/























































































































































































































































Author: Клевезаль Г.А
Tags: mammalia млекопитающие биология зоология охота
ISBN: 978-5-87317-355-6
Year: 2007




Text
Г.А. Клевезаль
ПРИНЦИПЫ И МЕТОДЫ
ОПРЕДЕЛЕНИЯ ВОЗРАСТА
МЛЕКОПИТАЮЩИХ
RUSSIAN ACADEMY OF SCIENCES
Koltzov Institute of Developmental Biology
G.A. Klevezal
PRINCIPLES AND METHODS
OF AGE DETERMINATION OF MAMMALS
KMK Scientific Press Ltd.
Moscow ❖ 2007
РОССИЙСКАЯ АКАДЕМИЯ НАУК
Институт биологии развития им. Н.К. Кольцова
Г.А. Клевезаль
ПРИНЦИПЫ И МЕТОДЫ ОПРЕДЕЛЕНИЯ
ВОЗРАСТА МЛЕКОПИТАЮЩИХ
Товарищество научных изданий КМ К
Москва ❖ 2007
G.A. Klevezal. Principles and methods of age determination of mammals. Moscow:
KMK Sci. Press Ltd. 2007. 283 p.
In the book a reader will find principal methods of age determination of mammals
allowing to estimate the age of an animal with necessary precision using the most
simple appropriate method. The book includes two parts. In the first part description
of different methods with indication of their resolution capacity, advantages and
disadvantages and area of application is given. Special attention is paid to aging living
animals. Specific data on using different methods for aging more than one hundred
species of Insectivores, Chiropterans, Rodents, Carnivores and Ungulates can be
found in the second part of the book.
The book is intended for zoologists, wild-game biologists, ecologists, archaezoologists
and conservation biologists.
Principal editor — Mikhail V. Mina, Doctor of Science (Biology).
The publication is supported by Presidium of RAS, the Program of fundamental
investigations «Biological Resources of Russia: Fundamental Backgrounds
of Rational Management»
©G.A. Klevezal, 2007
© KMK Sci. Press Ltd., 2007
ISBN 978-5-87317-355-6
УДК 599
ББК У 28.693.36
К 48
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих.' М.:
Т-во научных изданий КМК. 2007. 283 с.
Книга знакомит читателей с основными методами определения возраста млеко-
питающих и дает возможность любому исследователю определить возраст объекта
его исследования с необходимой точностью максимально простым и нетрудоем-
ким методом. Книга состоит из двух частей. В первой части дано описание всех
основных методов определения возраста с указанием разрешающей способнос-
ти, достоинств, недостатков и области применения каждого. Особое внимание
уделено методам прижизненного определения возраста животных. Во второй части
приведены конкретные данные, позволяющие определять разными методами воз-
раст более 100 видов из отрядов насекомоядных, рукокрылых, зайцеобразных,
грызунов, хищных, парно- и непарнокопытных.
Книга рассчитана на широкий круг зоологов, охотоведов, специалистов по ох-
ране природы, экологов, морфологов и популяционных биологов. Она будет слу-
жить пособием при оценке ресурсов как промышляемых и охраняемых видов
млекопитающих, так и видов, численность которых необходимо регулировать.
Ответственный редактор — д.б.н. М.В. Мина
Издание осуществляется при поддержке
Программы фундаментальных исследований Президиума РАН
«Биологические ресурсы России:
Фундаментальные основы рационального использования»
© Г. А. Клевезаль, 2007
© Товарищество научных изданий КМК, издание, 2007
ISBN 978-5-87317-355-6
СОДЕРЖАНИЕ
ПРЕДИСЛОВИЕ..........................................................18
Часть I. Методы определения возраста.................................20
Глава 1. Общие принципы............................................20
1.1. Понятия и термины.........................................20
1.2. Способы обработки и представления данных..................21
1.2.1. Выделение возрастных групп способами, не требующими
сложных вычислений..........................................21
1.2.2. Регрессионный анализ.................................24
1.2.3. Дискриминантный анализ...............................24
1.2.4. Кластерный анализ....................................26
1.2.5. Сравнение традиционных и многомерных методов.........26
1.2.6. Выбор конкретного метода для оценки возраста.........26
Глава 2. Определение возраста по признакам скелета.................28
2.1. Размеры черепа............................................28
2.2. Зарастание швов черепа....................................28
2.2.1. Область применения...................................28
2.2.2. Морфологическое обоснование..........................29
2.2.3. Разрешающая способность..............................29
2.2.4. Методика.............................................29
2.2.5. Изменчивость показателей.............................29
2.3. Форма черепа..............................................32
2.4. Развитие гребней..........................................33
2.5. Слияние эпифизов с диафизом костей конечностей............34
2.5.1. Область применения...................................34
2.5.2. Морфологическое обоснование..........................35
2.5.3. Разрешающая способность..............................36
2.5.4. Методика.............................................36
2.5.5. Изменчивость показателей.............................37
2.6. Состояние позвоночника и костей таза......................38
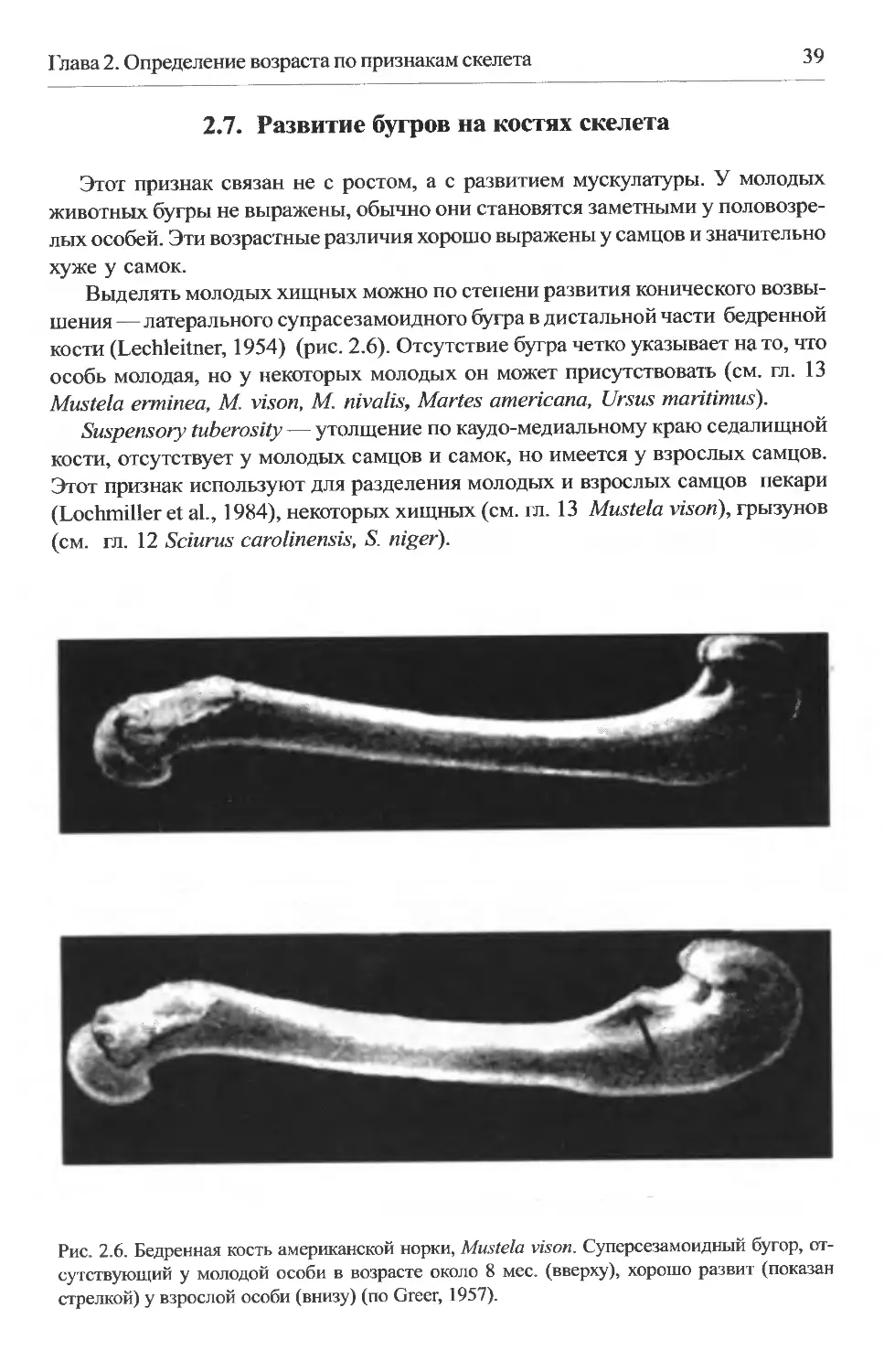
2.7. Развитие бугров на костях скелета.........................39
2.8. Размеры и форма кости полового члена (os penis, baculum)..40
2.8.1. Область применения...................................40
2.8.2. Морфологическое обоснование..........................40
2.8.3. Разрешающая способность..............................41
2.8.4. Методика.............................................41
2.8.5. Изменчивость показателей.............................43
2.8.6. Особенности использования метода в отдельных группах.43
2.8.6.1. Хищные..........................................43
2.8.6.2. Грызуны.........................................43
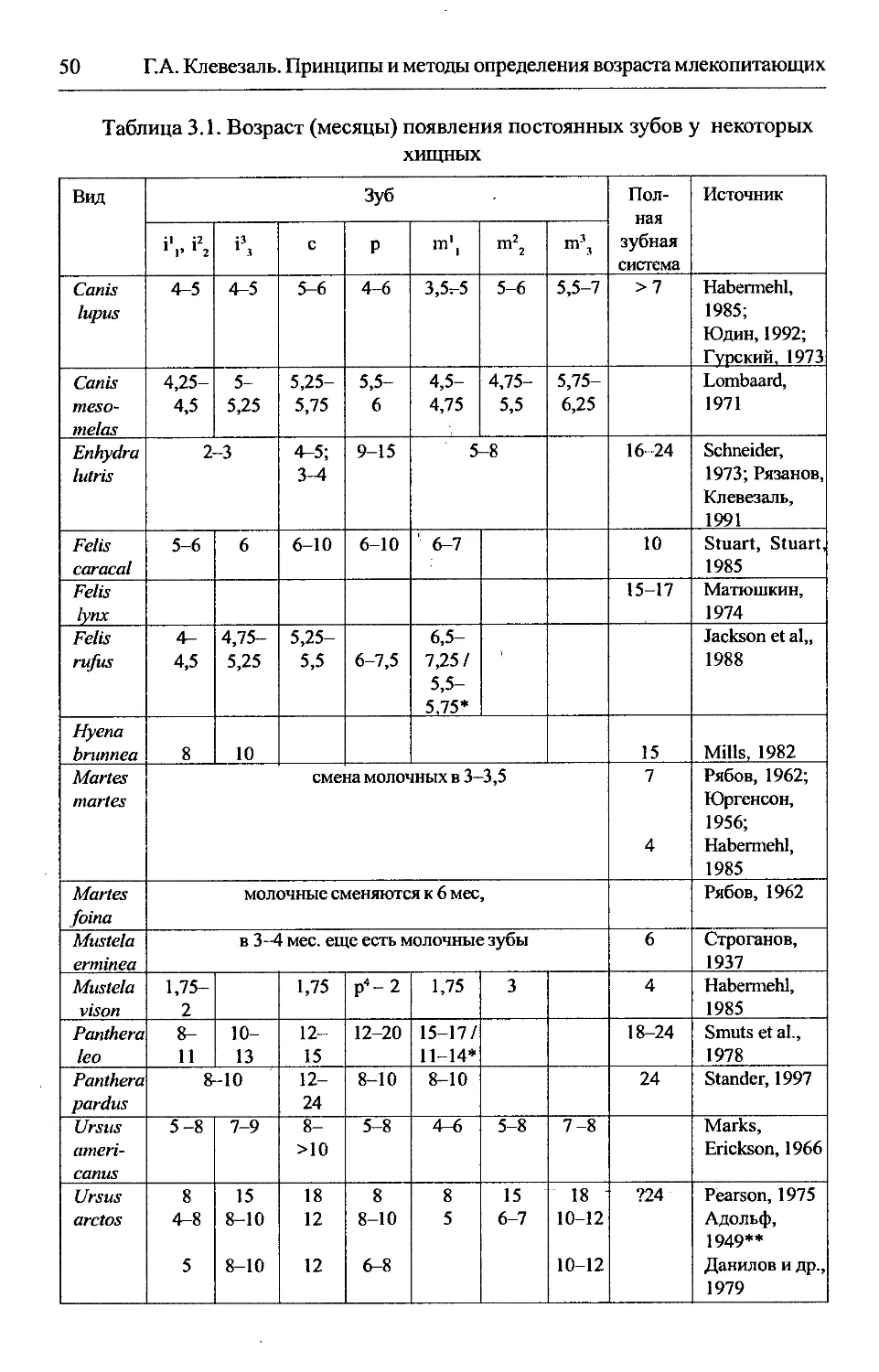
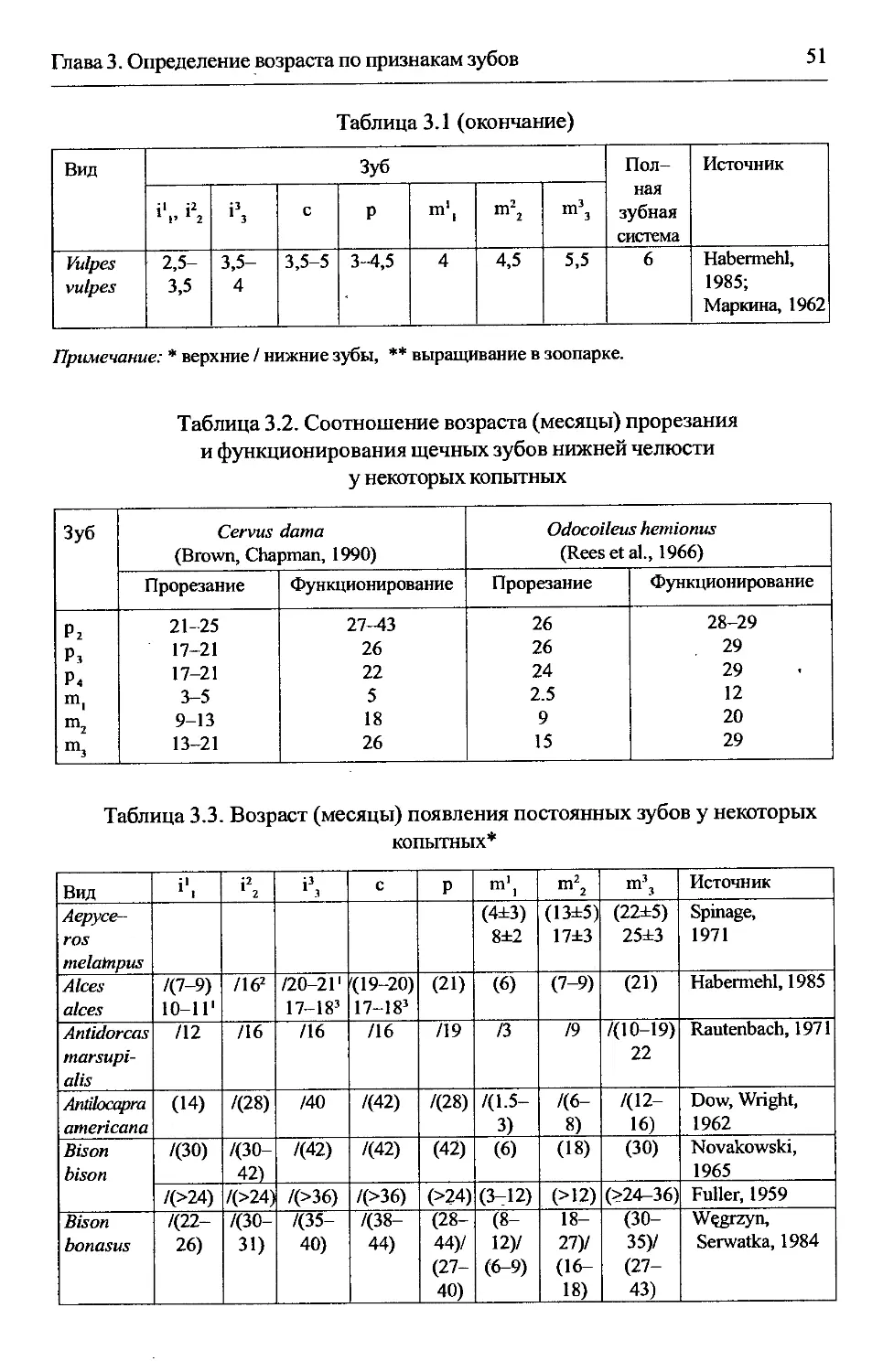
Глава 3. Определение возраста по признакам зубов...................45
Терминология...................................................45
3.1. Прорезание постоянных зубов...............................45
Содержание
7
3.1.1. Область применения...................................45
3.1.2. Морфологическое обоснование..........................46
3.1.3. Разрешающая способность..............................46
3.1.4. Методика.............................................47
3.1.5. Изменчивостьпоказателей..............................48
3.1.6. Особенности использования метода в отдельных группах.49
З.1.6.1. Хищные..........................................49
3.1.6.2. Парнокопытные...................................49
3.1.7. История применения...................................54
3.2. Рост зубов.................................................54
3.2.1. Рост зуба в длину....................................55
3.2.2. Замыкание апикального отверстия корня................55
3.2.3. Положение шейки зуба относительно альвеолы...........56
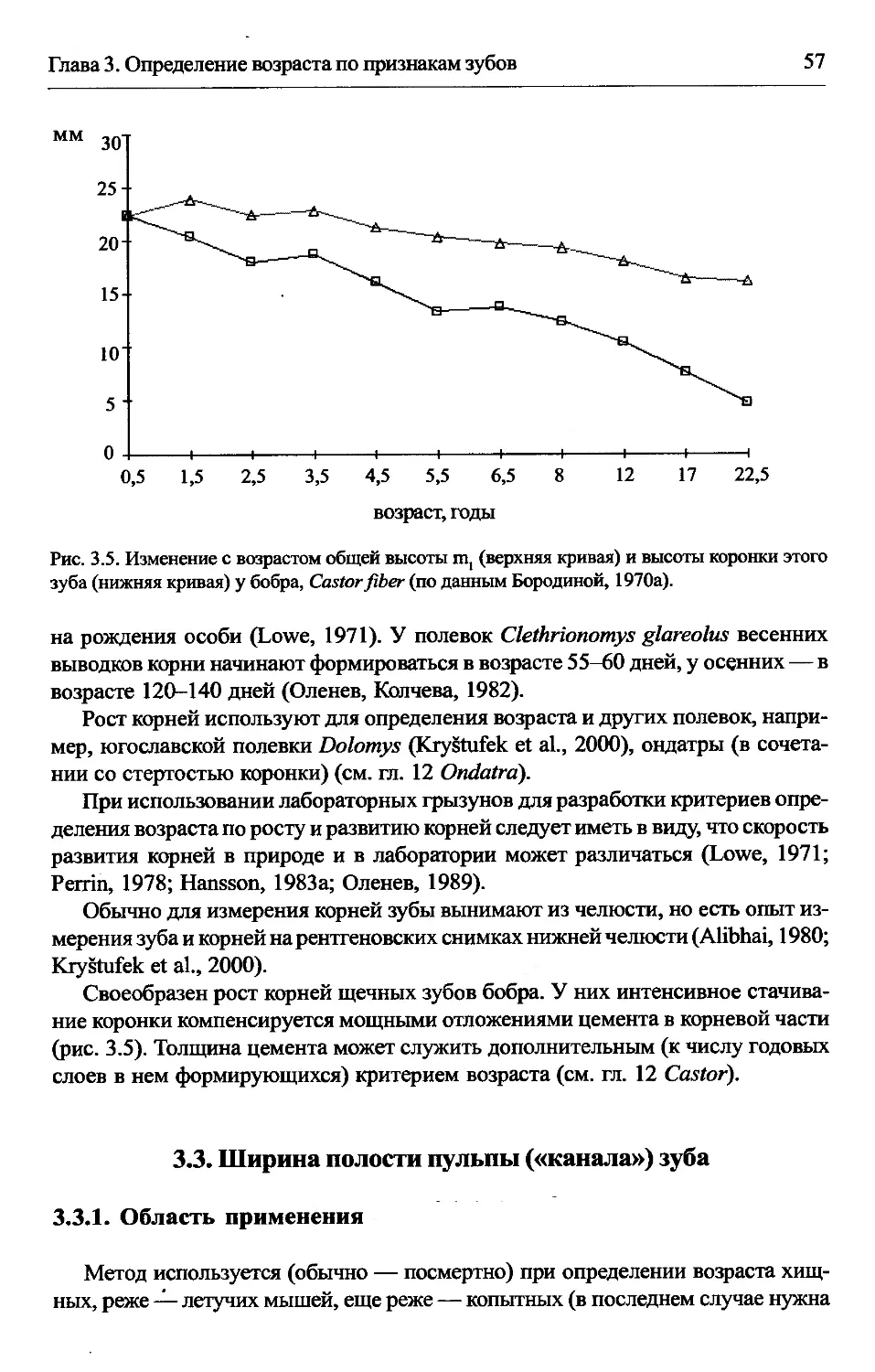
3.2.4. Рост корней и нарастание цемента.....................56
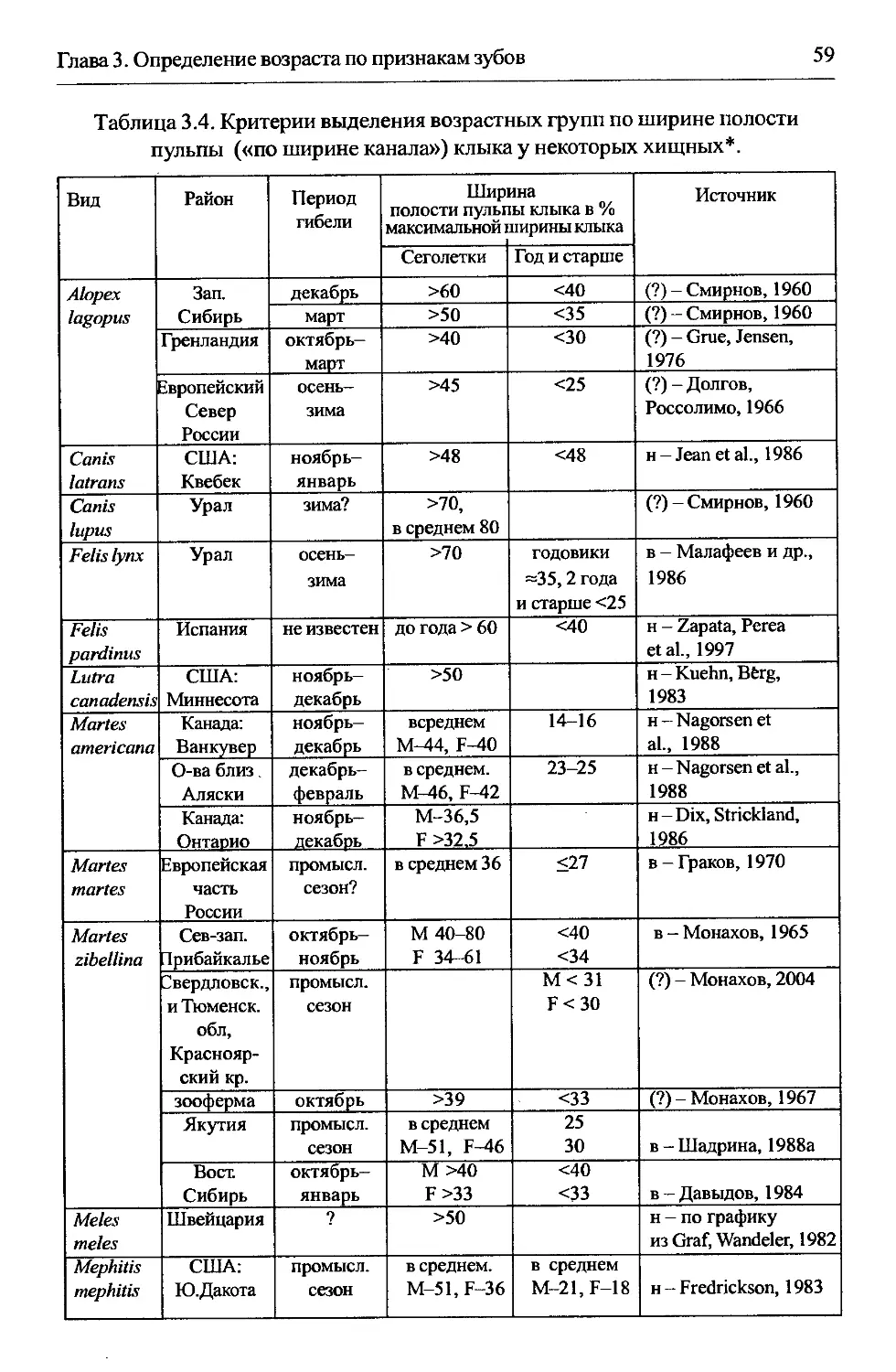
3.3. Ширина полости пульпы («канала») зуба......................57
3.3.1. Область применения...................................57
3.3.2. Морфологическое обоснование..........................58
3.3.3. Разрешающая способность..............................58
3.3.4. Методика.............................................61
3.3.5. Изменчивость показателей.............................62
3.3.6. Использование метода при работе с копытными..........64
3.3.7. История применения...................................64
3.4. Стачивание зубов с возрастом...............................64
3.4.1. Область применения...................................64
3.4.2. Морфологическое обоснование..........................65
3.4.3. Разрешающая способность..............................65
3.4.4. Методика.............................................66
3.4.5. Изменчивость показателей.............................67
3.4.6. Особенности использования метода в отдельных группах.67
3.4.6.1. Хищные..........................................67
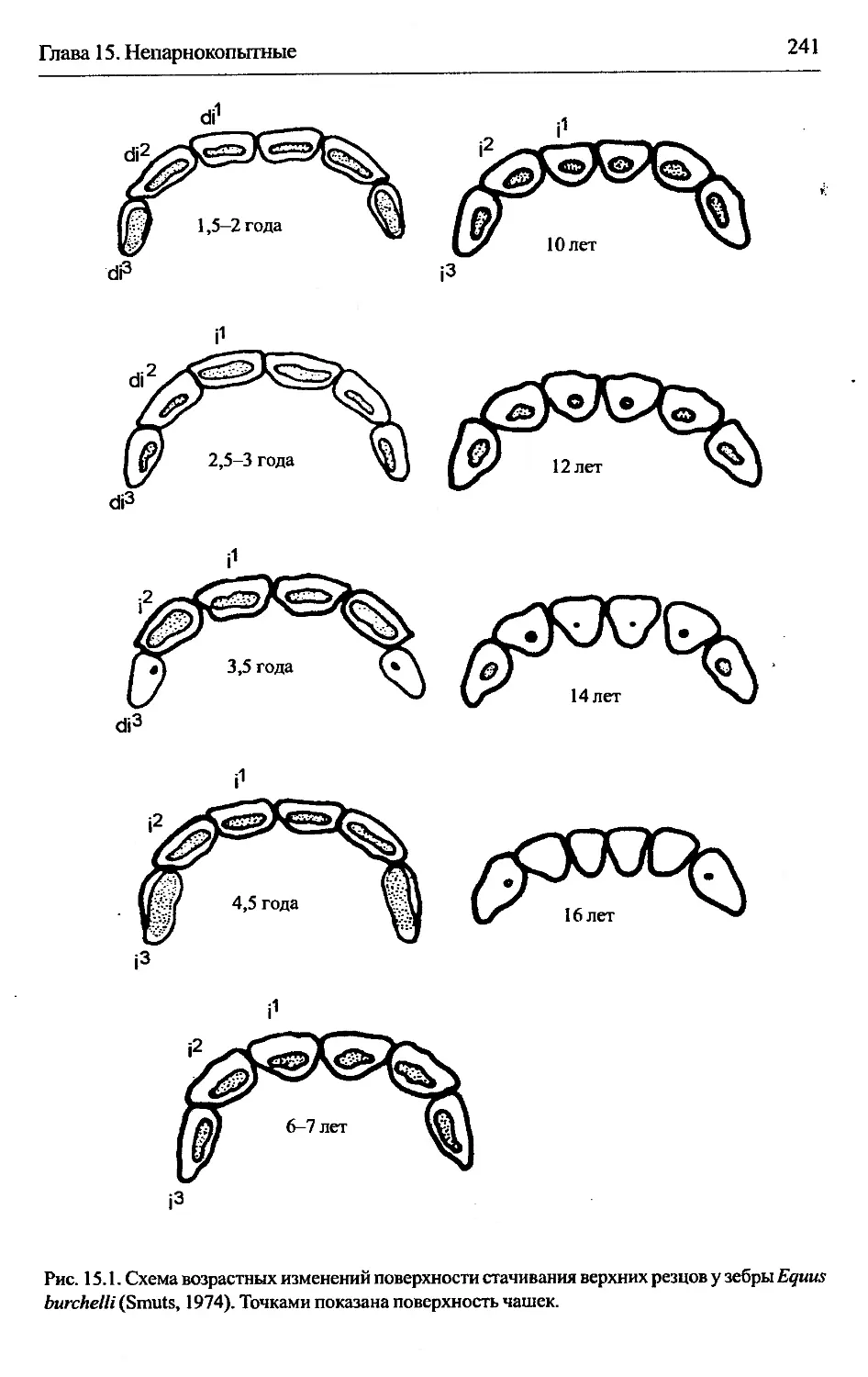
3.4.6.2. Непарнокопытные.................................69
3.4.6.3. Парнокопытные...................................69
3.4.6.4. Грызуны.........................................75
3.4.6.5. Насекомоядные и рукокрылые......................77
3.4.7. История применения...................................78
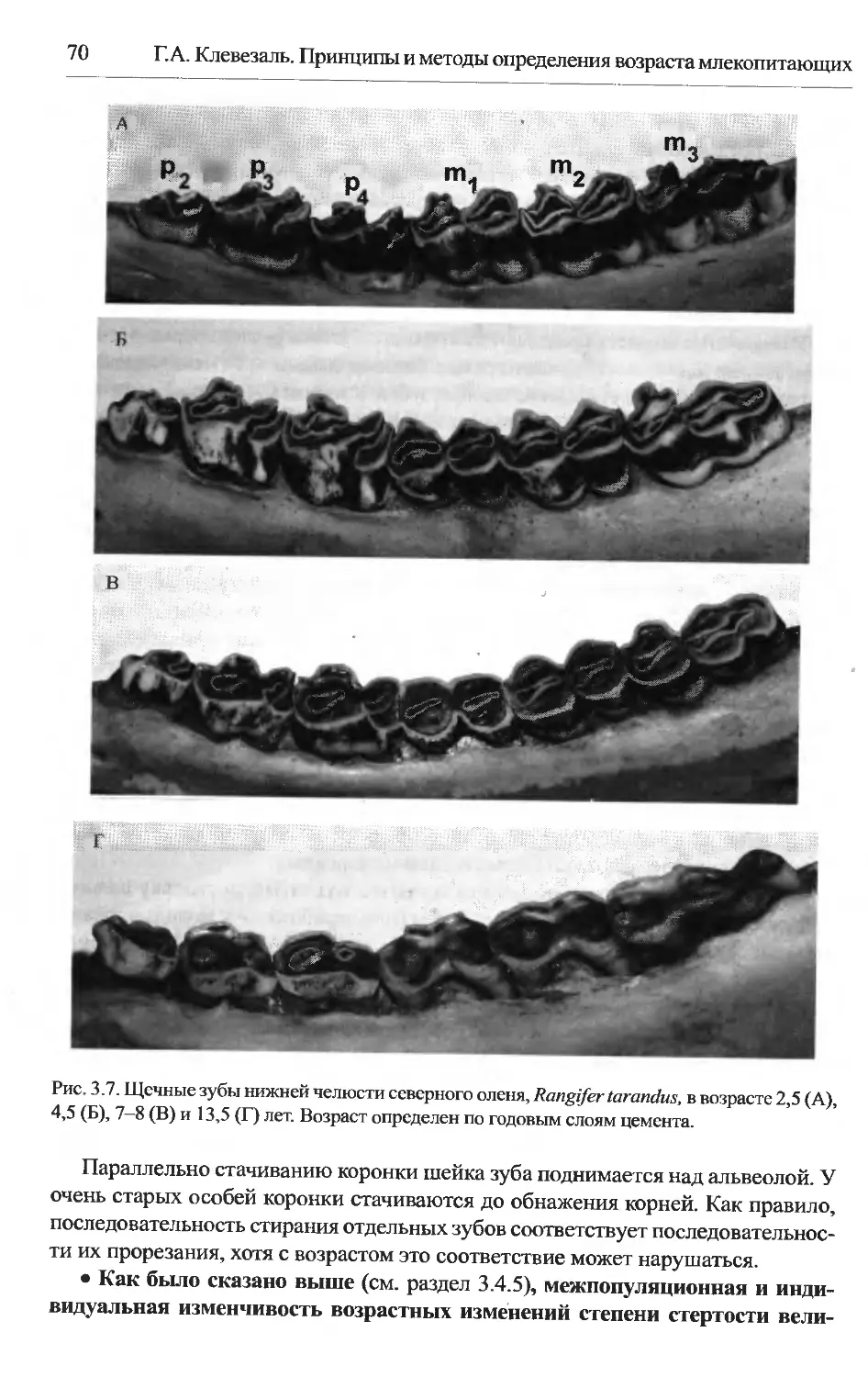
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости.79
4.1. Область применения.........................................79
4.2. Морфологическое обоснование...............................79
4.3. Разрешающая способность....................................82
4.4. Методика...................................................82
4.5. Изменчивость показателей...................................85
4.6. Особенности использования метода в отдельных группах.......86
4.6.1. Насекомоядные.....................................86
4.6.2. Рукокрылые........................................86
4.6.3. Зайцеобразные.....................................88
4.6.4. Грызуны...........................................88
8
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
4.6.5. Хищные...........................................89
4.6.6. Копытные.........................................91
4.7. История применения........................................92
Глава 5. Определение возраста по признакам рогов..................93
5.1. Рога полорогих.....................................93
5.2. Рога оленей........................................95
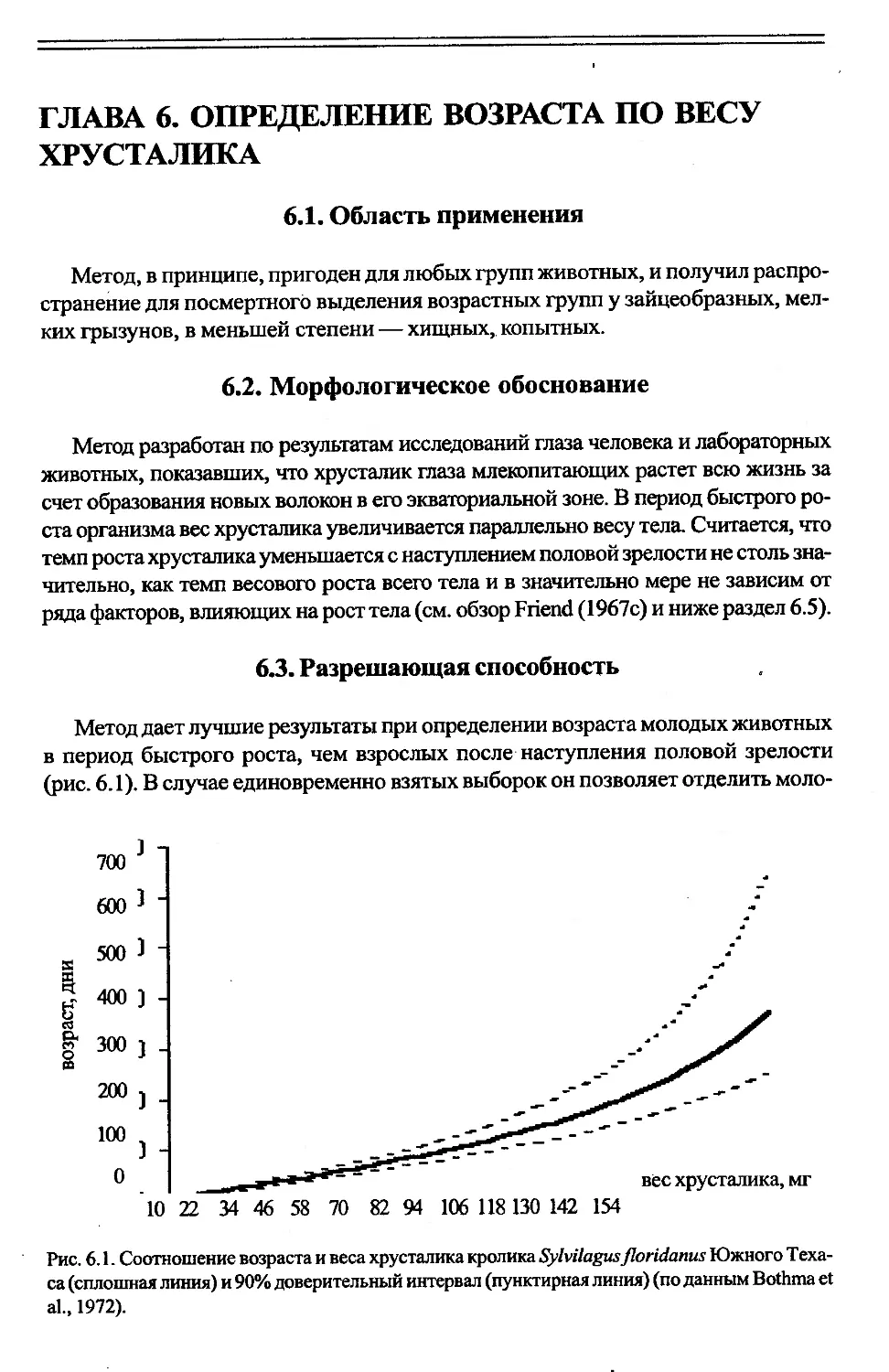
Глава 6. Определение возраста по весу хрусталика..................97
6.1. Область применения........................................97
6.2. Морфологическое обоснование...............................97
6.3. Разрешающая способность...................................97
6.4. Методика..................................................98
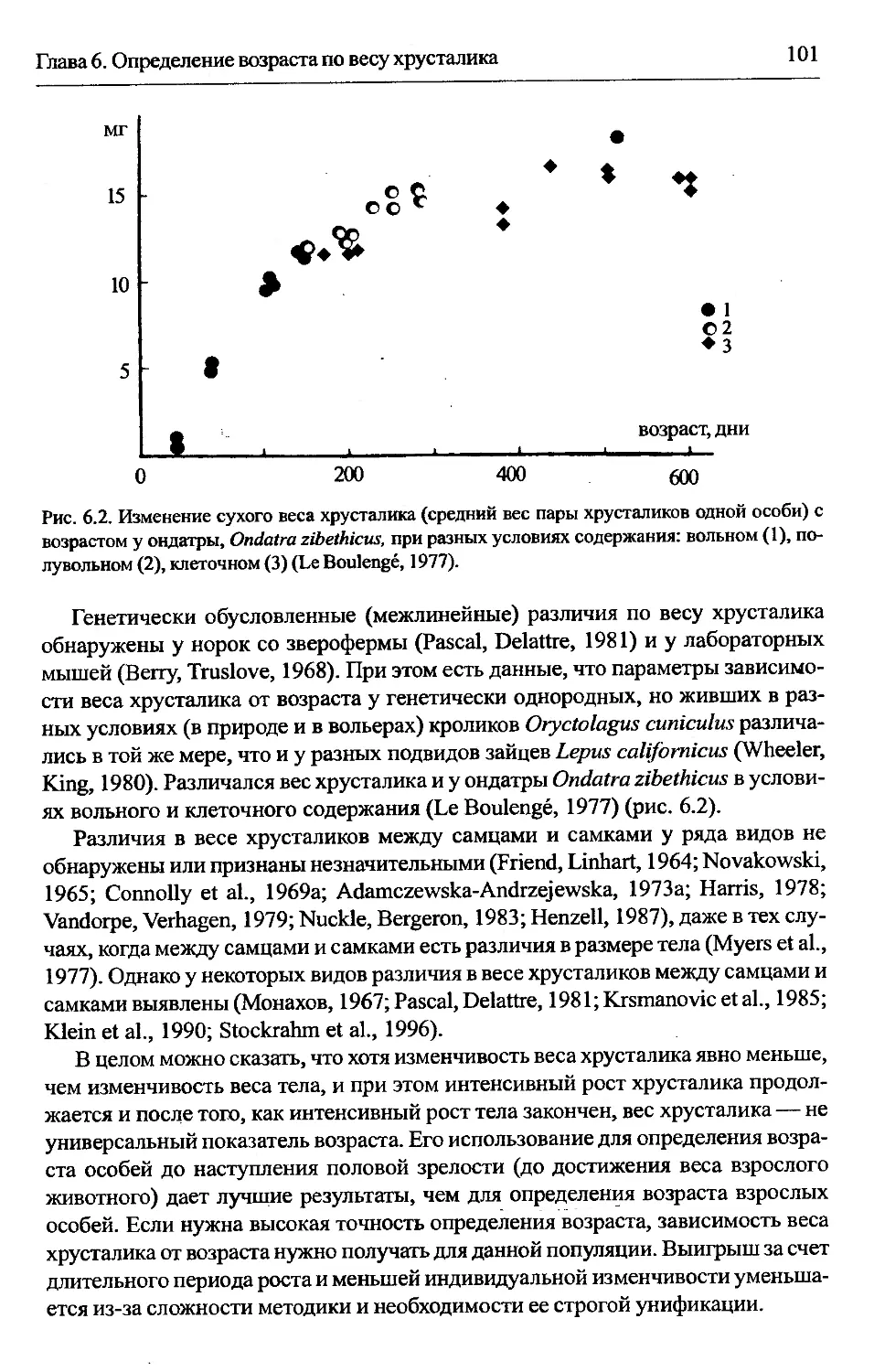
6.5. Изменчивость показателей.................................100
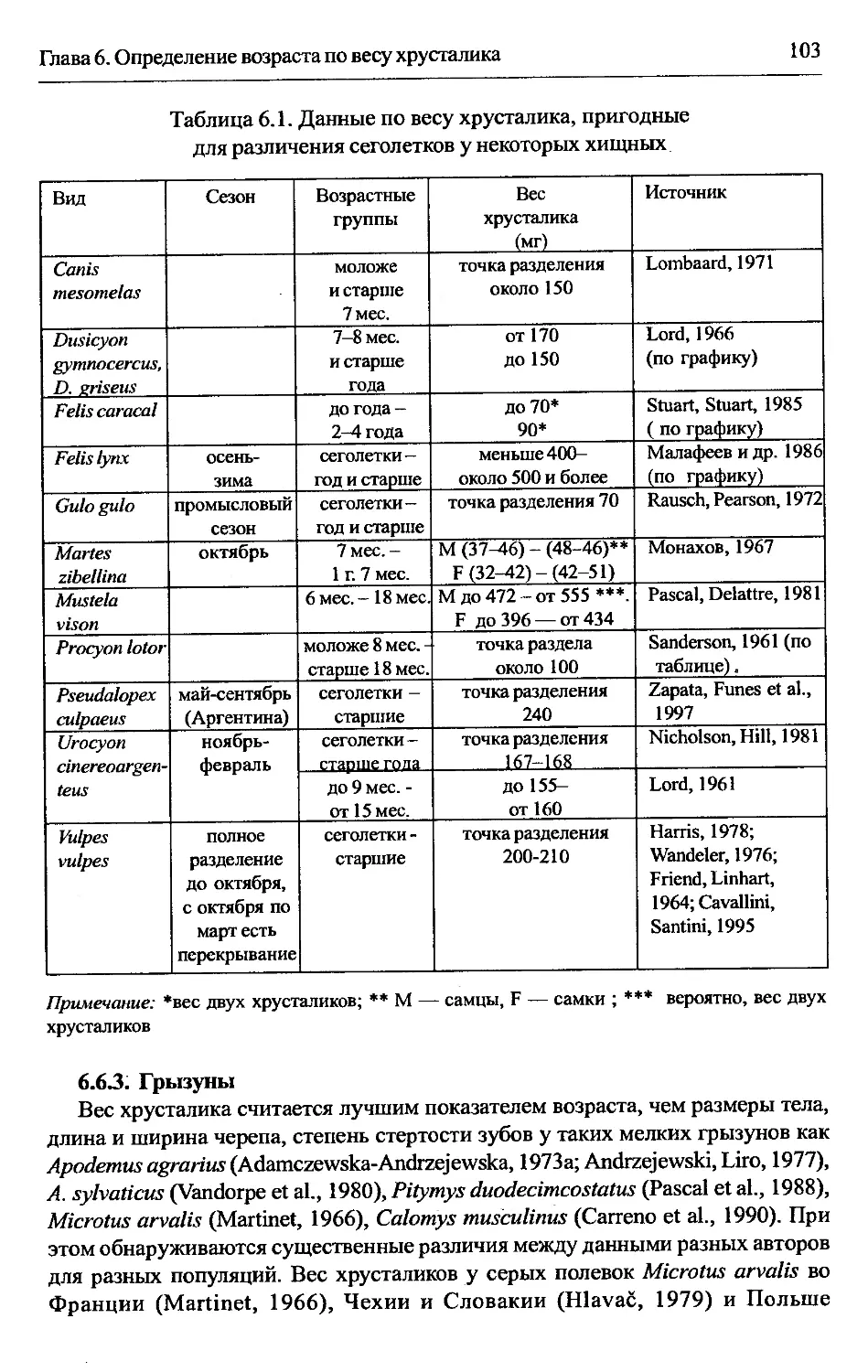
6.6. Особенности использования метода в отдельных группах.....102
6.6.1. Хищные..........................................102
6.6.2. Зайцеобразные...................................102
6.6.3. Грызуны...............:.........................103
6.6.4. Копытные........................................106
6.6.5. Насекомоядные...................................107
6.7. История применения.......................................107
Глава 7. Редко используемые методы...............................108
7.1. Изменение состояния сосудов сердца и вилочковой железы (тимуса).
.......................................................108
7.2. Содержание воды в организме..............................108
7.3. Содержание Sr-90.........................................108
7.4. Рацемизация аспарагиновой кислоты........................109
7.5. Содержание белка в хрусталике............................109
Глава 8. Методы прижизненного определения возраста...............111
8.1. Размеры тела и его частей.................................111
8.2. Окраска. Состояние меха..................................112
8.3. Возможности прижизненного использования других методов....112
8.3.1. Рога: форма, размер, годовые кольца................112
8.3.2. Зубы: прорезание, стертость, окраска...............114
8.3.3. Годовые слои в дентине и цементе зуба...............115
8.3.4. Годовые слои в фалангах пальцев....................115
8.3.5. Форма кости полового члена..........................115
8.3.6. Скелет: окостенение эпифизов.......................115
Часть II. Применение методов. Определение возраста представителей разных
систематических групп млекопитающих................................116
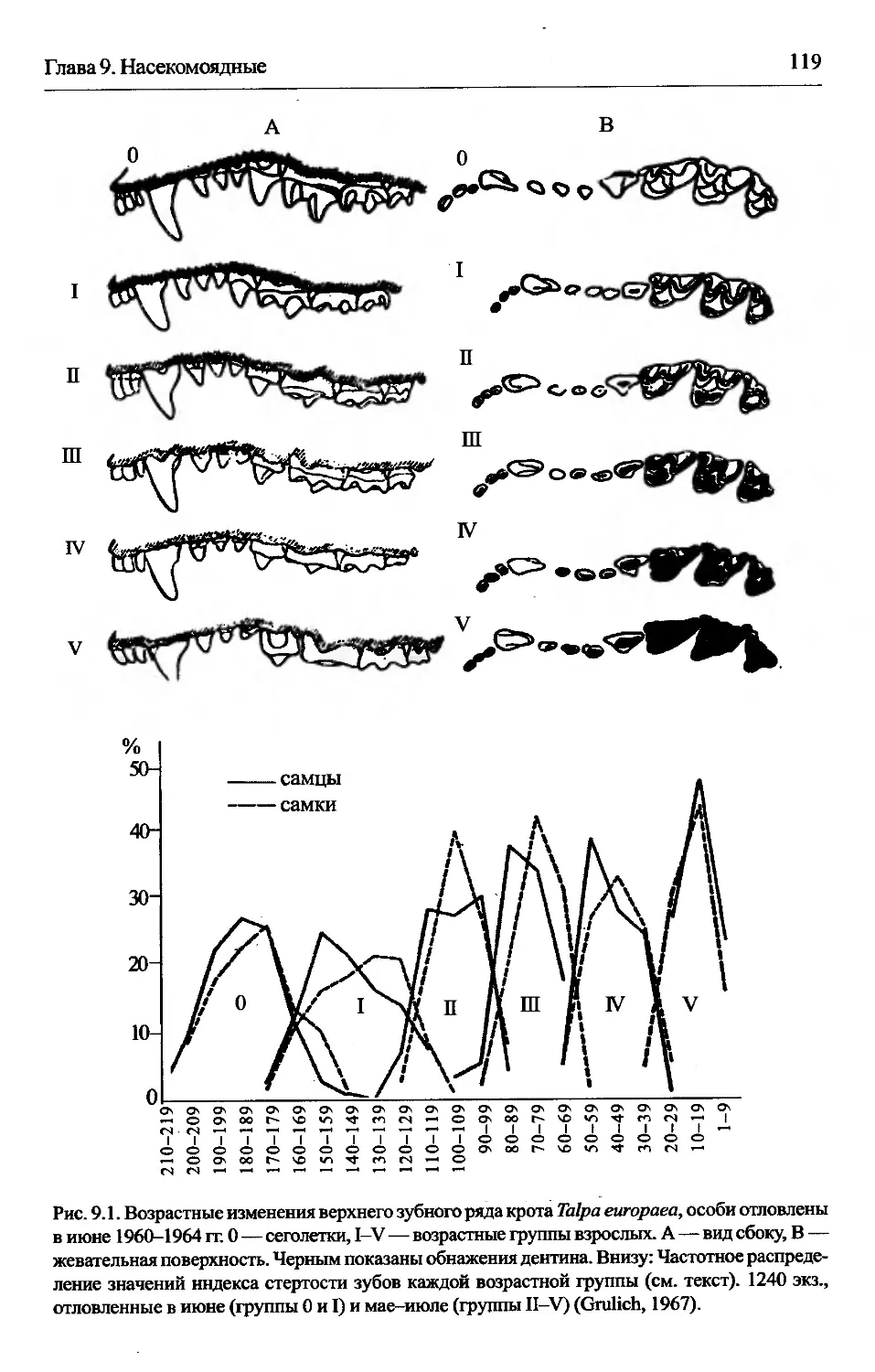
Глава 9. Насекомоядные...........................................116
Blarina brevicauda — короткохвостая бурозубка..........116
Crocidura — белозубки..................................116
Erinaceus europaeus — обыкновенный еж..................117
Galemys pyrenaicus — пиренейская выхухоль..............117
Sorex araneus — обыкновенная бурозубка.................117
Talpa еигораеа — обыкновенный крот.....................118
Содержание
9
Talpa micrura (Mogera robusta) — могера......................120
Глава 10. Рукокрылые. Летучие мыши..................................121
Внешний вид, окраска.........................................121
Кости крыловой перепонки.....................................121
Зубы. Стачивание.............................................123
Полость пульпы............................................. 124
Годовые слои.................................................124
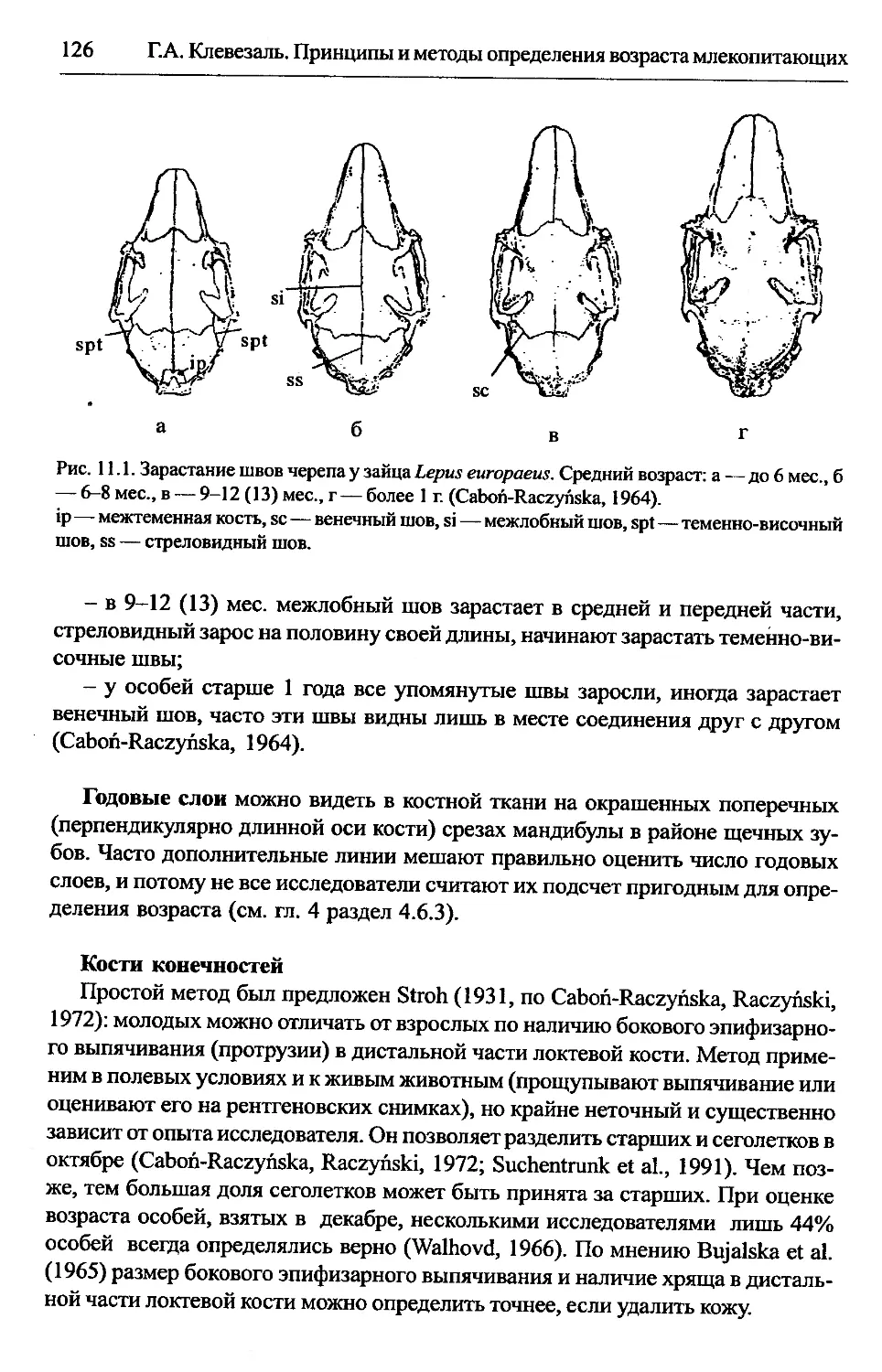
Глава 11. Зайцеобразные.............................................125
Lepus californicus — калифорнийский заяц.....................125
Lepus europaeus — заяц-русак.................................125
Lepus timidus — заяц-беляк...................................127
Ochotona — пищухи............................................127
Oryctolagus cuniculus — дикий кролик.........................128
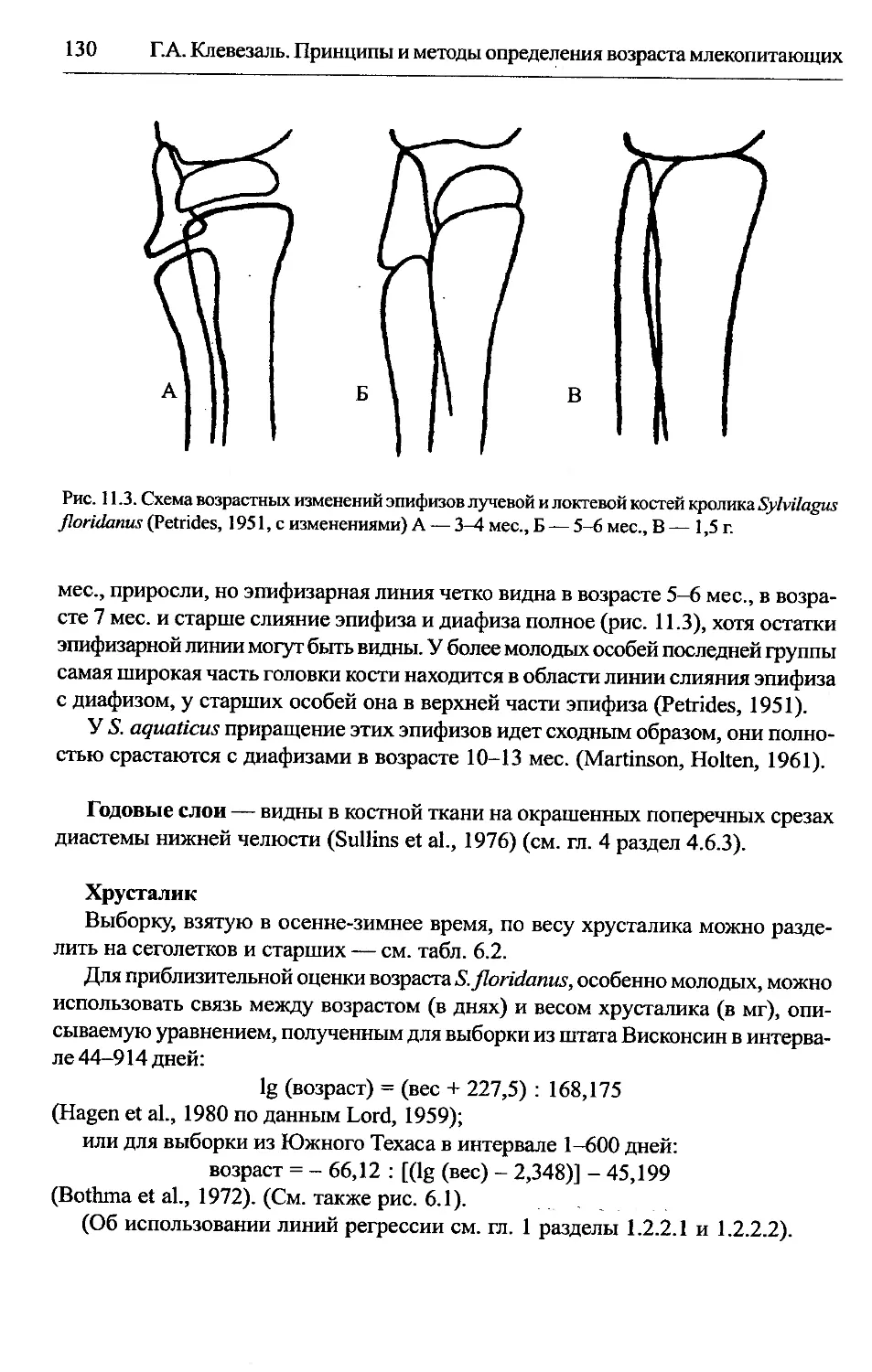
Sylvilagus — американские кролики............................129
Глава 12. Грызуны...................................................131
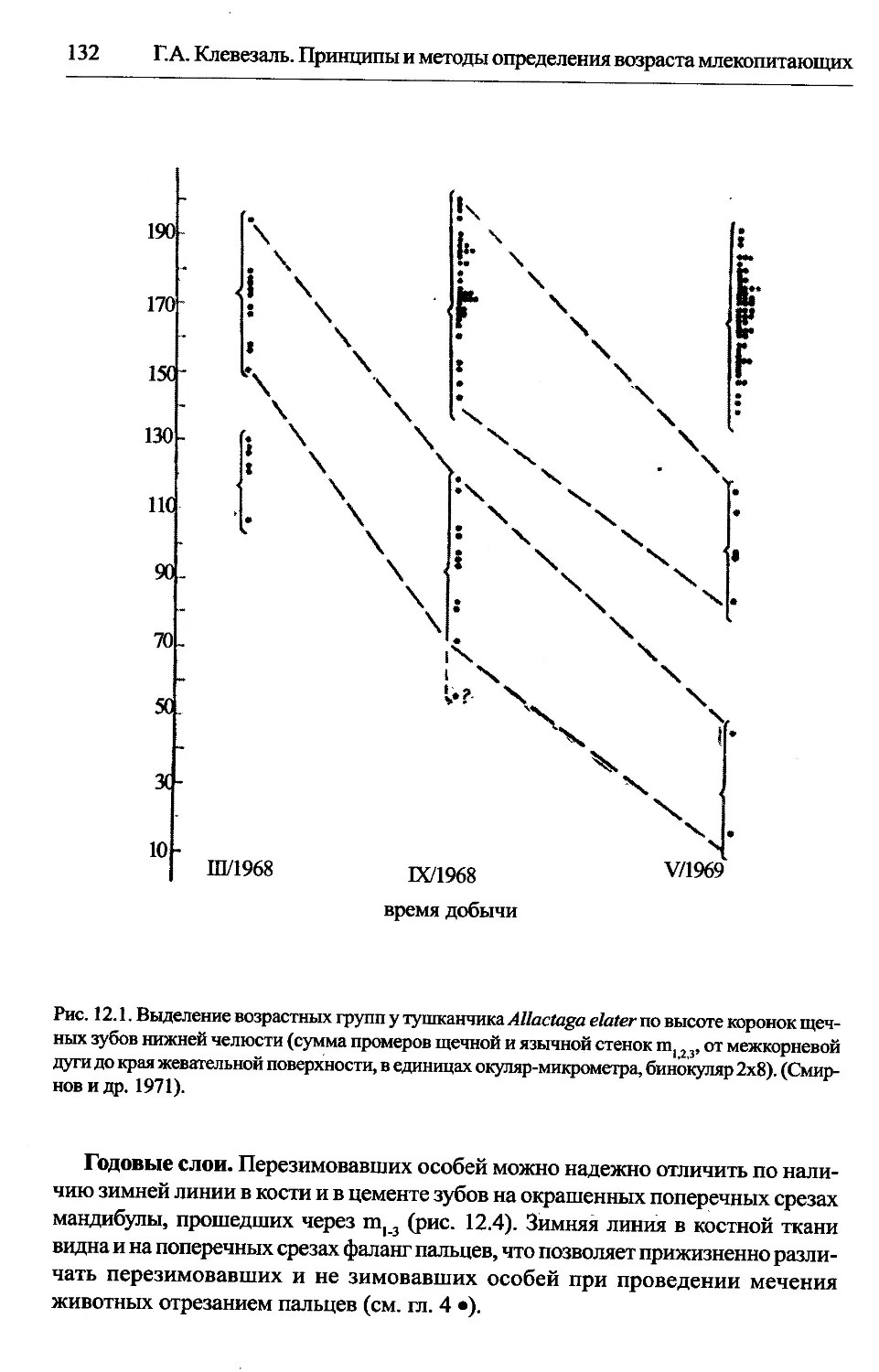
Allactaga и др. Dipodidae — тушканчики.......................131
Apodemus agrarius — полевая мышь.............................131
Apodemus Jlavicollis — желтогорлая мышь......................136
Apodemus sylvaticus — лесная мышь............................136
Arvicola terrestris— водяная полевка.........................137
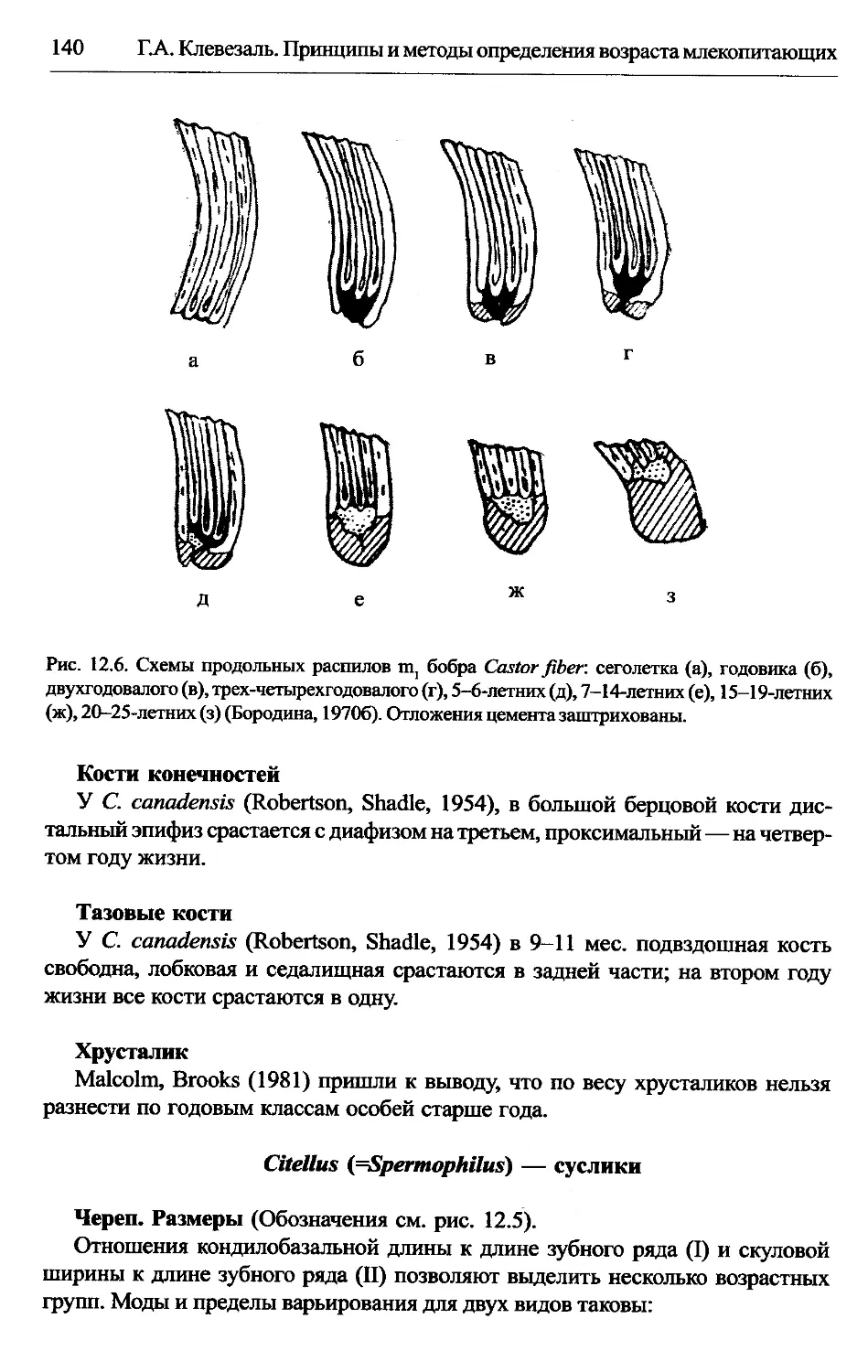
Castor — бобры...............................................137
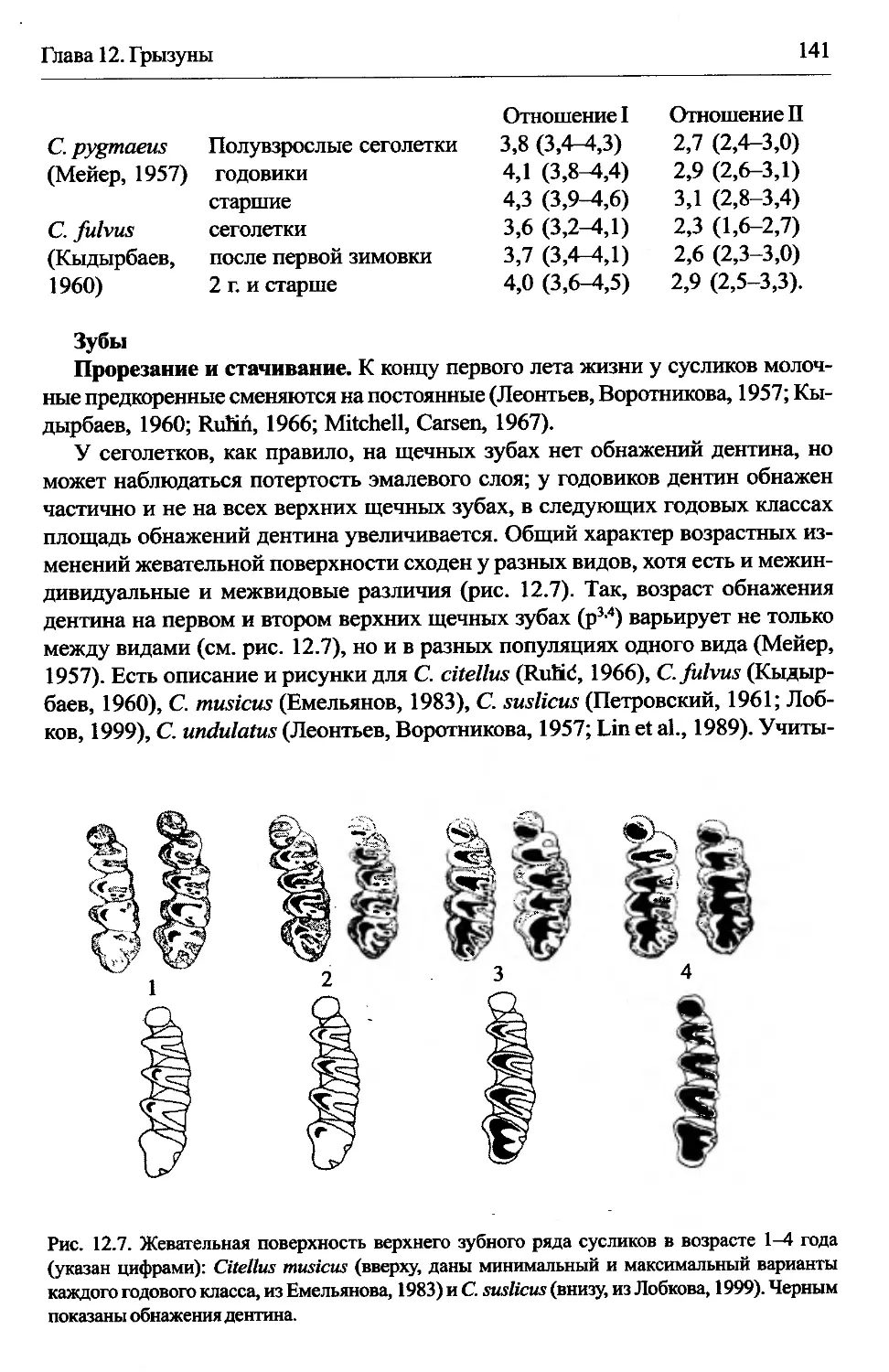
Citellus (ySpermophilus) — суслики...........................140
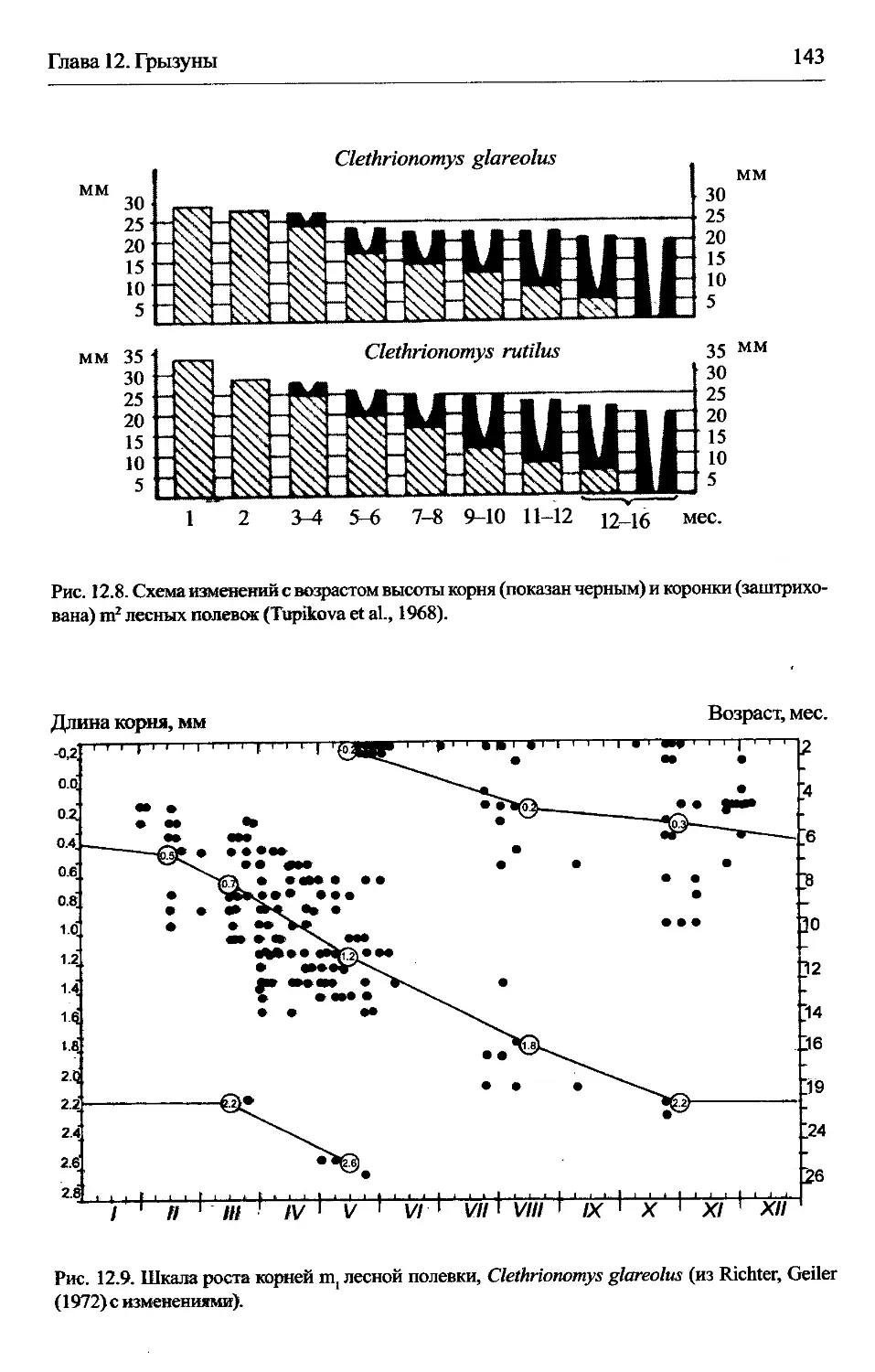
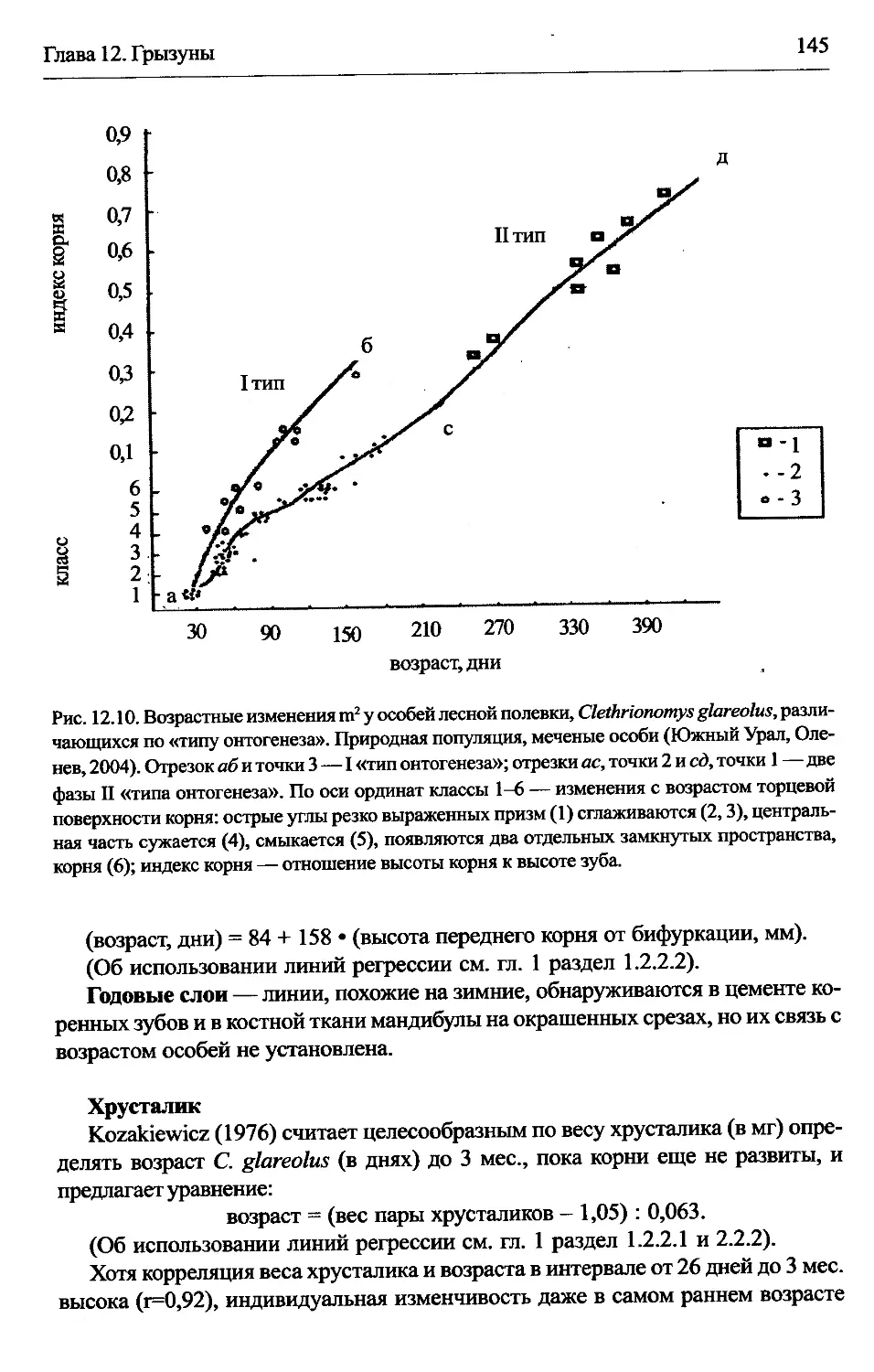
Clethrionomys —лесные полевки................................142
Cricetus cricetus — обыкновенный хомяк.......................146
Cynomys ludovicianus — чернохвостая луговая собачка..........147
Dicrostonyx torquatus — копытный лемминг.....................148
Dryomis nitedula — лесная соня...............................148
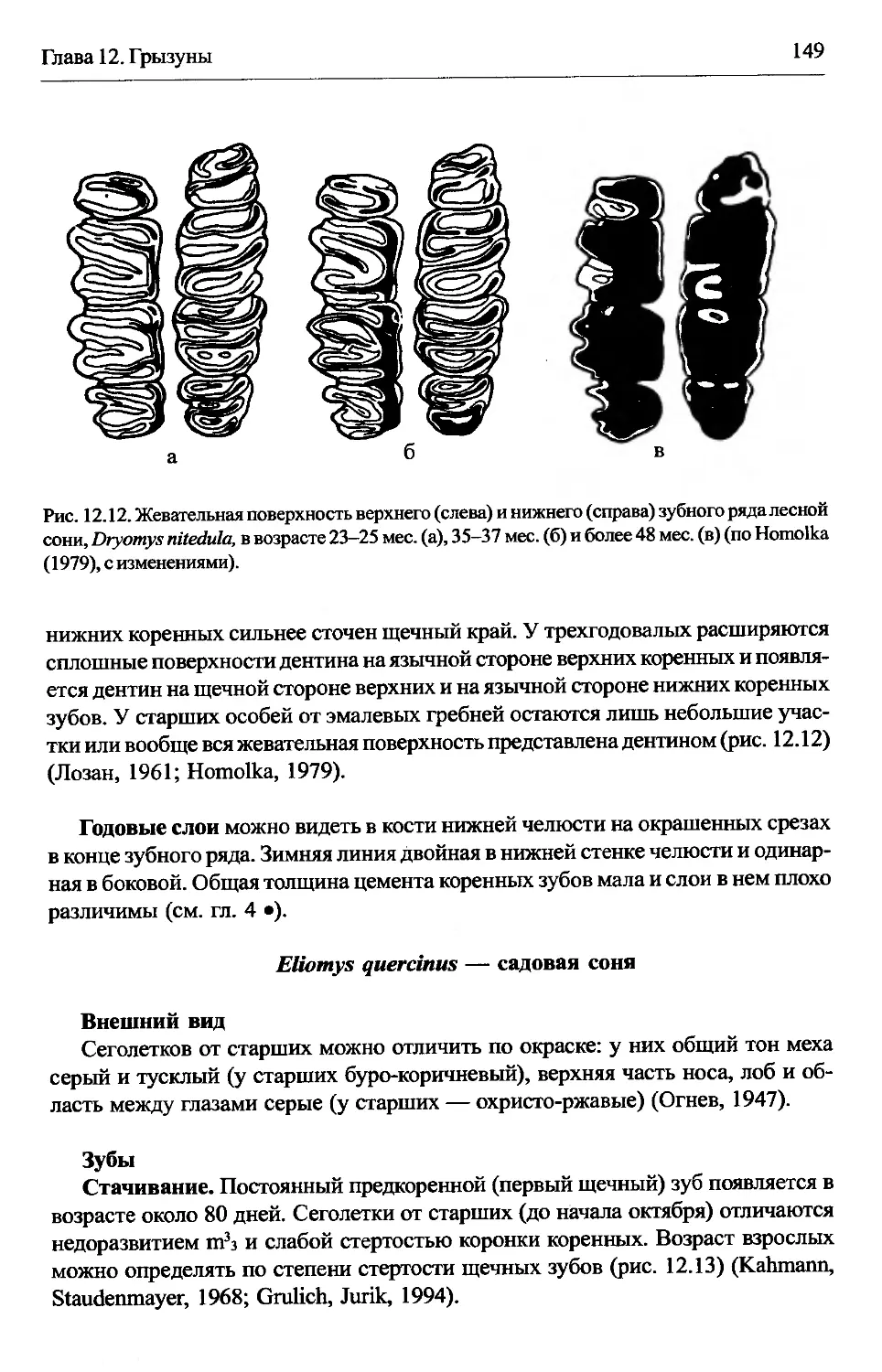
Eliomys quercinus — садовая соня.............................149
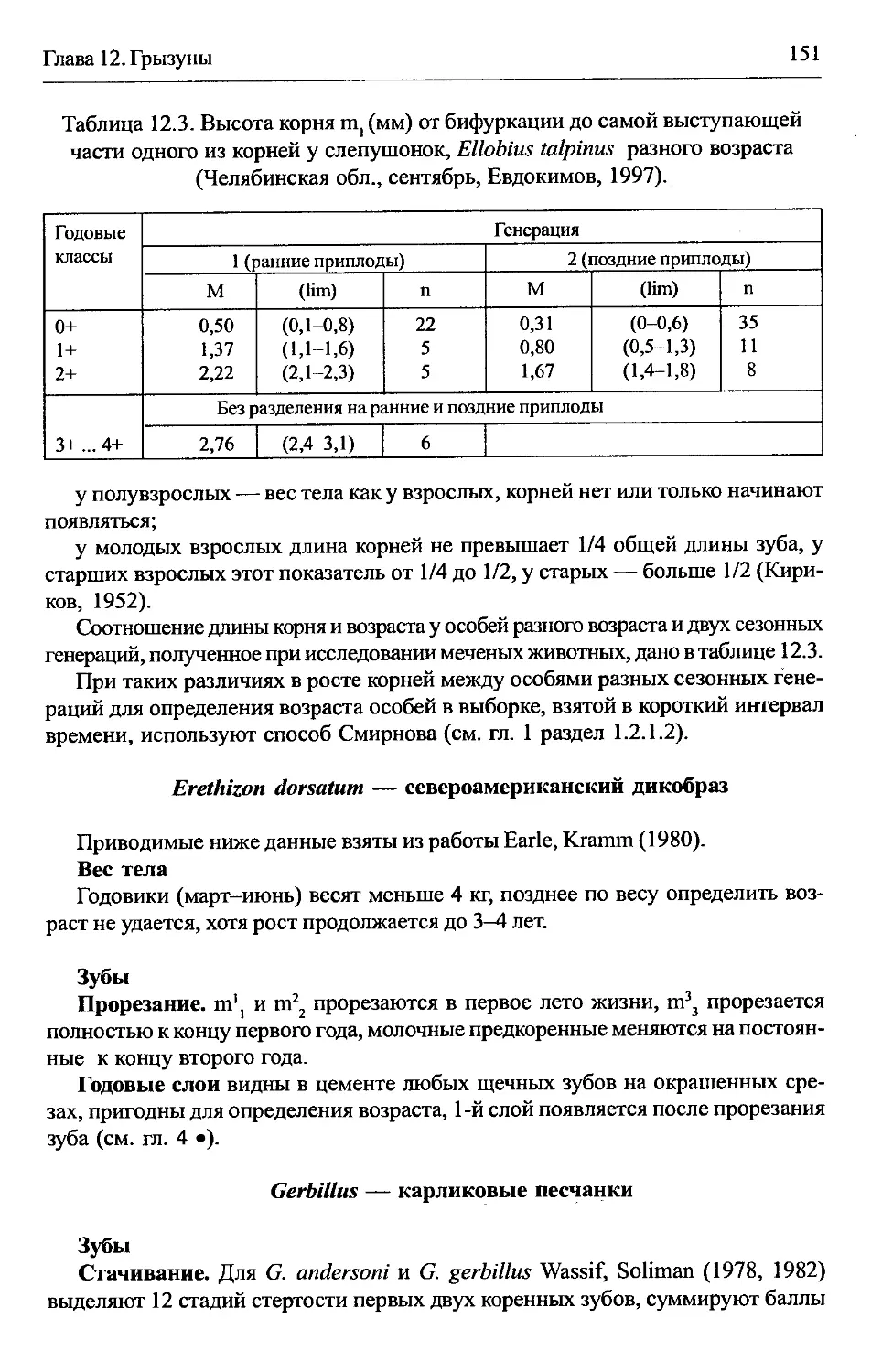
Ellobius talpinus — обыкновенная слепушонка..................150
Erethizon dorsatum — североамериканский дикобраз.............151
Gerbillus — карликовые песчанки..............................151
Glis glis — соня-полчок......................................152
Lagurus lagurus —- степная пеструшка.........................152
Lernmus lemmus — норвежский лемминг..........................153
Marmota baibacina, М. bobac, М. menzbieri, М. sibirica— сурки.154
Marmota топах — лесной сурок.................................156
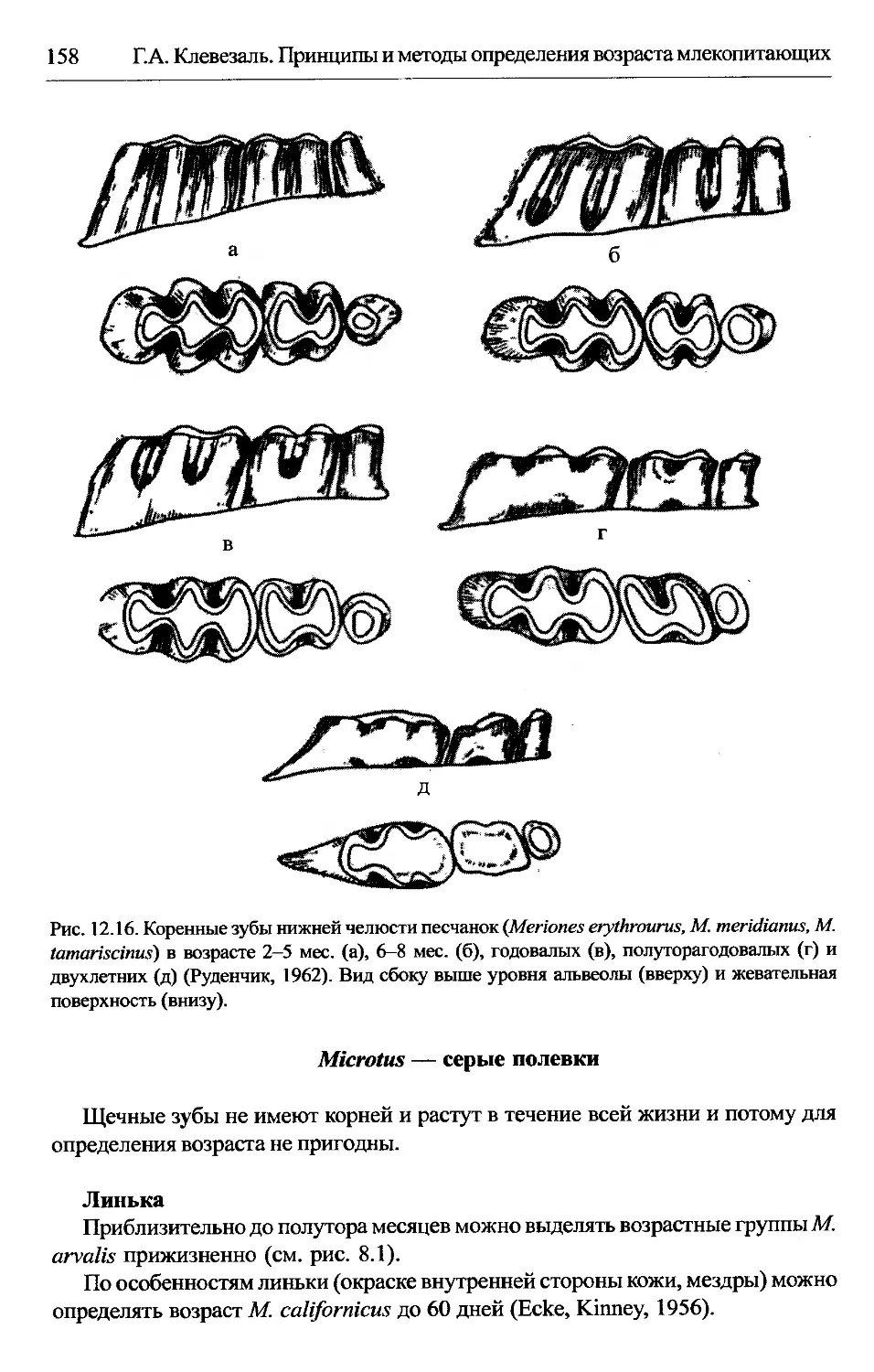
Meriones — малые песчанки....................................157
Microtus — серые полевки.....................................158
Mus musculus — домовая мышь..................................160
Muscardinus avellanarius — орешниковая соня..................161
Myocastor coypus — нутрия....................................161
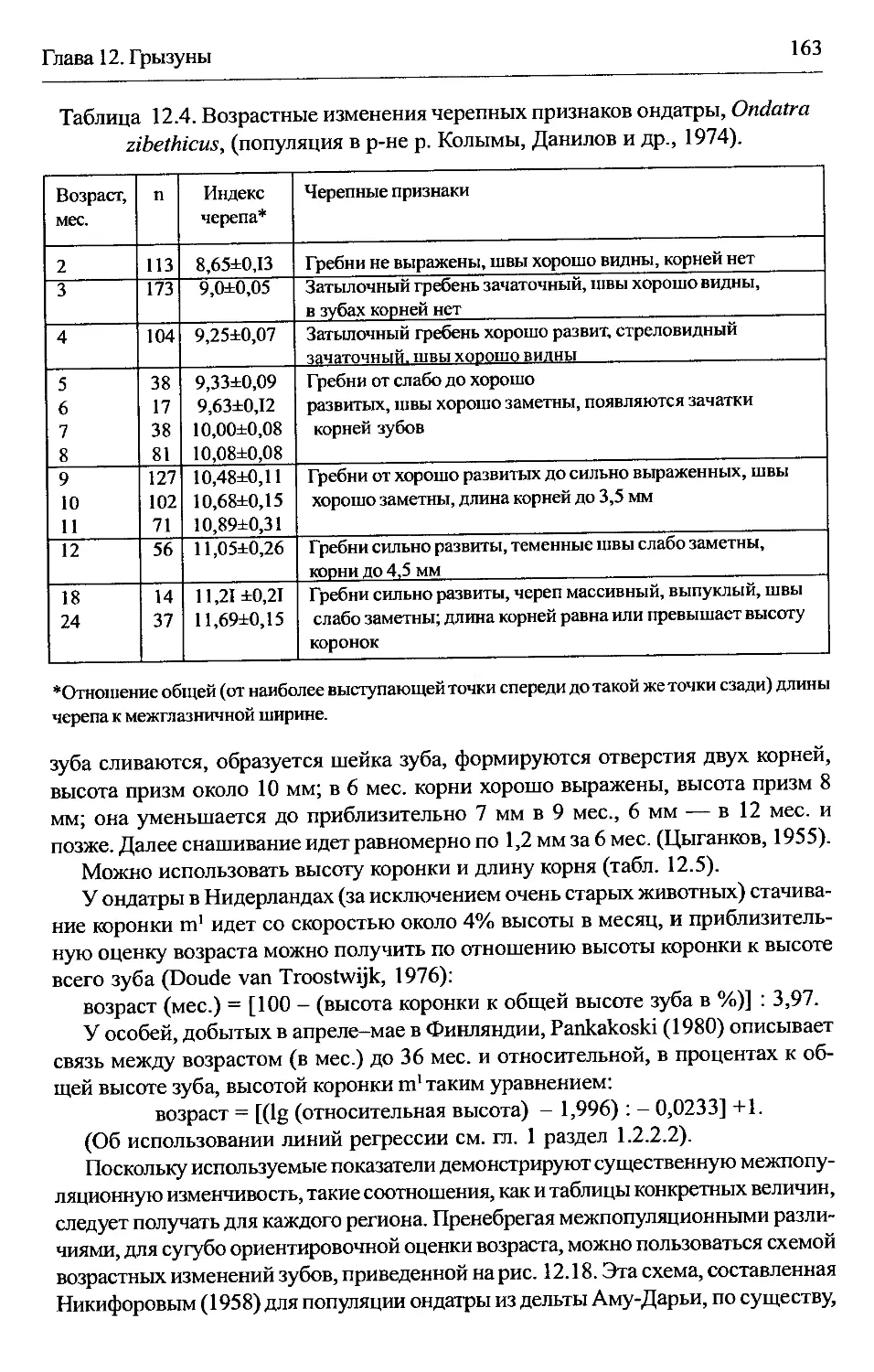
Ondatra zibethicus — ондатра.................................162
Rattus — крысы...............................................165
Rhombomys opimus — большая песчанка..........................165
Sciurus carolinensis, S. niger — каролинская и черная белки..166
Sciurus vulgaris — обыкновенная белка........................167
10
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Sicista — мышовки..............................................169
Sigmodon hispidus — колючий хлопковый хомяк....................170
Spalax microphtalmus — обыкновенный слепыш.....................170
Tamias (= Eutamias) — бурундуки................................171
Tamiasciurus hudsonicus — красная белка........................172
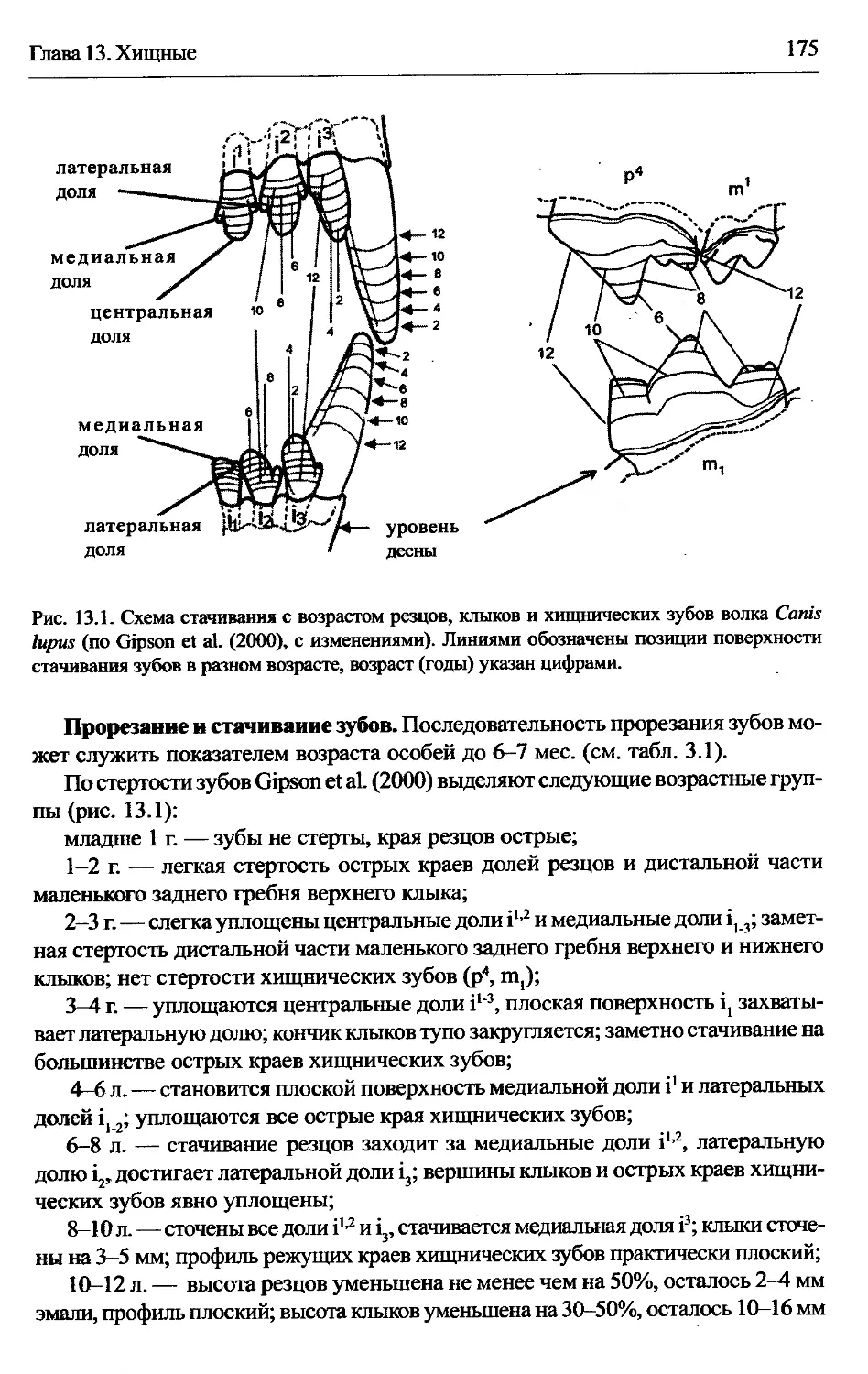
Глава 13. Хищные..................................................... 173
Alopex lagopus — песец.........................................173
Cants latrans — койот..........................................173
Cants lupus — волк.............................................174
Canis mesomelas — чепрачный шакал..............................176
Crocuta crocuta — пятнистая гиена..............................177
Enhydra lutris — калан.........................................177
Felts caracal — каракал........................................178
Felis catus, F.sylvestris — одичавшая и дикая кошки............179
Felts (=Рита) concolor — пума..................................179
Felis lynx — рысь..............................................180
Felis rufus — красная рысь.....................................181
Gulo gulo — росомаха...........................................181
Lutra canadensis — канадская выдра.............................182
Lutra lutra — выдра............................................182
Martes americana — американская куница.........................184
Martesfoina, M.martes — каменная и лесная куницы...............184
Martes pennanti — куница-рыболов...............................185
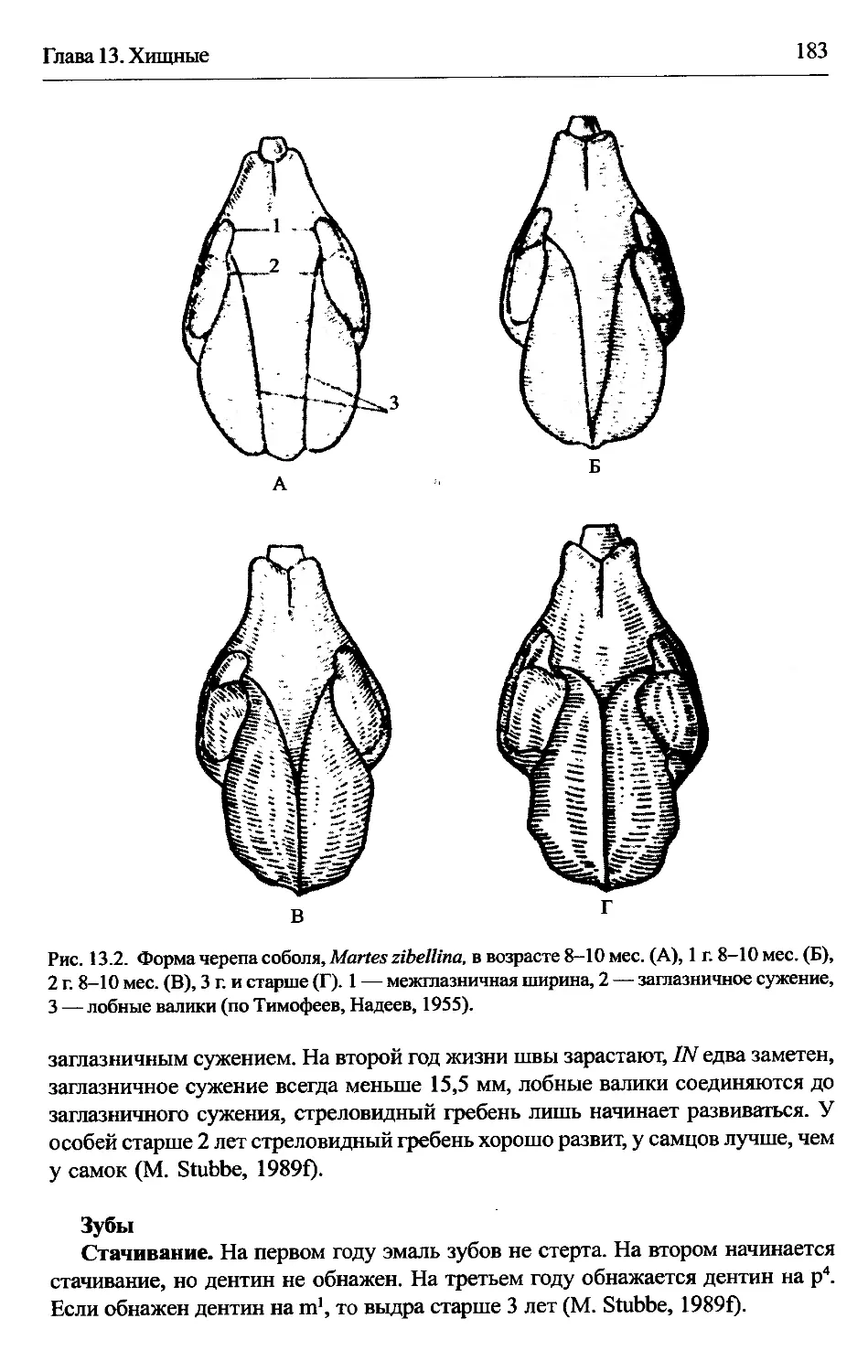
Martes zibellina — соболь......................................186
Meles meles — барсук...........................................188
Mustela erminea — горностай....................................189
Mustela nivalis — ласка........................................190
Mustela putorius — лесной хорек................................191
Mustela sibirica — колонок.....................................192
Mustela vison — американская норка.............................192
Nyctereutes procyonoides — енотовидная собака..................193
Panthera pardus — леопард......................................194
Procyon lotor— енот-полоскун...................................194
Urocyon cinereoargenteus — серая лисица........................196
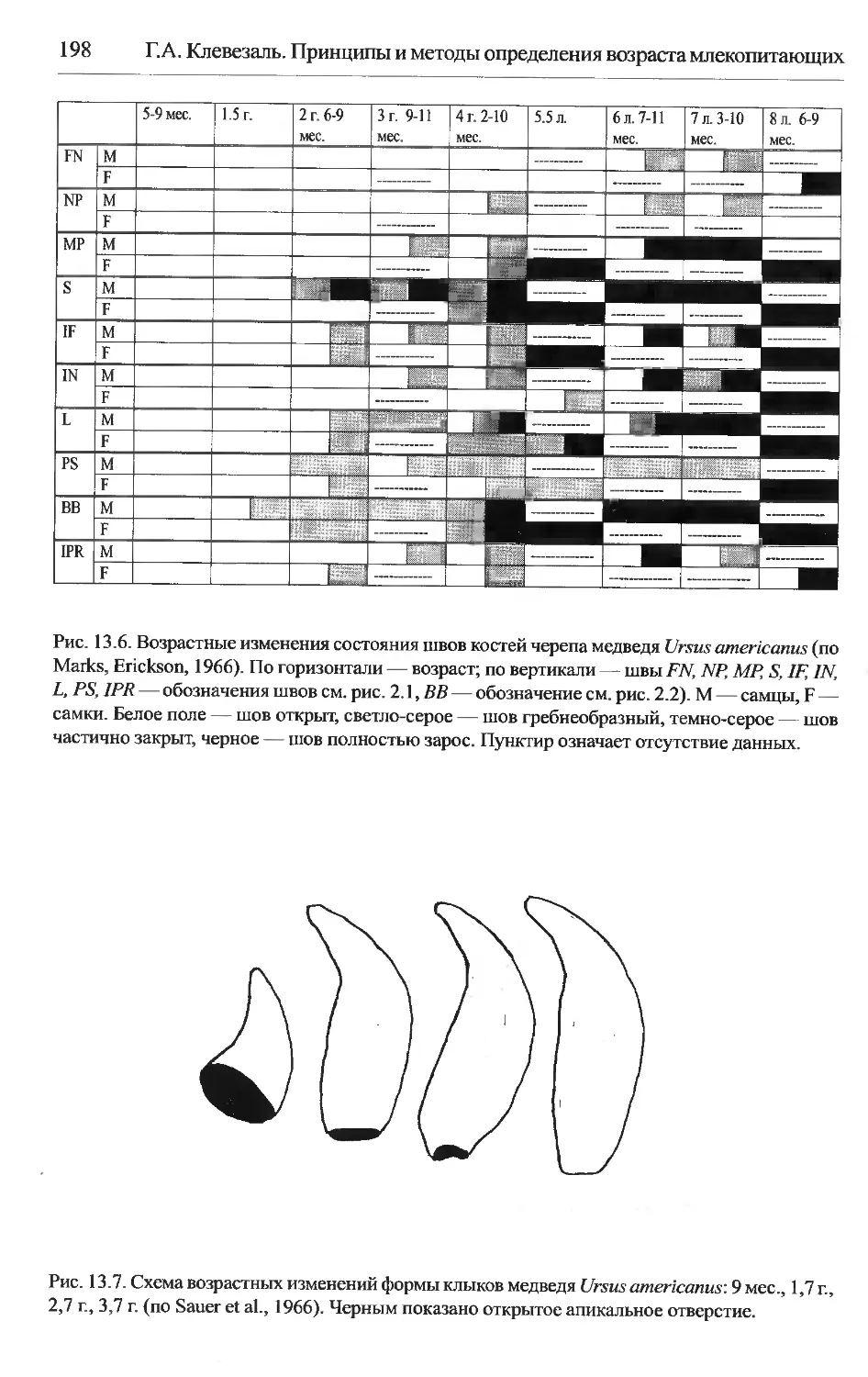
Ursus americanus — черный медведь..............................197
Ursus arctos — бурый медведь...................................199
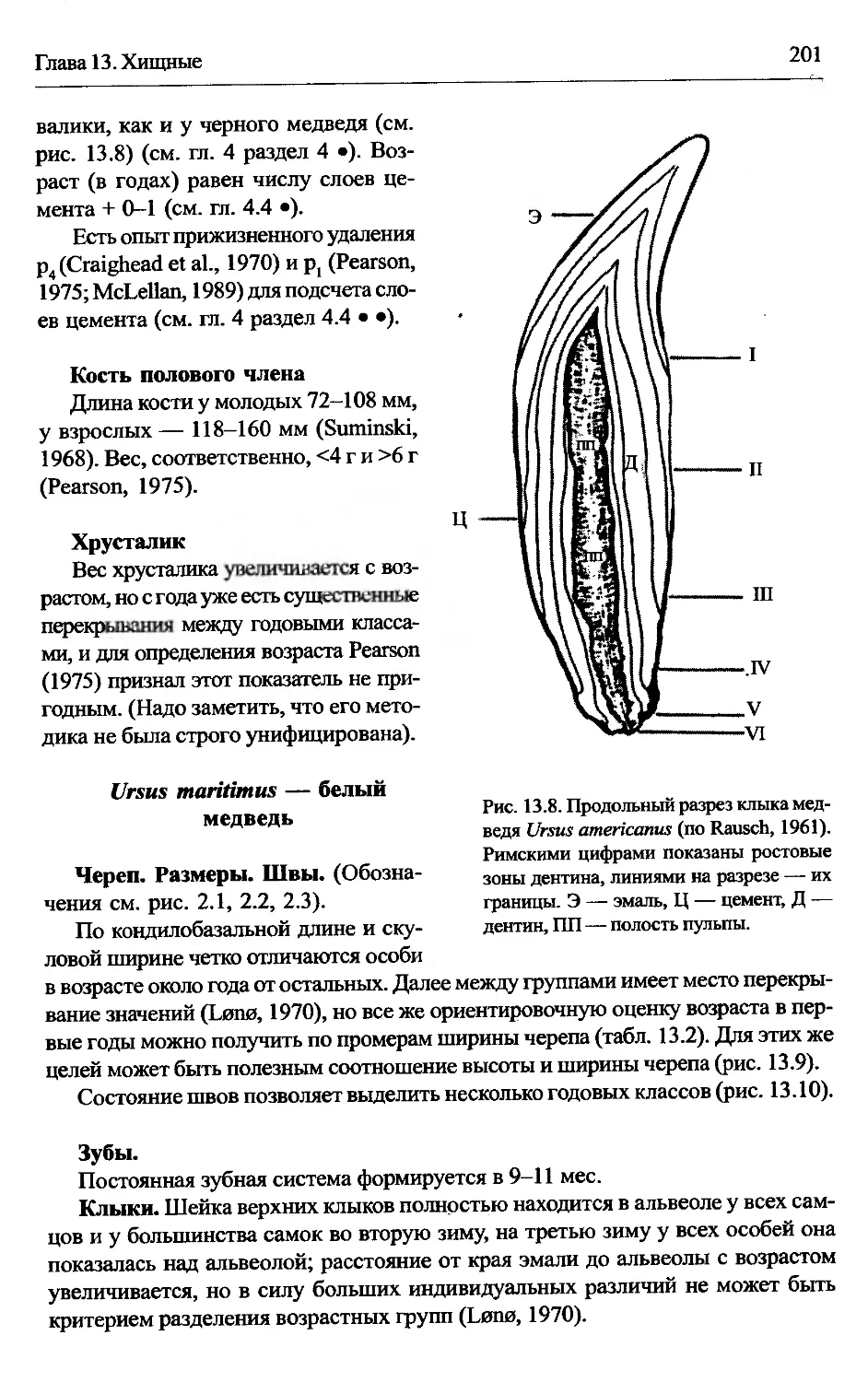
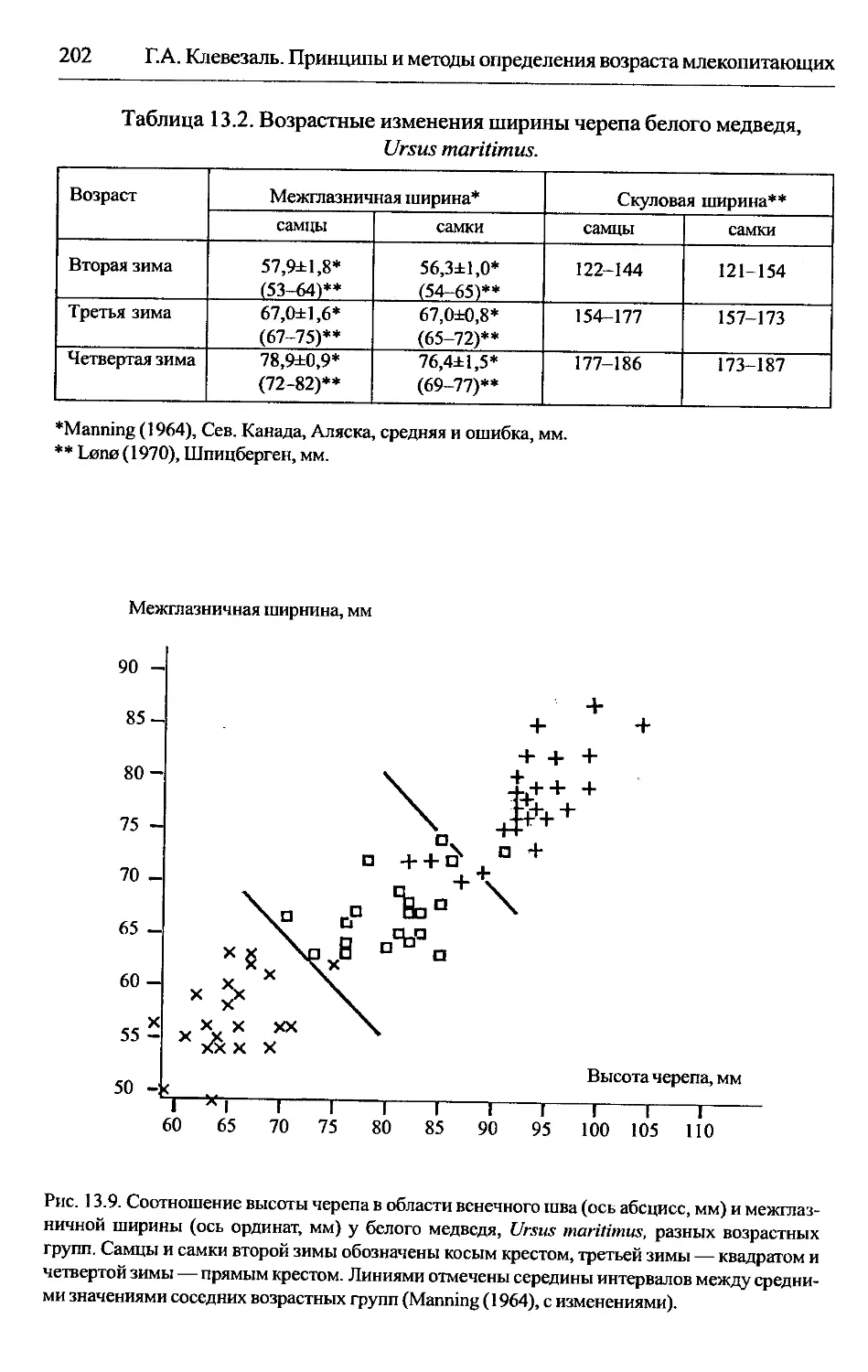
Ursus maritimus — белый медведь................................201
Vulpes vulpes — лисица.........................................204
Глава 14. Парнокопытные...............................................207
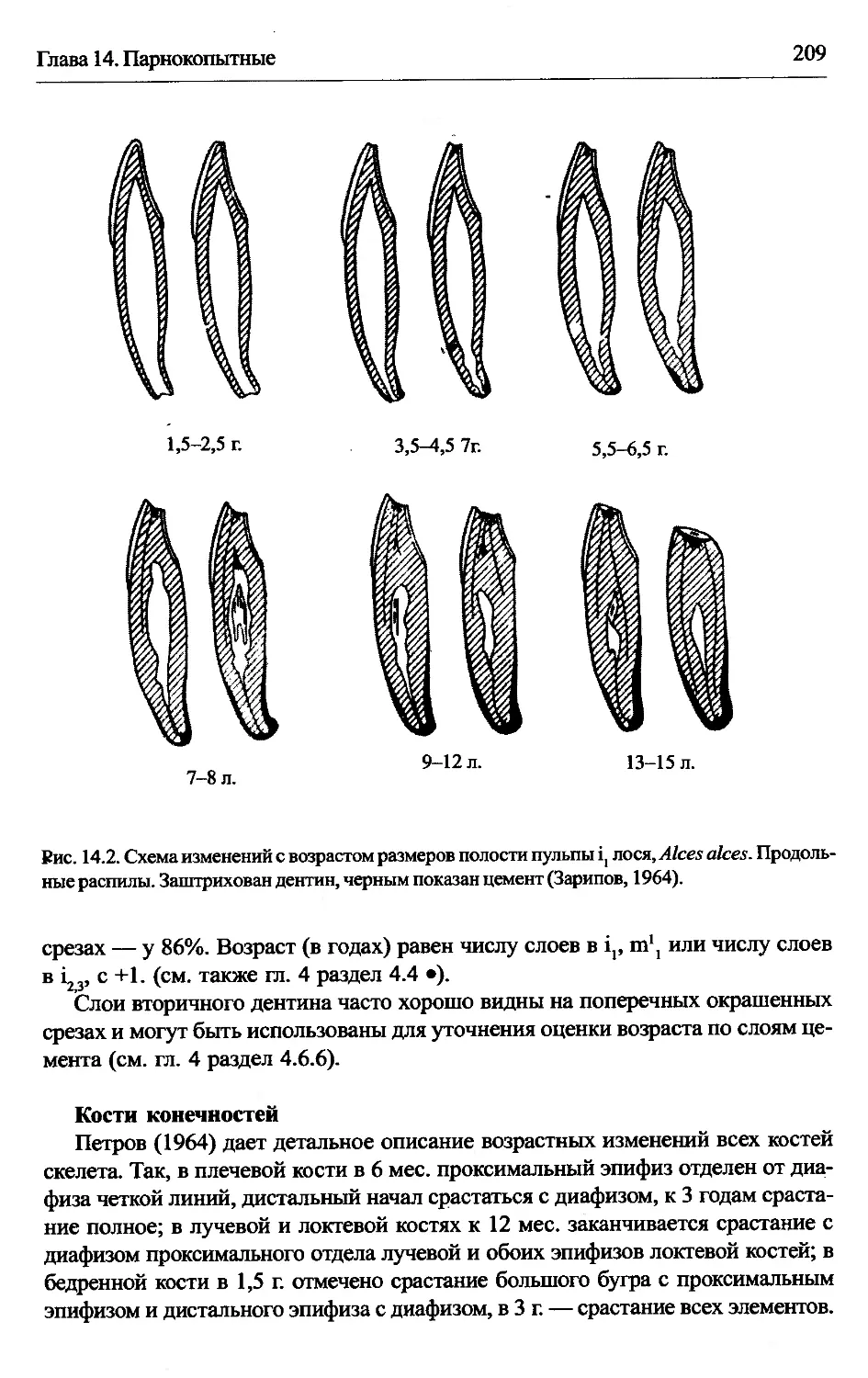
Alces alces — лось.............................................207
Antidorcas marsupialis — спрингбок.............................210
Antilocapra americana — вилорог................................210
Bison bison — бизон............................................211
Bison bonasus — зубр...........................................212
Capra — горные козлы...........................................213
Capreolus capreolus — косуля...................................214
Содержание U
Capricornis crispus — японский сероу..........................217
Cervus dama — лань............................................218
Cervus elaphus — благородный олень............................218
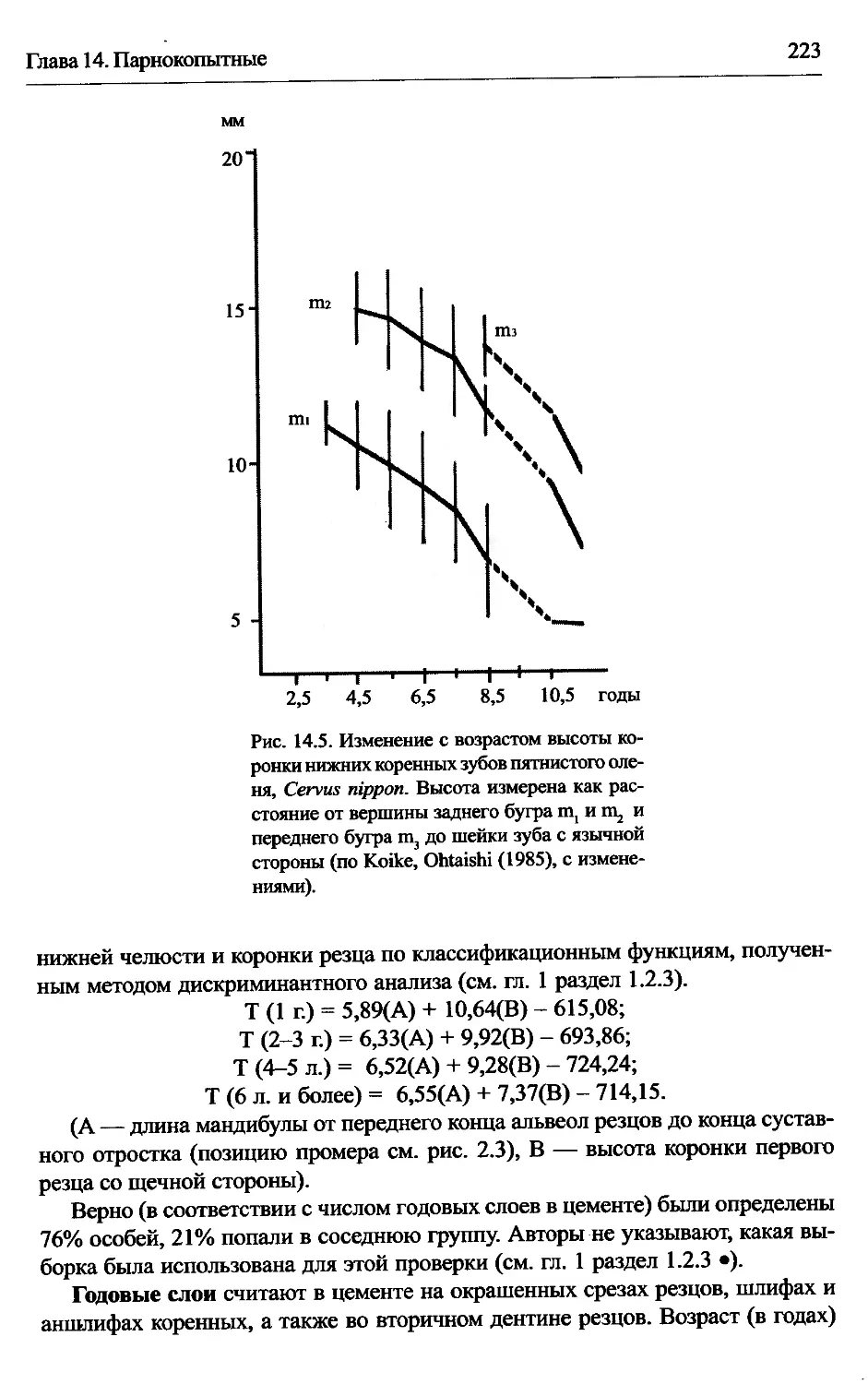
Cervus nippon — пятнистый олень...............................222
Giraffa Camelopardalis — жираф................................224
Nemorhaedus goral — горал.....................................225
Odocoileus hemionus — чернохвостый олень......................225
Odocoileus virginianus — белохвостный олень...................226
Oreamnos americanus — снежная коза............................227
Ovibos moschatus — овцебык....................................228
Ovis ammon — горный баран.....................................229
Ovis canadensis (Ovis dalli) — снежный баран..................230
Phacochoerus aethiopicus — бородавочник.......................230
Rangifer tarandus — северный олень............................231
Rupicapra rupicapra — серна...................................233
Saiga tatarica — сайгак.......................................233
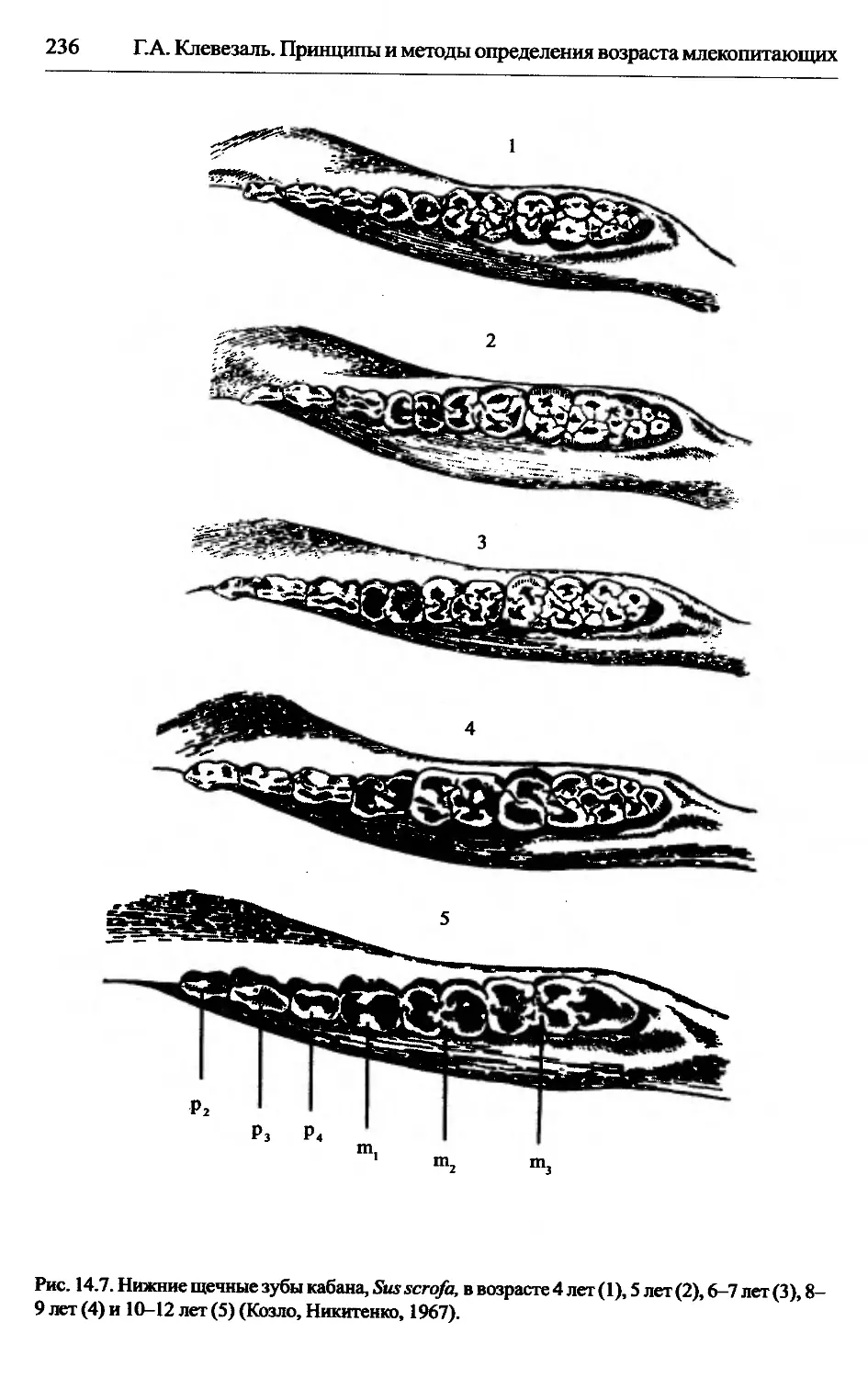
Sus scrofa — кабан............................................234
Tragelaphus scriptus — бушбок.................................238
Глава 15. Непарнокопытные...........................................239
Ceratotherium simum — белый носорог...........................239
Equus burchelli, E. zebra — зебры.............................240
Заключение ............................................................243
Литература ............................................................245
Указатель латинских названий животных..................................280
CONTENTS
Introduction....................................................................18
Part I. Methods of age determination............................................
