодолев до 250 км к концу мая, сайгаки тогда заселили Приманычские степи, Ергени и частично Сальские степи. В засушливые 1957 и 1959 гг. отмечалось расширение ареала в западном и юго-западном направлениях, где имелись лучшие условия водопоя и питания. В дождливые годы, например 1958 г., таких больших летних перемещений не бывает. Основное поголовье остается в пределах Прикаспийской низменности. Только часть популяции перемещается в Кума-Манычские степи и на Ергеня. Словом, места летовок меняются широко, но в каждом случае наблюдается тяготение к более увлажненным районам. При этом, как отмечал Л.В.Жирнов (1977), обычно во всех географических районах отдельные группировки сайгаков имеют свои излюбленные места летних выпасов. Однако сайгаки в летний период чаще, чем в другие сезоны года меняют пастбища, и их можно встретить в разных местах. Не прослеживается также строгой локализации районов их летнего обитания по годам.
О дальности летних кочевок можно судить по результатам мечения. Так, в засушливый 1959 г. из 8 меток лишь 2 возвращены из Прикаспийской низменности, тогда как 6 меток относились к западным районам: Сальские степи, Манычская впадина и южные Ергени. Самым западным пунктом добычи меченых сайгаков является с. Киста Апанасенского района Ставропольского края (43° в.д.). Протяженность-от мест мечения до места добычи сайгаков 170—420 км (Банников и др., 1961).
С наступлением осени и соответственно относительно прохладной и влажной погоды сайгаки постепенно перемещаются в места зимовки, расположенные в полупустыне. Они занимают пастбища с прутняком, эфедрой, разными солянками. В связи с холодными ветрами и снегопадами в конце ноября — начале декабря сайгаки формируют многотысячные стада, которые перемещаются в районы зимовок — 'Черные земли", Кума-Манычские степи. В пределах районов зимовок сайгаки не остаются оседлыми. Они совершают местные (кормовые) кочевки в зависимости от доступности кормов, погодных условий. Нередко во время снегопадов сайгаки уходят на юг, а при таянии снегов во время оттепели возвращаются обратно в районы зимовок. Так, по данным А.Г.Банникова и др. (1961), с декабря по февраль 1958—89 г. сайгаки на юг перемещались четыре раза и трижды возвращались обратно. В этом периоде наблюдался максимальный размах осенне-зимних кочевок, составляющий около 350—450 км в меридиональном направлении. При сильных холодах как в декабре 1959 г„ сопровождающихся ветрами, сайгаки откочевывают далеко на юго-запад. Кочующие стада в то время преодолевали густонаселенные районы, пересеченный рельеф, за 3—4 дня прошли 120—150 км и достигли окрестности Пятигорска и Железноводска. В некоторых пунктах они подошли к Тереку (ст.
395
Сайгак
Червленная) и т.д. Дальние кочевки, как это имело место в 1959 г., отмечались и ранее (например, в 1953/54 г.). В настоящее время в связи с увеличением населенных пунктов, оросительно-обводнительных каналов, распашки земель осенне-зимние кочевки сайгаков на юго-запад осложнены. Более того, все чаще слышатся сообщения, что в ряде мест сайгаки вынуждены вести лишь местные кочевки и лишены возможности совершать исторически сложившиеся миграции и обречены на гибель (Максимук и ДР-, 1989).
В заключение отмстим, что сайгакам, как многим стадным животным, высокая подвижность и смена местообитания необходимы для нахождения достаточного количества корма в условиях аридных зон с двумя периодами покоя вегетации травянистой растительности. Именно благодаря подвижности сайгаки обеспечивают себя полноценным кормом, что нейтрализует, например, отрицательное влияние избытка бора в биогеохимической провинции Прикаспия. Отсутствие такой способности у домашних овец приводит к частому появлению у них летней пневмонии и гастроэнтеритов (Лихачев, 1967).
Убежища, лежки. Сайгаки не имеют специальных убежищ, однако в снежные бураны и метели они укрываются в высоких зарослях тростника, в прибрежных кустарниках и другой гшрофильной растительности. Сайгаки часто спасаются от холодных и сильных ветров в бугристых песках, иногда заходят в населенные пункты, фермы и другие места, где отрицательное действие холодных ветров несколько снижается.
Сайгаки устраивают лежки. Перед тем как лечь на отдых животное выбивает копытами землю, топчет ее и обнюхивает и лишь потом ложится. Видимо, сайгаки предпочитают устраивать такие лежки на сусликовинах, о чем говорят разрытые ими кучки земли рядом с норой. Такие лежки известны как "родовые ямки", но редко в таких ямках встречаются сайгачата. Подобные лежки сайгаки делают и на снегу. Диаметр летних лежек около 30 см.
Сезонная и суточная активность. Об изменении активности сайгаков по сезонам года можно судить по степени миграционных движений, а также местных кочевок. Что касается суточной активности, то судя по нашим полевым исследованиям и данным Н.Н.Бакеева и А.Н.Формозова (1955), АТ.Банникова с соавторами (1961) и др., она подвержена значительным сезонным колебаниям. Весной сайгаки активны с утра и в течение всего дня с небольшими перерывами пасутся весь день. Ночью отдыхают. Летом, когда повышается температура, суточная активность приобретает двухфазный характер: пасутся утром и вечером, а в жаркое время отдыхают. Чем выше дневная температура, тем больше сдвигается утренняя активность на ранние часы. В разгар лета сайгаки пасутся примерно до 8 ч утра, а после дневного отдыха возобновляют выпас с 17 ч. Зимой они активны в течение всего дня. Ночная активность отмечается во время гона.
Определенный отпечаток накладывают на суточную активность кочевки, совершаемые чаще ночью. Однако кочевки могут быть в любое время суток.
396
Saiga tatarica Linnaeus, 1766
Поведение (рис. 51). Формирование сайгаков как типичного вида открытых пространств аридной зоны наложило глубокий отпечаток на все стороны жизни, в особенности на поведение. Они отличаются способностью к быстрому бегу, развивая скорость до 70—80 км/ч, хорошим слухом и зрением, дополненные привычкой делать "смотровые прыжки", хорошо плавают, они способны образовывать большие стада, что создает высокую вариабельность структуры их популяции. Уместно вспомнить, что в больших стадах сайгаков могут выжить ослабленные, даже слепые животные.
В стаде сайгаков иерархия наиболее четко выражена во время размножения, когда наиболее сильный самец становится обладателем "гарема". Он направляет движение своих самок во время пастьбы, отдыха, защищает от покушения других самцов и т.д. Вне брачного периода у сайгаков нет общепризнанных вожаков-лидеров, хотя самцы пользуются определенными преимуществами перед самками. В частности, самка в любое время года мирно уступает свою лежку самцу, демонстрирующему путем скребущего движения передней ноги свое намерение занять готовую лежку. Роль лидера обычно выполняют нацболее сильные, чаще взрослые самцы, способные прокладывать маршрут движения остальных. За ним следует ядро стада, которое включает также сильных животных, а за ними идут другие. Этологический механизм поддержания стадности недостаточно изучен, но, видимо, зрительная и акустическая коммуникации играют очень важную роль. Очевидно, такая существенная черта поведения сайгаков, как "смотровые прыжки", служит сигналом тревоги и мобилизуют других сайгаков к активным действиям. Как правило, напуганный сайгак делает 2—3 прыжка и быстро уходит от опасности. За ним следуют другие, нередко повторяя прыжки, что обеспечивает эстафетную передачу сигнала тревоги.
О высоком уровне развития зрительной коммуникации можно судить по тому, как трудно человеку подойти незамеченным к пасущимся или отдыхающим сайгакам. При этом достаточно, чтобы один сайгак обнаружил приближение опасности, как это станет достоянием других. Впоследствии они держат в поле зрения противника, сохраняя определенную дистанцию. Очевидно, зрительная коммуникация поддерживает единство пасущегося стада сайгаков. Бегущий сайгак воспринимается как сигнал тревоги не только внутри своего стада, но бег целого стада воспринимается другим стадом как сигнал тревоги и соответственно оно тоже пускается в бег.
В процессе становления отношения мать-детеныш у сайгаков велика роль химической коммуникации. С первых же часов жизни малыша оба они познают запахи друг друга. Они обнюхивают друг друга, мать после рождения сайгачонка тщательно облизывает его. Без этих процедур мать не оставляет своего детеныша и не уходит на пастьбу.
В поведении сайгачонка характерна способность затаиваться при приближении опасности. А.Г.Банников и др. (1961) отмечают, что сайгачата затаиваются настолько крепко, что к ним можно подходить, брать в руки и они не шевелятся. Четырех-пятидневные малыши при опасности бегут за матерью некоторое время, но затем со всего разбега падают и затаиваются. Иногда даже трудно заметить место, где неожиданно "исчез" сайгачонок.
397
Сайгак
Рис. 51. Сайгак
398
Saiga tatarica Linnaeus, 1766
Ряд исследователей (Петрищев, 1982, 1983; Хахин, 1983) свидетельствуют, что в поведении сайгачат встречаются генетически закрепленные типы оборонительного поведения. Так, Г.В.Хахин (1983) выделяет 5 этологических групп, которые отражают разнообразие отношения детенышей к человеку после мечения. Нет сомнений, что среди новорожденных имеются и спокойные и беспокойные, а также отвергающие или не отвергающие контакты с человеком. Этологическая разнокачественность молодняка бесспорна, но этот вопрос нуждается в дальнейшей разработке.
Все сайгачата независимо от их принадлежности к той или иной этологической группе игривые, уже в первые недели жизни совершают "смотровые прыжки", резво бегают за матерью.
Линька. Наши полевые наблюдения и многие литературные данные свидетельствуют, что рассматриваемому виду свойственны две линьки в году — весенняя и осенняя. Продолжительность линьки довольно растянута. По АТ.Банникову и др. (1961), в Западном Прикаспии она составляет весной 20—30 дней, а осенью около 2,5 мес. В.Д.Рашек (1966) считает, что’ на о-ве Барсакельмес длительность периода весенней линьки сайгака достигает приблизительно 100—ПО дней.
В зависимости от погодных условий весенняя линька сайгака в Калмыкии начинается в первой—третьей декаде апреля и заканчивается в начале или середине мая (Банников и др., 1961). На о-ве Барсакельмес линяющие самки были добыты еще 20—25 марта (Рашек, 1966). В 1990 г. весенняя линька сайгаков была запоздалой. Все самцы и самки, изученные нами 19—20 мая этого года, были линяющими. Если учесть, что соотношения площадей тела с новыми и старыми волосами было примерно равными, то можно полагать, что эта смена волос закончилась не ранее первой декады июня. Осенняя линька начинается обычно в первой декаде октября и длится до конца ноября или начала декабря.
Первая линька молодых начинается с двухнедельного возраста и тянется до конца июля или начала августа. За это время ювенильные волосы, обычно волнистые, постепенно сменяются волосами, характерными для летнего наряда взрослых. Осенняя линька молодых, в том числе прошлогодних, проходит позже взрослых. По ВЛ.Рашеку (1966), у последних она запаздывает более чем на месяц. .
По данным А-Г.Банникова и др. (1961), а также ВЛ.Рашека (1966), последовательность смены волос на теле сайгака довольно сложная. В общих чертах при весенней линьке первыми линяют волосы на конце морды, вокруг глаз и губ, на внутренней поверхности дистальных частей конечностей. Затем линяет верхняя поверхность морды, лба и спины, а впоследствии линька распространяется на бока, шею, брюхо и проксимальные отделы конечностей. Осенняя линька также начинается на конце морды, затем на маклоках, над хвостом и на внутренней поверхности конечностей. После этого линяют передняя часть спины, бока и шея. Затем линька распространяется на брюхо и наружную поверхность ног (рис. 52).
Разница в последовательности смены волос у самок и самцов не отмечалась (Рашек, 1966).
399
Сайгак
Рис. 52. Схема последовательности весенней линьки у сайгаков 1 — 6— стадии линьки (Рашек, 1966)
Питание. Сведения о питании сайгаков в Западном Прикасапии содержатся в ряде работ (Адольф, 1954; Бансев, .Формозов, 1955; Банников и др., 1961; Жирнов, 1977, 1982; Абатуров, 1984; Абатуров и др., 1987; и др.).
Сайгак, по А.Г.Банникову и др. (1961), поедает 67 видов растений, что в несколько раз меньше соответствующих показателей питания косули, оленя, тура и т.д. Видимо, это объясняется не только относительным однообразием растительности аридных зон равнины по сравнению с горами, но и определенной избирательностью питания сайгаков. В пище этих животных представлены растения 25 семейств, из которых на долю злаковых приходится 12 видов, маревых — 10, крестоцветных — 7 и сложноцветных — 6 видов. В качественном составе кормов сайгака характерно наличие ядовитых растений (более 13%), которые не поедаются или плохо поедаются скотом. Так, по данным многих авторов, скот фактически не поедает ана-базис, щирицу, бурачки, касатик, додарцию, зопник, тиллея и некоторых других, когда как в желудочке сайгака они попадаются в разное время года и притом некоторые их них довольно часто. АТ.Банников и др. (1961) относят к основным кормам (14,4—73,2% встреч) 7 видов: прутняк (73,2%), черную и белую полыни (49,1%), эфедру (33,7%) лишайники (25%), ана-базис и птичий горец (по 21%), пырей (14,4%). Группу второстепенных, т.е. со встречаемостью 3,1—8,7%, составляют 13 видов. Среди них особо выделяются костер (8,7%), малая полевичка (8,3%), мятлик (7,9%), камфоросма (7,0%), курай (6,6%). Из редких растений (частота встреч 0,4—2,6%) чаще других отмечались белая марь, двувильчатая лапчатка, колючий зопник, молочай, керемск, мясистая солянка и 1рудница. В пищевом рационе сайгаков характерно также наличие рудеральных видов, поедаемых животными. Как справедливо отмечает Б.Д.Абатуров (1984), это экологическое свойство способствует разобщению на одних и тех же пастбищах и снижению конкуренции между сайгаками и домашними животными. Сайгаки, использующие большее количество видов, в том числе ядовитых и сорных, оказываются в более выгодном положении, чем домашний скот.
Наряду с широким спектром и избирательностью поедаемых видов растений для сайгаков характерна избирательность в поедании наиболее пита-
400
Saiga tatarica Linnaeus, 1766
Таблица 77. Общее значение различных групп кормов в питании сайгака ____________________(по: Банников и др., 1961)________________
Растение Частота встреч в желудках Объем массы растений
абсолютная %
Злаки 172 75,4 32
Маревые 175 76,7 19
Прутняк 167 73,2 30
Солянки 83 36,4 17
Эфедра 77 33,7 27
Полынь 112 49,1 12
Эфемеры 43 18,8 17
Разнотравье 93 40,8 13
Лишайники 47 25,0 8
тельных и сочных частей растений, таких, как бутоны соцветий, семена, плоды, корневища и луковицы (Абатуров, 1984). В Западном Казахстане летом содержимое желудка сайгака на 80% состояло из таких частей растений (Раков, 1956).
Для оценки значения различных ipynn кормов А.Г.Банников и др. (1961) использовали условный показатель "значение", представляющий собой производное процентов частоты встреч и объема данного растения в желудках (табл. 77). Судя по этой таблице, основу питания сайгаков составляют прутняк и солянки, эфедра, полыни, а также злаки.
При оценке предпочитаемости и значения тех или иных групп растений интерес представляют результаты сопоставления данных по питанию в природе и в неволе, проведенное Б.Д.Абатуровым (1984). Оказалось, что результаты не соответствуют друг другу. В частности, злаки, как отмечено выше, относятся в природе к труппе предпочитаемых. В неволе, где предоставляется широкий выбор кормов, злаки уступают другим видам и не могут быть отнесены к предпочитаемым. Сайгаки не брали сочных и нежных всходов посевного ячменя, предпочитая другие виды растений. Наоборот, лебеда, молочай, кермек, грудница и др., которые в природе поедаются редко, в неволе оказались для них привлекательными. Это позволило автору заключить, что фактическая поедаемость тех или иных видов в природе определяется не только привлекательностью, но и их обилием. К этому уместно добавить, что в экстремальных условиях сайгаки, оказавшись в барханах, поедают крупные злаки (Elymus) и кустарники, в том числе те-рескен, тамариск, лох и др., а по долинам рек — тростник, рогоз и др. (Банников и др., 1961).
Значение различных ipynn растений в питании сайгака меняется в широких пределах по сезонам года (рис. 53). Значение злаков в ранневесеннем периоде велико, однако оно снижается почти в 3 раза весной и летом, особенно резко падает осенью. В позднеосеннем питании (Банников и др., 1961) злаков почти в 3 раза меньше, чем весной и летом, почти в 9 раз меньше, чем ранней весной (в марте). Зимой значение злаков минимально. Летом резко возрастает роль разнотравья, а зимой — лишайников. Словом, в питании сайгаков наблюдается четкая смена кормов в течение года.
26 В.Е.Соколов, А.К.Темботов
401
Сайгак
Рис. 53. Смена значения различных кормов по сезонам: 1 — злаки; 2 — прутняк; 3 — солянки;
4 — эфедра; 5 — полынь; 6 — эфемеры; 7 — разнотравье; 8 — лишайники (Банников и др., 1961)
Суточная потребность в корме составляет около 11% от массы тела сайгака, т.е. в сутки съедается от 3 до 6 кг кормов (Жирнов, 1977). По Б.Д.Абатурову и др. (1982), суточное потребление одной особью составляет (в килограммах сухого веса): 0,7 — с декабря по март, 1,0 — в апреле и ноябре, 1,6 — с мая по октябрь, 0,3 — для молодняка с июня по авхуст.
Потребность в водопое сильно меняется по сезонам года. При наличии сочной растительности, например весной, когда влажность потребляемых растений высока, сайгаки могут не посещать водопой. Как известно, весной злаки и эфемеры, составляющие основу питания сайгаков, содержат до 90% влаги. Летом, когда эти растения усыхают (влажность их падает до 30—35%), сайгаки нуждаются в водопоях, хотя они переходят на питание прутняком, солянками и разнотравьем. Значение водопоев особо возрастает в сухие лета, которые периодически отмечаются 2—3 раза в каждые 5—10 лет. В такие засушливые годы сайгаки начинают посещать водопои с 10—15 мая и длится это до конца засухи (Жирнов, 1977).
Воспроизводство и структура популяции
Размножение. Сайгаки относятся к полигамам. Размножение у них начинается с формирования "гаремов", которое приходится на декабрь. Оно сопровождается ожесточенными драками взрослых половозрелых самцов, в результате которого наиболее сильный рогач отбивает себе от 2—3 до 25—26 самок (Банников и др., 1961). Такие драки могут кончиться гибелью одного из дерущихся самцов. Ни в драках, ни в размножении самцы моложе 1,5 лет участия не принимают. Они не проявляют признаков половой активности, столь характерных для взрослых с половой активностью самцов: увеличение семенников (с 20—20,6 до 35 г в среднем), функционирование подглазничных желез и выделение сильно пахнущего секрета, набухание хобота, отрастание шерсти под глазами (бакенбарды), удлинение шерсти по бокам шеи (грива), имеющей 1рязно-бурую окраску. Рогачей с половой активностью легко отличить по этой окраске как от самок, так и молодых самцов. В результате формирования "гаремов" стадо распадается
402
Saiga tatarica Linnaeus, 1766
на мелкие труппы, в которых могут находиться, кроме самок, и молодые неполовозрелые самцы. Обладатель гарема ревностно охраняет свой гарем от других половозрелых самцов, но безразличен к самцам-сеголеткам. Он направляет движение группы, не позволяет самкам отлучаться от труппы и т.д. Самцы в это время фактически не питаются, но часто едят снег, сильно худеют, после гона они настолько истощены, что нередко становятся жертвой хищников. Спаривание происходит ночью и основная масса самок оплодотворяется в течение примерно 10 дней (обычно в середине декабря).
Беременность длится около 5 мес. Для ягнения сайгаки, преодолевая значительные расстояния, концентрируются на небольших участках полупустыни, например в окрестностях Сарпинских озер. Такие участки обычно имеют ровную поверхность, нередко здесь растительность разрежена и скудна, расположены они вдали от водопоев и населенных пунктов. При этих условиях лучше обеспечивается защита взрослых и молодняка. Подобные места известны как "родильные дома" сайгаков, и действительно плотность животных здесь достигает 700—1000 особей на 1 км2, а вся площадь, занимаемая таким домом составляет около 10 тыс. км2. Количество молодняка также высоко — на 1 га до 15—20 сайгачат, в среднем 5—6 (Жирнов, 1977, 1982). Рождение молодняка приходится на апрель-май, что зависит от погодных и иных факторов периода спаривания. Массовый окот обычно проходит, как и гон и беременность, всего за 3—4 дня. Так, в 1955 г. массовое размножение происходило со 2 по 5 мая (Даль, 1958). В 1990 г., по нашим наблюдениям, оно было в середине мая.
Плодовитость. Видоспецифической особенностью сайгака является высокая репродуктивная способность, которая восполняет смертность при различных обстоятельствах. Она достигается различными механизмами. У самок старше года, как правило, бывает по два детеныша, а до года — по одному. Так, у вскрытых зимой 1958/59 г. в Калмыкии 293 самок старше года и 140 самок до года по два зародыша обнаружено у 73,8% старых и 5% молодых самок; соответственно 26,2% старых и 95% молодых самок имели по одному эмбриону. Иногда встречаются и тройни. Среди 14 тыс. новорожденных, осмотренных в 1957 и 1958 гг., в четырех случаях было обнаружено по три сайгачонка, лежавшие вместе (Банников и др., 1961). Характерно высокое участие самок в размножении — за исключением лет с чрезвычайно суровыми климатическими условиями, в размножение вовлекаются 90—100% половозрелых животных (Жирнов, 1982). Не менее существенно то, что самки становятся половозрелыми в полугодовалом возрасте (самцы на год позже).
Рост и развитие. Сайгачата рождаются хорошо сформированными. Масса тела новорожденного в среднем около 3,5 кг, длина тела 60 см. Самцы тяжелее на 200 г, чем самки, а их длина тела больше на 4 см, в холке они на 1,7 см выше.
Сайгачата растут и развиваются очень интенсивно. По Г.В. Хайну (1983), в Калмыкии зарегистрирован случай, когда сайгачонок через 3—5 мин после рождения пытался встать, а через 45 мин начал сосать мать. Спустя час он ушел с лежки на 15 мин, а потом вернулся, сделал смотровой
26
403
Сайгак
Рис. 54. Изменения длины тела (сплошная линия) и скорости роста (пунктир) с возрастом у сайгака (Банников и др., 1961)
прыжок и бегал вокруг матери. Через 3 ч после рождения он спокойно шел за пасущейся матерью. Об активном перемещении молодняка в первые часы постнатальной жизни говорят итоги мечения. Все родившиеся помеченные в утренние часы сайгачата уже в тот же день покидают место окота. Помеченные вечером— утром уже не бывают на месте рождения. По А.Г.Банникову и др. (1961), сайгачата в возрасте 5—6 дней могут бежать 27—30 мин со скоростью 30—35 км/ч. В десятидневном возрасте малыши не отстают от взрослых.
О темпе роста размеров тела можно судить по кривой скорости роста, представленной на рис. 54. Она очень высока в первые 3—4 мес, а затем резко замедляется. Что касается массы тела, то она также интенсивно увеличивается с возрастом. За месяц она растет с 3 кг в среднем у новорожденного до 6—9 кг. В двухмесячном возрасте масса тела увеличивается в 2,5—3 раза, а к 6 мес она уже составляет в среднем 19,5 кг у самцов и 18,1 кг у самок (Присяжнюк, Проняев, 1988). Рост самок прекращается к 20 мес, а самцов — к двум годам.
Самцы в месячном возрасте обладают рожками черного цвета высотой примерно около 1 см. У полугодовалого они достигают около 10 см. Они сохраняют черный цвет. В возрасте 13—14 мес длина их 17—23 см, и они приобретает светлую окраску.
По А.Г.Банникову с соавторами (1961), в постнатальном онтогенезе сайгаков можно выделить три периода роста и развития.
Первый период, составляющий первые семь месяцев постнатальной жизни, соответственно падает на летне-осенний сезон, когда животные в силу высокой подвижности находят хорошую кормовую базу. В это время наблюдается наивысшая скорость увеличения размеров и массы тела.
. Второй период приходится на зимнее время, когда в природе бескормица, животные испытывают влияние неблагоприятных погодных условий и недостаток корма. У самцов этот период длится 3—3,5 мес, т.е. до весны, а у самок — 5,5—6,0 мес, что связано с беременностью и лактацией. Фактически в это время животные не растут.
Третий период у перезимовавших самцов начинается с весны (с апреля) и длится 7—8 мес (до ноября-декабря), а у самок — с лета (июнь-июль) и заканчивается в возрасте 18,5—19 мес (в ноябре-декабре).
404
Saiga tatarica Linnaeus, 1766
Таблица 78. Возрастной и половой состав стад сайгаков в декабре—январе и ____________ в мае—июне (Банников и др., 1961)___________________
Возрастная группа Пол Декабрь—январь (2828 экз.) Май—июнь следующего года (1223 экз.)
% соотношение полов % соотношение полов
Взрослые Самцы Самки 18,3 46,8 1:2,6 11,9 46,1 1:3,8
Молодые Самцы Самки 14,6 20,3 1:1,4 14,7 27,3 1:1,8
Максимальная продолжительность жизни самцов и самок в Западном Прикаспии составляет 9—10 лет, но основная масса живет до 4—5 лет (Банников и др., 1961).
Структура популяции. Все имеющиеся данные говорят о том, что сайгаки Северо-Западного Прикаспия представлены одной крупной популяцией, которая в отличие от казахстанской не образует территориальных подразделений. В этом отношении весьма убедительны географические карты Л.В.Жирнова (Банников и др., 1961), отражающие размещения сайгаков калмыцкой популяции по сезонам года и направление маршрутов кочевки в годы подъема численности (1957—1959 гг.). Эти данные и анализ результатов мечения позволили заключить, что в связи с номадным образом жизни и особенностями ландшафтного покрова полупустынь и сухих степей правобережья Волги регулярно происходят миграции сайгаков и перемешивания различных групп. Этому процессу способствует сезонная динамика стадности, в основе которой лежит взаимное тяготение особей вида. Типичной формой организации сайгаков является стадо среднее по величине — от 20 до 200 особей. Частота встреч таких стад не имеет резких сезонных колебаний (Банников и др., 1961). Объединение средних стад приводит к образованию более крупных, в том числе скоплений. Скопления особенно характерны в период размножения на тех участках, где происходит ягнение ("родильные дома”). Крупные и очень крупные стада образуются при кочевках зимой, весной и осенью. Наоборот, в период гона (декабрь) и летом, когда пастбища выгорают, характерно образование мелких стад, что также носит адаптивный характер.
Если оценить весь спектр изменчивости количества особей в стаде, то ее размах составит 1—2 порядка величин.
Возрастной состав популяции сайгака в Северо-Западном Прикаспии характеризуется высокой лабильностью, что зависит от многих факторов, в том числе от факторов среды, эпизоотии, наличия хищников и т.д. Так, засуха, периодически повторяющаяся в Прикаспии, уносит за лето до 80% молодняка. В благоприятные годы смертность за весь год составляет 61% (Банников и др., 1977; Жирнов, 1982; и др.). Соответственно меняется возрастная структура популяции. Массовый отход старых, особенно самцов, имеет место после гона. Как видно на табл. 78, в декабре-январе они соста-
405
Сайгак
Таблица 79. Гельмингофауна сайгака, интенсивность и экстенсивность инвазии в Калмыкии (по: Банников и др., 1961)
Вид гельминта Экстенсивность, % Интенсивность, экз. Локализация Авторы
Nematoda Trichostrongylus 5,8 62 Сычуг, тонкие Матевосян и др., 1959;
axci 8 127—600 КИШКИ Голубовский Ю.П.
Т. colubriformis 17,6 (370) 1-16 То же Матевосян и др., 1959;
Т. pr.obolurus 88,2 1-278 Слепая кишка, сы- Соколова, Лаврова, 1956;
Т. vitrinus 5,8 47 чуг, тонкие кишки Сычуг Матевосян и др., 1959 Матевосян и др., 1959;
Ostertagia 11,7 4—65 Сычуг, тонкие Соколова, Лаврова, 1956
circumcincta О. occidentalis 64,7 1-78 кишки Сычуг, тонкие и Соколова, Лаврова, 1956;
32 42-315 толстые кишки Матевосян и др., 1959;
Marshallagia 82,3 (59) 2-806 Сычуг, тонкие, Голубовский Ю.П. Матевосян и др., 1959;
marshalli 48 27-241 толстые и слепая Голубовский Ю.П.
M. dentispicul- 23,5 (82) 16-806 2-32 кишки Сычуг, тонкие Соколова, Лаврова, 1956 Матевосян и др., 1959;
aris Haemonchus 17,6 3-23 кишки Сычуг, тонкие Соколова, Лаврова, 1956;
contortus 32-232 кишки Матевосян и др., 1959;
Nematodirus 20 5,8 (52) 1 Тонкие, толстые и Голубовский Ю.П. Соколова, Лаврова, 1956;
abnormalis 20 3-24 (5) слепая кишки Матевосян и др., 1959;
N. oiratinus 17,6 5-349 Тонкие кишки Голубовский Ю.П. Матевосян и др., 1959
N. spathigcr 23,5 1-132 Тонкие кишки, Соколова, Лаврова, 1956;
Chabertia ovina 35,3 1-6 сычуг Толстые кишки Соколова, Боев, Бондырев, 1949; Матевосян и др., 1959 Соколова, Лаврова, 1956;
32 1-36 (5) Матевосян и др., 1959; Голубовский Ю.П.
вили 18,3% популяции, а в мае-июне их доля снизилась до 11,9%. Высокая смертность взрослых самцов отмечается во время джутов, приуроченных ко второй половине зимы. Ослабевшие во время гона самцы в это время гибнут в массе. Большой отход взрослых самцов к весне отмечается после суровой зимы. Тогда их доля в популяции падает до 3—5% (Банников и др., 1961).
406
Saiga tatarica Linnaeus, 1766
Таблица 79 (окончание)
Вид гельминта Экстенсивность, % Интенсивность, экз. Локализация Авторы
Oesophagosto- 11,7 1 То же Матевосян и др., 1959
mum Columbia-
num •
O. venulosum 5,8 4 То же
Nematodirella 5,8 667 Тонкие кишки
cameli — — Тонкие и толстые Соколова, Лаврова, 1956;
КИШКИ
N. longiasime- 16 9-32 Соколова, Боев,
spiculata Бовдырева, 1949;
Голубовский Ю.П.
N. gazelli 23,5 2-224 Тонкие кишки Матевосян и др., 1959;
Trichocephalus 41,2 1-3 Толстые и слепая Матевосян и др., 1959;
ovis КИШКИ
T. skrjabini 48 1-4 То же Голубовский Ю.П.
47,0 1-7 Соколова, Лаврова, 1956;
56,0 1-8 Соколова, Боев,
Бовдырева, 1949
— — Тонкая, слепая и Матевосян и др., 1959;
Skrjabinema ovis 84 47—4000 толстая кишки,
Cestoda (193) сычуг Голубовский Ю.П.
Moniezia 1-7 Толстые кишки Соколова, Лаврова, 1956;
exp ansa 41,2 1-3 Матевосян и др., 1959;
28 Голубовский Ю.П.
Thysaniczia 1-2 — Матевосян и др., 1959;
giardi 17,6 1 Голубовский Ю.П.
Taenia hydotige- 8 1-2 Полость тела Соколова, Лаврова, 1956;
na (Cystocercus 11,7 2-10 Печень, сальник, Матевосян и др., 1959
tennuicollis) 28 брыжжейка
Multiceps
multiccps
(Coenutus 5,8 1 Мозг Матевосян и др., 1959;
cerebralis) 8 1 - Голубовский Ю.П.
Результаты многочисленных исследований свидетельствуют, что у сайгаков Северо-Западного Прикаспия соотношение полов при рождении составляет 1:1. Но уже к 8—9-му мес жизни молодняка доля самок возрастает и это соотношение становится 1:1,4. Среди взрослых преобладание самок еще выше — в декабре-январе соотношение полов 1:2,6, а в мае-июне еще более увеличивается количество самок в популяции (1:3,8). Как отмечено выше, сдвиги в соотношении полов объясняются избирательной элиминацией самцов. При полигамии сайгаков эта диспропорция не сказывается отрицательно на воспроизводстве популяции. В "гаремах" отмечалось от 2 до 26 самок, в среднем 4—6 самок.
407
Сайгак
Высокая плодовитость и соответственно высокая смертность лежит в основе быстрого (за 3—4 года) обновления состава популяции сайгака Северо-Западного Прикаспия (Жирнов, 1977). Это позволило названному автору считать сайгака "эфемером" среди копытных аридных зон Евразии.
Враги, паразиты, конкуренты, смертность. У сайгака много врагов, но основным считается волк. Н.Я.Динник (1910) писал, что волки "подкрадываются к ним против ветра и душат их, прежде чем сайгаки успевают вскочить с лежки" (с. 214). А.Г.Банников и др. (1961, с. 277) отмечают, что "одиночный волк не может догнать здорового сайгака, но самцов, истощенных за период гона, самок перед родами и новорожденных волки режут в очень большом количестве". Волки догоняют сайгаков при многоснежье, удачны охоты волков труппами.
В настоящее время роль волков сведена к минимуму в связи с уничтожением их почти на всей территории Северо-Западного Прикаспия. Однако вместо волков появились домашние собаки, в том числе бродячие, которые наносят урон сайгакам. Новорожденные могут страдать от лисицы, степного орла, беркута, ворона.
Паразитофауна сайгака разнообразна (табл. 79), однако наиболее ощутима роль гельминтов, зараженность которыми сайгаков очень велика. Они встречаются почти у каждого сайгака, что объясняется общностью гельминтов домашних животных и сайгаков (67—84,4%) и пастбищ (Банников и др., 1961; Жирнов, 1982 и др.). Особенно часто встречается ценуроз. Очень патогенен телязиоз, вызывающий слепоту. В прошлом сайгаки сильно страдали от кожного овода (Pallasiomyia antilopum), который в массе откладывал личинки, паразитирующие под кожей. Животные изнурялись из-за них. В 20-х годах, как отмечают названные выше авторы, их не стало в связи с резким снижением численности сайгаков.
Существенные конкуренты сайгаков в Северо-Западном Прикаспии домашние животные, но тем не менее совместное их существование реально и даже экологически оправданно (Дежкин, 1986).
Смертность. Как отмечено выше, сайгаки в большом количестве гибнут от стихийных бедствий. В литературе приводятся факты массовой гибели сайгаков Северо-Западного Прикаспия в суровые зимы 1948/49, 1950/51, 1953/54 гг. Значительное количество сайгаков, особенно молодняка и кормящих самок, гибнет летом в засушливые годы. Заметный урон наносят оросительные каналы. В 1977 г. в Яшкульском распределителе погибло более 14 тыс. сайгаков (Максимук и др., 1987).
Численность и се динамика. Сайгаку, как типичному представителю стадных копытных равнин свойственны большие колебания численности. Об этом свидетельствуют многочисленные источники прошлых и нынешнего столетий. Учитывая, что этот вопрос обсуждался рядом специалистов (см. обзоры: Банников и др., 1961; Жирнов, 1982; и др.), здесь мы ограничимся лишь немногими иллюстрациями общих закономерностей динамики численности.
Еще в середине прошлого века сайгак был обычным видом Северо-Западного Прикаспия, но к началу текущего столетия численность его резко
. 408
Saiga tatarica Linnaeus, 1766
Таблица 80. Плотность населения сайгаков в Западном Прикаспия ______ по данным авиаучетов (по: Банников и др., 1961)__________
Год Месяц учета Плотность на 1 км2 Численность, ТЫС. голов
limit х
1950 Август 1,3-7,4 3,4 100
1952 0,18-43,0 5,6 180
1954 Июль 0,9-3,2 1,8 100
1955 Сентябрь 0,77-8,0 4,8 270
1958 Апрель 0,2-14,0 7,5 540
1959 0,9-9,5 5,9 348
1960 Май 0,09-15,0 2,8 186
сократилась. Глубокая се депрессия приходится на 20-е годы. Основная причина — истребление сайгаков человеком. Благодаря охране и ряду других благоприятных обстоятельств сайгаки стали успешно размножаться и к 1940—1941 гг. достигли уровня середины XIX в. в ряде районов Северо-Западного Прикаспия. Через 7—8 лет они заняли территорию, которую они населяли в то время. Первый авиаучет был проведен в августе 1950 г. — общая численность составила около 100 тыс. голов (табл. 80). Через 2 года она увеличилась до 180 тыс. голов, т.е. за два периода размножения прирост стада составил 80 тыс. особей. Спад численности в 1954 г. до уровня 100 тыс. голов (уровень 1950 г.) был обусловлен суровой и многоснежной зимой 1953/54 г., вызвавшей массовую гибель сайгаков. Количество животных, погибших в ту зиму, оценивается в 150 тыс. голов. Достаточно сказать, что в Караногайской степи, куда откочевали сайгаки, в конторы Заготжив-сырья было сдано 66 719 шкур дохлых животных.
Потери сайгаки восстановили быстро, с 1954 по 1958 г. численность сайгаков увеличилась в 5 раз. Ранней весной учтено было 540 тыс. голов; если учесть молодняк, появившийся этой весной, то летом 1958 г. численность сайгаков Северо-Западного Прикаспия составляла не менее 800 тыс. голов (Банников и др., 1961). По мнению названных авторов, стремительный рост поголовья сайгаков в 1954—1958 гг. обусловлен в первую очередь благоприятными условиями зимовки в течение всего периода. Эпизоотия ящура, имевшая место летом 1957 и 1958 гг., не оказала значительного влияния на динамику численности. В последующие годы отмечалось прогрессирующее снижение численности, что было вызвано многими неблагоприятными факторами. Так, сухое лето 1959 г. унесло значительное количество молодняка, а в декабре наступило резкое похолодание с последующими снегопадами. Сайгаки вынуждены были мигрировать, как и в 1953/54 г., до южных районов Ставрополья, Чечено-Ингушетии и Дагестана. К неблагоприятным погодным условиям и малопригодным местам жизни прибавилось массовое браконьерство, приведшее к гибели сайгаков в большом количестве. В 60-х годах численность сайгаков сократилась до 180 тыс. голов, что явилось следствием неумеренного и неумелого промысла и браконьерства (Темя-нова, Любаев, 1986). После введения дополнительных мер охраны численность стала восстанавливаться, но уровня 50-х годов не достигла. В
409
Сайгак
настоящее время в связи с распашкой целинных и залежных земель, с вводом в строй ряда оросительных систем и дальнейшим расширением пахотных и орошаемых площадей исключается возможность восстановления былой численности и распространения сайгака в Северо-Западном При-каспии. Более того, несмотря на снижение промысловой нагрузки и запрет промысла в 1984 и 1986 гг., численность сайгаков в последние 5 лет не увеличивается (Максймук и др., 1989). Сейчас речь, видимо, должна идти о том, чтобы определить пути разумного сочетания сайгаководства и сельскохозяйственного производства в аридной зоне страны. Прав В.В.Дежкин (1986), который пишет: "Необходимо срочно провести дополнительные эколого-экономические исследования с целью уточнения путей и методов поддержания промысловой плотности и эксплуатации калмыцкой популяции сайгака в условиях продолжающейся интенсификации животноводства и выработки компромиссных методов ведения в интересах охраны природы и охотничьего хозяйства".
Практическое значение. Сайгак является одним из промысловых животных на территории бывшего СССР. Лицензионный отстрел в Северо-Западном Прикаспии начат в 1951 г., за 5 лет добыто около 200 тыс. голов. С 1956 г. начат промысел и в том же году добыто 106 тыс., в 1957 г. — 128 тыс., а в 1958 г. — 200 тыс. сайгаков. В последующие годы объем заготовок резко упал. Всего за период с 1951 по 1985 г. в пределах Северо-Западного Прикаспия заготовлено более 2 млн сайгаков, что оценивается более чем в 30 млн руб. (Максймук и др., 1987). По данным Л.В.Жирнова (1977), в этом районе стоимость всей продукции сайгачьего промысла за 1957—1967 гг. составила в среднем 2700 руб. на 100 км2, а в годы наиболее развитого промысла (1957—1959 гг.) она составила 3500—5300 руб. на 100 км2. Такая высокая рентабельность была даже при наличии существенных недостатков в промысле путем отстрела. По мнению П.В.Житенко (1986), рациональное использование продуктов охотничьего промысла даст возможность увеличить прибыль до 3,5—4 руб. на каждую голову.
Рога сайгаков высоко ценятся как лекарственное сырье. Они пользовались большим спросом в Китае и странах Юго-Восточной Азии. Наша отечественная фармацевтика всерьез обратила внимание на изучение этого оригинального лекарственного сырья лишь в последние десятилетия. Результаты исследования подтверждают высокую ценность рогов сайгаков, реальность производства из него высокоэффективных лекарственных препаратов (Размахин, 1986).
Сайгаки могут причинить некоторый ущерб полеводству, особенно в сухие годы. В это время от них страдают посевы пшеницы, кукурузы, проса, ячменя, люцерны.
На естественных пастбищах имеет место конкуренция между сайгаками и овцами, однако в силу высокой подвижности сайгаков пастбищная нагрузка от них невелика. Достаточно сказать, что средние величины изъятия не превышают 5% урожая надземной фитомассы в год (Абатуров и др., 1987). К этому надо добавить, что значительную часть кормов сайгака овцы не употребляют.
410
Gazclla Blalnvillc, 1816
Что касается эпизоотического значения, то сайгаки не являются первопричиной вспышки ящура или бруцеллезной инфекции. Однако во время кочевок сайгаки могут распространять заболевания домашних животных.
Выше отмечено, что коэффициент общности гельминтов сельскохозяйственных животных и сайгаков высок, однако и здесь первоисточником являются домашние животные. При содержании сайгаков в загонах, исключающих контакты с овцами и другими домашними копытными, сайгаки полностью избавляются от паразитов (Звягинцев, Треус, 1986).
Род Газели — Gazclla Blainville, 1816
Антилопы средних и мелких размеров. По В.Е.Соколову (1979), длина тела 85—170 см, хвоста — 15—30 см, высота в холке 50—110 см. Крестец немного выше холки. Масса тела 12—85 кг. Самцы крупнее самок. Телосложение легкое и изящное. Ноги тонкие и высокие. Шея относительно длинная, а голова небольшая. Уши заостренные и сравнительно короткие. Глаза крупные и красивые. Между ноздрями небольшой голый участок кожи. Хвост средней длины, покрыт волосами, за исключением внутренней части основания^ на конце кисточка из удлиненных волос. II и V пальцы развиты нормально, средние копыта небольшие и заостренные. Рога у самцов и самок (за исключением джейрана) имеют лировидную форму, покрыты волнообразно изогнутыми кольцами, лишь верхушечная часть рогов гладкая. В поперечном сечении овальные или округлые.
Тело покрыто короткими и густыми волосами, но с небольшим количеством пуха. Спина и бока светло-желтые или коричневые, а низ тела, шеи и внутренние стороны ног белые. Окраска верха тела иногда неоднородна. По бокам головы, от уха до кончика морды, тянутся светлые или темные полосы. Хорошо развито белее околохвостовое "зеркало", которое обычно распространяется выше основания хвоста. Окраска волосяного покрова слабо различается по полу и возрасту. Характерно высокое развитие специфических кожных желез: пред глазничные, паховые, запястные, межпальцевые. Сосков одна пара.
В черепе хорошо развита мозговая часть, составляющая 1/3—1/2 длины лицевого отдела. Лобная поверхность между глазницами сильно вдавлена. В слезных костях расположены крупные ямки для предглазничной железы. Имеются этмоидальные отверстия. По В.Г.Гептнеру и др. (1961), характерно, что лицевая часть не удлинена и не увеличена, профиль лобно-носовой кости относительно прямой, межчелюстные кости обычно не соприкасаются с носовыми. Последние имеют вырезку на переднем крае. Щечные зубы брахиодонтные.
Зубная формула
I-C-P - М - = 32. 3 13 3
В роде диплоидное число хромосом варьирует от 30 до 58.
4 1 1
Джейран
Ареал рода обширный, он включает Северную Африку, северную часть Восточной Африки, весь Аравийский полуостров, Иран, Восточное Закавказье, Среднюю Азию, Афганистан, Монголию, Внутреннюю Монголию и Северный Тибет — в Китае, Северо-Западную, Северную и Среднюю Индию (Гептнер и др., 1961; Соколов, 1979). Ареал рода большей частью лежит в пределах аридных зон. Многие из газелей являются типичными представителями пустынь, полупустынь и лесостепей. Иногда проникают в горы до 2—3,5 тыс. м над уровнем моря.
Антропогенные изменения ареала рода и отдельных видов очень значительны, что обусловлено сельскохозяйственным и животноводческим освоением целинных и залежных земель, браконьерством с широким использованием современного транспорта и нарезного оружия. Для большей части видов сельхозугодия неблагоприятны,и они вынуждены покидать их. В итоге ряд видов стали редкими, оказались под угрозой исчезновения и занесены в "Красную книгу" (Соколов, 1979, 1986).
Газели встречаются поодиночке, парами, семейными группами, а в осенне-зимнее время объединяются в крупные стада, включающие сотни и тысячи голов. В ряде мест совершают миграции. При высыхании растительности нуждаются в водопоях. Активны летом, как правило утром и вечером, а зимой — в течение светлого периода суток. Питаются травянистой растительностью. Размножение в умеренных широтах и субтропиках приурочено к концу осени — началу зимы, а в тропиках сильно растянуто. В период гона самец занимает сравнительно небольшую территорию и активно ее охраняет. Продолжительность беременности 5—7 мес, количество детенышей 1—2, редко 3, совсем редко 4. Лактация длится около 6 мес. Половозрелость наступает в 1—1,5 года. Мелкие виды живут до 6—8, а крупные до 10—12 лет.
Газели относятся к ценным охотничьим животным, в недавнем прошлом добывались в большом количестве. В настоящее время в большинстве мест охота на газелей запрещена.
По современным данным в роде 12—13 видов, из которых на Кавказе и в б. СССР в целом один вид — джейран (Gazella subgutturosa Guld.).
Джейран — Gazella subgutturosa Giildenstaedt, 1780 t Рис. 55
Antilope subgutturosa, Guldenstaedt, 1778; Antilope dorcas,
Gray, 1843
Коллекционный материал. ЗИН РАН: 6 черепов, 6 рогов, 1 скелет; МГУ: 2 черепа.
Диагноз. Размеры средние, длина тела до 116 см, наибольшая длина черепа до 224 мм, длина хвоста более 12 см. Лировидные рога черного цвета, белое околохвостовое "зеркало" не заходит выше основания хвоста. Носовые кости с вырезкой, на переднем крае Р4 имеет открытую переднюю долинку.
