/





































































































































































































































































































































Text
ФАУНА
СССР
МЛЕКОПИТАЮЩИЕ
I
выпуск
3
ЗООЛОГИЧЕСКИЙ ИНСТИТУТ АКАДЕМИИ НАУК СССР
НОВАЯ СЕРИЯ № 71
МЛЕКОПИТАЮЩИЕ
Том I, вып. 3
И. И. СОКОЛОВ
КОПЫТНЫЕ ЗВЕРИ (ОТРЯДЫ PERISSODACTYLA
и ARTIODACTYLA)
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
МОСКВА • 1959 • ЛЕНИНГРАД
Главный редактор
Директор Зоологического института АН CCCI
акад. Е. Н. Павловский
Редакционная коллегия:
Б. Е. Быховский, | Б. С. Виноградов], А. А. Стрелков (редактор выпуска
А. А. Штакельберг (редактор серии)
ФАУНА СССР. МЛЕКОПИТАЮЩИЕ, Т. I, ВЫП. 3.
ИВАН ИВАНОВИЧ СОКОЛОВ. КОПЫТНЫЕ ЗВЕРИ
Утверждено к печати
Зоологическим институтом Академии наук СССР
Технический редактор А. В. Смирнова
Корректоры В. Ф. Виноградова, Е. В. Русакова и Л. В. Семенова
Сдано в набор 20/XI 1958 г. Подписано к печати 6/II 1959 г. РИСО АН СССР № 66-52 Р,
Формат бумаги 70 х 108"/,в. Бум. л. 20. Печ. л. 40 = 54.8 усл. печ. +1 вкл. Уч.-иад. л. 56.64 +
+ 1 вкл. (0.06). Изд. № 654. Тип. зак. № 861. М-22066. Тираж 1800, Цена 41 руб. 35 коп.
Ленинградское отделение Издательства АН СССР. Ленинград, В-164, Менделеевская лин., д. 1
1-я тип. Издательства АН СССР, Ленинград, В-34, 9-я линия, д. 12.
ПРЕДИСЛОВИЕ
Копытные — одна из важнейших в хозяйственном отношении групп
млекопитающих. Все копытные в той или иной мере используются чело-
веком. Подавляющее большинство их являются объектами промысловой
или спортивной охоты. Виды малочисленные и редкие служат для обо-
гащения фауны охотничьих угодий и заповедников, для украшения при-
городных парков. Наконец, копытные являются родоначальниками и цен-
нейшим материалом для совершенствования и выведения новых пород
наиболее важных видов домашних животных. Поэтому всестороннее
изучение этой группы животных весьма желательно.
Запасы большинства копытных нашей страны в дореволюционные
годы вследствие нерационального промысла и отсутствия мер охраны
сильно истощились. Некоторые виды находились на грани полного исчез-
новения. За последние десятилетия положение изменилось в лучшую
сторону. В результате принятых правительством СССР мер охраны,
установления сроков или полного запрета добычи многие виды копытных
значительно увеличили свою численность, расширили ареал, появились
в районах, где они в течение ряда десятилетий совсем не водились.
Особо бюлыпую роль в сохранении и увеличении численности копыт-
ных сыграли наши государственные заповедники. Помимо непосред-
ственной охраны от истребления редких видов (горала, пятнистого оленя,
кулана), заповедниками проводятся опыты по акклиматизации и реак-
климатизации копытных (зубра, лани, пятнистого оленя). Работниками
заповедников выполнен ряд ценных работ по экологии копытных (К. Г. Аб-
рамов, 1939, 1954; В. В. Дмитриев, 1938; С. С. Донауров и В. П. Теплое,
1938; М. А. Заблоцкий, 1939, 1948; А. А. Насимович, 1938, 1939, 1955).1
Однако сводок по всей группе копытных в целом в нашей литературе до
сих пор нет. Известная в свое время книга Н. В. Туркина и К. А. Сату-
нина «Звери России» (1902, 1904) безнадежно устарела, а кроме того,
обнимает даже не всех парнопалых. Только одной группы жвачных ка-
сается и недавно опубликованная в серии «Большой» фауны СССР прекрас-
ная монография К. К. Флерова «Кабарги и олени», к тому же уже исчез-
нувшая в продаже. Монография И. И. Соколова «Опыт естественной клас-
сификации полорогих (Bovidae)», помимо того, что касается только одного
семейства, посвящена в основном вопросам надвидовой систематики
и филогении.
Попытку в какой-то мере восполнить пробел в сводках по одной из
важнейших и интересной теоретически группе млекопитающих представ-
ляет предлагаемая вниманию читателей книга.
1 На стр. 20 приведен список только главнейшей литературы, касающейся боль-
ших групп копытных. Более подробные списки даются перед описанием отдельных
групп н видов.
4
ПРЕДИСЛОВИЕ
Мною была поставлена перед собой весьма трудная для одного лица,
к тому же в основном морфолога и систематика- задача попытаться
дать сводку главнейших данных по морфологии, систематике,
биологии и хозяйственному значению представителей этой группы оте-
чественной фауны.
Стремление осуществить исторический подход побудило включить
в сводку, кроме современных, два вида копытных, некогда входивших
в состав нашей фауны и вымерших в уже историческое время: европей-
скую дикую лошадь — тарпана и первобытного быка — тура. Отчасти
этой же причиной, а также большим научным интересом объясняется
включение в нее еще двух современных видов, обитающих на смежной
с Советским Союзом территории: лошади Пржевальского и дикого дву-
горбого верблюда. В результате получилась сводка по фауне копытных
млекопитающих не только СССР, но и Монгольской Народной Республики.
Помимо определительных таблиц, даются довольно подробные морфо-
логические характеристики всех систематических групп от видов до
отрядов и надотряда Ungulata включительно. Это я считал необходи-
мым ввиду того, что в отечественной литературе характеристика надвидо-
вых групп отсутствует. Материалом для этой части работ в основном
послужили черепа, скелеты, шкуры коллекционных фондов Зоологическо-
го института Академии наук СССР (в тексте сокращенно: ЗИН АН СССР).
Характеристики отдельных видов даются применительно к взрослым
особям; взрослыми считаются особи, у которых полностью развились и
начали стираться все постоянные зубы. При вычислении краниологических
индексов бралось не менее чем по пять черепов взрослых самцов и самок.
Для каждой группы и вида приводятся соображения об их филоге-
нии. Подвиды, за исключением некоторых заведомо спорных случаев,
просто перечислены без критического пересмотра, так как для оленей
и кабарог это недавно обстоятельно проделано К. К. Флеровым (1952),
а по полорогим составляет задачу выполняемой мною в настоящее время
отдельной работы.
Синонимика названий систематических групп и видов не приводится,
так как номенклатура подавляющего большинства наших копытных
достаточно - прочно установилась и обычно недоразумений не вызывает.
Очерки географического распространения для зарубежных стран
даются в обобщенном виде, для Советского Союза и Монголии — более
подробно.1 Особое внимание уделяется данным последних лет. Карты
географического распространения составлены так, чтобы отражали не
только современное, но и былое распространение видов.
Биология отдельных видов дается на основе критического сопостав-
ления и использования литературных данных, а также личных наблюде-
ний автора во время его поездок в разные районы Советского Союза
(Крайний Север европейской части СССР, Алтай, Хакассия, Молда-
вия, Заволжье, Сальские степи, Каракумы, Бет-Пак-Дала, остров
Барса-Кельмес, горный Таджикистан и др.).
Считая, что задуманная работа должна служить как бы введением
в изучение копытных и представлять основные исходные данные для же-
лающих углубленно изучать ту или иную их группу, большое значение
придается ссылкам на литературу. Помимо списка главнейшей литера-
1 Приведение данных по Монгольской Народной Республике значительно облег-
чилось выходом в свет капитальной сводки по млекопитающим этой страны А. Г. Бан-
никова (1954). '
ПРЕДИСЛОВИЕ
5
туры по копытным в общей части, перед описанием отдельных групп
и видов приводится перечень основной литературы, касающейся
данной группы или вида, а также литературы, на которую делаются
ссылки в тексте. За небольшими исключениями приводится лишь литера-
тура, вышедшая до 1955 г.
Приводим список сокращений в аннотациях, раскрывающих содержа-
ние цитируемой литературы:
акклим. — акклиматизация ох. — охота
ан ат. — анатомия охр. — охрана
биол. — биология параз. — паразиты
бол/ — болезни первооп. — первоописание
гибр. — гибридизация пит. — питание
изобр. — изображение прир. — приручение
ист. — история разв. — разведение
классиф. — классификация размн. — размножение
мигр. — миграции распр. — распространение
морф. — морфология синон. — синонимика
общ. — общая сист. — систематика
одомашн. — одомашнивание содерж. — содержание
опис. — описание стац. — стации
опр. возр. — определение возраста хоз. зная. — хозяйственное значение
Раздел Tylopoda (мозоленогие) написан по просьбе автора специалистом
по этой группе Я. И. Хавесоном. Автором раздела «Восстановление пого-
ловья зубров в СССР и за границей» (стр. 593) является М. А. Заблоцкий.
Пользуюсь случаем выразить этим лицам мою благодарность.
Рисунки зверей выполнены художником Е. Я. Захаровым. Авторами
рисунков черепов, их деталей и рогов являются художники Т. Ф. Бело-
цветова, В. Н. Ляхов и В. К. Азаров.
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ ВИДОВ КОПЫТНЫХ
ФАУНЫ СССР
Надотряд Ungulata — Копытные
Отряд Perissodactyla — Непарнопалые
Семейство Equidae — Лошадиные
Род Е q u u s Linnaeus — Лошади
Е. (Asinus) hemionus Pallas — Кулан ......................... 33
Е. cdballus gmelini Antonius — Тарпан.......................... 45
E. przewalskii Poljakov — Лошадь Пржевальского................. 52
Отряд Artiodactyla — Парнопалы e
Подотряд Nonruminantia — Нежвачные
Семейство Suidae — Свиные
Род Sus Linnaeus — Свиньи
<S. scrofa Linnaeus — Кабан ................................. 72
Подотряд Tylopoda — Мозоленогие
Семейство Camelidae — Верблюдов ые
Подсемейство Camelinae
Род G a m е 1 us Linnaeus — Верблюды
С. bactrianus ferns Przewalski — Дикий двугорбый верблюд...... 103
Подотряд Ruminantia — Жвачные
Семейство Moschidae — Кабарговые
Род М о s с h u s Linnaeus — Кабарги
М. moschiferus Linnaeus — Кабарга ........................... 122
Семейство Cervidae — Олени
Подсемейство Cervinae — Настоящие олени
Род Capreolus Gray — Косули
С. capreolus Linnaeus — Косуля ............................... 145
Род С е г v u s Linnaeus — Настоящие олени
С. (Dama) dama Linnaeus — Лань................................ 176
С. nippon Temminck — Пятнистый олень.......................... 183
•Ш. eiapfiu^ Linnaeus — Благородный олень .................... 204
СИСТЕМАТИЧЕСКИЙ УКАЗАТЕЛЬ
Род А 1 с е s Gray — Лоси
A. alces Linnaeus — Лось ................................................. 234
Подсемейство Neocervinae — Американские олени
Род Rangifer Н. Smith — Северн ые олени
К. tarandus Linnaeus — Северный олень .................................... 266
Семейство В о v i d а е — Полорогие
Подсемейство Gazellinae— Газелевые
Род G a z е 1 1 a Blainville — Газели
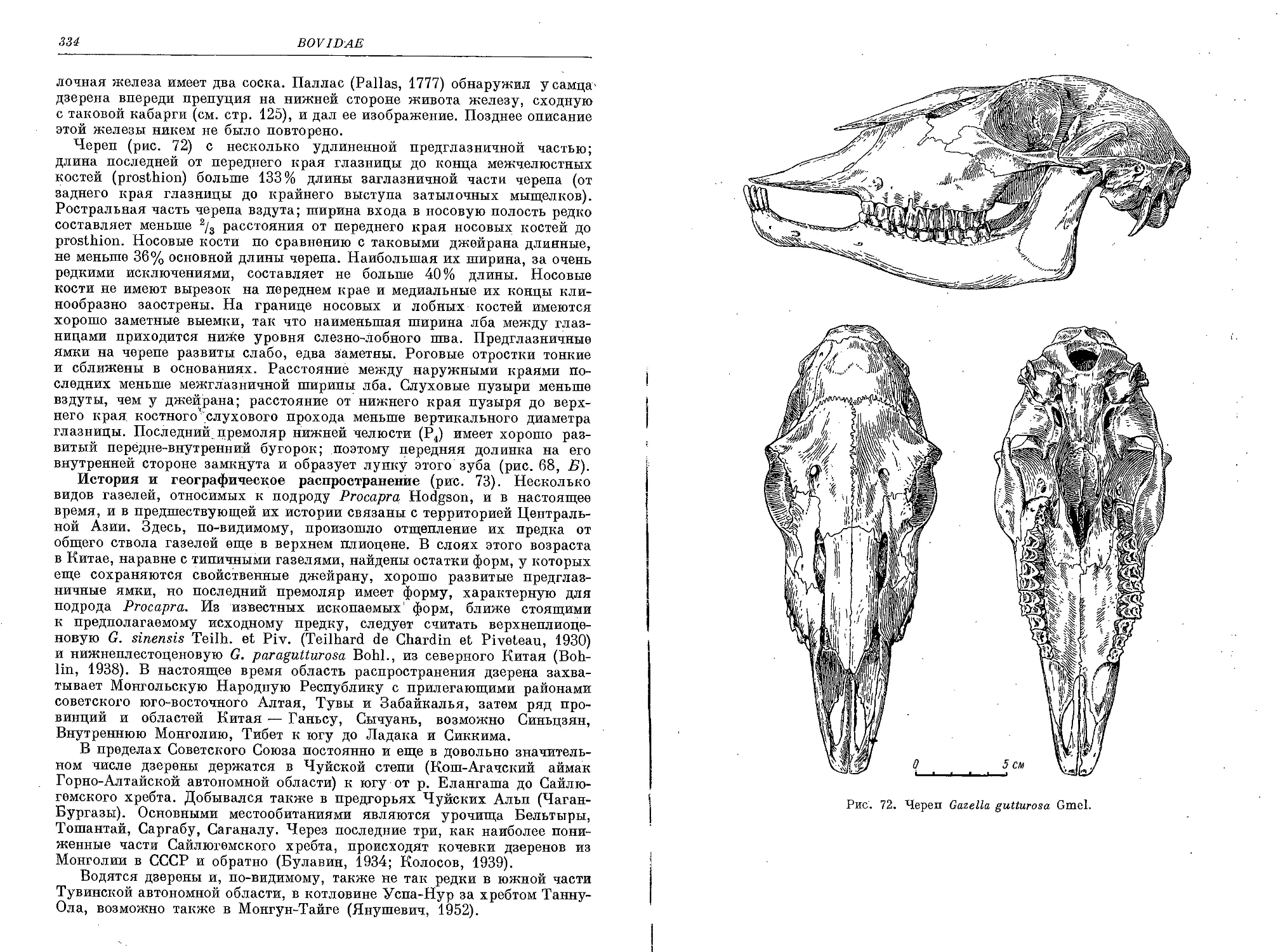
G. subgutturosa Giildenstaedt —Джейран ................................... 308
G. (Procapra) gutturosa Gmelin — Дзерен................................... 331
Подсемейство Caprinae — Козлообразные
Род Saiga Gray — Сайга
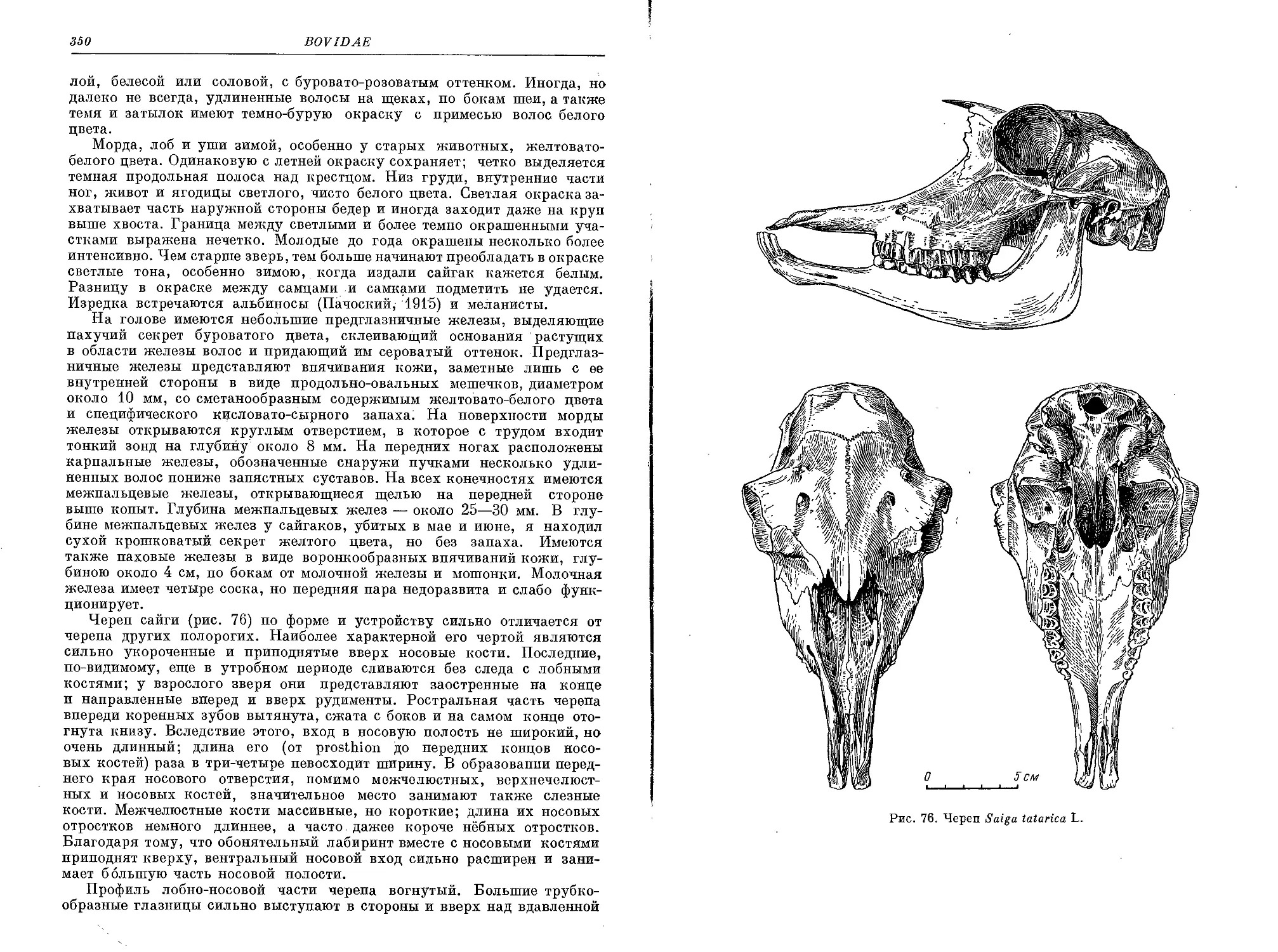
S', tatarica Linnaeus — Сайга............................................. 345
Род Nemorhaedus Flower et Garson — Горалы
.V. caudatus Milne-Edwards — Амурский горал .............................. 375
Род Rupicapra Blainville — Серны
R. rupicapra Linnaeus — Серна ........................................... 384
Род Capra Linnaeus — Козлы
C. aegagrus Erxleben — Безоаровый козел................................... 404
C. (Ibex) sibirica Meyer —Сибирский козерог .............................. 416
C. (Orthaegoceros) falconeri Wagner — Винторогий козел.................... 442
C. (Turus) caucasica Giildenstaedt — Кавказский каменный козёл . . . . . 455
Род О v i s Linnaeus — Бараны
О. musimon Pallas — Европейский муфлон................................. 482
О. orientalis Gmelin — Азиатский муфлон ................................. 497
О. аттоп Linnaeus — Архар ................................................ 516
О. (Pachyceros) nivicola Eschscholtz — Снежный баран.................... 544
Подсемейство Bovinae — Быковые
Род Bison Н. Smith — Бизоны
В. bonasus Linnaeus — Зубр ............................................... 568
Род Bos Linnaeus — Б ы к и
В. primigenius Bojanus — Первобытный бык, или тур......................... 601
ВВЕДЕНИЕ
Термином «копытные» (Ungulata Линнея) охватывается обширная
группа млекопитающих, к которой из современной фауны, кроме «на-
стоящих копытных», т. е. имеющих на концах пальцев копыта, относят
также слонов (Proboscidea), сиреновых (Sirenia), жиряков, или даманов
(Hyracoidea), а иногда также и трубкозубов (Tubulidentata). Вся группа
большинством зоологов определяется как надотряд.
Слоны, сирены и жиряки представляют пережиток некогда богатой
формами древней африканской группы прлукопытных (Subungulatd),
отошедшей от общего ствола раньше, чем предки остальных ветвей копыт-
ных.
Но даже без Subungulata «настоящие копытные», о которых главным
образом только и будет идти речь в дальнейшем, представленные двумя
отрядами — парнопалых (Artiodactyla) и непарнопалых (Perissodaclyld).,1
не являются с точки зрения филогенетической систематики естественной
группой; хотя они произошли, возможно, и от общих предков, но, по-ви-
димому, независимо друг от друга и не в одно время.
ХАРАКТЕРИСТИКА НАДОТРЯДА КОПЫТНЫХ
В подавляющем большинстве случаев крупные плацентарные млеко-
питающие (обычно величиною не меньше козы) на высоких ногах. Конеч-
ные фаланги пальцев лопаткообразно расширены или имеют форму треуголь-
ника, одеты со всех сторон в роговой башмак, копыто ((исключение —
семейство верблюдовых (Camelidae)). Отсутствует ключица. Число паль-
цев — от пяти до одного, но сильнее других развиты или один, третий,
или два — третий и четвертый, несущие на себе основную тяжесть тела.
Плечевая кость не имеет энтэпикондилярного отверстия (в отличие от
отряда хищных). В запястье запястная лучевая кость (scaphoideum)
и запястная промежуточная (lunare) всегда раздельны (не сливаются
вместе); отсутствует центральная кость запястья (carpi centrale); запяст-
ная лучевая (scaphoideum) сочленяется с третьей запястной (magnum).
Кости запястья (carpalia) и заплюсны (tarsalia) расположены по чередую-
щемуся типу, т. е. таким образом, что проксимальный конец пястной кости
каждого функционирующего пальца упирается не меньше, чем в две
кости дистального ряда запястья. На таранной кости (astragalus) имеется
закругленная верхняя поверхность с глубокой бороздой для сочлене-
ния с большой берцовой костью. Локтевая (ulna) и малоберцовая (fibula)
кости всегда в той или иной степени редуцированы, часто слиты с сосед-
ней (лучевой или большеберцовой) костью. Но проксимальные и дисталь-
1 В отечественной литературе часто применяются термины парнокопытные (Ai -
tiodactyla) и непарнокопытные (Perissodactvla}.
10
ВВЕДЕНИЕ
ные концы этих костей всегда сохраняются, служа местом прикрепле-
ния мускулатуры.
Глазницы черепа незамкнутые, но отделены от височных впадин по-
верхностной костной перемычкой из отростка лобной и скуловой или
чешуйчатой костей, образующей костное кольцо орбиты (исключение —
свиньи (Suina)). Зубы гетеродоптные и дифиодонтные. Типичная зубная
формула I®, С|, Р|, М®_ 441, но верхние резцы и клыки часто отсут-
ствуют. Коренные зубы бунодонтные, селенодонтные или лофодонтные.
Между переднекоренными (Р) и клыками обычно имеется лишенный
зубов промежуток (диастема). Анальные железы отсутствуют. Семен-
ники у взрослых самцов расположены вне брюшной полости. Os penis
отсутствует. Матка двурогая. Плацента диффузная или поликотиледон-
ная.
Географическое распространение, эволюция и морфологические адап-
тации. Копытные распространены по всем материкам земного шара,
кроме Антарктики, Австралии и Новой Зеландии. В последнюю они,
впрочем, завезены человеком. В Австралии существуют лишь в домашнем
состоянии.
В фауне СССР в настоящее время представлены оба отряда настоящих
копытных в числе 23 видов (не считая одомашненных и содержимых в зоо-
парках и зоосадах), что составляет около 7—8% мировой фауны этих
отрядов. Из этого числа подавляющее большинство (22 вида) относятся
к парнопалым, главным образом к семействам оленей (Cervidae) и поло-
рогих (Bovidae).
Копытные в широком смысле слова известны с начала третичного пери-
ода. В меловых отложениях они еще не найдены, во, уже начиная с па-
леоцена, на территории Европы, Азии и Америки были широко распро-
странены многочисленные группы так называемых первичнокопытных
(Protoungulatd), большая часть которых, как например обширная группа
южноамериканских копытных, A7otoungulata, вымерла, не оставив в со-
временной фауне потомков. Если не все, то большая часть копытных,
в том числе и отряды Perissodactyla и Artiodactyla, по современным пред-
ставлениям, ведет свое начало от одной из групп первичнокопытных,
кондилартр (Condylarthra). Примитивные представители последних
трудно отличимы от некоторых архаических хищных креодонтов (Сгео-
donta). Это и дало основание Г. Симпсону (Simpson, 1945) надотряды
хищных и копытных, как филогенетически наиболее близкие, объеди-
нить в одну когорту Ferungulata (=Ferae-\~Ungulata Линнея). Расхождение
стволов хищных и копытных имело место, вероятно, еще в конце мезо-
зоя.
Общее направление эволюции копытных находит наиболее полное
выражение в современных отрядах Perissodactyla и Artiodactyla. Все
строение их конечностей указывает на приспособленность к энергичным
поступательным движениям. Уже у самых ранних, а из современных
даже у самых примитивных форм, отсутствует ключица. Редукция ее
связана с ограничением движений конечностей в стороны; у подавляю-
щего большинства копытных, если сохраняются в какой-то мере движения
отведения и приведения конечностей (абдукция и аддукция), то враща-
тельные функции (пронация и супинация), равно как и хватательные,
практически отсутствуют.
1 Объяснение формулы см. стр. 17.
ХАРАКТЕРИСТИКА КОПЫТНЫХ
11
Редукция ключицы ведет также к ликвидации жесткого крепления
передней конечности; последняя связывается с туловищем только при
помощи мускулатуры, что способствует смягчению толчков при быстром
беге; смягчение толчков весьма важно, учитывая то, что основная функ-
ция передних конечностей — принятие на себя тяжести тела, оттолк-
нутого задними ногами.
С ликвидацией вращательных и хватательных функций связана редук-
ция локтевой и малоберцовой кости, от которых в крайних случаях
остаются только проксимальные и дистальные концы. Лучевая и больше-
берцовая кости соответственно развиваются сильнее. Ограничение боко-
вых движений компенсируется усилением функций сгибания и разги-
бания, причем последнее происходит не только путем усиления соответ-
ствующей мускулатуры, но и путем смены функций отдельных мышц
и суставов конечностей.
Второй характерной чертой в эволюции конечностей, связанной с адап-
тацией к энергичным поступательным движениям, является упрощение
строения дистальных участков. Предки всех филогенетических ветвей
копытных были, несомненно, пятипалыми животными. В процессе эво-
люции шла независимо в разных филогенетических ветвях постепен-
ная редукция и исчезновение боковых пальцев, в первую очередь I, II
и V; в результате конечность превращалась в четырех-, трех-, двупалую
и даже однопалую. Параллельно с сокращением числа пальцев происхо-
дила перестройка в соотношениях между карпальными и тарзальными
элементами с одной стороны, и пястными и плюсневыми костями функцио-
нирующих пальцев — с другой.1 Те группы, в которых достигнутый
уровень организации не позволял возможности согласованной эволю-
ции обоих сегментов конечностей, были обречены на вымирание (инадап-
тивная редукция, по В. О. Ковалевскому).
Упрощение конечностей в процессе эволюции сопровождалось пере-
ходом от стопохождения к пальце- и фалангохождению. а также удлине-
нием конечностей, особенно дистальных их звеньев. Наиболее ранние
из известных представителей копытных были не только пятипалыми,
но и стопоходящими формами, с короткими пястными и плюсневыми
костями. С развитием приспособления к быстрым движениям метапо-
дии приподнялись над землей; конечность стала пальцеходящей. С даль-
нейшим увеличением скорости движения первые фаланги пальцев также
приподнялись и животное стало опираться только на ногтевую фалангу.
Поднятие стопы, сопровождаемое удлинением метаподий, привело к тому,
что боковые пальцы совсем перестали касаться почвы, в результате чего
редукция последних происходила еще быстрее.
Конечные фаланги одного или двух средних пальцев, вынужденные
нести всю тяжесть обычно крупного тела при быстрых передвижениях
по твердому грунту, нуждались в защите. С этим связано развитие ко-
пыт — наиболее характерного признака группы. Впрочем, копыта,
несомненно, имелись у пятипалого и пальцеходящего представителя
архаических копытных, нижнеэоценового Phenacodus Соре. Строение
копыта может быть выведено из когтя. Расширенная ногтевая фаланга
спереди и с боков окружалась роговой стенкой, соответствующей когтю
других млекопитающих, а утолщенные и ороговевшие мякиши пальцев
превратились в подошву копыта, защищающую мягкие ткани снизу.
1 Запястье и заплюсна испытывали расширение, слияние и исчезновение отдель-
ных костей.
12
ВВЕДЕНИЕ
Не менее важной чертой эволюции копытных, нашедшей свое выра-
жение и в морфологии, является приспособление к питанию расти-
тельной пищей, к пережевыванию грубого травянистого корма. В связи
с этим трехбугорчатые зубы предков копытных становятся четырех- и
шестибугорчатыми с плоской жевательной поверхностью. Бугорки при-
тупляются, возникает бунодонтный тип зубов, лучше приспособленный
к перетиранию растительной пищи; этот тип зубов в несколько изменен-
ной форме из современных копытных сохранился лишь у свиней (Suina).
Дальнейшее увеличение жевательной поверхности шло путем образова-
ния на зубе гребней и складок: основные четыре бугорка или вытяги-
вались и изгибались, приобретая форму полулуний (селенодонтный тип),
или же соединялись продольными и поперечными гребнями (лофодонт-
ный тип). Захватывание и пережевывание травы вместе с твердыми
частицами почвы и связанное с этим усиленное изнашивание зуба вызы-
вало и необходимость увеличения крепости зуба, усиления его перетираю-
щей способности, продления срока службы. Последнее достигалось тем,
что первоначально низкокоронковые (брахиодонитные) зубы увеличи-
вались в высоту, приобретая вид призм (гипсодонтизм), что само по
себе значительно увеличивало срок изнашивания зуба. Кроме того це-
мент, первоначально покрывающий лишь корень зуба, распространяется
на коронку, заполняя все пространство между складками и гребнями
эмали. При изнашивании зуба на жевательной поверхности выступают
одновременно все три его элемента: эмаль, дентин (внутри эмали) и цемент
(снаружи); благодаря неодинаковой их твердости поверхность зуба
в процессе изнашивания получается неровной, что повышает ее перети-
рающую способность.
Адаптацией к питанию грубой растительной пищей являются также
некоторые особенности строения и функций желудочно-кишечного тракта.
Длинный кишечник приспособлен к перевариванию (с помощью специ-
альной кишечной микрофлоры) и усвоению клетчатки. У большей части
современных копытных желудок сложный, состоящий из 2—4 отделов.
Экологически копытные определяются, прежде всего, как раститель-
ноядные формы. Поэтому из элементов окружающей среды они теснее
всего связаны с растительностью, в меньшей степени — с климатом,
рельефом и животным миром. Лишь группа свиней (Suina) может быть
в известной мере названа всеядной; ио и для них части растений состав-
ляют основную пишу. Остальные же члены отрядов парно- и непарно-
палых являются типичными травоядными и древесноядными животными.
Поэтому, если отвлечься от влияния человеческой культуры, для ко-
пытных, как крупных форм млекопитающих, основным лимитирующим
распространение фактором является характер растительности и нали-
чие достаточной растительной массы. Узкая пищевая специализация,
за очень редкими исключениями (северный олень, некоторые виды тропи-
ческих копытных), в группе не имеет места; пищей могут служить все
части растений, за исключением самых грубых частей стволов деревьев,
включая в некоторых случаях (верблюд) даже колючки.
С приспособлением к максимальному использованию питательных
веществ грубого растительного корма связаны отмеченные выше особен-
ности устройства желудочно-кишечного тракта.
Климатические факторы, в частности температура, имеют меньшее
значение для копытных, благодаря наличию хорошего термо регуляци-
онного аппарата. Влияние элементов климата — температуры, влажности,
осадков — сказывается в первую очередь через растительность, отра-
ЧЕРЕП КОПЫТНЫХ
13
жаясь в сезонных колебаниях продуктивности естественных пастбищ.
Для некоторых видов в зимнее время, вследствие большой глубины снеж-
ного покрова, сильно затрудняется добывание корма и передвижение
(Формозов, 1946; Насимович, 1955). Возможно, что с этим связано то
обстоятельство, что основная масса фауны копытных в своем распростра-
нении ограничена зоной южных и средних широт. Лишь наиболее прогрес-
сивные (первобытный зубр, бараны) и специализированные формы (се-
верный олень, мускусный бык) сумели приспособиться к условиям Запо-
лярья.
Копытные служат добычей крупным хищникам: эволюция последних
была в значительной мере сопряжена с эволюцией копытных. Различ-
ные виды головного вооружения (рога, бивни, клыки, иногда резцы)
частично служат целям защиты от хищников, но защитная роль этих
образований сравнительно невелика. Основное средство спасения от
врагов, помимо органов чувств, — быстрый бег, в приспособлении к ко-
торому, вместе с необходимостью длительных переходов в поисках корма
и воды, заключается биологический смысл морфологических преобра-
зований конечностей копытных.
Большая часть филогенетических ветвей некогда обильной группы
копытных вымерли, не оставив потомков. В первую очередь исчезли
гигантские формы, от которых в современной фауне остались жалкие
остатки. В расцвете эволюции находится лишь одна из групп Ungulata —
это жвачные (Ruminanlia'). Несмотря на это, копытные до сих пор
имеют представителей почти во всех биотопах суши земного шара, вклю-
чая обитателей скал, деревьев (жиряки) и даже ведуших полуводный
образ жизни бегемотов и некоторых африканских полорогих.
Хозяйственное значение. Человек, самый мощный фактор в новей-
шем отрезке истории копытных, до последнего времени играл только
отрицательную роль. Но его разумная воля, при активном вмешатель-
стве в природу и переделке ее в интересах человечества, может и должна
способствовать процветанию данной группы. Хозяйственное значение
копытных очень велико. Едва ли можно найти другую группу наземных
позвоночных, которая приносила бы человеку большую пользу. Подав-
ляющее большинство копытных служит объектом спортивной или про-
мысловой охоты. Промысел копытных составляет в ряде районов важную
отрасль хозяйства и существенную часть бюджета местного населения.
При правильном планировании и организации промысла мясо диких
копытных может играть очень большую роль в создании местной продо-
вольственной базы. Но еще большую роль копытные сыграли в истории
человеческой культуры. Почти все одомашненные и прирученные млеко-
питающие принадлежат к этой группе. Характерно, что, по-видимому,
легче всего поддались приручению и наибольшее породное многообразие
дали формы (крупный рогатый скот, овцы, лошадь), принадлежащие
к наиболее прогрессивным и, вероятно, биологически наиболее пластич-
ным, филогенетически молодым ветвям копытных — семейства Equidae
и Bovidae (Соколов, 1955).
ЧЕРЕП КОПЫТНЫХ
Строение черепа. Наряду с особенностями наружной морфологии,
посткраниального скелета, зубной системы и внутренних органов в си-
стематике млекопитающих, в том числе и копытных, широко исполь-
зуются краниологические (черепные) признаки. Причиной этого является
14
ВВЕДЕНИЕ
сравнительная портативность, простота сбора, препаровки и хранения
черепов, по сравнению, например, с остальным скелетом. В коллекциях
музеев и научных учреждений черепа, наравне со шкурами, полнее
всего представляют ту или иную группу. Кроме того череп, как наиболее
сложно устроенная часть скелета млекопитающих, дает наибольшее
число признаков, которые могут быть использованы при выяснении от-
личий и сходств групп разного таксономического ранга.
Поскольку в дальнейшем краниологические особенности будут при-
водиться в определительных таблицах и в морфологических характери-
стиках отдельных групп и видов, целесообразно дать краткий очерк стро-
ения черепа и методику его измерений. Скелет головы, или череп (cranium,
рис. 1а) состоит из: а) собственно черепа (calvarium), б) прикрепляющейся
к последнему нижней челюсти (mandibula) и в) подъязычной кости (hyoi-
deum). В собственно черепе различают мозговой и лицевой отделы. Гра-
ницей между ними при измерениях принимается обычно линия, каса-
тельная к задним краям глазниц.
Мозговая часть черепа образует костные стенки мозговой полости,
вместилища головного мозга. В состав ее входят кости:
1) Непарная затылочная (occipitale), образованная тремя (вернее
четырьмя), рано сливающимися элементами: а) непарным телом, или
основной частью (basioccipitale), составляющей заднюю половину осно-
вания черепа; б) парными боковыми частями (exoccipitalia), несущими
суставные мыщелки для сочленения с первым шейным'позвонком (атлан-
том) и яремные отростки; в) непарной чешуей затылочной кости (supraoc-
cipitale, или squama occipitis), расположенной над боковыми частями.
Все четыре сегмента формируют большое затылочное отверстие (foramen
magnum).
2) Парная теменная кость (parietale), составляющая верхнюю крышу
мозговой коробки.
3) Небольшая непарная межтеменная кость (interparietale), обычно
рано сливающаяся с соседними костями (чешуей затылочной и теменными).
4) Лежащая впереди теменных парная лобная кость (frontale), уча-
ствующая в образовании крыши мозгового черепа и стенки глазницы
и несущая по бокам скуловые или глазничные отростки, а у большин-
ства оленей и полорогих также роговые отростки.
5) Парная чешуйчатая кость (squamosum), или чешуя височной кости
(squama temporis), образующая в основном боковые стенки мозговой
коробки; в стороны и вперед от чешуйчатых костей отходят скуловые
отростки; на нижней стороне оснований последних имеются суставные
площадки, с помощью которых к черепу причленяется нижняя че-
люсть.
6) Расположенная по бокам основания черепа между затылочной,
чешуйчатой и основной клиновидной костями парная каменистая кость
(os petrosum) или каменистая часть височной кости (pars petrosum ossis
temporis), составляющая слуховую класть черепа (regio tympanica).
Каждая каменистая кость состоит из трех частей: а) обращенной внутрь
мозговой коробки скалистой (petrosum), внутри которой заключен пере-
пончатый слуховой лабиринт, б) срастающейся с нею сосцевидной или
мастоидной (mastoideum), расположенной позади и снаружи и в) бара-
банной (tympanicum), которая у копытных не срастается так плотно
с остальными частями и представляет в различной степени вздутый кост-
ный пузырь (bulla ossea), заключающий внутри полость среднего уха
с тремя слуховыми косточками: молоточком, наковальней и стремеч-
ЧЕРЕП КОПЫТНЫХ
15
ком; от бокового отверстия барабанной кости в сторону отходит костный
слуховой проход (meatus auditorius externus).
7) Парная скуловая кость (zygomaticum, jugale), образующая вместе
со скуловым отростком чешуйчатой кости скуловую дугу черепа.
8) Непарная основная клиновидная кость или базисфеноид (basi-
sphenoideum), лежащая впереди тела затылочной кости, состоит из тела,
двух направленных в стороны и вверх височных или больших крыльев
(alisphenoidea) и двух направленных вниз крыловидных отростков.
9) Непарная передняя клиновидная кость или пресфеноид (praesphe-
noideum), составляющая переднюю часть основания черепа; от ее тела
отходят в стороны и вверх глазничные или малые крылья (orbitosphe-
noidea).
10) На границе мозгового и лицевого отделов черепа, отделяя мозго-
вую полость от носовой, расположена сложно устроенная непарная ре-
шетчатая кость (ethmoidale), состоящая, в основном, из системы тонких
костных пластинок, образующих завитки обонятельного лабиринта.
В состав лицевого черепа, кроме передней части лобной кости, вхо-
дят кости:
1) Парная носовая (nasale) в форме обычно вытянутой пластинки,
образующая верхнюю стенку носовой полости.
2) Парная верхнечелюстная (maxilla), образующая боковые и нижнюю
стенки носовой полости и основу твердого нёба. Кроме того, верхнече-
люстные кости несут глубокие альвеолы пли лунки для укрепления в них
верхних коренных зубов и клыков.
3) Относящаяся по происхождению к мозговому отделу черепа парная
слёзная кость (lacrimale), расположенная впереди глазницы между лоб-
ной, носовой, верхнечелюстной и скуловой костями; на лицевой поверх-
ности слёзных костей у многих форм имеются предглазиичные ямки для
кожных предглазничных желез.
У многих копытных между носовыми, лобными, слёзными и верхне-
челюстными костями имеются неправильной формы этмоидальные щели,
сообщающиеся с обонятельным лабиринтом носовой полости.
4) Прилегающая сзади к верхнечелюстным костям на нижней стороне
черепа парная нёбная кость (palatinum), образующая остов заднего
края твердого нёба.
5) Парная крыловидная кость (pterygoideum), тонкая пластинка,
прилегающая к крыловидным отросткам предыдущей кости с внутренней
стороны хоан.
6) Парная межчелюстная кость (intermaxilla, или premaxilla), приле-
гающая к верхнечелюстной, а иногда и к носовой кости, составляющая
переднюю оконечность черепа и участвующая в образовании переднего
края костного нёба. Передний край межчелюстных костей несет альвеолы
для укрепления резцовых зубов; но у жвачных последние отсут-
ствуют.
7) В средней части дна носовой полости лежит непарный сошник (vo-
mer), продольно изогнутая в виде желоба пластинка для закрепления
хрящевой носовой перегородки.
8) Парная нижняя челюсть (mandibula) на переднем резцовом крае
имеет альвеолы для резцов и клыков, а на теле — для нижних коренных
зубов. Загибающаяся вверх ее задняя часть, ветвь нижней челюсти,
несет два отростка: расположенный впереди более высокий венечный
и задний, более низкий, суставной, с поперечным суставным валиком
для сочленения с черепом.
16
ВВЕДЕНИЕ
У свиней между передними концами межчелюстных и носовых ко-
стей имеется еще небольшая непарная хоботная кость (os rostri), состав-
ляющая костную основу «пятачка», или рыла.
Срединные швы между одноименными парными костями (носовыми,
лобными и т. д.) носят название сагиттальных швов соответствующих
костей. Шов между теменной и затылочной костью называется ламбдо-
видным, между лобными и теменными — венечным. По бокам дорзаль-
ной (верхней) поверхности теменных костей идут в различной степени
выраженные, иногда переходящие и на задние участки лобных костей
теменные гребни, у непарнопалых и верблюдов в задней половине они
соединяются вместе и образуют сагиттальный теменной гребень. Гра-
ницу между дорзальной поверхностью черепа и плоскостью затылка обра-
зует (исключение — быки) поперечный затылочный гребень (crista occi-
pitalis), который по бокам переходит в боковые височные гребни, идущие
к основаниям скуловых отростков чешуйчатой кости. Выступающая
назад срединная точка затылочного гребня носит название затылочного
бугра (protuberantia occipitalis). Теменные, затылочный, височные гребни
и скуловая дуга образуют границу парных височных ям или впадин,
прилегающих к боковым стенкам мозговой коробки. Впереди височных
впадин лежат глазные впадины, или орбиты, образованные лобными,
скуловыми, верхнечелюстными, слёзными и передними клиновидными ко-
стями. Задний край наружного кольца орбит у копытных, за исключе-
нием свиней, замкнутый; в образовании его принимают участие скуловой
отросток лобной и лобный отросток скуловой, а у непарнопалых, вместо
последнего — скуловой отросток чешуйчатой кости. Орбита у копыт-
ных, как и у большинства других млекопитающих (кроме приматов),
сообщается с височной впадиной; на границе их находится крыловидный
гребень (crista pterygoidea), в области которого различные типы сочета-
ния костей мозгового черепа (лобных, теменных, чешуйчатых, али- и
орбитосфеноида) составляют в некоторых группах важный системати-
ческий признак. Внутри носовой полости находятся верхняя и нижняя
носовые раковины (naso- и maxilloturbinalia) в виде тонких, продыряв-
ленных мелкими отверстиями и трубкообразно свернутых костных пла-
стинок, составляющих костную основу носового лабиринта. Носовая
полость открывается позади хоанами, стенки которых образованы нёбными,
крыловидными костями и крыловидными отростками основной клино-
видной кости. Часть лицевого отдела черепа впереди коренных зубов
носит название ростральной (rostrum). Продольная ось черепа копытных
не представляет прямой линии. Оси мозгового и лицевого его отделов
сходятся под тем или иным углом, называемым углом излома оси черепа,
неодинаковым у отдельных групп надотряда.
На поверхность черепа открывается большое число отверстий, форма
и положение которых в отдельных случаях могут иметь система-
тическое значение. Ниже перечисляются некоторые из этих отвер-
стий.
Между передними частями межчелюстных костей образуется непар-
ное резцовое отверстие (for. incisivi) или резцовая вырезка. Между нёб-
ными отростками межчелюстных и верхнечелюстными костями в перед-
ней части костного нёба имеется парная нёбная щель (fissura palatina),
часто имеющая форму овального отверстия. На задней половине нёба,
на поверхности нёбной кости открываются отверстия нёбных каналов
(for. palatina). На боковой поверхности верхней челюсти в области передне-
коренных зубов (премоляров) открывается подглазничное отверстие (for.
ЧЕРЕП КОПЫТНЫХ
17
infraorbitale). На лобной кости в области орбит располагается пара над-
глазничных отверстий (for. supraorbitalia).
Измерения черепа. Единой стандартной схемы и методики измерения
черепов млекопитающих для систематических целей нет. Каждая группа
имеет свои краниологические особенности, которыми отличаются отдель-
ные ее члены и которые отличают ее от смежных групп равного ранга.
Поэтому ниже приводится методика взятия лишь тех наиболее общепри-
нятых промеров черепов, которые применимы и для выявления краниоло-
гических отличий отдельных групп копытных (рис. 1а); после наимено-
вания промера указываются точки, между которыми производится изме-
рение наикратчайшего расстояния по прямой (за исключением особо
оговоренных случаев).
1. Основная длина черепа: от нижнего края затылочного отверстия
(basion) до средины линии, соединяющей наиболее выступающие вперед
точки межчелюстных костей (prosthion); зубы во внимание не принимаются.
2. Наибольшая длина черепа: от средины линии, соединяющей наи-
более выступающие назад точки черепа (без рогов) (opisthokranion)
до prosthion.1
3. Наибольшая ширина черепа: измеряется в наиболее широком
месте (обычно на уровне орбит или скуловых дуг).
4. Длина мозговой части черепа: от basion до средины линии, соеди-
няющей задние края орбит.
5. Длина лицевой части черепа: от prosthion до средины линии, сое-
диняющей задние края орбит.
6. Межглазничная ширина черепа: в самом узком месте между орбитами.
7. Мастоидная, или затылочная, ширина черепа: измеряется или
между наружными краями наружных слуховых проходов или между
наружными выступами сосцевидных костей.
8. Длина ряда коренных зубов: от переднего края альвеолы перед-
него зуба (Р| или Р|) до заднего края альвеолы ’последнего коренного
(М|). Таким же образом измеряется отдельно длина ряда переднекорен-
ных (Р{ — Р|) и заднекоренных (М1 — М|).
9. Угол излома оси черепа: определяется дополнительный угол к углу,
образованному пересечением линии, проходящей через prosthion и точку
слёзно-скулового шва на краю глазницы и линии, касательной к верх-
нему краю орбиты и наружного отверстия слухового прохода. Обе линии
определяются в проекции на плоскость, параллельную сагиттальной
плоскости черепа (рис. 1а, 8).
Зубные формулы. Зубной формулой сокращенно обозначается число
зубов у животных на одной стороне. При этом резцы обозначаются бук-
вой I, клыки — С, переднекоренные (премоляры) — Р, заднекоренные
(моляры) — М,1 2 молочные премоляры — Pd. Рядом с обозначением рода
зубов ставится дробью число последних (числителем — в верхней челю-
сти, знаменателем — в нижней). Иногда в конце формулы обозначается
общее число зубов. Зубная формула зубра при таком обозначении будет
такой: I -2, С , Р М ^-=32; марала: I -2 , С 4-, Р М 4 = 34; ло-
о 1 о и 3 2 3 3
т 3 0—t 3—4 3 . . т 3 г, 1 п 4 ц 3 /,
шади: I -к-, С —?, Р -х—7 , М -5-=36—44; свиньи: I —, С -у, Р -=, М -х- =44.
О V х 4 3 3 1 4 3
1 В. дальнейшем под термином «длина черепа» подразумевается наибольшая его
длина.
2 Премоляры иногда еще называются «предкоренными» или «ложнокоренными»,
а моляры — просто «коренными» или «истинными коренными».
2 Фауна СССР, И. И. Соколов
Рис. la. Череп копытного (баран).
Схема строения: Ъ — барабанные кости; Ьо — основная часть за-
тылочной кости; bsph — базисфеноид; со — суставные мыщелки; с.pt — кры-
ловидные гребни;/ — лобные кости; I — слёзные кости; т—нижняя че-
люсть; max — верхнечелюстные кости; mst — сосцевидная (мастоидная)
кость; п — носовые кости; о — затылочная кость; р — теменные кости;
pal — нёбные кости; ртах — межчелюстные кости; ро — затылочный бугор;
psph — пресфеноид; so — чешуя затылочной кости; t — височные кости;
z — скуловые кости. Схема измерений черепа: 1 — основ-
ная длина; ?— наибольшая длина; 3 — наибольшая ширина; 4—длина
лицевой части; 5—межглазничная ширина; 6 — мастоидная ширина;
7 —длина ряда коренных зубов; 8 — угол излома оси черепа.
ИЗМЕРЕНИЕ ТЕЛА
19
ИЗМЕРЕНИЕ ТЕЛА ЖИВОТНЫХ
Ниже приводится методика взятия наиболее употребительных про-
меров копытных млекопитающих (рис. 16). Там, где особо не оговорено,
измерения производятся мягкой сантиметровой лентой по изгибу.
1. Длина тела: от переднего края верхней губы до корня хвоста.
о
Рис. 16.
Схема измерений тела копытных млекопитающих:
1 — длина тела; 2 — длина туловища; 3 — обхват туловища; 4 — высота в хол-
ке; <5— высота в крестце; 6—длина головы; 7—длина уха; 8—длина
хвоста; 9 — длина передней ноги; 10 — длина задней ноги; 11 — длина кисти;
12 — длина стопы; 13 — обхват пясти и плюсны; 14 —длина пальца; 15 —дли-
на копыта; 16 — высота копыта; 17 —длина таза.
2. Длина туловища (называется еще косой длиной туловища): от
переднего выступа плечелопаточного сустава до заднего выступа седа-
лищного бугра.
3. Обхват туловища (иначе — обхват груди): производится позади
лопаток.
4. Высота в холке: от высшей точки холки до кончиков копыт передней
ноги при вытянутой ноге по прямой. Иногда — от высшей точки холки
до нижнего края пяточных мякишей, что менее точно.
5. Высота в крестце: от высшей точки крестца до кончиков копыт
задней ноги при естественном ее полусогнутом положении по прямой.
6. Длина головы: от переднего края верхней губы до выступа заты-
лочного бугра.
2*
20
ВВЕДЕНИЕ
7. Длина уха: от нижнего края ушной вырезки до конника уха (без
волос) по прямой.
8. Длина хвоста: от корня до конника, без волос.
9. Длина передней ноги: от вершины локтевого бугра до кончиков
копыт при вытянутой ноге по прямой.
10. Длина задней ноги: от выступа тазобедренного сустава (прихо-
дится предварительно нащупывать его пальцами) по изгибу ноги при
естественном полусогнутом ее положении до кончиков копыт.
И. Длина кисти: от средины тыльной (дорзальной) поверхности за-
пястного сустава до кончиков копыт при вытянутых пальцах.
12. Длина стопы: от верхнего края пяточного бугра до копчиков
копыт при вытянутых пальцах по прямой.
13. Обхват пясти (и плюсны): в самом тонком месте.
14. Длина пальца (одинаково и на передних и на задних ногах): от
средины путового сустава до кончика копыт третьего пальца.
15. Длина копыта: от заднего края пятки копыта до его кончика
(отдельно третьего и четвертого пальцев). Измеряется штангенциркулем.
16. Высота копыта: от границы рога и кожи на переднем крае до кон-
чика копыта. Измеряется штангенциркулем.
17. Толщина копыта: в самом толстом месте в области пятки. Измеряется
штангенциркулем.
18. Длина таза: от переднего выступа моклока до заднего выступа
седалищного бугра.
19. Ширина таза: между наружными выступами моклоков. Измеряется
штангенциркулем^
Примечание. Приведенная методика отличается от общепринятой мето-
дики измерений домашних животных с учетом того, что у диких зверей, как правило,
измерения производятся на трупе.
ОСНОВНАЯ ЛИТЕРАТУРА ПО КОПЫТНЫМ
Абрамов К. Г. 1954. Копытные звери Дальнего Востока и охота на них. Ха-
баровск, Книжн. издат. : 1—128.
Антипин В. М. 1941. Млекопитающие Казахстана, III. Копытные. Алма-Ата,
Казахск. гос. изд. : 1—107
Банни к о в А. Г. 1954. Млекопитающие Монгольской Народной республики.
М., изд. Акад, наук СССР (Тр. Монгольск. комисс., вып. 53) : 1—669.
Баранчеев Л. М. 1955. Парнокопытные животные Верхнего Приамурья (Амур-
ской Области). Зап. Амурск, обл. муз. краевед, и общ. краевед., 3 : 200—218.
Б и хне р Е. А. 1902. Млекопитающие. СПб. :1—-XII Ц- 1—866.
Бобринский Н. А., Б. А. Кузнецов и А. П. Кузякин. 1944. Опре-
делитель млекопитающих СССР. М., изд. «Сов. наука» : 1—440.
Данилов Д. Н. 1945. Основные способы промысла диких копытных животных.
М., Заготиздат : 1—31.
Д ин н и к Н. Я. 1910. Звери Кавказа, I. Зап. Кавказск. отд. Русск. географ, общ.,
XXVII : 1—247.
Дмитриев В. В. 1938. Копытные звери Алтайского заповедника и прилежащих
мест. Тр. Алтайск. гос. заповеди., I : 171—262.
Жарков И. В. 1952. Основные методы учета диких копытных. Сборн. «Методы
учета численности и географического распределения наземных позвоночных».
М-, изд. Акад, наук СССР : 214—238.
Залесский П. М. 1934. Проппое и современное распространение копытных
в Западно-Сибирском крае. Охотник и пушник Сибири, № 7 8: 26—28.
Кириков С. В. 1952. Птицы и млекопитающие в условиях ландшафтов южной
оконечности Урала. М., изд. Акад, наук СССР : 1—412.
Кириков С. В. 1952—1953. Исторические изменения животного мира нашей
страны в XIII—XIX веках. Изв. Акад, наук СССР, сер. географ., 1952, № 6 :
31—48; 1953, № 4 : 15—27.
ЛИТЕРАТУРА
21
(Ковалевский В. О.) Kowalewskie W. 1873—1874. Monographic der
Gattung Anthracotherium. Guv. und Versuch einer natiirlichen Glassification der
fossilen Hufthiere. Palaeontographic.a, XX, 3—5 : 131—146.
Копылов И. П. 1950. Дикие копытные животные Иркутской области. Иркутск,
Облиздат, 1950 : 1—76.
Корнеев А. П. 1953. История промысла диких зверей на Украине. Киев, изд.
Киевск. универе. : 1—38.
Кулагин И. М. 1940. Фауна БССР, т. I, вып. IV. Копытные. Минск, изд. Акад,
наук БССР :1—61.
Л у с Я. Я. 1938. Современное состояние отдаленной гибридизации животных и пер-
спективы дальнейшей работы. Изв. Акад, наук СССР, сер. биол., 4 : 775—850.
Насимович А. А. 1938; К познанию минерального витания диких животных
Кавказского заповедника. Тр. Кавказск. гос. заповеди., I : 103—150.
Насимович А. А. 1939. Зима в жизни копытных западного Кавказа. Вопр.
эколог, и биоцонолог., 7 : 3—90.
Насимович А. А. 1955. Роль снежного покрова в жизни копытных животных
на территории СССР. М., изд. Акад, наук СССР : 1—402.
Рухлядев Д. П. 1939. К эпизоотологии случаев сибирской язвы у диких копыт-
ных животных Крымского заповедника. Науч.-метод, зап. Главн. управл.
по заповеди., IV : 154—165.
Саблина Т. Б. 1955. Копытные Беловежской пущи. Тр. инет, морфол. животн.
им. А. Н. Северцова, 15 : 1—191.
Слудский А. А. 1953. Джуты в пустынях Казахстана. Тр. Инет. зоол. Акад,
наук Казахск. ССР, II : 3—30.
Слудский А. А. 1954. Роль диких млекопитающих в распространении инфек-
ционных заболеваний домашних животных. Сборн. «Природная очаговость
заразных болезней в Казахстане», 2 : 69—98.
Соколов П. И. 1953. Опыт естественной классификации полорогих (Bavidae).
Тр, Зоол. инет. Акад, наук СССР, XIV, М.—Л., изд. Акад, наук СССР : 1—
295.
Соколов И. И. 1955. Биологические особенности домашних животных и их диких
предков. Почему до сих пор только немногие виды животвых одомашнены. При-
рода, 3 : 48—55.
Те плов В. П. 1954. К вопросу о соотношении полов у диких млекопитающих.
Зоол. жури., XXXIII, 1 : 174—179.
Туркин Н. В. и К. А. Сатунин. 1902. Звери России. Cervidae. М. : 1—506.
Туркин Н. В. и К. А. С а т у н и н. 1904. Звери России. Bovidae. М. : 491—794.
Флеров К. К. 1934. О некоторых географических и исторических изменениях
евразиатских копытных. Докл. Акад, наук СССР, IV : 167—171.
Флеров К. К. 1935а. Копытные звери (Ungulata) Таджикистана. В кн. : Б. С. Ви-
ноградов, Е. И. Павловский и К. К. Флеров. Звери Таджикистана. Л., изд.
Акад, наук СССР : 93—130.
Флеров К. К. 19356. Копытные (Ungulata) Арктических стран. В кн. : Г. П. Ад-
лерберг, Б. С. Виноградов, И. А. Смирнов, К. К. Флеров. Звери Арктики,
Л., изд. Главсевморпути : 105—263.
Флеров К. К. 1952. Кабарги и олени. Фауна СССР, т. I, вып. 2, М.—Л., изд. Акад,
наук СССР : 1—256.
Формозов А. Н. 1935. Колебания численности промысловых животных. М.,
КОИЗ : 1—108.
Формозов А. Н. 1946. Снежный покров в жизни млекопитающих и птиц. М., изд.
Моск. общ. игпыт. прир. : 1—141.
Шапошников Ф-Д. 1938. О «солонцевании» диких копытных животных в горно-
таежном Алтае. Билл. Моск. общ. испыт. прир., Отд. биол., LVIII, 1 : 3—10.
Antonins О. 1951. Uber Artbastarde bei Saugetieren. Verb. Zool.-Bot. Geseilsch.
Wien, 92 : 106—115.
Brehms Tierleben. 4 Aufl., Leipzig, 1925, B.d. 12 : I—XVI-|-1—722; Bd. 13 : I—
XX-H— 714.
В r uh in II. 1953. Zur Biologie der Stirnaufsatze bei Huftieren. Inauguraldissertation,
Physiologia Comparata et Oecologia, III, 1 : 63—92; 2 : 93—127.
Ellerman J. R. and T. C. Morrison-Scott, 1951. Checklist of palaearctic
and indian mammals 1758 to 1946. London, Brit. Mus. (Katur. hist.) : 1—810.
Flower S. 1931. Contribution to our knowledge of the duration of life in vertebrate
animals, V. Mammals. Proc. Zool. Soc. London : 145—234.
Flower W. H. and R. L у d e k k e r. 1891. Mammals living and extinct. London:
I—XVI4-1—763.
22
ВВЕДЕНИЕ
Frechkop S. 1936. Sur la classification des Ongules. Mammalia, I, 2 : 37—48.
Frechkop S. 1949a. La classification des Ongules actuels. G. R. XIII Congr. Inter-
nal. de ZoOl., Paris : 379—383.
Frechkop S. 1949b. Le replacement des dents chez les Ruminants. G. R. XIII
Gongr. Internet, de Zool., Paris : 408.
Friant M. 1954. Les principales caracteristiques du cerveau des ordres actuels
d'Ongules (Ungulate sensu lato): Proboscidea, Hyraeoidea, Perissodactyla, Arlio-
dactyla, Sirenia. C. R. Seances Acad. Sci. Paris, vol. 239, N 16 : 1004—1006.
Gray A. 1954. Mammalian hybrids. A checklist with bibliography. London—Edin-
burg : 1—144.
Gregory W. K. 1910. The orders of mammals. Bull. Amer. Mus. Natur. Hist, XXVII:
1—5'24.
Harper F. 1945. Extinct and vanishing mammals of the Old World. N. Y. : 1—850.
Keilbach R. 1953—1954. Vergleichend-anatomische Studien fiber die Saugernase
mit besonderer Beriicksichtigung des Knorpelskelettes. Wissenschaftl. Zeitschr.
d. Universit. Greifswald, III, Math.-naturwiss. Reihe, 4/5 : 201—244.
Lederer G. 1953. Ein Beitrag zur Ernahrung, Haltung und Zucht pflanzenfressender
Wildtiere in Gefangenschaft, I Teil. Futtermittel. Zool. Garten, Bd. 20, N 1 :
32-44.
Lydekker R. 1898. The deer of oil lands. London, Rowl. Ward : I—XX-|-1—329.
Lydekker R. 1913—1916. Catalogue of the Ungulate mammals in the British Mu-
seum (Natur. hist.). London; vol. I, 1913 : I—XVI-pl—249; vol. II, 1913 : I—
XVII-H—295; vol. Ill, 1914 : I—XV-pl—283; vol. IV, 1915 : I—XXI-f-l—438;
vol. V, 1916 : I—XLV+1—207.
Mochi U. and T. Garter. 1953. Hoofed mammals of the world. N. Y. : 1—94,
40 pls (silhouettes).
Romer A. 1946. Vertebrate paleontology. Chicago—Illinois : I—VIII + 1—687.
S с 1 a t e r Ph. and О. T h о m a s. 1894—1900. The book of antelopes. London; vol. I,
1894—1900 : I—XXXV+ 1—220; vol. II, 1896—1897 : I—V4-1—194; vol. Ill,
1897—1898 : I— V-f-1—245; vol. IV, 1899—1900 : I—V4-1—242.
Simpson G. 1945. The principles of classification and a classification of mammals.
Bull. Amer. Mus. Natur. History, 85 : I—XVI4-1—350.
V i 11 о z R. 1954. Considerations pratiques sur le role des animaux sauvages dans ]a
transmission des maladies contagieuses et la prophilaxie de selles-ci dans se Sud-
Est Asiatique. Bull. Office internat. epizooties, t. 42 : 206—212.
Von Antelope n, Ziegen und Schafen, den Rindern und Walen (Lebensbilder aus
der Tierwelt Europas. Saugetiere, Bd. IV), Leipzig, 1922 : 1—231.
Weber M. 1928. Die Saugetiere. 2 Aufl., Bd. II, Jena : I—XXIV+1—898.
СИСТЕМАТИЧЕСКАЯ ЧАСТЬ
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ОТРЯДОВ
КОПЫТНЫХ МЛЕКОПИТАЮЩИХ
1 (2). Число пальцев на задних конечностях — один или три. Сильнее
других развит средний (третий) палец, несущий основную тяжесть
тела. Через него проходит ось конечности. В образовании заднего
края орбиты принимают участие лобная и височная кости. Корен-
ные зубы лофодонтного типа ..............................
.............. Perissodactyla — Непарнопалые (стр. 23).
2 (1). Число пальцев на конечностях — два или четыре. В последнем
случае третий и четвертый пальцы развиты сильнее других и несут
на себе основную тяжесть тела. Между третьим и четвертым паль-
цами проходит ось конечности. Если орбита замкнутая, то в обра-
зовании ее заднего края принимают участие лобная и скуловая
кости. Коренные зубы селенодонтного или бунодонтного типа . .
........................ Artiodactyla — Парнопалые (стр. 63).
Отряд PERISSODACTYLA - НЕПАРНОПАЛЫЕ
Ковалевский В. О., 1873, Остеология Anchitherium aurelianense Cuv.
Киев : 1—86 (морф., эвол.); Lydekker R., 1916, Catal. Ungulate mammals Bri-
tish Mus., V, London : 1—58 (морф., сист.); Brehms Tierleben, 1925, 4 Aufl., 12,
Leipzig : 598—708 (общ., изобр.); Weber M., 1928, Die Saugetiere, II, Jena : 631 —
686 (морф., эвол., сист.); Wood H., 1937, J. Mammal, vol. 18, N 1 : 106 (классиф.);
Simpson G., 1945, Bull. Amer. Mus. Natur. Hist., 85 : 136—143-|-252—258 (клас-
сиф.).
Характеристика отряда. Копытные с числом пальцев на передних
конечностях 1, 3 или 4, на задних 1 или 3. Третий палец развит силь-
нее других, несет на себе основную тяжесть тела животного и через него
проходит ось симметрии конечности. Независимо от числа пальцев (три
или четыре), второй и четвертый симметричны по отношению к третьему
и в одинаковой степени недоразвиты. Третья фаланга функционирующих
пальцев уплощена сверху вниз и на свободном переднем крае симметрично
закруглена. На бедренной кости (рис. 2, Б) имеется третий вертел. Шейка
бедра выражена очень слабо. Межвертельный гребень соединяет большой
и третий вертел. Таранная кость (astragalus) имеет блоковидную поверх-
ность для сочленения с большой берцовой, но дистальная ее поверх-
ность уплощена и имеет большую поверхность для сочленения с ладье-
видной (naviculare), чем с кубовидной (cuboideum). Пяточная кость,
calcaneus не сочленяется с малой берцовой. Сумма числа грудных и пояс-
ничных позвонков не меньше 22 (у современных форм чаще 23).
24
PERISSODACTYLA
В черепе лицевой отдел длиннее мозгового. Характерно наличие ка-
нала (can. pterydoideus s. alaris), прободающего основание крыловид-
ных отростков базисфеноида. Хоаны передним краем доходят до уровня
второго или даже первого заднекоренного (М2 или М1). Суставная по-
верхность для причленения нижней челюсти поперечно вытянутая, с вы-
пуклой суставной поверхностью, позади имеет высокий засуставной
отросток (proc, postglenoideus), благодаря чему боковые движения ниж-
ней челюсти очень ограничены. Небольшая круглая tympanicum обра-
зует только наружную стенку барабанной полости (и то не всегда); внут-
реннюю стенку последней составляет petrosum. Между каменистой костью
petrosum и телом затылочной кости (basioccipitale) образуется большое
разорванное отверстие (lor. lacerum magnum), соответствующее ряду
отверстий основания черепа других млекопитающих. Носовые кости
расширены в задней половине. Правая и левая половины нижней челю-
сти срастаются без образования шва. Восходящие ветви нижней челю-
сти очень широки.
Коренные зубы лофодонтного, или селено-лофодонтного типа, с попе-
речными гребнями, всегда образуют замкнутый ряд. Переднекоренные
(премоляры), кроме переднего, сходны по форме и величине с заднекорен-
ными (молярами). Последний заднекоренной зуб нижней челюсти (М3)
состоит из двух долей; если же имеется третья, то таковая отсутствует
на последнем молочном премоляре (Pd4). Молочных переднекоренных —
четыре; постоянных — только 3. Длинная диастема отделяет коренные-
зубы от клыков и долотовидных или конусовидных резцов. Последние-
могут сильно редуцироваться, но не исчезают совсем. Губы хвататель-
ного типа, подвижные, иногда (у тапиров) вместе с другими мягкими
частями передней части головы превращены в недлинный хобот. Желу-
док простой, но на значительном протяжении от входа выстланный сли-
зистой оболочкой с многослойным плоским эпителием, лишенной пище-
варительных желез. Слепая кишка (соесшп) и ободочная (colon) длин-
ные и очень объемистые. Последняя имеет вид двойной петли, оба колена
которой плотно прилегают друг к другу. Продольные мускульные во-
локна слепой и ободочной собраны в укороченные, по сравнению с длиной
кишки, пучки (тении), благодаря чему в наполненном виде стенка кишеч-
ника между тениями образует ряды выпячиваний в виде карманов. Желч-
ный пузырь отсутствует. Боковое выпячивание стенки входа в носовую
полость образует так называемый носовой барабан, или дивертикул
(diverticulum nasi). Слизистая оболочка евстахиевых труб образует на
внутренней стороне выпячивания, так называемые воздухоносные или
заглоточные мешки.
Молочная железа двухдольчатая, расположена в паховой области.
Плацента всегда диффузная. Плод всегда один.
Географическое распространение. Африка, Передняя, Средняя, Цент-
ральная и Южная Азия с Малайским архипелагом; южная часть Север-
ной и северная половина Южной Америки (тапиры). В Европе в диком
состоянии обитали до конца XIX в. Отсутствуют в Австралии и на Мада-
гаскаре. В домашнем состоянии распространены по всему земному шару..
Эволюция непарнопалых. Непарнопалые представляют одну из много-
численных ветвей копытных. Общее направление эволюции последних
у Perissodactyla может находить наиболее яркое выражение. В процессе-
исторического развития имеет место переход от стопо- и пальцехождения
(которое из современных еще сохраняется у тапиров) к фаланго- или
копытохождению, редукция локтевой и малоберцовой кости, умень-
PERISSODACTYLA
25
шение числа лучей стопы и кисти. Нов противоположность парнопалым,
эволюция Perissodactyla шла в направлении образования трехпалой и
однопалой конечности с перенесением основной нагрузки на третий палец.
Даже в тех случаях, где, как и у многих парнопалых, сохраняется четыре
пальца (на передней конечности у тапиров), конечность сохраняет «меза-
Рис. 2. Бедро зубра (А) и лошади Пржевальского (Б) с
плантарной стороны.
а — третий вертел; б — малый вертел; в — межвертельный
гребень; г — вертлужная впадина.
ксонический» характер и ось ее проходит всегда через третий палец,
а второй и четвертый сохраняют по отношению к последнему симметрич-
ное расположение. При уменьшении числа лучей в стопе и кисти в пер-
вую очередь исчезает первый палец. Затем редуцируется пятый и в послед-
нюю очередь параллельно уменьшаются и исчезают второй и четвертый.
Своеобразное направление эволюции находит выражение и в зубной
системе. Исходный шестибугорчатый тип коренных зубов превращается
в лофодонтный. Происходит это путем попарного слияния внутренних
с наружными бугорками в поперечные гребни (передний — протолоф,
26
EQUIDAE
задний — метал оф), а также наружных бугорков в продольный гребень
(эктолоф). В процессе эволюции в ряде случаев имел место процесс
дополнительной складчатости основных гребней и образование новых
бугорков и гребней. В результате у лошадей, например, образуется очень
сложный рисунок жевательной поверхности зуба. Очень характерен
для непарнопалых процесс так называемой моляризации переднекорен-
ных зубов (премоляров), приобретения ими, в противоположность парно-
палым, формы и строения заднекоренных (моляров); даже у наиболее
примитивных из современных представителей отряда, тапиров, поляри-
зация не распространяется только на передний зуб Р|. Удлинение зуб-
ного ряда влечет за собой удлинение в черепе лицевого отдела.
Имея своими предками одну из древних групп первичнокопытных
(Condylarthrd), непарнопалые в самой ранней их истории представлены
рядом филогенетических ветвей, из которых в современной фауне сохра-
нились лишь три хорошо разграниченные группы, или семейства: одно-
палые лошади (Equidae), трехпалые носороги (Rhinocerotidae) и четырех-
палые (на передних конечностях) тапиры (Tapiridae); последние две
считаются стоящими ближе друг к другу, чем к семейству лошадиных.
Сохранившиеся до наших дней представители отряда являются жалкими
остатками некогда обильной группы, имевшей расцвет в средине и в конце
третичного периода. Вместо 158 родов, известных в ископаемом состоя-
нии, в настоящее время отряд насчитывает всего 6, бедных по количеству
видов и малочисленных по числу особей. Из вымерших групп, относя-
щихся к отряду непарнопалых, можно назвать более близких к лошадям
титанотериев, или бронтотериев (Т itanotheriidae s. Brontotheriidae), хали-
котериев (Chalicotheriidae), палеотериев (Palaeotheriidae) и несколько
мелких групп, родственных тапирам и носорогам. В фауне СССР непарно-
палые представлены только семейством лошадей (Equidae).
Семейство EQUIDAE — ЛОШАДИНЫЕ
Selve J., 1912, Коп. Svenska Vetenskapakad. Handl., Stockholm, XLVIII,
6 : 1—185 (морф., эвол.); Osborn H., 1918, Mem. Amer. Mas. Natur. Hist., n. s.,
2 : 1—217 (морф., эвол.); Loomis F., 1926, The evolution of the horse, Boston :
I—XVI-J-1—233; Matthew W., 1926, Quart. Rev. Biol., 1:139—185 (эвол.);
Matthew W. and R. S t i r t о n, 1930, Univers. California Publ., Dspartm. Geol.
Sci., 29 : 349—396 (морф., эвол.); Haughton S., 1931, Ann. South.-Afric. Mus.,
28 : 407—427 (морф., эвол.); S t i r t о n R., 1940, Univers. California Publ., Departm.
Geol. Sci., 25 : 16)—198; Громова В. II., 1949, История лошадей (рода Equus)
в Старом Свете, М.—Л., изд. Акад, наук СССР (Тр. Палеонтол. инет., XVII, 1, 2),
ч. I : 1—374, ч. II : 1 —163 (морф., эвол., сист.); Громова В. И., 1952, Гиппарионы
(род Hipparion), М.—Л., изд. Акад, наук СССР (Тр. Палеонтол. инет., XXXVI) : 1—
478 (морф., эвол.).
Характеристика семейства. Число пальцев и на передних и на зад-
них конечностях у современных форм — один. Конечности длинные.
Кожа покрыта плотным волосяным покровом, удлиненным на верхней
стороне шеи и на части или по всей поверхности хорошо развитого хво-
ста. В черепе у современных форм задний край глазниц отделен от височ-
ных впадин костным мостиком, образованным лобной и чешуйчатой
костями. Носовые отростки межчелюстных костей длинные, соединяю-
щиеся с носовыми костями. Барабанная часть каменистой кости (tympa-
nicum) образует костный слуховой проход. Proc, postympanicus не сли-
вается с яремным отростком (proc, paraoccipitalis). Число позвонков:
грудных 18—19, поясничных 5—6, крестцовых 5—6. Семенники у взрос-
EQUUS
27
дых самцов располагаются в специальном выпячивании брюшной стенки,
мошонке.
Распространение. Африка, Передняя, Средняя и Центральная Азия.
Некогда обильное по числу форм семейство лошадиных, насчитывавшее
свыше двадцати родов, вместе с вымершими палеотериями, халикотериями
и титанотериями, образует подотряд Hippomorpha. В современной фауне
представлено одним родом Equus L.
Род EQUUS LINNAEUS — ЛОШАДИ
Lydekker R., 1912, The horse and its relatives, London : I—ХП-j-1—286
'(общ., изобр.); Loomis F., 1926, The evolution of the horse, Boston : I—XVI+
+ 1—233; Ma tthe w W., 1926, Quart. Rev. Biol., I : 139—185(эвол.); Ф леровК. К.,
1931, Докл. Акад, наук СССР : 269—272 (морф.); Antonins О., 1937, Proc. Zool.
Soc., London, 107, ser. В: 557—564 (распр.); Гром о'в а В. И., 1941, Тр. Зоол. инет.
Акад, наук СССР, V :92—157 (эвол., сист., морф.); S t i г t о n R., 1942, J. Paleontol.,
vol. 16, № 5 : 627—637 (эвол., сист.); Развитие жизни на Земле, 1947,
Альбом наглядн. пособ., М., Госкультпросветиздат : 1—47, табл. 59 (эвол.); Bout-
del 1 е Е., 1948, С. R. XIII Congr. Internal, de Zool., Paris : 383—387 (распр.); Гро-
мова В. И., 1949, История лошадей (род Equus) в Старом Свете, М.—Л., изд. Акад,
наук СССР (Тр. Палеонтол. инет., XVII, 1 и 2), ч. I : 1—374, ч. II : 1—163 (морф.,
эвол., сист.); Bourdelle Е. etS. Frechkop, 1950, Mammalia, XIV, 4 : 126—
139 (классиф.).
Морфологическая характеристика. Диагностические признаки
рода — общие со всем семейством.
Общее направление эволюции непарнопалых в роде лошадей (Equus)
находит свое крайнее выражение. Это стройные высоконогие животные,
вся организация которых приспособлена к быстрому передвижению по
твердому грунту. Удлинение конечностей идет в первую очередь за счет
дистальных звеньев, тогда как проксимальные участки, плечо и бедро,
становятся относительно более короткими. Редукция числа лучей в стопе
и кисти достигает своей кульминации в линии лошадей уже к концу
плиоцена. От боковых (II и IV) метаподий остаются рудименты в про-
ксимальной части в виде так называемых грифельных косточек. У совре-
менных форм фаланги боковых пальцев в виде самостоятельных костей
не сохраняются.1 Сильной редукции подвергается локтевая и малобер-
цовая кости. От первой сохраняется лишь ее проксимальная (верхняя)
часть с локтевым отростком. Малоберцовая кость также представлена
очень тонким рудиментом проксимальной половины; но здесь сохраняется
отделенный от остальной части кости самый дистальный ее участок, рано
срастающийся с большой берцовой и образующий латеральную лодыжку
последней. В заплюсне сливаются в одну кость cuneiforme I и II.
Имеет место окончательный переход к «копытохождению», когда ко-
нечность и при ходьбе и стоянии опирается о почву только третьей фа-
лангой пальца. В связи с этим сагиттальный гребень на дистальном
конце третьей пястной кости не ограничивается вентральной ее поверх-
ностью, в противоположность пальцеходящим копытным, а распростра-
няется на всю поверхность суставного блока, увеличивая прочность со-
членения с первой фалангой. В связи с увеличением нагрузки третья
фаланга (копытная кость) утолщается и расширяется, приобретая форму
полукруга. Она одета в прочный роговой башмак—копыто, предназна-
ченное для смягчения толчков и для защиты лежащих в глубине мягких
1 Утолщения на дистальных концах грифельных костей, по воззрениям некоторых
исследователей, представляют рудименты скелета пальцев.
28
EQUIDAE
тканей от повреждений. Копыто (рис. 3) представляет собою видоизме-
ненный коготь (или ноготь) других млекопитающих. Охватывающая спе-
реди и боков конечную (третью) фалангу толстая роговая стенка копыта
соответствует ногтевой пластинке. Позади копытная стенка делает пово-
рот в сторону подошвы, образуя так называемые пяточные или заворот-
ные углы, и продолжается по бокам стрелки. Роговая стенка состоит
из двух основных слоев: 1) внутреннего, листочкового, более светлого,
образованного производящим слоем эпидермиса, одевающего листочко-
вый же слой основы кожи «мясной» стенки копыта, и 2) лежащего поверх
очень толстого и прочного трубчатого слоя, более темного цвета. Образо-
вание и рост трубчатого рога происходит от производящего слоя, эпидер-
миса, покрывающего сосочки основы кожи венчика на границе между
копытом и нормальной (не видоизмененной) кожей. Трубчатый слой вы-
полняет основную защитную и опорную функцию роговой стенки копыта.
Кроме того, снаружи копыто покрыто тонким слоем слабо ороговевших
клеток, называемым глазурью; образуется и растет глазурь от погра-
ничного между копытом и остальной кожей участка каймы рога. Назна-
чение этого слоя — предохранение трубчатого и листочкового рога от
высыхания. Венчик и кайма копыта соответствуют когтевому (ногтевому)
жолобу и валику других млекопитающих. Нижнюю (волярную или план-
тарную) поверхность копыта составляет копытная подошва, соответст-
вующая участку когтя (ногтя) между его стенкой и краем пальцевого
мякиша. Наружный слой ее образует несколько вогнутая роговая по-
дошва, выполняющая пространство между нижним (дистальным) краем
копытной стенки и стрелкой. Основой ее является трубчатый рог, образуе-
мый эпителием, выстилающим сосочки основы кожи («мясной» подошвы).
В прочности рог подошвы значительно уступает рогу копытной стенки.
Очень важной функционально частью копыта является его стрелка,,
представляющая сильно видоизмененный пальцевый мякиш. Она имеет
форму треугольника, со срединной бороздой, обращенного вершиной
вперед и вклиненного между заворотными частями стенки копыта и в сре-
динную часть копытной подошвы. Основу стрелки составляет сильно раз-
витый в этой части, в противоположность стенке и подошве, подкожный
слой в виде плотной упругой соединительнотканной мякишной подушки.
Поверх подушки мякиша лежит тонкий слой основы кожи («мясная»
стрелка), покрытый слоем производящего эпителия. Последний продуци-
рует толстый, но мягкий роговой слой, образующий роговую стрелку,
особенно массивную в передней части. Подкожный мякиш вместе с копыт-
ными хрящами,1 прикрепленными к боковым ветвям копытной фаланги,
и с роговой стрелкой образуют прекрасно приспособленный к выполне-
нию функции смягчения толчков рессорный механизм копыта. При опоре
о почву основная нагрузка падает на свободный (нижний) край копытной
стенки и на стрелку. Подошва касается почвы только на мягком грунте.
Подошвенные (пястные) мякиши сохраняются лишь в виде рудиментов,
так называемых «шпор» — бородавчатых разрастаний кожи в области
задней (волярной, плантарной) поверхности первой фаланги.1 2 Одно из
характерных для лошадей образований — «каштаны», возможно, являются
остатками запястных и заплюсневых мякишей. Скорее же всего, судя ш>
местоположению, они представляют собою видоизмененные кожные железы,
1 Являющимися также видоизмененными частями подкожного слоя этой части
копыта.
2 Впрочем, существует мнение, что шпоры представляют рудименты боковых
пальцев.
EQUUS
20
свойственные многим другим копытным. На передних конечностях каштаны,
в виде довольно крупных, плоских бляшек, расположены на внутренней
стороне несколько выше запястного сустава, а на задних (имеются не
у всех видов) — несколько ниже заплюсневого (скакательного).
В осевом скелете для лошадей, помимо перечисленных в диагнозе осо-
бенностей, характерно наличие на черепе сагиттального теменного гребня
(в противоположность носорогам), слияние tympanicum с perioticum
(в противоположность тапирам). Резко выражена опистоцельность шейных
Рис. 3. Копыто лошади. (По Ellenherger—Baum, 1943).
А — сагиттальный разрез (схема): пек — подкожный слой кожи;
ок — основа кожи и покрывающий ее волос; ср — сухожилие разгибателя пальцев;
к — кайма; в — венчик; окмс — основа кожи «мясной» стенки; л — листочки основы
кожи; и —надкостница; реп—роговая стенка; бл — белая линия; окп —основа кожи
подошвы с сосочковым слоем; оке — основа кожи стрелки с сосочковым слоем; рп —
роговая подошва; рстр — роговая стрелка; п — соединительнотканная подушка
(подкожный слой) мякиша и стрелки; I, II, III — фаланги пальцев. Стрелки указы-
вают направление роста рога. Б — вид с вентральной сторон ы: п — пя-
точные части; рстр — роговая стрелка; сб — ее срединная борозда; пк — подошвен-
ный край роговой стенки; бл — белая линия; рп — роговая подошва; зу — заворот-
ный угол роговой стенки; бб —• заворотная часть стенки и боковая борозда стрелки.
позвонков, возрастающая от третьего к седьмому; вместе с их значительной
длиной это обеспечивает большую подвижность шеи и способность ее к вра-
щательным движениям.
Особенности зубной системы отражают эволюцию в направлении пере-
хода к питанию жесткой травянистой пищей, собираемой непосредственно
с почвы. Лошади должны после одного пережевывания использовать ту
пищу, которая жвачными перетирается дважды с промежуточным мацери-
рованием в преджелудках. Число зубов — 44, но первый переднекоренной
(Р, и Р1) рудиментарен, иногда рано выпадает, находясь на пути к полному
исчезновению; при этом не ясен вопрос: является ли этот зуб у лошадей
молочным, не имеющим замены (Pd {), или он относится к серии постоянных,
предполагая, что молочный в процессе эволюции был утрачен раньше.
Кроме того, клыки у самок отсутствуют совсем (чаще) или рудиментарны.
Резцы, приспособленные к захвату и срыванию травы, в числе трех пар
хорошо развиты и в верхней и в нижней челюстях. Особенностью их у лоша-
30
EQUIDAE
дей являются карманообразные впячивания эмали на верхушках, так
называемые лунки или чашки,1 заполненные цементом. В процессе изна-
шивания зуба внешние очертания лунок меняются, что служит основным
показателем для определения возраста животных.
Коренные зубы достигают высокой степени гипсодонтии. Постоянный
рост зубов, как это имеет место у грызунов, здесь не происходит, нокоронка
зуба получается очень высокой. Складчатость эмали находит крайнее
выражение. Образуется эктолоф (наружный продольный гребень) и ряд
дополнительных стилей, увеличивающих жевательную поверхность зуба.
Зубной цемент покрывает коронку зуба до вершины включительно, выпол-
няя все углубления между складками эмали, гребнями и бугорками.
В процессе изнашивания зуба на жевательной его поверхности одновремен-
но выступают дентин, эмаль и цемент. Благодаря неодинаковой прочности
и неравномерности стирания этих трех элементов, жевательная поверхность
получается неровной, чем повышается ее перетирающая способноость.
Переднекоренные зубыкрупнее (несколько длиннее и шире) заднекоренных.
Третья доля на последнем нижнем заднекоренном (М3) не развита.
Из других характерных особенностей лошадей можно назвать следую-
щие. Пищевод при входе в желудок имеет особый клапан («заслонку»),
препятствующий обратному поступлению в пищевод из желудка пищевых
масс и газов. Поэтому рвота и отрыгивание пищевых масс и газов у лоша-
дей невозможны. Слепая кишка (в противоположность всем остальным
непарнопалым) верхушкой обращена вперед. Молочная железа двухдоль-
чатая, каждый сосок открывается наружу двумя каналами (в противопо-
ложность носорогам), ведущими в две самостоятельные молочные цистерны.
Почки гладкого однососочкового типа, но (в противоположность тапирам
и носорогам) концевые ходы почечной лоханки не ветвятся. Носонёбный
(стенонов) канал и сошниково-носовой (якобсонов) орган не имеют выхода
в ротовую полость.
Эволюция лошадей. Палеонтологическая история семейства Equidae
изучена едва ли не лучше, чем какой-либо другой группы животных.
«Ряд лошадей» (рис. 4) обычно служит образцом построения филогенети-
ческого древа. Общим предком всех лошадиных (Hippomorpha), очень близ-
кими таковому для всего отряда непарнопалых, в настоящее время считается
нижнеэоценовый род Hyracotherium Owen, найденный в Европе и почти
не отличимый от одинакового с ним возраста североамериканского Eohippus
Marsh. Это было животное величиною с небольшую собаку, с бугорчатыми
зубами, незначительной диастемой, незамкнутой орбитой; переднекорен-
ные зубы имели только три бугорка. Число лучей на передних конечно-
стях было четыре, на задних — только три; от двух боковых (I и V) оста-
вались только грифельные косточки; конечные фаланги пальцев были уже
снабжены копытами. Средне- и верхнеэоценовые потомки этих форм, севе-
роамериканские роды Orohippus и Epihippus Marsh, отличались тем, что
обладали несколько большей величиной и окончательной утратой на зад-
них конечностях следов грифельных косточек I и V лучей. Кроме того,
последние 2 переднекоренных зуба приобрели форму моляров.
В олигоцене предки современных лошадей представлены опять-таки
североамериканскими родами Mesohippus Marsh и Miohippus Marsh,
величиною с крупную собаку. Конечности, особенно метаподии, более длин-
ные. И на задней, и на передней число пальцев только три; третий развит
1 В полной мере на всех зубах развитые только у настоящих лошадей (подрод
Equus s. str.).
EQUUS
31
значительно сильнее других, но. о землю опирались все три пальца. Корен-
ные зубы стали ясно лофодонтными, образовался эктолоф. Переднекорен-
ные моляризовались за исключением Р}. Увеличилась диастема. Сохранив-
шаяся брахиодонтия указывает на питание в основном еще мягкой расти-
тельностью. Олигоценовые лошадиные дали начало ряду боковых миоце-
новых ветвей (Anchitherium Meyer, Archaeohippus Gidley, Hipohippus
Leidy), достигавших большой величины, но сохранивших примитивное
устройство конечностей и зубов и не оставивших в современной фауне
потомков. Часть их мигрировала в Евразию, где некоторые существовали
до нижнего плиоцена.
. Линия, ведущая к современным лошадям, и в миоцене продолжала свою
эволюцию в Новом Свете. Из форм этого периода’должны быть названы
Parahippus Leidy и Merychippus Leidy. Первый сохранил низкие коронки
зубов, еще не всегда замкнутую глазницу, но Merychippus имел уже гипсо-
32
EQUIDAE
донтные зубы с образовавшимися петлями эмали1 и хорошо развитым це-
ментом; образовался замкнутый костный мостик заднего края орбиты.
У обоих родов диастема стала длинной, удлинился лицевой отдел черепа;
боковые (II и IV) пальцы укоротились и, вероятно, при быстром беге уже
не касались земли. В лице Merychippus мы имеем, очевидно, качественно
новый этап эволюции ствола лошадиных, связанный, вероятно, с переходом
от лесного образа жизни к жизни на открытых пространствах с более жест-
ким травянистым покровом.
Merychippus дал начало, с одной стороны, группе гиппарионов, до
недавнего времени принимавшихся, по-видимому, неправильно за предков
однопалых лошадей, а с другой стороны, непосредственному предку совре-
менной лошади — роду Pliohippus Marsh. Гиппарионы проникли на тер-
риторию Старого Света на грани миоцена и плиоцена и на всем протяжении
последнего (а в Африке даже в нижнем плейстоцене) были широко распро-
странены, составляя характерный элемент нижнеплиоценовых «гиппарио-
новых фаун».
Нижнеплиоценовый род Pliohippus величиною с крупного пони харак-
теризовался тем, что у некоторых из его видов боковые пальцы уже исчезли,
но зубы при гипсодонтии еще сохраняют упрощенный, по сравнению
с Equus, рисунок. В верхнем плиоцене Америки появилась форма Plesip-
pus Matthew, которая является переходной к современным лошадям,
включается даже иногда в род Equus, отличаясь лишь некоторой упрощен-
ностью строения зубной системы. В верхнем плиоцене имело место послед-
нее, третье по счету, вселение лошадей в Евразию, а оттуда в Африку,
тогда как в самойАмерике лошадипо неизвестным пока причинам(вероятно,
вследствие истребления человеком) вымерли в доисторическую эпоху.
При открытии Америки европейцами лошадей там не было, хотя вновь
завезенные из Старого Света нашли все условия для своего процветания.
В настоящее время дикие лошади представлены тремя подродами:
1) собственно лошадей (Equus s. str.), одноцветных, с каштанами обычно
на всех четырех конечностях; 2) полосатых зебр (Hippotigris), населяющих
Африку, и 3) ослов и полуослов (Asinus), не имеющих поперечной полоса-
тости в окраске, но с каштанами только на передних конечностях и длин-
ным волосом только на дистальной части хвоста.
В фауне СССР род лошадей представлен только домашней лошадью
(Equus caballus L.) и куланом (Equus hemionus Pall.); в конце прошлого
столетия в степях юга СССР был истреблен близкий к домашней лошади
тарпан (Е. caballus gmelini Ant.). Для истории фауны СССР представляет
также интерес единственный в настоящее время дикий .вид настоящих
лошадей, лошадь Пржевальского (Е. przewalskii Poljak.), обитающая
в Джунгарии и западной Монголии.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА EQUUS ФАУНЫ
СССР И МОНГОЛИИ
1 (2). Уши длинные, не менее 170 мм. Хвост тонкий, покрыт длинным
волосом только в дистальной трети. Костные наружные слуховые
проходы длинные, более 5.5% основной длины черепа, направлены
косо назад и вверх, при взгляде на череп сверху значительно высту-
пают в стороны за контуры височных гребней. Тонкие длинные конеч-
1 Карманообразные углубления между складками эмали на трущейся поверхности
зуба. .... ........
EQUUS
33
ности и высокие узкие копыта; ширина плюсневой кости в средней
части не превышает 10 % ее длины; ширина третьей (копытной) фаланги
передней конечности не больше 137% ее длины по переднему
краю .... Equus (Asinus) hemion us Pall. — Кулан (стр. 33).
2 (1). Уши длиною не больше 170 мм. Хвост толстый, покрыт грубым
длинным волосом, не меньше чем на половину длины. Костные слу-
ховые проходы короткие (не более 5.5% основной длины черепа),
при взгляде на череп сверху концы их едва выступают за контуры
височных гребней. Ширина плюсневой кости в средней части больше
10% ее длины, ширина третьей фаланги передней конечности состав-
ляет больше 137 % длины ее по переднему краю.
3 (4). Масть у диких особей мышино-серая («мышастая»). На макушке
головы между ушами пук длинных волос, свешивающийся на лоб
(«челка»). Хвост несет длинные волосы на всем протяжении, кроме
самого корня. Каштаны имеются только на передних конечностях.
Длина ряда коренных зубов верхней челюсти меньше 37 % основной
длины черепа. Ширина второй фаланги передней конечности состав-
ляет 125—126% ее длины................................................
................Equus caballus gmelini Ant.—Тарпан (стр. 45).
4 (3). Масть с преобладанием красновато-желтых тонов («каурая» или
«гнедо-саврасая»). Челка на голове не развита. Хвост несет длинные
волосы только в дистальной половине. Каштаны имеются на всех
четырех конечностях. Длина ряда коренных зубов верхней челюсти
больше 37% основной длины черепа. Ширина второй фаланги конеч-
ности составляет не свыше 102% ее длины...............................
.......Е. przewalskii Poljak. — Лошадь Пржевальского (стр. 52).
Equus (Asinus) chemionus Pallas — Кулан или онагр« Джигетай
(рис. 5—8)
Pallas Р., 1775, Nov. Comm. Acad. Sci. Petropol, XIX : 394—417, pl. VII
(первооп.); Эве р с м а н н Э., 1850, Естеств. история Оренбургск. края, II, Казань :
229 (общ.); R a d d е G., 1861, Beitr. z. Kenntn. d. Russ. Reich., 23 : 430—433 (ox.);
R a d d e G., 1862, Reisen im Suden v. Ost. Sibirien, I, St.-Retersb. : 293—295 (сист.,
распр.); Карелин Г. С., 1875, Tp. СПб. общ. естествоисп., VI : 267—268 (распр.);
Пржевальский Н. М., 1875, Монголия и страна тангутов, I, СПб. : 282—285
(биол.); Radde G. u. Walter А., 1889, Zool. Jahrh., Abt. Syst., IV : 993—1094
(морф., распр.); Варенцов П., 1894, в кн. «Фауна Закасп. обл.», Ашхаб. : 1—38
(прир.); Леваневский М., 1894, Землевед., IV : 121—123 (опис.); Степняк,
1896, Природа и охота, 3 : 69—73 (ох.); Словцов И., 1897, Зап.-Сиб. отд. Русск.
географ, общ., XXI ; 131—132 (распр., биол.); Lydekker R., 1904, Novit. zool.,
XI : 583—596 (сист., изобр.); Lydekker R., 1904, Proc. Zool. Soc. London : 431 —
432 (сист., изобр.); Lydekker R., 1912, The horse. London: 176—186 (общ., изобр.);
Lydekker R., 1916, Catal. Ungulate Mammals Brit. Mus. (natur., hist.), V : 8—15
(сист., изобр.); Andrews R., 1924, Natur. hist., 24 (2) : 153—155 (общ., изобр.);
Brehms Tierleben, 1925, 12 : 668—675 (общ., изобр.); Schwarz E., 1929, Zool.
Gart., 2 : 85—94, (сист., изобр.); СелевинВ. A., 1932, Охотник и рыбак Сибири,
4 : 27 (распр.); Флеров К. К., 1932, Тр. Совета по изуч. производ. сил, сер. турк-
менск., 2 : 252—253 (сист., распр.); Шатерников а Т. М. и Н. В. Румян-
цев, 1934, Коневодство, 2 : 66—70 (общ., изобр.); Розанов М. П., 1937, сборн.
«Экспедиции Акад, наук СССР 1935 года», М., изд. Акад, наук СССР : 272—280 (распр.);
Шнитников В. Н., 1936, Млекопит. Семиречья, М.—Л., изд. Акад, наук СССР :
148—150 (распр., биол.); Селевии В. А., 1937, Бюлл. Среднеазиатск. гос. унив.,
22 : 299—301 (ист., распр.); Слудский А. А., 1939, Сборн. тр. Зоол. муз. Моск,
гос. унив., V : 24—25 (распр.); Allen, 1940, The mammals of China a. Mongolia,
2, N. Y. : 1281—1284 (общ., изобр.); Harper F., 1940, J. Mamm., 21 (2) : 197—201
(сист., распр.); Антипин В. М., 1941, Млекопит. Казахстана, III, Алма-Ата,
Каз. обл. гиз. : 93—96 (распр.); Громова В. И., 1941, Тр. Зоол. инет. Акад,
наук СССР, VI : 92—157 (морф., сист., ист.); И пг у н и и Г. И. и Е. П. К о р о в и н,
3 Фауна СССР, И. И. Соколов
34
EQUIDAE
1945, Природа, 4 : 47—58 (охр.); Harper F., 1945, Extinct a. vanishing Mammals,
N. Y. : 353—359 (общ.); Гептнер В. Г., 1948, Охр. прир. „2 : 50—60 (общ.); Гро-
мова В. И., 1949, Тр. Палеонтол. инет., XVII, 1 : 216—225, 2 : 3—162 (морф.,
эвол.); ПГарлемань Н.В., 1949, Охр. прир., 9 : 88—90 (ист.); КлюшкинЕ. А.,
1950, Изв. Туркменок, фил. Акад, наук СССР, 2:34—40 (охр.); Мурзаев Э.,
1950, Непроторенными путями, М. : 235—241 (распр.); Щекин В. А. и
А. В. Ш к у р г и н, 1950, Тр. Всес. научно-иссл. инет, конев., 18 : 106—118 (гибр.
с лошадью); Ц а лк ин В. И., 1951, Докл. Акад, наук СССР, н. с., LXXXI, 2 :
289—291 (распр.); Слудский А. А., 1953, Тр. Инет. зоол. Акад, наук Казахск.
ССР, II : 10—11 (биол.); Банников А. Г., 1954, Тр. Монг, комисс., 53 : 146—
159 (общ., изобр.); Д а л ь С. К., 1954, Изв. Акад, наук Армянск. ССР, VII 2 : 65—66
(ист.); Соколов И. И., 1958, Охота охотн. хоз., 9:21—24 (акклим.).
Кулан— единственный дикий представитель рода Equus в фауне СССР,
принадлежит к подроду ослов (Asinas). К последнему, кроме Е. hemio-
nus, относятся еще один или два вида африканских серых, или настоящих
ослов, от которых ведет свою родословную и современный домашний
осел. Кроме окраски, африканские ослы отличаются от азиатских на-
личием поперечных черных полос, идущих от холки по бокам плеч до
уровня локтевых суставов, длинными ушами и очень узкими передними
копытами.
Вместе с тибетским киангом и группой онагров Передней Азии по
морфологическим особенностям кулан занимает как бы промежуточное
положение между настоящими ослами и лошадьми, откуда все они по-
лучили название полуослов.
Описание. Высота в холке около 125 (от НО до 140) см, длина туло-
вища 175—200 см. Голова длиной около 50 см, кажется большой по сравне-
нию с общей величиной животного. Уши длиной 24—25 см, несколько
длиннее, чем у лошадей, но короче, чем у настоящих ослов. Общий тип
сложения легкий; поджарое туловище покоится на высоких тонких ко-
нечностях с узкими вытянутыми копытами. Передние копыта все же,
в отличие от африканских ослов, несколько шире задних. Каштаны
имеются только на передних конечностях, при этом, в отличие от лошадей,
они большие, расположены выше и представляют гладкие, лишенные
волоса участки кожи. Хвост тонкий, от основания покрыт коротким,
в летней шерсти плотно прилегающим к коже тонким волосом, и только
дистальная его треть несет кисть из длинных грубых волос. Длина хвоста
вместе с кистью составляет 60—80 см. Волосы на гриве короткие, торча-
щие вверх. Характерный для лошадей, особенно домашних, пучок волос
между ушами на макушке головы, так называемая челка, не раз-
вит.
Общий тон окраски верхней части' туловища, шеи, головы, а также
наружной поверхности ног до запястных и заплюсневых суставов от
светлого песчано-желтого, даже палевого, до красновато-коричневого,
в зимнем мехе иногда с сероватым оттенком. Вдоль средней линии спины
от гривы до начала длинных волос на хвосте тянется темно-бурая полоса
шириной в несколько сантиметров, сопровождаемая иногда по бокам
узкой светлой оторочкой. Грива, концы ушей и узкая полоса непосред-
ственно над копытами темно-бурые. Кисть из грубых волос на конце
хвоста, черная. Нижняя сторона туловища и шеи, конец морды, внутрен-
няя поверхность и нижние части конечностей и ушных раковин, а также
задние части бедер по сторонам хвоста светлые, от желтовато-сероватого
до чисто белого цвета. В летний период волосяной покров короткий,
плотно прилегает к коже, зимой более длинный (3—4 см) и волни-
стый.
Equus
35
Череп (рис. 6) с высоким лицевым отделом. Высота его впереди Р2
составляет не меньше 21.5 %1 основной длины. Расстояние от заднего
края сошника до заднего края костного нёба обычно больше расстояния
от заднего края сошника до нижнего края затылочного отверстия. Сошни-
ковый индекс (отношение второго промера к первому в процентах) низкий,
составляет от 84.5 до 113.4% (в среднем 99.8%). Глоточные бугорки не
заходят вперед дальше уровня большого разорванного отверстия (for.
lacerum). Костные слуховые каналы длинные, составляют не меньше
Рис. 5. Кулан (Equus hemionus Pall.).
5.5% основной длины черепа, направлены косо назад и вверх. Задний
край носовой вырезки (между носовой и межчелюстной костями) располо-
жен, как правило, на уровне заднего края Р2. У нижнего края входа
в носовую полость межчелюстные кости обычно образуют гребневидные
бугорки. Симфиз нижней челюсти (длина участка срастания левой и пра-
вой ее половины), по сравнению с лошадьми, короткий; длина его не
превышает 20% длины челюсти. Пространство между ветвями нижней
челюсти образует перед их слиянием с нижней стороны расширение
с закругленным передним и боковым краем. На крайних нижних резцах
(13) чашечка в той или иной степени недоразвита.
Число грудных и поясничных позвонков — 23. Конечности длинные,
особенно их дистальные звенья. Длина скелета свободной передней ко-
нечности (плечевая -(-предплечье-(-пястная) составляет больше 75%
длины туловища (грудной+поясничный+крестцовый отдел позвоноч-
ника). Длина пястной кости (metacarpale III) составляет не меньше 28%
длины всего скелета передней конечности. Пястные, плюсневые и кости
1 Все цифры по В. И. Громовой (1941, 1949).
3*
36
EQUIDAE
фаланг очень тонкие. Ширина пястной кости (metacarpale III) в средней
части не превышает 12% ее длины; тот же индекс для плюсневой кости
не свыше 10%. Ширина копытной (третьей) фаланги к ее длине по перед-
нему краю на передней конечности составляет меньше 140% (от 132
до 137%). Срединный гребень дистального суставного блока метаподий,
по сравнению с лошадьми, более низкий и закругленный.
История и географическое распространение
(рис. 7). Азиатские ослы ведут свою историю от верхнеплиоценовых,
уже однопалых лошадей типа Е. sanmeniensis Chard, et Piv. (подрод Al-
lohippus или Piesippus). Родиной их следует считать Центральную или
южную Азию, где в нижнем или среднем плейстоцене Индии обитал
их вероятный прямой предок Е. namadicus Falc. Форма, почти ничем
не отличимая от современного Е. hemionus, найдена в верхнем плейсто-
цене Китая. Территорию Европейского континента полуослы заселяют
лишь по окончании ледникового периода, в эпоху бронзы и неолита.
До этого здесь с нижнего и до верхнего плейстоцена обитала не имеющая
прямого отношения к современным ослам очень длинноногая и тонко-
ногая форма Е. hidruntinus.
В, настоящее время область географического распространения азиат-
ских ослов охватывает Переднюю, Среднюю и Центральную Азию, начи-
ная с Сирии, через Ирак, Иран, Афганистан, Среднюю Азию, Пакистан,
Тибетское плоскогорье и пустыню Гоби до Южного Забайкалья (Даурии)
и, возможно, северо-восточного Китая. Восточная граница распростране-
ния не установлена.
На территории СССР кулан прежде был распространен неизмеримо
шире, чем в настоящее время. Остатки, несомненно принадлежащие ку-
лану, найдены в слоях римского времени на территории Южного Крыма
(Симферополь). Есть указания (Шарлемань, 1949), что вплоть до XVI—
XVII в. он водился даже на территории Украины не только в степной
полосе, но и в лесостепи от побережья Черного моря, лимана Днепра
и Буга до широты Киева. С первых веков и до XII—XIII в. н. э. куланы
водились в Закавказье в долине среднего течения р. Аракса (Даль, 1954).
В конце XVIII и даже в середине XIX в. западной границей служила,
по-видимому, река Урал или Волга, а к северу, в районе р. Иртыша,
доходила до 52° с. ш. По сообщению Г. С. Карелина (1875), куланы, хотя и
не ежегодно, появлялись на левой стороне Урала против форта Редутского.
Экземпляр кулана был добыт им в 1855 г. между р. Сагизом и Эмбой.1
Есть сведения, что куланы в XVIII в. водились даже в Барабинской
степи (Селевин, 1932).
В середине прошлого века они были многочисленны по берегам Каспий-
ского и Аральского морей в районе Устюрта и Мангышлака, на севере
доходили до подножий Мугоджарских гор. В 1839 или 1840 г. «дикая
лошадь», судя по описанию — несомненный кулан, была поймана в 65 вер-
стах от бывшего Локтевского завода у западной окраины Алтая (Селе-
вин, 1937). Если не водился кулан постоянно, то в свое время имели
место регулярные его забеги из Монголии в степи южного Забайкалья
(Radde, 1861, 1862).
Еще в конце XIX в. куланы были широко распространены по равнин-
ным степям и пустыням Казахстана, Туркмении и Узбекистана. В началь-
ном периоде постройки Закаспийской железной дороги многочисленные
1 Согласно описи его коллекций, хранящейся в Зоологическом институте Акаде-
мии наук СССР.
3S
EQUIDAE
табуны их показывались около Казанджика, а также между Душаком
и Кары-Бендом. В 80-х годах А. Вальтер (Radde и Walter, 1889) в большом
числе встречал на холмах между реками Тедженом и Мургабом и в част-
ности в районе Ислам-Чешме и Акрабат. Изредка встречались они даже
на территории нынешней Павлодарской области Казахской ССР (Анти-
пин, 1941). В начале 80-х годов куланов ловили по левобережью Иртыша
недалеко от Усть-Каменогорска и Семипалатинска (Селевин, 1932).
В 1897 г. И. Словцов писал: «Кулан еще недавно заходил до р. Кулан-
утмес, впадающей с ю.-в. в озеро Денгиз, что лежит на юг от г. Атбасара,
и до притоков реки Терс-Аккана, впадающей с левой стороны в р. Ишим
близ того места, где последняя делает поворот, огибая урочище Джар-
гаин-Агач. Лет сто назад их встречали под 52° с. ш. на берегу Иртыша
против Коряковской столицы. . . В настоящее время, по рассказам кирги-
зов-охотников, находят табуны куланов на северном берегу Балхаша,
от р. Баканас и притока ее Денгильде на запад через горы Кызылрай
(красная трава), урочища Джелпаккалп и Бештюки до Актау. Позднею
осенью они удаляются отсюда южнее к р. Чу. Весною, в марте месяце,
идут с юго-запада к северо-востоку по северному берегу Балхаша, пере-
ходят пески Аккум и Сарыкум, останавливаясь по речкам Джемши и
Токрау». Разные источники подтверждают обитание многочисленных
табунов этого зверя в 80-х годах как в северном, так и в южном Прибал-
хашье. Огромные табуны их совершали кочевки между Балхашем и
хребтом Чингиз-Tay, в низовьях р. Аягуза. Обычным зверем кулан
был в Бет-Пак-Дале, где исчез в самом конце прошлого века. А. Брэм
в 70-х годах охотился на куланов в Зайсанской котловине. На Алакуль-
ской равнине куланы исчезли как будто в 80-х годах прошлого века. Охот-
ники-казахи передавали А. А. Слудскому (1939), что еще в самом конце
прошлого столетия куланы водились по правому берегу р. Сарысу в урочи-
щах Аяк-Касун, Орта-Касун и в степях Бике-Сары.
С начала 90-х годов ареал кулана начал катастрофически сокра-
щаться. По свидетельству В. Н. Шнитникова (1936), в период 1908—
1917 гг. куланы в незначительном числе сохранялись еще на пространстве
между р. Или и Караталом от Балхаша до предгорий и, возможно, между
Караталом и Аксу.
В литературе известно довольно большое число случаев появления
отдельных животных и небольших табунов куланов в разных местностях
Казахстана вплоть до 30-х годов этого века.1 Так, отмечались они для от-
дельных пунктов Прибалхашья, Бет-Пак-Далы, устья Аягуза, в Алакуль-
ской котловине и даже в центральном Казахстане. Однако подавляющее
большинство приводимых случаев основано на расспросах местных
жителей и охотников и поэтому не всегда достоверно. Два последние
более или менее достоверные указания на встречи следов пребывания
куланов в юго-восточном Казахстане относятся к 1936—1937 гг.: один —
к низовьям р. Или на протоке Топара-Кургак-Топаре (Слудский, 1939),
а второй — к району левобережья Или между государственной границей,
хребтом Турайгыр и р. Чарын (Селевин, 1937). В настоящее время в пре-
делах Казахстана кулан, как постоянный обитатель, отсутствует, но
возможны отдельные его заходы из Китая через Джунгарские ворота.
На территории СССР кулан сохранился лишь на самом юге Туркмении
в западной части Теджено-Мургабского междуречья, в районе между
1 Перечисление этих случаев в работах А. А. Слудского (1939) и В. М. Антипина
(1941).
EQU US
39
Кушкой и Серахсом. К середине 30-х годов нашего столетия небольшое
число куланов более или менее постоянно обитало в узкой полосе вдоль
границы Советского Союза с Ираном и Афганистаном от аула Миона на
западе до поста Ислам-Чешме на востоке (Шатерникова и Румянцев,
1934). В наибольшем числе они встречались в районе Акар-Чешме (хре-
бет Гёзгельды) и в котловине соленых озер Ер-Ойлан-Дуз. Как выяснил
М. П. Розанов (1937), весной и в начале лета табуны куланов с жеребя-
тами держатся севернее и северо-восточнее Акар-Чешме близ колодца
и хребта Элли-Бир. Осенью, вытесняемые стадами домашних овец, они
спускаются в долину Куллели-Дуз у самой афганской границы и к реке
Рис. 7. Распространение Equus hemionus Pall.
1 — в настоящее время; 2 — в первой четверти XX в.; 3 — северная гра-
ница в XIX в.; 4 —северная и западная границы в XVIII в. и раньше.
Эгригёк, около 70 км к востоку от Акар-Чешме, а зиму проводят в котло-
вине озер Ер-Ойлан-Дуз.
К западу от только что перечисленных мест кулан, как постоянный
обитатель, отсутствует. Однако, по свидетельству К. К. Флерова (1932),
небольшие табуны в 5—10 голов их появляются в окрестностях поста
Чильдухтер, куда они забегают из соседних районов Афганистана. Реже
наблюдали их вблизи долины Мургаба к востоку от погранзаставы Меру-
чак. Весной 1930 г. табунок из 4 голов наблюдался к востоку от аула
Сары-Чоп.
В СССР кулан взят под государственную охрану. Охота на него по-
всеместно и строжайше запрещена. На юге Туркмении учрежден спе-
циальный Бадхызский куланий заповедник, где в настоящее время число
куланов исчисляется уже сотнями голов и где проводится работа по его
одомашнению.
С 1953 года проводится работа по акклиматизации кулана в заповед-
нике Барса-Кельмес на Аральском море (Соколов, 1958). Издавна
в условиях полувольного паркового содержания куланы существуют на
юге Украины в Аскании-Нова (рис. 8).
. На территории Монгольской Народной Республики область распро-
странения кулана значительно шире, чем в Советском Союзе, но и здесь
40
EQUIDAE
она за последнее столетие сильно сократилась, занимая, в основном, лишь
юго-западную и южную части республики и Гоби. Северная граница
в Монголии, по А. Г. Банникову (1954), проходит с запада на восток по
нижнему течению р. Булугун, затем вдоль Гобийского и Монгольского
Алтая, до хребта Баянгииннуру (около 100° в. д.). Отсюда граница за-
гибает к северу и захватывает котловину озер Ногон-Нур, Бон-Цаган-
Нур и Бигэр-Нур. К востоку от котловины Бон-Цаган-Нур северная
граница распространения кулана проходит через нижнее течение рек
Байдарик, Туин-Гол, Тацын-Гол, а дальше, повернув на юго-восток и
обойдя с юга оз. Улан-Нур и пос. Далан-Дзадаган, мимо хребта Хурху
на 108° в. д., выходит за пределы монгольской государственной границы.
В некоторые годы, кроме того, куланы через Шаргиин-Гоби и Хуйсиин-
Гоби проникают в котловины озер Дургэ-Нур и Хара-Ус-Нур. Отдельные
забеги куланов наблюдаются в районе гор Шилин-Богдо, Богдо-Ула
и в окрестности Алтан-Обо, до 116° в. д. К югу от только что очерченной
северной границы куланы распространены не повсеместно. Нет их, на-
пример, в Аджи-Богдо, Нэмэгэту, Тосту, Гурбан-Сайхан и некоторых
других хребтах и отдельных поднятиях Гобийского и Монгольского
Алтая.
Подвиды. Систематика азиатских ослов крайне запутана. Описано
большое число форм (видов и подвидов), часто по одному-двум экземпля-
рам, главным образом по признакам окраски шерстного покрова, являю-
щимся показателями сезонного и полового диморфизма или индивидуаль-
ной изменчивости. Для многих форм не указаны даже хоть сколько-
нибудь точные их ареалы. По-видимому, все они должны быть отнесены,
к одному виду Equus hemionus Pall. В качестве подвидов могут быть
признаны лишь четыре.
1) Е. hemionus kiang Moorer. —Кианг. Размеры больше, чем у других
подвидов; высота в холке — около 130—140 см. Общий оттенок окраски
красновато-коричневый; характерно усиление интенсивности окраски
по направлению книзу, благодаря чему граница между светло- и темно-
окрашенной зоной туловища и шеи обозначена резко. Белая окраска
нижней стороны туловища вдается клином вверх по бокам лопаток.
Темная полоса вдоль хребта узкая и доходит на хвосте до основания длин-
ных волос; светлые полосы по бокам ее не развиты. Сезонный диморфизм
в окраске выражен не резко. Распространение: Ладак и Тибет.
2) Е. hemionus hemionus Pallas — Джигетай или Кулан. Рост немного
меньше, чем у кианга, но больше, чем у онагра; высота в холке состав-
ляет около 115—125 см. Как в летнем, так и в зимнем мехе красновато-
коричневый оттенок заменен рыжевато-песочным; при этом граница
между темно- и светлоокрашенной частью туловища не резко выражена.
Светлоокрашенная часть туловища и ног имеет желтовато-сероватый
оттенок и не образует выступа вверх в области лопаток; нижняя сторона
шеи окрашена почти в одинаковый тон с верхней. Темная полоса вдоль
хребта, как у кианга, но по бокам ее, особенно в задней части туловища,
следы беловатого цвета оторочки. Сезонный диморфизм в окраске слабый.
Распространение: Монголия, Средняя Азия, Китайский Турке-
стан, возможно северные районы Ирана и Афганистана.
3) Е. hemionus onager Boddaert — Онагр. Еще более мелкий, чем Е. h. he-
mionus. Высота в холке составляет около 110 см. Наиболее светлоокрашен-
ная форма. Общий фон окраски верхней части туловища палевый или
светлый, песчано-желтый. Светлая зона нижней части туловища распро-
страняется на заднюю часть крупа, образуя большое белое «зеркало»
Рис. 8а. Самка кулана. О-в Барса-Кельмес. Июнь 1957 г. Видна темная полоса
г> си т?° светлои оторочкой вдоль средппы спины. Фото автора
Рис. 86. Куланы на пастбище. О-в Барса-Кельмес. Mali 1957 г. Фото автора.
I
42
EQUIDAE
по бокам хвоста. Граница между светло- и темноокрашенной зоной ясно
выражена, но светлая зона не вдается снизу в область лопаток. Темная
полоса вдоль хребта более широкая, чем у кианга и джигетая, иногда не
доходит до конца хвоста и сопровождается по бокам ясными белыми
полосами, переходящими в околохвостовое «зеркало». Почти вся передняя
часть головы впереди глаз белая. Сезонный диморфизм в окраске ясно
выражен. Распространение: западная Индия, Пакистан, ве-
роятно Афганистан, Иран; на северной границе распространения, по-
видимому, незаметно переходит в предыдущую форму.
4) Е. hemionus hemippus I. Geoffroy—Сирийский онагр. Наиболее
мелкая форма из всех азиатских ослов. Высота в холке около 100 см.
Общий фон окраски более темный, коричневатый, особенно в задней
части, с возрастом приобретающий сероватый оттенок. Характерны
короткие уши и вздутая на конце морда. Возможно, заслуживает вы-
деления в самостоятельный вид. Находится на грани исчезновения,
быть может даже совершенно истреблен. Распространенная в пределах
Средней Азии популяция диких ослов ближе стоит к джигетаю (Е. h. he-
mionus Pall.), но обладает признаками, переходными к персидскому онагру
(Е. h. onager Pall.). Распространение: Сирия, Ирак; точные
границы распространения не установлены.
Биология. Типичными стациями для кулана являются различного
типа пустыни и полупустыни. Эволюция этой ветви семейства лошадиных
шла в направлении адаптации к условиям сухих ландшафтов. • В прежнее
время частично заселял и степи, но с освоением последних и усилением
преследования все больше удалялся в область трудно доступных пустынь.
Предпочитает места с плотным грунтом. Больших песчаных пространств
избегает. В летнее время косяки куланов иногда поднимаются в горы,
но все же предпочитают открытые равнины. Лишь при преследовании
куланы ищут укрытия в ущельях и оврагах. В спокойной же обстановке
даже отдых проводят на открытом месте.
Биология кулана изучена еще недостаточно. Пищу составляет травяни-
стая и кустарниковая растительность пустынь, включая горькие травы
и колючки. Организм кулана приспособился к нормальному существова-
нию на таких пастбищах, которые, на первый взгляд, не могут обеспечить
даже поддерживающего корма. Между тем, в условиях, например, мон-
гольских пустынь, не взирая на скудную растительность, животные
уже в июне—июле достигают хорошей упитанности. Могут пить соленую
воду, по-видимому, вообще могут долго обходиться без воды. Все же
последняя является основным лимитирующим фактором в засушливый
период года. В нормальных условиях в летнее время не уходит на рас-
стояние более чем 25—30 км от источника водопоя. В поисках корма могут
совершать длинные кочевки, но чаще причиной исчезновения куланов
в том или ином районе является лишение их водопоя, что имеет место
при выпасе в пустыне стад домашних животных.
Держатся, как правило, небольшими косяками, от 3 до 20 голов,
во главе которых находится взрослый самец. Но в местах, где куланы
многочисленны (Монголия), после ожеребления, а особенно к зиме, могут
собираться в стада до 1000 голов и больше. Самцы, изгнанные из стад
более сильными соперниками, пасутся одиночно, а иногда образуют от-
дельные косяки. На более мелкие группы, иногда на пары (самец и самка),
разбиваются перед ожереблением. Аллен приводит случай, когда самец
неохотно покинул застреленную самку. Молодые находятся в косяке
при матери до 2, а по некоторым данным даже до 3—4 лет. Часто выпа-
1
EQUUS
43
саются вместе с другими животными (джейран, дзерен, сайга, иногда
даже бараны). В Монголии джейранов вместе с джигетаями почти всегда
можно видеть и при пастьбе и при отдыхе (Allen, 1940).
В быстроте бега кулаи из копытных уступает лишь джейрану; бы-
строта сочетается с необычайной выносливостью. Опытами преследования
на автомобиле установлено, что на коротких дистанциях он может раз-
вивать скорость до 65—70 км в час. Куланы очень осторожны и пугливы.
При преследовании стараются бежать против ветра.
Кулан более молчалив, чем настоящий осел. Голос его сходен с криком
осла, но отдельные приступы рева не так длинны. Линька проходит в ап-
реле или мае. К средине июня все животные уже имеют полностью летний
волосяной покров. Период спаривания наступает в августе—сентябре,
тянется около месяца. Между самцами часто происходят жестокие драки
за обладание косяком самок. Молодые родятся в июне—июле; на за-
паде, по-видимому, раньше (март—май). Срок беременности около 11 меся-
цев. Самка приносит всегда одного жеребенка; для ожеребления выби-
рает открытое место, вероятно для лучшей защиты от хищников. Жеребе-
нок через час может стоять на ногах, но все же первые дни лежит спрятан-
ным в неровностях почвы. В трехнедельном возрасте бегает так же быстро,
как взрослые, только менее вынослив.
Из естественных врагов единственным является волк. Но, по-види-
мому, и он, как вредитель, имеет не слишком большое значение. Замет-
ный вред волк может принести главным образом недавно родившемуся
молодняку. Взрослые животные сравнительно легко спасаются бегом.
Главной причиной сокращения ареала и уменьшения численности ку-
лана является прямое и косвенное истребляющее влияние человека.
Применение усовершенствованного огнестрельного оружия явилось, на-
пример, причиной, вероятно, полного истребления сирийского онагра.
Освоение и использование полупустынных и пустынных пастбищ для
выпаса домашних животных вынуждало пугливого кулана отступать
в наиболее труднодоступные, не освоенные человеком места и вело к не-
уклонному сокращению ареала. Особенно большое значение, как спра-
ведливо отмечали В. Г. Гептнер (1948) и А. Г. Банников (1954), имело
лишение куланов открытых водоемов.
А. А. Слудский (1953) одной из основных причин исчезновения ку-
лана в степях и полупустынях Казахстана считает «джуты», стихийные
бедствия среди копытных млекопитающих, наступающие в неблагоприят-
ные по климатическим условиям суровые зимы. Уже при глубине снега,
немного превышающей 10 см, добыча корма для кулана становится за-
трудненной. При еще большей глубине животное должно затрачивать
значительную энергию на откапывание корма, особенно если к тому
же снег является уплотненным. Предельной глубиной, свыше которой
добыча корма становится практически невозможной, этот автор считает
40 см. Такое же значение имеет образовавшаяся после оттепели корка
наста свыше двух сантиметров или гололедица, образование на поверх-
ности почвы корки льда при быстрой смене дождливой погоды морозом.
Наступившая вследствие этих причин бескормица, особенно если она
продолжается достаточно длительный период (до двух недель и больше)
и сопровождается сильными морозами и буранами, влечет массовые
падежи животных. Губительным для куланов в Казахстане был джут
в зиму 1879/80 года. Это же явление, будто бы, явилось причиной оконча-
тельного исчезновения с зимы 1891/92 года куланов на Устюрте, в ни-
зовьях Сарысу и в некоторых других местах Казахстана.
44
EQUIDAE
Паразитофауна кулана остается почти не изученной. Известно только,
что в нем паразитирует желудочный овод, Gastrophilus pecorum F. (сообще-
ние К. Я. Грунина).
Хозяйственное значение. В прошлом кулан имел немалое значение
в качестве промыслового животного. Мясо и особенно жир высоко цени-
лись у местного населения. В ряде случаев его мясо предпочиталось
лучшему бараньему; жиру кулана приписывались целебные свойства.
Ценилась и кожа, как очень ноская для одежды; кроме того, она шла
иногда на выделку ценного цветного сафьяна. Все это служило причиной
интенсивного преследования кулана.
В настоящее время в пределах СССР кулан интересен главным обра-
зом как редкое животное, абсолютно запрещенное к добыче и взятое под
государственную охрану.
Данные о возможностях его приручения противоречивы. Кулан —
сообразительное, но своенравное, упрямое и в известных условиях свире-
пое животное. В зоологических садах иногда размножается. Пойманный
жеребенком, легко привыкает и приручается к человеку, но многочислен-
ные попытки использовать кулана в качестве верхового или упряжного
животного в большинстве кончались полной неудачей. Последние годы
в питомнике при Бадхызском заповеднике ставились опыты по прируче-
нию кулана. Удалось пойманных молодыми животных приучить к седлу
и упряжи и совершать на них поездки на расстояние 10—15 км (Плюш-
кин, 1950).
В Монголии имеют место случаи вольного скрещивания кулана с мест-
ной домашней лошадью. Гибриды внешностью похожи на куланов, но
имеют, подобно лошадям, длинную гриву и покрытый на всем протяже-
нии длинным волосом хвост. По характеру очень дики и не поддаются
дрессировке. В Узбекской ССР в течение ряда лет проводились успеш-
ные опыты по скрещиванию кулана-самца с- домашними ослицами и
кобылами. Четыре гибрида, куланоида, в 1940 г. экспонировались на
Всесоюзной сельскохозяйственной выставке. Двух осло-куланов я ви-
дел в Заповеднике Барса-Кельмес в 1957 г.
Гибриды лошадь X кулан обладали низкой по сравнению с лошадью
холкой, короткой гривой, не имели челки, репица (мясистая часть хвоста)
только с половины длины покрыта длинным волосом; каштаны на задних
конечностях не развиты; темный ремень вдоль хвоста в окраске сохра-
няется. По работоспособности они превосходили мулов, но применение
их ограничено вследствие «строгости» характера. Все до сих пор получен-
ные гибриды коне-куланы и осло-куланы оказались бесплодными.
Охота. Из способов охоты наиболее обычным было подкарауливание
на путях к водопою. Основан этот способ на том, что куланы пользуются
одним и тем же путем при ходьбе на водопой. Здесь их подкарауливали
охотники, расположившиеся цепью вдоль тропы, и зачастую расстрели-
вали все стадо беспорядочно мечущихся взад и вперед, обезумевших от
выстрелов животных. Иногда подошедших к водопою куланов загоняют
в воду; по выходе из воды Животные, утомленные непривычным для них
плаванием, убиваются поджидающими на берегу охотниками. Иногда
охотятся за куланом скрываясь за маневрирующим верблюдом, или под-
манивают с помощью домашней лошади, одинаковой с куланом масти.
Очень трудна и поэтому редко практикуется групповая охота с преследо-
ванием до полного утомления куланов.1
1 Преследование на автомобилях как хищнический способ, не заслуживает рас-
смотрения.
EQU US
45
Equus caballus gmelini Antonias — Тарпан, европейская дикая лошадь
(рис. 9 и 10)
Рычков П. И., 1762, Топография Оренбургская, СПб. : 290 (опис., распр.);
Г мел ин С., 1771, Путешествие от Петербурга до Черкас, СПб. : 68—73 (опис.,
изобр.); Pallas Р. S., 1776, Reise d. versch. Prov. d. Russ. Reich., I : 211, 272,
II : 662, III : 510—512, VIII (опис., изобр.); Pallas P. S., 1811, Zoogr. Rosso-
Asiat., I : 260 (опис.); Боплан, 1832, Описание Украины, СПб. : 92—93 (распр.,
хоз. знач.); Эверсманн Э. А., 1850, Естеств. история Оренбургск. края, II:
215—216 (опис., распр.); Шатилов И. Н., 1861, Акклим., II, 11 : 514—517 (опис.);
Brehms Tierleben, 1877, Grosse Ausg., 2 Aufl., Ill, Leipzig : 5—8 (опис., ист.,
изобр.); Брандт А. Ф., 1884, Нива, 38:906—910 (общ.); Шатилов И. Н-,
1884, Сообщ. о тарпанах, М. : 1—8 (опис., изобр.); Черский И. Д., 1891, Опис.
колл, послетрет. млекоп., СПб. : 412—431 (морф.); К е ппен Ф. П., 1896, К ист.
тарпана в России, Спб. : 1—78 (общ.); Анучин Д. Н., 1896, К вопросу о диких
лошадях и их прируч. в России, СПб. : 1—75 (критика статьи Кеппеяа); Етаг t J.C.,
1905, Proc. Zool. Soc., Edinburg, XXVI (общ.); Antoni us O., 1912, Verh. zool.-
botan. Gesellsch., Wien, 62 : (64)—(78) (сист.); Antonins О., 1912, Naturw. Wochschr.,
N. F. XI, 33 : 5'13—517 (ист.); Antonins O., 1913, Verh. zool.-botan. Gesellsch.,
Wien, 63 : 235—240 (опис., изобр.); Falz-Fein F., 1919, Sitzungsber. Gesellsch.
naturforsch. Freunde, Berlin, 5/6 : 196—206 (ист.); Браунер A. A., 1923, Сельско-
хоз. зоол., Одесса : 91—98 (общ.); Vetulani Т., 1928, Bull, internat. Acad.
Polon., Sci. et Lettr., Cracovie, ser. B, 7 : 75—78 (ист., восстанови.); V etulani T.,
1933, Zeitschr. f. Saugetierk., 8:281—282 (ист.); Гептнер В. Г., 1934, Zeitschf.
1. Saugetierk., 9:431—433 (ист.); Helzheimer M., 1935, Naturschutz, 7:154—
160 (ист., изобр.); Румянцев Б. Ф., 1936, Изв. Акад, наук СССР, сер. биол.,
2/3 : 415—444 (ист.); Северцов С. А., 1940, Природа, 9 : 92—94 (ист., восстановл.);
Громова В. И., 1941, Тр. Зоол. инет. Акад, наук СССР, VI : 92—157 (морф.);
Janicowski Т., 1942, Nature, 150 (3815) : 681—682 (опис., изобр.); Н а г р е г F.,
1945, Extinct a. vanishing mammals of Old World., N. Y. : 330—334 (ист.); Vetu-
lani T., 1947, Bull, internat. Acad. Polon. Sci. et Lettr., Cracovie ser. В, II (1—8) :
1—22, 7 pls (восстановление тарпана в Беловежской пуще, изобр. польских «коников»);
Громова В. И., 1949, Тр. Палеонт. инет., XVII, 1 : 1—374, 2 : 1—163 (морф.,
ист.); Vetulani Т., 1949, Med. Veteryn., V, 9 : 679—687 (ист.); Громова В. И.,
1950, Матер, по четвертичн. периоду СССР, 2 : 70—80 (морф., ист.); ВетуляниТ.,
1952, Зоол. журн., XXXI, 5 : 727—735 (ист., восстановл.); Кириков С. В., 1952,
Изв. Акад, наук СССР, сер. географ., 6 : 34 (ист.); Heck Н., 1952, Oryx, I, 7 : 338—
342 (восстанови.); Гептнер В. Г., 1955, Зоол. журн. XXXIV, 6 : 1404—1423 (общ.,
изобр.).
Еще лет 150—200 тому назад в степях юга СССР и даже в лесостепной
полосе в сравнительно большом числе водились дикие лошади, или тарпаны,
как их называло местное население, Происхождение тарпана и его связи
с другими дикими и домашними лошадьми — один из наиболее спорных
вопросов истории фауны СССР. Многими оспаривался исконно дикий ха-
рактер этой лошади, признаваемой лишь за потомка одичавших домашних
животных. Фактические данные, по которым можно выявить особенности
тарпана, по сравнению с другими дикими и домашними представителями
семейства лошадиных, очень скудны. В настоящее время тарпан истреблен.
Сохранился единственный в мире полный скелет и еще один череп, храня-
щиеся в коллекциях Зоологического института Академии наук СССР и
Зоологического музея Московского университета. Кроме того, имеется
подробное описание животного, которому принадлежал последний череп.
Следует отметить, что чистокровность и типичность этих «шатиловских»
тарпанов (по имени доставившего их автора) 1 подвергается большому
сомнению, хотя оба животные были пойманы на воле. К тому же оба они
были кастрированными самцами.
Несомненно, что с самых ранних времен появления в районах распро-
странения тарпана домашних лошадей происходило смешение дикой и
1 Подробнее см. стр. 51.
46
EQUIDAE
домашней популяций. Уже в первых достоверных известиях о тарпанах
и все время в дальнейшем сообщаются сведения о том, что дикие жеребцы
отбивали из домашних стад кобыл в свои косяки. Скрещивание с домаш-
ними лошадьми в течение веков не могло не сказаться на внешнем облике
дикого тарпана. Тип его менялся. Тем не менее, едва ли могут быть сомне-
ния в том, что это было исконно дикое животное, обитавшее с доисториче-
ских времен на обширной территории Европы, а возможно и Азии. За это
говорят, помимо памятников искусства древнего человека, ряд типичных
признаков внешней морфологии, как окраска, стойко сохраняемых даже
в помесях с домашними лошадьми и отмечаемых подавляющим большин-
ством лиц, видевших тарпана живым в естественной обстановке. Местное
население всегда прекрасно отличало в косяках тарпанов не только оди-
чавших домашних кобыл, но и помесей последних с дикими жеребцами.
Описание. По сохранившимся описаниям и изображениям внешний вид
тарпана может быть охарактеризован следующими чертами. Это сравни-
тельно небольшая лошадь, высотой в холке от 107 до 136 см и с длиной туло-
вища иногда меньше, иногда несколько больше высоты. Голова относитель-
но большая, грубоватая, с широкими ганашами (межчелюстным простран-
ством), с короткой, недовольно толстой и высокой мордой, выпуклым теме-
нем и несколько вдавленным между глазами лбом. Профиль головы у
корня носа вогнутый. Морда иногда слегка горбоносая. Уши острые, стоя-
чие, длиною около 17 см.1 В области ганашей удлиненные волосы часто
образовывали подобие бороды. Шея прямая или сверху вогнутая («оленья»),
толстая и короткая; длина ее по верхнему краю около 56 см. Голова дер-
жалась высоко. Для более ранних тарпанов типична короткая, стоячая
или полустоячая грива, но у последних описанных («шатиловских») экземп-
ляров она была длинная, до 50 см, свешивалась на обе стороны тела.Челка,
в отличие от полуослов и лошади Пржевальского, хорошо развита. Хвост
короче, чем у домашних лошадей, обычно на всем протяжении покрыт
длинным волосом, но иногда у корня на некотором расстоянии волос корот-
кий. Удлинение гривы и хвоста, возможно, является или результатом влия-
ния условий содержания в домашней обстановке, или показателем прилития
крови домашней лошади. Грудь широкая, ребра крутые; обхват груди за
лопатками у последнего «шатиловского» тарпана составлял 167 см. Спина
и поясница прямая, холка удовлетворительно развитая. Ширина таза
в моклоках 48 см, уже* чему Е. przewalskii Pol. Круп свислый. Конечности
короткие, но «сухие», с ясно обозначенными сухожильями и мускулами;
плечо прямое. Как на передних, так и на задних ногах хорошо развиты
«щетки» (пучки длинного грубого волоса на задней стороне путовых суста-
вов).1 2 Копыта высокие, «стаканообразные», как и у всех представителей
подрода Equus s. str. широкие, округлые; передние заметно шире задних.
Каштаны, в противоположность лошади Пржевальского, имеются только
на передних конечностях.3
Характерной окраской шерсти (мастью) тарпана была темная мышино-
серая («мышастая»), на нижней стороне более светлая («пепельная»), иногда
с подпалинами (рыжеватым оттенком) в пахах. Челка, грива, длинные
волосы на хвосте, ноги ниже запястных и голеностопных (скакательных)
суставов темно-бурые, почти черные. На передних ногах слабо заметная
1 По Э. Эверсманну (1850), край ушей «перегнут назад».
2 Суставов между запястной или плюсневой костью и первой фалангой.
з Признак этот проверен только на двух «шатиловских» тарпанах и, возможно,
не является для него типичным; известно, что и у некоторых пород домашних лошадей
каштаны на задних конечностях имеются не всегда.
EQU US
47
нолосатость. Очень характерна такого же цвета срединная узкая полоса
вдоль спины от конца гривы до начала длинных волос на хвосте. К зиме
волосяной покров становился густым и длинным, приобретал пепельный
оттенок. У жеребят шерсть слегка волнистая, с рыжеватым оттенком.
Уклонение во взрослом состоянии от типичной мышастой окраски несом-
ненно указывало на примесь крови домашней лошади.
Для черепа тарпана характерна по сравнению с Е. przewalskii меньшая
общая величина (основная длина не больше 470 мм), короткомордость
Рис. 9. Тарпан [Е quits caballus gmelini Ant.) (из Brehm, 1877).
выпуклость мозговой коробки и большая вдавленность лобно-лицевого
профиля, несколько относительно более широкая резцовая часть нижней
челюсти, относительно более короткий ряд коренных зубов верхней челю-
сти (36.5% основной длины черепа). Слуховые проходы короткие, не более
5% основной длины черепа, направлены почти прямо в стороны; при
взгляде на череп сверху концы их едва выступают за контуры височных
гребней. Высота лицевой части черепа впереди Р2 составляет не больше
21 % основной его длины.1 Сошниковый индекс, по сравнению с полуослами,
высокий; расстояние от basion до заднего края сошника не меньше 125%
расстояния от заднего края костного нёба до той же точки. Глоточные
бугорки расположены впереди разорванного отверстия. Задний край носо-
вой вырезки расположен не дальше, чем на уровне средины Р2. У нижнего
края входа в носовую полость парные гребневидные бугорки на межче-
люстных костях выражены очень слабо. Симфиз нижней челюсти длинный,
больше 20% ее длины. Пространство между ветвями нижнечелюстной
кости суживается постепенно и не образует расширения в передней части.
Чашечки развиты на всех резцах.
1 Все цифры по В. И. Громовой (1941, 1949).
49
EQUIDAE
Число грудных-j-поясничных позвонков — 24. Длина скелета свобод-
ной передней конечности (плечевая+предплечье+пястная) составляет
меньше 75% (74.6%) длины туловища (грудной-|-поясничный-|-крестцо-
вый отдел позвоночника). Длина пястной кости (metacarpale III) составляет
меньше 26% длины передней конечности. Ширина пястной кости в средней
части составляет около 16% ее длины; тот же индекс плюсневой кости
около 12 (11.9)%. Ширина копытной (третьей) фаланги на передней конеч-
ности составляет около 138% длины ее по переднему краю. Срединный
гребень дистального блока метаподий, по сравнению с полуослами, более
высокий и острый.
Отличиями в скелете от Е. przewalskii являются: 1) относительно более
длинные конечности, особенно задние (89% длины туловища); 2) более
короткие метаподии и, наоборот, более длинные плечевая и бедренная
кости (метакарп составляет 81.4% длины плеча; соответствующий индекс
плюсна—бедро равен 73.8%); 3) массивность первой и второй фаланг
(срединная ширина II фаланги передней конечности составляет 126%
ее длины).
История тарпана. Ранняя история тарпана в настоящее время не
совсем ясна. Дикие лошади обитали на территории Евразии в течение
всего ледникового и послеледникового периода. Однако сравнение их ске-
лета с единственным сохранившимся скелетом «шатиловского» тарпана
не позволяет ставить последнего в прямую генетическую связь с плейсто-
ценовыми лошадьми Европы. Тарпан обладает некоторыми особенностями
зубов (короткость протокона на верхних коренных) и конечностей (не-
обычайная массивность фаланг при тонкости метаподий), которые не согла-
суются с общим направлением эволюции, характерным для лошадей Евро-
пы, если его считать прямым потомком последних. Это обстоятельство за-
ставляет высказать предположение, что он представлял потомка лошадей,
вселившихся в Европу откуда-то извне уже в послеледниковый период
(В. И. Громова).1 Сравнение тарпана с лошадьми послеледниковой эпохи
не может быть проведено из-за неизученности доместикационных измене-
ний в скелете и невозможности, вследствие этого, определить, имеем ли
мы дело с остатками диких или одомашненных животных.
Остатки лошади, почти не отличимой от тарпана, найдены в одной из
стоянок человека (с. Погореловка Черниговской области) эпохи, пере-
ходной от палеолита к неолиту, когда лошадь, вероятно, не была еще
одомашнена. Пещерные изображения, относящиеся к палеолиту, говорят
о том, что лошадь, очень близкая по типу к тарпану, была распространена
в это время на обширном пространстве Европы и Азии от Атлантического
до Тихого океана.
Первые известия о диких лошадях в историческую эпоху относятся
к V в. до н. э. В это время они отмечаются для Испании, Полесья и вер-
ховьев р. Буга. Многочисленны сведения, относящиеся к средним векам
Западной Европы, Литвы и Польши. В Прусии, например, дикие лошади
существовали до конца XVI в.
На территории древней Польши, в Беловежской пуще, темно-серые
дикие лошади в начале XV в. (1409 г.) служили предметом королевской
охоты. Здесь они существовали до средины XVIII в., когда последние из
них были перемещены в охотничий парк одного из польских графов
1 Если только особенности «шатиловских» тарпанов не являются индивидуаль-
ными уклонениями или не могут быть отнесены за счет сомнительной типичности и
чистокровности данных экземпляров.
Тарпан, европейская дикая лошадь (Equus caballus qmelini Ant.)
(Рис. худ. В. Н. Ляхова)
I
EQU US
49
в Замостье около Билгораи (Люблинское воеводство). Конец существова-
нию тарпанов в Польше положила суровая зима 1812/13 года, когда уце-
левшие в парке графа Замойского от падежа животные были розданы окру-
Рис. 10. Череп Equus caballus gmelini Ant.
жающему населению и растворились в популяции местных домашних ло-
шадей. Сохранившиеся описания говорят за то, что эти «лесные тарпаны»,1
как их теперь называют, мало отличались от южнорусских степных со-
1 Equus gmelini silvaticus Vetulani.
4 Фауна СССР, И. И. Соколов
50
EQUIDAE
братьев. По мнению некоторых, собравших о них сведения лиц (Vetulani,
1928, 1933), они представляли лишь несколько измельчавших и приспо-
собившихся к лесной жизни степных тарпанов.
Тип «лесного тарпана» хорошо сохранился в примитивных домашних
лошадях некоторых районов Польши, в частности в районе Билгораи;.
на основе их предпринимаются попытки «восстановления»1 его в Бело-
вежской пуще (Ветуляни, 1952). По собранным С. В. Кириковым (1952)
данным, в литовских и белорусских пущах дикие лошади удерживались
до середины XVIII в.
Дольше всего тарпан сохранялся на территории юга европейской части
СССР, но и здесь его постигла общая участь диких лошадей Европы.
Вкратце история сокращения ареала и исчезновения этого вида в наших
пределах рисуется в следующем виде. Из «Поучения» Владимира Моно-
маха, который «по Роси ездя имал есмь своима рукама те же кони дикие»,
узнаем, что в XI—XII вв. тарпаны водились в Киевской и Черниговской
областях. Бойлан (1832), сочинение которого было впервые издано в 1650 г.,
писал: «По степям украинским... дикие лошади ходят табунами от 50 до
60-ти голов; нередко они заставляли нас браться за оружие: издали мы
принимали их за татарскую конницу». Об обилии диких лошадей в то время
говорит тот факт, что мясо их продавалось в «мясном ряду» наравне
с говядиной и бараниной. «Мясо их, — пишет Боплан, — чрезвычайно
вкусно и даже нежнее телятины, впрочем на мой вкус не так приятно».
Даже в начале XVIII в. тарпаны были еще многочисленны в южно-
русских степях, в частности в Молдавии и Приазовье. В 1768 г. С. Гмелин
лично охотился на тарпанов около г. Боброва Воронежской губ. и дал
первое удовлетворительное их описание и изображение. Современник
Гмелина Паллас говорит о распространении диких лошадей от Днепра
до Алтая, далее в Центральной Азии и к северу до 54—55 параллели.
Достоверные сведения о существовании тарпана в Оренбургском крае,
помимо Палласа, сообщают П. И. Рычков (1762) и Э. Эверсман (1850),
причем за столетие, разделявшее наблюдения этих авторов, северная гра-
ница его распространения сдвинулась к югу от широты Куйбышева до
Саратова.
Не ясен вопрос о типе диких лошадей, водившихся к востоку от Волги
и в Сибири. Ни изображений, ни удовлетворительного описания их не
сохранилось. Есть предположение, что это были животные, ближе стояв-
шие к лошади Пржевальского (Equus przewalskii Polj.), и что границу
между областями распространения этой последней и европейского тарпана
составлял водораздел Дона и Волги. За такое предположение говорят
отрывочные сведения о масти заволжских диких лошадей. Так, Э. Эверс-
ман (1850) называл ее серой или песочной, а П. И. Рычков (1762) — сав-
расой и «голубой». Нужно, однако, иметь в виду, что последний, помимо
упоминания о встречающейся голубой (иначе мышастой) масти, говорит
также о «западинах» на лбу, т. е. об одном из характерных признаков
южнорусского тарпана.
Сведения о сибирских диких лошадях очень скудны и, весьма вероятно,
относятся скорее к лошади Пржевальского. Э. Эверсман (1850) писал, что
они встречаются «всюду в степях на восток от Каспийского моря». Изве-
1 О подлинном восстановлении вымершего вида, конечно, речи быть не может.
Задача заключается в том, чтобы получить животных, дающих в какой-то мере пред-
став лев ие о дикой лесной лошади и обладающих некоторыми ее ценными в хозяйствен-
ном отношении свойствами.
EQUUS
&1
стно, что огромное число их погибло во время эпизоотии 1785 года, унес-
шей также около 85 000 домашних лошадей местного населения. Имеются
сведения о существовании тарпанов в низовьях р. Урала в начале прошлого
века (Кеппен, 1896).
Дольше тарпаны сохранились на еще мало в то время освоенных Хер-
сонских и Таврических степях между Днепром и Доном. В 30-х годах
XIX в. они были многочисленны по всему Азово-Черноморскому побережью
от Кубани и Маныча до низовьев Буга. Водились они также в глухих ме-
стах, в лесах и болотах, на границе Воронежской губернии и Донской
области. Но с этого времени область их распространения стала быстро
сокращаться. К 60-м годам сохранились лишь отдельные косяки в преде-
лах Херсонской и Таврической губерний. Из этих косяков в 1854-м и
в 1866 гг. были выловлены и И. Н. Шатиловым доставлены в Академию
наук и Московский Зоологический сад два (первые и последние), ставшие
достоянием науки, тарпана, очевидно с заметной примесью крови домашней
лошади, о черепах и скелете которых упоминалось раньше. Считалось, что
последняя самка тарпана была уничтожена в 1879 г. (Falz-Fein, 1919).
Но в 1934 г. появилась заметка (Гептнер, 1934) о том, что очень старый, но
по описанию типичный самец в 1914—1918 гг. доживал свой век в одном
из частновладельческих хозяйств — «Дубровке», около г. Миргорода Пол-
тавской губернии; животное было выловлено жеребенком из истреблен-
ного немцами-колонистами косяка диких лошадей. Таким образом, исчез-
новение тарпана на воле следует отнести на самый конец XIX в.
Причинами постепенного сокращения ареала и уменьшения числен-
ности тарпана, вплоть до полного исчезновения, явилось прямое истребле-
ние человеком и сельскохозяйственное освоение целинных степей — ос-
новных его стаций.
В средние века мясо диких лошадей употреблялось в пищу, и это слу-
жило одним из стимулов к преследованию животных. Не меньшую роль
в истреблении тарпана сыграл тот вред, который приносили дикие лошади
хозяйству человека. Помимо уничтожения ими запасов сена, о чем писал
уже Гмелин, причиной интенсивного преследования тарпанов был широко
распространенный угон дикими жеребцами в свои косяки домашних кобыл,
откуда последние никогда не возвращались. Поэтому местное население
при появлении вблизи поселений косяка диких лошадей старалось его
прогнать или уничтожить. Способы охоты на диких тарпанов были разные.
Применялся трудный способ преследования до полного утомления на ло-
шадях; для этого требовались, как правило, сменные, подставные кони,
но и в этих случаях загнать удавалось лишь отбитую от остального косяка,
чаще беременную, самку или жеребенка. Практиковалась описанная
С. Гмелиным поимка в расставленные сети. При этом, так как тарпаны
держались косяками от 6—7 до 30 голов под предводительством взрослого
жеребца, охота была более удачной, когда первым уничтожался вожак;
тогда мечущиеся в беспорядке животные легче попадали в сети. Иногда
тарпанов зимою загоняли на гладкий лед или в глубокие засыпанные сне-
гом овраги, где беспомощных животных связывали или убивали.
Немалую роль в исчезновении тарпана сыграло сельскохозяйственное
освоение степей и полупустынь. Дикие лошади с трудом выносили не-
посредственную близость поселений и с распашкой степей все больше
удалялись в необжитые места вплоть до несвойственных им болот и лесов.
Приручение тарпанов удавалось только при поимке их в возрасте жере-
бят. Но и в этом случае животные обладали беспокойным характером и
могли применяться главным образом лишь для верховой езды. Самцы
4*
52
EQUIDAE
должны были обязательно подвергаться кастрации для укрощения их
буйного нрава. Как уже отмечалось, тарпаны свободно скрещивались
с домашними лошадьми и давали, вероятно, плодовитое потомство. Помеси
ценились за быстроту бега и большую выносливость в работе.
Дикие лошади типа тарпана, вероятно, являются предками некоторых
групп домашних лошадей. Так, упомянутые польские коники даже по
величине и окраске представляют мало изменившихся потомков «лесных
тарпанов», или лошадей, изображенных палеолитическим человеком.
Характерную для коников «бороду» мне приходилось в свое время наблю-
дать у местных лошадей в Новгородской области. Тип тарпана внешне
и по краниологическим признакам проявляется в ряде отродий примитив-
ных лошадей Галиции, Боснии, Буковины, Восточных Карпат. Б. Ф. Ру-
мянцев (1936) пришел к заключению о том, что тарпан является предком
всех групп домашних лошадей северного типа.
j После того как настоящая работа была написана, появилась интерес-
ная статья В. Г. Гептнера «Заметки о тарпане» (1955).
В вопросе о том, был ли южнорусский тарпан исконно диким видом
или ^одичавшей домашней лошадью, наши мнения сходятся, так как мы
анализировали, и при этом без тенденциозности, по существу одни и те же
материалы. Наши точки зрения сходятся также и в том, что в последние
десятилетия его существования в популяции тарпана имелась и была зат
метна примесь крови местных домашних лошадей.
Однако я не согласен о В. Г. Гентнером в систематической трактовке
тарпана и лошади Пржевальского. Объединение этих двух форм в один вид
не оправдано. Во-первых, он сам неоднократно подчеркивает, что это
«резкие расы», что морфологически они «полярны». Во-вторых, обвиняя
В. И. Громову (1949) в тенденции к узкой трактовке вида, сам уважаемый
автор обнаруживает односторонний, сугубо морфологический подход
к этому вопросу.
Две популяции животных, хотя бы и весьма близкие по происхождению,
но переставшие в силу географической, физиологической изоляции или
иных причин скрещиваться в естественной обстановке 1 и в силу независи-
мого пути эволюции приобревшие после расхождения реальные морфоло-
гические отличия, должны относиться, по моему мнению, к разным видам.
Подобный пример мы и имеем в случае с лошадью Пржевальского и
тарпаном.
Не кажутся убедительными и номенклатурные изыскания автора
«Заметок о тарпанах». Научные названия обеих форм достаточно прочно
установились в литературе и общеизвестны, менять их, мне кажется, не
имеет никакого смысла.
Equus przewalskii Poljakow — Лошадь Пржевальского
(рис. И, 12, 13, 14)
Пржевальский Н. М., 1875, Монголия и страна тангутов, I, СПб., изд.
Русск. географ, общ. : 299 (первые сведения); П о л я к о в И. С., 1881, Изв. Русск.
географ, общ., XVII, 1 :1—19 (первооп., изобр.); Пржевальский Н. М.,
1883, Из Зайсана через Хами в Тибет к верховьям Желтой реки, СПб., изд. Русск.
географ, общ. : 40—42 (опис., изобр.); Черский И. Д., 1891, Опис. колл, после-
третичн. мпекопит., СПб. : 402—411 (морфол.); Грум-Гржимайло М. Е.,
1892, Нива, 17 : 374—382 (общ.); Грум-Гржимайло Г. Е. иМ. Е., 1896,
Опис. путешеств. в запади. Китай, I, СПб., изд. Русск. географ, общ. : 188—211 (общ.,
1 Я не имею в виду межвидовые и даже межродовые скрещивания, происходящие
ири содействии человека.
EQUUS
53
изобр.); Анучин Д. Н., 1896, К вопросу о диких лошадях и их приручении в Рос-
сии, СПб. : 62—75 (общ.); Тихомиров А. А., 1898, Естествозн. и геогр., 4 : 1—21
(опис., изобр.); К о з л о в П. К., 1899, Тр. эксп. Русск. географ, общ. по Центр. Азии,
II : 259 (прир.); Заленский В., 1902, Науч, результ. путеш. Н. М. Пржеваль-
ского, отд. зоол., т. I, ч. II, вып. 1 : 1—.76 (морф., сист., изобр.); N о а с k Th., 1902,
Zool. Anz., XXV, 663 : 135—145 (общ.); Hagenbeck, 1902, Proc. Zool. Soc. bond.,
I : 137—138, pl. XIII (распр., изобр.); Б и x н e p E. A., 1903, Лошадь Пржевальского,
СПб. : 1—40 (критика работы В. Заленского); Клеменц Д., 1903, С.-Петербургск.
Ведомости, № 186, от И (24) июля : 2—3 (ист., ох., хоз. зн.); Matschie Р., 1903,
Naturwiss. Wochschr., N. F., II, 49:581—583 (сист.); Кулагин Н. М., 1904,
Изв. Моск, сельскохоз. инет., 1 :1—31 (распр., биол.); Казнаков А. Н., 1907,
Монголия и Кам, II, 1 : 24 (распр.); Кащенко Н. Ф., 1907, Ежегодн. Зоол. музея
Акад, наук, XII : 177—194 (распр., морф., сист.); Hilzheimer М., 1909, Natur-
wiss. Wochschr., N. F., VIII, 51 : 810—812 (сист.); Antonins О., 1912, Verb,
zool.-botan. Gesellsch., 62 : 64—78 (ист.); Lydekker R., 1912, The horse, London :
: 87—92, pl. VIII, IX (опис., изобр.); Brehms Tierleben, 1926 , 4 Aufl., 12, Leipzig :
676—678, Taf. V (общ., изобр.); Spottel W., 1926, Kuhn-Archiv, XI : (общ.);
Antonins O., 1929, Zool. Gart, N. F., 1 : 87—98 (морф., сист., изобр.); Rey-
mond, M., 1932, Bull. Mus. Hist. Natur., ser. 2, t. 4, № 7 : 807—809 (расп.); P y-
мян дев Б. Ф., 1936, Изв. Акад, наук СССР, 2/3 : 415—444 (ист., изобр.); Anto-
nins О., 1937, Proc. Zool. Soc., London, 107, ser. В : 557—564 (распр.); Allen G.
1940, The mammals of China a. Mongolia, 2, N. Y. : 1285—1287 (общ.); Громов а В. И.,
1941, Тр. Зоол. инет. Акад, наук СССР, VI : 92—157 (морф., сист.); Harper F.,
1945, Extinct and wanishing mammals of Old World, N. Y. : 322—328 (распр., изобр.);
Bourdelle E., 1948, C. R. Congr. XIII Internal, de Zool., Paris : 383—387 (распр.);
Громова В. И., 1949, Тр. Палеонт. инет., XVII, 2 : 129, 149—151 (морф., эвол.);
Громова В. И., 1950, Матер, по четвертичн. периоду СССР, 2 : 70—80 (сист.);
Кравченко П., 1951, Огонек, 51 : 6—8 (прир.); Банников А. Г., 1954,
Млекопит. Монгольск. Народи. Республ., М., изд. Акад, наук СССР : 160—167
(общ., синон., изобр.); Доржин Э. Д., 1954, Природа, 9:105—106 (распр.,
биол.); Tong Е., 1954, Zoo life, vol. 9, № 2 : 58 (разв.; изобр.).
Лошадь Пржевальского, названная в честь открывшего ее в 1879 г.
знаменитого исследователя Центральной Азии Н. М. Пржевальского,
в настоящее время является единственным диким представителем настоя-
щих лошадей (подрод Equus s. str.). Первоописание Е. przewalskii было
сделано по шкуре и черепу трехлетнего самца, добытого в 1878 г. в песках
Джунгарии и подаренного Тихоновым на Зайсанском посту Н. М. Прже-
вальскому. Находка была столь неожиданна, что первое время вызывала
сомнения в принадлежности ее не гибриду и не одичавшему^ а настоя-
щему дикому животному. Однако дальнейшие исследования русских
путешественников (братьев Г. Е. и М. Е. Грум-Гржимайло, М. В. Пев-
цова, П. К. Козлова, В. И. Роборовского и др.) подтвердили существование
диких лошадей в западной части Центральной Азии. В начале 900-х годов
в Европу (в том числе в Асканию-Нова на юге России) были доставлены
первые партии живых животных. Все известные экземпляры по своему
сложению и окраске настолько однотипны, что исконно дикий характер
лошади Пржевальского в настоящее время никем не отрицается.
Описание. Небольшая лошадь, высотою в холке 124—140 см, с широ-
ким туловищем, на коротких, сухих, но более толстых, чем у джигетая,
ногах, с тяжелой, относительно большой головой (длиною у взрослых
животных 550—600 мм), короткой массивной шеей; длина последней,
по В. Заленскому (1902), меньше длины головы. Общий характер склада
и пропорции туловища сходны с примитивными породами азиатских
лошадей. Длина туловища приблизительно равна или несколько превос-
ходит высоту в холке. Копыта, в противоположность полуослам, широкие,
круглые, с хорошо развитой стрелкой. Каштаны имеются как на перед-
них, так и на задних конечностях в виде выступающих над поверхностью
кожи плоских шероховатых пластинок (в противоположность гладким
54
EQUIDAE
у джигетая) и располагаются на передних конечностях ближе к запяст-
ному суставу (у полу ослов ближе к грудной клетке). Уши относительно
короткие, в среднем около 135—140 мм, острые, с легкой вырезкой на
переднем крае около вершины.
Грива короткая, прямостоячая, несколько более длинная в средней
части (около 14—18 см); начинается несколько впереди ушей и идет до
холки. Сваливающийся у домашней лошади и тарпана на лоб пучок длин-
ных волос (челка) у лошади Пржевальского отсутствует. Хвост представ-
ляет нечто среднее между таковым Е. caballus и ослов. Репица (мясистая
часть хвоста) толще, чем у.последних, но настоящий длинный волос
начинается приблизительно с ее половины. Проксимальная половина
хвоста, отступя немного от его корня, по бокам несет толстый грубый
хвостовой волос, удлиняющийся по направлению к дистальной половине.
У самого корня хвоста и на дорзальной стороне проксимальной его части
волос в летнем наряде такой же, как на туловище; к зиме граница между
волосом боковых и дорзальной стороны хвоста выражена не резко; хвост
в целом по сравнению с домашней лошадью жидок, но длинен (около
90—100 см), спускается значительно ниже скакательного (голеностопного)
сустава. Вдоль нижнего края челюстей волос удлинен, но не грубый,
образует подобие «подвеса». В зимнее время на щеках он также становится
более длинным («бакенбарды»). Щетки на конечностях имеются, но раз-
виты не сильно.
Окраска лошади Пржевальского с трудом может быть подведена под
какую-либо из мастей, установленных для домашних лошадей. По-види-
мому, ближе других она может быть определена как каурая (Г. и М. Грум-
Гржимайло, 1896), но с темными, нетипичными для последней гривой,
хвостом и ногами. Иногда она определяется как саврасая (Поляков, 1881),
гнедо-саврасая (Тихомиров, 1898), светло-гнедая (в зимней шерсти;
М. Грум-Гржимайло, 1892). Вообще по окраске Е. przewalskii очень сходна
с джигетаем, от которого на не очень близком расстоянии ее с трудом
отличает даже местное население. Наиболее существенное отличие —
темная окраска нижних частей конечностей, светлых у Е. hemionus.
Общий фон окраски верхней части туловища, шеи и ног в летнем наряде
песчано-желтый, иногда с красноватым или буроватым оттенком. Вдоль
спины и дорзальной части хвоста до начала длинных волос идет темно-
коричневая с красноватым оттенком узкая, не свыше 10 мм, полоса (ре-
мень) без светлой по ее краям оторочки. Более интенсивная окраска верх-
ней части туловища книзу светлеет и без резкой границы переходит в жел-
товато-белую окраску живота. Светлые «подпалины» ясно выступают
в пахах и позади лопаток. Шея окрашена однотонно. Окраска верхней
части головы и щек более темная, кирпично-красного цвета. Конец морды
и губы серовато-белые. Вокруг ноздрей обычно имеется кольцо светло-
коричневых волос, соединяющееся с кольцом другой стороны коричне-
вой же перемычкой на уровне нижних краев носовых отверстий. Ноги
от копыт до путовых (пястно-фаланговых и плюснево-фаланговых)
суставов включительно и щетки (пучки волос позади суставов)
черно-коричневого с красноватым оттенком цвета; такую же окраску
имеет передняя (дорзальная) сторона конечностей до запястных и боковых
сторон скакательных суставов включительно. Задняя, наружная и внут-
ренняя сторона конечностей имеет желтовато-серую окраску с попереч-
ными коричневыми полосами; число последних (от 2 до 8) и степень вы-
раженности варьируют, иногда они совсем незаметны. Основная масса
волос гривы имеет также темно-коричневую окраску, но по бокам ее обрам-
EQUUS
55
ляют пучки менее грубых и более коротких волос одного с туловищем
цвета. Проксимальная половина хвоста имеет одинаковую с туловищем
окраску, хотя по бокам к желтому волосу примешиваются волосы черно-
коричневого цвета, а также волосы, имеющие зональную окраску. Длин-
ный волос конца хвоста черно-коричневого, почти черного цвета. Наруж-
ная сторона ушей одного цвета с туловищем, но концы имеют коричневую
окраску; внутренняя сторона ушей светло-желтого, иногда почти белого
цвета.
Рис. 11. Лошадь Пржевальского (Equus przewalskii Pol.).
Молодые имеют более светлую окраску с сероватым оттенком. Места,
окрашенные в коричневый тон, более бледны. Спинной ремень и попереч-
ные полосы на конечностях могут совсем отсутствовать. Летний волос
у взрослых животных короткий и гладкий, плотно прилегает к коже.
В зимнее время, а также у жеребят, он более длинный и волнистый, свет-
лый, приобретая ясно сероватый оттенок. Последнее, впрочем, имеет
место не всегда. Линька имеет место один раз в году — весною или в на-
чале лета. Переход к зимнему наряду происходит путем отрастания лет-
него волоса, начиная обычно с октября. Темный спинной ремень в зимней
шерсти сохраняется лишь в задней половине, и то не всегда.
По сравнению с тарпаном, кроме более низкого сошникового индекса,
в черепе лошади Пржевальского характерны: большая общая величина
(основная длина больше 470 мм), более широкая мозговая коробка и
яснее вдавленный лоб, в среднем более длинный лицевой отдел; более длин-
ный ряд коренных зубов (от 37.9 до 39.8 основной длины черепа),
частая недоразвитость чашечки на крайних нижних резцах (13) и не-
которые другие (Громова, 1949, 1950),
56
EQUIDAE
Костные слуховые проходы не больше 5.5% (от 4.4 до 5.5%) основной
длины черепа,1 направленные почти прямо в стороны. При взгляде
на череп сверху концы их едва выступают за контуры височных гребней.
Высота лицевой части черепа впереди Р2 составляет не свыше 21% (от
18.6 до 20.6%) основной его длины. Сошниковый индекс выше, чем у полу-
ослов, но ниже, чем у тарпана; расстояние от basion до заднего края
сошника составляет от 106.4 до 123% расстояния от последней точки до
заднего края костного нёба (в среднем 113.1%). Глоточные бугорки рас-
положены впереди разорванного отверстия. Задний край носовой вырезки
расположен не дальше, чем на уровне средины Р2. Парные гребневидные
бугорки на межчелюстных костях у нижнего края входа в носовую по-
лость выражены очень слабо или совсем отсутствуют. Пространство между
ветвями нижней челюсти суживается постепенно и не образует резко выра-
женного расширения в передней части. Симфиз нижней челюсти состав-
ляет 19.1 до 22.1% ее длины;
Число грудных-(-поясничных позвонков равно 24. Длина скелета
свободной передней конечности (плечевая -|- предплечье -(-пястная кость)
составляет 69—73.6% длины туловища (грудной-(-поясничный-(-крестцо-
вый отдел позвоночника). Длина метакарпа составляет от 26.4 до 27.7%
длины конечности и 85.9—91.1 % плеча. Длина задней конечности состав-
ляет от 78.4 до 85.9% длины туловища. Ширина пястной кости в средней
ее части составляет от 13 до 15.1 % ее длины; тот же индекс для плюсневой —
от 10.6 до 12.1%. Ширина копытной (третьей) фаланги передних конеч-
ностей составляет не меньше 140% (от 140.7 до 150%) длины ее по перед-
нему краю. Срединный гребень дистального блока метаподий высокий
и острый. Плюсневая кость составляет 76.8—79.3% длины бедра. Ширина
второй фаланги переднего пальца составляет от 93.3 до 101.2% ее длины.
Таким образом, отличия от тарпана в скелете конечностей сводятся
к более коротким конечностям, но с удлиненными и более тонкими мета-
подиями, к более тонким первым и вторым, но более широким третьим)
фалангам.
История и географическое распространение. Вопрос о былом рас-
пространении и палеонтологической истории Е. przewalskii не совсем
ясен. Достоверно относящиеся к ней остатки в ископаемом состоянии
не известны. Есть мнение, что лошади, по типу близкие к этому виду,,
еще во времена Палласа населяли область между Волгой и Уралом,
а также всю западную Сибирь (Hilzheimer, 1909). По некоторым данным
(Antonins, 1912) изображения лошадей художников французского палео-
лита должны быть также отнесены к этому типу. Предлагалось даже наи-
менование Е. przewalskii заменить на Е. equiferus, данное Палласом
для диких лошадей Заволжья.
Лошадь Пржевальского сохранила большое число архаических черт
(тонкость метаподий, длинная лицевая часть черепа, относительно мелкие
заднекоренные зубы, при одновременном относительном увеличении всего’
ряда коренных зубов, недоразвитые чашечки на нижних резцах и некото-
рые другие), которые не позволяют ее отнести к одной филогенетической
ветви с лошадьми Европы и северной Азии, в том числе и с тарпаном. По>
мнению В. И. Громовой (1949), лошадь Пржевальского представляет по-
томка самостоятельной центральноазиатской ветви лошадей, ведущей
свое начало от формы, близкой к верхнеплиоценовой Е. sanmeniensis
Chard, et Piv. Благодаря сравнительно большому постоянству климати-
1 Все цифры, относящиеся; к черепу и скелету,— по В. И. Громовой (1941, 1949)»
EQUUS
57
ческих условий в течение четвертичной эпохи, в этой части Азиатского
континента имела место задержка эволюции этой филогенетической ветви.
Крупные зубы, слабая их складчатость, вместе с длинными тонкими мета-
подиями являются приспособлением к условиям засушливой обстановки.
Рис. 12. Череп Equus przewalskii Pol.
Вопрос об участии лошади Пржевальского в образовании пород домаш-
них лошадей, в том числе и домашней монгольской, в настоящее время
решается в отрицательном смысле (Румянцев, 1936).
58
EQUIDAE
Область современного распространения лошади Пржевальского очень
незначительна, ограничиваясь Джунгарией и прилегающими частями
западной Монголии. Грубо схематично она представляет четырехуголь-
ник, заключенный между 42—47° с. ш. и между 86—100° в. д.1 Из отдель-
ных мест ее нахождения необходимо отметить следующие. Экземпляр,
послуживший типом для описания Е. przewalskii, был добыт в песках
Канабо (Ханобо). Примерно в этих же местах (между Булугуном и Бар-
кулем) встречал диких лошадей сам Н. М. Пржевальский (1883). В конце
Рис. 13. Лошадь Пржевальского (Equus przewalskii Pol.). Жеребец «Орлик».
Зоопарк Аскании-Нова. (Фот. А. И. Попова).
прошлого столетия в осенние месяцы дикие лошади были довольно много-
численны в районе урочища Гашун (Ачик-Су, около 65 км к северо-
востоку от Гучена) на восток до ур. Сыджира (около 30 км от Гашуна).
Здесь первыми из европейцев охотились на них братья Г. Е. и М. Е. Грум-
Гржимайло (1896). М. В. Певцов указывал в качестве области распростра-
нения Е. przewalskii южную часть пустыни между р. Манасом и восточ-
ной оконечностью Тянь-Шаня. Наиболее обычны дикие лошади, по его
данным, в саксауловой пустыне, начинающейся к востоку от меридиана
г. Гучена и до р. Манаса. Нередко встречаются они также к северу от
Гучена, а зимой посещают окраину Гученского леса (Анучин, 1896).
Г. Потанин (цит. по А. Банникову, 1954) указывал местонахождение
лошадей у северо-западного подножья Аджи-Богдо. Партии диких лоша-
дей были доставлены в Европу в начале 900-х годов из трех районов,
лежащих к югу и юго-западу от Коб до: 1) из равнины, ограниченной
щвостока Монгольским Алтаем, с запада оз. Улунгур, с севера р. Куи-
1 Совершенно неверная карта распространения у Аллена (Allen- 1940) основана,
вероятно, на незнании литературы.
EQUUS
59
Куиус и с юга р. Урунгу; 2) из предгорной равнины Эктаг-Алтая (Мон-
гольского Алтая), около 322 км к югу от Кобдо, и 3) из окрестностей
из. Цаган-Нур, уже к северу от Алтайского хребта.1 В то же время живот-
ные в этих районах не были редкостью и водились «тысячными» (?) табу-
нами (Нагрет, 1945).
Д. Клеменц (1903) писал, что лошади Пржевальского водятся к югу
и юго-востоку от хребта Байтаг-Богдо (пограничный хребет между Мон-
гольской Народной Республикой и Китаем). Местные жители сообщали
Рис. 14. Современный ареал Equus przewalskii Pol.
1 — граница постоянного ареала; 2 — места встреч и отдельных
заходов.
А. Н. Казнакову (1907) о том, что летом лошади водятся у подножья Шара-
Нуру (Тахиин-Шара-Нуру?), но на зиму куда-то исчезают.
В коллекциях ЗИН АН СССР, кроме упомянутого экземпляра, послу-
жившего типом для описания Е. przewalskii, и сборов братьев Грум-
Гржимайло, имеются экземпляры из следующих пунктов: колодец Эби
по дороге из Кобдо в Баркуль; местность около 53 км к востоку от Гашуна
между урочищами Нюрсу и Симигендзе; за горами Байтаг-Богдо (уро-
чище Харамелехстей); окрестности г. Гучена. Томскому университету
два экземпляра лошади Пржевальского были доставлены из района около
160 км к северу от Баркуля, из урочища Хони-Усу на р. Бижчин-Гол
у южных склонов Монгольского Алтая (Кащенко, 1907).
Ф. Харпер (Harper, 1945) называет еще следующие места встречи
лошадей Пржевальского: на равнинах у подножья южных склонов Мон-
гольского Алтая между 96 и 100° в. д.; в районе колодца Кайнарбулак;
па границе Монгольской республики и Китая, приблизительно около
400 км к юго-западу от горы Ихэ-Богдо; в Kapa-Гоби, западнее Эдзин-Гол
1 Вероятно, оз. Шаргын-Цаган-Нур в Шаргын-Гоби.
60
EQUIDAE
на 42° с. ш. и 98° в. д.; в местности около 20 км к западу от колодцев
Hou-Hung Chuan (42° с. ш. и 96° в. д.); в районе пункта Шиншинся на
границе между Синьцзяном и Ганьсу (на дороге Суну—Аньси—Хами).
В качестве наиболее восточного места нахождения лошади Пржеваль-
ского тот же автор отмечает пункт на границе Монгольской Народной
Республики и Китая на 42° с. ш. и 105° 30' в. д.
Основанное на расспросах местного населения утверждениеН.М. Прже-
вальского (1875) о том, что дикие лошади многочисленны в окрестностях
Лобнора, при фантастическом описании их внешности (грива «почти до
земли»), было им самим опровергнуто (1883).
Высказываемое иногда предположение (Antonius, 1937), что лошадь
Пржевальского в ее естественном ареале уже истреблена, лишено осно-
ваний. Имеются сведения самого недавнего времени о том, что по крайней
мере в юго-западной части Монгольской Народной Республики они со-
хранились до настоящего момента. По собранным в последние годы
А. Г. Банниковым (1954) сведениям, Е. przewalskii «обитает от нижнего
течения р. Булугун на западе, до хребта Аджи-Богдо на востоке. На се-
вере этот вид изредка доходит до подножий Монгольского Алтая. В насто-
ящее время регулярно встречается только у хребта Байтаг-Богдо и на
востоке до хребта Тахиин-Шару-Нуру». В этих районах были пойманы
живые экземпляры, которых А. Г. Банников видел в Улан-Баторе и на
животноводческой станции. А. Юнатов встретил табун лошадей Прже-
вальского в пустынной степи к северу от хребта Тахиин-Шара-Нуру.
По свидетельству местного уроженца Доржин Эрегдэн Дагва (1954),
обширная котловина Хонин Усан Гоби (Джунгарская пустыня), лежащая
между хребтами Тахиин-Шара-Нуру, Буга-Хайрханом, Их Хавтаг и
Бага Хавтаг, представляет благоприятные условия для существования
копытных. По его словам, еще лет десять назад дикие лошади Пржеваль-
ского встречались здесь табунами до 50 и даже до 100 голов.
Систематика. Доставленные Гагенбеком в Европу лошади Пржеваль-
ского из трех районов Джунгарии несколько отличались по окраске. В свое
время это послужило поводом рассматривать их как самостоятельные
расы (Matschie, 1903; Antonius, 1929). Однако приводимые отличия, оче-
видно, не выходят за пределы возрастной и индивидуальной изменчивости
и не дают основания на дробление вида со столь ограниченным ареалом.
Отмечаемая некоторая неоднородность среди животных даже из одного-
района (Кащенко, 1907) может объясняться примесью крови домашних
лошадей, скрещивание с которыми в пределах ареала Е. przewalskii
происходит почти повсеместно.
Биология. Сведения об образе жизни и привычках дикой лошади
Монголии немногочисленны и противоречивы, так как основаны главным
образом ца расспросных данных. Известно, что лошадь Пржевальского
является обитателем равнинных, преимущественно песчаных пустынь и
опустыненных степей. В области своего распространения распределена
неравномерно, предпочитая наиболее глухие, не посещаемые человеком
места. В горах поднимается на высоту до 2400 м. По М. В. Певцову (из
Анучина, 1896), дикие лошади обитают в саксаульных песках южной
части пустыни, расположенной между Манасом и меридианом восточной
оконечности Тянь-Шаня. Питаются они здесь камышами и солянками,
растущими в понижениях, жажду утоляют, выбивая в этих понижениях
копытами глубокие ямы, заполняемые солоноватой водой. Живут дикие
лошади здесь круглый год, но нередко из этих песков животные выходят
в расположенную по соседству «щебне-дресвяную пустыню, покрытую-
EQUUS
в1
местами тощим кипцом». Если лежит снег, то лошади задерживаются на
этих пастбищах дольше; в противном случае они вынуждены вскоре
возвращаться обратно в пески к своим водопоям. Заходят они из песков
и к югу в лиственный лес, расположенный, вероятно, в северных
предгорьях Тянь-Шаня. Здесь находят они обильный корм и воду,
ио в летнее время избегают его из-за обилия насекомых-кровосо-
сов.
В окрестностях урочища Гашун, где лошадей наблюдали и охотились
за ними братья Г. Е. иМ. Е. Грум-Гржимайло (1896), животные держатся
в щебенистой пустыне и среди бугристых песков, на пастьбу и водопой
в ночные часы они выходят к ключам, расположенным в понижении среди
зарослей камыша, дэриса и терескена. На день, часов с 5 утра, вновь
уходят в пустыню до захода солнца, где прекрасно маскируются благо-
даря защитной желтой окраске. В этом отношении они представляют, по
словам Г. Е. Грум-Гржимайло, противоположность куланам, находя-
щим убежище в горах, а не в пустыне.
По сообщенным А. Юнатовым сведениям, «в местах, где держатся
лошади Пржевальского, участки пустынных степей, образованные ковыль-
ком и солянками, чередуются с типичными пустынями: саксауловыми —
по тельвегу депрессий, терескеново-полынными и солянковыми (нанофи-
тоновыми)» (Банников, 1954).
Летом дикие лошади ведут более оседлый образ жизни, будучи
привязаны к местам водопоев. Зимой же, утоляя жажду снегом, не
привязаны к определенным местам обитания и широко кочуют (Клеменц,
1903).
Держатся лошади обычно группами, не превышающими 15—20 голов,
хотя раньше наблюдались табуны их до 50 и даже до 100 голов. Старые
самцы иногда держатся одиночно. Во главе табуна всегда находится
взрослый самец, проявляющий заботу о животных табуна, не исключая
и жеребят-сеголеток (Грум-Гржимайло, 1892; Клеменц, 1903).
Пасутся лошади всегда отдельно от куланов, не смешиваются с кося-
ками последних. В отличие от куланов, лошади Пржевальского имеют
очень характерную для них привычку ходить и даже бежать при пре-
следовании гуськом; следствием этого являются глубокие тропы, пере-
секающие во всех направлениях местности, где держатся эти животные.
Пищей диким лошадям служат различные пустынные травы: житняк,
гобийский ковылек, полыни, монгольский многокорешковый лук, вострец,
ревень, терескен, а также листья чия блестящего, камыша, баглура,
веточки саксаула. Зимой с помощью копыт добывают корм из-под снега;
На водопой дикие лошади ходят раз в 2-4 дня. Кормятся с рассвета до
полудня, после чего отдыхают, стоя или лежа на боку. Любят греться на
солнце. В непогоду укрываются в ложбинах, за скалами или в ущельях
(Доржин, 1954).
О времени спаривания в литературе сведений нет. Известно (Доржин,
1954), что самцы собирают гаремы по 10 и более кобыл. Между самцами
происходят ожесточенные драки, сопровождающиеся иногда увечьями
обрывом ушей, поломкой ребер и ног, что впрочем, по-видимому, слу-
чается не только в период гона.
Ожеребление кобыл происходит в период с конца апреля до 20-х чисел
мая. Приносят по одному жеребенку. Самки в это время придерживаются
определенных мест, богатых водой и кормом, чем обычно и пользуются
люди при поимке живых животных. На отдых самки с жеребятами также
приходят на одни и. те же места, результатом чего бывают наблюдавшиеся
62
EQUIDAE
Г. E. и M. Е. Грум-Гржимайло большие скопления кала жеребят в одном
месте.
Голос Е. przewalskii —звонкое ржание, сходное с таковым домашней
лошади, так же как и фырканье и храп. Дикая лошадь обладает острым
слухом и обонянием, может учуять человека на расстоянии 150—200 м,
но распространенное мнение о ее осторожности, как будет показано ниже,
по-видимому, преувеличино.
Паразитофауна и болезни лошади Пржевальского не изучены. Имеются
только указания, что у диких лошадей Монголии паразитирует желу-
дочный овод, Gastrophilus pecorum F. (сообщение К. Я. Грунина).
Хозяйственное значение. Дикая лошадь Пржевальского сохранилась
в таком незначительном числе, что имеет только научное значение, как
единственный дикий представитель настоящих лошадей. Большая часть
попыток приручать ее, даже пойманную в жеребячьем возрасте и воспи-
танную под домашней кобылой, оканчивалась неудачей. Животные со-
храняют дикость и на очень близкое расстояние человека не допускают.
Известен, впрочем, случай, когда пойманная жеребенком лошадь Прже-
вальского стала пригодной под верховую езду (Козлов, 1899 : 259).
Д. Клеменц (1903) также сообщал о попытках объезжать под седлом
воспитанных в неволе чистокровных пржевальцев и их гибридов с домаш-
ними монгольскими лошадьми, но опыты эти были неудачны. Не удава-
лось получить ни быстрого бега, ни красивых аллюров. Побежку их он
называет «незавидной». Кроме того, при выучке лошади Пржевальского
были дики, упрямы и бестолковы.
При скрещивании с домашней лошадью лошадь Пржевальского дает
вполне плодовитое потомство, но вследствие дикости животного и особенно
злобности самцов, скрещивание встречает большие затруднения. Легче
удается получать помеси между дикой кобылой и домашним жеребцом.
При скрещивании с английскими скаковыми жеребцами в Аскании-Нова
помеси унаследовали от отца формы тела, а от Е. przewalskii дикость,
стремление убегать и большую самостоятельность на пастбище, чем у
домашних южно-степных лошадей. В 1929 г. в Институте гибридизации,
в Аскании-Нова, получен единственный в мире, но бесплодный гибрид
между зеброй Чепмана [Equus (Hippotigris) chapmani Lag. ] и самцом
лошади Пржевальского. Общий тон окраски гибрид унаследовал от
отца; полосатость значительно уменьшена. Помеси с ослами получались, но
неизменно были бесплодны. Скрещивание с домашней монгольской ло-
шадью в области распространения Е. przewalskii, по-видимому, происхо-
дит не так редко и местные домашние лошади, вероятно, носят в себе
известную долю крови диких. Немногочисленные данные о быстроте и
выносливости лошади Пржевальского свидетельствуют, что правильная
постановка скрещивания с нею может значительно содействовать улучше-
нию рабочих качеств домашнего конского поголовья (Кравченко,
1951).
Поимка диких лошадей для целей приручения возможна только в же-
ребячьем возрасте. С этой целью в места, где имеют обыкновение держаться
самки с новорожденными жеребятами, отправляется группа всадников, имея
при себе лактирующих домашних кобыл. Встретив табун диких лошадей
с жеребятами, его начинают быстро преследовать до тех пор, пока послед-
ние, как менее выносливые, не начнут отставать. Отставших жеребят
ловят, накидывая на шею петли на длинных палках, и сразу же подсажи-
вают под домашних кобыл, у которых отнимают собственных жеребят.
Таким образом, были выловлены и с помощью домашних «кормилиц»
ARTIODACTYLA
63
воспитаны те партии живых лошадей Пржевальского, которые были
доставлены в Европу (Клеменц, 1903).
Охота на диких лошадей мало распространена из-за трудностей,
связанных с нею, а главное, вероятно, из-за малочисленности животных,
держащихся в самых отдаленных глухих местах. Стреляют их, под-
карауливая у троп, на водопоях. Распространенное мнение о невозмож-
ности охоты на них скрадом из-за их крайней чуткости и осторожности
преувеличено и опровергается хотя бы тем, что М. Е. и Г. Е. Грум-
Гржимайло успешно охотились на лошадей Пржевальского именно этим
способом. Мясо Е. przewalskii считается менее вкусным, чем у джи-
гетая.
Отряд ARTIODACTYLA — ПАРНОПАЛЫЕ
Соре Е., 1887, Proc. Amer. Philosoph. Soc., 24 : 377—400 (эвол., классиф.);
Weber М., 1928, Saugetiere, 2 Aufl., II, Jena : 517—625 (морф., классиф.); Mat-
thew W., 1929, Bull. Geol. Soc. Amer., 40 : 403—408 (классиф.); Matthew W.,
1934, I Mammal., 15 : 207—209 (эвол., классиф.); Simpson G., 1945, Bull. Amer.
Mus. Natur. Hist., 85 : 143—162+258—272 (эвол., классиф.); Romer A., 1946,
Vertebrate palaeontology, Chicago : 442—474 (эвол.); Schaeffer B., 1947, Amer.
Mus. Novit., 1356 : 1—24 (морф.); Frechkop S., 1948, C. R. XIII Congr. Internal,
de Zool., Paris : 379—383 (классиф.); Schaeffer B., 1948, Evolution, II, 2 : 164—
175 (эвол.).
Характеристика отряда. Копытные с числом пальцев как на перед-
них, так и на задних конечностях, два или четыре. Сильнее других раз-
виты третий и четвертый пальцы. Между ними проходит ось симметрии
конечности, и эти два пальца несут на себе основную тяжесть тела жи-
вотного. Второй и пятый пальцы в той или иной степени недоразвиты,
иногда совсем отсутствуют. Гомологичные фаланги функционирующих
пальцев имеют зеркальное сходство (как бы отражение одна другой).
Третья фаланга сжата с боков и имеет несимметричную треугольную форму.
Третий вертел (trochanter tertius) на бедре отсутствует (рис. 2, Л). Шейка
бедра ясно отделяет суставную головку от тела кости. Межвертельный
гребень идет от большого к малому вертелу и ограничивает с латераль-
ной и дистальной стороны вертельную ямку. Таранная кость (astragalus)
имеет два суставные блока: проксимальный для сочленения с костями го-
лени и дистальный — для naviculare с cuboideum. Пяточная кость (cal-
caneus), помимо таранной, всегда сочленяется также с малой берцовой
или ее рудиментом.
Сумма числа грудных и поясничных позвонков 19—20, крестцовых —
типично 4.
В черепе характерно отсутствие can. pterygoideus в основаниях крыло-
видных отростков базисфеноида. Хоаны передним краем редко заходят
вперед дальше второго заднекоренного зуба. Суставная ямка для причле-
нения нижней челюсти по сравнению с непарнопалыми менее вытянута
в поперечном направлении, более широкая (у верблюдовых даже круглая);
засуставный отросток (proc, postglenoideus) невысокий, или совсем от-
сутствует; поэтому нижняя челюсть может совершать движения не только
боковые, но иногда и в переднезаднем направлении. Tympanicum имеет
форму в той или иной степени вытянутого и вздутого пузыря (bulla),
образует не только наружную, но и большую часть внутренней стенки
барабанной полости. Характерна незначительная величина каменистой
части (petrosum), которая, как и у непарнопалых, не срастается с дру-
64
ARTIODACTYLA
гими костями. Всегда имеется хорошо развитый костный слуховой про-
ход, расположенный между proc, postglenoideus и proc, posttympanicus
чешуйчатой кости. Вместо общего большого разорванного отверстия по
бокам основания черепа остается лишь небольшой величины заднее (for.
lacerum posterior), тогда как for. ovale, rotundum и opticum разделены.
Сагиттальный гребень (crista sagittalis) имеется только в черепе Tylopoda
и Tragulidae. Носовые кости редко бывают сильно расширены в задней
половине. Если глазница имеет замкнутое кольцо, то задний край его
образуется только лобным отростком скуловой и скуловым отростком
лобной кости. Чешуйчатая кость участия в образовании заднего края
глазницы не принимает.
Коренные зубы селенодонтного или бунодонтного типа, в отдельных
случаях не образуют замкнутого ряда. Постоянные переднекоренные
(премоляры), даже последние из них, никогда не приобретают формы
заднекоренных (моляров) и устроены значительно проще. Последний
молочный премоляр (Pd4), а также последний моляр (М3) нижней
челюсти всегда состоят из трех долей.1 В отдельных случаях четыре пре-
моляра сохраняются и в серии постоянных зубов. В этих случаях диа-
стема может отсутствовать. Резцы и клыки в верхней челюсти часто
сильно редуцированы или совсем отсутствуют. Клыки нижней челюсти
часто имеют форму соседних резцов.
Губы разнообразной формы и устройства. Передняя часть головы
иногда приобретает характер недлинного хобота. Желудок у всех со-
временных форм в той или иной степени усложненный и состоит из 2—4
в различной мере отделенных друг от друга камер. Слизистая оболочка
с плоским многослойным эпителием, лишенная пищеварительных желез,
выстилает значительную, обычно большую часть желудка. Слепая кишка
(coecum) и ободочная (colon) менее объемисты, чем у непарнопалых,
обособленные тении и карманообразные выпячивания на них имеются
только у нежвачных. Ободочная кишка образует спираль в виде конуса
или диска. Желчный пузырь, за исключением сем. Cervidae, имеется.
Носовые барабаны и заглоточные воздухоносные мешки не развиты.
Молочные железы двух- или четырехдольчатые, паховые; реже (у не-
жвачных) множественные, расположенные на вентральной стороне жи-
вота. Плацента диффузная или котиледонная. Детенышей в одном помете
может быть несколько (у домашних свиней до 23).
Географическое распространение. Материк и прилегающие острова
Европы, Азии, Африки и Америки. Акклиматизированы в Новой Зелан-,
дии. В домашнем состоянии распространены по всему земному шару.
Эволюция парнопалых. Как и другие ветви копытных, парнопалые
ведут свое начало от примитивных палеоценовых форм одной из групп
первичнокопытных, кондилартр (Condylarthrd). Некоторые представи-
тели последних (например род Hyopsodus Leidy) по строению зубов и
пальцев почти не отличаются от ранних парнопалых. Почти одно-
временно в нижнем и среднем эоцене Европы и Америки появляются
роды Diacodexis Соре, Homacodon Marsh и Dichobune Cuvier, форма та-
ранной кости которых с двойным суставным блоком не оставляет сомне-
ния в принадлежности их к отряду парнопалых. Это были небольшой
величины животные, на низких ногах с четырьмя функционирующими
1 За очень редкими исключениями (род Neotragus Н. Smith из семейства Во-
vidae).
ARTIODACTYLA
65
пальцами, а на передних конечностях у некоторых сохранялся, по-видш.
мому, и небольшой первый палец. Низкий вытянутый череп не имел
замкнутого кольца глазницы. Мастоидная часть каменистой кости выхо-
дила на лицевую поверхность черепа. Зубы образовывали сплошной ряд
без диастемы. Строение зубов указывает на то, что эти животные не были
чисто растительноядными, а питались смешанной пищей. Коренные
зубы некоторых форм из них имели еще трехбугорчатое строение с при-
тупленными вершинами бугорков. Эта группа (инфраотряд Palaeodonta)
должна считаться исходной для всех последующих ветвей, в том числе
и современных, Artiodactyla. Уже к верхнему эоцену и нижнему олиго-
цену число групп парнопалых возросло (гигантские двупалые энте-
лодоны, аноплотерии, антракотерии х, ранние трагулиды, верблюды
и другие), большая часть которых вымерла, не оставив в современной
фауне потомков. Представители современных свиней, жираф, оленей
и полорогих появляются лишь в верхнем олигоцене — нижнем мио-
цене.
Парнопалые в процессе эволюции во многом развивались параллельно
пепарнопалым. Как у непарнопалых, общее направление эволюции —
адаптация к быстрым поступательным движениям и питанию раститель-
ной пищей. С этим связана в различной степени выраженная редукция
локтевой и малоберцовой кости, редукция и уменьшение числа боковых
лучей кисти и стопы, удлинение метаподий и фаланг пальцев, переход
от стопохождения к пальце- и фалангохождению, усложнение жеватель-
ной поверхности коренных зубов, развитие гипсодонтизма. Из современ-
ных форм наиболее близки к исходному состоянию свиньи й бегемоты.
Мозоленогие (Tylopoda) при полной редукции боковых лучей сохранили
неполное пальцехождение (опираются на вторую и третью фаланги)
н коготь вместо копыта на последних фалангах.
Своеобразное направление филогенетической ветви парнопалых на-
ходит выражение прежде всего в том, что у самых ранних представите-
лей отряда конечности, даже при сохранении первого пальца, имеют
«параксонический» характер, т. е. ось конечности проходит между третьим
и четвертым пальцем. В связи с этим увеличенную функциональную
нагрузку и усиленное развитие получает не один, а два названных луча
(III и IV). Еще у бегемотов третий палец несколько длиннее четвертого.
У остальных они уже одинаковы. Фаланги пальцев этих лучей приобре-
тают зеркальное сходство, а метаподий обнаруживают склонность сли-
ваться и образовывать функционально единую кость — цевку (os cannon).
Боковые лучи (II и V) подвергаются редукции, а их пальцы, по мере
удлинения центральных, перестают касаться почвы. В крайне выражен-
ных случаях они исчезают совсем или сохраняются в виде рудиментов,
лишенных костной основы. Наиболее быстро происходил и наибольшее
выражение нашел процесс усиления центральных и редукции боковых
лучей у форм, приспособленных к быстрому бегу и к обитанию на твер-
дом грунте. Передние конечности в этом отношении несколько отставали
от задних. Слияние центральных метаподий и исчезновение боковых
пальцев в филогенезе всех групп парнопалых происходили в первую
очередь в стопе, а не в кисти.
Второй специфической чертой парнопалых с самой ранней их.истории
является образование на таранной кости (астрагале) двойного суставного
1 Аноплотерии и некоторые антракотерии сохраняли следы непарнопалости:
третий палец у них был развит сильнее четвертого, а второй сильнее пятого.
5 Фауна СССР. И. И. Соколов
66
ARTIODACTYLA
блока. В процессе эволюции направление оси сустава между астрагалом
и пяточной костью (calcaneus) из косого превратилось в перпендикуляр-
ное оси конечности и параллельное оси голеностопного сочленения.
Образовавшийся тройной сустав с параллельными осями способствовал
увеличению размаха флексорно-экстензорных движений (сгибания и раз-
гибания), но почти нацело исключал вращательные (пронацию и супи-
нацию). У верблюдов, например, при сгибании конечности в голеностоп-
ном суставе во время лежания голень и плюсна приобретают почти
параллельное друг другу положение. Дистальный суставный блок "астра-
гала, вероятно, способствовал прыжкам — одному из обычных
способов передвижения некоторых примитивных современных (мел-
кие антилопы, оленьки), а также ранних представителей парнопа-
лых.
Исходная зубная формула 1-|-, Су, Р-^-, М-|- из современных форм
сохраняется только у свиней (Suidae). В остальных группах происходит
редукция переднекоренных (в первую очередь Рх и Р1, а у верблюдов
и Р2 и Р2) й, особенно, верхних резцов и клыков. Параллельно с редук-
цией резцов происходит и редукция межчелюстных костей. В нижней
челюсти клыки часто приобретают форму резцов и примыкают к послед-
ним, оставляя между собою и переднекоренными диастему. Исходный
трехбугорчатый тип коренных зубов в процессе эволюции превращается
в четырех-, пяти- и даже шестибугорчатый. Бугорки или округляются,
превращая зуб в бунодонтный (свиньи, бегемоты), или вытягиваются
в продольные изогнутые полулунные гребни, обусловливающие особен-
ности зубов селенодонтного (лунчатого) типа у жвачных и верблюдов.
Зубы первого типа приспособлены к всеядному питанию или питанию
мягкими, сочными частями растений. Селенодонтия же связана с адапта-
цией к пережевыванию жесткой травянистой пищи. Некоторые из вымер-
ших групп (например, антракотерии, аноплотерии) имели зубы смешан-
ного, буноселенодонтного типа. Широкая суставная поверхность для
сочленения с нижней челюстью, допускающая боковые движения послед-
ней, создает благоприятные условия для более совершенной функции
жевательного аппарата.
Приспособление к питанию травянистой пищей в эволюции парно-
палых копытных сопровождалось усложнением желудка. У свиней (Sus L.)
и бегемотов намечается еще не резко обособленное левостороннее выпя-
чивание стенки желудка; у пекари (Tayassu Fisch.) желудок состоит
уже из трех отделов. Наибольшего же усложнения он достигает в группе
жвачных (см. ниже, стр. 117).
Классификация отряда парнопалых. Классификация группы пар-
нопалых, охватывающей в современной фауне свиней, пекари, бегемотов
(гиппопотамов), верблюдов, оленьков, жираф, кабарог, оленей, вило-
рогов и полорогих («антилопы», быки, козлы, бараны), основывается
па строении зубов, конечностей и височно-затылочной области черепа.
Современные представители отряда без особого труда могут быть разде-
лены на три группы или подотряды: 1) нежвачные, Nonruminantia, или
Sutform.es — семейства свиней (Suidae'), пекари (Tayassuidae) и беге-
мотов (Hippopotamidae)', 2) мозоленогие, Tylopoda — семейство вер-
блюдовых (Camelidae)-, 3) жвачные, Ruminantia— семейства
оленьков (Tragulidae), кабарог (Moschidae), оленей (Cervidae), жи-
раф (Giraffidae), вилорогов (Antilocapridae) и полорогих (Bovi-
Аав). -
NONRUMINANTIA
67
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДОТРЯДОВ ПАРНОПАЛЫХ
КОПЫТНЫХ ФАУНЫ СССР И МОНГОЛЬСКОЙ НАРОДНОЙ
РЕСПУБЛИКИ
1 (2). Морда вытянутая, конусовидная, оканчивается обращенным впе-
ред круглым «пятачком» с выступающими краями. Волосяной покров
состоит, в основном, из грубой толстой щетины. Верхние клыки
выступают из-под верхней губы и вершинами загнуты вверх. В ске-
лете конечностей третьи и четвертые метаподий сохраняются в виде
самостоятельных неслившихся костей. В черепе лобная и скуловая
кости не соприкасаются между собою и кольцо орбиты в задней
части остается незамкнутым (рис. 16). Коренные зубы бунодонтные,
с коронками, несущими несколько тупых закругленных бугорков
(рис. 16). Число коренных зубов в каждой челюсти семь......
........ Nonruminantia (=Suiformes) — Нежвачные (стр. 67).
2 (1). Морда не имеет форму конуса и оканчивается губами или неболь-
шим мягким хоботом. Кожа покрыта мягким нещетинистым, хотя
иногда и ломким волосом. Верхние клыки если выступают из-под
верхней челюсти, то вершинами направлены вниз. В скелете конеч-
ностей третьи и четвертые метаподий слиты в одну кость (цевку).
В черепе лобная и скуловая кости соприкасаются между собою
и кольцо орбиты замкнуто (рис. 86). Коренные зубы селенодонтные,
коронки заднекоренных состоят из четырех дугообразно изогнутых
долей, слитых в основаниях (рис. 86). Число коренных в каждой
челюсти не больше шести.
3 (4). На спине один или два жировые горба. Верхняя губа раздвоена.
Конечности имеют только по два пальца, заканчивающиеся ногте-
видными копытами без подошвы, но с сильно развитыми мякишами.
При ходьбе и стоянии животное опирается на весь палец. Дисталь-
ные суставные блоки метаподий имеют срединные гребни только'
на задней стороне. Череп с сильно развитым сагиттальным гребнем.
В верхней челюсти имеется одна пара резцов. Нижние клыки отстоят
на некоторое расстояние от резцов и имеют заостренные вершины
..............................Tylopoda — Мозоленогие (стр. 98)..
4 (3). Жировые горбы на спине отсутствуют. Верхняя губа не раздвоена..
Кроме двух средних (третьего и четвертого) пальцев, имеются
в той или иной мере недоразвитые боковые (второй и пятый) пальцы.
Копыта имеют развитую подошву. При ходьбе и стоянии на твер-
дом грунте животное опирается только на копыто. Дистальные
суставные блоки метаподий на всей поверхности несут выступающие
срединные гребни, соответствующие углублениям на проксималь-
ных суставных поверхностях первых фаланг пальцев. Череп без
сагиттального гребня. В верхней челюсти резцов нет. Нижние
клыки имеют форму соседних резцов и сидят вплотную к последним
............................ Ruminantia — Жвачные (стр. 115).
Подотряд Nonruminantia (= Suiforrnes) —
Нежвачные
Характеристика подотряда. Коротконогие животные с четырьмя
(редко с тремя) пальцами на всех конечностях. Метаподий полностью
не сливаются и не образуют os cannon. Все кости запястья и зацлюсны
5*
68
ARTIODACTVLA
остаются самостоятельными. Локтевая и малая берцовая, как правило,
не срастаются с соседними костями (лучевой и большой берцовой). Ниж-
няя дуга первого шейного позвонка (атланта) на каудальном крае имеет
выступающий назад клювовидный отросток. Зубовидный отросток вто-
рого шейного позвонка (эпистрофея) имеет заостренную конусовидную
форму.
Мастоидная часть каменистой кости прикрыта чешуйчатой костью
и не выходит на наружную поверхность черепа. Рога отсутствуют. Глаз-
ница незамкнутая.1 Заднекоренные зубы бугорчатые (бунодонтные).
Клыки сильно развиты; нижние имеют открытый корень и обладают
постоянным ростом.
Кожа или почти совсем лишена волос, кроме вибрисс на передней
части головы, или покрыта редким волосяным покровом, состоящим,
в основном, из грубой толстой щетины, и лишь у некоторых видов (Sus
scrofa L.) имеется и подшерсток. Имеется склонность к образованию
толстого подкожного слоя жировой ткани. Желудок в большинстве слу-
чаев простой, не имеет ясного разделения на отделы и не приспособлен
к отрыгиванию пищи; поэтому вторичное пережевывание ее (жвачка)
не имеет места. Плацента диффузная.
Эволюция и систематика. Бунодонтные парнопалые с закрытой
мастоидной областью ведут происхождение от примитивных нижнеэоце-
новых палеодонтов. Однако прямые предки современных семейств с до-
стоверностью не установлены. Уже в верхнем эоцене и олигоцене Евразии
существовало значительное число групп Sutformes, большая часть которых
не оставила в современной фауне потомков. В эоцене и олигоцене полу-
чило расцвет, а в начале миоцена вымерло семейство энтелодонов (Ente-,
lodontidae). Эти «гигантские свиньи» с высокой холкой и черепом длиною
до 90 см имели бунодонтные, просто устроенные четырехбуг'орчатые
коренные зубы. В отношении же конечностей они были более специали-
зированы, по сравнению даже с современными свиньями. Лучевая и локте-
вая кости были слиты. Метаподии III и IV лучей хотя и не образовывали
os cannon, но боковые (II и V) были сильно редуцированы, и конечности
функционально были двупалые. Судя по характеру зубов и некоторым
особенностям скелета (длинные остистые отростки спинных позвонков,
форма затылочной и скуловой области черепа), энтелодоны питались кор-
нями растений, разрывая клыками почву. В прямой генетической связи
с современными Suidae они не стояли. Некоторые общие с последними
черты обусловлены, очевидно, сходным образом жизни.