Chapter 1. General principles................................................
1.1. Concepts and terms..................................................
1.2. Procedures of processing and presentation of data...................
1.2.1. Discrimination of age groups without complex calculations......
1.2.2. Regression analysis............................................
1.2.3. Discriminant analysis..........................................
1.2.4. Claster analysis...............................................
1.2.5. Comparison of multivariate methods with traditional ones.......
1.2.6. Choice of particular methods of age determination..............
Chapter 2. Age determination from skeleton characters........................
2.1. Size of skull.......................................................
2.2. Obliteration of skull sutures.......................................
2.2.1. Area of application...........................................
2.2.2. Morphological background......................................
2.2.3. Resolution capacity...........................................
2.2.4. Technique.....................................................
2.2.5. Variability of indices........................................
2.3. Form of skull.......................................................
2.4. Development of skull crests.........................................
2.5. Ossification of epiphyses of long bones.............................
2.5.1. Area of application...........................................
2.5.2. Morphological background......................................
2.5.3. Resolution capacity...........................................
2.5.4. Technique.....................................................
2.5.5. Variability of indices........................................
2.6. Vertebrae and pelvis ...............................................
2.7. Development of tubers on bones......................................
2.8. Size and form of baculum (os penis).................................
2.8.1. Area of application...........................................
2.8.2. Morphological background......................................
2.8.3. Resolution capacity...........................................
2.8.4. Technique.....................................................
2.8.5. Variability of indices........................................
2.8.6. Peculiarities of application for concrete groups of mammals...
2.8.6.1. Carnivores................................................
2.8.6.2. Rodents...................................................
Chapter 3. Age determination from teeth characters...........................
Terminology...........................................................
3.1. Permanent teeth eruption............................................
3.1.1. Area of application...........................................
3.1.2. Morphological background......................................
Contents 13
3.1.3. Resolution capacity............................................46
3.1.4. Technique..................................................... 47
3.1.5. Variability of indices.........................................48
3.1.6. Peculiarities of application for concrete groups of mammals....49
3.1.6.1. Carnivores................................................49
3.1.6.2. Artiodactyla..............................................49
3.1.7. History of using...............................................54
3.2. Teeth growth.........................................................54
3.2.1. Teeth growth in length.........................................55
3.2.2. Closure of apical foramen......................................55
3.2.3. Position of a tooth neck in relation to alveolus...............56
3.2.4. Root growth and cementum apposition............................56
3.3. Width of pulp cavity.................................................57
3.3.1. Area of application............................................57
3.3.2. Morphological background.......................................58
3.3.3. Resolution capacity............................................58
3.3.4. Technique......................................................61
3.3.5. Variability of indices.........................................62
3.3.6. Application for ungulates......................................64
3.3.7. History of using...............................................64
3.4. Teeth wear...........................................................64
3.4.1. Area of application............................................64
3.4.2. Morphological background.......................................65
3.4.3. Resolution capacity.......................................... 65
3.4.4. Technique......................................................66
3.4.5. Variability of indices.........................................67
3.4.6. Peculiarities of application for concrete groups of mammals....67
3.4.6.1. Carnivores................................................67
3.4.6.2. Perissodactyla........................................... 69
3.4.6.3. Artiodactyla..............................................69
3.4.6.4. Rodents...................................................75
3.4.6.5. Insectivores and Chiropterans.............................77
3.4.7. History of using...............................................78
Chapter 4. Age determination from annual layers in teeth and bone............79
4.1. Area of application..................................................79
4.2. Morphological background.............................................79
4.3. Resolution capacity..................................................82
4.4. Technique............................................................82
4.5. Variability of indices...............................................85
4.6. Peculiarities of application for concrete groups of mammals..........86
4.6.1. Insectivores................................................86
4.6.2. Chiropterans................................................86
4.6.3. Lagomorphs..................................................88
4.6.4. Rodents.....................................................88
4.6.5. Carnivores..................................................89
4.6.6. Ungulates...................................................91
4.7. History of using.....................................................92
Chapter 5. Age determination from horns’ characters..........................93
14
Klevezal G. A. Principles and methods of age determination of mammals
5.1 .Bovidae................................................................93
5.2 . Deers.................................................................95
Chapter 6. Age determination from a crystalline weight.........................97
6.1. Area of application....................................................97
6.2. Morphological background...............................................97
6.3. Resolution capacity....................................................97
6.4. Technique..............................................................98
6.5. Variability of indices................................................100
6.6. Peculiarities of application for concrete groups of mammals...........102
6.6.1. Carnivores......................................................102
6.6.2. Lagomorphs......................................................102
6.6.3. Rodents.........................................................103
6.6.4. Ungulates.......................................................106
6.6.5. Insectivores....................................................107
6.7. History of using......................................................107
Chapter 7. Rarely used methods................................................108
7.1. Age dependent changes of heart vessels and thymus..................108
7.2. Content of water......................................................108
7.3. Content of Sr-90......................................................108
7.4. Aspartic acid razemisation............................................109
7.5. Content of crystalline proteins.......................................109
Chapter 8. Methods of aging living animals....................................Ill
8.1. Size of body and its parts............................................Ill
8.2. Color. Pelt condition.................................................112
8.3. Possibilities of application of other methods for living animals.......112
8.3.1. Horns: form, size, annual rings.................................112
8.3.2. Teeth: eruption, wear, color....................................114
8.3.3. Annual layers in dentin and cementum of teeth...................115
8.3.4. Annual layers in phalanges......................................115
8.3.5. Form of baculum.................................................115
8.3.6. Skeleton: ossification of epiphyses.............................115
Part II. Application of the methods. Age determination of mammals of different systemat-
ic groups .........................................................................116
Chapter 9. Insectivora........................................................116
Blarina brevicauda...................................................116
Crocidura............................................................116
Erinaceus europaeus..................................................117
Galeniys pyrenaicus.............................................................................. 117
Sorex araiieus.......................................................117
Talpa europaea.......................................................118
Talpa micrura (Mogera robusta).......................................120
Chapter 10. Chiroptera........................................................121
Appearance and color........................................................121
Bones of patagium...........................................................121
Teeth. Wear.................................................................123
Pulp cavity.................................................................124
Annual layers...............................................................124
Contents
15
Chapter ll.L agomorpha.................................................125
Lepus californicus...............................................125
Lepus europaeus..................................................125
Lepus timidus....................................................127
Ochotona.........................................................127
Oryctolagus cuniculus............................................128
Sylvilagus.......................................................129
Chapter 12. Rodentia....................................................131
Allactaga and other Dipodidae....................................131
Apodemus agrarius................................................131
Apodemus flavicollis.............................................136
Apodemus sylvaticus..............................................136
Arvicola terrestris..............................................137
Castor...........................................................137
Citellus (=Spe rmophilus)........................................140
Clethriononiys...................................................142
Cricetus cricetus................................................146
Cynomys ludovicianus.............................................147
Dicrostonyx torquatus............................................148
Dryomis nitedula.................................................148
Eliomys quercinus............................................... 149
Ellobius talpinus................................................150
Erethizon dorsatum...............................................151
Gerbillus........................................................151
Glis glis........................................................152
Lagurus lagurus..................................................152
Lemmus lemmus....................................................153
Marmota baibacina, M. hobac, M. menzbieri, M. sibirica...........154
Marmota тонах....................................................156
Meriones.........................................................157
Microtus.........................................................158
Mus musculus.....................................................160
Muscardinus avellanarius.........................................161
Myocastor coypus.................................................161
Ondatra zibethicus...............................................162
Rattus...........................................................165
Rhombomys opimus............................................... 165
Sciurus carolinensis, S. niger...................................166
Sciurus vulgaris.................................................167
Sicista..........................................................169
Sigmodon hispidus................................................170
Spalax microphtalmus.............................................170
Tamias (= Eutamias)..............................................171
Tamiasciurus hudsonicus......................................... 172
Chapter 13. Carnivora...................................................173
Alopex lagopus...................................................173
Canis latrans....................................................173
Canis lupus......................................................174
16
Klevezal G.A. Principles and methods of age determination of mammals
Canis niesotnelas................................................116
Crocuta crocuta...................................................1П
Enhydra lutris....................................................YU
Fells caracal....................................................178
Felis catus, F. sylvestris.......................................179
Fells (=Puma) concolor...........................................179
Felis lynx...................................................... 180
Felis rufits.....................................................181
Gulo gulo........................................................181
Lutra canadensis.................................................182
Lutra lutra......................................................182
Martes americana.................................................184
Martesfoina, M.martes............................................184
Martes pennanti..................................................185
Martes zibellina.................................................186
Meles meles......................................................188
Mustela erminea..................................................189
Mustela nivalis..................................................190
Mustela putorius.................................................191
Mustela sibirica.................................................192
Mustela vison....................................................192
Nyctereutes procyonoides.........................................193
Panthera pardus..................................................194
Procyon lotor....................................................194
Urocyon cinereoargenteus.........................................196
Ursus americanus.................................................197
Ursusarctos......................................................199
Ursus maritimus..................................................201
Vulpes vulpes....................................................204
Chapter 14. Artiodactyla...............................................207
Alces alces......................................................207
Antidorcas marsupialis...........................................210
Antilocapra americana............................................210
Bison bison......................................................211
Bison bonasus....................................................212
Capra............................................................213
Capreolus capreolus..............................................214
Capricornis crispus..............................................217
Cervus dama......................................................218
Cervus elaphus...................................................218
Cervus nippon....................................................222
Giraffa Camelopardalis...........................................224
Nemorhaedus goral................................................225
Odocoileus hemionus..............................................225
Odocoileus virginianus...........................................226
Oreamnos americanus..............................................227
Ovibos moschatus.................................................228
Ovis ammon......................................................“229
Contents
17
Ovis nanadensis (Ovis dalli)......................................230
Phacochoerus aethiopicus..........................................230
Rangifer tarandus.................................................231
Rupicapra rupicapra...............................................233
Saiga tatarica....................................................233
Sus scrofa........................................................234
Tragelaphus scriptus..............................................238
Chapter 15. Perissodactyla..............................................239
Ceratotheriian sinium.............................................239
Equus burchelli, E. zebra.........................................240
Conclusion .................................................................243
Bibliography ...............................................................245
Index of Latin names........................................................280
ПРЕДИСЛОВИЕ
Необходимость оценки возраста животных при проведении разного рода ис-
следований, причем не только зоологических, не нуждается в обосновании. В
прошлом веке поискам точных методов определения возраста было посвящено
много работ, но направление поисков резко сузилось после обнаружения у мор-
ских, а затем и у наземных млекопитающих годовых слоев в кости и тканях
зуба. Анализ этих слоев стал преобладающим методом определения возраста, и
много работ было посвящено оценке точности этого метода и анализу воспроиз-
водимости оценок возраста отдельных видов. Оказалось, что хотя во многих
случаях анализ годовых и внутригодовых слоев открывает более широкие воз-
можности, нежели просто определение числа прожитых лет, есть немало приме-
ров, когда метод этот ненадежен или вообще не пригоден для определения воз-
раста. Кроме того, в ряде конкретных исследований достаточно выделить возра-
стные группы или даже просто разделить выборки на молодых и взрослых, а
иногда необходимо оценить, хотя бы приблизительно, возраст конкретной особи
при наличии лишь каких-то отдельных частей тела, где нет годовых слоев. Во
всех этих случаях приходится обращаться к старым, традиционным приемам,
изложенным в работах по биологии конкретных видов. Обзоры таких работ были
даны в статье Morris (1972) и в предназначенных главным образом охотоведам
сводках Larson, Taber (1980) и Habermehl (1985). Многие работы по тем или
иным причинам в эти сводки не вошли.
Цель данной книги, во-первых, познакомить заинтересованных читателей с
основными методами определения возраста млекопитающих, с возможностями
и ограничениями использования этих методов, а во-вторых, предоставить спра-
вочные данные для определения возраста разных видов, чтобы исследователь
мог судить, на какую точность оценки возраста особей данного вида он может
рассчитывать при анализе имеющегося у него материала. Рассмотрение ограни-
чено, за редким исключением, млекопитающими умеренной зоны. Не рассмат-
риваются морские млекопитающие (китообразные и ластоногие), для определе-
ния возраста которых используют, главным образом, годовые слои.
Книга состоит из двух частей. В первой части дано описание методов, во
второй части — приведены критерии для определения возраста особей конк-
ретных видов разными методами. Исследователю, который нашел во второй
части конкретные критерии оценки возраста интересующего его вида методом,
применимым к его материалу, рекомендуется ознакомиться с плюсами и мину-
сами этого метода, описанными в части первой. В тех случаях, когда такое
ознакомление просто необходимо, во второй части даны ссылки на соответ-
ствующие разделы первой части. Особо важные положения в главах первой
части выделены жирным шрифтом. Кроме того, черным кружком в начале аб-
заца отмечены те положения, которые читателю необходимо учитывать при ра-
боте с конкретными видами. Соответствующие ссылки на эти положения даны
в части II.
Предисловие
19
Виды, по определению возраста которых я смогла найти лишь отрывочные
данные, не включены в часть II, но большинство из них упоминается в части 1,
их можно найти по латинскому названию вида.
Указывая видовые названия, разные авторы пользуются разными таксономи-
ческими справочниками. Как правило, я даю то название, которое приводит
автор используемой работы. Однако, в отдельных случаях я полагала необходи-
мым дать синонимы, следуя сводкам Соколова (1977, 1979).
Я признательна всем сотрудникам лаборатории постнатального онтогенеза,
помогавшим мне в работе. Я крайне благодарна Н.Л. Крушинской, без постоян-
ной помощи которой книга не могла бы быть написана, а также М.В. Мине и
А.Б. Савинецкому за ценные замечания по рукописи. И большое спасибо со-
трудникам библиотеки Отделения общей биологии, особенно Л.Н. Митрохиной
и М.П. Токаренко, за большую помощь в получении литературы.
ЧАСТЬ I. МЕТОДЫ ОПРЕДЕЛЕНИЯ ВОЗРАСТА
ГЛАВА 1. ОБЩИЕ ПРИНЦИПЫ
1.1. Понятия и термины
Определение возраста понимается как любое суждение о возрасте особи. Если
мы ранжируем особей выборки от наиболее молодых до наиболее старых, мы
определяем относительный возраст этих особей. Если мы относим особей вы-
борки к возрастным группам и для каждой группы указываем возрастной интер-
вал в абсолютном выражении, мы тем самым определяем абсолютный возраст
особей. При этом речь может идти о возрастных группах и о годовых классах.
Возрастная группа объединяет особей того возраста, который соответствует
декларируемой оценке. Эту оценку задает исследователь. Например, взятую зи-
мой выборку животных, размножающихся весной, можно разделить на 2 возра-
стные группы: сеголетков и особей не моложе года, или на 3 возрастные группы:
сеголетков, годовиков и особей двух лет и старше. Годовой класс объединяет
особей, возраст которых различается менее чем на один год, например, особей,
достигших 5 лет, но не достигших 6 лет.
Следует различать правильность (верность, надежность) и точность оп-
ределения возраста. Точность—это декларированная степень приближения оценки
к абсолютному истинному возрасту особей. Метод определения возраста по го-
довым слоям называют точным, поскольку считается, что он позволяет опреде-
лить возраст особей с точностью до года или даже до месяца, если период раз-
множения короткий и дата гибели известна. Правильность — это мера того, на-
сколько велика вероятность ошибочного отнесения особи к возрастной группе
или годовому классу. Она определяет пригодность метода. Например, метод,
позволяющий разделить выборки на две возрастные группы по какому-то при-
знаку нельзя назвать точным, поскольку старшая группа включает особей разно-
го календарного возраста. Однако это разделение может быть правильным (на-
дежным), если все сеголетки попадают в первую группу, а все старшие особи
— во вторую, или может быть не вполне надежным, если лишь большинство
особей попадает в свою возрастную группу. Определение возраста по годовым
слоям, будучи точным методом по определению, может дать неправильную оцен-
ку возраста, оказаться ненадежным, например, если есть опасность принять не-
которые дополнительные слои за годовые.
Методы определения возраста обычно делят на две группы: 1 — связанные с
ростом (увеличением размеров особи) и 2 — базирующиеся на изменениях, обус-
ловленных длительностью периода от рождения особи и не зависящих от роста.
Методы первой группы используют показатели размеров тела и его частей, око-
стенение костей скелета, рост зубов и др. В отдельную подгруппу следует выде-
лить метод определения возраста по регистрирующим структурам (рогам, тка-
ням зуба и кости), где образуются и сохраняются следы сезонных изменений
Глава 1. Общие принципы
21
темпа роста. Ко второй группе относятся методы определения возраста по степе-
ни стертости зубов, изменению соотношения изомеров аспарагиновой кислоты,
накоплению в организме радионуклидов.
Все критерии возраста, связанные с ростом, имеют следующие общие черты:
— они позволяют разграничить возрастные группы до наступления половой
зрелости, т.е. до того времени, когда быстрый рост заканчивается; после наступ-
ления половой зрелости межиндивидуальные различия показателей становятся
больше, чем различия между возрастными группами и надежность показателя
возраста резко уменьшается;
- межпопуляционные различия в применимости показателей возраста соот-
ветствуют межпопуляционным различиям в темпе роста всего организма, и на-
дежность этих показателей различна для северных и южных популяций: в юж-
ных популяциях период быстрого роста заканчивается, как правило, в более ран-
нем возрасте, чем в северных, и соответственно надежность показателя возраста
уменьшается в более раннем возрасте.
Помимо того, методы можно разделить на: 1 — макроморфологические (ис-
пользующие внешние признаки, признаки скелета, характер зубной системы,
вес хрусталика и т.д.), 2 — микроморфологические (использующие годовые
слои в зубах и кости), 3 — биохимические (использующие содержание белка в
хрусталике, соотношение изомеров аспарагиновой кислоты, накопление радио-
нуклидов). Можно классифицировать методы и по системам органов или от-
дельным органам. Последняя классификация представляется наиболее удобной
для практического использования и потому принята ниже при описании методов
(главы 2-6).
1.2. Способы обработки и представления данных
При использовании большинства показателей возраста надежность выделе-
ния возрастных групп тем больше, чем короче как период размножения особей
данной популяции в течение года, так и период сбора материала.
1.2.1. Выделение возрастных групп способами, не требующими
сложных вычислений
1.2.1.1. Если период размножения и период сбора материала не велики, вы-
борку удается разделить на возрастные группы, построив кривую распределения
особей по величине признака и определив границы между возрастными группа-
ми по минимумам кривой распределения. Этот прием удобен для количествен-
ных, реже — для качественных признаков (если оценки выражены в баллах).
Для сглаживания мелких изменений частот и более четкого выявления границ
между группами можно использовать «метод скользящей средней»: последова-
тельно вычисляют средние частоты для трех (или пяти) соседних интервалов при-
знака, для получения каждой следующей средней сдвигаясь на один интервал
22
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
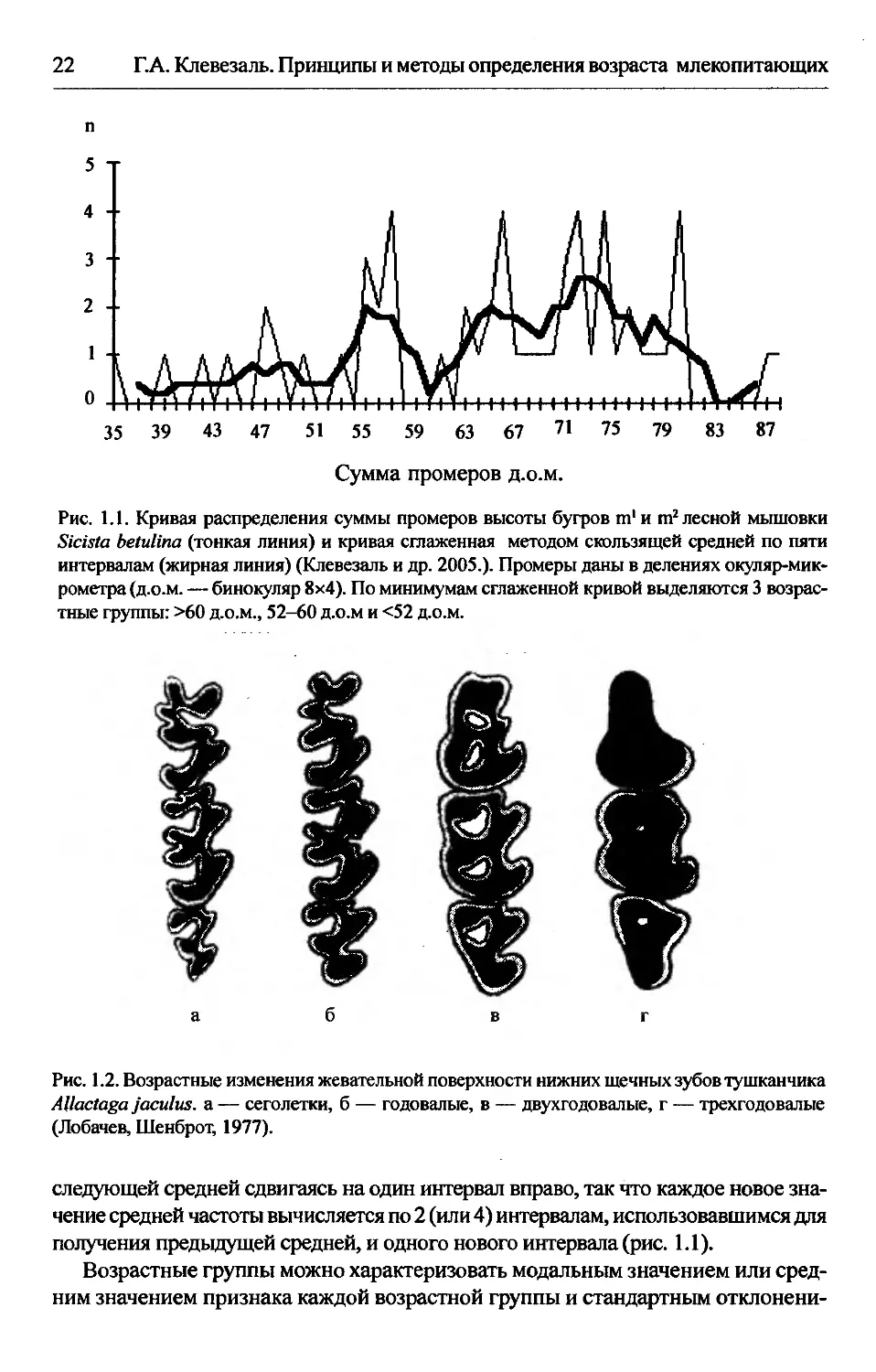
Рис. 1.1. Кривая распределения суммы промеров высоты бугров т1 и т2 лесной мышовки
Sicista betulina (тонкая линия) и кривая сглаженная методом скользящей средней по пяти
интервалам (жирная линия) (Клевезаль и др. 2005.). Промеры даны в делениях окуляр-мик-
рометра (д.о.м. — бинокуляр 8x4). По минимумам сглаженной кривой выделяются 3 возрас-
тные группы: >60 д.о.м., 52-60 д.о.м и <52 д.о.м.
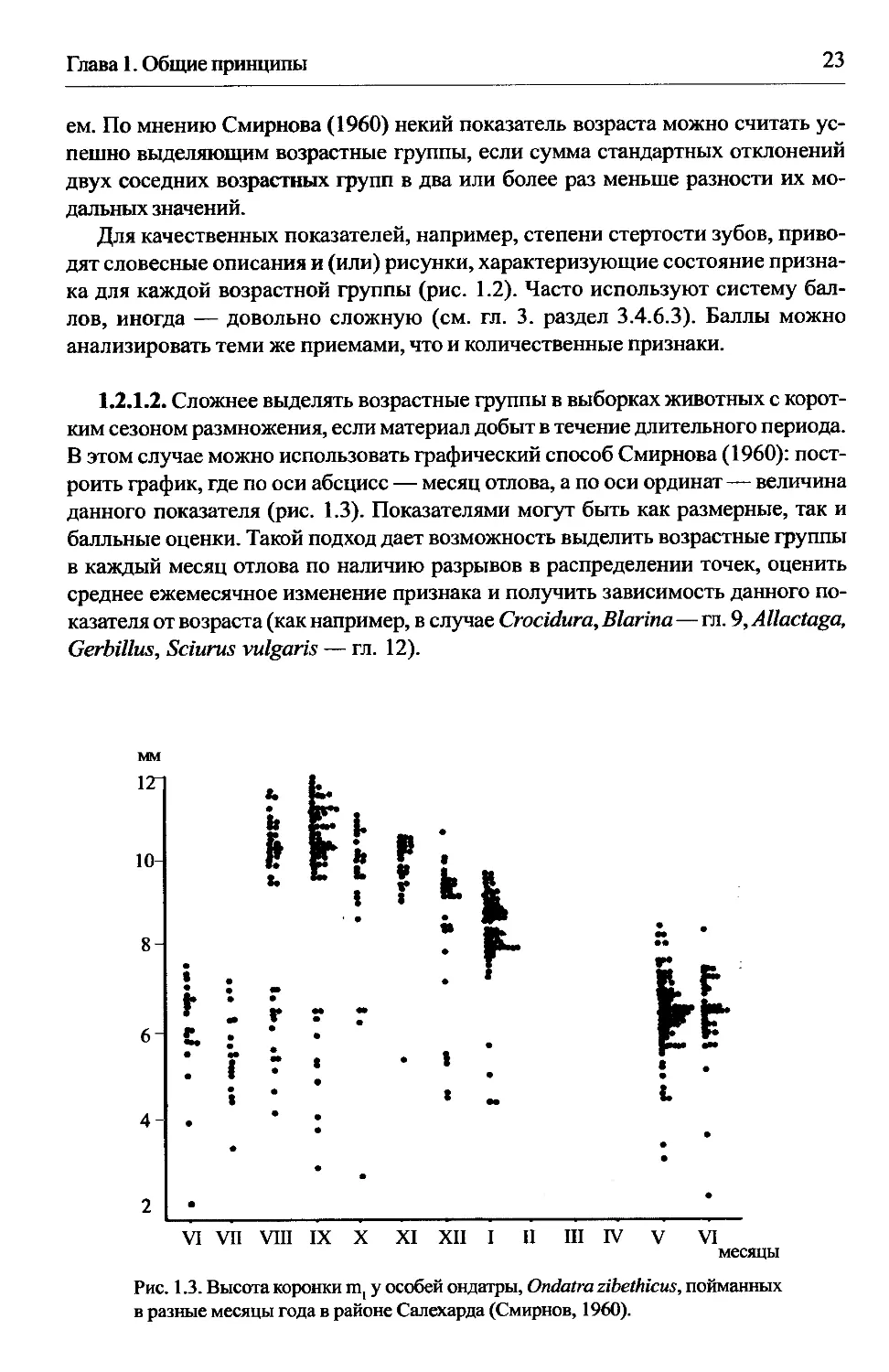
Рис. 1.2. Возрастные изменения жевательной поверхности нижних щечных зубов тушканчика
Allactaga Jaculus. а — сеголетки, б — годовалые, в — двухгодовалые, г — трехгодовалые
(Лобачев, Шенброт, 1977).
следующей средней сдвигаясь на один интервал вправо, так что каждое новое зна-
чение средней частоты вычисляется по 2 (или 4) интервалам, использовавшимся для
получения предыдущей средней, и одного нового интервала (рис. 1.1).
Возрастные группы можно характеризовать модальным значением или сред-
ним значением признака каждой возрастной группы и стандартным отклонени-
Глава 1. Общие принципы
23
ем. По мнению Смирнова (1960) некий показатель возраста можно считать ус-
пешно выделяющим возрастные группы, если сумма стандартных отклонений
двух соседних возрастных групп в два или более раз меньше разности их мо-
дальных значений.
Для качественных показателей, например, степени стертости зубов, приво-
дят словесные описания и (или) рисунки, характеризующие состояние призна-
ка для каждой возрастной группы (рис. 1.2). Часто используют систему бал-
лов, иногда — довольно сложную (см. гл. 3. раздел 3.4.6.3). Баллы можно
анализировать теми же приемами, что и количественные признаки.
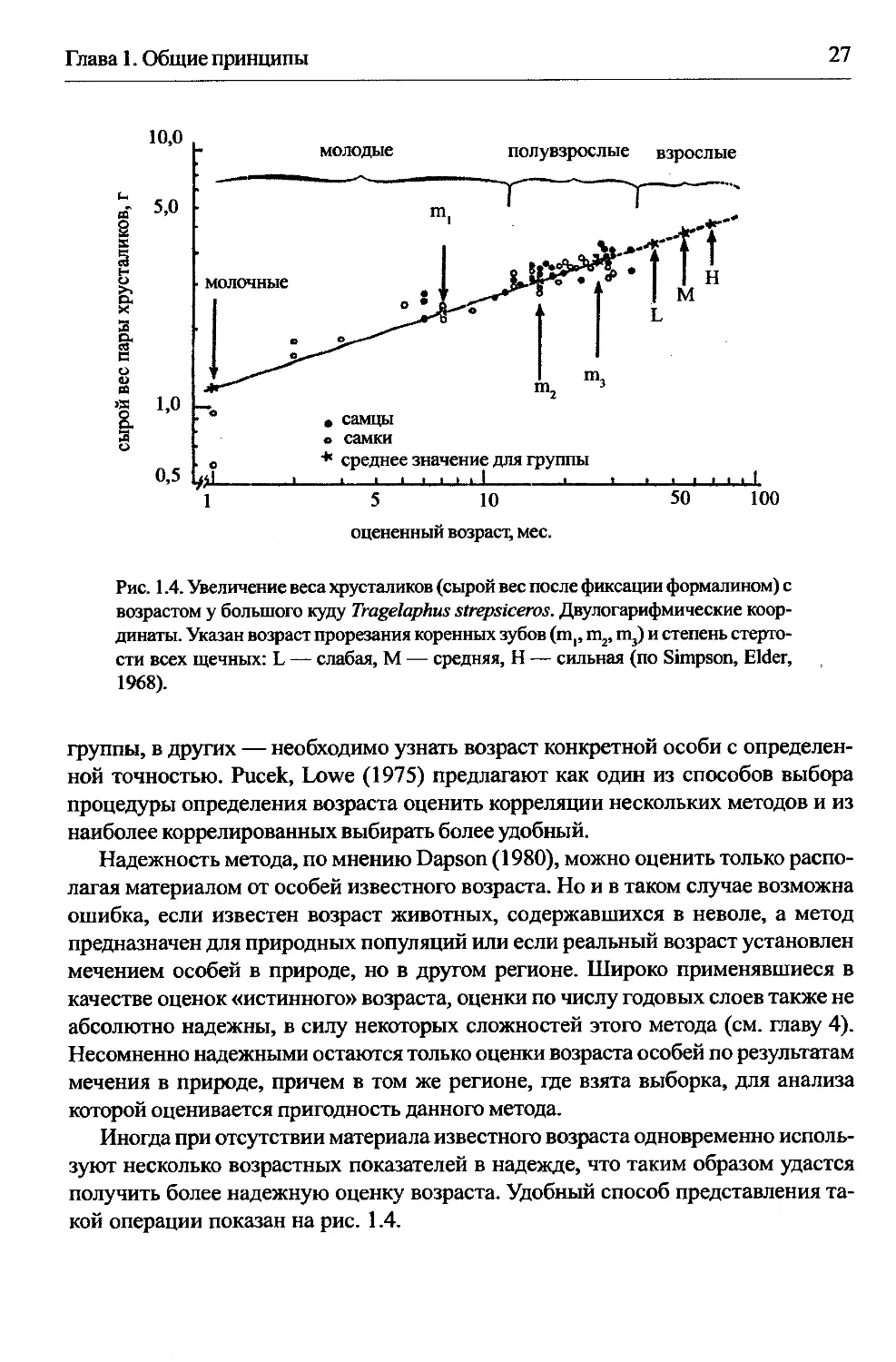
1.2.1.2. Сложнее выделять возрастные группы в выборках животных с корот-
ким сезоном размножения, если материал добыт в течение длительного периода.
В этом случае можно использовать графический способ Смирнова (1960): пост-
роить график, где по оси абсцисс — месяц отлова, а по оси ординат — величина
данного показателя (рис. 1.3). Показателями могут быть как размерные, так и
балльные оценки. Такой подход дает возможность выделить возрастные группы
в каждый месяц отлова по наличию разрывов в распределении точек, оценить
среднее ежемесячное изменение признака и получить зависимость данного по-
казателя от возраста (как например, в случае Crocidura, Blarina — гл. 9, Allactaga,
Gerbillus, Sciurus vulgaris — гл. 12).
Рис. 1.3. Высота коронки m, у особей ондатры, Ondatra zibethicus, пойманных
в разные месяцы года в районе Салехарда (Смирнов, 1960).
24
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
1.2.2. Регрессионный анализ
Получение уравнения регрессии какого-то показателя (например, веса хрус-
талика глаза) по возрасту — удобный прием для оценки возраста особей.
1.2.2.1. При построении линии регрессии, характеризующей зависимость меж-
ду у и х, не безразлично, какая из двух переменных принимается за независи-
мую. Dapson (1980) специально разбирает этот вопрос, рекомендуя использо-
вать в качестве независимой переменной показатель менее изменчивый и/или с
меньшей ошибкой измерения, применительно к методам оценки возраста — воз-
раст. Большинство исследователей так и делают. Величина признакау представ-
ляется как функция от возраста х, у = f(x). Поскольку, однако, требуется, как
правило, определить возраст, т.е. считать х по значению у, необходимо произвес-
ти алгебраическое преобразование уравнения. Для уравнения прямолинейной рег-
рессии вместо у = ах + b получим х — (у- Ь)/а. Доверительный интервал при
этом будет шире, но ошибка все же меньше, чем для линии регрессии х по у,
если за независимую переменную брать величину признака у и использовать
возраст как зависимую переменную х (Dapson, 1980).
Мы проверили это положение при построении зависимости высоты коронки
зубов северного оленя, Rangifer tarandus, от возраста, определенного по годо-
вым слоям, т.е. на примере, когда трудно было сопоставить величину ошибок из-
мерения хну (поскольку при определении возраста по слоям тоже может быть
ошибка, см. главу 4). Действительно, и в этом случае ошибка оценки возраста
оказалась меньше при использовании алгебраического преобразования уравне-
ния, в котором возраст —независимая переменная, чем при использовании урав-
нения, где возраст был зависимой переменной (Клевезаль, Соколов, 2004).
• Для удобства пользователя, в главах части II, приводя уравнение
регрессии, которое в оригинальной работе описывается зависимостью у
от х, я преобразую его, представляя х как функцию у.
1.2.2.2. Поскольку параметры зависимости возрастного показателя от возра-
ста могут меняться (и часто меняются) с возрастом, уравнение регрессии можно
использовать только для того интервала возраста, на котором оно получено.
Для проверки пригодности полученного уравнения следует брать не те дан-
ные, по которым уравнение построено. Хорошее совпадение, полученное по тем
же данным, не свидетельствует о пригодности уравнения для оценки возраста
особей другой выборки даже из той же популяции.
1.2.3. Дискриминантный анализ
Этот прием, во-первых, дает возможность из множества признаков выделить
те, что наилучшим образом различают возрастные группы, а во-вторых, может
позволить определять возраст отдельных особей при использовании нескольких
показателей с большей надежностью, чем при использовании каждого из этих
показателей в отдельности. Для его применения, однако, необходимо иметь
Глава 1. Общие принципы
25
выборку, уже разделенную на возрастные группы. Используя такую выборку, с
помощью дискриминантного анализа получают уравнения классификационных
функций, число которых зависит от числа выделяемых возрастных групп. Для
определения принадлежности отдельной особи к какой-то возрастной группе
нужно подставить ее параметры в каждое из этих уравнений и отнести животного
к той группе, для которой результат (Т) будет наибольшим.
Например, MacCracken, Van Ballenberghe (1987) предложили распознавать
молодых и взрослых лосей, A. alces, зимой по экскрементам, проведя дискри-
минантный анализ размеров «орешка»: объема (длина, умноженная на наиболь-
шую ширину, умноженная на ширину при повороте на 90° от наибольшей, мм3)
и отношения первой и второй ширины. Они получили такие классификационные
уравнения:
Т (взрослые самцы) = 0,0066 • объем + 3093,499 • отношение - 1621,945;
Т (взрослые самки) = 0,0025 • объем + 3120,612 • отношение - 1616,589;
Т (сеголетки) = — 0,0006 • объем + 3342,846 • отношение - 1835,457.
При проверке оказалось, что таким методом возрастные группы выделялись
надежнее, чем только по объему «орешка».
По данным Castro-Campillo et al. (1993) у гоферов Thomomys utnbrinus четыре
возрастные группы по промерам черепа выделяются дискриминантным анали-
зом лучше, чем по степени закрытия черепного шва. При этом для самцов и для
самок наилучшими дискриминантами оказались разные признаки.
Разделение итальянской выборки лисиц V vulpes, взятой в январе-апреле, на
сеголетков и старших, без перекрывания не получавшееся ни по весу хрустали-
ка (у, мг), ни по относительной ширине полости пульпы верхнего клыка (х, доли
общей ширины клыка), было надежным при использовании дискриминантного
анализа. Классификационные уравнения:
Т (сеголетки) = 136,5 х + 0,909 у - 124,9;
Т (взрослые) = 61,6 х + 1,073 у - 136,8 (Cavallini, Santini, 1995).
• При проверке классификационных функций важно, чтобы набор про-
верочных данных был иным, чем тот, который использовали для получе-
ния функций. Об этом пишут все руководства, но мало кто этому правилу
следует. Хорошая дискриминация, полученная при проверке теми же дан-
ными, не гарантирует хорошей дискриминации при использовании друго-
го набора данных. Для выяснения, насколько хорошо работают классификаци-
онные функции, их проверку следует проводить на других экземплярах.
Например, классификационные функции были получены для определения
возраста лесной мышовки Карелии, Sicista betulina, по комплексу признаков
стертости верхних коренных зубов (Клевезаль и др., 2005). При проверке этих
функций по той же самой выборке 98% особей оказались в своих возрастных
группах, при проверке по выборке из той же популяции, но взятой в другой
год — только 79%.
Если в работе не указано, какую выборку использовали для проверки, то
вывод авторов о преимуществе данного метода определения возраста перед дру-
гими, нельзя считать вполне обоснованным.
26
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
1.2.4. Кластерный анализ
Этот прием целесообразно использовать в ряде случаев для разделения возраст-
ных групп по сумме признаков (Пузаченко, 1991). При разделении возрастных групп
лесных мышовок (сеголетки, годовики и старшие) по оцениваемым в баллах при-
знакам стертости бугров и боковых гребней коренных зубов (Клевезаль и др., 2005)
этот метод дал лучшие результаты, чем распределение особей по каждому из этих
признаков (85% правильно определенных по сравнению с 75-78% по каждому
из признаков). Существенное достоинство кластерного анализа состоит в том,
что при его применении не встает вопрос, в какую группу включать «промежу-
точных» особей. Некоторые сложности возникают в связи с определением соот-
ветствия кластеров возрастным группам. Их, однако, легко избежать, построив
кривую распределения особей по сумме оценок признаков и сопоставив поло-
жение одних и тех же особей на дендрограмме и на кривой распределения.
1.2.5. Сравнение традиционных и многомерных методов
Не всегда методы многомерного анализа дают лучший результат, чем одномер-
ные. Например, разделение самцов лесной куницы, М. martes, на две (сеголетки и
старшие) и три (сеголетки, годовики и старшие) возрастные группы только по пози-
ции лобных валиков и разделение на основании кластерного анализа четырех при-
знаков (с добавлением состояния межносового шва, состояния стреловидного гребня
и стертости резцов) дали практически одинаковый результат (Клевезаль и др., 2005).
Goodyear (1983) при исследовании мышей Arvicanthis abyssinicus оценил степень
стертости каждого зуба, проанализировал данные методами факторного, клас-
терного и дискриминантного анализов и получил возрастные группы, сходные с
выделенными традиционным методом визуального осмотра зубов.
Yu, Lin (1999) использовали дискриминантный анализ для выделения возрас-
тных групп в популяции крыс Niviventer coxingi, где размножение шло в течение
всего года и материал собирали круглый год. По развитию и степени стертости
зубов они выделили пять возрастных групп, выбрали семь признаков размеров
тела и черепа для дискриминантного анализа, сравнили дистанции Махаланоби-
са между выделенными группами и сделали вывод, что разрывы есть между
первыми четырьмя группами, но не между четвертой и пятой. Результат много-
мерного анализа в данном случае свидетельствует, что средние показатели пер-
вых трех групп различаются и отличаются от показателей четвертой, но не дает
информации о степени перекрывания значений. Вероятно, для зоологического
анализа было бы более информативным представить графически распределе-
ние материала по выделенным возрастным группам.
1.2.6. Выбор конкретного метода для оценки возраста
Выбор должен определяться задачей исследования и наличием соответствую-
щего материала. В одних случаях достаточно разделить выборку на возрастные
Глава 1. Общие принципы
27
Рис. 1.4. Увеличение веса хрусталиков (сырой вес после фиксации формалином) с
возрастом у большого куду Tragelaphus strepsiceros. Двулогарифмические коор-
динаты. Указан возраст прорезания коренных зубов (тр т2, т3) и степень стерто-
сти всех щечных: L — слабая, М — средняя, Н — сильная (по Simpson, Elder,
1968).
группы, в других — необходимо узнать возраст конкретной особи с определен-
ной точностью. Pucek, Lowe (1975) предлагают как один из способов выбора
процедуры определения возраста оценить корреляции нескольких методов и из
наиболее коррелированных выбирать более удобный.
Надежность метода, по мнению Dapson (1980), можно оценить только распо-
лагая материалом от особей известного возраста. Но и в таком случае возможна
ошибка, если известен возраст животных, содержавшихся в неволе, а метод
предназначен для природных популяций или если реальный возраст установлен
мечением особей в природе, но в другом регионе. Широко применявшиеся в
качестве оценок «истинного» возраста, оценки по числу годовых слоев также не
абсолютно надежны, в силу некоторых сложностей этого метода (см. главу 4).
Несомненно надежными остаются только оценки возраста особей по результатам
мечения в природе, причем в том же регионе, где взята выборка, для анализа
которой оценивается пригодность данного метода.
Иногда при отсутствии материала известного возраста одновременно исполь-
зуют несколько возрастных показателей в надежде, что таким образом удастся
получить более надежную оценку возраста. Удобный способ представления та-
кой операции показан на рис. 1.4.
ГЛАВА 2. ОПРЕДЕЛЕНИЕ ВОЗРАСТА
ПО ПРИЗНАКАМ СКЕЛЕТА
2.1. Размеры черепа
По размерам черепа, как и по любому признаку, непосредственно связанно-
му с ростом особи, можно определять относительный возраст особей или выде-
лять возрастные группы среди растущих животных в период до достижения по-
ловой зрелости и отделять молодых от взрослых. У многих видов млекопитаю-
щих рост продолжается, хотя и в более низком темпе, и после достижения поло-
вой зрелости. Так, череп пумы, Felis concolor, растет в течение большей части
жизни: у самцов до 7-9 лет, у самок до 5-6 лет (Gay, Best, 1996). Однако, ме-
жиндивидуальные различия размеров у одновозрастных половозрелых особей,
как правило, превышают изменения размеров черепа с возрастом, что не дает
возможности выделять возрастные группы.
В ряде исследований было показано, что возрастные группы выделяются луч-
ше, если использовать не длину черепа (общую или кондилобазальную), а его
ширину (см. гл. 13 Lutra canadensis, Ursus americanus, U. arctos, U. maritimus),
соотношение длины и ширины черепа (см. гл. 13 Felis caracal) или отношение
ширины разных частей черепа (см. гл. 13 Martes zibellina, Mustela nivalis, Enhydra
lutris).
По величине отношения кондилобазальной длины или скуловой ширины к дли-
не зубного ряда выделяют несколько возрастных групп у сусликов (см. гл. 12
Citellus). Маркарян (1989) таким способом сумел выделить 5 возрастных групп до
возраста 30 мес. у хомяка Mesocricetus brandti. При таком подходе уменьшается
ошибка за счет индивидуальной изменчивости размеров одновозрастных особей,
поскольку длина зубного ряда достигает окончательных размеров после появле-
ния постоянной зубной системы и далее с возрастом не увеличивается.
Как межиндивидуальные, так и межпопуляционные различия размеров чере-
па очень велики, поэтому эти показатели возраста крайне не точны, и могут слу-
жить лишь как дополнительные.
Чаще, однако, при оценке возраста используют не размеры черепа, а степень
зарастания черепных швов и общую скульптуру поверхности черепа (см. ниже).
2.2. Зарастание швов черепа
2.2.1. Область применения
Метод пригоден для любых видов млекопитающих, но чаще используется
для хищных, а также для зайцеобразных (см. гл. 11 Lepus europaeus и Sylvilagus)
и грызунов, имеющих все зубы с постоянным ростом, например, для гоферов
Глава 2. Определение возраста по признакам скелета
29
Thomomys bottae (Daly, Patton, 1986). Его можно рекомендовать для выделения
сеголетков, лучше — в сочетании с другими методами.
2.2.2. Морфологическое обоснование
Метод использует показатель, изменяющийся с увеличением размера черепа,
которое, в свою очередь, связано с увеличением размера тела.
Интенсивное увеличение длины и ширины черепа возможно потому, что ко-
сти черепа у молодых особей не сращены друг с другом, шов — место их со-
единения — открыт. По мере увеличения размеров костей они срастаются, шов,
сначала четко видный, в процессе замедления и прекращения роста становится
менее заметным и наконец полностью зарастает, становится невиден. Разные швы
зарастают в разном возрасте.
2.2.3. Разрешающая способность
Метод позволяет выделить особей в фазе активного роста—молодых. Одна-
ко надежность его низкая ввиду большой индивидуальной изменчивости при-
знака. Все же можно считать, что наличие всех (или большинства) открытых
швов черепа — несомненный признак молодого животного. Иногда выделяют
несколько возрастных групп, используя сочетания нескольких швов (см., на-
пример, гл. 13 Procyon lotor, Ursus americanus).
2.2.4. Методика
На очищенном от мягких тканей черепе рассматривают соединение костей и
выделяют стадии зарастания швов, чаще всего три: 1 — шов открыт (контакт
между костями не полный), 2 — шов закрыт (кости в полном контакте), но еще
виден, 3 — шов полностью зарос и практически не виден (облитерирован). Вы-
бирают такое сочетание нескольких швов, которое позволяет выделить сеголет-
ков в том возрасте, когда их размеры близки к размерам взрослых, а иногда и
отделить группу более молодых взрослых от старших (см., например, гл. 13
Gulo, Lutra canadensis, Vulpes, Ursus maritimus). В данном тексте использованы
латинские обозначениями швов и их сокращения (рис. 2.1, 2.2).
2.2.5. Изменчивость показателей
Индивидуальная изменчивость очень велика. Нередки случаи, когда шов на
обеих или на одной половине черепа остается четко видным даже у старых
особей. Так, швы IN и FN были открыты у отдельных самцов медведей Ursus
arctos в возрасте 23 лет (Pearson, 1975), а шов РВ у некоторых волков Canis
lupus закрывается в возрасте 3,8 лет, но у других может полностью не зарас-
тать в течение всей жизни (Landon et al., 1998). У зубра, Bison bonasus, шов С
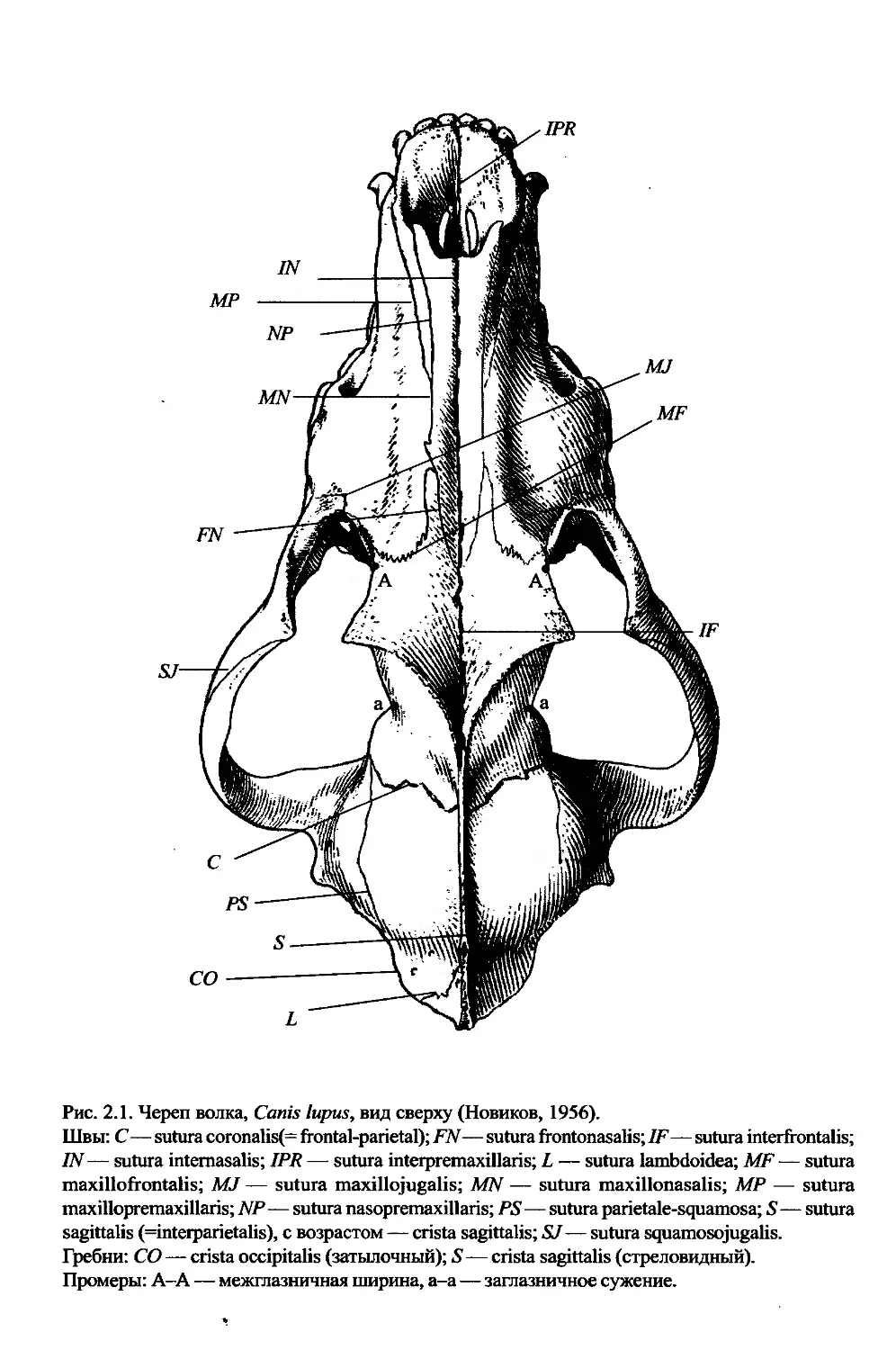
Рис. 2.1. Череп волка, Canis lupus, вид сверху (Новиков, 1956).
Швы: С — sutura coronalis(= frontal-parietal); FN— sutura frontonasalis; IF— sutura interfrontalis;
IN— sutura intemasalis; IPR — sutura interpremaxillaris; L — sutura lambdoidea; MF — sutura
maxillofrontalis; MJ — sutura maxillojugalis; MN — sutura maxillonasalis; MP — sutura
maxillopremaxillaris; NP — sutura nasopremaxillaris; PS—sutura parietale-squamosa; 5—sutura
sagittalis (=interparietalis), с возрастом — crista sagittalis; SJ — sutura squamosojugalis.
Гребни: CO — crista occipitalis (затылочный); 5—crista sagittalis (стреловидный).
Промеры: A-A — межглазничная ширина, a-a — заглазничное сужение.
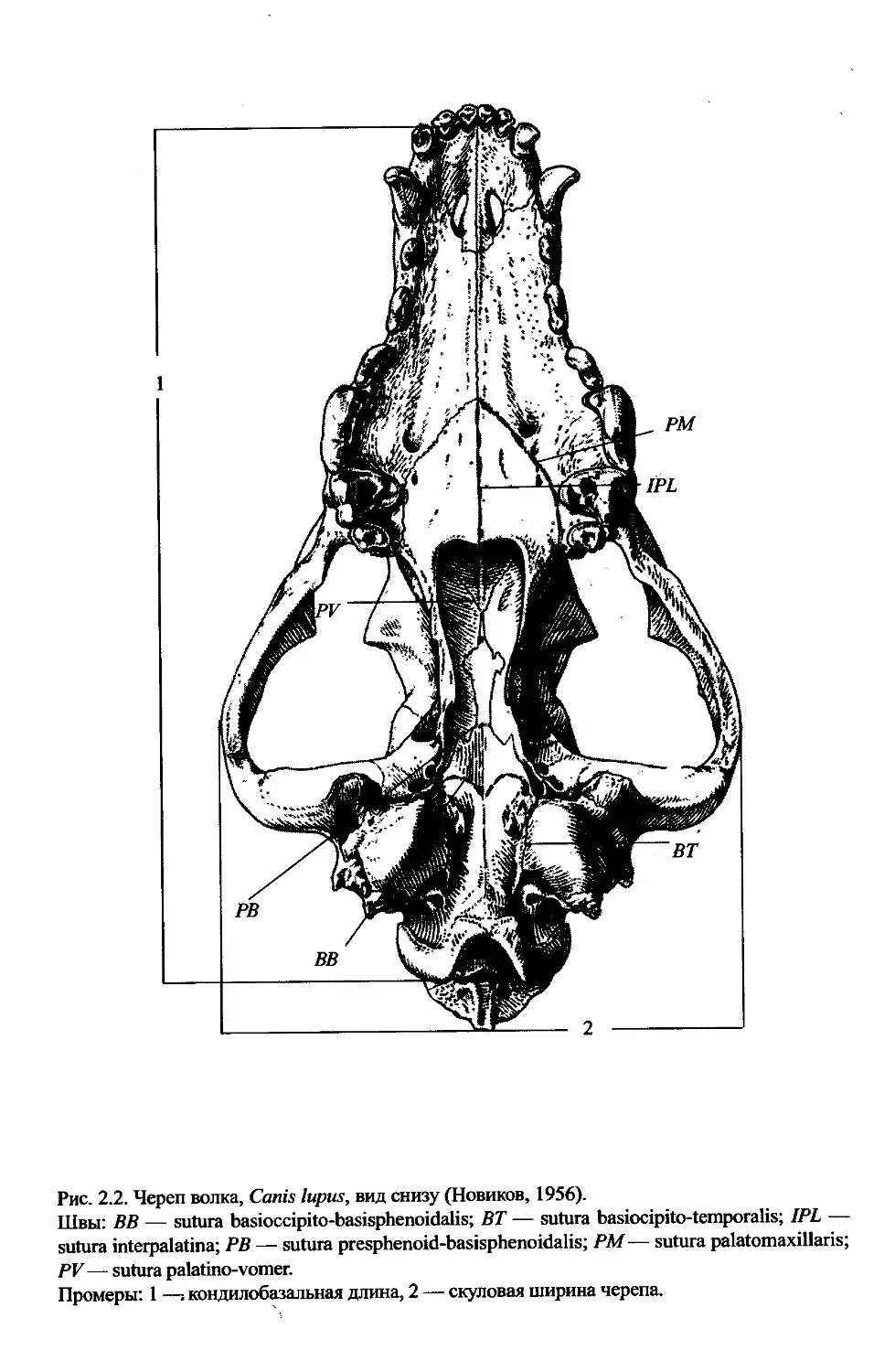
Рис. 2.2. Череп волка, Canis lupus, вид снизу (Новиков, 1956).
Швы: ВВ — sutura basioccipito-basisphenoidalis; ВТ — sutura basiocipito-temporalis; 1PL —
sutura interpalatina; PB — sutura presphenoid-basisphenoidalis; PM— sutura palatomaxillaris;
PV — sutura palatino-vomer.
Промеры: 1 —. кондилобазальная длина, 2 — скуловая ширина черепа.
32
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 2.1. Возраст (годы) зарастания швов у львов Panthera leo
(Smuts et al., 1978)
Швы* Самцы Самки
5 1,8-2,16/2,16-2,3** 1,75-2,25/2,25-2,3
IF 5/6 4,5-5/4,5-5
ВВ 3-7/4-7,5 4,5-5/4,5-5
PS 5-8/6,5-9 4,5-5 /4,5-5
MP 6,5-9/7,5-9 6-14/9-14
IN 8-16/никогда (?) 9-14 / никогда (?)
NP 14—16 / никогда (?) 9-14/никогда (?)
FN 14-16 / никогда (?) 9-14/9-14
♦Обозначения швов см. рис. 2.1.
** Шов частично закрыт / полностью зарос.
в средней своей части зарастает в возрасте 5—6 лет (но был открыт у 12-летней
самки), боковые его части — в возрасте 9-27 лет (Empel, 1962).
Если есть различия между самцами и самками по размерам тела и черепа и
скорости роста, то возраст зарастания швов также различен (см. гл. 13 Ursus
arctos, U. maritimus). Величину различий по возрасту зарастания между разны-
ми швами одной особи, между особями и между самцами и самками характери-
зует таблица 2.1.
По причине такой высокой индивидуальной изменчивости этот метод уступа-
ет методу определения возраста по окостенению эпифизов (см. ниже). Если не
окостеневшая эпифизарная пластинка (метэпифизарный хрящ) — несомненный
признак молодой особи, то один открытый шов таковым не является.
Внутривидовая изменчивость скорости зарастания швов может быть суще-
ственной, если есть внутривидовые различия в скорости роста или длительности
периода размножения. Такие различия описаны у скунса Spilogaleputorius (Mead,
1967): например, шов ВВ (рис. 2.2) полностью закрыт у всех особей старше 5
мес. одного подвида, и не закрыт у некоторых 8-месячных особей другого под-
вида.
Pucek, Lowe (1975) считают, что наличие незакрытых швов у взрослых мел-
ких млекопитающих может быть связано с кислотностью среды обитания.
2.3. Форма черепа
Форма черепа с возрастом несколько меняется у большинства млекопитаю-
щих: у молодых череп более округлый, крыша черепа выпуклая, у старших —
крыша уплощается, череп более угловатый. Этот признак можно использовать
как дополнительный критерий возраста (см. например, гл. 12 Glis, Arvicola,
Глава 2. Определение возраста по признакам скелета
33
Microtus, Lemmus). В некоторых группах млекопитающих особенно заметны
изменения формы с возрастом. У всех куньих заглазничное сужение с возрас-
том становится меньше. Его измеряют по отношению к межглазничной ширине
(рис. 2.1) (см. гл. 13 Enhydra, Martes zibellina, Mustela erminea, M. nivalis),
реже — по отношению к кондилобазальной длине черепа (см. гл. 13 М. martes).
Тимофеев и Надеев (1955) полагали, что этот «орбитальный индекс» может слу-
жить самостоятельным показателем возраста соболя, хотя Монахов (1967) счи-
тал его менее надежным, чем ширина канала клыка или вес хрусталика (см. гл.
13 Martes zibellina). Шадрина (1988а) сравнила результаты определения возраста
по орбитальному индексу и по годовым слоям соболя, Martes zibellina, и горно-
стая, Mustela erminea, (взятых в промысловый период). Она получила 70% со-
впадения при выделении сеголетков соболя и 90% при выделении сеголетков
горностая и только около 30% при выделении соболей в возрасте 1 г. 8 мес.,
причем в большинстве расхождений возраст соболя по орбитальному индексу
был завышен.
Наши данные по кунице М. martes (Клевезаль и др. 2005) показывают, что рас-
пределения самцов по величине обоих показателей заглазничной ширины (отноше-
нию к межглазничной ширине и отношению к кондилобазальной длине) не обнару-
живают минимумов, которые можно было бы использовать для разделения возрас-
тных групп. Не помогли выделению возрастных групп ни анализ распределения зна-
чений первой главной компоненты, ни кластерный анализ этих показателей, а диск-
риминантный анализ показал, что эти признаки выделяют возрастные группы хуже,
чем такие качественные признаки, как развитие гребней и стертость резцов.
Таким образом, как и в случае большинства признаков, непосредственно
связанных с ростом особи, по форме черепа можно выделить сеголетков, но не
всех. С возрастом межиндивидуальные различия в скорости роста делают метод
все менее надежным.
2.4. Развитие гребней
У всех молодых животных череп гладкий, с возрастом появляются бугры и
шероховатости, затем развиваются гребни.
У многих хищных как возрастной признак используют развитие гребней: стре-
ловидного (crista sagittalis), который появляется между теменными костями и с
возрастом «продвигается» вперед, и затылочного (crista occipitalis) появляющего-
ся по верхнему краю затылочной кости (рис. 2.1, 2.3). Высота и длина гребней с
возрастом увеличиваются. На первых стадиях своего развития гребень, уже до-
стигший определенного размера, может быть не плотным, спонгиозным. У са-
мок гребни развиваются в более позднем возрасте и не так хорошо выражены,
как у самцов.
У куньих (см. гл. 13 Martes zibellina, М. martes,) и некоторых грызунов (см.
например, гл. 12 Arvicola, Glis) в качестве критерия возраста используют положе-
ние лобных валиков (лобных гребней) на крыше черепа (см. рис. 12.5 и 13.2).
34
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
2
Рис. 2.3. Череп волка, Canis lupus, вид сбоку (Новиков, 1956).
Гребни : 5 — crista sagittalis (стреловидный), СО — crista occipitalis (затылочный).
Зубы: i — резцы, с — клыки, р - предкоренные, m — коренные.
Промеры: 1 — кондилобазальная длина черепа; 2 — наибольшая (общая) длина черепа; 3 —
длина нижней челюсти.
По всем этим признакам имеет место большая индивидуальная изменчивость,
поэтому, например, развитие стреловидного гребня является плохим показате-
лем возраста Spilogale putorius (Mead, 1967) и F. lynx (Kvam, 1983), однако они
часто полезны в сочетании с другими критериями возраста.
Для грызунов, у которых коренные зубы не имеют корней и потому постоян-
но растут и не пригодны для определения возраста, характер поверхности черепа
(в частности, развитие гребней) и форма черепа являются теми показателями,
которые дают ключ к разделению на возрастные группы, хотя бы и без привязки
к конкретному возрасту (см. гл. 12, Microtus, Arvicola, Lemmus).
2.5. Слияние эпифизов с диафизом костей конечностей
2.5.1. Область применения
Метод пригоден для определения возраста молодых животных до достиже-
ния дефинитивных размеров, в принципе любых систематических групп, но обыч-
но его используют для выделения сеголетков у хищных среднего размера, ко-
пытных, зайцеобразных, и для определения возраста рукокрылых. Полезен в
Глава 2. Определение возраста по признакам скелета
35
сочетании с другими методами, например, с определением возраста по слоям,
чтобы отличить явных сеголетков и не делать лишних срезов (например, Jensen,
Nielsen, 1968).
Метод посмертный, но может быть использован и прижизненно (при рентге-
новском исследовании костей или «на ощупь», например, у зайцев).
2.5.2. Морфологическое обоснование
Рост длинных костей конечностей в длину в постнатальный период обеспечи-
вается ростовыми зонами (метэпифизарными хрящевыми пластинками, которые
называют часто просто эпифизарными), находящимися между диафизом кости
и ее дистальным и проксимальным эпифизами. По мере замедления роста зона
становится уже и окостеневает, на ее месте остается линия слияния. С возрастом
линия исчезает, эпифиз сливается с диафизом (рис. 2.4). У большинства млекопи-
тающих рост длинных костей скелета, в основном, заканчивается, и эпифизы
сливаются с диафизами в период или вскоре после наступления половой зрело-
сти. У некоторых грызунов, однако, полного слияния эпифизов и диафизов длин-
ных костей не наблюдается даже у старых животных. Так, у полевок Lagurus,
Stenocranius в возрасте 9-12 мес. (практически предельный возраст) еще не было
полного слияния с диафизом проксимальных эпифизов большой и малой бер-
цовых костей (Тарасов, 1965), у лабораторных мышей (разных линий) в возра-
сте 550 дней и лабораторных крыс (разных линий) в возрасте 775 дней не было
полного слияния дистальных эпифизов локтевой, лучевой, бедренной костей и
проксимальных эпифизов берцовых костей (Dawson, 1934, 1935).
Иногда для определения возраста используют, вместе с другими критериями,
размер костей конечностей и размер костно-мозговой полости трубчатых костей
(см. тл. 12 Microtus califormcus).
Рис. 2.4. Состояние проксимального эпифиза плечевой кости черного медведя, Ursus americanus,
(Sauer, 1975). 0 — эпифиз и диафиз разделены хрящевой пластинкой, 1 — начинается сраста-
ние, видна эпифизарная линия, 2 — полное срастание эпифиза с диафизом.
36
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
2.5.3. Разрешающая способность
Метод позволяет выделить из всей выборки только активно растущих осо-
бей. Используется для различения сеголетков у хищных, зайцеобразных, и для
точного определения возраста молодых летучих мышей. У некоторых копыт-
ных он дает возможность выделить сеголетков, а иногда и особей нескольких
первых годовых классов (см. гл. 14 Capreolus, Odocoileus hemionus, Cervus
elaphus), но чаще межиндивидуальные различия не позволяют выделять годо-
вые классы. У мелких грызунов этим методом можно разделять на возрастные
группы и половозрелых размножающихся животных (см. гл. 12 Lagurus). При
исследовании выборок зайцеобразных и среднего размера хищных, взятых в
осенне-зимний период, по не приросшему эпифизу узнают сеголетка, но уже
с середины зимы приросший эпифиз не обязательно указывает на то, что эта
особь старше года.
При определении возраста зайцеобразных этот метод выделения сеголет-
ков уступает методу определения возраста по весу хрусталика (Wight,
Conaway, 1962; Connolly et al., 1969b; Rogers, 1982). Успех использования
зависит как от размеров животных (точнее от скорости их роста в первый год
жизни), так и от правильного выбора кости. Так, степень сращения эпифи-
зов лучевой и локтевой костей у ряда хищных среднего размера позволяет
выделять сеголетков зимой - ранней весной надежнее, чем относительная
ширина полости пульпы (см. гл. 13 Nyctereutes procyonoides), и по крайне
мере не хуже, чем вес хрусталика (см. гл. 13 Vulpes). Однако степень прира-
щения дистальных эпифизов бедренной кости у Martes americana уже осенью
позволяет выделить лишь малую долю сеголетков, и при том только самцов
(Dagg et al., 1975).
2.5.4. Методика
Обычно кости очищают от мягких тканей (вывариванием или с помощью ли-
чинок жуков-кожеедов — сем. Dermestidae) и исследуют область соединения
эпифиза и диафиза. Поскольку у молодых животных диафиз отделен от эпифиза
хрящевой пластинкой, при вываривании костей есть опасность потери эпифиза у
очень молодых особей, когда пластинка полностью хрящевая. Следующая ста-
дия окостенения ростовой зоны — образование бороздки (линии) между эпифи-
зом и диафизом, которая хорошо видна. Когда бороздка между эпифизом и диа-
физом зарастает, какое-то время остается ее след, который можно уловить, если
вести острие иглы вдоль кости, поперек бывшей бороздки. Последняя стадия —
полное слияние эпифиза с диафизом (рис. 2.4). Поскольку степень сращения
эпифиза с диафизом может быть разной, в работах обычно указывают, как опре-
деляют полное срастание. Различие в методике может существенно сказаться на
результате (см. гл. 11 Oryctolagus).
Используют и рентгеновские снимки костей как живых, так и мертвых жи-
вотных. При этом важно, чтобы кость лежала плоско и потому живых живот-
Глава 2. Определение возраста по признакам скелета
37
ных лучше обездвижить. На рентгеновском негативе широкая хрящевая плас-
тинка между эпифизом и диафизом выглядит широкой светлой тенью, эпифи-
зарная линия — тонкой светлой тенью. Если произошло полное слияние эпи-
физа с диафизом между ними не видно ни линии, ни тени. Некоторые зоологи
(например, Dagg et al., 1975) предпочитали давать рентгеновские снимки очи-
щенных костей опытным радиологам для оценки степени приращения эпифи-
зов к диафизам.
В принципе оба способа, и визуальный, и рентгеновский, дают одинаковый
результат при исследовании хищных среднего размера (Harris, 1978), но рентге-
новский метод менее надежен при исследовании мелких млекопитающих. Более
правильно выявляет степень слияния эпифизов с диафизом окраска очищенных
костей на наличие хряща (Thomsen, Mortensen, 1946), но трудоемкость этого
способа вряд ли оправдана результатом.
У летучих мышей (см. гл. 10) степень окостенения ростовых зон в местах
соединения пястных костей и пальцев крыловой перепонки можно определять на
просвет и даже измерять длину хрящевой зоны (Kunz, Anthony, 1982). Степень
окостенения ростовой зоны локтевой кости у зайцев определяют на ощупь (см.
гл. 11 Lepus europaeus).
2.5.5. Изменчивость показателей
Будучи типичным ростовым, этот признак подвержен влиянию всех факто-
ров, влияющих на рост организма в целом. Имеет место межпопуляционная и
индивидуальная изменчивость, а также различия между самцами и самками.
Истощение животного может задерживать окостенение эпифизов; так у исто-
щенных оленей Odocoileus hemionus было отмечено запаздывание примерно на
год (Lewall, Cowan, 1963).
Влияют на окостенение эпифизов и сезонные изменения темпа роста, поэтому у
животных с растянутым периодом размножения, например, у грызунов, отделе-
ние сеголетков от старших возможно не в любое время года. Рожденные в конце
лета - осенью особи развиваются медленно в зимний период и лишь во второй
половине следующего лета обгоняют быстрорастущих особей из ранних, весен-
них, выводков. Во второй половине зимы можно выделить сеголетков поздних
выводков, но у ранне-весенних выводков к этому времени исчезает хрящевая
пластинка и их можно принять за особей старше года (см. гл. 12 Sciurus nigef).
Слияние эпифизов с диафизами разных костей одной особи происходит в
разном возрасте, и исследование максимального числа разных костей может
дать более надежную информацию о возрасте особи. На практике, однако, для
определения возраста выбирают одну или несколько костей, обычно длинные
кости конечностей.