412
Gazclla subgutturosa Guldcnstacdt, 1780
Особенности морфологии и морфофизиологии
Размеры тела, черепа в отдельных элементов посткраниального скелета. Масса тела самцов 23—33 кг, самок 18—20 кг. Длина тела соответственно 110—116 и 93—110 см. Высота в холке самцов 64—75 см, самок 60—68 см, хвост 16—20 см, ухо 8—13 см (Гептнер и др., 1961). Туловище относительно короткое, ноги тонкие и длинные, копыта узкие и маленькие, что отражает в целом адаптированность к жизни в пустыне с твердым субстратом. Шея тонкая и длинная, на ее нижней стороне у самцов и самок ясно выражена гортань. Голова относительно маленькая, но глаза большие, выпуклые, черного цвета.
Рога в типичном виде свойственны только самцам, имеют длину 27—41 см, четко лировидные. Располагаясь почти над глазами, они у основания сближены, примерно до середины слабо расходятся и загибаются назад, но потом изгибаются в стороны, а концы сближаются и приподнимаются вверх. За исключением этой приподнятой концевой части, рога покрыты волнистыми кольцами в количестве до 26—28 шт. Как исключение у самок могут быть небольшие рожки длиной до 5 см.
Длина черепа небольшая: 172—224 мм у самцов, 182—208 мм у самок. В структуре черепа, по И.И.Соколову (1959), характерна недлинная предглаз-ничная часть, узкий вход в носовую полость, сильное развитие предглаз-ничных ямок, а также относительно короткие и широкие носовые кости с вырезкой на всю ширину и сильно вздутые слуховые пузыри. По мнению В.Г.Гептнера и др. (1961), в строении черепа джейрана типичны последние два признака. На наш взгляд, показательно мощное развитие слуховых капсул, отражающее, как и особенности строения конечностей, длительную и тонкую адаптацию к жизни в открытых пространствах пустынь.
Зубная система мало оригинальна. Наиболее видоспецифичной особенностью выделяется структура последнего предкоренного зуба нижней челюсти (Р4). Он имеет хорошо развитый лишь передне-наружный бугорок зуба. В связи с этим передняя долинка на внутренней его стороне остается открытой (Соколов, 1959).
В посткраниальном скелете, -как справедливо указывает Н.К.Верещагин (1939), весьма показательно максимальное увеличение длины метатарзальных и метакарпальных костей при уменьшении их толщины.
Окраска тела. Многие специалисты обоснованно обращали внимание на устойчивость и видоспецифичность окраски джейрана (Динник, 1910; Верещагин, 1939; Соколов, 1959; Гептнер и др., 1961; и др.). Те внутрипопу-ляционные сдвиги в окраске тела, которые наблюдаются в связи с возрастом и сезоном года, в основном определяются полосатостью и степенью изношенности отдельных волос. На верхней стороне тела окрашенный волос может иметь до 4 полос: у основания светло-бурая, нередко голубоватая, затем идет более темная полоса, которая сменяется выше желтой, а самый верхний кончик почти черного цвета. В зависимости от сохранности ломкой верхушечной части волос общая окраска верха тела рыже-
413
Джейран
414
Gazella subgutturosa Guldenstaedt, 1780
Рис. 55. Распространение джейрана
1 — окр. Тбилиси (Верещагин, 1959; ЗИН);
2 — Арарат (Динник, 1910); 3 — Вашлованский заповедник (Соколов, Сыроечковский, 1990); 4 — Владикавказ (Алекперов, 1977); 5 — Мингечаури (Алекперов, 1977); 6 — Евлах (Динник, 1910); 7 — Агдаш (Верещагин, 1947, 1959); 8 — Агджабсди (Верещагин, 1947, 1959); 9 — Геворкован (ЗИН); 10 — Физули (Алекперов, 1966); 11 — Мартуни (Алекперов, 1966); 12 — Имишли (Алекперов, 1977); 13 — Кюрдамир (Алекперов, 1977); 14 — Апшеронский п-ов (Динник, 1910, Верещагин, 1959); 15— о-в Булла (Сафаров, 1961); 16 — Бяцдованский заказник (Хагин, 1986); 17 — Ширванский заповедник (Соколов, Сыроечковский, 1990); 18 — Пушкино (Сатунин, 1909); 19 — устье р. Куры (Верещагин, 1959); 20 — Кызыл-Агачский заповедник (МГУ);
21 — Ленкорань (ЗИН)
вато-бурая, песочно-бурая, иногда рыжевато-серая, напоминающая, по Н.Я.Диннику (1910), зимнюю окраску косули. На нижней стороне тела окраска светлая благодаря белым волосам, покрывающим ее. На боках, где сменяется окраска верха на светлую низа, расположена полоса из темных волос. Аналогичная полоса окружает околохвостовое "зеркало", которое в отличие от ряда других видов не заходит выше основания хвоста. На этом фоне хорошо выделяется хвост с удлиненными волосами бурого или черно-бурого цвета. Окраска головы неоднородна. Здесь довольно сложно сочетаются участки с различной окраской: кончик и низ морды белые; темя и средняя часть лба темные, буроватые; щеки рыжевато-бурые; рыжеватая полоса тянется по середине
415
Джейран
Таблица 81, Промеры кожи джейрана
Показатель Толщина
КОЖИ, мм эпидермиса, мкм рогового слоя, мкм дермы, мм СОСОЧКОВОГО СЛОЯ, мм
Загривок № 1, зима 1,35 34 11 1,32 0,66
№ 2 1,46 39 22 1,43 0,77
№ 3 1,35 34 20 1,32 0,66
№ 4, лето 4,12 28 6 4,10 2,10
Крестец № 2, зима 1,13 33 20 1,10 0,66
№ 4, лето 1,56 28 11 1,54 1,10
№ 5 1,87 17 5 1,86 1,20
Грудь № 1, зима 1,07 34 17 1,04 0,66
№ 2 1,26 61 44 1,20 0,82
№ 3 1,62 _ 84 62 1,54 0,88
№ 4,лето 3,00 56 39 2,95 1,65
№ 5 — — — 1,90 1,10
Показатель Толщина сетчатого слоя, мм Диаметр мышц, поднимающих волосы, мк Размеры сальных желез, мкм Диаметр потовых желез, мкм Сумма диаметров потовых желез на протяжении 1 см препарата, мм
Загривок
№ 1, зима 0,66 55 22 х 121 88 2,7
№ 2 0,66 60 39 х 123 90 2,6
№ 3 0,66 50 44 х 140 100 2,4
№ 4, лето 2,00 НО 80 х 330 187 2,4
Крестец
№ 2, зима 0,44 44 39 х 140 88 1,8
№ 4, лето 0,44 60 66 х 330 120 3,4
№ 5 0,66 44 55 х 165 130 3,3
Грудь
№ 1, зима 0,38 40 56 х 145 90 2,6
№ 2 0,38 45 50 х 224 100 2,0
№ 3 0,66 40 55 х 275 121 1,6
№ 4, лето 1,30 40 120 х 330 ПО 2,2
№ 5 0,80 35 — НО —
носа, а темные — от предглазничных ямок к углам рта. Старые особи теряют описанный рисунок, и вся передняя часть головы становится почти белой. Передняя и наружная стороны ног в тон с верхом тела, но в области расположения выходов межпальцевых желез (над копытами), карпальных желез (ниже запястных суставов), а также главными и боковыми копытами, окраска сгущается, становится темнее. Обычно в перечисленных местах, как в области предглазничных желез, волосы грубее и длиннее,чем вокруг них.
416
Gazclla de Blainvillc, 1816
Рис. 56. Строение кожи загривка джейрана
1 — эпидермис; 2 — дерма; 3 — сальные железы; 4 — потовые, ув. 32 х 10
Кожный покров. Материал. Кожа загривка, груди и крестца (табл. 81) 5 джейранов, добытых в январе; № 1 — самка, длина тела 104 см; № 2 — самец, длина тела 101 см; № 3 — самец, длина тела — 10 см; в августе; № 4 — самец, длина тела 120 см и № 5 — самка, длина тела 113 см.
Летом толщина кожи джейрана больше на загривке и груди и меньше на крестце. Эпидермис и роговой слой наибольшей толщины достигают на груди. Роговой слой плотный. Эпидермис не пигментирован. В сосочковом слое дермы располагаются огромные корни растущих волос. Пучки коллагеновых волокон в самых верхних отделах сосочкового слоя образуют довольно плотное переплетение, проходя в различных направлениях. В средних и глубоких отделах они располагаются преимущественно вдоль корней волос. Эластиновые волокна многочисленны. Они залегают параллельно поверхности кожи. В сетчатом слое дермы преобладают горизонтальные пучки коллагеновых волокон, образующие средней плотности вязь. Эластиновые волокна в этом слое редки. Только по нижней границе сетчатого слоя идет узкая зона (примерно 80 мкм/толщиной), богатая сплетениями эластиновых волокон. В сетчатом слое кожи крестца встречаются группы жировых клеток (28 х 40, 34х 45 мкм). В коже груди в верхних отделах сетчатого слоя коллагеновые пучки проходят преимущественно в различных восходящих направлениях. Подкожная жировая клетчатка толщиной 0,5 мм имеется только в коже груди джейрана № 2. Размеры жировых клеток в ней 50 х 90, 70х 80 мкм.
27 В.Е.Соколов, А.К.Темботов
417
Джейран
Рис. 57» Строение остевых волос загривка джейрана
Наиболее крупные сальные железы находятся в коже груди. Потовые железы имеют преимущественно мешковидные, слегка извитые секреторные отделы (рис. 56), по Колесникову (1948), они ампулообразные, как правило, без изгибов. Длинная ось секреторных отделов в большинстве случаев проходит параллельно волосяным сумкам. В коже труди потовые железы образуют небольшие извивы, доходящие до волосяных луковиц. На крестце потовые железы наиболее многочисленны.
Волосы загривка подразделяются на три порядка одной категории остей; их длина 11,7; 6,6 и 3,4 мм; толщина 124; 90 и 47 мкм. По форме они одинаковые, отличаются только размерами. Наиболее толстое место у волос находится в нижней трети. У волос всех трех порядков хорошо развит сердцевинный слой (у остей первого порядка 85,0%). Стержни волос дугообразно загнуты назад; извилин они не имеют. Пучков или групп волос нет. Волосы растут слабо намеченными рядами, проходящими поперек тела. Ряды могут переходить один в другой или прерываться. Остевые волосы первого порядка наиболее многочисленны. Форма их поперечника плосковыпуклая, причем выпуклая сторона обращена к голове. В боковых отделах
418
Gazella de Blainville, 1816
корковый слой незначительно толще, чем в передне-задних. Поперек волоса насчитывается 2—4 кутикулярные клетки высотой 15 мкм. Очертания кутикулярных клеток остей второго и третьего порядков такие же, как и у остевых волос первого порядка. Высота их клеток 9 мкм. На 1 см2 поверхности кожи насчитывается 3499 волос всех порядков (рис. 57).
Волосы груди на категории и порядки не подразделяются. Все они сходны между собой, за исключением того, что часть из них растущие, без ножки, а другие — выросшие с ножкой. Их длина 6,9 мм, толщина 149 мкм. К вершине и основанию волосы утоньшаются. Наибольшая их толщина в нижней трети или половине стержня.
Сердцевина (84,8%) проходит по всему волосу, за исключением основания (у нерастущих волос) и вершины. В боковых частях волоса корковый слой немного толще, чем в передних и задних. Форма поперечника волоса близка к овалу с почти уплощенной (иногда слегка вогнутой) одной из сторон. Групп или пучков волосы не образуют. На 1 см2 поверхности кожи насчитывается в среднем 1333 выросших и 500 растущих волос.
Зимой мальпигиев слой эпидермиса 2—3-зарядный. Роговой слой слаборыхлый. Эпидермис не пигментирован. В дерме загривка сосочковый слой одинаковой толщины с сетчатым или немного толще его. В коже крестца и груди толщина сосочкового слоя больше сетчатого. Строение дермы такое же, как у летних зверей. Подкожная жировая клетчатка толщиной 0,75 мм обнаружена лишь в коже груди джейрана № 3. Кожные железы такие же, как летом.
Запястные железы были изучены у джейранов № 2 и 3 (оба самцы), добытых в январе, № 4 (самец) и № 5 (самка), добытых в августе. Сезонной и половой изменчивости не обнаружено. Снаружи железы вьщеляются более длинными и грубыми волосами темного цвета, склеенными у основания секретом желез. Кожа в местах этих образований утолщяется до 2,9 мм по сравнению с 1,9 мм в соседних участках. Утолщение тянется вдоль верхнего отдела запястья на 4,5—5 см и поперек на 2,2—3 см. По соседству с железой луковицы волос идут в глубину максимум до 0,8 мм от поверхности кожи; мышцы, поднимающие волосы, имеют в поперечнике 60 мкм; размеры сальных желез до ПО х 330 мкм; потовые железы невелики. В железе можно выделить две зоны: центральную и краевую. В центральной зоне луковицы волос находятся на глубине 2,2 мм от поверхности кожи. Вся ткань кожи почти нацело вытеснена охромными многодольчатыми сальными железами. Они простираются почти до подкожной мускулатуры; их размеры 1,0х 2,4 мм. В краевой зоне корни волос короче; сальные железы меньшей величины (0,9 х1,6 мм); трубчатые железы образуют клубки, но они немногочисленны; мышцы, поднимающие волосы, очень большие — в поперечнике до 350 мкм (рис. 58).
Паховая железа. В паховой области у самцов и самок расположены своеобразные кожные карманы. Н.Я.Динник (1910) ошибочно указывает, что они имеются только у самцов. У самки № 4, добытой 7 августа, размеры карманов были 5 см- в глубину и 3 см в поперечнике, у самца № 2 — 3,4 х2,6 см (выводное отверстие кармана 7 мм). Сезонных и половых 27*
419
Джейран
Рис. 58i Строение волос запястной железы джейрана (А—Г)
различий не установлено. Внутренняя поверхность паховых карманов покрыта редкими волосами. В волосяные влагалища открываются очень крупные многодольчатые (12—15 долек) сальные железы. Размеры желез возрастают от краев карманов к средним частям, а затем вновь уменьшаются на "дне" карманов, где они вытесняются потовыми железами. Наиболее крупные размеры сальных желез — ,660х 900, 660 xllOO мкм. В средних частях карманов сальные железы примыкают друг к другу, образуя сплошной железистый слой. Трубчатые железы, относительно слабо развитые в краевых частях карманов, достигают необычайно мощного развития на дне карманов. Здесь они образуют гигантские сильно завитые клубки размером 900 Х1300 мкм. Соседние клубки плотно примыкают друг к другу. Диаметр секреторных отделов некоторых желез достигает 70 мкм.
Ближе к средней линии от внутреннего угла глаза расположено небольшое кожное впячивание — предглазничная железа размером 0,8x1,5 см (самка № 5, добытая 7 августа); 2,4 х 2,9 см (самец № 4, добытый 2 августа); 1 х2 см (самец № 2, добытый 23 января). Внутренняя поверхность впячивания покрыта редкими волосами, по большей части склеенными густым выделением желез. Они образуют крупные сильно извитые клубки.
420
Gazclla de Blainville, IS 16
Зимой диаметр секреторных отделов достигает 110 мкм. Летом размеры трубчатых желез несколько возрастают и диаметр секреторных отделов увеличивается до 130 мкм. Протоки желез открываются в волосяные воронки. Сальные железы огромные. Каждая из них имеет своеобразное древовидное строение и представлена многочисленными протоками в общий секреторный выводной проток, который, в свою очередь, открывается в волосяное влагалище. Размеры сальных желез примерно 1,5x2,2 мм. Летом величина сальных желез немного возрастает.
Передние межпальцевыс железы представлены тонкостенными кожными мешками длиной 1,5—2,4 см и шириной 1,5—2,0 см. Внутренняя поверхность мешков покрыта редкими волосами. В волосяные влагалища открываются некрупные сальные железы, состоящие из нескольких долек общим размером у летнего джейрана до 165 хЗЗО мкм, и. трубчатые железы, извитые в небольшие клубки с диаметром секреторных отделов до 60 мкм. Никаких крупных железистых скоплений, подобных, например, предглаз-ничным, здесь нет. Зимой у джейрана размеры сальных желез увеличены до 400 х500 мкм, а величина трубчатых желез уменьшается (диаметр секреторных отделов 45 мкм).
Задние межпальцевые железы длиной 1,5—2,1 см и шириной 2,0—2,6 см, отличаются от передних только размерами. Сальные железы в них у джейрана летом имеют более крупные размеры, чем в коже передних межпальцевых органов (220—550 мкм), трубчатые железы образуют клубки размером до 400x550 мкм с диаметром секреторных отделов до 60 мкм. У джейрана зимой размеры сальных желез 330x550 мкм, клубков трубчатых желез — 250x660 мкм, диаметр секреторных отделов трубчатых желез достигает 60 мкм.
Мышцы, поднимающие волосы, хорошо развиты. Они начинаются почти от самой луковицы и идут наискось к верхней части волосяной сумки сзади стоящего волоса. Среди волос загривка можно выделить всего одну категорию остевых волос четырех размерных порядков. Их длина 38,8; 22,9; 12,3 и 3,5 мм, толщина 128; 81; 41 и 28 мкм. Групп и пучков волосы не образуют. Волосы растут слабо намеченными рядами. У всех хорошо развиты сердцевинные слои (у остей первого порядка 88,7%). Форма кутикулярных клеток у волос всех порядков сходна между собой. Остевые волосы первого порядка имеют, как правило, незначительное расширение в верхней части стержня. В поперечнике они бобовидной формы — выпуклые спереди и слегка вогнутые сзади. Сердцевина проходит по всему волосу, за исключением вершины и ножки. Поперек волоса насчитывается от 2 до 4 клеток кутикулы. Форма остевых волос второго порядка такая же, как и первого. Поперек волоса располагаются от 2 до 4, а у остей третьего порядка от 2 до 3 кутикулярных клеток. Общее число всех волос на 1 см2 поверхности кожи достигает 6000.
Среди волос груди можно выделить также лишь одну категорию остей, представленную тремя порядками. Их длина 43,9; 24,8 и 23 мм, толщина 279; 153 и 42 мкм. Волосы утоньшены в основании и к вершине и расширены в средней части. Сердцевина хорошо развита у всех волос (у остей первого
42 1
Джейран
порядка 91,1%). Форма поперечника волоса бобовидная с вогнутостью на каудальной стороне. Боковые отделы коркового слоя толще, чем передние и задние. Общее число волос на 1 см2 поверхности кожи достигает 2333.
Система крови. Сведения о крови джейранов содержатся в работе Н.Н.Булатовой (1953), согласно которой количество эритроцитов у самцов составляет 12,50 млн, а у самок 13,90 млн. Сопоставимое количество эритроцитов на Кавказе имеют лишь архар, кавказский тур, косуля. Объем эритроцита джейрана составляет 29,5 мкм3, что несколько больше, чем у кавказского тура (20,9 мкм3), но меньше, чем у косули (32,5 мкм3) и архара (31,5 мкм3). Показатели гематокрита: 38,7% у джейрана, 42,4% у архара, 37,8% у козла винторогого — рекордсмена среди позвоночных по количеству эритроцитов (25 млн). Что касается концентрации гемоглобина, то джейран по этому показателю мало отличается от названных выше копытных: у архара и винторогого козла он составляет 17,1, а у джейрана — 16,6%.
Несмотря на отрывочность приведенных данных, они прямо указывают на очень высокую кислородную емкость крови джейрана и дополняют такие адаптивные морфологические признаки, как большие глаза и уши, сильно вздутые слуховые капсулы, высокие и тонкие ноги и т.д.
Индексы внутренних органов нами не изучены, и соответствующими литературными данными мы не располагаем.
Кариотип. В диплоидном наборе 30 хромосом у самок и 31 хромосома у самцов. Пол определяется системой XY| XY2. Количество плеч аутосом, как и у других видов рода, 56 (Wurster, 1972).]
Структура и динамика ареала
Ареал вида (восстановленный) занимает равнинные, предгорные и низкогорные ландшафты Передней Азии от Аравийского полуострова до Белуджистана (Пакистана), Восточного Закавказья, Средней Азии, Южного Казахстана, Джунгарии, Кашгарии, Цайдамии, Северного Тибета, Гоби на север до Южной Монголии, Алашаня, Ордоса (Гептнер и др., 1961; Harrison 1968; Слудский, 1977; Corbert, 1978; и др.).
Былое распространение джейрана в СССР, также обширное, включало аридные зоны Восточного Закавказья, Средней Азии и Казахстана. Еще в 40—50-х годах XX в. западная граница азиатской части ареала простиралась до восточного берега Каспия, северная — до верховья р. Иргиз, Сарысу (48° с.ш.), северного берега Балхаша, а восточная — до оз. Алаколь. Встречались они также в Зайсанской котловине и отрогах Тарбагатая. Южную границу в то время составляли равнинно-предгорные районы Средней Азии. Джейраны проникали в горы по широким горным долинам и сыртам до 1500—3000 м над ур. моря.
В настоящее время, как свидетельствуют материалы совещания по джейрану (Москва, 1985) и наши данные, резко сократились распространение и численность джейрана по всей стране. Ареал вида не только
422
Gazclla de Blainyille, 1816
сокращается неуклонно, но и распадается на большие и меньшие очаги (Флинт, Присяжнюк, 1986).
Распространение на Кавказе. В прошлом столетии джейран, как ландшафтный вид полупустынь бородачевых степей, населял всю Кура-Арак-синскую низменность. До начала нашего столетия он встречался почти на всем Апшеронском полуострове, восточная граница простиралась до самого побережья Каспия, а на Запад по Куре — до окрестностей Тбилиси включительно и по долине Аракса — до пос. Арарат. На этой значительной территории джейраны попадались всюду, где были подходящие условия жизни. По Н.Я.Диннику (1910), до 70-х годов прошлого столетия за джейранами охотились в 30—40 верстах восточнее Тбилиси. В те же времена базары Тбилиси зимой были завалены этой дичью. По данным названного автора, до начала XX в. джейраны попадались в степях около Евлаха, но западнее Ширанской степи уже не встречались. В прошлом столетии джейрана не стало во всей Армении.
По Н.К.Всрсщагину (1942, 1959), в начале нашего столетия джейраны были выбиты на всем Апшеронском полуострове, к 20-м годам — в степях Шираки, в 30-х годах почти уничтожен в северной части Ширванской степи. Только у станции Карасу браконьеры добывали более 500 джейранов в год. Аналогичное положение было в Мильских степях и других районах. В результате безудержного браконьерства не только сократились численность и распространение, но ареал приобрел пятнистый характер. К 1937 г. он состоял из 8 фрагментов в пределах от 27 до 500 м над ур. моря: 1) Карталинскос плоскогорье; 2) Ширакская и Эльдарская степи; 3) долина р. Кюрокчай; 4) Аджиноурская и Турут-Сарыджинская степи; 5) Мильско-Карабахская степь; 6) Мутанская степь; 7) Ширванская степь; 8) Кабристан. Общая площадь, занятая джейранами, в то время составляла всего 8200 км2.
Сокращение ареала и его дизъюнктивности продолжались еще как минимум два десятилетия. Так, к 40-м годам джейраны были вытеснены севернее р. Сумгаит и отдельные особи встречались не ближе 25—30 км от Баку (в районах Карадага и Шахи-Кая), в Кабристане они оказались загнанными в малодоступные полупустынные увалы (Верещагин, 1959), нс стало их также в Ширакской и Гардабанской степях, исчезли они и на всей территории Грузии (Арабули, 1970).
15—24 июня 1960 г. были проведены авиаучеты (Сафаров, 1961) в Трут-Сарыджинской степи с охватом равнин и боздагов правого берега р. Куры и Мингечаурского водохранилища, между реками Шамхорчай, Кюрокчай (1) и Джейранчел (2), в Карабахской (3), Мильской (4), Ширванской (5) степях, юго-восточной Ширвани (6) и Муганской степи (7). Учеты нс проводились в Кобыстане и предгорьях Малого Кавказа между Мартуни й Агдамом. На всей обследованной территории общей общей площадью в 14 324 км2 зарегистрирован всего 131 джейран. Они были размещены изолированно в первых шести районах, а в Муганской степи их не нашли. Животные попадались по 1—2 головы, и лишь в районе мыса Бяндован и оз. Шор встречены группы по 7—9 животных. Между тем в начале текущего столетия в степи Джейранчел ("степь джейранов") встречались стада по
423
Джейран
100 голов и более, а во время авиаучетов 1960 г. здесь было встречено всего 7 голов. По Н.Я.Диннику (1910), больше всего джейранов в пределах Кавказа водилось в Карабахской и особенно в Муганской степях. Весной 1907 г. стада джейранов в 50—70 голов встречались в центре Муганской степи.
По данным М.А.Сафарова (1961), кроме перечисленных шести участков джейраны в количестве 17 голов были отмечены на о-ве Булла в начале июля 1960 г. (они были выпущены для акклиматизации первый раз в 1950 г.). Однако, судя по всем данным, опыт оказался неудачным.
Снижение численности и уменьшение области распространения джейранов в Закавказье продолжалось и в начале 60-х годов. По Е.А.Котлярову (1975), они встречались лишь в Ширванском заповеднике, Бяндованском, Керчайском и Апшеронском заказниках. При этом общее их поголовье в Азербайджане составляло всего 130 особей.
Организация Ширванского заповедника, Кировабадского и Апшеронского заказников и другие практические шаги по охране джейрана в Азербайджане дали ощутимый положительный эффект. В 1972 г. количество этих животных возросло в 5 раз, в 1983 г. достигнут уровень 40-х годов, а площадь, занятая ими, составила 0,3% ареала вида. По учетным данным, полученным в том же 1983 г., основная часть поголовья находилась на территории Ширванского заповедника и рядом лежащего Бандованского заказника. Это единая популяция и, видимо, самая крупная группировка в СССР (Шадилов, 1986). Кроме того, джейраны были учтены в Апшеронском и Керчайском заказниках, Ханларском госохотхозяйстве. Отдельные животные или небольшие группы зарегистрированы в Гобустанс, в Аджиноурской степи и в ряде других мест.
Современные тенденции антропогенных изменений ареала. В последние десятилетия все больше внимания привлекает проблема сохранения джейрана в природных экосистемах аридной зоны страны, в том числе Восточного Закавказья. Сложность проблемы усугубляется как особенностями биологии джейрана, так и региональными факторами. Как справедливо отмечают многие специалисты, для нормальной жизни джейрана нужны сухие субтропические ландшафты, ибо очень высокая нагрузка на опорную поверхность конечностей (у самцов 491, у самок 434 и у прибылых 349 г/см2) (Слудский, 1977) практически исключает возможность их существования в снежных районах или на сельскохозяйственных угодиях, особенно орошаемых. Поэтому мы разделяем точку зрения Н.К.Верещагина (1959) о том, что орошение и распашка Кура-Араксинской низменности приведут в ближайшем будущем к полному вытеснению джейранов из Восточного Закавказья, если не будет создан степной заповедник и организована действенная охрана этого животного. Однако при высокой плотности населения людей и дефиците сельхозугодий, как это наблюдается на Кавказе, проблему не разрешить созданием заповедников и совершенствованием охраны. Нужен целый комплекс мероприятий, чтобы местный генофонд джейрана сохранился. А.Н.Ахундов (1989) попытался свести их в систему и еще раз подтвердил, что перспектива сохранения полноценного генофонда джейрана
424
Gazella de Blainville, 1816
неоднозначна. Из многочисленных факторов, определяющих судьбу джейрана в Закавказье, особое внимание должны заслуживать совершенствование системы землепользования и выработка такой, которая позволяла существовать джейрану в природе. Создание сети заповедников и питомников мы рассматриваем лишь как часть этой системы. Поэтому, разделяя мнение В.Е.Флинта и В.Е.Присяжнюка (1986) и имея в виду, что заметное увеличение численности джейрана при стабилизации области распространения отмечены только в Азербайджане, следует подчеркнуть, что положение вида в Закавказье далеко не благополучно и не стабильно.
Систематика и изменчивость
Систематическое положение. По мнению В.Г.Гептнера и др. (1961), вид описан по экземпляру из Закавказья, вероятно даже из окрестностей Тбилиси; указание на типичную местность "Персия" или "Персия, вероятно Бус-сора" неверно, потому что там Гюльденштедт не был и не имел возможности получать оттуда коллекционный материал. В настоящее время уточнить происхождение типового экземпляра, очевидно, невозможно, тем более что и в Закавказье, и в Передней Азии встречается номинальная форма.
Реальность существования вида Gazella Subgutturosa Guld признается ведущими систематиками мира давно. В числе спорных вопросов остается надвидовая и внутривидовая систематика. О внутривидовой изменчивости будет сказано ниже; здесь отметим, что наши ведущие систематики В.Г.Гептнер и И.И.Соколов высказывали противоположные мнения. Так, И.И.Соколов (1959) в род Gazella включал, кроме джейрана, еще дзерена "Gazella gutturosa". В.Г.Гептнер и др. (1961), признавая систематическую близость обоих видов (безрогость самок, в той или иной степени редукция желез и др.), считали целесообразным относить дзерена к роду Ргосарга. Эта точка зрения имеет больше сторонников и мы ее признаем более обоснованной (Соколов, 1979). Наряду с другими систематическими признаками мы придаем большой систематике-таксономический вес различиям в структуре и степени развития целого ряда кожных желез.
Изменчивость. Для анализа различных аспектов изменчивости джейрана Закавказья мы располагали мизерным материалом. Весьма скудны также литературные сведения. Между тем целый ряд авторов высказывал неудовлетворенность состоянием изученности данного вопроса (Соколов, 1959; Гептнер и др., 1961; Слудский, 1977; и др.). При таком положении анализ имеющихся данных представляет определенный интерес.
Индивидуальная изменчивость. О размахе индивидуальной изменчивости массы тела и меристических параметров джейрана Восточного Закавказья можно судить по данным, приведенным выше. Более подробный анализ провести невозможно из-за отсутствия данных. На наш взгляд, интерес в плане индивидуальной изменчивости представляют этологические наблюдения, иллюстрирующие глубокую разнокачественность группы и популяции в целом. Так, в условиях питомника О.Б.Переладова (1986) выделила три группы детенышей джейрана, отличающиеся по ряду параметров поведения:
425
Джейран
отрицательно-социальные, нейтральные или пассивно-социальные и активно-социальные. Первой группе свойственны нервозность, возбудимость, склонность к панике у самок, агрессивность у самцов и т.д. Для животных второй группы более характерна реакция затаивания при опасности, отсутствие агрессивности, пассивность и малая подвижность. Третья группа отличается сильно выраженной ориентировочно-исследовательской реакцией, способностями к лидированию и т.д. Справедливо, очевидно, мнение автора о том, что индивидуальные особенности социальной активности джейранят не могут быть сведены лишь к одному воспитанию и соответственно должны быть учтены при организации работы в питомниках.
Возрастная изменчивость. Данные по разным аспектам возрастной изменчивости содержатся в ряде работ, в том числе общие вопросы в работах М.И.Исмагилова (1954), А.А.Слудского (1956), А.М.Мамбетжумаева (1970), В.В.Жевнерова (1984). Изменение экстерьера в условиях питомника изучено В.Е.Присяжнюком и Н.В.Солдатовой (1985); возрастные изменения зубной системы — Доан Чонг Бинем (1963) (табл. 82); последовательность дифференцирования конечностей рассмотрена И.М.Джафаровым (1982).
Масса тела джейрана, по всем имеющимся данным, характеризуется максимальным темпом возрастной изменчивости. Достаточно сказать, что к полутора годам она возрастает в 10—12 раз и приближается к размерам взрослых животных. По отношению к показателю полуторалетнего масса тела молодняка в полгода составляет 63—69%, годовалого — 76—98%. Интенсивность роста самок выше самцов, соответственно самки к полутора годам очень мало отличаются по массе от взрослых, а самцы отстают примерно на 1/4.
Возрастные изменения по меристическим показателям у джейрана, как следует из данных названных авторов, происходят не синхронно, что характерно не только для копытных, но и для различных групп млекопитающих, в том числе мелких землероек (Соколов, Темботов, 1989). Раньше bcqx, фактически в течение первого полгода постнатального периода, завершается формирование длины головы, высоты уха, длины хвоста, длины кисти, длины стопы, обхват пясти, длины шеи, ширины труди, ширины таза в седалищных бутрах, ширины переднего и заднего копыта. К концу первого года жизни заканчивается рост длины тела, длины передней конечности, глубины труди, длина переднего и заднего копыта, высоты в крестце и ширины в маклоках. В полтора года заканчивают в основном свое формирование косая длина туловища, обхват груди, косая длина таза, длина задней конечности и высота в холке. Некоторые из них сохраняют тенденцию к росту и в последующем. Интерес представляют возрастные изменения индексов телосложения, приведенные на рис. 59.
Возрастные изменения окраски тела у джейрана выражены слабо, и по ним выделять возрастные группы невозможно. Основные особенности окраски сводятся к тому, что у новорожденного верх головы, спины и хвоста светло-бурый, остальные волосы почти черные или темно-бурые. Такая окраска ягнят хорошо гармонирует с фоном глинисто-щебнистой
426
Gazclla de Blainvillc, 1816
Таблица 82. Возрастные изменения зубной системы джейрана (Доан Чонг Бинь, 1963)
Резцы и клыки Коренные зубы Воз] эаст
На нижней челюсти ^Имеются все молочные предкоренные До 1 мес
имеются все молочные резцы и клыки, иногда рудиментарные верхние клыки Начинают снашиваться Р^, рЗ и Р4. 1-2 мсс
Прорезались через кость передние лопасти
Nfl и через десну передние лопасти Ml
Иногда сохраняются Прорезались через десну передние лопасти 2-3 мсс
рудиментарные Nfl и задние лопасти Ml
клыки Стираются все молочные предкоренные. Задние лопасти прорезались через десну, а передние лопасти М2 — через кость Стираются задние лопасти M^i 3-4 мес
Прорезались через кость задние лопасти М2-Передние (у самки) или задние (у самца) 4-5 МСС
лопасти М^ прорезались через кость, а передние лопасти М2 — через десну Прорезались через десну передние лопасти 5-6 мсс
М^ и задние лопасти М2 6-7 мсс
Жевательная поверхность передней лопасти Р4 сглажена. Прорезались через десну задние лопасти М^_ Стираются передние лопасти М2- Сильно стерты р2, рЗ и р4 Стираются задние лопасти M2j. Прорезались через 7-8 9-10 мсс
кость передние лопасти М3. Передние лопасти МЗ3 в верхней челюсти мсс
прорезались только через кость, а в нижней и через десну. 12-13 мсс
Сменяются Прорезались через десну передние лопрети
молочные зацепы М3 и задние лопасти М3 Сглажена жевательная поверхность р2, рЗ и Р4. Между корнями молочных 13-14 мес
предкоренных видны прорезавшиеся через кость верхушки Р2, рЗд, р4д Задние лопасти М3 прорезались через десну, а
Сменяются на талонид — М3 — через кость Сменяются на постоянные все 14-15 мсс
постоянные вторые и предкоренные у самок. Прорезался через
третьи резцы (I2, I3) у самок десну талонид М3 15 —16 мес
427
Джейран
Таблица 82 (окончание)
Резцы и клыки Коренные зубы Возраст
Сменяются на Сменяются на постоянные все
постоянные вторые и предкоренные у самцов. Стираются задние 16—18 мес
третьи резцы (I2,13) лопасти М^З
у самцов Сменяются на Стираются рЗ^рЗ, р4. 18—19 мес
постоянные клыки.
Стираются 11 Стираются I2, 12 Стираются все предкоренные , стирается талонид М3 2—2,5 года
Стираются 11, 1?С Вершина задней стенки м3 начинает
стираться
Начинают стираться требни 12,С. Просвечивает дентин на 11 Сильно стерты р2 2,5—3 года 3—3,5 года 3,5—4,5 года
Просвечивает дентин След коронковой чашечки М i в виде
на I2 и стерты в верхней части черточки
Просвечивает дентин Жевательная поверхность Р^ сглажена.
на I3, гребни I2,13, След коронковой чашечки передней
С стерты и в нижней лопасти Мр виде пятнышка 1,5—5,5 лет
части
Появляются на 11 Почти сглажена жевательная поверхность
зубные звездочи в рз 5,5—6,5 лет
виде черточек Сглажена жевательная поверхность рЗ-След коронковой чашечки в виде пятнышка. Сглажена поверхность М1И талонида М3 6,5—7,5 лет
Зубные звездочки на След коронковой чашечки Р^ в виде
11 овальной формы пятнышка 7,5—8,5 лет
Зубные звездочки на Сглажена жевательная поверхность Ml
11 имеют передней лопасти М2«След коронковой 8,5—9,5 лет
треугольную форму чашечки М3 в виде пятнышка.
Жевательная Сглажена жевательная поверхность
поверхность и М22 и М3 Более 10
зубные звездочки на -11 округлые Резцы и клыки стерты почти до десны лет
полупустыни. Для полувзрослых и взрослых характерен темный рисунок на лбу. Старые животные, как отмечено выше, сильно светлеют.
В росте и развитии рогов в отличие от козлов отсутствует цикличность, и по ним определять возраст животных невозможно. Выпуклые бутры,
428
Gazclla de Blainvillc, 1816
Рис. 59. Изменения пропорций телосложения, джейрана в онтогенезе (Присяжнюк, Солдатова, 1985) Индексы:
А— большеголовости (1), длинноголовости (2), массивности (3), перерослости (4);
Б — грудной (ширина труди в плечелопаточных сочленениях — глубина груди) (5); формата (6), сбитости (7);
В — костистости (8); глубина груди (высота в холке (9); длина уха) длина хвоста (10); Г — длина передней конечности: косая длина (11); тазогрудной (ширина груди в плечелопаточных сочленениях) ширина таза в маклоках (12)
опоясывающие роговые чехлы, являются годовыми кольцами. Это не означает, что структура рогов не зависима от возраста. Рога становятся заметными в месячном возрасте, имея длину 1—1,5 см. В три месяца они увеличиваются до 6 см в среднем, а в полуторагодовалом возрасте они составляют 8,5—30 см (х = 17,3).
Анализ постнатального развития зубной системы позволяет (Доан Чонг Бинь, 1963) различать до 25 возрастных групп. Однако на практике, особенно при систематико-таксономических и полевых экологических исследованиях, пользоваться таким количеством возрастных групп трудно, практически невозможно. Поэтому мы свели его до 5 групп, положив в основу градации основные этапы формирования экстерьера в постнатальном периоде (см. выше).
429
Джейран
I группа — возраст около 6 мес. Прорезались через десну передние и задние лопасти М22-
II труппа — возраст около года. Сильно стерты все молочные предкоренные и задние лопасти М^; передние лопасти второго коренного прорезались через кость, а в нижней — через десну.
III группа — возраст около полутора лет. Сменяются на постоянные резцы, клыки, все предкоренные, стираются задние лопасти М33, а затем Р2, Р3, Р44.
IV .труппа — возраст 2 года и старше. Вначале стираются все предкоренные, а затем — коренные. Степень их стертости отражает возраст животных.
V группа — возраст более 10 лет. Резцы и клыки стерты до основания, поверхность Р4, М22, М3 сглажена.
Половые различия. У взрослых джейранов хорошо выражен половой диморфизм по массе тела — самцы превосходят самок более чем на 10 кг, т.е. тяжелее почти в 1,5 раза. В этом мнении специалисты сходятся, но различны суждения по молодняку. Так, А.А.Слудский (1956, 1977) считает, что у новорожденных телят самцы весят на 100—200 г больше, чем самки. Иными словами, у телят половой диморфизм выражен даже больше, чем во взрослом состоянии. В.В. Жевнеров (1984), проводивший исследования природных популяций на о-ве Барсакельмес, и В.Е.Присяжнюк, Н.В.Солда-това (1985) в Бухарском питомнике, пришли к мнению, что у джейранят отсутствуют половые различия не только по массе, но и по размерам тела. Различия появляются с возрастом и в связи с неодинаковыми темпами роста и развития.
Что касается окраски тела, то половой диморфизм отсутствует, видимо, на всех этапах постнатального развития. Существенным и очевидным показателем полового диморфизма считается наличие хорошо развитых рогов у самцов и комолость самок, что относится к числу примитивных признаков джейрана как вида. Наличие иногда рогов у самок в пределах от 5% (Слуд-ский, 1977) до 20—25% (Лобачев, Смирин, 1970) не противоречит изложенному выше. Рога самок небольшие (от 5 до 10 см), тонкие и неправильной формы. Их спутать с рогами самцов невозможно.
Из остеологических признаков пола, на наш взгляд, внимания заслуживают тазовые кости (Джафаров, Агаев, 1986). По данным названных авторов, целый ряд признаков тазовых костей позволяет на 100% диагностировать половую принадлежность. К таким признакам относятся особенности строения крестцового отростка, большой седалищной вырезки, ушковидной суставной поверхности, подвздошно-лонного возвышения, лонного бутра, лонного требня, седалищной дуги, наличие у самок отверстия в области симфиза, апофиза и др.
Половые различия в структуре черепа наиболее значительны в области мозговой части и обусловлены наличием у самцов роговых стержней. Половой диморфизм четко выражен и по меристическим показателям, что
430
Gazella de Blainvillc, 1816
связано с существенным расхождением между полами в массе и размерах тела.
Географическая изменчивость. Небольшой фактический материал, изученный нами, свидетельствует о том, что в пределах Закавказья обитает одна, довольно монолитная форма. Это связано, видимо, с высокой подвижностью животных, периодическими миграциями и колебаниями численности в очень широких пределах (до недавнего времени), а также со строгой приуроченностью джейранов к однообразным ландшафтам аридной зоны Закавказья. В наши дни в связи с восстановлением джейранов на отдельных изолированных участках Закавказья возможно появление новых территориальных группировок с определенными морфологическими, экологическими и этологическими параметрами. При этом нельзя не учитывать высокую пластичность и не менее высокую плодовитость джейранов. С таким прогнозом микроэволюционных процессов в новых, глубоко измененных ландшафтно-экологических условиях Закавказья согласуются некоторые этологические наблюдения. Так, по данным В.В.Жевнерова (1984), на о-ве Барсаксльмес поедание последа самкой джейрана не свойственно. Это объясняется отсутствием на острове волков. По той же причине при случайном появлении на острове волка у джейранов наблюдается шоковое состояние. В тех местах, где волки встречаются часто, такого состояния джейранов не отмечается. Поедание самкой последа при наличии волков — явление закономерное, свойственно многим видам копытных. Биологический смысл очевиден — чем меньше следов оставляет родившая самка, тем больше осложняется нахождение теленка хищником. Эта биологическая особенность регулярно отмечалась даже в условиях питомника (Пе-реладова, Перепадов, 1986).
Что касается положения джейранов Закавказья среди других внутривидовых форм, то они, как и псреднеазиатские, отличаются более длинными рогами и относительно мелкими размерами тела. Закавказские джейраны, например, уступают сородичам из пустынь Бетпак-Далы (Слудский, 1977) по длине тела примерно на 10 см, по высоте в холке — 5 см. Еще И.И.Соколов (1959) справедливо обращал внимание на не слишком резко выраженную географическую изменчивость окраски. В частности, западные популяции окрашены более -интенсивно, сильнее развиты бурые или сероватые тона. К востоку идет посветление, усиливается желтовато-рыжеватый тон. В Центральной Азии развивается даже розоватый оттенок.
Ряд териологов, в том числе И.И.Соколов (1959), В.Г.Гептнер и др. (1961), А.А.Слудский (1977) и др., высказывали неудовлетворенность изученностью географической изменчивости джейрана и состоянием внутривидовой систематики. Полагают, что джейраны Закавказья, изолированные от других популяций, вида, составляют отдельный подвид. Однако для разрешения этого вопроса пока нет достаточного материала.
Джейраны Закавказья относятся к одному подвиду — Gazella subgutturosa subgutturosa Guld.
431
Джейран
Экология
Ландшафтная приуроченность. Биотопы. Восстановленный ареал вида указывает на то, что джейран в прошлом занимал широкий круг ландшафтов аридной зоны Восточного Закавказья. Об этом свидетельствуют немногочисленные литературные данные, характер современного распространения в регионе и в других частях ареала.
К.А.Сатунин (1906) встречал джейранов в значительном количестве в глубине Муганских степей. Степь Джейранчель была местом концентрации этих копытных. Н.Я.Динник (1910) писал, что, подобно сайгакам, джейраны обитают в степях, но не в такой мере избегают горного рельефа, как они. В 30-х годах текущего столетия Н.К.Верещагин (1942) регистрировал их в Турут-Сарыджинской и Мильско-Карабахской степях. Они занимали полынную и полынно-солянковую полупустыню и "полустепи" с ровным, но иногда и с холмистым рельефом. Высотные пределы распространения в Закавказье составили от 27 до 500 м над ур. моря. Если судить по границам восстановленного ареала, то джейран в прошлом веке имел более широкие пределы высотного распространения от Каспия до 1000 м над ур. моря. На Иранском плоскогорье, где обитает тот же номинальный подвид, что и в Закавказье, они населяют горные ландшафты до высоты 2000 м над ур. моря. На том же уровне этот вид был обычен в Иссыккульской котловине (Янушевич и др., 1972). Однако как в Закавказье, так и в Средней Азии джейран проникает в горы по соответствующим ландшафтам — полупустынным и горным степям.
Современные антропогенные изменения природы аридной зоны Закавказья, как и в других частях ареала, очень сильно сказались на ландшафтной и биотопической приуроченности джейрана. В первую очередь они были вытеснены из тех мест, где возможно было вести орошаемое земледелие, ибо джейраны не приспособлены жить на пахоте, тем более орошаемой. По Н.К.Версщагину (1959), на Аджинаурском плоскогорье они отмечались только на богарных посевах ячменя и пшеницы. На невспаханных ровных местах джейраны не смогли удержаться; при всей их подвижности они становились легкой добычей моторизованных браконьеров. Спасение они находили в местах трудно проходимых автотранспортом — бугристых и грядовых песках, закрепленных в той или иной степени, боздагах, пересеченных местах и т.д. В годы минимальной численности и распространения джейрана в нашем столетии (начало 60-х годов) М.А.Сафаров (1961), обследовавший на самолете значительную территорию, находил скопление по 7—9 джейранов только в двух местах — в районе мыса Бяндован (побережье Каспия) и вблизи камышовых зарослей оз. Шор. Оба места были трудно доступны для автомобильной охоты. Во всех других местах, в том числе в самом их излюбленном месте в прошлом — Джейранчеле, где встречались стада по 100 голов и более, попадались лишь единицы. Джейраны, которые сохранились в то время, держались далеко от населенных пунктов, придерживались камышовых зарослей, подножий боздагов. Мы,
432
Gazclla de Blainville, 1816
обследовав эту Кура-Араксинс кую низменность неоднократно, в открытой степи встретили лишь однажды одного джейрана.
Сходные антропогенные изменения в распространении джейрана произошли в Средней Азии и Казахстане. Как отмечено выше, джейран широко населял указанные районы и был многочислен во многих местах, в том числе на плато Устюрт. Однако он стал здесь редким и сохранился лишь вдоль обрывов западного и южного чинков плато. Единичные особи и группы по 3—7 голов встречаются в районах песков Сам и Асманбай, Матай, соленой грязи Барсакельмес и Сарыкамыской котловины. Животные в этих местах не доступны для автомобильной охоты (Мамбетжумаев, 1970).