Другой обширной группой третичных нежвачных парнопалых были
верхнеэоценовые и олигоценовые ацтракотерии (сем. Anthracotheriidae).
Это были также крупные животные на коротких конечностях с четырьмя
хорошо развитыми и функционировавшими пальцами; иногда сохра-
нялся даже рудимент первого пальца. В противоположность другим
группам подотряда, у некоторых антракотериев наружные бугорки
пятибугорчатых брахиодонтных коренных зубов начали приобретать
полулунную форму. Антракотерии, по-видимому, вели' полуводный
образ жизни и были, вероятно, предками гиппопотамов (род Мегусо-
potamus Falc.).
Современные N onruminantia делятся на три семейства: 1) свиные
(Suidae), обитатели Старого Света; имеют вытянутую конусовидную
морду, просто устроенный желудок, самостоятельные третью и четвертую
1 За редким исключением у гиппопотамов.
SUIDAE
69
метаподии, хорошо развитые, но укороченные второй и пятый пальцы,
загнутые в стороны и вверх клыки верхнего зубного ряда; 2) южно-
и центральноамериканские пекари (Tayassuidae) с трехкамерпым желуд-
ком, начавшими сливаться в проксимальной части метаподиями, срос-
шимися локтевой и лучевой костями, сильно редуцированным пятым
пальцем и направленными вертикально вниз верхними клыками, и 3) рас-
пространенные в настоящее время только в Африке бегемоты, или гиппо-
потамы (Hippopotarnidae)', тяжелые, неуклюжие, полуводные животные
с почти лишенной волос кожей, расширенной на конце мордой, простым
желудком, самостоятельными несросшимися метаподиями, четырьмя функ-
ционирующими пальцами и постоянно растущими нижними резцами и
и клыками.
Первые две группы стоят друг к другу ближе, чем к бегемотам.
В фауне СССР имеется только представитель семейства свиней {Suidae) —
Sus scrofa L.
Семейство SUIDAE — СВИНЫЕ
Stehlin H. G., 1899—1900, Abhandl. Schweiz. Palaontol. Gesellsch., XXVI,
1 : 1—336; XXVII, 1 : 337—-527 (ист., сист.); Lydecker R., 1915, Gatal. Ungulate
mammals Brit. Mus., IV : 305—374 (сист.)', P о с о c k R. J., 1923, Proc. Zool. Soc.,
London : 531—549 (морф.); Brehms Tierleben, 4 Aufl., Bd. 13, Saugetiere, Bd. 4,
Berlin, 1925 : 2—31 (общ., изобр.); Б p эм А. Э., 1941, Жизнь животных, V, М.,
изд. Учпедгиза : 524—536 (общ., изобр.); Pilgrim G. Е., 1926, Palaeontol. indica,
vol. 4, № 1 : 1—65 (ист.); Pearson Н., 1927, Phylosoph. trans. Roy. Soc. London,
ser. B, 215 : 389—450 (морфол., эвол.); Pearson H.; 1928, Palaeontol. Sinica, ser.
G, 5 : 1—75 (морф., эвол., сист.); Weber M., 1928, Saugetiere, П, Jena : 547—554
(морф., сист., эвол.); Colbert Е. Н., 1935, Amer. Mus. Novit., 799 : 1—24 (эвол.).
Характеристика семейства. Животные средней величины. Редкий
волосяной покров состоит главным образом из грубых волос (щетины),
расположенных, по крайней мере на некоторых частях тела, группами
по три; между последними у многих видов в зимнее время отрастает
более тонкий подшерсток. Передняя часть головы вытянута в длинное
конусовидное рыло, оканчивающееся круглым, плоским, лишенным
волос, подвижным «пятачком». На поверхности последнего открываются
ноздри. Остов пятачка составляет непарная хоботковая кость (os rostri),
расположенная между передними краями межчелюстных и носовых костей.
В конечностях развиты все четыре луча, но боковые (II и V) значи-
тельно короче средних, и поэтому пальцы этих лучей только касаются
почвы; опирается на них животное лишь при ходьбе по мягкому грунту.
Метаподии и на передних и на задних конечностях не обнаруживают
даже склонности к слиянию. Локтевая и малая берцовая кости остаются
самостоятельными. Конечные фаланги пальцев (в противоположность
гиппопотамам) одеты настоящими копытами. Позвонков: грудных 14,
поясничных 5—6, крестцовых 4—5.
В черепе характерна плоская постепенно поднимающаяся в аборальном
(назад) направлении лобнотеменная поверхность, сходящаяся под острым
углом с плоской или вогнутой затылочной поверхностью. Соответственно
длине морды, лицевая часть черепа вытянута, с прочным срастанием
отдельных костей. Имеется, в противоположность другим парнопалым,
хорошо выраженная крыловидная ямка (fossa pterygoidea). Глазница
в задней половине широко открыта. Слуховые пузыри (bullae) вытянуты,
сжаты с боков и сильно выступают в вентральном направлении; внутрен-
няя их полость заполнена перегородками из тонких костных пластинок.
10
SUIDAE
Слёзная кость занимает значительное место на лицевой поверхности
черепа и имеет двойное отверстие носослёзного канала.
Полная зубная формула I ~, С-^-, Р М -|-=44 зуба из современных
Suidae сохранилась только у рода свиней (Sus L.); у остальных боковые
резцы или передние премоляры, а иногда и те и другие, исчезают. Резцы
имеют закрытые корни; у клыков же корни открыты, и эти трехгранного
сечения зубы обладают постоянным ростом. Верхние клыки загнуты
в стороны и вверх, а у бабирусс — вверх и назад. В месте выхода клыков
верхнечелюстные кости имеют утолщения или гребни. Заднекоренные
зубы, особенно последний, — со сложным бунодонтным рисунком. Кроме
основных, имеется иногда значительное количество дополнительных
бугорков, особенно в задней удлиняющей зуб части (гипоконе и гипоко-
ниде).
Желудок имеет левостороннее выпячивание в кардиальной части,
но в целом еще простой, не разделенный на отделы. Число сосков — не
меньше четырех. Число детенышей в помете — четыре-восемь, а у до-
машних форм еще больше.
Географическое распространение. Африка с Мадагаскаром; умерен-
ные и южные широты Европы и Азии, включая острова Малайского
архипелага.
Эволюция и классификация. Корни Suidae ведут, вероятно, к верхне-
эоценовым формам вроде родов Helohyus Marsh, и Choeropotamus Cuv.
(из группы палеодонтов), имевших еще сравнительно короткую лицевую
часть и просто устроенные четырехбугорчатые коренные зубы. Первые,
достоверно относящиеся к этому семейству ископаемые формы, появляются
только в олигоцене. Нижнеолигоценовый род Propalaeochoerus Stehlin
имел короткие верхние и лишь слегка удлиненные нижние клыки. В ли-
нии, ведущей к верхнеолигоценовым и миоценовым Palaeochoerus Pomel
и Hyotherium Meyer, а также в линии современных свиней происходило
превращение брахиодонтных клыков в постоянно растущие и, параллельно,
усложнение устройства коренных зубов. Начиная с нижнего плиоцена,
в Евразии существовало значительное число родов Suidae, в том числе
современный род Sus L. Большая часть их вымерла, не оставив потомков.
На территорию Нового Света эта группа никогда не проникала.
В современной фауне семейство Suidae представлено пятью родами.
1. Род Sus L. — Собственно свиньи, с полной зубной формулой I v,
О
14 3
С р Р р М у=44 зуба и слабо развитыми костными наростами над лун-
ками верхних клыков. Распространены в Европе, Азии и Африке (глав-
ным образом в северной ее части).
2. Род Potamochoerus Gray — Кистеухие или речные свиньи, имеющие
пучки волос на кончиках ушей и две пары костных наростов верхней
челюсти над клыками у самцов. Обитают в Африке к югу от Сахары и на
о. Мадагаскаре.
3. Род Hylochoerus Thos.—Лесные свиньи, с одним видом — Н. mei-
nertzhageni Thos., из зоны африканских экваториальных лесов. Резцов
в верхней челюсти имеется только одна пара, а переднекоренных в ниж-
ней — только две.
4. Род Phacochoerus Cuv. — Бородавочники. Характерные особен-
ности: мощные бородавчатые разрастания кожи по бокам морды впереди
глаз и сильно развитый последний заднекоренной зуб, в процессе роста
SUS
71
постепенно вытесняющий расположенные впереди него зубы. Распростра-
нены в Африке к югу от Сахары.
5. Род Babirussa Rafin. — Бабируссы. Отличается от других родов
том, что у самцов лунки верхних клыков открываются вверх. Растущие
вверх клыки прободают кожу, загибаются дугообразно назад и вниз
в надглазничную область. В противоположность другим родам семейства,
новорожденные поросята не имеют полосатой окраски. Распространение
ограничивается островами Борнео и Целебесом.
В фауне СССР представлен только род Sus L.
Род SUS LINNAEUS —СВИНЬИ
Muller S. u. Н. Schlegel, 1842, Zoogdig. van den Indisch. Archip., I:
169—182 (сист., изобр.); Fit zinger L. J., 1864, Sitzber. math.-natnrwiss. KI.
Acad. Wiss. Wien, L : 383—434 (сист.); Major F., 1883, Zool. Anzeig., 6 : 295—300
/сист.); Ly decker R., 1915, Catal. Ungulate mammals Brit. Mus., IV : 307—344
{сист.); Pilgrim G., 1926, Palaeont. indica, vol. 4, № 1 : 1—65 (ист.); Адлер-
6 e p г Г. IL, 1933, Тр. Лабор. генет. 1 : 185—207 (морф., сист.); С о 1 b е г t Е. Н.,
1935, Amer. Mus. Novit., 799 : 1—24 (ист.); Amon R., 1938, Ztschr. f. Zuchtung,
Reihe B., 40 : 49—88 (ист., сист.); E 11 e r m a n J. R. and T. G. Morrison-
Scott, 1951, Checklist of palaearctic and indian mammals, London : 344—348 (сист.).
Характеристика рода. За одним исключением,1 крупные или средней
величины животные. Зубная формула: I С -р Р-|, М 44 зуба,
но крайняя пара резцов и первый верхний премоляр иногда уже у молодых
животных выпадают. Первый переднекоренной не имеет молочного пред-
шественника и прорезывается одновременно с последним заднекоренным.
Стирание в процессе изнашивания зуба захватывает целиком вершины
верхних клыков (в противоположность Hylochoerus, Phacochoerus и Ba-
birussa). Череп длинный и узкий, но сагиттальный гребень (в противо-
положность Babirussa) не образуется. Поверхность лобно-теменной зоны
носит продольно выпуклый характер. На наружной стенке альвеол
верхних клыков имеется костный гребень, но мощного выроста здесь,
как и на границе с носовыми костями, верхнечелюстная кость не обра-
зует. Молодые первое время после рождения имеют продольно полосатую
окраску (исключение представляет S. verrucosus Mull, et Schleg.).
Географическое распространение — см. выше (стр. 70).
Эволюция и классификация. Род свиней в ископаемом состоянии
известен с верхнего миоцена или нижнего плиоцена. Произошел, вероятно,
от примитивных миоценовых видов рода Dicoryphochoerus Pilgr. Виды
S. advena Pilgr., S. adolescens Pilgr., S. comes Pilgr. из Индии имели
еще короткие и широкие заднекоренные зубы. Более поздние формы
(S. peregrinus Pilgr., S. strozzii Mencgh., S. namadicus Pilgr. и др.) харак-
теризуются постепенным усложнением коренных зубов и прогрессирую-
щими размерами клыков.
Классификация рода свиней, даже видовая, разработана до сих пор
неудовлетворительно. Описано большое число форм, иногда по одному-
двум экземплярам; видовая самостоятельность некоторых из них весьма
сомнительна. Бесспорно самостоятельными видами являются, видимо, лишь:
1. Sus barbatus Mull. — Бородатая свинья. Характеризуется необы-
чайно длинным вытянутым черепом, особенно в лицевой части, и формой
1 N. (Porcula) salvanius Hodgson из Непала и Бутана.
72
SUIDAE
нижних клыков, у которых задняя (аборальная) поверхность уже боко-
вой (латеральной). Распространена на островах: Борнео, Суматре и Фи-
липпинах. Возможно, к этому же виду, как подвиды, должны быть Отне-
сены: a) S. celebensis Miill. et Schleg. с Целебеса и Филиппин с прилегаю-
щими островами; отличается меньшей величиной и короткими волосами
на конце хвоста; б) S. verrucosus Miill. et Schleg. с о. Явы, отличительную
черту которого составляют три пары бородавчатых выростов кожи по
бокам головы. Эта группа по ряду особенностей черепа стоит ближе
других к исходным формам рода.
2. Sus cristatus Wagn. От других видов свиней отличается более высо-
кой и вытянутой мозговой частью черепа, уплощенными в аборальной
части носовыми костями, а также совершенно лишенным подшерстка
волосяным покровом.1 Распространена в Индии, Бирме, Индокитае
и Малайском п-ве.
3. Sus vittatusMvAX. et Schleg. Краниологически очень близка к S. scrofa.
Отличается окраской с огненно-рыжеватым оттенком, темной полосой
вдоль хребта и красновато-бурой вдоль средней части морды. Распростра-
нена на островах Зондского архипелага и, возможно, на крайнем юге
Малайского п-ва.
4. Sus scrofa L. — Обыкновенный кабан, распространенный в север-
ной Африке, южной и средней Европе, на Кавказе, в Передней, Средней,
Центральной и восточной Азии и на некоторых островах Японии (харак-
теристику см. ниже, стр. 72). К этому же виду, как подвид, должна быть
отнесена отличающаяся несущественными признаками S. leucomystax
Temm.,— форма с островов Японии и Тайваня, а также S. falzfeini Mat-
schie из Белоруссии.
Ю. Филипченко (1933) без достаточных оснований выделял в качестве
отдельного вида S. orientalis, кабанов Китая, Центральной Азии и Даль-
него Востока, которые несомненно относятся к типичному S. scrofa.
5. Sus salvanius Hodgs.—Карликовая форма из южных предгорий
восточных Гималаев, Непала и Бутана. Имеет только три пары сосков,
рудиментарный хвост и редуцированный последний коренной зуб верхней
челюсти. Выделяется обычно в особый род или подрод Porcula Hodgson.
Ряд форм, описанных с Новой Гвинеи и некоторых прилегающих
островов юго-восточной Азии (5. papuensis Less, et Garn., S. timoriensis
Miill. et Schleg.),' являются, вероятно, одичавшими потомками домашних
S. scrofa или S. vittatus, проникших на острова при посредстве человека.
Sus scrofa Linnaeus — Кабан. Дикая свинья (рис. 15, 16, 17, 18)
Eichwald, 1830, Naturhistorische Skizze von Lithauen, Volhynien and Podo-
lien, Wilna : 239 (распр.); Кесслер К. Ф., 1851, Естеств. ист. губерн. Киевск.
учебн. окр. : 82—83 (распр.); В е Ike G., 1866, Bull. Soc. natur. Moscou, XXXIX,
1 : 492 (распр.); R ad de G. und A. Walter, 1889, Zool. Jahrb., Abt. Syst., IV :
993—1094 (распр. и биол.); Г а а к e В., 1901, Животный мир, I, СПб. : 32—38 (общ.,
изобр.); Врублевский К., 1908, Архив ветеринарн. наук, 10 : 943—959 (бол.);
Greve К., 1909, Saugetiere Kur-, Liv-, Estlands, Riga : 133—135 (распр.); Алфе-
p а к и, 1910, Семья охотн. : 123—129 (распр.); Д и н н и к Н. Я., 1910, Звери Кав-
каза, I, Тифлис : 23—53 (общ.); Thomas О., 1912, Proc. Zool. Soc. London : 390—
393 (сист.); Lydecker R., 1915, Catal. Ungulate mammals Brit. Mus., IV : SOT-
SIS (сист.); Matschie P., 1918, Sitzber. Geselsch. naturforsch. Freunde, Berlin,
1 Нужно отметить, что ряд указываемых Г. Адлербергом (1933) краниологических
черт (например, форма лобной поверхности, ширина между теменными гребнями,
форма глазницы), как показал просмотр коллекционных фондов ЗИН АН СССР,
не являются специфическими для 5. cristatus и A. vittatus.
SUS
73
VIII : 300—306 (сист.); Sc hr 6te г H., 1922, Zool. Jahrb., Abt. Syst., XLVI : 303—
367 (морфол., сист.); Толстой H. Н., 1922, Охота на Кавказе, М. : 60—84 (ох.);
Браунер А. А., 1923, Сельскохозяйственная зоология, Одесса : 87 (ист., распр.);
Хранения В.,1925, Нарис фавни Поддшля, I, Вшниця : 26—27 (распр.);
Тассовский Г. Н., 1927, Произв. силы Дальн. Вост., IV : 471—570 (распр.);
Смирнов, 1928, Охотник и пушник Сибири, 6 : 15—16 (бол.); Шил лин-
г е р Ф. Ф., 1928, Кабан, М. : 1—38 (общ.); Раевский В. и Л. Капланов,
1929, Zool. Anzeig, Bd. 84, № 5/6 : 140—151 (распр.); Федюшин А. В., 1929,
Динамика и географии, распростран. охотн. фауны БССР, Минск : 28—32 (распр.);
Адлерберг Г. П., 1930, Докл. Акад, наук СССР : 91—96 (сист.); Селевин В. А.,
1930, Охотник, 4:14—15 (распр.); Flower S., 1931, Proc. Zool. Soc. London:
223—224 (продолж. жизни); Каверзнев В., 1932, Охота на кабанов, М. : 1—56;
Марков Е. Л., 1932, Дикие свиньи Закавказья, Тифлис : 1—40 (общ., изобр.);
Флеров К. К., 1932, Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. турки.,
2 : 253—255 (распр., биол., изобр.); Адлерберг F. П., 1933, сборн. «Проблемы
происх. дом. животн.», I : 185—207 (морф., сист., происх. дом. свиней); Филип-
пенко Ю. А., 1933, сборн. «Проблемы происх; дом. животн.», I : 157-—179 (морф.,
сист., происх. дом. свиней); Залесский П. М., 1934, Охотн. и рыбак Сибири,
7/8 : 26 (распр.); Афанасьев А. В., 1935, Млекоп. сев.-зап. части Малого Хин-
гана, Л. : 1^-4 (распр.); Флеров К. К., 1935, В кн. : Виноградов Б. С.,Е. Н. Пав-
ловский и К. К. Флеров, Звери Таджикистана, М.—Л. : 94—98 (общ., изобр.); Зо-
лотарев Н. Т., 1936, Млекопит. бассейна р. Имана, Л., изд. Акад, наук : 101—102
(распр., биол.); Логинов В. В., 1936, Бюлл. Моск. общ. испыт. прир., Отд. биол.,
XLV, 1 : 10—20 (биол.); Подаревский В. Б., 1936, Пробл. охотхозяйств,
акклиматиз. в вост. Сибири, Иркутск : 46—47 (распр.); Строганов С. У., 1936а,
Зоол. журн., XV, 3 : 541 (распр.); Строганов С. У., 19366, Сборн. Науч.-иссл.
инет. зоол. Моск. гос. унив., 3 : 108 (распр.); Шнитников В. Н., 1936, Мле-
копит. Семиречья, М.—Л. : 144—148 (распр., биол.); Новиков Г. А., 1937,
Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. вост.-сиб., 4 : 233 (распр.);
Птушенко Е.С., 1937, сборн. «Памяти акад. Мензбира», М., изд. Акад наук СССР:
409—462 (распр.); Amon R., 1938, Z itschr. f. Ziichtung., Reihe В. : 49—88 (ист.,
сист.); Анучин А. В., 1938, Тр. Зоол. сектора Груз. фил. Акад, наук СССР, II :
5—92 (сист.); Д о н а у р о в С. С. и В. П. Т е п л о в, 1938, Тр. Кавк, заповеди.,
I : 191—224 (общ.); Мигулин О. О., 1938, 3Bipi УРСР, Кшв, вид. Акад, наук
УРСР : 1—426 (распр.); Ляпунов И., 1939, Боец-охотник, 11 : 53—55 (акклим..);
Насимович А. А.., 1939, Вопр. экол. и биоценол., 7 : 72—80 (распр., биол.);
Кулагин Н. М., 1940, Фауна БССР, т. I, вып. I, Минск : 53—60 (общ.); Ант и-
пин В. М., 1941, Млекопитающие Казахстана, III, Алма-Ата, Каз. ОГИЗ : 8—16
(общ.); Бажанов В. С., 1945, Изв. Акад, наук Казахск. ССР, 5 : 75—76 (распр.,
биол.); Формозов А. Н., 1946, Снежный покров в жизни млекопит. и птиц,
М., изд. Моск. общ. испытат. прир. : 118—124 (распр., биол.); Верещагины. К.,
1947, Охотн. и промысл, животные Кавказа, Баку, изд. Азербайджанок, фил. Акад,
наук СССР : 43—45 (распр., хоз. знач.); Данилов Д. Н., 1947, сборн. «Очерки
природы Подмосковья и Московской обл.», М., изд. Моск. общ. испытат. прир. : 85—
95 (распр.); Громов В. И., 1948, Тр. Инет. геол, наук, 64 : 440—441 (ист.); Куз-
нецов Б. А., 1948, Млекопит. Казахстана, М., изд. Моск. общ. испытат. прир. :
142—143 (распр.); Скворцов Б. П., 1948, Тр. Воен.-мед. акад., XLIV : 173—
175 (распр.); Данилович А. П., 1949, Охрана природы, 8:79—80 (распр.);
Насимович А. А., 1949, сборн. «Пушные богатства СССР», 1 : 148 (распр.);
Страутман Ф. И. и К. А. Татаринов, 1949, Наук. зап. Львовск. державн.
универе., XVI, 5 : 26 (распр., хоз. знач.); Штегман Б. К., 1949, Охрана при-
роды, 8 : 64—70 (распр., биол.); Даршкевич Я., 1950, Птицы и звери Чкалов-
ской обл., Чкалов, Обл. изд. : 94—95 (распр.); Калниньш А. И., 1950, Охота
и охотничье хозяйство Латвийской ССР, Рига, Латгосиздат : 94—104 (распр., биол.);
Копылов И. П., 1950, Дикие копытные животные Иркутской обл., Иркутск,
Обл. изд. : 54—57 (распр., биол., изобр.); Осадчий П. и Б. Баврин, 1950,
Беловежская пуща, Минск, Госизд. БССР : 45—47 (распр., биол.); Сысоев В.,
1950, Охота в Хабаровском крае, Хабаровск, Дальгиз : 91—97 (распр.); Граве Г. Л.,
М. А. Емельянов и др., 1951, Животный мир Смоленской обл., Смоленск, Обл.
изд. : 155—156 (распр.); Дементьев Г. П., 1951, Изв. Турки, фил. Акад, наук
СССР, 1:58—63 (распр.); Исаков Ю. А., 1951, сборн. «Заповедники СССР»,
I : 317—340 (распр., биол.); О 1 о f f Н., 1951, Beitr. Tierk. u. Tierzucht, 2 : 1—92
(биол., изобр.); Пидопличко И. Г., 1951, О ледниковом периоде, II, Киев,
изд. Акад, наук УССР : 86—87 (ист.); Федосов А. В. и К. Н. Никитин, 1951,
Животный мир Брянской обл., Брянск, Обл. изд. : 10—11 (распр., биол.); Eller-
m a n J. R. and Т. С. М о г г i s о n-S с о 11, 1951, Checklist of palaearctic and in-
74
SUIDAE
dian mammals, London : 344—348 (сист.); Тассовский Г. H., 1952, Уч. зап.
Кишиневск. гос. увив., IV : 39 (распр.); Дубинин В. Б., 1952, Тр. Ленингр.
общ. естествоиспыт., LXXI, 4 : 73—81 (параз.); Залетаев В., 1952, Природа,
10 : 115 (распр.); К ириков С. В., 1952, Изв. Акад, наук СССР, сер. географ.,
6 : 37 (ист., распр.); Колесников И. И., 1952, Тр. Среднеазиатск. гос увив.,
XXXII : 20 (распр.); Корнеев О. П., 1952, Визначник зв!р1в УРСР, Кшв, вид.
«Радяньска школа» : 201—203 (распр.); Кузнецов Б. А., 1952, Изв. Молдавск.
фил. Акад, наук СССР, 4/5 : 141—142 (распр.); Линг X. И. и К. П а а в е р, 1952,
Список млекопитающих Эстонской ССР, Тарту, изд. общ. естествоиспыт. : 13 (распр.);
«Р у х л я д е в Д. П., 1952, Тр. Гельминтопогич. набор. Акад, наук СССР, VI : 331—
333 (бол.); Сокур I. Т., 1952, 3Bipi Радянських Карпат, Ки1в, вид. Акад, наук
УРСР : 54—56 (распр., биол.); Строганова А- С., 1952, Тр. Зоол. инет. Акад,
наук СССР, XI : 231 (распр.); Янушевич А. И., 1952, Фауна позвоночных Ту-
винской обл., Новосибирск, изд. Зап.-Сиб. фил. Акад, наук СССР : 61 (распр.);
Афанасьев А. В. и др., 1953, Звери Казахстана, Алма-Ата, изд. Акад, наук
Казахск. ССР : 455—459 (общ., изобр.); Дубинин В. Б., 1953, сбора. «Преобра-
зов. фауны позвон. нашей страны», М., изд. Акад, наук СССР : 27—36 (охр.); К и-
р и к о в С. В., 1953, Изв. Акад, наук СССР, сер. географ., 4 : 21—22 (ист., распр.);
Мадаминов С. Ю., 1953, Уч. зап. Ленинабадск. пед. инет., II : 121—122 (распр.,
хоз. знач.); Самородов А., 1953, Изв. Акад, наук Туркменск. ССР, 6 : 47 (распр.,
хоз. знач.); Абрамов К. Г., 1954, Копытные звери Дальнего Востока, Хабаровск,
Книжн. изд. : 11—27 (общ., изобр.); Банников А. Г., 1954, Млекопит. Мон-
гольск. Народи. Республ., М,—Л., изд. Акад, наук СССР : 167—177 (общ., синон..,
изобр.); Gray А., 1954, Mammalian hybrids, Edinburg:94 (гибр.); Кузнецов Б. А.,
1954, Бюлл. Моск. общ. испыт. природы, Отд. биол., LIX, 4 : 15 (распр., биол.);
Momberg Н., 1954, Forst u. Jagd, 4 : 158 (бол.); R о b 1 Н., 1954, Forst u. Jaga,
5 : 191—192 (терминол.); Фоканов В. А., 1954, Тр. Зоол. инет. Акад, наук СССР,
XVI : 134 (распр.); Ц алкин В. И., 1954, Сов. археол., XX : 253—287 (ист.);
Баранчеев Л. М., 1955, Зап. Амурск, обл. музея, 3 : 215—218 (распр:, биол.);
Насимович А. А., 1955, Роль режима снежн. покрова в жизни копытных на
территории СССР, М., изд. Акад, наук СССР : 288—297 (распр., биол.); Саблина Т. В.,
1955, Копытные Беловежской пущи, М., изд. Акад, наук СССР : 115—147 (биол.);
Сержанин И. Н., 1955, Млекопит. Белорусск. ССР, Минск, изд. Акад, наук
БССР : 243—253 (общ., изобр.); Сысоев В., 1955, Охотничьи богатства нашей
области, Владимир, Книжн. изд. :1—95 (распр.); Szaflarski J., 1955, Med.
Weteryn., Bd. 11, № 1 : 20 (бол.); Слудский А. А., 1956, Кабан, Алма-Ата, изд.
Акад, наук Казахск. ССР : 1—220 (общ., изобр.).
Описание. Животные крупной или средней величины. Высота в холке
взрослых самцов кавказских кабанов в среднем 103 см, с колебаниями
от 93 до 120 см, у самок — в среднем 75 см (61—96 см). Длина тела у сам-
цов — от 150 до 205 см, у самок — от 129 до 169 см (в среднем 144 см).
Общая величина является показателем расовых различий. Кабаны запад-
ной Европы и западных районов СССР мельче, чем кабаны Кавказа и Сред-
ней Азии. Для самцов из Германии приводятся цифры длины туловища
168 см и высоты в холке 89 см. Самыми крупными являются кабаны Даль-
него Востока, но в Забайкалье и Монголии обитает более мелкая раса.
Живой вес взрослых самцов из окрестностей Кавказского заповедника
колеблется от 64 до 178 кг, самок — от 48 до 109 кг (в среднем 68 кг, —
Донауров и Теплов, 1938). Как видно, самцы значительно крупнее самок.
Средняя величина животных в той или иной популяции в сильной степени
зависит от условий существования и от степени преследования человеком.
Еще в начале настоящего столетия, когда на них охотились меньше, на
Кавказе встречались животные весом до 250—300 кг (Марков, 1932)
и с длиной туловища без хвоста около 2 м (Динник, 1910). С увеличением
промысла предельного возраста достигает незначительная часть животных.
В районе г. Орджоникидзе, где на них интенсивно охотятся, средний
и максимальный вес кабанов меньше, чем в районах смежных с Кавказским
заповедником, где они подвергаются преследованию в значительно мень-
шей мере (Донауров и Теплов, 1938).
SUS
75
Особенностями сложения дикого кабана по сравнению с домашней
свиньей является большая голова с длинной вытянутой мордой и мощно
развитыми у взрослых самцов клыками, а также относительно короткое
и сжатое с боков уплощенное туловище на высоких крепких ногах. Ха-
рактерно для дикой S. scroja то, что высота в холке заметно превосходит
высоту в крестце (высокопередость). Вообще передняя часть туловища
производит впечатление более мощно развитой по сравнению с задней.
Рис. 15. Кабан (£us scro/a L.).
Длина головы у крупных экземпляров может доходить до 60 см. Обхват
груди у взрослых в среднем около 145 см. Хвост длиною около 24—25 см
(максимально 32 см), но, в противоположность домашней свинье, не за-
кручен в виде спирали, а прямой; при беге он поднимается вертикально.
На морде нет бородавчатых выростов кожи, как у S. verrucosus.
«Пятачок» на конце морды имеет форму поперечного овала с выпук-
лыми и наружными и верхним краями. Высота его составляет около
3/4 наибольшей ширины. Верхняя половина поверхности пятачка голая,
влажная; нижняя усажена очень редкими короткими волосами. Отвер-
стия ноздрей круглые; расстояние между ними составляет около т/4 ши-
рины пятачка. Края пятачка выступают несколько за уровень соседних
участков покрытой волосом кожи морды. Уши стоячие с заостренными
концами.
Одной из достопримечательных особенностей взрослых самцов кабанов
является так называемый «калкан». Последний представляет собою утол-
76
SUIDAE
щение соединительнотканного слоя кожи (cutis) по бокам задней части
груди и шеи. Наибольшей толщины, до 4 см, он достигает в области плеч
и лопаток, постепенно утончаясь в направлении к спине, голове и животу.
Калкан настолько плотен, что с трудом режется острым ножом даже
в свежем виде. На разрезе он имеет вид и консистенцию мозоли или
волокнистого хряща (Динник, 1910). Утверждение, что калкан представляет
слой смолы на поверхности кожи в результате трения кабана о деревья
(Шиллингёр, 1928) основано на недоразумении. У самок калкан не
развивается. У самцов от становится особенно толстым в период течки.
Тело, как и у других видов свиней, покрыто щетиной, между которой
в холодное время года расположен густой, довольно грубый, но все же
характера пуха извитой подшерсток (у южных рас может совсем от-
сутствовать). На нижней стороне шеи и задней части живота волос на-
правлен вперед (к голове), на остальных частях тела — назад. Длина
остевых волос на туловище около 6—7 см. На затылке, дорзальной части
шеи и холке щетина удлинена до 12—13 см, но не образует выдающейся
гривы или гребня. Концы волос, образующих щетину, обычно расщеплены
на 3—6 более тонких, обычно загнутых в стороны щетинок. Волосы щетины
тоньше у самок по сравнению с самцами, а также, по-видимому, тоньше-
у западных кабанов по сравнению с восточными. На голове, ушах, конеч-
ностях ниже скакательного и запястного сустава волос короче и, кроме
того, концы щетин не расщеплены. На конце хвоста грубые волосы обра-
зуют кисть длиною до 20 см.
Общая окраска кабана в зимнее время бурая с различными оттенками
от почти черного до серого или желтого цвета. Дикие свиньи западной
части ареала имеют более темную окраску. Светлее окрашены кабаны
Кавказа и Средней Азии.1 Помимо географической изменчивости, окраска
подвержена также изменчивости индивидуальной; последнее.' особенно
ярко проявляется у кабанов Дальнего Востока. Волосы щетины черно-
бурого или черного цвета, но расщепленные концы их имеют более свет-
лую окраску, от желтоватой до серой, почти белой. Подшерсток имеет
светло-бурый или темно-каштановый цвет, на нижних частях туловища
светлее. Летом он короток, иногда может совсем отсутствовать. Отличия
в оттенках окраски кабанов разных районов и на отдельных частях ту-
ловища у одного животного зависят от размеров посветлевших концов
щетины, степени их посветления, окраски и густоты подшерстка. Более
короткий и почти на всем протяжении светлый волос обусловливает
беловатую окраску конца морды и светлые полосы по ее бокам, на щеках
и горле^ особенно ясно выраженные у кабанов Дальнего Востока. Ясно
отграниченных от соседних участков белых пятен и полос при этом, не
образуется. Окраска лба иногда светлее, чем туловища, иногда, наоборот,,
темнее (у кабанов восточной Сибири и Дальнего Востока). Характерна
зональность окраски отдельных волос на лбу; светлый участок занимает
не конец волоса, а среднюю часть, тогда к’ак основание его и вершина
имеют черную окраску. Зональность окраски нам пришлось наблюдать
•также и на туловище у отдельных экземпляров кабанов Монголии и За-
байкалья. Общая окраска в этих случаях приобретает черноватый оттенок.
Встречаются отдельные экземпляры с почти сплошной серо-белой или
соломенно-желтой окраской туловища. Короткий, сплошь черно-бурый
волос обусловливает темную окраску окружности пятачка, губ, подбо-
родка. Сплошную темную окраску, иногда с огненно-рыжеватым оттен-
1 Подробности см. при перечислении подвидов.
SUS
77
ком, имеют также конечности ниже запястного и скакательного сустава
и кисть волос на конце хвоста. Основная часть хвоста имеет одинаковую
с туловищем окраску. Копыта, губы и лишенный волоса пятачок темно-
бурого или свинцово-серого цвета. Летняя окраска, благодаря отсутствию
или слабому развитию подшерстка и более короткой щетине, характери-
зуется более светлым бурым цветом, иногда с сероватым оттенком. Моло-
дые поросята первые два-три месяца после рождения имеют светлые
продольные полосы по рыжевато-бурому фону.
Череп S. scrofa (рис. 16) имеет умеренно развитую в длину по срав-
нению с другими видами лицевую и мозговую часть. Длина черепа
у мелких рас — от 345 до 375 мм, у крупных превосходит 400 мм, а у
самцов может достигать 490 мм. Некоторые особенности черепа (характер
лобно-лицевого профиля, форма и пропорции слезных костей, относи-
тельная длина лицевой части) являются отличиями подвидов. Из резцов
сильнее развиты две первые (средние) пары; третья пара недоразвита.1
В верхней челюсти резцы широкие, искривленные и отставлены друг от
друга, особенно последняя (третья) пары; первая и вторая пары направ-
лены вниз и навстречу одноименным зубам другой стороны. Узкие
долотовидные резцы нижней челюсти направлены почти прямо вперед,
расположены вплотную друг к другу; лишь альвеолы последней (третьей)
пары иногда отделены от соседних, так же как и от клыков, промежутком
в 2—3 мм. Между резцами и клыками в верхней челюсти имеется более
значительный беззубый промежуток длиною в 2—3.5 см. Длина нижних
клыков у взрослых самцов 6—10 см. Их задняя грань, в противополож-
ность некоторым другим видам свиней, шире наружной и стирается о пе-
реднюю поверхность загнутых в стороны и вверх верхних клыков.
Поверхность стирания и на нижних и на верхних клыках захватывает
и вершину зуба. Это обусловливает, с одной стороны, постоянную
остроту, отточенность, а с другой, ограничивает их рост, особенно верхних,
в длину. В редких случаях, когда стирание не захватывает вершину
верхних клыков, последние продолжают расти и, загибаясь кольцом вверх
и внутрь, могут насквозь прободать носовые кости. Эти случаи чрезмер-
ного отрастания клыков .следует, впрочем, отнести к аномалиям, а не
к норме. Из коренных зубов лучше всего развиты последние заднекоренные
(М3 и М3). Бугорки на задней части .этих зубов (гипоконе) обычно
образуют дополнительный ряд; особенно хорошо гипокон развит у ка-
банов западной части ареала. Расположенные впереди от последнего
заднекоренного зуба постепенно убывают в размерах. Самыми незначи-
тельными являются первые переднекоренные (Р1 и Pj или так назы-
ваемые «волчьи зубы»). Рано выпадая, у взрослых животных они могут
изредка вообще отсутствовать. Особенностью передних премоляров
является также то, что, в противоположность остальным переднекоренным,
они не имеют молочных предшественников и не сменяются в течение
всей жизни животного. В верхней челюсти Р1 почти всегда сидят вплот-
ную к следующим за ним премолярам и, обычно, на некотором расстоянии
от клыков; в нижней же челюсти они отставлены от остальных коренных
и расположены ближе к клыкам.
История и географическое распространение (рис. 17). Предком совре-
менного дикого кабана Палеарктики является, вероятно, S.. prisons
Serr. из верхнего плиоцена. Наиболее ранние, относящиеся к S., scrofa
остатки известны из раннечетвертичных слоев Сирии и Британских остро,-
В верхней челюсти изредка вообще, отсутствует дли выпадает. ;
18
SUIDAE
bob, а в плейстоцене кабан населял умеренные и теплые области южной,,
западной и восточной Европы и по крайней мере Средней Азии (Amon,.
1938; Пидопличко, 1951). В настоящее время область распространения
этого вида простирается от Атлантического до Тихого океана и охваты-
вает северную Африку, среднюю, южную и восточную Европу, а также
Малую, Среднюю, Центральную и восточную Азию к северу от Гима-
лаев, до южной Сибири, Забайкалья, Дальнего Востока и некоторых
островов Японии включительно. В прежнее время ареал был еще шире
и, кроме Британских островов, включал также южную часть Скандинав-
ского полуострова, где в настоящее время кабан в диком состоянии от-
сутствует. Некогда сплошной ареал S. scrofa оказался сравнительно
недавно (вероятно в середине XVIII или в начале XIX в.) разорванным
в европейской части Советского Союза.
На территории СССР ископаемые остатки дикого кабана впервые
обнаруживаются в хозарских толщах среднего плейстоцена (миндель—
рисе). Известны они в немногих палеолитических стоянках, из которых
наиболее обычны, в крымских. Редкость находок кабана в палеолите
В. И. Громов (1948) объясняет предпочтением, которое отдавал человек
этого периода открытым безлесным пространством при выборе мест для
поселения. Число находок увеличивается к концу палеолита, а в неолитиче-
ских стоянках (голоцен) остатки кабана встречаются очень часто.
На территории СССР область распространения кабана значительно-
сократилась уже в историческое время. Во времена Новгородского кня-
жества, например, кабанов было много у самого Новгорода,1 в XIII в.
даже в 60 верстах к северу от последнего. В Костромском наместничестве
дикие кабаны водились в конце XVIII в. (Кириков, 1953). А. Н. Формо-
зов (1946) северную границу распространения S. scrofa связывает с ли-
нией среднего максимума глубины снежного покрова 30—40 см. Помимо-
глубины снежного покрова, несомненно важную лимитирующую роль,
играет степень промерзания грунта (т. е. зимние минусовые температуры),,
затрудняющая рытье почвы в поисках пищи.
Не совсем ясны границы современного распространения кабана в при-
балтийских республиках. В Литовской ССР он, по-видимому, встречается
во всех подходящих биотопах. По сведениям, полученным от Управления
по делам охоты Литовской ССР, из 87 районов республики кабаны обитают
в 70-ти. В феврале 1954 г. было учтено 1950 штук.
В Латвии и Эстонии кабан, некогда многочисленный, резко уменьшил
численность с конца XVIII в., а временами исчезал совершенно.
В начале 90-х годов кабанов добывали, впрочем, даже под Тарту (Greve,
1909). В Латвийской ССР численность их стала возрастать с 1910 г., но-
резко колеблется по годам, в зависимости от климатических и кормовых
условий (Калниньш, 1950). На 1 марта 1954 г. в республике было учтено-
кабанов 1083 штуки, причем в наибольшем числе они зарегистрированы
в Лиепайском, Слатском, Екабпилсском, Тукумском, Вентспилсском.
и Добельском лесхозах (данные Республиканского управления охотничь-
его хозяйства). Значительно увеличились в числе дикие свиньи за послед-
ние годы также в Эстонской ССР. Здесь .они местами являются обычными
обитателями, особенно в центральных и восточных районах (Линг и
Паавер, 1952).
За последние годы отмечаются неоднократные встречи не только
отдельных животных, но и целых стад диких свиней (до 15—20 голов).
1 Подробности прежнего ареала S. scrofa с картой см. у А. Н. Формозова (1946)..
Рис. 16. Череп Sus scrofa L.
80
SUIDAE
в южных и западных районах Ленинградской области (Кингиссепском,
Волосовском и Гатчинском). Идет ли в данном случае речь о случайных
заходах, или о закономерном расширении ареала — сказать пока трудно.
Известный боровичский краевед С. Н. Поршняков сообщал автору
сведения, относящиеся к 1953—1954 гг., о двух заходах кабанов, вероятно
случайных, в пределы Новгородской области: одного в Валдайский район
(окрестности с. Шуя на тракте Боровичи—Валдай), а другого в Мошен-
ской, в окрестности дер. Середки.
В Белорусской ССР кабан распространен по всей республике до север-
ных и восточных областей. Наиболее плотна населенность им южных
и юго-западных районов. Издавна славится обилием этого зверя Беловеж-
ская пуща. В настоящее время в наибольшем числе кабаны встречаются
в областях Гомельской и Брестской. Обычными членами фауны они
являются в большинстве районов Гродненской, Минской, Молодеченской,
Витебской и Могилевской областей. При этом почти повсеместно отме-
чается увеличение их численности за последние годы (Сержанин, 1955).
Из прилегающих районов Латвии и Белоруссии кабаны, вероятно,
проникают, по крайней мере в западные и южные районы Псковской
и Великолукской областей, но точными данными по этому вопросу мы не
располагаем. Имеется лишь указание С. У. Строганова (1936а) о том,
что в районе оз. Селигер (Пеновский район Великолукской области)
в начале 30-х годов был убит случайно забежавший кабан.
Известны отдельные забеги кабанов в Калининскую область (Раев-
ский и Капланов, 1929). В 1935 г. партия кабанов, взятых из-под Наль-
чика, была завезена в Завидовское охотхозяйство Калининской области.
Здесь звери успешно акклиматизировались и к настоящему времени
число их значительно возросло (Ляпунов, 1939; Насимович, 1955).
В Подмосковье кабаны были обычны еще в XVIII в. До середины
прошлого века наблюдались отдельные их забеги, а к концу они исчезли
совершенно. Вновь кабаны появились в пределах Московской области
незадолго перед Великой Отечественной войной в связи с акклиматиза-
цией их в Завидовском охотхозяйстве, а в послевоенные годы, по-види-
мому, в результате естественного расширения ареала. К концу 40-х годов
кабаны встречались в Высоковском, Лотошинском, Клинском, Волоко-
ламском и Шаховском районах Московской области (Данилов, 1947).
Н. Сысоев. (1955) приводит кабана в списке охотничьих животных уже
Владимирской, области.
Почти исчезнувшие к началу 90-х годов в Смоленской области кабаны
вновь увеличили численность за годы Великой Отечественной войны.
В настоящее время они встречаются группами в 3—6 штук в Демидовском,
Понизовском, Велйжском, Слободском, Пречистенском, Холмжирков-
ском, Глинковском, Кордымовском и некоторых других районах области.
Совершенно отсутствуют, возможно, только в восточной ее части. Увели-
чение численности диких свиней в послевоенные годы отмечается также
для лесных районов Брянской области. Здесь они, по последним данным,
отсутствуют только в Бытожском, Дятьковском, Севском, Климовском,
Клинцовском и Стародубском лесхозах. Средняя плотность опреде-
ляется в одну голову на 2000 га леса; в некоторых районах она доходит
даже до 4.2 на 1000 га лесопокрытой площади (Федосов и Никитин, 1951).
Что касается территории Украинской и Молдавской ССР, то еще
в 30-х годах прошлого века кабан был частым зверем во всех лесах Волыни
и Подолии (Eichwald, 1830). Кроме того, он не только водился в плавнях
больших рек, но заходил даже в степь по долинам мелких речек. В средине
Рис. 17. Распространение Sus scrofa L.
XX в. (области постоянного обитания); 2 — XIX в. (места находок и предполагаемая северная граница
распространения); 3 — XVIII в. и раньше; 4 — отдельные заходы в настоящее время; 5 — места
акклиматизации.
6 Фауна СССР, И. И. Соколов
82
SUIDAE
прошлого столетия он был обычным зверем в северных частях Киевской
и Черниговской губерний. Даже в окрестностях Киева, по свидетельству
Кесслера, неоднократно убивали кабанов весом до 200—225 кг. Г. Белке-
(Belke, 1866) в списке млекопитающих округа Радомышля приводил
и S. scrofa. В низовьях Днепра кабан был окончательно истреблен громад-
ным наводнением 1845 г. (Браунер, 1923). Перед Великой Отечественной
войной, до воссоединения Западной Украины, дикие свиньи были редкими
животными в Винницкой, Каменец-Подольской, Киевской и Чернигов-
ской областях (Мигулин, 1938). Как и в других областях европейской
части СССР, в послевоенные годы на Украине наблюдается увеличение-
их численности и появление в районах, где они давно не водились. Наи-
большим распространением кабан пользуется в горных и низинных райо-
нах Западной Украины, в областях: Закарпатской, Станиславской,.
Черновицкой, Дрогобычской, Львовской и Тернопольской (Сокур, 1952).
В Одесской области он держится в плавнях Днестра, по которым подни-
мается вверх вплоть до Винницкой и Черниговской областей. Большое-
количество диких свиней в 1949 г. наблюдалось в районах Украинского-
Полесья: Лугинском, Словечанском и Олевском Житомирской области
(Данилович, 1949). *
Отмечается кабан теперь также в Кировоградской, Киевской и даже-
Полтавской областях, где свиней не было уже в средине прошлого века
(Корнеев, 1952). В восточных областях Украинской ССР в исторические
времена кабан, по-видимому, не водился. С незапамятных времен исчез-
он и в Крыму, хотя остатки его составляют на этой территории один из;
обычных компонентов фауны верхнепалеолитических стоянок. Несом-
ненные остатки кабана найдены при раскопках Неаполя Скифского в слоях,,
датируемых II в. до н. э. (Цалкин, 1954).1
На территории Молдавской ССР, кроме плавней Днестра и Прута,
дикие свиньи, по собранным мною в 1949 и 1950 гг. сведениям, нередки
в широколиственных лесах Кодр. В лесах северной части республики
бывают только заходом (Кузнецов, 1952).
К северу от УССР, в западной части Курской области кабаны были
многочисленны и служили предметом охоты около полутора столетий
тому назад; теперь они там отсутствуют, если только не появились вновь,
в послевоенные годы из соседних областей Украины (Птушенко, 1937).
В последние годы кабаны появились на территории Воронежского госу-
дарственного заповедника (Богданов и Куражсковский, 1955, Природа,.
9 : 120).1
Таким образом, имеет место сокращение былого ареала кабана в запад-
ной половине европейской части СССР и еще больше уменьшение его чис-
ленности, произошедшее за последние 100—150 лет. С другой стороны,,
несомненно увеличение его численности за последнее десятилетие и стрем-
ление населить территории, с которых он был вытеснен раньше. Является
ли последнее явление следствием применяемых мер охраны, или в основе-
его лежат причины иного порядка, сказать пока трудно.
К западу от только что перечисленных районов происходит разрыв,
ареала S. scrofa. Внойь на территории европейской части Союза кабан
появляется только на Северном Кавказе. Разъединение некогда сплош-
ного ареала кабана в восточной Европе произошло, как мы уже отмечали,
вероятно, в течение последних двух-трех столетий. В верхней частй
1 В 1957 году выпущена партия дальневосточных кабанов в б. Крымский запо-
ведник .
SUS
83
Дона он, впрочем, в исторические времена, вероятно, вообще не водился,
тогда как в низовьях исчез лишь в начале прошлого века.1
На Кавказе и область распространения, и численность кабана даже за
последние десятилетия сильно сократились. В 70-х годах он еще изредка
встречался в лесах под Ставрополем, но уже в начале XX в. из этих райо-
нов исчез совершенно (Динник, 1910). Северная граница распространения
в настоящее время проходит через плавни Кубани, верхнее течение р. Бе-
лой, Лабы, Урупа, Кубани, Терека (северные склоны Главного Кавказ-
ского хребта), долину реки Кумы до побережья Каспийского моря и по
последнему до низовьев Волги. На юге Кавказского перешейка кабан
распространен вплоть до государственной границы и дальше •— в Турции
и Иране. Плотность заселения кабаном отдельных частей территории
Кавказа неравномерна. Выше всего она до сих пор на Куба-Хачмасской
равнине, в бассейне р. Самура, на Закатало-Исмаиллинской, Куринской
и Ленкоранской низменностях. Истреблен почти полностью кабан на Ка-
рабахском нагорье и северных склонах Малого Кавказа вплоть до Грузии.
Сильно уменьшилась численность его в западной части Закавказья —
в Колхидской низменности, Аджарии, Имеретии, Абхазской АССР.
По прибрежным зарослям камышей область распространения кабана на
Северном Кавказе непосредственно смыкается с частью его ареала в низо-
вьях Волги. По долине последней дикие свиньи некогда заходили выше
Сталинграда, почти до Камышина, но уже в 60-х годах редко забегали
выше Астрахани и ограничивались в основном дельтой. В дореволюцион-
ное время и здесь кабаны были сильно истреблены. Только с организацией
в 1919 г. Астраханского государственного заповедника и с установлением
охраны численность их быстро возросла настолько, что в районах, приле-
гающих к заповеднику, стало возможным разрешение на него охоты (Иса-
ков, 1951). К востоку от Волги кабаны держатся по рекам Малому и Боль-
шому Узеням и степным озерам Западно-Казахстанской области. По-види-
мому, с этими районами связаны недавние их забеги на Волгу в районы
г. Вольска Саратовской области (Залетаев, 1952). Изредка заходят в район
долины Еруслана и Волги на севере Сталинградской области (Строганова,
1952). В средине прошлого столетия кабаны изредка водились по рекам
Ику и Сакмаре, и даже будто бы доходили до широкой излучины р. Белой,
т. е. до 53° с. ш. (Кириков, 1952). Но в 90-х годах в пределах Оренбург-
ского края и Башкирии их уже не было. Есть, впрочем, указание, что
в последнее время дикие свиньи вновь начали появляться в южных (Бу-
ранном) и восточных (Адамовском) районах Чкаловской области, заходя,
вероятно, из прилегающих районов Казахстана (Даршкевич, 1950).
В пределах Средней Азии кабан населяет главным образом долины
крупных рек и ббльшую часть горных районов. В Туркменской ССР он
занимает долины рек Атрека, Теджена, Мургаба, Аму-Дарьи, а также
горную область Копет-Дага. Особенно много кабанов в предгорьях
Серахского района, в районах Адам-Элена, Акар-Чешме, Зульфагара,
а также в долине р. Кашана (Флеров, 1932; Скворцов, 1948). По послед-
ним данным (Самородов, 1953), многочисленны они также в гребенчуко-
вых зарослях долины р. Атрека и в камышах озер Делили. В низовьях
Атрека область распространения кабана смыкается через южное побережье
Каспийского моря с ареалом его в восточной части Закавказья. Много-
1 Последние кабаны в гирлах Дона погибли от наводнения 1812 г. (Алфераки,
6*
S4
SUIDAE
численны кабаны до последних лет в западной части Узбоя, в местах,
где сохранились заросли камыша или кустарников. Как далеко отсюда
идет кабан на запад по долине Узбоя и на север по побережью Каспий-
ского моря — не ясно. Нет его на Больших Балханах.
Распространен кабан по всему Узбекистану, за исключением Кызыл-
кумов, где появляется он только в зимний период. В Таджикской ССР
населяет долины всех более или менее значительных рек, за исключением
полупустынных районов, а также горные области, кроме Памира (Флеров,
1935).
К областям почти сплошного распространения кабана необходимо
отнести Киргизскую ССР, но и здесь плотность заселения им не одинакова.
Сильнее он истреблен в северной половине республики, лучше сохранился
в южной. Многочисленны кабаны в долине р. Чаткала, в окрестностях
оз. Сары-Чилек. Обычным зверем кабан является в долине рр. Чу и Коч-
кора, на южных склонах Таласского Алатау и северных склонах Алай-
ского хребта. На северных склонах Таласского Алатау бывает только
заходом с южной стороны хребта. Отсутствует, видимо, в большей части
Алайской и Таласской долин. Мало его на северных склонах центральной
части Киргизского хребта (Кузнецов, 1948). Полностью истреблен кабан
в ряде районов Иссык-Кульской области (Шнитников, 1936). Споради-
чески встречается в районе Хан-Тенгри.
Сложно и недостаточно известно распространение диких свиней на
территории Казахской ССР. К востоку от Волги по побережью Каспий-
ского моря кабаны распространены до низовьев р. Урала. По долине по-
следнего поднимаются до Хорькина и изредка даже до Уральска (Фока-
нов, 1954). В междуречье Урал—Волга держатся на Камыш-Самарских
озерах, а также по Большому и Малому Узеням. Отмечались встречи
на оз. Челкар.
По побережью Каспия кабаны доходят также до устья Эмбы. По
последней водятся между Доссором и Косчагылом (Афанасьев и др.,
1953). Южнее кабаны, хотя и редки, встречаются на полуострове Бузачи,
Мангистау и у Западного чинка Устюрта. Попутно следует отметить,
что И. И. Колесников (1952) находил лежки кабанов на Южном Устюрте
во впадине Барса-Кельмес. В Казахстане, к востоку от указанных ранее
районов, кабаны водятся в Приаральских Каракумах, в районе оз. Чел-
кар-Тениз, по Иргизу и в низовьях Тургая. По Тургайской долине про-
никают до оз. Аксуат и даже до Семиозерного района Кустанайской об-
ласти. Заходят также на оз. Шубар-Тениз. По побережью Аральского
моря встречаются только между дельтами Аму-Дарьи и Сыр-Дарьи.
В долине последней наиболее многочисленны в районе нижнего течения.
Зимой кабаны довольно обычны в Кызылкумах, откуда проникают иногда
по льду даже на остров Уялы в Аральском море.
Из Сыр-Дарьинской низменности ареал кабана простирается в долину
Сарысу. Здесь кабаны известны в районе оз. Ащиколь и в Моинкумах
(Бажанов, 1945).
В центральном Казахстане кабаны обитают в районе озер Тенгиз
и Кургальджин, откуда иногда заходят к северу до линии железной до-
роги. Далее к востоку в Казахском нагорье кабаны, по-видимому, отсут-
ствуют. В северном и западном Прибалхашье встречаются в небольшом
числе по заливам Балхаша, поросшим тростником (Афанасьев и др., 1953).
По-видимому, изолированный очаг обитания кабанов находится в го-
рушках Джамбул, в юго-восточной части пустыни Бет-Пак-Далы. Здесь
во время экспедиции 1956 г. мы находили их лежки и копанки, хотя
SUS
85
самих зверей видеть и не удалось. Следует предполагать, что они есть
и в соседних горках Байгара, так как, вероятно отсюда, однажды наблю-
дался их заход вглубь пустыни, к ключам у урочища Тюкэн.
В южном Казахстане распространение кабана захватывает долины
рек Чу и Таласа, берега оз. Байликоль в Джамбульской области. Мно-
гочисленны кабаны в южном Прибалхашье, населяя долины рек Или,
Каратала, Аксу, Аягуза и их притоков. Обычным зверем они являются
в районе озер Алаколь, Сасыкколь и Зайсан. Распространены также по
всем горным хребтам юга и юго-востока республики: Таласском Алатау,
Каратау, Киргизском хребте, Чу-Илийских горах, Заилийском, Кунгей
и Джунгарском Алатау, Тарбагатае и Сауре. Много кабанов в Нарын-
кольском районе Алма-Атинской области.
К востоку от Иртыша в Восточно-Казахстанской области, особенно
в западных отрогах южного Алтая, кабаны очень редки. Здесь отмечаются
лишь отдельные заходы их из района оз. Зайсан и Черного Иртыша (Ан-
типин, 1941). К западу от Иртыша кабаны отмечались в Калбинском
хребте. По долине Иртыша и через ленточные боры у Семипалатинска
они проникают в южную часть Павлодарской области, доходя иногда
до самого Павлодара (Селевин, 1930).
В пределах Горно-Алтайской автономной области кабан как постоян-
ный обитатель сейчас отсутствует, но на берегу Телецкого озера был
найден его череп. Кроме того, до сих пор дикие свиньи изредка заходят
в юго-восточную часть области, в Кош-Агачский район, из Монголии.
Одиночные животные появлялись в юго-восточной части Усть-Коксин-
ского района, в верховьях р. Аккема (Залесский, 1934; Насимович, 1949).
Еще в средине и конце прошлого столетия кабаны, местами в значи-
тельном количестве, водились в южной части Курганской и Омской обла-
стей до широты самого Омска, а также по озерам Кулундинской и Бара-
бинской степей (в окрестностях оз. Чаны). Но с конца прошлого столетия
даже случайные заходы их на территорию западной Сибири, кроме Алтая,
ио-видимому, не имеют места. Нет кабана как постоянного обитателя
в пределах Красноярского края. Однако отмечались отдельные его заходы
в район притоков р. Кунгус, по-видимому из прилежащих районов Тувы
или Иркутской области (газ. «Красноярский рабочий» от 16 октября
1948 г.).
В Тувинской автономной области кабан встречается по всему хребту
Танну-Ола. Много кабанов в Восточно-Тувинском нагорье между реками
Хамсараи, Азас и особенно у оз. Успа-Нур (Янушевич, 1952).
В пределах Иркутской области кабан в настоящее время населяет
полосу предгорий Восточных Саян и Хамар-Дабанских гор к югу от линии
железной дороги вплоть до южной оконечности Байкала (Копылов, 1950).
В западной части этой зоны, по рекам Тагулу и Туманшету редок. Обычен
по рекам Уде, Ие, Оке, Большой и Малой Белой, Китою и Иркуту. Повы-
шенная, против небольшой средней, плотность кабана наблюдается
местами на р. Ие, в междуречье Китой—Иркут (р. Тойсук) и в треуголь-
нике Ангара—Иркут—Байкал (рр. Лона, Байсик и др.). Имеются непро-
веренные сведения о наличии кабана в верховьях Лены. В XVIII в.
кабаны водились по Ангаре в районе порога Долгого, около 57° с. ш.
(Насимович, 1955). В Бодайбинском районе, где еще недавно кабаны
держались около оз. Нечатка, они исчезли с 1922 г.
В Бурят-Монгольской АССР кабан обычен в аймаках Окинском, Тун-
кинском и Закаменском (Копылов, 1950), а также в районах юго-восточ-
ного Забайкалья, в частности в районе р. Селенги. Водится по рр. Турка,
86
SUIDAE
Гусиха и изредка даже заходит в долину Баргузина (Новиков, 1937).
С верховьев Баргузина северная граница распространения кабана
проходит к Витиму и правому притоку последнего, Калару, на 56-й па-
раллели, севернее которого в настоящее время кабана нет. В Читинской
области этот зверь обитает «по бассейнам рек Юмурген, Каренга, Нерча,
Джилинда, а также левой стороне Витима. Очень редок в урочище Гуленга
(Канарского района). Обычен в долине р. Урюмкан, протекающей по Га-
зимуро-Заводскому району. В Нерчинско-Заводском и Усть-Карийском
районах встречается реже, появляясь главным образом периодически,
заходя из-за Аргуни» (Подаревский, 1936). В 30-х годах много кабанов
было, по данным того же автора, в Красночикойском, Кыринском и Уле-
товском районах.
В Амурской области ареал кабана занимает широкую полосу вдоль
левого берега Амура; максимальной численности он достигает в Хин-
гано-Архаринском районе (Баранчеев, 1955). Северная граница его рас-
пространения в области проходит немного южнее 55° с. ш., по долинам
рр. Ольдоя и Уркана (Гассовский, 1927). Однако известны отдельные
случаи захода кабанов в верховья р. Тимптона, уже в пределы Якутской
АССР (Насимович, 1955).
Распространение кабана на Дальнем Востоке, по последним данным
К. Г. Абрамова (1954), рисуется в следующем виде: «Кабан распространен
в южной и средней части хребта Сихотэ-Алинь. По побережью он встре-
чается до реки Самарги и изредка севернее ее, до рек Ботчи и Коппи
включительно. Большая плотность кабанов наблюдается по среднему
течению Хора и Анюя, но в самые верховья этих рек животное не заходит.
Более редок кабан по нижнему и среднему течению реки Сунгари, где
встречается не выше стойбища Кун. В левобережной части бассейна
Амура он изредка попадается в районе реки Боктор, в ее нижнем течении,
и иногда заходит на реку Горин. Далее к западу он более обычен в кедро-
вых лесах по Куру и Урми и снова становится многочисленным по хребту
Шухи-Поктою и северным отрогам Малого Хингана, в Еврейской авто-
номной области. В Приморье кабанов более всего по рекам Иману и Би-
кину, в Хабаровском — по Хору и Подхоренку, а затем в истоках реки
Биджана». В числе млекопитающих зоны смешанных лесов верховьев
рек Бурей и Амгуни (Буреинский и Дуссень-Алиньский хребты) А. Афа-
насьев (1935) называет и S. scrofa. Отсутствует в настоящее время кабан
на Сахалине, хотя ископаемые его остатки, по сообщению К. Г. Абрамова
(1954), там встречаются довольно часто.
В пределах Монгольской Народной Республики область постоянного
обитания кабана занимает северные и частично центральные районы
страны. Некоторые детали его распространения здесь, по суммирован-
ным А. Г. Банниковым (1954) данным, рисуются в следующем виде.
Кабаны обычны по лесистым склонам хребта Танну-Ола и Хан-
Хухэя. В некоторые годы появляются в окрестностях оз. Убсу-Нур.
Многочисленны также по берегам оз. Хара-Ус-Нур и в низовьях р. Кобдо.
Вверх по р. Кобдо распространены вплоть до оз. Ачит-Нур.
По опросным данным, встречаются в небольшом числе по р. Хунгуй.
Далее к югу кабаны многочисленны в ряде оазисов у южных склонов
Монгольского Алтая (Дзакой, Дзарм, Ихэ-Толи и Дзулугутай). В не-
большом числе встречаются у оз. Шаргиин-Нур. Н. М. Пржевальский
встретил кабанов на р. Булугун, но водятся ли они теперь здесь, равно
как и на восточной оконечности Монгольского Алтая, А. Г. Банникову
выяснить не удалось.
SUS
87
Восточнее кабаны повсеместно распространены, начиная от северного
Хангая и Прикосоголья к северу до государственной границы с СССР.
Нет их только в широких степных долинах среднего и нижнего течения
рр. Селенги, Орхона, Идера, отчасти Тэса и Дельгер-Мурэна. В цент-
ральном и южном Хангае в подходящих стациях кабаны также много-
численны, но распространены не повсеместно.
Областью сплошного распространения кабана является хребет Хэнтэй
и прилегающие к нему районы северо-восточной Монголии. Наиболее
многочисленны здесь дикие свиньи в верхнем течении Керулена и его при-
токов и особенно в верховьях рр. Хара и Иро. К югу по Хэнтэю доходят
.вплоть до окрестностей Улан-Батора и горы Богдо-Ула.
На востоке МНР кабаны встречаются и весьма обычны по лесистым
склонам Большого Хингана, в бассейнах рр. Нумургиин-Гол и Халхин-
Гол. Далее А. Г. Банников называет ряд пунктов в южной части Мон-
гольской республики, куда они забегают из прилегающих районов Китая:
родники Мин-Булак, пустыня севернее Гашун-Нура и некоторые оазисы
Заалтайской Гоби.
Подвиды. Внутривидовая систематика S. scrofa L. и объем вида до
сих пор окончательно не установлены. Ряд морфологических особенно-
стей, (характер профиля лобно-лицевой части черепа, пропорции слёзной
кости, устройство последнего моляра, детали окраски волосяного покрова)
закономерно изменяется в направлении с востока на запад. При этом,
исходя из данных палеонтологии и онтогенеза, следует считать, что вос-
точные формы кабана стоят ближе к исходному для вида типу, тогда как
в направлении к западу популяции приобретают ряд морфологических
черт, имеющих прогрессивный характер.
Однако вывод Ю. А. Филипченко (1933) о видовой самостоятельности
восточных форм, выделяемых им в вид S. orientalis Fil., лишен оснований.
Западные и восточные кабаны морфологически связаны рядом незаметных
переходов; кроме того, они образуют практически сплошной ареал,
начиная с западной Европы до берегов Тихого океана. Необходимо иметь
в виду также большую однотипность окраски, строения черепа и коренных
зубов палеарктических кабанов с одновременной значительной индиви-
дуальной изменчивостью тех же признаков. Лишено основания и выделе-
ние двух типов (Amon, 1938) или экологических форм кабанов — гор-
ного и низинного (Динник, 1910). К. Флеров (1935) совершенно спра-
ведливо указывает, что в этих случаях идет речь о возрастных и половых
группах, обитающих в разной обстановке.
На территории СССР рыделяется пять подвидов S. scrofa, хотя гра-
ницы ареалов некоторых из них точно не установлены:
1) S. scrofa scrofa L. Средней величины, с вогнутым лобно-лицевым
профилем черепа и круто приподнятой теменной его частью. Слёзная
кость вытянутая; длина ее по нижнему краю превосходит высоту (ши-
рину). Последний верхний коренной зуб имеет хорошо развитый талон
(пятку). Окраска темная, с темными же ушами и более светлым рыжева-
тым оттенком на голове. Распространение: западная Европа;
в северной части ареала восточной границей считается Беловежская
пуща.
2) S. scrofa attila Thomas. Более крупная по размерам форма, с длин-
ной лицевой частью черепа. Профиль черепа самцов и слёзная кость,
как у типичной формы; у самок профиль более прямой. Окраска темная,
но несколько светлее, чем у S. s. scrofa. Распространение:
Венгрия, Румыния, Балканский п-ов, европейская часть СССР к востоку
88
SUIDAE
от Беловежской пущи, Кавказ, Закавказье и северный Иран. Кабаны
Белоруссии и западных областей европейской части СССР к востоку от
Беловежской пущи по величине занимают промежуточное положение
между типичной S. s. scrofa и формой attila с Северного Кавказа и ни-
зовьев Волги.
3) S. scrofa nigripes Blanford. Крупной величины, но лицевая часть-
черепа несколько короче, чем у предыдущей расы. Профиль черепа пря-
мой или слегка выпуклый в лобной области, с высоко приподнятой темен-
ной частью. Слёзная кость разнообразной формы. Окраска типично более
светлая, чем у других подвидов, варьируя от светло-бурой до соломенно-
желтой. Уши и ноги темно-бурые с рыжеватым оттенком. Такого же цвета
кисть волос на хвосте. Распространение: среднеазиатские
республики, Синьцзян, северо-западная Монголия, возможно южный
Иран. И по ареалу, и по ряду морфологических особенностей представляет
промежуточную форму между европейскими и восточноазиатскими каба-
нами.
4) S. scrofa raddeanus Adlerberg. Более мелкая, даже по сравнению-
с типичной, раса. Череп короткий, высокий с почти прямым или слегка
выпуклым в лобной части профилем. Слёзная кость короткая и высокая.
Типично длина ее по нижнему краю не превышает или меньше высоты
у переднего края глазницы. Талон на последнем заднекоренном слабо-
развит. Окраска более светлая, чем у S. s. scrofa, но темнеющая по бокам,
на голове и ушах. Пятно на горле и вымя серебристо-серое. Распро-
странение: южное Забайкалье, северная Монголия, к югу до-
пустынь центральной Монголии и к востоку до Большого Хингана.
5) S. scrofa continentalis Nehring. Одна из самых крупных рас S. scrofa.
Череп, достигающий длины 490 мм, широкий в скуловых дугах, типично-
имеет прямой профиль. Слёзная кость короткая и широкая. Длина ее
по нижнему краю меньше или равна высоте у переднего края глазницы.
Окраска сильно варьирует, иногда не отличаясь от S. s. scrofa. Распро-
странение: бассейн Амура от Большого Хингана до Тихоокеан-
ского побережья, Приморский край.
За пределами .СССР, как особый подвид, S. s. leucomystax Temm.,.
следует считать кабанов с островов Японии. Кроме того, описаны как
подвиды или виды, но мало изучены: S. s. meridionalis Major с Корсики
и Сардинии, S. s. castilianus Thom, из северной и S. s. boeticus Thom,
из южной Испании, S. s. reiseri Bolk. из Югославии, S. s. majori de Beam
et Festa из Италии, S. s. barbarus Sclat. из Марокко и Алжира, S. s. sen-
nariensis Fitz, из Сеннара и Кордофана, S. s. lybicus Gray из Малой Азии
И" Сирии, S. s. taivanus Swinh. с о. Тайваня, S. s. chirodontus Heude из
южного Китая, 5. s. coreanus Heude из Кореи и, наконец, S. s. moupinen-
sis Milne-Edw. из центрального и северного Китая. Внутривидовая систе-
матика S. scrofa настоятельно требует ревизии. Некоторые формы не-
заслуживают выделения в качестве подвидов. В частности, весьма сомни-
тельна целесообразность дифференцировки кабанов Кавказа, низовьев-
Волги и Средней Азии.
Биология. Места обитания кабана разнообразны и зависят в значи-
тельной мере от природных условий того или иного района. Он можег
населять долины и дельты больших и малых рек, приморские низменности,
леса, горы, вплоть до альпийской зоны. В определенные сезоны не избе-
гает даже пустынных ландшафтов. Характерна все же для диких свиней
склонность придерживаться сырых болотистых мест поблизости водоемов,,
там, где можно найти лужи с грязью, в которых они так любят купаться..
SUS
89-
Сезонность местообитаний определяется в сильной степени наличием
и доступностью кормовой базы. Необходимым условием является также
наличие в районе обитания надежных укрытий. В качестве последних
кабану служат густые заросли камыша, колючих и переплетающихся
кустарников, высокого бурьяна, заломы, молодая поросль хвой-
ного леса. Кабан свободно не только проходит, но и мчится в таких местах,
где не только человеку, но и собаке пройти почти невозможно. К .пере-
движению в этих условиях приспособлено все тело животного обтекае-
мой, сжатой с боков формы, на коротких ногах, с конусовидной головой,
глубоко сидящими маленькими глазами.
В западных областях европейской части СССР (в Беловежской пуще,
в лесах Белоруссии, Украинском Полесье, в Смоленской и Брянской
областях) излюбленными местообитаниями кабана являются пониженные
заболоченные участки смешанного и широколиственного леса. В густа
населенных районах держатся в наиболее глухих участках леса, окола
речек и ручьев с зарослями высокого тростника. Осенью и зимой, осо-
бенно в годы обильного урожая желудей, типичными местообитаниями
являются дубовые рощи. В восточных Карпатах в летний период кабаны
подымаются в горы выше зоны криволесья и ночами пасутся на откры-
тых полянах.
На Кавказе дикие свиньи обитают как в низинных местностях, так
и в поясе гор. Излюбленными стациями их являются камышевые заросли
в плавнях больших рек (Кубани, Терека, Кумы, Куры и др.), а также
сырые болотистые низины до самого побережья, Черного и Каспийскога
морей. В дневные часы кабаны скрываются в зарослях камыша, протап-
тывая многочисленные расходящиеся, в о всех направлениях тропы. Ночью
выходят для кормежки на более открытые места — луга, поля и даже на
огороды. В горах кабаны придерживаются, в основном, лесной зоны.
В местах интенсивного преследования дневные часы проводят в наиболее
«крепких» (трудно проходимых) местах: в зарослях рододендрона, терна,
самшита, густого мелкого дубняка, ельника, колючих кустарниках.
Сезонные особенности в размещении кабанов обусловливаются кормовой
базой, а в зимнее время, кроме того, характером залегания снежнога
покрова. Лето значительная часть кабанов (самки с поросятами, старые
самцы) проводит в нижнем поясе леса, в культурной зоне; часть популя-
ции (молодые самцы, подсвинки, холостые свиньи) подымается в горы,
выходя часто в зону альпийских лугов до 2500 м над ур. м., а изредка
даже в места обитания тура и серны (Динник, 1910 : 35). С конца лета и
всю осень основная масса зверей концентрируется в рощах диких фрук-
товых (яблонь, груш, алычи) и орехоплодных деревьев (дуба, бука,
каштана, чинара). От наличия опавших желудей и орехов зависит в зна-
чительной мере местопребывание зверей и в зимний период. Однако ли-
митирующим фактором является в это время также глубина снежного
покрова. При глубине снега в 60—80 см сильно затрудняется передвиже-
ние и добыча пищи даже для крупных животных (Насимович, 1939).
В низовьях Волги и на территории среднеазиатских республик излюб-
ленными стациями кабана являются тростниковые заросли и тугаи в до-
линах и по берегам рек и озер. В пустыню заходят лишь в зимнее время
и весною и поисках подземных частей луковичных растений. В долине
Сыр-Дарьи, например, нормальные зимы кабаны проводят на островах
роки или в тростниках дельты, но в годы больших снегопадов мигрируют
в Кызылкумы, откуда возвращаются в долину реки ко- времени созре-
вания культурных злаков (Бажанов, 1945). Как и на Кавказе, в горной
90
SUIDAE
полосе Средней Азии кабаны в летнее время подымаются до альпийских
лугов и зоны криволесья, к зиме спускаясь к нижней границе леса на
поля и в долины. В Копет-Даге, а также местами в горной зоне Таджи-
кистана кабацы охотно посещают фисташковые рощи.
На Дальнем Востоке кабан обитает в лесах манчжурского типа, осенью
придерживаясь более возвышенных мест, а зимой, весной и летом — долин
речек, пойменных лесов и островов на крупных реках (Золотарев, 1936).
В некоторых случаях кабаны не избегают и близости человеческих
.поселений. Широко известен их вред, наносимый сельскохозяйственным
культурам, расположенным даже на приусадебных участках. В ряде
местностей в зимнее время дикие свиньи держатся вблизи стогов сена,
которые для них служат и защитой от холода и источником корма.
Все представители семейства свиней, в том числе и кабан, — всеяд-
ные животные. Наравне с составляющей их основной рацион раститель-
ной пищей дикие свиньи охотно поедают доступные им животные про-
дукты, начиная с дождевых червей и кончая трупами птиц и крупных
млекопитающих.
Состав растительных кормов зависит от природных условий района
обитания и колеблется по сезонам. Постоянной составной частью пищи
кабана, особенно при отсутствии или недостатке плодоносящих деревьев,
являются травянистые растения как в виде подземных их частей (корне-
вища, клубни, луковицы), так и надземных. В ряде районов Средней
.Азии корневища и побеги тростника, рогоза и других прибрежных расте-
ний составляют, если ire, считать животной пищи, почти единственный
источник существования кабанов в течение всего года. Надземные зеленые
части травянистых растений (злаки, разнотравье) наибольшее значение
в пище кабанов имеют весною и в начале лета. В Кавказском заповед-
нике к числу растений, у которых они поедают надземные части, принад-
.лежат черемша, купена мутовчатая, ятрышник, щавель, сердечник, ман-
жетка и некоторые другие (Донауров и Теплов, 1938). В низовьях Волги
излюбленную пищу кабанов составляют плоды водяного ореха (чилима).
Удельный вес травянистых растений в рационе кабана в лесных районах
сильно снижается, начиная с конца лета, когда созревают и опадают
фрукты, а позднее орехи. Из диких фруктов пищей кабана на Кавказе
являются черешня, кизил, алыча, яблоки, груши. Наибольшее предпо-
чтение отдается последним. Вместе с мякотью поедаются и косточки фрук-
тов, предварительно раздробленные коренными зубами. В течение значи-
тельной части года, иногда шести-семи месяцев с сентября по апрель,
•основной пищей кабана в лесных районах являются плоды орехоносов —
дуба, каштана, бука, грецкого ореха, чинары, фисташки, реже лещины.
Наибольшее значение имеет дуб, широко распространенный в европей-
ской части ареала S. scrofa. Желуди служат пищей Диким свиньям иногда
даже весной, в проросшем состоянии. В дельте р. Или (Казахская ССР)
-существенную часть рациона кабана в годы урожая составляют ягоды
селитрянки (Nitraria schoberi L.). Из культурных растений кабан пред-
почитает початки кукурузы и просо. Иногда вредит рисовым, а также и
другим зерновым посевам. По утверждению Динника (1910), на Кавказе
дикие свиньи картофель не едят, чему противоречат сведения по этому
поводу из Закарпатской Украины.
Животная пища кабана чрезвычайно разнообразна. Одно из первых
мест занимают дождевые черви и обитающие в земле личинки насе-
комых (хрущей, чернотелок). При случае охотно едят и взрослых насе-
комых, особенно крупных жуков, а в годы массового размножения
SUS
91
саранчу (Штегман, 1949). Питаются также улитками, ловят лягушек.
При случае раскапывают норы мышевидных грызунов, остатки которых
нередко находят в их желудке. В дельте Или основной пищей кабанов
в летнее время является, по сообщению Б. К. Штегмана (1949), рыба,
остающаяся после спада весеннего паводка в замкнутых усыхающих
озерках по берегам русел.
Максимально поедаемое взрослым кабаном за одну кормежку коли-
чество пищи определяется в 2—3 кг; Динник (1910) в желудке убитого
им кабана обнаружил полведра пережеванных желудей. В случае не-
Рис. 18. Порой кабана. Кавказский заповедник. (Фот. И. В. Жаркова);
достатка или трудности добывания корма (зимою) поедают грибы, корни,
кору и даже ветви деревьев, мох, сухие листья, гнилую древесину. Не
гнушаются трупами животных. Разыскивая корни, луковицы и дожде-
вых червей, кабаны разрывают почву прекрасно приспособленным для этой
цели рылом, «вспахивая» иногда сплошь целые гектары. Эти «копанки»,
или «порой» (рис. 18) служат верным признаком наличия кабанов в дан-
ной местности.
Держатся кабаны, как правило, небольшими группами, редко более
10—20 голов, хотя в Уссурийской тайге изредка встречаются стада свыше
100 животных. Обычно группа состоит из самки и ее потомства. Молодые
держатся с матерью до полутора-двух лет, так что с нею обычно ходят два
поколения — текущего и предшествующего года. Несколько самок со
своими поросятами могут объединяться в один гурт; при этом они не
только ходят, но и на лежку устраиваются совместно. Самцы с возраста
1%—2 лет, как правило, ведут одиночный образ жизни, присоединяясь
к стадам самок лишь на период спаривания.
Образ жизни кабана, его сезонный и суточный цикл в сильной сте-
пени зависят от природных условий, урожая кормов, степени преследо-
92
SUIDAE
вания человеком. Сезонность размещения особенно резко выражена,
в горных районах.
Летом часть животных, как уже отмечалось, подымается в горы до
альпийской и субальпийской зоны. В зимний период снежный покров
вынуждает бблыпую часть популяции концентрироваться в зоне широко-
лиственных лесов, которая наиболее благоприятна в этот период и в кор-
мовом отношении (Донауров и Теплов, 1938). В лесной полосе европей-
ской части ареала летом кабаны предпочитают поросли молодого леса,,
тростниковые болота и берега рек; осень же и зиму проводят в дубовых
рощах, обеспечивающих в годы урожая желудей лучшую кормовую базу.
О сезонных миграциях кабанов в пустынных районах нами уже упоми-
налось. Если кабанов не преследуют, то они могут выходить на жировку
днем, а отдыхать устраиваются вблизи места кормежки. Однако в боль-
шинстве районов они вынуждены в дневное время скрываться в «крепких»
местах, а кормиться только с наступлением темноты или в ранние утрен-
ние часы. При этом свиньи нередко вынуждены совершать переходы
к месту кормежки в 15—20 км. Большую амплитуду суточные передви-
жения имеют в период массового созревания фруктов и орехов, а также
в период гона; сокращаются же они в зимнее время вследствие глубины
снега и морозов. Сравнительно невелики передвижения кабанов в дель-
тах и долинах рек. Обычно здесь они бродят в камышах, вырывая из земли
корневища, дождевых червей, откусывая зеленые побеги растений, по
ночам же выходят на соседние поляны и посевы. Только высокие паводки
заставляют животных уходить из поймы и переселяться иногда на
довольно значительные расстояния.
Бблыпая часть кабанов (как самцы, так и самки) устраивает так на-
зываемые лежки, или гнезда. В простейших случаях лежка представ-
ляет небольшое углубление в почве. В холодное же время года живот-
ное сгребает или натаскивает в одно место хворост, папоротник, сухую
траву и листья, в результате чего образуется своеобразная постель вы-
сотою иногда почти в полметра. По краям, после обминания средней:
части, получается нечто вроде вала, а сверху из наломанных веток иногда
образуется как бы крыша. Такие кучи хвороста и бурьяна служат не-
только для согревания, по и для защиты от жалящих насекомых (гнуса}
в период линьки. Две самки с поросятами могут устраивать совместную
лежку. Ложатся свиньи, особенно в холодное время года, вплотную друг
к другу, головами навстречу ветру. Лежки располагаются скрытно под
деревьями, у камней или в лесной чаще, а в дельтах рек и болотах —
среди камышей на возвышенном сухом месте. Более или менее продолжи-
тельное время одной лежкой кабаны пользуются только зимою, когда
их подвижность становится меньше. В южных районах Средней Азии
лежки служат кабанам защитой от летнего зноя и песчаных бурь. В этих
случаях они представляют ямы, вырытые свиньями в земле под берего-
выми обрывами, под корнями деревьев, в ущельях и достигающие иногда
глубины 1 м. В Латвии кабаны зимою иногда забираются в стога сена.
(Калниньш, 1950).
Характерной особенностью биологии кабанов является потребность
в регулярных грязевых ваннах. Несомненно, с этим связана их привер-
женность к сырым болотистым местам. Купанье в ямах с липкой грязью
или в илистых ключах производится с ранней весны, до декабря вклю-
чительно; особенно массовый и регулярный характер оно носит в осен-
ние месяцы (октябрь и ноябрь). Купаются даже в лужах, покрытых тол-
стой, какую только может пробить кабан, коркой льда. Купанье происходит
SUS
93
обычно утром или вечером, реже днем. Грязевые ванны, вероятно, являются
одним из средств избавиться от эктопаразитов. Привычкой кабанов
является также чесание о деревья, с которых они иногда стирают кору
до древесины.
Голос кабана сходен с голосом домашней свиньи и состоит в основном
из хрюканья и визга. При нападении или испуге свиньи могут издавать
короткие звуки вроде «ду-ду-ду» или «о-о-о» («гудеть»), а самцы сопеть
или реветь. В общем же, даже раненые, они очень молчаливы. Из чувств
у кабана хорошо развиты слух и обоняние. По ветру он чувствует чело-
века иногда за 350—400 м. Но зрение у него плохое (Динник, 1910).
Способностью к быстрому бегу кабан не обладает. На ровном месте его
легко догоняют собаки и верховая лошадь. Плавает хорошо, легко пере-
правляется вплавь через широкие реки и заплывает при необходимости
на километр и больше вглубь моря.
Линька у диких свиней начинается в апреле. На Кавказе к концу
мая—началу июня старая щетина и пух совсем выпадают и животные
становятся почти голыми. Сильно страдая от укусов насекомых-крово-
сосов, кабаны залезают в густой кустарник, в натаскиваемые для этой
цели кучи хвороста и бурьяна или в лужи с грязью, образующей на их
теле временный защитный панцирь. Отрастание щетины начинается
к концу июня, а в сентябре она становится уже длинной. Пух полного
развития достигает лишь к ноябрю.1
Половой зрелости кабаны достигают к полуторагодовалому возрасту,
по значительная часть их, поздно родившиеся или в неблагоприятные по
кормовым условиям годы, начинают размножаться лишь на третьем
году. Половой сезон (спаривание) охватывает период с ноября по январь.
Начало и конец его колеблются не только из года в год, в зависимости
от кормовых и климатических условий, но не одинаковы даже для раз-
ных районов на сравнительно небольшой территории в течение одного
года (Донауров и Теплев, 1938). У молодых самок половая охота и спа-
ривание происходят в более поздние сроки. В этот период самки держатся
сравнительно крупными группами, до 8—10 голов, по возможности в уда-
ленных от человеческих поселений местах. Ведут себя свиньи во время
гона беспокойно, много двигаются. Самцы сильно возбуждены, мало при-
нимают пищи. Между ними происходят жестокие турнирные бои с по-
мощью клыков, иногда кончающиеся смертью или тяжелым увечьем одного
из участников поединка. В этих условиях приобретает значение калкан,
защищающий от глубоких повреждений подвергающиеся ударам части
тела. Наиболее уязвимыми и опасными в отношении повреждений яв-
ляются стенки живота, пахи и задние конечности, не имеющие утолщен-
ной кожи. Самыми острыми клыками обладают и поэтому наиболее
опасны самцы в возрасте около 4—6 лет, называемые секачами. У ста-
рых клыки, хотя по размерам больше, но не так страшны, так как их
острые концы становятся больше загнутыми не в стороны, а внутрь.
Продолжительность беременности — около четырех месяцев.1 2 Мо-
лодые родятся с марта по май, основная масса — в апреле. Число поросят
1 Не верно утверждение Динника (1910 : 50), что пух начинает появляться лишь
в сентябре. На коллекционных материалах ЗИН АН СССР он заметен уже на июль-
ских шкурах.
2 У домашних свиней беременность длится 114—116 дней, при этом имеют место
случаи сложной беременности, когда самка вынашивает плоды разного возраста;
в этом случае и опорос бывает двойным, с промежутками в 2—4 недели (Б. В о »
к о п я л о в. Свиноводство. Сельхозгиз, М., 1950). Бывают ли случаи двойного опло-
дотворения у диких свиней —не известно.
94
SUIDAE
в помете колеблется от 3-х до 10-ти в зависимости от возраста самки и
условий предшествующей осени и зимовки. Средней цифрой в условиях
Кавказа в настоящее время является 4—5 поросят. У молодых самок
число детенышей в помете меньше, чем у взрослых. Перед опоросом самка
или несколько их совместно устраивают в укромном месте толстую постель,
(гнездо) с высокими краями, в которой и происходят роды. Родятся по-
росята беспомощными и в течение первой недели не покидают гнезда.
Свинья — хорошая мать, защищает своих детей, иногда бросается даже
на человека (Динник, 1910).
Родится на свет больше самок, но соотношение полов в дальнейшем
в результате гибели части популяции выравнивается и у взрослых оказы-
вается почти одинаковым (48% самцов и 52% самок, по Донаурову и
Теплову, 1938).
Считается, что в естественных условиях кабан может жить до 15—
20, а в исключительных случаях даже до 30 лет. Точные данные по этому
вопросу отсутствуют. Максимальная продолжительность жизни в неволе
(в Лондонском зоологическом саду) установлена в 19 лет 6 месяцев и
6 дней (Flower, 1931).
Численность кабанов в одном и том же районе может резко меняться
год от года. Колебания ее обусловливаются неравномерностью урожая кор-
мов и разной их доступностью в наиболее трудный зимний период, а также
гибелью животных от хищников, болезней и стихийных бедствий. Пло-
хой урожай кормов, глубокий снег и сильные морозы являются причиной
массового падежа диких свиней от голода. Неоднократные случаи этого,
явления имели место в Беловежской пуще, Латвии, на Кавказе, Карпатах
и в Средней Азии. При глубине снега свыше 55 см добывание корма для
свиней очень затруднено. Такие же последствия имеет образование после
оттепели наста, а в безлесных местностях промерзание почвы, когда жи-
вотные сильно ранят себе рыло и ноги, но корма достать не могут. Го-
лодовка сказывается не только в непосредственной гибели животных, но.
отражается также на количестве и качестве приплода. Только высокая
плодовитость свиней позволяет сравнительно быстро восстанавливать
их численность после стихийной гибели животных. Вследствие недостатка
кормов кабаны иногда откочевывают в другие места и на ряд лет могут
исчезать из того или иного района.
Врагами кабана из хищников являются волк, тигр и изредка барс.
В нормальных условиях взрослого самца кабана волк одолеть не может
не только в одиночку, но даже стаей. Известны случаи, когда нападающий
волк погибал от клыков кабана (Штегман, 1949). Жертвой волков в боль-
шом числе становятся молодые свиньи, подсвинки и поросята. Взрослые
кабаны гибнут от этого хищника лишь в глубокоснежные зимы и во.
время голодовок, когда могут уничтожаться целыми гуртами. Леопард
в горах часто нападает на кабанов; вследствие же редкости самого хищ-
ника наносимый им вред не играет существенной роли (Донауров и Теп-
лов, 1938).
В Средней Азии и на Дальнем Востоке кабаны в значительном числе
истребляются тигром. Недаром последнего называют в Приморье «па-
стухом» кабаньих стад. Нападения на кабана других хищников носят
случайный характер.
В дельтах и долинах рек причиной гибели только что родившихся
поросят являются пожары в прошлогоднем тростнике или высокие й
длйтельцые паводки; от последних в отдельные годы погибает не только
весь приплод, но и значительная часть взрослых зверей, не успевших
SUS
95-
перейти в верхние части дельты и оставшихся на узких незатопляемых
гривах (Исаков, 1951). В Астраханской! заповеднике с успехом приме-
няются для защиты кабанов от паводков искусственные «кочки». Послед-
ние представляют земляные насыпи, укрепленные бревнами на возвы-
шенных частях затопляемых островов (Дубинин, 1953).
Из эпизоотических заболеваний среди диких кабанов чаще других
наблюдаются ящур и чума. Наибольшее значение имеет чума, которая
иногда уносит значительную часть популяции зверя (Врублевский, 1908;.
Донауров и Теплов, 1938; Дубинин, 1952; Momberg, 1954; Саблина, 1955)..
Источником эпизоотии чаще всего являются домашние свиньи, а при
ящуре — и крупный рогатый скот. В Сибири наблюдалась массовая ги-
бель кабанов от геморрагической септицемии домашних свиней (Смирнов,
1928). В Чехословакии в 1950 г. имела место среди диких кабанов эпизо-
отия так называемой «тешенской болезни» вирусного происхождения.
В 1954 г. эпизоотия повторилась, па этот раз в пограничных с Чехосло-
вакией районах Польши (Szaflarski, 1955). Известны отдельные случаи
туберкулеза и сибирской язвы, но практического значения они не имеют.
У диких кабанов зарегистрировано около двух десятков 'видов пара-
зитических червей. Некоторые из ни?. {Gasterodiscoides hominis Lew. et
MacGonn., Gnathostoma hispidum Fedt.) могут паразитировать в массовых
количествах в течение круглого года и являться причиной истощения
животных. Круглые черви — метастронгилюсы {Metastrongylus elongatus-
Duj. и другие), паразитирующие в бронхах, вызывают тяжелые бронхо-
пневмонии, иногда со смертельным исходом (Дубинин, 1952; Рухлядев,.
1952). Заражение метастронгилюсом происходит при поедании свиньями
дождевых червей, являющихся промежуточными хозяевами этих пара-
зитов. Эпидемиологическое значение имеет сосальщик толстого кишеч-
ника {Gasterodiscoides hominis), паразитирующий и у человека, а также
нередкий среди диких кабанов низовьев Волги финноз (поражение мышц,
личиночной стадией ленточного червя Taenia solium L.) и трихинеллез
(возбудитель — мышечная трихина {Trichinella spiralis Ow.). Поедание
недостаточно проваренного или прожаренного мяса кабанов, поражен-
ного последними паразитами, может явиться причиной тяжелых заболе-
ваний человека.
Хозяйственное значение. Кабан ценен как мясное животное..
Выход мяса, в зависимости от упитанности животного, составляет около 55—
60% живого веса. Взрослый самец может дать, следовательно, свыше
100 кг мяса; но крупные звери попадаются теперь сравнительно редко-
и средний вес туши на Кавказе при заготовках определяется в 50 кг; основ-
ную массу при этом составляют полугодовалые и полуторагодовалые звери.
Лучшей упитанности кабаны достигают в ноябре. В это время взрослый
кабан, весом в 160 —180 кг, может дать около 18—20 кг внутреннего и
30—40 кг подкожного сала (Верещагин, 1947). Самцы быстро худеют
в период гона. Самки упитанность сохраняют дольше и теряют жир
только перед опоросом. Товарный выход мяса в большинстве районов
пока незначителен, но при правильной организации промысла кабанина
может играть весьма существенную роль в создании местной продоволь-
ственной базы. В некоторых районах Приморского края дикие свиньи
издавна являются источником мяса для русского населения, заготов-
ляющего его впрок путем засолки. Вкусовые и питательные качества,
мяса кабана по сравнению с другими дикими копытными очень высоки.
Только мясо самцов в период течки имеет специфический запах и при-
вкус.
$6 SUIDAE
Кроме мяса и сала, используется кожа и щетина. Первая может под-
вергаться, как и шкуры домашних свиней, фабричной выделке. Кроме
того, местное население на Кавказе шьет из нее прочную обувь — поршни
или каламани (Марков, 1932). Лучшая по упругости, чем у домашней
свиньи, щетина (около 350—400 г с головы) употребляется в шорном и
щеточном производстве. Более тонкий волос и пух годны для набивки
матрацев и мягкой мебели. Клыки взрослых самцов используются в ка-
честве украшения. Пойманные в молодом возрасте поросята диких свиней
легко привыкают к человеку и становятся ручными, но случаи размно-
жения диких кабанов в домашней обстановке нам не известны. В местах
обитания кабанов обычны помеси их с домашними свиньями. Так, счи-
тается, что кахетинские домашние свиньи, выпасаемые в дубовых и бу-
ковых лесах, являются продуктом такой метизации. Практическое зна-
чение одомашнение кабана и метизация его с домашними могут иметь
при улучшении существующих и создании новых, приспособленных
к местным условиям пород домашних свиней. Известны плодовитые гиб-
риды европейского кабана с бородатой свиньей (б1, barbatus Miill.,
Gray, 1954).
. Известную пользу кабан приносит истреблением вредных насекомых
и их личинок. Однако пользу эту превосходит вред, причиняемый истреб-
лением играющих видную роль в почвообразовании дождевых червей и
рытьем почвы. Иногда «вспахиваются» сплошь целые гектары, истреб-
ляются всходы и молодая поросль деревьев (Донауров и Теплов, 1938),
нарушается целостность растительного покрова, портятся сенокосы.
Большой вред дикие свиньи, наносят сельскохозяйственным культурам.
Особенно страдают, иногда нацело уничтожаемые, посевы проса и куку-
рузы. Могут повреждаться также посевы пшеницы, корнеплодов. Ф.Страут-
ман и К. Татаринов (1949) приводят случай, когда на одном хуторе в районе
Восточных Карпат кабанами были уничтожены за одну ночь все посевы
картофеля. В Средней Азии дикие свиньи сильно вредят бахчевым куль-
турам (Самородов, 1953). Вред при этом выражается не только в поедании,
но и во взрывании, вытаптывании почвы, исключающем часто дальней-
шую уборку урожая. В некоторых местностях дикие свиньи из-за этого
становятся настоящим бедствием, вынуждая организовывать планомер-
ную борьбу с ними. Кроме того, как пишет С. Мадаминов (1953), кабаны
разрывают и разрушают ирригационные системы, нанося, огромный
вред полям колхозов и совхозов.
В охотничьих хозяйствах кабаны могут наносить вред истреблением
яиц и молоди птицы. В Беловежской пуще известны даже случаи нападе-
ния кабанов на молодняк крупных зверей (Каверзнев, 1932).
Охота. Наиболее распространенными способами охоты на кабана
являются скрадывание, подкарауливание, охота с собаками и облава.
Скрадывание представляет один из наиболее трудных способов добычи
этого зверя. Оно возможно главным образом лишь в тех местностях, где
кабаны сравнительно мало преследуются человеком и выпасаются в свет-
лые часы суток. Скрадывают зверей на местах кормежки. Главное вни-
мание должно быть обращено на то, чтобы зверь не учуял раньше времени
человека; поэтому подход должен производиться против ветра, а не на-
оборот. Одежда и обувь охотника не должны издавать резкого запаха.
Требуется также строгое соблюдение тишины при подходе; излишняя же
маскировка не является необходимой. При спокойной кормежке кабан
все время помахивает хвостом, но при малейшем беспокойстве и насторожен-
ности животного, если оно даже продолжает кормиться, хвост перестает
SUS
97
двигаться. Поэтому для крадущегося охотника он является верным ин-
дикатором поведения зверя, сигнализирующим о необходимости сделать
остановку (Каверзнев, 1932).
Подкарауливание кабанов производится на посевах и бахчах, куда
кабаны приходят, обычно, в ночное время. Подкарауливают их также
па местах кормежки под фруктовыми и орехоплодными деревьями или
па тропах, ведущих от лежек к местам жировок, в местах, где звери
валяются в грязи. Во всех этих случаях охотник должен выбирать место
аасидки в укрытии камышей, деревьев, больших камней и т. п. и обяза-
тельно так, чтобы ветер дул не от него, а ему навстречу. Так как охота
с подкарауливанием производится в ночное время, то охотник должен
приходить на место засидки до захода солнца. По понятным причинам
для охоты выбирают светлые лунные ночи.
Охота с собаками требует значительного числа последних, к тому же
хорошо притравленных и злобных. Заключается она в том, что спущен-
ные собаки разыскивают, останавливают и задерживают кабана до под-
хода охотника.1 На долю последнего остается лишь прикончить зверя,
иногда с помощью рогатины или кинжала. Годны для этой охоты гончие,
по чаще применяются местные беспородные, специально притравленные
собаки. От хорошей кабаньей собаки требуется смелость, злобность и
ловкость, уменье хватать зверя за те места, где он не может достать ее
клыками. Большой процент собак при этих охотах гибнет от клыков
разъяренного зверя. Охотник также должен соблюдать осторожность при
подходе к задержанному собаками зверю; последний, увидев прибли-
жающегося прямо человека, может броситься на него, не взирая на со-
бак, и искалечить; поэтому рекомендуется подходить незаметно сбоку
пли сзади.
Облавная охота на кабана может быть очень добычливой (Марков,
1932). По своей технике она мало отличается от облавных охот на дру-
гих крупных зверей и заключается в том, что группа загонщиков, обло-
жившая участок леса, направляет зверей на линию стрелков. И в этом
случае охотники должны стоять против ветра и соблюдать абсолютную
тишину. И на Кавказе, и в Средней Азии практикуется охота на кабанов
верхом. На хорошей лошади догнать его не представляет труда. Важно
лишь заставить зверя выйти на открытое место и отрезать отступление
в заросли или скалистые горы. Загнанного кабана стреляют с седла или
прирезывают ножем.
Изредка практикуется «прочесывание» с собакой камышей и стрельба
подвернувшихся при этом зверей. Другие способы добычи кабанов (на-
пример ловля ямами, пастями ит. п.) носят случайный характер и большого
практического значения не имеют.
Охота на кабана связана с известным риском. Случаи неспровоцирован-
ного нападения его на человека не известны, и даже раненый кабан
чаще всего старается скрыться. Тем не менее, раненый, а особенно при-
веденный собаками в ярость зверь может броситься на охотника и на-
нести ему тяжелые увечья. Самцы наносят резкие короткие удары клы-
ками снизу вверх. Самки, наоборот, стараются сбить с ног противника
ударом и затем рвут его зубами, подобно собаке. Самцы этого никогда
не делают. Лучший способ избавиться от удара кабана — отскочить
в сторону или за дерево; зверь несется прямо и, раз промахнувшись, уже
нс возвращается обратно.
1 Подробное описание этого способа см. у Н- Н. Толстого (1922).
7 Фауна СССР, И. И. Соколов
98
TYLOPODA
Подотряд Tylopoda— Мозоленогие1
liliger J. К., 1811, Prodromus systematis mammalium et avium, Berolini :
102—103 (сист.); Matthew W. D., 1929, Bull. Geol. Soc. of America, 40 : 403—408
(сист.); Romer A. S., 1929,J. Geol., Chicago, 37 : 261—267 (эвол.); Scott W. B.,
1937, A history of land mammals in the Western Hemisphere, revised edition, N. Y. :
327—344 (сист., ист., реконструкции, изобр.); Simpson G. G., 1945, Bull. Amer.
Mus. Nat. Hist., 85 : 149—150, 258—260, 264—265 (сист.); Хавесон Я. И., 1954,
Труды Палеонтол. инет., 47 : 100—162 (сист., ист., изобр.).
Характеристика подотряда. Ноги опираются на фаланги; третью,
вторую и дистальную часть первой, а не на одну третью фалангу. Копыта
когтеобразные. Подошвы конечностей мозолистые. Trapezoideum и
magnum, а также cuboideum и naviculare раздельные.
Шейные позвонки, от эпистрофея до шестого включительно, характерны
тем, что у них межпоперечные отверстия имеют каудальный выход в по-
звоночный канал, и притом далеко от каудального края тела позвонка.
Седьмой шейный позвонок может иметь межпоперечные отверстия с пра-
вой и левой сторон, либо одно из них может отсутствовать, либо могут
отсутствовать оба; если данные отверстия имеются, то они не выходят
в позвоночный канал.
Нижняя челюсть массивна, и (в отличие от Ruminantia) обе ее половины
вполне слиты между, собою. Имеется ангулярный отросток, как у хищных.
Рога отсутствуют. Имеется сагиттальный гребень черепа, в разной мере
выраженный у отдельных групп. Строение дорзальной и затылочной
частей мозгового отдела черепа обнаруживает большое сходство с Car-
nivora.
Моляры селенодонтны. Премоляры редуцируются в направлении ис-
чезновения Р|, а затем Р3.1 2 Первые премоляры (Р|), имеющие клыко-
образную форму, редуцированы лишь у некоторых поздних групп (Ca-
melops Leidy, Lama Cuv.), сохраняясь даже у современных Camelus L.
Клыки хорошо развиты; у самцов они значительно крупнее, чем у самок.
Верхние резцы клыкообразны, причем у современных форм I1 и I2 реду-
цированы, существуя лишь в молочной генерации; I3 развит хорошо.
Эволюция и систематика. Группа Tylopoda четко прослеживается
в отложениях верхнего эоцена Северной Америки (Scott, 1937; Simpson,
1945). По Г. Симпсону, она может быть смутно определена и в отложениях
среднего эоцена. Представители верхнеэоценовой группы Protylopus
Wortman были несколько крупнее кролика; они обладали полным зуб-
ным рядом и совершенно раздельными лучевой и локтевой костями.На-
чиная с олигоценовых групп, локтевая кость срастается с лучевой,
образуя у современных форм кость, на которой сохраняются лишь прокси-
мальная и дистальная части локтевой кости. Малая берцовая (fibula)
редуцируется, как у Ruminantia, образуя лодыжковую кость (malleo-
lare). Метаподий у более поздних групп образуют os canon с характерной
широко раздвоенной дистальной частью; однако вплоть до верхнего мио-
цена были группы с раздельными III и IV метаподиями (например Oxyda-
ctylus Peters, из нижнего миоцена, Protolabis Соре из среднего и верх-
него миоцена). Рудименты II и V метаподий исчезли в олигоцене.
1 Раздел Tylopoda (стр. 98—115) написан Я. И. Хавесоном.
2 Описана также полная редукция Р 3 и частичная редукция Р4 у четвертичного
американского верблюда Eschatius Соре. У позднейших форм наблюдается сильная
моляризация Р4.
CAMELIDAE
99
Эволюция Tylopoda в основном происходила одновременно с эволю-
цией лошадей на американском континенте. В Северной Америке обе
эти группы исчезли (геологически совсем недавно), но зато в Южной
Америке доныне уцелела (вселившаяся туда по образовавшемуся, при-
близительно в начале плейстоцена, вулканическому Панамскому пере-
шейку) ветвь мозоленогих, объединяемая в род Lama. Североамерикан-
ские верблюды рода Camelops вымерли в постплейстоцене (Romer, 1929),
по-видимому, уже после вселения человека, обладавшего ко времени своего
появления на континенте Америки довольно совершенными орудиями,
заведомо не палеолитического типа.
Миграция мозоленогих в Старый Свет (через Аляско-Чукотское сое-
динение) произошла, по всей вероятности, в среднем плиоцене, когда
в Восточное полушарие вселились верблюды рода Paracamelus Schl.
(Хавесон, 1954).
Расцвет Tylopoda длился от олигоцена до верхнего плиоцена вклю-
чительно.
По В. Скотту (Scott, 1937), мозоленогие образовали четыре группы:
во-первых, основной ствол, доходящий до верблюдов и лам, а также до
крупных ламоподобных форм плиоцена и плейстоцена; во-вторых, пи-
тавшихся листвой деревьев «жирафообразных» верблюдов (в частности
известна очень крупная форма Alticamelus altus (Marsh) из среднего
миоцена Колорадо), причем эта ветвь возникла в олигоцене, достигла
расцвета в среднем и верхнем миоцене и угасла в нижнем плиоцене;
в-третьих, мелких «газелеобразных» верблюдов, ранним представителем
которых считается род Stenomylus Peters, из нижнего миоцена, а поздней-
шим — род Rakomylus Frick из нижнего плиоцена; наконец, в-четвер-
тых, олигоценовую группу Pseudolabis Matthew.
В. Мэтью (Matthew, 1929) отнес к мозоленогим открытую Кювье
группу ксифодонтов («мечезубых») из среднего эоцена—нижнего олиго-
цена Европы, на основании чего Г. Симпсон (1945) представил подотряд
Tylopoda состоящим из двух семейств: Xiphodontidae Flower, 1884, и
Camelidae Gray, 1821.
Следует, однако, подчеркнуть, что положение ксифодонтов недоста-
точно выявлено, так что отнесение их к Tylopoda представляется спорным.
Начиная с миоцена, появляется ряд крупных и очень крупных форм
мозоленогих. С другой стороны, доныне дожили и относительно мелкие
формы — представители рода Lama.
Семейство CAMELIDAE — ВЕРБЛЮДОВЫЕ
Gray J. Е., 1821, Lond. med. repository, 15 : 296—310 (сист.); Z i 11 е 1 К. A.,
1893, Handbuch derPalaeontologie, Abt. I.Palaeozoologie, IV. Miinchen ; XI-|-799 (сист.,
ист.); Wortman J. L., 1898, Bull. Amer, Mus. Nat. Hist., 10 (7) : 93—142 (сист.,
изобр.); Scott W. В., 1937, A history of land mammals in the Western Hemisphere,
N. Y. : 327—344 (сист., ист., реконструкции, изобр.); Simpson G. G., 1945, Bull.
Amer. Mus. Natur. Hist., 85 : 149—150, 258—260, 264—265 (сист.).
Приведенная выше характеристика подотряда Tylopoda (liliger, 1811;
emend. Simpson, 1945) относится также к семейству Camelidae, так как
включение в данный подотряд семейства Xiphodontidae требует точных
доказательств, без чего диагноз группы, объединяющей оба упомянутых се-
мейства, сформулировать невозможно. Пока можно лишь утверждать, что
подотряд содержит по крайней мере одно семейство, четко прослежи-
ваемое с верхнего эоцена.
7*
100
CAMELIDAE
Семейство Camelidae было разделено Г. Симпсоном на пять подсемейств:
1) Poebrotheriinae Zittel, 1893 (верхний эоцен—средний олигоцен);
2) Camelinae Zittel, 1893 (верхний олигоцен—современность); по
Г. Симпсону подсемейство Camelinae включает также подсемейство
Protolabinae Zittel, 1893;
3) Pseudolabidinae Simpson, 1945 (верхний олигоцен);
4) Alticamelinae Simpson, 1945 (средний олигоцен—нижний плиоцен),
«жирафообразные» верблюды;
5) Stenomylinae Frick, 1937 (нижний миоцен—нижний плиоцен), «газе-
леобразные» верблюды.
Деление Camelidae на подсемейства, предложенное Г. Симпсоном на
основании признания перечисленных выше четырех групп, указанных
В. Скоттом (Scott, 1937), преимущественно по работам В. Мэтью, а также
Ч. Фрика, и дополнения этих групп пятой за счет выделения архаической
группы Poebrotheriinae, представляется вполне логичным. Однако от-
сутствие диагнозов у Г. Симпсона делает все эти построения недостаточно
определенными. Например, в таблице родов (стр. 150) Г. Симпсон, следуя
В. Мэтью, отнес к новой группе Alticamelinae, кроме родов Alticamelus
Matthew, 1901, и Оху dactylus Peterson, 1904, еще два рода — Paratylopus
Matthew, 1904 (средний олигоцен—нижний миоцен), и Miolabis Нау, 1899
(средний миоцен). В тексте же (стр. 265) отмечается, что оба эти рода не
могут являться представителями Alticamelinae. Во избежание подобных
противоречий необходимо установить четкие отличия между Camelinae,
Stenomylinae, Alticamelinae и Pseudolabidinae. Относящиеся сюда мате-
риалы могут быть выявлены по разрозненным описаниям американских
мозоленогих. Однако поскольку современные формы верблюдовых отно-
сятся к одному и тому же подсемейству Camelinae, то в данной работе
приводятся особенности только Camelinae в широком смысле этого слова,
т. е. здесь приходится рассматривать это подсемейство как неогеновую
(в основном) группу, восходящую к полностью палеогеновой группе
Poebrotheriinae.
Подсемейство CAMELINAE
Zittel К. А., 1893, Handbuch der Palaeontologie, Abt. I, Palaeozoologie, IV,
Munchen : XI4-799 (сист., анат., ист.); Bogdaschew N., 1930, Anat. Anzeig., 70 :
143—154 (микроск. анат. костей); Авербург В. В., 1931, Практич. ветеринария,
И : 52—58 (анат. желудка); Авербург В. В., 1933, Арх. анат., гист. и эмбр.,
т. 12, вып. 2 : 295—317 (анат. желудка); Авербург В. В., 1940, Тр. Угб. с.-х.
инет., Самарканд, 2 : 37—44 (анат. желудка); Simpson G. G., 1945, Bull. Amer.
Mus. Natur. Hist., 85 : 150 (сист.); Никитин В. H., 1949, Атлас клеток крови
сельскохозяйственных и лабораторных животных, М. : 48-}—70 цв. табл, (кровь); Ха-
ве с о н Я. И., 1954, Тр. Палеонтол. инет., 47 : 100—162 (сист., ист., изобр.).
Характеристика. Высоконогие, с длинной шеей мозоленогие, с двумя
пальцами на всех конечностях. Глазница замкнутая.
Нижеуказанные особенности, непосредственно не относящиеся к ске-
лету, достоверны для групп, начиная с нижнего плиоцена, когда разо-
шлись предки современных родов Camelus L. и Lama Cuv.,h весьма ве-
роятны для более ранних представителей Camelinae. Бедренная часть
задней конечности не включена в контур туловища, так что конечность
на некотором протяжении выше колена охвачена со всех сторон кожным
покровом. При лежании тело опирается на запястья, локти, грудь и ко-
лени, причем все эти семь участков безволосы и покрыты особыми мо-
золистыми образованиями. Верхняя губа раздвоена. Желудок сложный,
CAMELUS
101
многокамерный, причем не только сами камеры, но и их гистологическое
строение сильно отличаются от того, что характерно для Ruminantia.1
Только рубец и сычуг могут быть сопоставляемы с соответствующими от-
делами желудка последних. В противоположность жвачным, рубец
у Tylopoda весьма развит с первых дней жизни. Желчного пузыря нет.
11лацента диффузная. Двойни не известны. Препуций направлен каудальио.
Эритроциты овальные, чего нет ни у какой иной группы современных
млекопитающих. Шерстный покров своеобразный: вокруг более мощного
волоска группируются более тонкие шерстинки. Для трубчатых костей
характерна очень густая сеть гаверсовых каналов (Bogdaschew, 1930).
Современные Camelinae делятся на два рода — Camelus Старого Света
и Lama горной Южной Америки, которые следует рассматривать как
относящиеся к разным трибам—Camelini и Camelopsini.1 2
Триба Camelini Havesson (1954): клыкообразные премоляры почти
у всех самцов и у значительной части самок имеются; затылочный, са-
гиттальный и теменные гребни выражены резко; в трибу входят три
рода — Gigantocamelus Barbour et Schultz (1939), Paracamelus Schlosser
(1903) и Camelus Linnaeus (1758).
Триба Camelopsini Havesson (1954): клыкообразные премоляры сов-
сем отсутствуют; затылочный, сагиттальный и теменные гребни выражены
слабо; в трибу входят два рода — Camelops Leidy (1854) и Lama Cuvier (1800).
Род CAMELUS LINNAEUS — ВЕРБЛЮДЫ
Linnaeus С., 1758, Syst. Natur., ed. X, 1 : 65 (сист.); Falconer H.,1868,
Fauna Antiqua Sivalensis, London: 227—242 (ист.); P о m e 1 A., 1893, Cameliens et Cer-
vidcs, Alger: 5—22 (ист.); Nehring A., 1901, Globus, Braunschweig, 80,12 : 188—189
(ист.); Lesbre F. X., 1903, Arch. Mus. Hist. Natur. de Lyon, 8 : 1—195 (анат.);
I.eche W., 1904, Scientific results of a journey in Central Asia, 6, I, Stockholm :
I — 69 (анат.); Boule M. et Teilhard de Chardin P., 1928, Arch. Inst.
Paleont. Hum., 4 : 27—102 (ист.); Weber M., 1928, Die Saugetiere, Jena, II : XXI V-j-
+898 (сист., ист.); Боголюбский С. Н., 1929, Происхождение верблюдов,
Алма-Ата : 1—48+2 табл, (анат., ист.); Пхакадзе Г. М., 1932, Тр. Лабор. генет.
Акад, наук СССР, 9 : 243—249 (микр. анат.); Берггрин А. П,, 1934, сб. «Верб-
людоводство», Алма-Ата : 9—56 (анат.); Colbert Е. Н., 1935, Trans. Amer. Phi-
los. Soc., n. s., 26 : X+401 (ист.); Бабушкина В. A., 1938, сб. «Верблюдоводство
и СССР», М. : 208—230 (биол. размнож.); Барминцев IO. Н., 1939, Коневодство,
12, I : 42 (биол. размнож.); Новиков И. И., 1940, Тр. Инет, генет. Акад, наук
СССР, 13 : 285—296 (сперматогенез); Хавесон Я. И., 1940, сб. Акад, наук СССР
«Пробл. происхожд., эволюции и породообразования дом. животных», М. : 291—358
(карта распростр.); Буйновский Н. А. и Я. И. Хавесон, 1953, Бюлл,
Комисе, по изуч. четвертичн. периода Акад, наук СССР. 19 : 79—81 (ист.); Хаве-
сон Я. И., 1954а, Тр. Палеонтол. инет., 47 : 100—162 (сист., ист.); X а в е с о н Я. И.,
19546, Докл. Акад, наук СССР, т. 98, № 3 : 475—478 (ист.).
1 В. В. Авербург (1931, 1933, 1940), изучая двугорбого верблюда, установил пять
отделов желудка: рубец (rumen), выстланный безжелезистой слизистой оболочкой,
покрытой плоским многослойным эпителием; двойной дивертикул (diverticulum
dextra et sinistra) ho обе стороны пищеводного желоба рубца, разделенный на боль-
шое число ячей, выстланных слизистой оболочкой, содержащей простые трубчатые
железы и покрытой цилиндрическим эпителием; губка (myospongiosa) с карманооб-
разными ячейками, покрытыми слизистой с трубчато-разветвленными железами и
цилиндрическим эпителием; труба (tubus) со складками слизистой с простыми трубча-
тыми железами, покрытой цилиндрическим эпителием; сычуг (abomasus) с кардиаль-
ными, фундальными и пилорическими железами.
2 Названия приводятся впервые; диагнозы триб соответствуют характеристикам
групп I и III на стр. 124—125 (см. также схему и пояснения на стр. 129—130) в ра-
боте Хавесона (1954).
102
C AM ELI DAE
Характеристика рода. Верхние моляры не обладают хорошо раз-
витыми передней и срединной складками на наружных стенках (парасти-
лем и мезостилем), а также отчетливо выраженным срединным ребром на
латеральной стороне каждой доли. Третий нижний премоляр отсутствует,
сохраняясь лишь в виде молочного зуба. Орбита расположена полностью
над зубным рядом: передний край — над передней долей М2, задний —
над срединой М3.
Эволюция. Род Camelus, возникший, по-видимому, в верхнем плио-
цене, разделился около среднего плейстоцена (или несколько ранее)
на две ветви, давшие одногорбых и двугорбых верблюдов. Эти две ветви
отличаются, в частности, тем, что у одногорбых четвертый нижний пре-
моляр имеет совершенно открытую переднюю долинку, у двугорбых же
эта долинка почти закрыта или же совершенно закрыта (Хавесон, 1954а).
Систематика. К одногорбым верблюдам относятся: 1) С. thomasi
Pomel (1893) из среднего плейстоцена Алжира; 2) С. sivalensis Falconer
et Cautley (1836) из плейстоцена Сиваликских холмов на севере Индии
(Colbert, 1935); 3) современный С. dromedarius Linnaeus (1758), известный
и в ископаемом состоянии, начиная с верхнего плейстоцена Алжира.
К двугорбым относятся: 1) С. knoblochi Nehring (1901) из среднего
плейстоцена Поволжья; к этой же форме относят находки из плейсто-
цена Алтайского края (Буйновский и Хавесон, 1953) и северного Китая
(Boule et Teilhard de Chardin, 1928); 2) современный C. bactrianus Lin-
naeus (1758).
Приручение одногорбого верблюда С. dromedarius L., по-видимому,
связано с древними культурами Ирана.1 На территории СССР была най-
дена нижняя челюсть дикой формы одногорбого верблюда в отложениях
оз. Севан в Армении, получившая название С. dromedarius dahli Haves-
son (Хавесон, 19546). Эта форма, по всей вероятности, существовала, на-
чиная с верхнего плейстоцена,и вымерла не позже III тысячелетия до н. э.
Скорее всего, ископаемый верхнеплейстоценовый С. dromedarius из Уэд
Сеген в Алжире был гораздо ближе к верблюду Даля, чем к С. thomasi.
Дикий двугорбый верблюд С. bactrianus ferus Przewalski дожил до
современности в Синьцзяне и в Заалтайской Гоби.
Домашние одногорбые и двугорбые верблюды столь явственно отли-
чаются друг от друга, что К. Линней присвоил им особые видовые обозна-
чения. Тем не менее, ряд авторов пытался оспаривать видовую самостоя-
тельность данных форм, выдвигая представление, что соответствующие
различия возникли якобы после одомашнения. Так, М. Вебер (Weber,
1928) утверждал, что С. dromedarius является лишь расой С. bactrianus,
развившейся в домашнем состоянии.
Подобные заключения не выдерживают критики, так как наряду
с такими, казалось бы, «зоотехническими» отличиями, как более грубый
и менее обильный шерстный покров у С. dromedarius по сравнению с С. ba-
ctrianus, более высокая молочная продуктивность и меньшая жирномолоч-
ность у одногорбого, необходимо отметить: меньший срок плодоношения,
в среднем около 388 дней у одногорбого (Бабушкина, 1938) и 411 дней
у двугорбого (Барминцев, 1939); большее развитие нёбной занавески
у одногорбого; существенные особенности в отношении не только на-
личия одного или двух горбов, но и самого положения горбов (в частности,
1 Южную Аравию и восточную Африку следует рассматривать лишь как вто-
ричные очаги возникновения многообразия домашних форм, но не как места первичного
одомашнения, вопреки не вполне точной формулировке, допущенной мною (Я. X.)
на экспликации к карте распространения .верблюдовых (Хавесон, 1940).
CAMELUS
103
у дромедара холка не закрыта, а у бактриана закрыта основанием горба);
иной характер оброслости отдельных частей тела (например: у бактриа-
нов предплечья имеют обильную опушку из более длинных волос, а лопатки
оо не имеют; у дромедаров же на лопатках оброслость выражена сильнее,
и на предплечьях опушки нет; оброслость нижнего края шеи у обеих
форм совершенно различная).
Структурные отличия между бактрианами и дромедарами хорошо про-
слеживаются и на черепе, в частности на нижней челюсти. Так, у дро-
медаров нёбная вырезка U-образная, а у бактрианов V-образная; венеч-
ный отросток нижней челюсти у дромедаров направлен вертикально,
у бактрианов же имеет аборально загнутое окончание; промеры, характе-
ризующие у взрослых особей расстояние от аборального края симфиза
до зубного ряда, у любых (крупных и мелких) дромедаров меньше, чем
у бактрианов; имеется также ряд трансгрессирующих отличий в отно-
шении краниометрических индексов (Lesbre, 1903; Leche, 1904; Боголюб-
с.кий, 1929; Хавесон, 19546).
Особое же значение (наряду с указанной выше особенностью строе-
ния четвертого нижнего премоляра) следует придать тому, что наблю-
даются отклонения от нормального сперматогенеза у гибридов первой
генерации между бактрианами и дромедарами (Пхакадзе, 1932; Новиков,
1940). Более того, вековой опыт среднеазиатских животноводов говорит,
что гибридные верблюды второй генерации, как правило, представляют
собою посредственных, либо совершенно малоценных животных. Гиб-
риды же первого поколения, наоборот, выделяются своей крепостью и
хозяйственной ценностью, связанной с проявлением пышного гетеро-
зиса. Таким образом, факт относительно давнего расхождения обеих
форм, совершенно независимого от воздействия одомашнения, выявляе-
мый наличием некоторой физиологической изоляции и морфологическими
особенностями, вполне очевиден.
Представители обоих видов верблюдов в состоянии поедать такие ко-
лючие растения, которых избегают решительно все копытные. Кроме
того, они хорошо приспособлены выносить без ущерба более длительное
отсутствие воды и более высокую концентрацию солей в корме и воде,
чем другие травоядные. Зато верблюды очень чувствительны к сырости,
укусам насекомых и инвазиям. Дромедары — в основном обитатели за-
сушливой жаркой зоны; бактрианы — жители прохладных сухих стран,
по боящиеся даже суровых морозов, например на плато Памира, а также
при практиковавшихся бурятами Забайкалья перевозках грузов по за-
мерзшим рекам на золотых приисках в Якутии.
Camelus bactrianus ferus Przewalski — Дикий двугорбый верблюд,
или хабтагай (рис. 19, 20, 21)
Potocki J., 1829, Voyage dans les steps d’Astrakhan et du Caucase, Paris.
1:1—361 (ист.); Ritter C., 1832, Die Erdkunde, Berlin, 2:1—1143 (pacnp.,
ист.); Ritter G., 1847, Die geographische Verbreitung des Kameels in dor Alien Welt.
Die Erdkunde, Berlin, 13; Anhang II : 609—759 (распр., ист.); Пржеваль-
c кий H. M., 1875, Монголия и страна тангутов, СПб., I : 1—381 (опис.); Прже-
вальский Н. М., 1878, Изв. Русск. географ, общ., т. 13, вып. 5 (1877) : 263—329
(опис.); Певцов М. В., 1879, Зап. Зап.-сиб. отдела Русск. географ, общ., 1 : 13—
14, 44 (распр.); Пржевальский Н. М., 1883, Из Зайсана через Хами в Тибет
и на верховья Желтой реки, СПб. : 1—473 (опис., изобр.); Пржевальский Н. М.,
1888, От Кяхты на истоки Желтой реки, исследование северной окраины Тибета и
путь через Лоб-Нор по бассейну Тарима, СПб. : 1—536 (распр.); Blanford W. Т.,
1892, Proc. Zool. Soc., London : 370—371 (сист.); Littledale G., 1894, Proc.
104
CAMELIDAE
Zool. Soc. London : 446—448 (распр., ox., изобр.); Грум-Гржимайло Г. E.,
1896, Описание путешествия в западный Китай, СПб., I : 1—547 (распр.); Pousar-
gues Е. de, 1898, Mem. de la Soc. Zool. de France, 11 : 126—224 (распр., сист.);
Hedin S., 1898, Through Asia, London, vol. I : 1—663, vol. II : 665—1278 (опис.);
Козлов П. К., 1899, Тр. эксп. имп. Русск. географ, общ. по Центральной Азии,
совершенной в 1893—1895 гг. под начальством В. И. Роборовского, СПб., 2 : 1—296
(опис., ох.); Ладыгин В. Ф., 1900, Изв. Русск. географ, общ., т. 36, вып. 2 : 169—
197 (распр., ох.); РоборовскийВ. И., 1900, Тр. эксп. имп. Русск географ, общ.
по Центральной Азии, совершенной в 1893—1895 гг., СПб., I : 1—610 (распр., ох.);
L е с h е W., 1904, Scientific results of a journey in Central Asia (1899—1902),Stockholm,
6, I : 1—69 (анат., изобр.); H e d i n S., 1905, Lop-Nor, Scientific results etc., 2 :1—717
(изобр., ox.); H e d i n S., 1905, North and East Tibet, Scientific results
etc., 3 : 1—593 (опис., распр.); Pira A., 1909, Zool. Jahrb., Supplem. X (2) : 233—
426 (морф.); Браунер A. A., 1922, Породы сельскохозяйственных животных.
Рогатый скот. Одесса : 1—116 (морф.); Boule М. et Teilhard de С h a r-
d i n P., 1928, Arch. Inst. Paleont. hum., 4 : 27—102 (ист.); Lattimore O., 1928,
The desert road to Turkestan, London:!—331 (распр.); Силуков А. Д., 1928,
Хозяйство Монголии, Улан-Батор, 1 (8):86—99 (биол., ох.); Боголюбский С. Н.,
1929, Происхождение верблюдов, Алма-Ата : 1—48 (анат., ист.); Громова В. И.,
1932, Тр. Комисе, по изуч. четвертичн. периода, 2 : 69—184 (ист.); ХавесонЯ. И.,
1933, сборн. «Проблемы происхождения домашних животных», (I) : 313—373 (морф.);
Симуков А. Д., 1938, Современная Монголия, Улан-Батор, I (26) : 31—43 (биол.,
распр.); Allen G., 1940, The mammals of China and Mongolia, New York, Natural
History of Central Asia, 11, (2) : XXVIII+621—1350 (общ.); Хавесон Я. И.,
1940, сборн. «Проблема происхождения, эволюции и породообразования домашних
животных», I : 291—358 (морф.); Н е d i n S., 1944, The Sino-Swedish expedition,
Stockholm, publ. 25, III (1933—1935) : 1—346 (распр., биол.); Банников А. Г.,
Э. M. M у p з a e в и А. А. Ю н а т о в, 1945, Изв. Всесоюзн. географ, общ., т. 77,
вып. 3 : 127—144 (распр.); Банников А. Г., 1945, Зоол. журн., 24 (3) : 190—200
(распр., биол.); Банников А. Г., 1947, Докл. Акад, наук СССР, т. 55, № 7 : 673—
676 (биол.); Мурзаев Э. М., 1948, Непроторенными путями, М. : 1—223 (распр.,
биол.); Хавесон Я. И., 1948, Докл. Акад, наук СССР, т. 60, № 6 : 1093—1096
(морфол., сист.); Б у й н о в с к и й Н. А. и Я. И. X а в е с о н, 1953, Бюлл. Комисе,
по изуч. четвертичн. периода, 19:79—81 (ист.); Банников А. Г., 1953, Тр.
Монгольск. комисс., 51 : 1—111 (сист.); Банников А. Г., 1954, Тр. Монгольск.
комисс., 54 : 178—196 (сист., распр., биол.); Д а г в а Д. Э., 1954, Природа, 9 : 105—
106 (биол.); Хавесон Я. И., 1954а, Тр. Палеонтол. инет., 47 : 100—162 (морфол.);
Хавесон Я. И., 19546, Докл. Акад, наук СССР, т. 98, № 3 : 475—478 (морфол.).
Описание. Дикий верблюд настолько мало отличается от домашнего
двугорбого, что в сводке Г. Аллена (Allen, 1940) о млекопитающих Китая
и Монголии автор ограничился оговоркой о возможной одичалости этого
животного, не приводя его в списке видов. Де-Пузарг (de Pousargues,
1898) в своем очерке жвачных Центральной Азии пришел к заключению,
что сомнительно, чтобы зоологи смогли когда-либо решить вопрос об
исконной дикости или же одичалости данной формы, «даже имея в своем
распоряжении все желаемые материалы для изучения, а также полные
серии черепов и костяков».1
Наружные признаки дикого верблюда по сравнению с признаками
домашнего в основном сводятся к несколько меньшему росту и к лепто-
сомности (узкотелости) телосложения, так что, по утверждению мон-
голов, можно даже отличить след хабтагая от более широкого следа до-
машнего верблюда (Симуков, 1938; Дагва, 1954; Банников, 1953).
Горбы у диких определенно меньше, чем у домашних. Эту особенность,
отметил еще историк и лингвист Ян Потоцкий (Potocki, 1829) со слов
посещенного им в 1797 г. буддийского священника в Астраханской степи,
ходившего в Тибет и видевшего этих зверей вблизи горы Богдо-Ола. Впо-
следствии об этом же упоминал ряд авторов, непосредственно видевших
1 Однако краниометрическая методика, как мы увидим ниже, оказалась'достаточно
чувствительной, чтобы обнаружить характерные особенности данной формы.
CAMELUS
105
дикого верблюда либо слышавших о нем от местных жителей Синьцзяна
и западной Монголии (Пржевальский, 1878, 1883; Певцов, 1879; Cum-
berland, 1891 (цит. по Blanford, 1892); Hedin 1898 : 828; Lattimore, 1928,
со слов монголов вблизи Эдзин-Гола; Симуков, 1928; Дагва, 1954). О не-
больших размерах горбов дикого верблюда свидетельствуют также ри-
сунок В. И. Роборовского (в книге Пржевальского, 1883) и фотографии
С. Хедина (Hedin, 1905, II, фиг. 68 и 69; Leche, 1904, фиг. 60 и 61), изо-
Рис. 19. Дикий верблюд {Camelus bactrianus ferus Przew.). (Рис. В. Н. Ляхова),
бражающие в разных положениях дикого верблюда, добытого в феврале
1901 г. у Алтимиш-Булака (около 41° с. ш. и 90° в. д.).
Н. М. Пржевальский (1875) упомянул со слов монголов Цайдама,
что шерсть диких верблюдов имеет сероватый цвет. В другом описании
(1878) Н. М. Пржевальский отметил, уже на основании собственных
впечатлений, что «цвет шерсти у всех диких верблюдов один и тот же —
красновато-песчаный; морда у дикого серее». В. Дакке (Leche, 1904)
обнаружил, что у экземпляра, добытого С. Хедином, шерстинки на туло-
вище у основания серо-коричневые, у вершины красновато-коричневые.
Впрочем, подобная окраска нередко встречается и у домашнего двугор-
бого верблюда, в особенности в Кашгарии и других странах Центральной
Азии. Поэтому еще Н. М. Пржевальский (1878) указал на необходимость
специальных краниометрических изысканий для выявления особенностей
дикой формы бактриана.
Первая попытка подобного рода была предпринята И. С. Поляковым
(1847—1887), написавшим соответствующую статью около 1879 г. Хотя
эта статья и осталась неопубликованной, но благодаря bL М. Пржеваль-
106
CAMELIDAE
скому (1883 : 43—44) можно получить представление о материале, ис-
пользованном Поляковым для изучения, а также о характере получен-
ных им выводов.
Оказывается, И. С. Поляков по ошибке привлек в качестве материала
по диким не только черепа, привезенные Н. М. Пржевальским, но и
черепа домашних верблюдов, доставленные в 1878 г. под видом диких
охотниками из Зайсана, посланными начальником местного пограничного
поста за китайскую границу добыть диких лошадей и верблюдов.
Имея под видом диких смешанный материал из диких и домашних,
к тому же очень малочисленный и включавший черепа неполновозрастных
экземпляров, И. С. Поляков, конечно, не смог получить каких-либо
определенных выводов.
Более успешными оказались изыскания В. Лэкке (Leche, 1904) и
С. Н. Боголюбского (1929). Первый располагал тремя черепами дикого
верблюда, один из которых был от неполновозрастного индивида. Один
из черепов взрослых экземпляров был от самца, добытого Н. М. Прже-
вальским, другой от самца, добытого С. Хедином. Всего было взято
14 промеров черепа. Индексы не вычислялись. Пользуясь, хотя и не-
большим, но достоверно идентифицированным материалом, В. Лэкке
получил ряд выводов, часть которых была подтверждена впоследствии.
Так, он обратил внимание на то, что край подглазничного отверстия рас-
положен у домашнего бактриана аборальнее, чем у дикого. Иначе го-
воря, аборальная часть лицевого отдела черепа короче у домашней формы.
Было отмечено также, что нижняя челюсть у домашних выше, чем
у диких.
С. Н. Боголюбский (1929) промерил 4 черепа взрослых диких бактриа-
нов, в том числе три, добытых экспедицией В. И. Роборовского и П.К.Коз-
лова 1893—1895 гг. Впервые применив морфологические индексы для
изучения черепа верблюдов, С. Н. Боголюбский отчетливо выявил отно-
сительное укорочение лицевой части у домашних по сравнению с дикими.
Таким образом, ему удалось несколько уточнить выводы В. Лэкке.
Я. И. Хавесон (1940, 1948, 19546), сравнивая различные формы вер-
блюдов, представил таблицы морфологических индексов, выявляющие
краниометрические различия между дикими и домашними двугорбыми
верблюдами. Работа 1948 г., специально посвященная этим отличиям,
базируется на изучении семи черепов взрослых индивидов дикого вер-
блюда Синьцзяна, хранящихся в Ленинграде (6 экз.) и в Стокгольме
(1 экз.). Эти черепа были доставлены Н. М. Пржевальским (2 экз.),
В. И. Роборовским и П. К. Козловым (4 экз.) и С. Хедином (1 экз.). Та-
ким образом, в отношении музейного материала не были использованы
только хранящиеся в Лондоне черепа диких верблюдов, добытые в Каш-
гарии Кемберлендом (Cumberland) и Г. Литльдейлом (Littledale) в 90-х
годах прошлого века. Первый доставил один череп, а второй четыре;
часть последних, вероятно, не от вполне взрослых особей. Эти черепа
доныне никем не описаны.
Из семи черепов диких бактрианов, изучавшихся Я. И. Хавесоном
(1948), два были от самок и пять от самцов. Следует отметить, что у всех
верблюдов половой диморфизм в отношении черепа выражен очень слабо.
У самцов клыки и клыкообразные премоляры развиты, однако, сильнее;
общие размеры черепа в среднем крупнее.
Наиболее показательные различия между дикими и домашними двугор-
быми верблюдами выявились в работе 1948 г. по 26 индексам, подавляю-
щее большинство которых трансгрессирует.
CAMELUS
107
Высота нижней челюсти в венечном, суставном и ангулярном отростках
ко отношению к базальной и мандибулярной длине у Camelus bactrianus
/crus определенно меньше, чем у домашней формы. Высота челюсти в су-
Рис. 20. Череп Camelus bactrianus ferus Przew.
ставном и ангулярном отростках по отношению к высоте в венечном
у диких меньше.
Лицевая часть у диких несколько более вытянута по отношению к ба-
зальной длине; соотношение между осью мозгового отдела черепа и осью
108
CAMELIDAE
лицевого отдела у них меньше. Ширина черепа у дикой формы меньше
как по отношению к базальной длине, так и по отношению к длине ли-
цевой части. Ширина нижней челюсти по отношению к базальной и
мандибулярной длине у диких меньше.
Общая высота черепа с нижней челюстью у диких относительно меньше.
У дикой формы длина аборальной части лицевого отдела черепа по
отношению к ширине черепа больше, чем у домашней. Удлиненность
аборальной части лицевого отдела черепа хорошо выявляется по отно-
шению к общей длине лицевого отдела: 28.6—33.1% у диких и 27.1 —
29.3% у домашних; отношение общей амплитуды (6.0) к величине за-
хождения (0.7) превышает 8, тогда как из цитируемой работы (1948) сле-
дует, что и при отношении между общей амплитудой и захождением (транс-
грессией), равном 2 или даже несколько меньше, трансгрессирующие
ряды уже оказываются достаточно характерными для каждой из сопо-
ставляемых форм.
В другой статье (19546) Я. И. Хавесон привел четыре дополнительных
индекса промеров высоты и продольных осей нижней челюсти, по которым
дикие и домашние двугорбые верблюды разделяются настолько хорошо,
что на использованном в этой статье материале (по 6 черепов каждой
формы) трансгрессии не было. Речь идет о соотношениях: (1) высоты
суставного отростка над горизонтальной плоскостью к расстоянию от
аборального края симфиза до наиболее глубокой точки вырезки между
суставным и ангулярным отростками; (2) той же высоты к расстоянию
от аборального края симфиза до орального края foramen mandibulae;
(3) высоты над горизонтальной плоскостью наиболее глубокой точки вы-
резки между венечным и суставным отростками к расстоянию от абораль-
ной точки симфиза до указанной точки вырезки между суставным и ангу-
лярным отростками; (4) той же высоты к расстоянию, принятому знаме-
нателем второго соотношения.
Следующая табличка представляет эти индексы:
Номера соотношений С. bactrianus ferus С. bactrianus domesticus
(n=6) (n=6)
(1) 49.3—53.5 53.9—58.1
(2) 57.8—61.7 62.8—68.2
(3) 46.6—48.4 49.4—53.4
(4) 54.3—55.9 57.8—62.1
Почти все перечисленные различия имеют явно доместикационный
характер, проявляя замечательный параллелизм с изменениями, обнару-
женными, в частности, при сопоставлении черепов кабанов и домашних
свиней (A. Pira, 1909; Я. И. Хавесон, 1933), тура и серого степного скота
(А. А. Браунер, 1922).
Изменчивость черепа у диких верблюдов меньше, чем у домашних,
что легко проследить по пределам колебаний индексов.
Для суждения об отличиях посткраниального скелета у диких и до-
машних бактрианов материал еще более скуден, чем для выявления кра-
ниологических отличий. В Зоологическом институте Академии наук СССР
имеются два скелета и в Стокгольме один. Кроме того, один скелет был
доставлен в Лондон из Кашгарии и один в Улан-Батор из юго-западной
Монголии. Последние два экземпляра до сих пор не описаны.
Все же можно отметить отчетливо выраженную легкость сложения
(точнее, лептосомность, т. е. «узкотелость» конституции) у дикой формы.
CAMELUS
109
Например, у крупного дикого экземпляра (ЗИН, № 11173) наибольшая
длина предплечья 613 мм, у мелкого домашнего из Кашгарии (ЗИН,
№ 24637) 536 мм, а у крупного домашнего в музее Ленинградского вете-
ринарного института (ЛВИ) 581 мм. Зато ширина дистальной суставной
поверхности предплечья у дикого только 82.5 мм, тогда как у мелкого
домашнего 83, а у крупного домашнего 93. Наибольшая длина бедренной
кости у № 11173 552 мм, у № 24637 496 мм, у экземпляра ЛВИ 517 мм;
в то же время наибольшая ширина дистальной части femur у первого
116 мм, у мелкого домашнего 121 мм, а у крупного домашнего 128 мм.
I Гаиболыпая длина tibia у № 11173 495 мм, у № 24637 431 мм, у экземпляра
Л ВИ 451 мм; ширина же дистального конца этой кости у первого 87 мм,
у второго 86 мм и у третьего 92 мм. Аналогичные соотношения легко
усмотреть в промерах метаподий. Астрагал у дикого верблюда менее
массивен, чем у домашнего. Например, длина между точками сближения
седловин проксимальной и дистальной суставных поверхностей астрагала
у крупного дикого № 11173 60 мм, у мелкого синьцзяньского домашнего
№ 24637 60 мм, у крупного домашнего (ЛВИ) 63 мм; наибольшая длина
латерального края астрагала у № 11173 75 мм, у мелкого домашнего 77 мм,
у крупного домашнего 81 мм. Промеры отдельных костей для экземпляров
№ 11173 (ЗИН) и из ЛВИ приведены в сравнительных таблицах у Я. И.Ха-
несона (1954а).
История и географическое распространение (рис. 21). Ископаемых
остатков собственно Camelus bactrianus /erus пока не обнаружено. По-види-
мому, к рассматриваемой форме близок верхнеплейстоценовый верблюд, от
которого сохранилась первая фаланга (№ 3568, ЗИН), добытая из зо-
лотоносных песков речки Узунжул, в бассейне Енисея, сравнительно
недалеко от Минусинска. Узупжулский верблюд (С. bactrianus aff.
I eras') является элементом фауны, в состав которой входили «сайга, шер-
стистый носорог и мамонт; таким образом, мы должны ее отнести к плей-
стоцену и притом к наиболее позднему отделу его» (Громова, 1932). Уточ-
ненные промеры этой фаланги опубликовал Я. И. Хавесон (1954а). Ока-
палось, что данная кость не массивнее, чем у домашнего бактриана, как
полагала В. И. Громова, но действительно более массивна, чем у скелета
крупного экземпляра дикого бактриана.
Нет оснований сомневаться в том, что дикий бактриан восходит к форме,
достаточно близкой к плейстоценовому Camelus knoblochi Nehring. Однако
чрезвычайно массивный С. knoblochi из среднего плейстоцена бассейнов
Волги и Урала обнаруживает некоторые черты специализации, исключаю-
щие возможность того, что дикий бактриан является потомком этого вида.
Скорее всего, С. bactrianus ferus происходит от какой-то несколько ме-
нее крупной формы плейстоценового верблюда из внутренней Азии. Круп-
ный верблюд был открыт и в плейстоцене Шара-Оссо-Гол в Северном Ки-
тае. Последняя форма была отнесена М. Булем и П. Тейлар де Шардэном
(Boule et Teilhard de Chardin, 1928) к найденному на Волге виду С. kno-
blochi. Очень вероятно, что то был особый подвид, доныне не обозна-
ченный.
Особенно интересна находка черепа плейстоценового верблюда из
района, примыкающего к Кулундинской степи в южной Сибири (Буйнов-
г-кий и Хавесон, 1953). Будучи значительно крупнее современных двугор-
бых верблюдов, у которых базальная длина черепа максимально состав-
ляет около 520 мм (Хавесон, 1948), череп приалтайского верблюда, имею-
щий базальную длину 555 мм, определенно уступает по величине С. kno-
blochi из бассейнов Волги и Урала с базальной длиной около 610 мм.
по
GAM ELI DAE
Еще в самом начале XX в. убежищем дикого верблюда в Китае явля-»
лась часть пустынного высокогорья, ограниченная хребтом Алтынтаг
и некоторыми другими смежными с ним горными цепями системы Нань-
шаня, а также гораздо более низкие горы Куруктаг и Чолтаг. Оттуда
в прохладное время года этот зверь проникал в пески Такла-Макан и
Кумтаг, а также на западную окраину Цайдама. Очаг обитания диких
бактрианов имелся и в зоне хребтов Бэйшаня, западнее низовьев Эдзин-
Гола, вблизи района, где хозяйствовали торгоуты, непосредственно не
преследовавшие диких верблюдов. Отмеченное поголовье было более
или менее изолировано от основного ареала диких верблюдов большой
дорогой из Ганьсу в Джунгарию. По-видимому, оно было связано с по-
головьем этих зверей в Заалтайской Гоби в Монголии.
Вблизи Лобнора (на Алтынтаге и в низовьи Тарима) нанятые охот-
ники добыли для Н. М. Пржевальского трех диких бактрианов (экспе-
диция 1876—1878 гг.). Координаты места добычи не известны. Впослед-
ствии Н. М. Пржевальский видел следы этих зверей южнее главного
хребта Алтынтага, на высоте более 3000 м, в пункте около 38°30' с. ш.
и 90 в. д. (Пржевальский, 1888).
Охотники, посланные экспедицией братьев Грум-Гржимайло 1889—
1890 гг., убили дикого верблюда в пустыне между Чолтагом и Курукта-
гом вблизи урочища Палуан-Булак (около 41°30' с. ш. и 91° в. д.).
В. И. Роборовский добыл два экземпляра в декабре 1893 г. вблизи дороги
от Хами на Ga-Чжоу в районе около 41°50' с. ш. и 94°20' в. д. По опре-
делению Я. И. Хавесона, эти два экземпляра, по-видимому, были есте-
ственными гибридами между домашней и дикой формами. В ноябре 1893 г.
местные охотники убили для П. К. Козлова самца дикого верблюда
в Куруктаге, юго-восточнее оазиса Кызыл-Сыныр, у ключа Ногусту
(около 40°50' с. ш. и 89°36' в. д.). В январе 1894 г. П. К. Козлов добыл
самку с почти доношенным эмбрионом в пустыне Кумтаг (урочище Туя,
около 40°15' с. ш. и 91°50' в. д.), а затем еще двух самок в той же пустыне
вблизи урочища Тограк-Худук (около 40°30' с. ш. и 92°31' в. д.). В фев-
рале 1896 г. люди, нанятые С. Хедином (экспедиция 1893—1897 гг.),
убили трех диких верблюдов в пустыне севернее р. Керии, пропадающей
в песках Такла-Макан; однако черепа не были взяты. Зато в экспедицию
1899—1902 гг. С. Хедину удалось взять в коллекцию (в феврале 1901 г.)
Рис. 21. Распространение дикого двугорбого верблюда. (По Свену Хедиву (в Китае;
в начале XX в.) и по Э. М. Мурзаеву (в пограничной зоне между Китаем и Монголией;
по экспедиции 1943 г.), а также по материалам различных исследователей. Составил
Я. И. Хавесон, 1956 г.).
1 — Алтимиш-Булак, 41 ° с. ш. и 90° в. д. (II 1901, добыт экземпляр № 3613; S. Hedin).
2 — хребет Алтынтаг, высота 3000 м, 38°30' с. ш. и 90° в. д; (были замечены следы;
Пржевальский, 1888 : 279). 3 — низовья р. Кара-Муран, около 38° с. ш. и 84°30' в. д.
(общее указание, там же : 377). [? Я. X.]. 4 — севернее городов Керия и Хотан (общее
указание, там же : 471). (?? Я. X.]. 5 — пустыня между Чолтагом и Куруктагом,
урочище Палуан-Булак, около 41°30' с. ш. и 91° в. д. (убит экземпляр; Грум-Гржи-
майло, 1896, I : 417). 6 — 41°50' с. ш. и 94°20' в. д. (XII 1893, добыт гибридный эк-
земпляр № 8676; Роборовский). 1 — 40°15' с. ш. и 91°50' в. д., урочище Туя (I 1894,
добыт экземпляр № 7937; Козлов). 3 — 40°30' с. ш. и 92°30' в. д., урочище Тограк-
Худук (I 1894, добыт экземпляр № 8673; Козлов). 9 —• 40°50' с. ш. и 89°36' в. д., уро-
чище Ногусту; XI 1893, добыт экземпляр № 8672; Козлов). 70—41 °20' с. ш. и 94°8' в. д.
(видели во время пути; S. Hedin, 1944 : 250). 11 — 40°30' с. ш. и 92°30' в. д. (видели
следы; S. Hedin, 1944 : 257). 12 — 42° с. ш. и 98° в. д. (указание со слов проводников
на места водопоя диких верблюдов; Lattimore, 1928 : 219). 1-1 — 38°30' с. ш. и 95° в. д.
(указание на охраняемое обычаем убежище самок и молодняка диких верблюдов;
Роборовский, 1900 : 471).
Рис. 21.
112
CAMELIDAE
полный скелет самца из Куруктага (урочище Алтимиш-Булак, около
41° с. ш. и 90° в. д.). Много позднее, в трудах шведско-китайской экспе-
диции 1927—1935 гг. отмечено, что в ноябре 1934 г. к востоку-северо-
востоку от Лобнора (около 41°20' с. ш. и 94° 8' в. д.) С. Хедин видел,
как он считал, дикого верблюда, который, пустившись в бег, быстро
исчез на севере. Несколько западнее члены экспедиции увидели «шесть
диких верблюдов». Около 40°30' с. ш. и 92°30' в. д. «следы диких верблю-
дов пересекали местность во всех направлениях».
Таким образом, после 1901 г., когда был добыт упомянутый сейчас
экземпляр на Алтимиш-Булаке, конкретные сведения относительно су-
ществования данного зверя в Синьцзяне как бы внезапно оборвались.
Зато появилось несколько сообщений о диком верблюде на юго-за-
падной окраине Монголии.
В ноябре 1899 г. В. Ф. Ладыгин, участник Тибетской экспедиции
П. К. Козлова, видел свежие следы диких верблюдов вблизи меридиана
города Сучжоу в горной пустыне к северу от монголо-китайской границы.
О. Лэттимор (Lattimore, 1928), пересекая в 1926 г. степь к западу от
Эдзин-Гола, собрал сведения о том, что дикие верблюды посещают группу
ключей в районе около 42° с. ш. и 98° в. д., т. е. вблизи монгольской
границы.
В 1927 г. монгольский краевед А. Д. Симуков убил на юго-западной
окраине Монголии, вблизи нагорья Цаган-Богдо, крупного самца ди-
кого (как он полагал) верблюда, причем полный скелет животного был
доставлен в Улан-Батор. К сожалению, этот скелет до сих пор никем не
описан. Судя по дневнику А. Д. Симукова, добытый верблюд имел до-
вольно большие горбы (высотою в 32 см), тогда как у диких, по Н. М. Прже-
вальскому, их высота оказывается меньше 20 см; на основании этого
можно предполагать, что данный экземпляр, хотя и был диким по образу
жизни, но содержал в основном кровь домашнего.
Летом 1943 г. географ Э. М. Мурзаев, ботаник А. А. Юнатов и зоолог
А. Г. Банников, совершая экспедиционную поездку по Заалтайской Гоби,
подтвердили наличие там таких верблюдов, которых местные монголы
считают дикими. Однако из числа виденных ими экземпляров ни один
не попал в коллекцию. Наконец, в 1947 г. около хребта Эдэрэнгин-Нуру
в Заалтайской Гоби были добыты самец и самка с почти доношенным
эмбрионом. Череп самки впоследствии попал в остеологическое храни-
лище Московского университета. По определению Я. И. Хавесона, этот
череп принадлежал особи естественного гибрида дикого верблюда с до-
машним, причем доля крови домашнего не велика. В этом отношении
можно усмотреть аналогию для монгольского дикого бактриана и ди-
кого коня тарпана южнорусских степей, последние представители ко-
торого имели (по исследованию проф. Браунера) явно гибридное про-
исхождение.
Что касается раннеисторического и доисторического ареала дикого
бактриана, то он, по-видимому, простирался далеко на восток в сторону
Ордоса. К. Риттер (Ritter, 1832, 1847) полагал, что в упомянутом районе
некогда, но притом чуть ли не в историческое время, произошло одомаш-
нение двугорбых верблюдов. В пользу высокой степени вероятности по-
добного высказывания в отношении локализации одомашнения говорят
и современные культурно-исторические представления. Однако сообра-
жения К. Риттера должны рассматриваться лишь как догадка, очень
правдоподобная в отношении протяженности древнего ареала, но едва ли
удачная в смысле установления времени одомашнения дикого бактриана.
CAMELUS
113
Торгоуты и другие монголы мало преследовали диких верблюдов,
хотя и не мешали охотиться другим. Кое-где они, по-видимому, про-
пилили некоторую заботу о воспроизводстве поголовья этих зверей. Во
исяком случае, В. И. Роборовский (1900) сообщил, что урочище Куку-
Сай на плато Цайдама, вблизи северо-западной оконечности хребта Рит-
тера (около 38"30' с. ш. и 95° в. д.), являлось убежищем для самок и
молодняка диких верблюдов, где охота на них порицалась обы-
чаем.
Преследование этих животных практиковалось преимущественно охот-
никами из мусульманских народностей Синьцзяна. В XIX в. оно уси-
лилось благодаря росту населения. Освоение горных и пустынных паст-
бищ, лишающее зверей водопоев, а также проникновение современного
оружия в самые глухие районы Центральной Азии, по-видимому, уже
а первом десятилетии XX в. настолько подорвали поголовье данного
зверя, что он оказался на грани исчезновения. В. И. Роборовский (1900)
отметил, что охотники из Люкчуна убивали за зиму до десятка диких
верблюдов в пустыне, примыкающей к Турфанской котловине. В. Ф. Ла-
дыгин (1900) сообщил некоторые подробности о беспорядочном истребле-
нии монгольских хабтагаев охотниками чаньту, т. е. мусульманами Ха-
мийского оазиса.
Поэтому самый факт существования в настоящее время диких бактриа-
нов в Синьцзяне вызывает сомнение, разрешить которое может только
специальное обследование так называемой Гашуньской Гоби, Алтынтага
и смежных с ними хребтов системы Куньлуня и Наныпаня, а также Бэй-
нншя и песков Такла-Макан. Популяция же хабтагая в юго-западной
Монголии, по-видимому, состоит из естественных гибридов настоящих
диких с домашними бактрианами. Тем не менее и эта гибридная форма
представляет выдающийся интерес как зоологический и зоотехнический
объект и требует всемерной охраны.
Биология дикого верблюда не может считаться сколько-нибудь
удовлетворительно изученной, и значительная часть того, что известно
и этом отношении, связана с рассказами местных охотников, проверить
которые не легко.
Несомненно, что для данного зверя характерны сезонные миграции.
В Кашгарии диких верблюдов видели (преимущественно летом) на вы-
сокогорье северной окраины Тибетского плато. Н. М. Пржевальский
(1878) был изумлен, многократно встречая в этой части Центральной Азии
следы пребывания верблюдов в очень тесных ущельях и на таких скло-
нах, по которым с трудом взбираются охотники. Там следы верблюдов
встречались вместе со следами архаров и куку-яманов (Pseudois nahoor\
llodgs.). В более прохладное время года верблюды устремлялись в не-,
счаную страну Кумтаг и даже в окруженные многочисленными, плотно
населенными оазисами, но крайне малодоступные для людей пески гро-
мадной пустыни Такла-Макан. Впрочем, в настоящее время трудно себе
представить, чтобы дикие верблюды могли регулярно мигрировать
а пустыни великой котловины Кашгарии, минуя возделываемую зону,
опоясывающую значительную часть границы этих пустынь.
В северной части ареала кашгарского дикого верблюда наблюдались
сезонные передвижения этого зверя к большому пресному оз.Баграшкуль
(Грум-Гржимайло, 1896). Несомненно, что эти миграции были связаны
с недостатком водопоев во время летней жары.
С. Хедин (Hedin, 1905, III) перечислил шесть более широких и от-
крытых ущелий, точнее проходов Алтынтага, по которым, согласно ука-
8 Фауна СССР, И. И. Соколов
114
CAMELIDAE
заниям местных охотников, дикие верблюды ежегодно устремлялись из
пустынь восточнее Лобнора в горы и обратно.
В Заалтайской Гоби дикие верблюды пасутся летом в более высоких
частях страны, а зимою кормятся в расположенной южнее пустыне, со-
стоящей из многочисленных невысоких каменистых гряд. Э. Дагва (1954)
считает, что в большинстве дикий верблюд Заалтайской Гоби уходит
к зиме за пределы Монголии. Однако А. Д. Симуков, оставивший очень
интересные записи об этом звере в Монголии (1928, 1938; см. также
опубликованные А. Г. Банниковым в 1954 г. обширные извлечения из
дневников путешествия А. Д. Симукова на запад Монголии в 1927 г.,
а также из его рукописи «Западная Гоби», составленной в 1935 г.), со-
мневался, чтобы популяция хабтагаев Монголии имела контакт с попу-
ляцией диких верблюдов пустынь, окружающих Лобнор.
А. Г. Банников (1947) характеризовал дикого верблюда как обитателя
кустарниковых пустынь.
А. Д. Симуков (1935) считал, что дикий верблюд характерен для щеб-
нистой пустыни равнин Западной Гоби. При этом в поисках корма и
воды зверь «заходит в горы, держится в мелкосопочнике и холмах, но
настоящее его место —широкие пустынные равнины и долины, где его
рост и острое зрение позволяют видеть опасность с весьма большого рас-
стояния». «Несомненно, что дикий верблюд знает водопои на огромном
пространстве, а кроме того, чует воду на весьма большом расстоянии,
так как обоняние у него прекрасное». Вообще, следует отметить, что
у верблюда нервная система и органы чувств достигают значительного
развития. По А. П. Берггрину (1934), у домашнего бактриана головной
мозг достигает веса в 730 г, причем большой мозг обладает глубокими бо-
роздами и более развит, чем у рогатого скота и у лошади.
Относительно пищи дикого верблюда Э. М. Мурзаев (1948) писал,
что эти звери «пасутся в пустынях, где растут парнолистники, саксаул,
солянки, луки, ковыльки: в оазисах они охотно едят листья тополя».
Э. Дагва (1954) несколько уточнил это указание, отметив, что в юго-
западной Монголии «летом дикий верблюд поедает гобийский ковыль,
многокорешковый и монгольский лук, пижму тысячелистную, песчаную
и холодную полынь, баглур, камыш, дерис, солянку воробьиную, ревень
белокорневой, а также ветки и листья караганы, саксаула и тополя евфрат-
ского».
Дикий верблюд кормится, по-видимому, утром и вечером, а в полдень
и ночью лежит (Симуков, 1935). О полуденном отдыхе этого животного
писали также Э. М. Мурзаев (1948) и Э. Дагва (1954).
Гон происходит в начале февраля (Дагва). Н. М. Пржевальский
(1878) писал, что «время течки у дикого верблюда бывает зимою — с по-
ловины января почти до конца февраля» (по старому стилю!), т. е. до
второй декады марта. Э. М. Мурзаев (1948) указал на январь—февраль.
Молодняк рождается в марте. Двойни не известны. Самки оплодотво-
ряются на третьем году жизни (Пржевальский, 1878).
Ряд авторов писал о свирепых драках между самцами во время яри,
но решительно никто из писавших не наблюдал подобных столкновений,
хотя все это и правдоподобно, судя по тому, что самцы домашних бактриа-
нов кусают друг друга во время случного сезона. Рассказывают также,
что одиночные дикие самцы нападают на группы домашних верблюдов,
жестоко избивают домашних самцов и отбивают самок, так что «нередки
случаи скрещивания диких самцов с домашними верблюдицами» (Дагва,,
1954).
RUMINANTIA
116
Дикие верблюды держатся небольшими косяками или даже парами,
редко собираясь в группы более десяти-пятнадцати голов. Во главе
косяка, по утверждению рассказчиков, находится осторожный старый
самец, зорко осматривающий окружающую местность. Считается, что
зти звери обладают исключительно тонким зрением, слухом и обонянием.
Поэтому охотники обыкновенно подкарауливали зверей на водопоях.
Однако, судя по описаниям В. И. Роборовского и П. К. Козлова (1899,
1900), при наличии хорошего оружия, тренированные охотники добывали
диких верблюдов и с подхода, причем эта охота не оказывалась особенно
утомительной. В. Ф. Ладыгин (1900), со слов проводника, обрисовал
картину не то что охоты, а избиения монгольских хабтагаев людьми,
прибывавшими зимою из Хамийского оазиса в Заалтайскую Гоби на
арбах, запряженных лошадьми или быками. Впрочем, возможно, что
хабтагаи, будучи до некоторой степени смешанного, происхождения,
не столь сторожки, как собственно дикие верблюды Алтынтага, Куруктага
п Кумтага.
Хозяйственное значение. В начале XX в. дикие верблюды стали
настолько редкими, что теперь не приходится говорить об их хозяйствен-
ном значении. Во всяком случае, А. Д. Симуков (1938) писал: «В былое
время некоторые охотники прилегающих к Западной Гоби хошунов
(мелкая территориальная единица в Монголии,— Я. X.) специально
занимались охотой на дикого верблюда. Эта охота, несмотря на кажу-
щуюся выгоду (большое количество прекрасного мяса и жира, ценная
своею прочностью шкура), требовала столько времени, сил и средств,
что, несомненно, была скорее романтическим спортом, нежели экономи-
чески целесообразным предприятием. Охотники выезжали верхом на
лучших верблюдах поздней осенью, снаряжаясь на срок от двух недель
до полутора-двух месяцев и рыскали по безводным просторам, отыскивая
свежие следы желанного зверя». Проводник А. Д. Симукова говорил,
что бывали трагические случаи, когда увлекшийся охотник, истомив
верхового верблюда, оставался в пустыне пешим и без воды.
Совершенно очевидно, что необходимо эффективно оградить дикого-
верблюда от грозящего ему истребления. Б Монгольской Народной
Республике охота на хабтагая запрещена.
Подотряд —Жвачные
Ковалевский В. О., 1875, Изв. Общ. любит, естествозн,, антроп. и. этно-
граф., XVI, 1 : 1—59 (морф., эвол.); Schlosser М., 1887, Morphol. Jahrbuch,
XII : 1—36 (эвол., сист.); Р о с о с k R., 1910, Proc. Zool. Soc. London : 840—896
(морфол.); P о с. о с к R-, 1918—1919, Ann. Magaz. Natur. Hist. (ser. 9), I ; 426—435;
ll : 125—144, 214—225, 367—374, 440—459; III : 103-114 (морф.); Matthew W.
nod W. Granger, 1925, Amer. Mus. Novitates, 195:1—12; 196 : 1—11. (эвол.);
Colbert E., 1941, Amer. Mus. Novitates, 1135 : 1—24 (морф., эвоп.); Pilgrim _G.,
1041, Biol. Review, vol. 16, No. 2:134—163 (ист.); Simpson G., 1945, Bull. Amer.
Mus. Natur. Hist., 85 : 143—162-J-265 —272 (эвол., классиф.).
Характеристика подотряда. Высоконогие, в большинстве случаев
стройные животные. Число пальцев — два или четыре, но функционально
конечность всегда двупалая, так как боковые пальцы, если и имеются,-
то недоразвиты и в нормальных условиях при ходьбе обычно не ка-
саются почвы. Метаподий боковых лучей стопы и кисти в той или иной
степени редуцированы и не сочленяются с костями заплюсны и запястья;
от боковых метаподий сохраняются обычно только проксимальные или»
8*
116
RUMIN ANTIA
дистальные рудименты; часто, особенно на задних конечностях, они ис-
чезают вовсе. Метаподии средних (III и IV) лучей, как правило, слиты
и образуют непарную кость (os cannon). Локтевая кость в дистальной
и средней части значительно редуцирована, часто срастается с лучевой.
Еще большей редукции подвергается малая берцовая кость; от нее в ка-
честве небольшой самостоятельной кости сохраняется только дистальный
конец, так называемая лодыжковая кость (os malleolare), сочленяющаяся
с большой берцовой, пяточной (calcaneus) и таранной костью (astragalus)
и функционально составляющая часть заплюсны. Исключение составляют
члены семейства оленьковых (Tragulitiae), у которых малая берцовая
кость сохранилась полнее и в нижней половине сливается с большой бер-
цовой. В запястье малая многоугольная кость (trapezoideum) сливается
с головчатой (capitaturn s. magnum) или рудиментарна (у оленьковых);
большая многоугольная кость (trapezium) исчезает или сливается с пре-
дыдущими костями. В заплюсне характерно для всех групп жвачных
слияние кубовидной кости (cuboideum) с ладьевидной (naviculare). Сли-
ваются в одну также вторая и третья клиновидные кости (cuneiforme
II и III). На дистальном суставном блоке средних метанодий имеется
в той или иной мере выраженный срединный гребень. Основания попереч-
ных отростков шейных позвонков прободаются каналом для прохожде-
ния позвоночных артерий.
В отличие от мозоленогих, конечные фаланги пальцев одеты настоя-
щими копытами. Вместо клювовидного отростка нижняя дуга атланта
несет на вентральной поверхности лишь слегка выступающий вниз буго-
рок. Зубовидный отросток второго шейного позвонка (эпистрофея) имеет
форму полого полуцилиндра. Грудных позвонков тринадцать, редко че-
тырнадцать.
Сосцевидная (мастоидная) часть позади чешуйчатой кости выходит
на наружную поверхность черепа. Глазница всегда замкнута. Лобные
кости обычно несут той или иной формы выросты, рога. Сагиттальный
стреловидный гребень на черепе не развит, хотя бы теменные гребешки
обеих сторон и соприкасались друг с другом. Суставная ямка для сочле-
нения с нижней челюстью и суставной мыщелок последней имеют попе-
речно вытянутую форму. Лицевая и глазничная часть слезной кости
развиты равномерно. На лицевой поверхности ее часто имеется предглаз-
ннчная ямка для предглазничных кожных желез. Между слезными, но-
совыми, лобнымй и верхнечелюстными костями у многих форм имеются
так называемые этмоидальные щели.
Резцы в верхней челюсти отсутствуют. В нижней они имеют лопатко-
видную или долотовидную форму. Верхние клыки также могут исчезать,
но у безрогих форм они, наоборот, получают сильное развитие и высту-
пают из ротовой полости вниз (оленьки, кабарга). Клыки нижней челюсти
примыкают к резцам и приобретают форму последних. Заднекоренные
зубы лунчатые (селенодонтные). У некоторых групп развивается гипсо-
донтия (рис. 22, В). Переднекоренные (премоляры) образуют сплошной
ряд с заднекоренными. Первый премоляр (Р1 и PJ не развивается. Вто-
рой премоляр не имеет форму клыка, как у верблюдов. Между клыками
И коренными зубами имеется значительный беззубый промежуток (диа-
стема).
Кожа имеет нормальный волосяной покров, состоящий из более тонкой,
чем у свиней, ости и тонкого, нежного пуха (подшерстка). Образование
толстого подкожного слоя жировой ткани не имеет места. Помимо свой-
ственных всем млекопитающим молочных, сальных и потовых желез,
RUMINANTIA
117
в кожном покрове большинства жвачных образуется ряд специальных,
свойственных только им, кожных желез. Главные из них:
1) межкопытные, или межпальцевые в виде мешкообразного или бутыл-
кообразного впячивания кожи, открывающегося или между основаниями
копыт, или несколько выше их на передней стороне конечностей;
2) предглазничные железы различной величины и формы, расположен-
ные в соответствующих углублениях на поверхности слезных костей
черепа;
3) карпальные железы, внешне выступающие в виде подушки или
пучка волос на передней (дорзальной) стороне конечностей, пониже за-
пястного сустава (имеются только у некоторых полорогих, Bovidae)',
4) тарзальные (предплюсневые) и метатарзальные (плюсневые) железы,
также имеющие вид подушек или пучков выступающего волоса; первые
расположены на внутренней (медиальной) стороне скакательного (голено-
стопного) сустава, а вторые — ниже, на внутренней стороне плюсны;
5) паховые железы — мешкообразные впячивания кожи в задней части
живота по бокам от молочной железы (имеются только у некоторых
цолорогих, Bovidae).
Кожные железы выделяют различной консистенции и запаха секрет,
служащий, вероятно, для целей распознавания и нахождения животными
друг друга по следу. Функция некоторых желез связана с половой дея-
тельностью. Наличие или отсутствие отдельных желез в ряде случаев
является систематическим признаком той или иной группы.
Желудок сложный, разделен на ясно отграниченные четыре (редко три)
отдела: рубец, сетку, книжку и сычуг. Собственно желудок, пищевари-
тельную его часть, представляет лишь последний из названных отделов.
В процессе пищеварения имеет место отрыгивание заглоченной в первый
отдел желудка пищи и ее вторичное пережевывание (жвачка). Плацента
множественно котиледонная, за исключением оленьковых. Молочная же-
леза двух- или четырехдольчатая, расположена в области задней части
брюшной стенки.
Эволюция и классификация. Жвачные появились на геологической
сцене в эоцене в виде мелких форм, занимавших по сравнению с нежвач-
ными незначительное место в фауне той эпохи. В настоящее время они
представляют наиболее прогрессивную и многочисленную группу, копыт-
ных, еще не пережившую своего расцвета. Эволюция жвачных шла в на-
правлении приспособления к питанию исключительно растительной пищей
и быстрому бегу как средству спасения от врагов и способу использо-
вания обширных, но скудных и лишенных водопоя кормовых площадей.
С этим связаны: форма лунчатых коренных зубов, приспособленных к пе-
режевыванию жесткой растительной пищи, удлинение средних и редукция
боковых лучей четырехпалой конечности, функционально превращающейся
в двупалую, усиление центральных лучей (III и IV) и слияние их мета-
подий в одну непарную кость, повышающее крепость конечности. Услож-
нение желудка также связано с приспособлением к питанию труднопере-
варимой, богатой клетчаткой, растительной пищей и с защитой от воз-
можных врагов. Объемистый первый отдел желудка, рубец, позволяет
животному наскоро заглатывать большое количество слабо или вовсе
пепережеванной пищи и перерабатывать ее в укрытии, при спокойной
обстановке. Под действием слюны и расщепляющих клетчатку микро-
организмов (инфузорий) пища в рубце мацерируется и небольшими пор-
циями отрыгивается для вторичного пережевывания в ротовую полость.
Вторично пережеванная, она поступает для дальнейшей переработки
118
RUMINANTIA
пищеварительными соками и бактериями в следующие отделы желудка
и кишечник. Это направление эволюции позволило малочисленным вна-
чале жвачным стать победителями в жизненной борьбе и вытеснить боль-
шую часть остальных, менее приспособленных к меняющимся условиям
среды групп копытных.
Как и другие группы парнопалых, жвачные ведут свое начало от при-
митивных нижне- или среднеэоценовых палеодонтов (Palaeo donta). Самые
ранние их представители появились во второй половине эоцена.1 Лучше
других известен верхнеэоценовый род Archaeomeryx Matthew and Gran-
?er из Монголии — животное величиной с лисицу (Matthew and Granger,
925). Судя по строению заплюсны и селенодонтным коренным зубам,
это был типичный представитель жвачных, но многие черты примитив-
ности указывают на то, что хотя он и не мог быть прямым предком всех
последующих Ruminantia, однако стоял весьма близко к таковому. Зад-
ние конечности у Archaeomeryx были значительно длиннее передних, ука-
зывая на то, что передвигался он прыжками. Боковые лучи (II и V) стопы
и кисти сохраняли полностью и метаподий и фаланги. Средние метаподий
и на передних и на задних конечностях были еще самостоятельными.
Рогов не было. Нижние клыки уже приняли форму резцов и заняли
место рядом с последними, но зато в верхней челюсти еще сохранялись
резцы. Archaeomeryx вместе с рядом других примитивных олигоценовых
и нижнемиоценовых форм Северной Америки образуют семейство Нурег-
tragulidae, особенностью которого являлась клыкообразная форма увели-
ченных и отставленных от других зубов передних премоляров нижней
челюсти (PjhPj).
Морфологически близким и, весьма вероятно, прямым предком совре-
менных высших жвачных (Ресога) был род Gelocus Aymard из нижнего
олигоцена Европы. Верхние резцы у Gelocus были утрачены, передние
премоляры не имели формы и положения клыка. На задних конечностях
средние метаподий уже слились в одну кость, но на передних были еще
раздельны. Близок современным оленькам (Tragulidae) и иногда вклю-
чается с ними в одно семейство.1 2 Сам Gelocus может рассматриваться
как один из непосредственных предков полорогих (Bovidae). Начавшаяся
рано в группе Gelocidae дивергенция привела к появлению форм (роды
Lophiomeryx, Prodremotherium и некоторые другие), послуживших исход-
ными для других семейств Ресога.
Из других вымерших групп древних жвачных следует упомянуть
протоцератид (Protoceratidae) — вероятных потомков гипертрагулид, суще-
ствовавших с нижнего олигоцена до нижнего плиоцена на территории
Северной Америки. У представителей этой группы впервые в истории
парнопалых появились рога. Последние представляли две или три пары
костных выростов на верхнечелюстных, носовых и лобных костях, по-
крытых, вероятно, кожей с волосом, как у современных жираф. В совре-
менной фауне протоцератиды не оставили потомков.
Современные жвачные составляют пять или шесть семейств.
1) Оленьковые (Tragulidae) — наиболее примитивная группа, сохра-
нившая большое число архаических черт, свойственных общим предкам
подотряда. Рогов нет. Полностью сохранились, хотя и в более слабой
1 В основание ствола Ruminantia ставят эоценовую группу Amphime.rycidae,
известную по отдельным костям конечностей и нижним челюстям, в которых впервые
появляются некоторые признаки жвачных.
2 Чаще Gelocus с рядом близких к нему родов выделяется в отдельное семейство
Gelocidae (Schlosser, 1887; Simpson, 1945).
RUMINANTIA
119
Рис. 22. Верхний заднекоренной
зуб.
А — оленя (брахиодонтный),
Б — козла (гипсодонтный).
степени развитые, локтевая, малая берцовая, а также кости боковых
лучей запястья. Метаподий центральных лучей полностью слились только
на задних конечностях; на передних же они остаются или совсем самосто-
ятельными, или сливаются лишь частично. В желудке развиты только
три отдела, книжка остается в зачаточном состоянии. Плацента диффуз-
ная. Включает всего два современные рода: Tragulus Brisson из юго-
восточной Азии и Hyemoschus Gray из Экваториальной Африки.
Все остальные, так называемые высшие жвачные, имеют вполне разви-
тую цевку на всех конечностях, четырехраздельный желудок, множественно
котиледонную плаценту, и обычно объединяются в надсемейство (или ин-
фраотряд) Ресога, включающий остальные
пять семейств.
2) Кабарги (Moschidae), особенностью
которых является отсутствие рогов, вме-
сто которых имеются сильно развитые у
самцов верхние клыки. Часто включается
в состав следующего семейства.
3) Олени (Cervidae), в подавляющем
большинстве имеющие рога у самцов и
лишенные выдающихся клыков. Рога оле-
ней представляют выросты лобных ко-
стей, ежегодно сбрасываемые и в период
их роста одетые покрытой волосом кожей.
4) Жирафовые (Gira'ffidae) — современ-
ные обитатели африканских лесов и са-
ванн. От других жвачных отличаются тем,
что рога их представляют несменяемые
выросты лобных костей, в течение всей
жизни покрытые нормального строения
кожей с волосами; вторую особенность представляют уплощенные, но
раздвоенные коронки нижних резцов.
5) Вилороги (Antilocapridae), имеющие рога в виде постоянных не-
сменяемых выростов лобных костей, одетых роговым чехлом из видоиз-
мененного эпидермального слоя кожи, ежегодно сбрасываемым и заме-
няемым новым. Единственный в современной фауне представитель се-
мейства вилорог (Antilocapra americana Ord.) живет в прериях Северной
Америки.
6) Полорогие (Bovidae) — группа, охватывающая так называемых анти-
лоп, быков, козлов и баранов. Отличительной особенностью этого семей-
ства являются постоянные несменяемые рога, одетые в также несменяемый
роговой чехол, представляющий видоизмененный эпидермальный слой
кожи.
Последние два семейства филогенетически более близки друг к другу,
чем к другим жвачным и поэтому иногда объединяются в одно семейство
или надсемейство Bovoidea.
В фауне СССР имеются представители семейств кабарог, оленей и
полорогих.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ СЕМЕЙСТВ ЖВАЧНЫХ
ФАУНЫ СССР
1 (2). Размеры небольшие; длина тела от конца морды до корня хвоста
меньше .120 см; длина черепа меньше 170 мм. Ни у самцов, ни у са-
мок рогов нет. На хвосте имеется железа, открывающаяся на верх-
120 MOSCHIDAE
ней его стороне. У самцов длинные, саблевидные верхние клыки
сильно выступают вниз из-под верхней губы. У самок клыки ко-
роче, но все же их длина не меньше высоты коронок коренных зубов.
В черепе длинные верхние отростки межчелюстных костей дости-
гают середины длины носовых. Теменные гребешки сильно сближены
в средней части, соединяются друг с другом, а если не соединяются,
то расстояние между ними меньше длины последнего коренного
зуба (МР)...............Сем. Moschidae — Кабарги (стр. 120).
2 (1). Размеры средние или большие; длина тела от конца морды до корня
хвоста не менее 120 см; длина черепа не меньше 170 мм. Самцы всегда,
а часто и самки, имеют рога. Железы на хвосте отсутствуют. Верхние
клыки короткие, меньше высоты коренных зубов. Верхние отростки
межчелюстных костей не достигают уровня середины длины носовых.
Наименьшее расстояние между теменными гребешками больше длины
последнего заднекоренного зуба (М3).
3 (4). Рога, если имеются, то ветвистые (у взрослых), состоящие во вполне
развитом состоянии из ничем не прикрытой кости. Они ежегодне
сбрасываются и вырастают вновь. Во время роста рога покрыты
, кожей с волосами. Зубы брахиодонтные (рис. 22, Л). Отверстие,
ведущее из глазницы в носослезный канал, двойное. Желчный пу-
зырь отсутствует............Сем. Cervidae — Олени (стр. 137).
4 (3). Рога, если имеются, то неветвистые и представляют постоянные,
не спадающие выросты лобных костей, покрытые в течение всей жизни
несменяемыми роговыми чехлами. Зубы гипсодонтные (рис. 22, Б).
Отверстие, ведущее из глазницы в носослезный канал, одно. Желч-
ный пузырь имеется..........Сем. Bovidae — Полорогие (стр. 295).
Семейство MOSCHIDAE —КАБАРГОВЫЕ
Литература — см. описание вида Moschus moschiferus (стр. 122).
Характеристика семейства. Как самцы, так и самки лишены рогов.
Роль вторично полового признака и турнирного оружия у самцов играют
длинные, саблевидно изогнутые верхние клыки, обладающие длительным
ростом и выступающие из-под верхней губы ниже подбородка. На черепе
предглазничные ямки отсутствуют. Теменные гребешки обеих сторон
сильно сближены, иногда даже соприкасаются.
Задние конечности значительно длиннее передних, вследствие чего>
пояснично-крестцовая часть у живого животного много выше холки.
Боковые (II и V) пальцы хорошо развиты. Их копыта по величине не-
много уступают копытам средних (III и IV) пальцев. От метаподий боко-
вых лучей сохраняются лишь дистальные концы, сочленяющиеся с пер-
выми фалангами соответствующих пальцев. Пальцы способны сильна
раздвигаться, увеличивая опору тела.
Предглазничные кожные железы и железы на конечностях отсутствуют;
зато у самцов имеется свойственная только данному семейству особая
мускусная железа (кабарожья струя), расположенная на вентральной
стенке живота между препуцием и пупком. Имеется также железа на
хвосте. При печени имеется желчный пузырь. Penis с нитевидным отрост-
ком, на конце которого открывается отверстие мочеиспускательного ка-
нала. Молочная железа двудольчатая. Слизистая оболочка матки, «будучи
расположена шестью узкими продольными складками очень небольшой
глубины, проходящими почти по всей длине рожков матки и местами
MOSCHUS
121
прерывающимися, никогда не имеет на свободных краях этих складок
язычкообразных выростов, которые образуют котиледонные папиллы
у Cervidae или характерные продольные ряды возвышений у Bovidae»
(Garrod, 1877 : 291).
Географическое распространение. Гималаи, Восточная Азия, вклю-
чая Индокитай; Алтай, таежная зона восточной и северо-восточной Си-
бири и Дальнего Востока до Сахалина включительно. Отсутствуют на
Камчатке.
История. Предшествующая история семейства Moschidae. не ясна.
Все известные ископаемые остатки датируются не раньше голоцена.
Кабарга обнаруживает признаки сходства с одной из древних групп
оленей — с подсемейством Palaeomerycinae. Но некоторые примитивные
черты, сближающие ее с оленьками, заставляют предполагать независи-
мое происхождение кабарог от общего с другими семействами высших
жвачных (Ресога) предка из олигоценовых трагулид Старого Света.
И в современной фауне, и в ископаемом состоянии семейство Moschi-
dae представлено одним родом кабарог — Moschus L.
Род MOSCHUS LINNAEUS-КАБАРГИ
Характеристика и географическое распространение рода — те же,
что семейства.
Классификация. Обширный ареал кабарги естественно распадается
на две части: северную и южную. Первая охватывает всю Сибирь, север-
ную Монголию и Дальний Восток. Южная часть включает Гималаи,
восточный Тибет, Китай и некоторые области юго-восточной Азии. Обе
половины сообщаются на востоке полосой вдоль побережья Желтого и
Японского морей, хотя возможно, что в настоящее время в области се-
верного Китая они уже разобщены (Allen, 1940). Как из северной, так
и из южной части ареала описано большое число видов и подвигов ка-
барги, зачастую на основании несущественных признаков, являющихся
лишь показателями индивидуальной или возрастной изменчивости. Несом-
ненно,' однако, наличие существенных морфологических отличий между
двумя основными типами кабарги — северной и южной.
Основная часть популяции кабарги из Китая по сравнению с си-
бирской имеет более крупные размеры, в частности большую длину
туловища и черепа, но зато и абсолютно, и относительно короткие конеч-
ности. Характерна для китайской кабарги также несколько более светлая
штриховатая окраска. Характер местообитаний, а также, вероятно, и
образ жизни в северной и южной части, по-видимому, различны. В свете
этих данных видовая самостоятельность сибирской и китайской кабарог
едва ли подлежит сомнению.
На территории провинции Сычуань и южной части провинции Ганьсу
в Китае, наряду с обычной Moschus sifanicus Przew., обитает другая,
сильно отличающаяся карликовая форма кабарги, по-видимому, спра-
ведливо выделенная К. К. Флеровым в качестве самостоятельного вида —
Moschus berezovskii Fler.
Таким образом, следует принимать наличие трех видов кабарги, от-
личия между которыми заключаются в следующих чертах:
1) Moschus moschtferus L. — Обыкновенная, или сибирская кабарга.
Средней величины; туловище на высоких ногах; длина пястных костей
не менее 126 мм, плюсневых — не менее 167 мм. Окраска сравнительно
темная (более подробная характеристика приводится ниже). Рас-
122
MOSCHIDAE
простран ение: Алтай, Саяны, горнолесные области восточной
Сибири, Дальний Восток, северная Монголия, возможно Дунбэй (Маньч-
журия) и Корея.
2) Moschus sifanicus Przew. — Западно-китайская кабарга. Имеет
в среднем более крупное туловище на коротких ногах; длина пястных
костей от 112 до 119 мм, плюсневых — от 143 до 148 мм. Череп имеет
вытянутую лицевую часть; длина последней, как правило, больше поло-
вины длины черепа. Слезные кости ясно вытянутые; длина их больше
ширины. Распространение: Гималаи, восточный Тибет, страна
Кам, западный и южный Китай; не ясно, как далеко заходит распростра-
нение этого вида в Индокитае.
3) Moschus berezovsk.it Fler. — Кабарга Березовского. Отличается
незначительной величиной. Длина черепа, по имеющимся данным, не
превышает 149 мм; длина пястных костей не более 89 мм, длина плюсне-
вых — не более 120 мм. Распространение: пока установлено
лишь в провинциях Сычуань и Ганьсу в Китае. Возможно, встречается
в Корее и Индокитае,
Moschus moschiferus Linnaeus — Кабарга1 (рис, 23—25)
Linnaeus С., 1758, Syst. Natur., ed. X : 66 (первооп.); Pallas P., 1779, Spicil.
zool., fasc. XIII : 2—45, tab. IV—V (морф.); E sc h ho 1 z F., 1830, Isis : 606
(сист.); Brandt F., 1833, Medic. Zool., vol. 1 : 341—351, vol. 2 : 347—348 (морф.);
Hodgson, 1839, J. Asiat Soc., Bengal, VIII : 203 (сист.); Milne-Edwards A.,
1834, Ann. sci. nat. zool. (ser. 5), II : 55—79 (морф., сист.); FlowerW., 1875,
Proc. Zool. Soc., London : 159—190 (анат.); G a r ro d A., 1877, Proc. Zool. Soc.,
London, 287—292 (анат.); Никольский A. M., 1883, Тр. СПб. общ. естество-
йсп., XIV : 182 (распр.); Buchner Е., 1890, Melanges Biol., XIII : 162—163
(сист.); Кащенко Н. Ф., 1899, Результ. Алтайск. зоол. эксп. 1898 г., По-
звоночные, Томск : 40—41 (распр.); Туркин Н. В. и К. А. Сатунин, 1902, Звери
России, Cervidae : 42—43Д-491—499 (общ., изобр.); Hollister, 1911, Proc.
Biol. Soc., Washington, XXIV : I (сист.); Соловьев Д. К., 1920, Саянский про-
мыслово-охотн. район, Пг., Госизд. : 245—248 (распр., ox.); Pocock В., 1923,
Proc. Zool. Soc., London : 181—207 (морф.); Доппельмаир Г. Г., 1926, Собо-
линый промысел на северо-вост, побережье Байкала, Верхнеудинск—Ленинград, изд.
Госплана Бур.-Монг. АССР : 193—195 (распр., биол., ох.); Тассовский Г. Н.,
1927, Произв. силы Дальн. Вост., IV : 534 (распр.); Емельянов А. А., 1927,
Произв. силы Дальн. Вост., IV : 257—266 (распр., хоз. знач.); Флеров К. К.,
1928, Докл. Акад, наук СССР (А), 24 : 515—519 (сист.); Флеров К. К., 1930,
Ежегодн. Зоол. муз. Акад, наук СССР, XXXI : 1—20 (сист., синон.); Яковлев Е.,
1930, Тр. Зоол. секц. Средне-Сибирск. отд. Русск. географ, общ., I : 58 (распр.); Нау-
мов С. П., 1931, Тр. Полярн. комисс., 4 : 42—43 (распр., враги); Белополь-
ский Л. О., 1932, Тр. Зоол. инет. Акад, наук СССР, 1 : 183 (распр.); Козлова-
Пушкарева Е. В., 1933, Тр. Монг, комисс., 10:12—13 (биол.); А ф а н а-
сьев А. В., 1934, Тр. Сов. по изуч. произв. сил, сер. дальневост., 2 : 254—256 (распр.,
биол ); 3 о л о т а р е в Н. Т., 1934, Тр. Сов. по изуч. произв. сил, сер. дальневост.,
3 : 156—161 (распр., биол.); Наумов Н. П., 1934, Тр. П >лярн. комисс., 17 : 66
(распр.); Salesski Р., 1934, Ztschr. f. Saugetierkunde, 99:369—370 (распр.);
Тугаринов А. Я., Н. А. Смирнов и А. И. Иванов, 1934, Птицы и
млекопит. Якутии, М.—Л., изд. Акад, наук СССР : 1—67 (распр.); Флеров К. К.,
.1935, В кн.: Адлерберг Г. П., Н. А. С м и р н о в иК. К. Флеров, «Звери
Арктики», М.—Л., изд. Главсевморпути: 195—223 (общ., синон., изобр.); Золота-
рев II. Т., 1936, Млекопит. бассейна р. Имана, М.—Л., изд. Акад, наук СССР :
101—102 (распр., биол., параз.); Подаревский В. Б., 1936, Пробл. охотхоз.
акклим. в Воет. Сибири, Иркутск, ОГИЗ : 33—34 (распр.); Новиков Г. А., 1937,
1 В качестве terra typica для Moschus moschiferus следует принимать Алтай, как
это делал вначале Флеров (1930), а за ним Цалкин (1947). Поэтому это видовое назва-
ние следует сохранять за популяцией из Сибири и Дальнего Востока. Мне непонятна
столь резкая и, по существу, ничем не обоснованная перемена точки зрения К. К. Фле-
ровым (1952), относящим теперь к М. moschiferus кабаргу южной части ареала, а си-
бирскую к другому виду — Moschus sibiricus Pall.
MOSCHUS
123
Тр. Сов. по изуч. произв. сил, сер. вост.-сиб., 4 : 233—234 (распр., биол., ох.); Юрген-
соя П. Б., 1938, Тр. Алтайск. гос. запов., I : 157—159 (распр., биол.); Allen G.,
1940, Mammals of China and Mongolia, N. Y. : 1129—1137 (общ., синон.); Анти-
пин В. М., 1941, Млекопит. Казахстана, т. III, Копытные, Алма-Ата, ГИЗ : 16—
19 (общ.); Скалой В., И. Щербаков и М. Базы кин, 1941, Природа и
соц. хозяйство, VIII : 276—277 (распр.); Ца лк ин В. И., 1947, в кн.: Гент-
нер В. Г. и В. И. Ц а л к и н, Олени СССР, М., изд. Моск. общ. испыт. прир. :
120—175 (сист., синон., изобр.); Грунин К. Я., 1949, Энтомол. обозр., XXX,
3/4 : 440—442 (параз.); Насимович А. А., 1949, сборн. «Пушн. богатства СССР»,
I ; 145—147 (распр., биол., изобр.); Павлов Е. II., 1949, Промысл, звери Читинск.
области, Чита, Обл. изд. : 82—85 (общ., изобр.); Копылов И. П., 1950, Дикие
копытные Иркутской обл., Иркутск, Облиздат : 49—54 (распр., биол., изобр.); М a t-
Рис. 23. Кабарга (Moschus moschiferus L.).
he so n C., 1950, J. Soc. Preserv. Faun. Emp., London, 61 : 26—31 (мускус, изобр.);
Черкасов A. A., 1950, Записки охотника Вост. Сибири, Иркутск, Облиздат :
201—207 (биол., ох.); Судариков В. Е. иК. М. Рыжиков, 1951, Тр. Гель-
минтол. лабор., V : 53—58 (параз.); Сысоев В. П., 1952, Охота в Хабаровском
крае, Хабаровск, Крайизд. : 112 (распр., ох., изобр.); Флеров К. К., 1952, Ка-
барги и олени, М.—Л., изд. Акад, наук СССР (Фауна СССР, и. с., № 55) : 25—58 (общ.,
синон., изобр.); Янушевич А. И., 1952, Фауна позвоночных Тувинской обл.,
Новосибирск, изд. Зап.-Сиб. фил. Акад, наук СССР : 61 (распр.); Щербаков А. Н.,
1953, Кабарга, ее экол. и хоз. использов., автореферат, М., изд. Моск, области, педагог,
инет. : 1 — 11 (общ.); Лаптев И. П., 1955, Заметки по фауне и флоре Сибири, 18 :
4—5 (распр.); Насимович А. А., 1955, Роль режима снежн. покрова в жизни
копытных. М., изд. Акад, наук СССР : 287—288 (распр., биол.).
Описание. Общие размеры сибирской кабарги по сравнению с другими
копытными СССР небольшие. Длина тела взрослых экземпляров состав-
ляет 85—100 см, редко превышая последнюю цифру. Высота в холке
колеблется от 56 до 67 см. Живой вес, в зависимости от упитанности,
составляет 10—17 кг; у самцов, обычно, несколько больше, чем у самок.
Характерную особенность внешнего Сложения кабарги Представляет
слабое развитие передней части туловища, а также и передних конеч-
ностей по сравнению с задними. Крестец приподнят значительно выше
холки и задняя часть тела много шире и массивнее, чем плечи и грудная
клетка.
124
MOSCHIDAE
Голова небольшая, равномерно суживается в лицевой части, но у сам-
цов морда несколько расширена в области клыков. Голое пространство
(«носовое зеркало») на верхней губе охватывает и окружность ноздрей.
По средней линии оно разделено неглубокой бороздкой. Зрачки круглые,
но при ярком свете могут сжиматься в узкую щель. Радужина глаз зо-
лотистая, темно-коричневая или темно-желтая (Туркин и Сатунин, 1902).
Хвост короткий, длиной 4—6 см, скрыт в волосах, уплощен сверху вниз,
у самок и молодых покрыт волосами, у самцов голый, лишь с пучком
волос на конце.
Копыта главных (III и IV) пальцев узкие, сжатые с боков, заостренные;
на боковых пальцах (II и V) они сравнительно хорошо развиты, по длине
(30—45 мм) лишь немного уступают средним; при стоянии животного
касаются почвы. Цвет копыт коричневый, разной интенсивности оттенков.
Шея, туловище и верхние части конечностей кабарги покрыты густым,
грубым и ломким волосом с пустотами в сердцевинной части. Мех у М.
moschiferus по сравнению с китайским видом более густой и длинный;
на крестце он может доходить до 85—95 мм. И длина и грубость волоса
возрастают от головы к крупу; наиболее длинные и толстые волосы нахо-
дятся на огузке, вокруг хвоста. Ближе к животу волосы несколько утон-
чаются; наиболее мягкие волосы находятся в подмышках и пахах, на
вымени, мошонке, а также на внутренней стороне ушей. Волосы вокруг
отверстия мускусной железы у самцов удлинены и направлены концами
к его средине. Основная масса волосяного покрова туловища состоит
из волнообразно изогнутых волос, постепенно утончающихся кверху и
имеющих резко заостренную, иногда нитевидную верхушку. Голова и конеч-
ности ниже запястных и скакательных суставов покрыты значительно более
тонким, коротким и почти лишенным волнистости волосом. Подшерсток
между основным, кроющим волосом практически отсутствует.
Окраска волосяного покрова в деталях значительно варьирует, но
основной фон у сибирской кабарги создают коричневые тона от светлого
рыжеватого до очень темного, черно-коричневого. Детали расцветки обу-
словливаются распределением и количественным соотношением различно
окрашенных отдельных волос. Основание волоса всегда светлое, чисто
белое или светло-бурое. К вершине идет постепенное потемнение, и ин-
тенсивность потемнения верхушечных частей налегающих друг на друга
волос создает основной фон окраски меха. Большая или меньшая часть
волос в верхней трети имеет хорошо отграниченное от ниже и выше ле-
жащих темных участков светлое, белое или рыжеватое кольцо. Если
волосы этого типа в большом числе распределены равномерно среди
остальных, то получается равномерная штриховатая окраска, типичная
для китайской кабарги. У сибирской же собранные группами волосы
с белым кольцом образуют резко отграниченные светлые пятна на ко-
ричневом общем фоне. Пятна расположены часто рядами или, сливаясь,
образуют полосы. Пятнистость сильнее выражена у молодых животных,
у взрослых же она сосредоточена, в основном, по бокам задней части
туловища (на пояснице и бедрах); у самцов с возрастом она может почти
вовсе исчезнуть. Иногда, впрочем редко, светлые участки преобладают,
создавая основной фон, на котором выступают темные неправильной фор-
мы полосы и пятна, Волосы на нижней части груди и животе имеют светло-
бурый цвет с белыми верхушками, обусловливая сероватую мелкоштрихова-
тую окраску этих частей тела. Голова светлая, в средней части буровато-
коричневая, по бокам буровато-серая-благодаря наличию широкого свет-
лого кольца на предверхушечной части бурых волос. Такую же окраску
MOSCHUS
126
имеет наружная сторона ушей; внутренняя сторона последних покрыта
светлыми волосами; концы ушей черно-бурые. Верхние части конечно-
стей окрашены одинаково с туловищем, ниже запястных и скакательных
суставов буровато-коричневые с сероватым налетом. Характерную осо-
бенность кабарги составляют парные белые полосы по бокам нижней
стороны шеи и головы до передних конечностей, теряющиеся на внутрен-
ней стороне последних. Полосы эти образованы белыми верхушками во-
лос, в основной части имеющих светло-бурую окраску. С течением времени
волос выцветает, и подрастающая молодая шерсть всегда окрашена за-
метно темнее старой. Половой диморфизм в окраске выражен слабо,
у самок лишь несколько сильнее развита пятнистость.
Молодые до шести месяцев имеют ярко выраженную пятнистость
на общем коричневом фоне. Многочисленные пятна рыжевато-желтого
и белого цвета образуют поперечные и продольные полосы. Последние
в числе нескольких пар, начинаясь от холки и лопаток, сходятся к осно-
ванию хвоста.
Продольные складки кожи на хвосте образуют железу, выделяющую
пахучий секрет (Рососк, 1910). На нижней стороне живота у взрослых
самцов, между пупком и препуцием, расположена (из всех копытных
имеющаяся только у кабарги) мускусная железа, или так называемая
«кабарожья струя». Она представляет собою мешетчатое впячивапие кожи
длиною около 6 см с расположенными в его стенках железами, выделяю-
щими студнеобразный, резко пахучий секрет; открывается мускусный
мешочек двумя отверстиями: одним на наружной поверхности живота
и другим в полость препуция. Наличие этой железы только у самцов
ставит ее в связь как будто с половой функцией. Но судя по тому, что
ни запах, ни количество мускуса не меняется по сезонам, это заставляет
подозревать ее иное назначение.
Подобно оленькам и некоторым полорогим, половой член самцов на
конце имеет нитевидный придаток. Молочная железа, расположенная
в задней части брюшной стенки между бедер, имеет два близко располо-
женных друг к другу соска.
Общая длина черепа (рис. 24) у взрослых экземпляров колеблется
от 144 до 163 мм.1 Лицевая часть черепа сравнительно короткая, но вы-
сокая. Длина ее2 редко более 50%, а высота за клыками не менее 14%
общей длины черепа (Цалкин, 1947). Слезные кости короткие и широкие;
их наибольшая ширина обычно не уступает длине.
Коренные зубы не высокие, брахиодонтные. Резцы и прилегающие
к ним нижние клыки лопаткообразные, симметричные; самые крупные —
средняя пара (IJ, остальные постепенно убывают в ширине, самые уз-
кие — крайние (клыки). Верхние клыки у самцов длинные, до 70 мм
(а по Флерову — даже до 100 мм), саблевидно изогнуты назад и выступают
из-под верхней губы ниже подбородка. Корни их глубоко сидят в верх-
ней челюсти, имеют открытую пульпу, и поэтому эти зубы обладают
постоянным ростом. Переднюю стенку альвеол клыков образуют носовые
отростки межчелюстных костей. Поперечное сечение верхних клыков
самцов сильно сжатое с боков с закругленным передним и заостренным
задним краем и вершиной. У живого животного клыки сидят в альвеолах
слегка подвижно. У самок верхние клыки небольшие, круглого сечения
1 У М. sifanicus от 157 до 172 мм.
3 От prosthion до переднего края глазницы.
126
MOSCHIDAE
л не выступают из-под верхней губы. У старых самок они иногда вообще
отсутствуют (выпадают).
Сибирская кабарга по сравнению с китайской высоконога. Длина
метаподий у нее на передних конечностях составляет 126—148 мм, на
задних — от 167 до 189 мм.1 Соответственно, ширина дистального сус-
тавного блока пястной составляет не более 14.5%, а у плюсневой — не
более 12.5% всей длины соответствующей кости.
Географическое распространение (рис. 25). В общих чертах ареал
М. moschiferus охватывает Алтай, Саяны, восточную Сибирь (кроме Кам-
чатки), Дальний Восток, Сахалин, северную Монголию, вероятно север-
ный Китай и Корею. Точные южные границы распространения вида пока
не ясны. Только в пределах современного ареала и только из поздне-
четвертичных отложений (неолит) кабарга известна и в ископаемом со-
стоянии.
В пределах СССР западной границей распространения кабарги яв-
ляется южный Алтай. В пределах Казахстана она указывается для Кур-
чумского и Сарымсактского хребтов, Саурских гор, для окрестностей
Катон-Карагая, верхнего течения р. Бухтармы (выше с. Алтая).1 2 Далее
к востоку кабарга является обычным зверем в большей части хребтов
центрального и северного Алтая почти до 52° с. ш. (Кащенко, 1899).
Многочисленна в Катунских горах, по северным склонам Чуйских Альп,
по р. Аргуту и ее притокам (Коксу и др.), в хребте Сайлюгем. Приво-
дился в качестве места обитания также водораздел Коксу, Урсула и Бух-
тармы. Более редка в западной части Горно-Алтайской автономной об-
ласти по рекам Коксе и Чарышу. Широко распространена кабарга в меж-
дуречьях Катуни и Башкауса, а также между Башкаусом и Чулышма-
ном (Salesski, 1934).
Сведения о современном распространении кабарги в центральном
Алтае (Усть-Коксинском районе Горно-Алтайской области) приводит
А. А. Насимович (1949). В настоящее время, пишет он, «кабарга очень
редка в верховьях Катуни (она более обычна ниже впадения Бирюксы);
ничтожно количество ее на северных склонах хребта Листвяга и в сосед-
них участках Холзуна — в верховьях Красноярки. Всюду здесь выпа-
дают глубокие снега. Следуя вниз по течению правым берегом Катуни,
можно заметить, что кабарги ничтожно мало по реке Верхней Зайчихе;
она обычна у Тэкэли, в окрестностях Сахсабая, по р. Чернушке у Верх-
него Уймона, и среднему течению Большого Скола; ее много по рр. Мульте,
Нижнему Курачану, Кочурле и, в особенности, в горах по Аккему и Ар-
гуту, на Ак-Чане ее значительно меньше. В системе Теректинского хребта
кабарги меньше. Здесь она обычна на рр. Громатухе, Черной Терехте,
Чендеку, в верховьях рр. Катандинок и в горах близ Тюнгура. На р. Бы-
струхе кабарги нет, на Бирюксе ее мало. В системе р. Коксы больше
всего ее по притокам Карагаю, Банной и Хайдуну; по р. Айле кабарги
нет».
Обычна и местами многочисленна кабарга на территории бывшего
Алтайского заповедника, к востоку от р. Чулышмана, особенно между
Телецким озером и верховьями Большого и Малого Абакана (Юргенсон,
1938). Нет ее в безлесных районах Алтая, например в Чуйской степи.
1 Соответственные цифры для М. sifanicus и М. berezovskii см. раньше, стр. 122.
2 В коллекциях ЗИН АН СССР хранятся два черепа кабарги, добытые в 1929 г.
в окрестностях с. Аргаты по р. Крутой, левому притоку Бухтармы (коллектор В. До-
ценко) и один череп из окрестностей Катон-Карагая.
MOSCHUS
127
Область сплошного распространения кабарги представляют Саяны;
особенно многочисленна она в узле Западных и Восточных Саян. В ле-
вобережье Енисея много этого зверя к западу от р. Кантегира, в южной
Рис. 24,. Череп Moschus moschiferus L.
части Хакасской автономной области. В Западных Саянах к востоку
от Енисея северная граница распространения проходит, по-видимому,
в среднем течении р. Ои (Кебежа). Отсутствует в Хакасско-Минусинской
котловине. Обыкновенна во всех горных елово-кедровых лесах Тувинской
автономной области (Янушевич, 1952).
128
MOSCHIDAE
В Восточных Саянах кабарга держится в верхнем и среднем течении
рр. Маны и Кана и доходит до Красноярска. Известны места обитания
в таежных частях Новоселовского и Балахтинского районов, в бассейнах
рр. Базаихи и Рыбной ( в пределах Уярского района), а также повсе-
местно в подходящих местах в Агинском районе (Подаревский, 1936).
В окрестностях Красноярска многочисленна на территории заповедника
«Столбы». Здесь она переходит на левый берег Енисея, но как далеко
распространяется на запад — не ясно. В Кузнецком Алатау кабарга
отмечается для бассейна рр. Большого и Малого Июса. Известен случай
захода беременной самки кабарги в 1952 г. в Григорьевский сельсовет
Туганского района Томской области (Лаптев, 1955). В Горной Шории,
по-видимому, отсутствует.