В разных ipynnax млекопитающих порядок слияния эпифизов с диафизами
разных костей не одинаков, но у близких видов различия незначительны (Тара-
сов, 1965, 1966), что дает основание использовать данные, полученные для од-
ного вида, при определении возраста близких видов.
38
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
2.6. Состояние позвоночника и костей таза
Позвонки растут, в принципе, так же как и кости конечностей, но их исполь-
зуют для определения возраста значительно реже. С возрастом уменьшается тол-
щина эпифизов и хрящевой межпозвонковой прослойки и увеличивается длина
тела позвонка. Степень срастания можно также определять по наличию линии
между эпифизом и диафизом, но Trout, Lelliott (1992) считают этот метод более
субъективным, чем проверка на отламывание эпифиза от диафиза (см. гл. 11
Oryctolagus cuniculus).
Hagen (1956) предложила определять относительный возраст мелких грызу-
нов по индексу — отношению длины двух соседних хвостовых позвонков (а) к
толщине их эпифизов (б) (рис. 2.5), значение которого увеличивается с возрас-
том. При исследовании короткохвостых грызунов ею были выбраны 8 и 9 по-
звонки (считая от последнего хвостового). У полевки Microtus arvalis это —
примерно середина хвоста. При исследовании длиннохвостых — 15 и 16-й по-
звонки. Для полевок Microtus arvalis и Clethrionomys glareolus и домовой мыши
Mus musculus (лабораторные мыши) индексы в возрасте 2 мес. — около 5, в 6
мес. — 7, в 9 мес. — 10.
Есть примеры использования для оценки возраста степени сращения тазовых
костей. Например, у пекари Tayassu tajacu швы между подвздошной, седалищ-
ной и лобковой костями зарастают в возрасте 8-12 мес.; у годовалых они едва
видны (Lochmiller et al., 1984). У некоторых хищных критерием относительного
возраста может быть степень зарастания тазового шва (симфиза) (см. гл. 13
Gulo, Lutra canadensis, Martes pennantl).
Рис. 2.5. Схема измерения хвостовых позвонков мелких грызунов. Хвостовые позвонки полев-
ки Microtus arvalis в возрасте примерно 2 мес. (1) и 12 мес. (2) (Hagen, 1956). Пояснения в
тексте.
Глава 2. Определение возраста по признакам скелета
39
2.7. Развитие бугров на костях скелета
Этот признак связан не с ростом, а с развитием мускулатуры. У молодых
животных бугры не выражены, обычно они становятся заметными у половозре-
лых особей. Эти возрастные различия хорошо выражены у самцов и значительно
хуже у самок.
Выделять молодых хищных можно по степени развития конического возвы-
шения — латерального супрасезамоидного бугра в дистальной части бедренной
кости (Lechleitner, 1954) (рис. 2.6). Отсутствие бугра четко указывает на то, что
особь молодая, но у некоторых молодых он может присутствовать (см. гл. 13
Mustela етйпеа, М. vison, М. nivalis, Martes americana, Ursus maritimus).
Suspensory tuberosity — утолщение по каудо-медиальному краю седалищной
кости, отсутствует у молодых самцов и самок, но имеется у взрослых самцов.
Этот признак используют для разделения молодых и взрослых самцов пекари
(Lochmiller et al., 1984), некоторых хищных (см. гл. 13 Mustela vison), грызунов
(см. гл. 12 Sciurus carolinensis, S. niger).
Рис. 2.6. Бедренная кость американской норки, Mustela vison. Суперсезамоидный бугор, от-
сутствующий у молодой особи в возрасте около 8 мес. (вверху), хорошо развит (показан
стрелкой) у взрослой особи (внизу) (по Greer, 1957).
40
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
2.8. Размеры и форма кости полового члена (os penis, baculum)
2.8.1. Область применения
Метод пригоден для оценки возраста самцов тех групп млекопитающих, у
которых есть поддерживающая кость полового члена. Его часто применяют для
оценки возраста хищных, в отдельных случаях — грызунов. Обычно метод по-
смертный. Иногда используется прижизненно, когда можно на ощупь оценить
размер и форму этой кости.
Метод целесообразно использовать (1) для прижизненной приблизительной
оценки особи как молодой или взрослой на ощупь, если это трудно сделать по
размерам тела, (2) как дополнительный критерий возраста, (3) когда нет других
возможностей разделить самцов на молодых и взрослых, например, при нали-
чии неполных останков животных.
2.8.2. Морфологическое обоснование
В кости полового члена различают тело (обычно, вытянутый стержень), осно-
вание (утолщенную проксимальную часть) и тонкий дистальный конец (рис. 2.7
А). У многих грызунов дистальный конец несет пальцевидные отростки. Разме-
ры и форма кости — ростовой признак, однако связан в большей мере с поло-
Рис. 2.7. Строение кости полового члена (на примере кости взрослого черного хорька, Mustela
putorius) — А и кости молодых (слева) и взрослых (справа) особей — Б (по Walton, 1968).
Глава 2. Определение возраста по признакам скелета
41
вым созреванием, чем с ростом как таковым. С возрастом, по мере полового
созревания, увеличивается вес, длина и толщина кости, меняется ее форма. У
хищных до наступления половой зрелости вес увеличивается линейно, затем теми
увеличения снижается, но вес продолжает увеличиваться практически в течение
всей жизни (Marks, Erickson, 1966; Walton, 1968; Habermehl, 1985; King, 1991).
Несколько меняется форма кости. У молодых проксимальная часть почти такой
же толщины, что и тело кости. По мере полового созревания она утолщается
(рис. 2.7Б). Это утолщение проксимальной части — хороший визуальный крите-
рий взрослой особи, иногда его можно выявить на ощупь у обездвиженных жи-
вотных.
2.8.3. Разрешающая способность
Метод пригоден для разделения самцов на молодых и взрослых в выбор-
ках, взятых до начала полового созревания молодых. Он мало пригоден в пе-
риод, когда у молодых начинается половое созревание. Например, наибольшая
индивидуальная изменчивость веса кости у барсука обнаружена у особей, воз-
раст которых приближается к году, потому что в этом возрасте часть животных
уже достигает половой зрелости (Ahnlund, 1976). В результате у большинства
тех хищных, которые могут стать, но могу! и не стать половозрелыми в возра-
сте около года, отделение сеголетков от годовалых в зимний период трудно. В
это время размеры кости рано созревающих сеголетков могут быть сходны с
таковыми половозрелых годовиков, поэтому метод пригоден для выделения
сеголетков у хищников мелких и средних размеров во второй половине лета —
осенью (Mead,1967; Walton, 1968; Harris, 1978). В выборке, взятой в это вре-
мя, сеголетки от старших разделяются с заметным хиатусом. У крупных хищ-
ников можно выделить несколько возрастных групп (см. гл. 13 Ursus americanus,
U. maritimus).
Некоторые исследователи считают, что у хищных разделять самцов на моло-
дых и взрослых проще по прорезанию зубов, зарастанию швов черепа, прира-
щению эпифизов костей конечностей, весу хрусталика (например, см. гл. 13
Canis mesomelas). Выделение сеголетков по кости полового члена оказалось столь
же надежным, как и по каналу клыка у колонка Mustela sibirica (Панкратьев,
1971), и более надежным, чем по внешнему виду черепа и стертости зубов у
куницы М. martes (Граков, 1962).
2.8.4. Методика
При посмертном определении очищают кость от мягких тканей и оценивают
сухой вес или длину. Вес является лучшим показателем возраста, чем длина: в
приводимых авторами данных часто между группами молодых неполовозрелых
и половозрелых нет перекрывания по весу, но есть по длине (табл. 2.2). Это,
видимо, связано с утолщением проксимального конца кости при наступлении
половой зрелости, при котором вес увеличивается больше, чем длина кости.
42
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 2.2. Вес (W, г) и длина (L, мм) кости полового члена у некоторых
хищных
Вид Lutra canadensis Молодые (неполовозрелые) W—1,6 (0,7-2,8) L — 69 (57-82) Взрослые (половозрелые) 3,6-8,4 81-107 Источник Stephenson, 1977
W — до 4,7 L — до 87 от 4,8 от 87 Friley, 1949
Lutra lutra W — 0,9 (до 1 года) L —47 (-«-) L < 50 1,75-2,6 54-61 >50 Вшивцев, 1972 M.Stubbe, 1989f
Martes foina W —0,22-0,32 L —51-58 0,38-0,80 56-67 Рябов, 1962
Martes martes W —0,16(0,13-0,19) L — 37 (35-38) L — 35-40 0,29 (0,2-0,35) 42 (40-46) 39-46 Попов, 1943; Рябов, 1962; Suminski, 1968
Martes pennanti W<1,2 Wright, Coulter, 1967
Martes zibellina W —0,050-0,093 (октябрь) большинство > 0,15 Монахов, 1967
Meles meles W <0,5 L<65 1,26±0,21 74-81 Graf, Wandeler, 1982; Suminski, 1968
Mustela erminea W<0,03 L —как правило <23 >0,04 >23 King, 1991; Van Soest, VanBree, 1970; Попов, 1943; Fog, 1969
Mustela eversmanni W —0,2(0,13-0,28) L —37 (34-41) 0,44 (0,37-0,53) 39 (37-42) Попов, 1943
Mustelafrenata W —0,014-0,029 0,053-0,101 Wright, 1947 (no Petrides, 1950)
Mustela lutreola W — 0,12 (0,10-0,13) L — 34 (33-35) 0,29 (0,23-0,38) 36 (35-37) Попов, 1943
Mustela nivalis W <0,01 L<13 W <0,025 0,025 16 >0,03 Habermehl, 1985 Fog, 1969
Mustela putoris W —0,15(0,08-0,24) L — 34(31-37) 0,34 (0,28-0,47) 36 (33-38) Попов, 1943
L — 31-38 W< 0,425 42-50 >0,425 Suminski, 1968; Hartung, 1980 (noM,Stubbe, 1989c)
Mustela sibirica W<0,15 L < 34 >0,20 >35 Попов, 1943; Войлоч- ников, Михайловский, 1970
W<0,13 Синицын, 1982
W —0,077-0,230 L —31-38 0,249-0,350 34-39 Панкратьев, 1971
Mustela vison W —0,17(0,13-0,21) L —42(38-45) 0,34 (0,24-0,45) 45 (40-50) Попов, 1943; Petrides, 1950; Lechleitner, 1954
Nyctereutes procyonoides L<38 58-72 Suminski, 1968
Глава 2. Определение возраста по признакам скелета
43
Таблица 2.2 (окончание)
Вид Молодые (неполовозрелые) Взрослые (половозрелые) Источник
Procyon lotor W —0,2 0,3 L — 40- 50 (до 90) 1,5-3,0 80-95 (больше 90) Habermehl, 1985 (Grummt, 1989)
Taxidea taxus L —J86 >93 Petrides, 1950
Urocyon cinereoargenteus W - 0,280±0,062 0,528±0,1 Petrides, 1950*
Vulpes vulpes fulvus W - 0,342±0,7 0,492±0,103 Petridcs, 1950*
Vulpes vulpes W < 0,27 L —40-50 до 0,6 52-66 Habermehl, 1985 Harris, 1978
W <0,4 (до 6 мес) Hartung, 1982 по M. Stubbe, 1989a
L —44-52 52-66 Suminski, 1968
Примечание: * — выборки малы.
Кость крупных животных можно очищать вывариванием или с помощью дер-
местид. Отделение кости полового члена грызунов требует специальной методи-
ки: взятый от тушки половой член помещают в 1N или 2N раствор КОН на срок
от 1 ч. (Microtus) до 4 ч. (Sciuridae), затем под бинокуляром удаляют мягкие
ткани, споласкивают в 5%-ном р-ре уксусной к-ты и в воде и, если необходимо,
окрашивают ализарином (на 2000 см3 2%-ного р-ра КОН 0,1 г ализарина крас-
ного) (Friley, 1947).
2.8.5. Изменчивость показателей
Индивидуальная и межпопуляционная изменчивость размеров этой кости,
очевидно, коррелированна с возрастом полового созревания. Например, у скун-
са Spilogale putorius некоторые особи подвида S', р. latifrons размножаются на
первом году жизни, в возрасте 5 мес., другие — на следующий год. У другого
подвида, S', р. interrupta, все особи размножаются в возрасте 1 года. У первого
подвида обнаружена большая индивидуальная изменчивость веса и формы кости
у сеголетков, чем у второго (Mead, 1967).
2.8.6. Особенности использования метода в отдельных группах
2.8.6.1. Хищные
Для отнесения отдельных особей хищных к категории молодых или взрос-
лых могут быть использованы данные таблицы 2.2.
2.8.6.2. Грызуны
Есть примеры использования этой кости для выделения сеголетков у грызу-
нов в тех случаях, когда размножение сезонное и сеголетки не достигают поло-
44
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
вой зрелости в первое лето жизни. У белки и ондатры сеголетков от перезимо-
вавших можно отличить осенью и в начале зимы (см. гл. 12 Sciurus vulgaris,
Ondatra). У бурундука Tamias ruficaudus (Beg, Hoffmann, 1977) уже после июля-
августа не было различий между сеголетками и взрослыми по размерам кости.
У лемминга Myopus schisticolor (Artimo, 1969) после наступления половой зре-
лости не было корреляции между размером кости и хронологическим возрастом
особи; в июне-октябре по длине тела кости можно было отличить сеголетков от
старших, но столь же четкие различия были и по длине и по весу тела.
Видимо, нет смысла использовать кость полового члена для определения воз-
раста грызунов, имеющих несколько пометов в год и рано достигающих поло-
вой зрелости. Например, Chen et al. (1988) выделили 5 возрастных групп по
длине этой кости у крыс Rattus norvegicus, но распределение особей по этому
параметру практически не показывает границ между группами. Именно потому,
что рост кости полового члена связан не с возрастом как таковым, а с наступле-
нием половой зрелости, в случае мелких грызунов, как считают Pucek, Lowe
(1975), данный критерий возраста имеет ограниченную ценность, поскольку у
этих грызунов более, чем у млекопитающих других групп, наступление половой
зрелости зависит от динамики численности и других внутрипопуляционных про-
цессов.
ГЛАВА 3. ОПРЕДЕЛЕНИЕ ВОЗРАСТА
ПО ПРИЗНАКАМ ЗУБОВ
Терминология
У млекопитающих различают резцы (i), клыки (с), предкоренные, или премо-
ляры, (р) и коренные, или моляры, (т) (см. рис. 2.3). В каждой половине верх-
ней и нижней челюсти может быть несколько резцов, предкоренных и коренных
и не более 1 клыка. Каждый зуб, кроме клыков, имеет свой порядковый номер,
который ставится после обозначения зуба как верхний или нижний индекс в за-
висимости от того, о зубе из верхней или нижней челюсти идет речь. Например,
нижний коренной т3, верхний — т3. Для обозначения молочного зуба ставят
впереди d — di, и т.д. Часто для совместного обозначения предкоренных и ко-
ренных зубов используют термин «щечные» зубы.
Исходным у плацентарных млекопитающих считается наличие с каждой стороны
верхней и нижней челюсти по 3 резца, 1 клыку, 4 предкоренных и 3 коренных. У
млеиэпитающих многих групп число зубов сокращается и при их обозначении мо-
жет возникнуть путаница. Так, у грызунов число предкоренных зубов редуцируется
до 0-2, у оленей — до 3. Одни авторы дают им нумерацию в соответствии с их
положением в челюсти (например, рр р2, р3), другие нумеруют их в соответствии с
исходной зубной формулой млеиэпитающих как р2, р} и р4. В таких случаях, чтобы
избежать разночтений, кроме буквенного обозначение зуба я указываю его по-
ложение в челюсти (например, у бобра — первый нижний щечный р4).
У большинства млекопитающих имеется две смены зубов — молочные и
постоянные. Молочные прорезаются до или вскоре после рождения и функцио-
нируют в течение периода быстрого роста организма и, соответственно, черепа и
зубного ряда. Молочных предшественников имеют резцы, клыки и предкорен-
ные зубы. Принято считать, что у всех млекопитающих, кроме человека, корен-
ные зубы не имеют молочных предшественников. В одонтологии человека при-
нята другая терминология (Зубов, 1968): сменяющиеся щечные зубы называют
молочными молярами, а сменяющие их постоянные — премолярами.
Зубы чаще, чем любые другие органы, используют для определения возраста.
Можно выделить такие критерии возраста: прорезание и смена зубов, рост зуба в
длину, зарастание полости пульпы (канала) зуба, стачивание зубов, годовые слои в
тканях зуба. Эти критерии используют как порознь, так и в сочетании друг с другом.
3.1. Прорезание постоянных зубов
3.1.1. Область применения
Метод применяют для оценки возраста средних и крупных хищных, копыт-
ных, слонов, обычно — посмертно, а когда удается заставить животное «пока-
46
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
зать зубы» или у обездвиженных особей, то и прижизненно, чтобы отделить мо-
лодых особей от старших, выделить возрастные группы среди молодых живот-
ных или уточнить возраст формирования первого годового слоя при определе-
нии возраста по годовым слоям в дентине или цементе.
3.1.2. Морфологическое обоснование
Постоянные зубы, имеющие молочных предшественников, появляются, как
правило, на месте молочных; вырастая под молочными они как бы выталкивают
молочные в полость рта. Иногда постоянный зуб (например, клык хищных) по-
является немного сбоку, и какое-то время в челюсти могут быть видны и молоч-
ный, и постоянный зубы.
Первый коренной зуб прорезается обычно еще до окончания смены всех мо-
лочных зубов, последний коренной — самым последним. С его появлением за-
канчивается увеличение в длину зубного ряда.
После прорезания зуба в течение какого-то времени он растет в высоту, ко-
ронка полностью видна над десной и зуб начинает функционировать—приходит
в соприкосновение с соответствующим зубом другой челюсти.
Особая ситуация имеет место у слонов и сирен (ламантинов, дюгоней). У них
щечные зубы не делятся на молочные и постоянные, и смена зубов происходит
не в вертикальном, а в горизонтальном направлении. Новые зубы появляются в
заднем конце челюсти и продвигаются вперед, а на переднем конце челюсти ста-
рые щечные зубы выпадают. У слонов зубы пластинчатые и пластины поочеред-
но начинают функционировать и со временем выпадать так, что одновременно
могут функционировать задние пластины предыдущего зуба и передние — пос-
ледующего.
У бородавочника сходным образом ведет себя третий коренной; постепенно
продвигаясь вперед он со временем вытесняет сточенные первый и второй ко-
ренные (см. гл. 14 Phacochoerus).
Постоянно растущие (постоянно прорезающиеся) зубы — это резцы у зай-
цеобразных и грызунов и коренные у полевок и зайцеобразных. Эти зубы не
имеют настоящих корней и постоянный рост в корневой части компенсирует ин-
тенсивное стачивание коронки. Их рост и стачивание не используют как крите-
рий возраста.
3.1.3. Разрешающая способность
Прорезание зубов коррелированно с увеличением размеров тела и хорошо
«работает» до достижения дефинитивных размеров тела.
Для тех млекопитающих, у которых смена молочных дубов на постоянные
растянута во времени, этот способ пригоден для выделения возрастных групп
среди молодых животных. У некрупных хищных (шакал, лисица, енотовидная
собака, куницы) смена молочных на постоянные заканчивается к первой зиме и,
следовательно, по этому признаку летом и осенью можно выделить сеголетков.
Глава 3. Определение возраста по признакам зубов
47
Чем дольше период активного роста, тем медленнее происходит появление по-
стоянных зубов, и тем выше ценность этого признака как критерия возраста. У
крупных копытных можно выделить 2-5 годовых классов.
У слонов возрастные группы выделяют в течение всей жизни, судя по тому,
какой из шести поочередно функционирующих щечных зубов функционирует в
данное время (см. ниже, раздел 3.1.4).
3.1.4. Методика
Замену молочных зубов на постоянные и появление постоянных зубов опре-
деляют осмотром челюстей, что иногда возможно и у живых особей. Различают
альвеолярное (зуб виден выше края альвеолы) и десневое (выше края десны)
прорезание, последнее происходит позже, но на фоне индивидуальной изменчи-
вости этой разницей во времени можно пренебречь. Строго говоря, при описа-
нии следует указывать, что (какую стадию) автор называет прорезанием зуба.
Например, Hemming (1969) пишет: «зуб считали прорезающимся, если какая-
нибудь его часть была видна выше уровня альвеолы».
КАК ОТЛИЧИТЬ МОЛОЧНЫЕ ЗУБЫ ОТ ПОСТОЯННЫХ: молочные замет-
но меньше по общему размеру, по высоте коронки, у них четко выражена шейка
зуба (рис. 3.1), корни уплощены, у щечных зубов — «растопырены». У оленей
последний молочный предкоренной имеет 3 корня, а замещающий его постоян-
ный — 2 корня (рис. 3.2). Если вынуть функционирующий молочный зуб из
а
б
Рис. 3.1. Резцы зубра, Bison bonasus, в возрасте 1 года (а), 2 лет (б), 4 лет (в), 8 лет (г) и более
10 лет (д) (по Krasinska, Krasiiiski, 2004).
а — все резцы молочные, б — оба i, постоянные, остальные молочные, в-д — постоянные
резцы с разной степенью стертости: поверхность стачивания постоянных резцов продольно-
овальная — в, трапециевидная — г и почти округлая — д.
48
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 3.2. Нижние предкоренные зубы северного оленя, Rangifer
tarandus. Молочные предкоренные у полуторагодовалого оленя
(вверху) и постоянные у оленя в возрасте 11 лет.
альвеолы, то задолго до полного его выпадения на корне видна резорбция, а на
дне альвеолы — вершинка постоянного зуба.
В некоторых случаях различить молочные и постоянные зубы не легко. На-
пример, Wilson et al. (1984) полагают, что поскольку у дукера Sylvicapra grimmia
трудно отличить молочные резцы от постоянных, для определения возраста луч-
ше использовать щечные зубы.
При оценке возраста слонов (Elephantidae) важно определить, какие пласти-
ны какого зуба функционируют в данное время. Зубы у них различаются разме-
рами и числом пластин, но индивидуальная изменчивость обоих параметров при-
водит к тому, что легко распознать только первые 2—3 зуба и для оценки возраста
приходится разрабатывать довольно сложные схемы (Sikes, 1967a,b; Roth,
Shoshani, 1988).
3.1.5. Изменчивость показателей
Последовательность прорезания зубов может быть разная у близких видов,
есть межпопуляционные и межиндивидуальные различия. Например, у лани Cervus
dama постоянные нижние зубы обычно появляются в такой последовательности:
Глава 3. Определение возраста по признакам зубов
49
m^-i -т2-12-ц-очп3-р4-(р3, р2), но у некоторых особей т3 и р4 прорезались одно-
временно или р4 раньше, чем (Chapman, Chapman, 1970).
Имеются межпопуляционные и межиндивидуальные различия и по воз-
расту прорезания постоянных зубов. Изменчивость возраста прорезания раз-
ных зубов одной и той же особи может быть разной. В целом, чем позже
появляется зуб, тем шире диапазон изменчивости возраста прорезания
(сравните, например, возраст прорезания первого и третьего коренных зубов
у С. capreolus, Cervus darna, Capricornis crispus, Ranfiger tarandus, Saiga
tatarica — см. табл. 3.3). При этом есть значимая корреляция возраста про-
резания одного зуба с возрастом прорезания другого зуба у той же особи,
например, для и!] и т3 у оленей Rangifer tarandus г = 0,7; р<0,001 (Клеве-
заль, Соколов, 1997).
3.1.6. Особенности использования метода в отдельных группах
З.1.6.1. Хищные
Говоря о смене зубов, обычно имеют в виду выпадение молочного и появле-
ние постоянного зуба. У хищных, в большинстве случаев, зуб начинает функци-
онировать вскоре после прорезания. Только клыки крупных хищных долго рас-
тут до начала функционирования.
В целом, как можно видеть из таблицы 3.1, у разных хищных первыми сме-
няются первые два резца и одновременно или чуть раньше прорезается первый
коренной.
З.1.6.2. Парнокопытные
У копытных от начала прорезания зуба (зуб становится виден в полости рта)
до начала его функционирования проходит определенное время (табл. 3.2).
У зубра Bison bonasus, по данным W^grzyn, Serwatka (1984), коронка m^
достигает полного размера через 5-12 мес., т22 — через 5-10 мес. т33 — через
10-13 мес. после начала десневого прорезания.
Этот период длиннее, если начало прорезания пришлось на осень, чем в тех
случаях, когда оно приходится на весну, поскольку все ростовые процессы, в
том числе и рост зубов, зимой идет медленнее. Зимнее замедление роста зубов
отмечали многие авторы (например, Соколов, 1956).
Последний, третий, коренной имеет обычно три доли, первая доля начинает
стачиваться тогда, когда последняя еще не достигла окончательной высоты, по-
этому если указывается возраст полного прорезания зуба, то для третьего корен-
ного этот период сильно растянут (табл. 3.3).
Растянутый период прорезания разных зубов копытных дает возможность
детально оценивать возраст молодых животных. Brown, Chapman (1991a,b) при
исследовании рентгеновских снимков нижних челюстей выделили 10 стадий
формирования зуба (от зубного зачатка, видного на снимке как темная округлая
лакуна, до стадии, когда закрывается апикальное отверстие корня), присвоили
по 1 баллу каждой стадии и по сумме баллов всех щечных зубов предложили
50
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 3.1. Возраст (месяцы) появления постоянных зубов у некоторых
хищных
Вид Зуб Пол- ная зубная система Источник
i1 i2 1 1» 1 2 Р3 С Р т1, ш22 т’3
Canis lupus 4-5 4-5 5-6 4-6 3,5-5 5-6 5,5-7 >7 Habermehl, 1985; Юдин, 1992; Гуоский. 1973
Canis rneso- melas 4,25- 4,5 5- 5,25 5,25- 5,75 5,5- 6 4,5- 4,75 4,75- 5,5 5,75- 6,25 Lombaard, 1971
Enhydra lutris 2-3 4-5; 3-4 9-15 5 -8 16-24 Schneider, 1973; Рязанов, Клевезаль, 1991
Felis caracal 5-6 6 6-10 6-10 • 6-7 10 Stuart, Stuart, 1985
Felis lynx 15-17 Матюшкин, 1974
Felis rufus 4,5 4,75- 5,25 5,25- 5,5 6-7,5 6,5- 7,25/ 5,5- 5.75* Jackson et al„ 1988
Hyena brunnea 8 10 15 Mills, 1982
Martes martes смена молочных в 3- -3,5 7 4 Рябов, 1962; Юргенсон, 1956; Habermehl, 1985
Martes foina молочные сменяются к 6 мес, Рябов, 1962
Mustela erminea в 3-4 мес. еще есть молочные зубы 6 Строганов, 1937
Mustela vison 1,75- 2 1,75 р<-2 1,75 3 4 Habermehl, 1985
Panthera leo 8- 11 10- 13 12- 15 12-20 15-17/ 11-14* 18-24 Smuts et al., 1978
Panthera pardus 8 -10 12- 24 8-10 8-10 24 Stander, 1997
Ursus ameri- canus 5-8 7-9 8- >10 5-8 4-6 5-8 7-8 Marks, Erickson, 1966
Ursus arctos 8 4-8 5 15 8-10 8-10 18 12 12 8 8-10 6-8 8 5 15 6-7 18 10-12 10-12 ?24 Pearson, 1975 Адольф, 1949** Данилов и др., 1979
Глава 3. Определение возраста по признакам зубов
51
Таблица 3.1(окончание)
Вид Зуб Пол- ная зубная система Источник
i‘ i2 1 Р 1 2 i’s С Р m'|
Vulpes vulpes 2,5- 3,5 3,5- 4 3,5-5 3-4,5 4 4,5 5,5 6 Habermehl, 1985; Маркина, 1962
Примечание: * верхние / нижние зубы, ** выращивание в зоопарке.
Таблица 3.2. Соотношение возраста (месяцы) прорезания
и функционирования щечных зубов нижней челюсти
у некоторых копытных
Зуб Cervus dama (Brown, Chapman, 1990) Odocoileus hemionus (Reeset al., 1966)
Прорезание Функционирование Прорезание Функционирование
Р2 21-25 27-43 26 28-29
Р3 17-21 26 26 29
Р« 17-21 22 24 29
Hl, 3-5 5 2.5 12
т2 9-13 18 9 20
т3 13-21 26 15 29
Таблица 3.3. Возраст (месяцы) появления постоянных зубов у некоторых
копытных*
Вид i'. i’a c P га1. m22 т’з Источник
Aepyce- ros melampus (4±3) 8±2 (13±5) 17±3 (22±5) 25±3 Spinage, 1971
Alces alces /(7-9) 10-1Г /162 /20-21' 17—18’ (19-20) 17-18’ (21) (6) (7-9) (21) Habermehl, 1985
Antidorcas marsupi- alis /12 /16 /16 /16 /19 /3 /9 /(10-19) 22 Rautenbach, 1971
Antilocapra americana (14) /(28) /40 /(42) /(28) /(1.5- 3) /(6- 8) /(12- 16) Dow, Wright, 1962
Bison bison /(30) /(30- 42) /(42) /(42) (42) (6) (18) (30) Novakowski, 1965
/(>24) /(>24) /(>36) /(>36) (>24) (3-12) (>12) (>24-36) Fuller, 1959
Bison bonasus /(22- 26) /(30- 31) /(35- 40) /(38- 44) (28- 44)/ (27- 40) (8- 12)/ (6-9) 18- 27)/ (16- 18) (30- 35)/ (27- 43) Wegrzyn, Serwatka, 1984
52
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 3.3 (продолжение)
Вид *’i *’з c P m1, m22 m3 Источник
Capra ibex /(15) /(28- 30) /(36- 40) /(44- 46) (28— 30) (4- 6) (12- 15) (28- 35) Habermehl, 1985
Capra pyrenaica /(15) /(30) /(35- 46) /(48- 57) (25- 33) (1-2) (13) (25) Vigal, Machordom, 1985
Capreolus capreolus /(5-6) /(7-9) /(8- 12) /(9- 12) (12-) 13 (3-4) (4-6) 9- 12 Соколов 1956; Habermehl, 1985
Capricornis crispus /(10- 22) /(17- 24) /(го- 31) /(26- 36) /(19- 31) /(5-6) /(4- 12) /(12- 21) Miura, Yasui, 1985
Cervus elaphus /(14- 15) /(16- 17) /(го- 21) (14- 15)/ (22- 23) (26- 28) (4-6) (Il- IS) (20- 30) Шостак, 1988
/(14- 17) (19) (>24) (4) (10) (21) Briedermann etal., 1989
Cervus dama /(7-9) /(13) /(13- 17) /(13- 17) /(17- 25) /(3- 5) /(9- 13) /(13- 24) Chapman, Chapman, 1970
Cervus nippon /19 /28 /(3-4) 6 /17 /(18- 20) 28 Ueckermann, Scholz, 1971
/(4) /(12) /(20) Koike, Ohtaishi, 1985
Damaliscus dorcas /18 /33 /33 /33 (24) 6 33 (12) 18 (18- 24) 33 Olivier, Greyling, 1991
Damaliscus korrigum /18 /24 /30 />30 /24- 30 /8- 10 /18 /24 Mertens, 1984
Giraffa Camelopar- dalis /48 /(60) 66 /66 /(72) 78 /(54) 66 /(до 10- 12) /(18- 30) /(42) Hall-Martin, 1976
Muntiacus reeves i /(14) 14.5 /(16,5) 18.5 /(18,5) 20 M** (4)/ (18,5) 20 p** (14) 15/ (18,5) 20 (20) 21/ (18,5) 20 (<0,25) 5 (6) 11,5/ (5) 10 (8) 19- 23/ (13) 21 Chapman et al., 1985
Nemorhae- dus goral /(16) 24 /24 /30 36 24 (0,5- 3) 6 (6) 12 (16) 24 Бромлей, 1956; Соломкина, Хохряков, 1993
Odocoileus hemionus /12- 13 /16 /17- 18 18- го 28- 29 12 20 29 Rees et al., 1966
Odocoileus virginianus /8- 10 /10- 12 /12- 18 12- 18 (=19) (2,5-4] 4-6 (6-8) 8- 10 (10- 18) Brokx, 1972
/6-7 /(lO- ll) 11-12 /(lO- ll) 11-12 /(lO- ll) 11-12 /(18- 19) /(3) 4 /(7) 9- 10 /(11- 13)18 Severinghaus, 1949
Глава 3. Определение возраста по признакам зубов
53
Таблица 3.3 (продолжение)
Вид c P Источник
Oreaninos americanus /(15- 16) 23 /(26- 29) 38-40 /(38- 40) 48 /(48) (23-29) 38-40 /(6- 10) 15- 16 /(10- 16) 23 /(15- 29) 38- 40 Brandborg, 1955 (no Larson, Taber, 1980)
Oryx leucoryx 24-26 27-30 31-3'4 34-35 27-42 6-7 19-23 31-42 Ancrenaz, Delhomme, 1997
Ovibos moschatus /(24— 30) /(30- 36) /(48) (48- 54) (36- 48) (6) (12) (30) Henrichsen, Grue 1980 (no Vigal, Machordom, 1985)
Ovis ammon /(17- 19) /(24— 27) /(32- 34) (42- 45) (35- 50'1*** (24- 27) (5) (10- 14) (32- 34) Habermehl, 1985
Ovis canadensis /(12) 16 /(36) 42 /48 /48 /(30- 36)42 /(6) 12 /(12) 16 /(30- 36) 42 Larson, Taber, 1980
Ovis canadensis (Ovis dalli) /(13- 16) /(25- 28) /(33- 36) /(45— 48) /(25- 32) /(1- 4) /(8- 13) /(22- 40) Hemming, 1969
Rupicapra rupicapra /(15- 17) (18- 19, 24)*** /(27- 30) /(36) (30- 44)*** (38- 44) (30- 40)*** (27- 30) (28- 44)*»* (4-6) (3- 4)*** (15- 19) (15- 16)*** (27- 31) (27- 32)*** Habermehl, 1985
(13- 15) (25- 29) (36- 39) (36- 46) (25- 29) (2-3) (9-13) (22-25) Perez-Barberia, Mutuberria, 1996
Rangifer tarandus /9- 13 ZU- 15 /11- 15 /11- 16 /21- 28 /3-5 /10- 15 /15- 28 Miller, 1974
/(4) 6 /(6- 15)15- 16 /(15- 27) 27-28 Клевезаль, Соко- лов, 1997
Saiga tatarica /(13- 14) /(15) /(16- 18) (17- 19) (12- 20) (2-3)/ (1-2) 2-3 (5-7)/ (4) (15)/ (10- lD Соколов, Рашек, 1961
/(13- 14) /(17) /(19) (15) (2-3)/ 3 (7-9) (4) /7-9 (15- 17) Банников и др., 1961
Sus scrqfa (12- 14) (22- 27) /(19- 22) (7- 10) (7- H) (14- 18) (4-6) (12- 14) (26 33)/ (22- 26) Baubet et al., 1994; Matschke, 1967
(12- 14) (20- 24) (6-9) (10-12) (15-18) (4) (11-12) (го- 36) Genov, 1992 (Алжир)
Sylvicapra grimmia /(34- 37) /(34- 37) /(34- 37) (22- 25) 26 (1-4) 4-7/ (1-3) 4 (8- 10) 10-18 /(8-10) 15-20 (18-25) 26/ (15-23) 22-25 Wilson et al., 1984
54
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 3.3(окончание)
Вид i', c P m22 m3 Источник
Tayassu tajacu (10,5- 15) /(10- 13,5) (13- 18) /(13- 15) /(9- H) (7- 10) (16,5- 21) (4-6) (9- 12,5) (18,5- 23,5) Kirkpatrick, Sowls, 1962
Taurotra- gus oryx /24- 27 /28- 29 /30- 31 /32- 35 /36- 40 /9- 13 /24- 27 /28- 29 Jeffery, Hanks, 1981
Trage- laphus scriptus (21) (24- 26) (4,5) 7 (8,5- 10) 14 (19- 21) 26 Simpson, 1973
Примечание: * В скобках указан возраст, когда зуб находится в стадии прорезания, в осталь-
ных случаях — возраст начала функционирования; косой чертой разделены данные для вер-
хних и нижних зубов; если не указано, то либо возраст практически одинаков, либо у авторов
дано без указания, какие зубы, верхние или нижние авторы имели в виду. ** М — самцы, F —
самки. *** —данные разных авторов.
' Зарипов (1964); 2 Кнорре, Шубин (1959); ’Белейшие (1970).
оценивать с точностью до месяца возраст оленей Cervus dama до возраста 38
мес. и С. elaphus до 40 мес.
Возраст прорезания одноименных зубов в верхней и нижней челюстях может
различается, но как правило эти различия значительно меньше, чем межиндиви-
дуальные.
Существенные различия в данных разных авторов по прорезанию зубов у
некоторых копытных (Odocoileus virginianus, Sus scrofa—табл. 3.3) можно объяс-
нить, по крайней мере отчасти, межпопуляционной изменчивостью.
3.1.7. История применения
Метод известен давно. Для определения возраста домашних копытных при-
менялся еще во времена Аристотеля (Корневен, Лесбр, 1929).
3.2. Рост зубов
После прорезания зуб некоторое время растет в длину за счет нарастания
дентина, при этом в апикальной части корня канал зуба (полость пульпы) открыт.
По мере замедления роста зуба в длину и нарастания цемента на корне апикаль-
ное отверстие закрывается (рис. 3.3). Нарастание цемента на корне идет парал-
лельно со стачиванием коронки в результате чего шейка зуба выдвигается из
альвеолы. Иногда нарастание цемента столь мощное, что существенно компен-
сирует стачивание коронки. Все эти особенности можно использовать для оцен-
ки возраста.
Глава 3. Определение возраста по признакам зубов
55
3.2.1. Рост зуба в длину
Этот признак используется для опреде-
ления возраста лишь отдельных видов, на-
пример, рост клыка в длину у бородавоч-
ника Phacochoerus aethiopicus (см. гл. 14).
Как и у любого ростового признака, инди-
видуальная изменчивость велика.
3.2.2. Замыкание апикального
отверстия корня
Большинство постоянных зубов быстро
достигают дефинитивной высоты, и апи-
кальное отверстие корня замыкается рано.
Исключение составляют клыки хищных и
некоторые зубы копытных. Степень зарас-
тания апикального отверстия зуба легко оп-
ределить визуально, когда зуб вынут из аль-
веолы (см. рис. 3.3).
Открытый апикально канал клыка у хищ-
ных среднего размера (шакалов, песцов,
лисиц) несомненный признак сеголетка.
Рис. 3.3. Схема строения зуба.
Следует помнить, что все особи с открытым каналом — сеголетки, но не
все сеголетки имеют открытый канал. Причиной тому — межиндивидуаль-
ные различия в темпе роста особи (и зуба) и времени рождения особи. Имеет
место и межпопуляционная изменчивость. У волка на Дальнем Востоке апикаль-
ное отверстие клыка закрывается у сеголетков уже в ноябре - начале декабря
(Юдин, 1992), в Пермской обл. и Удмуртии оно еще не закрыто в январе (Вега-
нов, 1984). Поэтому у мелких и средних хищных по относительной ширине по-
лости пульпы клыка (см. гл. 3.3) удается распознать больше сеголетков в выбор-
ке, чем по наличию открытого апикального отверстия.
У крупных хищных рост клыков медленный и апикальное отверстие закрыва-
ется постепенно в течение нескольких первых лет жизни (см. гл. 13 Ursus
americanus). К трем годам закрывается апикальное отверстие у льва Panthera
leo, у самок раньше, чем у самцов, в нижних клыках раньше, чем в верхних
(Smuts et al., 1978). По величине апикального отверстия клыка медведей можно
различить те же возрастные группы, что и по ширине полости пульпы: сеголетки,
годовалые и двухгодовалые и старше (см. гл. 13 Ursus americanus). У крупных
млекопитающих (крупнее волка и рыси) выделять два первые годовых класса
проще по величине апикального отверстия, чем по ширине полости пульпы.
По зарастанию апикального отверстия можно выделить несколько возраст-
ных групп и у некоторых копытных (см. гл. 14 Sus scrofa, Cervus elaphus).
56
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
3.2.3. Положение шейки зуба относительно альвеолы
У молодых особей, как правило, весь корень зуба находится в альвеоле. С
возрастом, по мере нарастания цемента на корне, корень как бы выдвигается из
альвеолы и этот видимый участок корня (расстояние от края альвеолы до шейки
зуба) увеличивается. Есть примеры использования этого признака для оценки
возраста (см. гл. 9 Crocidura, гл. 13 Canis latrans, Urocyori). У хищных общая
тенденция увеличения этого параметра с возрастом хорошо выражена, но инди-
видуальная изменчивость велика. В осенне-зимний период по этому признаку
можно выделить сеголетков у хищных, но не всех, у части сеголетков этот пара-
метр такой же, как и у старших особей. Из-за индивидуальной изменчивости не
удалось использовать этот параметр для выделения возрастных групп медведя
Ursus maritimus (Lone, 1970).
3.2.4. Рост корней и нарастание цемента
У некоторых грызунов формирование и рост корней происходит в течение
длительного периода, что позволяет использовать длину корней для определения
возраста. Начиная с работ Кошкиной (1955) и Mazak (1963), появление и длина
корней стали популярными признаками для определения возраста лесных по-
левок (рис. 3.4) (см. гл. 12 Clethrionomys}. Следует отметить, что у размно-
жающихся несколько раз в году грызунов скорость роста корней зависит от сезо-
Ш IV
Рис. 3.4. Условные возрастные группы рыжей полевки, Clethrionomys glareolus, выделяемые по
характеру пц в естественных популяциях (по Башениной, 1981).!—сам< кгтоятельный молодняк,
II—молодые в возрасте примерно 1,5-3 мес., III—сеголетки в возрасте около 3-4 мес., достиг-
шие размеров взрослых, IV — взрослые, перезимовавшие. Корни начинают формироваться в
группе III и хорошо развиты и высота их увеличивается с возрастом в группе IV.
Глава 3. Определение возраста по признакам зубов
57
Рис. 3.5. Изменение с возрастом общей высоты т, (верхняя кривая) и высоты коронки этого
зуба (нижняя кривая) у бобра, Castor fiber (по данным Бородиной, 1970а).
на рождения особи (Lowe, 1971). У полевок Clethrionomys glareolus весенних
выводков корни начинают формироваться в возрасте 55-60 дней, у осенних — в
возрасте 120-140 дней (Оленев, Каляева, 1982).
Рост корней используют для определения возраста и других полевок, напри-
мер, югославской полевки Dolomys (Krystufek et al., 2000), ондатры (в сочета-
нии со стертостью коронки) (см. гл. 12 Ondatra).
При использовании лабораторных грызунов для разработки критериев опре-
деления возраста по росту и развитию корней следует иметь в виду, что скорость
развития корней в природе и в лаборатории может различаться (Lowe, 1971;
Perrin, 1978; Hansson, 1983а; Оленев, 1989).
Обычно для измерения корней зубы вынимают из челюсти, но есть опыт из-
мерения зуба и корней на рентгеновских снимках нижней челюсти (Alibhai, 1980;
Krystufek et al., 2000).
Своеобразен рост корней щечных зубов бобра. У них интенсивное стачива-
ние коронки компенсируется мощными отложениями цемента в корневой части
(рис. 3.5). Толщина цемента может служить дополнительным (к числу годовых
слоев в нем формирующихся) критерием возраста (см. гл. 12 Castor).
3.3. Ширина полости пульпы («канала») зуба
3.3.1. Область применения
Метод используется (обычно — посмертно) при определении возраста хищ-
ных, реже — летучих мышей, еще реже — копытных (в последнем случае нужна
58
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
поправка на возраст прорезания постоянного зуба). Для оценки возраста живых
животных он применим, если удается получить рентгеновские снимки зубов обез-
движенных особей.
3.3.2. Морфологическое обоснование
Однокорневые зубы к началу функционирования имеют полностью сформи-
рованные коронку и часть корня. Ширина функционирующего зуба в самой ши-
рокой его части (близ шейки зуба или в верхней части корня) меняется незначи-
тельно, поскольку отложения цемента в этой области малы. В начале функциони-
рования таких зубов, как клыки хищных, не подверженных интенсивному стачи-
ванию, внутренняя полость (полость пульпы, «канал») большая, а стенки зуба
тонкие. С возрастом происходит отложение дентина со стороны тканей пульпы, и
ширина полости уменьшается, а стенки зуба утолщаются. Со временем, ско-
рость этого процесса резко уменьшается. В клыках многих хищных это умень-
шение совпадает с замедление роста всего животного в связи с наступлением
половой зрелости. Позже межиндивидуальные различия в темпе заполнения по-
лости пульпы существенно перекрывают возрастные.
3.3.3. Разрешающая способность
Метод удобен для отделения сеголетков от старших в одномоментно взятой
выборке. В случае хищных, размножающихся на втором году жизни, надежность
разделения выше осенью — в начале зимы, чем в конце зимы — весной. При растя-
нутом периоде сбора материала этим методом можно выделить явно молодых и
явно взрослых с большой переходной группой. Приблизительная оценка возраста
отдельной особи (как сеголетка или старшего) возможна, если относительная
ширина полости пульпы у нее равна или больше среднего для сеголетков данного
вида или равна или меньше среднего для взрослых (табл. 3.4).
При работе с мелкими и средними хищными, когда материал взят в промыс-
ловый сезон (при анализе возрастного состава пушных зверей добытых в осен-
не-зимний период) метод позволяет отделить явных сеголетков, чтобы возраст
остальных определять по годовым слоям. Хотя при этом могут быть выделены не
все сеголетки, такой прием позволяет сэкономить время и деньги.
Для выделения сеголетков крупных хищных этот метод не имеет преимуществ
по сравнению с использованием диаметра апикального отверстия клыка (см.
выше, раздел 3.2.2).
При определении возрастного состава выборок волка Canis lupus и рыси Felis
lynx этот метод (в модификации Смирнова, см. ниже, раздел 3.3.4) был признан
не уступающим по надежности методу определения возраста по годовым слоям,
поскольку ошибка индивидуального определения возраста по ширине полости
пульпы клыка была случайной (т.е. завышение и занижение оценок равновероят-
ны) и потому возрастной состав существенно не отличался от определенного по
числу годовых слоев (Смирнов, 1983, Малафеев и др. 1986). Монахов (2004) на
Глава 3. Определение возраста по признакам зубов
59
Таблица 3.4. Критерии выделения возрастных групп по ширине полости
пульпы («по ширине канала») клыка у некоторых хищных*.
Вид Район Период гибели Ширина полости пульпы клыка в % максимальной ширины клыка Источник
Сеголетки Год и старше
Alopex Зап. декабрь >60 <40 (?)-Смирнов, 1960
lapopus Сибирь март >50 <35 (?)-Смирнов, 1960
Гренландия октябрь- март >40 <30 (?) - Grue, Jensen, 1976
Европейский Север России осень- зима >45 <25 (?) - Долгов, Россолимо, 1966
Canis latrans США: Квебек ноябрь- январь >48 <48 н-Jeanetal., 1986
Canis lupus Урал зима? >70, в среднем 80 (?)-Смирнов, 1960
Felis lynx Урал осень- зима >70 годовики =35,2 года и старше <25 в - Малафеев и др., 1986
Felis pardinus Испания не известен до года > 60 <40 н - Zapata, Perea etal., 1997
Lutra canadensis США: Миннесота ноябрь- декабрь >50 н - Kuehn, Btrg, 1983
Martes americana Канада: Ванкувер ноябрь- декабрь всреднем М-44, F-40 14-16 н - Nagorsen et al., 1988
О-ва близ. Аляски декабрь- февраль в среднем. М-46, F-42 23-25 н - Nagorsen et al., 1988
Канада: Онтарио ноябрь- декабрь М-36,5 F >32.5 н - Dix, Strickland, 1986
Martes martes Европейская часть России промысл, сезон? в среднем 36 <27 в - Граков, 1970
Martes zibellina Сев-зап. Прибайкалье октябрь- ноябрь М 40-80 F 34-61 <40 <34 в-Монахов, 1965
Звердловск., и Тюменск. обл, Краснояр- ский кр. промысл, сезон М< 31 F < 30 (?) - Монахов, 2004
зооферма октябрь >39 <33 (?) - Монахов, 1967
Якутия промысл, сезон в среднем М-51, F-46 25 30 в - Шадрина, 1988а
Вост. Сибирь октябрь- январь М >40 F >33 <40 <33 в-Давыдов, 1984
Meles meles Швейцария ? >50 н - по графику из Graf, Wandeler, 1982
Mephitis mephitis США: Ю.Дакота промысл, сезон в среднем. M-51,F-36 в среднем M-21,F-18 н - Fredrickson, 1983
60
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 3.4 (окончание)
Вид Район Период гибели Относительная ширина полости пульпы клыка в % Источник
Сеголетки Год и старше
Mustela erminea Тюменская обл. промысл, сезон М >20 F>16,5 <20 <16,5 в-Копеин, 1967
Mustela sibirica Сибирь промысл, сезон >25 <20 в - Войлочников, Михайловский, 1970
Зап. Сибирь промысл, сезон ? М >28 F >21 в-Синицын, 1982
Дальний Восток России? промысл, сезон? М >30 F >19 <24 <16 (?) - Панкратьев, 1971
Nyctereu- tes procyo- noides Финляндия сентябрь- ноябрь >50 <50 н - Kauhala, Helle, 1990
Дальний Восток России осенне- зимний сезон М >44 <39 в-Юдин, 1977
Pseud- alopex culpaeus Аргентина май- сентябрь >50 <50 (?) - Zapata, Funes et al., 1997
Taxidea taxus США: Ю. Дакота осень в среднем 60 в среднем М-35, F-29 н - Fredrickson, 1983
Urocyon cinereo- argenteus США: Висконсин ноябрь- январь >40 <38 в - Root, Payne, 1984
Vulpes vulpes fulvus США: Миннесота промысл, сезон >35 <35 н - Simon, Frydendall, 1981
Vulpes vulpes Урал промысл, сезон >41 (?) Корытин, Ендукин, 1982
Япония январь ноябрь- >33 <33 1990 (?) - Uraguchi et al.,
Дальний Восток промысл, сезон М >45 F >38 <40 <35 (?)-Юдин, 1986
России Швейцария июль- сентябрь >54 <44 н-Kappeler, 1985
Италия октябрь- декабрь >40 <40 H-Kappeler, 1985
январь- март >36 <30 h —Kappeler, 1985
январь- февраль F > 40 <30 в - Cavallini, Santini, 1995
Примечание: * М — самцы, F — самки; если не указано, то вместе для самцов и самок;
в — верхний клык, н — нижний, (?) — не указано.
Глава 3. Определение возраста по признакам зубов
61
большом материале из разных регионов Сибири сопоставил разделение на сего-
летков и взрослых в промысловых выборках соболя Martes zibellina по ширине
полости пульпы и по годовым слоям в цементе. Ошибка (принимая число слоев
за оценку истинного возраста) составила в среднем 5% (в разных выборках от
3,8 до 7,3%) и была не направленной, так что несовпадающие оценки взаимно
компенсировались.
3.3.4. Методика
На распиле или на рентгеновском снимке клыка измеряют ширину полости
пульпы и общую ширину клыка в самой широкой его части. Чтобы нивелировать
индивидуальную изменчивость размеров зуба, ширину полости пульпы выража-
ют в процентах общей ширины зуба.
Такое представление результата позволяет несколько варьировать место изме-
рения зуба. Измерения в области шейки или немного ниже дают примерно одина-
ковое значение относительной ширины полости пульпы, поскольку форма полости
пульпы на этих участках практически повторяет наружную форму зуба.
Не следует брать клыки, если коронки повреждены так, что обнажена полость
пульпы; Grue, Jensen (1976) показали, что в этом случае результат определения
возраста будет неверным.
Измерения, как правило, проводят по большой оси поперечного разреза клы-
ка, поскольку это технически более удобно. Кроме того, как было показано при
исследовании колонка, Mustela sibirica (Войлочников, Михайловский, 1970),
измерение по малой оси дает менее четкое разделение возрастных групп.
На рентгенограммах зубов полость пульпы выглядит темной, стенки зуба —
более светлыми. Рентгеновские снимки не следует использовать при работе с
мелкими зубами. Так, Fredrickson (1983) затруднялся отличать на рентгеновских
снимках полость пульпы от дентина в мелких зубах самок скунса, Mephitis
mephitis.
У летучих мышей оказалось возможно видеть полость пульпы под бинокуля-
ром, когда целый клык погружен в глицерин (Baagoe, 1977).
Используют либо нижние, либо верхние клыки; было показано, что выбор
клыков на результат существенно не влияет. Например, у сеголетков енотовид-
ной собаки, Nyctereutesprocyonoides, максимальная относительная ширина поло-
сти пульпы верхних клыков 44%, нижних — 41% (Юдин, 1977). Различия отно-
сительной ширины полости пульпы разных клыков одной особи песца Alopex
lagopus, как правило, не превышали 5% (Grue, Jensen, 1976).
Для уточнения разделения возрастных групп Смирнов (1983, 1988) предло-
жил брать логарифм произведения оценок относительной ширины (в процентах)
полости пульпы, полученных при измерении по малой и большой осям попереч-
ного разреза зуба. Этот прием позволил выделить несколько возрастных групп у
волков и рысей (см. гл. 13 Canis lupus, Felis lynx).
Jenks et al. (1986), определяя точку разделения возрастных групп как полу-
сумму средних величин соседних возрастных групп (см. гл. 13 Martespennanti),
62
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
выделили 4 возрастные группы и получили возрастные распределения выборок,
не отличающиеся от таковых, полученных при определении возраста по годовым
слоям цемента.
Для оценки возраста отдельных особей предлагают уравнения регрессии, что
может быть полезно, если материал собран в течение всего года. Например,
Thomson, Rose (1992), используя в качестве показателя не относительную ши-
рину полости пульпы, а обратную величину — отношение ширины зуба к шири-
не полости пульпы (измерено на рентгеновских снимках на уровне десны), на
возрастном интервале от сеголеток до 54 мес. дают такие уравнения для оценки
возраста динго, Canis farniliaris dingo'.
верхние клыки: In (возраст, мес.) = 0,232 • соотношение + 1,865;
нижние клыки: In (возраст, мес.) = 0,183 • соотношение + 1,898.
(См. также гл. 13 Canis lupus, гл. 14 Sus scrofa).
3.3.5. Изменчивость показателей
Изменение скорости заполнения полости пульпы в клыках хищных, видимо,
соответствует общему изменению темпа роста организма. У размножающихся
на втором году жизни хищных полость пульпы заполняется быстро в первое лето
жизни и ее заполнение не прекращается зимой, хотя темп его явно снижается.
Полость пульпы клыков песца Alopex lagopus уменьшается с октября по март
примерно на 15% (Смирнов, 1959), клыков соболя Martes zibellina с октября по
январь — на 10% (Давыдов, 1984).
Межиндивидуальные различия относительной ширины полости пульпы в од-
ной и той же возрастной группе велики. Например, у сеголетков песца, добытых
в декабре, ширина полости пульпы клыка варьирует от 60 до 90% (Смирнов,
1960). Эти различия определяются как различиями в темпе роста, так и различи-
ями в сроках рождения особей. Даже у сезонно размножающихся хищных осо-
би, рожденные в начале и в конце сезона размножения, существенно различают-
ся по темпу роста. Например, в начале августа относительная ширина полости
пульпы клыков у барсука М. meles, родившегося в конце марта, была 30%, а у
родившихся на 10 дней позже — 19-20% (Клевезаль и др., 1991).
Межиндивидуальные различия в темпе роста и снижение темпа роста с на-
ступлением половой зрелости приводят к тому, что у хищных, размножаю-
щихся на втором году жизни, даже в выборке, взятой в сжатые сроки, но в
конце зимы - начале весны, не удается безошибочно определить особей преды-
дущего года рождения. Выборки соболя, Martes zibellina, по относительной ши-
рине полости пульпы можно было четко разделить на сеголетков и старших, если
они были взяты в октябре-ноябре, но не в феврале-марте (Монахов, 1965). У
лисицы, V. vulpes, Швейцарии разделение сеголетков и старших, безошибочное
осенью (п = 634), дало ошибку в декабре 0,4% (п = 231), в январе 2% (п = 156)
и 5,5% (п = 345) в феврале-марте (Kappeler, 1985), а в выборке енотовидной
собаки, Nyctereutesprocyonoides, взятой в феврале-марте, этим методом практи-
чески не удалось выделить молодых (Kauhala, Helle, 1990).
Глава 3. Определение возраста по признакам зубов
63
Как и по любому ростовому признаку, по относительной ширине полости
пульпы могут иметь место различия между самцами и самками у видов, у кото-
рых скорость роста самок и самцов различается. Например, у колонков, Mustela
sibirica, добытых зимой, относительная ширина полости пульпы клыков самцов
сеголетков в среднем на 10% больше, чем у самок (Войлочников, Михайловс-
кий, 1970). Такие же различия, на 10%, обнаружены и у горностая, Mustela
erminea, Новой Зеландии в возрастной группе до 4 мес. (King, 1991).
Есть и межпопуляционные различия скорости заполнения полости пульпы.
Небольшие (в среднем на 2-3%), но статистически значимые различия были
выявлены на большом материале у куниц Martes americana, добытых в ноябре-
декабре на о-ве Ванкувер и в штате Онтарио (Nagorsen et al., 1988). В случае
межпопуляционных различий справедлива общая для ростовых признаков зави-
симость размеров и скорости роста от абиотических факторов среды (Мина,
Клевезаль, 1976). Огрубляя, можно сказать, что если исследуемая популяция
обитает в более мягком климате, чем референтная, можно ожидать, что данная
относительная ширина полости пульпы будет достигнута раньше.
Межгодовые различия ширины полости пульпы могут даже превышать меж-
популяционные. У добытых в один и тот же месяц сеголетков енотовидной соба-
ки Финляндии межгодовые различия достигали в среднем 10% (Kauhala, Helle,
1990), у соболя Прибайкалья — 8% (Монахов, 1965). При таких различиях, ви-
димо, нет смысла пытаться найти точные границы разделения выборки на моло-
дых и взрослых, характерные для данной популяции вне зависимости от условий
года.
На фоне всей описанной выше изменчивости все-таки проявляется некото-
рая общая закономерность скорости зарастания полости пульпы. Если суммиро-
вать данные по разным видам разных авторов, то можно выделить как бы 4
группы хищных:
(1) крупные — Canis lupus, Felis lynx, Ursus;
(2) средние псовые — Canis latrans, Nyctereutes procyonoides, Alopex lagopus,
V. vulpes, и крупные куньи — Meles, Lutra;
(3) средние куньи — Martes;
(4) мелкие куньи — Mustela sibirica, M. erminea.
В первой группе у сеголетков зимой ширина полости пульпы в среднем не
менее 60-70% общей ширины клыка, во второй — не менее 50%, в третьей —
30-40%, в четвертой — 20-30%.
Смирнов (1960) связывал скорость зарастания полости пульпы с общими
размерами животного. Однако различия в размерах тела и черепа между вол-
ком, Canis lupus, и рысью, Felis lynx, относящимся к одной группе, больше, чем
различия между рысью и койотом, Canis latrans, которые относятся к разным
группам. Возможно, что скорость заполнения полости пульпы связана с разме-
ром тела не прямо, а опосредованно, через возраст достижения размеров тела
взрослых. Волк и рысь, как правило, приносят потомство (и, следовательно,
достигают размеров тела взрослых) в 2-годовалом возрасте, а койот, как и ос-
тальные виды второй группы, в годовалом.
64
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Конкретные данные по относительной величине полости пульпы клыков хищ-
ных разных видов в разные сезоны приведены в таблице 3.4. Если нужного вида
в таблице нет, то можно воспользоваться следующим огрублением, сделанным
с учетом всего спектра описанной выше изменчивости.
Данную особь осенью - в начале зимы можно уверенно считать сеголетком,
если относительная ширина полости пульпы клыка больше 70% у животных раз-
мера не меньше рыси, больше 60% у животных размера лисицы или барсука,
больше 50% у особей размера куницы, и больше 40% у особей размера колонка
или горностая.
Следует помнить, что все выделенные по такому критерию сеголетки, несом-
ненно, являются таковыми, но не все сеголетки будут таким образом выделены.
3.3.6. Использование метода при работе с копытными
По относительной ширине полости пульпы резцов можно оценить возраст
кабана (см. гл. 14 Sus scrofa). Визуальную оценку ширины полости пульпы пер-
вого резца, вероятно, можно использовать как дополнительный критерий возра-
ста многих копытных (см. рис. 14.2).
Kleymann (1972) предложил определять возраст косули, С. capreolus, по пло-
щади полости пульпы щечных зубов, в частности р3, определяемой на рентгено-
вских снимках мандибулы в сагиттальной плоскости. Судя по приведенным им
графикам, по этому признаку годовики четко отличаются от 2-годовалых, разни-
ца между остальными годовыми группами не столь очевидна. Он полагал, что
этот метод может быть использован при определении возраста других копытных,
но метод не получил распространения.
3.3.7. История применения
Метод впервые предложен и обоснован В.С. Смирновым (1959, 1960) и по-
лучил широкое распространение при определении возраста промысловых хищ-
ных мелких и средних размеров.
3.4. Стачивание зубов с возрастом
3.4.1. Область применения
Метод наиболее пригоден для оценки возраста тех видов животных, у которых
возрастные изменения степени стертости существенно больше межиндивидуальных
различий. Этому требованию в наибольшей мере отвечают зубы с большой функцио-
нальной нагрузкой, в частности, зубы копытных, грызунов и насекомоядных. В ряде
случаев он успешно используется при исследовании хищных. Он не годится для
групп, имеющих все зубы с постоянным ростом (зайцеобразные, многие полевки).
Обычно этот метод используют для посмертного определения возраста. Для при-
жизненного определения он может быть полезен в тех случаях, когда есть возмож-
Глава 3. Определение возраста по признакам зубов
65
ность получить отпечаток жевательной поверхности, измерить или оценить на глаз
высоту коронки или площадь жевательной поверхности (см. ниже, раздел 3.4.4).
Метод применяется для оценки относительного возраста мелких грызунов, гру-
бого разделения хищных на несколько возрастных групп, выделения годовых клас-
сов копытных, долго живущих грызунов и насекомоядных. Для одномоментно
взятых выборок этот метод используют совместно с определением возраста по
годовым слоям: по степени стертости материал разделяют на дискретные возраст-
ные группы, по годовым слоям в зубах «модальных особей» определяют возраст
каждой группы (так, например, поступил Simpson (1973) при работе с бушбоком
Tragelaphus scriptus), но иногда определяют возраст и «промежуточных» особей.
3.4.2. Морфологическое обоснование
У хищных, копытных, насекомоядных, рукокрылых и многих грызунов полнос-
тью прорезавшийся, но еще не функционирующий зуб имеет острые эмалевые вер-
шины или бугорки. По мере функционирования эти вершины постепенно стираются,
и когда эмаль на определенных участках снашивается полностью, появляются обна-
жения дентина, по цвету несколько отличающегося от эмали. Далее, с возрастом
стачиваются и бугорки, поверхность зубов уплощается, еще позже уменьшается
высота всей коронки. В процессе стачивания меняется и форма зубов.
У некоторых млекопитающих (см. гл. 12 Castor, Ondatra, Clethrionomys) ду-
ющая поверхность коренных зубов плоская изначально; по мере стачивания ри-
сунок поверхности практически не меняется, но высота коронки существенно
уменьшается.
В ряде случаев принимают, что скорость стачивания отдельных зубов посто-
янна на больших возрастных интервалах (см. гл. 13 Crocuta, гл. 14 Bison bison,
Girajfa), но чаще при детальном анализе обнаруживается изменение скорости
стачивания с возрастом. Solounias et al. (1994) полагали, что скорость стачива-
ния зуба высока в раннем возрасте, уменьшается в среднем и в конце жизни
вновь увеличивается, потому что в раннем возрасте мала поверхность стачива-
ния, а в позднем нарушается соотношение эмали и дентина. Уменьшение скоро-
сти стачивания с возрастом отмечено для разных типов зубов и для разных групп
млекопитающих: для многих копытных (Spinage, 1973: Соколов и др., 1996; Кле-
везаль, Соколов, 2004), ондатры (Смирнов, Шварц, 1959), бурозубок (см. гл. 9
Blarina) и, вероятно, имеет место у большинства млекопитающих. Однако уве-
личение скорости стачивания у старых особей отмечено даже не во всех популя-
циях одного вида (Клевезаль, Соколов, 2004).
3.4.3. Разрешающая способность
Метод связан не с увеличением размера тела, а с прожитым интервалом вре-
мени. Он «работает» от начала функционирования постоянных зубов до конца
жизни особи. Стачивание продолжается в течение всей жизни и, в отличие от
ростовых признаков, нет остановки или замедления процесса в зимний период.
66
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
При работе с взятыми в короткие сроки выборками размножающихся раз в
году долгоживущих млекопитающих этот метод дает возможность выделить не-
сколько возрастных групп и даже годовые классы. При этом выделение возрас-
тных групп затруднено у старших особей, когда зубы сильно стерты. У коротко
живущих грызунов этим методом можно выделить несколько возрастных групп
в пределах одного года.
3.4.4. Методика
Предварительно надо тщательно выбраковывать особей с поврежденными
зубами. Это особенно важно при работе с материалом капканного промысла
хищных. По данным Григорьева, Попова (1952) у песцов, добытых капканным
промыслом, около 70% особей имели механическое повреждение зубов. Далее
ранжируют материал по степени стертости на основании визуального осмотра и
(или) измерения высоты коронки зуба (или зубов). Выборку, взятую в течение
короткого периода, делят на возрастные группы на основании «разрывов» в из-
менении рисунка поверхности стачивания или высоты коронки. Для размножаю-
щихся раз в году долгоживущих млекопитающих каждую последующую возра-
стную группу считают старше предыдущей на год. При этом следует иметь в
виду, что индивидуальная изменчивость степени стертости с возрастом увеличи-
вается, а надежность метода уменьшается.
Возрастные группы характеризуют описанием и (или) рисунками степени стер-
тости зубов. Если измеряют высоту коронки зубов, то приводят конкретные оценки
высоты для возрастных групп или уравнение регрессии высоты коронки по воз-
расту на определенном возрастном интервале. Можно характеризовать стертость
в баллах и далее работать с этими данными как с размерными признаками. Слож-
ную систему балльных признаков иногда используют при работе с копытными
(см. ниже, раздел 3.4.6.3).
Надежность выделения возрастных групп увеличивается, если есть эталон-
ные черепа от особей известного возраста (см. ниже, раздел 3.4.6.1). В идеале
эталонные особи должны быть из того же региона, откуда взята исследуемая
выборка. Можно калибровать выделенные возрастные группы, определив воз-
раст особей из каждой возрастной группы по годовым слоям в цементе или
дентине. Следует, однако, осторожно относится к материалу известного возрас-
та, полученному при содержании животных в лаборатории или меченых в при-
роде, но не в районе исследования.
При необходимости прижизненного определения возраста можно оценить на
глаз степень стертости или получить отпечатки зубов живых животных (см. гл.
12 Cynomys, гл. 13 Pantherapardus, Crocuta, гл. 14 Capreolus, гл. 15 Equus burchelli,
а также гл. 8).
Если период размножения растянут и (или) если материал собран в течение
длительного периода, можно использовать графический способ Смирнова (1960)
(см. гл. 1 раздел 1.2.1.2). Этот способ был успешно применен для выделения
возрастных групп насекомоядных и грызунов, с использованием в качестве по-
Глава 3. Определение возраста по признакам зубов
67
казателей, как высоты коронки зуба, так и степени стертости, оцененной в бал-
лах (см. гл. 9 Crocidura, Blarina, гл. 12 Gerbillus, Allactaga).
3.4.5. Изменчивость показателей
Скорость стачивания зубов с возрастом подвержена индивидуальной и меж-
популяционной изменчивости как в силу влияния качества корма на стачивание,
так и в силу индивидуальных особенностей твердости эмали и дентина. Микро-
твердость эмали, определяющая скорость ее стачивания, может быть различна не
только у близких видов, но даже у особей, рожденных в разные годы, принадле-
жащих разным генерациям одного года и даже у одновозрастных особей (Adam-
czewska-Andrzejewska, 1966; Kierdorf, Becher, 1997).
Географическая изменчивость не дает возможности дать характерную для вида
схему определения возраста по стачиванию зубов. Например, существенные раз-
личия в темпе стачивания зубов европейских, сибирских и кавказских популя-
ций кротов, Talpa еигораеа, сделали невозможным создание единой для вида
схемы определения возраста (Депарма, 1954). Межпопуляционные различия как
в скорости стачивания, так и в начальной высоте зубов являются основным ис-
точником ошибок при оценке возраста (Hewison et al., 1999).
Другим источником ошибок является индивидуальная изменчивость твердо-
сти тканей зуба и предпочтения кормов, приводящая к существенным различиям
в скорости стачивания зубов. Так, питавшиеся относительно мягкими кормами
годовалые белки Sciurus vulgaris в июле-августе имели стертость зубов, харак-
терную для сеголетков (Егоров, 1958). У сурков Marmota топах известного
возраста степень стертости зубов некоторых 2-годовалых особей была выше,
чем 4-годовалых (Davis, 1964).
У некоторых копытных отмечены различия в скорости стачивания зубов меж-
ду самцами и самками (Erickson et al., 1970; Mitchell, 1971; Miller, 1974; Thomas,
Bandy, 1975; Anderson, 1986; Kerwin, Van Deelen et al., 2000).
3.4.6. Особенности использования метода в отдельных группах
З.4.6.1. Хищные
В большинстве случаев по степени стертости легко отделить молодых от взрос-
лых, иногда удается выделить и старшие возрастные группы (см. гл. 13 Vulpes,
Meles), особенно если есть возможность использовать для сравнения черепа
животных известного возраста. По стертости резцов, клыков и хищнических
зубов, сравнивая со стертостью зубов особей известного возраста, удалось пра-
вильно распределить по двухлетним возрастным группам 15 из 18 волков Canis
lupus известного возраста (от 1 до 14 лет) (Gipson et al., 2000). Однако, у мелких
куньих, например, у Mustela sibirica, добытых в промысловых сезон, на основа-
нии стертости зубов трудно отличить даже сеголетков от старших. В силу разли-
чий в сроках рождения молодняка и индивидуальной изменчивости стачивания
описание изменений стертости годовых классов иногда ограничивают самыми
68 Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
общими указаниями (см. гл. 13 Mustela putorius). Межпопуляционные различия,
однако, могут быть и небольшими. Например, у волков Gipson et al. (2000) не
выявили различий в характере стачивания зубов между особями трех популяций
(Аляска, Миннесота, Онтарио) и между особями из неволи и из природы. В от-
дельных случаях трудности выделения возрастных групп связаны с различной
стертостью зубов правой и левой половинок челюсти, как например, у лисиц
Urocyon cinereoargenteus (Root, Payne, 1984).
Как правило, первыми начинают стачиваться те постоянные зубы, которые
первыми прорезаются. Как следует из раздела 3.1 («Прорезание зубов»), у боль-
шинства хищных первыми прорезаются постоянные первые два резца ит1,. На
основании имеющихся данных можно сделать такое грубое обобщение:
1) у хищных средних размеров {Meles, Vulpes, Lutra, Martes) заметные обна-
жения дентина на т1 появляются в возрасте около 2-3 лет (рис. 3.6);
2) поверхность стачивания нижних резцов у молодых взрослых имеет попе-
речно-овальную форму, у взрослых среднего возраста — округлую или треу-
гольно-округлую, у старых — продольно-овальную (рис. 3.6).
При сравнении оценок возраста по степени стертости и по числу годовых
слоев в зубах оказалось, что по степени стертости можно надежно выделить
только сеголетков, причем часть фактических сеголетков может попасть в стар-
шие возрастные группы. В старших возрастных группах возраст, определенный
по степени стертости, оказывался, как правило, завышен. Ошибка больше в тех
случаях, когда выборка взята в течение всего года. Так, в выборке М. meles,
взятой в течение всего года, в группе, которую по степени стертости следовало
отнести к сеголеткам, оказалось равное число особей без годовых слоев и с
одним годовым слоем, а у особей самой старшей (выделенной по степени стертос-
ти) группы было от 2 до 8 годовых слоев (Van Вгее et al., 1974).
Рис. 3.6. Возрастные изменения жевательной поверхности m1 (I—IV) и нижних резцов (a-d)
лисицы, Vulpes vulpes. I — 8-10 мес., II—20-22 мес., Ill — 32-34 мес., IV—старше 3 лет (М.
Stubbe, 1965). По уточнению М. Stubbe (1989а) — III—IV — старше 2 лет.
с
Глава 3. Определение возраста по признакам зубов
69
3.4.6.2. Непарнокопытные
Для определения относительного возраста непарнокопытных широко исполь-
зуют степень стертости резцов. У непарнокопытных есть верхние резцы и стачи-
вание всех резцов более закономерно, чем у жвачных парнокопытных, которые
верхних резцов не имеют. Привлекает также то, что степень стертости резцов
можно определить у живых животных (см. гл. 15).
3.4.6.3. Парнокопытные
У нежвачных возраст определяют по степени стертости резцов и клыков (см. гл.
15 Sus scrofa). У жвачных стачивание резцов более вариабельный и менее надежный
критерий возраста, чем стачивание щечных зубов. У косули С. capreolus, например,
вообще нет закономерности в стирании резцов (Соколов, 1956). Это, видимо, связа-
но с отсутствием резцов в верхней челюсти: при смыкании челюстей нижние резцы
не имеют твердого упора в верхней челюсти. Тем не менее, форма поверхности
стачивания резцов может служить дополнительным критерием: с возрастом из попе-
речно-овальной она становится трапециевидной, треугольной или округлой (Соко-
лов, Рашек, 1961; Шостак, 1988) (см. рис. 3.1). Этот показатель привлекателен тем,
что дает возможность оценить возраст (хотя и приблизительно) у живых особей.
Mertens (1984) в качестве вспомогательного критерия для оценки возраста
антилопы Damaliscus korrigum предложил использовать длину и ширину повер-
хности стачивания резца, первая с возрастом уменьшается, вторая — увеличива-
ется. Вспомогательным критерием может служить обнажение дентина на повер-
хности стачивания.
Обычно у жвачных парнокопытных критерием возраста считают стачивание
всех щечных зубов, как предкоренных, так и коренных. Однако Brown, Chapman
(1990), используя сложную балльную оценку степени стертости зубов лани Cervus
dama, пришли к выводу, что стачивание предкоренных зубов является менее
надежным критерием возраста, чем стачивание коренных.
Предкоренные и коренные зубы складчатые, лунчатые, поскольку щечные
(наружные) и язычные (внутренние) бугры (или «гребни» — у молодых живот-
ных) коронки зуба изогнуты. У молодого зуба острые бугры разделены зубной
воронкой (чашкой), имеющей вид глубокой щели, которую часто называют также
лункой и которая открыта в передней и задней части. В процессе стачивания
постепенно уменьшается высота бугров, сначала появляются обнажения эма-
ли, затем — обнажения дентина, но лунки еще открыты с одной или с обеих
сторон (как у р3 4и т3на рис 3.7А). Далее площадь обнаженного дентина увели-
чивается, дентин язычного и щечного бугров в передней и задней части зуба
образует единое поле и лунка приобретает вид окантованного эмалью овала
или полумесяца (рис. 3.8). В зависимости от числа бугров в зубе, лунок может
быть 1, 2 и 3. В четырехбугорных зубах (таких как и т2) лунок две, дентин
щечных и язычных бугров сливается в передней и задней части зуба, а также
между лунками. Далее по мере стачивания лунки становятся мелкими и исче-
зают (рис. 3.7В, Г). Когда дентин коронки сточен до обнажения вторичного
дентина, его можно видеть на поверхности зуба как «глазки» (рис 3.8).
70
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Б
Рис. 3.7. Щечные зубы нижней челюсти северного оленя, Rangifer tarandus, в возрасте 2,5 (А),
4,5 (Б), 7-8 (В) и 13,5 (Г) лет. Возраст определен по годовым слоям цемента.
Параллельно стачиванию коронки шейка зуба поднимается над альвеолой. У
очень старых особей коронки стачиваются до обнажения корней. Как правило,
последовательность стирания отдельных зубов соответствует последовательнос-
ти их прорезания, хотя с возрастом это соответствие может нарушаться.
• Как было сказано выше (см. раздел 3.4.5), межпопуляционная и инди-
видуальная изменчивость возрастных изменений степени стертости вели-
Глава 3. Определение возраста по признакам зубов
71
Рис. 3.8. Жевательная поверхность т2 северного оленя, Rangifer tarandus, возраст 8 лет (воз-
раст определен по годовым слоям). Стрелками указаны «глазки» вторичного дентина. Э —
эмаль, Д — дентин, Л — лунки.
ка и не позволяет дать такое описание, которое было бы надежным для
определения возраста особей какого-то вида в целом. Во многих работах по
оценке возраста особей конкретных популяций приводятся описания и ри-
сунки или фотографии поверхности стачивания зубов в разном возрасте. Во
II части данной книги в повидовом описании указано, в каких работах та-
кие описания и рисунки приведены. Если исследователь имесг большую
выборку, взятую в течение одного сезона, он сам разделит ее на возраст-
ные группы, различающиеся по степени стертости зубов. Если же этим
методом необходимо оценить приблизительный возраст отдельных осо-
бей, следует найти в литературе описание, сделанное для выборки особей
того же вида, наиболее сходной с исследуемой по условиям обитания.
Хотя по степени стертости зубов пытаются выделять не только возрастные
группы, но и годовые классы, из-за большой индивидуальной изменчивости стер-
тости зубов одновозрастных особей в большинстве случаев авторы затрудняют-
ся выявить различия степени стертости зубов между соседними годовыми клас-
сами взрослых особей и потому часто объединяют годовые классы по два- три
(например, см. гл. \4 Antidorcas marsupialis). Aitken (1975) отмечает, что у косу-
ли после 4-5 лет можно говорить лишь о картине, видной у большинства особей
данного годового класса, и даже при этом он не дает характеристику годовых
классов после 7 лет.
Кроме описания и рисунков для выделения возрастных групп используют
балльную систему. Соколов и др. (1996) сравнили оценки возраста оленя Rangifer
72
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
tarandus по степени стертости всех щечных зубов нижней челюсти, оцененной
по 5-балльной системе, и по годовым слоям цемента и обнаружили, что степень
стертости в 3 балла в трех популяциях была модальной как у особей 4-6, так и у
особей 7-9 лет. Этого можно было ожидать, учитывая упоминавшуюся выше
высокую индивидуальную изменчивость степени стертости.
Более сложная и более объективная система балльных оценок разработана
для домашних копытных (Payne, 1973 и Grant, 1982 по Hillson, 1986), при ее
использовании основное внимание уделяется площади обнаженного дентина и
характеру лунок и в баллах оценивается стертость каждого зуба. Payne (Payne,
1973 по Hillson, 1986) сначала предложил схематическое изображение вариантов
стертости (см. рис 3.9), а затем присвоил балл каждому варианту (Payne, 1987).
На этих принципах Brown, Chapman (1990,1991с) для зубов лани, Cervus darna,
и благородного оленя, Cervus elaphus, и Lubinski (2001) для зубов вилорога,
Antilocapra americana, разработали свои системы балльных оценок степени стер-
тости зубов. Каждому участку каждого зуба присваивается балл в зависимости
от стертости эмали и обнажения дентина, баллы суммируются для каждого зуба.
По системе Brown, Chapman (1990) учитывались даже мелкие изменения в стер-
тости разных участков зуба. Например, стертость п^ в возрасте 7 мес. оценива-
ется в 4 балла, 8 мес. — 10 баллов, 11 мес. — 13 баллов, в 91 мес.— 34 балла.
Сумма баллов стертости всех щечных зубов дала возможность авторам полу-
чить линию регрессии стертости зубов по возрасту. Схема Lubinski (2001) была
несколько проще: 1 балл присваивается обнажению дентина на какой либо из
двух сторон каждого бугра зуба (в т1 — 4 бугра и 8 сторон, максимально 8
баллов), 1 балл — каждому слиянию полей дентина между буграми в щечно-
Рис. 3.9. Нижние коренные зубы пятнистого оленя, Cervus nippon, в возрасте 3 лет (А 1) и 9 лет
(Б 1), 2—схематическое изображение их жевательной поверхности (черным показан обнажен-
ный дентин) и 3 — условное изображение по схеме, предложенной Payne (1973 по Hillson,
1986).
Глава 3. Определение возраста по признакам зубов
73
язычном направлении (в mt их может быть 3) и 2 балла — исчезновению каждой
лунки (в т, их две); таким образом, максимальная стертость т, оценивается в 15
баллов. Например, сумма баллов стертости всех щечных зубов нижней челюсти
Antilocapra americana в возрасте 3,5 лет — 57, а у особей старше 9 лет — 76-81.
Чаще, однако, получают уравнения регрессии высоты конкретного зуба по
возрасту (см. гл. 14 Capreolus, Bison bison, Giraffa Camelopardalis, Rangifer).
Этот прием пригоден для приблизительной оценки возраста отдельных особей, в
идеале, той же популяции, для которой уравнение получено. Например, по урав-
нению регрессии высоты коронки по возрасту, полученному для одной выборки
северного оленя, Rangifer tarandus, (см. гл. 14) удалось определить возраст 75%
особей другой выборки той же популяции с ошибкой не более 1 года, и 46% с
точностью до года, но возраст особей другой популяции того же вида был опре-
делен с ошибкой не более 1 года лишь в 39% случаев (Клевезаль, Соколов,
2004).
Была попытка оценивать возраст по отношению площади поверхности стачи-
вания зуба к высоте коронки (Erickson et al., 1970), однако, по мнению самих
исследователей, этот способ в силу его сложности не имеет существенных пре-
имуществ перед визуальной оценкой степени стертости.
Spinage (1971, 1976) на примере газели Гранта, Gazella grand, и импалы,
Aepyceros melampus, апробировал общую формулу зависимости стирания зубов
отвозраста: ,,,, ,,
У = Yfl - (t/nf), откуда t = п (1- Y/Ytfk
где Y — высота зуба в возрасте t, Yn — предполагаемая начальная высота (до
начала стачивания), t — возраст, п — возраст, при котором высота зуба стачи-
вается до нуля. У газели и импалы расчетная константа к была определена как
0,5. Spinage измерял высоту коронки вместе с межкорневой цементной подуш-
кой, и не по буграм, а по впадине между ними. При таком способе измерения
возрастные изменения сглаживаются. Во-первых, не учитывается быстрое ста-
чивание бугров. Во-вторых, толщина цементной подушки с возрастом увели-
чивается, и измеряемая высота уменьшается медленнее, чем высота одной только
коронки.
По мнению Spinage (1971, 1976), определение возраста таким способом не
надежнее, чем визуально по степени стертости. Возможно, однако, что он не
прав и эту формулу можно считать более универсальной и (при измерении высо-
ты именно коронки) пригодной для ориентировочной оценки возраста отдельных
особей других видов копытных, если есть возможность определить константу
(к) на основании фактических данных. При исследовании зубов северного оле-
ня было показано, что ошибка определения возраста отдельных особей по этой
формуле для средних возрастных групп меньше, чем по общей линии регрес-
сии, и что константа к, рассчитанная для одной популяции, пригодна для другой
популяции того же вида (Клевезаль, Соколов, 2004).
Для тех случаев, когда нет возможности определить эмпирическое соотноше-
ние между высотой коронки и возрастом зуба, Klein et al. (1981) предлагают
использовать такую формулу:
74 Г, А. Клевезаль. Принципы и методы определения возраста млекопитающих
At = Ат - [(Ат - Ae)/He]Ht,
где At— возраст особи в данный момент, Ат — предельная продолжительность
жизни для особей данного вида, Ае — возраст при прорезании зуба, Не — высо-
та коронки до начала стачивания, Ht — фактическая высота коронки зуба в дан-
ный момент. Авторы протестировали формулу на разных щечных зубах северо-
американского благородного оленя, Cervus elaphus, и пришли к выводу, что она
обеспечивает разумную точность выделения возрастных групп и потому пригод-
на для копытных, имеющих сходный тип зубов. По-видимому, эту формулу сто-
ит использовать, когда нет других возможностей. Следует помнить, что при этом
делаются два весьма сомнительных допущения: равномерное стачивание зуба в
течение жизни и полное стачивание коронки данного зуба в возрасте, макси-
мально возможном для особей данного вида. Об изменении скорости стачива-
ния с возрастом упомянуто выше (в разделе 3.4.2. Морфологическое обоснова-
ние). Скорость стачивания разных щечных зубов также не одинакова и высота
коронок в предельном возрасте может существенно различаться (Клевезаль, Со-
колов, 2004).
Для оценки надежности определения возраста по стертости зубов были про-
ведены многочисленные сравнения определения возраста по степени стертости и
по годовым слоям. Для косули, С. capreolus, ( Aitken, 1975) и чернохвостого
оленя, Odocoileus hemionus (Erickson et al., 1970) точное соответствие было от-
мечено у 63-73% экземпляров, с ошибкой не более, чем в 1 год — у 90%. Для
лося, A. alces, полное соответствие было лишь у 17% особей (Dzieciolowski,
1976). Для благородного оленя, Cervus elaphus, выделение годовых классов по
степени стертости 11 операторами в среднем было ошибочным в 50%. Когда
годовые классы объединили по три, ошибка снизилась до 16% (Keiss, 1969).
При сравнении двух методов выяснилось, что возможны разные варианты:
- возраст, определенный по степени стертости зубов, чаще завышен, чем за-
нижен (С. capreolus — Szabik, 1973; Cervus nippon — Hrabe et al., 1989; Odocoileus
virginianus — Lockard, 1972);
— чаще занижен (Odocoileus hemionus — Erickson et al., 1970; A. alces —
Dzieciolowski, 1976; Odocoileus virginianus — Gilbert, Stolt, 1970; Lapierre, 1976);
- занижен в молодом возрасте (2-4 года) и завышен в старшем (Antilocapra
americana — Kerwin, Mitchell, 1971; Odocoileus hemionus — Thomas, Bandy, 1975).
Следует отметить, что при сравнении оценок возраста по степени стертости и по
числу годовых слоев нужно учитывать возможную ошибку при подсчете слоев
(см. гл. 4), поэтому реальную оценку надежности метода может дать только срав-
нение с особями известного (например, в результате мечения) возраста. Такое срав-
нение показало, что по стертости зубов возраст оленей Odocoileus virginianus в
возрасте 3-12 лет (с точностью до 1 года) был верно определен лишь у 20%
особей (Gee et al., 2002), Cervus elaphus в возрасте 1-7 лет — у 43% (Hamlin et
al., 2000), Odocoileus hemionus в возрасте 1-7 лет — у 62% (Hamlin et al., 2000).
Доля верно определенных особей С. capreolus ни в одном годовом классе от 2
лет и старше не превышала 50% (Hewison et al., 1999). При оценке по стертости
операторы завышали возраст молодых и занижали возраст старых особей.
Глава 3. Определение возраста по признакам зубов
75
Gee et al. (2002) оценивают методику в целом очень низко: у оленей Odocoileus
virginianus по степени стертости (и прорезанию) можно надежно выделить лишь
такие группы: сеголетки, годовики и взрослые (а среди последних — молодые
взрослые и старые). Если говорить об определении с точностью до года, то они
правы, но для приблизительной оценки возраста метод вполне годится и удобен
своей простотой. Использование эталонных челюстей от особей известного воз-
раста улучшает результат определения возраста всей серии. Например, когда
имелись в распоряжении эталонные челюсти косуль С. capreolus из южной Шве-
ции известного в результате мечения возраста (годовые классы 1—9), возраст
косуль по степени стертости опытный оператор определил верно у 68% особей, с
ошибкой ±1 год у 91% особей, причем в группе 3-9 лет верно было определено
47% и с ошибкой ± 1 год — 80% особей (Cederlund et al., 1991). Даже когда
введение эталонных образцов лишь немного улучшало общую надежность опре-
деления возраста, оно уменьшало расхождение между операторами, имеющими
разный опыт (Hewison et al., 1999).
3.4.6.4. Грызуны
У грызунов, имеющих зубы с дефинитивным ростом и бугорчатой жеватель-
ной поверхностью, возрастные группы выделяют по степени стертости эмали и
обнажения дентина, данные представляют в виде рисунков (например, рис. 3.10).
Несмотря на субъективность оценок степени стертости этот метод дает хороший
результат для многих видов мелких грызунов (Pucek, Lowe, 1975). В зубах со
складчатой жевательной поверхностью рисунок поверхности у молодых живот-
ных сложный, с возрастом происходит его упрощение (рис. 3.11).
Для количественной оценки используют систему баллов (см. гл. 12 Gerbillus,
Spalax). Для объективизации оценок предложены разные приемы: оценивать от-
ношение площадей поверхности стачивания, занимаемых дентином и эмалью
(Freudenthal et al., 2002) или отношение площади обнажения дентина к площади
жевательной поверхности (см. гл. 12 Tatnias sibiricus), определять высоту коро-
нок коренных зубов (см. гл. 12 Sciurus vulgaris, Spalax).
У грызунов, имеющих зубы с дефинитивным ростом, но с плоской жеватель-
ной поверхностью, возраст определяют по высоте коронки, соотношению высо-
ты коронки и длины корня (см. гл. 12 Castor, Clethrionomys, Ondatra). У бобров
стачивание коронки существенно компенсируется мощными отложениями це-
мента на корне, количество которого служит дополнительным критерием возра-
ста (см. гл. 12 Castor).
Возможный источник ошибки при определении возраста грызунов, особенно
— живущих несколько лет, это увеличение индивидуальной изменчивости сте-
пени стертости в старших возрастных группах. При разделении выборки на воз-
растные группы есть опасность из одного (старшего) годового класса сде-
лать два, поэтому оценка возраста старших особей по степени стертости может
быть завышена. Например, при сравнении возраста белки Sciurus vulgaris, опре-
деленного по числу годовых слоев в цементе и по степени стертости щечных
зубов, совпадение было в 55% случаев, оценка по степени стертости была завы-
76
Г, А. Клевезаль. Принципы и методы определения возраста млекопитающих
I П Ш IV I П Ш IV
Рис. 3.10. Схема изменения с возрастом жевательной поверхности коренных зубов мышей
разных видов (Тупикова, 1964). Левые 4 столбца — верхняя челюсть, правые — нижняя
челюсть, а — Mus musculus, б — Apodemus flavicollis, в — Apodemus agrarius, г — Micromys
minutus. Возраст: I — 1-1,5 мес., II — 5-10 мес., Ill — 12-16 мес., IV — 1,5-2 года. Серым
показано обнажение дентина.
Глава 3. Определение возраста по признакам зубов
77
Рис. 3.11. Возрастные изменения жевательной поверхности верхних коренных зубов обыкно-
венного слепыша, Spalax microphtalmus. 1,2 — молодые и полувзрослые, 3,4 — взрослые, 5-
7 — старые особи (из Топачевский, 1969).
шена в 2 раза чаще, чем занижена (Шадрина, 19886). При этом из 14 особей,
возраст которых по стертости был оценен в 3-6 лет, у одной совпали оценки
обоими методами, у всех остальных возраст по стертости оказался завышен.
3.4.6.5. Насекомоядные и рукокрылые
Возраст определяют по высоте зуба (или зубов) или по площади стертой по-
верхности (рис. 3.12). Поскольку эмаль бурозубок окрашена в бурый цвет, а
дентин светлый, легко отличить перезимовавших особей, у которых эта эмаль
практически стерта, от сеголетков. У живущих несколько лет насекомоядных по
степени стертости коренных зубов легко выделить годовые классы, если особи
пойманы в одно и то же время года, но как и в других группах, возникают
трудности при определении возраста старых особей.
78
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 3.12. Характер стачивания ш1 малой бурой ночницы, Myotis lucifugus, типичный для раз-
ных возрастных групп (Stegeman, 1956). Верхний ряд — вид сбоку, нижний ряд — жеватель-
ная поверхность. Черным показано обнажение дентина.
3.4.7. История применения
Метод старый как мир. Возраст лошадей по степени стертости резцов арабы
определяли, по крайней мере, уже в I веке н.э. (Корневен, Лесбр, 1929).
ГЛАВА 4. ОПРЕДЕЛЕНИЕ ВОЗРАСТА ПО
ГОДОВЫМ СЛОЯМ В ТКАНЯХ ЗУБОВ И КОСТИ
• Разработке, применению и оценке этого метода посвящено очень
много исследований и упоминать их все здесь нет возможности. Подроб-
ное описание метода и ссылки на работы даны в книге Клевезаль (1988) и
в дополненной английской версии (Klevezal, 1996), ниже дан лишь крат-
кий обзор. Если в этой главе или в сооэ ветствующих разделах II части нет
указаний на источник данных, его можно найти в упомянутых книгах.
4.1. Область применения
Метод пригоден для всех млекопитающих, живущих более года, у которых
выражена сезонность роста и имеются регистрирующие структуры с длитель-
ным (в идеале — пожизненным) периодом регистрации и сохранения записи. В
большинстве случаев, метод используется для определения возраста мертвых
животных, но иногда — живых (см. гл. 8.3)
4.2. Морфологическое обоснование
Метод использует факт наличия у млекопитающих так называемых регистри-
рующих структур (дентина и цемента зубов, костной ткани), которые в процессе
своего роста реагируют на изменение физиологического состояния организма
изменением своей морфологии, причем раз возникшие изменения сохраняются
длительное время.
Сезонные изменения скорости роста организма и его тканей приводят к
тому, что в дентине и цементе зубов и в костной ткани образуются ростовые
слои. Каждый слой состоит из относительно широкого элемента, соответ-
ствующего периоду интенсивного роста весной-летом, и узкого, соответ-
ствующего периоду остановки или существенного замедления роста в зим-
ний период. Зимний узкий (основной) элемент выглядит в цементе как ли-
ния или полоска, в дентине — как узкая полоска, в кости — как линия (рис.
4.1, 4.2).
В дентине зубов слои образуются со стороны полости пульпы, которая посте-
пенно заполняется. Первый по времени образования слой — наружный, после-
дний — внутренний (см. рис. 4.1,1). Когда полость пульпы заполнена, формиро-
вание слоев прекращается. Сформированные слои сохраняются в течение всей
жизни особи, если нет патологического изменения дентина.
Дентин, формирующийся в начальный период утолщения стенки зуба, назы-
вается первичным. Когда стенка зуба достигла определенной толщины, полость
пульпы и следовательно поверхность отложения дентина уменьшается, отложе-
ние дентина замедляется и начинает формироваться вторичный дентин, отделен-
ный от первичного зоной, где направление дентиновых канальцев меняется.
80
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 4.1. Схема корня зуба и участки поперечного среза резца лося, Alces alces (I) и продоль-
ного среза клыка лисицы, Vulpes vulpes (II). Д — дентин, (Д1 — первичный ДП— вторичный),
Ц — цемент, ПП — полость пульпы. Точками обозначены годовые слои. Срезы окрашены
гематоксилином.
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
81
Часто (например, в клыках хищных) слои во вторичном дентине более четкие,
чем в первичном.
В щечных зубах грызунов и в резцах копытных слой вторичного дентина в
коронке откладываются параллельно жевательной поверхности, предохраняя по-
лость пульпы от обнажения при интенсивном стачивании зуба. Их число можно
использовать для определения возраста, если первый слой не сточен и если из-
вестно, в каком возрасте он начал формироваться (см. гл. 14 Cervus elaphus, С.
dama).
Для определения возраста по числу слоев дентина обычно используют зубы,
не несущие большой механической нагрузки, полость пульпы в которых закры-
вается в позднем возрасте, например, клыки. Ростовые слои, формирующиеся
после наступления половой зрелости, обычно более четкие, чем формирующие-
ся у молодых животных.
В зубах, корень которых растет в течение нескольких лет, таких как клыки
медведей, формирование нескольких первых годовых слоев дентина сопровож-
дается образованием ростовых зон, ограниченных валиками, на наружной по-
верхности корня. Эти зоны видны до тех пор, пока они не сглажены отложением
цемента (см. гл. 13 Ursus americanus и рис. 13.8).
В цементе слои образуются со стороны покрывающих корень тканей, пер-
вый по времени образования слой находится близ границы с дентином, пос-
ледний слой — наружный (см. рис. 4.1, II). В большинстве случаев слои в
Рис. 4.2. Годовые слои в костной ткани. Участок поперечного среза щечной стенки нижней
челюсти бобра, Castor fiber. Окраска — гематоксилин. ПК — периостальная костная ткань,
Ост — остеоны, точками показаны годовые слои.
82
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
цементе формируются в течение всей жизни особи, хотя есть и отдельные ис-
ключения (см. ниже, раздел 4.4). Слои, в принципе, сохраняются пожизненно,
но в некоторых случаях (например, в резцах многих копытных) даже в норме
имеет место их частичная резорбция.
В кости слои формируются в периостальной костной ткани со стороны по-
крывающего кость периоста, последний по времени образования слой — наруж-
ный (рис. 4.2) и сохраняются до тех пор, пока не резорбируются при замене
пластинчатой периостальной ткани остеонами. У копытных это происходит в ран-
нем возрасте, у грызунов — в позднем, у некоторых из них слои могут сохра-
няться пожизненно.
Тот факт, что ростовые слои в зубах и в кости являются годовыми, подтвер-
ждено многочисленными исследованиями животных разных видов известного
возраста. Учитывая морфологические особенности тканей, для определения воз-
раста по годовым слоям в первую очередь можно рекомендовать цемент, затем —
дентин, и в последнюю очередь — костную ткань.
4.3. Разрешающая способность
Метод позволяет определить возраст с точностью до года, а у животных с
коротким сезоном размножения, если известно время гибели, и с большей точ-
ностью. Точность метода была проверена многочисленными исследованиями
особей известного возраста, его надежность оценена независимыми параллель-
ными подсчетами слоев разными исследователями.
4.4. Методика
Ростовые слои можно выявить на окрашенных тонких срезах, на тонких шли-
фах или на аншлифах. Считают их под микроскопом или бинокуляром.
Пригоден как хранящийся в сухом виде, так и фиксированный материал.
Длительное, в течение нескольких десятков лет, хранение материала в сухом виде
не мешает приготовлению окрашенных препаратов дентина и цемента, но часто
не дает возможности исследовать слои костной ткани. Мало пригоден для выяв-
ления слоев в кости и в цементе музейный материал, если кости и зубы были
отбелены химическим способом.
Слои дентина, формирующиеся по всей поверхности полости пульпы, напри-
мер в простых конических зубах, таких как клыки, обычно выявляют на попе-
речных срезах, проходящих через середину зуба (или верхнюю часть корня, см.
рис 4.1). Слои вторичного дентина, формирующиеся в коронке параллельно же-
вательной поверхности зуба, можно выявить только на продольных срезах или
шлифах. Слои в цементе считают на продольных срезах, прошедших через сере-
дину полости пульпы, реже — на поперечных срезах через середину корня. Слои
в костной ткани обычно считают на поперечных (перпендикулярной длинной оси
кости) срезах челюсти или костей конечностей.
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
83
Для получения окрашенных препаратов зуб или кость декальцинируют, про-
мывают, режут на микротоме и срезы окрашивают гистологическими красителя-
ми. Декальцинировать можно в 4-5%-ном растворе азотной кислоты или любом
другом декальцинате. Время декальцинации зависит от размера объекта и от спо-
соба его хранения. В растворе азотной кислоты декальцинация хранившегося в
сухом виде материала может длиться от полусуток (челюсти мышевидных гры-
зунов) до нескольких суток (зубы копытных). Фиксированный в формалине ма-
териал перед декальцинацией промывают и он декальцинируется дольше, чем не
фиксированный, хранившийся сухим.