В связи с организаций заповедников и других охраняемых территорий в Восточном Закавказье происходят положительные' сдвиги в ландшафтном и биотопичсском размещении джейранов. Однако Лучшие местообитания все еще остаются недоступными джейранам, их занимают домашние животные и эти места в перспективе могут быть распаханы и превращены в орошаемые поля. В ШирванскОхМ заповеднике и Бяндованском заказнике наиболее предпочитаемыми биотопами для джейранов являются бугристые участки полынной, солянковой и полынно-солянковой полупустынь. Они составляют около 40% всей территории заповедника и заказника (Шадилов, 1986).
Миграции. Территориальная организация популяции джейрана подвержена значительным сезонным колебаниям, что зависит от состояния кормовой базы, наличия водопоев, режима погоды, особенно снежного покрова и т.д. Как известно, все эти факторы глубоко региональны в Восточном Закавказье и определяют многие особенности поведения джейрана. В Кура-Араксинской низменности с сухим субтропическим климатом обычно нс бывает глубокого снежного покрова, превышающего критический для джейрана уровень (более 10 см). Как правило, выпавший -снег к вечеру растаивает. Поэтому из-за снега джейраны не мигрируют, нс образуют скопления и т.д. В Казахстане, например, джейраны, спасаясь от холодных ветров и снега, мигрируют из более северных районов на юг. Протяженность пути по прямой линии к местам лстовок и обратно здесь достигает 700 км (Слудский, 1977). В Закавказье иногда бывают климатические отклонения от нормы, что сопровождается выпадением большого снега и снижением температуры. В этом случае джейраны оказываются в катастрофическом положении, так как они нс способны добывать корм под глубоким снегом, нс могут уходить по такому снегу от хищников, не имеют также возможности мигрировать, ибо с трех сторон их местообитание окружено горами, где выпадает снега больше, чем на равнине. Четвертая сторона — ворота Кура-Араксинской низменности "заперты" Каспийским морем. Джейраны оказываются в безвыходном положении и в большом количестве гибнут (Верещагин, 1959).
У джейранов, как и сайгаков, наблюдаются миграции, кочевки и другие перемещения в связи с выгоранием растительности и увеличением потребности в воде. В Восточном Закавказье летние перемещения джейранов также небольшие; здесь немало возможности для утоления жажды —
28 В.Е.Соколов, А.К.Темботоп _
Джейран
близость Каспия, Куры и Аракса, многочисленные старицы, озера, оросительные каналы и т.д. По Н.К.Верещагину (1942), для посещения водопоя джейран пробегает 15—20 км по ночам (Юго-Восточная Ширвань — берег Каспия; Мильская степь — озера по Куре и Мильские каналы). Изучение водного обмена у населяющих аридные области копытных показали, что такие дальние пробеги на водопой маловероятны. Проблема водопоя в летнюю жару смягчается еще микромозаикой растительного покрова почти на всей территории Восточного Закавказья. Наряду с выжженными землями, нередки различные оазисы с вегетатирующей растительностью в течение всего лета. Джейраны в аналогичных условиях обходятся влагой, содержащейся в таких растениях. Как отмечено выше, учеты, проведенные в июне 1960 г., показали, что летом джейраны скапливались вблизи берега моря и камышовых зарослей озер, а единичные особи — у подножия боз-дагов, где в силу особенностей рельефа сохранялись лучшие кормовые и защитные условия. Сходное поведение джейрана отмечено в Казахстане. Так, на Устюрте джейраны летом живут по окраинам саксауловых зарослей, по обрывам чинков плато (Слудский, 1956, 1977; Мамбетжумаев, 1977; и др.); в пустынях Сары-Ишикотрау они заселяют закрепленные гряды и бугристые пески и такыровидные равнины с саксаульниками (Бекенов, Бежанов, 1989).
Сезонная и суточная активность. Поведение. Данные по Закавказью фрагментарны (Верещагин, 1942; Алекперов, 1966; наши наблюдения), но они согласуются с относительно полной информацией по Казахстану и Средней Азии (Слудский, 1956, 1977; Мамбетжумаев, 1972; и др.). Все эти сведения говорят о том, что подвижность джейранов меняется в широких пределах по сезонам года. Летом, когда наступают жаркие дни, у джейранов четко выражена двухфазная активность — утренняя и вечерняя. На пастбищах они появляются с рассвета и с небольшими перерывами пасутся до 9—10 ч. Возобновляется пастьба в 18—20 ч и с такими же перерывами длится до поздней ночи. Продолжительный дневной отдых животные проводят в более прохладных и обдуваемых местах (в тени под кустами древовидной солянки, на бугре, под кустарниками и т.д.). Осенью в связи с приближением гона и наступлением прохлады заметно повышается активность — сокращается продолжительность дневного отдыха, больше времени они проводят на пастьбе, играют, дерутся, учащаются ухаживания самцов за самками и т.д. Зимой они активны в основном в светлые часы суток, дневной перерыв резко сокращается и фактически суточная активность -становится однофазной. Ночью они находятся на лежках, где меньше ветра и более тепло. В Закавказье зимние лежки Н.К.Верещагин находил на кучках колосовых чешуек злаков, у муравейников. С наступлением весны и повышением температуры воздуха джейраны начинают пастись с рассвета до полудня с некоторыми перерывами. Отдыхают в промежутках от 12 до 15 ч, а потом возобновляют пастьбу и активны до полной темноты. Выбор места отдыха весной более разнообразен, ибо они широко используют открытые и незащищенные от солнца места — открытые солончаки, песчаные островки, лишенные соли и др.
434
Gazclla de Blainvillc, 1816
НЛ.Динник (1910, c. 218) писал, что джейрана "надо считать за животное высокоодаренное. Кроме того, он необыкновенно подвижен, проворен, чрезвычайно миловиден и обладает способностью необыкновенно быстро бегать" (рис. 60). По его мнению, нрав у джейранов очень хороший, они легко и быстро привыкают к человеку, и потому их часто приручали и держали во дворах. Такие прирученные джейраны часто встречались раньше, их можно было видеть свободно бегающими на улицах, они сами возвращались во двор своего хозяина. Такие джейраны легко уходили от собак, сохраняли легкость и грациозность движения. Н.Я.Динник обращал внимание на высокоразвитые органы чувств: острое зрение, тонкий слух и превосходное обоняние. В.Г.Гептнер и др. (1961), признавая тонкий слух джейрана, считают, что у них слабо развито зрение и еще хуже обоняние. Очевидно роль последних двух органов чувств принижена в названной работе. Об уровне развития обоняния можно судить не только по морфологии кожных желез (см. выше), но и по тому, какое место занимает в воспроизводстве популяции химическая и зрительная коммуникация’ (Бланк, 1985; Мармазинская, 1990). В поведении половозрелого самца выделен целый комплекс территориальности, в том числе оставление мочи и экскрементов ("гоиные уборные"), мечение растений секретом предглазничной железы (сигнал о занятости территории), принятие различных поз (голова и шея вытянуты вперед, уши прижаты назад, хвост поднят). Все эти и другие элементы полового поведения воспринимаются органами зрения и обоняния. В социальном поведении немало элементов, направленных для зрительного восприятия. То же самое можно сказать и в отношении оборонительного поведения. По А.А.Слудскому (1977), джейраны при обнаружении хищника чихают, бьют передними копытами, часто принимают своеобразные позы ("мимические сигналы"); заметив это, другие животные настораживаются и убегают. При этом делают, как и сайгаки, большие своеобразные прыжки вверх ("сигнальный прыжок"). То же самое можно сказать и в отношении пищевого поведения. Некоторые аспекты поведения затронуты еще в соответствующих разделах.
Линька. По литературным данным, которыми мы располагаем (Верещагин, 1938, 1942; Мамбетжумаев, 1955, 1970; Слудский, 1956; Гептнер и др., 1961), джейран в году линяет один раз. В Азербайджане это приходится на период с середины марта до конца апреля — начала мая. Линька начинается с щеки, ушей, зеркала и хвоста. От щек она переходит на темя, верх шеи и на ее бока, а затем на спину. От зеркала линька распространяется на бедра и дистальные части задних ног, затем на бока тела. В последнюю очередь линяют грудь и брюхо, которые соприкасаются с холодной весенней почвой дольше всех других частей тела. Осенью, видимо, происходит лишь подрастание шерсти, которое отмечается с октября по ноябрь — начало декабря.
Лежки. Характерной особенностью экологии джейрана является строительство лежек, представляющих собой небольшие углубления в земле. Выбивает джейран ее передними копытами, обычно располагает под древовидной солянкой, у муравейников, на пухлых солянках, по окраинам
28*
435
Рис. 60. Джейраны
436
Gazclla de Blainville,. 1816
барханов и т.д. По мнению А.А.Слудского (1977), чаще всего лежки располагаются под одиночными растениями саксаула, гребенщика или джуз-гуна. Нередко под одним кустом расположены до 4 лежек, используемых в жаркие дни по очереди в зависимости от освещения. Зимой под одним кустом Г.Н.Сапожников (1976) находил на лежке трех джейранов. Покидая лежку, джейран оставляет помет и мочу, выполняющие, видимо, функции территориального мечения.
Питание. По Н.К.Верещагину (1939, 1942), джейраны в Азербайджане питаются злаковым эфемеретумом (Aegilaps sguarrosa, Lolium rigidum, Роа bul-bosa, Aeluropus repens, A. littoralis), каперцами (Capparis spinosa), всходами ячменя, пшеницы, хлопка. В четырех вскрытых Х.М.Алекперовым (1966) желудках джейрана из юго-восточной части республики, кроме названных видов, в большом количестве обнаружены полынь, солянка, культурные злаки. Поедание всходов и листьев хлопка не наблюдалось. Эта скудная информация несколько дополнена Е.А.Котляровым (1975); согласно его данным, животные поедают все доступные растения, в том числе тростник, кермек каспийский, латук татарский, ячмень дикий, ячмень заячий, костер краснеющий, рожь дикую, тамариск и др. Наиболее распространенными кормовыми растениями являются песчаная и метельчатая полыни. Их листьями и молодыми побегами охотно кормятся джейраны. Для более полной характеристики питания джейрана ниже приводятся данные А.А.Слудского (1977), полученные на о-ве Барсакельмес. Из 216 видов высших растений, произрастающих на этом острове, животные поедают в течение года 70 видов. В зимнее время пищевой рацион включает 37 видов, в том числе два вида древесных, 16 кустарников и полукустарников, 7 кустарничков и 12 многолетних и однолетних трав. При этом джейраны предпочитают два вида хвойника, курчавку шиповатую, два вида лебеды, биюр-гуну и терескену серую, саксаулы черный и белый, франкению, гребенщик и полынь белоземельную. Поедается также выброшенный на берег волнами малый взморник (растет в морской воде на глубине 8—9 м). При недостатке кормов это растение может составлять 50—80% содержимого желудка джейранов. Весной и летом список предпочитаемых растений включает хвойник (ветки с листьями и костянками), два вида пырея, два вида мор-тука, мятлик луковичный, Осоку разночешуйчатую, один вид камыша, лук песчаный, один вид тюльпана, ежовник безлистый, марью городскую и др.
В питании джейрана, как и сайгака, примечательно, что они поедают ядовитые для домашнего скота растения, в том числе гармалу, парнолистник туркменский, солянку Рихтера, хвойники шишконосный, двухколосковый и средний, ежовники лиловый, однолетний и безлистный, додарцию восточную и др. На материке, в пустыне Бетпак-Дала, А.А.Слудский (1977) зарегистрировал поедание джейраном в весенне-летний период 47 видов растений. В их числе 7 видов злаков, 3 вида ковыля, 2 вида осоки, 3 вида лука.
Итак, можно заключить, что состав кормов характеризует джейрана как высокопластичный вид. Это положение следует учитывать при оценке со
437
Джейран
стояния кормовой базы и емкости угодий джейрана в новых, глубоко измененных человеком условиях аридной зоны Восточного Закавказья.
Воспроизводство и структура популяции
Размножение. За последние десятилетия накопились данные по воспроизводству популяции джейрана в различных ландшафтных районах страны. К сожалению, мы не располагаем наблюдениями по Закавказью, чтобы выявить региональные особенности этой стороны экологии джейрана.
Самец ухаживает за самкой в течение всего года, физиологическая активность семенников и яичников зарегистрирована весной, летом и зимой (Цаплюк, 1972). В поведении самца наиболее значительна половая активность с середины лета, когда половозрелый самец на путях ежедневных перемещений самок учащает выбивание копытами участков земли, мочится на них и на следах мочи оставляет кучи помета. Посещая их заново или чужую "гребку", он копытами разбрасывает помет и оставляет свой. В итоге такие участки превращаются в "гонные уборные", выполняющие функции маркера индивидуальной территории размножающегося самца (площадь около 50—80 га). В период гона, что приходится в Закавказье на середину декабря (Верещагин, 1942), а в большинстве районов Средний Азии на конец ноября — начало декабря (Бланк, 1985), постепенно возрастает территориальность самцов, его активность по охране индивидуальной территории и агрессивность по отношению к другим половозрелым самцам. Наличие таких строго охраняемых индивидуальных территорий отмечали многие зоологи (Слудский, 1956; Бланк, 1985; и др.). Но по Ю.К.Горелову (1972), в Бадхызе (юго-восточная Туркмения) территориальность самцов выражена слабо (что касается рытья гребок, то это, видимо, лишь проявления "экологического рудимента"). Думается, что факты, приведенные в перечисленных выше работах, достаточно убедительны в том, что поведение джейрана по время гона в условиях Бадхыза нетипично для вида в целом.
Все специалисты утверждают, что самки своих территорий н^имеют.
Джейраны — полигамы, но эта особенность проявляется своеобразно. При одном рогаче обычно находится несколько самок, он ими владеет, оберегает от других самцов, вступая с ними в единоборство. Однако как только какой-то самке удается пересечь границу территории соседнего самца, то на этом прекращается преследование со стороны первого самца. Она становится достоянием соседа (Бланк, 1985). Во время гона самец мало пасется, активен почти круглосуточно и охраняет свой табунок (обычно 2—5 самок), соответственно худеет, теряет осторожность, гибнет чаще самок от хищников и стихийных бедствий (Верещагин, 1938). Нетерриториальные самцы менее агрессивны, имеет место ухаживание нескольких таких самцов за одной самкой, меняющих друг друга в погоне за ней (Бланк, 1985). Отмечено также, что в одном стаде из 17 самок имелось 3 самца и они относились терпимо друг к другу (Горелов, 1972).
В ряде работ, в том числе А.А.Слудского (1977), показана вариабельность сроков гона — их зависимость от географического положения района, по
438
Gazella de Blainvillc, 1816
годы, от участия самок прибылых (вступающих в размножение позже, чем взрослые самки).
Продолжительность беременности, по В.Г.Гептнеру и др. (1961) и А.А.Слудскому (1956), составляет 5—5,5 мес, по О. Б. Пер слад о вой и С.В.Пе-реладову (1986) — 160—164 дня, по В.ВЖевнерову (1984) — 148—152 дня, по Н.К.Верещагину (1942) — 5 мес. Изменения продолжительности беременности отмечены даже в одном и том же питомнике Сюнт-Хасардагского заповедника (Персладова, Перепадов, 1986).
В зависимости от сроков спаривания и беременности молодняк появляется в разное время весны: на Аравийском полуострове — в марте-апреле (Harrison, 1968), В Бадхызе — с середины апреля до конца мая (Горелов, 1972), в Закавказье — в начале мая (Верещагин, 1942), в Илийской и Сары-тогайской долине (900 м над ур. моря) — 20 апреля — 29 июня (Слудский, 1977).
По наблюдениям Д.А.Бланка (1985), беременная самка остается в группе (взрослые самки, молодняк) до последнего часа перед родами. Плод выходит легко и быстро, самки рожают в положении лежа или "вприсидку", обычно в местах более или менее защищенных (под кустами, в лощинах и т.д.). В годы высокой численности джейранов отмечались "родильные дома” (Гептнер и др., 1961). В 50-х годах такой "родильный дом", состоящий из 500 взрослых и родившихся джейранов, был отмечен в Западном Казахстане (Горбунов, Зархидзе, 1986).
Сразу после родов самка облизывает детеныша, кормит его молоком. Повторив эти процедуры 2—3 раза, она возобновляет пастьбу. Однако первые недели не уходит далеко (не более 0,5 км), лишь при водопое удаляется до 4—5 км. Время между приемами молока детеныш, затаившись, лежит на земле, вытянув шею и голову вперед; больше всего его в это время выдают большие темные и выразительные глаза: он их не закрывает и при испуге (Динник, 1910).
Плодовитость. У джейрана в году бывает один помет. Наличие 2—3 периодов подъема половой активности, изредка весеннее, летнее (кроме продуктивного зимнего), видимо, указывает на возможность существования в прошлое геологическое время и большего количества пометов в году.
Ягнят в помете обычно у молодых самок — 1, у взрослых — 2, редко 3, исключительно редко 4. В Бадхизс, когда отмечался подъем численности (1975—1979 гг.), среди самок с потомством двойни оказались у 63% (Кузнецов, 1986). При стечении благоприятных условий резко падает яловость. Так, по А.А.Слудскому (1977), в Бетпак-Дале из 36 исследованных взрослых и полувзрослых самок яловыми оказались всего 4, что автор связывает с благопритной зимовкой предыдущего года.
Рост и развитие. Ягнята рождаются хорошо сформировавшимися. При появлении на свет открываются глаза, тело покрыто густым покровом волос, ягненок очень быстро становится на ноги: по В.Г.Гептнеру и др. (1961), через несколько часов, а по А.А.Слудскому (1977), через 1—2 ч. Непосредственный хронометраж поведения джейраненка в первый час жизни, проведенный Д.А.Бланком (1985) в Илийской долине, показал: не более,
439
Джейран
чем через 11 мин после рождения он ходил и даже пытался пробежать и прыгать, спустя еще несколько минут совершил первое сосание молока. Первые 7—10 дней джейраненок лежит на месте, определенном матерью. Потом он сам выбирает себе это место. При наличии двух ягнят самка укладывает их врозь, кормит в разное время и переводит по очереди на новые места. Сводит их через 3—4 дня, после чего они уже сосут одновременно и лежат рядом. Джейраненок в первую неделю, следуя за матерью, обнюхивает те растения, которые она поедает, а потом самостоятельно различает растения и начинает скусывать листья и жевать их. Спустя 15—17 дней он уже пасется, но недолго и нуждается в молоке матери в течение 3—3,5 мес.
Молодые набирают массу тела очень быстро. Так, по данным А.А.Слудского (1977), при средней массе взрослого самца 30 кг и самки 24 кг, годовалый самец весит в среднем 24 кг. Особенно высок темп роста в первые месяцы жизни. Если новорожденный весит в среднем 2,4 кг, то к месяцу — 7,1—9,5 кг, к 3 мес — 13,5 кг, к 6 мес — 17,5—19,5 кг.
Развитие зубной системы проходит сложно, основные его этапы отражены в табл. 82.
Структура популяции. Сведения о половом и возрастном составе популяции джейрана Восточного Закавказья в годы его высокой численности мы не имеем. Некоторые данные, полученные М.А.Сафаровым (1961), относятся к периоду минимальной численности вида в регионе. В трех районах (Джейранчел, близ устья Шамхорчай-Кюрокчай и Ширванская степь) отмечено всего 23 джейрана, из которых взрослых 10 самцов, 6 самок, а остальные 7 животных сеголетки. По остальным районам, где учтено 108 голов, данных о составе популяции нет.
Многолетние данные, полученные А.А.Слудским (1977) в Бстпак-Дале, показали значительную динамику соотношения полов. За несколько лет среди взрослых самцы составляли от 5,4 до 21,1%, тогда как самки — от 78,9 до 94,6%. На одного самца в среднем приходилось соответственно от 3,5 до 17,5 самок. Автор считает, что выборка данных была достаточной и, следовательно, исключался недоучет одного из полов. Соотношения полов эмбрионов и новорожденных (л = 59 экз.) в 1955 г.: самцы составляли 47,4%, самки — 52,6%.
Возрастная структура, по данным названного автора, меняется широко не только по годам, но и в течение одного года за счет высокой смертности молодняка. В среднем за три года на одну самку приходится 1,7 ягненка, а в сентябре — 1,1, т.е. сохранилось около 60% родившихся. В благоприятные годы поголовье возрастает за счет прибылых на 85—90%, а при неблагоприятных (джуты) — всего на 54,2% (1950/51 г.).
Враги, паразиты, конкуренты, смертность. У джейрана в Восточном Закавказье много врагов среди хищных зверей и птиц. По данным Ю.М.Шади-лова (1986), все еще много волков, лисиц и шакалов в Ширванском заповеднике и Бяндованском заказнике, где в настоящее время обитает самая крупная популяция джейрана. Здесь также много собак, в том числе бродячих и одичавших. По данным названного автора, на площади 69 тыс.га
440
Ovis Linnaeus, 1758
одних только пастушьих собак бывает около 400. Роль хищников увеличивается в связи с мероприятиями по восстановлению джейрана в ряде районов Восточного Закавказья. Известно, что синантропизация волка сопровождается увеличением их численности на охраняемых территориях (Бараташвили, 1989). Немало данных свидетельствует о том, что при уничтожении волков резко возрастает численность шакалов, бродячих собак.
У джейранов найдено около двух десятков гельминтов, но патогенное значение их неясно (Гептнер и др., 1961). По Б.Е.Курашвили и др. (1985), в Закавказье зарегистрированы гельминтозы: дикроцеллиоз, хаберциоз, трихо-стронгилидоз. На джейранах паразитируют 6 видов клещей, один вид вшей, один вид пухоеда. Джейраны страдают от кровососущих насекомых, а также оводов (особенно Pavlavskiata subgutturosae и Crivellia corinnae).
В Закавказье серьезными конкурентами джейранов являются домашние животные, особенно овцы. Смертность джейрана в Закавказье очень высока в многоснежные зимы, которые бывают раз в 15—20 лет. Последний раз такая высокая смертность была зарегистрирована в 1950 г.
Численность и ее динамика. В 1960 г., когда джейран находился на храни гибели во всем Закавказье, на территории Азербайджана (без Гобустана) учтен всего 131 джейран (Сафаров, 1961). С конца 60-х годов и по настоящее время происходит увеличение численности, что можно проиллюстрировать примером охраняемой территории Ширванской степи (здесь обитает основное поголовье). Если в 1960 г. на территории Бяндованского заказника обитало 77 джейранов (Сафаров, 1961), то в 1969 г. их стало 450 голов. В 1980 г. в Ширванском заповеднике и южной части названного заказника обитало 3600 джейранов; в 1982 г. 4000, а в 1983 г. — 4500 (Хахин и др., 1986).
Практическое значение. В Восточном Закавказье джейран имел важное экономическое значение. Ради очень вкусного мяса его добывали в большом количестве. Еще в 1932—1937 гг. добыча составляла до 2000 голов в год (Верещагин, 1942). При живой массе самцов до 32 кг и самки 28 кг, по данным названного автора, убойный вес составляет до 60%. Шкурки в зимнем мехе используются в качестве ковриков.
Джейран может быть одомашнен и представляет интерес в селекции новых пород домашних животных. Ущерб полевым культурам от джейранов незначителен. Неизвестны случаи, когда джейраны были истчниками инфекции для домашних животных.
Род Горные бараны — Ovis Linnaeus, 1758
Бараны мелких и средних размеров. Длина тела ПО—200 см, длина хвоста 3—13 см, высота в холке 65—125 см. Масса тела 20—230 кг. Телосложение плотное, но стройное. Удлиненное тело на относительно высоких и стройных ногах, хвост покрыт снизу волосами, короче длины ушей, не имеет утолщения. Горные бараны по экстерьеру резко отличаются от домашних овец. Конец морды опушенный, лишь по середине верхней губы и
441
Горные бараны
по краю ноздрей проходит узкая полоска голой кожи. У самцов рога большие, длиной 50—190 см, а у самок значительно меньше — 10—30 см. Структура рогов сильно изменчива, но довольно своеобразна, что существенно отличает представителей рода Ovis от рода Capra. В поперечном сечении рог имеет обычно треугольную форму благодаря наличию на его поверхности трех ребер: передне-внутреннего, передне-наружного и заднего. Иногда у отдельных форм и особей, в том числе закавказских баранов, слабо развито передне-наружное ребро. Соответственно различаются три грани: наружная, внутренняя и лобовая. В зависимости от степени развития ребер может меняться форма поперечного сечения рога от треугольной до линзообразной. Рога самок не только короче, но и тоньше.
Большое систематико-таксономическое значение придается характеру за-крученности рогов у самцов. В связи с разработкой хромосомной изменчивости диких баранов складывается мнение, что фенотип рогов соответствует кариотипу (Воронцов и др., 1972; Сопин, Харрисон, 1986). Поэтому во всех работах, посвященных систематике и изменчивости диких баранов, большое место занимает описание рогов самцов. Рога самок в этом отношении не представляют интереса. Очень подробный анализ всех аспектов изменчивости рогов диких баранов провел В.И.Цалкин (1951). Здесь мы отметим, что широкий спектр географической изменчивости можно свести к трем типам. Первый, или гомонимный, тип: рога сначала загибаются вверх, назад, а затем наружу, вниз, вперед и наружу. Второй, или перверти-рованный тип: рога с изгибом вверх, назад, наружу, а затем в концевой половине загибаются вниз и внутрь к шее. Третий, или гетеронимный, тип: сначала вверх, назад, наружу, а в концевом отделе идут более или менее горизонтально к шее.
В отличие от козлов на поверхности рогов нет бугров или поперечных валиков, однако поверхность их морщинистая; в той или иной степени хорошо выражены годовые сегменты. Они отделены друг от друга поперечными бороздами. Ширина и длина годичных сегментов подвержены сезонной и географической изменчивости, что зависит от ряда факторов. Волосяной покров, состоящий из ости и подшерстка, относительно низкий, лишь у некоторых форм .развит подвес из длинных, жестких волос, занимающих низ шеи и отчасти труди. В отличие от коз "борода" отсутствует. Окраска тела одноцветная, но варьирует от белой, беловато-серой и желтой до коричневой и черной. Окраска низа светлее боков и спины. Смена волосяного покрова происходит один раз в год — весной. Хорошо развиты предглазничные. паховые и межпальцевые железы. Подхвостовая железа в отличие от коз обычно отсутствует. Нет также запястных желез. Сосков одна пара.
Череп относительно укороченный, сильно пневматизированный, особенно его зароговая часть. Изгиб лобно-теменной кости значительный, особенно у взрослых и старых самцов, обладающих большими рогами. Носовые кости развиты нормально. Верхние отростки межчелюстных костей обычно длинные и вдаются между носовыми и верхнечелюстными костями, реже укорочены и не доходят до носовых костей. Носовые отверстия
442
Ovis Linnaeus, 1758
нормального размера. Этмоидальные отверстия чаще отсутствуют или развиты слабо. Слезная кость имеет хорошо развитую ямку для предглазнич-ной железы. Костные слуховые барабаны небольшие. Характерны гипсодонтные зубы. Резцы долотовидной формы.
Зубная формула:
0 0 3 3
I-C - Р - М - = 32.
3 13 3
В посткраниальном скелете специалисты (Громова, 1953; Соколов, 1959; и др.) обращают внимание на такие особенности, как более короткая пястная кость, чем плечевая или предплечевая кости, более длинные и узкие метаподии, по сравнению с козами. Длина пястной кости не менее 78% длины лучевой, плюсневой — не менее 62% длины берцовой и т.д.
Диплоидное число хромосом у горных баранов труппы vignei (уриалов) составляет 58, группы poll (архары) — 56, труппы musimon (муфлоны) — 54.
Ареал рода обширный, охватывает некоторые острова Средиземноморья, Переднюю, Среднюю, Центральную, Северо-Восточную и частично Северную Азию и запад Северной Америки от Аляски до Мексики.
Места обитания связаны с открытыми горными ландшафтами, имеющими сглаженный рельеф; в отличие от козлов горные бараны избегают скальных участков, но нередко выбирают пологие пастбища, чередующиеся ущельями и скалами, где находят в случае необходимости защиту от врагов, благоприятный микроклимат в жару или в холод. Могут занимать различные высоты, начиная от низменности (ниже уровня Мирового океана) до 5000 м над ур. моря, что обеспечивается высокой кислородной емкостью крови.
В питании горных баранов основное значение имеют злаки, некоторые виды разнотравья, в высокогорьях — осоки, в полупустынях — солянки, полыни. Зимой животные поедают высохшую траву, ветки кустарников и листья. Однако веточный корм и вообще древесно-кустарниковая растительность не играет той роли, что у коз. Летом бараны активны обычно в ранние утренние часы, а зимой преимущественно днем. Стадные животные; группировки в сотни и более' голов образуются осенью и зимой. Нередко бараны совершают миграции, вызванные недостатком пищи или появлением глубокого снегового покрова.
Гон происходит в октябре-ноябре, занимает 3—6 нед. "Гаремы" включают 1—2 самцов и от 5 до 25 самок, их формирование сопровождается драками, но не ожесточенными. Беременность около 5 мес, приносят 1—2, редко 3 ягнят в период с марта до середины июня. Для ягнения самка уединяется, выбирая укромное место (пещеры, навесы камней и др.). Половозрелость наступает в 1,5—3 года, но самцы, как правило, участвуют в размножении на 4—5-м году жизни, так как не выдерживают конкуренции более старших самцов.
Бараны относятся’к важным промысловым животным. От 54-хромо-сомных горных баранов (группы musimon) произошли все домашние поро
443
Азиатский муфлон
ды. Успешно используются для гибридизации с домашними породами в целях выведения новых пород (горный меринос, архаромеринос).
Систематика рода нуждается в коренной ревизии с привлечением комплексного материала. 54-хромосомных диких баранов Закавказья, Северо-Западного Ирана и Малой Азии мы рассматриваем в качестве самосто-ятельнго вида Ovis orientalis Gmelin.
Азиатский муфлон — Ovis orientalis Gmelin, 1774
Рис. 61
Ovis ammon, Linnaeus, 1758; Цалкин, 1951; Гептнер и др., 1961; Ovis orientalis, Gmelin, 1774; Бобринский, 1944; Ellerman et Morrison-Scott, 1951; Соколов, 1959, 1963; Ovis gmclini, Bluth, 1841; Динник, 1910; Musimon ana-tolica, Северцов, 1873; Ovis ophion, Насонов, 1923.
Коллекционный материал. ЗИН РАН: 25 черепов с роговыми чехлами, 16 скелетов, 11 роговых чехлов; МГУ: 2 черепа с роговыми чехлами.
Диагноз. Бараны средних размеров. Масса тела самцов до 95 кг, самок — 75 кг. Наибольшая длина черепа самцов до 264 мм, самок — 238 мм. Рога гетеронимные, длиной до 67 см. На верхней части боков имеется большое светлое пятно различной формы.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. В Закавказье масса тела самцов в среднем 65—75 кг, самок — 50—60 кг (Саркисов, 1939, 1950; Явруян, 1970). По Н.Я.Диннику (1910), длина тела самцов до 150 см, высота в холке 84—92 см; по Э.Г.Явруяну (1970), она составляет в возрасте старше 5 лет соответственно 121—148 и 88—93 см. У самок того же возраста длина тела 93—96 см, высота в холке 72—76 см.
При сравнении этих параметров с таковыми европейских муфлонов на западе и туркменскими уриалами на востоке нетрудно заметить промежуточное положение закавказских баранов между ними как по массе, так и размерам тела. Этот факт указывает ещё раз на очень высокую вариабельность экстерьера диких баранов й на малую значимость для систематики массы и размеров тела. Зона перекрывания этих показателей у закавказских муфлонов (2п = 54) и туркменских уриалов (2п = 58) настолько значительна, что трудно их использовать для раз деления этих видов.
По размерам и структуре череп закавказских баранов относится к западному, или типу муфлона (Цалкин, 1951; Гептнер и др., 1961). Череп средней величины. По данным Э.Г. Явруяна (1969), общая длина черепа составляет 228—264 мм (х = 244,4), высота черепа 117—139 мм (х' = 128,7), ширина черепа по задним стенкам орбит 117—154 мм (х - 131,2), длина мозговой коробки 88—108 мм (х=94,7), длина предкоренных зубов 18—32 - мм (Х= 25,1), длина коренных зубов 40—53 мм (х— 47,6) (табл. 83).
По величине черепа закавказские бараны занимают также промежуточное положение между европейскими муфлонами и туркменскими аргали;
444
Ovis orientalis Grnclin, 1774
Таблица 83. Основные промеры черепа азиатского муфлона из Закавказья.
__________________________Самцы (п = 46)__________________________
Признак limit X
Общая длина черепа 228-264 244,4
Кондилобазальная длина черепа 216-256 231,7
Базилярная длина 198-236 213,1
Длина лицевой части 174-197 185,2
морды 130-150 140,1
мозговой коробки 88-108 94,7
носовых костей 71-100 88,5
ложнокоренных зубов 18-32 25,1
настоящих коренных 40—53 47,6
Высота черепа 117-139 128,7
Длина межчелюстной кости . 70-97 84,9
Межглазничная ширина 81-102 90,1
Ширина черепа по задней стенке орбит 117-154 131,2
Ширина черепа в области слуховых отверстий 73-91 81,2
Ширина мозговой коробки 63-79 72,4
хорошо выражена зона перекрытия по всем изученным измерениям черепа как с западными, так и восточными популяциями.
Как справедливо отмечали предыдущие исследователи (Цалкин, 1951; Гептнер и др., 1961), кроме относительно небольших размеров, для закавказских баранов характерен ряд особенностей. Так, костные стержни рогов тонкие и длинные (обхват основания составляет около 60% длины); высшая точка лобных костей находится над орбитами или немного позади; затылочная область .выдастся за линию задних краев оснований костных стержней рогов; лобная кость сравнительно невелика и слабее изогнута в вертикальном направлении; теменная кость относительно велика; сильно изогнута верхняя Затылочная кость; морда довольно низкая, узкая и короткая (до 60% общей длины черепа); межчелюстные кости короткие и обычно не выдаются между носовыми и верхнечелюстными костями; глазницы сравнительно крупные. Такой тип черепа имеют, кроме баранов Закавказья, популяции, населяющие острова Средиземного моря, Малую Азию, Иран, Афганистан, Туркмению, Устюрт и Кызылкумы и далее на восток до северо-западной Индии (Цалкин, 1951; Гептнер и др., 1961).
Длина рогов по лобной поверхности у взрослых самцов (п = 46) составляет 370—784 мм (х = 588,7), обхват у основания рогов 215—270 мм (X = 238,1), а длина 4 сегментов рогов 212—652 мм (х = 472). Длина костного стержня рогов тех же самцов 200—450 мм (х = 342), расстояние между наружными краями стержней рогов 27—36 мм (х = 31,4). Рога самок небольшие, длина по лобной поверхности 90—152 мм (х = 125), обхват у основания 83—108 мм (х= 94).
По структуре рога закавказских баранов составляют определенный закавказский тип. Это гстсронимные рога, небольшие и тонкие у основания. От черепа они отходят вверх и в стороны, а затем идут назад и внутрь, навстречу друг другу; на самом конце они могут быть слегка загнуты вверх. В
445
Азиатский муфлон
Рис. 61. Распространение азиатского муфлона
446
Ovis orientalis Gmelin, 1774
1 — Кедский р-н (Арабули, 1969);
2 — Мастара (Саркисов, 1941, 1944;
Даль, 1954; Верещагин, 1959);
3 — Алагяз (Верещагин, 1959; ЗИН);
4 — Арагац (Насонов, 1923);
5 — Арегунский хребет (Ананян, 1962);
6 — Арташат (Красная книга СССР, 1984);
7 — Веди (Даль, 1944);
8 — Арарат (Марков, 1924, Верещагин, 1959);
9 — Хосровский заповедник (Соколов, Сыроечковский, 1990);
10 — нижнее течение Арпачая (Сатунин, 1912);
11 — Хачик (Даль, 1948., Красная книга СССР, 1984);
12 — Хидзорут (Даль, 1948; Красная книга СССР, 1984);
13 — Азизбеков (Явруян, 1968);
14 — Заритап (Красная книга СССР, 1984);
15 — Шахбуз (Верещагин, 1959;
ЗИН);
16 — верховья р. Нахичеваньчай (Явруян, 1968);
17 — Сисиан (Явруян, 1970);
18 — Горис (Явруян, 1970);
19 — Кафан (Явруян, 1970);
20 — Шикахохский заповедник (Соколов, Сыроечковский, 1990);
21 — Мегри (Красная книга СССР, 1984);
22 — горы между Мегри и Ордубадом (Марков, 1924);
23 — Ордубад (Верещагин, 1959;
Красная книга СССР, 1984)
качестве индивидуальной изменчивости встречаются отдельные особи со слабо первертированным типом — концевые части рогов направляются вниз, а не внутрь. Закавказский тип рогов свойствен в пределах СССР только баранам
447
Азиатский муфлон
Закавказья, а за рубежом они развиты у баранов Кипра, западных частей Ирана. Типичные первертированные рога имеют животные из Передней Азии (ирано-афганской части ареала рода). Как видно, гетеронимные рога имеют 54-хромосомные бараны Запада, а первертированные — как 54-, так и 58-хромосомные бараны.
Изучению посткраниального скелета диких баранов посвящен ряд работ, обзор которых с дополнением новых данных приводится в статье Ф.Б.Чср-нявского (1964). Оказалось, что у представителей подсемейства Caprinae 30—31 позвонков в позвоночном столбе; вариация количества происходит за счет крестцового отдела, включающего 4—5 позвонков. Шейных 7, грудных 13 и поясничных 6 позвонков. Длина названных отделов по отношению к общей длине позвоночника (6%) у О. orientalis и О. ammon сходна, но существенно отличается от О. nivicola, живущего в скалах высокогорья. Так, длина шейного отдела у самцов (нет половых различий) трех названных видов составляет: 31,1; 30,2; 28,7%, а поясничного отдела соответственно 21,6; 21,5; 23,5. Длина передних конечностей к длине позвоночного столба составляет 58,4; 59,2; 57,8%, длина задних конечностей: 72,0; 72,2; 70,4%. Особенности первых двух видов автор связывает со способностью их к быстрому бегу в условиях сглаженного горного рельефа. Как отмечалось ранее, у типично горных копытных (туры, безоаровые козлы) ярко выражены приспособления к высокогорью, и по этим признакам они ближе стоят к О. nivicola, чем к другим видам баранов. Из других частей посткраниального скелета, рассмотренных Ф.Б.Чернявским, обратим внимание лишь на ширину лопатки по отношению к ее длине — признак, который отражает степень развития мышц передних конечностей. Она оказалась значительно больше у толсторогов, чем у О. orientalis и О. ammon. В связи с обсуждаемым вопросом интерес представляет сравнение мышц конечностей закавказских баранов и кавказских туров, проведенное Н.М.Ахмедовым (1973). Мускулатура передних конечностей составляет у баранов Кавказа 6,95 % от массы тела, у туров — 7,00%, а у задних конечностей соответственно 17,22 и 14,84%. Как видно, мышцы задних конечностей закавказских баранов лучше развиты, чем у туров, а передних — уступают турам лишь немного.
Окраска тела, по мнению В.Г.Гептнера и др. (1961), будучи строго локализованной географически, у диких баранов представлена четырьмя различными типами. Одним из них является тип закавказского барана, свойственный популяциям Малой Азии, Северо-Западного Ирана и Закавказья. Общий тон верха зимнего покрова, как еще отмечал Н.Я.Динник (1910), рыжевато-желтый, рыжевато-бурый, каштаново-рыжий различных оттенков. У самцов имеются резко выраженные белые поля на боках. Форма их различна, на спине они могут сливаться в виде седла. Иногда к белым волосам пятен могут примешиваться волосы основного тона туловища и тем самым затушевывать интенсивность пятна. У отдельных особей эти поля могут почти полностью отсутствовать.
Белые или беловатые волосы покрывают брюхо, заднюю сторону ляжек ("зеркало"), конец морды, нижнюю губу, внутренние и нижние стороны
448
Ovis oricntalis Gmclin, 1774
ног. Окраска головы серовато-бурая или рыжевато-желтая. Характерно наличие хорошо развитого темного (темно-коричневого или коричневатобурого) подвеса; длина составляющих волос 8—20 см. Подвес обычно начинается в 12—17 см ниже горла и тянется, постепенно расширяясь и удлиняясь, до начала груди. Летняя окраска самцов бледнее, чем зимняя, светлого поля нет, слабо развит подвес, длина волос не более 8 см. Окраска тела самок зимой и летом почти одинакова, отличается тем, что более тусклая, отсутствуют боковые пятна, а также подвес.
Окраска тела закавказских баранов сходна с типом муфлона, свойственного баранам Корсики и Сардинии, но существенно отличается от туркменских. В частности, у последних нет белых пятен, зеркало невелико и слабо выражено, подвес сильно развит (до 20 см) и длинными волосами начинается при основании головы в виде, двух пучков по бокам и потом соединяются на горле. Такая борода переходит в подвес. В области бороды обычно волосы белые, лишь к труди увеличивается число темных волос в той или иной степени.
При оценке экстерьерных признаков закавказских баранов и родственных им популяций Северо-Западного Ирана и Малой Азии обратим внимание, что, по Л.В.Сопину (1983), ведущими признаками горных баранов являются: тип рогов, окраска тела, форма и размеры околохвостового пятна, форма и цвет подвеса.
Кожный покров. Морфологический материал по кожному покрову архара из Закавказья добыть не удалось. Однако исследования близких видов рода Ovis позволяют выделить некоторые общие характерные черты.
Кожа имеет среднюю толщину, изменяющуюся по сезонам: по-видимому, она толще летом, чем зимой. Непигмснтированный или слабо пигментированный эпидермис кожи туловища тонкий на спине и утолщен на груди, с хорошо развитым роговым слоем, иногда содержит прерывистый зернистый слой (на крестце) с ячеистой внутренней поверхностью (на труди). Сосочковый слой дермы развит лучше сетчатого. Подкожная жировая клетчатка обнаружена на труди. Обычные сальные железы остевых волос однодольчатые или двух-трехдольчатые, лежат по одной или по две с обоих сторон волоса, иногда в два яруса. Сальные железы пуховых волос простые мешковидные, располагаются по одной у каждого волоса. Волосы на загривке растут густо, подразделяются на остевые (иногда выделяются промежуточные) и пуховые. Пуховые волосы растут пучками. Мышцы, поднимающие волосы, хорошо различимы. Из специфических кожных желез обнаружены: мейбомиевые, носового зеркала, предглазничные, паховые, цир-куманальные, межпальцевые, половых туб. Возможно присутствие подхвостовых, препуциальных и желез наружного слухового прохода.
Система крови. В работе Н.Н.Булатовой (1962) содержатся сведения по крови архаров, всего 12 взрослых самок, изученных с 29 октября по 9 ноября; уриалов, всего 5 особей (2 самца и 3 самки) возрастом от 1 до 4 лет (с 28.VI по 18.VII) и муфлона, одной самки в возрасте 1,5 года (19.VI). У архаров количество эритроцитов составило 10,38—18,60 млн/мм3 (Г = 13,5), диаметр одного эритроцита 4,61—5,22 мкм (X = 4,95), объем одного
29 В.Е.Соколов, А.К.Тсмботов
449
Азиатский муфлон
эритроцита 30—34,4 мкм3 (д’ = 31,5), концентрация гемоглобина в эритроцитах 38,2—43,5% (лГ = 40,4), гематокрит 16,6—19,0 г% (х =17,9).У трех самцов архара, возрастом 2—3 года, с 4 ноября по 7 декабря (масса тела 51—70 кг) изучен объем циркулирующей лдзови. Отношение массы крови к-массе тела составило 8,8—10,8% (х = 9,9). У уриалов количество эритроцитов было 13,4—18,7 млн/мм3 (х = 14,67), диаметр одного эритроцита 5.03—5,28 мкм (х= 5,15), объем одного эритроцита 26,2—28,0 мкм3 (х= 27,4), гемоглобина 15,3—18,7 г% (х= 16,9). У одного трехлетнего самца с массой тела 30 кг изучен 9.VI объем циркулирующей крови. Отношение массы крови к масе тела 9,63%. У полуторагодовалой самки муфлона, массой тела в 25 кг, изученной 19.VI, оказалось эритроцитов 12,22 млн/мм3, диаметр одного эритроцита 5,16 мкм, объем одного эритроцита 34,2 мкм3, гемоглобина 16,6 г%. Как видно, кислородная емкость крови всех изученных диких баранов высокая. Аналогичные результаты получены по муфлонам зоопарков Великобритании и Нидерландов (Hawkey et al., 1984), гривистым баранам Лондонского зоопарка (Hawly et al., 1984), толсторогам Нельсона (McDonald et al., 1981), баранам Далла с Аляски. В условиях мягкого климата Великобритании, близких к средиземноморским, количество эритроцитов 13,0 млн/мм3 у самцов и 11,9 млн/мм3 у самок, лейкоцитов 7,8— 9,1 тыс./мм3. В суровых условиях Аляски у барана Далла число эритроцитов 13,8 млн/мм3, лейкоцитов 7,6 тыс./мм3, гемоглобина 14,3 г%, гематокрит составляет 46%, общий протеин — 6,8 г%, в том числе альбумина 3,3 г%, глобулина 3,5 г%. Кальция содержится 9,6 мг%, фосфора — 4,6 мг%.
У муфлона из зоопарка в Великобритании выявлена динамика гематологических показателей в связи с возрастом, полом, сезоном и содержанием витамина Е в рационе. Так, с возрастом увеличивается количество эритроцитов: при рождении — 8,2 млн/мм3, 1,5 нед — 11,8, старше — 13 млн/мм3; лейкоцитов соответственно 4,0 тыс/мм3, 9,8 и 7,8—9,1. Изменения показателей лейкоцитов происходит за счет нейтрофилов и лимфоцитов. Как отмечено выше, эритроцитов у самцов больше, чем у самок. Минимальное число эритроцитов и наибольшие их размеры приходятся на период отела и лактации (февраль-июнь), что, очевидно, связано с повышенной деятельностью эритропоэза в это время. Аналогичное явление нами обнаружено у ряда видов мелких млекопитающих (Темботова, 1974; Темботова и др., 1979, 1980, 1987; и др.). Обнаружены некоторые малосущественные различия, связанные с характером местообитания. Добавление к естественному корму витамина Е не вносит существенных изменений в гематологические показатели.
Четкие возрастные изменения, согласующиеся с изложенной выше динамикой, выявлены также у гривистого барана Лондонского зоопарка. Изучены новорожденные, шестинедельные, годовалые и животные старше 1 года. С возрастом увеличивается в 2 раза число эритроцитов, но также вдвое сокращается их объем, в 1,5—2 раза возрастает количество нейтрофилов, в 5—10 раз — количество лимфоцитов, в 1,5—2 раза растет число тромбоцитов и содержание фибриллогенов.
450
Ovis orientalis Gmelin, 1774
Индексы внутренних органов закавказских баранов не изучены.