По Енисейскому кряжу кабарга распространена до низовьев р. Под-
каменной Тунгуски. Бассейн последней является одним, из наиболее
плотно заселенных ею районов Сибирского плоскогорья. К северу кабарга
встречается значительно реже. Так, по данным Е. Яковлева (1930), на
Нижней Тунгуске она является «редкостью, проникая в систему ее юж-
ных притоков (рр. Учами, Таймура, Нидыма) через водораздел с Под-
каменной Тунгуски. Случаи более северного ее обитания известны лишь
для верховьев рр. Тембенчи и Кочечумы». Однако, по Н. П. Наумову
(1934), кабарга изредка встречается еще у Осиновского и Большого по-
рога, у оз. Неконгда, в устье р. Виви. В конце прошлого века, в промыс-
ловом количестве (т. е. служила предметом регулярного промысла) встре-
чалась в горах Сыверма, уже за полярным кругом. В 30-х годах этого
столетия следы кабарги отмечались по соседству с Сыверма, в верховьях
р. Кочечумы (Наумов,. 1934). Наиболее северной точкой енисейского
севера, где отмечался этот вид, являются окрестности оз. Есей, уже к се-
веру от 68-й параллели (Флеров, 1935). Изредка кабарга появляется и на
левобережье нижнего Енисея. С. П. Наумов (1931) сообщает о случае
добычи ее в 20 км к западу от д. Алинской (около 63.5° с. ш.).
Почти повсеместно, но неравномерно распространена кабарга, на тер-
ритории Иркутской области и Бурят-Монгольской АССР (Подаревский,
1936; Копылов, 1950). Наиболее многочисленна она в Восточных Саянах
и Хамар-Дабанских горах. В Тофаларии, например, по численности
кабарга занимает первое место среди копытных. Обычна в южной части
Тайшетского района и в Слюдянском районе. В бассейне Ангары чаще
всего встречается по р. Ерейке и в верховьях Илима. Многочисленна
в горах западного Прибайкалья, в верховьях Лены и по всему течению
Киренги. К западу и северу от этих мест, в районах Жигалевском, Усть-
Кутском и Киренском, она значительно более редка, а местами вообще
отсутствует. То же относится к значительной части Усть-Илемского
района. В Катангском районе более многочисленна в южной части. Срав-
нительно многочисленна в местности между верхними течениями Нижней
Тунгуски и Чоны к югу от Ербогачена. Крайние пункты уже единичных
заходов кабарги на севере Иркутской области расположены на 62—63° с. ш.
(рр. Иритка, левый приток Н. Тунгуски). Распространена кабарга по всему
Бодайбинскому району, но наиболее многочисленна по р. Б. Патону
и притокам Витима — Синюче, Эмалыку и Нерпо, а также в районе оз.
Орон.
В Бурят-Монгольской республике кабарга обычна по всем хребтам
восточного побережья Байкала. Нет ее только в борах нижних течений
рр. Сосновки и Таркулика (Доппельмаир, 1926). Наиболее многочисленна
в бассейнах рр. Урбыкана, Ирингды и Яксакана, в верховьях Баргузина,
MOS CH US
129
в районе мыса Понгонье. В районе северной оконечности Байкала много
кабарги в бассейне р. Котери, тогда как по верхней Ангаре, особенно
в нижнем течении, она сравнительно редка (Г. Новиков, 1937). Много-
численна по всему Баунтовскому району, но малочисленна в Кабанском.
В южной части Бурятии обычна в Окинском, Тункинском и Закамен-
ском аймаках; встречается также всюду в районах юго-восточного При-
байкалья.
В Читинской области более многочисленна в северной половине. В бас-
сейне Калара и Калакана по численности стоит на первом месте среди
Рис. 25. Распространение Moschus moschiferus L.
1 —современный ареал; 2 —-места отдельных заходов.
других копытных. Обычна по среднему течению Олекмы и ее притоку
Тунгиру. Сравнительно редка по рр. Каренги и Нерчи. Водится по р. Ин-
годе и служит предметом промысла в Красночикойском районе.
В Амурской области кабарга распространена, по-видимому, также
во всех местах, где находит подходящие условия для существования.
Отмечается она, например, для бассейна р. Нюкжи, Олекмы, Гилюя,
Могота, Тынды, левых притоков Уркана, Токо, Оконона (район истоков
рр. Зеи и Маи), для южных склонов Станового хребта, хребта Тукурингра.
Обычным зверем она является в районе верхнего течения р. Селемджи
(Тассовский, 1927; Подаревский, 1936).
Недостаточно выяснено распространение кабарги на территории
Якутии. Несомненно, обычным животным она является по Чаре, Олекме,
в верхнем течении Алдана и Амги, в Становом хребте и на Алданском на-
горье. Наиболее многочисленна, по Скалону (1941), по рр. Чаре и Токко.
На Лене распространена от южных границ республики до Олекминска
и несколько ниже. Встречается в промысловом количестве даже в преде-
лах, бывшего Якутского округа (Тугаринов, Смирнов и Иванов, 1934),
9 Фауна СССР, И. И. Соколов
130
MOSCHIDAE
в верхнем течении Вилюя, изредка до самого Вилюйска, в горах по р. Чоне.
В Верхоянском хребте и Верхоянском округе вообще, по Скалону (1941),
в противоположность другим литературным данным, редка. В коллекциях
ЗИН АН СССР имеется экземпляр кабарги, добытый в 460 км к северу
от Верхоянска (гора Тупаях, или Сироты). Встречается, но сравнительно
редка, на пространстве между верхними течениями Индигирки и Колымы;
на правобережье Индигирки севернее р. Момы вообще не заходит. Отсут-
ствует кабарга, по Скалону (1941), к западу от Лены, севернее Олекминска,
а также в центральной части республики в районах Западно-Кангалах-
ском, Мегино-Кангалахском, Чурапчинском, Усть-Алданском, Таттин-
ском и Амгинском.
Как широко распространена она к востоку и югу от Колымы не ясно.
Известно, что встречается кабарга по рр. Омолону и Коркодону (пра-
вые притоки Колымы), а также на восточном и южном склонах Колым-
ского хребта. Западная граница ее здесь несколько не доходит до р. Пен-
жины (Белопольский, 1932). Вероятно водится в пределах Ольского,
Сеимчанского и других районов к северу от Охотского побережья. Отме-
чается для хребта Джугджур. Отсутствует в пределах большей части
Чукотского и Корякского национальных округов к северу и востоку от
водораздела Пенжины и Омолона. Нет ее также и на Камчатке.
Обычным зверем кабарга является в южной части Хабаровского края,
в Буреинском и соседних с ним хребтах Дуссе-Алинь и Ям-Алинь. Здесь
ее ареал смыкается с ареалом в Амурской области. Особенно многочис-
ленна кабарга в верховьях Бурей, Селемджи, Уда, Амгуни. Как близко
кабарга подходит с севера к Амуру в его нижнем течении — не известно.
По утверждению Сысоева (1952), в южной части Амурской области и в Би-
робиджане она отсутствует.
В Приморском крае кабарга распространена по всему хребту Сихотэ-
Алинь от южной его части 1 до низовьев Амура. В нижнем течении рек,
стекающих с западных склонов Сихотэ-Алиня (например, Имана), она.
значительно более редка. Широко до сих пор распространена во всех
гористых лесах о-ва Сахалина, особенно в его южной половине.
В пределах Монгольской Народной Республики кабарга распростра-
нена главным образом в северной части, в районах, прилегающих к Во-
сточным Саянам и Забайкалью. Обычна, а местами и многочисленна,
в Прикосоголье, т. е. в горах, окружающих оз. Хубсугул. Обычна кабарга,
по последним данным А. Г. Банникова (1954), по всему Хэнтэю вплоть
до Улан-Батора. Много следов кабарги и самого зверя он отмечал в за-
поведнике Богдо-Ула к югу от города. В Хангае распространение ее
ограничивается Хантай-Нуру, верховьями р. Орхона, а также по
хребтам Тарбагатай и Бургутани. В качестве редкого зверя, по словам
того же автора, она отмечалась в верховьях р. Урду-Тамир. Следы ее от-
мечались в лесочках по Цаган-Турту. В конце прошлого столетия она во-
дилась на хребте Хан-Хухэй, но сохранилась ли до настоящего времени —
не известно. А. Г. Банников подчеркивает, что на Монгольском Алтае
и в предгорьях Большого Хингана кабарги, по-видимому, нет.
Подвиды. Внутри М. moschiferus L. в нашем понимании выделялись
следующие подвиды:
1) М. moschiferus moschiferus L. Длина черепа 140—164 мм; носовые
кости сравнительно широкие, несколько суженные посредине. Общий фон
окраски коричневый. Пятнистость выражена хорошо. Светлая полоса на.
1 В коллекциях ЗИН АН СССР имеются экземпляры из Ольгинского района.
MOSCHUS
131
внутренней стороне ног выражена не резко. Распространение:
горы западной и средней Сибири — Алтай, Саяны, Танну-Ола, северо-
западная Монголия, междуречье Енисея и Лены.
2) М. moschiferus sibiricus Pallas. От типичной формы отличается не-
сколько меньшей величиной (длина черепа от 140 до 159 мм), более уз-
кими носовыми костями, более вытянутой ростральной частью черепа.
Окраска сероватого тона, пятнистость развита слабее, светлая полоса
на внутренней стороне конечностей выражена более ясно. Распро-
странение: восточная Сибирь — Забайкалье, Яблоновый и Стано-
вой хребты, северо-восточная Монголия, Якутия к северу до 62° с. ш.
3) М. moschiferus arcticus Flerow. Меньшей, в среднем, величины по
сравнению с типичной формой (длина черепа от 140 до 157 мм). Череп
имеет укороченную лицевую часть (не свыше 50% длины всего черепа),
длинные по сравнению с другими подвидами клыки и короткие, не имею-
щие сужения в средней части носовые кости. Окраска темная, до коричне-
во-черного тона. Пятнистость слабая, но светлая полоса на внутренней
стороне конечностей выражена очень резко. Распространение:
северо-восточная Сибирь к востоку от р. Лены до Колымы и к югу до
60-й параллели.
4) М. moschiferus turowi Zalkin. От типичной формы отличается не-
сколько меньшими размерами и более интенсивной, яркой темно-коричне-
вой окраской. Длина черепа у взрослых от 144 до 157 мм. Лицевая часть
черепа не превосходит 50% всей его длины. Широкая мозговая коробка.
От М. т. arcticus отличается меньшей длиной зубного ряда, меньшими
размерами коренных зубов и, как следствие этого, большей длиной без-
зубого края верхней челюсти. Распространение: Амурская
область, Приморский край, возможно северо-восточный Китай.
5) М. moschiferus parvipes Hollister. От других рас отличается темной
окраской, меньшими размерами, короткими и тонкими метаподиями.
Распространение: Корея; Флеров к этому подвиду в свое время
относил такжу кабаргу восточной части Маньчжурии и Приморья.
6) М. moschiferus sachalinensis Flerow. По размерам сходна с кабаргой
материка Дальнего Востока. Отличается от других подвидов тем, что.
глазницы сближены внутренними краями и открываются косо вбок и вверх.
Межглазничная ширина меньше ширины мозговой коробки непосредственно
за глазницами или равна ей. Метаподий короткие. Распростране-
ние: о-в Сахалин.
К. К. Флеров в своей последней работе (1952) сохраняет в качестве
подвида лишь сахалинскую кабаргу (Af. т. sachalinensis).
Биология. Биология кабарги изучена недостаточно. Приводимые раз-
ными лицами сведения противоречивы. На территории своего ареала
кабарга распространена неравномерно, так как отличается сравнительно
узкими требованиями к условиям внешней обстановки.
Распространение кабарги тесно связано с горной тайгой. Типичные
для нее стации.— темнохвойные леса на крутых склонах гор по соседству
с выходами скал и каменистых россыпей. Скалы и россыпи используются
как место отдыха и убежище в случае необходимости скрываться от вра-
гов. Чем круче скалы, тем больше удобств представляют они для кабарги;
но за пределы лесной растительности, в альпийскую зону кабарга не за-
ходит, как никогда не выходит и на открытые увалы. В Саянах, например,
типичными местами обитания ее являются утесистые берега горных ре-
чек, недалеко от верхней границы леса. Гораздо реже и, по-видимому,
только в летнее время кабарга держится в равнинной тайге, у подножья
9*
132
MOSCH I DAE
гор, в борах по речным долинам. По сообщению Подаревского (1936),
в Баунтовском районе Бурят-Монгольской АССР в значительном числе
населяет даже заболоченные увалы лиственничной тайги, но и эти стации
не принадлежат к типичным. В условиях Алтая кабарга придерживается
темнохвойной тайги склонов северной экспозиции; на солнцепечных
встречается также, но реже. В другие стации заходит редко и, как правило,
только в тех случаях, когда по соседству имеются типичные для нее угодья
(Юргенсон, 1938). Чаще населяет кедровые, пихтовые, елово-кедровые
насаждения или чистые ельники; лиственничных, особенно разреженных
лесов, избегает. «В тех местах, — пишет Золотарев (1934), — где нагор-
ные насаждения представлены в основном лиственничными или листвен-
ными лесами, кабарга держится в узких полосах ельников или елово-
пихтовых насаждений, тянущихся хотя бы узкой ленточкой вдоль горных
ключей. Там, где нет этих условий, нет и кабарги».
Одним из важных, если не необходимых, условий местообитания кабарги
является также наличие сплошного напочвенного мохового покрова.
Основу питания кабарги составляют различные лишайники и мхи.
В летнее время поедаются виды Cladonia, Cetraria, Marschantia, гипно-
вые мхи. Кроме них, пищей в этот период служат листья и тонкие ветви
ольхи, осины, хвощи, а также некоторые травянистые растения, как
василистник, герань, чина. Осенью поедаются даже кедровые орехи
(Копылов, 1950). Зимою мхи и лишайники местами составляют почти
единственную пищу. Вследствие затрудненной добычи корма из-под
снега огромное значение в это время приобретают свисающие с деревьев
бородатые лишайники Usnea. В дополнение к ним поедаются высохшие
части («ветошь») травянистых растений: кипрея, мышиного горошка,
частично зимнезеленые растения. К зимнему корму относятся также
тонкие ветви жимолости, ивы, осины, молодая кора и хвоя цихты, сосны
и сибирского кедра (Pious sibirica Мауг.). Хвоя и побеги пихты (Abies
sibirica Led.) составляли основную массу содержимого желудка кабарог,
добытых в период февраль—апрель в северо-восточном Забайкалье (Доп-
пельмаир, 1926). При этом заглатывались непережеванными одревес-
невшие побеги пихты длиной в 1—3 см и толщиной до 2.5 мм, вместе с си-
дящей на них хвоей. Побеги хвойных деревьев иногда употребляются
кабаргой в пищу и в летнее время.1 На Алтае посещает солонцы вместе
с другими видами копытных.
В отличие от ряда других копытных, кабарга ведет оседлый образ
жизни и не совершает сколько-нибудь значительных суточных или сезон-
ных миграций. «Если не тревожить ее, — пишет Е. В. Козлова-Пушка-
рева (1933), — а только наблюдать за ней, то можно легко убедиться
в том, что одна и та же кабарожка встречается всегда в одном излюбленном
ею месте». Есть, правда, указания, что осенью и зимой кабарга предпо-
читает держаться в наиболее трудно доступных для человека скалистых
участках горной тайги. В летнее время ее чаще можно встретить у под-
ножья склонов, в долинах горных речек (Черкасов, 1950). В этих слу-
чаях, вероятно, высокая температура воздуха и перегретый солнцем
каменистый субстрат заставляют животных искать убежища под покро-
вом густого леса в низинах. Однако В. Н. Скалой (1941) утверждает, что
в Бодайбинском районе Иркутской области самки с молодняком прово-
1 Вызывает сомнение цитируемое иногда сообщение П. С. Далласа (Pallas, 1799)
о том, что в желудке кабарги он находил остатки корневищ болотных трав, когда из-
вестно, что ни конечности, ни зубы ее не приспособлены к роющей деятельности.
MOSCHUS
133
дят лето, наоборот, на высоких скалистых местах («отстоях»), а самцы —
в истоках речек. В зимнее время кабарга, естественно, выбирает места,
где .снег сдувается ветром или тает на солнцепеках.
Держится кабарга, как правило, одиночно, за исключением периода
спаривания, когда самец собирает вокруг себя несколько самок, или,
наоборот, за одной самкой ухаживает несколько самцов. По-видимому,
даже прошлогодний молодняк редко выпасается вместе с матерью. G дру-
гой стороны, указание, что при охоте подражанием голосу детеныша на
зов часто прибегает не только самка, но и самец (Сысоев, 1952), говорит,
как-будто, за то, что, по крайней мере в этот период, звери живут парами,
а не одиночно. Е. В. Козлова-Пушкарева (1933) сообщает, что изредка
кабарги пасутся парами, но зато несколько лежек рядом никогда не
бывает. Обитая в течение длительного времени в одних местах, кабарги
протаптывают тропы, которые используются промышленниками для рас-
становки различного рода ловушек. Кормятся обычно по утрам или ве-
черам. Днем, как правило, отдыхают. Для отдыха и лежки выбирают
небольшие горизонтальные площадки на обрывистых склонах или недо-
ступных выступах скал, покрытые лишайниками или мхами.
Обычный способ передвижения — прыжки. Есть указания, что даже
в неволе кабарга передвигается прыжками (Туркин и Сатунин, 1902).
На быстром ходу ее бег несколько напоминает бег зайца. Передвижение
прыжками, с помощью которых кабарга может резко изменять направление
бега, представляет адаптацию к жизни в условиях скалистых ландшафтов.
С этим, несомненно, связаны особенности пропорций конечностей (более
мощное развитие задних по сравнению с передними), изгиб позвоночника,
форма и пропорции таза. При всякой опасности и испуге кабарга бросается
в россыпи и скалы, где с необычайной легкостью, делая большие прыжки,
мчится по таким кручам, где никакой преследователь за ней угнаться не
может. С помощью широко раздвигаемых копыт она цепляется за самые
незначительные выступы и неровности камня, взбирается по отвесной
стене на какую-либо неприступную скалу и замирает там неподвижно
«на отстое». На ровном месте кабарга может быстро бежать только на ко-
ротком расстоянии, вскоре утомляется, и в этих условиях ее быстро до-
гоняет собака. При необходимости хорошо плавает, даже через горные
реки.
В отношении развития органов чувств сведения противоречивы.
По одним данным, хорошо развиты обоняние и слух, хуже зрение (Павлов,
1949); по другим, наоборот, хуже развито обоняние при отличном зре-
нии и слухе (Черкасов, 1950).
Голос кабарги напоминает хриплое «чиф-фый» или мягкое шипение
вроде «ф ш ш ш» (Козлова-Пушкарева, 1933). Этот звук кабарги издают
во время гона, испуга и вообще при всякого рода возбуждении. Раненые,
по словам Афанасьева (1934), издают резкий протяжный крик; такой же
крик, но более отрывистый, кабарги, якобы, издают, перекликаясь в тайге
друг с другом. «Часто по вечерам, — пишет он, — мы слышали их далеко
разносящиеся своеобразные крики». Голос молодых кабаржат похож не-
сколько на голос козленка, но более нежный и протяжный.
Слабо изучен у кабарги процесс линьки шерсти. Не ясно даже, линяет
ли она один или два раза в году. Если линька однократная, то, по-ви-
димому, очень растянутая, возможно в связи с возрастом и полом. По
Флерову (1952), самцы линяют в феврале и марте, самки — с конца июня
по начало сентября, причем имевшие кабаржат, линяют позднее, чем
холостые; иногда линька у них продолжается еще в сентябре. Молодые
134
MOSCHIDAE
животные линяют несколько раньше. В коллекциях Зоологического
института АН СССР имеются шкуры кабарог, находящиеся в стадии
смены волосяного покрова, с датами добычи с 26 июня по конец фев-
раля. У добытой Афанасьевым (1934) 30 августа в бассейне р. Бурей самки
большая часть волоса еще не сменилась. Линька начинается с головы
и нижних частей конечностей, затем постепенно распространяется на бока
и шею. В последнюю очередь и почти одновременно смена волоса происхо-
дит на спине и животе. Кроме более интенсивной насыщенной окраски
молодого волоса, других отличий, например в густоте или структуре,
между ним и старым мехом не заметно.
Половой зрелости самцы достигают к ЗУг годам; к этому возрасту
у них вырастают клыки1 и развивается мускусная железа, содержимое
которой начинает издавать специфический запах. Есть сведения, что
самки способны оплодотворяться уже на втором или третьем году жизнщ
Половой сезон у кабарги начинается иногда в конце ноября, чаще
в декабре и длится около месяца. В этот период самец собирает вокруг себя
несколько самок; иногда, наоборот, за самкой ходят несколько самцов.
Между самцами происходят поединки; турнирным оружием служат
выступающие верхние клыки, которыми звери наносят противникам
удары сверху вниз, главным образом по шее и верхним частям туловища.
Есть указание, что поединок иногда кончается тяжелыми ранениями
и даже смертью одного из противников. Как долго длится период течки
у отдельных животных — не известно. Самец все время преследует и го-
няет самку; даже самый акт покрытия происходит, говорят, на ходу.
По окончании периода гона самец и самка расходятся и вновь ведут оди-
ночный образ жизни.
Беременность длится пять или шесть месяцев. Роды происходят в мае
или июне. Приносит самка одного или двух детенышей. Специального
логова для новорожденных мать не устраивает, выбирая для родов лишь
наиболее укромные места под навесом или в расселинах скал. Рождаю-
щиеся беспомощными кабаржата первое время лежат и прячутся в укры-
тиях непосредственно на земле или камнях. Мать навещает их только
для кормления. До средины лета, как сообщает Черкасов (1950), самок
вместе с молодыми не встречают, а находят последних только в логовищах.
Кабарга — хорошая мать. В случае опасности она пытается отвлечь
на себя внимание и увести охотника или другого врага подальше от места
нахождения детенышей. По данным Е. В. Козловой-Пушкаревой (1933),
самка кормит молоком детей не более двух месяцев, а позднее, якобы,
детеныши ходят уже одни. Этим сведениям противоречат факты охоты
с манком на кабаргу до самого сентября.
Продолжительность жизни кабарги не известна.
Кабарга имеет многочисленных врагов. Преследуют ее все хищники,
начиная с тигра и медведя и кончая, как говорят, даже соболем и куницей.
Особенно много гибнет молодняка в первые недели его жизни. Кабаржата
гибнут и от хищных птиц, например от ястреба-тетеревятника или фи-
лина. Главным врагом является россомаха, на Дальнем Востоке — харза,
в меньшей степени — рысь. Известны случаи, когда даже взрослые ка-
барги становятся добычей лисиц (Доппельмаир, 1926; Наумов, 1931).
Из эктопаразитов кабарги известна муха Cordilobia inexpectata
Grun. из семейства Calliforidae в личиночной фазе паразитирующая под
кожей, главным образом на спине. Поражаются этим паразитом как
1 Полного развития клыки достигают только к 4х/2 годам.
MOSCHUS
135
взрослые, так и молодые животные в массовом количестве: до 100 особей
паразита на 1 см2 и почти до 2000 паразитов на одном животном. «В пе-
риод развития личинок, — пишет К. Я. Грунин (1949), — кабарга сильно
истощена и теряет свою обычную чуткость и осторожность до такой сте-
пени, что к ней часто можно подойти почти вплотную». Как следствие
поражения паразитами имеет место задержка линьки. Пока этот пара-
зит установлен только на кабарге из Ситотэ-Алиня и в горах Кентея
(Монголия). От кабарги из Забайкалья описан новый вид паразитирую-
щей в печени трематоды Dicrocoelium orienlalis Sud. et Riz. (Судариков
и Рыжиков, 1951). У добытой Н. Т. Золотаревым (1936) на р. Имане ка-
барги на морде, ушах и в нахах имелось большое количество клещей
Ixodes persulcalus Р. Sch.
Хозяйственное значение. Главным продуктом, ради
которого промышляют кабаргу, является мускусная железа, или «каба-
рожья струя», взрослых самцов, о которой упоминалось раньше. Содер-
жимое мускусного мешочка — мускус, в свежем виде представляет студ-
невидную массу красновато-коричневого цвета, жирную на ощупь, обла-
дающую специфическим запахом и, по Туркину и Сатунину, острым,
горьким вкусом. Мускус кабарги считается лучшим видом натурального
мускуса животного происхождения и находит применение главным об-
разом в парфюмерной промышленности.1 Его действующее пахучее на-
чало — мускон — представляет жидкость, кипящую при температуре
142—143°, и с химической стороны относится к группе циклических
кетонов. В парфюмерном производстве мускус имеет значение как фик-
сатор, т. е. средство, удерживающее запах духов.2 В настоящее время
находят применение синтетические вещества, обладающие сходными
с натуральным мускусом свойствами, но полностью его не заменяют.
Поэтому спрос на последний со стороны парфюмерной промышленности
остается прежним, несмотря на высокую цену (Matheson, 1950).
Вырезанный целиком у убитого самца мускусный мешочек высу-
шивается на воздухе и в таком виде сдается на заготовительные пункты.
Качество мускуса, по-видимому, не зависит от сезона, в который добыта
кабарожья струя, и поэтому промысел самцов кабарги может произво-
диться в течение всего года.
По сообщению К. К. Флерова (1935), для получения мускуса нет
необходимости убивать животное. Мускус легко можно извлечь, вставив
в отверстие железы трубочку и надавливая рукой на мускусный мешочек.
Этот способ мог бы открыть перспективы для развития кабарговодства.
Однако случаев содержания кабарги с этой целью мне не известно.
Мясо кабарги, как пищевой продукт, большого значения не имеет
в силу незначительного его выхода по сравнению с другими дикими
копытными. Кроме того, оно жестко, не вкусно, а у самцов имеет мус-
кусный привкус. Часто поэтому, особенно в глубине тайги, при добыче
кабарги тушка просто выбрасывается. Шкура также не имеет большой
цены, вследствие грубости и ломкости волоса. Лишь иногда из нее делают
коврики для сиденья. Снятый со шкуры волос может служить для набивки
матрацев или мягкой мебели. Кожа без волоса выделывается на замшу,
из которой шьют кисеты, рукавицы, брюки, а иногда даже мягкую обувь 1 2
1 Применение мускуса в качестве лекарственного средства в настоящее время
почти не имеет места и, кроме того, лишено научных оснований.
2 Большая советская энциклопедия, 2-е изд., 1954, т. 28, стр. 583.
136
MOSCHIDAE
(Емельянов, 1927). Клыки кабарги в некоторых местах употребляются
как шило или в качестве украшения, а из кожи лапок шьют
шапки.
Охота. Способы охоты на кабаргу разнообразны. В прежнее время
ее в больших количествах промышляли с помощью кулём, пастей, петель
и других самоловных орудий. С этой целью перегораживался склон горы,
по которому происходит передвижение кабарги. В изгороди, представ-
ляющей, обычно, простой залом из подручного материала, оставляются
один или несколько проходов, в которых и расставляют ловушки.
Петли, а также капканы ставят на тропах, особенно в зимнее время.
Эти хищнические приемы промысла, при которых большое число живот-
ных (самки, молодняк) гибнет бесцельно, в настоящее время повсеместно'
запрещены.
Основными способами добычи являются охота с собакой и на пищик-
манок. Первый способ заключается в том, что, напав на след кабарги,
собака начинает ее преследовать с лаем. Кабарга, как и всегда при воз-
никновении опасности, бросается к скалам. Несколькими гигантскими
прыжками она взбирается на какой-либо отвесный выступ, куда собака
проникнуть не может, и замирает неподвижно «на отстое». Собака, про-
должая облаивать снизу или сверху неподвижно стоящего зверя, отвле-
кает на себя ее внимание и дает возможность охотнику подойти к ней на
верный выстрел.
Следует отметить, что кабарга — доверчивый зверь. Е. В. Козлова-
Пушкарева (1933) приводи! случай, когда группа кабарог жила длитель-
ное время по соседству с лагерем экспедиции, не смущаясь близостью
людей, рубкой леса, частыми выстрелами и лаем собак. При встрече с чело-
веком без собаки кабарга, издав свое шипящее «чиф-фый», делает несколько
прыжков и метрах в двухстах останавливается, повернувшись в сторону
охотника. При приближении к ней она вновь сделает несколько прыж-
ков и будет делать такие перебежки, оставаясь в пределах одного и того
же склона горы (Сысоев, 1952).
Основной промысел на кабаргу совершается в зимнее время, так как
летом она скрывается в наиболее глухих, труднодоступных местах и в лес-
ных чащах. В Забайкалье она добывается главным образом в апреле
(Новиков, 1937).
В летнее время практикуется охота на кабарог с манком. Последний,
по словам Сысоева (1952), представляет сложенную вдвое пластинку из
бересты величиной не более трехкопеечной монеты. С помощью этого
приспособления охотник издает резкий вибрирующий звук, подражаю-
щий голосу детеныша кабарги. На этот звук выбегают взрослые звери
(как уже отмечалось, часто выбегает и самец, особенно во второй поло-
вине лета). Влечение на зов бывает так велико, что по зверю можно сде-
лать не один выстрел. На Саянах охота с манком производится с конца
мая до самой осени.
Из других, менее распространненых способов охоты на кабаргу,сле-
дует назвать скрадывание — трудный способ благодаря скалистому
рельефу мест обитания. Менее чутки кабарги во время гона, когда самцы
иногда сами выбегают на охотника, приписывая производимый последним
шум приближающемуся сопернику или самке (Черкасов, 1950). При
охоте с собакой поднятая кабарга также иногда набегает на охотника,
так как имеет привычку возвращаться на старый след. Кабарга крепка на
рану: даже тяжело раненная в живот, она может уйти и пропасть для
охотника бесследно.
CERVIDAE
137
Семейство CERVIDAE — ОЛЕНИ
S с 1 a t e r Ph., 1870, Proc. Zool. Soc., London : 114—117 (сист., распр.); Bro-
oke V., 1878, Proc. Zool. Soc., London : 883—928 (сист.); Dombrowski R.,
1885, Geweihe und Gehorne, Die Geweihbildung der europaischen Hirscharten, Wien.
1—132, Taf. I—XL; Lydekker R., 1898, The deer oi all lands, London : I—XX-|-
1—329 (общ.); Туркин H. В. и К. А. Сатуиин, 1902, Звери России, Gervidae,
М. : I—XI4- 1—506 (общ.); Р о с о с k R., 1910, Proc. Zool. Soc., London : 939—972
(морф.); Lydekker R., 1915, GataL Ungulate mammals Brit. Mus., vol. IV, Lon-
don : I—XXI-J-1—438 (сист.); M a c e w e n W., 1920, The growth and schedding of
the antler of the deer, Glasgow : 1—109; Macewen W., 1923, Brit. Medic. Journ.,
London, 2 : 91—93 (рога); P о с о с к R., 1923, Proc. Zool. Soc., London : 181—207
(морф., сист.): Matthew W. and W. Granger, 1924, Amer. Mus. Novit., 105 :
1—7 (опис. Eumeryx); Noback Ch. and W. Model!, 1928—1933, Zoologica,
XI : 19—30 (рога); P о с о с к R., 1933, Proc. Zool. Soc., London : 377—406 (рога);
P о с о с к R., 1935, Proc. Zool. Soc., London : 179—194 (зубы); R о-m e r A., 1946,
Vertebrate paleontology, Chicago-Illinois : 468—469, 621 (ист.); Waldo С. и др.,
1949, Amer J. Anat., v. 84, No I : 27—61 (рога); ФлеровК. К., 1950, Матер, по четверт.
периоду, 2 : 50—69 (ист.); Северцов С. А., 1951, Пробл. экол. животн., М., изд.
Акад, наук СССР: 61—91 (рога); Флеров К. К., 1952, Кабарги и олени, М.—Л.,
изд. Акад, наук СССР (Фауна СССР, Млекопит., I, 2) : 1—256 (общ.); W i 1 s с k i В.
and Ch. Waldo, 1953, Anat. Rec., vol. 117, No 3 : 353—375 (рога).
Характеристика семейства. За исключением одного рода (Hydro-
potes S winhoe) самцы имеют рога.1 Последние во вполне развитом со-
стоянии представляют костные, обычно ветвящиеся образования, сидя-
щие на концах выростов лобных костей (пеньков). Рога у оленей ежегодно
сбрасываются и вырастают вновь. Выросты лобных костей (пеньки)
остаются постоянными. На их верхушках, через определенный проме-
жуток после сбрасывания старого рога, начинается рост нового. Расту-
щий рог последовательно проходит стадии соединительнотканную, хря-
щевую и костную; снаружи он одет кожей с короткими тонкими волосами
(«бархат»). Процесс окостенения начинается от основания и по перифе-
рии стержня, постепенно распространяясь внутрь и к вершине. В отли-
чие от полорогих (Bovidae) рост рога происходит не в основании, а на
вершине. Последняя до окончания формирования рога сохраняет упру-
гую соединительнотканную консистенцию. На верхушке происходит
и образование всех отростков, как правило дихотомически ветвящегося
рога. Повреждение верхушки растущего рога влечет неправильное его
формообразование только в течение данного года. Если же травма захва-
тывает венчик или вершину пенька (несменяемой части), то неправиль-
ный, уродливый рост может наблюдаться и в дальнейшем. По окончании
процесса окостенения покрывающая рог кожа отмирает и спадает. Окон-
чательно сформировавшиеся рога представляют кость, не прикрытую
ни кожей (как у жираф), ни роговым чехлом (как у полорогих). Место
будущей резорбции кости обозначается неровным вздутым кольцом (ро-
зеткой) в основании спадающей части рога.
От основного ствола (штанги) рога обычно отходит различное число
отростков. Самый нижний, отходящий над розеткой отросток носит
наименование первого надглазничного (рис. 26), следующий за ним —
второго надглазничного; третий отросток, отходящий от ствола ближе к се-
редине последнего, называется средним, а расположенные над ним — верху-
шечными или венечными. Концы отростков, как и вершина самой штанги, У
У северных оленей (род Rangifer Н. Smith) рога имеют и самки.
138
CERVIDAE
могут вторично ветвиться или приобретать уплощенную форму. Наличие
или отсутствие, число, направление и форма отростков могут служить
важными диагностическими признаками для отдельных групп оленей.
Поверхность штанги и главных отростков у большинства видов имеет
многочисленные продольные желобки, следы кровеносных сосудов, про-
ходящих под кожей в период роста рога.
В черепе характерно наличие двойного отверстия носослезного ка-
нала, расположенного вблизи или на самом крае глазницы. На наруж-
ной поверхности слезных костей всегда имеются предглазничные ямки.
Теменные гребешки отставлены друг от друга и не образуют сагитталь-
ного гребня. Носовые отростки межчелюстных костей короткие, и если
касаются носовых костей, то на незначительном протяжении. Щель или
овальное (резцовое) отверстие между межчелюстными костями всегда
имеется.
0 13 3
Зубная формула в типичных случаях ly, Cyi М —• Премоляров
всегда три. Верхние клыки, лишь у самцов немногих форм оленей, длинные,
выступающие вниз из-под верхней губы. Обычно они не выше осталь-
ных зубов, а иногда отсутствуют вовсе. Коренные зубы в подавляющем
большинстве случаев брахиодонтные (рис. 22, Л).
Длина задних конечностей значительно преобладает над длиной перед-
них только у примитивных форм оленей. Боковые (II и V) пальцы со-
храняют костный скелет, но развиты слабее по сравнению с кабаргами.
В запястье сращены (слились) головчатая кость (capitatum s. magnum)
с малой многоугольной (trapezoideum), а в заплюсне — кубовидная
(cuboideum) с ладьевидной (naviculare) и вторая клиновидная с третьей
(cuneiforme II и III). От метаподий боковых лучей сохраняются лишь
верхние или нижние концы, реже те и другие.
В коже развивается большое число специальных желез: предглаз-
ничные, межпальцевые, железы на внутренней стороне плюсны и скака-
тельного (голеностопного) суставов, иногда и на лбу. Зато свойственная
самцам кабарог мускусная железа на нижней части живота у оленей
отсутствует. Отсутствуют также свойственные многим полорогим карпаль-
ные и паховые железы. Наличие или отсутствие тех или иных желез,
их местоположение и строение являются важными признаками в систе-
матике Cervidae. Желчный пузырь отсутствует. Penis не имеет на конце
нитевидного отросгка. Молочная железа четырехдольчатая. Матка имеет
хорошо развитые котиледоны, но число последних меньше, чем у поло-
рогих.
Географическое распространение. Весь материк Евразии с прилегаю-
щими островами от Шпицбергена и Новосибирских островов до остро-
вов Средиземноморья, Малайского архипелага, Филиппин и Японии;
Северная и Южная Америка; Северная Африка. Отсутствуют в Африке
к югу от Сахары, в Австралии, Новой Зеландии,1 на островах Тихого
океана.
Эволюция (рис. 27). Корни семейства оленей ведут к верхнеэоцено-
вым трагулидам (Tragalidae). Флеров (1950) считает весьма близким
к основанию ствола Cervidae род Lophiomeryx Pomel из олигоцена Европы
1 В Повой Зеландии акклиматизирован благородный олень (Cervas elaphus L.)
и лань (Cervus dama L.), а на островах Антарктики — северный олень (Rangifer tu-
rundas L.).
CERVIDAE
139
и Азии. Lophiomeryx turgaicus Fl. имел еще короткие, функционально
четырехпалые конечности с неслившимися метаподиями, первый ниж-
ний премоляр и длинные верхние клыки; переднекоренные зубы у него
еще не имели селенодонтных складок. Формой, отвечающей требованиям
к идеальному предку всех последующих оленей и уже относимой к этому
семейству, является нижне- и среднеолигоценовый Eumeryx Matthew
and Grangen из Монголии. Он имел уже слившиеся центральные мета-
подии, но кили на их суставных блоках развиты еще слабо; сохранялся
первый премоляр нижней челюсти.
Род Eumeryx вместе с рядом других, главным образом миоценовых,
примитивных оленей Евразии и Америки, в большинстве лишенных ро-
гов, но имевших длинные изог-
нутые верхние клыки, образует
подсемейство Palaeomerycinae. Не-
которые формы этой группы дали
начало современным подсемей-
ствам Cervidae. Есть предположе-
ние, что палеомерицины являются
предками не только оленей, но и
жирафе кабаргами (Romer, 1946).
Наиболее характерной чертой
Cervidae является своеобразное
направление эволюции рогов.
Предки и наиболее ранние пред-
ставители семейства были безро-
гими. На этой стадии эволюции
из современных остановился ки-
тайский водяной олень {Hydro-
poles inermis Swinh.). В своей
эволюции рога оленей прошли
стадию простых выростов лобной
кости, в течение всей жизни не-
сменяемых и покрытых кожей с
Рис. 26. Рог благородного оленя {Cervus
elaphus L.) и его отростки.
1 — первый надглазничный; 2 — второй
надглазничный; 3—средний; 4—венеч-
ные.
волосами, как у современных
жираф. В миоцене и плиоцене Северной Америки существовала
своеобразная боковая ветвь оленей (подсемейство Dromomerycin.ae'),
не оставившая в современной фауне потомков. Несменяемые ко-
стные выросты у представителей этой группы достигали большой ве-
личины и вырастали не только на лобной, но и на затылочной кости.
В ходе эволюции остальных оленей на кончиках этих выростов обра-
зовывались вначале, вероятно, спадающие роговые колпачки, а затем
периодически сбрасываемые и вновь вырастающие небольшие костные
образования. В дальнейшем несменяемые выросты, ставшие пеньками,
укорачивались, а сменяемая часть, наоборот, удлинялась и приобрела
дихотомическое ветвление. Большую часть этих стадий можно просле-
дить и среди современных форм семейства. Первоначально, а у многих
оленей и сейчас, рога играют роль вторичнополового признака, украше-
ния самцов и турнирного оружия. Значение рогов как орудия защиты
очень невелико, тем более, что в течение большей части года они или
совсем отсутствуют, или находятся в состоянии, исключающем воз-
можность их применения для этой цели.
Основное направление эволюции филогенетической ветви оленей —
то же, что и у других жвачных. Это — приспособление к окончательному
140
CERVTDAE
переходу на питание растительной пищей, в противовес еще в известной
мере всеядным оленькам. С этим связана дальнейшая дифференцировка
отделов желудка, повышение степени гипсодонтии, моляризация перед-
некоренных зубов. Повышение уровня ор1аиизации, по сравнению с олень-
ками, выражается также в большей дифференцированности извилин и
борозд на полушариях головного мозга. Однако во всех этих отношениях
олени значительно еще уступают полорогим (Bovidae). Зубы у них, в, це-
лом, еще брахиодонтны и не приспособлены к пережевыванию жестких
травянистых растений. Большая часть оленей питается листвой, корой
и тонкими ветками кустарников и деревьев, т. е.. сравнительно мяг-
кой пищей. Травянистая растительнссть, за исключением мягкого
разнотравья, не составляет у них существенной доли пищевого
рациона.
В ходе эволюции оленей имел место также переход от крадущейся
походки и передвижения прыжками к быстрому бегу рысью. В связи
с этим исчезает резкая непропорциональность развития передних и зад-
них конечностей, происходит общее их удлинение, особенно дистальных
звеньев (метаподий и пальцев), усиление центральных и редукция боко-
вых лучей кисти и стопы.
Характерной, но пока трудно объясняемой с биологической точки
зрения чертой в эволюции Cervidae является переход от пятнистой
к более однотонной окраске волосяного покрова. У примитивных из
современных и, вероятно, у всех предковых форм пятнистая окраска
наблюдается и у молодых, и у взрослых. У многих современных оленей
пятнистость сохраняется только у молодых особей, тогда как взрослые
имеют однотонную окраску. Наконец, у наиболее специализирован-
ных (например у северного оленя) и молодые родятся в одноцветной
шерсти.
Классификация. 11 родов и 48 видов современных оленей К. К. Фле-
ров (1952) делит на 4 подсемейства.
1) Cervulinae Sci. (=Muniiacinae Pocock) — Мунтжаки. Мелкие при-
митивные олени, стоящие близко к исходным формам семейства, высо-
той в холке не более 60 см и с длиною черепа не свыше 25 см. Несменя-
емые выросты лобных костей (пеньки рогов) длинные; их длина от зад-
него края глазницы до вершины больше длины верхнего зубного ряда.
Спадающая часть рога, наоборот, очень мала. И у самцов, и у самок
вдоль наружных краев лобных костей проходят продольные гребни.
Включает два современных рода: 1) Muniiacus Rafinesque — мунтжаки
и 2) Elaphodus Milne-Edw. — хохлатые олени. Распростране-
ние: Индия, п-ов Индокитай, Малайский архипелаг, о. Тайвань,
южный и средний Китай.
2) Hydropotinae Truess. — Водяные олени. Также мелкие животные,
высотой в холке не более 60 см и с длиною черепа менее 20 см. От всех
других оленей отличаются тем, что ни у самок, ни у самцов нет рогов.
Взамен последних у самцов, как у кабарги, мощно развитые верхние
клыки, сильно выступающие вниз из-под верхней губы. Единственный
вид, Hydropotes inermis Swinh. — китайский водяной олень, рас-
пространен в Восточном Китае от долины р. Янцзы к северу
до Кореи.
3) Cervinae Baird — Настоящие олени. Животные от мелких (высотою
в холке немного более 60 см) до очень крупных (свыше 2 м). Несменя-
емые выросты лобных костей (пеньки рогов) короткие; их длина всегда
менее длины зубного ряда верхней челюсти. Спадающая часть рога,
CERVTDAE 141
наоборот, велика и имеет не менее трех отростков. Включает современ-
ные роды; 1) Capreolus L. — косули, 2) Cervus. L — благородные олени,
3) Elaphurus Milne-Edw. — олень Давида, 4) Alces Gray — лоси. Распро-
странение: северная Африка, Европа и Азия с прилегающими
островами (кроме зоны тундры).; Северная Америка на юг до Мексики.
4) Neocervinae Caret. ( = Odocoileinae Pocock) — Американские олени.
Размеры от очень мелких до крупных (высота в холке от 34 до 130 см).
Пеньки рогов короткие; их длина всегда короче длины зубного ряда;
спадающая часть рога бывает или незначительной, или, наоборот, мощно
развита. Наиболее существенный признак, отличающий эту группу от
Рис. 27. Схема филогении семейства оленей (Cervidae).
(По К. К. Флерову, 1950).
всех других подсемейств оленей, — хоаны, полностью разделенные вы-
ступающим далеко назад сошником. Задний край сошника достигает
и касается основной клиновидной кости, (базисфеноида). Включает со-
временные роды: 1) Mazama Rafinesque — мазамы, 2) Pudu Gray —пуду,
3) Odocoileus Rafinesque — виргинские пампасовые олени и гуэмалы,
4) Rangtfer Н. Smith — северные олени. Р а с и ространение:
северная часть Европы и Азии , (в последней к югу до 50—45° с. ш.),
Северная и Южная Америка с некоторыми прилегающими остро-
вами.
Представители первых двух подсемейств в современной фауне СССР
отсутствуют. Подсемейства Сervinae и Aeocervinae представлены родами
Capreolus, Cervus, Alces и Rangijer.
142
CERVIDAE
ОПРЕДЕЛИТЕЛЬНАЯ ТАБЛИЦА РОДОВ СЕМЕЙСТВА ОЛЕНЕЙ
(CERVIDAE) ФАУНЫ СССР
1 (2). Рога, за крайне редкими исключениями, имеют и самцы и самки.
Все пространство верхей губы между ноздрями покрыто волосами.
Зароговая часть черепа укорочена; расстояние от заднего края
черепа (затылочного бугра) до переднего края теменных костей
меньше расстояния между срединой линии, соединяющей задние
края глазниц, и линией, соединяющей задние выступы лобных
костей. Сошник срастается с небными костями и делит хоаны на
две половины. Передние края глазниц расположены позади уровня
заднего края зубного ряда......................................
.............. Rangifer Н. Smith — Северные олени (стр. 265).
2 (1). Рога имеют только самцы. На передней стороне верхней губы
между ноздрями имеется хотя бы небольшое пятно голой, лишен-
ной волоса кожи. Зароговая часть черепа хорошо развита. Рас-
стояние от заднего края черепа (затылочного бугра) до переднего
края теменных костей больше расстояния между срединой линии,
соединяющей задние края глазниц, и линией, соединяющей задние
концы лобных костей. Задний край сошника не срастается с нёб-
ными костями и не делит хоаны на две половины. Передний край
глазниц расположен впереди уровня заднего края зубного ряда.
3 (4). Размеры очень большие. Высота взрослых зверей в холке более
170 см. Длина черепа свыше 500 мм. Верхняя губа сильно увели-
чена и нависает над нижней. Кожа на нижней стороне головы под гор-
лом образует отвисающий вырост («серьгу»). Вся верхняя губа,
за исключением небольшого голого пятна треугольной или оваль-
ной формы, покрыта волосами. Волосы задней части крупа имеют оди-
наковую с остальным туловищем окраску. В окраске молодых пят-
нистость отсутствует. Линия затылочного профиля при взгляде
на череп сверху имеет вогнутую форму (рис. 53). Длина носовых
костей значительно меньше расстояния от их передних концов
до переднего края межчелюстных костей. Межчелюстные кости
длинные, значительно длиннее ряда верхних коренных зубов, но
не соприкасаются с носовыми костями............................
......................... Alces Gray — Лоси (стр. 234).
4 (3). Размеры не очень большие. Высота в холке менее 170 см. Длина
черепа не превышает 480 мм. Верхняя губа не увеличена и не на-
висает над нижней. Кожа на нижней стороне головы не образует
выроста или складки. Всё или почти всё пространство верхней
губы между ноздрями покрыто голой, не имеющей волос кожей.
Волосы на задней части крупа окрашены светлее соседних участ-
ков и образуют вокруг или по бокам хвоста «зеркало». Молодые
первые месяцы жизни имеют пятнистую окраску. Линия затылоч-
ного отверстия, при взгляде на череп сверху, имеет прямую или
выпуклую форму. Длина носовых костей больше расстояния от
их передних концов до переднего края межчелюстных костей.
Межчелюстные кости короче ряда коренных зубов верхней челю-
сти, но соприкасаются с носовыми костями.
5 (6). Предглазничные железы отсутствуют. Зрачки глаз поставлены
косо. Хвост снаружи не виден; длина его не более 3 см. Рога не
имеют надглазничных отростков. Длина лицевой поверхности слез-
CERVIDAE
143
ной кости менее передне-заднего диаметра глазницы. Передний край
нёбных костей находится на уровне передней половины первого
верхнего моляра (М1). В мыщелковую ямку открывается только от-
верстие подъязычного канала . Capreolus Gray — Косули (стр. 144).
6 (5). Предглазничные железы имеются и хорошо развиты. Зрачки глаз
имеют форму расположенного горизонтально овала. Хвост снаружи
ясно виден, длиною свыше 3 см. Рога имеют один или два надглазнич-
ных отростка. Длина слезной кости не меньше передне-заднего
диаметра глазницы. Передний край нёбных костей не заходит вперед
дальше уровня задней половины первого верхнего моляра. В мыщел-
ковую ямку открываются два отверстия: подъязычное отверстие
и отверстие, ведущее в мыщелковый канал..........................
.................. Cervus L. — Настоящие олени (стр. 173).
Подсемейство CERVINAE — НАСТОЯЩИЕ ОЛЕНИ
Характеристика подсемейства.1 Олени от мелких до очень крупных
размеров. Высота в холке от 60 до 235 см. Рога у самцов всегда имеются,
у самок отсутствуют; пеньки их короткие; расстояние от заднего края
глазницы до вершины пенька обычно меньше длины ряда коренных зу-
бов верхней челюсти. Спадающая часть рогов, наоборот, велика и у взрос-
лых животных имеет не менее трех отростков. В скелете конечностей
от боковых метаподий сохраняются или верхние, или нижние концы
(рудименты). Боковые пальцы и копыта сравнительно хорошо развиты
(в отличие от подсем. Cervulinae).
В черепе, в отличие от Cervulinae, лобные кости в передней части не удли-
нены и поэтому задние концы носовых костей достигают уровня передних
краев глазниц, а если не достигают, то на расстояние не больше длины
последнего верхнего моляра. В отличие от Cervulinae, шов между лоб-
ными и слезными костями сравнительно короткий, меньше длины сво-
бодного края слезной кости вдоль этмоидальных щелей. Сошник своим
вентральным краем не достигает и не касается заднего края нёбных ко-
стей и не делит хоаны на две половины (в противоположность Neocer-
vinae). Задний край сошника не достигает и не соприкасается с базис-
феноидом. Слуховые пузыри небольшие, угловатые, не выступают или
лишь едва выступают ниже вентральной поверхности basioccipitale
(в отличие от Hydropotinae). В отличие от Cervulinae и Hydropotinae,
края глазниц сильно выступают в стороны. Ширина черепа на уровне
их нижних краев больше ширины на уровне задних краев скуловых ко-
стей. Верхние клыки и у самцов, и у самок не велики, не выступают выше
уровня остальных зубов, а иногда даже отсутствуют вовсе.
Географическое распространение и история. Северная Африка,
Европа и Азия и часть прилегающих к ним островов. Два вида распро-
странены в Северной Америке к югу до Мексики. Отсутствуют в зоне аркти-
ческих тундр.
Корни подсемейства Cervinae ведут к какой-то из примитивных форм
мунтжаков (Dicrocerus Lartet и другие), в миоцене широко распростра-
ненных по Азии и Европе. Уже в верхнем миоцене и начале плиоцена
появляется ряд видов, обнаруживающих черты сходства с современными
косулями и объединяемых в род Procapreolus Schl. В основание другой
1 В основном по К. Флерову (1952).
144
CERVIDAE
ветви оленей должен быть положен также верхнемиоценовый и нижне-
плиоценовый потомок Dicioeerus, род Cervavitus КЬош.,еще сохраняющий
черты сходства с мунтжаками, но уже имевший рога типа настоящих
оленей. Потомки Cervavitus, по-видимому, дали начало не только группе
благородных оленей (род Cervus L.), но и лосю (Флеров, 1950).
Классификация. Подсемейство включает четыре современ-
ные рода.
1) Capreolus Gray — Косули. Животные сравнительно небольшой
величины. Длина черепа не превосходит 250 мм. Слезные кости корот-
кие, меньше передне-заднего диаметра глазницы. Предглазничные ямки
имеют вид пологого, неглубокого вдавления. Предглазничные железы
рудиментарные. Носовое зеркало занимает все пространство между ноз-
дрями. Хвост короткий, скрыт в окружающих его волосах. Рога не имеют
надглазничного отростка. Их ствол в начальной (проксимальной) части
усажен множеством резко выступающих бугорков или тонких отростков.
2) Cervus L. — Настоящие олени. Животные средней или крупной
величины. Длина черепа от 250 до 500 мм. Слезные кости не меньше
передне-заднего диаметра глазницы, но не превышают его больше чем
в 1 /4 раза. Предглазничные ямки глубокие. Носовое зеркало большое,
занимает все пространство между ноздрями. Хвост не длинный, но ясно
выступает, иногда несколько длиннее уха. Рога всегда имеют один или
два надглазничных отростка. Ствол рогов над розеткой имеет шерохо-
ватую поверхность, но без резко выступающих бугорков.
3) Elaphurus Milne-Edw. — с единственным видом Е. davidianus Milne-
Edw. — оленем Давида, обитавшим, вероятно, в Китае. В настоящее время
уже не встречается в диком состоянии. Размеры довольно крупные. От
других оленей отличается тем, что череп (длиною около 420 мм) имеет
вытянутую предглазничную часть. Слезные кости более чем в 2 раза
превышают диаметр глазницы. Длинный хвост (с волосами почти дости-
гает скакательного сустава). Длинные боковые копыта; кожа на окружаю-
щем их пространстве до пяточных частей главных копыт лишена волоса.
Все отростки рогов, из которых самый длинный нижний, направлены
назад.
4) Alces Gray — Лоси. Очень крупные животные. Длина черепа
у взрослых самцов более 500 мм. Очень короткие носовые кости, в про-
тивоположность другим оленям, более чем вдвое короче длины перед-
него носового входа (от prosthion до передних концов носовых костей).
Необычайно развитая вздутая верхняя губа нависает над нижней. Но-
совое зеркало представляет лишь небольшое голое пятно в центре по-
крытого волосами пространства между ноздрями.
Иногда выделяют в качестве самостоятельного рода Duma Frisch—
ланей, отличающихся,, помимо некоторых других признаков (в част-
ности, отсутствием верхних клыков), уплощенными и лопаткообраз-
ными, расширенными в верхней части рогами.
В фауне СССР имеются представители всех перечисленных родов,
кроме Elaphurus.
Род CAPREOLUS GRAY — КОСУЛИ
Характеристика рода — см. выше.
Род включает один вид — Capreolus capreolus L. — обыкновенную
косулю.
CAPREOLUS
145
Capreolus capreolus Linnaeus. — Косуля, или дикая коза
(рис. 28—33)
Linnaeus С., 1758, Syst. nat., ed. X, 1 : 68 (первооп.); П а л л а с П. С.,
1773, Путешеств. по разным провинциям Российск. импер., т. 1 : 151, Прибавл. :
4—2 (распр.); Богданов М., 1871, Птицы и звери черноземн. полосы Поволжья,
Казань : 177 (распр.); Сабанеев Л. П., 1874, Позвоночные Средн. Урала, М. :
1—2 (распр.); Сабанеев Л. П., 1875, Природа, IV : 1—21 (распр., биол., ох.,
изобр.); Dombrowski R., 1876, Das Reh, Wien : 1—91, Taf. I—XIV (биол.,
ox.); Туркин H. В. и К. А. Сатунин, 1902, Звери России, Cervidae, М. :
399—490 (общ., изобр.); Разевиг В. А., 1904, Псовая и ружейн. охота, 4 : 25—
38; 5 : 82—95, 6 : 38—54 (общ., изобр.); Sakurai Т., 1906, в кн. К е i Ь е 1, Nor-
mentafeln zur Entwicklungsgeschichte d. Wirbeltiere, Jena : 6 (размн.); К руликов-
c к и й П., 1909, Календарь и памяти, книжка Вятск. губ. :5 (распр.); Дин ник Н. Я.,
1910, Звери Кавказа, ч. 1, Тифлис : 64—93 (общ.); М ii 1 1 е г-L iebenswalde J.,
1910, Lebensbilder aus der Tierwelt, Bd. II, Folge I, Saugetiere, II : 91—165 (общ.,
изобр.); Pocock R., 1910, Proc. Zool. Soc. London : 957—958 (морф.); Врублев-
ский К., 1912, Архив ветер, наук, 8 : 756 (пит.); В рауне р А. А., 1915, Зап.
Крымск, общ. естественен., 5 : 112—114 (сист.); Lydekker R., 1915, Catal. Ungu-
late Mammals Brit. Mus., IV, London : 218—228 (сист., синон.); Федорович Ф. Ф.,
1915, Тр. Пензенск, общ. любит, естествозн., 11 : 51—52 (распр.); Браунер А. А.,
1923, Сельскохоз. зоол., Одесса, Госизд. Украины : 83—85 (распр.); Туров С. С.,
1924, Тр. Общ. иссл. Рязанск. края, III : 61 (распр.); Мил Д., 1926, Охотник, 1 :
20 (распр.); Тассовский Г. Н., 1927, Произв. силы Дальн. Вост., III : 534
'(распр.); Кузнецов Б. А., 1929, Тр. по лесному опытн. делу, VI : 103—104 (распр.,
биол., ох.); Лавров Н. П., 1929, Тр. по лесн. опытн. делу, VI : 49—81 (распр.,
биол., хоз. знал.); Раевский В. В. и Л. Г. Капланов, 1929, Zool. Anzeig.,
84, № 5 : 151 (распр.); Федюшин А. В., 1929, Динамика и геогр. распр. охотн.
фауны BGGP, Минск, изд. «Паляуничи беларуси» : 24—28 (распр.); Даль С. К.,
1930, Зап. Крымск, общ. естествоисп. и любит, прир., XII : 64—127 (сист., биол.,
изобр.); Л а в р о в А., 1930; Охотник, 1 : 14 (распр.); Лентовский М. Н., 1930,
Вести. Центр, муз. Казахстана, 1:109—110 (распр.); Шиллингер Ф. Ф.,
1930, Европейский олень, лань и косуля в охотхозяйстве, Харьков, изд. ЦС ВУСОР :
54—76 (общ., изобр.); Яковлев Е., 1930, Тр. Зоол. секц. Среднесибирск. отд.
Русск. географ, общ., 1 : 58 (распр.); Скалой В. Н., 1931, Тр. по защите раст.,
1, 8 : 152 (хоз. знач.); Jacobi А., 1932, Jenaische Ztschr. f. Naturwissensch., 67 :
326—345 (морф., сист.); Дементьев Г. П., 1933, Козуля, М., КОИЗ : 1—51 (общ.,
изобр.); Шухов И. Н., 1933, Охотничий промысел Приенисейского края, Красно-
ярск : 1—24 (распр.); Афанасьев А. В., 1934, Тр. Совета по изуч. произв. сил
Акад, наук СССР, сер. дальневост., 2 : 259—260 (распр.); Золотарев Н. Т.,
1934, Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. дальневост., 3 : 134—
135 (распр.); Тугаринов А. Я., Н. А. Смирнова и А. И. Иванов, 1934,
Птицы и млекопит. Якутии, Л., изд. Акад, наук СССР : 49, 57, 64 (распр.); Фле-
ров К. К., 1935, в кн.: Виноградов Б. С., Е. Н. Павловский и
К. К. Флеров, Звери Таджикистана. М,—Л., изд. Акад. наук. СССР : 106—
109 (сист., распр.); Подаревский В. Б., 1936, Пробл. охотхоз. акклиматиз.
:в вост. Сибири, Иркутск, ОГИЗ : 43—46 (распр.); Строганов С. У., 1936, Зоол.
журн., XV, 3:542—543 (распр.); Шнитников В. Н., 1936, Млекопитающие
Семиречья, М.—Л., изд. Акад, наук СССР : 140—144 (общ.); Новиков Г. А.,
1937, Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. вост.-сиб., 4 : 234—236
(распр.); Птушенко Е. С., 1937, Сборн. «Памяти акад. Мензбира», М., изд. Акад,
наук СССР : 450—451 (распр.); Дмитриев В. В., 1938, Тр. Аптайск. гос. заповеди.,
1 : 226—229 (распр.); Куклин С. А., 1938, Звери и птицы Урала, Свердловск,
Облгиз : 122—124 (распр., биол., изобр.); ГО р г е н с о н П. Б., 1938, Тр. Алтайск.
гос. запов., 1 :159—160 (распрА; Рухлядев Д. П., 1939, Науч.-метод. зап. Комит.
но заповеди., III : 133—140 (бол.); Кулагин Н. М., 1940, Фауна БССР, т. 1,
вып. IV, Минск, изд. Акад, наук БССР : 40—53 (общ., изобр.); Антипин В. М.,
1941, Млекопитающие Казахстана, т. III, Алма-Ата, Каз. обл. гос. изд. : 18—28 (общ.).
С е в е р ц о в С. А., 1941, Сов. охотник, 1 : 19 (распр.); Скалой В. Н., И. Щ ер-
бак о в иМ. Базыкин, 1941, Природа и соц. хозяйство VIII : 277—278 (распр );
Афанасьев А. В., 1945, Изв. Акад, наук Казахск. ССР, 5 : 29—31 (распр.);
Бажанов В. С., 1945, Изв. Акад, наук Казахск. ССР, 5 : 75—78 (распр.); Ска-
лой В. Н., 1946, Природа, 6:73 (распр.); Формозов А. Н., 1946, Снежн.
покров в жизни млекопит. и птиц, М., изд. Моск. общ. испыт. прир. : 114—117 (распр.);
Барабаш-Никифоров И. И., 1947, Сборн. «Воронежск. гос. заповедник»,
Ю Фауна СССР, И. И. Соколов
146
CERVIDAE
Воронеж, Обл. изд. : 31—32 (распр.); Фетисов А. С., 1947, Сезонные корма си-
бирской косули, Иркутск, Облиздат: 1—12; Формозов А. Н., 1947, Сборн.
«Природа гор. Москвы и Подмосковья», М.—Л., изд. Акад, наук СССР : 303—304
(распр.); Иваненко Б. И., 1948, Тр. Крымск, гос. заповеди., III : 87—94 (хоз.
знач.); Кузнецов Б. А., 1948, Звери Киргизии, М., изд. Моск. общ. испыт. прир. :
109—112 (распр., биол.); Марков Е. Л., 1948, Тр. Тбилисск. зоопарка, И : 43—
48 (биол., размн.); Аверин Ю. В., 1949, Тр. Ильменск. гос. заповеди., 4 : 9—62
(общ.); Штегман Б. К., 1949, Сборн. «Охрана природы», 8 : 71—72 (биол.); Д а р щ-
к е в и ч Я., 1950, Птицы и звери Чкаловск, обл., Чкалов, Облиздат : 88—91 (распр.,
изобр ); Е л п а т ь е в с к и й В. С., 1950, Уч. зап. Саратовск. гос. унив., сер. биол.,
26 : 44 (распр.); Калниньш А. И., 1950, Охота и охотн. хоз. Латвийской ССР,
Рига, Латгосиздат : 122—138 (общ., изобр.); Копылов И. П., 1950, Дикие копытн.
животн. Иркутской обл., Иркутск, Облиздат : 28—37 (общ., изобр.); Stive Н.,
1950, Ztsc.hr. mikrosk.-anat. Forsch., Leipzig, 5 : 427—530 (размн.); WettsteinO.,
1951, Osterr. Weidwerk, 2 : 13—14 (определ. возр.); Граве Г. Л., 1951, Животн.
мир Смоленск, обл., Смоленск, Облиздат : 156—157 (распр., изобр.); Райский А. П.
1951, Сборн. «Очерки физич. геогр. Чкаловск, обл.», Чкалов, Облиздат : 151—202
(распр.); Скалов В. Н., 1951, Бюлл. Моск. общ. испыт. прир., LVI, 1:27—32
(распр.); Судариков В. Е. и К. М. Ры ж и ков, 1951, Тр. Гельминтол. лабор.,
V : 53—58 (параз.); Т е g п е г Н., 1951, The roe deer, London : 1—176 (общ., изобр.);
Федосов А. В. и К. Н. Никитин, 1951, Животн. мир. Брянск, обл., Брянск.,
изд. «Брянский рабочий» : 10 (распр., изобр.); Е ] 1 е г m a n J. and Т. Morrison-
Scot t, 1951, Checklist of palaearctic and Indian mammals, London : 371—373
(сист., синон.); Верещагин H. К., 1952, Млекопитающие Кавказа. Диссертация
(рукопись), Ленинград : 894—900 (распр.); Jaczewski Z., 1952, Fragm. faunis-
tica Mus. Zool. Polonici, VI, 12 : 199—207 (биол.); Кириков С. В., 1952, Птицы
и млекопит. в условиях ландшафтов южн. оконечн. Урала. М., изд. Акад, наук СССР :
268—270 (распр., биол.); Корнеев О. П,, 1952, Визначник 3Bipie УРСР, Ки1в,
изд. «Радянська школа» : 206—208 (распр.); Кузнецов Б. А., 1952, Изв.Молдавск.
фил. Акад, наук СССР, 4/5 : 40—41 (распр.); Пастернак Ф. А., 1952, Монография
косули, автореферат, М. : 1—9 (общ.); Сысоев В. П., 1952, Охота в Хабаровском
крае, Хабаровск, Дальгиз : 108—111 (ох.); Флеров К. К., 1952, Кабарги и олени,
М.—Л., изд. Акад, наук СССР : 92—ИЗ (общ., синон., изобр.); Янушевич А. И.,
1952, Фауна позвоночн. Тувинск. обл., Новосибирск, изд. Западносибирск, фил. Акад,
наук СССР : 61 (распр.); Янушевич А. И. и И. Благовещенский, 1952,
Промысл, звери и птицы запади. Сибири, Новосибирск, Облиздат : 130—133 (распр.,
изобр.); Афанасьев А. В. и др., 1953, Звери Казахстана, Алма-Ата, изд. Акад,
наук Казахск. ССР : 464—468 (общ., изобр.); Лаптев И. П., 1953, Охотн. хоз.
Томской обл., Томск, изд. Общ. по распр. полит, и научи, знаний : 11 (распр.);
Ралль Ю. М., 1953, Уч. зап. Ростовск. н/Д. унив., XIX, 3 : 115—126 (распр.);
Тимофеев В. В., 1953, Тр. Всесоюзн.: инет, охотн. промысла, XII : 203—206
(биол.); Томилин А. Г., 1953, Тр. Моск, пушно-мех. инет., IV : 32 (распр.); Фе-
тисов А. С., 1953, Козуля в вост. Сибири, Иркутск, изд. Иркутск, гос. унив. : 1—
76 (общ., изобр.); Абрамов К. Г., 1954, Копытные звери Дальн. Востока, Хаба-
ровск, Книжн. изд. : 35—43 (общ., изобр.); Асадов С. М., 1954, Изв. Акад, наук
Азербайдж. ССР, 9:33—46 (параз.); Банников А. Г., 1954, Тр. Монгольск.
комисс., 53 : 200—208 (общ., изобр.); Berger К., 1954, Forst u. Jagd, 5 : 197—198
(морф.); Попов В. А. и др., 1954, Тр. Казанск. фил. Акад, наук СССР, сер. биол.
наук, 3:190—191 (распр.); Слудский А. А., 1954, Сб. «Природн. очаговость
заразных болезней в Казахстане», 2 : 69—98 (бол.); Асадов С. М., 1955, Докл.
Акад, наук Азербайдж. ССР, т. И, № 1 : 39—42 (параз.), Баранчеев Л. М.,
1955, Зап. Амурск, обл. музея, 3 : 209—212 (распр., биол.); Лаптев И. П., 1955,
Заметки по фауне и флоре Сибири, 18 : 5 (распр.); Насимович А. А., 1955,
Роль снежн. покрова в жизни копытных, М., изд. Акад, наук СССР : 1—402 (распр.,
биол.); Пастернак Ф. А., 1955, Уч. зап. Моск. гор. педагогии, инет., XXXVIII :
29—140 (морф., стад., пит.); Саблина Т. Б., 1955, Копытные Беловежской пущи,
М., изд. Акад, наук СССР : 90—114 (биол.); Соколов И. И., 1956, Зоол. журн.,
XXXV, 8 : 1238—1249 (определ. возр.).
Описание. Животные средней величины. Высота в холке у взрослых
самцов составляет от 65 до 95—100 см. Длина тела от 100 до 150 см. Вес
од 20 до 60 кг. Размеры самок несколько меньше. Длина черепа 190—250 мм.
Высоконоги. Высота в холке приблизительно равна длине туловища.
CAPREOLUS
14T
Высота в крестце сантиметра на два-три превышает высоту в холке.
Поэтому линия спины несколько наклонена вперед.
Голова постепенно суживается к концу морды и не производит впе-
чатления длинной или тяжелой. Уши длиною 10—15 см, составляют
больше половины длины головы, имеют заостренные верхушки. Носовое
зеркало темного цвета, занимает почти все пространство между ноздрями
и срединную часть верхней губы до ее нижнего края. Верхняя его гра-
Рис. 28. Косуля {Capreolus capreolus L.).
ница в виде прямой или слегка вздутой линии проходит ниже верхних
краев ноздрей. Полоска лишенной волоса кожи тянется также вдоль
нижнего, а иногда и наружного их края. Глаза имеют темно-коричневую
радужную оболочку и косо поставленные зрачки.
Хвост короткий, не длиннее 2—3 см, скрыт в окружающих его воло-
сах ягодиц и крупа. Копыта узкие, заостренные; боковые — примерно
вдвое короче главных и при ходьбе, обычно, не касаются почвы. Цвет
копыт черный.
Рога длиною до 40 см, дихотомически ветвящиеся, нормально имеют
каждый по три остростка, при этом надглазничные отростки всегда от-
сутствуют, а средние направлены вперед. Изредка число отростков может
доходить на каждом роге до четырех и даже пяти. На месте отхождения,
среднего отростка вершина рога отогнута назад и вновь ветвится так,
что передний из конечных отростков направлен почти вертикально.,,
10*
148
CERVIDAE
Поперечное сечение рога круглое, но поверхность главного ствола от
основания и иногда почти до последней развилки усажена множеством
выростов (перлов, или жемчужин); некоторые из последних достигают
высоты свыше 1 см. Розетки рогов у косули, по сравнению с другими
оленями, очень широки и могут превосходить диаметр ствола рога в 1 % или
2 раза. Главные стволы рогов наклонены слегка назад, лирообразно рас-
ходятся в стороны, но вновь сближаются в вершинах, особенно концами
задних отростков.
У самок рога в норме отсутствуют, но на месте их часто имеются не-
большие бугорки на лобных костях. В отличие от самок рода Cervus L.,
бугорки эти у косуль расположены не по краям боковых лобных гребней,
а на задних выступах лобных костей, и направлены не в стороны, а вер-
тикально. В виде редкого исключения встречаются рогатые самки.
В полном зимнем наряде волосяной покров туловища и шеи состоит
из редкого, тонкого, сильно извитого пуха и густой, волнообразно извитой
грубой и ломкой ости, длиной на спине и крупе до 50—55 мм. На животе
в окружности препуция длинные волосы у самцов образуют свисающий
пучок. На голове, наружной стороне ушей и на конечностях, ниже ска-
кательного сустава, волос прямой, короткий, менее ломкий и плотно
прилегающий к коже. На лбу он несколько приподнят и слегка завит.
Внутренняя сторона ушей покрыта редкими, но длинными и мягкими во-
лосами. Более редкий и мягкий волос растет также в нахах и в подмышках.
Общий тон зимней окраски у взрослых животных серый разного от-
тенка — от бурого до рыжеватого. Окраска, более темная на спине и
крупе, на боках и животе постепенно светлеет. Подшерсток светло-бу-
рого цвета. Волосы ости от основания светло-серые, постепенно темнеют
к вершине; в верхней трети волоса темно-бурое кольцо сменяется светлым,
а заостренная верхушка вновь приобретает темную, почти черную
окраску. Чередование в верхушечной части светлых и темных участков
обусловливает штриховатость зимней окраски косули. Посветление
окраски на боках и нижней части туловища вызвано тем, что исчезает
нижнее темное кольцо, а светлое становится шире. Основная же часть
волоса, равно как и верхушка, не меняют своего цвета. Длинные мягкие
волосы в задней части живота, нахах и подмышках имеют более или
менее равномерную светлую, рыжевато-белую окраску. Тот или иной
оттенок окраски меха придает светлое кольцо в верхней трети остевых
волос: если оно белого цвета, то окраска имеет характер чисто серой;
обычно наблюдающаяся рыжеватость обусловлена соответствующей
окраской светлой кольцевой зоны. Конечности ниже запястных и скакатель-
ных суставов имеют однотонную светлую, рыжеватую окраску. Наруж-
ная сторона ушей имеет одинаковую с верхней частью туловища окраску,
но концы ушей обычно темные. Внутренняя их сторона грязно-белого
цвета. Верхняя губа по бокам от носового зеркала, средняя часть нижней
губы и подбородка белые. Узкая полоска выше носового зеркала, полоса
от краев ноздрей к углам рта и боковые части нижней губы темно-корич-
невые. Белые волосы на ягодице по бокам от хвоста образуют резко от-
граниченное от соседних темноокрашенных участков «хвостовое зеркало».
У косули оно сравнительно не велико, не подымается выше основания
хвоста и почти не заходит на стороны крупа.
Летний мех состоит из почти прямых коротких остевых волос, более
тонких, чем зимою, и менее ломких. Подшерсток практически отсутствует.
Окраска туловища и шеи летом однотонная рыжая, только на боках и
особенно на животе более светлая. На голове, особенно на морде, сохра-
Рис. 29. Череп Capreolus capreolus L.
150
CERVIDAE
няется, хотя и со значительной примесью рыжего, серый оттенок. Конец
морды и ушей сохраняет темную окраску. Околохвостовое зеркало ста-
новится светло-рыжим, почти неотличимым’от цвета соседних участков.
Молодые родятся пятнистыми. На общем рыжеватом фоне распо-
ложены с каждой стороны три ряда пятен желтовато-белого цвета. Ме-
диальный из них тянется от затылка к основанию хвоста; остальные
начинаются в области лопаток и теряются на крупе. Пятнистость, выра-
женная вначале отчетливо, к месячному возрасту обычно исчезает. Из-
редка встречаются полные или частичные альбиносы, а также пестро-
окрашенные косули,
Предглазничные железы рудиментарные, нефункционирующие. И на
передних, и на задних^ конечностях на передней стороне выше копыт
имеются межпальцевые железы: на задних — в виде глубокого мешочка
с узким выходным отверстием, на передних —в виде пологого вдавления.
Железы выделяют секрет, обладающий специфическим запахом. Имеются
метатарзальные железы; последние представляют небольшие участки
утолщенной кожи на внутренней стороне плюсны, покрытые более
длинными и выступающими над общей поверхностью волосами, скле-
енными в основаниях. Поверхность кожи здесь выделяет черноватый, не
обладающий запахом секрет (Рососк, 1910).
^Ширина черепа (рис. 29) на уровне скуловых дуг составляет менее
половины, но более 40% его длины. Лицевая часть черепа умеренной
длины, не производит впечатления вытянутой. Носовые кости прибли-
зительно в 1 Yz раза превосходят длину носового отверстия (от переднего
края межчелюстных до передних концов носовых костей). Межчелюстные
кости не всегда соприкасаются с носовыми. Длина их короче длины верх-
него ряда коренных зубов, а передние концы не образуют расширения
с приостренными наружными краями, как это имеет место у рода Cervas.
Глазницы выступают в стороны сравнительно мало. Наиболее глубокое
вдавление лба между ними приходится немного впереди надглазничных
отверстий. Слезные кости широкие и короткие; длина их лицевой поверх-
ности меньше передне-заднего диаметра глазницы. Предглазничные ямки
на лицевой поверхности слезных костей не глубокие, иногда едва заметны.
Средняя часть шва между нёбными и верхнечелюстными костями имеет
прямую или выпуклую вперед форму и достигает уровня передней поло-
вины первого заднекоренного зуба. Линия профиля поперечного заты-
лочного гребня при взгляде на череп сверху имеет форму угла, обращён-
ного вершиной назад. На границе между глазницей и височной впадиной,
в области крыловидного гребня, с теменной костью соприкасаются и
али-, и орбитосфеноид (рис. 30, 4). Лобная кость, за крайне редкими исклю-
чениями, алисфеноида не достигает. Височные гребни позади скуловых
дуг развиты слабо, и при взгляде на череп сверху костные слуховые про-
ходы хорошо видны. В мыщелковую ямку (углубление между мыщелком
и яремным отростком), открывается только отверстие (иногда двойное)
подъязычного канала. Имеющееся у благородного оленя отверстие, ве-
дущее в мыщелковый канал, у косули не развивается.
Клыки в верхней челюсти, как правило, отсутствуют; изредка встре-
чаются у молодых самцов, но вскоре выпадают. Резцы асимметричные;
средняя их пара сильно расширена по сравнению с остальными, наруж-
ные углы ее оттянуты в сторону. Вторая пара значительно уже, а третья
и клыки представляют тонкие изогнутые наружу штифты. Коренные
зубы брахиодонтные; коронки их суживаются от основания к вершине
зуба. Между передней и задней половиной заднекоренных зубов, на внут-
CAPREOLUS
151
ренней стороне у верхних и на наружной у нижних, у основания коро-
нок расположены зачаточные бугорки; на верхних зубах они могут отсут-
ствовать. Передне-внутренняя долька последнего нижнего премоляра
Рис. 30. Характер сочетания костей в области крыловидных
гребней.
А — Capreolus capreolus L.; Б — Cervus nippon Temm. as — алис-
феноид; / — лобная кость; os — орбитосфеноид; р — теменная
кость; t — височная кость.
•отделена от передне-наружной почти до основания коронки; обе задние
дольки соединены друг с другом и с передне-наружной уже у вершины
зуба (рис. 31, Л). Второй нижний пре-
моляр (Р3) вместо передне-внутренней
дольки обычно имеет складки на сре-
дине и впереди главной (передне-наруж-
ной) доли зуба. На переднем нижнем
премоляре (Р2) имеются только слабо
развитые задние и передняя складки.
История и географическое распро-
странение (рис. 32). Корни рода Ca-
preolus Gray ведут к миоценовым мунт-
жакам (подсемейства Cervulinae'), Уже
в верхнем миоцене и нижнем плиоцене
и в Европе, и в Азии жила группа
форм, сходных по ряду признаков с
•современными косулями и объединяе-
мых в род Procapreolus Schloss. Еще бо-
лее близок к ним среднеплиоценовый
род Pliocervus Hilzh. Род Capreolus, по
К. К. Флерову (1952), ведет свою исто-
рию с верхнего плиоцена или нижнего
плейстоцена, а вид Capreolus capreolus
с достоверностью установлен лишь в
конце ледниковой эпохи (рисе—вюр-
Рис. 31. Перёднекоренные зубы
(премоляры) правой половины ниж-
ней челюсти.
А — Capreolus capreolus L.; Б —
Cervus nippon Temm.; В — Alces
alces L.; Г — Rangifer tarandus L.
ме). Ареал ископаемых косуль существенно не отличался от такового
•современных,
В общих чертах современный ареал косули охватывает всю западную
и восточную Европу к северу до 60° с. ;ш., Британские о-ва, Сицилию,
152
CERVIDAE
южную часть Скандинавского полуострова до 65° с. ш. на западном и
до 61° на восточном побережье, южную Финляндию, Переднюю Азию
на юг до Месопотамии и Палестины, Кавказ с Закавказьем, Ирак, Иран,
Сибирь до 58—60° с. ш. и до Дальнего Востока включительно, горы Сред-
ней Азии до Памирского нагорья, северные и восточные окраины Цент-
ральной Азии, северо-восточный и центральный Китай с восточным
Тибетом, Корею.
В сравнительно недалеком прошлом ареал косули, по крайней мере
в умеренных широтах, был сплошным. А. Н. Формозов (1946) северную
его границу связывает с линией среднего максимума глубины снежного-
покрова в 50 см. Зона максимальной численности этого зверя охватывает
области, где глубина снега не превышает 10—20 см. Вследствие хищни-
ческого истребления в предреволюционные годы ареал распался на не-
сколько частей; лишь в результате принятых мер за последние годы
косуля вновь начала заселять районы, в которых отсутствовала уже в те-
чение ряда десятилетий.
Современное распространение ее на территории СССР рисуется в сле-
дующем виде.
Всюду в подходящих местах косуля водится в Эстонии, Латвии, Литве.
В Эстонии, однако, по данным С. А. Северцова (1941), косуль мало
и встречаются они главным образом в граничащих с Латвией районах.
Впрочем, в коллекциях ЗИН АН СССР хранится экземпляр шкуры самца,,
добытого в 1948 г. в окрестностях г. Таллина.
До сих пор держатся косули в западных и южных районах Ленин-
градской области вплоть до Ломоносовского района. В качестве северной
границы до недавнего времени указывались р. Нева и южный берег Ладож-
ского озера, а в качестве восточной — р. Сясь. Однако в послевоенные
годы косуля проникла на территорию Карельского перешейка и в неболь-
шом числе постоянно держится в районах Приморском, Лесогорском,
Выборгском и Приозерском.1 В Новгородской области очень редка, по-
встречается вплоть до северных районов, возможно заходит даже в юго-
западные районы Вологодской области. В 1913—1914 гг. две косули были
убиты в лесах недалеко от впадения р. Колодней в Мологу (58.7° с. ш.,
36° в. д.). В 1924 г. встречались дважды в окрестностях ст. Пестово. Не-
большая группа косуль держалась зимою 1921 г. около д. Пелюшни Боро-
вичского района (58.9° с. ш., 33° в. д.) (Мил, 1926). В Боровичском крае-
ведческом музее находится чучело косули, загнанной волками несколько-
лет назад в д. Семерицы Опеченского района (на границе Новгородской
и Калининской областей). Здесь на водоразделе рр. Меты и Березайки
издавна существует очаг постоянного обитания косуль. В последние
перед Великой Отечественной войной годы численность и ареал косуль
в Новгородской области увеличились. По сведениям, полученным от
известного боровичского краеведа G. Н. Поршнякова, стайки косуль
не так редко встречались в окрестностях с. Кончанского-Суворовского,
но за последние годы вновь стали редки. При посещениях Боровичского
района Новгородской области в 1951—1957 гг. автору приходилось слы-
шать о редких встречах косули в Зехновском сельсовете. О косуле в Кре-
стецком уезде Новгородской губернии упоминал в конце прошлого сто-
летия Л. П. Сабанеев (1875). С. У. Строганов (1936) называет косулю не
очень редким зверем для Валдайского района; здесь она еще в 30-х годах
настоящего столетия встречалась группами до 15—20 голов.
’ Устные сообщения Г. В. Сердюковой и А. А. Гуреева.
Рис. 32. Распространение Capreolus capreolus L.
XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше; 4 — места случайных заходов в настоящее
время; 5 — места акклиматизации.
154
CERVIDAE
В Калининской области косуля отмечалась в окрестностях оз. Сели-
гер, в Осташковском, Кировском, Болотовском, Вышневолоцком районах
и даже в окрестностях г. Калинина. Встречается также в верховьях р. Мо-
лоти, в Бежецком и Весьегонском районах (Строганов, 1936). В 1945 г.
одиночки и даже небольшие группы косуль отмечались в районе
Дарвинского заповедника на юге Вологодской области (Насимович, 1955).
По-видимому, повсеместно водится косуля в Псковской и Велико-
лукской областях (Лавров, 1930). По устному сообщению О. В. Егорова,
держится в довольно значительных количествах по восточному побе-
режью Чудского озера»
Еще в XVI в. косуля была многочисленна на территории Москов-
ской области до самого Подмосковья, но к концу прошлого века была
полностью истреблена. Вновь завезенная в предвоенные годы, успешно
размножилась и в настоящее время довольно широко распространена
на западе и севере области (Формозов, 1947). Заходы отдельных неболь-
ших стад в период 1920—-1940 гг. отмечались на восток до Ярославля,1
Переяславля-Залесского, г. Балахны. В конце XIX в. постоянно води-
лась в Костромской и западной части Нижегородской губернии, откуда,
по словам Сабанеева (1874), битых косуль ежегодно привозили в Москву.
К областям почти сплошного распространения косули следует отне-
сти территорию Белорусской ССР. В наибольшем числе она держится
в западной половине, тогда как в районах к востоку от Днепра местами
исчезла почти совершенно. В Смоленской области косуля обитает во всех
лесных районах, наиболее обыкновенна в северных и восточных — Ве-
лижском, Слободском, Демидовском, Пречистенском, Знаменском, Всход-
ском, Ершичском и других. В Брянской области не многочисленна и
сохранилась в небольшом числе на территории Новозыбковского, Кли-
мовского, Клетнянского, Гаваньского и Брянского лесхозов. Достовер-
ные сведения о современном обитании косули в Калужской, Тульской1 2
и Орловской областях отсутствуют, но еще в конце прошлого столетия,
по Сабанееву (1875), она здесь водилась. Изредка встречается в преде-
лах Рязанской области, в частности указывается для Касимовского,
Чаплыгинского и Спасского районов. В последнем еще сравнительно
недавно на косуль устраивались охоты (Туров, 1924).
На территории Украинской ССР еще в XVII в. косуля была широко
распространена не только в лесах, но и в степях и плавнях, совершая
регулярные кочевки летом из лесов в степи, зимой — в обратном направ-
лении (Браунер, 1923). До сих пор косули отмечаются для всех областей
Украины к западу от Днепра, но сохранились, в основном, лишь в лес-
ных и лесостепных районах. Больше всего их в районах Карпат и Закар-
патской Украины. Обычны в Житомирском Полесье. В коллекциях ЗИН
АН СССР хранится череп косули, добытой в окрестностях Киева в 1902 г.
По-видимому, косули отсутствуют в Одесской и Николаевской областях.
В Левобережной Украине косули до сих пор держатся в лесах и лесо-
степных районах областей Черниговской, Сумской, Полтавской, Харь-
ковской, Днепропетровской и в северной части Запорожской (Корнеев,
1952). Отсутствуют, по-видимому, в Ворошиловградской, Херсонской и
Сталинской областях, хотя в последней водились еще в конце XVIII в.
(Формозов, 1946). В настоящее время проводится реакклиматизация косули
1 В Ярославской губернии (Данилевский уезд) косуля изредка встречалась и
в конце прошлого века (Сабанеев, 1874).
2 В списке животных заповедника Тульские засеки косули нет.
CAPREOLUS
155
в районах Украины, где она раньше водилась, но была вытеснена или
истреблена.
На территории Молдавской ССР, по собранным мною в 1949—1950 гг.
сведениям, косуля сохранилась еще в довольно значительном числе в ле-
сах Молдавских К одр и в прибрежных лесах по рр. Днепру и Пруту.
По данным Б. А. Кузнецова (1952), заходом появляется в Бельцской
степи.
Изолированный очаг распространения косули представляет горный
Крым. Раньше она была многочисленна во всей лесной его части, вплоть
до окрестностей Севастополя, Симферополя, Ялты и Судана. Теперь со-
хранилась главным образом на северном склоне Крымского хребта между
Чоргу-Чоргунем и Старым Крымом. Больше всего косуль на территории
Крымского государственного заповедника, откуда они расселяются и
в соседние районы.
К северу от Украины косули в небольшом числе сохранились в Кур-
ской области, особенно в южных и западных районах, например в Дмит-
риевском и Льговском (Птушенко, 1937). До сих пор некоторое их число
встречается на территории учлесхоза Ленинградского университета «Лес
на Ворскле» Борисовского района Белгородской области и в Борисов-
ском лесхозе.
В XIV в. косули были многочисленны на берегах Дона. К 20-м го-
дам текущего столетия они сохранились отдельными группами в круп-
ных лесных массивах Воронежской области (Усманском, Шиповом, Хре-
новском). Около десятка голов их насчитывалось в 1947 г. на территории
Воронежского заповедника (Барабаш-Никифоров, 1947). Косуля как по-
стоянный обитатель отсутствует в Ростовской области. До последних
лет наблюдаются ее заходы в районы Каменской области — Каменский,
Басковский и Тарасовский (Ралль, 1953).
Нет сведений о наличии косули в Тамбовской области. В Пензенской
области в прошлом столетии водилась в пределах Городищенского уезда;
рог косули был найден также в Чембарском районе (Федорович, 1915).
Дальше к востоку имеет место разрыв некогда сплошного ареала ко-
сули, произошедший, вероятно, сравнительно недавно. Вновь этот вид
появляется на Северном Кавказе и в Заволжье. Кавказская часть ареала
сливалась с украинской, вероятно, по зарослям кустарников и камышей
долин Кубани и Дона. При раскопке крепости Саркел в нижнем течении
Дона остатки косули обнаружены в слоях, относящихся к X—XIII вв.
н. э. ;
На Кавказе косуля обыкновенна в нижней трети лесов северного
и южного склонов Главного хребта, особенно в западной его половине.
Северная граница ее распространения в настоящее время на западе про-
ходит, по Верещагину (1952), приблизительно на линии 45° с. ш. от Ново-
российска параллельно Главному хребту до нижнего течения Терека
(Кизляра). В лесах под Ставрополем исчезла в 20-х годах настоящего столе-
тия. Мало косуль в широколиственных лесах между Ходыженском и Май-
копом, лишь изредка заходят они в Пятигорье. Нет их в сухих долинах
Кабарды и Дигории, а также в долине р. Сунжи. По Тереку водятся до
Кизляра и по всему Сулаку, кроме низовьев. Сохранились косули в уце-
левших лесах предгорных увалов Грозненской области. Отсутствуют
в центральном Дагестане, хотя в нижнем течении Самура водились еще
в 20-х годах этого века. Под Махачкалой бывают лишь как редкость.
В незначительном числе косули держатся в предгорьях Кусарской рав-
нины, в окрестностях города Кубы,, с. Кусары и урочища. Сувал. . >т
156
CERVIDAE
He повсеместно держится и не везде многочисленна косуля и в Закав-
казье. Водится в болотистых лесах Колхидской низменности, например,
около г. Поти. Мало ее в нижнем поясе лесов Абхазии и Грузии, а в верхо-
вьях р. Ингура косуля, вследствие глубоких снегов, отсутствует вовсе.
Наиболее обычна на южном склоне Главного хребта в Юго-Осетии, Ка-
хетии, Азербайджане. На Малом Кавказе косули держатся всюду, где-
сохранились лиственные леса. В Карабахе косули еще водятся в буковых
лесах Тертер-чая. Нет их в южной части Армянской ССР, мало и на Та-
лыше. Отсутствует косуля в Прикуринской низменности.
В среднем Заволжье косуля была многочисленна в конце XVIII в..
В то время она водилась и в Правобережье (Паллас, 1773) 1 и к востоку
за Волгой по рр. Соку, Кинели и Самаре. В небольшом числе в первой
четверти прошлого века держалась в лесах Сингилеевского уезда Симбирской
губернии, но позднее исчезла (Богданов, 1871). В начале 30-х годов,
этого столетия появилась и за последние годы значительно увеличила
свою численность на территории Бузулукского бора. Имеются указания
о случайных заходах косули в пределы Татарской АССР, в низовья р. Камы
(Попов и др., 1954).
Не ясен вопрос о наличии и современном распространении косули
в Пермской области,* 2 но в прошлом столетии она водилась в северной
части Кунгурского уезда Пермской губернии (Сабанеев, 1874). По-види-
мому, не так давно встречалась в Коми-Пермяцком округе (Томилин, 1953).
До сих пор косуля является обычным зверем в пределах Сред-
него и Южного Урала, в областях Свердловской, Челябинской и в Баш-
кирской АССР. Западная граница ее распространения здесь проходит
по предгорным лесам Башкирии, по рр. Белой и Уфе (подробности см.
Лавров, 1929 и Кириков, 1952), затем поднимается к г. Красноуфимску,,
а от последнего, уклоняясь несколько на восток, на севере доходит до-
c. Ивдель (около 60.5° с. ш.). Далее к востоку северная ее граница про-
ходит по р. Лозьве, Тавде и Тоболу до широты Тобольска (Куклин, 1938)..
Известны отдельные заходы к западу и северу от этой границы. Так, она
отмечалась в бассейне Северной Сосьвы и Ляпина до 64° с. ш. (Насимо-
вич, 1955). Имели место неоднократные заходы на территорию Печорско-
Илычского заповедника (Коми АССР), до устья р. Илыч, но здесь косули,,
как правило, зиму не переживают (Формозов, 1946).
К югу и востоку от очерченной линии косуля наиболее многочисленна,
и в прошлом и сейчас между 57—55° с. ш., в Нязепетровском, Уфалей-
ском, Кыштымском и особенно Каслинском Урале (Туркин и Сатунин,.
1902; Куклин, 1938). Весьма обыкновенна в Южном Урале, в частности
на территории бывшего Башкирского и Ильменского заповедников. Го-
раздо малочисленнее косуля в степных предгорьях Южного Урала, но-
еще встречается в Губерлинском мелкосопочнике уже на территории
Чкаловской области.
В последней, кроме Бузулукского бора, по данным Я. Даршкевйча
(1950), по-видимому требующих уточнения, присутствие косули уста-
новлено в большинстве северных районов, а также в районах Буртин-
,ском, Краснохолмском, Илекском и даже Ак-Булакском. Более обыкно-
венна она в восточных районах области, в частности в Орском, Кваркен-
ском и Адамовском (Райский, 1951).
, 1 Имеются требующие проверки сведения (П. Козлов, по Елпатьевскому, 1950)
О том, что в 1943 г. кбсуля появилась в лесах Вольского района Саратовской области.
2 В соседней с ней Удмуртской АССР были найдены в земле рога косули на терри-
тории б; Сарапульского уезда (Круликовский, 1909).
CAPREOLUS
157
В западной Сибири косуля водится в Тюменской, Курганской, Омской
и Новосибирской областях, а также в Алтайском крае. Северная граница
-ее распространения в Тюменской области проходит в бассейне р. Конды,
-севернее Шаима, до 61° с. ш., дальше идет на северо-восток до Оби, до-
стигая почти 62-й параллели (Скалой, 1946). По Иртышу косуля, как
уже отмечалось, встречается до широты Тобольска. Далее к востоку
граница проходит севернее Омска, на широте Барабинска и Томска на-
правляется к Красноярску. В левобережье Енисея теперь она достигает
58° с. ш. (Фетисов, 1953). В последнее время И. П. Лаптев (1955) указал
ряд дальних заходов косули в бассейне Оби вплоть до района Сургута.
Еще лет двадцать-тридцать назад в лесостепной зоне западной Си-
'бири косуля была малочисленна и водилась лишь местами, напрймер
в юго-западной части Тарского района и в Ировских лесах Омской
•области. В результате запрета промысла и мер охраны к настоящему
моменту размножилась и является, по словам А. И. Янушевича и И. Бла-
говещенского (1952), обычным зверем даже в Барабинской и Кулундин-
ской степях вплоть до Салаирского кряжа.
Еще недавно разорванный здесь ареал косули в настоящее время вновь
сомкнулся с ее ареалом в южной и восточной Сибири. Область практи-
чески сплошного распространения косули представляет Алтай. Однако
заселена его территория этим зверем неравномерно. В отдельных районах,
например в верховьях Большого Абакана и безлесных частях Чуйской
степи, ее нет. В наибольшем числе косуля водится вШебалинском, Онгудай-
ском и Чемальском аймаках Горно-Алтайской автономной области, в верх-
них течениях рр. Семы, Урсула, Песчаный, Чарыша, а также в бассейне
левых притоков рр. Коксу и Катуни. Второй очаг высокой плотности попу-
ляции этого зверя представляет бассейн и междуречье Чулышмана и
Башкауса. На территории б. Алтайского заповедника к востоку от Чу-
лышмана держится в меньшем числе, хотя во многих участках и не пред-
ставляет редкости (Дмитриев, 1938).
Многочисленна косуля во всей лесной зоне Тувинской автономной
области вплоть до лесов речных долин и Тункинского нагорья (Януше-
вич, 1952).
В небольшом числе косуля держится по всей Хакассии, кроме цент-
ральных безлесных районов, а также к северу до Сибирской магистрали
на пространстве между Абаканской железнодорожной веткой и Енисеем.
Около 25 лет назад, по Лаврову (1929), водилась только к югу от г. Ста-
лине, но в настоящее время, по-видимому, распространена по всей Кеме-
ровской области, так как летом заходит даже в горные районы соседней
Томской области (Лаптев, 1953).
Из Южного Урала и лесостепной зоны западной Сибири ареал косули
распространяется в северную часть Казахстана, в области Западно-Ка-
захстанскую, Актюбинскую, Кустанайскую, Северо-Казахстанскую,
Кокчетавскую и Акмолинскую. В западном Казахстане косуля встречается
в окрестностях с. Приурального и в пойменных лесах Урало-Илекского
речного узла (Афанасьев и др., 1953). В Актюбинской области косуля
отмечена для Хобдинского района (Антйпин, 1941), тогда как на Мугод-
жарах ее, по-видимому, нет. Кроме того, изолированный очаг обитания
имеется в небольшом лесном острове близ Джуруна. В Кустанайской
области распространение косули ограничивается также северной частью,
по с 1928 г. она появилась в Аман—Карагайском бору, а за послед-
ние годы проникла на территорию Наурзумского заповедника. Держится
косуля, по-видимому, и до настоящего времени в большинстве горно-
158
CERVIDAE
лесных районов Кокчетавской области. Отсюда она проникает в зону
лесостепи, в частности в Пресновский район Северо-Казахстанской
области и Пресногорьковский Кустанайской области (Лентовский, 1930).
Нет сведений о деталях распространения косули в Северо-Казахстанской
и северной части Павлодарской областей, но считается (Афанасьев и др.,
1953), что южная граница ее распространения от Улутау (где этот вид.
отмечен Н. Павловым), огибая с севера Казахское нагорье (в котором
этого зверя нет), выходит на Иртыш в районе Павлодара. Впрочем А. Се-
дельников (цит. по Антипину, 1941) в начале этого столетия установил
наличие косули в районе Баян-Аула и Каркаралинска. Необходимо'
отметить, что косуля, вообще малочисленная во всех перечисленных об-
ластях Казахстана, за последние десятилетия, в связи с охраной или
в силу других причин, начала увеличивать свою численность и появляться'
в районах, где она ранее не водилась.
Более многочисленна косуля в восточном Казахстане, в Восточно-
Казахстанской и Семипалатинской областях. Здесь ареал ее смыкается
с ареалом остального Алтая и охватывает верховья Бухтармы, хребты
Южный Алтай, Саур, Тарбагатай, Калбинский, окрестности оз. Зай-
сан. Во многих из этих районов косуля является обычным зверем. Но
в наибольшем числе она водится в южных областях Казахской ССР.
Многочисленна в хребтах Каратау, Таласском Алатау, Киргизском, Чу-
Илийском, Заилийском, Кунгей и Джунгарском Алатау, в хребте Кет-
мень. Весьма обычна также в области Семиречья, в частности в тугаях
рр. Чу, Или, Каратала, Аксу и Лепсы, западной части котловины озер-
Сасыкколь и Алаколь. В бассейне Сыр-Дарьи косули нет (Антипин,
1941; Афанасьев и др., 1953).
Практически косуля отсутствует на территории Туркменской и Узбек-
ской ССР; но имеются требующие проверки данные, будто бы она изредка
появляется на Больших Балханах (Бажанов, 1945)1 и в тугаях левобе-
режья Аму-Дарьи (Лавров, 1929). Косуля встречается местами в Ферган-
ской области.
В пределах Таджикистана распространение косули ограничивается^
хребтами Гиссарским, Алтайским и Зеравшанским (Флеров, 1935).
Почти повсеместно распространена, а местами весьма многочисленна
косуля в Киргизской ССР. К районам, наиболее богатым ею, относятся'
верховья р. Чаткала, Алайский, Киргизский хребты, хребты Кунгей'
и Терскей Алатау, горы к востоку от р. Тон. Многочисленна на Сусамыр-
ском плато и в горах, окружающих Джумгальскую долину. Сравнительно-
редка косуля в Кочкорском, почти безлесном районе и на северном склоне-
Таласского Алатау. Нет ее в Алайской долине (Б. Кузнецов, 1948).
Переходим к распространению косули в восточной Сибири. В заени-
сейской части Красноярского края она водится в основном к югу от Си-
бирской магистрали. В умеренном количестве встречается по р. Ба-
заихе, Мане, в Уярском, Манском, Канском и других смежных районах.
Многочисленна косуля, вероятно, благодаря охране, на территории за-
поведника «Столбы» в окрестностях Красноярска. На севере, по И. Шухову
(1933), доходит до р. Чуны, а по Е. Яковлеву (1930), до верховьев Подка-
менной Тунгуски. В коллекциях ЗИН АН СССР имеется экземпляр ко-
сули, добытый в 1914 г. в Тасеевской волости Канского уезда в окрестно-
стях д. Глинной. Если и встречается, то очень малочисленна в безлесной
1 Посетивший в 1952 г. Большие Балханы О. В. Егоров здесь о косуле ничего не:
слышал.
CAPREOLUS
159
части Минусинской котловины, по в южных подтаежных и таежных
районах к югу от Минусинска весьма обычна. Многочисленным и важ-
ным промысловым зверем косуля является на всем пространстве Запад-
ных и Восточных Саян В Иркутской области косуля распространена
от южных границ до 57—58° с. ш. на западе и до 60° с. ш. на востоке.
Однако плотность популяции ее не равномерна. Нет ее вовсе в пределах
Тофаларии, кроме северной окраины; больше всего — в полосе, приле-
гающей с севера и юга к железнодорожной магистрали между Нижне-
удинском и Иркутском, а также в Усть-Ордынском районе и Бурят-
Монгольском национальном округе. Менее многочисленна косуля в Тай-
шетском и Шиткинском районах. По Ангаре она доходит до широты
Нижне-Илимска и даже несколько к северу, а на Лене — до Усть-Кута.
В бассейне Киренги держится только в верхнем течении до с. Карала.
В северных районах — Киренском и Катангском, по крайней мере в на-
стоящее время, косуля отсутствует (Копылов, 1950). По данным В. Н. Ска-
лона (1951), относящимся к 1943 г., косуля водится, хотя и не везде,,
в большей части Бодайбинского района, но более поздние сведения го-
ворят о том, что здесь она имеется в небольшом числе лишь в северо-
восточной части, по р. Жуе.
Обычным, а местами весьма многочисленным зверем косуля является
в пределах Бурят-Монгольской АССР. Более редка она севернее р. Верх-
ней Ангары и по Муе (Новиков, 1937), а также в самом подлеморье
к северу от Чивыркуйского залива. Зато в Баунтовском районе (Витим-
ское плоскогорье) она достигает, по Подаревскому (1936), наиболее вы-
сокой численности: при. учете здесь приходится в среднем 30—40 встреч
косуль за один день. Многочисленна также в Закаменском районе (южное
Забайкалье), в долинах Баргузина и Большого Чивыркуя. Обычным
зверем она является на обоих склонах Яблонового хребта.
Область распространения косули в Якутии захватывает только юж-
ные и юго-восточные ее районы. Многочисленна косуля по рекам, стекаю-
щим с Патомского нагорья, по Чаре, Жуе, Токко. По Олекме обычна
до самого устья и, по-видимому, постоянно держится даже несколько
севернее по левобережью и правобережью Лены. Встречается на Алдан-
ском плоскогорье, в районах Томмотском, Тимптонском, а также в Учур-
ском, на водоразделе Гонама и Алгана (Скалой и др., 1941). Г. П. Демен-
тьев (1933) считает северной границей распространения здесь р. Алдан,
но изредка косуля встречается, по-видимому, и севернее, в среднем тече-
нии Амги, т. е. на широте 60-й параллели.1 За последнее время отмечены
случаи появления косули под Якутском.1 2 Имеется старое указание Р. Ма-
ака о том, что косуля изредка забегает с Лены на р. Вилюй в район р. Сун-
тар. Известны такие случаи и за последние годы. В 1934 г., например,
косуля добыта в Мархинском наслеге, около 64° с. ш. (Скалой, 1941,
1946). В коллекциях 'ЗИН АН СССР имеется череп косули, переданный
в 1927 г. В. Дорофееву жителем урочища Кюпчкюй Первого Бордонского
наслега Нюрбинского улуса Вилюйского округа.
Широко косуля распространена в Читинской области. Здесь она
встречается даже по островкам леса, балкам и оврагам Даурских степей.
Много косули на пространстве между верхними течениями Витима и
Олекмы, в долинах Нерчи и Каренги, а также в Красночикойском и Сре-
1 Пара рогов косули из этого района (с р. Тенгутта, левого притока Амги) достав-
лены в коллекцию ЗИН в 1927 г. Л. Бианки.
2 Устное сообщение Н. К. Верещагина.
160
CERVIDAE
тенском районах. Гораздо меньше на Тундаке, редка она по Калакану,
а на Каларе встречается лишь единично. Добывается в долине Конды,
Чары, а также в верховьях Олекмы и по Тунгиру (Подаревский, 1936).
Многочисленна в районе Джалинды.1
В Амурской области косуля широко распространена как в лесной,
так и в лесостепной зонах. Несколько меньше ее плотность лишь в се-
верных районах — Джелтулакском и Зейском (Баранчеев, 1955).
В пределах Хабаровского края косуля в очень небольшом количестве,
не имея уже никакого промыслового значения, еще встречается в верх-
нем течении р. Уды, но доходит ли до побережья Удской губы — не из-
вестно. Южнее, в верхнем течении Селемджи и в хребте Ям-Алинь ее
нет (Золотарев, 1934). Более многочисленна косуля в бассейне р. Бурей
до с. Чекунда. Выше становится все реже, а на восточной стороне хребта
Дуссе-Алинь отсутствует вовсе. Нет ее и в верхнем течении Амгуни (Афа-
насьев, 1934). Больше всего косуль в Хабаровском крае, по В. П. Сы-
соеву (1952), в Еврейской автономной области и в Комсомольском районе.
В южной части Дальнего Востока, по К. Г. Абрамову (1954), косули
-«много от южного Приморья до р. Великой Кемы, откуда северная гра-
ница ее ареала поворачивает на запад, удаляясь от побережья, перехо-
дит Сихотэ-Алинь и на р. Бикине проходит по Улунгинской поляне.
Далее она идет к северо-западу и проходит по нижним притокам Хора.
Здесь северная граница несколько отклоняется на северо-восток и, про-
ходя параллельно Амуру, заходит в низовья р. Анюя. От низовьев Анюя
северная граница распространения косули переходит на левобережье
Амура и достигает Эворон-Чукчагирской низменности. Но встречается
здесь это животное только в отдельных участках. Также только в отдель-
ных участках водится косуля в северо-западной части Чумиканского
района, где она есть в верховьях р. Уды, по р. Шевли, в окрестностях
золотого прииска Баладека».
Отсутствует косуля на о. Сахалине.
В Монгольской Народной Республике ареал косули захватывает се-
верную и, частично, центральную ее части. Известно, что этот вид встре-
чается на южных (монгольских) склонах хребтов Сайлюгем и Танну-
Ола. Много косуль, по последним данным, в Монгольском Алтае, в част-
ности на поднятии Хархира-Тургень, хребте Цаган-Шибэту, на север
от оз. Урюк-Нур и в верхнем течении р. Булугун. Известны ее места
нахождения на хребте Хан-Хухей. Далее косуля широко распростра-
нена по всему Прикосоголью и Хангаю; в последнем она отсутствует
только в степных долинах и на безлесных Хангайских горах. Водится
этот вид в горах бассейна Селенги и ее притоков (Орхона, Эгин-Гола),
вдоль границы с советским Прибайкальем. Широко распространена по
всему Хэнтэю от горы Богдо-Ула до Угтум-Хан, Тэмэн-Чулу, Согод и
Ихэ-Гадзар к югу от р. Ульдзя. На западе республики косуля встре-
чается в предгорьях Большого Хиигана до рр. Дэгэ-Гол и Мухурин—
Гол (Банников, 1954 : 200—208).
Подвиды. Описано большое число местных форм, принимаемых раз-
ными авторами то как подвиды, то как самостоятельные виды. В настоя-
щее время более общепринятой считается точка зрения, рассматриваю-
щая все местные формы рода Capreolus как подвиды одного вида.
В отношении числа подвидов также нет единого мнения. Некоторыми
•принимается свыше пятнадцати подвидов (Пастернак, 1952). Более пра-
1 Устное сообщение С. М. Друри.
CAPREOLUS
161
Пильной следует считать точку зрения К. Флерова (1952), сводящего
число их к четырем.
1) С. capreolus capreolus L. — Европейская косуля. Размеры сравни-
тельно небольшие; длина тела около 125 см, высота в холке около 80 см;
длина черепа от 190 до 216 мм; живой вес до 41 кг. Общий фон зимней
окраски серовато-бурый, более темный, чем у других рас, особенно на
задней части спины и на крестце. В летней шерсти окраска головы серая
или бурая, резко отличающаяся от окраски спины и боков. Основания
волос до половины длины серо-бурые или темно-бурые. Слуховые пузыри
на черепе небольшие. Рога тонкие, обычно не длиннее 30 см; сильно сбли-
жены основаниями, так что розетки часто соприкасаются друг с другом
(рис. 33, А). Стволы рогов от оснований направлены вверх почти парал-
лельно, иногда даже с наклоном внутрь. Жемчужины на них развиты
слабо. Распространение: западная Европа (включая Британ-
ские острова и Скандинавский полуостров), европейская часть СССР
до Волги и Кавказа, Крым, Закавказье, Малая Азия, Палестина, Иран.
2) С. capreolus pygargus Pallas — Сибирская косуля. Размеры круп-
ные; длина тела около 140 см, высота в холке до 90 см и более; длина
черепа 215—250 мм; живой вес до 65 кг. Окраска зимой серая, на спине
буроватая с примесью рыжеватых тонов. В летней окраске голова одно-
тонная со спиной и боками. Волосы на всем теле, кроме хребта, имеют
белые основания. Слуховые пузыри на черепе крупные, вздутые. Рога
(рис. 33, Б) длиной до 40 см и более, часто имеют по 4 и более отростков,
широко расставлены в основаниях; расстояние между венчиками почти
равно диаметру рога, а то и больше. Стволы рогов уже от основания на-
правляются в стороны и вверх. Жемчужины на них развиты сильно и
иногда приобретают вид коротких отростков. Распространение:
восточные районы европейской части СССР за Волгой, Кавказ, Средняя
Азия, Урал, Сибирь до Забайкалья и Якутии включительно, западный
Китай (Синьцзян), северная и северо-западная Монголия.
3) С. capreolus bedfordi Thomas — Маньчжурская косуля. Размеры
крупные, но несколько мельче предыдущей формы; длина черепа 211—
215 мм. Зимняя окраска серовато-рыжая, зеркало со слабым рыжеватым
оттенком. Голова более рыжая и бурая, чем все тело. Летняя окраска
интенсивно рыжая, иногда буреющая на верхней стороне тела. Пропор-
ции черепа, как у С. с. pygargus. Распространение: Хабаров-
ский и Приморский края, северный и северо-восточный Китай, Корея.
4) С. capreolus melanotis Miller — Сычуанская косуля. Сходна с сибир-
ской и маньчжурской расой, но несколько меньше; наибольшая длина
черепа — от 207 до 223 мм. Окраска в зимнем меху буровато- или ры-
жевато-серая, голова ржаво-буроватая с темным лбом. Уши более буро-
ватые, чем голова. Летний мех рыжий. Слуховые пузыри более вздуты,
чем даже у двух предыдущих подвидов. Рога более сближены,
в противоположность другим подвидам почти не имеют лирообразного
раскида, тонкие и длинные. Распространение: Китай — вос-
точный Тибет, провинции Ганьсу, Сычуань, Наныпань на север до Гоби,
Кам.
Биология. На обширной территории своего ареала косуля обитает
в неодинаковой обстановке. В целом этот зверь — обитатель несплош-
ных лесов и лесостепи. Современное ее распределение по стациям, осо-
бенно в европейской части ареала, в значительной мере обусловлено
развитием сельскохозяйственной культуры й прямым истребляющим
влиянием человека. Еще в XVII в., как отмечалось, косуля во множестве-
11 Фауна СССР, И. И. Соколов
162
CERVIDAE
населяла целинные степи Украины и совершала по ним регулярные пере-
кочевки. В настоящее же время в степной полосе она держится только
там, где имеются островные леса, степные колки или заросли кустарни-
ков по оврагам, балкам и долинам степных речек.
Типичными для косули стациями в равнинных местностях являются
участки леса, перемежающиеся с открытыми лесными полянами и лу-
гами. И в европейской части СССР и в Сибири косуля избегает сплошных
массивов тайги, особенно темнохвойной. Если и встречается в больших
лесах, то придерживается главным образом опушек по соседству с поля-
нами или лугами. Явное предпочтение отдается лиственным и смешан-
ным лесам, особенно с наличием хорошо развитого лиственного подлеска
и травянистого покрова. Лес в первую очередь служит местом отдыха
и укрытия от врагов, тогда как открытые участки являются, обычно,
местами жировок. В летние месяцы косули не избегает болотистых мест,
поросших высокими травами, камышами или редким лесом («кардашёй»
на Урале). Последние предпочитаются самками в период рождения и
вскармливания молодняка.
В лесостепной зоне Сибири и северного Казахстана типичными ста-
циями являются островные леса и березовые колки. В условиях Средней
Азии косуля, кроме горных еловых лесов и ленточных боров северного
Казахстана, держится в кустарниках предгорий, тугаях речных долин,
по балкам ковыльной степи и даже в полупустынных предгорьях. В дельте
Или косуля не придерживается тугаев и вообще не связана с древесной
растительностью; лежки устраивает в зарослях чия, среди солянок,
в кустах тамарикса, у края тростниковых зарослей. Последними косуля
пользуется в качестве укрытия от преследующих врагов (Штегман, 1949).
В восточной Сибири одной из излюбленных летних стаций являются
болотистые леса с травянистым покровом из трехлистной вахты. Во всех
случаях необходимо наличие в районе обитания водопоя, в виде ручья
и речки, хотя, по свидетельству Лаврова (1929), при нужде косуля пьет
и стоячую болотную или дождевую воду.
Область вертикального распространения косули охватывает мест-
ности от покрытых лесом приморских равнин до высот в две-три тысячи
метров над уровнем моря и даже до альпийских лугов, на которые иногда
выходит пастись летом. В Крыму в наибольшем числе косули держатся
в буковых лесах северных склонов Южного хребта и в смешанных моло-
дых насаждениях с густым подлеском, особенно удобных для косуль
в первое время после рождения молодняка. Излюбленные места зимой
приурочены к редколесью южных склонов гор, вероятно в связи с от-
сутствием на них стойкого снегового покрова (Лавров, 1929; Даль. 1930).
На Кавказе распространение косуль связано с лесами нижнего и
среднего пояса гор. Выше верхней границы леса косуля, как правило,
не заходит. Больших сплошных лесов она и здесь избегает, и «чем больше
в лесу полян, тем они больше нравятся козам» (Динник, 1910). Особенно
многочисленны косули по краям заросших бурьяном полян и опушек
лесов. Любят поросшие молодым лесом старые лесосеки, но сплошных
непролазных чащ, в противоположность кабанам, избегают. На отдых
и зимой, и летом охотно располагаются в «редкодубах» — редкостойных
дубовых лесах с невысоким травянистым покровом, обеспечивающих
широкий кругозор отдыхаюшим животным.
На Алтае излюбленные стации косуль — пологие склоны гор без вы-
ходов коренных пород, поросшие редким лиственничным лесом с под-
леском из березы, осины, жимолости, красной смородины и других кус-
CAPREOLUS
16-3
тарниковых и древесных пород. Сплошной почвенный покров в таких
местах образован злаками и широколистными травами высотой до 50—
70 см. Наиболее часто косуль можно встретить здесь поблизости стекаю-
щих с гор ручьев и речек, в местах, где участки густых зарослей кус-
тарников чередуются с открытыми полянами или редким древостоем
без подлеска. Охотно держатся также в межгорных долинах, особенно
если по соседству имеются хотя бы небольшие участки леса или кустар-
Рис. 38. Рога европейской (Л) и сибирской (Б) косули.
никовая поросль. Как и всюду, косуля избегает и здесь россыпей;и скал
и не живет в темнохвойной тайге.1 Небольшие сосновые боры в устьях
рек, изреженные сосняки по склонам и лиственное берегово-осиновое
редколесье с болыперазнотравьем и разнотравьем являются обычными
местообитаниями косуль в районе Прителецкого Алтая (Юргенсон, 1938).
Если косуля не подвергается усиленному преследованию, она не из-
бегает и близости человеческих поселений. В охотничьих хозяйствах,
при условии строгой охраны, косуля превращается в постоянного оби-
тателя полей и лугов, изредка даже причиняя некоторый вред культур-
ным посевам.
Пищу косуль составляют разнообразные травы (главным образом
из двудольных), листва, побеги, кора и тонкие ветви кустарников и де-
ревьев. Видовой состав и количественное соотношение составных частей
рациона зависят от сезона и условий, в которых живет косуля. Летом
1 Личные наблюдения в центральном Алтае летом 1930 г.
....................................................................11*
164
CERVIDAE
основным кормом служат травы и зеленая листва кустарников и деревьев.
Зимой основу питания составляют почки, ветки и кора, в первую оче-
редь осины и ивы, а в дополнение к ним травяная ветошь, неопавшая
листва и зимнезеленые растения, добываемые из-под снега. Впрочем,
веточный корм, как показало исследование содержимого желудка крым-
ских косуль, играет заметную роль и в летнее время (Даль, 1930).
Из травянистых растений, в большом количестве поедаемых в усло-
виях Крыма, на первом месте стоит горечавка (Gentiana cruciata L.).
Охотно употребляются в пищу косулями также: тысячелистник (Achi-
llea millefolium L.), подмаренник (Gallium hybridum L.), горькуша (Gen-
taurea jacea L.), душица (Origanum vulgare L.), клевер (Trifolium me-
dium L.), зверобой (Hypericum hirsutum L.), ластовник (Gynanchum
scandens Kuns.); у последних двух объедаются не только листья и вер-
хушки, но и плоды (Лавров, 1929). Хорошо поедаются листья липы (Ti-
lia cordata Mil.), вяза (Ulmus glabra Schmalh.), ясеня (Fraxinus oxicarpa
Wild, и F. excelsior L.), а также молодые побеги ежевики (Rubus сае-
sius L.). В список особо охотно поедаемых и лакомых для косули расте-
ний Даль (1930) включает плющ (Hedera helix L.) и омелу (Viscum al-
bum L.). Осоки и злаки охотно поедаются ранней весной., когда еще нет
другой зелени, и осенью, благодаря более позднему сохранению ими
листьев и стеблей в зеленом состоянии (Пастернак, 1955). В горах Крыма
и Кавказа пищей служат также опавшие плоды яблонь, груш, кизила,
каштаны, буковые орехи, желуди. Излюбленными растениями, по Са-
банееву (1875), являются дягиль, или медвежья дудка (Archangelica sp.),
а осенью грибы, в первую очередь грузди и опенки. Во многих местах
восточной Сибири основным летним кормом является вахта (Мепуап-
thes trifoliata L.).
Интересно отметить, что косуля без вреда поедает ряд растений, ядо-
витых для домашних травоядных животных, даже такие, как чемерица
и пострел (Фетисов, 1953). Из культурных растений косули ощипывают
листочки и головки клевера, а также ботву картофеля (Лавров, 1929).
На Урале кормами, по Л. И. Сабанееву (1875), служат также ягоды ма-
лины, брусники, морошки и княженики, а на Дальнем Востоке даже
клюквы.
Зимой заметную роль в пищевом рационе косули играют древесные
лишайники, а при недостатке других кормов — трутовики и мхи, выка-
пываемые из-под снега. В хвойном лесу в пищу идут хвоя и концы вет-
вей сосен и можжевельника; при этом могут откусываться побеги тол-
щиною до 7 мм.1 Наконец, всюду, где только это возможно, косули зи-
мой очень охотно держатся и питаются сеном из стогов, не вывезенных
с лесных сенокосов. Разрывая сено, косули в первую очередь выбирают
мелколистное разнотравье (Фетисов, 1953).
Как и другие виды оленей, испытывая недостаток в минеральных
солях, косули посещают естественные и специально устраиваемые искус-
ственные солонцы. Здесь животные лижут, а иногда и выгрызают почву,
содержащую повышенное, по сравнению с другими местами, количество
солей, необходимых для организма. Особо регулярный характер посе-
щение солонцов носит весной и в начале лета с марта по июнь и осенью
с половины августа и в сентябре. При этом установлено (Пастернак,
1955), что весной солонцы посещаются почти исключительно самками,
1 О громадных количествах молодых еловых побегов, находимых в желудках ко-
суль из Беловежской лущи, упоминает К. Врублевский (1912).
CAPREOLUS
16-j
тогда как осенью, особенно в период гона, на них чаще можно встретить
самцов. В средине лета (июль—начало августа) косуль на солонцах ви-
дят редко, а зимой, вероятно вследствие замерзания, они ими не посе-
щаются вовсе.
Образ жизни косули в разных областях ее обитания и в разные сезоны
не одинаков. В теплое время года косуля всюду расселяется более ши-
роко; зимой концентрируется на ограниченных территориях, обычно
более благоприятных в кормовом и климатическом отношении. Во мно-
гих районах сезонные перемещения приобретают характер регулярных,
нередко массовых миграций. Сезонные перекочевки косули отмечены
на Урале (Сабанеев, 1875). По окончании течки, с выпадением первых
снегов, обычно в октябре, косули, до того державшиеся небольшими
группами, начинают собираться в табуны по 20—40 голов и постепенно
передвигаются со всех прилегающих районов в пределы Каслинского
Урала, где зимой концентрируется значительное число животных. Эти
перекочевки из года в год, как правило, происходят по одним и тем же
путям и вызваны тем, что в районе зимовок глубина снежного покрова
значительно меньше, чем в соседних, северном, южном и особенно запад-
ном склоне Урала.
Регулярные кочевки косуль имеют место также в Красноярском крае,
Иркутской области, Забайкалье, Дальнем Востоке и в ряде других об-
ластей Сибири. В восточной Сибири различают «местных» и «ходовых»
косуль. Первые в течение всего года. обитают в ограниченном районе,
меняя лишь стации по сезонам. «Ходовая» же косуля не живет в одном
районе и ежегодно предпринимает длительные перекочевки из одной
местности в. другую (Фетисов, 1953). И здесь основным фактором, вызы-
вающим миграции, является глубина снежного покрова и условия кормо-
добывания в зимний период. В равнинных районах европейской части
СССР они не известны, возможно в связи с малочисленностью сохранив-
шегося поголовья этого зверя.1
Несколько иной характер сезонные миграции косуль носят на Кав-
казе и Алтае. Здесь с наступлением теплого времени не все, но значитель-
ная часть их, поднимается в горы к верхней границе леса, даже выходят
на альпийские луга, где встречаются до сентября. С наступлением осени
косули вновь спускаются в нижний пояс гор и в долины. Причиной и
здесь является характер снежного покрова. Снег глубиной более 50 см
делает существование косули почти невозможным (Дмитриев, 1938).
Известны также массовые непериодические переселения косуль, вызы-
ваемые, вероятно, причинами стихийного порядка. В годы обильных
снегопадов, например, имеют место массовые перекочевки косуль из се-
веро-восточного Китая через Амур в прилегающие районы Хабаровского
края (Сысоев, 1952).
Зиму косули проводят табунками в 10—12, редко до 18 голов, состоя-
щими из зверей разного возраста и пола. Во главе табунка стоит вожак,
обычно старая самка, реже самец. Пасутся в течение всей светлой части
суток. Лежки устраивают, выкапывая ногами снег до земли или твер-
дого наста. Несколько животных не устраивают одной общей лежки,
но на отдыхе располагаются зимой друг к другу ближе, чем летом. Лежка
обычно устраивается около дерева, куста, пня или другого предмета.
Лежит косуля на груди и животе, поджав под себя ноги; на лежке часто
1 В XVII в. происходили ежегодные массовые перекочевки косуль на Украине —
весной из лесов в открытые степи, а осенью в обратном направлении (Браунер, 1923).
166
CERVIDAE
мочится и по следу от мочи зимой иногда можно отличить лежку самца
от самки. В сильные метели и морозы косули забираются в чащу и от-
стаиваются там кучкой. В тихую погоду придерживаются малоснежных
участков, а в горах — солнцепечных склонов, где раньше, чем в других
местах, появляется и первая зелень.
С февраля—марта табунки начинают разбиваться; первыми отделя- '
ются и держатся группами или в одиночку самцы. В апреле отделяются
беременные самки. В апреле обычно начинается обратная перекочевка
косуль к местам летовок. В апреле и мае большая часть животных дер-
жится одиночно, только самцы иногда держатся парами или небольшими
группами, вместе с холостыми самками и молодыми. Самки время отела
проводят в самых глухих местах; только после того как молодняк ок-
репнет, они появляются на лугах и опушках. Начиная с весны и все лето,
косули пасутся главным образом вечером и по утрам, часов до шести
или семи. Кормятся на опушках, лесных лужайках, по берегам речек,
по краям болот. Дневные часы проводят в лежках, устраиваемых в те-
нистых местах, на дне оврагов, вблизи рек и ручьев, в которых охотно
купаются в жаркое время. Лишь в наиболее глухих местах и там, где
их охраняют и мало беспокоят, косули пасутся иногда в дневные часы
и летом. В гористых местностях косули придерживаются по возможности,
северных склонов, как дающих лучшую защиту от зноя; кроме того,
кормовые условия здесь более благоприятны вследствие раннего выго-
рания травы на южных склонах.
Переходы от мест отдыха к местам кормежки, водопоя и к солонцам
совершаются по определенным маршрутам, на которых косули вытап-
тывают тропы, особенно хорошо заметные в зимнее время. Табунок в
спокойной обстановке движется гуськом. Обычный способ передвижения
косули — шагом, но она способна двигаться крупными, высокими прыж-
ками. Преследуемая или испуганная косуля делает прыжки до 8 м в длину,
свободно перепрыгивает овраги, кусты высотою до 2 м, поваленные де-
ревья. На ровном месте за ней не может угнаться собака; правда, ко-
суля утомляется при этом очень быстро. К бегу рысью она не приспособ-
лена, в тростниках, где вследствие множества острых торчащих стеблей
прыжки не возможны, передвигается недостаточно быстро, с шумом,
и часто становится добычей волка (Штегман, 1949). Хорошо плавает;
известны случаи, когда косули переплывали озера шириною до 7 км.
Преодолевает такую крупную и быструю реку, как Ангара (Фетисов,
1953). Но на гладком, не прикрытом снегом льду скользит, падает и быстро
подняться не может. Раненная или преследуемая собаками косуля де-
лает круги, петли, броски в сторону, в воду, для того чтобы сбить пре-
следователей со следа (Фетисов, 1953).
Из органов чувств у косули хорошо развиты слух и, особенно, обо-
няние. Последнее особенно обострено в ветренную сырую погоду. Че-
ловека, по свидетельству G. К. Даля (1930), косуля может учуять по
ветру за полкилометра. Обонянием она пользуется не только для свое-
временного обнаружения опасности, но и при розыске пищи под снегом.
Зрение, наоборот, развито гораздо хуже, и на него косуля, по-видимому,
полагается гораздо меньше, чем на другие чувства.
Голос косули представляет подобие хриплого отрывистого лая боль-
шой собаки («гау-гау» или «гоф-гоф»; отсюда и выражение: «козел гав-
кает»). По силе звук не соответствует величине животного и иногда
его сравнивают с ревом медведя. Чаще всего косуля «гавкает» при
испуге, как во время бега, так и стоя на месте. Следующие часто
CAPREOLUS
167
друг за другом звуки указывают на сильный испуг зверя. Тот же звук,
ло-видимому, служит для переклички и во время турнирных поединков
(Кузнецов, 1929). Чаще кричат косули весной по зорям, и почти не кри-
чат днем и зимой. Голос самки более отрывистый и грубый. Во время
гона косули, кроме рева, издают особые отрывистые звуки, трудно пе-
редаваемые словами; по словам Г. П. Дементьева — что-то среднее между
визжанием и ревом. Кроме рева, иногда можно слышать звук, похожий
на писк, протяжный и нежный, издаваемый при закрытом рте; писком
является и голос теленка, высокий и протяжный при голоде, отрывистый
и резкий при испуге. Писком созывает самка своих детей. Пойманная
раненной косуля издает резкий дребезжащий звук («верещанье»).
Смена волосяного покрова у косули происходит в году дважды.
Однако настоящая линька с полным выпадением всего старого волоса
имеет место только весною. Сроки, быстрота процесса и окончание зави-
сят от климатических условий и состояния организма зверя. В средних
широтах, Крыму и на Кавказе линька начинается в марте или даже в
конце февраля. В это время под покровом старой шерсти начинает от-
растать короткий летний волос. С наступлением теплой погоды зимняя
шерсть вылезает клочьями, оставаясь на лежках, кустах и деревьях,
о которые трутся животные, ощущая, по-видимому, раздражение кожи.
В первую очередь волос выпадает на боках, у лопаток; затем процесс
переходит на голову и конечности; последними вылинивают спина и круп.
Заканчивается весенняя линька в мае или начале июня. Молодые выли-
нивают несколько позднее взрослых. Задерживается также линька у
зверей больных и очень старых. Осенняя линька заключается в основ-
ном, в отрастании пуха и зимнего грубого волоса. Уже в конце августа
сквозь короткую летнюю шерсть начинает пробиваться зимний волос,
придающий сероватый оттенок рыжему фону. Еще раньше, вскоре после
весенней линьки, начинает отрастать пух (подшерсток). В сентябре пре-
обладает серая окраска, но с рыжеватыми летними тонами, околохвосто-
вое зеркало белеет. Есть требующее детальной проверки предположение
(Динник, 1910; Дементьев, 1933), что летний мех к зиме, по крайней мере
целиком не выпадает, а лишь отрастает и превращается в остевой волос.
Порядок отрастания зимней шерсти — обратный весенней линьке: на-
чинается со спины и боков и заканчивается на голове и конечностях.
Полного развития зимний мех достигает в ноябре. Уже с января волос
становится ломким и непрочным, темные кончики волос снашиваются
и окраска животных становится более бледной, грязно-серой.
Нормально рога у косуль имеют только самцы, но известно много
случаев появления их у самок, особенно престарелых. Однако типичных
для самцов размеров и формы рога у самок не приобретают и представ-
ляют почти всегда небольшие уродливые образования.1 Нередко и у сам-
цов, вероятно, в результате травмы, рога также имеют уродливую не-
правильную форму или увеличенное число отростков. Изредка встре-
чаются самцы с тремя и даже четырьмя рогами. Рост рогов у црибылых
самцов (сеголеток) начинается в трех-четырехмесячном возрасте. Уже
в начале сентября можно прощупать бугорки с торчащими волосами
на месте будущих рогов. В октябре покрытые кожей рожки уже ясно
выступают, а к декабрю имеют вид простых тонких стержней («спичек»)
1 В коллекциях ЗИН АН СССР имеется череп взрослой самки косули из Германии
с рогами длиной 17 см. Рога имеют хорошо развитые розетки, но представляют простые
«спички» без развилок. Оба рога очищены от кожи, а один из них или во время убоя
или препаровки отделился от пенька.
168
CERVIDAE
без розеток и отростков, длиной до 5—7 см. С января по март—апрель
темп роста рогов замедляется. Вновь усиленно рога начинают расти в
апреле. При благоприятных условиях существования уже первые рога
косули достигают к годовалому возрасту полного развития и имеют по 3
конца на каждом (Соколов, 1956). Но часто рога годовалых зверей пред-
ставляют форму «вилок» с раздвоением на верхушках или даже простых
спичек.1 Кожа с первых рогов сбрасывается в мае и июне, а сами они
спадают и заменяются новыми в декабре или январе.
У взрослых самцов рога сбрасываются после спаривания в период
с октября, а иногда даже с сентября, по декабрь. Недели через три-четыре
после этого начинается рост новых рогов, вначале имеющих форму про-
стых шишек. Окончательную форму, но еще покрытые кожей и мягкие
на концах, рога приобретают обычно к марту—апрелю. С апреля по май
происходит очистка кожи с рогов путем трения о кусты, тонкие деревья
и другие предметы, вероятно вследствие раздражения, ощущаемого
животным от подсыхающей и омертвевающей на рогах кожи. У молодых
время сбрасывания, роста и очистки рогов от кожи запаздывает по срав-
нению со взрослыми, в среднем, на месяц. Кроме того, сроки этих про-
цессов, как и линьки, зависят от климатических условий местности и со-
стояния организма животного.1 2 * * * 6 Только что очищенный от кожи рог имеет
белый цвет, но в дальнейшем приобретает темно-коричневую окраску.
Спадение рога происходит вследствие резорбции (рассасывания) кости
на границе между пеньком рога и розеткой, отпадающей вместе с рогом.
Спадает рог вследствие собственной тяжести или при случайных ударах
и толчках о твердые предметы и сотрясениях. После спадения кровото-
чащая некоторое время ранка вскоре затягивается кожей, под которой
начинается рост нового рога.
Есть указания, что кастрация или повреждение семенников приоста-
навливает или нарушает процесс роста рогов. Развившиеся до кастрации
рога, после нее уже не сбрасываются. Опыты, поставленные в Польше
(Jaczewski, 1952), показали, что односторонняя кастрация не оказывает
никакого влияния на рост и сроки сбрасывания рогов у косули.
Способность к оплодотворению (половая зрелость) у косуль в благо-
приятных условиях наступает на втором году жизни, но обычно самки
приносят впервые детей только в возрасте трех лет. Самцы практически
начинают принимать участие в гоне также в возрасте не ранее трех лет,
когда становятся способными принимать участие в турнирных боях за
обладание самкой.
Период гона, даже в одной местности, растянут, обычно, на срок не
менее двух месяцев. Начало и конец его зависят от широты местности,
высоты над уровнем моря, от кормовых и климатических условий. В боль-
шинстве равнинных местностей у взрослых зверей он начинается в начале
августа или даже в конце июля; разгар падает на конец августа—начало
1 Это явилось причиной господствовавшего в течение свыше полустолетия не-
правильного представления о том, что якобы по числу отростков на рогах косули
можно определять ее возраст до 4 лет (Туркин и Сатунин, 1902; Дементьев, 1933;
Флеров, 1952, и др.).
2 В Джунгарском Алатау, например, по данным А. В. Афанасьева (1945 : 29—
30), сбрасывание рогов происходит только в конце марта (последнего самца со старыми
рогами он видел 20 марта); рост новых начинается с конца апреля, а окостеневают и
очищаются от кожи рога только к концу июня. В коллекциях Зоологического инсти-
тута Академии наук СССР хранится череп самца косули, добытый в Таласском Алатау
6 декабря с шестиконечными рогами, покрытыми шерстью, и экземпляр с такими же
рогами, добытый 28 августа на Талыше (Закавказье).
CAPREOLUS
169
сентября, а заканчивается гон обычно в октябре. В горах течка начи-
нается позднее, чем в низинах. В Средней Азии, по И. А. Долгушину
(Антипин, 1941) и Б. К. Штегману (1949), гон происходит в сентябре.
Молодые животные приходят в охоту позднее старых, но и заканчивается
она у них также с запозданием.
Самцы в период течки сильно возбуждены, мало едят, по следу ра-
зыскивают самок, теряют обычную чуткость и осторожность. Найдя
самку, козел преследует ее, перегоняя с места на место. Часто на полянах
и опушках, гоняясь друг за другом по кругу вокруг одного дерева или
куста, косули выбивают растительность до земли, образуя так называемые
«точки», которых может быть несколько на сравнительно небольшом
участке. Кроет самец самку на ходу, причем несколько покрытий могут
следовать с короткими промежутками друг за другом (Даль, 1930). Один
самец за сезон может оплодотворить до 6 самок; вокруг самца держатся
несколько самок, но иногда, наоборот, за одной самкой гоняется несколько
самцов. В последнем случае между самцами происходят жестокие по-
единки, нередко заканчивающиеся смертью одного из соперников (Кириков,
1952). Известны случаи (Даль, 1930), когда погибают оба самца, крепко
сцепившись во время драки рогами. Иногда самец наносит удары рогами
и уклоняющейся от покрытия самке.
Считается, что беременность у косули длится около 9 месяцев, но
в течение первых четырех оплодотворенное яйцо развивается чрезвы-
чайно медленно и только с декабря зародыш начинает развиваться более
быстро (Sakurai, 1906). Исследования последних лет (Stieve, 1950), однако,
показали, что период замедленного развития имеет место не всегда. Ока-
залось, что у косули течка бывает в году дважды: от середины июля до
конца августа (главная) и от конца ноября до середины или конца декабря
(вторая, дополнительная). Самец может оплодотворять с мая по январь.
Если самка оплодотворяется во время главной течки, то беременность
длится от 9 до 10 месяцев, причем 5—6 месяцев приходится на первую
ее часть. В это время плоды развиваются очень медленно, и лишь в конце
декабря начинается вторая половина вынашивания, длящаяся 5 месяцев.
Если яйца оплодотворяются во время второй течки, то первая часть бе-
ременности отпадает и молодые рождаются через 5—6 месяцев после
оплодотворения.
Отел самок в большинстве районов в массе происходит во второй по-
ловине мая—первой половине июня, но, в соответствии с растянутыми
сроками гона, тянется с последних чисел апреля до июля включительно.
Перед отелом самка удаляется в глухие места, где в зарослях кустарника,
в высокой густой траве или в другом укрытии приносит детенышей, число
которых в помете обычно два, реже три; четыре бывают как исключение.
Молодые, особенно первотелки, приносят, как правило, одного детеныша.
Детеныши родятся весом около 2—2.5 кг, беспомощны, не могут даже
стоять и первые 6—8 дней лежат, притаившись в высокой траве. В это
время их легко можно взять руками. Косуля —- хорошая мать. Она па-
сется поблизости, подходит к детенышу на его писк, когда он голоден
или испуган, делает попытки защищать от врагов и даже следует неко-
торое время за уносящим козленка человеком. С семидневного возраста
косулята могут следовать за матерью и даже сносно бегать, но до 15 дней,
особенно в случае опасности, еще прячутся, вытянув шею и голову с при-
жатыми ушами. В возрасте месяца с небольшим в беге не уступает взрос-
лым. Мать кормит детенышей молоком в продолжение около месяца.
К этому времени косулята начинают поедать зеленый корм, появляется
no
CERVIDAE
жвачка (Марков, 1948). В период гона прибылые уходят от матерей и ве-
дут самостоятельную жизнь, но по окончании его вновь возвращаются
и держатся вместе с ними до появления следующего помета.
Родятся косулята со всеми молочными резцами, клыками и премоля-
рами. Смена резцов и клыков начинается с пятимесячного возраста и за-
канчивается к году. Первыми сменяются внутренние резцы (I т), а послед-
ними наружные (13) и клыки. В 13—14 месяцев выпадают молочные
и вырастают постоянные премоляры. Заднекоренные прорезываются,
начиная с передних (М! и М1), в трехмесячном возрасте. Окончательного
развития зубная система косули достигает к возрасту 15—16 месяцев,
когда прорезались и достигли уровня остальных зубов задние верхние
моляры (М3). По развитию, смене и стиранию жевательной поверхности
коренных зубов можно определять возраст косули до полутора лет с точ-
ностью до одного месяца (Соколов, 1956).
Продолжительность жизни косули в естественной обстановке не
установлена, но, судя по состоянию зубов на коллекционных экземпля-
рах, она не превышает 10—12 лет.
Главными врагами косуль являются волки, от которых гибнут и взрос-
лые звери, и молодняк. Особенно большой ущерб наносят волки в конце
зимы, когда тонкий слой непрочного наста поверх глубокого снега вы-
держивает хищника, но проламывается под острыми копытами косули.
Зимою волки часто загоняют косуль на гладкий лед, где последние бе-
жать не могут, а летом — в скалы, с которых неприспособленная к ним
косуля срывается и разбивается о камни (Динник, 1910). В лесных райо-
нах косули становятся иногда жертвой росомах и рысей, подкараули-
вающих их на деревьях около троп. В Крыму главным врагом косуль
является лисица, выслеживающая молодняк, преследующая иногда даже
взрослых животных. Много гибнет от хищников молодняка в первые
недели жизни. В этом возрасте козлята становятся легкой добычей не
только волка, но и медведя. С притаившимся косуленком расправляется
не только лисица, но и дикая кошка, куница и даже хорек. Молодые
косули гибнут иногда также от крупных хищных птиц, таких, как бер-
кут и филин (Фетисов, 1953).
Косуля является хозяином эндо- и эктопаразитов. Из числа послед-
них в первую очередь следует назвать кожных оводов {Hypoderma diana
Вт. и дальневосточный вид Н. capreoli Rubzow), а также носового овода
{Cephenomyia stimulator Cl.).1 В Средней Азии (Штегман, 1949) косули
от оводов страдают очень сильно, но на Кавказе, по Диннику (1910),
на них не паразитирует ни кожный, ни носовой овод. Беспокоят косуль
и мешают нормальному нагулу слепни, комары, оленья муха-крово-
соска {Lipoptena cervi L.), иксодовые клещи.
В Крымском заповеднике зарегистрированы неоднократные случаи
падежа косуль от тейлериоза, вызываемого кровепаразитом Theileria
capreoli Ruehl. (Рухлядев, 1939). Переносчиком последнего являются
иксодовые клещи {Ixodes ricinusL., Haemaphysalis otophila P. Sell, и др.).
Кроме того, косуля является хозяином ряда паразитических сосальщиков,
ленточных и круглых червей (Судариков и Рыжиков, 1951; Асадов, 1954,
1955). Численность косуль зависит и от имеющих место в районах их
массового распространения эпизоотий. Последние отмечались в Закав-
казье (Лавров, 1929) и Средней Азии (Шнитников, 1936); причины этих
1 По сообщению К. Я. Грунина, изредка на косулях находят единичные личинки
другого вида носоглоточного овода — Pharyngomyia picta Meyg.
CAPREOLUS
111
эпизоотий не всегда ясны. Известно, что у косуль наблюдались отдельные
случаи заболеваний и вспышки пастереллеза, ящура, сибирской язвы
и даже чумы крупного рогатого скота (Слудский, 1954). Местами косули
страдают от периодических бескормиц вследствие гололедицы, наста или
глубокого снега. Такие случаи наблюдались в Баунтовском районе Бурят-
Монгольской АССР (Тимофеев, 1953) и в Башкирском заповеднике (Ки-
риков, 1952). В этих случаях истощенные косули гибнут не столько от
голода, сколько от волков, бродячих собак и браконьеров.
Хозяйственное значение. Косуля имеет большое значение как про-
мысловый мясной зверь. Во многих районах Сибири, Алтая и Дальнего
Востока промысел косуль является важным дополнительным источником
питания местного населения. Мясо довольно вкусное и не жесткое, хотя
редко бывает жирным и по вкусовым качествам уступает говядине и сви-
нине (даже мясу дикого кабана). На Алтае из мяса косули делаются
вкусные пельмени, незаменимые в замороженном виде в зимних поездках
на далекие расстояния. Лучшую упитанность косуля имеет в августе
перед началом гона. Самцы во время гона сильно худеют, и их мясо ста-
новится невкусным. Взрослая европейская косуля дает чистого мяса
15—18, редко 25 кг. Сибирская косуля гораздо крупнее и туша уже 2—3-лет-
них зверей весит около 32 кг, а старых самцов еще больше — до 50 кг
и выше. От хорошо упитанной косули получается около 2—2.5 кг внут-
реннего сала (Фетисов, 1953).
Шкуры косуль не представляют большой ценности, так как мех на
них летом короткий, а зимой грубый и ломкий. Из шкур, добытых осенью,
когда зимний волос еще не потерял блеска, более прочен и не слишком
длинен, в Сибири и на Алтае шьют так называемые «бардовые» дохи. Эти
дохи очень теплы, но требуют особого ухода. Их нельзя, например, вно-
сить с мороза в теплое помещение. И все же, даже при этом условии,
остевой волос выпадает с них все время в значительном числе. Шкуры
косуль обычно употребляются еще в виде ковриков под ноги и для сиденья,
а на Кавказе из них делают потники под седло шерстью вниз, предо-
храняющее, благодаря упругости меха, спину лошади от набоев и ссадин.
Из выделанной по типу замши кожи иногда шьют брюки и куртки, режут
ее на ремни. У самцов в период гона кожа на шее утолщается; эта так
называемая «кукуя» употребляется даже на подошвы (Дементьев,
1933).
Несмотря на то, что местами, особенно в азиатской части СССР, до-
бывается весьма значительное количество косуль, большая часть и мяса,
и шкур от них употребляется на месте; товарный выход этой продукции
до последнего времени был ничтожен.
Рога косули употребляют в качестве украшений и для разных поде-
лок, таких, как рукоятки к ножам, вилкам и т. п., и местами служат
предметом сбыта.
Пойманные на воле в молодом возрасте косули легко привыкают
к человеку и становятся совершенно ручными. Известны неоднократные
случаи размножения их в неволе, но молодняк, обычно, в этих случаях
нежизнеспособен. и рано или поздно погибает. Одомашнению косуль
препятствует неуживчивый нрав взрослых зверей, особенно самцов. По-
следние, когда имеют уже окостеневшие рога, набрасываются не только
на своих собратьев, но и на человека.
Благодаря способности жить в различной обстановке й сравнительно
высокой плодовитости, косуля представляет; большие персцективы в смысле
акклиматизации и разведения её в охотничьих хозяйствах, средней и юж-
172
CERVIDAE
ной полосы Союза. В Сибири же и на Дальнем Востоке при надлежащей
охране и рациональной организации промысла она может играть суще-
ственную роль в создании продовольственной базы для местной промыш-
ленности.
Вред, приносимый косулей сельскохозяйственным культурам и лесу
ощипыванием верхушек ветвей и сдиранием коры с тонких деревьев,
в большинстве районов ничтожен. Мне известен из литературы единствен-
ный случай, сообщаемый В. Н. Скалоном (1931), когда, спасаясь от гнуса,
косули выходили из леса на открытые места в посевы хлебов и сильно
вытаптывали их. Интересно, что звери не ели колосьев или других частей
хлебных растений, а пользовались ими как щеткой для сбивания с кожи
кровососущих насекомых. Однако подобные случаи, по-видимому, уни-
кальны и могут иметь место лишь в районах высокой численности ко-
сули.
Более ощутимым вред косули может, вероятно, стать в районах лес-
ных полезащитных и других посадок. Здесь она, как в основном дре-
весноядный вид, может сильно вредить молодым деревьям и подлеску.
В Крымском госзаповеднике косуля, наравне с благородным оленем, иг-
рает наибольшую роль в повреждении подроста бука и других деревьев
(Иваненко, 1948).
Охота. Способы охоты на косуль разнообразны. С собаками охо-
тятся редко, так как косуля, благодаря быстрому бегу, уводит их слиш-
ком далеко. В Средней Азии, на Кавказе и, особенно, в восточной Сибири
охотятся облавой или загоном, но этот способ требует большого числа
участников и не всегда оправдывается результатом (описание его см. Фе-
тисов, 1953).
Чаще охотятся с подхода по вечерам и утрам, когда звери находятся
на жировке. Этот способ бывает удачен только по чернотропу, когда нет
снега, хрустящего под ногами. Требуется большая выдержка и осторож-
ность при подходе (обязательно против ветра) к очень чуткому зверю.
Малейшая неосторожность, хруст сучка под ногой охотника сводит на
нет затраченные усилия и время..
Подкарауливают косуль у источников водопоя и особенно у солонцов,
иногда специально устраиваемых для этой цели.1 В районах, где косули
держатся в значительном числе, добывают их капканами, используя
привычку их ходить на жировку и водопой по постоянным тропам. Этот
способ практикуется главным образом зимою, когда звери придерживаются
более стойко троп, протоптанных в снегу. Капканы применяются типа
волчьих; иногда к капкану привязывают за веревку обрубок дерева,
мешающий попавшему зверю уйти, далеко.
Раньше в местах переходов косуль устраивали замаскированные ямы
с приманкой из лакомого корма (Сабанеев, 1875). Практиковавшееся
в дореволюционное время массовое истребление косуль путем преследо-
вания по непрочному насту или глубокому снегу в настоящее время за-
прещено. Запрещенной является также охота «на пищик» путем подра-
жания голосу косуленка, на который выбегает и становится жертвой охот-
ника самка. На рану косуля не крепка. Раненый самец редко, но иногда
пытается ударить рогами подошедшего охотника; сильный удар может
быть нанесен также задними ногами.
1 Этот способ, как приходилось наблюдать, является обычным при добыче косуль
на Алтае летом для удовлетворения текущих потребностей семьи охотника в мясе.
CERVUS
173
Род CFRVUS LINNAEUS —НАСТОЯЩИЕ ОЛЕНИ
Характеристика рода. Животные средней или крупной величины.
Высота в холке от 60 до 165 см. Длина черепа взрослых самцов от 250
до 500 мм.1 Высота в крестце не меньше высоты в холке. Конец морды
не увеличен. Профиль морды в носовой части прямой. Верхняя губа не
увеличена и не нависает над нижней. Носовое зеркало занимает все про-
странство между ноздрями (исключение — подрод Dama Frisch). Пред-
глазничные железы хорошо развиты. Глаза с горизонтально располо-
женным зрачком. Светлое околохвостовое зеркало хорошо развито и обычно
ясно отграничено от окружающей окраски.
Хвост длиной более 3 см, всегда виден, но не достигает уровня ска-
кательных суставов.
Рога имеют один или два надглазничных отростка, более или менее
круглого сечения (лишь у подрода Dama верхняя половина рогов упло-
щена в виде лопат).
Поверхность ствола и нижних отростков шероховатая, но резко вы-
ступающих наростов («жемчужин»), как у косули, нет. Розетки выступают
в стороны от основания ствола не более чем на 1 см. Копыта относительно
более широкие, чем у лося (род Alces Gray) и косули {Capreolus Gray),
но уже, чем у северных оленей (Rangijer Н. Smith).
Череп с умеренно развитой лицевой частью. Наибольшая его ширина
составляет более 40% длины. Межчелюстные кости не превышают (а если
превышают, то немного) длину ряда коренных зубов верхней челюсти
и всегда соприкасаются с носовыми костями. Передние концы их уплощены
и имеют приостренные края. Носовые кости не укорочены; длина их
больше длины переднего носового отверстия (от prosthion до переднего
края носовых костей) и не меньше длины верхнего зубного ряда. Гребешки
на беззубом крае верхнйх’челюстей (между Р1 2 и С) сближены не сильно;
расстояние между ними в самом узком месте больше длины Р2. Глазницы
выступают в стороны не сильно. Вдавление лба между ними расположено
на уровне или немного впереди надглазничных отверстий. Длина слез-
ных костей не менее передне-заднего диаметра глазницы, но не превышают
его более чем в 1 % раза. Предглазничные ямки глубокие. Шов между
нёбными и верхнечелюстными костями не заходит вперед дальше уровня
задней половины М1.
Профиль поперечного затылочного гребня при взгляде сверху прямой
или выпуклый.
По краям лобных костей на месте рогов у самок имеются ясно высту-
пающие в стороны бугорки. На границе глазницы и височной впадины
лобная кость соединяется с алисфеноидом.2 Орбитосфеноид на черепах
взрослых животных не соприкасается с теменной костью (рис. 30, Б).
Височные гребни позади скуловых дуг развиты хорошо и при взгляде
на череп сверху костные слуховые проходы не видны. В мыщелковую
ямку, кроме подъязычного отверстия (часто двойного), открывается от-
верстие, ведущее в мыщелковый канал. Слуховые пузыри вздутые, вы-
ступают значительно ниже засуставных отростков.
Верхние клыки имеются (исключение — подрод Dama). Коронки
резцов асимметричные; средняя их пара сильно расширена, по сравне-
1 Последняя цифра относится к оленям из американской части ареала.
2 Исключение иногда представляют подроды Axis Н. Smith и Rusa Н. Smith.
174
CERVIDAE
нию с остальными, наружные углы оттянуты в стороны; остальные резцы
изогнуты наружу и постепенно убывают в ширине от 12 к С. Коренные
зубы менее брахиодонтные, чем у других родов Cervidae. Добавочные
бугорки или столбики на внутренней стороне верхних и наружной стороне
нижних заднекоренных имеются, но на последних не всегда. Передне-
внутренняя и обе задние дольки Pt связаны с передне-наружной долей
почти до самой вершины зуба. Второй нижний премоляр (Р3) вместо
передневнутренней дольки имеет складки на середине и впереди главной
(передне-наружной) доли зуба. На переднем нижнем премоляре (Р2)
только слабо развитые задние и передняя складки.
От боковых (II и V) пястных костей (метакарпалий) сохраняются
лишь дистальные рудименты (рис. 31, Б).
История и географическое распространение. Корни рода Cervus
ведут к потомкам миоценовых мунтжаков — роду Cervavitus Khom.
Несколько видов этого рода, по К. Флерову (1950), были распростра-
нены с верхнего миоцена до нижнего плиоцена в умеренной полосе Европы
и Азии. У них были большие рога с 3—4 отростками, но сравнительно
примитивные брахиодонтные зубы; конечности имели сросшиеся централь-
ные (III и IV), полные боковые (II и V) метаподии и хорошо развитые
боковые пальцы. К концу нижнего плиоцена в Индии, а со среднего
плиоцена и в Европе, появляются олени, принадлежащие к современному
роду Cervus (подроды Axis и Ruso).
В настоящее время распространение рода включает Европу и Азию
к северу до 60-й параллели, северо-западную Африку и Северную Америку
от 60-й параллели на севере до Мексики на юге.
Классификация. По современным представлениям (Флеров,
1952) 16 видов рода Cervus делятся на 5 подродов.
1) Axis Н. Smith — Аксисы, или свиные олени. Рога с 3 концами,
причем второй отросток отходит от ствола назад. Хвост длинный, пре-
восходит длину уха, носовые кости короткие, не достигают задними кон-
цами уровня передних краев глазниц. Слуховые пузыри очень крупные,
выступают значительно ниже уровня basioccipitale. Четыре вида этого
рода распр остранены в Индии, Пакистане, о. Цейлоне, Бирме,
Таиланде, Аннамё, западных Филиппинах и о. Бевин (между островами
Борнео и Явой).
2) Rusa Н. Smith — Замбары. Сходен с предыдущим; отличается тем,
что второй отросток рога отходит от главного ствола не назад, а вперед.
Слуховые пузыри менее вздутые и выступают сравнительно мало, ниже
уровня basioccipitale. Семь видов этого подрода распространены
в южной, юго-восточной Азии: Индии, Индокитае с прилегающими ост-
ровами и в Китае на север до Сычуани.
3) Przewalskium Flerow — Олень Пржевальского. От других подро-
дов отличается длинными узкими ушами, коротким хвостом (короче
уха), высокими и широкими копытами, а также белой окраской носа,
губ и подбородка до горла. Рога с 5 концами, из которых один является
надглазничным отростком. Череп с очень глубокими предглазничными
ямками и длинными носовыми костями; задние концы последних распо-
ложены позади уровня передних краев глазниц. Задняя расширенная
половина носовых костей имеет по бокам четырехугольные выступы,
образующие внутренние края этмоидальных щелей. Наибольшая ширина
носовых костей больше передне-заднего диаметра глазницы.. Отверстия
носослезных каналов расположены на внутренней стенке глазниц. Един-
ственный вид С. (Przewalskium) albirostris Fl. распространен
CERVUS
175
в восточном и северном Тибете, Наныпане, Южно-Кукунорском хребте
и в верховьях р. Янцзы.
4) Dama Frisch — Лани. От других подродов отличается прежде всего
формой рогов, которые на вершинах уплощены и иногда образуют лопа-
тообразные расширения. Хвост длиннее уха. Задние края носовых костей
расположены приблизительно на уровне передних краев глазниц. Наи-
большая ширина носовых костей меньше передне-заднего диаметра глаз-
ницы. Коренной областью распространения двух видов ланей
являются средиземноморские страны, Малая Азия и Иран. В диком со-
стоянии сохранилась лишь в Иране, юго-западном побережье Мрамор-
ного моря и в Малой Азии. Акклиматизирована в умеренных областях
большинства стран Европы.
5) Подрод Cervus Linnaeus •— Настоящие олени. Рога имеют не менее
4 отростков и не уплощены. Хвост короткий, не превышает длину уха.
Задние концы носовых костей расположены на одном уровне или несколько
позади передних краев глазниц. Наибольшая ширина носовых костей
меньше, равна или больше передне-заднего диаметра глазницы. Рас-
пространение: западная Европа, включая Британские о-ва,
Корсику, Сардинию и южную часть Скандинавского полуострова, юг
европейской части СССР, Кавказ, Малая Азия, Иран, Афганистан, Каш-
мир, горы и тугаи Средней Азии, южная Сибирь, Забайкалье до Даль-
него Востока включительно, горы северной Монголии, Китай на юг до
23° с. ш., включая Кашгарию, восточный Тибет и о. Тайвань, острова
Японии (включая Рюкю), Гималаи, северная Африка (Алжир, Тунис,
Сенегамбия), Северная Америка от 59—60° на севере до Мексики на юге.
В фауне СССР представлены только подроды Duma (акклиматизиро-
ван) и Cervus.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА CERVUS I,
ФАУНЫ СССР
1 (4). Размеры средние; высота в холке (плечах) не больше 120 см; наи-
большая длина черепа меньше 350 мм. Окраска взрослых животных
пятнистая. Пятнистость доходит до корня хвоста, а также захва-
тывает лопатки и плечи. Развитых надглазничных отростков на
каждом роге имеется только по одному. Слуховые пузыри вздутые,
гладкие; при горизонтальном положении черепа они выступают
ниже уровня средней части основной затылочной кости (basioc-
cipitale).
2 (3). Вершины рогов у взрослых самцов уплощены в виде лопат; от
верхнего и заднего края лопат отходят несколько (более трех)
пальцевидных отростков. Голое пространство на конце морды (но-
совое зеркало) сравнительно небольшое; средняя его часть едва
поднимается выше уровня нижних краев ноздрей. Хвост длиннее
уха. Наименьшая ширина лба между глазницами и рогами на че-
репе больше ширины черепа в надушных буграх. Подъязычное
отверстие черепа расположено впереди переднего края суставной
поверхности на основной затылочной кости. Верхние клыки от-
сутствуют ................Cervus (Dama) dama L. — Лань (стр. 176).
3 (2). Вершины рогов не уплощены и имеют один или два отростка.
Голое пространство на конце морды (носовое зеркало) большое;
верхний его край расположен немного ниже уровня верхних углов
ноздрей. Хвост не длиннее уха. Наименьшая ширина лба между
116
CERVIDAE
глазницами и рогами меньше ширины черепа в надушных буграх.
Подъязычное отверстие черепа расположено позади уровня перед-
него края суставной поверхности на основной затылочной кости.
Верхние клыки имеются ........................................
............Cervus nippon Temm. — Пятнистый олень (стр. 183).
4 (1). Размеры крупные; высота в холке более 120 см; наибольшая длина
черепа более 350 мм. Окраска взрослых животных не пятнистая.
Если иногда у самок в летней шерсти имеется неясно выраженная
пятнистость, то она не доходит до корня хвоста и не распростра-
няется на лопатки и плечи. У вполне взрослых самцов число над-
глазничных отростков на каждом роге равно двум. Слуховые пу-
зыри небольшие, угловатые; при горизонтальном положении черепа
они не выступают ниже уровня средней части основной затылочной
кости .... Cervus elaphus L. — Благородный олень (стр. 204).
Cervus (Dama) dama Linnaeus — Лань, или даниэль (рис. 34—36)
Linnaeus С., 1758, Syst. Natur., ed. X, 1 : 67 (первооп.); Туркин Н. В.
и К. А. С а т у н и н, 1902, Звери России, Cervidae, М. : 37, 393—397 (общ., изобр.);
Greve К., 1909, Saugetiere Kur-Liv-Esthlands, Riga : 1—183 (акклим.); Вруб-
левский К., 1912, Архив ветер, наук, 8 : 749 (распр., пит.); W i m a n s W.,
1913, Deer breeding, London : 47—63 (общ., изобр.): Завадовский М. М., 1926,
Тр. Лабор. эксперим. биол. Московок, зоопарка, 1 : 19—22 (кастрация); Шиллин-
г е р Ф. Ф., 1930, Европейский олень, лань и косуля в охотхозяйстве, Харьков, изд.
ВУСОР : 45—54 (общ., изобр.); Житков Б. М., 1940, Науч.-методич. зап.
Главн. управл. по заповеди., VII : 100 (акклим.); Успенский Г. А., 1950,
Аскания-Нова, М., Сельхозгиз : 78 (акклим.); Кириков С. В., 1952, Изв.
Акад, наук СССР, сер. географ., 6 : 31—48 (ист.); Флеров К. К., 1952, Кабарги
и олени, М.—Л., изд. Акад, наук СССР (Фауна СССР, Млекопит., 1,2): 128—134
(общ., изобр.); Buxton А., 1954, Oryx, vol. 2, No. 6 : 392 (морф., изобр.); Куз-
нецов Б. А., 1954, Бюлл. Моск. общ. испытат. прир., отд. биол., LIX, 4 : 14 (ист.,
распр.); R i п е у Th., 1954, Trans. Roy. Soc., New Zealand, vol. 82, No. 2 : 569—578
(рога); На r r i s о n R. and A. H у e 1 1, 1954, J. Anatomy, vol. 88, pt. 3 : 338—355
(размн.); Саблина T. Б., 1955, Тр. Инет, морфол. животн. им. А. Н. Северцова,
15 : 16 (ист.); Сержанин И. Н., 1955, Млекопит. Белорусск. ССР, Минск, изд.
Акад, наук Белорусск. ССР : 223—226 (общ.).
Описание. Олень средней величины. Высота в холке у взрослых сам-
цов колеблется от 85 до 100 см. Длина тела около 140 см. Длина черепа
254—290 см. Живой вес редко превосходит 100 кг, обычно меньше. Самки
несколько мельче самцов.1 Высота в крестце на несколько сантиметров
превосходит высоту в холке.
Профиль спины прямой, поднимающийся кзади или слегка выпуклый.
Голова сравнительно широкая в заглазничной части, имеет сильно су-
женное рыло. Уши сравнительно длинные; будучи отогнуты вперед, они
достигают предглазничных ямок; верхушки их заострены. Носовое зеркало
небольшое, занимает лишь среднюю часть верхней губы; верхняя его
граница едва заходит выше уровня нижних краев ноздрей и только по
бокам тянется вдоль внутренних и наружных краев последних. Пред-
глазничные железы небольшие. Радужная оболочка глаз коричневая.
Рога у самцов в основания х и в средней части круглого или продольно
овального сечения. От главного ствола отходят вперед один надглазнич-
ный и один средний отросток, загнутые верхушками вверх. Надглазнич-
ный отросток по длине не уступает среднему. Второй надглазничный
1 В Шлезвиг-Голщтинии и Ютландии разводится никем до сих пор не описанная
крупная раса или популяция лани (Buxton, 1954).
CERVUS
in
отросток отсутствует. Выше среднего отростка главный ствол уплощается
и имеет вид лопаты, от верхнего и заднего края которой отходят 7—9'
небольших пальцеобразных или тоже уплощенных отростков; при этом
самый задний и нижний отросток наиболее длинный и отставлен дальше
от других отростков, являясь, очевидно, гомологом заднего отростка
у остальных представителей рода Cervus L.1 Главные стволы рога выше
надглазничных отростков расходятся назад и в стороны, делая резко
выраженный лирообразный изгиб; передний и задний отростки лопат
Рис. 34. Лань (Cervus dama L.).
направлены внутрь, навстречу одноименным отросткам противополож-
ного рога. Хвост длиной около 18—20 см, длиннее уха. Средние копыта
удлиненные, с заостренными концами. Копыта боковых пальцев слабо раз-
виты, обычно не превышают 73 длины главных и при ходьбе не касаются
почвы.
Общий фон зимней окраски верхней части туловища, шеи и головы
типично темно-бурый с желтоватым или рыжеватым оттенком. Наиболее
темно окрашена верхняя часть спины и крестца, нередко черно-бурого
цвета. По бокам и книзу окраска постепенно светлеет, приобретает ясно
серый оттенок. Серый оттенок имеют также шея, бока и низ головы, тогда
1 Близкий к обыкновенной лани вид — лань месопотамская (Cervus (Dama) те-
sopotamica Brooke), помимо более круппых размеров, отличается тем, что надглазнич-
ный отросток слабо развит, рог уплощен в основании среднего отростка, тогда как
лопатообразное уплощение вершины рога отсутствует.
12 Фауна СССР, И. И. Соколов
MS
CERVIDAE
как верхняя часть последней (лоб и нос) окрашена одинаково с верхом
туловища — коричнево-бурая. Вокруг глаз светлое кольцо. Внутренняя
сторона ушей серовато-белая, наружная коричнево-бурая. Конец
морды по краям рта беловатый. Низ туловища и внутренняя поверхность
ног белого цвета, иногда с рыжеватым или сероватым оттенком. Конеч-
ности ниже запястных и скакательных суставов светлые, спереди рыже-
ватые, сзади и с боков почти белые. Хвост белый, но по средней линии
его проходит черно-бурая полоса. Околохвостовое зеркало небольшое,
почти не выходит на бока бедер и не поднимается выше основания хвоста,
сверху и с боков окаймлено узкой полосой черно-бурого цвета, сливаю-
щейся с темной окраской верха крестца. По верхней части, по бокам
туловища и на крупе разбросаны едва заметные в зимней шерсти светлые
круглые или овальные небольшие пятна. Копыта и носовое зеркало
черно-бурого цвета. Общий фон летней окраски буровато-рыжий или
чисто рыжий. Распределение темно- и светлоокрашенных участков то же,
что и зимой. Пятнистость выражена ярко, захватывая не только спину,
бока и бедра, но и лопатки до шеи. По бокам туловища пятна, сливаясь,
образуют почти сплошные продольные белые полосы. Окраска ланей,
особенно в условиях паркового, полувольного содержания, подвержена
изменчивости. Часто встречаются альбиносы, пятнистые и сравнительно
редко меланисты («черная лань»).
Зимний волосяной покров состоит из буроватого тонкого подшерстка
(пуха), более толстой ломкой ости и из редко разбросанных грубых,
длинных направляющих волос. На лицевой части головы ость короткая,
упругая, на лбу и шее она мягкая.1 На верхней стороне туловища она
более грубая, чем на шее, но самый грубый и толстый волос находится
на боках и животе. Свойственная самцам других видов оленей грива на
нижней стороне шеи у самцов ланей не развивается. Волос вдоль верх-
него края шеи вершинами направлен вперед. Характерен для ланей
пучок длинных волос в области половых органов (у самцов вокруг отвер-
стия препуциального мешка). Отдельные волосы верхней половины
туловища имеют зонарную расцветку; от основания они светло-бурые,
кверху постепенно темнеют, но перед верхушкой имеют светло-рыжее или
белое кольцо; кончик волоса черно-бурый, почти черный. Волосы вдоль
средней линии спины и по краям околохвостового зеркала обычно лишены
предвершинного светлого кольца, обусловливая равномерную темную
окраску этих частей тела. Серый оттенок окраски боков туловища, шеи
и головы обусловлен более светлой, белой окраской предвершинных
колец.
Половой диморфизм в окраске и характере волосяного покрова
у ланей практически отсутствует. Молодые всегда имеют пятнистую
окраску.
Длина черепа колеблется от 254 до 290 мм. Череп (рис. 35) относительно
широкий в лобноглазничной части, клинообразно суживается по на-
правлению к рылу и не образует расширения на границе верхнечелюст-
ных и межчелюстных костей. Ширина лба в самом узком месте между
рогами и глазницами больше ширины черепа в надушных буграх. Задний
край носовых костей расположен приблизительно на уровне передних
краев глазниц. Слуховые пузыри умеренно вздутые, с гладкой поверх-
ностью, выступают несколько ниже уровня basioccipitale. Подъязычные
1 У одной исследонанной нами шкуры лани из Италии волос на шее имеет харак-
тер мягкой шерсти.
Рис. 35. Череп Cervus dama L.
12*
180
CERVIDAE
отверстия расположены впереди переднего края суставной поверхности
затылочных мыщелков.
Первый переднекоренной зуб верхней челюсти (Р2) имеет внутреннюю
долю из двух половинок, разделенных вертикальной бороздой. Клыки
в верхней челюсти отсутствуют.
История и географическое распространение. Происхождение под-
рода Dama Frisch еще не выяснено. Есть предположение, что он пред-'
ставляет боковую ветвь плиоценового рода Eucladocerus Falc. (Флеров,
1952). Близкие к современным виды известны из плейстоценовых отло-
жений Европы и Азии.
Коренным ареалом лани считаются страны Средиземноморья: Порту-
галия, Италия, Греция, о-ва Родос и Сардиния, северная Африка, южная
часть Малой Азии, Палестина. В диком состоянии она сохранилась на
юго-западпом побережье Мраморного моря, вдоль южного берега Малой
Азии на восток до Аданы, включая горы Тавра и в небольшом числе
в северо-западной Африке (Флеров, 1952). В полудомашнем состоянии
содержится в большинстве стран Европы, включая Англию и южную часть
Скандинавского полуострова. Принято считать, что в Среднюю и Север-
ную Европу лань завезена с юга. Однако судя по тому, что уже в плей-
стоцене на территории Германии существовала форма лани (С. dama fossi-
lis auct.), почти не отличимая от современной, можно предполагать, что
ареал ее раньше был значительно шире и что сохраняемое в парках по-
головье представляет прямых потомков аборигенных обитателей средней
Европы. Известно, например, что во времена Великого княжества Ли-
товского (XIII—XVI вв.) лани были не редки на территории современной
Польши и Литвы. В начале XVIII в., по-видимому, еще местная лань
обитала в Беловежской пуще (Кириков, 1952). К XIX в. она здесь была,
вероятно, истреблена, так как вновь начала завозиться в пущу неболь-
шими партиями лишь с 1860 г. В Беловежской пуще лани жили вплоть до
Первой мировой войны. В 1912 г. их здесь было 1250 голов (Врублев-
ский, 1912). В годы хозяйничания немцев (с 1'915 г.) лани были истреблены,
и с 1920 г. в Беловежской пуще ланей нет (Саблина, 1955). В дореволю-
ционное время лань содержалась в некоторых частновладельческих охот-
ничьих парках в Прибалтике, в угодьях царской охоты под Петербургом,
в бывшем Боржомском отводе в Закавказье, в некоторых центральных
и южных губерниях. Так, свыше столетия лани жили в лесах одного
из поместий Сычевского уезда Смоленской губернии, на границе с нынеш-
ней Калининской областью. Они держались не только около усадьбы,
где их подкармливали, но и в окружающих имение лесах без специаль-
ной охраны. Лани сильно одичали, были крайне осторожны, жили на
естественных кормах и хорошо переносили местные зимы. Еще около
1910 г. здесь на них неоднократно охотился покойный Б. М. Житков
(1940). Жили акклиматизированные лани и под Москвой (Туркин и Са-
тунин, 1902; Greve, 1909; Шиллингер, 1930).
На территории СССР в настоящее” время небольшое число ланей
сохранилось на территории Литвы,. Белоруссии и на Украине. В Литов-
ской ССР, куда лани были завезены около 60 лет назад (Кузнецов, 1954),
они обитают на территории лесхозов Юрваркского, Куршепского и Шау-
ляйского. По данным учета, в феврале 1954 г. здесь насчитывалось 112
ланей.
В Белорусской ССР некоторое количество уцелевших после Великой
Отечественной войны потомков от завезенных еще в 1932 г. из Германии
8 ланей сохранилось на территории Вяловского заповедника (Молодеч-
CERVUS
181
неясная область, Воложинский и Ивенецкий районы). В 1949 г. их
насчитывалось 13 голов (Сержанин, 1955).
На Украине лань издавна разводится в зоопарке Аскания-Нова
(Херсонская область). Некоторое количество их выпущено па о. Хортица
на Днепре (территория бывшей Запорожской сечи). В 1952 г. 10 голов
ланей из Аскании-Нова выпущено в госохотзаказнике Сухолесы Узинского
района Киевской области. Лапи здесь успешно прижились и дали при-
плод.1
В 1949 г. некоторое количество ланей также из Аскании-Нова за-
везено в охотничий заповедник в Колхиде (Успенский, 1950), но каковы
результаты этого опыта акклиматизации — не известно.
Опыт показал, что в наших условиях лань удовлетворительно аккли-
матизируется в местностях до 50° с. ш. В более северных районах она
трудно переносит зимы и требует устройства на этот период крытых
помещений.
Биология. В естественной обстановке и при полувольном содержании
на достаточно обширных пространствах лань предпочитает смешанные
или лиственные леса, чередующиеся с открытыми полянами, на которых
она в летние месяцы предпочитает кормиться. Сплошных, особенно хвой-
ных, лесов она избегает. Охотно держится в густых зарослях кустарника,
лесного подроста, по густым опушкам леса. Рельеф местности предпочи-
тает ровный или слегка всхолмленный.
Пищу ланей составляют в летний период травы, а также листва и мо-
лодые побеги кустарников и деревьев. Зимой основу питания, при отсут-
ствии подкормки, составляет веточный корм. Поедают заготовляемое
для них сено, жолуди, каштаны, буковые орехи, а также картофель и
корнеплоды (Шиллингер, 1930). Охотно пользуются устраиваемыми для
них искусственными солонцами. ............
Держатся лани группами, состоящими из нескольких самок с при-
плодом и молодняка прошлых лет. Самцы большую часть года держатся
отдельно, присоединяясь к стадам самок лишь на период гона и зимой.
,В смешанном табунке самец обычно является вожаком.
Сколько-нибудь значительных миграций лани не совершают; незна-
чительные передвижения в- пределах обитаемой территории связаны
с кормовыми и защитными условиями. Кормятся не только по ночам и
утром, но и в дневные часы. В условиях паркового содержания лани менее
пугливы, чем, например, благородный олень, проявляют признаки любо-
пытства по отношению к незнакомому предмету, смело подходят к местам
подкормки, заходят даже в специально устраиваемые для них навесы
и сараи, легко привыкают к человеку, идут на зов или сигнал, берут из
рук корм. Однако полного одомашнения не бывает, и животные обычно
не позволяют себя удерживать руками или погладить (Туркин и Сатунин,
1902). Там, где лань попадает па полную свободу, она быстро дичает;
становится очень пугливой и осторожной. Из органов чувств хорошо
развит слух.
Передвигается лань шагом, рысью и прыжками. В быстроте бега
уступает благородному оленю. Прыжки может совершать высотою до
2 м (Шиллингер, 1930). Хорошо плавает.
Первые рога у самцов начинают расти в шестимесячном возрасте
в виде простых спичек, кожа с которых сходит в августе следующего года.
1 Сведения по Литовской и Украинской ССР получены от Управлений охотни-
чьего хозяйства этих республик в октябре 1954 г. 1
182 CERVIDAE
Эти первые рога сбрасываются в июле третьего года жизни. В процессе
развития рогов последовательно появляются надглазничный и средний
отростки. В последнюю очередь развиваются лопаты и задние отростки
(рис. 36). Полного развития рога достигают к пятилетнему возрасту.
Взрослые самцы сбрасывают рога в апреле или мае. Рост новых начи-
нается дней через восемь после сбрасывания старых и заканчивается в ав-
густе или сентябре. Рост и развитие рогов у лани зависит от функции'
половых желез. Кастрация самцов в раннем возрасте, до начала заметного
роста рогов, ведет к тому, что рога совсем не развиваются или на месте
их образуются шишкообразные выросты хрящевой консистенции, покры-
тые кожей и не сменяемые в течение всей жизни. Кастрация взрослых
самцов с вполне развитыми и очистившимися от кожи рогами влечет
преждевременное их сбрасывание. Отрастающие в дальнейшем рога
лишены или имеют недоразвитые отростки и, не сменяясь всю жизнь,
остаются покрытыми кожей (Завадовский, 1926).
Половой сезон (период гона) у ланей протекает в октябре. В это время
они удаляются там, где это возможно, на постоянные «токовища», распо-
ложенные в глубине леса. Здесь самцы собирают вокруг себя гаремы
из возможно большего числа самок. В возбуждении они выбивают ногами
в земле по нескольку углублений, расположенных одно поблизости от дру-
гого (Шиллингер, 1930). Между самцами нередко происходят ожесточенные
драки. В парковых хозяйствах рекомендуется на каждые 60 голов дер-
жать 7—8 самцов (Туркин и Сатунин, 1902). Беременность самок длится
около 8 месяцев. В июне самка приносит одного, редко двух телят. Перед
отелом она удаляется от стада и присоединяется к нему вновь обычно
в августе. На второй день после рождения теленок способен следовать
за матерью, но первое время лежит спрятанным в траве. Мать кормит
молоком детеныша долго, чуть ли не до следующего отела, но уже с четы-
рехнедельного возраста теленок начинает щипать траву (Туркин и Са-
тунин, 1902).
Сведения о продолжительности жизни ланей противоречивы. Туркин
и Сатунин (1902) определяют его в 20 лет, тогда как Шиллингер (1930)
называет маловероятную цифру в 70 лет и указывает на случаи, когда
40—50-летние самки регулярно приносили телят.
Врагами ланей являются главным образом волк и рысь; для молодых
первое время после рождения опасна лисица. Злом в условиях паркового
содержания могут быть бродячие собаки.
Из болезней среди ланей наблюдался пастереллез. Греве (Greve,
1909) из числа паразитов называет 10 видов червей, один вид власоеда
{Trichodectes longicornis Ntz.) и оленью кровососку (Lipoptena cervi L.).
В западной Европе на ланях зарегистрированы два вида носоглоточных
оводов: Cephenomyia miltispinosa Ulr. и изредка Pharyngomyia picta
Meig.1
Хозяйственное значение. Благодаря способности легко акклимати-
зироваться и переходить в полудомашнее состояние, лань представляет
большой интерес для нашего охотничьего хозяйства. Она с успехом,
вероятно, может быть акклиматизирована в ряде заповедников и охотни-
чьих хозяйств, при условии надлежащей охраны от хищников, бродя-
чих собак и браконьеров. В средней полосе потребуется заготовка сена
на зимний период, но в Закавказье лань в течение круглого года может
1 Данные К. Я. Грунина.
CERVUS
183
находиться на подножном корме.1 При правильном ведении хозяйства она
может давать значительное количество вкусного мяса, шкуру, годную
для выделки замши, и рога, которые находят применение в качестве
настенного украшения и для различных поделок.
Рис. 36. Возрастные изменения рогов у лани (Cervus dama L.).
(По Ф. Шиллингеру, 1930).
Cervus nippon Temminck-—Пятнистый олень (рис. 37—40)
Temminck, 1837, Fauna japonica, Introduct. : XXII (первооп.); M а а к P.,
1861, Путешеств. по долине р. Уссури, т. 1, СПб. : 136—137 (распр.); Прже-
вальский Н. М., 1870, Путешеств. в Уссурийском крае, СПб. : 265—266
(биол.); Туркин Н. В. и К. А. Сатунин, 1902, Звери России, Cervidae, М. :
28—36, 363—392 (общ., изобр.); Lydekker R., 1914, Catal. Ungulate mammals
Brit. Mus., IV, London. : 106—116 (сист., синон.); Абрамов К. Г., 1930, Пятни-
стый олень, Владивосток, изд. акц. общ., «Приморский зоопитомник» : 1—75 (общ.,
изобр.); Дорогостайский В., 1930, Охотник, 2:12—14 (акклим., пит.,
изобр.); Менард Г. А., 1930, Пантовое хозяйство, М.—Л., Госторгиздат : 1—167
(общ., изобр.); Мей се ль М. Н. и Т. А. Григорьева, 1933, Вести. Дальне-
вост. фил. Акад, наук СССР, 1, 2, 3 : 155—123 (панты); Граменицкий М. И.
и И. И. С и в е р ц о в, 1934, Вести. Дальневост. фил. Акад, наук СССР, 1 (8) : 64—
66 (панты); Мейсель М. Н., 1934, Вести. Дальневост. фил. Акад, наук СССР,
1 (8) : 61—63 (панты); М иро любое И. И., 1934а, Сборн. «Вопросы пантового
оленеводства», II : 1—22 (бол.); М иро любов И. И., 19346, Сборн. «Вопросы
пантового оленеводства», II : 31—45 (определ. возр.); Рябова Т. И., 1935, Вести.
Дальневост. фил. Акад, наук СССР, 14 : 175—178 (пит.); Миро любов И. И.,
1936, Вести. Дальневост. фил. Акад, наук СССР, 16 : 155—174 (биол.); Пав-
ленко С. М., 1936, сборн. «Пантокрин (хуалукрин)», 2 : 3—9 (панты); Т э в и А. С,.,
1936, сборн. «Пантокрин (хуалукрин)», 2 : 49—60 (панты); Рябова Т. И. и
Н. П. С а в е р к и н, 1937, Тр. Дальневост. фил. Акад, наук СССР, сер. ботан, II :
533—673 (пит.); Венд панд О. В., 1938, Вести. Дальневост. фил. Акад, наук
СССР, 28:134—140 (пит.); Абрамов К. Г., 1939, Научно-метод. зап. Главн.
управл. по заповеди., III : 160—163 (распр., стац); Даршкевич Я., 1939, На-
учно-метод. зап. Главн. управл. по заповеди., IV : 144—153 (акклим.); Козлов В. В.
1939, Научно-метод. зап. Главн. управл. по заповеди., V : 201—202 (акклим.); Сар-
кисов А. А., 1944, Тр. Ереванск. зоол. парка, I—II : 91—97 (акклим.); Аве-
рин Ю. В. и С. Л. У ш к о в, 1947, Тр. Ильменск. гос. заповеди., Ill, 1 : 161—185
(акклим.); Миролюбов И. И., 1948, Каракулеводство и звероводство, 6 : 73—
75 (панты); Миролюбов И. И. иЛ. П. Рященко, 1948, Пятнистый олень,
Владивосток, изд. Дальневост, звероводческ. треста : 1—114 (общ., изобр.); Арсе-
ньев В. А., 1949, Охрана природы, 7 : 79—96 (акклим.); Миролюбов И. ИГ.,
1949, Каракулеводство и звероводство, 1 : 74—75 (гибр.); Митюшев П. В.,
1 Подробности о технике содержания ланей в охотничьих хозяйствах можнанайти
в брошюре Ф. Шиллингера (1930).
184
CERVIDAE
M. П. Любимов и В. К. Новиков, 1950, Пантовое оленеводство и болезни
пантовых оленей, М., изд. «Международн. книга» : 1—249; Бромлей Г. Ф., 1951,
сборн. «Заповедники СССР», II : 220 (распр.); Лихачев А. И., 1951, Тр. Ново-
сибирск. с.-х. инет., VIII : 117—132 (акклим.); Ellerman J. and Т. Morri-
son-Scott, 1951, Checklist of palaearctic and indian mammals, London : 364—366
(сист., синон.); Ленский A. A., 1952, Каракулеводство и звероводство, 4:78
(нит.); Рященко Л. П., 1952, Каракулеводство и звероводство, 4 : 50—52 (разв.);
С а л ганский А. А., 1952, Каракулеводство и звероводство, 5 : 52—58 (акклим.,
разв., содерж.); Флеров К. К., 1952, Кабарги и олени, М.—Л., изд. Акад, наук
СССР (Фауна СССР, Млекопит., I, 2) : 138—147 (общ., синон., изобр.); Р я-
щ е н к о Л. II., 1953, Каракулеводство и звероводство, 4 : 54—59 (разв.); Абра-
мов К. Г., 1954, Копытные звери Дальн. Вост., Хабаровск, Книжн. изд. : 44—55
(общ., изобр.); Козлов В. В., 1954, Зоол. журн., XXXIII, 4 : 927—933 (акклим.);
Рященко Л. П., 1954, Каракулеводство и звероводство, 6 : 39—42 (разв.); Бром-
лей Г. Ф. и 3. И. Г у т н и к о в а, 1955, Супутинский заповеди., Владивосток,
Книжн. изд. : 24—26 (биол.); Двойченко Г., 1955, Ставропольский край, Став-
рополь, Книжн. изд. : 69 (распр.); Ильина Г. И., 1955, Экологические особен-
ности пяти, оленя и перспективы его акклим. в европейской части СССР, Авторефе-
рат, М. : 1—15 (акклим.); Насимович А. А., 1955, Роль режима снежн. покрова
в жизни копытных, М., изд._Акад. наук СССР : 215—217 (распр., биол.).
Описание. Олень средних размеров. Высота в холке у взрослых сам-
цов колеблется в пределах 85—118 см, косая длина туловища от 90 до
118 см. Длина черепа 265—335 мм, живой вес составляет 93 до 148 кг.
Самки несколько мельче самцов (живой вес меньше примерно на 20%).
Высота в крестце несколько больше высоты в холке. Самцы, по данным
И. Миролюбова и Л. Рященко (1948), в среднем, более низкопереды.
Небольшая голова с узкой мордой, сравнительно длинная шея и тонкие
ноги придают пятнистому оленю стройный вид. Уши длинные, заостренные,
будучи загнуты вперед, они достигают вершинами предглазничных ямок,
или, даже, заходят вперед от последних. Носовое зеркало большое, зани-
мает средину верхней губы, все пространство между ноздрями и распро-
страняется на наружные края последних. Предглазничные железы боль-
шие, глубокие. Радужная оболочка глаз коричневая, с горизонтально
расположенным зрачком.
Рога типично имеют по 4 конца: надглазничный, средний и два кон-
цевых. Надглазничный отросток отходит от ствола рога выше, чем у
благородного оленя и под углом, близким к прямому, но верхушка его,
как и у среднего отростка, загнута вверх. Второй надглазничный от-
росток отсутствует или представлен рудиментом. В тех случаях, когда
рога имеют по 5 концов, дополнительным является третий концевой
отросток; при этом вместо простой вилки вершина рога приобретает
вид чаши из трех отходящих от общего корня отростков. Главные стволы
рогов образуют лировидный изгиб и вершинами обычно направлены
навстречу друг другу. Сечение ствола рога и отростков круглое или
овальное. Хвост длиннее, чем у благородного оленя, и лишь немного
уступает длине уха. Боковые копыта составляют около половины длины
главных, при наступании или ходьбе по мягкому грунту касаются
почвы.
Зимний волосяной покров состоит из тонкого извитого подшерстка
и грубой ломкой ости. Длина волос на спине в это время около 5—6 см.
Морда впереди глаз и конечности ниже запястных и скакательных суста-
вов покрыты коротким, плотно прилегающим к коже волосом. Волосы
на плюсневых железах образуют подобие упругой подушки, выступаю-
щей над общей поверхностью окружающей кожи. Хвост покрыт волосом,
одинаковой с туловищем длины, не образующим на конце кисти. Внутрен-
няя (вентральная) поверхность хвоста волоса не имеет. У самцов
CERVUS
185
волосы на шее, особенно на нижней стороне, удлинены до 9—10 см и
образуют подобие гривы.
Общая окраска туловища в зимнем меху буровато-серая, исчерченная,
иногда с рыжеватым оттенком. По средней линии шеи и передней части
спины идет более или менее ясно выраженная темная полоса. На боках
шерсть несколько светлее; в подмышках, пахах, на внутренней стороне
Рис. 37. Пятнистый олень (Cervus nippon Temm.).
бедер у живота и на животе в области половых органов окраска светлая,
почти белая; передняя часть живота, особенно пятно непосредственно
впереди отверстия полового члена у самцов темные, почти черные. Шея
темно-серого цвета, верхняя часть головы и наружная сторона ушей
рыжая, морда сверху и с боков рыжевато-желтая. Конечности ниже
запястных и скакательных суставов светлые рыжевато-желтые с темной
розоватой полосой на передней стороне. Белое околохвостовое зеркало
занимает внутреннюю сторону ягодиц ниже основания хвоста и внизу
сливается со светлой окраской живота. По бокам от хвоста, а иногда ц
сверху, оно резко отграничено от остальной окраски туловища узкой
полосой черных волос. Волосы по краям хвоста белые, сверху одинако-
вого цвета с туловищем и темной полосой по средней линии; кончик
хвоста у самцов белый, у самок рыжий. В зимнем меху на спине, боках,
186
CERVIDAE
лопатках и бедрах проступают неясные беловатые пятна. Иногда они
почти не заметны. Самки окрашены светлее самцов. Низ живота у них
сплошь светлый, почти белый; грива на шее не развивается.1
Подшерсток (пух) светло-бурого цвета. Отдельные волосы ости со
спины и боков туловища имеют зонарную окраску: в основной части
светло- или темно-бурые, к вершине постепенно темнеют, но в пред-
вершинной части имеется светлое (от чисто белого до рыжего) кольцо,
а самый заостренный кончик волоса — черного цвета. От степени потем<
нения основной части волоса, от размера и цвета светлого предвершин-
ного кольца зависит интенсивность и оттенок окраски. Волосы, образую-
щие темную полосу вдоль спины, хвоста и сверху хвостового зеркала,
сплошь темные, иногда черные без светлой предверхушечной зоны. Ко-
роткие волосы на морде и нижних частях конечностей равномерно желто-
ватые, за исключением темных верхушек. Околохвостовое зеркало и
пятна на туловище образованы белыми волосами без темных верхушек.
С течением времени темно окрашенные верхушки остевых волос обламы-
ваются и вследствие этого к концу зимы окраска оленя становится более
светлой и тусклой.
Летом туловище как у самцов, так и у самок ярко-рыжее, кроме темной
полосы вдоль шеи, спины и крестца и многочисленных белых пятен на
спине, боках и задней половине шеи. Пятна в летнем меху чисто белые,
ясно отграниченные от основного рыжего фона, располагаются 5—6
продольными рядами. В нижних рядах пятна крупнее, почти сливаются
друг с другом и образуют продольные полосы по бокам живота. Шея и
конец морды сероватые. Лоб, бока головы и ноги желтовато-песчаного
цвета Низ живота белый, с рыжеватым оттенком. Хвостовое зеркало
белое. Летний волос более короткий, длиной на спине 2—3 см. Под-
шерсток в летнем меху отсутствует. Молодые имеют пятнистую окраску.
Кожа на носовом зеркале и копыта темно-бурого, почти черного цвета.
На черепе (рис. 38) ширина лба в самом узком месте между глазницами
и рогами меньше ширины черепа в надушных буграх. Задние края носовых
костей расположены на уровне передних краев глазниц или несколько
впереди последних. Слуховые пузыри вздутые, гладкие, выступают ниже
уровня средней части basioccipitale. Подъязычные отверстия расположены
позади уровня переднего края суставных мыщелков.
Внутренняя доля переднего премоляра верхней челюсти (Р2) сплошная;
вертикальная борозда на ее медиальной поверхности имеется, но не делит
ее на две половины, как у С. dama. Клыки в верхней челюсти имеются.
История и географическое распространение (рис. 39). В ископаемом
состоянии пятнистый олень не известен, но корни его, по-видимому, ведут
к плиоценовому виду С. magnus Zdansky из Китая, уже имевшему на рогах
четыре отростка (Флеров, 1952). В настоящее время распространение
С. nippon ограничено восточными провинциями Китая, Кореей, южной
частью Дальнего Востока, а также островами Японии и Тайванем.
В пределах СССР пятнистый олень в диком состоянии водится в При-
морском крае к югу от 46-й параллели. В прошлом он занимал всю об-
ласть восточных склонов Сихоте-Алипя в этих пределах. Теперь же
распространен крайне неравномерно. Причиной является отсутствие во
многих районах благоприятных условий или вытеснение и истребление
1 Кастрированные самцы (если даже кастрация была произведена по достижении
половой зрелости) имеют характер волосяного покрова и окраску, сходную с самками.
В частности, у них не развивается на шее грива (Миролюбов, 1948).
CERVUS
187
человеком. В средине прошлого столетия, по Мааку (1861), пятнистый
олень встречался в долине р. Уссури от верховьев до устья Нора. На
восточном склоне хребта Сихотэ-Алинь северной границей в то время
Рис. 38. Череп Cervus nippon Ternm.
была долина Бикина. В настоящее время его там нет, если не считать
изредка содержимых в неволе. Отдельные очаги пятнистого оленя в 1928 г.
сохранялись по рр. Амагу, Великой Кеме и Колумбе.
188
CERVIDAE
До настоящего времени в незначительном количестве он держится
в южной части Сихоте-Алиньского заповедника к югу от Куруминского
хребта (Бромлей, 1951). В прошлом пятнистые олени были многочисленны
между бухтой св. Владимира и р. Тапоузой. В настоящее время вдоль
большей части побережья Японского моря они встречаются лишь местами
в крайне незначительном числе. На пространстве от р. Пхусун до р. Ку-
руму (левый приток р. Иобзыхе) отсутствуют вовсе (Абрамов, 1939).
В качестве отдельных пунктов, где в 20-х годах настоящего столетия еще
сохранялось незначительное количество этого зверя, называются вер-
ховья рр. Тетюхе, Ахобе и район мыса Низменного. Область более или
менее сплошного распространения пятнистого оленя представляет южная
оконечность Приморского края к югу от р. Пхусун до Посьетского рай-
она включительно. В последнем он отмечается в вершинах рр. Рязановой,
Адими, Сидими, Брусья, в пределах заповедника Кедровая падь. Далее
к востоку дикий пятнистый олень водится в Суйфунском, Шкотовском и
Сучанском районах, в вершинах рр. Майхе, Сицы, Мамонтовой пади,
Бепги, Попугоу, Стеклянухи, в верхних притоках р. Сучана, по всему
побережью от бухты Восток (Абрамов, 1930). Наибольшее количество
сохранилось в прибрежной части Судзухинского заповедника. Общее
поголовье диких оленей в Приморском крае в несколько раз меньше
числа их, содержимых в домашней обстановке.
Подвиды. В настоящее время общепринято выделение четырех под-
видов пятнистого оленя.
1) С. nippon nippon Temminck. Мелкая раса; высота в холке около 85 см.
От других рас, по Флерову (1952), отличается ясно выраженной пят-
нистостью в зимней шерсти,1 белой окраской волос, покрывающих плюс-
невые железы, и большими размерами околохвостового зеркала, захо-
дящего па боковую часть бедер и ограниченного черной каймой и с бо-
ков, и сверху. Распространение: острова Японии, включая
Рюкю.
2) С. nippon taiouanus Blyth. Размеры средние; высота в холке
около 98 см. Туловище относительно более длинное, а конечпости более
короткие, чем у типичного подвида. Пятнистость зимой выражена ясно.
В летнем меху выделяется ярко-рыжий оттенок задней части шеи. Во-
лосы на плюсневых железах буроватые. Темная оторочка околохвосто-
вого зеркала ясно выражена только сверху. Распространение:
о-в Тайвань.
3) С. nippon kopschi Swinhoe. Размеры крупные; высота в холке
около ПО см. Пятнистость в зимнем меху почти незаметна. От следую-
щего подвида (С. п. hortulorum) отличается более яркой темной полосой
вдоль спины и более бледной пятнистостью верхней стороны спины летом.
Распространение: Китай к югу от р. Хуанхэ.
4) С. nippon hortulorum Swinhoe. Размеры как у предыдущего вида.
От последнего отличается отсутствием или слабым развитием темной по-
лосы вдоль спины, более яркой пятнистостью верхней стороны спины и
распространением пятнистости на плечи и бедра. Распростране-
ние: Китай к северу от р. Хуанхэ, Корея, южная часть Приморского
края СССР.
К. К. Флеров (1952) высказывает весьма вероятное предположение
об идентичности двух последних подвидов.
1 По Лайдекеру (Lydekker, 1914 : 108), наоборот, пятнистость зимой почти не
заметна. Проверить это, за отсутствием материала, нам не было возможности.
Рис. 39. Распространение Cervus nippon Temm.
— коренной ареал; 2 — места акклиматизации.
190
CERVIDAE
Биология. Распространение дикого пятнистого оленя приурочено
к зоне широколиственных лесов и смешанной уссурийской тайги с пре-
обладанием монгольского дуба, клена, липы, ясеня, маньчжурского ореха,
бархата и других растений маньчжурской флоры. Рельеф местности райо-
нов обитания представляет горные склоны с каменистыми выходами и
россыпями, большим числом долин (падей), ручьев и речек. Предпочтение
отдается местам, где сплошные леса чередуются с травянистыми полянами,
излюбленными кормовыми угодьями оленей в летнее время. Пятнистый
олень охотно селится в зарослях молодого леса, выросшего на месте
былых гарей.
Фактором, играющим важнейшую роль и лимитирующим распростра-
нение пятнистого оленя, является глубина снежного покрова. Он не спо-
собен доставать корм из-под глубокого снега, и высота снежного по-
крова в 20—30 см является крайним пределом. Не меньшее значение имеет
продолжительность зимы. На западных склонах Сихотэ-Алиня средняя
глубина снега меньше, чем на восточных, но так как здесь он залегает
сплошным покровом и держится дольше, пятнистый олень на большей
части их, по-видимому, никогда не водился (Абрамов, 1939). Основная
масса его поголовья держится вблизи побережья Японского моря, где
зима малоснежна, а выпадающий снег быстро тает. Чем дальше от моря,
тем оленей становится меньше.
Сколько-нибудь значительных миграций пятнистые олени не совер-
шают и в основном каждое стадо придерживается одного обжитого уча-
стка. В Судзухинском заповеднике, например, где сохранилась наиболее
многочисленная популяция этого зверя, олени занимают малоснежные
солнечные склоны хребта Тачиньчжан вблизи морского побережья; здесь
они проводят и отел, и период гона (Бромлей, 1951). Могут иметь место
лишь местные передвижения, сводящиеся к тому, что летом звери более
широко и равномерно расселяются по занимаемой территории, придер-
живаясь тенистых участков, а в зимнее время концентрируются на бес-
снежных или малоснежных склонах. Летом олени часто держатся непо-
средственно на побережье, где прохладный сырой воздух служит защитой
от жалящих насекомых, а также представляется возможность питаться
морскими водорослями и глотать соленую морскую пену.
Пятнистые олени живут группами в 7—10 голов, изредка собираются
в табуны по нескольку десятков, а в парковых хозяйствах даже по не-
скольку сотен. Табуны состоят из животных различного возраста и пола.
В период роста рогов (пантов) самцы обычно держатся одиночно; самки
на время отела и первых недель лактации также уединяются от табунка,
к которому присоединяются лишь тогда, когда оленята достаточно окреп-
нут. Во главе табунка находится, по Миролюбову (1936), самка. В слу-
чае опасности последняя издает резкий свист с добавлением низкого звука.
Испуганные олени несутся крупными прыжками. При этом белые волосы
в окружности хвоста, по-видимому, рефлекторно подымаются и около-
хвостовое зеркало сильно увеличивается в размерах, помогая бегущим
животным не терять друг друга из вида.
По характеру пятнистые олени мирные животные. Драки между ними
(если не считать самцов в период гона) бывают редко. В качестве орудий
защиты и нападения самцам служат окостеневшие рога. В период же
роста последних самцы, как и самки, дерутся зубами или передними
ногами, нанося острыми концами копыт быстрые и резкие удары вперед.
Пятнистый олень прекрасный пловец. Известны многочисленные случаи
переплывания им проливов шириною свыше 10 км для переселения с ма-
CERVUS
191
терика на острова или обратно, и при этом не только в теплое время года,
но и зимой.
При передвижениях в пределах занимаемых территорий пятнистые
олени имеют привычку придерживаться определенных путей, на которых
вытаптываются тропы. Эта привычка раньше использовалась промышлен-
никами для установки изгородей с ловчими ямами (лудев), капканов и
других ловушек.
Дневные часы пятнистые олени проводят в зарослях под прикрытием
кустарников и деревьев. На открытые поляны для пастьбы выходят ве-
чером, рано утром или в ночное время.
Пищей пятнистым оленям в естественной обстановке служат растения,
начиная с осок и кончая листвой, побегами, корой, и даже относительно
толстые (до 1.5 см) ветви ряда деревьев. При этом состав кормовых расте-
ний меняется по сезонам. В весенний период основную пищу составляют
молодые осоки, а также разнообразные злаки, зонтичные, сложноцвет-
ные и другие травянистые растения. Летом излюбленным и основным кор-
мом являются листья и побеги винограда (Vitis amurensis Rupr.), леспе-
деци (Lespedeza bicolor Turcz.), плектрантуса (Plectranthus exisus Max.),
а местами также липы (Tilia amurensis Кот. и др.) и монгольского дуба
(Quercus mongolica Fisch.) (Рябова, 1935). Дуб предоставляет пятнистому
оленю корм не только в виде листьев и побегов, но и желудей, являющихся
в годы богатого урожая почти исключительным осенне-зимним кормом
(Вендланд, 1938). В числе излюбленных пятнистым оленем растений
Т. И. Рябова (1935) называет один из видов травянистых лиан (Smilax
oldhami Miq.). Зимой основу питания составляет древесный корм и лишь
в дополнение к нему некоторые зимне-зеленые растения. Из древесных
пород наиболее охотно в это время объедаются ветви бархата (Phelloden-
dron amurense Rupr.), маньчжурского ореха (Juglans manshurica Max.)
и аралии (Aralia manshurica Rupr.); у последней поедаются не только
побеги, но и кора, что нередко служит причиной гибели деревьев. При
недостатке других кормов поедаются побеги липы, ясеня (Fraxinus rhyn-
chophylla Hance), клена (Acer pseudosieboldianum Korn.). Охотно поедае-
мым зимним кормом являются побеги и молодая кора ильма (Ulmus
montana With.).
На взморье олени поедают водоросли и особенно охотно выброшен-
ную прибоем на берег морскую капусту. Попутно глотают морскую пену,
вероятно, удовлетворяя потребность организма в соли. Естественные со-
лонцы пятнистые олени посещают, но не столь регулярно, как другие
Cervidae. В питомниках охотно лижут предлагаемые куски соленого мор-
ского льда. И в парковых условиях, и на воле имеет место обгладывание
сброшенных рогов, вероятно в связи с недостатком в корме кальциевых
солей. При случае пятнистые олени посещают посевы культурных расте-
ний, из которых особенно охотно поедаются листва и стручки фасоли и
сои. Представление о том, что пятнистый олень разборчив в пище и при-
способлен к питанию лишь ограниченным кругом растений, опровер-
гается практикой паркового содержания на площадях с недостаточной кор-
мовой базой, а также акклиматизацией его в районах с совершенно от-
личной от дальневосточной флорой.
У пятнистого оленя хорошо развиты слух и обоняние, тогда как зрение
не является острым. Вообще этот зверь гораздо менее чуток, и под-
красться к нему легче, чем, например, к изюбрю (Пржевальский, 1870).
Голос у самок пятнистого оленя представляет громкий отрывистый
писк. Самцы в период гона ревут; рев приблизительно может быть выражен
192
CERVIDAE
протяжным звуком «и-о-у», начинающимся на высоких и обрывающимся
на низких нотах.1
Линька у пятнистого оленя происходит дважды в году. Весной олени
начинают линять в марте, изредка даже в феврале. Разгар линьки падает
на апрель, но у больных и истощенных животных она запаздывает, и
нередко в июне или, даже, июле можно видеть отдельных зверей с невы-
лезшими клочьями старой шерсти. Линька происходит быстро, старая
зймняя шерсть вылезает клочьями вместе с пухом, обнаруживая уже под-
росший под ее покровом короткий летний волос. У отдельного животного
линька начинается и заканчивается в 3—4 недели. Осенняя линька начи-
нается в августе и, как правило, заканчивается в октябре. Лишь истощен-
ные звери приобретают зимний мех на месяц-полтора позже. Самки пере-
линивают несколько позднее самцов. Летние волосы в процессе линьки,
длящейся у них также 3—4 недели, выпадают; только на шее часть их
остается и лишь меняет желтый пигмент на темный (Миролюбов, 1936).