Срезы, толщиной примерно 20 мкм, можно получать на замораживающем
микротоме, это не занимает много времени, поскольку не требуется заключать
материал в блок. Окрашивают срезы разными гистологическими красителями:
гематоксилином Эрлиха или другим гематоксилином, толуидиновым синим, по
Гимза и др. Заключают срезы в глицерин или синтетические смолы. Детали мож-
но найти в руководствах по микроскопической технике, в сводке Клевезаль (1988)
или в работах по использованию годовых слоев для определения возраста конк-
ретных видов (см. часть II, повидовое описание). Под микроскопом или биноку-
ляром узкие зимние элементы годового слоя выглядят темно-окрашенными (см.
рис. 4.1, I, II).
Иногда слои видны на зашлифованных половинках зуба (аншлифах) или на
тонких шлифах. В отраженном свете, на аншлифах или толстых шлифах, зимние
элементы выглядят темными (рис 4.3), в проходящем свете на тонких шлифах —
прозрачными, светлыми. В некоторых
случаях у копытных в межкорневой це-
ментной подушке щечных зубов годо-
вые слои выявляются лучше, чем на ок-
рашенных препаратах, поскольку менее
заметны дополнительные (см. ниже) эле-
менты.
При исследовании крупных зубов для
увеличения контрастности слоев можно
зашлифованную поверхность зуба обра-
ботать слабым раствором кислоты, на-
пример, 5%-ной муравьиной кислотой и
затем промыть и высушить. Годовые слои
на такой обработанной поверхности вы-
являются как валики или гребни.
Годовые слои обычно считают по чис-
лу узких (зимних, основных) элементов
слоя. Следует помнить, что формирова-
ние зимнего элемента слоя оканчивается
раньше и следующего за зимним эле-
ментом широкого слоя начинается вес-
ной также раньше того, как особи,
Рис. 4.3. Зашлифованная поверхность ниж-
ней части нижнего коренного зуба бобра,
С as tor fiber. Продольный распил. Отражен-
ный свет. Видны слои в цементе. Ц— це-
мент.
84
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
рожденной весной, исполнится очередной год. Следовательно, число основных
элементов (узких полос или линий) является, строго говоря, показателем числа
полных прожитых зим, а не полных годов.
• При определении возраста по числу слоев в цементе тех зубов, кото-
рые прорезаются не в первый год жизни особи (у копытных, крупных хищ-
ных), следует установить возраст появления первого слоя цемента и вно-
сить соответствующую поправку. Часто имеются разногласия относительно
поправки при использовании зубов, прорезающихся в конце первого лета жизни
(например, m1 , копытных, клыки некоторых хищных). Поскольку имеет место
межпопуляционная изменчивость как возраста прорезания зуба, так и скорости
его роста, вопрос о поправке должен решаться для каждой популяции отдельно.
Можно руководствоваться следующим правилом. Если в зубе, прорезающимся
в конце лета - осенью, первая узкая (зимняя) линия видна близ дентина, то она
сформирована в первую зиму существования зуба, а если между ней и денти-
ном видна широкая полоса цемента, ее следует считать линией второй зимы пе-
риода существования зуба. Например, у волка Canis lupus первая видимая зим-
няя линия может быть линией как первой, так и второй зимы. Корытин, Габченко
(1988) предлагают такое решение: если первый слой значительно (в 2 раза) шире
последующего, к числу слоев нужно прибавить 1, если первый видимый слой
не шире последующего, возраст равен числу слоев.
• • Следует иметь в виду, что в отдельных случаях ежегодное формиро-
вание цемента может нарушаться. Это было документировано для зубра Bison
bonasus: в it годовые слои образуются ежегодно в течение всей жизни лишь на
язычной стенке, а на щечной после 7 лет регулярность нарушается, хотя отложе-
ние цемента не прекращается (Klevezal, Pucek, 1987). По-видимому, это связано
с характером механической нагрузки на зуб; в резцах зубра она существенно
больше на язычную стенку. Вероятно, то же имеет место и в ряде других случа-
ев. McLaughlin et al. (1990), и Keay (1995) у медведей Ursus americanus, и Arthur
et al. (1992) у куниц Martes pennanti обнаружили, что возраст, определенный по
годовым слоям цемента первого предкоренного зуба, соответствовал реальному
у молодых и средневозрастных особей, а у старших (6 лет и старше у медведей
и 3 г. и старше у куниц) он был занижен. Вероятно, в этих, практически рудимен-
тарных, предкоренных зубах, не имеющих существенной механической нагруз-
ки, отложение цемента с возрастом становится не регулярным. Тенденция к за-
нижению на 1—2 года оценки возраста, полученной по слоям цемента первого
предкоренного, по сравнению с таковой, полученной по слоям в клыке, была
обнаружена также и у волков, Canis lupus, старше 6 лет (Ballard et al., 1995).
При определении возраста по годовым слоям в кости выбирают такую кость и
такой ее участок, где аппозиция слоев периостальной костной ткани продолжает-
ся долго, а темп резорбции первых сформированных слоев низкий. Обычно ис-
пользуют нижнюю челюсть или середину диафиза длинных костей конечностей.
Срез должен пройти перпендикулярной длинной оси кости. Например, у бобра
Castor fiber и сурка Marmota baibacina годовые слои хорошо видны в щечной
стенке челюсти за концом зубного ряда, причем у бобров старше 11 лет первый
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
85
слой может резорбироваться, а у сурков он может быть частично резорбирован
уже в 3 года, а после 5 лет площадь отложения новых годовых слоев уменьша-
ется.
4.5. Изменчивость показателей
Годовые слои могут быть очень четкими, когда зимний элемент контрастный
(см. рис. 4.1) и легко отличается от дополнительных. Бывает, однако, что основной
элемент не столь контрастен, а дополнительные линии и полоски относительно чет-
кие и многочисленны, и тогда разграничить годовые слои трудно (рис. 4.4). Есть
межвидовые и межпопуляционные различия по степени четкости слоев. В ряде
случаев уменьшение четкости ростовых слоев у особей одного вида происходит
параллельно уменьшению степени континентальности климата района обитания, но
есть и явные нарушения этого соответствия. Межпопуляционными различиями чет-
кости слоев в значительной мере объясняются разногласия исследователей отно-
сительно пригодности ростовых сло-
ев для определения возраста особей
одного и того же вида.
Вследствие межвидовых разли-
чий в четкости слоев надежность
метода различна для разных видов.
Так, на большом материале от оле-
ней известного возраста было ус-
тановлено, что один и тот же иссле-
дователь (Gary Matson, Matson’s
Laboratory, Montana, USA) правиль-
но определил 97% (из 111) Cervus
elaphus canadensis, 93% (из 108)
Odocoileus hemionus и 85% (из 74)
О. virginianus из одного и того же
региона (Hamlin et al., 2000).
На практике полезно сопровож-
дать оценку возраста каждой осо-
би баллом ее надежности: от 1 до 5
по возрастающей (Клевезаль и др.,
1981), от 0 до 3 по возрастающей
(Gasaway et al., 1978), от А до С по
убывающей (Matson, 1981) или от
1 до 4 по убывающей (Norton,
Fairall, 1991). Оценки с низкой сте-
пенью надежности следует исполь-
зовать только в сочетании с оценка-
ми возраста, полученными други-
ми методами.
Рис. 4.4. Участок продольного окрашенного
среза корня р( горала, Nemorhaedus goral.
Нечеткие слои в цементе. Д — дентин, Ц —
цемент, ПП — полость пульпы.
86
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
У ряда млекопитающих слои не обнаружены (Aepyceros melampus, Spinage,
1971), или признаны малопригодными (Sylvicapra grimmia, Wilson et al., 1984),
или их рекомендуется использовать с осторожностью, поскольку трудно разли-
чить основные и дополнительные (например, у Tragelaphus angasi, Anderson, 1986).
4.6. Особенности использования метода в отдельных группах
4.6.1. Насекомоядные
Годовые слои формируются и сохраняются в костной ткани, в дентине и в
цементе зубов. Слои во всех трех тканях можно видеть на окрашенных срезах,
если поперечный срез мандибулы в районе коренных зубов прошел таким обра-
Рис. 4.5. Окрашенный поперечный срез ниж-
него резца ушастого ежа, Hemiechinus auritus.
Д—дентин, Ц—цемент, ПП—полость пуль-
пы. Видны слои в цементе.
Фото препарата С.И. Исакова.
зом, что на нем представлен и про-
дольный срез зуба через полость
пульпы. Однако не всегда удается
выявить слои во всех трех тканях.
Так, у ежей Hetniechinus auritus и
Н. hypomelas (Исаков, 1984) слои в
цементе (на поперечных срезах кор-
ней коренных зубов) были четкие
(рис. 4.5) и число их совпадало с
числом слоев в кости на попереч-
ных срезах мандибулы и середины
диафиза бедренной кости, но в ден-
тине слои были нечеткие.
Для определения возраста лучше
считать слои в цементе, корректируя
подсчет по слоям в кости и, если они
видны, в дентине. Это помогает отли-
чить основные линии от дополнитель-
ных, поскольку в разных тканях чет-
кость дополнительных линий может
быть разной. Для таких видов, как
выхухоль Desmana moschata, у кото-
рой не легко выделить годовые слои и
в цементе, и в дентине, и в кости, хо-
роший результат может дать исследо-
вание всех трех тканей каждой особи.
4.6.2. Рукокрылые
Годовые слои образуются в дентине и в цементе зубов летучих мышей, а в
костной ткани их формирование, по-видимому, прекращается в первые годы.
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
87
Рис. 4.6. Участок окрашенного гематоксилином поперечного среза коронки ггц рыжей вечер-
ницы, Nyctalus noctula. Д — дентин, ПП — полость пульпы, цифрами показаны годовые слои
дентина.
Общие наслоения цемента у большинства видов не велики, слои в них считать
трудно и для определения возраста их используют редко. В большинстве слу-
чаев возраст определяют по числу годовых слоев дентина, которые у большин-
ства видов хорошо видны на окрашенных поперечных срезах клыков или ко-
ренных зубов в районе шейки (рис. 4.6); в резцах слои видны плохо. Полость
пульпы клыков и крупных щечных зубов заполняется дентином медленно; у
Myotis lucifugus в дентине обнаружено 17 слоев, у Eptesicus fuscus — 19
(Schowalter et al., 1978), у Eptesicus serotinus — 21 слой (Рахматулина, 1984).
У разных видов четкость слоев дентина различна. В ряде случаев авторы не
упоминают никаких проблем при определении возраста по слоям дентина, на-
пример, Myotis blythi, Nyctalus noctula, Vespertilio pipistrellus (Бутовский, Шай-
марданов, 1988), Murina leucogaster (Тиунов, 1988). Для некоторых видов, осо-
бенно относительно мелких {Myotis daubentoni, Barbastella harbastellus, Myotis
nattereri), метод признан ненадежным (Batulevicius et al., 2001).
У представителей подотряда крыланов, Pteropus alecto и P poliocephalus, бо-
лее крупных, чем летучие мыши, Cool et al. (1994) выявили ростовые слои и в
дентине, и в цементе, и в кости, но только слои цемента, по их мнению, можно
использовать для оценки возраста. По их данным, и толщина цемента, и число
слоев в нем увеличиваются с возрастом, однако ежегодно формируется не один,
а в среднем 1,4 слоя. Иной результат получен Divljan et al. (2006), которые пока-
зали четкое соответствие числа слоев цемента числу прожитых лет у Pteropus
poliocephalus, нарушавшееся только у особей старше 10 лет.
88
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
4.6.3. Зайцеобразные
У зайцеобразных и резцы, и щечные зубы обладают постоянным ростом, а
потому ни дентин, ни цемент не имеют годовых слоев. Годовые слои можно
найти только в кости. Они видны на окрашенных поперечных (перпендикулярно
жевательной поверхности зубов) срезах или на тонких шлифах середины тела
нижней челюсти, диастемы нижней челюсти, середины диафиза костей конечно-
стей. Число слоев в середине тела нижней челюсти и в диастеме пищухи Ochotona
pricei было одинаковым, при этом проще было делать срезы диастемы (Крыло-
ва, 1973). Часто зимние линии не просто отличить от дополнительных. При ис-
следовании особей известного возраста разных видов примерно в 85% случаев
подтвердилось, что слои годовые, но определение возраста по их числу не про-
сто и требует определенного опыта.
4.6.4. Грызуны
У большинства грызунов зимние линии можно видеть в цементе и вторичном
дентине щечных зубов, а также в кости или только в цементе и в кости. В посто-
янно растущих резцах годовых слоев нет. Для разных видов традиционно ис-
пользуют одну из регистрирующих структур (например, только цемент — см. гл.
12 Castor), но в трудных случаях правильное решение дает исследование и ден-
тина, и цемента, и кости (см. гл. 12 Citellus).
У долгоживущих грызунов линия первой зимы в цементе часто образуется
только в верхней части корня и расположена очень близко к границе цемента и
дентина, поскольку к первой зиме корень еще не полностью вырос. Все слои при
этом видны только на продольных срезах корня. Поскольку часто на поперечных
срезах слои видны более четко, можно считать их на поперечном срезе через
середину корня, добавляя 1 год (см. гл. 12 Marmota).
При определении возраста большинства мелких грызунов удобно делать по-
перечные срезы нижней челюсти так, чтобы на срезе оказались продольные сре-
зы т2или т3 (см. гл. 12 Apodemus). Тогда на одном препарате можно видеть
слои в костной ткани, в цементе, а иногда и во вторичном дентине. Однако у
многих живущих несколько лет грызунов слои цемента лучше видны на попе-
речных срезах нижней половины корня, чем на продольных в нижней части кор-
ня (например, у Meriones, см. гл. 12).
Вторичный дентин может формироваться не по всей поверхности полости
пульпы (см. гл. 12 Marmota).
Годовые слои часто видны и на поперечных срезах диафизов трубчатых
костей (см. например, гл. 12 Tarnias) и фаланг пальцев. У сеголетков мож-
но видеть линию, обычно несколько извилистую, отделяющую наружную
периостальную зону кости и не считающуюся годовой (рис. 4.7). Наличие
слоев костной ткани в диафизах фаланг пальцев дает возможность прижиз-
ненно определять возраст особей у ряда видов (см. гл. 12 Apodemus agrarius,
Citellus).
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
89
Рис. 4.7. Поперечный срез второй фаланги третье-
го пальца перезимовавшей пашенной полевки,
Microtus agrestis. Стрелкой указана зимняя линия,
двойная в нижней части среза, лр—линия резорб-
ции, отделяющая наружную периостальную зону
кости.
У грызунов, имеющих все зубы с постоянным ростом, например, у серых
полевок Microtus, годовые слои можно найти только в костной ткани. Из-за на-
личия двойных и дополнительных линий срезы нижней челюсти вряд ли можно
рекомендовать для выявления перезимовавших особей у этих коротко живущих
грызунов. Однако, для прижизненного выявления перезимовавших можно ис-
пользовать фаланги пальцев, которые ампутируют при мечении грызунов. На их
поперечных срезах у перезимовавших особей в периостальной зоне видна зим-
няя линия, отсутствующая у сеголетков (рис. 4.7).
• У зимоспящих грызунов зимняя линия в кости становится заметной
только тогда, когда рост очередного слоя кости уже начался, поэтому она
видна не у всех особей, пойманных весной сразу после выхода из спячки.
4.6.5. Хищные
Хотя годовые слои можно видеть и в дентине, и в цементе, а в ряде случаев и
в периостальной костной ткани, для определения возраста обычно используют
только цемент. Чаще всего слои в цементе считают на окрашенных срезах клы-
ков (рис. 4.8), но можно использовать и предкоренные и коренные зубы. На
аншлифах и шлифах, как правило, слои видны хуже. Специального внимания
требует вопрос о времени формирования первого слоя цемента и длительности
90
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 4.8. Нижняя часть корня клыка енотовидной собаки, Nyctereutes procyonoides. Продоль-
ный срез, окрашенный гематоксилином. Д—дентин, Ц — цемент, точками показаны годовые
слои цемента.
формирования регулярных слоев в предкоренных, не несущих существенной
механической нагрузки (см. выше раздел 4.4 • и •• ).
Для большинства видов хищных (за исключением Lutra, Gulo и Ursus
maritimus) подсчет слоев цемента признан точным и надежным методом опре-
деления возраста. Кроме видов, упомянутых в главе 13, этот метод признан
пригодным для определения возраста Mephitis mephitis, Canis aureus, Taxidea
taxus, Panthera leo.
Для экономии времени и средств при определении возраста особей в боль-
ших выборках отделяют как несомненных сеголетков особей, имеющих откры-
тое апикальное отверстие корня или широкую полость пульпы клыка (см. гл. 3
разделы 3.2.2 и 3.3), а возраст остальных определяют по годовым слоям (напри-
мер, Шиляева и др., 1975).
У белого медведя Ursus maritimus слои в цементе очень нечеткие. Hensel,
Sorensen (1980) сообщают, что только у 32-45% особей известного возраста
три оператора верно определили возраст по слоям цемента на окрашенных сре-
зах рудиментарных Р] 3 Однако при исследовании нескольких тщательно при-
готовленных окрашенных срезов предкоренных зубов большой серии медве-
дей известного возраста (5-18 лет), в 90% случаев три опытных оператора
правильно определили возраст и в 95% — двое из этих троих (Calvert, Ramsay,
1985).
Во многих случаях слои хорошо видны во вторичном дентине и их подсчет
можно использовать для корректировки подсчета слоев цемента. Для этого нуж-
но делать поперечные срезы в районе шейки зуба или чуть ниже (чтобы выявить
Глава 4. Определение возраста по годовым слоям в тканях зубов и кости
91
Рис 4 9 Поперечный срез корня i, лося, Alces alces. Окрашен гема-
токсилином. Видны слои в цементе и в дентине. Д — дентин (Д1 —
первичный, ДП— вторичный), Ц — цемент, ПП — полость пульпы.
слои дентина) и продольные срезы корня (чтобы выявить все слои цемента).
Первый слой вторичного дентина образуется на втором году жизни или позже, в
зависимости от скорости роста и времени полового созревания особей.
4.6.6. Копытные
Годовые слои видны в цементе и во вторичном дентине зубов на окрашенных
срезах (рис. 4.9), шлифах и аншлифах. В кости слои не образуются или быстро
резорбируются. Хотя слои формируются во всех зубах, для определения возра-
ста чаще берут резцы. Следует помнить, что по числу слоев определяют возраст
зуба и для получения возраста особи необходимо ввести поправку на возраст
появления зуба. При этом надо иметь в виду индивидуальную и межпопуляцион-
ную изменчивость роста зуба (см. выше, раздел 4.4 • ).
Четкость слоев цемента бывает различна у разных видов и в разных популя-
циях одного вида. Например, у косули С. capreolus слои нечеткие, хотя и не во
всех регионах. Cederlund et al. (1991) получили лучшее соответствие известно-
му возрасту при определении возраста косуль из Швеции по степени стертости
(при наличии эталонов от меченых животных для каждого годового класса),
чем по числу слоев цемента на шлифах шг
Слои во вторичном дентине часто более четкие, чем в цементе, однако пер-
вый слой вторичного дентина может формироваться не в первый год существо-
вания зуба, и число слоев вторичного дентина может быть равно, а может быть
меньше числа слоев цемента. Тем не менее, число слоев вторичного дентина
92
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
можно использовать для корректировки подсчета слоев цемента, поскольку оно
является минимально возможной оценкой возраста данной особи.
Минимально возможной оценкой возраста может служить и число слоев вто-
ричного дентина, которые видны в коронке резцов оленей на продольных шли-
фах (срезах) и формируются параллельно поверхности стачивания зуба, предох-
раняя полость пульпы от обнажения при стачивании. Образование первого слоя у
благородного оленя Cervus elaphus происходит в 3 года, а в возрасте 10-12 лет
он может быть сточен (Eidmann, 1932).
4.7. История применения
Данные о наличии слоев в зубах и кости некоторых млекопитающих появля-
лись с конца XIX в. Eidmann (1932) обнаружил слои вторичного дентина в рез-
цах оленей и предложил их использовать для определения возраста. Однако только
после того, как годовые слои дентина были описаны у морских млекопитающих
и использованы для определения возраста (Scheffer, 1950; Laws, 1952), на них
обратили внимание и исследователи наземных видов (Sergeant, Pimlott, 1959;
Sergeant, 1967; Клевезаль, Клейненберг, 1967) и метод получил широкое рас-
пространение. К 1987 г. он был признан пригодным для определения возраста
112 видов морских и наземных млекопитающих разных отрядов (Клевезаль, 1988).
ГЛАВА 5. ОПРЕДЕЛЕНИЕ ВОЗРАСТА
ПО ПРИЗНАКАМ РОГОВ
Есть два типа рогов: роговые, постоянные (у полорогих) и костные, ежегодно
сменяющиеся (у оленьих). Возможности их использования для определения воз-
раста различны и потому рассматриваются отдельно.
5.1. Рога полорогих
Рога полорогих — это постоянные (несменяемые) выросты лобных костей,
покрытые роговым чехликом; рог растет от своего основания. Рост рогового
чехлика неравномерный, у многих видов ежегодно рост рога начинается весной
и заканчивается осенью, зимой имеет место остановка роста. В результате на
роговом чехлике образуются кольцевые бороздки, «кольца», определяющие гра-
ницы между соседними годовыми приростами — годовыми «сегментами» (рис.
5.1). И у козлов, и у баранов самым большим является прирост второго года
жизни (второй годовой сегмент), далее с возрастом ширина сегментов умень-
шается, у баранов быстрее, чем у козлов (Цалкин, 1950). В отдельных случаях (у
баранов с некрупными рогами — реже, а у баранов с крупными рогами — чаще)
самым большим бывает прирост третьего года (Цалкин, 1948). Ширина годового
прироста зависит не только от возраста, но, как и рост организма в целом, от
ряда других факторов. Так, зависимость ширины годового прироста от темпера-
туры среды и фенологии растений весной выявлена у горных козлов Capra
(Giacometti et al., 2002) и от климатических условий предыдущего года — у
серны Rupicapra (Perez-Barberia et al., 1996).
Такая регистрация сезонных изменений темпа роста рога и длительное сохра-
нение в виде годовых колец и годовых приростов (сегментов) позволяет опреде-
лять возраст многих видов с точностью до года, а если дата гибели известна, то
идо месяца (см. гл. 14 Ovis canadensis, Oreanmos americanus, Capra, Capricornis
crispus, Rupicapra).
Годовые сегменты или годовые кольца можно считать на поверхности рога
при осмотре трупа, живого пойманного животного, а иногда даже при наблюде-
нии в природе. Годовые приросты хорошо выявляются на продольном распиле
рога (Miura, 1985).
Рост очередного годового сегмента, как правило, начинается весной до оче-
редного дня рождения особи, и осенью сегмент очередного года полностью сфор-
мирован. Весной особь неполных трех лет имеет четыре сегмента, последний в
самом начале его формирования (рис. 5.1), осенью, при возрасте около 3,5 лет,
у нее четыре полностью сформированных годовых сегмента.
Необходимо отличать основные (годовые) кольца от дополнительных неровно-
стей поверхности сегментов. Годовые кольца — глубокие бороздки с выраженны-
ми краями, дополнительные — более мелкие бороздки и валики с закругленными
краями (Hemming, 1969). С возрастом ширина годовых приростов уменьшается,
94
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 5.1. Схема роста рога козлов рода Capra (по Couturier, 1961). Стрелками показаны основ-
ные («зимние») кольца, разделяющие годовые сегменты. Пунктиром указано состояние рога в
конце полного очередного года жизни особи.
дополнительные кольца труднее отделить от основных и надежность определения
возраста снижается. В первую зиму жизни особи зимняя задержка роста, в том
числе и роста рогов, может быть плохо выражена, поэтому первое годовое коль-
цо может быть нечетким.
Koubek, Hrabe (1983) обнаружили, что у сеголетков серны Rupicapra, у кото-
рых первое годовое кольцо нечеткое, в ноябре-декабре длина рога была суще-
ственно меньше, чем ширина первого годового сегмента у взрослых, и следова-
тельно, рост рога должен был продолжаться в зимний период. Есть данные, что
у серны рост рогов в зимний период происходит в течение не только первой, но
и второй зимы жизни особей (Perez-Barberia et al., 1996).
У старых животных годовой сегмент первого года жизни может исчезнуть в
процессе стачивания рога.
У взрослых и старых самок многих видов полорогих годовые кольца выра-
жены значительно хуже, чем у самцов, и определение возраста менее надежно.
Яркий пример — шотландские домашние козы, уже столетие живущие на сво-
боде. У самцов и молодых (до двух лет) самок число колец на рогах соответ-
ствует возрасту, у старших самок — не соответствует (Bullock, Pickering, 1984).
Вероятно, подсчет колец пригоден для определения возраста Ammotragus
lervia (Gray, Simpson, 1985), Hemitragus jemlahicus (Caughley, 1965). Однако
Глава 5. Определение возраста по признакам рогов
95
для определения возраста ряда видов метод признан непригодным, несмотря
на то, что число колец увеличивается с возрастом. Такая ситуация обнаружена
в Новой Зеландии и Австралии у Capra hircus (Rudge, 1972; Henzell, 1987), у
африканских Damaliscus dorcas (Olivier, Greyling, 1991), Antidorcas marsupialis
(Rautenbach, 1971), Aepyceros melampus (Spinage, 1971), Tragelaphus angasi
(Anderson, 1986), Gazella grand (Spinage, 1976), Kobus defassa (Spinage, 1967),
а также у Bison bison (Fuller, 1959), Saiga tatarica и Nemorhaedus goral (см. гл.
14). При этом иногда удается выделить несколько возрастных групп по размеру
и форме рогов. Так, возрастные группы особей, не достигших половой зрело-
сти, можно выделять у Taurotragus oryx (Jeffery, Hanks, 1981). По форме рогов
у самцов можно визуально выделить возрастные группы в 6 мес., 9 мес., 1 г.,
1,5 г., 2,5 г. и старших у Tragelaphus angasi (Anderson, 1986), группы в 6 мес.,
1 г., 2 г., 6 л., 10 л. у самцов и самок Gazella grand (Spinage, 1976). По размеру
и форме рогов можно различать несколько возрастных групп у Bison bison,
Ovibos moschatus, Antidorcas marsupialis, Ovis canadensis, Ovis amrnon, Saiga
tatarica (см. гл. 14).
5.2. Рога оленей
У оленей сезонность роста рогов проявляется не только в ежегодном сбрасы-
вании весной старых рогов и отращивании новых, но и в морфологии пеньков.
Banfield (1960) на срезах пеньков рогов северного оленя Rangifer tarandus обна-
ружил кольца, судя по результатам сравнения со степенью стертости зубов —
годовые. Однако вероятно в силу вторичной перестройки костного материала,
было обнаружено не более 4 колец даже у старших особей.
Хотя число отростков рогов с возрастом увеличивается, оно не является чет-
ким критерием возраста из-за большой индивидуальной изменчивости. У благо-
родного оленя Cervus elaphus по числу отростков можно лишь сказать, молодой
олень, среднего возраста или старый (Habermehl, 1985). У лося (см. гл. 14 Alces
и табл. 14.1) в возрасте 6-7 лет может быть от 2 до 9 отростков, но все же можно
выделить несколько возрастных групп, используя число отростков в сочетании с
диаметром основания рога. У косули (см. гл. 14 Capreolus) три возрастные груп-
пы можно выделить по внешнему виду и размеру рогов, а также по диаметру
основания рога. Приблизительно возраст по форме рогов можно, видимо, опре-
делить у лани (см. гл. 14 Cervus dama).
В охотничьей практике определению возраста оленей по рогам уделяется боль-
шое внимание, для этой цели используют как промеры пенька, так и промеры
рога. Хороший обзор разных методов, пригодных, однако, лишь для приблизи-
тельной оценки возраста, дает Habermehl (1985) для благородного оленя. В час-
тности, поверхность сбрасывания рога, выпуклая у молодых, с возрастом упло-
щается. Примером того, с какой степенью точности можно определять возраст
по размерам пенька, может служить рис. 5.2.
Lockow (1987) предлагает номограмму для определения приблизительного
возраста благородного оленя (от 2 до 10 лет) по окружности рога в нижней его
96
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
возраст, лет
Рис. 5.2. Изменение с возрастом диаметра (средняя и пределы варьирования) пенька рога
благородного оленя, Cervus elaphus (поданным Schreiber, 1993).
части и длине штанги рога. По его данным, ошибка определения возраста по
этой номограмме лишь у 6% особей превышала 2 года по сравнению с возрас-
том, определенным экспертами по степени стертости зубов. Преимущество это-
го подхода в том, что он дает возможность, хотя и приблизительно, оценить воз-
раст оленей по сброшенным рогам.
Несмотря на межпопуляционные различия, вес рогов, который увеличивается
с возрастом, может быть критерием приблизительной оценки возраста (см. гл.
14 Cervus elaphus).
ГЛАВА 6. ОПРЕДЕЛЕНИЕ ВОЗРАСТА ПО ВЕСУ
ХРУСТАЛИКА
6.1. Область применения
Метод, в принципе, пригоден для любых групп животных, и получил распро-
странение для посмертного выделения возрастных групп у зайцеобразных, мел-
ких грызунов, в меньшей степени — хищных, копытных.
6.2. Морфологическое обоснование
Метод разработан по результатам исследований глаза человека и лабораторных
животных, показавших, что хрусталик глаза млекопитающих растет всю жизнь за
счет образования новых волокон в его экваториальной зоне. В период быстрого ро-
ста организма вес хрусталика увеличивается параллельно весу тела. Считается, что
темп роста хрусталика уменьшается с наступлением половой зрелости не столь зна-
чительно, как темп весового роста всего тела и в значительно мере не зависим от
ряда факторов, влияющих на рост тела (см. обзор Friend (1967с) и ниже раздел 6.5).
6.3. Разрешающая способность
Метод дает лучшие результаты при определении возраста молодых животных
в период быстрого роста, чем взрослых после наступления половой зрелости
(рис. 6.1). В случае единовременно взятых выборок он позволяет отделить моло-
700
600
500
400
300
200
100
0
Рис. 6.1. Соотношение возраста и веса хрусталика кролика Sylvilagus floridanus Южного Теха-
са (сплошная линия) и 90% доверительный интервал (пунктирная линия) (по данным Bothma et
al., 1972).
98
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
дых животных от взрослых и выделить несколько годовых групп, если период
роста растянут на несколько лет. Удобен для построений линий регрессии при-
знака по возрасту для получения приблизительной оценки возраста отдельных
особей.
6.4. Методика
Методика подробно описана в ряде работ (Friend, 1967а; Morris, 1972; Hagen
et al., 1980). Глазные яблоки (целиком, мелкие можно проткнуть, крупные —
надрезать с задней стороны) фиксируют как можно скорее после гибели живот-
ного в 4%-ном нейтральном растворе формальдегида в дистиллированной воде.
Этот же раствор иногда называют 10%-ным формалином: 1 часть стандартного
продажного (40%) формальдегида на 9 частей воды. Длительность фиксации
влияет на окончательный вес хрусталика и потому должна быть стандартной для
всей выборки, не менее 1 и не более 40 недель (Friend, 1967а, 1968).
Без серьезных причин не следует использовать иные фиксаторы. Смена фик-
сатора может существенно повлиять на результат. Например, когда разные хрус-
талики одной и той же особи скунса Spilogale putorius фиксировали в разных
фиксаторах, вес их различался в среднем на 13-17% (Mead, 1967).
Фиксированные хрусталики, не промывая, осторожно препарируют (следует
брать неповрежденные хрусталики с гладкой сферической поверхностью) и вы-
сушивают до постоянного веса при 60-80 °C. Время высушивания зависит от
размера хрусталика. Например, для высушивания при 80 °C до достижения по-
стоянного веса хрусталиков лисицы потребовалось 48 ч. (Friend, Linhart, 1964),
а хрусталиков бобра — 56 ч. (Malcolm, Books, 1981). Предложены некоторые
изменения схемы высушивания: предварительное высушивание при 40-80 °C и
досушивание при 100 °C, для ускорения — при низком давлении и в присут-
ствии гигроскопического материала (Cabon-Raczynska, Razynski, 1972) или вы-
сушивание в течение 24 ч. при 100 °C (Suchentrunk et al., 1991).
В принципе, можно высушивать и свежий хрусталик, но из-за его хрупкости
с ним труднее манипулировать, чем с фиксированным. Кривые изменения с воз-
растом сухого веса высушенных свежих и предварительно фиксированных хру-
сталиков были практически одинаковы (Friend, 1967а). Если материал был фик-
сирован не свежим, вес хрусталика уменьшится по сравнению со свежефикси-
рованным.
Свежий (но не подпорченный!) материал можно замораживать до фиксации,
но затем его нужно фиксировать замороженным, без предварительного оттаива-
ния. Предварительное замораживание существенно не влияет на вес (Millar,
Iverson, 1976), хотя отмечено снижение веса при замораживание хрусталиков на
срок более одного дня (Montgomery, 1963).
Снижался вес и при фиксации оттаявших хрусталиков, особенно большие
потери веса имели место при неоднократном оттаивании и замораживании
(Rongstad, 1966; Ostbye, Semb-Johansson, 1970). По среднему весу и пределам
варьирования веса фиксированных таким образом хрусталиков росомах G. gulo
Глава 6. Определение возраста по весу хрусталика
99
Аляски, полученных от охотников, не было выявлено различий даже между се-
голетками и годовиками, хотя по весу хрусталиков того же вида с Юкона, замо-
роженных свежими, имелись четкие различия между этими двумя группами
(Rausch, Pearson, 1972).
При взвешивании сухих хрусталиков нужно свести до минимума контакт с
атмосферным воздухом, поскольку высушенные хрусталики гигроскопичны. При
хранении в сухом прохладном месте хрусталик пригоден для дальнейшей обра-
ботки в течение одного-двух дней. Следует избегать высокой температуры и влаж-
ности и не хранить хрусталики в полиэтилене.
Различия веса правого и левого хрусталиков, как правило, незначительные и
ненаправленные (Lombaard, 1971; Sanderson, 1961; Ashby, Henry, 1979; Myers et
al., 1977), поэтому если в качестве показателя используют вес одного хрустали-
ка, то берут или среднее значение веса обоих хрусталиков, или вес наиболее
тяжелого или наименее поврежденного. Если возможно, лучше взвешивать оба
хрусталика одной особи и оперировать со средним весом (Morris, 1972). Со
средним весом или весом одного хрусталика, как правило, оперируют при ис-
следовании хищных, копытных, средних и крупных грызунов и зайцеобразных.
При исследовании мелких грызунов взвешивают одновременно оба хрусталика
одной особи.
• Чтобы использовать данные других авторов для определения возра-
ста особей исследуемой популяции, нужно строго следовать их методи-
ческим указаниям, т.к. во многих случаях имеют место разные отступле-
ния от «классической» прописи.
Когда вместо сухого веса использовали сырой вес нефиксированных хруста-
ликов, общая тенденция изменения веса с возрастом сохранялась, но конкрет-
ные значения веса в данном возрасте различались. Сравнение динамики сухого
и сырого веса фиксированных хрусталиков крыс Rattus fuscipes и R. villocissimus
(Myers et al., 1977) показало, что высушивание уменьшает ошибку оценки воз-
раста по весу хрусталика, но уменьшение незначительное и не оправдывает до-
полнительные затраты на процедуру высушивания. Сходные данные были полу-
чены и для песчанок Tatera indica (Jain, Prakash, 1981). В интервале 5-150 дней:
(вес пары сухих хрусталиков, мг) = 1,4450 + 0,72111 • возраст, дни;
г = 0,9905;
(вес пары сырых хрусталиков, мг) = 9,8328 + 1,2835 • возраст, дни);
г = 0,9793.
Вес свежего, не высушенного, хрусталика используют редко, поэтому в гла-
вах части II, если не указано иначе, все приведенные данные относятся к сухому
весу хрусталика.
Часто приводят уравнения регрессии веса хрусталика по возрасту и 95%-ный
доверительный интервал, позволяющий оценить надежность данного уравнения (см.
га. 1 1.2.2.1 и 1.2.2.2). Поскольку скорость увеличения веса хрусталика с возрастом
уменьшается, а вариабельность увеличивается, зависимость не линейна и довери-
тельный интервал с возрастом увеличивается (рис. 6.1). Для более точного опи-
сания такой зависимости используют либо систему полу- или двулографми-
100
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
ческих координат (см., например, гл. 12 Apodemus sylvaticus), либо выделяют
линейные участки и приводят несколько уравнений, каждое для своего возраст-
ного интервала, либо сочетают оба приема (см. гл. 12 Microtus).
• • При использовании уравнений регрессии для оценки возраста по
весу хрусталика следует помнить, что они дают лишь приблизительную
оценку возраста и пригодны только для оценки возраста на том возраст-
ном интервале, для которого они получены.
Для более точного описания зависимости веса хрусталика от возраста
Dudzinski, Mykytowycz (1961) предложили выражать возраст как 1/(возраст +
к), где к — показатель, определяемый посредством перебора до получения ли-
нейности в полулогарифмических координатах. Того же типа уравнения исполь-
зовали Myers, Gilbert (1968) и Connolly et al. (1969a) (см. гл. 11 Oryctolagus
cuniculus, Lepus californicus, гл. 14 Odocoileus hemionus).
6.5. Изменчивость показателей
Есть данные, что индивидуальная изменчивость веса хрусталика меньше, чем
веса тела. Так, изменения в питании в постнатальный период, влияющие на вес
тела, не влияли на вес хрусталика; не было корреляции веса хрусталика с весом
тела у особей одного возраста и пола (Friend, 1967b). Не выявлено влияния из-
менения темпа роста особи в целом на вес хрусталика (Henzell, 1987) и влияйия
упитанности на зависимость веса хрусталика от возраста (Gosling et al., 1980).
Вес хрусталика менее подвержен влиянию внешних факторов, чем вес тела (Myers,
Gilbert, 1968; Malcolm, Brooks, 1985). При этом, однако, не отрицалось суще-
ственное влияние на вес хрусталика внешних факторов в период внутриутробно-
го развития (Friend, Severinghaus, 1967).
Можно было надеяться, что межиндивидуальные различия веса хрусталика у
особей одной возрастной группы меньше различий между смежными возрастны-
ми группами, и следовательно, этот показатель окажется хорошим критерием воз-
раста. Эта надежда оправдалась не полностью. Большинство работ по диким мле-
копитающим подтверждает; что вес хрусталика увеличивается долгое время после
наступления половой зрелости, когда темп роста тела резко уменьшается. Однако
при этом подчеркивается, что период относительно быстрого роста хрусталика
сменяется периодом медленного увеличения веса и что у половозрелых особей
межиндивидуальные различия веса перекрывают возрастные (см. ниже раздел 6.6).
Есть примеры как большого сходства возрастных изменений хрусталика
даже у разных подвидов (см. гл. 14 Capreolus), так и существенных межпопу-
ляционных различий. У кроликов Sylvilagus floridanus вес хрусталика в возрас-
те 46 дней в четырех разных штатах США был от 42 до 60 мг., а вес в 64 мг
соответствовал возрасту 52 дней у особей из Висконсина и 78 дней у особей из
Юж. Техаса (Lord, 1959; Bothma et al., 1972). При весе хрусталика 50 мг по урав-
нению для калифорнийской популяции зайцев Lepus californicus возраст был рас-
считан как 13 дней (95%-ный интервал 10-16 дней), а по уравнению для канзасской
популяции—как 25 дней (95%-ный интервал 20-31 дней) (Connolly et al., 1969b).
Глава 6. Определение возраста по весу хрусталика
101
Рис. 6.2. Изменение сухого веса хрусталика (средний вес пары хрусталиков одной особи) с
возрастом у ондатры, Ondatra zibethicus, при разных условиях содержания: вольном (1), по-
лувольном (2), клеточном (3) (Le Boulengc, 1977).
Генетически обусловленные (межлинейные) различия по весу хрусталика
обнаружены у норок со зверофермы (Pascal, Delattre, 1981) и у лабораторных
мышей (Berry, Truslove, 1968). При этом есть данные, что параметры зависимо-
сти веса хрусталика от возраста у генетически однородных, но живших в раз-
ных условиях (в природе и в вольерах) кроликов Oryctolagus cuniculus различа-
лись в той же мере, что и у разных подвидов зайцев Lepus califomicus (Wheeler,
King, 1980). Различался вес хрусталика и у ондатры Ondatra zibethicus в услови-
ях вольного и клеточного содержания (Le Boulenge, 1977) (рис. 6.2).
Различия в весе хрусталиков между самцами и самками у ряда видов не
обнаружены или признаны незначительными (Friend, Linhart, 1964; Novakowski,
1965; Connolly et al., 1969a; Adamczewska-Andrzejewska, 1973a; Harris, 1978;
Vandorpe, Verhagen, 1979; Nuckle, Bergeron, 1983; Henzell, 1987), даже в тех слу-
чаях, когда между самцами и самками есть различия в размере тела (Myers et al.,
1977). Однако у некоторых видов различия в весе хрусталиков между самцами и
самками выявлены (Монахов, 1967; Pascal, Delattre, 1981;Krsmanovicetal., 1985;
Klein et al., 1990; Stockrahm et al., 1996).
В целом можно сказать, что хотя изменчивость веса хрусталика явно меньше,
чем изменчивость веса тела, и при этом интенсивный рост хрусталика продол-
жается и после того, как интенсивный рост тела закончен, вес хрусталика — не
универсальный показатель возраста. Его использование для определения возра-
ста особей до наступления половой зрелости (до достижения веса взрослого
животного) дает лучшие результаты, чем для определения возраста взрослых
особей. Если нужна высокая точность определения возраста, зависимость веса
хрусталика от возраста нужно получать для данной популяции. Выигрыш за счет
длительного периода роста и меньшей индивидуальной изменчивости уменьша-
ется из-за сложности методики и необходимости ее строгой унификации.
102
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Только для зайцеобразных и мелких грызунов этот метод признан превосхо-
дящим другие, более простые. Для хищных технически проще использовать раз-
меры полости пульпы клыка, для копытных — смену и развитие зубов.
6.6. Особенности использования метода в отдельных группах
6.6.1. Хищные
Метод признан пригодным только для выделения сеголетков у тех хищных,
которые достигают половой зрелости на втором году жизни (табл. 6.1). Он столь
же точен, как и методы, использующие степень сращения эпифизов с диафиза-
ми, замещение и стирание зубов, размер и форму кости полового члена. Однако
Sanderson (1961) полагает, что у сеголетков енота Procyon lotor по весу хруста-
лика можно определять даже месяц рождения.
В применении к крупным хищным метод не получил высокой оценки. Из-
за большой индивидуальной изменчивости у рысей Felis lynx, например, не
удается отличить даже годовалых особей от двухгодовалых (Малафеев и др.,
1986). Существенное перекрывание значений между годовыми классами от-
мечено у медведя Ursus arctos уже с первых лет жизни (Pearson, 1975). (В
последнем случае, правда, по признанию автора, методика не была унифици-
рована).
6.6.2. Зайцеобразные
В этой группе метод получил наибольшее распространение, поскольку, как
правило, позволял четко разделить особей, добытых в промысловый сезон,
на сеголетков и старших. В выборках, взятых в разные годы, точка разделе-
ния сеголетков и старших зайцев смещалась незначительно (рис. 6.3), по хру-
сталику удавалось выделить на 15% больше молодых, чем методом пальпа-
ции конечностей (см. гл. 11 Lepus europaeus) (Cabon-Raczynska, Razynski, 1972;
Semizorova, 1975). Однако граница разделения молодых и взрослых в одном
районе может несколько смещаться от осени к зиме (см. табл. 6.2, Sylvilagus).
Этот метод выделения сеголетков кроликов считается более надежным, чем
исследование эпифизов конечностей (Connolly et al., 1969b; Rogers, 1982).
Есть, примеры и неудачного использования метода. Так, Hell (1967), который
отступил от классической методики (см. раздел 6.4) и высушивал хрусталики
при комнатной температуре, не получил четкого разделения сеголетков и стар-
ших зайцев Lepus europaeus, добытых в ноябре-декабре.
По мнению большинства исследователей, этот метод годится лишь для разде-
ления сеголетков и старших в силу существенной индивидуальной изменчивос-
ти веса хрусталиков. Однако Keith, Сагу (1979) сочли возможным у зайцев Lepus
americanus выделять три группы (0,5-1,5 г., 1,5-2,5 г. и старшие), a Hearn, Mercer
(1988) — даже 4 группы (сеголетки, годовики, двухгодовалые и старшие) у
размножающегося 1 раз в году зайца Lepus arcticus.
В ряде случаев для оценки возраста отдельных особей приводят линии рег-
рессии (см. гл. 11 Lagomorpha).
Глава 6. Определение возраста по весу хрусталика
103
Таблица 6.1. Данные по весу хрусталика, пригодные
для различения сеголетков у некоторых хищных
Вид Сезон Возрастные группы Вес хрусталика (мг) Источник
Canis mesomelas моложе и старше 7 мес. точка разделения около 150 Lombaard, 1971
Dusicyon gymnocercus, D. griseus 7-8 мес. и старше года от 170 до 150 Lord, 1966 (по графику)
Felis caracal до года - 2-4 года до 70* 90* Stuart, Stuart, 1985 ( по графику)
Felis lynx осень- зима сеголетки— год и старше меньше 400- около 500 и более Малафеев и др. 1986 (по графику)
Gulo gulo промысловый сезон сеголетки- год и старше точка разделения 70 Rausch, Pearson, 1972
Martes zibellina октябрь 7 мес. - 1 г. 7 мес. М (37^46) - (48-46)** F (32-42)-(42-51) Монахов, 1967
Mustela vison 6 мес. - 18 мес. М до 472 - от 555 ***. F до 396 — от 434 Pascal, Delattre, 1981
Procyon lotor моложе 8 мес. - старше 18 мес. точка раздела около 100 Sanderson, 1961 (по таблице).
Pseudalopex culpaeus май-сентябрь (Аргентина) сеголетки - старшие точка разделения 240 Zapata, Funes et al., 1997
Urocyon cinereoargen- teus ноябрь- февраль сеголетки- старше года точка разделения 167-168 Nicholson, Hill, 1981
до 9 мес. - от 15 мес. до 155- от 160 Lord, 1961
Vulpes vulpes полное разделение до октября, с октября по март есть перекрывание сеголетки - старшие точка разделения 200-210 Harris, 1978; Wandeler, 1976; Friend, Linhart, 1964; Cavallini, Santini, 1995
Примечание: *вес двух хрусталиков; ** М — самцы, F — самки ; *** вероятно, вес двух
хрусталиков
6.6.3. Грызуны
Вес хрусталика считается лучшим показателем возраста, чем размеры тела,
длина и ширина черепа, степень стертости зубов у таких мелких грызунов как
Apodemus agrarius (Adamczewska-Andrzejewska, 1973а; Andrzejewski, Lire, 1977),
A. sylvaticus (Vandorpe et al., 1980), Pitymys duodecimcostatus (Pascal et al., 1988),
Microtus arvalis (Martinet, 1966), Calomys musculinus (Carreno et al., 1990). При
этом обнаруживаются существенные различия между данными разных авторов
для разных популяций. Вес хрусталиков у серых полевок Microtus arvalis во
Франции (Martinet, 1966), Чехии и Словакии (Hlavac, 1979) и Польше
104 Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
вес хрусталика, мг
Рис. 6.3. Частотное распределение по весу хрусталика зайцев Lepus europaeus, добытых в
Польше в ноябре-январе в разные годы (Cabon-Raczynska, Razynski, 1972).
Глава 6. Определение возраста по весу хрусталика
105
Таблица 6.2. Вьщеление возрастных групп зайцеобразных
по весу хрусталика
Вид Регион, сезон Возрастные группы Вес хрусталика (мг) в области разделения возрастных групп Источник
Lepus americanus США: Альберта, январь-июнь июль-декабрь особи предыдущего лета рождения - старшие 192 204 Keith, Сагу, 1979 (меченые)
Lepus europaeus Разные районы Европы, октябрь-март сеголетки- старшие 270 Cabon-Raczynska, Razynski, 1972
Р-н Праги, охотничий сезон сеголетки - старшие ’ 250-280 Semizorova, 1975
Дания, октябрь-декабрь сеголетки- старшие около 2/0 Walhovd, 1966
Дания, ноябрь сеголетки - старшие 230 Andersen, Jensen, 1972
Австрия, осень сеголетки - старшие 275 Suchentrunk et al., 1991
Голландия, октябрь-декабрь сеголетки- старшие 280-290 Broekhuizen, Maaskamp, 197.9
Sylvilagus floridanus США: Огайо, ноябрь. сеголетки - старшие 200-210 Edwards, 1962
Орегон, август-сентябрь. сеголетки - старшие <168 >185 Sullins et al., 1976
Миссури, ноябрь-январь сеголетки - старшие 180190 200-230 Wight, Conaway, 1962
Sylvilagus nuttalli США: Орегон <177дней >291 дней <150 >177 Sullins et al., 1976
Sylvilagus aquaticus США: Миссури, декабрь-март сеголетки - старшие <205 >212 Martinson et al., 1961
(Adamczewska-Andrzejewska, 1973а) в одном и том же возрасте различался по-
чти вдвое. Для объяснения этого Adamczewska-Andrzejewska (1973а) выдвигает
три возможные причины: подвидовые различия, различия в методике, различия
в условиях обитания (лабораторная или природная популяции).
Данные о соотношении скорости роста хрусталика в лаборатории и в природе
разноречивы. Различия не выявлены у мышей Mus musculus (Rowe et al., 1985) и
Apodemus agrarius (Косой, Косая, 1989). У лесных полевок Clethrionomys glareolus
во всех возрастных группах вес хрусталика в природной выборке был меньше,
чем в лабораторной, у серых полевок Microtus agrestis соотношение было обрат-
ным (Hansson, 1983а), что может быть связано со спецификой питания. У крыс
Rattus vilosissimus (Myers et al., 1977) в природе хрусталик увеличивался с воз-
растом медленнее, чем в лаборатории.
106
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Учитывая разноречивость данных, следует указывать, для какой выборки
получены приводимые результаты. Так, Millar, Iverson (1976) указывают, что связь
между возрастом (в днях) двух подвидов Peromyscus и весом хрусталика (сред-
нее значение веса обоих хрусталиков одной особи, мг):
Р leucopus от 18 до 148 дн.: In (возраст) = (вес + 7,3099) : 3,6679;
Р. maniculatus от 26 до 80 дней: In (возраст) = (вес + 6,1478) : 2,9854;
получены для первого поколения грызунов, выращенных в неволе.
Используя этот показатель для определения возраста грызунов, особенно
мелких, следует помнить, что будучи ростовым признаком, темп роста хруста-
лика может различаться у особей разных сезонных генераций. Такие различия
не обнаружены у мышей (Vandorpe, Verhagen, 1979), но были найдены у неко-
торых полевок. У Microtus oeconomus и M.(Stenocranius) gregalis в возрасте
90-100 дней вес хрусталиков у особей весенней (март, май) генерации был зна-
чимо больше, чем у полевок позднелетней (июль) генерации (Schwarz et aL,
1964; Добринский, Михалев, 1966).
Различия в темпе роста хрусталиков у грызунов из выводков разной величи-
ны были выявлены у полевок Microtus montanus и М. pinetorum (Gourley, Janett,
1975) и не найдены у Clethrionomys glareolus (Hansson, 1983b).
Используют вес хрусталика и для определения возраста грызунов, живущих
несколько лет. Хотя у белок рода Sciurus индивидуальная изменчивость веса хру-
сталика велика (Карпухин, Карпухина, 1971), все же по этому показателю моле-
но осенью выделить сеголетков с той же точностью, что и по сращению эпифи-
зов с диафизами костей конечностей (Beale, 1962; Fisher, Perry, 1970; Degn, 1973)
и точнее, чем по величине кости полового члена (Degn, 1973). Выделять среди
сеголетков особей первого и второго помета, видимо, лучше по хрусталику (Beale,
1962). По мнению Fisher, Perry (1970) по весу хрусталика можно выделить воз-
растные группы среди особей старше года и этот метод дешевле и проще, чем
рентген эпифизов. У белок Tamiasciurus hudsonicus, судя по данным Rusch et al.
(1982), по весу хрусталика можно определять возраст в течение первых 5-6 мес.
и отделить особей второго года жизни от старших.
Для крупных грызунов {Castor, Marmota) вес хрусталика плохой показатель
для определения возраста особей старше года.
6.6.4. Копытные
Большинство исследователей считают, что вес хрусталика пригоден для
выделения особей младших возрастных групп, но для этих целей проще
использовать прорезание и стертость зубов (Novakowski, 1965; Connolly et
al., 1969a; Rautenbach, 1971; Hall-Martin, 1976; Ashby, Henry, 1979; Feldhamer,
Chapman, 1980; Mason, 1984). По весу хрусталика лучше, чем по развитию
зубов, определять возраст оленят моложе года (Connolly et al., 1969а). По
мнению Klein et al. (1990), изучавших материал от меченых Sus scrofa, меж-
популяционные различия велики, но вес хрусталика все же можно исполь-
зовать как критерий возраста, имея дело с особями одной и той же популя-
ции.
Глава 6. Определение возраста по весу хрусталика
107
Таким образом, по весу хрусталика у копытных можно отделить молодых от
взрослых в единовременно собранных выборках (если нет возможности иссле-
довать зубы), но перекрывания между старшими годовыми группами велики.
Этот показатель можно использовать для определения возраста особей в тече-
ние первого лета жизни, но кривую (таблицу) зависимости веса от возраста сле-
дует получать для каждой конкретной популяции.
6.6.5. Насекомоядные
Вес хрусталика мало используется для определения возраста насекомоядных.
Gliwicz, Jancewicz (2001) пришли к выводу, что это — хороший показатель воз-
раста бурозубок, поскольку вес увеличивается в течение всей жизни особей. По
суммарному весу двух хрусталиков они выделили 3 возрастные группы среди
молодых Sorex araneus и две — среди молодых 5. minutus.
6.7. История применения
Первым вес хрусталика для определения возраста использовал Lord (1959).
На основании исследования особей известного возраста, он предложил таблицу
и кривую изменения веса сухого хрусталика глаза кролика Sylvilagus floridanus
от 1 до 914 дней, что позволяло определять месяц рождения сеголетков кроли-
ков и год рождения кроликов старше года. После этого метод стали применять к
другим млекопитающим. В обзоре 1967 г. указаны 20 видов, на которых был
апробирован этот метод (Friend, 1967с), а в обзоре 1995 г. — более 100 видов
(Stubbe et al., 1995).
ГЛАВА 7. РЕДКО ИСПОЛЬЗУЕМЫЕ МЕТОДЫ
Параллельно с развитием и совершенствованием традиционных методов шли
поиски новых методов определения возраста. Некоторые из них заслуживают
упоминания, хотя они и не стали распространенными.
7.1. Изменение состояния сосудов сердца
и вилочковой железы (тимуса)
У копытных в фиброзном кольце аорты образуются сердечные кости. С возрас-
том их размеры увеличиваются, что может служить критерием относительного воз-
раста (Habermehl, 1985), хотя в самых старших возрастных группах размеры этих
костей несколько уменьшаются. Например, у лани Cervus dama кость в основании
аорты очень мала в 6 мес., в возрасте 2 лет достигает длины 12 мм, в возрасте 7-
12 лет — 20 мм и уменьшается до 15 мм в возрасте 14 лет (Habermehl, 1985).
Степень развития вилочковой (зобной) железы одно время была популярным
критерием относительного возраста грызунов, в частности серых полевок Microtus.
Эта железа, крупная у молодых в период быстрого роста, уменьшается вплоть
до полного исчезновения у взрослых. При этом, однако, следует учитывать се-
зонные изменения железы, которые различны у разных сезонных генераций гры-
зунов (Оленев, 1967).
Вероятно, таким образом, т.е. в качестве дополнительных критериев возрас-
та, можно использовать и другие внутренние органы, состояние которых с воз-
растом меняется.
7.2. Содержание воды в организме
Fedyk (1974a,b) показала, что содержание воды в организме рыжих полевок
Clethrionomys glareolus коррелированно с возрастом, и до возраста 90 дней этот
показатель можно использовать как критерий возраста особей как весенних, так
и осенних генераций. Содержание воды определяли по разнице веса тушки (без
пищеварительного тракта) до и после высушивания.
7.3. Содержание Sr-90
Rosenthal (1968) предложил метод оценки возраста по относительному содер-
жанию в зубах Sr-90. Метод основан на том, что (1) эмаль и основная масса
дентина коронки постоянных щечных зубов большинства крупных млекопитаю-
щих формируются в ранний и относительно короткий период жизни особи и (2) в
течение этого периода при кальцинации тканей зуба в них включается Sr-90 и оста-
ется там навсегда (если пренебречь его естественным распадом). Этот метод вклю-
чает следующие этапы. Во-первых, нужно получить данные по выпадению и на-
Глава 7. Редко используемые методы
109
коплению Sr-90 в районе обитания исследуемой популяции. Их можно найти в
отчетах учреждений, ведущих мониторинг радиационного загрязнения среды. Да-
лее, по соотношению содержания Sr-90 в среде и в диете изучаемого вида нужно
построить график изменения содержания Sr-90 в диете на протяжении ряда лет
Показатель включения в зуб Sr-90, получаемого с пищей принимается, на основа-
нии экспериментальных данных, равным 0,3. Наконец, определяют относительное
(на грамм кальция) содержание Sr-90 в коронке зуба, делят на показатель 0,3,
получают содержание Sr-90 в диете в год формирования зуба, а затем по графику
изменения концентрации Sr-90 в диете по мере увеличения загрязнения среды
определяют хронологический год формирования зуба, на основании чего и уста-
навливают год рождения особи. Rosenthal (1968) тестировал этот метод на мар-
тышках Alouatta сагауа и оценил его точность как ±2 года. Метод предполагает
целый ряд допущений, но при определенных условиях может быть использован
как ориентировочный для оценки возраста крупных млекопитающих.
7.4. Рацемизация аспарагиновой кислоты
Метод (Helfman, Bada, 1976) основан на том, что аминокислоты включаются
в белки организма в виде L-изомера и постепенно идет процесс их перехода в D-
изомер, накапливается рацемат—смесь изомеров. Процесс этот зависит от тем-
пературы. С наибольшей скоростью идет рацемизация аспарагиновой кислоты.
При температуре тела млекопитающих скорость достаточна для того, чтобы в
метаболически инертных белках долгоживущих млекопитающих количество D-
формы можно было измерить. В дентине зубов в год переходит из одной формы
в другую 0,1% кислоты (Helfman, Bada, 1976).
Возраст долгоживущих млекопитающих оценивали по соотношению этих изоме-
ров аспарагиновой кислоты в дентине зубов или хрусталике глаза (Bada et al., 1980,
1983). При этом было выяснено, что скорость рацемизации в молочных и постоян-
ных зубах различается примерно вдвое (Ohtani et al., 1996), и что данные разных
лабораторий по скорости этого процесса в постоянных зубах существенно различа-
ются (Waite et al., 1999). Метод оценивается как перспективный применительно к
человеку, но требующий строгой унификации методики (Waite et al., 1999; Ritz-Timme
et al., 2000); следует учитывать не только условия, при которых проводится анализ,
но и условия, в которых находился анализируемый орган в организме (Ohtani,
Yamamoto, 2003). Вероятно, какими-то пока неизвестными причинами, влияющими
на результат, можно объяснить то, что при определении этим методом по хрусталику
глаза возраст 48 особей гренландского кита Balaena mysticetus George et al. (1999)
для трех особей получили крайне маловероятный возраст 135, 159 и 211 лет.
7.5. Содержание белка в хрусталике
Были попытки использовать для определения возраста не вес хрусталика, а
содержание в нем белка. Количество нерастворимых белковых фракций в хрус-
талике глаза увеличивается с возрастом и зависит от возраста более, чем от
по
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
каких-либо иных факторов (Kauffman et al., 1967; Dapson et al., 1968; Otero,
Dapson, 1972; Dapson, Irland, 1972) .
Показателем служит содержание в хрусталике тирозина. Кратко процедура
такова: свежий хрусталик гомогенизируют, центрифугируют (если для цетрифу-
гирования материала мало, используют фильтрацию), добавляют реагенты для
получения окрашенного комплекса с тирозином, определяют содержание коло-
риметрически и сравнивают с таковым от особей известного возраста.
Зависимость содержания тирозина от возраста, полученная для хомячков
Peromyscuspolionotus от 45 до 752 дней, оказалась криволинейной:
1g (вес тирозина в микрограммах) = 1,624 + 0,491 1g (возраст в днях)
(Dapson, Irland, 1972).
Точность метода, по мнению авторов, высокая, 95% особей с весом тирозина
404 микрограмма имеют возраст от 70 до 147 дней, с весом 797 микрограмм —
285-583 дней. Для мелких грызунов в период после окончания быстрого роста
этот метод точнее, чем метод определения возраста по весу хрусталика, (см. гл.
12 Sigmodon hispidus, Microtuspennsylvanicus), но исследователь должен решить,
стоит ли эта точность тех затрат времени и сил, которые требует данный метод.
Для долгоживущих млекопитающих его преимущество не очевидно. Grau et
al. (1970), исследовавшие енота Procyon lotor, пришли к выводу, что по содер-
жанию белка (оцененному по азоту) в хрусталике можно отличить сеголетков от
перезимовавших, но рекомендовать этот метод нет смысла, поскольку это же
можно сделать и по весу хрусталика, что проще.
У кабана Sus scrofa выявлено увеличение содержание азота в хрусталике с
возрастом до 320 дней, причем разный уровень кормления практически не влиял
на содержание белка (Kauffman et al., 1967). Ludwig, Dapson (1977) полагают,
что для определения возраста копытных, например, оленя Odocoileus virginianus,
этот метод более точный, чем определение возраста по степени стертости. Его,
однако, вряд ли стоит рекомендовать для широкого применения, учитывая слож-
ность процедуры и влияние на содержание белка замораживания хрусталиков,
чего трудно избежать.
ГЛАВА 8. МЕТОДЫ ПРИЖИЗНЕННОГО
ОПРЕДЕЛЕНИЯ ВОЗРАСТА
Методы определения возраста, не требующие умерщвления животных, стано-
вятся все более популярны в связи с уменьшением промысла и охраной многих
видов млекопитающих и возрастающим интересом к методам исследований, наи-
менее травмирующих животных. Большинство прижизненных методов позволя-
ют отделить молодых от взрослых или определить примерный возраст взрослых
особей. Основные из них следующие.
8.1. Размеры тела и его частей
Во всех группах млекопитающих детенышей и очень молодых особей от взрос-
лых можно отличить по размеру тела. К возрасту достижения половой зрелости
фаза быстрого роста заканчивается, но в разных группах млекопитающих к это-
му периоду особи достигают разной доли от окончательных размеров (см обзор.
Мина, Клевезаль, 1976), и потому для разных групп размеры играют разную
роль как вспомогательный критерий при определении возраста.
Размеры, форма тела и поведение позволяют оценить примерный возраст не-
которых копытных при слежении за ними на расстоянии (см. гл. 14 Capreolus,
Cervus elaphus, Odocoileus hemionus, Oreamnos americanus, Ovibos moschatus, Ovis
amnion).
Размеры могут помочь в ряде случаев, когда другие показатели возраста дают
нечеткий результат. Многие исследователи приводят данные по возрастным изме-
нениям размеров тела в качестве вспомогательного критерия (см. гл. 11 Lepus
europaeus, гл. 12 Cricetus, Erethizon, Tamias, гл. 13 Enhydralutris, Ursus americanus,
гл. 14 Bison).
Размеры тела, как никакой другой показатель, подвержены индивидуальной и
межпопуляционной изменчивости. При работе не с той популяцией, для которой
имеются проверенные данные, следует помнить об очень большой межпопуляци-
онной изменчивости размеров, в том числе и об увеличении размеров в более
холодных условиях обитания (правиле Бергмана).
Кроме того, самые старые животные не всегда самые крупные. Есть много
данных, свидетельствующих, что до предельного возраста доживают относительно
мелкие, медленно растущие особи.
При оценке возраста мелких грызунов иногда используют зависимость веса
тела от возраста. Однако вывод, сделанный на основании исследования лесных
полевок Clethriononiys gapperi (Fuller, 1988), сводящийся к тому, что нельзя су-
дить о возрастной структуре популяции, если вес особи — единственный крите-
рий возраста, можно отнести и к другим грызунам.
Размеры отдельных частей тела также могут служить прижизненным кри-
терием возраста, если можно осмотреть и измерить обездвиженных животных.
Длина задней лапы позволяет отделить молодых особей от взрослых и иногда
112
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
выделить несколько возрастных групп среди молодых (см. гл. 11 Lepus europaeus,
Sylvilagus, гл. 12 Castor, Cricetus, гл. 13 Ursus arctos, U. americanus). Возраст
носорога прижизненно можно определять по размерам рога (см. гл. 15
Ceratotherium simum).
У большинства видов млекопитающих молодых, неполовозрелых самок от
взрослых, размножавшихся, можно отличить по размеру и внешнему виду со-
сков: у молодых они маленькие, слабо заметные. Однако следует помнить, что
не все самки могут вступить в размножение сразу после достижения половой
зрелости.
Следы, при наличии четких отпечатков, отражают размер лапы и позволяют
отличить молодых от взрослых или выделить несколько возрастных групп (см.
гл. 12 Castor, гл. 14 Alces).
По длине отпечатка задней конечности слона Loxodonta africana можно оце-
нивать возраст самцов и самок моложе 15 лет (Western et al., 1983).
Экскременты, Размер и форма экскрементов связаны с размером тела, но
эти показатели менее точные, чем размеры тела. По экскрементам удается разли-
чать молодых и взрослых лосей, а иногда и отличать самцов и самок среди
взрослых (см. гл. 14 Alces). Размер экскрементов используют для оценки возра-
стного состава группировок слонов Elephas maximus (Reilly, 2002).
8.2. Окраска. Состояние меха
У многих видов млекопитающих молодые особи отличаются от старших ок-
раской тела (см. гл. 9 Talpa, гл. 10, гл. 12 Apodemus, Lemmus, гл. 13 Enhydra
lutris, гл. 14 Capreolus). У мелких грызунов смена волосяного покрова на раз-
ных участках позволяет определять возраст молодых; ход линьки можно уви-
деть, если раздувать мех и на месте раздвинувшихся волос видеть «щетку» но-
вой шерсти (Тупикова, Каледа 1957) (рис. 8.1). Следует отметить, что указанный
на рисунке 8.1 возраст разных стадий линьки, следует принимать как сугубо
ориентировочный, в других условиях он может быть несколько иным. У некото-
рых видов окраска, опушенность и форма хвоста и/или лап позволяют отличить
молодых от взрослых (см. гл. 9 Sorex, гл. 12 Marmota baibacina, М. bobac, М.
menzbieri, Sciurus carolinensis, S. niger).
8.3. Возможности прижизненного использования
других методов
8.3.1. Рога: форма, размер, годовые кольца
Форма и размер рогов у оленьих и некоторых полорогих позволяют отделить
молодых от взрослых и иногда выделить еще несколько возрастных групп при
слежении за животными на расстоянии (см. данные в разных разделах гл. 14, а
также гл. 5).
Глава 8. Методы прижизненного определения возраста
113
I
Ш
IV
Рис. 8.1. Первая линька у мелких грызунов разного возраста (на основании наблюдений в
неволе) (по Тупикова, Каледа, 1957, с изменениями). I-V — стадии линьки. Белое поле —
ювенильный наряд, точки — участки, где при раздувании меха видна «щетка» новой шерсти,
черное поле — наряд взрослого зверька.
Cricetulus triton-. 25-30 дней—la; 35 дней—Па; 40 дней—П1а; 45 дней — IVa; 50 дней—Va.
Cricetulus migratoriusi 27-30 дней—la; 32-34 дня—Па; 35-38 дней—Ша; 40 дней—IVa; 45
дней—Va.
114
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Rhombomys opimus'. 30 дней — la; 37 дней — Па; 45 дней — Ша; 52 дня — IVa; 60 дней — Va.
Nesokia indica: 40 дней —16; 45 дней — Па; 50 дней — Ша; 55 дней — IVa; 60 дней — Va.
Apodemus agrarius: 30-33 дня—1в; 35 дней — Пб; 38 дней — Шб; 42 дня—IVa; 45 дней—Va.
Apodemus sylvaticus: 30-32дня 1в; 35 дней—Пб; 40 дней—Шб; 43 дня—IVa; 45-50дней—Va.
Apodemusflavicollis: 30 дней —- 1в; 35 дней — Пб; 40 дней—Шб; 45 дней — IVa; 50 дней—Va.
Apodemus speciosus'. 30 дней—1в; 35 дней—Пб; 40 дней — Шб; 50 дней—IVa; 60 дней—Va.
Mus musculus-. 28-30дней—1в; 32-35 дней—Пб; 35-38 дней—Шб; 40 дней—IVa; 40-45 дней—Va.
Lagurus lagurus-. 25 дней — Ir; 27 дней — Пб; 30 дней — Шб; 33 дней — IVa; 35 дней — Va.
Microtus arvalis: 26-28 дней — Ir; 30-32 дня — Пв; 34-35 дней — Шв; 37-39 дней — IV6; 40-
42 дня — Va.
Dicrostonyx torquatus: 28-30дней—1д; 32 дня—Пг; 35 дней—Шг; 40 дней—I Vb; 45 дней—V6.
По кольцам на рогах некоторых полорогих (см. гл. 5.1) можно определять
возраст особей с точностью до года у обездвиженных особей.
8.3.2. Зубы: прорезание, стертость, окраска
По прорезанию и стачиванию зубов можно выделять несколько возрастных
групп, если удается получить слепок или отпечаток жевательной поверхности
или просто заглянуть в пасть усыпленного или физически обездвиженного жи-
вотного. Gipson et al. (2000) предложили схему стачивания зубов волка (см. гл
13 Canis lupus), позволяющую прижизненно определить возраст особи «в преде-
лах двухлетнего и интервала его фактического возраста» (с. 757).
У млекопитающих среднего размера и крупных, раздвинув челюсти, мож-
но осмотреть зубы с помощью фонарика и зубного зеркала (см. гл. 12 Cynomys,
гл. 13 Panthera pardus и гл. 14 Capreolus), а также получить отпечатки зубов, а
затем и их слепки — с помощью материалов и оборудования, предлагаемого
для дантистов (см. ш. 13 Crocuta, гл. 14 Odocoileus hemionus). Malcolm (1992),
работавший со щетинистыми крысами Proechimys, получал отпечатки зубов
живых животных следующим образом: на узкую алюминиевую пластинку на-
носил пластилин толщиной 2 мм и, держа ее за свободный от пластилина ко-
нец, вводил в пасть и прижимал к зубному ряду. Отпечатки жевательной по-
верхности резцов копытных можно сделать на копировальной бумаге (см. гл.
15 Equus burchelli). /
По отпечаткам на пластилине всех зубов, включая щечные, у обездвиженных
животных, возраст 76% особей Kobus defassa был определен с ошибкой не бо-
лее года (по сравнению со слоями цемента в первом резце) (Spinage, 1967).
Есть опыт прижизненного измерения отдельных зубов. Например, у канны,
Taurotragus oryx, у живых животных мерили высоту коронки ii и ширину (в на-
правлении соседних зубов) его поверхности стачивания (Jeffery, Hanks, 1981)
(см. также гл. 13 Crocuta).
Окраска зубов
У молодых и старых особей некоторых видов она различается, что позволяет
их разделять довольно точно (см. гл. 9 Sorex, гл. 12 Marmota топах).
Глава 8. Методы прижизненного определения возраста
115
8.3.3. Годовые слои в дентине и цементе зуба
Накоплен большой опыт удаления зуба у обездвиженного животного для пос-
ледующего анализа (см. гл. 13 Enhydra, Martes pennanti; Ursus arctos; U. ameri-
canus; гл. 14 Rangifer, Capreolus; Odocoileus virginianus). Во многих случаях
были произведены повторные отловы, показавшие, что рана быстро заживает
(Bergerud, Russel, 1966.; Блажис, 1971; Машкин, 1979; Garshelis, 1984) и через
несколько лет не выявляется отрицательного влияния удаления зуба на состояние
животного (Craighead et al., 1970; Pearson, 1975).
8.3.4. Годовые слои в фалангах пальцев
У грызунов при мечении обычно отрезают вторую и третью фаланги. По на-
личию на поперечном срезе фаланги зимней линии склеивания можно отличить
сеголетков от перезимовавших (см. рис. 4.7), а у некоторых видов выделить и
следующий годовой класс (Клевезаль, 1988).
8.3.5. Форма кости полового члена
У хищных с возрастом меняется и размер, и форма это кости. У молодых
кость тонкая, проксимальная часть практически такой же толщины, что и тело
кости. По мере полового созревания тело кости увеличивается и проксимальная
часть утолщается. Утолщение проксимальной части можно прощупать у обезд-
виженного животного.
8.3.6. Скелет: окостенение эпифизов
У обездвиженных мелких млекопитающих с помощью рентгенографии мож-
но определить приблизительный возраст по окостенению тел позвонков или эпи-
физов конечностей. У зайцев степень окостенения эпифизов оценивают также и
прощупыванием кости (см. гл. 11 Lepus europaeus). У летучих мышей прижиз-
ненно измеряют длину предплечья и состояние ростовой зоны фаланг пальцев
(см. гл. 10).
ЧАСТЬ II. ПРИМЕНЕНИЕ МЕТОДОВ.
ОПРЕДЕЛЕНИЕ ВОЗРАСТА ПРЕДСТАВИТЕЛЕЙ
РАЗНЫХ СИСТЕМАТИЧЕСКИХ ГРУПП
МЛЕКОПИТАЮЩИХ
В этой части книги приведены фактические данные, дающие возможность
определять относительный или абсолютный возраст особей отдельных видов од-
ним или несколькими методами. Виды одного отряда размещены в алфавитном
порядке по их латинским названиям. В основном это—виды, распространенные
в умеренной и приполярной зонах Северного полушария. Другие виды включе-
ны в том случае, когда они служат хорошей иллюстрацией материала, изложен-
ного в части I. Не включены виды, по оценке возраста которых имеются лишь
отрывочные данные с использованием только одного из методов. Они, однако,
могут упоминаться при описании соответствующего метода и ссылку можно
найти по названию вида в указателе.
ГЛАВА 9. НАСЕКОМОЯДНЫЕ
Blarina brevicauda — короткохвостая бурозубка
Зубы
Стачивание. Стертость зубов можно оценивать по индексу стачивания-—сумме
промеров (окуляр-микрометром под микроскопом) высоты коронок по одному
внутреннему и двум наружным буграм (протокониду, паракониду и метакониду)
nij и ш2 и внутреннему бугру (протокониду) mr Построение таблицы частоты встре-
чаемости разных значений индекса в разные месяцы круглогодичного отлова (ана-
лог способа Смирнова — см. гл. 1 раздел 1.2.1.2) дает возможность в каждый
месяц отлова, хотя и несколько субъективно, отделить сеголетков от старших и
построить график зависимости темпа стачивания от возраста. Эта зависимость не
прямолинейна (с возрастом темп стачивания уменьшается) (Dapson, 1968).
Crocidura — белозубки
Череп. Зубы
Стачивание. У С. guldenstaedti, относя к группе «молодых» (juvenis) только
детенышей гнездового периода, можно выделить группу полувзрослых (subadultus,
от 1 до 5 мес.), не имеющих гребней на черепе, с двумя подгруппами (с нестертыми
и со слабо стертыми промежуточными1 зубами), группу взрослых (от 5 до 13 мес.)
с заметно стертыми промежуточными зубами и затылочными гребнями на черепе и
1У землероек промежуточными называют сходные по форме резцы, клыки и предкоренные.
Глава 9. Насекомоядные
117
группу старых (от 13 мес. и старше) с резко выраженными гребнями черепа и
промежуточными зубами, стертыми более чем на две трети (Темботова, 1986).
Сеголетков от старших особей С. suaveolens можно отличить по длине види-
мой части корня первого верхнего резца, от края альвеолы до шейки зуба. Этот
параметр увеличивается с возрастом. Используя графический способ Смирнова
(гл. 1 раздел 1.2.1.2) можно выделить эти возрастные группы в каждый конкрет-
ный месяц сбора, несмотря на большой разброс значений, связанный с наличи-
ем нескольких пометов за лето (Vohralik, 1988).
Erinaceus europaeus — обыкновенный еж
Зубы
Стачивание. В выборке, взятой в мае-июле, по обнажению дентина на жева-
тельной поверхности Skoudlin (1976) выделяет 5 годовых классов (не считая
сеголетков), приводя в подтверждение распределение особей по промерам зу-
бов (см. ниже Talpa).
Годовые слои видны на окрашенных срезах в нижней стенке мандибулы за
концом зубного ряда, в цементе и во вторичном дентине щечных зубов. Годовой
слой может включать несколько сближенных зимних линий (см. гп. 4 •).
Кости конечностей
По рентгенограммам передних лап у живых и мертвых ежей можно отличить
особей в возрасте около года и моложе от старших: у молодых есть эпифизарный
хрящ в дистальной части метакарпальных костей. Дистальные эпифизы лучевой и
локтевой костей сливаются с диафизами, вероятно, на 4-е лето жизни (Morris, 1971).
Galemys pyrenaicus — пиренейская выхухоль
Зубы
Gonzalezesteban et al. (2002) полагают, что можно определять приблизитель-
ный возраст особей по стачиванию коронки верхнего клыка. Сравнение с оцен-
ками возраста по слоям цемента на продольных срезах клыка и первого предко-
ренного показало, что в 1-й год коронка не сточена, на 2-й год сточена на 1/4, на
2-3-й годы — на 1/2, на 3-5-й— на 3/4, на 3-6-й годы — полностью сточена,
видны 2 корня.
Sorex araneus — обыкновенная бурозубка
Внешний вид
У перезимовавших, в отличие от сеголетков, ушные раковины и подошвы
лап лишены волос, хвост кажется голым (Дунаева, 1955).
Зубы
Стачивание. Зубы у молодых острые с хорошо заметной бурой эмалью, у
перезимовавших они стертые, белые, могут иметь лишь остатки желтоватой эма-
ли (Дунаева, 1955).
118
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Викторов (1967,1994) предлагает использовать промеры поперечника повер-
хности стачивания первого нижнего промежуточного (соседнего с крупным рез-
цом) зуба (мерить под бинокуляром). У особей из Тверской обл. этот промер до
2 мес. не больше 100 мкм, в 2-4 мес. — 100-150 мкм, в 4-6 мес. — 150-200
мкм, в 6-8 мес. — 200-250 мкм, в 8-10 мес. — 250-300 мкм, в 11 и более —
больше 300 мкм. При выделении последней возрастной группы дополнительным
критерием служит отношение высоты данного зуба к его продольному диаметру,
которое менее 50-52%.
Годовой слой — четкий, у перезимовавших особей виден в нижней стенке
мандибулы и в цементе зубов на окрашенных срезах (см. гл. 4 •)
Talpa еигораеа — обыкновенный крот
Внешний вид
До осени сеголетки легко отличаются от старших серебристым мехом и
темной (почти черной) пигментацией кожи подошв конечностей. К концу
сентября подошвы белеют, мех темнеет еще раньше (Башкиров, Жарков,
1934). По данным Grulich (1967), только в течение первых двух месяцев
жизни подошвы темные у всех особей, позже у части особей пигментация
исчезает.
Зубы.
Стачивание. Перезимовавших кротов от сеголетков, по мнению Кирилова
(1952), можно отличить по тому, что у первых хорошо видны корни верхних
клыков и отчетливое стачивание р4. Однако по данным Skoczen (1966), корни
клыков становятся видны после второй зимовки.
По степени стертости эмали и обнажению дентина щечных зубов выделяют
5-6 возрастных (годовых) групп (Депарма, 1954; Skoczen, 1966; Grulich, 1967).
Grulich (1967) разделил выборку, взятую в короткий срок в летний период, на
6 возрастных групп, и не выявил переходов между ними (рис. 9.1). Дополни-
тельным критерием, по его мнению, может быть распределение величины пока-
зателя стертости в каждой возрастной группе. Как показатель стертости он ис-
пользовал сумму высот бугра р4 и обоих внутренних бугров ш'~3 (от вершины до
шейки зуба), отнесенную к кондилобазальной длине черепа и умноженную на
100 (рис. 9.1). Деление на кондилобазальную длину введено потому, что выяв-
лена положительная корреляция высоты бугров с кондилобазальной длиной че-
репа у сеголетков с нестертыми зубами.
Годовые слои — хорошо видны на окрашенных поперечных срезах нижней
челюсти, прошедших через коренные зубы: в кости челюсти, в цементе и во
вторичном дентине. Число четких слоев костной ткани в альвеоле зуба служит
оценкой минимального возможного возраста особи, поскольку с возрастом пло-
щадь аппозиции кости уменьшается, и срез может пройти не через все слои. В
некоторых популяциях у сеголетков в цементе видна ювенильная (не годовая)
темно-окрашенная линия близ границы с дентином (см. гл. 4 •).
Глава 9. Насекомоядные
119
Рис. 9.1. Возрастные изменения верхнего зубного ряда крота Talpa еигораеа, особи отловлены
в июне 1960-1964 гт. 0 — сеголетки, I-V—возрастные группы взрослых. А — вид сбоку, В —
жевательная поверхность. Черным показаны обнажения дентина. Внизу: Частотное распреде-
ление значений индекса стертости зубов каждой возрастной группы (см. текст). 1240 экз.,
отловленные в июне (группы 0 и I) и мае-июле (группы II-V) (Grulich, 1967).
120 Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Talpa micrura (=Mogera robusta) — могера
Зубы
Стачивание. Охотина (1966) по степени стертости выделяет 4 группы и, кро-
ме описания стертости зубов, характеризует группы величиной отношения дли-
ны верхнего зубного ряда к максимальной высоте коронки ш1 со щечной сторо-
ны. С мая по ноябрь это отношение у кротов в возрасте 1-6 мес. составляет 5,9-
7,8, в возрасте 1 г. — 8,6-13,9, 2 г. — 17,5-19,6, 3 г. — 24,0-26,9, 4 г. — 29,4.
ГЛАВА 10. РУКОКРЫЛЫЕ. ЛЕТУЧИЕ МЫШИ
Набор методов определения возраста летучих мышей невелик и не видоспе-
цифичен. Поэтому для этой группы не дано повидовое описание. Обзоры см.
Baagoe (1977) и Anthony (1988).
Внешний вид, окраска
Молодые растут быстро и в большинстве случаев достигают размера взрос-
лых уже летом. Так, у Eptesicus fuscus это происходит в июле и с середины
августа по размерам нельзя отличить сеголетков от взрослых (Christian, 1956).
У молодых особей многих видов мех короче и другого цвета, чем у взрос-
лых. У М. myotis молодые — серо-коричневые, взрослые — коричневые с от-
тенком желтизны на беловато-палевой брюшной части (Mazak, 1965). Молодые
Nyctalus noctula остаются более серыми, чем взрослые вплоть до осени, до ухо-
да в спячку (Kleiman, 1969). У Myotis daubentoni, осмотренных в апреле, более
однотонно окрашенным и серым был мех у особей, по состоянию эпифизов —
молодых (Baagoe, 1977). Однако по мнению Anthony (1988) эти различия в ок-
раске невелики и в силу индивидуальных вариаций как у молодых, так и у взрос-
лых различать эти возрастные группы только по окраске непросто.
Молодые Myotis daubentoni, по данным Richardson (1994), осенью и зимой
имеют черное пятно, заходящее с нижней губы на подбородок, хорошо заметное,
когда рот открыт. Весной оно становится более размытым. Большинство стар-
ших особей его не имеют вообще, у некоторых оно сохраняется еще несколько
лет как серо-коричневое, размытое.
Кости крыловой перепонки
Абсолютный возраст молодых особей определяют по длине предплечья, которая
увеличивается в первые недели жизни. Измерения можно делать прижизненно. По
длине предплечья можно определять возраст молодых Vespertilio murinus до 20 дней
(Швецов, 1966), Pipistrellus kuhli до 24 дней (Шагоян, Явруян, 1974), Myotis lucifugus
в первые 11 дней (Kunz, Anthony, 1982), Carollia perspicillata до 20 дней (Cosson et
al., 1993), до 21 дня у листоносых мышей Hipposideros terasensis (Cheng, Lee, 2002).
Старый классический способ — отличать молодых особей от взрослых по от-
сутствию слияния эпифиза и диафиза в местах соединения пястных костей с фа-
лангами пальцев при рассматривании на просвет расправленной крыловой пере-
понки (для мелких — под бинокуляром в проходящем свете) или на ощупь.
Таким способом можно отличать молодых М. myotis до возраста 4 мес., а
Rhinolophus hipposideros до 80 дней, и вводя «индекс оссифйкации», определять
возраст в течение этого периода с точностью до 5 дней (Rybar по Baagoe, 1977).
Baagoe (1977), кроме этих приемов, использовал еще и рентгеновские сним-
ки дистальной части пястных костей и проксимальной части III—V пальцев,
122
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 10.1. Разные стадии (см. текст) слияния с диафизами эпифизов дистального конца пястной
кости и проксимального конца первой фаланги третьего пальца летучих мышей Eptesicus
serotinus и Myotis daubentoni. Позитив рентгеновского снимка (Baagoe, 1977).
полученные с помощью мягкого рентгеновского облучения живых или коллек-
ционных Myotis daubentoni, и коллекционных Р. pipistrellus, Nyctalus noctula,
Eptesicus serotinus. Он выделил 5 стадий (рис. 10.1): (I) — эпифизарная линия
четко видна как полоска (на негативе темная, на позитиве светлая) у дистальных
эпифизов лучевой и пястных костей и проксимальных эпифизов первой фаланги
пальцев, (II) эпифизарная линия становится нечеткой у эпифиза лучевой кости,
(III) эпифизарных линий не видно ни у одного из эпифизов, но видна неровная
нечетко очерченная переходная зона между эпифизом и диафизом, по крайней
мере в дистальной части третьей пястной кости, (IV) более субъективно выделя-
емая на большом материале группа, четко промежуточная между (III) и (V), (V)
Глава 10. Рукокрылые. Летучие мыши
123
переходная (ростовая) зона не прослеживается, вся кость в этой зоне однородно
белая. Этим способом молодые особи могут быть отделены от старших в течение
более долгого периода, чем без рентгеновских снимков. Часть молодых Nyctalus
noctula и Р pipistrellus, по-видимому, не отличима от взрослых уже зимой. Мо-
лодых (прошлогодних) (стадии III и IN) Myotis daubentoni можно было отличить
от взрослых еще и весной, в апреле мае, но не позже, a Eptesicus serotinus —
даже в июне-августе.
Под бинокуляром или на рентгеновских снимках можно прижизненно изме-
рять длину хрящевой прослойки в соединении пястной кости с фалангой пальца.
Этот параметр сначала увеличивается, а затем уменьшается. У Pipistrellus mimus
длина предплечья линейно увеличивается в первые 20 дней, длина хряща в со-
единении четвертой пястной кости с фалангой пальца линейно увеличивается в
первые 2 недели, затем линейно уменьшается до 80 дней, когда происходит око-
стенение (Isaac, Marimuthu, 1996). По сочетанию длины предплечья и длины хря-
щевой прослойки можно определять возраст с точностью до нескольких дней у
Myotis lucifugus до возраста 32 дня (Baptista et al., 2000) и приблизительный
возраст до 2 мес. (Kunz, Anthony, 1982), у Eptesicus fuscus — до 43 дней (Burnett,
Kunz, 1982), у М. myotis—до 75-76 дней (de Paz, 1986), у Carolliaperspicillata —
до 60 дней (Cosson et al., 1993), у Pipistrellus subflavus — до 45 дней (Hoying,
Kunz, 1998), у Megaderma lyra — до 34 дней (Rajan, Marimuthu, 1999) у
Hipposideros terasensis — до 44 дней (Cheng, Lee, 2002).
Зубы. Стачивание
Молочные зубы у летучих мышей функционируют первые недели жизни, по-
том начинают сменяться. У Myotis lucifugus последними, приблизительно в
возрасте 54 дней, меняются верхние резцы (Stegeman, 1956). Позже как
критерий возраста используют стертость зубов. Christian (1956) пришел к
выводу, что хорошее разделение на возрастные группы летом дает ширина
сточенной площадки кончика верхнего клыка; можно выделить 5 возрастных
групп, соответствующих годовым классам, где первая—это сеголетки перед
спячкой. Использование клыков для определения возраста казалось очень
перспективным, поскольку по стертости только этих зубов можно было бы
оценивать возраст летучих мышей прижизненно. Поэтому Twente (1955)
сделал попытку оценивать возраст Myotis velifer (для части особей был
известен минимальный возможный возраст) по стертости верхних клыков.
Он выделял 6 возрастных групп: от «1 - кончик клыка не сточен, острый» до
«6 - клык сточен полностью до уровня десны», и пришел к выводу, что четкие
годовые возрастные группы выделить нельзя. Stegeman (1956) разделил
большую выборку Myotis lucifugus, собранную в один день в июле, на годовые
классы по степени стертости т1 (см рис. 3.12); несмотря на большую
изменчивость степени стертости внутри классов, между классами были
заметные разрывы.
124
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Sluiter (1961) для М. myotis в качестве индекса стачивания использовал сумму
промеров высоты (до кольцевого валика) верхнего и нижнего клыков, бугров р33,
ш3, двух наружных (видных со щечной стороны) бугров 2 и трех внутренних
(с язычной стороны) бугров т1,2 с правой и левой стороны, всего 30 промеров,
и имея небольшой материал от особей, минимальный возможный возраст кото-
рых был известен, вынужден был сделать заключение, что в целом метод для
определения возраста мало пригоден. Показатель не меньше 51,3 мм был свой-
ственен особям первого года жизни, 51,3-48,4 мм — первого или второго года,
48,4—47,5 мм — 1-3-го годов, 47,5-43,9 — 2-4-го (или даже 5-го) годов и т.д.
Baagee (1977) измерял высоту обоих наружных бугров (до нижнего края коль-
цевого валика) ш1 и использовал как индекс стертости сумму высот, отнесен-
ную к длине зуба (измеренной с язычной стороны). Он показал, что есть корре-
ляция этого индекса со степенью окостенения ростовой зоны костей конечнос-
тей, но индивидуальная изменчивость большая и, судя по его графику, особи
Myotis daubentoni с индексом более 1,65 — все молодые, с индексом 1,3 и
меньше — все старшие, в промежуточной области могут быть и те, и другие.
Таким образом, в целом надежность выделения годовых классов по степени
стертости не велика. Это подтвердили и данные Hall et al. (1957) на материале от
меченых особей Myotis lucifugus и М. keenii.
Полость пульпы
При определенном навыке полость пульпы верхнего клык можно разглядеть
под микроскопом в проходящем свете, если зуб поместить в глицерин (Baagoe,
1977). Отношение ширины полости пульпы к общей ширине зуба в прокси-
мальной части «воротничка» больше 60% характерно для молодых особей
Myotis daubentoni, меньше 45% — для старших, в промежутке могут быть
и молодые, и взрослые. Перекрывание возрастных групп такое же, как и по
степени стертости зубов.
Годовые слои
У многих видов летучих мышей четко видны в дентине на окрашенных попе-
речных срезах клыков. Зимняя линия первой зимы жизни может быть нечеткой.
Если полость пульпы закрыта, число слоев в дентине показывает минимальный,
а не фактический возраст особи. Слои в цементе выявлены у летучих лисиц
Pteropus, крупных представителей рукокрылых (детали см. гл. 4 раздел 4.6.2).
ГЛАВА 11. ЗАЙЦЕОБРАЗНЫЕ
У зайцеобразных не только резцы, но и все щечные зубы не имеют настоящих
корней и растут постоянно, поэтому они не пригодны для определения возраста.
Lepus californicus — калифорнийский заяц
Кости конечностей
В течение 1-6 мес. ростовая зона между диафизом и проксимальным эпифи-
зом плечевой кости зарастает до стадии бороздки, в 6-15 мес. между эпифизом
и диафизом видна линия, у особей старше 15 мес. срастание полное, линии не
видно (Tiemeier, Plenert, 1964).
Хрусталик
Связь между возрастом (в днях) и весом хрусталика (в мг) в интервале от О
до 1120 дней описывается уравнением:
100/(возраст + 53) = 4,375 - 1,686 • 1g (вес одного хрусталика) (Connolly et
al., 1969b).
(Что такое 53 и почему инвертирован возраст см. гл. 6 раздел 6.4, поел, абзац).
Проверка по данным, использованным для построения зависимости, показа-
ла, что при весе хрусталика в 110 мг (известный возраст 56 дней) — ошибка в 2
дня, при весе в 300 мг (возраст 350 дней) — ошибка 100 дней. (Об использова-
нии линий регрессии см. гл. 1 разделы 1.2.2.1. и 1.2.2.2).
Lepus europaeus — заяц-русак
Размеры тела и его частей
По весу тела можно определять примерный возраст до 4 мес. (вес 3 кг). Веса
взрослой особи (около 4 кг) зайцы достигают в среднем в 8 мес., но иногда и в
5 мес. В зимний период имеет место уменьшение веса, особенно заметное у
особей на первом году жизни (Pielowski, 1971; Broekhuizen, Maaskamp, 1979).
По длине задней лапы, измеренной от кончика наиболее выступающего когтя
до конца пяточной кости, можно дать приблизительную оценку возраста: при длине
менее 10,5 см — возраст, вероятно, меньше 1 мес., при длине менее 12 см —
меньше 8 мес. (Broekhuizen, Maaskamp, 1979).
Череп. Швы
По состоянию швов крыши черепа выделяют несколько возрастных групп
(рис. 11.1):
- у особей в возрасте до 6 мес. все швы хорошо видны, межтеменная кость
четко обозначена;
- в возрасте 6-8 мес. межтеменная кость различима с трудом или вообще не
различима, задняя часть стреловидного шва зарастает;
126
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 11.1. Зарастание швов черепа у зайца Lepus europaeus. Средний возраст: а—до 6 мес., б
— 6-8 мес., в — 9-12 (13) мес., г — более 1 г. (Cabon-Raczynska, 1964).
ip—межтеменная кость, sc—венечный шов, si — межлобный шов, spt—теменно-височный
шов, ss — стреловидный шов.
- в 9-12 (13) мес. межлобный шов зарастает в средней и передней части,
стреловидный зарос на половину своей длины, начинают зарастать теменно-ви-
сочные швы;
- у особей старше 1 года все упомянутые швы заросли, иногда зарастает
венечный шов, часто эти швы видны лишь в месте соединения друг с другом
(Cabon-Raczynska, 1964).
Годовые слои можно видеть в костной ткани на окрашенных поперечных
(перпендикулярно длинной оси кости) срезах мандибулы в районе щечных зу-
бов. Часто дополнительные линии мешают правильно оценить число годовых
слоев, и потому не все исследователи считают их подсчет пригодным для опре-
деления возраста (см. гл. 4 раздел 4.6.3).
Кости конечностей
Простой метод был предложен Stroh (1931, по Cabon-Raczynska, Raczynski,
1972): молодых можно отличать от взрослых по наличию бокового эпифизарно-
го выпячивания (протрузии) в дистальной части локтевой кости. Метод приме-
ним в полевых условиях и к живым животным (прощупывают выпячивание или
оценивают его на рентгеновских снимках), но крайне неточный и существенно
зависит от опыта исследователя. Он позволяет разделить старших и сеголетков в
октябре (Cabon-Raczynska, Raczynski, 1972; Suchentrunk et al., 1991). Чем поз-
же, тем большая доля сеголетков может быть принята за старших. При оценке
возраста особей, взятых в декабре, несколькими исследователями лишь 44%
особей всегда определялись верно (Walhovd, 1966). По мнению Bujalska et al.
(1965) размер бокового эпифизарного выпячивания и наличие хряща в дисталь-
ной части локтевой кости можно определить точнее, если удалить кожу.
Глава 11.Зайцеобразные
127
По данным Broekhuizen, Maaskamp (1979) на рентгеновских снимках костей
выпячивание перестает быть заметным у особей старше 5 мес.; следы срастания
дистальных эпифизов лучевой и локтевой костей с диафизами еще заметны в
возрасте 7 мес. При визуальном исследовании или на рентгенограммах пере-
дней конечности взрослых особей от молодых можно отличать по полному сли-
янию их проксимального эпифиза плечевой кости с диафизом (Walhovd, 1966).
Хрусталик
Вес хрусталика дает возможность довольно четко отделить сеголетков от стар-
ших, добытых в охотничий сезон (см. табл. 6.2); разделение старших на возрас-
тные группы проблематично.
Для приблизительной оценки возраста отдельных особей можно использо-
вать уравнение регрессии. В интервале от рождения до 450 дней:
возраст, дни = 0,0007646 вес, мг в степени 2,2958 или
1g (возраст, дни) - -3,12 + 2,2958 • 1g (вес, мг);
95% доверительный интервал при весе 300 мг ± 100 дней (Suchentrunk et
al.,1991). (Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Lepus timidus — заяц-беляк
Кости конечностей
В возрасте до 3 мес. эпифизы бедренной кости легко отделяются от диафизов
при вываривании, в 3-4 мес. они начинают срастаться, но шов виден до февраля,
постепенно становясь менее заметным. В марте, в возрасте 8-9 мес. происходит
полное срастание эпифизов с диафизом, шов у проксимального эпифиза не виден,
у дистального может еще быть заметен у отдельных особей (Егоров и др., 1957).
Кости таза
В возрасте 1-2 мес. кости таза легко отделяются друг от друга, их слияние
происходит в возрасте 3-4 мес. (в сентябре), но швы еще заметны, в октябре
швы исчезают. В возрасте 4-5 мес., в октябре, виден шов на месте прирастания
седалищного бугра, он прослеживается у всех молодых вплоть до мая (Егоров и
др., 1957).
Годовые слои — видны в нижней челюсти на поперечных окрашенных сре-
зах или шлифах в верхней части язычной стенки в районе начала зубного ряда.
Хорошее соответствие числа слоев известному возрасту получил lason (1988),
считавший слои в этом участке на аншлифах нижней челюсти. От годовых ли-
ний следует отличать линию резорбции (внутреннюю) и дополнительные линии.
Ochotona — пищухи
В конце лета - осенью сеголетков от старших можно отличить по состоянию
половой системы (поскольку они начинают размножаться лишь на следующий
128
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
год), и по скульптуре поверхности нижней челюсти: у взрослых хорошо выра-
жены гребни для прикрепления жевательной мускулатуры, у молодых они не
развиты (Хмелевская, 1961).
Развитие гребней, в частности гребня на угловом отростке нижней челюсти,
замещение молочных предкоренных на постоянные и характер последнего предко-
ренного позволяют оценить возраст пищух до четырех месяцев (Lissovsky, 2004).
Годовые слои — видны в костной ткани на окрашенных поперечных срезах
нижней челюсти, в наружной стенке середины тела челюсти или в диастеме (см.
гл. 4 раздел 4.6.3).
Oryctolagus cuniculus — дикий кролик
Кости конечностей и тазовые кости
У кроликов из Новой Зеландии в возрасте до 10 мес. проксимальный эпифиз
большой берцовой кости не сращен с диафизом; в 10-25 мес. эпифизы этой
кости срослись с диафизом, все задние эпифизы крестцовых позвонков не сра-
щены; в 26-30 мес. задний эпифиз седьмого крестцового позвонка сращен с
диафизом, шестого — еще нет; у особей старше 33 мес. сращен и задний эпифиз
шестого позвонка. Есть срастание или нет, определяли по наличию или отсут-
ствию видимой эпифизарной линии (Taylor, 1959).