Кариотип. В ряде работ анализирован кариотип рода Ovis, в том числе рассматриваемого вида (Nadler et al., 1971; Кулиев, 1974; Valdez et al., 1978; и др.). Согласно этим данным, в хромосомном аппарате 2п = 54, NF = 60, среди которых шесть крупных метацентрических аутосом, крупная акроцентрическая Х-хромосома и мелкая субметацентрическая У-хромосома. При дифференциальной окраске удастся идентифицировать все элементы, но с высокой точностью различаются первые три пары двуплечих хромосом как по количеству полос, так и их локализации.
В пределах закавказской части ареала кариотип мономорфный, хотя, видимо, нельзя исключать возможности обнаружения других кариотипических форм, имея в виду обнаружение полиморфной популяции на Центральном Эльбрусе, включающей особей с 2л — 54, 55, 56, 57, 58 (Nadler et al., 1971; Воронцов и др., 1972; Valdez et al., 1978).
К 54-хромосомным баранам относятся, кроме популяции Закавказья, также бараны Северо-Западного Ирана (в том числе западной части Эльбруса), Малой Азии, островов Средиземноморья (Ovis musimon и О. orientalis). Восточный Иран, включая восточную часть Эльбруса на западе до Ишкашима на востоке, и Малый Тибет населяют бараны с 2л = 58 (Nadler et al., 1971; Воронцов и др., 1972; Valdez et al., 1978).
Структура и динамика ареала
Ареал вида ограничен Восточным Закавказьем, Северо-Западным Ираном и Малой Азией. Более конкретно очертить его затруднительно. Северная граница проходит по Малому Кавказу, восточная — по линии, соединяющей Центральный Эльбурс с Ларистаном. К востоку от этой линии располагается гибридная зона между 54- и 58-хромосомными видами, ширина которой определяется примерно в 150 км (Valdez et al., 1978). В этой зоне обнаружены разные кариотипические формы, в том числе 54- и 55-хромо-сомные формы, обладающие смешанными морфологическими признаками обоих видов. Мало современных данных и по южной границе. Получение однозначных сведений осложняется тем фактом, что дикие бараны Юго-Восточной Аравии относятся к другому виду — Capra vignei (Сопин, Харрисон, 1986). Пока трудно сказать, где кончается ареал одного вида и начинается распространение другого. Или имеет место налегание ареалов двух разных видов. Западная граница закавказского барана, по К.А.Сатунину (1912), доходит до Сарыкамыша, много этих баранов было на Зорских горах, южнее с. Йгдыр, оседло живут они на Арарате. В.И.Цалкин (1951) проводит западную границу восточнее Карского плато и оз. Ван. А.А.Сар-кисов (1944) полагал, что эти бараны не доходят до Карского плато.
На территории б. СССР азиатский муфлон встречается только в Закавказье. Азиатский муфлон, видимо, в историческое время никогда не имел широкого распространения на Кавказе. В начале текущего столетия, как писал К.А.Сатунин (1910), он встречался только в немногих местах, "а
29
451
Азиатский муфлон
именно в самой южной части его, на горах, находящихся в Эриванской губернии и Карской области вблизи траниц России с Персией и Турцией" (с. 159). Обобщая данные К.А.Сатунина, Г.И.Радде сообщал, что муфлоны живут в горах по нижнему течению Арпачая, часто встречаются недалеко от Ордубада, во время суровых зим спускаются в долину Аракса, оседло живут на Арарате и в южной части Нахичевани. Видимо, азиатский муфлон в то время встречался в Кедском районе Аджарии, о чем свидетельствует современное его обитание на пограничной с Турцией полосе (Арабули, 1989). На карте Н.К.Верещагина (1959) эта точка отсутствует.
В пределах Восточного Закавказья в середине и, видимо, в конце XIX в. северная граница, вероятно, простиралась до Арйгаца (Насонов, 1923). Об этом свидетельствует коллекционный материал ЗИН РАН, происходящий из Алагеза (4095 м над ур. моря, 1849 г.) и окрестностей с. Мастара (1911 г.) той же горы. В 30—40 годах нашего века муфлонов там уже нс было (Саркисов, 1941, 1944; Даль, 1954). Есть указание, что лет 300—350 тому назад муфлоны жили на Арегунском хребте, к северу от оз. Севан (Ананян, 1962). Э.Г.Явруян (1969) считает, что это маловероятно, ибо там в прошлом росли большие леса, непригодные для жизни муфлонов. По данным Э.Г.Яв-руяна с 50-х годов текущего столетия эти животные уже не появляются в районе Джсмурука. Словом, северная граница ареала постоянно смещалась на юг, сокращалась площадь, занятая дикими баранами повсеместно. Распространение в наши дни (в 60-х годах) отражено на карте Э.Г.Явруяна (1969), где указано всего 5 точек постоянного обитания диких баранов. Они расположены в следующих местах: 1) ущелье Арацо Урцского хребта; 2) ущелье Багсадзор, расположенное на западе Нахичевани; 3) Айоцдзор-ский хребет, ущелье Каракуш; 4) хр. Гапу-Даш-Кара в верховьях Нахиче-ваньчая; 5) Южно-Зангезурский хребет. В различных исследованиях неоднократно упоминались эти места нахождения муфлонов. В частности, на обитание баранов наУрцскомхребте указывали почти все, кто занимался этими животными в Закавказье (Саркисов, 1941, 1944; Даль, 1944, 1951; Явруян и др., 1971; и др.). Заповедник, созданный там, видимо, обеспечит сохранение муфлона в Армении. На карте Э.Г.Явруяна (1969) северную траницу ареала в настоящее время составляет Ероносский хребет, она проходит севернее Ехегнадзора, в окрестностях Азизбекова, восточнее которого по Северо-Зангезурскому хребту делает большую петлю, достигая почти границы Азербайджана. Далее на вбеток, вновь по Баргуштанскому хребту образует узкий клин, занимающий почти весь названный боковой отрог Зангезурско-го хребта. Муфлон занимает также Мсгринский и Хуступ-Катарский хребты. Южная граница проходит по склонам хребтов и отрогов, обращенных к р. Араке, не достигая ее на 20—30 км. Лишь в районе Ордубада, по отрогам Зангезурского хребта граница близко подходит к Араксу. Однако по сей день дикие бараны пересекают Араксинскую долину и саму реку, совершая митрации с севера на юг и обратно. О таких переходах писали А.А.Саркисов (1941, 1944), С.К.Даль (1944), Э.Г.Явруян (1969) и др.
Сокращение распространения азиатского муфлона за пределами Армении, в том числе в Грузии, было интенсивным. По данным А.Б.Арабули
452
Ovis orientalis Gmelin, 1774
(1989), он сохранился лишь в одной точке и то на нейтральной зоне, на границе с Турцией, в Кедском районе Аджарии. Здесь в 1985 г. насчитывалось всего 40 особей. Хотя они защищены от браконьерства в силу пограничного режима, но лишены возможности расширения своего распространения.
Современные тенденции антропогенных изменений ареала. Факты, которыми мы располагаем, свидетельствуют о том, что за последние 100— 150 лет ареал азиатского муфлона в Закавказье сократился как минимум в 2 раза. Немало оснований утверждать, что уменьшение распространения сопровождалось интенсивным увеличением спорадичности поселений. Однако нет достаточно данных, чтобы определить приблизительно темп процесса.
Все исследователи, занимавшиеся муфлонами Закавказья, сходятся в том, что основными причинами столь катастрофического сокращения ареала являются антропогенные факторы — прямое преследование и косвенное влияние путем изменения типичных местообитаний. К сожалению, обе группы факторов и по сей день сохранили свою решающую силу, и сообщения, поступающие в печать, говорят об этом. Так, в феврале-марте 1968 г. проводился учет муфлонов в Армении, охватив Сисианское, Горис-ское и Кафанское охотхозяйства, Зангезурский и Бартазский заказники, а также Абовянский, Веденский, Ехегнадзорский, Азизбековский районы Армении и северную часть Нахичевани. Учетом было охвачено около 70% наиболее типичных местообитаний этих животных. Немногочисленные стада были обнаружены лишь на склонах Урцского, Айоцдзорского и Бар-гуштанского хребтов — всего 4 группы численностью в 16 голов (5 самцов и 11 самок). При оценке этих данных нужно учесть, что работа велась зимой, когда основное поголовье мигрирует, на самолете АН-2, уступающем маневренными возможностями вертолету. Однако перемещения муфлонов в Закавказье в связи с многоснежьем не выходят за пределы б. СССР, для них практически непреодолимы пограничные ограждения по долине Арак-са. Поэтому мы разделяем общее заключение Явруяна с соавторами (1974, с. 54): "Учет копытных животных в южных районах Армении, с какими бы большими пропусками он ни проводился, показывает, что численность баранов и безоаровых козлов очень ограничена, и это вызывает тревогу за их дальнейшую судьбу". Данные последующих лет не внесли утешений: "...несмотря на более чем 30-летнюю охрану этих видов и организацию в 1958 г. Хосровского заповедника, основной задачей которого являлась охрана и воспроизводство муфлонов и козлов, положение с безоаровым козлом остается малоутешительным, а с муфлоном безнадежным" (Айрумян, Гаспарян, 1973, с. 6). С таким мнением в целом согласуются и другие литературные данные (Алекперов и др., 1973; Явруян, 1975; Арабули, 1989).
Систематика и изменчивость
Систематическое положение. По сей день нет единого мнения о месте закавказских диких баранов в системе рода Ovis. Достаточно сказать, что в
453
Азиатский муфлон
таких крупных сводках, как работы В.И.Цалкина (1951), В.Г.Гептнера и др. (1961), признается подвид Ovis ammon gmelini Blyth, в синоним с ним сводят О.a. armeniana Nasonov. Этот подвид, по мнению названных авторов, населяет Закавказье, Восточную Турцию и Северо-Западный Иран. В других сводках, как, например, И.И.Соколова (1959, 1963), за баранами Закавказья сохраняется самостоятельный таксон Ovis orientalis armeniana Nasonov, отделяя их тем самым как от других подвидов Малой Азии, так и от вида островов Средиземноморья. Оба взгляда имеют глубокие корни, а также последующее развитие.
Как известно, О.а. gmelini описан Е. Блайсом в 1840 г. по экземпляру из Эрзерума. Реальность существования таксона признавали такие известные систематики, как Р. Лидеккер (Lydekker, 1913, 1915), Н.В.Насонов (1909) и др. По их мнению, этот подвид населял значительную территорию Малой Азии и Закавказье, тогда как в горах Эльбруса обитал О.о. erskinei, а на о-ве Коюндаг, на оз. Урмии — О.о. urmiana. Н.В.Насонов (1919) на основании сравнительного изучения диких баранов Армении пришел к выводу, что они отличаются от типового экземпляра и заслуживают таксономического обозначения, и соответственно предложил новый подвид — О.о. afmeniana. По его мнению, для популяции Эрзерума характерны в изгибе рогов слабая перверсия и хорошо выраженное наружное ребро. Он полагал, что закавказские бараны имеют только гетеронимные рога со слабо выраженным наружным ребром. Исследования, проведенные В.И.Цалки-ным (1951), показали, что систематико-таксономический вес указанных Н.В.Насоновым признаков очень низок, и они не могут служить основанием для отделения Закавказских баранов от эрзерумских. Среди закавказских баранов, как справедливо отмечал В.И.Цалкин, имеются особи, у рогов которых четко выражено наружное ребро, есть и экземпляры, почти лишенные такого ребра. Таково положение и с перверсией. Тем не менее некоторые исследователи до сих пор признают О.о. armeniana (Соколов, 1959, 1963).
В системе Н.В.Насонова для нас представляет интерес то, что, выделяя в самостоятельный вид Ovis gmelini, он в состав этого вида включал 3 подвида: O.g. erskinei (Эльбрус), O.g. urmiana (о-в Коюндаг на оз. Урмии) и O.g. isphaganica (Юго-Западный Иран). Небезынтересен и тот факт, что Северный и Северо-Восточный Иран, по мнению Н.В.Насонова, населяет другой политипический вид — Ovis orientalis, который образует самостоя-j тельные подвиды: О.о. orientalis на Восточном Эльбрусе, О.о. dolgopolovi в горах между Эльбрусом и Копетдагом. Как видно, по мнению Н.В.Насо-нова, на сравнительно небольшой территории Закавказья, Эльбруса и Ко-петдага обитают два вида диких баранов с четырьмя подвидами. Как отмечает В.И.Цалкин (1951), предложенная Н.В.Насоновым система имела немало сторонников. Однако в последующие годы был высказан ряд дополнений и уточнений, анализ которых проведен детально в названной выше монографии. Мы здесь остановимся на некоторых фундаментальных работах, появившихся с 50-х годов. В первую очередь следует отметить крупную сводку по горным баранам Европы и Азии, подготовленную В.И.Цал-
454
Ovis orientalis Gmelin, 1774
киным (1951). Комплексный анализ систематике-таксономического веса многих параметров диких баранов позволил автору заключить, что многочисленные формы, населяющие территорию от островов Средиземноморья до Центральной Азии, Южной Сибири и Забайкалья, не являются группой видов, но единым политипическим видом, образующим большое количество локальных форм. Существенно, что изучение биологии рассмотренных баранов не противоречит такому суждению, соответственно все ранее описанные формы этой территории составляют один вид — Ovis ammon L. На другой части ареала рода Ovis в Северо-Восточной Сибири представлен самостоятельный вид Ovis canadensis Shaw. Значит, по его мнению, дикие бараны Европы и Азии принадлежат к двум аллопатрическим видам: Ovis ammon и Ovis canadensis.
Что касается внутривидовой систематики О. ammon, В.И.Цалкин отмечал ее неудовлетворительность и полагал, что она будет подвергаться изменениям, направленным главным образом в сторону сокращения числа рас. Он сохранил 24 подвида, в число которых входят закавказский баран О. ammon gmelini (Закавказье, Восточная Турция и Северо-Западный Иран), урмийский баран О. ammon urmiana (о-в Коюндаг на оз. Урмии), эльбрусский баран О. ammon orientalis (горная система Эльбруса, возможно и прилегающие районы Северного Ирана). Иные подвиды населяют Южный, Юго-Западный Иран (О. ammon isphaganica, О. ammon laristanica), а также южную часть Малой Азии (О. ammon anatolica), Кипр (О. ammon ophion), Сардинию и Корсику (О. ammon musimon).
Взгляд В.И.Цалкина (1951) на систему рода Ovis разделяют многие специалисты, в том числе В.Г.Гептнер и др. (1961). Однако количество автохтонных подвидов на территории б. СССР, по его мнению, не более 7.Ре-альность существования закавказского подвида О. ammon gmelini у них не вызывает сомнения. В качестве его синонима рассматривают О. ammon аг-meniana, что ближе к истине.
Из противоположных взглядов на систематику рода Ovis укажем на мнение И.И.Соколова (1959, 1963), согласно которому названный род понимается очень широко и включает 4 подрода; число видов на территории б. СССР 4: Ovis musimon (акклиматизирован), О. orientalis, О. ammon, О. nivicola. Реальность существования названных видов по И.И.Соколову диктуется не только с морфологических, но и с биологических позиций. Закавказских баранов он относит к О. orientalis, выделяя их в самостоятельный подвид О. orientalis armeniana. В его ареал этот автор включает, кроме Закавказья, Восточную Турцию и Северо-Западный Иран. Малообоснованным он считает отделение подвида О. orientalis anatolica, обитающего в южных районах Малой Азии. Признавая и другой подвид — О. orientalis urmiana, он допускал идентичность и этого подвида с О. orientalis armeniana. В области горных систем Эльбруса И.И.Соколов различает два подвида: О. orientalis orientalis (восточная часть Эльбруса) и О. orientalis erskinei (Эльбрусский хребет).
Изучение систематики р. Ovis значительно продвинулось благодаря ка-риологическим исследованиям, проводимым в последние десятилетия
455
Азиатский муфлон
(Nadler et al., 1971; Воронцов и др., 1972; Valdez et al., 1978; Орлов, 1982; Сопин, Харрисон, 1986; и др.).
На наш взгляд, эти данные позволяют сделать ряд предварительных заключений, имеющих прямое отношение к систематическому положению диких баранов Закавказья.
1. Ovis ammon в понимании В.И.Цалкина (1951), В.Г.Гептнера с соавторами (1961) и других исследователей широкой концепции вида, очевидно, состоит из 2—3 самостоятельных видов.
2. Дикие бараны, населяющие Закавказье, Северо-Западный Иран и Малую Азию имеют один и тот же кариотип с 2п = 54, NF = 60.
3. В связи с тем, что на этой территории популяции имеют сходный фенотип, а он связан с кариотипом, то целесообразно объединение их в один вид.
4. Учитывая, что terra typica orientalis расположено близ Решта в провинции Гилян, т.е. в западной части Эльбруса (Воронцов и др., 1972), где обитают 54-хромосомные бараны, мы считаем обоснованным мнение именовать баранов Закавказья, Северо-Западного Ирана и Малой Азии Ovis orientalis Gmelin.
Изменчивость. Литературные сведения по изменчивости диких баранов Закавказья немногочисленны, что объясняется рядом объективных причин. В частности, закавказская популяция занимает небольшую территорию периферии ареала вида. Соответственно численность ее невелика и с глубокой древности она испытывает мощный антропогенный фактор, отрицательное влияние которого усилено из-за сравнительно низкогорного и сильно пенепленизированного рельефа. Этим можно объяснить малочисленность коллекционного материала — В.И.Цалкин (1951) располагал всего 19 экз. разных возрастов. В других центральных хранилищах научной коллекции б. СССР (ЗИН РАН и Зоомузей МГУ) по сей день имеется всего 38 экз. баранов из Закавказья. Если к ним прибавить и коллекцию баранов на местах, и тогда общее количество будет меньше полсотни. Полнота анализа изменчивости ограничена еще и тем, что распространение диких баранов в Закавказье приурочено лишь к одному джавахетско-армянскому варианту поясности. Как известно (Соколов, Темботов, 1989), экологические условия этого варианта менее разнообразны, чем в других районах Кавказа, что несомненно не может не сказаться на изменчивости местных баранов.
Индивидуальная изменчивость. Мнения исследователей по этому вопросу сходны — во всех популяциях баранов, в том числе и Закавказье, размах индивидуальной изменчивости значителен. Однако проиллюстрировать это, в сущности справедливое, положение возможно не по всем параметрам вида. Фактически нет данных по массе тела баранов Закавказья. Этот пробел в какой-то степени можно восполнить по материалам В.И.Цалкина (1951), полученным им из Копетдага, где обитает сходная форма. В январе-марте среди шестилетних самцов масса тела (л = 6 экз.) составляла 56— 79 кг, пятилетних (л = 6 экз.) — 54—66 кг, четырехлетних (л = 4 экз.) — 48—58 кг. Размах вариации массы тела у взрослых самцов составляет от 10
456
Ovis orientalis Gmclin, 1774
до 23 кг. Индивидуальная изменчивость самок, очевидно, выражена в меньшей степени, чем у самцов. У самок старше 2 лет (п = 13 экз.) масса тела составила от 30,0 до 46 кг, т.е. различие составляет 16 кг.
Менее изменчивыми являются размеры тела — в трех ранее рассмотренных возрастных группах самцов разница в высоте в холке варьирует в пределах от 2 до 6 см, а разница в длине тела — от 7 до 15 см.
В черепе, как справедливо отмечал В.И.Цалкин (1951), очень высокой вариабельностью характеризуются длина носовой кости, ее форма, длина предальвеолярной части верхней челюсти, длина лобных костей, межглазничная ширина и др. Менее изменчивы общая и кондилобазальная длина черепа, длина морды, длина мозговой коробки, высота черепа. Большой изменчивостью отличаются очертания и размеры межчелюстной и слезной костей. Исключительной вариабельностью характеризуются размеры и форма рогов. По этому поводу В.И.Цалкин (1951) писал, что "при изучении серии рогов из строго определенных частей ареала обнаруживается, как правило, крайне пестрая картина. Этим же объясняется существование значительной трансгрессии в размерах рогов, с которой приходится постоянно сталкиваться даже при сравнении относительно далеких друг от друга рас" (с. 31). Наши и литературные данные (Цалкин, 1951; Манасерова, 1988) свидетельствуют о высокой индивидуальной изменчивости архитектоники поверхности роговых чехлов. Это относится в равной степени к размерам одних и тех же годовых сегментов, степени развития наружного ребра, морщин и бороздок; хотя наиболее типичны для закавказских баранов гетеронимные рога, но у некоторых животных наблюдается слабо первер-тированный тип. Из всех перечисленных признаков, видимо, больше всего варьирует длина годовых сегментов — у разных особей размеры одного и того же годового сегмента могут колебаться более чем в 3 раза. Более стабильным оказался обхват основания рогов, что соответственно придает этому параметру высокий систематике-таксономический вес.
По индивидуальной изменчивости окраски меха наиболее полные данные приводит В.И.Цалкин (1951), согласно которому у баранов Северо-Восточной Турции, Армении и Северо-Западного Ирана основной тон окраски спины, шеи и боков весьма изменчив, варьирует от яркого красновато-рыжеватого до рыжеватого или темно-коричневого. Столь же сильно варьирует светлое пятно по бокам. У экземпляра из Эрзерума пятна большие, распространяясь на спину, сливаются. Экземпляр из Нахичевани (окрестности с. Джамалдин) имеет небольшое и неконтрастное светлое пятно. Среди сборов из Закавказья есть экземпляр, у которого совсем не развито светлое пятно. Индивидуальная изменчивость четко проявляется и в других деталях окраски меха, а также в степени развития подвеса. Словом, систематико-таксономический вес окраски меха невелик.
Возрастная изменчивость. Новорожденные дикие бараны довольно мелкие. По В.И.Цалкину (1951), в условиях Центрального Копетдага недавно родившийся ягненок (самец) имел массу тела 2,8 кг, длину тела 56 см, высоту в холке 45 см. Они растут очень интенсивно, достигая в возрасте 9 мес (в феврале) массы от 20 до 22 кг, длины тела до 104 см, в холке до
457
Азиатский муфлон
75 см. В возрасте 2 лет, по данным названного автора, самец имел массу 34 кг, длину тела 120 см, высоту в холке 84 см. По достижении 3 лет их масса составляет от 38 до 46 кг, длина тела от 126 до 134 см, высота в холке от 89 до 94 см. У четырехлетних животных масса возрастает значительно, составляя 48—58 кг, длина тела 132—139 см, высота в холке 92—94 см. В дальнейшем с возрастом продолжает расти масса (у семилетних — 98—101 кг), но рост размеров тела идет менее интенсивно, и в силу высокой индивидуальной изменчивости особи старших возрастов по размерам имеют широкую трансгрессию. Что касается возрастных изменений массы и размеров тела самок, то результаты исследований не столь однозначны, как по самцам. Известно, что уверенно возраст самок можно определить до 2 лет. Группа животных старше 2 лет имела массу от 30 до 46 кг, длину тела от 121 до 136 см, В холке — от 77 до 85 см. Следовательно, у самок, как справедливо заметил В.И.Цалкин (1951), масса и размеры тела формируются быстрее, чем у самцов. Очевидно, к трем годам самки достигают своих полных размеров, тогда как самцы — лишь в четырехлетием возрасте. Полную массу тела самцы приобретают, вероятно, только в 7 лет, тогда как самки — в 3 года.
Возрастные изменения волосяного покрова наиболее значительны у самцов, что связано с появлением подвеса на 2-м или 3-м году жизни, который достигает своего полного развития только у взрослых животных в зимний период. Летом он развит слабо.
Сведения по возрастной изменчивости окраски тела в условиях Закавказья отрывочные. Очевидно, смена ювенильного наряда происходит осенью, тогда сеголетки становятся похожими на самок. После зимовки появляется боковое пятно.
Череп и рога претерпевают очень большие возрастные изменения. Подробный анализ их выходит за рамки настоящей работы. Обратим внимание на те стороны вопроса, которые, на наш взгляд, необходимо учитывать при решении систематике-таксономических проблем. Как известно, ягнята рождаются с относительно крупным черепом, особенно мозговой частью. Соответственно у них относительная длина морды и связанных с ней отдельных костей невелика. Как следствие, в ходе постнатального развития интенсивность роста различных костей черепа осуществляется различно. Лишь к двум годам жизни происходит выравнивание скорости роста костей черепа и стабилизируются пропорции черепа. Например, если относительная длина морды к общей длине черепа увеличивается очень быстро в первые два года, то ее рост потом происходит одинаково с ростом общей длины черепа. Словом, по В.И.Цалкину (1951), "череп самцов к двухлетнему возрасту оказывается сформированным во всех своих основных пропорциях, и в дальнейшем отчетливо выраженных возрастных изменений уже не наблюдается" (с. 85). Что касается роста роговых стержней, то увеличение их длины продолжается в течение всей жизни, однако пик его приходится, судя по всем коллекционным материалам, на первые 3—5 лет. У животных старше 6 лет резко падает интенсивность роста стержня в длину. В этом плане особый интерес представляет тот факт, что обхват
458
Ovis orientalis Gmelin, 1774
стержня у основания стабилизируется, как и у бородатого козла, к 4-му году жизни (Цалкин, 1950, 1951). Соответственно мы разделяем точку зрения названного автора, "что в целях систематического исследования, черепа самцов от четырех лет и старше могут быть объединены в одну общую группу, не требующую дополнительного элиминирования" (с. 92). Что касается самок, однозначных данных все еще мало. Вероятно, в трехлетием возрасте размеры и структура черепов мало отличимы от более старших. Если они имеются, то укладываются в пределах индивидуальной изменчивости черепов взрослых животных.
Зубная система диких баранов в постнатальный период претерпевает сложные и глубокие изменения. Анализ коллекционного материала, сопоставление количества годовых сегментов и показателей зубной системы свидетельствует о том, что полная смена молочных зубов на постоянные и соответственно формирование зубной системы у диких баранов завершаются лишь в четырехлетием возрасте. В последующие годы возрастные изменения зубной системы проявляются в прогрессирующем увеличении стертости зубов. Комплекс качественных-и количественных изменений в зубной системе позволяет определять возраст диких баранов с определенной уверенностью до восьмилетнего возраста. Вслед за В.И.Цалкиным (1951) при работе с коллекционным материалом мы рекомендуем руководствоваться следующими изменениями зубной системы, которые отражают возраст животного.
Молочные зубы от резцов до предкоренных появляются в первые недели жизни. Первая пара коренных имеется у животных шестимесячного возраста, вторая пара — в годовалом возрасте, а третья — к двум годам жизни.
Первая и вторая пары резцов и все молочные предкоренные сменяются на постоянные к 2 годам, а третья пара молочных резцов — к 3 годам жизни. Лишь на четвертом году осуществляется замена молочных клыков на постоянные.
Стертость становится заметной в 3 года на первом коренном в виде слабой притупленности. В 4 года совершенно стерт первый предкоренной, сильно сношен второй предкоренной, притуплены третий предкоренной и первый коренной. Пятилетние характеризуются сильной стертостью всех предкоренных и сношенностью первых коренных, а вторые коренные притуплены. У шестилетних гребни зубов совсем стерты на первом коренном, сильно притуплены на втором коренном. В 7—8 лет гребни зубов сношены на всех коренных.
Половые различия. Во всех исследованиях, посвященных закавказским диким баранам, указывалось на наличие целого ряда показателей, по которым различаются самцы и самки.
По данным А.А.Саркисова (1939), Э.ГЛвруяна (1970), в Закавказье максимальная масса тела самцов превосходит таковую самок более чем на 25%. Близкие результаты получаются и при сравнении средних показателей массы тела. У баранов Копетдага половой диморфизм по массе тела выражен еще резче — самцы тяжелее самок в среднем на 75 % (Цалкин,
459
Азиатский муфлон
1951). Если судить по данным этого автора, легко заметить, что, чем крупнее форма, тем резче выражен половой диморфизм по массе тела.
Самцы всегда рогатые, тогда как среди самок баранов Закавказья примерно половина не имеет рогов. У тех самок, у которых имеются рога, они небольшие, не имеют резко выраженных ребер, годовых сегментов, морщинистость слабая. Отсутствие или слабое развитие рогов существенно сказывается на размерах и структуре черепа самок, что обусловливает в конечном итоге половой диморфизм по многочисленным краниологическим показателям.
Обратим внимание на некоторые из них. Так, размеры черепа и роговых стержней у самцов значительно крупнее, чем у самок, у основания стержней лобная кость самцов сильно выпуклая и соответственно зароговая часть лобных костей резче спадает, лобно-носовой профиль имеет более заметную вогнутость и т.д. Фактический материал, которым мы располагаем, свидетельствует о том, что все 19 признаков краниологических различий между самцами и самками диких баранов Копетдага, выявленных В.И.Цалкиным (1951), справедливы и по отношению к популяции баранов Закавказья.
По экстерьеру половой диморфизм, согласно данным Э.Г.Явруяна (1970), выражен четко. Так, самцы отличаются от самок относительно более длинным туловищем, более глубокой и широкой грудью, а также более длинным и широким тазом; голова самцов длиннее и шире, чем самок. По окраске тела оба пола во многом сходны, но у самок она несколько светлее. У самцов с возрастом появляются светлые пятна на боках, иногда соединяющиеся на спине. Кроме того, у самцов на груди и нижней части шеи со временем появляется подвес, окрашенный интенсивно в темный или даже черный цвет. В отличие от самцов у самок нет ни светлых пятен на боках, ни подвеса на груди и шее.
Географическая изменчивость. Этому вопросу много внимания уделяли ведущие специалисты страны и за рубежом. Чтобы оценить уровень разработанности проблемы, достаточно напомнить, что в их числе были Н.А.Се-верцов (1873), РЛидсккср (Lydekker, 1898, 1913, 1915), Н.В.Насонов (1911, 1923), В.И.Цалкин (1951) и многие другие ведущие специалисты своего времени. Видимо, долго еще останется непревзойденной монография В.И.Цалкина ‘Торные бараны Европы и Азии" (1951), где различные аспекты изменчивости, в особенности географической, подвергнуты всестороннему анализу. Закавказские дикие бараны, на наш взгляд, в этой сводке достаточно четко "вписаны" в общую картину географической изменчивости р. Ovis Евразии. Ряд специальных работ (Явруян, 1970; Манасерова, 1988) лишь подтвердили общие положения, высказанные В.И.Цалкиным по оценке морфологических особенностей диких баранов Закавказья. Более существенные дополнения сделаны в связи с изучением хромосомного набора различных рас, обитающих в Армении и прилегающих районах Турции и Ирана (Valdez et al., 1978). Новые данные получены ио диким баранам Армении в связи с их разведением на экспериментальной базе Зоологи-' ческого института Армении.
460
Ovis orientalis Gmclin, 1774
На основе анализа всего этого материала можно высказать наше отношение к данному вопросу. На наш взгляд, морфологические параметры закавказских баранов, какими являются средние размеры тела, гетероним-ные рога, светлое пятно на боках у самцов, детали структуры черепа, вполне укладываются в общую закономерность географической изменчивости вида. Они соответствуют также географическому положению вида в области его распространения (северное, внутриматериковое, континентальный климат и т.д.) и ландшафтной структуре гор, занимаемых этими животными. Как справедливо отмечает В.И.Цалкин (1951), увеличение массы и размеров тела с запада на восток и с юга на север, четко выраженное у дикого барана, объяснимо с позиции теории Бергмана. Существует немало данных, в том числе по рассматриваемому виду, свидетельствующих о справедливости этой теории и в отношении горных популяций широко распространенных равнинных видов. В плане адаптивных реакций к экстремальным условиям жизни объяснимо также положение, по которому указанным градиентам соответствует географическая изменчивость размеров рогов. Такое соответствие объяснимо, если учесть роль костных стержней рогов как дополнительного органа кроветворения. Думается, что с такой концепцией согласуется не только географическая изменчивость рогов самцов, но и самок. В самом деле, на островах Средиземноморья самки комолые, а в высокогорьях Таджикистана самки встречаются только рогатые с относительно крупными рогами. Между этими крайними вариантами можно найти целый ряд переходных форм. Словом, значительная доля комолых самок (до 50%) в популяциях Закавказья соответствует географическому положению их в пределах единого ареала вида.
Особенности окраски волосяного покрова закавказских баранов также укладываются в общую закономерность изменения ее в связи с широтным градиентом климатических факторов. Так, по Э.Г.Явруяну (1970), у более южных популяций, как урмийская, окраска тела менее интенсивная, желтовато-серая. Слабее, чем у закавказских, выражены у них светлые пятна на боках тела. Более того, светлые пятна могут почти отсутствовать, тогда как у закавказских они настолько развиты, что могут слиться на спине.
В связи с географической изменчивостью естественно возникает вопрос о том, насколько выражено влияние высотной и горизонтальной неоднородности горных ландшафтов на морфологические признаки дикого барана в пределах закавказской части ареала. Постановка такого вопроса правомочна, если учесть характер изменчивости других копытных Кавказа, например кавказского тура. Все имеющиеся данные говорят о том, что территориальных различий по морфологическим признакам среди диких баранов Закавказья нет. Во всяком случае, коллекционный материал, которым в настоящее время мы располагаем, не позволяет выявить такие различия. Видимо, сказывается не только ограниченность территории и ее однородность, но и высокая подвижность диких баранов, меньшая, чем у туров, привязанность к определенным районам. Судя по литературным данным, дикие бараны Закавказья совершают не только высотные миграции, до и горизонтальные. По А.А.Саркисову (1941), известны пункты
461
Азиатский муфлон
перехода диких баранов через р. Араке из Турции и Ирана в Армению и обратно.
Исходя из изложенного выше, мы считаем более обоснованной точку зрения, согласно которой Дикие бараны Закавказья, Восточной Турции и Северо-Западного Ирана объединяются в один подвид — Ovis orientalis gmelini Blyth (syn. armeniana).
Экология
Ландшафтная приуроченность. Биотопы. По азиатскому муфлону мало данных, в том числе палеозоологических, археологических и летописных. Поэтому однозначно сказать, при каких ландшафтных условиях формировался вид, какие экосистемы для него являются первичными, а какие — вторичными, затруднительно. Между тем нынешняя территория ареала азиатского муфлона освоена человеком очень давно и плотность населения людей здесь высока. Соответственно человек охотится на муфлонов с незапамятных времен, с ним конкурируют домашние животные, вытесняя их с лучших пастбищ. Существенно, что здесь не действовало ограничение со стороны религии, как это имело место по отношению к кабанам. С другой стороны, высокая пластичность муфлонов, в том числе большая кислородная емкость крови, позволяла им осваивать горы до больших высот, хотя условия жизни там явно экстремальны.
Если исходить из характера распространения в последнее столетие, по которому имеются некоторые литературные данные, то азиатского муфлона правомочно рассматривать как представителя нагорных степей. А.А.Саркисов (1941), специально изучавший азиатского муфлона, отмечает, что в области распространения этого копытного в Закавказье представлены полынная полупустыня, солянковая полупустыня из более галофит-ных солянок (преимущественно Salsola crassa, Gamantus pilosus, Petrosi-monia brachoata и др.), солянковая полупустыня на предгорных склонах (Artemisieto-salsoletum nodulosae), ксерофитная растительность со злаковым дерном и большой примесью ксерофитов, а также субальпийская и альпийская растительность. Муфлоны, по мнению этого автора, более подвижные животные, чем бородатые козлы. Они часто перемещаются, но обычно держатся в районах горно-степной и ксерофитной растительности. Они, как правило, избегают глубоких ущелий, скал, субальпийских и альпийских лугов. Это не означает, что муфлоны не заселяют субальпийский и альпийский пояса, особенно в наши дни, когда количество отар домашних овец резко возросло и для муфлона не остается места в пастбищный сезон. А.А.Саркисов справедливо отмечает, что в районах лиственных лесов, таких, как Мегри, муфлоны не появляются. О том, что они избегают сплошные леса, писали почти все, кто занимался изучением диких баранов. Так, Н.К.Верещагин (1959) отмечал, что распространение муфлона в настоящее время в Закавказье связано с нагорными степями с преобладанием группировок плотнокустовых злаков и зиббальдии с формацией колючих астрагалов на склонах средней полосы гор. Он также
462
Ovis orientalis Gtnelin, 1774
указывал на долину Аракса с солянково-полынной группировкой, как на область распространения муфлонов. По мнению В.Г.Гептнера и др. (1961), вид в целом характеризуется как житель в основном открытых пространств с мягким горным рельефом: плато, пологих склонов гор, извилистых предгорий и т.д. Мы склонны рассматривать нынешнее распространение азиатского муфлона в субальпийском и альпийском поясах вторичным, связанным с деятельностью человека или с погодными условиями. Еще Г.И.Радде (1899) писал, что в жаркие дни лета муфлоны поднимаются до вечных снегов. Прямое браконьерство вытеснило муфлонов со всей долины Аракса, единичные особи, пытающиеся перейти с берега на берег, становятся, как правило, жертвой человека (Явруян, 1969). Что касается среднегорий, являющихся исконными местами обитания муфлонов, то их они вынуждены покинуть с появлением многочисленных отар в пастбищный сезон.
Митрации — характерная черта экологии диких баранов, очевидно, во многих местах, в том числе в Закавказье. О сезонных перемещениях писал еще Н.Я.Динник (1910), отмечая, что во время суровых зим в Армении бараны спускаются с гор в долину Аракса. Ссылаясь на К.А.Сатунина, он говорил, что они живут оседло только на Арарате и южной части Нахичевани, а на всем протяжении от Арарата до Сарыкамыша бывают только зимой. То же подтверждает и А.А.Саркисов (1941) — летом, в знойную пору животные поднимаются в высокогорье, а зимой спускаются в предгорья, нередко используя при этом озимые поля в качестве временного пастбища. Из-за глубоких снегов муфлоны иногда подходят близко к населенным пунктам, при этом бывали случаи, когда они заходили в села и города или присоединялись к стадам домашних овец.
По мнению А.А.Саркисова, азиатские муфлоны Закавказья регулярно совершают кочевки сезонного характера, вызванные более суровыми зимами Армении и Нахичевани, чем в Турции и Иране. Массовые кочевки через р. Араке он наблюдал в двух пунктах: 1) недалеко от с. Джейранлы, где муфлоны переправляются через р. Араке в Иран и двигаются на Дарашанские горы (Карадаг); 2) переправляясь через р. Араке, частью попадают на Каре-инское плато (Иран, Макинский район), частью направляются на южные склоны горы Арарат и на равнины Баязетского Санджака. Аналогичные кочевки этот автор регистрировал и в пределах Ирана и Турции. Э.Г.Яв-руян (1969) приводит 10 таких миграционных путей, по которым муфлоны переходят р. Араке из Закавказья в Иран и Турцию и обратно, но вряд ли при наличии пограничного режима такими миграционными путями животные могут пользоваться регулярно.
Сроки кочевок зависят от погодно-климатических факторов; обычно массовая миграция наблюдается после выпадения больших снегов в горах (Насимович, 1955). Пути следования животных относительно постоянные, реки они переходят в одних и тех же местах; они идут, как правило, более или менее крупными группами (Саркисов, 1941, 1944; Насимович, 1955 и др.). Судя по учетным данным Э.ГЛвруяна (1971), не все дикие бараны зимой перекочевывают из Армении в Нахичеванские районы. Зима 1967/68 г. была очень снежной, холодной и неблагоприятной для баранов, но
463
Азиатский муфлон
отдельные группы их не покинули летних местообитаний и держались на высоте около 2800 м над ур. моря (недалеко от с. Лернашен), обычно на южных склонах и выдувах.
Высотные пределы территориального размещения азиатского муфлона в Закавказье очень широкие. По А.А.Саркисову (1941), верхняя хранила распространения проходит на уровне вечных снегов. Он находил этих животных на высоте до 3856 м над ур. моря (склоны г. Ячлудара). Г.И. Радде (1899) указывал, что муфлоны заходят в область вечных снегов. Н.Я.Динник (1910) утверждает, что в Зорских горах муфлоны поднимаются до 4575 м. Очевидно, в этих данных нет преувеличения, если учесть, что дикие бараны Средней Азии могут жить от высот ниже уровня мирового океана (Каспийское побережье) до 4500—5000 м (см. обзор: Гептнер и др., 1961).
Убежища. Лежки. Для избегания или смягчения влияния неблагоприятных факторов бараны используют различные убежища. От людей и хищников уходят в горы, в скалы. Спасаясь от жары или кровососов, занимают более обдуваемые и тенистые места — под навесами камней, на карнизах, среди кустарников или под отдельными деревьями. Лежки свои устраивают обычно недалеко от места пастьбы, иногда они уходят выше или ниже по склону на сотни метров и там устраиваются на отдых. Судя по многочисленному помету, удобными лежками бараны пользуются долго. Такие места со временем выбиваются и углубляются. Такие выбитые места можно встретить под деревьями и кустарниками. Лежки были обнаружены даже на сельскохозяйственных угодьях или вблизи построек человека. Так, Э.Г.Явруян (1969) в сентябре 1967 г. встретил взрослых самцов в 200—300 м от пустовавшей овчарни с. Арени, а их лежки на противоположном склоне холма в 80—100 м. Другой раз две взрослые самки и три годовалых самца были вспугнуты на кукурузной плантации.
Сезонная и суточная активность. В зависимости от состояния организма (пол, возраст, упитанность и т.д.), конкретных условий жизни, отношения к животным и человеку в широких пределах меняется сезонная ритмика. Тонкости этой ритмики у диких баранов Закавказья не изучены. Имеются лишь отдельные наблюдения, отражающие сезонные сдвиги в активности муфлонов в связи с размножением, кочевками, защитой от кровососов. Половой цикл вносит резкие изменения в ритм жизни половозрелых самцов. Видимо, ритм активности в связи с кочевками и защитой от кровососов сходен у разных полов и возрастов.
Суточная активность летом двухфазная, пики приходятся на вечерние и утренние часы, иногда муфлоны активны ночью. Отдыхают они примерно с 10 до 18 ч, выбирая для этого тени кустов и скал. Зимой активны днем, ночью отдыхают. В пасмурную и дождливую погоду суточная ритмичность нарушается. Когда идет сильный дождь, бараны не пасутся, а когда дуют сильные и холодные ветры, они уходят в скалы и выстаивают в подветренных местах. Аналогично поступают зимой при обильном снегопаде и сильном ветре. Иногда они ложатся в глубокий снег. В суточную активность большие изменения вносят люди, находящиеся долгое время в горах
464
Ovis orientalis Gmelin, 1774
(скотоводы, геологи, туристы и др.). Днем, когда скот и пастухи пребывают на пастбищах, муфлоны держатся высоко в горах, занимая обычно скалистые места, а ночью спускаются на выпасы, освободившиеся от домашних животных.
Поведение (рис. 62). Наблюдения за архарами в Бадхызе (Баскин, 1976, 1986), за муфлонами в Малых Карпатах (Ccrvan, Duha, 1987) и некоторые наши и литературные сведения (Явруян, 1969) свидетельствуют о том, что для диких баранов характерна сложная система поведения — широкий спектр социальных взаимодействий, способы коммуникации, демонстрация и восприятие сигналов органами чувств и т.д. Особенно яркое внешнее проявление отмечено при опасности и в связи с формированием гаремов. Заметив опасность, они издают свист как сигнал опасности, при этом стучат копытами. Заметив человека, муфлоны нс выпускают его из поля зрения, уходят от него, но нс отрываются, чтобы не терять из вида врага. В период гона и подготовки к нему половозрелые самцы проявляют агрессивность к соперникам, бьются мощными рогами, издавая сильйый шум. Эти Черты сохранились и у домашних овец. Полевые наблюдения свидетельствуют о том, что благодаря хорошо развитым органам чувств они свободно ориентируются в природе, умело выбирают свои миграционные пути, места безопасные для отдыха, проявляют к ним определенную привязанность. Нет сведений о том, что во время отдыха у муфлонов бывают сторожа, но в их стаде имеются вожаки, видимо наиболее опытные самцы или самки.
Муфлоны относятся к стадным копытным, состав и размер групп которых меняется широко в течение года. Перед гоном начинается укрупнение стад, в составе которых в этот период насчитывается до 12—15 голов (Явруян, 1969). Такие стада наблюдаются в Закавказье обычно в конце ноября — начале декабря и держатся примерно до апреля, когда наступает период ягнения. В это время стада покидают беременные самки; самцы тоже отделяются в самостоятельные группы. Летом самки с ягнятами держатся отдельно. Самцы самостоятельными группами пасутся недалеко от самок и ягнят до начала следующего периода гона и образования смешанных групп. В прошлом, когда численность закавказских баранов была высока, встречались стада в 20Q—300 голов (Динник, 1910).
Линька четко выражена у баранов Закавказья, но конкретных сведений о ней очень мало. Известно, что, как и у других видов баранов, линька в полном ее объеме происходит один раз в году — обычно начинается с конца марта и завершается в первых числах июня (Явруян, 1969). Согласно данным названного автора, смена волос начинается с лопаток, таза и по хребту. Затем интенсивно сменяются волосы подвеса, шеи и боков тела. На остальной части тела выпадение волос увеличивается после полного выпадения длинных волос подвеса. Отрастание зимних волос (удлинение остевых, появление пуха) начинается с конца лета и заканчивается в основном к началу гона, т.е. в конце ноября — начале декабря.
Питание. В ряде литературных источников содержатся сведения о питании азиатского муфлона в Закавказье, но наиболее полные они у Э.Г.Яв-
30 В.Е.Соколов, А.К.Темботов
465
Азиатский муфлон
Рис. 62. Азиатский муфлон
466
Ovis orientalis Gmclin, 1774
руяна (1969). Перечень растений, поедаемых муфлоном, составленный им, содержит большой и довольно разнообразный набор видов, но явное преобладание травянистой растительности очевидно. Обращает внимание и тот факт, что перекрывания набора кормов азиатского муфлона и бородатого козла нет; видно, что у первого меньшую роль играют древесно-кустарниковые растения, чем у второго. Характерно и то, что сезонная динамика состава и соотношения различных видов растений значительна. Особенно велика роль травянистой растительности в период весенней вегетации, который обычно тянется с марта до конца июня. В это время муфлоны поедают, по данным упомянутого автора, преимущественно злаки, осоки, зо-зимию, лук, маки, беллевилию, спаржу, василистник, гладиолус, марь, хохлатку, желтушник и др. Здесь уместно отметить, что на значительной части ареала рода Ovis наибольшую роль играют злаки, в том числе ряд видов рода Festuca, особенно типчак, пырей Agropyrum и др. (Гептнер и др., 1961; Rodriguez et al., 1987; Heroldova, 1988;. Martinez, 1988; и др.). В летний период, когда выгорает травянистая растительность на значительной части среднегорья, закавказские бараны питаются преимущественно эспарцетом, резедой, молоканом, бинерцией, синеголовником и др. Возрастает доля веточных кормов за счет миндаля, вишни, шиповника и др. Осенью, особенно в период с сентября по ноябрь, в связи с полным выгоранием травянистой растительности заметна роль в питании азиатского муфлона древесно-кустарниковой растительности. Поедаются ветки и листья вишни, миндаля, ивы, шиповника, можжевельника, барбариса, каркаса. В зимний период муфлоны испытывают большие затруднения в связи с ухудшением кормовой базы, нередко из-за малой доступности корма под снегом, низкой калорийностью пожухлой травы, которой им приходится питаться. В связи с этим они вынужденно перемещаются на выдувы и другие места, где меньше снега и доступнее трава; питаются ветками деревьев и кустарников, а также опавшими листьями. В отличие от коз муфлоны кору не гложут.