Первые рога у самцов пятнистого оленя вырастают лишь на втором
году жизни. В возрасте около 9—10 месяцев у них под кожей лба начи-
нают образовываться вздутия лобных костей — будущие пеньки рога
(«стаканчики» по терминологии оленеводов) высотой 2.5—3 см. В течение
последующих 2—3 летних месяцев на стаканчиках вырастают хрящевид-
ной консистенции рога в виде простых спичек 1 2 длиной 20—30 см, которые
осенью окостеневают и освобождаются от покрывающей их кожи. На гра-
нице пенька и стержня рога уже намечается слабо выраженная розетка
(венчик). Эти первые рога-спички сбрасываются в двухлетнем возрасте
(в мае—июне). На 3-м году вырастают рога с тремя концами, и обычно лишь
после 4-летнего возраста развивается полное число отростков. С возра-
стом пеньки постепенно укорачиваются и одновременно утолщаются,
розетка становится более выпуклой, а сами рога увеличиваются в весе
и размерах, сохраняя прежнюю форму. С 12—14 лет ежегодно отрастаю-
щие панты постепенно начинают уменьшаться в размерах.
Взрослые, нормально упитанные самцы сбрасывают старые рога
в апреле и мае. Подобно линьке, сбрасывание рогов происходит позднее
у молодых, истощенных и больных животных (у последних иногда в июле).
На 5—7-й день после сбрасывания молодая кожица закрывает костную
поверхность пенька на месте былого соединения его с рогом, а еще через
5—6 дней здесь образуется упругое вздутие — «шишка». Последняя
растет и на 20—25-й день на ее вздутой вершине начинается раздвоение.
Передняя половина последнего превращается в надглазничный отросток,
задняя же половина, продолжая расти, образует ствол, а на конце после-
довательно вторые и третьи утолщения, также развивающиеся и дающие
начало остальным отросткам рога и его вершине (рис. 40).
Формирование хрящевого рога заканчивается в 3 недели, но рост
концов отростков продолжается несколько дольше. Еще до сформирования
рога начинается его окостенение. Последнее, начинаясь с основания, за-
канчивается к 4—5-му месяцу с начала роста рога.
Покрывающая рог кожа, лишенная питания, вследствие запустения
кровеносных сосудов, подсыхает. По-видимому, ощущая от нее раздраже-
ние, олень начинает тереться рогами о стволы и ветви деревьев, ударяет
1 По словам Абрамова (1930 : 10), рев пятнистого оленя даже отдаленно не на-
поминает рева изюбря и скорее может быть сравнен с криком ишака.
2 Лишь изредка у лучше развитых самцов уже эти первые рога на верхушках
образуют раздвоение («вилку»).
CERVUS
193
ими о кусты и т. п. В результате омертвевшая кожа очищается с поверх-
ности окостеневших рогов, а сами рога под влиянием дубильных веществ
растений, о которые они терлись, окрашиваются в буровато-коричневый
тон.1 Процесс очистки рогов заканчивается в августе или в начале сентября
перед началом полового сезона.
Рис. 40. Стадии роста и развития рогов у пятнистого оленя
(Cervus nippon, Temm.). (По Г. Менарду, 1930).
Как вторичнополовой признак, развитие рогов у самцов пятнистых
оленей связано с функцией половых желез. При полной кастрации в ран-
нем возрасте, до начала роста первых рогов, образование последних
вообще не происходит; если же рост рогов уже начался, то после кастрации
наблюдается недоразвитие их и уродливое формирование. Полная кастра-
ция половозрелых самцов в стадии роста рогов влечет в первые два-три
года нарушение нормального хода формирования, окостенения и сроков
смены. В частности, с них совершенно не счищается кожа, окостеневший
рог имеет пористую структуру, и на распиле он темный (вместо нормально
белого). Имеет место недоразвитие, уменьшение числа отростков, а спустя
2—3 года после кастрации рога перестают расти вовсе. Односторонняя
1 У оленей, содержащихся в неволе и не имеющих возможности тереться рогами
о деревья, потемнение рогов, по К; Абрамову (1930), не происходит.
13 Фауна СССР, И. И. Соколов
194
CERVIDAE
кастрация взрослых самцов в период роста рогов никакого влияния на
развитие их не оказала (Миролюбов, 1948).
Повреждение рога в период роста, если не затронуты пенек и розетка,,
не отражается на росте их в последующие годы. Если же травма захваты-
вает пенек или розетку, то в последующие годы рога будут стойко разви-
ваться в уродливой форме. Если растущий рог удалить (спилить), не по-
вреждая розетки, до начала образования «вторых шишек», то часто он вы-
растает в тот же год вторично («отава»), но полного развития, как правило,,
уже не достигает.
Полного физического развития пятнистый олень достигает к 6—7-лет-
нему возрасту, когда наблюдается не только расцвет воспроизводительной
функции, но и наиболее мощный рост рогов. Половое же созревание насту-
пает раньше.1 Уже на втором году жизни, в возрасте около 16—18 месяцев,,
и самцы и самки могут участвовать в размножении, а двухлетние самки
обычно приносят первый приплод.
Половой сезон (гон) у пятнистых оленей протекает в период с сентября
по ноябрь включительно. Самый разгар гона имеет место в октябре. В это
время самцы собирают вокруг себя гаремы из самок. Считается, что один
самец в течение полового сезона может оплодотворить 5—7, изредка ДО'
10 и даже более самок. Олени держатся в это время группами по 10—30
голов (включая молодняк). Самки внешне не проявляют признаков воз-
буждения. Самцы же, в обычное время очень мирные, в период гона сильно'
возбуждены, ревут, выкапывают в земле ямки и валяются в грязи. Между
ними, как и у других оленей, происходят турнирные бои, причем звери
могут наносить друг другу рогами тяжелые раны. Известны случаи, когда
соперники во время борьбы сцеплялись рогами и погибали (Менард, 1930).
Побежденный самец или совсем уходит из табуна в поисках нового гарема
или держится от победителя на почтительном расстоянии. К концу гона
самцы сильно худеют. Самка кроется самцом с промежутками несколько'
раз. Продолжительность и цикличность течки точно не известны, но
в случае неоплодотворения в первую течку, последняя наблюдается пов-
торно. Продолжительность беременности установлена точными наблюде-
ниями и составляет немного менее 7 % месяцев. Отел нормально начинается
с конца мая и тянется до половины июля. Но в условиях полувольного'
паркового содержания, при нарушении элементарных условий существо-
вания, у отдельных животных может иметь место позднее наступление
половой охоты, и, соответственно, поздние отелы. В этих случаях приплод
бывает обычно не жизнеспособным.
Самка пятнистого оленя, как правило, приносит по одному олененку;
двойни очень редки. Перед отелом она удаляется из табуна и рождает
детеныша в укрытом месте. Роды проходят в течение 1 %—2 часов и сравни-
тельно легко. Вес телят в условиях домашнего или полувольного содер-
жания колеблется от 4.75 до 7 кг (в среднем 5.5 кг); разница в весе между
самцами и самками почти не заметна. Родится примерно одинаковое число
самцов и самок. Олененок сразу после рождения беспомощен, но уже на
вторые сутки может довольно быстро бегать. Все же первые дни обычно
лежит спрятанным в траве, самка же далеко не уходит. Кормление моло-
ком вначале производится до 10—12 раз в день, затем постепенно реже.
Лактация тянется обычно около 8—10 месяцев, но иногда теленок сосет
почти до года. Молоко содержит от 13 до 30% (к концу лактации) жира.
С матерями оленята держатся всю зиму, а иногда и лето следующего года.
1 Раздел о размножении написан, в основном, по И. И. Миролюбову (1936, 1948).
CERVUS . 195
Первые месяцы жизни они растут быстро и к шестимесячному возрасту
достигают веса 25—35 кг. Первая линька происходит в те же сроки, что и
у взрослых самок. В зимний период рост замедляется, но усиленно олени
растут до двухлетнего возраста, достигая к этому времени около 80% веса
взрослых. Максимальный вес, по И. Миролюбову, у самцов наблюдается
в возрасте 7—10 лет, у самок в возрасте 4—6 лет. Дряхление организма,
если судить по степени развития рогов, начинается с возраста 12 лет.
Предельный возраст, до которого может доживать пятнистый олень в усло-
виях паркового полувольного содержания, 20—25 лет; известны случаи,
когда самки в питомниках давали приплод до 18—20-летнего возраста.
Показателем возраста у оленей, как и у других жвачных, служит
развитие и стирание зубов. Родятся оленята, имея все молочные резцы
и премоляры. Смена молочных резцов на постоянные происходит в возрасте
от 1 до 1 % лет, к двухлетнему возрасту они достигают полного развития,
и их режущие края начинают стираться. Смена молочных премоляров
происходит в возрасте от 1 года 8 месяцев до 2 лет почти одновременно.
Заднекоренные (моляры) прорезываются в следующие сроки (в нижней
челюсти несколько раньше, чем в верхней): первые в возрасте 2—3 месяцев,
вторые в возрасте 6—8 месяцев, третьи в возрасте от1 года 8 месяцев до 2 лет.
К трем годам постоянные зубы достигают полного развития и начинают
стираться. В дальнейшем возраст животных определяется главным обра-
зом по величине стертой поверхности на резцах и нижних клыках.1
Из врагов пятнистого оленя должен быть назван в первую очередь
серый волк {Canis lupus L.) и в меньшей мере красный волк {Суоп alpinus
Pall.). Волки приносят колоссальный вред и свободно живущим оленям
и содержащимся в парковых хозяйствах. Известны случаи, когда про-
бравшаяся в парк стая волков в один прием уничтожала несколько десят-
ков оленей. Нападают волки группами, устраивая подобие облав, когда
отдельные хищники забегают вперед наперерез гонимому остальной стаей
табуну, или же загоняя оленей на лед у ключей и речек, где те, скользя,
быстро двигаться не могут и легко становятся добычей. Летом волки под-
карауливают оленей на постоянных тропах.
Врагом пятнистого оленя является также леопард {Fells pardus L.), но
он держится только в самой южной части Приморья и вообще очень редок.
Кроме того, леопард, охотясь в одиночку при нападении ограничивается
одним оленем и массового уничтожения, как волки, не производит.
Изредка нападает рысь, которая может справиться даже со взрослым
пятнистым оленем. Молодняк, особенно только что родившиеся оленята,
может стать жертвой диких кошек, лисиц, барсука. Утаскивают оленят
также белоплечий и белохвостый орланы.
В тайге пятнистые олени подвергаются нападению клещей (до 8видов),
мух-кровососок и других кровососущих насекомых. Однако кровепарази-
тарных заболеваний, переносчиками которых являются клещи, среди пят-
нистых оленей не наблюдалось.
В носоглотке и лабиринте решетчатой кости паразитирует личинка
носоглоточного овода {Pharyngomyia picla Meig.). Подкожные овода
нормально у пятнистого оленя не паразитируют, но К. Я. Грунин 1 2нашел
у оленей из питомника на р. Билимбе мертвых недоразвившихся личинок
подкожного оленьего овода {Hypoderma diana Вт.), обычного паразита
водящегося в той же местности изюбря. У пятнистых оленей, содержимых
1 Подробности см. Миролюбов и Рященко (1948).
2 Устное сообщение.
13*
196
CERVIDAE
в домашней обстановке, зарегистрировано около 20 видов паразитических
червей (Миролюбови Рященко, 1948), но какие из них являются исконными
паразитами и какие приобретены оленем в домашнем состоянии — пока
не известно.
Хозяйственное значение. От пятнистого оленя получают вкусное мясо,
имеющую применение кожу и ряд других побочных продуктов, но основ-
ным и наиболее ценным продуктом, ради которого охотятся и разводят
этих зверей в неволе, являются панты.
Панты представляют собою растущие и еще не окостеневшие рога этого
зверя, издавна находившие применение и очень ценимые в восточной меди-
цине. Несмотря на значительно меньшие размеры, панты пятнистого оленя
всегда ценились гораздо выше находивших то же применение пантов
сибирского марала {Cervus elaphus canadensis Erxl.) и дальневосточного
изюбря (С. е. xanthopygus Milne-Edw.). Особо высоко ценились так назы-
ваемые лобовые панты, снятые с убитого самца вместе с основаниями и
прилегающим участком лобной кости. Цена пары лобовых пантов в доре-
волюционные годы доходила до 500 рублей.1 Это в сильной степени стиму-
лировало промысел и явилось одной из основных причин сильного истреб-
ления пятнистого оленя. Лучшие панты получаются в тот момент, когда
надглазничный отросток разовьется, а на конце продолжающего рост
главного ствола образуется утолщение («вторые шишки»), но образование
среднего отростка еще не началось, т. е. в возрасте около 48—55-го дня
с момента сбрасывания старых рогов и начала роста новых. Поэтому сезон
пантовки (промысла зверей для получения лобовых пантов) происходил
обычно в июле. Уже на «втором раздвое», т. е. когда начал формироваться
третий отросток, ценность пантов снижается и они относятся ко второму
сорту.
При гистологическом исследовании пантов установлено, что кожа
содержит большое число крупных сальных желез; в эпидермисе наблю-
дается значительное развитие тонофибриллей и клеток Лангерганса, а
в соединительнотканном слое — крупных фибробластов. Находящийся
под кожей промежуточный слой состоит из соединительной ткани, богатой
веретенообразными фибробластами, и содержит многочисленные кровенос-
ные сосуды. Центральная (мозговая) часть панта представляет эмбрио-
нальную соединительную ткань с развивающимися внутри ее пятнами хря-
щевой ткани и пронизанную многочисленными кровеносными лакунами и
сосудами. В этой же части, в непосредственной близости к сосудам, рас-
полагаются очаги кроветворения. Образование костной ткани в роге про-
исходит за счет окостенения хряща (Мейсель и Григорьева, 1933).
Панты пятнистого оленя от пантов изюбря отличаются, во-первых,
более высокой постановкой надглазничного отростка, затем коротким,
ровным, нежным бархатистым волосом и в свежем виде светлой желтой
окраской, иногда с красноватым оттенком. Первосортные панты должны
иметь не более двух концов — надглазничного отростка и ствола, причем
последний на вершине не должен иметь даже признаков раздвоения, а
надглазничный отросток должен быть не менее половины длины ствола.
Не допускаются в первом сорте даже признаки окостенения. Большое зна-
чение при определении сортности пантов имеет степень их симметрии.
Всякого рода дефекты от механических повреждений также снижают их
1 В книге К. Абрамова «Пятнистый олень» (1930 : 65) приведена фотография
лобовых пантов, оцененных по прейскуранту 1929 г. в 1300 рублей.
CERVUS
197
ценность. Чем массивнее и толще панты (при условии отсутствия переро-
слости), тем они более ценны.
Снятые сочные панты быстро подвергаются разложению и порче. Для
предотвращения последней производится консервировка пантов. Сущест-
вует несколько способов консервировки,1 но суть их одна и заключается
в том, что только что снятые панты подвергаются варке в кипящей или
в близкой к точке кипения воде. Варка производится путем многократного
опускания их на короткое время в горячую воду с краткими же промежут-
ками для остывания. Иногда после первой и единственной варки на другой
и в последующие дни производится сушка пантов в русской печи при тем-
пературе около плюс 100—110° С с соблюдением определенного режима
(русский способ). В других случаях производят, с промежутками от не-
скольких часов до суток, несколько (4—6) варок, но последующая сушка
производится не в печи, а на воздухе при комнатной или уличной темпера-
туре (китайский способ). Во время консервировки все время следят
и исправляют возникающие дефекты (лопанье кожи и трещины, сморщивав
ние верхушек и т. п.). В процессе консервировки, вероятно, происходит
отдача тканью пантов излишней влаги и свертывание тканевых белков.
Готовность пантов определяется твердой на ощупь консистенцией и посту-;
киванием пантов друг о друга, при котором должен издаваться звук сухого
дерева (Миролюбов и Рященко, 1948). Хорошо консервированные панты
имеют коричневую окраску и теряют в весе, вследствие отдачи влаги,
до 70%.
Химический состав консервированных (сухих) пантов пятнистого оленя
по исследованию Н. В. Хрустова и Е. Д. Иваненко (Мейсель, 1934), сле-
дующий:
Влаги ..............................................................12.54%
Общего азота ..................................................... 10 36%
Из него белкового азота........................................... 9.87%
Жира................................................................ 1.90%
Холестерина ........................................................0.173%
Золы ........................................................... 45.42%
Фосфора в виде Р2О.->.............................................. 17.95%
Кальция .............................................................4.68%
Качественные реакции на углеводы давали отрицательные результаты.
В течение ряда столетий панты марала и пятнистого оленя находили
применение в китайской и тибетской медицине в качестве лечебного сред-
ства при самых разнообразных заболеваниях. Китайская фармакопея
приписывала им общее тонизирующее действие, особенно на мужскую
половую сферу, а также стимулирование функций костного мозга и крове-
творения (Мейсель и Григорьева, 1933). Однако целебное действие пантов
научно было не обосновано, явно переоценивалось, а порою оценка при-
обретала мистический характер: панты одним своим присутствием якобы
предохраняют владельца их от всякого рода заболеваний и несчас-
тий.
Советскими учеными проведено всестороннее изучение пантов, в ре-
зультате чего получило научное обоснование применение их в качестве
лекарственного средства. Было доказано экспериментально, что водные и,
особенно, спиртовые вытяжки пантов оказывают возбуждающее действие
1 Некоторые из них описаны И. Миролюбовым и Л. Рященко (1948), К. Абрамо-
вым (1930), Г. Менардом (1930) и другими.
198 CERVIDAE
на работу сердца; особенно сильно проявлялось это действие на предва-
рительно ослабленную сердечную мышцу (Граменицкий и Сиверцов, 1934)?
Коллективом сотрудников пантовой лаборатории под руководством
йроф. С. М. Павленко был приготовлен из пантов особый высокоактивный
препарат «Пантокрин» (хуалукрин). Состав пантокрина и химическая
сущность его действующих составных частей еще не известны. Но экспери-
ментальная проверка на животных и применение его в ряде клиник при
лечении разного рода заболеваний заставляют рассматривать терапевти-
ческий эффект пантокрина как совокупное действие многих действующих
начал.
Как показали эксперименты, пантокрин активно воздействует на азо-
тистый и углеводный обмен, сердечно-сосудистую систему, органы пищева-
рения, снимает утомление с экспериментально утомленной мышцы, усили-
вает выделительную функцию почек. Под его влиянием резко повышаются
регенеративные процессы в застарелых язвах и вяло заживающих ранах.
«Необходимо отметить, — пишет С. М. Павленко (1936), — исключитель-
ный терапевтический эффект от применения впантокрина“ при функцио-
нальных неврозах, вызванных переутомлением и истощением нервной
системы, при понижении половой функции эндокринного происхождения,
при явлениях овариальной недостаточности, особенно в период климакса,
при инфицированных, вяло текущих гнойных язвах и ранах и пр. Глав-
нейшая и исключительно важная терапевтическая особенность „панто-
крина “, отмечаемая всеми клиницистами, испытавшими его в своей лечеб-
ной практике, проявляется в его изумительной способности резко повы-
шать общий тонус организма больного».
Одним из основных действующих начал пантов является, по-видимому,
мужской половой гормон. Проведенные в связи с этим сравнительные ис-
следования активности действия экстракта показали, что панты северногр
оленя (Rangifer tarandus L.) не содержат полового гормона; наиболее
активное действие оказывает экстракт из консервированных пантов пят-?
нистого оленя, тогда как активность экстракта свежих пантов марала
(Cervus elaphus sibiricus Sev.) оказалась заметно более низкой (Тэви,
1936).
Как в природе, так и икусственным путем легко получаются межви-
довые гибриды между пятнистым оленем и изюбрем (Cervus elaphus xantho-
pygus Milne-Edw.). Они известны у местного населения под названием
«чин-да-гуйза». По размерам, складу и окраске гибриды имеют промежуточ-
ный характер между исходными видами. Околохвостовое зеркало у них
рыжеватого, но более светлого, чем у изюбря, цвета и не окаймлено черными
волосами. Замечательно, что ареал и время гона у изюбрей и пятнистых
оленей совпадают, гибриды получаются вполне плодовитыми, и тем не
менее массовая гибридизация не имеет места, а видовая самостоятельность
обеих форм сохраняется очень стойко. Панты гибридов по размерам и
форме близки к пятнистому оленю, в частности имеют один надглазничный
отросток, но поставленный низко, как у изюбря. Панты гибридов расцени-
ваются низко, так как имеют темный цвет, плоскую форму, покрыты густым
длинным («собачьим») волосом, и не столь «сочны», как у пятнистого оленя
(Миролюбов, 1949). Поэтому с хозяйственной точки зрения гибридизация
пятнистого оленя и изюбря не целесообразна. Известны также плодовитые
гибриды между пятнистым и кавказским благородным оленем (Саркисов,
1944).
Охота, на пятнистого оленя в пределах Советского Союза строжайше
воспрещена.
CERVUS
199
Пантовое оленеводство. Высокая стоимость пантов и быстрое
уменьшение числа диких оленей вызвали к жизни новую отрасль животно-
водства — пантовое оленеводство.
Вполне ручным пятнистый олень становится только тогда, когда взят
от матери в первые дни жизни и воспитался в непосредственном и постоян-
ном общении с человеком. Но и в этом случае олень обычно не позволяет
совершать с ним ряда манипуляций, как например, зоотехнического или
ветеринарного осмотра, а самцы, даже самые ручные и ласковые в осталь-
ное время, в половой сезон становятся агрессивными и опасными для уха-
живающего персонала (Миролюбов и Рященко, 1948). В условиях же обыч-
ного их полувольного содержания результаты одомашнения в характере
оленя сказываются лишь в том, что в период зимней подкормки олени пере-
стают дичиться человека, подходят в присутствии его к кормушкам, а не-
которые животные даже принимают корм из рук ухаживающих за ними
людей.
Пантовое оленеводство в Приморье возникло в конце 60-х годов прош-
лого столетия на базе хищнического вылавливания в тайге с помощью
лудев диких самцов и передерживания их в загородках до момента, когда
разовьются панты, с последующим забоем. Пионерами настоящего олене-
водства были русские переселенцы, которые, наравне с самцами, стали со-
держать и выловленных самок, с целью получения и выращивания припло-
да. Почти одновременно была открыта возможность, вместо забоя самцов,
производить срезку (спиливание) пантов у живых оленей, что обеспе-
чивало ежегодное получение пантовой продукции от одних и тех же живот-
ных и намного повышало доходность оленеводства. Спиленные сантиметра
на 1.5—2 выше коронки панты на следующий, а иногда и в тот же год
вырастают вновь. В настоящее время твердо установлено, что ежегодная
срезка пантов ни в какой мере не отражается отрицательно ни на самом
животном, ни на размерах и качестве пантов, отрастающих в последующие
годы (Миролюбов и Рященко, 1948).
Наряду с этими домашними оленниками, в которых животные в течение
всего года находились на искусственном кормлении и были лишены необ-
ходимого моциона, уже с конца прошлого столетия стали организовываться
крупные помещичьи оленеводческие хозяйства паркового типа. В этих
хозяйствах пятнистые олени содержались на больших обнесенных оградой
площадях — парках, где пользовались подножным кормом в течение всего
года. Лишь в суровые зимы или при ограниченности площади и недостатке
естественных кормов производилась подкормка животных сеном. Олени
оставались почти такими же дикими, как на воле, и хозяйство базирова-
лось на отстреле самцов с целью получения лобовых пантов. Самки и в
парках, и в домашних оленниках содержались вместе с самцами; племен-
ная зоотехническая работа практически не велась; даже наоборот, часто
практиковался отстрел лучших самцов из соображений высокой стоимости
пантов. В результате имело место вырождение оленей, выражавшееся
в уменьшении живого веса, ухудшении качества и размеров пантов.
Усиленное развитие и качественно иное направление пантовое олене-
водство получило в советское время. Наша страна является единственной
в мире, где эта отрасль животноводства получила надлежащее развитие.
Сохранившееся поголовье дикого пятнистого оленя взято под строгую
охрану. Охота на него запрещена в течение круглого года. Уже в ближай-
шие годы после установления на Дальнем Востоке Советской власти было
приступлено к реорганизации домашнего оленеводства на социалисти-
ческой основе. Ведущей организационной формой были признаны совхозы,
200
CERVIDAE
в которых в настоящее время сосредоточено основное поголовье пятнистых
оленей.
Принятая в совхозах система содержания — парковая, с выловом
и содержанием в течение зимы в оленниках самцов и оленят приплода
минувшего года. Парки устраиваются на территориях, включающих как
древесную растительность, так и травянистые поляны, из расчета 1—2 га
на голову оленей (Митюшев, Любимов и Новиков, 1950). Парк обносится
оградой высотою в 2.5 м из укрепленной на столбах металлической сеткп
с ячеями И—13 см и толщиной проволоки 2.0—2.5 мм. На территории
парка устраиваются, в зависимости от его размеров, один или несколько
Олейников. Последние представляют систему сооружений, включающую
кормовой и сортировочный двор, двор для доращивания пантов, панто-
резный сарай с денниками и панторезным станком, а также несколько
выгулов. Обычно по соседству с оленником устраивается изолятор для
больных и подозрительных по заболеванию олепей. Рогачей (самцов) по,
окончании сезона спаривания переводят в оленник, заманивая подкормкой
или вылавливая с помощью специальной ловушки. Здесь самцы содер-
жатся всю зиму и следующую весну вплоть до времени срезки пантов.
После срезки пантов самцы, признанные годными в качестве производите-
лей, выпускаются в парк к самкам, а остальные содержатся отдельно.
В оленниках же всю зиму до весны выдерживается молодняк последнего
года. Самки и молодые олени старше года все время находятся на выпасах,,
лишь подкармливаясь, в случае необходимости, зимой. При такой системе
обеспечивается, с одной стороны, максимальное использование пастбищных
ресурсов парка и необходимый моцион животных, а с другой — ежегодная
съемка высококачественных пантов.
Советское пантовое оленеводство развивается на научной основе.
Созданная на Дальнем Востоке в первые годы организации совхозов
Научно-исследовательская станция по пантовому оленеводству разраба-
тывает рациональные приемы ведения хозяйства, в частности разведения
и кормления, изучает биологию и болезни пятнистых оленей, способы
повышения их продуктивности, улучшенные способы консервировки
пантов. В настоящее время разработаны инструкции по бонитировке оле-
ней и ведению племенного дела, рационы для различных групп оленей,
меры борьбы с важнейшими заболеваниями этих животных. В результате
поголовье пятнистых оленей за советский период возросло в 6 раз, а про-
дукция пантового оленеводства — в 10 раз (Миролюбов и Рященко, 1948).
Улучшаются породные качества оленей и их пантов. Так, если в 1902 г.
средний вес срезных пантов с о. Аскольд составлял 372 г, то в 1940 г. по
совхозам он уже достиг 1080 г, т. е. превысил прежнюю цифру в 3 раза;
передовая бригада совхоза «Майхэ» в 1938 г. добилась увеличения среднего
веса пантов до 1214 г (Митюшев, Любимов и Новиков, 1950 : 10—13).
Побочной продукцией пантового оленеводства является мясо, шкуры,
хвосты, пенисы, зародыши и сухожилья.
Мясо пятнистых оленей обладает хорошими вкусовыми качествами.
Сейчас оно получается лишь от случайно заболевших неизлечимыми болез-
нями и от выбракованных животных. Выбраковку и убой самцов в хозяй-
ствах производят в период пантовки с целью получения лобовых пантов.
Самки выбраковываются осенью перед постановкой на зимовку. Мяса от
взрослого самца (вес туши) получается в среднем около 60 кг, максимально
до 103 кг; от самок несколько меньше.
Кожи пятнистого оленя используются в кожевенном производстве.
Отрезанные, особым образом обработанные и высушенные хвосты и пенисы
CERVUS
201
(половые члены) с семенниками идут на экспорт. Экспортируются и, по-
видимому, также ценятся китайской медициной высушенные или заморо-
женные эмбрионы («лу-тай») в возрасте от 2 до 3—4 месяцев.
При содержании в домашней обстановке у оленей появляется ряд забо-
леваний, не свойственных или встречающихся очень редко у их диких
собратьев. Из числа их следует упомянуть такие, как некробациллез,
геморрагическая септицемия, туберкулез — инфекционные заболевания,
часто уносящие значительное число животных. Большой вред могут нано-
сить глистные инвазии, если не проводятся надлежащие профилактические
мероприятия. Массовый характер при скученном содержании самцов может
приобретать воспаление препуция (крайней плоти) не инфекционного
происхождения: от попадания загрязненной почвы в его полость; оно часто
ведет к некрозу полового члена, закупорке мочеиспускательного капала
и гибели оленя от разрыва мочевого пузыря. При нарушении кормового
режима, особенно при неправильном скармливании концентрированных
кормов, бывает вздутие рубца (тимпанит).
Акклиматизация пятнистого оленя в европейской части СССР
и в Сибири. Первый, более или менее удачный опыт акклиматизации
небольшой группы пятнистых оленей за пределами коренного ареала их
обитания был предпринят в 1926 г. в Забайкалье (Дорогостайский, 1930).
Был создан специальный питомник, но чем в конце концов закончился
этот опыт — не известно. С 30-х годов начались плановые опыты акклима-
тизации их в других районах Европейской и Азиатской частей Союза,
притом в более крупных масштабах. Акклиматизация преследовала:
1) сохранение пятнистого оленя как исчезающего вида; 2) обогащение
охотничьей фауны и 3) расширение базы пантового оленеводства. Первая
партия пятнистых оленей была завезена в 1933 г. на Алтай в Шебалинский
мараловодческий совхоз, где до того времени разводились только маралы
(Cervus elaphus sibiricus Sev.). Второй завоз пятнистых оленей туда же
был произведен в 1935 г. Олени успешно акклиматизировались в условиях
Алтая и настолько размножились, что оказалось возможным перевести
часть поголовья в два другие совхоза. В самом Шебалинском совхозе
к 1948 г. пятнистые олени по численности сравнялись с маралами и в связи
с большей ценностью их пантов стали основой хозяйственной деятельности
совхоза. В последние предвоенные годы прирост поголовья пятнистых
оленей на Алтае был выше, чем на их родине и чем у местных маралов
(Лихачев, 1951).
В 30-х же годах со стороны Центрального совета Всесоюзного военно-
охотничьего общества предпринимались попытки акклиматизации пятни-
стых оленей в некоторых охотничьих хозяйствах. Но они окончились
неудачей, так как завезенные в незначительном числе и сразу выпущенные
на волю олени быстро рассредоточивались и исчезали, погибая, вероятно,
от хищников, браконьеров, а зимой от истощения.
В 1938 г. сравнительно большая партия пятнистых оленей (около
240 голов) была завезена из пантовых совхозов Дальнего Востока и груп-
пами по 27—53 головы размещена с целью акклиматизации в семи заповед-
никах: Тебердинском (Грузинская ССР), Ильменском (Челябинская об-
ласть), Мордовском (Мордовская АССР), Окском (Рязанская область),
Куйбышевском (Куйбышевская область), Хоперском (Воронежская об-
ласть), Бузулукском (Чкаловская область).
После содержания в течение 1—2 лет в загонах олени всюду были вы-
пущены на свободу. В большинстве мест выпуска, благодаря надлежащей
охране и организации зимней подкормки, они прижились, увеличили
202
CERVIDAE
численность и в ряде случаев расселились далеко за пределы заповедников
(Козлов, 1954). Имеются, например, сведения (Двойченко, 1955), что пят-
нистые олени, выпущенные в Тебердинском заповеднике, появились в
Зеленчукском районе Ставропольского края.
Основные стации, которые пятнистые олени занимают в новых местах,—
смешанные леса с преобладанием лиственных пород. Особенно охотно они
выбирают участки лиственных лесов с богато развитым подлеском, чере-
дующиеся с травянистыми полянами. Чистых спелых сосновых насажде-
ний, например на территории Бузулукского бора, избегают. В Байкальском
оленьем питомнике охотно поедали листву березы, осины и ивы; излюб-
ленным кормовым растением оказалась сильно ядовитая для многих дру-
гих жвачных чемерица (Veratrum album L.; Дорогостайский, 1930).
Как и на родине, основу питания их в новых районах составляют
в летнее время травы, листва и побеги кустарников и деревьев, а зимой —
древесно-веточный корм. Однако видовой состав кормов стал иным.
Ряд растений, которые на Дальнем Востоке совсем не употреблялись
в пищу или поедались плохо, в условиях акклиматизации стали осно-
вой питания. Так, охотно поедается бересклет бородавчатый, рябина,
листья и молодые побеги клена. На Урале излюбленным кормом служит
черемуха, у которой поедаются не только цветы, листья и побеги, но и
сдираемая зубами кора (Аверин и Ушков, 1947). Олени могут вредить
подросту сосны, у которой не только объедают побеги, но и сгрызают
до древесины молодую сочную кору.
Сроки отела и гона у пятнистых оленей в районах акклиматизации
существенно не отличаются от таковых на их родине, но, по данным
Г. И. Ильиной (1955), чаще наблюдаются отдельные случаи запоздалого
рева и, соответственно, деторождения. Вследствие неблагоприятных
в большинстве районов условий зимовки, несколько запаздывает ве-
сенняя линька. Сбрасывание рогов происходит в среднем на месяц позднее,
чем на Дальнем Востоке. Но появление зимнего волосяного покрова,
а также отрастание и развитие новых рогов, благодаря хорошей летней
кормовой базе, происходят своевременно и не уступают дальневосточным
оленям. Что касается влияния измененных условий существования на каче-
ство пантов, то имеются лишь требующие проверки указания на то, что
экстракт из пантов пятнистого оленя Мордовского заповедника менее
активен, чем из пантов дальневосточных оленей (Арсеньев, 1949).
Главными врагами акклиматизированных пятнистых оленей оказы-
ваются волки. При этом тактика нападения последних иная, чем на Даль-
нем Востоке: преследуя оленей стаей, они стараются загнать их не на
лед (см. стр. 195), а в глубокий снег, где копытные также лишаются воз-
можности быстрого бега.
В целом опыт акклиматизации пятнистых оленей в европейской части
Союза ССР нельзя назвать удачным. Как показало обстоятельное исследо-
вание этого вопроса Г. И. Ильиной (1955), первые годы, благодаря по-
стоянному наблюдению, подкормке зимой и истреблению волков, числен-
ность оленей всюду возрастала. Однако в годы Великой Отечественной
войны она сильно сократилась. В послевоенные годы наблюдается возра-
стание численности, но темпы его совершенно недостаточны. В некоторых
заповедниках (Ильменском, Тебердинском, в ликвидированных Бузу-
лукском и Куйбышевском) оленей осталось или меньше, чем было выпу-
щено в 1938 г., или число их едва превышает первоначальную цифру.
Основными факторами, препятствующими успешной акклиматизации
пятнистого оленя, оказываются неблагоприятные условия зимовки и
CERVUS
203‘
волки. Самым трудным сезоном для акклиматизированных в европей-
ских заповедниках пятнистых оленей является зима. Пятнистые олени
оказались способными переносить низкие температуры вплоть до ми-
нус 40° С, — в сильные морозы от холода погибали только истощенные
животные и неокрепший молодняк. Гораздо же чувствительнее они к глу-
бине и продолжительности снежного покрова. При глубине снега, пре-
вышающей 30 см, пятнистые олени не в состоянии добывать травянистый
корм и вынуждены довольствоваться грубым малопитательным веточным
кормом. В некоторых районах акклиматизации в многоснежные зимы
снег лежит слоем 70—80 см, при котором становятся недоступными не
только травянистые, но и веточные корма. Кроме того, глубокий снег
сильно затрудняет передвижение. В связи с этим, почти повсюду основная
масса даже далеко разошедшихся оленей к зиме собирается и держится
в районе расположения подкормки в виде выставляемых стогов сена.
Однако значительная часть их не подходит к подкормочным пунктам
и в течение круглого года живет на подножных кормах. В этих случаях
звери выбирают в качестве зимовочных мест сильно обдуваемые участки
с неглубоким снегом. Поэтому, так же как и на востоке, наблюдается
не равномерное распределение оленей на занимаемой территории, а в виде
отдельных очагов, более или менее удаленных друг от друга.
Не пользующиеся подкормкой олени к весне сильно худеют, слабеют,
легко становятся добычей волка; среди них имеют место случаи запозда-
лой течки и отела; поздно родившиеся телята не успевают к зиме окреп-
нуть и в большинстве погибают. В результате имеет место значительное
снижение интенсивности размножения.
Не меньшим злом являются волки. Гибель от волков составляет почти
половину всех потерь пятнистых оленей в заповедниках. Волки страшны
не только непосредственным истреблением оленей, но и тем, что раз-
гоняют их с удобных мест зимовок, усиливают истощение, рассредото-
чивают и без того малочисленное поголовье.
Не случайно, по данным Г. И. Ильиной (1955), только в Хоперском
заповеднике число пятнистых оленей со времени выпуска увеличилось
в 7й раз: здесь в течение последних лет снежный покров не превышал
допустимого максимума, нет волков, а кроме того, благодаря особому
положению угодий заповедника, оленям некуда разбегаться.
В условиях европейской части СССР олени, предоставленные самим
•себе, без подкормки и охраны быстро исчезают. Ликвидация Бузулукского
и Куйбышевского заповедников повела к почти полному исчезновению
выпущенных на их территории оленей.
«Анализ результатов акклиматизации пятнистого оленя в европей-
ских заповедниках показывает, что обогащение охотничьей фауны за
счет пятнистого оленя в других районах европейской части Союза не-
рентабельно и бесперспективно» (Ильина, 1955).
Издавна пятнистые олени акклиматизированы на юге Украины в за-
поведнике Буркуты Херсонской области (филиал зоопарка Института
гибридизации и акклиматизации животных Аскании-Нова). За последние
годы здесь успешно проводятся первые в истории пантового оленеводства
опыты по выпасному содержанию оленей без изгородей. Путем специально
разработанной системы воспитания и тренировки создано стадо пятнистых
оленей свыше сотни голов, выпасаемое под присмотром пастуха, подобно
другим домашним животным, в плавнях Днепра, среди березовых кол-
ков, перемежающихся с дюнами и участками травянистой и кустарнико-
вой степи (Салганский, 1952). В настоящее время этот положительный
204
CERVIDAE
опыт внедряется в практику пантовых совхозов Дальнего Востока (Ря-
щенко, 1953, 1954).
Автору известно, что с 1952—1954 гг. проводятся опыты акклиматиза-
ции пятнистых оленей в Закавказье — в Армении и Хизинском районе
Азербайджанской ССР. Олени в обоих местах находятся в условиях
полуволыюго паркового содержания. Окончательные результаты этих
опытов еще не ясны.
Cervus elaphus Linnaeus — Благородный олень (рис. 41—50)
L i n n а е u s С., 1758, Syst. Natur.,ed. X, 1 : 67 (первооп.); Blasius J., 1857,.
Naturgeschichte d. Saugetiere Deutschlands, Braunsihweig : 539—553 (общ.); Саба-
неев Л. П., 1868, Тр. Ярославск. губ. статистич. комит., 4 : 265 (распр.); М и д-
д е н д о р ф А. Ф., 1869, Путешеетв. на север и восток Сибири, II, 5 : 1—310 (распр.);
Север цов Н. А., 1873, Изв. Общ. любит, естествозн., антроп. и этногр., VIII,
2 : 103—107 (сист., распр.); Никольский А. М., 1883, Тр. СПб. общ. естество-
исп., XIV : 178—182 (распр., биол. разв.); Черский II. Д., 1891, Опис. колл,
послетретичн. млекопит., собранных Новосибирск, эксп. 1885—1886 гг., СПб., изд.
Акад, наук : 309—355 (морфол., сист.); Кузнецов А., 1898, Зап. Читинск, отд.
Приамурск. отд. Русск. географ, общ., III : 80—173 (общ., ох., развод.); Брау-
нер А. А., 1900, Зап. Новороссийск, общ. естествоисп., XII, 2 : 1—21 (сист.); Тур-
кин Н. В. и К. А. С а т у н и н, 1902, Звери России, Cervidae, М. : 2—34-197—
362 (общ., разв. изобр.); St га hl Н., 1906, Anat. Hefte, 31 : 199—218 (анат.);
Greve К., 1909, Siiugetiere Kur-I.iv-Estlands, Riga : 151—152 (распр.); Дин-
ни к Н. Я., 1910, Звери Кавказа, ч. I, Тифлис : 93—137 (общ.); Врублевский К.,
1912, Архив ветерин. наук, 8 : 746—778 (пит., хоз. знач.); W i m a n s W., 1913,.
Deer breeding, London : 1—105 (разв., ox.); A n t о n i u s H., 1920, Proc. Koninkl.
Akad. Wetenschap. Amsterdam, XXIII : 37—43 (сист., морф.); Соловьев Д. К.,.
1920, Саянский промысл.-охотн. р-н, Птг. гос. изд. : 222—238 (распр., биол., ох.,
изобр.); Толстой Н. Н., 1922. Охота на Кавказе, М. :5—114 (ох.); Kiesse-
1i п g W., 1925, Der Rothirsch., Neumann-Neudamm (цит. no Linke, 1954); Форту-
натов Б. К., 1925, сборн. «Аскания-Нова», М., Госизд. : 324—329 (гибр.); Д о п-
п е л ь м а и р Г. Г., 1926, Соболиный промысел на северо-восточном побережье
Байкала, Верхнеудинск—Ленинград, изд. Госплана Бур.-Монг. АССР : 177—188-
(биол.); Завадовский М. М., 1926. Тр. Либор, эксперим. биол. Моск, зоопарка,
I : 33—39 (кастр.); Тассовский Г. Н., 1927, Произв. силы Дальн. Востока, IV :
528—531 (распр., биол.); Доппельмаир Г. Г., 1927, сборн. «Якутия», Л.,
изд. Акад, наук СССР : 15 (распр.); Емельянов А. А., 1927, Произв. силы Дальн.
Вост. IV : 263 (распр.); Смирнов, 1928, Охотник и путник Сибири, 6:15 (бол.),
Шиллингер Ф. Ф., 1930, Европ. олень, лань и косуля в охотхоз., Харьков,
изд. ЦС ВУСОР : 10—44 (общ., разв., изобр.); Соколов И. II., 1932а, Союзпуш-
нина, 15/16:43—44 (определ. возр.); Соколов И. И., 19326, Союзпушнина,
19/20 : 43—48 (разв.); Соколов И. И., 1932в, Союзпушнина, 17/18 : 34—37 (разв.);
Eidmann Н-, 1932, Mitteil. aus Forstwirtsch. u. Forstwiss., 2 (зубы) (цит. па
Linke, 1954); Бобринский H. A., 1933, Зоол. журн., XII, 4 : 77—85 (распр.);
Шухов И. Н., 1933, Охотничьи промыслы Приенисейск. края, Красноярск : 8—
9 (распр.); Флеров К. К., 1935, сборн. «Матер, по паразитол. и фауне южн. Тад-
жикист.», X : 315—343 (общ., изобр.); Heck L., 1935, Der Deutsche Edelhirseh,
Parey—Berlin : 1—187 (общ., изобр.); Боев С. Н., 1936, Тр. Казахст. научно-иссл.
ветер, инет., I (бол.); Котовщикова М. А., 1936, Научн. тр. гос. заповеди.,
сер. II, Крымск, гос. заповеди., I : 105—130 (биол.); Лаптев М. К., 1936, Определ.
позвон. животы. Туркменск. ССР, вып. V, Млекопит., Баку, Госиздат : 90 (распр.);
Подаревский В. Б., 1936, Пробл. охотхозяйств, акклиматиз. Вост. Сибири,
Иркутск, Огиз : 40—42 (распр., стан.); Т э в и А. С., 1936, сборн. «Пантокрин»,
2 : 49—60 (панты); В е n i n d е J., 1937, Zentralbl. f. Kleintierzucht und Pelztierzucht,
XIII, 10 : 1—223 (общ., изобр.); Новиков Г. А., 1937, Тр. Совета по изуч. произв.
сил, сер. вост.-сиб., 4 : 237—242 (биол., ох.); Дмитриев В. В., 1938, Тр. Алтайск.
гос. заповеди., I : 171—226 (распр., биол.); Колосов А. М., 1939, Уч. зап.Моск,
гос. унив., 20 : 169—170 (распр.); Насимович А. А., 1939, Вопр. эколог, и био-
ценол., 7 : 44—53 (биол.); Рухлядев Д. П., 1939, Науч.-метод, зап. Комит.
по заповеди., III : 133—140 (бол.); Eidmann Н-, 1939, L'ntersuch. am Gebi^
des Rothirsches, Hannover (определ. возр.) (цит. no Linke, 1954); Житков Б. М.,
1940, Тр. Моск. Зоопарка, I : 50—57 (сист.); Насимович А. А., 1940, Науч.-
метод. зап. Глав. упр. по заповеди., VI : 149—150 (пит,); Скалой В. Н., 1941,
CERVUS
205
Прир. и соц. хоз., сб. VIII, ч. II : 278 (распр.); Ц а л к и и В. И., 1944, Зоол. журн.,
XXIII : 369—376 (размн.); Бажанов В. С., 1945, Изв. Акад, иаук Казахск. ССР,
5 : 75 (распр.); Ц а лк ин В. И., 1945, Зоол. журн., XXIV, 4 : 224—234 (рога);
Формозов А. Н., 1946, Снежный покров в жизни млекопитающих и птиц, М.,
изд. Моск. общ. испыт. прир. : 111—114 (распр.); Ц а лк ин В. И., 1946, Зоол.
журн., XXV, 2 : 157—166 (линька); В ер е щ а г и н Н. К., 1947, Охотн. и промысл,
животн. Кавказа, изд. Акад, наук Азербайдж. ССР : 54—55 (распр., изобр.); Г е п т-
нер В. Г. иЦалкин В. И., 1947, Олени СССР, М., изд. Моск. общ. испыт. прир :
5—119 (морф., сист., распр., изобр.); Кузнецов Н., 1947, Звери и птицы Ярославок,
обл., Ярославль, Облизд : 5 (распр.); Vogt Т., 1948, Das Rotwild, Vienna:
I—Vlll-f-l—207 (пит., рога); Капланов Л. Г., 1948, Тигр, изюбрь, лось, М.,
изд. Моск. общ. испыт. прир. : 50—78 (общ., изобр.); Кузнецов Б. А., 1948,
Звери Киргизии, М., изд. Моск. общ. испыт. прир. : 107—109 (распр.); Клюш-
к и н Е. А., 1949, Охрана природы, 7 : 71—78 (распр., биол.); Калниньш А. И.,
1950, Ох. и охотн. хоз. Латвийск. ССР, Рига, Латгосизд. : 111—122 (общ.); Копы-
лов И. П., 1950, Дикие копцтн. Иркутск, обл., Иркутск, Облизд. : 18—28 (распр.,
биол., изобр.); Куражсковский Ю. Н., 1950, Охрана природы, 10 : 115—
130 (акклим.); М и т ю ш е в П. В., М. П. Любимов и В. К. Новиков, 1950,
Пантовое оленевод, и болезни пантовых оленей, М., изд. «Междунар. книга» : 1—240;
Осадчий П. иВ. Баврин, 1950, Беловежская пуща, Минск, Госизд. : 41—
43 (общ.); Черкасов А. А., 1950, Записки охотн. вост. Сибири, Иркутск, Облизд. :
176—200 (биол., ох.); Мертц П. А., 1951, Бюлл. Моск. общ. испыт. прир., Отд.
биол., LVI, 5 : 31—44 (стац., изобр.); Murie О., 1951, The Elk of North America,
Harrisburg : 1—176 (общ.); Swanson C., 1951, Murrelet, 32 : 19—22 (опр. возр.);
Скалой В. H., 1951, Бюлл. Моск. общ. испыт. прир., Отд. биол., LVI, 1 : 28—29
(распр.); Ц а лк ин В. И., 1951, Докл. Акад, наук СССР, LXXVII, 3:521—523
(ист., распр.); Е 1 1 е г m a n J. and Т. Morrison-Scott, 1951, Checklist of
Palaearctic and Indian mammals, London : 367—370 (сист., синон.); Жарков И. В.,
1952, сборн. «Методы учета численности и география, распределения наземп. позво-
ночных», М., изд. Акад, наук СССР : 231; Со кур I. Т., 1952, 3Bipi Радянських
Карпат, Киев, изд. Акад, наук УССР : 56—58 (распр., биол.); Сысоев В. П.,
1952, Охота в Хабаровск, крае, Дальгиз : 98—105 (распр., биол., ох.); Флеров К. К.,
1952, Кабарги и олени, Л., изд. Акад, наук СССР (Фауна СССР., Млекопит., I, 1) :
147—181 (общ., изобр.); Якушев ич А. И., 1952, Фауна позвон. Тувинск. обл.
Новосибирск, изд. Западно-Сибирск. фил. Акад, наук СССР : 61—62 (распр.); Яну-
ше в ич А. И. и И. Благовещенский, 1952, Промысл, звери и птицы за-
пади. Сибири, Новосибирск, Облизд : 128—130 (распр.); Афанасьев А. В.
В. С. Б а ж а н о в и др., 1953, Звери Казахстана, Алма-Ата, изд. Акад, наук
Казахск. ССР : 472—477 (распр., изобр.); Зайцев С. В., 1953, Каракулеводство
и звероводство, I : 61—63 (панты); Мертц П. А., 1953, Тр. Воронежск. заповеди.,
4:123—132 (пит.); Цагарев П., 1953, Газ. «Сов. Киргизия», 27/11 (распр.);
Шапошников Ф. Д., 1953, сборн. «Преобраз, фауны позв. животных», М.,
изд. Моск. общ. испыт. прир. : 97—100 (пит.); Шарлемань Н. В., 1953, При-
рода, 4 : 103—104 (распр.); Банников А. Г., 1954, Млекопит. Монгольск. Пар.
Республ., М.—Л., изд. Акад, наук СССР : 208—216 (общ.); Казновский П. Ф.,
1954, Природа, 2:110—112 (акклим.); Linke W., 1954, Der Rothirsch, Witten-
berg Luterstadt : 1—94 (общ., изобр.); Jaczewski Z., 1954, Folia biologica, Wars-
zawa, II, 2:140—141 (рога); Насимович A. A., 1955, Роль режима снежн.
покрова в жизни копытных, М., изд. Акад, наук СССР : 199—215 (распр., биол.);
Саблина Т. В., 1955, Копытные Беловежской пущи, М., изд. Акад, наук СССР :
20—89 (биол.); Я н у ш к о П. А., 1957а, Тр. Крымск, гос. заповеди., IV : 139—155
(размн.). Я н у ш к о П. А., 19576, Тр. Крымск, гос. заповеди., IV : 107—138 (биол.,
хоз. знач.).
Описание. Олень крупных, реже средних размеров. Самые мелкие
островные расы (с о-вов Корсики и Сардинии) имеют высоту в холке
около 87 см и вес около 80 кг. Сибирские благородные олени (маралы)
ростом до 150—155 см и живым весом до 320 кг. Еще крупнее американ-
ские вапити, отдельные экземпляры которых достигают высоты свыше
160 см и до 500 кг живого веса (Murie, 1951). Длина тела от конца морды
до конца хвоста у самцов европейского благородного оленя колеблется
в пределах 185—215 см. Самки мельче самцов; их живой вес на 20—25%
меньше.
206
CERVIDAB
Высота в крестце приблизительно равна высоте в холке. Профиль-
спины прямой или слегка вогнутый, с несколько выступающими холкой
и крестцом. Туловище покоится на высоких тонких ногах. Шея, особенно
у самок, длинная, тонкая; у самцов она сильно утолщается в период гопа.
Голова производит впечатление легкой, узкая, с вытянутой мордой и
прямым профилем носа. Уши длинные; отогнутые вперед, они достигают
заострёнными верхушками предглазничных ямок или даже заходят вперед
от последних. Носовое зеркало большое, занимает почти все пространство
между ноздрями от их верхних углов до края верхней губы. Предглазничные
железы большие и глубокие. Радужная оболочка коричневого цвета,
Зрачек в форме горизонтального овала.
Рога с числом концов на каждом роге у взрослых самцов не менее пяти,
обычно больше. Надглазничных отростков типично два, расположены
поблизости друг к другу. Нижний надглазничный отросток отходит от
ствола рога почти сразу над розеткой. Средний отросток один. Верхушка
вполне развитого рога несет не меньше двух, часто большее число отро-
стков. В одних случаях верхушечные отростки расходятся из одной точки
в разные стороны кустом, образуя «чашу», или «венец» на вершине рога.
В других случаях они дихотомически ветвятся и при этом расположены
приблизительно в одной плоскости. Максимально ветвистая пара из из-
вестных рогов имела 66 концов (Шиллингер, 1930). Обычно же число по-
следних на одном роге не превышает 8—9. Главные стволы имеют ясно
выраженный лирообразный изгиб и вершинами направлены несколько
навстречу друг другу. Сечение ствола и отростков округлое. Поверхность
ствола и оснований отростков темпо-коричневая, шероховатая, с про-
дольными бороздками. Концы отростков гладкие, белого или желто-
ватого цвета. Форма изгиба стволов, число и расположение отростков,
наравне с индивидуальной, обнаруживают ясно выраженную закономер-
ную географическую изменчивость и поэтому служат одним из основных
систематических признаков при определении подвидов. Как и у большинства
других оленей, самки рогов не имеют.
Хвост длиною от 5 до 16 см, как правило, короче уха. На конце
хвоста под кожей (и у <3 и у есть железа без выводных протоков
неизвестного назначения. Копыта овальной формы, с закругленным
наружным краем. Боковые копытца небольшие, не больше половины
длины главных, при ходьбе по твердому грунту не касаются почвы.
Волосяной покров в зимний период состоит из грубой ломкой ости и
тонкого извитого подшерстка. Длина зимней ости на спине составляет
около 4 см. Морда впереди глаз и конечности ниже запястных и скакатель-
ного суставов покрыты толстым, но коротким и упругим волосом, плотна
прилегающим к коже. Волосы на местах плюсневых желез выступают вад
остальной поверхностью кожи и в основаниях склеены темным густым
секретом. Хвост покрыт волосом одинакового с туловищем типа и на
конце не имеет кисти. На нижней стороне шеи у самцов, а часто и у са-
мок, волосы длиною до 8—10 см образуют гриву, не одинаково разви-
тую у разных рас оленей.
На огромной территории ареала благородных оленей окраска волося-
ного покрова значительно варьирует от песчано-серой до интенсивно-
коричневой с красноватым оттенком или рыжей. Основу ее все же состав-
ляют бурый и коричневый тона, к которым присоединяются оттенки того
или иного цвета. При этом в изменениях окраски наблюдается хорошо вы-
раженная географическая закономерность. Наиболее темно окрашенные
олени населяют западную и северную часть ареала. Посветление окраски
CERVUS
207
идет в направлении с запада на восток и с севера на юг. Наиболее ярко
окрашенные формы населяют крайний восток и крайний запад. Серые
и тускло окрашенные занимают среднюю часть ареала. Наиболее светло
окрашены олени Средней и Центральной Азии. Голова окрашена,
обычно, несколько светлее туловища. Подбородок, а иногда также во-
лосы вдоль краев губ и кольцо вокруг глаз имеют беловатую окраску.
Рис. 41. Европейский благородный олень (Cervus elaphus elaphus L.).
По бокам нижей губы вблизи углов рта с обеих сторон имеется по темному
пятну. Нижняя сторона живота и шеи, а также конечности у одних рас
окрашены светлее, а у других, наоборот, темнее, чем спина и бока. Более
темную окраску имеют также бедра и плечи. По передней стороне ног
от запястных и скакательных суставов и почти до копыт идет неясная
темная полоса, тогда как боковые и задняя сторона заметно светлее.
Вдоль средней линии шеи и спины тянется узкая полоса черно-бурого
цвета, которая, впрочем, часто не заходит назад дальше передней части
спины и холки. Пятнистость в окраске у взрослых, как правило, отсут-
ствует. Лишь иногда у самок, и только в летней шерсти, она заметна
в виде неясных размытых пятен рыжеватого цвета по бокам хребта или
208
CERVIDAE
в задней части туловища и на бедрах. На лопатки, плечи и шею пятни-
стость никогда не заходит. Околохвостовое зеркало больших размеров,
захватывает область на крупе выше хвоста. Цвет его от белого и кремового
до ржаво-рыжего. Иногда оно окаймлено сверху и с боков узкой полосой
волос темного цвета, иногда незаметно сливается с окраской окружающих
частей тела. По средней линии околохвостовое зеркало часто разделено
рыжевато-бурой полосой, переходящей на хвост. Светлая его окраска
распространяется на промежность и внутренние стороны бедер, посте-
пенно сливаясь с окраской смежных частей тела. Хвост или одноцветный
с околохвостовым зеркалом, или сверху более темный, рыжевато-бурый.
Отличия в зимней и летней окраске выражены в разной степени. В летнем
меху преобладает рыжеватый тон, а в зимнем сероватый или бурый.
Самки более одноцветны по сравнению с самцами и не имеют ясно вы-
раженных контрастов между светло- и темно-коричневыми местами. На-
равне с формой рогов и особенностями окраски, форма и размеры около-
хвостового зеркала служат основанием для выделения географических
рас или подвидов (см. ниже, стр. 216). Молодые первые месяцы после
рождения имеют рыже-бурую, резко пятнистую окраску. Кожа на но-
совом зеркале, на голых краях губ, а также копыта черно-бурого
цвета.
Череп (рис. 42) имеет длину от 325 мм у мелких рас до 490 мм у самцов
из американской части ареала (Флеров, 1952). Ширина лба в самом узком
месте между рогами и глазницами у взрослых самцов меньше ширины
черепа в надушных буграх. Задние концы носовых костей расположены
впереди уровня передних краев глазниц. Слуховые пузыри небольшие,
угловатые, почти никогда не выступают ниже уровня basioccipitale. Подъ-
язычные отверстия расположены позади уровня передних краев суставных
поверхностей затылочных мыщелков. Внутренняя доля переднего пре-
моляра верхней челюсти (Р2) иногда сплошная, иногда разделенная вер-
тикальной бороздой на две половины, как у Cervus dama L. Верхние клыки
имеются как у самцов, так и у самок, но к старости часто выпадают.
История и географическое распространение (рис. 43). Корни со-
временных благородных оленей ведут к общему предку с пятнистым оле-
нем типа Cervas perrieri Croiz. et Job. или C. etuerianus Croiz. et Job. и
близких к ним форм из верхнего плиоцена западной Европы. На грани
верхнего плиоцена и нижнего плейстоцена (гюнц—миндель) в Европе жил
первый представитель благородных оленей, С. elaphus antiquus Pohl.,
имевший рога с простой вилкой на вершине, но уже со вторым надглаз-
ничным отростком. В нижнем плейстоцене (миндель) впервые-появляются
и олени, имевшие не только два надглазничных отростка, но и развет-
вленную в виде кроны вершину. Современные расы благородного оленя
известны в Европе, Азии и Америке с верхнего плейстоцена (Флеров, 1952).
В настоящее время область распространения благородных оленей
охватывает северную Африку, западную и юго-восточную Европу (вклю-
чая Британские острова, Корсику и Сардинию), южную часть Скандинав-
ского полуострова, Прибалтийские республики, западную часть Бело-
русской ССР, Крым, Кавказ, Малую, Среднюю и Центральную Азию,
южную и восточную Сибирь с Дальним Востоком, а также умеренную
зону Северной Америки. В западной части Скандинавского полуострова
северная граница распространения лежит выше 60-й параллели. Южная
граница доходит приблизительно до 30° с. ш. Некогда, по-видимому,
сплошной ареал С. elaphus L. в настоящее время разорван и состоит из
отдельных изолированных частей.
CERVUS
209
В СССР остатки благородного оленя встречаются, начиная с низов
четвертичного периода, при этом в ряде местностей, где он в исторические
времена уже не водился. Остатки, не вызывающие сомнения в принадлеж-
Рис. 42. Череп Cervus elaphus L.
ности к этому виду, известны даже с низовьев р. Яны и о. Ляхова (Чер-
ский, 1891). По А. Н. Формозову (1946), естественная северная граница
ареала С. elaphus, если принимать во внимание и области, где он истреб-
лен человеком, совпадает с линией среднего годового максимума снего-
14 Фауна СССР И. И. Соколов
210
CERVIDAE
вогб покрова в 40—50 см; области с глубиной снега не свыше 20—30 см
совпадают с зоной максимальной численности благородных оленей.
Распространение благородного оленя в пределах СССР и изменения
ареала в историческое время представляются в следующем виде. На тер-
ритории прибалтийских республик, в частности в Латвии и Литве, он был
истреблен уже к началу XIX в. Только перед первой мировой войной,
вновь завезенные в охотничьи хозяйства из Беловежской пущи, олени
размножились не только в парках, но и на воле.
В Литве по учету в феврале 1954 года насчитывалось 254 оленя,
большая часть которых обитала в северной части республики, в Кур-
шенском и Шяуляйском лесхозах. Около 40 оленей учтено в запад-
ных лесхозах — Юрбарском и Таурагском. В Латвии оленей еще больше.
На 1 марта 1954 г. по республике числилось 836 оленей, распространенных
в большинстве лесхозов. Наибольшее число оленей, согласно учету,
обитает в Вентспилсском, Угальском, Добельском, Алсунгском, Кул-
дигском и Бауском лесхозах.1
На территории самой Беловежской пущи исконная популяция оленей
также была истреблена. Вновь олени начали завозиться в пущу из Гер-
мании и Польши для реакклиматизации начиная с 1865 г. (Врублевский,
1912). В настоящее время число их в советской части Беловежа превы-
шает 400 голов. На остальной части Белорусской ССР благородный олень
отсутствует. Любопытно, что аборигенный олень западных областей СССР,
был, по-видимому, значительно крупнее западно-европейской расы (Цал-
кин, 1951).
Некогда благородный олень был широко распространен на Украине.
Об этом свидетельствуют не только данные археологических раскопок,
но и воспоминания очевидцев (Шарлемань, 1953). Последние достоверные
сведения об обитании их в левобережной Украине относятся к концу
XVIII в. (Формозов, 1946).
В настоящее время в диком состоянии олень сохранился на территории
Украинской ССР в горных лесах Карпат, в областях Закарпатской,Чер-
новицкой, Станиславской и Дрогобычской. Наиболее многочисленны
олени в Раховском округе Закарпатской области (Сокур, 1952). Единич-
ные экземпляры изредка появляются также в Львовской и даже в Волын-
ской и Ровенской областях. С конца прошлого столетия благородные
олени акклиматизированы в Аскании-Нова. Пять оленей, завезенных
в 1925 г. на о. Бирючий в Азовском море, размножились к 1951 г. до 100
экземпляров. Одичавшие потомки бежавших из Аскания-Нова оленей
держатся в плавнях нижнего течения Днепра. До Великой Отечественной
войны стадо около 400 голов, произошедшее от завезенных из Аскания-
Нова 26 оленей, жило в Печенежском охотничьем хозяйстве Харьков-
ской области. За годы Великой Отечественной войны это стадо было
уничтожено. В 1952 г. в Печенежское охотхозяйство вновь было завезено
по семь голов оленей из Крымского и Воронежского заповедников. Ре-
зультаты их акклиматизации пока не известны. Завезенные из Аскании-
Йова олени живут в настоящее время в бывшем охотничьем хозяйстве
Гавриловское в районе Большая Лепетиха Херсонской области.1 2 Аска-
нийское стадо и связанные с ним популяции оленей имеют метисный ха-
1 Сведения получены от Управлений охотничьего хозяйства Литовской и Латвий-
ской ССР, начальникам которых — товарищам В. Бергасу и Н. Вульфу и звероводу
Охотуправления Латвийской ССР тов. К. П. Гринбергу автор пользуется случаем
выразить благодарность.
2 Сведения получены от Управления охотничьего хозяйства Украинской ССР.
Рис. 43. Распространение Cervus elaphus L-
XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше; 4 — отдельные заходы в настоящее
время; о — места акклиматизации.
14*
212
CERVIDAE
рактер, так как произошли от скрещивания (в значительной степени бес-
системного) европейского благородного оленя с кавказским оленем и си-
бирским. маралом и поэтому в настоящее время известны больше под име-
нем асканийского марала.
Имеются сведения (Туркин и Сатунин, 1902) о том, что еще в прошлом
столетии благородные олени водились в некоторых местах Московской,
Ярославской, Нижегородской и даже Вологодской губерний, но насколько
эти сведения достоверны — судить трудно. Л. П. Сабанеев в 1868 г. отно-
сительно Ярославской губернии писал, что благородные олени составляли
«исконных обитателей лесов по р. Пушме в Мологском уезде. В других
уездах они замечены были.-только в Романовском около с. Шолыпи
(осенью 1866 г.)».
В XI—XV вв. олени водились в нынешней Курской, а в XVIII — в Во-
ронежской (Кириков, 1952а) областях. В Воронежскую губернию (имение
Рамонь) небольшое число оленей было завезено в 70—80-х годах прош-
лого века из Германии. Разбежавшиеся после Революции и Гражданской
войны остатки этого стада были взяты под охрану на территории Воронеж-
ского государственного заповедника (недалеко от Воронежа) в Усманском
лесу. К 1951 г. число их превышало 500 голов; часть поголовья рассели-
лась по смежным с заповедником лесным массивам (Мертц, 1951). В пред-
военные годы сибирские маралы были завезены с целью акклиматизации
в Мордовский и Башкирский заповедники. Завезенная в последний в 1941 г. 1
полсотня голов к 1953 г. размножилась в десять раз. В настоящее время
основная часть стада концентрируется в западной части заповедника,
в бассейнах рр. Кагана и Южного Узяна. На севере эти маралы посте-
пенно заселяют Средний Кракк, а на юге доходят до р. Бетери (Казнев-
ский, 1954).
Изолированный очаг обитания благородного оленя расположен в гор-
ной части Крыма. Многочисленный в прошлом столетии и встречавшийся
даже в лесах под Севастополем и Бахчисараем, олень после Революции
сохранился лишь на территории нынешнего Крымского государственного
заповедника. Благодаря принятым морам охраны, число оленей неуклонно
возрастает. Число их с 729 голов в 1945 г. увеличилось к 1957 г. до 2387 .
(данные Крымского заповедника). Есть сведения, что олени в Крыму раз-
множились настолько, что начали расселяться за пределы заповедника
(Котовщикова, 1936; Жарков, 1952).
Область распространения С. elaphus на Кавказе является продолже-
нием малоазиатской части ареала. Распространен олень здесь не повсе-
местно и неравномерно. Северная граница здесь проходит по линии южнее
Краснодара, Майкопа, Черкесска, Нальчика и идет по среднему течению
Терека до Кизляра. По Н. А. Диннику (1910), изолированные участки
обитания его в начале этого столетия были на правом берегу Кубани
у Краснодара (в Красном Лесу — сюда олени вновь завезены недавно)
и ь лесах около Горячего Ключа. В свое время они водились даже в Став-
ропольских островных лесах, откуда исчезли в начале XIX в. В Закав-
казье олень еще в начале этого столетия был распространен, хотя и не
повсеместно, от Батуми на западе до Талышинских гор на востоке. На
Талыше их сейчас уже нет. Истреблены они в этом столетии и в районе
Малого Кавказа к северо-востоку от оз. Севана. Более всего в настоящее
время оленей на территории Кавказского заповедника (в западной части
Главного Кавказского хребта). Много их также в заповедниках Боржом-
скомиЗакатальском и в низовьях р. Сулака (Дагестанская АССР; Вере-
щагин, 1947).
CERVUS
213
Есть косвенные доказательства (Формозов, 1946), что ареал оленя не-
прерывно продолжался в Заволжье и на Урале. В качестве мест обитания
в XVIII в. указываются окрестности Уфы, а в XIX в. — район Сверд-
ловска и г. Иремель на Южном Урале. Кости и рога оленей находились
неоднократно в пляжных отложениях среднего течения р. Урала.
В пределах Средней Азии обитают две расы благородных оленей: бу-
харский олень (хангул) и марал. Для юго-западной части Туркмении
Рис. 44. Марал (Cervus elaphus sibiricus Sev.).
(Копет-Даг к югу от Чандыра) Лаптевым (1936) указывается кавказская
раса; но сохранился ли здесь этот олень в настоящее время — не из-
вестно.
Область распространения бухарского оленя (хангула, С. е. bactria-
nus Lyd.) в СССР в прежнее время захватывала большую часть долин
Аму-Дарьи и Сыр-Дарьи с их притоками. Еще в начале XX в. за ними
охотились на Сыр-Дарье выше Кзыл-Орды (Бажанов, 1945). Не так давно
он водился в долине Теджена. В настоящее время на Аму-Дарье хангул
держится лишь в дельте (Бобринский, 1933) и в тугаяхДарган-Атинского
района (Туркменская ССР), но число их здесь не велико, по данным Плюш-
кина (1949) всего около 60 голов. Есть сведения, что в небольшом числе
хангул еще недавно держался в Кызыл-кумах между нижним течением
Аму-Дарьи и Сыр-Дарьи, (Бобринский, 1933), но сохранился ли он там
сейчас — не известно. Единственное место в наших пределах, где бухар-
ский олень и сейчас держится в более или менее значительном числе, —
это долина Пянджа с его притоками в Таджикской ССР. По данным Фле-
%14
CERVIDAE
рова (1935), подтвержденным опросами на месте в 1951 г., в Таджикистане
олень многочислен в тугаях окрестностей Пархара. По Пянджу держится
от Кафирнигана на западе до Чубека на востоке. По Кафирнигану доходит
вверх до Кабадиана, по Вахшу от устья до урочища Устатаймас. По
Кызылсу встречается почти до Куляба. Об изменении ареала в этой части
последние годы сведений в литературу не поступало.
Системы гор Тянь-Шаня, Джунгарского Алатау, Тарбагатая, Алтая
и Саян представляют область распространения другого подвида благо-
родного оленя — марала (С. elaphus sibiricus Sev.).
В Таджикистане марал встречается лишь в северной части, в хребтах
Алайском и Зеравшанском. Последний является, по-видимому, западной
его границей.
Еще не так давно обычный по всей Киргизии, марал в настоящее
время сохранился здесь главным образом в северных районах. Встре-
чаются маралы до сих пор в ельниках Кавактау, Сарыджаз, в урочищах
Джаны-Джер и Улан. Особенно многочисленны в двух последних. За-
ходят иногда в пределы Талды-Суйского и Тюпского районов (Цагарев,
1953). По Кузнецову (1948) есть они до сих пор в хребтах Алайском,
Кунгей и Терскей Алатау, а также в долине р. Нарын.
В Казахстане современная область распространения марала охваты-
вает Заилийский и Джунгарский Алатау, Кегенский хребет, Кетмень
(Нарынкольский р-н) и Саур, где марал, по-видимому, еще сравнительно
многочислен. В Тарбагатае редок. Почти истребленный в Восточно-Ка-
захстанской области (на Алтае), в результате мер охраны увеличил чис-
ленность, встречаясь, например, в верхнем течении Бухтармы. Еще
в прошлом столетии маралы водились по лесам Каркаралинских гор и
Чингиз-тау, около Баян-Аула (Северцов, 1873), а также по ленточным
борам северного Казахстана, о чем свидетельствуют находки сброшенных
ими рогов (Афанасьев и др., 1953). Г. Карелин (по Формозову, 1946)
находил маралов в Актюбинской области на Мугоджарах.
Естественное продолжение современного ареала марала в Казахстане
представляет сибирский Алтай. Однако и здесь в ряде районов за послед-
нее столетие он или исчез, или сильно уменьшил свою численность. Так,
практически он отсутствует к западу от р. Катуни, если не считать неболь-
шого участка на Терехтинском хребте. Зато к востоку от Катуни, начи-
ная с ее верховьев, марал распространен почти повсеместно за исключе-
нием верхнего безлесного пояса гор и участков степей.1 На севере грани-
цей распространения является линия между с. Чемал и северной оконеч-
ностью Телецкого озера (Дмитриев, 1938). На востоке область распро-
странения переходит в Туву и Саяны.
В остальной части западной Сибири между Уралом и Обью в настоя-
щее время марал отсутствует, но есть основания предполагать, что по лен-
точным борам Казахстана и южной Сибири область его современного рас-
пространения на Алтае в свое время смыкалась с европейской частью
ареала. Так, имеются достоверные сведения, что в XVIII в. марал встре-
чался у оз. Кончи и в Куртамышском районе Курганской области (Фор-
мозов, 1946), а в начале XIX в., по-видимому, заходил даже в Барабин-
скую степь (Миддендорф, 1869).
Сохранился до сих пор марал в Кемеровской области, в Горной Шории
(Янушевич и Благовещенский, 1952), но подробности его распространения
здесь не ясны.
1 Хотя в конце прошлого века встречался даже в центре Чуйской степи (Колосов,
1939).
CERVUS
215
В Хакасской автономной области, по собранным мною в 1942—1945 гг.
сведениям, в очень небольшом числе марал сохранился главным образом
в южных п западных горно-лесных районах. До широты Ачинска он,
по-видимому, в настоящее время уже не доходит. В Тувинской области
марал является до сих пор обычным зверем во всех горных лесах (Януше-
вич, 1952).
Область сплошного распространения марала представляют Западные
и Восточные Саяны. Здесь он весьма обычен по течению рр. Уса, Кызыр-
Сука, Кебежа, Казыра и Кизира (Соловьев, 1920).
По Восточным Саянам доходит до Красноярска, встречаясь по Мане,
Базаихе и Кану. Переходит южнее Красноярска даже на левый берег
Енисея до Чулыма. Встречается на южных отрогах Енисейского кряжа
по р. Кану.1
Многочисленны до сих пор маралы в узле Западных и Восточных
Саян, а также в бассейнах рр. Бирюсы и Гутары. В пределах Иркутской
области марал, кроме Восточных Саян, повсеместно и в значительном
числе водится в хребтах западного Прибайкалья (Копылов, 1950), к се-
веру до правых притоков Киренги — Мини и Черепанихи (выше север-
ной оконечности Байкала). В меньшем количестве и не повсеместно он
встречается в междуречьях к северу от главной Сибирской магистрали,
доходя до 56—57° с. ш. Нет его в Катангском районе, и только случайно
заходит он в Нижне-Илимский район. Очаг обитания марала, изолирован-
ный от южной части ареала, имеется на севере Бодайбинского района
по р. Б. Патоме и Чаре (Скалой, 1951).
К востоку от оз. Байкал начинается область распространения восточ-
ного подвида благородного оленя— изюбря (С. elaphus xanthopygus
Milne-Edw.), который распространен почти повсеместно в Бурят-Монголь-
ской АССР, начиная от Саян и Хамар-Дабанского хребта до северной
оконечности Байкала и бассейна Верхней Ангары. Нет его только
в южных остепененных районах. Особенно много изюбря в Баунтовском
районе, а также в Яблоновом хребте и его отрогах. Северная граница
здесь приблизительно совпадает с горной границей Бодайбинского района
Иркутской области, поднимаясь на севере в верхнее течение Чары около
58° с. ш. (Подаревский, 1936).
Точные сведения о распространении изюбря в Якутии отсутствуют.
В начале этого столетия он служил объектом промысла под Олекминском
(Доппельмаир, 1927). В настоящее время держится в среднем и нижнем
течении р. Чары и Токко, а также изредка и случайно появляется между
верховьями р. Амги и Алданом (Скалой, 1941, 1951).
Повсеместно, кроме степных районов, обычным зверем изюбрь является
в Читинской и Амурской областях. Особенно многочислен он, кроме
Яблонового хребта, на севере в междуречье Витима и Олекмы (Подарев-
ский, 1936). Много изюбрей в районе хребта Тукурингра, в бассейнах
рр. Ольдоя, Гилюя и Уркана, а также в прилегающих районах к югу
от железнодорожной магистрали, а по последним данным также в север-
ной части Хингано-Архаринского района Амурской области. Становой
хребет является северной границей распространения изюбря, но в не-
большом числе он встречается и к северу, в верховьях р. Нюкжи (Гас-
совский, 1927).
1 Устное сообщение К. А. Юдина. Насколько достоверны сведения, которыми
располагал Подаревский (1936), проводивший северную границу по р. Чуне и через
Енисей около 58° с. ш. к верхнему течению р. Кети, сказать трудно.
216
CERVIDAE
В пределах Хабаровского края северной границей распространения
изюбря является Тугуро-Чумиканский район. Северная граница здесь
проходит в верхнем течении рр. Зеи, Уды, по среднему течению Левой
Бурей, верховьям Амгуни, по Амуру около 52° с. ш. и выходит на побе-
режье около р. Самарги (Капланов, 1948). В наибольшем количестве
изюбри водятся в районах Бикинском, Вяземском, им. Лазо и северной
части Еврейской автономной области (Сысоев, 1952).
В Приморском крае область сплошного распространения изюбря пред-
ставляет Сихотэ-Алинь, но и здесь он встречается не повсюду. Много
изюбрей в районах Судзухинского и Супутинского заповедников, но уже
в заповеднике Кедровая Падь и около Владивостока его нет. Многочислен
изюбрь по всему южному и среднему Сихотэ-Алиню, а также по рр. Хору,
Подхоренку и Анюю (Капланов, 1948). Отсутствует благородный олень на
Сахалине.
В пределах Монгольской Народной Республики С. elaphus распро-
странен спорадически в горнолесной и лесостепной полосах, прилегаю-
щих к границе с Советским Союзом. Областями его обитания являются:
западная часть Монгольского Алтая к востоку приблизительно до 93° в. д.,
затем бассейн р. Коб до, район озер Ачит-Нур и Ургэ-Нур, хребет Хан-
Хухей, верховья р. Тес-Хема, Прикосоголье, местность к западу, востоку
и югу от оз. Хубсугул от 50° с. ш., хребты Хангай и Хэнтэй, среднее те-
чение рр. Эгин-Гола, Селенги и Орхона, а также предгорья и отроги
Большого Хингана к востоку до 113° в. д. (Банников, 1954).
Подвиды. Вид С. elaphus в настоящее время большинством исследо-
вателей принимается в широком объеме. Ряд форм (сибирский марал,
изюбрь, бухарский олень (хангул), американский вапити и др.), ранее
считавшихся за самостоятельные виды, теперь рассматриваются лишь
как подвиды благородного оленя. Число последних разными авторами
принимается различно. Наиболее общепринятыми являются:
1) С. elaphus elaphus L.— Европейский благородный олень (рис. 41).
Размеры средние; высота в холке 120—125 см, длина черепа 403—440 мм.
Окраска насыщенная рыже-бурая или серовато-рыжая, зимой иногда
серая. Грива не темнее туловища. Пространство между ветвями нижней
челюсти светлее щек. Околохвостовое зеркало средних размеров, буро-
ржавого цвета с неясной темной полосой по средней линии, лишь немного
светлее боков и постепенно сливается с окраской спины, благодаря от-
сутствию сверху темного окаймления. Конечности, а также плечи и бедра
у самцов часто окрашены темнее остального туловища. Конечные отростки
на рогах, число которых иногда велико, расходятся кустом и образуют
крону («чашу»). Средний (третий) отросток рога длиннее четвертого. Глав-
ный ствол рога в верхней части не отогнут назад. Р а с п р о с т-р а н е-
н и е: западная Европа (за исключением Скандинавского полуострова,
к северу от 60-й параллели, южной Испании и о-вов Корсики и Сардинии),
прибалтийские республики СССР, Беловежская пуща, Карпаты,
Крым.
2) С. elaphus maral Oglby — Каспийский благородный олень. Раз-
меры более крупные; высота в холке около 135 и до 150 см; длина черепа
до 480 мм. Окраска менее насыщена красно-бурыми тонами, преобладают
желтоватые и серые. Плечи и бедра более темные. Пространство между
ветвями нижней челюсти серовато-белое. Околохвостовое зеркало не-
большое, ржаво-кремовое, наверху сливающееся с окраской спины,
по бокам окаймленное размытой темной полосой. Крона на конце рогов
образуется не всегда и имеет обычно меньшее число отростков. Рас-
CERVUS
пространение: Кавказ, северный Иран, быть может, вся Малая
Азия, юго-восточная Туркмения.
3) С. elaphus bactrianus Lydekker — Бухарский олень, или хангул
(рис. 45). Размеры несколько меньше, чем у типичного подвида; высота
в холке около 120 см; длина черепа до 406 мм. Окраска более однородная,
светлая, песчано-серая, особенно в зимнем меху; книзу еще более свет-
леет; от холки по спине тянется буроватая полоса, сливающаяся с темнс-
Рис. 45. Бухарский олень, или хангул (Cervus elaphus bactrianus Lyd.)..
ватой окраской бедер. Ноги светлее спины, но по передней их стороне
идет узкая темноватая полоса. Передняя часть морды беловатая, кроме
носового зеркала и темных пятен на нижней губе у углов рта. Простран-
ство между ветвями нижней челюсти светлее щек. Околохвостовое зер-
кало сливается с окраской спины, по бокам окаймлено узкой черно-бу-
рой полосой. Рога, как правило, пятиконцевые; крону на вершине почти
никогда не образуют. Главный ствол рога в проксимальной половине
прогнут назад, а около среднего отростка делает изгиб внутрь. Р а с-
про ст.ранение: бассейн Аму-Дарьи (в недалеком прошлом и Сыр-
Дарьи) в Туркмении и Таджикистане, северный Афганистан.
4) С. elaphus sibiricus Severzov1 — Марал (рис. 44). Размеры крупные;
высота в холке до 150—155 см; длина черепа до 480 см. Окраска зимой
серовато-бурая, шея, низ живота и конечности более темные. Летом
в окраске имеются рыжеватые или коричневые тона и нет потемнения
живота и конечностей. Самки окрашены и зимой и летом более однотонно.
1 См. примечание к подвиду С. elaphus canadensis Erxleben.
218
CERVIDAE
Пространство между ветвями нижней челюсти такого же темного цвета,
как и щеки. Зеркало больших размеров, высоко заходящее на круп, от
оранжеватого до соломенного цвета, хорошо ограниченное и с боков и
сверху. Рога массивные, но число отростков обычно шесть-семь, при этом
четвертый отросток, как правило, длиннее среднего (третьего). Конечные
отростки расположены почти в одной плоскости и никогда не образуют
кроны («чаши»). Главный ствол рога в месте отхождения четвертого от-
ростка заметно отогнут назад. Распространение: горы Сред-
ней Азии и южной Сибири от Тянь-Шаня до западного Прибайкалья вклю-
чительно, северная Монголия, Большой Хинган.
5) С. elaphus xanthopygus Milne-Edwards — Изюбрь. Отличается от
марала несколько меньшей величиной, яркой красновато-рыжей окраской
летней шерсти и более темным цветом зимою. Летом околохвостовое зер-
кало почти сливается с окраской спины. Зимою оно остается рыжего
цвета. Рога сходны с маралом, но четвертый отросток развит слабее.
Распространение: к востоку от Байкала, бассейн Амура, Си-
хотэ-Алинь, северный Китай к югу от р. Хуанхэ.
6) С. elaphus corsicanus Erxleben — Корсиканский, или варварийский
олень. Самая мелкая раса благородных оленей; высота в холке около
80 см; длина черепа около 340 мм. Рога не образуют кроны. Отсутствует,
как правило, второй надглазничный отросток. Распространение:
о-ва Корсика и Сардиния, южная Испания, северная Африка (С. elaphus
barbarus Bennett).
7) С. elaphus) affinis Hodgson — Шу, или кашмирский хангул. Близок
к С. elaphus bactrianus, отличается лишь некоторыми особенностями
окраски. Распространение: Кашмир, Гималаи, западный Китай,
восточный Тибет, Кам на юг до Юнани.
8) С. elaphus wallichi Cuvier — Мансароварский олень. Отличается
большим белым, резко отграниченным околохвостовым зеркалом и белым
хвостом. Распространение: юго-западный Тибет.
9) С. elaphus yarkandensis Blanford — Яркендский олень. Крупный
олень; характерная особенность — один изгиб ствола рога на месте
среднего отростка, выше и ниже этого места рог прямой. На конце рога
образуется чаша из 3—5 концов. Распространение: бассейн
Тарима, оз. Лобнор.
10) С. elaphus alashanicus Bobrinskoi et Flerow. — Алашанский ва-
пити. От С. elaphus sibiricus и С. elaphus xanthopygus отличается более
мелкими размерами и одноцветностью окраски в зимнем меху. Рас-
пространение: Алашанский хребет (юго-восточная Монго-
лия).
И) С. elaphus canadensis Erxleben — Вапити. Самая крупная раса
благородных оленей, высотою в холке до 165 см с мощно развитыми
рогами, по форме сходными с С. elaphus sibiricus.1 Окраска темная, но
несколько светлее, чем у марала. Слабо выражен сезонный и половой
диморфизм. Распространение: Северная Америка от 60° с. ш.
до Калифорнии и Аризоны на юге.
Без достаточных оснований выделяются и у нас и в Северной Америке
еще ряд подвидов, как например С. elaphus hippelaphus Kerr, (среднеевро-
1 К. Флеров (1952) и объединяет его в один подвид с последним, с чем нельзя
согласиться. Популяции сибирского марала и американского вапити слишком давно
стали на независимые пути эволюции, для того, чтобы могли приниматься не только
ла один подвид, но даже за один вид. Черты их сходства — проявление параллелизма.
CERVU.S
219
пейский олень). С. brauneri Charlemagne (крымский), С. elaphus songari-
cus Severtzov (тяншанский марал) и некоторые другие (Гептнер и Цалкин,
1947; Флеров, 1952).
Биология. На огромной территории своего ареала благородный
олень обитает в разнообразной обстановке, Его можно встретить от при-
морских низин до альпийской зоны и от типичной тайги до лесостепи
и даже пустыни. В летнее время олени, если не преследуются человеком,
хищниками или гнусом, занимают практически все стации в районе оби-
тания; только степень встречаемости в разных его частях будет раз-
личной.
Все же можно определить некоторые характерные особенности мест
обитания оленей, свойственные большинству подвидов во всех частях
ареала. Это — наличие леса и гористый или хотя бы холмистый характер
рельефа. В совершенно безлесных местностях благородные олени не
живут; равным образом, только в крайнем случае мирятся с низинным
и особенно болотистым ландшафтом.
Однако темнохвойной тайги и густого леса олени, как правило, избе-
гают. Предпочтение всегда отдается смешанным и разреженным лесным
насаждениям с хорошо развитым подростом или подлеском. Густые хвой-
ные леса служат местом обитания лишь тогда, когда они перемежаются
с безлесными травянистыми полянами или граничат с лугами, например
в субальпийской зоне, в долинах рек и речек. В этих случаях лес служит
местом отдыха и укрытия от хищников и жалящих насекомых. Повсюду
излюбленными стациями оленей являются гари или места порубок с за-
рослями кипрея, порослью лиственных древесных и кустарниковых пород.
В Крымском заповеднике наибольшая плотность оленей в течение всего
года наблюдается в полосе дубовых лесов. Здесь летом обитает около 70%,
а зимой даже до 96% всего поголовья оленей. Наименьшее значение для
оленей здесь имеют сосновые леса, в которых обитает не больше 2%°/о всего
стада (Янушко, 19576).
В отношении рельефа благородный олень также отдает явное предпоч-
тение горному, но спокойному рельефу. Крутого скалистого ландшафта
и каменистых россыпей избегает. «Независимо от экспозиции склона и
абсолютной высоты, все крутые скалистые и богатые россыпями места
неохотно посещаются маралами; следы их здесь объясняются случайными
заходами или тем, что данное место находится на пути из одной основной
стации в другую» (Дмитриев, 1938 : 199).
Места обитания в большинстве районов, особенно в Азиатской части
ареала, не остаются постоянными в течение всего года. Эти сезонные
передвижки, иногда достигающие значительных масштабов, в зимнее
время, в основном, обусловливаются характером залегания и глубиною
снежного покрова. Олени свободно достают корм и даже опавшие желуди
и фрукты из-под снега глубиною не свыше 25—30 см. При глубине снега
свыше 50—60 см затрудняется не только добыча корма, но и передвижение
(рис. 46). Только сильные самцы могут еще более или менее свободно
двигаться в метровом снегу (рис. 47). Неглубокий, но плотный или по-
крытый коркой наста снег также заставляет оленей нередко откочевывать
в другие места: наст не только затрудняет добычу корма, но и представ-
ляет прямую угрозу для оленей в случае нападения хищников, так как
не выдерживает тяжести бегущих животных и потому создает большие
помехи при быстром беге. В летнее время стационарное распределение
обусловливается, в основном, кормовыми условиями и возможностью спа-
саться от кровососущих насекомых. Сезонные миграции более ясно вы-
220
CERVIDAE
ражены у взрослых самцов, как у более сильных и подвижных животных,
вообще более склонных к бродяжнической жизни.
В равнинных местах, как в Беловежской пуще, в лесах Воронежской
области, олени летом держатся преимущественно в участках, богатых
лиственным подростом в дубовых и смешанных лесах. Дубняки, а на Кав-
казе и заросли диких фруктовых деревьев, являются также местами
осенней концентрации этих животных, привлекаемых обилием излюблен-
ного корма — опавшими желудями и яблоками. Заболоченных низин
избегают. Зимою, как и везде, площади, занимаемые оленями, сильно
сокращаются; животные концентрируются в основной своей массе в мало-
снежных и кормовых участках леса.
В горных районах Карпат, Кавказа и Сибири в летнее время излюб-
ленными стациями являются верхний пояс леса, зона субальпийских и
даже альпийских лугов. Олени поднимаются до высот в 3000 м (Динник,
1910). Основная масса летует на высотах от 100—1200 до 2500 м. Побу-
ждающим мотивом является обилие гнуса (кровососущих насекомых)
в нижнем и среднем поясе гор и отсутствие его в верхнем, прохладном
и обдуваемом ветрами. Особенно чувствительны к укусам насекомых
самцы с их нежными, растущими в это время рогами. В жаркое время года
оленей можно встретить даже по соседству с вечными снегами, но, как
правило, далеко от верхней границы леса они не уходят. Ельники на гра-
нице с альпийскими лугами, по Н. А. Северцову (1873), являются основ-
ными местообитаниями марала и на Тянь-Шане. На альпийских лужай-
ках и травянистых склонах между еловыми рощами маралы пасутся,
но в ельники возвращаются для отдыха и пережевывания жвачки.
В зависимости от высоты местности и климатических условий эти
перекочевки на летние пастбища начинаются уже в конце апреля и за-
канчиваются обычно в мае или начале июня. Иногда летние выпасы рас-
положены далеко от мест зимовок и поэтому олени дважды в год выну-
ждены совершать большие переходы. Места летовок и зимовок в пределах
одного района постоянны. Постоянны и пути, по которым совершаются
сезонные перекочевки, представляя нередко хорошо пробитые тропы на
удобных перевалах и других местах, по которым вынуждена проходить
значительная часть животных. На постоянстве путей миграции основаны
способы поимки живых оленей ловчими изгородями для целей разведения.
Первыми начинают кочевки взрослые самцы. Последними движутся
беременные самки. Самки вообще более равномерно распределены по району
обитания. Летом их можно встретить от гольцовой зоны до предгорных
лесов и речных долин, но в условиях Алтая, например, они преимуще-
ственно держатся по пологим склонам смешанно-хвойных насаждений
среднего пояса лесной зоны. Защиту от насекомых самки ищут в лесных
чащах, чего не могут делать самцы с их растущими, чувствительными ко
всякого рода повреждениям рогами. Избегают маралы большетравных
участков, так как при травостое свыше 60—70 см сильно сужается круго-
зор зверя (Дмитриев, 1938). Самки с телятами иногда предпочитают ши-
рокие долины нижнего пояса лесной зоны.
С исчезновением насекомых и когда окостеневшие рога самцов стано-
вятся менее чувствительными, что наступает обычно с половины августа,
начинаются обратные откочевки оленей с мест летовок в зону широколи-
ственных и смешанных лесов. Первыми в этом случае трогаются молодые
и самки. Распределение по территории в осенний период зависит от кор-
мовых условий и наличия .мест, удобных для гона (проведения полового
сезона). ...
CERVUS
221
Зимою в горных районах Крыма и Кавказа олени спускаются в нижний
пояс лесов, в предгорья, а иногда остаются на средних высотах (Динник,
1910; Янушко, 19576). На Алтае основными зимними стациями являются
долины рек и ключей на высоте от 1400 до 1800 м над уровнем моря
в средней полосе леса, где снег сравнительно неглубок и- имеется в изо-
билии один из излюбленных зимних кормов марала —- ивняки. Есте-
ственно,-предпочитаются места, защищенные от ветра. Охотно, особенно
ближе к весне, олени держатся на солнечных, обогреваемых склонах,
Рис. 46. Самка марала в снегу глубиной 100—110 см. Долина р. Малый
Шалтан (бывш. Алтайский заповедник). (Фото Ф. Д. Шапошникова).
где и снег не так глубок, и раньше, чем в других местах, появляется
зелень. Часть животных, в первую очередь крепкие быки, вероятно из
соображений безопасности, остаются зимовать в высоких горах, особенно
если зима малоснежна (Сокур, 1952), или задерживаются там до декабря,
даже января, пока глубокие снега не заставят их спуститься ниже.
На Дальнем Востоке, в Сихотэ-Алине лучшими стациями изюбря
являются старые гари и прибрежные дубняки; из хвойных лесов пред-
почитаются кедровники. Сезонные миграции здесь не велики и несколько
отличны от тех, которые были упомянуты раньше. На обнаженных вер-
шинах и хребтах изюбри спасаются от мошек только в первую, более
прохладную половину лета. С наступлением жары во второй половине,
когда появляется большое количество слепней, звери спускаются в урему,
в прохладные сырые места, ольховые заросли и хвойные насаждения,
появляются на берегу моря (Капланов, 1948).
Особые стации занимает бухарский олень (хангул). Его основные
места обитания — тугайные заросли в долине Аму-Дарьи, Пянджа и его
притоков. Предпочитаются тугаи по соседству с участками пойменных
222
CERVJDAE
лесов из разнолистного тополя и джиды (Флеров, 1935). В нижнем тече-
нии Аму-Дарьи хангул предпочитает заросли пятты (Populus pruinosa
Schrenk) с покровом из чаира (Aeluropus litoralis Pari.) (Клюшкин, 1949).
Иногда олени выходят из тугаев в пустыню; но и в этом случае они при-
держиваются саксаульников и порослей тамариска. Сезонных миграций
в настоящее время хангулы не совершают.
По характеру потребляемой пищи благородного оленя следует счи-
тать не столько травоядным, сколько древесноядным животным. Иссле-
дуя содержимое желудка у зубров и оленей Беловежской пущи в летнее
время, Врублевский (1912) установил, что у первых пища состояла,
в среднем, из 80% травянистого и только на 20% из древесного корма.
У оленей же наоборот — травы составляли всего около 10%, а 90% со-
держимого состояло из остатков древесного корма. Близкие к этим дан-
ные получил П. А. Янушко (19576), изучавший питание крымского благо-
родного оленя. Оказалось, что и здесь «основным кормом в течение всего
года является древесная и кустарниковая растительность, остатки ко-
торой в содержимом желудков оленей составляют по весу от пищевой
массы 60—85%». Травы, очевидно, имеют существенное значение в лет-
ний период, но зимой олени живут, если только не получают искусствен-
ной подкормки, почти исключительно на веточном корме.
Видовой состав употребляемых в пищу растений во многом зависит
от характера растительности района обитания и различается, естественно,
в одном и том же районе по сезонам года. Есть ряд растений, поедаемых
оленями с большой охотой во всех районах. Так, излюбленным кормовым
растением повсюду является кипрей, или иван-чай (Epilobium angusti-
folium L.), ради которого олени так охотно посещают гари. Повсеместно
охотно поедаются листья и ветви различных видов ив. Излюбленную пищу
оленей и хороший нажировочный корм составляют желуди,1 буковые
орехи, плоды яблонь, груш, кизила, собираемые зверями даже из-под
снега. В западной части ареала (Беловежская пуща, Крым, Кавказ,
Воронежский заповедник) к числу основных древесных кормов относятся
листва и ветки дуба, липы, клена, ясеня, бука, тиса, ильма и некоторых
других. Интересно отметить, что на Кавказе и Алтае береза также отно-
сится к одному из основных кормов (Дмитриев, 1938; Насимович, 1939);
в Беловежской же пуще, по свидетельству Врублевского (1912), олени
ее не трогают, так же как ель и ольху. На Дальнем Востоке излюбленным
веточным кормом служит также бархат, аралия, леспедеция и осина
(Капланов, 1948). В Крыму и на Кавказе охотно поедаются паразитиче-
ские растения — омела (Viscum album L.) и плющ (Hedera helix L.).
Из травянистых растений по вкусовым качествам и значению в рационе
оленя на первом месте, кроме кипрея, следует поставить виды разно-
травья: белокопытник (Petasites officinalis Moench.), калужницу (Caltha
palustris L.), борщевник (Heracleum pubescens MB), гречишник (Polygonum
sp.) и некоторые другие. Хуже поедаются, даже молодые, злаки и папорот-
ники. В Забайкалье изюбри охотно поедают хвощ (Equisetum scirpoides
Michx.) (Новиков, 1937).
Передвигаясь по пастбищу., олень не срывает траву подряд, как это
делает, например, крупный рогатый скот. Он выбирает и схватывает от-
дельные растения или даже только части их: листочки, верхушки, бу-
1 В Воронежском заповеднике желуди играют столь видную роль в питании
благородных оленей, что одну зиму имел место падеж последних от истощения в ре-
зультате неурожая и сбора желудей для семенных целей (Мертц, 1953).
CERVUS
223
тоны соцветий. У древесных растений летом объедаются листья, побеги
этого года, тонкие ветви. Осенью пока древесина не замерзла, а иногда и
летом, олени срывают и поедают с ряда деревьев (осины, липы, ильма, ив)
кору, или сгрызая ее резцами, или сдирая в виде лыка. Зимою олени
объедают с деревьев, помимо верхушек, довольно толстые ветки (превра-
щая их зубами в мелкие опилки; Врублевский, 1912), почки, сережки
берез, листву зимнезеленых кустарников и деревьев. При возможности
откапывают из-под снега зеленую траву; не брезгают и торчащими из
Рис. 47. Взрослый самец марала на зимовке в лесу. Окрестности Телецкого
озера. (Фото Ф. Д. Шапошникова).
снега верхушками высохших растений. Очень охотно поедают зимою дре-
весные лишайники (Usnea), собираясь иногда с этой целью на лесосеках
у поваленных деревьев. Всюду, где это возможно, благородные олени под-
ходят и едят из стогов сено (Шапошников, 1953). В европейской части
ареала лакомством оленя считаются грибы, но на Алтае, по словам Дмит-
риева (1938), маралы их не особенно любят.
Пища тугайного оленя (хангула) состоит из зеленых частей растущих
в тугаях растений. Излюбленным кормом являются сочные листья пятты
(Populus pruinosa Schrenk.), верхней части кендыря (Apocynum venetum L.),
а также чаир (Aeluropus litoralis Pari.) (Клюшкин, 1949). Весной хангулы
едят зеленые части эфемеров. Выходя в пустыни, ощипывают побеги сак-
саула, тамарикса; не отказываются, в случае нужды, и от сухой травы.
Иногда благородные олени делают потравы зерновых, огородных и
бахчевых культур. В частности, известны случаи поедания_капусты на
огородах при охотничьих хозяйствах и в заповедниках.
Для удовлетворения потребностей в минеральном питании благород-
ные олени посещают, как и другие копытные, солонцы. Усиленное солон-
224
CERVIDAE
цевание начинается весною, с переходом на питание зеленью, и продол-
жается до июля (Дмитриев, 1938). После этого, очевидно в связи с тем,
что потребность в солях с окончанием роста рогов и обогащением мине-
ральными веществами кормовых растений уменьшилась, посещение со-
лонцов носит случайный характер. Вновь оно усиливается с конца августа
до октября (Дмитриев, 1938). Наравне с сухими солонцами, посещаются
«оленые источники, а в Приморье изюбри спускаются пить соленую мор-
скую воду. На Пяндже и Вахше хантулы лижут напочвенную соль в за-
рослях тамарикса.
Большую часть года благородные,олени живут небольшими группами,
крупных стад не образуют. Перед отелом самки уединяются и, пока оленята
не окрепнут, держатся в-одиночку. Бйвает так, что две самки с телятами
пасутся'вместе. Иногда к ним присоединяется приплод прошлого года,
особенно самки. С августа в группе самок можно встретить и взрослого
самца. Самцы отделяются от самок в конце зимы и держатся чаще в оди-
ночку или'группами по 2-—3 головы,, а старые быки почти круглый год
(за исключением полового сезона) ведут одиночный образ жизни.
По окончании периода спаривания олени начинают табуниться и
образуют несколько большие, чем в остальное время, группы, до 10 п
более голов в каждой.1 В состав последних входят животные всех поло-
возрастных групп. Табунки распадаются в начале весны перед весенними
миграциями и подготовкой к отелу. "
В пределах избранного местообитания, если кормовые условия благо-
приятны и оленей не тревожат, они ведут почти оседлый образ жизни,
особенно зимою. Дневные часы, как правило, они отдыхают и пережевы-
вают жвачку.;1 Для отдыха выбираются разреженные леса. Иногда олень
ложится у небольшой скалы, у корней поваленного дерева так, что имеет
перед собой хорошее поле зрения, а сам мало заметен. Густых зарослей
для мест лежек олени избегают. Летом пасутся, в основном под вечер и
рано утром, а также, особенно там, где их беспокоят, и ночью. В Приморье
изюбри день проводят в ropai, а на ночь спускаются и пасутся в зарослях
по долинам рек и ключей (Капланов, 1948). Зимою, вследствие затрудне-
ний в добыче пищи, часто кормятся и в дневные часы. Солонцы посещают
после заката солнца или даже глубокой ночью. После солонцевания звери
обычно направляются к водопою.
Передвигается олень шагом, рысью, галопом или прыжками. Рысью,
впрочем, двигается мало; будучи потревожен, быстро переходит на галоп.
Во время бега легко перепрыгивает большие камни, овраги, поваленные
деревья; сравнительно легко (хотя и далеко уступает в этом отношении
козлам) спускается по крутым каменистым склонам. Испуганный делает
сильные и в то же время необычайно мягкие пружинистые прыжки.1 2
У благородного оленя хорошо развито обоняние; человека марал мо-
жет по ветру учуять за несколько сот метров. Зрение, возможно, менее
-остро, чем слух и обоняние (Дмитриев, 1938). Там, где оленей преследуют,
1 Динник (1910 : 122) указывает, что зимою табуны могут достигать 40 штук.
2 В 1930 г. мне удалось наблюдать на Алтае случай, когда выпущенный после
осмотра в станке домашний марал-рогач сделал прыжок около 1.5—2 м в высоту и
длиною не менее 6—7 м. Тогда же пришлось быть свидетелем другого, не менее инте-
ресного и почти невероятного случая, иллюстрирующего не только силу, но и ловкость
прыжка. Направленный в панторезный станок с опущенным колпаком, взрослый ро-
гач перед самым станком (длиною 3 м) сделал прыжок, перемахнул станок вдоль и
через оконце в передней части колпака размером 45X45 см (описание станка см. Со-
колов, 1932в), которое не успели закрыть, выскочил наружу, не сделав ни единой ца-
рапины на несрезанных еще пантах.
CERVUS
225
они очень скрытны и осторожны. Однако опыт организации заповедни-
ков и охотничьих хозяйств показал, что олени не всегда так пугливы.
К хангулу на Вахше можно подойти метров на 60—70, а иногда и ближе
(Флеров, 1935). Самки вообще более доверчивы, чем самцы.
При быстром беге голова и шея вытянуты так, что у самцов рога за-
кинуты назад и стволами почти касаются спины. Прекрасно плавают.
Переплывают бурные горные потоки, а также могут заплывать в открытое
море на несколько километров (Капланов, 1948).
Большой смышленностью благородные олени не обладают. Ухаживаю-
щего за ними человека от других не отличают. Бывают очень злобны, осо-
бенно самцы. Известны случаи неспровоцированного нападения на лю-
дей. Рассерженный олень морщит верхнюю губу, шипит, скрипит зубами,
топает передней ногой, мочится, на губах появляется пена. Защищаясь
или нападая, старается нанести резкий удар сверху вниз передней ногой,
для чего последнюю подымает вперед и вверх.1
В обычной обстановке олени, особенно самцы, молчаливы. Потрево-
женные, они иногда издают предостерегающий крик, весьма отдаленно
сходный с мычанием, только более отрывистый и короткий. Теленок
с матерью перекликается особым непередаваемым «писком».
Линька у благородного оленя в году бывает дважды. Видимое начало
весенней линьки — апрель, в мае она обычно заканчивается; однако
первые признаки в виде выпадающих отдельных волос можно наблюдать
уже в марте. И начало, и конец линьки зависят от состояния организма
зверя. У здоровых, крепких животных она начинается и заканчивается
раньше. У больных и истощенных можно видеть большие клочья не-
вылинявшей шерсти еще в июле. Индивидуальная продолжительность
весенней линьки — от 2 до 3% месяцев (Цалкин, 1946). Взрослые самцы
вылинивают раньше. Последними заканчивают линьку молодняк и бе-
ременные самки. Весною линька начинается с Пог и головы, затем ли-
няет туловище, дольше всего удерживается зимняя шерсть около хвоста
и на шее. Старая свалявшаяся шерсть отпадает большими клочьями,
зацепляясь за кустарники и деревья. Испытывая раздражение, олени
во время линьки забираются в воду, валяются в грязи, оставляя иногда
в ней целые пласты старой шерсти.
Летнюю, короткую и лишенную подшерстка, шерсть олень носит
не долго. Уже в половине августа начинается осенняя линька. В сен-
тябре олени покрыты, хотя и коротким, но уже зимним мехом, полное
развитие которого заканчивается обычно только в ноябре. Весенняя
линька начинается со спины, ягодиц и боков, распространяясь постепенно
книзу. Она не носит такого бурного характера, как весною, и, возможно,
вообще выпадает не весь летний волос.
В условиях зоопаркового содержания южные расы оленей (хангул и
кавказский олень) весеннюю линьку заканчивают раньше, а осеннюю
начинают позднее и поэтому в летнем наряде ходят на 16—35 дней дольше
(Цалкин, 1946).
Оленята родятся безрогими. В сентябре, а иногда и раньше на месте
будущих рогов видны завихряющиеся пучки волос, а в декабре прощупы-
ваются возвышения лобных костей (начало роста пеньков). К годовалому
возрасту пеньки достигают высоты 5—7 см; вскоре после этого кожа
1 О силе удара можно судить по тому, что брезентовый плащ, нечаянно уронен-
ный в тесный загон к рассерженному маралу, в течение l1^—2 минут ударами копыт
был превращен в лохмотья.
15 Фауна СССР. И. И. Соколов
226
CERVIDAE
на верхушке пенька лопается (Митюшев и др., 1950) и начинается рост
сменяемой части рога. Первые рога, окостеневающие обычно к сентябрю—
октябрю (т. е. к 15—16-месячному возрасту), имеют вид простых шпилек,
длиною от 30 до 50 см; у хорошо развитых оленят иногда на вершине
рожек появляется «вилка» (раздвоение).
Вторые рога, развивающиеся на третьем году жизни, имеют обычно
надглазничный и средний отростки (т. е. три, а иногда и четыре конца).
Трехлетние самцы имеют уже и второй надглазничный отросток. В по-
следующие годы число отростков увеличивается за счет разветвления
вершины главного ствола рога.1 Крона образуется в возрасте 4—5 лет.
Число отростков увеличивается до 6 или 7 лет. В дальнейшем, с 7—8
лет, когда рога достигают уже полного развития, увеличение веса идет
не за счет роста числа концов, а лишь за счет утолщения главного ствола
и основных отростков (рис. 48). Пеньки рогов с каждым годом становятся
все ниже и толще, так что у старых самцов рога кажутся сидящими не-
посредственно на лобных костях. Длина главного ствола по изгибу у бла-
городного оленя может доходить до 120 см, а вес до 18.8 кг, но, как пра-
вило, значительно меньше (5—8 кг; Туркин и Сатунин, 1902).
Намеченная схема и сроки развития рогов и сама их форма у оленей
подвержены сильной изменчивости и зависят в значительной мере от
условий жизни животного, в первую очередь от питания. Изредка уже
первые рога имеют надглазничные отростки; иногда на них появляются
дополнительные отростки или наблюдается вторичное ветвление концов
кроны. Чаще, однако, бывает запаздывание или даже недоразвитие числа
отростков. Иногда, даже у вполне взрослых самцов, отсутствует второй
надглазничный отросток.1 2 Поэтому предложенные методы определения
возраста благородного оленя по рогам (Blasius, 1857; Туркин и Сатунин,
1902) практически, как и у косули, мало пригодны.
С возраста И—12 лет наблюдается обратное развитие рогов: умень-
шаются их размеры, убывает число отростков (в первую очередь исче-
зает опять же второй надглазничный отросток), вплоть до того, что ос-
тается лишь один надглазничный.
Ежегодное сбрасывание рогов у взрослых самцов происходит в боль-
шинстве районов в период с половины марта до половины—конца апреля.
В западной Европе (Blasius, 1857) и в Крыму (Котовщикова, 1936; Янушко,
19576) оно происходит раньше — в феврале или начале марта. Молодые
сбрасывают рога позднее, иногда уже в мае. Позднее (в апреле—мае)
сбрасывание их происходит также у бухарского оленя (Флеров, 1935;
Цалкин, 1945). Спадают оба рога редко одновременно, — обычно же
с промежутком в 1—2 дня.
Сразу после сбрасывания старых рогов начинается рост новых. На
месте сброшенного рога на вершине пенька образуется кровяной струп.
По краям последнего кожа припухает в виде синевато-серого кольца,
которое, постепенно расширяясь и утолщаясь, наплывает на средину
пенька, приподымая струп кверху. Уже дней через 7—10 растущие рога
представляются в виде вздутых, покрытых кожей с нежным волосом
мягких шишек. Через некоторое время наверху шишек намечается по-
перечная бороздка отделяющая переднюю их треть — зачаток первого
1 Порядок появления отростков в течение индивидуальной жизни оленей (в онто-
генезе) отражает последовательность появления их в истории данной формы (в фило-
генезе).
2 Как отмечалось, этот признак является даже расовой особенностью корсикан-
ского оленя (С. elaphus corsicanus Erxl.).
CERVUS
227
надглазничного отростка (первый раздвои). В процессе развития (рис. 49)
таким же путем, последовательно и параллельно с ростом главного ствола
образуются второй надглазничный, средний и концевые отростки.
Для марала в условиях
зоопаркового содержания
Цалкин (1945) приводит
следующие сроки образова-
ния раздвоев (зачатков от-
ростков) после сбрасывания
старого рога: первый на
13—17-й день, второй на
23—30-й, третий на 46—
52-й, четвертый на 66—75-й,
пятый на 84—90-й день.
Окончательную форму ра-
стущие рога приобретают у
взрослых на 105—115-й день,
у молодых несколько раньше.
Кроме того, на рост пантов
Рис. 48. Возрастные изменения рогов у благород-
ного оленя {Cervus elaphus elaphus L.).
(По Ф. Шиллингеру, 1930).
несомненное влияние оказывает погода. Наступившие вскоре после
сброса старых рогов холода задерживают рост новых. Окостенение рогов
начинается с оснований; завершается оно обычно к концу июля (по Цал-
Рис. 49. Стадии роста и развития рогов у марала {Cervus elaphus sibiricus
Sev.). (Из В. И. Цалкина, 1945).
кину, у взрослых на 140—148-й день с начала роста). Внешне окостене-
ние выражается в подсыхании (омертвении) покрывающей рог кожи и
приостренности концов отростков.
15*
228
CERVIDAE
Испытывая, вероятно, раздражение, олень трет и ударяет рогами
о кусты или деревья и сдирает таким образом с них кожу, которую иногда
съедает (Динник, 1910).
Первое время после очистки рога имеют белый цвет, иногда с полос-
ками крови. Только потом, от трения о кору и древесину они приобретают
свойственную им темную окраску со светло-желтыми или белыми кон-
цами отростков. Чистка происходит с конца июля (у крымских и западно-
европейских) до сентября (у хангула). У отдельных животных она может
тянуться от 6 до 20 дней. Молодые чистят несколько позже; «перво-
рожки» — обычно в сентябре.
Повреждение розетки и прилегающей к ней ткани пенька растущего
рога влечет нарушение процесса его формирования не только в этом,
но и в следующие годы (Соколов, 19326). Срезанный без повреждения
розетки растущий рог, нередко регенерирует, образуя в том же году
вторые недоразвитые рожки («отаву»).
Рост рогов у благородного оленя в сильной степени зависит от функ-
ции половых желез. Полная кастрация самцов в раннем возрасте, до
начала видимого роста рогов, ведет к тому, что рога не развиваются.
Полная кастрация зверей с растущими или уже заканчивающими развитие
рогами ведет к различного рода нарушениям их дальнейшего развития, —
неодинаковым, в зависимости от стадии развития рогов и возраста самца,
в котором произведена кастрация. Односторонняя кастрация на рост и
формирование рогов заметного влияния, по последним данным, не имеет
(Завадовский, 1926).
На периодичность роста и сбрасывание рогов у благородного оленя
оказывают влияние также условия освещения. В результате опытов,
проведенных в Польше (Jaczewski, 1954), установлено, что после искус-
ственного сокращения дневного освещения, с последующим внезапным
его продлением, во время роста рогов, самцы теряют их в год дважды.
Половое созревание, при нормальных условиях развития, у благо-
родных оленей наступает в возрасте около 15—16 месяцев. Но самки чаще
впервые оплодотворяются на третьем году жизни, а самцы в естествен-
ной обстановке практически участвуют в размножении с 4—5 лет
(Янушко, 1957а).
Половой сезон, как правило, падает на сентябрь—октябрь и про-
должается около 1 — 1Уг месяцев. У самцов он сопровождается сильным
возбуждением и ревом. Если олени не подвергаются преследованию, то для
рева и гона они ежегодно избирают одни и те же места. Уже в конце ав-
густа самцы, закончившие к тому времени очистку рогов, становятся
беспокойными, много движутся, ударяют рогами о кусты и деревья,
в определенных местах, обычно на открытых лужайках, устраивают так
называемые «точкй», участки с выбитой ими наголо копытами и рогами
почвой размером до 15—16 м2, посещаемые ими ежедневно (Динник,
1910). Один самец устраивает иногда по нескольку таких точкбв непо-
далеку один от другого. В это время у самцов сильно утолщается шея
и широко открываются отверстия предглазничных желез. Животные из-
дают специфический запах.
Иногда с конца августа же, а чаще с начала или даже середины сен-
тября, самцы начинают реветь, сначала негромко, редко и только по
утренним и вечерним зорям. Самый разгар рева происходит во второй
половине сентября или начале октября. Только у хангулов он и начи-
нается и кончается недели на 2—3 позднее. В это время громкий рев
слышится почти целые сутки, затихая, и то не всегда, лишь на несколько
CERVUS
229
часов днем. Для рева бык выбирает гребни и вершины холмов, с которых
его голос хорошо слышен, иногда за несколько километров.1 Самец, вы-
тянув голову и шею и закинув на спину рога, начинает рев на низких
тонах, постепенно повышает их до высоких и иногда кончает вновь низ-
ким придыхательным звуком. Отдельный рев тянется около 8—10 секунд
и следует один за другим с разными интервалами, в зависимости от ин-
тенсивности гона. Самцы часто мочатся; быть может в связи с этим стоит
появление почти черного пятна на животе впереди отверстия препуция.
В это время они мало едят и к концу гона сильно худеют. Больше, чем
в другое время, самцы во время гона любят валяться в грязи и лужах.
У самок течка наступает недели через две после начала у самцов рева
(по наблюдениям В. И. Цалкина (1944) в зоопарке). Они собираются,
по-видимому привлекаемые звуками, вокруг ревущих самцов, но ведут
себя пассивно, очень спокойно и поэтому упитанности во время гона
не теряют. Самцы создают вокруг себя гаремы от 3 до 10 самок, ревниво
оберегая их от посягательства других самцов. Между самцами происхо-
дят, как и у других оленей, поединки рогами, которые иногда заканчи-
ваются увечьем или смертью одного из соперников. Известны случаи,
когда от ударов ломались стволы рогов и отростки диаметром в 5 см (Дми-
триев, 1938). Изредка оба самца погибают, намертво сцепившись рогами
или сорвавшись с края скалы в пропасть. Молодые и слабые самцы бес-
пощадно изгоняются; поэтому гаремы создают и практически участвуют
в размножении обычно самцы в возрасте старше 4—5 лет. Но, так же
как и телята текущего года, молодые самцы держатся скрытно около
гаремов, и иногда им удается покрыть отдельных самок. Акт совокупле-
ния длится весьма короткое время. Самец кроет, по наблюдениям в ма-
ральниках и зоопарках, одну самку несколько раз.
Уже в начале октября гон и возбуждение самцов постепенно затихают,
но отдельные, обычно более молодые, самцы ревут еще в ноябре.
Период беременности длится около 34 недель (Митюшов и др., 1950).
Отел начинается во второй половине мая и, соответственно гону, тянется
1—I1/? месяца. Разгар его падает на конец мая—начало июня. Отдельные,
обычно более молодые, самки телятся в конце июня и даже в июле.1 2
Перед отелом самка удаляется от табуна, телится в уединенном глу-
хом месте прямо на земле, не устраивая никакой подстилки. Число оленят
в помете один, очень редко два. Теленок родится беспомощным; лишь
через несколько, 4—5 часов, едва подымается, чтобы пососать молока.
Первые 7—10 дней почти все время лежит спрятанным в кустах или
высокой траве, вытянув по земле голову и шею. До месячного или даже
полуторамесячного возраста самка с теленком живут почти на одном ме-
сте. При малейшей опасности и предостерегающем звуке матери олененок
припадает к земле и лежит неподвижно. С месячного возраста он стано-
вится настолько крепким, что следует с матерью повсюду и не отстает
от нее при беге. С этого же приблизительно времени он начинает поне-
многу питаться зеленью, появляется жвачка. Пользуется молоком матери
почти до следующего ее отела. Сосут, первое время, 4—6 раз в день (Цад-
кин, 1944). На время гона теленок отгоняется самцом, но держится часто
неподалеку, а по окончании спаривания вновь возвращается к матери.
1 В Шебалинском совхозе Горно-Алтайской области я его отчетливо слышал за
7 км.
2 Капланов (1948) сообщал о случае отела 25 ноября.
230
CERVIDAE
Телята-самцы покидают мать весною следующего года; самки же дер-
жатся с нею и дольше, пока сами не становятся матерями.
Новорожденные оленята маралов, по взвешиваниям в маральниках,
весят 12—14кг (Митюшов и др., 1950). Окраска их коричневая, разных
оттенков, с резко выраженной пятнистостью. Пятнистость держится до
сентября, когда начинается первая линька, и к октябрю обычно совсем
исчезает. Первый зимний наряд теленок получает окончательно в октябре.
К шестимесячному возрасту они достигают около 35% веса взрослых
животных. Полного физического развития достигают к возрасту 6—7 лет,
но, как уже говорилось, к размножению способны гораздо раньше.
Продолжительность жизни благородных оленей в зоопарках и ма-
ральниках максимально 20—25 лет; но в природе они обычно до этого
возраста не доживают, а в домашнем состоянии также выбраковываются
раньше. Определение возраста марала с известной точностью можно про-
изводить по резцам (Соколов, 1932а). Родится мараленок со всеми мо-
лочными резцами. Первая пара их сменяется в годовалом возрасте или
немного позднее, вторая — в двухгодовалом, третья — в трех- и клыки
нижней челюсти — в четырехгодовалом. С 6 лет начинается стирание
режущего края резцов. Дентин обнажается на первой паре в 7 лет, на
второй в 8, на третьей в 9 и на клыках в 10 лет. В дальнейшем о возрасте
судят приблизительно по величине и форме стертой поверхности тех же
зубов. До верхушек корней зубы снашиваются к возрасту 20—23 лет.
Врагами оленя является ряд хищников. На первом месте по значению
стоят волки, от которых гибнет значительное количество оленей всех
возрастов. Наиболее опасны волки зимою, в период глубокого снега и
наста, когда они легко догоняют проваливающегося и режущего себе
ноги оленя. Охотятся волки стаями, загоняя оленей на лед, где послед-
ние не могут быстро двигаться или падают и становятся добычей хищни-
ков. В горах же иногда загоняют их на кручи, с которых олени срываются
и разбиваются, также становясь добычей волка (Динник, 1910). На Даль-
нем Востоке врагом, хотя и меньшим, чем волк, является тигр. От тиг-
ров гибнут бухарские олени в Таджикистане (Флеров, 1935).
Гораздо реже погибают благородные олени от рыси, росомахи и лео-
парда. Совсем редко, вероятно только случайно, они становятся добычей
медведя, хотя такие случаи и известны (Дмитриев, 1938). От лис и других
мелких хищников даже телята практически не гибнут, так как самка
их легко отгоняет.
Сильно страдают олени от кровососущих насекомых, особенно самцы
в период роста рогов. Спасаются, откочевывая в верхний прохладный
пояс гор или в густую лесную чащу. Иногда с этой же целью принимаются
быстро бегать (Динник, 1910).
Нападают на оленей во множестве клещи, являясь иногда переносчиком
некоторых паразитов крови, например возбудителей тейлериоза (Рухля-
дев, 1939). Паразитируют на благородных оленях кожный овод (Hypo-
derma diana Вт.), носовой овод (Pharyngomyia picta Meiger), а также муха-
кровососка (Lipoptena cervi L.), вылет которой в Крыму наблюдается
в октябре, после чего она обламывает крылья и поселяется на олене
(Котовщикова, 1936). В западной Европе на благородном олене заре-
гистрированы еще два вида оводов — подкожный (Hypoderma acteon Вг.)
и носоглоточный (Cephenomyia auribarbis Meig.).1 Существенного вреда
эктопаразиты оленям, по-видимому, не приносят.
1 Устное сообщение К. Я. Грунина.
CERVUS 231
Наблюдаются среди диких оленей некоторые заразные болезни, пере-
даваемые от домашних животных. В литературе упоминаются эпизоотии
пастереллеза, сибирской язвы (Кузнецов, 1898), ящура (Динник, 1910),
чумы крупного рогатого скота (Смирнов, 1928), некробациллеза («копыт-
ной болезни») и других, еще по известных заболеваний, принимающих
иногда массовый характер.
Хозяйственное значение. Благородный олень является ценным мяс-
ным зверем. В европейской части СССР охота на него запрещена, но в не-
которых районах Сибири и на Дальнем Востоке промысел на него ради
этой цели имеет большое значение для местного населения. От взрослого
самца кавказского оленя получают тушу мяса весом около 140—150 кг,
а от крупного зверя даже больше; туша самки, по Диннику (1910), весит
100—130 кг; от добытой нами на Кавказе самки вес туши был 88.5 кг.
Приблизительно такие же цифры (137—180 кг) приводятся для самцов
дальневосточного изюбря (Капланов, 1948). Крупнее сибирский марал,
дающий иногда свыше 200 кг мяса. Мясо маралов имеет своеобразный
красно-кирпичный оттенок и «дичный» привкус. У молодых, особенно
телят, и у самок оно нежнее. Наиболее жирны олени в конце августа, когда
у них под кожей на крупе слой жира может достигать 4—5 см. Внутрен-
него и подкожного сала иногда снимается около 30 кг (Капланов, 1948).
Мясо самцов во время гона в пищу не годится, так как имеет неприятный
специфический запах.
Кожа годна для выделки замши; но обычно ее выделывают домашним
способом и употребляют на пошивку брюк и курток. Режут ее также на
ремни для веревок и изготовления упряжи. Дохи из шкур взрослых оленей
слишком тяжелы. Весенняя шкура мало ценится, так как часто содер-
жит много отверстий, проделанных оводами. Очень прочная, покрытая
коротким гладким волосом кожа с ног ниже запястного и скакательного
суставов (камысы) идет на изготовление зимней обуви (унтов). Зимняя,
очень ломкая, но упругая шерсть могла бы служить для набивки мебели
или . матрацев.
Окостеневшие очищенные рога служат украшением. Раньше на Кав-
казе существовал даже промысел по сбору сброшенных оленями рогов.
Рога служат также для поделок (рукояток ножей, настенных украшений
и т. п.).
Самой ценной продукцией благородных оленей являются неокостенев-
шие растущие рога — панты. Значение и применение их то же, что и у пят-
нистого оленя (см. стр. 196), но, как показали исследования (Тэви,1936),
действующее начало пантов марала менее активно, чем у пятнистого оленя.
Внешне панты марала и изюбря также заметно отличаются от таковых
пятнистого оленя. Во-первых, они более значительных размеров. Вес сред-
ней величины пары только снятых и еще не консервированных мараловых
пантов составляет от 4—5 до 6—7 кг; имеются указания (Никольский,
1883), что в исключительных случаях даже срезанные панты достигают
24 кг веса. Во-вторых, различна у того и другого их форма. Если панты
пятнистого оленя имеют всего один или два отростка, то у марала и изюбря
их обычно не меньше четырех. Надглазничный отросток отходит непосред-
ственно у основания главного ствола. Волос, покрывающий кожу пантов,
изюбрей и маралов более длинный, густой и грубый от почти черного до
рыжеватого и светло-серого цвета. Лобовые панты, как и у пятнистого
оленя, на заграничном рынке ценятся дороже, чем срезные. Лучшими счи-
таются панты с четырьмя концами и закругленной плотной вершиной. Не-
пременное требование к лобовым пантам — симметричность. Лучший сезон
232
CERVIDAE
для пантовки (добычи пантов) дикого марала и изюбря — июнь; панты,
добытые даже в начале июля, часто имеют излишнее окостенение и идут
более низким сортом.
Говоря о хозяйственном значении, следует сказать несколько слов
и о вреде, наносимом благородным оленем. Он выражается иногда в унич-
тожении подроста древесных пород, поедаемых зверем, что является
препятствием к нормальному возобновлению леса. Это может иметь зна-
чение главным образом при парковом содержании и при перенаселении
занимаемой территории, как это имело место последние годы в Крымском
заповеднике (Янушко, 1957). Известно, какую отрицательную роль в свое
время сыграли сильно размножившиеся олени в Беловежской пуще,
когда в некоторых ее кварталах совершенно прекратилось возобновле-
ние леса. Интересно, что в этих условиях имела место пищевая конкурен-
ция между оленями, с одной стороны, и между лосями и зубрами — с
другой (Врублевский, 1912). Наличие пищевой конкуренции между оле-
нями и косулями в Крымском заповеднике отмечает П. А. Янушко (19576).
Отрицательное влияние на лесовозобновление может сказаться, в пер-
вую очередь, в местах концентрации оленей на зимовках. В большинстве
районов вольного существования благородного оленя в СССР это вред-
ное его влияние не сказывается или оно крайне ничтожно.
Охота. Способы охоты на благородных оленей разнообразны (Тур-
кин и Сатунин, 1902; Кузнецов, 1898; Соловьев, 1920). Весьма распро-
страненный способ — скрадывание. Применяется оно круглый год, но
чаще осенью во время рева, когда бдительность самцов сильно понижена.
Особенности его те же, что и при скрадывании других диких копытных
(см. стр. 96). С большими трудностями применяется этот способ летом,
когда скрадывают, обычно на зорях, пасущихся на полянах животных.
Самый распространенный способ добычи оленей с целью получения
пантов — подкарауливание на солонцах. Используются или естествен-
ные солонцы, или устраиваются искусственные в удобных для этой цели
местах. В последнем случае соль или просто рассыпается по земле, или
засыпается в пробитые в земле глубокие ямки. Охотник должен, соблюдая
все меры предосторожности, придти к солонцу и засесть в укрытии (в
кустах, вырытой яме или на специально устроенном «лабазе») до заката
солнца, так как олени приходят солонцеваться в сумерках, а иногда
еще и раньше, после вечерней пастьбы. Главное требование при этой
охоте — соблюдение абсолютной тишины и отсутствие запаха от обуви и
одежды. При выборе места для засидки (укрытия) важно учитывать на-
правление ветра. Олень очень осторожен при подходе к солонцу, но,
увлекшись солонцеванием, часто теряет осторожность (Дмитриев, 1938).1
Подкарауливают оленей также на водопоях, пастбищных лужайках и
других местах, куда олени приходят регулярно.
В Сибири широко применяется также способ охоты на самцов в период
рева с трубой («охота на рев»). Заключается в подражании древесинной или
берестяной трубой голосу ревущего самца. Последний, принимая услы-
шанный звук за голос соперника, или спешит навстречу охотнику и по-
падает под пулю, или, стоя на месте и откликаясь на звук трубы, под-
пускает к себе охотника на верный выстрел. Охота производится вечером
или рано утром, а также в светлые лунные ночи. Успех ее зависит от
1 В настоящее время в государственных заповедниках разработаны рациональные
методы устройства искусственных солонцов, конечно не для целей охоты, а для ми-
неральной подкормки животных.
CERVUS
233
1922), в настоящее время, в связи
районах, отошли в историю. Повсе-
Рис. 50. Пант марала I сорта. Начало
образования четвертичного отростка.
(По П. Митюшову, 1950).
качества трубы и уменья охотника подражать голосу зверя. Иногда на
звук трубы выбегают не только самцы, но и самки.
Реже применяется охота с собаками, загоняющими оленя на непри-
ступную скалу. Поставленный «на отстой» олень удерживается до под-
хода охотника собакой, изредка и осторожно подлаивающей, чтобы от-
влечь внимание, но не испугать и не заставить зверя уйти раньше вре-
мени. Парфорсная охота с гончими и охоты облавой, широко применяв-
шиеся раньше в западных областях Европейской России и на Кавказе
(Туркин и Сатунин, 1902; Толстой,
с запретом охоты на оленей в этих
местно запрещены хищнические
способы добычи по насту, глу-
бокому снегу, ямами, самострела-
ми и т. п., в свое время сыграв-
шие большую отрицательную роль
в уменьшении поголовья зверя.
Особой опасности охота на
оленей не представляет; лишь
при подходе к раненому зверю
необходимо соблюдать осторож-
ность, так как он, собрав послед-
ние силы, может ударить ногой
или рогами (Динник, 1910 ).
Мараловодство. Еще раньше,
чем разведение пятнистого оленя,
возникло мараловодство и изюб-
реводство. Впервые оно появилось
на Алтае и в Забайкалье в 40-х
годах прошлого века (Кузнецов,
1898) и развилось из первона-
чально единичных случаев поим-
ки и содержания зверей в тес-
ных загородках для доращивания ш
цу XIX в. разведение маралов и изюбрей приняло более правильный
характер и явилось самостоятельной отраслью хозяйства в ряде районов
Сибири и Забайкалья. Однако подлинная реконструкция мараловодства
была осуществлена только в Советское время. Был создан ряд мараловод-
ческих совхозов, существенным образом изменена система содержания,
разведения и эксплуатации оленей. Маралы и изюбри содержатся на
огороженных площадях паркового типа из расчета 1—2 га на голову оле-
ней в полувольном состоянии. Площадь хозяйства делится на ряд заго-
нов для содержания разных половозрастных групп оленей; кроме того,
там имеется целая система дворов и коридоров, заканчивающихся панто-
резным станком (Соколов, 19326). В летнее время животные находятся на
подножном корме. Зимою пользуются подкормкой заготовленным сеном
и концентрированными кормами. Воспитанные в неволе с раннего воз-
раста, олени привыкают к человеку, берут из рук кОрм, позволяют себя
гладить, но полного одомашнения не происходит.
Изгородь, которою обносится площадь для содержания оленей, и
внутренние перегородки имеют высоту около 2.5 м и делаются из нетол-
стых бревен. Для срезки пантов в настоящее время применяются такого
же типа станки, что и для пятнистого оленя, только, соответственно
величине марала, значительно массивнее и больше (Соколов, 1932в).
и последующего забоя. К кон-
234
CERVIDAE
Срезка пантов у марала производится тогда, когда они имеют 4 конца
и начинает намечаться четвертый раздвои (рис. 50), и проводится, в основ-
ном, с десятых чисел июня до первых чисел июля. Но не так редки случаи,
когда у молодых или слабых животных панты снимаются в конце июля.
Иногда при ранней срезке вырастает и «отава». Срезать панты марала
несколько труднее, чем у пятнистого оленя, но правила и предосторож-
ности соблюдаются те же. Особо требуется предохранять от повреждений
розетку рога. В качестве кровоостанавливающего и дезинфецирующего
средства применяется смесь квасцов и нафталина, втираемая в поверхность
среза.
В тот же день срезанные панты подвергаются консервировке. При-
менялся до последнего времени китайский способ консервировки (см.
стр. 197), заключающийся в многократной, не менее 5—6 раз, варке
по 15—30 минут в воде при температуре 95—98° и последующей дли-
тельной (около 15 дней) сушке пантов на открытом воздухе (Митюшев
и др., 1950).
Раньше, при изготовлении пантов для китайского рынка, в воду
для варки добавляли для придания им цвета чай. В настоящее время
и для пантов марала разработаны более совершенные методы комби-
нированной консервировки (варки и жаровой сушки) (Зайцев, 1953).
При консервировке панты убывают в весе, вследствие потери влаги,
на 60—7О°/о (в среднем на 64%).
Как и у пятнистого оленя, при убое марала и изюбря используются
и идут на экспорт хвосты, семенники с половыми членами и зародыши
(«лу-таи»).
При содержании в домашней обстановке у маралов и изюбрей на-
блюдается ряд инфекционных и инвазионных болезней, не свойственных
их диким сородичам или встречающихся у последних крайне редко (ту-
беркулез, ящур, некробациллез, эмфизематозный карбункул, ряд парази-
тарных заболеваний). Некоторыми из них олени заражаются и от других
домашних животных (Митюшов, Любимов и Новиков, 1950).
Род ALCES GRAY — ЛОСИ
Характеристика рода — см. выше (стр. 144).
Род включает всего один вид — Alces alces L.
Alces alces Linnaeus — Лось, или сохатый (рис. 51—56)
Linnaeus С., 1758, Syst. Natur., ed. X, 1 : 66—67 (первооп.); Brook C.,
1827, A winter in Lapland, London : 192—193 (одом.); Северцов H. A., 1854,
Вести, естеств. наук, 19 : 289—300 (общ., изобр.); Похилевич Л., 1864, Сказа-
ние о населенных пунктах Киевский губ., Киев : 11 (ист.); Brandt J., 1870, Mem.
1’Acad. Sci. St. Petersb. (ser. VII), XVI, 5 : 1—84 (ист., распр.); Morrisson W.
and A. Young, 1879, J. Linn. Soc., London, XIV : 371—390 (анат.); Носи-
л о в К., 1890, Природа и охота, окт. : 30—39 (распр., биол., ox.); Lydekker R.,
1898, The Deer of all lands, London : 52—60 (общ., изобр.); Нарышкин Д. Н.,
1900, Охота на лосей, СПб. : 1—112 (общ., изобр.); Lydekker R., 1902, Proc.
Zool. Soc., London107—109 (сист.); Туркин H. В. и К. А. С а т у н и н, 1902,
Звери России, М. : 8—10+119—196 (общ., изобр.); Bley F., 1910, Das Elch (Le-
bensbilder aus der Tierwelt, Bd. II, Erste Folge, Saugetiere, II), Leipzig : 189—224
(общ., изобр.); P о с о c k R., 1910, Proc. Zool. Soc., London : 958—960 (морф.);
Zukowsky L., 1910, Wild und Hund, 16 : 807 (сист.); Горбачев С. H.,
1915, Матер, к позн. природы. Орловск. губ., 21 : 8—10 (распр.); Lydekker R.,
1915, Catal. Ungul. Mamm. Brit. Mus. (Natur. Hist.), IV : 230—238 (сист., синон.);
Федорович Ф. Ф., 1915, Тр. Пензенск. общ. любит, естествозн., II : 51 (распр.);
ALCES
235
ZukowskyL., 1915, Arch. f. Naturgesch., 80 A, 9 : 33—45 (сист.); Толстов С. M.
1916, Наша охота, 18 : 13—14 (распр.); J а с о b i А., 1919, Anat. Anzeig., Bd. 52,
Н. 1/2 — : 41—46 (анат.); Соловьев Д. К., 1920, Саянский промыслово-охотн.
район и соболиный промысел в нем, Птг., Госиздат. : 246—248 (биол., распр.);
Туров С. С., 1924, Тр. Общ. иссл. Рязанск. края, III : 61 (распр.); О m s t е d А.,
1925, Der Heger, 1 (общ., изобр.); Храневич В., 1925, Нарис фавни Подшля,
I, В1нниця : 98 (распр.); Бутурлин С. А., 1928, Охотник, 6:21—22 (пит.);
Schulz Н., 1929, Wild und Hund, 45 (цит. по Юргенсону, 1935, ох, изобр.); Б а-
жанов В. С., 1930, Бюлл. Средне-Волжск. краев, станции защ. раст. за 1926—
1928 гг., Самара : 65 (распр.); v. Osternburg Walter, 1930, Von deutschen Elch
in Ostpreussen, Berlin (распр.); Яковлев E., 1930, Тр. Зоол. секц. Ср.-Сиб. отд.
Русск. географ, общ. : 58 (распр.); Flower S., 1931, Proc. Zool. Soc., Lon-
don : 220 (продолж. жизни); Белопольский Л. О., 1932, Тр. Зоол. инет. Акад,
наук СССР, 1,3/4 : 182—183 (распр.); Кулагин Н. М., 1932, Лоси СССР, М.,
изд. Акад, наук СССР : 1—120 (общ.; литерат. сводка); Бутурлин С. А., 1934,
Лоси, М., КОИЗ : 1—67 (общ., изобр.); Залесский П. М., 1934, Охотник и пуш-
ник Сибири, 7/8 : 27 (распр.); Lubicz-Niezabitowski Е., 1934, Ztschr.
f. Saugetierk., 9 : 195 (распр.); Наумов Н. П., 1934, Тр. Полярной комисс., 17 :
: 64—65 (распр., биол.); Подсосов Л. А., 1934—1935, Бюлл. зоопарков и зооса-
дов, 6/7 : 35—37, 10—11 (размн.); Капланов Л. Г., 1935, в кн. Юргенсон, Капла-
нов и Книзе «Лось и его промысел», М., изд. Главпушнины : 103—124 (общ.); П ту-
га е н к о Е. С., 1937, сборн. «Памяти акад. Мензбира», М., изд. Акад, наук СССР :
451—452 (распр.); Дмитриев В. В., 1938, Тр. Алтайск. заповеди., 1 : 229—242
(распр., биол.); Eidmann Н., 1938, Mitt. Forstwirtsch. : 1—55 (морф.); К о s t-
г о п К., 1938, Bull. Inst. Natur. Agron., Brno, D. 25 : 1—99 (общ., изобр.); К н о p-
р е Е. П., 1939, Науч.-метод, зап. Главн. управл. по заповеди., IV : 100—117 (одом.);
Лихачев Г. Н., 1939, Тр. по лесн. опытн. делу Тульск. засек, III : 243—257
(пит.); Попов С. П., 1939, Тр. Науч.-иссл. инет, полярн. земледел., животнов.
и промысл, хоз., сер. оленеводство, 6 : 75—102 (одом.); Б о р о д ин Л. П., 1940,
Тр. Окск. гос. заповеди., 1 : 129—149 (стац., пит.); Портенко Л. А., 1941,
Фауна Анадырск. края, III : 87—89 (распр.); Северцов С. А., 1941, Советск,
охотник, 1 : 19 (распр.); Скалой В., И. Щербаков и М. Базыкин, 1941,
Прир. и соц. хоз., VIII : 270 (распр.); Пузанов И. И. и др., 1942, Звери, птицы,
гады и рыбы Горьковской области, Горький, Облизд. : 131—134 (общ.); Дани-
лов Д. Н., 1945, Основн. способы промысла диких копытных, М., Заготизд. : 4—13
(ох.); Куклин С. А., 1946, Лоси Молотовской обл., Молотов, изд. Обл. НИТОЛес :
1—64 (общ., изобр.); Формозов А. Н., 1946, Снежн. покров в жизни млекоп.
и птиц, М., изд. Моск. общ. испыт. прир. : 110—111 (распр.); Громов В. И.,
1948, Тр. Инет. геол, наук, 64 : 439 (ист.); Капланов Л, Г., 1948, Тигр, изюбрь,
лось, М., изд. Моск. общ. испыт. прир. : 79—128 (общ., изобр.); Семенов-Тян-
Шанский О. И., 1948, Тр. Лапландск. гос. заповеди., 11 : 91—162 (распр., биол.,
изобр.); Т е п л о в В. П., 1948, Зоол. жури., XXVII, 1 : 75—78 (причины гибели);
Абрамов К. Г., 1949, Бюлл. Моск. общ. испыт. прир., Отд. биол., LIV, 1 : 17—
28 (биол., ох.); Данилов Д. Н., 1949, Охр. прир., 7 : 60—70 (распр., числ.);
Данилович А. П., 1949, Охр. прир., 8 : 80 (распр.); Кнорре Е. П., 1949,
Науч.-метод зап. Глав, управл. по заповеди., XIII : 105—114 (биол., одом.); S k и п-
с к е F-, 1949, Algen, studier. jakt och ward, Stockholm : 1—400 (общ., изобр.); Д a p-
шкевич Я., 1950, Птицы и звери Чкаловск, обл., Чкалов, Облизд. : 92—93
(распр.); Калниньш А. И., 1950, Охота и охотничье хоз. Латвийск. ССР, Рига,
Латгосизд. : 104—111 (распр., биол., ох.); Копылов И. П., 1950, Дикие копытп.
животн. Иркутск, обл., Иркутск, Обл. изд. : 10—20 (общ., изобр.); М а ч у л ь-
•с к и й С. Н., 1950, Докл. Акад, наук СССР, LXXIII, 6 : 1313—1315 (паразиты);
Peterson R., 1950, Occas. papers Roy. Ontario mus. zool., 9 : 1—7 (сист.); Сер-
жанин И., 1950, Сборн. научн. тр. Инет. биол. Акад, наук Белорусск. ССР, 1 :
141—146 (распр., биол.); Флеров К. К., 1950, Матер, по четверт. периоду, 2 :
61 (эвол.); Черкасов А. А., 1950, Записки охотн. вост. Сибири, Иркутск, Обл.
изд. : 142—175 (биол. ох.); Данилов Д. Н., 1951, Охр. прир., 13 : 93—101 (распр.)
Karpinski I., 1951, Chronmy przyrode, ojezasta, 11/12 : 40—48 (ист., распр.);
Мартынов Н. М., 1951, Охр. прир., 13:71—74 (распр., биол.); Строга-
нова А. С., 1951, Природа, 6 : 53—54 (распр.); Федосов Л. В. и К. Н. Н и-
китин, 1951, Животн. мир Брянской обл., Брянск, изд. «Брянский рабочий» :
6—8 (распр.); Findley, 1951, J. Mammal., vol. 32, N 1 : 116 (быстрота бега);
Шапошников Л. В., 1951, Охр. прир., 13 : 75—92 (пит.); Шварц С. С.,
В. Н. Павлинин и Н. Н. Данилов, 1951, Животный мир Урала, Свердловск,
Обл. издат. : 103—104 (распр.); Ellerman J. and Т. Morrison-Scott, 1951,
Checklist of palaeartic and Indian mammals, London : 373—374 (распр., синон.);
236
CERVIDAE
Bromee-Skuncke F., 1952, Ark. f. zool., Bd. 4, H. 1:1—43 (зубы,
продолж. жизни); Верещагин Н. К. и И. М. Громе в, 1952, Тр. Зоол. инет.
Акад, наук СССР, IX : 1226—1229 (распр.); Казаченко Б., 1952, газ. «Сталин-
ское знамя», Пенза, 17 VIII : 3 (распр.); К и р и к о в С. В., 1952а, Изв. Акад, наук
СССР, сер. географ., 6 : 31—48 (ист., распр.); К и р и к о в С. В., 19526, Птицы и
млекопит. в условиях ландшафтов южной оконечности Урала, М., изд. Акад, наук
СССР : 276—280 (распр., стац.); Корнеев О. П., 1952, Визначник зв!р!в УРСР,
Ки1в, вид. «Радянська школа» : 204—205 (распр.); Лаптев И. П., 1952, Тр. Томск,
гос. унив., 118, сер. биол. : 91—100 (количеств, учет с самолета); Peterson R.,
1952, Roy. Ontario mus. zool., Contribut., 34 : 1—30 (сист., синон.); Сысоев В. П.,
1952, Охота в Хабаровском крае, Хабаровск, Дальгиз : 93—98 (распр., ох.); Ф ле-
ров К. К., 1952, Кабарги и олени (Фауна СССР, 1,2), М.—Л., изд. Акад, наук СССР :
185—212 (общ., синон., изобр.); Янушевич А. И., 1952, Фауна позвоночн.
Тувинск. обл., Новосибирск, изд. Зап.-Сиб. фил. Акад, наук СССР : 61 (распр.);
Янушевич А. И. и И. Благовещенский, 1952, Промыслов, звери и
птицы запади. Сибири, Новосибирск, Облизд. : 127—128 (распр., биол.); Аге-
енко П. Д.иК.Р. Ергольский, 1953, сборн. «Рационализация техники охотн.
промысла», 2 : 60—62 (учет); Altmann М., 1953, Animal kingdom, v. 56, No. 3 :
66—72 (биол.); Г а в e м а н А. В., 1953, Природа, 12 : 116 (наблюдения с самолета,
стац.); Г о дне в Е. Д., 1953, Бузулукский бор, ,М.—Л., Гослесбумизд. : 23—24
(хоз. знач.); Долгушин Л. Д., 1953, Изв. Акад, наук СССР, сер. геогр., 5 : 64—
69 (распр.); Казаченко Б., 1953, Охотничья фауна Пензенск. обл., Пенза, Книжн.
изд. : 19—21 (распр.); Кнорре Е. П., 1953а, сборн. «Преобразов, фауны позво-
ночн. животн.», М., изд. Моск. общ. испыт. прир. : 191—200 (одом.); Кнорре Е. П.,
19536, Зоол. журн., XXXII. I : 140—149 (физиол., одом.); Леб ле Б. Б., 1953,
Пушные звери Арханг. обл., Архангельск, Книжн. изд. : 71—74 (распр.); Наза-
рова И. В., 1953, сборн. «К изучению животн. мира Куйбышевск. водохранилища»,
Казань, Татгосизд. : 36—43 (хоз. знач.); Осокорев С., 1953, газ. «Тамбовская
правда», 13 (6277) : 4 (распр.); Пидопличко И. Г. и В. А. То пачев ский,
1953, Природа, 7 : 101—103 (ист., распр.); Слудский А. А., 1953, Бюлл. Моск,
общ. испыт. прир., отд. биол., LVIII, 2 : 14—18 (распр.); Туров И. С., 1953, Зоол.
журн., XXXII, 5 : 886—892 (вредит.); Erdbrink D., 1954, Geol. en mijnbouw,
v. 16, No. 7 : 301—309 (ист.); Жарков И. В., 1954, Бюлл. Моск. общ. испыт. прир.,
отд. биол., LIX, 5 : 3—8 (пит.); К н о р р е Е. П., 1954, Изв. Коми фил. Всес. географ,
общ., 2 : 53—59 (мигр.); Лихачев А. И., 1954, Молочные железы лосей и их про-
дуктивность, изд. Новосибирск, с.-х. инет. : 1—4; Попов В. А. и др., 1954, Тр.
Казанск. фил. Акад, наук СССР, сер. биол. наук, 3 : 191—194 (биол., хоз. знач.);
Слудский А. А., 1954, сборн. «Природн. очаговость заразных болезней в Ка-
захстане», 2:69—98 (бол.); Верещагин Н. К., 1955, Зоол. журн., XXXIV,
2 : 460—463 (ист.); Егоров Н. Н., 1955, Природа, 1 :111—112 (распр.); Красов-
ский В. П., 1955, Природа, 1 : 119 (распр.); Лаптев И. П., 1955, Заметки по
фауне и флоре Сибири, 18 : 5 (распр.); Лихачев А. И., 1955а, Тр. Томск, гос. унив.,
131 : 269—282 (размн.); Л и х а ч е в А. И., 19556, Новый источник мясной продукции,
изд. Новосибирск, с.-х. инет. : 1—4 (хоз. знач.); Макридин В. П., 1955, При-
рода, 1 : 119 (распр.); Мартынов Н. М., 1955, Природа, 9 : 104—106 (распр.);
Н а с и м о в и ч А. А., 1955, Роль режима снежн. покрова в жизни копытных, М.,
изд. Акад, наук СССР : 1—102 (распр., биол.); Саблина Т. Б., 1955, Копытные
Беловежской пущи,|М., изд. Акад, наук СССР : 148—159 (биол.); Шарлемань Н. В.,
1955, Природа, 4 : 119—120 (распр.); Кнорре Е. П., 1956, Опытная лосеферма,
Сыктывкар, Коми книжн. изд. : 3—34 (одомашн.).
Описание. Животное очень крупной величины. Высота в холке от
170 до 235 см. Длина тела от конца морды до хвоста от 220 до 300 см.
Косая длина туловища составляет около 160 см (Капланов, 1948). Наи-
большая длина черепа взрослых самцов свыше 500 мм (от 540 до 620 мм).
Живой вес взрослых самцов колеблется от 350 до 570 кг.1 Самки несколько
меньшей величины и легче сложены, чем самцы; живой вес их редко
бывает больше 400 кг.
1 Вес отдельных крупных экземпляров доходит до 655 кг (Бутурлин, 1934). В При-
морье лоси более мелки, иногда меньше 300 кг.
ALCES
237
В телосложении обращает внимание длинноногость, высокопередость
и длинная тяжелая голова. Высота в холке значительно (иногда на 15 см)
превосходит высоту в крестце. Холка высокая, производит впечатление
горба. Конечности, особенно передние, длинные, в сочетании с ними
туловище кажется коротким. Шея короткая, массивная.
Рис. 51. Лось (Alces alces L.).
Голова длинная, составляет меньше 1/3, но больше 1/4 длины всего
тела, сильно горбоносая; впереди глаз она суживается, но конец морды
вновь расширен и вздут. Верхняя губа сильно увеличена, спереди четырех-
угольная, мясистая, нависает над нижней губой не только спереди, но
и по бокам. Почти все пространство между ноздрями и до края верхней
губы покрыто волосами. Остатки носового зеркала представлены неболь-
шим овальным или треугольным голым пятном темного цвета на средине
верхней губы. У самцов иногда имеется еще по небольшому пятну около
внутренних краев обеих ноздрей. Предглазничные железы небольшие,
но функционирующие. Глаза с радужиной коричневого цвета и гори-
зонтально расположенным зрачком. Уши длинные, 25—30 см, около
73 длины головы, с заостренными вершинами, снаружи и внутри покры-
тые волосами. На нижней стороне головы, в области гортани и у самцов,
и у самок имеется в различной степени развитый нижний вырост («серьга»,
238
CERVIDAE
или «борода») длиною 20—35 см, покрытый более длинными, чем на со-
седних участках, волосами. Лучше всего «серьга» развита в возрасте
4—5 лет. В дальнейшем она с верхушки подсыхает, крошится и поэтому
становится меньше.
Спина позади холки прямая или вогнутая. Круп сильно свислый.
Хвост короткий, не более 10—15 см. Копыта узкие и длинные, у самцов более
широкие и с более закругленными концами. Боковые копыта сравнительно
хорошо развиты, на передних ногах составляют около половины длины
главных, и при ходьбе по мягкому грунту касаются почвы. Как на перед-
них, так и на задних конечностях имеются межпальцевые железы. На
задних расположены также предплюсневые и плюсневые железы, обозна-
ченные снаружи темными пучками волос, склеенных в основаниях.
Пеньки рогов, отходя от черепа, направляются в стороны перпенди-
кулярно продольной его оси и почти в одной плоскости со лбом. В этой
же плоскости расположены и основания самих рогов, но затем рога за-
гибаются вверх. Типично у взрослых самцов уже вблизи оснований рога
расширяются и образуют широкую плоскую лопату с значительным
числом (до 8 и даже 16 на каждом роге) отходящих от нее в стороны и
вверх пальцевидных отростков. Нормально развитая лопата обычно
состоит как бы из двух частей: 1) передней, или нижней, меньшей, с не-
большим числом отростков и представляющей разветвленный и уплощен-
ный надглазничный отросток, и 2) задней, или верхней, большей, постав-
ленной более вертикально и имеющей большое число отростков. Отростки,
отходящие от лопаты, в отличие от лани, направлены не назад, а вперед,
в стороны и вверх. Вес пары рогов крупного самца может доходить до
15—20 кг, обычно же меньше (у уссурийского лося, например, всего
5—6 кг). Степень развития рогов подвержена сильной индивидуальной,
возрастной и географической изменчивости (рйс. 52). У молодых, а также
у престарелых лосей и у лосей, живущих в неблагоприятных условиях,
лопата может совсем не развиваться. , Отсутствие лопаты характерно
для уссурийского лося; в последнем случае число отростков на каждом
роге обычно четыре: надглазничный, средний и два концевых; если
имеется пятый, то он представляет разветвление первого (надглазнич-
ного) отростка.
Волосяной покров зимой состоит, как и у других оленей, из грубой
ломкой ости и тонкого извитого, но редкого подшерстка. Длина зимней
шерсти на спине около 6—8 см. Волосы ости имеют слабую волнистость
в средней части и утончены как в основании, так и к вершине. Голова,
а также ноги, ниже середины предплечья, и голени покрыты коротким
изогнутым, но не волнистым волосом, плотно прилегающим к коже.
Только пучки волос иа местах плюсневых желез подымаются над общим
покровом. Хвост покрыт одинаковым с туловищем волосом и на конце
не имеет кисти. Волосы на «серьге» и нижней части шеи удлинены до
12—15 см, а по верхнему краю шеи и на холке они образуют подобие
гривы, поднимающейся при возбуждении зверя. Мягче остевой волос
на внутренней стороне ушей, на задней части живота и в пахах.
Общий фон окраски лося темно- или черно-бурый, на боках иногда
почти черный. Вдоль средней линии спины идет черная полоса. Характер-
ного для многих оленей светлого околохвостового зеркала у лося нет.
Голова окрашена одноцветно с туловищем, но конец морды, а часто й
лоб, светлого рыжеватого или серого тона; верхняя губа имеет белесо-
вато-серую окраску. Ноги, начиная с запястий и нижних частей голеней,
светлые с желтым или сероватым оттенком. Передняя их сторона, особенно
ALCES
23!)
над копытами, более темная. Задняя часть живота и внутренняя сторона
ног окрашены светлее туловища, иногда почти белые. Светло окрашен-
ные места ясно отграничены от темных.
Летняя окраска мало отличается от зимней, лишь иногда несколько
светлее. Рыжеватый или серый оттенок, особенно на голове, выражен
сильнее. Кроме того, ость летом короче, а пуховые волосы отсутствуют
вовсе. Пятнистость в окраске молодых отсутствует. Копыта, голое про-
странство между ноздрями (носовое зеркало) и голые места губ черно-
бурого цвета. Половой диморфизм и географическая изменчивость в окраске
выражены слабо.
В черепе (рис. 53) обращает внимание сильно вытянутая ростральная
часть и укороченные носовые кости. Наибольшая ширина черепа меньше
40% его длины. Межчелюстные кости длинные, значительно больше
Рис. 52. Рога европейского лося без лопат (Л) ис лопатами (В).
(Из С. А. Бутурлина, 1934).
длины зубного ряда верхней челюсти (Р2—М3) и не соприкасаются с носо-
выми костями. Передние их концы не образуют расширений. Носовые
кости короткие; их длина менее длины верхнего зубного ряда. Расстояние
между передними концами межчелюстных костей (prosthion) и носовыми
костями больше чем в 2 раза превышает длину последних. Гребешки на
беззубном крае верхних челюстей сближены; расстояние между ними
в самом узком месте меньше длины Р2. Глазницы выступают в стороны
не сильно. Задняя часть шва между лобными костями вздута в виде
бугра; впереди него лобные кости образуют глубокое вдавление, располо-
женное позади уровня надглазничных отверстий. Слёзные кости длиннее
передне-заднего диаметра глазницы. Предглазничные ямки небольшие.
Шов между нёбными и верхнечелюстными костями расположен на уровне
передней половины М1. Линия профиля поперечного затылочного гребня
при взгляде сверху имеет вогнутую форму. По краям лобных костей
позади глазниц, на месте рогов, у самок имеются ясно выступающие
в стороны костные бугорки. Кроме того, в противоположность другим
родам, и у самцов, и у самок на границе лобных и теменных костей имеются
ясно выступающие костные бугорки, достигающие высоты 1 см, как бы
зачатки второй пары рогов. На границе височной впадины и глазницы
в области крыловидного гребня лобная кость входит в контакт с алисфе-
ноидом. Орбитосфеноид не соприкасается с теменной костью. Височные
гребни позади скуловых дуг развиты хорошо, но наружные концы костных
слуховых проходов, при взгляде на череп сверху, видны. В мыщелко-
вую ямку открывается только подъязычное отверстие. В стенку послед-
него открывается и отверстие мыщелкового канала, Слуховые пузыри
240
CERVIDAE
маленькие, угловатые и, если не считать выдающихся на их поверхности
острых шипов, совсем не выступают ниже уровня засуставных отростков.
Коренные зубы брахиодонтные, имеют суживающиеся к вершинам
коронки. Как на верхних, так и на нижних заднекоренных имеются
незначительные добавочные бугорки.1 Задние концы внутренних полу-
луний верхних заднекоренных, как правило, раздвоены, образуя так
называемую «палеомериксову складку».1 2 На наружной стороне задней
доли М3 имеется характерная карманообразная складка. Передне-наруж-
ная и задне-внутренняя дольки последнего, нижнего премоляра (Р4)
образуют одну плотную лопасть, от которой почти до основания кармана
отделены передне-внутренняя и задне-наружная дольки (рис. 31, В).
На втором нижнем премоляре (Р3) хорошо развита передне-внутренняя
доля. Передний нижний премоляр (Р2) имеет на внутренней стороне
хорошо развитые среднюю и переднюю складки. Клыки в верхней че-
люсти отсутствуют как у самцов, так и у самок. Резцы имеют форму
симметричных лопаток и постепенно убывают от внутренней пары (IJ
к клыкам (G).
От боковых пястных (II и V) костей сохраняются только дистальные
концы.
Из анатомических особенностей лося наиболее примечательно устрой-
ство носа с необычайно сильно развитыми и своеобразно устроенными
хрящами. (Jacobi, 1919).
История и географическое распространение (рис. 54). Корни рода
Alces L. ведут, по Флерову (1950), к общему предку с настоящими оле-
нями — нижнеплиоценовому роду Cervavitus Khomenko. Потомками
последнего является средне- и верхнеплиоценовый род Eucladocerus
Falc., отдельные виды которого (например, Е. pliotarandoides Aless.)
обнаруживают в строении лицевого черепа и зубов черты сходства с ло-
сями. Представители рода A Ices впервые обнаруживаются в верхнем плио-
цене или нижнем плейстоцене Тираспольского гравия (Молдавская ССР),
в нижнечетвертичных отложениях р. Ишима в Сибири и в ряде других
мест. В доледниковый период и позднее, вплоть до начала миндель—
рисса в Евразии существовал огромных размеров широколобый лось
(Alces lattfrons Johns.).3 Современный вид, Alces alces L., появляется
на геологической сцене со среднего плейстоцена (миндель—рисе) и весьма
обычен в фауне верхнепалеолитических и, особенно, неолитических
стоянок (Громов, 1948; Пидопличко и Топачевский, 1953).
Современный ареал лося охватывает почти всю таежную зону Европы,
Азии и Северной Америки, местами вклиниваясь в лесостепь и северное
криволесье (лесотундру). В результате интенсивного преследования и
вследствие ряда других, еще невыясненных причин, и северная, и, осо-
бенно, южная границы ареала неоднократно и сильно менялись уже в исто-
рическое время (Бутурлин, 1934). В западной Европе, если не считать
Польши (Lubicz-Niezabitowski, 1934), лось сохранился в ничтожном
числе. Обычным зверем он является до сих пор на большей части Сканди-
навского полуострова и в Финляндии.
В пределах СССР за последние 20—25 лет, в результате принятых
мер по охране сильно истребленного в дореволюционное время лося, имеет
1 Отсутствуют иногда на М2.
2 Признак примитивности.
3 Самая ранняя из известных находок ископаемого лося в западной Европе (Ни-
дерланды) датируется рисской эпохой (Erdbrink, 1954).
ALCES
241
место не только почти повсеместное увеличение численности, но и заселе-
ние этим зверем новых районов, в которых он в течение последних де-
сятилетий совсем не водился (Данилов, 1951). В южной Сибири и Казах-
Рис. 53. Череп Alces alces L.
стане, например, с 30-х годов настоящего столетия граница распростране-
ния лося переместилась к югу на 500—800 км (Слудский, 1953). То же
самое имело место на севере Карелии, Мурмана и на Урале, только
в обратном направлении.
16 Фауна СССР, И. И. Соколов
242
CERVIDAE
На Кольском полуострове северная граница лося в настоящее время
проходит в западной части на широте около 68.5—69° с. ш., от Нотозера
на Пул озеро, дальше через среднее течение р. Вороньей и по Поною.
Изредка появляются лоси даже на побережье Кольского залива. Лось-
является обитателем Кольского полуострова с четвертичного периода;,
однако в конце прошлого столетия он был здесь, как и в северной Карелии,
крайне редок; только за последние 55 лет увеличил свою численность,,
а во многих южных районах стал весьма обычным зверем (О. Семенов-Тян-
Шанский, 1948).
Распространен лось по всей Карелии и лесной зоне Архангельской
области. В последней он держится в бассейнах рр. Онеги, Северной Двины,.
Мезени, доходит до низовьев последней; известны даже случаи захода
лосей на северную часть п-ва Канин (Бутурлин, 1934; Лебле, 1953).
В целом же северная граница леса здесь является и пределом постоянного’
обитания этого вида.
В Коми АССР лоси многочисленны в южной половине республики..
В бассейне самой Печоры они до недавнего времени были сравнительно’
редки; только с организацией Печоро-Илычского заповедника стали уве-
личиваться в числе. Еще недавно считалось, что на Урале северной гра-
ницей постоянного обитания лося считается 65-я параллель. Последние-
данные (Долгушин, 1953) свидетельствуют о том, что даже на западном
склоне они теперь регулярно встречаются до 66° с. ш. (р.Кожима, урочища
Пяседа), а на восточном — еще выше, в значительном числе выходят
в тундру до р. Щучьей, около 67°40' с. ш.; изредка добываются даже на
берегу Карского моря. О появлении лосей в Малоземельской и Болыпе-
земельской тундрах сообщил недавно В. П. Макридин (1955). По данным
этого автора, обычны они на р. Суле и ее притоке р. Сойме; несколько ран
он наблюдал их на Печоре в районе с. Великовисочного; неоднократно
наблюдались зимой 1952 г. в районе г. Нарьян-Мара; обычны, по его сло-
вам, они также по рр. Шапкиной, Куе и Колве.1 В феврале 1952 года
В. П. Макридин обнаружил лося в типичной тундре, более чем в 80 км
к северо-западу от пос. Хоседа-Хард (между 67—68° с. ш. и 58° в. д.).
Зоной сплошного, хотя и не повсеместного распространения лося,,
являются все западные и центральные области европейской части СССР —
от прибалтийских республик до северного Заволжья. Южная граница за
последние 20—25 лет в большинстве районов заметно сдвинулась к югу
(Данилов, 1951). На западе Белоруссии в настоящее время она проходитна
широте 52-й параллели через Брестскую область, где лось еще встречается
в Антопольском и Коссовском районах,1 2 равно как и в Пинском. В Полес-
ской области граница проходит вдоль р. Угорть до Припяти, от которой, не
доходя километров 40 до г. Мозыря, направляется на Жлобин. Далее гра-
ница идет к северу по Днепру и, не доходя 20—30 км Орши, поворачивает
на г. Рославль Смоленской области. В районы Полесской, Гомельской и
Могилевской областей, лежащие к востоку от указанной границы, лоси
заходят только случайно. Больше всего лосей в Белоруссии в областях
Витебской, Пинской и Полесской (Сержанин, 1950).
В свое время сравнительно широко лоси были распространены на
Украине. Остатки их найдены в низовьях Днепра и Буга, в слоях, относя-
1 В бытность мою в этих местах в 1933 г. слышать о лосях здесь в то время не-
приходилось.
2 По данным И. Сержанина (1950), в пределах Брестской области лоси сохрани-
лись только в Беловежской пуще; нет их также в Гродненской и б. Барановичской
•областях.
Рис. 54. Распространение Alces alces L.
XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше; 4 — отдельные заходы в настоящее время.
244
CERVIDAE
щихся к началу нашей эры (Пидопличко и Топачевский, 1953). Во времена
Владимира Мономаха на лосей охотились в Черниговских лесах, а в XV в.
мясом добытых в окрестных лесах лосей кормились отряды войск в г. Луцке
(Шарлемань, 1955). Еще в XVII—XVIII вв. лоси водились в верховьях
Днестра, в некоторых лесах Молдавии и Украины (Кириков, 1952а).
В 60-х годах прошлого столетия лоси держались в лесах около местечка
Дымера, в 40 км к северу от Киева (Похилевич, 1864). Истребленные
к началу XX в., лоси вновь начали размножаться в северных частях
Украины в годы Советской власти. В настоящее время постоянно они дер-
жатся в районах Ровенской и Житомирской областей к северу от 51-й па-
раллели и к востоку до р. Уборть. Заходят иногда в северные части Киев-
ской, Черниговской и Волынской областей, а за последние годы их наблю-
дали в окрестностях Киева, в Хмельницкой, Сумской и, даже, в Винницкой
и Полтавской областях (Храневич, 1925; Корнеев, 1952; Шарлемань,
1955). Весьма вероятно, что многие пункты, из обозначенных на приводимой
карте (рис. 54) как места случайных заходов в пределы Украины, следует
рассматривать как южную границу их современного постоянного ареала.
В Смоленской области лоси распространены во всех районах, кроме
самых южных, куда они заходят только случайно. Населяют лоси также
Брянские леса, особенно в северной и восточной половине этой области.
При этом граница постоянного их обитания проходит, по Д. Н. Данилову
(1951), восточнее г. Унечи к г. Середина-Вуда, отсюда на восток по краю
лесного массива в бассейнах рр. Севы и Навли. Больше всего лосей в лесах
Клетнянского, Жуковского, Дятьковского, Брянского, Карачевского,
Трубчевского и некоторых других районов (Федосов и Никитин, 1951).
В Орловской области лось в былые времена, по-видимому, обитал повсе-
местно (Горбачев, 1915), но в настоящее время постоянно держится только
в северных лесных районах — Хотынецком, Знаменском и Волховском
и только изредка заходит в Дмитровский и Мценский. В Курской области
в настоящее время лосей нет, но еще в конце прошлого и в начале этого
столетия единичные особи встречались в лесах по р. Свапе в пределах
б. Дмитриевского уезда, а раньше, судя по находкам рогов, обитали по
всему краю (Птушенко, 1937 ). В Тульской области лось распространен
главным образом в северо-западной, лесной части; южная граница его,
распространения проходит здесь по линии Болохово—Венев (Данилов,
1951). В Рязанской области постоянно держится в районах к северу от
Оки, а также в восточных, начиная с б. Сапожковского уезда (Туров, 1924).
Регулярные выходы лосей наблюдаются в пределы Ряжского лесхоза.
В Тамбовской области водятся в районах вдоль р. Хны — в Тамбовском,
Рассказовском, Лысогорском, Бондарском, Пичаевском, Моршанском
и Алгасовском, а иногда также заходят в северную часть Знаменского
района. Последние годы они размножились настолько, что появляются
даже в окрестностях Тамбова (Осокорев, 1953). G 1950—1951 гг. неболь-
шое число лосей появилось на территории Воронежского госзаповедника.1
В пределах б. Пензенской губернии лоси водились во всех уездах,
кроме Пензенского и Чембарского (Федорович, 1915), но были весьма мало-
численны. В последние годы они вновь размножились и сейчас в пределах
области насчитывается около 800 голов. Больше всего их в северо-западной
и восточной частях области, в районах Салтыковском, Шемышейском.
Бессоповском, Терновском, Кузнецком, Болыпе-Вьясском, Сосновобор-
ском, Городищенском, Лунинском и Николо-Пестравском. Но проходом
лоси бывают также в Кучкипском, Телегинском и Колышлейском, уже
степных районах (Казаченко, 1953).
ALCES
24S
В Саратовской области лоси впервые появились в 70-х годах прошлого
века, к настоящему времени населяют, хотя и не повсеместно, большую
часть правобережной ее половины. Южную границу их распространения
П. М. Мартынов (1951) проводит от г. Ртищева на восток к р. Медведице,
отсюда вниз по течению последней до р. Широ-Карамыша; далее граница
идет к Татищеву и выходит к Волге на широте Саратова и г. Энгельса.
В наибольшем числе лоси держатся в районах Вольском, Хвалынском,
Черкасском и Б. Карабулакском. Изредка показываются даже в районе
г. Балашова. В 1953 г. лоси зарегистрированы в числе 550—570 голов в 35
районах Саратовской области, появившись в водораздельных лесах Балан-
динского, Лысогорского, Татищевского и других районов, где их раньше
не было вовсе. По-видимому, проникнув из Саратовской области, в этом же
году лоси появились в лесах Фрунзенского, Фроловского и некоторых
других районов Сталинградской области (Мартынов, 1955), а В. П. Кра-
совский (1955) сообщил о заходе пары лосей в Хоперский заповедник
в Новохоперском районе Балашовской области. В левобережной части
Саратовской области лоси держатся главным образом в пойме р. Волги.
Говоря о европейской части ареала, следует упомянуть о Кавказе, где
лося сейчас нет, но где, судя по целому ряду доказательств, он был с конца
плейстоцена и до конца XVIII или начала XIX в.1 Остатки лося, датируе-
мые VIII—XIII в. нашей эры, известны с нижнего Дона, особенно много-
численны среди кухонных остатков древней крепости Саркел (Белая
Вежа) близ ст. Цимлянской (Верещагин, 1955).
В Заволжье южная граница распространения лося, если не считать
узкой приречной полосы в Саратовской области, проходит в средних частях
Куйбышевской и Чкаловской областей. В первой лоси обитают постоянно
вдоль право- и левобережья Волги и к северу от р. Самары, в лесных и
лесостепных районах. Сильно размножились (до 300 голов) лоси в Бузулук-
ском бору, где даже наносят вред лесным культурам (Годнев, 1953).
Неоднократно отмечались они, как случайные звери, по р. Иргиз (Марты-
нов, 1951; Бажанов, 1930) и в других степных районах. Последние годы
имели место заходы их по долине Волги в полупустынные районы
Сталинградской области, в долину р. Еруслана (Строганова, 1951).
В Чкаловской области лоси держатся, кроме Бузулукского, в северных
и северо-западных районах, а также в степных колках бассейна р. Сак-
мары. Южную границу в пределах области Данилов (1951) проводит от
г. Бузулука до с. Сакмары, в 30 км к северо-востоку от Чкалова. В 1950 г.
мною получены достоверные сведения о встречах лосей по островкам леса
в Люксембургском районе. Даршкевич (1950) указывает их даже для Соро-
чинского и Буртинского районов.
По Уральскому хребту, согласно последним данным С. В. Кирикова
(1952), лоси доходят в районе Губерлинской до р. Урала.
В западной Сибири северная граница распространения лося идет от
названного ранее пункта на Северном Урале к Оби и пересекает послед-
нюю севернее Салехарда. По последним данным (Лаптев, 1955), лоси встре-
чаются в бассейне рр. Полуя и Надыма. Дальше граница выгибается
дугой к югу до 62-й параллели и вновь поднимается к северу в среднем
течении Таза (Яковлев, 1930).
Южная и одновременно восточная граница в Зауралье проходит вдоль
верхнего течения р. Уралак оз. Миасс и далее по линии Миасс — Кыштым —
1 Указание на остатки лося в нижне-четвертичной фауне Таманского полу-
острова (Верещагин, 1955) оказалось ошибочным. (
246
cervidae
Касли—Багаряк и по р. Синаре (Данилов, 1951).1 В Челябинской области
лоси держатся в северо-западной части, а А. А. Слудский (1953) считает
их обычными зверями в Троицкой лесостепи. В Курганской области они
доходят до Звериноголовского района на границе с Казахстаном (Слуд-
ский, 1953).
В Омской и Новосибирской областях лоси населяют не только таеж-
ную зону, но и лесостепье, встречаются южнее оз. Чаны. В значительном
числе последние годы появились в районах Омска и Новосибирска (Яну-
шевич и Благовещенский, 1952). С 1948—1949 гг. лоси появились в лен-
точных борах степной части Алтайского края, между Барнаулом и Семи-
палатинском, в частности в Алеусском бору, а также в Ребрихинском и
Ракитовском лесхозах. Имеются, впрочем, неоспоримые данные, что лоси
здесь водились и раньше (Егоров, 1955). Граница распространения лося
в остальных частях севера Алтайского края нуждается в уточнении. Из
южных областей западной Сибири и Урала ареал лося проникает в запад-
ные и северные районы Казахстана. Здесь, во-первых, наблюдаются от-
дельные заходы лосей по пойме Урала до широты Чапаевска и южнее, а
также в окрестности Актюбинска и даже до Джурупа в верховья р. Эмбы.
По последним данным (Слудский, 1953), лоси встречаются почти во всех
районах Кустанайской области. С 1938 г. являются постоянными обитате-
лями островных боров и колков Северо-Казахстанской области и ряда
районов Кокчетавской. В последней лоси доходят до заповедника Боровое.
В Акмолинской области встречаются на юг до Атбасара. Отмечено неодно-
кратное появление лосей в правобережных лесных массивах Павлодарской
области. Еще в XVIII—XIX вв. лоси были постоянными обитателями Вос-
точно-Казахстанской области в пределах Южного Алтая. В настоящее
время их здесь нет и известен лпшь единичный случай захода в 1949 или
1950 г. в район с. Верхубинки (Слудский, 1953).
На Алтае изолированные очаги обитания лося сохранились к востоку
от Катуни, в верховьях ее притока Кадрина, по р. Пыже (приток Бии),
в междуречье Башкауса и Чулышмана, на территории б. Алтайского запо-
ведника (около Телецкого озера) и далее в верховьях Большого и Малого
Абаканов (Дмитриев, 1938). Раньше он встречался, по-видимому, в большей
части горного Алтая. В небольшом числе лоси держатся в горно-лесных
районах Хакассии, а также в Кемеровской области. В Тувинской авто-
номной области лоси обычны в Восточно-Тувинском нагорье и в восточной
части хребта Танну-Ола, тогда как в западной их нет (Янушевич, 1952).
На Енисее северная граница лося, по Н. Наумову (1934), проходит на
широте с. Ярцева,1 2 идет по водоразделу Верхней и Подкаменной Тунгуски до
94—95° в. д., затем круто поворачивает на север и проходит по течению
р. Кочечумы к верховьям Курейки и Хеты до 70° с. ш. Имели место отдель-
ные случаи еще более дальних заходов, например севернее Дудыпты и до
с. Носок на Енисее (Яковлев, 1930).
На Хатанге, как постоянный обитатель, лось встречается до устья Хеты,
а иногда до 73° с. ш. (Толстов, 1916). На севере Якутии граница лося,
в общих чертах, совпадает с таковой леса. На Анабаре он доходит до 70°
(Скалой и др., 1941) (отдельные выходы даже до 72°). Известен случай
добычи лося на одном из островов дельты Лены (Насимович, 1955). На
1 С. С. Шварц с соавторами (1951) южную границу лося на Урале проводят выше—
через Сысерть, Камышлов и Ялуторовск, что едва ли соответствует истинной картине
в настоящее время.
2 А. И. Янушевич и И. Благовещенский (1952) проводят ее значительно севернее,
немного южнее впадения Нижней Тунгуски.
ALCES 247
Колыме, по С. А. Бутурлину (1934), обыкновенен до устья р. Омолона,
встречаясь, по-видимому, и в самой дельте. К востоку от Колымы в качестве
постоянного обитателя лось водитсявбассейнахБольшогоиМалого Анюев.
Крайним районом на северо-востоке, где еще в настоящее время изредка
появляются лоси, являются верховья р. Анадыря (Белопольский, 1932).
Имеются сведения, что в XVIII в. лосей добывали около с. Марково; не
исключена возможность, что в свое время они доходили и до устья Ана-
дыря (Портенко, 1941).
Восточная граница ареала проходит по восточным склонам Колымского
хребта. Изредка лоси выходят на Охотское побережье в районе с. Гижиги;
.далее к югу держатся по всему Охотскому побережью. Иногда их добы-
вают даже в Пенжинском районе (Белопольский, 1931). На Камчатке
лосей нет. В Бурят-Монгольской АССР, Читинской и Амурской областях
лось доходит до государственных границ СССР. Здесь южная граница его
ареала уходит в пределы Монголии и северо-восточного Китая; нет лосей
только в южных степных районах. В Приморском крае южная граница
распространения лося в настоящее время доходит до 44°45' с. ш. В неболь-
шом числе он встречается в верхнем течении р. Имана и Тетюхе.В прошлом
•столетии доходил до верховьев р. Суйфуна, но в настоящее время лосей
здесь уже нет (Капланов, 1948).
В пределах Монгольской Народной Республики распространение лося
ограничивается северными горно-таежными районами Хэнтэя и Прикосо-
голья. В Хэнтэе этот зверь обычен в верховьях рр. Онона, Керулена,
Худэри, Тола, Тэрэлдж, Иро, Меньзя, к югу встречается вплоть до г. Богдо-
Ула. В Прикосоголье лоси встречаются по р. Ури-Гол, особенно много-
численны в левобережье р. Эгин-Гол и в Дархаттах, доходя до восточной
части Танну-Ола. В Хангае встречаются не южнее хр. Хантай-Нуру.
Изредка лоси появляются на правом берегу Нумэргин-Гола, в предгорьях
Б. Хингана. Нет лосей на Монгольском Алтае (Банников, 1954).
В пределах очерченных границ лось распространен не повсюду и весь-
ма неравномерно. Так, на юге европейской части ареала плотность заселе-
ния им даже лесных территорий незначительна. Более высока она в сред-
ней полосе, но к северу вновь убывает (Данилов, 1949). Мало или только
случайно заходят лоси в степные районы Забайкалья (в Даурские степи).
Нет лосей в степях Хакасско-Минусинской котловины, в центральных
районах Якутии (Скалой и др., 1941), а также в некоторых частях Хабаров-
ского края (в южной части Амурской области, в Бикинском и Вяземском
районах; Сысоев, 1952). Нет и, вероятно, никогда не было лося на Саха-
лине.
Подвиды. Более общепринятым является выделение трех подвидов
лося.
1) A. alces alces Linnaeus — Европейский лось. Более мелкая раса.1
Высота в холке до 216 мм. Вес до 500 кг. Окраска туловища сравнительно
однотонная коричнево-бурая. Чисто черные тона отсутствуют. Нижние
части боков, перед живота и верхние части ног только немного темнее шеи
и спины. Сезонный диморфизм в окраске выражен слабо. Голое пятно на
верхней губе (носовое зеркало) близко к ромбической или эллиптической
форме. Нёбные отростки межчелюстных костей черепа впереди сошника
узкие, образуют вместе желобок; их совместная ширина меньше верти-
кального отверстия for. infraorbitale. Восходящие (носовые) отростки меж-
челюстных костей вверху лопастеобразно расширены и заканчиваются
1 Признаки даются по К. Флерову (1952).
248
CERVIDAE
тупо. Передний край хоан закруглен. Носовое отверстие сравнительно'
короткое. Длина его от переднего конца носовых костей до переднего конца
межчелюстных (prosthion) равна или меньше расстояния от переднего конца
носовых костей до средины заднего края черепа. Распростра-
н е н и е: Европа, западная Сибирь к востоку до Енисея, Алтай.
2) A. alces americanus Clinton — Восточно-сибирский, или американ-
ский лось. В целом — более крупная раса (если не считать лосей из южных
областей ареала); высота в холке до 235 см, весдо570кг. Окраска — сильно
отличающаяся на разных частях тела. Шея и верхняя часть туловища
ржаво-бурая или серо-бурая с черным налетом, как бы чепраком охваты-
вающая чисто черный цвет нижней части боков, живота и верхних частей
ног. Разнотонность окраски сильнее всего выражена в конце зимы, слабее
в осеннем меху. Голое пятно на верхней губе (носовое зеркало) грушевид-
ное, Т-образное. Нёбные отростки межчелюстных костей черепа впереди
сошника широкие, плоские; их совместная ширина больше вертикального-
диаметра for. infraorbitale. Восходящие отростки межчелюстных костей
кверху сужены и заострены. Передний край хоан клинообразно заострен..
Носовое отверстие очень длинное. Расстояние от переднего конца носовых
костей до переднего конца межчелюстных (prosthion) обычно значительно-
превышает расстояние от переднего края носовых костей до средины зад-
него края черепа. Распространение: восточная Сибирь, Дальний
Восток, Северная Америка.
3) A. alces bedfordi Lydekker — Уссурийский лось. Характеризуется
незначительными размерами, — высота в холке около 180 см, живой вес
не свыше 400 кг (Капланов, 1948), — укороченной носовой областью
черепа, слабо развитыми рогами без лопатообразных расширений и насы-
щенной темной окраской. Распространение: к югу от Амура,,
в южной части Хабаровского и в Приморском краев.
Биология. Лось — типично лесной зверь. Если заходит на севере-
в тундру, а на юге в степную полосу, то далеко от границы леса не уходит
и придерживается участков, где имеется растительность древесного или
кустарникового типа.
В отличие от благородного оленя, лось, как правило, избегает расчле-
ненного рельефа, отдавая явное предпочтение равнинным ландшафтам и
низинам. В то время как благородный олень предпочитает сухие, возвы-
шенные места, лось, наоборот, любит, особенно летом, сырые, заболочен-
ные участки с замедленным водостоком, по соседству с озерами, медленно-
текущими реками и старицами. В гористых местностях (Алтае, Саянах,
Туве, Приморском крае) придерживается предгорных лесов, пологих
лесных склонов среднего пояса гор, не свыше 1900—2000 м над уровнем
моря, с мягким рельефом, болотами и лесными озерками, широких забо-
лоченных и заросших кустарниками долин речек по соседству с лесом
(Дмитриев, 1938). Если в некоторые сезоны лоси поднимаются в горы выше
пределов лесной растительности, то держатся лишь там, где много болот
и воды, главным образом, по плоскогорьям альпийской зоны. Крутых
и скалистых склонов избегают.
Стационарное размещение лосей меняется не только в зависимости от
природных условий местности, но и по сезонам. Условиями, от которых
зависит выбор животными типов местообитаний в разные сезоны, являются
наличие кормов, глубина и характер залегания снежного покрова,
а также хищники и кровососущие насекомые. ’-м
В зимний период наибольшее значение имеет снег. Благодаря длинным
конечностям, лось свободно передвигается по45—50-сантиметровому снегур
ALCES
249
но уже при глубине 60 см должен применять заметные усилия при беге,
а при 75—80 см передвигается медленно и с трудом. Критической для
лося, по-видимому, является глубина снега 80—90 см, свыше которой,
особенно при недостаточно хорошей кормовой базе, он существовать долго
не может и вынужден переселяться в менее снежные районы (Семенов-Тян-
Шанский, 1948). Вероятно, с этим связано отсутствие лося в ряде северных
районов западной Сибири и на Камчатке, где максимальная глубина
снежного покрова превышает 80 см, хотя остальные условия благоприят-
ствовали бы его существованию.
Плохо переносят лоси высокие температуры воздуха. Поэтому в летнее
время выбор местообитаний, помимо обеспеченности кормами, обусловли-
вается необходимостью спасаться от вредного действия жары и крово-
сосущих насекомых.
В средней полосе европейской части Союза лоси летом, как правило,
держатся в глухих пойменных лесах, по низинам и болотам, за-
росшим сосной и ивняком, по берегам глухих озер и речек по соседству
с лесом. Забираясь в воду и проводя в ней целые часы, лоси не только
находят прохладу, но и корм в виде сочных листьев и корневищ водных
растений. Очень охотно посещают они летом старые вырубки и гари, где,
помимо лиственного древесного подроста, находят излюбленный корм
в виде кипрея (иван-чая) и других сочных растений. Восстановление моло-
дого лиственного леса на местах обширных пожаров считают одной из
причин расширения ареала лося на Урале. Зиму лоси в равнинных райо-
нах проводят также в болотах с сосной и ивняком или в осинниках и берез-
няках, где снег хотя и глубже, но зато кормами звери обеспечены здесь
гораздо лучше, чем в спелых лесных насаждениях. В последние они бы-
вают вынуждены переходить в периоды глубокого снегопада, так как
в лесу снег лежит более тонким слоем. Наблюдения с самолета показали,
что в Калининской области зимой лоси концентрируются в осиновых мо-
лодняках и зарослях ивняка по окраинам болот. При этом в открытые
места не выходят, если их не выгонят хищники, также избегают и густых
ельников (Гавеман, 1953).
На Кольском полуострове летние стации лося в общем те же, что и
в средней полосе Союза: и здесь предпочитаются болота, заросли ивняков,
реки, озера, гари, участки с высоким разнотравьем. Охотно держатся они
также в участках ельников и березняков с широколиственным разно-
травьем; зато ягельных боров и ельников с чернично-вороничным покро-
вом избегают. Зиму лоси проводят в сосновых борах и речных долинах,
так как глубина снега в ельниках больше. При этом они совершают, хотя
и не слишком дальние, но все же регулярные перекочевки к местам зимо-
вок и обратно (Семенов-Тян-Шанский, 1948).
В равнинной тайге западной Сибири, в бассейне р. Демьянки, лоси
болот, по Л. Капланову (1935, 1948), не посещают. Летом они держатся
в лесной полосе вдоль рек, вблизи озер и стариц, в которых проводят зна-
чительную часть суток. Осенью откочевывают к югу и зиму проводят
в светлых лиственных лесах, на участках свежих гарей в мелком осиннике,
березняке и по низинам, поросшим ивняком. Темнохвойных лесов избе-
гают. Масштабы кочевок значительны (до 300 км) и обусловливаются,
в основном, большой глубиной снега в районе летовок и лучшей кормовой
базой на зимовках.
На Алтае и летние, и зимние стации лосей остаются почти одни и те же,
но летом самки живут, в основном, около лесных озер и кустарниковых
долин, тогда как самцы подымаются до гольцовой зоны, где держатся так-
250
CERVIDAE
же по болотам и озерам. Зимой и самцы, и самки концентрируются в ерни-
ковых долинах в прилегающих к ним опушках леса с подлеском из кустар-
никовой березки (Дмитриев, 1938). В Саянах и Восточной Сибири лето
лоси проводят в кедровой тайге, а к зиме опускаются в зону подтайги,
в гривы, поросшие березняком, осиной и'рябиной.
Несколько отличное стационарное распределение по сезонам наблю-
дается в Приморском крае и в Приамурье.
В целом распространение лосей здесь связано с распространением
охотской флоры хвойных лесов, тогда как в районах преобладания расти-
тельности манчжурского типа их мало. В летнее время лоси населяют
разнообразные стации, но все же основная масса концентрируется по
пониженным, заболоченным участкам с обилием воды и сочной раститель-
ности. В Сихотэ-Алине лоси держатся во влажных елово-кедрово-листвен-
ных лесах и гарях по ним, у озерных заболоченных впадин, в поймах рек
с кустарниковыми зарослями, высокогорных ельниках, высокотравных
лугах. По заболоченным долинам иногда доходят до морского побережья,
где ищут защиту от гнуса. Зимой лоси в Приморье сосредоточиваются на
вершинах хребтов и на плоскогорьях. При этом Уссурийский лось, в от-
личие от европейского, по словам Л. Капланова (1948), предпочитает тем-
нохвойные и хвойные леса старым гарям с лиственным мелколесьем.
Излюбленными зимними стациями здесь являются лиственничные, елово-
лиственные и темнохвойные леса с лиственными породами во втором
ярусе, а также прибрежные кустарниковые заросли в верхних частях
долин западных склонов Сихотэ-Алиня.
На крайнем северо-востоке, в бассейне притоков Колымы и Анадыря,
лоси зимой держатся по заросшим тальником островам рек, а летом выхо-
дят отстаиваться от комаров на речные косы (Портенко, 1941). В тундро-
вой полосе Северного Урала стациями их являются островки леса в защи-
щенных долинах, а также заросли ивняков по оврагам и поймам речек
(Долгушин, 1953).
Места зимовок и летовок лосей зачастую отстоят далеко друг от друга,
и животные во многих районах вынуждены дважды в году совершать пере-
кочевки на десятки и даже сотни километров. Так, в Печорской тайге имеют
место ежегодные перекочевки лосей осенью в юго-западном направлении,
а весной в обратном направлении. Наблюдается также ежегодная откочев-
ка лосей на зиму с западного склона Урала на восточный, менее снежный
(Кнорре, 1954). В других местах они переходят лишь из одной стации
в другую в пределах сравнительно небольшого района. В пределах средней
равнинной полосы европейской части Советского Союза значительных
регулярных сезонных миграций лосей не отмечалось. Однако в последние
(послевоенные) годы ежегодно в июне—июле наблюдаются передвижки
лосей в пригородах Ленинграда с заходами в черту города.1 Направление
и причины кочевок лосей здесь пока не ясны; одни годы они обнаружива-
ются в южной части города, а в 1955 г. большая часть их наблюдалась на
северном берегу Невы — на Выборгской стороне и в Новой Деревне.
Сезонные перекочевки лосей совершаются, как правило, по определен-
ным и постоянным из года в год путям. В гористых районах эти пути идут
вдоль речных долин, через невысокие перевалы. В местах, богатых лосями,
они обозначаются тропами. Как правило, кочевки совершаются неболь-
шими группами животных и в большинстве случаев проходят незаметно.
Иногда передвижки на зимние угодья начинаются рано, в сентябре, но
1 Только благодаря этим заходам кочевки лосей и стали известны.
ALCES
251
основная масса зверей откочевывает по окончании гона в октябрей ноябре,
когда глубина снега достигает 25—50 см (Капланов, 1948; Семенов-Тян-
Шанский, 1948; Кнорре, 1954). Время обратных миграций зависит от метео-
рологических условий. Нередко первые подвижки к местам летовок заме-
чаются уже в феврале или марте, но заканчиваются не раньше мая или
июня.
По характеру питания лось еще больше, чем благородный олень,
является древесноядным зверем. Части кустарников и деревьев, начиная
с листвы и кончая корой и древесиной, служат основой питания почти
в течение всего года. Даже в летний период, когда в большом количестве
потребляются в пищу сочные травянистые растения, древесно-веточный
корм занимает видную долю в рационе лося.
Видовой состав растений, употребляемых лосем в пищу, и предпочте-
ние, отдаваемое тому или иному виду, зависят от характера растительности
района его обитания, от доступности кормов, и, само собою разумеется,
меняется по сезонам. Повсеместно излюбленными кормовыми растениями,
особенно в зимний период, являются различные виды ив и осина, но послед-
няя в местах обитания лося на Алтае отсутствует или редка, и основу
питания зимой здесь, наряду с ивами, составляет кустарниковая береза
(Дмитриев, 1938). К числу основных зимних кормов относятся также сосна,
береза, черемуха, тополь.
На Кольском полуострове сосновый подрост, береза и можжевельник
составляют вместе около 80% зимнего корма, но поедаются главным обра-
зом в глубокоснежный период, при недостатке более предпочитаемых ив,
черемухи, осины и рябины (Семенов-Тян-Шанский, 1948). В Бузулукском
бору кроме сосны, зимой пищей служат ветви дуба, бересклета бородав-
чатого, ильма и некоторых степных кустарников (Кнорре, 1949). В других
местах Поволжья поедаются также клен, желтая акация, липа (Мартынов,
1951). В Тульских засеках основу питания составляют дуб, осина, ивы,
тогда как рябина, ясени, клены, липа и другие породы играют второстепен-
ную роль (Лихачев, 1939). Дуб, как и липа с кленом, имеет второстепенное
значение в составе лосиных кормов в Белоруссии, а в списке кормовых
растений лося на Дальнем Востоке Л. Капланов (1948) его совсем не при-
водит. Первостепенную роль в зимнем корме уссурийского лося, помимо
осины, нескольких видов ив и берез, играют таволга иволистная, рододен-
дрон, желтый и зеленокорый клены, а также бородатый древесный лишай-
ник (Usnea barbata L.). В лесах европейской части СССР последний яв-
ляется вынужденным кормом при недостатке прочих кормов (Семенов-
Тян-Шанский, 1948).
У деревьев зимой обламываются и поедаются ветки толщиной до 1 см
и больше, предпочитаются прошлогодние побеги. Зубы лося прекрасно
приспособлены для размалывания древесины, и желудок лося зимой часто
представляет, по выражению С. А. Бутурлина (1934), «мешок с опилками».
У сосны и можжевельника поедаются преимущественно верхушечные
побеги с хвоей, для чего лось нагибает тяжестью туловища стволы тех
деревцов, которые выше роста, и, пропуская их между ног, начисто объ-
едает. Хвое сосны и можжевельника отдается предпочтение, тогда как ело-
вая употребляется в пищу очень редко.
Кроме ветвей, пищей служит также кора. Охотнее всего лоси обгры-
зают и сдирают резцами кору у осины, ивы, рябины, черемухи, менее охот-
но едят кору молодых сосен. С этой целью лоси часто посещают лесосеки,
разбрасывают штабели дров, обгрызают поваленные бобрами осины (Сер-
жанин, 1950). Кора служит пищей поздней осенью, пока не замерзла, и
252
CERVIDAE
весной, когда она оттает, так как мерзлую даже крепкие резцы лося,,
действующие как стамеска, отодрать от древесины не могут. Поздней
осенью и ранней весной, а также и зимою, при неглубоком снеге, лоси
охотно объедают кустики черники.
Основу летнего питания повсеместно составляет сочная травянистая
растительность со значительным участием в рационе листьев с молодыми
побегами кустарников и деревьев. На первом месте среди них и летом стоят
ивы, осина, рябина и некоторые другие лиственные породы, сосна же летом
поедается редко.
Из трав употребляются виды высокого сочного разнотравья. При нужде
лось поедает пырей и полынь, например в степном Поволжье (Мартынов,
1951), но весь склад животного с короткой шеей и длинными ногами, а
также толстая верхняя губа, являются помехой при сборе низко растущих
растений. Для того, чтобы достать мордой до земли, лось вынужден обычно
широко раздвигать передние ноги.1 Даже высокие травы лоси не поедают
целиком, а обрывают лишь верхушки, сощипывают цветочные бутоны,
отдельные листья. К излюбленным растениям относятся кипрей, ради
которого лоси так охотно посещают вырубки и гари, затем таволга вязо-
листная, белокрыльник, медвежья дудка. Объедают кусты смородины,
иногда малины, черники, багульника, голубики. На болотах одним из
важнейших кормовых растений является вахта (Menyanthes trifoliata L.),
различные виды хвощей, пушица, калужница и некоторые другие. Заби-
раясь в воду, лоси в большом количестве поедают рогоз, тростник, кубыш-
ку, аир, кувшинку, рдесты, стрелолист и другие водяные растения; при
этом в пищу идут не только надводные части, но и мясистые корневища,
ради добычи которых лоси погружают голову в воду до дна. К. Г. Абрамов
(1949) включает в список кормов уссурийского лося даже морскую водо-.
росль — ламинарию японскую. Осоки поедаются только на ранней стадии
вегетации.
В Жигулях из летних травянистых кормов лося на первом месте по,
частоте поедаемости стоят сныть, скерда сибирская, володушка золотистая,
кипрей, таволги, два вида злаков и лесной купырь (Жарков, 1954).
Сведения в отношении поедаемости грибов противоречивы. По Туркину
и Сатунину (1902), лоси их не едят также как и ягоды, но на Дальнем Вос-
токе грибы якобы являются даже нажировочным кормом (Абрамов, 1949).
Сена дикие лоси, по-видимому, не едят; иногда лишь разрывают стоги,
выбирая попавшие в сено древесные ветки и отдельные лакомые растения
(Бутурлин, 1934). Посевов культурных растений, как правило, не трогают,
хотя и имеются отдельные указания на потравы лосями овса в стадии нали-
вания зерна и озимей (Нарышкин, 1900).
На Урале, Алтае, в Восточной Сибири и Дальнем Востоке лоси, как,
и другие копытные, посещают соленые источники, а также естественные
и искусственные солонцы. Предпочтение, по-видимому, отдается первым.
Солонцы посещаются с весны и до начала зимы, пока почва не замерзнет
настолько, что звери не могут пробить замерзшую корку копытом. В рав-
нинной части Сибири и в европейской части СССР солонцевание лосей
не известно. Попытки приучить их пользоваться искусственными солон-
цами в б. Лапландском заповеднике окончились неудачей — звери прохо-
дили их мимо, не обращая внимания на них (Семенов-Тян-Шанский, 1948).
Большую часть года лоси держатся небольшими группами в 3—5 голов,
состоящими из 1—2 взрослых самок и их потомства: очень редко, особенно
1 Это хорошо видно на фотографии лося в работе Л. Г. Капланова (1948 :108).
ALCES
253
в летнее время, можно встретить в подобной группе и взрослого самца.
Самцы вне периода гона держатся отдельными группами по 2—3, а еще
чаще — поодиночке. Лишь зимою по окончании гона лоси иногда соби-
раются в небольшие табунки по 7—10 голов. Самые большие группы, кото-
рые удается наблюдать зимой в богатых лосями местах, 15—18 голов
(Бутурлин, 1934).
Лось по натуре мало подвижное животное. Если он в достаточном коли-
честве находит пищу и никем не тревожим, то может долго держаться на
одном месте. Особо оседлый образ жизни лоси ведут зимой, в период глубо-
коснежья. Известны многочисленные примеры, когда группа лосей прово-
дила почти всю зиму на весьма ограниченном участке — на лесном острове
в болоте, в долине речки. О. И. Семенов-Тян-Шанский (1948) приводит
случай, когда лось в течение 5 дней держался на площади в 10 га. По-
движность несколько повышается в период гона, а также в сезоны осенних
и весенних перекочевок. Осенью лоси делают переходы по 5—6 км в сутки,
зимой гораздо меньше — всего 0.7—0.9 км, но зато сильнее объедают ку-
старники и деревья (Семенов-Тян-Шанский, 1948).
Значительную, а зимой даже большую часть суток лоси проводят
в лежке. Принимаются за еду 5—6 раз, чаще всего по утренним и вечерним
зорям, а также ночью. Отдыхать ложатся также по нескольку раз в сутки
или в тенистой чаще по соседству с местами жировок, или на возвышенном
месте, откуда кругом хорошо видно. Летом, спасаясь от жары и жалящих
насекомых забираются в воду, нередко по шею, и кормятся водными расте-
ниями или отдыхают, лежа на мелководье. В жаркие дневные часы часто
выходят на обдуваемые ветром речные косы и даже морские побережья.
В Приморье лоси ходят определенными тропами, иногда за несколько
километров, на водопои и к солонцам. На последние они приходят перед
закатом солнца и нередко задерживаются до 8—10 часов утра. Хорошо
протоптанные «лосиные дороги» известны и в других богатых лосями ме-
стах Сибири (Капланов, 1935, 1948).
В спокойной обстановке лоси передвигаются шагом. Преследуемые же
или испуганные бегут рысью 1 со скоростью 15—16 км в час (Бутурлин,
1934): Известен случай, когда лосиха, преследуемая на автомобиле, пробе-
жала за час 22 мили (около 35.3 км; Findley, 1951). На галоп переходят
обычно на исходе сил, при сильном утомлении. И при ходьбе и на бегу лось
высоко поднимает ноги. Во время бега слышится легкий треск или пощел-
кивание, по-видимому, происходящее в суставах пальцев. Длинные конеч-
ности, снабженные к тому же хорошо развитой кожной складкой между
копыт (Рососк, 1910), позволяют лосю преодолевать не только снег свыше
метра глубиною, но и топкие болота, по которым даже человек часто пройти
не может. Несмотря на кажущуюся неуклюжесть сложения, лось легко
перепрыгивает через рвы, поваленные деревья, высокие кочки и другие
препятствия на пути.
Прекрасно плавает, легко переплывает большие реки и озера шириною
в 1—2 км.
Из внешних чувств у лося лучше всего развит слух. Даже осторожно
идущего человека он может услышать за километр. И во сне с закрытыми
глазами лось обычно все время поводит ушами (Семенов-Тян-Шанский,
1948). Обоняние и зрение, по-видимому, развиты слабее (Бутурлин, 1934).
1 Утверждения Н. Нарышкина (1900) и С. А. Бутурлина (1934) о том, что лоси бе-
гают только иноходью, в опытах по одомашниванию не подтвердились (Попов, 1939;
рисунок в работе Кнорре, 1939).
254
CERVIDAE
. Там, где их преследуют, лоси чрезвычайно боязливы и осторожны. Но
многочисленные сообщения за последние годы о случаях заходов их в пре-
делы даже таких крупных населенных пунктов, как Москва и Ленинград
(рис. 55), свидетельствуют о том, что пугливость и осторожность не явля-
ются прирожденными свойствами лося и всецело зависят от отношения
к нему человека.
За исключением периода гона, лоси молчаливы, самка подзывает
теленка коротким, отрывистым, часто повторяющимся стоном (Капланов,
1948). Голос, издаваемый новорожденным «ммя-ммя». По Нарышкину
(1900), лоси перекликаются звуком, похожим на спаренное «у-о» с выдохом
на последнем слоге. Испуганные издают хриплые звуки напоминающие
грубое взлаивание (Абрамов, 1949). Самцы во время гона издают громкий
рев, иногда слышимый по ветру за 2 км. Смертельно раненый лось не кри-
чит, а стонет.
Линька у лосей происходит один раз в году. Первые признаки ее заметны
уже в апреле, разгар падает на май и первую половину июня, а на севере,
например в Мурманской области, на 2—3 недели позднее. Первыми выпили-
вают лучше упитанные взрослые самцы, за ними молодняк, а последними
беременные самки. В массе линька заканчивается в большинстве районов
к концу июня, но истощенных зверей с клочьями старой шерсти можно
видеть и в конце июля. Старая шерсть выпадает в первую очередь на голове
и ногах, дольше удерживается на спине и крупе. Вновь отрастающая шерсть
вначале редкая и очень темного цвета. К сентябрю она подрастает и ста-
новится гуще, появляется подшерсток. Полного развития зимний мех
достигает в октябре.
В большинстве районов рога взрослыми самцами сбрасываются во
второй половине ноября и в декабре. Молодые лишаются их позднее —
в январе или феврале. В восточной Сибири, по И. П. Копылову (1950 : 16),
быки начинают терять рога не ранее февраля или марта.1 В противопо-
ложность благородным оленям, новые рога начинают отрастать не сразу
после спадения старых, а позднее, начиная приблизительно с марта.
Полного развития рога достигают в конце июля—начале августа. В ав-
густе же происходит очистка окостеневших рогов от кожи, хотя у молодых
быков можно видеть их в коже еще в сентябре. Испытывая раздражение,
лоси ударяют и трутся рогами о кусты и деревья, сдирая при этом большими
лоскутами кожу. Первые 2—3 дня после освобождения от кожи на белых
рогах видны полосы крови, но вскоре они приобретают типичную коричне-
вую окраску.