Trout, Lelliott (1992) оценивали в баллах степень приращения эпифизов семи
крестцовых позвонков другим способом: если эпифиз отламывался целиком, он
считался не сращенным (балл 0), и если его нельзя отломить — сращенным
(балл 1). У кроликов из Англии до 8 мес. сумма баллов — 0, до 14 мес. — не
выше 7, до 20 мес. — не выше 10, с 26 мес. — 14 (что соответствует макси-
мальной сумме баллов для обоих эпифизов семи крестцовых позвонков). Trout,
Lelliott (1992) полагали, что этот метод более объективный, чем оценка наличия
линии между эпифизом и диафизом.
Существенные различия в скорости окостенения ростовой зоны позвонков у
кроликов из Новой Зеландии и из Англии могут быть объяснены, кроме межпо-
пуляционных различий, и существенными различиями в оценке процесса окос-
тенения.
Годовые слои — можно видеть в костной ткани в диастеме нижней челюсти
(Henderson, Bowen, 1979) (см. гл. 4 раздел 4.6.3).
Хрусталик
Для кроликов из Австралии связь между возрастом (в днях) и весом хруста-
лика (в мг) описывается такими уравнениями:
возраст = 181,4 : In (314: вес) - 57,
стандартное отклонение 11 %, зависимость калибрована до 250 мг, когда особям
(возраст был известен) было 24 мес. (Myers, Gilbert, 1968);
(возраст + 41) = 60,1912 : [2,4598 - 1g (вес)],
Глава 11. Зайцеобразные
129
зависимость дана для интервала 0-800 дней, но надежно можно оценивать воз-
раст до 150 дней (Dudzinski, Mykytowycz, 1961).
(Что такое 314 и 41 и почему инвертирован возраст в обоих уравнениях см.
гл. 6 раздел 6.4.4, последний абзац).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2)
Sylvilagus — американские кролики
Размеры частей тела
У S. floridanus в течение первых 3 мес. жизни можно определять возраст по
длине задней лапы (от кончика наиболее выступающего когтя до конца пяточной
кости, мм):
в возрасте 1 день — 20-23; в возрасте 1 мес. — 51-56;
5 дней — 26-33; 2 мес. — 70-75;
10 дней — 35-41; 3 мес. — 82-90;
21 дней — 43-53; 4 мес. — 95 и более.
(Hale, 1949; Bothnia et al., 1972).
Череп. Швы
У S', floridanus по зарастанию швов в затылочной области можно выделить
несколько возрастных групп среди особей до 5-6 мес. (рис. 11.2).
Кости конечностей
У S. floridanus дистальный эпифиз плечевой кости срастается с диафизом в
возрасте 4-5 мес., проксимальный — в 8-9 мес., у особей старше 10 мес. про-
ксимальный эпифиз слит с диафизом (Bothnia et al., 1972).
Дистальные эпифизы лучевой и локтевой костей у S. floridanus отделены от
диафиза на 1мм и более в возрасте 2 мес., менее чем на 1 мм — в возрасте 3-4
Рис. 11.2. Возрастные изменение в затылочной области черепа кролика Sylvilagus floridanus.
Шов между наружными и верхней затылочными костями (указан стрелкой): 1 — открыт (пред-
полагаемый возраст до 92 дней), 2—частично зарос, но все еще явно виден (93-105 дней), 3 —
виден только под лупой (106-169 дней), 4 — полностью зарос (от 170 дней) (Hoffrneister,
Zimmerman, 1967).
130
Г.А. Клевезаль, Принципы и методы определения возраста млекопитающих
Рис. 11.3. Схема возрастных изменений эпифизов лучевой и локтевой костей кролика Sylvilagus
floridanus (Petrides, 1951, с изменениями) А — 3-4 мес., Б — 5-6 мес., В — 1,5 г.
мес., приросли, но эпифизарная линия четко видна в возрасте 5-6 мес., в возра-
сте 7 мес. и старше слияние эпифиза и диафиза полное (рис. 11.3), хотя остатки
эпифизарной линии могут быть видны. У более молодых особей последней группы
самая широкая часть головки кости находится в области линии слияния эпифиза
с диафизом, у старших особей она в верхней части эпифиза (Petrides, 1951).
У S. aquaticus приращение этих эпифизов идет сходным образом, они полно-
стью срастаются с диафизами в возрасте 10-13 мес. (Martinson, Holten, 1961).
Годовые слои — видны в костной ткани на окрашенных поперечных срезах
диастемы нижней челюсти (Sullins et al., 1976) (см. гл. 4 раздел 4.6.3).
Хрусталик
Выборку, взятую в осенне-зимнее время, по весу хрусталика можно разде-
лить на сеголетков и старших — см. табл. 6.2.
Для приблизительной оценки возраста S. floridanus, особенно молодых, можно
использовать связь между возрастом (в днях) и весом хрусталика (в мг), опи-
сываемую уравнением, полученным для выборки из штата Висконсин в интерва-
ле 44-914 дней:
1g (возраст) = (вес + 227,5) : 168,175
(Hagen et al., 1980 по данным Lord, 1959);
или для выборки из Южного Техаса в интервале 1-600 дней:
возраст = - 66,12 : [(1g (вес) - 2,348)] - 45,199
(Bothma et al., 1972). (См. также рис. 6.1).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
ГЛАВА 12. ГРЫЗУНЫ
Allactaga и другие Dipodidae — тушканчики
Зубы
Стачивание. По степени стертости верхних коренных зубов Сабилаев (1969)
у A. severtzovi выделял следующие возрастные группы:
1-1,5 мес. — режущие края эмалевых петель стерты незначительно, пятна
обнажения дентина изолированные, высота коронки т1 — >3 мм;
2-5 мес. — эмалевые петли несколько стерты, пятна дентина увеличены и
некоторые соединены между собой, высота коронки т1 до 2,5 мм;
6 мес. - 1 год — эмалевые петли на жевательной поверхности сглажены,
закруглены, площадь жевательной поверхности расширена, обнажения дентина
увеличены, высота коронки ш' достигает 2 мм;
старше 1 года—эмалевые петли стерты почти до основания углублений между
ними, пятна дентина достигают максимального размера, высота коронки гл' —
<1,5 мм.
Возрастные (годовые) группы можно выделять и по суммарной высоте коро-
нок коренных зубов (рис. 12.1) или по сочетанию высоты коронок и характера
жевательной поверхности (рис. 12.2).
Годовые слои — описаны в кости на окрашенных поперечных срезах нижней
челюсти в нижней стенке и в цементе ш?. В костной ткани дополнительные линии
мешают выделять годовые слои, в цементе годовые слои диагностируются луч-
ше. Возраст, оцененный по числу слоев, в целом соответствует возрастным груп-
пам, выделенным по степени стертости и по весу хрусталика (Шенброт, 1984).
Apodemus agrarius — полевая мышь
Внешний вид
Прижизненно молодых мышей от взрослых можно отличить по весу тела и
по окраске. Молодые имеют сероватую окраску спины с менее яркой черной
полоской, взрослые рыже-бурую окраску — с яркой черной полоской. Прибли-
зительный возраст первой линьки — см. рис. 8.1.
Зубы
Стачивание. Общая схема стачивания представлена на рис. 3.10. Adamczew-
ska-Andrzejewska (1973b) выделила 6 возрастных групп, отметив большую ин-
дивидуальную изменчивость. Группы в целом сходны с описанными ею (1967)
для A. flavicollis и Keller (1974) для Mus musculus (см. рис. 12.3), стертость
зубов в 5-й и 6-й группах соответствует стертости зубов Mus musculus в возрас-
те 10-11 мес. и старше. Мыши с такой степенью стертости встречаются в популя-
ции редко. Так, из 142 полевых мышей, пойманных нами в апреле-октябре в
Волгоградской обл. только восемь имели стертость, соответствующую пятой груп-
пе и две — шестой.
132
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.1. Выделение возрастных групп у тушканчика Allactaga elater по высоте коронок щеч-
ных зубов нижней челюсти (сумма промеров щечной и язычной стенок го, 2 3, от межкорневой
дуги до края жевательной поверхности, в единицах окуляр-микрометра, бинокуляр 2x8). (Смир-
нов и др. 1971).
Годовые слои. Перезимовавших особей можно надежно отличить по нали-
чию зимней линии в кости и в цементе зубов на окрашенных поперечных срезах
мандибулы, прошедших через т( 3 (рис. 12.4). Зимняя линия в костной ткани
видна и на поперечных срезах фаланг пальцев, что позволяет прижизненно разли-
чать перезимовавших и не зимовавших особей при проведении мечения
животных отрезанием пальцев (см. гл. 4 •).
Глава 12. Грызуны
133
,80f.
160
140
190
100
П
Рис. 12.2. Высота коронок (сумма промеров щечной и язычной стенок го, 2 3) в единицах оку-
ляр-микрометра (бинокуляр 8x1), число зимних линий в кости (а—нет линий, б — 1 линия,
в — 2 линии) и характер поверхности стачивания нижних коренных зубов у тушканчиков
Alactagulus pygmaeus (I), взятых в конце июня - начале июля, и Scirtopoda telum (II), взятых в
мае в Приаральских Каракумах. Точками обозначены особи с данной высотой коронок, звез-
дочкой — особи, у которых было определено число линий в кости. (Лобачев, Шенброт, 1977).
Хрусталик
По весу можно выделить несколько возрастных групп.
У мышей Польши, вес пары хрусталиков (в мг):
в возрасте <2 мес. — 1,9-5,5; в возрасте 9-10 мес. —11,4-12,2;
3-4 мес. — 5,6-8,5; 11-12 мес. — 12,3-13,0;
5-6 мес. — 8,6-10,3; >12 мес. - >13,1
7-8 мес. — 10,4-11,3;
(Adamczewska-Andrzejewska, 1973а).
Когда эта шкала была проверена на особях известного возраста (мечение в
природе), из 31 особи (в возрасте 8-12 мес.) у 9 возраст был определен верно с
точностью до месяца, у остальных оценка по хрусталику была занижена в сред-
нем на 1,6±1,2 мес. (Andrzejewski, Lire, 1977) (см. гл. 6 раздел 6.4 •).
134
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.3. Схема стертости верхних коренных зубов мышей Apodemus flavicollis (A.f.) (по Adam-
czewska-Andrzejewska, 1967) и Mus musculus (M.m.) (no Keller, 1974). Возраст: A — 3-4
недели (tn3 еще не прорезался), В — около 2 мес., С — около 5 мес., D — около 10 мес., Е —
11 мес. и старше.
Косой и Косая (1989) дали такую шкалу для оценки возраста полевых мышей
Дальнего Востока России. Вес одного хрусталика (в мг):
в возрасте до 20 дней — <1,8;
в 20-30 дней — 1,8-2,4;
с 1 до 2 мес. — 2,5-3,7;
с 2 до 3 мес. — 3,8-4,6;
с 3 до 4 мес. — 4,7-5,1;
с 4 до 6 мес. — 5,2-5,7;
с 6 до 8 мес. — 5,8-6,2;
с 8 до 10 мес. — 6,3-6,6;
с 10 до 12 мес. — 6,7-6,9;
>12 мес. — >7,0.
Глава 12. Грызуны
135
Рис. 12.4. Поперечный срез нижней челюсти мыши рода Apodemus.
А — схема среза, прошедшего через mr В — нижняя часть корня m, Apodemus sylvaticus, d —
дентин, С— цемент, стрелкой указана зимняя линия в цементе; окраска гематоксилин. С —
участок нижней челюсти в конце зубного ряда Apodemus agrarius, стрелкой указана зимняя
линия.
136
Г, А. Клевезаль. Принципы и методы определения возраста млекопитающих
Их данные существенно отличаются от приведенных выше данных Adam-
czewska-Andrzejewska (1973a). Может быть различия связаны с тем, что поле-
вые мыши Дальнего Востока более крупные, чем европейские.
Apodemus flavicollis — желтогорлая мышь
Внешний вид
Прижизненно молодых мышей от взрослых можно отличить по весу тела и
по окраске. Молодые имеют сероватую окраску спины и брюха, горловое пятно
выражено слабо, у взрослых охристо-ржавая окраска спины резко отличается
от светлого брюха, горловое пятно яркое. Приблизительный возраст первой линь-
ки — см. рис. 8.1.
Зубы
Стачивание. Возрастные группы выделяют по степени стертости корен-
ных: см. рис. 12.3 и рис. 3.10. Для привязки степени стертости к возрасту в
конкретной популяции можно использовать способ Смирнова (см. гл. 1 раздел
1.2.1.2).
Годовые слои — см. Apodemus agrarius
Хрусталик
Для ориентировочной оценки возраста можно использовать данные табл. 12.1.
Таблица 12.1. Вес пары хрусталиков (в мг) у желтогорлых мышей,
Apodemus flavicollis, разного возраста (Haferkom, 1995)
Возраст п Средний вес Стандартное отклонение
11 нед. 22 4,56 1,17
4 мес. 12 8,69 0,756
6 мес. 10 10,98 0,882
8 мес. 7 15,07 0,916
10 мес. 10 19,0 1,339
16 мес. 9 22,4 2,214
18 мес. 3 23,43 1,25
Apodemus sylvaticus — лесная мышь
Внешний вид
Прижизненно молодых мышей от взрослых можно отличить по весу тела и
по окраске. Молодые имеют сероватую окраску спины и брюха, у взрослых
светлое брюхо отличается от рыжевато-охристой спины. Приблизительный воз-
раст первой линьки — см. рис. 8.1.
Глава 12. Грызуны
137
Зубы
Стачивание. Общие изменения степени стачивания с возрастом такие же,
как и у других мышей — см. рис. 12.3 и рис. 3.10. Для привязки степени стер-
тости к возрасту в конкретной популяции можно использовать способ Смирнова
(см. гл. 1 раздел 1.2.1.2).
Годовые слои — см. Apodemus agrarius
Хрусталик
Vandorpe, Verhagen (1979) на материале из природы и из лаборатории, не об-
наружив ни сезонных различий, ни различий между самцами и самками в темпе
роста хрусталика, описали связь возраста (в днях) и веса хрусталиков (в мг) до
возраста 6 мес. таким уравнением:
In (возраст) = (вес пары хрусталиков + 15,213) : 6,568.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Это уравнение дает возможность оценивать возраст мышей по данным, приве-
денным Gurnell, Клее (1984) для первого лабораторного поколения мышей, в це-
лом не хуже, чем приведенные этими авторами множественные регрессии, в кото-
рых кроме веса хрусталика использован вес тела или индекс стачивания зубов.
Arvicola terrestris — водяная полевка
Щечные зубы не имеют корней и растут в течение всей жизни и потому для
определения возраста не пригодны.
Череп. Форма (Обозначения см. рис. 12.5).
Возрастные группы выделяют по скульптуре черепа, по развитию гребней (Ку-
черук, Кулик, 1955; Пантелеев, 1966; Терехина, 1973). С возрастом уплощается
крыша черепа, развиваются затылочный, теменные и лобные (межглазничные)
гребни (аналогичные возрастные изменения претерпевает череп Lemmus, см. ниже,
рис. 12.14). В возрасте 1-1,5 мес. крыша черепа выпуклая, гребней нет. В возра-
сте 2-2,5 мес. крыша черепа уплощается, лобные гребни не сомкнуты, в возра-
сте 3-4 мес. они смыкаются. Сеголетки весенней и летней генераций к осени
имеют либо не сомкнутые лобные гребни, либо длина смыкания не более 4 мм.
У перезимовавших длина сомкнутого гребня увеличивается в течение лета, при-
чем у летней генерации она в апреле-мае менее 4 мм, в июле не превышает 5 мм,
у весенней генерации весной она более 4 мм, а в июле более 5 мм. Осенью,
однако, значения для разных генераций перекрываются. Этот показатель уже осе-
нью и весной второго года жизни более 6-7 мм.
Castor — бобры
Размеры частей тела
По длине задней лапы, можно определять возраст С. fiber по следам на грязи
или мокром снегу: сеголетки — 9-15 см; годовики — 15-17 см; 2 г. и старше —
16-19 см (Лавров, 1953).
138
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.5. Схематический рисунок черепа грызуна (Rattus norvegicus, по Гамбарян, Дукельс-
кая, 1955).
1 — лобные (межглазничные) гребни, 2 — теменные гребни, 3 -— затылочный гребень, 4 —
стреловидный гребень, 5 — верхнезатылочная кость, 6 — затылочные мыщелки, 7 — венеч-
ный шов, 8 — кондилобазальная длина, 9—межглазничная ширина (межглазничное расстоя-
ние), 10 — скуловая ширина, 11 — длина зубного ряда. В боковой проекции скуловая кость
удалена, чтобы показать зубной ряд.
Глава 12. Грызуны
139
Зубы
Резцы. По кривизне резцов С. fiber, как верхних, так и нижних, можно выде-
лить три возрастные группы. Если положить сухой резец на бумагу, обвести на-
ружный и внутренний контуры, затем нарисовать окружности, части которых
составляют эти контуры, и определить радиусы этих окружностей — наружной
(нрад.) и внутренней (врад.), то средние оценки для групп (мм) таковы
(Troszynski, 1977):
до года 1-3 г. более 3 лет
верхи, резец нрад. 21,2 27,8 31,2
врад. 13,9 18,7 20,7
нижн. резец нрад. 43,8 54,7 58,8
врад. 36,8 45,7 48,5
Прорезание и стачивание. По прорезанию можно определять возраст боб-
ров в течение первых полутора лет. У С. fiber m, появляется в возрасте 2 недель,
полностью вырастает в 4 недели; т2 появляется в 6 недель и полностью выраста-
ет в возрасте 2 мес., т3 — соответственно, в 4 и 5 мес., первый постоянный
щечный зуб (р4) сменяет молочный в 10-12 мес., полностью вырастает в 12-16
мес. (Piechocki, Stiefel, 1977). По данным Бородиной (1970а) у бобрят в февра-
ле-марте р4 уже постоянный. У С. canadensis р4 полностью прорезается в возра-
сте около года (Van Nostrand, Stephenson, 1964).
У бобров стачивание коронки существенно компенсируется мощными отло-
жениями цемента на корне (см. гл. 3 раздел 3.2.4). Бородина (19706) предложи-
ла надежный способ приблизительной оценки возраста по характеру базальной
части зуба и соотношению высоты коронки и толщины отложений цемента, вид-
ных на продольном распиле зуба (рис. 12.6).
Количественно возрастные изменения отношения толщины цемента ш3 к вы-
соте зуба Борисов (1988) выразил таким образом:
в возрасте 3 г. — 1/6, в возрасте 7-9 л. — 1/3-1/2,
4 г. — 1/5-1/4, 10-14 л. — 1/2-2/3,
5-6 л. — 1/5-1/3, 15-20 л. — 2/3—4/5
У С. canadensis (судя по описанию Van Nostrand, Stephenson, 1964) отверстия
корней зубов с возрастом закрываются в том же темпе, что и у европейского
бобра, и видимо, одну и ту же схему (рис. 12.6) можно использовать для опре-
деления возраста обоих видов.
Годовые слои — в цементе видны на продольных аншлифах коренных зубов
обоих видов (см. рис. 4.3). Иногда дополнительные линии мешают выделению
основных. Первые 2-3 годовых слоя, образующиеся только в боковых частях
корня, тонкие и плохо заметны, поэтому возраст молодых особей определяют по
состоянию полости пульпы (рис. 12.6), а с 3-4 лет — по числу годовых слоев
цемента, формирующихся по всей поверхности корня. При необходимости мож-
но использовать годовые слои костной ткани, которые хорошо видны на окра-
шенных срезах мандибулы в щечной стенке за концом зубного ряда (см. рис.
4.2), у особей старше 11 лет первый слой может резорбироваться.
140
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.6. Схемы продольных распилов бобра Castor fiber, сеголетка (а), годовика (б),
двухгодовалого (в), трех-четырехгодовалого (г), 5-6-летних (д), 7-14-летних (е), 15-19-летних
(ж), 20-25-летних (з) (Бородина, 19706). Отложения цемента заштрихованы.
Кости конечностей
У С. canadensis (Robertson, Shadle, 1954), в большой берцовой кости дис-
тальный эпифиз срастается с диафизом на третьем, проксимальный—на четвер-
том году жизни.
Тазовые кости
У С. canadensis (Robertson, Shadle, 1954) в 9-11 мес. подвздошная кость
свободна, лобковая и седалищная срастаются в задней части; на втором году
жизни все кости срастаются в одну.
Хрусталик
Malcolm, Brooks (1981) пришли к выводу, что по весу хрусталиков нельзя
разнести по годовым классам особей старше года.
Citellus (=Spertnophilus) — суслики
Череп. Размеры (Обозначения см. рис. 12.5).
Отношения кондилобазальной длины к длине зубного ряда (I) и скуловой
ширины к длине зубного ряда (II) позволяют выделить несколько возрастных
групп. Моды и пределы варьирования для двух видов таковы:
Глава 12. Грызуны
141
С. pygmaeus
(Мейер, 1957)
С. fulvus
(Кыдырбаев,
1960)
Полувзрослые сеголетки
годовики
старшие
сеголетки
после первой зимовки
2 г. и старше
Отношение I
3,8 (3,4—4,3)
4,1 (3,8-4,4)
4,3 (3,9-4,6)
3,6 (3,2-4,1)
3,7 (3,4-4,1)
4,0 (3,6-4,5)
Отношение II
2,7 (2,4-3,0)
2,9 (2,6-3,1)
3,1 (2,8-3,4)
2,3 (1,6-2,7)
2,6 (2,3-3,0)
2,9 (2,5-3,3).
Зубы
Прорезание и стачивание. К концу первого лета жизни у сусликов молоч-
ные предкоренные сменяются на постоянные (Леонтьев, Воротникова, 1957; Кы-
дырбаев, 1960; Rulin, 1966; Mitchell, Carsen, 1967).
У сеголетков, как правило, на щечных зубах нет обнажений дентина, но
может наблюдаться потертость эмалевого слоя; у годовиков дентин обнажен
частично и не на всех верхних щечных зубах, в следующих годовых классах
площадь обнажений дентина увеличивается. Общий характер возрастных из-
менений жевательной поверхности сходен у разных видов, хотя есть и межин-
дивидуальные и межвидовые различия (рис. 12.7). Так, возраст обнажения
дентина на первом и втором верхних щечных зубах (р3-4) варьирует не только
между видами (см. рис. 12.7), но и в разных популяциях одного вида (Мейер,
1957). Есть описание и рисунки для С. citellus (Rufid, 1966), С. fulvus (Кыдыр-
баев, 1960), С. musicus (Емельянов, 1983), С. suslicus (Петровский, 1961; Лоб-
ков, 1999), С. undulatus (Леонтьев, Воротникова, 1957; Lin et al., 1989). Учиты-
Рис. 12.7. Жевательная поверхность верхнего зубного ряда сусликов в возрасте 1-4 года
(указан цифрами): Citellus musicus (вверху, даны минимальный и максимальный варианты
каждого годового класса, из Емельянова, 1983) и С. suslicus (внизу, из Лобкова, 1999). Черным
показаны обнажения дентина.
142
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
вая большую изменчивость данного признака, лучше для каждой популяции
разрабатывать свою схему, при этом можно оценивать степень стачивания в
баллах и для выделения возрастных групп использовать способ Смирнова (см.
гл. 1 раздел 1.2.1.2).
Для всех сусликов стертость щечных зубов весной соответствует таковой в
предыдущую осень перед залеганием в спячку.
Годовые слои — видны в костной ткани (нижней стенке мандибулы), в це-
менте (в боковых стенках) и во вторичном дентине (в боковых стенках корня и в
дне коронки) коренных зубов на окрашенных поперечных срезах нижней челю-
сти, прошедших через коренные (по типу рис. 12.4А). Часто дополнительные
линии мешают разграничению годовых слоев, поэтому для получения более на-
дежного результата лучше использовать все три регистрирующие структуры. По
числу слоев в костной ткани на окрашенных поперечных срезах первой и второй
фаланг третьего пальца задней лапы можно выделить сеголетков (нет слоев),
годовиков (1 слой) и старших особей (2 слоя) (см. гл. 4 • и 4.6.4 •).
Кости конечностей
Слияние дистальных эпифизов плечевой кости у С. beecheyi происходит по-
степенно в возрасте между двумя и тремя годами (Tomich, 1962).
Хрусталик
По весу хрусталика С. suslicus можно отличать молодых от взрослых в пер-
вое лето жизни и в марте-апреле следующего года (Лобков, 1999).
Clethrionomys — лесные полевки
Большинство исследований выполнено на С. glareolus, но в целом сходные
возрастные изменения можно ожидать у С. rutilus и С. rufocanus.
Внешний вид
В возрасте около месяца тусклый ювенильный наряд меняется на более яркий
взрослый (Тупикова, 1964). У молодых С. glareolus окраска спины и брюха бо-
лее темная, чем у взрослых, рыжины на спине практически нет; у С. rutilus мо-
лодые от взрослых отличаются только темной окраской брюха (Кириков, 1952).
Зубы.
Рост корней (см. также гл. 3 раздел 3.2.4). Наиболее распространенный ме-
тод определения возраста. Основан на том, что корни коренных зубов появляют-
ся приблизительно с 3-месячного возраста, и далее их рост идет параллельно со
стачиванием коронки (рис. 12.8). Обычно используют mi или т2. В mi корни
увеличиваются, в среднем, по 0,15 мм в мес., в т2 — по 0,1-0,2 мм (Pucek,
Zejda, 1968; Tupikova et al., 1968; Тупикова и др., 1970).
Как и любой ростовой признак, рост корней существенно различен у особей
разных сезонных генераций. Так, корни Ш] начинают развиваться в среднем в
Глава 12. Грызуны
143
мм
Clethrionomys glareolus
30
25
20
15
10
5
мм
Рис. 12.8. Схема изменений с возрастом высоты корня (показан черным) и коронки (заштрихо-
вана) т2 лесных полевок (Tupikova et al., 1968).
Длина корня, мм Возраст, мес.
/ 1 « 1 ш ' iv ' v ] Vi 1 у//1 viii ' ix ' х ' Xi ' хн
Рис. 12.9. Шкала роста корней т, лесной полевки, Clethrionomys glareolus (из Richter, Geiler
(1972) с изменениями).
144
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
возрасте 8 недель у особей весенней генерации, в 12 недель — у летней и в 14
недель — у особей осенней генераций. Интенсивный рост корней у особей ве-
сенней генерации происходит в возрасте 2-3 мес., у особей летней и осенней
генераций — после зимовки (Zejda, 1977). В соответствии с этим, в зависимости
от времени года и от принадлежности особи к той или иной сезонной генерации,
корни пн увеличиваются со скоростью от 0,05 до 0,55 мм/мес. (Lowe, 1971).
Zejda (1977) взял за основу полученную им зависимость возраста начала
формирования корней от месяца рождения и сконструировал масштабную ли-
нейку, используя которую по длине корней и дате поимки можно определить
ориентировочно дату рождения. Richter, Geiler (1972) использовали графичес-
кий способ Смирнова (см. гл. 1 раздел 1.2.1.2) для выделения каналов измене-
ния высоты корней ш] в течение года (рис. 12.9).
Учитывая различия сезонных генераций и межпопуляционные различия, Ба-
шенина (1981) предложила для практических целей выделять 4 возрастные груп-
пы (по характеру т(): у самых молодых призмы видны по всему зубу; в возра-
сте около 1,5-3 мес. в основании зуба первые призмы сливаются, первая бороз-
дка укорочена; приблизительно в возрасте 3-4 мес. корневые части призм сли-
ты, видна выемка, соответствующая будущему межкорневому своду; у старших
перезимовавших особей корни развиты, коронка стерта в разной степени (см.
рис. 3.4).
Оленев (1989), на основании исследования меченых в природе особей,
предложил свой способ выделения возрастных групп. У молодых особей
возраст оценивают по форме базального отверстия т2, позже — по индексу
корня (отношению высоты корня к высоте коронки). Учитывая дату поимки,
вес тела и состояние репродуктивной системы он выделил «типы онтогене-
за»: I — быстро растущие, рано вступающие в размножение и короткоживу-
щие особи и II — медленно растущие, поздно вступающие в размножение и
долго живущие. На практике, по индексу корня отделяют перезимовавших, у
которых индекс уже весной более 0,2, от сеголетков. Далее сеголетков раз-
деляют по «типу онтогенеза»: набирающие вес и в генеративной системе име-
ющие признаки вступления в размножение — тип I, с малым весом, без при-
знаков размножения—тип II. Затем по характеру корней определяют возраст
по схеме, показанной на рис. 12.10. Эта схема должна хорошо работать в
популяциях, где исключено выживание рожденных весной особей до следу-
ющей весны.
Поскольку различия в развитии корней у особей разных сезонных генераций
большие, нет смысла пытаться получить уравнение зависимости высоты корня
от возраста для определения возраста всех европейских лесных полевок, но это
можно сделать для оценки возраста особей конкретной популяции с не очень
растянутым периодом размножения. Perrin (1978) на основании исследования
развития корней ш, у лабораторной колонии С. gapperi, у которых корни появля-
ются в возрасте 80 дней и сезонный период размножения короткий, привел такое
уравнение, предназначенное для определения возраста особей из природной по-
пуляции до возраста 380 дней:
Глава 12. Грызуны
145
Рис. 12.10. Возрастные изменения т2 у особей лесной полевки, Clethrionomys glareolus, разли-
чающихся по «типу онтогенеза». Природная популяция, меченые особи (Южный Урал, Оле-
нев, 2004). Отрезок аб и точки 3—I «тип онтогенеза»; отрезки ас, точки 2 и сд, точки 1 — две
фазы II «типа онтогенеза». По оси ординат классы 1-6 — изменения с возрастом торцевой
поверхности корня: острые углы резко выраженных призм (1) сглаживаются (2,3), централь-
ная часть сужается (4), смыкается (5), появляются два отдельных замкнутых пространства,
корня (6); индекс корня — отношение высоты корня к высоте зуба.
(возраст, дни) = 84 + 158 • (высота переднего корня от бифуркации, мм).
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Годовые слои — линии, похожие на зимние, обнаруживаются в цементе ко-
ренных зубов и в костной ткани мандибулы на окрашенных срезах, но их связь с
возрастом особей не установлена.
Хрусталик
Kozakiewicz (1976) считает целесообразным по весу хрусталика (в мг) опре-
делять возраст С. glareolus (в днях) до 3 мес., пока корни еще не развиты, и
предлагает уравнение:
возраст = (вес пары хрусталиков - 1,05) : 0,063.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.1 и 2.2.2).
Хотя корреляция веса хрусталика и возраста в интервале от 26 дней до 3 мес.
высока (г=0,92), индивидуальная изменчивость даже в самом раннем возрасте
146
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
так велика, что кажется неоправданным применение столь трудоемкого метода.
Проще использовать размеры черепа или вес тела.
Хвостовые позвонки — см. гл. 2.6.
Cricetus cricetus — обыкновенный хомяк
Размеры тела и черепа
Для приблизительной оценки возраста можно использовать данные, получен-
ные при содержании хомяков в неволе (табл. 12.2).
Зубы
Стачивание. Развитие коренных заканчивается к возрасту 50 дней. По сте-
пени стертости ш1 летом-осенью можно отделить сеголетков от перезимовав-
ших: у сеголетков, если и есть обнажения дентина, то их полоски со щечной и
язычной сторон в заднем конце зуба не замкнуты, у перезимовавших — замкну-
ты. Высота (с язычной стороны) среднего бугра т’ у сеголетков (возраст 2—5
мес.) 0,57 мм, ширина обнажения дентина на средней паре бугров (измерено
поперек зуба от крайней точки с язычной до крайней точки со щечной стороны)
до 1,10 мм. В возрасте 13-17 мес. высота бугра — 0,18-0,48, обнажение денти-
на — 1,18-1,58; в возрасте от 25 мес., соответственно, 0-0,3 и 1,41—1,76 мм
(Vohralik, 1975).
Поскольку хомяки имеют несколько пометов в год, для выделения возраст-
ных групп, видимо, имеет смысл применять способ Смирнова (см. гл. 1 раздел
1.2.1.2), используя либо приведенные выше промеры, либо оценку стертости в
баллах.
Таблица 12.2. Возрастные изменения веса тела и размеров черепа
обыкновенного хомяка, Cricetus cricetus (Vohralik, 1975)
Воз- раст, мес. Вес тела, г Кондилобазальная длина черепа*, мм Длина нижней челюсти*, мм
самцы самки самцы самки самцы самки
1 75-114 66-106 35,1-36,7 34,0-35,2 23,2-24,0 22,9-23,4
2 137-240 115-207 39,6-42,7 39,6-41,6 25,6-27,3 24,9-26,0
4-5 210-356 179-310 41,9-46,5 40,0-46,0 27,2-29,6 25,4-29,3
13-17 280-515 215-390 47,6-51,1 43,9-48,6 30,8-33,1 28,6-31,9
>25 405-515 228-420 49,4-52,9 47,7—49,6 32,2-34,9 30,7-32,6
'схему измерения см. на рис. 12.5 и 2.3.
Глава 12. Грызуны
147
Годовые слои можно видеть на окрашенных срезах нижней челюсти в
цементе коренных зубов и в костной ткани нижней челюсти в конце зубного
ряда. Первая зимняя линия в цементе расположена близко к дентину (см. гл.
4«).
Хрусталик
Вес двух хрусталиков (мг) югославской популяции в апреле-октябре менял-
ся с возрастом (возраст оценен приблизительно) следующим образом (Krsmanovic
et al., 1985):
1 мес. 2 мес. 3-9 мес. 12-20 мес. > 20 мес.
самцы 2,3-16,8 16,0-18,5 24,5-28,0 36,4-40,6 43,1 46,3
самки 7,8-9,9 13,9-16,5 21,2-23,9 29,8-33,6 37,1-44,3.
Cynomys ludovicianus — чернохвостая луговая собачка
Зубы
Прорезание. Стачивание. ш33 полностью прорезаются в возрасте 80-100
дней, dp44 полностью сменяется на р44 (второй щечный верхний и первый щеч-
ный нижний зубы) в возрасте около 100 дней (Stockrahm, Seabloom, 1990).
Hoogland, Hutter (1987) прижизненно определяли высоту бугров нижних ко-
ренных на глаз как высокие (около 3 мм) — у сеголетков, средние (около 2
Рис. 12.11. Вид спереди нижней челюсти и первый нижний щечный зуб луговой собачки,
Cynomys ludovicianus (по Сох, Franklin (1990), с изменениями). Стрелками указаны бугры,
между которыми измеряли расстояние (см. текст).
148
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
мм) — в возрасте 1,5 года и низкие (меньше 1 мм) — в 2,5 года и старше. 86%
особей известного возраста, были определены правильно.
Сох, Franklin (1990) измеряли штангенциркулем на живых животных рассто-
яние между щечным и язычным буграми (протоконидом и параконидом) первого
нижнего щечного зуба (рис. 12.11). Челюсти раздвигали круглыми щипцами. В
среднем это расстояние в возрасте 20 недель около 1,8 мм, в 1,5 г. — 2,3 мм, в
2,5 г. — 3,1 мм, в 3,5 г. — 3,6 мм, в 4,5 г. — 4,8 мм. Этот промер до 3 лет
увеличивается прямолинейно. Доверительные интервалы годовых классов не пе-
рекрываются при уровне вероятности 75%.
Хрусталик
По данным Stockrahm et al. (1996) в мае-июле сухой вес 1 хрусталика (мг)
имеет следующие значения:
сеголетки годовики двухгодовалые 3 г. и старше
самцы 5,4-11,0 10,8-16,6 16,4-20,6 20,8-22,0
самки 4,2-11,4 10,2-17,0 15,6-21,0 19,2-22,4.
Dicrostonyx torquatus — копытный лемминг
Щечные зубы не имеют корней и растут в течение всей жизни, потому для
определения возраста не пригодны.
Вес тела
По весу тела весной в первой половине лета можно отличить перезимовав-
ших особей от особей весенней генерации, а во второй половине лета выделить
особей летних выводков. Так, на о. Врангеля до 20-х чисел июля с вероятностью
95% самцы весом не более 60 г и самки весом менее 80 г были особями весен-
ней генерации, а во второй половине лета все зверьки весом менее 45 г — осо-
бями летней генерации (Травина, Стишов, 1996, по результатам мечения).
Dryomys nitedula — лесная соня
Внешний вид
Окраска сеголетков тусклее и серее, чем старших, мех мягче, ровнее и коро-
че (Огнев, 1947).
Зубы
Стачивание. У сеголетков летом эмалевые гребни щечных зубов острые, осе-
нью перед спячкой и весной следующего года гребни начинают притупляться. У
годовиков летом эмаль гребней стирается, появляются небольшие обнажения ден-
тина особенно на двух последних коренных зубах. У двухгодовалых летом эмаль
сильно стерта, обнажения дентина видны на всех зубах, по язычному краю верх-
них коренных обнаженный дентин образует общие небольшие поверхности, на
Глава 12. Грызуны
149
Рис. 12.12. Жевательная поверхность верхнего (слева) и нижнего (справа) зубного ряда лесной
сони, Dryomys nitedula, в возрасте 23-25 мес. (а), 35-37 мес. (б) и более 48 мес. (в) (по Homolka
(1979), с изменениями).
нижних коренных сильнее сточен щечный край. У трехгодовалых расширяются
сплошные поверхности дентина на язычной стороне верхних коренных и появля-
ется дентин на щечной стороне верхних и на язычной стороне нижних коренных
зубов. У старших особей от эмалевых гребней остаются лишь небольшие учас-
тки или вообще вся жевательная поверхность представлена дентином (рис. 12.12)
(Лозан, 1961; Homolka, 1979).
Годовые слои можно видеть в кости нижней челюсти на окрашенных срезах
в конце зубного ряда. Зимняя линия двойная в нижней стенке челюсти и одинар-
ная в боковой. Общая толщина цемента коренных зубов мала и слои в нем плохо
различимы (см. гл. 4 •).
Eliomys quercinus — садовая соня
Внешний вид
Сеголетков от старших можно отличить по окраске: у них общий тон меха
серый и тусклый (у старших буро-коричневый), верхняя часть носа, лоб и об-
ласть между глазами серые (у старших — охристо-ржавые) (Огнев, 1947).
Зубы
Стачивание. Постоянный предкоренной (первый щечный) зуб появляется в
возрасте около 80 дней. Сеголетки от старших (до начала октября) отличаются
недоразвитием т3з и слабой стертостью коронки коренных. Возраст взрослых
можно определять по степени стертости щечных зубов (рис. 12.13) (Kahmann,
Staudenmayer, 1968; Grulich, Jurik, 1994).
150
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.13. Желательная поверхность верхнего (слева) и нижнего (справа) зубного ряда садовой
сони, Eliomys quercinus, в возрасте 90 дней (а), 1 г. (б), 2 г. (в), 3 г. (г) и 4 г. (д). Черным показаны
обнажения дентина. Дентин окрашен азотнокислым серебром (Kahmann, Staudenmayer, 1968).
Ellobius talpinus — обыкновенная слепушонка
Зубы
Рост корней. Стачивание. Выделить нескольких групп, без строгой привяз-
ки к возрасту можно по отношению высоты корней к общей высоте ш2в сочета-
нии с весом тела:
у молодых вес тела меньше веса взрослых, корней нет;
Глава 12. Грызуны
151
Таблица 12.3. Высота корня т, (мм) от бифуркации до самой выступающей
части одного из корней у слепушонок, Ellobius talpinus разного возраста
(Челябинская обл., сентябрь, Евдокимов, 1997).
Годовые Генерация
классы 1 (ранние приплоды) 2 (поздние приплоды)
М (Нт) п М (Нт) п
0+ 0,50 (0,1-0,8) 22 0,31 (0-0,6) 35
1+ 1,37 (1,1-Е6) 5 0,80 (0,5-1,3) 11
2+ 2,22 (2,1-2,3) 5 1,67 (1,4-1,8) 8
Без разделения на ранние и поздние приплоды
3+... 4+ 2,76 (2,4-3,1) 6
у полувзрослых — вес тела как у взрослых, корней нет или только начинают
появляться;
у молодых взрослых длина корней не превышает 1/4 общей длины зуба, у
старших взрослых этот показатель от 1/4 до 1/2, у старых — больше 1/2 (Кири-
ков, 1952).
Соотношение длины корня и возраста у особей разного возраста и двух сезонных
генераций, полученное при исследовании меченых животных, дано в таблице 12.3.
При таких различиях в росте корней между особями разных сезонных гене-
раций для определения возраста особей в выборке, взятой в короткий интервал
времени, используют способ Смирнова (см. гл. 1 раздел 1.2.1.2).
Erethizon dorsatum — североамериканский дикобраз
Приводимые ниже данные взяты из работы Earle, Kramm (1980).
Вес тела
Годовики (март-июнь) весят меньше 4 кг, позднее по весу определить воз-
раст не удается, хотя рост продолжается до 3—4 лет.
Зубы
Прорезание, т’ и т22 прорезаются в первое лето жизни, т33 прорезается
полностью к концу первого года, молочные предкоренные меняются на постоян-
ные к концу второго года.
Годовые слои видны в цементе любых щечных зубов на окрашенных сре-
зах, пригодны для определения возраста, 1-й слой появляется после прорезания
зуба (см. гл. 4 •).
Gerbillus — карликовые песчанки
Зубы
Стачивание. Для G. andersoni и G. gerbillus Wassif, Soliman (1978, 1982)
выделяют 12 стадий стертости первых двух коренных зубов, суммируют баллы
152
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
стертости двух верхних и двух нижних каждой особи, и используя графический
способ Смирнова (см. гл. 1 раздел 1.2.1.2), по соотношению сезона размноже-
ния и степени стертости определяют средний балл стертости в месяц и отсюда—
индивидуальный возраст каждой особи. Хотя эти виды и симпатрические, харак-
тер стачивания их зубов несколько различен.
Glis glis — соня-полчок
Гребни. (Обозначения см. рис. 12.5).
Зубы
Стачивание. Возрастные группы можно выделять по степени стертости ко-
ренных зубов и развитию гребней черепа.
У сеголетков к осени сменяется молочный предкоренной на постоянный, вы-
растает ш33, лобные гребни черепа тонкие, низкие и не доходят до венечного
шва, теменные гребни отсутствуют.
У годовалых в первой половине лета есть потертость эмали на гребнях зубов,
обнажений дентина либо нет, либо они появляются местами на т2 и т3, лобные
гребни и боковые затылочные гребни черепа хорошо заметны.
У годовалых в конце летнего сезона и двухлетних в начале летнего сезона
есть обнажения дентина в виде тонких полосок на вершинах широких эмалевых
гребней зубов, хотя и не на всех зубах.
У двухгодовалых в конце летнего сезона и трехгодовалых в начале четверто-
го летнего сезона сплошное обнажение дентина вдоль эмалевых гребней зани-
мает значительную часть площади гребней; лобные и теменные гребни черепа
соединяются с боковыми затылочными, лобные сближены в области заглазнич-
ного сужения.
У старых (4 г. и старше) особей зубы сильно стерты, гребни зубов очень
низкие и обнажения дентина видны между ними, гребни черепа резко выражены,
череп более угловатый, чем у особей предыдущих групп, лобные гребни соеди-
нены в области заглазничного сужения. (Попов, 1938 по Огнев, 1947; Алекпе-
ров, Грекова, 1969; Homolka, 1979).
Годовые слои кости видны в нижней стенке нижней челюсти в районе т3. Их
не было видно у сеголетков осенью, но они были обнаружены у перезимовав-
ших. Вероятно, слои годовые (Krystufek et al., 2005).
Lagurus lagurus — степная пеструшка
Щечные зубы не имеют корней и растут в течение всей жизни и потому для
определения возраста не пригодны.
Внешний вид
Прижизненно зверьков до месяца можно узнать по характеру смены волося-
ного покрова — см. рис. 8.1.
Глава 12. Грызуны
153
Скелет
Рентгеновские снимки целого зверька в двух проекциях: дорсо-вентральной
и боковой позволяют выделить следующие возрастные группы (Тарасов, 1966):
1 мес. — все вторичные очаги окостенения (эпифизы и апофизы) четко отгра-
ничены от диафизов, начало сращения апофиза пяточной кости;
2 мес. — сращение костей таза, эпифизов грудных и поясничных позвонков и
апофиза локтевого отростка;
3 мес. — слияние с диафизами всех эпифизов позвонков, дистальных эпифи-
зов плечевой и большеберцовой костей, проксимальных эпифизов лучевой и
бедренной костей;
4 -5 мес. — в скелете туловища и конечностей массовые сращения, кроме
проксимальных эпифизов плечевой и большеберцовой костей;
6 -7 мес. — сужение зон соединения эпифизов и диафизов плечевой и боль-
шеберцовой костей;
8-9 мес. — сращение с диафизом проксимального эпифиза плечевой кости;
10-12 мес. — проксимальный эпифиз большеберцовой кости не сращен с
диафизом, у отдельных особей полное слияние апофиза пяточной кости.
Кость полового члена
В возрасте 2 мес. на рентгеновских снимках всего скелета в дорзо-вентраль-
ной проекции видно тело кости и три точечные тени отростков; в 3 мес. отростки
становятся пальцевидными, центральный длиннее и шире боковых, в 4-7 мес.
основание принимает ромбовидную форму с закругленными краями, отростки
полностью окостеневают, основание центрального утолщено; к 8—9 мес. форма
основания возвращается к округлой, тело становится тоньше, общая длина с
отростками 3,5 мм. Позже с возрастом тело кости становится еще тоньше (Тара-
сов, 1984).
Lemmus lemmus — норвежский лемминг
Щечные зубы не имеют корней и растут в течение всей жизни и потому для
определения возраста не пригодны.
Внешний вид
Молодые до месяца отличаются от старших тусклой, без блеска окраской спи-
ны и серым (а не светлым, как у старших) брюшком (Кошкина, Халанский, 1961).
Коропеп (1970) разработал систему оценки линьки (и по характеру волосяно-
го покрова, и по пигментным пятнам на мездре), позволяющую определять воз-
раст полувзрослых и молодых взрослых особей, в сложных случаях — с при-
влечением данных по размеру тела и состоянию половой системы.
Череп. Форма (Обозначения см. рис. 12.5).
По форме черёпа и развитию гребней можно выделить несколько возрастных
групп. До месяца крыша черепа выпуклая, гребни не развиты; с 1,5 мес. крыша
154
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.14. Возрастные изменения формы черепа лемминга Lemmus lemmus.
I — от 15 до 25-30 дней, II — от 25-30 дней до 1,5-2 мес., III — от 1,5-2 мес. до 3-3,5 мес.,
IV — от 3-3,5 мес. до 5-6 мес. V—старше 5-6 мес. (Кошкина, Халанский, 1961).
уплощается, начинают развиваться гребни, межглазничные (лобные) гребни сбли-
жаются; после 3 мес. они начинают смыкаться, скуловая ширина увеличивается,
все отростки и гребни становятся более выпуклыми (Кошкина, Халанский, 1961)
(рис. 12.14).
Хрусталик
Связь между возрастом (в днях) и весом хрусталика (в мг) от 20 до 200 дней
описывается уравнением (Hagen et al. (1980) по данным Ostbye, Semb-Johansson
(1970) для лабораторных особей):
1g (возраст) = (вес пары хрусталиков + 1,378) : 2,075.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Однако после 100 дней рассчитанная оценка возраста оказывается занижен-
ной.
Marmota baibadna, М. bobac, М. menzbieri, М. sibirica — сурки
Внешний вид.
Прижизненно можно отличать годовалых от старших у М. baibadna, М. bobac,
М. menzbieri по равномерному у первых и неравномерному у вторых опушению
хвоста (Машкин, Колесников, 1990).
Зубы
Прорезание и стачивание. У всех этих видов возрастные изменения зубов
в целом сходны.
прорезаются в возрасте около 40 дней; т33 начинают прорезаться в 75
дней и достигают полной высоты в 100-110 дней, предкоренные прорезаются в
конце осени и достигают полной высоты в возрасте 13-14 мес. В двухлетнем
возрасте на первых коренных обнажения дентина в виде подковки. С возрастом
площадь обнажения дентина увеличивается (рис. 12.15), на верхних зубах несколько
быстрее, чем на нижних. У особей в возрасте от 6 до 8-10 лет поперечные гребни
Глава 12. Грызуны
155
Рис. 12.15. Возрастные изменения жевательной поверхности верхних (слева) и нижних (справа)
зубов сурков, типичные для Marmota baibacina, М. bobac, М. menzbierv. сеголетков (а), пере-
зимовавших одну (б), две (в), три (г), четыре (д), пять (е), шесть (ж), семь (з) и восемь и более
(и) зим (Машкин, Колесников, 1990).
156
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
на зубах могут быть еще заметны; у старших эмаль остается только в виде по-
лоски по краю зубов, жевательная поверхность представлена только дентином
(Петров, 1961; Леонтьев, Пешков 1966; Червякова, Червяков, 1973; Машкин,
Колесников, 1990).
Для всех сурков стертость щечных зубов весной соответствует таковой в
предыдущую осень перед спячкой.
Годовые слои хорошо видны на окрашенных срезах в цементе и вторичном
дентине коренных зубов, в костной ткани нижней челюсти за концом зубного
ряда.
Возраст лучше определять по числу слоев цемента на поперечных срезах се-
редины корня. Возраст (в годах) равен числу слоев +1. (Линия первой зимы в
цементе видна лишь в верхней части корня и на такой поперечный срез не попа-
дает). На таком срезе видны и слои вторичного дентина, которые можно исполь-
зовать для корректировки. У М. baibacina в т3 первая зима отмечена в дентине
светлоокрашенной полосой (на окрашенных препаратах), линия второй зимы
отделяет первичный дентин от вторичного и далее слои формируются только на
одной стороне полости пульпы.
В нижней челюсти после 5 лет площадь аппозиции костной ткани уменьшает-
ся. До 4 лет можно определять возраст и по числу годовых слоев кости на попе-
речных срезах середины диафиза второго пальца (см. гл. 4 •).
У М. menzbieri Машкин (1979) удалял первый верхний щечный зуб у живых
сурков для последующего определения возраста по слоям.
Marmota топах — лесной сурок
Череп (Обозначения см. рис. 12.5).
В сентябре можно отличить сеголетков от старших по наличию венечного
шва и слабо развитому затылочному гребню (Hamilton, 1934)
Зубы
Резцы. У сеголетков и у особей весной после первой зимовки резцы не ок-
рашены, в отличие от старших особей. Этот признак легко использовать для
оценки возраста прижизненно (Davis, 1964; Nuckle, Bergeron, 1983).
В сентябре можно отличить сеголетков от старших по наличию молочных
предкоренных зубов (Hamilton, 1934).
Хрусталик
Для популяции Квебека в интервале 2-48 мес. связь между возрастом (в
мес.) и весом хрусталика (в мг) описывается уравнением:
In (возраст) = (вес одного хрусталика — 7,724) : 5,060.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
95% доверительный интервал линии регрессии очень большой: 5,6-27,0 мг
для возраста 12 мес. и 11,1-22,3 мг для 48 мес. (Nuckle, Bergeron, 1983).
Глава 12. Грызуны
157
Этому уравнению вполне удовлетворяют данные, приведенные для сурков
Пенсильвании (добытых в марте-мае, за исключением сеголетков):
вес хрусталика (мг, средняя ± стандартное отклонение, в скобках число осо-
бей) сеголетков 12,32±2,8 (34); годовиков 21,78±1,7 (52); взрослых 28,53±4,5
(170) (Davis, 1964).
Кости конечностей
В сентябре можно отличить сеголетков от старших по неполному срастанию с
диафизами эпифизов лучевой, локтевой и плечевой костей (Hamilton, 1934).
Meriones — малые песчанки
Зубы.
Прорезание и стачивание. Характер возрастных изменений зубов сходен у
М. erythrourus, М. tamariscinus, М. meridianus (Гинтлис, 1959; Руденчик, 1962)
(рис. 12.16) и М. unguiculatus (Крылова, 1978).
В возрасте 1-1,5 мес. т33 не дорос до уровня других коренных или имеет
неоформленную жевательную поверхность;
в 2-5 мес. — т33 дорос до уровня других коренных и имеет оформленную
жевательную поверхность, основания призматических (боковых) бороздок на-
ходятся в альвеоле; коронки заметно расширяются к основанию;
в 6-8 мес. — основания призматических бороздок подняты над краем альве-
ол, особенно у первого коренного, зубы менее конусовидны;
в 1 г. — призматические бороздки составляют примерно половину высоты
коренных зубов, углы эмалевых петель закруглены;
в 1,5 г. — призматические бороздки составляют не более трети высоты ко-
ренных зубов, обнажены основания корней (у гребенчуковой — еще нет);
в 2 г. — коронки почти стерты, бороздки отсутствуют или едва заметны, ос-
нования корней обнажены у всех песчанок, эмалевые петли местами исчезают.
Годовые слои видны на окрашенных срезах (позиция среза — как и у
мышей, см. рис. 12.4) в цементе коренных зубов и в костной ткани нижней
стенки мандибулы под т2 3. Зимние линии в кости могут быть двойными или
состоять из группы сближенных линий. Слои цемента часто более четко вид-
ны на поперечных срезах нижней половины корня, чем на продольных, а на
поперечных срезах корня ближе к шейке можно видеть слои вторичного ден-
тина, число которых в т3 у М. libycus было равно числу слоев цемента (см.
гл. 4 •).
Хрусталик
Связь между возрастом (в днях) и весом хрусталика (в мг) особей лабора-
торной колонии М. shawi в интервале 21-752 дней описывается уравнением:
In (возраст) = (вес одного хрусталика + 68,88) : 28,16 (Rodel, 1987).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
158
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
д
Рис. 12.16. Коренные зубы нижней челюсти песчанок (Meriones erythrourus, М. nieridianus, М.
tamariscinus) в возрасте 2-5 мес. (а), 6-8 мес. (б), годовалых (в), полуторагодовалых (г) и
двухлетних (д) (Руденчик, 1962). Вид сбоку выше уровня альвеолы (вверху) и жевательная
поверхность (внизу).
Microtus — серые полевки
Щечные зубы не имеют корней и растут в течение всей жизни и потому для
определения возраста не пригодны.
Линька
Приблизительно до полутора месяцев можно выделять возрастные группы М.
arvalis прижизненно (см. рис. 8.1).
По особенностям линьки (окраске внутренней стороны кожи, мездры) можно
определять возраст М. californicus до 60 дней (Ecke, Kinney, 1956).
Глава 12. Грызуны
159
Размеры тела и черепа
Для определения возрастал/, californicus до 100 дней и от 100 дней до 1 года
по нескольким промерам черепа и длине тела Lidicker, MacLean (1969) предло-
жили уравнения множественной регрессии, пригодные, видимо, для оценки воз-
раста особей только исследованной ими популяции.
Череп. Форма. (Обозначения см. рис. 12.5).
Возраст М. arvalis Башенина (1953) предлагает определять по форме черепа и
развитию гребней. Возрастные изменения этих признаков сходны с таковыми у
леммингов — см. рис. 12.14, но скорость развития иная:
в возрасте дух недель верхнезатылочная кость в боковой проекции выступает
назад дальше затылочных мыщелков;
в 2-4 мес. — верхнезатылочная кость и мыщелки на одном уровне, хорошо
выражены боковые затылочные и теменные гребни;
в 5-6 мес. — затылочные мыщелки выступают дальше верхнезатылочной кости,
крыша черепа плоская; заметные, сходящиеся межглазничные (лобные) гребни;
в 7-10 мес. — межглазничные гребни полностью сомкнулись в середине,
сзади они образуют с теменными костями ясно очерченное пространство, впере-
ди расходятся.
Тот же подход использовали Лаврова и др. (1960) при определении возраста
М. oeconomus:
черепная коробка гладкая, кондилобазальная длина черепа (Lc) не более 23 мм —
полувзрослые; на лобных и теменных костях намечаются выступы и шероховатости,
Lc — 24-25 мм — молодые взрослые; рельеф на поверхности черепной коробки
хорошо заметен, Lc — 26-27 мм — средние взрослые; на лобных, теменных и заты-
лочных костях выражены гребни, Lc—28 мм и более — старшие взрослые и старые.
Хрусталик
Adamczewska-Andrzejewska (1973а) предлагает три уравнения, описывающие
связь между возрастом (в неделях) и весом хрусталика (в мг), для разных пери-
одов жизни М. arvalis'.
4-12 недель: In (возраст) = [In (вес пары хрусталиков) + 0,4595] : 0,6724;
12-23 недели: In (возраст) = [In (вес пары хрусталиков) + 0,0070] : 0,4788;
23-58 недель: In (возраст) = [In (вес пары хрусталиков) - 0,0659] : 0,4507.
Точность оценки возраста (с 95% вероятностью) — две недели для особей в
возрасте до 20 недель, три недели для особей до 30 недель и 14 недель и более
для особей от 58 недель (см. также М. arvalis в гл. 6).
Для М. oeconomus в интервале 21-365 дней:
1g (возраст, дни) = (вес пары хрусталиков, мг + 1,729) : 2,799.
При возрасте 50 дней, 95% доверительный интервал 39-64 дней,
при возрасте 300 дней — 236-381 дней (Hagen et al.,1980).
Для особей лабораторной колонии М. pennsylvanicus (Thomas, Bellis, 1980)
связь между возрастом (в днях) и весом хрусталика (в граммах) в интервале
20 до 120 дней описана уравнением:
160
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
(возраст) = 1: [0,0982 - 0,0508 • 1g (вес пары хрусталиков *104)] - 6.
Уравнение наиболее пригодно в интервале веса хрусталиков от 0,0001 до 0,0065 г.
При весе 0,0052 г возраст 88,9 дней, 95%-й интервал 61,6-152,7 дней.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Stump, Anthony (1983) предложили для М. pennsylvanicus несомненно более
точный, но более сложный способ определения возраста по содержанию нера-
створимых белков в хрусталиках (см. гл. 7 раздел 7.5). В интервале 18-400 дней
(лабораторная колония) связь возраста (в днях) с содержанием белков (среднее
содержание белков двух хрусталиков одной особи в микрограммах) описана
уравнением:
возраст = [0,580 + 0,054 • (белок одного хрусталика)]2.
При оцененном возрасте в 100 дней 95%-й доверительный интервал 92-109
дней, в 400 дней — 367-434 дня.
Кости конечностей
Окостенение костей конечностей у М. (=Stenocranius) gregalis сходно с тако-
вым Lagurus lagurus (см. выше) (Тарасов, 1966).
На рентгеновских снимках всей особи (мертвой или усыпленной) М.
californicus под микроскопом Myers (1978) измерял длину, толщину стенки кос-
тей конечностей, ширину костномозговой полости, степень сращения с диафиза-
ми эпифизов костей конечностей и хвостовых позвонков (всего 9 параметров),
получены уравнение множественной регрессии для самцов и самок и совместно
для особей до 100 дней. Ошибка не более 10 дней. По-видимому, такой метод
имеет смысл использовать только, если с данной популяцией планируется мно-
голетняя работа.
Хвостовые позвонки — см. гл. 2 раздел 2.6.
Mus musculus — домовая мышь
Зубы
Стачивание. Возрастные изменения стертости эмали и обнажения дентина,
описываемые Варшавским (1950) и Keller (1974) сходны (см. рис. 12.3). Первый
выделяет девять возрастных групп до группы «2,5 г.», второй — пять возрастных
групп до «старше 11 мес.».
Хвостовые позвонки — см. гл. 2 раздел 2.6.
Хрусталик
Связь между возрастом (в днях) и весом хрусталиков (в мг) в интервале
3-72 недели описывается уравнением (Rowe et al., 1985):
самцы: 1g (возраст + 20) = 1,019 + 0,175* вес пары хрусталиков;
самки: 1g (возраст + 20) = 1,004 + 0,182 • (вес пары хрусталиков, мг).
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Глава 12. Грызуны
161
Muscardinus avellanarius — орешниковая соня
Зубы
Стачивание. У сеголетков летом эмалевые гребни щечных зубов острые,
осенью перед спячкой и весной следующего года гребни начинают притуплять-
ся. У годовиков летом эмаль гребней всех щечных зубов стирается, появляются
небольшие обнажения дентина. У двухгодовалых летом эмаль сильно стерта,
обнажения дентина видны между гребнями, особенно в т2 и т3. У старших осо-
бей гребни стираются и обнаженный дентин образует связанные между собой
поверхности (Лозан, 1961; Homolka, 1979).
Myocastor coypus — нутрия
Зубы
Прорезание. Первый коренной начинает стачиваться в возрасте 1 мес., вто-
рой прорезается в 3 мес. и начинает стачиваться в 4 мес., третий прорезается в 7
мес., достигает полной высоты в 9 и начинает стачиваться в 10 мес.
Постоянный нижний предкоренной (первый щечный) зуб есть уже у ново-
рожденных, но без корней. Корни едва достигают 1/4 высоты всего зуба к
началу второго мес., 1/3 — в 3-9 мес., и половины к 10-12 мес. (Habermehl,
1985).
Резцы. Наибольший диаметр резцов позволяет дать приблизительную оцен-
ку возраста. Jentzsch (1956, по Habermehl, 1985) дает такие размеры наибольше-
го диаметра резцов (мм):
Возраст до 1 мес. 1-2 мес. 2-3 мес. 3-4 мес. 4-9 мес. 9-12 мес. 1г.
и старше
верхний <2,2 2,2-3,4 3,5-3,7 3,8-5,0 5,1-6,0 6,1—6,5 от6,5
нижний <1,9 1,9-2,5 2,6-3,2 3,3^1,0 4,1-5,0 5,1-5,5 от5,5
По данным Алиева (1954) диаметр верхнего резца в 2 мес. — 4,4 мм, в 5-7 мес. — 5,0-
5,2 мм, в 12 мес. — 6,4 мм, в 72 мес. — 8,5 мм.
Годовые слои в цементе корней и межкорневой подушки всех щечных зу-
бов видны на окрашенных срезах, они нечеткие и среди дополнительных выде-
лить годовые не просто. В костной ткани нижней челюсти и бедренной кости
картина еще более сложная (см. гл. 4 • ).
Хрусталик
Связь между возрастом (в мес.) и весом одного хрусталика (в мг) от рожде-
ния до 6 лет, выявленная при исследовании особей известного возраста при воль-
ном и клеточном содержании, описывается уравнением:
162
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
1g (возраст + К) = 0,511 + 0,013 • вес одного хрусталика,
где К = 4,34 мес. — период беременности (Gosling et al., 1980).
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Ondatra zibethicus — ондатра
Линька
В 90% случаев молодых от взрослых в осенне-зимний (промысловый) сезон
можно отделить по характеру линьки — рисунку светлых и темных участков на
внутренней стороне шкуры, хорошо видному на снятой шкуре. Темные участки
могут занимать разную площадь, но у сеголетков они сплошные и симметричны
относительно продольной оси, а у старших имеют вид разбросанных пятен с
неправильными очертаниями (Слудский, 1948; Shanks, 1948; Корсаков, 1949)
(рис. 12.17). Признак пригоден до окончания линьки в феврале-марте (Larson,
Taber, 1980).
Череп. Размеры. Форма. (Обозначения см. рис. 12.5).
Для выделения возрастных групп можно использовать комплекс признаков
(табл. 12.4).
Зубы
Рост корней. Стачивание. С возрастом уменьшается высота коронки при-
зматических коренных зубов и формируются и вырастают корни.
В возрасте меньше 2,5-3 мес. ш1 состоит только из призм, каждая заканчива-
ется в основании отдельным отверстием; в 2,5-3 мес. края призм в основании
Рис. 12.17. Внутренняя сторона шкуры (спина и брюхо) ондатры, Ondatra zibethicus, в сентяб-
ре с пятнами линьки. 1 — молодая самка, 2 — старый самец (Корсаков, 1949).
Глава 12. Грызуны
163
Таблица 12.4. Возрастные изменения черепных признаков ондатры, Ondatra
zibethicus, (популяция в р-не р. Колымы, Данилов и др., 1974).
Возраст, мес. п Индекс черепа* Черепные признаки
2 113 8,65±0,13 Гребни не выражены, швы хорошо видны, корней нет
3 173 9,0±0,05 Затылочный гребень зачаточный, швы хорошо видны, в зубах корней нет
4 104 9,25±0,07 Затылочный гребень хорошо развит, стреловидный зачаточный, швы хорошо видны
5 38 9,33±0,09 Гребни от слабо до хорошо
6 17 9,63±0,12 развитых, швы хорошо заметны, появляются зачатки
7 38 10,00±0,08 корней зубов
8 81 10,08±0,08
9 127 10,48±0,11 Гребни от хорошо развитых до сильно выраженных, швы
10 102 10,68±0,15 хорошо заметны, длина корней до 3,5 мм
11 71 10,89±0,31
12 56 11,05±0,26 Гребни сильно развиты, теменные швы слабо заметны, корни до 4.5 мм
18 14 11,21 ±0,21 Гребни сильно развиты, череп массивный, выпуклый, швы
24 37 11,69±0,15 слабо заметны; длина корней равна или превышает высоту коронок
*Отношение общей (от наиболее выступающей точки спереди до такой же точки сзади) длины
черепа к межглазничной ширине.
зуба сливаются, образуется шейка зуба, формируются отверстия двух корней,
высота призм около 10 мм; в 6 мес. корни хорошо выражены, высота призм 8
мм; она уменьшается до приблизительно 7 мм в 9 мес., 6 мм — в 12 мес. и
позже. Далее снашивание идет равномерно по 1,2 мм за 6 мес. (Цыганков, 1955).
Можно использовать высоту коронки и длину корня (табл. 12.5).
У ондатры в Нидерландах (за исключением очень старых животных) стачива-
ние коронки т1 идет со скоростью около 4% высоты в месяц, и приблизитель-
ную оценку возраста можно получить по отношению высоты коронки к высоте
всего зуба (Doude van Troostwijk, 1976):
возраст (мес.) = [100 - (высота коронки к общей высоте зуба в %)] : 3,97.
У особей, добытых в апреле-мае в Финляндии, Pankakoski (1980) описывает
связь между возрастом (в мес.) до 36 мес. и относительной, в процентах к об-
щей высоте зуба, высотой коронки ш1 таким уравнением:
возраст = [(1g (относительная высота) - 1,996) : - 0,0233] +1.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Поскольку используемые показатели демонстрируют существенную межпопу-
ляционную изменчивость, такие соотношения, как и таблицы конкретных величин,
следует получать для каждого региона. Пренебрегая межпопуляционными разли-
чиями, для сугубо ориентировочной оценки возраста, можно пользоваться схемой
возрастных изменений зубов, приведенной на рис. 12.18. Эта схема, составленная
Никифоровым (1958) для популяции ондатры из дельты Аму-Дарьи, по существу,
164
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 12.5. Средние показатели размеров зубов (мм) у ондатры,
Ondatra zibethicus, Юж. Богемии (Tmkova, 1966)*
Возрастной интервал (мес.)
2,5-3 3-4,5 4,5-6 6-9 9-12 12-18 18-24
т2 8,46/- 8,16/0,67 7,54/1,10 6,52/1,76 5,15/2,79 3,12/3,58 1,67/4,93
т, 8,14/- 7,80/0,97 7,08 / 1,48 6,07/2,28 4,79/3,15 2,88/4,09 1,84/5,53
* Высота призматической части коронки / длина корня.
коронка стерта, в
альвеолах остались
одни корни
около 3, 5 лет
Рис. 12.18. Схема для определения возраста ондатры, Ondatra
zihethicus, по форме ш1. (Никифоров, 1958).
не отличается от таковой, полученной на меченых особях в Новосибирской обл.
(Корсакова, 1970).
Выборку, взятую в марте-апреле можно разделить на три возрастные группы
и по соотношению края наружной первой бороздки т1 и альвеолы:
Глава 12. Грызуны
165
бороздка глубоко в альвеоле — молодые (рожденные в конце сезона, воз-
раст около 7 мес.);
бороздка подходит к краю альвеолы — полувзрослые (рожденные в начале
предыдущего сезона размножения, возраст около 10 мес.);
бороздка поднимается над краем альвеолы—взрослые (старше 1 года) (Olsen, 1959).
Смирнов (1960) предложил графический способ определения возраста он-
датр, пойманных в разное время года, по высоте коронки т1 (см. рис. 1.3 в гл. 1).
Этот прием может быть использован для определения возраста ондатры из любо-
го региона по высоте коронки любого зуба (например, Ларин, Мохов (1969)
использовали ш').
Кость полового члена
В декабре у особей в возрасте 6-8 мес. пальцевидные отростки хрящевые, у
особей старше 15 мес. они окостеневшие, но не приросшие к телу кости. Этот
критерий дал возможность точнее разделить выборку на молодых и взрослых, чем
состояние зубов (Elder, Shanks, 1962). (О технике обработки см. гл. 2 раздел 2.8.4).
Хрусталик
По весу можно отличить молодых от взрослых (см. рис. 6.2 в гл. 6).
Rattus — крысы
Зубы
Стачивание. Возрастные группы у R. rattus и R. norvegicus можно выделить
по степени стертости щечных зубов. Карноухова (1971) выделила 7 возрастных
групп (последняя группа — особи старше 3,5 лет) у серых и черных крыс, от-
ловленных в городских и портовых строениях г. Владивостока. Однако лишь
первые 4 группы соответствовали возрасту серых крыс, отловленных на рисо-
вых полях и в населенных пунктах Кубани (Рыльников, 1990).
Годовые слои — ростовые слои видны в костной ткани и в цементе щечных
зубов, но их пригодность для определения возраста сомнительная (см. гл. 4«).
Хрусталик
На материале от первого лабораторного поколения диких R. norvegicus полу-
чено следующее уравнение, описывающее связь между возрастом (в днях) и
весом хрусталиков (в мг) до возраста 74 недели:
1g (возраст + 22) = 1,313 + 0,021* вес пары хрусталиков (Hardy et al., 1983).
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Rhombomys opimus — большая песчанка
Хрусталик
По весу можно, вплоть до декабря, отличать сеголетков от перезимовавших
особей.
166
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Вес одного хрусталика (мг) в возрасте 4-5 мес. — 15,6 (13,0-17,6); в 6-7
мес. — 18,2 (16,6-20,4); в 8-9 мес. — 20,2 (18,7-22,9); в 12-18 мес. — 27,9
(25,1-31,6); в 19-24 мес. — 30,0 (28,0-34,5); у особей старше 24 мес. — 33,7
(30,3—42,9) (Корнеев и др., 1976).
Sciurus carolinensis, S. niger — каролинская и черная белки
Внешний вид
В октябре-ноябре сеголетков от взрослых S. carolinensis и S. niger можно
отличить по форме хвоста: у молодых он заостренный, треугольный, стороны
его не параллельны; у взрослых он прямоугольный, стороны параллельны или
почти параллельны (Uhlig, 1956 по Larson, Taber, 1980).
В промысловый сезон сеголетки 5. carolinensis отличаются от взрослых тем,
что на брюшной поверхности хвоста на красноватом фоне четко видны две-три
продольные темные полосы. У взрослых они маскируются вторичным опушени-
ем (Sharp, 1958).
Осенью перелинявших S. carolinensis можно разделить на 3 группы по состо-
янию волосяного покрова боковой части бедра: у сеголетков летних выводков
нет желтой каймы близ основания черных волос подпуши, большинство окайм-
ленных остевых волос имеют черный кончик; у сеголетков весенних выводков
окаймление волос подпуши отсутствует или нечеткое, но кончики остевых волос
белые; у взрослых бело-желтое окаймление волос подпуши четкое, кончики ос-
тевых волос белые (Barrier, Barkalow, 1967).
Зубы
Прорезание и стачивание. У молодых (до 140 дней) 5. carolinensis есть
молочные предкоренные, или если предкоренные уже постоянные, то они не
полностью прорезаны, у полувзрослых (до 400 дней) все постоянные зубы
полностью прорезаны, на поверхности стачивания т3 нет обнажения дентина, у
взрослых (490 дней и старше) на т3 есть обнажения дентина (Hench et al.,
1984).
Годовые слои видны в цементе на окрашенных продольных срезах корен-
ных зубов (Fogl, Mosby, 1978).
Кости конечностей
На рентгеновских снимках передних лап дистальные эпифизы лучевой и лок-
тевой костей у S. carolinensis и 5. niger отделены от диафиза широкой щелью у
особей от рождения до 18 недель, отделены узкой щелью — в возрасте 19-33
недель у самцов S. carolinensis и в 19-39 недель у самцов и самок У niger, и
отделены темной полоской — в 33-46 недель у самцов 5. carolinensis, и 39-50 (в
среднем 54) недель у самцов и самок S. niger. Сеголетков от старших надежно
можно определять с июля по декабрь у S. niger и с февраля по ноябрь у S.
carolinensis. Осенью можно различать сеголетков, принадлежащих к весенним
или к летним выводкам (Carson, 1961).
Глава 12. Грызуны
167
Тазовые кости
У самцов по каудо-медиальному краю седалищной кости с возрастом разви-
вается бугристость, достигающая максимальных размеров уже в возрасте 8-
11,5 мес. у S. carolinensis и 10—13,5 мес. у S. niger. У самок эта бугристость
отсутствует (Colburn, 1986).
Хрусталик
Вес хрусталика может быть использован для разделения выборки на возрас-
тные группы. Возрастные группы у взятых в сентябре-октябре S. carolinensis впол-
не надежно можно выделить, приняв сухой вес 1 хрусталика в 32 мг как точку
разделения сеголетков весенних и летних выводков и в 37,5 мг — как точку
разделения всех сеголетков и старших (Fisher, Perry, 1970). У S', niger в октябре-
ноябре сеголетки весеннего выводка имеют вес 1 хрусталика 29-39 мг с модой
35 мг, летнего выводка — 22-29 мг с модой 25, особи старше года — от 40 мг
(Beale, 1962).
Sciurus vulgaris — обыкновенная белка
Зубы.
Прорезание и стачивание. Постоянные зубы достигают нормальной высоты
в возрасте 6-9 мес.
Кирис (1937) в выборке, взятой осенью, по прорезанию и росту постоянных
зубов выделял 4 возрастные группы в течение первого года жизни и далее 6
годовых классов по степени стертости зубов. Однако, выделенные им старшие
возрастные группы, видимо, не соответствуют реальному возрасту (см. гл. 3
раздел 3.4.6.4).
Более четкие результаты деления на возрастные группы особей, добытых в
течение года, были получены графическим способом (Смирнов, 1960) по сред-
ней высоте коронок 8 щечных зубов (рис. 12.19). Средняя высота щечных зубов
за год уменьшается приблизительно на 0,3 мм.
Годовые слои видны в кости и в цементе щечных зубов на окрашенных
срезах нижней челюсти (позицию срезов см. рис. 12.4). По данным Шадриной
(19886), возраст лучше определять по слоям в цементе нижнего предкоренного,
по слоям в кости оценка может быть занижена. Цемент под коронкой и на корнях
наслаивается не равномерно и потому может быть виден не на всех продольных
срезах зуба. Слои цемента можно считать и на поперечных срезах корней. Слои
вторичного дентина образуются со второго года и их число можно использовать
для корректировки подсчета слоев в цементе как оценку минимального возмож-
ного возраста (см. гл. 4 •).
Кости конечностей
Дистальный эпифиз бедренной кости в возрасте 0-5 мес. отделен глубокой
бороздой от диафиза (при вываривании костей особей в первые 3 мес. жизни
эпифиз отваливается); в 6-10 мес. эпифиз отделяет борозда с фронтальной
168
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 12.19. Средняя высота коронки 8 щечных зубов белок Sciurus
vulgaris (Курганская обл.), добытых в течение года (Смирнов, 1960).
(Какие зубы исследовали и как измеряли высоту коронки не указано).
стороны и линия сращения с противоположной; приблизительно в 11-15 мес. —
линия сращения с обеих сторон; от 16 мес. — линия не видна или очень слабая
(Lemnell, 1974).
В октябре-декабре сеголетки первого помета отличаются от старших по степе-
ни сращения дистального эпифиза бедренной кости и проксимальных эпифизов
большой берцовой и плечевой костей с диафизами: у сеголетков отчетливо виден
шов — линия сращения, у зверьков старше года он не виден (Егоров, 1958)
В июле-сентябре по отсутствию сращения дистального эпифиза лучевой кости
с диафизом визуально (на очищенных костях) можно четко отличить сеголетков от
старших (Degn, 1973).
Глава 12. Грызуны
169
Кости таза
Седалищный бугор безымянной кости у самцов выражен только у особей
старше года (Егоров, 1958).
Кость полового члена
У 92% сеголетков в августе-декабре вес кости меньше 20 мг, у 88% старших —
больше. В январе-феврале этот критерий разделения молодых и взрослых уже
не годится (Degn, 1973).
Хрусталик
В возрасте 1 мес. вес одного хрусталика — 11,2 мг, в 2-4 мес. — 20,0 (16,9-
23,9), в 6-9 мес. — 27,4 (19,9-23,0), в 1 г. — 29,1 (26,5-30,6) мг (Карпухин,
Карпухина, 1971).
В сентябре-ноябре вес одного хрусталика у сеголетков 23 (16-28) мг, у стар-
ших— 30 (25-38) мг, приблизительная граница разделения возрастных групп—
29 мг (Швецов, 1975). В октябре-декабре — граница разделения сеголетков и
старших — 30 мг (Degn, 1973).
Sicista — мышовки
Зубы
Стачивание. У сеголетков S. параеа в августе - начале сентября щечные
зубы не стерты; на второе лето — бугорки зубов стерты не более чем наполови-
ну; на третье лето бугорки почти исчезают; на четвертое лето зубы стерты и име-
ют форму чаши со слабо выступающими краями (Зимина, Меркова, 1960).
По нашим данным, у добытых в течение летнего периода особей S. betulina в
возрасте 0-3 г. (возраст был определен по годовым слоям, см. ниже) по степени
стертости удалось выделить только 3 возрастные группы: сеголетки, годовики и
старшие (Клевезаль и др., 2005). Корреляция с возрастом оценок степени стер-
тости зубного ряда в целом была существенно ниже, чем балльных оценок от-
дельно бугров и боковых гребней (межбугорных перегородок) ш1,2 и стертости
ш3 и суммы всех баллов. В среднем, у сеголетков бугры ш1-2 не сточены или
сточены с внутренней стороны без обнажения дентина, боковые гребни не сточе-
ны и в ш3 бугры или не сточены или один, самый высокий, чуть затронут стачи-
ванием. У годовиков видно слабое обнажение дентина на буграх т1-2, заметно
стачивание их боковых гребней и стачиванием затронуты все бугры ш3. У стар-
ших особей бугры и боковые гребни всех зубов сточены сильно или очень силь-
но. По распределению особей по этим признакам стертости 78% были отнесены
в свою возрастную группу (особи с промежуточными показателями считались
неверно определенными). Кластерный анализ по этим признакам (см. гл. 1 раз-
дел 1.2.4) дал возможность более точного разделения на возрастные группы за
счет распределения по группам промежуточных вариантов.
Годовые слои видны в костной ткани нижней стенки мандибулы, в цементе
и во вторичном дентине коренных зубов на окрашенных срезах. Слои в разных
170
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
тканях можно видеть на одном срезе, если поперечный срез челюсти прошел
продольно через полость пульпы т2 или тг (см. гл. 4 • и 4.6.4 •)
Sigmodon hispidus — колючий хлопковый хомяк
Череп. Размеры (Обозначения см. рис. 12.5).
Скуловая ширина — лучший показатель возраста из всех промеров, она ме-
нее 17,5 мм в возрасте 1 мес., до 18,7 мм — в возрасте 2-3 мес., не более 19,8 мм
у большинства 3-4-месячных, и более у большинства старших (Chipman, 1965).
Зубы
Прорезание. В возрасте 1 мес. проходит альвеолярное, но не десневое проре-
зание т33, в возрасте 2 мес. эти зубы достигают полной высоты. (Chipman, 1965).
Кости конечностей
Дистальные эпифизы пястных костей начинают сращиваться с диафизами в
возрасте 2 мес. и полностью сращены в возрасте 3 мес. и старше. Проксималь-
ный и дистальный эпифизы локтевой и дистальный эпифиз лучевой костей начи-
нают сращиваться с диафизом в возрасте 2-3 мес., полностью сращены в возра-
сте 4 мес., но линия шва видна и у старших особей (Chipman, 1965).
Хрусталик
Связь между возрастом (в днях) и весом хрусталика (в мг) в интервале 10-130
дней для особей из лабораторной колонии описывается уравнением (Birney et al., 1975):
возраст=(вес одного свежего не фиксированного хрусталика -9,194) : 0,222.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Как показали данные другой лабораторной колонии, вес 2 высушенных хру-
сталиков в возрасте 1 мес. — 10-14 мг, 2 мес. — 17-20 мг, 3 мес. — 20-25 мг,
4-5 мес. — 25-33 мг, 6-12 мес. — от 33 мг (Chipman, 1966).
Сравнение возрастных изменений веса хрусталика и содержания нерастворимых
белков (количество тирозина) (Birney et al., 1975) показало, что до 130 дней оба
показателя одинаково пригодны для определения возраста особей, позже определе-
ние белка дает более надежную оценку возраста в силу меньшей индивидуальной
изменчивости. Вес свежего хрусталика быстро увеличивается до возраста 130 дней
(в среднем по 0,22 мг/день) и затем более медленно (по 0,03 мг/день) по крайней
мере до возраста 600 дней. Нерастворимая белковая фракция увеличивается быстро
до 160 дней (7,24 микрограмма/день), потом медленно (2,97 в день) до 600 дней.
Spalax microphtaltnus — обыкновенный слепыш
Зубы
Стачивание. Рисунок жевательной поверхности меняется с возрастом (см.
рис. 3.11). У особей, добытых в апреле-июне, Пузаченко (1991) оценивал
стертость каждого верхнего и нижнего коренного зуба в баллах (0-5) и при
Глава 12. Грызуны
171
сопоставлении с возрастом, определенным по числу годовых слоев в кости по-
лучил соотношение:
(возраст, годы) = 0,193 • сумма баллов стертости - 0,903.
Возраст 13,6% особей был определен с ошибкой на 1 год.
Определение возраста по высоте коронок коренных зубов оказалось менее
надежным:
(возраст, годы) = 6,753 - 0,236 • произведение высот т* и т3.
Возраст 26,6% особей был определен с ошибкой, не превышающей 1 год.
Кластерный анализ (см. гл. 1 раздел 1.2.4) (метод «до ближайшего соседа») дал
20% ошибочных определений (ошибки на 1 год).
Годовые слои можно считать в костной ткани, во внешней (щечной) стенке
нижней челюсти на поперечных срезах через последний коренной зуб, много
дополнительных линий, но выделяются группы, соответствующие годовым сло-
ям (Пузаченко, 1991).
Tamias (=Eutamias) — бурундуки
Размеры тела и черепа
По длине тела и черепа можно отличать сеголетков от старших Т. ruficaudus,
на 95% доверительном уровне. Длина тела сеголетков самцов не позже августа
и самок не позже июля не более 120 мм, наибольшая длина черепа не позже
августа — не более 34 мм (Beg, Hoffmann, 1977).
Зубы
Прорезание и стачивание. У Т. ruficaudus в возрасте 39-45 дней т3 не проре-
зан, 45-67 дней коренные прорезаны, есть еще молочные предкоренные, в 79 дней
и старше предкоренные постоянные. В возрасте 10-16 мес. р3 (первый щечный,
мелкий) не сточен; в 30-40 мес. р3 явно стачивается; в 46-52 мес. поверхность
зубов уплощается, площадь эмали резко уменьшена (Beg, Hoffmann, 1977).
У добытых весной Т. sibiricus Шубин (1965) определял отношение площади
обнажения дентина к площади жевательной поверхности т2. В выборке одного
района выделены 3 группы — 0,1-5% (годовалые), 6,1-10% (двухгодовалые),
более 12,1-18% — трехгодовалые и старше. В выборке из другого района раз-
рывов в распределении особей по этому признаку (см. гл. 1 раздел 1.2.1.1) меж-
ду возрастными группами не было.
Годовые слои Barker et al. (2003) выявили в костной ткани у Т. amoenus на
окрашенных поперечных срезах бедренной кости, их число соответствовало воз-
расту, если не считать первую линию, видную у сеголетков (очевидно, линию
резорбции — см. гл. 4 раздел 4.6.4).
Хрусталик
У Т amoenus в сентябре у сеголетков вес обоих сухих хрусталиков был ме-
нее 12 мг, у старших — более 13 мг (Barker et al., 2003).
172
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Tamiasciurus hudsonicus — красная белка
Приводимые ниже данные взяты из работы Davis, Sealander (1971/
Кости конечностей
Бороздка между проксимальным эпифизом и диафизом плечевой кости вид-
на до 7 мес.; в возрасте 7-10 мес. она зарастает латерально, но еще открыта
медиально, у особей старше 10 мес. видна только тонкая линия на месте бороз-
дки. Бороздка между дистальным эпифизом и диафизом бедренной кости четкая
до 8 мес.; в возрасте 8-18 мес. она закрыта вся, кроме медиальной ее части; в
возрасте 21-24 мес. тонкая линия в медиальной части бывшей бороздки улавли-
вается, если провести иглу вертикально от эпифиза к диафизу.
Хрусталик
Вес одного хрусталика, мг (лабораторная колония):
25 дней — 6,5; 3 мес. — 14,7; 7 мес. — 19,3; 10-12 мес. — 22,1-22,2;
48 дней — 10,5; 4-5 мес. — 17,2-17,5; 8 мес. — 20,2; 14 мес. — 22,8;
57 дней — 11,4; 6 мес. — 18,4, 9 мес. — 21,6; 16-24 мес. — 23,0-23,8;
26-36 мес. — 24,6-25,9.
ГЛАВА 13. ХИЩНЫЕ
Alopex lagopus — песец
Череп. Швы. (Обозначения см. рис. 2.1, 2.2)
В возрасте 6-9-10 мес. все швы открыты кроме ВВ',
в 10-12 мес. в стадии зарастания находятся швы РВ, S;
в 1,5-2 г. швы РВ, S зарастают полностью;
у особей старше 2 лет зарастают PM, IPR и большинство других швов (Дол-
гов, Россолимо, 1966).
Зубы
Смена молочных зубов на постоянные начинается в возрасте 3 мес. и закан-
чивается через 1-2 мес. (Данилов и др. 1979).
Клыки. Сеголетков в возрасте до 4-5 мес. можно отличить по не замкнуто-
му апикальному отверстию корня клыка (Смирнов, 1960), а сеголетков и стар-
ших осенью-зимой можно различать по относительной ширине полости пульпы
(«канала») клыка — см. табл. 3.4.
Можно использовать и абсолютную ширину полости пульпы верхних клыков
(в сагиттальном (передне-заднем) направлении): в ноябре-декабре у сеголетков
она более 2,5 мм, у старших — менее 1,5 мм, но в январе-феврале у некоторых
особей моложе года полость пульпы уже такой же ширины, как у старших (Bradley
et al., 1981).
Годовые слои видны в цементе, часто и во вторичном дентине клыков на
окрашенных срезах. Поперечные срезы, прошедшие чуть ниже шейки зуба, удобно
использовать для подсчета слоев дентина, а продольные срезы остальной части
корня — для подсчета слоев цемента. Слои, как правило, четкие. Возраст (в
годах) равен числу слоев в цементе (см. гл. 4 • и 4.4 •).
Кость полового члена
Отношение веса кости к ее длине:
в возрасте 6-10 мес. — 2,51-4,87; в 10-12 мес. — 4,62-6,31; в 1,5-2 г. — 5,77-
6,38; у особей старше 2 лет—7,48-11 (Долгов, Россолимо, 1966). В возрасте 1-1,5
лет кость достигает окончательной формы, длина около 60 мм (Тарасов, 1984).
Canis latrans — койот
Череп. Швы. (Обозначения см. рис. 2.2)
Осенью сеголетков можно различать по открытому шву РВ (Linhart, Knowlton,
1967).
Зубы
Клыки. Животное с открытым апикально каналом клыка является, несом-
ненно, сеголетком, поскольку апикальное отверстие корня клыка замыкается к
174
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
возрасту 9 мес. (Linhart, Knowlton, 1967). Возрастную группу сеголетков можно
выделить и по относительной ширине полости пульпы клыка (см. табл. 3.4).
Nellis et al. (1978) нашли возможным определять возраст (в мес.) до 60 мес.
по расстоянию (в мм) от края эмали верхнего клыка до края альвеолы (точнее,
до линии соединяющей края альвеолы двух соседних с клыком зубов):
возраст = 8,9 + 5,56 • указанное расстояние
Стертость зубов дает возможность выделять группы сеголетков и годови-
ков. Использование этого критерия для выделения старших возрастных групп,
как правило, занижает оценку возраста (Bowen, 1982).
Годовые слои видны на окрашенных срезах в цементе клыков. Возраст (в
годах) равен числу слоев + 0-1 (см. гл. 4 • и 4.4 •).
Canis lupus — волк
Череп. Швы. (Обозначения см. рис. 2.2)
У волков из разных штатов Сев. Америки шов ВВ зарастает (при этом линия
шва может быть видна) не раньше 3 и не позже 4 лет (см. также гл. 2 раздел
2.2.5) (Landon et al., 1998).
Зубы
Клыки. Осенью-зимой сеголетков можно отличить от старших по относи-
тельной ширине полости пульпы клыка (см. табл. 3.4)
Сеголетков можно узнать по открытому апикальному отверстию корня, кото-
рое закрывается у волков Квебека примерно в возрасте 9 мес. (О межпопуляци-
онной изменчивости этого показателя см. гл. 3 раздел 3.2.2). Зимой сеголетки с
уже закрытым апикальным отверстием корня клыка и старшие могут быть разде-
лены по толщине дентина (на уровне десны): при максимальной толщине менее
3,5 мм особь можно смело отнести к сеголеткам, более 4 мм — к старшим
(Parker, Maxwell,1986).
Landon et al. (1998) приводят уравнение регрессии для определения возраста
волка (в годах) до 7 лет по относительной толщине стенки клыка, измеренной в
сагиттальной плоскости в районе шейки (общая ширина зуба минус ширина по-
лости пульпы, отнесенная к общей ширине, в процентах):
возраст= 1,181’ относительная толщина: (94,359-относительная толщина).
Однако уже со второго годового класса значения перекрываются.
Смирнов (1983) предложил использовать как показатель возраста сумму деся-
тичных логарифмов относительной ширины (в процентах) полости пульпы, изме-
ренной по обеим осям поперечного сечения зуба. Значения для разграничения
соседних годовых классов таковы: 1,5; 1,1; 0,925; 0,75; 0,575; 0,45; 0,325; 0,2.
Превышение этих значений позволяет отнести животное к младшей из двух со-
седних групп. При этом Смирнов отмечает, что только первый класс (сеголетки)
отделен от старших четким разрывом.
Глава 13. Хищные
175
Рис. 13.1. Схема стачивания с возрастом резцов, клыков и хищнических зубов волка Canis
lupus (по Gipson et al. (2000), с изменениями). Линиями обозначены позиции поверхности
стачивания зубов в разном возрасте, возраст (годы) указан цифрами.
Прорезание и стачивание зубов. Последовательность прорезания зубов мо-
жет служить показателем возраста особей до 6-7 мес. (см. табл. 3.1).
По стертости зубов Gipson et al. (2000) выделяют следующие возрастные груп-
пы (рис. 13.1):
младше 1г. — зубы не стерты, края резцов острые;
1-2 г. — легкая стертость острых краев долей резцов и дистальной части
маленького заднего гребня верхнего клыка;
2-3 г. — слегка уплощены центральные доли i1,2 и медиальные доли i] 3; замет-
ная стертость дистальной части маленького заднего гребня верхнего и нижнего
клыков; нет стертости хищнических зубов (р4, ш();
3—4 г. — уплощаются центральные доли i1’3, плоская поверхность i, захваты-
вает латеральную долю; кончик клыков тупо закругляется; заметно стачивание на
большинстве острых краев хищнических зубов;
4-6 л. — становится плоской поверхность медиальной доли i1 и латеральных
долей i ; уплощаются все острые края хищнических зубов;
6-8 л. — стачивание резцов заходит за медиальные доли i1,2, латеральную
долю i2, достигает латеральной доли i3; вершины клыков и острых краев хищни-
ческих зубов явно уплощены;
8-10 л.—сточены все доли i1-2 и i3, стачивается медиальная доля i3; клыки сточе-
ны на 3-5 мм; профиль режущих краев хищнических зубов практически плоский;
10-12 л. — высота резцов уменьшена не менее чем на 50%, осталось 2-4 мм
эмали, профиль плоский; высота клыков уменьшена на 30-50%, осталось 10-16 мм
176
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
эмали; высота хищнических зубов уменьшена не менее чем на 30%, задний бу-
гор т1 сточен почти до десны;
> 13 л. — резцы сточены до десны, некоторые утеряны; на клыках осталось
менее 10 мм эмали, кончик их тупой; хищнические зубы уплощены, задние буг-
ры р4 и Ш] видны почти у десны (см. также гл 3 раздел 3.4.6.1).
Годовые слои видны в цементе клыков на окрашенных срезах. Возраст (в
годах) равен числу слоев + 0-1 (см. гл. 4 • и 4.4 •).
Кость полового члена
Длина у молодых 93-100 мм, взрослых — 103-117 мм (Suminski, 1968).
Canis mesomelas — чепрачный шакал
Приводимые ниже данные взяты из работы Lombaard (1971).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до 6 мес. (см. табл. 3.1). По стертости можно
выделить следующие группы:
1 г. — бугорки жевательной поверхности i* и i2 не сточены;
2 г. — средний бугорок стачивается в i' и начинает стачиваться в i2;
3 г. — стачивается краевой-медиальный бугорок, щель между ним и средним
бугром исчезает в i1, но еще видна в i2;
4 г. — в i* не видно ни бугров, ни щелей, в i2 латеральная щель еще видна;
5 л. — с этого возраста и далее прогрессирует стачивание резцов, и годовые
классы плохо разграничиваются.
Годовые слои видны в цементе на продольных окрашенных срезах клыка.
Возраст (в годах) равен числу слоев + 1 (см. гл. 4 •).
Кости конечностей
Дистальные эпифизы лучевой и локтевой костей срастаются с диафизами
в возрасте 11 мес., проксимальный эпифиз лучевой кости — в 12 мес.; нали-
чие бороздки между эпифизом и диафизом (см. гл. 2 раздел 2.5.4) плечевой
кости — признак полувзрослых, полное окостенение имеет место в 12-13
мес.
Кость полового члена
В возрасте 12 недель длина кости 24 мм, в 23 недель длина кости 48 мм, вес
114 мг. Уже после 5 мес. возрастные группы по весу кости перекрываются. По-
скольку до 5 мес. легко определить возраст по прорезанию зубов, определять
его по кости полового члена не имеет смысла.
Хрусталик
По весу хрусталика можно опознать молодых особей (до 7 мес.) (см. табл.
6.1).
Глава 13. Хищные
177
Crocuta crocuta — пятнистая гиена
Зубы
Клыки. Ширина полости пульпы клыков, измеренная на рентгеновских сним-
ках (на уровне нижнего края эмали с вентральной стороны) у особей до 3 лет
5,9±0,6 мм для верхнего клыка и 5,О±О,5 мм для нижнего, у особей старше
3 лет 2,7±0,1 мм и менее для верхнего и 2,2±0,1 мм и менее для нижнего клыка
(Lindeque, Skinner, 1984).
Прорезание и стачивание зубов. До 6,5—7,5 мес. имеются только молоч-
ные зубы, в возрасте 7,8-10 мес. появляются постоянные первый и второй рез-
цы, полная постоянная зубная система формируется в 13-18 мес. (Van Hom et al.,
2003).
Jaarsveld et al. (1987) в Южной Африке брали отпечатки р3 у обездвиженных
животных, по ним делали гипсовый слепок и на нем измеряли высоту и площадь
поверхности стачивания зуба. По этим признакам они предложили приблизи-
тельно оценивать возраст отдельных особей (до 18 лет):
возраст (число годовых слоев в дентине нижнего клыка) = (21,4 - высота
зуба) : 0,39;
(8 из 22 точек лежат вне 95% доверительного интервала).
Van Hom et al. (2003) прижизненно измеряли зубы у гиен известного возраста
из восточной Африки (Кения). Они проверили возможность оценки возраста по
вышеприведенному уравнению и получили ошибку до 10 лет.
По их данным для определения возраста (в мес.) взрослых гиен практически
на протяжении всей жизни более пригодны следующие уравнения.
Для самок (или если пол не известен):
возраст = 34,281 + 15,423 • длина поверхности стачивания р3 - 2,473 • мини-
мальная высота коронки нижнего клыка;
если клык сломан, то:
1g (возраст) = 1,004 + 0,116 • длина поверхности стачивания р3;
ошибка до 5 лет в первом и до 7 во втором случае;
для самцов:
1g (возраст) = 1,237 - 0,070 • длина поверхности стачивания р3 + 0,040 • (дли-
на поверхности стачивания р3)2 - 0,002 • (длина поверхности стачивания р3)3;
ошибка до 1 года.
Годовые слои в дентине, видные на травленых кислотой и окрашенных про-
дольных аншлифах клыков, более надежный критерий возраста, чем слои в цемен-
те. Первый слой образуется на 2-й год жизни (Jaarsveld et al., 1987) (см. гл. 4 •).
Enhydra lutris — калан
Размеры тела и внешние признаки
Прижизненно можно выделить несколько возрастных групп по размеру и
окраске. В возрасте около 1 мес. самки и самцы весят 2,2 кг, 2 мес. — 3,9 кг.
Самцы в возрасте 1-2 г. весят 15-25 кг, окраска меха головы темная; в 3-5 лет
178
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
весят 25-33 кг, окраска головы темная или промежуточная между темной и бе-
лой; от 6 лет весят более 30 кг, окраска головы белая. Возрастные группы самок
по весу тела различаются плохо, окраска головы темная в 1-6 лет, промежуточ-
ная — от 7 лет, белая — от 9 лет (Garshelis, 1984).
Череп. Комплекс признаков
Schneider (1973) дает такую схему для приблизительной оценки возраста:
1 мес. — заглазничное сужение черепа (3Q больше межглазничной ширины
(МИГ) (положение промеров см. рис. 2.1), зубы нижней челюсти молочные;
2-3 мес. — резцы и первый предкоренной постоянные, ЗС = МИГ,
4-5 мес. — прорезаются постоянные клыки;
5-8 мес. — клыки прорезались почти полностью, прорезаются коренные;
9-15 мес. — коренные по высоте достигли уровня молочных предкоренных,
ЗС < МИГ,
16-24 мес. — все зубы постоянные;
25-34 мес. — формируется затылочный гребень, на верхней челюсти выше
альвеолы клыков образуется небольшой гребень;
33-36 мес. — образуется стреловидный гребень, затылочный гребень отчет-
ливый, шов ВВ зарастает (обозначения см. рис. 2.2, 2.3);
3 г. и старше — оба гребня отчетливы, шов ВВ зарос, у старших животных
стачиваются зубы.
Зубы
Клыки. Апикальное отверстие постоянных верхних клыков закрывается в воз-
расте около года; возраст молодняка можно определить по числу суточных слоев,
которые следует считать на окрашенных препаратах или тонких шлифах между ба-
зальным концом неонатальной линии и полостью пульпы (Рязанов, Клевезаль, 1991).