Азиатские муфлоны в теплое время года регулярно посещают солонцы. Такой солонец, по Э.Г.Явруяну (1969), расположенный на Урцском хребте близ с. Араздаян (Таки ахпур), имеет вид открытых узких пластов соленосных пород. В окрестностях с. Хнздорут находится родник Try джур, содержащий слабо ощутимый на вкус раствор соли. Этим родником муфлоны пользуются часто, к нему подходят хорошо проторенные тропы. Соленые и обычные источники воды муфлоны посещают в жаркое время года рано утром и поздно вечером. В ненастное весеннее и осеннее время, когда они пасутся и днем и в сумерках водопой посещают один раз.
Воспроизводство и структура популяции
Размножение. По данным Н.Я.Динника (1910), гон у закавказских баранов начинается в декабре. А.А.Саркисов (1944) указывает на более ранний срок начала гона у муфлона-Армении — 20-е числа ноября. Э.ГЛвруян (1969) считает, что сроки спаривания приходятся на конец ноября — начало декабря. Очевидно, сроки гона сдвигаются в ту или иную сторону в зави
31*
467
Азиатский муфлон
симости от конкретных условий года. Продолжительность гона растягивается до 1—1,5 мес; раньше спариваются самки старших возрастных групп, а потом молодые самки.
Перед гоном смешанное и укрупненное стадо разбивается на мелкие группы, включающие несколько самок, 1—2 самцов и несколько молодых. За владение таким "гаремом" самцы часто дерутся между собой. Нередко схватки между самцами бывают ожесточенные (Саркисов, 1944), но, видимо, судя по данным ряда авторов (см. обзор: Гептнер и др., 1961), не сопровождаются гибелью одного из соперников. В отличие от ряда копытных взрослые половозрелые самцы терпимо относятся к молодым самцам, находящимся в стаде. Молодые самцы не участвуют в размножении до 4—5-летнего возраста.
Продолжительность беременности, по Н.Я.Диннику (1910), составляет около пяти месяцев, что подтверждено последующими исследованиями (Саркисов, 1944; Явруян, 1969). Молодые рождаются в конце апреля или в мае. Перед этим самка покидает труппу, уходит в скалы и выбирает место отела так, чтобы была защита от дождя и окружающая местность обозреваема.
Плодовитость. О количестве ягйят в одном помете данные в литературе противоречивы. Н.Я.Динник (1910), А.А.Саркисов (1944) считают, что в помете азиатского муфлона чаще всего два ягненка, редко один или три ягненка. По Э.Г.Явруяну (1969), молодых в помете один-два, редко три. В.Г.Гептнер и др. (1961) не разделяют мнение, что в условиях Закавказья два ягненка в помете — явление обычное.
Мы не располагаем данными, позволяющими определить какое количество самок в популяции приносит потомство, какое остается яловым.
Рост и развитие. Ягнята рождаются слабыми, но хорошо сформировавшимися. Чутко реагируют на сигналы матери, которая находится вблизи от ягненка 2—3 дня. Часто самка облизывает ягненка, приучает его к своему голосу. А сама самка, судя по ее поведению, привыкает к запаху своего ягненка. Хотя малышу нужно всего 2—3 ч, чтобы встать на ноги и 7—8 ч для сравнительно быстрого передвижения,1 он остается на месте, забившись в траве, достаточно долго. По наблюдениям Э.Г.Явруяна (1969) за 3-4-дневным ягненком в течение 4,5 ч (с 12 ч 30 мин до 17 ч), малыш прикладывался к вымени матери девять раз и сосал каждый раз 2—5 мин. Ягнята (п — 4) возрастом около одного месяца за 2 ч (с 9 ч 15 мин до 11 ч 15 мин) сосали своих матерей всего один раз в течение 1—2 мин. Указание В.Г.Гептнера и др. (1961) о том, что ягнята получают молоко от матери в основном утром и вечером, видимо, относится к хорошо подросшим. А.А.Саркисов (1944) Отмечает, что первые 2 дня после рождения молодняка самка фактически не потребляет пищу. Через 6—9 дней самка с хорошо окрепшими ягнятами возвращается в стадо. В составе стада самка ревностно охраняет своего ягненка, отгоняя других от него, но через некоторое время нужда в этом отпадает. Однако рефлекс материнства сохраняется долго — самка кормит молоком ягненка около 5 мес, молодняк остается при ней до года; хотя часть молодых переходит с осени к самостоятельной
468
Ovis orientalis Gmelin, 1774
жизни. Нередко после ягнения к самке вновь возвращаются годовалые ягнята и держатся вместе с ней долго, даже до второго ягнения. Молодые очень рано начинают употреблять растительную пищу — через 7—8 дней после рождения они пытаются щипать зеленую траву, наравне со взрослыми они пасутся уже в возрасте 1 мес.
По наблюдениям за живыми баранами, а также судя по результатам сравнительного анализа длины годовых колец, молодые животные интенсивно растут первые годы жизни. Самки достигают полной массы к 3 годам, а самцы значительно позже — к 7 годам. Дальнейший рост сравнительно незначительный и притом преимущественно за счет жировых отложений (Цалкин, 1951). Самки половозрелыми становятся к 3 годам. Видимо, и самцы в это время созревают, но в размножении могут участвовать и отбивать самок у более взрослых самцов в возрасте не ранее 4—5 лет.
Возрастные изменения экстерьерных показателей наиболее полно изучены у горных баранов Копетдага (Цалкин, 1951), близких к Закавказью территориально и по типу поясности. В силу этого сходства и в связи с недостатком данных по Закавказью ниже приводятся данные названного автора.
Развитие черепа идет со значительными структурными изменениями. Так, череп самца в возрасте 2—3 нед имеет узкую и короткую морду, занимающую около половины общей длины черепа. Лобно-носовой профиль слабо вогнутый, свод черепа в виде правильной сферы, костные стержни рогов отсутствуют. В трехмесячном возрасте увеличиваются относительная длина морды и носовых костей, а мозговой коробки — уменьшается (43,5% против 45% в 2—3 нед); появляются костные стержни рогов (около 10 мм длиной). Ягнята в возрасте 9 мес характеризуются увеличением длины морды до 57% и носовых костей до 37% от общей длины черепа. Костные стержни рогов увеличиваются до 35—45 мм. Сокращается относительная длина мозговой части черепа (41,5—42,5%). В возрасте двух лет относительная длина морды достигает 60%, носовых костей — 40%, длина стержней рогов — до 160—245 мм. Лобно-носовой профиль сильно вогнут. Относительная длина мозговой коробки составляет около 40% общей длины черепа. Резко возрастает относительная высота черепа. Трехлетние самцы имеют череп, сходный с двухлетними, но прогиб лобных костей у них сильнее, а профиль зарогового отдела круто согнут. В четыре года череп по конфигурации сходен с трехлетним, лишь в абсолютных размерах увеличен и больше швов сливается. Дальнейшие возрастные изменения мало заметны. Не вдаваясь в подробное описание развития черепа самок, отметим, что интенсивный рост черепа у них происходит в первые 2 года, сильно замедляется на 3-м году, а на 4-м череп достигает полного размера. Значительно раньше (примерно в 2 года) формируются основные пропорции черепа, и в дальнейшем отчетливо выраженных возрастных изменений уже не отмечается.
Формирование зубной системы происходит следующим образом. У новорожденных в первые же недели их жизни появляются все три пары
469
гХзиатский муфлон
молочных резцов, молочные клыки, три пары молочных предкоренных. К шестимесячному возрасту появляются первые коренные на обеих челюстях. К году появляется вторая пара коренных. К двум годам сменяются на постоянные I и II пары резцов,'а также все предкоренные; вдобавок появляется III пара коренных. К 3 годам на постоянные сменяется III пара молочных резцов. На 4-м году жизни сменяются молочные клыки. Таким образом, к 4 годам заканчивается формирование зубной системы, а дальнейшие возрастные изменения сводятся к износу зубов.
Рост и развитие рогов легко и достаточно точно прослеживаются по сегментации роговых чехлов. В отличие от восточносибирских форм у азиатского муфлона, как справедливо отмечали предыдущие исследователи, рога интенсивно растут в 1-й и 2-й годы, а начиная с 3-го года жизни их рост замедляется, что хорошо видно по постоянному сокращению длины сегментов. Максимальный рост рогов характерен для 2-го года, а минимальный — для последнего года жизни. В Закавказье азиатский муфлон живет, очевидно, 13—15 лет (Явруян, 1969). По мнению В.И.Цалкина (1951), предельный возраст баранов составляет 12—13 лет.
Структура популяции. Очень мало данных по этому вопросу можно найти не только в старых литературных источниках, но и в современных, что, видимо, объясняется низкой численностью данного вида на всем протяжении закавказской части ареала. Имеются лишь отдельные фрагменты. Так, в феврале-марте 1968 г. наУрцском хребте учтено 16 баранов, среди которых было 5 самцов и 11 самок (Явруян, 1961).
Уместно отметить, что для нормальной и продуктивной жизни муфлонов в ГДР планировалось формирование стада в следующей структуре: ягнята — 40%, молодняк — 20%, до 2 лет и старше — 40%. Оптимальным соотношением полов считается 1:1 (Bottcher, 1988).
Враги, паразиты, конкуренты, смертность. Хищные млекопитающие, которые могли бы влиять на численность диких баранов (волк, леопард, рысь) стали настолько редкими, что уже не играют какую-либо заметную роль в регуляции численности копытных. Самым опасным был и остается браконьер, вооруженный современной техникой и оружием. Что касается хищных птиц, которые якобы причиняют ущерб молодняку (Саркисов, 1944), то мы не разделяем эту точку зрения. Во всяком случае нужны дополнительные данные с учетом катастрофического сокращения на Кавказе крупных хищных птиц.
Болезни и паразиты азиатского муфлона в Закавказье обсуждались в ряде работ (Саркисов, 1939, 1944; Григорян, 1949; Явруян, 1969, 1975; и др.). Установлено, что дикие бараны страдают всеми болезнями, которые зарегистрированы у домашних овец (некробациллез, сибирская язва, пастереллез, инфекционная плевропневмония, паратиф, пироплазмоз и др.). Они нередко погибают от кокцидиоза, глистных инвазий. В кишечнике почти всех взрослых баранов встречаются гельминты из сем. Strongilidae. Интенсивность инвазии у диких по сравнению с домашними овцами невысокая, но средняя величина инвазии желудочно-кишечными нематодами
470
Rupicapra Blainvillc, 1816
равна 242 экз., а максимальная — 601 экз. На диких баранах зарегистрированы также клещи, паразитирующие на домашних животных.
Численность и ее динамика. Н.Я.Динник (1910) писал, что в прежние времена бараны встречались на Кавказе и соседних с ним странах, как говорят, стадами в сотню голов и более, но теперь это, может быть, случается только в самых глухих местах Турции. О стадах диких баранов в 300 и более голов, в недалеком прошлом обитавших в горах Армении, писали также Е.Л.Марков (1924), А.А.Саркисов (1944). По К.А.Айрумяну и К.М.Гаспаряну (1977), в этой республике еще в 1975 г. встречались стада в 200 голов, а в последние годы, по их мнению, максимальный размер стада не превышает^ голов. Сходные данные приводил Э.Г.Явруян (1970). Что касается современного состояния численности азиатского муфлона в Закавказье, литературные сведения весьма противоречивы. По данным Э.Г.Явруяна (1970), К.А.Айрумяна и К.М.Гаспаряна (1973), в Армении и Нахичевани обитает не более 400 муфлонов. В том же сборнике "Редкие млекопитающие фауны СССР” (1973) рядом со статьей упомянутых выше авторов, Х.М.Алекперов и др. (1973) пишут, что азиатский муфлон в Азербайджане "распространен только в горах Нахичеванской АССР. В 30—40-х годах здесь численность муфлона была невысокой. В настоящее время благодаря строгой охране его поголовье составляет 1000—1200 особей" (с. 34). В Грузии, на границе с Турцией, в нейтральной зоне, недалеко от Кедского района Аджарии в 1985 г. зарегистрировано до 40 особей (Арабули, 1989).
Фактором катастрофического сокращения численности азиатского муфлона в Закавказье и в целом диких баранов является антропогенный пресс — как прямое истребление в прошлом, так и в настоящее время, а также вытеснение их домашним скотом с помощью человека. О ведущей роли в регуляции численности диких баранов человека свидетельствуют и положительный опыт в Западной Европе и отрицательный в нашей стране (Яв-руян, 1969, 1975; Айрумян и др., 1973; Выпарев, 1980; Rodriguez et al., 1987; Schiftze, 1987; Федосенко, 1988; Dubray, 1988; Арабули, 1989; и др.).
Практическое значение. Как объект охоты азиатский муфлон исключительное животное. Мясо обладает высокими вкусовыми качествами. При разумном сочетании интересов охотничьего хозяйства с горным животноводством значительно повысится выход продукции с единицы площади гор Закавказья. Нам представляется исключительно важным дальнейшее использование муфлонов для получения новых пород домашних овец. Такой аспект значения рассматриваемого вида усилился в связи с кариологи-ческими исследованиями, показавшими идентичность кариотипа муфлонов и всех домашних овец.
Род Серны — Rupicapra Blainvillc, 1816
Размеры небольшие; длина тела 125—135 см, длина хвоста 5—8 см, высота в плечах 70—80 см, а в тфестце выше на 4—5 см. Масса гсца, по В.Е. Соколову (1979), составляет у самцов 30—50 кг, у самок 25—42 кг. У карпатских серн, по И.И.Соколову (1959), масса тела достигает 60 кг.
471
Серна
Телосложение плотное, туловище укороченное, профиль спины немного выпуклый. Ноги длинные и массивные, средние копыта большие, но сужены, способны раздвигаться широко. Боковые копыта сильно развиты. Голова сравнительно небольшая, легкая, суживается к концу морды, лобо-носовой профиль резко вогнут. Ноздри щелевидные, направлены вверх, между их внутренними краями имеется небольшое носовое зеркало. Уши относительно длинные, а хвост короткий, с голой нижней поверхностью. Рога имеются как у самцов, так и у самок. Их строение весьма оригинально. Они поставлены вертикально, но их вершины загнуты назад и вниз крючком, поверхность у них гладкая, годовые кольца выражены. Цвет обычно черный или почти черный, в поперечном сечении круглые или овальные. Такое строение рогов среди семейства полорогих свойственно только сернам (Гептнер и др., 1961). Глаза хорошо развиты, выпуклые и значительно выступают в стороны, что увеличивает обозреваемое животным пространство. Волосяной покров относительно грубый, но четко подразделяется на ость и подшерсток. Особенно длинные остевые волосы наблюдаются зимой вдоль хребта. Окраска летнего волосяного покрова шеи и спины ржаво-желтая или желтая, отграниченная от боков широкой черно-бурой или серо-бурой полосой. Грудь темно-бурого цвета. Голова более светлая, чем туловище и шея, но ясно выражена неоднородность ее окраски (узор из светлых и темных участков). Ноги темные; задние светлее передних. Околохвостового зеркала и "бороды" на морде нет. Окраска волосяного покрова мало подвержена географической изменчивости, тогда как сезонный диморфизм в окраске резко выражен. Сходное явление наблюдается также в густоте и длине волос. Хорошо развита зароговая кожная специфическая железа, функционирующая активно в период гона. Имеются межпальцевые железы на всех конечностях, но отсутствуют предглазничные, паховые и запястные железы. Млечная железа имеет две пары сосков. Половой диморфизм выражен слабо — самки немного меньше по массе и размерам тела, слабее также и их рога.
В черепе лицевая часть сужена, основания роговых стержней вздуты. Лобная поверхность имеет впадину между глазницами. Последние хорошо развиты; трубковидные. Ширина черепа на уровне глазниц больше половины основной длины черепа. Кости черепа слабо пневматизированы, но лобные кости имеют небольшие пазухи, переходящие в полости роговых стержней. В слезной кости нет ямки для пр ед глазничной железы. Этмоидальные отверстия узкие, с возрастом они вовсе исчезают. Костные слуховые барабаны маленькие. Верхнечелюстные кости соприкасаются с носовыми. По И.И.Соколову (1959), характерно, что передний край хоан не достигает уровня заднего края верхнего зубного ряда и боковых выемок заднего края костного неба. В отличие от козлов и баранов коренные зубы менее гипсодонтные. На долю длины ряда предкоренных зубов верхней челюсти приходится 34,8—36,7% длины всего ряда коренных зубов. Резцы брахиодонтные; четко выражен перехват на границе между коронкой и корнем зуба. В ряду нижних резцов выделяются более крупными размерами средние резцы, вторые коренные. Задние коренные зубы
472
Rupicapra Blainville, 1816
верхней челюсти имеют 4 эмалевые трубки, а нижние — 5 (три наружные и две внутренние).
Зубная формула:
I — С — Р - М - = 32. 3 13 3
Посткраниальный скелет характеризуется мощным развитием грудной клетки; в скелете конечностей пястная кость меньше плечевой и составляет около 29,5% суммы длины плечевой, лучевой и пясти (Соколов, 1959). Спинных позвонков 38.
Диплоидное число хромосом серн Западной Европы 58 при количестве плеч 60 (Gropp et al., 1970).
Серны, по палеонтологическим данным, формировались в условиях гор и предгорий Западной Палеарктики. Костные остатки плейстоценового и голоценового возраста обнаружены в пещерах горных областей Италии, Франции, Бельгии, Германии, Западного Закавказья и др. Словом, серны являются типично западнопалеарктическими (европейскими) автохтонами и палеоэндемиками.
Ареал рода охватывает горы Центральной и Южной Европы от Пиренеев до Балкан и Карпат, Малой Азии и Кавказа. В б. СССР встречается лишь на Главном Кавказском хребте и горах Малого Кавказа. В настоящее время распространение серн сокращено человеком, приобрело многократные разрывы даже в одной горной области.
Места обитания связаны с горами, хотя высотные пределы распространения серн охватывали пространство от предгорий', в сущности равнин Западной Европы (Гептнер и др., 1961), до высокогорья Кавказа. Однако, видимо, оптимальными местообитаниями следует считать полосу верхней границы лесов и субальпийского криволесья, где имеются крутые скалистые участки, чередующиеся пологими склонами с луговой растительностью. Такие условия обеспечивают серн кормовой базой и убежищами. Питаются различными травянистыми растениями, в первую очередь злаками и разнотравьем. Зимой, кроме злаков, серны поедают листья, побеги и ветви кустарников. Активны в светлый период времени, но ритмика активности выражена слабо. Пасущиеся животные регистрировались в разных горных областях в течение всего дня. Однако летом серны наиболее активны утром и вечером. Гон в разных областях происходит с середины октября до начала января с разгаром в ноябре. Полигамы, с одним самцом бывает несколько самок или с 2—3 самцами 5—6 самок. Беременность длится 160—190 дней, рождает самка одного, реже двух и совсем редко трех детенышей с конца апреля до начала июня. Лактация длится до 6 мес. Половозрелыми самки становятся в 1,5—2,5, а самцы в 2,5—3,5 года. Продолжительность жизни до 12—20 лет. К основным врагам серн относятся волк и рысь. На детенышей часто охотятся лисица, беркут, ворон. Служат объектом охоты, в основном спортивной. В прошлом много серн добывалось ради получения безоаровых шаров (безоары).
Систематика рода в настоящее время не вызывает дискуссии и признается лишь один вид в роде — Rupicapra rupicapra L. В недавнем прош
473
Серна
лом насчитывалось несколько видов серн. Количество подвидов, очевидно, около 10.
Серна — Rupicapra rupicapra Linnaeus, 1758 Рис. 63
Rupicapra tragus, Gray, 1843; Lydekker, 1908, 1910; Динник, 1910; Caprella rupicapra, Динник, 1896.
Коллекционный материал. ЗИН РАН: 36 черепов с рогами, 2 черепа без рогов 37 пар рогов, без черепа; МГУ: 14 черепов с рогами, 2 пары рогов без черепа.
Диагноз. Мелкие копытные с вертикальными рогами, концы которых загнуты назад и вниз в виде крючков. Лобно-носовой профиль вогнут. Верхнечелюстные кости соприкасаются с носовыми.
Особенности морфологии и морфофизиологии
Размеры тела, черепа и отдельных элементов посткраниального скелета. На Западном Кавказе, по данным В.А.Котова (1964), масса тела взрослых самцов в среднем 36,2 кг, самок — около 30 кг. Длина тела взрослых самцов 115—132 см (х = 120,7). Примерно такие же показатели и у самок. Высота в холке у взрослых самцов в среднем 88 см, у самок — 77 см. Высота в крестце соответственно 95 и 84 см, обхват груди за лопатками у самцов 90, у самок 80 см. По данным И.Ф.Попковой (1967), длина шеи 21—30 см, обхват горла у головы около 31 см.
Литературные сведения по размерам черепа очень фрагментарны. Доступный нам коллекционный материал мы свели в табл. 84—85. Эти данные относятся к Западному Кавказу, где, на наш взгляд, находится оптимум ареала серны в регионе. У взрослых самцов (п = 14) основная длина черепа составляет 182,3—195,0 мм (х = 190,6), максимальная ширина черепа 104,0—118,0 мм (х= 111,5), межглазничная ширина 72,6—82,0 мм (х = 77,2), длина лицевой части 122,0—133,0 мм (х = 127,7), длина верхнего ряда зубов 53,5—63,5 мм (Х = 58,6). Размеры черепа взрослых самок немного меньше, чем самцов соответствующего возраста.
Рога серн, характеризующиеся, как отмечено выше, глубоким своеобразием, подробно описаны еще Н.Я.Динником (1910). Он располагал значительной выборкой (около полусотни). По его данным, средняя длина рогов по внешнему изгибу у взрослых старых самок 23 см. Наши данные по размерам рогов серны Западного Кавказа отражены в табл. 84—85.
Посткраниальный скелет, как отмечено в обзоре рода, имеет глубокие адаптивные черты. Не менее показательны особенности строения копыт. Согласно данным И.Ф.Попковой (1967), на Кавказе у серны угол передней стенки копыта составляет 40°, тогда как у кавказского тура этот угол равен 60°. Более пологий угол копыта серн указывает, по мнению автора, что этому животному чаще, чем туру, приходится передвигаться по мягкому грунту. Характерно также наличие на передней части когтевой пластинки плотного гребня длиной 45 мм. Бока ее, как и вся подошва, мягкая и
474
Rupicapra rupicapra Linnaeus, 1758
шероховатая, что усиливает сцепление конечностей с грунтом. Кроме того, на задней стороне подошвы имеется небольшой выступ, выполняющий роль тормоза, особенно когда серна спускается вниз. Дополнительную опорную функцию выполняют увеличенные боковые пальцы. Кроме того, раздвижение копыта (до 86 мм) и наличие щетин между пальцами сокращает удельное давление с 0,330 до 0,236 кг на 1 см2.
Удельное давление туров на мягком грунте почти в 3 раза выше, чем у серн (Попкова, 1967). Имеющиеся данные позволяют характеризовать серну как представителя горных районов с влажным климатом.
Окраска тела серны, как справедливо отмечал еще Н.Я.Динник (1910), на Кавказе сохраняет свою видоспецифичность, но имеет существенные региональные особенности. Летом серна выглядит ярко-рыжей или даже красновато-рыжей. Этот цвет особенно интенсивно развит по верхней части туловища, контрастирующий с черно-бурым ремнем на шее и спине. Ширина ремня от 1,5 до 4 см, и он преимущественно состоит из блестящих черно-бурых или черных волос. По мере перехода от спины к нижней части тела интенсивность окраски снижается. По боку тела от локтя до паха, а затем по передней стороне задней ноги тянется широкая полоса серобурого или черно-бурого цвета. Сходную окраску имеет и нижняя сторона груди. Волосы паха беловатые. Из общего фона верха тела выделяется задняя часть ляжек, имеющая более светлую окраску. Шея, наоборот, того же цвета, что и туловище. Хвост у основания рыжий, а на конце рыжебурый, иногда черно-бурый. Окраска головы сложена из нескольких разноцветных участков. Лоб, верхняя часть морды, нижняя челюсть, подбородок и горло ржаво-рыжие или даже красновато-рыжие. По сравнению с туловищем голова окрашена светлее. Характерно, что темно-бурая полоса в 3—4 см шириной тянется от угла рта через глаз и бока головы, а затем сливается с темной полосой шеи. Уши на внешней стороне серо-бурые, темнеющие на конце; внутри волосы грязно-белые. Ноги по окраске резко отличаются от туловища. Передние от локтевого сустава и почти до конца черно-бурые, лишь у самых копыт, особенно по бокам, появляется рыжеватый цвет. В темно-бурый или черно-бурый цвет окрашена также передняя часть задней ноги начиная от коленного сустава и до копыт.
В зимней окраске серны хорошо развиты черно-бурые волосы; ости почти черные, примерно на 7 см длиннее других волос. Они сильно удлин-нсны (до 15—17 см) на спине и холке. Здесь кончики этих волос бывают белыми. В целом серна зимой черно-бурая, особенно черный цвет развит на лопатках, бока головы черно-бурые, грудь снизу светло-бурая. Затылок и прилегающие к нему верхняя часть шеи и горло светло- или желтовато-рыжие. Задняя сторона бедер и живот ярко-рыжие.
Кожный покров. Волосы. Изучен волосяной покров загривка взрослой самки, добытой в августе в Кавказском заповеднике. Он представлен двумя категориями волос: остевыми и пуховыми. Остевые волосы на размерные порядки не подразделяются. Их длина колеблется в пределах от 13,2 до 38 мм. Волос волнист по всей длине стержня. В прикорневой зоне слабо
475
Серна
476
Rupicapra rupicapra Linnaeus, 1758
Рис. 63. Распространение серны
1 — Гайдук (Динник, 1910);
2 — Новороссийск (Гинеев и др., 1988);
3 — Кабардинка (Динник, 1910; Насимович, 1949; Верещагин, 1959);
4 — Адербиевка (Гинеев, 1988);.
5 — Эриванская (Гинеев и др., 1988);
6 — верховья р. Абин (Гинеев и др.
1988);
7 — верховья р. Убинка (Гинеев и др., 1988);
8 — верховья р. Ппгада (Гинеев и др., 1988);
9 — верховья р. Афипс (Гинеев и др., 1988);
10 — верховья р. Шебш (Гинеев и др., 1988);
11 — Крепостная (Гинеев и др., 1988);
12 — Дефановка (Гинеев и др., 1988);
13 — перевал Гойтх (Динник, 1910;
Верещагин, 1959);
14 — окрестности Майкопа (ЗИН);
15 — Даховская (Гинеев и др., 1988);
16 — Гузсрипль (Динник, 1910);
17 — Кавказский заповедник, гора Пше-киш (МГУ);
18 — гора Бамбак (ЗИН);
19 — гора Фишт (Динник, 1910);
20 — Верховья р. Белая (Динник, 1910);
21 — верховья р. Сочи (Динник, 1910; Насимович, 1949; Верещагин, 1959);
22 — окрестности Адлера (Динник, 1910; Насимович, 1949; Верещагин, 1959);
23 — Гагринский хребет (Капанидзс, 1973);
24 — Рицинский заповедник (Соколов, Сыроечковский, 1990);
25 — верховья р. Малая Лаба (Динник, 1910; Верещагин, 1959);
26 Пссбай (ЗИН);
27 — Соленое (Гинеев и др., 1988);
28 — верховье р. Уруп (Динник, 1910);
29 — Бзыбский хребет (Капанидзе, 1972);
30 — Псху-Гумистинский заповедник (Соколов, Сыроечковский, 1990);
31 — верховье р. Малый Зеленчук (Динник, 1910);
32 — гора IIIисс (Динник, 1910);
477
Серна
33 — Абхазский хребет (Капанидзе, 1972);
34 — Марухский перевал (ЗИН);
35 — Теберда (Тарасов, 1977); ,
36 — Тебердинский заповедник (хребет Гоначхир (МГУ);
37 — Кодорский хребет (Верещагин, 1959; Капанидзе, 1972);
38 — верховье р. Кубань (Динник, 1910);
39 — Батуми (Динник, 1910);
40 — Чохатаури (Капанидзе, 1974);
41 — Хуло (Капанидзе, 1974);
42 — Абастумани (Капанидзе, 1974);
43 — Боржомский заповедник (Соколов, Сыроечковский, 1990);
44 — верховья р. Малка (ЗИН);
45 — Тырныауз (Темботов, 1972);
46— верховье р. Чегем (Верещагин, 1959; Темботов, 1972);
47 — верховье р. Черек (Верещагин; 1959; Темботов, 1972);
48 — верховье р. Урух (Верещагин, 1959; Темботов, 1972);
49 — верховье р. Рио ни (Динник, 1910);
50 — верховья р. Ардон (Верещагин, 1959; ЗИН);
51 — Северо-Осетинский заповедник (Соколов, Сыроечковский, 1990);
52 — Лиахский заповедник (Соколов, Сыроечковский, 1990);
53 — Казбегский заповедник (Соколов', Сыроечковский, 1990);
54 — Ахметский р-н (Капанидзе, 1974);
55 — Ахметский заповедник (Соколов, Сыроечковский, 1990);
56— Ахметский заповедник, Тушетский участок (Соколов, Сыроечковский, 1990);
57 — Ведено (Динник, 1910);
58 — Кварельый!й р-н (Капанидзе, 1974);
59 — Советский р-н (Точиев, 1975);
60 — Тляратинский р-н (Точиев, 1975);
61 — Гутонский заказник (Маломагомедов, 1967);
62 — Лагодехский заповедник (Соколов, Сыроечковский, 1990);
63 — Лагодехи (ЗИН);
64 — Закатальский заповедник (Верещагин, 1959; Соколов, Сыроечковский, 1990);
65 — Закаталы (Марков, 1924);
66— Илисунский заповедник (Соколов, Сыроечковский, 1990);
67 — Алагез (Динник, 1910);
68 — Дилижан (Динник, 1910);
69 — верховье р. Шамхор (Динник, 1910);
70 — Исмаиллинский заповедник (Соколов, Сыроечковский, 1990);
71 — Пиркулинский заповедник (Соколов, Сыроечковский, 1990).
выражена "ножка”. Наибольшая толщина (120—165 мкм) приходится на нижнюю треть стержня. К вершине стержень незначительно сужается. Сверху вниз от основания к вершине стержень немного сплющивается. Поперечные срезы у основания имеют правильную округлую форму (рис. 64, А). Корковый слой равномерно развит вокруг стержня. В верхней половине стержня поперечные срезы вытянутой бобовидной формы: с дорсальной и вентральной сторон проходят слабо выраженные желобки.
478
Rupicapra rupicapra Linnaeus, 1758
Таблица 84. Размеры черепа и рогов серны Западного Кавказа (в мм).
Взрослые и старые
Признак Самцы
п limit х ± ш о Су
Максимальная длина черепа 14 210,5-222,0 216,7+0,95 3,41 1,6
Кондилобазальная длина 14 196,0—212,0 206,3+1,22 4,39 2,1
Основная длина 14 182,3-195,0 190,6± 1,03 3,71 1,9
Максимальная ширина черепа 14 104,0-118,0 111,5+0,95 3,41 3,1
Скуловая ширина 15 84,6-95,5 89,1+0,78 2,91 3,3
Межглазничная ширина 15 72,6-82,0 77,2+0,75 2,81 3,6
Длина лицевой части 14 122,0+133,0 127,7+0,92 3,31 2,6
Максимальная длина носовых костей 15 63,0-78,0 72,9+1,23 4,60 6,3
Длина верхнего ряда зубов 15 53,5-63,5 58,6+0,80 2,98 5,1
Максимальная ширина мозговой капсулы 15 60,0-68,0 63,2+0,62 2,32 3,7
Длина нижней челюсти 12 169,0-176,3 172,1+0,73 2,41 1,4
Длина нижнего ряда зубов 12 58,0-67,0 62,4+0,96 3,19 5,1
Длина диастемы нижней 12 48,0-53,0 49,8+0,43 1,42 2,8
челюсти
Расстояние между внутренни- 15 10,0-20,0 13,5+0,62 2,35 17,4
ми сторонами осевых стержней рогов Расстояние между внешними сторонами осевых стержней 15 46,7-61,0 54,5+0,84 3,14 5,8
рогов Максимальная длина рогов 14 185,5-237,0 207,8+3,48 12,55 6,0
Максимальное расстояние между рогами 12 55,0-99,0 77,2+4,06 13,47 17,4
Обхват рогов у основания 15 62,0-89,0 81,5+1,96 7,32 9,0
Желобок с дорсальной стороны глубже, чем с вентральной. Корковый слой неравномерно развит вокруг стержня. На боковых закругленных сторонах он почти в 1,5—2 раза толще, чем с дорсальной и вентральной сторон (рис. 64, Б).
Сердцевинный канал проходит по всему стержню. Сердцевина в основании развита лучше, чем в верхней половине стержня. В основании она занимает 92% диаметра волоса. Здесь она крупноячеистая. Сердцевинные клетки неправильной многоугольной формы, немного сплющены сверху вниз. По периферии располагается ряд мелких уплощенных клеток (рис. 64, В). При большом увеличении сердцевины видно,’ что радиальные межклеточные перегородки часто укреплены волосковидными выростами, вдающимися в полость клетки (рис. 64, Б). В верхней половине стержня сердцевина также крупноячеистая, но составляет 83% его диаметра. Здесь стенки сердцевины клеток имеют многочисленные волосковидные выросты. В основании выроста располагаются ''опоры”, поддерживающие его. Такая структура сердцевины обеспечивает, вероятно, дополнительную прочность волоса.
479
Серна
Таблица 84 (окончание)
Признак Самки
П‘ limit х + т О Су -
Максимальная длина черепа 5 211,5-214,0 212,6+0,54 1,08 0,5
Ковдилобазальная длина 5 201,0-204,7 202,8±0,90 1,80 0,9
Основная длина 5 185,5-189,3 187,0±0,81 1,62 0,9
Максимальная ширина черепа 5 105,0-107,3 106,6+0,48 0,96 0,9
Скуловая ширина 5 83,4-89,0 86,5+1,07 2,13 2,5
Межглазничная ширина 5 70,5-77,4 73,5+1,58 3,15 4,3
Длина лицевой части 5 124,6-126,0 125,1±0,26 0,52 0,4
Максимальная длина носовых 5 70,0-76,0 72,3±1,25 2,49 3,4
костей Длина верхнего ряда зубов 5 54,0-60,5 56,9+1,55 3,09 5,4
Максимальная ширина 5 61,0-62,5 61,8±0,29 0,57 0,9
мозговой капсулы Длина нижней челюсти 4 169,0-175,0 172,6+1,47 2,54 1,5
Длина нижнего ряда зубов 4 58,0-60,5 58,9±0,66 1,14 1,9
Длина диастемы нижней 4 49,0-57,0 52,8—2,23 3,86 7,3
челюсти Расстояние между внутренни- 5 10,5—18,0 15,2±1,40 2,80 18,4
ми сторонами осевых стержней рогов Расстояние между внешними 5 45,5-52,6 48,6±1,28 2,56 5,3
сторонами осевых стержней рогов Максимальная длина рогов 5 176,0-199,0 193,4+4,93 9,86 5,1
Максимальное расстояние . 4 59,0-84,0 72,0+5,91 10,23 14,2
между рогами Обхват рогов у основания 5 61,0-88,0 70,4+5,13 10,26 14,6
Рисунок кутикулы мало меняется вдоль стержня. Чешуйки вытянуты поперек его длинной оси. Свободные края относительно ровные и плотно прилегают к корковому слою (рис. 64, Д).
Пуховые волосы немногочисленные, сильно извитые, не подразделяются на размерные порядки. Их длина не превышает 14,5 мм. Толщина (15 мкм) не меняется. Поперечные срезы округлой формы. Сердцевина отсутствует. Рисунок кутийулы кольцевидного типа и не меняется вдоль стержня.
Толщина кожи на груди у самки равна в среднем 3,85 мм. Эпидермис довольно толстый — 22—28 мкм (25 мкм) и непигментирован. Сосочковый слой дермы значительной толщины, в области загривка он составляет в среднем 1,5 мм, а на груди — 1,7—1,8 мм. Видимо, это связано с сильным развитием корней растущих волос. Сальные железы крупных размеров на загривке (192,5 х512,5 мкм) и чуть меньше в области груди (110 х415 мкм). По данным В.Е.Соколова (1973), сальные железы серны на загривке наиболее крупные по сравнению с другими изученными представителями подсемейства козлиных.
480
Rupicapra rupicapra Linnaeus, 1758
Таблица 85. Размеры черепа и рогов серны Западного Кавказа (в мм).Самцы, 2 года
Признак Самцы
п limit X + т G Q
Максимальная длина черепа 5 184,0-206,6 199,02±4,56 9,12 4,6
Кондилобазальная длина 5 175,0-197,5 189,6±4,37 8,74 4,6
Основная длина 5 160,0-184,0 174,08±4,54 9,07 5,2
Максимальная ширина черепа 5 97,3-110,5 101,9+2,56 5,12 5,0
Скуловая ширина 4 78,5-91,4 82,9±3,35 5,81 7,0
Межглазничная ширина 5 70,0-77,3 72,7±1,44 2,87 3,9
Длина лицевой части 5 106,5-124,5 117,0±3,38 6,76 5,8
Максимальная длина носовых 5 55,0-69,0 64,2±2,82 5,63 8,8
костей Длина верхнего ряда зубов 3 51,0-61,0 54,7±13,90 5,51 10,1
Максимальная ширина 5 56,0-62,8 59,1±1,27 2,53 4,3
мозговой капсулы Длина нижней челюсти 5 146,3-167,0 156,9±3,82 7,63 4,9
Длина нижнего ряда зубов 3 51,0-67,0 56,3±6,53 9,24 . 16,4
Длина диастемы нижней 4 43,0-46,5 45,1±0,89 1,55 3,4
челюсти Расстояние между внутр енни- 4 14,5-15,5 15,0+0,33 0,58 3,9
ми сторонами осевых стержней рогов Расстояние между внешними 4 48,0-54,7 50,7±1,79 3,10 6,1
сторонами осевых стержней рогов Максимальная длина рогов 4 142,0-207,0 168,0±16,31 28,25 16,8
Максимальное расстояние 4 49,0-80,0 61,8±7,91 13,07 21,1
между рогами Обхват рогов у основания 4 70,0-85,0 75,8+3,72 6Д5 8,5
Потовые железы с длинными трубчатыми, сильно извитыми секреторными отделами. Предглазничные железы, как отмечено при характеристике рода, отсутствуют. Межпальцевые имеются на передних и задних конечностях и представляют собой неглубокие вдавливания кожи, покрытые внутри редкими волосами. Они открываются треугольными отверстиями на передней стороне ног, выше копыт. Паховые железы отсутствуют. Как самцы, так и самки имеют своеобразные, свойственные, кроме серн, еще американской снежной козе и горалу, зароговые, или затылочные, железы. В период полового сезона железы вздуваются, сильно выступают в виде прилегающих друг к другу и к основаниям рогов багровых наростов, поверхность которых пересекается несколькими глубокими бороздами, напоминающими извилины головного мозга (Hcssling, 1855 г.; Schick, 1913 г.; Рососк, 1910, 1918 гг.; Schattcr, 1940 г., по: Соколов, 1973). По данным Соколова (1959), животные в это время издают сильный, напоминающий козлов запах, обусловленный секретом зароговых желез. Для кожи туловища характерны крупные мышцы, поднимающие волосы (100—165 мкм на загривке и 50—70 мкм на груди).
31 В.Е.Соколов, А.К.Темботов
481
Серна
Рис. 64. Строение остевых волос затривка взрослой самки серны
А — поперечный срез в основании, ув. х 500; Б — то же в верхней половине стержня, ув. х 600; В, Г, Е, — сердцевина, ув.хЗОО и 2000; Д — кутикула волоса, ув. х 1000
482
Rupicapra rupicapra Linnaeus, 1758
Система крови у серны, насколько нам известно, на Кавказе не изучена.
Индексы внутренних органов остаются также мало изученными. Вся известная нам информация ограничивается фрагментарными данными И.Ф.Попковой (1967) по отдельным органам. Согласно этим данным, индекс сердца (h — 6 экз.) у серны составляет 6,9—9,7%о (Г = 8,1), тогда как у кавказского тура, по сведениям того же автора, значительно меньше — 4,3—7,5%о (х = 5,7). Серна по индексу сердца ближе к джейрану, у которого он составляет в среднем 9,2%о. Такое положение автор связывает со способностью серны и джейрана к быстрому бегу на длинные дистанции, тогда как обитание туров в сильно скалистых и расчлененных местах сделало невозможным бег на большое расстояние. С таким выводом согласуется и положение серны по быстроаллюрности между джейраном и кавказским туром. Кроме того, по легкости костяка серны занимают также промежуточное положение между этими видами.
Индекс печени у серны составляет (h — 3 экз.) 14,8—27,0%о (X — 19,6), что очень близко к соответствующему показателю кавказского тура. Сходны также индексы легких (п = 6 экз.) у серны 15,2—33,3%о (£ — 27,7), а у тура — 16,2—26,3%о (х= 20,0).
Кариотип. Данными по хромосомному набору кавказской серны мы не располагаем. В Западной Европе (Швейцария) кариотип серны включает 58 хромосом с числом плеч 60. Х-хромосома — акроцентрик среднего размера, У-хромосома точечная, морфология ее не определена (Gropp et al., 1970). Сведений по дифференциальной окраске хромосом серны у нас нет.
Структура и динамика ареала
Ареал вида Rupicapra rupicapra совпадает с распространением монотипического рода Rupicapra. Что касается распространения в б. СССР, то восстановленный его ареал, кроме гор Большого и Малого Кавказа, видимо, включал юго-западную часть Карпат, прилегающую к Румынии (Гептнер и др., 1961). В настоящее время серна в Карпатах отсутствует.
Распространение в горах Кавказа в прошлом веке было обширным, и все имеющиеся литературные и архивные данные свидетельствуют о том, что серна входила в группу фоновых видов копытных горно-лесных и горно-луговых ландшафтов региона (Динник, 1893—1910; Шильдер, 1895—1902; Туркин, Сатунин, 1902; Верещагин, 1947, 1959; и др.). Судя по этим данным, в конце прошлого и самом начале текущего столетия западная граница ареала простиралась до гор окрестностей пос. Кабардинка ”...но сюда, вероятно, загоняют их лишь в исключительных случаях глубокие снега, выпадающие на более высоких местах. Начиная же от перевала Гоитх... на восток по Главному хребту и его высоким отрогам серны встречаются везде в Кубанской области" (Динник, 1910, с. 229). Северо-восточная граница, как пишет названный автор, занимала горы Скалистого хребта, встречаясь спорадично, но "прежде же попадались здесь в очень многих местах. На горе Герпегеж около Псебайской и Бесланеевской станиц они и теперь живут постоянно в небольшом количестве. Попадаются серны нередко на
31
483
Серна
горе Шиссе недалеко от Зеленчукской и Сторожевой станиц” (Там же, с. 230). На Центральном Кавказе распространение серны в рассматриваемый период уже было сужено, однако еще в 80-х годах XIX в., по данным К.Н.Россикова (1887), на горах Балк-Баши и Ташлы-Сырт, в верховьях р. Малка серна встречалась "очень часто на самых недоступных скалах и нередко далеко выше снеговой линии". Смещение распространения серны в высокогорья эльбрусского варианта имело, на наш взгляд, антропогенную основу. Зольские пастбища, занимающие обширные плато, интенсивно используются со дня окончания кавказских войн в качестве летних пастбищ. Здесь уместно привести данные Н.Я.Динника (1910) о том, что серны избегают высоких гор и мрачных, безлесных ущелий, любимых турами. Этой особенностью поведения он объяснял отсутствие серны в Девдоракском ущелье, спускающемся с Казбека. Между тем, как пишет сам автор, здесь раньше серна была отстреляна местным охотником. На южном макросклоне Центрального Кавказа, где лучше сохранялись кормовые и защитные условия, серны встречались часто. Сам Н.Я.Динник их видел много и часто в верховьях Риона и на горе Шод. В Сванетии они были, по его данным, очень обыкновенны. В Дагестане серны встречались в то время значительно реже, чем в бывших Кубанской и Терской областях. Однако нередкие поселения регистрировались в б. Андийском и Гунибском округах, на обоих склонах Главного Кавказского хребта, но на местах, удаленных от этого хребта, серны попадались очень редко. Отмечены также в б. Самурском округе, но их уже не было в б. Темир-Хан-Шуринском, Аварском, Даргинском и Кайтаго-Табасаранском округах. То же самое можно сказать и в отношении Кюринского и Казикумухского округов.
Распространение серн на Малом Кавказе на рубеже XIX и XX вв. было мало известно. Н.Я.Динник (1910) располагал сведениями о наличии серн лишь в нескольких точках: горы недалеко от Батуми и Артвина, окрестности Боржоми и Белого Ключа на Триалетском хребте, верховья Шам-хорчая и Дзсгемчйя до с. Барсуна. Видимо, серны встречались на горе Арагац. Мы разделяем мнение В.Г.Гептнера и др. (1961), что серны на Малом Кавказе были распространены в то время значительно шире, чем представляется это в наши дни. По данным названных авторов, серны по северному склону Шахдагского хребта (бассейн Шамхора, Акстафы) также спускались очень низко. Тем не менее нет оснований считать, что эти копытные в обозримое прошлое обитали в Талыше, Зангезурских горах, в Карабахе и на восточной части Армянского нагорья. К 30-м годам текущего столетия серны перевелись на Шахдагском и Муров-Дагском хребтах. В те же годы сокращение и дробление некогда сплошного ареала серн происходило на Большом Кавказе, в том числе на Западном Кавказе, где расположен оптимум ареала. Такие негативные сдвиги ряд авторов (см. обзор: Котов, Рябов, 1963) связывают с быстрым увеличением населения людей в горах и сопутствующим этому освоением новых территорий, а главное в связи с хищнической охотой. По мнению Н.К.Верещагина (1959), исчезновение серн на Карабахе, как и на всем Малом Кавказе, нельзя свести лишь к одному антропогенному фактору, ибо по Муров-Дагскому
484
Rupicapra rupicapra Linnaeus, 1758
хребту и в ряде других горных районов сохранилось немало глухих лесов, в которых уцелели крупные животные (бородатый козел). Надо полагать, что влияние человека имело больше отрицательных последствий на юго-востоке Кавказа, где представлены худшие природные условия для серн. К тому же здесь поселение человека в горах наиболее древнее и густое. В 50-х годах XX в. ареал серны стад еще более узким, смещенным выше в горы и сильно пятнистым. Серны оказались выбитыми почти повсеместно на периферии ареала,-на многих легко доступных браконьерам горных урочищах (Гецтнср, Формозов, 1941; Насимович, 1955; Темботов, 1960; Котов, Рябов, 1963; и др.). Так, на Западном Кавказе в 30-х годах серны встречались в окрестностях с. Соленое, но потом были выбиты. В 1953 г. две серны обитали в окрестностях Псебая, однако вскоре были отстреляны. В последующие годы здесь уже серны не встречались. Те, которые вынужденно оказываются в низкогорьях или предгорьях, как это было в мае 1956 г. в черте Майкопа, обычно погибают.