Половой зрелости лоси, по данным Е. П. Кнорре (1956), достигают, как
правило, на третьем году жизни, реже в 3 % года и совсем редко в воз-
расте 1 % лет. Самцы фактически начинают участвовать в размножении
не раньше, чем на четвертом году, так как до этого возраста отгоняются
взрослыми, более сильными соперниками. Начало полового сезона у лосей
обнаруживается ревом самцов. Первых ревущих самцов можно слышать
уже во второй половине августа, но, как правило, начало рева падает на
первые числа сентября. В европейской части ареала самцы ревут по опуш-
кам леса, на лесных полянах, в островах леса среди болот. Рев скорее
напоминает стон, трудно выражаемый звуками человеческого голоса; одни
передают его как «иох-иох» (Абрамов, 1949), другие сравнивают с корот-
ким отрывистым рявканьем, мычанием или со звуком удара обуха топора
1 Сведения требуют проверки, так как А. А. Черкасов (1950 : 144) указывает
сроки, не отличающиеся от других районов.
ALCES
255
о древесину (Туркин и Сатунин, 1902); ни одно из этих сравнений нельзя
назвать удачным. Ревут чаще по вечерам и утром, а иногда даже и ночью.
Следует отметить, что рев не везде и не всегда выражен ярко. Например,
нет его в Печорской тайге (Теплов, 1948); слабо и вовсе не наблюдается
в Саянах (Соловьев, 1920).
Самки приходят в охоту, по-видимому, несколько позднее самцов и на
этот период покидают своих телят. Находящаяся в охоте лосиха отвечает
на рев самца своеобразным мычаньем или храпом. В отличие от благород-
Рис. 55. Молодой лось на окраине Ленинграда, сентябрь 1955 г. (Фото автора).
пых оленей лоси — моногамные животные. Большей частью самца в поло-'
вой сезон можно видеть с одной самкой. Но моногамия эта довольно условна.
По наблюдениям Кнорре (1949), изредка (в 10% наблюдаемых случаев)
можно видеть около одного самца несколько самок. В домашней обстановке
один молодой самец покрыл за сезон трех самок. Не исключена возмож-
ность, что и в природе пары могут распадаться и сильный самец последо-
вательно оплодотворит несколько лосих.
Самцы в период гона сильно возбуждены, ударяют рогами о кусты и
деревья. У них сильно утолщается шея. Они обнюхивают самок; на том
месте, где мочилась самка, выбивают копытами ямку, сами в нее мочатся,
иногда валяются в этой грязи (Бутурлин, 1934), лижут мочу самок, вытя-
гивая голову и оттопыривая верхнюю губу (Капланов, 1948). Самцы в этот
период, вероятно вследствие усиленного секретирования кожных желез,
издают резкий специфический запах. Последний ощущается даже на сле-
дующий день на том месте, где были лоск. При встрече двух самцов, между
256
CERVIDAE
ними происходят поединки рогами, иногда кончающиеся увечьем или
смертью (Теплов, 1948).1 Самцы в период гона совсем мало едят и за это
время сильно худеют. Самки ведут себя гораздо спокойнее и заметно упи-
танности не теряют. Спаривание происходит быстро, но самец кроет самку
несколько раз.
По наблюдениям на Дальнем Востоке гон у лосей протекает в более
сжатые сроки, чем у изюбрей (Капланов, 1948). Разгар его падает на вто-
рую половину сентября и начало октября, а заканчивается гон к середине
или концу октября. Позднее ревут лишь отдельные молодые самцы.
Гон из года в год происходит в одних и тех же местах.
Беременность у лосих длится, по наблюдениям в зоопарках, около
240 дней с уклонениями до 10 дней в обе стороны (Подсосов, 1934, 1935).
Отел в большинстве районов происходит в мае и июне, но в Саратовской
области, по Мартынову (1951), самки начинают телиться в начале апреля,
а заканчивается отел к концу мая. На Мурмане, наоборот, он начинается
только в конце мая итянетсяиногдадо началаиюля (Семенов-Тян-Шанский,
1948). Перед тем как родить, самка удаляется из табуна, но специального
места для родов не избирает, и последние проходят там, где придется.
Новорожденный лосенок имеет короткое туловище и необычно длин-
ные ноги. Длина тела около 70 см, высота в холке 70—90 см. Вес новоро-
жденных европейских лосей от 6 до 16 кг, средний вес нормального
теленка 9—12 кг (Кнорре, 1949, 1956). Число телят в помете, как и вес,
зависит от условий эмбрионального развития. В европейской части СССР
и западной Сибири взрослые самки чаще приносят по два'теленка, молодые
обычно по одному. На Мурмане и в Бузулукском бору двойню родят меньше
половины лосих (Семенов-Тян-Шанский, 1948). В Печоро-Илычском запо-
веднике, после многоснежных зим, большой процент самок имеет только
по одному теленку. На Колыме лосихи нормально приносят по два теленка
но в восточной Сибири и на Дальнем Востоке, за редкими исключениями,
только по одному. По некоторым наблюдениям, самцов рождается несколь-
ко больше, чем самок.
Родится лосенок слабым и беспомощным; он едва подымается через
несколько часов, чтобы пососать молока. В течение первых 5—7 дней он
лежит, затаившись в кустах, и легко может быть пойман руками; но уже
с 7—10-дневного возраста человеку догнать его не всегда удается (Кнорре,
1953а). В противовес общепринятым представлениям, самка не пытается
защищать теленка от человека. Приблизительно до месячного возраста
лосенок большую часть времени лежит, а мать кормится неподалеку.
Только с месячного возраста теленок всюду следует за матерью и не от-
стает от нее в беге. На Дальнем Востоке самки с телятами начинают появ-
ляться на солонцах с конца июня (Капланов, 1948). Зеленый корм начи-
нают принимать понемногу с двухнедельного возраста, а окончательно
переходят на растительное питание с сентября или октября, когда мать
покидает детей на период гона.
Лактирующая способность самок лосей, принимая во внимание их
крупную величину, не высокая. Как и у других оленей, молоко выде-
ляется из всех четырех сосков, но, как установлено, из задней пары оно
выделяется в значительно большем количестве, чем из передней. По опы-
там на Печорской лосеферме (Кнорре, 1956), суточные удои подопытных
лосих не превышали 3 л. Пара лосят высасывала естественным путем
1 Прекрасная фотография двух сцепившихся рогами и погибших самцов имеется
в статье Горбачева (1915).
ALCES
257
у матери до 3.8 л молока в сутки. У молодых лосих первого отела молоч-
ность раза в два уступала таковой взрослых самок.
Лосиное молоко белого (а по А. И. Лихачеву (1954) кремового) цвета,
гуще коровьего, без запаха, но специфического терпко-солоноватого
вкуса. Прибавленное к сладкому чаю или кофе, оно производит впечат-
ление коровьих сливок. Жирность молока высокая, колеблется в раз-
ные периоды лактации от 6—8 до 19% (в среднем, по-видимому, около
12%). Химические анализы показали, что оно содержит сухого вещества
около 18%, белков 9—15%, молочного сахара свыше 5%, зольных веществ
1—1.5%. Плотность в градусах ареометра —43 (Лихачев, 1954; Кнорре,
1956).
Продолжительность лактации в норме 3—4 месяца, но если не после-
довало оплодотворения в очередной половой сезон, может продолжаться
до 7—9 месяцев.
До 3—4 месяцев лосята имеют однотонную буровато-рыжую окраску,
которая сереет к осени, а зимой делается сходной со взрослыми лосями.
Как уже говорилось, на период гона самка покидает телят, но затем
они вновь разыскивают мать и держатся с ней по крайней мере до следую-
щего отела. Нередко с лосихой можно встретить приплод не только этого,
но и предшествующего года.
Развитие молодых идет быстро. Уже к шестимесячному возрасту евро-
пейские лоси достигают живого веса 120—130 кг, а отдельные экземпляры
160 кг. Средний вес полуторагодовалых составляет 250, а в ЗУг года
300 кг (Кнорре, 1949, 1956).
. У новорожденных самцов на месте рогов имеется только завихрение
волос. В возрасте около 6 месяцев под кожей на голове прощупываются
бугорки — начало роста костных пеньков. Последние продолжают расти;
весной кожа на их верхушках лопается и начинается рост сменяемой
пасти рога (Туркин и Сатунин, 1902). Первые рога в виде простых спичек
длиной 20—30 см вырастают и очищаются от кожи в возрасте 16—17 ме-
сяцев. На третьем году формируются рога, с двумя концами каждый,
так называемые «вилки». В начале четвертого года рога имеют по три
конца и в этом возрасте намечается образование лопаты в виде легкого
уплощения у оснований отростков. На пятом году, наряду с появлением
четвертого отростка, имеется хотя и небольшая, но ясно выраженная
лопата. Вполне развитая лопата у европейского и американского лося
образуется, когда животное достигнет полных пяти лет.1 В этом же воз-
расте вырастает и пятый отросток (Бутурлин, 1934). От этой общей схемы
имеются многочисленные уклонения, зависящие от условий индивиду-
ального развития животных, в частности от условий питания (Капла-
нов, 1948). После шести лет число отростков продолжает увеличиваться,
но закономерность нарушается и вес рогов возрастает главным образом
за счет их утолщения. К старости рога дегенерируют, — не только умень-
шаются их размеры, но и число отростков. При кастрации лосят до на-
чала роста у них рогов последние не развиваются вовсе (Кнорре, 1953а).
Возрастные изменения в зубах и методы определения возраста лося
по ним изучены очень слабо. Известно, что лосенок родится с тремя па-
рами резцов, но без клыков (Семенов-Тян-Шанский, 1948) и с двумя
из молочных премоляров. В годичном возрасте происходит смена двух
внутренних пар резцов (I г и 12), но премоляры сохраняются молочные.
Моляры начинают вырастать с восьми месяцев. Полное развитие зубов,
1 Как известно, уссурийский лось лопаты на рогах вообще не имеет.
17 Фауна СССР, И. И. Соколов
258
CERVIDAE
в том числе смена молочных резцов, клыков и премоляров, заканчи-
вается к возрасту 16 месяцев (Bromee-Skuncke, 1952).
Продолжительность жизни лося — в среднем 15-—16 лет, макси-
мально 23—25 лет (Flower, 1931; Bromee-Skuncke, 1952).
Врагами лося в первую очередь являются крупные хищники, но он
страдает от них, по-видимому, меньше, чем другие копытные звери. Изве-
стны случаи, когда лось убивал передними ногами не только волка, но
и медведя. Волки отваживаются нападать на взрослого лося только
стаей, загоняя его на лед или гоняя по насту и глубокому снегу. В боль-
шинстве малоснежных районов они, как вредители лося, серьезного
значения не имеют. Более серьезным врагом является медведь, в пита-
нии которого, например на Кольском полуострове, лось играет видное
место. В свое время в Центральном лесном заповеднике (Калининская
область) прирост популяции лосей был приостановлен вследствие чрез-
мерного истребления медведями (Семенов-Тян-Шанский, 1948). Большой
ущерб лосиному поголовью медведи приносят также на Дальнем Востоке.
Они подкарауливают лосей, догоняют по насту, глубокому снегу, и даже
по чернотропу, в последнем случае пользуясь тем, что лоси плохо пере-
носят жару и скоро утомляются от быстрого бега (Абрамов, 1949). От
волка и от медведя гибнет больше молодых лосей, чем взрослых. Тигры
в Приморском крае на лосей нападают редко, так как основу их питания
составляют кабаны. На Печоре вредителем является также росомаха,
от которой в многоснежные годы гибнет лосей больше, чем от медведя
(Теплов, 1948). Однако, с крупным зверем росомаха справляется редко.
Жертвой рыси и других мелких хищников могут быть только телята,
и то случайно.
Сильно страдают лоси от кровососущих насекомых — слепней, до-
ждевок, комаров, мошек. Местами на них нападают в большом количестве
клещи. В первую очередь кровососы садятся на места, покрытые корот-
кими волосами: на ноги, особенно выше запястных и скакательных су-
ставов, на уши, растущие рога. Спасаясь от них, лоси вынуждены скры-
ваться в тени, отстаиваться на хорошо обдуваемых ветром открытых ме-
стах, даже забираться в воду.
Вредителем лося является носоглоточный овод Cephenomyia ulrichii Br.,
паразитирующий в личиночной стадии и приносящий животным беспо-
койство, главным образом весной и в первую половину лета. Поселяясь
в большом числе в носовой полости и носоглотке, личинки вызывают
воспаление и набухание слизистой оболочки, затрудняют дыхание, являясь
причиной кашля и исхудания животных. Распространен этот вид овода
по всей Палеарктике и паразитирует только на лосе. Редким, случай-
ным паразитом является второй вид носоглоточного овода — Pharyngo-
myia picta Meig. Паразитирует на лосях в некоторых районах (по Черка-
сову, например, в Забайкалье и на Дальнем Востоке) подкожный овод
изюбря, Hypoderma diana Вт.1 В большом числе находят на убитых лосях
также оленью муху-кровососку — Lipoptena cervi L. Из клещей на лосях
зарегистрированы, по данным Г. В. Сердюковой (устное сообщение),
два вида — Ixodes persulcatus Sch. (вероятно нападает и I. ricinus L.)
и Dermacentor pictus Herm. Внутренние паразиты лося изучены слабо.
1 По устному сообщению специалиста по оводам К. Я. Грунина, имеющиеся в ли-
тературе данные о паразитировании на лосе нескольких видов подкожных оводов
(Kostron, 1938) не соответствуют действительности. К. Я. Грунин даже не уверен в том,
что подкожный овод изюбря способен проходить полный цикл развития на лосе.
ALCES
259
G. H. Мачульский (1950) у лосей из Бурят-Монгольской АССР обнару-
жил 10 видов паразитических червей.
Лоси поражаются рядом эпизоотических заболеваний, свойственных
домашним животным. Наблюдались массовые падежи от сибирской язвы,
в результате которых численность лосей резко снижалась (Капланов,
1935; Слудский, 1954). По данным последнего автора, наблюдались слу-
чаи заболевания лосей ящуром1 и чумой крупного рогатого скота. Есть
указания также на некробациллез, поражающий копыта (Туркин и Сату-
нин, 1902), а также на эмфизематозный карбункул (Кнорре, 1949).
Хозяйственное значение. Лось прежде всего ценен как крупный
мясной зверь. При правильном планировании и надлежащей организа-
ции промысла лосиное мясо может иметь большое значение в создании
продовольственной базы промышленности в ряде районов, особенно азиат-
ской части ареала.1 2
По С. А. Бутурлину (1934) и Кнорре (1939), туша и пригодные в пищу
внутренности лося составляют около 60 % живого веса зверя, а по А. И. Ли-
хачеву (19556) — в период лучшей упитанности даже до 64—70%. Следо-
вательно, от средней величины самца можно получить 250—280 кг чистого
мяса.3
Годовалый лось из Приамурья дает тушу около 70 кг (Абрамов, 1949),
а европейский — не менее 90—100 кг. Самцы лучшую упитанность имеют
в конце августа перед началом гона, а самки в сентябре и октябре. От
хорошо упитанного лося получается свыше 30 кг внутреннего и подкож-
ного сала. Слой последнего на крупе может превышать 5 см (Туркин и
Сатунин, 1902; Капланов, 1935).
Вкус мяса лосей приятный, за исключением периода гона, когда у сам-
цов оно имеет неприятный специфический запах. По исследованиям,
произведенным в лаборатории Новосибирского сельскохозяйственного
института, отмечается большое содержание в нем белков (22.0—24.5%)
и высокая калорийность (до 2148 калорий). Цвет жира белый, а в расплав-
ленном состоянии желтоватый. Характерна его твердость и тугоплавкость.
По данным той же лаборатории, точка плавления внутреннего жира 50.6°,
подкожного 45.2° и околохвостового 45.4°. Вес костей в туше лосиного
мяса не превышает 8—9% (Лихачев, 19556). Особым лакомством среди
охотников считается верхняя губа в отваренном виде или в виде студня.
Ценным продуктом, получаемым от убитого лося, является кожа.
Из нее при выделке получается лучшая, чем из оленьей кожи, замша.
Из выделанной домашним способом лосиной замши шьют куртки, брюки,
рукавицы. Легкие шкуры молодых лосей употребляются на изготовление
дох и другой зимней одежды. Из шейной части выделывают даже подошвен-
ную кожу. Кожа, особенно с ног, идет также на ремни для конской упряжи.
1 Ящур среди лосей наблюдался в Ленобласти в 1957 г.
2 В годы Великой Отечественной войны через г. Абакан Хакасской автономной
области транспортировалось значительное количество лосиных и кабаньих туш, за-
готовленных в Минусинской тайге и Тувинской области.
3 Произведенное нами взвешивание по частям взрослого лося самца, добытого
15 марта 1949 г. во Всеволожском районе Ленинградской области, дало следующие
результаты: вес туши мяса без нижних частей конечностей (убойный вес по боенской
терминологии) 207 кг, вес кожи 30.3 кг, головы (без кожи и рогов) 13 кг, печени 6.5 кг,
сердца 2.6 кг, почек 1.2 кг, легких 3.5 кг, языка 1.1 кг, желудка (без содержимого)
7.8 кг, селезенки 2.4 кг (переполнена кровью), головного мозга 410 г, семенников
80 г, тонкого кишечника (без содержимого) 1240 г, толстого кишечника (без содержимого
и без прямой кишки) 1440 г, длина тонкого кишечника 38.25 м. Аналогичные данные по
западносибирским и уссурийским лосям имеются в работе Л. Г. Капланова (1935, 1948).
17*
260
CERVIDAE
Из толстой кожи с нижних частей ног (камысов) шьют весьма прочную
обувь (унты и др.) волосом кверху. Кожа с камысов употребляется также
на подбивку лыж. Из сухожилий ног делаются прочные, не боящиеся
сырости нитки, а кожа с головы на Дальнем Востоке идет на шитье кра-
сивых ковриков — камаланов (Сысоев, 1952).
Длинная зимняя шерсть лося, получающаяся при выделке кожи,
годится для набивки мебели, седел, матрацев. Рога имеют больший спрос
как украшение. Могут они идти также на различные поделки и выварку
клея.
Лось может приносить и вред в условиях культурного лесоводства.
Объедая верхушечные части деревьев, например сосны, сдирая кору, он
препятствует нормальному возобновлению леса и губит молодые посадки.
Такие случаи известны за границей (Bley, 1910) и у нас в Бузулукском
бору, где лоси не только «подстригают» верхушки молодых сосен, но и
губят иногда тополевые посадки (Годнев, 1953). О существенном вреде
от лосей сосновым посадкам в Татарии сообщалось в самые последние
годы (Назарова, 1953; Попов и др., 1954). В связи с тем, что в настоящее
время имеет место заселение лосями лесостепных районов, необходимо
считаться с вредом, который они могут нанести полезащитному лесо-
разведению. В большинстве же областей ущерб, приносимый лосем лесу,
настолько ничтожен, что ни в какой мере не может идти в сравнение
с его промысловым значением.
Охота. Способы добычи лосей многообразны; некоторые из применя-
емых на промысле марала и благородного оленя практикуются и при
охоте на лося. Подкарауливают их на солонцах, водопоях или у озер
и рек, в которые звери забираются, спасаясь от насекомых. К находя-
щимся в воде лосям можно, соблюдая осторожность, подъезжать на лодке.
Лучше, если последняя замаскирована по бортам тростником или вет-
ками кустарников, растущих в окружении. Иногда догоняют и бьют
плывущих лосей с лодки, так как скорость их на воде не превышает 2 км.
Там, где наблюдается достаточно интенсивный рев быков, применяется
охота «на трубу», или «вабу», заключающаяся в подражании голосом
или с помощью трубы реву зверей. В принципе эта охота не отличается
от таковой на благородного оленя (см. стр. 232). Успех ее зависит от ис-
кусства охотника подражать лосю и от уменья соблюдать необходимые
меры1 предосторожности при подходе. Облавные охоты—весьма распростра-
ненный способ, но применяются главным образом охотничьими коллекти-
вами, так как требуют сравнительно большого числа участников.
Почти повсеместно, где в более или менее значительном числе дер-
жатся лоси, распространена охота на них с лайками. Иногда достаточно
одной хорошей собаки, но чаще двух-трех и более. Пущенные по следу,
собаки догоняют лося, забегают вперед, лаем и хватками за морду останав-
ливают зверя. Лось, отбиваясь от нападающих собак, сравнительно
легко подпускает на выстрел спешащего на лай охотника. Успех охоты
зависит от злобности собак, уменья их задержать зверя, а также и от
охотника, который и в этом случае должен соблюдать необходимую скрыт-
ность. Легче и скорее останавливаются старые самцы, но от ударов их
копыт гибнут нередко и опытные собаки. Охота с гончими на лосей теперь
применяется редко.
Обычным способом является охота «скрадом», применяемая с начала
зимы и до глубокого снега. Найдя след лося, охотник «скрадывает» его,
что легче удается в пасмурную и ветреную погоду. Основное правило
при этой охоте — не идти по следу зверя, так как, поняв, что его
ALCES
261
преследуют, и обладая прекрасным слухом, лось охотника никогда
не подпустит. Следует идти стороной, обходить кругом или вперед, стараясь
выходить в места, которыми может пройти лось. Успех охоты всецело
зависит от осторожности охотника, знания и уменья его использовать
привычки зверя.
Охота нагоном производится двумя или несколькими охотниками
и также, обычно, по неглубокому снегу. Выследив или обойдя лосей, один
или часть охотников занимают места, где могут пройти звери, а осталь-
ные, зайдя в круг, неторопливо направляют их и гонят на затаившихся
стрелков.
Нерациональные и хищнические способы добычи лося (преследо-
вание по насту и глубокому снегу, ловля в ямы, расстановка самостре-
лов и т. п.) сейчас применяются крайне редко и запрещены законом.
Неспровоцированные нападения лосей на человека, хотя и известны,
но довольно редки (Бутурлин, 1934). Раненый лось очень опасен и при
неосторожном подходе к нему может убить ударом передней ноги охот-
ника наповал. Преследование не слишком тяжело раненного рекомен-
дуется возобновлять не в тот же день, так как сгоряча лось может уйти
далеко, а оставленный в покое вскоре ложится, ослабевает и легче по-
зволяет взять себя на следующий день. Следует отметить, что па терри-
тории Европейской части СССР и многих областях Сибири охота на лосей
или вовсе запрещена, или производится по лицензиям, выдаваемым охот-
ничьим коллективам.
Одомашнивание лося. Мысль об одомашнивании и использовании
лося в качестве транспортного животного, благодаря его величине и силе,
весьма заманчива и не нова. Попытки приручения лосей издавна пред-
принимались во многих местах. Имеются сведения о том, что будто еще
во времена Карла IX в Швеции ездили на лосях (Brook, 1827).
Многочисленные сообщения о приручении и использовании лосей
появлялись и в нашей, еще дореволюционной литературе. Однако во
всех описываемых случаях речь шла о единичных, чисто любительских
попытках именно приручения, а не об одомашнивании лося.
В более широких масштабах, в плановом порядке и на научной основе
эти работы стали проводиться в Советском Союзе. Опыты по одомашнива-
нию лося были поставлены на Якутской опытной станции Института Поляр-
ного земледелия (Попов, 1939) и в ряде государственных заповедников
(Кнорре, 1939, 1949, 1953, 1956). При Печорско-Илычском заповеднике
организована первая опытная лосиная ферма. Результаты ее работ, равно
как и весь предшествующий опыт, позволяют сделать следующее заклю-
чение. Выловленный до месячного возраста, вскормленный и воспитан-
ный в постоянном общении с людьми лосенок легко привыкает к человеку,
перестает дичиться, позволяет обращаться с собою, как с домашним живот-
ным, легко поддается тренировке в упряжи, верхом, под вьюком. Чем
моложе теленок взят от матери, тем он легче приручается и поддается
выучке в работе. Лосята, выловленные в месячном возрасте, сохраняют
следы нрава диких зверей и в прирученном состоянии (Попов, 1939).
Вскармливаются выловленные телята коровьим молоком, но с первых
дней рекомендуется предлагать им зелень в виде веточного корма. На
выпойку тратится 150—200 л молока влечение 3—3% месяцев. С раннего
возраста необходимо предоставлять теленку выпас.
В настоящее время, благодаря работам Печорской опытной лосефермы,
в положительном смысле решен основной вопрос при доместикации каж-
дого животного — размножение лосей в домашней обстановке. Полу-
262
CERVIDAE
чено несколько поколений вполне жизнеспособных животных от лосих,
родившихся и выращенных в неволе. Десятилетняя одомашненная ло-
сиха Милка за 7 отелов принесла 12 лосят. По физическому развитию
выращенные в домашней обстановке лоси второго поколения не отли-
чаются от диких (Кнорре, 1956).
Стойловое содержание лоси выносят плохо даже зимою. Домашние
лоси в зимний период пользуются свободным выпасом, без пастуха. По-
следний лишь раз в сутки совершает контрольные обходы района выпаса
для того, чтобы вернуть отдельных животных, вышедших за пределы
контрольной круговой тропы. Раза два-три в месяц лосей пригоняют
на ферму или они приходят сами, чтобы полакомиться картофелем или
корнеплодами.
Пока остается неразрешенным вопрос о свободном летнем выпасе
лосей, так как в этот период они склонны расходиться и пастись в оди-
ночку, а кроме того, обнаружить их пребывание в бесснежный период
почти невозможно. Поэтому на Печорской опытной лосеферме пока
практикуется летнее содержание лосей в больших загонах. Вольный
выпас летом, предоставляется только самкам, имеющим лосят, содер-
жимых в загонах, куда матери приходят их кормить (Кнорре, 1956).
В течение всего года домашние лоси находятся на подножном корме.
Подкормка концентрированными кормами им не требуется, и даже пере-
носят они ее плохо. И взрослым лосям, и телятам осенью рекомендуется
давать ботву овощей и корнеплодов, а зимой — и сами корнеплоды или
картофель в количестве 2—4 кг на голову в сутки. Сено едят только
молодые.
Лось — очень скороспелое животное. Прирост живого веса за первые
шесть месяцев жизни может доходить до 830 г в сутки, превосходя в этом
отношении крупный рогатый скот (Кнорре, 1939). Не требуя заготовки кор-
мов и находясь в течение круглого года на подножном корме, на пастби-
щах, непригодных для других домашних животных и сенокоса, одомаш-
ненный лось создает перспективы вовлечения в круг хозяйственного
использования «неудобных» земель таежной зоны.
Беговые качества лося еще не изучены. На коротких дистанциях он,
по-видимому, может развить довольно большую скорость (Попов, 1939).
Но, с другой стороны, несмотря на присущие ему экстерьерные особен-
ности, лось — животное по преимуществу шагового аллюра. Вследствие
плохой способности к терморегуляции, увеличение скорости передвижения
в санной упряжи с 4 до 8 км в час уже при незначительной нагрузке
вызывает иногда резкое повышение температуры тела (Кнорре,
19536).
По-видимому, лучше лось приспособлен к работе под вьюком. Мак-
симальная грузоподъемность двухлетней самки под вьюком на Якут-
ской опытной станции оказалась 135 кг. С вьюком в 120 кг в начале марта
она прошла 7 км за 67 минут без заметного утомления. Нормальная нагрузка
в санной упряжи для той же самки оказалась 500—600 кг (Попов, 1939).
Е. П. Кнорре (1956) наибольшей полезной нагрузкой под вьюком для
взрослого кастрированного самца считает летом 60, а осенью 100 кг.
Полезную нагрузку саней в условиях пересеченной местности тот же
автор определяет в 350 кг, а по ровной, хорошо наезженной зимней дороге
500 кг.
С вьюком лось способен проходить по таким местам (бурелому, болотам),
где другие домашние животные применяться не могут. Поэтому большее
значение он может иметь в качестве транспортного животного при охот-
NEOCERVINAE
263
ничьем промысле в равнинных таежных районах. Лучшие результаты
получаются при использовании кастрированных самцов.
Отрицательным качеством лося, как транспортного животного, яв-
ляется то, что, во-первых, он ленив, во-вторых, иногда отказывается
идти или свертывает в сторону с дороги (Попов, 1939). Кроме того, лось
плохо переносит жару, и летом на нем можно работать только в ночные
часы, и то не всегда. Вопрос об использовании лося в качестве транспорт-
ного животного пока нельзя считать вполне решенным.
Рис. 56. Лоси в упряжке. Печоро-Илычский заповедник.
(Фото О. И. Семенова-Тян-Шанского).
Весьма ограничены перспективы использования молочной продуктив-
ности домашних лосей. Лосиное молоко обладает высокой калорийностью
и неплохими вкусовыми качествами (см. стр. 257), но молочность лосих
не высокая. При опытах на Печорской лосеферме удои за весь период
лактации, не превышавший, по-видимому, двух-трех месяцев, в луч-
шем_случае достигали 150 л (Кнорре, 1956).
Подсемейство NEOCERVINAE—АМЕРИКАНСКИЕ ОЛЕНИ
Характеристика подсемейства.1 Олени от очень мелких до крупных
размеров. Высота в холке от 34 до 150 см. Рога у самцов имеются всегда,
иногда (род Rangifer) они имеются и у самок. Пеньки рогов короткие;
расстояние от заднего края глазницы до вершины пенька короче зубного
ряда верхней челюсти. Спадающая часть рогов развита в разной степени.
В скелете конечностей сохраняются нижние концы боковых метаподий.
Боковые пальцы и копыта хорошо развиты.
1 По К. Флерову (1952).
264
CEBVI DAE
В черепе лобные кости не удлинены; задние концы носовых костей
достигают уровня передних краев глазниц, а если не достигают, то не-
много больше, чем на длину последнего верхнего моляра. Боковые лоб-
ные гребни (в отличие от Cervulinae) отсутствуют. Шов между слёзными
и лобными костями редко превышает длину свободного края слёзной
кости вдоль этмоидальных щелей. В отличие от других подсемейств Cer-
vidae, сошник своим вентральным краем срастается с нёбными костями
до их заднего края и разделяет хоаны на две половины. Задний край
сошника достигает и соприкасается на довольно значительном расстоя-
нии с базисфеноидом. Слуховые пузыри небольшие, угловатые; лишь
отростками, и то не всегда, выступают ниже уровня basioccipitale. Края
глазниц иногда сильно выступают в стороны, иногда же почти не выхо-
дят за контуры остального черепа (Mazama Rat., Pudu Cray). Передние
края Межчелюстных костей имеют выемки, так что самые кончики их
(prosthion) образуют угловой выступ. Верхние клыки как у самцов,
так и у самок не велики, не выступают выше уровня остальных зубов;
иногда они совсем отсутствуют.
Географическое распространение. Тундровая и таежная зоны Европы
и Азии, Северная и Южная Америка с некоторыми из прилегающих остро-
вов.
Классификация. Подсемейство включает четыре современных рода.
1) Магата Rafinesque —Мазамы. Размеры сравнительно небольшие;
длина черепа, около 220 мм. Лицевая часть черепа укороченная,
а мозговая, наоборот, удлинена; расстояние от переднего края глазницы
до конца межчелюстных костей (prosthion) лишь немного больше
расстояния от заднего края глазницы до заднего края черепа. Рога
только у самцов, в виде простых спичек, на голове волосы удли-
нены в виде пучка. Верхние клыки у самцов обычно имеются. Рас-
пространение: Центральная Америка и тропическая часть Южной
Америки.
2)Pudu Gray — Пуду. Размеры очень маленькие; длина черепа не
больше 150 мм. Череп с укороченной лицевой частью, как у предыду-
щего рода, но отличается от последнего очень глубокими предглазнич-
ными ямками, расширенными в задней части и частично прикрываемыми
краем слёзной кости. Рога в виде простых коротких спичек имеют только
самцы. Верхние клыки всегда отсутствуют. Распространение:
Чили и, возможно, Эквадор (Lydekker, 1915).
3) Odocoileus Rafinesque — Виргинские пампасовые олени, гуэмалы.
Размеры средние; длина черепа около 270 мм. Череп с хорошо развитой
лицевой частью. Расстояние от переднего края глазницы до переднего
края межчелюстных костей (prosthion) намного превосходит расстояние
от заднего края глазницы до заднего края черепа. Рога имеются только
у самцов, но, в отличие от двух предыдущих родов, имеют ветвистый вид
или, по крайней мере, вид двухконцовых вилок. Верхние клыки имеются
не всегда. Распространение: Северная, Центральная и Южная
Америка.
4) Rangifer Н. Smith — Северные олени. Сравнительно большой вели-
чины; длина черепа почти никогда не бывает меньше 300 мм. Лицевая
часть черепа еще длиннее, чем у предыдущего рода. Хорошо развитые
ветвистые рога имеют как самцы, так и самки. И самцы, и самки имеют
клыки, но последние скрыты под слизистой оболочкой и видны только на
препарированных черепах. Отличием от остальных родов подсемейства яв-
ляется отсутствие носового зеркала. Верхняя губа сплошь покрыта во-
RANGIFER
265
л осами. Распространение: тундровая и лесная зона Восточ-
ного и Западного полушария, включая ряд островов Полярного бассейна,
а на юге доходит местами до 48° с. ш.
Род RANGIFER Н. SMITH - СЕВЕРНЫЕ ОЛЕНИ
Lydekker R., 1915, Catal. Ungul. mammals Brit. Mus., IV, London : 238—256
(сист. синон.); Wollebaek A., 1926, Result, staats Spitsbergenexpedit., I, 4:
1—71 (общ. изобр.); J а с о b i A., 1931, Das Renntier, Zool. Anzeig., Erganzungsband
zu Bd. 96 : 1—264 (общ., синон., изобр.); M urie О., 1935, Alaska-Youkon caribou
(North Amer. Fauna, 54) : 1—93 (общ., изобр.); G o колов И. И., 1937, Сов. олене-
водство, 9 : 9—101 (морф., сист., синон.); Флеров К. К., 1952, Кабарги и олени,
М.—Л., изд. Акад, наук СССР (Фауна СССР, Млекопит., I, 2) : 222—247 (общ., изобр.).
Характеристика рода — см. выше.
Классификация северных оленей производится по-разному. Одними
исследователями (Lydekker, 1915; Флеров, 1952) все формы с необычайно
обширного ареала распространения объединяются в один вид — Rangifer
tarandus L. Другими (Jacobi, 1931) признается существование ряда само-
стоятельных видов. По моему мнению (Соколов, 1937), несомненно за-
служивает выделения в качестве отдельного вида Rangifer platyrhynchus
Vrol., северный олень Шпицбергена, так как он обладает целым рядом
резких морфологических отличий, самостоятельным, давно изолирован-
ным ареалом и особенностями биологии (Wollebaek, 1926), Все осталь-
ные формы рода Rangifer Евразии должны рассматриваться принадле-
жащими к одному виду — Rangifer tarandus L.
Главнейшие краниологические отличия видов следующие.
1) Rangifer tarandus L. Размеры сравнительно крупные; длина черепа
у самцов не менее 330 мм. Череп имеет хорошо развитую лицевую часть;
расстояние от переднего конца черепа (prosthion) до.переднегокрая глазницы
составляет не менее 140 % расстояния отпереднего края глазницы до нижнего
края затылочного отверстия (basion). Ширина черепа над клыками больше
ширины его на уровне задних концов верхних отростков межчелюстных
костей. Передние концы носовых костей не расширены. На их переднем
крае имеется выступающий вперед медиальный зубец. Расстояние между
наружными краями затылочных мыщелков составляет не менее 74%
ширина затылка.
2. Rangifer platyrhynchus Vrol. Размеры меньше; длина черепа у сам-
цов редко превышает 345 мм. Череп имеет укороченную лицевую часть;
расстояние от переднего конца черепа до переднего края глазницы состав-
ляет не более 135% расстояния от переднего края глазницы до нижнего
края затылочного отверстия. Череп в ростральной части вздут;-ширина
его над клыками меньше ширины на уровне задних концов верхних отрост-
ков межчелюстных костей. Плоские носовые кости образуют резкое рас-
ширение в передней половине. Передние их концы вместо медиального
зубца образуют направленную вершиной назад вырезку. Расстояние
между наружными краями затылочных мыщелков составляет не более
70% ширины затылка.
Систематика северных оленей американской части ареала требует
капитальной ревизии. Описано свыше десятка «видов», в большинстве
случаев без достаточных оснований. Но, по-видимому, по крайней мере
некоторые формы следует рассматривать как отдельные от евразиатских
виды, изменяющиеся во многом параллельно последним (Seton, 1939;
Murie, 1935).
266
CERVIDAE
Rangifer tarandus Linnaeus — Северный олень (рис. 57—59)
Linnaeus С., 1758, Syst. natur., ed. X, vol. I : 67 (первооп.); В и л ь д е р-
м е т Г., 1838, Леев, жури., III : 259 (распр.); Врангель Ф., 1841, Путешеств.
по северн. берегам Сибири, СПб. : 79,88 (хоз. знач., ох.); Эверсманн Э., 1850,
Естеств. история Оренбургск. края, II, Казань : 249—252 (распр.); Аргентов А.,
1860, Акклим., I, 1 : 20—33 (опис., биол.); М и г г а у А., 1866, Geographic, distribut.
of mammals., London : 153, 155, 334 (сист.); Сабанеев Л. П., 1868, Тр.
Ярославок, статист, комит., 4 : 263—264 (распр.); Миддендорф А. Ф., 1869,
Путешеств. на север и восток Сибири, ч. II, отд. 5 : 170—183 (общ.); Гримм О,
1870, Тр. Вольного экономил, общ., I, 3 : 215—219 (распр.); Богданов М., 1871,
Птицы и звери черноземной полосы Поволжья, долины средней и нижней Волги, Ка-
зань : 176—177 (распр.); Никольский А. М., 1887, Остров Сахалин и его фауна
позвон. животн., СПб., изд. Акад, наук : 155—158 (распр.) Майде ль Г., 1896,
Путешеств. по северо-вост, части Якутской обл., СПб., изд. Акад, наук : 201—204
(хоз. знач., ox.); Lydekker R., 1898, Deer of all lands, London : 33—49 (обш-,
изобр.); Александров Л., 1899, Природа и охота, I : 1—6 (распр.); Добро-
творский М. И., 1902, Тр. Общ. ветеринарн. врачей при Казанск. ветеринарн.
инет. : 19—28 (распр.); Круликовский Л., 1902, Зап. Уральск, общ. любит,
естествозн., XXIII : 109—118 (распр.); Lydekker R, 1902, Proc. Zool. Soc. Lon-
don, II : 360—362 (сист.); Туркин H. В. и К. A. Ca ту н ив, 1902, Звери Рос-
сии, Cervidae, М. : 4—7+45—118 (общ., изобр.); Roister R., 1909, Anat. Hefte,
38 : 101—192 (размн.); Lonnberg Е., 1909, Ark. Zool. 4 : 10 (сист.); P о с о c k R.,
1910, Proc. Zool. Soc. London : 960—962 (морф.); T a n d 1 e г J., 1910, Anzeig. Kais.
Akad. d. Wissenseh., mathem.-naturwiss. KI., 47 : 252—257 (кастрация); Hollis-
ter N., 1912, Smithson Miscell. Coll., vol. 56, Ks 35 : 6 (сист.); Hornaday, W.,
1912, Our wanishing wild life, N. Y. (бол.); L у d e k k e r R., 1915, Catal. Ungulate
mammals in the Brit. Mus., IV, London : 238—256 (сист., синон.); Соловьев Д. К.,
1920, Саянский промыслово-охотничий район и соболиный промысел в нем, Птг,
Госизд. : 248—250 (распр., биол.); Carette Е., 1922, Revista d. Museo de La Plata,
Bu nos-Aires, XXVI : 393—472 (морф., сист., эвол.); P о с о c k R., 1923, Proc.
Zool. Soc. London : 204—206 (морф.); Щеголев В. Н., 1925, Фауна, биол. и эконом,
знач. млекопит. Череповецк. губ., Череповец : 1—37 (распр.); ДоппельмаирГ. Г.,
1926, Соболиный промысел на сев.-вост, побер. Байкала, Л., изд. Госплана Бур.-
Монг. АССР : 192 (распр., стац.); Wollebaek А., 1926, Res. Staats Spitsbergen-
expedit., I, 4 : 1—71 (общ., изобр.); Тассовский Г. Н., 1927, Произв. силы
Дальн. Вост., IV : 530—532 (распр., биол.); Емельянов А. А., 1927, Произв.
силы Дальн. Вост., IV : 257—266 (распр.); Куроптев А., 1927, сборн. «Природа
и люди Севера», I, Архангельск : 34 (распр., ох.); Максимов А. Н., 1928, Уч.
зап. Ивет, истории, VI : 3—37 (одомашн.); Шухов И., 1928, Охотник, 7 : 21—23
(распр.); Зубаровский М., 1929, Охота и природа, 9 : 9 (распр.); Кузне-
цов Б. А., 1929, Тр. по леей, опытн. делу, VI : 102—103 (распр.); Лобачев С. В.,
1930, Тр. Центр, лесн. опытн. станц., Отд. биол. и промысл, охоты, VII: 63—64 (распр.);
Троицкий В. П., 1930, Изв. Сибирск. краев, науч.-промысл, станц., I : 25—26
(распр.); Башкиров И. и Н. Григорьев, 1931, Работы Волжско-Камск.
краев, пром.-биол. станц., I : 39 (распр.); Jacobi А., 1931, Das Renntier, Zool.
Anzeig., Erganzungsband zu Bd. 96 : 1—264 (общ., синон., изобр.); Наумов С. П.,
1931, Тр. Полярн. комис., 4 : 37—42 (распр., биол.); Flower S., 1931, Proc. Zool.
Soc. London : 145—234 (продолж. жизни); Боль Б. К. и Л. Д. Николаевский,
1932, сборн. по оленеводству, тундров. ветерин. изоотехн., М., изд. «Власть Советов» :
258—274 (биол.); Ф л е р о в К. К., 1932, Тр. Совета по изуч. произв. сил, сер. Якутск.,
4 : 1—15 (сист., синон.); Н a d w е n S., 1932, Trans. Roy. Soc. of Canada, ser. 3,
vol. XXVI, sect. V : 237—256 (сист.); Белопольский Л. О., 1933, Тр. Зоол.
инет. Акад, наук СССР, I, 3/4 : 181—182 (распр.); Наумов Н. П., 1933, Дикий
северный олень, М., КОИЗ : 1—72 (общ., изобр.); Соколов И. И., 1933а, Сов.
оленеводство, 3 : 1—88 (библиогр.); Соколов И. И., 19336, Карело-Мурманск.
край, 9/10:69—-72 (быстрота бега); Соколов И. И., 1933в, сборн. «Arctica»,
I : 115—129 (распр., разв.); Flerow С., 1933, J. Mammal., 14, 4 : 328—338 (сист.,
морфол.); Золотарев Н. Т., 1934, Тр. Совета по изуч. произв. сил, сер. дальне-
вост., 3 : 136 (распр.); Наумов Н. П. 1934, Тр. Полярн. комис., 17 : 61—64 (pacnj).
биол.); Palmer L., 1934, Reising reindeer in Alaska, Unit. Stat. Departm. of Agri-
cult., Misc. Publ. № 307 : 1—41 (разв.); Сергеев A. M., 1934, Зоол. журн. ХШ,
4 : 760 (распр.); М u г i е О., 1935, Alaska-Youkon caribou, North Amer. Fauna, 54 :
12—93 (общ., изобр.); Сдобников В. М., 1935, Тр. Арктич. инет., XXIV :
5—66 (биол.); Соколов И. И., 1935, Тр. Арктич. инет. XXIV : 67—128 (морф.,
биол., разв.); Флеров К. К., 1935. В кн.: Адлерберг Г. П., Б. С. Виноградов,
RANGIFER
267
H. А. Смирнов, К. К. Флеров. «Звери Арктики», Л., изд. Главсевморпути : 145—182
(общ., изобр.); Формозов А. Н., 1935, сборн. «Природа Горьковск. и Кировск,
краев», Горький, Госиздат : 159—161 (распр.); Боль Б. К. и Л. Д. Н и к о л а е в-
с к и й, 1936, Советск, оленеводство, 8 : 45—60 (рога); Гептнер В. Г., 1936,
сборн. тр. Гос. Зоол. муз. Моск. гос. унив., III : 19—21 (распр., биол.); К о л ra-
nt е в И. И., 1936, Тр. Биол. науч.-иссл. инет. Томск, гос. унив. II : 308 (распр.,
морфол., сист.); Подаревский В. Б., 1936, Пробл. охотхозяйств, акклим.
в вост. Сибири, Иркутск, Огиз : 34—36 (распр.); Формозов А. Н. и Л. Н. Фор-
мозова, 1936, сборн. науч.-иссл. инет. зоол. Моск. гос. унив., 3 : 115—116 (пит.);
Новиков Г. А., 1937, Тр. Совета по изуч. произв. сил, сер. вост.-сиб., 4 : 236—
237 (распр., биол.); Соколов И. И., 1937, Сов. оленеводство, 9 : 9—101 (морф.,
сист., синон.); Брюн Л. А., 1938, Природа, 7/8:197—198 (распр.); Дми-
триев В. В., 1938, Тр. Алтайск. гос. заповеди., I : 243—248 (распр., биол.); Кук-
лин С. А., 1938, Звери и птицы Урала, Свердловск, Обл. издат. : 119—122 (распр.,
биол.); Акаевский А. И., 1939, Анатомия сев. оленя, Л., изд. Главсевморпути :
1—323; Горбунов Е. И., 1939, Тр. Инет, полярн. земл., животн. и промысл,
хоз., сер. «Оленеводство», 5 : 101—142 (размн.); Исаков Ю. А., 1939, Бюлл.
Моск. общ. испыт. прир., Отд. биол., XLVIII, 2/3 : 37—49 (распр.); Саморо-
дов А. В., 1939, сборн. тр. Гос. зоол. муз. при Моск. гос. унив., V : 18 (распр.);
Seton Th. 1939, Live of game animals, vol. 2, pt. 1/2, Garden City : I—XX+413—
780 (общ., изобр.); Александрова В. Д., 1940, Тр. Инет, полярн. земл., жи-
вотн. и промысл, хоз., сер. «Оленеводство», И : 9—96 (пит.); Машковцев А. А.,
1940, Докл. Акад, наук СССР, I : 78—80 (морф.); Cabrera А., 1941, Publ. Field
Mus Natur. Hist., zool. ser., 27 : 125—135 (сист.); Портенко Л. A., 1941, Фауна
Анадырск. края, ч. Ill, Л., изд. Главсевморпути : 77—87 (распр., биол.); Ска-
лой В., И. Щербаков и М. Б азыкин, 1941, сборн. «Природа и соц. хоз.»,
VIII : 270—276 (распр., миграции); Dechambre Е., 1942, Mammalia, VI, 2:
74—81 (одомашн.); Не г re W., 1943, Anatom. Anzeig., 141 : 196—214 (одомашн.);
Бобринский Н. А. и др., 1944, Определитель млекопит. СССР, М., изд. «Сов.
наука» : 236—238 (сист., распр.); Аверин IO. В., 1948, Тр. Кроноцк. гос. заповеди.
I : 190—199 (распр., биол.); Громов В. И., 1948, Тр. Инет. геол. Акад, наук
СССР, 64 : 439—440 (ист.); Северное оленеводство, М., Сельхозгиз,
1948 : 1—367;Семенов-Тян-Шанский О. И., 1948, Тр. Лапландск. гос.
заповеди., II : 3—90 (общ., биол., изобр.); Д р у'р и И. В., 1949а, Дикий северный
олень, М.—Л., изд. Главсевморпути : 1—77 (общ.); Д р у ри И. В., 19495, в кн.:
Доброхотов А. Ф. «Частное животноводство», М., Сельхозгиз : 805—830 (разв.);
Кузнецов Б. А., 1949, сборн. «Пушные богатства СССР», 1:158 (акклим.);
Ку ражсковс кий Ю. Н., 1949, Охрана природы, 7 : 50—59 (распр., биол.);
Павлов Е. И., 1949, Промысл, звери Читинск. обл., Чита, Облиздат. : 105—109
(распр., изобр.); Копылов И. П., 1950, Дикие копытн. животн. Иркутск, обл.,
Иркутск, Облиздат : 37—49 (общ., изобр.); L a n t i s М., 1950, Arktic, vol. 3, № 1 :
27—44 (разв.); Марвин М. Я., 1950, Промысл, звери и птицы Карело-Финск.
ССР, Петрозаводск, Госизд. : 46—47 (распр.); Флеров К. К., 1950, Матер,
по четверт. периоду в СССР, 2 : 50—69 (эвол.); Николаевский Л. Д-, 1951,
Некробациллез северн. оленей, М., Сельхозгиз:!—98 (бол.); Скалой В. Н.,
1951, Бюлл. Моск. общ. испыт. прир., Отд. биол., LVI, 1 : 27—28 (распр.); Ш а-
потников Ф. Д., 1951, Вопр. геогр. Сибири, 2 : 299—304 (распр.); Е 1 Ге г т а п
andT.M orr iso n Scott, 1951, Checklist of palaearctic and Indian mammals, London:
375—376 (сист., синон.); Br о me e -Skuncke F., 1952, Ark. f. zool., Stockh.,
Bd. 4, № 1 : 1—43 (определ., возр.); Гребельский С. Г., 1952, Борьба с кожи,
оводом северн. оленя, М., Сельхозгиз : 1—88; К и р и к о в С. В., 1952, Птицы и
млекопит. в условиях южн. оконечн. Урала, М., изд. Акад, наук СССР : 268 (распр.);
Мишин И. П., 1952, Природа, 1 : 120 (распр., стац.); Флеров К. К., 1952,
Кабарги и олени, М.—Л., изд. Акад, наук СССР (Фауна СССР, Млекопит., I, 2) :
222—247 (общ., изобр.); Янушевич А. И., 1952, Фауна позвон. Тувинск. обл.,
Новосибирск, изд. Зап.-Сиб. фил. Акад, наук СССР : 62 (распр.); В о u с h и d J.,
1953, Mammalia, XVII, 1:29—49 (морфол. зубов); Верещагин Н. К., 1953,
Бюлл. Комисе, по изуч. четверт. периода, 18 : 47 (распр.); Кириков С. В., 1953,
Изв. Акад, наук СССР, сер. география., 4 : 24 (ист.); Л е б ле Б. Б., 1953, Пушные
звери Арханг. обл., Архангельск. Книжн. изд. : 74—75 (распр.); М и л ь к о в Ф. Н.,
1953, Среднее Поволжье, М., изд. Акад, наук СССР : 151 (распр.); Пидоплич-
к о И. Г. и В. А. Т о п а ч е в с к и й, 1953, Природа, 7 : 101—103 ^ист.); Сдоб-
ников В. М., 1953, По арктической тундре, М., Географгиз : 39—65 (биол., ох.);
Томилин А. Г., 1953, Тр. Моск, пушно-мехов. инет., IV : 31—42 (распр.); Фе-
тисов А. С., 1953, Тр. Иркутск, гос. унив., VII, 1/2:34 (распр.); Банни-
ков А. Г., 1954, Млекопит. Монгольск. Народи. Республ., М., изд. Акад, наук СССР;
268
CERVIDAE
219—220 (морф., сист., распр.); Баранчеев Л. М., 1955, Зап. Амурск, обл.
музея краеведен., 3 : 200—218 (распр.); Насимович А. А., 1955, Роль режима
снежи, покрова в жизни копытных, М., изд. Акад, наук СССР : 249—287 (распр., биол.);
Шапошников Ф. Д., 1955, Зоол. журн., XXXIV, 1 : 191—207 (морф., биол.).
Описание. Сравнительно крупный представитель своего семейства, хотя
намного уступает лосю и некоторым расам благородного оленя. Высота
в холке составляет от 80 до 140 см, длина тела 150—210 см, живой вес
от 80 до 180 кг. Общее телосложение менее стройное и легкое, чем у бла-
городных оленей. Туловище покоится на сравнительно коротких, не-
сколько утолщенных в суставах ногах. Его косая длина составляет
около 105% высоты в холке (данные по домашнему оленю; Соколов, 1935).
Крестец одинаковой высоты с холкой или немного выше. Линия спины
прямая или слегка вогнутая в передней части. Холка обычно в той
или иной мере выступает. Характерна некоторая согнутость (саблистость)
задних ног в скакательных суставах и сближенность последних при
взгляде сзади («коровий постав»). Шея, по сравнению с лосем, длинная,
но не длиннее, чем у благородного оленя.
Голова сравнительно длинная, с несколько вытянутой и немного
утолщенной на конце мордой. Профиль носа vneraa выпуклый, реже
прямой. Носового зеркала (голого пятна на конце морды), свойственного
всем другим Cervidae, у северного оленя нет; все пространство между
ноздрями до самого края верхней губы покрыто волосами.1 Предглаз-
ничные железы хорошо развиты. Глаза с радужной оболочкой коричне-
вого цвета и горизонтально расположенным овальным зрачком. Уши
длиною около 15—18 см с закругленными концами.
Кроме нредглазничных желез, у северного оленя имеется «синус
преддверия носа», расположенный у основания крыловидной складки
носа и представляющий кожное выпячивание глубиною 15—20 мм со
щелевидным входом. Полость его заполнена жироподобным секретом
желтоватого цвета (Акаевский, 1939).
Хвост короткий, обычно не длиннее уха. Копыта главных пальцев,
в противоположность другим оленям, широкие, округлые и изогнуты так,
что их верхушки направлены навстречу друг другу; со стороны подошвы
копыт края обоих вместе образуют фигуру более или менее правильного
круга. Боковые копыта, особенно на передних ногах, хорошо развиты,
немного уступают по длине главным и при ходьбе всегда касаются почвы.
И главные, и боковые пальцы способны очень широко раздвигаться.
Межкопытные железы развиты хорошо лишь на задних конечностях.
На передних они представлены в виде едва заметных углублений на перед-
ней стенке межпальцевого пространства. Плюсневых желез нет, но. на
внутренней стороне скакательных суставов имеются предплюсневые
железы, обозначаемые снаружи пятнами более длинных, чем на окружаю-
щих участках, грубых волос; основания последних иногда склеены желто-
ватым секретом.
Рога имеют не только самцы, но, за редкими исключениями, и самки.
У последних они короче и тоньше. Комолые самки встречаются, но редко
(Соловьев, 1920; Соколов, 1935; Машковцев., 1940). Поверхность рогов
более гладкая, чем у других оленей; сосудистые желобки на ней едва
заметны. Главные стволы образуют дугообразный изгиб и вершинами
загнуты вперед и слегка внутрь. Поперечное сечение у оснований круглое
или слегка уплощенное с боков. Длина рога по изгибу может доходить
1 Приспособление к низким температурам полярной зоны.
RANGIFER
269
до 130—140 см. От главного ствола в типичных случаях вперед отходят
только два надглазничных отростка. Средний отросток, как и все верху-
шечные, отходят от задней стороны ствола и направлены назад. Над-
глазничные отростки лопаткообразно расширены и, в свою очередь,
на вершине несут несколько пальцевидных концов. Первый надглазнич-
ный отросток отходит у самого основания рога и направляется несколько
внутрь и вниз параллельно лбу зверя. Второй надглазничный отросток,1
Рис. 57. Дикий северный олень (JRangifer tarandus L.) .
отходя от ствола, направляется обычно несколько в сторону и вперед,
вершиной загибается внутрь, навстречу другому отростку. Вершины
главных стволов уплощены, но не в такой степени, как у лани или лося.
От этого основного типа строения рогов имеются разнообразные уклоне-
ния. Часто надглазничные отростки на концах не уплощаются, а раз-
дваиваются; иногда второй надглазничный отросток вообще отсутствует.
Изредка, наоборот, на передней стороне рога имеется третий отросток.
Волосяной покров северного оленя состоит из длинной грубой и лом- -
кой ости и топкого извитого подшерстка. Толстые, слабо волнистые
волосы ости имеют в сердцевинной части воздушные полости, благодаря
сильной вакуолизации клеток. Длина зимнего меха может доходить до
8—9 см. На нижней стороне шеи удлиненные до 20—25 см волосы и у сам-
цов, и у самок образуют длинную гриву. На голове и нижних частях
конечностей волосы короткие, более плотно прилегающие к коже. Уши по-
крыты длинной, но более мягкой шерстью. Мягкая шерсть также в пахах.
Удлиненные и грубые, но упругие волосы вокруг копыт образуют густую
плотную щетку, увеличивающую площадь опоры при ходьбе по рыхлому
1 Иногда называется «ледяным», так как считалось, что в помощью его олень
достает корм из-под снега.
210
CERVIDAE
снегу и мягкому грунту. Длинный тонкий подшерсток хорошо развит.
Вместе с очень густым остевым мехом он создает надежную защиту от
холода в зимнее время. Сразу после линьки летний волос короток, всего
около 2—2.5 см; подшерсток в летнем меху отсутствует и начинает раз-
виваться только с августа месяца.
Общий фон летней окраски от светло-коричневого до темно-коричне-
вого или бурого. Наиболее интенсивно окрашены лоб и морда, особенно
позади ноздрей. От средней части спины книзу окраска светлеет, но на
уровне локтевого сустава по бокам тянутся темные полосы от локтя до
колен. Живот светлый, иногда белый. Шея буровато-серого цвета, осо-
бенно к концу лета. Белый волос на ягодицах вокруг хвоста, а также на
нижней стороне последнего образует небольшое «зеркало». Верхняя
сторона хвоста коричневая. Передняя сторона ног темная, с боков и по-
зади они более светлые. Светлая, иногда белая кайма окружает копыта.
Пространство между ноздрями до края верхней губы чисто белого цвета.
Отдельные волосы летней ости от основания имеют светло-бурый или
даже белый цвет, но к вершине постепенно темнеют. Характерного для
многих оленей, предвершинного светлого кольца волосы северного оленя
не имеют. От степени потемнения вершин зависит интенсивность окраски.
С конца лета происходит постепенное посветление окраски. Начи-
нается она с шеи и холки, затем распространяется на бока, в последнюю
очередь на голову и ноги.1 В конце концов к зиме коричневая окраска
обычно сохраняется лишь на местах, более темно окрашенных и летом,
т. е. на голове, спине, на ногах. Все остальные части тела приобретают
светло-серую, иногда почти белую окраску. Посветление происходит
благодаря тому, что, начиная с конца лета, сквозь летний бурый волос
во все возрастающем количестве прорастают более длинные волосы бе-
лого цвета, перекрывающие летнюю окраску. От смешения белых и
бурых волос зависит переходная серая окраска осеннего меха.
Половой диморфизм в окраске отсутствует. Молодые первые месяцы
жизни имеют однотонную коричневую, иногда с рыжеватым оттенком
окраску; живот и у них несколько светлее туловища. Пятнистость в ок-
раске молодых обычно отсутствует, но изредка наблюдаются едва замет-
ные размытые пятна.
Череп (рис. 58) характеризуется укороченностью зарогового отдела.1 2
Расстояние от заднего выступа затылочного бугра до переднего края
теменных костей меньше расстояния от средины линии, соединяющей
задние края глазниц, до средины линии, соединяющей задние выступы
лобных костей позади рогов. При положении черепа на горизонтальной
поверхности передние края глазниц расположены позади уровня зад-
него края зубного ряда. Глазницы трубкообразные, сильно выступающие
в стороны. Межглазничная ширина превосходит ширину черепа в щечных
буграх.
Носовые кости в передней части узкие, позади сильно расширены и в
виде лопастей вдаются между лобными и верхнечелюстными костями.
Межчелюстные кости образуют на переднем крае выступ в виде угла; их
верхние концы не достигают и не соприкасаются с носовыми костями. Пред-
глазничные ямки и этмоидальные щели хорошо развиты. Хоаны сдвинуты
назад. Их передний край расположен заметно позади уровня заднего
1 Вариации и сезонные изменения окраски у домашних оленей, см.: Соколов,
1935.
2 Подробное описание черепа северного оленя см.: Соколов, 1937.
RANGIFER
271
края зубного ряда. Вентральный край сошника срастается с нёбными
костями и разделяет хоаны на две половины, а задний край достигает
основной клиновидной кости и прилегает к ней. Слуховые пузыри неболь-
Рис. 58. Череп Rangifer tarandus L.
шие, угловатые, почти не выступают ниже уровня basioccipitale. Диа-
стема нижней челюсти длинная, обычно больше длины соответствующего
ряда коренных зубов, а если короче, то не больше, чем на половину дли-
ны последнего нижнего моляра (М?).
272
CERVIDAE
Коренные зубы брахиодонтныё. Добавочный бугорок выражен, и то
не всегда, только на первом и втором верхних молярах. Передне-наружная
и задне-внутренняя дольки второго и последнего премоляров нижней
челюсти образуют одну сплошную лопасть. Задне-наружная долька на
обоих зубах отделена от нее почти до основания, тогда как передне-
внутренняя уже вблизи вершины соединяется с главной лопастью
и в процессе стирания образует внутреннюю стенку лунки зуба. Перед-
ний нижний премоляр имеет хорошо развитые складки и иногда даже
образуется передняя луночка этого зуба (рис. 31, Г). Клыки в верхней
челюсти имеются как у самцов, так и у самок, но в течение всей жизни
животного покрыты слизистой оболочкой. Резцы имеют форму почти
симметричных лопаток и равномерно убывают от средней пары к клыкам.
В скелете конечностей сохраняются дистальные рудименты боко-
вых пальцев.
История и географическое распространение. Род. Rangifer Н. Smith
происходит от общего корня с американскими оленями, потом-
ками палеомерицин (Blastomeryx Соре и близкие к нему роды). Формирова-
ние его произошло, по-видимому, в умеренной зоне Северной Америки
в нижнем плейстоцене (Carette, 1922; Флеров, 1950). В среднем плейстоцене
северные олени проникли на территорию Европы и Азии и широко рас-
селились в пределах тундровой и лесной зон. В плейстоцене распростра-
нение доходило до южной Франции. В пределах СССР остатки северного
оленя начинают встречаться с начала верхнего плейстоцена (рисе), напри-
мер в Крыму (Громов, 1948). В начале послеледникового периода северный
олень встречался в долине Днестра, Южного Буга, Днепра и Донца.
Обломок рога северного оленя найден на берегу Бугского лимана в слоях,
относящихся к II—III в. нашей эры (Пидопличко и Топачевский, 1953).
Эволюция северных оленей шла в направлении приспособления к жизни
в условиях полярной зоны. В связи с этим стоит ряд уже упомянутых
морфологических адаптаций (строение нижней части конечностей, меха,
конца морды). Но как в Старом, так и в Новом Свете они живут не только
в тундре, но и в лесной полосе и физически развиваются здесь даже лучше.
В противоположность другим представителям семейства Cervidae снег
не является фактором, лимитирующим распространение северного оленя.
Гораздо большее значение в этом отношении имеет наличие в районе
обитания основного корма, наземных и древесных лишайников, без уча-
стия которых в рационе, хотя бы в незначительном количестве, этот
зверь, по-видимому, существовать не может.1
Область распространения северных оленей уже в последние тысяче-
летия была шире. Есть основания предполагать, что во времена Юлия
Цезаря (I в. до нашей эры) они водились на территории Германии
и Польши (Пидопличко и Топачевский, 1953). В настоящее время ареал
вида охватывает всю тундровую и часть таежной зоны материка Европы
и Азии, начиная со Скандинавского полуострова и кончая Камчаткой
и Сахалином, а также ряд островов- Полярного бассейна (Новая Земля,
Вайгач, Белый, Диксон, Северная Земля, некоторые острова из группы
Новосибирских) (рис. 59). '
Так же как и у лося, у северного оленя имеют место периодические
колебания южной границы распространения, исчезновение и появление
1 В. М. Сдобников (устное сообщение) придерживается иной точки зрения, так
как по его наблюдениям на северном Таймыре олени практически живут без лишай-
ников.
RANGIFER
273
его в ряде районов, которое нельзя объяснить только истребляющей
или охраняющей деятельностью человека. Численность и плотность
популяции северных оленей на обширной территории их ареала также
неравномерна и непостоянна. Даже в пределах тундры во многих местах
северные олени крайне малочисленны или исчезли совершенно. Несомненно,
что главную роль в данном случае сыграли развитие оленеводства и, в связи
с этим, вытеснение с лучших пастбищ дикого оленя домашним. С другой
стороны, несомненно и то, что популяции дикарей пополняются за счет
ухода из стад и одичания домашних оленей, как это имеет место, например
в Северной Карелии и на Кольском полуострове. И в настоящее время, и
в прошлом районы наибольшей численности оленей расположены в Си-
бири к востоку от Енисея.
В пределах СССР распространение северного оленя и изменения
его ареала в историческую эпоху рисуются в следующем виде.
Северный олень населяет как северный, так и южный остров Новой
Земли, хотя в текущем столетии он уже не так многочислен как раньше.
По-видимому, до сих пор сохранился на Вайгаче. На о. Колгуеве водился
в середине XVIII в., но, в силу неизвестных причин, в течение нескольких
лет вымер (Соколов, 1933в).
По сообщению И. В. Друри (1949), имеются дикие олени на Соло^
вецких островах, но нет уверенности, что это не потомки завезенных еще
в XVI в. домашних (Добротворский, 1902).
На Мурмане, вследствие чрезмерного истребления диких оленей в до-
революционные годы и развития оленеводства, к 30-м годам настоящего
столетия дикари сохранились в небольшом количестве, главным образом
в западной половине. В результате принятых мер охраны, в последние
десятилетия дикие олени на Кольском полуострове вновь начали увеличи-
вать свою численность и снова появились в районах к востоку от желез-
ной дороги в центральной части Кольского полуострова (Друри, 1949).
На территории Лапландского заповедника число их только за 12 лет
(с 1929 по 1941 г.) возросло в десять раз (Семенов-Тян-Шанский, 1948).
В Карельской АССР в настоящее время они держатся главным образом
в северных районах: Кестеньгском, Лоухском, Кемском, Калевальском,
Беломорском, Тунгудском, Ругозерском, Сегозерском, а также в части
Медвежьегорского и Пудожского районов (Марвин, 1950).1 Аспирантом
Карело-Финского филиала Академии наук СССР И. П. Волковой два
диких оленя были добыты в сентябре 1951 г. в окрестностях д. Пирти-
Губа Калевальского района. Шкуры и скелеты этих экземпляров хра-
нятся в коллекциях Зоологического института АН СССР в Ленинграде.
Между Ладожским и Онежским озерами в прошлом столетии олени
к югу доходили до берегов р. Свири.
Вероятно, сохранился дикий олень в большинстве лесных районов
Архангельской области. С достоверностью он указывается для верховьев
Пинеги и Северной Двины, для бассейна Мезени и ее притоков — Пижмы
и Пезы, далее для Шенкурского и Красноборского районов, а также
для междуречья Пеши и Цильмы (Куроптев, 1927; Зубаровский, 1929;
Сергеев, 1934; Куражсковский, 1949; Лебле, 1953). По-видимому, с давних
времен вытеснен дикий олень из тундр Архангельской области и Коми
1 Таким образом, утверждение Ю. А. Исакова (1939) о резком сокращении ареала
северного оленя в Карелии не находит подтверждения. С другой стороны, при расспро-
сах в 1939 г. жители Ругозерского и Ребольского районов называли мне олеНя, как
известного им, но крайне редкого зверя.
18 Фауна СССР, И. И. Соколов ,
274
CERVIDAE
АССР. Лесная полоса Коми АССР представляет зону почти сплошного
распространения дикого оленя; в наибольшем числе он здесь встречается,
с одной стороны, в области Тиманского кряжа, а с другой — в районах
Полярного Урала. В последнем северные олени доходят до верхнего
течения р. Усы в устьях ее притоков Косью и Роговой (Куражсковский,
1949). Сравнительно не редки в верхнем течении Печоры на территории
Печеро-Илычского заповедника. В небольшом числе северные олени
сохранились в северной половине Кировской области, в районах Омут-
нинском, Подосиновском, Халтуринском, Слободском и в некоторых
других. В наибольшем числе держатся в правобережье Камы, по ее при-
токам Чус и Кым (Лобачев, 1930). Как о нередких зверях в южных
уездах Вятской губернии упоминал о них в 1902 г. Круликовский. Не-
сомненно, сохранился олень в лесных районах Пермской области; однако
сведениями о распространении его здесь за последние годы, кроме дан-
ных А. Г. Томилина (1953) по Коми-Пермяцкому национальному округу,
где олень водится к югу до 60° с. ш., мы не располагаем. На Урале юж-
ная граница распространения проходит около 59° с. ш. в верховьях,
р. Косьвы (Куклин, 1938).
Мало данных о распространении северных оленей в Вологодской
области. В конце прошлого столетия они держались в окрестностях са-
мой Вологды (Александров, 1899). Вероятно, кое-где в глухих, мало-
доступных местах они обитают и в настоящее время. Достоверные сведения
об обитании оленей в Кирилловском и Устюженском районах Воло-
годской области, относящиеся к 20-м годам нашего столетия, приводит
В. Щеголев (1925). Южную границу современного ареала его в Европей-
ской части СССР обрисовать трудно. Туркин (Туркин и Сатунин, 1902)
проводил ее по восточному побережью Ладожского оз. к Тихвину. Даже
в начале зтого столетия северный олень водился на о Валааме. А. Ф. Мид-
дендорф (1869), со слов охотников, указывал его для болот между Волховом
и Сясью до Ильменя. Сейчас его здесь нет, но имеются сравнительно
недавние данные о том, что в небольшом числе олени держатся на востоке
Ленинградской области в Ефимовском районе, в междуречье р. Лидь
иКолпь (Брюн, 1938). По сведениям, полученным автором от известного
Боровичского краеведа С. Н. Поршнякова, последний табун оленей в Хвой-
нинском районе Новгородской области был уничтожен в 1917 — 1918 гг.
Будто бы не редки северные олени были еще в средине XIX в. в Валдай-
ском уезде (Кириков, 1953). По сведениям этого автора, в XV в. олени
водились во владениях Бежецкого князя, а в XVIII в. они упоминаются
при описании Клинского уезда.
До конца 60-х годов прошлого столетия охотились за оленями в Весь-
егонском и Вышневолоцком уездах Тверской губернии. Убивали их даже на
оз. Селигер. В настоящее время северных оленей в пределах Калинин-
ской области нет. Нет их и в Ярославской области, хотя по Л. П. Саба-
нееву (1868), олени встречались здесь в уездах Мологском, Угличском
и Пошехонском до 57° с. ш. Есть указания, что нередко они доходили до
границ Владимирской и Московской губерний (Туркин и Сатунин, 1902).
О появлении оленей в окрестностях г. Владимира сообщалось в «Губерн-
ских ведомостях» за 1856 г. (№ 25 : 193—194). В 40-х годах XIX в.
в большом числе водились они в северной Заволжской части нынешней
Горьковской области, равно как и в прилежащих районах Костромской;
в последней, в 90-х годах олени удерживались еще по р. Керженцу,
в Усть-Керженском лесничестве, а также в Шарьинском и Макарьевском
уездах. Последние олени в Макарьевском уезде были убитыв 1917— 1918 гг.
Рис. 59. Распространение Rangifer tarandus L.
XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше; 4 — отдельные заходы в на
стоящее время.
18*
!
276
CERVIDAE
Возможно, они сохранились в некоторых районах Костромской области,
но в Горьковской их в настоящее время уже нет (Формозов, 1935).
В Кировской области южная граница современного распространения
проходит по истокам рр. Малой Влосницы и Лупьи (притока Камы)
около 58—59° с. ш. (Лобачев, 1930).
Рог северного оленя найден в культурных слоях городища X—XIV вв.
около г. Чебоксары (Мильков, 1953). В пределах Татарской АССР в настоя-
щее время северный олень не водится, но упоминания о добыче его встре-
чаются в старой литературе неоднократно (Богданов, 1871; Добротвор-
ский, 1902).1 В 1863 г. олень был убит под Казанью, а в 20-х годах этого
столетия в Мамадышском кантоне (Башкиров и Григорьев, 1931).
Нет северного оленя и на территории Марийской АССР, хотя в про-
шлом столетии, по Эверсманну (1850), он здесь водился.
В Свердловской области, как указывалось, северные олени встреча-
ются по Уралу не южнее 59° с. ш. Кроме того, по Бобринскому (1944),
небольшая изолированная популяция сохранилась около Красноуфимска.
Во время путешествий Палласа и Эверсманна северные олени встречались
будто'бы около Оренбурга и даже заходили южнее до.46° с. ш. В настоящее
время в списках млекопитающих Чкаловской области северного оленя
уже не приводят. В конце прошлого и начале нынешнего столетия он
указывался для Златоуста, Верхнеуральска и некоторых других мест
Южного Урала. Отдельные забеги до 53° с. ш. наблюдались и в последние
годы (Туркин и Сатунин, 1902; Кирилов 1952).
В музее г. Уральска (Западно-Казахстанская область) хранится
чучело северного оленя, добытого в окрестностях города в средине про-
шлого века (Верещагин, 1953).
В пределах западной Сибири дикий северный олень является сейчас
не столько тундровым, сколько таежным зверем. На Ямале и Гыданском
полуострове он распространен только в северных частях, ближе к побе-
режью. Южнее встречается лишь в пределах лесотундры и лесной зоны.
В южных частях тундр он давно вытеснен стадами домашних оленей
(Наумов, 1931). Держится дикий олень на о-вах Белом, Оленьем и Сиби-
рякова (И. Друри, 1949). Южная граница к востоку от Урала проходит
на широте 58—59° с. ш. Есть дикие северные олени в верховьях Сосьвы,
в бассейнах Лозьвы и Тавды, на водоразделе Тавды и Туры, к северу от
Тюмени, в лесных дачах около Тобольска (Куклин, 1938). Далее к востоку
они еще встречаются в бассейне р. Демьянки, в верховьях Васюгана,
Парабеля, Тары и даже Оми к югу от 56° с. ш. (Друри, 1949). Во время
путешествия Миддендорфа (1869) зимою регулярно появлялись в Барабин-
ской степи на широте Омска. В конце 20-х годов нашего века были много-
численны в Васюганье (Шухов, 1928). К востоку от Оби южная граница
распространения северного оленя проходит по водоразделу рр. Кети
и Чулыма и пересекает Енисей южнее г. Енисейска (Друри, 1949).
К югу от очерченной границы северные олени постоянными обита-
телями являются, хотя и в незначительном количестве, в Кузнецком
Алатау, на водоразделе рр. Кии и Томи и по долине первой до границы
с лесостепью. В Горной Шории они известны на хребте Мустаг и по
рр. Кондоме и Мрассу. Встречаются также в Абаканском хребте, в вер-
ховьях Большого и Малого Абаканов. На Алтае в пределах СССР обитают
к востоку от верховьев р. Бии, Телецкого озера и о. Чулышмана, на се-
1 В коллекциях Зоологического института АН СССР хранится череп самки
R. tarandus из Казанской губернии без точного указания места нахождения.
RANGIFER
277
вере доходят до р. Лебедь, правого притока Бии (Шапошников, 195.1),
а на юге почти до 50° с. ш.
Из Алтая область распространения дикого северного оленя пере-
ходит и охватывает Саяны, где в наибольшем числе он держится в узле
Западных и Восточных Саян. По восточным Саянам к северу доходит до
верховьев рр. Маны и Кана. Имеется олень в Тувинском нагорье (Яну-
чпевич, 1952), но детали его распространения здесь не известны. А. Г. Бан-
ников (1954) указывает в качестве мест обитания верховья рр. Эрсын-
Гол и Наран-Гол в восточной части Танну-Ола.
Почти повсеместно распространены олени, начиная с низовьев Енисея
и Таймыра на западе, до Колымы на востоке, а также на прилегающих
островах Полярного бассейна. Регулярно заходят они с Таймыра на
о. Диксон, на острова архипелага Норденшельда. Повсеместно встре-
чаются на о. Большевик (южный остров Северной Земли) Являются оби-
тателями большинства Новосибирских островов: Большого и Малого
Ляховских, Котельного, Фаддеевского, Новой Сибири и даже о. Беннета.
По крайней мере до 20-х годов нашего, века заходили и удерживались
на Медвежьих островах, в частности на Четырехстолбовом (Друри,
1949). На материке наибольшая концентрация поголовья дикого оленя
имеет место на Таймыре, в низовьях Лены и междуречье Яны и Инди-
гирки. На территории Эвенкийского национального округа в наиболь-
шем количестве встречается в трех районах: 1) на водоразделе Ангары
и Подкаменной Тунгуски, 2) на водоразделе Подкаменной и Нижней
Тунгусок и 3) на водоразделе Нижней Тунгуски, Вилюя и Котуя (На-
умов, 1934). В Якутии северный олень распространен от побережья до
южных границ республики, кроме центральных, густо населенных
районов.
Еще около 100 лет назад был сказочно многочислен на Колыме и
к востоку от последней, на территории нынешнего Чукотского националь-
ного округа. В настоящее время, в результате развития оленеводства,
а еще больше вследствие нерационального, хищнического промысла,
дикий северный олень и в этих районах стал редким зверем, а местами
исчез совершенно. До сих пор в качестве постоянного обитателя дер-
жится в верховьях Большого и Малого Анюев, в бассейне верхнего те-
чения Анадыря до устьев Майны и Белой, в хребтах Колымском, Русском
и Щучьем. Местами сохранился на р. Омолоне. Изредка встречается на
Анадырском хребте до района к северу от бухты Крестовой (Белополь-
ский, 1933; Портенко, 1941; Друри, 1949). Нет дикого оленя на полу-
острове Тайгоносе, а также в Олюторском районе (Корякский нацио-
нальный округ); в пределах последнего, по словам местного населения,
оленя не было и раньше (Самородов, 1939).
На Камчатке область распространения дикого северного оленя лежит
к югу от 58° с. ш. По последним данным (Друри, 1949), северная граница
проходит здесь по рр. Уке и Хайлюму. Отсюда дикий олень распростра-
нен до южной оконечности полуострова, но очень неравномерно; в ряде
районов его совсем нет, вследствие большой глубины снежного покрова.
В общем же олени на Камчатке многочисленны; на территории Кроноц-
кого заповедника, например, до сих пор встречаются табуны их до 1500—
2000 голов в каждом (Аверин, 1948).
В восточной Сибири южная граница распространения дикого северного
оленя проходит южнее низовьев Верхней Тунгуски (Ангара) по р. Тасее-
вой (в коллекциях Зоологического института АН СССР имеется череп
R. tard.nd.us, добытый в 1914 г., в 25 верстах от с. Тасеево), затем в бассейне
278
CERVIDAE
Чуны, не заходя к югу и западу за р. Ону. Известен олень в Нижнеудин-
ском районе (Троицкий, 1930). Далее к востоку встречается в верховьях
р. Илима, Киренги и Лены (Копылов, 1950).
Из Восточных Саян ареал северного оленя переходит в Южное При-
байкалье. 'Водится в Хамар-Дабане и других хребтах вдоль восточного
побережья Байкала вплоть до бассейна Верхней Ангары и Муи (Новиков,
1937; Фетисов, 1953). Обычен северный олень в хребтах Баунтовского
района Бурят-Монгольской АССР.1 2 По-видимому, только через Забай-
калье ареал северного оленя на Алтае и Саянах смыкается с таковым
северной части Иркутской области, Красноярского края и западной
Сибири. Прямых доказательств смыкания ареала к западу от Байкала
в литературе встретить не удается.
Не редок северный олень в бассейне Витима. Заходит на восточные
склоны Яблонового хребта. Встречается в верховьях Нерчи и Олекмы,
а также по рр. Нюкже, Тунгиру, Каренге (Подаревский, 1936; Павлов,
1949). К югу от этих мест в пределах Бурят-Монгольской АССР и Читин-
ской области, по Б. Кузнецову (1929), не встречается.
В пределах Амурской области дикий северный олень обитает на южных
склонах Станового хребта, например в верховьях р. Ольдоя, по рр. Гат-
кану, Сигикте, Моготу (правые притоки Гилюя), а также по высоким
предгорьям к югу от хребта Тукурингра (Гассовский, 1927). Дальше
к востоку держится в верхнем течении рр. Зеи, Селемджи и Бурей (Зо-
лотарев, 1934). По последним• данным (Баранчеев, 1955), максимальная
численность дикого оленя наблюдается на северо-западе области по
р. Олекме, на северо-востоке по р. Мае и в самой северной части Селем-
джинского района по южным склонам хребта Джагды между 131.5 и
133° в. д. В коллекциях ЗИН АН СССР имеется экземпляр северного оленя,
добытый в низовьях Нимана, правого притока Бурей.
В Хабаровском крае, кроме уже упомянутых ранее северных райо-
нов, R. tarandus распространен по всему Охотскому побережью до Амура
и Сихотэ-Алиня. Нет его только в Хабаровском, Бикинском, Вяземском
районах и в Еврейской автономной области. По Сихотэ-Алиню к югу
доходит приблизительно до 48° с. ш.; А. А. Емельянов (1927) обнаружил
следы его пребывания в бассейне рр. Ботчи и Копи.
На Сахалине, еще в конце прошлого столетия, северные олени были
распространены по всему острову (Никольский, 1887). В настоящее
время населяют северную его часть от полуострова Шмидта до побережья
залива Терпения. Стада их в зимнее время достигают здесь 50—100 го-
лов и даже больше (Мишин, 1952). В свое время несколько оленей было
завезено японцами на о. Сансиру (Курилы); здесь они размножились и
держатся в горах северной его части (Кузнецов, 1949).
В пределах Монгольской Народной Республики, по А. Г. Банникову
(1954), имеется несколько изолированных друг от друга очагов обита-
ния северных оленей. Во-первых, они иногда заходят из ВосточнойТанну-
Ола с верховьев рр. Эрсын-Гол и Наран-Гол (правые притоки р. Тэс).
Далее, дикие олени обычны в Дархатах по верхнему течению рр. Тэнгэ-
син-Гол и Шишхид-Гол, в гольцах Хойт-Агуй-Ула и Хильцзар-Дубху-
Ула, на границе с Тувинской автономной областью. Наконец, в неболь-
шом числе они встречаются к востоку от оз. Хубсугул в верховьях р. Ури-
Гол, на гольцах Хас-Ихэ-Ундур и Умухэй-Ула.
1 По В. Б. Подаревскому (1936), нет оленя в бассейнах рр. Уды (речь, вероятно,
идет о верховьях), Гутары и Такула, а на побережье Байкала — севернее Култука.
2 Устное сообщение С. М. Друри.
RANGIFER
279
Подвиды. Вир, Rangifer tarandus L. в том объеме, как он принимается
в настоящей работе, включает четыре подвида, отличающиеся не только
морфологическими особенностями, но и биологией.
1) R. tarandus tarandus L. — Тундровый олень Европы и Азии. Раз-
меры сравнительно небольшие; длина черепа у самцов от 348 до 387 мм
(в среднем 368 мм), у самок от 309 до 352 мм (в среднем 335 мм). Окраска
в зимнем меху светлая, но все же темно окрашенные места, особенно
полосы по бокам туловища, на голове и на спине, ясно выражены; во-
лосы зимнего меха очень длинные. Череп с умеренно развитой лицевой
частью; расстояние от переднего края глазницы до переднего конца че репа
(prosthion) обычно не больше, чем в 1% раза превышает расстояние от
переднего края глазницы до нижнего края затылочного отверстия (Ьа-
sion). Наибольшая ширина черепа составляет редко меньше 44% его
длины. Длина лобных костей не больше их ширины в области глазниц.
Затылок неширокий; ширина его редко превышает 55% наибольшей
ширины черепа. Носовые кости в передней части сравнительно узкие;
их ширина в самом узком месте составляет от 37 до 56% их наибольшей
ширины. Продольно они слабо выпуклы и слабо выступают над верхне-
челюстными костями. Лоб между глазницами сильно вдавлен, края глаз-
ниц сильно приподняты. При горизонтальном положении черепа (ли-
нии hasion — prosthion) затылочный гребень расположен выше уровня
ростральной части черепа. Распространение: тундровая зона
Европы и Азии с прилегающими островами Полярного бассейна азиат-
ской части СССР.
2) R. tarandus pearsoni Lydekker — Новоземельский олень. Размеры,
в среднем, меньше, чем у типичной расы; длина черепа у самцов от 312
до 329 мм (большею частью не более 357 мм). Окраска в зимнем меху
очень светлая, почти белая, иногда с розоватым оттенком. Темная полоса
по бокам туловища, свойственная другим подвидам северного оленя,
отсутствует. Волосы зимнего меха очень длинные. Краниологически
сходен с R. t. tarandus, но отличается более короткими носовыми костями,
длина которых, в среднем, не превышает 31% основной длины черепа.
Передние концы носовых костей уплощены и не выступают, при взгляде
на череп сбоку, над верхнечелюстными костями. Распростране-
ние: Новая Земля.
3. R. tarandus valentinae Flerow — Лесной северный олень. Размеры,
по сравнению с типичной расой, более крупные; длина черепа у самцов
от 355 до 405 мм (в среднем 382 мм). Окраска в зимнем меху темнее, чем
у типичной расы, обычно буровато-серая, часто с рыжеватым оттенком.
Темные участки на голове, спине и по бокам туловища имеются, но
слабо отграничены и постепенно переходят в светлую окраску соседних
частей тела. Волосы зимнего меха более коротки. Череп с несколько
удлиненной лицевой частью; расстояние от переднего края глазницы до
переднего края черепа обычно не менее чем в 1 % раза превышает расстоя-
ние от переднего края глазницы до нижнего края затылочного отверстия.
Длина лобных костей может быть больше или меньше их ширины в об-
ласти глазниц. Ширина носовых костей в самом узком месте редко бы-
вает меньше 41% их наибольшей ширины (в среднем у самцов около 45%,
у самок около 44.5%). Носовые кости большею частью имеют хорошо
выраженный аркообразный изгиб передних концов и заметно поднимаются
над верхнечелюстными костями. В остальном сходен с R. t. tarandus.
Распространение: лесная зона восточной Европы (?) и Сибири,
Алтай, Саяны; на юго-востоке граница не заходит южнее северной око-
280
CERVIDAE
нечности Байкала и Станового хребта; во время летних кочевок может
доходить до Таймыра и тундровой зоны Якутии.
4) R. t. phylarchus Hollister — Охотский северный олень. Размеры
крупные; длина черепа у самцов от 392 до 438 мм (в среднем 415 мм),
у самок от 344 до 377 мм. Окраска темнее, чем у типичной расы, темные
и светлые участки хорошо отграничены. Длина меха — средняя между
тундровым и лесным. Череп узкий, вытянутый, особенно в лицевой части.
Расстояние от переднего края глазницы до переднего конца черепа со-
ставляет обычно не меньше 155% расстояния от переднего края глаз-
ницы до нижнего края затылочного отверстия. Наибольшая ширина
черепа обычно меньше 44% его длины. У самцов длина лобных костей
всегда больше их ширины. Затылок широкий: ширина его у самцов со-
ставляет редко меньше 59% наибольшей ширины черепа. Носовые кости
широкие; ширина их в самой узкой части составляет от 46 до 66% наи-
большей их ширины. Передние концы носовых костей сильно аркообразно
изогнуты и, как правило, значительно выступают над верхнечелюстными
костями. Основание ростральной части черепа приподнято, межглазнич-
ное вдавление не глубокое; края глазниц почти не поднимаются выше
уровня лба. При горизонтальном положении черепа затылочный гребень
расположен ниже уровня ростральной части. Распространение:
северо-восточное побережье Байкала, бассейн р. Амура, северная часть
хребта Сихотэ-Алинь, Камчатка и Сахалин; западная граница распро-
странения подвида требует уточнения.
Как показали краниологические исследования (Соколов, 1937) и
принимая во внимание большую изменчивость окраски, нет оснований
для обособления европейских и сибирских тундровых оленей; поэтому
сохраняемый К. К. Флеровым (1952) в качестве самостоятельного под-
вида сибирский тундровый олень (R. t. sibiricus Murray) мною относится
к типичному подвиду. По тем же основаниям должен быть объединен
с охотским подвидом выделенный К. К. Флеровым (1932) в качестве само-
стоятельного вида R. angustirostris Flerow 1 — баргузинский (забайкаль-
ский) олень.
Биология. По образу жизни северные олени делятся на тундровых,
лесных и горных.
Тундровые олени весну, лето и начало осени проводят в открытой
тундре, доходя до самого побережья Ледовитого океана. Здесь они на-
ходят в достаточном количестве корм, а благодаря более низкой темпера-
туре воздуха и ветрам, меньше страдают от кровососущих и паразити-
рующих насекомых. С наступлением осени, с исчезновением насекомых
и увяданием зеленой растительности, основная масса оленей покидает
тундру и откочевывает в зону криволесья (лесотундры). Здесь они про-
водят зиму, а весной вновь начинают передвижку в обратном направле-
нии. В лесу снег не так плотен и не представляет сильных затруднений
в добыче корма. Кроме обычно хорошо развитого ягельного покрова,
в лесу олени находят дополнительный источник питания — древесные
лишайники, в изобилии растущие на деревьях. Наконец, и климатиче-
ские условия зимою в лесу более благоприятны — олени не так сильно
страдают от сильных морозов и снежных буранов.
Но, как правило, за южную границу лесотундры тундровые олени
заходят зимой редко, так как при глубоком, хотя бы и рыхлом, снеге
затрудняется не только добыча корма, ..но и передвижение. Свободно
1 Позднее Флеров (1952) стал рассматривать его лишь как подвид.
RANGIFER
281
олень достает корм из-под снега глубиною не больше 50 см (Семенов-
Тян-Шанский, 1948), критической же является глубина снега около
80 см, при которой добыча корма практически не возможна (Наумов,
1933). В глубокоснежные зимы олени вынуждены выходить даже обратно
в южную полосу тундры. В лесную зону заходят лишь более сильные
самцы, способные доставать корм из-под глубокого снега.
На севере Ямала и Гыданского полуострова дикие олени вынуждены
зимовать в тундре, так как лесотундра и север лесной полосы заняты
зимующими стадами домашних оленей (Наумов, 1934). Значительное
количество диких оленей, возможно, приходящих на материк с островов
Ледовитого океана, остается зимовать на северном побережье Таймыра.1
Не имеют возможности переселяться к югу северные олени некоторых
островов, например Новой Земли. Условия существования оленей в тундре
зимой более трудны, чем в лесной зоне. Снег хотя обычно и не ложится
глубоким слоем, но настолько уплотняется ветрами, что олени с трудом
могут откапывать из-под него пищу. Еще страшнее гололедица, покры-
вающая сплошь поверхность снега ледяной коркой и совершенно исклю-
чающая возможность добычи корма. В этих условиях гибнут не только
дикие, но и домашние олени. Зимующие в тундре олени придерживаются
малоснежных районов. На Гыданском полуострове и Таймыре они дер-
жатся ближе к побережью и только в случае гололедицы уходят в глубь
тундры. На Новой Земле зимовки диких оленей расположены на восточ-
ной гористой стороне и северной оконечности острова, где больше обна-
женных от снега участков, тогда как на юге снеговые условия менее благо-
приятны. В годы широкого распространения гололедицы олени пытаются
перекочевать с Новой Земли на о. Вайгач и даже на Ямал (Друри, 1949).
Лесные олени зиму и лето проводят в лесной зоне. Однако больших
сплошных массивов леса избегают и предпочитают местности, где имеются
хотя бы небольшие участки разреженного древостоя или открытые про-
странства. Лето лесные олени проводят чаще на открытых моховых,
хорошо обдуваемых болотах, где меньше досаждают насекомые-крово-
сосы и, кроме того, где звери обеспечены лучшим кругозором. Осенью
держатся не только в болотах, но и в лесах, привлекающих, вероятно,
обилием грибов. Зимой предпочитают ягельные боры, где не так глубок
снег и имеется основной зимний корм — ягель. Живут также по болотам
и редколесью, где снег не так глубок и имеется корм в виде древесных
лишайников и сохраняющихся под снегом зеленых частей растений. На
севере Сибири летующие в лесотундре олени оттесняются приходящими
из тундры на зимовку домашними стадами и уходят вглубь лесной зоны.
Часть из них направляется не на юг, а на север (Наумов, 1931). Одним
из факторов, лимитирующих распространение дикого оленя зимою в лес-
ной полосе, является глубина снежного покрова; поэтому восточные мало-
снежные районы Заенисейской Сибири всегда были и местами максималь-
ной концентрации оленей.
Сезонное размещение по стациям оленей в горнолесных областях не
одинаково и зависит от местных условий. В малоснежных районах, на-
пример на Саянах и в восточной Сибири, лето северные олени проводят
в высокогорье, в гольцовой зоне, по соседству со снежными забоями,
где находят условия, близкие к тундровым. На зиму они спускаются
в лесную зону и живут здесь в тех же условиях, что и лесные олени.
В многоснежных районах смена мест обитания носит обратный характер.
1 Устное сообщение В. М. Сдобникова.
282
CERVIDAE
На Кольском полуострове, например, летом дикие олени держатся и
в горной тундре, и в лесу. В начале зимы, пока снег неглубок,излюблен-
ными местообитаниями являются чернично-ягельные боры. С углубле-
нием снегового покрова олени переселяются в тундру, где неглубокий
и плотный снег предоставляет свободу передвижения, а ягельники обе-
спечивают кормом. В тундре олени проводят весь остаток зимы до на-
чала массового таяния снега (Семенов-Тян-Шанский, 1948).1 В Кроноц-
ком заповеднике, на Камчатке, олени дважды в году подымаются
в горы: с декабря по май, когда в лесу лежит глубокий снег, и с июня
по сентябрь, спасаясь от жары и жалящих насекомых (Аверин, 1948). На
Алтае северные олени лето и зиму проводят в гольцовой зоне, выше границы
леса на высотах от 1500 до 2000 м; в горах снег сдувается ветром, легко
добывать корм, а кроме того, почти нет хищников. Только в бассейне р. Ле-
бедя олени держатся в пихтово-кедровых насаждениях, так как здесь
хотя и глубокий, но плотный снег позволяет свободно передвигаться
и кормиться лишайниками, обильно растущими на деревьях (Дмитриев,
1938).
В связи с сезонной сменой мест обитаний стоят миграции диких се-
верных оленей выраженные у них сильнее, чем у других представителей
семейства Cervidae. Наибольший размах миграции приобретают у тун-
дровых оленей восточной Сибири и Якутии, места зимовок которых не-
редко отстоят от мест, где звери проводят лето, на сотни километров.
Направление миграций более или менее постоянно, но пути, по которым
движутся животные, могут год от года несколько, а иногда и весьма суще-
ственно меняться, в зависимости от конкретной обстановки.1 2 Отмечается
даже периодичность в смене путей кочевок, зависящая, вероятно, от
стравливания оленями пастбищ (Наумов, 1933).
Весною движение оленей с мест зимовок к северу у тундровых оленей
начинается еще до начала таяния снега и продолжается иногда не менее
месяца. На Таймыре на побережье олени появляются с конца марта—
начала апреля (Гептнер, 1936); заканчиваются перекочевки обычно
к концу мая. Время массового прибытия на летовку совпадает с началом
появления зелени в тундре. Первыми перекочевывают беременные самки,
позднее молодняк; последними в тундре появляются взрослые быки.
Двигаются иногда небольшими группами в 10—12 голов, но в некоторых
районах на общих путях собирались двигавшиеся сплошной лавиной стада
в несколько тысяч животных. При массовых переправах мигрирующих
оленей через реки производились знаменитые, ныне сохранившиеся
лишь в немногих местах охоты «на плавях». Скорость передвижения при
миграциях различна (вероятно, в зависимости от состояния пастбищ на
пути миграций и погоды), но вообще довольно значительна, особенно
осенью (до 20 км в сутки). Кормятся животные только урывками, на
ходу. На местах летовок происходят менее заметные местные передвижки
оленей, в зависимости от кормов и погоды. На побережье Таймыра,
у Диксона например, олени появляются дважды: весной в апреле—мае и
вторично, ненадолго, в первых числах сентября (Гептнер, 1936). Обрат-
ные перекочевки тундровых оленей к югу в лесотундру начинаются в за-
1 Семенов-Тян-Шанский (1949) считает причиной ухода оленей из глубокоснеж-
ных лесов не столько трудность добывания корма, сколько отсутствие свободы пере-
движения по глубокому рыхлому снегу, с чем едва ли можно согласиться хотя бы по-
тому, что движется олень зимою, в основном, в поисках пищи.
2 Более подробно о причинах, путях и масштабах миграций северных оленей
см.: Наумов, 1933; Друри, 1949, а по Северной Америке — Murie, 1935, Seton, 1939.
RANGIFER .
283
висимости от местных условий уже в августе, а в массе происходят
в сентябре и октябре. Двигаются осенью олени смешанными стадами —
и самцы, и самки вместе. Во время осенних перекочевок происходит и
гон у северных оленей.
В европейской части СССР и в западной Сибири, а также во всех горно-
таежных районах, сезонные перекочевки оленей носят местный характер
и обычно не превышают нескольких десятков километров. Только на
Северном Урале имеют место сравнительно дальние перемещения в сен-
тябре с западных многоснежных склонов и припечорских низменностей
на восточные малоснежные склоны и низменности западной Сибири,
а весною в обратном направлении (Куражсковский, 1949). В равнинных лес-
ных районах олени только меняют стации по сезонам, выбирая к зиме
более кормные и малоснежные места, шире расселяясь в бесснежное время
года. В противоположность другим оленям, северные олени зимой, даже
в лесной полосе, если благоприятствуют снеговые условия, ведут до-
вольно подвижный образ жизни и за зиму проделывают путь иногда до
сотни километров (Наумов, 1934). Незначительные, и при этом уже не
широтные, а вертикальные миграции северных оленей имеют место также
в горно-лесных районах. Осенние откочевки в гольцы и горную тундру
происходят, когда снег в лесу станет глубоким настолько, что затрудняет
передвижение, обычно в период с ноября по январь. Спускаются с гор
весною с началом массового таяния снега, обычно в мае. Переход на лето
в высокогорья от границы лесов, где олени держатся до наступления жары,
происходит обычно в июле. Никаких постоянных путей перекочевок при
вертикальных миграциях, по-видимому, нет (Семенов-Тян-Шанский, 1948).
Пищей северному оленю служит подавляющее большинство растений,
произрастающих в районах его обитания. Растений, непоедаемых со-
вершенно из-за плохих вкусовых качеств или ядовитых свойств, очень
немного. Определенные виды растений оленями поедаются с большим
аппетитом, другие служат пищей только при отсутствии более желатель-
ных кормов, но в общем основа питания определяется не столько пред-
почтением, отдаваемым тому или иному корму, сколько составом и до-
ступностью в данный сезон пастбищных растений.
Характерная биологическая особенность северного оленя — употреб-
ление в пищу в течение всего года наземных и древесных лишайников.
Способность оленя, в противоположность другим жвачным животным,
переваривать этот вид пищи является одним из физиологических при-
способлений к обитанию в условиях тундры. Лишайники не являются
полноценным кормом, обеспечивающим организм необходимыми пита-
тельными веществами. Как показали химические исследования (Формо-
зов и Формозова, 1936; Александрова, 1940), большинство лишайников
богаты углеводами, из которых специфическим для них является лихенин,
но содержат мало минеральных и азотистых веществ. Вследствие этого,
при обильном питании одними лишайниками создается отрицательный
азотистый баланс, наступает белковое голодание и происходит потеря
в весе. Кроме того, лишайники почти не содержат витаминов. Вероятно,
результатом не только недостаточного количественного, но и качественно
неполноценного питания является наблюдающееся к концу зимнего
периода исхудание, сопровождающееся глубокими дистрофическими
процессами в организме оленя (Боль и Николаевский, 1932). Вопрос
о том, насколько лишайники необходимы для северного оленя и может
ли он существовать без них, не ясен. Среди оленеводов распространено
мнение, что олени хорошо нагуливаются только тогда, когда наравне
284
CERVIDAE
с зеленым кормом на пастбищах имеется в достаточном количестве ягель.1
Во всяком случае, даже и летом, когда имеются в избытке другие корма,
олень поедает, хотя бы в небольшом количестве, и его.
Ягель является основной пищей оленя зимою. В это время лишайники,
откапываемые им из-под снега, составляют до 80% содержимого желудка.
Но, как теперь установлено (Александрова, 1940), непременной частью «
рациона и в зимний период являются сохраняющиеся под снегом зеленые
части некоторых травянистых растений (осок, пушиц, хвощей), а иногда
и высохшие их части. Из лишайников, в первую очередь и наиболее
охотно, поедаются кустистые из рода Cladonia (С. alpestris L. и С. гап-
giferina L. и. др.). Важную роль играют также виды листоватых и труб-
чатых лишайников (роды Getraria, Stereocaulon, Sphenophorus). Дре-
весные лишайники (Usnea, Bryogopon) служат пищей, когда наземного
корма мало или он не доступен вследствие наста, гололедицы или глу-
бокого снега.
Весной, с появлением первой зелени, олени с жадностью набрасы-
ваются на сочные ростки пушиц (род Eriophorum), осок и других одно-
дольных растений и разнотравья. Наравне с зеленью, за недостатком
последней, весною олени в большом количестве поедают и ягель.
Летом зеленый корм составляет до 80% содержимого желудка. Основу
питания в это время в тундре составляют листья различных видов ив и
карликовой березки (Betula папа L.). Из травянистых растений отдается
явное предпочтение разнотравью. Однодольные растения видное участие
в рационе оленей принимают лишь при отсутствии или недостатке кустар-
никового корма и разнотравья. В лесной полосе летом северные олени
питаются, кроме листвы ив и берез, болотным разнотравьем: калужницей,
морошкой, гравилатом, а также хвощами, в меньшей мере осоками и
пушицей. Лишайники летом уже не являются основным кормом, но все же
поедаются, особенно во влажном состоянии, и составляют до двадцати,
а иногда и больше процентов рациона (Александрова, 1940).
Осенью, по мере огрубения и отмирания зеленых растений, лишай-
ники вновь занимают видное место в питании оленя. Травянистые расте-
ния олени не поедают целиком, а, захватывая губами, срывают отдель-
ные листочки, верхушки и цветочные бутоны. У наземных кустистых
лишайников зимой также обрываются верхушки. Летом же вырываются
часто целые кустики. В сухом виде олени ягеля не едят, вероятно вслед-
ствие грубости, а также потому, что он при схватывании крошится на
мелкие частицы.
Любимым лакомством оленей осенью являются грибы, ради которых
они спускаются на осенние месяцы с гор в полосу леса.
Летом олень, при наличии некоторого количества ягеля, обеспечен
полноценным питанием. Но уже осенью начинает сказываться солевое
и азотистое голодание. В связи с этим стоят общеизвестные факты ловли
оленем леммингов, рыбы, обгладывания сброшенных рогов, поедания
сброшенной с рогов кожи (Семенов-Тян-Шанский, 1948). Этим же сле-
дует объяснять пристрастие и диких, и домашних оленей к человеческой
моче (вернее к снегу, смоченному последней) в зимнее время. А. Н. Фор-
мозов (1936) наблюдал поедание оленями весной птичьих яиц, но уже
с начала июля, вероятно в связи с исчезновением потребности организма
в дополнительном белковом питании, они не обращали ни на яйца, ни
1 Под общим названием «ягель» подразумевают всю совокупность наземных
лишайников, служащих северному оленю пищей.
RANGIFER
285
да птенцов никакого внимания. В восточной Сибири дикие северные олени
посещают не только естественные, но и искусственные солонцы, особенно
осенью, после гона (Наумов, 1934; Скалой, 1951). На Сахалине олени, по
сообщению А. М. Никольского (1887), выходят лизать соль на морское
побережье. Сена дикие олени не едят.
Северный олень — типично стадное животное. Встречаются и одиночно
живущие экземпляры, главным образом взрослые самцы, но эти случаи
составляют не правило, а исключение. Размеры стад сильно варьируют,
от единиц до сотен и даже тысяч голов в каждом, в зависимости от коли-
чества оленей в данном районе и от сезона.
Зимние смешанные стада распадаются перед отелом важенок, которые
прогоняют от себя прошлогодних телят. В течение лета олени держатся
группами не свыше десятка голов, а чаще по 3—5 животных. При этом
молодняк с яловыми самками, взрослые быки и важенки с телятами
образуют обособленные группы. В таежной полосе восточной Сибири и
на Сахалине в комариную пору самцы чаще, чем в другое время, живут
одиночно. Самки с телятами держатся парами или объединяются по не-
скольку голов вместе. Гораздо реже образуются летом смешанные стада
из важенок с телятами и молодняка обоего пола.
К осени табунки объединяются в создаваемые самцами на период гона
таремы. В состав последних входят один взрослый сильный самец, не-
сколько молодых самцов и 8—13 самок с приплодом. Редко, но встре-
чаются в это время стада по нескольку десятков голов или, наоборот,
группы, состоящие из самца и 1—2 самок.
По окончании гона самцы отделяются от самок и держатся группами
отдельно. Остальные олени образуют к зиме смешанные стада от 15—
20 до нескольких десятков и даже сотен голов в каждом. Иногда к этим
стадам присоединяются и самцы. На путях массовых миграций, особенно
осенью, собираются тысячные, собственно уже не стада а массы движу-
щихся лавиной оленей, распадающиеся на более мелкие группы по при-
бытии к местам зимовок. Еще более укрупняются стада оленей в конце
зимы. На Камчатке, например, наблюдались табуны до 2000 голов (Аве-
рин, 1948).1 Такие большие стада не выпасаются плотной массой, как
домашние олени, а широко рассеиваются небольшими группами по вы-
пасному участку.
Северный олень — весьма подвижное животное. Кормится на ходу,
почти непрерывно передвигаясь с места на место. Особенно велика под-
вижность зимою, но и летом олени редко задерживаются подолгу на
одном месте. Спасаясь от гнуса, они бегают взад и вперед, нередко вы-
таптывают тропы, которые особенно хорошо бывают заметны в лесу и
в горах. Тропами животные пользуются также при переходе из одного
выпасного участка в другой, а в некоторых районах тропы являются
показателями путей сезонных миграций. При переходах небольшими
группами олени следуют друг за другом гуськом, но при массовых мигра-
циях двигаются широким фронтом.
Кормятся как в ночные, так и дневные часы суток, хотя отдыхающими
оленей чаще можно видеть днем, чем ночью. В высокогорье от жары
спасаются, кроме бега, тем, что держатся вблизи снеговых забоев, ло-
жатся даже на снег или становятся на обдуваемые ветром хребты и вер-
шины. От морозов северные олени не страдают, но во время пурги лежат
1 Самое большое стадо, которое наблюдал в апреле Семенов-Тян-Шанский (1948)
в б. Лапландском заповеднике; состояло из 832 оленей.
286
CERVIDAE
спиной к ветру и повертывают голову на подветренную сторону тела
(Семенов-Тян-Шанский, 1948).
Передвигается северный олень в спокойной обстановке шагом; испу-
ганный же бежит рысью или вскачь; таких прыжков, какие делает бла-
городный олень, у него наблюдать не приходилось. При движениях
северного оленя, в суставах нижних частей его конечностей происходит
характерный треск, слышимый хорошо за 20—40 м. Из многих предло-
женных объяснений происхождения этого звука (Jacobi, 1931) ни один
не может быть назван удачным. При беге голова оленя поднята кверху
так, что рога ложатся на спину. Хвост при этом держится вертикально
и его белая вентральная сторона увеличивает размеры и без того расши-
ренного во время бега околохвостового зеркала. Северный олень хорошо
плавает, легко переплывает широкие реки и небольшие озера; когда
плывет, хвост держит также поднятым кверху.
У северного оленя лучше всего развито обоняние. Человека по ветру
может учуять по крайней мере за 1—2 км (Дмитриев, 1938). С помощью
обоняния обнаруживает даже ягель под покровом снега (Семенов-Тян-
Шанский, 1948). Хорошо слышит, но острым зрением не обладает. «На
открытом месте на фоне снега олень видит другого оленя или человека
примерно за километр, но замечает его только в движении. . . Глаз
оленя охватывает широкий угол, в пределах которого он замечает все
перемены в положении предметов, но рассмотреть что-либо в деталях
он не может» (Семенов-Тян-Шанский, 1948).
Смышленностью олень не обладает; хозяина и ухаживающего за ним
человека от посторонних не отличает. Нрав имеет сравнительно кроткий;
драки между животными, по крайней мере у домашних, за исключением
самцов в период гона, бывают очень редко. Дикие олени очень осторожны
и пугливы; однако осторожность не является врожденной и вырабаты-
вается в результате общения с человеком. Известны случаи, когда, впер-
вые увидев человека, олени свободно подпускали охотников на близкий
выстрел. Домашних оленей не только избегают, но при виде их, обычно,
обращаются в паническое бегство. Только самцы в период гона, находясь
в возбужденном состоянии, часто забегают в стада домашних оленей.
Голос взрослых северных оленей отдаленно напоминает негромкое
низкое «хрюканье», которым важенки (самки) перекликаются с теля-
тами. Теленок ищет мать высоким звуком, по моему мнению не совсем
удачно изображаемым О. И. Семеновым-Тян-Шанским как «еа. . . еа. . .
еа. . .» или «кау. . . кау. . .». Самцы во время гона также «хрюкают»,
только еще более низким и грубым, чем самки, тоном.
Смена волосяного покрова происходит один раз в году. Линька на-
чинается выпадением отдельных волос, обычно в мае. Разгар ее падает
на июнь и июль. В первую очередь вылинивают взрослые самцы и яло-
вые самки. На истощенных животных еще в августе1 можно изредка
видеть остатки старой шерсти.
В августе сквозь короткую летнюю буро-коричневую ость начинают
пробиваться более длинные и светлые направляющие волосы зимнего
меха, придающие седоватый оттенок окраске зверя. Одновременно про-
должается рост летней ости и развитие подшерстка.
Полного развития зимний мех достигает в большинстве районов к концу
сентября или к началу октября.
1 И. И. Колюшев (1936) август считает нормальным сроком окончания линьки.
RANGIFER
287
Рога взрослые самцы сбрасывают вскоре после окончания периода
гона, в ноябре или декабре, молодые самцы несколько позднее старых.1
Яловые самки сбрасывают их одновременно с самцами или несколько
позднее. Остальные важенки теряют рога через несколько дней после
отела, т. е. в мае—июне; но известны редкие случаи, когда беременные
важенки сбрасывали рога в марте.1 2 В мае же сбрасывают первые рога
дикие олени-годовики. Начало роста рогов у самцов, по О. И. Семенову-
Тян-Шанскому (1948), падает на апрель, но, по расспросным сведениям,
у некастрированных самцов домашних оленей в Болыпеземельской тундре
они начинают отрастать уже через две недели после сбрасывания старых.3
Вначале рога растут медленно, но с наступлением тепла темпы нарастают.
Формирование новых рогов заканчивается к концу июля—началу ав-
густа. Вскоре после потери старых начинают отрастать рога и у важенок.
Быстрота и степень развития рогов зависят от упитанности оленей, усло-
вий погоды, количества «гнуса». У больных и истощенных животных
окостенение рогов происходит рано, но они недоразвиваются и достигают
лишь незначительных размеров (Наумов, 1934). Несколько позднее
формируются рога у самок.
Самцы начинают очищать рога от кожи с конца августа и, по иашим
наблюдениям в Болыпеземельской тундре, в основной массе заканчивают
этот процесс уже в начале сентября. Почти одновременно с самцами очи-
щают рога от кожи яловые самки., У самок, имеющих телят, этот процесс
начинается позднее, во второй декаде сентября, и в двадцатых числах
этого месяца неочищенные рога можно видеть только у единичных живот-
ных. Испытывая раздражение от начинающей омертвевать и подсыхать
кожи, олени ударяют рогами о кусты, трутся ими о попадающиеся на
пути предметы, оставляя на них лоскутья и полосы содранной и часто
еще окровавленной кожи.4 5 Процесс чистки у отдельного животного длится,
обычно, не больше 1—2 дней. Телята-первогодки чистят рога с конца
сентября до начала ноября. Сбрасывание ими первых рогов производится
в мае следующего года (Семенов-Тян-Шанский, 1948).
Половой зрелости северные олени, как правило, достигают в возрасте
около 16—17 месяцев; однако не так редки случаи, особенно среди до-
машних оленей, когда рано родившиеся и развивавшиеся в благоприят-
ных кормовых и климатических условиях самочки оплодотворяются
уже в первый год своей жизни, в пяти-шестимесячном возрасте, и сле-
дующей весной приносят жизнеспособных телят. Самцы практически на-
чинают принимать участие в размножении не раньше двух лет, так как
раньше этого отгоняются более сильными соперниками.
Период гона (спаривания) у северных оленей в большинстве райо-
нов происходит в период с сентября по ноябрь. Чаще начало его падает
на вторую половину сентября,8 но иногда признаки полового возбужде-
ния у отдельных, хорошо упитанных взрослых самцов проявляются
1 Сообщение С. II. Наумова (1931) о том, что некастрированные самцы на Гыдан-
ском полуострове сбрасывают рога несколько позднее кастрированных (для которых
сроки протекания этого процесса им указываются с начала февраля до десятых чисел
мая), очевидно, основано на недоразумении.
2 Наблюдения автора (Соколов, 1935) над домашними оленями в Большеземель-
ской тундре.
3 По-видимому, на неверных расспросных данных основано утверждение И. И. Ко-
люшева (1936) о том, что на сибирском севере образование новых рогов у самцов и хо-
лостых самок начинается в июне.
4 Гистогенез рогов у северного оленя см.: Боль и Николаевский, 1936.
5 На Алтае, по Д. К. Соловьеву (1920), несколько позднее, чем у марала.
288
CERVIDAE
уже в последних числах августа. У самок течка наступает несколько
позднее. У диких оленей начало гона определяется образованием сме-
шанных стад. Разгар гона происходит в конце сентября и в первой поло-
вине октября, но отдельных гоняющихся оленей можно наблюдать и
в ноябре. На Камчатке, по некоторым данным, и отел и гон проходят
недели на две или на три позднее. В Кроноцком заповеднике, например,
разгар гона наблюдается в средине ноября (Аверин, 1948). На севере
Якутии, кажется, наоборот, имеет место некоторое ускорение.
Самцы в период гона сильно возбуждены и драчливы, теряют свою
обычную осторожность, нередко подбегают к домашним оленям даже
в присутствии человека (Наумов, 1934; Семенов-Тян-Шанский, 1948).1
В это время у них утолщается шея, набухают брови, слизистая оболочка
глаз краснеет («глаза наливаются кровью»), а от тела исходит специфи-.
ческий запах, из-за которого мясо их становится на некоторое время
непригодным в пищу. Возбужденный самец издает особый храп или хрю-
канье, на который отвечает его соперник. Каждый самец стремится соз-
дать гарем из 10—15 самок, который он ревниво охраняет от соперников
и, бегая вокруг, не позволяет им расходиться. Между самцами происхо-
дят драки не только рогами, которые иногда даже ломаются из-за этого,
но и передними и задними ногами. Серьезные повреждения при этом,
по-видимому, наносятся редко. Самец во время гона ест мало, сильно
худеет и нередко теряет гарем в результате поединка с менее утомлен-
ным конкурентом. Самки ведут себя более спокойно, но порой вынуждены
много бегать, спасаясь от настойчиво преследующего самца. В противо-
положность другим оленям, телята не покидаются на период гона и все
время находятся с матерями. Самец ходит или бегает за находящейся
в охоте самкой, вытянув голову, и время от времени как бы приседает
задом. Неоплодотворенная самка вторично приходит в охоту через 11 —
22 дня.
Беременность, по последним данным, полученным на домашних
оленях, в среднем равна 7% месяцам, но колеблется от 192 до 246 дней
(Горбунов, 1939). Единичные случаи рождения молодых бывают уже
в конце апреля, но массовый отел в большинстве районов происходит
во второй половине мая.1 2 Отдельные поздние отелы, по крайней мере
у домашних, известны не только в июне, но и в июле. Так как на севере
в период массового отела снег сошел еще не везде, то для родов самка
выбирает проталины и более сухие возвышенные места. Самка приносит,
как правило, одного теленка, но у домашних оленей иногда бывают и
двойни.
Теленок встает сосать мать через 2—3 часа после рождения, но пер-
вые дни беспомощен, больше лежит и начинает следовать за матерью
лишь на пятый или шестой день. Оленуха — не очень хорошая мать и
в случае опасности нередко бросает теленка (Семенов-Тян-Шанский,
1948). Уже с двухнедельного возраста теленок начинает понемного щипать
зеленый корм. Молоком мать кормит его до наступления полового се-
зона, но ходят оленята с матерями до следующего отела, а иногда даже
и все следующее лето.
Родятся оленята с живым весом в среднем около 6.5 кг (от 4.1 до 9.2 кг)
(сборн. «Северное оленеводство», 1948 : 212), к осени достигают 60%
1 Помеси между дикими и домашними оленями всюду ценятся за их высокие
ездовые качества.
2 По сообщению В. М. Сдобйикова, на Таймыре и гон, и отел у диких оленей про-
текают на месяц позднее.
RANGIFER
289
веса матерей. Самцов родится, судя по домашним оленям, несколько
больше, чем самок, но в стадах диких оленей взрослых самцов раза в три
или четыре меньше. Объясняется последнее большей гибелью самцов
с двухлетнего возраста вследствие их меньшей осторожности, особенно
во время гона.
Рога у оленят начинают расти уже с месячного возраста. К осени
они имеют вид простых, несколько изогнутых вершинами вперед спичек,
но иногда, особенно у домашних, уже в первый год образуется развилка
на вершине рога или вырастает небольшой первый надглазничный от-
росток. Как уже отмечалось, в октябре—ноябре с этих первых рогов
сбрасывается кожа, а в апреле—мае следующего года сбрасываются
и сами рога. Вторые рога обычно имеют только один надглазничный от-
росток, у третьих вырастает и второй («ледяной»); но в силу большой
изменчивости и зависимости от условий жизни животного, целям опреде-
ления возраста рога северного оленя служить не могут.
Кастрация не приостанавливает роста рогов, но искажает ход и сроки
окостенения, очистки и смены. Если кастрация самца произведена в пе-
риод, когда рога покрыты кожей, то сдирать последнюю олень начинает
только в конце сентября или в октябре, а рога сбрасывает в конце марта—
первой половине апреля. Если же к моменту кастрации олень уже начал
чистить или очистил рога, то он сбрасывает их через 10—15 дней после
операции, иногда вместе с кожей, и до конца апреля ходит безрогим.
В последующие годы очистка кожи у кастратов происходит, начиная
с конца сентября—октября, но процесс этот идет вяло, у большей части
полного освобождения рогов от кожи не происходит, и последние сбра-
сываются в марте—апреле вместе с остатками кожи.
Возрастные изменения в зубной системе представляются в следую-
щем виде (Соколов, 1937). При рождении у северного оленя имеются
все три пары молочных резцов и нижние клыки. Премоляры еще покрыты
слизистой оболочкой и обнажаются через несколько дней. В годовалом
возрасте молочные резцы сменяются постоянными и появляются пер-
вые моляры. В 2 года развиты все моляры, а в начале 3-го года про-
исходит выпадение молочных премоляров. Постоянные премоляры до-
стигают полного развития к 3 годам. К этому же возрасту становится
заметно стирание внутренней пары резцов (1Х); в 4 года стирается уже
вторая, а в 5 лет третья пара.1
Предельную продолжительность жизни домашнего северного оленя
определяют в 25—28 лет, но, как на исключительный случай, мне ука-
зывали в Болыпеземельской тундре на важенку, прожившую якобы
около 35 лет. Самки нормально плодоносят до 15 лет и дольше этого воз-
раста домашних оленей содержат редко. Продолжительность жизни
дикого оленя в естественной обстановке, вероятно, еще меньше; в кол-
лекциях черепа зверей старше 13 лет встречаются очень редко (Вгошёе-
Skuncke, 1952).
Из врагов северного оленя на первом месте по значению стоит волк.
От волков на севере гибнет огромное количество и диких, и домашних
оленей. Олени в тундровой зоне составляют основной источник питания
волков и численность обоих видов находится в тесной зависимости друг
от друга. Волки совершают такие же сезонные миграции, как олени,
почему и получили местами прозвище «пастухов» оленьих стад. Сильно
1 Сходные, но несколько отличные данные о сроках появления и смены зубов
у оленей Скандинавского полуострова приводит Броме-Скунке (Bromee-Skuncke,
1932).
19 Фауна СССР, И. И. Соколов
290
CERRIDAE
размножившиеся волки могут совершенно приостанавливать естествен-
ный рост популяции диких оленей в том или ином районе (Наумов, 1933;
Семенов-Тян-Шанский, 1948). Гибнут от волков олени всех возрастов
и во все времена года. Но, как показал учет в б. Лапландском заповед-
нике (Семенов-Тян-Шанский, 1948), самцов гибнет в несколько раз
больше, чем самок. Охотятся волки большей частью стаями, окружая
или заганивая оленей. Поэтому зимой, когда снег более глубок, к тому же
часто покрыт настом, не выдерживающим оленя, а сами олени, в силу
неполноценного питания, слабы, ущерб, наносимый им волками, имеет
наибольшие размеры.
От медведя и росомахи гибнет сравнительно немного оленей, главным
образом телят в первые дни после отела. Взрослого оленя ни тот, ни дру-
гая догнать не могут, и поэтому последний только случайно может стать
их добычей. Случайный характер носят и заметного влияния на числен-
ность оленей не оказывают потери от мелких хищников и птиц (Сдоб-
ников, 1935).
Кроме непосредственного истребления, вред от волков увеличивается
тем, что они разгоняют стада, лишают зверей нормального пастбищного
режима и препятствуют восстановлению и поддержанию упитанности
животных.
Сильно страдают северные олени от кровососущих насекомых (мошек,
комаров, слепней) и оводов, нападающих на них в несметных количествах
в теплое время года, с конца июня по август. Насекомые причиняют
оленям сильное беспокойство, лишают их нормального отдыха и пита-
ния. Необходимость уменьшить себе страдания от насекомых являетса
одной из причин сезонных миграций северных оленей. Олени спасаются
от насекомых на хорошо обдуваемых ветром и прохладных морских по-
бережьях, вершинах горных хребтов, в альпийской зоне вблизи снега.
Так как эти возможности имеются не везде, то олени спасаются иногда,
бегая быстро взад и вперед по прямой линии или кругу. При беге соз-
дается встречный ток воздуха, отгоняющий насекомых, но олени затра-
чивают колоссальное количество энергии в ущерб состоянию организма..
Из наружных паразитов северного оленя наибольшее значение имеют
овода: кожный (Oedemagena tarandi L.) и носовой (Cephenomyia trompe-
Modeer), паразитирующие в личиночной стадии. Личинки первого посе-
ляются под кожей оленя, иногда в числе нескольких сотен экземпляров,,
и пробуравливают насквозь кожу. Вокруг них происходит реактивное-
воспаление соседних тканей с нагноением, что не может не отразиться
вредно на организме зверя. У домашних оленей снятая в конце зимы
или весною кожа, на спине и боках сплошь усеяна большим числом от-
верстий, через которые в мае—июне происходит выпадение личинок..
Еще больший вред северным оленям причиняет носовой овод. Помимо
беспокойства и панического страха, которые вызывает у оленя прибли-
жение взрослого насекомого, выбрызгивающего ему в носовую полость
живых личинок, присосавшиеся в большом числе к слизистой оболочке
гортани и носоглотки личинки могут иногда вызвать удушение (Семенов-
Тян-Шанский, 1948). Дикие олени, в силу своей большей подвижности,
от оводов страдают меньше.
Из эпизоотических болезней у диких оленей бывает некробациллез
(«копытная болезнь», см. ниже), в Северной Америке известны падежи
диких северных оленей от сибирской язвы (Hornaday, 1912), наблюда-
лись заболевания чесоткой (Наумов, 1933). Во всех этих случаях источ-
ником инфекции, по-видимому, являлись стада домашних оленей.
RANGIFER
291
Хозяйственное значение. Дикий северный олень необычайно ценен,
прежде всего как мясное животное. Промысел его во многих районах
имеет важное значение как дополнительный источник питания, а в до-
революционное время на Колыме и Анадыре он служил основой благо-
состояния оседлого населения. Неудачная охота на «дикаря» нередко
ставила жителей в крайне тяжелое положение, влекла за собой голо-
довку (Врангель, 1841). При рациональной постановке промысел ди-
кого оленя и в настоящее время может играть весьма существенную роль
в создании местной продовольственной базы для промышленности Край-
него Севера.
Взрослый олень-самец может дать при забое тушу весом от 45 до 100
и больше килограммов хорошего мяса (Друри, 1949; Аверин, 1948).
Лучшей упитанности и наиболее жирны олени осенью перед началом
гона. Аверин (1948) приводит случай, когда от не очень крупного самца
было получено 12.2 кг подкожного и 5.6 кг внутреннего сала. Толщина
слоя подкожного жира на крестце достигала 8 см.
Не меньшую ценность представляет кожа северного оленя. Если
шкура не продырявлена, из нее получается при выделке известная своим
прекрасным качеством оленья замша. Шкурка убитого в первые недели
жизни теленка дает ценный и красивый мех «пыжик», употребляемый
на изготовление меховых пальто и шапок. Из осенних, имеющих еще
недлинный мех шкурок шьют различного рода одежду, от брюк и руба-
шек до пальто или малиц. Зимние шкуры менее ценны, так как имеют
пышный, но очень ломкий волос. Они идут на изготовление верхней
одежды типа дох (совиков, кухлянок). Оленьи меха, благодаря их струк-
туре и характеру волоса, очень теплы. В оленьей дохе можно спать на
открытом воздухе при тридцатиградусном морозе. Широкое применение
на Крайнем Севере зимние оленьи шкуры находят в качестве постелей,
ковров, спальных мешков и тому подобных изделий.
Особо прочная кожа с нижних частей ног, так называемые «камысы»,
идет на пошивку зимней обуви, а также на подбивку лыж.
Сухожилья с ног употребляются как очень крепкие и не боящиеся
сырости нитки при шитье обуви и одежды. Рога идут на всевозможные
поделки.
Охота. Способы добычи диких северных оленей разнообразны.1
Некоторые из них, как например, скрадывание, подкарауливание на
кормежках и местах, куда звери приходят спасаться от насекомых, те
же, что и на других оленей и уже описаны раньше. Следует отметить,
что охота с лайками, практикуемая при промысле кабарги, изюбря и
лося, на северного оленя не применима, так как он не выдерживает го-
лоса собаки и быстро уходит.
В районах, лежащих к востоку от Енисея, в прежние годы широко
практиковался ныне запрещенный способ коллективной массовой их
добычи «поколкой на плавях». Этот способ был основан на свойстве се-
верных оленей придерживаться из года в год более или менее постоян-
ных путей сезонных миграций. Зная места переходов мигрирующих мас-
сами оленей через реки, охотники группами подкарауливают их в засаде
на берегу, и когда звери входят в воду и начинают плыть, на легких лод-
ках преграждают им путь и, врезываясь в середину плывущего стада,
копьями убивают их.1 2 Применяемая осенью поколка на плавях сыграла
1 Описание главнейших из них см.: Сдобников, 1953.
2 Хорошее описание охоты на плавях дали Матюшкин (Врангель, 1841) и Майдель
(1896). .
19*
292
CERVIDAE
не малую роль в уменьшении общей численности оленей. Истреблялось
огромное количество животных без разбора возраста и пола. В короткое
время убивались сотни и даже тысячи оленей; не всегда успевали даже
справиться с обработкой и вывозкой из тундры добычи. Большое число
подранков и убитых, но своевременно не вытащенных из воды животных
пропадало бесполезно. Однако этот способ добычи, при правильном пла-
нировании и организации, мог бы стать одним из основных при плано-
вых заготовках оленьего мяса, так как позволяет в короткий срок и без
большой затраты труда заготовить значительное количество требуемого
продукта. Промысел можно усовершенствовать применением так назы-
ваемых «маховок». Последние представляют собой загоны из двух посте-
пенно сближающихся к выходу в русло реки рядов вбитых на некото-
ром расстоянии друг от друга кольев с привязанными к их вершинам
гусиными крыльями («маховками») или лоскутками цветной ткани. Стадо
диких оленей группой загонщиков осторожно направляется в загон,
а у узкого выхода из последнего скрытно располагаются стрелки. При
выходе из загона может быть растянута сеть, в которой запутываются
звери (Наумов, 1933).
Из индивидуальных способов охоты на северного оленя во многих
районах распространена охота с «подводчиком». Принцип ее заключается
в том, что охотник со специально обученным и сходным с диким по ок-
раске домашним оленем, самцом или самкой («подводчиком»), направ-
ляется в места, где водятся дикие олени, и под прикрытием управляемого
с помощью длинного ремня подводчика подкрадывается к последним
на выстрел. Успех зависит от выучки подводчика и осторожности охот-
ника. Этим способом можно охотиться в течение круглого года. Но наи-
более успешна охота в период гона, когда дикий самец принимает спе-
циально обученного домашнего самца, называемого «манщиком» или
«манчиком», за соперника и сам бежит к нему навстречу для поединка.
В этом случае манщика иногда пускают одного с петлей на рогах, в ко-
торой запутывается дикарь, вступая в драку с домашним оленем.
В тундре применяется охота с подъезда на домашних оленях. Охот-
ник кружит, стараясь объехать диких оленей с наветренной стороны,
а так как те, в свою очередь, стараются встать с подветренной, то, делая
перебежки, могут набежать на выстрел (Наумов, 1933).
При скрадывании оленей в тундре пешком применяют щиток с про-
резью для стрельбы, обтянутый чем-нибудь белым и толкаемый охот-
ником впереди себя на лыжах или санках. Миддендорф (1869) в числе
способов охоты упоминал подманивание диких самцов подражанием
их голосу во время гона. Во времена Врангеля (1841) на севере Якутии
летом иногда загоняли диких оленей с помощью собак в воду, где дого-
няли медленно плывущих зверей на лодках и убивали.
Широко применявшиеся раньше способы добычи самострелами, ямами,
капканами, устанавливаемыми на тропах, а также преследование по
насту и глубокому снегу, в настоящее время запрещены законом.
Оленеводство. Человек издавна и независимо в нескольких местах
приручил и сделал домашним животным северного оленя. Время одомаш-
нения точно не известно, но, по-видимому, начало его относится к периоду
не позднее V в. нашей эры (Максимов, 1928). Однако условия существо-
вания домашнего оленя слишком мало изменились по сравнению с диким,
чтобы вызвать существенные изменения в его организме. Признаки домести-
кации находят выражение лишь в повышенной изменчивости окраски
домашних оленей. Наравне с типичной для большинства домашних оле-
RANGIFER
293
ней «дикой» окраской, у них часто встречаются уклонения от нее в сто-
рону как потемнения до почти черной, так и посветления до полного
или частичного альбинизма (Соколов, 1935; Dechambre, 1942; Herre,
1943). У местного населения всегда существовал повышенный спрос на
белый, а также на темный мех для пошивки нарядной одежды, и поэтому
как особо темные, так и белые олени в первую очередь оставлялись на
племя. Селекция по другим признакам, кроме величины и крепости жи-
вотных, не производилась. По этой причине среди домашних оленей не
создалось четкого деления на породы, как у других домашних живот-
ных. Однако и среди них ясно намечаются два основных типа: более
мелкий тундровый и рослый лесной, отличающиеся не только общей
величиной, но и пропорциями телосложения, а также краниологической
структурой. Краниологические исследования (Соколов, 1937) показали,
что географическое распространение тех и других не повторяет распро-
странения соответствующих форм диких оленей и что отличия между
лесными и тундровыми домашними оленями связаны не только с разными
условиями существования, но и с их различным происхождением.
Значение домашнего оленя огромно. Во многих местностях Крайнего
Севера жизнь человека была бы без оленя невозможна. Домашний олень
дает человеку не только пищу в виде мяса, одежду и обувь в виде шкуры,
но служит незаменимым транспортным животным, так как питается и
зимой, и летом подножным кормом, а в пути не требует везти для себя
корм, как это бывает при езде на собаках. Особенно ценно то, что олень
в условиях северного бездорожья проходит почти везде и даже там, где
порой никакое другое животное пройти не может. Олень переплывает
реки в упряжке, свободно проходит по болотам, где даже человек без
риска провалиться в трясину ступить не может.
От оленьего транспорта до сих пор зависит успешность охотничьего
промысла во многих районах тайги и тундры.
В тундре как зимой, так и летом оленей запрягают в сани (нарты),
так как езда на колесах при полном отсутствии дорог не возможна. В
таежной полосе и в горах, где и нарты не применимы, оленей используют
для верховой езды и перевозки грузов вьюками.
В санной упряжи зимой на каждого оленя можно грузить, в среднем,
около 100 кг. Испытания на коротких дистанциях показали, что в отдель-
ных случаях олени могут свезти груз до тонны и даже больше. Летом
нагрузка уменьшается, в зависимости от состояния пути. Под вьюком
средняя нагрузка оленя 30—40 кг, но отдельные сильные быки поды-
мают гораздо больше. Для верховой езды пригодны лишь крупные лес-
ные олени, разводимые в некоторых районах Сибири (например, в Сая-
нах, Якутии) и на Дальнем Востоке. Для перевозки грузов зимой за-
прягают в нарты по два, а летом, по три оленя.
Налегке, при удовлетворительном состоянии снежного покрова,
зимой ездят со скоростью в 9—10 км в час, но в случаях особо быстрой
езды на небольших расстояниях на хороших оленях за час проезжают
свыше 20 км. При испытаниях на Ленинградском ипподроме зимою
1932/33 года олени проходили дистанцию в 1600 м по снеговой дорожке
в среднем за 3 мин. 27.1 сек., а лучшие упряжки за 2 мин. 39 сек. (Со-
колов, 19336).
Для легковой езды запрягают в санки от 3 до 5 оленей. Для перевозки
тяжестей употребляются большею частью кастрированные самцы, а для
езды налегке иногда и важенки (самки), а также и некастрированные
самцы (хоры).
294
CERVIDAE
В некоторых районах самок оленей доят. Оленье молоко содержит
много молочного сахара (свыше 2%) и до 20% жира, а поэтому обладает
высокими вкусовыми и питательными качествами. Однако, в силу незна-
чительной молочности оленух, молоко целиком потребляется внутри
хозяйства. Оленуха дает в сутки 200—300 гг; изредка удой составляет
700—800 г. За сезон лактации с одной самки обычно получают молока
не более 50 л (Друри, 1949).
В Советском Союзе произведена реконструкция оленеводства на со-
циалистической основе. Оно является ведущей отраслью народного хо-
зяйства Крайнего Севера и важнейшим средством вовлечения в сферу
хозяйственного использования пастбищных ресурсов тундровой зоны.
С 1930 г. начали издаваться оленеводческие совхозы и колхозы. В кол-
хозах поголовье оленей за период с 1933-го по 1946 г. возросло почти
в 10 раз (сборн. «Северное оленеводство», 1948), а общее поголовье оле-
ней на севере СССР за тот же период увеличилось почти в 1^ раза. Ог-
ромное значение и сейчас имеет' оленеводство в осуществлении на севере
грузоперевозок. Тысячекилометровые перевозочные трассы обслужи-
ваются оленьим транспортом. Не меньшую роль играет оленеводство
в' создании продовольственной базы для развивающейся промышленности
Крайнего Севера.
Разработаны новые организационные формы колхозного и совхоз-
ного оленеводства. Основной производственной единицей является
стадо, состоящее в тундровой полосе из 1200—1500 голов, а в таежной
из 500—600 голов, обслуживаемое пастушеской бригадой (Друри, 19496).
Стада совершают сезонные перекочевки — весною на север в тундру,
осенью обратно в лесотундру или лесную зону. Благодаря проведенному
повсеместно землеустройству совхозов и колхозов, масштабы кочевок
за последние годы значительно сокращены. Осуществляется плановый
пастбищеоборот, предупреждающий чрезмерное и преждевременное
стравливание ягельных пастбищ.
Усовершенствованы приемы ухода и содержания оленей. Повсеместно
внедряются стационарные и переносные изгороди (корали), облегчающие
проведение работ по ветеринарно-зоотехническому обслуживанию и об-
работке стад без лишней гонки и беспокойства стада. Созданы усовер-
шенствованные типы упряжи, вьючных и верховых седел для оленя,
значительно повышающие его рабочую продуктивность (сборн. «Север-
ное оленеводство», 1948).
Значительно улучшены обработка-и использование продукции олене-
водства и повышена его товарность. В районах оленеводства оборудована
сеть убойных пунктов и комбинатов, построены оснащенные современ-
ной техникой замшевые заводы. Массовый забой оленей на мясо и на
шкуры для замши производится, в основном, в возрасте 4)4—5 месяцев,
так как до этого времени идет наиболее интенсивный рост животных.
К осени первого года жизни телята достигают 50—60 кг и даже до 80 кг
живого веса, а при убое дают тушу мяса весом около 27 кг. Из мяса из-
готовляются консервы, копчености и колбасные изделия. Используются
и второстепенные продукты убоя оленей: кровь для выработки альбу-
мина, рога и копыта для выварки клея, кишки в колбасном производ-
стве, железы внутренней секреции в производстве эндокринных препа-
ратов. Собираемая весною шерсть-линька и снимаемая при выделке
замши шерсть используются для набивки мягкой мебели, матрацэв, седел,
спасательных кругов и тому подобных вещей. Из кожи можно из-
готовлять не только замшу, но и хром, юфту и другие сорта, при-
BOVIDAE
295
годные для пошивки кожаных пальто, дамской обуви и кожгаланте-
реи.
Успешно решаются вопросы борьбы с потерями оленеводства от бо-
лезней. Бич оленеводства в дореволюционные годы, сибирская язва, бла-
годаря плановым профилактическим прививкам практически полностью
ликвидирована. Наибольшее значение в настоящее время имеет некро-
бациллез, или так называемая «копытная болезнь», поражающая не
только нижние фаланги конечностей, но и внутренние органы оленя.
Вызывается особым микробом — палочкой некроза (Proactinomyces
necrophorus (Flugge) Krassilnikov) и сопровождается рядом других ми-
кробов: стафилококков, стрептококков и других, подготавливающих
почву для развития основного возбудителя болезни. Внешне заболева-
ние выражается хромотой, исхуданием и часто кончается гибелью жи-
вотных. Радикального способа лечения этого заболевания пока не най-
дено. Иммунитета (невосприимчивости к повторному заболеванию) не
образуется и поэтому предохранительные прививки невозможны. Бла-
годаря исследованиям советских ученых по выяснению условий возник-
новения и развития болезни, в последние годы намечаются мероприятия
по предупреждению и лечению этого заболевания (Николаевский, 1951).
От завезенных в конце прошлого столетия из восточных областей
Сибири домашних оленей ведет свою историю оленеводство в США на
Аляске (Lantis, 1950).*
Семейство BOVIDAE—ПОЛОРОГИЕ
Riitimeyer L., 1867—1868. Neue Denkschr. allgem. Schweiz. Gesellsch. Na-
turwissensch., XXII : 1—102, XXIII : 1—175 (морф., сист.); Ковалев ский В. О.,
1875, Изв. Общ. любит, естествозн., антроп. и этнограф., XVI, 1 : 1—59 (эвол.);
Riitimeyer L., 1877—1878, Abhandl. Schweiz. Palaontol. Gesellsch., IV : 1—72,
V : 73—203 (морф., сист.); Flower W. and R. Lydekker, 1891, Mammals
living and extinct, London : 334—367 (сист.); S с 1 a t e г P. and 0. Thomas,
1894—1900, The book of antelopes, London, vol. I : I—XXXV+1—220, vol. II : I—IV+
+1—194, vol. Ill : I—IV+1—245, vol. IV : I—IV+1—242 (общ. изобр.); Lydek-
ker R., 1898, Oxen, sheep and goats of all lands, London : I—XIV-j-1—318 (общ.
изобр.); Lonnberg E. 1903, Nova Acta Soc. Sci. Upsal., (ser. 3), XX : 1—61 (анат.);
Major F., 1904, Proc. Zool. Soc. London : 419—422 (зубы); Туркин H. В. и
К. А. С а т у н и н, 1904, Звери России, Bovidae, М. : 491—794 (общ., изобр.); Р о-
с о с k R., 1910, Proc. Zool. Soc. London : 845—935 (морф.); Lydekker R., 1913—
1914, Catal. Ungulate mammals Brit. Mus., London, vol. I : I—XVI+1—249, vol. II :
: I—XVII+1—295, vol. Ill : I—XV+1—283 (сист., синон.); P о cock R., 1918—
1919, Ann. and Magaz. Natur. Hist. (ser. 9), I : 426—435, II : 125—144, 214—225,
367—374, 440—459, III : 103—114 (морф.); Zitte 1 K., 1923, Grundzuge d. Pala-
ontol., II, Miinchen—Berlin : 587—596 (сист., эвол.); Diierst J., 1926, Denkschr.
Schweiz. Naturforsch. Gesellsch., LXIII, 1 :1—180 (рога); Weber M., 1928, Die
Saugetiere, Bd. II, 2 Aufl., Jena : 587—596 (сист.); Schwarz, E., 1937, Wissensch.
Ergebn. d. Oldoway Expedit., N. F., 4 : 7—90 (сист.); Pilgrim G., 1939, Paleont.
Indica, n. s., XXVI, 1 : 1—356 (сист., эвол.); Benson В., 1943, Amer. Midi. Natur.,
vol. 30, № 3 : 786—789 (зубы); Simpson G., 1945, Bull. Amer. Mus. Natur. Hist.,
85 : 157—162+270—272 (сист.); Соколов И. И., 1949, Докл. Акад, наук СССР,
т. 67, № 1 : 1101—1104 (эвол.); Северцов С. А., 1951, Пробл. экол. животн.,
М., изд. Акад, наук СССР : 81—94 (рога); Соколов И. И., 1952, Тр. Палеонтол.
инет., XLI : 155—158 (происх.); Соколов И. И., 1953, Опыт естественной клас-
сификации полорогих, М.—Л., изд. Акад, наук СССР (Тр. Зоол. инет., т. XIV) : 1—295
(морфол., сист., эвол.).
Характеристика семейства. Самцы, а большей частью и самки, имеют
рога.1 Рога полорогих представляют постоянные, несменяемые выросты
1 Отсутствие рогов (комолость) у самцов наблюдается иногда как доместикацион-
яый признак.
296
BOVIDAE
лобных костей, одетые снаружи роговыми чехлами из видоизмененного
эпидермального слоя кожи.
В отличие от семейства вилорогов (Antilocapridae), роговые чехлы
не спадают и не меняются в течение всей жизни животного.1 Рост рога,
в противоположность оленям (Cervidae), происходит не на вершине, а в ос-
новании; верхушка же представляет наиболее старую его часть, обра-
зовавшуюся в первые стадии формирования. Характерно периодическое
усиление и замедление роста рогов, выражающееся в образовании на
поверхности роговых чехлов колец и связанное, очевидно, с цикличностью
функции половой системы.
Форма рогов весьма разнообразная, но никогда не бывает ветвистой-
Рога могут иметь вид простых спичек; бывают дугообразно загнутые
вперед или назад; улиткообразные; свернутые или скрученные спиралью;
прямые, стоящие вертикально или направенные назад. Скручивание и свер-
тывание рогов может быть гомонимное или гетеронимное.1 2 Длина рогов
может быть небольшой, не превышающей половины длины черепа или,
наоборот, превосходить последнюю в несколько раз.
В черепе отверстие, ведущее из полости глазницы в носослёзный канал,
одно. Предглазничные ямки на наружной поверхности слёзных костей
и этмоидальные щели иногда имеются, но часто отсутствуют. Теменные
гребешки широко расставлены, сагиттальный гребень на черепе никогда
не образуется. Носовые отростки межчелюстных костей сравнительно
короткие и если и касаются носовых костей, то обычно на незначительном
расстоянии. Щель, или овальное отверстие между межчелюстными ко-
стями (резцовое отверстие) на нёбной поверхности черепа всегда имеется.
Длина задних конечностей заметно преобладает над длиной передних
только у примитивных членов семейства. В запястье сращены (слились)
головчатая кость (magnum, s. capitatum) с малой многоугольной (trape-
zoideum), а в заплюсне—кубовидная (cuboideum) с ладьевидной (naviculare)
и вторая клиновидная с третьей (cuneiforme II и III). От боковых метапо-
дий (II и V) сохраняются только проксимальные рудименты, которые на
задних конечностях обычно сливаются с цевкой (os cannon). Боковые
пальцы сильно редуцированы, иногда совсем исчезают (например, род
Neotragus Н. Smith). В скелете боковых пальцев сохраняются, и то не
всегда, только рудименты одной или двух фаланг. Зубная формула ти-
0 0 3 3
пична: I-^-, С-р P-у, М-р, но часто премоляров в нижней, а изредка и
в верхней, челюсти только два, так как Р 2 и Р2 или совсем не развиваются,
или рано выпадают. Верхние клыки развиваются лишь, как аномалия,
крайне редко (Major, 1904; Benson, 1943). Коренные зубы иногда брахио-
донтные, но у большинства полорогих коронки их выше, чем у оленей и
кабарог, в отдельных случаях достигают высокой степени гипсодонтии
(рис. 22, Б). Постоянным ростом зубы, однако, у Bovidae не обладают,
так как имеют замкнутые корни.
1 Несменяемость роговых чехлов у Bovidae, впрочем, понятие относительное.
У некоторых представителей семейства роговые отростки в начальном периоде их
роста покрыты так называемым «гиалиновым» рогом, вскоре сменяемым настоящим
роговым чехлом (Duerst, 1926).
2 Гомонимными рогами называются такие, у которых направление изгиба идет
внутрь, навстречу рогу противоположной стороны, а гетеронимными — те, у которых
правый рог свернут или изогнут вправо, а левый влево; если рога от основания изог-
нуты гомонимно, а к вершине изгиб принимает противоположное направление (как
например, у дагестанского тура, Capra caucasica cylindricornis Blyth), то они носят
название первертированных.
BOVIDAE
297
Из кожных желез, имеющихся у оленей и кабарог, у полорогих часто
отсутствуют предглазничные, межпальцевые, плюсневые и предплюсневые.
Зато многие из них имеют железы, не свойственные оленям: паховые,
карпальные, зароговые, околохвостовые. У некоторых групп полорогих
имеются железы на животе или у добавочных копыт (Рососк, 1910, 1918-—
1919). Наличие или отсутствие тех или иных кожных желез, так же как
форма, размеры и положение рогов, могут служить ценными диагности-
ческими и филогенетическими признаками отдельных видов и групп по-
лорогих. Желчный пузырь имеется, за исключением рода Cephalophus
Н. Smith (Lonnberg, 1903). Penis у многих видов (но не у всех) на конце
с нитевидным отростком. Молочная железа двухдольчатая или четырех-
дольчатая. Матка имеет большое число хорошо развитых котиледо-
нов.
Географическое распространение. Европа, Азия, Африка, Северная
Америка и прилегающие к ним острова. Отсутствуют в Австралии, Южной
Америке, на Мадагаскаре и на Сахалине. В Новой Зеландии акклимати-
зированы. В домашнем состоянии распространены по всему свету.
Эволюция полорогих. Семейство Bovidae филогенетически самая мо-
лодая и наиболее многочисленная из современных групп копытных, еще
не пережившая эпохи своего расцвета. Корни полорогих ведут к нижне-
олигоценовым оленькам (Tragulidae). Непосредственный их предок или
исходная форма не известны, но, вероятно, весьма близким к таковому
являлся род Gelocus Aymard, живший на территории Европы в нижнем
олигоцене. Gelocus не имел рогов, локтевая кость у него была самостоя-
тельна, но малая берцовая сильно редуцирована. Боковые пальцы при
ходьбе, вероятно, касались земли. На передних конечностях центральные
(III и IV) метаподий были раздельны, но на задних соответствующие кости
слились и образовали цевку (os cannon). От боковых метаподий сохранялись
и проксимальные, и дистальные рудименты. Коренные зубы были крайне
брахиодонтного типа, сохранялись верхние саблевидные клыки, но верх-
ние резцы уже исчезли, а клыки нижней челюсти функционально были
резцами. Премоляры имели крайне примитивное устройство, причем пер-
вый из них в верхней челюсти уже исчез, тогда как в нижней еще сохра-
нялся (Ковалевский, 1875).
Формы, промежуточные между оленьками и настоящими полорогими,
пока не известны. В среднем миоцене Европы жили антилопы, обладавшие
несменяемыми рогами, но еще с крайне примитивно устроенными брахио-
донтными коренными зубами черепа и длинной, горизонтально располо-
женной зароговой частью. Они могли бы считаться исходными формами
для всех последующих полорогих. Но в слоях того же возраста в Европе
и еще более раннего в Монголии найдены уже сравнительно высоко спе-
циализированные представители семейства, которые заставляют пред-
полагать, что отхождение предков полорогих от общего ствола Ресога
произошло не позднее верхнего или даже среднего олигоцена (Соколов,
1949, 1952, 1953). Родиной полорогих следует считать Евразийский кон-
тинент, где на стыке его с Африкой лежал первичный центр расселения
данной группы. Вторичными центрами являлись, с одной стороны, Цен-
тральная Азия, а с другой — области по соседству с Индией, к западу
от последней (Pilgrim, 1939; Соколов, 1953).
Характерная черта полорогих — покрытые несменяемым чехлом рога —
в истории данной группы появилась, по-видимому, не сразу. Исходные
формы, вероятно, рогов не имели или обладали небольшими выростами
лобных костей, покрытых периодически сбрасывавшимися колпачками
298
BOV Г DAE
ороговевшей кожи.1 Первоначальное назначение рогов — украшение
самцов и турнирное оружие. В качестве орудия защиты от врагов и на-
падения они начали служить позднее.
Общее направление эволюции полорогих во многом сходно с таковым
оленей. Это — окончательный переход на питание исключительно расти-
тельной пищей, приспособление к быстрому бегу. Однако в некоторых
отношениях они определили оленей. Судя по наиболее примитивным из
современных, предки и ранние представители полорогих были, по всей
вероятности, мелкими лесными формами или обитателями кустарниковых
зарослей и лесных опушек, питались плодами, ягодами и частично листвой
кустарников и деревьев. В ходе эволюции они приспособились к питанию
более жесткой и грубой травянистой пищей. Полорогие собирают корм боль-
шею частью прямо с земли, вместе с частицами почвы, песка и пыли, вы-
зывающими быстрое изнашивание зубов. В связи со всем этим выработалась
гипсодонтия. Коронки коренных зубов становятся не только высокими,
но благодаря усилению эмалевого слоя и развитию гребней и эмалевых
складок увеличивают также свою крепость. Усиление складчатости и
отложение на поверхности зуба и между складками эмали цемента спо-
собствуют также увеличению перетирающей поверхности зуба, а чередо-
вание слоев эмали, дентина и цемента, обладающих разной степенью
прочности, делает трущуюся поверхность неровной, повышая функцио-
нальную способность зуба. В процессе эволюции у многих форм проис-
ходит увеличение размеров заднекоренных зубов, как наиболее важных
при пережевывании корма, в силу близкого их положения к точке при-
ложения силы жевательной мускулатуры. Становящиеся менее важными
переднекоренные зубы (премоляры), наоборот, постепенно редуцируются,
и у многих форм передний коренной зуб нижней челюсти (Р2), а изредка
и верхней исчезают.
Желудок, слабо разделенный у примитивных представителей полоро-
гих, у прогрессивных достигает высокой степени дифференцировки, по-
вышая способность к усвоению грубого растительного корма.
У полорогих происходит еще более полное слияние центральных ме-
таподий, повышающее крепость конечностей. От боковых метаподий и
боковых пальцев остаются ничтожные, почти не имеющие функциональ-
ного значения рудименты, а в некоторых случаях и они исчезают.
Характерным для направления эволюции ряда филогенетических
ветвей Bovidae является переход от пассивных к активным способам за-
щиты. Роль орудия защиты и нападения приобретают рога, увеличиваю-
щие свои размеры и прочность. В связи с этим рога, первоначально имев-
шиеся только у самцов, в филогенезе многих форм приобрели и самки.
В связи с преобразованием рогов важные изменения претерпевает
череп. У наиболее древних из ископаемых и у примитивных из современных
форм рога расположены на черепе над или непосредственно за глазни-
цами;1 2 длинная зароговая часть черепа расположена почти горизонтально;
крыша мозговой коробки образована в основном теменными костями.
Так как применение рогов в качестве оружия более эффективно, если
они расположены ближе к основанию черепа, затылочной его части, где
прикрепляется основная масса двигающей его мускулатуры, то в про-
цессе эволюции происходит неуклонное сдвигание рогов вместе с задними
1 Отголоском этой стадии, общей с американским вилорогом, являются упомя-
нутые раньше чехлики на начинающих расти рогах некоторых современных полоро-
гих, позднее замещаемые чехлом из настоящего рога.
2 Имеется в виду свободное положение черепа на горизонтальной поверхности.
BOVГDAE
299
участками лобных костей к вершине черепа. Зароговая часть при этом
постепенно укорачивается, теменные кости оттесняются назад, вместо
горизонтального принимают наклонное назад положение, в крайних слу-
чаях становятся в одну плоскость с затылком или даже, как например
у быков, нависают над последним. Крыша мозговой коробки при этом
образуется уже не теменными, а только лобными костями.
Параллельно с преобразованием зарогового отдела происходит увели-
чение излома оси черепа, улучшающее эффективность применения рогов
в качестве колющего и ударного оружия при защите и нападении. В ходе
эволюции полорогих имеет место также пневматизация лобных (вклю-
чая роговые стержни), а иногда также теменных и затылочных костей,
т. е. развитие в них внутренних полостей,, способствующих облегчению
черепа без ущерба прочности. Прочность черепа даже возрастает благо-
даря более тесному соединению или полному слиянию отдельных костей,'
особенно в зароговом отделе.
Описанное общее направление эволюции, происходившее под воздей-
ствием изменений в окружающей обстановке, осуществлялось неодинако-
выми темпами в разных филогенетических ветвях Bovidae. Наряду с весьма
прогрессивными представителями, как быки и бараны, до сих пор суще-
ствуют примитивные группы, во многих отношениях недалеко ушедшие
от исходных форм своего семейства. У отдельных членов и групп в ходе
эволюции выработались приспособления к частным условиям среды,
способствовавшие усиленной, по сравнению с другими парнопалыми,
адаптивной радиации общего ствола полорогих. Среди них до сих пор
сохранились обитатели умеренных и тропических лесов, степей, пустынь
п полупустынь. Некоторые представители, и при этом назависимо из
разных филогенетических ветвей, приспособились к жизни в горах.
Есть даже полуводные обитатели прибрежных зарослей, находящие
в воде не только пищу, но и защиту от врагов, например африканские
так называемые водяные козлы (роды Kobus A. Smith, Adenota Gray) и
ситатунга (Tragelaphus spekei Sci.). Наконец, мускусный бык (Ovibos
moschatus Zimm.) и снежный баран (Ovis nivicola Eschsch.) приспособились
к суровым условиям Заполярья.
В связи с этим внешний вид полорогих также весьма разнообразен.
Среди них имеются животные, величиной не превышающие кролика,
например африканские карликовые антилопы (N eotragini), и гиганты
вроде дикого буйвола (Bubalus bubalis L.) или вымершего не так давно
тура (Bos primigenius Boj.). Наряду со стройными высоконогими газелями
(род Gazella De Blainville) есть коротконогий неуклюжий мускусный
бык (Ovibos moschatus Zimm.) или гималайский такин (Budorcas taxico-
lor Hodgs).
Классификация полорогих. Существовавшее со времен Палласа
деление полорогих на быков, козлов, баранов и антилоп не соответствует
современным представлениям об их филогенетических отношениях (рис.
60) и поэтому большинством зоологов в настоящее время оставлено.
Явно искусственная группа «антилоп» в системе ликвидирована, так как
многие из них генетически стоят ближе к быкам или козлам с баранами,
чем к другим антилопам. Однако согласие в вопросе об отношениях друг
к другу отдельных групп Bovidae и связанном с этим-делении всего се-
мейства на соподчиненные группы отсутствует, а классификация его
проводится по-разному (Lydekker, 1913—1914; Schwarz, 1937; Pilgrim,
1939; Simpson, 1945). Мною (Соколов, 1953) принимается деление поло-
рогих на шесть подсемейств.
300
BOVIDAE
1) Gazellinae — Газелевые. Рога, за немногими исключениями, име-
ются только у самцов; основаниями расположены над глазницами, мас-
сивные, без внутренних полостей внутри стержней. Ноздри расположены
близко друг к другу; расстояние между ними не больше высоты верхней
губы (от ее нижнего края до ноздрей). Молочная железа имеет четыре соска
(исключение — роды Gazella De Blainville и Antilope Pall.). Зароговая часть
черепа длинная, больше длины лба. Лобные кости короткие, не больше
36% основной длины черепа. Слуховые пузыри вздутые. Средняя пара
резцов сильно расширена по сравнению с другими и имеет форму асим-
метрических лопаток (рис. 61, Л). Распространение: Африка,
Передняя, Средняя, Центральная и Южная Азия, некоторые районы
южной Сибири (Алтай, Тува, южное Забайкалье). Объем подсемейства
см. стр. 304.
2) Cephalophinae — Дукеры. Рога часто имеются и у самок, массивные,
без внутренних полостей внутри стержней. Ноздри расположены близко
друг к другу, расстояние между ними не больше высоты верхней губы
от ее нижнего края до ноздрей. Молочная железа имеет четыре соска.
В отличие от других полорогих, предглазничные железы расположены
на средине расстояния между ноздрями и глазами и открываются серией
линейно расположенных мелких отверстий на лишенном волос участке
кожи. Отличительными чертами в черепе являются также очень большие
предглазничные ямки, в образовании которых принимают участие сильно
расширенные в задней половине носовые кости, и сдвинутые далеко за
глазницы основания роговых отростков, не выходящих при этом в стороны
за границы мозговой коробки. Зароговая часть черепа значительно ко-
роче длины лба. Лобные кости длинные, больше 36% основной длины
черепа. Слуховые пузыри вздутые. Средняя пара резцов сильно расши-
рена по сравнению с другими и имеет форму асимметричных лопаток.
Распространение: Африка к югу от северного тропика. Свыше
30 видов дукеров морфологически близки друг к другу и обычно объ-
единяются в один род Cephalophus Н. Smith.
3) Hippotraginae — Лошадиные антилопы. Рога имеют как самцы,
так и самки, причем у последних они обычно не меньше, чем у самцов;
основаниями рога расположены над или сразу же за глазницами, мас-
сивные, без внутренних полостей внутри стержней. Ноздри расположены
близко друг к другу; расстояние между ними меньше высоты верхней
губы от нижнего ее края до ноздрей. Очень характерна своеобразная
расцветка лицевой части головы с чередующимися светлыми и темными
полосами и черным пятном различной величины посредине лба и носа.
Молочная железа имеет четыре соска. Зароговая часть черепа несколько
укорочена и немного короче лобной зоны. Лобные кости составляют от
28 до 40% основной длины черепа. Слуховые пузыри умеренно вздуты
и сжаты с боков. Средняя пара резцов сильно расширена по сравнению
с другими и имеет форму асимметричных лопаток. Характерную отли-
чительную черту данного подсемейства представляет чрезвычайно силь-
ное развитие как на верхних, так и на нижних заднекоренных зубах
(молярах) добавочных столбиков, приобретающих Т-образную форму.
Распространение: Африка к югу от Сахары, Месопотамия
и Аравия. Подсемейство включает три современных рода: Hippolragus
Sundevall (лошадиные антилопы), Oryx De Blainville (сернобыки) и A Idax
Rafinesque (мендес).
4) Alcelaphinae — Бубалы, или коровьи антилопы. Рога имеют и
самцы, и самки, причем у последних они немного меньше, чем у самцов.
Рис. 60. Схема предполагаемой филогении и классификации семейства полорогих
(Bovidae).
Рис. 61. Типы резцов полорогих.
А — Gazella subgutturea Piild.; Б —Rupicapra rupicapra L.; В — Bison bonasus L.;
Г — Capra caucasica Giild.
302
BOVIDAE
Основаниями рога расположены позади глазниц; в дистальной части
роговых отростков имеются полости, но высоко в роговые стержни они
не заходят. В отличие от других полорогих, морда расширена и уплощена
на конце. Сильно складчатые ноздри усажены по краям длинными щети-
нистыми волосами. Расстояние между ноздрями меньше высоты верхней
губы от ее нижнего края до ноздрей. Молочная железа имеет два соска.
Характерной отличительной чертой в черепе является необычайно длинная
лицевая часть, не меньше чем в два раза превышающая длину мозговой
части. Зароговая часть черепа короче лобной зоны (исключение —
род Damaliscus Sclater and Thomas). Лобные кости длинные, не меньше
37% основной длины черепа. Слуховые пузыри вздутые. Средняя пара
резцов сильно расширена по сравнению с другими й имеет форму асим-
метричных лопаток. Распространение: вся Африка как к се-
веру, так и к югу от Сахары. Подсемейство включает четыре современ-
ных рода: Alcelaphus Blainville, Damaliscus, Gorgon Gray, Connochaetes
Licht.
5) Caprinae — Козлообразные. Рога, за очень редкими исключениями,
имеют и самцы и самки, но у последних они по сравнению с самцами
развиты очень слабо. Основания рогов лежат над глазницами, но иногда
немного сдвинуты назад за края последних. Роговые стержни содержат
внутренние полости, часто распространяющиеся до верхушек. Ноздри
расположены близко друг к другу; расстояние между ними не больше
высоты верхней губы от ее нижнего края до ноздрей. Молочная железа
имеет два или четыре соска. Длина зароговой части черепа может быть
меньше или больше длины лба. Лобные кости средней длины или длинные;
их длина редко составляет меньше 36 % основной длины черепа. В черепе
характерной отличительной чертой является короткая, уплощенная, почти
квадратной формы основная затылочная кость (basioccipitale); ширина ее
в передней части, за одним исключением (род Budorcas Hodgson), не меньше,
чем в задней. Слуховые пузыри небольшие, мало или совсем не вздуты,
всегда сжаты с боков. Все резцы узкие, равномерно убывающие в ширине
от средней пары к клыкам (рис. 61, Б, Г). Добавочных столбиков и бу-
горков на коренных зубах нет. Распространение: Европа,
Азия, Северная Америка с прилегающими арктическими островами,
северная Африка. В домашнем состоянии — по всем материкам земного
шара. Объем подсемейства см. стр. 343.
6) Bovinae — Быковые. Рога у большинства видов имеют и самцы,
и самки, причем у последних они немного меньше, чем у самцов. Осно-
вания рогов в большинстве случаев сдвинуты в той или иной степени за
глазницы, иногда располагаются над самим затылком. Роговые стержни,
за немногими исключениями, содержат внутренние полости, нередко
доходящие до вершины. Ноздри широко расставлены друг от друга;
расстояние между ними больше (или во всяком случае не меньше)
высоты верхней губы от верхнего ее края до ноздрей. Характерно отсут-
ствие предглазничных 1 и межпальцевых желез, но зато у ряда видов
имеются не свойственные представителям других подсемейств железы
около добавочных копыт. Молочная железа всегда имеет четыре соска.
Зароговая часть черепа в типичных случаях сильно укорочена, иногда
совсем оттесняется назад и становится в одну плоскость с затылком.
Длина лобных костей составляет от 35 до 59% основной длины черепа.
1 Исключение — примитивные индийские роды Boselaphus Blainville и Tetra-
cents Leach.
BOVIDAE
303
Основная затылочная кость сильно расширена в задней и заметно сужена
в передней части. Слуховые пузыри сильно вздуты и сжаты с боков.
Резцы или широкие, но равномерно убывающие в ширине от средней пары
к клыкам (рис. 61, В), или передняя их пара имеет форму асимметричных
лопаток, а остальные резко убывают в ширине от первой пары к клыкам.
Заднекоренные (по крайней мере нижние) имеют добавочные бугорки
или столбики. Распространение: Европа, Азия, Африка,
Северная Америка. В домашнем состоянии распространены по всем ма-
терикам земного шара. Объем подсемейства см. стр. 565.
В фауне СССР представлены подсемейства газелевых (Gazellinae),
козлообразных {Caprinae) и быковых {Bovinae).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ПОДСЕМЕЙСТВ ПОЛОРОГИХ
(BOVIDAE) ФАУНЫ СССР
1 (4). Размеры средние; длина тела от конца морды до корня хвоста не
больше 200 см. Основная длина черепа не больше 350 мм. Рога
расположены над глазницами или непосредственно позади послед-
них; расстояние от края глазницы до основания рогового чехла
(или основания рогового стержня) меньше вертикального диаметра
глазницы. Ноздри сближены; расстояние между их внутренними
. углами меньше расстояния от края верхней губы до ноздри. Верх-
няя губа покрыта волосами, кроме узкой вертикальной полоски
на ее средине. Хвост короткий, не достигает концом скакательного
сустава. Заднекоренные зубы не имеют ни добавочных столбиков,
ни бугорков.
2 (3). Межпальцевые железы имеются на всех четырех конечностях,
открываются в виде вертикальных щелей на передней стороне
выше копыт. На ягодницах и иногда на крупе вокруг хвоста уча-
сток кожи с волосами чисто белого цвета, резко отграничен темной
полоской от окраски смежных частей тела. Передняя пара резцов
сильно расширена в виде асимметричных лопаток; остальные
резцы резко убывают в ширине от первой пары к клыкам (рис. 61, А).
Пястная кость длиннее лучевой1. . Gazellinae— Газелевые (стр. 304).
3 (2). Межпальцевые железы если имеются, то открываются круглым
отверстием на передней стороне ног выше копыт. Ягодницы и круп
окрашены однотонно с остальным туловищем, а если около хвоста
окраска более светлая, желто- или грязно-белая, то она нерезко
отграничена от окраски соседних частей тела. Передняя пара рез-
цов имеет форму долот или узких, почти симметричных лопаток.
Остальные резцы постепенно убывают в ширине от первой пары
к клыкам (рис. 61, Б, Г). Пястная кость короче лучевой.........
....................... Caprinae — Козлообразные (стр. 342).
4 (1). Размеры крупные; длина тела от конца морды до корня хвоста
не меньше 230 см; основная длина черепа не меньше 420 мм. Рога
расположены позади глазниц; расстояние от края глазницы до
основания рогового чехла (или рогового стержня) не меньше верти-
кального диаметра глазницы. Ноздри широко расставлены; рас-
стояние между ними больше расстояния от края верхней губы до
ноздри. Хвост длинный, с кистью на конце, достигает скакательных
1 Здесь, как и всюду в дальнейшем, имеется в виду физиологическая длина труб-
чатых костей, т. е. расстояние между наиболее выступающими точками проксимальной
и дистальной суставных поверхностей.
304
BOVIDAE
суставов. Конец морды голый; безволосый участок кожи (носовое
зеркало) занимает переднюю сторону верхней губы и все простран-
ство между ноздрями. Заднекоренные зубы имеют добавочные
столбики: верхние на внутренней, а нижние на наружной сто-
роне ............................Bovinae — Быковые (стр. 564).
Подсемейство GAZELLINAE — ГАЗЕЛЕВЫЕ
Характеристика подсемейства. Рога у большей части видов под-
семейства имеют только самцы (исключение — некоторые виды газелей).
Основания рогов расположены над глазницами. Роговые стержни не имеют
внутренних полостей. Форма рогов разнообразная: в виде простых спичек,
лировидные, дугообразно загнутые вперед или свернутые в плотную
гетеронимную спираль; на поверхности рогов нет продольных ребер (ки-
лей).
Ноздри расположены близко друг к другу; расстояние между ними не
больше (обычно заметно меньше) расстояния между краем верхней губы и
ноздрей. Белые волосы на ягодицах, а иногда и на крупе вокруг хвоста
образуют резко отграниченное от смежных, иначе окрашенных участков
«околохвостовое зеркало». Предглазничные и межпальцевые железы,
хотя бы в редуцированном виде, обычно имеются. Хвост короткий, не
достигает скакательных суставов. Сосков 4 (исключение — роды Ga-
zella De Blainville и Antilope Pall.).
Череп имеет хорошо развитую зароговую часть. Длина ее превосхо-
дит длину лба. Лобные кости короткие; их длина вдоль сагиттального
шва редко превышает 36% основной длины черепа. Глазницы передними
краями расположены на уровне коренных зубов верхней челюсти. У боль-
шинства представителей подсемейства на черепе имеются предглазнич-
ные ямки и этмоидальные щели. Основная затылочная кость (basioccipi-
tale) уже в передней части, чем в задней, и никогда не бывает в форме
уплощенного квадрата. Передний край хоан расположен впереди уровня
заднего края верхних моляров. Слуховые пузыри вздутые, выступают
ниже уровня основной затылочной кости.
Средняя пара резцов сильно расширена по сравнению с другими
и имеет форму асимметричных лопаток. Остальные резко убывают в ши-
рине, а крайний резец и клык имеют форму тонких изогнутых штифтов.
Последний премоляр нижней челюсти (Р4.) имеет открытой переднюю
или заднюю долинку. Добавочные столбики на заднекоренных зубах
имеются или отсутствуют.
Географическое распространение. Африка, Передняя, Средняя, Цен-
тральная и южная Азия, Закавказье, некоторые районы южной Сибири
(Алтай, Тува, южное Забайкалье).
Классификация. Подсемейство Gazellinae включает в себя разнообраз-
ных по внешнему виду и биологии животных, начиная с карликовых
антилоп — левреток (род Madoqua Ogilby) и кончая крупными водяными
козлами (род Kobus A. Smith). Есть среди них обитатели кустарниковых
степей, пустынь, прибрежных зарослей и травянистых лужаек и даже
высокогорий. Но все они характеризуются в целом как наиболее при-
митивная группа своего семейства. Морфологическими показателями
примитивности являются длинная зароговая часть черепа, просто устро-
енные рога, которые у большинства видов имеют только самцы, и неко-
торые другие. Роды данного подсемейства группируются в четыре трибы.
GAZELLA
305
1) Neotragini — Карликовые антилопы. Размеры небольшие; основ-
ная длина черепа у самок меньше 130 мм.1 Рога в виде почти прямых
спичек. Носовое зеркало хорошо развито (за исключением рода Madoqua').
Предглазничные и межпальцевые железы имеются. Кожа меж копыт
покрыта волосом. Большинству представителей свойственны также кар-
пальные железы. Молочная железа, за исключением одного случая (Mado-
qua megalotis Meng.), четырехдольчатая, имеет четыре соска. Наибольшая
ширина черепа не меньше 50% основной его длины. Распростра-
нение: Африка к югу от Сахары. Включает роды: Neotragus Н. Smith,
Ourebia Laurill., Raphicerus H. Smith, Madoqua Ogil. и Oreotragus A. Smith.
2) Gazellini — Газелиные. Размеры средние; основная длина черепа
от 155 до 240 мм. Рога лировидные или свернутые в штопорообразную
спираль. Носовое зеркало слабо развито; верхняя губа почти сплошь
покрыта волосами. Предглазничные и межпальцевые железы, за немно-
гими исключениями (Aepyceros melampus Licht.), имеются. Кожа меж
копыт покрыта волосом. Молочная железа двухдольчатая или четырех-
дольчатая. Наибольшая ширина черепа составляет от 43 до 58% основ-
ной его длины. Распространение: то же, что для всего под-
семейства. Включает роды: Gazella Blainv., Lithocranius Kohl., Aepy-
ceros Sund. и Antilope Pall.
3) Reduncini — Водяные и болотные козлы. Размеры средние и круп-
ные; основная длина черепа от 190 до 330 мм. Лировидные или дугообразно
загнутые вперед рога. Носовое зеркало хорошо развито, передняя часть
верхней губы голая (исключение — род Ammodorcas Thom.). Предглазнич-
ные железы рудиментарные или вовсе отсутствуют. Нет межпальцевых
желез. Кожа меж копыт голая, не покрыта волосами. Молочная железа
четырехдольчатая. Ширина черепа составляет меньше половины основ-
ной его длины. Отличительными чертами от всех других триб являются
также сильно развитые мускульные бугорки (tubercula muscularia) на
переднем крае basioccipitale и наружные складки на переднем крае нижних
заднекоренных зубов. Распространение; Африка к югу от
Сахары (в ископаемом состоянии также в Индии). Включает роды: Kobus
A. Smith, Adenota Gray, Redunca H. Smith и Ammodorcas Thom.
4) Peleini — Антилопа-косуля. Размеры средние или небольшие; ос-
новная длина черепа около 190 мм. Рога почти прямые, лишь слегка
загнутые вершинами вперед, носовое зеркало хорошо развито. Предглаз-
ничные железы отсутствуют, но межпальцевые хорошо развиты. Кожа
меж копыт голая, не имеет волос. Молочная железа четырехдольчатая.
Наибольшая ширина черепа составляет около 50% его основной длины.
В отличие от других триб, срединная длина лобных костей больше 40%
основной длины черепа. Отличительной чертой является также шерсти-
стый характер волосяного покрова, напоминающего мех кролика или
зайца. Распространение: южная Африка. Триба включает
один род и вид — Pelea capreolus Bechst.
В фауне СССР представлен только один род данного подсемейства —
Gazella.
Род GAZELLA BLAINVILLE — ГАЗЕЛИ
Blainville М., 1816, Bull. Sci. Soc. Philomatique, Paris : 75 (первооп.);
Hodgson В., 1846, J. Asiat. Soc. Bengal, XV : 334; Brooke W., 1873, Proc.
Zool. Soc. London : 535—554 (сист.); Lat aste M., 1885, Act. Soc. Linn., Bor-
1 Размеры и индексы черепа у полорогих всюду, где особо не оговорено, приво-
дятся для самок.
20 Фауна СССР, И. И. Соколов
306
BOVIDAE
deaux, XXXIX : 129 (сист.); S с 1 a t e r Ph. and О. Thomas, 1897—1898, Book
of antelopes, III, London : 53—216 (общ., изобр.); P о с о с к В., 1910, Proc. Zool.
Soc. London : 887—890 (морф.); Борисяк А. А., 1914, Тр. Геол, комит., н. сер.,
87:1—104 (эвол.); Lydekker JEU, 1914, Catal. Ungulate mammals Brit, mus.,
Ill, London : 28—113 (сист.); P о с о c k B., 1918, Ann. Mag. Natur. Hist. (ser. 9),
II : 125—129 (морф.); Schwarz E., 1937, Wissensch. Ergebn. Oldoway Expedit.,
N. F., 4 : 7—90 (сист.); В о h 1 i n B., 1939, Bull. Geol. Inst. Univers. Upsala, 28 :
79—122 (эвол.); Pilgrim G., 1939, Paleont. indica, n. ser., XXVI, 1 : 32—39 (эвол.);
Саркисов A. A., 1945, Тр. Ереванск. зоопарка, IV :5—20 (сист.); E 11 e r m a n J.
and T. M о r r i s о n-S с о 11, 1951, Checklist of palaearctic and indian mammals,
London : 387—394 (сист., синон.); Вялов О. С. и К. К. Ф л e p о в, 1952, Бюлл.
Моск. общ. испыт. прир., Отд. геол., XXVII, 5 : 80—88 (ист.); Соколов И. И.,
1953, Тр. Зоол. инет., XIV : 234—238 (сист., эвол.).
Характеристика рода. Животные очень легкого, стройного тело-
сложения. Рога лирообразно изогнутые, от основания более или менее
параллельные, к вершинам расходящиеся в стороны, с концами, загну-
тыми вперед или внутрь. Поверхность роговых чехлов почти на всем
протяжении, кроме гладких верхушек, покрыта ясно выступающими
отставленными друг от друга поперечными кольцами. Поперечное сечение
рогов в основаниях продольно-овальное. У большинства видов они имеются
не только у самцов, но и у самок.
Окраска буровато-песчаная разных оттенков; низ живота, груди,
шеи и внутренние стороны ног белые. Характерно чередование темных
и светлых полос на морде и боках, а также белое околохвостовое зеркало.
Хвост средней длины или короткий, далеко не достигает концов Скакатель-
ных (голеностопных) суставов. Предглазничные железы, за одним исклю-
чением (Gazella picticaudata Hodgs.), имеются, но иногда очень слабо
развиты. Межпальцевые железы открываются в виде щелей выше копыт.
У большинства видов имеются паховые и карпальные железы (исклю-
чение— G. marsupialis Zimm, и G. picticaudata Hodgs.). Молочная железа
двухдольчатая ( в противоположность родам Lithocranius Kohl, и Aepyceros
Sund.).
В черепе мозговая часть составляет редко меньше 65% длины лицевой.
Поверхность лба сильно вдавлена между глазницами. Предглазничные
ямки имеются, но иногда очень мелкие. Этмоидальные щели имеются,
за исключением одного вида (G. marsupialis Zimm.). Межчелюстные кости
на переднем конце не образуют лопаткообразных расширений, характер-
ных для большинства полорогих.
Коренные зубы гипсодонтные, крупные; длина беззубого края ниж-
ней челюсти редко превышает 65% длины ряда Р2—М3.
История и географическое распространение. Род Gazella — самый
древний из современных полорогих. Наиболее ранние остатки газелей
известны уже в верхнем миоцене (сармате) и в нижнем плиоцене (понте).1
Это были мелкие формы с брахиодонтными зубами и слабо изогнутыми
рогами (Борисяк, 1914; Bohlin, 1939). В миоцене и плиоцене газели на-
селяли не только Африку (куда проникли только в верхнем плиоцене)
и Азию, но и южную половину Европы, вплоть до Англии. Близкие
к современным виды Gazella появились в верхнем плиоцене.
В настоящее время область распространения газелей включает се-
верную, восточную и южную Африку, Переднюю, Среднюю и Центральную
Азию (включая восточное Закавказье), Индийский полуостров.
1 Следы (отпечатки копыт) газелей обнаружены даже в нижнемиоценовых
слоях Предкарпатья (Вялов и Флеров, 1952).
GAZELLA
307
Классификация. Многочисленные формы газелей, насчитывающие по
каталогу Лайдекера (Lydekker, 1914) 27 видов и еще большее число под-
видов, различающихся главным образом особенностями окраски и формы
рогов, довольно естественно распадаются на три подрода:
1) Gazella s. str. Хорошо развитые предглазничные ямки и этмоидаль-
ные щели. Носовые кости с вырезкой на передних концах. Последний
нижний премоляр (Р4) имеет открытую переднюю долинку (рис. 68, Л).
Хвост сравнительно длинный. Подрод включает 23 вида, распростра-
ненных в северной и восточной Африке, в Закавказье, Передней, Средней,
Центральной и южной Азии.
2) Procapra Hodgson. Предглазничные ямки едва заметны. Носовые
кости клинообразно заостренные на переднем конце. Последний нижний
премоляр (Р4) имеет замкнутую переднюю долинку (рис. 68, Б). Хвост
короткий, почти не виден среди окружающих волос. Включает 3 центрально-
азиатских вида: G. gutturosa Gmel., G. przewalskii Buchn. и G. picticau-
data Hodgs.
3) Antidorcas Sundev. От остальных газелей отличается отсутствием
паховых и карпальных желез, этмоидальных щелей в черепе, а также
наличием на спине железистой складки кожи с длинными белыми воло-
сами. Включает один вид G. marsupialis Zimm., антилопу-прыгуна,
распространенный в Африке к югу от р. Замбези.
В качестве подрода Nanger Latast. иногда выделяются три крупные
газели восточной Африки — G. soemmeringi Cretz., G. granti Br. и G.
dama Pall., которые, помимо величины, отличаются от других Gazella s. str.
длинными рогами и большими размерами околохвостового зеркала.
Без достаточных оснований выделяется иногда в подрод Prodorcas
Pocock дзерен (G. gutturosa Gmel.) из-за не проверенного впоследствии
никем указания Палласа на наличие у него препуциальных желез.
Эллерман и Моррисон-Скотт (Ellerman and Morrison-Scott, 1951)
в качестве подрода Trachelocele выделяют джейрана (G. subgutturosa Guld.),
отличающегося от других видов Gazella s. str. наличием у самцов во время
течки зобовидного вздутия гортани.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ РОДА GAZELLA
ФАУНЫ СССР
1 (2). Размеры сравнительно небольшие. Основная длина черепа меньше
210 мм. Длина хвоста больше 120 мм. Белое околохвостовое зеркало
не заходит на круп выше корня хвоста. Волосы на участке между
пяточными мякишами главных копыт и добавочными копытами
имеют темно-бурую окраску. Носовые кости с вырезкой на перед-
нем крае. Расстояние между наружными краями оснований ро-
говых отростков больше межглазничной ширины черепа. Послед-
ний премоляр нижней челюсти (Р4) имеет открытую переднюю
долинку (рис. 68, А) .............................................
............ Gazella subgutturosa Giild. — Джейран (стр. 308).
2 (1). Размеры больше. Основная длина черепа взрослых животных
больше 210 мм. Хвост короткий, меньше 120 мм. Белое околохво-
стовое зеркало заходит на круп заметно выше корня хвоста. Волосы
на участке между мякишами главных копыт и добавочными копы-
тами имеют светлую окраску. Носовые кости без вырезки на перед-
нем крае. Срединные их концы клинообразно заострены. Рассто-
яние между наружными краями оснований роговых отростков
20*
308
BOVIDAE
меньше межглазничной ширины черепа. Последний премоляр
нижней челюсти (Р4) имеет замкнутую переднюю долинку (рис. 68, Б)
......... Gazella (Procapra) gutturosa Gmel. — Дзерен (стр. 331).
Gazella subgutturosa Giildenstaedt — Джейран (рис. 62—70)
Giildenstaedt A., 1780, Acta Acad. Sci. Petrop., t. II, pt. 1:251—274
(первооп.); Nordmann, 1840, Fauna pontique, Paris : 61 (распр.); Эверсман Э.,
1850, Естествен, ист. Оренбургск. края, II, Казань : 262—263 (распр.); Brooke W.,
1873, Proc. Zool. Soc. London : 545—546 (сист.); R adde G. und A. Walter, 1889,
Zool. Jahrb., Abt. System., IV : 1060—1064 (распр., биол.); H e u d e P., 1894, Mem.
Hist. Nat. Emp. Chinois, II : 245—246 (сист.); S elater P. and 0. Thomas,
1897—1898, Book of antelopes, III, London : 89—94 (синон., сист., изобр.); Lydek-
ker В., 1900, Great und small game of India, London : 184 (общ., изобр.); С а т y-
н ин К. А-, 1905—1906, Изв. Кавк. муз. II, 3—4 : 212—213 (распр., морф.); Д и н-
ник Н. Я., 1910, Звери Кавказа, I, Тифлис : 215—220 (общ.); Lydekker R.,
1914, Catal. Ungulate mammals Brit. Mus., Ill : 42—46 (сист., синон.); Boule M.
et P. T e i 1 h.a r d, 1928, Arch, de 1’inst. Paleontol. Hum., 4 : 66 (эвол.); В и т о-
в ич В. В., 1928, Охотник, 6 : 18 (распр., биол.); X а х л о в В. А., 1928, Зайсан-
ская котловина и Тарбагатай, Томск : 93 (распр.); Flower S., 1931, Proc. Zool.
Soc. London : 214 (продолж. жизни); Флеров К. К., 1932, Тр. Совета по изуч.
произв. сил Акад, наук СССР, сер. туркменок., 2 : 255—256 (распр., биол., изобр.);
Каверзнев В. Н., 1933, Полорогие фауны СССР и их добывание, М., КОИЗ :
21—26 (общ., изобр.); Флеров К. К. иИ. М. Г ромов, 1934, Тр. Совета по изуч.
произв. сил Акад, наук СССР, сер. туркменок., 6 : 369 (распр.); Дементьев Г. П.,
1935, Дзерен, джейран, сайга, М., КОИЗ : 15—30 (общ., изобр.); Флеров К. К.,
1935, в кн.: Павловский Е. Н., Б. С. Виноградов и К. К. Флеров, «Звери Таджики-
стана», М.—Л., изд. Акад, наук СССР : 109—112 (общ,, синон., изобр.); Фле-
ров К. К., и И. М. Громов, 1935, Тр. Тадж. компл. экспед. 1932 г., X : 281—284
(распр., биол., изобр.); Витович В. В., 1936, Боец-охотник, 9 : 56—57 (распр.);
Шнитников В. Н., 1936, Млекопитающие Семиречья, М.—Л., изд. Акад. Наук
СССР : 127—128 (распр., биол.); Богачев В. В., 1938, Тр. Азербайдж. фил. Акад,
наук СССР, геол, сер., IX : 14—17 (ист.); Верещагин Н.К., 1939, Тр. Зоол. инет.
Азербайдж. фил. Акад, наук СССР, X : 89—90 (распр.); Салихбаев X., 1939,
Тр. Узбекск. зоол. сада, I : 118—119 (распр., хоз. знач.); Соколов С. С., 1939,
Зоол. журн., XVIII, 3 : 449 (распр., биол.); Султанов Г., 1939, Тр. Узбекист.
зоол. сада, I : 103 (распр., биол., хоз. знач.); Allen G., 1940, Mammals of Chine
and Mongolia, II, N. Y. : 1223—1227 (общ., изобр.); Огнев С. И., 1940, Матер,
к позн. фауны и флоры СССР, н. с., отд. зоол., 3 (XVIII) : 78 (распр.); Анти-
пин В. М., 1941, Млекопит. Казахстана, III, Алма-Ата, ОГИЗ : 47—54 (общ., изобр.);
Бобринский Н. А. и др., 1944, Определит, млекопит. СССР, М., изд. «Сов.
наука» : 246 (распр.); М ин е р в ин В. Н., 1944, Тр. Туркменок, фил. Акад, наук
СССР, V : 227—241 (биол.); Бажанов В. С., 1945, Изв. Акад, наук Казахск. ССР,
5 : 75—76 (распр.); Дементьев Г. П., 1945, Учен. зап. Моск. гос. унив., 83,
биол. : 76—78 (распр., биол.); Ишунин Г. И. и Е. П. Коровин, 1945, При-
рода, 4 : 52 (биол.); Деревягин П. Я., 1947, Вести. Акад, наук Казахск. ССР,
4 : 33—38 (мигр.); Эверт В. К., 1947, Изв. Акад, наук Казахск. .ССР, 6 : 131—133
(биол.); Г р о м о в В. И., 1948, Тр. Инет. геол, наук, 64 : 436 (ист.); Зверев М. Д.,
1948, Тр. Алма-Атинск. заповеди., VII : 145—147 (быстрота бега); Соколова И. Б.
и В. И. Бондарева, 1948, Изв. Акад, наук Казахск. ССР, сер. паразитол., 6 :
110—112 (параз.); Грунин К. Я., 1949, Докл. Акад, наук СССР, LXIV, 4 : 603—
606 (параз.); Басенко Е. П., 1950, Тр. Гос. заповеди. Барса-Кельмес, 1:104
(акклим.); Рустамов А. К., 1950, Изв. Туркменок, фил. Акад, наук СССР, 2 :
45 (распр.); Станюкович К. В., 1950, Сообщ. Таджикск. фил. Акад, наук СССР,
XXIV : 19—22 (биол.); Дементьев Г. П., 1951, Изв. Туркменок, фил. Акад,
наук СССР, I : 58—59 (распр., биол.); Е 1 1 е г m a n J. and Т. Morrison-Scott,
1951, Checklist of palaearctic and indian mammals, London : 390—391 (сист., синон.);
Асадов С. M., 1952, Изв. Акад, наук Азербайдж. ССР, 12 : 11—18 (параз.); К о-
лесников И. И., 1952, Тр. Среднеазиатск. гос. унив., 32, биол. науки, 2 : 13—
26 (распр.); Афанасьев А. В., В. С. Бажанов и др., 1953, Звери Казахстана,
Алма-Ата, изд. Акад, наук Казахск. ССР : 484—488 (общ., изобр.); Слудский А. А,,
1953, Тр. Инет. зоол. Акад, наук Казахск. ССР, II : 15—16 (биол.); Банников А. Г.,
1954, Млекопит. Монгольск. Народи. Республ., М.—Л., изд. Акад, паук СССР : 250—
258 (общ., изобр.); Д у б и н и н В. Б., 1954, Тр. Инет. зоол. и паразитол. Акад.
GAZELLA
309
наук Узбекск. ССР, 3 : 178 (распр.); Исмагилов М. И., 1954, Бюлл. Моск. общ.
испыт. прир., Отд. биол., ЫХ, 6 : 3—9 (размн.); Кульпин П. И., 1954, Природа,
4 : 102—104 (распр., биод.); Костин В. П., 1955, Тр. Инет. зоол. Акад, наук Ка-
захск. ССР, IV : 67—82 (биол.); Грунин К. Я., 1956, Энтомол. обозр., XXXV,
3 : 716—722 (параз.); И ш у н и н Г., 1956, Охота и охотн. хозяйство, 8 : 21—23
(общ.); Слудский А. А., 1956, Тр. Инет. зоол. Акад, наук Казахск. ССР, VI :
78—108 (размн.); Маленков И. К., 1958, Зоол. ж., 37 : 629 (распр.).
Описание. Животные в пределах своего семейства небольшой или
средней величины. Длина тела от конца морды до корня хвоста от 94 до
Рис. 62. Джейран {Gazella subgutturosa Giild.), самец.
115 см; высота в холке от 67 до 80 см. Живой вес самцов от 29 до 33 кг,
самок от 24 до 29 кг.1 Основная длина черепа от 165 до 205 мм. Телосложе-
ние стройное и очень легкое. Короткое узкое туловище покоится на длинных
тонких ногах. Высота в холке обычно немного меньше высоты крестца.
Голова легкая с заостренной мордой. Верхняя губа покрыта волосами, от
носового зеркала сохраняется лишь узкая полоска голой темной кожи
между нижними углами ноздрей. Зрачок глаз слегка продольноовальный,
почти круглый. Радужная оболочка темно-коричневая. Уши средней длины,
около 12—16 см, с заостренными верхушками.
Рога, как правило, имеются только у самцов, длиной от 27 до 41 см.
Основаниями они расположены почти целиком над глазами. Форма рогов
типично лировидная. В основаниях они сближены, часто почти параллель-
ные, но вскоре сильнее расходятся в стороны и одновременно дугообразно
1 Размеры и вес — по материалам автора из Бет-Пак-Далы.
310
BOVIDAE
загибаются назад, а вершинами внутрь и вперед (рис. 63, Л). Цвет рогов
черно-бурый или серый. Поверхность роговых чехлов состоит из большого
числа поперечных, сильнее развитых на передней стороне колец; у осно-
вания рогов кольца расположены близко друг к другу; в средней части
к вершинам более редко. Загнутые концы рогов на протяжении 8—9 см
от верхушек гладкие. В виде исключения рога бывают и у самок, но дли-
ною не больше 5 см.1
Шея длинная и тонкая. На нижней ее стороне и у самцов, и у самок
ясно выступает гортань (рис. 64). Конечности тонкие. Копыта длинные, но
узкие, с сильно заостренными верхушками. Длина передних копыт 49—
56 мм, задних на 1—6 мм меньше. Высота передних копыт по переднему
краю 28—35 мм, задних обычно на 2—3 мм меньше, реже равна или меньше,
чем передних. Небольшие добавочные копыта, длиной 10—12 мм, распо-
ложены высоко над мякишами копыт средних пальцев. Хвост по сравнению
с дзереном длинный, от 17 до 25 см, а с волосами до 30 см, снизу на две
трети от основания голый.
Волосяной покров по сравнению, например, с дикими козлами и осо-
бенно оленями довольно мягкий и упругий, но на верхушках волос все же
ломкий. Ясное разделение на остевой волос и подшерсток отсутствует.
Зимний мех густой и плотный; длина волос на спине до 4—5 см. Летом
волосы более редкие и короткие, всего 1—1.5 см. На голове, кроме лба,
и ногах волос всегда более короткий, плотно прилегающий к коже. На
животе он, наоборот, зимой длиннее. У самок на месте рогов имеются не-
большие пучки удлиненных волос. Отдельные волосы верхней части ту-
ловища и шеи от основания светло-бурые или голубоватые, выше тем-
неют, а к вершинам желтые. Самые кончики волос тонкие, заострен-
ные (если не обломаны), темно-бурые, почти черные. Волосы на светло
окрашенных местах от основания до вершин белого цвета, а на хвосте
темно-бурого.
Свойственная большинству других видов газелей резкая разграничен-
ность темно окрашенных и светлых участков у джейрана выражена срав-
нительно слабо. У нестарых экземпляров средняя часть лба и темя темного,
бурого цвета. По средней линии носа тянется более светлая рыжеватая
полоса, которая, не доходя до ноздрей, заканчивается, хотя и не всегда,
темным бурым пятном. Конец морды, кроме темных безволосых участков
верхней губы, белый. От предглазничных ямок к углам рта тянутся темные,
бурого цвета полосы. Последние отделены от срединной рыжеватой по-
лосы на носу белыми полосами, сливающимися со светлой окраской конца
морды. Щеки светлого рыжевато-бурого тона. Низ головы белый. С возрастом
окраска головы светлеет и у старых экземпляров становится белой, за
исключением неясных темных полос перед глазами. Наружная сторона
ушей песчано-серая, кончики иногда черноватые. Волосы на внутренней
стороне ушей светлые, почти белые.
Общая окраска верхней части туловища и шеи песчано-серая и серо-
вато-желтая. Общий фон окраске придают предвершинные желтые участки
волос. По бокам туловища, на границе с белой окраской живота, немного
выше локтевого и коленного суставов тянется полоса более темных, бурых
волос. Такая же полоса обрамляет с боков белое околохвостовое зеркало.
Последнее не заходит выше основания хвоста. Хвост темно-бурый, из более
длинных, чем на остальных частях туловища, и более грубых волос, со-
бранных на верхней его стороне в виде гребня. Передняя и наружная сто-
1 Хорошо выраженные бугорки на месте рогов на черепах самок имеются всегда.
GAZELLA
311
роны ног обычно одноцветны с туловищем, но участки на передней стороне
непосредственно над копытами (вокруг отверстий межпальцевых желез),
а также позади между главными и боковыми копытами более темного,
буро-коричневого цвета (рис. 65). Живот, нижняя сторона груди, шеи
и внутренние части ног чисто белые.
Половой возрастной и сезонный диморфизм в окраске выражен слабо,
но зимой она становится обычно более светлой и тусклой вследствие обла-
мывания темно окрашенных верхушек волос. Имеет место хотя и не слиш-
ком резко выраженная географическая изменчивость окраски. Так, джей-
Д 5.
Рис. 63. Рога джейрана (Л) и дзерена (Б).
раны западной части ареала окрашены более интенсивно, с большим уча-
стием бурых или сероватых тонов, К востоку идет посветление, с преобла-
данием желтовато-рыжеватых, а в Центральной Азии даже розоватых от-
тенков,
Имеются хорошо развитые предглазничные железы в виде образован-
ных складками кожи вертикальных щелей или ямок. На передних ногах
чуть ниже запястных суставов расположены так называемые карпальные
железы, представляющие утолщение кожи с заложенными в ней железами,
выделяющими желтый, на ощупь жирноватый секрет. Снаружи эти железы
выделяются более длинными и грубыми по сравнению с соседними участ-
ками пучками волос темного цвета (рис. 65); основания волос склеены
секретом желез. На всех конечностях имеются межпальцевые железы,
открывающиеся на передней стороне бабок выше копыт продольной щелью.
Как у самцов, так и у самок 1 имеется по одной паре паховых желез в виде
узких карманов (глубиной около 3—5 см), расположенных по соседству
1 В противоположность неверному указанию Динника (1910) о том, что паховые
.железы имеются только у самцов.
312
BOVIDAE
с молочной железой (рис. 66). Молочная железа двухдольчатая, имеет два
основных и иногда еще два, расположенных спереди от основных, нефунк-
ционирующих соска. Вымя, кроме самих сосков, покрыто довольно длин-
ными мягкими волосами.
Череп (рис. 67) имеет сравнительно недлинную предглазничную часть;
длина последней от переднего края глазницы до конца межчелюстных
костей (prosthion) составляет меньше 133% длины заглазничной части
черепа от заднего края глазницы до крайнего выступа затылочных мы-
щелков. Вход в носовую полость узкий; ширина его не больше половины
Рис. 64. Самка джейрана (Gazella subgutturosa Giild.), центральная Бет-
Пак-Дала (Фото автора).
расстояния от prosthion до передних концов носовых костей. Носовые кости
сравнительно короткие и широкие; длина их составляет не больше 35%
основной длины черепа, а наибольшая ширина — не меньше 40% собствен-
ной длины. Передние концы носовых костей имеют вырезки во всю ширину
кости, причем латеральные их отростки лишь немного уступают по длине
медиальным. На границе носовых и лобных костей профиль черепа прямой
или даже вогнутый. Внутренние края глазниц имеют вырезки выше
слезно-лобного шва, на уровне их приходится и наименьшая ширина
лба между глазницами. Предглазничные ямки на черепе хорошо развйты.
Роговые отростки массивные; расстояние между наружными краями их
оснований больше межглазничной ширины лба. Слуховые пузыри сильно
вздутые; расстояние от нижнего края пузыря до верхнего края костного
слухового прохода обычно равно или больше вертикального диаметра
глазницы. Последний премоляр нижней челюсти (Р4) имеет хорошо
развитым только передне-наружный бугорок зуба; поэтому передняя
долинка на внутренней его стороне остается открытой (рис. 68, 4).
История и географическое распространение (рис. 69). Вследствие
неизученности остеологических отличий между видами подрода Gazella
GAZELLA
313
s. str., точное время появления на геологической сцене G. subgutturosa
определить трудно. Но, судя по ряду находок в Европе и Китае (Boule
et Teilhard de Chardin, 1928; Богачев, 1938), близкие к современным
формы жили уже во второй половине плиоцена. В отложениях четвертич-
ного периода остатки этого вида известны, кроме областей современного
распространения, лишь в фауне палеолитических стоянок Забайкалья
(Громов, 1948).
Современный ареал вида захватывает Закавказье, Переднюю, Среднюю,
и Центральную Азию — Иран, Афганистан (возможно и часть Пакистана),
Рис. 65. Молодая самка джейрана (Gazella subgutturosa Gold.), центральная
Бет-Пак-Дала. (Фото автора).
Джунгарию, Гоби, северную часть Тибета, Алашань и Ордос. В пределах
СССР джейран в настоящее время распространен в восточном Закавказье,
а также в пустынной и полупустынной зонах среднеазиатских республик
(Туркменской, Узбекской, Таджикской, Киргизской) и Казахстана.
В Закавказье еще около 100 лет назад он встречался повсеместно
в степях от Баку до Тбилиси и даже к северу от последнего, около Авчалы.
В 70-х годах прошлого века на джейранов охотились в долине Куры, кило-
метрах в 30—40 ниже Тбилиси (Радде, 1899, цит. по Диннику, 1910).
В настоящее время ни в одном из этих пунктов их нет, и современную
западную границу распространения этого вида на Кавказе Н. К. Вереща-
гин (1939) проводит в Порском плоскогорье, километрах в 60 к востоку
от Тбилиси. В 30-х годах прошлого столетия джейраны водились на тер-
ритории Эриванской губернии между Араратом и Алагезом (Nordmann,
1840), но в начале этого века, по свидетельству К. А. Сатунина (1905—
1906), здесь от них не осталось и следа.
Современный ареал джейрана в Закавказье представлен рядом изоли-
рованных очагов, что зависит как от наличия подходящих стаций, так
и от прямого или косвенного истребляющего влияния человека. В ка-
честве районов обитания Н. К. Верещагин перечисляет следующие:
1) междуречье Поры и Куры; в 80-х годах джейран служил предметом
314
BOVIDAE
охоты около Караязы (50—60 км от Тбилиси вниз по долине Куры);
в настоящее время западная граница проходит на долготе Казаха; более
многочисленны в восточной части междуречья; 2) центральная часть
Ширакской степи (междуречье Иоры и Алазани), где в начале этого
столетия распространение джейрана доходило до ее северо-западного
угла; 3) долина Сарыджа, степи Аджиноур и Турут — пространство,
лежащее в углу, образованном левобережьями Куры и Алазани; 4) до-
лина Кюрок-Чая и Боздаг; в незначительном числе к востоку от Киров-
абада, больше между Боздагом и Кюрок-Чаем к северо-востоку от Евлаха;
Рис. 66. Паховые и молочные железы самки джейрана (Gazella subgutturosa
Giild.), центральная Бет-Пак-Дала. (Фото автора).
5—6) Мильско-Карабахская и Муганская степи;1 в последней джейраны
держатся главным образом между Азизбековским каналом им.
В. И. Ленина, а также вдоль иранской границы от Белясувара до Аракса;
7) юго-восточная Ширвань, в районах вдоль каспийского побережья от
Пирсагата до низовьев Куры включительно; 8) Ширванская степь в про-
странстве междуречья Аксу—Пирсагат к северу и югу от железной до-
роги; за последнее десятилетие здесь сильно истреблены (Витович, 1928,
1936) и сохранились главным образом близ озер Караса и Аджикабула;
9) Кабристан, районы вдоль побережья Каспийского моря, непосред-
ственно прилегающие к Апшеронскому полуострову.
Сто с небольшим лет назад распространение джейрана доходило
до Баку, тогда как теперь этот вид встречается лишь километрах в пят-
надцати к западу от последнего. Северная граница проходит здесь около
станций Килязи и Насосная. Общее количество джейранов, обитавших
1 По устному сообщению X. М. Алекперова, в Карабахской степи к западу от
Карягино и Агдама в настоящее время джейран отсутствует. По-видимому, уже в по-
следние десятилетия он вытеснен с территории между Агдамом и Евлахом, около ко-
торого, по Н. Я. Диннику (1910), джейраны попадались до начала XX в. Отсутствует
ранее водившийся джейран также в степях и полупустынях Зангеланского и Джеб-
раильского районов Азербайджана.
GAZELLA
315
в Закавказье на конец 30-х годов нынешнего века, Н. К. Верещагин
(1939) определял в 5730 голов. В последующие годы шло дальнейшее
прогрессирующее сокращение ареала и снижение численности джейрана
Рис. 67. Череп Gazella subgutturosa Giild.
в результате не только прямого истребления браконьерами, но и освое-
ния степей и полупустынь под посевы сельскохозяйственных культур
и под выпасы для домашнего скота.
316
BOVIDAE
Область почти сплошного распространения джейрана представляют
пустынная и полупустынная зоны, а также предгорья Средней Азии.
Нет его в высокогорных областях, но в широких горных долинах и сыр-
тах этот вид может обитать до высот 1500—3000 м. Современная северная
граница распространения джейрана проходит в Казахстане. На западе
она начинается у побережья Каспийского моря на широте около 46° с. ш.
«Проходя на восток севернее чинка Устюрта,1 она поднимается до 47° с. ш.
близ Аральского моря и следует к среднему течению р. Сарысу, проходя
несколько южнее Джезказгана. Далее она идет по северной окраине пустыни
Бет-Пак-Дала к оз. Балхашу. Джейраны встречаются также в Алакуль-
ской котловине и очень редко по западным и южным отрогам Тарбага-
тая. Сравнительно недавно они обитали в Зайсанской котловине» (Афа-
насьев и др., 1953). В последней джейраны исчезли в 90-годах прошлого
века (Хахлов, 1928).