Прорезание зубов. Последовательность прорезания зубов может служить
показателем возраста особей до 1,5-2 лет (см. табл. 3.1).
Годовые слои видны на продольных аншлифах клыков в цементе: в 84-88%
случаев расхождение оценок 2 операторов не более 1 года (Рязанов, Маминов,
1996). На аншлифах и шлифах клыков слои цемента видны хуже, чем на окра-
шенных препаратах (Клевезаль, Мараков, 1966). Возраст (в годах) равен числу
слоев в клыке +0-1 (в зависимости от времени рождения, см. гл. 4 раздел 4.4 •).
Слои нечеткие.
Есть опыт прижизненного удаления р] для подсчета слоев цемента (Garshelis, 1984).
Felis caracal — каракал
Приводимые ниже данные взяты из работы Stuart, Stuart (1985).
Череп. Размеры (Позицию промеров см. рис. 2.2, 2.3).
По соотношению общей длины черепа и его скуловой ширины можно выде-
лить несколько возрастных групп в пределах первого года жизни и отличить их
Глава 13. Хищные
179
от старших: у особей в возрасте до 10 мес. общая длина до 120 мм, скуловая
ширина до 80 мм.
Зубы
Последовательность прорезания зубов может служить показателем возраста
особей первого года жизни (см. табл. 3.1).
Клыки. Наибольший диаметр апикального отверстия верхнего клыка в возрасте
10 мес. (при прорезании) 3,4 мм, в 1г. 3 мес. — 1,6 мм, в возрасте 2 г. оно закрыто.
Годовые слои — вероятно, как у F. catus, Е sylvestris (см. ниже).
Хрусталик
По весу (см. табл. 6.1) можно различить особей до года и особей от 2 лет.
Felis catus, Е sylvestris — одичавшая и дикая кошки
Зубы
Клыки. У кошек Е sylvestris из Германии апикальное отверстие верхнего клыка
закрывается в возрасте 6-8 мес. у самок и в 7-10 мес. у самцов (Piechocki, Stiefel,
1988), а у кошек из Испании—в возрасте с 6 до 18 мес. (Garsia-Perea, Baquero, 1999).
Годовые слои видны на окрашенных срезах в цементе клыков или корен-
ных. Возраст (в годах) равен числу слоев +0-1 (см. гл. 4 • и 4.4 •) (Piechocki,
Stiefel, 1988; Garsia-Perea, Baquero, 1999).
Хрусталик
Перекрывания веса соседних годовых классов по весу хрусталика отмечено
у самок Е catus со второго года, у самцов с третьего года жизни. Средний сухой
вес одного хрусталика у самцов и самок сеголетков, соответственно, 90 и 70 мг,
годовалых — 235 и 233 мг, двухгодовалых — 279 и 255 мг, трехгодовалых —
307 и 282 мг (Pascal, Castanet, 1978).
Кости конечностей
У Е sylvestris эпифизы бедренной и большой берцовой костей не срослись с
диафизом у самцов и самок в возрасте 5-15 мес., срослись, но эпифизарные
линии видны у самцов в возрасте 18-20 мес., у самок в возрасте 8-17 мес.,
полное слияние может иметь место с 18 мес. у самцов и 8-10 мес. у самок.
Следовательно, самцы с не приросшими эпифизами—не старше 15 мес., самки —
7 мес. (Piechocki, Stiefel, 1988).
Felis (-Рита) concolor — пума
Зубы
Стачивание (Gay, Best, 1996):
2 г. — полная постоянная зубная система, верхние клыки белые, у самцов
резцы и клыки не стерты, у самок — слегка стерты;
180
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
3-4 г. — клыки слегка окрашены, кончики клыков слегка стерты у самцов и
на 2 мм у самок, у самцов видна потертость самых высоких бугров i1 и Р, Р едва
стерт или не стерт, у самок Р стерт, но по уровню выше, чем Р и Р;
5-6 л. — клыки умеренно окрашены, кончики стерты на 3 мм у самцов и на
3-5 мм у самок, Р стерт, но выше Р и Р;
7-9 л. — клыки темно окрашены, кончики уплощены до округлых, Р стерт до
уровня Р и Р;
10 л. и старше—резцы стерты до десны, клыки стерты до округлости, тупые.
Годовые слои видны в цементе предкоренных на окрашенных срезах, нечет-
кие, но пригодны для определения возраста (Matson, 1993).
Felis lynx — рысь
Череп. Швы. Гребни (Обозначения см. рис. 2.1, 2.2, 2.3).
Шов ВВ открыт у большинства самцов и самок в возрасте до 37 мес. и зак-
рыт у большинства старших, но у некоторых самцов может быть открыт до 60
мес. Этот шов открыт почти у половины самок в возрасте 37-48 мес., закрыт у
большинства самок в возрасте 49-60 мес. и у всех старших.
Стреловидный гребень не развит у самцов-сеголетков, у старших развивается
и увеличивается с возрастом. Возраст самого молодого самца, имевшего гре-
бень, был 15 мес., а самого старого без гребня — 72 мес. Для самок этот крите-
рий не годится, половина самок старше 72 мес. не имела гребня (Kvam, 1983).
Зубы
Клыки. Относительная ширина полости пульпы клыка (см. табл. 3.4) позво-
ляет разделить сеголетков, годовиков и старших. Для ориентировочного выде-
ления старших групп предлагается использовать индекс Смирнова (см. Canis
lupus) (Малафеев и др., 1986):
в возрасте 1 г. индекс 0,850 и более; 4 г. — 0,550-0,635;
2 г. — 0,722-0,849; 5 л. — 0,464-0,549;
3 г. — 0,636-0,721; 6 л. и старше — 0,463 и менее.
У рысей, обитающих в Испании, апикальное отверстие клыка закрывается в
возрасте 12-18 мес. (Zapata, Perea et al., 1997), у рысей на Урале диаметр апи-
кального отверстия клыка (по большому эллипсу поперечного сечения) у осо-
бей моложе года в октябре — 8,2 мм, в марте — 5,2 мм (Малафеев и др., 1986).
Годовые слои как правило, хорошо видны в цементе на окрашенных продоль-
ных срезах клыков. Возраст (в годах) равен числу слоев + 0-1 (см. гл. 4 • и 4.4 •).
Есть опыт прижизненного удаления резца у анестезированных животных для
подсчета слоев цемента (Breitenmoser et al., 1993 — к сожалению, авторы не
приводят никаких подробностей, просто упоминают этот факт).
•Хрусталик
По весу хрусталика сеголетки четко отличаются от старших возрастных групп
(см. табл. 6.1), но различить возрастные группы годовиков и двухгодовалых
уже не удается (Малафеев и др., 1986).
Глава 13. Хищные
181
Felis rufus — красная рысь
Череп. Швы (Обозначения см. рис. 2.2).
Шов ВВ полностью зарастает не ранее 3-летнего возраста (Garsia-Perea, 1996).
Зубы
Клыки. По данным Crowe (1972) апикальное отверстие клыка закрывается
в возрасте 13-18 мес. Поскольку некоторые особи с открытым корнем могут
быть старше 12 мес., Johnson et al. (1981) предлагают такой критерий: рысь
можно считать старше 12 мес., если отношение общей длины клыка к макси-
мальной ширине апикального отверстия корня равно или больше 10 у самок и
8 у самцов.
Прорезание. Последовательность прорезания зубов может служить показа-
телем возраста особей до полугода (см. табл. 3.1).
Годовые слои видны в цементе на окрашенных срезах клыков. Возраст (в
годах) равен числу слоев +1 (Crowe, 1972). (См. гл. 4 • и 4.4 •)
Gulo gulo — росомаха
Череп. Швы. Гребни (Обозначения см. рис. 2.1, 2.2, 2.3).
У всех сеголетков, добытых осенью - в начале зимы еще открыты швы IN и
MN. Они могут быть закрыты у некоторых самцов весной. У половозрелых сам-
цов стреловидный гребень (5) развит в дорзальном направлении дальше, чем у
неполовозрелых. Для определения, насколько этот гребень у самцов выступает
дорзально, надо из общей длины черепа вычесть кондилобазальную длину. У 7
взрослых самцов средняя величина этого показателя 14,9 мм, у 12 неполовозре-
лых — 11,6 мм (Wright, Rausch, 1955).
Rausch, Pearson (1972) полагают, что по зарастанию швов IN и SJ в сочета-
нии с прирастанием эпифизов костей конечностей (см. ниже), можно надежно
отличить особей моложе 10-11 мес.
Зубы
Годовые слои видны в цементе на окрашенных срезах клыков и щечных
зубов, нечеткие (см. гл. 4 •).
Кости конечностей
В ноябре-декабре у сеголетков, в отличие от старших, дистальные эпифизы
лучевой и локтевой костей или еще не срослись с диафизом или только что
срослись (видна бороздка) (Rausch, Pearson, 1972).
Тазовый пояс
Тазовый шов (лобковое сращение) полностью открыт осенью - ранней вес-
ной у всех особей моложе года, и хотя бы частично зарос дорзально или вент-
рально у всех особей старше года (Wright, Coulter, 1967).
182
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Кость полового члена
На второй год жизни созревает только часть самцов, поэтому между сеголет-
ками, годовалыми и двухлетними есть перекрывания предельных значений раз-
меров кости. В среднем, вес кости у сеголетков до 1-1,2 г, у годовиков 1,6-1,8
г, у двухгодовалых 1,8-2,0 г (Rausch, Pearson, 1972). У взрослых кость длиной
80-90 мм, вес 1,8-2,9 г (Wright, Rausch, 1955).
Хрусталик
По весу хрусталика (см. табл. 6.1) можно отличить сеголетков от остальных.
Lutra canadensis — канадская выдра
Череп. Размер. Швы (Обозначения см. рис. 2.1, 2.2).
По скуловой ширине можно отличить особей моложе года от старших в но-
ябре-марте:
самцы < 1г. 65,6±0,3 мм, >1 г. от 69,5±0,3 мм;
самки < 1г. 63,0±0,4 мм, >1 г. от 66,8±0,4 мм (Stephenson, 1977).
В ноябре-марте у особей моложе года открыты, а у старших закрыты швы
РМ, В В, FN, SJ (Hamilton, Eadie, 1964; Stephenson, 1977).
Зубы
Клыки. Если ширина полости пульпы больше половины ширины зуба, то
возраст особи меньше года (измеряли на рентгеновских снимках, Kuehn, Berg
(1983)).
Годовые слои видны в цементе на окрашенных срезах клыков и щечных
зубов, нечеткие, но их можно использовать для оценки возраста (См. гл. 4 •).
Тазовый пояс
Тазовый шов (лобковое сращение, симфиз) хрящевой в возрасте 8-9 мес., с
явным швом — в 11—12 мес., заметным швом — в 20-21 мес., с возраста 35 мес.
шва не видно (Hamilton, Eadie, 1964).
Кость полового члена
По длине и весу кости можно отличить молодых (см. табл. 2.2). В возрасте
8-9 мес. кость тонкая, проксимальный конец ее лишь чуть толще; в 11-12 мес.
проксимальный конец увеличивается, с 20-21 мес. форма кости как у взрослых,
проксимальный конец отделен от тела кости небольшим боковым гребнем
(Hamilton, Eadie, 1964).
Lutra lutra — выдра
Череп. Швы. Форма черепа. Гребни (Обозначения см. рис. 2.1, 2.2, 2.3).
У сеголетков швы видны, особенно IN, загпазничное сужение лишь в ред-
ких случаях меньше 15,5 мм, лобные валики (см. рис. 13.2) соединяются за
Глава 13. Хищные
183
Рис. 13.2. Форма черепа соболя, Martes zibellina, в возрасте 8-10 мес. (А), 1 г. 8-10 мес. (Б),
2 г. 8-10 мес. (В), 3 г. и старше (Г). 1 — межглазничная ширина, 2 — заглазничное сужение,
3 — лобные валики (по Тимофеев, Надеев, 1955).
заглазничным сужением. На второй год жизни швы зарастают, IN едва заметен,
заглазничное сужение всегда меньше 15,5 мм, лобные валики соединяются до
заглазничного сужения, стреловидный гребень лишь начинает развиваться. У
особей старше 2 лет стреловидный гребень хорошо развит, у самцов лучше, чем
у самок (М. Stubbe, 19891).
Зубы
Стачивание. На первом году эмаль зубов не стерта. На втором начинается
стачивание, но дентин не обнажен. На третьем году обнажается дентин на р4.
Если обнажен дентин на т1, то выдра старше 3 лет (М. Stubbe, 1989f).
184
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Годовые слои видны в цементе на окрашенных срезах клыков и щечных
зубов, нечеткие, но их можно использовать для оценки возраста. Возраст (в
годах) равен числу слоев (см. гл. 4 •). По мнению Heggberget (1984) слои, по-
видимому, годовые, но считать их трудно.
Кости конечностей
У особей до года не срослись с диафизами проксимальный эпифиз плечевой
кости, дистальные эпифизы лучевой, локтевой и бедренной, проксимальный и
дистальный эпифизы большой и малой берцовых костей (Zeiler, 1988).
Кость полового члена
Молодых можно отличить по весу и длине кости (см. табл. 2.2).
Martes americana — американская куница
Череп. Гребни. (Обозначения см. рис. 2.1, 2.2, 2.3)
Распределение куниц, добытых зимой, по длине стреловидного гребня дву-
вершинное. Самцы с гребнем менее 20 мм и самки без гребня — неполовозре-
лые, самцы с гребнем более 20 мм и самки с гребнем — половозрелые (Marshall,
1951).
Зубы
Клыки. По относительной ширине полости пульпы клыка (см. табл. 3.4) мож-
но отличить особей моложе года от старших осенью-зимой.
Годовые слои видны в цементе на окрашенных срезах клыков, слои средней
четкости (Matson, 1995). Matson (1993) рекомендует использовать р4 (верхний
или нижний, не указано).
Кости конечностей
Самцов моложе года в ноябре-феврале можно отличить по отсутствию лате-
рального супрасезамоидного бугра бедренной кости (см. гл. 2.7); для самок этот
метод менее надежен. Отсутствие бугра — признак молодого животного, но у
некоторых молодых он может присутствовать (Leach et al., 1982).
Martes foina, M. martes — каменная и лесная куницы
Череп. Швы. Гребни (Обозначения см. рис. 2.1, 2.3).
В возрасте 9-12 мес. сохраняются только швы IN (Рябов, 1962), у М. martes
они зарастают в 1,5-2 г. (Юргенсон, 1956). По нашим данным (Клевезаль и др.,
2005), у добытых в осенне-зимний сезон самцов только полностью открытый
шов является несомненным признаком особи моложе года, но он был обнару-
жен лишь у 19% (из 37) таких особей. Полностью заросший шов — признак
особи старше года. Он был найден у 22% (из 45) годовиков и у половины (из 38)
старших. Самцы с частично заросшим швом были во всех возрастных группах.
Глава 13. Хищные
185
Стреловидный гребень начинает развиваться в конце первого года жизни, у
самок позже, чем у самцов. У М. martes гребень хорошо выражен с конца вто-
рого года, у М. foina — на третью зиму (Рябов, 1962; Habermehl, 1985). Полно-
стью не выраженный гребень у добытых осенью-зимой самцов—несомненный
признак особи моложе года, но у некоторых молодых особей (примерно у 20%)
он может быть слабо или даже хорошо выражен; сильно развитый гребень —
несомненный признак особи не моложе года (Клевезаль и др., 2005).
Соединение правого и левого лобных валиков с возрастом смещается к затлаз-
ничному сужению. Судя по данным Habermehl (1985), их положение у М. foina
соответствует позиции «А» (рис. 13.2) в возрасте около 9-21 мес. у самок и 9 мес.
у самцов, позиции «Б» в возрасте около 33 мес. у самок и 21 мес. у самцов, пози-
ции «В» в возрасте 44-57 мес. у самок и 33 мес. у самцов и позиции «Г» в возрасте
44—57 мес. у самцов. По нашим данным у М. martes в осенне-зимний период пози-
цию «А» имели 70% самцов моложе года, 70% годовиков имели позицию «В», 80%
особей в возрасте 2 г. и старше—позицию «Г» (Клевезаль и др., 2005). Используя
распределение особей по позиции лобных валиков нам удалось правильно разде-
лить 90% самцов (121 экз.) на особей моложе года и старших. Это подтверждает
мнение Гракова (1962), что даже по комплексу признаков (внешнему виду черепа,
стертости зубов, ширине канала клыка) как самцов, так и самок лесных куниц про-
мысловой выборки можно надежно разделить лишь на сеголетков и старших.
Зубы
Прорезание и стертость. Возраст появления постоянных зубов см. табл. 3.1.
В возрасте 6 мес. зубы не стерты; в 9 мес. уплощается и исчезает зубчатый
край i остальные зубы не стерты; в 21 мес. (вторая зима) заметно незначитель-
ное стачивание эмали i1-2 и почти всегда слабое округление кончиков клыков и
эмалевых бугорков предкоренных зубов; в 33 мес. видно обнажение дентина на
верхних резцах, стачивание эмали на хищнических зубах и сильное округление
бугров предкоренных и коренных; в 44-46 мес. исчезают лопасти на коронке 3-
го резца, клыки сточены на треть их первоначальной длины и бугры на р2 4 и Ш]
почти исчезли (Habermehl, 1985). По нашим данным (Клевезаль и др., 2005) у
добытых осенью-зимой самцов лесной куницы верхние резцы были не стерты
или чуть стерты только у особей моложе года (65% из 37), а стерты с обнажени-
ем дентина — только у особей старше года (80% из 84).
Годовые слои видны в цементе на окрашенных срезах клыков. Возраст (в
годах) равен числу слоев (см. гл. 4 •).
Кость полового члена
Молодых от взрослых можно отличить по весу и длине кости (см. табл. 2.2).
Martes pennanti — куница-рыболов
Череп Швы Гребни (Обозначения см. рис. 2.1, 2.2, 2.3).
Осенью-зимой самцы-сеголетки отличаются от взрослых открытыми или, по
186
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
крайней мере, видимыми швами SJ, MN, IN, FN. У самок отделить сеголетков от
старших можно по открытому шву PM (Wright, Coulter, 1967).
У половозрелых самцов стреловидный гребень выдается в дорзальном на-
правлении дальше, чем у неполовозрелых. Если у добытых осенью-зимой раз-
ница между общей и кондилобазальной длиной черепа 6 мм и более — это взрос-
лый самец, если менее 6 мм — молодой самец (Wright, Coulter, 1967).
У самок осенью-зимой сеголетков от старших отличают по отсутствию стре-
ловидного гребня (Eadie, Hamilton, 1958 по Kuehn, Berg, 1981).
Зубы
Клыки. До декабря-января включительно, молодых можно опознать по от-
крытому апикальному отверстию корня клыка (Kuehn, Berg, 1981).
У добытых в октябре-ноябре особей Jenks et al. (1984) измеряли ширину
полости пульпы на рентгеновских снимках приклеенных на лист картона клы-
ков. По относительной ширине полости пульпы («канала») клыка Jenks et al.
(1986) сочли возможным в составе выборок выделять 4 возрастные группы:
сеголетки, годовики, двухгодовалые и старшие. Границы разделения соседних
групп по полусумме средних величин показателей таковы: самцы — 0,45,0,24 и
0,18, самки — 0,36,0,20 и 0,15. Они получили возрастные распределения выбо-
рок, не отличающиеся от таковых, полученных при определении возраста по го-
довым слоям цемента.
Годовые слои видны в цементе на окрашенных срезах клыков и предкорен-
ных. Слои четкие (Matson, 1995). Matson (1993) рекомендует использовать р4.
Есть опыт прижизненного удаления р(, (Strickland et al., 1982; Arthur et al., 1992)
для подсчета слоев цемента. (Связанные с этим проблемы см. гл. 4 раздел 4.4 ••).
Кости конечностей
См. М. americana
Тазовый пояс
Осенью-зимой - ранней весной у молодых животных тазовый шов (лобковое
сращение) открыт, у особей старше года он зарос, хотя бы частично (Wright,
Coulter, 1967).
Кость полового члена
Осенью и зимой самцов-сеголетков можно отличить по весу этой кости (см.
табл. 2.2). С февраля значения веса у особей моложе года и старших уже пере-
крываются (Wright, Coulter, 1967).
Martes zibellina — соболь
Мышцы головы
По данным Тимофеева и Надеева (1955) возрастные группы можно выделить
по положению жевательных мышц на голове после снятия шкурки (рис. 13.3).
Глава 2. Определение возраста по признакам скелета
187
Рис. 13.3. Голова соболя, Martes zibellina, после снятия шкурки у особей в возрасте 8-10 мес.
(А), 1 г. 8-10 мес. (Б) и 2 г. 8-10 мес. (В). Видно изменение расположения жевательной муску-
латуры (Тимофеев, Надеев, 1955).
Череп. Форма
Возрастные группы можно выделить по положению слияния лобных валиков
(см. рис. 13.2). Возможно, что как и у куницы (см. выше М. martes), у самок
продвижение места слияния фронтально происходит позже, чем у самцов.
По орбитальному индексу—отношению межглазничной ширины к заглазнич-
ному сужению (см. рис. 13.2) — Тимофеев и Надеев (1955) выделяли 4 возраст-
ные группы (табл. 13.1). Видимо, более или менее успешно этот прием можно
использовать лишь для выделения сеголетков (критику см. гл. 2 раздел 2.3).
Зубы
Клыки. По относительной ширине полости пульпы клыка (см. табл. 3.4) в
промысловых сезон можно отличать сеголетков от старших.
Годовые слои видны в цементе на окрашенных срезах клыков (см. гл. 4 •).
Возраст (в годах) равен числу слоев.
Таблица 13.1. Возрастные изменения орбитального индекса черепа соболя,
Martes zibellina (Тимофеев, Надеев, 1955)
Возрастные группы Самцы Самки
8-10 мес. 1,06(0,84-1,07) 1,06 (0,78-1,14)
1 г. 8-10 мес. 1,10 (1,04—1,22) 1,11 (1,04-1,24)
2 г. 8-10 мес. 1,20 (1,09-1,37) 1,17 (1,08-1,33)
старшие 1,30(1,14-1,55) 1,24 (1,17-1,43)
188
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Кость полового члена
По весу кости (см. табл. 2.2) можно осенью отличать молодых от взрослых.
Хрусталик
По весу хрусталика (см. табл. 6.1) можно отличать 7-месячных от особей
старших на год и более.
Meles tneles — барсук
Череп. Швы. Гребни. (Обозначения см. рис. 2.1, 2.2, 2.3).
Сеголетков от старших можно отличать по открытым швам до следующей
весны, когда почти все швы не видны.
Стреловидный гребень выражен у особей в 1,5 года, но он еще спонгиозный
(рыхлый) (Ahnlund, 1976).
Зубы
Клыки. Относительные размеры полости пульпы клыка (см. табл. 3.4) по-
зволяют отличать сеголетков от старших.
Стачивание зубов. М. Stubbe (1989d) предложил следующую схему опре-
деления возраста по стертости зубов:
6-9 мес. — начинает стачиваться эмаль 2 и ш1;
второй год — дентин на поверхности ш1 еще не слит в одну ленту, поверх-
ность стачивания резцов нижней челюсти поперечно-овальная;
старше 2 лет — поверхность стачивания нижних резцов более или менее ок-
руглая, позже — продольно-овальная; на всех буграх ш1 видны обнажения ден-
тина, позже — широкая сплошная лента дентина (последовательность стачива-
ния см. рис. 13.4).
Когда выборка особей известного (в результате мечения) возраста одной
популяции была использована для обучения оператора определять возраст по
степени стертости, возраст (годовые классы до 6 лет и далее 7+) особей дру-
гой популяции (также известного возраста) по степени стертости коренных,
особенно последнего коренного, был определен с точностью ± 1 год у 89-98%
особей, с точностью до года -— у 53-74% (в разных популяциях) (Harris et al.,
1992).
Годовые слои видны в цементе (на продольных) и в дентине (на поперечных)
окрашенных срезах клыков. Слои четкие. Возраст (в годах) равен числу слоев в
цементе или числу слоев в дентине +1 (см. гл. 4 •).
Кости конечностей
В первое лето жизни оба эпифиза большой берцовой кости не сращены с
диафизом, дистальный срастается в первую зиму, в возрасте около 10 мес. Про-
ксимальный не сращен в течение второго лета жизни и срастается осенью-зимой
(Ahnlund, 1976).
Глава 13. Хищные
189
Рис. 13.4. Стадии стачивания m1 (I—VI) и нижних резцов (а-f) барсука, Meles meles (Stubbe,
1965). I — 7-9 мес., II— 19-21 мес., Ill — 31-33 мес., IV-VI — старше 3 лет.
Тазовый пояс
Тазовый шов начинает зарастать на третьем году (Stubbe, 1965 noHabemjehl, 1985)
Кость полового члена
По весу и длине кости (см. табл. 2.2) можно отделить молодых, неполовозре-
лых от взрослых. Особи старше 6 мес. разделяются на возрастные группы плохо
(Graf, Wandeler, 1982).
Mustela ertninea — горностай
Череп. Швы. Форма (Обозначения см. рис. 2.1).
У самцов шов IN открыт у сеголетков, совсем не виден у старших, но стадия
«шов зарастает, но виден» встречается в обеих возрастных группах. У самок-
сеголетков встречается также стадия, когда шов не виден (King, 1991).
По орбитальному индексу (межглазничная ширина деленная на заглазничное
сужение) можно отделить молодых от взрослых, у последних он больше 1,1 у
самок и 1,15 у самцов (Grue, King, 1984).
Зубы
Клыки. По относительной ширине полости пульпы клыка (см. табл. 3.4) можно
отличать сеголетков от старших. Копеин (1967) считал возможным выделять среди
последних две группы: 1,5-2 г. (относительная ширина канала верхнего клыка у
самцов 19,5-10,5%, у самок 16,0-9,5%) и 2,5 г. и старше (соответственно, ме-
нее 10 и 9%). Учитывая существенную индивидуальную изменчивость этого по-
казателя, вряд ли такое выделение надежно.
190
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Стачивание зубов. Строганов (1937) дал ключ для определения возраста по
степени стертости до 6-7 лет:
3-4 мес. — есть молочные зубы;
6 мес. — все зубы постоянные, не стерты;
около 1г. — эмаль верхнего хищнического1 (р3) стерта;
около 2 л. — на р3 есть обнажения дентина на вершине, профиль зуба не
изменен;
около 3 л. — обнажения дентина на р3 увеличены, профиль зуба имеет форму
арки с полукруглым сводом;
около 4 л. — профиль р3 — гребень с сильно притупленной вершиной, задняя
лопасть этого зуба сглажена почти до основания коронки;
около 5 л. — полосы дентина на двух задних зубах верхней челюсти слились,
образуя общую полоску.
Поскольку индивидуальная изменчивость этого показателя велика, следует
признать более реалистичным выделение только трех возрастных групп (М. Stubbe,
1989b):
(1) сеголетки — резцы почти не стерты, на хищническом зубе р3 может быть
видна узкая полоска дентина;
(2) особи на втором году жизни — обнажения дентина на резцах и широкая
полоска дентина на р3;
(3) старше 2 лет—прогрессирующее стачивание зубов, угол стачивания глав-
ного бугра р3— от прямого до тупого.
Годовые слои видны в цементе на окрашенных продольных срезах клыков и
mr Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кости конечностей
Отсутствие супрасезамоидного бугра бедренной кости (см. гл. 2 раздел 2.7)
надежный признак молодой (до 5 мес.) особи, но некоторые молодые самки
могут его иметь. У молодых (до 5 мес.) эпифизы бедренной кости не срослись
или почти срослись с диафизом (Van Soest, Van Bree, 1970; King, 1991).
Кость полового члена
Вес и длина кости (см. табл. 2.2) позволяют отличить молодых, неполовозре-
лых особей от половозрелых.
Mustela nivalis — ласка
Череп. Швы. Форма (Обозначения см. рис. 2.1).
Шов IN открыт у особей моложе 3 мес., зарос, но заметен в 3-6 мес., не
заметен у особей старше 6 мес. (King, 1980).
1 В зубной формуле куньих рода Mustela р1 отсутствует, поэтому хищническим зубом, последним
верхним предкоренным, является р3.
Глава 13. Хищные
191
Орбитальный индекс (межглазничная ширина деленная на заглазничное су-
жение) —хороший критерий возраста. По зависимости орбитального индекса от
возраста (до 20 мес.) можно дать приблизительную оценку возраста отдельных
особей (King, 1980):
самцы: возраст (мес.) = (орбитальный индекс - 0,8) : 0,028;
самки: возраст (мес.) = (орбитальный индекс - 0,783) : 0,011.
(Об использовании зависимости см. гл. 1 раздел 1.2.2.2)
Оценить относительный возраст, те. ранжировать особей по увеличению возраста,
как и у других куньих, можно по характеру слияния лобных валиков (см. рис. 13.2).
Зубы
Клыки. По относительному диаметру полости пульпы клыка (измеренному
по рентгеновским снимкам нижней челюсти) в выборке, взятой в разные сезоны
года, можно сказать только, что особи моложе 3 мес. имеют относительную
ширину полости пульпы не менее 20% у самок и 25—30% у самцов, а особи
старше 1 года — не более 20% для самок и 30% для самцов. С относительной
шириной в 20% для самок и 30% для самцов могут быть особи и моложе 3 мес.,
и старше года (Pascal et al., 1990).
Годовые слои видны в цементе на окрашенных продольных срезах клыков.
Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кости конечностей
Отсутствие супрасезамоидного бугра бедренной кости (см. гл. 2 раздел 2.7) —
надежный признак сеголетка, но начиная с октября у некоторых молодых он
может уже быть (King, 1980).
Кость полового члена
Вес кости более надежный показатель возраста, чем длина.
Связь между возрастом (в мес.) и весом (в мг) до 10 мес. описывается урав-
нением (King, 1980), которое можно использовать для приблизительной оценки
возраста:
1g (возраст) = [1g (вес) - 0,614] : 0,915
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Если выборка, взята в разные сезоны года, можно только сказать, что при
весе кости до 10 мг особь, очевидно, не старше 3 мес., при весе 34 мг — старше
года (Pascal et al., 1990).
Mustela putorius — лесной хорек
Череп. Швы. Форма (Обозначения см. рис. 2.1, 2.3).
У особей в возрасте 7-9 мес. отчетливо виден шов IN, он зарастает в 19-22
мес. (Habermehl, 1985).
На первом году жизни стреловидный гребень еще плоский, грубый и рых-
лый. Лобные валики (см. рис. 13.2) соединяются за заглазничным сужением. Не
192
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
полностью развитый стреловидный гребень—признак молодых. На втором году
лобные валики соединяются впереди (или у самок — в области) заглазничного
сужения (М. Stubbe, 1989с).
Зубы
Стачивание. У сеголетков может быть заметно стачивание верхнего и ниж-
него хищнических зубов (р3 и и начало стачивания резцов; на втором году-
сильное стачивание и даже выпадение зубов (М. Stubbe, 1989с).
Годовые слои видны в цементе на окрашенных продольных срезах клыков.
Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кость полового члена
По весу и длине кости (см. табл. 2.2) можно отличить молодых особей от
половозрелых.
Mustela sibirica — колонок
Зубы
Клыки. По ширине полости пульпы клыка (см. табл. 3.4) можно отличить
сеголетков от старших в промысловый сезон.
Стертость зубов. По этому признаку в выборке, взятой в промысловый се-
зон, трудно отличить даже сеголетков от взрослых (Войлочников, Михайловс-
кий, 1970).
Годовые слои видны в цементе на окрашенных продольных срезах клыков.
Возраст (в годах) равен числу слоев (см. гл. 4 •)
Кость полового члена
По весу и длине кости (см. табл. 2.2) можно отличить молодых особей от
половозрелых. В ноябре-январе, ширина проксимальной части («основания»)
кости у сеголетков 0,83 (0,5-1,05 мм), у взрослых 2,36 (1,78-3,49 мм) (Войлоч-
ников, Михайловский, 1970).
Mustela vison — американская норка
Череп. Швы. Форма (Обозначения см. рис. 2.1, 2.3).
У сеголетков (возраст около 8 мес.) в отличие от взрослых (1 г. 8 мес. и
старше) четко виден шов SJ (Greer, 1957). В выборке, взятой в ноябре-марте,
доля особей моложе года, выделенная по этому признаку, значимо не отлича-
лась от таковой, выделенной по годовым слоям в цементе клыка (Askins, Chapman,
1984).
Lechleitner (1954), однако, не обнаружил открытых швов у особей, добытых
в осенний промысловый сезон, но нашел, что всех особей без стреловидного
гребня можно смело считать сеголетками, особи с гребнем могут быть любой
возрастной группы, в том числе и сеголетками.
Глава 13. Хищные
193
Заглазничное сужение составляет у самок 0,95 межглазничной ширины в
возрасте 8 мес., 0,85 — в возрасте 20 мес., 0,78 и 0,72 в возрасте 32 и 44 мес.
У самцов — 0,85 в 8 мес., 0,74 в 20 мес. и 0,55 в 32 мес. (Rottcherr, 1965, по М.
Stubbe, 1989е).
Зубы
Возраст появления постоянных зубов см. табл. 3.1.
Годовые слои видны в цементе на окрашенных продольных срезах клыков.
Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кости конечностей
По развитию супрасезамоидного бугра бедренной кости (см. гл. 2 раздел 2.7)
Greer (1957) верно разделил 93% особей (всего около 300) на молодых (около 8
мес.), у которых бугор отсутствовал или был маленький, и взрослых (1 г. 8 мес.
и старше), у которых бугор был средний или большой. По этому признаку разде-
ление выборки, взятой в ноябре-марте, на особей моложе года и старших значи-
мо не отличалось от разделения на эти две группы по годовым слоям в цементе
клыка (Askins, Chapman, 1984).
Кости таза
Молодых самцов можно отличить от взрослых по характеру заднего края
седалищной кости: у молодых он гладкий, у старших — бугристый (Greer, 1957).
Кость полового члена
По весу и длине кости (см. табл. 2.2) можно отличить молодых особей от
половозрелых.
Морфология кости более надежный критерий разделения, чем вес: у особей
старше года (в осенний промысловый сезон) кость более массивная, чем у мо-
лодых, с хорошо выраженным основанием (проксимальным концом) и четким
гребнем в месте прикрепления пещеристого тела (Lechleitner, 1954).
Хрусталик
По весу хрусталика (см. табл. 6.1) можно отличать 6-месячных от 1,5-годо-
валых.
Nyctereutes procyonoides — енотовидная собака
Зубы
Смена молочных резцов начинается в возрасте 2-3 мес., полная постоянная
зубная система функционирует с возраста 6 мес. (Habermehl, 1985).
Клыки. По относительной ширине полости пульпы клыка (см. табл. 3.4) осе-
нью-зимой можно отличать сеголетков от старших.
Годовые слои видны в цементе на окрашенных продольных срезах клыков.
Возраст (в годах) равен числу слоев (см. гл. 4 •).
194
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Кости конечностей
До конца ноября сеголетков можно безошибочно отделить от старших по на-
личию линии между дистальными эпифизами лучевой и локтевой костей и диа-
физами. Ошибка такого разделения увеличивается с 2% в декабре до 17% в
апреле-мае. Все же с декабря-января и до апреля-мая по этому признаку надеж-
нее отделять взрослых от особей моложе года, чем по ширине полости пульпы
клыка.
Осенью в ряде случаев сеголетков можно определить и прижизненно прощу-
пыванием выпячивания дистального эпифизарного хряща локтевой кости (Kauhala,
Helle, 1990).
Panthera pardus — леопард
Приводимые ниже данные взяты из работы Stander (1997)
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до 2 лет (см. табл. 3.1). Далее возраст можно
оценить по стертости зубов:
2 ,5—3 г. — слабое, затем заметное стачивание резцов, высоких бугров р3 и
р42, клыков, продольного гребня клыка, в 3 г. — легкая желтизна предкоренных
и коренных;
4 г. — стачивание видно на всех зубах, в i’, может быть видна полость пуль-
пы, продольный гребень на клыках сточен по всей длине, все зубы желтые;
5-6 л. — полость пульпы видна на всех резцах, один-два резца могут быть
утеряны, кончики клыков сточены до плоских, бугор на р3ч 4 округло сточен, все
зубы желтые;
7-10 л. — зубы сильно сточены, резцы короткие и плоские с обнаженной
полостью пульпы на поверхности стачивания, некоторые утеряны, клыки сточе-
ны до обнажения полости пульпы, бугор на р33 4 сточен плоско, все зубы интен-
сивно желтые.
Визуальное исследование состояния зубной системы позволяет дать прибли-
зительную оценку возраста живых (усыпленных) особей.
Procyon lotor — енот-полоскун
Череп. Швы
По сочетанию степени зарастания разных швов Grau et al. (1970) выделяли 7
возрастных групп у самцов. Учитывая большую индивидуальную изменчивость,
для определения возраста можно принять общую для самцов и самок схему воз-
растных изменений зарастания швов по Junge, Hoffmeister (1980) — рис. 13.5.
2 В зубной формуле леопарда р1 и р2 отсутствуют, р2 очень мал.
Глава 13. Хищные
195
Рис. 13.5. Возрастные изменения состояния швов костей черепа у енота, Procyon lotor, из
Иллинойса (по табл ице из Junge, Hoffmeister, 1980). Белое поле—более чем 95% шва открыто,
кости в контакте, но шов ясно виден; черное поле — шов зарос и не виден; серое поле — шов
находится в промежуточной стадии, виден частично. По горизонтали возраст в месяцах, по
вертикали швы: MP, S, С, IF, MJ, PS, SJ, FN, NP, L, IN— обозначения швов см. рис. 2.1; ВВ, РМ,
IPL, — обозначения см. рис. 2.2.
Зубы
Клыки. Особь моложе года можно опознать по открытому апикальному от-
верстию клыка, оно закрывается примерно в возрасте 12 мес. (Grau et al., 1970).
Стачивание зубов. В возрасте 0-14 мес. зубы не стерты;
в возрасте 15-38 мес. начинает стираться щечная сторона т, и задне-щечная
т2, но бугры на обоих зубах присутствуют;
в 39-57 мес. передне-щечный бугор Ш] и бугры на щечной поверхности пт,
сточены плоско, появляются первые признаки стачивания предкоренных;
в 58 86 мес. ш2 сточен плоско, в ш, остались два островка эмали, остальная
поверхность сточена плоско, передне-язычный бугор р4 сточен полностью, а пе-
редне-щечный еще имеется;
у особей старше 86 мес. Ш] и т2 сточены плоско, передне-щечный бугор р4
сточен полностью (Grau et al., 1970).
Годовые слои видны в цементе на окрашенных срезах клыков, предкорен-
ных и первого коренного. Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кость полового члена
По весу и длине кости (см. табл. 2.2) можно отличить молодых от взрослых.
По форме осенью можно выделить 3 группы: в 5-6 мес. кость г-образная, в 1,5 г.
кроме резко изогнутой дистальной части кость имеет изгиб и на несколько утол-
щенном (проксимальном) конце в 2 г. и старше — кость S-образной формы
(Руковский, 1970).
Хрусталик
По весу хрусталика можно определять месяц рождения особей моложе года.
Для разделения выборки на две возрастные группы — до года и старше года
196
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
(см. табл. 6.1) проще использовать срастание эпифизов с диафизами или кость
полового члена (Sanderson, 1961).
Urocyon cinereoargenteus — серая лисица
Череп. Швы (Обозначения см. рис. 2.2).
Большинство швов закрываются к январю первого года жизни особи кроме
шва РВ, который закрывается на 10-11 месяце жизни. Сошник достигает полно-
го развития на втором году жизни (Wood, 1958).
Зубы
Клыки. Часть сеголетков можно опознать по открытому апикальному отверстию
корня клыка (в ноябре 65% сеголетков имели открытое апикальное отверстие, в конце
декабря—только 30%), а оставшихся—по относительной ширине полости пульпы
клыка (на уровне шейки): у сеголетков она 40% и больше (Root, Payne, 1984).
Можно выделить сеголетков по расстоянию от края эмали верхнего клыка до
линии соединяющей края альвеол двух соседних с клыком зубов: с начала нояб-
ря по конец января все особи, у которых это расстояние было не более 2 мм,
были моложе года, но величина в 3-3,5 мм была обнаружена у особей до года и
у особей в возрасте 1-2 года, (Root, Payne, 1984)
Стачивание зубов. Wood (1958) выделял возрастные группы по степени стертос-
ти внутренних бугров т1 (не стерты в течение первого года жизни, слегка стерты—на
втором году, стерты до появления двух пятен обнажения дентина — на третьем, на
четвертом году пятна сливаются в одну полоску, которая дальше с возрастом, расши-
ряется). Nicholson, Hill (1981) использовали этот метод для разделения на возрастные
группы выборки, взятой зимой. Все особи, определенные по степени стертости как
сеголетки, не имели годового слоя цемента. Однако не имели годового слоя (т.е. по
цементу были определены как особи моложе года) и более половины особей, по
стертости отнесенных к годовалым, и часть особей, отнесенных к старшим группам.
Годовые слои видны в цементе на окрашенных срезах клыков. Возраст (в
годах) равен числу слоев (см. гл. 4 •).
Кости конечностей
Приращение дистального эпифиза лучевой кости к диафизу у сеголетков на-
чинается с декабря; в ноябре сеголетки хорошо отличаются от старших по этому
признаку (Sullivan, Haugen, 1956).
Кость полового члена
Особи с весом кости до 400 мг — несомненно молодые, с весом от 700 мг —
взрослые (Wood, 1958).
Хрусталик
Сравнение веса хрусталика с числом годовых слоев показало, что по весу
хрусталика можно отделять сеголетков от старших (см. табл. 6.1).
Глава 13. Хищные
197
Ursus americanus — черный медведь
Размеры тела
Первые два годовые класса можно ориентировочно выделить по размерам.
Средние значения таковы (Sauer, 1975):
Показатели возраст самцы самки
общая длина тела* (см) 0,5 г. 79 76
1,5 г. 123 115
2,5г. 138 132
ширина задней ступни (см) 0,5 г. 6 6
1,5 г. 9 8
2,5г. 9,5 8,6
живой вес (кг) 0,5 г. 15 13
1,5 г. 48 38
2,5 г. 72 58
* до кончика хвоста
Череп. Размеры. Швы (Обозначения см. рис. 2.1, 2.2, 2.3).
У самок длина черепа достигает значений, характерных для взрослых особей в
3,5 г., ширина — в 4,5 г. Средние величины в разном возрасте таковы (Sauer, 1975):
Показатели возраст самцы самки
Наибольшая длина (мм) 1,5 г. 203 180
2,5 г. 237 206
3,5 г. 240 214
Скуловая ширина (мм) 1,5 г. 130 116
2,5 г 152 135
3,5 г. 159 139
4,5 г. 161 145
Последовательность зарастания швов с возрастом показана на рис. 13.6. По
мнению Sauer (1975), этот признак является ненадежным критерием возраста из-
за большой индивидуальной изменчивости.
Зубы
Клыки. Клыки растут в длину в течение первых нескольких лет жизни (рис.
13.7). Апикальное отверстие корня закрывается в возрасте 3—4 лету медведей из
штатов Нью-Йорк и Мичиган (Sauer et al., 1966; Marks, Erickson, 1966), на 6-е
лето жизни — у медведей из Аляски (Rausch, 1961).
Судя по данным Sauer et al. (1966), по ширине полости пульпы нижних клы-
ков можно различить особей 1,75 г. и моложе (ширина более 3,5 мм) и 2,75 г. и
старше (менее 3,5 мм).
198
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 13.6. Возрастные изменения состояния швов костей черепа медведя Ursus americanus (по
Marks, Erickson, 1966). По горизонтали — возраст; по вертикали — швы FN, NP. MP, S, IF, IN,
L, PS. IPR — обозначения швов см. рис. 2 1, ВВ — обозначение см. рис. 2.2). М — самцы, F —
самки. Белое поле — шов открыт, светло-серое — шов гребнеобразный, темно-серое — шов
частично закрыт, черное — шов полностью зарос. Пунктир означает отсутствие данных.
Рис. 13.7. Схема возрастных изменений формы клыков медведя Ursus americanus: 9 мес., 1,7 г.,
2,7 г., 3,7 г. (по Sauer et al., 1966). Черным показано открытое апикальное отверстие.
Глава 13. Хищные
199
У медведей из Аляски по размерам корня и числу границ годовых зон денти-
на (валиков) на корне клыка Rausch(1961) различал 6 годовых групп (рис. 13.8);
на корне Р, который формируется раньше клыка, можно видеть до трех годовых
зон. Но у медведей, обитающих на территориях штатов Нью-Йорк и Мичиган,
число наружных валиков на клыках оказалось ненадежным критерием возраста
(Marks, Erickson, 1966).
Прорезание зубов. Постоянные зубы появляются в течение первого года жиз-
ни (см. табл. 3.1).
Годовые слои видны в цементе на окрашенных срезах клыков, резцов, ко-
ренных и предкоренных. Возраст (в годах) равен числу слоев в клыках + 1. На
большом материале от особей известного возраста метод признан надежным
(Harshyne et al., 1998).
Есть опыт прижизненного удаления рр р2, для подсчета слоев; число слоев в це-
менте соответствовало возрасту у животных не старше 6 лет, у старших число слоев
было меньше числа прожитых лет (McLaughlin et al., 1990) (см. гл. 4 раздел 4.4 ••).
Кости конечностей
У самок эпифизы длинных костей конечностей начинают срастаться с диафиза-
ми в 2,5 г. и полностью срастаются не ранее 8,5 лет. Раньше всех, в 2,5 г. у самок
и в 3,5 г у самцов, срастаются дистальные эпифизы лучевой и плечевой костей,
позже всех—проксимальный эпифиз плечевой кости (в 8,5 л. у самок) (Sauer, 1975).
Кость полового члена
У сеголетков вес кости (г) 0,84 (0,6-1,4), длина (см) 6,86 (4,5-8,6)
в возрасте 1-2 г. 2,32 (1,4-3,6) 9,98 (8,4-11,5)
у особей старше 2 л. 3,2—16,8 11,6-19
(Marks, Erickson, 1966).
Ursus arctos — бурый медведь
Размеры тела
По ширине передней лапы (измеренной позади пальцев) летом можно разли-
чить три возрастные группы. Для медведей из Якутии этот показатель такой (сред-
няя, пределы варьирования, п):
сеголетки — 6,8 см (6,0-7,5 см, 22);
особи прошлого года рождения — 8,5 см (8,0-9,5 см, 36);
взрослые самки — 12,3 см (11,0-13,5 см, 40);
взрослые самцы — 14,1 см (13,0-17,0 см, 192) (Тавровский и др., 1971).
У медведей из Карелии такая ширина передней лапы: сеголетки — 6-9 см,
годовалые — 9,5-11,5 см, взрослые — более 12 см (Данилов и др., 1979).
Череп. Размеры. Швы. Форма (Обозначения см. рис. 2.1, 2.2, 2.3).
Для самцов от 1 до 23 лет Pearson (1975) приводит два разных уравнения
связи между возрастом (в годах) и скуловой шириной черепа (в мм) для двух
разных районов провинции Юкон (Сев. Америка):
200
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
1g (возраст) = 4,510008 • 1g (ширина) - 9,29264;
1g (возраст) = 3,95651 »lg (ширина) - 8,10864;
и для самок общее:
1g (возраст) = 5,50499 »lg (ширина) - 11,39523.
Для медведей, обитающих на Балканах (Sladek, 1992), эта связь возраста (в
годах) и скуловой ширины черепа (в мм) для самцов от 2 до 18 лет:
In (возраст) = (ширина - 110,560) : 37,186;
для самок от 2 до 11 лет:
In (возраст) = (ширина - 123,916) : 24,113.
До трех лет швы не зарастают. С четырех лет начинается зарастание швов L,
IF, MN, PM, IPL (IF полностью или частично зарастает почти у половины пяти-
летних самцов и у всех 14-летних; у самок на пятом году он зарастает у 80%, и
на седьмом году у 100%). IN у самцов начинает зарастать в шесть лет, он
может быть полностью заросшим у некоторых особей в 15 лет, но у некоторых
виден и после 20 лет. У самок начало зарастания IN отмечено у 5-летней, пол-
ное — у 14-летней. От 7 до 10 лет зарастает MF. В последнюю очередь, в возра-
сте 15-18 лет, зарастает SJ (Завацкий, 1978; Sladek, 1992).
По мнению Pearson (1975) использование зарастания швов как критерия возрас-
та ограничено большой индивидуальной изменчивостью. В среднем возраст сам-
цов с закрытыми швами IN и FN можно считать не менее 12 лет, самок —10 лет,
но у отдельных самок в 16 лет и самцов в 23 г. эти швы были открыты.
Длина стреловидного гребня у самцов, и в меньшей степени у самок, в сред-
нем увеличивается с возрастом, но индивидуальная изменчивость очень велика.
У самцов гребень появляется в возрасте трех лет, начинает заходить за венечный
шов в возрасте 6-8 лет, но может оставаться коротким даже у старых особей
(Sladek, 1992).
Зубы.
Смена молочных на постоянные и прорезание постоянных даны в табл. 3.1.
Клыки. Перед первой зимовкой (возраст 8-9 мес.) клыки находятся в аль-
веолах, их коронки сформированы. На второе лето жизни нижние клыки самцов
вырастают до 50 мм и более, у самок — не более 46 мм, шейка в альвеоле.
Длина верхних клыков несколько меньше. На третье лето развитие верхних и
нижних клыков сходно, шейка находится у края альвеолы или даже слегка выс-
тупает за край. На четвертое лето шейка выступает над альвеолой (на 1-5 мм), у
отдельных особей видны первые следы стачивания. На 5-6-е лето апикальное
отверстие корня клыка закрывается. С девяти лет идет прогрессирующее стачи-
вание (Rausch, 1969).
Стертость. Резцы начинают стачиваться в возрасте 2-3 лет, сильно стачива-
ются в 12-15 лет у самцов и 6-7 лет у самок; т3 начинает стачиваться в 8-9 лет
у самцов и 6-7 лет у самок, сильно сточен в 21—22 г у самцов и 11-12 лет у
самок (Sladek, 1992).
Годовые слои видны в цементе и в дентине всех зубов на окрашенных срезах.
Границы первых пяти годовых слоев дентина на клыках видны как наружные
Глава 13. Хищные
201
валики, как и у черного медведя (см.
рис. 13.8) (см. гл. 4 раздел 4 •). Воз-
раст (в годах) равен числу слоев це-
мента + 0-1 (см. гл. 4.4 •).
Есть опыт прижизненного удаления
р4 (Craighead et al., 1970) и р( (Pearson,
1975; McLellan, 1989) для подсчета сло-
ев цемента (см. гл. 4 раздел 4.4 • •).
Кость полового члена
Длина кости у молодых 72-108 мм,
у взрослых — 118-160 мм (Suminski,
1968). Вес, соответственно, <4 г и >6 г
(Pearson, 1975).
Хрусталик
Вес хрусталика увеличивается с воз-
растом, но с года уже есть существенные
перекрывания между годовыми класса-
ми, и для определения возраста Pearson
(1975) признал этот показатель не при-
годным. (Надо заметить, что его мето-
дика не была строго унифицирована).
Рис. 13.8. Продольный разрез клыка мед-
ведя Ursus americanus (по Rausch, 1961).
Римскими цифрами показаны ростовые
зоны дентина, линиями на разрезе — их
границы. Э — эмаль, Ц — цемент, Д —
дентин, ПП — полость пульпы.
Ursus maritimus — белый
медведь
Череп. Размеры. Швы. (Обозна-
чения см. рис. 2.1, 2.2, 2.3).
По кондилобазальной длине и ску-
ловой ширине четко отличаются особи
в возрасте около года от остальных. Далее между группами имеет место перекры-
вание значений (Lona, 1970), но все же ориентировочную оценку возраста в пер-
вые годы можно получить по промерам ширины черепа (табл. 13.2). Для этих же
целей может быть полезным соотношение высоты и ширины черепа (рис. 13.9).
Состояние швов позволяет выделить несколько годовых классов (рис. 13.10).
Зубы.
Постоянная зубная система формируется в 9-11 мес.
Клыки. Шейка верхних клыков полностью находится в альвеоле у всех сам-
цов и у большинства самок во вторую зиму, на третью зиму у всех особей она
показалась над альвеолой; расстояние от края эмали до альвеолы с возрастом
увеличивается, но в силу больших индивидуальных различий не может быть
критерием разделения возрастных групп (Lono, 1970).
202
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 13.2. Возрастные изменения ширины черепа белого медведя,
Ursus maritimus.
Возраст Межглазничная ширина* Скуловая ширина**
самцы самки самцы самки
Вторая зима 57,9±1,8* (53-64)** 56,3±1,0* (54-65)** 122-144 121-154
Третья зима 67,0±1,6* (67-75)** 67,0±0,8* (65-72)** 154-177 157-173
Четвертая зима 78,9±0,9* (72-82)** 76,4±1,5* (69-77)** 177-186 173-187
♦Manning (1964), Сев. Канада, Аляска, средняя и ошибка, мм.
** Leno (1970), Шпицберген, мм.
Рис. 13.9. Соотношение высоты черепа в области венечного шва (ось абсцисс, мм) и межглаз-
ничной ширины (ось ординат, мм) у белого медведя, Ursus maritimus, разных возрастных
групп. Самцы и самки второй зимы обозначены косым крестом, третьей зимы — квадратом и
четвертой зимы — прямым крестом. Линиями отмечены середины интервалов между средни-
ми значениями соседних возрастных групп (Manning (1964), с изменениями).
Глава 13. Хищные
203
3-я зима* 4-ая зима* 5-ая зима* 6-ая** зима* 7-ая зима* Старшие
ВВ М ммц
F —
МР М 1..
F — —
NP М
F I.S — —
FN М
F — —
IN М
F — —
С М г
F —
IPL М
F lilo — —
* — включая осенние месяцы и март
** — для самок — особи старше 5 лет
Рис. 13.10. Возрастные изменения состояния швов костей черепа у белого медведя, Ursus
maritimus (по Leno, 1970). Белое поле — шов открыт, черное поле — шов закрыт, серое
поле — закрыта часть шва. Разные поля в одной клетке означают наличие разных вариантов.
Прочерк — нет данных.
По горизонтали — возраст; по вертикали — швы; MP, NP, FN, IN,С— обозначения швов см.
рис. 2.1; ВВ, IPL — обозначения см. рис. 2.2; М — самцы, F — самки.
У самок по ширине полости пульпы нижних клыков (измерено на рентгено-
вских снимках зубов) четко отделяются только годовики, далее есть перекрыва-
ние между группами. Во вторую зиму ширина ровна 10-12 мм, на третью — 3-
5 мм, на 4-ю зиму — 2-3 мм, на 5-ю зиму — 2 мм. В целом, в возрасте 2 года и
старше ширина меньше 5 мм (Lono, 1970).
Годовые слои видны в цементе на окрашенных срезах предкоренных и ко-
ренных, очень нечеткие, особенно в первые четыре года (см. гл. 4 • и гл. 4
раздел 4.6.5). Для правильного выделения годовых слоев следует использовать
дополнительные методы из описанных для этого вида. Есть опыт прижизненного
удаления р( для подсчета слоев цемента (Hensel, Sorensen, 1980), возраст в го-
дах равен числу слоев в р, 3 +1.
Кости конечностей
Эпифизы бедренной кости (головки, большого вертела и берцовых мыщел-
ков) у самцов начинают прирастать к диафизу на 6-ю зиму и полностью сраста-
ются через полгода, у самок на два года раньше.
204
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Латеральный супрасезамоидный бугор (см. гл. 2 раздел 2.7) у самцов плос-
кий в первые четыре года, бугристый у пятигодовалых (на шестую зиму) и хоро-
шо выражен у старших, у самок картина менее четкая, бугор плоский у всех
годовалых и двухгодовалых и большинства трехгодовалых (Lono, 1970).
Кость полового члена
Размер кости можно использовать для выделения нескольких первых годо-
вых классов. По данным Leno (1970) во вторую зиму вес кости 1,4-1,8 г, длина
84-95 мм (п = 5), кость цилиндрическая; на третью зиму вес кости 3,3-5 г и
длина 118-134 мм (п = 4), появляется утолщение дистального конца; на четвер-
тую зиму эти показатели, соответственно, 7,1-8,7 г и 144—166 мм (п=3); на пя-
тую зиму — 9,9-13,8 г, и 115-172 мм (п = 7); у особей старше шести лет —
14,5-22,1 г и 168-194 мм (n=15). Dyck et al. (2004) исследовали большой мате-
риал и определяли возраст особей по годовым слоям цемента в нижнем предко-
ренном. По их данным есть существенные перекрывания показателей между воз-
растными группами: во вторую зиму вес и длина, соответственно, 1,2-5,2 г и
78-133 мм (п=34); на третью зиму — 2,1-7,6 г и 100 151 мм (n= 109); на четвер-
тую зиму — 2,5-13,3 г и 108-178 мм (п = 158); у особей старше шести лет —
11,3-31,9 г и 151-222 мм (п = 325). Причиной такого перекрывания могут быть
как индивидуальные различия, выявленные при исследовании большого матери-
ала, так и ошибки при определении возраста по числу годовых слоев цемента
(см. выше и гл. 4 раздел 4.6.5).
Vulpes vulpes — обыкновенная лисица
Череп. Швы. Гребни (Обозначения см. рис. 2.1, 2.2, 2.3)
В возрасте 5-6 мес. затылочные гребни отсутствуют, все швы заметны; с 6
мес. до 1 г. гребни стреловидный и поперечный затылочный хорошо развиты,
первый короткий, все швы хорошо различимы, кроме ВВ; у старших все швы
срослись, стреловидный гребень доходит до венечного шва С (Маркина, 1962).
Открытый шов ВВ в октябре-декабре—признак сеголетка, в декабре шов начи-
нает закрываться. Шов РВ открыт почти у всех сеголетков в декабре, закрывает-
ся на второй год жизни (Harris, 1978; Churcher, 1960; Johnston, Beauregard, 1969).
Зубы
Возраст появления постоянных зубов см. табл. 3.1.
Клыки. По относительной ширине полости пульпы клыка можно отличать
сеголетков от старших в промысловый сезон (см. табл. 3.4).
Хорошее разделение на сеголетков и старших было получено методом диск-
риминантного анализа по весу хрусталика и ширине полости пульпы (классифи-
кационные уравнения см. гл. 1 раздел 1.2.3).
Стертость. Сеголетков в сентябре легко отличить от старших по отсутствию
обнажений дентина (Kappeler, 1985). На втором году жизни обнажения дентина
Глава 13. Хищные
205
Рис. 13.11. Последовательность стачивания верхних резцов у добытых зимой лисиц Vulpes
vulpes (Harris, 1978). Цифрами указаны годовые классы.
появляются на резцах, но форма поверхности стачивания еще поперечно-оваль-
ная, после двух лет она становится сначала округлой, а затем продольно-оваль-
ной (М. Stubbe, 1989а), заметны обнажения дентина на т’ (см. рис. 3.6). Этот
критерий возраста удобен для выборок, взятых в промысловый сезон. В выбор-
ке, взятой в октябре- марте, Harris (1978), используя свою схему стачивания вер-
хних резцов (рис. 13.11), правильно определил 73% особей в возрасте до года,
65 % особей второго года, 44% третьего и 40% — четвертого года. В большин-
стве неверных определений ошибка была на год.
В выборке, полученной в течение всего года, Van Bree et al. (1974) выделили
4 возрастные группы по степени стертости резцов и коренных, лишь 35—47%
были определены правильно (за истинный возраст принимали возраст, опреде-
ленный по числу годовых слов в зубах).
Годовые слои видны в цементе на окрашенных продольных срезах клыков,
обычно четкие. Возраст (в годах) равен числу слоев (см. гл. 4 •).
Кости конечностей
Сеголетков можно выделить по не сросшимся с диафизами:
проксимальным эпифизам большой и малой берцовых костей, не позже ок-
тября (Harris, 1978); проксимальным эпифизам лучевой кости, с октября по се-
Таблица 13.3. Изменение веса хрусталика лисицы, Vulpes vulpes,
в первые два года жизни (Bromel, Zettl, 1974)
Возраст (мес.) Вес одного хрусталика (мг) Возраст (мес.) Вес одного хрусталика (мг) Возраст (мес.) Вес одного хрусталика (мг)
2 50-80 6 160-180 10 195-205
3 80-105 7 165-185 11 200-215
4 105-125 8 170-190 12 210-230
5 125-170 9 185-200 1-2 года 220-250
206
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
редину января (Jensen, Nielsen, 1968); дистальным эпифизам лучевой и локтевой
костей, до возраста 8-9 мес. (Sullivan, Haugen, 1956).
Кость полового члена
По мнению Harris (1978), по размеру кости (см. табл. 2.2) разделение сего-
летков и взрослых возможно только до конца сентября.
Хрусталик
По весу хрусталика до осени-зимы можно различать сеголетков и старших
(см. табл. 6.1). При выделении сеголетков в сентябре-октябре полное соответ-
ствие было получено между определениями по весу хрусталика и по окостене-
нию проксимального эпифиза плечевой кости (Friend, Linhart, 1964).
По мнению Bromel, Zettl (1974) до 2 лет точность определения возраста по
весу хрусталика (табл. 13.3) такая же, как по замещению и стиранию зубов.
Хорошее разделение на сеголетков и старших было получено методом диск-
риминантного анализа по весу хрусталика и ширине полости пульпы (классифи-
кационные уравнения см. гл. 1 раздел 1.2.3).
ГЛАВА 14. ПАРНОКОПЫТНЫЕ
Alces alces — лось
Следы
При наличии четких отпечатков можно выделить три возрастные группы:
0,5 г. — длина 90 мм ширина 65 мм; 1,5 г. — соответственно 115 мм и 85 мм;
>2,5 г. — 130 мм и ПО мм (Язан, 1961).
Экскременты (для оценки возраста пригодны только в осеннее-зимний пе-
риод).
У сеголетков форма «орешка» удлиненная, длина 23 25, ширина 10—15 мм;
в 1,5-3,5 г. — удлиненная форма, длина 30-35, до 40, ширина 15-20 мм;
в старшей группе «орешки» неправильной формы, смяты с боков, длина 25-
30, ширина 20-25мм (Язан, 1961).
MacCracken, Van Ballenberghe (1987) предложили использовать объем экск-
рементов (длина, умноженная на наибольшую ширину, умноженная на ширину
при повороте на 90° от наибольшей, мм3): у сеголетков объем 4000-5100; у
взрослых самцов —- 8200-11800; у взрослых самок — 4900-8500. По этому
критерию были идентифицированы правильно 91% кучек экскрементов той же
выборки и 69% выборки, взятой в другую зиму в том же районе. Несколько
более точное различение было получено с помощью дискриминантных функций
(см. гл. 1 раздел 1.2.3).
Рога —- самцы (самки рогов не имеют)
Число отростков в среднем увеличивается с возрастом, но у старых особей
уменьшается. Как следует из табл. 14.1, оно может дать лишь приблизительную
оценку возраста.
По мнению Язана (1961), по сочетанию данных о числе отростков и толщине
рога можно выделить возрастные группы 1,5 г., 2,5 г., 3,5 г., 4,5—5,5 л. и стар-
шие. Дицевич и др. (1983) считали, что у лосей из Восточной Сибири эти пока-
затели увеличиваются с возрастом лишь до 6,5-7,5 лет.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до двух лет (см. табл. 3.3). Позже стертость
щечных зубов позволяет приблизительно выделять годовые классы или возрас-
тные группы (см. гл. 3 раздел 3.4.6.3 •). Рисунки и описания стертости зубов у
особей разных возрастных групп даны в работах: Кнорре, Шубин (1959), Зари-
пов (1964), Херувимов, Орлов (1967), Habermehl (1985).
Приблизительную оценку возраста можно получить по обнажению дентина
на поверхности стачивания резцов (рис. 14.1) и ширине полости пульпы (рис.
14.2).
208
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 14.1. Возрастные изменения признаков рогов лося, Alces alces, севе-
ро-востока Европы (Язан, 1961) и средние значений этих показателей
(в скобках) для лосей из Восточной Сибири (Дицевич и др., 1983).
Показатель Возрастные группы
1,5 2,5 3,5 4,5-5,5 старшие
Число отростков 1/1-2/2* 1/2-3/3 2/2-474 474-7/7 2/2**-9/9
(одного / другого рога) (1,0) (2,0) (3,0) (4,2) (3,7-4,5)
Окружность рога 85-100 110-125 135-140 145-175 160-195
у розетки (мм) (68,6) (125,0) (141,0) (132,4) (151,5-165,3)
* только 1 из 15 имел 3/3,
** 2 из 12.
Рис. 14.1. Схема возрастных изменений резцов лося, Alces alces. 1— возраст 6 мес., 2—9 мес.,
3 — 1 г. 7 мес., 4 — 1 г. 9 мес., 5 — 2,5 г., 6 — 3,5 г., 7 — 4,5 г., 8 — 5,5 л., 9 — 6,5 л., 10 — 7,5 л.
(Зарипов, 1964).
Годовые слои — видны в цементе резцов и коренных, на аншлифах и окра-
шенных срезах. У особей из Восточной Европы и Сибири слои, как правило,
четкие (см. гл. 4 •). Gasaway et al. (1978), считавшие слои на шлифах резцов
особей известного врзраста (2-11 лет), получили совпадение числа слоев с воз-
растом только у 56% особей, Matson (1991), считавший слои на окрашенных
Глава 14, Парнокопытные
209
Вис. 14.2. Схема изменений с возрастом размеров полости пульпы i1 лося, Alces alces. Продоль-
ные распилы. Заштрихован дентин, черным показан цемент (Зарипов, 1964).
срезах — у 86%. Возраст (в годах) равен числу слоев в ip m'j или числу слоев
в i23, с +1. (см. также гл. 4 раздел 4.4 •),
Слои вторичного дентина часто хорошо видны на поперечных окрашенных
срезах и могут быть использованы для уточнения оценки возраста по слоям це-
мента (см. гл. 4 раздел 4.6.6).
Кости конечностей
Петров (1964) дает детальное описание возрастных изменений всех костей
скелета. Так, в плечевой кости в 6 мес. проксимальный эпифиз отделен от диа-
физа четкой линий, дистальный начал срастаться с диафизом, к 3 годам сраста-
ние полное; в лучевой и локтевой костях к 12 мес. заканчивается срастание с
диафизом проксимального отдела лучевой и обоих эпифизов локтевой костей; в
бедренной кости в 1,5 г. отмечено срастание большого бугра с проксимальным
эпифизом и дистального эпифиза с диафизом, в 3 г. — срастание всех элементов.
210
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Antidorcas marsupialis — спрингбок
Приводимые ниже данные взяты из работы Rautenbach (1971).
Рога
По форме и общему размеру рогов можно выделить 5 возрастных групп,
старшая — 21 мес.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей почти до двух лет (см. табл. 3.3). По стертос-
ти выделены такие группы:
22 мес. - 3,5 г. — слабая стертость зубов, лунки (см. гл. 3 раздел 3.4.6.3)
видны на всех коренных, небольшая стертость может быть видна на жевательной
поверхности р4 (третий щечный зуб), Ш] и на последней доле т3;
3,5-5,5 л. — умеренная стертость, лунки исчезают на тр но видны на осталь-
ных коренных;
5,5-7,5 л. — сильная стертость, исчезают лунки на всех коренных.
Самыми стертыми были зубы у 8-летних (возраст определен по слоям цемен-
та), р4 и были стерты до корней.
Годовые слои видны в цементе i] на окрашенных срезах. В год образуется
широкий светлоокрашенный и узкий темноокрашенный слои. Возраст (в годах)
равен числу годовых слоев + 1 (см. также гл. 4 раздел 4.4 •).
Antilocapra americana — вилорог
Рога
Особь с рогами, вершины которых выше ушей — взрослый самец (Larson,
Taber, 1980).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей в первые три года жизни (см. табл. 3.3).
Постоянные щечные зубы цилиндрические, корни или не развиваются, или
развиваются в позднем возрасте, коронка долго растет в высоту, компенсируя
стачивание, и потому высота коронки является ненадежным показателем возрас-
та. В молодом зубе лунки (см. гл. 3 раздел 3.4.6.3) (= «зубные чашки») очень
глубокие, по мере стачивания зуба глубина их уменьшается, и они исчезают. Это
может служить ориентировочным показателем возраста. С четырех лет исчезают
обе лунки в тр задняя лунка в т3, с пяти лет — передняя лунка в ш2 и задняя
лунка в р4 (третьем щечном зубе), в возрасте шести лет уже может не быть также
двух задних лунок в р4 и обеих лунок в m, (Dow, Wright, 1962).
Lubinski (2001) предложил оценивать стертость каждого зуба в баллах и вы-
делять возрастные группы по числу баллов (см. гл. 3 раздел 3.4.6.3).
Глава 14. Парнокопытные
211
Годовые слои видны в цементе на окрашенных срезах ip возраст (в годах)
равен числу слоев +1 (см. также гл. 4 раздел 4.4 •).
Bison bison — бизон
Размеры
По весу тела без шкуры можно отличить сеголетков и полуторагодовалых;
начиная с 2,5 лет пределы варьирования веса особей разных годовых классов
перекрываются. Несмотря на это Novakowski (1965) дает таблицу значений для
веса особей разных годовых классов до 18 лет.
Рога
Не пригодны для оценки возраста, поскольку годовые кольца трудно разли-
чить (Fuller, 1959).
Череп (Обозначения см. рис. 2.1).
Самцов 5,5 лет и старше от более молодых можно отличить по заросшим
швам С и IF. Слияние черепных швов у самок не является надежным критерием
возраста (Shackleton et al., 1975).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе трех лет (см. табл. 3.3). Темп проре-
зания зубов бизонов из разных районов совпадает до 2,5 лет, в более позднем
возрасте межпопуляционные различия возраста прорезания одних и тех же зу-
бов могут достигать 12 мес. (Shackleton et al., 1975).
Frison, Reher (1970) дают следующую схему определения возраста:
0,5 г. — резцы и предкоренные молочные, Ш] прорезан, но не сточен;
1,5 г. — i( прорезается, остальные молочные, сточены, видны их корни, т(
умеренно сточен, ш2 не сточен;
2,5 г. — ij прорезан и стачивается, i2 в стадии прорезания, остальные молочные
резцы сильно сточены, р2 и р3 (первые два щечных зуба) в стадии прорезания, dp4
сильно стерт, Ш], т2 умеренно стерты, у т3 прорезались первые две доли, но не третья;
3,5 г. — ip i2 стачиваются, i в стадии прорезания, р2, р3 умеренно стерты, р4
частично или полностью прорезан, третья доля т3 прорезана, но не стерта;
4,5 г. —- все зубы постоянные, вторая и третья доли т3 еще не стерты;
5,5-9,5 л. — зубы стерты, эмаль видна по краю зубов, лунки еще видны;
10,5-13,5 л. — зубы сильно стерты, лунок не имеет как минимум тр эмаль
видна лишь на небольшом участке, часто коронки зубов стерты целиком, видны
лишь корни (См. также гл. 3 раздел 3.4.6.3 •).
Весьма приблизительно можно определить возраст отдельных особей по про-
мерам зубов. По высоте переднего внутреннего бугра т1 (измеренной от повер-
хности стачивания до нижней границы эмали, в мм) в интервале 2-16 лет можно
оценить возраст (в годах), используя зависимость (Haynes, 1984):
212
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
возраст = (45,3 - высота) : 1.61.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Можно использовать промеры поперечной (губно-язычной) ширины поверх-
ности стачивания (мм) (Novakowski, 1965):
в 3,5 г. ширина 0,9-1,4; в 9,5 л. — 6,4-6,7;
4,5 г. — 2,0-2,6; 12,5 л. — 6,4-6,9;
5,5 л. — 3,1-4,7; 16,5 л. — 6,9-7,4;
6,5 л.—5,4-5,6; 18,5 л. — 7,6-8,3.
8,5 л. — 6,0-6,3;
Годовые слои видны в цементе резцов и щечных на аншлифах и окрашен-
ных срезах. Для оценки возраста к числу слоев нужна поправка, зависящая от
возраста прорезания используемого зуба (см. также гл. 4 раздел 4.4 •).
Novakowski (1965) прибавлял четыре года к числу слоев на протравленном
аншлифе р4, Haynes (1984) считал возможным определять возраст в годах с
точностью ±2 г. по числу слоев цемента в межкорневой подушке Ш] на про-
травленных аншлифах.
Bison bonasus — зубр
Размеры тела
Вес тела и длина тела (измеренная от кончика носа до основания хвоста по
прямой) могут быть приблизительным критерием возраста в течение первых 5
лет у самок и 7 лет у самцов. У самок до 60 мес.:
(возраст, мес.) = (вес тела, кг - 55,647) : 6,157;
In (возраст, мес. + 7,751) = (длина тела, см) : 54,480.
У самцов до 84 мес.:
(возраст, мес.) = (вес тела, кг - 53,254) : 6,956;
In (возраст, мес. + 5,063) = (длина тела, см) : 59,321.
(Krasinska, Krasinski, 2002).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Рога
У телят в возрасте двух мес. длина рога 2 см. и до 12 мес. она увеличива-
ется примерно на 5 см. каждые три месяца. В 6 мес. рога прямые, а в 12 мес.
слегка загнуты к середине головы. В возрасте двух лет размеры рогов у сам-
цов больше, чем у самок, у последних они больше загнуты к середине. У
молодых зубров поверхность рога матовая, у старших — блестящая. Длина
рога по внешней кривизне около 10 см в возрасте 6 мес., 20 см — в 12 мес.,
30 см у самцов и 27 у самок в возрасте 2 г, и 39 см у самцов и 30 см у самок
в 3 г. После 3 лет рост рогов замедляется, окончательных размеров рога дости-
гают у 7-8-летних. У самцов старше 10 лет концы рогов стерты и закруглены
(Krasinska, Krasinski, 2004).
Глава 14. Парнокопытные
213
Череп
Череп (включая нижнюю челюсть) достигает размеров, характерных для взрос-
лых особей, в 5 лет (Empel, 1962)
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе четырех лет (см. табл. 3.3). По стер-
тости резцов выделяют несколько возрастных групп: в и i2 поверхность стачи-
вания выглядит тонкой линией у самцов и самок в 3—4 г, толстой линией — в 4—
5 лет, приобретает поперечно-овальную форму в 5-6 лет у самцов и 6-7 лет у
самок, губно-язычная (поперечная) ширина овала увеличивается в 7—9 лет у сам-
цов и 8-11 лет у самок, форма становится треугольной со сглаженными углами
у самцов в возрасте 10-11, самок — 14-15 лет, и вытянутой в губно-язычном
направлении в 14-16 лет у самцов и 18-19 у самок (Wegrzyn et al., 1994). (См.
также рис. 3.1 и гл. 3 раздел 3.4.6.3 •).
Годовые слои видны в цементе резцов и коренных на аншлифах и окрашен-
ных срезах. Подсчету годовых слоев мешают дополнительные линии. Возраст (в
годах) равен числу слоев в ij + 2 (Klevezal, Pucek, 1987) (см. также гл. 4 разделы
4.4 • и 4. 4 ••).
Позвоночник
Срастание эпифизов позвонков всех участков позвоночника начинается у самцов и
самок в возрасте шести лет и заканчивается в возрасте восьми лет (Roskosz, 1962).
Кости конечностей
В длинных костях эпифизы срастаются с диафизами после 6 лет и полностью
сливаются после восьми лет (Krysiak, Swiezynski, 1967).
Capra — горные козлы
Рога
Число колец на рогах С. aegagrus строго соответствует возрасту, хотя с 15
лет годовые приросты становятся очень узкими (Couturier, 1961) (см. рис. 5.1).
У С. ibex у самцов годовые кольца на рогах так хорошо выражены, что можно
определять возраст у живых животных, у самок годовые приросты после третье-
го малы, что не дает возможности определять возраст живых самок. У мертвых
особей можно по рогам определять возраст и самцов, и самок. У самок с 8-го,
а у самцов с 12-го года годовые приросты становятся очень узкими («миллимет-
ровые кольца» у самок и «сантиметровые» кольца у самцов). Если в процессе
снашивания рогов первые, широкие, кольца видны плохо, можно подсчитать эти
узкие и прибавить 7 для самок и 11 для самцов (Onderscheka, Hartl, 1989).
На рогах самцов С. sibirica годовые кольца четкие, ширина сегментов с воз-
растом уменьшается, но даже у 9-10-летних особей ширина сегментов 4-9 см
(Цалкин, 1950).
214
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
У обитающих во Франции С. ibex можно выделять возрастные группы (для
самцов — первые 4 годовые класса и >5 лет; для самок — годовалые и >2 г.) по
размерам тела, расстоянию между кончиками рогов, окружности основания рога.
Однако полученные показатели годятся только для оценки возраста особей дан-
ной популяции (Michallet et al., 1994).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе 3—4 лет (см. табл. 3.3). Стачивание,
главным образом резцов, у С. aegagrus становится очевидным с 8 лет, с 15 лет
зубы сильно стерты, в 20 лет коронки резцов сточены полностью, коронки щеч-
ных зубов — сильно редуцированы (Couturier, 1961). Фотографии нижнего щеч-
ного зубного ряда разных возрастных групп (до 20 лет) С. ibex даны у Habermehl
(1985). (См. также гл. 3 раздел 3.4.6.3 •).
Годовые слои считают в цементе первого резца, возраст (в годах) равен
числу слоев + 1 (см. также гл. 4 раздел 4.4 •) (Habermehl, 1985).
Кости конечностей
У испанских С. pyrenaica дистальный эпифиз плечевой кости полностью сра-
стается с диафизом уже у годовалых самок, а у самцов в этом возрасте еще
видна бороздка. Дистальный эпифиз бедренной кости не сращен с диафизом в
возрасте 1—2 лет, начинает срастаться на 3-м году у самок и 4-м году у самцов и
полностью сращен на 5—7-м году у самок и с 7 лет у самцов. Проксимальный
эпифиз начинает срастаться на 4-7-м году у самок и окончательно сращен на 5-
8-м году у самок и в возрасте 7-9 лет у самцов. Метод определения возраста по
окостенению эпифизов этих и других костей конечностей уступает по разрешаю-
щей способности всем другим методам (Serrano et al., 2004).
Хрусталик
Для С. pyrenaica в интервале от 3 мес. до 19 лет связь между возрастом (в
мес.) и весом хрусталика (в мг) описывается уравнением:
1g (возраст) = (вес одного хрусталика - 79,36) : 259,94.
Начиная с двух лет не удается выделять годовые классы на 95% доверительном
уровне. Метод полезен для оценки в месяцах возраста сеголетков, в остальных
случаях лучше использовать стертость зубов и кольца на рогах (Vigal, Machordom,
1988). (Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Capreolus capreolus — косуля
Внешний вид
Косули родятся пятнистыми, пятна у сеголетков сохраняется до августа (Смир-
нов, 1979).
У самцов европейской косули в мае-сентябре у 2-летних имеется белое пятно
над носом и черное или черно-бурое на лбу, у 3—4-летних белое пятно занимает
Глава 14. Парнокопытные
215
почти половину морды, у пятилетних оно седеет и расплывается по лбу, в 6-8 лет
лоб и нос седые, в 9 лет вся голова седая (Дык, 1965).
У сибирской косули самцы сеголетки и самки всех возрастов имеют бурый
лоб и спинку носа, рыжеватые или желтоватые брови; у большинства самцов
старше года лоб и спинка носа темно-пепельные или желтовато-серые, брови
бледно-серые или желтовато-серые (Смирнов, 1977).
Рога — самцы (самки рогов не имеют).
У сибирской косули в возрасте от года до двух лет вершины рогов редко
выступают за кончики ушей, они без отростков и без розетки; в 2-7 лет вершины
рогов выше ушей с отростками и перлами розетки; наиболее мощные рога в
возрасте 3—5 лет; у особей старше 7 лет они становятся непропорционально, по
сравнению с пеньками, тонкими, перлы мельчают (Смирнов, 1977, 1979).
Для европейской косули Habermehl (1985) приводит следующие данные по
изменению толщины пенька рога с возрастом:
годовики — 8-10 мм., 2 г. — 13-14 мм, 3 г. — 16-17 мм, 4-5 л. — 18-20 мм,
после 10 лет — 25-27 мм.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе года (см. табл. 3.3). Сеголетки в пер-
вую зиму имеют молочный (трехбугорчатый) последний предкоренной (третий
щечный зуб), последний коренной может отсутствовать. У годовалых зимой все
зубы постоянные, нет обнажений дентина на внутренних буграх последнего пред-
коренного, стертость т3 незначительна, если вообще имеется. В следующих го-
довых классах стертость всех зубов увеличивается. Cicognani, Monti (2000) по-
лагают, что по этим признакам в выборках, взятых охотниками, можно надежно
различать первые два годовые класса и группу старших, последнюю по нали-
чию обнажений дентина на внутренних буграх последнего предкоренного.
Описание годовых классов по степени стертости и рисунки до семи лет есть
в работах Aitken (1975) и Смирнова (1977). Koubek, Hrabe (1988), взяв за основу
схему Aitken (1975), добавили описание возрастных групп 9-12 лет. Habermehl
(1985) приводит фотографии нижнего зубного ряда годовых классов вплоть до
возрастной группы «11 лет и старше». Однако Aitken (1975) отмечает, что после
4-5 лет можно говорить лишь о картине, характерной для большинства особей
данного годового класса. (См. также см. гл. 3 раздел 3.4.6.3 •).
Ashby, Henry (1979) в выборках, взятых в мае-августе (самцы) и в ноябре-
феврале (самки) выделяли возрастные группы следующим образом. У особей в
возрасте один год зубная система полная и зубы мало сточены; в возрасте 10 лет
(предельный возраст) зубы максимально сточены; остальные группы выделяли
исходя из предположения, что стачивание в течение всего периода было равно-
мерным. Сравнение результатов такого визуального определения возраста с оцен-
ками возраста по числу годовых слоев и по высоте коронки т, подтвердило его
надежность.
216
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Высота коронки щечной стенки т, (определена как средняя из четырех про-
меров от линии десны высоты двух бугров правого и левого зубов) линейно
уменьшается с 1,5 до 6 лет (Ashby, Henry, 1979, Англия):
(возраст, годы) = (7,854 — высота коронки шр мм) : 0,881.
Большой разброс значений в каждом классе делает оценку возраста сугубо
приблизительной.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
С. Stubbe (1989) дает номограммы изменений с возрастом высоты коронки
п^ и средней высоты коронки всех нижних щечных зубов и диаметра основания
рога. По его мнению, по таким номограммам можно оценить возраст самцов
точнее, чем по степени стертости.
Прижизненно по прорезанию и степени стертости зубов нижней челюсти
можно (подтверждено на меченых особях во Франции) выделять три группы:
(1) моложе года (зубы молочные, о чем свидетельствует трехлопастной тре-
тий предкоренной), (2) 1-6 лет (все зубы постоянные, третий предкоренной1
двухлопастной, бугорки предкоренных острые или тупые с обнажением ден-
тина в виде пятен), (3) 7 лет и старше (щечные зубы стертые, плоские, у
особей старше 10 лет резцы или коренные могут выпадать) (Van Laere et al.,
1989).
Была попытка выделять возрастные группы по размеру полости пульпы по-
стоянных зубов (см. гл. 3 раздел 3.3.6). Метод позволял отделять годовиков от
двухгодовалых, но не получил распространения.
Годовые слои исследуют на окрашенных срезах (реже на шлифах) в
цементе резцов и коренных. Возраст (в годах) равен числу слоев в ц , т, 2
или числу слоев в i3, р, т3 +1 (см. также гл. 4 раздел 4.4 •). По мнению
Ashby, Henry (1979) слои лучше считать на окрашенных срезах i чем на
аншлифах шр
Определение возраста косуль из многих районов вызывает затруднение из-за
наличия дополнительных слоев (см. гл. 4 • и гл. 4 раздел 4.6.6).
Есть опыт прижизненного удаления нижнего клыка для подсчета слоев це-
мента (Блажис, 1971).
Кости конечностей
Эпифизарные хрящи дистальной части плюсневых (метатарзальных) и пяст-
ных (метакарпальных) костей исчезают в возрасте 14-16 мес. Визуальное иссле-
дование очищенных от кожи и мягких тканей пястных костей позволяет с октяб-
ря по февраль отделить сеголетков от особей старше года так же точно, как и
исследование состояния зубов (Navarre, 1993).
Хрусталик
Для приблизительной оценки возраста можно воспользоваться таблицей
14.2.
1 Третий щечный зуб.
Глава 14. Парнокопытные
217
Таблица 14.2. Вес сухого хрусталика (мг, средняя и пределы) у косуль,
Capreolus capreolus, разного возраста.
Возраст Забайкалье, октябрь- декабрь (Смирнов, 1977) Австрия (Maringgele, 1979)
<0,5 г. <190 п=14
0,5 г. 175(152-196) п=9
1 г. 223-233 п=4
1.5 г. 240 (201-278) п=18 234-277 п=4
2 г. 303-330 п=2
2,5 г. 272 (247-298) п=9
3 г. 301-302 п=3
3-4 г. 332 (302-408) п=14 314-340 п=4
Capricornis crispus — японский сероу
Приводимые ниже данные взяты из работ Miura (1985); Miura, Yasui (1985).
Рога
Возраст можно определять по числу колец на рогах самцов и самок. В пер-
вую зиму вместо кольца образуется утолщение. Первое четкое кольцо образует-
ся во вторую зиму, когда животному 1,5 г. Годовые кольца отличаются от допол-
нительных четким краем по всей окружности кольца (у дополнительных края
сглажены). У старых кончики могут быть сношены, но не было случаев, чтобы
снашивание затрагивало кольцо второй зимы.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе 34 мес. (см. табл. 3.3). По степени
стертости можно выделить 8 возрастных групп:
2,5 г. — зубы не сточены;
3,5 г. — есть легкая стертость р3 и р4 (второго и третьего щечных зубов);
4,5 г. — задняя лунка (см. гл. 3 раздел 3.4.6.3) на р4 стерта не полностью;
6,5 лет—задняя лунка на р4 стерта полностью;
9,5 лет — стерта полностью задняя лунка в тр
11—13 лет — стерта полностью передняя лунка в Шр
15 лет -— в т2 и т3 стерты по одной лунке;
17-19 л. — в т2 и т3 стерты по крайней мере две лунки.
Годовые слои можно считать в цементе на окрашенных срезах гар первая
темная полоса образуется в первую зиму жизни (см. гл. 4 • и гл. 4 раздел 4.4 •).
218
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Cervus dama (=Dama dama) — лань
Рога — самцы (самки рогов не имеют).
Первые рога появляются на втором году жизни, позже индивидуальные вари-
ации, по мнению Chaplin, White (1969), делают определение возраста по рогам
ненадежным. Однако рога пригодны для определения приблизительного и отно-
сительного возраста. Habermehl (1985), суммируя данные немецких исследова-
телей, пишет, что на третьем году жизни появляются два отростка и заметное
расширение верхушки рога. На четвертом году «лопата» рога уже хорошо за-
метна. Далее с возрастом «лопата» расширяется, диаметр пенька рога (в середи-
не его высоты) увеличивается от 24-32 мм на четвертом году до 39-46 мм на
13-14-м году. Вес рога увеличивается от менее чем 0,6 кг на третьем году жизни
до 2 кг на 7-8-м году. Максимальный вес рогов, зарегистрированный у ланей в
Германии, 3,5-4 кг (Ueckermann, Hansen ,1983).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей моложе двух лет (см. табл. 3.3). Позже воз-
растные группы можно выделять по степени стертости: см. гл. 3 раздел 3.4.6.3 •.
Описание и таблицы приведены у Chaplin, White (1969), таблица и рисунки у
Ueckermann, Hansen (1983), Habermehl (1985).
Brown, Chapman (1990) предложили и опробовали довольно сложную балльную
систему оценки степени стертости для выделения возрастных групп (см. гл. 3 раздел
3.4.6.3) и получения связи возраста и стертости, характеризуемой суммой баллов.
Годовые слои. Возраст (в годах) равен числу слоев вторичного дентина в
коронке на продольных аншлифах if+l (если первый слой начал стачиваться,
число слоев отражает минимальный возраст) или числу слоев цемента на шли-
фах m] (Ueckermann, 1978).
Хрусталик
Из 31 особи моложе 6 мес. 29 имели вес хрусталика меньше 350 мг (Stubbe
et al., 1995).
Cervus elaphus — благородный олень
Внешний вид
По внешнему виду можно различать сеголетков, особей второго лета жизни
и взрослых (рис. 14.3).
Околохвостовое «зеркало» остроконечное у особей до 5 лет, у старых (10 и
более лет) — прямоугольное, у средневозрастных (5-8 лет) — переходного типа
(Шостак, 1988).
Рога — самцы (самки рогов не имеют).
Вес рога увеличивается с 1,2-1,4 кг у особей в возрасте двух лет до 2,4-3 кг
у особей в возрасте четырех лет, 3,4-4,2 кг — в 6 лет, достигая 4,9-5,5 кг в 10-
Глава 14. Парнокопытные
219
б
Рис. 14.3. Самки и самцы благородного оленя, Cervus elaphus: а—сеголетки, б—второе лето
жизни, в — взрослые (Шостак, 1988).
11 лет (Drechsler, по Habermehl, 1985; Schreiber, 1993). (Другие возможности
оценки возраста по рогам см. в гл. 5 раздел 5.2).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до 2,5 лет (см. табл. 3.3): до 12 мес. резцы,
клыки и предкоренные молочные, появляется т,; в 12-24 мес. предкоренные
еще молочные, появляются постоянные резцы, т, и т2, постоянный верхний
220
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
клык; в 24-36 мес. заканчивается смена предкоренных (в начале этого периода),
появляется т3, его третий бугор даже в конце этого периода белый и без потерто-
сти эмали (Quimby, Gaab, 1957; Mitchell, 1967). Старшие возрастные группы
выделяются по степени стертости, учитывая стачивание эмали, обнажение денти-
на и характер лунок (рис 14.4). См. гл. 3 раздел 3.4.6.3 •. Описание и рисунки
степени стертости зубов разных возрастных групп европейских оленей есть у
Lowe (1967), Habermehl (1985), Шостака (1988), северо-американских — у
Quimby, Gaab (1957).
Lowe (1967) определял возраст оленей до восьми лет (с ошибкой не более
года) по такой зависимости:
(возраст, мес.) = 11,7076 Н/М + 1,88683LA - 88,76;
(Н/М — высота мандибулы под задним гребнем т, деленная на минимальную
высоту коронки п^ между буграми со щечной стороны; LA—расстояние между
задним краем альвеолы т3 и задним краем ангулярного отростка).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
По состоянию верхних постоянных клыков северо-американских оленей Greer,
Yeager (1967) выделяют возрастные группы:
1,5 г. — коронки прорезаны на четверть у самцов и от четверти до половины
у самок, корень сформирован не полностью у самцов, но иногда полностью у
самок, полость пульпы занимает 75-95% зуба у самцов — 70-90% у самок;
2,5 г. — коронки прорезаны на половину, апикальное отверстие корня откры-
то у самцов и полностью или частично закрыто у самок, полость пульпы занима-
ет 30-70% у самцов и почти заполнена у самок;
3,5 г. — коронки прорезаны более чем наполовину, апикальное отверстие
частично или полностью закрыто, полость пульпы занимает менее 30% у самцов;
к 6 г. — коронка прорезана полностью.
Стачивание начинается в 2,5 г у самцов и немного раньше у самок, когда
прорезание коронки еще не закончено.
У большинства самцов и самок европейских оленей в возрасте 2—4 лет вер-
хние клыки не сточены, но у отдельных особей они были не сточены и в возрасте
6-7 лет. Сильное стачивание, когда коронка сточена наполовину, в большинстве
случаев наблюдалось в 10-14 лет у самцов и в 9-20 у самок, но менее сильное
можно было видеть у многих самцов в возрасте 7-14 лет и самок в возрасте 8-
19 лет (d’Errico, Vanhaeren, 2002).
Годовые слои считают в цементе на окрашенных срезах и аншлифах. Возраст
(в годах) равен числу слоев в i.+0-l, в mI 2 +1, в р, т3+2 (см. гл. 4 раздел 4.4 •).
Подсчет слоев в межкорневой цементной подушке на аншлифах mp Mitchell
(1967) признал методом надежным, aLowe (1967) - не надежным. Quere, Pascal
(1983) отмечали, что на окрашенных срезах i] слои выявляются лучше, чем на
шлифах коренных.
При определении возраста по числу слоев вторичного дентина на продоль-
ных аншлифах i( к числу слоев нужно прибавить 2-3 (по оценкам разных авто-
ров, см. Habermehl, 1985); если первый слой начал стачиваться, число слоев
отражает минимальный возможный возраст.
Глава 14. Парнокопытные
221
Рис. 14.4. Рисунки жевательной поверхности (a-в) и всего нижнего зубного ряда (г-е) северо-
американского благородного оленя, Cervus elaphus canadensis, в возрасте трех (а), четырех (б),
пяти (в), семи (г), девяти (д) и пятнадцати (е) лет (по Quimby, Gaab, 1957).
222
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Кости конечностей
По размерам, состоянию эпифизов и скульптуре (бугристости) дистальной
части лучевой и локтевой костей североамериканских оленей Knight (1966) вы-
делял телят, годовиков и двухгодовалых; эти показатели в старших возрастных
группах перекрываются. У сеголетков и годовалых особей эпифизы не сращены
с диафизами, между ними видна щель или трещина. В лучевой кости диафиз не
сращен с эпифизом у половины двухлетних самок, всех двухлетних самцов;
срастание отмечено у всех самок старше двух лет и самцов старше четырех лет.
В локтевой кости срастание имеет место у всех самок старше трех лет и самцов
старше четырех лет.
Хрусталик
По предварительным данным Maringgele (1978/1979) для европейских оленей
вес одного хрусталика (мг) в возрасте 5-11 мес. — 220-341, 16-18 мес. — 595,
в 3 г. — 551-675, 4 г. — 634, 681, 5 лет — 715, 855, 6-9 лет — 629, 642, 13-14
лет — 759 мг.
Cervus nippon — пятнистый олень
Рога — самцы (самки рогов не имеют).
Число отростков по одному на каждом роге у годовалых, у двухгодовалых -
чаще по 3 (от 2 до 4), у старших — чаще всего по 4 (от 3 до 4). Длина (вдоль
рога, до самой верхней точки) менее 100 мм у годовалых, в среднем 300 мм у
двухгодовалых и в среднем 400 мм у старших (Zejda, Babicka, 1983).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до 2,5 лет (см. табл. 3.3). Позже возрастные
группы выделяют по стертости — см. гл. 3 раздел 3.4.6.3 •. Описание стертости
резцов и щечных зубов оленей разных возрастных групп и рисунки дают Миро-
любов, Рященко (1948), Habermehl (1985), Ohtaishi (1980).
Ueckermann, Scholz (1971) пришли к выводу, что для определения возра-
ста пятнистых оленей можно использовать таблицы степени стертости зубов
лани.
Приблизительно возраст можно оценить по высоте коронки коренных зубов
(рис. 14.5) и по форме поверхности стачивания она узко-овальная в возрасте
2,5-4 лет у самцов и 3-5 лет у самок, широко-овальная в 4-6 лет у самцов и 6-
9 у самок, почти квадратная (площадь квадрата с возрастом уменьшается) в 6-
14 лет у самцов и в 8-16 у самок, треугольная у самцов в 12-21 год и самок в
12-20 лет и округлая у самок в 16-24 г. (Ohtaishi, 1980).
Hrabe et al. (1989) сравнили результаты определения возраста по стертости
щечных зубов и по числу годовых слоев в : полное совпадение было в 50%
случаев, в 85% ошибка не превышала года. Они пришли к выводу, что более
надежно выделять возрастные группы не по степени стертости, а по промерам
Глава 14. Парнокопытные
223
Рис. 14.5. Изменение с возрастом высоты ко-
ронки нижних коренных зубов пятнистого оле-
ня, Cervus nippon. Высота измерена как рас-
стояние от вершины заднего бугра пц и и
переднего бугра гл, до шейки зуба с язычной
стороны (по Koike, Ohtaishi (1985), с измене-
ниями).
нижней челюсти и коронки резца по классификационным функциям, получен-
ным методом дискриминантного анализа (см. гл. 1 раздел 1.2.3).
Т (1 г.) = 5,89(A) + 10,64(B) - 615,08;
Т (2-3 г.) = 6,33(A) + 9,92(B) - 693,86;
Т (4—5 л.) = 6,52(A) + 9,28(B) - 724,24;
Т (6 л. и более) = 6,55(A) + 7,37(B) - 714,15.
(А — длина мандибулы от переднего конца альвеол резцов до конца сустав-
ного отростка (позицию промера см. рис. 2.3), В — высота коронки первого
резца со щечной стороны).
Верно (в соответствии с числом годовых слоев в цементе) были определены
76% особей, 21% попали в соседнюю группу. Авторы не указывают, какая вы-
борка была использована для этой проверки (см. гл. 1 раздел 1.2.3 •).
Годовые слои считают в цементе на окрашенных срезах резцов, шлифах и
аншлифах коренных, а также во вторичном дентине резцов. Возраст (в годах)
224
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
равен числу слоев в цементе т'р или в i( + 1 (см. также гл. 4 раздел 4.4 •), числу
слоев вторичного дентина на продольных аншлифах i,-*-l (до начала стачивания
первого слоя) (см. гл. 4 •).
НгаЬё et al. (1989) сравнили число слоев цемента на окрашенных срезах i и в
цементной подушке на аншлифах Расхождение не более чем на один слой
было найдено в 75% случаев. При использовании межкорневой цементной по-
душки обычно трудно найти слой первой зимы, который очень тонок и виден
лишь в центральной части (Koike, Ohtaishi, 1985).
Хрусталик
Связь между возрастом (в годах) и весом хрусталика (в мг) в интервале от
сеголетка до 9 лет описывается уравнением:
1g (возраст + 1) = (вес одного хрусталика - 266,46) : 390,38.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Перекрывания значений веса в ноябре-декабре нет только между группами
«телята» и «годовалые» (Feldhamer, Chapman, 1980).
Giraffa Camelopardalis — жираф
Приводимые ниже данные взяты из работы Hall-Martin (1976).
Зубы
Прорезание и стачивание. По прорезанию (см. табл. 3.3) и стертости зу-
бов можно выделять годовые классы до 15 лет. Несмотря на вариабельность
степени стертости у особей старшего возраста, ошибка при определении не
превышает двух лет (за истинные были принятые оценки возраста по годовым
слоям).
Связь между возрастом (в годах) и высотой коронки т1 (в мм, средняя
максимальной высоты переднего и заднего бугров язычной стороны правого и
левого зубов) до возраста 20 лет описывается уравнением:
возраст = (20,7986 - высота коронки ш1) : 0,9558.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Годовые слои видны в цементе на окрашенных продольных срезах ш1. Не-
смотря на наличие двойных и дополнительных, годовые слои цемента выделить
можно. Возраст (в годах) равен числу слоев +1.
Хрусталик
По весу хрусталика можно определять возраст неполовозрелых особей, у
взрослых — большой разброс значений, хотя отмечено практически линейное
увеличение веса хрусталика с 17 мес. до 12 лет. Связь между возрастом (в
годах, определен по слоям цемента) и весом хрусталика (в мг) на этом возраст-
ном интервале описывается уравнением:
возраст = 0,0199 • вес одного хрусталика - 10,9.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Глава 14. Парнокопытные
225
Nemorhaedus goral — горал
Приводимые ниже данные взяты из работ: Бромлей (1956); Соломкина, Хох-
ряков (1993).