На южном макросклоне Западного Кавказа, в Геленджикском, Туапсинском, Лазаревском и Адлерском районах серны встречаются в основном по Главному Кавказскому хребту. По Н.К.Верещагину (1959), в 1955 г. несколько серн обитало северо-западнее Туапсе, на скалах Джубги. Отмечались серны также в верховьях рек Пшада, Псекупс, Аше и др. По данным В.А.Котова й Л.С.Рябова (1963), на Западном Кавказе граница проходила по линии населенных пунктов: Шапсугская (западный пункт ареала) — севернее Адербиевки — южнее Михайловского Перевала — севернее Пшада — южнее Дефановки и Подхребетового — Георгиевское — Солох-Аул — Четвертая — Чвежипсе. Северная граница южнее линии Эриванская — Крепостная — Безымянное — Самурская — Даховская — Бугунжа — Соленое. В пределах очерченной территории более или менее широко серны распространены в Псебайском, Тульском, Апшеронском, Адлерском и Лазаревском районах. По свидетельству названных авторов в западной части ареала распространение носит островной характер. На территории Тебер-динского заповедника серна обычна в лесистых отрогах Главного Кавказского хребта, а на расположенных выше 2300 м скалистых альпийских лугах отмечались только изредка (Эквтимишвили, 1952; Аренс, 1957). В 50-х годах распространение серны в эльбрусском варианте еще больше сократилось, отступив до Бокового хребта. Ни на Меловом, ни на Скалистом хребтах нами серны не обнаружены, однако, по словам скотоводов, отдельные животные спускаются по лесистым скалам глубоких ущелий до Долины нарзанов. В бассейне Терека, где возобновляется пояс широколиственных лесов, а рельеф типично альпийский, ареал серны более широкий, занимает наряду с Главным Боковой и Скалистый хребты. В Баксанском ущелье мы ее регистрировали от г. Тырныауза до верховья Баксана, в Чегемском — от с. Булунгу до верховья Чегема: в Черекском — в верховьях одноименной реки. В Северной Осетии, по свидетельству И.В.Наниева, серна есть на Скалистом и Главном хребтах в лесных ландшафтах (Насимович, 1955). Сведениями о распространении серны в Чечено-Ингушетии рассматриваемого периода мы не располагаем. Видимо,
485
Серна
серны здесь встречались широко и довольно часто, что объясняется малочисленностью людей в горах в то время и благоприятными природными условиями. Эта область, по Н.Я.Диннику (1910), В.Г.Гептнеру и А.Н.Фор-мозову (1941), отличалась высокой численностью серн. В 30-х годах, по В.Г.Гептнеру и А.Н.Формозову (1941), серны обитали в высокогорьях Андийского хребта; спорадично в б. Гунибском округе, на высоких горах водораздела Кара-Койсу и Аварского Койсу; редко на западном массиве Богосского хребта, но часто в той части хребта, которая примыкает к аулу Хорда. Больше всего серн было на склонах Главного хребта, но на местах, удаленных от него, их видели очень редко. Словом, еще в 30-х годах в Дагестане распространение серны было приурочено к высокогорью. Они существовали отдельными изолированными колониями с низкой численностью. По мнению Н.К.Всрещагина (1959), вскоре серн не стало в центральном безлесном Дагестане и только на восточном склоне горы Шах-Даг оставалось небольшое число серн.
На южном макросклоне Большого Кавказа их было немало в Абхазии и Сванетии (колхидский вариант). Они занимали крутые склоны с пихтой, елью и кленом в верховьях Кодора, Ингура, Риона. Очень широким размахом высотного распространения отличались стада в горной системе Рачинского, хребта, выступающего в качестве климатораздела между западнозакавказскими и восточнозакавказскими вариантами поясности. По Н.К.Ве-рещагину (1959), серны редко встречались в Южной Осетии и Кахетии. В пределах Азербайджана в значительном количестве серны, по данным того же автора, держались в березовых ущельях Кахетичая, Шинчая и в верховьях Туряньчая. Много серн было в Закатальском заповеднике, а далее на восток они отмечались вблизи Курбан-Эфснд Исмаиллинского района. Восточной границей являлась гора Бабадаг в Азербайджане.
Современный ареал серны на Кавказе имеет вид узкой полосы с многочисленными разрывами. Самый северный участок обитания, по данным А.М.Гинеева и др. (1988), находится у ст. Гайдуки на горе Различная. Следующее поселение обнаружено в верховьях р. Пшада на Хр. Подкова, горах Обляго и Папай. Далее от линии, соединяющей горы Шесси и Б. Псеушхо, серны занимают все крупные хребты, подходя вплотную к поселкам Гузе-рипль, Кардану, Черноречье и горе Маркопидже. Современное распространение серны в верховьях Зеленчука и Кубани изучено слабо. В пределах Тебердинского заповедника их регистрировали в верхней полосе лесов, субальпийском и альпийском поясах (Тарасов, 1977; Бобырь, 1980).
В Кабардино-Балкарии, по нашим данным, серны изредка встречаются в высокогорье (верховья рек Малка, Баксан, Чегем, Черек, Урух). Очень мало данных по Северной Осетии и Чечено-Ингушетии. По Т.Ю.Точиеву (1975), серны обычны в ущельях Шони, Аши, Гулой-Лом. По А.М.Абдурах-манову и Т.Х.Спасской (Гинеев и др., 1988), в Дагестане этот вид встречается в пределах Главного Кавказского хребта Тляратинского, Чародин-ского, Советского, Агульского, Цумадинского и Цунтинского районов. Изредка серна появляется в соседних горах.
486
Rupicapra rupicapra Linnaeus, 1758
По Закавказской части ареала свежих материалов мало. А.Б.Арабули (1989) считает, что в Грузии серна распространена в наши дни в верхней полосе лесов, а также в безлесном субальпийском поясе. На охраняемых территориях (Лагодехский и Боржомский заповедники, в Саирме, Окуни) они спускаются до нижнего лесного пояса. В Азербайджане, по Х.М.Алск-перову и др. (1973), серны встречаются главным образом в пределах Большого Кавказа и в незначительном количестве на Муровдагском хребте Малого Кавказа. С.М.Кулиев (1989) сообщает, что в Исмаиллинском заповеднике и заказнике (восточная часть Большого Кавказа) они обитают в субальпийском и альпийском поясах до 2500—2800 м над ур. моря. Зимой спускаются почти до предгорий.
Итак, в настоящее время серна на Кавказе имеет весьма О1раниченное распространение, приуроченное в основном к высокогорью; лишь на охраняемых территориях заселяет лесные пояса до предгорьсв. За исключением отдельных участков Кавказского заповедника, всюду встречается спорадично, и дать детальную картину современного распространения пока невозможно.
Современные тенденции антропогенных изменений ареала. За последнее столетие распространение и численность серны на Кавказе неуклонно сокращались. Этот процесс имел место как в оптимуме ареала, каким является Западный Кавказ, так и на периферии, в том числе в восточной части Закавказья, где природные условия менее благоприятны для серн. В итоге из широко распространенного и фонового вида она превратилась в довольно редкое и спорадично распространенное животное. Более того, растет угроза значительного обеднения генофонда местной и глубоко автохтонной формы с высоким таксономическим рангом. На наш взгляд, стало очевидным, что без специальных мероприятий по сохранению, увеличению численности и распространению серны трудно предотвратить, полное уничтожение данного вида на Кавказе. По оценке причин, обусловивших такое положение кавказской серны, общепринятого суждения нет. Многие исследователи не без основания считают таковыми антропогенные факторы: изменения ландшафтных условий, кормовой и защитной базы, прямое преследование, вытеснение их домашними животными, занос последними различных возбудителей болезни (например, ящура), фактор беспокойства в связи с ростом туризма и т.д. С таким суждением согласуются результаты работы некоторых заповедников, особенно Кавказского. Существует и другое мнение, не столь популярное, как первое, но, на наш взгляд, принципиально важное. Суть его в том, что исчезновение серны в ряде районов Кавказа нельзя объяснить только антропогенными факторами. Положение дел с воспроизводством кавказской серны осложняется особенностями биологии, в том числе высокой избирательностью мест обитания. На основе обоих мнений, на их синтезе возможно продолжить мероприятия для сохранения и воспроизводства кавказской серны на значительной территории, чтобы она вновь стала объектом спортивной охоты.
487
Серна
Систематика и изменчивость
Систематическое положение. Проблема "вид или подвид" в отношении кавказской серны в настоящее время не обсуждается. Мнения различных систематиков по этому вопросу сходятся — кавказская серна составляет хорошо выраженный подвид R.r. caucasica. Несмотря на это, имеется определенная неудовлетворенность состоянием изученности серн Кавказа в систематическом плане. Более того, мы разделяем мнение Н.К.Всрещагина (1959, с. 363), о том, что "говорить о расселении серны в четвертичном периоде из Малоазийско-Кавказских гор в европейские Альпы или, наоборот, из Европы на восток через Балканы и Малую Азию нет оснований. Кавказскую серну следует, в сущности, считать не подвидом швейцарской, а самостоятельным видом. По сравнению с альпийской у кавказской серны имеются не только известные морфологические отличия, но и иной голос — длинный шипящий звук, отмеченный еще Динником (1896)". О наличии у кавказской серны целого комплекса морфологических признаков Н.Я.Динник писал неоднократно, в том числе в известной монографии (Динник, 1910). Это касается окраски тела, отдельных волос (зональности их), расцветки головы, размеров тела и черепа и т.д. Не все морфологические признаки, указанные им, выдержали ревизию. Но пока нет работ, основанных на более значительном коллекционном материале.
Межу тем изучение серны Западной Европы в последние годы (Blahout, 1971; Hartl, 1986; Miller, Hartl, 1987) показало, что генетическое разнообразие у серны значительно выше, чем у подавляющего большинства крупных млекопитающих. Межпопуляционные генетические дистанции у альпийских серн очень значительны (от 0,0031 до 0,0169). Вероятно, сказывается разнообразие условий жизни и значительная привязанность их к местам обитания. Ведь у серны наблюдаются, как правило, только высотные перемещения и притом лишь в тех районах, где выпадает большой снег. Эти и другие доводы не могут быть отброшены при ревизии систематики рода Rupicapra, которую мы считаем актуальной.
Индивидуальная изменчивость. Для анализа размаха индивидуальной изменчивости экстерьера кавказской серны очень мало данных. Н.Я.Динник (1910) приводит промеры всего 6 разновозрастных самцов и самок, В.А.Котов (1964) — 8 самцов и 3 самок, И.В.Попкова (1967) — 6 разновозрастных зверей без разделения по полу. При этом животные добыты в разные сезоны года и соответственно с различной упитанностью. К сожалению, мы тоже не располагаем соответствующими данными.
Более полные сведения мы имеем по размерам черепа, которые иллюстрируют определенную индивидуальную изменчивость.
У взрослых самцов максимальная величина коэффициента вариации свойственна длине носовых костей (см. табл. 84). У взрослых самок этот показатель также очень сильно варьирует. Как справедливо подметил И.И.Соколов (1959), диагностического значения не может иметь и форма носовых костей. В частности, отсутствие или наличие боковых отростков в задней половине носовых костей не является устойчивым признаком
488
Rupicapra rupicapra Linnaeus, 1758
кавказской серны, как это полагал Р. Лидеккер (Lydekker, 1913). Широко изменяется также длина зубных рядов, что обусловлено, видимо, степенью их стертости. У самцов и у самок сильно колеблются размеры скуловой и межглазничной ширины, а у самцов — также максимальная ширина черепа. Между тем как у самцов, так и у самок высокой стабильностью .характеризуется максимальная, основная длина черепа и длина нижней челюсти.
Индивидуальная изменчивость особенно велика по размерам рогов как самцов, так и самок, что говорит о небольшом систематике-таксономическом весе этого показателя у серны. Относительно низкой вариабельностью отличается максимальная длина рогов. В равной степени это свойственно и самцам и самкам. Как видно, прав был РЛидеккер (Lydekker, 1913), когда он придавал длине рогов значительный систематико-таксономический вес. Однако этого нельзя сказать в отношении толщины рогов, ибо этот показатель высоко изменчив.
Как справедливо отмечал еще НЛ.Динник (1910), многие детали окраски кавказской серны отличаются высокой индивидуальной изменчивостью. Так, черный ремень на спине может иметь различную ширину, протяженность на шее и на огузке, а у некоторых особей он вовсе не выражен. Однако при всей вариации деталей окраски тела у кавказской серны все же сохраняются особенности расцветки головы, горла и других частей тела.
У кавказской серны не изучена генетическая изменчивость, но если судить по результатам аналогичных исследований по альпийским сернам, то она, видимо, значительна. Достаточно сказать, что при анализе биохимической изменчивости белков, кодируемых 42 локусами у 136 особей, для всех пяти изученных “популяций" оказались полиморфными 10 локусов (Miller, Hartl, 1987).
Возрастная изменчивость. Литературные сведения по этому вопросу фрагментарны, нередко противоречивы, так как фактических данных по Кавказу очень мало. По В.А.Котову (1964), кавказская серна на Западном Кавказе к годовалому возрасту достигает размеров, близких к размерам взрослых. Самка в возрасте 9—10 мес, добытая 14 января, весила 21,3 кг; длина тела 103 см, высота в холке 73, высота в крестце 75 см. В условиях загона самец и самка к концу октября первого года жизни имели массу тела 17,2 и 21,2 кг соответственно. К концу мая следующего года — 25,4 и 26,0 кг, к полутора годам — около 30 кг. Масса тела самца кавказской серны в возрасте трех лет достигает 38—43 кг; масса тела самки такого же возраста немного меньше. Как видно, экстерьер кавказской серны формируется быстро, в течение первых трех лет жизни они набирают массу и размеры тела, очень близкие к максимальным показателям. В соответствии с возрастной изменчивостью массы и размеров тела происходит рост рогов. В первую зиму у серн имеются совсем небольшие рожки, слабо выдающиеся из волос. К концу лета следующего года они достигают длины до 12—14 см. На втором году жизни концы рогов начинают загибаться, в два года длина рогов 15—17 см. На третьем году жизни рога увеличиваются в
489
Серна
среднем на 3,4 см, на четвертом — 1,7 см, на пятом — до 0,5 см, а в последующие годы прирост рогов составляет лишь 0,2—0,1 см (Динник, 1910; Попкова, 1967). Аналогичные результаты получены и по европейской серне, что свидетельствует о видоспецифичности характера возрастной изменчивости рогов у серн — интенсивный рост в первые 3—4 года, а затем резкое торможение (Couturier, 1938; Blahout, 1976; и др.). Как видно, наличие рогов как у самцов, так и у самок, а также хорошо заметные годовые сегменты у тех и у других делают роговые чехлы удобным показателем для определения возраста серн. Возрастные изменения окраски сводятся лишь к замене на втором году жизни нарядом взрослого.
Формирование зубной системы и возможность использования зубов для определения возраста серны, судя по имеющимся у нас данным и литературным источникам (Cauturier, 1938; Blahout, 1976; и др.), одинаковы в различных частях ареала вида. Полная замена молочных зубов на постоянные завершается, как правило, на третьем году жизни. На четвертом году жизни у серн происходит окостенение черепа.
Половые различия. По данным В.А.Котова (1964), на Западном Кавказе среди взрослых животных, как правило, самцы тяжелее самок примерно на 6 кг. При очень сходной длине тела самцы выше самок в крестце и в холке в среднем на 11 см, а обхват груди за лопатками у самцов больше в среднем на 10 см. Половой диморфизм по размерам и форме роговых чехлов слабо выражен. Рога самок лишь немного уступают самцам в длине и толщине. В нашей выборке максимальная длина рогов по внешнему изгибу в группе взрослых у самцов в среднем больше на 1,4 см, чем у самок, толщина рогов у основания — на 1,1 см. По размерам черепа за редким исключением самцы превосходят самок, по максимальной длине всего на 0,4 см, ковдилобазальной длине — на 0,55 см, максимальной ширине черепа — на 0,49 см, межглазничной ширине — на 0,37 см и т.д. По нашим данным, самцы уступают самкам по длине нижней челюсти и длине диастемы в нижней челюсти. Достоверность различий по ряду показателей высокая.
Географическая изменчивость. В пределах видового ареала, как отмечено выше, описано большое число форм, но степень морфологического различия между ними в ряде случаев невелика или очень мала (Гептнер и др., 1961). Однако, судя по результатам электрофоретических исследований белков (Hartl, 1986; Miller, Hartl, 1987), генетическая изменчивость серн в связи с геотрафическими изоляциями может быть значительной. Однако по кавказской серне такими данными мы не располагаем. Нет также достаточной выборки коллекционного материала, который позволил бы с использованием математических методов провести анализ изменчивости кавказской серны в пределах Кавказа и оценить ее положение в кругу других форм вида. Поэтому мы вынуждены ограничиться лишь некоторыми замечаниями, почерпнутыми из литературы и просмотра небольшого коллекционного материала.
Н.Я.Динник (1910) обращал внимание, что в Закавказье около Абас-Тумана добытая зимой серна имела общий цвет не черно-бурый, а
490
Rupicapra rupicapra Linnaeus, 1758
каштаново-бурый. В.Н.Гсптнср и др. (1961) допускали, что серны Малого Кавказа идентичны популяции Малой Азии, выделяемой в качестве самостоятельного подвида R.r. asiatica. Его распространение, видимо, простиралось через Аджаро-Имсртинский хребет по всему Малому Кавказу. При таком положении возможно, как отмечают названные авторы, форма caucasica существует только на Большом Кавказе, а в Закавказье, по крайней мере Западном, жила R.r. asiatica. "В связи с уничтожением закавказской серны вопрос о се систематическом положении уже, по-видимому, не разрешим" (Гептнер и др., 1961, с. 507). Однако взаимодействия между соседними подвидами могли произойти через Лихский хребет, выполняющий своеобразный мост между горными системам Закавказья и Большого Кавказа. В связи с этим, на наш взгляд, было бы интересно провести сравнения по всем доступным параметрам, в том числе биохимическим, западнокавказских (Кавказский заповедник) и восточнозакавказских (Лаго-дехский и Закатальский заповедники) серн. По данным И.Ф.Попковой (1967), длина рогов по изгибу несколько больше у западнокавказских серн (23,6 см), чем у восточнокавказских (20 см). Эти данные основаны на мизерной выборке и мало что проясняют.
Итак, мы считаем, что систематическое положение кавказской серны недостаточно разработано. До получения новых данных мы выделяем на Кавказе один подвид R.r. caucasica Lydekker, 1910.
Экология
Ландшафтная приуроченность. Биотопы. Все имеющиеся у нас сведения, в том числе данные по распространению в прошлом и настоящем, по морфологии и морфофизиологии, а также палеозоологический материал однозначно указывают на становление серны как горного лесолугового вида окраинно-материковых типов поясности Средиземноморского бассейна. Мы разделяем мнение Н.К.Всрещагина (1959), что серна приспособлена к мезофитным участкам горных поднятий Средиземноморья. Связи ее с лесом и мезофитной луговой растительностью прочнее, чем у козлов. Н.Я.Динник (1910), характеризуя местообитания серн, отмечал, что излюбленными местами обитания являются скалистые, более или менее высокие горы, поблизости от которых растет лес. Не меньше предпочитаются, по его мнению, крутые, обрывистые скалы, поросшие редким лесом, состоящим из сосен, елей или берез. Очень часто серны посещают также субальпийское криволесье. В большом количестве серны встречались Н.Я.Диннику на всех безлесных лугах Западного Кавказа, где их не тревожили. Они обычно не встречаются на очень высоких горах и в мрачных, безлесных ущельях, любимых турами (Диннику, 1910). А.А.Насимович (1938), изучавший длительное время серн на Западном Кавказе, считал, что серны, будучи привязанными к скалистым ландшафтам, в отличие от туров избегают высокогорных скал. Поэтому они малочисленны там, где много туров, а где мно'го серн — мало туров.
491
Серна
На основе многолетних полевых наблюдений, проведенных в Кавказском заповеднике, составлена характеристика ландшафтных условий, предпочитаемых кавказскими сернами: задернованные склоны крутизной в 20—40°; скалы с карнизами и нишами, необходимые в качестве убежища от врагов, непогоды и отдыха; достаточная кормовая база, особенно зимой; различные солонцы. Если отсутствует какой-либо из перечисленных компонентов, то привлекательность территории как местообитания серны резко падает (Дубень, 1985). Сходное отношение к выбору ландшафтов подмечено и у европейских серн. Так (Blahout, 1985), в Татрах сернам необходим наряду с горными лугами для пастьбы крутой, сильнопересеченный скалистый рельеф в качестве убежища. Большое значение этим автором придается температуре, ветру, снегу, гололедице, дождю, которые при жестких режимах исключают возможность существования серн, вынуждают их к смене местообитания. При благоприятных условиях на Кавказе диапазон высотного размещения серн может колебаться от 150—200 до 4000 м над ур. моря. Максимальная высота местообитания, известная науке, составляет у альпийских серн 4750 м на Монблане (Cautuirc, 1938).
На Западном Кавказе, где расположен оптимум ареала, летом серны встречаются во всех поясах Кавказского заповедника, но больше всего на высоте 1700—2500 м — по альпийским и субальпийским лугам и соседним участкам верхней ч^сти лесного пояса (Насимович, 1955). Сходная ландшафтная приуроченность наблюдается и на Восточном Кавказе, где представлены иные типы поясности. По И.Ф.Попковой (1967), в Закатальском заповеднике, где больше всего сохранилось кавказских серн на Восточном Кавказе, эти животные, концентрируясь на мезофитных лугах, расположенных близ крутых осыпей и скал, обильно заселяют верхние и средние части боковых отрогов Главного хребта. При этом серн больше всего в местах с относительно спокойным рельефом, Хорошей облесенностыо и хорошо развитыми лугами, имеющими богатое увлажнение. Что касается обитания серн круглогодично в высокогорьях Дагестана, верховьях Арагвии или Малки, то это следствие преследования человеком или вытеснения с пастбищ домашним скотом. То же самое имеет место, когда серны вынуждены спасаться в лесном поясе (Северная Осетия и др.). Изменение местообитания серн наблюдается и в тех районах, где браконьерство уже не выступает как лимитирующий фактор. В Татрах, например, его заменил фактор беспокойства со стороны туристов. В результате серны меняют свои излюбленные местообитания на менее благоприятные, нередко весьма опасные для их жизни (лавиноопасные места) (Blahout, 1985).
Миграции. Кавказские серны, как отмечено выше, крепко привязаны к излюбленным участкам и покидают их лишь вынужденно. Для них не характерны такие дальние и регулярные перемещения в разные сезоны года, как у азиатского муфлона в Закавказье. Там, где в течение года сохраняются защитные и кормовые условия, серны могут жить в течение всего года. Так, в Кавказском заповеднике часть серн живет оседло в лесном поясе; иногда их регистрируют и в альпийском поясе, где животные занимают малоснежные участки (Динник, 1910; Насимович, 1955; Котов, Рябов,
492
Rupicapra rupicapra Linnaeus, 1758
1963). На западном пределе своего распространения (район горы Фишт и др.), где высота местности невелика, серны практически живут оседло. Однако значительная часть поголовья серн совершает сезонные кочевки. Протяженность их наиболее значительна в Западном Закавказье, где снежный покров в горах достигает 2 м и более. По данным А.А.Насимовича (1955), размах сезонных кочевок по вертикали достигает 1500, даже 2000 м, а протяженность — 20—25 км. Такие большие сезонные кочевки чаще наблюдаются в верховьях рек Головинки, Сочи и Белой. На южном макросклоне Западного Кавказа серны зимуют, по данным названного автора, в поясе гор от 500 до 1200 м. На северном макросклоне Главного Кавказского хребта, где выпадает меньше снега, чем на южном, зимовки серн находятся ближе к местам летовок, примерно на расстоянии в 5—10 км. Те серны, которые живут в течение года в поясе лесов, кочуют еще меньше, занимая ограниченную территорию в течение года (около 8—10 км2);
В Закатальском заповеднике, по данным И.Ф.Попковой (1967), серны в течение года почти одинаково используют все высотные пояса гор в пределах 600—3000 м над ypV моря. Их территориальное размещение определяется в основном наличием хороших пастбищ вблизи скал — надежных убежищ. В силу более теплого климата и значительно меньшего количества годовых осадков, в том числе снега, размах пространственных перемещений по вертикали составляет до 300—1500 м, а протяженность переходов — не более Ю-15 км. Время наступления сезонных кочевок зависит от погодных условий; в каждом районе имеются свои сроки. В Кавказском заповеднике серны обычно спускаются в лесной пояс в октябре, а возвращаются в высокогорье с конца апреля или начала мая (Насимович, 1955; Жарков 1959). В Закатальском заповеднике время нахождения зимой в лесном поясе значительно сокращено — серны спускаются позже, а возвращаются в высокогорья на месяц раньше (Попкова, 1967). Что касается суточных перемещений, то они у серны незначительны — на Западном Кавказе в альпийском поясе не более 4—5 км в длину по прямой, а в лесном еще меньше (Насимович, 1949), в Закатальском заповеднике — весной 2—3 км, летом из-за кровососов достигает 4—5 км.
Убежища. Лежки. Серну спасает способность быстро уйти в скалы и преодолевать, казалось бы, совершенно недоступные кручи. Они не боятся ни скалистых стен, ни холодных рек, они могут, как пишет Н.Я.Динник (1910), спасаясь от врагов, бросаться с отвесных скал и, цепляясь за самые незначительные выступи их, спускаться в страшные пропасти, взбираться на отвесные скалы. По М.Благоут (Blahout, 1985), серны могут преодолевать скалы трудностью даже II и III степени альпийской квалификации. Она прыгает с места на высоту до 180 см, а в длину до 560 см.
Для отдыха серны используют различные места. Мы их наблюдали летом на снегу, где они лежали и отдыхали часами. Об этом пишет также Н.Я.Динник (1910). По его данным, на Западном Кавказе серна нередко отдыхает днем в пещерах, имеющих широкие отверстия и протяженность до 4—6 м. Они залезают туда глубоко и лежат, пока не спадет полуденная жара. В местах, где часто тревожат серн, они уходят высоко в скалы и
493
Серна
отдыхают на выступах, позволяющих обозревать подходы к ним. Нередко они спускаются вниз, в густые леса, чаще березняки и ельники, и в тени отдыхают. В условиях неволи летом серны также отдыхают в полдень в тени или в сарае (Инякова, 1957). В зимнюю стужу или при продолжительных дождях серны находят укрытия также в скалах под навесами камней, нишах, пещерах, под деревьями.
Сезонная и суточная активность. Специальных исследований по сезонной ритмике кавказской серны не проводилось. Однако фрагментарные полевые данные свидетельствуют, что периоды размножения, как и сезонных кочевок, характеризуются повышенной активностью. Низкая температура и глубокий снег зимой, наоборот, снижают ее. Суточная активность серн изучена лучше. Литературные и наши данные говорят о том, что кавказская серна ведет дневной образ жизни, как правило с двумя пиками активности летом, а зимой они слабо выражены. На характер дневной активности влияет много факторов. По данным Н.Я.Динника (1910), в совершенно безлюдных местах пасущихся серн можно было видеть в течение всего дня, притом на сравнительно небольшой территории. Среди ненапуганного стада серн встречались свободно пасущиеся животные, другие лежали, пережевывая жвачку, третьи играли и т.д. В течение дня они неоднократно менялись ролями. Там, где серн часто пугают, они обычно активны рано утром и поздно вечером. В альпийском поясе Кавказского заповедника летом серны начинают пастись с рассветом, к 10—11 ч большинство прекращают пастись, а возобновляют в 17—18 ч. Однако и в жаркий полдень можно видеть отдельных пасущихся животных. В пасмурные дни на открытых местах серны активны в течение дня. Под лесным пологом у большинства серн полуденный отдых не наблюдается. Ритмичная смена отдыха и пастьбы нарушается зимой, осенью и в пасмурную погоду. В такие дни пасущихся животных можно видеть в течение всего светлого периода суток. В условиях загона в Тебердинском заповеднике серны сохраняли аналогичный суточный ритм.
Поведение (рис. 65). Н.Я.Динник (1910), разделяя точку зрения Брэма, отмечал, что серны могут быть эмблемой осторожности. В равной степени у них хорошо развиты зрение, слух, обоняние. Если что и подводит серну, то это "консерватизм" (привязанность к местам обитания, путям подхода к солонцам и т.д.) и любопытство (желание узнать источник опасности). У серны нрав крутой, в неволе она очень агрессивна по отношению к человеку. В раннем возрасте, по данным А.П.Иняковой (1957), серна довольно дружелюбна к человеку, позволяет трогать, подходить, взвешивать, но с возрастом дичает, а во взрослом состоянии готова нападать на человека. Такое же отношение сернят к другим видам животных. Однако, как отмечает М.Благоут (Blahout, 1985), иерархия, свойственная сернам, устанавливается не силой, ибо рога у них страшное оружие. Вместо поединка широко используются запугивающие позы — позы нападения, наступления, угрожающий прыжок, выбивание передними копытами почвы и т.д. Покорность и подчинение демонстрируют тем, что животные сутулятся, самки мочатся, отворачиваю! голову, удаляются, или ложатся при нападении.
494
Rupicapra rupicapra Linnaeus, 1758
Рис. 65. Серна
495
Серия
В период гона, по данным названного автора, у самцов оптические и химические виды коммуникации таковы: гримасничание, отряхивание, маркирование секретом пахучих желез, рытье земли рожками. Во время гона осуществляется контроль течки, ухаживание, высовывание языка, преследование. Беден набор акустической сигнализации и ее проявления — блеяние разных полов и возрастов, предупреждающий свист. От боли стонет очень редко.
Линька. Специфические особенности сезонной смены волос были замечены и обсуждены еще Н.Я.Динником (1910). Как он справедливо писал, линька проходит медленно и постепенно, почти в течение всего лета можно встретить линяющих животных. На сроки линьки влияют возраст (раньше линяют взрослые и старые, чем молодняк), упитанность (худые позже, чем хорошо упитанные), состояние организма (здоровые раньше, чем больные) и т.д. Несмотря на все эти привходящие факторы, которые имеют место среди копытных, растянутость линьки — характерная черта вида.
Процесс смены волос у кавказской серны хорошо прослежен названным выше автором и его данные вошли в современные сводки (Соколов, 1959; Гептнер и др., 1961; Попкова, 1967); в их объективности мы убеждались при просмотре коллекционного материала и полевых данных (пролиняв-шис волосы на пастбищах, визуальные наблюдения и т.д.). В соответствии с этими данными линька весной начинается с посветления зимних волос, потом в апреле на Западном Кавказе, а в марте в Восточном Закавказье начинается их выпадение. Одновременно взамен старых вырастают короткие рыжие волосы будущего летнего наряда. Разгар линьки наблюдается в мае, однако линяющих животных можно встретить в июне и июле. В самом конце июля Н.Я.Динник (1910) застрелил трех животных, у которых много зимней шерсти было в пахах, по бокам туловища, на внутренней поверхности задних ног и на задней стороне бедер. Смена волос весной проходит раньше на голове, шее, а затем на лопатках, бедрах, боках, крестце, а завершается на животе и конечностях.
Осенняя линька в строгом ее понимании не выражена; с конца августа летние волосы постепенно увеличиваются в длине, изменяется их цвет, все больше приближаясь к зимней, черно-бурой окраске. Одновременно подрастает пух и становится гуще.
Питание. В Кавказском заповеднике зарегистрировано 203 вида растений, поедаемых кавказской серной (Насимович, 1949; Жарков, 1959; Дубень, 1985). Этот список в Закатальском заповеднике включает 109 видов (Попкова, 1967). В их составе много видов травянистых растений (разнотравье, осоковые, злаки, мхи, лишайники и др.), деревья и различные кустарники, бобовые и др. Очевидно, что набор кормовых растений кавказской серны можно расширить, если будет изучено питание этого вида в различных частях ареала. Достаточно напомнить, что флора Кавказа, как и фауна глубоко эндемична, многие виды узкоареальные, викарирующие и могли по этой лишь причине не оказаться в списке. Нужно иметь в виду то, что трудно определить, каким видам растений серна отдает предпочтение (Blahout, 1985). Этот автор в качестве такого предпочитаемого растения
496
Rupicapra rupicapra Linnaeus, 1758
называет только лишайники, которые серна ищет даже в тех случаях, когда в природе имеется достаточное количество другого корма.
Все имеющиеся данные свидетельствуют, Что соотношение фитомассы тех или иных растений, потребляемых кавказской серной, определяется фитосредой, в которой оказывается она в каждом сезоне года. Как отмечено выше, летом основная часть популяции сосредоточивается в субальпийском и альпийском поясах, и животные пасутся на злаково-разнотравных лугах. Основу питания, по данным А.А.Насимовича (1949), И.В.Жаркова (1959), И.Ф.Попковой (1967) в это время составляет сочное и цветущее разнотравье. Меньше потребляются осоки, злаки и бобовые. При этом в начале лета серна щиплет листья, молодые побеги, распускающиеся бутоны таких растений, как подорожника, анемона, манжетки, прострел, тмин, бутень и др. Начиная с июля в Кавказском заповеднике значительно увеличивается количество природных концентрированных кормов в виде соцветий и бутонов. Эти части растений оказываются откушенными серной у конского щавеля, водосбора, мытника, различных сложноцветных, колокольчиков и др. В августе их доля в питании серны увеличивается, особенно за счет сложноцветных. В Закатальском заповеднике, где климат суше и соответственно раньше огрубевают листья и стебли растений, серны вынуждены подниматься выше в горы в поисках цветущего колокольчика, цефалярии, щавеля, одуванчика и т.д. В августе, видимо по той же причине, серны нередко предпочитают сырые ложбины и затененные участки карнизов с более сочной растительностью. И в этих условиях животные чаще всего поедают верхушки растений с завязями, таких, как борщевик, золотистый бутень, кавказский тмин, порезник и другие зонтичные. Из сложноцветных часто поедают различные васильки, пупавки, одуванчики. В рационе нередко оказывались щавель, лютики, кавказский клевер, колокольчики и т.д. В течение всего лета, когда в природе богатая и разнообразная растительность, серны потребляют мало веточного корма. Однако зимой и ранней весной его значение в жизни серн велико. Они часто поедают в это время ветки, побеги и почки рябины, ивы, березы, бука, клена, дуба; хорошо поедают чернику, ежевику, кавказскую жимолость, шиповник и др. Из травянистых растений привлекательны те, которые остаются зелеными (например, горная овсяница). Вынужденно поедают в большом количестве и высохшие злаки, щиплят хвою сосен, елей и пихт, а также лишайников.
В жизни кавказских серн, как и других копытных, важное значение имеет минеральное питание. В бесснежные периоды года они охотно посещают солонцы, минеральные источники, часто поедают моренные глины. Охотно посещают искусственные солонцы, что нередко человек использует для привлечения серн. Солонцующйхся животных чаще можно встретить весной, когда в пище серн много сочной растительности, а также осенью, когда растения высыхают и выщелачиваются. Зимой потребность в минеральных веществах компенсируется веточным кормом.
32 В.Е.Соколов, А.К.Темботов
497
Серна
Воспроизводство и структура популяции
Размножение. Сведения о размножении кавказской серны содержатся_в ряде работ, в том числе Н.Я.Динника (1910), А.А.Насимовича (1939), В.Г.Гептнера с соавторами (1961) и др. Они выполнены в основном по наблюдениям на Западном Кавказе, преимущественно в Кавказском заповеднике. В Закатальском заповеднике (Восточное Закавказье) аналогичные исследования проведены И.Ф.Попковой (1967). Данные по размножению европейской серны в Татрах имеются в монографии М.Б.Благоут (Blahout, 1985).
По нашим наблюдениям и литературным данным половая активность серн замечается еще со второй половины августа, когда половозрелые самцы постепенно отделяются от стад из самок и молодняка. Они потом груп-пируются в 2—3 особи. У самок распад крупного стада на мелкие в 2—3 особи или формирование из отдельных особей таких же мелких групп наблюдается в октябре. Не группируются в основном лишь животные, недостигшие половой зрелости. После формирования новых группировок все чаще замечаются агрессивные действия самцов — они становятся беспокойными, драчливыми, часто бегают и кричат, создаются гаремы (один самец и несколько самок) или смешанные группы из 2—3 самцов, 5—6 взрослых самок и нескольких молодых самцов и самок. Количество животных в группе может сильно меняться. Спаривание начинается в Кавказском заповеднике в первых числах ноября, а в Закатальском — с 20-х чисел октября. Длится как на Кавказе, так и в Татрах несколько недель и соответственно завершается в конце ноября или начале декабря. Продолжительность беременности 160—170 дней; наиболее раннее появление козлят в Закатальском заповеднике 25 апреля 1957 г. (Попкова, 1967); на Западном Кавказе также изредка новорожденные наблюдаются в конце апреля, но массовый отел обычно приходится на май и начало июня; отдельные случаи были до середины июня (Динник, 1910). Для рождения детеныша самка уходит в глухие, редко посещаемые места, где имеются малодоступные скалы среди лесов, обычно расположенные невысоко в горах, а, наоборот, ниже, в лесном поясе.
По И.Ф.Попковой (1967), все обнаруженные места отела отвечали таким требованиям, как редкая посещаемость людьми, наличие или близость надежных убежищ в виде скал и осыпей, хорошей кормовой базы и минеральных источников.
Плодовитость. Кавказская серна, как правило, приносит одного детеныша, редко двух. Троих никогда не встречали, нет также и в литературе таких фактов.
Рост и развитие. Детеныш серны рождается хорошо сформировавшимся. Сразу после появления мать приступает к облизыванию, очищая и массажируя его. Детеныш встает на ноги через два часа после рождения. По М.Благоут (Blahout, 1985), в первые дни жизни детеныш сосет молоко до 30 раз в день. Количество приема молока потом снижается до 2—3 раз в день к концу лактации, которая продолжается до конца лета. Пастьбу
498
Rupicapra rupicapra Linnaeus, 1758
начинают, по В.Г.Гептнеру и др. (1961), в месячном возрасте. В Татрах, согласно данным М. Благоут, они пасутся уже на второй неделе жизни. В условиях вивария Тебердинского заповедника сернята начинали поедать зеленый корм в месячном возрасте (Инякова, 1957). Молодняк серны растет довольно быстро, через два месяца они имеют больше половины веса взрослых животных. Если в недельном возрасте их масса тела всего около 3 кг, то в полгода — около 20 кг. Как отмечено выше, в августе появляются маленькие черные рожки в виде конуса высотой около 2 см. Наиболее интенсивный рост организма и соответственно роговых чехлов наблюдается в первые три года, потом прирост рогов резко замедляется. То же самое можно сказать в отношении роста массы тела — в трехлетием возрасте самец кавказской серны весит до 38—43 кг, а самка несколько меньше по массе и некоторым показателям размеров тела (Гептнер и др., 1961). Увеличение массы тела и величины рогов фактически завершается к 4 годам (Попкова, 1967) , Череп серны растет дольше, даже после. 5,5 лет продолжает расти, но незначительно (Koubck et al., 1989). Формирование зубной системы кавказской серны проходит также, как у альпийской серны (Coutuier, 1938). По данным этого автора, предельный возраст, известный по западноевропейской серне, составляет 15—18 лет, очень редко 25 лет.
Половозрелыми серны становятся в 1,5 года, но в гоне участвуют с 2,5 лет; первое потомство самка приносит, по данным Н.Я.Динника (1910), на третьем году жизни.
Структура популяции. Наиболее полные данные по структуре популяции серн получены в Кавказском заповеднике и они неоднократно обсуждались в трудах его сотрудников, в том числе А.В.Дубня (1985). В таблице этого автора приводятся результаты учета численности и процентное соотношение молодняка за 1960—1981 гг. Работа проводилась преимущественно летом, в июне-июле. Лишь в 1975 г. она велась в августе, а в 1978 и 1980 гг. .— в течение года. Нет данных за 1962, 1964, 1967 и 1970 гг. Количество учтенных животных за год в среднем превышало тысячу, в пределах от 370 особей (1969 г.) до 2601 особи (1968 г.). За весь период численность молодняка в популяции колебалась в пределах от 13,2% в 1981 г. до 26,5% в 1963 г. Сравнительно низкой численностью молодняка выделяется 1976 г., когда на него приходилось 14,9% от 5,3 тыс. учтенных серн. Низка была численность молодняка также в 1977, 1978, 1981 гг., а сравнительно высокой х. 1960, 1961, 1963 гг. Годы низкой численности сеголеток характеризовались холодными зимами, прохладными веснами, резкими колебаниями температур в период рождения детенышей. Высокий процент сеголеток приходился на годы с ранними теплыми веснами и жарким летом. По мнению автора, уровень сеголеток в 15% и ниже, как и высокий процент взрослых, указывает на неблагополучие, депрессию популяции. Уровень в 16—19% свидетельствует о стабильном состоянии численности, тогда как превышение этого уровня — о росте численности популяции. Соотношение полов с 1978 по 1981 г. составило 1:2,5—4, в
32*
499
Серна
среднем 1:3,3 в пользу самок. Между тем, как и у других копытных, при рождении детенышей серны соотношение полов 1:1.
В Закатальском заповеднике, по данным И.Ф.Попковой (1967), за 1953— 1958 гг. численность молодняка составила от 14,0 до 21,8%. Минимальный показатель 14,8% зарегистрирован в сентябре 1957 г., а максимальный 21,8% — в июле 1953 г. По мнению автора, полученные данные характеризуют стабильное состояние популяции серны в Закатальском заповеднике. Для такого утверждения, как нам представляется, учетных данных у автора было мало.
Враги, паразиты, конкуренты, смертность. Кавказские серны в наши дни оказываются жертвами волка, рыси и медведя, а в прошлом — еще и леопарда. Однако все имеющиеся у нас данные свидетельствуют, что взрослая и здоровая серна обычно уходит от них свободно, в мало доступные для хищников скалы. Больший урон могут они понести в многоснежные зимы от рыси. От названных хищников гибнет в значительном количестве молодняк. По А.А.Насимовичу (1949), в Кавказском заповеднике количество молодняка с 17% от всего поголовья в июне снижается зимой до 6%, и основную причину он усматривает в деятельности волка и рыси. Тем не менее годовые колебания численности серны в Кавказском и Закатальском заповедниках, где сохранилось основное поголовье этого копытного, определяются не хищниками (Попкова, 1967; Дубень, 1985). Во многих работах, посвященных питанию хищников в горах Большого Кавказа, приводятся высокие показатели (до 20%) встречи остатков серны в желудках или экскрементах волка и рыси (см. обзор: Гептнер и др., 1961; Попкова, 1967; и др.). Справедливо их рассматривать как остатки, чаще всего, от погибших в лавинах и других стихийных явлениях, а также больных и ослабевших животных.
Кавказские серны очень часто подвержены инвазиям гельминтов. Д.П.Рухлядев (1964) в Кавказском заповеднике у серн обнаружил 16 видов гельминтов: 1 вид трематоды, 3 — цестоды и 12 видов нематод. Интенсивность инвазии по 14 вскрытиям 100%, а экстенсивность заражения составляла 606 гельминтов на одну особь. Инвазированные гельминтами серны нередко гибнут. Из 16 видов гельминтов, выявленных у серн, 15 общие с домашними животными. В Азербайджане, преимущественно в Закатальском заповеднике, С.М.Асадовым (см. обзор: Рухлядев, 1964) обнаружено у серн 26 видов гельминтов, в том числе три новых вида трихостронгилид и один новый вид протостронгилид. Инвазированность серн гельминтами и на Восточном Кавказе составляла 100%, но экстенсивность значительно ниже (в среднем 10,4 экз. на особь), чем на Западном Кавказе. Объясняется это относительно сухим климатом Восточного Кавказа.
Серны подвержены целому ряду болезней (ящур, сибирская язва, бруцеллез, некробациллез, туберкулез, кератоконъюнктивит, кокцидоз). Известен случай массовой гибели серн на Западном Кавказе от ящура в результате их заражения от домашнего скота (Динник, 1910).
В горах Большого Кавказа конкурентами серн являются кавказские туры. Как отмечалось выше, обычно эти виды исключают друг друга, что,
500
Rupicapra rupicapra Linnaeus, 1758
видимо, объясняется значительным сходством кормов и налеганием биотопов, населенных ими. Несмотря на это, высокая численность как серн, так и туров на Западном Кавказе отмечалась неоднократно. Значительно сложнее конкуренция серн и домашних животных, особенно коз и овец. Она, как правило, кончается тем, что серны оказываются вытесненными из лучших пастбищ в малокормные, селе- и лавиноопасные места. В таких экстремальных условиях серны часто гибнут.
Численность и ее динамика. В специальном исследовании А.В.Дубеня (1985) численность серн на Западном Кавказе в середине прошлого века определена в 40—50 тыс. особей, а в настоящее время — в 5—6 тыс. особей. Как видно, сокращение почти десятикратное. Обусловлено оно в первую очередь антропогенными факторами — вырубкой леса, выпасом скота, необузданным браконьерством и т.д.
В наши дни больше серн сохранилось в Кавказском заповеднике. В 1960—1981 гг. общая численность там определялась, в пределах 5—6 тыс. особей, т.е. была довольно стабильной и высокой. Однако в последние годы отмечается постепенное падение численности, и пока трудно однозначно определить его причины. За пределами Кавказского заповедника серна редка. На всем протяжении гор Северного Кавказа их поголовье не превышает 3,4—3,7 тыс. особей (Гинеев и др., 1989). В Грузии, по А.Б. Арабули (1989), серн, в настоящее время не более 5 тыс. особей; причины падения численности — многоснежные зимы и сходы лавин, а также браконьерство. Катастрофическое падение численности отмечается не только на неохраняемых территориях, но и в заповедниках. Так, в Лагодехском заповеднике серн было много до 50-х годов, а к середине 80-х их осталось не более 300 особей. Тенденция снижения и сейчас сохраняется. В Азербайджане численность серн в 70-х годах определялась в 700—800 особей (Алекперов, 1979). О современном состоянии данных нет. Как видно, численность серн на всем Кавказе интенсивно сокращается (на наш взгляд, в основном в связи с антропогенным процессом).
Практическое значение. Хотя по вкусовым качествам мясо и жир серны уступают таковым других копытных, в том числе туру, однако они издавна служат объектом охоты. Ежегодно добывалось около 1,5 тыс. голов (Верещагин, 1947). Выход мяса составляет около 60% от живого веса. Используются шкуры в качестве ковриков, иногда для получения меха. Ставились опыты по разведению в неволе (Инякова, 1957), скрещиванию серны с овцами и козлами, но желаемых результатов получено не было (Лопырин, 1960).
При надлежащей охране и воспроизводстве серна может вновь стать обычным копытным во многих районах Кавказа.
501
Литература
Абатуров Б.Д. Млекопитающие как компонент экосистем. М.: Наука, 1984. 288 с.
Абатуров Б.Д., Субботин А.Е., Джапова Р.Р. Использование сайгаками запасов пастбищной растительности в Северо-Западном Прикаспии // Экология. 1987. № 5. С. 93—95.