Обыкновенны джейраны до сих пор по всему Устюрту, особенно в юж-
ной его части, на территории впадины Барса-Кельмес и причинковых
тугаях левобережья низовьев Аму-Дарьи (Колесников, 1952; Костин,
1955), на полуострове Мангышлаке. Обычным зверем они являются
в Приаральских Каракумах, в долине Сыр-Дарьи, на всем пространстве
Кызылкумов между Сыр-Дарьей и Аму-Дарьей (Бажанов, 1945). Джей-
раны обычны до настоящего времени в пустыне Бет-Пак-Дала, в пе-
сках Муюнкумах и на обширной территории Сары-Аркинской степи
к северу от Бет-Пак-Далы (Кульпин, 1954). В северном Прибалхашье
они встречаются, но очень редки (Антипин, 1941). По данным Д. Н. Каш-
карова (Антипин, 1941), только изредка появляются в районе озер Ащи-
коль и Бийликоль. В Семиречье (в Алма-Атинской и Талды-Курган-
ской областях Казахской ССР) больше всего этого зверя в бассейне
р. Или к востоку от Заилийского Алатау и хребта Кетмень, в частности
в нагорных долинах Сюгаты и Джаланаш (Огнев, 1940). Обычен в пред-
горьях Джунгарского Алатау (Катутау,. Ортатау, Дулантау, в долине
между Катутау и Алтынэмельским хребтом). Как уже отмечалось, встре-
чаются джейраны в Алакульской котловине, но, по В. Н. Шнитникову
(1936), отсутствуют в степной полосе западной части б. Лепсинского и
восточной части Канальского уездов.
В 1929 г. 9 голов джейранов (7 самок и 2 самца) были завезены на
остров Барса-Кельмес в Аральском море, где они, по-видимому, никогда
до этого не водились. Джейраны очень хорошо акклиматизировались
на этом острове, начали быстро увеличивать свою численность. По архив-
ным данным Барса-Кельмесского заповедника, поголовье их к 1947 г.
исчислялось в 2300 голов, а к 1948 г. будто бы возросло до 4650 голов
(хотя последняя цифра и вызывает сомнения). Суровая зима 1948/
49 года с глубоким плотным снегом и гололедицей свела на нет резуль-
таты акклиматизации джейранов на Барса-Кельмесе. В 1949 г. было
учтено только 64 живых и 1807 трупов джейранов; к концу этого года,
в результате продолжавшегося отхода истощенных животных, их оста-
лось еще меньше. С тех пор, несмотря на нормальные в среднем условия,
по не выясненным пока причинам, джейраны, в противоположность сай-
гакам, не могут восстановить свою численность и в 1957 г., когда я был
на острове, количество их не превышало два-три десятка.
1 По-видимому, много севернее Устюрта джейран едва ли заходил и раньше
(Эверсманн, 1850).
GAZELLA
317
В равнинных и предгорных районах Туркмении и Узбекистана джей-
ран распространен почти повсеместно, за исключением районов интенсив-
ного земледелия и поэтому густо населенных. Однако сведения о сравни-
тельной численности его в отдельных районах крайне не-
полны. Водится джейран в Ферганской долине (Мален-
ков, 1958). Встречаются джейраны в Хавастском районе
Ташкентской области (Дубинин, 1954), равнинной и буг-
ристой частях Гузарского района Кашка-Дарьинской
области (Салихбаев, 1939) и Ширабадского района Сур-
хан-Дарьинской области (Султанов, 1939). Еще в 80-х го-
дах прошлого века они были весьма многочисленны
в южной Туркмении, в частности между Мургабом
и Тедженом (Radde und Walter, 1889). В долине Мур-
габа джейраны и сейчас весьма обыкновенны, но о
прежнем изобилии, по-видимому, теперь не может идти
Рис. 68. Перед-
некоренные зу-
бы (премоляры)
нижней челю-
сти.
А — Gazella sub-
gutturosa Gii Id.;
Б—Gazella gut-
turosa Gmel.
речи (Флеров, 1932). Обычным, но уже немногочислен^
ным зверем являются они в юго-западной Туркмении, в
рацоне Гасан-Кули, Кизыл-Атрека, Мадау, Мессериана.
Численность их здесь уменьшается с каждым годом, что
Г. П. Дементьев (1945, 1951) ставит в связь со все уси-
ливающейся охотой с автомобиля. Лет двадцать назад,
а возможно и сейчас, джейран был обыкновенным зве-
рем в полосе горных склонов юго-западного Копет-Дага,
в бассейне Сумбара и Чандыра (Флеров и Громов,
1934). Повсеместно, хотя и в небольшом числе, встречается на Узбое
и в центральных Каракумах (Дементьев, 1951). Автору неодно-
кратно приходилось видеть свежие следы джейранов осенью 1949 г.
Рис. 69. Распространение Gazella subgutturosa Giild.
1 — XX в. (области постоянного обитания); 2 — XIX в.
на территории совхоза Уч-Аджи в юго-восточных Каракумах. Повсе-
местно встречаются они в северо-западной Туркмении, в частности в окрест-
ностях колодца Чагыл, особенно на такырах и глинисто-полынных рав-
нинах (Рустамов, 1950).
318
BOVIDAE
В Таджикистане область распространения джейрана занимает юго-
восточную часть республики. Наиболее многочислен он в долинах ниж-
него течения Вахша, Кафирнигана, смыкаясь на западе с ареалом его
в Узбекистане. В 1951 г. мне довольно часто удавалось наблюдать следы
и видеть живых джейранов в Дангаринском районе Кулябской области
по долинам и пологим склонам гор в бассейнах рр. Таирсу и Кызылсу.
На востоке границу распространения джейрана в Таджикистане, по
Флерову (1935), составляют предгорья Памира, а на севере — хребты
северной части республики. По-видимому, едва ли встречается этот вид
севернее южной оконечности Вахшского хребта.
В Киргизии единственным очагом распространения джейрана является
Иссык-Кульская котловина. Прежде он водился, по-видимому, по всему
северному побережью Иссык-Куля и здесь ареал его сливался с ареалом
в юго-восточном Казахстане. В настоящее же время сохранился лишь
у западной его оконечности, на Семисбельском плато (между р. Чу
и оз. Иссык-Куль). К северу здесь за последние десятилетия не заходил
дальше окрестностей поселка Рыбачьего (Дементьев, 1935). В коллекциях
ЗИН АН СССР имеется экземпляр джейрана, добытый в 1913 г. на
юго-западном берегу оз. Иссык-Куль, в урочище Агален.
В Монгольской Народной Республике распространение джейрана
охватывает южную часть страны, пустыню Гоби, долину между хребтом
Хангай и Монгольским Алтаем и котловину Больших озер до оз. Убсу-
Нур. Северная граница, по А. Г. Банникову (1954), с востока на запад
проходит так: «От опустыненной равнины Эргиин-Тала, примерно у пункта
пересечения государственной границы (Монгольской Народной Респуб-
лики, — И. С.) с 112° в. д., джейран поднимается к северу в направлении
Байшинту. Здесь по опустыненным сухим долинам газели регулярно
проникают до 46° с. ш. у Тубшин-Ширэ сомона. В отдельные годы в этом
районе они заходят и дальше к северу до мелких озер: Усуни-Цаган,
Улан-Гобиини Гобиин-Цаган, вплоть до урочища Шара-Уху. Как исключе-
ние, по опросным сведениям, отмечены забеги в урочища Салхитуин-Тала.
«Далее, к западу граница распространений джейрана, следуя при-
мерно 46° с. ш., проходит к Чойрэну, затем опускается к Мандал-Гоби
и хр. Дельгер-Хангай. Пересекая р. Онгин-Гол вблизи Хущу-Хид, граница
опять поднимается по котловине Гобийских озер. Здесь северная граница
обитания джейрана пересекает низовья рр. Тацын-Гол, Туин-Гол, Нарин-
Гол и Байдариг и по южным предгорьям Хангая вновь языком подни-
мается к северу, доходя до южной излучины р. Дзабхан. Обойдя хр. Та-
шири-Сэрхэ с юга через котловину оз. Бигэр-Нур и узкую степную пере-
мычку урочища Халюн, где джейраны бывают редко, граница уходит
в Шаргын и Хуйсиин-Гоби, следуя подножьям хр. Тайшири, Хасагт-
Хайрхан. В котловине больших западных озер, обходя барханные пески
нижнего течения р. Дзабхан с запада, граница подходит к восточной
окраине котловины оз. Хиргис-Нур. В котловине оз. Хиргис-Нур джей-
раны бывают непостоянно. Так, после гибельной для копытных зимы
1935 г. они там не отмечались. До указанного года, по опросным сведен-
ниям, как редкость отмечались забеги джейрана еще севернее, в котловину
оз. Убсу-Нур. В котловине озер Хара-Ус-Нур, Хара-Нур (Кобдосский)
и Дургэ-Нур, начиная с южных и юго-восточных склонов горы Хара-
Аргалинту, джейраны бывают постоянно. По долине р. Кобдо джейраны
поднимаются довольно высоко до линии гор Цаст-Ула-—Алтан-Хухэй.
В отдельные годы бывают забеги и выше, вплоть до Боро-Бургасу, Суока,
оз. Ачит-Нур и нижнего течения р. Буху-Мурэн.
GAZELLA
319
«К юго-востоку от озер Хара-Ус и Дургэ-Нур встречаются джейраны
в котловине оз. Дзэргэн-Цаган-Нур, в котловине между хребтами
Бумбат-Хайрхан и Баян-Дарбиин-Нуру.
«Огибая Монгольский Алтай, ио склонам магистрального хребта и
далее по хребтам Хара-Адзирга, Бургах-Будда, Баян-Цаган граница
распространения джейрана опускается к югу, в перемычку между горами
Цэцэн-Хайрхан и Бахар, в котловину оз. Ногон-Нур. В котловину оз. Ху-
тун-Нур джейраны заходят редко. Мимо восточной оконечности хребтов
Гичгэнэ-Нуру и Байбаян-Ундур северная граница распространения
вдоль южных склонов Монгольского Алтая проходит на нижнее течение
р. Булугун.
«Южнее джейран отсутствует по значительным хребтам и поднятиям,
таким, как Аджи-Богдо, Хубчи, скалистая часть Эдэрэнгин-Нуру, гор-
ной группы Цаган-Богдо-Ула, Ихэ-Богдо, Нэмэгэту, Тост-Нуру, Арца-
Богдо, Гурван-Сайхан, Дзблен и некоторым другим».
Подвиды. В настоящее время признается наличие трех подвидов
джейрана.
1) G. subgutturosa subgutturosa Guldenstadt. Размеры сравнительно не-
большие. Высота в холке не превышает 67 см. Рога длинные, до 40 см.
Распространение: Сирия, Ирак, Иран, Афганистан, средне-
азиатские республики СССР, Закавказье, Казахстан.
2) G. subgutturosa sairensis Lydekker. Размеры больше, чем у типичной
расы. Высота в холке до 70 см, но рога более короткие, до 27 см, редко
больше. Распространение: Джунгария (западный Китай).
3) G. subgutturosa reginae Adlerberg. Размеры сравнительно крупные,
но рога недлинные, длина по прямой линии от основания рогового чехла
до вершины не больше 30 см. Главное отличие от других подвидов — очень
бледный общий тон окраски, песчаный, с легким желтовато-рыжеватым
оттенком. Распространение: Цайдамское плоскогорье до
Алтынтага на север, к югу до Куньлуня; на востоке доходит до оз. Кукунор.
Д. Эллерман и Т. Моррисон-Скотт (Ellerman and Morrison-Scott, 1951)
включают в качестве подвидов джейрана яркендскую газель (G. yarkandensis
Blanf.) из Китайского Туркестана и сеистанскую (G. seistanica Lydd.),
рассматриваемых Р. Лайдекером (Lydekker, 1954) как самостоятельные
виды, отличающиеся большей величиной и ясно выраженной зональностью
окраски морды. Кроме того, как подвид выделяется G. s. hillieriana Heude
из Монголии (Allen, 1940). Систематическое положение последней и ее
отношение с G. s. reginae требуют уточнения.
Биология. Хотя джейран в ряде областей является обычным зверем,
биология его изучена недостаточно, а по некоторым сторонам образа жизни
сведения противоречивы.
В общем джейран — типичный представитель фауны пустынных и
полупустынных ландшафтов. Обитает практически во всех типах пустынь
и полупустынь Азии. Наиболее характерными местообитаниями являются
полынно-солянковые пустыни, полынные и полынно-мятликовые полупу-
стыни.
Основные районы распространения джейрана представляют открытые
равнины, но, в отличие от двух других азиатских антилоп — дзерена и
сайги, этот вид легче мирится с пересеченным рельефом местности, а в не-
которых случаях отдает явное предпочтение полузакрытым участкам
ландшафтов.
В Закавказье, Казахстане и в Средней Азии джейраны заселяют не
только открытые равнины, но и пологие предгорья, а также горные долины.
320
BOVIDAE
В Таджикистане держатся на пологих склонах хребта Каратау, Сарса-
риака и других гор вдоль нижнего течения Вахша и Пянджа. В Дангарин-
ской впадине джейранов чаще можно встретить не на равнине, а на полу-
пустынных увалах соседних невысоких хребтов. В ряде районов они даже
откочевывают на лето из открытых пустынь в предгорья, привлекаемые
наличием на последних зеленой ксерофитной растительности и меньшим
количеством кровососущих насекомых. В горах Богуты (восточная оконеч-
ность Заилийского Алатау) живут и зимой на обнаженных от снега солнце-
печных склонах (Соколов, 1939). В отрогах Джунгарского Алатау джей-
раны подымаются до 1500 м (Антипин, 1941), а в Монголии и Иранском
нагорье встречаются даже на высотах около 2000 м (Дементьев, 1935;
Банников, 1954).
Предел вертикального распространения кладет не сама высота местно-
сти, а характер связанной с нею растительности и рельеф. Высокогорий
и скалистых участков, даже невысоких гор джейраны, не приспособлен-
ные к передвижению в этих условиях, тщательно избегают.
Джейраны обитают как в глинистых и каменистых, так и в песчаных
пустынях, но предпочитают местности с плотным грунтом. Явно избегают,
особенно летом, слабо закрепленных и барханных песков, равно как и
открытых площадей с сильно каменистой почвой (Костин, 1955). Излюб-
ленными стациями, по данным последнего автора, являются закрепленные
бугристые пески с разреженной кустарниковой растительностью, а также
редкая саксауловая поросль, обеспечивающая защиту как от зноя, так и от
непогоды в холодное время года. Впрочем, как густых кустарников, так
и зарослей саксаула джейраны, вероятно из инстинкта безопасности, избе-
гают. Летом посещают солончаки.
В восточном Азербайджане летом держатся в каменистых россыпях
известковых скал, где, благодаря наличию более разнообразной расти-
тельности, лучше обеспечены свежим зеленым кормом (Верещагин, 1939).
Участки песчаной пустыни чаще служат местами обитания в зимний период,
когда грунт становится более уплотненным.
На южном Устюрте основными стациями джейрана, по крайней мере
в летнее время, являются обширные впадины с такыровидными почвами.
Однако джейранов привлекают сюда не особенности рельефа и почвы,
а наличие во впадинах долго сохраняющейся сочной зеленой раститель-
ности, дающей возможность животным длительное время существовать
•без водопоя. На возвышенных частях плато джейраны отсутствуют или
появляются очень редко (Минервин, 1944).
В Бет-Пак-Дале в мае нам чаще удавалось встречать и добывать джей-
ранов в участках глинисто-каменистой пустыни с редкими кустами сак-
саула, спириантуса и других древесных растений. В противоположность
обитающему в тех же районах сайгаку, наличие редких кустов, по-види-
мому, является непременным условием мест, пригодных для джейрана.
В кустах джейран не только спасается от жары, отдыхает днем и прячет
детенышей, но в случае опасности и убегает туда, прячась от врагов.
В Монголии А. Г. Банников (1954) чаще встречал джейранов в ковыль;
ково-луковой полупустыне, а также в солянковых (балгуровых) пустынях.
На большей части ареала имеют место сезонная смена мест обитания
и связанные с ней миграции джейранов. Основной причиной миграций
является, по-видимому, неодинаковая обеспеченность кормами в разные
сезоны. В недалеком прошлом, а возможно и в настоящее время происхо-
дили массовые перекочевки джейранов из Туркмении в Иран и Афганистан
через плоскогорья Парапамиза между Аму-Дарьей и Тедженом (Дементьев,
GAZELLA
321
1935). Ежегодные перемещения их осенью, в ноябре, к югу, а весной, в мае,
в обратном направлении до сих пор имеют место в Каракалпакской части
Устюрта (Костин, 1955). Как отмечалось, из-за Сыр-Дарьи на зиму звери
перекочевывают в Кызылкумы, а ранней весной, еще до ледохода, пере-
правляются через реку в обратном направлении. Из района Карсакпая
джейраны переселяются осенью в низовья Иргиза и Тургая, где зимы не
так многоснежны. Из равнинной части междуречья Или-Каратал в южном
Прибалхашье джейраны ежегодно на зиму в массе откочевывают в долину
Калкан-Матая в Мын-Булак (юго-западные отроги Джунгарского Алатау;
Деревягин, 1947). О массовых миграциях джейранов осенью из Бет-Пак-
Далы на юг, в пески Муюнкумы, а ранней весной в обратном направлении»
совершающихся ежегодно, сообщает П. И. Кульпин (1954). В последнем
случае протяженность миграционных путей исчисляется, по-видимому,
сотнями километров и обусловлена лучшими защитными условиями зимой
(бугристый рельеф, наличие саксаульников). Откочевки на юг в Бет-Пак-
Дале начинаются с выпадением дождей и снега (конец ноября—декабрь),
по р. Чу переходят только по льду.
В ряде мест известны миграции противоположной направленности.
Так, в Семиречье значительная часть самок весной для деторождения под-
нимается в горы юго-западной части Джунгарского Алатау, где происхо-
дит отел и сохранение молодняка, а для кормежки звери спускаются
в прилегающие долины.
Чисто местные, а иногда и далекие, но нерегулярные миграции могут
происходить вследствие выгорания растительности и высыхания источни-
ков водопоя, наступления гололедицы, выпадения глубокого снега, а также
из-за вытеснения зверей отарами домашних овец.
Суточный цикл джейранов не сложен. В основном — это дневные жи-
вотные. Но в лунные, светлые ночи, особенно если нет условий для спокой-
ного выпаса в дневные часы суток, выходят кормиться и ночью. Обычно же
ночью отдыхают; летом выпасаются с рассвета до 10—11 часов и с 17—18 ча-
сов до заката солнца и дольше. В жаркое время дня лежат и пережевы-
вают жвачку. Места для лежек, как правило, выбирают в тени: под обры-
вом в овраге или в высохшем русле реки, в тени куста. Если нет тенистого
укрытия, располагаются на открытом, обдуваемом ветром месте.
Лежки джейранов представляют небольшие яйцевидные углубления
в земле, выбиваемые ногами. Иногда под одним кустом располагается
несколько лежек, которыми, как полагают, зверь пользуется последова-
тельно, по мере перемещения тени. Зимой джейраны разгребают копы-
тами снег и устраивают лежки в понижениях или с подветренной сто-
роны кустиков и холмов. Удобными для отдыха, безопасными местами
джейраны могут пользоваться долго. Такие места узнаются по скоплению
около них помета.
В холодное время года джейраны пасутся весь день, лишь на время
прекращают пастьбу для отдыха и пережевывания жвачки. На водопой
обычно ходят на рассвете и перед закатом или в сумерках. В жару нередко
посещают водопои и днем. Места водопоев более или менее постоянны.
Так как число удобных для питья мест в пустыне ограничено, то на водо-
поях часто собирается довольно большое число животных.
Места таких водопоев со множеством следов джейранов автору при-
шлось наблюдать в Таджикистане, на рр. Кызылсу и Таирсу. Для подхода
к питьевым источникам джейраны пользуются определенными путями, где
образуются тянущиеся на несколько километров тропы. На пути постоян-
ных переходов и вблизи водопоев пастбища иногда так выбиваются, что
21 Фауна СССР. И. И. Соколов
322
BOVIDAE
растительный покров нарушается и образуются подвижные пески (Станю-
кович, 1950).
Основную пищу джейранов составляют травянистые растения, произ-
растающие в районах обитания. Обычно — это виды разнотравья и солянки,
но в ряде случаев имеется несомненная избирательность видового со-
става кормовых растений. Так, в Мильско-Карабахской степи излюблен-
ным кормом являются ксерофитные злаки (Верещагин, 1939). В юго-запад-
ной Туркмении, наравне с полынью, охотно поедаются мятлик, осочки,
пыреи (Дементьев, 1945). В Монголии, по А. Г. Банникову (1954), из 13 по-
едаемых джейраном видов растений основу питания составляют, по су-
ществу, только четыре: гобийский ковылек (Stipa gobica Boshev.), баглур
(Anabasis brevifolia С. A. M.), монгольский лук (Allium mongolicum Begel)
и галогетон (Halogeton sp.), тогда как полыни по крайней мере летом, по-
видимому, совсем не употребляются в пищу. В предгорьях Джунгарского
Алатау основными кормовыми растениями являются таволга, подмареник
желтый, биюргун (Anabasis salsa Benth.), головки лука (Allium sp.),
тырса, типчак, курчавка (Atraphaxis frutescens L.) и даже тростник
(Phragmites sp.) (Антипин, 1941). «На острове Барса-Кельмес, — пишет
Е. ,И. Страутман (Афанасьев и др., 1953), —джейраны охотно поедают
весной и летом пырей (Agropyrum sibiricum Eichw. и A. desertorum Fisch.),
белую полынь (Artemisia terrae albae H. Krasch.), биюргун (Anabasis
salsa Benth.), солянку (Salsola crassaM. В.)и др. Поздней осенью, зимой и
ранней весной джейраны питаются молодыми побегами саксаула (Haloxy-
lon aphyllum Mink, и H. persicum Bge.), сухими листьями ревеня (Rheum
tataricum L.). Основными нажировочными кормами являются полыни
и солянки, в том числе биюргун (Anabasis salsa Benth.), особенно его се-
мена». В желудках убитых весной в Бет-Пак-Дале джейранов, среди остат-
ков других растений, нам часто приходилось видеть листочки бо-
ялыча.
Как показали наблюдения и исследования В. Н. Минервина (1944),
летом питание джейрана зависит также от обеспеченности животных питье-
вой влагой. Так, на южном Устюрте, где звери лишены источников для
питья, они совершенно не едят ни полыней, ни солянок, во многих других
частях ареала джейрана составляющих основу его питания. Летом здесь
этот зверь держится по впадинам и питается зелеными частями произра-
стающих в них растений, содержащих значительное количество влаги:
в первую очередь — это листья, верхушки побегов и плоды капорца (Cappa-
ris spinosa L.), хотя последний и не является массовым растением; в мень-
шей мере — плоды янтака (верблюжьей колючки, Alhagi camelorum Fisch.),
и совсем мало — плоды соленосного кустарника (Reaumuria fruticosa
Bge.) и побеги тамарикса. Интересно отметить, что, в противоположность
домашним овцам, джейраны не трогают зеленых частей янтака, тогда как
без вреда поедают ядовитый для овец и коз капорец (Capparis spinosa
L.).
На южном Мангышлаке, в районе колодца и мыса Суе, джейраны имеют
возможность пить морскую воду и, кроме того, утоляют жажду за счет
поедания сочных водорослей на берегу моря. Вероятно, поэтому основу
их питания составляет наземный лишайник чоп-кна (Theloschystes lacuno-
sus Rupr.), по виду напоминающий олений мох (Cladonia). Несмотря на то,
что летом этот лишайник очень сух, ломок и при растирании между паль-
цами превращается в порошок, до 75% содержимого желудка убитых джей-
ранов, по В. Н. Минервину (1944), составляли остатки этого растения.
Звери его много и охотно поедают, вероятно, благодаря его высокой пита-
GAZELLA
323
тельной ценности,1 а сухость компенсировалась наличием в достаточном
количестве питьевой влаги. Чоп-кна встречается и на Устюрте, но вслед-
ствие сухости и отсутствия источников водопоя, совсем не поедается здесь
джейраном.
В культурной полосе джейраны посещают посевы сельскохозяйствен-
ных растений. В Закавказье, например, зимой пасутся на полях ячменя
и пшеницы.1 2 Имеются сообщения, что иногда приходят кормиться на
всходы хлопка (Дементьев, 1935; Верещагин, 1939). В Сюгатинской до-
лине (Казахстан) отмечались случаи поедания листьев кукурузы (Афана-
сьев и др., 1953). В современных условиях, однако, едва ли может идти
речь о серьезном вреде от джейрана сельскохозяйственным культурам.
Спорен вопрос о водопойном режиме джейранов. По одним данным,
они не могут обходиться без питья и будто бы за десятки километров
совершают переходы к местам водопоев. По другим, наоборот, организм
этого зверя приспособлен к пониженному водному обмену, так что он
может долго жить в условиях полного отсутствия влаги. По-видимому,
и то и другое утверждение верно только в известной мере.
Как показали наблюдения В. Н. Минервина (1944), на южном Устюрте
джейраны месяцами существуют в самое жаркое время года при полном
отсутствии питьевой воды без вреда для организма. Потребность в воде
в данном случае, как уже отмечалось, удовлетворяется за счет поедания
сочных зеленых растений. Никаких, даже незначительных миграций
в поисках воды джейраны южного Устюрта не предпринимают, зато
вынуждены отказываться от очень питательных и массовых, но сухих
кормов и довольствоваться колючими, сочными капорцами. Аналогичное
явление отмечает А. Г. Банников (1954) для антилоп Монголии. Совер-
шать более или менее значительные переходы или даже вовсе покидать
известный район джейраны вынуждены в тех случаях, когда нет побли-
зости водоемов для питья и в то же время ощущается недостаток в сочном
зеленом корме. Последнее обычно случается в засушливые годы.
При наличии удобных водопоев джейраны посещают их ежедневно
по два и даже по три раза. Охотнее пьют пресную воду, но при недостатке
последней не отказываются и от горько-соленой из пересыхающих луж
или из моря. Известны случаи посещения джейранами бассейнов, устраи-
ваемых для водопоя домашних животных.
Весной и летом джейраны держатся по 2—8 штук, представляющих
часто семейные группы. Весной в Бет-Пак-Дале нам они встречались
парами, реже больше. Самый крупный табунок состоял из 12 самок.
В одиночку встречаются обычно самки перед и первое время после отела,
а также самцы. Последние в течение большей части года держатся от-
дельно от молодняка и самок, хотя изредка даже весной нам приходилось
видеть самцов и самок вместе. Самостоятельные группки образуют также
яловые самки и молодняк прошлого года. Последний иногда, впрочем,
держится с матерями. Ближе к осени и к зиме джейраны объединяются
в более крупные смешанные табуны, насчитывающие по нескольку де-
сятков, иногда до 150—200 голов. Особо большие скопления джейраны
образуют при перекочевках. С. И. Билькевич (цит. по Г. Дементьеву,’
1935) наблюдал весной 1919 года в Иране шедшее сплошной лавиной
стадо джейранов, определяемое им в 10 тысяч голов. Это было пересе-
1 Чоп-кна является, по данным того же автора, прекрасным нажировочным кор-
мом домашних коз и овец.
2 Устное сообщение X. М. Алекперова.
21*
324
BOVIDAE
ление зверей из песчаных пустынь на летовки в предгорья Копет-Дага.
Распадение табунов на более мелкие группы происходит в марте—начале
апреля. Вожаков и специальных сторожей в табунах, по-видимому, нет.
В спокойной обстановке, при пастьбе джейраны передвигаются шагом.
Вспугнутые делают крупные прыжки и вначале несутся вскачь, время
от времени делая остановки и озираясь. Преследуемые обычно бегут
рысью или галопом. Бег джейранов необычайно изящен, особенно по срав-
нению с водящейся в тех же местностях сайгой. Слегка закинув голову
назад и подняв вертикально хвост, несется джейран мягкой пружинистой
рысью. Сильно испуганный, он делает со всех четырех ног прыжки про-
тяженностью 2—3 м. Настойчиво преследуемый, переходит на галоп и
как бы стелется по земле.
Спасаясь, джейраны бегут цепочкой и рассыпаются в стороны метров
за 100—150 от настигающей машины. К преследуемому табуну часто
присоединяются другие. Характерно стремление бежать наперерез и пере-
секать путь непосредственно впереди машины, но в местах усиленного
преследования джейраны выработали новые рефлексы и оставили
губительную для них привычку. Раненные и загнанные, часто ложатся
и затаиваются, плотно прилегая к земле.
Насколько незаметен джейран, лежащий на земле и сливающийся
с окружающим фоном благодаря песчано-рыжей окраске, настолько
бросается в глаза стайка убегающих и мелькающих белыми задами жи-
вотных. Поднятый хвост делает еще более заметным яркое белое около-
хвостовое зеркало, вероятно служащее зверям при групповом беге ночью
хорошим ориентиром.
Бег джейрана считается классическим образцом не только изящества,
но и быстроты. Однажды в Бет-Пак-Дале мы преследовали на автомашине
двух (молодую и взрослую беременную) самок. Первая легко вырвалась
вперед, перебежав дорогу перед автомобилем, и быстро ушла. Вторая,
заведомо беременная самка в течение 5 минут мчалась параллельно авто-
машине, шедшей со скоростью 60 км, и в конце концов, также перерезав
наш путь почти перед колесами машины, ушла в сторону и скрылась
в кустах. В другой раз на такыре самка с перебитой передней ногой
успешно конкурировала с автомобилем, развившим скорость 60 км, и спас-
лась в зарослях боялыча, где машина на большой скорости следовать
за ней не могла. При этом джейрануха, держа перебитую конечность
на весу, бежала так легко, как будто четвертая нога ей была совсем не
нужна. Нам не удавалось долго преследовать джейранов, но В. П. Костин
(1955) утверждает, что со скоростью 60 км в час они могут бежать от 20
до 90 минут, в жару, впрочем, утомляясь очень быстро. По данным
М. Д. Зверева (1948), в исключительных случаях скорость джейранов
доходит до 65 км.
Джейран очень осторожный и чуткий зверь. Зрение, слух и обоняние,
по-видимому, развиты превосходно.1 В местах, где их часто преследуют,
джейраны редко подпускают на открытом месте человека ближе 1.5—2 км.
Там же, где их не пугают, они доверчивы, проявляют даже признаки
любопытства, и известны случаи, когда на виду можно было прибли-
зиться к ним на 50—60 шагов (Шнитников, 1936).
Наблюдения за джейранами в Бет-Пак-Дале, где их теперь довольно
часто беспокоят, создали у меня впечатление, что это зверь осторожный,
1.Хотя, по П. И. Кульпину (1954), чутье будто бы развито слабо.
GAZELLA
325
но не пугливый. Джейраны редко подпускали грузовую машину с людьми
ближе, чем на 300—400 м, но панического страха при этом не проявляли.
Однажды к нашему лагерю подошла метров на 500 самка, долго и напря-
женно смотрела в нашу сторону, пока ее не отогнал выстрел. Уходя в сто-
рону от машины, джейраны бегут медленной спокойной рысью. Даже
после неудачного выстрела они очень часто, сделав несколько прыжков,
тут же останавливаются, давая возможность выстрелить повторно, а по-
том убегают, время от времени останавливаясь и поворачивая голову
в сторону, откуда появилась опасность. Раза два мне удавалось наблюдать
за самками, по-видимому, имевшими где-то поблизости детей. Они заме-
чали меня, подпускали метров на 250—200, но при дальнейших моих
попытках сблизиться медленно уходили, все время наблюдая за мною
и двигаясь по кругу, где-то внутри которого, вероятно, находились джей-
ранята.
В спокойной обстановке хвост у джейрана опущен вниз. Поднятый
вертикально хвост служит признаком настороженности или беспо-
койства зверя. Сигналом тревоги для табунка, по В. П. Костину (1955),
служат удар копытом о землю первым заметившим опасность зверем или
тревожный писк находящихся по соседству песчанок, крики и взлеты
птиц.
Джейран — молчаливый зверь. Мне ни разу не удалось услышать
его голос в обычной обстановке. Только смертельно раненный, при прибли-
жении человека он иногда издает хриплые звуки, отдаленно напоминаю-
щие блеяние овцы. Ягнячьим, но более грубым голосом закричал и пойман-
ный нами джейраненок (рис. 70).
Линька у джейрана происходит в году, по-видимому, дважды. Первый
раз линяют весной, начиная со второй половины февраля. Тянется линька
долго и заканчивается в мае, а у молодых еще позднее. Даже у здоро-
вых, нормально развитых джейранов остатки старой шерсти на боках
и около границы околохвостового зеркала нами наблюдались до конца
второй декады мая. Больные и истощенные звери линяют позднее здоровых.
У добытых 21 и 22 мая в Бет-Пак-Дале истощенных самок остатки старой
шерсти еще сохранялись на затылке, шее, боках и наружной стороне
голеней. Выпадение старой шерсти происходит постепенно, а так как
сезонный диморфизм в окраске у джейрана выражен слабо, то линька
проходит как бы незаметно. Даже в хороший бинокль нельзя увидеть
на джейране висящие клочья старой шерсти, как это приходится нередко
наблюдать, например, у линяющих баранов, и закончил ли джейран
линьку или нет, обычно удается установить только на трупе. Осенняя
линька происходит с сентября по октябрь включительно.
Большая часть джейранов, по-видимому, уже с полуторалетнего воз-
раста активно участвует в размножении. Самки, по словам А. Слудского
(1956), иногда приносят детей в возрасте 1 года. Течка начинается с ноября,
а по некоторым данным (Афанасьев и др., 1953) даже с конца октября
и тянется по декабрь включительно.
Г. П. Дементьев (1935) называет джейранов моногамами. Однако по
Е. И. Страутману (Афанасьев и др., 1953), в период гона они держатся груп-
пами по 3—5 штук во главе со взрослым самцом, прогоняющим молодых, так
что едва ли в этом случае может идти речь о моногамии. В это время у джей-
ранов набухают паховые железы, выделяющие большее, по сравнению
с обычным временем, количество пахучего секрета. Самцы приходят в воз-
буждение, у них еще больше вздувается гортань, и за время гона они
худеют. Между самцами происходят столкновения (турнирные поединки),
326 BOVIDAE
не носящие, однако, той ожесточенности, как у других копытных.1 Самки
ведут себя спокойно и упитанности заметно во время течки не снижают.
Рождение молодых происходит в период с конца апреля до конца
мая. В Бет-Пак-Дале первую родившую самку мы добыли 14 мая. Бывают
отдельные случаи запоздалой беременности; В. П. Костин (1955) 29 июня
наблюдал беременную самку и новорожденных джейранят. В Монголии,
по данным А. Г. Банникова (1954), сроки размножения у джейранов
запаздывают по сравнению со среднеазиатскими на целый месяц и даже
больше. В норме взрослая самка приносит двух детенышей, молодые
по одному.1 2 Изредка, говорят, бывают тройни и даже, как исключение,
по четыре у одной самки.
Перед отелом самки обычно покидают табунки, уединяются и родят
где-нибудь в кустарнике, под обрывом, на дне старого русла и в других
укромных местах. Послед после родов самка съедает. Остатки его мы
часто находили в содержимом желудков убитых самок.
Вес новорожденных джейранов, по нашим данным из Бет-Пак-Далы,
составляет от 2.7 до 3.1 кг. Первые несколько дней джейранята беспо-
мощны, лежат, плотно прижавшись к земле, с подогнутыми ногами и вытя-
нутой вперед шеей (рис. 70). В это время они легко могут быть взяты
руками. Мать обычно пасется поблизости, но в предгорьях Джунгарского
Алатау самки на день покидают детенышей и выходят пастись в межгор-
ные долины (Антипин, 1941). Через 3—4 дня джейраненок начинает хо-
дить, а с 10—15-дневного возраста не отстает от матери и способен про-
бежать за ней значительное расстояние. Первых самок с детенышами
на ногах мы встретили 18 мая, но, как сообщил мне А. А. Слудский, самок
с молодыми можно встретить на пастбище редко раньше 15 июня, так
как они долго прячут детенышей, если даже последние уже могут бегать
за ними. С этого времени самки с молодыми вновь собираются в небольшие
группки.
Молоко у джейранов густое, сладковатого вкуса, по данным Г. Йшу-
нина (1956), содержит до 39% жира. Кормит самка молодых до конца
августа, но иногда можно встретить брошенных матерями джейранят уже
в конце июля. Растут джейранята сравнительно быстро и уже к июлю
могут достигать половинного роста взрослых. Рога у самцов начинают расти
ввозрастеоколо месяца. Молодые с матерями держатся иногда до 1—1 % лет.
Продолжительность жизни джейрана в естественных условиях не из-
вестна. В Лондонском зоологическом саду самка прожила 9 лет 4 месяца
и 2 дня (Flower, 1931).
Из врагов в первую очередь следует назвать волка. Волк не может
догнать быстроногого джейрана; поэтому обычный прием охоты этого
хищника — подкарауливание и неожиданный прыжок на жертву у водопоя
или на путях переходов, а также скрадывание из-за прикрытий при пере-
сеченном рельефе. В Таджикистане на Вахше и Пяндже джейран на во-
допоях иногда становится добычей подстерегающего его тигра (Флеров,
1935). Молодые джейранята гибнут от орлов, лисиц и даже хорьков.
Известны случаи нападения орлов на годовалых джейранов (Верещагин,
1939).
1 А. А. Слудский (1953), впрочем, сообщает о случаях серьезных повреждений и
даже гибели самцов после драк во время гона.
2 В Монголии или резко понижена плодовитость самок джейрана, или слишком
велика смертность молодняка, так как только 2.6% встреченных А. Г. Банниковым
(1951) самок имели по два джейраненка.
GAZELLA
327
Джейран является хозяином ряда внутренних и наружных napa3HTOBf
Из наружных паразитов следует отметить, во-первых, два вида подкож-
ных оводов: Pavlovskiata subgutturosae Grunin и Crivellia corinnae Grunin
(Грунин, 1949, 1956). Далее, с достоверностью установлено паразитиро-
вание пяти видов клещей: Rhipicephalus turanicus Pom., R. rossicus Jak.
et K.-Jak., R. pumilio Sch.. Hyalomma asiaticum Sch. et Schl. и H. plum-
beum Panz.1
Рис. 70. Новорожденный джейран (Gazella subgutturosa Giild.), центральная Бет-Пак-
Дала. (Фото автора).
Из прочих эктопаразитов на джейране найдены вошь, Linognathus
tibialis Pg., и пухоед, Damalinia appendiculata Pg.,1 2 паразитирующие и
на некоторых других копытных, в частности на африканских антилопах.
Из внутренних паразитов у джейрана установлено наличие около
двух десятков видов гельминтов (Соколова и Бондарева, 1948; Асадов,
1952). Наибольшее число их паразитирует в желудке и кишечнике, мень-
шее — в печени (печеночная двуустка, Fasciola hepatica L.), в брюшной
полости, подкожной клетчатке. Подавляющее большинство видов гель-
минтов, найденных у джейрана, являются общими с домашними жвачными
животными. Специфичных для него установлено только четыре вида.
О частоте встречаемости эндопаразитов можно судить по тому, что из
10 зверей, вскрытых И. Б. Соколовой и В. И. Бондарцевой (1948) методом
полных гельминтологических вскрытий, свободным от паразитов не ока-
залось ни одного.
Ни один из обнаруженных до сих пор у джейрана экто- и эндопара-
зитов серьезного патогенного значения для него, по-видимому, не пред-
ставляет.
1 Сведения, любезно сообщенные Г. В. Сердюковой.
2 Данные, любезно сообщенные Д. И. Благовещенским.
328
BOV I DAB
Иногда джейраны поражаются инфекционными заболеваниями от до-
машних животных через пастбища, водопои и воздух. В 40-х годах этого
столетия в Дангаринском районе Таджикистана джейраны поражались
и гибли в большом числе от ящура, свирепствовавшего среди домашних
овец. По сообщению ветеринарного врача овцесовхоза Дангара (Таджик-
ская ССР) тов. Полутона, болели и гибли джейраны в годы Великой
Отечественной войны также от овечьей оспы.
Однако наибольшим бедствием для джейранов, как и для других
копытных Средней Азии, является джут. Нередко среди зимы наступает
потепление, к тому же иногда сопровождаемое выпадением осадков.
Внезапно наступающее затем похолодание сковывает оттаявший снег,
покрывает его непроницаемой для копыт животных, особенно для таких
как джейран, коркой. Слоем льда и сосульками покрываются и торчащие
из-под снега растения. Животные оказываются лишенными корма и об-
речены на голодную смерть, если не сумеют быстро откочевать в местность
не подверженную джуту. Положение усугубляется, кроме холода, тем,
что образовавшаяся корка наста нередко не выдерживает тяжести жи-
вотных; последние, проваливаясь сквозь нее, не могут быстро передви-
гаться, режут острыми краями льда ноги, обессиленные или гибнут от
холода и голода, или легко становятся добычей волков. Случаи массовой
, гибели джейранов по этой причине общеизвестны. Они наблюдались
в южной Туркмении (Ишунин и Коровин, 1945). Зимой 1945/46 года
во время джута джейраны сотнями гибли в низовьях р. Или (Эверт,
i 1947).
Преследуемые голодом, звери заходилив населенные пункты в поисках
корма. Сильно истощенных (живой вес не превышал 12—17 кг), с изранен-
ными ногами, их давили бродячие собаки, расстреливали почти в упор
охотники и даже прирезывали ножом на лежках под кустами. Весной
десятками находили скелеты погибших от голода и холода живот-
ных.
К явлениям джута относятся и случаи гибели джейранов вследствие
глубокого снегопада. Джейраны не способны к тебеневке (добыче корма
из-под снега); уже снег в 20 см сильно затрудняет добывание пищи, осо-
бенно если снегопад сопровождается сильным ветром. О массовой гибели
джейранов во время снежного бурана на о. Барса-Кельмес сообщает
А. А. Слудский (1953). На следующий после бурана день находили де-
сятки трупов; некоторые из них были найдены среди кустов в позе стояния.
Известны также случаи гибели джейранов при переправе через реки по
неокрепшему льду.
Хозяйственное значение. Джейран в первую очередь ценен как
мясное животное. Мясо его считается вкусным, бывает жирным, ино-
гда не уступая мясу сайги. При правильной организации промысла
и охраны от истребления джейран может стать одним из средств исполь-
зования кормовых ресурсов пустынной и полупустынной зоны важным
дополнительным источником получения мяса. Еще в довоенные и годы
и особенно в годы Великой Отечественной войны были предприняты попыт-
ки плановых заготовок мяса джейранов (Зверев, 1948; Шнитников, 1936).
Из кожи джейрана выделывают замшу. Шьют из нее также одежду
и летнюю обувь. Однако главное ее применение, по В. П. Костину (1955), —
это изготовление бурдюков для воды, айрана и кумыса. Помимо того
что при этом экономятся шкуры домашних овец и коз, джейраньи бур-
дюки отличаются легкостью, эластичностью и не придают напиткам
неприятного запаха. Из шкур молодых джейранов изготовляются ко-
GAZELLA
329
жаные ведра, очень удобные при разъездах. Рога самцов находят при-
менение в качестве украшения.
Джейран, пойманный молодым, легко привыкает к человеку, обла-
дает кротким нравом и делается совсем ручным. Известны многочислен-
ные случаи содержания прирученных джейранов на свободе. Однако раз-
множение в домашней обстановке, по-видимому, не происходило и практи-
ческого значения разведение и содержание этого зверя не имеет.
Охота. Способы добычи джейранов разнообразны. Обычное скрады-
вание благодаря открытым местообитаниям и высокой чуткости зверя
почти не возможно. В некоторых местностях практикуют подход к джейрану,
прячась за верблюдом, вид которого не внушает зверю особого подозрения.
Один из распространенных способов охоты — подкарауливание зверя и
стрельба на водопое и тропах. На привычке джейранов ходить к водопою
определенными путями основана добыча их с помощью расставляемых
на тропах капканов (Колесников, 1952).
Травят джейранов с помощью среднеазиатских борзых (тазы). Нам
показывали в юго-восточных Каракумах тазы, с помощью которой за один
год ее владельцем было взято пятнадцать джейранов. Так как даже луч-
шая борзая редко может догнать джейрана, обычно применяют сменных со-
бак; с этой целью охоту организуют так, чтобы уже утомленного травлей
зверя направить к запасной группе собак, которая без труда его дого-
няет (Дементьев, 1935).
Местами в советской Средней Азии, но особенно, по Г. П. Дементьеву,
в Индии и Афганистане, еще сохранился способ охоты с помощью спе-
циально с этой целью содержимых и обученных хищных птиц — беркутов,
крупных ястребов и соколов. Подъехав насколько возможно на лошади
к джейрану, охотник пускает в угон ему птицу. Задача последней —
догнать, схватить и удержать до прибытия охотника зверя. Дрессировка
крупных соколов-балобанов и ястребов ведется так, чтобы хищник бил
зверя в голову и старался клевать его в глаза, так как удержать
схваченного за спину джейрана даже крупный балобан не в состоя-
нии.
Из групповых способов охоты на джейрана распространенным яв-
ляется охота нагоном. Найдя табун зверей, охотники делятся на две пар-
тии. Одна прячется с ружьями за какими-либо прикрытиями, а другая
очень осторожно и не спеша старается объехать с противоположной сто-
роны джейранов и направить их на притаившихся стрелков.
Г. П. Дементьев (1935) описывает варварский способ заганивания
джейранов в воронкообразный с широким входом и суженным выходом
загон, стороны которого составляют кучки земли, пугающие животных.
Группа всадников с криками гонит зверей к широкой стороне загона.
Попав в воронку, джейраны мчатся к узкому выходу на противополож-
ном конце и с разбега натыкаются на расставленные за ним в поперечный
ряд концами навстречу бегущим животным острые сухие камышины.
Бесспорно много животных гибло при этом бесполезно.
Не менее хищнический способ охоты, который развился и в Средней
Азии, и в Закавказье за последние десятилетия, — это преследование и
стрельба с автомобиля. Загнанные до полусмерти звери расстреливаются
почти в упор, нередко прирезываются ножами или давятся колесами
автомашины. Большое число уцелевших, но загнанных животных при
этом гибнет позднее в результате переутомления сердечной мышцы. Хотя
охота с автомобилей повсеместно запрещена, но, к сожалению, кое-где
она до сих пор практикуется почти открыто.
330
BOVIDAE
Позволю себе более подробно описать еще один, лучше известный
мне способ добычи джейрана также с помощью автомашины, но менее
вредный. Это — ночная охота с фарой. Применение ее основано на том,
что ослепленные направленным на них потоком яркого света (для этой
цели обычно приспосабливается автомобильная или мотоциклетная фара)
животные не в состоянии от него оторваться, как бы парализуются и под-
пускают на очень близкое расстояние подходящего из темноты стрелка.
В полной темноте (в лунную ночь охота не бывает удачной) на машине,
кроме шофера, выезжают два человека — фарящий и стрелок. Доехав
до места, где держатся джейраны, фары автомашины выключаются и
включается фара (рефлектор) в руках напарника стрелка. Машина дви-
гается малой скоростью, а фарящий плавно водит по сторонам и впереди
фарой, стараясь поймать в поле зрения джейранов. Заметить последних
довольно легко. Глаза их, обращенные в сторону света, сами начинают
светиться и обнаруживают животных.
Очень эффектное зрелище представляет стайка попавших в полосу
света джейранов. Вначале замечаешь несколько пар горящих фосфори-
ческим блеском глаз. Потом различаешь силуэты животных. Как зачаро-
ванные, смотрят они на приближающийся к ним источник света, пытаясь,
но зачастую не будучи в силах от него оторваться. В идеальных случаях
джейраны остаются неподвижны, пока не грянет выстрел, но обычно они
пытаются вырваться из полосы света, и иногда им это удается. Фарящий,
-заметив зверей, начинает «играть» фарой, слегка трясти ее, чтобы мигаю-
щим светом еще больше ослепить животных. Условным световым сигналом
он указывает шоферу направление, в котором должна двигаться машина,
и момент, когда она должна остановиться, в то же время стараясь не упу-
стить из поля зрения джейранов. В нужный момент, когда машина оста-
новится, а фарящий продолжает усиленно играть фарой, стрелок, на-
ходящийся в темноте, незаметно выскакивает с ружьем из машины и,
•ни в коем случае не попадая в полосу света, стороной бежит к зверям.
Последние подпускают стрелка очень близко, на несколько десятков
щагов, на верный выстрел картечью.
Успех охоты с фарой зависит, помимо обилия джейранов в районе
охоты, в первую очередь от искусства фарщика, от умения его вовремя
обнаружить зверя и удержать его в полосе света до подхода на должную
дистанцию автомобиля и стрелка. Большое значение имеет автомашина.
Лучше всего для этой охоты подходит бесшумная легковая или полу-
тонная машина. Тяжелая, сильно тарахтящая отпугивает большую часть
джейранов задолго до того, как на них скажется парализующее действие
•света.
Нужно сказать, что эта охота лишена высокого спортивного интереса.
При недостаточно высокой культуре охотников, она может превратиться
в простое избиение животных, нередко допускающих расстреливать себя
почти в упор. Но она имеет то важное преимущество, что при умелой
стрельбе при ней почти не получается подранков и загнанных до полусмерти
зверей. Кроме того, охота с фарой бывает весьма добычлива и поэтому,
при правильной ее организации и регулировании, может стать основным
Способом планового отстрела или заготовок мяса и шкур джейранов.
Джейран очень крепок на рану. В этом мы имели неоднократную
возможность убедиться во время экспедиции в Бет-Пак-Далу, где про-
водился отстрел джейранов для научных целей.
Даже при стрельбе из трехлинейной винтовки с телескопичес-
ким прицелом, к сожалению, много уходило подранков. Я уже
GAZELLA
331
упоминал о джейране, бежавшем со скоростью 60 км в час на трех
ногах. С перебитой брюшной аортой джейрануха пробежала около 100 м.
Пробитая пулей насквозь от промежности до основания шеи другая
самка, прежде чем свалиться, пробежала около 10 м. Еще более порази-
тельный случай произошел с самцом: после нескольких выстрелов, с ви-
сящими из разрыва брюшной стенки внутренностями, он продолжал дви-
гаться и ушел от нас, так как ни пешком, ни на медленном ходу машины
его догнать было нельзя, а условия местности (заросли боялыча, неров-
ности почвы) не допускали быстрого движения автомашины.
Gazella (Procapra) gutturcsa Gmelin — Дзерен, или зобатая газель
(рис. 71—73)
Gmelin J., 1760, Novi Comment. Petropol., V : 347—351, Tab. IX, X (первооп.,
изобр.); Pallas P. S., 1777, Spicilegia zoologica, XII : 46—60, tab. II, III (опис.,
изобр.); В a dde G., 1862, Reisen im Siiden von Ost-Sibirien, I, St.-Petersb. : 254—
261, Taf. XI, Fig. I (опис., биол., изобр.); Brooke V., 1873, Proc. Zool. Soc. Lon-
don : 546—547 (сист., опис.); Пржевальский H. M., 1875, Монголия и страна
тангутов, I, СПб., изд. Русск. географ, общ. : 18—21 (биол.); Черкасов А. А.,
1884, Записки охотника вост. Сибири, СПб., изд. А. Суворина : 517—521 (биол., ох.);
S с 1 a t е г Ph. and О. Thomas, 1897—1898, The book of antelops, III, London :
S3—88 (общ., синон., изобр.); К о п ы p и н С., 1907, Природа и охота, I : 1—15 (ох.);
Р о с о с k R., 1910, Proc. Zool. Soc. London : 892 (морф.); Hollister N., 1913,
Smithson Misc. Collect. LX, 19 : 531—532+1—2 (сист.); Lydekker R., 1914,
Catal. Ungulata mammals Brit. Mus. Ill : 34—40 (сист.,синон., изобр.); Рос ос к R.,
1918, Ann. Mag. Natur. Hist., II, 8 : 128 (морф.); Andrews R., 1924, Natur Hist.,
24, 2 : 155—159 (биол.); Andrews R., 1926, On the trail of ancient man, N. Y. —
London (цит. no Allen, 1940); Л у к а ш к и н А. С., 1927, Тр. Общ. изуч. Маньчжурск.
края, Харбин, Зоология, I : 8—19 (биол., ох.); Формозов А. Н., 1929, Матер.
Комисс. по исследов. Монгольск. и Танну-Тувинск. Респ., 3 : 97—98 (биол.); Те i 1-
hard de Chardin P. etR. Piveteau, 1930, Ann. Paleontol., 19 : 3—132
(эвол.); Каверзнев В. H., 1933, Полорогие фауны СССР и их добывание, М.,
КОИЗ : 17—21 (общ., изобр.); Булавин А., 1934, Охотник Сибири, 3 : 25 (распр.);
Дементьев Г. П., 1935, Дзерен, джейран, сайга, М., КОИЗ : 3—15 (общ., изобр.);
Лодаревский В. Б., 1936, Пробл. охотхозяйственн. акклим. в вост. Сибири,
Иркутск, ОГИЗ : 48 (распр.); В о h 1 i п В., 1938, Nova Acta Soc. Sci. Upsal. (ser. 4),
Xl, 2 : 1—54 (эвол.); Колосов A. M., 1939, Уч. зап. Моск, унив., 20 :171—174
(распр., биол.); Allen G., 1940, The mammals of China- and Mongolia, 2, N. Y. :
1210—1216 (общ., синон.); Мачульский С. Н., 1941, Тр. Бурят-Монгольск.
зооветинст., II : 120 (параз.); Фетисов А. С. и В. Н. Хрусцеле вский,
1948, Тр. Иркутск, гос. унив., сер. биол., III, 3 :14 (распр.); П а в л о в Е. И., 1949,
Промысловые звери Читинской обл., Чита, Обл. йздат. : 109—112 (общ., изобр.);
Скалой В. Н., 1949, Бюлл. Московск. общ. испыт. прир., Отд. биол., LIV, 3 :
7 (биол.); Грунин К. Я., 1950, Докл. Акад, наук СССР, LXXIII, 4 : 861—864
(параз.); Банн, и к о в А. Г., 1951, Бюлл. Московск. общ. испыт. прир., Отд. биол.,
LVI, 3:21—41 (распр., биол.); Янушевич А. И., 1952, Фауна позвоночн. Тувинск.
автономн. обл., Новосибирск, изд. Зап.-сиб. фил. Акад, наук СССР:62 (распр.);
Банников А. Г., 1954, Млекопит. Монгольск. Народи. Республ., М., изд. Акад,
наук СССР (Тр. Монгольск. комисс., 53) : 232—250 (общ., изобр.); Слудский А. А.,
1954, сборн. «Природн. очаговость заразных болезней в Казахстане», 2 : 81 (бол.).
Описание. Средней величины животное, но крупнее джейрана. Длина
тела от конца морды до корня хвоста у самцов от 105 до 148 см, высота
в холке 62—84 см (Банников, 1954). Самки несколько меньше (длина
тела 100—121 см, высота в холке 54—74 см). Основная длина черепа от 215
до 255 мм. Живой вес 16—24 кг, максимально до 30—32 кг (приведенные
цифры по литературе вызывают сомнение — вероятно они преуменьшены).
Телосложение легкое, стройное; ноги тонкие. Высота в крестце на 2—3 см
превышает высоту в холке. Голова производит впечатление более грубой,
чем у джейрана; передняя часть морды несколько расширена и вздута.
Верхняя губа покрыта волосами; от носового зеркала сохраняется лишь
332
BOVIDAE
узкая полоска голой кожи между нижними углами ноздрей и вдоль сред-
ней линии верхней губы. Радужная оболочка глаз темно-коричневая.
Уши сравнительно короткие, всего 9—12 см, с заостренными верхушками.
Рога имеются только у самцов; по сравнению с джейраном рога дзе-
рена более тонкие и короткие; длина их по изгибу не больше 25—28 см.1
Основаниями рога почти целиком расположены над глазами. Форма
рогов лировидная, но характер изгиба несколько иной, чем у G. subgut-
turosa Guld. На протяжении всей дистальной половины они сближены,
почти параллельные и только в вершинах образуют довольно крутой
изгиб в стороны (рис. 63, Б). Концы рогов загнуты несколько внутрь
и вверх, но не вперед, как у G. subgutturosa Guld. Цвет рогов светлый,,
грязно-серый. Поверхность роговых чехлов состоит из отставленных друг
от друга поперечных колец, более тесно расположенных у основания
рога. Только концы рогов на 5—6 см от вершин гладкие.
Шея не длинная, но тонкая. У самцов на нижней стороне шеи заметно
выступает сильно развитая гортань, от которой зверь получил прозвище
«зобатая газель». Тонкие конечности заканчиваются узкими копытами
с заостренными верхушками. Длина копыт по переднему краю: передних
33—39 мм, задних на 1—2 мм меньше. Длина копыт от заднего края пя-
точных мякишей до верхушек на передних ногах 45—53 мм, на задних
также на несколько миллиметров меньше. Хвост короткий от 7 до 11.5 см.
Добавочные (боковые) копыта длиною 12—15 мм, расположены высоко
над пяточными мякишами средних пальцев.
Волосяной покров мягкий, но на верхушках волос ломкий. Ясное
разделение его на остевой волос и подшерсток отсутствует. Зимний мех
густой и плотный; длина волос на спине достигает 5 см. Летом мех
более редкий и короткий, обычно около 2—3 см. Лоб и темя по-
крыты более длинным волосом, чем нос и щеки. Более длинные упругие
волосы по бокам верхней губы, у ноздрей, оттопыриваясь и загибаясь
концами книзу, образуют подобие усов и усиливают впечатление взду-
тости передней части морды. В задней части живота, в области вымени,
мошонки и препуция, волосы очень редкие, летом сквозь них просвечи-
вает кожа. Нижняя сторона хвоста и пространство вокруг заднепроход-
ного отверстия голые. Отдельные волосы на верхней части туловища
от основания светло-бурые, выше темнеют, но ближе к вершине обра-
зуют ясно отграниченное желтое кольцо; тонкие заостренные кончики
волос темно-бурого цвета. Волосы на светло окрашенных местах сплошь,
белого, а на темно окрашенных буро-коричневого цвета.
Свойственная большинству других видов газелей резкая разграни-
ченность светло и темно окрашенных участков на голове и боках туло-
вища у дзерена отсутствует. Верхняя часть носа от передних углов глаз
до ноздрей буро-коричневая. Лоб и темя желтовато-рыжие. Щеки свет-
лые, желто-песчаные, книзу постепенно светлеют. Средняя часть верх-
ней губы, нижняя губа, низ головы и шеи белые, с легким желтоватым
или сероватым оттенком. Уши снаружи одноцветны со лбом или немного
светлее, внутри желтовато-белые. Верх шеи желто-песчаный или рыже-
ватый, бока шеи более темные, с сероватым оттенком. Общая окраска
верха туловища, как и у джейрана, песчаная, но более светлая, в средней
части спины и крупа с сероватым оттенком, и темнеет по направлению
к хвосту. Бока туловища книзу светлеют и приобретают желтоватую
1 Аллен (Allen, 1940) почему-то приводит более высокие, но мало вероятные
цифры (ог 33 до 40 см).
GAZELLA
333
окраску. Свойственная многим газелям темная бурая полоса на границе
с белой окраской живота отсутствует. Нижняя сторона груди, живот,
нахи и внутренние стороны ног белые, иногда с желтоватым оттенком.
Околохвостовое зеркало большое, заходит на стороны ягодиц и на круп
выше основания хвоста; отграничивающей его с боков от окраски
туловища темной полосы нет. Передняя сторона ног коричнево-бурая
до самых копыт, но характерная для джейрана темная окраска простран-
ства между добавочными копытами и пятками копыт средних пальцев
Рис. 71. Дзерен {Gazella gutturosa Gmel.).
у дзерена отсутствует; этот участок у него одинакового с окружающими
частями, светлого желтоватого цвета. Хвост белый, кроме буро-корич-
невого конца. Часто, но не всегда, темная верхушка хвоста соединяется
с основной окраской туловища узкой рыжеватой полоской, разделяющей
в этих случаях околохвостовое зеркало в средней части на две половины.
К концу зимы вследствие выгорания и обламывания верхушек во-
лос окраска становится более светлой и тусклой, иногда с серо-голу-
боватым оттенком. Ясный половой диформизм в окраске отсутствует.
У молодых животных на первом году жизни серый оттенок верхней части
туловища выражен более интенсивно.
Предглазничные железы рудиментарны, а иногда, по-видимому, от-
сутствуют вовсе. Карпальные железы также лишь едва заметны в виде
слегка удлиненных и более грубых волос на передней стороне запястных
суставов. Межпальцевые железы имеются, но отличаются ли они суще-
ственно от таковых у других газелей не известно. Недостаточно ясно,
имеются ли у дзерена паховые железы; по некоторым, не совсем убеди-
тельным данным (Рососк, 1910, 1918; Lydekker, 1914; Allen, 1940) они
имеются, но на коллекционных шкурах их обнаружить не удается. Мо-
334
BOVIDAE
лочная железа имеет два соска. Паллас (Pallas, 1777) обнаружил у самца
дзерена впереди препуция на нижней стороне живота железу, сходную
с таковой кабарги (см. стр. 125), и дал ее изображение. Позднее описание
этой железы никем не было повторено.
Череп (рис. 72) с несколько удлиненной предглазничной частью;
длина последней от переднего края глазницы до конца межчелюстных
костей (prosthion) больше 133% длины заглазничной части черепа (от
заднего края глазницы до крайнего выступа затылочных мыщелков).
Ростральная часть черепа вздута; ширина входа в носовую полость редко
составляет меньше 2/3 расстояния от переднего края носовых костей до
prosthion. Носовые кости по сравнению с таковыми джейрана длинные,
не меньше 36% основной длины черепа. Наибольшая их ширина, за очень
редкими исключениями, составляет не больше 40% длины. Носовые
кости не имеют вырезок на переднем крае и медиальные их концы кли-
нообразно заострены. На границе носовых и лобных костей имеются
хорошо заметные выемки, так что наименьшая ширина лба между глаз-
ницами приходится ниже уровня слезно-лобного шва. Предглазничные
ямки на черепе развиты слабо, едва заметны. Роговые отростки тонкие
и сближены в основаниях. Расстояние между наружными краями по-
следних меньше межглазничной ширины лба. Слуховые пузыри меньше
вздуты, чем у джейрана; расстояние от нижнего края пузыря до верх-
него края костного' слухового прохода меньше вертикального диаметра
глазницы. Последний премоляр нижней челюсти (Р4) имеет хорошо раз-
витый передне-внутренний бугорок; поэтому передняя долинка на его
внутренней стороне замкнута и образует лунку этого зуба (рис. 68, Б).
История и географическое распространение (рис. 73). Несколько
видов газелей, относимых к подроду Procapra Hodgson, и в настоящее
время, и в предшествующей их истории связаны с территорией Централь-
ной Азии. Здесь, по-видимому, произошло отщепление их предка от
общего ствола газелей еще в верхнем плиоцене. В слоях этого возраста
в Китае, наравне с типичными газелями, найдены остатки форм, у которых
еще сохраняются свойственные джейрану, хорошо развитые предглаз-
ничные ямки, но последний премоляр имеет форму, характерную для
подрода Procapra. Из известных ископаемых’ форм, ближе стоящими
к предполагаемому исходному предку, следует считать верхнеплиоце-
новую G. sinensis Teilh. et Piv. (Teilhard de Chardin et Piveteau, 1930)
и нижнеплестоценовую G. paragutturosa Bohl., из северного Китая (Boh-
lin, 1938). В настоящее время область распространения дзерена захва-
тывает Монгольскую Народную Республику с прилегающими районами
советского юго-восточного Алтая, Тувы и Забайкалья, затем ряд про-
винций и областей Китая — Ганьсу, Сычуань, возможно Синьцзян,
Внутреннюю Монголию, Тибет к югу до Ладака и Сиккима.
В пределах Советского Союза постоянно и еще в довольно значитель-
ном числе дзерены держатся в Чуйской степи (Кош-Агачский аймак
Горно-Алтайской автономной области) к югу от р. Елангаша до Сайлю-
гемского хребта. Добывался также в предгорьях Чуйских Альп (Чаган-
Бургазы). Основными местообитаниями являются урочища Бельтыры,
Тошантай, Саргабу, Саганалу. Через последние три, как наиболее пони-
женные части Сайлюгемского хребта, происходят кочевки дзеренов из
Монголии в СССР и обратно (Булавин, 1934; Колосов, 1939).
Водятся дзерены и, по-видимому, также не так редки в южной части
Тувинской автономной области, в котловине Успа-Нур за хребтом Танну-
Ола, возможно также в Монгун-Тайге (Янушевич, 1952).
Рис. 72. Череп Gazella gutturosa Gmel.
33 в
BOVIDAE
Третье и последнее место в Советском Союзе, где более или менее
Постоянно обитают дзерены, является крайний юго-восток Забайкалья,
Борзинский и Быркинский районы Читинской области. По Фетисову
и Хрусцелевскому (1948), северо-западная граница здесь «проходит
сейчас по линии: северные склоны горы Чир-Абогайтуйский (Аргунский
хребет), падь Николай-Торм, окрестности Березовой гривы, ст. Мациев-
ская, оз. Чиндант, Федькин брод на р. Борзе, окрестности с. Кулусутай
и с. Баин-Цаган». Кроме того, ежегодно большие стада дзеренов на зиму
заходят из Монголии в Даурские степи между рр. Снопом и Аргунью,
занимая пространство до 50.5° с. ш.; часть из них остается здесь и на
лето. Е. И. Павлов (1949) встречал летом самок с молодыми в окрест-
ностях оз. Хара-Нур. Появляются иногда даже в Агинской степи (По-
даревский, 1936; Павлов, 1949). В 1911 г. случайно забежавший дзерен
был убит под Иркутском (Лукашкин, 1927). По свидетельству Палласа,
в конце XVIII в. область распространения доходила якобы до побережья
Тихого океана; в настоящее же время Большой Хинган является восточ-
ным пределом обитания дзерена (Alien, 1940).
В пределах Монгольской Народной Республики дзерен населяет
обширные пространства в центральной и восточной частях страны. Рас-
пространение его здесь очень сложно. Нет его в юго-восточных и север-
ных районах республики, к югу от Монгольского Алтая, а также в Хан-
гае. Крайне неравномерно и далеко не повсеместно встречаются дзе-
рены в северо-западной части МНР, хотя и доходят до государственной
границы с СССР в Сайлюгемском хребте. Южная граница массового рас-
пространения в республике с востока на запад идет приблизительно по
линии Дариганг—Алтан-Ширэ—Мандал—Гоби—южные склоны Хангая
и Монгольского Алтая. На севере ареал дзерена из Монголии заходит
в пределы советского Алтая и Тувинскую автономную область. Далее
к востоку северная граница его распространения проходит несколько
южнее 50° с. ш. от верхнего течения р. Тэс к г. Мурэн, захватывает верх-
нее течение р. Идэра, огибает с юга Хангай, около 103° с. ш. вновь резко
поворачивает к северу, по остепненной долине р. Орхона и его притоков
поднимается почти до 49-й параллели. Далее к востоку по правобережью
р. Толы на широте Улан-Батора проходит через верхнее течение р. Ке-
рулен и огибает с юга и востока хребет Хэнтэй и, пересекая верхнее те-
чение р. Ульдзи близ впадения в последнюю р. Дзучи, уходит на север
в пределы СССР приблизительно на 112° в. д. В зимнее время ареал дзе-
рена в Монголии несколько расширяется как в горной, так и в северной
его части. В районах верхнего течения Селенги, Орхона (севернее 49° с. ш.),
Онона и Керулена наблюдаются лишь отдельные забеги дзеренов (Бан-
ников, 1954).
Подвиды. Внутривидовая систематика G. gutturosa не разработана.
До сих пор, кроме типичной расы, описана в качестве подвида лишь
G. gutturosa altaica Holl, из Монгольского Алтая (Hollister, 1913), отли-
чающаяся якобы большими размерами околохвостового зеркала, широ-
ким черепом и более широко раскинутыми на концах рогами.
Биология. В противоположность джейрану, обитателю пустынных
и полупустынных ландшафтов, дзерен больше связан со степями; явно
избегает пустынь, особенно каменистых. В пределах коренного ареала,
в Монголии, как отметил еще Н. М. Пржевальский (1875), дзерены за-
нимают «лучшие пастбища пустыни». А. Г. Банников (1954) в летнее
время чаще всего встречал их в ковыльных степях и ковыльковых полу-
пустынях (около 75% встреч). Значительно реже дзерены встречались
GAZELLA
337
в ковыльно-вострецовых и злаково-солянковых степях, равно как явно
они избегают степей полынных, горных и разнотравных. Необходимо,
однако, иметь в виду, что на Алтае дзерены держатся как раз в горных
степях с разнотравной растительностью, а в Забайкалье населяют разное
травные Даурские степи. В районе Хайлара (северо-восточный Китай,
к западу от Большого Хингана) стациями дзеренов являются песчаные
степи, растительность которых состоит из вострецов, ковыля, полыни
и гречихи, а в понижениях и на дюнах произрастают еще кое-где верба,
шиповник, яблонька и черемуха (Лукашкин, 1927). Разнотравные степи,
вплоть до южного лесостепья, дзеренами используются в Монголии и За-
байкалье главным образом зимой.
Рис. 73. Распространение Gazella gutturosa Gmel.
1 — постоянный ареал; 2 — отдельные забеги.
Рельеф местообитаний предпочитается равнинный, но не избегают
дзерены пологих склонов и широких долин. Для Монголии А. Г. Банни-
ков (1954) определяет амплитуду колебания высоты мест обитания от
600 до 1900 м. На Алтае, как уже отмечалось, дзерены живут в условиях
степных горных плато, в при сезонных миграциях переходят даже через
наиболее пологие места Сайлюгемского хребта (Колосов, 1939). Кустар-
ников и высокотравной степи дзерены, за исключением самок в период
отела, избегают.
Выбор мест обитания в зимний период определяется наличием корма
и характером снежного покрова. В Монголии, по А. Г. Банникову (1954),
первую половину зимы дзерены держатся в ковыльных и ковыльково-
ковыльных степях. По обеднении этих пастбищ от усиленного выпаса
и с выпадением глубокого снега, звери перемещаются в малоснежные
и более богатые кормами районы. Часть их откочевывает на север в по-
лосу разнотравных степей, например в Даурские степи 'Забайкалья;
часть, наоборот, подается к югу в полосу южных полупустынь и даже
пустыню, куда летом эти звери никогда-не заходят (Банников, 1954).1
Около Хайлара в суровые зимы дзерены предпочитают держаться в кот-
ловинах с высокими песчаными дюнами и даже в сосновых рощах,
спасаясь от ветров и буранов (Лукашкин, 1927). О заходах зимою в по-
1 Г. Радде (Radde, 1862) причину откочевок дзеренов из Монголии в степи Забай-
калья видел в недостатке питьевой воды вследствие замерзания источников и от-
сутствия снега. А. Г. Банников (1954) считал несостоятельным подобное объяснение.
22 Фауна СССР, И. И. Соколов
. 338
BOVIDAE
лосу лесостепья и изредка даже леса сообщалось и из других мест, но,,
как правило, это вынужденные стации, где дзерену трудно спасаться от
своего главного врага — волка. В исключительных случаях, когда на
равнинах лежит глубокий снег, дзерен, в нормальных условиях избе-
гающий хоть сколько-нибудь пересеченного рельефа, вынужден под-
ниматься в горы для того, чтобы прокормиться на обдуваемых ветром
склонах. Регулярные миграции дзеренов весною, в июне—начале
июля из Монголии в Чуйские степи, а осенью еще по чернотропу обратно,
имеют место и на советском Алтае (Колосов, 1939).
В некоторых случаях протяженность сезонных перемещений дзеренов
исчисляется, по-видимому, сотнями километров. В большинстве райо-
нов такие перемещения проходят спокойно и почти незаметно, но известны
и массовые миграции этих животных. Картину весенней миграции много-
тысячного стада дзеренов в Монголии наблюдал Эндрюс (Andrews, 1926).
Массовый характер зимние перемещения зверей носят в годы джута
(см. стр. 328). Громадные табуны дзеренов вынуждены в этих случаях
делать длинные и порой беспорядочные переходы в поисках мест с до-
ступным кормом. Именно в такие годы наблюдаются наиболее много-
численные скопления дзеренов в северо-восточном Китае и в Забайкалье.
Стадное чувство у дзерена развито еще сильнее, чем у джейрана.
Как и у последнего, наибольшая концентрация стад имеет место зимой.
Летом дзерены держатся группами от одного-двух до нескольких десят-
ков экземпляров. Размер как летних, так и зимних стад зависит от общей
численности животных в данном районе. На Алтае, например, летом
дзерены держатся группами всего около 10 голов (Колосов, 1939); не-
больших размеров достигают летние табунки также в окрестностях Хай-
лара и в Забайкалье. Средний размер табуна для этого периода в усло-
виях Монголии А. Г. Банников (1954) определяет в 26 голов, максималь-
ный — не более 120 голов. Близкие к этим цифры в свое время приводил
Н. М. Пржевальский (1875). Летние табунки состоят из самок, сеголеток
и приплода предшествующего года. Взрослые самцы держатся самостоя-
тельными группами и присоединяются к остальным зверям только осенью
перед началом гона.
К осени, с сентября, дзерены начинают табуниться. Максимальных
размеров стада, по-видимому, достигают к декабрю. Средний размер
зимних табунов в Монголии, по А. Г. Банникову (1954), около 100 голов,
но очень часто стада насчитывают несколько сотен, а то и тысяч живот-
ных. Особенно бывают велики скопления дзеренов в неблагоприятные
по кормовым условиям моменты, когда десятитысячные стада их вынуж-
дены концентрироваться на участках с более или менее доступным кор-
мом. Зимние стада — смешанные, состоят из самцов и самок. Крупные
табуны удерживаются, по-видимому, всю зиму и весну, распадаются
только перед самым отелом. Эндрюс (Andrews, 1924) наблюдал в июне
стада дзеренов, насчитывавшие от шести до восьми тысяч голов.1
Соотношение самцов и самок в популяции дзерена равно приблизи-
тельно один к одному.
Суточный цикл дзеренов известен плохо. Ночью, по-видимому, от-
дыхают. Выпасаются, как и джейраны (по крайней мере летом), в утрен-
ние и дневные часы. В жаркое время дня лежа пережевывают жвачку.
1 Этот автор, впрочем, считает, со слов местных жителей, что дзерены дважды
собираются крупными стадами — в сентябре перед гоном и весной перед отелом. По
окончании брачного сезона будто бы происходит распадение крупных стад на более
мелкие группы.
GAZELLA
339
На водопой ходят в конце ночи или на рассвете. Лежки устраивают на
открытом месте, вероятно в целях обеспечения необходимого кругозора.
Лежат группами неподалеку друг от друга. Зимой, прежде чем лечь,
разрывают копытами снег до земли. В холодную погоду и в бураны сби-
ваются в кучу и ложатся вплотную друг к другу (Лукашкин, 1927).
Кормятся, находясь все время в движении. В Монголии иногда за-
ходят в отары домашних овец. А. С. Лукашкин (1927) наблюдал их в со-
обществе дроф, спокойно разгуливавших среди пасущихся дзеренов.
Пищей дзерену служат травянистые растения, произрастающие в ме-
стах их обитания.
В противоположность многим другим копытным, состав пищи срав-
нительно мало меняется по сезонам. Летом основу питания, по наблю-
дениям и исследованиям А. Г. Банникова (1954) содержимого желуд-
ков, составляют разнообразные злаки: ковыль, ковылек, вострец. В же-
лудках большей части зверей в значительном количестве находились
также остатки многокорешкового лука (Allium polyrrhizum Turcz.).
Вострец составляет основную пищу дзерена в степях северо-западного Ки-
тая (Лукашкин, 1927). По-видимому, довольно охотно, но в меньшем
количестве поедаются полыни, солянки и некоторые виды разнотравья —
бобовые и их семена, тонконог, карагана, прутняк, пижма и другие.
В центральной части ареала ковыль составляет главную пищу дзерена
и в зимний период, но в северных степях в это время большее участие
в питании этого зверя принимают виды разнотравья.
При наличии на пастбище достаточного количества зеленого корма
дзерены, по-видимому, подолгу могут обходиться без питья. А. Г. Бан-
ников (1954) часто встречал их в местах, расположенных за десятки и даже
за сотни километров от открытых водоемов. При наличии последних
дзерены не отказываются от их посещения.
У дзерена хорошо развиты как зрение и слух, так и обоняние. На
открытом месте, по Н. М. Пржевальскому (1875), подкрасться даже к не-
пуганому табуну ближе чем на 500 шагов очень трудно. Развитой смыш-
леностью дзерен не обладает. Напуганные выстрелами, подряд несколько
раз, могут набегать на стрелков, от пуль которых только что погибли
их соседи. Испуганный бежит, делая время от времени крупные прыжки
высотой до 2 м, а длиной до 5 м и больше. В быстроте бега и выносливости
превосходит даже джейрана. Как установлено преследованиями с авто-
мобиля, дзерен может развивать скорость до 75 км в час, причем не сдается
даже после часового бега (Лукашкин, 1927). Как й у джейрана, харак-
терна привычка бежать некоторое время параллельно, как бы вперегонки,
а затем пересечь путь впереди, иногда почти перед самой машиной.
Голос дзерена слышен редко и, по Н. М. Пржевальскому (1875), со-
стоит из довольно громкого отрывистого рявканья.
Весенняя линька у дзеренов протекает в мае—июне. Зимпий наряд
звери получают в ноябре.
Время наступления половой зрелости не известно, но, вероятно, оно
то же, что и у джейрана, т. е. в возрасте около 1 /4 лет. Спаривание про-
исходит в период с конца ноября по первую половину января. Самцы
в это время держатся с самками и гоняются эа ними. Во время гона они
сильно возбуждены, издают громкий рев, слышимый чуть ли не за 2 км.
Между самцами часто происходят драки, но ожесточенного характера,
по-видимому, не носят. Имеются сообщения о том, что самец будто бы
кроет только одну самку (Лукашкин, 1927). Однако, это мало вероятно,
принимая во внимание стадный образ жизни этого вида.
22*
340
BOVIDAE
Время отела, по А. М. Колосову (1939) и А. С. Лукашкину (1927),
конец мая—июнь, но А. Г. Банников (1954) встречал дзеренов с новорож-
денными не раньше первых чисел июля.
По Эндрюсу (Andrews, 1924), громадные стада дзеренов перед самым
отелом широко рассыпаются по равнине с широким горизонтом, но даже
во время родов самки держатся группами и рожают неподалеку друг от
друга. По другим данным, самки для отела выбирают, наоборот, участки
с более пересеченным, чем в обычное время рельефом, мелкосопочники,
места с зарослями высокой травы дырсуна (Пржевальский, 1875). Новоро-
жденные помещаются в понижениях под кустами или в зарослях дыр-
суна. Противоречивы данные, касающиеся плодовитости дзерена. По
А. С. Лукашкину (1927) и Е. И. Павлову (1949), самки приносят в норме
по два, реже по одному, иногда по три детеныша. По А. Г. Банникову же
(1954), в Монголии двойни являются не правилом, а исключением. Ана-
логичные сведения А. М. Колосов (1939) приводит для Алтая. Вес ново-
рожденного, по данным последнего автора, около 3 кг. У добытого им
экземпляра этого возраста прорезавшимися зубами были только четыре
резца (по-видимому, две первые пары). Первые дни после рождения мо-
лодые лежат в укрытии, плотно прижавшись к земле и вытянув по ней
шею. Через 4—8 дней они начинают следовать за матерью и уже в этом
возрасте могут бежать со скоростью 25 миль в час (Andrews, 1924). Самки
кормят детенышей молоком, по-видимому, до брачного сезона.
Развиваются молодые дзерены сравнительно быстро. В возрасте 4—5
месяцев у самцов начинают расти рога.1 К годовалому возрасту почти
достигают размеров взрослых.
Главным и почти единственным врагом дзерена является волк. Так
как догнать и взять на открытом месте здорового дзерена волк не в со-
стоянии, то добычей последнего являются главным образом больные,
ослабевшие и отставшие звери. Однако Н. М. Пржевальский (1875)
упоминает о групповых облавах волков на дзеренов. Легче жертвой
подкарауливающих волков дзерены становятся в мелкосопочниках и ку-
старниковых или высокотравных степях. Молодые за первые дни жизни
погибают иногда также от лис, корсаков и крупных хищных птиц.
По-видимому, бывают массовые падежи дзеренов от повальных ин-
фекционных заболеваний, но последние еще совсем не изучены. Н. М. Прже-
вальский (1875) упоминает о падеже дзеренов зимою 1871 г., не ука-
зывая причины. Возможно, что это был один из многочисленных падежей
в период джута, происходящих при таких же обстоятельствах, что и у джей-
рана. В Монголии во время ящурных эпизоотий ящуром поражаются
и дзерены (Слудский, 1954).
Из наружных паразитов дзерена известен, во-первых, подкожный
овод (Przevalskiana aenigmatica Grun.), паразитирующий в личиночной
стадии в подкожной клетчатке животного на спине по обеим сторонам
хребта. Далее, на дзеренах из Монголии найден упоминаемый еще Палла-
сом (Pallas, 1777) носоглоточный овод (Pharyngomyia dzerenae Grun.; Гру-
нин, 1950). А. М. Колосов (1939) на волосах добытого им молодого дзе-
рена обнаружил в большом количестве муху-кровососку (Melophagus sp.).
Из внутренних паразитов у дзеренов пока зарегистрированы в За-
байкалье эхинококки (Echinococcus granulosus Batsch) в печени, сердце
и легких; затем на печени встречены личинки тонкошейной финны (Cys-
ticercus tenuicollis Rud.); в скелетной мускулатуре, при сильных степе-
1 По коллекционным материалам ЗИН АН СССР.
GAZELLA
341
нях поражения и в сердечной мышце встречаются личинки бычьего со-
литера (Cysticercus bovis Cob.; Мачульский, 1941). Патогенное значение
для животного могут иметь лишь мухи-кровососки и сильная степень
пораженности эхинококкозом.
Хозяйственное значение. В Монгольской Народной Республике
хозяйственное значение дзерена велико. Благодаря высокой численности
он является основным промысловым видом копытных. В годы Великой
Отечественной войны проводились плановые заготовки мяса дзеренов
(Банников, 1954). Переработка их туш проводилась, по-видимому, еще
до войны на мясокомбинате в Улан-Удэ Бурят-Монгольской АССР (Ма-
чульский, 1941). Мясо дзерена обладает хорошими вкусовыми качествами.
По словам А. С. Лукашкина (1927), оно «больше напоминает баранину,
чем косулю». В отличие от джейрана, у дзерена откладывается довольно
большое количество подкожного жира. Особенно жирны дзерены бывают
поздней осенью перед началом брачного сезона.
Шкура дзерена в выделанном виде идет на изготовление дох, которые
шьются мехом наружу и отличаются, по А. С. Лукашкину (1927), лег-
костью и сравнительно теплы. Из выделанной в виде замши кожи мон-
голы шьют верхнюю одежду, а кроме того, замша идет на пошивку пере-
метных сум, кисетов, чехлов для трубок и тому подобных изделий. Рога
в свое время находили спрос в Китае и употреблялись в качестве рукоя-
ток для особых ножей, служивших одновременно и шилом.
Охота. Способы добывания дзеренов разнообразны. Скрадывание
хотя и применяется, но редко и трудно вследствие высокой чуткости
зверя и открытому характеру мест обитания. Иногда практикуется скра-
дывание дзеренов под прикрытием верблюда, к виду которого они более
доверчивы и привычны (Пржевальский, 1875). Применяется охота с лов-
чими птицами, в принципе ничем не отличающаяся от таковой на джей-
рана (см. стр. 329). В прежнее время монголы для добычи применяли
борзых собак.
В местах, где дзеренов еще много и они не сильно напуганы человеком,
одними из распространенных и добычливых способов до последнего вре-
мени являлись описанные А. С. Лукашкиным(1927) охоты в наездку и на-
гоном. Основаны они целиком на использовании сильно развитого стад-
ного инстинкта дзерена. «Завидев вдали пасущееся стадо, охотники едут
рысцой, как бы мимо его. Не доезжая до табунка метров 400, они круто
поворачивают и во весь опор несутся к дзеренам. Последние в это время
удивленно разглядывают надвигающуюся лавину. Охотники же, при-
близившись на незначительную дистанцию, останавливаются и начинают
обстреливать стадо. Увидев такую развязку, дзерены бросаются в сторону,
но отбежав сотни две-три метров, они останавливаются и ждут повторе-
ния того же приема. После двух-трех таких обстрелов стадо разбегается
в стороны. Другой способ охоты несколько разнится от первого тем, что
участвуют обязательно две партии, нагоняющие друг на друга встре-
чающиеся табуны дзеренов. В то время, когда первая партия налетает
на табун, вторая уже прячется в засаде на пути следования дзеренов,
убегающих от выстрела первой партии. Вторая партия обстреливает
„навернувшийся" на нее табун и своими выстрелами заворачивает его
на первую, а та в свою очередь на вторую. Так повторяется несколько
раз, пока дзерены сообразят, что есть и другие направления для отступ-
ления».
Своеобразный способ охоты нагоном, практикуемый в Чуйской степи
на Алтае, описывает А. М. Колосов (1939). В основе его лежит привычка
342
BOVIDAE
дзеренов, спасаясь, бежать вереницей по кругу и возвращаться на преж-
нее место. Поэтому один из группы охотников на коне, не слишком быстро,
едет позади уходящего стада, а другие прячутся за какими-либо укры-
тиями около того места, где по их расчетам должны пройти, сделав круг,
звери. При всякой охоте нагоном важно уменье очень осторожно,
не напугав, направить табун в сторону залегших стрелков, а для по-
следних умело рассчитать направление и место, где должны пройти
звери. Облавную охоту на дзеренов в Забайкалье описал С. Копырин
(1907).
В Забайкалье прежде, в годы массового зимнего нашествия дзеренов
из Монголии, группы охотников, оцепив стадо, старались загнать его
на лед, на котором звери не могут двигаться, и без единого выстрела
вырезывали их ножами.
В последние десятилетия всюду, в том числе и в Монголии, все большее
применение находит незаконная охота на дзеренов с автомобиля. Не-
допустимость этого способа была указана при перечислении способов охоты
на джейранов. При охоте на дзерена он, по существу, носит еще более
истребительный характер, так как благодаря крепости этого зверя на
рану большое количество подранков погибает бесполезно.
[Подсемейство CAPRINAE —|КОЗЛООБРАЗНЫЕ
Характеристика подсемейства. Рога, за редкими исключениями,
имеют и самцы, и самки, но у последних они по сравнению с самцами
развиты слабо. Основания рогов расположены над глазницами или не-
много сдвинуты назад за края последних. Роговые стержни содержат
внутренние полости, часто распространяющиеся до верхушек. Форма
рогов различная: серповидные, дугообразные, свернутые улиткообразно,
гетеронимно скрученные, иногда с двойным изгибом или в виде загнутых
на вершинах крючков. Поверхность рога несет часто от одного до трех
продольных ребер (килей).
Носовое зеркало, если имеется, небольшое. Верхняя губа покрыта
волосами, за исключением узкой вертикальной полоски на ее средине.
Ноздри расположены близко друг к другу. Расстояние между ними
меньше расстояния между краем верхней губы и ноздрей.
Предглазничные и межпальцевые железы имеются или отсутствуют.
Хвост короткий, концом не достигает скакательных суставов. Молочная
железа с 2 или 4 сосками.
Длина зароговой части черепа может быть больше или меньше длины
лба. Лобные кости средней длины или длинные; их длина вдоль сагит-
тального шва редко составляет меньше 36% основной длины черепа.
Передние края глазниц нередко располагаются позади уровня корен-
ных зубов. Предглазничные ямки и этмоидальные щели часто отсутствуют.
Основная затылочная кость (basioccipitale) короткая, широкая, упло-
щенная, почти квадратная; в передней части обычно не уже, чем в зад-
ней (рис. 76). Передний край хоан расположен впереди или позади уровня
заднего края верхних моляров. Слуховые пузыри небольшие, мало или
совсем не вздутые, всегда сжаты с боков.
Резцы узкие, равномерно убывающие в ширине от внутренней пары
к клыкам (рис. 61, Б, Г). Добавочные бугорки и столбики на задне-
коренных зубах (по крайней мере на нижних) отсутствуют. Последний
CAPRINAE
343
переднекоренной зуб нижней челюсти (Р4) имеет замкнутую переднюю
долинку (рис. 68, Б) (исключение — род Saiga Gray).
Географическое распространение. Европа, Азия, северная
Африка и Северная Америка с прилегающими арктическими остро-
вами.
Классификация. Подсемейство Caprinae включает средних и круп-
ных в пределах семейства Bovidae животных. Для большинства их харак-
терно крепкое тяжелое сложение, короткость плюсны и пясти; в ске-
лете передних конечностей метаподий уступают по длине не только пред-
плечью (лучевой), но у большинства и плечевой кости. По образу жизни
Caprinae — жители по преимуществу горных ландшафтов. Исключение
представляют сайга (Saiga tatarica Ь.)иоронго (Paniholopshodgsoni Abel.),
приспособившиеся к обитанию на равнинах. В связи с этим последние
сохранили ряд примитивных особенностей (длинные метаподий, длин-
ный зароговый череп), но приобрели ряд черт своеобразной специали-
зации. Современные роды подсемейства Caprinae мною (Соколов, 1953)
группируются в пять триб.
1) Saigini — Сайга. Наиболее характерным отличием от всех других
триб является преобразованный в виде короткого хобота конец морды
с трубкообразными ноздрями (рис. 75). Самки рогов не имеют. Имеются
предглазничные, межпальцевые, паховые и карпальные железы. Молоч-
ная железа имеет четыре соска. В черепе сильно укорочены носовые
кости, еще в эмбриональный период сливающиеся без следа с лобными
костями. В связи с этим вход в носовую полость резко увеличен; в об-
разовании его переднего края принимают участие слезные кости (рис. 76).
Последний нижний премоляр (Р4) имеет открытой переднюю и замкнутой
заднюю долинки. Распространение: восточная Европа, Сред-
няя и Центральная Азия. Триба включает один монотипический род Saiga
Gray.
2) Pantholopini — Оронго, или чиру. Отличительными от других
триб признаками являются длинные, значительно превосходящие длину
головы вертикальные рога и наличие в верхней челюсти только двух
премоляров. Самки рогов не имеют. Межпальцевых, предглазничных и
карпальных желез нет, но паховые железы имеются. Молочная железа
имеет два соска. Носовые кости короткие, но не редуцированы; длина
их не превышает 36% основной длины черепа. Вход в носовую полость
несколько расширен, но в образовании его переднего края слезные кости
участия не принимают. Последний нижний премоляр (Р4) имеет замкну-
тую переднюю долинку. Распространение: Тибет. Триба вклю-
чает один род и вид — Panthalops hodgsoni Abel.
3) Nemorhaedini — Горалы и cepao. Характерной отличительной
чертой от всех остальных триб подсемейства является выпуклый (реже
прямой) профиль черепа на границе носового и лобного отдела, а также
мало, по сравнению с другими родами, выступающие в стороны глаз-
ницы. Самки имеют рога. Межпальцевые и предглазничные железы
имеются, но последние иногда недоразвиты. Паховых и карпальных
желез нет. Молочная железа имеет четыре соска. Носовые кости не ре-
дуцированы; длина их редко составляет меньше 35% основной длины
черепа. Вход в носовую полость иногда несколько увеличен, но слезные
кости и в этом случае лишь на очень небольшом протяжении принимают
участие в образовании его переднего края. Последний премоляр нижней
челюсти (Р4) имеет замкнутую переднюю долинку. Распростра-
нение: восточная и юго-восточная Азия, включая южную часть Даль-
344
BOVIDAE
него Востока. Триба включает роды Nemorhaedus Н. Smith (горалы) и
Capricornis Og. (серао).
4) Caprini — Козлы и бараны. Рога (за редкими и случайными исклю-
чениями у некоторых видов рода Ovis) имеют и самцы, и самки. Форма
рогов разнообразная, но если они длинные, то не стоят вертикально.
Предглазничные, межпальцевые и паховые железы иногда имеются, но
часто отсутствуют. Карпальных желез нет. Молочная железа, за редкими
исключениями, имеет два соска. Носовые кости не укорочены; длина их
обычно не меньше 36% основной длины черепа. Вход в носовую полость
не расширен и слезные кости участия в образовании его переднего края
не принимают. Передний премоляр нижней челюсти (Р4) имеет замкну-
тую переднюю долинку. Распространение: средняя и южная
Европа, Азия, Северная Америка. Триба включает роды: Capra L.
(козлы), Ovis L. (бараны), Hemitragus Hodgs. (тары), Rupicapra Blainv,
(серна) и Oreamnos Rafin. (американская снежная коза).
5) Ovibovini — Овцебыки и такины. От всех других триб подсемей-
ства отличаются, прежде всего, большей величиной; основная длина
черепа больше 350 мм. Рога есть и у самцов, и у самок с своеобразным
двойным изгибом. Предглазничные железы имеются, но межпальцевые,
паховые и карпальные отсутствуют. Молочная железа имеет 4 соска.
В черепе характерна сильная укороченность теменной части чешуи
затылочной кости, представленной на дорзальной поверхности черепа
узкой полоской впереди затылочного гребня, а также узость мозговой
коробки. Ширина последней составляет не свыше 25% основной длины
черепа (соответствующие показатели у всех других родов подсемейства
больше). Носовые укорочены, но не редуцированы, длина их составляет
около 34—35% основной длины черепа. Вход в носовую полость несколько
расширен, но слезные кости лишь передне-верхним углом, и то не всегда,
на очень небольшом протяжении принимают участие в образовании его
переднего края. Последний нижний премоляр (Р4) имеет замкнутую перед-
нюю долинку. Распространение: крайний север Америки с при-
легающими островами; восточная Азия (восточные Гималаи, Юньнанщ.
Сычуань, Ганьсу и Шэньси в Китае). Триба включает роды Ovibos Blainv.
(овцебык) и Budorcas Hodgs. (такины).
В фауне СССР представлены роды: Saiga, Nemorhaedus, Rupicapra,,
Capra и Ovis.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ ПОДСЕМЕЙСТВА
КОЗЛООБРАЗНЫХ (CAPRINAE) ФАУНЫ СССР И МОНГОЛЬСКОЙ
НАРОДНОЙ РЕСПУБЛИКИ
1 (2). Передняя часть головы (морда) сильно горбоноса и вздута. Конец
ее вытянут в виде небольшого хобота с открывающимися на его
конце круглыми, тесно сближенными ноздрями (рис. 75). У самцов
рога желто-белого, воскового цвета. Самки безрогие и имеют четыре
хорошо развитые соска. Носовые кости очень короткие и без следа
слиты с лобными костями. Носовой вход на черепе очень длинный;
расстояние от переднего края межчелюстных костей до переднего кон-
ца носовых не меньше чем в три раза превосходит ширину носового
входа. В скелете пястная кость длиннее плечевой 1.........
...............................Saiga — Сайга (стр. 345).
1 О способе измерения плечевой кости см. стр. 352.
SAIGA
345
2 (1). Конец морды не вздут и не образует на конце хобота. Ноздри
щелевидные. Рога серо-бурого и темно-рогового цвета. Самки имеют
рога, а если безрогие, то имеют только два развитые соска. Носовые
кости не укорочены и имеют видимый шов с лобными костями.
Расстояние от переднего края межчелюстных костей до передних
концов носовых значительно меньше тройной ширины носового
входа. Пястная кость короче плечевой.
3 (6). Рога круглого или продольно-овального сечения и на поверхности
не имеют продольных ребер (килей). Самки имеют хорошо развитые
четыре соска. Резцы брахиодонтные; на них хорошо заметна шейка
на границе корня и коронки (рис. 61, Б).
4 (5). Рога вертикальные или наклоненные немного вперед. На концах
загнуты назад в виде крючков (рис. 82). Позади рогов имеются
шишковидные железы. Верхний профиль черепа в предроговой
части ясно вогнутый. Верхнечелюстные кости соприкасаются с носо-
выми ................................Rupicapra — Серна (стр. 384).
5 (4). Рога загибаются назад и в стороны, почти не поднимаясь над
уровнем лба и не загнуты на концах в виде крючков (рис. 79). Же-
лез позади рогов нет. Верхний профиль черепа в предроговой
части прямой или выпуклый. Верхнечелюстные кости не сопри-
касаются с носовыми.............Nemorhaedus — Горалы (стр. 374).
6 (3). Рога у самцов треугольного, грушевидного или линзовидного се-
чения, на поверхности имеют от одного до трех острых или закруг-
ленных ребер (килей). Самки имеют только два хорошо развитых
(функционирующих) соска. Резцы гипсодонтные, долотовидные;
граница между корнем и коронкой (шейка) на них не заметна
(рис. 61, Г).
7 (8). Самцы, а часто и самки, имеют на нижней стороне головы бороду
(пучок удлиненных волос). Нет предглазничных, паховых и меж-
пальцевых желез (по крайней мере на задних ногах). Самцы имеют
у основания хвоста снизу железы, придающие животным специфи-
ческий запах. На черепе не имеется ни предглазничных ямок, ни про-
дольного гребешка на лицевой поверхности слезных костей. Рас-
стояние между теменными гребешками в самом узком месте не пре-
вышает 16% основной длины черепа . . Capra — Козлы (стр. 400).
8 (7). Пучка удлиненных волос на нижней стороне головы (бороды) нет.
Предглазничные и паховые железы, а также межпальцевые железы
на всех четырех конечностях имеются. Желез около хвоста ни
у самок, ни у самцов нет. На черепе имеются в той или иной мере
развитые предглазничные ямки и продольный гребешок на лицевой
поверхности слезных костей. Расстояние между теменными гребеш-
ками в самом узком месте больше 16% основной длины черепа.
......................................... Ovis— Бараны (стр. 475).
Род SAIGA GRAY —САЙГА
Родовые признаки — см. выше (стр. 343).
В современной фауне род представлен одним видом Saiga tatarica L.
Saiga tatarica Linnaeus — Сайга, или сайгак (рис. 74—78)
Gmelin J., 1760—1761, Nov. Comm. Acad. Petropol., V (1760) : 345—346,
VII (1761) : 39—47 (опис., изобр.); Linnaeus C., 1766, Syst. Nat., ed. 12, I : 97
(опис.); П а л л а с П. C., 1773—1786, Путешествие по разным провинциям России-
346
BOVIDAE
ской Империи, т. I (1773) : 318—319+331, т. II, кн. 2 (1786) : 117—118 (распр.);
Т| о лето й| А., 1842, Жури, коннозаводства и охоты, II, 5 : 51—75 (ох.); Э в ер-
с м а п н Э., 1850, Естеств. ист. Оренбургск. края, II, Казань : 257—262 (распр.,
биол.); G 1 i t s с h L., 1865, Bull. Soc. Imp. Natur. Moscou, XXVIII : 207—245 (опис.,
биол.); Murie J., 1870, Proc. Zool. Soc. London : 451—503 (анат.); G 1 i t s c h L.,
1873, Verhandl. Leop. Carol. Deutsch. Akad. Naturforscher., XXXVI : 1—21, Taf. I—
III (анат.); Анучин Д. H., 1875, Природа, 3 : 201—222 (общ.); Б о п л а н Г.,
1890, Описание Украины, сборн. «Мемуары, относящиеся к истории южной Руси»,
вып. I, Киев : 365 (ист., распр.); N е h г i n g А., 1891, Neuen Jahrb. Mineral., Geol.,
Palaontol., 2 : 107—155 (эвол.); Черский И. Д., 1891, Опис. коллекц. послетре-
тичн. млекопит., собранных Новосибирск, эксп., СПб., изд. Акад, наук : 263—274
(ист., распр.); S с 1 a t е г Ph. and О. Thomas, 1897—1898, Book of Antelopes, III :
29—41 (общ., синон., изобр.); G r e v ё С., 1898, Korrespondenzblatt Naturfor. Vereins,
Riga, XLI : 109—112 (распр.); Туркин H. В. и К. А. Сатун ин, 1904, Звери
России, Bovidae, М. : 491—499+583—591 (общ., синон., изобр.); Бер г Л. С., 1905,
Природа и охота, 5 : 1—5 (распр., биол.); Гербер штейн С., 1908, Записки о мо-
сковитских делах, перев. А. И. Малеина, СПб. : 175 (ист., распр.); АлферакиС. Н.,
1910, Семья охотников, 5 : 128 (распр.); Динник Н. Я., 1910, Звери Кавказа, I,
Тифлис : 206—215 (общ.); Lydekker R., 1914, Catal. Ungulate Mammals Brit.
Mus., Ill, London : 12—16 (сист., синон., изобр.); Хоменко И. П., 1914, Тр.
Ставропольск. общ. изуч. Сев.-Кавказск. края, III, 1 :19—29 (эвол.); Пачоский И.,
1915, Бюлл. Харьковск. общ. любит, прир., 1 : 71 (альбинизм); So ffe 1 К., 1922,
в кн. «Lebensbilder aus d. Tierwelt Europas», IV : 1—12 (общ., изобр.); В о-
р о б ь е в К. А., 1929, Охотник, 2 : 30 (распр., биол.); Плигинский В. Г.,
1929, Изв. Курск, общ. краевед., 1/2 : 42—43 (распр.); Бажанов В. С., 1930,
Бюлл. Средне-Волжск. краев, станц. защиты раст. за 1926—1928 гг., Самара : 65
(распр.); DegerbolM., 1932, Medd. Dansk. Geol. Foren, 8 : 175—184 (ист.); Глад-
ков H. А. и Г. В. Никольский, 1933, Сборн. тр. Гос. зоол. муз. при МГУ,
II : 15 (распр.); Каверзнев В. Н., 1933, Полорогие фауны СССР, М.—Л., КОИЗ :
9—16 (общ., изобр.); Орлов Е. И. и Г. А. К а й з е р, 1933, Уч. зап. Саратовск.
гос. унив., X, 2 : 142 (распр.); Беляева Е. И., 1935, Тр. Комисе, по изуч. четвер-
тичн. периода, IV : 305 (ист.); Дементьев Г. П., 1935, Дзерен, джейран, сайга,
М.—Л., КОИЗ : 30—44 (общ., изобр.); Шнитников В. Н., 1936, Млекопит.
Семиречья, М.—Л., изд. Акад, наук СССР : 121—127 (общ., изобр.); Громова В. И.,
1937, Докл. Акад, наук СССР, XVI, 6 : 351—352 (ист.); Liester С. W., 1937,
Bull. Zool. Soc. N.Y., 40:23—24 (общ., изобр.); Frick Ch., 1937, Bull. Amer.
Mus. Natur. Hist., 69 : 546 (ист.); П i д о и л i ч к а I. Г., 1938, Матер, до вивчення
минулих фаун УРСР, I, Ктв, вид. Акад, наук УРСР : 122, 126, 127, 141 (ист.); Б е-
ляева Е. И., 1939, Бюлл. Моск. общ. испыт. прир., Отд. геол., XII, 6 : 88—89
(ист.); К о л о с о в А. М., 1939, Уч. зап. Моск, унив., 20:174 (распр.); Rey-
nolds S., 1939, Monograph Brit, pleistocene mamm., vol. Ill, pt. VI, London : 1—
65 (ист.); Слудский A. A., 1939, Сборн. тр. Гос. зоол. муз. при Моск. гос. унив.,
V : 22—24 (распр.); Доброхотов В. И., 1940, Астраханск. гос. заповеди.,
М., изд. Главн. управл. по заповеди. : 71 (распр.); Туркевич Б. Г., 1940, Тр.
Узбекск. гос. унив., нов. сер., 2 : 1—13 (анат.); Антипин В. М., 1941, Млекопит.
Казахстана, III, Алма-Ата, Казогиз : 54—64 (общ., изобр.); Гептнер В. Г. и
А. Н. Формозов, 1941, Сборн. тр. Гос. зоол. муз. Моск. гос. унив., VI : 70—72
(хоз. знач.); Mohr Е., 1943, Zool. Garten, 15 : 175—185 (общ., изобр.); Гюльм-
с и р я н II. Г., 1944, Тр. Ереванск. зоопарка, I—II : 75—81 (биол.); Harper F.,
1945, Extinct and vanishing mammals of Old world, N. Y. : 677—682 (ист., распр.);
Банников А. Г., 1946, Докл. Акад, наук СССР, LI, 5 : 397—399 (сист.); А д о-
л ь ф Т. А., 1948, Охрана природы, 2 : 61—93 (распр.); Б а б е н ы ш е в В. П., 1948,
Природа, 6 : 63—64 (распр.); Б е р г г р и н А. П., 1948, Тр. Алма-Атинск. ветзоо-
инст., V : 333—345 (морф.); Гептнер В. Г., 1948, Докл. Акад, наук СССР, LX,
4 : 709—712 (синон.); Громов В. И., 1948, Тр. Инет. геол, наук, 64 : 435 (ист.);
Зверев М. Д., 1948, Тр. Алма-Атинск. гос. заповеди., VII : 145—147 (бег.); Ц а л-
к и н В. И., 1948, Докл. Акад, наук СССР, LIX, 3 : 605 (ист.); Грунин К. Я.,
1949, Докл. Акад, наук СССР, LXIV, 4 : 603—606 (параз.); Соколова И. Б.
и др., 1949, Изв. Акад, наук Казахск. ССР, сер. паразитол., 7 : 91—94 (параз.);
А д о л ь ф Т. А., 1950, Охрана природы, 10 : 73—82 (биол. распр.); В асенкоЕ. П.,
1950, Тр. Гос. заповеди. Барса-Кельмес, I : 38—115 (общ.); Колесников И. М.,
1950, Природа, 10 : 64—67 (биол.); Лавровский А. А., 1950, Охрана, природы,
10 : 83—87 (биол., распр.); Карачаровский В. В., 1951, Мат. по археол.
СССР, 21 : 250 (ист.); Пидопличко И. Г., 1951, О ледниковом периоде, II,
Киев, изд. Акад, наук УССР : 63—66 (ист., распр.); Соколов С. С., 1951, Вести.
Акад, наук Казахск. ССР, 3 : 143—151 (биол.); Формозов А. Н., 1951, Пробл.
SAIGA
347
физич. геогр., XVI : 152—160 (хоз. знач.); Адольф Т. А., 1952, сборн. «Методы
учета числ. наземн. позвоночн.», М., изд. Акад, наук СССР : 245—249 (учет); Ива-
нов В. В., 1952, Природа, 8 : 120—121 (распр., биол.); К и р и к о в С. В., 1952,
Изв. Акад, наук СССР, сер. географ., 6 : 31—48 (ист.); Л оды женская В. И.,
1952, Учен. зап. Карело-Финск. гос. унив., IV, 3:17—41 (анат.); Алекпе-
рова Н. А., 1953, Изв. Акад, наук Азербайдж. ССР, 4 : 65—70 (эвол., ист.); Афа-
насьев А. В. и др., 1953, Звери Казахстана, Алма-Ата, изд. Акад, наук Казахск.
ССР : 479—484 (общ., изобр.); Р а л л ь Ю. М., 1953, Учен. зап. Виол-почв, факульт.
Ростовск. н/Д. гос. унив., XIX, 3 : 115—126 (распр.); Слудский А. А., 1953,
Тр. Инет. зоол. Акад, наук Казахск. ССР, II : 11—15 (биол.); Соколов И. И.,
1953, Тр. Зоол. ипст. Акад, наук СССР, XIV : 184—187 (морф., эвол.); Читан С. А.,
1953, Изв. Акад, паук Армянск. ССР, биол. и сельскохоз. науки, т. 6, № 8 : 87—91
(анат.); Адольф Т. А., 1954, Учен. зап. Московск. гос. педагог, инет. им. Потем-
кина, XXVIII : 247—256 (биол.); Банников А. Г., 1954, Млекопит. Монгольск.
Народи. Республ., М., изд. Акад, наук СССР (Тр. Монгольск. комисс., 53) : 220—232
(общ., изобр.); КульпинП. И., 1954, Природа, 4 :102—104.(распр., биоп.); Стро-
ганова А. С., 1954, Тр. Зоол. инет. Акад, наук СССР, XVI : 83—84 (распр.);
Воршев В. Н., 1955, сборн. «Рационализ. охотн. промысла», 4 : 44—46 (учет,
хоз. знач.); Слудский А. А., 1955, Тр. Инет. зоол. Акад, наук Казахск. ССР,
IV : 18—55 (распр., биол., хоз. знач., изобр.); Даль С. К., В. М. Гусев и
С. Н. Бедный, 1956, Тр. Научно-исслед. противочумн. инет. Кавказа и Закав-
казья, I : 430—437 (распр., гибель); Соколов И. И., 1958, Ох. и ох. хоз., 9 : 21—24.
Описание. Животное средней величины. Длина тела от 104 до 144 см.
Высота в холке у самцов 70—-83 см, у самок 63—74 см. Живой вес самцов
осенью 37—60 кг, самок 29—37 кг. Основная длина черепа от, 190 до
235 мм.
Телосложение несколько неуклюжее, напоминающее домашнюю овцу.
Массивное бочкообразное туловище покоится на невысоких, тонких
ногах. Профиль спины слегка выпуклый. Высота в крестце приблизи-
тельно равна высоте в холке. Впечатление неуклюжести усиливается
непропорционально большой головой необычной формы. Последняя
по длине (29—45 см) может превосходить даже длину шеи. Передняя
часть головы (морда) сильно горбоноса и вздута. Конец ее вытянут
в виде срезанного впереди небольшого хобота. На конце последнего
открываются широкие круглые, тесно сближенные ноздри. В спокой-
ном состоянии хобот у старых животных может свешиваться вниз на
несколько сантиметров, а при ходьбе и беге пассивно раскачиваться вправо
и влево. При обнюхивании и беспокойстве хобот напрягается, вытяги-
вается еще больше, наверху его при этом ясно обозначается продольная
бороздка (рис. 75). Обладая большой подвижностью, он часто поводится
из стороны в сторону, то вытягивается, то втягивается; в последнем
случае кожа на нем сморщивается в складки. Основу вздутой части морды
и хобота составляют мягкие ткани: сильно развитые носовые хрящи,
соединительная ткань, мускулатура.1 Вся наружная поверхность верх-
ней губы (кроме очень узкой срединной полоски голой кожи) и края
ноздрей покрыты волосами. Большие, с сильно выступающими вверх
надбровными дугами глаза имеют темно-коричневую радужную оболочку.
Зрачок глаз продольно-овальный.
Уши короткие, длиной 7—8 см, на концах закруглены.
Рога имеются только у самцов, длиной у взрослых экземпляров от
28 до 37 см. На черепах самок, на месте рогов имеются лишь небольшие,
не выше 5 мм костные бугорки, покрытые сверху кожей. Основания
рогов расположены почти целиком над глазницами и отставлены друг
от друга на несколько сантиметров. Форма рогов неправильно лировид-
1 Анатомические особенности носа сайги см. в работах В. И. Ладыженской (1952)
и С. А. Читина (1953), а из старых — в работе К. Глича (Glitsch, 1873).
348
BOVIDAE
ная. Они слабо наклонены назад; отходя от черепа до половины длины
расходятся слегка в стороны, затем начинают сближаться, перед верши-
нами направляются параллельно друг другу или даже вновь расходятся;
а кончиками загнуты несколько внутрь. Характерна асимметрия рогов;
изгиб одного рога редко является зеркальным отображением другого,
особенно в верхней половине. Цвет рогов у взрослых самцов желтовато-
белый, восковой. У оснований он имеет буроватый оттенок, а самые
кончики, особенно у молодых, иногда темнее. Поверхность роговых чех-
Рис. 74. Сайга (Saiga tatarica L.), самец.
лов несет до 20—22 сильно выступающих колец, отставленных друг от
друга на 1—2 см. Только верхняя треть или четверть рогов гладкая, без
колец, да верхние кольца представлены лишь поперечными бугорками
на передней стороне рога. Поперечное сечение рогов почти круглое, но
продольный диаметр несколько больше поперечного.
Шея не длинная, но тонкая. В спокойном состоянии она держится
горизонтально и голова не поднимается выше, уровня спины и холки,
усиливая сходство сайги с домашней овцой. Только при возбуждении
и настороженности сайгак поднимает голову кверху, но и тогда она не
держится так гордо и высоко, как у джейрана. Конечности тонкие, зад-
ние с саблистым поставом (подогнутые в скакательных суставах). Копыта
низкие, но длинные, с сильно развитыми мякишами; высота копыт, й
передних и задних, по переднему краю от 32 до 39 мм; длина копыт от
заднего края пяточных мякишей до верхушек на передних ногах от 55
до 66 мм, на задних на 3—10 мм меньше. Добавочные копыта небольшие,
около 10—13 мм. Хвост тонкий и короткий, около 13—18 см, снизу го-
лый. .7 ' ' . ’.1 .
SAIGA
349
Летний волосяной покров короткий (длина волос 1.5—2 см), плотно
прилегающий к коже, лоснящийся, упругий. Зимний мех значительно
гуще и длиннее, состоит из довольно грубых волнообразно извитых
волос; длина их на спине в это время может достигать 5—6 см, а по
Е. П. Басенко (1950) даже 10 см. Кончики волос заострены, но как в лет-
нем, так и в зимнем меху вследствие хрупкости волоса часто обломаны.
Ясное разделение волос на ость и подшерсток отсутствует. На морде и
ногах волос короткий; по сравнению со спиной он несколько короче
и мягче также на животе. В зимнем меху волосы удлинены на щеках
Рис. 75. Сайга (Saiga tatarica L.), самка в летней шерсти.
(Фото В. И. Ерещенко).
ниже глаз, а также на нижней стороне шеи, где они достигают 10—15 см
и образуют подобие гривы от подбородка до грудной клетки. Несколько
удлинены (до 8 см) и собраны в виде торчащего вверх гребешка волосы
на конце хвоста. Кожа на нижней стороне хвоста, вокруг анального
отверстия и в пахах голая и зимой и летом.
Общая окраска верха и боков тела летом вскоре после линьки яркая
желто-рыжая, иногда с буроватым оттенком. Отдельные волосы от осно-
вания светло-бурые, в средней части несколько темнеют, а к вершинам
желтеют; тонкие заостренные их верхушки темно-бурого цвета. Бока
головы, ноги снаружи и хвост окрашены одноцветно с туловищем. Верх
носа и конец морды более светлые, у старых животных даже белые, а лоб
и темя буровато-коричневые со значительной примесью волос белого
цвета. Вдоль средней линии спины от холки до корня хвоста
тянется узкая темная буровато-коричневая полоса. Последняя, впрочем,
всегда ясно выражена лишь в области поясницы и крестца, где более
длинные, по сравнению с окружающими участками, волосы при испуге
и возбуждении животного могут подниматься дыбом. Нижняя сторона
шеи, груди, живот, внутренние стороны ног, а также промежность и
ягодицы желтовато-белого цвета.
К зиме окраска постепенно светлеет, приобретая бледный, буровато-
песчаный оттенок. К концу зимы, в результате выцветания волос и обла-
мывания темно окрашенных верхушек она становится еще более свет-
350
BOVIDAE
лой, белесой или соловой, с буровато-розоватым оттенком. Иногда, но
далеко не всегда, удлиненные волосы на щеках, по бокам шеи, а также
темя и затылок имеют темно-бурую окраску с примесью волос белого
цвета.
Морда, лоб и уши зимой, особенно у старых животных, желтовато-
белого цвета. Одинаковую с летней окраску сохраняет; четко выделяется
темная продольная полоса над крестцом. Низ груди, внутренние части
ног, живот и ягодицы светлого, чисто белого цвета. Светлая окраска за-
хватывает часть наружной стороны бедер и иногда заходит даже на круп
выше хвоста. Граница между светлыми и более темно окрашенными уча-
стками выражена нечетко. Молодые до года окрашены несколько более
интенсивно. Чем старше зверь, тем больше начинают преобладать в окраске
светлые тона, особенно зимою, когда издали сайгак кажется белым.
Разницу в окраске между самцами и самками подметить не удается.
Изредка встречаются альбиносы (Пачоский, 1915) и меланисты.
На голове имеются небольшие предглазничные железы, выделяющие
пахучий секрет буроватого цвета, склеивающий основания растущих
в области железы волос и придающий им сероватый оттенок. Предглаз-
ничные железы представляют впячивания кожи, заметные лишь с ее
внутренней стороны в виде продольно-овальных мешечков, диаметром
около 10 мм, со сметанообразным содержимым желтовато-белого цвета
и специфического кисловато-сырного запаха. На поверхности морды
железы открываются круглым отверстием, в которое с трудом входит
тонкий зонд на глубину около 8 мм. На передних ногах расположены
карпальные железы, обозначенные снаружи пучками несколько удли-
ненных волос пониже запястных суставов. На всех конечностях имеются
межпальцевые железы, открывающиеся щелью на передней стороне
выше копыт. Глубина межпальцевых желез — около 25—30 мм. В глу-
бине межпальцевых желез у сайгаков, убитых в мае и июне, я находил
сухой крошковатый секрет желтого цвета, но без запаха. Имеются
также паховые железы в виде воронкообразных впячиваний кожи, глу-
биною около 4 см, по бокам от молочной железы и мошонки. Молочная
железа имеет четыре соска, но передняя пара недоразвита и слабо функ-
ционирует.
Череп сайги (рис. 76) по форме и устройству сильно отличается от
черепа других полорогих. Наиболее характерной его чертой являются
сильно укороченные и приподнятые вверх носовые кости. Последние,
по-видимому, еще в утробном периоде сливаются без следа с лобными
костями; у взрослого зверя они представляют заостренные на конце
и направленные вперед и вверх рудименты. Ростральная часть черепа
впереди коренных зубов вытянута, сжата с боков и на самом конце ото-
гнута книзу. Вследствие этого, вход в носовую полость не широкий, но
очень длинный; длина его (от prosthion до передних концов носо-
вых костей) раза в три-четыре певосходит ширину. В образовании перед-
него края носового отверстия, помимо межчелюстных, верхнечелюст-
ных и носовых костей, значительное место занимают также слезные
кости. Межчелюстные кости массивные, но короткие; длина их носовых
отростков немного длиннее, а часто дажее короче нёбных отростков.
Благодаря тому, что обонятельный лабиринт вместе с носовыми костями
приподнят кверху, вентральный носовой вход сильно расширен и зани-
мает большую часть носовой полости.
Профиль лобно-носовой части черепа вогнутый. Большие трубко-
образные глазницы сильно выступают в стороны и вверх над вдавленной
Рис. 76. Череп Saiga tatarica L.
352
BOVIDAE
между ними плоскостью лба. Внутренние полости (пазухи) в лобных ко-
стях развиты очень слабо и даже в основания роговых отростков не за-
ходят. Предглазничные ямки на черепе имеются, но неглубокие и с по-
логими краями. Этмоидальные щели отсутствуют. Характерно для черепа
сайги наличие выступающих гребешков вдоль задней части сагитталь-
ного лобного и венечного швов. Передний край хоан расположен позади
уровня боковых выемок заднего края костного нёба и уровня заднего
края верхнего зубного ряда.
Заднекоренные зубы сильно гипсодонтные, верхние — с узкими сти-
лями на наружной поверхности. Переднекоренные сильно редуцированы;
в нижней челюсти их обычно только два. Длина ряда переднекоренных
верхней челюсти составляет не больше одной трети длины всего зубного
ряда. Последний премоляр нижней челюсти имеет открытую переднюю
долинку. Резцы гипсодонтные, долотовидные; граница между корнем и
коронкой заметна слабо.
В скелете конечностей длина пястной кости превосходит длину пле-
чевой кости (от вершины суставной головки до средины дистального
суставного блока).
История и географическое распространение (рис. 77). Происхо-
ждение и ранние этапы эволюции сайги не известны. Остатки, несомненно
принадлежащие этому роду, появляются только в плейстоцене. На тер-
ритории СССР наиболее ранние находки обнаружены в составе хазарской
фауны Нижнего Поволжья (средний плейстоцен), в средне-и верхнепалео-
литических стоянках Украины и Крыма, Северного Кавказа и Сибири
(Громов, 1948). Судя по степени примитивности в ряде особенностей,
отхождение предка сайги от общего ствола подсемейства Caprinae должно
было произойти еще в третичный период (в плиоцене или даже миоцене).
Современный ареал сайги занимает часть европейской территории
Советского Союза к востоку от правобережных степей низовьев Волги,
полупустынную зону Казахстана и Средней Азии до северо-западной Мон-
голии включительно, Прежде он был гораздо шире и включал всю степ-
ную полосу Евразии от Атлантического до Тихого океана. В плейстоцене
сайга доходила до юго-западной Франции, Бельгии, Польши. Остатки
ее найдены даже на Британских островах (Greve, 1898; Хоменко, 1914;
Reynolds, 1939; Mohr, 1943). На севере Европы она доходила до низовьев
Камы (Адольф, 1948), а в Азии обнаружена в составе верхнеплейстоце-
новой фауны Новосибирских островов. Бесспорно принадлежащие сайге
остатки найдены на Аляске (Frick, 1937).
Степная зона европейской части СССР не так давно составляла часть
коренного ареала сайги. Остатки ее найдены во многих местах юга Украины:
в среднечетвертичных отложениях (палеолит) на берегу Куяльницкого
лимана, там же при раскопках трипольского поселения около с. Усатого
(первое тысячелетие до нашей эры), на месте греческой колонии Ольвия
(Бугский лиман) в слоях, датированных первыми веками нашей эры,
в торфяниках около Херсона и в других местах (Пидопличко, 1938,
1951). Широко сайга была здесь распространена уже в историческое время.
В Молдавии, например, она была истреблена в начале XVIII в. (Harper,
1945). Вконце XVIII в. сайгаки водились в степях между Днепром и Южным
Бугом.
Судя по находкам в древних городищах, в XIII в. встречались около
Киева и Харькова (Пидопличко, 1951). В середине XVI в., по С. Гербер-
штейну (1908), они обитали на степных равнинах по Днепру, Дону и
Волге. В приднепровских степях встречал сайгаков в первой половине
SAIGA
353.
XVII в. Боплан (1890), В середине XVII в. сайгак в числе других зве-
рей указывается для Чугуевского и Змиевского лесостепных районов под
Харьковом. Водились будто бы они еще в первой половине XVIII в. даже
в Воронежской губернии (Плигинский, 1929) и даже Курской (Greve,
1898), но, по-видимому, их там не было уже к концу XVIII в. В начале
XVIII в. сайгаки «еще встречались возле Умани, Новой Сечи, в бассейнах
рр. Тащлыка, Синюхи, Мертвой Воды, Ингула, Ингульца, Кожной,
Анчахрака, Карачахрака» (Кириков, 1952).1
Рис. 77. Распространение Saiga tatarica L.
1 — XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше;
4 — отдельные заходы в настоящее время; 5 —места акклиматизации.
Область распространений сайги в свое время включала и Крым. По-
мимо уже упоминавшихся палеолитических находок (Громов, 1948),
остатки сайгака были найдены при раскопках Неаполя Скифского,
в слоях II в. до и. э.—II в. н. э. (Цалкин, 1948). О существовании этого
вида в Крыму в более поздние исторические времена ничего не известно.
К началу XIX в. к западу от р. Дона сайга была вытеснена или истреб-
лена.1 2
Междуречье Волги и Дона в их нижнем течении стало крайним запад-
ным районом постоянного ее обитания. В доисторические времена сайга
была распространена не только на Северном Кавказе, но и в Закавказье.
Четвертичные ее остатки найдены при раскопках палеолитической стоянки
1 Подробные данные о былом распространении сайги в степях европейской части
Союза имеются в статьях К. Греве (Greve, 1898) и Т. А. Адольф (1948). К сожалению,
некоторые из приводимых ими данных,, по-видимому, не вполне достоверны.
2 Уже несколько десятков лет сайга акклиматизирована в зоопарке Аскания-
Нова, где содержится в большом загоне вместе с рядом других копытных.
23 Фауна СССР, И. И. Соколов
354
BOVIDAE
Ильской к юго-западу от Краснодара (Громова, 1937), а также в биту-
минозных отложениях Бинагадов около Баку (Алекперова, 1953).
В 60-х годах прошлого столетия, по Л. Гличу (Glitsch, 1865), ареал
сайги в Волго-Донском междуречье образовывал треугольник, вершина
которого находилась около Царицына, стороны образовывали берега Дона
и Волги, а основанием служил Маныч. Сальские и Манычские степи
были местами концентрации сайгаков на зимовках, откуда летом они
распространялись к северу почти до Сарепты, а иногда и Царицына. В 80-х
годах сайгаки изредка показывались поздней осенью и зимою в гирлах
Дона (Алфераки, 1910).
В самом конце прошлого и быть может в первых годах этого столетия
сайгаки были обычны в Кумо-Манычских степях к югу от Маныча по
р. Кугульте, — например, добывали их в окрестностях Ставрополя
(Динник, 1910).
В начале нынешнего столетия область обитания сайги в правобережье
нижней Волги к атастрафичесви сокращалась и к 20-м годах ограничивалась
Прикаспийской низменностью к востоку от Ергеней и к северу от р. Кумы
до северных границ Астраханской губернии. Благодаря принятым при
Советском строе мерам охраны с полным запретом вылова и отстрела за
последние десятилетия численность поголовья сайги здесь сильно воз-
росла; она стала массовым зверем. Последние годы в Астраханской
области проводится организованный промысел сайгаков; только
в ноябре и декабре 1958 г. здесь заготовлено около 150 тыс. туш сайга-
ков. Одновременно вновь расширяется и ее ареал. Сайга появилась
в тех местах, где ее не было уже в течение ряда десятилетий. В насто-
ящее время она обитает, по Т. А. Адольф (1950), во всех районах
Астраханской области (только в более густо населенном Прикаспийском,
сайга бывает набегом). Распространена сайга в настоящее время в ряде
районов Сталинградской области: в Малодербентовском, Сарпинском,
Ворошиловском, Котельниковском. Северную границу распространения
здесь Т. А. Адольф проводит по 48.5° с. ш., но отдельные забеги наблю-
даются вплоть до Сталинграда. В наибольшем числе сайгаки держатся
в северной части Астраханской и южных районах (в первую очередь,
по Лавровскому (1950), в Мало-Дербентовском) Сталинградской области.
К югу численность зверей, вследствие бедности кормов и недостатка воды,
заметно меньше. К берегам Волги сайгаки, вследствие постоянного при-
сутствия там людей, не подходят ближе 40—70 км. На территории Астра-
ханского заповедника появляются только при случайных заходах (Доб-
рохотов, 1940).
В смежном с Астраханской областью Ставропольском крае сайгаки
в настоящее время держатся в северных и восточных районах: Степном,
Апанасенковском, Арзгирском, Туркменском и Левокумском (Бабенышев,
1948; Адольф, 1950). В Ростовской области они появились в 1947 г. и по-
следние годы стали нередкими в Заветинском и Ремонтненском районах
(Ралль, 1953). Несколько позднее стали встречаться в Зимовниковском,
Орловском. В отношении последнего я располагаю сведениями, собран-
ными на месте в 1953 г. Сайгаки появились здесь после суровой зимы
1949/50 года и с тех пор встречаются в большем или меньшем числе и
летом, и зимой. Держатся главным образом в восточной части района,
в частности на территории племовцесовхоза Орловский. Встречаются
табунами от 30 до 300 голов, но в 1951 г. наблюдались тысячные стада,
пришедшие, по словам местных жителей, вероятно, с Черных Земель,
в связи с занятием последних под зимовки домашних овец. Западная гра-
Рис. 78а. Самка сайги. О-в Барса-Кельмес. Июнь 1957 г. (Фото автора).
Рис. 786. Сайгаки на водопое. О-в Барса-Кельмес. Июнь 1957 г. (Фото автора).
23*
356
BOVIDAE
ница распространения сайги в Ростовской области и Ставропольском
крае в настоящее время проходит около 42—43° в. д. По последним дан-
ным (Адольф, 1950), сайгаки после длительного отсутствия вновь появились,
хотя и в небольшом числе, в Каясулинском, Ачикулакском, Карано-
гайском и Тарумовском районах Грозненской области, достигая на юге
44-й параллели. Имеются сведения о встречах сайгаков около Кизляра,
а в годы джута они изредка заходят даже в степную часть Дагестана до
Хасавюрта (Колесников, 1950).
В Закавказье в исторические времена сайга не водилась. В 1949 г.
была предпринята попытка акклиматизировать ее на о. Булла у западного
побережья Каспийского моря в Азербайджане (газ. «Бакинский рабочий»
от 16 XI 1949), куда группа сайгаков была завезена из Астраханской
области. Однако попытка эта окончилась неудачей: как сообщило в письме
Управление по делам охотничьего хозяйства Азербайджанской ССР,
в одну из ближайших суровых зим «с большим ледоставом и глубоким
снегом» все сайгаки на о. Булла погибли.
Заволжье и Южный Урал в свое время также входили в коренной
ареал Saiga tatarica. Четвертичные ее остатки найдены на Волге в устье
р. Камы, на полуострове Тунгуз (ниже устья р. Черемшана) в составе
«хазарской фауны» (средина плейстоцена), на р. Чагре, севернее Малого
Иргиза (Адольф, 1948; Беляева, 1935, 1939), а также в верхнепалеоли-
тических, неолитических и более поздних слоях пещер Южного Урала
(Карачаровский, 1951).
В 60-х годах XVIII в., по свидетельству Лапласа (1773), «многочислен-
ные стада» сайгаков появлялись на р. Бузулуке (ныне Куйбышевская
область). В большом числе встретил он их летом 1769 г. в степи у р. Са-
мары близ Новосергиевска. Многочисленны сайгаки были в то время
в районе Уральска. Еще в средине прошлого столетия будто бы были
обычны в Николаевском уезде Самарской губернии (ныне Пугачевский
район Саратовской области; Бажанов, 1930), а изредка, говорят, показы-
вались и у Бузулука (Анучин, 1875). В это же время они не составляли
редкости в Новоузенских степях (Адольф, 1948).
В левобережье Волги сайга держится и в настоящее время; однако
современная граница ее распространения здесь не достаточно ясна.
Можно предполагать, что она проходит где-то на широте Сталинграда
или немного южнее и в связи с общим увеличением за последние 15—20
лет численности этого зверя, по-видимому, прогрессивно сдвигается
к северу и в этой части ареала.
Кроме того, и в прошлом столетии, и в настоящее время наблюдаются
многочисленные заходы сайги в районы, далеко отстоящие от границ
постоянного обитания. В 20-х годах, например, сайгаки появлялись
в Пугачевском уезде Саратовской губернии, на рр. Малом Иргизе (с. Ива-
новка, 52°10' с. ш.) и Б. Чалыкле (около сел. Бобров-Гай и Жестянка;
Бажанов, 1930). В эти районы сайгаки заходят из района Синих гор,
на границе с Казахстаном, где в районе станций Озинка и Семиглавый
Мар они в небольшом числе до сих пор держатся постоянно (Адольф,
1948). В 1911 г. сайгаки отмечались в Приерусланских степях (Орлов
и Кайзер, 1933). О появлении сайгаков в 1949—1952 гг. в Палласовском,
Гмелиновском и Старо-Полтавском районах Сталинградской области,
в Приерусланских полупустынных степях сообщает А. С. Строганова
(1954). Сайгаки в эти годы держались здесь группами по 8—10 голов
не только зимой, но и летом. Идет ли в данном случае речь лишь о вре-
менных заходах, связанных с неблагоприятными условиями этих лет
SAIGA
357
в Прикаспии, или имеет место естественное расширение основного ареала
должны показать последующие годы.1 В прошлом столетии имели место
отдельные заходы сайги и в правобережные районы Средней Волги
(Greve, 1898), но теперь таких случаев давно не отмечалось.
В середине прошлого столетия, по Э. Эверсманну (1850), сайгаки
не переходили р. Урала в районе Уральска и Оренбурга, но южнее
бесчисленные стада их ежегодно переплывали эту реку.
В Азии в далеком прошлом ареал сайги охватывал не только степную,
но и часть современной таежной зоны. И. Д. Черский (1891) перечисляет
следующие места проверенных им лично ископаемых находок этого вида
в Сибири, большая часть которых, по-видимому, относится к верхнему
плейстоцену и голоцену: 1) Окрестности Свердловска, Березовские руд-
ники, 2) около г. Тюмени с р. Салаирки, 3) окрестности г.Омска,
4) около Салаирского рудника (ныне Кемеровская область), 5) речка
Узун-жул Минусинского округа (53° с. ш., 90° в. д.), 6) окрестности
Красноярска, 7) Нижнеудинская пещера Иркутской губернии (54° с. ш.,
98.5° в. д.), 8) р. Вилюй на 63°45' с. ш. и 121°42' в. д., 9) устье р. Оле-
нека, 10) Ляховские острова у северного побережья Якутии. Еще
в XVIII в. сайгаки были многочисленны в Барабинской степи
(Паллас, 1786 : 117).
Основная часть современного ареала Saiga tatarica расположена
в степной и полупустынной зонах Казахстана.1 2
Как и в других местах, область распространения и численность этого
вида здесь с конца прошлого века, вследствие хищнического промысла
и джутов, резко сократились. К 20-м годам текущего столетия сайгаки
в небольшом числе сохранились лишь в наиболее глухих, трудно доступ-
ных углах республики.
Однако за последние 20—25 лет, в результате полного запрета добычи,
благоприятных климатических условий и ряда других причин, если не
ареал, то былая численность этого вида в Казахстане восстановилась.
Особо сильное возрастание численности имело место в годы Великой
Отечественной войны, когда некоторые глубинные районы перестали
посещаться охотниками и стадами домашних животных. По приблизи-
тельным подсчетам запасы этого зверя в Казахстане к 1954 г. определялись
около 900 тысяч голов (Слудский, 1955).
Северная граница встреч сайги, по А. А. Слудскому (1955), в настоя-,
щее время проходит «через Уральск—ст. Казахстан—пос. Чилик—Орск—
Атбасар—оз. Кургальджин—пос. Киевку на р. Нуре, затем поворачи-
вает к югу, проходя через Кент, западные предгороя хр. Чингиз-Тау
(пос. Чубартау) и отсюда на восток через Аягуз и северное побережье
оз. Зайсан. Далее указанная граница уходит в Китай».
Практически повсеместно распространена в настоящее время сайга
в междуречье Урал—Волга. В последние годы здесь наблюдались стада
до 2 тысяч голов. Обычным, хотя и не столь многочисленным зверем
она является на пространстве между Уралом и Эмбой, от низовьев до
верхнего течения последней. Сайга отмечается теперь во всех районах
Западно-Казахстанской и Гурьевской областей. В последней она наи-
1 Следует упомянуть о сообщенном автору В. А. Поповым случае захода перед
Великой Отечественной войной сайгака в Нурлатский район Татарской АССР, веро-
ятно в связи с имевшим место в эти годы джутом в низовьях Волги.
2 Дается в основном по А. А. Слудскому (1955), В. М. Антипину (1940) и А. В. Афа-
насьеву с соавторами (1953).
358
BOVIDAE
более многочисленна на северном побережье Каспия. Местами зимней
концентрации сайгаков являются также низовья Эмбы, район оз. Сам,
полуостров Бузачи и южная половина Устюрта.1
В суровую зиму 1949/50 года группа в 20—30 сайгаков, гонимая ветром,
перешла по льду с полуострова Мангышлака или Бузачи на о. Кулалы
в Каспийском море (около 50 км от ближайшего пункта материка). Здесь
они весной 1950 г. дали приплод. Какова дальнейшая их судьба на острове
не известно.1 2 На самом Мангышлаке сайгак очень редок; южная граница
постоянного его пребывания здесь, по-видимому, проходит на широте
форта Шевченко.
Есть требующие проверки данные о том, что сайга в последние годы
появилась на Устюрте в пределах Каракалпакской АССР и в северных
районах Туркмении, в частности около побережья Кара-Богаз-Гола и
в северной части Каракумов. Отмечались случаи забегов ее к югу до
линии железной дороги на широте Ашхабада (Адольф, 1948).
В Актюбинской области Казахской ССР сайга встречается почти во
всех районах, вплоть до Новороссийского, Степного и Карабутакского;
заходит даже в южную часть Орских степей.3 Однако в значительном
числе держится лишь в районах Байганинском, Челкарском и Иргиз-
ском.
Область сплошного распространения сайги включает также северное
и восточное Приаралье. Она встречается во всех районах Кзыл-Ордин-
ской области. Здесь в полосе от Казалинска на западе до ст. Соло-Тюбе
на востоке расположены одни из самых крупных зимовок сайгаков в Ка-
захстане. Часть сайгаков, переходя Сыр-Дарью, зимует по окраинам
Кызылкумов. В последних, по Н. Гладкову и Г. Никольскому (1933),
они доходят до дельты Аму-Дарьи, но как далеко распространяются
в южном и восточном направлениях не известно. В 80-х годах прошлого
века сайга в большом числе встречалась в левобережьи Аму-Дарьи на
пространстве между Турткулем и Чарджоу в пределах теперешних Узбек-
ской и Туркменской ССР (Адольф, 1948).
Возвращаясь к Казахстану, следует сказать, что еще в начале этого
столетия сайга обитала на некоторых островах Аральского моря: о. Воз-
рождения и Барса-Кельмес (Берг, 1905). К настоящему времени сохра-
нилась только на последнем, и то лишь благодаря заповедному режиму
и двукратному дополнительному завозу небольших партий с материка.
По данным учета (Соколов, 1958), к маю 1957 г. на острове было
1167 сейгаков (учет проводился до окота и поэтому приплод 1957 г.
в зту цифру не вошел).
В Кустанайской области сайга в значительном количестве обитает
лишь в Тургайском и Амангельдинском районах, в долине Тургая и его
притоков. Только летом появляется в северной половине области, заходя
иногда в Семиозерный и Тарановский районы до Наурзума.
Также только летом в небольшом числе наблюдаются сайгаки в Акмо-
линской области, доходя на север до оз. Кургальджин и Атбасара.
Весьма многочисленна сайга во многих местах центрального Казах-
стана. Крупнейшим районом летовок этого вида является Карагандин-
1 К. Я. Грунин сообщил мне, что в Мангистауском районе Гурьевской области
сайгаки очень редки.
2 Газета «Туркменская искра», от 16 VIII 1950.
“•> Во время своего путешествия Паллас большое число сайгаков встречал на пути
между Орском и Губерлинском. В районе Губерлинских гор в 40-х годах прошлого
столетия охотился на них А. К. Толстой (1842).
SAIGA
359
ская область, особенно территория к югу от железной дороги Жарык—
Байконур и к западу от магистрали Караганда—Балхаш. К северу и
востоку от указанных районов она встречается во всей области, но зна-
чительно более редка, а в Тельманский и Нуринский районы заходит,
и то лишь изредка, летом.
Весьма многочисленны сайгаки в Бет-Пак-Дале, где до сих пор не
составляют редкости стада, насчитывающие тысячи животных. На юге
здесь распространение доходит до предгорий Каратау. Пески Муюнкумы
служат местами постоянных зимовок сайги, причем иногда она появляется
в саксаульниках в 100 км к северу от г. Джамбула (Соколов, 1951) и
в районе оз. Бийликоль. Отмечалась она также для низовьев р. Чу (Шнит-
ников, 1936), в Чуйском и Курдайском районах Джамбульской области
(Слудский, 1955) и недалеко от Алма-Аты, в предгорьях Анархая (Зве-
рев, 1948).
К востоку от Караганды сайга обитает в северном Прибалхашье.
Здесь она держится главным образом летом; на зиму откочевывает к за-
паду или по льду к югу, после замерзания Балхаша, в южное Прибал-
хашье. Имеются указания (Антипин, 1941), что в начале нынешнего
столетия сайгаки встречались в окрестностях Баян-Аула и будто бы до-
ходили даже до Иртыша.
В настоящее время в Павлодарской области, по-видимому, сайги нет
совсем, а в Семипалатинской она отмечается только для районов Абра-
линского, Абайского, Аягузского и Чубартауского, причем в первых
двух бывает только заходом.
В южном Прибалхашье областью, где сайгаки бывают не только на
зимовках, но и летом, является равнинная часть между рр. Или и Кара-
талом, южная оконечность оз. Балхаш, окраины песков Таукум, восточ-
ный шлейф Чу-Илийских гор. В суровые зимы заходят в Джамбульский
и Илийский районы Алма-Атинской области. Постоянными обитателями
сайгаки являются в междуречье Каратал—Аксу к югу до предгорий
Джунгарского Алатау; особенно многочисленны в низовьях этих рек.
Кроме того, встречаются у невысоких горок Ушкара, Аркарлы и Арга-
наты (у восточного побережья Балхаша). До сих пор сайга обитает
в Алакольской котловине, где сохранилась главным образом в южной
части побережья Алаколя.
В XVIII в. Гмелин большое число сайги наблюдал в районах Усть-
Каменогорска и Семипалатинска, по рр. Ульбе, Шульбе и Алею, где
в настоящее время их нет и в помине (Адольф, 1948). В прошлом столе-
тии сайга водилась в Зайсанской котловине и южных предгорьях Тар-
багатая (Антипин, 1941). Еще в 1925 г. она изредка попадалась по юго-
западному побережью Зайсана. Позднее она здесь исчезла, вновь по-
явилась только в 1948 г. и теперь держится в небольшом числе на се-
верном побережье Зайсана (Слудский, 1953).
Появившиеся в литературе сведения о наличии сайги в Чуйской степи
на Алтае в дальнейшем не подтвердились (Колосов, 1939).
В Монголии, по А. Г. Банникову (1954), изолированный очаг обита-
ния сайги расположен в районе Больших Западных озер Хиргис-Нур,
Хара-Нур, Хара-Ус-Нур, Дургэ-Нур и к югу от них в Хуйсиин-Гоби
и Шаргын-Гоби. «На севере граница проходит по южным склонам хр. Хан-
Хухей, на западе — по предгорьям Хархира-Алтанхухей, т. е. примерно
по меридиану 92° до нижнего течения р. Кобдо. Отсюда юго-западная,
а затем южная граница ареала уходит в Хуйсиин-Гоби и далее в Шаргын-
Гоби. . . Восточная граница идет по склонам Хасагту-Хайрхан, далее
360
BOVIDAE
на нижнее течение р. Дзабхан и затем еще севернее к склонам Хан-Хухея.
Отдельные забеги известны в районе озер Битер-Нур, Бон-Цаган-Нур,
Орок-Нур и даже Убсу-Нур, недалеко от границы Тувы. Кроме того,
из пограничных в Джунгарии районов ареал сайги заходит в юго-запад-
ную часть Монголии, где она постоянно обитает у хребта Байтаг-Богдо-
к югу от Монгольского Алтая».
Подвиды. Кроме типичного подвида (5. t. tatarica L.), А. Г. Бан-
ников (1946) описал как самостоятельный вид, а позднее (1951) стал счи-
тать подвидом: S. tatarica mongolica Bannicov — Монгольская сайга.
Отличается от типичного подвида меньшими размерами (длина тела не
свыше 120 см, наибольшая длина черепа до 216 мм), слабо развитыми ро-
гами, меньшей степенью горбоносости, серо-бурым тоном зимней окраски,
а также тем, что у большей части индивидуумов в нижней челюсти
имеются по три премоляра. Распространение: котловина
Больших Западных озер Монголии — Хиргис-Нур, Хара-Нур, Хара-Ус-
Нур, Дургэ-Нур и далее к югу от них в Хуйсиин-Гоби и Шаргын-
Гоби.
Биология. В настоящее время сайга является, в основном, обитателем
полупустынь; на зимовки во многих местах заходит и вглубь пустын-
ной зоны. В прежние, уже исторические времена в значительном числе
населяла также типчаково-ковыльные степи. Причиной сокращения
ареала, как отмечалось, было не только непосредственное преследование
й истребление человеком, но и вытеснение вследствие освоения обшир-
ных пространств целинных степей под выпас домашних животных и под
сельскохозяйственные культуры.
Летними стациями сайги в большинстве районов служат полынно-
типчаковые полупустыни, а на юге Сталинградской области и в централь-
ном Казахстане она заходит также в полосу сухих полынно-злаковых
степей. Поздней осенью и зимой сайгаки концентрируются в районах
полынно-солянковых полупустынь и в глинистых солянковых пустынях.
В Ростовской области они и зимой придерживаются сохранившихся мас-
сивов целинной полынно-типчаково-ковыльной степи. В Монголии основ-
ными местообитаниями являются злаково-солянковые, ковыльковые и
ковыльково-луковые полупустыни (Банников, 1954).
Еще в большей степени, чем джеран и дзерен, сайга является типич-
ным обитателем открытых равнин с плотным грунтом. Избегает она
холмистого рельефа, кустарниковых зарослей, а тем более тугаев.
В этом я имел возможность неоднократно убеждаться при посещении
Бет-Пак-Далы и о. Барса-Кельмес. Если джейранов чаще можно-
было встретить на участках с редким саксаулом, спириантусом и дру-
гими высокими, одиноко стоящими кустарниками, то сайгаков мы
встречали, как правило, на самых ровных открытых участках полынно-
боялычевой и полынно-биюргуновой пустыни, а еще чаще по краям боль-
ших такыров. На такырах сайгаки часто и отдыхают, великолепно гар-
монируя с ними по своей глинисто-рыжей покровительственной окраске.
Избегает сайга и песчаного грунта, поэтому держится только по окраи-
нам песчаных пустынь и то лишь в зимнее время. В саксаульниках и бар-
ханах нам сайгаки встречались лишь на переходах к водопою и с одного
участка пастбищ на другие. Предельная высота мест обитания сайги
в Монголии 1600 м, но и при этом она придерживается больших озерных
котловин.
Важным фактором, играющим роль в распространении и стадиальном
размещенйй сайги, является глубина и характер залегания снежного-
SAIGA 361
покрова. Снег глубиною уже 15—20 см создает большие трудности не
только для добывания пищи, но и для передвижения.1
В связи с сильно выраженной зависимостью сайги от рельефа и усло-
вий кормодобывания стоят имеющие место в большинстве районов се-
зонные миграции этого зверя, достигающие в ряде случаев протяжен-
ности в сотни километров. В пределах Украины в свое время зимой сАйга,
по-видимому, концентрировалась на побережье Черного и Азовского
морей, а на летовки откочевывала в северные разнотравные степи, До-
ходя до современной Воронежской области. В правобережье Волги и
в настоящее время основная масса поголовья сайги зимует в малоснежных
Черных Землях на юге Астраханской области, заходя частично в пре-
делы Грозненской области и Ставропольского края. Ранней весной, ещё
до отела, многочисленные табуны сайгаков из Черных Земель прикоче-
вывают в степи южных районов Сталинградской области, где проводит
отел и летовки. Значительная часть животных впрочем и летом остается
в Астраханских степях и полупустынях. Интересно, что, по весьма убе-
дительным данным С. К. Даля с соавторами (1956), причиной миграций
является не только глубокий снег, но и сильные морозы.
В прошлом столетии огромные табуны сайги, зимущей в районах
Устюрта и в Приаралье ежегодно уходили на север до Уральска, массами
переплывали р. Урал в нижнем течении и расселялись по всему Волго-
Уральскому междуречью. Осенью они уходили обратно в бесснежные
полупустыни к востоку от Каспийского моря (Эверсманн, 1850).
В настоящее время массовый характер носят сезонные миграции сайги
в центральном Казахстане. Основные места зимовок здесь расположены
в Бет-Пак-Дале к северу от р. Чу. Значительная часть животных по
льду переходит эту реку, особенно в суровые зимы, и расселяется по
Муюнкумам. Весною Имеет место обратное движение сайги в северные
районы, в Сары-Аркинские степи. Дальше всего на север заходят самцы,
тогда как значительная часть самок и лето проводит в северных частях
пустыни (Соколов, 1951). Более или менее ясно выраженные сезонные
миграции имеют место и в других районах Казахстана (Афанасьев и
др., 1953).
Миграции часто протекают незаметно, но местами приобретают мас-
совый характер и внушительные размеры. Во время наблюдавшейся
А. А. Слудским (1953) в центре Бет-Пак-Далы осенней миграции сайгаков
в конце октября 1947 г. сотенные табуны их медленно следовали один
за другим, причем в течение одного дня проходили тысячи животных.
Весной 1948 г. в течение дня мимо одного пункта крупными табунами
прошло около 12—13 тысяч сайгаков, а в последующие дни следовали
лишь отдельные небольшие группы.
Сроки осенних и весенних миграций зависят от географического по-
ложения, а также от кормовых и климатических условий. На правобережье
Волги сайгаки подходят к местам отела в средине марта (Лавровский,
1950). В районе Черных Земель, по собранным мною сведениям, массовое
появление сайги осенью наблюдается в ноябре. В центральном Казах-
стане откочевка сайги на летние пастбища начинается в марте. Основ-
ная масса мигрирующих на север весной сайгаков проходит через уро-
чище Когашик (Бет-Пак-Дала) в конце апреля или начале мая. Обрат-
ное движение на зимовки в нормальные годы происходит в сентябре или
1 В связи с этим вызывает сомнение сообщение С. К. Даля с соавторами (1956)
о том, что сайга может добывать корм из-под снега глубиною до 40 см.
362
BOVIDAE
октябре. В засушливые же годы, когда рано выгоревшая скудная расти-
тельность не обеспечивает зверей кормом, уже в августе наблюдаются
откочевки сайгаков в южные районы. Время откочевок сайги из север-
ного в южное Прибалхашье на зимовки и весною в обратном направле-
нии по льду определяется сроками начала и конца ледостава.
Помимо регулярных сезонных миграций, имеют место местные не-
регулярные, но иногда приобретающие массовый характер передвиже-
ния сайги. Летом они вызываются недостатком корма или воды вслед-
ствие засухи, а зимой образованием ледяной корки, наста или глубокого
снега. В поисках лучших пастбищ сайгаки предпринимают массовые
перекочевки нередко в далеко расположенные районы, не подвержен-
ные засухе и зимнему джуту. А. А. Слудский (1953) наблюдал в бассейне
р. Сары-Су в июне передвижение множества табунов сайги числен-
ностью от 50 до 2000 голов каждый, двигавшихся в одном направлении.
Чаще в таких случаях кочевки носят беспорядочный характер. Так,
в суровую зиму 1949/50 года сайгаки из Черных Земель уходили не
только к югу, но и к северу, появляясь в далеко отстоящих пунктах
среднего Поволжья. В годы джута, говорят, направление движения
стад сайгаков указывает скотоводам путь, в каком следует искать не
пораженных стихийным бедствием пастбищ.
Сайга — дневное животное. Ночью она отдыхает. Выпасается в лет-
ний период с утра до вечера, делая перерыв для отдыха через каждые
3—4 часа,1 в жаркие дневные часы лежит на обдуваемых ветром участ-
ках на такырах, берегах водоемов. Ложась, вытаптывает копытами
небольшую лежку. Когда лежит, морду иногда засовывает между подо-
гнутых под себя ног или утыкает в землю, по-видимому, в целях защиты
широких носовых отверстий от насекомых. Подолгу на одном месте не
лежит; через каждые 40—50 минут встает, возможно в связи с перегре-
ванием, и, походив минут десять, ложится на новом месте снова. На Барса-
Кельмесе в самое жаркое время дня некоторые сайгаки имеют обыкно-
вение ложится на отдых в теневой стороне кустов тамарикса или сак-
саула. При этом звери, по-видимому, по мере движения тени меняют под
одним и тем же кустом место лежки. Там довольно часто попадаются
одиноко стоящие кусты, у основания которых почва с северной и восточ-
ной стороны сильно уплотнена, лишена растительности и усеяна пометом
сайгаков. Особенно часто и охотно лежат в тени кустов сайгачата. В про-
хладные дни сайгаки активны в течение всего дня, пасутся и ложатся
для отдыха и пережевывания жвачки по мере насыщения. Зимой также
пасутся весь короткий день, лишь изредка ложатся. Благодаря густому
волосяному покрову даже во время непогоды сайга не нуждается в укры-
тии и ложится на открытом месте, не разгребая снега (Слудский, 1953).
На лежке или около нее часто можно видеть и помет сайгаков. Однако
мне пришлось на Барса-Кельмесе видеть большие скопления помета,
которые никак не могли принадлежать одному животному и были рас-
положены вне места лежек. В данном случае речь идет о своего рода
«общественных уборных», которые наблюдаются иногда и у джейранов.
В пределах избранного местообитания сайга, по наблюдениям
Е. П. Басенко (1950), придерживается определенных мест отдыха и жи-
ровок. При переходах к местам кормежки и водопоя пользуется одними
и теми же путями, в результате чего может протаптывать хорошо замет-
ные тропы, используемые промышленниками для расстановки капканов.
1 Мне приходилось видеть отдыхающих сайгаков в 7—8 часов утра.
SAIGA
363
Питание сайги определяется, в основном, составом растительности
в местах ее обитания. Однако имеет место и несомненная избирательность
в выборе кормовых растений. А. Г. Банников (1954) подчеркивает: «Мно-
гие виды растений, встречающиеся в значительном количестве там, где
обитает сайга, совсем или почти совсем не поедаются этими животными».
Е. П. Басенко (1950). на основе собственных наблюдений и исследований
содержимого желудков составила список кормовых растений сайги,
включающий около 30 видов. Основу питания на Барса-Кельмесе со-
ставляют солянковые, злаки, сложноцветные и, в меньшей степени,
бобовые растения. Из злаков наибольшее значение имеют произрастаю-
щие на острове в значительном количестве пырей и мятлик, поедаемые
в течение всего года, за исключением лета. Из солянок на первом месте
по пищевому значению стоит биюргун (Anabasis salsa Benth.), поедаемый
сайгаками в течение всего года, особенно зимой. В некоторые годы большое
кормовое значение приобретает полынь, (которую, по нашим наблюде-
ниям, сайгаки едят и в июне), но, по-видимому, меньшее, чем солянки.
Из бобовых весною пищей служат астрагалы, ас августа до конца осени —
листья и молодые побеги верблюжьей колючки (Alhagi pseudoalchagi
Desv.). На Барса-Кельмесе я наблюдал, как сайгаки объедали белый
саксаул.
В степях и полупустынях Казахстана основу летнего питания сайга-
ков составляют разнотравье и злаки. У самок, летующих в северной
части Бет-Пак-Далы, основную роль в питании играют, по С. С. Соко-
лову (1951), разные виды солянок, тогда как самцы в этих же условиях
поедали преимущественно солодку (Glyzyrrhiza aspera Pall.). Интересны
сообщения этого автора о том, что в летние месяцы главную пищу сам-
цов в Тургайских степях составляли два вида лишайников из рода
Aspicilium, остатки которых составляли до 90% содержимого желудков.
По Л. Гличу (Glitsch, 1865), в состав летнего корма, помимо полыни
и солодки, в значительном количестве входит лебеда (Atriplex sp.).
Зимний корм сайги в условиях Казахстана, в основном, составляют
полыни и солянки. Из последних на первом месте стоит биюргун. У по-
лыней поедаются главным образом веточки с почками, содержащими соч-
ные, еще не распустившиеся листочки. Поедание зимою в значительном
количестве солянок объясняется тем, что к этому времени происходит
выщелачивание отмерших частей растений, уменьшается содержание
солей, повышается, наоборот, содержание белка и жира. Исследование
содержимого желудков показало, что на 60—70% оно состояло из раз-
ных видов полыни и до 30% составляли остатки биюргуна (Соколов,
1951).
В монгольских полупустынях основу питания сайги, по А. Г. Бан-
никову (1954), в течение всего года составляют гобийский ковылек (Stipa
gobica Roshev.), луки (Allium), а летом, кроме того, баглур (Anabasis
brevifolia С. А. М.). Еще в старой литературе (Glitsch, 1865) встречаются
указания на поедание сайгаками молодых всходов культурных растений.
Случаи повреждения ими посевов хлебов отмечаются и за последние
годы. Только крайняя необходимость заставляет сайгаков приближаться
и искать защиты у населенных пунктов. В суровую зиму 1949/50 года
сайгаки в Ерусланских степях собирались и, вероятно, кормились у сто-
гов сена (Строганова, 1954).
Утверждения о том, что сайгаки длительное время могут обходиться
без питья, по-видимому, лишены основания. Правда, Е. П. Васенко
(1950) сообщает, что по ее наблюдениям сайгаки иногда, когда мало воды,
364
BOVIDAE
пьют с интервалом в трое-четверо суток, но в нормальных условиях при-
ходят на водопой ежедневно, а иногда даже под два раза. А. А. Лавров-
ский (1950) сообщает, что в правобережье Волги во время засухи 1949 г.
после прошедших грозовых дождей дороги в степях были испещрены
множеством следов сайгаков, приходивших утолять жажду в грязных
дождевых лужах.
Пьют сайгаки как пресную, так и морскую воду. Морскую, по нашим
наблюдениям на Барса-Кельмесе, пьют даже при наличии пресной, но
последнюю явно предпочитают. На о. Возрождения сайгаки пили во-
обще только морскую воду, так как источников пресной там в то время
не имелось; правда, Л. С. Берг (1905), сообщавший об этом, подчерки-
вал слабую соленность воды Аральского моря. Специальные неоднократ-
ные наблюдения, проведенные нами на Барса-Кельмесе, показали, что
сайгаки, если их не тревожат, на водопой приходят в разное время,
практически весь день, но максимум посещений падает на время от 7 до
9 часов утра и после полудня между 15 и 16 часами. Сайгаки для того,
чтобы напиться, не боятся заходить в воду по запястья и глубже (рис. 786).
С самками в воду иногда заходят и сайгачата. Процесс питья длится
не больше 5 минут. В воду опускается только рот и верхняя губа; ши-
роко раскрытые ноздри всегда остаются поверх воды.
Считается, что потребность в минеральном питании у сайги удовлет-
воряется пастбищными кормами, содержащими большое количество солей.
Мне солонцевания сайгаков тоже ясно видеть не удалось, но однажды
издали я наблюдал, как сайгаки, низко наклонив головы к земле, как
будто обнюхивая ее, что-то делали на совершенно голом такыре. Не-
сколько раз пришлось замечать взрыхленную в виде узких полосок по-
верхность пухлых солончаков (кебиров), на которых часто держались
сайгаки. Наконец, в желудке (книжке и сычуге) одного убитого сайгака
было обнаружено некоторое количество довольно крупного гравия,
который случайно, Вместе с травой едва ли мог быть заглочен. В неволе
сайгаки охотно лижут каменную соль.
Сайга — типичное стадное животное. Стадное чувство у нее развито
сильнее, чем у джейрана и дзерена. В этом отношении сайга сходна
с домашней овцой. Если одно или несколько потревоженных живот-
ных побежит в сторону, за ними бросается все стадо. Если табун
сайгаков переправляется через реку, и передние уже достигли противо-
положного берега, то даже нападение на плывущих зверей не всегда и не
сразу заставит их повернуть обратно.
Только в период отела, и то не всегда, самки держатся некоторое
время в одиночку. В остальное же время сайга держится табунами разной
величины в зависимости от численности ее в том или ином районе и
сезона года. После отела, когда молодняк немного окрепнет, самки со-
бираются в стада, насчитывающие иногда тысячи животных (Кульпин,
1955) и состоящие из самок с приплодом текущего года. Обычно, как
показали наши наблюдения на Барса-Кельмесе, в состав табуна входят
и один или несколько самцов; однако последние чаще держатся и даже
мигрируют отдельными, сравнительно небольшими группами, включаю-
щими нередко и холостых самок. Самый большой табун самцов, который
мы встретили в мае в Бет-Пак-Дале, состоял из 70 рогачей.
В некоторых районах летом наблюдаются стада сайги, достигающие
крупных размеров. Выше отмечалось наблюдавшееся А. А. Слудским
передвижение в начале июня стад, доходивших до двух тысяч. С. С. Со-
колов (1951) 1 июня видел табун в несколько сотен самок с молодыми,
SAIGA
365
направлявшийся через р. Сарысу. Образование крупных табунов летом,
как и летние кочевки, возможно, связаны с неблагоприятными условиями
и поисками лучших пастбищ и водопоев. Больших размеров табуны до-
стигают в июле—ноябре. Однако в период гона сайгаки держатся та-
бунками по 20—26 голов, представляющими гаремы из одного самца и
самок с приплодом текущего года.
Максимальную численность табуны сайги имеют зимой после гона.
Взрослые самцы по окончании гона отделяются и держатся в одиночку
или группами по 3—10 голов (Слудский, 1955). Крупные стада распа-
даются весной, с апреля. Однако П. И. Кульпин (1954) 14 мая 1953 г.
наблюдал массовый отел сайги на ограниченной площади в центральной
части Бет-Пак-Далы, когда, следуя на машине, он попал в полосу, где
«территория в 6—8 км по фронту и 2—2.5 км вглубь представляла собой
живое желтое поле. Здесь собрались десятки тысяч самок для окота.
Значительная чдсть их уже паслась с новорожденными сайгачатами.
Большинство сайгачат еще не успело обсохнуть, и, не научившись дер-
жаться на собственных ногах, лежало на земле, вытянув передние ножки
вперед и спрятав голову между ними. . . Шоферу поминутно приходилось
объезжать густо расположенных на земле, большею частью попарно,
маленьких обитателей пустыни, а несколько раз мы вынуждены были
проезжать, оставляя их в колее, между колесами машины».
Старые указания о том, что в стадах сайги имеются сторожа и вожаки
(по одним данным самцы, по другим самки), наблюдениями последних
лет не подтвердились. За сторожей принимали, по-видимому, случайно
стоящих зверей, какие всегда найдутся в большом отдыхающем стаде.
В современных условиях существования сайга является осторожным
зверем. Однако осторожность является, очевидно, свойством, приобре-
тенным в результате постоянного преследования человеком. Непуганая
сайга гораздо менее осторожна. Когда на о. Николая в Аральском море
(ныне о. Возрождения) впервые высадились люди, они были встречены
сайгаками. Звери, ранее не видевшие людей, с любопытством смотрели
на них и даже от выстрелов не разбегались (Берг, 1905). После того, как
на о. Барса-Кедьмес был учрежден заповедник и сайга взята под охрану,
звери стали уже через несколько лет менее осторожны. Оци перестали
избегать близости человеческих поселений, бывают на отдыхе и жировках
вблизи центральной усадьбы, иногда подходят к ней даже для водопоя.
И вообще они менее осторожны, чем джейраны. По свидетельству
Е. П. Басенко (1950), при подходе к находящимся по соседству табункам
сайгаков и джейранов, последние, будучи потревожены, убегали раньше.
Однако с мнением Е. П. Васенко о большой доверчивости сайгаков на
Барса-Кельмесе я не могу согласиться. Они там не пугливы, но доста-
точно осторожны. Перед окотом и в период последнего они редко под-
пускали нас даже на лошади и верблюде ближе чем на 200—300 м. Когда
с матерями стали ходить сайгачата, по непонятной нам причине самки
стали вести себя гораздо более доверчиво; но и тогда на открытом месте
в полный рост ближе, чем на 100 м подойти к ним не удавалось. Только
един раз, по-видимому, стимулируемая жаждой, одинокая самка метрах
в пятидесяти от нас, занятых погрузкой на повозку убитого сайгака, про-
шла к водопою, на наших глазах принялась пить и дважды позволила
сделать с нее метрах в тридцати фотографические снимки.
У сайги хорошо развиты слух и обоняние, хотя, по-видимому, и не
в такой мере, как у большинства других диких копытных. Еще видимо
слабее у нее зрение. Неподвижно сидящего человека сайга, по нашим
366
BOVIDAE
наблюдениям, замечает метров за 30—40, но распознать, по-видимому,
не может. Нам неоднократно приходилось подкарауливать с фотоаппара-
том сайгаков у водопоя, не спрятавшись, а лишь прижавшись к кусту
тамарикса или саксаула на той стороне, откуда подходили звери. Заме-
тив неподвижно сидящего фотографа, сайгак настороживается, но с оста-
новками, напряженно всматриваясь в фигуру человека, продолжает
двигаться в нужном ему направлении, иногда подходит метров на 8—10 и
обычно лишь движение руки при наводке на резкость или при переводе
пленки заставляет зверя в испуге шарахнуться в сторону, если только
еще раньше этого попутный ветерок не нанесет на него запаха человека.
Что касается слуха, то должен сказать, что на звук щелкающего
затвора фотоаппарата даже вблизи сайгаки не реагировали вовсе. Спря-
тавшись в кустах, мы с компанионом могли негромко переговариваться,
не вызывая беспокойства пьющих метрах в 40—50 сайгаков. На звук
хлопка в ладоши, по наблюдениям Е. П. Басенко (1950), сайгаки реаги-
руют за 150—200 м; громкий крик заставляет в тихую погоду сайгаков
поднять головы, но определить — откуда идет звук, на этой дистанции
они, по-видимому, еще не могут, тогда как метров за 100—150 они по-
ворачивают голову сразу в сторону звука, хотя бы человека им и не было
видно. Еще лучше, очевидно, развито обоняние. Если ветерок тянул
от сидящего в засидке охотника, то охота ни с ружьем, ни с фотоаппара-
том почти никогда не была удачной.
Сайгаки отличаются спокойным нравом, за исключением самцов во
время гона. Пойманные в молодом возрасте легко становятся ручными и
ходят на свободе. Еще П. Паллас (1773) сообщал о многочисленных слу-
чаях приручения сайгаков в Заволжье. Пойманные сайгачата уже через
несколько дней начинают привыкать к ухаживающим за ними людям,
бросаются к ним навстречу, трутся о ноги, хватают за одежду. Колхоз-
ники по р. Сарысу выкармливают молодых сайгачат, подпуская к коровам,
и был случай, когда пойманный ослабленным взрослый сайгак стал вы-
пасаться вместе со скотом (Соколов, 1951). Практического значения раз-
ведение сайгаков до сих пор не имело, так как большинство приручен-
ных зверей в домашней обстановке в силу невыясненных причин погибало.
Сайга является, по-видимому, самым быстрым из современных наших
копытных. Взрослого здорового сайгака не может догнать самая резвая
борзая. За последние годы имеются более точные данные о быстроте этого
зверя, полученные при преследованиях на автомобилях со спидометром.
М. Д. Зверев (1948) приводит максимально развитую самками скорость
при десятиминутном преследовании 72 км в час. А. Г. Банников (1954)
утверждает, что сайга со скоростью 60—70 км пробегает легко расстояние
25—30 км. Еще более высокие цифры приводит С. С. Соколов (1951).
По данным этого автора, сайгаки выдерживали преследование в течение
15 минут со скоростью 80 км, а в исключительных случаях километр
пробегали за 40 секунд. При преследовании на автомобиле со скоростью
80 км в течении 5 минут, впереди оказывались молодые двухмесячные
ягнята, за ними самки, а позади взрослые самцы. Во время экспедиции
в Бет-Пак-Далу нам пришлось однажды преследовать одну, по-видимому
беременную, самку. Она бежала по полынно-боялычевой равнине парал-
лельно автомашине, развившей скорость 50 км, в продолжении 5 минут,
но затем заметно стала сдавать и, повернув, ушла в сторону.
По аллюру и общему характеру движений сайга сильно отличается от
обитающего в тех же местностях джейрана. При движении шагом заме-
чается некоторая угловатость. И на шагу, и при беге голова держитсяг
SAIGA
367
опущенной вниз, отчего внешний вид зверя не вяжется с его быстротой.
Особенно характерен и своеобразен бег сайги. В противоположность
джейрану, бегущему пружинистой рысью и карьером, сайга, обычно
быстро перебирая ногами и опустив голову, как бы стелется по земле
плавной рысью. Отдельные животные время от времени делают забавные
прыжки «свечой» вверх, отталкиваясь задними ногами. Эти прыжки,
вероятно, дают возможность животному, движущемуся с опущенной го-
ловой, увеличивать кругозор во время бега.
Большинством автором аллюр сайгака определяется как иноходь
(одновременное выбрасывание передней и задней ног одной стороны),
чего нельзя, впрочем, сказать при рассматривании моментальных фото-
графий, снятых с самолета.1 Правильнее его назвать рысью. На Барса-
Кельмесе мне удавалось видеть и сайгаков, бегущих галопом; но этот
аллюр не является типичным, и движутся им лишь не испуганные и не
слишком спешащие звери. Если сайга вынуждена спасаться быстрым
бегом, то бежит только плавной стелющейся рысью. Останавливаются
сайгаки во время бега, в противоположность джейрану, редко, особенно
если движутся табуном.
Как показали исследования А. П. Берггрина (1948), особености мор-
фологии опорно-двигательного аппарата сайги, в частности устройство
тазобедренного сустава, имеющего блоковидную форму, свидетельствуют
о резко выраженной специализации в направлении быстрых поступатель-
ных движений. Поэтому сайга бежит обычно по прямой, не делает петель
и резких поворотов, как это имеет место у джейрана,1 2 по пересеченной
местности и в кустарнике ей бежать трудно.
Сайга хорошо плавает. Преследуемая заплывает от берега на пол-
километра и больше, без труда переплывает широкие реки с быстрым
течением (Берг, 1905; Воробьев, 1929).
Взрослый сайгак вне периода размножения — зверь молчаливый.
Лишь иногда, говорят, издает имеющие предостерегающее значение глу-
хие хрюкающие или храпящие звуки. В период же окота самки и сайга-
чата громко и часто перекликаются друг с другом. Голос самки в это
время представляет что-то среднее между хрюканьем и низким хриплым
блеянием, больше все же приближаясь к первому. Голос сайгачат также
является хрюканьем, только более звонким и нежным. Голоса перекли-
кающихся матерей и сайгачат в начале лета нередко заранее предупреж-
дали нас, сидящих в засидке у водопоя с целью наблюдений, о прибли-
жении табуна сайгаков. В местах же Массового окота звуки хрюкающих
тысяч сайгаков будто бы напоминают звук морского прибоя. Взрослые
самцы во время гона, когда дерутся, по Е. П. Васенко (1950), «хрюкают,
храпят и кричат».
Линька у сайги происходит в году дважды — весной и осенью. Ве-
сенняя линька в условиях Барса-Кельмеса начинается, по Е. П. Васенко,
около середины апреля, а в раннюю весну даже во второй половине марта.
Однако нельзя согласиться с этим автором в том, что в первой половине
мая линька уже заканчивается. В 1957 г., который по климатическим
условиям был нормальным и благоприятным, первый из отстрелянных
нами сайгаков, крупный самец (весом 48 кг), 12 мая находился в стадии
1 См., например, журнал «Огонек» № 50 за 1950 г.
2 М. Зверев (1948), впрочем, приводит случай, когда преследуемые на автомобиле
сайгаки свернули резко в сторону, а П. Кульпин (1954) указывает на привычку сайги,
подобно джейрану, перебегать дорогу перед машиной.
368
BOVIDAE
интенсивной линьки; старая шерсть у него еще сохранялась на нижней
стороне головы, на спине, боках, верхней стороне крупа, на ягодицах
и бедрах. Не вполне вылинявших зверей нам пришлось видеть в табунах
до середины июня. Первый вполне вылинявший самец был добыт лишь
7 июня. Как и у других зверей, в первую очередь линяют хорошо упи-
танные самцы и холостые самки. Задерживается линька у подсосных ста-
рых маток и у истощенных больных животных. В Бет-Пак-Дале первого
вполне вылинявшего самца мы в 1956 г. встретили 19 мая, самки же в се-
редине мая еще имели остатки старой шерсти. Четко выраженной после-
довательности в топографии линьки нам подметить не удалось; несомненно
только, что в первую очередь старая шерсть выпадает на голове и ниж-
них частях ног; остатки старой шерсти дольше всего удерживаются на
боках, подгрудке и на хвосте.
Осенняя линька происходит путем не подрастания, а смены старого
волоса на новый; начинаясь в начале октября, она тянется свыше двух
месяцев и заканчивается только к середине декабря. Смена волосяного
покрова осенью начинается с конца морды, с пространства вокруг осно-
вания хвоста и внутренней стороны ног. Затем процесс линьки после-
довательно распространяется на голову, круп, верх спины и бока ту-
ловища. В последнюю очередь старая шерсть выпадает на низу живота и
на наружной стороне ног (Васенко, 1950).
Половой зрелости самцы сайгака достигают на втором году жизни.
Часть самок, по сообщению А. А. Слудского, становится половозрелыми
в возрасте 7 месяцев. Половой сезон (спаривание) имеет место поздней
осенью или в начале зимы, Начало его падает на вторую половину
ноября, разгар — на декабрь. Заканчивается гон в конце декабря, реже
к середине января. Самцы в это время присоединяются к самкам и
создают гаремы по 10—12, изредка до 25 самок с приплодом текущего года
(Васенко, 1950). Самцы сильно возбуждены, строго охраняют свой та-
бунок, не позволяют самкам уходить далеко в сторону от своей группы
и не подпускают близко других самцов. Между ними в это время про-
исходят турнирные бои, приобретающие нередко ожесточенный характер..
Известны не единичные случай обламывания одного или обоих рогов,
повреждения мягких тканей и даже костей черепа. Правда, по сообще-
нию Е. П. Васенко, утомленные получасовой борьбой соперники часто
тут же на месте ложатся отдыхать по соседству друг с другом, но зато
известны случаи гибели одного из борющихся во время поединка или
вскоре после него.
Самцы в период гона едят очень мало и к концу его сильно худеют.
Самки ведут себя в это время спокойно и упитанности не снижают.
Продолжительность беременности сайги около 5—5% месяцев.1
Массовый окот в Казахстане и Монголии протекает во второй и тре-
тьей декаде мая, у молодых и престарелых несколько позднее. Первый
случай окота на Барса-Кельмесе в 1957 г. отмечен 13 мая, 17 мая мы ви-
дели уже бегающих с матерями сайгачат. Вообще окот у сайгаков про-
текает очень дружно, основная их масса приносит детей в несколько дней
третьей декады мая. Но 4 июня нами была добыта старая самка с эмбрио-
ном, который, очевидно должен был родиться не раньше, чем через две
недели. 12 июня были встречены совсем маленькие, едва бегающие сай-
1 Е. П. Васенко (1950) приводит единственный, точно зафиксированный случай
беременности сайги в Алма-Атинском зоопарке продолжительностью ровно 6 меся-
цев.-
SATGA
369
гачата. На Барса-Кельмесе известен случай окота в августе. В право-
бережье Волги, по А. А. Лавровскому (1950), и гон, и окот проходят на~
1—2 недели раньше.
Самки, как уже отмечалось, перед окотом иногда удаляются из та-
бунов. Для окота выбирают равнины или понижения. Подавляющее
большинство самок приносят по два детеныша, хотя в Монголии, по учетам
А. Г. Банникова (1954), с двойнями летом встречается только 53.8% са-
мок. По одному сайгачонку приносят чаще самки-первоокотки. Изредка
бывает по три детеныша в одном помете (Васенко, 1950; Строганова, 1954).
Первые 2—3 дня сайгачата слабо стоят на ногах, через 4—5 дней
довольно хорошо бегают, но, пробежав 30—40 м, ложатся и затаи-
ваются на земле. Ложась на землю, они прикладывают уши, вытяги-
вают голову и шею, плотно прижимаются к земле и благодаря
хорошо выраженной покровительственной окраске сливаются с нею.
«Рыжевато окрашенный сайгачонок, — пишет Е. П. Васенко (1950),—
настолько напоминает по цвету желтеющие к этому времени листья ре-
веня (Rheum tataricum L.), что заметить его крайне трудно, даже если
знаешь, где он лежит. На расстоянии 10—12 шагов от наблюдателя сай-
гачонок совершенно сливается с общим фоном». Мать от новорожденного
обычно далеко не уходит; в случае опастности она бегает широкими кру-
гами вокруг того места, где лежит сайгачонок, чем иногда обнаруживает
его и облегчает поимку.
К 7—8-дневному возрасту сайгачата обычно крепнут настолько, что,
по сообщению Э. Эверсманна (1850), их не может догнать верховая ло-
шадь. Молодые сайгачата — забавные и очень подвижные существа;
испугавшись человека, они стремглав срываются с места, мчатся быстрее
взрослых зверей, и уже с первых дней делают характерные для сайга-
ков «свечки». Однако если матери остаются более или менее спокойными,
сайгачата останавливаются, некоторое время напряженно смотрят на
человека и обычно даже начинают медленно двигаться ему навстречу,
вероятно для того, чтобы рассмотреть получше. С этого времени сайгачата
начинают ходить с матерями, хотя мне неоднократно пришлось наблю-
дать их, лежащими метров за 500—700 от пасущихся или ушедших на
водопой маток. Последние, после того как малыши достаточно окрепли,
вновь начинают объединяться в группы. По Е. П. Васенко (1950), сай-
гачиха кормит детенышей в день дважды — утром и под вечер; но мне
приходилось на Барса-Кельмесе видеть сосущих сайгачат в разное время
светлой части суток. Акт сосания очень короток, длится не дольше 1 ми-
нуты, обычно гораздо меньше. Сосут и по одному и сразу оба сайгачонка.
Высасывает сайгачонок за один прием, по-видимому, не менее 150—-200 г
(Гульмисирян, 1944). Из молочной железы беременной последние дни
самки нами было собрано молозива около 500 г. Лактация продолжается
до сентября, но зеленым кормом сайгачата начинают понемногу пи-
таться с возраста 2—3 недели.
Родятся сайгачата весом, в среднем, около 3.1 кг (от 2.8 до 4 кг). Ра-
стут довольно быстро; к пятимесячному возрасту достигают половины
веса взрослых (Васенко, 1950). В месячном возрасте у самцов начинают
расти рога, приблизительно по 1 см в месяц. Первые рога темного цвета,
бурого. Особенностью сайгака является то, что образовавшиеся в первый
год жизни роговые чехлы слущиваются и заменяются новыми.1 Как уже
сказано, первые роговые чехлы темные. На их поверхности растут редкие
1 Факт, в литературе почему-то совсем не отмеченный.
24 Фауна СССР, И. И. Соколов
370
BOVIDAE
грубые волоски. Уже в возрасте около 7 месяцев сквозь старые чехлы
начинают прорезываться верхушки дефинитивных (постоянных) рогов.
Старые чехлы после этого одевают в виде трубки основание рогов, по-
степенно отслаиваются и отпадают. У годовалых самцов остатки первых
чехлов обычно еще охватывают кольцом основание рогов поверх дефи-
нитивных. У 15-месячных старые чехлы совсем сброшены, но до двухлет-
него возраста поверхность проксимальной части дефинитивного рога
еще шелушится и совершенно светлой, гладкой становится лишь на тре-
тьем году. У годовалых самцов годовые чехлы еще не плотно прирасли
к стержням и при ощупывании ощущается их подвижность.
Родятся сайгачата желтовато-рыжими. На втором-третьем месяце
у них проходит первая линька, когда впервые появляется темная полоса
на хребте. Осенью того же года, одновременно со взрослыми, они ли-
няют второй раз.
Соотношение полов при рождении приближается один к одному;
по данным С. С. Соколова (1951), имеется даже перевес в сторону самцов.
Однако вследствие повышенной смертности последних (см. ниже) с вбз-
растом соотношение полов меняется и в Монголии, например, по А. Г. Бан-
никову (1954), оно равняется 1 : 3.7 в пользу самок.
Продолжительность жизни в естественных условиях не известна.
А. А. Слудский (1955) сообщает, что по утверждениям опытных охотников,
при общей продолжительности жизни сайги в 10—12 лет, самцы редко
живут больше половины этого срока. Я сомневаюсь в возможности до-
стижения 12-летнего возраста не только самцами, но и самками. У за-
ведомо (благодаря кольцеванию в ягнячьем возрасте) 3-летнего самца,
добытого нами на Барса-Кельмесе, зубы были настолько изношены, что
едва ли он мог нормально пережевывать пищу, и, следовательно,нормально
питаться еще больше чем 2 года.
Из врагов сайги по значению на первом месте стоит волк. Так как
последний не может состязаться в скорости бега с сайгой, то в одиночку
применяет охоту подкарауливанием или скрадом. Обычно волки караулят
сайгаков на пути их постоянных переходов, на местах водопоев, следуют,
прячась, позади табунов и, улучив удобный момент, схватывают отставших
или ослабевших животных. Подкрадываются они к сайгакам в жаркие
дни на лежках, когда, уткнувшись мордой в землю, звери менее
чутки.
Зимой волки чаще охотятся на сайгаков стаей. В это время особенно,
много гибнет взрослых самцов, сильно ослабевших после гона. Если
здоровая самка может бежать со скоростью до 70—75 км, то скорость
бега истощенных самцов не превышает таковую волка (40—45 км). Волки
десятками режут ослабевших сайгаков, сразу не съедают их трупы п
кормятся ими всю последующую часть зимы. В богатых сайгой районах,
трупы сайгаков после гона находят в большом числе. А. А. Слудский
(1955) полагает, что «ежегодно более половины всех взрослых самцов
гибнет от противников и волков без всякой пользы для человека и вос-
производства стада». Молодые сайгачата в первые дни после рождения
могут стать жертвой лисиц, крупных хищных птиц, даже чаек, как
я мог убедиться в этом на Барса-Кельмесе.
В летнее время сайгаки страдают от кровососущих насекомых (гнуса),
которые не только наносят укусы, но и забираются в их широкие ноздри,
отчего звери вынуждены в ущерб собственной безопасности прятать
конец морды. Особенно много терпят сайгаки летом вследствие очень
короткого волоса от укусов слепней (Т abanidae).
SAIGA 371
Из наружных паразитов наибольшее значение до недавнего времёнй
имел подкожный овод Pallasiomyia anlilopum Pall. (Грунин 1949). Мнрго-'
численные личинки последнего нередко сплошь изрешечивали на верх-
ней стороне спины сайги кожу и, естественно, сильно изнуряли живот-
ных. Интересно отметить, что в период последнего резкого сокращения
мысленности сайги (20-е годы текущего столетия) этот паразит, по край-
ней мере в Казахстане и Поволжье, по-видимому, вымер. Несколько
десятков сайгаков, добытых А. А. Слудским (1955) й его сотрудниками
в Бет-Пак-Дале и в Волжско-Уральских степях, были свободны от ли-
чинок оводов. То же подтвердили ему местные охотники, добывавшие
сотни сайгаков в разных районах Казахстана. Е. П. Васёнко (1950) упо-
минает о паразитирующих изредка на сайге клещах, которые к тому же'
могут быть переносчиками кровепаразитарных заболеваний. Однако мне
на Барса-Кельмесе и Бет-Пак-Дале в мае и июне присосавшихся клещей
находить не удавалось.
Внутренние паразиты сайги изучены еще недостаточно. У сайгаков,
добытых на о. Барса-Кельмес и в пустыне Бет-Пак-Дала, обнаружено 12 ви-
дов паразитических червей. Патогенное значение имеют два вида — Skrja-
binodera saiga Gned. et Vsevol. и Parajilaria antipini Buehl., паразити-
рующие в глазном яблоке и могущие быть причиной слепоты животных
(Соколова и др., 1949). На. Барса-Кельмесе взрослые сайгаки на 100%
поражены нематодой(пока не определенной), в большом числе парази-
тирующей в межмышечной ткани и под серозными оболочками брюшной
полости. Мясо при большом числе Этих паразитов, обнаруживающихся
при его разрубке, приобретает непривлекательный вид.
Наибольший ущерб численности сайги наносит гибель от неблаго-
приятных климатических условий, особенно в зимний период. В годы
джута гибнут тысячи сайгаков и резко снижается их поголовье.
В правобережье Волги зима 1949/50 года была очень суровой, сопро-
вождалась буранами, сильными снегопадами, лишившими зверей воз-
можности добывать пищу. Сайгаки вынуждены были искать лучших
мест, уходили на юг и на север. Много их гибло в пути, а также и на но-
вых местах, где они часто также не находили необходимых условий.
В эту зиму в некоторых местах находили группы по 10—15 голов замерз-
ших и погибших от голода животных (Колесников, 1950; Лавровский,
1950). Подобная же картина в Поволжье и западном Прикаспии повто-
рилась в суровую зиму 1953/54 года. При этом сайгаки гибли не только
от голода, но и от мороза. С. К. Даль (1956) находил весной во множестве
трупы взрослых самцов, павших при наполненном пищей желудке. Самки
и молодые самцы гибли также от переедания на богатых кормом пастби-
щах при перекочевке на них после длительной голодовки.
Не меньшее значение, чем холода и глубокий снег, имеет гололедица
и наст, если они продолжаются дольше недели (Слудский, 1953). Звери
и в этом случае не могут достать себе корма, а при насте, к тому же, часто
лишены и возможности передвигаться в поисках лучших пастбищ. Корка
льда, когда звери проваливаются сквозь наст, режет кожу на ногах,
вызывая глубокие повреждения, надолго лишающие животных возмож-
ности передвижения даже тогда, когда стихийное бедствие миновало.
Ослабевшие животные в первую очередь становятся добычей
волка.
В Казахстане десятки, а быть может и сотни тысяч сайгаков погибли
от бескормицы и волков в суровые зимы 1940/41, 1941/42, 1948/49, 1950/51
и 1953/54 годов. Только в Западно-Казахстанской и Гурьевской областях
24*
372
BOVIDAE
после джута 1949/50 года численность сайгаков, по данным А. А. Слуд-
ского (1955), сократилась в 2 раза.
Губительными для сайгаков могут стать и летние засухи. В засушливое
и жаркое лето 1949 г. на правобережье Волги, вследствие раннего выго-
рания растительности, сайгаки были лишены не только корма, но и воды
в результате пересыхания водоемов. Звери были вынуждены широко
кочевать, многие ослабевшие животные становились жертвой появившихся
в большом количестве крупных хищных птиц (белоголовых сипов и бу-
рых грифов; Лавровский, 1950).
Следует отметить, что сайга после джутов быстрее восстанавливает
численность по сравнению с джейраном. Так, во время суровой зимы
1948/49 года на Барса-Кельмесе погибло почти все поголовье сайгаков и
джейранов. И вот с того времени, меньше чем за десять лет, сайга прак-
тически восстановила былую численность, тогда как число джейранов,
количество которых к 1948 г. было около четырех тысяч, в настоящее
время едва насчитывает несколько десятков.
Наиболее высока смертность от всех указанных причин среди самцов;
и от хищников, и во время джута в первую очередь гибнут взрослые самцы,
истощенные гоном. Из зарегистрированных Е. П. Васенко 39 случаев
падежа сайгаков на о. Барса-Кельмес только четыре приходилось на
долю самок и два на долю молодых самцов. Остальными павшими зверями
были взрослые самцы, погибшие главным образом во вторую половину
зимы, после гона.
Из случайных причин можно упомянуть о гибели сайгаков при пере-
правах через реки. П. И. Кульпин (1954) сообщил о случае гибели це-
ликом табуна сайгаков в 500—700 голов при переправе по неокрепшему
льду через р. Чу во время осенних перекочевок.
Хозяйственное значение. Сайга ценна, прежде всего, как мясной
зверь. Вес туши взрослого, нормально упитанного самца осенью, со-
ставляя в среднем 30 кг, может достигать 36 кг, а самок около 18 кг.
Вес туши взрослых самцов, убитых в мае—июне на о. Барса-Кельмесе,
составлял от 20 до 28 кг, самок — от 14 до 18 кг. По собранным
Ф. Б. Чернявским в 1958 г. данным, средний вес выпотрошенной
(без желудка и кишечника) туши в шкуре сайгаков из Астраханского
Охотпромхоза составил для самцов 34.2 кг. (от 29.3 до 39.5 кг),
для самок—-21.3 кг (от 19.7 до 23.2 кг). Особенностью сайги по срав-
нению с джейраном и дзереном является то, что при благоприят-
ных условиях существования она способна к быстрому ожирению. От
убитого поздней осенью сайгака получают, в среднем, 3.5—4.5 кг сала,
по качеству сходного с бараньим, а от крупного, хорошо упитанного
самца, по А. А Слудскому (1953, 1955), до 8—12 кг. Слой жира на спине
достигает иногда нескольких сантиметров. Даже весной, в мае, нам уда-
валось собрать от самцов и имевших детей самок до 2.8 кг подкожного
жира.
Мясо сайгаков обладает неплохими вкусовыми качествами, особенно
у молодых зверей, по вкусу несколько напоминает баранину.
Наиболее ценным продуктом, получаемым от сайги, до последних
лет были рога. Они высоко ценятся и находят применение в китайской
медицине. В начале 90-х годов в Казахстане цена на сайгачьи рога дохо-
дила до 20—25 рублей за пару, что в то время равнялось стоимости хоро-
шей лошади или верблюда (Слудский, 1955). Даже в 1926 г. в Астрахани
и Махач-Кале они скупались по 100—150 рублей (Гептнер и Формозов,
1941). Высокая стоимость рогов была важнейшей причиной хищнического
SAIGA
373
истребления сайги. В дореволюционные годы ежегодно экспортировались
из России в Китай десятки тысяч пар сайгачьих рогов.
Шкура сайгака не представляет особой ценности. Из зимних шкур
шились дохи (ергаки), которые, впрочем, из-за ломкости волоса, проч-
ностью не отличались. Непораженные подкожным оводом шкуры, вероятно,,
годятся для выделки хрома или замши.
Несколько слов о возможном вреде сайгака. По сообщению В. В. Ива-
нова (1952), «работники многих совхозов жаловались на то, что сайгаки
наносят значительный вред хозяйствам, уничтожая стога сена в степи».
А. Н. Формозов (1951) отмечал случаи поедания сайгой побегов молодых
деревьев в полезащитных полосах и проса на посевах. В последние годы
вновь усиленно раздаются голоса о вреде от сайгака посевам. Насколько
справедливы эти жалобы, пока сказать трудно (Воршев, 1955). Ставится
вопрос и о конкурентной роли сайгаков в овцеводстве на Черных Землях.
Во всяком случае эти соображения играли не последнюю роль в вопросе
о разрешении организованного промысла сайгаков в Астраханской об-
ласти, который осуществляется в последние годы.
В большинстве других областей едва ли может серьезно идти речь
о вреде от сайгака по сравнению с пользой, которую можно извлечь при
рациональной охране и промысле этого зверя.
Охота. Способы охоты и добычи сайги многообразны. Многие из
применяющихся ранее хищнических способов ее промысла (Адольф,
1948) в настоящее время имеют лишь исторический интерес и в дальней-
шем безусловно применяться не будут. Большинство их основано на
использовании сильно развитого у сайги стадного чувства. Так, широко
практиковался способ загона сайгаков в воронки из куч земли с расстав-
ленным в их узком конце барьером из острых камышин, какой приме-
нялся раньше и при добыче джейрана (см. стр. 329). Кроме того, загоняли
большие табуны сайгаков на предварительно скошенный на некоторой
высоте от земли острый камыш, на который натыкались обезумевшие от
крика и преследования звери. Помимо нерационального истребления
большого числа животных без разбора пола и возраста, масса их беспо-
лезно погибала позднее от полученных ран. Применялось не менее хищни-
ческое истребление сайгаков, загнанных на гладкий лед или в глубокий
снег, где беспомощные звери убивались палками или закалывались но-
жами. В прежнее время практиковалась охота при переправах через
реки, основанная на том, что если переправились передовые звери,
за ними следует весь табун, несмотря на избиение.
Из способов добычи сайгаков, не носящих истребительного харак-
тера и которые будут, вероятно, допустимы при разрешении промысла
в дальнейшем, могут быть названы следующие:
а) Скрадывание. Способ трудный, вследствие открытого ха-
рактера местообитаний сайги, но возможный благодаря несколько мень-
шей ее чуткости и более слабому зрению, чем у других копытных. Против
ветра можно подойти шагов на двести, особенно летом в жару, когда
сайгаки находятся на лежках, уткнув в землю морду.
б) Охота нагоном. Заключается она в том, что партия охот-
ников, найдя табун сайгаков, разделяется на две группы. Одна залегает
с ружьями на пути, по которому должны по расчетам пройти звери,
а вторая, обойдя табун кругом, старается осторожно направить его
в сторону притаившихся стрелков. Успех охоты при этом способе зависит
от того,, насколько удачно выбрано место для стрелков, а также от уменья
и выдержки загонщиков.
374
BOVIDAE
в) Подкарауливание на тропах у водопоев.
Этот способ применялся не так часто вследствие дневного образа жизни
сайги и открытого характера мест, избираемых зверями для водопоя.
г) Расстановка капканов на тропах, по которым
сайгаки ходят на водопой и жировки. В Семиречье устраивают подобие
длинных изгородей из разложенного на земле саксаула. В центре изго-
роди оставляется узкий просвет для прохода сайгаков, в котором и рас-
ставляются капканы (Шнитников, 1936).
д) Охота с борзыми. Увлекательный, но трудный способ.
Так как даже самая резвая среднеазиатская борзая (тазы) не может
догнать здорового сайгака, то охотятся только группами, имея сменных
собак и лошадей. При этом охотники расставляются так, что одни из них
гонят зверя в направлении остальных, спускающих свежих собак на
смену утомленным (Динник, 1910). Успех и в этом случае зависит в пер-
вую очередь от уменья направить ход зверя и от умелой расстановки
запасных охотников со свежими собаками и лошадьми.
В. Н. Шнитников (1936) описывает способ преследования сайгаков
на лошадях по кругу, рассчитанный, впрочем, на добычу только молодых
и слабых животных.
Сайгаков, как и джейранов, можно добывать с помощью автомобиля
на скорости около 70 км на ровном и твердом грунте. Однако в силу наз-
ванных ранее причин (см. стр. 329), этот способ, как хищнический, за-
прещен.
Практикуется уже описанный ранее (см. стр. 330) способ охоты на
автомобиле с фарой. Достоинства и недостатки его те же, что и при охоте
на джейрана. Этот способ, по-видимому, будет в дальнейшем основным
при плановых заготовках мяса сайгаков и джейранов.
Род NEMORHAEDUS FLOWER et GARSON — ГОРАЛЫ
Heude Р., 1894, Mem. Hist. Natur. Emp. Chinois, II : 234—245 (морфол.,
сист); Pocock R., 1998, Proc. Zool. Soc. London : 19Q—202 (сист., изобр.); Ly-
dekker R., 1913, Catal. Ungul. mamm. Brit. Mus., vol. I, London : 202—210
(сист., синон.); Pocock R., 1913, J. Bombay Natur. Hist. Soc., XXII, 2 : 311 —
317 (сист., синон., изобр.); Schaub S., 1928,Ecl. Geol. Helvet., vol. 21, № 1 : 260—
266 (эвол.); Adlerberg G., 1932, Изв. Акад, наук СССР, Отд. матем. и естеств.
наук : 259—285 (сист., синон.); Е Herman J. and Т. Morriso n-S с о t t,
1951, Checklist of palaearctic a. indian mamm., London : 401—402 (сист., синон.);
Соколов И. И., 1953, Тр. Зоол. инет. Акад, наук СССР, XIV: 162—167 (морфол.,
эвол.).
Характеристика рода. Животные средней величины. Рога круглого
сечения, слабо расходящиеся в стороны, иногда почти параллельные,
дугообразно загнутые назад, почти не подымаются над поверхностью
лба; имеются как у самцов, так и у самок. Носовое зеркало хорошо раз-
вито, занимает почти все пространство между ноздрями до их верхних
углов. Предглазничные железы рудиментарные.' Межпальцевые железы
есть, паховые отсутствуют. Молочная железа имеет четыре соска.
Профиль лобно-лицевой части черепа прямой или выпуклый. Края
глазниц слабо выступают в стороны. Носовые кости несколько укоро-
чены, но составляют все же редко меньше 36% основной длины черепа.
Лобные кости вздутые, поверхность лба поднимается выше краев глаз-
ниц. Основания роговых отростков расположены недалеко за глазницами.
Этимоидалъные щели отсутствуют. Резцы брахиодонтные; граница между
корнем и коронкой зуба хорошо заметна в виде перехвата. Последний
нижний премоляр имеет замкнутую переднюю долинку.
NEMORHAEDUS
3Z5
От очень близкого рода Capricornis Ogilby (серао) горалы отличаются
меньшей величиной, слабым развитием предглазничных желез. Основ-
ными отличиями в черепе являются: а) отсутствие предглазничных ямок,
б) сближенность теменных гребешков, наименьшее расстояние между
которыми редко бывает больше срединной длины теменной поверхности
затылочной кости, в) укороченность теменных костей, длина которых
по средней линии менее 60% ширины мозговой коробки, г) увеличенные
размеры костного носового входа; верхнечелюстные кости не соприка-
саются с носовыми; с последними также лишь передним уголком, и то
не всегда, соприкасаются слезные кости.
В скелете передних конечностей пястная кость короче и локтевой,
и плечевой костей.
История и географическое распространение. В настоящее время
область распространения рода Nemorhaedus включает южную часть
Дальнего Востока, Корею, большую часть провинций Китая (в том числе
Тибет), Бирму, Ассам, Непал, Пенджаб, Кашмир. В далеком прошлом
ареал был гораздо шире. Остатки, не вызывающие сомнения в принад-
лежности к роду горалов, найдены в верхнеплейстоценовых отложениях
Франции и Италии (Schaub, 1928). В области современного распростра-
нения , они обнаруживаются не раньше плейстоцена (Соколов, 1953).
Классификация. Систематика рода Nemorhaedus разработана неудо-
влетворительно и настоятельно требует ревизии. Без достаточных осно-
ваний описано большое число «видков» (Heude, 1894; Lydekker, 1913).
Необосновано и объединение всех форм в один вид (Ellerman and Morri-
son-Scott, 1951). По-видимому, самостоятельными видами следует счи-
тать:
1) Nemorhaedus goral Hardwicke — Гималайский горал. Имеет срав-
нительно короткий хвост, длиною без волос, около 7.5 см. Темная окраска
дорзальной стороны передних ног простирается почти до главных копыт.
Распространение: Тибет, Кашмир, Пенджаб, Непал, Сикким.
2) Nemorhaedus caudatus Milne-Edwards — Амурский, или китайский
горал. Хвост болре длинный, около 12.8 см. Темная окраска дорзальной
стороны передних ног от запястных суставов переходит на латеральную
сторону нижних частей конечностей до добавочных копыт. Распро-
странение: юг Дальнего Востока, Корея, Китай, Бирма.
Nemophaedus caudatus Milne-Edwards — Амурский, или китайский ,
горал (рис. 79—81)
Schrenk L., 1858, Reisen u. Vorschungen im Amurlande, Bd. 1, Lief. 1, St.-
Petersb. : 158—160 (распр.); Radde G., 1862, Reis, im Siiden v. Ost-Sibirien, I,
St.-Petersb. : 262—270 (опис., изобр., биол.); M i 1 n e-E d war ds M., 1868, Rec-
berches pur servir & 1’bist. natur. des mammiferes, t. I—II, Paris : 186—189 (сист., изобр.);
Туркин H. В. иК. А. Сатунин, 1905, Звери России, Bovidae, М. : 523—525
(опис.); Evans G., 1905, Proc. Zool. Soc. London : 311—314 (опис., биол.); Po-
cock R., 1908, Proc. Zool. Soc. London : 196 (сист., синон., изобр.); P о с о с k R.,
1910, Proc. Zool. Soc. London : 854—855 (морф.); Lydekker R., 1913, Catalogue
Ungul. mammals Brit. Mus. London : 206—210 (сист., синон., изобр.); Pocock R.,
1913, Bombay Natur. Hist. Soc., XXII, 2 : 314—317 (сист., синон.); Pocock R.,
1918, Ann. Mag. Natur. Hist. (ser. 9), II : 135—136 (морф.); Таежник, 1930, Охот-
ник, 1 : 15 (биол., хоз. знач.); Flower S., 1931, Proc. Zool. Soc. London : 211 (npo-
долж. жизни); Adlerberg G., 1932, Изв. Акад, наук СССР, Отд. матем. и естеств.
наук 2, : 264—267 (сист., синон.); Каверзнев В. Н., 1933, Полорогие фауны
СССР, М., КОИЗ : 35—37 (общ., изобр.); Золотарев Н. Т., 1936, Млекопит.
басе. р. Имана, М.—Л., изд. Акад, наук СССР : 108—110 (распр.); Абрамов К. Г.,
1939, Научи.-метод, зап. Главн. управл. по заповеди., IV : 198—201 (распр., биол.);
Allen G., 1940, Mammals of Chine and Mongolia, Pt. 2, N. Y. : 1240—1244 (сист.,
376
BOVIDAE
синон., биол.); Щульц Р. С. и А. Н. К а денаци, 1950, Тр. Гельминтол. лабор.
Акад, наук СССР, III : 152—160 (параз.); Соколов И. И., 1953, Тр. Зоол. инет.
Акад, наук СССР, XIV : 167 (эвол.); Абрамов К. Г., 1954, Копытн. звери Дальн.
Востока, Хабаровск, Книжн. издат. : 103—112 (опис., распр., биол., хоз. знач., изобр);
Насимович А. А., 1955, Роль режима снежн. покрова в жизни копытных, М.,
изд. Акад, наук СССР : 165—166 (распр., биол.).
Описание. Животные средней величины; длина тела от 100 до 130 см,
высота в холке 70—90 см. Основная длина черепа от 172 до 214 мм. Вес
до 48 кг.
Телосложение неуклюжее, несколько напоминающее козлов. Массив-
ное, покрытое длинным мохнатым волосом туловище покоится на корот-
ких, довольно толстых ногах. Профиль спины прямой или выгнутый;
иногда крестец держится ниже холки. Голова не тяжелая, конусообразно
суживающаяся впереди глаз, но Притупленная на конце. Носовое зер-
кало (голое пятно на конце морды) лучше развито, чем у козлов и бара-
нов; оно занимает большую часть пространства между ноздрей, доходит
до верхних краев последних, а также тянется в виде узкой вертикальной
полоски посредине верхней губы. Волосистая кожа вдается Сверху между
ноздрями лишь в средней части. Цвет носового зеркала черно-бурый.
Верхняя губа, кроме узкой срединной полоски, покрыта волосами. Глаза
небольшие, слабо выступающие в стороны; радужная оболочка темно-
коричневого цвета. Уши длинные, 12—14 см, не менее половины длины
головы.
Рога имеют и самцы, и самки. Основания их расположены непосред-
ственно позади глазниц, форма — дугообразно загнутая назад. Попереч-
ное сечение — круглое, с обхватом в основаниях около 8—11 см; к вер-
хушкам рога конусообразно заостряющиеся. Расстояние между их основа-
ниями не превышает 1—1.5 см. Длина рогов у самцов от 16 до 19—20 см;
у самок они немного короче и тоньше. Цвет темно-бурый, почти черный.
Поверхность роговых чехлов на две трети от оснований состоит из мелких,
расположенных почти вплотную друг к другу колец; только верхушка
рога гладкая. Число колец с возрастом увеличивается, но точным кри-
терием возраста они служить не могут.
Шея короткая и благодаря длинной шерсти кажется толстой; дер-
жится в спокойном состоянии горизонтально. Конечности короткие,
особенно ниже запястных и скакательных суставов.. Высота копыт по
переднему краю на передних конечностях от 33 до 40 мм, на задних на
2—5 мм меньше; длина от заднего края пяточных мякишей до верхушек
на передних ногах 47—58 мм, на задних 42—52 мм. Добавочные копыта
довольно крупные, около 20—25 мм. Хвост сравнительно с другими
видами подсемейства длинный; длина его без волос 11—19 см, с волосами
до 46 см; волосами он покрыт и сверху, и снизу.
Волосяной покров у амурского горала зимой густой и длинный, но
мягкий. Только на передней части головы он более короткий. Волосы
не ломкие и крепко сидят на коже. Очень хорошо выражено деление
волосяного покрова на ость и пух, или подшерсток. Длина остевых волос
на туловище зимой доходит до 8—9 см. Подшерсток очень густой, длиной
до 3.0—3.5 см. И остевые волосы, и волосы подшерстка извиты, что вместе
с длиной и мягкостью придает волосяному покрову несколько взлохма-
ченный характер. Более грубые и упругие, направленные назад волосы
на верхней стороне шеи и холке образуют подобие гривы высотой 13—
14 см, подымающейся, когда зверь находится в возбужденном состоянии.
Волосы на хвосте также более грубые, постепенно удлиняются от корня
NEMORHAEDUS
377
к вершине и на конце достигают длины 26—27 см. Удлиненные, но мягкие
волосы на лбу перед основаниями рогов образуют - подобие вихра или
челки.