Рога
Длина рогов (по лобной поверхности) 10-20 мм в возрасте 3 мес., 100-160
мм — в 12 мес., 150-180 мм — в 3 г. и 180-220 мм в 14-15 лет. Число колец на
рогах увеличивается с возрастом, но они не годовые: 2-4 кольца в возрасте 12
мес., 9-11 возрасте 3 лет, 12-17 в 5 лет и 21-24 кольца в возрасте 14-15 лет.
Рога у молодых темно-оливкового цвета, а в возрасте 16—18 мес. ювениль-
ный рог меняется на постоянный, становится чисто черным.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до трех лет (см. табл. 3.3). У горала в возрас-
те трех лет все зубы постоянные, но не стерты, зубы заметно стерты в 8 лет и
щечные стерты почти до десен в 14-15 лет. (См. также гл. 3 раздел 3.4.6.3 •).
Годовые слои видны в цементе на окрашенных срезах резцов и предкорен-
ных, считать их трудно из-за многочисленных дополнительных (см. гл. 4 •).
Odocoileus hemionus — чернохвостый олень
Внешний вид
Прижизненно по окраске, размерам, форме тела и поведению могут быть опоз-
наны сеголетки, годовики и старшие, самцы и самки (детальная таблица для О.
h. columbianus дана у Dasmann, Taber, 1956).
Рога — самцы (самки рогов не имеют).
У сеголетков с декабря видны шишковидные выступы, у годовиков рога прямые
не ветвистые, у взрослых с двумя или более отростками (Dasmann, Taber, 1956).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до двух лет (см. табл. 3.3). Позже возрастные
группы выделяют по степени стертости — см. гл. 3 раздел 3.4.6.3 •.
Есть опыт прижизненного получения слепков щечных зубов у не усыплен-
ных оленей с использованием арсенала средств зубного протезирования (Barnes,
Longhurst, 1960). Метод оказался точнее использовавшегося до этого прижиз-
ненного определения возраста по осмотру зубов и дал возможность получить
материал для последующего сравнения стертости зубов меченых особей при
повторном их отлове.
Годовые слои видны в цементе на окрашенных срезах i] и mr Возраст (в
годах) равен числу слоев в m( или в ц + 1 (см. гл. 4 • и гл. 4 раздел 4.4 •).
226
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 14.3. Возраст (мес.), при котором активный рост в длину разных
костей чернохвостого оленя, Odocoileus hemionus, прекращается, но линия
слияния эпифиза с диафизом еще видна (Lewall, Cowan, 1963).
Кость Проксимальный эпифиз Дистальный эпифиз
Плечевая 52/35-60* 14/17
Лучевая 14/14 34/35-60
Локтевая 34/35 52/35-60
Бедренная 24/29 52/35
Б. берцовая 34/35-60 15/14
Фаланги 14/ 14
Пяточная 27/29-35
Плюсна 29/29-35
Пясть 29/29
* самцы / самки
Кости конечностей
По рентгеновским снимкам очищенных (вывариванием или с помощью ли-
чинок жуков-кожеедов — сем. Dermestidae) костей можно определять возраст
до 72 мес. с разумной точностью по окостенению эпифизов костей конечностей
(табл. 14.3).
Если есть возможность использовать лишь несколько костей, рекомендуется
брать проксимальный эпифиз плечевой и дистальный лучевой костей.
Хрусталик
У О. h. columbianus до 100 мес. связь между возрастом (в мес.) и весом
хрусталика (в мг) описывается уравнением:
возраст = {100 : [61,6584 - 21,8917 • 1g (вес одного хрусталика)]} - 6;
(см. гл. 6 раздел 6.4 ••).
Метод дает более точные, чем прорезание зубов, оценки возраста особей до
12 мес. При возрасте 18 мес. 95% доверительный интервал 14-24 мес. (Connolly
et al., 1969а).
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Odocoileus virginianus — белохвостый олень
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может
служить показателем возраста особей до 1,5 лет (см. табл. 3.3). Далее возрас-
тные группы выделяют по степени стертости — см. гл. 3 раздел 3.4.6.3 •. Де-
тальное описание характера стертости и рисунки по возрастным группам дает
Severinghaus (1949).
Годовые слои — в цементе, лучше видны на окрашенных срезах ip чем на
аншлифах tnr Возраст (в годах) равен числу слоев (см. гл. 4 • и гл. 4 раздел 4.4 •).
Глава 14. Парнокопытные
227
Таблица 14.4. Определение возрастных групп снежной козы, Oreamnos
americanus, с ноября по май (Smith, 1988).
Ростральная длина* (мм) Длина бороды (мм) Длина рога** (мм) ДР:ДУ***
Сеголетки (зимой) <120 <90 45-90 0,71 (с осени до ранней весны)
Годовики 130-155 Около 100 125-165 (осенью) 1,3-1,5
Двухгодовалые* * * * 159-171 115-135 165-219 1,66
Взрослые 171 222 155-180 205-260
*от кончика носа до переднего угла глаза;
** по наружной кривизне;
*** отношение длины рога к длине уха;
**** переходная группа, наиболее трудно различимая. В двух последних группах толщина и
кривизна рогов позволяют отличить самцов от самок.
Есть опыт прижизненного удаления ц для подсчета слоев цемента (DeYoung,
1989).
Хрусталик
Вес одного хрусталика (мг) в возрасте 4-6 мес. — 183-218, в 6-8 мес. —
226-275, в 8-10 мес. — 257-302, в 10-12 мес. — 286-322 и в 12-18 мес. —
308-398 (Brokx, 1972, Венесуэла).
Oreamnos americanus — снежная коза
Внешний вид
По размерам частей головы, рогов и бороды можно выделить несколько воз-
растных групп (табл. 14.4).
Рога
Кольца на рогах годовые, но точность оценок возраста по числу этих колец
уменьшается после 5-7 лет; первое четкое годовое кольцо образуется на вторую
зиму (Stevens, Houston, 1989).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до четырех лет (см. табл. 3.3). Проверка этого
метода на материале от особей известного возраста показала, что в 12 из 15
случаев возраст был определен верно, причем верной была оценка для всех
шести годовиков и двухгодовалых, ошибки были в оценках возраста трех- и
четырехгодовалых (Stevens, Houston, 1989). Позже возрастные группы можно
выделять по стертости зубов — см. гл. 3 раздел 3.4.6.3 •.
Годовые слои — видны в цементе на окрашенных срезах i,. Возраст (в го-
дах) равен числу слоев + 1 (см. гл. 4 •).
228
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Ovibos moschatus — овцебык
Внешний вид (Olesen, Thing, 1989).
Телята: рогов не видно, зимой они закрыты белой шерстью на лбу; нет свет-
лоокрашенного «седла» на спине, которое есть у годовиков и старших; остевые
волосы не развиты.
Годовики: рога короткие прямые, слега наклонены вперед, длина видимой
части рога зависит от степени развития белых курчавых волос на лбу; чернова-
тые остевые волосы на туловище достигают нижней части живота, но не покры-
вают конечности.
Двухлетние: у самок рога во фронтальной плоскости проецируются вбок пря-
мо, в профиль они изогнуты вперед и слегка вверх; у самцов они толще, более
конические, проксимальные две трети — под углом 46° к поверхности головы,
их дистальная часть резко искривлена вперед; лоб покрыт белой шерстью, осно-
вания рогов не видны; на туловище остевые волосы достигают дистального сус-
тава конечностей.
Трехлетние: у самок рога достигают полной длины, тонкие, их кривизна вид-
на и анфас и в профиль; лоб еще покрыт шерстью кремового оттенка, покрыва-
ющей основания рогов; у самцов проксимальная часть рогов опущена вниз,
дистальная изогнута вверх; остатки шерсти на лбу закрывают основания рогов
дорсо-латерально таким образом, что видны в виде двух пушистых образований
в проксимальной части рогов; остевые волосы на туловище могут закрывать
половину нижней части конечностей.
У самок в 7-10 лет белая шерсть на лбу частично исчезает, обнажая основа-
ния рогов, шириной до 6-7 см; у очень старых самок остевые волосы туловища
снашиваются и становятся видны конечности кремовой окраски.
У самцов на пятое лето и осень шерсть на лбу исчезает и становится виден
массивный лобный выступ. Он достигает почти полных размеров у самцов в
возрасте пяти лет, остается только узкая, в палец шириной, оволосненная по-
лоска посередине. Лобный выступ может быть до 20 см шириной, светлого
цвета. У особей старше 8-9 лет рога снашиваются и обламываются. После 10
лет остевые волосы туловища обламываются, обнажая кремового цвета конеч-
ности.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до четырех лет (см. табл. 3.3). Позже возрас-
тные группы можно приблизительно выделить по степени стертости — см. гл. 3
раздел 3.4.6.3 •.
Годовые слои видны в цементе на окрашенных срезах и на шлифах i Слои
нечеткие, метод не надежен (Gronquist, Dinneford, 1984). На тонких шлифах в
проходящем свете слои видны в средней и верхней части корней предкоренных и
коренных зубов (Savelie, Beattie, 1983).
Глава 14. Парнокопытные
229
Ovis аттоп — горный баран
Внешний вид
Светлая лицевая маска отсутствует у ягнят и появляется и увеличивается с
возрастом. Этот признак подвержен большой индивидуальной изменчивости. Так,
у О. musimon (= О. аттоп) маска может появляться у некоторых самок уже на
втором году жизни, а у некоторых трех-четырехлетних отсутствовать, хотя у че-
тырехлетних встречается часто (Bousses, Reale, 1994). Поэтому Prien et al. (1989)
считают возможным выделять возрастные группы (у самцов: первый год, вто-
рой, 3-5-й год, 5-6-й год, 7-й год и старше; у самок: первый, второй, 5-6-й год,
7-9-й год, старше 9 лет) только по совокупности внешних признаков и особен-
ностям поведения.
Рога
Habermehl (1985) в качестве вспомогательных и сугубо ориентировочных
предлагает следующие критерии определения возраста самцов. Рога, при рас-
смотрении сбоку:
у ягнят составляют 1/6 дуги окружности по нижней поверхности рога, кончик
рога направлен вверх-назад;
у годовалых — 1/2 окружности, кончик смотрит вниз-назад;
у двухгодовалых — 2/3 окружности, кончик смотрит вниз-вперед;
у трех-четырехгодовалых—4/5 окружности, кончик смотрит вперед и вверх-
вперед;
у пятигодовалых и старше — полная окружность, кончик смотрит сначала
вверх, позже вверх-назад.
Можно определять возраст по годовым кольцам на рогах, после 5 лет кольца
становятся узкими (Habermehl, 1985).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей почти до четырех лет (см. табл. 3.3). Далее
для выделения возрастных групп можно использовать стачивание зубов — см.
гл. 3 раздел 3.4.6.3 •. Фотографии нижних челюстей разных годовых классов до
15 лет дает Habermehl (1985).
Хрусталик
Связь между возрастом (в мес.) и весом хрусталика (в сантиграммах) в
интервале от 3 мес. до 3 лет описывается уравнением (Сапожников, Усманов,
1967):
1g (возраст) = 2,30* 1g (вес одного хрусталика,) - 2.66.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
При оценке по этой формуле возраста до года ошибка равна 0,5 мес., далее —
не более 4 мес.
230
Г,А. Клевезаль. Принципы и методы определения возраста млекопитающих
Ovis canadensis (=Ovis dalli) — снежный баран
Рога
Молодые самцы имеют менее 3/4 завитка рогов, двухгодовалые и старше —
3/4 завитка и больше (Jones et al., 1954 по Larson, Taber, 1980).
Число годовых колец на рогах — точный критерий возраста самцов до 7-9
лет, позже появляются дополнительные кольца, которые нужно отличать от ос-
новных. У особей старшего возраста первый годовой сегмент может быть сто-
чен. Его исчезновение можно обнаружить по размерам годовых сегментов: если
рог не сточен, второй сегмент в два раза больше первого а далее ширина сег-
ментов постепенно уменьшается.
Число годовых колец, по крайней мере у самцов, можно считать прижизнен-
но (Geist, 1966; Hemming, 1969).
Возраст старых (12-13 лет, по данным мечения в природе) самок по кольцам
на рогах оценили 12 операторов и все 12 его занизили (Hoefs, Konig, 1984).
Череп. Швы (Обозначения см. рис. 2.1).
При большой индивидуальной изменчивости, в среднем у самцов в 5-6 лет
зарастают швы IF, в 6-7 лет — С, FN, в 8-9 лет — MF, MJ, в 9-10 лет — SJ, IPL
(Cowan, 1940).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей почти до четырех лет (см. табл. 3.3). Далее
возрастные группы можно выделять по степени стертости зубов — см. гл. 3
раздел 3.4.6.3 •.
Годовые слои видны на тонких шлифах и на окрашенных срезах i Возраст
(в годах) равен числу слоев + 1 (см. гл. 4 • и гл. 4 раздел 4.4 •).
Кости конечностей
В возрасте 3 г. 5-6 мес. все эпифизы длинных костей конечностей слиты с
диафизами и линии слияния не видны (Walker, 1987).
Phacochoerus aethiopicus — бородавочник
Внешний вид
У годовиков верхние клыки едва видны над губой, их длина 1-3 мм; в 24
мес. клыки выступают над краем губы на 1,5-2,3 см у самок и 2-3 см у самцов
(Mason, 1984).
Зубы
Прорезание и стачивание. Рост и смена зубов происходит не так, как у боль-
шинства других млекопитающих. Клыки меняются и т1, прорезается в возрасте от
полугода до года, резцы и предкоренные меняются в возрасте от года до двух, т22
Глава 14. Парнокопытные
231
начинают прорезаться в возрасте 9-11 мес., т3— в 17-20 мес. Коренные зубы
имеют своеобразное строение. Третьи коренные состоят в среднем из 21-22 стол-
биков дентина, окруженных эмалью и скрепленных между собой цементом. Про-
резание начинается с передних столбиков и продолжается в течение нескольких
лет параллельно со стачиванием зуба. При этом длина горизонтальной жеватель-
ной поверхности увеличивается, а высота передних столбиков оказывается ниже,
чем высота задних. Продвигающийся вперед третий коренной давит на второй и
первый при фиксированной позиции предкоренных. В результате этого давления и
стачивания первый коренной выпадает в возрасте около 3—4 лет, второй коренной
обычно сохраняется до 7-8 лет, предкоренные выпадают в то же время, что и
второй коренной или несколько позже. Продолжительность существования зу-
бов подвержена существенной индивидуальной изменчивости (Mason, 1984).
До 10 лет можно оценивать возраст по отношению длины поверхности смы-
кания т3с зубами нижней челюсти к его высоте в передней части:
(возраст, годы) = (отношение + 0,72) : 0,699.
После 10 лет зависимость величины отношения от возраста перестает быть
линейной (Spinage, Jolly, 1974).
(Об использовании линий регрессии см. гл. 1.2.2.2).
Приблизительную оценку возраста в течение первых нескольких лет жизни дает
процентное отношение длины верхнего клыка (от основания на уровне губы до кон-
чика по прямой) к ширине рыла (максимальное расстояние между губами на уровне
клыков). Уравнения регрессии, позволяющие оценивать возраст, получены отдель-
но для самцов и самок и для возрастных групп 12-36 и 36-60 мес. (Mason, 1984).
12-36 мес., самцы: возраст (мес.) = (отношение - 5,98) : 0,91;
самки: возраст (мес.)= (отношение + 9,83) : 0,92;
36-60 мес., самцы: возраст (мес.)= (отношение - 3,71): 0,59;
самки: возраст (мес.)= (отношение - 0,98) : 0,53.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Годовые слои в цементе на окрашенных срезах i( пригодны для определения
возраста взрослых особей (хотя у трети особей слои были нечеткие) (Mason, 1984).
Хрусталик
Вес хрусталика достаточно надежно выделяет два первые годовые класса, но
проще выделить эти годовые классы по состоянию зубной системы (Mason, 1984).
Связь между возрастом (в мес.) и весом хрусталика (в мг) до возраста 36 мес.
описывается уравнением:
возраст = (вес одного хрусталика - 94,7337) : 2,8264.
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Rangifer tarandus — северный олень
Зубы
Прорезание и стачивание._Последовательность прорезания зубов может слу-
жить показателем возраста особей почти до 2,5 лет (см. табл. 3.3). Позже возра-
232
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
стные группы выделяют по степени стертости (см. гл. 3 раздел 3.4.6.3 •). Под-
робное описание возрастных групп и фотографии дает Miller (1974).
Соколов (1937) приводит такое описание:
2 г. — предкоренные не достигли уровня коренных, все коренные прорезались,
нет признаков стирания на резцах, но есть на mp п^, и на передней половине т3;
3 г. — стирание заметно на всех щечных зубах, кроме р2 (первого щечного
зуба) и, возможно, i,;
4 г. — стирание заметно на первых двух резцах и всех щечных зубах;
5 л. — стирание видно на всех резцах.
Связь между возрастом (в мес.) и высотой коронки т( (измеренной по щеч-
ной стороне от прикорневой части эмали до поверхности стачивания переднего и
заднего гребней, взято среднее, мм) оленей-карибу Каминуриакской популяции
в интервале 34-181 мес. Morrison, Whitridge (1997) описывают уравнениями:
самцы: возраст = -10,20 • высота коронки + 160,25;
самки: возраст = -14,53 • высота коронки + 209,26;
для тех и других вместе: возраст = -12,56 • высота коронки + 186,97.
(Об использовании линий регрессии см. гл. 1 раздел 1.2.2.2).
Для таймырской популяции связь между возрастом (в годах) и высотой ко-
ронки т( (измеренной как и в выше описанном случае, мм) для самцов и самок
в интервале 2,5-12,5 лет:
возраст = (13,74 - высота коронки) : 0.87.
По этой зависимости можно определять возраст особей другой выборки из
той же популяции, но не особей другой популяции (Клевезаль, Соколов, 2004).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
При определении возраста отдельных особей по формуле Спинэджа (см.
Spinage (1971, 1976) в гл. 3 раздел 3.4.6.3) правильно, с точностью до года, был
определен возраст большего числа особей, чем по уравнению регрессии. Кон-
станта к, полученная для расчета по высоте щечной и язычной стенок коронки
nij была равна, соответственно, 0,79 и 0,87. Она оказалась пригодной для расче-
та возраста не только оленей другой выборки той же популяции, но и оленей
другой популяции (Клевезаль, Соколов, 2004).
Годовые слои видны в цементе на окрашенных срезах резцов и щечных
зубов. Возраст (в годах) равен числу слоев в mf или равен числу слоев в i + 0-
1. (см. гл. 4 • и 4.4 •).
Есть опыт прижизненного удаления i( и i2 для подсчета слоев цемента (Bergerud,
Russell, 1966).
Кости конечностей
К возрасту три года происходит срастание с диафизом дистальных эпифизов
большой берцовой и плечевой костей, к четырем годам — проксимального эпи-
физа бедренной кости, к пяти годам — проксимального и дистального эпифизов
локтевой кости, проксимального большой берцовой и дистального эпифизов лу-
чевой и бедренной костей, к шести годам — проксимального эпифиза плечевой
кости (Егоров, 1971).
Глава 14. Парнокопытные
233
Rupicapra rupicapra — серна
Рога
Число годовых колец является хорошим показателем возраста. Первое годо-
вое кольцо нечеткое и исчезает (сшелушивается) с возрастом. Последующие
сохраняются пожизненно (Bargagli, Lovari 1984; Perez -Barberia, Fernandez-Lopez,
1996). С шестого года годовые сегменты становятся очень узкими («миллимет-
ровые кольца»). При определении возраста можно сосчитать эти узкие кольца и
прибавить пять (Briedermann, 1989).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до трех лет (см. табл. 3.3).
По стертости можно определять возраст не старше восьми лет, далее метод не
надежен (описание годовых классов см. Perez-Barberia, 1994). См. также гл. 3
раздел 3.4.6.3 •.
Годовые слои видны в цементе на окрашенных срезах i1. Их число — надеж-
ная оценка возраста особей не старше 10 лет (Bargagli, Lovari 1984). Все же
подсчет колец на рогах проще и точнее (Perez-Barberia, Fernandez-Lopez, 1996).
Saiga tatarica — сайгак
Рога — самцы (самки рогов не имеют).
Первичный роговой чехлик темно-бурого цвета сшелушивается в течение
первых двух лет жизни и заменяется постоянным с блестящей светлой поверх-
ностью (иногда с темным концом). По этому признаку можно опознать особей
старше двух лет. Ни длина рогов, ни число колец сами по себе для определения
возраста служить не могут (Соколов, Рашек, 1961).
Череп. Швы (Обозначения см. рис. 2.1, 2.2).
Шов 5 — зарастает полностью в возрасте 2 мес., В В — ясно виден до 13
мес., в 14-20 мес. он полностью зарастает (Соколов, Рашек, 1961).
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей в первые полтора года (см. табл. 3.3).
Возраст старших определяют по степени стертости:
2 г. — цис имеют потертости только по верхнему краю, на р4 (третьем щеч-
ном зубе) исчезли лунки (см. гл. 3 раздел 3.4.6.3) или остались точками, на т,
сохраняются только задние лунки, они исчезают в начале третьего года;
3 г. — резцы и клыки сильно стерты, исчезли передние а иногда и задние
лунки на т’,, передние лунки на пц;
4 г.—резцы стерты настолько, что поверхность стачивания вместо овальной
становится треугольной, иногда исчезают задние лунки на пц и передние на пц.
234
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
5 л. — от резцов и клыков остаются только корни, исчезли лунки на т2;
6 л. — сильно изношены верхние коренные, от предкоренных (а иногда и от
ш’) остаются лишь слабо держащиеся корни;
7 л. — видимо предельный возраст в природе; от резцов, клыков и предко-
ренных (кроме р4) остаются только корни, исчезли лунки на ш2 и т3 (Соколов,
Рашек, 1961).
Годовые слои видны в цементе на окрашенных срезах i(, Возраст (в годах)
равен числу слоев + 0-1 (см гл. 4 раздел 4.4 •); слои нечеткие (Груздев, Проня-
ев, 1994).
По данным Груздева, Проняева (1994) оценки возраста по числу слоев и по
степени стертости совпадали у 60% особей, занижений по стертости было боль-
ше, чем завышений. По данным Lundervold et al. (2003) оценки возраста по чис-
лу слоев и по степени стертости хорошо согласовались друг с другом.
Sus scrofa — кабан
Зубы
Прорезание. Последовательность прорезания зубов может служить показа-
телем возраста особей до двух лет (см. таб. 3.3).
Клыки. У самцов возраст можно приблизительно оценивать по форме и ста-
чиванию клыков (рис. 14.6). До четырех лет корень нижнего клыка суживается
к коронке, у старших он по всей длине одинаковой ширины. Чем больше разли-
чия в ширине между вершиной и основанием корня клыка, тем моложе самец.
Измеряют ширину корня (на 10 мм выше апикального отверстия) и ширину ко-
ронки у нижней части поверхности стачивания. У молодых, до четырех лет, сам-
цов второй промер существенно меньше первого, к 8 годам оба промера при-
мерно равны. По отношению ширины корня к ширине коронки Brandt (1961)
выделяет возрастные группы:
2-3 г. — отношение 1,5; 6-7 л. — отношение 1,08;
3-4 г,— 1,3; 7-8 л,— 1,05;
4-5 л.— 1,2; 8 л. и старше — 1,03-1,00.
5-6 л. — 1,12;
По мнению Hell (1976) даже до пяти лет ошибка определения возраста по этим
показателям ± 1 год.
Приблизительно оценить возраст самцов можно по длине поверхности стачи-
вания нижних клыков: 10 мм в двухлетнем возрасте, 50 мм — в возрасте около
6 лет, и с возраста 8 лет максимальная длина 70 мм (Habermehl, 1985).
У самок апикальное отверстие нижних клыков шириной 11 мм в возрасте 12-
14 мес., 10 мм — в возрасте 21-24 мес., 6 мм — в 28-32 мес., 3 мм — в 36-42
мес. и 1 мм — в 48-60 мес. (Habermehl, 1985).
Резцы. У самцов и самок кабана выделяют несколько возрастных групп по
ширине апикального отверстия первых двух нижних резцов:
в возрасте 2—3 лет открыты апикальные отверстия обоих резцов, в возрас-
те 4—6 лет в первом резце отверстие закрыто, во втором — открыто, в возрасте
Глава 14. Парнокопытные
235
Рис. 14.6. Клыки самцов кабана, Sus scrofa, верхней (А) и нижней (Б) челюсти в возрасте 14-15
мес. (1), 22-24 мес. (2), 3 г. (3), 4 г. (4), 5 лет (5), 7-8 лет (6), более 10 лет (7) (Козло, Никитенко,
1967).
236
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Рис. 14.7. Нижние щечные зубы кабана, Sus scrofa, в возрасте 4 лет (1), 5 лет (2), 6-7 лет (3), 8-
9 лет (4) и 10-12 лет (5) (Козло, Никитенко, 1967).
Глава 14. Парнокопытные
237
7-9 лет апикальное отверстие закрыто и во втором резце (Iff, 1976 по Habermehl,
1985).
Для ориентировочной оценки возраста можно использовать длину плоскости
стачивания первых двух нижних резцов. Для второго резца этот показатель в 2 г. —
О мм, в 3 г. — 4-5 мм, в 4 г. — 10-12 мм, в 5 л. — 13,0-13,5 мм, в 6 л. — 14-
15 мм (Hell, 1976).
По ширине полости пульпы (в самой широкой ее части) резцов (ip i2),
отнесенной к ширине зуба, без перекрывания отличаются полуторагодова-
лые (соотношение больше 0,45) от старших (отношение меньше 0,45). (Из-
мерения были сделаны на рентгеновских снимках). Приблизительный воз-
раст до 108 мес. можно оценить по этому отношению для i, используя урав-
нение:
возраст, мес. = 2,25 - 27,65 • In (отношение,) (Saez-Royuela et al., 1989).
(Об использовании линий регрессии см. гл. 1 разделы 1.2.2.1 и 1.2.2.2).
Стачивание щечных зубов
В 3 г. начинают стираться вершины предкоренных; полностью сношены бугры
у тр исчезли складки эмали кроме отделяющей передние и задние доли; у т2
бугры стерты, но эмалевые складки заметны; задняя доля т3 проходит десневое
прорезание.
В 4-5 л. стерты вершины и в виде узкой полоски обнажен дентин на р3 и р4, в
mp т2 обе доли гладкие, есть остатки эмалевых складок, стерты бугры и начина-
ет обнажаться дентин всех долей т3.
В 6—7 л. отмечаются отчетливые полоски дентина на предкоренных, более или
менее сильное углубление жевательной поверхности долей т, и т2, бугры на т3
стерты, но границы между ними видны.
В 8 л. и старше широкие полоски дентина видны на предкоренных, на
коренных расширяется площадь дентина; высота nij уменьшена так, что по-
верхность стачивания может быть ниже десны; в т3 эмалевые складки посте-
пенно исчезают, обнажения дентина видны как отдельные пятна или занимают
всю поверхность (Козло, Никитенко, 1967; Козло, 1975; Moller, 1984) (рис.
14.7).
Годовые слои видны в цементе на окрашенных срезах i, 2 и тр они нечет-
кие. Clarke et al. (1992) делали аншлифы подушки т, 2, травленые и окрашен-
ные, у половины особей слои цемента нечеткие, но в целом их число соответ-
ствовало возрасту особей известного возраста. Возраст (в годах) равен числу
слоев в mt или числу слоев в m2+l. Hell (1976) считал слои в цементе на попе-
речных аншлифах корня i2(l см ниже альвеолы). Возраст (в годах) равен чис-
лу слоев + 1,5.
Хрусталик
В 4,5-8,5 мес. вес одного хрусталика до 177 мг (вероятность 0,91),
в 9-11 мес. — 177-208 мг (вероятность 0,76),
в 11,5-20,5 мес. — 208-254 мг (вероятность 0,8),
в 21-47 мес. — больше 254 (вероятность 0,97) (Klein et al., 1990).
238
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Tragelaphus scriptus — антилопа бушбок
Приводимые ниже данные взяты из работ Simpson, Elder (1969); Simpson (1973)
Череп
По длине черепа и нижней челюсти можно оценить возраст примерно до трех
лет.
Зубы
Прорезание и стачивание. Последовательность прорезания зубов может слу-
жить показателем возраста особей до 30 мес. (см. табл. 3.3). Позже группы
выделяют по степени стертости щечных зубов: слабая (возраст около 37 мес.),
средняя (64 мес.), сильная (114 мес.) с промежуточными градациями (у указан-
ных выше авторов даны описание и рисунки).
Годовые слои видны в цементе на аншлифах и окрашенных срезах р4 (тре-
тьем щечном зубе). Зимние линии двойные, вероятно для определения возраста
пригодны. Возраст (в годах) равен числу слоев + 2.
Хрусталик
Средний (сырой, после фиксации формалином) вес пары хрусталиков (в грам-
мах):
в возрасте 1 мес. — 0,7 (0,4-1,1); в возрасте 37 мес. — 1,7 (1,3-2,2);
6 мес. — 0,9 (0,5-1,3); 52 мес. — 1,7 (1,5-2,0);
12 мес. — 1,3 (1,0-1,7); 64 мес. — 1,9 (1,5-2,4).
22 мес. — 1,6 (1,2-2,1);
ГЛАВА 15. НЕПАРНОКОПЫТНЫЕ
Относительный возраст непарнокопытных традиционно определяют по стачи-
ванию резцов. На жевательной поверхности резцов у Equidae имеются выст-
ланные эмалью углубления — чашки, идущие внутрь зуба на разную глубину.
Например, у зебры средняя глубина чашки в i, 31 мм (Smuts, 1974).
Общую картину возрастных изменений резцов лошадиных можно видеть на
примере приведенного ниже детального описания изменения резцов у зебры. То
же происходит и у лошадей (Корневен, Лесбр, 1929): форма поверхности стачи-
вания сначала уплощенная в губно-язычном (поперечном) направлении, потом
овальная, округлая, треугольная и сплющенная с боков прямоугольная. Соответ-
ственно меняется и форма чашки. Сначала в процессе стирания чашка из удли-
ненной становится овальной, потом округлой и потом стирается полностью и
исчезает (рис. 15.1).
Кроме поверхности стачивания резцов, глубины и формы чашки, при опреде-
лении возраста используют бороздку Гальвэйна—боковую бороздку на третьем
резце, которая у лошадей появляется в 10-летнем возрасте, видна на половине
коронки в возрасте 15-16 лет; достигает верхнего края коронки в возрасте 21 г.,
начинает исчезать в 25 лет и полностью исчезает к 30 г. (Spinage, 1973). Как крите-
рий возраста используют также появление на поверхности стачивания резцов об-
нажения вторичного дентина в виде желто-коричневых пятен («зубные звезДы»).
Последний признак, считающийся не очень надежным для лошадей (Корневен и
Лесбр, 1929), оказался более надежным для определения возраста шетландских
пони и ослов: у последних зубная звезда на поверхности стачивания появляется
в возрасте 3,5-4 лет, i2 — 4—4,5 лет, i3 — 5,5-7 лет (Muylle et al., 1999).
Ceratotherium simum — белый носорог
Приводимые ниже данные взяты из работы Hillman-Smith et al. (1986)
Внешний вид
Прижизненно можно определять приблизительный возраст по длине рога
относительно длины уха (табл. 15.1).
Зубы
Прорезание. Стачивание. Типичная зубная формула: молочные — di 0/0
dcO/O dp 4/4, постоянные — i0/0 сО/О рЗ/З m3/3, dp1 (не замещается постоянным,
но может сохраняться долго после смены остальных предкоренных.
Первый постоянный предкоренной (р2) появляется в возрасте 3-4 лет, полная
смена предкоренных происходит к 8 годам, т1, появляется в возрасте около 3
лет, т22 — в 4-7 лет, т33 — в 8-16 лет.
Как и при прорезании имеет место большая индивидуальная изменчивость
стачивания, особенно нижних зубов. По мере стачивания увеличивается пло-
щадь обнажения дентина, внутренние эмалевые складки становятся мелкими и
замкнутыми и к возрасту 35-40 л. исчезают на р4 (третьем щечном зубе) и т1.
240
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Таблица 15.1. Размеры рогов белого носорога, Ceratotherium simum, в разном
возрасте
Возрастной интервал (мес.) Передний рог Длина заднего рога (см)
длина (см) окружность у основания (см)
0-2 шишка на черепе нет
2-4 небольшой вырост нет
4-9 0,25 длины уха различим
9-18 0,5 длины уха шишка на черепе
18-30 0,75-1 длины уха, 23-30 42-48 округлый вырост, 7-9
30-72 1-1,5 длины уха, 34-17/27—11* 45-60/46-54* округлый вырост, 7-16
72-120 1,5-2 длины уха, 41-57/43-52* 67-72/60* 0,2-0,5 длины уха, 15-21/15-17*
120-480 1,5-4 длины уха, 43-69/52-82* 70-79/65-68* 0,25-1,25 длины уха, 16-39/16-26*
* Самцы/самки
Высота коронки т1 (измеренная как расстояние от бифуркации корней до
выемки между буграми со щечной стороны) в 3-4 г. — в среднем 9,3 см, в 7-
9 лет — 7,9 см., в 10-15 лет — 6,4 см., в 14-20 лет — 5,3 см., в 25-32 г. — 3,1 см,
в 30-38 лет — 1,6 см.
В уравнении Spinage (см. гл. 4 раздел 4.3.6.3 )у0= 11см., п = 42 г., к 0,6 и
следовательно
возраст, годы = 42 • - высота коронки см : И)
или 1g (возраст, годы) = 1,6232 + lg(l - высота коронки см : 11): 0,6
Годовые слои видны в отраженном свете в цементе на продольных (в щеч-
но-язычном направлении) аншлифах т1 в межкорневой цементной подушке, в
год образуется одна темная и одна светлая полоса, но считать слои трудно. Воз-
раст (в годах) равен числу слоев +3.
Equus burchelli, Е. zebra — зебры
Приводимые ниже данные взяты из работ Smuts (1974), Penzhom (1982).
Зубы
Прорезание и стачивание. Характер поверхности стачивания резцов у жи-
вых Е. burchelli оценивали по отпечаткам на копировальной бумаге: квадрат
копировальной бумаги 10x10 см укрепляли на куске плотной белой бумаги
того же размера и оборотной стороной копировальной бумаги плотно прижи-
мали к верхним резцам. Поскольку твердая эмаль стачивается медленнее ден-
тина, на отпечатках хорошо видна форма поверхности стачивания зубов и фор-
ма чашек.
Возраст прорезания практически одинаков для верхних и нижних зубов.
Глава 15, Непарнокопытные
241
Рис. 15.1. Схема возрастных изменений поверхности стачивания верхних резцов у зебры Equus
burchelli (Smuts, 1974). Точками показана поверхность чашек.
242
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
3-6 мес. — di3 явно прошел альвеолярное прорезание и начинает прорезаться
сквозь десну, т1 приближается к краю альвеолы, пт2 часто виден в альвеоле;
6-9 мес. — di3 прорезан, но не стачивается, di1-2 стачиваются, т1 выходит за
край челюсти и чувствуется сквозь десну, т2 виден в альвеоле;
9-12 мес. — di3 только начинает стачиваться, т1 начинает десневое прореза-
ние, т2 приближается к краю кости;
1-1,5 г. — di3 сточен наполовину, т1 начал стачиваться, т2 прошел альвеоляр-
ное прорезание и прощупывается через десну, ш3 может быть виден в альвеоле;
1,5-2 г. — di3 стерт на три четверти, di1 — пенек, под которым в альвеоле
виден i’, m2 прорезан и начал стачиваться, т3 виден в альвеоле;
2-2,5 г. — i* заместил di1, чашка di2 почти сточена, т2 стачивается, ш3 подхо-
дит к краю альвеолы, но не прошел альвеолу, р2 и р3 (первые два щечных зуба)
видны под пеньками dp2 и dp3;
2,5-3 г. — i1 начал стачиваться, т3 прощупывается сквозь десну или начал
прорезаться, р2 и р3 заменили dp2 и dp3, с виден в альвеоле (у самцов), т3 выше
уровня альвеолы, р4 виден под обломком dp4;
3,5 г — i2 стачивается, от di3остался короткий пенек, с прорезается у самцов,
р4 заместил dp4, m3 начал стачиваться, i3 виден в альвеоле позади di3;
4 г.—i3 прорезан, но не стачивается, все предкоренные и коренные стачиваются;
5 л. — i3 стачивается, чашки i1-3 продольно-овальные (см. рис. 15.1);
6-7 л. —чашки i1*3 продольно-овальные;
8-9 л. — чашка i1 овальная, чашки i2 3 продольно-овальные;
10-11 л. — чашка i1 овальная или кругло-овальная, i2 — овальная, i3 — про-
дольно-овальная;
12-13 л. — чашка i1 круглая, i2 — кругло овальная, i3 — овальная;
14—15 л. — чашка i1 точечная или отсутствует, i2 — круглая, i3 — кругло-
овальная или круглая, поверхность i1 —равносторонний треугольник;
16-17 л. — чашки i2, i3 становятся едва заметны и исчезают, поверхность i1 -
удлиненный треугольник, i2—равносторонний треугольник;
18-20 л. — поверхность i’ — прямоугольник, i2 — удлиненный треугольник
или прямоугольник, i3 — неправильный прямоугольник.
У Е. zebra чашки в резцах сохраняются до более позднего возраста.
Годовые слои видны в цементе на тонких шлифах i1 в проходящем свете,
возраст (в годах) равен числу слоев +2,5 (Е. burchelli) или на продольных окра-
шенных срезах резцов, возраст (в годах) равен числу широких полос +3 (Е.
zebra).
Хрусталик
По весу хрусталиков можно выделить лишь несколько первых возрастных
групп: у молодых до полугода Е. burchelli вес пары высушенных хрусталиков
менее 500 мг, в 0,5-2 г. — 500-700 мг, в 2-3 г. — 700-800 мг. Позже индивиду-
альная изменчивость веса хрусталика у одновозрастных особей перекрывает
возрастные изменения.
ЗАКЛЮЧЕНИЕ
На протяжении всей истории разработки методов определения возраста зоо-
логи старались найти метод, позволяющий получить объективную оценку ка-
лендарного возраста особей. После описания годовых слоев в тканях зуба и
кости казалось, что такой метод наконец-то найден. На какой-то период все ранее
использовавшиеся методы оценки возраста отошли на второй план. Однако при
долговременном мониторинге определенных популяций исследователи предпо-
читают использовать оценки возраста по годовым слоям лишь для проверки и
корректировки выделения возрастных групп, выделенных менее трудоемкими
методами. Например, по мнению Cicognani, Monti (2000) для правильного веде-
ния охотничьего хозяйства (речь идет о косуле) важно четко определять лишь
группу годовиков. Они используют оценки возраста по годовым слоям в цемен-
те зубов для проверки надежности выделения трех возрастных групп в промыс-
ловой выборке: сеголетков (по прорезанию зубов), годовиков (по отсутствию
обнажения дентина на некоторых зубах) и старших (по наличию обнажений ден-
тина). Учитывая значительную индивидуальную и межпопуляционную изменчи-
вость большинства показателей возраста, следует признать такой подход разум-
ным. При этом, однако, необходимо помнить, что оценка возраста по годовым
слоям может иметь свою ошибку, определяемую как четкостью слоев, так и
опытом исследователя. Следовательно, по-прежнему актуальным остается тес-
тирование разных методов оценки возраста на материале от особей из природ-
ных популяций, возраст которых точно известен в результате мечения.
В соответствии со степенью интереса зоологов и охотоведов к тому или ино-
му виду методы определения возраста разных видов разработаны в разной сте-
пени. Как можно видеть во второй части книги, для одних, «популярных», видов
есть данные, позволяющие выделять возрастные группы почти всеми распрост-
раненными методами, для других же видов опубликованных данных крайне мало
и они отрывочны. Хотя авторы современных работ по этологии, динамике попу-
ляций и другим популярным проблемам не могут обойтись без оценки возраста
отдельных особей, что необходимо, в частности, для выяснения возрастного со-
става выборки, определение возраста не считается серьезной задачей, и в тексте
соответствующих публикаций ей, как правило, уделяют минимальное внимание.
Например, Breitenmoser et al. (1993) в работе посвященной состоянию реинтро-
дуцированной популяции рыси, указывают, без всяких пояснений, что возраст
определяли по числу годовых слоев в прижизненно удаленных резцах. Между
тем, заинтересованный читатель хотел бы знать, какие резцы удаляли, какую ане-
стезию использовали, как животные переносили процедуру. Эти сведения край-
не важны, поскольку данных по прижизненному удалению зубов не так уж мно-
го и ценен опыт каждого исследователя. Но даже при использовании многократ-
но апробированных методов посмертного определения возраста желательно при
изложении полученных результатов приводить данные, позволяющие судить об
эффективности данной методики в применении к конкретному виду и конкретной
популяции.
244
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Наконец, развитие неинвазивных исследований млекопитающих в природе
требует расширения набора методов прижизненного определении возраста осо-
бей. Следует подчеркнуть, что не все методы прижизненного определения возра-
ста являются неинвазивными в строгом смысле слова, то есть позволяют оце-
нить возраст животного без отлова, визуально, на расстоянии, как, например,
определяют возраст копытных по размерам тела, окраске, поведению, форме
рогов и, в отдельных случаях, по числу колец на рогах. Методы же прижизнен-
ного определения возраста по отпечаткам жевательной поверхности зубов или
по числу годовых слоев в удаленном зубе и т.п. нельзя считать неинвазивными.
Они в разной степени травмирующие, но при этом имеют общее достоинство: их
использование не требует умерщвления животного и не создает опасности для
его жизни. Необходимо совершенствовать старые методы, отыскивая возмож-
ность применять методы, ранее использовавшиеся только посмертно, для опре-
деления возраста живых животных и уменьшая травмирующий эффект уже су-
ществующих методов прижизненного определения возраста. При этом, конечно,
актуальной задачей остается разработка новых методов прижизненного опреде-
ления возраста, минимально травмирующих животное.
ЛИТЕРАТУРА
Адольф ТА. 1949. Рост и развитие медвежат по наблюдениям в Московском
зоопарке в 1940 г. // Тр. Московского зоопарка. Вып. 4. С. 78-99.
Алекперов А.М., Грекова В.Х. 1969. К возрастной изменчивости и линьке полч-
ка (Glis glis L.) в северо-западной части Кавказа // Уч. зап. Азербайджанс-
кого гос. ун-та. Сер. биол. наук. Вып. 3. С. 53-59.
Алиев Ф.Ф. 1954. Возрастная изменчивость в зубной системе нутрии (Myocastor
coypus Molina) // Докл. АН Аз.ССР. Т. 10. Вып. 4. С. 291-295.
Балейшис Р. 1970. Определение возраста лосей по слоистой структуре корнево-
го цемента резцов и клыков И Зоол. журн. Т. 49. Вып. 5. С. 778-780.
Банников А.Г., Жирнов Л.В., Лебедева Л.С., Фандеев А.А. 1961. Биология сай-
гака. М.: Изд-во сель-хоз. лит-ры. 336 с.
Башенина Н.В. 1953. Об определении возраста обыкновенной полевки (Microtus
arvalis Pall.) И Зоол. журн. Т. 32. Вып. 4. С. 730-743.
Башенина Н.В. 1981. Онтогенез И Европейская рыжая полевка. М.: Наука. С.
211-226.
Башкиров И.С., Жарков И.В. 1934. Биологии и промысел крота в Татарии // Уч.
зап. Казанского ун-та. Т. 94. Вып. 8. Ч. 3. 125 с.
Блажис Ф.С. 1971. Метод прижизненного извлечения клыка у косули (Capreolus
capreolus) для определения возраста // Зоол. журн. Т 50. Вып. 6. С. 926-929.
Борисов Б.П. 1988. О некоторых возрастных признаках европейских бобров И
Научно-технический прогресс в практику перестройки охотничьего х-ва /
Научная конференция. М., ноябрь 1988. С. 41-43.
Бородина М.Н. 1970а. Возрастная изменчивость некоторых морфологических
признаков бобров мокшанской популяции // Тр. Мордовского гос. заповед-
ника. Вып. 5. С. 91-130.
Бородина М.Н. 19706. Справочные таблицы для определения возраста речных
бобров // Тр. Мордовского гос. заповедника. Вып. 5. С. 131-136.
Бромлей Г.Ф. 1956. Горал (Nemorhaedus caudatus raddeanus Heude, 1894) // Зоол.
журн. Т. 35. Вып. 9. С. 1395-1405.
Бутовский П.М., Шаймарданов З.Т 1988. Структура и балансы популяций трех
видов рукокрылых юго-восточного Казахстана И Рукокрылые (морфология,
экология, эхолокация, паразиты, охрана). Киев: Наукова Думка. С. 116-120.
Варшавский С.Н. 1950. Определение возраста курганчиковой и домовой мышей //
Бюл. Моск, об-ва испыт. природы. Отд. биол. Т. 55. Вып. 6. С. 21-34.
Викторов Л.В. 1967. Определение возраста обыкновенной бурозубки (Sorexaraneus
L.) И Бюл. Моск, об-ва испыт. природы. Отд. биол. Т 72. Вып. 6. С. 151-152.
Викторов Л.В. 1994. Определение возраста обыкновенной бурозубки // Фауна и
экология животных Верхневолжья. Сб. научных трудов Твер. гос. ун-та. Тверь.
С. 70-88.
Войлочников А.Т, Михайловский Б.А. 1970. Определение возраста колонков//
Сб. научн.-технич. информации ВНИИОЗ. Охота, пушнина и дичь. Вып. 29.
Киров. С. 3-12.
246
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Вшивцев В.П. 1972. Выдра Сахалина. Новосибирск: Наука. Сиб. отд. 106 с.
Гамбарян П.П., Дукельская Н.М. 1955. Крыса: Сов. Наука. 254 с.
Гинтлис Р.Б. 1959. К вопросу об определении возраста краснохвостой песчанки //
Бюл. Моск, об-ва испыт. природы. Отд. биол. Т. 64. Вып. 6. С. 23-30.
Граков Н.Н. 1962. К методике определения возраста лесной куницы // Проблемы
зоологич. исследований в Сибири. Горно-Алтайский книгоиздат. С. 55-57.
Граков Н.Н. 1970. Опыт определения возраста по годичным слоям в цементе
клыка лесной куницы // Оптимальная плотность и оптимальная структура
популяций животных. Информационные материалы. Вып. 2. Свердловск. С.
193-194.
Григорьев Н.Д., Попов В. А. 1952. К методике определения возраста материкового
песца // Изв. Казан, фил. АН СССР сер. биол. и с/х наук. Вып. 3. С. 207-215.
Груздев А.Р., Проняев А.В. 1994. Определение возраста сайгака (Saiga tatarica)
по слоям цемента зубов // Зоол. журн. Т. 73. Вып. 7,8. С. 223—226.
Гурский И.Г. 1973. Определение возраста волка (Canis lupus L.) по черепу //
Вести, зоологии. № 3. С. 55-59.
Давыдов Н.П. 1984. К методике определения возраста соболей // Тез. докл. Все-
союзной конф. Регистрирующие структуры и определение возраста млеко-
питающих. М. С. 10-11.
Данилов Г.Д., Луковцев Ю.С., Попов М.В. 1974. Обзор и анализ различных
способов определения возраста ондатры // Бюл. научн.-техн. информ. Якутск,
фил. Сиб. отд. АН СССР. Якутск. С. 4-10.
Данилов П.И., Русаков О.С., Туманов И.Л. 1979. Хищные звери Северо-запада
СССР. Л.: Наука. 162 с.
Депарма Н.К. 1954. К методике определения возраста кротов // Бюл. Моск, об-
ва испыт. природы. Отд. биол. Т. 59. Вып. 6. С. 11-26.
Дицевич Б.Н., Свиридов Н.С., Дюба Г.П. 1983. Возрастные особенности лося
Прибайкалья // Биол. и хоз. использование промысловых зверей и птиц Си-
бири и Дальнего Востока. Иркутск. С. 52-70.
Добринский Л.Н., Михалев М.В. 1966. К методике определения возрастной
структуры популяций животных // Тр. Ин-та биологии УФАН СССР. Т. 51.
С. 107-115.
Долгов В.А., Россолимо О.Л. 1966. Возрастные изменения некоторых особен-
ностей строения черепа и бакулюма хищных млекопитающих и методика
определения возраста на примере песца (Alopex lagopus L.) // Зоол. журн. Т.
45. Вып. 7. С. 1074-1080.
Дунаева Т.Н. 1955. К изучению биологии размножения обыкновенной бурозуб-
ки (Sorex araneus L.) // Бюл. Моск, об-ва испыт. природы. Отд. биол. Т. 60.
Вып. 6. С. 27-43.
Дык В. 1965. Как мы определяли возраст европейской косули // Охота и охотни-
чье хозяйство № 5. С. 55.
Евдокимов Н.Г. 1997. Методика определения возраста обыкновенной слепушон-
ки Ellobius talpinus (Rodentia, Cricetidae) // Зоол. журн. Т. 76. Вып. 9. С.
1094-1101.
Литература
247
Егоров О.В. 1958. К методике определения возраста белки // Научные сообщ.
Якутского фил. АН СССР. Вып. 1. С. 104-108.
Егоров О.В. 1971. Морфо-физиологические особенности диких северных оле-
ней {Rangifer tarandus) Якутии. Биологические проблемы Севера // Тр. Сев-
Вост. компл. ин-та ДВЦ АН СССР. Вып. 42. С. 37-47.
Егоров О.В., Попов М..В., Лабутин Ю.В. 1957. К методике определения возраста
зайца-беляка Lepus timidus gihganus II Докл. на восьмой научн. сессии Якут-
ского фил. АНСССР. С. 124-134.
Емельянов П.Ф. 1983. Определение возраста горного суслика // Вести, зооло-
гии. № 3. С. 68-71.
Завацкий Б.П. 1978. Краниометрическая характеристика енисейской популяции
бурого медведя {Ursus arctos) И Зоол. журн. Т. 57. Вып. 2. С. 308-311.
Зарипов Р.З. 1964. К методике определения возраста лосей// Природные ресур-
сы Волжско-Камского края. Животный мир. Вып. 1. М.: Наука. С. 30-45.
Зимина Р.П., Меркова М.А. 1960. Экология Тянь-Шаньской мышовки в север-
ном Тянь-Шане И Фауна и экология грызунов. М.: Изд-во М ГУ. Вып. 6. С.
183-207.
Зубов А.А. 1968. Одонтология. М.: Наука. 199 с.
Исаков С.И. 1984. Использование регистрирующих структур для определения
возраста ежей Hemiechinus auritus и Paraechinus hypomelas И Регистр, струк-
туры и определение возраста млекопитающих. Тезисы докл. Всес. конф. 22-
25 мая. М. С. 19-20.
Карноухова Н.Г. 1971. Определение возраста серых и черных крыс // Экология
№ 2. С. 71-76.
Карпухин И.П., Карпухина Н.М. 1971. Вес хрусталика глаза как критерий возра-
ста белки обыкновенной (Sciurus vulgaris) И Зоол. журн. Т. 50. Вып. 2. С.
274-277.
Кириков С.И. 1952. Птицы и млекопитающие в условиях ландшафтов южной
оконечности Урала. М.: Изд-во АН СССР. 411 с.
Кирис И.Д. 1937. Методика и техника определения возраста и анализ возрастно-
го состава популяции белки // Бюл. Моск, об-ва испыт. природы. Отд. биол.
Т. 46. Вып.1. С. 36-42.
Клевезаль Г. А. 1988. Регистрирующие структуры млекопитающих в зоологичес-
ких исследованиях. М.: Наука. 285 с.
Клевезаль Г.А., Клейненберг С.Е. 1967. Определение возраста млекопитающих
по слоистым структурам зубов и кости. М.: Наука. 142 с.
Клевезаль Г.Н., Мараков С.В. 1966. О возрасте калана // Сб. научно-техн, ин-
формации ВНИИЖП. Вып. 16.
Клевезаль Г.А., Соколов А.А. 1997. Оценка выживаемости поздно рожденных или
отставших в развитии северных оленей {Rangifer tarandus) по результатам
анализа регистрирующих структур // Зоол. журн. Т. 76. Вып. 7. С. 865-871.
Клевезаль Г.А., Соколов А.А. 2004. Возрастные и межпопуляционные особеннос-
ти скорости стачивания щечных зубов северных оленей, Rangifer tarandus И
Зоол. журн. Т. 83. Вып. 6. С. 745-756.
248
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Клевезаль Г.А., Груе X., Мина М.В. 1981. Способ оценки пригодности регист-
рирующих структур для подсчета слоев при определении возраста животных//
Зоол. журн. Т. 60. Вып. 12. С. 1869-1877.
Клевезаль Г.А., Мина М.В., Крушинская Н.Л. 2005. Использование методов
многомерного статистического анализа при определении возраста млекопи-
тающих (на примере лесной мышовки, Sicista betulina и лесной куницы,
Martes martes) //Зоол. журн. Т. 84. Вып. 11. С. 1389-1401.
Клевезаль Г.А., Юдин В.Г., Суховская Л.И. 1991. Рост клыков и формирование
слоев дентина у хищных // Зоол. журн. Т. 70. Вып. 11. С. 110-119.
Кнорре Е.П., Шубин Г.Г. 1959. Определение возраста лося // Тр. Печоро-Илыч-
ского гос. заповедника. Вып. 7. С. 123-132.
Козло П.Г. 1975. Дикий кабан. Минск: Ураджай. 222 с.
Козло П.Г., Никитенко М.Ф. 1967. Методика определения возраста дикого каба-
на И Экология млекопитающих и птиц. М.: Наука. С. 209-221.
Копеин К.И. 1967. Анализ возрастной структуры популяций горностая // Тр.
МОИП. Вып. 25. С. 33-39.
Корневен С., Лесбр М. 1929. Распознавание возраста по зубам и другим произ-
водным кожи лошади, коровы, овцы, свиньи, верблюда, собаки, кошки и
домашних птиц. М.: Новая деревня. 383 с.
Корнеев А.П., Карпов А.А., Пейсахис А.Л. 1976. Возможности использования
веса хрусталика глаза в качестве критерия возраста большой песчанки —
Rhombomys opimus // Зоол. журн. Т. 55. Вып. 3. С. 443—444.
Корсаков Г.К. 1949. Опыт организации учета ондатры. М.: Гос. изд-во техн, и
эконом, лит-ры по вопросам заготовок. 32 с.
Корсакова И.Б. 1970. Приемы определения возраста ондатры по эталонам зубов И
Тр. Кировского сель-хоз. ин-та. Т. 22. Вып. 52. С. 85-91.
Корытин Н.С., Габченко В.Н. 1988. Анализ изменчивости размеров годичных
слоев в цементе клыка у волка//Вопросы динамики популяций млекопита-
ющих. Свердловск. С. 32-33.
Корытин Н.С., Ендукин А.Ю. 1982. Сравнение двух методов определения возра-
ста у красной лисицы И Вопросы экологии животных. Информ, мат-лы Ин-
та экологии растений и животных. Свердловск. С. 17.
Косой М.Е., Косая О.И. 1989. Возрастные изменения массы хрусталика глаза у
полевой мыши Apodemus agrarius // Соврем, подходы к изуч. изменчивос-
ти. Владивосток: АНСССР, ДВО, Биол.почв. ин-т. С. 106-115.
Кошкина ТВ. 1955. Метод определения возраста рыжих полевок и опыт его
применения // Зоол. журн. Т. 34. Вып. 3. С. 631-639.
Кошкина Т.В., Халанский А.С. 1961. Возрастная изменчивость черепа норвеж-
ского лемминга и анализ возрастного состава популяции этого вида // Бюл.
Моск, об-ва испыт. природы. Отд. биол. Т. 66. Вып. 2. С. 3-14.
Крылова ТВ. 1973. Возрастной состав популяции монгольской пищухи (Ochotona
pricei) И Зоол. журн. Т. 52. Вып. 9. С. 1422-1425.
Крылова ТВ. 1978. Возрастная структура природной популяции монгольской пес-
чанки (Meriones unguiculatus) II Зоол. журн. Т. 57. Вып. 12. С. 1842-1847.
Литература
249
Кучерук В.В., Кулик И.Л. 1955. Опыт анализа популяции водяной крысы по кра-
ниологическим признакам // Бюл. Моск, об-ва испыт. природы. Отд. биол. Т.
60. Вып. 4. С. 45-52.
Кыдырбаев X. 1960.0 возрастных особенностях желтого суслика (Citellus fulvus
oxianus Thom) // Тр. Ин-та зоологии АН Каз. СССР. Вып. 13. С. 160-169.
Лавров Л.С. 1953. Определение возраста у речных бобров // Тр. Воронежского
гос. заповедника. Вып. 4. С. 77-84.
Лаврова М.Я., Прохорова Е.В., Лист Л.В. 1960. Некоторые особенности биоло-
гии полевок-экономок и течение лептоспирозной эпизоотии среди них на
западе Московской области И Бюл. Моск, об-ва испыт. природы. Отд. биол.
Т. 65. Вып. 2. С. 13-24.
Ларин Б.А., Мохов Н.Г.1969. Сравнительная оценка способов определения воз-
раста ондатры // Тр. Кировского сель-хоз. ин-та. Т. 21. Вып. 46. С. 106-118.
Леонтьев А.Н., Воротникова Л.М. 1957. Определение возраста длиннохвостого
суслика И Известия Иркутского гос. науч. иссл. противочумного ин-та Си-
бири и Дальнего Востока. Вып. 16. С. 60-68.
Леонтьев А.Н., Пешков Б.И. 1966. К определению возраста тарбагана // Изв.
Иркутского н-и противочум. ин-та Сибири и Дальнего Востока. Вып. 26. С.
175-185.
Лобачев В.С., Шенборт Г.И. 1977. Определение возраста и возрастная структура
популяций тушканчиков И Бюл. Моск, об-ва испыт. природы. Отд. биол.. Т.
82. Вып. 2. С. 18-23.
Лобков В.А. 1999. Крапчатый суслик северо-западного Причерноморья. Одес-
са: Астропринт. 270 с.
Лозан М.Н. 1961. Определение возраста лесной и орешниковой сонь И Зоол.
журн. Т. 40. Вып. 11. С. 1740-1743.
Малафеев Ю.М., Кряжимский Ф.В., Добринский Л.Н. 1986. Анализ популяции
рыси Среднего Урала. Свердловск: УНЦ АН СССР. 115 с.
Маркарян Р.Е. 1989. Способ определения возраста закавказского хомяка
(Mesocricetus brandti Nehring) И Вклад ученых по защите растений в научно-
техн. прогрессе. Ереван. С. 83-85.
Маркина А.Б. 1962. К вопросу о возрастной краниологии лисицы И Тр. Биол.
ин-та Сиб. отд. АН СССР. Вып. 8. С. 171-179.
Матюшкин Е.Н. 1974. Рысь. М.. Р. Лесная промышленность. 62 с.
Машкин В.И. 1979. Прижизненное определение возраста сурка Мензбира // Тр.
Кировского сель-хоз. ин-та. Вып. 63. С. 14-19.
Машкин В.И., Колесников В.В. 1990. Определение возраста сурков (Marmota,
Sciuridae) по рисунку стертости жевательной поверхности зубов // Зоол.
журн. Т. 69. Вып. 6. С.124-131.
Мейер М.Н. 1957. О возрастной изменчивости суслика (Citellus pygmaeus) //
Зоол. журн. Т. 36. Вып. 9. С. 1393-1402.
Мина М.В., Клевезаль Г.А. 1976. Рост животных. М.: Наука. 291 с.
Миролюбов И.И., Рященко Л.П. 1948. Пятнистый олень. Владивосток: МВТ-СССР,
Дальневосточный звероводческий трест. 115 с.
250
Г. А. Клевезаль, Принципы и методы определения возраста млекопитающих
Монахов В.Г. 2004. Оценка возрастной структуры выборок соболей с примене-
нием трех методов определения возраста // Экология. № 6. С. 430-435.
Монахов Г.И. 1965. Определение возраста соболей по относительной ширине
канала клыка И Сб. научно-техн, информ. ВНИИЖП. Охота, пушнина, дичь.
Вып. 11. С. 46-53.
Монахов Г.И. 1967. Сравнительная оценка некоторых методов определения воз-
раста соболей // Сб. научно-техн, информ. ВНИИЖП. Охота, пушнина и дичь.
Вып. 19. С. 35-46.
Неганов В.Г. 1984. Об определении возраста волка по слоистой структуре це-
мента клыка // Регистр, структуры и определение возраста млекопитающих.
Тезисы докл. Всес. конф. 22-25 мая. Москва. С. 46-47.
Никифоров Л.П. 1958. Опыт определения возраста ондатры в полевых условиях И
Организация охотничьего промысла. М.: Изд. Центрсоюз. Вып. 7. С. 137-141.
Новиков Г.А. 1956. Хищные млекопитающие фауны СССР. М.-Л.: Изд-во АН
СССР. 294 с.
Огнев С.И. 1947. Звери СССР и прилежащих стран. Том V. Грызуны. М.-Л.:
Изд-во АН ССР. 809 с.
Оленев В.Г. 1967. Сезонные и возрастные изменения веса зобной железы у гры-
зунов // Тр. Моск, об-ва испыт. природы. Вып. 25. С. 67-77.
Оленев Г.В. 1989. Функциональная детерминированность онтогенетических из-
менений возрастных маркеров грызунов и их практическое использование
в популяционных исследованиях // Экология. № 2. С. 19-31.
Оленев Г.В. 2004. Функционально-онтогенетический подход в изучении популя-
ций цикломорфных млекопитающих. Автореф. дисс. на соиск. уч. ст. докто-
ра биол. наук. Екатеринбург. 47 с.
Оленев Г.В., Колчева Н.Е. 1982. Динамика возрастных изменений зубов корне-
зубых полевок и их использование при определении абсолютного возраста
грызунов // Вопросы экологии животных. Информ, мат-лы Ин-та экологии
растений и животных. Свердловск. С. 21.
Охотина М.В. 1966. Дальневосточный крот и его промысел. М.: Наука. 136 с.
Панкратьев А.Г. 1971. Определение возраста колонка по признакам кости при-
апа // Биологические и медицинские исследования на Дальнем Востоке. Вла-
дивосток. С. 77-84.
Пантелеев П.А. 1966. Опыт определения возраста водяных полевок (Arvicola
terrestris L.) в осенней популяции И Бюл. Моск, об-ва испыт. природы. Отд.
биол. Т. 71. Вып. 4. С. 20-26.
Петров А.К. 1964. Окостенение скелета в индивидуальном развитии лося // Зоол.
журн. Т. 43. Вып.12. С. 1837-1847.
Петров Б.М. 1961. К систематике и возрастной изменчивости сурка Мензбира
(Marmota menzbiri Kaschk.) // Зоол. журн. Т. 40. Вып. 1. С. 93-105.
Петровский Ю.Т. 1961. Особенности экологии крапчатого суслика в Белорус-
сии // Зоол. журн. Т. 40. Вып. 5. С. 736-748.
Попов В.А. 1943. Новый показатель возраста куницевых (Mustelidae) // Докл. АН
СССР. Т. 38. Вып. 8. С. 289-292.
Литература
251
Пузаченко Ю.А. 1991. Определение возраста обыкновенного слепыша Spalax
microphthalmus (Rodentia, Spalacidae) // Зоол. журн. Т. 70. Вып. 12. С.
113-124.
Рахматулина И.К. 1984 Возрастная структура скоплений самок позднего кожана //
Тез. докл. Всесоюзной конф. Регистрирующие структуры и определение
возраста млекопитающих. Москва. С. 53—54.
Руденчик Ю.В. 1962. Определение возраста полуденной, гребенчуковой и крас-
нохвостой песчанок (род Meriones) по степени стертости коренных зубов //
Узб. биолог, журн. Вып. 4. С. 58-62.
Руковский Н.Н. 1970. О прижизненном определении возраста самцов енота-по-
лоскуна // Сб. научно-техн. инф. ВНИИОЗ. Вып. 29. С. 10-13.
Рыльников В.А. 1990. Размножение, возрастной состав и смертность // Серая
крыса. / В.Е. Соколов, Е.В. Карасева (ред.). М.: Наука. С. 181-229.
Рябов Л.С. 1962. Некоторые возрастные особенности кавказских лесных и ка-
менных куниц // Зоол. журн. Т. 41. Вып. 11. С. 1731-1738.
Рязанов Д.А., Клевезаль Г.А. 1991. Развитие верхних клыков калана Enhydra
lutris и некоторые замечания относительно определения возраста особей //
Зоол. журн. Т. 70. Вып. 1. С. 121-128.
Рязанов Д.А., Маминов М.К. 1996. Определение пола и возраста по клыкам у
калана (Enhydra lutris) II Зоол. журн. Т. 75. Вып. 4. С. 593-601.
Сабилаев А.С. 1969.Экология тушканчика Северцова (Allactaga severtzovi) в
Каракалпакии // Зоол. журн. Т. 48. Вып. 6. С. 902-911.
Сапожников Г.Н., Усманов З.Д. 1967. Определение возраста Ovis аттоп
bocharensis Nasonov по весу хрусталика глаза И Доклады АН Таджик. ССР.
Т. 10. Вып. 11. С. 64-67.
Синицын А.А. 1982. К методике определения возраста колонка (Mustela sibirica
Pall.) // Бюл. Моск, об-ва испыт. природы. Отд. биол. Т. 87. Вып. 4. С. 32-35.
Слудский А.А. 1948. Ондатра и ее акклиматизация в Казахстане. А.-Ата.: Изд-во
АН Каз. ССР. 181с.
Смирнов В.С. 1959. Определение возраста и возрастная структура популяций
песца на Ямале // Тр. Салехардского стационара Урал. Фил. АН СССР. Вып.
1. С. 220-228.
Смирнов В.С. 1960. Определение возраста и возрастные соотношения у млеко-
питающих на примере белки, ондатры и пяти видов хищников // Тр. Ин-та
биологии УФАН. Вып. 14. С. 97-112.
Смирнов В.С. 1983. Сравнение двух методов определения возраста, используе-
мых для анализа возрастной структуры волка И Исследование актуальных
проблем териологии. Ин-т экологии растений и жив. УНЦ АН СССР. Сверд-
ловск. С. 79-82.
Смирнов В.С. 1988. Исследование динамики численности волка с использова-
нием двух методов определения возраста // Анализ размерно и возрастной
структуры популяций позвоночных. Свердловск. С. 114-132.
Смирнов В.С., Павленко ТА., Покровский А.В. 1971. Метод анализа возрастной
структуры популяции малого тушканчика // Экология. № 4. С. 88-90.
252
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Смирнов В.С., Шварц С.С. 1959. Сравнительная эколого-физиологическая ха-
рактеристика ондатры в лесостепных и приполярных районах // Вопросы
акклиматизации млекопитающих на Урале. Тр. Ин-та биологии УФАН. Вып.
18. Свердловск.
Смирнов М.Н. 1977. К методике определения возраста косуль по комплексу мор-
фологических признаков И Сб. научно-тех. инф. ВНИИОЗ. Вып. 5. С. 34-39.
Смирнов М.Н. 1979. Как определить пол и возраст косули на расстоянии//Охота
и охотничье хозяйствово. № 1. С. 22-23.
Соколов А.А., Груздев А.З., Проняев А.В. 1996. Особенности стачивания щеч-
ных зубов у северных оленей {Rangifer tarandus) разных популяций И Зоол.
журн. Т. 75. Вып.10. С. 1573-1580.
Соколов В.Е. 1977. Систематика млекопитающих. Отряды зайцеобразных, гры-
зунов. М.: Высшая школа. 494 с.
Соколов В.Е. 1979. Систематика млекопитающих. Отряды китообразных, хищ-
ных, ластоногих, трубкозубых, хоботных, даманов, сирен, парнокопытных,
мозоленогих, непарнокопытных. М.: Высшая школа. 528 с.
Соколов И.И. 1937. Половозрастная и расовая изменчивость в черепе дикого и
домашнего северного оленя// Сов. оленеводство. № 9. С. 2-102.
Соколов И.И. 1956. К методике определения возраста косули {Capreolus capreolus) И
Зоол. журн. Т. 35. Вып. 8. С. 1238-1249.
Соколов И.И., Рашек В.Л. 1961. Развитие зубов и черепа как показатель возра-
ста у сайги {Saiga tatarica) И Бюл. Моск, об-ва испыт. природы. Отд. биол.
Т. 66. Вып. 6. С. 77-98.
Соломкина Н.В., Хохряков С.А. 1993. Половой диморфизм и возрастные измене-
ния черепа амурского горала, Nemorhaedus caudatus (Artiodactyla, Bovidae) И
Зоол. журн. Т. 72. Вып. 3. С. 153-159.
Строганов С.У 1937. Методика определения возраста и анализ возрастного состава
популяций горностая {Mustela erminea L.) И Зоол. журн. Т. 16. Вып. 1. С. 113-128.
Тавровский В. А., Егоров О.В., Кривошеев В.Г., Попов М.В., Лабутин Ю.В. 1971.
Млекопитающие Якутии. М.: Наука. 670 с.
Тарасов С.А. 1965. Особенности развития скелета полевок {Lagurus lagurus,
Stenocranius gregalis) И Зоол. журн. Т. 44. Вып. 9. С. 1376-1381.
Тарасов С.А. 1966. Определение возраста полевок рентгенологическим мето-
дом // Зоол. журн. Т. 45. Вып. 8. С. 1247-1250.
Тарасов С.А. 1984. Возрастные особенности бакулюма некоторых млекопитаю-
щих // Зоол. журн. Т. 63. Вып. 1. С. 120-125.
Темботова Ф.А. 1986. Половозрастная структура кавказской белозубки {Crocidura
guldenstaedti Pall., 1811) И Экология. № 6. С. 35—41.
Терехина А.Н. 1973. Изменчивость водяной полевки {Arvicola terrestris L.) в Волго-
Ахтубинской пойме И Бюл. Моск, об-ва испыт. природы. Отд. биол. Т. 78.
Вып. 4. С. 13-26.
Тимофеев В.В., Надеев В.Н. 1955. Соболь. Заготиздат. 404 с.
Тиунов М.П. 1988. Численность, возрастной состав и особенности зимовки боль-
шого трубконоса Murina leucogaster на юге Дальнего Востока // Рукокры-
Литература
253
лые (морфология, экология, эхолокация, паразиты, охрана). Киев: Наукова
Думка. С. 120-122.
Топачевский В.А. 1969. Слепышовые (Spalacidae) // Фауна СССР. Млекопитаю-
щие. Т. 3. Л.: Наука. 247 с.
Травина И.В., Стишов М.С. 1996. Об определении возраста по весу тела у ко-
пытного лемминга (Dicrostonyx torguatus Vinogradovi Ognev) // Экология. №
1. С. 40-43.
Тупикова Н.В. 1964. Изучение размножения и возрастного состава популяции
мелких млекопитающих // Методы изучения природных очагов болезней
человека / П.А.Петрищева, Н.Г.Олсуфьев (ред.). М.: Медицина. С. 154-208.
Тупикова Н.В., Каледа Л.В. 1957. Определение возраста грызунов // Фауна и
экология грызунов. М.: Изд-во М ГУ Вып. 5.
Тупикова Н.В., Сидорова Г.А., Коновалова Э.Ф. 1970. Определение возраста
лесных полевок // Фауна и экология грызунов. М.: Изд-во МГУ. Вып. 9. С.
160-167.
Херувимов В.Д., Орлов И.И. 1967. Определение возраста лосей (Alces alces) из
южных областей Европейской части СССР // Зоол. журн. Т. 46. Вып. 8. С.
1229-1233.
Хмелевская Н.В. 1961. О биологии алтайской пищухи // Зоол. журн. Т. 40. Вып.
10. С. 1583-1585.
Цалкин В.И. 1948. О росте рогов и строении черепа у диких баранов // Зоол.
журн. Т. 27. Вып. 5. С. 445-460.
Цалкин В.И. 1950. Сибирский горный козел. М.: Изд-во МОИП, 118с.
Цыганков Д.С. 1955. Методика определения возраста и продолжительности жиз-
ни ондатры // Зоол. журн. Т. 34. Вып. 3. С. 640-651.
Червякова В.Б., Червяков В.Д. 1973. Возрастные изменения в черепе серого сус-
лика (Marmota baibadna)!I Зоол. журн. Т. 52. Вып. 7. С. 1055-1062.
Шагоян В.А., Явруян Э.Г. 1974. К биологии средиземного нетопыря (Pipistrellus
kuhli Natt.) в Армении // Материалы Первого Всес. совещ. по рукокрылым.
Л. С. 96-98.
Шадрина Е.Г. 1988а. Сравнительная характеристика методик определения возра-
ста соболя (Martes zibellina) и горностая (Mustela erminea) // Зоогеография
и экологические исследования териофауны Якутии. Якутск. С. 76-83.
Шадрина Е.Г. 19886. К методике определения возраста белки обыкновенной
(Sciurus vulgaris) II Зоол. журн. Т. 67. Вып. 8. С. 1229-1237.
Швецов А.И. 1975. К методике определения возраста белок по весу хрусталика //
Сб. научн.-техн. информ. ВНИИОЗ. Вып. 47—48. С. 92—96.
Швецов Ю.Г. 1966. О возрастных изменениях обыкновенного двуцветного ко-
жана (Vespertilio murinus L.) в Прибайкалье // Научн. докл. высшей школы.
Биол. науки. Вып. 3. С. 44-45.
Шенброт Г.И. 1984. Определение возраста, продолжительность жизни и динами-
ка возрастной структуры популяции тушканчика Северцова // Регистрирую-
щие структуры и определение возраста млекопитающих. Тезисы докл. Всес.
конф. 22-25 мая 1984 г. М. С. 61-62.
254
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Шиляева Л.М., Морозова О.И., Леушина Н.С. 1975. Сравнительная оценка ме-
тодов и техника определения возраста песца // Сб. научн-техн. информ. ВНИ-
ИОЗ. Вып. 49-50. С. 110-117.
Шостак С.В. 1988. Определение возраста европейского благородного оленя.
Минск: Ураджай. 61 с.
Шубин Н.Г. 1965. К методике определения возраста бурундука И Уч. зап. Томс-
кого гос. ун-та. Вып. 51. С. 89-91.
Юдин В.Г. 1977. Енотовидная собака Приморья и Приамурья. М.: Наука, 162 с.
Юдин В.Г., 1986. Лисица Дальнего Востока СССР. Владивосток. 280 с.
Юдин В.Г. 1992. Волк Дальнего Востока России. Благовещенск: ДО РАН. 31с.
Юргенсон П.Б. 1956. Определение возраста у лесной куницы // Зоол. журн. Т.
35. Вып. 5. С. 781-783.
Язан Ю.Н. 1961. Биологические особенности и пути хозяйственного освоения
популяции мигрирующих лосей Печорской тайги // Тр. Печоро-Илычского
гос. заповедника. Вып. 9. С. 114-201.
Adamczewska-Andrzejewska К.А. 1966. Variations in the hardness of the teeth of
Sorex araneus Linnaeus, 1758 // Acta Theriol. Vol. 11. No. 3. P. 55-69.
Adamczewska-Andrzejewska K.A. 1967. Age reference model for Apodemus flavi-
collis (Melchior, 1834) // Ekol. Polska. Vol. 15. No. 41. P. 787-790.
Adamczewska-Andrzejewska K.A. 1973a. The lens weight as indicator of age in the
wildMicrotus arvalis population // Bull. Acad. Pol. Ser. Sci. Biol. Vol. 21. No. 5.
P. 331-336.
Adamczewska-Andrzejewska K.A. 1973b. Growth, variations and age criteria in
Apodemus agrarius (Pallas,1771) // Acta Theriol. Vol. 18. No. 19. P. 353-394.
AhnlundH. 1976. Age determination in the European badger, Meles meles LI I Ztschr.
Saugetierk. Bd. 41. No. 2. S.120-125.
Aitken R.J. 1975. Cementum layers and tooth wear as criteria for ageing Roe deer
{Capreolus capreolus) // J. ZooL Vol. 175. No. 1. P. 15-28.
Alibhai S.K. 1980. An X-ray technique for ageing Bank voles {Clethrionomys glareo-
lus) using the first mandibular molar // J. Zool. Vol. 191. No. 3. P. 418-423.
Ancrenaz M., Delhomme A. 1997. Teeth eruption as a means of age determination in
captive Arabian oryx, Oryx leucoryx // Mammalia. Vol. 61. No. 1. P. 135-138.
Andersen J., Jensen B. 1972. The weight of the eye lens in the European hares of
known age// Acta Theriol. Vol. 17. No. 1-11. P. 87-92.
Anderson J.L. 1986. Age determination of the nyala Tragelaphus angasill South Afr.
Wildl. Res. Vol. 16. No. 3. P. 82-90.
Andrzejewski R., Liro A. 1977. Effect of different kinds of feed on wear of the teeth
in individuals of a field mouse population И Acta Theriol. Vol. 22. No. 20-29. P.
393-395.
Anthony E.L.P. 1988. Age determination in bats // Ecological and behavioral methods
for the study of bats. Ed. T.H.Kunz. Wash. L.: Smithsonian Ins. Press. P. 47-58.
Arthur S.M, Cross R.A., Paragi T.F., Krohn W.B. 1992. Precision and utility of ce-
mentum annuli for determining ages of fishers // Wildl. Soc. Bull. Vol. 20. No. 4.
P. 402-405.
Литература
255
Artimo A. 1969.The baculum in the wood lemming, Myopus schisticolor (Lilljeb.), in
relation to sexual status, size and age// Ann. Zool. Fenn. Vol. 6. No. 4. P. 335-344.
Ashby K.R., Henry B.A.M. 1979. Age criteria and life expectancy of Roe deer (Capre-
olus capreolus) in coniferous forest in north-eastern England // J. Zool. Vol. 189.
P. 207-220.
Askins G.R., Chapman J.A. 1984. Age determination and morphological characteris-
tics of wild mink from Maryland, USA // Ztschr. Saugetierk. Bd. 49. No. 3. S.
183-189.
Baagoe H. J. 1977. Age determination in bats (Chiroptera) // Vid. medd. Dan. naturhist.
Foren. Vol. 140. P. 53-92.
Bada J.L., Brown S., Masters P.M. 1980. Age determination of marine mammals
based on aspartic acid racemization in the teeth and lens nucleus// Age determi-
nation of toothed whales and sirenians // W.F.Perrin, A.C.Myrick (eds.). IWC
Cambridge. P. 113-118.
Bada J.L., Mitchell E., Kemper B. 1983. Aspartic acid racemization in narwhal teeth //
Nature. Vol. 303. P. 418-420.
Ballard W.B., Matson G.M., Krausman P.R. 1995. Comparison of two methods to age
gray wolf teeth // Ecology and conservation of wolves in a changing world /
L.N.Carbyn, S.H. Fritts, D.R.Seip (eds.). Occas. publ, ser. No. 35. P. 455-^459.
Banfield A.W.F. 1960. The use of caribou antler pedicels for age determination // J.
Wildl. Manag. Vol. 24. No. 1. P. 99-102.
Baptista T.L., Richardson C.R., Kunz T.H. 2000. Postnatal growth and age estima-
tion in free-ranging bats. A comparison of longitudinal and cross-sectional sam-
pling methods // J. Mammal. Vol. 81. No. 3. P. 709-718.
Bargagli R., Lovari S. 1984. Age correlation between horn segments and tooth ce-
mentum annuali in the chamois (Rupicapra rupicapra ornata) // Saugetierk. Mitt.
Bd. 31. No. 2-3. S. 179-183.
Barker J.M., Boonstra R., Schulte-Hostedde A.1.2003. Age determination in yellow-
pine chipmunks (Tamias amoenus): a comparison of eye-lens masses and bone
sections // Canad. J. Zool. Vol. 81. No. 10. P. 1774-1779.
Barnes R.D., Longhurst W.H. 1960. Techniques for dental impressions, restraining
and embedding markers in live-trapped deer // J. Wildl. Manag. Vol. 24. P. 224-
226.
Barrier M.J., Barkalow F.S.Jr. 1967. A rapid technique for aging gray squirrels in
winter pelage // J. Wildl. Manag. Vol. 31. No. 4. P. 715-719.
Batulevicius D., Pauziene N. and Pauza D. H. 2001. Dental incremental lines in some
small species of the European vespertilionid bats // Acta Theriol. Vol. 46. P. 33-42.
Baubet E., Brandt S., Jullien J. M., Vassant J. 1994. Valeur de 1’examen de la denture
pour la determination de 1’age chez le sanglier (Sus scrofa) // Gibier Faune Sau-
vage, Game Wildl. Vol. 11. P. 119-132.
Beale D.M. 1962. Growth of the eye lens in relation to age in fox squirrels// J. Wildl.
Manag. Vol. 26. No. 2. P. 208-211.
Beg M.A., Hoffmann R.S. 1977. Age determination and variation in the red-tailed
chipmunk, Eutamias ruficaudus // The Murrelet. Vol. 58. P. 26-36.
256
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Bergerud A., Russell H.L. 1966. Extraction of incisors of Newfoundland caribou // J.
Wildl. Manag. Vol. 30. No. 4. P. 842-843.
Berry R.J., Truslove G.M. 1968. Age and eye lens weight in the House mouse // J.
Zool. Vol. 155. No. 2. P. 247-252.
Birney E.C., Jenness R.B., Baird D.D. 1975. Eye lens proteins as criteria of age in
cotton rats I I J. Wildl. Manag. Vol. 39. No. 4. P. 718-728.
Bothma J. du P., Teer J.G., Gates Ch.E. 1972. Growth and age determination of the
cottontail in south Texas // J. Wildl. Manag. Vol. 36. No. 4. P. 1209-1221.
Bousses P., Reale D. 1994. Valeur du masque facial comme critere d’age des femelies
de mouflons de Corse (Ovis musimon) dans la population de Kerguelen // Mam-
malia. Vol. 58. No. 2. P. 301-304.
Bowen W.D. 1982. Determining age of coyotes, Canis latrans, by tooth sections and
tooth-wear patterns // Canad. Field-Natur. Vol. 96. No. 3. P. 339-341.
Bradley J.A., Secord D., Prins L. 1981. Age determination in the arctic fox (Alopex
lagopus) // Canad. J. Zool. Vol. 59. P. 1976-1979.
Brandt E. 1961. Der Wert der Keilerwaffen als Altersweiser// Tragungsberichte No.
37. Beitrage zur Jagd- und Wildforsch. Bd. 1. S. 53-77.
Breitenmoser U., Kavczensky P., Dotterer M., Breinetmoser-Wursten C., Capt S.,
Bemhart F., Liberek M. 1993. Spatial organization and recruitment of lynx (Lynx
lynx) in re-introduced population in the Swiss Jura Mountains // J. Zool. Vol.
231. No. 3. P. 449-464.
Briedermann L. 1989. Gamswild Rupicapra rupicapra (L) // Buch der Hege. Bd. 1.
Haarwild. VEB Deutscher Landwirdschaftverlag / H.Stubbe (ed). Berlin. S. 2-56.
Briedermann L., Dittrich G., Lockow K.-W. 1989. Rotwild Cervus elaphus L. 11 Buch
der Hege. Bd. 1. Haarwild / H.Stubbe (ed). VEB Deutscher Landwirdschaftver-
lag. Berlin. S. 344-382.
Broekhuizen S., Maaskamp F. 1979. Age determination in the European hare (Lepus
europaeus Pallas) in The Netherlands // Zeitschrift Saugetierk. Bd. 44. S. 162-175.
BrokxP.A. 1972. Age determination of Venezuelan white-tailed deer//J. Wildl. Man-
ag. Vol. 36. No. 4. P. 1060-1067.
Bromel J., Zettl K. 1974. Beitrag zur Altersbestimmung beim Rotfuchs (Vulpes vulpes
L., 1758) H Ztschr. Jagdwiss. Bd. 20. No. 2. S. 96-104.
Brown W.A.B., Chapman N.G. 1990. The dentition of fallow deer (Dama dama): a
scoring scheme to assess age from wear of the permanent molariform teeth// J.
Zool. Vol. 221. P. 659-682.
Brown W.A.B., Chapman N.G. 1991a. Age assessment of fallow deer (Dama dama):
from a scoring scheme based on radiographs of developing permanent molari-
form teeth // J. Zool. Vol. 224. No. 3. P. 367-379.
Brown W.A.B., Chapman N.G. 1991b. Age assessment of red deer (Cervus elaphus) :
from a scoring scheme based on radiographs of developing permanent molari-
form teeth // J. Zool. Vol. 225. No. 1. P. 85-97.
Brown W.A.B., Chapman N.G. 1991c. The dentition of red deer (Cervus elaphus ). A
scoring scheme to assess age from wear of the permanent molariform teeth // J.
Zool. Vol. 224. No. 4. P. 519-536.
Литература
257
Bujalska G., Caboii-Raczinska К., Raczinski J. 1965. Studies on the European hare. 4.
Comparison of different criteria of age // Acta Theriol. Vol. 10. No. 1-5. P. 1-9.
Bullock D. J., Pickering S.P. 1984. The validity of horn ring counts to determine the
age of Scottish feral goats (Capra domestica)H J. Zool. Vol. 202. No. 4. P. 561-
564.
Burnett Ch.D., Kunz T.H. 1982. Growth rates and age estimation in Eptesicus fuscus
and comparison with Myotis lucifugus // J. Mammal. Vol. 63. No. 1. P. 33-41.
Cabori-Raczynska K. 1964. Studies on the European hare. III. Morphological varia-
tion of the skull // Acta Theriol. Vol. 9. No. 17. P. 249-285.
Cabon-Raczynska K., Raczynski J. 1972. Methods for determination of age in the
European hare // Acta Theriol. Vol. 17. No. 1-11. P. 75-86.
Calvert W., Ramsay M.A. 1985. Accurate aging of polar bears using tooth cementum
incremental lines // IV Intern. Theriological Congr. Abstr. Pap. and Posters. Ed-
monton; Alberta (Canada). No. 0094.1.
Carreno N.B., Brigada A.M., Rosi M.I., Casto V.A. 1990. Estimating ages of com
mice (Colomys musculinus) I I J. Mammal. Vol. 71. No. 3. P. 468-470.
Carson J.D. 1961. Epiphyseal cartilage as an age indicator in fox and grey squirrels // J.
Wildl. Manag. Vol. 25. No. 1. P. 90-93.
Castro-Campillo A., Gonzalez-Robles O., Pulido J.R. 1993. Determination of age
groups in Thomomys umbrinus (Rodentia: Geomydae) // Tex. J. Sci. Vol. 45. No.
3. P. 217-230.
Caughley G. 1965. Hom rings and tooth eruption as criteria of age in the Himalayan
thar, Hemitragus jemlachicus // N.Z.J. Sci. Vol. 8. No. 3. P. 333-351.
Cavallini P., Santini S. 1995. Age determination in the red fox in a Mediterranean
habitat // Ztschr. Saugetierk. Bd. 60. No. 3. S.136-142.
Cederlund G., Kjellander P., Stalfelt F. 1991. Age determination of the roe deer by
tooth wear and cementum layers - tests with known age material // Trans 20 th.
Congr. Int. Union Game Biol., Godollo, Aug 21-26, Pt. 2-Godolo. P. 540-545.
Chaplin R.E., White R.W.G. 1969. The use of tooth eruption and wear, body weight
and antler characteristics in the age estimation of male wild and park Fallow deer
(JDama dama) H J. Zool. Vol. 57. P. 125-132.
Chapman D.I., Chapman N. 1970. Development of the teeth and mandibles of fallow
deer // Acta Theriol. Vol. 15. No. 7. P. 111-131.
Chapman D.I., Chapman N.G., Colles C.M. 1985. Tooth eruption in Reeves muntjac
(Muntiacus reevesi) and its use as a method of age estimation // J. Zool. Vol. 205.
No. 2. P. 205-221.
Chen Ronghai, Dong Zhigang, Yang Chunwen. 1988. Использование бакулюма для
определения возраста самцов серой крысы // Шоулэй сюэбао-Acta Theriol.
Sin. Vol. 8. No. 4. P. 288-293.
Cheng H.C., Lee L.L. 2002. Postnatal growth, age estimation, and sexual maturity in
the formosan leaf-nosed bat Hipposideros terasensis 11 J. Mammal. Vol. 83. No.
3. P. 785-793.
Chipman R.K. 1965. Age determination of the cotton rat (Sigmodon hispidus) 11 Tu-
lane Studies in Zoology. Vbl.12. No. 2. P. 19-38.
258
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Chipman R.K. 1966. Cotton rat age classes during a population decline // J. Mammal.
Vol. 47. No. 1. P. 138-140.
Christian J.J. 1956. The natural history of a summer aggregation of the big brown
bat, Eptesicus fuscus I I Amer. Midland Natur. Vol. 55. No. 1. P. 66-95.
Churcher C.S. 1960. Cranial variation in the North American red fox // J. Mammal.
Vol. 41. P. 349-360.
Cicognani L., Monti F. 2000. Testing a quick and reliable method to discriminate age
classes in roe deer Capreolus capreolus on the basis of tooth wear // Hystrix. Vol.
11. No. 2. P. 127-130.
Clarke C.M.H., Dzieciolowski R.M., Batcheler D., Frampton C.M. 1992. A compar-
ison of tooth eruption and wear and dental cementum techniques in age determi-
nation of New Zealand feral pigs // Austral. Wildl. Res. Vol. 19. No. 6. P. 769-
777.
Colburn M.L. 1986. Suspensory tuberosities for aging and sexing squirrel // J. Wildl.
Manag. Vol. 50. No. 3. P. 456-459.
Connolly G.E., Dudzinski M.L., Longhurst W.M. 1969a. An improved age-lens weight
regression for black-tailed deer and mule deer П J. Wildl. Manag. Vol. 33. No. 3.
P. 701-704.
Connolly G.E., Dudzinski M.L., Longhurst W.M. 1969b. The eye lens as an indicator
of age in the black-tailed jack rabbit // J. Wildl. Manag. Vol. 33. No. 1. P. 159—
164.
Cool S.M., Bennett M.B., Romaniuk K. 1994. Age estimation of Pteropodid bats
(Megachiroptera) from hard tissue parameters I I Austral. Wildl. Res. Vol. 21. P.
353-364.
Cosson J.F., Rodolphe E, Pascal M. 1993. Determination de Page individuel, cr-
oissance post-natale et ontogenese precoce de Carollia perspicillata (L. 1758)
(Chiroptera, Phyllostomidae) // Mammalia. Vol. 57. No. 4. P. 565-578.
Couturier M. A. J. 1961. Determination de Г age du bouquetin des alpes {Capra aegagrus
ibex ibex) a 1’aide des dents et des comes // Mammalia. Vol. 25. No. 4. P. 453-461.
Cowan I.M. 1940. Distribution and variation in the native sheep of North America //
Amer. Midland. Natur. Vol. 24. No. 3. P. 505-580.
Сох M.K., Franklin W. L. 1990. Premolar gap technique for aging live black-tailed
prairie dogs // J. Wildl. Manag. Vol. 54. No. 1. P. 143-146.
Craighead, J. J., Craighead F.C., McCutchen H.E. 1970. Age determination of grizzly
bears from fourth premolar tooth sections // J. Wildl. Manag. Vol. 34. No. 2. P.
353-363.
Crowe D.M. 1972. The presence of annuli in bobcat tooth cementum layers // J.
Wildl. Manag. Vol. 36. No. 4. P. 1330-1332.
Dagg A.I., Leach D., Sumner-Smith G. 1975. Fusion of the distal femoral epiphysis
in male and female marten and fisher // Canad. J. Zool. Vol. 53. P. 1514-1518.
Daly J.C., Patton J.L. 1986. Growth, reproduction, and sexual dimorphism in Tho-
momys bottae pocket gophers // J. Mammal. Vol. 67. No. 2. P. 256-265.
Dapson R.W. 1968. Reproduction and age structure in a population of short-tailed
shrews, Blarina brevicauda // J. Mammal. Vol. 49. P. 205-214.
Литература
259
Dapson R.W. 1980. Quidelines for statistical usage in age-determination techniques // J.
Wildl. Manag. Vol. 44. P. 541-548.
Dapson R.W., Irland J.M. 1972. An accurate method of determining age in small
mammals // J. Mammal. Vol. 53. No. 1. P. 100-106.
Dapspn R.W., Otero J.G., Holloway W.R. 1968. Biochemical changes with age in
lenses of white mice // Nature. Vol. 218. No. 5141. P. 573.
Dasmann R.F., Taber R.D. 1956. Determining structure in Columbian black tailed
deer populations // J. Wildl. Manag. Vol. 20. No. 1. P. 78—83.
Davis D.E. 1964. Evaluation of characters for determining age of woodchucks // J.
Wildl. Manag. Vol. 28. No. 1. P. 9-15.
Davis D.W., Sealander J.A. 1971. Sex ratio and age structure in two red squirrel
populations in Northern Saskatchewan // Canad. Field-Natur. Vol. 85. No. 4. P.
303-308.
Dawson A.B. 1934. Further studies on epiphyseal union in the skeleton of the rat //
Anat. Rec. Vol. 60. P. 83—86.
Dawson A.B. 1935. The sequence of epiphyseal union in the skeleton of the mouse
with special reference to the phenomenon of «lapsed» union // Anat. Rec. Vol.
63. P. 93-99.
Degn H.J. 1973. Systematic position, age criteria and reproduction of Danish red
squirrels (Sciurus vulgaris L.) // Danish Rev. Game Biol. Vol. 8. No. 2. P. 3—24.
DeYoung Ch.A. 1989. Aging live white-tailed deer on southern ranges // J. Wildl.
Manag. Vol. 53. No. 3. P. 519-523.
d’Errico F., Vanhaeren M. 2002. Criteria for identifying red deer (Cervus elaphus)
age and sex from their canines. Application to the study of Upper Palaeolithic and
Mesolithic ornaments // J. Archaeol. Sci. Vol. 29. P. 211-232.
Divljan A., Parry-Jones K., Wardle G.M. 2006. Age Determination in the Grey-Head-
ed Flying Fox // J. Wildl. Manag. Vol. 70. No. 2. P. 607-611.
Dix L.M., Strickland M.A. 1986. Use of tooth radiographs to classify martens by sex
and age // Wildl. Soc. Bull. Vol. 14. P. 275-279.
Doude van Troostwijk W.J. 1976. Age determination in muskrats, Ondatra zibethicus
(L.), in the Netherlands // Lutra. Vol. 18. No. 3. P. 33-43.
Dow S. A., Wright P.L. 1962. Changes in mandibular dentition associated with age in
pronghorn antelope // J. Wildl. Manag. Vol. 26. No. 1. P. 1-18.
Dudzinski M.L., Mykytowycz R. 1961. The eye lens as an indicator of age in the wild
rabbit in Australia // Austral. Wildl. Res. Vol. 6. No. 2. P. 156-159.
Dyck M.G., Bourgeois J.M., Miller E.H. 2004. Growth and variation in the bacula of
polar bears (Ursus maritimus) in the Canadian Arctic // J. Zool. Vol. 264. No 1.
P. 105-110.
Dzieciolowski R. 1976. Age appraisal in moose // Acta Theriol. Vol. 21. No. 23. P.
307-310.
Earle R.D., Kramm K.R. 1980. Techniques for age determination in the Canadian
porcupine // J. Wildl. Manag. Vol. 44. No. 2. P. 413-419.
Ecke D.H., Kinney A.R. 1956. Aging meadow mice, Microtus califomicus, by obser-
vation of moult progression // J. Mammal. Vol. 37. No. 2. P. 249-254.
260
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Edwards W.R. 1962. Age structure of Ohio cottontail populations from weight of
lenses // J. Wildl. Manag. Vol. 26. No. 2. P. 125-132.
Eidmann, H. 1932. Alterbestimmungen am Gebiss des Rothirches (Cervus elaphus
L). Mitt. Forstwirsch., Forstwiss. Bd. 3. No. 2. S. 291-341.
Elder W.H., Shanks C.E. 1962. Ages changes in tooth wear and morphology of the
baculum in muskrats // J. Mammal. Vol. 43. No. 2. P. 144-150.
Empel W. 1962. Morphologic des Schadels von Bison bonasus И Acta Theriol. Vol. 6.
No. 4. P. 53-111.
Erickson J.A., Anderson A.E., Medin D.E., Bowden D.C. 1970. Estimating ages of
mule deer— an evaluation of technique accuracy // J. Wildl. Manag. Vol. 34. No.
3. P. 523-531.
Fedyk A. 1974a. Gross body composition in postnatal development of the bank vole. I.
Growth under laboratory conditions // Acta Theriol. Vol. 19. No. 26. P. 381-401.
Fedyk A. 1974b. Gross body composition in postnatal development of the bank vole.
III. Estimating age H Acta Theriol. Vol. 19. No. 26. P. 429-440.
Feldhamer G.A., Chapman J.A. 1980. Evaluation of the eye lens method for age
determination in sika deer // Acta Theriol. Vol. 25. No. 14-21. P. 239-244.
Fisher E.W., Perry A.E. 1970. Estimating ages of gray squirrels by lens-weight // J.
Wildl. Manag. Vol. 34. No. 4. P. 825-828.
Fog M. 1969. Studies on the weasel (Mustela nivalis) and the stoat (Mustela er-
minea) in Denmark // Danish. Rev. Game Biol. Vol. 6. No. 2. P. 1-14.
Fogl J.G., Mosby H.S. 1978. Aging grey squirrels by cementum annuli in razor-
sectioned teeth // J. Wildl. Manag. Vol. 42. No. 2. P. 444 448.
Fredrickson L. 1983. Use of radiographs to age badger and striped skunk // Wildl.
Soc. Bull. Vol. 11. No. 3. P. 297-299.
Freudenthal M., Martin-Suarez E., Bendala N. 2002. Estimating age through tooth
wear. A pilot study on tooth abrasion in Apodemus (Rodentia, Mammalia) //
Mammalia. Vol. 66. No. 1. P. 275-284.
Friend M. 1967a. Some observations regarding eye-lens weight as a criterion of age
in animals // N. Y. Fish and Game J. Vol. 14. No. 2. P. 91-121.
Friend M. 1967b. Relationship between eye-lens weight and variation in diet// N.Y.
Fish and Game J. Vol. 14. No. 2. P. 122-151.
Friend M. 1967c. A review of research concerning eye-lens weight as a criterion of
age in animals // N.Y. Fish and Game J. Vol. 14. No. 2. P. 152-165.
Friend M. 1968. The lens technique H Transactions of Thirty-Third North American
Wildl. Conference. P. 279-298.
Friend M., Linhart S.B. 1964. Use of the eye lens as an indicator of age in the red fox //
N.Y. Fish and Game J. Vol.ll. No. 1. P. 58-66.
Friend M., Severinghaus C.W. 1967. Influence of nutrition on eye-lens weight in deer //
N.Y. Fish and Game J. Vol. 14. No. 2. P. 166-175.
Friley C.E. 1947. Preparation and preservation of the baculum of mammals // J.
Mammal. Vol. 28. No. 4. P. 395-397.
Friley C.E. 1949. Age determination by use of the baculum in the River Otter (Lutra
c. canadensis Schreber) H J. Mammal. Vol. 30. No. 2. P. 102-110.
Литература
261
Frison G.C., Reher Ch. A. 1970. Age determination of buffalo by teeth eruption and
wear // Plains Anthropologist. Vol. 15. No. 50. Part 2. P. 46-50.
Fuller W.A. 1959. The horns and teeth as indicators of age in bison // J. Wildl. Manag.