Абатуров Б.Д., Холодова М.В., Субботин А.Е. Потребление и перевариваемость кормов у сайгаков // Млекопитающие: Докл. 3-го съезда ВТО. М.: Наука, 1982. Т. 1. С. 150-151.
Абдурахманов М.Г. Задачи охраны дагестанских туров // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 145—147.
Абдурахманов М.Г. Распространенность и численность туров // 2-я Даг. респ. науч.-техн. конф, по охране природы. Махачкала, 1971. С. 27—29.
Абдурахманов М.Г. Экология, охрана, использование туров Дагестана: Автореф. дис. ... кавд. биол. наук. Махачкала, 1973. 24 с.
Абдурахманов М.Г. Восточнокавказский тур // Охота и охотничье хоз-во. 1979. № 8. С. 10-12.
Абдурахманов М.Г. Современное состояние популяции кавказских туров и задачи их охраны // Копытные фауны СССР: Тез. докл. М.: Наука. 1980. С. 225—226.
Абдурахманов М.Г. Некоторые особенности возрастной и половой структуры туров Дагестана // Экология и охрана горных видов млекопитающих. М.: Наука, 1987. С. 3-4.
Адлерберг Г.П. Предварительный обзор русских и монгольских кабанов // Докл. АН СССР. 1950. Т. 70, С. 91-96.
Адольф Т.А. Некоторые данные по биологии сайгака в Астраханских степях // Уч.зап. МГПИ им. В.П. Потемкина. Зоология. 1954. Т. 28, вып. 2. С. 247—256.
Айрумян В.А. К вопросу об акклиматизации пятнистого оленя в Армении // Изв. АН АрмССР. Биол. науки. 1962. Т. 15, № 11. С. 69—78.
Айрумян В.А., Гаспарян К.М. Редкие копытные и хищные млекопитающие Армении Ц Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 6-7.
Айрумян К.А., Гаспарян К.М. Редкие копытные и хищные Армении // Редкие млекопитающие фауны СССР. М., 1977. С. 35—42.
Айунц К.Р. Роль особенностей высокогорья в жизни Кавказского тура (Capra caucasica Guld) // 8-я Всесоюз. зоогеогр. к’онф.: Тез. докл. Л., 1985, С. 263—264.
Алекперов Х.М. Млекопитающие юго-западного Азербайджана. Баку: АН АзССР, 1966. 144 с.
Алекперов Х.М., Ерофеева С.Н., Рахматулина И.К. Современное состояние некоторых видов млекопитающих Азербайджана в связи с необходимостью их охраны и рационального использования // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 8—10.
Алекперов Х.М., Кулиев С.М. Результаты акклиматизации пятнистого оленя в Гпр-канском заповеднике // Млекопитающие: Докл. 3-го съезда ВТО. М.: Наука, 1982. Т. 2. С. 195.
Алекперов Х.М., Кулиев С.М. Некоторые данные по экологии кабана и косули в Ку-ба-Хачмасской зоне Азербайджана // Экология и охрана горных видов млекопитающих. М.: Наука, 1987. С. 8—9-
Александров В.Н. К изучению естественного питания зубров в Кавказском заповеднике //Тр. Кавказ, гос. заповедника. 1958. Вып. 4. С. 37—55.
502
Литература
Александров В.Н. Кавказский государственный заповедник и охрана природы Северо-Западного Кавказа // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 10-12.
Александров В.Н. Экология кавказского оленя // Тр. Кавказ, гос. заповедника. 1968. Вып. 10. С. 95—200.
Алиев Ф.Ф. Акклиматизация и реакклиматизация млекопитающих в Азербайджанской ССР // 1-е Всесотоз. совещ. по млекопитающим: Тез. докл. М.: Изд-во МГУ, 1961. Т. 3. С. 17-18.
Алиев Ф.Ф. Современное распространение и динамика численности диких копытных АзССР // Там же. С. 77-79.
Алиев Ф.Ф. Олени Азербайджанской ССР // Тр. Ин-та зоологии АзССР. 1965. Т. 25. С. 175-202.
Арабули А.Б. Косуля в Восточной Грузии: Автореф. дис. ... кавд. биол. наук. Тбилиси, 1966. 22 с.
Арабули А.Б. Распространение и численность некоторых охотничье-промысловых млекопитающих в Восточной Грузии // Зоол. журн. 1970. Т. 9/11, вып. 3. С. 418—421.
Арабули А.Б. Дикий кабан (Sus scrofa attila Thomas) и европейская косуля (С. capreolus capreolus L.) в Колхидской низменности // Материалы к фауне Грузии. Тбилиси: Мецниереба, 1973. Вып. 3. С. 288—291.
Арабули А.В. Влияние волка на численность кавказского оленя в Боржомском заповеднике // Копытные фауны СССР: Экология, морфология, использование, охрана. М.: Наука, 1975. С. 189-190.
Арабули А.Б. Современное состояние редких видов копытных Грузии // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 1. С. 28—29.
Аренс Л.Е. О влиянии заповедности на охотничье-промысловую фауну Тебердинского заповедника и прилегающих районов // Тр. Теберд. гос. заповедника. Ставрополь, 1957. Т. 1. С. 113-140.
Асадов С.М. Гельм и нто фауна серны в Азербайджане // Изв. АН АзССР. 1959. № 5. С. 95-96.
Ахмедов Н.М. О влиянии условий обитания на характер организации скелетной муску-латуры некоторых диких и домашних копытных Азербайджана // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 105-106.
Ахмедов Н.М., Керимова А.Ю. К сравнительной морфологии скелетной мускулатуры диких и домашних коз // Экология и охрана горных видов млекопитающих. М.: Наука, 1987а. С. 9—10.
Ахмедов Н.М., Саркисов Л.М. Возрастные особенности линейных промеров посткраниального скелета диких (Capra cylindricornis Blyth., Capra aegagrus Erx.) и домашних (Capra hircus L.) коз // Там же. 19876. С. 11—13.
Ахундов А.Н. Формирование популяций копытных, адаптированных к антропогенному ландшафту при помощи системы отбора, выращивания, обучения и реинтродукции животных с последовательным использованием "эффекта основателя" (на примере джейрана) // Экология, морфология, использование и охрана диких копытных. М.: ВАСХНИЛ, 1989. С. 252-254.
Ахундов А.Н. Стратегия охраны копытных в Азербайджане на примере джейрана // Там же. 19896. С. 255-257.
Байдавлетов Р.Ж. Экология и поведение лося в Западном Алтае //Тр. Ин-та зоологии АН КазССР. 1988. Т. 44. С. 7-36.
503
Литература
Байдавлетов Р.Ж. Численность лося в Казахстане и причины ее изменения // Всесоюз. совещ. по проблемам кадастра и учета животного мира: Тез. докл. Уфа, 1989. Ч. 2. С. 132-134.
Бакеев Ю.Н. Миграция лосей на Кавказе // Бюл. МОИП. Отд. биол. 1980. Т. 85, № 2. С. 31-42.
Бакеев Н.Н., Формозов А.Н. Распространение и некоторые черты экологии сайги в западной части Прикаспийской низменности // Материалы по биогеографии СССР. М., 1955. Вып. 2. - .
Балейшис Р. Определение возраста лосей по слоистой структуре корневого цемента ,_резцов и клыков // Зоол. журн. 1970. Т. 19, вып. 5. С. 778—780. ____
Балейшис Р.М. Различия в строении рогов благородных оленей Литвы //Тр. АН
ЛитССР. 1988. № 4. С. 87-93. _____________ ......,
'Балейшис Р.М., Блузма П.П., Прусайте Я.А. Предпочитасмость й значимость разных компонентов зимних пастбищ лосей // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 121-122.
Банников А.Т. Новый вид сайги из Монголии // Докл. АН СССР. 1946. Т. 51, № 5. С. 397-400.
Банников А.Г. Материалы к познанию млекопитающих Монголии. V. Антилопы // Бюл. МОИП. 1951. Т. 56, вып. 3. С. 21-41.
Банников А.Г., Жирнов Л.В., Лебедева Л.С., Фандеев А.А. Биология сайгака // М.: Сельхозиздат, 1961. 336 с.
Банников А.Г. Жизнь животных. Млекопитающие, или звери. М.: Просвещение, 1971. Т. 6. С. 526-527.
Бараташвили Т. Дикие копытные Боржомского заповедника // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 62—63.
Бараташвили Т.К. Синантропные волки Грузии: Опыт контроля, охотничье поведение // Экология, поведение и управление популяциями волка. М., 1989. С. 17—24.
Баскин Л.М. О питании благородных оленей // Зоол. журн. 1965. Т 44, вып. 3.
С. 465 - 469. — -.. .... • ..........-...-..-_________
Баскин Л.М. Поведение копытных животных. М.: Наука, 1978. 295 с?
Баскин Л.М. Экология и поведение зубра // Зубр: Морфология, систематика, эволюция, экология. М.: Наука, 1979. С. 442—462.
Баскин Л-М- Этология стадных животных. М.: Знание, 1986. 189 с.
Батхиев А.М. Распределение и численность безоаровых козлов в Чечено-Ингушетии // Фауна, экология и охрана животного мира Северного Кавказа. Нальчик, 1980. С. 46-56.
Батхиев А.М. Безоаровый, или бородатый, козел в горной Чечено-Ингушетии (экологический анализ географического распространения) // Природа и хозяйство Чечено-Ингушской АССР. Грозный, 1989. № 5. С, 103—109.
Башкиров И., Заблоцкий М.А. Кавказский зубр. М., 1939. 144 с.
Бекетов А., Бежанов Б. Современное состояние популяций джейрана в песках Сары-Ишикотрау // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 1. С. 257—259.
Белов С.Н., Юдин Б.С. Зимнее распределение и плотность населения копытных в бассейне р. Ларь-Еган // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 124-126.
Белянин В.Н. Материалы по смертности копытных в Жигулевском заповеднике // Там же. С. 122-123.
Беме Л.Б. Дикие звери Северо-Кавказского края. Пятигорск: Севкавгиз, 1936. 167 с.
504
Литература
Бланк Д.А. Особенности социального и репродуктивного поведения джейрана (Gazella subgutturosa) в Илийской долине // Зоол. журн. 1985. Т. 64, вып. 7. С. 1059—1069.
Бобринский Н.А., Кузнецов Б.А., Кузякин А.П. Определитель млекопитающих СССР. М.: Сов. наука, 1944. 440 с.
Бобринский Н.А., Кузнецов Б.А., Кузякин А.П. Определитель млекопитающих СССР. М.: Просвещение, 1965. 381 с.
Бобырь Г.Я. Взаимоотношения бурого медведя с копытными в горах Северного Кавказа // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 126 —127.
Богачев А.С. Пятнистый олень в современной фауне. Уссурийск, 1989. 44 с.
Богомолова Е.М., Курочкин ТО.А. Роды у лосих: Поведение лосихи и новорожденного лосенка // Зоол. журн. 1984. Ni> 11. С. 1713- 1724.
Богомолова Е.М., Курочкин Ю.А., Никольский А.А. Звуковые сигналы в коммуникативном поведении лося (Alces alces) // Там же. № 12. С. 1872—1882.
Брагин А.Б. Влияние частного скотоводства на состояние Чечене-Ингушской популяции зуброй // Влияние антропогенной трансформации ландшафта на поселение наземных позвоночных животных: Тез. Всесоюз. совещ. М., 1987. Ч. 1. С. 185— 186.
Бромлей Г.Ф. Экология дикого пятнистого оленя в Приморском крае // Сб. материалов по результатам изучения млекопитающих в гос. заповеднике. М., 1956. С. 5- 19.
Булатова Н.Н. Некоторые особенности дыхательной функции крови высокогорных животных: Автореф. дис. ... канд. биол. наук. М., 1953. 24 с.
Булатова Н.Н. Особенности крови высокогорных животных // Тр. Ин-та морфологии и физиологии животных. 1962. Вып. 41. С. 11—46.
Бурчак-Абрамович Н.И. К истории наземных позвоночных Азербайджана // Животный мир Азербайджана. Баку: Изд-во АН АзССР, 1944. С. 7—33.
Бурчак-Абрамович Н.И., Ширвапптдзе Л.А. Лось в Абхазии // Биология и промысел лося. М.: Россельхозиздат, 1967. Сб. 3. С. 38—47.
Варзиев А.Б., Курятников Н.Н. Итоги реакклиматизации зубров в Северо-Осетинской АССР // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 229—230.
Варнаков А.П. Особенности питания лося в Волгоградской области // Там же. 1980а. С. 130-131.
Варнаков А.П. Сравнительная морфологическая характеристика лося //Там же. 19806. С. 129-130.
Вейнберг П.И. К поведению дагестанского тура во время гона // Там же. 1980. С. 284- 285.
Вейнберг П.И. Попытка относительного учета туров в условиях сложного рельефа высокогорья // Эколого-фаунистические исследования в заповедниках. М., 1981. С. 69-74. /
Вейнберг П.И. Дагестанский тур (Capra cylindricornis Blyth) на Центральном Кавказе (экология, поведение и охрана): Автореф. дис. ... канд. биол. наук. М., 1981. 24 с.
Вейнберг П.И. К познанию брачного и агонистического проведения дагестанского тура // Млекопитающие: Докл. 3-го съезда ВТО. М.: Наука, 1982. Т. 2. С. 118—119.
Вейнберг П.И. Внутривидовые различия в экологии и сегрегация взрослых самцов от самок с молодняком у дагестанского тура // Там же. Т. 1. С. 167—168.
Вейнберг П.И. Дагестанский тур. М.: Наука, 1984. 88 с.
Вейнберг П.И. О формах рогов рода Сарга // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 1. С. 47-48.
505
Литература
Вейнберг П.И. Годовая динамика возрастной структуры популяции тура в Северо-Осетинском заповеднике // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 2. С. 237—238.
Верещагин Н.К. Дагестанский тур в Азербайджане // Тр. Зоол. ин-та АН АзССР. 1938. Т. 9, вып. 45. С. 1—70.
Верещагин Н.К. Джейран в Азербайджане // Там же. 1939. Т. 10. С. 109—122.
Верещагин Н.К. Каталог зверей Азербайджана. Баку: Изд-во АН АзССР, 1942. 95 с.
Верещагин Н.К. Охотничьи и промысловые животные Кавказа. Баку: Изд-во АН АзССР, 1947. 144 с.
Верещагин Н.К. Лось Alces sp. как недавно вымершее животное // Докл. АН АзССР. 1948. Т. 4, № 3. С. 124-126.
Верещагин Н.К. К истории и систематическому положению лося на Кавказе // Докл. АН СССР. 1949. Т. 1, № 3. С. 491-493.
Верещагин Н.К. Млекопитающие Кавказа. М.; Л.: Изд-во АН СССР, 1959. 704 с.
Верещагин Н.К., Наниев В.И. Прежнее и современное распространение копытных в Северной Осетии // Зоол. журн. 1949. Т. 28, вып. 3. С. 277—280.
Виноградов А.Ф. (Bison europaeus) Северо-Западного Кавказа // Тр. 2-го съезда естест-воиспытателей и врачей. М., 1870.
Виноградов А.Ф. Зубр Северо-Западного Кавказа // Журн. охоты и коннозаводства. 1872. № 7/8.
Воронцов Н.Н., Коробицына К.В., Надлер Ч.Ф. и др. Цитогенетическая дифференциация и границы видов у настоящих баранов Палеарктики // Зоол. журн. 1972. Т. 51, вып. 8. С. 1109-1124.
Вырыпаев В.А. Факторы, определяющие численность архара в Центральном Тянь-Шане, и меры по охране // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 233-234.
Газалиев Н.А. Дагестанская популяция сайгаков и перспективы ее рационального использования // Биологическая продуктивность ландшафтов Дагестана. Махачкала, 1980а. № 3. С. 33-40.
Газалиев Н.А. Об экологии и продуктивности дагестанского тура // Там же. 19806. С. 54-57.
Галкин В.С. Факторы доместикации и изменения пантовых оленей при одомашнивании // Проблемы доместикации животных. М.; 1987. С. 76—85.
Гамбарян П.П. Итоги акклиматизации уссурийского пятнистого оленя в Хосровском лесу // Изв. АН АрмССР. Биол. науки. 1959. Т. 12, № 5. С. 85—92.
Гамбарян П.П., Гаспарян К.М., Григорян В.Г., Айрумян В.А. Учет копытных с вертолета в условиях сильно пересеченной местности // Ресурсы фауны промысловых зверей в СССР и их учет. М.: Изд-во АН СССР, 1963. С. 104—109.
Гаспарян К.М. К кормовому режиму безоаровых коз на Урцском (Сарайбулагском) хребте // Изв. АН АрмССР. Биол. науки. 1964. Т. 17, № 2. С. 85—100.
Гаспарян К.М. Экология безоарового козла // Фауна позвоночных животных АрмССР: Зоол. сб. Ереван, 1975. Вып. 16. С. 78—106.
Гаспарян К.М., Межлумян С.К. О прошлом и настоящем распространении безоарового козла в Армении // Тез. докл. на юбилейной сессии по фауне республики, посвящ. 25-летию АН АрмССР. Ереван, 1969. С. 29—32.
Гептнер В.Г., Формозов А.Н. Млекопитающие Дагестана // Сб. тр. Зоол. музея МГУ. 1941. Т. 6. 74^ с. .
Гептнер В.Г., Насимович А.А., Банников А.Г. Млекопитающие Советского Союза. М.: Высш, шк., 1961. 776 с.
506
Литература
Гинеев А.М. Численность и охрана диких парнокопытных в Северном Дагестане // 1-я Даг, респ. науч.-техн. конф, по охране природы: Сб. тр. Махачкала, 1971. С. 30—32.
Гинеев А.М. Расселение лосей в Краснодарском крае // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 116.
Гинеев А.М. Лоси на Кавказе // Копытные фауны СССР: Тез. докл. М.: Наука, 1975. С. 74-75.
Гинеев А.М. Динамика численности копытных в плавнях Терека // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 1. С. 40-42.
Гинеев А.М., Абдурахманов М.Г., Спасская Т.Х. и др. К современной численности и распространению некоторых редких и исчезающих млекопитающих Северного Кав-каза // Ресурсы животного мира Северного Кавказа: Тез. докл. Ставрополь, 1988. С. 42-49.
Горбунов А.В., Зархидзе В.А. Многолетние изменения численности, современное распространение и перспективы восстановления популяций джейранов в Западной Туркмении // Охрана и перспективы восстановления численности джейрана в СССР: Сб. науч. тр. М., 1986. С. 22-28.
Гордиюк Н.М. Факторы смертности лося и марала в Башкирском заповеднике // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 140—141.
Горелов Ю.К. Размножение джейрана (Gazella subgutturosa Guld.) и проблема восстановления его численности в Бадхызе (Юго-Восточная Туркмения) // Териология. Новосибирск: Наука, 1972. Т. 1. С. 420—424.
Грачев И.И., Галанцев В.П. Физиология лактации. Л., 1973. 590 с.
Григорян Г.А. К изучению фауны паразитических червей диких жвачных Армении и их роли в распространении гельминтозов среди домашних овец и коз // Тр. НИИ встеринарии^АрмССР. 1949-Вып. 6. С. 188.
Григорян Г.А. Паразитические черви безоаровых коз Армении // Там же. С. 151—158.
Громов В.И. Первобытный зубр (Bison priscus Bojanus)B СССР // Тр. Зоол. ин-та АН СССР. 1935. Т. 2. С. 2-204.
Громов В.С. О различиях в окраске тела у европейской (Capreolus capreolus) и сибирской (С. pygargus) косуль // Зоол. журн. 1986. Т. 65, вып. 11. С. 1693—1703.
Громов В.С. Морфологическая изменчивость, поведение и систематика косули: Авто-реф. дне. ... канд. биол. наук. М., 1986. 24 с.
Громов В.С. Закономерности роста европейских (Capreolus capreolus) и сибирских (С. pygargus) косуль//Зоол. журн. 1988. Т. 67, вып. 9. С. 1381 — 1392.
Громов И.М., Гуреев А.А., Новиков Г.А. и др. Млекопитающие фауны СССР. М.; Л.: Наука, 1963. Ч. 1/2. 639 с.
Громов В.С., Скулкин В.С. Географическая изменчивость неметрических признаков черепа косули (Capreolus capreolus) // Зоол. журн. 1986. Т. 65, вып. 6. С. 911—923.
Гунчак Н.С. Морфологическая характеристика кабана (Sus scrofa) Украинских Карпат //Там же. 1978. Т. 57, вып. 12. С. 1870-1877.
Даль С.К. Позвоночные животные Сарайбулагского хребта // Зоол. сб. АН АрмССР. 1944. С. 5-46.
Даль С.К. Позвоночные Памбакского хребта // Там же. 1948. Вып. 5. С. 42—56.
Даль С.К. Позвоночные животные прибрежной полосы оз. Севан и изменение их группировок в связи со спуском водоема //Там же. 1950. Т. 7. С. 5—51.
Даль С.К. Данные по биологии, распространению и количественному соотношению в стадах безоаровых коз на Урцском хребте // Изв.АН АрмССР. Сер. биол. и с.-х. наук. 1951. Т. 1, вып. 4. С. 33—40.
507
Литература
Даль С.К. Очерк позвоночных животных Баргуштанского и Мергинского горных массивов // Материалы по изучению фауны АрмССР: Зоол. сб. 1953.
Даль С.К. Животный мир Армянской ССР. Ереван: Изд-во АН АрмССР, 1954, 415 с.
Даль С.К., Гусев В.М., Бедный С.Н. Влияние зимы 1953/54 г. на состояние, численность и распространение сайги в Западном Прикаспии // Тр. Науч.-исслед. противочум. ин-та Кавказа и Закавказья. Ставрополь, 1956. Вып. 1. С. 430—440.
Даль С.К., Гусев В.М., Бедный С.Н. Об экологии и размножении сайги // Зоол. журн. 1958. Т. 37, вып. 3. С. 447-456.
Данилкин А.А. Структура ареала косуль в СССР и факторы, определяющие его границы// Экология. 1989. № 4. С. 46—51.
Двойченко Г.Г. Пушные и промысловые звери Ставрополья. Ставрополь, 1955. 88 с.
Дежкин В.В. О комплексном использовании пастбищных угодий Калмыцкой АССР в интересах сохранения сайгака и ведения промыслового сайгаководства в условиях интенсификации сельскохозяйственного производства // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 144-145.
Дементьев Г.П. Косуля. М.; Л.: КОИЗ, 1933. 52 с.
Демьянова О.М., Любаев В.Л. Проблемы и перспективы охраны сайгака в Калмыкии // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 145—146.
Джанатпвили А. Млекопитающие Тславского района // Тр. Тславского гос. пед. ин-та им. Гогебашвили. 1947.
Джанашвили А.Г. К вопросу об одомашнивании туров // Тр. Тбилис. зоопарка. 1948. Вып. 1.
Джафаров И.М. Последовательность дифференцировки суставов конечностей джейрана // Млекопитающие: Докл. 3-го съезда ВТО. М.: Наука, 1982. Т. 2. С. 30—31.
Джафаров И.М., Агаев И.Г. Признаки пола на тазовых костях джейрана // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 2. С. 34—35.
Дзуев Р.И. Редкие виды млекопитающих Кабардино-Балкарской АССР и проблемы их охраны //Ресурсы животного мира Северного Кавказа. Ставрополь, 1988. С. 61-64.
Динник Н.Я. О кавказских горных козлах //Тр. СПб. о-ва естествоиспытателей. 1882. Т. 13. С. 73-91.
Динник Н.Я. Несколько слов о кавказском зубре // Естествознание и география. 1899. № 2.
Динник Н.Я. По Чечне и Дагестану. Тифлис, 1906. 78 с.
Динник Н.Я. Истребление дичи в горах Кубинской области // Природа и охота. 1909. С. 69-78.
Динник Н.Я. Звери Кавказа. Китообразные и копытные // Зап. Кавказ, отд. Рус. геогр. о-ва. 1910. Т. 27, вып. 1, ч. I. 246 с.
Динник Н.Я. Звери Кавказа // Там же. 1914. Т. 28, вып. 1. С. 247—536.
Доан Чонг Бинь. Возрастная изменчивость зубной системы джейрана (Gazclla subgut -turosa Guld., 1780) // Зоол. журн. 1963. Т. 42, вып. 6. С. 932—943.
Донауров С.С., Теплов В.П. Кабан в Кавказском заповеднике //Тр.Кавказ, гос. заповедника. 1938. Вып. 1. С. 191—226.
Дормидонтов Р.В. Пятнистый олень // Копытные звери. М.: Лесн. пром-сть, 1977. С. 10—28.
Дубень В.В. Численность и структура популяции серн во взаимосвязи с некоторыми экологическими факторами // Экологические исследования в Кавказском биосферном заповеднике. Ростов н/Д: Изд-во Рост, ун-та, 1985. С. 31—49.
508
Литература
Дуров B.B. Размещение, миграции и численность кабана в Красноярском крае // Вопросы экологии позвоночных животных. Краснодар: Кубан. ун-т, 1973. Вып. 163. С. 99-136.
Дуров В.В. Динамика численности и сезонное размещение косули в Кавказском заповеднике М.: Лесн. пром-сть, 1982. 224 с.
Дуров В.В. Кабан Западного Кавказа: Автореф. дис. ... кацд. биол. наук. М., 1987. 20 с.
Дуров В.В., Александров В.Н. Размеры и вес кабанов в Кавказском заповеднике // Тр. Кавказ, гос. заповедника. М.: Лесн. пром-сть, 1968. Вып. 10. С. 294—301.
Евтушевский Н.Н. Некоторые итоги акклиматизации оленя пятнистого (Cervus nippon hortulorum Sw.) в Черкасской области // Вести, зоологии. 1977. Т. 1. С. 7—11.
Егоров О.В. Дикие копытные Якутии. М.: Наука, 1965. 259 с.
Елфимова С.С., Граевская Б.М., Сипко Т.П., Буневич А.Н. Характеристика метаболизма зубров разных регионов // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 2. С. 220—221.
Емельянов П.Ф., Рыбалов В.Е. Реакклиматизация косули (Capreolus capreolus L.) в Ставропольском крае // Зоол. журн. 1967. Т. 46, вып. 6. С. 966—968.
Емельянов П.Ф., Рыбалов В.Е. Косули в Ставрополье // Охота и охотничье хоз-во. 1968. № 8. С. 13.
Енукидзе Г.П. К фауне млекопитающих Лагодехского гос.заповедника // Тр. Ин-та зоологии АН ГССР. 1953. Т. 12. С. 169-182.
Жарков И.В. Результаты учета животных в Кавказском заповеднике в 1939 г. // Науч.-метод. зап. комитета по заповедникам. 1940. Т. 7. С. 138—150.
Жевнеров В.В. Джейран острова Барса-Кельмес. Алма-Ата: Наука, 1984. 145 с.
Жирнов Л.В. О сезонных кочевках сайгаков в Западном Прикаспии // Бюл. МОИП. Т. 64, вып. 5. 1959. С. 134.
Жирнов Л.В. Сайгак // Редкие животные СССР: Копытные звери. М.: Лесн. пром-сть, 1977. С. 79-118.
Жирнов Л.В. Возвращение к жизни (экология, охрана и использование сайгаководства в условиях интенсификации сельскохозяйственного производства) // 4-й съезд ВТО: Тез. докл. М.: Наука. 1986. Т. 3. С. 144—145.
Житенко П.В. Рациональное использование продуктов сайгачьего промысла // Там же. С. 147-148.
Журавлева К.В., Раушенбах Ю.О. Рекогносцировочное гельминтологическое исследование как первый этап изучения гельминтозов копытных Западного Кавказа // Науч.-метод. зап. гл. упр. по заповедникам. 1939. Вып. 5. С. 87—103.
Заблоцкий М.А. Некоторые биологические особенности зубра и их изменение в условиях загонного содержания //Тр. Приокско-Террасского заповедника. 1957. Т. 1.
Заблоцкий М.А., Заблоцкая М.М. Реакклиматизация зубра на Кавказе и ее насущные проблемы /^ Экология и охрана горных видов млекопитающих. М.: Наука, 1987. С. 64-66.
Закариев А.Я. К экологии безоарового козла в Дагестане // Млекопитающие: Докл. 3-го съезда ВТО. М.: Наука, 1982. Т. 1. С. 202-203.
Залиханов М.Ч. Распространение и динамика численности туров в Кабардино-Балкарии // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 117-118.
Залиханов М.Ч. Туры в Кабардино-Балкарии. Нальчик: Эльбрус, 1967. ПО с.
Звегинцова Н.С., Треус М.Ю. Зараженность сайгаков гельминтами в условиях полу-вольного содержания в Аскании-Нова // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 148-149.
Зубр: Морфология, систематика, эволюция, экология. М.: Наука, 1979. 496 с.
509
Литература
Зуев Е.Я. Проблемы охраны сайгаков Калмыкии // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 150.
Ильина Г.И. Экологические особенности пятнистого оленя и перспективы его акклиматизации в европейской части СССР // Уч. зап. МГПИ им. В.П.-Потемкина. 1956. Т. 61, вып. 4/5. С. 3—90.
Ильинский В.О., Ладова Л.А. Опыт комплексного ведения лесного и охотничьего хозяйства. М.: Лесн. пром-сть, 1976. 121 с.
Инякова А.П. К вопросу численности и распределения охотничье-промысловых животных в Тебердинском гос. заповеднике (1953—1955 гг.) // Тр. Теберд. гос. заповедника. 1957. Т. 1. С. 249—257.
Инякова А.П. Опыт содержания серн Rupicapra rupicapra caucasica Lyd. в неволе // Там же. С. 257—267.
Инякова А.П. К вопросу размножения западнокавказского тура (Capra caucasica Guld.) в неволе // Там же. С. 269—270.
s Иржак Л.И., Гладилов В.В. Возрастная характеристика сродства гемоглобина к / кислороду у лося Alces alces // Журн. эволюц. биохимии и физиологии. 1981. Т. 17, Na 1. С. 66-69.
Иржак Л.И., Качмарчик Э.В. Типы гемоглобина у растущих лосей // Журн. общ. биологии. 1971. Т. 32, № 3. С. 377-380.
Исмагилов М.И. К экологии джейрана на о-ве Барса-Кельмес // Бюл. МОИП. Отд. биол. 1954. Т. 60. вып. 6. С. 3—9.
Калецкий А.А. Корма лосей в зимний период и общегодовой объем потребления кормов // Биология и промысел лося. М.: Россельхозиздат. 1967. Вып. 3. С. 221—247.
Калецкий А.А. Состав кормов и количественная характеристика питания лосей: Автореф. дис. ... кацд. биол. наук. М., 1968. 22 с.
Калугин С.К. К восстановлению зубров на Кавказе // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 137—138.
Калугин С.Г. Восстановление зубра на Северо-Западном Кавказе // Тр. Кавказ, гос. заповедника. 1968. Вып. 10. С. 1—94.
Калугин С.Г. Зубры на Северном Кавказе // Копытные фауны СССР: Экология, морфология, использование, охрана. М.: Наука. 1975. С. 99—101.
Камьянов Б.Т. Сравнительное изучение морфологических показателей кавказского зубра в условиях Северного Кавказа: Автореф. дис. ... канд. биол. наук. Нальчик, 1969. 24 с.
Камьянов Б.Т., Нусов Н.И. Возрастные изменения трубчатых костей по морфологическим и интерьерным показателям у кавказского зубра // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 165—169.
Капанадзе А.Д. Новые данные о распространении серны (R. rupicapra caucasica Lyd.) в Грузии // Сообщ. АН ГССР. 1972. Т. 65, № 3. С. 705-707.
Капанадзе А.Д. Серна на Малом Кавказе // Материалы к фауне Грузии. Тбилиси, 1974. Вып. 4. С. 479.
Кириков С.В. Распространение зубра на территории Советского Союза в XI—XX вв. // Зубр. М.: Наука, 1979. С. 476.
Кнорре Е.П. Экология лося в связи с его одомашниванием // Науч.-метод. зап. гл. упр. по заповедникам. 1949. Вып. 13. С. 222—224.
Кнорре Е.П. Лорь в СССР: Автореф. дис.... кавд. биол. наук. М., 1953, 24 с.
Кнорре Е.П., Кнорре Е.К. Закономерности роста и сезонных изменений живого веса лося И Зоол. журн. 1956. Т. 35, вып. 8. С. 1229—1236. ..._____________
Кнорре Е.П., Шубин Г.Г. Определение возраста лося //Тр. Печоро-Илычского гос. заповедника. 1959- Вып. 7. С. 123—132. -------
510
Литература
Князев С.П., Тихонов В.Н. Краниологические особенности европейских и азиатских диких кабанов и их гибридов с домашними свиньями // Морфология и генетика кабана. М.: Наука, 1985. С. 33—49.
Кожухов М.В. О репродуктивном потенциале лося (Alces alces) в условиях одомашнивания // Зоол. журн. 1989. Т. 68, вып. 5. С. ,150—152.
Козло П.Г., Никитенко М.Ф. Методика определения возраста дикого кабана// Экология млекопитающих и птиц. М.: Наука, 1967. С. 209—221.
Ткозло П.Г. Дикий кабан. Минск: Ураджай, 1975. 223 с. "
Козлов Д ГГ., Назарова Н.С. Гельминтофауна зубра и пути ее формирования // Зубр. М.: Наука, 1979. 471 с.
Коржуев П.А. Гемоглобин. М.: Наука, 1964. 287 с./
Коржуев П.А., Тимошевская Г.В., Кадыкова И.А., Спасская Г.Х. Рога как форма адаптации дагестанского тура к высокогорным условиям// Экология. 1978. № 2. С. 100-102.
Коряков Б.Ф. Лоси на Северном Кавказе // Охота и охотничье хоз-во. 1965. № 6. С. 16-17.
Костоглад В.Е. Кабан в Приазовских плавнях // Сб. науч.-техн. информации ВНИИЖП. 1969. Вып. 24. С. 214-217.
Котляров Е.А. Условия обитания джейрана на заповедных территориях АзССР // Копытные фауны СССР. М.: Наука, 1975. С. 266—267.
Котов В.А. Питание рыси в Кавказском заповеднике //Тр. Кавказ, гос. заповедника. 1958. Вып. 4. С. 214-217.
Котов В.А. Количественный учет серн в Кавказском заповеднике // Там же. 1960. Вып. 6. С. 185-191.
Котов В.А. Отлов и мечение туров (Capra caucasica) на Западном Кавказе // Бюл. МОИП. Отд. биол. 1964а. Т. 69, вып. 2. С. 54-60.
Котов В.А. Некоторые данные по морфологии западнокавказского тура // 2-я науч, конф, зоологов пед. ин-тов РСФСР: Тез. докл. Краснодар, 19646. С. 180—181.
Котов В.А. К экологии западнокавказского тура: Автореф. дис. ... канд. "биол. наук. Воронеж, 1966. 24 с.
Котов В.А. Вес и размеры кубанских туров // Зоол. журн. 1966. Т- 45, вып. 8. С. 1229- 1334.
Котов В.А. Роль Кавказского заповедника в увеличении численности кубанских туров // Природа Северного Кавказа и ее охрана: Тез. докл. Нальчик, 1967. С. 16—17.
Котов В.А. Кубанский тур, его экология и хозяйственное значение // Тр. Кавказ, гос. заповедника. 1968. Вып. 10. С. 201- 293.
Котов В.А., Рябов А.С. Промысловые и ценные млекопитающие предгорных и горных районов Краснодарского края // Там же. 1963. Вып. 7. С. 239.
Кочанов Н.Е., Симакова Т.В. Содержание марганца, кобальта и меди в кормах и крови лосей // Тр. Псчоро-Илычского гос-заповедника. 1976. Вып. 13. С. 127—129.
Кудрявцев А.А., Кудрявцева П.А. Клиническая гематология животных. М.: Колос, 1975.399 с.
Кузнецов В.Ш. Джейраны в районе Бадхыза // Охрана и перспективы восстановления численности джейрана в СССР: Сб. науч. тр. М., 1986. С. 31—37.
Кузякин В.А. Ресурсы основных видов охотничьих млекопитающих в РСФСР // Всссоюз. совет, по проблеме кадастра и учета животного мира. Уфа: Башк. кн. изд-во, 1989. С. 119-161.
Кулиев С.М. К распространению и экологии Capra aegagrus (Artiodactyla) Нахичеванской АССР // 1-й Мсждунар. конгр. по млекопитающим: Рсф. докл. М., 1974. Т. 1. С. 328.
5 1 1
Литература
Кулиев С.М. Безоаровый козел в дагестанский тур Азербайджана (биология, охрана и хозяйственное значение): Автореф. дис. ... канд. биол. наук. М., 1981. 24 с.
Кулиев С.М. Распространение и численность дагестанского тура в Закатальском заповеднике // Тез. Всесоюз. совещ. по пробл. кадастра и учета живота, мира: Тез. докл. Уфа, 1989. Ч. 2. С. 232-233.
Кулиев Г.К., Мамедов Т.О. Изучение хромосомного набора безоарового козла (Capra aegagrus End.) и дагестанского тура (С. cylindricornis) // Изв. АН АзССР. Сер. биол. 1974. № 5/6. С. 88—93.
Кулиев Г.К., Мамедов Т.О. Изучение хромосомного набора Ovis orientalis Gmcl. с помощью дифференциальной окраски // 1-й Междунар. териол. конгр.: Реф. докл. М., 1974. Т. 1. С. 327.
Курашвили Б.Е. Охрана и рациональное использование животного мира Грузии. М.: Наука, 1985.^143 с. (Человек и окружающая среда).
Курашвили Б.Е., Кохия С.С., Арабули А.Б. и др. Млекопитающие животные Грузии: Атлас. Тбилиси: Мсцнисреба, 1981. С. 94—95.
Курносов К.М., Петрищев Б.И. Эмбриогенез кабана (Sus scrofa): Состояние внешних морфологических признаков у эмбрионов кабана разных стадий развития // Зоол. журн. 1977. Т.6, вып. 3. С. 412— 419.
Курятников Н.Н. Особенности стадного поведения туров на-Центральном Кавказе // Материалы III Всесоюз. конф, по поведению животных. М.: Наука. 1983. Т. 2. С. 154-155.
Лавров Н.П. Географическое распределение, биология и хозяйственное значение косули в СССР // Тр. по лесному опытному делу Центр, лесн. опыт, станции. 1929. Вып. 6. С.49-82. .............
Лавровский В.В. Поведение стаи волков при охоте на лося // Экология, поведение и ' управление популяциями волка. М., 1989. С. 75—86.
Липкович А.Д. Современное состояние и перспективы восстановления чистокровных зубров на Северном Кавказе // Ресурсы животного мира Северного Кавказа: Ставрополь, 1988. С. 101-104.
Литвинов В.П. Волк (Canis lupus) и кабан (Sus scrofa) в Кызыл-Агачском заповеднике // Зоол. журн. 1981. Т. 60, вып. 10. С. 1588-1591.
Литвинова Н.А. К питанию зубров в Цейском заказнике // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 177—179.
Лихачев А.И. Адаптированность сайга к суровым условиям полупустынь Прикаспия // Материалы 3-й зоол. конф. под. ин-тов РСФСР. Волгоград, 1967. С. 501—503.
Лобачев В.С., Смирин В.М. Рога у самок сайги и джейрана // Зоол. журн. 1970. № 9.
С. 1421-1423. /
Лобашев М.Е., Ватта К.В., Тихомирова М.М. Генетика с основами селекции. М., 1970. 432 с.
Логинов В.В. Новые данные к познанию биологии кавказского кабана // Бюл. МОИП. Отд. биол. 1936. Т. 15, вып. 1. С. 10—21.
Максимук А.В. Промысел сайгака в Северо-Западном Прикаспии // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 156-157.
Максимук А.В., Петрищев Б.И., Хахин Г.Б. Динамика численности сайгака Северо-Западного Прикаспия // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 2. С. 272—273.
Максимук А.В., Хахин Г.В., Позняк В.Г. Ирригационное строительство и проблема сохранения популяции сайгака в Северо-Западном Прикаспии // Животные водных и околоводных биогеоценозов полупустыни. Элиста, 1987. С. 81—87.
512
Литература
Маломагомедов М.А. Некоторые данные о промысловых животных Гутонского заказника Дагестанской АССР // Материалы 3-й зоол. конф. пед. институтов РСФСР. Волгоград, 1967. С. 507.
Мамбетжумаев А.М. О численности и размещении джейрана (Gazella subbutturosa) на плато Устюрт и Кызылкумах // Зоол. журн. 1970. № 3. С. 431—434.
Мамукелашвили Г.В. Сезонное использование пастбищ кавказским оленем в Лагодсх-ском госзаповедникс и его кормовой ассоартимент // Заповедники Грузии (Тбилиси). 1987. Т. 6. С. 215-233.
Мамукешвили Г.В. Кавказский олень — украшение заповедника "Лагодехский заповедник". Тбилиси: Сакартвело, 1989. № 2. С. 15—16.
Манасерова Н.С. Морфологическая характеристика черепа арменийского муфлона // Биол. журн. Армении, 1988. Т. 41, № 8. С. 668 -675.
Марков Е.Т. Охотничье хозяйство Закавказья. Тифлис, 1931. 221 с.
Марков Е.Л. Дикие свиньи Закавказья. Тифлис, 1932. 38 с.
Мармазинская Н.В. Социальное поведение джейранов в период гона // 5-й съезд ВТО: Тез. докл. М. Наука, 1990. Т. 3. С. 32-33.
Межлумян С.К., Гукасян Р.Х. Динамика ареала и эколо-го-морфологические особенности кавказского благородного оленя // Зоол. сб. АН АрмССР. 1986. Т. 20. С. 99—127.
Миддендорф А.Ф. Путешествие на север и восток Сибири // Сибирская фауна. Отд. 5. 1869. Ч. 2. 310 с.
Мойсеенко Н.А., Мочалов Н.Н. Эколого-физиологическая характеристика красной крови и энерготраты у лосей в раннем постнатальном онтогенезе // Тр. Коми науч, центр АН СССР. 1987. № 89. С. 135-144.
Наниев В.И. Некоторые изменения ареалов и видового состава млекопитающих СО АССР // Уч. зап. Сев.-Осет. пед. ин-та им. С. Орджоникидзе. 1956. Вып. 20. С. 283—298.
Наниев В.И. Кавказский зубр — Домбай // Там же. С. 249—276.
Наниев В.И. Некоторые изменения ареала и видового состава млекопитающих СО АССР // Изв. Сев.-Осет. пед. ин-та. 1956. Т. 17. С. 283-298.
Насимович А.А. Динамика запасов благородного оленя в Кавказском заповеднике // Тр. Кавказ, гос. заповедника. 1936. Т. 1. С. 3—33.
Насимович А.А. Учет туров в Кавказском заповеднике в июне 1935 г. // Материалы Кавказ, гос. заповедника. Майкоп, 1936. № 2. С. 18—21.
Насимович А.А. О некоторых закономерностях зимнего распределения копытных в горах Западного Кавказа // Бюл. МОИП. Отд. биол. 1936. Т. 45, № 1. С. 3—9.
Насимович А.А. К познанию минерального питания диких животных Кавказского заповедника //Тр. Кавказ, гос. заповедника. 1938. Вып. 1. С. 103—152.
Насимович А.А. Зима в жизни копытных Западного Кавказа // Экологии и биоценологии. 1939. Вып. 7. С. 3—91.
Насимович А.А. К методике количественного учета поголовья тура // Науч.-метод, зап. гл. упр. по заповедникам. 1940. Вп. 7. С. 211 — 227.
Насимович А.А. К методике количественного учета благородного оленя в Кавказском заповеднике // Там же. 1941. Вып. 8. С. 173—177.
Насимович А.А. Очерк экология западнокавказского тура // Тр. Кавказ, гос. заповедника. 1949а. Вып. 3. С. 5—38.
Насимович А.А. Новые данные по биологии серны на Западном Кавказе // Там же. 19496. Вып. 3. С. 51-64.
Насимович А.А. Об организации заповедника высокогорной природы Центрального Кавказа // Охрана природы. 1950. № 11. С. 72 — 79.
33 В.Е.Соколов, А.К.Темботов
5 1 3
Литература
Насимович А.Л. Роль режима снежного покрова в жизни копытных животных на территории ёССР. М.: Изд-во АН СССР, 1955. С. 403.________ _____________
Насонов" Н?В. Муфлоны и близкие к ним формы диких баранов”// ИзвГ Имп. Рос. акад- наук. 1911. С. 1267-1296.
Насонов Н.В. Географическое распространение диких баранов Старого Света. Пг., 1923. 254 с.
Немцев А.С. Формы социальных группировок зубров Кавказского заповедника // Копытные фауны СССР: Тез. докл. М.: Наука, 1980а. С. .300—301.
Немцев А.С. Учет численности горных зубров во время яра // Там же. 19806. С. 36— 38.
Немцев А.С. Морфоэкологические особенности черепа восстановленных горных зубров // Экология горных млекопитающих. Свердловск, 1982. С. 85—87.
Немцев А.С. Происхождение популяции кавказского гибридного зубра и предпосылки ее сохранения // Популяционная изменчивость вида и проблемы охраны генофонда млекопитающих. М.: Наука, 198.3. С. 73—74.
Немцев А.С. Динамика населения зубров Кавказского заповедника// Экологические исследования в Кавказском биосферном заповеднике. Ростов н/Д: Изд-во Рост, ун--та, 1985. С. 49-63.
Немцев А.С. Демографическая структура популяции гибридных горных зубров // Экология и охрана горных видов млекопитающих. М.: Наука, 1987а. С. 122—124.
Немцев А.С. Проблемы восстановления зубров на Северном Кавказе // Там же. 19876. С. 125-127.
Немцев А.С. Особенности биологии, охрана и пути использования популяции горных зубров Северо-Западного Кавказа: Автореф. дис. .. кавд. биол. наук. М., 1988а. 19 с.
Немцев А.С. Демографический анализ популяции горных зубров // Ресурсы животного мира Северного Кавказа: Тез. докл. Ставрополь, 19886. С. 129—131.
Немцев А.С. Перспективы и особенности охоты на зубров в Краснодарском крае // Там же. 1988в. С. 131-133.
Немцев А.С., Киселева Е.Г. Сравнительное морфоэкологическос изучение горных и равнинных зубров // Экология и охрана горных видов млекопитающих. М.: Наука, 1987. С. 127-129.
Пинов Л.К., Хрусталева И.В. Морфология скелета домашних и диких свиней в постнатальном онтогенезе // Морфология, генетика кабана. М.: Наука, 1985. С. 122-130.
Оноприенко Л.Г., Хохлова З.И. Размещение и численность лося на Ставрополье // Ресурсы животного мира Северного Кавказа: Тез. докл. Ставрополь, 1988.
Оноприенко Л.Г., Хохлов А.Н. Современная численность охотничье-промысловых животных Ставропольского края // Всесоюз. совещ. по пробл. кадастра и учета животного мира: Тез. докл. Уфа, 1989. Ч. 2. С. 263- 266.
Орлов В.Н., Чудиновская Г.А. Хромосомный набор зубра и других быков трибы Во-vinii // Зубр. М.: Наука, 1979. С. 435—442.