Остевые волосы имеют ясно выраженную зональность окраски. От
оснований они обычно светло-бурые, кверху постепенно темнеют, затем
резко обрываются беловатым широким кольцом, а верхушки на значи-
тельном протяжении вновь приобретают черно-бурую или черную окраску.
Волосы подшерстка одноцветные буро-коричневые.
Рис. 79. Горал {Nemorhaedus caudatus Milne-Edw.).
Общий тон окраски амурского горала желтовато-бурый или серовато-
желтый, но варьирует от грязного желто-белого до серого и темно-бурого,
почти черного. У отдельных экземпляров наблюдается явное преоблада-
ние коричневого оттенка. Верхняя сторона носа, лоб, темя и затылок
темно- или даже черно-бурые. Хохол на лбу имеет иногда примесь
белых волос. Бока головы и губ желто-бурые или желтовато-серые.
Иногда над глазами и впереди них имеется более светлая желтоватая
полоса. Передний край верхней и нижней губ беловатые. Волосы
на внутренней стороне ушей светлые, почти чисто белые; наружная их
сторона имеет одинаковую с туловищем окраску, кончики черные. Под-
бородок черный. На нижней стороне головы и шеи в области гортани
большое белое пятно, иногда с легким рыжеватым налетом. Удлиненные
волосы гривы, а также узкая полоса вдоль спины, крестца и тыльной
поверхности хвоста черно-бурого, почти черного цвета. Темная полоса
378
BOVIDAE.
вдоль спины развита не одинаково и иногда бывает почти незаметна'
Окраска шеи, туловища, бедер и низа живота желтовато-серая или серо-
бурая, местами очень темная, почти черная. Оттенок и интенсивность
окраски того или иного участка тела зависят от размеров темно и светло
окрашенных участков волос, густоты кроющего волоса и подшерстка.
Пахи, область молочной железы, наружных половых органов и про-
межность белого цвета. Иногда по сторонам от хвоста на ягодицах заметно
очень небольшое околохвостовое белое зеркало. Нижняя сторона хвоста
белая. Кисть на его конце состоит из смеси длинных волос темно-бурого
и белого цвета. Наружные стороны передних и задних ног до запястных
и скакательных суставов имеют одинаковую с туловищем окраску. По
передней стороне предплечий сверху до запястий тянутся темно-бурые
полосы; ниже запястий, передняя и внутренняя стороны ног имеют
ржаво-рыжую окраску; вдоль наружной стороны тянется темная полоса,
захватывающая и область добавочных копыт сзади. Задние ноги книзу
от скакательных суставов спереди светлые, ржаво-желтые, сзади темно-
бурые. Книзу от путовых суставов и передние, и задние ноги до копыт
светлые, беловатые, с легким буроватым оттенком.
Самцы и самки по окраске отличаются мало.
Возрастные и сезонные изменения окраски до сих пор не изучены.
По-видимому, не так редко встречаются звери белой окраски, альбиносы
(Абрамов, 1954; Таежник, 1930).
Предглазничные железы у горалов рудиментарные, но имеются и пред-
ставляют небольшой, почти лишенный волос участок кожи с утолщением
соединительнотканным слоем. Межпальцевые железы имеют характер
довольно узких изогнутых каналов, расположены на уровне первых
и вторых фаланг; внутренние стенки их покрыты редкими волосами,
склеенными секретом. Открываются железы круглыми отверстиями на
передней стороне ног повыше копыт (Рососк, 1910).
Характеристика черепа (рис. 80) дана при описании рода.
История и географическое распространение, (рис. 81). В ископаемом
состоянии остатки, достоверно относящиеся к современному горалу, до
последнего времени не были известны. Видовая, а иногда и родовая при-
надлежность плейстоценовых находок из Китая не установлена. По полу-
ченным автором от Амурского областного музея краеведения сведениям,
в последние годы при раскопках древних поселений кости горала найдены
в горных районах верхнего Амура — Хингано-Архаринском и Маза-
новском. К сожалению, возраст слоев или поселений, где были сделаны
эти находки, мне не известны.
В настоящее время ареал N. caudatus включает южную часть Дальнего
Востока, Корею, а также западные, центральные и северо-восточные
провинции Китая: Сычуань, Юнань, Шэньси, Шаньси, а также Бирму
до Аракана.
В пределах СССР, еще в прошлом столетии, горал был распространен
шире, чем в настоящее время. Шренк (Schrenk, 1858) застал этого зверя
на нижнем течении Амура, причем устье этой реки он считал северной
границей его распространения. Существование горала в области левых
притоков нижнего течения Амура категорически отрицалось; Уссури
считалась западной границей его распространения в Приморье. Однако
Г. Радде (Radde, 1862) во время путешествия по восточной Сибири через
несколько лет после Шренка получил экземпляр горала из южной части
Буреинского хребта, добытого на высотах, спускающихся к Лагару,
левому притоку Амура. Правда, уже тогда зверь в тех местах был очень
Рис. 80. Череп Nemorhaedus caudatus Milne-Edw.
380
BOVIDAE
редок и, кроме названного пункта’ встречался лишь’ на северо-западной
оконечности Буреинского хребта; следы его Г. Радде находил также
в верховьях р. Дичуна (приток Амура). В коллекциях Зоологического
института АН СССР имеется шкура и череп горала, полученные из Па-
рижского музея естественной истории с обозначением места добычи:
«Никольск. Граница с Маньчжурией». Можно предполагать, что речь
идет о с. Екатеринино-Никольское Еврейской авт. области, т. е. неда-
леко от районов, указываемых Г. Радде. Дата добычи этого экземпляра
не известна.
В настоящее время горала ни в Буреинском хребте, ни на Амуре
в пределах СССР, по-видимому, нет.1 Северным пределом в наши дни
следует считать верховья р. Бикина к югу от Хоро-Бикинского водораз-
дела.
Современный ареал в Приморье ограничивается южной половиной
Сихотэ-Алиня и разобщен на ряд далеко отстоящих друг от друга изоли-
рованных местообитаний (Абрамов, 1939, 1954). Около трех четвертей
сохранившегося в Приморье поголовья сосредоточено на территории
Судзухинского заповедника. Размножившиеся здесь под охраной звери
расселились с прибрежных сопок Горал и Туманная по прилегающим
горам Тачин-Джана, Тачин-Гуана и по Алексеевскому хребту. Проникли
горалы также на гору Павлины и даже на гору Три Сестры в Чугуевском
районе. «Помимо территории бывшего Судзухинского заповедника и со-
седних с ним участков местности, горал есть в Приморье в Хасанском
районе, на горе Синий Утес, в Буденновском — на горе Чинда-Лаза,
в Сучанском — на отрогах Сихотэ-Алиня по р. Малазе, в окрестностях
селений Молчановки и Сергеевки. Много горалов и по скалистым берегам
р. Ванцин, у ключей Фасольего и Бараньего. Встречаются эти животные
по р. Пхусуну, близ впадения ее притоков Сяо-Вангоу и Та-Вангоу,
на так называемых Бараньих Скалах.
«Севернее горал есть около Тетюхе, на лазе Китовое ребро, на мысе
Абрек, в приморской части Сихотэ-Алиньского заповедника и дальше на
север до устья р. Пайна. Самая северная точка, где сейчас встречаются
горалы, находится, по-видимому, в верховьях Бикина, на речных щеках
р. Килоу. В 1947 г. удэгеец Гамбу случайно добыл там, около устья
Улунги, одного самца горала.
«На западных склонах Сихотз-Алиня горал есть в верховьях Тудагоу
(даубихинской) и по р. Даубихэ, выше с. Виноградовки. В Чугуевском
районе горал недавно встречался в предгорьях Лабо-Лазы и Лаоне-Лазы.
Далее на север он обитает до р. Имана, где сохранился близ с. Лаулю
на скалах сопки Чичивезы и около с. Вахумбо на скале Сяо-Лаза.
«В 1935 г. в Санчихезе мы видели шкуру горала у П. М. Слесарева,
получившего ее из рук удэгейца, добывшего этого зверя на Сяо-Лазе.
Теперь животные в этих местах уже не водятся» (Абрамов, 1954). Н. Золо-
тарев (1936) указывал горала также для районов бухты Терней, для
верховьев р. Моно, притока Бикина, и для Ольгинского района.
Подвиды. Описаны три формы горалов, которые следует рассматри-
вать как подвиды Nemorhaedus caudatus Milne-Edwards.
1) N. caudatus caudatus Milne-Edwards. Длинный, мягкий и пушистый
волосяной покров. Окраска ног ниже запястных и скакательных суставов
более или менее однотонная, светлого желтовато-коричневого цвета.
1 По словам Н. Т. Золотарева (1936), горал известен в окрестностях с. Помпе-
евки на Амуре, но после него эти сведения никем не были подтверждены.
NEMORHAEDUS
381
Конец хвоста черный, темнее его основания и срединной полосы на спине.
Распространение: западный, центральный и северный Китай.,
2) N. caudatus raddeanus Heude. Длинный, мягкий и пушистый воло-
сяной покров. Наблюдается резко выраженный контраст между светлой
окраской ног ниже путовых суставов и выше расположенными, темно
окрашенными частями. Верхняя сторона хвоста темно-бурая, однотонная
со срединной полосой на спине. Судя по размерам черепа, отличается
Рис. 81. Распространение Nemorhaedus caudatus
Milne-Edw. 1 — XX в.; 2 — XIX в.
от других рас более крупной величиной. Распространение:
Дальний Восток, Корея, вероятно также северо-восточный Китай.
3) N. caudatus griseus Milne-Edwards. От двух других рас отличается
меньшей величиной, более коротким и грубым волосяным покровом,
темной и сверху, и снизу окраской хвоста, а также ясно выраженной
желтизной светлого пятна на горле. Распространение: южный
и западный Китай (провинции Сычуань и Юньнань), Бирма.
Биология. Биология горала изучена слабо, вследствие его малочис-
ленности и обитания в трудно доступных местах. Распространение этого
зверя неразрывно связано со скалистым ландшафтом. В Китае горал
населяет наиболее высокие вершины (Allen, 1940). Чем круче и недоступ-
нее скалы, тем они больше предпочитаются и в первую очередь заселяются
горалом. Только рано утром и поздно вечером отваживаются горалы
покидать на короткое время скалистые уступы, чтобы покормиться на
травянистых площадках, поблизости от своих убежищ (Evans, 1905).
382
BOVIDAE
В Приморье, по данным К. Г. Абрамова (1939, 1954), горалы обитают
В нижнем и среднем поясах гор, не свыше 500 м. Важным фактором ста-
диального размещения является глубина снега; при глубине его свыше
35 см они могут ходить только по тропам. По этой причине основная
масса горалов придерживается юго-восточных склонов Сихотэ-Алиня,
где выпавший снег сохраняется недолго или не задерживается вовсе.
Наиболее типичными местообитаниями являются приморские крутые
обрывы, а в стороне от моря — каменистые россыпи, перемежающиеся
со скалами, поросшими широколиственной растительностью с преобла-
данием дуба и примесью кедра, а также склоны ущелий рек, так называе-
мые речные щеки.
«Наиболее удобны для жизни горалов приморские скалистые участки,
сохранившие местное название „лазы“, что значит скалы. Здесь, в этих
нагромождениях скал горалы находят наилучшие условия жизни. Зимой
среди обрывов имеются бесснежные площадки и солнцепеки с рано зеле-
неющей ланцетовидной осокой. Есть надежные места, недоступные для
волка и трудно доступные для охотников. Здесь хорошо сохраняется
молодняк, здесь спасаются от преследования врагов и взрослые особи.
«Сохранившиеся вдали от побережья гористые участки менее удобны
для горалов. Скалы там постепенно разрушаются, рельеф сглаживается,
„стареет”, и сюда легче проникает волк. Таким образом, наилучшие угодья
горалов в настоящее время расположены вдоль побережья Японского
моря» (Абрамов, 1954).
Пища амурского горала разнообразна. С ранней весны и до мая основу
питания составляют лесные осоки, из которых, по К. Г. Абрамову (1954),
ланцетовидная осока имеет наибольшее значение. Летом горалы поедают
овсяницу, ясьменник, зубровку, плейкрантус, вику и некоторые другие
травы, а также полынь, леспедецу, листья и побеги древесных растений:
амурского винограда, дуба, липы, маньчжурского ясеня и других. Осенью
питаются желудями, опавшими листьями деревьев, засохшей травой.
Засохшая трава вместе с древесно-веточным кормом составляют основу
и зимнего питания. Из других кормов горала называют древесные лишай-
ники и морские водоросли.
В связи с сезонностью кормовой базы наблюдаются перекочевки гора-
лов. Ранней весной они поднимаются выше на солнечные склоны, где
зелень появляется раньше. Осенью же, когда в горах растительность
отмирает, звери выходят кормиться на приморские склоны, где зеленый
корм сохраняется дольше.
Держатся горалы небольшими группами или в одиночку; но в Бирме,
по словам Г. Эванса (Evans, 1905), живут группами по 4—6 и даже до ^го-
лов вместе.
Суточный цикл жизнедеятельности весьма не сложен. На кормежку
утром выходят иногда затемно и кормятся часов до 10 утра. Днем отды-
хают. Около 4 часов пополудни вновь отправляются на пастбище, где
проводят время до сумерек. Весной на молодой зелени пасутся круглые
сутки. Выходят днем на пастьбу иногда также в дождлйвые дни и в пас-
мурную холодную погоду (Evans, 1905). Дневные часы, как правило,
проводят поблизости от мест кормежки в скалах. Для отдыха ложатся
в тени под навесом йли на выступе скалы, в таком месте, где обеспечен
достаточный кругозор и возможность наблюдать за всем происходящим.
Лежащий или стоящий па фоне темных скал горал, удивительно сход-
ный с ними по тону окраски, становится почти незаметным. Известен
случай, когда одним выстрелом были убиты два горала, причем охотником
NEMORHAEDUS
383
второй зверь был обнаружен только при падении (Allen, 1940). На водо-
пой спускаются после утренней кормежки и сразу же после этого ухо-
дят в скалы.
Раз избрав место жительства, горал придерживается его стойко,
и если его здесь никто не тревожит, то быстро его не меняет. Места лежек
также постоянны и узнаются по так называемым «уборным» — кучкам
помета, напоминающего козий (Абрамов, 1939). Переходы от мест отдыха
на пастбища, к водопою и обратно горалы совершают по определенным
тропам. В тихую ясную погоду в Приморье любят отдыхать на скалистых
обрывах в непосредственной близости от моря.
Горал хорошо приспособлен к передвижению по крутым скалам,
не уступая в этом отношении козлам и серне. С этим связана короткая
у него пясть и плюсна, как и у других горных копытных. В скалах горал
чувствует себя в родной стихии, лучше, чем на ровном месте. Прекрасно
и лазает, и прыгает по самым незначительным выступам скал. Даже тяжело
раненный быстро и подымается, и спускается почти по отвесным склонам,
исчезая бесследно и от охотника, и от собаки. На ровном же месте горал
становится беспомощным, передвигается короткими неловкими прыжками
и легко догоняется собакой, если не успевает скрыться в скалах.
У горала хорошо развиты, по Абрамову (1939), слух и обоняние,
тогда как зрение довольно слабо. Даже от стоящего открыто человека,
если тот неподвижен, горал не бросится прочь, а сначала будет пытаться
разобраться в чем дело. Эванс (Evans, 1905) утверждает обратное и гово-
рит, что этот зверь больше полагается на зрение, чем на слух и обоняние.
При возбуждении или испуге горал поднимает хвост кверху, подобно
козе; приподнимаются при этом также и длинные волосы на гриве.
Голос горала при испуге Абрамов (1939) передает, как тревожное
«чек-чек-чек-чек-чек...»; Эванс (Evans, 1905) сравнивает его с коротким,
повторяющимся несколько раз подряд отрывистым шипением или чиха-
нием.
О размножении горала почти ничего не известно. Самцы и самки парами
держатся в сентябре. Возможно в это время происходит и гон. На Даль-
нем Востоке самки приносят чаще одного, реже двух горалят в конце
мая или в первой половине июня. Для отела выбираются наиболее непри-
ступные места в скалах. Продолжительность жизни на воле не известна.
В Лондонском зоологическом саду самец горала прожил 17 лет, 7 месяцев
и 23 дня (Flower, 1931).
Из врагов горала на первом месте по значению в Приморье стоит
волк, который приносит вред не только непосредственным уничтожением,
но и тем, что разгоняет зверей с обжитых участков. С тех пор как на терри-
тории б. Судзухинского заповедника появились в большом количестве
волки, горалы были вынуждены покинуть пологие склоны Сихотэ-Алиня,
где кормовые условия для них, вероятно, значительно более благо-
приятны, и переместиться в наиболее трудно доступные скалы (Абрамов,
1939). Не будучи, вероятно, в состоянии преследовать в горах, волки
подкарауливают зверей на постоянных тропах при переходах к местам
кормежек и на водопои. Большой вред приносит рысь, которая чаще
давит горалов на лежке. Молодых давит харза, а также уносят орланы.
У амурских горалов, содержавшихся в Московском зоопарке, при
вскрытиях обнаружено 26 видов внутренних паразитов-гельминтов, из
которых четыре явились новыми, а два специфичными для N. caudatus
(Шульц и Каденаци, 1950). Фауна эктопаразитов не изучена, но из-
вестно, что оводами, ни подкожными, ни носоглоточными, горалы в совет-
384
BOVIDAE
ском Приморье не поражены.1 Иногда на них нападают клещи и муха-
кровососка.
Хозяйственное значение. Количество сохранившихся в советском
Приморье горалов столь невелико, что практического значения в настоя-
щее время они совершенно не имеют. Охота на них запрещена повсе-
местно. При увеличении их численности в результате охраны промысел
горала может стать дополнительным источником получения шкур и мясд.
Мясо его обладает хорошими вкусовыми качествами. Шкура с ее мягким
и прочным мехом может использоваться для пошивки верхней меховой
одежды.
Вероятно была бы успешна акклиматизация горала в некоторых
горных районах, например на Кавказе или в горах Средней Азии.
Благодаря хорошим качествам меха следовало бы предпринять по-
пытки к одомашнению горала. Опыт Судзухинского заповедника показы-
вает, что горалы хорошо выносят содержание в неволе при надлежащем
уходе и учете специфики кормового режима (Абрамов, 1939).
Продукты убоя горалов находят применение в китайской медицине.
Наиболее ценится кровь, особенно из «сердечных сумок» (имеется в виду,
вероятно, предсердие). Кровь промышленники собирают и высушивают
вместе с сердцем. Китайцы, говорят, отличают кровь горала от крови
других животных даже в высушенном состоянии. Употребляется она при
малокровии, ушибах и даже переломах костей. Кроме крови, исполь-
зуется как лекарственное средство и вся туша горала, оцениваемая
в связи с этим, по словам К. Г. Абрамова (1939) до 1000 рублей. «Ки-
тайцы, — пишет Таежник (1930), — будто бы кладут очищенную от
внутренностей тушку с кожей и шерстью в котел и варят до тех пор, пока
тушка не разваривается совершенно, после чего содержимое котла про-
цеживается сквозь сито и остуживается. Таким образом изготовляется
холодное, которое дается людям, больным малокровием». Высокая оценка
горала китайской медициной явилась, вероятно, причиной быстрого хищ-
нического его истребления в прошлое время.
Охота. Охотились на горалов главным образом скрадом или
с подъезда на лодке к прибрежным скалам, на которых держатся звери.
Собак при охоте не применяют, так как они не могут преследовать и оста-
навливать зверя в крутых диких скалах. Известно (Абрамов, 1954),
что горал очень крепок на рану. В настоящее время охота на горала
строго запрещена.
Род RUPICAPRA BLAINVILLE —СЕРНЫ
Отличительные признаки рода см. выше (стр. 345).
[Rupicapra rupicapra Linnaeus — Серна (рис. 82—84)
Linnaeus С., 1758, Syst. Natur., ed. X, vol. 1:68 (первооп.); Bonaparte
Ch., 1844, Atti Riun. Sci. Ital. : 337 (сист.); Mann C., 1867, Zool. Garten, VII : 275—
276 (гибр.); Чуди, 1873, Альпийский мир, СПб.: 421—470 (биол., ох., изобр.); Gir-
tanner Н., 1880, Zool. Garten, XXI : I—II-1-44—48 (прир., содерж.); Keller F.,
1887, Die Gemse, Klagenfurt:!—VIII-J-l—515 (общ., изобр.); Васильев Я.,
1896, Природа и охота, 2 : 14—15 (распр.); Динник Н. Я., 1896, Природа и охота,
2 : 36—59 (биол.); Neumann О., 1899, Ann. Mus. Genova, ser. 2, vol. XX : 347
(сист.); Туркин H. В. и К. А. Сатунин, 1904, Звери России, Bovidae, М. :
506—522-J-595—607 (общ., изобр.); Lydekker R., 1908, Field., GXII : 104 (сист.);
Boule М., 1910, Les grottes de Grimaldi, I, 3 :159—236 (ист.); Динник H. Я., 1910,
1 Устное сообщение К. Я. Грунина.
RUPICAPRA
385
Звери Кавказа, I, Тифлис : 220—246 (общ.); Cabrera А., 1910, Proc. Zool. Soc.
London : 998—999 (сист.); Meerwarth H., 1910, В кн. «Lebensbilder aus der
Tierwelt, Bd. II, F. I, Saugetiere, II», Leipzig : 1—69 (общ., изобр.); P о -
cock R., 1910, Proc. Zool. Soc. London : 849—851 (морф.); Lydekker R.., 1913,
Catal. Ungulate mammals, Brit. Mas., I, London : 179—186 (морф., сист., синон.);
Рос ос k R., 1918—1919, Ann. Mag. Natur. Hist., ser. 9, 11:131—132 (морф.);
M e r k-B u c h b e r g M., 1922, В кн. «Lebensbilder aus der Ticrwelt Europas, Sau-
getiere, IV», Leipzig : 13—33 (биол., изобр.); В о 1 k a у St., 1925, Novit. Mus. Sarae-
voensis, I : 15—16 (сист.); Brehms Tierleben, 1925, 4, AufL, Bd. 13, Leipzig :
231—240 (общ., изобр.); Koller О., 1929, Zool. Anzeig., LXXXIII, 1—4 : 46 (сист.);
Flower S., 1931, Proc. Zool. Soc. London : 211 (продолж. жизни); Каверз-
нев В. Н., 1933, Полорогие фауны СССР, М.—Л., КО113 : 26—35 (общ., изобр.);
Zedtwitz F., 1937, Gams in ihrer Bergheimat, Berlin—Lichterfelde:!—60
(общ., изобр.); Couturier M., 1938, Le chamois, Rupicapra rupicapra (L.), Gre-
noble : 1—855 (общ., синон., изобр.); Насимович, А. А., 1939, Воир. эколог, и
биоцеполог., 7 : 53—59 (распр., биол.); Гептнер В. Г. и А. Н. Формозов,
1941, Сборн. тр. Гос. зоол. муз. Моск. гос. унив., VI : 72—73 (распр., биол.); Н а-
симович А. А., 1941, Научно-метод. зап. Главн. управл. по заповеди., VIII :
178—181 (биол.); Саркисов А. А., 1944, Тр. Ереванск. зоопарка, 1/2 : 139—142
(распр., стац.); Т h о ring ton J., 1944, Bull. Hist. Medic., t. 15, № 1 : 65—78 (хоз.
знач.); Верещагин H.K., 1947, Охотничьи и промысл, животные Кавказа, Баку,
изд. Акад, наук Азербайдж. ССР : 48—50 (распр.); Whitehead G., 1947, Country
Life, London, 102 : 1214—1215 (ox.); Cobb E., 1948, Tield, London, 192 : 553 (ак-
климат., изобр.); Whitehead G., 1949, Country Life, London, 105 : 668—669 (ox.);
Couturier M., 1949a, Mammalia, XIII, 1/2 : 49—51 (рога); Couturier M.,
19495, Sur le traces de mes 500 chamois de France, Paris : 1—273 (ox.); Насимо-
вич A. A., 1949, Тр. Кавказск. гос. заповеди., 3 : 51—64 (биол.); Y a r h a m E. R.,
1949, Zoo life, London, vol. 4, № 2 : 60—62 (изобр.); Couturier M., 1951, Mamma-
lia, XV, 3 : 73—83 (морф.); E Iler ma n J. and T. Morrison-Scott, 1951,
Checklist of palaearctic and indian mammals, London : 397—398 (сист., синон.);
Верещагин H. К., 1952, Млекопитающие Кавказа, Диссерт. : 915—921 (распр.,
ист.); Эквтимишвили 3. С., 1952, Сообщ. Акад, наук Грузинск. ССР, 8 : 477—
484 (стац.); Соколов И. И., 1953, Тр. Зоол. ипст. Акад, наук СССР, XIV : 157—
158 (морф., эвол.); Эквтимишвили 3. С., 1953, Сообщ. Акад, наук Грузинск.
ССР, XIV, 3:163—166 (стац.); Эквтимишвили 3. С., 1953, Сообщ. Акад,
наук Грузинск. ССР, XIV, 5 : 303—308 (стадность); Асадов С. М., 1954, Докл.
Акад, наук Азербайдж. ССР, X, 10 : 643—647 (параз.); Gray А., 1954, Mamma-
lian hybrids, London—Edinburg : 81 (гибр.); D a ne k Z., 1954, Wszechswiat., I : 5—
11 (общ., изобр.); Насимович A. A., 1955, Роль режима снежного покрова
в жизни копытных, М., изд. Акад, наук СССР : 157—165 (распр., биол.).
Описание. Животные сравнительно небольшой или средней вели-
чины; высота в холке от 71 до 86 см; длина тела от 102 до 119 см. Основ-
ная длина черепа 169—201 мм. Живой вес в пределах 25—45 кг, редко
больше.1 Средний вес самца около 30—36 кг (Динник, 1910).
Телосложение в общем легкое. Сравнительно короткое туловище
с широкой и глубокой грудной клеткой покоится на довольно толстых
высоких ногах. Профиль спины несколько выпуклый; высота в крестце
на 4—6 см превосходит высоту в холке. Голова легкая, с узкой, суживаю-
щейся к концу мордой. Носовое зеркало небольшое, захватывает лишь
пространство между внутренними углами ноздрей, но иногда вдоль
внутренних краев доходит почти до верхних углов последних. Вся верх-
няя губа, кроме узкой срединной полоски, покрыта волосами. Цвет но-
сового зеркала черно-бурый. Глаза не очень большие, но выпуклые,
заметно выступают в стороны. Радужная оболочка глаз желто-бурая.
Уши сравнительно длинные, около 12—14 см, обычно не меньше по-
ловины длины головы.
Рога имеют и самцы, и самки; у последних они сравнительно хорошо
развиты, лишь немного короче и тоньше, чем у самцов. Основания ро-
1 Как рекорд, указывается вес старого самца из Карпат в 62 кг (Couturier, 1938).
25 Фауна СССР, И. И. Соколов
386 BOVIDAE
гов расположены над глазницами. Характерна их форма в виде почти
параллельных, вертикально поставленных, а у живого животного даже
часто несколько наклоненных вперед и правильно загнутых назад на
вершинах крючков. Поперечное сечение рогов круглое или овальное;
обхват в основаниях около 7—8 см. Расстояние между основаниями
редко превосходит 10—15 см. Длина рогов по изгибу от 17—18 см у са-
мок, до 24—24 см у самцов.1 Цвет рогов серовато-бурый или темно-бу-
рый. Поверхность роговых чехлов несет большое число мелких попереч-
ных колец с бороздками между ними. У оснований рогов кольца распо-
ложены вплотную друг к другу и более узкие, ближе к вершинам ши-
рокие, сглаженные, с заметными интервалами между ними. Загнутые
концы рогов поперечной кольчатости не имеют. Кроме того, на всем
протяжении от оснований до верхушек на рогах заметна мелкая, но хо-
рошо выраженная продольная исчерченность.
Шея серны короткая, но сравнительно тонкая. Голова даже в спо-
койном состоянии держится приподнятой кверху. Ноги, как уже отме-
чалось, высокие и довольно толстые. Обращает внимание большая в
сравнении с общими размерами животного величина копыт, способных
к тому же сильно раздвигаться. Длина их от заднего края пяточных мя-
кишей до верхушек на передних конечностях от 66 до 73 мм, на задних
от 62 до 68 мм. Высота копыт по переднему краю на передних ногах 43—
47 мм, на задних на 1—2 мм меньше. Характерная особенность копыт
серны заключается в том, что их толстая роговая стенка загибается
внутрь и образует непосредственно позади пяточных мякишей хорошо
заметный выступ. Этот выступ, по-видимому, играет роль тормоза или
зацепа при спуске зверя по крутым каменистым склонам. Добавочные
копыта расположены высоко, но хорошо развиты (длиною около 34 и
шириною до 22—23 мм) и, вероятно, также играют немалую роль в ка-
честве тормозов при передвижениях в скалах. Цвет копыт темно-
бурый.
Хвост у серны короткий (около 12 см), даже с волосами лишь немного
превосходит длину уха.
Волосяной покров более грубый, чем у горала, и ясно делится на
ость и тонкий подшерсток. Длина меха в полном зимнем наряде на туло-
вище может доходить до 8—9 см, а наиболее удлиненные остевые волосы
вдоль средней линии спины и шеи достигают, по Н. Я. Диннику (1910),
даже 15—47 см. На морде и нижних частях конечностей волосы и зимой,
и летом более коротки по сравнению с другими частями тела. Волосы
на хвосте, наоборот, несколько длиннее, хотя и не образуют сколько-
нибудь заметной кисти. Нижняя поверхность хвоста, кроме концевой
части, волос не имеет. Как остевые волосы, так и волосы подшерстка
имеют легкую волнистость.
Зональность окраски отдельных волос выражена сравнительно слабо.
Остевые волосы летнего меха от основания светло-бурые, затем их окраска
переходит в желтую, занимающую большую часть волоса, и только са-
мые верхушки вновь несколько темнеют, приобретая темно-бурый отте-
нок. Более длинные волосы вдоль средней линии шеи и спины темно-
‘бурые, но кончики их имеют яркую желтовато-рыжую окраску.
Окраска серны довольно однотипная, подвержена незначительной
индивидуальной изменчивости, но сезонный диморфизм выражен сильно.
1 Известны рога серны длиною 28.6 и 33.7 см (Couturier, 1938, 1949).
RUPICAPRA
387
Общий тон окраски летом ржаво-рыжий или желтовато-рыжий,
иногда с легким буроватым налетом, в зависимости от числа волос с бу-
рыми кончиками и степени их побурения.
Голова светлее туловища и шеи. Верхняя сторона носа желтая, лишь
иногда с легким буроватым оттенком; последний становится более ясным
на лбу. Темя ржаво-рыжее, одного тона с боками туловища и шеи. Ко-
нец морды, особенно передний край верхней губы, желтовато-белый.
Рис. 82. Серна (Rupicapra rupicapra L.).
Вдоль нижнего края глаз и вперед до верхних губ и ноздрей тянется
коричнево-бурая или темно-бурая полоса. Бурая полоса обрамляет ле-
том обычно и надбровье. Низ головы светлый, песчано-желтый. Более
светлая, песчано-желтая окраска захватывает также переднюю часть
шеи, образуя характерное горловое пятно, но резкая граница между
ними и прилегающими частями шеи в летней окраске отсутствует. Внут-
ренняя сторона ушей желтовато-белая; наружная серо-желтая, состоя-
щая из смеси коричнево-бурых и желтых волос; кончики ушей коричнево-
бурые. Вдоль средней линии шеи и спины тянется, но несколько не до-
ходит до корня хвоста, узкая, шириною 2—3 см, темно-коричневая или
темно-бурая полоса, исчерченная желто-рыжими кончиками волос. Бока
шеи и спины ржаво-рыжие, несколько светлее нижележащих участков.
По бокам живота от локтевых к коленным суставам тянется неясная
размытая коричнево-бурая полоса, переходящая на переднюю сторону
25*
388
BOVIDAE
задних ног. Круп желто- или ржаво-рыжий; задняя сторона бедер не-
сколько светлее.
Передняя и нижняя сторона груди коричнево-бурая. Живот более
светлый, песчано-желтый. Подмышки, нахи, область мошонки, вымени
и промежность грязно-белые с желтоватым оттенком. Хвост темно-бурый,
почти черный. Передние ноги темные, коричнево-бурые, сзади немного
светлее. Волосы вокруг добавочных копыт и по сторонам пальцев ниже
путовых суставов имеют золотисто-желтую окраску. Задние ноги свет-
лее передних благодаря примеси желтых волос.
С середины или даже с начала августа начинается побурение окраски.
Общая окраска вполне развитого зимнего меха коричнево-бурая. По-
всеместно, особенно на голове, усиливаются контрасты между светло и
темно окрашенными участками тела. Верхняя сторона и конец морды
становятся желто-белыми. Темные полосы по бокам головы тянутся впе-
реди от оснований ушей до ноздрей и губ. Беловатое кольцо окружает
глаза сверху и сзади. Весь низ головы от основания ушей и темных бо-
ковых полос, а также горловое пятно светлые, желто-белые. Светлая
окраска горлового пятна резко отграничена от обрамляющих ее черно-
бурых полос, идущих книзу от оснований ушей и сливающихся с темно-
бурой окраской нижней стороны шеи. К белизне горлового пятна на гра-
нице с темными полосами примешивается ржаво-рыжий оттенок. Темя,
затылок и наружная сторона ушей коричнево-бурые, одноцветные с ту-
ловищем. Внутренняя сторона ушей грязно-белая.
Более светлые боковые стороны шеи заметно отделяются от темно
окрашенного низа и черно-бурой полосы по ее верхнему краю. Темная
полоса вдоль средней линии спины и поясницы заметна не всегда. Жи-
вот в зимнем меху желто-рыжий. Задняя сторона бедер более светлая,
чем спина, рыжеватая. Грудь, полосы по бокам туловища на границе
со светлой окраской живота и ноги еще более темные, чем летом, темно-
бурые. Золотисто-желтая окраска волос вокруг добавочных копыт и по
бокам пальцев сохраняется и в зимней шерсти. Хвост у основания имеет
одинаковую с туловищем окраску, а в остальной части черно-бу-
рый.
Самки по окраске не отличаются сколько-нибудь заметно от самцов.
Окраска молодых на первом году жизни имеет серо-бурый оттенок
и может быть названа рыжевато-серой. Вдоль спины у них также тянется
срединная темно-бурая полоса, но живот покрыт серовато-белой, а грудь
светло-рыжей шерстью. Ноги серо-бурого или шоколадно-бурого цвета.
Волосяной покров у молодых серн более мягкий и пушистый, чем у взрос-
лых.
Предглазничные железы у серны отсутствуют. Межпальцевые име-
ются на всех четырех конечностях и представляют неглубокие вдавле-
ния кожи, покрытые внутри редкими волосами. Открываются межпаль-
цевые железы треугольными отверстиями на передней стороне ног выше
копыт. Паховые железы отсутствуют. Как самцы, так и самки имеют
своеобразные, свойственные, кроме серн, еще американской снежной
козе (Oreamnos americanus Ord.) зароговые, или затылочные железы.
Последние в обычное время представляют расположенные позади рогов
участки утолщенной и сморщенной в складки кожи. В период полового
сезона железы вздуваются, сильно выступают в виде прилегающих друг
к другу и к основаниям рогов багровых наростов, поверхность которых
пересекается несколькими глубокими бороздами, напоминающими из-
вилины головного мозга (Pocock, 1910, 1918). Животные в это время из-
Рис. 83. Череп Rupicapra rupicapra L.
390
BOVIDAE
дают сильный, напоминающий козлов запах, обусловленный секретом
зароговых желез.
Молочная железа имеет четыре соска.
Череп (рис. 83) серны характеризуется суженностью и приплюсну-
тостью лицевой части, а также резко выступающими трубкообразными
глазницами. Ширина его на уровне глазниц редко составляет меньше
55% основной длины черепа. Носовые кости средней длины, от 35 до
39% основной длины черепа. Предглазничные ямки на слезных костях
отсутствуют. Профиль черепа на границе носовых и лобных костей резко
вогнутый. Зароговая часть черепа (мозговая коробка) круто спадает
назад и поставлена к плоскости лба под более острым углом, чем к плос-
кости затылка. Поверхность лба заметно вдавлена между глазницами.
Лобные кости содержат небольшие внутренние полости (пазухи), кото-
рые распространяются и в роговые отростки до трети высоты последних.
Передний край хоан расположен впереди уровня заднего края верхнего
зубного ряда и впереди уровня боковых выемок заднего края костного
нёба
Коренные зубы по сравнению с козлами и баранами менее гипсо-
донтные. Длина ряда переднекоренных верхней челюсти составляет
от 34.8 до 36.7% длины всего ряда коренных зубов. Резцы брахиодонт-
ные; граница между коронкой и корнем зуба ясно заметна в виде пере-
хвата (рис. 61, Б).
В скелете конечностей пястная кость короче плечевой кости и состав-
ляет около 29.5% суммы длин плечевой, лучевой и пясти.
Из других особенностей серны Динник (1910) отмечает сильное раз-
витие у нее соединительной ткани под кожей на нижней стороне гру-
дины, которое этот автор справедливо связывает с необходимостью ле-
жания животного на голых камнях. Соединительная ткань при этом
играет роль эластической подушки, подобно аналогичному образованию
у козлов и верблюдов (см. стр. 100).
История и географическое распространение (рис. 84). Происхож-
дение рода Rupicapra Blanville до сих пор не ясно. В верхнем плио-
цене на территории Франции и Италии обитала форма, по направлению
специализации весьма близкая к серне, но имевшая изгиб верхушек
рогов не назад, а вперед.
Все достоверные находки Rupicapra относятся к периоду, начиная
со среднего или даже верхнего плейстоцена. В плейстоцене этот род
распространен был гораздо шире не только на юг, но и на север.
Остатки серны, например, найдены на территории Бельгии (Boule,
1910).
В настоящее время серна обитает в Кантабрийских горах (Испания),
Пиренеях, Французских, Итальянских, Баварских, Швейцарских и
Австрийских Альпах, на Апеннинах (область Абруцци в Италии), на Кар-
патах в пределах Чехословакии, южной Польши и Румынии, в некото-
рых горах Югославии, Албании и Греции, в горах восточной части Ма-
лой Азии, Главном Кавказском хребте и в Закавказье (Couturier, 1938).
Современные разрозненные ареалы серны представляют остатки некогда
сплошной области распространения этого вида.
Во второй половине прошлого столетия дважды пробовали пересе-
лить серну в горы Норвегии; но обе попытки окончились неудачей, так
как завезенные животные через короткие сроки, в силу невыясненных
причин, погибали. С 1911 г. удачно акклиматизирована в Чешских Су-
детах. В 1907 г. завезено и выпущено 8 штук серн в Новой Зеландии
RUPICAPRA
391
(горы Кока), где число их к 1920 г. возросло до 70 голов (Cobb, 1948;
Danek, 1954).
В советских Карпатах серны нет. В пределах Советского Союза она
распространена в Главном Кавказском хребте и на некоторых горах
Закавказья. На западе Большого Кавказа они изредка добывались около
с. Кабардинки, километрах в 15—20 от Новороссийска, куда серны, по-
видимому, заходят лишь случайно в многоснежные зимы. Изредка встре-
чаются они в верховьях Пшады, а также в окрестностях Геленджика.
Область более или менее сплошного распространения серны, по Н. Я. Дин-
Рис. 84. Современный ареал Rupicapra rupicapra L. (По Н. К. Верещагину (1947)
с дополнениями).
нику (1910), начинается от Гойтхского перевала, неподалеку от кото-
рого на горе Индык (в верховьях р. Пшиша) серны живут постоянно.
Начиная от этого места, они по обеим сторонам Главного Кавказского
хребта встречаются вплоть до Бабадага и восточных склонов Шахдага
включительно (Верещагин, 1947).
На северной стороне в наибольшем числе до настоящего времени
серны обитают в верховьях рр. Белой, Уруштена и Малой Лабы, на тер-
ритории Кавказского госзаповедника (Насимович, 1939, 1941). Много
их до сих пор в районе гор Фишта и Оштена. В конце прошлого и начале
нынешнего столетия были еще не так редки во многих местах Черных
Гор, расположенных километрах в 65 к северу параллельно Главному
хребту, но теперь их там почти уже нет. За пределами Кавказского
заповедника, к западу и северу от него, серна в настоящее время
распространена спорадически, гнездами, отделенными одно от дру-
гого пространством порой в несколько десятков километров (Насимо-
вич, 1939, 1949). По-видимому, наиболее северными пунктами обита-
ния серн являются горы: Матаза (в верховьях р. Цица), Корыта (на
р. Белой) и Герпегеж в районе Псебая. В самом конце прошлого столетия
верховья рр. Теберды, Аксаута, Зеленчука, Кяфара, Урупа, Большой
392
BOVIDAE
и Малой Лабы, по выражению Я. Васильева (1896), были «переполнены
этой чудесной дичью», которую автор встречал сотенными стадами. На
территории Тебердинского заповедника серны в довольно значительном
числе живут и в настоящее время (Эквтимишвили, 1953).
В центральной части Большого Кавказа, по Н. К. Верещагину (1952),
встречаются, хотя и не так многочисленны, на склонах Эльбруса, в вер-
ховьях рр. Баксана, Чегема и Черека; но на передовой цепи Скалистого
хребта их уже нет. Дальше на восток серны живут в горах Осетии
и Дигории, но и здесь они обитают не повсеместно.
Живет серна в верховьях Уруха и Ардона, но, по Верещагину (1952),
«нет ее на хребтах, пересекающих внутреннюю сухую зону продольных
долин». По крайней мере, в начале этого столетия встречалась она в верх-
нем течении Аргуна, в частности на горе Даргендук, а также в районе
с. Ведено, Зиберлихи и некоторых других местах (Динник, 1910).
В северном и центральном Дагестане, в районах Хасавюртовском,
Буйнакском и смежных с ними серн нет в силу отсутствия подходящих
стаций. Во внутреннем Дагестане они сохранились лишь отдельными
изолированными очагами и весьма немногочисленны. Так, есть они в
пределах бывшего Андийского округа, спорадически в Гунибском (в
частности, вблизи Вантляшетского перевала Аваро-Кахетинской дороги).
По сравнению с турами редки, но встречаются серны в Богосском хребте.
В. Г. Гептнер и А. Н. Формозов (1941) предполагают, что должны они
быть также на водораздельных хребтах между Самуром, Аварским и
Казикумухским Койсу. По данным этих авторов, держатся серны в го-
рах на водоразделе рр. Каракойсу и Аварского Койсу (над аулом Карда),
в верховьях Самура и, может быть, Ахтычая. Изредка попадается в рай-
оне гор Базар-Дюзи, Шалбуздага и Шахдага.
На горных склонах западной части Главного Кавказского хребта
серна в незначительном числе встречается на горах: Аутль, Хуко, Бзныге
и Ануке, в верховьях рр. Шахе, Сочи, Мацесты, Дагомыса, а также
в хребтах по нижнему и среднему течению Мзымты (Насимович, 1939).
Верховья Мзымты, Псоу, Хошупсе и Жоупсе считались лучшим местом
по числу серн в бывшем Черноморском округе (Васильев, 1896).
Далее, к востоку она еще в значительных количествах держится в
горах Абхазии и Сванетии, в верховьях рр. Кодара, Ингура и Риони,
где много их видел Н. Я. Динник (1910) на горе Щоде. В Южной Осетии
и Кахетии, по Н. К. Верещагину (1952), серн мало. В значительном
числе они держатся еще в березовых ущельях Кахетичая, Шинчая и
в верховьях Турьянчая в пределах Азербайджанской ССР. Много серн
на территории Закатальского госзаповедника. Далее, к востоку они
держатся еще близ Курбан-Эфенд в Исмаилинском районе.
Отдельные очаги обитания серны, как следы былого соединения с
ареалом этого вида в Малой Азии, сохранились на территории Малого
Кавказа. Еще в начале этого столетия они жили в горах недалеко от Ба-
туми (Динник, 1910). В это же время встречались около Боржоми, Аба-
стумани, а в правобережье Куры до сих пор обитают на Триалетском
хребте в районе Белого Ключа. А. А. Саркисов (1944) имел экземпляры
серн из района Бахмаро (Аджаро-Имеретинский хребет). Встречались
недавно серны на Алагезе, но теперь, по данным этого автора, их там
уже нет.
В Армении серны водились, по-видимому, еще в начале нынешнего
столетия в бывших Дорийском и Дилижанском уездах (Динник, 1910),
на скалах Трет около разъезда Кобер, но во всех этих местах их теперь
RUPICAPRA
393
уже нет. Далее, не так давно серны держались в Шахдагском хребте
(к северо-востоку от Севана), в верховьях рр. Дзегамчая и Шамхорчая.
На последней они встречались будто бы до с. Барсум (Динник, 1910).
В настоящее время серны в пределах Армянской ССР нет.1 В 30-х го-
дах этого столетия серны исчезли также в хребте Муров-Даг (Саркисов,
1944).
Подвиды. К настоящему моменту описаны 10 подвидов (географи-
ческих рас).
1) R. rupicapra rupicapra L. Рога почти вертикальные, отставленные
друг от друга настолько, что остается плоское пространство на лобных
костях между ними. Основная длина черепа 190—207 мм. Распро-
странение: Альпы в пределах Франции, Италии, Швейцарии,
Австрии, Баварии, северная часть Югославии, Тироль, Трансильва-
ния, Карпаты.
2) R. rupicapra ornata Neumann. От типичной расы отличается блед-
ной окраской верхней стороны шеи, ясно выраженными темными поло-
сами по бокам шеи от оснований ушей до грудины, между которыми вкли-
нивается светлая окраска низа головы и горла. Основная длина черепа
до 190 мм. Распространение: область Абруццы в Италии.
3) R. rupicapra pyrenaica Bonaparte. От типичной расы отличается
более светлой зимней окраской. Окраска шеи одинаковая с R. г. ornata.
Рога сильно сближены в основаниях, так что поверхность лобных костей
между ними слегка вогнута. Основная длина черепа до 182—190 мм.
Распространение: Пиренеи.
4) R. rupicapra parva Cabrera. Размеры небольшие, еще меньшие,
чем у предыдущего подвида (кондилобазальная длина черепа около
184 мм). От последнего отличается также более темной окраской и слабо
выраженным пятном на горле, которое лишь немного светлее, чем общая
окраска тела. Распространение: Кантабрийские горы (Испания).
5) R. rupicapra caucasica Dinnik. Описание окраски дано выше (стр. 386).
От других рас, по Лайдекеру (Lydekker, 1913), отличается более корот-
кими, толстыми рогами, светлым, почти белым низом головы и пятном
на горле, а также особенностями окраски головы. Указываемые этим ав-
тором отличия в форме носовых костей (отсутствие боковых отростков
в их задней половине), при просмотре серий черепов в коллекциях ЗИН
АН СССР, не подтвердились. Распространение: горы Кавказа
и Закавказье.
6) R. rupicapra asiatica Lydekker. От остальных рас отличается тем-
ной, дымчато-бурой окраской, с черно-бурыми конечностями и шеей,
а также незначительными размерами светлой зоны на лице. Распро-
странение: горы Малой Азии.
7) R. rupicapra balcanica Bolkay. Отличается некоторыми особенно-
стями черепа: сближенными основаниями роговых отростков, плоским
широким лбом и большой длиной верхнего зубного ряда. Распро-
странение: страны Балканского полуострова, Югославия, Алба-
ния, Болгария и Греция.
8) R. rupicapra olympica Koller. Описана по единственному черепу
с горы Олимп в Греции, на основании несущественных отличий в изгибе
рогов. Вероятно, ничем не отличается от предыдущей расы.
9) R. rupicapra cartusiana Couturier. Несколько большей, чем типич-
ная раса, величины. Кроме того, отличается заметно уплощенными и до-
1 Во всяком случае С. К. Даль в его монографии по животному миру Армении
(1954) в списке современных млекопитающих серн не приводит.
394
BOVIDAE
вольно сильно расходящимися к вершинам рогами, а также увеличен-
ной длиной верхнего и нижнего зубного ряда. Распространение:
горы Шартр (Дофине) во Франции.
10) R. rupicapra carpatica Couturier. Наиболее крупный подвид. Об-
щая длина черепа достигает 220 мм, живой вес до 60 кг. Окраска во все
сезоны темнее, чем у других подвидов, летом каштановая, зимой черная.
Рога длинные, до 33 см. И абсолютно, и относительно длинный зубной
ряд. Коренные зубы не только длинные, но и широкие. Распростра-
нение: Карпатские горы, в частности Трансильванские Альпы, по
черепам из которых была описана эта форма.
Биология. Серна — типичное горно-лесное животное. Крутые ска-
листые участки гор с наличием леса являются непременным условием
существования серны. Область ее вертикального распространения рас-
положена от нижней границы леса, иногда на высотах всего 600—700 м,
до субальпийской и альпийской зон, до 3000 м и больше. Как скалы,
так и лес служат в первую очередь местами убежищ. На последний серна
полагается, по-видимому, даже больше и в случае опасности скорее бро-
сается в густой лес, если он имеется поблизости, чем в скалы. В лесу
она заметит человека и скроется от него раньше, чем тот ее обнаружит
(Динник, 1910). Впрочем, в местах, где серн мало тревожат, они охотно
пасутся и в безлесных горах, и на открытых пастбищах, особенно если
по соседству имеются участки скал и других убежищ (Динник, 1910;
Насимович, 1949).
Подобно кавказскому козлу (туру, Capra caucasica Guld.), обитающему
на Кавказе в тех же районах, серна—жительница скалистых ландшаф-
тов, однако, в противоположность туру, особо «тяжелых» скальных мест
избегает. В связи с этим А. А. Насимович (1939) отмечает одну особен-
ность распространения этих видов в пределах западного Кавказа: на
хребтах, где много туров, серны сравнительно редки, и наоборот — в
местах, где много серн, туры часто отсутствуют вовсе. Объяснение этому
автор находит в тяготении зверей к разным стациям. Серна в высоко-
горье отдает предпочтение открытым спокойным стациям — плато, дну
широких трогов, перемежающихся со скалами, осыпями, крутыми скло-
нами. Тур же предпочитает наиболее пересеченные скалистые участки
с небольшими участками зеленых пастбищ. В лесной полосе гор встре-
тить козлов и серн в одних и тех же местах можно чаще. По Н. Я Дин-
нику (1910), серны живут в горах в среднем выше, чем туры, а в горном
Дагестане они держатся в крайнем высокогорье, что, помимо отсутствия
на более низких горах подходящих стаций, объясняется, вероятно, не-
померным преследованием их со стороны человека (Гептнер и Формо-
зов, 1941).
Ясно выраженных сезонных миграций, как у некоторых других ко-
пытных, у серн не наблюдается; в течение круглого года их можно встре-
тить от нижней зоны леса до высокогорья. Меняется лишь степень встре-
чаемости их на разных высотах и местообитаниях. Летом основная масса
зверей держится на высотах от 1700 до 2500 м, в верхней полосе леса,
в субальпийской и альпийской зонах. Излюбленными местообитаниями
в это время являются обрывистые, скалистые места вблизи леса, крутые
лесистые склоны, поросшие редким сосняком, пихтой, елью и березой.
Охотно серна держится также на субальпийских и альпийских лугах
по соседству с густыми зарослями березняка, где звери скрыва-
ются в случае опасности и отдыхают в жаркие дневные часы су-
ток.
RUPICAPRA
395
С конца октября начинается перемещение серн в ниже расположенные
части горных склонов, увеличивается встречаемость в средней полосе
леса (Эквтимишвили, 1952, 1953). Зиму проводят в полосе темнохвойных
лесов на высотах 1000—1500 м. В противоположность летнему периоду,
в большем числе серны встречаются в нижней зоне широколи-
ственного леса. В субальпийской полосе остаются единичные экзем-
пляры.
Масштабы осенне-зимних перемещений серн не одинаковы в разных
местах и в разные годы и определяются, в основном, глубиной снежного
покрова. На южном склоне западной части Большого Кавказа, где снега
выпадает много, расстояние между местами летовок и зимовий достигает
25 км (Насимович, 1939). В пределах зимних мест обитания серны вы-
бирают крутые склоны гор с выходами скал, образующих навесы и пе-
щеры. Выбор таких мест, помимо наличия укрытий на случай непогоды,
обусловлен, вероятно, и меньшей глубиной снега. Предпочтение зимой
отдается солнечным склонам, но в противоположность турам, серны не
избегают и тенистых мест, если защитные и кормовые условия на них
достаточно благоприятны. Обратные перемещения серн в верхнюю по-
лосу леса и субальпийское криволесье происходят по мере таяния снега,
появления зеленого корма и начинаются обычно в апреле (Насимович,
1939).
В пределах избранных мест обитания серны ведут сравнительно
оседлый образ жизни. Масштабы суточных передвижений по сравнению,
например, с теми же турами незначительны; район кочевок определен-
ной группы серн часто ограничивается площадью 8—10 км2, а по прямой
4—5 км. При этом животные стойко придерживаются определенных мест
обитания, очень неохотно их меняют и, по словам А. А. Насимовича
(1949), «даже там, где их преследуют, они продолжают оставаться в ста-
рых районах, лишь меняя дневной образ жизни на ночной». В определен-
ные часы они появляются в разных частях своего микроареала, пере-
двигаются одними и теми же путями, на которых вытаптываются ясно
обозначенные тропы.
В нормальных условиях серны летом кормятся в утренние и вечерние
часы. В жаркие солнечные дни пасутся до 10—11 часов, затем ложатся от-
дыхать на снег, если он имеется по соседству, или ищут прохлады в тени
деревьев и под навесами скал. Очень охотно забираются на день в неглу-
бокие пещеры. Иногда местами отдыха служат высокие выступы скал
с хорошим кругозором (Динник, 1910). Вновь выходят на пастьбу летом
после 5—6 часов вечера, причем время жировки захватывает и часть ночи.
В средине ночи, по крайней мере в нормальных условиях, серны отдыхают.
Правильная смена часов отдыха и пастьбы нарушается в пасмурную,
прохладную погоду, осенью и зимой, а также и летом в полосе леса, где
звери находят достаточно тени. В этих случаях и в середине дня можно
встретить большое число жирующих зверей, а отдыхает большая часть во
второй половине дня, под вечер. В непогоду серны проводят целые дни под
навесами скал, в пещерах или забираются под деревья, объедая низко
свисающие ветви.
Серна — стадное животное. Размеры и состав стад изменчивы даже
в пределах одного местообитания и сезона. Потревоженное стадо серн не
бежит в одном направлении, а бросается врассыпную и вновь собирается
в случайные группы уже иного состава (Насимович, 1949). Кроме того,
наблюдаемые размеры стад зависят в сильной степени также от общей
численности серн в том или ином районе. В местностях, где серны мало-
396
BOVIDAE
численны они держатся по 2—3 штуки. В прежнее время на западном Кав-
казе нередки были стада в сотню экземпляров. Н. Я. Динник (1910) часто
встречал группы в 15—20 голов; по А. А. Насимовичу (1949), средний
размер стада в Кавказском заповеднике в 1924 г. составлял всего 7 голов,
а в 1937 г., когда поголовье их в результате охраны увеличилось, он опре-
делялся уже цифрой 16—17 голов. Максимальные из наблюдавшихся этим
автором стада в 88 и 93 головы представляли, по-видимому, временные,
случайные скопления.
В некоторых случаях образование больших стад, по-видимому, свя-
зано с ограничением мест возможного обитания серн вследствие вытесне-
ния стадами домашних животных.
Более крупные стада серн наблюдаются в августе и сентябре.1 В про-
тивоположность многим другим копытным, к зиме и особенно весной раз-
меры стад постепенно снижаются, и только летом после окота серны вновь
начинают табуниться.
Состав стад чаще бывает смешанным, из зверей всех возрастов и обоего
пола. Но иногда взрослые самцы и самки с молодыми образуют самостоя-
тельные группы.
Пищей серн в летний период являются, в основном, различные травя-
нистые растения. Составленный А. А. Насимовичем (1949), по наблюдениям
в субальпийской и альпийской зонах, список их летнего корма включает
33 вида трав и 8 видов кустарников и деревьев. Из травянистых растений
по числу видов превалирует разнотравье, но в списке особо охотно поедае-
мых кормов на первом месте стоят злаки — овсяница и мятлик, а за ними
следуют мяун, колокольчики, анемона, лютики, копеечник, белокопытник
и борщевник. У древесно-кустарниковых пород поедаются листья и моло-
дые побеги, а у пихты хвоя.
Основу зимнего питания составляет, помимо сухих трав, добываемых
на бесснежных и малоснежных участках, древесно-веточный корм — ветви
и кора ивы, бука, рябины. В большом количестве поедается овсяница,
листья и побеги зимне-зеленых растений: ожины, омелы, лавровишни.
Употребляются в пищу также мох, древесные лишайники и даже хвоя сос-
ны, пихты и ели. По некоторым данным, охотно поедаются сернами плоды
каштана. Случаев поедания заготовленного в стогах сена, как это будто бы
наблюдается у европейской серны (Чуди, 1873), на Кавказе не известно.
По-видимому, в связи с недостатком минеральных веществ в пище,
серны посещают естественные и искусственные сухие солонцы, пьют воду
из соленых источников, а также поедают перемешанную с песком глину,
осаждающуюся по берегам вытекающих из-под ледников горных ручьев
и речек (Динник, 1910). Наиболее часто и регулярно серны посещают
солонцы весной и летом, реже осенью. О солонцевании их в зимний период
ничего не известно.
Серна — очень подвижное животное. В пределах своего небольшого
сезонного местообитания она много двигается. Даже в часы отдыха, она
то ложится, то, спустя короткое время, встает, чтобы снова немного покор-
миться или перейти на лежку в другое место.
На ровном месте серна быстро бежать не может; бег ее на рав-
нине — неуклюжий голоп. Зато она превосходно приспособлена к
передвижению по скалам и крутым каменистым склонам. «Наблю-
дающий серн даже не первый раз, — пишет Н. Я. Динник (1896),—
нередко бывает буквально поражен их способностью карабкаться по ска-
1 Приведенные ранее цифры относятся к августу.
RUPICAPRA
397
лам. Иногда видишь серну над отвесной стеной в сотню сажен высоты и не
допускаешь мысли, чтобы она отважилась спуститься с нее; но вдруг серна
испугавшись ли чего-нибудь, или по какой-либо другой причине, бросается
с кручи и, цепляясь за незаметные издали неровности скалы, смелыми раз-
машистыми прыжками начинает спускаться в пропасть. Временами она
скользит и катится, широко расставив задние ноги, но, встретив незначи-
тельный выступ, ухитряется встать на него всеми четырьмя ногами и задер-
жать падение своего тела. . . Наблюдая серну, когда она спускается с от-
весной стены в пропасть, я несколько раз был не в состоянии словами или
каким-нибудь восклицанием не обнаружить своего страха за участь серны,
которая, как мне казалось, должна была неминуемо сорваться со скалы
и разбиться вдребезги. Также жутко смотреть на нее, когда она несется по
узкому, почти незаметному карнизу высокой отвесной скалы. По осыпям,
представляющим самый ужасный хаос из камней всевозможных величин,
серны бегают с изумительным проворством и ловкостью». Легко и без вред-
ных последствий спрыгивают вниз с высоты 10—15 м.
Большую роль в способности серн карабкаться и спускаться по крутым
каменистым склонам, по-видимому, играют отмеченные ранее (стр. 386)
особенности строения копыт, к тому же обладающих способностью широко
раздвигаться.
В местностях, где на серн не охотятся (например, в заповедниках),
они не слишком пугливы и осторожны. При встрече с человеком не стре-
мятся в таких случаях быстро скрыться, нередко подпускают шагов на две-
сти триста, а иногда обнаруживают даже признаки явного любопытства.
Там же, где они подвергаются систематическому преследованию, серны
становятся очень осторожным зверем. У них хорошо развиты как зрение
и слух, так и обоняние. Однако на слух, как думал Н. Я. Динник (1896),
серна в естественных условиях полагается меньше, чем на чутье и зрение,
почему и позволяет иногда приблизиться к ней под прикрытием на 20—30 м.
По ветру она нередко чует охотника за полкилометра, равно как на откры-
том месте издали не только заметит, но и различит от окружающих предме-
тов даже неподвижно стоящего человека. Панического страха, впрочем,
серна при виде подозрительного предмета не проявляет и, насторожив-
шись, не обращается в бегство, пока не убедится в грозящей ей опасности,
а убегая, время от времени оглядывается назад, используя для этого
выступающие высокие камни или скалы.
Специальных сторожей в табунах серн, по-видимому, не бывает, так
как даже в отдыхающем стаде всегда находится несколько бодрствующих
животных.
Голос серны, который иногда называют свистом, в действительности,
по Н. Я. Диннику (1896), представляет что-то среднее между настоящим
свистом и шипением, довольно протяжный, длящийся около двух секунд.
Свист является предупреждающим сигналом, издаваемым зверем в случае
опасности и тревоги. Другие звуки сернами издаются очень редко. Голос,
которым перекликаются мать и дитя, говорят, напоминает блеяние коз.
Смена волосяного покрова у серн происходит в году дважды. Сроки
весенней линьки установить трудно, так как этот процесс сильно растянут
и происходит столь постепенно, что трудно, говорят, добыть серну в чисто
летней или чисто зимней шерсти.
Исследование коллекционных шкур показывает, что в конце апреля
на голове и шее под длинным зимним волосом пробивается короткая летняя
шерсть. Экземпляры, добытые 18 июня, еще не закончили линьку. Осо-
бенно затягивается линька у кормящих самок, у истощенных животных,
398
BOVIDAE
а также у молодых. Н. Я. Динник (1910) находил остатки зимней шерсти
даже у серн, добытых в конце июля старого стиля. Обычно уже в начале
августа сквозь рыжую летнюю шерсть начинают пробиваться темные осте-
вые волосы и бурый подшерсток, а в начале или середине сентября рыжая
летняя шерсть выпадает и заменяется более длинным зимним волосом тем-
ного цвета.
Половой зрелости серны достигают и начинают участвовать в размно-
жении на третьем году жизни. В трехлетием возрасте обычно приносят
первых ягнят. Период гона и спаривания падает на время с конца октября
по начало декабря. В это время более крупные стада серн разбиваются на
небольшие группы по 3—5, максимально до 10 голов. Самки в этот период
ведут себя спокойно, но самцы приходят в сильное возбуждение, много
двигаются, перебегают от одного стада к другому, в результате чего сильно
худеют. Между самцами происходят турнирные поединки. Случаев по-
вреждений и ран от ударов во время драк не известно, но иногда один самец
сталкивает другого в пропасть, что может сопровождаться увечьем или
смертью. Во время гона самцы часто кричат, издавая звуки, отдаленно
напоминающие блеяние овцы, но более глухие (Динник, 1910).
Продолжительность беременности у серн — около шести месяцев.
Первые ягнята, по А. А. Насимовичу (1949), появляются в конце апреля.
Массовый окот происходит в мае, но отдельные случаи более поздних
родов наблюдаются, по-видимому, до июня.
Как и у других копытных, перед окотом самки удаляются из своих
групп, уходят в глухие скалистые места в средней или верхней полосе
леса, где и приносят по одному, реже по два ягненка. Достоверных случаев
троен у серн в условиях Кавказа не наблюдалось. Уже вскоре после ро-
ждения молодые могут следовать за матерью и не хуже нее карабкаться по
скалам. Первый месяц или два самки с детенышами держатся одиноко
в скалистых, поросших лесом местах и только позднее, в июле—августе
выходят в более открытые участки и присоединяются к общим смешанным
группам. К 5-месячному возрасту молодые достигают 50% веса взрослых.
Добытая нами 17-месячная самка весила 28.5 кг, 21/2-летняя — 34.6 кг.
Новорожденные имеют бледную рыжеватую окраску; позднее окраска
становится рыжевато-серой.
Рога у серн начинают отрастать на третьем-четвертом месяце жизни.
В начале они имеют вид черноватых шишек, а потом тонких спичек. Заги-
баться на верхушках в виде крючка начинают только на втором году жизни.
Приводимые в литературе данные о продолжительности жизни серны
в естественных условиях до 20—25 лет основаны на предположениях или
мало достоверных сведениях, полученных от охотников. Ф. Цедтвич (Zedt-
witz, 1937) на основании исследования степени стирания зубов предпола-
гает, что максимальная продолжительность жизни серны едва ли может
быть больше 20 лет. Самому ему удавалось добывать лишь 14—15-летних
самцов. С. Флауэр (Flower, 1931) приводит максимальные цифры продол-
жительности жизни серн в зоологических садах 15 лет 8 месяцев 1 день и
16 лет 10 месяцев 19 дней.
Из врагов серны, по степени опасности для нее, на первом месте следует
поставить леопарда, но, ввиду малочисленности этого хищника в настоя-
щее время, практическое его значение не велико. По-видимому, не так редко
серна становится жертвой рыси. И барс, и рысь скрадывают и подкараули-
вают серн на путях переходов, кидаясь прыжком на свою жертву. Изве-
стны, впрочем, случаи, когда рысь гонялась за сернами, загоняла их на
скалы, с которых те падали и разбивались. Волк опасен сернам, по-види-
RUPICAPRA
399
мому, лишь зимой, в глубокоснежных местах. В условиях же бесснежного
скалистого ландшафта, этот хищник преследовать серну не в состоянии.1
То же самое, но еще в большей степени относится к медведю. Известны
случаи, когда серны спокойно паслись в 30—40 шагах от медведя (Дин-
ник, 1896). Сообщения о том, что врагами крупных взрослых серн являются
хищные птицы, основаны на легкомысленных охотничьих рассказах.
Не меньше, чем от хищников,гибнут серны от снежных лавин и обвалов,
погребающих иногда целые табуны этих животных (Насимович, 1949).
В горах западной Европы известны массовые падежи серн от парши,
а также от эпизоотий, переходящих на них с домашних животных (Brehm,
1925). Неопределенные сообщения о массовых падежах серн от эпизоотий
домашних животных, в частности от ящура, поступали и с Кавказа, но
достоверно никем не подтверждались.
Паразитофауна серны в условиях Кавказа почти совсем не изучена.
С. М. Асадов (1954) описал из сычуга серны новый, до того не известный
вид нематоды семейства трихостронгилид — Marshallagia skrjabini Asad.
На сернах найден один вид клеща — Ixodes ricinus L. 1 2
Хозяйственное значение. Мясные качества серны не так высоки. Вес
туши мяса взрослого самца достигает 20—25 кг, самки около 15—21 кг.
Вкусовые качества мяса не очень высокие: оно темное и довольно
жесткое, уступает в этом отношении мясу других копытных, в том числе
мясу кавказского тура. Несколько мягче оно становится после того, как
полежит в течение одних или двух суток. Наиболее жирны серны осенью,
но жир у них твердый, тугоплавкий (температура плавления около 37°),
и поэтому, застывая во рту, он производит непрятное ощущение (Динник,
1896, 1910).
Снятые шкуры серн употребляются на коврики, зимние изредка идут
на воротники. Иногда их режут на тонкие, но прочные ремни. Рога служат
для различных поделок: ручек для тростей и ножей, вешалок и т. п., а
также в качестве украшений.
Интересно отметить, что различным частям и даже испражнениям сер-
ны, средневековой медициной приписывалось целебное действие. Особенно
в этом отношении ценились находимые иногда в желудке и кишечнике
серны «безоары». Это круглые, величиной с орех или куриное яйцо шары,
состоящие из смеси непереваренных частей пищи и заглоченной шерсти,
склеенных слизью (Thorington, 1944).
Пойманные в молодом возрасте серны привыкают к человеку довольно
легко, однако по-настоящему ручными становятся редко. Вследствие,
вероятно, отсутствия свободы для движения развиваются в неволе медлен-
но, хиреют и долго обычно не живут. Даже выращенные с раннего возраста,
во взрослом состоянии проявляют нередко дикий характер и становятся
опасными для человека (Чуди, 1873; Girtanner, 1880). Размножаются
в неволе с трудом, хотя случаи подобного рода за последние десятилетия
известны (Brehm, 1925).
Имеющиеся в старой литературе сообщения о получении якобы гибри-
дов между серной и домашней козой (Mann, 1867; Чуди, 1873; Girtanner,
1880) мало достоверны. Теоретически получение подобных гибридов,
учитывая сравнительно большую генетическую разобщенность козлов и
серн (разные не только роды, но и подтрибы), мало вероятно. Научно до-
1 Следует оговориться, что, по данным А. А. Насимовича (1949), в Кавказском
заповеднике волк является главным врагом серн и притом, по-видимому, больше ле-
том, чем зимою.
2 Устное сообщение Г. В. Сердюковой.
400
BOVIDAE
стоверных случаев этого рода не известно, и описания гибридов козы
с серной до сих пор никем не дано (Gray, 1954).
Охота. Наиболее распространенным способом охоты на серн на Кав-
казе, является охота с подхода. В принципе она мало отличается от охоты
этого рода на других копытных. Задача заключается в том, чтобы, пользуясь
прикрытиями в виде выступов скал и камней, бесшумно подойти против
ветра к зверю. От охотника требуется, кроме силы и выносливости, необ-
ходимой при передвижении в горных условиях, большая выдержка, на-
ходчивость, хорошее знание местности и уменье ориентироваться в горной
обстановке. Если приходится в силу необходимости подходить к сернам на
открытом месте, больше шансов на успех будет при движении на четверень-
ках или ползком, когда серна не всегда отличает человека от животных.
Стрелять приходится в этих условиях, даже при подходе против ветра,
редко ближе чем на 300—500 шагов, так что требуется, помимо прочего,
хорошее оружие (лучше всего винтовка с телескопическим прицелом)
и большая меткость.
Иногда применяется охота нагоном, когда часть охотников, после того
как серны обнаружены, располагается на пути, по которому должны прой-
ти потревоженные звери (обычно ниже по склону), а остальные, обходя
серн кругом, стараются заставить их идти в нужную сторону. В этом слу-
чае, помимо знания местности, важнейшее значение имеет знакомство
с привычками зверя, уменье предугадать путь, по которому легче всего
гнать зверя.
Из других способов добычи серн сравнительно легким является практи-
куемое часто подкарауливание у солонцов, мест водопоя или на пастбищах,
основанное на использовании склонности этого зверя стойко, придержи-
ваться установившегося суточного цикла.
Род CAPRA LINNAEUS — КОЗЛЫ
Linnaeus С., 1758, Syst. Natur., ed. 10, 1 : 68 (первооп.); Scbinz H., 1838,
Neue Denkschrift Schweiz. Gesellsch. Naturwissensch., II : 3—25 (сист., биол.); Hod-
gson В., 1847, J. Asiat. Soc., Bengal, XVI : 700 (сист.); Ne wtonE., 1880, Geol.
magaz. : 447—452 (ист.); Sc la ter Ph., 1886, Proc. Zool. Soc. London : 314—318
(сист.); Lydekker R., 1898, Wild oxen, sheep and goats of all lands, London:
239—296 (общ., изобр.); Lydekker R., 1913, Catal. Ungulate mammals, Brit. Mus.,
London, 1 : 123—173 (сист., синон., изобр.); A d a m e t z L., 1915, Mitt, landwirt'sch.
Lehrkanzel d. Hochschule f. Bodenkultur in Wien., Ill, 1 : (цит. по В. Громовой, 1940)
(ист.); Brehms Tierleben, 1925, 4. Aufl., Bd. 13, Leipzig: 272—298 (общ., изобр.);
H ilz heimer M., 1933, Arch. f. Tierernahrung u. Tierzucht, VIII : 323—371 (сист.
гибр.); Громова В. И., 1935, Докл. Акад, наук СССР, 4 (XI), 1/2 : 97—99 (ист.);
Schwarz Е., 1935, Ann. Mag. Natur. Hist., 16 : 433—437 (сист.); В о h 1 i n В.,
1937., Palaeont. Sinica, ser. C. vol. 14, fasc. 1 : 1—111 (эвол.); Громова В. И.,
1940, Пробл. происх., эвол. и породообразов. дом. животных, 1 : 63—120 (морф.);
К г е t z о i М., 1942, Foldtani Kozlony, v. 72, No. 4/12 : 353—356 (ист.); M a x i m J.,
1948, Bull. Soc. Sti. Cluj, t. 10, No. 1 : 67—76 (ист.); de В e a u x O., 1949, Atti Soc.
Ital. Sei. Natur., Milano, 88, No. 1/2 : 17—20 (сист.); Пидопличко И. Г., 1951,
О ледниковом периоде, II, Киев, изд. Акад, наук УССР : 59—61 (ист.); Е 1 1 е г m a n J.
and Т. Morrison-Scott, 1951, Checklist of palaearctic and indian mammals,
London : 404—409 (сист., синон.); Громова В. И., 1953, Тр. Комисе, по изуч.
четверт. периода, X, 1 : 1—122 (морф.); Соколов И. И., 1953, Тр. Зоол. инет.
АН СССР, XIV : 145—147 (морф., сист., ист.).
Характеристика рода. Животные средней величины, плотного, не-
сколько неуклюжего сложения. Массивное туловище покоится на сравни.
1 Описание охот на серн в горах западной Европы см.: Чуди, 1873; Keller, ,1887;
Couturier, 1849.
CAPRA
Ml
тельно коротких и толстых ногах. Копыта низкие, но длинные и массивные.
Добавочные копыта хорошо развиты, короткие и широкие, почти квадрат-
ные. Хвост треугольный, снизу голый. Рога имеют и самцы, и самки.
У самцов они длинные, значительно превосходят длину головы. Форма
рогов разнообразная: дугообразно загнутые назад, иногда при этом слегка
изогнутые гомонимно или гетеронимно; прямые, направленные назад
и в стороны, в этом случае одновременно свернутые или скрученные в што-
порообразную гетеронимную спираль; иногда рога первертированные.
Поперечное сечение рогов от линзообразного или грушевидного до тре-
угольного и четырехугольного, сжатого с боков или почти круглого (кав-
казские туры). У взрослых самцов рога имеют более или менее выраженные
2—3 продольные ребра (киля) и, соответственно, две или три грани. По-
верхность роговых чехлов у самцов, помимо мелкой поперечной морщини-
стости, на передней стороне несет отставленные друг от друга узлы (бугры),1
или сильно выступающие поперечные валики. У самок рога тоньше и ко-
роче, обычно не превышают длину головы и не имеют на передней стороне
ни валиков, ни узлов.
Верхняя губа покрыта волосами, за исключением узкой вертикальной
полоски голой кожи по средней линии и узких полосок по краю ноздрей.
Характерно для козлов, особенно для самцов, наличие на нижней стороне
головы пучка длинных волос, так называемой «бороды». Волосяной покров
в развитом состоянии ясно дифференцирован на ость и подшерсток.
Предглазничные, паховые и межпальцевые железы отсутствуют.1 2
Молочная железа имеет два функционирующих и иногда еще два недораз-
витых соска. У самцов около хвоста расположены особые железы, усиленно
функционирующие в период полового сезона и обусловливающие специфи-
ческий запах козлов.
Череп имеет хорошо развитую зароговую часть; длина последней
(от заднего края затылочного бугра до среднероговой точки, т. е. точки на
средине между основаниями роговых отростков) больше длины лба от
среднероговой точки до передних концов лобных костей.3 Теменные гре-
бешки сближены; расстояние между ними не превышает 16% основной
длины черепа и, за исключением одного вида (Capra falconeri Wagner),
менее длины теменных костей по средней линии. Предглазничные ямки на
черепе отсутствуют, равно как и продольный гребешок на лицевой поверх-
ности слезной кости. Этмоидальные щели всегда имеются. Межчелюстные
кости соприкасаются с носовыми, глубоко вклиниваются между носовыми
и верхнечелюстными. Роговые отростки содержат внутренние воздушные
полости, доходящие до верхушек; стенки роговых отростков состоят из
плотного костного вещества.
Зубы гипсодонтные. Особенно сильно выражена гипсодонтия резцов,
которые приобретают вытянутую долотовидную форму (рис. 61, Г).
Отличиями в скелете козлов от близкого рода баранов являются укоре-
ненность и относительно большая ширина метаподий; ширина верхнего
края пястной кости составляет не менее 20% ее наибольшей длины. Длина
пястной кости не превышает 73% длины лучевой, а длина плюсневой — не
свыше 60—61% длины большой берцовой. Длина лучевой кости равна
или меньше наибольшей длины плечевой (В. Громова, 1953).
1 Исключение представляет винторогий козел (Capra falconery Wagn.), у которого
слабо развитые узлы имеются на ребре, идущем по заднему краю рога.
2 У домашних коз иногда имеются слабо развитые межпальцевые железы йа
передних конечностях.
3 По крайней мере у самок.
26 Фауна СССР. И. И. Соколов
402
BOVIDAE
История и географическое распространение. Корни рода Capra ведут
к нижеплиоценовому роду Tossunnoria Bohlin из Монголии (Bohlin, 1937),
совмещавшему в себе признаки козлов и баранов. Остатки, уже заведомо
относящиеся к роду козлов, появляются только в плейстоцене. Из них
лучше других известна так называемая первобытная коза — С. prisca
Woldr. (Adametz, 1915; Maxim, 1948), найденная в верхнеплейстоценовых
отложениях Галиции, Австрии, Румынии и у нас в верхнепалеолитических
стоянках Крыма (пещере Аджи-Коба; Громова, 1935). С. prisca является
одним из наиболее вероятных предков домашней козы (С. hircus L.).
В настоящее время ареал диких козлов охватывает ряд горных обла-
стей Средиземноморья и средней Европы: Испанию, Швейцарию, Авст-
рию, северную Италию, ряд островов Греческого архипелага, северо-вос-
точную Африку, Кавказ, Переднюю, Среднюю и Центральную Азию,
Алтай и Саяны, а также северную Индию и часть Пакистана (западный
Синд).
Современные разрозненные ареалы отдельных видов представляют
остатки некогда обширной и сплошной области распространения от Атлан-
тического океана до Индии и от Средиземноморья до Англии и Ирландии
включительно (Newton, 1880; Пидопличко, 1951).
Козлы — типично горные животные. Эволюция их, по-видимому,
с ранней истории шла в направлении адаптации к обитанию в условиях
скалистых ландшафтов. Вся их организация приспособлена не к быстрому
бегу, а к передвижению, лазанию по крутым скалистым склонам. Чертами
адаптации к жизни в горах являются увеличенные, широко раздвигающиеся
в стороны копыта с крепкими стенками, короткие толстые ноги с особенно
укороченными и широкими метаподиями, повышающие устойчивость жи-
вотного при передвижении в условиях сильно пересеченного рельефа.
С этим же связана более шаровидная по сравнению с баранами и газелями
головка бедренной кости, допускающая отведение в сторону ноги при
ходьбе вдоль крутых склонов, а также некоторые другие особенности
в устройстве опорно-двигательного аппарата (Громова, 1953).
Классификация. Около 10 видов современных козлов довольно хорошо
группируются в четыре подрода.
1) Capra s. str.— Безоаровые козлы. Дугообразно развитые рога сам-
цов имеют линзообразную форму поперечного сечения. На передней сто-
роне рога имеется только одно острое ребро и не развита передняя (лобо-
вая) грань рога. Распространение: острова Греческого архипе-
лага, Кавказ, Малая Азия, Иран, Белуджистан, на восток до западного
Синда. В современной дикой фауне один вид — С. aegagrus Erxl.; к этому
же подроду должны быть отнесены плейстоценовая С. prisca Woldr. и рас-
пространенная по всем материкам домашняя коза С. hircus L.
2) Ibex Hodgs. — Козероги. Рога у самцов дугообразные, с двумя
хорошо развитыми на передней стороне килями и передней (лобовой)
гранью между ними. Роговые чехлы на передней стороне несут сильно
выступающие поперечные валики. Распространение: ряд гор-
ных областей средней Европы и северной Италии, горы Средней Азии
(Гиссарский хребет, Памир, Тянь-Шань) и южной Сибири (Алтай, Саяны),
Монголия, западные провинции Китая, Верхний Египет, Судан, Абисси-
ния, Синай, Палестина, Сирия, Аравия, Афганистан, Кашмир, северный
Пенджаб. Подрод включает виды: 1) С. (Ibex) ibex L., европейский, или
альпийский козерог; 2) С. (Ibex) sibirica Meyer, сибирский козерог, или тэк;
3) С. (Ibex) nubiana F. Cuv., нубийский козел, населяющий африканскую
часть ареала рода, кроме Абиссинии; 4) С. (Ibex) sinaitica Ehrend., аравий-
CAPRA
403
ский козерог, обитающий в Синае, Сирии, Палестине и на Аравийском
полуострове (обычно объединяется с предыдущим видом); 5) С. (Ibex)
wait Rupp., абиссинский козел, сохранившийся кое-где в горах Абиссинии.
3) Orthaegoceros Trouessart. — Винторогие козлы. Рога у самцов
с двумя (передним и задним) килями, причем последний, в противополож-
ность другим подродам, наиболее острый. Лобовая грань рога не развита,
наружная поверхность рога сильно выпукла по сравнению с внутренней.
Форма рогов или прямая, но скрученная гетеронимно в виде штопора, или
свернутая в открытую гетеронимную спираль. Распространение:
Пиренеи, горы южной части Узбекистана и юго-западного Таджикистана,
Афганистан, Кашмир, Пенджаб, Белуджистан. Включает два вида:
С. (Orthaegoceros) pyraenaica Schinz., испанского козла, и С. (Orthaegoceros)
falconeri Wagn., винторогого козла Средней и южной Азии. Испанский
вид иногда выделяется в самостоятельный род Turocapra (de Beaux, 1949).
4) Turns Hilzheimer — Кавказские каменные козлы, или туры. Харак-
теризуются приближающимся к круглому сечением рогов у самцов,
и склонностью к перверзии или гетеронимному их изгибу. Распро-
странение: Главный Кавказский хребет. Один вид — С. caucasica
Guld. Подрод в целом является как бы связующим подроды Ibex и Orthae-
goceros. Тур Северцова (С. caucasica sever tzowi^enxli.) обнаруживает несом-
ненные черты сходства с козерогами, а изгиб рогов С. caucasica cylindri-
cornis несколько напоминает таковой у испанского козла (С. pyrenaica).
В Фауне СССР представлены все четыре подрода.
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ КОЗЛОВ (РОД CAPRA)
ФАУНЫ СССР
1 (2). Рога свернуты в виде штопора или винта, правый рог вправо, левый
влево, и имеют на каждом роге два приостренные ребра — переднее
и заднее. У самцов на нижней стороне шеи и груди имеется подвес
из длинных волос (рис. 92). В черепе теменная кость укорочена;
длина ее по средней линии меньше расстояния между теменными
гребешками...................................................
Capra (Orthaegoceros) falconeri Wagner—Винторогий козел (стр. 442).
2 (1). Рога дугообразно загнуты назад или образуют пологий спирале-
видный изгиб наружу, назад, вниз и концами внутрь. Заднее ребро
на рогах отсутствует или сильно закруглено. Подвес из длинных волос
на нижней стороне шеи и груди отсутствует. Теменная кость не уко-
рочена; длина ее по средней линии не меньше расстояния между те-
менными гребешками в самом узком месте.
3 (4). Рога расходятся в основаниях под углом не менее 45° (рис. 98).
Лобовая грань на рогах у самцов широкая, лишь немного уступает в
ширине боковым. Все ребра на рогах закруглены и нечетки, а грани
сильно выпуклы, особенно внутренняя. Поэтому поперечное сече-
ние рогов у оснований имеет округло-треугольную, иногда почти
круглую форму..........................Capra (Tunis) caucasica
Guldenstaedt — Кавказский каменный козел, или тур (стр. 455).
4 (3). Рога расходятся в основаниях под углом меньше 45°. Лобовая
грань или совсем отсутствует, или сравнительно узкая, значительно
уступающая по ширине боковым. На передней стороне рогов имеется
по одному или по два резко обозначенных ребра. Передняя (лобовая)
грань рога если имеется, то плоская; боковые также слабо выпуклы.
Поэтому поперечное сечение рогов в основаниях ясно сжатое с боков,
2S*
404
BOVIDAE
линзовидное, грушевидное или .четырехугольное с закругленной
задней гранью.
5 (6). На передней стороне рогов имеется по одному передне-внутреннему
острому ребру с расположенными на нем сжатыми с боков буграми
(рис. 85). Лобовая грань рога отсутствует. Поперечное сечение рогов
линзовидное или грушевидное. Лоб узкий; межглазничная его ши-
рина на черепе меньше расстояния от переднего края хоан до нижнего
края затылочного отверстия (basion). Длина ряда переднекоренных
зубов в верхней челюсти (Р2— Р4) составляет не менее 33% длины
всего соответствующего ряда коренных зубов (Р2—М3)..................
Capra aegagrus Erxleben — Безоаровый козел (стр. 404).
6 (5). На передней стороне рогов развито по два четко обозначенных ребра.
Между ними имеется плоская лобовая грань с расположенными на
ней отставленными друг от друга поперечными валиками (рис. 88).
Поперечное сечение рога ближе к вершине треугольное, у оснований
сжатое с боков, четырехугольное с выпуклой задней гранью. Лоб
широкий; межглазничная его ширина на черепе не меньше расстоя-
ния от переднего края хоан до нижнего края затылочного отверстия
(basion). Длина ряда переднекоренных зубов в верхней челюсти
(Р2—Р1) не больше 33% длины всего соответствующего ряда корен-
ных зубов (Р2—М3) ..................................................
Capra (Ibex) sibirica Meyer. — Сибирский козерог (стр. 416).
Capra aegagrus Erxleben — Безоаровый козел (рис. 85—87)
Erxleben F., 1777, Syst. Regne Anim., 1 : 260 (первооп.); Blanford W.,
1875, J. Asiat. Soc. Bengal : 12—20 (сист., синон.); Danford C., 1875, Proc. Zool.
Soc. London : 458—468 (общ.); R a d d e G. and A. Walter, 1889, Zool. Jahrb.,
Abt. System., IV : 1064—1065 (распр.); Lydekker R., 1898, Wild oxen, sheep
and goats of all lands, London : 260—266 (общ., синон., изобр.); I v г e a, 1899, Proc.
Zool. Soc. London : 599 (сист.); Lorenz—Lib urn a u L., 1899, Wissensch.
Mitteil. aus Bosnia u. Herzog., VI : 851—886, Taf. XXVI—XVIII (сист., изобр.,);
Туркин H. В. и К. А. Сатунин, 1904, Звери России, Bovidae, М. : 527—529
(общ., синон., изобр.); Динник Н. Я., 1910, Звери Кавказа, 1, Тифлис : 162—168
(общ.); Lydekker R., 1913, Catal. Ungulate mammals Brit. Mus., I, London:
156—161 (сист., синон., изобр.); Пфиценмайер Е. В., 1915, Изв. Кавказок,
муз., VIII : 251—266 (гибр.); Билькевич С., 1918, Изв. Закаспийск, муз., 1 :
12 (сист.); Brehms Tierleben, 1925, 4. AufL, 13, Leipzig : 285—288 (общ.,
изобр.); Огнев С. И. и В. Г. Гептнер, 1929, Тр. Научно-иссл. инет. зоол.
Моск. гос. унив., Ill, 1 : 143—145 (распр., биол.); Flower S., 1931, Proc. Zool.
Soc. London : 209—210 (продолж. жизни); Флеров К. К., 1932, Тр. Совета по изуч.
произв. сил Акад, наук СССР, сер. туркменск., 2 : 256—258 (морф., распр.); Н i 1 z-
heimer М., 1933, Arch. f. Tierern. u. Tierzncht, VIII : 327—371 (сист., гибр.);
Лаптев M. К., 1934, Изв. Туркменск. междуведомств. комит. по охране природы,
1 : 134—135, 187 (распр., биол.); Флеров К. К. и И. М. Г р о м о в, 1934, Тр.
Совета по изуч. произв. сил Акад, наук СССР, сер. туркменск., 6 : 369 (распр.); W о 1 f В.,
1938, Fauna fossilis cavernorum. I. Anitnalia, pars 82 : 1—192, 1—96; pars 89 : 193 —
240, 97—208 (ист.); Гептнер В. Г. и A. H. Формозов, 1941, Сборн. тр. Гос.
зоол. муз. Моск. гос. унив., VI : 64—65 (распр., стад.); Даль С. К., 1944, Зоол.
сборн. Зоол. инет. Акад, наук Армянок. ССР, 3 : 39—41 (биол.); Саркисов А. А.,
1944, Тр. Ереванск. зоол. парка, 1/2 : 133—144 (бол.); Дементьев Г. И., 1945,
Учен. зап. Моск. гос. унив., вып. 83, биол. : 76 (распр.); Даль С. К., 1948, Зоол.
сборн. Акад, наук Армянок. ССР, 5 : 57 (распр.); Скворцов В. П., 1948, Тр.
Воен.-мед. акад., XLIV : 176 (распр.); Григорян Г. А., 1949, Тр. Армянск.
ваучно-иссл. ветеринарн. инет., VI : 151—158 (параз.); Ц а лк ин В. И., 1950,
Докл Акад, наук СССР, LXX, 2 : 323—325 (сист.); Butler R., 1951, Fild, Lon-
don, 197 : 128 (охр., изобр); Даль С. К., 1951, Изв. Акад, наук Армянск. ССР,
IV, 1 : 33—39 (биол.); Пидопличко И. Г., 1951, О ледниковом периоде, II,
Киев, изд Акад, наук УССР : 61 (ист); Е 1 1 е г m a n J. and Т. М orrison-Scott,
1951, Checklist of palaearctic and indian mammals, London : 405 (сист., синон.); В e-
CAPRA
405
рещагинН. К., 1952, Млекопит. Кавказа, Диссерт., Л. (распр.); G а р к и с о в А. А.-,
1953, Природа, 3 : 113 (гибр.); Эквтимишвили 3. С., 1954, Тр, Инет... зоол.
Акад, наук Грузинск. ССР, XIII ; 96—97 (размн.); Насимов ич А. А., 1955,
Роль снежного покрова в жизни копытных, М., изд. Акад, наук СССР : 140 (распр.,
биол.).
Описание. Животные меньшей по сравнению с другими дикими коз-
лами величины. Длина тела до 150 см; высота в холке 80т—90 и до 95 см.
Основная длина черепа у самцов до 255 мм, у самок от 185 до 222.мм. Живой
вес, по А. А. Саркисову (Даль, 1951), самцов 35—38 кг, самок 26—30 кг.
Рис. 85. Безоаровый козел (Capra aegagrus Erxl.).
Телосложение также несколько более легкое, чем у других видов рода
Capra L. Высота в кресте на 1—2 см превосходит высоту в холке. Голова
с прямым носом, но с заметно выпуклым лбом, особенно у самцов. Уши
длиной 10—И см, с заостренными верхушками. Как и у других козлов,
остатки носового зеркала представлены полосками голой кожи вдоль верх-
них краев ноздрей, между внутренними краями последних и по средней
линии верхней губы. Радужная оболочка глаз коричневая. Шея у самцов
короткая, толстая; у самок она производит впечатление, особенно летом,
более длинной и тонкой. Линия спины прямая, круп заметно свислый.
Хвост длиной вместе с волосами 17—20 см, без волос 12—13 см. Копыта
длинные, но низкие, с тупыми закругленными концами. Длина передних
копыт от пятки до верхушки у взрослых самцов 67—72 мм, задних от 56 до
60 мм; высота копыт — передних 33—38 мм, задних‘на 1—2 мм меньше.
На передней стороне запястных суставов и у самцов, и у самок имеются
406
BOV I DAU
лишенные волос, довольно толстые роговые мозоли круглой формы раз-
мерами с трех-пятикопеечную монету.
Характерной особенностью рогов безоарового козла является наличие
острого ребра (киля) вдоль передне-внутреннего края и отсутствие перед-
ней, лобовой грани. Форма поперечного сечения рога сильно сжатая с бо-
ков, линзовидная или грушевидная; слабо намеченное заднее ребро рога
закруглено. Из двух боковых граней рога наружная более выпуклая.
Вдоль переднего ребра рога у самцов расположен ряд бугров на расстоянии
8—14 см один от другого, сжатых с боков и отчетливо выступающих над
поверхностью рога. У самок бугры лишь намечаются, а чаще отсутствуют
вовсе. От основания и почти до самых верхушек поверхность рога несет
мелкую поперечную морщинистость и отставленные друг от друга более
глубокие поперечные борозды (кольца), отграничивающие участки годич-
ного прироста рога. Как раз на пересечении переднего ребра с кольцами
годичного прироста расположены бугры по переднему ребру рога.1 Цвет
рогов от серовато-бурого до почти черного.
Рога дугообразно или саблевидно загнутые назад, не сильно, под углом
35—40° расходящиеся в стороны; сравнительно тонкие, но широкие;
передне-задний их диаметр в основаниях у самцов может доходить до 10.7 см,
при поперечнике около 5.5 см. У взрослых самцов рога несколько сходятся
вершинами, так что наибольшее расстояние между рогами имеет место
в предвершинной части. Изредка концы рогов даже перекрещиваются.
В основаниях рога сильно сближены; расстояние между передними их
краями часто не превосходит 1 см. Длина рогов самцов в несколько раз
превосходит длину черепа, достигая в рекордных случаях 140 см по изгибу
(Lydekker, 1898).
У самок рога короче, от 18 до 39 см, и редко превосходят длину черепа
больше чем в полтора раза. Переднее ребро рога у них менее острое, а
основания рогов расставлены несколько шире, на 2—3 см. Динник (1910)
отмечает изредка встречающуюся комолость самок.
Изгиб рогов назад, в начальной половине, пологий и равномерный,
к вершинам становится крутым; у самцов концы их обычно направлены
вниз и вперед.
Волосяной покров безоарового козла состоит из упругих, слабо вол-
нистых, сравнительно негрубых волос и тонкого извитого подшерстка.
Длина остевых волос в зимнем меху составляет 5—6 см. Несколько дли-
нее волосы на нижней и верхней стороне шеи и вдоль средней линии
спины. В летнем меху подшерсток отсутствует, остевые волосы упругие,
плотно прилегают к коже. Зимой мех становится более пушистым и
благодаря наличию подшерстка на ошупь более мягким. Борода длин-
ная, заостренная на конце, состоит из толстых грубых волос длиной до
20 см, у взрослых самцов она занимает весь низ головы, кроме нижней
губы и самого подбородка. У самок борода нормально отсутствует, по
иногда развивается, хотя и не в такой степени, как у самцов (Динник,
1910). Волосы на конце хвоста длиннее, чем у основания, но сколько-
нибудь заметной кисти не образуют.
Подшерсток у безоарового козла имеет дымчато-серую окраску. Во-
лосы ости на боках туловища и спине от основания светло-серые, выше
буреют, затем следует желто-рыжее предвершинное кольцо, а заострен-
ный кончик волоса имеет вновь темно-бурую окраску. Иногда, вместо
1 Число бугров, поэтому, может служить критерием возраста зверя, хотя возможно,
что на границе прироста первого года жизни узла не образуется. Этот вопрос требует
уточнения.
CAPRA
407
рыжего, предвершинное кольцо бывает белым. Темно окрашенные кон-
чики волос легко обламываются уже в конце лета и осенью. От интен-
сивности окраски различных зон остевых волос, от их протяженности,
числа волос с необломанными верхушками зависит общий тон окраски.
Летом окраска красноватая или рыжая с буроватым оттенком. На
боках шеи, по сторонам предплечий и бедер, она светлее, чем на остальном
туловище. Верх головы, верхняя и нижняя губа буро-коричневые.
Края губ белые. Вока головы обычно одноцветны с туловищем, в отдель-
ных случаях бурые, но более светлые, чем верхняя сторона; иногда от
переднего угла глаз тянутся не доходящие до углов рта темные полосы,
отделенные от темного верха носа светлой шерстью. Борода темно-бурая,
почти черная, но обычно содержит незначительную примесь белых
волос. Уши снаружи желтовато-серые, внутри серовато-белые. Концы
их черные. Вдоль верхнего края шеи и по средней линии туловища тя-
нется узкая буро-коричневая полоса, сильнее всего развитая на холке
и постепенно сходящая на нет к корню хвоста. В некоторых случаях
в срединной бурой полосе заметна незначительная примесь белых волос.
Хвост черно-бурый или черный, но у основания часто имеет примесь
рыжих, а на конце — светлых волос. В типичных случаях от холки вдоль
переднего края лопаток тянется различной ширины темно-бурая
полоса, охватывающая наподобие хомута основание шеи. Темно-бурая
окраска «хомута» распространяется на переднюю сторону груди, а также
по всей нижней стороне шеи. Низ груди, живот, нахи, внутренняя сторона
ног и ягодиц белые, иногда с желтоватым или сероватым оттенком.
От локтевого к коленному суставу по бокам туловища тянутся темно-
бурые полосы, отграничивающие рыжевато-бурые бока от светлой окраски
живота. Передняя сторона ног темно-бурая, постепенно темнеющая, до
почти черной, книзу. Темные полосы передних ног в ясно выраженных
случаях сливаются с темной окраской хомута, охватывающего основание
шеи, на задних ногах служат продолжением темных полос по бокам живота.
Наружная, внутренняя и задняя стороны ног ниже запястных и скака-
тельных суставов белые с желтоватым или сероватым оттенком. Темная
продольная полоса передних ног, выше запястных суставов, прерывается
поперечной белой полоской, соединяющей светлую окраску внутренней
и наружной стороны ног.
К зиме окраска безоаровых козлов светлеет и вместо красновато-
рыжеватых начинают преобладать буровато-серые и белесые тона. Рас-
пределение светло и темно окрашенных участков остается прежним.
Общая окраска самок та же, что и у самцов, но все темные отметины
у них выражены слабо, а частично отсутствуют вовсе. Так, хомут у осно-
вания шеи у них, как правило, не заметен, равно как далеко не всегда
имеются и темные полосы по бокам живота. Темно-бурая полоса вдоль
хребта мало заметна. Темная окраска передней стороны ног ограничи-
вается областью ниже средины голени и предплечья и не достигает ин-
тенсивности ее у самцов. Молодые самцы окрашены сходно с самками;
полное развитие окраски, свойственной взрослым животным, достигается
на третьем-четвертом году жизни. Козлята в первые месяцы после рожде-
ния имеют более насыщенную желтовато-рыжую окраску, со слабо раз-
витыми темными участками только на верхней стороне головы и нижних
частях ног.
Имеют место индивидуальные колебания в особенностях окраски.
Так, темный хомут в основании шеи у некоторых взрослых самцов раз-
вит очень слабо, почти не заметен. У отдельных экземпляров отсутствуют
408
BOVIDAE
и темные полосы по бокам тела. Зимняя шкура взрослого самца с Кавказа
в коллекции ЗИН АН СССР при хорошо развитых темных участках имеет
серовато-белую с едва заметной рыжеватостью окраску; последняя обу-
словлена примесью к рыжим значительного количества белых с темными
кончиками волос.
В черепе безоарового козла (рис. 86) характерно, кроме формы рогов,
вздутие в виде бугра срединной части лба перед основаниями роговых
отростков. Особенно велик лобный бугор у самцов. У самок он менее
развит, но все же, как правило, ясно заметен в виде сходящихся на сред-
ней линии лба продольных валиков, идущих от оснований роговых
отростков. Поэтому поверхность лба в средней части расположена выше
уровня верхних краев глазниц. Лоб сравнительно узкий; межглаз-
ничная ширина его меньше расстояния от переднего края хоан до ниж-
него края затылочного отверстия. Роговые отростки мало сдвинуты назад;
передний край их основания расположен впереди или на одном уровне
с задним краем глазниц.
Носовые кости по сравнению с другими видами короткие и широкие;
их наибольшая ширина составляет от 32 до 49 % собственной длины.
Расстояние от переднего края межчелюстных костей (prosthion) до перед-
него края альвеолы Р2 превосходит расстояние от prosthion до передних,
концов носовых костей не более, чем на длину Р2, а иногда даже меньше.
Передний край хаон расположен, как правило, несколько впереди уровня
боковых выемок заднего края костного нёба.
Передние коренные (премоляры) по сравнению с другими видами
мало редуцированы; длина их ряда в верхней челюсти составляет не менее
трети (от 33 до 37%) длины всего соответствующего ряда коренных зубов
(Р2—М3).
История и географическое распространение (рис. 87)Capra aega-
grus — наиболее примитивный из современных видов козлов, сохранив-
ший ряд архаических черт, свойственных вероятному предку рода
Capra. Ввиду недостаточной четкости морфологических отличий от близ-
кой плейстоценовой формы, С. prisca Woldr., установить время появления
современного вида на геологической сцене довольно трудно. Остатки
козлов, сходных с С. aegagrus, найдены в пещерных отложениях Италии,
Ирландии, Франции, Болгарии (Wolf, 1938), а также в неолите восточ-
ного побережья Средиземного моря. В областях современного распро-
странения следы обитания этого вида отмечаются начиная с голоцена.
Современный ареал безоарового козла включает греческие острова:
Крит, Киклады (Иура, Эримомилос), затем Малую Азию, Кавказ, Иран,
южную Туркмению, Белуджистан, и на востоке доходит до западного
Синда. В пределах СССР С. aegagrus обитает в восточной части Главного
Кавказского хребта, в Закавказье, Копет-Даге и в Больших Балханах.
На Кавказе ареал безоарового козла представлен рядом изолированных
очагов обитания.1 Одним из крупных очагов является горный Дагестан.
Здесь по северному склону Главного Кавказского хребта этот вид более
или менее повсеместно встречается от верховьев Аргуна и Алазани па
западе (в пределах Грузии) до верховьев Гердыманчая, около с. Исмаиллы,
на востоке. На северо-западе граница распространения проходит, не до-
ходя 20—30 км Гуниба; раньше козлы встречались и около Ботлиха.
В прибрежных и равнинных районах Дагестана козлы отсутствуют.
1 Распространение на Кавказе дается по Н. Я. Диннику (1910), В. Г. Гептнеру
и А. Н. Формозову (1941), С. К. Далю (1951) и Н. К. Верещагину (1952).
Рис. 86. Череп Capra aegagrus Erxl.
410 BOVIDAE
Редки они и в средних, густо населенных горных районах области, а
местами нет их вовсе. Во времена Н. Я. Динника (1910) уже не было безо-
аровых козлов на безлесных горах водораздела Самура, Каракойсу и
Казикумухского Койсу. По данным Гентнера и Формозова (1941), они
встречаются в Лакском (Казикумухском) районе. В небольшем количестве
безоаровые козлы до сих пор сохранились в районах верхнего течения
Андийского и, особенно, Аварского Койсу, где Богосский хребет
считается одним из лучших охотничьих угодий Дагестана.
Н. Я. Динник встречал козлов в Тушетии, но держатся ли они там
в настоящее время — не известно. В верховьях Терека и Арагвы их
уже нет, как не встречались они и раньше к западу от Военно-Грузинской
дороги. Не встречал Н. К. Верещагин (1952) безоаровых козлов и на
южном склоне Большого Кавказского хребта от Лагодехи до Шемахи,
а по словам охотников, на южные склоны Большого Кавказа безоаровые
козлы вообще заходят крайне редко.
В Закавказье безоаровый козел распространен также отдельными
очагами. Встречается он до сих пор по западным отрогам Триалетского
хребта у Боржоми. В Армении, по Далю (1951), «в настоящее время безо-
аровые козлы постоянно встречаются в скалистых горах от долины р. Азат
до склонов Зангезурского, Мегринского иБаргушатского хребтов. На север
ареал этих животных простирается до южных склонов Гегамского и
Варденисского хребтов. Наибольшее количество безоаровых коз
известно на Урцском и Айцдзорском хребтах, а также на ближайших
к ним скалистых массивах. На юге безоаровые козлы местами встречаются
вплоть до Аракса».
В 70-х годах прошлого столетия безоаровые козлы отмечались в окрест-
ностях с. Еленовки на Севане, а в 90-х годах — в Шахдагском хребте
в районе Кедабека и Калакента. В 30—40-х годах нынешнего века встре-
чались от верховьев Шамхора до восточной оконечности Шахдагского
хребта (г. Кечал-Даг). Следы пребывания до 1941 г. отмечались также
к западу от Севана, в Памбакском хребте, но в настоящее время они, по-
видимому, там исчезли (Даль, 1951).
В пределах Нахичеванской АССР безоаровые козлы водятся на изо-
лированных хребтах Алинджа-Даг и Илянлу-Даг, а также по всему
Зангезурскому хребту вплоть до долины Аракса. По Араксу область
распространения С. aegagrus доходит до юго-восточных склонов Мегрин-
ского хребта вплоть до Охчичая. Как место, особо богатое козлами,
Н. К. Верещагин называет Неграмские горы между Джульфой и Орду-
бадом. Небольшое число их держалось в 1947 г. даже в окрестностях
Акулиса и Ордубада.
По-видимому, сохранились безоаровые козлы в том или ином числе
в Карабахском нагорье. Как крайнюю восточную точку их распростра-
нения в 30-х годах Н. К. Верещагин называет гору Кире в Азербайджане.
По данным X. М. Алекперова,1 на горе Кирсе (над Степанакертом) безо-
аровые козлы держались до 1944 г. По собранным им сведениям, они
водятся до сих пор в Карабахском хребте (урочище Гырх-Кыз), на ска-
листых вершинах Дели-Дага, Муров-Дага, Кечал-Дага. В небольшом
числе сохраняются на вершине Кошкара, на горе Кяпая у Гек-Геля.
Имеются козлы в ущелье Баргушатчая (близ с. Новлу, на границе с Ар-
менией), а также в районе Охчичая к востоку от Кафана.
1 Устное сообщение.
CAPRA
'III
В Талышинском хребте безоаровых козлов в настоящее время давно
нет, но в средине прошлого столетия они встречались на вершине горы
Кызюрду, представляя, вероятно, соверо-восточную границу иранского
ареала вида.
В пределах Туркменской ССР безоаровый козел населяет, по-видимому,
весь Копет-Даг и Большие Балханы (Лаптев, 1934). Распределение ого
здесь изучено недостаточно.
В центральном Копет-Даге безоаровые козлы, по С. И. Огневу и
В. Г. Гентнеру (1929), весьма обычны. Главная масса их держится в мас-
сиве Дуптака. В районе Фирюзы встречаются несколько реже, а местами,
Рис. 87. Распространение Capra aegagrus Erxl.
1 — XX в.; 2 — XIX в.; 3 — XVIII в. и раньше.
особенно вблизи некоторых населенных пунктов, их нет вовсе. Встре-
чаются безоаровые козлы в Гермабском ущелье и в ущельях к западу
от пос. Гермаба. Обыкновенны также в ущелье р. Мергень-Ули около
с. Дмитриевского (Куркулаба), но выше бывшего пограничного поста
Мергень-Ули, где рельеф становится ровным и спокойным, не подходя-
щим для обитателя резко скалистых ландшафтов, козлов уже нет. Много
козлов во всех ущельях, тяготеющих к пограничной вершине Гулюр
(Хайнилау, Тернаули, Сундукли, Миимли и др.).
В бассейне рр. Сумбара и Чандыра (юго-западный Копет-Даг, Кара-
Калинский район) безоаровые козлы держатся в безлюдных местах на
Иранской границе. Лет двадцать назад довольно много их было в окрест-
ностях пограничных застав Арваз и Кизыл-Имам (Флеров и Громов,
1934). М. К. Лаптев (1934) встречал козлов в горах между рр. Чандыром
и Атреком. В колекциях ЗИН АН СССР хранятся экземпляры С. aega-
grus из окрестностей ст. Джабель (Большие Балханы), Кошакуй, г. Ду-
шак и из ряда других пунктов Копет-Дага.
Восточная граница распространения безоарового козла в пределах
Советского Союза проходит в междуречье Теджена и Мургаба. Б. П. Сквор-
цов (1948) наблюдал его в ущелье Нурдыванлы. По словам местных охот-
ников, козлы добываются как в данном месте, так и восточнее, в районе
Зульфагара, встречаясь здесь, возможно, уже совместно с сибирским козе-
412
BOVIDAU
рогом. По собранным К. К. Флеровым (1932) данным, безоаровые козлы'
встречаются в Афганистане к югу от Кушки и в районе Рабат-Кашана,
но, как он сам пишет, в последнем случае речь идет, возможно, уже о си-
бирском козероге.
Подвиды. По-видимому, справедливым следует считать выделение
четырех подвидов:
1) С. aegagrus aegagrus Erxl. Размеры сравнительно крупные, рога
массивные; обхват последних в основаниях не менее 20 см. Распро-
странение: Малая Азия, Иран, Кавказ с Закавказьем.
2) С. aegagrus blythi Hume. Размеры меньше, чем у типичной расы.
От последней отличается также более тонкими рогами (обхват у основа-
ний не более 20.5 см) со слабым развитием бугров по переднему краю
и более светлой окраской волосяного покрова. Распространение:
западный Синд, Белуджистан, возможно также юго-восточный Иран
(Билькевич, 1918).
3) С. aegagrus turcmenicus Zalkin. По общим размерам не уступает
типичному подвиду, но отличается от него более тонкими, в среднем,
рогами; обхват последних в основаниях у самцов от 17 до 23 см. Окраска
сходная с С. aegagrus aegagrus. Распространение: Туркмено-
Хоросанские горы (в пределах СССР — Копет-Даг и Большие Балханы).
4) С. aegagrus cretica Schinz. Мелкая островная форма. От остальных
подвидов отличается, помимо меньшей величины, тем, что рога обнаружи-
вают в той или иной мере выраженный гомонимный, сходный с домаш-
ними козами, изгиб с загнутыми в стороны концами. Распростра-
нение: острова Греческого архипелага — Крит, Иуры, Эримомилос.
Иногда оспаривается исконно дикий характер коз, населяющих Крит п
острова Эгейского моря.
Биология. Образ жизни безоарового козла изучен слабо. Объяс-
няется это обстоятельство трудностями наблюдений, вследствие недоступ-
ности мест его обитания и относительной малочисленности этого зверя.
Безоаровый козел — типичный обитатель скалистых ландшафтов.
«Громадные ущелья, стены, отвесы и уступы их склонов с отдельными
участками, поросшими травой, иногда с лепящимися кое-где арчами, —
вот типичное местопребывание Capra aegagrus. Всякое другое животное
рискует сломать себе шею на этих головокружительных обрывах» (Огнев
и Гептнер, 1929). В отличие от другого обитателя гор —• серны, которая
в спокойных условиях существования требует наличия по соседству
со скалами достаточно обширных и открытых мест выпаса, безоаровый
козел довольствуется незначительными лужайками зелени у подножья
и между скал. Еще больше, чем для серны, скалы и ущелья важны для
него как убежища, с которыми безоаровые козлы расстаются очень редко.
Пределы вертикального распространения безоаровых козлов довольно
широки, во многом определяются наличием подходящих стаций, а также
степенью преследования со стороны человека. В обычных условиях они
обитают на высотах от 1000 до 2300—2500 м, но встречаются и ниже,
а на Больших Балханах спускаются нередко к самому подножью (Лаптев,
1934). Зато на Арарате они живут на высотах свыше 4000 м, а в Дагестане,
избегая преследования, поднимаются к подножью ледников и вечного
снега (Динник, 1910). Решающим в выборе местообитаний является не
высота над уровнем моря, а наличие крутых обрывистых скал и ущелий.
Важным условием существования безоаровых козлов является нали-
чие в местах обитания леса. В противоположность турам, на Кавказе
козлы предпочитают не голые утесы, а облесенные склоны, где скалы
CAPRA
413
сочетаются или чередуются с лесом. Придерживаясь, в основном, средней
пли верхней полосы леса, они чаще встречаются в глухих лесистых местах,
где вовсе не водятся туры. По-видимому, отдавая дань защитным свой-
ствам леса, в Дагестане безоаровые козлы иногда держатся в густых
зарослях даже на сравнительно пологих склонах (Гептнер и Формозов,
1941). Следует, однако, сказать, что в ряде местностей, как например
в южном Закавказье, около Сарыкамыша и Ордубата (Динник, 1910),
а также в Брлыпих Балханах и Копет-Даге, этот вид хорошо уживается
и в безлесных горах.
Сезонная смена мест обитания у безоаровых козлов выражена, по-
видимому, нечетко. Имеются указания, что на зиму козлы спускаются
ниже, чем летом, и держатся, в основном, по южным солнцепечным
склонам.
Живут безоаровые козлы, как правило, небольшими группами, редко
более 7—10 голов вместе. Около половины встреч, отмеченных С. К. Да-
лем (1944, 1951), приходилось на одиночных и державшихся парами
животных. Максимальные из наблюдавшихся этим автором стад состояли
в одном случае из 23, в другом из 31 штуки. Впрочем, размеры табунов
зависят в значительной мере от общей численности зверей в том или ином
районе; так, по словам Г. Радде и А. Вальтера (Radde a. Walter, 1889),
в 80-х годах прошлого века в Копет-Даге стада в 30—40 и даже 90 голов
не составляли большого исключения.
Состав стада безоаровых козлов не постоянен. Одиночками держатся
главным образом взрослые самцы, реже — самки (и то лишь перед око-
том). Вожаком в стадах чаще бывают старые самки. Потревоженное стадо
бежит, не рассыпаясь, в одном направлении. При ночлеге и отдыхе козлы
располагаются в одном месте, но на расстоянии нескольких метров один
от другого. Специальных сторожей, по-видимому, не выставляют, но
в табунке всегда находится на ногах часть животных (Даль, 1951).
Суточный цикл у безоаровых козлов не сложен. С рассвета до 9—10 ча-
сов утра они пасутся, не уходя далеко от места ночевок. Днем отдыхают
в тени деревьев, под навесами скал или в пещерах. Эти же места зимой
служат убежищем на время непогоды. Летом с 14—15 часов вновь отправ-
ляются на пастьбу, спускаясь на лужайки к основаниям скал, где иногда
собирается по нескольку стад вместе (Даль, 1944). Пасутся, по-видимому,
и часть ночи. Лежки располагаются в скалах, но следует подчеркнуть,
что они находятся недалеко от мест жировок; ущелья служат местами
не только отдыха и убежищ, но и кормежки.
О питании безоаровых козлов известны лишь отрывочные данные,
основанные на исследовании поедей на местах выпаса в Закавказье (Даль,
1944, 1951). Пищу составляют в летний период разнообразные травы,
а также листва и побеги кустарников и деревьев. С. К. Даль приводит
список из девятнадцати видов кормовых растений, но, вероятно, число
поедаемых видов значительно больше. Из трав сильнее всего на пастбищах
оказываются объеденными гиппомаратрум (Hyppomarathrum sp.), мала-
байла (Malabaila graveolens Hoffm.), подмаренник (Galium sp.), белльвалия
(Bellevalia sp.), синеголовник (Eryngium sp.), весною ковыль (Stipa sp.)
и некоторые другие, а из кустарников — Каракас (Celtis sp.) и крушина
(Ramnus sp.). Питание козлов в зимний период не изучено совершенно.
Известно, что безоаровые козлы, по крайней мере на Кавказе, охотно
посещают естественные солонцы.
В спокойной обстановке безоаровые козлы передвигаются шагом
Быстро бежать не могут. На ровном месте их легко догоняет собака.
414
BOVIDAE
Зато козлы прекрасно, еще лучше, чем серна, приспособлены к пере-
движениям в условиях сильно пересеченного скалистого рельефа. Безо-
аровые козлы прекрасно лазают по почти отвесным скалам, узким кар-
низам, в случае опасности делают прыжки свыше трех метров (Даль,
1951). В отношении способности лазать они мало уступают турам, подобно
последним «легко взбираются на отвесные скалы, смело спускаются
в страшные пропасти, пробираются по самым опасным тропинкам и де-
лают самые смелые прыжки» (Динник, 1910). Застигнутое врасплох стадо
козлов ищет спасение не в беге на ровном месте, а бросается вверх по
скалам или бежит вдоль узких карнизов ущелий.
В осторожности и сообразительности безоаровые козлы, по-видимому,
значительно уступают другим представителям рода. Н. Я. Динник (1910)
наблюдал их в полуверсте от жилья. В Копет-Даге близ Гермама козлы
встречаются на обрывах в нескольких сотнях шагов от домов над селе-
нием, а около Куркулаба иногда можно видеть козлов из самого поселка
(Огнев и Гептнер, 1929).
Безоаровые козлы обладают неплохим зрением, слухом и обонянием,
но и в этом отношении уступают кавказским турам. В закрытой местности
к ним можно подкрасться на 40—60 шагов, чего почти не случается
с туром (Динник, 1910; Туркин и Сатунин, 1904).
Голос безоарового козла сходен с блеянием домашней козы, только
более отрывистый и грубый.
Характер и порядок смены волосяного покрова у безоарового козла
почти не изучен, но, по-видимому, линька бывает в году дважды. Смена
зимнего волоса происходит в апреле и мае. Во всяком случае, в первой
половине мая козлы в Сарайбулагском хребте ходили еще в старой шер-
сти (Даль, 1944). В Ленинградском зоопарке в первой половине июня
1954 г. самец безоарового козла полностью приобрел летнее одеяние.
Линька из летного меха в зимний, по К. К. Флерову (1932), начинается
в августе или даже в конце июля. Уже в это время сквозь летнюю рыжую
шерсть начинает просвечивать серый зимний волос. Отрастает ли к зиме
и летний волос, или он выпадает — не известно.
Сроки размножения у безоаровых козлов не одинаковы в разных
частях ареала и варьируют по годам даже в пределах одного района
в зависимости от погодных условий.
Период гона и спаривания на Кавказе падает на время с середины
ноября по конец декабря. Самый разгар приходится на декабрь, но от-
дельные случаи спаривания бывают до конца января (Эквтимишвили,
1954). Самцы во время гона сильно возбуждены. Между ними часто бы-
вают драки.
Беременность самки длится около 5 месяцев. Окот начинается в сере-
дине или конце апреля. Большинство самок родят в мае, но отдельные
случаи запоздалого окота могут иметь место до конца июня. В более
южных широтах и гон, и окот происходят раньше. На самом юге Кавказа
молодые родятся в апреле (Динник, 1910); В. Г. Гептнер и С. И. Огнев
(1929) наблюдали в Копет-Даге появление новорожденных козлят в пер-
вой декаде мая, а в Больших Балханах, по М. К. Лаптеву (1934), окот
наблюдается в конце марта или начале апреля. Перед родами самки уда-
ляются от своих табунков, уходят в наиболее глухие места, где и при-
носят по одному или по два козленка. Тройни у безоаровых козлов пред--
ставляют редкое исключение. Первые дни козлята беспомощны и лежат
на земле между камнями. В возрасте до 3 дней их обычно ловят для зоопар-
ков (Саркисов, 1944). Самка привязана к козленку; в случае опасности
CAPRA
415
она старается отвлечь врага, рискуя собственной жизнью. Спустя несколько
дней козленок не только может следовать за матерью, но вскоре не хуже
нее карабкается по скалам. Когда козлята достаточно окрепнут, самки,
державшиеся до того отдельно, соединяются в табунки. Молодые дер-
жатся с матерью до следующего окота, а если она осталась почему-либо
холостой, то и дольше. Соотношение полов в стадах безоаровых козлов,
по исследованиям С. К. Даля (1951), близко один к одному.
Продолжительность жизни безоаровых козлов в естественных условиях
не известны. В зоопарках они живут в среднем от 8 до 10 лет, но известны
единичные случаи, когда самки доживали до 14 и даже до 17 лет (Flower,
1931).
Основными врагами безоаровых козлов из животного мира являются
леопард и рысь. Первый, в силу своей малочисленности, большого практи-
ческого значения, по-видимому, не имеет. И леопард, и рысь охотятся
на козлов скрадом. Рысь, вероятно, также может гоняться за ними по
скалам. Волк, как вредитель, едва ли имеет значение, так как не может
преследовать добычу в условиях сильно пересеченного рельефа.
О болезнях и эпизоотиях среди безоаровых козлов на воле ничего
не известно. Г. А. Григорян (1949), исследуя методом полных гельминто-
логических вскрытий 26 экземпляров безоаровых козлов из Армении,
обнаружил зараженность их 22 видами паразитических червей, в боль-
шинстве своем общими с домашними овцами и козами. Только два вида
нематод из сычуга и тонкого отдела кишечника являются специфическими
для безоарового козла. Патологическое значение, по полученным данным,
имеют только два вида протостронгилид {Protostrongylus davtiani Ssav.
и P. muraschinzewi Davt.), вызывающие очаги поражений в легких. Инте-
ресно отметить, что безоаровые козлы в Армении совершенно свободны
от диктиокаулеза, дикроцелиоза и фасциолеза, широко распространенных
у домашних овец и коз этих же районов. Меньше у диких козлов оказалась
по сравнению с домашними и степень зараженности трихостронгилидами.
Из эктопаразитов пока известен только один вид клеща — Hemaphy-
salts otophila Р. Sch., найденный на козле из Копет-Дага.1
Хозяйственное значение. Безоаровый козел — один из основных
родоначальников домашней козы, Capra hircus L. Поэтому он может
иметь значение при создании новых пород и при улучшении породных
качеств и конституциональной крепости существующих пород коз.
Известное практическое значение он имеет и как промысловый мясной
зверь. Мясо его по вкусовым качествам мало отличается от мяса домашней
козы. Шкуры до сих пор использовались большей частью в местном обиходе
на коврики, но безусловно годны для выделки из них ценных сортов
кожи, шевро или замши. Рога находят применение в качестве настенных
украшений.
При скрещивании диких безоаровых козлов с домашними козами
во всех комбинациях и поколениях родится вполне плодовитое потомство
(Hilzheimer, 1933). Подобные гибриды иногда получаются и естественным
путем в районах обитания диких козлов (Пфиценмайер, 1915). У гибри-
дов безоарового козла с домашней козой окраска и форма рогов насле-
дуются от дикой формы.
Известны случаи получения гибридного потомства от вольного скре-
щивания безоарового козла с кавказским туром в бывшем Боржомском
зверином отводе (Пфиценмайер, 1915). У гибридов этого рода наблю-
1 Устное сообщение Г. В. Сердюковой.
416
BOV I DAU
дается промежуточный характер наследования формы рогов и окраски.
Борода в одном случае была длинной, как у безоарового козла, но росла
не только на подбородке, но и на щеках, подобно туру. В другом случае
борода гибрида и по длине не отличалась от турьей. Приплод при скре-
щивании безоаровой козы с самцом гибридом от кавказского тура и до-
машнего козла был получен в Ереванском зоопарке (Саркисов, 1953).
О плодовитости гибридов безоарового козла с туром ничего не известно.
Наконец, следует упомянуть, что в зоопарках Германии в свое время
были получены плодовитые у обоих полов гибриды между безоаровыми
и винторогими козлами (Hilzheimer, 1933).
Охота. Из способов охоты на безоарового козла главными и почти
единственными являются скрадывание и охота с подхода. В принципе
она мало отличается от таковой на других горных копытных, например
серну, но в некоторых отношениях еще труднее. Правда, к безоаровому
козлу вообще легче подкрасться, благодаря его меньшей чуткости и осто-
рожности, но зато он обитает в еще более «тяжелых» местах, вследствие
чего от охотника требуется колосальная сила, ловкось, смелость и умение
лазать и цепляться за незначительные выступы скал и карнизы. Поэтому
настоящих охотников на диких козлов не так много.
Иногда охотятся на них группами нагоном (см. стр. 400), а Н. Я. Дин-
ник (1910) упоминает также об охоте с гончими собаками, но как исполь-
зуются при охоте на этого зверя собака — не ясно.
Capra (Ibex) sibirica Meyer — Сибирский козерог (рис. 88—91)
Meyer F., 1794, Zool. Annal., 1 : 397 (первэоп.); А л ф e p а к и С. В., 1880,
Природа и охота, V : 1—13 (общ.); Иванов Д., 1885, Природа и охота, 1 : 1—48
(биол., ох.); Г е р н Б. Ф., 1891, Семипалатинск, ведом., 15 : 3 (ист.); Ly dekke г R.,
1898, Wild oxen, sheep and goats of all lands, London : 275—285 (общ., синон., изобр.);
Головнин Д. Н., 1901, Природа и охота, 7 : 21—29, 8 : 1—14 (общ., изобр.);
Lydekker R., 1901, Proc. Zool. Soc. London, 1 : 91—94 (сист.); Noack Th.,
1902, Zool. Anzeig., XXV : 622—626 (сист.); N о a c k Th., 1903, Zool. Anzeig., XXVI :
381—390 (сист.); Туркин H. В. иК. А. Сатунин, 1904, Звери России, Bo-
vidae, М : 530—534-J-635—671 (общ., изобр.); Сатунин К. А., 1905, Природа
и охота, V : 26—27 (сист.); Leizewitz W., 1906, Zool. Anzeig., XXIX : 654—656
(сист.); Lorenz L., 1906, Denkschr. mathem.-naturwissensch. KI. K. Akad. d. Wis-
sensch., Wien, LXXX : 1—23 (сист.); Lydekker R., 1913, Catal. Ungulate mam-
mals Brit. Mus., I : 142—153 (сист., синон., изобр.); Rowland Ward, 1914,
Records of big game, London : 167—370 (рога); Дорогостайский В. Ч.,
1918, Ежегодн. Зоол. музея Акад, наук, XXIII, 1 : 32—43 (распр., биол.); Соло-
вьев Д. К., 1920, Саянский промыслово-охотничий район, Пгр., Гос. изд. : 250—
251 (распр., биол.); Дорогостайский В. Ч., 1925, Пушные и промысловые
звери Прибайкалья, Иркутск : 1—15 (распр.); Селевин В. А., 1925, Охотник,
6—7 : 28—29 (распр.); Северцов С. А., 1929, Землеведение, XXXI, 2—3 : 181 —
182 (распр., биол.); Скалой Н. В., 1929, Охрана природы, 3 : 75—77 (распр.);
Троицкий В. И., 1930, Тр. Общ. изуч. Сибири, 2 : 1—48 (распр.); Flower S.,
1931, Proc. Zool. Soc. London : 210 (продолж. жизни); Флеров К. К., 1932, Тр.
Совета по изуч. произв. сил, сер. туркменок., 2 : 258 (распр.); Булавин А.,
1934, Охотн. и рыбак Сибири, 3 : 24—25 (распр.); Залесский П. М., 1934, Охотн.
и пушник Сибири, 7—8 : 26—28 (распр.); Денисов В. Ф., 1935, Тр. Киргизок-
комилексн. экспед., IV, 3 : 81—ИЗ (морф., хоз. знач.); Розанов М. П., 1935,
сборн. «Матер, по млекопит. и птицам Памира», Л., изд. Акад, наук СССР : 34 (распр.,
биол.); Флеров К. К., 1935, в кн.: Виноградов Б. С., Е. Н. Павловский и К. К. Фле-
ров, Звери Таджикистана, Л., изд. Акад, наук СССР : 115—120 (общ., синон., изобр.);
Подаревский В. Б., 1936, Пробл. охотхозяйств, акклим. в вооточн. Сибири,
Иркутск, Огиз : 47—48 (распр.); Шнитников В. Н., 1936, Млекопит. Семире-
чья, М.—Л., изд. Акад, наук СССР : 129—134 (общ.); Гу реев iA. А., 1937, Тр.
Совета по изуч. произв.сил Акад, наук СССР, сер. туркменок., вып. 9:357—370 (распр.);
Селевин В. А., 1937, Бюлл. Средне-азиатск. гос. унив., 22 : 318 (распр,); Дмит-
риев В. В., 1938, Тр. Алтайск. гос. заповеди., 1 : 171—259 (библ., распр.); Ко-
CAPRA
417
л о с о в А. М., 1939, Учен. зап. Московск. гос. унив., 20 : 174—176 (распр., биол.,
ох.); Левиев П., 1939, Тр. Узбекист. зоол. сада, 1 : 133 (распр.); Разоре-
нова А. П., 1939, Сборн. тр. Гос. зоол. муз. Моск. гос. унив., V : 13 (распр.); Гро-
мова В. И., 1940, Пробл. происх., эвол. и породообразов. дом. животн., 1 : 89—93
(ист.); Машковцев А. А., 1940, Боец-охотник, 7 : 24—28 (распр.); О гн е в С. И.,
1940, Матер, к позн. фауны и флоры СССР, нов. сер., отд. зоол., 3 : 78—79 (распр.);
Антипин В. М., 1941, Млекопит. Казахстана, III, Копытные, Алма-Ата, Каз.
ОГИЗ : 65—72 (общ., изобр.); Афанасьев А. В., 1945, Изв. Акад, наук Казахск.
ССР, 5 : 29 (распр.); Микулин М. А. и А. Я. Исаева, 1945, Изв. Акад, наук
Казахск. ССР, 5 : 22 (распр.); Антипин В. М., 1946, Вести. Акад, наук Казахск.
ССР, 11 : 61—62 (биол.); Антипин В. М., 1946, Вести. Акад, наук Казахск. ССР,
7—8 : 49 (распр.); Б у т а р и и Н. С., 1947, Вести. Акад, наук Казахск. ССР, 1—
2 : 86—91 (гибр.); Демин В. С., 1947, Тр. Алма-Атинск. гос. заповеди., IV : 88—
89 (морф., сист.); Зверев М. Д. и А. А. Келейников, 1947, Тр. Алма-Атинск.
гос. заповеди., IV : 49—-59 (мигр.); Громов В. И., 1948, Тр. Инет. геол, наук,
64 : 434 (ист.); Зверев М. Д., 1948, Тр. Алма-Атинск. зоопарка, 1 : 94—116 (биол.);
Кузнецов Б. А., 1948, Звери Киргизии, М., изд. Моск. общ. испыт. прир : 112—
115 (общ.); Скворцов Б. И., 1948, Тр. Воен.-мед., акад., XLIV : 176 (распр.); Шна-
ревичИ. Д., 1948, Тр. Алма-Атинск. зоопарка, 1:73—93 (биол.); ШульпинЛ. А.,
1948, Изв. Акад, наук Казахск. ССР, сер. зоол., 7 : 65—67 (распр., биол.); Б о е в С. Н.
и И. Б. Соколова, 1949, Изв. Акад, наук Казахск. ССР, сер. паразитол.,
7 : 87—90 (параз.); Громова В. И., 1949, сборн, «Тешик-Таш», М., изд. Моск. гос.
унив. : 89—93 (ист.); Лев Д. Н., 1949, Тр. Узбекск. гос. унив., нов. сер.,
39 : 3—47 (ист.); Мекленбурцев Р. Н., 1949, Зоол. журн., XXVII, 5 : 482—
483 (распр., биол.); Насимович А. А., 1949, сборн. «Пушные богатства СССР»,
1 : 147—148 (распр.); Тимофеев В. В., 1949, Звери нашей области, Иркутск,
Обл. изд. : 17—18 (опис., распр., биол.); Ц а л к и н В. И., 1949, Бюлл. Моск. общ.
испыт. прир., отд. биол., LIV, 2 :3—21 (сист.); Адилдабеков К. А., 1950-,
Изв. Киргизск. фил. Акад, наук СССР, I—IX : 77 (распр., биол.); Копылов И. П.,
1950, Дикие животн. Иркутск, обл., Иркутск, обл. изд. : 57—-59 (общ., изобр.);
Петров О. В., 1950, Тр. Ленингр. общ. естествоиспыт., LXX, 4 : 155—193 (распр.,
стац.); Ц а л к и н В. И., 1950, Сибирский, горный козел, М., изд. Моск. общ. испыт.
прир. : 1—119 (общ., изобр.); Виноградов Б. С., 1951, сборн. «Ущелье Кон-
дара», М.—Л., изд. Акад, наук СССР : 107 (распр.); Ellerman J.R.a.T.C.Mor-
rison-Scott, 1951, Checklist of palaearctic and. Indian mammals, London : 406—
407 (сист., синон.); Янушевич А. И., 1952, Фауна позвон. Тувинск. обл., Но-
восибирск, изд. Зап.-сиб. фил. Акад, наук СССР : 62 (распр.); Афанасьев А. В.,
В. С. Бажанови др., 1953, Звери Казахстана, Алма-Ата, изд. Акад, наук Казахск.
ССР : 489—493 (общ., изобр.); Егоров О. В., 1953, Бюлл. Моск. общ. испыт. прир.,
отд. биол., LVIII, 3 : 3—13(размн.); Банников А. Г., 1954, Млекопит. Монгольск. На-
роди. Республ., М., изд. Акад, наук СССР : 258—266 (общ.); Б о е в С. Н., 1954, Зоол.
журн., XXXIII, 4:758 (параз.); Gray А., 1954, Mammalian hybrids, London-
Edinburg : 73 (гибр.); Дементьев Д. П. и П. С. Тюрин, 1954, Тр. Инет,
зоол. и паразитол. Киргизск. фил. Акад, наук СССР, II : 152—153 (распр., биол.);
Дубинин В. Б., 1954, Тр. Инет. зоол. и паразитол., Акад, наук Узбекск. ССР,
3 : 178 (распр.); Егоров О. В., 1955, Тр. Зоол. инет. Акад, наук СССР, XVII :
7—134 (морф., экол., изобр.); НаСимович А. А., 1955, Роль снежн. покрова
в жизни копытных, М., изд. Акад, наук СССР : 132—140 (распр., биол.); Л ялиц-
кая С., 1958, по Крымскому заповеднику, М., Географгиз: 1—87 (акклим.).
Описание. Козел крупной величины. Высота в холке у взрослых
самцов составляет в среднем около 92 см, в отдельных случаях доходит
до 105 см. Длина тела в среднем 142 см, максимально до 165 см (Демин,
1947). Живой вес взрослых, находящихся в хорошей упитанности сам-
цов составляет около 100—110 кг, а изредка, по-видимому, может дохо-
дить до 125 кг (Бутарин, 1947). Основная длина черепа у самцов от 230
до 273 мм. Самки значительно меньше самцов. Живой вес их составляет
50—ВО кг; длина черепа колеблется в пределах 220—229 мм.
Массивное и относительно длинное туловище покоится на коротких
толстых ногах. Ясно выражена низкопередость; высота в крестце на
3—5 см превосходит высоту в холке. Голова не производит впечатления
тяжелой. Профиль носа прямой, лоб менее выпуклый, чем у безоарового
козла. Длина ушей около 14—16 см. Шея у самцов относительно корот-
27 Фауна СССР, И. И. Соколов
418
BOVIDAE
кая и толстая; у самок она тоньше. Линия спины прямая или несколько
выпуклая в поясничной части. Круп заметно свислый. Хвост длиной
с волосами около 25 см, без волос 10—12 см. Копыта длинные и массив-
ные, но низкие с притупленными закругленными концами; длина их
у взрослых самцов на передних ногах от 70 до 80 мм, на задних от 60
до 70 мм; высота копыт на передних конечностях 37—45 мм, на задних
на 1—3 мм меньше. Характерные для безоарового козла круглые, ли-
шенные волос, роговые мозоли на тыльной поверхности запястных су-
ставов у сибирского козерога имеются не всегда.1
Рога сжаты с боков, у самцов с двумя хорошо развитыми ребрами
на переднем крае (передне-внутренним и передне-наружным) и широкой
плоской лобовой гранью между ними. Внутренняя грань рога слегка
выпукла, наружная более плоская, часто даже вогнутая. Передне-вну-
треннее ребро рога приостренное, наружное слегка закруглено, но
у вполне взрослых экземпляров все же даже несколько нависает над бо-
ковой гранью. Скоса наружу лобовой грани не заметно, и последняя
расположена под более или менее прямым углом к боковым граням и
к продольной оси рога. В верхушечной части ясно развито заднее ребро,
но у основания оно становится очень широким и закругленным. Поэтому
поперечное сечение рога, ясно треугольное ближе к вершинам, у осно-
ваний приобретает характер продольно вытянутого четырехугольника
с закругленной задней гранью.
Поверхность рога от основания и почти до самых вершин несет мел-
кую, сплошную поперечную морщинистость и ряд более глубоких, от-
ставленных друг от друга круговых борозд (колец) по границам участков
годичного прироста рога. Кроме того, на передней, лобовой грани рога
расположен ряд резко выступающих поперечных валиков, отставленных
друг от друга на 3—5 см и заходящих несколько на боковые грани. На
каждом участке годичного прироста рога расположено, как правило,
по два поперечных валика, редко по одному или по три; при этом, в про-
тивоположность Capra aegagrus, валики всегда расположены между коль-
цами годичного прироста. Длина рогов у вполне взрослых самцов по
изгибу составляет, в среднем, 100—110 см, в исключительных, рекорд-
ных случаях до 148 см (Rowland Ward, 1914). Продольный диаметр рога
у основания достигает 7.5—8 см при поперечнике около 6—7 см.
Рога дугообразно или саблевидно загнутые назад и не сильно, под
углом 35—42°, расходящиеся в стороны. Изгиб их, пологий у оснований
и более крутой к вершинам, происходит чаще всего в одной плоскости.
Концы рогов направлены вниз, а иногда одновременно несколько внутрь,
навстречу друг другу или, наоборот, слегка в стороны. Основаниями рога,
особенно у самцов, сближены, но расстояние между ними редко бывает
меньше 2—2.5 см.
У самок рога тоньше и короче; длина их не более 36—37 см и не пре-
вышает полуторную длину черепа. Основаниями рога самок расставлены
больше, чем у самцов, лобовая грань отсутствует; лишь иногда имеется
слабо развитое передне-внутреннее ребро. Поэтому поперечное сечение
рога имеет форму продольного овала. Поперечные валики и бугры на пе-
редней стороне рогов отсутствуют, но зато поперечная морщинистость,
имеющая характер круговых колец, у самок по сравнению с самцами
развита сильнее. Цвет рогов у сибирского козерога серовато-бурый.
1 На просмотренном коллекционном материале их удалось обнаружить только
на шкурах зверей из сибирской части ареала. У козлов из Памира и Тянь-Шаня мо-
золи на ногах, по-видимому, даже у старых животных не развиваются.
CAPRA
419
Волосяной покров состоит из слабо волнистой, грубоватой ости и
тонкого извитого подшерстка. Длина остевых волос на туловище в зим-
ней шерсти около 6—8 см; удлиненные до 10—12 см волосы по верхнему
краю шеи у самцов иногда образуют подобие гривы. Зимой мех взъеро-
шен, подшерсток очень густой. Летние, более короткие и упругие волосы
со слабо развитым пухом плотнее прилегают к коже. Борода у самцов
длинная (около 20 см), заостренная, занимает весь низ головы, кроме
Рис. 88. Сибирский козерог (Capra sibirioa Meyer).
самого подбородка. У самок она обычно отсутствует, но иногда пред-
ставлена небольшим тонким пучком длинных грубых волос, растущих
позади подбородка. Хвост от основания покрыт короткими, такими же,
как на туловище, волосами; на конце его волосы более грубы, удлинены
до 10—13 см, хотя кисти, как таковой, и не образуют.
Окраска сибирского козерога не одинакова в разных частях его
ареала; кроме того, она подвержена в значительной степени индивидуаль-
ной изменчивости. По сравнению с безоаровый козлом в окраске пре-
обладают не красновато-рыжие, а бурые и серые тона.
В полном зимнем меху взрослые самцы из южных областей Сибири
(Алтая, Саян) и прилегающих частей Монголии имеют общую окраску
туловища и шеи — светлую желтовато-белую с легким буроватым нале-
том, придающим ей грязноватый оттенок. Вдоль средней линии спины
от холки к корню хвоста тянется неширокая коричнево-бурая полоса.
27*
420
BOVIDAE
Плечи, лопатки, предплечья, а также передняя сторона бедер и голеней
буро-коричневые, заметно контрастирующие со светлыми боками. Та-
кую же окраску имеют низ шеи, грудь и полосы по бокам туловища от
локтевых до коленных суставов. Темная окраска лопаток с темной про-
дольной полосой вдоль хребта, в противоположность С. aegagrus, не сли-
вается. Затылок, темя, лоб и нос буро-коричневые. Темная окраска носа
переходит на щеки впереди глаз. Конец морды более светлый, бурова-
тый. Губы белые. Низ живота, пахи, промежность, внутренняя сторона
ягодиц по бокам от хвоста светлые, беловатые. Иногда, впрочем, низ
живота, наоборот, бывает более темным. Борода и хвост черно-бурые,
у старых зверей на конце желто-рыжие.
Зимняя окраска самцов из горных хребтов Средней Азии сильно от-
личается от таковой зверей Саян и Алтая. Общая окраска туловища у
них серовато-бурая. Коричневый тон, характерный для темно окрашен-
ных участков южносибирских козлов, здесь отсутствует и заменяется
чисто бурым. Бедра, наружная сторона голеней и предплечья более свет-
лые, желтовато-серые. Плечи, шея, нижняя сторона груди, наоборот,
более темные, но нет хоть сколько-нибудь заметного контраста между
темно и светло окрашенными участками тела. По передней стороне ног
от средины предплечий и голеней до копыт тянутся темно-бурые, почти
черные полосы, от которых на уровне путовых суставов отходят темные
же боковые полоски к добавочным копытам.1 Задняя сторона пясти и
плюсны светлая, желтоватая, иногда почти белая.
Довольно часто, но не у всех особей, на спине и шее имеются светлые
буровато-белые, так называемые седловидные пятна. Посредине они раз-
делены темной полосой, идущей вдоль хребта, а по краям седловидные
пятна обычно нерезко отграничены от соседних, более темных участков
кожи. Интенсивность и размеры седловидных пятен варьируют. Иногда
они занимают почти всю спину и шею, а седловидное пятно спины может
сливаться с таким же светлым пятном на шеё. В других случаях посвет-
ление на спине и шее отсутствует, верхняя сторона туловища даже бо-
лее темная, тогда как книзу, по бокам, окраска постепенно светлеет.
Темная продольная полоса вдоль хребта в этих случаях до корня хвоста
не доходит и сливается с темной окраской крестца и поясницы.
Темные полосы по бокам между локтевым и коленным суставами,
как правило, отсутствуют или едва заметны. Низ живота, пахи, промеж-
ность, внутренние стороны ягодиц по бокам от хвоста и задняя сторона
ног ниже запастных и скакательных суставов (кроме участка вокруг и
ниже добавочных копыт) светлые, разных оттенков, от грязноватого
-желто-серого до почти белого. Голова, обычно светлее туловища, буро-
вато-серая с желтоватым оттенком. Характерного для козерогов Алтая
и Саян потемнения переносья и щек впереди глаз здесь не наблюдается.
Уши снаружи одного цвета с головой, внутри грязно белые. Конец морды,
пространство между ноздрями и губы светло-бурые. Борода и хвост темно-
^бурые, обычно с примесью небольшого числа рыжеватых или белых во-
лос. Следует отметить, что в конце зимы, вследствие выцветания и обла-
мывания большей части темно окрашенных верхушек волос, окраска
и у самцов, и у самок становится более светлой, тусклой.
1 Эта особенность хорошо отличает сибирского козерога от безоарового козла,
у которого основания добавочных копыт всегда отделены от продольной темной по-
лосы на передней стороне ног светлой окраской.
CAPRA
421
Летом окраска у самцов во всех частях ареала более однотонная,
серовато-бурая, иногда с легким желтоватым или даже рыжеватым на-
летом, особенно на нижних частях бедер и на предплечьях. Ясно выра-
женных потемнений, кроме темно-бурой полосы вдоль хребта от затылка
до хвоста и темной передней стороны ног, нет; Светлые седловидные пятна
на спине и шее отсутствуют или едва заметны. Голова окрашена сходно
с туловищем, но более темная окраска переносья у козлов северной
Рис. 89. Сибирский козерог (Capra sibirica Meyer). Ленинградский зоопарк.
(Фото автора).
части ареала иногда сохраняется и летом. Низ живота, пахи, промеж-
ность, участки по бокам от хвоста и задняя сторона ног сохраняют бело-
ватую с разными оттенками окраску.
Самки окрашены более однотонно по сравнению с самцами. Даже
срединная полоса вдоль хребта у них имеется не всегда и выражена слабо.
В отдельных случаях можно видеть едва заметную темную полосу на гра-
нице светлой окраски живота и боков. Общая окраска самок серовато-
бурая, но желтовато-рыжий оттенок по сравнению с самцами выражен
сильнее, особенно в летней шерсти. Светлое седловидное пятно на спине
иногда имеется и у самок.1 Темно-бурая полоса на передних ногах у са-
мок из Саян и Алтая часто, хотя и не всегда, прерывается светлой по-
1 В противоположность утверждению В. И. Цалкина (1950), считающегося сед-
ловидные пятна вторичнополовым признаком самцов. Ясно видимое седловидное
пятно имеется, например, нашкуре у самки из восточного Памира (колл. ЗИН, №36116).
422
BOVIDAE
перечной полосой выше запястных суставов; у самцов, так же как и у
самок Средней Азии, судя по просмотренному материалу, она всегда
сплошная.
У молодых козлят первые месяцы жизни в окраске преобладают жел-
товато-рыжие тона. Полувзрослые самцы сходны с самками. Полного
развития окраска у них достигает, по-видимому, на четвертом или пятом
году жизни.
Окраска сибирских козлов подвержена и индивидуальной изменчи-
вости, даже в пределах сравнительно небольших участков ареала. На
Памире, например, даже в одном и том же стаде, наряду с темно окрашен-
ными, можно видеть «красных» и очень светлых «белых» козлов (Егоров,
1955). О светло-серых козлах Алма-Атинского заповедника писал В. Де-
мин (1947). К индивидуальным особенностям следует отнести случаи,
когда ноги ниже запястных и скакательных суставов и спереди, и с бо-
ков имеют сплошь светлую окраску. Встречается изредка также очень
светлая, почти чисто белая морда.
Волосы пуха чаще светло-бурые или дымчатые. Расцветка отдельных
волос ости разнообразна. Типично волос со спины и боков от основания
очень светло-бурый, кверху становится более темным, бурым, затем идет
не очень широкое предвершинное желтовато-рыжеватое кольцо, а тонкий
приостренный конец волоса, пока он не обломан, темно-бурый. Волосы
темной полосы вдоль хребта от основания до верхушек темно-бурые,
без предвершинной светлой зоны. Сероватый оттенок получается обычно
от примеси к бурым светлых дымчатых, иногда почти белых волос, а ры-
жеватый зависит от интенсивности окраски светлых предвершинных
колец. Светлые седловидные пятна на спине и шее образованы белыми
с желтоватым оттенком волосами, часть которых имеет темно-бурые за-
остренные верхушки.
Череп сибирского козерога (рис. 90) по сравнению с другими козлами
в частности с безоаровым (С. aegagrus Erxleb.), обладает следующими
отличительными чертами. Роговые отростки у самок слегка сдвинуты
назад; передние края оснований роговых стержней расположены, как
правило, несколько позади, редко на одном уровне с задними краями
глазниц. У самцов на передней стороне роговых стержней имеется ясно
видимая лобовая грань; характерна также широколобность. Межглаз-
ничная ширина черепа не менее (по крайней мере, у самок) расстояния
от переднего края хоан до нижнего края затылочного отверстия. Ни
у самцов, ни у самок нет предрогового вздутия на лбу, а если лоб в сред-
ней части выпуклый, то нет продольных валиков, идущих к нему от ос-
нований роговых отростков, как у С. aegagrus. Поверхность лба вдоль
сагиттального лобного шва обычно вогнута. У самцов она перед основа-
ниями роговых отростков шероховатая, с ясно отграниченным и при-
поднятым на границе с гладкой поверхностью кости передним краем.
Эта шероховатость, вероятно, вызвана утолщением и ороговением кожи;
здесь, возможно, происходит формирование поперечных валиков на ло-
бовой грани рога.
Носовые кости по сравнению с другими видами длинные; их наиболь-
шая ширина составляет от 28 до 40% собственной длины. Расстояние
от переднего конца носовых костей до переднего края межчелюстных
(prosthion), как правило, значительно меньше расстояния от prosthion
до переднего края Р2 (разница всегда превышает длину этого зуба). Пе-
редний край хоан расположен или впереди, или позади уровня боковых
выемок заднего края костного нёба.
Рис. 90. Череп Capra sibirica Meyer.
424
BOVIDAE
Переднекоренные (премоляры) сильно редуцированы; длина их ряда
в верхней челюсти не более 33% всего соответствующего ряда коренных
зубов (Р2 — М3).1
История и географическое распространение (рис. 91). Наиболее
древние остатки, достоверно относящиеся к сибирскому козерогу, из-
вестны из среднепалеолитической (мустьерской) стоянки неандертальца
в гроте Тешик-Таш в Отрогах Гиссарского хребта в Узбекистане (Громова,
1949). Ископаемые остатки С. sibirica найдены также в верхнепалеолити-
ческих стоянках у г. Красноярска и вверх от него по Енисею. Кроме
того, есть требующие уточнения указания на находки этого вида в сред-
нем и верхнем плейстоцене на Южном Урале (Громов, 1948).
В настоящее время область распространения С. sibirica включает
горные хребты южной Сибири, Средней и Центральной Азии: Алтай,
Саяны, Танну-Ола, Гобийский Алтай, систему Тянь-Шаня, Памиро-
Алая, Гиндукуш, Парапамиз, Куэнь-Лунь, Гималаи. Есть основания
предполагать, что еще в верхнем плейстоцене ареалы сибирского и аль-
пийского козерогов (С. ibex L.), Несомненно имеющих общее происхож-
дение, смыкались через юго-западную Азию или, судя по находкам остат-
ков козлов в Крыму, через юго-восточную Европу (Громова 1949).
В пределах СССР современный и былой ареал этого вида представ-
ляется в следующем виде.2
Где-то в районе Бадхыза (крайний юг Туркмении) имеет место стык
ареалов безоарового козла (С. aegagrus Erxl.) и сибирского козерога.
Заведомо относящиеся к этому виду экземпляры были добыты в се-
верном Афганистане, вблизи советской границы, в Баламургабе (Ly-
dekker, 1913).
Есть весьма неопределенные указания на обитание С. sibirica и в по-
граничных районах советского Бадхыза. К. К. Флеров (1932) склонен,
например, относить к этому виду козлов из окрестностей пограничной
заставы Рабат-Кашана (к западу от Кушки), о которых ему удалось
только слышать. Б. П. Скворцов (1948), со слов местных охотников,
сообщает о встречах козерога одновременно с безоаровым козлом в ущелье
Нурдыванлы (хребет Гез-Гельды) и в районе Зульфагара (на Теджене).
Местные охотники уверяли А. А. Гуреева (1937) в том, что сибирский
козерог встречается в горах Карлюка и Куйтана, т. е. в районе хребта
Кугитангтау на границе Туркмении и Узбекистана.
В хребте Байсунтау (юго-западное продолжение Гиссарского хребта
в Узбекистане) С. sibirica в настоящее время не водится, но в знаменитой
среднепалеолитической стоянке в гроте Тешик-Таш на южной оконеч-
ности этого хребта он по числу костных остатков, среди других млеко-
питающих, занимает первое место (Громова, 1949).
Областью сплошного распространения сибирского козерога являются
Гиссарский, Зеравшанский и Туркестанский хребты в Таджикистане.
По южным склонам Гиссарского хребта козлы проникают в северную
и западную часть Сары-Ассийского района Сурхандарьинской области
Узбекистана (Левиев, 1939). Остатки сибирского козерога найдены в 1 2
1 Ряд указываемых В. И. Цалкиным (1950) и В. И. Громовой (1940) отличий от
других козлов в пропорциях черепа (суженность лицевого отдела черепа, ширина
fossa mesopterygoidea, положение высшей точки верхнего профиля черепа и др.) верны
но, к сожалению, не абсолютны.
2 Сводку литературных данных по распространению и биологии сибирского ко-
зерога недавно дал В. И. Цалкин (1950). Часть материалов этой сводки используется
в настоящей работе.
CAPRA
425
мустьерской стоянке (средний палеолит) Аман-Кутан, на западной око-
нечности Зеравшанского хребта, в 45 км к югу от Самарканда (Лев, 1949).
По северным склонам Туркестанского хребта изредка и случайно козлы
заходят в предгорья южной части Хавастского района Ташкентской об-
ласти. В. Б. Дубинин (1954) видел здесь пару рогов козла, добытого
в 1943 г. в 15 км южнее кишлака Баландчакир.
Детали распространения козерога в Таджикистане не известны, но,
по-видимому, козлы многочисленны во всех высокогорных районах се-
Рис. 91. Распространение Capra sibirica Meyer.
1— XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше;
4 — места отдельных заходов.
верной и восточной части республики. Известно, что по южным скло-
нам Гиссарского хребта козлы спускаются зимой иногда в ущелье Дже-
ринодсой. Зимой 1944/45 года экземпляр козла был добыт в ущелье над
с. Такобом (Виноградов, 1951). Распространены сибирские козероги
в Заалайском, Дарвазском хребтах и на Памире. В более низких и поло-
гих хребтах южной части Таджикистана, в частности в хребтах Каратау
и Сарсариак, по собранным автором в 1951 г. сведениям, С. sibirica от-
сутствует.
Широко и практически повсеместно сибирский козел распространен
на Памире. Более многочислен в скалистых хребтах и ущельях запад-
ного Памира, тогда как сглаженных увалов восточной его половины,
по словам М. П. Розанова (1935), избегает. Р. Н. Мекленбурцев (1949)
встречал козлов в ущельях притоков р. Шахдары и в долине послед-
ней, а также в верховьях и в среднем течении Гурумды и в районе Ра-
426
BOVIDAE
бат-Чакобая, в 45 км к западу от г. Мургаба. Большое количество чере-
пов и скелетов козлов им было обнаружено в верховьях р. Башгумбеза.
В. И. Цалкин (1950) перечисляет пункты, в которых М. П. Розанов встре-
тил сибирских козлов; это — ущелье Маркансу, район Мургаба, ущелье
Хан-Юлы и район поселка Кызылрабата. Кроме того, М. П. Розанов
(1935) наблюдал их на северных склонах Заалайского хребта близ Бор-
Добы, уже в пределах Киргизии.
В Киргизской ССР сибирский козел распространен практически
повсеместно там, где имеется подходящая для него экологическая обста-
новка.1 Весьма многочисленны козлы по всему Киргизскому хребту
(Машковцев, 1940), особенно на его северных склонах. Наблюдались
они на очень крутом хребте Ичкелетау, а также в верховьях р. Тегерек.
Обычным зверем являются они в верхнем поясе гор Таласского Алатау
(Абдилдабеков, 1950). Многочисленны в заповеднике Джебаглы.
Д. Н. Кашкаров, а также Б. А. Кузнецов (1948) наблюдали козлов в
истоках р. Чаткала, в горах Кара-Току, Мустера и в окрестностях
оз. Сары-Чилек. По Чаткальскому хребту козлы водятся до западной
оконечности его, вплоть до верховьев р. Шавассай (северо-восточная
часть Ташкентской области в Узбекистане; Цалкин, 1950). В 10-х годах
этого столетия они были обычны в Пскемском и Угамском хребтах (Южно-
Казахстанская область). Нет достоверных данных, но вероятно, что
держатся козлы и по Кураминскому хребту. Находились они на пере-
вале Аир-Бель на Узунахматском хребте, а также в горах по верхнему
течению р. Узунахмата. Много козлов в горах, окружающих Кетмень-
Тюбинскую котловину. Н. М. Пржевальский встречал их по р. Караколу,
а сейчас они водятся, хотя и не так многочисленны, в Джумголтау, юж-
нее Киргизского хребта.
В большом числе водятся козлы в горах, окружающих оз. Сонкель,
на северных склонах Кавакского хребта и на водоразделе речек бассейна
Монкуш и Кара-Киче. Еще южнее они обитают в бассейне р. Нарына,
в частности в окрестностях пос. Атбаши, Нарына и в урочище Арпа.
Сравнительно мало козлов в окрестностях Арсланбоба; водятся в
горах восточной части Гульчинского района. Многочисленны в Алай-
ском и Заалайском хребтах. Встречаются в горах по р. Коксу, к востоку
от Алайской долины.
К северу от Иссык-Куля сибирские козлы населяют весь хребет Кун-
гей-Алатау, являясь здесь наиболее многочисленным видом из всех ко-
пытных (Петров, 1950; Дементьев и Тюрин, 1954). Водятся они и в горах
по северному побережью Иссык-Куля, в частности близ поселков Са-
зоновки, Чалпанаты, а также в горах к северу от Рыбачьего. Д. Н. Каш-
каров (цит. по Кузнецову, 1948) наблюдал козлов в Терскей-Алатау,
на Каракольских сыртах, в частности по рр. Оттуку и Куэлу. В Терскей-
Алатау вел изучение экологии сибирских козлов О. В. Егоров (1955).
Южнее Иссык-Куля козлы являются обычным зверем в бассейне
рр. Нарына, Атбаши, а также по Аксаю. На востоке Киргизии они во-
дятся в Джеты-Огузском районе, а также в районе Хан-Тенгри, где они
распространены спорадически; но там, где сибирские козлы имеются,
они встречаются сотнями голов (Антипин, 1946).
В пределах Казахской ССР сибирской козел держится, как уже от-
мечалось, в заповеднике Аксу-Джебаглы, в Пскемском и Угамском
хребтах, затем по северным склонам Киргизского хребта. Указание
1 Конкретные данные приводятся, в основном, по Б. А. Кузнецову (1948).
CAPRA
427
В. M. Антипина (1941) об обитании козлов в горах Каратау и в Чу-Илий-
ских горах, по-видимому, основано на недоразумении. Далее козлы на-
селяют Заилийский и Кунгей-Алатау. В числе мест, где они обычны,
С. И. Огнев (1940) называл Сюгатинское ущелье, Сарыбулак, Орта-Мерке,
Джаланашскую лесную дачу Алма-Атинского заповедника, а также
ущелья Куль-Су и Урюкты. Стада козлов в 100—200 и более голов на-
блюдались в долине р. Кебени. По словам В. Н. Шнитникова (1936),
в долине р. Большой Алмаатинки в 1933 г. наблюдались стада до 100
голов. В восточной части Заилийского Алатау они обитают в горах Тур-
айгыр, Большие и Малые Богуты, Алабайтал и Ортатау (Шнаревич,
1948). Многочисленны козлы в Нарынкольском районе Алма-Атинской
области в верховьях рр. Текес и Кегень, по-видимому, вплоть до хребта
Кетмень (Микулин и Исаева, 1945).
Обычны и за последние десятилетия, по-видимому, увеличили свою
численность козлы в Джунгарском Алатау. В. М. Антипин (1946) ука-
зывает их для юго-западной оконечности гор Чулак. По А. В. Афанасьеву
(1945), в пределах Джунгарского Алатау нет козлов в хребте Чаган,
но встречаются они в Кугалинском хребте, а особенно много в Матае.
Областью распространения С. sibirica является горная система
Алтая.
Еще совсем недавно, а возможно и сейчас, козлы держатся в неболь-
шом числе на долготе Семипалатинска. Во всяком случае, до 1925 г.
они встречались, хотя и очень редко (по В. А. Селевину, 1925), в горах
Коконь к юго-западу от Семипалатинска, а также к юго-востоку от пос-
леднего в Дельбегетейских горах. Тот же автор в 1937 г. называет козла
редким обитателем луго-степного пояса высот Сары-Арка. Есть указания
(Берн, 1891), что будто бы раньше он доходил к западу еще дальше, вплоть
до Ортау. В Южном Алтае в пределах Казахстана козлы обитают в Са-
рымсакты, Нарымском и Курчумском хребтах, Сауре и Тарбагатае
(Афанасьев и др., 1953).
Много козлов в горах по течению р. Сарымсака, стекающей с Нарым-
ского хребта возле оз. Катон-Карагай (Цалкин, 1950). В конце прошлого
столетия, а вероятно и до сих пор, водятся они в горах по р. Бухтарме.
Перечень мест в этой части Алтая, в которых в конце прошлого и начале
этого столетия встречались козлы, приводят Н. В. Туркин и К. А. Са-
тунин (1904).
Достаточно хорошо известно в настоящее время распространение
сибирского козерога в пределах центрального и юго-восточного Алтая
(Залесский, 1934; Дмитриев, 1938; Булавин, 1934; Колосов, 1939; Наси-
мович, 1949). Судя по карте в работе Дмитриева (1938), распространен
он здесь отдельными гнездами, не повсеместно. Хотя Ф. Геблер (1837;
цит. по Колосову, 1939) в свое время указывал козлов для устьев Коксу
до Каира и для южных склонов Яманушки, но, по-видимому, к западу
от верховьев Катуни их теперь нет. На хребте Холзуне они также от-
сутствуют. Указание П. М. Залесского (1934) на хребет Листвяга, как
на место их нахождения, не определенно. В левобережье Катуни в на-
стоящее время с достоверностью установлено наличие козлов только
на Теректинском хребте. Однако и здесь они обычны только в юго-восточ-
ной его части у Тюнгура. Много меньше их в верховьях Катандинок
и притока Катуни, Маргалы. По р. Теректе водились до 1940 г., а теперь
сохранились только на северном склоне Теректинского хребта в верховьях
Б. Яломана. В небольшом числе держатся еще козлы в верховьях Ку-
рунды и Тюгурука (на притоке последней Маргале).
428
BOVIDAE
Больше всего в пределах юго-восточного Алтая козлов в Катунском
хребте, по рр. Аккему и Кочурле. Обычны они в верховьях Нижнего
Курагана и на притоке последнего Ермаке. На склонах горы Белухи
их сравнительно мало из-за глубокоснежья, а к западу от нее, говорят,
козлов не было и раньше. Много до сих пор козлов по р. Аргуту.
В правобережье Катуни самым северным пунктом обитания козерога
П. М. Залесский (1934) считает среднее течение р. Немала, а В. В. Дмит-
риев (1938) — горы между устьями рр. Б. Сумульты и Эдигана. Дальше к се-
веро-востоку очаг обитания имеется в верховьях р. Уйменя. В среднем и
верхнем течении рр. Большой и Малой Сумульты, а также р. Кадрина козлов
нет. Крупная и по числу особей, и по занимаемой территории популяция
сибирских козлов обитает в нижнем течении р. Башкауса от устья до
р. Паспарту и в горах по всему течению р. Чебдара, левого притока Баш-
кауса. В бассейне р. Чулышмана козел распространен от низовьев до
впадения в него р. Шавлы. В прежние времена он водился, по словам
местных жителей, по всему западному берегу Телецкого озера, а по скло-
нам долины р. Ян-Чили доходил до вершины последней. Водятся козлы
в нижнем течении р. Чульчи, притока Чулышмана.
К востоку от Телецкого озера очаг обитания козлов находится на
пространстве между верховьями рр. Большого Абакана и Кыги. Далее
к юго-востоку козлы встречаются в верховьях рр. Гульчи, Шавлы и их
притоков (Кара-Кем, Ян-Сару, Узун-Карасу, Мейдугем), а также на
западных склонах Шапшальского хребта. Следы козлов наблюдались
в верховьях р. Абдулюгем и по левым притокам Чулышмана, выше устья
р. Пепши-Оюк.
В среднем течении р. Башкауса, по В. В. Дмитриеву (1938), козлов
нет; вновь они появляются выше впадения р. Арталаш и обычны до самых
ее верховий.
На самом юге Ойротии (Горно-Алтайская автономная область) си-
бирские козлы обычны в хребтах Курайском, где их много на склонах,
обращенных в сторону Чуйской степи, в Чуйских Альпах, в хребте Сай-
люгеме. В последнем, по словам А. Булавина (1934), козлов особенно
много в урочище Сартомар, а по А. М. Колосову (1939) — в урочищах
Кара-Алаха, Ака-Алаха, по р. Кара-Кему и в верховьях р. Аргута.
Известно, что много их в верховьях Чаган-Бургазы и Тархатты.
Сведения о распространении сибирского козла в Саянах менее полны.
По данным В. Н. Скалона (1929), козлы держатся, хотя и весьма не-
многочисленны, в хребтах Тепсель и Каранем. Водятся они в верховьях
р. Оны И' притока последней Каратоны.1
На восток они здесь распространены и обычны до р. Аксу, а на запад,
вероятно, до Малого Абакана.
По данным П. М. Залесского (1934), кроме р. Оны, козлы в запад-
ных Саянах встречаются в верховьях р. Джебаша (правый приток Аба-
кана), а также по обеим сторонам хребта Тепсель, особенно по рр. Боль-
шому и Малому Тепселю и Мандараку.
В Тувинской области, по В. Н. Скалону (1929), козлы встречаются
в хребте Танну-Ола и в верховьях р. Кемчик (южная часть Шапшаль-
ского хребта), а по В. Н. Троицкому (1930), они сохранились также по
хребту Каратыш и в горах вдоль бывшей границы СССР с Тувой между
рр. Малым Абаканом и Енисеем.
1 В Атласе Мира, издания 1954 г., обозначена, вероятно, как Карасума.
CAPRA
429
На Енисее северная граница распространения сибирского козла про-
ходит на широте р. Кызыр-Сука. В этих местах, по словам Д. К. Соловьева
(1920), козлы в то время были столь обыкновенны, что их можно было
наблюдать и даже стрелять, спускаясь на лодке по Енисею. К востоку
от с. Верхне-Усинского и до верхнего течения р. Морхой, по данным
того же автора, козлов нет вовсе. К востоку от Морхоя они попадаются
чаще, а в верхнем течении р. Уды их опять много. Здесь в узле западных
и восточных Саян, по-видимому, находит свой северный предел совре-
менное распространение сибирского козерога.
Далее к востоку козел, по данным В. Б. Подаревского (1936), зани-
мает на всем протяжении альпийскую зону восточных Саян, где мес-
тами его довольно много. В левобережье верховьев р. Уды, например
до устья р. Едене, встречаются стада более чем в 50 экземпляров. Много
козлов в Тункинских Альпах, в частности на хребте Мунку-Сардык.
По В. Ч. Дорогостайскому (1925), имеются козлы и в Китойских Голь-
цах (верховья р. Оки). Следует полагать, хотя прямые данные еще отсут-
ствуют, что козлы должны быть также и в восточной части Тувинской
области в верховьях Большого и Малого Енисея. Тункинские Гольцы,
вероятно, являются восточным пределом распространения сибирского
козерога в Советском Союзе, хотя И. П. Копылов (1950) и указывает,
что, кроме Тофаларии, Окинского и Тункинского районов, козлы довольно
большими табунами встречаются также в Закаменском аймаке Бурят-
Монгольской АССР. Определенно отсутствуют они в Хамар-Дабане (До-
рогостайский, 1918).
Сведения Подаревского (1936) о том, что очаг обитания козлов будто бы
имеется в Бодайбинском районе Иркутской области вокруг оз. Орон,
нуждаются в проверке.
Наконец следует отметить, что 7 штук С. sibirica в 1947 г. заве-
зены из Киргизии в Крым и выпущены в районе г. Четырдага.
В 1957 г. их здесь было около 20 голов (Лялицкая, 1958).
В пределах Монгольской Народной Республики С. sibirica населяет
все более высокие хребты системы Монгольского и Гобийского Алтая,
Хангая, горы к западу от оз. Хубсугул (Косогол) и возвышенности За-
алтайской Гоби. По А. Г. Банникову (1954), «весь магистральный хребет
Монгольского Алтая (Алтаин-Нуру) населен козлами, практически без
заметных разрывов, на восток до Цагай-Хайрхана и Бус-Хайрхана.
Далее, на юго-восток, этот вид с небольшими перерывами населяет хребты
как Монгольского, так и Гобийского Алтая, а именно: Хара-Адзирга,
Ихэ-Даян, Бурхан-Будда, Гичгэнэ-Нуру и Сэрхэ. Видимо, не ежегодно
заходят козлы и на Тайшири. С несколько большими разрывами рас-
пространен козел еще далее на юго-восток по хребтам Баянгин-Нуру,
Хара-Аргалинтэ, Нарин-Хара, Ихэ-Богдо, Бага-Богдо, Арца-Богдо,
Гурван-Сайхан, Баян-Ундэр, Буур, Номогон, Хурхэ. Последний хре-
бет (примерно 106° в. д.) — крайний юго-восточный предел распро-
странения вида в стране.
К югу от Гобийского Алтая, в Заалтайской и Западной Гоби, козел
весьма обычен на хребтах Байтаг-Богдо, Аджи-Богдо, Атас-Ула, Цаган-
Богдо-Ула, Эдэрэнгин-Нуру, Тост-Нуру, Нэмгэт-Ула-Дзолен, Ихэ-Арга-
линтэ и по ряду других мелких поднятий.
К северу от Монгольского Алтая горные козлы обычны на хребтах
Цаст-Ула, Алтан-Хухэй, Джиргаланту, Хархира-Нуру и Тургунь
и, далее к северу, на склонах хребтов Мунх-Хайрхана и Шибэту.
430
BOVIDAE
Что касается Хангая, то здесь козлы населяют весь магистральный
хребет — от верховий рр. Идэр и Охтон-Тэнгри на западе до верховий
Орхона на востоке. В северных хребтах Хангая (Болиай, Босхоту) коз-
лов, видимо, нет. Напротив, по хребту Хан-Хухей он обычен.
Далее, по Танну-Ола, сибирский козел проникает в Монголию на
Алтан-Хулмин-Нуру и в верховья р. Дэльгер-Мурэн. В Прикосоголье
этот вид держится от Улан-Тайги, Хоридул-Сардыка и Баян-Ула до
Мунку-Сардыка. В восточном Прикосоголье, видимо, отсутствует».
Подвиды. Систематика сибирских козлов очень запутана. Описано
большое число «видов» и подвидов, в ряде случаев по одной-двум шкурам
или паре рогов, не всегда даже точно выясненного происхождения. Лай-
декер (Lydekker, 1913) принимал число подвидов С. sibirica Меу., равное
тринадцати. К. К. Флеров (1935) число последних свел к двум, а Эллер-
ман и Моррисон-Скотт (Ellerman and Morrison-Scott, 1951). не только
аннулировали все выделяемые ранее подвиды, но и вообще сибирского
козерога рассматривают как подвид единого евразийского вида С. ibex L.,
к которому, кроме альпийского казерога (С. ibex ibex L.), относят также
западнокавказского тура (С. caucasica severtrzowi Menzb.).
По-видимому, наиболее приближающуюся к истинной картину си-
стематических взаимоотношений форм сибирских козерогов дал В. И. Цал-
кин (1949). Этот автор признает наличие пяти рас или подвидов С. sibirica.
1) С. sibirica sibirica Meyer — Типичная раса. Размеры сравнительно
небольшие; общая длина черепа у самцов в среднем 270 мм, максимально
293 мм. Рога у самцов более короткие и тонкие, длиною не свыше 100 см.
В окраске характерно общее посветление ее зимой у самцов и резко вы-
раженное контрастирование потемнений на плечах, бедрах, нижней сто-
роне шеи, груди и передней стороне ног.1 Распространение:
советский и монгольский Алтай, Саяны, северная Монголия.
2) С. sibirica hagenbeckii Noack. Краниологически и по размерам схо-
ден с типичной расой, но отличается от последней однотонной серовато-
коричневой зимней окраской самцов. Распространение: Го-
бийский Алтай, возвышенности Заалтайской Гоби.
3) С. sibirica alaiana Noack. Размеры сравнительно крупные; общая
длина черепа у самцов в среднем 292 мм, максимально 306 мм. Носовые
кости и абсолютно и относительно более длинные, чем у типичного под-
вида. Рога у самцов более толстые и длинные, длиною у старых зверей
больше 100 см. Общая окраска самцов зимой темная, коричнево-бурая.
На спине имеется светлое седловидное пятно.1 2 Распространение:
восточный и центральный Тянь-Шань, Памир, Алайский и Заалайский
хребты.
4) С. sibirica formosovi Zalkin. От предыдущего подвида отличается
только отсутствием в зимнем наряде взрослых самцов светлого седло-
видного пятна. Распространение: Киргизский хребет, Талас-
ский Алатау и другие хребты западного Тянь-Шаня. Целесообразность
выделения этого подвида вызывает серьезные сомнения, поскольку от-
сутствие седловидного пятна довольно часто наблюдается, например,
у козерогов Памира.
1 Следует отметить также наличие ороговевших мозолей на передней стороне
запястий и светлую поперечную полоску выше последних у самок (см. стр. 421).
2 Характерными признаками подвида, по-видимому, следует считать также от-
сутствие ороговевших мозолей на передней стороне запястий и сплошную темную
окреску передней стороны ног как у самцов, так и у самок.
CAPRA
431
5) C. sibirica dementievi Zalkin. От всех остальных рас отличается
незначительными размерами; общая длина черепа у взрослых самок
230—232 мм. Распространение: предположительно — запад-
ный Куньлунь, на восток до Карийского хребта (более точно не установ-
лено). Недостаточно изученная раса с невыясненным ареалом. Описана
по двум черепам самок.
Биология. Сибирский козерог еще больше, чем другие виды Capra,
связан в выборе мест обитания со скалистым ландшафтом высокогорья.
В некоторых районах, даже в зимний период, козлы не спускаются ниже
верхней границы леса. Однако это верно только в общей форме. Пределы
вертикального распространения у сибирского вида широки —от 500
до 4000 м над ур. м. и даже выше. В Саянах, по словам Д. К. Соловьева
(1920), козлы живут выше границы леса и в наиболее трудно доступных
скалистых местах с отвесными обрывами и каменистыми россыпями.
Так же, в общем, характеризует стации козлов в Семиречье В. Н. Шнит-
ников (1936), где, по словам этого автора, они не встречаются ниже 2500 м.
Но уже на Алтае этот вид в ряде мест, например в районах среднего
и нижнего течения рр. Башкауса, Чулышмана, Чебдара, Чульчи, Ка-
туни, занимает склоны глубоких долин, спускаясь до 500 м, и только ле-
том поднимается, и то в небольшом числе, выше границы леса (Дмитриев,
1938).
Одним из непременных условий местообитаний сибирского козла
является наличие поблизости скал и каменистых осыпей, имеющих,
по-видимому, большее значение, чем высота местности над уровнем моря.
В районе Хорога (Памир), где вершины хребтов сглажены, а средняя
полоса гор скалиста, большая часть козлов предпочитает держаться
в этой последней (Егоров, 1955). Скалы сами по себе не являются местом
постоянного пребывания козлов уже потому, что звери часто не находят
здесь достаточного количества корма. Они играют главным образом
защитную роль. Козел не может быстро бегать и в случае опасности
полагается не на быстроту своих ног, а на великолепную способность
карабкаться по самым крутым, неприступным каменистым склонам.
Скалы же служат местом укрытий в жаркие дневные часы и в зимнюю
непогоду. Далеко от них козлы, как правило, не уходят. В верхней части до-
лины р. Билянд-Киик на Памире, по О. В. Егорову (1955), довольно
крутые и изобилующие скалами склоны верховий совершенно не насе-
лены козлами по той причине, что нижняя граница скального пояса
отделена полосой лишенных растительности осыпей от роскошных аль-
пийских пастбищ, расположенных по отлогим склонам вдоль ру-
сла.
«Стацией козлов, — пишет Г. Н. Мекленбурцев (1949), — надо счи-
тать не только скалы, как это указывается у Иванова и Головина для
Памира и у многих авторов для других местанахождений этих зверей,
но целый ландшафтный комплекс, в котором главное место принадлежит
крупным скалистым массивам. Помимо последних, в этот комплекс должны
входить альпийские лужайки, служащие излюбленным пастбищем коз-
лов, ручьи, необходимые для водопоя, и небольшие участки между ска-
лами, покрытые твердой, мелкощебнистой почвой, на которой обычно
лежат козлы».
Определенно сибирские козлы избегают мест с мягким покатым релье-
фом (если поблизости нет скал), темнохвойной тайги, таежного высоко-
травья и сырых болотистых участков, даже если последние и расположены
в альпийской зоне. При прочих равных условиях они явно предпочитают
432
BOVIDAE
места с твердым грунтом, районы с наличием остепненной раститель-
ности и незначительным количеством выпадающих осадков.
Очень важным фактором в жизни сибирского козла, определяющим
не только стадиальное размещение, но порою и самую возможность оби-
тания в той или иной местности этого зверя, является глубина снежного
покрова. При большой глубине снега для козла затруднительна не только
добыча из-под снега корма, но и передвижение. Большая крутизна скло-
нов, при которой не образуется глубокой пелены снега, и сильные ветры,
сдувающие снег, являются залогом того, что в долинах Чулышмана,
Башкауса и некоторых других рек на Алтае становится возможным су-
ществование козлов в полосе леса в течение круглого года. В районе
Хантенгри, наоборот, козлы в течение круглого года живут в высоко-
горье на открытых склонах, где достаточно высокая инсоляция и ветры
обнажают почву от снега и не создают препятствий к добыче корма даже
в зимнее время (Антипин, 1946). В некоторых местах Восточного Памира,
как например в долине р. Каинды, несмотря на наличие прекрасных
альпийских пастбищ, козлы очень редки, а зимой отсутствуют вовсе,
вследствие большой глубины снежного покрова (Егоров, 1955).
Неравномерность залегания снежного покрова и необеспеченность
кормами в зимний период являются причиной, имеющей место во мно-
гих районах сезонной смены мест обитания и даже миграций этого, вообще
довольно оседлого зверя.
На Алтае в Чуйских Альпах в верховьях Башкауса, Чулышмана
и во многих других местах лето козлы проводят в высокогорье, зоне так
называемой каменистой тундры, или фигурных гольцов, значительно
выше верхней границы леса. Большую часть времени звери скрываются
в неприступных вершинах хребтов, а кормиться выходят на расположен-
ные по соседству альпийские лужайки. Зимой, когда здесь выпадает
глубокий снег и козлам становится трудно добывать пищу, они спус-
каются на крутые травянистые склоны, опушки леса и другие места юж-
ной экспозиции, где солнце и ветер препятствуют образованию глубо-
кого снежного покрова. В особо снежные и безветренные зимы козлы
заходят глубже в полосу леса, где снег не только мельче, но и рыхлее
(Дмитриев, 1938).
В пределах б. Алма-Атинского заповедника (Заилийский Алатау)
козлы на зиму перемещаются с северной на южную экспозицию «отщел-
ков» северных склонов (Зверев и Келейников, 1947). В особо снежные
зимы они здесь спускаются даже в тальниковые ельники нижней части
лесной зоны. В Терскей-Алатау из верхнего скалистого пояса гор, где
козлы проводят лето, осенью они спускаются на лишенные скал солнце-
печные склоны, а зимой нередко заходят даже в полосу леса (Егоров,
1955). Такова же в общем, по данным последнего автора, закономерность
сезонного перемещения козлов и на Памире. Здесь к зиме козлы, дер-
жавшиеся летом в верховьях боковых ущелий, спускаются в нижние
части долин, преимущественно на южные склоны, где снеговой покров
не глубок или отсутствует вовсе. В районе Хорога поздней осенью, зи-
мой и ранней весной козлы большими стадами выходят на посевы, вблизи
которых, даже при отсутствии поблизости скал, они иногда устраивают
лежки.
Масштабы сезонных кочевок сибирских козлов не велеки; даже по
горизонтали они редко превышают 7—10 км, а по вертикали составляют
иногда всего 200—300 м. Сроки миграций зависят от местных условий.
Осенью они начинаются в конце сентября или начале октября, но мае-
CAPRA
433
совые перемещения происходят поздней осенью после выпадения глубо-
кого снега. Первыми к местам зимовок откочевывают самки с козлятами
и молодняк; старые самцы уходят с летовок позднее. Обратные переме-
щения местами начинаются уже в марте или апреле, но О. В. Егоров
(1955) большую часть смешанных стад в Терскей-Алатау наблюдал на
местах зимовок еще в середине мая. Окончательная откочевка на летние
пастбища происходила только в середине июня. Весенние перекочевки
проходят теми же путями, что и осенью, только в обратном направлении.
Первыми весной откочевывают в горы старые самцы, а дольше всего за-
держиваются в среднем скалистом поясе гор самки. Старых сам-
цов, говорят, можно встретить у границ ледников уже в начале
июня.
Сроки перекочевок могут нарушаться преследованием со стороны
человека, приходом в места обитания диких козлов домашних стад и дру-
гими случайными причинами, не нарушающими общего сезонного харак-
тера миграций. Иногда наблюдаются временные нерегулярные переко-
чевки козлов летом вследствие нередкого в районах высокогорья выпаде-
ния глубокого снега. О подобных случаях для б. Алма-Атинского заповед-
ника сообщали М. Д. Зверев и А. А. Келейников (1947), а для Терскей-
Алатау О. В. Егоров (1955).
Сибирские козлы — стадные животные. В одиночку держатся крайне
редко. Размеры стад зависят, прежде всего, от общей численности коз-
лов в данном районе. На Алтае, например, по В. В. Дмитриеву (1938),
они даже осенью и зимой не образуют стад более чем в 30 голов. Зато
в районах высокой численности, например на Памире, и в июле можно
наблюдать табуны по 65—70 экземпляров.
Меняется стадность козлов и по сезонам. Взрослые самцы весной
отделяются от зимних смешанных стад и, уходя выше в горы, в течение
всего лета держатся небольшими группами по 4—6 голов в каждой. Самки
с приплодом текущего года и молодняк обоего пола в возрасте до 3—4
лет образуют отдельные стада, достигающие нескольких десятков экзем-
пляров. Беременные самки перед родами отделяются на короткое время
и держатся по одной-две отдельно. Молодые самцы на это время образуют
самостоятельные стада, в которых можно видеть также яловых самок.
Изредка и летом можно встретить среди смешанного стада одного-двух
старых самцов, но, как правило, последние вливаются в общие табуны
только осенью, незадолго до начала гона. Табуниться козлы начинают
уже в сентябре; на период гона стада вновь распадаются, так как самцы
отбивают себе гаремы. Максимальной численности табуны достигают
зимой по окончании гона. Образованию более крупных зимних стад спо-
собствует ограниченность кормовой базы, благодаря чему на малоснеж-
ных участках, обеспеченных зимними кормами, происходит концентра-
ция животных. В таких местах не составляют большой редкости стада
козлов в 100—150 и даже более экземпляров.
В стадах козлов имеются вожаки. Роль последних в обычное время
исполняет старая самка, а во время же гона — старый самец. Имеются
ли в стадах козлов специальные сторожа, не известно. Вероятнее всего,
роль последних выполняют случайно оторвавшиеся от пастьбы или бодр-
ствующие звери.
Для сибирского козла также характерна привязанность к постоян-
ному местообитанию. Даже напуганные и изгнанные охотниками из ка-
кого-либо ущелья, козлы через несколько дней вновь возвращаются
на старое место (Егоров, 1953).
28 Фауна СССР, И. И. Соколов
434
BOVIDAE
Суточный цикл жизнедеятельности С. sibirica в принципе не отлича-
ется от такового других козлов. Детали же его в разных районах опреде-
ляются местными условиями: состоянием погоды, степенью преследова-
ния со стороны человека, наличием в горах домашних стад и тому подоб-
ными причинами.
Летом козлы поднимаются с ночлега и начинают пастись еще до рас-
света. Медленно, кормясь на ходу, они двигаются вверх по склонам,
к полудню достигают мест дневного отдыха, где лежат и пережевывают
жвачку. Часов с четырех или пяти пополудни звери встают и начинают,
также не торопясь, спускаться вниз на пастбища. Пасутся не только
весь остаток дня, но и значительную часть ночи (Егоров, 1955). На ноч-
лег, если их не беспокоят, козлы устраиваются поблизости пастбищ,
а иногда и на местах кормежки. Дневные лежки обычно располагаются
или на открытых и обдуваемых ветром каменистых россыпях с широким
кругозором, или, наоборот, в закрытых местах, в тени, под выступами
скал, в нишах, реже в пещерах.
Протяженность суточных миграций летом не велика; по вертикали
она может доходить 600—700 м, а по горизонтали обычно не превышает
1.5—2 км.
В летние пасмурные дни, а также осенью и весной, сокра-
щается продолжительность дневного отдыха. Весной и осенью протяжен-
ность суточных миграций больше; иногда зверям приходится ежедневно
совершать путь в оба конца до 10—12 км. Суточные передвижения, если
зверей не беспокоят, происходят одними и теми же путями, на которых
часто образуются ясно обозначенные тропы.
Основную причину суточных миграций О. В. Егоров (1955) весьма
правдоподобно усматривает в температурных условиях. Весной и первую
половину лета звери, еще одетые зимним мехом, ищут прохладу в жаркие
дневные часы высоко в горах; осенью же, когда шерсть еще коротка,
а в горах начинают усиливаться заморозки, звери вынуждены на ночь
спускаться в более теплые места. Последним, по мнению этого автора,
объясняется тот факт, что в ряде долин Восточного Памира козлы выхо-
дят кормиться на участки с высохшей травой, когда на местах летовок
сохраняются еще непокрытые снегом участки зеленых альпийских паст-
бищ. Известную роль, как причина сезонных и суточных передвижений
и перемещений, играют насекомые-кровососы, спасаясь от которых козлы
нередко ложатся отдыхать прямо на снег.
Зимой козлы ведут более оседлый образ жизни. Двигаются мало.
Большую часть светлой половины суток проводят в пастьбе. Регулярных
суточных ; миграций не совершают.
На водопой, по мнению О. В. Егорова, в противовес утверждению
других авторов, козлы специально не ходят, довольствуясь питьем из
попадающихся ручейков или влагой, содержащейся в зеленом корме,
а также снегом.
К зимнему холоду козлы устойчивы, так как защищены длинным,
густым мехом. В тихую погоду часто лежат, полузанесенные снегом.
Нередко пасутся под проливным дождем. Укрытия ищут только во время
сильных буранов или дождя, сменяемого снегопадом. Постоянными
зимними укрытиями служат ниши и углубления под навесами скал;
эти места узнаются по значительному иногда скоплению на них помета.
На открытом месте козлы ложатся головами против ветра, вероятно
в целях большей безопасности, а также потому, что в этом случае холод-
ный воздух меньше проникает в толщу меха.
CAPRA
435
Пищей сибирского козерога является большинство травянистых
растений, произрастающих в районах его обитания. Видовой ее состав
зависит не только и, быть может, даже не столько от предпочтения,
отдаваемого зверем тому или иному растению, сколько от наличия его
в растительном покрове пастбищ. Предпочитаемость различных кормов
в сильной степени меняется и по сезонам. Зимой степень поедаемости
и видовой состав пищи в сильнейшей степени определяются доступностью
их, в зависимости от глубины и плотности снежного покрова.
Основу летнего питания составляет зеленая травянистая раститель-
ность. В противоположность многим другим копытным, предпочитающим
разнотравье, значительный удельный вес в питании козлов имеют
злаки и осоки, к пережевыванию которых хорошо приспособлена со-
вершенная зубная система козлов и баранов. В числе поедаемых коз-
лами летом на северных склонах Терскей-Алатау растений О. В. Егоров
(1955) называет мятлики, типцы, осоки, кобрезии, гречихи, лук, астра-
гал и некоторые другие. Основную кормовую базу козлов в восточном
Памире летом составляет растительность лугов альпийской зоны. В числе
кормовых растений, кроме тех же злаков и осок, что и в Терскей-Алатау,
О. В. Егоров (1955) называет еще горец, примулу, лисохвост и некото-
рые другие. Злаки имеют значение основного корма для козлов и в Илий-
ском Алатау; в число поедаемых растений здесь входят ковыль, дикий
овес. Кроме того, по И. Д. Шнаревичу (1948), и летом значительную
роль в питании будто бы играют побеги можжевельника и эфедры. На
Алтае во второй половине лета козлы пасутся на ягодниках и
особенно охотно поедают ягоды красной смородины (Дмитриев,
1938).
Осенью основу питания по-прежнему составляют злаки и разнотравье;
очень часто в это время козлы выпасаются на склонах, покрытых типча-
ковой степью. Видную роль осенью начинает играть в питании и дре-
весно-веточный корм: листья и побеги смородины и ивы, ягоды шипов-
ника, красной смородины, рябины. Основная масса содержимого желуд-
ков трех убитых О. В. Егоровым (1955) осенью на Памире козлов состояла
из семян и стеблей зонтичного растения варх (Prangos sp.) и гречихи
(Polygonum sp.) (Егоров, 1955). По данным И. Д. Шнаревича (1948),
в б. Алма-Атинском заповеднике основой осеннего питания козлов яв-
ляются побеги кустарников, составляющие по весу до 85% содержи-
мого всех исследованных желудков, причем свыше 50% приходится на
долю можжевельника и эфедры.
Зимним кормом козлов служат высохшие наземные части травяни-
стых растений на местах с неглубоким снежным покровом или вообще
лишенных такового. Они составляют основу зимнего питания козлов
в некоторых местностях на Алтае, например на склонах долин Баш-
кауса и Чулышмана (Дмитриев, 1938), на северных склонах Терскей-
Алатау и в некоторых местах восточного Памира. В Терскей-Алатау
у верхней границы леса козлами используются и веточные корма, осо-
бенно в глубокоснежные зимы; в наибольшем количестве поедаются по-
беги ивы. Основным зимним кормом в б. Алма-Атинском заповеднике
являются побеги различных кустарников, в первую очередь можжевель-
ника и эфедры, тогда как злаки имеют второстепенное значение (Шна-
ревич, 1948).
В восточном Памире основными зимними пастбищами козлов, по
О. В. Егорову (1955), являются пустыни субальпийского пояса с незна-
чительным снеговым покровом. Основными кормами служат здесь тере-
28 *
436
BOVIDAE
скен (Eurotia ceratoides G. A. M.) и полыни (Artemisia skorniakowii
Winkl. и др.) с примесью ковылей, мятлика и типца.
Л. А. Шульпин (1948) и И. Д. Шнаревич (1948) к числу кормовых
растений сибирских козлов относят арчу. О. В. Егоров (1955) поедание
козлами арчи в Терскей-Алатау, Памире и Кунгей-Алатау, по крайней
мере в летний период, отрицает. В Терскей-Алатау козлы добывались
им неоднократно, но побеги этого растения ни разу не находились в со-
держимом желудков.
Весеннее, как и осеннее питание сибирского козла носит смешанный
характер. Переход на зеленые травянистые корма совершается по мере
развития вегетации растений.
На Алтае козлы, по В. В. Дмитриеву (1938), посещают естественные
солонцы в виде выходов желтовато-белой или серо-белой глины, вые-
дая в ней целые пещеры шириной в метр и высотой 60—70 см.
На северных склонах хребта Терскей-Алатау солонцы в виде мелко-
зема пепельного цвета также посещались, но нерегулярно, не чаще од-
ного-двух раз в неделю. В восточном Памире, попутно с поеданием травы,
козлы заглатывают соли, выступающие в виде светлого налета на об-
ширных участках горных склонов (Егоров, 1955).
В спокойной обстановке сибирские козлы двигаются шагом. Походка
их медленная. Бег рысью медленный и неуклюжий. Бегущего козла
на ровном месте легко догоняет собака. Спасает зверя необычайная спо-
собность передвигаться по каменистым россыпям и скалам, куда они
при опасности бросаются быстрыми сильными прыжками. То, что гово-
рилось о передвижениях безоарового козла в горах, в одинаковой сте-
пени относится и к сибирскому козерогу. «Уменье их передвигаться по
скалам, — пишет В. Н. Шнитников (1936), — прямо изумительно, и
тому, кто не видел этого лично, трудно даже представить ту обстановку,
среди которой теке1 не только свободно и легко ходит, но и бежит пол-
ным аллюром в случае преследования. Также изумительны и его прыжки
вниз на камни с огромной высоты ... В районе обитания теков особенно
широко' развиты каменные осыпи из всевозможных размеров каменных
обломков. Ходьба по иным из таких осыпей является подчас особенно
трудной, а на более крутых и опасной, так как неустойчиво лежащие
камни Часто выскакивают из-под ног, катятся вниз, иногда увлекая и
соседние, и в результате целый участок осыпи может поползти вниз,
увлекая с собой идущего. Но теки ходят по всяким осыпям необычайно
легко и уверенно и как будто бы даже особенно любят ходить по ним».
Стараясь пройти... опасную крутую осыпь как можно быстрее, козлы,
тем не менее, замедляют шаг или останавливаются вовсе, если начинают
падать камни (Антипин, 1946). В спокойной обстановке козлы обычно
двигаются цепочкой друг за другом; испуганное же стадо бежит, как
попало, и нередко некоторые из зверей обгоняют вожака. При передви-
жении по снегу впереди идут обычно наиболее сильные старые звери.
При нужде неплохо плавают, могут переплывать довольно широкие
'реки (Цалкин, 1950).
В местах, где козлов преследуют, они очень чутки и осторожны. Там
же, где их мало тревожат, они могут подпускать человека близко, пас-
тись по соседству со стадами домашних животных. Из органов чувств
у козла, по В. Н. Шнитникову (1936), хорошо развиты как слух и зре-
1 Местное название сибирского козла в Семиречье.
CAPRA
437
ние, так и обоняние. Заподозрив опасность, козел может полчаса не спу-
скать глаз с подозрительного предмета.
Звуки сибирские козлы издают редко; чаще других можно услышать
предостерегающий свист. По Н. В. Туркину и К. А. Сатунину (1904),
самцы во время гона будто бы издают рев. Голос самок и козлят напоми-
нает таковой домашних коз, но более сдавленный и глухой (Цалкин, 1950).
Линька у сибирских козлов хорошо прослежена И. Д. Шнаревичем
(1948) и О. В. Егоровым (1955). Смена волосяного покрова происходит
один раз в году —• весною и в начале лета. Начало и продолжительность
весенней линьки варьируют в зависимости от условий местности и со-
стояния животных. Самцы и яловые самки линяют раньше. Позднее
начинается и более растянута она у детных самок, а также у истощенных
и больных животных. В Илийском Алатау выпадение старой шерсти
начинается уже в марте, а заканчивается линька у нормально упитан-
ных зверей в конце мая или начале июня. На северном склоне Терскей-
Алатау и на Памире, по личным наблюдениям О. В. Егорова (1955),
линька у козлов начинается не раньше середины мая, что он связывает
с более высоким расположением здесь мест обитания козлов над уровнем
моря. Заканчивается она у большинства в середине июля, но у отдельных
зверей, истощенных самок с козлятами, остатки зимней шерсти ему цри-
ходилось наблюдать до конца августа. В первую очередь выпадает под-
шерсток. Дольше удерживается более тонкая ость (так называемый пе-
реходный волос). Старая шерсть обычно свисает и выпадает клочьями.
Испытывая раздражение, звери трутся о выступы скал, о кусты.
В первую очередь шерсть выпадает на лопатках, плечах, бедрах и
боках головы, затем линька распространяется на бока и остальную часть
головы. Дольше всего старая шерсть удерживается, по И. Д. Шнаревичу
(1948), на животе и конечностях, а по О. В. Егорову (1955) — на верх-
ней части спины и шеи.
Параллельно с выпадением зимней шерсти идет отрастание летнего
волоса. Последний в августе достигает длины 2—3 см и тогда же заме-
чается появление подшерстка. В сентябре начинает расти зимняя ость,
которая вскоре обгоняет в росте летний волос, остающийся в виде про-
межуточного на всю зиму. Окончательного развития зимний волосяной
покров достигает к концу ноября.
Половой зрелости и самцы, и самки сибирского козла при благоприят-
ных условиях существования достигают на втором году жизни и в воз-
расте 1% лет могут оплодотворяться. Однако большая часть самок в
естественных условиях приносит первых козлят в возрасте не моложе
3 лет. Самцы же начинают участвовать в размножении еще позже, так,
примерно, не ранее 4—5 лет, так как до этого отгоняются более сильными
старыми козлами и могут покрыть самку только случайно.
Сибирские козлы.— типичные полигамы. Взрослые сильные самцы
отбивают себе гаремы из нескольких самок.- Число ходящих при одном
самце особей, включая и козлят-сеголеток, колеблется от 10 до 16 го-
лов (Шнаревич, 1948). По-видимому, нормой на 1 самца следует считать
5—6 самок. При большей нагрузке, которая наблюдается в районах
интенсивного преследования козлов, может иметь место яловость извест-
ной части самок (Егоров, 1953).
Сведения о сроках протекания гона противоречивы. На Алтае, по
В. В. Дмитриеву (1938), он начинается во второй половине ноября и тя-
нется около трех недель. Приблизительно такие же сроки указываются
для Заилийского и Таласского Алатау (Шнаревич, 1948; Шульпин, 1948).
438
'BOVIDAE
По заслуживающим доверия сведениям О. В. Егорова (1953), совпадаю-
щим с таковыми Р. Н. Мекленбурцева (1949), в Терскей-Алатау и на Па-
мире период гона падает на время со второй половины декабря по пер-
вую половину января.
Самцы в период гона сильно возбуждены и к концу его резко худеют.
Между ними происходят ожесточенные турнирные поединки. До смерти
или увечья дело доходит, по-видимому, редко, но известен случай, когда
трупы двух сцепившиеся рогами самцов были прибиты к берегу тече-
нием реки (Шнаревич, 1948). Молодые, отогнанные взрослыми соперни-
ками самцы бродят позади стад и, вероятно, при случае кроют отставших
от гарема самок.
Беременность у самок сибирских козлов, судя по наблюдениям в
зоопарках (Шнаревич, 1948) и по срокам окота в природе, равна 5%—6
месяцам. В Терскей-Алатау и на Памире, по О. В. Егорову (1955), рожде-
ние молодняка происходит в первой половине и в середине июня. Случаи
запоздалого окота имеют место до конца июня.
Перед окотом самки удаляются по одной-две из табунов и уходят
в глухие скалистые ущелья, где приносят одного, реже двух козлят.
Достоверные случаи рождения троен не известны. Двойни наблюдаются
чаще у вполне взрослых самок и в зоопарках, где животные содержатся
в условиях более благоприятного питания.
Вес новорожденного козленка колеблется, по данным О. В. Егорова
(1953), в пределах 2.8—4.8 кг. Уже в первый день послеутробной жизни
козленок может подниматься на ноги и немного ходить, но первые дни
очень слаб и лежит, затаившись между камней или в кустах. Спустя
несколько дней он не только следует за матерью, но не уступает послед-
ней в способности бегать и карабкаться по скалам. Молодые, имеющие
от рождения несколько месяцев, и полуторагодовалые козлы легче и
быстрее бегают в горах, чем вполне взрослые звери (Дмитриев,
1938).
Рога у козлят начинают расти вскоре после рождения и ясно заметны
уже на третьей-четвертой неделе. В природных условиях в течение пер-
вого года жизни рог вырастает на 10—12 см, зимой рост его замедляется,
иногда прекращаясь почти вовсе, а поперечные валики начинают обра-
зовываться лишь на втором году жизни (Егоров, 1938). У козлов в зоо-
парках, благодаря лучшему, чем в природной обстановке, и равномер-
ному питанию, рога продолжают расти и зимою, достигают к годовалому
возрасту длины 30 см, а на их лобовой грани появляются валики, число
которых может доходить до пяти (Шнаревич, 1948).
Молоком самки кормят детенышей до конца октября, а иногда и до
декабря. Зелень козлята начинают щипать на третьей-четвертой неделе,
а с месячного возраста травянистый корм принимает заметное участие
в их рационе.
К годовалому возрасту козлята достигают, по данным О. В. Егорова
(1953), около одной трети веса и около двух третей роста взрослых. Рост
у сибирских козлов продолжается долго, до 8—9 лет. Особенно долго
растут рога.
Соотношение полов у новорожденных точно не известно; в смешанных
стадах козлов преобладают самки, число которых составляет в среднем
57-60%.
Судя по рогам коллекционных экземпляров самцов, продолжитель-
ность жизни сибирского козла в естественной обстановке редко превы-
шает 10 лет, но в Лондонском зоологическом саду самка прожила 22
CAPRA
439
года 3 месяца и 19 дней, причем была убита, а не умерла естественной
смертью (Flower, 1931).
Из врагов сибирского козла на первом месте по значению стоят барс
(Felts uncia Schreb.) и серый волк (Cants lupus L.). Барс, будучи пре-
красно приспособленным к передвижению в горах, охотится на козлов
в течение круглого года. От него гибнут животные обоего пола от ново-
рожденных козлят до старых самцов. Охотится барс на козлов скрадом,
подкарауливает на местах отдыха и переходах, бросается на жертву
и догоняет ее несколькими крупными прыжками.
От волков много гибнет козлов в глубокоснежные зимы. Летом гибель
от них носит случайный характер: во-первых, козлы в это время дер-
жатся в скалистом высокогорье, где неумеющему бегать по скалам волку
невозможно догнать даже немного подросшего козленка; во-вторых, летом
волки охотятся в одиночку, а взять один на один взрослого козла волк
обычно не в состоянии. Зато зимой в некоторых районах козлы составляют
главную пищу волков. Охотясь стаями, волки легко догоняют козлов по
хоть сколько-нибудь глубокому снегу. Гибнут от волков в первую оче-
редь большерогие и тяжелые на бегу взрослые самцы, к тому же исто-
щенные бескормицей и предшествующим гоном (Егоров, 1955).
Из других хищных зверей на козлов нападают красные волки (Суоп
alpinus Pall.), которые в некоторых местностях, например в Саянах и
Семиречье, будто бы являются главными их врагами (Скалой, 1929;
Шнитников, 1936). Охотятся красные волки также стаями по 10—12
штук. Гибель от медведей носит случайный характер; жертвой их могут
стать только недавно родившиеся или раненные козлы. На недавно ро-
дившихся козлят нередко нападают лисица и рысь. О. В. Егоров (1955)
неоднократно находил в помете лисиц остатки шерсти козлят. По его
словам, «большой урон поголовью сибирского козерога причиняет бер-
кут. Однако им уничтожается исключительно молодняк, о чем можно
судить по наличию костей козлят и архарят под гнездами этих птиц».
Многими в качестве врага называется орел-бородач, от которого, вероятно,
гибнут также только козлята.
Массовые эпизоотические заболевания сибирских козлов не изучены,
но, по-видимому, иногда имеют место. Так, по Р. Н. Мекленбурцеву
(1949), на Памире очень распространена эпизоотическая пневмония
«кара-юпка» (черное легкое), которая, раз начавшись в каком-либо рай-
оне, губит всех обитающих в нем козлов. В. В. Дмитриев (1938) упоми-
нает о крупной эпизоотии, имевшей место среди козлов в 1917—1920 гг.
на Алтае. Заболевание было похоже на паршу и сопровождалось слепо-
той. Звери падали от болезни, их легко ловили собаки. Эпизоотия охва-
тила также домашних и некоторых других диких животных, включая
медведей. С этой эпизоотией связывают исчезновение козлов на запад-
ном побережья Телецкого озера и в долине р. Ян-Чили. В зоопарках
сибирские козлы болеют такими, свойственными домашним животным
заболеваниями, как некробациллез и катарральная горячка.
Из других причин убыли сибирских козлов следует отметить нередкие
случаи гибели целых групп их при снежных обвалах. Существенную
роль в снижении численности козлов играет гибель их вследствие недо-
статка корма и истощения в глубокоснежные зимы. Известны случаи,
когда ослабевших козлов ловили в снегу руками (Соловьев, 1920). Чаще
погибают от бескормицы уже истощенные предшествующим гоном самцы,
особенно старики с их сильно стертыми зубами, не пригодными для пе-
режевывания единственно доступного в такое время веточного корма.
440
BOVIDAE
Из эктопаразитов сибирского козла на первое место следует поста-
вить носового овода {Oestrus ovis L.), личиночной стадией которого в не-
которых местах заражены все козлы поголовно. Личинки носового
овечьего овода поселяются на слизистой оболочке носовой полости,
лобных пазух, даже в пазухах роговых стержней (Егоров, 1955). На
восточном Памире и Заилийском Алатау на козлах паразитирует другой
вид носового овода — Ое. caucasicus Grunin.1
Иногда на козлах в большом числе паразитируют мухи-кровососки
(Melophagus sp.). С крупного самца О. В. Егоровым (1955) было собрано
476 кровососок, не считая паразитов, свалившихся при транспортировке
зверя. У сильно пораженных кровососками козлов наблюдалось исто-
щение и задержка линьки. Известно паразитирование блох из родов
Xenopsylla и Vermipsilla. И. Д. Шнаревич (1948) у большого числа коз-
лов (до 30%), добытых им в Заилийском Алатау, находил вшей из рода
Linognathoides. Из иксодовых клещей на сибирском козле с достоверностью
зарегистрированы два вида — Dermacentor pavlovskyi 01. и Haemaphy-
salis walburtoni Nutt.1 2
Сибирский козерог является хозяином ряда паразитических червей.