Vol. 23. No. 3. P. 342-344.
Fuller W.A. 1988. Is weight a reliable index to age in microtine rodents? // Acta
Theriol. Vol. 33. No. 17. P. 247-261.
Garshelis D.L. 1984. Age estimation of living sea otters 11 J. Wildl. Manag. Vol. 48.
No. 2. P. 456-463.
Garsia-Perea R. 1996. Patterns of postnatal development in skulls of lynxes, genus
Lynx (Mammalia: Carnivora) H J. Morphol. Vol. 229. No. 3. P. 241-254.
Garsia-Perea R., Gaquero R.A. 1999. Age estimation in Iberian wildcats Felis silves-
tris, by canine tooth sections // Acta Theriol. Vol. 44. No. 3. P. 321-327.
Gasaway W.C., Harkness D.B., Rausch R.A. 1978. Accuracy of moose age deter-
mination from incisor cementum layers // J. Wildl. Manag. Vol. 42. No. 3. P.
558-563.
Gay S.W, Best T.L. 1996. Age-related variation in skulls of the puma (Puma con-
color) H J. Mammal. Vol. 77. No. 1. P. 191-198.
Gee K.L., Holman J.H., Kausey M.K., Rossi A.N., Armstrong J.B. 2002. Aging white-
tailed deer by tooth replacement and wear: a critical evaluation of a time-hon-
oured technique H Wildl. Soc. Bull. Vol. 30. No. 2. P. 387-393.
Geist V. 1966. Validity of horn segment counts in aging bighorn sheep // J. Wildl.
Manag. Vol. 30. No. 3. P. 634-635.
Genov P.V. 1992. Teeth eruption and their replacement in wild boar (Sus scrofa
Solater 1986) in Angeria // Acta Zool. Bulgarica. Vol. 43. P. 43-46.
George J.C., Bada J., Zeh J., Scott L., Brown S.E., O’Hara T, Suydam R. 1999. Age
and growth estimates of bowhead whales (Balaenoptera mysticetus) via aspartic
acid racemization // Canad. J. Zool. Vol. 77. P. 571—580.
Giacometti M., Willing R., Defila C. 2002. Ambient temperature in spring affects
horn growth in male alpine ibexes // J. Mammal. Vol. 83. No. 1. P. 245-251.
Gilbert F.F., Stolt S.L. 1970. Variability in aging Maine white-tailed deer by tooth-
wear characteristics // J. Wildl. Manag. Vol. 34. No. 3. P. 532-535.
Gipson P.S., Ballard W.B., Nowak R.M., Meeh L.D. 2000. Accuracy and precision
of estimating age of gray wolves by tooth wear // J. Wildl. Manag. Vol. 64. No.
3. P. 752-758.
Gliwicz J, Jancewicz E. 2001. Aging and kohort dynamics in Sorex shrews // Acta
Theriol. Vol. 46. No. 3. P. 225-234.
Gonzalezesteban J., Villate I., Castien E., Rey I., Gosalbez J. 2002. Age determination
of Galemys pyrenaicus 11 Acta Theriol. Vol. 47. No. 1. P. 107-112.
Goodyear J.J. 1983. A computer method of creating tooth wear classes for the study
of age structure in rodent population // Amer. Zoologist. Vol. 23. No. 4. P. 949.
Gosling L.M., Huson L.W., Addison G.C. 1980. Age estimation of coypus (Myocas-
tor coypus) from eye lens weight I I J. Appl. Ecol. Vol. 17. No. 3. P. 641-647.
Gourley R.S., Jannett F.J. 1975. Pine and montane vole age estimates from eye lens
weights // J. Wildl. Manag. Vol. 39. No. 3. P. 550-556.
262
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Graf М., Wandeler A.I. 1982. Altersbestimmung bei Dachsen (Meles meles) // Rev.
Suisse Zool. Bd. 89. No. 4. P. 1017-1023.
Grau G.A., Sanderson G.C., Rogers J.P. 1970. Age determination of raccoons // J.
Wildl. Manag. Vol. 34. No. 2. P. 364-372.
Gray G.G., Simpson C.D. 1985. Hom growth and aging of free-ranging Barbary
sheep (Ammotragus lervia) 11 Mammalia. Vol. 49. No. 1. P. 85-91.
Greer K.K. 1957. Some osteological characters of know-age ranch minks // J. Mam-
mal. Vol. 38. No. 3. P. 319-329.
Greer K.R., Yeager H.W. 1967. Sex and age indications from upper canine teeth of
elk (wapiti) // J. Wildl. Manag. Vol. 31. No. 3. P. 408-417.
Gronquist R.M., Dinneford W.B. 1984. Age determination of musk-oxes from dental
cementum annuli // Biol. Pap. Univ. Alaska. Spec. Rep. No. 4. P. 67-68.
Grue H., Jensen B. 1976. Annual cementum structures in canine teeth in Arctic foxes
(Alopex lagopus L.). from Greenland and Denmark // Danish Rev. Game Biol.
Vol. 10. No. 3. P. 3-12.
Grue H.E., King C.M. 1984. Evaluation of age criteria in New Zealand stoats (Muste-
la erminea) of known age // New Zealand J. Zool. Vol. 11. P. 437-443.
Grulich 1.1967. Zur Methodik der Altersbestimmung des Maulwurfs, Talpa europaea L., in
der Periode seiner selbstandigen Lebensweise // Zool. Listy Bd. 16. No. 1. S. 41-59.
Grulich I., Jurik M. 1994. A morphometric study of Eliomys quercinus from the
Slovakian Karst // Folia Zool. Vol. 43. No. 3. P. 209-218.
Grummt W. 1989. Waschbar, Procyon lotor (L.) H Buch der Hege. Bd. 1. Haarwild I
H.Stubbe (ed). VEB Deutscher Landwirdschaftverlag. Berlin. S. 410-416.
Gumell J., Knee Cl. 1984. Determining the age of wood mice (Apodemus sylvaticus) I I
Folia Zool. Vol. 33. No. 4. P. 339-348.
Habermehl K.H. 1985. Alterbestimmung bei Wild- und Pelztieren. Verlag Paul Parey-
Hamburg, Berlin. 223 S.
Haferkom J. 1995. Zur Alterbestimmung von Apodemus flavicollis anhand der Au-
genlinsentrockenmasse // Metoden feldokologischer Saugetierforschung. Hrsg.
Stubbe M., Stubbe A., Heidecke D. Halle/Saale. S. 143-145.
Hagen A., Stenseth N. Chr., Ostbye E., Skar H.J. 1980. The eye lens as an age
indicator in the root vole // Acta Theriol. Vol. 25. No. 4. P. 39-50.
Hagen В. 1956. Alterbestimmung an einiger Muriden-Arten // Ztschr. Saugetierk. Bd.
21. S. 39-43
Hale J.B. 1949. Aging cottontail rabbits by bone growth // J. Wildl. Manag. Vol. 13.
No. 2. P. 216-225.
Hall J.S., Cloutier R.J., Griffin D.R. 1957. Longevity records and notes on tooth
wear of bats 11 J. Mammal. Vol. 38. No. 3. P. 407-409.
Hall-Martin A. J. 1976. Dentition and age determination of the giraffe Giraffa Camel-
opardalis 11 J. Zool. Vol. 180. P. 263-289.
Hamilton W.J. 1934. The life history of the rufescent woodchuck, Marmota топах
rufescence Howell // Annals of the Carnegie Museum. Vol. 23. P. 85-178.
Hamilton W.J., Eadie W.R. 1964. Reproduction in the otter, Lutra canadensis II J.
Mammal. Vol. 45. No. 2. P. 242—252.
Литература
263
Hamlin K.L., Рас D.F., Sime С.A., DeSimone R.M., Dusek G.L. 2000. Evaluating the
accuracy of ages obtained by two methods for Montana Ungulates // J. Wildl.
Manag. Vol. 64. No. 2 . P. 441-449.
Hansson L. 1983q. Differences in age indicators between field and laboratory small
rodent populations // Mammalia. Vol. 47. No. 3. P. 371-376.
Hansson L. 1983b. Reproductive development related to age indicators in microtine
rodents 11 J. Wildl. Manag. Vol. 47. No. 4. P. 1170-1172.
Hardy A.R., Quy R.J., Huson L.W. 1983. Estimation of age in the Norway rat (Rat-
tus norvegicus Berkenhout) from the weight of the eye lens // J. Appl. Ecol. Vol.
20. No. 1. P. 97-102.
Harris S. 1978. Age determination in the red fox (Vulpes vulpes) — an evaluation of
technique efficiency as applied to a sample of suburban foxes // J. Zool. Vol.
184. P. 91-117.
Harris S., Cresswell W.J., Cheeseman C.L. 1992. Age determination of badgers (Me-
les meles) from tooth wear: the need for a pragmatic approach // J. Zool. Vol.
228. No. 4. P. 679-684.
Harshyne W.A., Diefenbach D.R., Alt G.L., Matson G.M. 1998. Analysis of error
from cementum annuli age estimates of known-age Pensylvania black bears // J.
Wildl. Manag. Vol. 62. No. 4. P. 1281-1291.
Haynes G. 1984. Tooth wear rate in northern bison // J. Mammal. Vol. 65. No. 3. P.
487-491.
Hearn B.J., Mercer W.E. 1988. Eye-lens weight as an indicator of age in Newfound-
land arctic hares // Wildl. Soc. Bull. Vol. 16. No. 4. P. 426-429.
Heggberget T.M. 1984. Age determination in the European otter Lutra lutra lutra 11
Ztschr. Saugetierk. Bd. 49. No. 5. S. 299-305.
Helfrnan P.M., Bada J.L. 1976. Aspartic acid racemisation in dentine as a measure of
ageing И Nature. Vol. 262. No. 5566. P. 279-281.
Hell P. 1967. Variabilita vahy ocych sosoviek vzhl’adom na urcovanie veku zajacov //
Lynx. Vol. 8 (N.S.). P. 3-6.
Hell P. 1976. Porovnanie urcovania veku diviacej zveri podl’a anul v zubnom cemente
a podl’a opotrebovania chrupu I I Folia Venator. Vol. 5-6. P. 57-66.
Hemming J.E. 1969. Cemental deposition, tooth succession, and horn development
as criteria of age in Dall sheep// J. Wildl. Manag. Vol. 33. No. 3. P. 552-558.
Hench J.E., Gordon L_, Kirkland Jr., Setzer H.W., Douglass L.W. 1984. Age classifi-
cation for the gray squirrel based on eruption, replacement, and wear of molar-
iform teeth // J. Wildl. Manag. Vol. 48. No. 4. P. 1409-1414.
Henderson A., Bowen H.M. 1979. A short note: Estimating the age of the European
rabbit, Oryctolagus cuniculus, by counting the adhesion lines in the periosteal
zone of the lower mandible // J. Appl. Ecol. Vol. 16. No. 2. P. 393-396.
Hensel, R.J., Sorensen F.E. 1980. Age determination of live polar bears 11 Bears —
their biology and management. Kalispel, Montana. (Bear Biol. Assoc. Conf. Ser. 3) /
C.J. Martinka , McArthur K.L. (eds.). P. 93-100.
Henzell R. 1987. Eye-lens weight as an indicator of age in Australian goats (Capra
hircus L.) // Austral. Wildl. Res. Vol. 14. No. 1. P. 69-78.
264
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Hewison Vincent J.P., Angibault J.M., Delorme D., Van Laere G., Gaillard J-M.
1999. Tests of estimation of age from tooth wear on roe deer of known age: variation
within and among populations // Canad. J. Zool. Vol. 77. No. 1. P. 58-67.
Hillman-Smith A.K.K., Owen- Smith N., Anderson J.L., Hall-Martin A. J., Selaladi J.P.
1986. Age estimation of the White rhinoceros (Ceratotherium simum) I I J. Zool.
Vol. 210. P. 355-379.
Hillson S. 1986. Teeth. Cambridge Univ. Press. Cambridge. 376 p.
Hlavac A. 1979. Contribution to the age determination of the Microtus arvalis (Pallas,
1779) by dry lens weight 11 Lynx. Vol. 20. P. 35-44.
Hoefs M., Konig R. 1984. Reliability of aging old Dall sheep eves by the horn annulus
technique 11 J. Wildl. Manag. Vol. 48. No. 3. P. 980-982.
Hoffmeister D.F., Zimmerman F.G. 1967. Growth of the skull in the cottontail (Syl-
vilagus floridanus) and its application to age-determination // Amer. Midland
Natur. Vol. 78. No. 1. P. 198-206.
Homolka M. 1979. Zur Alterbestimmung der Schlafer (Gliridae) H Folia Zool. Vol. 28.
No. 2. P. 103-114.
Hoogland J.L., Hutter J.M. 1987. Using molar attrition to age live prairie dogs H J.
Wildl. Manag. Vol. 51. No. 2. P. 393-394.
Hoying K.M., Kunz Т.Н. 1998. Variation in size at birth and post-natal growth in the
insectivorous bat Pipistrellus subflavus (Chiroptera: Vespertilionidae) // J. Zool.
Vol. 245. No. 1. P. 15-27.
НгаЬё V., Zejda J., Horakova M. 1989. Ageing of sika deer (Cervus nippori) H Folia
Zool. Vol. 38. No. 2. P. 97-107.
lason G.R. 1988. Age determination of mountain hares (Lepus timidus}. A rapid meth-
od and when to use it // J. Appl. Ecology. Vol. 25. No. 2. P. 389-395.
Isaac S.S., Marimuthu G. 1996. Postnatal growth and age estimation in the Indian
pygmy bat Pipistrellus mirnus // J. Mammal. Vol. 77. No. 1. P. 199-204.
Jaarsveld A.S. van, Henschel J.R., Skinner J.D. 1987. Improved age estimation in
spotted hyenas (Crocuta crocuta) // J. Zool. Vol. 213. No. 4. P. 758-762.
Jackson D.L., Gluesing E.A., Jacobson H.A. 1988. Dental eruption in bobcats // J.
Wildl. Manag. Vol. 52. No. 3. P. 515-517.
Jain A.P., Prakash I. 1981. Eye-lens as an indicator of age in the Indian gerbil, Tatera
indica // Growth. Vol. 45. No. 2. P. 119-122.
Jean Y., Bergeron J.M., Bisson S., Larocque B. 1986. Relative age determination of
coyotes, Canis latrans, from Southern Quebec // Canad. Field-Natur. Vol. 100.
No. 4. P. 483-487.
Jeffery R.C.V., Hanks J. 1981. Age determination of eland Taurotragus oryx (Pallas,
1766) in the Natal Highveld// S. Afr. Tydskr. Dierk. Vol. 16. No. 2. P. 113-122.
Jenks J.A., Bowyer R.T., Clark A.G. 1984. Sex and age-class determination for
fisher using radiographs of canine teeth // J. Wildl. Manag. Vol. 48. No. 2. P.
626-628.
Jenks J.A., Clark A.G., Bowyer R.T. 1986. Sex and age determination for fisher
using radiographs of canine teeth: a response // J. Wildl. Manag. Vol. 50. No. 2.
P. 277-278.
Литература
265
Jensen В., Nielsen N.B. 1968. Age determination in the red fox (Vulpes vulpes L.)
from canine tooth sections // Danish Rev. Game Biol. Vol. 5. No. 6. P. 1-15.
Johnson N.F., Brown B.A., Bosomworth J.C. 1981. Age and sex characteristics of
bobcat canines and their use in population assessment // Wildl. Soc. Bull. Vol. 9.
No. 3. P. 203-206.
Johnston D., Beauregard M. 1969. Rabies epidemiology in Ontario // Bull. Wildl.
Disease Assoc. Vol. 5. P. 367-370.
Junge R., Hoffmeister D.F. 1980. Age determination in raccoons from cranial suture
obliteration // J. Wildl. Manag. Vol. 44. No. 3. P. 725-729.
Kahmann H., Staudenmayer T. 1968. Zahnwachstum, Zahnwechsel und Zahnabnut-
zung bei dem Gautenschlafer Eliomys quercinus Linnaeus, 1766// Ztschr. Sauge-
tierk. Bd. 33. No. 6. S. 358-364.
Kappeler A. von 1985. Untersuchungen zur Altersbestimmung und zur Altersstruktur
verschiedener Stichproben aus Rotfuchs-Populationen (Vulpes vulpes L.) in der
Schweiz. Lizentiatsarbeit Zoologisches Institut der Universitat Bern. 129 S.
Kauffman R.C., Norton H.W., Harmon B.C., Breidenstein B.C. 1967. Growth of the
porcine eye lens as an index to chronological age // J. Anim. Sci. Vol. 26. No. 1.
P. 31-35.
Kauhala K., Helle E. 1990. Age determination of the raccoon dog in Finland // Acta
Theriol. Vol. 35. No. 3-4. P. 321-329.
Keay J.A. 1995. Accuracy of cementum age assignments for black bears // Calif.
Fish and Game. Vol. 81. No. 3. P. 113-121.
Keiss R.E. 1969. Comparison of eruption-wear patterns and cementum annuli as age
criteria in elk // J. Wildl. Manag. Vol. 33. No. 1. P. 175-180.
Keith L.B., Cary J.R. 1979. Eye lens weights from free-living adult snowshoe hares
of known age // J. Wildl. Manag. Vol. 43. No. 4. P. 965-969.
Keller A. 1974. Determination de 1’age de Mus musculus Linne par 1’usure de la dentition
// Rev. Suisse Zool. Bd. 81. 4. S. 839-844.
Kerwin M.L., Mitchell G.J. 1971. The validity of the wear-age technique for Alberta
pronghorns // J. Wildl. Manag. Vol. 35. No. 4. P. 743-747.
Kierdorf U., Becher J. 1997. Mineralization and wear of mandibular first molars in
red deer (Cervus elaphus) of known age // J. Zool. Vol. 241. No. 1. P. 135-143.
King C.M. 1980. Age determination in the Weasel (Mustela nivalis) in relation to the
development of the skull // Ztschr. Saugetierk. Bd. 45. S. 153-173.
King C.M. 1991. A review of age determination methods for the stoat Mustela er-
mineall Mamm. Rev. Vol. 21. No. 1. P. 31-49.
Kirkpatrick R.D., Sowls L.K. 1962. Age determination of the collared peccary by the
tooth-replacement pattern // J. Wildl. Manag. Vol. 26. No. 2. P. 214-217.
Kleiman D.G. 1969. Maternal care, growth rate, and development in the noctule
(Nyctalus noctula), pipistrelle (Pipistrellus pipistrellus), and serotine (Eptesicus
fuscus) bats // J. Zool. Vol. 157. P. 187-211.
Klein E, Breton D., Brandt S., Gaillard J.M. 1990. Appreciation de Page du sanglier
(Sus scrofa) a partir de la masse du cristallin et de la masse de 1’animal // Gibier
Faune Sauvage. Vol. 7. P. 39-51.
266
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Klein R. G., Wolf С., Freeman L.G., Allwarden К. 1981.The use of dental crown
heights for constructing age profiles of red deer and similar species in archaeo-
logical samples // J. Archaeol. Sci. Vol. 8. No. 1. P. 1-31.
Klevezal G.A. 1996. Recording structures of mammals. Determination of age and
reconstruction of life history. A.A.Balkema, Rotterdam. 274 p.
Klevezal G.A., Pucek Z. 1987. Growth layers in tooth cementum and dentine of
European bison and its hybrids with domestic cattle // Acta Theriol. Vol. 32. No.
9. P. 115-128.
Kleymann M. 1972. Altersbedingte Veranderungen des Pulpencavum beim Rehwild //
Ztschr. Jagdwiss. Bd. 18. No. 1. S. 36-39.
Knight R.R. 1966. Bone characteristics associated with aging in elk // J. Wildl. Man-
ag. Vol. 30. No. 2. P. 369-374.
Koike H., Ohtaishi N. 1985. Prehistoric hunting pressure estimated by the age com-
position of excavated sika deer (Cervus nippon) using annual layers of tooth
cement // J. Archaeol. Sci. Vol. 12. P. 443-456.
Koponen T. 1970. Age structure in sedentary and migratory populations of the Nor-
wegian lemming, Lemmus letnmus (L.), at Kilpisjarvi in 1960 // Ann. Zool. Fenn.
Vol. 7. No. 2. P. 141-187.
Koubek P., Hrabe V 1983. Dynamics of horn growth in the Jeseniki Mts population
of chamois, Rupicapra rupicapra rupicapra (CSSR) // Folia Zool. Vol. 32. No.
2. P. 97-107.
Koubek P., Hrabe V. 1988. Urcovani veku smcu {Capreolus capreolus) jihomoravske
populace //Folia Venator. Vol. 18. P. 83-97.
Kozakiewicz M. 1976. The weight of eye lens as the proposed age indicator of the
bank vole // Acta Theriol. Vol. 21. No. 23. P. 314-316.
Krasinska M., Krasinski Z. 2002. Body mass and measurements of the European
bison during postnatal development // Acta Theriol. Vol. 47. No. 1. P. 85-
106.
Krasinska M., Krasinski Z. 2004. Zubr. Monografia przyrodnicza. Warszawa-Bialow-
ieza. 312 p.
Krsmanovic L., Purger J. J., Mikes M. 1985. Some taxonomic characters and age of
the common hamster, Cricetus cricetus L., in the area of Vbjvodina (Yugoslavia)
// Глас. Прир. муз. Белград. Vol. 41. Р. 123-134.
Krysiak К., Swiezyhski К. 1967. The present status of research on the morphology
of the European bison // Acta Theriol. Vol. 12. No. 22. P. 339-348.
Krystufek B., Kolaric K., Paunovic M. 2000. Age determination and age structure in
Martino’s vole Dinaromys bogdanovi H Mammalia. Vol. 64. No. 3. P. 361-370.
Krystufek B., Pistotnik M., Casar K.S. 2005. Age determination and age structure in
the edible dormice Glis glis based on incremental bone lines // Mammal Rev. Vol.
35. No. 2. P. 210-214.
Kuehn D.W., Berg W.M. 1981. Use of radiographs to identify age-classes of fisher // J.
Wildl. Manag. Vol. 45. No. 4. P. 1009-1010.
Kuehn D.W., Berg W.M. 1983. Use of radiographs to age otters // Wildl. Soc. Bull.
Vol. 11. No. 1. P. 68-70.
Литература
267
Kunz Th. H., Anthony E.L.P. 1982. Age estimation and post-natal growth in the bat
Myotis lucifugus // J. Mammal. Vol. 63. No. 1. P. 23-32.
Kvam T. 1983. Age determination in European lynx Lynx I. lynx (L) based on cranial
development // Fauna Norv. Ser. A. Vol. 4. P. 31-36.
Landon D.B., Waite C.A., Peterson R.O., Meeh L.D. 1998. Evaluation of age determi-
nation techniques for gray wolves // J. Wildl. Manag. Vol. 62. No. 2. P. 674-682.
Lapierre L.E. 1976. Comparaison entre deux methodes d’estimation de 1’age chez le
cerf de Virginie (Odocoileus virginianus) // Natur. Canad. Vol. 103. No. 2. P. 73-75.
Larson J.R., Taber R.D. 1980. Criteria of sex and age// Wildlife management tech-
niques manual. 4 ed. Wildlife Soc. Washington. P. 143-181.
Laws R.M. 1952. A new method of age determination for mammals // Nature. Vol.
169. P. 972-973.
Le Boulenge E. 1977. Two ageing methods for muskrats: live or dead animals// Acta
Theriol. Vol. 22. No. 30-36. P. 509-520.
Leach D., Hall B.K., Dagg A.J. 1982. Aging marten and fisher by development of the
suprafabellar tubercle // J. Wildl. Manag. Vol. 46. No. 1. P. 246-247.
Lechleitner R.R. 1954. Age criteria in mink, Mustela vison // J. Mammal. Vol. 35. No.
4. P. 496-503.
Lemnell P.A., 1974. Age determination in red squirrels (Sciurus vulgaris L.) // 11-th
Int. Congr. Game Biol., Stockholm, 1973, Stockholm. P. 573-580.
Lewall E.F., Cowan I. McT. 1963. Age determination in black-tail deer by degree of
ossification of the epiphyseal plate in the long bones // Canad. J. Zool. Vol. 41. P.
629-636.
Lidicker W.Z., MacLean S.F. 1969. A method for estimating age in the California
vole, Microtus californicus И Amer. Midland Natur. Vol. 82. No. 2. P. 450-470.
Lin J., Zhang Y, Zhang L. 1989. Определение возраста и структура популяции
Citellus undulatus И Acta Theriol. Sin. Vol. 9. No. 3. P. 216-220.
Lindeque M., Skinner J.D. 1984. Size frequency analysis of tooth wear in spotted
hyaenas Crocuta crocuta // South Afr. J. Zool. Vol. 19. No. 4. P. 291-294.
Linhart S.B., Knowlton F.F. 1967. Determining age of coyotes by tooth cementum
layers // J. Wildl. Manag. Vol. 31. No. 2. P. 362-365.
Lissovsky A. A. 2004. Contribution to age determination of pikas (Lagomorpha, Ocho-
tonidae, Ochotona) II Russian J. Theriol. Vol. 3. No. 1. P. 43-48.
Lochmiller R.L., Helligren E.C., Grant W.E. 1984. Sex and age characteristics of the
pelvic girdle in the collared peccary // J. Wildl. Manag. Vol. 48. No. 2. P. 639-641.
Lockard G.R. 1972. Further studies of dental annuli for aging white-tailed deer // J.
Wildl. Manag. Vol. 36. No. 1. P. 46-55.
Lockow K.W. 1987. Neues zur Altersbestimmung des Rothirsches // Unsere Jagd.
Bd. 37. No. 8. S. 232-233.
Lombaard L.J. 1971. Age determination and growth curves in the black-backed jack-
al, Canis mesonielas Schreber, 1775 (Carnivora: Canidae) //Annals of Transvaal
Museum. Vol. 27. No. 7. P. 135-170.
Leno Odd. 1970. The polar bear (Ursus maritimus Ph.) in the Svalbard area // Norsk
Polarinst. Skrifter. Vol. 149. P. 1-103.
268
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Lord R.D. 1959. The lens as an indicator of age in cottontail rabbits // J. Wildl.
Manag. Vol. 23. No. 3. P. 358-360.
Lord R.D. 1961. The lens as an indicator of age in the gray fox // J. Mammal. Vol. 42.
No. 1. P. 109-111.
Lord R.D. 1966. Growth of the lens of the pampas gray fox {Dusicyon gymnocercus
antiquus) and the Patagonian gray fox {Dusicyon griseus griseus) 11 J. Mammal.
Vol. 47. No. 3. P. 536-538.
Lowe V.P.W. 1967. Teeth as indicators of age with special reference to Red Deer
{Cervus elaphus) of known age from Rhum // J. Zool. Vol. 152. No. 2. P. 137-
153.
Lowe V.P.W. 1971. Root development of molar teeth in the bank vole {Clethrionomys
glareolus) И J. Anim. Ecol. Vol. 40. No. 1. P. 49—61.
Lubinski P.M. 2001. Estimating age and season of death of pronghorn antelope {An-
tilocapra americana Ord) by means of tooth eruption and wear // Internat. J.
Osteoarchaeol. Vol. 11. No. 3. P. 218-230.
Ludwig J.R., Dapson R.W. 1977. Use of insoluble lens proteins to estimate age in'
white-tailed deer // J. Wildl. Manag. Vol. 41. No. 2. P. 327-329.
Lundervold M., Langvatn R., Milner-Gulland E.J. 2003. A comparison of age estima-
tion methods for the saiga antelope Saiga tatarica // Wildl. Biology. Vol. 9. No. 3.
P. 219-227.
MacCracken J.G., Van Ballenberghe V. 1987. Age and sex related differences in fecal
pellet dimensions of moose I I J. Wildl. Manag. Vol. 51. No. 2. P. 360-364.
Malcolm J.R. 1992. Use of tooth impressions to identify and age live Proechimys guyan-
nensis and P. curvieri (Rodentia: Echimyidae) // J. Zool. Vol. 227. No. 4. P. 537-546.
Malcolm J.R., Brooks R.J. 1981. Eye lens weight and body size as criteria of age in
beaver {Castor canadensis) И Canad. J. Zool. Vol. 59. No. 6. P. 1189-1192.
Manning TH. 1964. Age determination in the polar bear Ursus maritimus Phipps //
Canad. Wildl. Serv. Occas. Papers. Vol. 5. P. 3-12.
Maringgele F.J. von. 1978/1979. Aftersbestimmung beim Reh {Capreolus capreolus
[L.]) und beim Rothirsch {Cervus elaphus L.) mit Hilfe der Trockengewichts-
bestimmung der Augenlinse // Ztschr. Jagdwiss. Bd. 24. No. 4. S. 178-182.
Maringgele F.J. von. 1979. Aftersbestimmung beim Reh {Capreolus capreolus) mit
Hilfe der Trockengewichtsbestimmung der Augenlinse // Zool. Anz. Bd. 202.
No. 1-2. S. 29-32.
Marks S.T., Erickson A.W. 1966. Age determination in black bear//J. Wildl. Manag.
Vol. 30. No. 2. P. 389-410.
Marshall W.H. 1951. An age determination method for the pine marten // J. Wildl.
Manag. Vol. 15. No. 3. P. 276-283.
Martinet L. 1966. Determination de 1’age chez le campagnol des champs {Microtus
arvalis Pallas) par la pesee du cristallin // Mammalia. Vol. 30. P. 425-430.
Martinson R.M., Holten J.W 1961. Age criterion and population dynamics of the
swamp rabbit in Missouri // J. Wildl. Manag. Vol. 25. No. 3. P. 271-281.
Mason D.R. 1984. Dentition and age determination of the warthog Phacochoerus
aethiopicus in Zululand, South Africa // Koedoe. Vol. 27. P. 79-119.
Литература
269
Matschke G.H. 1967. Aging European wild hogs by dentition // J. Wildl. Manag. Vol.
31. No. 1. P. 109-113.
Matson G.M. 1981. Workbook for cementum analysis. Milltown: Matson.
Matson G. M. 1991. Age determination by tooth cementum analysis // Progress Re-
port. No. 12. Matson’s Laboratory. Milltown, Montana.
Matson G. 1993. Matson’s laboratory. Progress Report. No. 13. 12 p.
Matson G. 1995. Matson’s laboratory. Progress Report. No 14. 8 p.
Mazak V. 1963. Notes on the dentition in Clethrionomys glareolus Schreber, 1780 in
the course of postnatal life // Saugetierk. Mitt. Bd. 11. No. 1. S. 1-11.
Mazak V. 1965. Changes in pelage of Myotis myotis Borkhausen, 1798 (Mammalia,
Chiroptera) // Vestn. Cheskosl. Spol. Zool. Vol. 29. No. 4. P. 368-376.
McLaughlin, C.R., Matula G.J., Cross R.A., Halteman W.H., Caron M.A., Morris
K.I. 1990. Precision and accuracy of estimating age of Maine black bears by
cementum annuli // Int. Conf. Bear Res. and Manag. 8-th. P. 415-419.
McLellan B.N. 1989. Dynamics of a grizzly bear population during a period of indus-
trial resource extraction. 1. Density and age-sex composition // Canad. J. Zool.
Vol. 67. P. 1856-1860.
Mead R.A. 1967. Age determination in the spotted skunk // J. Mammal. Vol. 48. No.
4. P. 606-616.
Mertens H. 1984. Determination de 1’age chez le topi {Damaliscus korrigum Ogilby)
au Parc National des Virunga (Zaire)// Mammalia. Vol. 48. No. 3. P. 425-436.
Michallet J., Loison A., Gaillard J.M., Gauthier D. 1994. Valeur de criteres biometriques
extemes pour la determination de 1’age du bouquetin des Alpes {Capra ibex
ibex)\ role du sexe et de 1’habitat // Gibier Faune Sauvage, Game Wildl. Vol. 11.
P. 99-118.
Millar J.S., Iverson S.L. 1976. Weight of the eye lens as an indicator of age in
Peromyscus // Canad. Field-Natur. Vol. 90. No. 1. P. 37-41.
Miller F.L. 1974. Biology of the Kaminuriak population of barren-ground caribou.
Part 2. Dentition as an indicator of age and sex; composition and socialization of
the population // Canad. Wildlife Service. Report Series. No. 31. 87 p.
Mills M.G. L. 1982. Notes on age determination, growth and measurements of brown
hyaenas, Hyaena brunnae from the Kalahari Gemsbok National park // Koedoe.
Vol. 25. P. 55-61.
Mitchell B. 1967. Growth layers in dental cement for determining the age of red deer
{Cervus elaphus L.) // J. Anim. Ecol. Vol. 36. P. 279-293.
Mitchell O.G., Carsen R.A. 1967. Tooth eruption in the Arctic ground squirrel // J.
Mammal. Vol. 48. No. 3. P. 472-474.
Miura S. 1985. Hom and cementum annulation as age criteria in Japanese serow // J.
Wildl. Manag. Vol. 49. No. 1. P. 152-156.
Miura S., Yasui K. 1985. Validity of tooth eruption-wear patterns as age criteria in the
Japanese serow, Capricornis crispus // J. Mammal. Soc. Japan. Vol. 10. No. 4.
P. 169-178.
Moller D. 1984. Beitrage zur Altersbestimmung des Schwazwildes // Unsere Jagd.
Bd. 34. No. 8. S. 232-233.
270
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Montgomery G.G. 1963. Freezing, decomposition and raccoon lens weights // J.
Wildl. Manag. Vol. 27. No. 3. P. 481-483.
Morris P.A. 1971. Epiphyseal fusion in the forefoot as a means of age determination
in the hedgehog (Erinaceus europaeus) // J. Zool. Vol. 164. P. 254-259.
Morris P.A. 1972. A review of mammalian age determination methods // Mammal
Rev. Vol. 2. No. 3. P. 69-104.
Morrison D., Whitridge P. 1997. Estimating the age and sex of caribou from mandib-
ular measurements // J. Archaeol. Sci. Vol. 24. P. 1093-1106.
Muylle S., Simoens P, Lauwers H., Van Loon G. 1999. Age determination in Mini-
Shetland Ponies and Donkeys II J. Veter. Med. Ser. A- Physiol. Pathol. Clin. Med.
Vol. 46. No. 7. P. 421-429.
Myers K., Carstairs J., Gilbert N. 1977. Determination of age of indigenous rats in
Australia H J. Wildl. Manag. Vol. 41. No. 2. P. 322-326.
Myers K., Gilbert N. 1968. Determination of age of wild rabbits in Australia // J.
Wildl. Manag. Vol. 32. No. 4. P. 841-849.
Myers Ph. 1978. A method for determining the age of living small mammals // J.
Zool. Vol. 186. No. 4. P. 551-556.
Nagorsen D.W., Forsberg J., Giannico G.R. 1988. An evaluation of canine radio-
graphs for sexing and aging Pacific coast martens (Martes americana) // Wildl.
Soc. Bull. Vol. 16. P. 421-426.
Navarre P. 1993. Distinction des chevrillards et des chevreuils (Capreolus capreolus)
de plus d’un an par examen du metacarpe ou du metatarse // Gibier Faune Sau-
vage. Vol. 10. P. 135-142.
Nellis C.H., Wetmore S.P., Keith L.B. 1978. Age-related characteristics of coyote
canines // J. Wildl. Manag. Vol. 42. No. 3. P. 680-683.
Nicholson W.S., Hill E.P. 1981. A comparison of tooth wear, lens weights and ce-
mentum annuli as indices of age in the gray fox // Worldwide Furbearer Conf.,
Frostburg, Md. Aug. 3-11, 1980, Proc. Vol. 1. S. 1. P. 355-367.
Norton P.M., Fairall N. 1991. Mountain reedbuck Redunca fulvorufula growth and
age determination using dentition // J. Zool. Vol. 225. No. 2. P. 293-307.
Novakowski N.S. 1965. Cemental deposition as an age criterion in bison, and the
relation of incisor wear, eye-lens weight, and dressed bison carcass weight to
age // Canad. J. Zool. Vol. 43. P. 173-178.
Nuckle J., Bergeron J.M. 1983. Eye lens weight as a criterion for age classification of
wild woodchucks // J. Wildl. Manag. Vol. 47. No. 3. P. 846-852.
Ohtaishi, N. 1980. Determination of sex, age and death season of recovered remains
of sika deer (Cervus nippon) by jaw and tooth-cementum // Archaeol. and Nat.
Sci. Vol. 13. P. 51-74.
Ohtani S., Yamamoto I. 2003. Differences in the D/L aspartic acid ratios in dentin
among different types of teeth from the same individuals and estimated age //
Intern. J. Legal Medicine. Vol. 117. No. 3. P. 149-152.
Ohtani S., Kato S., Sugeno H. 1996. Changes in D-aspartic acid in human deciduous
teeth with age from 1-20 Years I/ Growth, Development and Aging. Vol. 60. P.
1-6.
Литература
271
Olesen C.R., Thing H. 1989. Guide to field classification by sex and age of the
muskox // Canad. J. Zool. Vol. 67. No. 5. P. 1116-1119.
Olivier G.C., Greyling F.J. 1991. Criteria for age assessment ofblesbok// S. Afric. J.
Wildl. Res. VoL 21. No. 1. P. 1-5.
Olsen P.F. 1959. Dental patterns as age indicators in muskrats // J. Wildl. Manag. Vol.
23. No. 2. P. 228-231.
Onderscheka K., Hartl G.B. 1989. Steinwild Capra ibex L. // Buch der Hege. Bd. 1.
Haarwild. /Н. Stubbe (ed). VEB Deutscher Landwirdschaftverlag. Berlin. S. 236-
248.
Ostbye E., Semb-Johansson A. 1970. The eye lens as an indicator in the Norwegian
lemming (Lemmus lemmus L.) // Norwegian J. Zool. Vol. 18. No. 2. P. 239-243.
Otero J.G., Dapson R.W. 1972. Procedures in the biochemical estimation of age of
vertebrates // Res. Popul. Ecol. Vol. 13. P. 152-160.
Pankakoski E. 1980. An improved method for age determination in the muskrat,
Ondatra zibethica (L) // Ann. Zool. Fenn. Vol. 17. No. 2. P. 113-121.
Parker G.R., Maxwell J.W. 1986. Identification of pups and yearling wolves by den-
tine width in the canine // Arctic. Vol. 39. No. 2. P. 180-181.
Pascal M., Castanet J. 1978. Methodes de determination de 1’ age chez le chat haret
des iles Kerguelen // Terre et Vie. Vol. 32. P. 530-555.
Pascal M., Delattre P. 1981. Comparaison de differentes methodes de determination
de Page individuel chez le vison (Mustela vison Schreiber) // Canad. J. Zool. Vol.
59. No. 2. P. 202-211.
Pascal M., Damange J.-P., Douville P, Guedon G. 1988. Recherche de critbres d’age
chez le campagnol proven^al, Pitymys duodecimcostatus (De Selys-Longchamps,
1839)// Mammalia. Vol. 52. No. 1. P. 85-91.
Pascal M., Delattre P., Damange J.P., Haffner P, Poser S. 1990. Identification des
classes d’age dans les populatoins de belettes (Mustela nivalis L. 1766) // Rev.
Ecol. Vol. 45. No. 2. P. 93-112.
Payne S. 1987. Reference codes for wear states in the mandibular cheek teeth of
sheep and goats// J. Archeol. Sci. Vol. 14. P. 609-614.
Paz O. de 1986. Age estimation and postnatal growth of the Greater Mouse bat Myo-
tis myotis (Borkhause, 1797) in Guadalajara, Spain // Mammalia. Vol. 50. No. 2.
P 243-251.
Pearson A.M. 1975. The northern interior grizzly bear Ursus arctos L. // Canadian
Wildlife Service. Report Series. No. 34. 86 p.
Penzhom B.L. 1982. Age determination in Cape mountain zebras Equus zebra zebra
in the Mountain zebra National park // Koedoe. Vol. 25. P. 89-102.
Perez-Barberia F.J. 1994. Determination of age in Cantabrian chamois (Rupicapra
parva) from jaw tooth-row eruption and wear // J. Zool. Vol. 233. No. 4. P. 649-
656.
Perez-Barberia F.J., Mutuberria G. 1996. Teeth eruption pattern in Cantabrian cham-
ois Rupicapra pyrenaica parva // Acta Theriol. Vol. 41. No. 2. P. 217-221.
Perez-Barberia F.J., Fernandez- Lopez J.M. 1996. Using cementum annuli to estimate
Cantabrian chamois age // J. Wildl. Manag. Vol. 60. No. 1. P. 62-67.
272
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Perez-Barberia F.J., Robles L., Nores С. 1996. Horn growth pattern in Cantaberian
chamois Rupicapra pyrenaica parva: Influence of sex, location and phenology //
Acta Theriol. Vol. 41. No. 1. P. 83-92.
Perrin M.R. 1978. Molar root-length as an indicator of age in Clethrionomys gapperi I I
Acta Theriol. Vol. 23. No. 19-30. P. 423-434.
Petrides G.A. 1950. The determination of sex and age ratios in fur animals // Amer.
Midland. Natur. Vol. 43. No. 2. P. 355-382.
Petrides G.A. 1951. The determination of sex and age ratios in the cotton tail rabbit //
Amer. Midi. Natur. Vol. 46. P. 312-336.
Piechocki R., Stiefel A. 1977. Zahndurchbruch und Zahnwechsel beim Elbebiber,
Castor fiber albicus // Anat. Anz. Bd. 142. S. 374-384.
Piechocki R., Stiefel A. 1988. Uber die Altersstruktur der Verluste der Wildkatze (Fe-
lis silvestris Schreber 1777) // Hercynia N.F. Bd. 25. No. 2. S. 235-258.
Pielowski Z. 1971. The individual growth curve of the hare // Acta Theriol. Vol. 16.
No. 5. P. 79-88.
Prien S., Peukert R., Telle R. 1989. Muffelwild Ovis ammon musimon (Pallas) //
Buch der Hege. Bd. 1. Haarwild / H. Stubbe (ed.). VEB Deutscher Landwird-
schaftverlag. Berlin. S. 172-208.
Pucek Z., Lowe VP. W. 1975. Age criteria in small mammals // Small mammals: their
productivity and population dynamics I F.B. Goleey, K. Petrusewicz, L. Rysz-
kowski (eds.). Cambridge Univer. Press. P. 55-72.
Pucek Z., Zejda J. 1968. Technique for determining age in the red-backed vole, Clethri-
onomys glareolus (Schreber, 1780) // Small mammals Newsletters. Vol. 2. No. 4.
P. 51-60.
Quere J.P., Pascal M. 1983. Comparaison de plusieurs methodes de determination de
1’age individuel chez le Cerf elaphe (Cervus elaphus L.) // Ann. Sci. Natur, Zool.
В Ser. Vol. 5. No. 4. P. 235-252.
Quimby D.C., Gaab J.E. 1957. Mandibular dentition as an age indicator in Rocky
Mountain elk // J. Wildl. Manag. Vol. 21. No. 4. P. 435-451.
Rajan K.E., Marimuthu G. 1999. Postnatal growth and age estimation in the Indian
false vampire bat (Megaderma lyrd) // J. Zool. Vol. 248. No. 4. P. 529-534.
Rausch R.A., Pearson A.M., 1972. Notes on the wolverine in Alaska and the Yukon
territory // J. Wildl. Manag. Vol. 36. No. 2. P. 249-268.
Rausch R.L. 1961. Notes on the black bear, Ursus americanus Pallas, in Alaska, with
particular reference to dentition and growth // Ztschr. Saugetierk. Bd. 26. No. 2.
S. 77-107.
Rausch R.L. 1969. Morphogenesis and age-related structure of permanent canine
teeth in the Brown Bear, Ursus arctos L., in Arctic Alaska H Ztschr. Morph. Tiere.
Bd. 66. S. 167-188.
Rautenbach I.L. 1971. Ageing criteria in the springbok, Antidorcas marsupialis (Zim-
mermann, 1780) (Artiodactyla: Bovidae) //Annals Transv. Mus. Vol. 27. No. 6.
P. 83-133.
Rees J.W., Kainer R.A., Davis R.W. 1966. Chronology of mineralization and eruption
of mandibular teeth in mule deer // J. Wildl. Manag. Vol. 30. No. 3. P. 629-631.
Литература
273
Reilly J. 2002. Growth in the Sumatran elephant (Elephas maximus sumatranus)
and age estimation based on dung diameter // J. Zool. Vol. 258. No. 2. P. 205-
213.
Richardson P. W. 1994. A new method of distinguishing Daubenton’s bats (Myotis daub-
entoni) up to one year old from adults // J. Zool. Vol. 233. No. 2. P. 307-309.
Richter H., Geiler H. 1972. Uber die Altersstruktur je einer Population der Gelbhals-
mauS Apodemus tauricus flavicollis (Melchior 1834) und der Rotelmaus,
Clethrionomys glareolus (Schreber 1780) im Rabenauer Grund und an den Weis-
seritztalhangen. (Informationsartikel mit Fachdepot) // Wiss. Z. Techn. Univ.
Dresden. Bd. 21. No. 6. S. 1225-1227.
Ritz-Timme S., Rochhoiz G., Schutz H.W., Collins M.J., Waite E.R., Cattaneo C.,
Kaatsch H.-J. 2000. Quality assurance in age estimation based on aspartic acid
racemisation // Intern. J. Legal Medicine. Vol. 114. No. 1-2. P. 83-86.
Rodel R. von 1987. Alter und Linsengewicht von Meriones shawi grandis (Cabrera,
1907) // Ztschr. Saugetierk. Bd. 52. S. 291-294.
Robertson R. A., Shadle A.R. 1954. Osteologic criteria of age in beaver // J. Mammal.
Vol. 35. No. 2. P. 197-203.
Rogers P.M. 1982. Reliability of epiphysial fusion as an indicator of age in rabbits //
Mammalia. Vol. 46. No. 2. P. 267-269.
Rongstad O.J. 1966. A cottontail rabbit lens-growth curve from southern Wisconsin //
J. Wildl. Manag. Vol. 30. No. 1. P. 114-121.
Root D.A., Payne N.F. 1984. Evaluation of techniques for aging gray fox // J. Wildl.
Manag. Vol. 48. No. 3. P. 926-932.
Rosenthal H.L. 1968. Chronological age determination as estimated from strontium-
90 content of teeth and bone // Biology of the howler monkey (Alouatta caraya) //
Bibl. Primatol. Vol. 7. P. 48-58.
Roskosz T. 1962. Morphologie der Wirbelsaule des Wisents, Bison bonasus (Linnae-
us 1758) // Acta Theriol. Vol. 6. No. 5. P. 113-164.
Roth V.L., Shoshani J. 1988. Dental identification and age determination in Elephas
maxmus // J. Zool. Vol. 214. No. 4. P. 567-588.
Rowe F.P., Bradfield A., Quy R.J., Swinney T. 1985. Relationship between eye lens
weight and age in the wild house mouse (Mus musculus) // J. Appl. Ecol. Vol. 22.
No. 1. P. 55-61.
Rudge M.R. 1972. Homs as indicators of age in goats (Capra hircus L.) // New
Zealand J. Sci. Vol. 15. No. 2. P. 255-263.
Rusch D.A., Reeder W.G., Rusch D.H. 1982. Eye lens, testes, and body weight
trends in Alberta red squirrels // J. Wildl. Manag. Vol. 46. No. 4. P. 1010—
1017.
Ruftc A. 1966. Odredivanje uzrasnih kategorija u populaciji tekunice Citellus citellus
L. // Arh. Biol. Nauka. Vol. 18. No. 1. P. 65-70.
Saez-Royuela C., Gomariz R.P., Telleria J.L. 1989. Age determination of European
wild boar // Wildl. Soc. Bull. Vol. 17. P. 326-329.
Sanderson G.C. 1961. The lens as an indicator of age in the raccoon // Amer. Midland
Natur. Vol. 65. No. 2. P. 481-Л85.
274
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Savelle J.M., Beattie В. 1983. Analysis of dental annuli in Muskoxen {Ovibos tnoscha-
tus) as an aid in the determination of archaeological site seasonality // Canad. J.
Anthropol. Vol. 3. No. 1. P. 123-129.
Sauer P.R. 1975. Relationship of growth characteristics to sex and age for black
bears from the Adirondack region of New York //N. Y. Fish and Game J. Vol. 22.
No. 2. P. 81-113.
Sauer PR., Free S., Browne St. 1966. Age determination in black bears from canine
tooth sections // N.Y. Fish and Game J. Vol. 13. No. 2. P. 125-139.
Scheffer V.B. 1950. Growth layers on the teeth of Pinnipedia as an indication of age //
Science. Vol. 112. P. 309-311.
Schneider K.B. 1973. Age determination of sea otter // Final Report Federal Aid in
Wildlife Restoration Projects W-17-4 and W-17-5, Job 8. 10R. 23 p.
SchowalterD.B., Harder L.D., Triechel B.H. 1978. Age composition of some vespertilio-
nid bats as determined by dental annuli // Canad. J. Zool. Vol. 56. No. 2. P. 355-358.
Schreiber R. 1993. Rosenstock und Geweihentwicklung im Mittelgebirge vom Alter
der Hirsche // Wild und Hund. Bd. 96. No. 16. S. 36-38.
Schwarz S.S., Pokrovski A.V., Istchenko V.G., Olenjev V.G., Ovtschinnikova N.A.,
Pjastolova A.0.1964. Biological peculiarities of seasonal generations of rodents,
with special reference to the problem of senescence in mammals // Acta Theriol.
Vol. 8. No. 2. P. 11-43.
Semizorova I. 1975. Hmotnost vysusenych ocnich cocek ukazatelem veku zajice
polniho {Lepus europaeus Pall.) // Lesnictvi. Vol. 21. No. 6. P. 553-560.
Sergeant D.E. 1967. Age determination of land mammals from annuli // Ztschr. Sauge-
tierk. Bd. 32. No. 5. S. 297-300.
Sergeant D.E., Pimlott D.H. 1959. Age determination in moose from section incisor
teeth // J. Wildl. Manag. Vol. 23. No. 3. P. 315-321.
Serrano E., Gallego L., Perez J.M. 2004. Ossification of the appendicular skeleton in
the Spanish ibex Capra pyrenaica Schinz, 1838 (Artiodactyla: Bovidae), with
regards to determination of age // Anat., Histol. Embriol. Vol. 33. P. 33-37.
Severinghaus C. W. 1949. Tooth development and wear as criteria of age in white-
tailed deer // J. Wildl. Manag. Vol. 13. No. 2. P. 195-216.
Shackleton D.M., Hills L.V., Hutton D.A. 1975. Aspects of variation in cranial char-
acters of plains bison {Bison bison bison Linnaeus) from Elk Island National
Park, Alberta // J. Mammal. Vol. 56. No. 4. P. 871-887.
Shanks Ch. E. 1948. The pelt-primeness method of aging muskrats // Amer. Midland
Natur. Vol. 39. No. 1. P. 179-187.
Sharp W.M. 1958. Aging gray squirrels by use of tail-pelage characteristics // J.
Wildl. Manag. Vol. 22. No. 1. P. 29-34.
Sikes S.K. 1967a. How to tell the age of an African elephant // Afric. Wild Life. Vol.
21. No. 3. P. 191-202.
Sikes S.K. 1967b. The African elephant, Loxodonta Africana: a field method for the
estimation of age // J. Zool. Vol. 154. P. 235-248.
Simon D.E., Fiydendall M.J. 1981. Age study of Minnesota red fox using cementum
annulae counts and tooth X-rays // J. Minnesota Acad. Sci. Vol. 46. No. 3. P. 2-5.
Литература
275
Simpson C.D. 1973. Tooth replacement, growth and ageing criteria for the Zambezi
bushbuck— Tragelaphus scriptus ornatus Pocock// Amoldia. Vol. 6. No. 6. P. 1-25.
Simpson C.D., Elder W.H. 1968. Lens weight related to estimated age in greater kudu
// J. Wildl. Manag. Vol. 32. No. 4. P. 764-768.
Simpson C.D., Elder W.H. 1969. Tooth cementum as an index of age in greater kudu //
Amoldia. Vol. 4. No. 20. P. 1-9.
Skoczen S. 1966. Age determination, age structure and sex ratio in mole, Talpa euro-
paea Linnaeus, 1758 populations // Acta Theriol. Vol. 11. No. 26. P. 523-536.
Skoudlin J. 1976. Zur Altersbestimmung bei Erinaceus europaeus und Erinaceus con-
color (Insectivora, Erinaceidae) // Vest. Os. SpoL Zool. Vol. 40. No. 4. P. 300-
306.
Sladek J. 1992. Moznosti rozlisovania pohlavia a veku medveja hnedeho podl’a
lebkovych a zubnych znakov// Folia Venator. Vol. 22. P. 175-190.
Sluiter F.W 1961. Abrasion of teeth in the connections with age in the bat Myotis
myotis // Proc. Koninkl. Nederl. Akad. Wet., Ser. C. Vol. 64. No. 3. P. 424-434.
Smith B.L. 1988. Criteria for determining age and sex of American mountain goats in
the field // J. Mammal. Vol. 69. No. 2. P. 395-402.
Smuts G.L. 1974. Age determination in Burchell’s zebra (Equus burchelli antiquo-
rum} from the Kruger National Park // J. Sth. Afr. Wildl. Mgmt. Ass. Vol. 4. No.
2. P. 103-116.
Smuts G.L., Anderson J.L., Austin J.C. 1978. Age determination of the African lion
(Panthera leo) // J. Zool. Vol. 185. No. 1. P. 115-146.
Solounias N., Fortelius M., Fruman P. 1994. Molar wear rates in ruminants: a new
approach // Ann Zool. Fenn. Vol. 31. No. 2. P. 219-227.
Spinage C.A. 1967. Ageing the Uganda defassa waterbuck (Kobus defassa Ugandae
Neumann) // East. Af. Wildl. J. Vol. 5. P. 1-17.
Spinage C.A., 1971. Geratodontology and horn growth of the impala (Aepyceros
melampus) // J. Zool. Vol. 164. No. 2. P. 209-225.
Spinage C.A. 1973. A review of the age determination of mammals by means of teeth,
with especial reference to Africa HE. Afr. Wildl. J. Vol. 11. P. 165-187.
Spinage C.A. 1976. Age determination of the female Grant’s gazelle // East. Afr. Wildl.
J. Vol. 14. No. 2. P. 121-134.
Spinage C.A., Jolly G.M. 1974. Age estimation of warthog // J. Wildl. Manag. Vol. 38.
No. 2. P. 229-233.
Stander PE. 1997. Field age determination of leopards by tooth wear // Afr. J. Ecol-
ogy. Vol. 35. No. 2. P. 156-161.
Stegeman L. 1956. Tooth development and wear in Myotis!! J. Mammal. Vol. 37. No.
1. P. 58-63.
Stephenson A.B. 1977. Age determination and morphological variation of Ontario
otters // Canad. J. Zool. Vol. 55. No. 10. P. 1577-1583.
Stevens V, Houston D.V 1989. Reliability of age determination of mountain goats//
Wildl. Soc. Bull. Vol. 17. No. 1. P. 72-74.
Stockrahm D.M.B., Seabloom R.W. 1990. Tooth eruption in black-tailed prairie dogs
from North Dakota // J. Mammal. Vol. 71. P. 105-108.
276
Г. А. Клевезаль. Принципы и методы определения возраста млекопитающих
Stockrahm Dickerson B.J., Adolf S.L., Seabloom R.W. 1996. Aging black-
tailed prairie dogs by weight of eye lenses // J. Mammal. Vol. 77. No. 3. P. 874-
881.
Strickland, M.A., Douglas C.W., Brown M.K., Parsons G.R. 1982. Determining the
age of fisher from cementum annuli of the teeth // N. Y. Fish and Game J. Vol.
29. No. 1. P. 90-94.
Stuart Ch.T., Stuart T.D. 1985. Age determination and development of foetal and
juvenile Felis caracal Schreber, 1776 // Saugetierk. Mitt. Bd. 32. No. 3. S. 217-
229.
Stubbe A., Stubbe M., Hamisch J. 1995. Die Augenlinse als Alterskriterium bei Sauge-
tieren // Metoden feldokologischer Saugetierforschung / Stubbe M., Stubbe A.,
Heidecke D. (Hrsg.). Halle/Saale. S. 103-142.
Stubbe С. 1989. Rehwild Capreolus capreolus (L.) // Buch der Hege. Bd. 1. Haarwild.
H. Stubbe (ed.).VEB Deutscher Landwirdschaftverlag. Berlin. S. 140-170.
Stubbe M. 1965. Zur Biologie der Raubtiere eines abgeschlossenen Waldgebietes //
Ztschr. Jagdwiss. Bd.ll. No. 2. S. 73-102.
Stubbe M. 1989a. Fuchs, Vulpes vulpes L. // Buch der Hege. Bd. 1. Haarwild / H.
Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin. S. 344-382.
Stubbe M. 1989b. Hermelin, Mustela erminea L. // Buch der Hege. Bd. 1. Haarwild
/ H. Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin. S. 514-527.
Stubbe M. 1989c. litis, Mustela putorius L. // Buch der Hege. Bd. 1. Haarwild / H.
Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin. S. 503-513.
Stubbe M. 1989d. Dachs, Meles meles L. // Buch der Hege. Bd. 1. Haarwild / H.
Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin. S. 456-477.
Stubbe M. 1989e. Amerikanischer Nerz, Mustela vison Schreber // Buch der Hege.
Bd. 1. Haarwild / H. Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin.
S. 541-549.
Stubbe M. 1989f. Fishotters (Lutra lutra L.) // Buch der Hege. Bd. 1. Haarwild / H.
Stubbe (ed.). VEB Deutscher Landwirdschaftverlag. Berlin. S. 550-575.
Stump W.Q., Anthony R.G. 1983. Use of eye lens protein for estimating age of
Microtus pensylvanicus H J. Mammal. Vol. 64. No. 4. P. 697-700.
Suchentrunk E, Willing R., Hartl G.B. 1991. On eye lens weights and other age
criteria of the Brown hare (Lepus europaeus Pallas, 1778) // Ztschr. Saugetierk.
Bd. 56. No. 6. S. 365-374.
Sullins G.L., McKay D.O., Verts B.J. 1976. Estimating age of cotton-tails by perio-
steal zonations //Northwest Sci. Vol. 50. No. 1. P. 17-22.
Sullivan E.G., Haugen A.O. 1956. Age determination of foxes by x-ray of forefeet //
J. Wildl. Manag. Vol. 20. No. 2. P. 210-214.
Suminski P.P. 1968. Der Penisknochen als Altersmerkmal des Raubwildes //Tagungsber.
Dtsch. Acad. Landwirtschaftswiss. Berlin. Bd. 104. S. 205-209.
Szabik E. 1973. Age estimation of roe-deer from different hunting grounds of south-
eastern Poland // Acta Theriol. Vol. 18. No. 12. P. 223-236.
Taylor R.H. 1959. Age determination in wild rabbits // Nature. Vol. 184. No. 4693. P.
1158-1159.
Литература 277
Thomas D.C., Bandy P.J. 1975. Accuracy of dental wear age estimates of black-
tailed deer//J. Wildl. Manag. Vol. 39. No. 4. P. 674-678.
Thomas R.E., Bellis E.D. 1980. An eye-lens weight curve for determining age in
Microtus pennsylvanicus II J. Mammal. Vol. 61. No. 3. P. 561-563.
Thomsen H.P., Mortensen O.A. 1946. Bone growh as an age criterion in the cotton-
tail rabbit H J. Wildl. Manag. Vol. 10. No. 2. P. 171-174.
Thomson P.C., Rose К. 1992. Age determination of dingoes from characteristics of
canine teeth // Austral. Wildl. Res. Vol. 19. P. 597-599.
Tiemeier O.W, Plenert M.L. 1964. A comparison of three methods for determining
the age of black-tailed jack rabbits // J. Mammal. Vol. 45. No. 3. P. 409-416.
Tomich P.Q. 1962. The annual cycle of the California ground squirrels, Citellus
beecheyi I I Calif. Univ. Pub. Zool. Vol. 65. No. 3. P. 213-281.
Tmkova E. 1966. К metodice urcovani stari ondatry pizmove, Ondatra zibethica
Linnaeus 1758 //Lynx. Vol. 6. P. 165-172.
Troszynski W. 1977. Changes in the form of incisors as an age criterion for beaver
(Castor fiber L.) // Zool. Pol. Vol. 26. No. 2. P. 167-175.
Trout R.C., Lelliott L.M. 1992. Ageing wild rabbits (Oryctolagus cuniculus) from
southern England by determining epiphyseal ossification in the lumbar
vertebrae // J. Zool. Vol. 228. No. 4. P. 653-687.
Tupikova N.V., Sidorova G.A., Konovalova E.A. 1968. A method of age determina-
tion in Clethrionomys H Acta Theriol. Vol. 13. No. 8. P. 99-115.
Twente J.W. 1955. Aspects of a population study of cavern-dwelling bats // J. Mam-
mal. Vol. 36. No. 3. P. 379-390.
Ueckermann E. 1978. Methoden zur Alterbestimmung beim Schalenwild // Wild und
Hund. Bd. 80. S. 1068-1070.
Ueckermann E., Hansen P. 1983. Das Damwild. Naturgeschichte, Hage, Jagd. 2. Aufl.
Verlag Paul Parley. Hamberg, Berlin. 336 S.
Ueckermann E., Scholz H. 1971. Zur Zahnentwicklung und Altersschatzung beim
Sikawild (Sika nippori) И Ztschr. Jagdwiss. Bd. 17. No. 2. S. 49-52.
Uraguchi K., Takahashi K., Maekawa K. 1990. The age structure of the red fox
population in Hokkaido Japan // Proc. 5-th Intern. Congr. of Ecology Aug.
Yokohama.
Van Bree P.J., Van Soest R.W.M., Stroman L. 1974. Tooth wear as an indication of
age in Badgers (Meles meles L.) and Red Foxes (Vulpes vulpes L) // Ztschr.
Saugetierk. Bd. 4. S. 243-248.
Van Deelen T.R., Hollis K.M., Anchor C., Etter D.R. 2000. Sex effects age determi-
nation and wear of molariform teeth in white-tailed deer // J. Wildl. Manag. Vol.
64. No. 4. P. 1076-1083.
Van Hom R.C., McElhinny T.L., Holekamp K.E. 2003. Age estimation and dispersal
in the spotted hyena (Crocuta crocuta) H J. Mammal. Vol. 84. No. 3. P. 1019-
ЮЗО.
Van Laere G., Boutin J.M., Gaillard J.M. 1989. Estimation de Г age chez le chevreuil
(Capreolus capreolus) par 1’usure dentaire.Test de fiabilite sur des animaux marques //
GibierFaune Sauvage. Vol. 6. P. 417-426.
278
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
Van Nostrand ЕС., Stephenson А.В. 1964. Age determination for beavers by tooth
development // J. Wildl. Manag. Vol. 28. No. 3. P. 430-434.
Van Soest R.V.M., Van Bree P.J.H. 1970. Sex and age composition of a Stoat popu-
lation {Mustela erminea Linnaeus, 1758) from a coastal dune region of the Neth-
erlands // Zool. Museum Univ. Amsterdam. Vol. 17. No. 226. P. 51-77.
Vandorpe E., Verhagen R. 1979. An age reference model for the wood mouse, Apode-
mus sylvaticus (Linnaeus, 1758), by use of the lens technique // Ann. Soc. Roy.
Zool. Belg. Vol. 109. No. 2-4. P. 133-140.
Vandorpe E.J.C., Verhagen R., Verheyen W.N. 1980. Comparison of different age
indicators for the wood mouse, Apodemus sylvaticus И Ann. Soc. Roy. Zool.
Belg. Vol. 110. No. 3-A P. 185-197.
Vigal C.R., Machordom A. 1985. Tooth eruption and replacement in the Spanish wild
goat // Acta Theriol. Vol. 30. No. 19. P. 305-319.
Vigal C.R., Machordom A. 1988. Evaluation de la methode de determination de 1’age
en fonction de la masse du cristallin chez le bouquetin {Capra pyrenaica) H
Canad. J. Zool. Vol. 66. No. 12. P. 2836-2839.
Vohralik V. 1975. Postnatal development of the common hamster Cricetus cricetus
(L.) in captivity // Rozpr. Ceskoslv. Acad. Ved. 48 p.
Vohralik V. 1988. Age determination and the population structure in Crocidura suave-
olens (Mammalia; Insectinora) in Prague, Czechoslovakia // Vestn. Cs. Spolec.
Zool. Vol. 52. No. 1. P. 63-74.
Waite E.R., Collins M.J., Ritz-Timme S., Schitz H.-W., Cattaneo C., Bornnan H.I.M.
1999. A review of the methodological aspects of aspartic acid racemization anal-
ysis for use in forensic science // Forensic Sci. Intemat. Vol. 103. P. 113-124.
Walhovd H. 1966. Reliability of age criteria for Danish hares {Lepus europaeus) II
Danish Rev. Game Biol. Vol. 4. No. 3. P. 105-128.
Walker D.N. 1987. Sequence of epiphysael fusion in the Rocky Mountain bighorn
sheep // Great Basin Natur. Vol. 47. No. 1. P. 7-12.
Walton K.C. 1968. The baculum as an age indicator in the polecat Putorius putorius 11
J. Zool. Vol. 156. No. 4. P. 533-536.
Wandeler A.I. 1976. Altersbestimmung bei Fuchsen // Revue Suisse Zool. Bd. 83. No.
4. S. 956-963.
Wassif K., Soliman S. 1978. Tooth wear and age in the gerbil, Gerbillus andersoni de
Winton (Rodentia, Cricetidae) from the Western Desert of Egypt // Bull. Zool.
Soc. Egypt. Vol. 28. P. 73-83.
Wassif K., Soliman S. 1982. Age estimation in Gerbillus gerbillus Olivier (Rodentia,
Cricetidae) from the Western Desert of Egypt // Bull. Zool. Soc. Egypt. Vol. 32.
P. 65-76.
Wegrzyn M., Serwatka St. 1984. Teeth eruption in the European bison // Acta Theri-
ol. Vol. 29. No. 9. P. 111-121.
Wegrzyn M., Malyska E., Rostokosz T., Serwatka St. 1994. Observations on the
wearing of the permanent incisors in European bison, Bison bonasus L. // Ann.
Warsaw Agr. Univ.-SGGW. Vet. Med. Vol. 19. P. 19-31.
Western D., David M.C., Georgiadis N. 1983. Age estimation and population age
Литература
279
structure of elefants from footprint dimensions // J. Wildl. Manag. Vol. 47. No.
4. P. 1192-1197.
Wheeler S.H., King D.R. 1980. The use of eye-lens weights for aging wild rabbits,
Oryctolagus cuniculus (L.), in Australia // Austral. Wildl. Res. Vol. 7. No. 1. P.
79-84.
Wight H.M., Conaway C.H. 1962. A comparison of methods for determining age of
cottontails // J. Wildl. Manag. Vol. 26. No. 2. P. 160-163.
Wilson V.J., Schmidt J.L., Hanks J. 1984. Age determination and body growth of the
Common duiker, Sylvicapra grimmia (Mammalia) // J. Zool. Vol. 202. No. 2. P.
283-287.
Wood J. 1958. Age structure and productivity of a gray fox population // J. Mammal.
Vol. 39. P. 74-86.
Wright P.L., Coulter M.W. 1967. Reproduction and growth in Maine fishers // J.
Wildl. Manag. Vol. 31. No. 1. P. 70-87.
Wright P.L., Rausch R. 1955. Reproduction in the wolverine, Gulo gulo 11 J. Mam-
mal. Vol. 36. No. 3. P. 346-355.
Yu Hon-Tsen, Lin Yao-Sung. 1999. Age, reproduction, and demography of the spini
rat (Muridae: Niviventer coxingi) in Subtropical Central Taiwan // Zoological
Studies. Vol. 38. No. 2. P. 153-163.
Zapata S.C., Funes M.C., Novaro A.J. 1997. Estimation de la edad en el zorro Colo-
rado Patagonico (Pseudalopex culpaeus) // Mastozoologia Neotropical. Vol. 4.
No. 2. P. 145-150.
Zapata S.C., Perea R.G., Beltran J.F., Ferreras P., Delibes M. 1997. Age determina-
tion of Iberian lynx (Lynx pardinus) using canine radiograph and cementum
annuli enumeration // Ztschr. Saugetierk. Bd. 62. S. 119-123.
Zeiler J.T. 1988. Age determination based on epiphyseal fusion in post-cranial bones
and tooth wear in Otters (Lutra lutra) I I J. Archaeol. Sci. Vol. 15. P. 555-561.
Zejda J. 1977.'A device serving to determine the birth of Clethrionomys glareolus by
the length of Mj roots // Folia Zool. Vol. 26. No. 3. P. 207-211.
Zejda J., Babicka C. 1983. Variability of antlers in a sika deer (Cervus nippon) popu-
lation in Czechoslovakia // Folia Zool. Vol. 32. No. 3. P. 225-240.
УКАЗАТЕЛЬ
Aepyceros melampus 51,73,86,95
Alces 95,112
-alces25,51,74,80,91,207-209
Allactaga 23,67,131
- elater VS2
-jaculus 22
-severtzovi 131
Alactagulus pygmaeus 133
Alopex lagopus 59,61-63,173
Alouatta caraya 109
Ammotragus lervia 94
Antidorcas marsupialis 51,71,95,210
Antilocapra americana 51,72-74,210,211
Apodemus 88,112,135
-agrariuslb,88,103,105,114,131-136
-flavicollislb, 114,131,134,136
-speciosus 114
-sylvaticus 100,103,114,135,136-137
Arvicanthis abyssinicus 26
Arvicola 32,34
-terrestris 137
Balaena mysticetus 109
Barbastella barbastellus 87
Bison 111
- bison 51,65,73,95,211-212
- bonasus 29,47,49,51,212-213
Blarina 23,65,67
— brevicauda 116
Calomys musculinus 103
Canis
— aureus 90
- familiaris dingo 62
- latrans 56,59,63,173-174
-lupus 29,50,58,59,61-63,67,84,114,
174-176
-mesomelas4\,50,103,176
Capra 93,94,213-214
- aegagrus213,214
— ibex 52,214
- hircus 95
- pyrenaica 52,214
— sibirica 213
Capreolus 36,66,73,95,100, 111, 112,
114,115
-capreolus 49,52,64,69,74,75,91,214—
217
Capricornis crispus 49,52,93,217
Carollia perspicillata 121, 123
Castor 57,65,75,88,106,112,137,139-
140
-canadensis 139,140
-fiber 57,81,83,84,137,139,140
Ceratotherium simum 112,239-240
Cervus
- dama 48,49,51,52,54,69,72,81,95,108,
218
- elaphus 36,52,54,55,72,74,81,92,95,
96,111,218-222
- canadensis 85,221
-nippon 52,72,74,222-224
Citellus 28,88,140-142
-beecheyi 142
-citellus 141
-fulvus 141
-musicus 141
—pygmaeus 141
—suslicus 141,142
- undulatus 141
Clethrionomys 56,65,75,142-146
-gapperi 111, 144
-glareolus 38,56,57,105,106,108,142,
143,145
— rufocanus 142
-rutilus 142,143
Cricetulus migratorius 113
Cricetulus triton 113
Cricetus 111, 112,
-cricetus 146-147
Crocidura 23,56,67,116
- guldenstaedti 116
— suaveolens 117
Crocuta 65,66,114
-crocuta 177
Cynomys 66,114
- ludovicianus 147-148
Damaliscus dorcas 52,95
Damaliscus korrigum 52,69
Desmana moschata 86
Указатель латинских названий животных
281
Dicrostonyx torquatus 114,148
Dolomys 57
Dryomis nitedula 148-149
Dusicyon gymnocercus 103
Dusicyon griseus 103
Elephas maximus 112
Eliomys quercinus 149-150
Ellobius talpinus 150-151
Enhydra 33,115
-lutris 28,50,111,112,177-178
Eptesicus
-fuscusSl, 121,123
-serotinus 87,122,123
Equus
— burchelli 66,114,240-242
-zebra 240-242
Erethizon 111
-dorsatum 151
Erinaceus europaeus 117
Eutamias — ng. Tamias
Felis
-catus 179
-sylvestris 179
-concolor28,179-180
-caracal 28,50,103,178-179
-lynx34,50,58,59,61,63,102,103,180
—pardinus 59
-rufus 50,181
Galemys pyrenaicus 117
Gazella granti 73,95
Gerbillus 23,67,75,151-152
— andersoni 151
-gerbillus 151
Giraffa Camelopardalis 52,65,73,224
Glis glis 152
Gulo 29,38,90
-gw/o 98,103,181-182
Hemiechinus
- auritus 86
- hypomelas 86
Hemitragus jemlahicus 94
Hipposideros terasensis 21,123
Hyena brunnea 50
Kobus defassa 95,114
Lagurus 35,36
-lagurus 114,152-153
Lemmus 34,112
-lemmus33, 153-154
Lepus
-americanus 102,105
-arcticus 102
-californicus 100,101,125
-europaeus 28,37,102,104,105,111,112,
115,125-127
— timidus 127
Loxodonta africana 112
Lutra 63,68,90
- canadensis 28,29,38,42,59, 182
- lutra 42,182-184
Marmota 88,106
-baibacina 84,112,154-156
-bobac 112,154-156
-menzbieri 112,154-156
-топахбТ, 114,156-157
-sibirica 154—156
Martes 63,68
- americana 36,39,59,63,184
-foina 42,50,184-185
-martes 26,33,41,42,50,59,184-185
-реииаий‘38,42,61,84,115,185-186
-zibellina 28,33,42,59,61,62,103,183,
186-188
Megaderma lyra 123
Meles 63,67,68
-wefey 42,59,62,68,188-189
Mephitis mephitis 59,61,90
MerionesBS, 157-158
-erythrourus 157-158
-libycus 157
— meridianus 157—158
- tamariscinus 157-158
-shawi 157
- unguiculatus 157
Mesocricetus brandti 28
Micromys minutus 76
Microtus 33,34,43,89,100,108,158-160
-agrestis 89,105
-arvalis 38,103,114,158-160
-californicus 35, 158-160
- montanus 106
— oeconomus 106,159
-pennsylvanicus 110,159-160
282
Г.А. Клевезаль. Принципы и методы определения возраста млекопитающих
-pinetorum 106
Mogera robusta - ng. Tapia micrura
Muntiacus reevesi 52
Murina leucogaster 87
Musmusculus 38,76,105,114,131,134,
160
Muscardinus avellanarius 161
Mustela
- erminea 33,39,42,50,60,63,189-190
- eversmanni 42
- frenata 42
- lutreola 42
- nivalis 28,33,39,42,190-191
-putorius 40,42,68,191-192
-sibirica 41,42,60-62,67,192,
- vison 39,42,50,103,192-193
Myocastor coypus 161-162
Myopus schisticolor 44
Myotis
- blythi 87
- daubentoni 87,121-124
- keenii 124
-lucifugus 78,87,121,123,124
-myotis 121,123,124
- natter eri 87
-velifer 123
Nemorhaedus goral 52,85,95,225
Nesokia indica 114
Niviventer coxingi 26
NyctalusnoctulaRT, 121-123
Nyctereutes procyonoides 36,42,60-63,
90,193-194
Ochotona 127—128
- pricei 88
Odocoileus
-Лепнопн^Зб, 37,51,52,74,85,100, 111,
114,225-226
- virginianus 52,54,74,75,85,110,115,
226-227
Ondatra 44,57,65,75,
-zibethicus 23, 101,162-165
Oreamnos americanus 53,93,111,227
Oryctolagus cuniculus 36,38,100,101,
128-129
Oryx leucoryx 53
Ovibos moschatus 53,95,111,228
Ovis
— аттоп53,95, 111,229
- nanadensis 53,93,95,230
-dalli 53,230
Panthera
-leo 32,50,55,90
-pardus 50,66,114,194
Peromyscus 106
-leucopus 106
-maniculatus 106
-polionotus 110
Phacochoerus aethiopicus 46, 55, 230-
231
Pipistrellus
— kuhli 121
— mimus 123
-pipistrellus 122,123
-subflavus 123
- duodecimcostatus 103
Procyonlotor 29,43,102,103,110,194-
196
Proechimys 114
Pseudalopex culpaeus 60, 103
Pteropus 124
— alecto 87
- poliocephalus 87
Rangifer 73,115
- tarandus 24,48,49,53,70,71,95,231-
232
Rattus 165
-fuscipes 99
— norvegicus 44,138,165
- rattus 165
- vilosissimus 99,105
Rhinolophus hipposideros 121
Rhombomys opimus 114,165-166
Rupicapra rupicapra 53,93,94,233
Saiga tatarica 49,53,95,233—234
Scirtopoda telum 133
Sciurus 106
- carolinensis 39,112,166-167
-niger 37,39,112,166-167
- vulgaris 23,44,67,75,167-169
Sicista 169-170
- betulina 22,25,169
-napaea 169
Sigmodon hispidus 110,170
Sorex 112,114
- araneus 107,117-118
— minutus 107
Spalax 75
- microphtalmus 77,170-171
Spermophilus - ng. Citellus
Spilogale putorius 32,34,43,98
Stenocranius 35
-gregalis 106,160
Sus scrofa 53-55,62,64,69,106,110,234-
237
Sylvicapra grimmia 48,53,86
Sylvilagus2%, 102,112,129-130
-aquaticus 105,130
-floridanus 97,100,105,107,129,130
-nuttalli 105
Talpa 112,117
-.europaea 67, 118-119
-micrura 120
Tamiasffi, 111, 171
— amoenus 171
-ruficaudus 44,171
-sibiricuslS, 171
Tamiasciurus hudsonicus 106,172
Tatera indica 99
Taurotragus oryx 54,95,114
Taxidea taxus 43,60,90
Tayassu tajacu 38,54
Thomomys bottae 29
Thomomys umbrinus 25
Tragelaphus
- angasi 86,95
-scriptus 54,65,238
- strepsiceros 27
Urocyon cinereoargenteus 43,56,60,68,
103,196
Ursus 63
- americanus 28,29,35,41,50,55,81,84,
111,112,115,197-199,201
-arctos 28,29,32,50,102,112,115,199-
201
-maritimus 28,29,32,39,41,56,90,201-
203
Vespertilio
— murinus 121
-pipistrellus 87
14^29,36,67,68
- vulpes 25,43,51,60,62,63,68,80,103,
204-206
---fulvus 43,60
Книги Товарищества научных изданий КМК
БИОЛОГИЯ
СЕРИЯ «ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ РОССИИ»
Флора Северо-Западного Кавказа. [Вып.7]. А.С. Зернов. 2006. 664 с. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 400 руб. — Флора Нижнего Повол-
жья. Том 1 (споровые, голосеменные, однодольные). [Вып.6]. А.К. Скворцов
(ред.). 2006. 435 с. Формат 170 х 240 мм. Те. перепл. — Цена 300руб. — Ивы
европейской части России. [Вып.5]. Е.Т. Валягина-Малютина. 2004. 217с.
Формат 170х 242 мм. Те. перепл. — Цена 150 руб. — Булавоусые чешуек-
рылые Северной Азии. [Вып.4]. Ю.П. Коршунов. 2002. 424 с. с портр., илл.
Формат 170x 244 мм. Те. перепл. — Цена 300руб.—Определитель сосуди-
стых растений севера Российского Причерноморья. [Вып.З]. А.С. Зер-
нов. 2002. 283 с., илл. Формат 170 х 244 мм. Те. перепл. — Цена 150 руб. —
Наземные звери России. Справочник-определитель. [Вып.2]. И.Я. Пав-
линов и др. 2002. 298 с. Формат 170x244 мм. Те. перепл. — Цена 150руб.
Планируется: А.Л. Львовский, Д.В. Моргун. Булавоусые чешуекрылые Восточ-
ной Европы (в печати). — Е.Т. Валягина-Малютина. Деревья и кустарники зи-
мой. 2-е изд. (подготовлено к печати). — 3.H. Рябинина, М.С. Князев. Определи-
тель сосудистых растений Оренбургской области.
ПРОЧИЕ ОПРЕДЕЛИТЕЛИ ПО ФЛОРЕ И ФАУНЕ
Растения средней полосы Европейской России. Полевой атлас. И.А.
Шанцер. 2-е изд. 2007.470 с., цветной фотоатлас. Бум. мелов. Формат 125
х 170 мм. Те. перепл. — Цена 300руб. — Иллюстрированный атлас беспоз-
воночных Белого моря. Н.Н. Марфенин, С.А. Белорусцева (ред.). 2006. 312
с., цветной фотоатлас. Бум. мелов. Формат 150 х 220 мм. Те. перепл. —
Цена 400 руб. — Иллюстрированный определитель сосудистых расте-
ний Ленинградской области. Л.В. Аверьянов и др. 2006. 799 с. Формат 170
х 240 мм. Те. перепл. — Цена 400 руб. — Флора средней полосы европей-
ской части России. 10-е изд. П.Ф. Маевский. 2006. 600с. Формат 21 Ох290
мм. Те. перепл. — Цена 400 руб. — Определитель сосудистых растений
Соловецкого архипелага. К.В. Киселёва и др. 2004. 175 с., цв. фото. Фор-
мат 170x 240мм. Те. перепл. — Цена 130руб. — Иллюстрированное руко-
водство для ботанических практик и экскурсий в Средней России. В.Э.
Скворцов. 2004. 506 с. Формат 170 х 240 мм. Те. перепл. — Цена 220 руб. —
Флора Восточной Европы. Том 11. Н.Н. Цвелев (ред.). 2004. 535 с. Формат
170 x240 мм. Те. перепл. — Цена 200руб. — Иллюстрированный определи-
тель растений Средней России. Том 3. И.А. Губанов и др. 2004. 520 с. Фор-
мат 21 Ох 295 мм. Те. перепл. — Цена 280руб. Том 2 (распродан) — Том 1.
2002. 526 с. с портр. — Цена 280 руб. — Флора мхов средней части Евро-
пейской России. Том 2. М.С. Игнатов, Е.А. Игнатова. 2004. С.609-944. Бум.
мелов. Формат 195 х 270 мм. Те. перепл. — Цена 300 руб. Том 1. 2003. С. 1-
608, илл. — Цена 500руб. — Определитель грибов России. Дискомицеты.
Вып.1. Копротрофные виды. В.П. Прохоров. 2004. 255 с. Формат 145x218
мм. Те. перепл. —Цена 120 руб.
Планируется: Е.А. Коблик, Е.Н. Курочкин. Атлас птиц запада России.
СЕРИЯ «РАЗНООБРАЗИЕ ЖИВОТНЫХ»
Мамонт [Вып.З]. А.Н. Тихонов. 2005. 90 с., цв. вкл. Формат 145 х 205 мм. —
Цена 50 руб. — Городские комары, или «дети подземелья» [Вып.2]. Е.Б.
Виноградова. 2004. 96 с., цв. вкл. Формат 145 х 205 мм. — Цена 50 руб. —
Гидра: от Абраама Трамбле до наших дней [Вып.1 ]. С.Д. Степаньянц и др.
2003. 101 с. + цв.вкл. Формат 145 х 205 мм. — Цена 50руб.
УЧЕБНИКИ ДЛЯ ВУЗОВ
Морская биогеоценология. И.В. Бурковский. 2006. 285 с. Формат 170x 240
мм. Те. перепл. — Цена 180 руб. — Основы микологии (морфология и сис-
тематика грибов и грибоподобных организмов). Л.В. Гарибова, С.Н.
Лекомцева. 2005. 220 с. Формат 170 x240 мм. Те. перепл. — Цена 180 руб. —
Малый практикум по зоологии беспозвоночных. И.А. Тихомиров и др. 2005.
304 с., 14 ч/б вкл. Формат 170 х 240 мм. Те. перепл. — Цена 200 руб. —
Основы биогеографии. В.Г. Мордкович. 2005. 236 с., 1 цв. вкл. Формат 170
х 240 мм. Те. перепл. — Цена 180руб. — Биология дрожжей. И.П. Бабьева,
И.Ю. Чернов. 2004. 221 с., бум. мелов. Формат 140х 203 мм. В обл. — Цена
100 руб. — Лекции о клеточном цикле. О.И. Епифанова. 2-е изд. 2003. 160
с. Формат 140х 200 мм. В обл. — Цена 70руб. — Развитие эволюционных
идей в биологии. Н.Н. Воронцов. 2-е изд. 2004. 432 с. Формат 145 х 210мм.
Те. перепл. — Цена 150 руб.
Планируется: В. Вестхайде, Р. Ригер (ред.). Руководство по зоологии. Беспозво-
ночные (пер. с нем.). —Д. Футуяма. Эволюционная биология (пер. с англ.) — И.А.
Тихомиров (СПбГУ). Малый практикум по зоологии беспозвоночных (часть 2). —
И.А. Жирков. Жизнь на дне. — К. Хаусман. Протистология (пер. с англ.).
СЕРИЯ «СОВРЕМЕННАЯ ОТЕЧЕСТВЕННАЯ БИОЛОГИЯ»
Избранные труды. В.В. Кучерук. 2006. 523 с. спортр. Формат 170 x 240 мм.
Те. перепл. — Цена 250 руб. — Избранные труды. Е.Н. Матюшкин. 2005.
658 с. с портр. Формат 170 х 240 мм. Те. перепл. — Цена 300руб. — Избран-
ные труды по эволюционной биологии. А.П. Расницын. 2005. iv + 347 с. с
портр., 16 фототаблиц. Формат 170 х 240 мм. Те. перепл. — Цена 200руб.
— Избранные труды. Организм, геном, язык. Б.М. Медников. 2005. 452 с. с
портр. Формат 170х 240 мм. Те. перепл. — Цена 220 руб.
Планируется: С.М. Разумовский. Избранные труды.
СПРАВО ЧНЫЕ ИЗДАНИЯ, ИСТОРИЯ БИОЛОГИИ
Морские биологические станции на Русском Севере (1881-1938). С.И.
Фокин и др. 2006.130 с., ил., бум. мелов., вобл. Формат 145 x210 мм. — Цена
120руб. — Виды и формы хвойных, культивируемые в России. Часть 1.
Juniperus, Cephalotaxus, Taxus, Тоггеуа. Д.Л. Матюхин, O.C. Манина, H.C.
Королева. 2006. 259с., бум. мелов., цв. фото, вобл. Формат 165x235мм. —
Цена 400руб. — Эрнст Майр и современный эволюционный синтез. Э.И.
Колчинский. 2006. 149 с., ч/б вкл., в обл. Формат 145 х 215 мм. — Цена 100
руб. —Александр Николаевич Формозов: Жизнь русского натуралиста.
А. А. Формозов. 2006. 208 с., в обл. Формат 135х 203 мм. — Цена 100 руб. —
Пока горит свеча... Очерки по истории кафедры зоологии беспозвоночных
МГУ. 2-е изд. В.В. Малахов.2006.153 с., бум. мелов., в обл. Формат 145x210
мм. — Цена 120 руб. — Морские и солоноватоводные брюхоногие мол-
люски России и сопредельных стран: иллюстрированный каталог. Ю.И.
Кантор, А.В. Сысоев. 2006. 371 с., 140 цв. табл., в те. перепл. Формат 210
х 290 мм. — Цена 900руб. — Конспект фауны земноводных и пресмыка-
ющихся России. С.Л. Кузьмин, Д.В. Семенов. 2006. 139 с., в те. перепл.
Формат 145х 210 мм. — Цена 150руб. — Список птиц Российской Федера-
ции. Е.А. Коблик и др. 2006. 281 с., бум. мелов., печать двухцветная, в обл.
Формат 145 х215 мм. — Цена 200руб. — Биологический факультет МГУ.
А.И. Нетрусов и др. (ред.). 2005. 242 с., в те. перепл. Формат 170 х 240 мм.
— Цена 180 руб. — Эволюционные факторы формирования разнообра-
зия животного мира. Э.И. Воробьева, Б.Р. Строганова (ред.). 2005. 308 с., в
те. перепл. Формат 170х 240 мм. — Цена 150 руб. — Каталог бесчелюст-
ных и рыб пресных и солоноватых вод России с номенклатурными и
таксономическими комментариями. Н.Г. Богуцкая, А.М. Насека. 2004. 389
с. с вложенным лазерным диском. Формат 170 х 240 мм. Те. перепл. — Цена
200 руб. — Каталог типовых образцов сосудистых растений Восточной
Азии, хранящихся в Гербарии Ботанического института им. В.Л. Кома-
рова (LE). часть 1 (Япония и Корея). В.И. Грубов (ред.). 2004. 188 с., ил.
Формат 140 х 205 мм. В обл. — Цена 80 руб. — Международный кодекс
зоологической номенклатуры. Изд. 4-е. 2-е изд. русск. пер. 2004. 223 с.
Формат 143x213 мм. — Цена 70руб. — Биология гидротермальных сис-
тем. А.В. Гебрук (ред.). 2002. 543 с. с цв. вкл., в те. перепл. Формат 210x260
мм. — Цена 300руб.
Планируется: Г.Ю. Любарский. Эволюция зоологии. История одного музея. —
Д.Л. Матюхин, О.С. Манина, Е.С. Сысоева. Виды и формы хвойных, культиви-
руемые в России. Часть 2. Picea A. Diet г., Thuja L.
НАУЧНЫЕ МОНОГРАФИИ
Новости систематики низших растений. Том 40. М.П. Андреев (отв. ред.).
2006. 339 с., ил. Формат 150х 220 мм. Те. перепл. — Цена 200руб. — Состо-
яние и проблемы продукционной гидробиологии. К 100-летию проф. Г.Г.
Винберга. 2006. 329 с., ил., портр. Формат 150 x 220мм. Те. перепл. — Цена
150 руб. — Микокалициевые грибы Голарктики. А.Н. Титов. 2006. 296 с.,
ил. + 40 с. цв. вкл. Формат 170 х 240 мм. Те. перепл. — Цена 200 руб. —
Наука о развитии жизни. Опыт теории эволюции. Ю.В. Чайковский. 2006.
712 с., ил. Формат 150 х 220 мм. Те. перепл. — Цена 300 руб. — Биология
морских нематод. А.В. Чесунов. 2006. 367с., ил. Формат 170 х 240 мм. Те.
перепл. — Цена 200 руб. — Пресноводные раковинные амебы. Ю.А. Ма-
зей, А.Н. Цыганов. 2006.300с., ил. Формат 170x240мм. Те. перепл. — Цена
180 руб. — Позвоночные животные северо-востока центрального регио-
на России. В.А. Зайцев. 2006. 513 с., ил., цв. вкл. Формат 170 х 240 мм. Те.
перепл. — Цена 300руб. — Семейство гвоздичные во флоре Кыргызста-
на. ГА. Лазьков. 2006.272 с., ил. Формат 170 х 240 мм. Те. перепл. — Цена
180 руб. — Новости систематики высших растений. Том 38. Т.В. Егорова
(отв. ред.). 2006. 377 с. Формат 148x213 мм. Те. перепл. — Цена 200руб. —
Принципы таксономии животных. Дж.Г. Симпсон, (пер. с англ.). 2006. хх +
293 с. Формат 145 х 210 мм. Те. перепл. — Цена 180 руб. — Экология рыб
Обь-Иртышского бассейна. Д.С. Павлов, А.Д. Мочек(ред.). 2006. 596с. Фор-
мат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Чума: происхождение и
эволюция эпизоотической системы. В.В. Сунцов, Н.И. Сунцова. 2006. 247
с. Формат 170 x240 мм. Те. перепл. — Цена 180 руб. —Адаптации хальци-
доидных наездников к паразитированию на ложнощитовках в услови-
ях различных широт. Е.С. Сугоняев, Н.Д. Войнович. 2006. 263 с. Формат
170 х 240 мм. Те. перепл. — Цена 180руб. — Гепатоидные кожные железы
млекопитающих. С.А. Шабадаш, О.Ф. Чернова. 2006. 217 с. Формат 170 х
240 мм. Те. перепл. — Цена 180 руб. — Эволюция биосферы и биоразно-
образия. К 70-летию А.Ю. Розанова. С.В. Рожнов (отв. ред.). 2006. 600 с. с
портр. Формат 170 х 240 мм. Те. перепл. — Цена 300 руб. — Флора субарк-
тических гор Евразии и высотное распределение её видов. В.Б. Куваев.
2006. 568 с., ч/б вкл. Формат 170 х 245 мм. Те. перепл. — Цена 280 руб. —
Пластинчатоусые жуки подсемейства Scarabaeinae фауны России и со-
предельных стран. О.Н. Кабаков. 2006. 374 с., цв. вкл. Формат 170 х 240
мм. Те. перепл. — Цена 270 руб. — Атлас-определитель усоногих раков
(CirripediaThoracica) надсемейства Chthamaloidea Мирового океана. О.П.
Полтаруха. 2006.198 с. Формат 170x240мм. Те. перепл. — Цена 200руб. —
Растения Центральной Азии. Вып.15. В.И. Гоубов (отв. ред.). 2006. 143 с.
Формат 164х 238 мм. В обл. — Цена 150руб. — Млекопитающие Вьетнама.
Г.В. Кузнецов. 2006.420 с. Формат 170 x 240 мм. Те. перепл. — Цена 270руб.
— Нематоды надсемейства Drilonematoidea - паразиты дождевых чер-
вей. С.Э. Спиридонов, Е.С. Иванова. 2005. 296 с. Формат 170 х 240 мм. Те.
перепл. — Цена 180 руб. — Травы на градиенте влажности почвы. С.Н.
Шереметьев. 2005. 271 с. Формат 170 x240мм. Те. перепл. — Цена 180 руб.
— Бделлоидные коловратки фауны России. Л.А. Кутикова. 2005. 315 с.
Формат 170х 240 мм. Те. перепл. — Цена 180руб. — Кариотипы паразити-
ческих перепончатокрылых. В.Е. Гэхман. 2005. 185 с., бум. офсетн. и ме-
лов. Формат 150 х 220 мм. Те. перепл. — Цена 120 руб. — Ископаемые
цветковые растения. Том 4. Nyctaginaceae - Salicaceae. Л.Ю. Буданцев
(ред.). 2005. 466 с., бум. офсетн. и мелов. Формат 228 х 295 мм. Те. перепл.
— Цена 400 руб. — Китайская восковая пчела на Дальнем Востоке Рос-
сии. В.Н. Кузнецов. 2005.111 с., бум. мелов., цв. фото. Формат 148x215 мм.
В обл. — Цена 100 руб.
Животное население почв бореальных лесов Западно-Сибирской рав-
нины. Б.Р. Строганова, Н.М. Порядина. 2005. 234 с. Формат 170 х 240 мм.
Те. перепл. — Цена 180руб. — Лишайники - индикаторы радиоактивного
загрязнения. Л.Г. Бязров. 2005. 476 с. Формат 150 х 220 мм. Те. перепл. —
Цена 200 руб. — Полорогие А.А. Данилкин. 2005. 550 с., цв. вкл. Формат 170
х 240 мм. Те. перепл. — Цена 250руб. — Земноводные российского Даль-
него Востока. С.Л. Кузьмин, И.В. Маслова. 2005.434 с., цв. вкл. Формат 170
х 240 мм. Те. перепл. — Цена 200руб. — Введение в современную филоге-
нетику. И.Я. Павлинов. 2005.391 с. Формат 148x220 мм. Те. перепл. — Цена
180руб. —Трансформационная типологическая систематика. Б.П. Заха-
ров. 2005. 164 с. Формат 145 х 210 мм. В обл. — Цена 60 руб. — Проблемы
эволюции и теоретические вопросы систематики. А.К. Скворцов. 2005.
293 с. с портр. Формат 148 x 220 мм. Те. перепл. — Цена 180 руб. — Анато-
мия коры розоцветных. Л.И. Лотова, А.К. Тимонин. 2005. 264 с. Формат
170 х 240 мм. Те. перепл. — Цена 150руб. — Биология возбудителя опис-
торхоза. С.А. Беэр. 2005. 336 с. Формат 170 x 240 мм. Те. перепл. — Цена
180руб. — Проблемы теоретической морфологии и эволюции растений.
Н.Н. Цвелев. 2005. 407 с. с цв. портр. Формат 145 х 218 мм. Те. перепл. —
Цена 200 руб. — Ветвистоусые ракообразные отряда Ctenopoda миро-
вой фауны. Н.М. Коровчинский. 2004. 410 с. Формат 170 х 240 мм. Те. пе-
репл. — Цена 250руб. — Атлас волос млекопитающих. Тонкая структура
остевых волос и игл в сканирующем электронном микроскопе. О.Ф. Чер-
нова, Т.Н. Целикова. 2004. 429 с., бум. мелов. Формат 170 х 240 мм. Те. пе-
репл. — Цена 300 руб. — Почвы, биогеохимические циклы и биосфера.
Развитие идей Виктора Абрамовича Ковды. К 100-летию со дня рожде-
ния. Н.Ф. Гпазовский (отв. ред.).2004. xii + 403 с. с портр. и ч/б фото. Фор-
мат 170 x 240 мм. Те. перепл. — Цена 250руб. — Зоогенная дефолиация и
лесное сообщество. Е.Н. Иерусалимов. 2004. 263 с. Формат 148 x213 мм.
Те. перепл. — Цена 120 руб. — Морфогенез и эволюция. В.Г. Черданцев.
2003. 360 с. Формат 145х 205 мм. Те. перепл. — Цена 150 руб. — Головохо-
ботные черви Мирового Океана. Л.В. Адрианов, В.В. Малахов.1999.328 с.,
бум. мелов. Формат 205 х 285 мм. В обл. — Цена 150руб. — Приапулиды:
строение, развитие, филогения и система. А.В. Адрианов, В.В. Малахов.
1996. 268с., бум. мелов. Формат 210x285мм. Вобл. — Цена 150руб.
Планируется: К).Б. Бызова. Дыхание почвенных беспозвоночных. — О. Т. Руси-
нек. Паразиты рыб озера Байкал. — А.М. Амирханов. Растительность Северо-
Осетинского заповедника. — А.М. Прокофьев. Морфология, систематика и про-
исхождение усатых гольцов. — В.А. Чащухин. Ондатра: причины и следствия
биологической инвазии. — Новости систематики высших растений. Том 39.—А.В.
Крылов (ред.). Сукцессионные изменения экосистемы малой реки. — И.Н. Ильин.
Экология океанического обрастания в пелагиали. — Е.Б. Поспелова, И.Н. Поспе-
лов. Флора Таймыра (высшие сосудистые растения). Часть 1. Аннотированный
список флоры и её общий анализ. — Е.О. Фадеева. Экология грача в антропоген-
ных ландшафтах Окско-Донского междуречья. — А.К. Маркова и др. Эволюция
экосистем Европы при переходе от плейстоцена к голоцену. — В.А. Паевский.
Демографическая структура и популяционная динамика певчих птиц.
Заказать эти и другие издания (геогарфия, медицина,история) изд-ва КМК можно
по адресу:
123100 Москва, а/я 16 изд-во КМК, Михайлову Кириллу Глебовичу
Комп, почта: kmk2000@online.ru
Интернет: http://webcenter.ru/~kmk2000 (аннотации изданных книг)
Факс: (495) 203-2717 Тел. (495) 692-5894 раб.
Научное издание
КЛЕВЕЗАЛЬ Г алина Александровна
Принципы и методы определения возраста
млекопитающих
М.: Т-во научных изданий КМК. 2007. 283 с.
Для заявок:
123100 Москва а/я 16
или:
kmk2000@online.ru
Отпечатано в ГУП ППП «Типография «Наука» АИЦ РАН
121099 Москва, Шубинский пер., 6.
Подписано в печать 23.03.2007. Заказ № 572
Формат 70x100/16. Объем 18 печ.л. Бум. офсетная. Тираж 1000 экз.