Орлов В.Н., Булатова Н.Ш. Сравнительная цитогенетика и кариосистематика млекопитающих. М.: Наука, 1983. 405 с.
Падайга В.Н. Значение косули в лесном хозяйстве и система мероприятий по защите от нес лесовозобновлений в Литовской ССР: Автореф. дис. ... кавд. биол. наук. Таллинн, 1965. 25 с.
Попкова И.Ф. Серна на южных склонах Главного Кавказского хребта // Тр. Теберд. гос. заповедника. М.: Лесн. пром-сть, 1967. Вып. 7. С. 160—211.
514
Литература
Пастернак Ф.А. Материалы к систематике и биологии косули // Уч. зап. МГПИ им. В.П. Потемкина. 1955. Т. 38, вып. 3. С. 29—140.
Переладова О.Б. Индивидуальные особенности социального поведения джейранов // Охрана и перспективы восстановления численности джейрана в СССР. М., 1986а. С. 74-84.
Переладова О.Б., Перепадов С.В. Размножение джейранов в вольерах питомника Сюнт-Хасардагского заповедника //Там же. 19866. С. 49—59.
Перерва В.И. Успешность размножения и иерархический статус зубров // Экология, морфология, использование и охрана диких копытных: Тез. докл. Всссоюз. совещ. М., 1989. 4.2. С. 228-229.
Перовский М.Д. История расселения лосей в южных районах европейской части России // Охотоведение. Использование и охрана лесных копытных. М., 1976. С. 5-2.3.
Петрищев Б.И. Линейно-весовой рост эмбрионов дикой свиньи // Вести, зоологии. 1980. Вып. 6. С. 63-67.
Петрищев Б.И., Бородин А.Д. Развитие эмбрионов кабана популяции широколиственных лесов Брянской области // Морфология и генетика кабана. М.: Наука, 1985. С. 87-104. ~
Петров А.К. Окостенение скелета в индивидуальном развитии лося // Зоол. журн. /
1964. Т. 43, Вып. 12. С. 1837-1847.__------------------------——---------'
/'ЙриедйтЛ^А.'А.’В^ШбСТяз^мёжду' весом тела и размерами рогов у лосей (Alces alces)/ i разного возраста // Там же. 1979. Т. 58, вып. 1. С. 105—110.
ХПрштуцкаГЛТТГТТТштГгаткягТГ^^
промысловых животных Дагестана // Ресурсы животного мира Северного Кавказа: Тез. докл. Ставрополь, 1988. С. 136—140.
Прилуцкая Л.И., Ппшванов Ю.В. Ареал и численность безоарового козла в Дагестане // Экология, мор'фология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 2. С. 243-245.
Присяжнюк В.Е. Вес и размеры оленей пятнистых (Cervus nippon Hort.) с о-ва Аскольд // Вести, зоологии. 1973. Т. 5. С. 18—23.
Присяжнюк В.Е. Морфометрические характеристики аборигенного пятнистого оленя (Cervus nippon Hort.) Приморья // Зоол. журн. 1981. Т. 60. С. 1817—1828.
Присяжнюк В.Е., Проняев А.В. Морфологический статус сайгаков Западного Прикаспия и Северйого Казахстана. Сообщ. 1. Описание экстерьера. М.: ВНИИ природы и заповедного дела, 1988. 28 с.
Присяжнюк В.Е., Проняев А.В., Каплун М.В. Краниометрическая характеристика сай-гаков Калмыкии и Северного. Казахстана // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т.З. С. 162-163.
Присяжнюк В.Е., Солдатова Н.В. Формирование экстерьерных признаков в онтогенезе (Бухарский заповедник) // Экологические особенности охраны животного мира: Сб. науч. тр. М., 1985. С. 84—99.
Проняев А.В. Сравнительное исследование черепов двух популяций сайгака: Автореф. дис. ... канд. биол. наук. М., 1985. 23 с.
Проняев А.В. Фенотипическая, генотипическая характеристики и современное состояние популяций сайгака: Автореф. дис. ... канд. биол. наук. М., 1985а. 23 с.
Проняев А.В. Фенотипическая характеристика популяций сайгака на основе количественных признаков экстерьера // Управление популяциями диких копытных .животных: Сб. науч. тр. М., 19856, С. 43—54.
Проняев А.В. Фенотипическая, генотипическая характеристики популяций сайгака // 4-й съезд ВТО; Тез. докл. М.: Наука, 1986. Т. 3. С. 163—165.
33*
515
Литература
Проняев А.В., Аксенов С.В. Фенотипическая характеристика поколений у сайгака Северо-Западного Прикаспия // Биол. основы охраны и воспроизводство охот, ресурсов. М., 1987. С. 82-89.
Проняев А.В., Рожков Ю.П. Генетический эффект действия стабилизирующего отбора в изолированной популяции сайгака (Saiga tatarica L.) // Генетика. 1988. № 7. С. 1256-1261.
Пфиценмайер Е.В. Несколько интересных ублюдков семейства полорогих из Закавказья // Изв. Кавказ, музея. 1914. Т. 8, вып. 3/4.
Равкин Е.С., Овчаренко Г.И. О турах, на Северном Кавказе // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 137-139.
Радде Г.И. Коллекции Кавказского музея // Зоология. 1899. Т. 1. 520 с.
Размахни В.Е. Рога сайгака — ценное лекарственное сырье // 4-й съезд ВТО: Тез. докл. М.: Наука, 1986. Т. 3. С. 165-167.
Размахин В.Е., Сабуров Д.Н. Рациональное планирование норм отстрела лосей // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 255—256.
Раков Н.В. Сайрак в Западном Казахстане // Тр. Ин-та зоологии АН КазССР. 1956. Т. 6. С. 28-60.
Раков Н.В. О географической последовательности проявления подвидовых признаков у лося в Амуро-Уссурийском крае // Внутривидовая изменчивость наземных позвоночных и микроэволюция. Свердловск, 1966а. С. 263—270.
Раков Н.В. О значении некоторых факторов изоляции в проявлении и сохранении подвидовых признаков у лосей в Амуро-Уссурийском крае // Там же. 19666. С. 271-278.
Ратеани Дж. Быть может, этим путем спасем... Тбилиси: Сакартвело, 1989. № 2. С. 17-18.
Рашек В.Л. Высота, окраска и линька зимнего волосяного покрова сайгаков // Зоол. журн. 1966. Т. 45, вып. 4. С. 624—628.
Рашкевич Н.А. Результаты реакклиматизации зубра в Чечено-Ингушской АССР // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 139-140.
Решетов В.Ю., Суханов В.Б. Посткраниальный скелет // Зубр. М.: Наука, 1979. С. 142-173.
Россиков К.Н. Обзор млекопитающих долины реки Малки // Зап. Акад. наук. Физ,-мат. отд. 1887. Т. 54. С. 1—97.
Россиков К.Н. В горах Северо-Западного Кавказа // Изв. Рус. геогр. о-ва. 1890. Т. 26, вып. 4.
Русаков О.С. Структура популяции лося на разных фазах динамики численности // Копытные фауны СССР: Экология, морфология, использование, охрана. М.: Наука, 1975. С. 21—22.
Русаков О.С., Тимофеева Е.К. Кабан. Л.: Изд-во ЛГЗ7, 1984. 205 с.
Рухлядев Д.П. /ельминтофауна диких парнокопытных животных Крыма и Кавказа в эколого-зоогеотрафическом освещении // Тр. Кавказ, гос. заповедника. Саратов: Изд-во Сарат. ун-та, 1964. 449 с.
Рухлядев Д.П. К гельмпнтофауне реликтовых животных. Паразитические черви зубров И Природа Северного Кавказа и се охрана: Тез. докл. Нальчик, 1967. С. 82—83.
Сапожников Г-Н. Дикие копытные Таджикистана. М.: Наука, 1976. С. 48—62.
Саркисов А.А. О географическом распространении Ovis ophion armenia Nas. // Сообщ. АН ГССР. 1941. Т. 2, № 3. С. 277-283.
516
Литература
Саркисов А.А. О копытных Ирана // Тр. Ереван, зоопарка, 1944а. Вып. 1. С. 39—59. Саркисов А.А. Армянский муфлон Насонова //Там же. 19446. Т. 1, вып. 2. С. 89—91. Саркисов А.А. Полорогие Армянского нагорья: Автореф. дис. ... кавд. биол. наук.
Ереван, 1950. 24 с.
Сатунин К.А. Кавказский зубр // Естествознание и география. 1898. Т. 2. С. 1—21.
Сатунин К.А. О млекопитающих степей Северо-Восточного Кавказа // Изв. Кавказ, музея. 1901. Т. 1. Вып. 4. С. 1—66.
Сатунин К.А. Обзор исследования млекопитающих Кавказского края // Зап. Кавказ, отд. Рус. геогр. о-ва. 1903. Т. 24, вып. 2. С. 1—63.
Сатунин К.А. Распределение млекопитающих Талыша и Мугани по зоологическим стациям. Систематический каталог млекопитающих Талыша и Мугани // Изв. Кавказ, музея. 1905/1906. С. 87—262.
Сатунин К.А. Судьба кавказского зубра // Охотн. вести. 1908. № 18. с. 292—295.
Сатунин К.А. Род Capra: Козлы // Природа и охота. 1909. № 8. С. 1—6.
Сатунин К.А. Некоторые соображения о происхождении фауны Кавказского края // Изв. Кавказ, отд. Рус. геогр. о-ва. 1909/1910. Т. 20, № 2. С. 1 — 12.
Сатунин К.А. Животный мир Мугани. Тифлис, 1912. С. 1—88.
Сафаров М.А. Результаты учета с самолета численности джейрана в Азербайджане // Изв. АН АзССР. 1961. № 8. С. 51-55.
Сафаров М.А. Морфологические особенности и систематическое положение дикого кабана Азербайджана // Фауна и экология наземных позвоночных Азербайджана. Баку: Изд-во АН АзССР, 1965. С. 3—68.
Сафронова Ж.И. О формировании современных ареалов копытных на Северном Кавказе // Науч. тр. Кубан. ун-та. 1973. Вып. 163. С. 81—83.
Сихарулидзе 3. Влияние популяции копытных на растительность лесных искусственных окон и леса сагурамского заповедника // Заповедники Грузии. Тбилиси, 1987. Т. 6. С. 257.
Слепов Е. Охотничье-промысловые звери и птицы Кубани. Краснодар: Кн. изд-во, 1956. 74 с.
Слудский А.А. Размножение джейрана // Тр. Ин-та зоологии АН КазССР. 1956. Т. 6. С. 24-143.
Слудский А.,А. Джейран //,Коды.тные.а.вери. М„ 1977. С. 28—6L ------------
/Соколов И.И. К методике определения возраста косули (Capreolus capreolus L.) // .
Зоол. журн. 1956. Т. 35, вып. 8. С. 1238—1249._
Соколов ТТ.№—Фауна—СССР; Млекопитающие. Копытные звери (отряды Perissodactylan Artiodactyla). М.; Л.: Изд-во АН СССР, 1959. Т. 1, вып. 3. 640 с. 9
Соколов В.Е. Кожный покров млекопитающих. М.: Наука, 1973; 487 с. "7
Соколов В.Е. Систематика млекопитающих. М.: Высш, шк., 1979. 528 с.
Соколов В.Е. Словарь названий животных: Млекопитающие. М.: Рус. яз., 1984. 350 с.
Соколов В.Е., Данилкин А.А. Сибирская косуля: экологические аспекты поведения. М.: Наука, 1981. 144 с.
Соколов В.Е., Громов В.С. Морфометрические исследования черепа разных подвидов косули (Capreolus capreolus) // Зоол. журн. 1985. Т. 64, вып. 5. С. 738—749-
Соколов В.Е., Громов В.С. Опыт по гибридизации европейских и азиатских косуль // Докл. АН СЙСР. 1985. Т. 285, № 4. С. 1022-1024.
Соколов В.Е., Громов В.С., Рутовская М.В. Звуковая сигнализация у европейской (Capreolus capreolus) и сибирской (С. pygargus) косуль // Зоол. журн. 1987. Т. 66, вып. 3. С. 430—443.
Соколов В.Е., Громов В.С. Новая таксономия современных косуль (Capreolus Grag, 1821) //Изв. АН СССР. Сер. биол. 1988. № 4. С. 485- 493.
517
Литература
Соколов В.Е., Данилкин А.А. Сибирская косуля: Экологические аспекты поведения. М.: Наука, 1981. 144 с.
Соколов В.Е., Орлов В.Н., Сыроечковский Е.Е. Заповедники СССР: Заповедники Кавказа. М.УМысль, 1990. 365 с.
Соколов В.Е., Орлов В.Н., Чудиновская Г.А., Данилкин А.А. Хромосомные различия двух подвидов косули Caprcolus capreohis capreohis L. и С. с. pygargus Pall. // Зоол. журн. 1978. Т. 57. № 7. С. 1109-1112.
Соколов В.Е., Темботов А.К. Позвоночные Кавказа: Насекомоядные. М.: Наука, 1989. 548 с.
Сопин Л.В. Фенетика горных баранов как основа их внутривидовой Таксономии // Популяционная изменчивость вида и проблемы охраны генофонда: Тез. докл. Всссоюз. совещ. М., 1983. С. 173—175.
Старкин А.В. Некоторые отличительные черты таранной кости зубра и бизона // Вести, зоологии. 1988. № 2. С. 77—79.
Стекленев Е.П. О сезонности размножения пятнистых оленей (Cerus nippon hortulorum Sw.) в условиях юга Украины // Вести, зоологии. 1978. Т. 3. С. 28—33.
Страйер Л. Биохимия. М.: Мир, 1984. Т. 1. С. 88—103.
Сухорослов М.С. Экологические особенности и хозяйственное значение кабана Центральной Черноземной области: Автореф. дис. ... канд. биол. наук. Воронеж, 1972. 17 с.
Сысоев Н.Д. К биологической характеристике и перспективам использования популяций лося Владимирской области // Исследования по фауне Советского Союза (млекопитающие). М., 1968. С. 121 — 129.
Тарасов М.А. Некоторые данные о динамике численности копытных в Тебердинском госзаповеднпке // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 147—148.
Тарасов М.А. Факторы, определяющие динамику численности копытных в Тсбердин-ском заповеднике и прилегающих районах // Редкие млекопитающие фауны СССР. М.: Наука, 1977. С. 156-165.
Телегин В.И., Ивлева Н.Г., Николаев В.В. Наблюдения за суточной активностью лосей зимой // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 204—205.
Темботов А.К. Млекопитающие Кабардино-Балкарской АССР. Нальчик: Эльбрус, 1960. 196 с.
Темботов А.К. К изучению географического распространения животных в горах. Нальчик: Эльбрус, 1970. 36 с.
Темботов А.К. География млекопитающих Северного Кавказа. Нальчик: Эльбрус, 1972. 245 с.
Темботов А.К. О современном распространении и состоянии охраны туров в Кабардино-Балкарской АССР // Редкие виды млекопитающих фауны СССР и их охрана. М.: Наука, 1973. С. 148-149.
Темботов А.К. О географической изменчивости туров (Cara caucasica Guld.) на Северном Кавказе // Фауна, экология и охрана животных Северного Кавказа: Сб. науч. тр. Нальчик, 1974. Вып. 2. С. 118—131.
Темботов А.К. Ресурсы живой фауны. Ч. 2. Позвоночные животные суши. Ростов н/Д.: Изд-во Рост, ун-та, 1982. 320 с.
Темботов А.К., Кетенчиев Х.А., Дзуев Р.И. Популяционная структура и состояние охраны генофонда кавказских туров // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 266 - 267.
518
Литература
Темботов А.К., Темботова Э.Ж. Параметры крови — основа прогнозирования динамики ареалов млекопитающих в горах Кавказа // Грызуны. М.: Наука, 1980. С. 290-291.
Темботов А.К., Шхашамишев Х.Х. Животный мир Кабардино-Балкарии. Нальчик: Эльбрус, 1984. 190 с.
Темботова Э.Ж., Василенко В.Н., Темботова Ф.А. Сезонные изменения гематологических показателей у Ар. sylvaticus в условиях равнины Кабардино-Балкарской АССР // Фауна, экология и охрана животных Северного Кавказа: Сб. науч. тр. Нальчик, 1979. Вып. 4. С. 119-129-
Темботова Э.Ж., Темботов А.К. Система крови полевой мыши и ее сезонные изменения в условиях лесостепного пояса Кабардино-Балкарской АССР // Фауна и экология млекопитающих Кавказа: Сб. науч. тр. Нальчик, 1979- С. 144—162.
Теплов В.А. Волк в Кавказском заповеднике // Тр. Кавказ, гос. заповедника. 1938. • Вып. 1. С. 366-443.
Точиев Т.Ю. Охотничье-промысловыс животные Чсчс но-Ингушской АССР: Автореф. дис. ... канд. бпол. наук. Баку, 1968. 22 с.
Точиев Т.Ю. Вопросы экологии и охраны кавказского оленя и безоарового козла в условиях Чечено-Ингушской АССР // Редкие виды млекопитающих фауны СССР и их охрана: Сб. материалов АН СССР. М.: Наука, 1973. С. 150—151.
Точиев Т.Ю. Редкие виды диких парнокопытных Чечено-Ингушетии и их охрана // Копытные фауны СССР. М.: Наука, 1975. С. 284—285.
Точиев Т.Ю. Охотничья фауна Чечено-Ингушской АССР // Ресурсы животного мира Северного Кавказа: Тез. докл. Ставрополь, 1988. С. 152—156.
Точиев Т.Ю., Бахтисв А.М. К вопросу о питании безоаровых коз в Чечено-Ингушской АССР // Фауна, экология и охрана Северного Кавказа: Сб. науч. тр. Нальчик, 1980. С. 57-64.
Трошина А.И., Тихонов В.Н. Цитологические особенности некоторых диких свиней Европы, Азии, Африки и Америки // Морфология и генетика кабана. М.: Наука, 1985. С. 17-28.
Туров С.С. Опыт систематического обзора млекопитающих Осетии (Северный Кавказ) // Учен. зап. Северо-Кавказского ин-та краеведения. Владикавказ, 1926. Т. 1. С. 311 -337.
Тимофее! Е.К. Лось (экология, распространение, хозяйственное значение). Л.: Изд-во ЛГУ, 1974. 168 с.
Тимофеева Е.К. Роль косули в биоценозе лесостепных дубрав // Комплексные исследования биогеоценозов лесостепных дубрав. Л.: Изд-во ЛГУ7, 1986. С. 105—113.
Тихонов В.Н., Горелов И.Г., Бобович В.Е. Иммунологические особенности некоторых форм диких свиней Европы, Азии, Африки и Америки // Морфология и генетика кабана. М.: Наука, 1985. С. 3 — 17.
Тихонов В.Н., Трошина А.И. Идентификация хромосом и их перестроек в кариотипах подвидов дикого кабана Sus scrofa методом дифференциальной окраски // Докл. АН СССР. 1974. Т. 214, № 4. С. 932-935.
Туркин В.Н., Сатунин К.А. Звери России. М., 1902. Т. 4. 506 с.
Унанян А.К. Акклиматизация пятнистого оленя в Хосровском заповеднике // Копытные фауны СССР: Экология, морфология, использование, охрана. М., Наука, 1975. С. 135-136.
Фадеев В.А., Иванов А.И; Численность сайгаков в Казахстане в зависимости от кормовых ресурсов // Тр. Ин-та зоол. АН КазССР. 1988. Т. 44. С. 37—47.
519
Литература
Федоров С.М. Млекопитающие (Mammalia) Ставропольского края: Видовой состав, значение и использование // Материалы по изучению Ставропольского края. Ставрополь, 1954. Вып. 6. С. 177—192.
Федосенко А.К. Современное состояние популяций архара в СССР // Редкие наземные позвоночные Сибири: Материалы совещ. Новосибирск, 1988. С. 251—256.
Филатов Д.П. Летняя и зимняя поездки в Северо-Западный Кавказ в 1909 г. для ознакомления с кавказским зубром // Ежегодн. Зоол. музея Акад. наук. 1910. Т. 15. С. 171-217.
Филатов Д.П. О кавказском зубре // Зап. Имп. Акад. наук. Физ.-мат. отд. 1912. Т. 30, № 8. 40 с.
Филонов К.П., Калецкая М.Л. Особенности воспроизводства популяции лося в Дарвинском заповеднике // Зоол. журн. 1984. № 8. С. 1232—1241.
Флеров К.К. Очерк фауны млекопитающих долины рек Чороха // Ежегодн. Зоол. музея АН СССР. 1927. Т. 28, вып. 1. С. 63-71.
Флеров К.К. О диагностических принципах рода Caprcolus // Докл. АН СССР. 1928. № 23. 479 c.z
Флеров К.К. Обзор диагностических признаков беловежского и кавказского зубров // Изв. АН СССР. Сер. 7. Отд-ние мат. и естеств. наук. 1932. Т. 10. С. 1579—1590.
Флеров К.К. О некоторых географических и исторических изменениях евразийских копытных//Докл. АН СССР. 1934. Т. 4, № 3. С. 167—169.
Флеров К.К. Фауна СССР. Млекопитающие. Кабарга и олени. М.; Л.: Изд-во АН СССР, 1952. Т. 1, вып. 2. 255 с.
Флеров К.К. Систематика и эволюция // Зубр. М.: Наука, 1979. С. 9—112.
флинт В.Е., Присяжнюк В.Е. Состояние популяции, охрана и перспективы восстановления джейрана в СССР // Охрана и перспективы восстановления численности джейрана в СССР: Сб. науч. тр. М., 1986. с. 3—8.
Формозов А.Н. Заметки о млекопитающих Северного Кавказа // Учен. зап. Северо-Кавказского ин-та краеведения. Владикавказ, 1926. Т. 1. С. 72—89.
Хахин П.В. Некоторые особенности поведения сайгаков // Прикладная этология: Материалы 1П Всесоюз. конф, по поведению животных. М., 1983. С. 89—92.
Хахин Г.В., Шадилов Ю.М., Кузьмин И.Ф. Опыт учета джейранов на охраняемых территориях Азербайджана // Охрана и перспективы восстановления численности _ джейрана в СССР: Сб. науч. тр. М., 1986. С. 258—259.
Херувимов В.Д., Орлов И.И. Определение возраста лосей (Alces alces L.) из южных областей европейской части СССР // Зоол. журн. 1967 Т. 14. вып. 8. С. 1229—1233.
Хихнева Т.Д. ОхОтничье-промысловые звери Дагестана, их зональное распределение и хозяйственное значение // 2-я Даг. респ. науч.-тех. конф, по охране природы. Махачкала, 1971. С. 18—20..
Хихнева Т.Д. Охотничье-промысловые млекопитающие Дагестана: Автореф. дис. ... кацд. биол. наук. Махачкала, 1972. 20 с.
Хохлов А., Дубень А. Сколько туров в Кавказском заповеднике? // Охота и охотничье хоз-во. 1977. № 12. С. 36-37.
Цалкин В.И. К биологии размножения настоящих оленей // Зоол. журн. 1944. Вып. 6. С. 369-376.
Цалкин В.И. Новый подвид безоарового козла из Туркмении // Докл. АН СССР. 1950. Т. 29, № 2. С. 323-325.
Цалкин В.И. Горные бараны Европы и Азии. М.: МОИП, 1951. 343 с.
Цалкин В.И. Изменчивость и систематика туров Западного Кавказа. (Caprinae, Artio-dactyla) // Бюл. МОИП. Отд. биол. 1955. Т. 60. вып. 4. С. 17—33.
520
Литература
Цаплюк О.Э. Возрастная и сезонная динамика половой активности джейранов // Изв. АН КазССР. Сер. биол. 1972. Вып. 3. С. 39-45.
Цаплюк О.Э. Возрастные и сезонные особенности биологии размножения косули (Capreolus capreolus) в Казахстане // Зоол. журн. 1977. Т. 56, вып. 4. С. 611—618.
Червонный В.В. Географическая изменчивость некоторых элементов экологической ниши лося в зимний период // Экология, морфология, использование и охрана диких копытных: Тез. Всесоюз. совещ. М., 1989. Ч. 1. С. 143—144.
Чернявская С.И. Сезонное размещение и кочевки диких копытных и медведя в районе Кавказского заповедника в связи с распространением урожая плодов фруктарников и орехоносов // Бюл. МОИП. Отд. биол. 1956. Вып. 4. С. 7—21.
Чернявский Ф.Б. О некоторых приспособительных особенностях строения посткраниального скелета снежного барана (Ovis nivicola Esch.) // Зоол. журн. 1964. Т. 13, вып. 2. С. 242-252.
Члаидзе З.И. Capra caucasica на южном склоне Главного Кавказского хребта // 1-й Междунар. териол. конгр.: Реф. докл. М., 1974а. Т. 2. С. 321.
Члаидзе З.И. Некоторые вопросы экологии тура на Рачинском, Сванетском и Абхазском Кавказе // Материалы к фауне Грузии. Тбилиси: Мецниереба, 19746. Вып. 4. С. 481-521.
Чхиквишвили И.Д. К фауне млекопитающих и птиц Абхазии // Материалы к фауне Абхазии. Тбилиси: Фан, 1939. С. 1—44.
Шадилов Ю.М. Джейран в Азербайджане // Охрана и перспективы восстановления численности джейрана в СССР: Сб. науч. тр. М., 1986. С. 41—43.
Шахмарданов З.А., Рамазанов З.М. Ареалы копытных Дагестана // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 113-114.
Шахмарданов З.А., Спасская Т.Х. Популяция и промысел кабана в Дагестане // Копытные фауны СССР: Экология, морфология, использование, охрана. М.: Наука, 1975. С. 141-142.
Шварц С.С. Эволюционная экология животных: Экологические механизмы эволюционного процесса // Тр. Ин-та экологии растения и животных. Свердловск, 1969. Вып. 65. С. 3-198.
Шидловский М.В. Млекопитающие фауны высокогорья Большого Кавказа в Грузии // Фауна высокогорья Большого Кавказа в пределах Грузии. Тбилиси, 1964, С. 175—198.
Шилвдер В. Кубанская охота Его Императорского Высочества Великого Князя Сергея Михайловпча в 1894 г. Олени // Природа и охота. 1895. С. 1—55.
Шильдер В. Кубанская охота Его Императорского Высочества Великого Княза Сергея Михайловича // Природа и охота. 1897. Т. 8. С. 56—64.
Шималов В.Т. К изучению гельминтофауны копытных Белоруссии // Копытные фауны СССР: Тез. докл. М.: Наука, 1980. С. 220—222.
Штуббе Г., Брухгольц 3. Опыты по гибридизации европейской и сибирской косуль (Capreolus capreolus L., 1758, Capreolus capreolus pygargus Rail., 1771) II Зоол. журн. 1979. T. 58, вып. 9. С. 1398-1403.
Шубин Г.Г., Язан Ю.П. Опыт организации и ведения лесопромыслового хозяйства // Тр. Печоро-Илычского гос. заповедника. 1959. Вып. 7. С. 231—240.
Шумов А.В. Хромосомы подрода Bos (Bison) и их гибридов с крупным рогатым скотом // Вопросы гибридизации копытных. Мд Наука, 1980. С. 91—94.
Эквтимишвили З.С. Вертикальное распространение некоторых видов копытных на южном склоне Кавказского хребта // Сообщ. АН ГССР. Тбилиси, 1952. Т. 13, вып. 6. С. 476-485.
521
Литература
Эквтимишвили З.С. Материалы к изучению размножения безоарового козла // Тр. Ин-та зоологии АН ГССР. 1954. Т. 13. С. 93-97.
Юргенсон П.Б. Охотничьи звери и птицы: Прикладная экология. М.: Лесн. пром-сть, 1968. 308 с.
Явруян Э.Г. Распространение дикого барана Ovis ammon Gmelin в Закавказье // Бюл. МОИ1Т Отд. биол. 1969. Т. 24(2). С. 26-32.
Явруян Э.Г. Закавказский дикий баран: Автореф. дис. ... канд. биол. наук. Ереван, 1970. 24 с.
Явруян Э.Г. Врага, болезни и паразиты закавказского горного барана // Копытные фауны СССР: Экология, морфология, использование, охрана. М.: Наука, 1975а. С. 211-212.
Явруян Э.Г. Численность и методы охраны дикого барана в Армении и Нахичеванской АССР Ц Там же. 19756. С. 285.
Явруян Э.Г. Аэро- и пеший учет некоторых копытных в горных районах Армении и Нахичеванской АССР // Там же. 1975в. С. 60-61.
Явруян Э.Г., Попков А.С., Степанян Ж.С. Учет промысловых животных с самолета в условиях сильно пересеченной горной местности (Армения) // Бюл. МОИП. Отд. биол. 1971. Т. 76, № 2. С. 52-55.
Язан Ю.П. Биологический особенности и пути хозяйственного освоения популяции мигрирующих лосей Печорской тайги // Тр. Печоро-Илычского гос. заповедника. 1961. Вып. 9. С. 114-201.
Язан Ю.П. Лось на Северном Кавказе // Бюл. МОЮТ. Отд. биол. 1966. Т. 71, вып. 4. С. 1(20^125.
Язан Ю.П. Охотничьи звери Печорской тайги: Биология популяций, механизмы регуляции численности. Киров: Волго-Вят. кн. изд-во, 1972. 384 с.
Янушевпч А.И., Айзин Б.М., Кыдыралиев А.К. и др. Млекопитающие Киргизии. Фрунзе: Илим, 1972. 463 с.
Andrzejewski R. Spotty mutation of the wild boar Sus scrofa Linnaeus, 1758 // Acta theriol. 1974. Vol. 19. № 1, pt 13. P. 159-163.
Andrzejewski R., Jezierski W., Kostelecka-Myrcha A., Myrcha A. Colour mutation in wild boars and the physiological causes of mortality of the muhants in the population I I Bull. Acad. Pol. Sci. Ser. Sci. Biol. 1975. Vol. 23, № 12. P. 793-797.
Bertram G. Nebenhierengewichte und corpora lutea bei Rehwild // Kith Intern. Congr. Game Biol. Stockholm, 1974. S. 125—131.
Blahout M. ZurTaxonomie der Population von Rupicapra rupicapra (Linne, 1785) im der Hohen Tatra // Zool. listy. 1971. Vol. 21, № 2. P. 115-132.
Blancou J., Barrat J., Devaud J.P., Vannier Ph., Gourreau J.M. Pathologic du sanglier (Sus scrofa L.) coonaissances actuelles en France // Gibier faune sauvage. 1987. Vol. 4, sept. P. 279-294.
Blasetti A., Boitani L., Riviello M.C., Visalbeighi E. Activity budgets and use of enclosed space by wild boars (Sus scrofa) in captivity // Zool. Biol. 1988. Vol. 7, № 1. P. 69—79.
Bottcher W. Selektionsabschluss bei weiblichem Muffelwildeine Notwendigkeite // Unsere Jagd. 1988. Bd. 38, № 10. S. 294-295.
Cervan D., Duha J. Niektore poznatky socialneho spravania druhu Ovis musimon musimon (Pallas, 1811) // Biologia (^SSR), 1987. Cv. 42, № 10. S. 991-998.
Chang Hong, Xiao Qianzhu. Выбор местообитания благородным оленем в районе Дайлин зимой // Шоулэй сюэбао = Acta theriol. sin. 1988. Vol. 8, № 2 P. 81—88.
Chaudhri S.S., Mandokhot V.M., Gupta R.P., Yadav C.L. Haematological and biochemical observations in buffaloes naturally infected with fasciola gigantica // Ind. Vet. J. 1988. Vol. 65, № 1. P. 23-27.
522
Литература
Corbet G.B. The mammals of the Palaearctic region: A taxonomic review // British museum (Natural History). L.; Ithaca: Cornell Univ, press, 1978. P. 3.14.
Corbet G.B., Hill J.E. A world list of Mammalian Species. L.: Brit. Museum. 1980. 254 p.
Couturier M.A. Le chamois (Rupicapra rupicapra L.). Grenoble, 1938. 455 p.
Dubray D. Abondance, structure et dynamique de la population de mouflons de Corse (Ovis amon musimon S.) du secteur est du massif du Cinto (Haute—Corse), et analyse du role de protection de la reserve de 1‘office national de la chasse d'Asco // Bull. ecol. 1988, Vol. 19, № 2/3. P. 439-450.
Duda H. Jednak podyskutujmy о zubrach // Przyr. pol. 1987. № 8. S. 10.
Ellerman J.R., Morrison-Scott T.C.S. Checklist of Palearctic and Indian Mammals 1758 to 1946. L., 1951. 811 p.
Franzmann A.W., Zeresche R.E. Alaskan moose blood Studies with emphasis on condition evaluation // J. Wildlife Manag. 1978. Vol. 42, № 2. P. 334—351.
Haines H., Chichester H.G., Landreth H.F. Blood respiratory properties of Bison bison // Respirat. Physiol. 1977. Vol. 30, № 3. P. 305-310.
Haltenorth T. Klassiflkation der Saugetiere: Artiodactyla 1(18) // Walter der Gruyter Handbuch der Zoologie. B., 1963.
Harrison D.L. The Mammals of Arabia. L., 1968. 381 p.
Hawkey C.M., Hart M.G., Fitzgerald A.K. Haematological values in mouflon (Ovis musimon): influence of age, sex, season and vitamin E status // Res. Vet. Sci. 1984. Vol. 36, № 1. P. 37-42.
Hawly C.M., Hart M.G., Fitzgerald A.K. Age related hematological changes in captive Barbary sheep (Ammotragus lervia) // Ibid. 1984. Vol. 36, № 1. P. 118—119.
Heroldova M. The diet of mouflon (Ovis musimon) outside the growing period 1983—1984 // Folia zool. 1988. Vol. 37, № 4. P. 309-319.
Herzog S. The karyotype of the red deer (Cervus elaphus I..) // Caryologia. 1987. Vol. 40. P. 299-305.
Hoffman M. Die Bisamratte. Leipzig: Geest und Portig., 1952. 43 S.
Honacki J.H., Kinman K.E., Koeppe J.M. Mammal species of the world. 1982.
Kent J.E., Chapman D.L, Chapman N.G. Scrum consituents of red deer (Cervus elaphus) // Res. Vet. Sci. 1980. Vol. 28, № 1. P. 55-57.
Kitchen H., Putham F.W., Taylor W.J. Hemoglobin Polymorphism: its relation to sickling of Erythrocytes in white-tailed deer // Science. 1964. Vol. 144. P. 1237—1239.
Konig K.P. Raum-Zeit-Verhalten von Rehwild in geschlossencn Waldrevieren // Ztschr. Jagdwidd. 1987. Bd. 33, № 3. S. 168-175.
Kostelecka-Myrcha A. Changes in haematological values in wild boar during postandtal development //Acta theriol. 1974. Vol. 19, № 1/13. P. 165—176.
Kubek P., Hrabe V., Hora'kova M. Postnatal development of the Skull of Rupicapra rupicapra (Mammalia, Bovidae) // Folia zool. 1989. Vol. 38, № 1. S. 31—44.
Lehmann E. von. Bemerkungen zu zwei Rehschadeln aus dem Elbrus—Gebirge, Iran // Bonn. zool. Bcitr. 1988. Bd. 39. N° 2/3. S. 229-235.
I/ickow K.-W. Neues zur Altersbestimmung des Rothirsches // Unsere Jagd. 1987. Bd. 37, № 8. S. 232-233.
Marler R.J. Some hematologic and blood chemistry values in two herds of American bison in Kansas // J. Wildlife Diseases. 1975. Vol. 11. P. 97-100.
Martinez T. Cotrfparacion de los habitos alimentarios de la cabra montes у de la oveja en la zona alpina de Sierra Nevada // Arch. Zootechn.. 1988. Vol. 37, № 137. P. 39—49.
Mayer J.J., Brisbin L.I. Sex identification of Sus scrofa based on canine morphology // J. Mammal. 1988. Vol. 69, № 2. P. 308-412.
523
Литература
Мауг В., Tesarik Е., Auer Н., Burger Н. Nucleolus-organizer regions and heterochromatin in three species of Bovidae // Genetica. 1987. Vol. 75, № 3. P. 207—212.
McDonald S., Paul S.R., Bunch Th.D. Physiologic and hematologic values in Nelson desert bighorn sheep //J. Wildlife Diseases. 1981.*Vol. 17, № 1. P. 131 — 134.
Mchrer C.F. Some hematologic values of bison from five areas of the United States // Ibid. 1976. Vol. 12, № 1. P. 7-13.
Meidel E. Abasen von Spitzentrieben durch Rehwild // Wild und Hund. 1987. Bd. 90, № 5. S. 18-19.
Melander Y. The mititic chromosomes of Some Covicorn mammals (Bos taurus L., Bison bonasus L., Ovis areis L.) // Hereditas. 1959. Vol. 45, № 4. P. 649—664.
Mcnetries El. Catalogue raisonne des objets de zoologie recuellis dansun voyage au Caucase et jusqu'aux frontieres actuelles de la Perse, entrepris par ordre de sa majeste I'empereur. 1832. Vol. 3g, 271 p.
Miller Ch., Hartl G.B. Genetische Variation bei Gemsen der Alpen (Rupicapra rupicapra L.) // Ztschr. Jagdwiss. 1987. Bd. 33, № 4. S. 220-227.
Muir P.D., Sykes A.R. Effect of winter nutrition on antler development in red deer (Cervus elaphus): a field study // N. Z. J. Agr. Res. 1988. Vol. 31, № 2. P. 145-150.
Nadler C.F., Lay D.M., Hassinger J.D. Cytogenetic analyses of wild sheep populations in northern Iran // Cytogenetics. 1971. Vol. 10. P. 317—328.
NeXas J. Srncfzver. Pr.: Statni zemed. naklad, 1975. 302 s.
Ondcrscheka K. Wildbiologjsche Studie belegt: OO-Raps verantwortlich fur Rehwildstcrben, nicht fflr Hasensterben // Wild und Hund. 1987. Bd. 90, № 7. S. 13.
Ondcrscheka K., Tataruch F., Steineck Th., Klansek E., Vodnansky M. Geh'aufte Rehwildver luste nach Aufnahme von Raps // Ztschr. Jagdwiss. 1987a. Bd. 33, № 2. S. 139-142.
Ondcrscheka K., Tataruch F., Steineck Th., Klansek E., Vodnansky M., Wagner J.,
Echsel H. GehSufte Rehwildverluste nach Aufnahme von OO-Raps // Ibid. 1987b. Bd. 33, № 3. S. 191-205.
Pauling L., Itang H.A., Singer S.J., Wells J.C. Sickle cell anemia: a molecular disease // Science. 1949. Vol. ПО. P. 543-548.
Pemberton J.M., Albon S.D., Guinness F.E., Clutton-Brock Т.Н., Berry RJ. Genetic variation and juvenile survival in red deer // Evolution. 1988. Vol. 42, № 5, P. 921—934.
Prossinagg H. Vom Waldreh zum Fcldrch // Osterr. Weidwerk. 1988. № 8. S. 25—29.
Raczynski I. Biologischc Grundlagen der Zuchtung und der Restitution des Wisents, Bison bonasus// Zool. Gart. 1980. Bd. 50, № 5. S. 311—316.
Renecker L.A., Hudson R.L. Seasonal activity budgets of moose in aspen-dominated boreal forests// J. Wildlife Manag. 1989. Vol. 53, № 2. P. 296-302.
Rodriguez P.J.C., Rodriguez L.J.L., Dominguez C.F. Dates sobre la alimentacion del muflon de Corcega (Ovis amnion musimon (Bovidae) en Tenerife, Islas Canarias // Uieraea. 1987. Vol. 17, № 1/2. JP. 11-18.
Schutze M. Bewirtschaftung des Muffelwildes wesentlich intensivieren // Unsere Jagd. 1987. Bd. 37, № U.S. 324-325.
Simpson G. The principles of classification and a classification of Mammals // Bull. Amer. Mus. Natur. Hist. 1945. Vol. 85. 350 p.
Stubbe C., Lockow K.-W., Stubbe W. Ergebnisse der Rehwildmarkierung im Wild-forschungsgebiet Hakel // Unsere Jagd. 1987. № 7. S. 196—198.
Tyler N.J.C. Rangifer tarandus platyrhynchus. Sexual dimoiphism in the pelvic bones of Svalbard reindeer, Rangifer tarandus platyrhynchus // J. Zool. 1987. Vol. 213, № 1. P. 147-152.
524
Литература
Valder R., Nadler C.F., Bunch T.D. Evolution of wild sheep in Iran // Evolution. 1978. Vol. 32, № 1. P. 56-72.
Williamson M.J., Pelton M.R. Some hematological parameters of European wild hogs (Sus scrofa) //J. Tenn. Acad. Sci. 1976. Vol. 51, № 1. P. 25—28.
Zhou Shiland, Wu Shanzhi. Изучение наследования количественных признаков у пятнистого оленя // Йчуань = Hereditas. 1988. Vol. 10, № 2. Р. 17—19.
Zhou Sh., Wu S. Излучение репродуктивных показателей и генетика количественных признаков пятнистого оленя // Дуньусюэ яньцзя = Zool. Res. 1988. Vol. 9, № 2. Р. 175-182.
SUMMARY
The moderri theriofauna of the Caucasus contains 12 ungulates species. Of these, Capra caucasica Guld. belongs to the Region's paleoendemics. There are 8 indigenous species: Sus scrofa L., Capreolus capreolus L., Cervus elaphus L., Capra aegagrus Erxl., Saiga tatarica L., Gazella subgutturosa Guld., Ovis orientalis Gmelin, Rupicapra euricapra L.
The remaining three species: Cervus nippon Tern., Alces'alces L. and Bison bonasus L. are recent newcomers.
All the species parameters under study, including morphological, morphophysiological (e.g. hematological), range structure and ecology are characterized by regional perculiaritics. Their development is essentially influenced by the factors associated with the three-dimensional situation of the mountain ecosystems and the history of the development of the altitudinal zonality of the Caucasian landscapes.
Оглавление
Предисловие .................................................... 3
Отряд Парнокопытные — Artiodactyla Owen, 1848 .................. 5
Семейство Свиные — Suidae Grey, 1821......;............................... 7
Род Кабаны — Sus L., 1758 ................................. 8
Кабан - Sus scrofa L., 1758 ............................ 10
Особенности морфологии и морфофизиологии ............ 10
Структура и динамика ареала.......................... 27
Систематика и изменчивость........................... 32
Экология ............................................ 51
Воспроизводство и структура популяции................ 59
Семейство Оленьи — Corvidae Gray, 1821 ...................... 74
Род Косули — Capreolus Gray, 1821 ........................ 76
Европейская косуля — Capreolus capreolus L., 1758 ...... 78
Особенности морфологии и морфофизиологии............. 78
Структура и динамика ареала.......................... 87
Систематика и изменчивость........................... 95
Экология .......................................... 106
Воспроизводство и структура популяции............... 115
Род Олени — Cervus Linnaeus, 1758........................ 120
Пятнистый олень — Cervus nippon Temmink, 1838 ......... 121
Особенности морфологии и морфофизиологии............ 123
Структура и динамика ареала......................... 138
Систематика и изменчивость.......................... 141
Экология ........................................... 151
Воспроизводство и структура популяции............... 156
Благородный олень — Cervus elaphus Linnaeus, 1758 ..... 160
Особенности морфологии и морфофизиологии............ 160
Структура и динамика ареала......................... 177
Систематика и изменчивость.......................... 183
Экология........................................... 188
Воспроизводство и структура популяции .............. 196
Род Лоси — Alces Gray, 1821 ............................. 205
Лось — Alces alces Linnaeus, 1758 ..................... 208
Особенности морфологии и морфофизиологии........... 208
Структура и динамика ареала......................... 217
Систематика и изменчивость.......................... 221
Экология............................................ 224
Воспроизводство и структура популяции............... 234
Семейство Полорогие — Bovidae Gray, 1821 ................... 243
Род Зубры — Bison Н. Smith,1827 ....................... 246
Зубр — Bison bonasus Linnaeus, 1758 ............... 248
Особенности морфологии и морфофизиологии............ 248
Структура и динамика ареала......................... 268
Систематика и изменчивость.......................... 274
Экология............................................ 280
Воспроизводство и структура популяции............... 293
Оглавление
Род Горные козлы Capra Linnaeus, 1758 .................... 301
Кавказский тур — Capra caucasica Guldcnstacdt et Pallas, 1783 ....................................... 303
Особенности морфологии и морфофизиологии............. 303
Структура и динамика ареала......................... 316
Систематика и изменчивость........................... 319
Экология............................................. 329
Воспроизводство и структура популяции................ 332
Бородатый козел — Capra aegagrus Erxlebcn., YT11 ................ 338
Особенности морфологии и морфофизиологии............. 338
Структура и динамика ареала.......................... 345
Систематик^ и изменчивость........................... 350
Экология............................................. 358
Воспроизводство и структура популяции................ 367
Род Сайгаки — Saiga Gray, 1843 ........................... 372
Сайгак — Saiga tatarica Linnaeus, 1766 ................ 373
Особенности морфологии и морфофизиологии ............ 373
Структура и динамика ареала.......................... 384
Систематика и изменчивость........................... 387
Экология .......................................... 393
Воспроизводство и структура популяции................ 402
Род Газели — Gazclla de Blainville, 1816.................. 411
Джейран — Gazclla subgutturosa Guldcnstacdt, 1780 ...... 412
Особенности морфологии и морфофизиологии............. 413
Структура и динамика ареала.......................... 422
Систематика и изменчивость........................... 425
Экология............................................. 432
Воспроизводство и структура популяции.............. 438
Род Горные бараны — Ovis Linnaeus, 1758 ................ 441
Азиатский муфлон — Ovis orientalis Gmelin, 1774 ........ 444
Особенности морфологии и морфофизиологии............. 444
Структура и динамика ареала......................... 451
Систематика и изменчивость........................... 453
Экология............................................. 462
Воспроизводство и структура популяции.............. 467
Род Серны — Rupicapra de Blainville, 1816................. 471
Серна — Rupicapra rupicapra Linnaeus, 1758 ............. 474
Особенности морфологии и морфофизиологии........... 474
Структура и динамика ареала.......................... 483
Систематика и изменчивость........................... 488
Экология......................................... 491
Воспроизводство и структура популяции................ 498
Литература...................................................... 502
Научное издание
Серия Позвоночные Кавказа
Соколов Владимир Евгеньевич
Темботов Асланби Казиевич
МЛЕКОПИТАЮЩИЕ
Копытные
Утверждено к печати
Институтом эволюционной морфологии и экологии животных им. А.Н. Северцева Российской академии наук
Редактор издательства Г.М. Орлова
Художник Ф.Н. Буданов
Художественный редактор Н.Н. Михайлова
Технический редактор З.Б. Павлюк
Подписано к печати 30.12.92
Формат 70 X 100 1/16
Печать офсетная
Усл. печ. л.42,57. Усл. кр.-отт. 42,57. Уч.-изд. л. 44,7
Тираж 350 экз. Тип. зак. ЗД^.
Ордена Трудового Красного Знамени
издательство "Наука”
117864 ГСП-7, Москва В-485
Профсоюзная ул., 90.
Ордена Трудового Красного Знамени
1-я типография издательства "Наука”
199034, Санкт-Петербург В-34,9-я линия, 12