В Казахстане, например, у него зарегистрировано свыше 20 видов гель-
минтов (Боев и Соколова, 1949). Только легочных нематод найдено 7
видов, из которых один (Cystocaulus vsevolodowi Boj.) является специ-
фичным для сибирского козерога (Боев, 1954). Наиболее распространен-
ными гельминтами являются дикроцелиоз и протостронгилидозы легких,
которыми поражены все без исключения исследованные экземпляры.
Практическое значение имеют ценуроз межмышечной ткани и легочно-
глистные инвазии, вызывающие бронхопневмонии.
Хозяйственное значение. Наиболее ценным продуктом, получае-
мым при убое сибирского козла, является мясо. Во многих районах,
как например в Киргизии, оно играет видную роль в питании населения,
а при рациональной организации промысла может играть заметную роль
в создании продовольственной базы местной промышленности и крупных
населенных пунктов.
Мясо сибирского козерога по вкусу несколько напоминает мясо до-
машнего козла; в отличие от последнего, оно лишено, за исключением
периода гона, специфического козлиного запаха, но зато обладает свое-
образным привкусом дичи (Егоров, 1955). У взрослых животных мясо
довольно жестко и трудно уваривается благодаря обилию межмускуль-
ной соединительной ткани. При химическом анализе трех образцов мяса
козлов, произведенном В. Ф. Денисовым (1935), сухое его вещество со-
держит 75.5 — 88% сырого протеина, до 14% жира и от 4 до 7% без-
азотистых экстрактивных веществ.
Вес туши мяса вполне взрослого самца составляет 36—61 кг, в сред-
нем 40—45 кг. У самок соответствующие цифры колеблются в пределах
30—36 кг (Егоров, 1955). Выход чистого мяса по отношению к живому
весу (убойный вес) сравнительно высок, у самок, по данным О. В. Его-
рова (1955), в среднем около 65%, у самцов несколько меньше — около
58—62%. У отдельных хорошо упитанных зверей вес туши мяса может
достигать 70% живого веса.
Осенью перед гоном козлы бывают очень жирны. В отличие от бара-
нов, основная масса жироотложений скапливается в брюшной полости,
1 Устное сообщение К. Я. Грунина.
2 Сведения, полученные от Г. В. Сердюковой.
CAPRA
441
в области почек, сальника и толстых кишок. От 8—9-летних самцов позд-
ней осенью, по словам О. В. Егорова, можно набрать около 4—5 кг сала.
Подкожные жироотложения, наоборот, развиты очень слабо. Жир коз-
лов твердый, по данным В. Ф. Денисова (1936), содержит повышенное,
даже по сравнению с диким бараном, количество тугоплавких (стеари-
новой и пальмитиновой) жирных кислот.
Ценным продуктом, получаемым от сибирского козла, является
шкура. У козлов она прочнее, чем у баранов. У взрослых зверей, осо-
бенно осенью и зимой, она сравнительно толста и пригодна для выделки
хрома. Из шкур молодых козлов можно изготовлять шевро. Из нее же
шьют теплые прочные шубы. Хорошо подобранный, «тау-течный» мех,
говорят, довольно красив (Туркин и Сатунин, 1904). Выделанные шкуры
козлов используются, как коврики для подстилки на пол; из них шьется
мягкая и довольно прочная обувь «чокои». Из козлиных же шкур ме-
стами делаются бурдюки для кумыса. Рога используются в качестве
настенного украшения и на мелкие поделки. Даже очищенный и высу-
шенный желудок иногда используется для хранения масла или сала
(Егоров, 1955).
Пойманные молодыми сибирские козероги легко привыкают к домаш-
ней обстановке и размножаются в неволе. Серьезных опытов по их одо-
машниванию, вероятно, в виду малой практической значимости этого
мероприятия, не предпринималось.
Известны многочисленные и успешные опыты скрещивания С. sibi-
rica с домашними козами. В Академии наук Казахской ССР с 1944 г.
ведутся плановые работы по гибридизации местных домашних коз с ди-
кими сибирскими козлами (Бутарин, 1947). Целью этих работ является
выведение новой породы коз, совмещающей высокую шерстную продук-
тивность домашней козы с хорошей приспособленностью дикого вида
к условиям высокогорья. Осуществление этой задачи позволило бы го-
раздо полнее использовать пастбищные ресурсы высокогорья Средней
Азии, чем это имеет место в настоящее время, вследствие недостаточной
приспособленности к этой обстановке домашних животных.
До сих пор скрещивались домашние самки с дикими самцами. Из
особенностей полученных гибридов обращает внимание, прежде всего,
гетерозис, увеличенные по сравнению с исходными формами размеры.
Общее сложение и окраска шерсти в одних случаях была сходной с до-
машними, в других — с дикими козлами. В опытах Н. С. Бутарина гиб-
ридная самочка имела пеструю окраску. Характер (структура) шерсти
сходен с диким видом. Гибрид от скрещивания домашней козы с сибир-
ским козерогом в Ленинградском зоосаду имел характерное для домаш-
него козла острое переднее ребро рога (Егоров, 1955).
Скрещивание домашних и диких сибирских козлов, по-видимому,
нередко происходит и в природной обстановке. О. В. Егорову (1955)
на Памире пришлось видеть домашних коз, гибридное происхождение
которых подтверждалось пастухами. Гибридные животные имели близ-
кую к диким окраску с темным ремнем вдоль спины и на передней сто-
роне ног. Они отличаются так же, как и при специально поставленных
опытах по гибридизации, более живым темпераментом, подвижностью,
лучше переносят бескормицу в многоснежные зимы.
Гибриды диких и домашних коз вполне плодовиты. В Лондонском
зоологическом саду были получены гибриды от сибирского козерога
и кавказского тура (Gray, 1954). О внешности и плодовитости этих гиб-
ридов ничего не известно.
«2
BOVIDAE
Охота. Способы добычи козлов довольно разнообразны. Наиболее
распространена охота скрадом, в принципе ничем не отличающаяся от
таковой на других горных копытных, например на безоарового козла
или серну. От охотника требуется большая сила, выносливость и вы-
держка, так как козлы могут учуять или увидеть человека, особенно если
их часто беспокоят, за 1—1.5 км. Иногда при этом способе берут с собой
собаку, которую, однако, пускают со сворки лишь для того, чтобы дог-
нать и остановить раненого зверя.
Практикуется охота нагоном, когда одна группа охотников медленно
двигается фронтом по обеим сторонам хребта на расстоянии 100—200 м
друг от друга и гонят козлов на другую группу, расположившуюся
в засаде. При другом способе охоты нагоном один или несколько охот-
ников, обнаружив на пастбище козлов, располагаются на вероятных
путях (лазах), по которым должны пройти звери, а один трогает стадо
и заставляет его бежать мимо засевших в засаде стрелков. Успех охоты
зависит от знания привычек зверя и от уменья выбрать место для засады.
При втором способе на Алтае часто применяют собак, которых загонщик
пускает со сворки только тогда, когда по его расчетам стрелки займут
свои места, а им обнаружены звери. От собак при такой охоте требуется
не чутье, а злобность и настойчивость («вязкость»). Иногда собакам уже
вскоре удается одного или нескольких зверей «поставить на отстой» на
каком-нибудь выступе скалы, и такой козел в первую очередь становится
добычей. Однако опытные охотники в таких случаях не спешат, так как
с отстоя козел скоро не уйдет, и стараются дождаться ходового зверя.
j,O. В. Егоров (1955) сомневается в успешности и целесообразности
охоты на козлов с собаками в расчете на добычу отстойного зверя. Он
неоднократно убеждался в этом сам, а по словам промышленников, козлы
становятся на отстой только после продолжительного преследования,
что бывает редко, так как собаке невозможно угнаться за ними по кру-
тым скалистым склонам.
Подкарауливают козлов на местах пастьбы и на переходах, пользуясь
их привычкой придерживаться одних и тех же участков пастбищ и путей
суточных миграций. На Алтае практикуется подкарауливание на солон-
цах (Разоренова, 1939; Колосов, 1939). На Тянь-Шане этот способ не при-
меняют ввиду нерегулярности посещения солонцов козлами.
Иногда расставляют капканы на тропах козлов, а для ловли живьем
применяют растягиваемые на местах переходов сети, в которых с раз-
бега запутываются напуганные преследованием звери (Егоров, 1955).
Во всех случаях охоты скрадом, нагоном и подкарауливанием чрезвы-
чайно важно учитывать направление не только ветра, но и слабой воз-
душной тяги.
Capra (Orthaegoceros) falconeri Wagner — Винторогий козел, или
мархур (рис. 92—95)
₽Wagner J., 1839, Mnnch. Gelehrt. Anzeig., 9 : 430 (первооп.); Hutton Th.,
1842, Calcutta, J. Natur. Hist., II : 535—542 (морф., сист.); Kinloch A., 1885,
Large game shooting of Tibet, Himalayas and Northern India, Calcutta : 136—
144 (биол., ox., изобр.); Lydekker R., 1898, Wild oxen, sheep and goats, Lon-
don : 286—296 (общ., синон., изобр.); Трубецкой П. С., 1910, Природа и охота,
1 : 41—48 (распр., бйол., изобр.); Lydekker R., 1913, Catal. Ungulate mammals
Brit. Mus., I, London : 161—173 (сист., синон., изобр.); Rowland War d's re-
cords of big game, 1914, London : 358—366 (рога); Brehms Tierleben, 1925, 4. Aufl.,
Bd. 13, Leipzig, : 297—298 (опис., изобр.); HilzheimerM., 1933, Arch. f. Tier-
ernahrung u. Tierzucht., VIII : 323—371 (гибр.); Флеров К. К., 1935, вкн.:
CAPRA
443
Виноградов Б. С., Е. Н. Павловский и К. К. Флеров, Звери Таджикистана, Л., изд.
Акад, наук СССР : 112—115 (общ., изобр.); Флеров К. К. и И. М. Громо в,
1935, Матер, по паразитол. и фауне южн. Таджикистана, X : 268—269 (распр.); Г у-
р е е в А. А., 1937,' Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. туркменск.,
9 : 370 (распр.); Султанов Г., 1939, Тр. Узбекск. зоол. сада, 1 : 102 (распр.,
биол.); Шапошников Л. В., 1939, Тр. Моск, зоопарка, 1 : 59—63 (морф.); Гепт-
нер В. Г., 1940, Тр. Моск, зоопарка, 1 : 39—40 (морф.); Harper F., 1945, Ex-
tinct a. wanishing mammals of Old World, N. Y. : 627—635 (распр., изобр.); Цал-
кин В. И., 1945а, Уч. зап. Моск. гос. унив., 83 : 5—30 (общ.); Цалкин В. И.,
19456, Докл. Акад, наук СССР, XLVI, 5 : 229—232 (сист.); Zoo life London, I, 1, 1946,
обложка (изобр.); Цалкин В. И., 1948, Охрана природы, 3 : 79—83 (общ.); Гро-
мова В. И., 1949, сборн. «Тешик-Таш», изд. Моск. гос. унив. : 8—99 (ист., распр.);
Cobb Е., 1949, Field, London, 193:190 (изобр.); Antonins О., 1951, Verb,
d. Deutsch, zool.-botan. Gesellsch., 92 : 109—110 (гибр.); Ellerman J. and
T. Morriso n-S с о t t, 1951, Checklist of palaeartic and indian mammals, Lon-
don : 408—409 (сист., синон.); Железняков Д. Ф., 1952, Тр. Средне а зиатск.
гос. унив., XXXII : 27—33 (распр., биол.); Богданов О. П., 1953, Природа,
7 : 112 (распр.); Султанов Г. С., 1953, Тр. Инет. зоол. и паразитол. Акад, наук
Узбекск. ССР, II : 3—27 (распр., биол., изобр.); Cray А., 1954, Mammalian hyb-
rids, London : 70 (гибр.); Насимович А. А., 1955, Роль снежного покрова в жизни
копытных, М., изд. Акад, наук СССР : 141 (распр., биол.).
Описание. Козел крупной величины. Длина тела у взрослых самцов
от 155 до 170 см. Высота в холке 85—100, максимально до 115 см (Brehm,
Рис. 92. Винторогий козел (Capra falconeri Wagn.).
1925). Основная длина черепа 231—255 мм. Живой вес 86—109 кг. Самки
значительно меньше самцов: длина тела у них (по В. И. Цадкину, 1945а)
около 145 см, живой вес 32—41 кг, основная длина черепа 196—207 мм.
Телосложение крепкое, но несколько более легкое, чем у сибирского
козерога. Туловище покоится на довольно толстых, средней длины но-
444
BOVIDAE
rax. Голова сравнительно тяжелая, иногда слегка горбоносая. Длина
ушей у самцов составляет 12—15 см. Массивность шеи самцов увеличи-
вается мощно развитым снизу подвесом. У самок шея тонкая, особенно
в летней шерсти. Линия спины прямая; круп заметно свислый. Хвост
у самцов длиной 12—15 см, у самок около 8 см. Копыта массивные, длин-
ные, но низкие, с закругленными концами; длина их у взрослых самцов
на передних конечностях 64—78 мм, на задних 60—64 мм; высота копыт:
передних 33—40 мм, задних на 1—2 мм меньше. На передней (тыльной)
поверхности запястных суставов и у самцов, и у самок имеются лишенные
волоса и утолщенные роговые мозоли круглой или овальной формы,
величиной с трех-пятикоопеечную монету.
На рогах имеется два хорошо развитые ребра — передне-внутреннее
и задне-внутреннее, и две грани — внутренняя и наружная. У молодых
зверей на первом году лучше развито переднее ребро; у взрослых же оно
слегка закруглено, тогда как заднее становится значительно более острым.
Из двух граней внутренняя плоская, иногда даже слегка вогнута, а на-
ружная сильно выпукла; поэтому поперечное сечение рога, кроме верху-
шечной уплощенной части, близко к полукруглому. Поверхность рога
от основания до самих верхушек несет мелкую сплошную поперечную
морщинистость и ряд отставленных друг от друга более глубоких кру-
говых борозд (колец) на границе участков годичного прироста рога.
В отличие от других видов козлов, совершенно отсутствуют бугры (утол-
щения) или поперечные валики на передней стороне рога. Слабо разви-
тые утолщения, и то далеко не всегда, имеются лишь на заднем ребре
рога. При этом, расположены они, подобно безоаровому козлу, на границах
годичного прироста рога.
Рога и у самцов, и у самок, в отличие от других козлов нашей фауны,
свернуты или скручены в гетеронимную спираль. Степень свертывания
и скручивания весьма различна, обнаруживая не только индивидуаль-
ную, но и ясно выраженную географическую изменчивость. В одних
случаях рога самцов образуют очень пологий изгиб, сначала сильно
расходясь в стороны, затем в предвершинной части вновь несколько
сближаясь, а концами обращены наружу или вверх. При этом располо-
женные на поверхности роговых чехлов ребра (кили) описывают один
полный поворот вокруг оси рога. В других случаях рога свернуты в более
плотную штопорообразную спираль, а ось рога и ребра описывают в про-
странстве два и даже три полные оборота. Наконец, для мархуров Сулей-
мановых гор характерна форма винтообразно скрученных рогов, когда
сама ось рога остается прямой, а ребра на поверхности роговых чехлов
описывают вокруг нее до двух и более оборотов. Степень расхождения ро-
гов в стороны подвержена сильной индивидуальной изменчивости. Степень
наклона их назад более или менее постоянна; у живого животного рога
почти не поднимаются выше профиля лба и носа. Длина рогов по изгибу
заднего киля у вполне взрослых самцов (старше 5 лет) составляет обычно
от 55 и до 110 см, по прямой же от основания переднего киля до верхушек —
от 45 до 73 см. Роуланд Вард (Rowland Ward, 1914) приводит максималь-
ную длину рогов по изгибу, равную 63 дюймам (161.3 см).
Рога сжаты с боков, хотя и не так сильно, как у Capra aegagrus Erxl.;
передне-задний диаметр рога в основании у крупного самца составляет
около 91—92 мм при поперечнике 56—60 мм и обхвате около 220 мм.
Основания рогов почти параллельные и тесно сближены; расстоя-
ние между ними, как правило, не превышает 1—2 см, а иногда даже
меньше.
CAPRA
445
У самок рога значительно тоньше и короче, обычно не более 15—20 см.
Скручивание не превышает половины оборота. Передний киль, в противо-
положность самцам, острее, чем задний. Основания рогов более широко
расставлены, чем у самцов. Внутреняя грань вогнутая, наружная вы-
пуклая, но рога сильнее сжаты с боков, особенно у верхушек. Цвет рогов
серовато- или темно-бурый.
Волосяной покров состоит из довольно длинной шелковистой ости
и тонкого, развитого только в зимнее время подшерстка. Длинная, до
30 см, заостренная борода занимает не только всю нижнюю сторону
головы, кроме самого подбородка, но и нижние части щек. Кроме того
Рис. 93. Винторогий козел (Capra falconeri Wagn.). Алма-Атинский
зоопарк. (Фото автора).
длинные, до 20 см, грубые волосы образуют подвес на нижней стороне шеи,
груди и плеч. Между бородой и верхним краем подвеса имеется разрыв
с более короткими волосами. Пучки удлиненных волос имеются часто
также на передней и боковой сторонах предплечий. Удлинены, но зна-
чительно меньше, волосы также на голенях ниже колен и на верх-
ней стороне шеи и на затылке. У самок борода в виде небольшого пучка
длинных волос имеется, но грива по верху шеи и подвесы на нижней
стороне отсутствуют. Хвост от основания покрыт волосами одинаковой
с туловищем длины, но на конце они слегка удлинены, хотя и не образуют
кисти.
Общая окраска туловища и шеи в зимнем меху серовато-белая, состав-
ленная йз белых с темными, буровато-коричневыми верхушками волос.
Иногда она бывает более темной, с преобладанием буровато-серых тонов
(Цалкин, 1945а). Голова окрашена темнее туловища. Низ живота, нахи,
внутренние части бедер, наоборот, заметно светлее. Передняя сторона
446
BOVIDAE
ног темная, буро-коричневая, причем на передних ногах темная окраска
резко обрывается поперечной светлой полосой выше запястных суставов.
Преобладающий тон окраски в летнем меху буровато-рыжий, довольно
изменчивый у отдельных животных. Основная масса волос туловища,
ног и боков шеи имеет белые основания, переходящие в буровато-корич-
невую окраску средней части; затем идет широкое предвершинное рыжее
кольцо и темно-бурый заостренный кончик. Часть волос сплошь буро-
коричневые, без рыжих предвершинных колец. Среди окрашенных волос
разбросаны чисто белые с темной верхушкой. От преобладания того или
иного типа волос, степени развития на них рыжей или буроватой зоны,
зависит общий тон окраски. Число белых волос, по-видимому, увеличи-
вается с возрастом; поэтому старые самцы могут иметь и летом светлую
белесоватую окраску. Вдоль средней линии спины до корня хвоста тянется
темная буро-коричневая полоса, с примесью небольшого количества
белых волос. У старых самцов эта полоса становится чисто белой. Из
смеси буро-коричневых и белых волос состоит имеющаяся у самцов и
в летнее время гривка наверху шеи. Низ живота, пахи, внутренние части
ягодиц и бедер, как и зимой, светлые, желтовато-белые. На границе
боков и живота от локтевых к коленным суставам иногда тянутся темные
неясные полосы, переходящие на переднюю сторону задних ног. Темя
и затылок буро-коричневые с примесью белых волос. Лоб, переносье,
бока головы имеют коричневато-серую окраску, составленную бурыми
со светлыми кончиками волосами. Конец морды более однотонный, бу-
роватый; края губ беловатые. Темная буровато-коричневая окраска
передних ног, как и в зимнем меху, прерывается поперечной желтовато-
белой полоской выше запястного сустава. Ниже запястий и скакательных
суставов темная окраска, постепенно слабеет и в общем эти части ног
становятся белесыми с примесью большего или меньшего количества
буро-коричневых волос. Борода как зимой, так и летом, в передней части
темно-бурая, почти черная (рис. 93) в задней половине грязно- или
желто-белая. Подвес на шее и груди желтовато-белого цвета, с примесью
черно-бурых волос. Хвост темно-бурый, почти черный, в основании
иногда рыжеватый.
Очень часто летняя окраска бывает более яркой рыжевато-песочной;
только голова и ноги немного темнее туловища, да затылок буроватого
или светло-коричневого тона. Потемнение на передней стороне задних
конечностей едва заметно и ноги до самых копыт имеют сходную с туло-
вищем окраску, кроме небольших размытых, буро-коричневых пятен
выше копыт. Темной полосы по бокам туловища в этих случаях нет,
а на боках по направлению книзу окраска заметно светлеет, хотя и ясно
отграничена от желтовато-белого живота.
Половой диморфизм практически отсутствует, и самки имеют такую же
окраску, как и самцы.
Окраска молодых летом однотонная, рыжевато-бурая, со светлым
животом и темной полосой вдоль хребта. Зимой она становится более
светлой, сероватой.
Череп винторогового козла (рис. 94) характеризуется, прежде всего,
гетеронимной скрученностью роговых отростков. Так же как и на роговых
чехлах, на них ясно видны передний и задний кили (ребра), внутренняя
почти плоская и более выпуклая наружная грани. У самцов основания
роговых отростков расположены частично над глазицами, у самок они цели-
ком сдвинуты за края последних. Лоб относительно узкий; межглазнич-
ная его ширина не больше расстояния от переднего края хоан до нижнего
Рис. 94. Череп Capra falconeri Wagn.
448
BOVIDAE
края затылочного отверстия.1 Поверхность лба и у самцов, и у самок равно-
мерно выпуклая в средней части, но без предрогового вздутия и идущих
к нему валиков от оснований роговых отростков. Свойственная черепам
самцов сибирского козерога шероховатость перед основаниями роговых
отростков у винторогого козла отсутствует, и поверхность соответствую-
щих участков лобных костей гладкая. Носовые кости умеренной ширины
и длины; их наибольшая ширина составляет от 26 до 33% собственной
длины. Расстояние от переднего края носовых костей до конца межчелюст-
ных (prosthion) не меньше или немного меньше расстояния от prosthion
до переднего края Р2 (разница не превышает длину этого зуба). Передний
край хоан расположен немного впереди уровня боковых выемок заднего
края костного нёба.
Характерной чертой мархура, в отличие от всех остальных видов
козлов, является укороченность теменной кости. Длина ее по средней линии
всегда меньше расстояния между теменными гребешками. Характерен
также вогнутый характер верхнего профиля зароговой части черепа на
границе лобных и теменных костей.
Переднекоренные зубы (премоляры) сравнительно мало редуцированы;
длина их ряда в верхней челюсти составляет более одной трети всего
соответствующего зубного ряда (Р2—М3).
История и географическое распространение (рис. 95). История и
происхождение группы винторогих козлов не известны.
В настоящее время ареал С. falconeri охватывает южные районы Сред-
ней Азии (Туркменской, Узбекской и Таджикской ССР), Афганистан,
Белуджистан, Балтистан, Кашмир, северную часть Пенджаба, западные
Гималаи. На восточные склоны Гималаев, по В. И. Цалкину (1945а),
этот вид не проникает. Восточной границей распространения является
верхнее течение Инда к востоку до Ронду (Harper, 1945). На юге водится
еще в Сулеймановых горах и на хребте Чиалтан в Белуджистане около
Кветты. Область распространения винторогого козла в Советском Союзе
представляет крайнюю северную часть его ареала.
Имеется три очага обитания мархура в пределах Средней Азии. Пер-
вый из них включает западные и юго-западные отроги Гиссарского хребта.
Здесь винторогие козлы держатся, по-видимому, еще в сравнительно
значительном числе, на хребте Кугитангтау, образующем естественную
границу на юге между Туркменской и Узбекской ССР. Группами по
нескольку десятков голов встречаются они до сих пор на восточном склоне
Кугитангтау (Железняков, 1952), а на севере в горах Танги-Дувал (хребты
Зиралик и Куюк; Султанов, 1939). На западной, туркменской стороне
хребта Кугитангтау мархуры держатся к северо-востоку от Карпюка и
Куйтана (Гуреев, 1937).
Правый рог винторогого козла в колекциях ЗИН АН СССР с обозна-
чением в качестве места нахождения Ширабада, вероятно, относится
к экземпляру, добытому в Кутитангтау.
Имеются сведения о существовании винторогих козлов в хребте Бай-
сун-Таге (Громова, 1949), но Г. Султанов (1953) существование здесь
этого вида севернее Кугитангтау, в том числе и в Байсунском районе,
на основании личных исследований, отрицает. Интересно отметить,
что остатков винторогого козла не обнаружено в мустьерской стоянке
неандертальцев в гроте Тешик-Таш, тогда как кости сибирского козе-
рога (Capra sibirica Meyer), ныне в этом районе отсутствующего, составляют
1 Этот признак мог быть проверен только на трех экземплярах.
CAPRA
449
основную массу костного материала (Громова, 1949). В. И. Громова
(1949) предполагает, что мархур проник в южные отроги Гиссарского
хребта после того, как стоянка в Тешик-Таше была покинута человеком.
Далее, имеются сообщения о встречах с козлами в районе Мачая Сур-
хан-Дарьинской области, а в последние годы окончательно подтверждено
наличие их в хребте Бабатаг, на границе Узбекистана и Таджикистана.
Здесь они держатся главным образом на западных склонах южной поло-
вины хребта; в качестве мест обитания в Бабатаге указываются родники
Кашка-Булак, Большая и Малая Турапга, Паталлы, Биес-Симас, Кара-
Рис. 95. Распространение
Capra falconeri Wagn.
Джумалак, Ит-Бутак, а также южная вершина хребта Беш-Арча (Богда-
нов, 1953).
Третий, более или менее крупный очаг распространения винторогого
козла в СССР представляет восточная часть Кулябской и южная часть
Гармской областей в Таджикистане. Здесь он встречается и, по-видимому,
не так редок, в горах между рр. Яхсу и Пянджем, приблизительно от
линии Куляб—Чубек на юго-западе и до Дарвазского хребта включи-
тельно на северо-востоке. В качестве местообитания в этой части ареала
перечисляются: Дарваз, Даштиджумский район Кулябской области,
горы Хазрат-Имам Аскари, километрах в 20 к востоку и километрах
в 50 к югу от Куляба, хребет Хозретиши и продолжение его к югу —
Кишваристан-Тау, ущелье Дагарион около бывшего пограничного поста
Хирманджая, горы Табакчи и Терекли. Указания П. С. Трубецкого
(1910) о том, что в небольшом числе мархуры встречаются около Бельджуана
(в западной части Кулябской области), подтверждается экземплярами
из гор Сарсаряк, добытыми Регелем в 80-х годах прошлого века. Об
этом же сообщает, со слов местных жителей, К. К. Флеров и И. М. Гро-
мов (1935). Однако при посещении мною этого района в 1951 г. слышать
о существовании здесь мархуров мне не приходилось. По словам П. С. Тру-
бецкого, рога их попадаются нередко на гробницах (мазарах) бывшего
Каратегинского бекства. Водятся ли винторогие козлы здесь в настоящее
время, не известно. Есть непроверенные указания Д. Н. Кашкарова
(цит. по Цадкину, 1945а) об обитании винторогих козлов в районах
29 Фауна СССР, И. И. Соколов
45 О
BOVIDAE
Гарма, никем не подтвержденные позднее. На Памире винторогого козла
нет.
Таким образом, западная часть ареала винторогого козла в СССР
изолирована от восточной, так же как и от очага обитания его в горах
Бабатаг.
Подвиды. Внутривидовая систематика винторогих козлов до сих пор
строится, в основном, на форме рогов без учета данных краниологии
и других морфологических признаков, вследствие трудностей добычи
и бедности материала в музеях. Поэтому правомочность выделения не-
которых подвидов вызывает серьезные возражения.
До сих пор описано семь подвидов С. falconeri.
1) С. falconeri falconeri Wagner. Рога очень широко расходятся в сто-
роны и описывают пологую спираль не более чем в полтора оборота.
Распространение: Астор, Балтистан, долина верхнего Инда.
2) С. falconeri cashmiriensis Lydekker. Рога образуют менее открытую
спираль, чем у предыдущей расы, и у крупных экземпляров делают два
полные оборота. Распространение: Кашмир, хребет Пар-Панд-
жал, возможно Гильгит.
3) С. falconeri megaceros Hutton. Отличается от предыдущих подвидов
еще более сильным скручиванием рогов, имеющих уже почти прямую,
но еще не винтообразную форму. Распространение: горы
северного Афганистана, Кандагар до Белуджистана.
4) С. falconeri jerdoni Hume. Рога конические, прямые, типично вин-
тообразные, с килями, описывающими вогруг оси до трех полных оборотов.
Общие размеры несколько меньше, чем у других подвидов, и более ко-
роткая борода. Распр остранение: Сулеймановы горы, заинд-
ские области Пенджаба.
5) С. falconeri chialtanensis Lydekker. Сходен с типичной расой
(С. falconeri falconeri Wagner), но отличается от нее более сближенными
концами рогов. Распространение: горы Чиалтан около Кветты
в Белуджистане.
6) С. falconeri heptneri Zalkin. Тяжелые массивные рога с крутыми
и относительно узкими штопорообразными завитками спирали, коли-
чество которых может достигать у взрослых экземпляров более двух.
Распространение: горы восточного Таджикистана, тяготею-
щие к правому берегу Пянджа.
7) С. falconeri ognevi Zalkin. От предшествующего подвида отличается
более широкими и пологими завитками спирали, количество которых
достигает двух. Распространение: хребет Кугитангтау, на
границе Туркмении и Узбекистана.
Биология. Биологические особенности винторогого козла изучены
недостаточно, а об образе жизни этого зверя в пределах Советского Союза
более или менее конкретные данные получены только в последние годы
(Султанов, 1953; Железняков, 1952). Подобно другим представителям
рода Capra L., мархур — житель скалистых гор. Пределы вертикального
его распространения — от нижней полосы арчевой зоны (около 1500 м
над ур. м.) до высот 3000 м и более. Однако винторогие козлы счита-
ются теплолюбивыми животными, и по сравнению, например, с сибир-
ским козерогом (С. sibrica Meyer) держатся, в общем, в горах заметно ниже.
П. С. Трубецкой (1910) охотился на мархуров ниже верхних пределов
культурной полосы гор, тогда как сибирских козлов на таких высотах
никогда не бывает. Известны даже случаи добычи и поимки мархуров
в населенных пунктах (Lydekker, 1898; Султанов, 1939).
CAPRA
451
В выборе предпочитаемых мест обитания отдельные расы обнаружи-
вают некоторые отличия, например: асторский подвид (С. falconeri fal-
coneri Wagn.) придерживается, в основном, более открытых безлесных
участков гор, скрываясь в лесных рощах лишь в жаркие дни, в целях
защиты от кровососущих насекомых; пар-панджалская же раса (С. /. са-
shmiriensis Lyd.), наоборот, является жителем скал, окруженных глу-
хим лесом, из которого звери отваживаются выходить на кормежку
только поздно вечером и рано утром.
В пределах Советского Союза мархуры большую часть года проводят
в лесной полосе и наиболее часто встречаются на высотах 1500—2500 м.
Однако густых арчевых зарослей в Кугитангтау, например, они избе-
гают и укрытия на случай опастности ищут не в арчевниках, а в скалах.
«Жизнь винторогого козла теснейшим образом связана со скалами, со-
ставляющими необходимое условие его существования. Козлы могут
временно покинуть скалы, но когда угражает опасность, быстро взби-
раются по скалам и спасаются в них от преследования. Большую часть
жизни мархуры проводят в скалах, в которых находят и укрытие от
врагов и пищу, и отдыхают. Здесь же проходит у них окот. Характерная
стация мархура — глубокие скалистые ущелья в поясе арчевого редко-
лесья, на склонах которых растут кустарники, встречаются отдельные
площадки с хорошо выраженным травяным покровом» (Султанов, 1953).
Значительных вертикальных и горизонтальных миграций винторогие
козлы не предпринимают, но имеет место ясно выраженная посезонная
смена мест обитания. Летом основная масса козлов живет в верхнем
поясе арчевой зоны, где на альпийских лужайках между скал звери
в изобилии находят достаточное количество сочного зеленого корма.
В Таджикистане самцы мархуров поднимаются в горы еще выше и дер-
жатся летом в альпийской зоне.
В Кугитангтау поздней осенью, когда высоко в горах выпадает глу-
бокий снег, винторогие козлы откочевывают на юг хребта или спускаются
в среднюю полосу арчевой зоны; здесь нет снега или он мелок, а кроме
того значительно теплее. Весной, когда в предгорьях начинается раз-
витие эфемеровой растительности, козлы спускаются к нижней границе
арчевого леса, где держатся до конца июня (Железняков, 1952). Обратное
перемещение в горы летом происходит по мере выгорания растительности
в нижних зонах. В горах Таджикистана мархуры на зиму спускаются
в скалы по р. Пянджу (Трубецкой, 1910).
Большую часть года винторогие козлы живут небольшими группами
в 3—5 голов. Взрослые самцы чаще держатся отдельно, тогда как группы
самок включают обычно приплод не только текущего, но и предшествую-
щего года, а нередко и молодых неполовозрелых самцов. Впрочем, не
составляют редкости встречи вместе с самками, даже и летом, взрослых
самцов. Живущие одиночками звери наблюдаются как среди самцов,
так и молодых самок. В период гона и зимой мархуры образуют более
крупные и смешанные стада, насчитывающие по нескольку десятков
голов. Максимальное по размерам стадо, которое пришлось наблюдать-
Г. Султанову (1953), состояло из 27 голов, а Д. Ф. Железнякову (1952):
сообщали о встрече в одном табуне около 50 животных. Более крупные
стада наблюдаются .чаще среди самок. Самцы группами более чем 10 го-
лов, по-видимому, не встречаются вовсе. В упомянутом выше табуне
из 27 голов, самцов было только 5 экземпляров.
Имеются ли в крупных стадах мархуров вожаки и сторожа, не изве-
стно.
29*
452
BOVIDAE
Летом суточный цикл жизнедеятельности мархуров, по имеющимся
данным, складывается так: утром с рассвета и часов до 10 утра козлы
пасутся. После этого, когда становится жарко, они уходят в теневые
места скал или в кустарник, где отдыхают и пережевывают жвачку.
Под вечер или на закате идут на водопой, а после этого на пастьбу. Па-
сутся почти всю ночь. На рассвете идут на водопой вторично и вновь
принимаются за пастьбу. В жаркие дни иногда пьют и в середине дня.
В Кугитангтау, где мест, пригодных для водопоя, не много, звери порою
вынуждены с этой целью проходить значительные расстояния. Зимой
мархуры используют для кормежки светлые часы суток, ночью же и
в непогоду скрываются в углублениях и под навесом скал. На лежки
устраиваются по нескольку особей вместе, особенно зимой. Перед тем
как лечь, мархур иногда предварительно очищает копытами место лежки
от крупных камней.
Не известно, насколько привержены винторогие козлы к местам
обитания и легко ли их меняют. Известно только, что места водопоев
постоянны и мархуры придерживаются их более или менее стойко.
Г. Султанов (1953) приводит список кормовых растений винторогого
козла в Кугитангтау, состоящий из 16 видов и, конечно, не охватывающий
всего разнообразия поедаемых растений. Основу летнего питания мархура
составляют разнообразные травы. Основными и, по-видимому, наиболее
излюбленными кормами являются зизифора (Ziziphora clinopodioides
Lam.), затем мятлик (Роа bulbosa L.) и пустынная осока (Carex pachy-
stylis Gay.), находимые постоянно в желудках убитых животных. Охотно
поедаются также прангос (Prangos pabularia Lindl.) и ревень (Rheum
maximowiczii A. Los.); молодые побеги последнего употребляются весной,
а засохшие листья служат зимним кормом.
Как показывают наблюдения (Трубецкой, 1910; Султанов, 1953), на-
равне с травой, мархуры летом не отказываются и от древесно-веточного
корма. Так, они охотно едят ветки с листьями жимолости и других кустар-
ников. Г. Султанов отмечал поедание мархурами молодых побегов на
богарных посевах пшеницы.
Зимой кормом служит большинство усохших травянистых растений,
которыми звери питались и летом. Однако основу питания в это время,
по-видимому, составляют различные виды полыни (Artemisia) и хвоя
арчи (Juniperus seravschanica Кот.). Хвоя и побеги арчи, даже от молодых
деревьев, являются, по-видимому, вынужденным кормом, поедаемым
лишь при недостатке другой пищи. Во всяком случае, пойманный только
что самец мархура из предложенной ему в неволе разнообразной пйщи
в первую очередь поедает травы, вплоть до верблюжьей колючки, и только
затем, с видимой неохотой, принимается жевать ветки арчи (Железня-
ков, 1952).
Воду винторогие козлы пьют не только из горных ручьев и источни-
ков, но и из хаков — чашеобразных углублений, где с весны скаплива-
ется вода от дождей и тающего снега. Зимой мархуры утоляют жажду
поеданием снега.
В отношении минерального питания винторогих козлов, в частности
об отношении их к естественным и искусственным солонцам, ничего не
известно.
В спокойной обстановке мархуры — флегматичные животные. На
пастбище и при переходах двигаются шагом. На ровном месте, особенно по
более или менее мягкому грунту, быстро и долго бежать не могут. Известен
случай (Султанов, 1953), когда козел, преследуемый собаками, едва
CAPRA
453
пробежав полкилометра, упал на землю обессиленный с высунутым языком.
Зато превосходно, как и другие козлы, они прыгают и карабкаются по
самым крутым скалам, непринужденно бегут по узким карнизам.
Г. Султанов (1939) приводит случай, когда две попавшие в тупик самки
мархура быстро выбрались по отвесным склонам, чего ни одно другое
животное из местной фауны, по его мнению, не в состоянии было бы
сделать. И в спокойной обстановке, и потревоженное стадо мархуров бежит
гуськом.
Винторогий козел, по-видимому, легче, чем другие его собратья,
мирится с близостью человека и вообще менее осторожен. Об этом свиде-
тельствует то, что в Кугитангтау, например, летом он держится в местах
по соседству и, даже вероятно, на одних выпасах с домашними стадами,
посещает, как уже отмечалось, богарные посевы. Для характеристики
отношения винторогого козла к людям интересен случай, о котором
сообщал О. П. Богданов (1953). Однажды в Бабатаге он встретил нахо-
дившихся почти рядом самку уриала (Ovis orientalis bochariensis Nas.)
и самку мархура. И при той, и при другой находились детеныши. «На наше
неожиданное появление, — пишет О. П. Богданов, —животные реагировали
по-разному. Самка уриала с детенышем и вслед за ней два молодых вин-
торогих козла бросились от нас вверх по склону. Самка же винторогого
козла, оставаясь на месте и смотря в нашу сторону, подпустила людей
на расстояние, примерно, в сто метров».
Быть может, эта особенность винторогого козла являлась залогом
того, что он явился одним из вероятных родоначальников домашней козы —
Capra hircus L.
Винторогие козлы — молчаливые звери. Только убегая, издают хрип-
лый носовой звук, который одновременно служит и сигналом тревоги.
Молодые кричат голосом, сходным с таковым домашнего козленка.
Смена волосяного покрова у винторогих козлов происходит один раз
в году — весною, в апреле и мае. Сначала от кожи отделяется пух, неко-
торое время задерживающийся между волосами ости. Ость выпадает
позднее. Линька начинается с головы и нижней части туловища. Дольше
всего старая шерсть удерживается на спине.
Половая зрелость у мархуров, по данным Г. Султанова (1953), насту-
пает на третьем году жизни, но молодые самцы, по-видимому, вследствие
соперничества с более взрослыми, начинают участвовать в размножении
позднее. Гон и спаривание протекает в период с середины ноября до
конца декабря. В этот период винторогие козлы, как уже отмечалось,
образуют более крупные, чем в обычное время, смешанные стада. Каждый
самец стремится отбить себе группу в несколько самок. Между ними
происходят довольно ожесточенные драки. Увидев друг друга, соперники
начинают рыть копытами землю, постепенно сближаются, и с разбега
в несколько шагов с силой ударяются основаниями рогов и лбами.
Вероятно, в целях усиления удара, перед нанесением его, звери припод-
нимаются на задние ноги и ударяют лбом немного сверху. Удары рогов
в период гона слышны на далекое расстояние. Они повторяются много
раз подряд, после чего, если один из соперников не отступил, звери просто
начинают толкать друг друга лбами. Достоверных случаев нанесения во
время драк серьезных повреждений не известно, но, вероятно, имеют
место случаи поломки рогов или сталкивания одного самца другим в про-
пасть. Такой самец с потерянным еще на воле одним рогом жил в Москов-
ском зоопарке (Шапошников, 1939). Побежденные соперники и молодые
самцы ходят на почтительном расстоянии позади стада и иногда, при
454
BOVIDAE
случае, кроют самок. Самцы в период гона сильно возбуждены, много
двигаются, утомляются в драках и сильно худеют. В это время они сами,
и особенно их моча, издают сильный специфический запах, по которому
опытные охотники узнают, не видя, о близком присутствии зверя. Самки
во время течки ведут себя спокойно и упитанность не теряют.
Беременность у самок мархуров длится около 5—5% месяцев. Окот
начинается в конце апреля и тянется до начала июня. Наибольшее число
окотов приходится на май. За несколько дней перед родами самка уда-
ляется из своего табуна и уходит для оката в самые глухие и трудно до-
ступные места. Взрослые самки приносят обычно двух козлят. По одному
чаще бывает у молодых (первоокоток). Случаев троен до сих пор не из-
вестно. Родятся козлята довольно беспомощными, но уже на второй день
могут следовать за матерью.
Молоком детеныши пользуются до начала следующей течки, но с ма-
терью держатся иногда до наступления половой зрелости.
Рога начинают расти вскоре после рождения и растут довольно быстро.
К годовалому возрасту нормально развивающиеся самцы имеют рога
с одним полным оборотом спирали (Шапошников, 1939).
Продолжительность жизни винторогих козлов в природных условиях
редко превышает 8—10 лет.
Естественных врагов у мархура, обитающего на территории СССР,
не много. Волк в Кугитангтау, например, очень редок. Орел-ягнятник,
помимо того, что также малочислен, нападает иногда только на молодых
козлят. Ни волки, ни хищные птицы существенной роли в динамике
численности винторогих козлов не играют.
Ничего не известно и о массовых заболеваниях среди мархуров. Па-
разитофауна совершенно не изучена. Г. Султанов (1953) под кожей до-
бытых им в ноябре мархуров находил личинок подкожного овода разме-
ром до 0.5 см. Видовая принадлежность последнего не известна. Большая
степень поражения оводами наблюдалась у молодых зверей и доходила
в отдельных случаях до 30—35 личинок на одном экземпляре. Выпадение
оводовых личинок из-под кожи происходит весной, так как в июне они
у убитых зверей уже не находились. На одной молодой самке Г. Султановым
были обнаружены иксодовые клещи.
Хозяйственное значение. В виду крайней ограниченности ареала и
незначительной численности винторогого козла охота на' него в СССР
с 1935-1936 гг. категорически воспрещена. Поэтому в настоящее время
этот вид в нашей фауне ценен, прежде всего, как редкое животное.
При увеличении его численности и расширении ареала, что при про-
ведении соответствующих мероприятий, по-видимому, вполне возможно,
винторогий козел может иметь и несомненное промысловое значение.
Мясо его по качеству, вероятно, не уступает мясу других диких козлов.
Правда, в период гона у самцов оно очень жестко, совершенно не содер-
жит жира, а кроме того, благодаря специфическому запаху, вообще
в пищу не пригодно.
Охотились на мархуров раньше скрадом или устраивали загоны.
Изредка, особенно в глубокоснежные зимы, травили собаками.
Интересен вииторогий козел и как один из вероятных родоначальников
домашней козы. Правда, с достоверностью участие мархура в ее проис-
хождении не установлено, так как при скрещивании его с домашними
козами получающиеся вполне плодовитые гибриды имеют всегда форму
рогов, свойственную мархуру (Antonins, 1951). Поэтому фалькоиероидный
изгиб рогов у некоторых пород домашних коз свидетельствует, возможно,
CAPRA
455
не о происхождении, а лишь о прилитии крови винторогого козла или
испанского тура. В Германии в свое время были получены плодовитые
гибриды между винторогими и безоаровыми козлами (Hilzheimer, 1933).
В Лондонском зоосаду в 1864 г. родился гибрид от скрещивания винторо-
гого козла и нубийского козерога {Capra nubiona F. Cuv.; Gray, 1954).
Capra (Turns) caucasica Giildenstaedt—Кавказский каменный козел,
или тур (рис. 96—100)
Giildenstaedt, 1779, Acta Acad. Sci. Petropol., Ill : 273—281 (первооп.);
В 1 у t h E., 1840, Proc. Zool. Soc. London : 62—69 (сист.); Реутт H., 1841, Русск.
вести., 6 : 724—727 (опис., биол.); Rouiller С., 1841, Bull. Soc. Imp. Natur.,
Moscou, XIV : 910—923 (сист.); Динник H. Я., 1879, Природа и охота, 1 : 1—12
(ох.); Динник Н. Я., 1880, Природа и охота, 6 : 1—22 (ох.); Динник Н. Я.,
1882, Тр. СПб. общ. естествоисп., XIII : 73—91 (общ., изобр.); В ii chner Е.,
1887, Mem. Acad. Sci. St.-Petersb., sir. VII, t. XXXV, 8 : 1—27, 1 Taf. (сист., морф,,
изобр.); Динник Н. Я., 1887, Природа и охота, 3 : 1—20 (ox.); Menzbier М.,
1887, Proc. Zool. Soc. London : 618—620 (сист.); Шильдер В., 1892, Боржомский
звериный отвод СПб. : 1—24 (акклим.); Lydekker R., 1898, Wild oxen, sheep a.
goats of all lands, London : 242—253 (общ., синон., изобр.); R a d d e G., 1899, Museum
caucasieum, I, Tiflis ; 75—77, Taf. XIII (сист., распр., изобр.); Matschie P.,
1931, Sitzber. Ges. Naturforsch. Freund., Berlin : 27—33 (сист.); P а з e в и г В. A.,
19Э4, Псовая и ру.-к. охота, 4 : 25—38, 5 : 82—95, 6 : 38—54 (общ., изобр.); Т у р-
к и н Н. В. и К. А. С а т у н и н, 1994, Звери России, Bovidae, М. : 537—544+607—
633 (общ., сини!., изобр.); Ендржеевский А., 1905, Псовая и руж. охота,
2 : 53—67 (биол., ох., изобр.); С ат у н ин К. А., 1905, Изв. Кавказск. муз., II,
1 : 63—67 (сист.); S a t u n i п К., 1995, Zool. Anzeig., XXIX, 11 : 336—344 (сист.);
Динник Н. Я., 1999, Матер, к познан, фауны и флоры Российской имп., IX : 1 —
47 (общ., изобр.); Сатунин К. А., 1999, Природа и охота, VIII, Иовейш. допол-
нения : 1—16 (сист.); Динник Н. Я., 1910, Звери Кавказа, I, Тифлис : 168—205
(общ., изобр.); Lydekker R., 1913, Catal. Ungulate mammals, British. Mus., I,
London : 131 —138 (сист., синон., изобр.); Rowland Ward, 1914, Records of big
Game, London : 381—384 (рога); Пфиценмайер E. В., 1915, Изв. Кавказск.
музея, VIII : 251—266 (гибр.); Brehms Tierleben, 1925, 4. Aufl., Leipzig, 13:
273—274 (морф., изобр.); Flower S., 1931, Proc. Zool. Soc. London : 210 (продолж.
жизни, гибр.); Шерешевский Э. И., 1931, Сборн. работпо изуч. фауны Крымск,
гос. залэвод!., М. : 63—74 (акклим.); Hilzheimer М., 1933, Arch. f. Tierer-
nahrung u. Tierzucht, 8 : 323—371 (гибр.); Верещагин H. К., 1938, Дагестан-
ский тур, Баку, изд. Азербайдж. фил. Акад, наук СССР : 1—69 (общ., изобр.); Н а-
с и м о в и ч А. А., 1938, Тр. Кавказск. гос. заповеди., 1 : 130—133 (пит.); Н а с и м о-
в и ч А. А., 1939, В >пр. экол. и биоценол., 7 : 59—68 (распр., биол.); Громова В. И.,
1940, Прэбл. прэисх., эволюции и породообразов. дом. животн., 1 : 63—120 (морф.);
Насимович А. А., 1940, Научн.-метод, зап. Главн. управл. по заповеди., 7 :
23—28 (учет); Гептнер В. Г. и А. Н. Формозов, 1941, Сборн. тр. Гос. зоол.
муз. Моск. гос. унив., VI : 65—70 (распр., биол.); Верещагин Н. К., 1947,
Охотн. и промысл, животные Кавказа, Баку, изд. АН Азербайдж. ССР : 45—47 (распр.,
гибр., хоз. знач.); Г р у н и н К. Я., 1948, Докл. Акад, наук СССР, LXI, 6 : 1125—
1127 (параз.); Джанашвили А. Г., 1948, Тр. Тбилисок. зоопарка, 1 : 46—49
(одом., гибр.); Насимович А. А., 1949, Тр. Кавказск. гос. заповеди., III : 5—
38 (распр., биол.); Джанашвили А. Г., 1950, Тур, Тбилиси, Гос. изд. : 1—32
(обц.); Е 1 1 о г m a n J. and Т. Morrison-Scott, 1951, Checklist of palaearctic
and Indian mammals, London : 407 (сист., синон.); Верещагин H. К., 1952,
Млекопитающие Кавказа, Диссертация, Ленинград : 576—985 (распр.); Т у р о в С. С.,
1952, Очерки охотника-натуралиста, М., изд. Моск. общ. испыт. прир. : 114—130
(ох.); Эквтимишвили 3. С., 1952, Сообщ. Акад, наук Грузинок. ССР, 8 : 477 —
484 (стац.); Саркисов А. А., 1953, Природа, 2 : 113 (гибр.); Эквтимиш-
вили 3. С., 1953а, Сообщ. Акад, наук Грузинок. ССР, XIV, 3 : 163—166 (стац.);
Эквтимишвили 3. С., 19535, Сообщ. Акад, наук Грузинок. ССР, XIV, 5 :
303—303 (стада.); Эквтимишвили 3. С., 1953в, Тр. Инет. зоол. Акад, наук
Грузинок. ССР, XI : 22—23 (пит.); Асадов С. М., 1954, Докл. Акад, наук Азер-
байд:к. ССР, X, 9 : 643—647 (параз.); Эквтимишвили 3. С., 1954, Тр. Инет,
зоол. Акад, наук Грузинок. ССР, XIII : 104—105 (размн.); Насимович А. А.,
1955, Роль режима снежного покрова в жизни копытных, М., изд. Акад, наук СССР :
125—132 (распр., биол.); Цалкин В. И., 1955, Бюлл. Моск. общ. испыт. прир.,
отд. биол., LX, 4 : 17—33 (морф., сист., распр., изобр.).
456
BOVIDAE
Описание. Козел средней или крупной величины. Высота в холке
у взрослых самцов составляет 79—99 см, но в отдельных случаях пре-
восходит 100 см. Длина тела около 130—155 см, в исключительных
случаях до 165 см.1 Основная длина черепа от 228 до 287 мм. Живой вес
от 64 до 96 кг. Самки заметно мельче самцов; основная длина черепа
у них колеблется в пределах 205—234 мм, а живой вес даже у семилет-
них экземпляров редко превосходит 60 кг (Верещагин, 1938).
Телосложение крепкое; короткое массивное туловище покоится на
сравнительно коротких и толстых ногах. Голова в сравнении с общим
сложением животного небольшая, не более 41 см, иногда горбоносая.
Уши длиной 10—14 см с заостренными верхушками. Высота в крестце
несколько (иногда на 10 см) превосходит высоту в холке. Хвост
длиной от 10 до 20 см. Копыта большие; длина их у взрослых сам-
цов на передних конечностях составляет 80—92 мм, на задних 70—76 мм;
высота копыт и на передних, и на задних ногах около 34—54 мм.
Характерные для безоарового и винторогого козлов роговые мозоли
на тыльной стороне запястных суставов у кавказского тура, за очень
редкими исключениями, отсутствуют; кожа здесь, хотя и утолщена, но,
как правило, сплошь покрыта волосами.
Рога самцов характеризуются наличием на их поверхности трех ребер
(килей): передне-внутреннего, передне-наружного и заднего; но все кили
сильно закруглены. Передняя (лобовая) грань развита, очень широкая,
слабо выпуклая, но, как и другие две, на углах закруглена. В противо-
положность другим видам Capra, сильно выпукла внутренняя (вернее,
задне-внутренняя) грань рога; на ее поверхности часто развивается до-
полнительное закругленное ребро. Если последнее располагается ближе
к передне-внутреннему килю, то последний оттесняется на поверхность
очень широкой лобовой грани и сохраняется в виде продольного слабого
гребешка, обозначенного более заметно выступающими на нем буграми
и зигзагообразным изгибом поперечной морщинистости рога. В этом
случае поперечное сечение в основании приобретает округленно-треуголь-
ную, иногда почти круглую форму. Если продольная выпуклость внутрен-
ней грани располагается ближе к ее заднему краю, то передне-внутреннее
ребро занимает обычное место, а поперечное сечение рога в основании
имеет неправильно-четырехугольную форму. Верхушки рогов всегда сжаты
с боков и уплощены.
Поверхность роговых чехлов несет хорошо выраженную поперечную
морщинистость с глубокими кольцевидными бороздами на границах участ-
ков годичного прироста рога. Поперечные валики на лобовой грани рога
имеются, но развиты в разной степени. У зверей из западной части ареала
они очень высокие и крутые, резко обособленные от остальной поверх-
ности рога. В направлении к востоку степень развития их постепенно
уменьшается и у туров из восточной части Главного Кавказского хребта
поперечные валики низкие, пологие, слабо дифференцированные среди
общей морщинистости рога, а иногда почти и вовсе не заметны. Степень
развития валиков связана также с возрастом; на сегментах прироста,
начиная с 3—4 года, они наиболее высоки и отчетливы, тогда как на
участках рога, образовавшегося после 7—8 лет, валики выражены обычно
значительно слабее. Число поперечных валиков на участке годичного
прироста рога колеблется от одного до трех, но чаще бывает два.
1 Добытый во время нашей поездки в Кавказский заповедник в 1958 г. взрос-
лый самец тура Северцова имел высоту в холке 109 см, длину тела — 156 см и
живой вес 148.4 кг.
CAPRA
451
Характер изгиба рогов у кавказских каменных козлов разный, причем
и в этом отношении наблюдается ясно выраженная географическая зако-
номерность. Общей особенностью тура, отличающей его от других видов
рода Capra L., является то, что рога, отходя от черепа, изгибаются не
вверх и назад, как у безоарового козла и сибирского козерога, а больше
вверх и в стороны. Бросается в глаза также заметно больший раскид
рогов; угол их расхождения1 у взрослых самцов не бывает менее 60°,
Рис. 96. Западнокавказский тур, или тур Северцова [Capra caucasica severtzowi
Menzb.).
в исключительных случаях становится почти прямым (80—83°). У туров
западного Кавказа рога, изгибаясь в стороны и назад, описывают дугу
почти в одной плоскости, вершинами направлены вниз и одновременно
слегка наружу или, наоборот, внутрь. Рога туров из восточной части
ареала делают очень пологий, но ясно выраженный гетеронимный изгиб;
отходя от черепа, они направляются в стороны и вверх, затем загибаются
назад и вниз, а вершины их обращены вверх и внутрь навстречу друг
ДРУГУ (рис. 97). Между этими двумя крайними типами изгиба существует
ряд переходов, особенно у зверей центральной части Большого Кавказа.
Основаниями рога сильно сближены; расстояние между внутренними
краями роговых чехлов обычно не превышает 1—2 см.
Длина рогов у взрослых самцов варьирует в широких пределах, от
48 см до 97 см, а по Р. Варду (Rowland Ward, 1914) — даже до 46 дюймов
(117.8 см). Обхват в основании колеблется от 26 до 30 см, в отдельных
случаях достигает 33—35 см.
1 Измеряется по передне-внутреннему килю на протяжении 10 см от оснований.
458
BOVIDAE
У самок рога гораздо короче и тоньше, чем у самцов, шире (на 3—4 см)
расставлены в основаниях. Длина их не более 22—25 см и только в исклЮ'
чительных случаях достигает 30 см. Обхват рогов в основаниях у самок
около И—13 см. Поперечные валики на передней стороне не развиты,
но общая поперечная морщинистость рогов у них выражена даже сильнее.
Степень расхождения рогов в стороны у самок слабее, но все же менее
45—50° не бывает и, наравне с более округлым их сечением в основаниях,
хорошо отличает самок кавказских туров от других видов Capra. Цвет
рогов и копыт темно-бурый, почти черный. Радужная оболочка глаз
темно-бурая.
Вполне развитый волосяной покров состоит из довольно мягкой, хотя
и ломкой ости и тонкого извитого подшерстка. Длина остевых волос
на туловище в зимнем меху составляет около 4—6 см. Удлинены (до 10—
11 см) и более грубы волосы у самцов на верхней стороне шеи. Более
длинные, чем на соседних частях головы, и упругие волосы образуют
иногда вихры на лбу и щеках, по бокам носа. Короткие, но очень грубые,
почти щетинистые волосы растут на нижней стороне груди. На хвосте
длина волос около 7—8 см. Борода у самцов из западной части ареала
хорошо развита, длиной около 17—48 см, но не всегда заострена и часто
снизу как бы подстрижена. У зверей восточных популяций борода широ-
кая, заходящая на щеки, но короткая, состоит из изогнутых жестких
волос длиной 8—10 см. У самок борода в виде небольшого тонкого пучка
длинных волос имеется, но не всегда.
Общая окраска туловища и шеи в зимнем меху варьирует, но пре-
обладают желто-серые и коричнево-бурые тона. Резко выраженная зо-
нальность в расцветке отдельных волос у туров отсутствует. Остевые
волосы от основания обычно светлые, выше постепенно буреют или жел-
теют; приостренные кончики их, пока не обломаны, всегда темно-бурые.
Волосы подшерстка светло-бурые. Верх головы, холка, плечи и круп
обычно темнее остального туловища. Стороны головы одного цвета с туло-
вищем. Конец морды более светлый, бурый или коричневатый. Подбородок
темно-бурый. Края губ грязновато-желтовато-белые. Уши снаружи одно-
тонные с туловищем, внутри грязно-белые.
Передняя сторона ног, начиная с запястных и скакательных суста-
вов и книзу темно-бурая, почти черная на самих запястьях и около копыт.
Темная, черно-бурая окраска захватывает также пространство вокруг
добавочных копыт, заднюю сторону бабок (фаланг пальцев), а на задних
ногах также пятки. Темная окраска на передней стороне ног выше за-
пястий и скакательных суставов постепенно сходит на нет и сливается
с окружающими частями.
Характерные для безоарового и винторогого козлов светлые пятна
на тыльной стороне передних ног выше запястных суставов у туров неиз-
менно отсутствуют. Задние стороны ног ниже запястий и скакательных
суставов обычно светлые, того или иного оттенка. Низ живота, пахи,
промежность, внутренние стороны ягодиц по сторонам от хвоста также
светлые.1 Хвост покрыт темно-бурыми, иногда с коричневатым оттенком
волосами. Борода бурая, часто с желтыми верхушками волос. Темная
полоса вдоль средней линии спины и шеи у кавказских туров, если имеется,
1 М. А. Мензбир (Menzbir, 1887) упоминает о светлом пятне на затылке у западно-
и центральнокавказских туров; при просмотре коллекционного материала в ЗИН
АН СССР установить наличие этого пятна не удалось. Не упоминает о нем и Н. Я. Дин-
ник (1910).
CAPRA
459
развита очень слабо, а часто вообще отсутствует. Лишь в редких случаях
можно заметить также слабые размытые темные полосы по бокам, на гра-
нице со светлой окраской пахов и живота. Зато нижняя сторона груди
часто темно-бурая, подмышки позади и внутрь от локтевых суставов
с ясно выраженным рыжеватым оттенком.
У зверей из западной части ареала окраска зимой серовато-бурая,
часто с более или менее выраженным желтоватым оттенком, иногда бывает,
наоборот, темной, почти чисто бурой. В более темный цвет, кроме перед-
ней стороны ног, окрашены темя и затылок, тогда как лоб и бока головы
имеют одноцветную и однотонную с туловищем окраску. Низ живота,
пахи, задняя сторона ног и другие светло окрашенные участки тела —
от буровато-серого цвета до грязно-белого с желтоватым оттенком.
Туры восточной половины Большого Кавказа окрашены значительно
темнее. Общая окраска их темно-бурая или каштаново-бурая. Только
пахи, внутренние части бедер и задние стороны нижних частей ног более
светлые, ржаво-бурые или белесоватые, да верхняя сторона шеи у сам-
цов желтовато-бурая, состоящая из бурых с рыжими верхушками волос.
Часто окраска бывает еще более темной и однотонной. Даже живот в этих
случаях не имеет посветления, а ноги ниже запястных и скакательных
суставов сплошь темно-бурые без светлой полосы на задней стороне.
К весне, вследствие обламывания темно окрашенных верхушек и
общего выцветания волос, окраска сильно светлеет, становится белесой.
Иногда выцветание становится столь сильным, что окраска приобретает
грязновато-желто-белый характер, за исключением небольших темных
пятен на боках и бурых полос на передней стороне ног. Особенно сильное
посветление наблюдается у старых животных.
460
BOVIDAE
Летом окраска туров рыжевато-серая или рыже-бурая разной интен-
сивности, немного светлеющая на боках. Как и зимой, верх головы темнее
туловища; конец морды и верхняя губа светлые, рыжевато-бурые. Перед-
няя сторона ног, низ груди, борода и хвост бурые. Живот, нахи, внутрен-
ние стороны бедер и ягодиц, а также задняя сторона ног ниже запястья
и заплюсны желто-белые с грязноватым оттенком. Туры восточной части
ареала и летом окрашены темнее. Потемнение с переходом от желтовато-
рыжеватых к бурым тонам начинается уже в сентябре, но полное развитие
зимний наряд получает только в декабре.
Резко выраженный половой диморфизм в окраске отсутствует; самки
обычно лишь немного светлее самцов. У молодых первые месяцы после
рождения окраска равномерно буровато-рыжая, позднее может становиться
голубовато-серой.
В черепе кавказского тура (рис. 98) характерно более широкое, по
сравнению с другими козлами, расхождение роговых отростков. Основа-
ния их у самок всегда заметно сдвинуты назад за глазницы. Характерен
также сильный излом профиля черепа между лобной и зароговой зоной;
угол схождения их, даже у самок, приближается к прямому и обычно
более острый, чем угол схождения затылочной и теменной поверхности
черепа.1 У взрослых самцов лобная и зароговая поверхности сходятся
под острым углом, и зароговой профиль черепа на границе между лобными
и теменными костями имеет ясно вогнутый характер. Лоб широкий; меж-
глазничная ширина его, за крайне редкими исключениями, превосходит
расстояние от переднего края хоан до нижнего края затылочного отверстия.
Поверхность лба у самок плоская или слегка выпуклая в средней части
перед рогами; но и в этом случае без шишкообразного вздутия и идущих
к нему валиков от оснований роговых отростков. Вдоль сагиттального
лобного шва обычно имеется легкая продольная вогнутость. У самцов
поверхность лба плоская или вогнутая. В том случае, если на передней
стороне рогов хорошо развиты поперечные валики, поверхность лба
перед основаниями роговых отростков шероховатая, отграниченная от
остальной гладкой поверхности лобных костей приподнятым краем. Носо-
вые кости умеренной длины или длинные; их наибольшая ширина со-
ставляет у самок от 25 до 34%, а у самцов от 31 до 41 % собственной длины.
Расстояние от переднего конца носовых костей до переднего конца меж-
челюстных (prosthion) меньше расстояния от prosthion до переднего края
альвеолы Р2, по крайней мере на длину этого зуба.
Переднекоренные зубы сильно редуцированы; длина их ряда в верх-
ней челюсти у вполне взрослых животных не более одной трети длины
всего ряда коренных зубов.
История и географическое распространение. Современное распро-
странение, а также, по-видимому, и вся предшествующая история кав-
казских каменных козлов связаны с Большим Кавказом. Здесь они,
вероятно, и произошли от какого-то весьма близкого к безоаровому козлу
(Capra aegagrus Erxl.), но еще более примитивного предка. Находок,
достоверно относящихся к этому виду и датируемых раньше позднего
голоцена, не известно. Весьма фрагментарные остатки, происходящие
из верхнеплейстоценовых отложений западного Закавказья (пещеры
в ущельях рр. Квирилы, Чхеримела, Риона, Куденста и Мзымты), вблизи
1 У других видов козлов углы равны или угол схождения лобной и зароговой
плоскостей, по крайней мере у самок, менее острый.
Рис. 98. Череп Capra caucasica Giild
462
BOV f DAE
границ современного ареала тура, отнесены к этому виду только про-
визорно (Верещагин, 1952).
Область современного распространения кавказского тура (рис. 99)1
простирается от 39°45' до 48°30' в. д., а в широтном отношении огра-
ничивается Главным Кавказским хребтом с его отрогами и прилегающими
непосредственно к нему параллельными цепями. В горах Малого Кав-
каза и вообще в Закавказье туры ни в настоящее время, ни в прошлом
не водились.
Крайним западным пунктом, где до последних лет держатся туры,
является гора Чугуш. В 80-х годах прошлого столетия, по Н. Я. Дин-
нику (1910), они встречались еще километров тридцать западнее, в районе
вершин Фишта и Оштена.1 2 Достоверно известны случаи добычи их также
на г. Абаго к северо-западу от Чугуша.
Одним из наиболее богатых турами мест, благодаря хорошо поставлен-
ной издавна охране, является территория Кавказского заповедника и
непосредственно прилегающие к нему районы. Здесь каменные козлы
распространены не только на Главном хребте и на его отрогах, но захо-
дят иногда и на вершины параллельного ему Скалистого хребта. В боль-
шом числе они держатся здесь в верхнем течении рр. Белой, Большой
и Малой Лабы и их притоков.
Областью обитания туров являются истоки и верхнее течение Большого
Зеленчука, Марухи, Аксаута, Теберды и Доута, но здесь их не так уж
много и на территории Тебердинского заповедника они держатся неболь-
шими группами голов по десять-пятнадцать. Во времена Н. Я. Динника,
да, вероятно, и сейчас, многочисленны туры на западных склонах Эльб-
руса.
Благодаря сравнительно незначительной высоте западной оконечности
Кавказского хребта туры свободно проникают на южные его склоны
и встречаются в верхнем течении Мзымты, Бзыби и Кодора.
Местности между Эльбрусом на западе и горой Шхара на востоке
являются областью распространения центральнокавказского тура или,
как эту расу еще называют, тура Гюльденштедта. До сих пор в довольно
значительном числе он держится здесь по хребтам меридионального
направления к северу от Эльбруса, в верхнем течении Малки, Баксана,
Западного и Восточного Череков. Имеются данные о том, что еще в конце
прошлого столетия козлы встречались в районе гор Бармамут и Кинжал,
а в начале XIX века даже на Бештау севернее Пятигорска. На южных
склонах центрального Кавказа туров очень мало в верховьях Ингура
и Мульхры (на южной стороне горы Тенгульды). Если и есть козлы,
то лишь в небольшом числе на расположенных к югу от Главного хребта
вершинах Шоде (верховья Риона) и Лайле (Сванетский хребет). Нет,
по Н. К. Верещагину, в настоящее время туров у Зекарского перевала,
хотя в конце прошлого, а быть может даже в начале нынешнего века, и
на южных склонах этой части Большого Кавказа они не были так редки
(Динник, 1910).
К востоку от истоков Балкарского Черека и Ингура идет, в основном,
область распространения восточнокавказского или дагестанского тура
(С. с. cylindricornis Blyth). Горы Дигории и Осетии были издавна богато
заселены козлами, о чем свидетельствуют огромные скопления черепов
и рогов этих животных в местных святилищах-пещерах. Обычны туры
1 Дается по Н. Я. Диннику (1910) и Н. К. Верещагину (1952).
2 А. А. Насимович (1949) это указание Н. Я. Динника подвергает сомнению.
CAPRA
463
также и в районе Казбека, в верховьях Терека, Чанты-Аргуна и Ш,аро-
Аргуна. Еще более многочисленны они в горном Дагестане, в частности
в бассейне Аварского и Андийского Койсу и в расположенном между
ними Богосском хребте. Много туров в Катехском ущелье и верховьях
Самура. Но местами, особенно богатыми этим зверем, до сих пор являются
районы гор Базар-Дюзи, Бабадаг и Шахдаг. Севернее многочисленны,
по Н. К. Верещагину, каменные козлы в верховьях Карачая и на верши-
нах, окружающих ущелье Кусарчая. В центральной части Дагестана
каменных козлов нет или они крайне редки, вероятно вследствие истреб-
ления. Так, издавна отсутствуют они в районе Ботлиха, Гуниба, как и
Рис. 99. Распространение Capra caucasica Giild.
1 — XX в. (области постоянного обитания); 2 —• XIX в.; 3 — XVIII в. и раньше.
вообще в среднем течении р. Каракойсу, хотя безоаровые козлы и серны
в некоторых глухих местах здесь еще сохранились. В Самурском и Лакском
районах, по Н. К. Верещагину, туры бывают лишь при случайных заходах.
На южном склоне восточной части Кавказского хребта мало каменных
козлов в верховьях рр. Арагвы и Алазани. Нет их вовсе в Телавском
хребте, хотя в прошлом они здесь, возможно, водились. Мало их в настоя-
щее время в пределах Лагодехского заповедника, а после неизвестной
эпизоотии, также в Беланухском и Мазымчайском ущельях Закатальского
заповедника. Число туров на территории последнего в настоящее время
Н. К. Верещагин определяет в 3500—4000 голов.Встречаются туры также
в Варташенском и Исмаиллинском районах, в верховьях Геокчая (Азер-
байджанской ССР). Много их в амфитеатрах ущелий Шинчая и Демира-
парончая. Группами в 15—20 голов встречаются они до сих пор в верховьях
Гильгинчая. В качестве крайнего восточного пункта распространения
тура в настоящее время принимается гора Айвазель, километрах в двад-
цати пяти к востоку от Бабадага. Таким образом, существенных изменений
ареала кавказского каменного козла за исторический отрезок времени,
да, по-видимому, и в течение всей его истории, не произошло. Уменьши-
лась только в ряде районов его численность, вследствие прямого истреб-
ления и вытеснения стадами домашнего скота.
464
BOVIDAE
Имеются указания на то, что в прошлом столетии туры водились
в горах к северу от Шемахи.
В конце 80-х годов прошлого столетия около десятка туров были за-
везены в Боржомский звериный отвод (охотничье хозяйство одного из
великих князей) и помещены в обширный огороженный загон (Шильдер,
1892). Обстановка для туров оказалась благоприятной; они успешно раз-
множались, скрещивались с сохранившимися там безоаровыми козлами
(Пфиценмайер, 1915). К 1917 г. число туров и их гибридов в отводе воз-
росло до 500 голов (Верещагин, 1938). Позднее они были частично выпу-
щены на волю, частью уничтожены в годы интервенции и гражданской
войны. В настоящее время здесь, по-видимому, не сохранилось от них
и следов.
Перед Первой мировой войной незначительное число кавказских ту-
ров содержалось в зверинце бывшей Крымской охоты, на территории
нынешнего Крымского заповедника, но, как и в Боржомском отводе,
в годы гражданской войны, они погибли (Шерешевский, 1931). Опыт
акклиматизации каменных козлов на территории горного Крыма следовало
бы повторить.
Подвиды. Систематика кавказских туров разработана неудовлетвори-
тельно и строится до сих пор главным образом на форме рогов. Описано
пять форм, принимаемых описавшими их авторами за самостоятельные
виды. Ареалы этих «видов» лишь частично налегают друг на друга, причем
популяции в районах соприкосновения по морфологическим особенностям
имеют явно промежуточный характер. Не замечается существенных от-
личий и в условиях местообитаний, равно как и в биологии животных
в разных частях ареала. Поэтому нельзя считать обоснованным призна-
ние за самостоятельные, реально существующие виды даже западно-
кавказского тура Северцова и сильно отличающегося от него дагестан-
ского. 1
Еще менее обосновано объединение западнокавказских туров в один
вид с альпийским и сибирским козерогом (ЕПегшан and Morrison-Scott,
1951) при одновременном выделении в особый, общий с дагестанским,
вид центральнокавказского тура, представляющего явно промежуточную
форму, более близкую к турам западного Кавказа.
Учитывая то, что кавказские каменные козлы образуют практически
сплошной ареал, а также то, что ареалы отдельных форм только частично
налегают друг на друга, что все они обнаруживают общность биологии,
существенное сходство в отношениях к внешней среде, свободно скрещи-
ваются и образуют промежуточные формы, более обосновано считать всю
популяцию туров от западной до восточной оконечности Кавказского
хребта принадлежащей к одному полиморфному виду, объединяющему
три или четыре подвида.
1) С. caucasica severlzowi Menzbir — Тур Северцова, или западно-
кавказский. Рога у самцов изогнуты дугообразно или серпообразно в од-
ной плоскости (рис. 96). Отходя от черепа, они направляются вверх и
в стороны, затем назад и в стороны, а концами загнуты вниз, иногда
несколько вперед и наружу. Расстояние между концами рогов редко
бывает менее 45 см, а в отдельных случаях (Динник, 1910) доходит до
90 см. Бугры или поперечные валики на передней стороне рога выражены
1 Такую точку зрения защищает в недавно опубликованной, весьма обстоятель-
ной работе по систематике западнокавказских туров В. Н. Цалкин (1955), подходя-
щий к вопросу о видовых критериях с чисто морфологических позиций.
CAPRA
465
обычно довольно ясно. Окраска сравнительно светлая, в зимнем меху
серовато-желтовато-бурая. Борода у самцов длинная, до 17—18 см, часто
клинообразно заостренная на конце. Распространение: запад-
ная часть Кавказского хребта к востоку до Эльбруса.
2) С. caucasica cylindricornis Blyth — Дагестанский, или восточнокав-
казский тур. Рога у самцов образуют ясно выраженный гетеронимный
изгиб (рис. 97). Отходя от черепа, они направляются вверх и в стороны,
затем назад и вниз, а концами загнуты внутрь навстречу друг другу и
немного вверх. Расстояние между концами рогов у взрослых самцов
очень редко бывает более 45 см. Бугры и поперечные валики на передней
стороне рога развиты очень слабо, иногда почти вовсе не заметны. Ок-
раска более темная, чем у других подвидов, в зимнем меху темно-бурая
или каштаново-бурая. Борода у самцов широкая, но короткая, обычно
около 7—8 см или короче и, как правило, не заострена. Распростра-
нение: восточная половина Кавказского хребта к востоку от р. Че-
река, но отдельные экземпляры проникают на запад до Теберды.
3) С. caucasica caucasica Guldenstaedt — Центральнокавказский тур,
или тур Гюльденштедта. Форма во всех отношениях промежуточная
между двумя предыдущими подвидами. Рога у самцов, отходя от черепа,
направляются вверх и в стороны, затем назад и в стороны, а концами
загибаются навстречу друг другу, но не так как у С. cylindricornis. Рас-
стояние между концами рогов около 50—52 см, изредка, по Н. Я. Дин-
нику (1910), до 60—62 см. Поперечные валики или бугры на.передней
стороне рогов иногда ясно выражены. Окраска более 'сходная с С. с.
severtzowi, но борода средней длины или короткая и широкая, как у С. с.
cylindricornis. Распространение: центральная часть Кавказского
хребта к востоку от Эльбруса и до горы Шхары; в меньшем числе встре-
чается и к востоку от последней, от Дигории и до Осетии включительно
{Верещагин, 1952).
4) С. caucasica dinniki Satunin — тур Динника. Изгиб рогов, сходный
с С. с. severtzowi, но в средней части рога имеют еще дополнительный
прогиб внутрь. Кроме того, рога отличаются тем, что коротки и, отно-
сительно собственной длины, толсты. На передней их стороне хорошо
развиты поперечные, отставленные друг от друга валики. В качестве
области распространения указывается западная и северо-западная часть
ареала вида: верховья рр. Белой, Уруштена, Малой и Большой Лабы
{Сатунин, 1909). Целесообразность выделения этого подвида подвергается
справедливым сомнениям. Возможно, что описание его основано на ин-
дивидуальных вариациях или на молодых экземплярах.
: На явном недоразумении основано выделение в качестве «вида» С. rad-
dei (Matschie, 1900) — козла из гор Сванетии (верховья р. Ингура).
Биология. Кавказские каменные козлы, как и другие виды рода
•Capra, типичные обитатели скалистых гор. Всюду они предпочитают
держаться в наиболее трудно доступных участках горных вершин и скло-
нов. Хоть сколько-нибудь открытых мест и пологого рельефа, в противо-
положность сернам, туры явно избегают. Однако в деталях их образ
жизни, в частности выбор местообитаний, сезонный и даже суточный цикл,
в разных частях ареала не одинаков и в сильной степени определяется
не только местными природными условиями, но и деятельностью человека.
Границы вертикального распространения туров расположены в пре-
делах высот от 1000 до 3300 м и даже выше, от средней полосы леса до
альпийской зоны, границ ледников и вечных снегов. В местах, где они
не вытесняются с исконных пастбищ стадами домашних животных и не
30 Фауна СССР, И. И. Соколов
466
BOVIDAE
преследуются человеком, как в заповедниках, наибольшее число туров
придерживается верхней полосы леса, субальпийской и альпийской зон
(Насимович, 1949; Эквтимишвили, 1952, 1953а). При этом верхняя и
нижняя границы вертикального распространения существенно не меняются
по сезонам; меняется только степень встречаемости на разных высотах.
В районах, где в альпийской зоне в широких масштабах производится
выпас домашних овец и коз, а звери усиленно преследуются вооружен-
ными огнестрельным оружием пастухами, туры вынуждены приспособ-
ляться к изменившейся обстановке. Частично они поднимаются в крайнее
высокогорье, на самые вершины и скалистые кручи, недоступные даже
домашним козам; при этом заходят иногда значительно выше нижней
границы снежной зоны, держась все светлые часы суток на выступающих
из-под снега скалах (Динник, 1910; Гептнер и Формозов, 1941). Значи-
тельная часть зверей, вытесняемых с альпийских пастбищ, наоборот,
спускается ниже, в пределы лесной полосы, в глубокие скалистые ущелья.
В связи с этим, в некоторых районах, например на южных склонах Кав-
казского хребта в пределах Азербайджана, у местного населения создалось
представление о наличии двух экологических форм — горного и лесного
тура (Верещагин, 1938).
Меняются местообитания туров и по сезонам. Летом подавляющая
часть зверей держится в субальпийской и альпийской зонах, на высотах
от 1500 м и выше. В пределах леса в этот период встречаются главным обра-
зом поздно родившие самки или звери, вытесненные с альпийских цастбищ
домашними стадами. Здесь туры придерживаются наиболее крутых ска-
листых участков склонов и ущелий. В альпийской зоне летом расселяются
широко, в наиболее жаркий период особенно предпочитают соседство
ледников и снега, где находят в изобилии сочный зеленый корм, необхо-
димую прохладу и защиту от насекомых. Особенно высоко поднимаются
взрослые самцы, тогда как самки и молодняк придерживаются более
низко расположенных участков.
В конце лета, когда в альпийской зоне трава в значительной мере
высыхает или грубеет, туры охотнее держатся в верхней части леса и
субальпийском криволесье, где по лесным лужайкам и тенистым местам
им легче находить зелень. По свидетельству А.А.Насимовича (1949),
в сентябре в альпийской полосе туров держится по крайней мере раза
в два-три меньше, чем в. июле.
Зимой большее число туров держится в лесной полосе, особенно молод-
няк и самки. Однако значительная часть популяции и в это время не
покидает субальпийской и альпийской зон, особенно в западной части
ареала. По-видимому, глубина снега около 40 см является пределом, огра-
ничивающим возможность добычи турами корма (Эквтимишвили, 1952). По-
этому большинство зимовок его на западном Кавказе расположено на более
малоснежном северном склоне Главного хребта (Насимович, 1949). По той
же причине нет туров на перевалах Главного хребта в пределах Азер-
байджана, а в многоснежные зимы они вообще держатся только в верх-
ней трети лесной зоны, спускаясь до высот в 1000 м. Звери, остающиеся ,
на зимовку в альпийской полосе, держатся на солнечных склонах южной
экспозиции и по выдувам. Здесь на обнаженных от снега участках они
находят необходимый для них минимум корма. В таких местах временами
концентрируется значительное число животных, нередко в сотни голов.
В лесной полосе туры зимой придерживаются мест с перемежающейся
снежностью, скалистых склонов, преимущественно южной экспозиции,
где снег мало, задерживается и быстрее тает под лучами солнца.
CAPRA
467
Зима — наиболее трудный период в жизни туров. За это время они
сильно худеют. В случаях глубоких снегопадов звери вынуждены пред-
принимать перекочевки с одного хребта на другой, иногда по глубокому
снегу. Протяженность этих чисто местных миграций не бывает, впрочем,
велика и обычно не превышает расстояния в 12—15 км. В восточной части
Большого Кавказа сезонные миграции принимают регулярный характер.
Так, большая часть каменных козлов южных склонов Главного хребта,
Рис. 100. Стадо восточнокавказских туров, Лагодехский заповедник.
(Фото И. А. Шавлиашвили).
напротив Лагодех и Закатал, перед началом гона откочевывает и проводит
зиму на северном склоне в Дагестане (Верещагин, 1938). Причину мигра-
ций усматривают в большом количестве снега, выпадающего на южном
склоне, и в недостатке здесь кормов, вследствие малого числа мест, удоб-
ных для зимовки.
Обратная передвижка туров с зимовочных мест к местам летовок про-
исходит по мере таяния в горах снега и появления свежего зеленого корма.
В основном, она падает на период с конца апреля до начала июня.
Н. К. Верещагин (1938) отмечает летние перекочевки части туров из Зака-
тальского заповедника на север в пределы Дагестана, происходящие в
июне и связанные с наличием в лесу и на альпийских лугах южного склона
большого числа кровососущих насекомых.1
Каменные козлы — типично стадные животные. Одиночно держа-
щиеся как самки, так и самцы составляют не правило, а исключение.
Размер стад не одинаков и зависит от сезона, условий места обитания, а
1 3. С. Эквтимишвили (1952) отрицал наличие миграции туров с южного склона
Главного хребта на северный и обратно.
30*
468
BOVIDAE
еще больше от общей численности данного вида в том или ином районе.
По словам Н. Я. Динника (1909), на Эльбрусе, в Дигории и Осетии, уже
в его время группы в 10—15 голов встречались редко, и даже на охраняе-
мой территории бывшей Великокняжеской охоты в верховьях Большой
и Малой Лабы стадо в 50 голов представляло редкое исключение. В области
восточной части Большого Кавказа, где эти звери более многочисленны,
стада дагестанских туров в несколько десятков голов и в настоящее время
не составляют исключения. Н. К. Верещагин (1938) наблюдал летом стадо
в 225 голов. Другие лица на территории Закатальского заповедника назы-
вали ему еще большие цифры — 500 и 400 голов, а в прежние годы осенью
перед гоном и перекочевкой на северные склоны козлы будто бы образовы-
вали табуны до 1000 штук или немного меньше.
В настоящее время средний размер стад туров в Кавказском заповед-
нике в летний период определяется в 11—12 голов (Насимович, 1949).
Близкие к этим цифры для Тебердинского заповедника приводит 3. С. Экв-
тимишвили (19536). Максимальные из наблюдаемых летом стад состояли
из 80—100 животных; только в одном случае А. А. Насимович встретил
табун, насчитывавший 152 экземпляра, а мне называли цифру 184.
Взрослые самцы и самки образуют отдельные, не смешивающиеся,
кроме периода гона, стада. Смешанные табуны наблюдаются редко й пред-
ставляют, вероятно, результат случайных встреч. Обычно, как отмечалось,
самцы держатся в верхней части альпийской зоны, тогда как самок чаще
можно встретить ближе к верхней границе леса. Иногда к группам взрос-
лых козлов присоединяются и молодые самцы. С самками, кроме приплода
текущего года, обычно держится и полувзрослый молодняк, в возрасте до
двух-трех лёт, обоего полд. Молодняк часто образует самостоятельные
стада. , .
Минимальный размер стад наблюдается в июне—июле, в период окота
и первое время после него, когда самки с новорожденными ведут одиноч-
ный образ жизни.: В лесной полосе туры держатся более мелкими груп-
пами, чем в альпийской зоне. Перед началом гона, а в некоторых местах и
раньше, ониначинают табуниться. К группам самок присоединяются самцы.
Максимальные размеры стад наблюдаются в период спаривания. По окон-
чании гона при благоприятных кормовых условиях происходит частичное
распадение крупных табунов на более мелкие группы. Обычно же увели-
ченные размеры стад остаются до весны, главным образом в силу того, что
число пригодных для зимовок мест и кормовых площадок ограничено,
особенно в альпийской зоне, и на них поэтому происходит концентрация
значительного числа животных. В лесной полосе и зимой чаще наблюдают-
ся более мелкие стада туров (Насимович, 1949).
Состав стад у туров в пределах одного сезона относительно постоянен.
Если их не преследуют, то в определенные часы суток изо дня в день в те-
чение длительного времени можно наблюдать в одном месте одну и ту же
группу животных. Вожаков и сторожей, как таковых, туры не имеют.
В отдыхающем стаде всегда находится одно или несколько случайно бодр-
ствующих и посматривающих по сторонам животных. Естественно, что
чаще, хотя и не всегда, первыми замечают опасность старые, более осторож-
ные звери, занимающие и при отдыхе возвышенные места с более широким
кругозором.
При переходах в спокойной обстановке туры двигаются гуськом, реже
плотной группой. Потревоженное или испуганное стадо, в противополож-
ность сернам, не бросается врассыпную, а бежит в одном направлении,
:хотя обычный строй при этом и бывает нарушен.
CAPRA
469
Суточный цикл жизни каменных козлов в значительной мере опреде-
ляется условиями существования. В нормальной, спокойной обстановке,
летом туры с рассвета и до 9—10 часов утра пасутся на альпийских лужай-
ках. Когда становится жарко, часть животных подымается выше, ближе
к ледникам, или устраивается на отдых в тени под навесом скал, в пещерах.
Часть зверей, наоборот, спускается ниже в полосу леса, где находит
прохладу в тени деревьев. В середине дня в альпийской зоне пасутся
только отдельные звери, равно как только изредка можно видеть в утрен-
ние часы отдыхающих туров. Спасаясь от назойливых насекомых и ища
прохлады, туры часто лежат часами на снегу, забираются даже в воду.
Иногда они располагаются на открытых, хорошо обдуваемых ветром скло-
нах, перевальных гребнях или на выступающих высоко отдельных скалах,
обеспечивающих зверям, наравне с прохладой, широкий кругозор и без-
опасность. В сильно изрезанной местности, наоборот, предпочитают отды-
хать под выступами скал и в пещерах, где иногда образуются так назы-
ваемые «стойла», с большим скоплением помета (Верещагин, 1938).
Вновь выходят на жировку туры часа за четыре перед заходом солнца;
пасутся не только весь остаток дня, но с небольшими промежутками и всю
ночь.
В местах, где звери подвергаются преследованию, а альпийские луга
используются под выпас домашних стад, туры почти всю светлую часть
суток вынуждены проводить в высокогорье среди ледников и неприступных
скал или уходить в полосу леса в глубокие ущелья. Только перед самым
закатом они выходят на прилегающие участки субальпийских лужаек, где
кормятся всю ночь до рассвета, с тем, чтобы рано утром вновь скрыться
до прихода людей со стадами.
В лесной полосе Азербайджана туры предпочитают и пастись, и рас-
полагаться на отдых на местах осыпей по склонам и на дне ущелий. В пас-
мурные дни, а также при постоянном обитании туров в лесной зоне нет
столь четко выраженной смены часов отдыха и кормежки. И пасущихся,
и отдыхающих зверей можно встретить в этих условиях в любое время
суток.
Иногда, в особенности в лесу, места отдыха располагаются недалеко
от мест жировок. В альпийской же полосе турам ежедневно приходится
совершать дважды в день переходы по нескольку километров (Насимович,
1949). При сильном ветре и отдыхают и пасутся в ущельях.
Водопоями, по наблюдениям А. А. Насимовича, туры пользуются чаще
в утренние часы, тогда как сухие солонцы посещают преимущественно во
второй половине суток.
В отношении выбора и перемены мест обитания каменные козлы кон-
сервативны. Временами, с удивительной настойчивостью они придержи-
ваются одних и тех же мест и посещают их изо дня в день, даже если их
беспокоят. При переходах от мест отдыха к местам кормежки, к водопоям
и солонцам пользуются из года в год одними и теми же путями. Последние
обозначаются не только глубоко вытоптанными тропами на мягкой почве,
но узнаются и по отшлифованному копытами камню, даже на голых ска-
лах (Гептнер и Формозов, 1941).
В зимний период туры ведут более оседлый образ жизни, чем летом.
Вследствие ограниченности мест кормежки, они еще более привязаны к ним
и далеко не уходят. Пасутся большую часть дня, лишь на короткое время
ложатся отдыхать на подветренных и солнцепечных склонах. Ночью,
по-видимому, не пасутся. От непогод спасаются под навесами скал, в пеще-
рах или под нависающими ветвями деревьев. Впрочем, туры очень легко
410
BOVIDAE
переносят климатические невзгоды. А. А. Насимовичу (1939, 1949) не раз
приходилось наблюдать их спокойно пасущимися и под проливным дож-
дем и.с шапками снега на спине. Лишь сильный буран с ветром заставляет
уходить с открытых мест и искать убежища под покровом леса. Лежки рас-
полагаются нередко прямо на снегу, иногда на пучках травы, между кор-
нями у стволов деревьев, для чего животные копытами разгребают неболь-
шие ямки. Места зимних лежек постоянны, и на них нередко скапливается
значительное количество помета (так называемые «стойла»).
Летней пищей турам служат, в основном, разнообразные травы, произ-
растающие в местах обитания. А. А. Насимович (1949) приводит список
летних кормов западнокавказских туров для альпийской зоны, включаю-
щий 90 видов, около 90% которых составляют травянистые растения.
Наиболее излюбленным из последних, судя по степени поедаемости, яв-
ляется ряд злаков (пахучий колосок, мятлик, лисохвост, вейник; овсяницы),
а также многие виды разнотравья — колокольчики, манжетка, анемона,
белокопытник, некоторые виды лютиков, гречишник и другие. Интересно,
что ряд из поедаемых турами видов растений считаются ядовитыми для
домашнего скота; например лютики, рододендрон. Н. Я. Динник (1910)
к числу пищевых растений туров относит также некоторые виды драб,
осок и ряд других растений, отсутствующих в приведенном предыдущим
автором списке. Близкий по составу и общему характеру список кормовых
растений дагестанского тура приводит Н. К. Верещагин (1938).
Из древесно-кустарниковых растений летом служат пищей листья и по-
беги карликовой ивы (Salix arbuscula L.), карликовой низкорослой альпий-
ской березки, почки и листья рододендрона (Rhododendron ponticum L.),
хвоя пихты и сосны, а также кора последней. В грибной сезон охотно
поедаются некоторые виды шляпочных грибов (сыроежки, рыжик, скри-
пица); А. А. Насимовичу пришлось видеть желудок убитого тура, напол-
ненный измельченными грибами.
Зимнее питание туров изучено слабо; В альпийской и субальпийской
зонах оно состоит почти целиком из засохших и незанесенных глубоким
снегом трав, служивших пищей и в летний период. Охотно, конечно, по-
едаются немногие зимнезеленые растения. В лесной полосе, кроме остатков
трав, основу питания составляют тонкие ветви ив, березы, других кустар-
ников и деревьев, а также хвоя пихты, ели, хвоя, кора и даже древесина
сосны (Насимович, 1949). В Закатальском заповеднике, кроме древесно-
веточного корма, в пищу употребляются листья зимнезеленой здесь еже-
вики и особенно плюща, составляющего в этот период один из излюблен-
ных и основных кормов тура (Верещагин, 1938).
При недостатке другого корма, пищей служат также свисающие с де-
ревьев бородатые лишайники (Usnea barbata L.). Чтобы достать последние,
туры нередко забираются на высокие пни и стволы поваленных деревьев.
Испытывая, вероятно, недостаток минеральных веществ в пище, туры
охотно посещают в течение всего года как естественные, так и искусствен-
ные солонцы. К последним они привыкают очень быстро, почему их из-
давна применяют для привлечения этих животных. Искусственным солон-
цам турами отдается даже предпочтение. Наиболее интенсивное солонце-
вание наблюдается в период с марта по июнь. Чаще посещают солонцы
самки и молодняк. Взрослых самцов, возможно в силу их большей осторож-
ности, на солонцах видят реже. Солонцуются даже молодые полутора-
двухмесячиые турята (Насимович, 1938, 1949).
Кроме посещения постоянных сухих и мокрых солонцов, наблюдается
выедание турами ямок в почве в самых разнообразных и как будто случай-
CAPRA
411
ных местах. Н. Я. Динник (1909) отмечал поедание перетертых частиц
горных пород, выносимых вытекающими из-под ледников ручьями. Охотно
грызут туры землю, смоченную человеческой мочой. Наконец/в Азербайд-
жане наблюдается посещение и питье из источников, вода которых издает
запах сероводорода (Верещагин, 1938). Хотя в местах обитания туров нет
недостатка воды, звери не только зимой, но часто и летом, заглатывают
комки снега.
На пастбище туры подвижны; при пастьбе на одном месте, особенно
если кругом достаточно корма, долго не задерживаются. С другой стороны,
они кажутся довольно флегматичными животными. Отдыхая, часами ле-
жат неподвижно или, встав и сделав несколько шагов, ложатся снова
(Динник, 1909). Однако эта флегматичность только кажущаяся. Почуяв
опасность или находясь в обстановке, где она может угрожать, туры момен-
тально теряют флегматичный вид и буквально «превращаются в слух
и зрение». На переходах передние животные часто останавливаются, ос-
матриваются кругом, втягивают воздух с целью вовремя обнаружить
опасность. В противоположность серне, сравнительно легко мирящейся
с близостью домашних стад, туры, как уже отмечалось, по крайней мере
на светлые часы суток, совершенно исчезают из мест, где появились люди
и овцы. Кстати сказать, с сернами на пастбище или на солонце туры почти
не встречаются вместе. Если на солонец пришли туры, серны быстро отсту-
пают (Насимович, 1949). Характер у туров драчливый. Довольно часто, :
говорят, можно видеть и в обычное время, и особенно во время гона бо-
дающихся ТУРОВ. . :
В спокойной обстановке туры передвигаются шагом. Потревоженные,:
особенно если уходят вниз по склону, бегут крупными прыжками. Пре-
красно, как и другие-козлы, приспособлены к быстрому передвижению
по узким карнизам и карабканию по крутым, неприступным скалам.
Каменному козлу ничего.не стоит сделать в случае необходимости прыжок
длиной 2—2.5 м, и даже молодой полуторамесячный козленок без особого
труда вспрыгивает на вертикальный уступ высотой около метра (Динник,
1909); По способности карабкаться и прыгать по скалам С. С. Туров (1952)
ставит каменных козлов на первое место среди других копытных, но
Н. Я. Динник (1909) утверждает, что туры в этом отношении уступают
сернам. . •
Из органов чувств у тура лучше развито обоняние. По ветру он может
учуять человека за полкилометра и более. Слух также не плохой, но, как
и многие другие горные животные, тур на него полагается меньше, так как
в горах вообще много звуков, к тому же усиливаемых резонансом. Зрение,
по-видимому, особой остротой не отличается. Человека издали туры рас-
познают, по-видимому, только стоящего или идущего во весь рост. Ползу-
щего Же или лежащего на земле не всегда узнают и на расстоянии 200—300 м.
Соблюдая необходимые меры предосторожности, к ним против ветра можно
подкрасться на несколько десятков шагов. У охотников (Динник, 1909;
Туров, 1952) даже создалось представление, что тур опасается не столько
вида человека, сколько его запаха. Молодые звери, быть может даже не
впервые видящие человека, при встрече с ним нередко проявляют не па-
нический страх, а любопытство.
Звук, который чаще всего можно услышать от тура, — это своеобраз-
ный свист, короткий и резко обрывающийся. Этот свист является предосте-
регающим, издается как в случае явной опасности, так и при подозрении
на опасность. Изданный вначале одним зверем, он повторяется часто и по
нескольку раз подряд другими. Иногда туры издают еще звуки, напоми-
412
BOVIDAE
нающие чихание, а убегая фыркают, что, по мнению Н. Я. Динника (1909),
служит выражением гнева. Самки подзывают козлят коротким тихим блея-
нием. Таким же голосом, но более нежным, отвечает и козленок.
Смена волосяного покрова у каменных козлов происходит один раз
в году, весной и в начале лета. К концу зимы мех сильно вынашивается
и выцветает. Весенняя линька у западнокавказских туров, судя по кол-
лекционным экземплярам, начинается в апреле, а у дагестанских, по
Н. К. Верещагину (1938), даже раньше — с первых чисел марта. Затяги-
вается она до конца июня или даже начала июля. В противоположность
многим другим копытным, взрослые самки вылинивают раньше, чем самцы
(Верещагин, 1938). Первым вылезает подшерсток, клочьями висящий неко-
торое время на удерживающих его волосах старой ости. В последнюю
очередь выпадает ость наверху спины и животе. Новый подшерсток начи-
нает отрастать в августе; в сентябре он уже хорошо развит. С сентября же
начинается удлинение летней ости, но полное развитие зимний волосяной
покров получает только в декабре.
Половой зрелости каменные козлы, по-видимому, достигают в возрасте
2J4 лет, на третьем году жизни. Самцы практически начинают участвовать
в размножении позднее, так как первые годы не допускаются к спариванию
более взрослыми козлами. Период гона и спаривания падает на время
с конца ноября по начало января включительно. Разгар его происходит
в декабре. Животные в этот период собираются в крупные смешанные стада,
состоящие из самцов и самок. Между самцами происходят ожесточенные
турниры, во время которых один из дерущихся иногда сталкивает своего
соперника в пропасть. При этом бывают случаи поломки рогов или даже
гибели одного из соревнующихся.
Продолжительность беременности у туриц около 5% месяцев (Вереща-
гин, 1938). Массовый окот происходит с середины мая до средины июня.
Отдельные случаи раннего окота наблюдаются в начале мая и даже в конце
апреля. Поздние окоты имеют место в июле, а в исключительных случаях
даже в начале августа (Эквтимишвили, 1954). Окот проходит, как правило,
в верхней полосе леса или субальпийской зоне. Самки перед родами уходят
из своих табунов в наиболее глухие, трудно доступные места, где приносят
обычно по одному козленку.
Первые два-три дня козлята беспомощны; в это время их можно поймать
руками. Но спустя несколько дней, они могут следовать за матерями
и вскоре не хуже их прыгают и карабкаются по скалам. Когда турята до-
статочно окрепнут, самки присоединяются к общим табунам, но некоторые
из них держатся со своим приплодом все лето отдельно. Турицы — хоро-
шие матери и не всегда оставляют своих детей даже при стрельбе (Гептнер
и Формозов, 1941).
Судя по тому, что самки, имевшие в вымени молоко, добывались зимой,
лактация и кормление молодняка продолжаются по крайней мере до на-
ступления следующей беременности или течки. В случае, когда самка не
оплодотворилась, иногда, по-видимому, пользуются молоком и годовалые
козлята (Насимович, 1949). Молоко туриц, по исследованиям, проведен-
ным в Тбилисском зоопарке, содержит свыше 5% жира, около 4% белка
и до 5.76% сахара (Верещагин, 1938).
Родятся турята, имея вес 5—6 кг. Интенсивный рост наблюдается в те-
чение первых трех-четырех лет, затем замедляется, но продолжается, по-
степенно затухая, почти до конца жизни. Рога у самцов начинают расти
уже в первые недели жизни и к шестимесячному возрасту вырастают до
8—10 см.
CAPRA
473
Продолжительность жизни каменных козлов в естественных условиях
ограничивается в среднем 14—15 годами. Известен случай добычи 22-лет-
него тура (Верещагин, 1938). Соотношение полов в стадах туров близко
один к одному.
Из врагов тура на первом месте по значению стоит волк. Больше стра-
дают от волков каменные козлы в зимний период и в лесной полосе, где
глубокий снег и концентрация зверей на ограниченных площадях делают
их более легкой добычей хищников. В альпийскую зону волки проникают
реже, главным образом весной, по насту. Волки охотятся на туров скрадом
или подкарауливают на местах переходов и на солонцах, так как пресле-
довать в угон в сильно пересеченной скалистой местности они не могут.
Опасным для туров высокогорным хищником является леопард; из тех
мест, где появляется эта кошка, туры иногда на некоторое время исчезают
вовсе. Но благодаря своей малочисленности практического значения как
вредитель леопард не имеет. Нападает на туров также рысь, для которой
тур в Кавказском заповеднике — один из основных объектов питания
(Котов, 1958, Тр. Кавк. Гос. запов., IV : 214—217). Медведь не в силах
догнать взрослого козла; поэтому не представляет большой редкости
видеть спокойно пасущихся туров недалеко от медведя. Случайной жерт-
вой последнего могут стать только недавно родившиеся турята. Н.К. Вере-
щагин (1938) наблюдал попытку нападения орла-ягнятника на четырех-
месячного туренка. Рано родившиеся козлята иногда гибнут от заморозков
и непогоды.
Не представляют редкости случаи гибели целых групп туров от частых
в горах снежных лавин и обвалов. Изредка гибнут они также, срываясь со
скал при неосторожных прыжках. Существенно отражаться на численности
вида эти случайные причины, конечно, не могут. В многоснежные и суро-
вые зимы туры, особенно самцы после гона, сильно худеют (Динник, 1909).
Однако достоверно установленных случаев гибели от истощения не из-
вестно.
Изредка наблюдались среди туров массовые заболевания. В одних
случаях они передавались им от домашних животных, как например ящур
или чума (Динник, 1910; Насимович, 1949). Но, по-видимому, изредка
имеют место эпизоотии, свойственные только турам. В 1932 г. на южных
склонах Кавказского хребта среди них наблюдалось какое-то кожное
заболевание, сопровождавшееся выпадением почти со всей поверхности
тела шерсти; от этой эпизоотии, по-видимому, вымерла большая часть
животных в Беланухском ущелье (Верещагин, 1938).
Из эктопаразитов на кавказских турах, кроме обыкновенного овечьего
овода (Oestrus ovis L.), найден носоглоточный овод (Oestrus caucasicusGrwo.',
Грунин, 1948), пока единственный, достоверно установленный представи-
тель африканского рода Oestrus в Азии и Европе. Из других эктопаразитов
известно нападение иксодового клеща (Ixodes ricinus L.) и пухоедов (Вере-
щагин, 1938). .
Эндопаразиты изучены очень плохо. С. М. Асадов (1954) описал из сы-
чуга тура новый вид трихостронгилид — Marshallagia scrjabini Asad.,
найденный им одновременно также и у серны.
Хозяйственное значение. Кавказский каменный козел ценен прежде
всего как мясной зверь. Даже в настоящее время на Кавказе ежегодно
добывается несколько тысяч голов туров. От убитого взрослого самца
получается мяса до 55 кг, от самки 25—30 кг. Убойный вес (выход чистого
мяса) составляет около 60% живого веса. Мясо по вкусовым каче-
ствам лучше, чем у серны, и не отличается существенно от домашней
414
BOVIDAE
козлятины. Мясо старых самцов более жестко и не так вкусно. Лучше
туры упитаны осенью перед началом гона. В это время слой под-
кожного жира может достигать сантиметра, а внутреннего (брыжеечного)
сала от трехлетнего зверя набирается несколько килограммов. От упо-
мянутого на стр. 456 добытого нами в октябре 1958 г. тура Северцова
туша мяса без внутреннего жира весила 89.2 кг; кроме того, от него
было собрано 12.1 кг(!) внутреннего жира. Жир тугоплавкий, поэтому
при употреблении в пищу жирной козлятины во рту получается неприят-
ное ощущение застывшего сала. Шкуры используются на месте. Из них
шьются мягкие башмаки чарыхи, вырезываются ремни. Употребляются
они также в качестве ковриков, для изготовления заплечных мешков,
обивки седел, а изредка даже на куртки и шубы. В Дагестане из зимнего
пуха (подшёрстка) тура изготовляют высоко ценимые, легкие и теплые
башлыки, а из выщипанных зимних шкур шьют папахи. Пух вычесывают из
шкур или собирают весной на ветках кустов в местах обитания туров.
От взрослого тура можно получить 330—350 г пуха (Гептнер и Формо-
зов, 1941; Верещагин, 1938).
Рога находят применение в качестве украшений. Оправленные в се-
ребро, они употреблялись как кубки. Идут рога также для разных по-
делок, вроде рукоятей к ножам, кинжалам и т. и.
Наибольшее значение промысел тура имеет для чабанов, выпасающих
высоко в горах овец, составляя нередко основу их питания и тем самым
способствуя сохранению поголовья домашних животных. Для прочих
групп охотников добыча туров имеет ограниченное значение, ввйду
трудности выноса убитых животных из. гор в низины.
Серьезные опыты по приручению и одомашниванию туров стали про-
водиться в последние годы в Тбилисском зоопарке (Джанашвили, 1948»
1950). Пойманный молодым, тур легко привыкает к домашней обстановке
и к ухаживающим за ним людям. Известны случаи, когда ручные туры
спокойно разгуливали на полной свободе в населенных пунктах.
В зоопарках легко переносят летнюю жару даже в низинах^ Свободно
размножаются в неволе. При скрещивании с домашними козами (Capra
hircus L.) дают вполне плодовитое потомство. Наследственность у гибри-
дов туро-коз носит Промежуточный характер. Рога у них имеют форму,
Свойственную домашней козе и безоаровому козлу. Шерсть длинная,
сходная с домашним видом. Характер телосложения — промежуточный
между козой и туром. Подучаются гибриды с живым весом во взрослом
состоянии до 120 кг.
Вопрос о практической целесообразности скрещивания туров с домаш-
ними козами остается открытым. Возможно, оно окажется полезным
в целях повышения конституциональной крепости домашних животных.
О гибридах туров с безоаровыми козлами (Capra aegagrus Erxl.) го-
ворилось раньше Дем. стр. 464). В Лондонском зоологическом парке
было получено большое число гибридов между турами и сибирским козе-
рогом (Capra sibir tea'Meyer). О .внешнем виде их и плодовитости ничего
не известно (Flower, 1931). .
Охота. Хорошо описана охота на туров Н. Я. Динником (1879, 1887),
В. А. Шильдером (Туркин и Сатунин, 1904) и С. С. Туровым (1952).
Основной способ добычи — охота скрадом с подхода, ничем не отличаю-
щаяся от таковой на серн и безоаровых козлов.
В Дагестане считается, что добыть тура легче, чем оленя, косулю,
серну или безоарового козла, возможно в связи с большей численностью
Этого зверя. Охотятся также нагоном группами по нескольку человек,
OVIS 475
когда одни из охотников гонят туров, а другие располагаются на пути,
по которому должны пройти звери. При этом способе требуется хорошее
знание местности и привычек зверя. Подкарауливают туров на тропах,
на солонцах, у серных источников. В последнем случае, если идет целое
стадо, опытный охотник старается пропустить первых животных; тогда
даже после выстрелов, повинуясь стадному инстинкту, все звери про-
несутся мимо стрелка и последний при сноровке сможет взять несколько
экземпляров.
В. Г. Гептнер и А. Н. Формозов (1941) упоминают об охоте в Даге-
стане на туров со стаей злобных притравленных собак. Последние, взяв
зверя по следу, молча гонят его, пока не поставят «на отстой» на какой-
нибудь выступающий скале и, давая знать охотнику лаем, удержат тура
до его подхода. Иногда ловят туров в расставленные на тропах капканы.
По сравнению с серной тур на рану не крепок и падает от ранения,
при котором серна легко и далеко убегает (Динник, 1910).
Род OVIS LINNAEUS —БАРАНЫ
Bly th Е., 1841, Journ. Asiat. Soc. Bengal, X : 858—888 (сист., распр.); Се-
верцов Н. А., 1873, Природа, 1:144—245 (общ., изобр.); Lydekker R.,
1898, Wild oxen, sheep and goats, London : 149—239 (общ., синон., изобр.); S h i t-
kow B. u. L. Sabanejew, 1910, Zool. Jahrb., Abt. Syst., XXVIII : 456—472
(морф.; сист.); Насонов H. В., 1911, Изв. Российск. Акад, наук : 1267—1296
(морф., сист.); Lydekker R., 1912, The sheep and its cousins, London : I—XV+
1—315 (общ., изобр.); Kowarzik R., 1913, Zool. Anzeig., XLI : 439—445 (сист.,
распр.); Lydekker R., 1913, Catal. Ungulate mammals Brit. Mus., I, London:
73—122 (сист., синон., изобр.); Kowarzik R., 1914, Petermann’s Geograph. Mit-
teil., I : 70—72 (сист., распр.); Millais J., 1914, bkh.: The gun at home and abroad.
The big game of Africa and Europe, London : 370—379 (общ.); Насонов H. В.,
1919, Изв. Российск. Акад, наук : 1215—4246 (морф.); Насонов Н. В., 1923,
Географич. распростр. диких баранов Старого Света, Птг., изд. Акад, наук : I—IV+
1—255 (сист., синон., распр.); В г е h m’s Tierleben, 1925, 4. Aufi., Bd. 13 Leipzig :
244—271 (общ., изобр.); Teilhard de Chardin P. et.I. Piveteau, 1930,
Ann. Paleontol., XIX : 3—132 (эвол.); Громова В. И., 1935, Докл. Акад, наук
СССР, IV (XI), 1—2 : 97—99 (ист.); Ф л е р о в К. К., 1935, в кн.: Виноградов Б. С.,
Е. Н. Павловский и К, К. Флеров, Звери Таджикистана, Л., изд. Акад, наук СССР :
127—130 (общ., изобр.); Gromova V., 1936, Neue Forsch. in Tierzucht und Ab-
stammungslehre (Festschr. z. 60. Geburtstag v. prof. J. U. Duerst), Bern : 81—91 (ист.);
К a p д ы м о в и ч Э. И., 1936, Успехи зоотехнич. наук, II, 3 : 397—417 (морф.);
В о h 1 i п В., 1937, Palaeontol. sinica, ser. С., XIV, 1,: 1—111 (эвол.); В о h 1 i n В.,
1938, Nov. Acta Soc. sci. Upsal., ser. IV, vol. XI, № 2 : 1—54 (эвол.); Л у с Я. Я.,
1938, Изв. Акад, наук СССР, сер. биол., 4 : 775—850 (гибр.); Teilhard de Char-
din Р. et М. Trassaert, 1938, Polaeontol. sinica (n. s. C.), 6 : 1—68 (эвол.);
Schaub S., 1943, Eclogue geol. Helvet., XXXVI, 2 : 270—289 (эвол.); Цал-
кин В. И., 1948, Охрана природы, 2 : 94—102 (ист., распр.); П и д о п л и ч ко И. Г.,
1951, О ледниковом периоде, II, Киев, изд. Акад.' наук УССР : 58—59 (ист.); Цал-
кин В. И., 1951, Горные бараны Европы и Азии, М., изд> Моск. общ. испыт. прир. :
1—343 (морф., сист., синон., распр., изобр.); Ellerman J. and Т. Morrison-
Scott, 1951, Checklist of palaearctic and indian mammals, London : 409—418 (сист.,
синон.); Громова В. И., 1953, Тр. Комисе, по изуч. четвертичн. периода, X,
1 : 1—121 (морф.); Соколов И. И., 1953, Тр. Зоол. ийст. Акад, наук СССР, XIV:
151—156 (морф., сист., эвол.); Herre W., 1953, Zool. Anzeig., Bd. 151, № 7/8 :'
204—209 (ист.).
Характеристика рода. Животные средней величины, плотного, но
несколько более легкого, чем козлы, сложения. Туловище покоится на
не тонких, но сравнительно высоких ногах. Хвост треугольный, обычно
со всех сторон покрытый волосами. Самки некоторых видов иногда не
имеют рогов. У самцов рога длинные, не менее чем в 2 раза, превышают
длину головы. Отходя от черепа, они направляются вверх и одновременно
загибаются назад и в стороны. Форма рогов у самцов разнообразная:
загнутые в виде кольца или круга с концами, обращенными вперед и не.
476
BOVIDAE
сколько внутрь или, наоборот, наружу; свернутые в улиткообразную
гомонимную спираль с концами, обращенными в стороны; реже гетеро-
нимно изогнутые с концами, обращенными позади головы внутрь на-
встречу друг другу, иногда первертированные.1 На поверхности рогов
имеется два или три продольных ребра — заднее, передне-внутреннее
и передне-наружное (последнее не всегда), и, соответственно, две или
три грани — внутренняя, наружная и иногда передняя, или лобовая.
Поперечное сечение рогов у самцов обычно треугольное, сильно сжатое
с боков у вершин. Если передне-наружное ребро и передняя грань не
развиты, то сечение приближается к линзовидному с плоской внутренней
и сильно выпуклой наружной гранью. У самок рога тоньше и короче,
чем у самцов, обычно не длиннее головы и, не образуя сложного изгиба,
дугообразно загнуты в стороны и назад; поперечное сечение рогов у са-
мок продольно-овальное, сильно сжатое с боков.
Характерные для козлов бугры и поперечные валики на передней
стороне рога у баранов отсутствуют, но зато на поверхности рогов имеется
сплошная поперечная морщинистость, особенно сильно развитая у сам-
цов. Кроме общей морщинистости, на поверхности рогов, как и у козлов,
ясно видны более глубокие круговые борозды (кольца) на границах
участков годичного прироста рога.
Верхняя губа покрыта волосами, за исключением узкой вертикальной
полоски по средней линии и узких полосок по краю ноздрей. Характерный
для козлов пучок длинных волос на нижней стороне головы («борода») у ба-
ранов отсутствует. Волосяной покров в развитом состоянии ясно диф-
ференцирован на ость и подшерсток.
Предглазничные, паховые и межпальцевые железы, как правило,
хорошо развиты, но свойственные самцам козлов околохвостовые (при-
анальные) железы отсутствуют или развиты очень слабо. Молочная же-
леза имеет два функционирующих и иногда еще два недоразвитых соска.
Зароговая часть черепа несколько укорочена; длина последней от
заднего края выступа затылочного бугра до среднероговой точки (т. е.
точки на средине между основаниями роговых отростков) меньше длины
лба от среднероговой точки до средины линии, соединяющей передние
края лобных костей. Теменные гребешки не сближены; расстояние между
ними в самом узком месте больше длины теменных костей по средней
линии и всегда превышает 16% основной длины черепа. Предглазничные
ямки на черепе большею частью хорошо развиты, равно как и гребешок
на лицевой поверхности слезной кости.1 2 Этмоидальные щели иногда
имеются, но чаще отсутствуют. Межчелюстные кости не всегда достигают
носовых костей и не всегда соприкасаются с ними, а если соприкасаются, то
редко глубоко вклиниваются между носовыми и верхнечелюстными костями.
Полости в роговых отростках не всегда доходят до верхушек последних.
Проксимальная часть роговых отростков, равно как и стенки пневма-
тизированной их части, состоят из пористого костного вещества. Поверх-
ностный слой компактного вещества на роговых отростках очень тонок
или совсем отсутствует.
Зубы гипсодонтные. Особенно сильно выражена гипсодонтия рез-
цов, имеющих вытянутую долотовидную форму.
В скелете конечностей пястная кость, за крайне редкими исключе-
ниями, короче и плечевой кости, и предплечья. Метаподии по сравнению
1 Объяснение терминов «гомонимный», «гетеронимный» и «первертированный»
см. примечание на стр. 296.
2 Исключение составляют подроды Ammotragus Blyth и Pseudois Hodgson.
OVIS
477
с козлами более длинные и узкие; длина пястной кости составляет не
менее 78% длины лучевой; а длина плюсневой — не менее 62% длины
берцовой. Ширина верхнего конца пястной кости не свыше 18—19%
ее наибольшей длины. Длина лучевой кости, за крайне редкими исключе-
ниями, больше длины плечевой (Громова, 1953).
История и географическое распространение. Род баранов филоге-
нетически весьма близок к роду козлов. Корни его ведут к общему с по-
следним предку — нижнеплиоценовому роду Tossunnoria Bohlin из Монго-
лии (Bohlin, 1937), совмещающему в себе признаки Capra L. и Ovis L.
Настоящие бараны на геологической сцене появились позднее. Наи-
более ранние остатки их найдены в слоях верхнеплиоценового возраста
и в Азии, и в Европе. Изображение фрагмента черепа из верхнего плио-
цена Сенеза во Франции (Schaub, 1943) по форме и постановке рогов
и теменной зоны не оставляет сомнений в принадлежности его к баранам.
В Азии наиболее ранними формами баранов являются Ovis zdanskyi
Bohl. (Bohlin, 1938) из верхнеплиоценовых отложений провинции Хэнань и
О. schantungensis Teilh. et Piv. из слоев Никована провинции Шаньси
в Китае (Teilhard de Chardin et Piveteau, 1930; Teilhard de Chardin e.t
Trassaert, 1938). Формирование и обособление филогенетической ветви
баранов произошло, вероятно, в среднем плиоцене. Предковая форма,
из которой могли бы быть выведены более специализированные современ-
ные и ископаемые бараны, не известна; следует, однако, полагать, что
она была морфологически близка к некоторым современным формам
муфлонов, вроде О. ophion Blyth. с еще удлиненной зароговой частью,
слабо развитым передне-наружным ребром на рогах и дугообразным
изгибом последних.
В настоящее время ареал диких баранов включает острова Средизем-
ного моря — Корсику, Сардинию, Кипр, а также северную Африку,
Малую, Среднюю, Центральную и северо-восточную Азию, запад Север-
ной Америки от Аляски до Мексики. На материке Европы в ряде стран
живут в качестве интродуцированного зверя в заповедниках и охотничьих
парках. В прошлом ареал был более широким и, в противоположность
современному, сплошным. Остатки баранов найдены, например, в палеоли-
тических отложениях Крыма (Громова, 1953), Англии и в ряде других
стран западной Европы (Пидопличко, 1951).
Большинство современных рас баранов являются горными живот-
ными, однако по сравнению с козлами они обитают, как правило, в усло-
виях более открытых ландшафтов и спокойного рельефа. По-видимому
и эволюция их шла в направлении выработки приспособлений не к лаза-
нию по скалам, а к быстрому бегу по сравнительно слабо пересеченной
местности и пологим горным склонам. Чертами морфологических адапта-
ций к этим условиям являются более легкое по сравнению с козлами сложе-
ние, сравнительно длинные конечности, менее короткие заостренные копыта,
приближающаяся к цилиндрической форма головки бедренной кости.
Классификация. Род Ovis мною принимается в широком объеме.
В него, кроме настоящих баранов, включаются также африканский гри-
вистый баран (О. lervia Pall.) и гималайский нахур, или голубой баран
(О. nahoor Hodgs.), часто выделяемые в качестве представителей самостоя-
тельных родов.1 Морфологически больше других видов выделяются также
толстороги, или снежные бараны северо-восточной Сибири и Америки.
Поэтому род Ovis делится на четыре подрода.
1 Основания для подобной трактовки систематических отношений баранов см. Со-
колов, 1953 : 154—155.
478
BOVIDAE
1) Ovis s. str. — Настоящие бараны. Характеризуются хорошо разви-
тыми предглазничными, паховыми железами и межпальцевыми железами
на всех четырех конечностях, а также наличием предглазничных ямок на
черепе. Распространение: острова Средиземного моря, Малая,
Передняя, Средняя и Центральная Азия, южная Сибирь.
2) Pachyceros Gromova — Снежные бараны, или толстороги. Предглаз-
ничные железы и соответствующие им ямки на черепе развиты очень слабо.
От других рас отличается также более слабо развитой морщинистостью
роговых чехлов, нависанием края лобовой грани рога над боковой, уко-
рененностью роговых отростков черепа и отсутствием иногда переднего
коренного зуба (Р2) в нижней челюсти. Подрод включает один или два
вида [О. (Pachyceros) canadensis Shawn О. (Pachyceros) nivicola Eschscholtz],
распространенных в северо-восточной Азии и Северной Аме-
рике.
3) Ammotragus Blyth —Гривистый баран. Отличается полным отсут-
ствием межпальцевых и предглазничных желез, а также предглазничных
ямок на черепе, сильным развитием рогов у самок, не уступающих по
размерам таковым у самцов, и наличием на нижней стороне шеи, груди
и передних ногах мощно развитого подвеса из длинных волос. Подрод вклю-
чает один вид — О. (Ammotragus) lervia Pallas, распространенный
в северной Африке от Рио-де-Оро и Мавритании через Марокко, Алжир, *
Тунис и Ливию до Египта, а на юге до Нигера, Асбена и Кордофана |
в Судане (Ellermann and Morrison-Scott, 1951).
4) Pseudois Hodgson — Haxyp, или голубой баран. Отличается отсут-
ствием предглазничных желез и ямок на черепе, отсутствием межпаль-
цевых желез на всех конечностях, своеобразной голубоватой окраской
самцов, а также резко выраженным первертированным изгибом рогов. Под-
род включает один вид — О. (Pseudois) nahoor Hodgson, распростра-
ненный в ряде провинций Китая, в Сиккиме и Непале до Кашмира. |
Общепризнанная классификация настоящих баранов (подрод Ovis s. I
str.) до сих пор также отсутствует. Объясняется последнее обстоятельство (
сильно выраженной в этой группе половозрастной, сезонной и индивидуаль-
ной изменчивостью морфологических признаков, а также параллелизмом |
в развитии отдельных форм. Систематика их, в большинстве случаев, до j
сих пор строится главным образом, если не исключительно, на форме рогов. I
Большинство местных форм баранов к настоящему времени достаточно 1
хорошо изучено и серьезных разногласий в отношении их морфологических ।
особенностей нет. Однако трактовка систематических взаимоотношений
и таксономического ранга отдельных рас различна и находится в зависи-
мости от понимания тем или иным исследователем объема и границ вида и
от подхода к вопросу о видовых критериях.
Н. А. Северцов (1873) делил всех диких баранов на две группы — арка-
ров и муфлонов, рассматривая их как отдельные роды. Н. В. Насонов
(1923), описавший ряд новых форм из различных мест обширного
ареала рода, сохранил деление их на муфлонообразных (Species
rn.uflonijorm.es) и аргалообразных (Species argaliformes), не при-
давая, впрочем, этим группам определенного систематического ранга.
Как и Н. А. Северцов, этот исследователь в пределах каждой из основных
групп различал ряд комплексов форм, которым придавал видовое значение.
Все многообразие форм диких баранов, число которых насчитывалось 36,
Н. В. Насонов сводил, не считая снежного и близких к нему американских
баранов, к 8 самостоятельным видам. Однако сам он должен был отметить,
что выделяемые им виды не имеют абсолютных отличий и отдельными при-
OVIS
479
знаками связываются друг с другом и что даже между группами mujloni-
formes и argaltformes невозможно провести границы (Насонов, 1923 : 140).
Позднее В. И. Громова (Gromova, 1936), основываясь на результатах кра-
ниологических исследований, пришла к тем же выводам об отсутствии ясно
выраженной обособленности муфлонообразных и аргаловидных баранов.
Подробный сравнительно-морфологический анализ современных форм
диких баранов недавно произвел В. И. Цалкин (1951).1 Этот автор также
пришел к выводу, что ни один из рассмотренных им признаков «не дает воз-
можности разделить муфлонов и аргали на четко разграниченные группы»
и что не только отдельные формы (расы) баранов, но и целые комплексы
их, которым предшествующими исследователями придавалось видовое
значение, «не изолированы ни морфологически, ни биологически и связаны
многочисленными формами, соединяющими в себе признаки различных
групп». В результате В. И. Цалкин высказал точку зрения, что «много-
численные формы диких баранов, распространенные от островов Среди-
земного моря на западе до Центральной Азии, южной Сибири и Забай-
калья на востоке, представляют собой не группу видов, как принималось
всеми нашими предшественниками, но единый политипический вид, насе-
ляющий огромную территорию и образующий большое количество локаль-
ных форм» (1951 : 217).
Действительно, все местные формы баранов представляют пеструю
мозаику сочетания отдельных признаков, каждый из которых встречается
в той или иной степени выраженности и в популяциях из других частей
ареала. Ни по одному из исследованных до сих пор признаков нельзя
провести четкой границы, отделяющей какую-либо расу или группу рас
от всех других. Даже комбинация ряда признаков не всегда дает возмож-
ность различать отдельные формы баранов. Если рассматривать расы бара-
нов только с морфологической точки зрения, как это делает В. И. Цалкин,
и требовать в качестве обязательного критерия вида наличие морфологи-
ческого хиатуса с другими близкими формами видового значения, то всех
евразийских, а быть может и американских баранов следует счи-
тать за один вид.
Однако возможна и другая точка зрения, если к представлению о виде
подходить не только с морфологических, но и с биологических позиций.
При этом следует принимать во внимание следующее.
1) Общий ареал баранов, относимых к подроду Ovis s. str., состоит из
двух, по-видимому в какой-то мере разобщенных частей — западной,
населенной мелкими муфлонообразными расами, и восточной, представ-
ляющей область распространения крупных аргали и архаров. Как устано-
вил еще в свое время Н. А. Северцов (1873), граница между этими частями
проходит по западным хребтам системы Памира и южным склонам Кара-
корума. Разрыв ареала между западными и восточными формами, неви-
димому, имеет место в восточном Таджикистане. Нет баранов на Дарваз-
ском хребте. В известной мне литературе нет сведений об обитании их (оче-
видно, нет и самих животных) на хребтах Петра Великого и Гиссарском.
К востоку от этих областей бараны обитают лишь на Памире, на
Заалайском и Алайском хребтах, в горах Киргизии, Казахском Каратау,
Чу-Илийских горах. По-видимому, не смыкаются также ареалы барана
Винье (О. orientalis vignei Blyth) и архара (О. аттоп poll Blyth) на
Памире. Первый из них встречается в юго-западной его части (Насонов,
1 Некоторыми его данными, в первую очередь цифровыми, я буду пользоваться
в дальвейшем.
480
BOVIDAE
1923), тогда как восточная половина Памира представляет область рас-
пространения архара (Флеров, 1935). Нет никаких давних, говорящих
за то, что ареал баранов в только что перечисленных областях был сплош-
ным когда-либо раньше.
Не совсем ясно, соприкасаются ли области распространения муфлонов
и аргали в Центральной Азии к югу и юго-востоку от Памира.
2) Если даже ареалы западных и восточных форм баранов и соприка-
саются, то смешения и скрещивания популяций в районах соприкоснове-
ния не происходит. Если бы последнее имело место, то, несомненно, именно
в этих пограничных областях должны были встречаться промежуточные
между теми и другими формы. Между тем этого нет; как легко можно ви-
деть и из приводимых в работе В. И. Цалкина (1951) данных, крайняя
восточная форма муфлонообразных баранов (О. orientalis vignei Blyth)
и по общим размерам, и по структуре черепа и рогов резко отличается от
смежных с нею географически форм аргаловидных баранов: О. аттоп
poll и О. аттоп karelini Sev., О. a. hodgsoni Blyth, О. a. dalailamae Przew.
3) До сих пор не доказана скрещиваемость диких муфлонообразных
с дикими же аргаловидными баранами и возможность получения от подоб-
ных скрещиваний вполне плодовитого потомства. Известна скрещивае-
мость тех и других с домашними овцами, но достоверные случаи получения
гибридов между муфлонами и аргали не известны (Лус, 1938).
Таким образом, утверждение В. И. Цалкина об отсутствии биологиче-
ской изоляции между mufloniformes и argaliformes не имеет под собой до-
статочно веских оснований.
4) Морфологически промежуточными и как бы связующими формами
между муфлонами и аргали являются расы cycloceros, arcar, severtzowi
и nigrimontana, сравнительно далеко расположенные от границ распростра-
нения типично аргаловидных баранов и которые, поэтому, не могут быть
рассматриваемы как продукт смешения последних с популяциями муфло-
нов. Еще Н. А. Северцов (1873) подчеркивал факты проявления элементов
относительно большого морфологического сходства у рас баранов не смеж-
ных, а более удаленных районов. То же самое, на примере европейского
муфлона (О. musimon Schr.) и снежного барана (О. nivicola Esch.), отметила
В. И. Громова (Gromova, 1936). Эти факты, как и промежуточный характер
некоторых, отмеченных выше форм аргаловидных и муфлонообразных
баранов, следует рассматривать не как явление незавершившегося про-
цесса морфологической дивергенции, а как проявление параллелизма,
независимого появления сходных черт строения в двух близких
филогенетически ветвях под влиянием сходных условий существова-
ния.
5) Следует также иметь в виду, что промежуточный характер некоторых
рас выражается лишь в немногих признаках, а кроме того, трансгрессив-
ность в отношении, например, размеров и пропорций черепа, так же как
и рогов, носит не массовый характер, а наблюдается буквально у единич-
ных животных. Во всяком случае, имея перед собою даже только шкуру
и череп с рогами взрослого самца, можно всегда без труда определить,
имеем ли мы дело с муфлоном или аргали.
Не следует упускать из вида и то обстоятельство, что не только особен-
ности биологии, отношения с окружающей средой, несомненно лежащие
в основе дивергентной эволюции животных, но даже морфология баранов
изучена еще недостаточно. Нельзя поручиться за то, что в дальнейшем не
будут найдены признаки, которые позволят четко диагносцировать от-
дельные их формы.
OVIS'
.481
Исходя из изложенного, видовая обособленность муфлонообразных
и аргаловидных баранов и трактовка их как самостоятельных видов пред-
ставляются более обоснованными, хотя с диагностической стороны и не сов-
сем удобными.
Как особый вид, должен рассматриваться европейский, или корсикан-
ский муфлон (О. musimon Schreb.), проделавший длительную эволюцию
в условиях островной изоляции, в некоторых отношениях параллельную
другим расам, например снежному барану (Gromova, 1936), и приобрет-
ший некоторые черты своеобразной специализации в черепе и окраске.
Под сомнением остается видовая обособленность кипрского муфлона
(О. ophion Blyth), наиболее примитивной расы из современных рас баранов.
Что касается материковых рас муфлонообразных баранов, тохотяареал
их в настоящее время частично разорван и некоторые из них изолированы
друг от друга, но, как будет показано ниже, еще в недалеком прошлом он
был сплошным. Кроме того, отдельные расы в пограничных областях дей-
ствительно обнаруживают наличие переходных особей от одной к другим, и
поэтому более правильным.представляется рассматривать всех их как под-
виды одного вида, в силу приоритета носящего название О. orientalis Gmelin.
То же самое следует сказать о всех аргалоподобных формах восточной
части ареала настоящих баранов, которые должны быть объединены в один
вид О. аттоп Linnaeus.
Поэтому в настоящей работе признается существование на территории
СССР четырех видов баранов: О. musimon Schreb., О. orientalis Gmel.,
О. аттоп L. и О. nivicola Eschs
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ ВИДОВ БАРАНОВ (РОД OVIS)
ФАУНЫ СССР
1 (2). В нижней челюсти с каждой стороны только два переднекоренных
зуба (премоляра) .... Ovis (Pachyceros) nivicola Eschscholtz —
..........................Снежный баран, или толсторог (стр. 544).
2 (1). В нижней челюсти с каждой стороны по три переднекоренных зуба.
3 (8). Окраска передней стороны ног ниже запястий и скакательных су-
ставов не темнее (обычно светлее) окраски туловища. У самцов наруж-
ный край передней (лобовой) грани не выступает над боковой поверх-
ностью рога. Наибольшая ширина черепа на уровне глазниц у самок
не больше расстояния от переднего конца межчелюстных костей
(prosthion) до заднего края верхнего зубного ряда. Длина роговых
стержней (без роговых чехлов) больше обхвата их в основаниях.
-4 (7). Размеры средние или небольшие. Длина тела у самцов меньше
160 см, у самок не свыше 90 см. Основная длина черепа у самцов
не больше 255 мм, у самок не больше 230 мм. Если длина черепа превы-
шает приведенные цифры, то у самцов на нижней стороне шеи имеется
сильно развитый подвес из длинных волос, доходящий до головы
и образующий у основания ее подобие бороды или бакенбардов, а
обхват рога у основания не превышает 28 см.
5 (6). Размеры мелкие; высота в холке меньше 80 см; общая длина черепа
не свыше 245 мм. Окраска темная, черноватая или красновато-ко-
ричневая. Вдоль средней линии шеи и спины идет узкая черно-коричне-
вая полоса. Подвес на нижней стороне шеи у самцов черный или черно-
коричневый, не доходящий до головы. Передний край хоан при
горизонтальном положении костного нёба находится на одном уровне
с передними краями глазниц.......................................
.... Ovis musimon Pall. — Европейский муфлон (стр. 482).
31 Фауна СССР, И. И. Соколов
482
BOVIDAE
6 (5). Размеры более крупные; высота в холке у самцов не меньше80 см;
общая длина черепа больше 245 мм, а если меньше, то подвесна
нижней стороне шеи спереди беловатый и доходит до головы. Темной
полосы на средней линии шеи и спины нет. Передний край хоан при
горизонтальном положении костного нёба расположен впереди
уровня передних краев глазниц ........................................
.........Ovis orientalis Gmelin — Азиатский муфлон (стр. 497).
7 (4). Размеры крупные. Длина тела у самцов больше 160 см. Высота в
холке у самцов не меньше 100 см, у самок свыше 90 см.Основная
длина черепа у самцов больше 255 мм, у самок больше 230 мм. Под-
веса из длинных свисающих волос на нижней стороне шеи и у осно-
вания головы ни у самок, ни у самцов нет..............................
................Ovis ammon Linnaeus — Архар (стр. 516).
8 (3). Окраска передней стороны ног ниже запястий и скакательных суста-
вов темнее туловища. У самцов наружный край передней (лобовой)
грани выступает в сторону над боковой поверхностью рога; на перед-
нем крае последней имеется продольный желобок вдоль выступаю-
щего передне-наружного ребра тела. Наибольшая ширина черепа на
уровне глазниц больше расстояния от переднего края межчелюст-
ных костей (prosthion) до заднего края верхнего зубного ряда.
Длина роговых стержней (без роговых чехлов) меньше обхвата их
в основаниях.......................................Ovis (Pachyceros)
nivicola Eschscholtz — Снежный баран, или толсторог (стр. 544).
Ovis musimon Pallas — Европейский муфлон (рис. 101—104)
Pallas Р., 1811, Zoographia Rosso-Asiatica, 1:230—231 (первооп.); Р о ru-
mp г о 1 F., 1879, Assoc. Franc, sur 1’avance de sci., C. R. de la 8-e session : 600 (ист.);
Ne bring A., 1891, Nene Jahrb. Mineral., Geol., Palaontol., II: 107—155 (эвол.,
ист.); Buxton E., 1893, Short stalks. London : 1—35 (биол., ox., изобр.); Lydek-
ker R., 1898, Wild oxen, sheep and goats of all lands, London : 154—159 (общ., синон.,
изобр.); Б и x н e p E., 1902, Млекопитающие, СПб. ; 599—601 (общ., изобр.); Koch А.,
1902, Foldtani kozloni, XXXIII : 346—350 (ист.); Toula F., 1909, Jahrb. Geol.
Reichsanst., LIII : 51 (цит. по H. В. Насонову, 1922) (ист.); Tesdorpf 0., 1910,
Einbiirgerung des Muffelwildes, Berlin : 9—57 (общ., изобр.); Насонов H. В.,
1911, Изв. Российск. Акад, наук : 1292—1295 (морф., сист.); Kowarzik R.,
1913, Zool. Anzeig., XLI, 10 : 439—445 (сист.); Lydekker R., 1913, Catal. Ungu-
late mammals Brit. Mus., I, London : 75—77 (сист., синон., изобр.); К owarzik R.,
1914, Petermann’s Geograph. Mitteil. I : 70—72 (сист.); Насонов H. В., 1922,
Изв. Российск. Акад, наук : 439—451 (морф., сист., изобр.); Tesdorpf О., 1922,
в кн.: Lebensbilder aus der Tierwelt Europas, Saugetiere. Bd. IV, Leipzig : 64—78 (общ.,
акклимат., изобр.); Насонов Н. В., 1923, Географии, распр. диких баранов Ста-
рого света, Птг., изд. Акад, наук : 60—62 (сист., синон., распр., изобр.); В г е h m’s
Tierleben, 1925, 4. Aufl., Bd. 13, Leipzig : 246—249 (общ., изобр.); Фортуна-
тов Б. К., 1925, сборн. «Аскания-Нова», М., ГИЗ : 322—323 (гибр.); -Р удин-
ский К., 1926, Украинский охотник и рыболов, 5:12—13 (биол., изобр.); Flo-
wer S., 1931, Proc. Zool. Soc. London : 207—208 (продолж. жизни); Heck L.,
1931, Sitzungsber. Naturforsch. Freunde. Berlin, 10/12 : 257—267 (биол., акклим.,
изобр.); Шерешевский Э. И., 1931, Сборник раб. по изуч. фауны Крымск,
заповеди., М. : 63—74 (акклим., биол.); Иванов М. Ф. и Л. К. Гребень, 1932,
Бюлл. зоотехнич. опытной и племенной станц., Гос. научно-иссл. степи, инет., Гос.
заповеди. Чапли (бывш. Аскания-Нова), 7 : 120—155 (гибр., изобр.); Schnei-
der К., 1932, Zool. Garten, Leipzig, 5 : 129—131 (размн.); Браунер А. А., 1935,
сборн. «Гибридизация и акклим. животн.», II, М. : 42—48 (ист., морфол., акклим.);
Gromova V., 1936, Neue Forsch. in Tierzucht. und Abstammungslehre (Festschr. z.
60. Geburtstag v. prof. J. U. Duerst), Bern : 81—91 (морф.); Паровщиков В. Я.,
1936, Боец-охотник, 4 : 54—55 (акклим.); Громова В. И. и В. И. Громов,
1937, Тр. Советск, секции Междунар. ассоц. по изуч. четвертичн. периода, I : 55—
59 (ист.); Рухлядев Д. П., 1939, Научно-метод. зап. Главн. управл. по заповеди.,
IV : 154—155 (бол.); Каденации А. Н., 1941, Научно-метод. зап. Главн. управл-
OVIS
483
по заповеди., VIII : 87—88 (параз.); Рухлядев Д. П., 1941, Научно-метод. зап.
Главн. управл. по заповеди., VIII : 81—82 (бол., параз.); Schaub S., 1943, Ecl.
geol. helvet., XXXVI, 2 : 282 (эвол.); Harper F., 1945, Extinct, and vanishing
mamm. Old World., N. Y. : 573—576 (ист., распр., изобр.); Громов В. И., 1948,
Тр. Инет. геол. Акад, наук СССР, 64 : 434 (ист.); Рухлядев Д. П., 1948, сборн.
«Паразитофауна и заболевания диких животных», М., изд. Главн. управл. по запо-
веди. :5—102 (параз.); Иванов М. Ф., 1949, Избран, соч., I, М., Сельхозгиз :
310—329 (гибр.); Цалкин В. И., 1951, Горные бараны Европы и Азии, М., изд.
Моск. общ. испыт. прир., 239—241 (морф., сист., распр., изобр.); Сарки-
сов А. А., 1953, Природа, 2 : ИЗ (гибр.); Н е г г е W., 1953, Zool. Anzeig., Bd.
151, № 7/8 : 204—209 (ист.); Gray А., 1954, Mammalian hybrids : 80 (гибр.); D у k V.,
1954, Ochrana prirody, 3 : 76—89 (морф., биол., акклим.); Ян ушко П. А., 1955,
Тр. Крымск, фил. Акад, наук УССР, IX : 133—150 (биол., акклим.).
Описание. Баран сравнительно небольшой величины. Высота в холке
у взрослых самцов составляет от 65 до 83 см; длина тела 113—444 см;
Рис. 101. Европейский муфлон (Ovis musimon Pall.).
основная длина черепа 202—225 мм; живой вес осенью около 40—50 кг,
но в отдельных случаях бывает, по-видимому, и больше. Самки заметно
мельче самцов; основная длина черепа у них составляет от 180 до 204 мм,
живой вес до 35—36 кг (Янушко, 1955).
Стройное туловище покоится на высоких и нетолстых ногах. Голова
небольшая, сидит на не слишком толстой и длинной шее. Профиль лицевой
части прямой. Уши сравнительно небольшие. Высота в крестце несколько
превосходит высоту в холке. Холка приподнята, так что линия спины
в передней части несколько вогнута. Хвост небольшой, длиною около
10 см. Копыта не длинные, но сравнительно высокие; длина их на передних
31*
484
BOVIDAE
конечностях составляет 57—63 мм, на задних 50—58 мм; высота копыт:
передних 34—38 мм, задних одинаковая с передними, иногда на 1—2 мм
больше или меньше.1
Рога у вполне взрослых самцов длиною по изгибу передней поверхности
от 58 см .до 75 см, очень редко больше. Длина четырех конечных сегмен-
тов (прирост второго, третьего, четвертого и пятого года жизни; см. Цад-
кин; 1951 : 14) составляет от 35 до 55 см. Относительно своей длины рога
тонкие; обхват их в основаниях от 20 до 23 см, редко больше, и составляет
,о.т 29.5,до 39.7% длины всего рога. Объем четырех тех же сегментов
в среднем составляет около 435 см3.1 2 Рога сравнительно круто поставлены
по отношению к продольной оси черепа. С профилем носовых костей их
основания образуют угол около 130—150° (Gromova, 1936). Изгиб рогов
разнообразный, чаще всего гомонимный, с концами, обращенными прямо
вперед по бокам головы или вперед и немного внутрь. Но нередко рога
первертированные или даже гетеронимные, обращенные концами позади
головы внутрь, навстречу друг другу. Поперечное сечение рогов в основа-
нии треугольное, с более острыми задним и передне-внутренним и сильно
закругленным передне-наружным ребрами. Продольный диаметр сечения
рога в основании от заднего до наиболее выпуклой точки передне-внутрен-
него ребра колеблется от 73 до 83 мм. Поперечный диаметр (от средины
внутренней грани до наиболее выступающей точки наружной3 грани) —
от 51 до 65 мм. Верхушки рогов сильно сжаты с боков и имеют только зад-
нее и переднее ребра.
Передне-наружное ребро обычно ясно выражено, но всегда сильно
закруглено,; особенно ближе к основанию рога. Передняя (лобовая) грань
плоская или (чаще) слегка выпуклая; выпуклость увеличивается с возра-
стом, поэтому у'взрослых зверей больше заметна у основания рога. Лобо-
вая грань всегда скошена наружу и поэтому угол, образуемый ею с внут-
ренней4 гранью, значительно более острый, чем с наружной. Внутренняя
грань в дистальной (верхушечной) части вогнутая, к основанию становится
1 Как и у других видов, цифры размеров копыт приводятся по измерениям на
коллекционных экземплярах.
2-Предложенный В. И. Цалкиным (1951 : 15) метод определения прибли-
зительного объема годовых сегментов рогов в редакции его автора заключается
в следующем: «Условно принимая поперечное сечение рога за треугольник и имея
возможность легко измерить величину основания и высоту его, можно, пользуясь
общеизвестной формулой площади треугольника, вычислить площадь поперечного
сечения в любой точке протяжения рога, например при основании и при вершине
каждого годового сегмента. Названные сегменты в схеме могут быть представлены
как усеченные пирамиды, объем которых должен определяться по обычной для этих
фигур геометрической формуле. Надо лишь учитывать, что каждый сегмент всегда
более или менее сильно изогнут и длина его по лобной поверхности всегда больше длины
по нижнему (заднему, по моей терминологии, — И. С.) ребру. Таким образом, для
определения высоты сегмента, рассматриваемого как усеченная пирамида, необхо-
димо вычислить полусумму длины его по лобной поверхности и нижнему ребру. Рас-
полагая всеми’этими исходными данными, мы исчисляем объем усеченной пирамиды
по формуле:.' • • •
Л
. V = уКВ + Ь + ' ВЪ),
где В, И' Ь — площади оснований, a h — высота.
«(Определенный таким образом объем сегмента, разумеется, не точен, всегда ока-
зываясь значительно меньше истинного и составляя от 60 до 80% последнего».
’ По'терминологии В. И. Цалкина (1951) —височной.
' ; *'По: терминологии В. И. Цалкина (1951) — затылочной.
OVIS
485
плоской или даже несколько выпуклой. Наружная грань выпуклая, но
в средней части рога, по соседству с задним ребром, обычно продольно вог-
нута. Поперечные складки (морщины) на роговых чехлах в дистальной
половине широкие и высокие, ближе к основаниям становятся более сгла
женными и мелкими. Цвет рогов и копыт серовато-бурый по сравнению
с другими баранами более темный.
Самки чаще всего комолые. По некоторым данным (Harper, 1945), па
о. Сардиния они обычно имеют короткие рожки, а на Корсике, как пра-
вило, безрогие.
Вполне развитый волосяной покров состоит из прямой, более грубой
ости и тонкого извитого подшерстка. Длина остевых волос в зимнем меху
на верхней стороне туловища около 3—4 см. Длинные грубые волосы на ниж-
ней стороне шеи у самцов образуют подвес длиною до 12—45 см, который,
за крайне редкими исключениями, не доходит до основания головы и не
образует на углах нижних челюстей подобия бакенбардов. Удлиненные до
7—9 см волосы по верхнему краю шеи, холки и передней части также вы-
глядят в виде невысокой гривки. Морда впереди глаз и ноги ниже запяст-
ных и скакательных суставов покрыты коротким, плотно прилегающим к
коже, блестящим волосом. Волосы на хвосте по толщине и длине не отли-
чаются заметно от таковых на туловище.
Общий фон окраски европейского муфлона заметно отличается от
остальных баранов и характеризуется преобладанием насыщенных черно-
ватых, коричневых и ржаво-рыжих тонов. Окраска обусловливается сме-
шением остевых волос двух типов: а) от основания светло-бурых и посте-?
пенно, без перерыва темнеющих до полной черноты к вершинам и б) светло-
бурых от основания, темнеющих в средней части и желто-рыжих в верхней
трети, с темной заостренной верхушкой. От преобладания волос того или
иного типа зависит общий оттенок окраски. Волосы подшерстка всегда1
желто-бурые. В полном зимнем наряде туловище с боков и сверху от рыже-
вато-коричневого до черновато-коричневого или черно-бурого цвета.
Вдоль средней линии спины и шеи, до хвоста включительно, тянется ко-
ричнево-черная полоса. Наиболее отчетлива и широка она на задней поло-
вине шеи и холке; на средине спины иногда едва заметна. Более темно
окрашены, иногда почти черные, область холки, лопатки и пространство
около хвоста на крупе. Несколько светлее задняя часть боков и бедра. По
бокам груди за лопатками с каждой стороны расположено по крупному
светлому, так называемому седловидному пятну, отделенному от такого
же пятна другой стороны срединной хребтовой полосой темного цвета;
Степень выраженности и размеры их не одинаковы: в одних случаях они
резко отграничены от соседней буро-коричневой окраски, в других неза-
метно переходят в нее; иногда они невелики; зато у отдельных животных
внизу сливаются со светлой окраской живота и как бы перепоясывают
туловище на его средине (Шерешевский, 1931). Окраска седловидных
пятен, образованных смесью белых и коричнево-бурых волос, от почти
чисто белого до белесоватого желто-коричневого цвета. От локтевых к ко-
ленным суставам тянутся темные черно-коричневые полосы, резко отграни-
чивающие светлую желто- или грязно-белую окраску живота от темных
боков. Иногда, впрочем, эти полосы не сплошные и ограничиваются участ-
ками непосредственно позади локтей и впереди коленных суставов. Свет-
лая окраска живота захватывает пахи, промежность, внутреннюю часть
бедер и выходит иногда немного на наружную сторону ягодиц; в этих
случаях образуется подобие зеркала по сторонам хвоста, обрамленного
черноватой, более темной, чем соседние участки крупа, полоской.
486
BOVIDAE
Шея по бокам ржаво-рыжая с буроватым оттенком. Удлиненные во-
лосы верхнего ее края черно-коричневого, иногда почти черного цвета.
Подвес на нижней стороне шеи и груди состоит из черных, лишь иногда
с незначительной примесью светлых волос.
Голова окрашена светлее туловища. Лоб и морда сверху желтовато-
бурые с коричневым оттенком. Темя и затылок желто-серые с буроватым
налетом. Стороны головы коричневые, постепенно темнеющие книзу.
Вокруг глаз светлое кольцо. Конец морды грязно-белый. Белая окраска
конца морды наверху доходит иногда до уровня глаз, а также переходит
в виде полосы на нижнюю сторону головы до гортани. Уши снаружи жел-
товато-серые, внутри грязно-белые. Хвост сверху и на конце черно-корич-
невый или черный, снизу и с боков белый.
Ноги выше запястных и скакательных суставов интенсивного шоко-
ладно-коричневого цвета, на передней стороне переходят в черноватую
окраску низа шеи и груди, а у задних ног — в черно-коричневые полосы
по бокам живота. По задней стороне ног тянутся узкие, резко отграничен-
ные от соседних коричневых частей белые полоски, сливающиеся наверху
с белой окраской промежности и живота. Ноги ниже запястных и скака-
тельных суставов белесые, с желтоватым или рыжеватым оттенком. По
передней их стороне обычно тянется неясная, размытая коричневая
полоска.
Летняя окраска самцов светлее зимней. В ней преобладают красно-
ватые тона, но все же и в этот сезон буровато-коричневый оттенок выражен
достаточно ясно. Общая окраска туловища рыжевато-бурого или красно-
вато-коричневого, более однотонного, чем зимой, цвета. При этом наблю-
даются более светло и более темно окрашенные экземпляры. У светлых на
первый план выступает рыжеватый, у темных — буро-коричневый оттенок.
Седловидные пятна иногда хорошо видны, иногда же представлены неболь-
шой примесью на их месте к шерсти основной окраски желто-белых волос.
Продольная полоса вдоль шеи и туловища имеется, но более узкая; только
на холке она немного шире. На уровне средины спины она иногда совсем
исчезает. Темные полосы по бокам живота заметны и летом, но часто не
сплошные, прерывающиеся в средней части. Живот, пахи, промежность
и ягодицы, как и зимою,, грязновато-желто-белые. Шея по бокам однотон-
ная с туловищем. Удлиненные волосы на верхней стороне шеи представ-
лены, и то не всегда, черно-бурым пучком на затылке. Подвес, как таковой,
на нижней стороне шеи.отсутствует, но волосы на его месте удлинены до
3—3.5 см и имеют, как и зимой, черную окраску. Голова летом коричневая,
только вокруг глаз иногда видно светлое кольцо, да конец морды грязно-
белой окраски, переходящей на подбородок и межчелюстное пространство.
Темя и затылок темно-коричневые. Уши снаружи желтовато-серые, внутри
грязно-белые. Верхние части ног до запястных и скакательных суставов
рыжеватые, с примесью белых и коричневых волос. Передняя сторона ног
иногда буро-коричневая, резко обрывающаяся при переходе в светлую
окраску ниже запястного сустава. Светлые полосы тянутся по задней
стороне предплечий и голеней, а иногда также по передней стороне послед-
них. Нижние части ног от запястий и скакательных суставов до копыт,
как и зимой, грязновато-желто-белые, обычно с размытой слабой коричне-
вой полосой на передней стороне.
Самки окрашены более тускло. Зимой у них, как и у молодых самцов,
вместо рыжевато-коричневых преобладают бурые тона. Но распределение
ш контрасты между светло и темно окрашенными местами те же, что и
у самцов. Часто отсутствует темная полоса вдоль шеи и туловища, а также
OVIS
481
нет свойственных самцам гривки по верхнему краю шеи и подвеса. Окраска
головы желто-серая. Почти никогда нет черной полосы на нижней стороне
шеи и не всегда имеются ясно выраженные седловидные пятна. Летом
Рис. 102. Череп Ovis musimon Pall.
у самок окраска туловища желтовато-коричневая или желтовато-бурова»
тая. Шея однотонна с туловищем или с более интенсивным рыжеватым
оттенком. Голова серовато-коричневая, более темная на лбу и затылке.
Уши желтовато-серые, внутри белые.
488
BOVIDAE
Молодые на первом году жизни желтовато-буровато-серые с серой голо-'
вой и ушами. К осени у них начинает проявляться местами черноватый
налет, срединная полоса в области холки и зачатки темных полос по бокам
живота.
В черепе европейского муфлона (рис. 102) характерна сравнительно
короткая лицевая и хорошо развитая мозговая часть. Длина мозговой
коробки (от заднего выступа затылочного бугра (acrocranion) до заднего
края глазницы) составляет от 43.5% до 47.3% основной длины черепа и
от 67.4% до 74% длины морды (от prosthion до переднего края глазницы).
Длина морды составляет от 62.2 до 66.8% основной длины черепа. Меж-
глазничная ширина черепа на уровне верхних краев слезных костей на
краю глазницы составляет не менее или лишь немного менее двух третей
длины морды от prosthion до точки слезно-скулового шва на краю глазницы.1
Зароговая часть черепа даже у самцов сильно выступает позади рогов и
слабо отогнута книзу. Наружные слуховые проходы (meatus auditorius
externus) расположены на уровне задних краев оснований роговых отрост-
ков или даже несколько позади последних. Длина роговых отростков зна-
чительно превосходит обхват их в основаниях. Носовые кости сравнитель-
но короткие и широкие; ширина их редко составляет меньше 38% (от 36
до 49%) собственной длины. Межчелюстные кости почти никогда не сопри-
касаются с носовыми. Костное нёбо сравнительно длинное; при горизон-
тальном его положеции передний край хоан расположен приблизительно
на одном уровне, с передними краями глазниц. В области крыловидного
гребня (crista pterygoidea), на границе височной впадины и глазницы,
теменная кость, за крайне редкими исключениями, входит в соприкоснове-
ние с алисфеноидом (рис. 103, Л). В области поперечного затылочного
гребня у взрослых самцов мастоидная кость соприкасается с теменной,
вклиниваясь между чешуей височной и затылочной кости; у самок же часто
наблюдаются обратные отношения (рис. 104).
История и географическое распространение. Ovis musimon Schr.
представляет одну из сравнительно примитивных рас баранов, хотя в не-
которых отношениях он более специализирован по сравнению не только
с другой островной формой — кипрским муфлоном (О. orientalis ophion
Blyth), но и с некоторыми из континентальных форм рода Ovis L. Муфлоно-
образные бараны появились на территории Европы позднее аргалоидных.
Остатки их начинают попадаться с верхнего плейстоцена и известны как
из ряда стран западной Европы — из Венгрии, Австрии, Франции, Голлан-
дии и др. (Koch, 1902; Toula, 1909; Nehring, 1901; Herre, 1953), так и с тер-
ритории Советского Союза. Если принять развиваемую В. И. Громовой
(Gromova, 1936) теорию вторичного мельчания плейстоценовых мле-
копитающих, то предков муфлонов следует усматривать в Ovis sp.
из верхнего плиоцена Франции (Schaub, 1943), в нижнеплейстоценовом
О. antiqaa Pom. (Pommerol, 1879) и в некоторых других формах верхне-
третичных и раннечетвертичных баранов (Насонов, 1922; Gromova, 1936).
Однако следует иметь в виду, что в Крыму почти одновременно обитали
вплоть до азиля (неолит) и аргаловидные бараны и муфлоны (Громова и
Громов, 1937). Остатки муфлонообразного барана найдены также в верхне-
палеолитической стоянке Мезин в Черниговской области (Громов, 1948).
В западной Европе кости диких муфлонов находились вместе с орудиями
палеолитического человека и костями домашних животных. Вероятно,
1 Указание Н. В. Насонова (1922 : 440) на то, что она никогда не бывает меньше
двух третей длины морды, не совсем точно.
OVIS
489
одной из причин исчезновения диких баранов на материке Европы явилось
истребление и вытеснение их человеком.
В настоящее время распространение европейского муфлона ограни-
чивается островами Корсикой и Сардинией. Сюда они, несомненно, про-
Рис. 103. Сочетание костей в области крыловидного гребня.
А —Ovis musimon Pall.; Б — Ovis аттоп L.; В — Ovis nivicola Eschsch.
as — алисфеноид; / — лобная кость; os — орбитосфеноид; p—теменная
кость; t — чешуя височной кости.
никли с материка до отделения от последнего названных островов, которое,
по мнению Н. В. Насонова (1922), произошло не ранее нижнего плейсто-
цена.
Рис. 104. Сочетание костей в области поперечного затылоч-
ного гребня у самцов (Л) и самок {Б) Ovis musimon Pall.
т— мастоидная кость; р— теменная кость; so — затылоч-
ная кость; t — височная кость.
Некогда весьма многочисленные муфлоны на этих островах за послед-
ние полтора столетия подверглись сильнейшему истреблению. Если рань-
ше они ходили стадами по 100—200 голов, то теперь группа в 10—15 голов
считается редкой. Еще в начале 900-х годов на Корсике насчитывалось около
490
BOVIDAE
4000 муфлонов (Brehm, 1925; Tesdorpf, 1922), но в 1927 г. число их в ре-
зультате отсутствия надлежащей охраны определялось цифрой порядка
350 животных (Heck, 1931). На Сардинии муфлонов, по-видимому, не мно-
гим больше, чем на Корсике, если только не меньше. Начиная с 40-х годов
прошлого века, европейский муфлон был завезен и акклиматизирован
в полудиком состоянии в ряде стран западной Европы — в Италии, Авст-
рии, Венгрии, Чехословакии, Германии, Англии (Tesdorpf, 1922; Heck,
1931), а также в России (см. ниже). В настоящее время число реакклимати-
зированных муфлонов на материке Европы, вероятно, превосходит коли-
чество их на островах Средиземного моря.
Подвиды. Коварцик (Kowarzik, 1913, 1914) описал, без достаточных
оснований, шесть географических форм диких баранов с островов Корсики
и Сардинии, которые Н. В. Насонов (1922) справедливо рассматривал,
как проявление высоко развитой у О. musimon индивидуальной изменчи-
вости.
Н. В. Насонов (1922) различал две «формы» европейского муфлона:
1) f. matschiei (Duerst) с гомонимными рогами, концы которых направлены
вперед и иногда в стороны, и с более коричневой окраской и 2) f. pallasii
(Nasonov) с перевертированными рогами, концы которых направлены
назад и внутрь, и с более рыжеватой окраской. Однако он не придавал этим
«формам» систематического ранга и рассматривал их, как типы индиви-
дуальных изменений.
Биология. На Корсике и Сардинии муфлоны населяют возвышенные
части островов. Однако они и здесь не являются типично горным, а скорее
горно-лесным зверем (Tesdorpf, 1922; Heck, 1931). В горы выше 2000 м
в нормальных условиях не подымаются, очень крутых склонов и скалистых
мест избегают. Зато наличие леса является непременным условием сущест-
вования европейского муфлона. Лето муфлоны проводят в полосе светлых
каштановых и дубовых лесов, а также в сосняках и на вырубках листвен-
ных насаждений, где звери находят не только корм, но также тень и защиту
в дневное время. На альпийские лужайки, расположенные по соседству
с лесом, выходят для пастьбы только в ночное время.
К зиме, когда в горах выпадает снег, муфлоны спускаются в более
низко расположенную зону «макквийи» (macquia), представляющую собою
густые низкорослые кустарнико-древесные поросли, перемежающиеся
•с открытыми полянами, служащими в летнее время местами выпаса домаш-
них стад. Часть зверей и на зиму остается в горных лесах, представляющих
большую защиту и пищу в виде лишайников ипобегов сосны (Tesdorpf, 1922).
Весной, когда на пастбищах появляются домашние стада, муфлоны из
макквийи вновь поднимаются выше в горы.
Преследование людьми и близость стад домашних овец вынуждают
часто диких баранов забираться в непроходимые чащи макквийи или ухо-
дить в скалистые высокогорья.
По образу жизни муфлон — ночное животное. Большую часть дня
он проводит скрытно в лесу. Только перед заходом солнца выходит на жи-
ровку, нередко отстоящую довольно далеко от мест дневных укрытий.
Постоянных путей переходов у муфлонов, по-видимому, нет. Пасутся всю
ночь, перед восходом солнца возвращаются в лес. Зимой для отдыха и
пастьбы выбирают теплые солнечные долины и склоны на солнцепеках,
а летом, в жаркие дневные часы ищут прохлады в тени кустарников и
деревьев.
Держатся муфлоны группами. Только перед самым окотом самки отде-
ляются от стад и первые дни после рождения молодняка держатся в оди-
OVIS
491
ночку. Когда ягнята подрастут, самки вновь соединяются по нескольку
вместе и образуют табунки по 10—12 голов. В состав табунка входят,
кроме самок с ягнятами, приплод предшествующего года. Взрослые самцы
иногда ведут одиночный образ жизни, но чаще образуют небольшие, до
3—4 голов, самостоятельные группы. Изредка вместе с самками и летом
держатся один или два самца, но чаще — это молодые одногодовалые или
.двухгодовалые звери. К осени размеры стад увеличиваются, доходят до
.20—30 голов, к ним присоединяются взрослые самцы. Как уже отмечалось,
прежде размеры стад были гораздо больше.
Группа муфлонов всегда имеет вожака. Таковым, как правило, бывает
старая самка. Но роль сторожа на отдыхе и жировке нередко играет
взрослый самец.
Пищей муфлонам летом служит разнообразная растительность мест
их обитания: травы, вереск, вегетативные части черники, листва кустар-
ников и деревьев (Brehm, 1925). У разводимых в Австрии муфлонов излюб-
ленным кормом являются молочаи (Рудинский, 1926). Осенью с жадностью
поедают желуди и буковые орехи. Зимой питаются надснежными частями
растений; выкапывать пищу из-под снега муфлоны, по-видимому, не могут.
В это время они едят торчащую из-под снега засохшую траву, тонкие ветки,
побеги сосны, а также древесные лишайники. В противоположность
многим другим копытным, обрывающим только верхушки трав, все бараны
благодаря устройству гипсодонтных резцов, подстригают траву почти
у самого корня (Шерешевский, 1931).
Голос муфлонов-ягнят, по словам О. Тесдорпфа (Tesdorpf, 1922), напо-
минает что-то среднее между блеянием козы и домашней овцы. Сходен
с этим и голос взрослой самки, только более низкий и короткий. Самцы
очень молчаливы. При тревоге издают звук, напоминающий не то шипение,
не то чихание, а при испуге нечто вроде свиста, похожего на свист серны.
Во время гона самцы, по Э. Шерешевскому (1931), издают «нечто сред-
нее между фырканьем, хрюканьем и блеянием».
Дикие муфлоны — очень подвижные, ловкие и быстрые звери. Они
•совершенно легко спускаются и поднимаются по крутым склонам, прыгают
с довольно высоких уступов. При спуске с крутых склонов широко рас-
ставляют передние и сближают задние ноги, приседая задом. При быстром
беге переходят на прыжки, сильно отталкиваясь задними ногами и, по-
добно домашним овцам, вскидывая задом. Зимой бегут цепочкой, а летом
более скученной группой. Весьма чутки и осторожны. Все чувства у муф-
лонов развиты хорошо, включая, по-видимому, и зрение. Охотника иногда
замечают за 1500—2000 м (Brehm, 1925). Однако полагаются больше на
обоняние. П. А. Янушко (1955) в Крымском заповеднике приходилось
наблюдать муфлонов с расстояния 20—25 м; они видели его, но особого
беспокойства не проявляли, если он не производил резких движений.
Но стоило зверям учуять запах человека, как, издав полушипящий-полу-
свистящий звук, они моментально исчезали.
При содержании в парках и зоосадах иногда перепрыгивают заборы
высотою до 2.2 м (Tesdorpf, 1922).
Особо осторожны и чутки самцы, за исключением периода полового
сезона. Они постоянно находятся настороже, молчаливы, прислушиваются
ко всякому шороху, с большой осторожностью выбирают места остановок
и отдыха, используют малейшие укрытия. В местах, где их часто тревожат,
муфлоны-самцы днем забираются в такую чащу, куда не только человек,
но и собака проникнуть не всегда может. Ложась на отдых в закрытой
пересеченной местности, муфлоны, будто бы (Tesdorpf, 1922), выбирают
492
BOVIDAE
место так, чтобы токи воздуха, завихряясь, приносили запахи со всех
направлений. Именно осторожностью самцов, долго принимающих уча-
стие в размножении и дающих крепкое полноценное потомство, объяс-
няют отсутствие у муфлонов Корсики и Сардинии, несмотря на остров-
ную изоляцию, следов вырождения (Tesdorpf, 1922).
Линяют муфлоны один раз в году — весной. Сроки и порядок прохож-
дения линьки на родине его не изучены. Осенью происходит лишь удли-
нение ости и вырастание подшерстка. По О. Тесдорпфу (Tesdorpf, 1922),
подшерсток появляется только в конце сентября, но в коллекциях ЗИНа
имеется шкура взрослого самца (№ 14615), добытого на о. Сардиния 20'
июля, на которой сквозь еще короткую ость ясно выступает плотный
густой подшерсток.
Половой зрелости самцы и самки муфлонов могут достигать в возрасте
полутора лет. Самки на втором году обычно и оплодотворяются, а в двух-
летнем возрасте приносят первого ягненка. Самцы же практически начи-
нают участвовать в размножении не раньше, чем на третьем или четвертом
году жизни, так как раньше отгоняются более сильными взрослыми
баранами. По наблюдениям П. А. Янушко (1955) в Крымском заповеднике,,
и самки не все созревают на втором году жизни.
Сведения о времени гона и спаривания у муфлонов противоречивы
Р. Лайдекер (Lydekker, 1898) со слов Букстона указывает, что драки между
самцами происходят в декабре и январе; по М. Хильцхеймеру (Brehm,
1925), на Сардинии течка протекает с августа по октябрь, на Корсике на
месяц позднее — с конца сентября по декабрь; по О. Тесдорпфу (Tesdorpf,
1922), она зависит от погодных условий и начинается с первыми осенними
дождями, в- октябре или ноябре, и тянется до конца декабря.
Самцы в это время разыскивают табуны самок, между ними происходят-
ожесточенные бои. Молодые удаляются без боя. Полуторагодовалые самцы,
как безопасные соперники, даже не всегда прогоняются от стада. Зато-
взрослые бараны бьются часами. Расходясь метров на двадцать, они с раз-
бега с силой ударяются лбами и рогами до тех пор, пока не дойдут до-
изнеможения или один из соперников не будет оглушен и повержен.
Известны случаи, когда бараны убивают друг друга насмерть или сталки-
вают в пропасть. Стук дерущихся муфлонов издали напоминает удары
бросаемых один на другой камней. Во время гона и разыскивания
самок самцы теряют обычную осторожность и чаще попадают под,
пулю.
Беременность у самок муфлона продолжается около пяти месяцев.
Роды происходят с конца марта по конец апреля, изредка в мае. Перед,
окотом, как уже говорилось, самка удаляется от стада и где-нибудь в укром-
ном месте приносит одного, реже двух ягнят (Schneider, 1932). Уже сразу
после рождения ягненок может стоять на ногах; в этот момент его еще-
можно поймать, но уже через несколько часов он может следовать за ма-
терью и поимка становится трудной (Heck, 1931). В отличие от многих
других копытных, ягнята диких баранов не затаиваются в случае опас-
ности, а всегда пытаются спастись бегством (Шерешевский, 1931). Через-
несколько недель, когда ягнята окончательно окрепнут, самки присоеди-
няются к табунам, с ними начинают ходить и ягнята прошлого года, но-
взрослых баранов самки избегают, так как те недружелюбно относятся
к ягнятам.
Если самка вскоре после родов почему-либо потеряла ягненка, она
может снова придти в охоту, быть покрытой самцом и осенью принест®
второго ягненка (Tesdorpf, 1922).
OVIS
493
Приблизительно с четырехмесячного возраста у самцов начинают расти
рога; форму, свойственную взрослым самцам, они приобретают на 4—5-м
году жизни.
Будучи пойманными в раннем возрасте, муфлонята легко привыкают
ж человеку и становятся вполне ручными. Однако подрастая, они стано-
вятся назойливыми, начинают проявлять дикий нрав и с полуторагодова-
лого возраста делаются опасными не только для домашних животных, но
и для человека (Tesdorpf, 1922).1
Продолжительность жизни европейского муфлона в естественных усло-
виях и при парковом полувольном содержании определяется в 7—8 лет;
но при содержании в зоопарках и садах при особо благоприятных условиях
.живут до 10—14 лет, а в исключительных случаях даже до 19 лет (Flower,
1931).
В естественной обстановке скрещивание домашних овец с муфлонами
происходит очень редко (Heck, 1931). Искусственно было получено боль-
шое число гибридов муфлона с разными породами домашних овец.
Гибриды вполне плодовиты. Шерстные качества носят промежуточный
между исходными формами характер. От диких родителей гибридное по-
томство наследует высокую живость и подвижность. Известны случаи
получения гибридов от скрещивания европейского и азиатского муфлонов
{Ovis orientalis Gmel.; Саркисов, 1953). Сохраняли ли гибриды плодови-
тость и в какой мере — не ясно. Имеются сведения о получении плодови-
того потомства от скрещивания муфлона с американским толсторогом, но
они требуют подтверждения (Gray, 1954).
Волки на Корсике и Сардинии отсутствуют. Практически единственный
враг муфлона на средиземноморских островах — человек. Одной из при-
чин уменьшения численности этого вида здесь, по О. Тесдорпфу (Tesdorpf,
1922), являются также различные болезни и эпизоотии, переходящие на
муфлонов с домашних овец. Особенно опустошительна повальная легоч-
ная болезнь, появление которой связывается с резкой переменой погоды.
Она выражается в параличе зада у животных и шатающейся походке.
Сильно страдают муфлоны на Корсике и Сардинии от бескормицы в много-
снежные и суровые зимы.
Мясо дикого муфлона обладает неплохим вкусом. Незначительный
специфический запах его у взрослых самцов легко исчезает при вымачи-
вании в простой воде. Наиболее упитаны муфлоны бывают осенью перед
гоном.
Акклиматизация европейского муфлона в СССР. На территории СССР
акклиматизация европейского муфлона началась в конце прошлого века.
В 1894 г. три муфлона были завезены в зоопарк Аскании-Нова. Здесь они
частично разводились в чистоте, а также скрещивались с мериносами ста-
рого типа. В 1898 г. в Асканию-Нова был завезен самец азиатского муф-
лона; принимал ли он участие в скрещивании с европейским муфлоном —
не известно (Браунер, 1935).
Смешанное стадо чистокровных и гибридных муфлонов частично со-
держалось в загоне, но большая часть жила в полудиком состоянии в степи
без пастуха. Чтобы бараны не разбегались, им ставилась подкормка из
зерен ячменя, которую муфлоны приходили поедать ежедневно. С целью
одомашнения новорожденные муфлонята подсаживались после рождения
1 Следует иметь в виду, что европейский муфлон является вероятным родоначаль-
ником некоторых пород домашних овец, в частности северных короткохвостых.
494
BOVIDAE
к недавно окотившимся и потерявшим своих ягнят самкам мериносов или
выкармливались искусственно.
Сохранившееся после Октябрьской революции и гражданской войны
асканийское стадо муфлонов было весьма пестрым. С 1921 г. началась
систематическая выбраковка из него всех явно гибридных и нетипичных
по фенотипу животных. В результате этой работы, к 1935 г. в Аскании-
Нова была создана популяция, по экстерьеру и краниологическим призна-
кам не отличимая от чистокровного европейского муфлона (Браунер,.
1935).
Уже в советское время в Аскании-Нова велись плановые работы по-
гибридизации муфлона с разными породами домашних овец и всестороннее
изучение полученных гибридов (Иванов и Гребень, 1932). Профессором
М. Ф. Ивановым на основе скрещивания муфлона с мериносом была выве-
дена новая порода овец — горный меринос, совмещающий в себе высокие
шерстные качества мериносов с приспособленностью муфлона к горным
условиям существования. Правда, насколько известно, широкого распро-
странения новая порода, вероятно вследствие наступившей вскоре Великой
Отечественной войны, не получила.1
Акклиматизацию муфлона в условиях сухих южных степей Аскании-
Нова нельзя назвать удачной; имели место большие потери зверей от ту-
беркулеза и воспаления легких.
Удачный опыт акклиматизации европейского муфлона проведен в гор-
ной полосе Крыма. В 1913—1914 гг. в бывшую царскую охоту в Крыму
было завезено 13 муфлонов: 3 из Аскании-Нова и 10 штук с о. Корсики.
Звери были помещены в обширный, в 2—3 га, обнесенный оградой «звери-
нец» на вершине горы Большая Чучель, где содержались вместе с некото-
рыми другими копытными, в частности с крымским оленем и дагестанским
туром. Звери нормально размножались и к 1917 г. муфлонов насчитыва-
лось около 30 голов (Шерешевский, 1931). Осенью 1917 г. звери были вы-
пущены на свободу и в последующие годы гражданской войны и разрухи
подверглись сильному истреблению. К 1923 г., когда на территории бывшей
царской охоты был учрежден Крымский государственный заповедник, муф-
лонов оставалось не больше 6—8 штук. Благодаря принятым мерам ох-
раны поголовье, хорошо приспособившееся за предшествующие годы
к горным условиям Крыма, начало быстро восстанавливать численность
и к 1927 году вместе с приплодом насчитывалось около 45 голов.
К 1941 г. численность муфлонов в Крымском заповеднике доходила
до 490 голов. В то время они обитали, главным образом, по склонам
хребта Синаб-Даг, заходя вниз по рр. Сырой и Сухой Альме до устья
последней и до Камышлов. Отдельные экземпляры встречались у перевала
Кепит-Богаз, на Монастырском хребте до Большой поляны и на хребте
Конек (Рухлядев, 1948). В годы Великой Отечественной войны, во время
оккупации немцами Крыма, муфлоны вторично пережили критический
период. Большая часть их была вновь перебита. После освобождения
Крыма и восстановления охраны, к концу 1946 г. их оставалось около
30—40 голов (Янушко, 1955). В послевоенные годы муфлоны вновь быстро
увеличивают свою численность: на начало июля 1955 г. количество их
1 Как сообщил автору б. заведующий зоопарком Аскании-Нова Г. А. Успенский,
от старого асканийского стада муфлонов после Великой Отечественной войны уце-
лели лишь два самца, павшие в последующие годы. Новая партия корсиканских
муфлонов завезена в 1950 г. К началу 1955 г. в Аскании имелось 8 чистокровных муф-
лонов и около полутора десятков гибридов с домашними овцами.
OVIS
49a
в Крымском заповеднике определялось примерно в 350 голов. Отдельные
довольно значительные группы обнаружены далеко за пределами заповед-
ника.
Условия в горной полосе Крыма для существования европейского муф-
лона оказались благоприятны. Основным местообитанием их является
горный хребет в центре заповедника, между горами Черной на севере и
Большая Чучель на юге, протяженностью около 3 км. Здесь в довоенные
годы муфлоны совершали правильные сезонные перемещения. Лето они
проводили на более ровном и открытом западном склоне, в его высокой
части (на высоте около 1000—1300 м), поросшей буковым и сосновым лесом,
чередующимся с буреломами, скалами, полянами. На зиму муфлоны
перемещались на восточный склон того же хребта, более закрытый со всех
сторон горами и прорезанный множеством балок и боковых хребтов, покры-
тых на южной стороне дубом, а на северной буком. Зимою муфлоны держа-
лись ниже, спускаясь до высот около 600 м (Шерешевский, 1931).
Перекочевки с летовок на зимние местопребывания происходят в конце-
ноября—начале декабря. Обратное перемещение имеет место в период,
со второй половины апреля по середину мая.
Теперь положение изменилось. На западном склоне Синаб-Дага муф-
лоны в настоящее время бывают, но постоянно не держатся, так как с ран-
ней весны до поздней осени здесь производятся дровозаготовки. Теперь,,
по данным П. А. Янушко (1955), «летом стадо самок сконцентрировано,,
в основном, на восточных склонах горы Большая Чучель и хребта Синаб-
Даг; днем они находятся внизу в долине р. Сарысу, а вечером поднимаются
на пастбища — вершины гор. Самцы летом, в основном, придерживаются
района горы Черной. Днем они лежат на северном и северо-восточном
склонах горы в верховьях р. Черной и в балках западного склона между
горой Черной и хребтом Синаб-Даг, а вечером выходят на безлесные-
пространства вершин горы Черной и хребтов Муфлоньего, Срединного и
Веселого, спускающихся в долину р. Сарысу».
Летом часть поголовья муфлонов расселяется также по Бабуган-
Яйле и Никитской яйле, куда до войны они не заходили.1 Два почти изо-
лированные очага обитания муфлонов находятся на безлесных вершинах
гор Саурка и Базмана на окраине Никитской яйлы.
Зимой область обитания муфлонов в Крымском заповеднике сужи-
вается и ограничивается, в основном, восточными склонами горы Си-
наб-Даг, где на южных склонах отрогов хребта, спускающихся в уро-
чище Центральная котловина, они находят достаточное количество корма.
Муфлоны, обитающие летом на Саурке и Базмане, на зимовку спускаются
на отрог Хасал-Сырт (между р. Кача и ее притоком Писсара).
Зиму в Крымских горах муфлоны переносят в общем легко. Снег
редко бывает глубже 20—25 см и не служит препятствием к передвиже-
нию. Затруднения муфлоны испытывают при глубине его свыше 30 см.
Кроме того, в горах всегда находятся малоснежные места, где звери
находят достаточно корма. Глубокоснежные зимы, когда муфлоны стра-
дают от недоедания и истощения, здесь сравнительно редки.
Суточный цикл муфлонов в Крыму тот же, что и на родине. Только
зимой, как и в ненастную погоду в любое время года, ночь они проводят
где-нибудь в защищенной от ветра балке, а на жировку выходят днем
на открытые места, где иногда на солнцепеке и отдыхают. При переходах
пользуются определенными путями, на которых вытаптывают тропы.
1 В августе 1958 г. я наблюдал муфлонов неоднократно около кордона Алпбпц
(г. Роман-Кош).
496
BOVIDAE
Зимой муфлоны держатся группами по 12—18 голов, летом же обычно
не больше 3—5 вместе. Самцы большую часть года проводят отдельно
от самок. Интересно, что одно время вожаком в стаде крымских муфло-
нов, по сообщению Э. Шерешевского (1931), была державшаяся вместе
с ними самка дагестанского тура.
Основу питания муфлонов в Крымском заповеднике в летний период
составляют различные травы. П. А. Янушко (1955) установил поедание
118 видов растений. Основным кормом является 21 вид, из которых
на первом месте по значению стоят герань кровяно-красная (Geranium
sanguineum L.), ежа сборная (Dactylis glomerata L.), дубровник пурпур-
ный (Teucrium chamaedrus L.), а по Э. Шерешевскому (1931) — также
асфоделина (Asphodeline lutea Rchb.) и типчак (Festuca ovina L.). Дре-
весно-веточный корм в летнем питании муфлонов имеет весьма подчи-
ненное значение и поедается в более или менее значительных размерах
лишь при выгорании травянистой растительности и недостатке другого
зеленого корма.
Зимой муфлоны в первую очередь и наиболее охотно. кормятся на
бесснежных и малоснежных местах травяной ветошью и листочками
зимнезеленых осок и злаков, таких, как ежа сборная, пырей собачий,
осока пальчатая и некоторые другие. Так как муфлоны не способны
копытить (доставать корм из-под снега), то важную роль в питании,
особенно в многоснежные зимы, играют тонкие ветки различных ку-
старников и деревьев вплоть до хвои можжевельника и сосны, а также
древесные лишайники и даже мхи (Янушко, 1955). Ежедневно, а иногда
и дважды в сутки, муфлоны посещают искусственные солонцы.
Линька проходит один раз в году, в апреле—мае. Самки линяют позд-
нее самцов. Зимняя шерсть отрастает в октябре; в ноябре все звери
уже ходят в зимнем наряде.
Муфлоны и в условиях Крымского заповедника сохраняют свою
обычную чуткость. По Э. Шерешевскому (1931), в смешанных стадах
из оленей, баранов и косуль первыми человека замечают муфлоны.
Однако они отнюдь не пугливы, а временами проявляют даже любопыт-
ство. Часто, отбежав несколько в сторону, звери останавливаются и на-
блюдают за человеком, а иногда, заметив проходящего вблизи человека,
если последний не делает резких движений, даже не трогаются с места.
По данным П. А. Янушко (1955), муфлоны даже менее осторожны, чем
благородные олени.
Гон у муфлонов в Крыму протекает с конца октября по ноябрь включи-
тельно. Взрослые бараны отгоняют молодых, но драк между ними не на-
блюдалось, возможно вследствие незначительного числа взрослых самцов.
Окот происходит, в основном, в апреле.1 Приносят по одному, но не так
редко и по два ягненка. Вес ягненка при рождении около 1.5—2 кг, но
уже к восьмимесячному возрасту он доходит до 17 кг, а в двухлетнем
приближается к весу взрослых. По наблюдениям П. А. Янушко (1955),
часть самок на втором году жизни еще не готова к размножению. Соотно-
шение полов 46.6 : 53.4 в пользу самок.
Волков в Крыму нет. Единственным хищником, от которого здесь
страдает муфлон, является лисица. От последней, впрочем, гибнут не
только ягнята, но и взрослые муфлоны, особенно в суровые многоснежные
зимы.
Приводимые Э. Шерешевским (1931) иные сроки, по-видимому, не верны.
OVIS
497
В редкие глубокоснежные зимы наблюдается массовое истощение
и даже гибель зверей от бескормицы. Такое положение наблюдалось,
например, в 1932 г., а также в январе 1940 г. (Рухлядев, 1941).
В Крыму муфлон является хозяином многочисленных паразитов
(Рухлядев, 1948). При полном гельминтологическом вскрытии муфлона
А. Н. Каденации (1941) обнаружил 10 видов паразитических червей,
большинство которых общи с домашними овцами, являющимися источни-
ком заражения и диких баранов. Согласно данным Д. П. Рухлядева
(1948), муфлоны поголовно заражены возбудителем дикроцелиоза (Dicro-
celium lanceatum St. et Hass.) и легочно-глистной болезни (Synthetocaulus
kochi Schulz). Эти паразиты, размножаясь в большом числе, вызывают
исхудание животных, которое в совокупности с неблагоприятными кормо-
выми условиями зимой может привести к смерти. Клещи Д. П. Рухляде-
вым реже находились на муфлонах, чем на косулях и оленях. В неболь-
шом числе обнаружились два их вида — Ixodes ricinus L. и Haemaphy-
salis caucasica Olen.
Позднее T. Г. Мельникова (цит. по Янушко, 1955) обнаружила еще
четыре вида — Haemaphy salts concinna Koch., H. inermis Bir., H. punctata
Can. и Dermacentor marginatus Schulz.1
Клещи вредят не только высасыванием крови, но могут являться
переносчиками кровепаразитарных заболеваний. Пока у муфлона в Крыму
обнаружен один вид кровепаразита — тейлерия (Theileria musimi Ruehl.).
Степень ее патогенности пока не ясна.
Отмечались единичные случаи падежа муфлонов в Крыму от сибир-
ской язвы, заражение которой происходило через пастбища от домашних
животных (Рухлядев, 1939).
В целом, акклиматизацию европейского муфлона в Крыму нужно
признать удачной. Доказательством этого следует считать хотя бы тот
факт, что крымские муфлоны крупнее, чем их собратья с Корсики и Сар-
динии. Вес самцов достигает 55 кг, а самок до 36 кг (Янушко, 1955).
Можно надеяться, что в недалеком будущем этот вид интересного
с точки зрения спортивной охоты зверя прочно войдет в состав дикой
фауны горного Крыма.
Перспективной, вероятно, была бы акклиматизация европейского
муфлона также в советских Карпатах.
Ovis orienfalis Gmelin — Азиатский муфлон (рис. 105—107)
Gmelin S., 1774, Reise durch Russland z. Untersuch. d. drei Natur-Reiche,
3:432+486—493 (первооп., изобр.); Bly th E., 1840, Proc. Zool. Soc. London:
62—81 (сист.); Hutton Th., 1842, Calcutt Journ. Natur. Hist., II : 514—521 (сист.);
Федченко А. П., 1873, Ежегодн. Туркестанск. края, II : 436 (распр.); Lydek-
ker R., 1898, Wild oxen, sheep and goats, London : 159—174 (общ., синон., изобр.);
Туркин Н. В. и К. А. С а т у н и и, 1904, Звери России, Bovidae, М. : 553—559
(морф., сист., изобр.); Динник Н. Я., 1910, Звери Кавказа, I, Тифлис : 158—
161 (общ.); Насонов Н. В., 1910, Изв. Росс. Акад, наук, IV : 681—710 (морф.,
сист., синон.); Насонов Н. В., 1911, Изв. Российск. Акад, наук : 1292—1295
(сист., синон., изобр.); Lydekker R., 1913, Catal. Ungulate Mammals Brit. Mus.,
I, London : 77—92 (сист., синон., изобр.); Насонов Н. В., 1913, Изв. Российск.
Акад, наук : 3—32, 14 рис. (сист., синон., биол., изобр.); Nasonov N., 1914,
Bull. Acad. Sci. St.-Petersbourg : 761—778 (морф., синон., сист., изобр.); Ewart J.,
1 Специфических для данного вида подкожных и носовых оводов у европейского
муфлона до сих пор не обнаружено. К. Я. Грунин (устное сообщение) находил иногда
под кожей крымских муфлонов личинок овода Hypoderma diana Вг., паразитирующего
в норме на благородном олене. Однако закончить развитие эти личинки в организме
муфлона, по-видимому, не могут.
32 Фауна СССР, И. И. Соколов
498 BO V ID A К
1914, Trans, Highl. agricult. Soc. Scotl., 26 : 74—101 (гибр.); Насонов H. В.,
1923, Географии, распростр. баранов Старого Света, Птг., изд. Акад, наук : 18‘—59
(морф., сист., синон., распр., изобр.); Марков Е. Л., 1924, Спорт и охота, 11
(одом., гибр.); Brehms Tierleben, 1925, 4. Aufl., Leipzig : 249—250 (общ., изобр.);
Огнев С. II. и В. Г. Г е п т и е р, 1929, Тр. Научно-иссл. инет. зоол. Моск. гос.
унив., Ill, 1 : 138—143 (распр., биол., сист.); Мориц Л., 1931, Охотник, 5 : 27,
6 : 27 (морф., биол.); Flower S., 1931, Proc. Zool. Soc. London : 208 (продолж.
жизни); Флеров К. К., 1932, Тр. Совета по изуч. произв. сил, сер. туркменок.,
2 258—261 (биол., сист.); Лаптев М. К., 1934, Изв. Туркменок, междуведомств.
комитета но охране природы, I : 135—136 (биол.); Флеров К. К., 1935, в кн.:
Виноградов Б. С., Е. Н. Павловский и К. К. Флеров, Звери Таджикистана, Л., изд.
Акад, наук СССР : 120—127 (общ., синон., изобр); Флеров К. К. и И. М. Гро мо в,
1935, Матер, но паразитол. и фауне южн. Таджикистана, X : 266—268 (распр., биол.);
G го m о v а V., 1936, Neue Forsch. in Tierzucht und Abstammungslchre (Festschr. z. 60.
Geburtstag v. Prof. J. U. Duerst), Bern : 81—91 (морф., сист.); Громов И. М., 1937,
Тр. Совета по изуч. произв. сил Акад, наук СССР, сер. туркменок., 9 : 353—354
(распр., биол.); Гуреев А. А., 1937, Тр. Совета по изуч. производ. сил Акад,
наук СССР, сер. туркменок., 9 : 370 (распр.); Me кленбу рцев Р. Н., 1937,
Тр. Среднеазиатск. гос. унив., сер. Villa, зоол., 26 : 46 (распр., биол.); Р о з а-
н о в М. П., 1937, сборн. «Экспедиции Акад, наук СССР 1935 г.», М.—Л., изд. Акад,
наук СССР : 277 (распр.); Султанов Г., 1939, Тр. Узбекск. зоол. сада, I : 102—
103 (распр.); Антипин В. М., 1941, Млекопит. Казахстана, III, Копытные, Алма-
Ата, Казахск. гос. изд. : 72—77 (общ., изобр.); Саркисов А. А., 1944, Тр. Ере-
ванок. зоол. парка, I—II : 89—90 (миграции); Саркисов А. А., 1944, Тр. Ере-
ванок. зоол. парка, III : 5—51 (общ.); Бажанов В. С., 1945, Извест. Акад, наук
Казахск. ССР, сер. зоол., 5 : 77—78 (распр., биол.); Дементьев Г. П., 1945,
Учен. зап. Моск. гос. унив., 83 : 76 (распр.); Скворцов Б. П., 1948, Тр. Воен.-
мед. акад., XLIV : 175—176 (распр.); Цалкин В. И., 1948, Бюлл. Моск. общ.
испыт. прир., отд. биол., LIII, 1 : 31—48 (распр., биол.); Рустамов А. К., 1950,
Изв. Туркменок, фил. Акад, наук СССР, 2 : 45 (распр.); Григорян Г. А., 1951,
Изв. Акад, наук Армянок. ССР, биол. и сельскохоз. науки : 821—826 (napasi); Гру-
нин К. Я., 1951, Энтомол. обозр., XXXI, 3—4:467—473 (параз.); Демен-
тьев Г. П., 1951, Изв. Туркменск. фил. Акад, наук СССР, I : 58 (распр.); Цал-
кин В. И., 1951, Горные бараны Европы и Азии, М., изд. Моск. общ. испыт. прир. :
241—255 (сист., сипон., изобр.); Elie rm an J. and Т. М о г г i s о п-S с о 11,
1951, Checklist of palaearctic and indian mammals, London : 416—418 (сист., синон.);
M а д а м и н о в С. Ю., 1953, Тезисы докл. на выездной сессии Акад, наук Таджикск.
ССР в Ленинабаде, Сталинабад : 175 (распр.); Саркисов А. А., 1953, Природа,
2:113 (гибр.); Gray А., 1954, Mammalian hybrids, London—Edinburg : 80—81
(гибр.); Д а л ь G. К., 1954, Животн. мир Армянок. ССР, I, Позвоночные животные,
Ереван, изд. Акад, наук Армянок. ССР : 157—158 (распр., биол.); Дубинин В. Б.,
1954, Тр. Пнет. зоол. и паразитол. Акад, наук Узбекск. ССР, 3 : 178 (распр.); Н а-
с и м о в и ч А. А., 1955, Роль снежн. покрова в жизни копытных, М., изд. Акад,
наук СССР : 144—157 (распр., биол.)
Описание. Баран средних размеров; длина тела взрослых самцов
от 132 до 147 см, высота в холке от 82 до 98 см.1
Основная длина черепа 205—253 мм, только в исключительных слу-
чаях до 265 мм. Живой вес от 53 до 79 кг, очень редко до 96 кг (Цалкин,
1951). Самки мельче самцов; длина тела у них составляет 121—133 см,
высота в холке 77—85 см, основная длина черепа от 183 мм до 235 мм и
живой вес от 36 кг до 46 кг, хотя известен случай добычи старой самки
весом в 56 кг (Цалкин, 1951).
Телосложение крепкое, но не тяжелое. Короткое туловище покоится
на довольно длинных ногах. Голова не тяжелая, профиль лицевой части
прямой. Уши длиной около 10—13 см. Радужная оболочка глаз желто-
коричневая. Шея умеренной длины и толщины, у самок тонкая. Высота
в крестце на 3—5 см превосходит высоту в холке. Линия спины прямая,
но несколько наклонная вперед. Длина хвоста 13—15 см. Копыта срав-
нительно длинные; длина их на передних конечностях составляет 64—
1 По Цалкину (1951) и Саркисову (1944).
OVIS
499
69 мм, на задних 55—58 мм; высота передних копыт от 33 до 39 мм, задних
на 1—3 мм меньше.
Рога у вполне взрослых самцов по размерам и форме сильно варьи-
руют. Длина их по изгибу передней поверхности колеблется в пределах
от 51 до 93 см. Длина четырех сегментов (прирост второго, третьего,
четвертого и пятого годов жизни) составляет от 42 до 82 см (Цалкин,
1951). Относительно своей длины рога, в общем, тонкие; обхват их в осно-
ваниях 21—30 см, составляя от 29.5 до 42.2% длины всего рога. Продоль-
Рис. 105. Азиатский муфлон (Ovis orientalis Gmel.).
иый диаметр сечения рога в основании (от заднего ребра до наиболее выдаю-
щейся точки передне-внутреннего) составляет 79—109 мм, наибольший
поперечный от 54 до 76 мм.
Средние величины объема четырех сегментов у разных рас варьируют
от 420 до 900 см3 (Цалкин, 1951). Рога круто поставлены по отношению
к продольной оси черепа; с профилем носовых костей они образуют угол
около 133—165°. Характер изгиба рогов разнообразный. У более при-
митивных и мелких западных рас он, как правило, гетеронимный, с кон-
цами, загнутыми назад и внутрь. У южных и восточных рас изгиб чаще
бывает первертированным; при этом, в зависимости от степени первер-
зии, вершины рогов могут быть обращены вниз и внутрь по бокам шеи,
вниз и слегка вперед или даже прямо вперед с самыми кончиками, сверну-
тыми внутрь. Наконец, для баранов Копет-Дага, Устюрта и Мангыш-
лака типичен гомонимный изгиб рогов, которые при этом нередко, описы-
вая в пространстве спираль в полный оборот, концами направлены вперед,
вверх, а иногда и наружу.
32*
5D0
BOVIDAE
Степень выраженности передне-наружного ребра также подвержена
ярко выраженной географической изменчивости, от нее зависит форма
передней (лобовой) грани и характер поперечного сечения рога. У мел-
ких, примитивных рас передне-наружное ребро нередко практически
•отсутствует, передняя и наружная грани рогов образуют общую вы-
пуклость на его месте. Внутренняя грань рога в верхней трети вогнутая,
в остальной части плоская или даже слегка выпуклая. Поперечное
сечение рога в основании получается линзовидное, с плоской внутренней,
сильно выпуклой наружной поверхностью и острыми углами на местах
передне-внутреннего и заднего килей. Иногда передне-наружное ребро
ясно выражено, но притуплено, и лобовая грань с наружной образует
значительно более тупой угол, чем с внутренней гранью. Лобовая грань
в этих случаях ясно выражена, плоская или, чаще, в той или иной мере
выпуклая, но всегда заметно скошена наружу. Внутренняя грань рога
у вершин вогнутая, к основаниям становится постепенно плоской или
слегка выпуклой. Наружная грань слегка выпукла, особенно в основании
рога. Поперечное сечение рога имеет, форму, приближающуюся к тре-
угольнику с более тупым углом, соответствующим передне-наружному
ребру. У рогов с ясно выраженным гомонимным Изгибом передне-на-
ружное ребро выражено, как правило, резко и по степени развития не
уступает передне-внутреннему. Лобовая грань в этом случае может ста-
новиться не только совершенно плоской, но даже слегка вогнутой. С на-
ружной гранью рога она образует приблизительно такой же угол, что
и с внутренней, а иногда производит впечатление даже скоса внутрь,
а не наружу. Внутренняя грань рога в верхней половине вогнутая, книзу
уплощается, изредка становясь слегка выпуклой. Наружная грань слегка
выпукла, но нередко при общей выпуклости образуется продольная
вогнутость перед нависающим над ней передне-наружным ребром рога.
Поперечное сечение гомонимных рогов приближается к форме равно-
бедренного треугольника с более острым углом, соответствующим зад-
нему ребру рога.
Верхушки рогов во всех случаях сильно сжаты с боков и имеют толйко
два ребра — переднее и заднее несоответственно, только внутреннюю
и наружную грани. Поперечные складки на рогах (морщины) в одних
случаях широкие и высокие, в других более мелкие, тесно сидящие друг
к другу и заметно сглаженные ближе к основанию рога.
Описанные типы рогов не являются обособленными друг от друга
и не свойственны только определенным расам О. orientalis. В пределах
одной и той, же популяции могут встречаться в качестве индивидуаль-
ных уклонений рога разной формы. Кроме того, степень выраженности
передне-наружного ребра лобовой грани и даже разные типы изгиба
рогов связаны рядом постепенных переходов. Поэтому выделение от-
дельных местных форм азиатских муфлонообразных баранов в качестве
самостоятельных, видов на основании особенностей формы рогов, как
это неоднократно имело место в истории систематики рода Ovis, лишено до-
статочных оснований.
; Самки в большинстве случаев имеют рога, но у западных рас часто
.комолые. Рога, самок короткие, как правило, не длиннее черепа, сжаты
О боков.. Из ребер более или менее выражены только заднее и передне-
внутреннее, оба закругленные. Поперечное сечение продольно-оваль-
ное,, с более плоской внутренней и выпуклой наружной гранью. Форма
.рогов дугообразная, с загнутыми иногда внутрь или, наоборот, наружу
верхушками. Морщинистость более мелкая, чем у самцов, но всегда
OVIS 5Q1
имеется. Цвет рогов и у самцов, и у самок желтоватый или желтовато^
серый, более светлый, чем у европейского муфлона.
Вполне развитый волосяной покров состоит из слегка волнистой,i
грубой, ломкой ости и тонкого извитого подшерстка. Длина остевых волос;
в зимнем мехе на верхней стороне туловища около 5—6 см. У самцов
на нижней стороне шеи длинные грубые волосы образуют подвес. Сте-
пень развития последнего у разных рас (подвидов) не одинакова. У од-
них он тонкий и короткий, даже зимой не превосходит 9—10 см и огра-
ничивается нижней и средней третью шеи, не достигая основания головы
на 10—15 см. У других рас подвес, наоборот, чрезвычайно густой и длин-1
ный; длина его иногда достигает 25—30 см. Длинные волосы в этих слу-
чаях занимают всю нижнюю сторону шеи, захватывают область углов
нижней челюсти и щек и образуют подобие широкой бороды или бакен-,
бард на границе головы и шеи. В летнем меху подвес развит слабее,
а иногда отсутствует вовсе. Волосы на верхней стороне шеи, хотя не-
сколько длиннее, чем на туловище (до 8—9 см), но верхней гривы не
образуют; только иногда на затылке имеется пучок особо длинных волос.
Морда впереди глаз, а также ноги ниже запястных и скакательных су-,
ставов покрыты коротким блестящим, плотно прилегающим к коже во-
лосом. Хвост покрыт одинаковыми с туловищем волосами. Кожа в пахах,<
по бокам мошонки и вымени покрыта редким мягким волосом, почти
голая. , 1
Окраска у отдельных рас О. orientalis несколько разниться и, кроме
того, подвержена индивидуальной изменчивости. В общем она произво-
дит впечатление блеклой и однотонной. Нет той насыщенности красно-
вато-коричневыми и черноватыми тонами, а также тех контрастов между;
светло и темно окрашенными участками тела, которые так характерны
для европейского муфлона. Преобладают желтовато-р.ыжие тона, иногда
с коричневым, реже буроватым оттенком. Волосы с боков туловища,
и шеи у оснований очень светлые, почти белые, выше становятся светло-
коричневыми, а в верхней четверти или трети приобретают желтовато^
рыжую окраску. Самые кончики остевых волос, пока не обломаны, темно-
коричневые. От степени пожелтения верхушечной части волос обычно
зависит тот или иной оттенок общей окраски. Подшерсток светло-коричнеР
вого или желтовато-белого цвета. ,
Общая окраска туловища и шеи зимой рыжевато-желтая с коричнев
вым оттенком. Верх спины, холка, иногда крестец и плечи немного'темнее..
Более темную коричневатую окраску нередко имеют также ягодицы
по соседству со светлыми участками по сторонам от хвоста. Шея, бока]
и бедра, наоборот, более светл:ые. Темная полоса вдоль средней линии,
спины и туловища, свойственная большинству особей О. musimon Schr.,
у азиатских муфлонов отсутствует. Только на холке иногда имеется:
едва заметное потемнение в виде продольного пятна. По бокам грудной
клетки за лопатками расположено иногда по светлому, так называемому
седловидному пятну, образованному смесью белых и коричнево-рыжих
волос. Размеры и цвет седловидных пятен зависят от преобладания волос
того или иного типа. Иногда они большие, почти белые и резко отграни^
ченные, иногда едва намечаются в виде незначительной примеси белых,
волос к основному фону, а часто отсутствуют вовсе. От локтевых к колен-,
ным суставам по бокам туловища обычно тянутся нерезко обозначенный
светло-коричневые полосы, отграничивающие окраску боков отщветлогц
низа живота. Нередко эти полосы не сплошные; прерваны в средней
части, а иногда представлены лишь.коричневыми пятнами позади локте^
502
BOVIDAE
на подмышках. В летней окраске они часто совсем не заметны. Нижняя
сторона груди, живот, пахи, промежность и ягодицы желто- или грязно-
белого цвета. Хвост сверху одного тона с туловищем, снизу и с боков
белый; кончик иногда коричневого цвета.
Верхняя сторона шеи одного цвета с верхом туловища. Подвес на ниж-
ней ее стороне спереди грязно- или желто-белый, в остальной части состоит
из смеси рыжих и коричневых волос, иногда сплошь белый, с отдельными
коричневыми волосами, но не бывает черно-коричневым или черным.
Голова блеклой желтовато-коричневой или желтовато-серой окраски.
Верхняя сторона носа светло-коричневая, лоб, как правило, темнее,
иногда с ясным буроватым оттенком, более темная окраска часто рас-
пространяется также на бока головы. Вокруг глаз светлое желтоватое
кольцо. Темя и затылок одноцветные с верхом туловища или более тем-
ные, коричневые. Конец морды грязно-белый; беловатая окраска захва-
тывает также низ головы до самого подвеса. Уши снаружи желто-серые
или серовато-коричневые, внутри грязновато-белые.
Ноги выше запястий и скакательных суставов окрашены в один тон
с туловищем, но иногда наружная сторона предплечий и голеней имеет
ясно выраженный коричневато-серый оттенок. Ниже запястных и ска-
кательных суставов ноги белесые, из смеси белых и рыжевато-коричне-
вых волос. Контраст между темным верхом и светлым низом ног выражен
слабее, чем у европейского муфлона.
На задней стороне ног имеются неясные беловатые полосы, соеди-
няющиеся со светлой окраской промежности и живота.
По передней стороне запястий и плюсен обычно проходят неясные
коричневые полосы. Волосы непосредственно над копытами имеют чисто
белую окраску.
Летняя окраска самцов светлее зимней, но более яркая, желтовато-
рыжая, изредка становится песчано-желтой. По нижней стороне шеи
на месте подвеса тянется черно-коричневая полоса. Седловидные пятна,
как правило, отсутствуют; коричневых полос по бокам живота или со-
всем нет, или они едва заметны.
Самки окрашены почти так же, как и самцы; можно говорить лишь
о некотором общем посветлении, о полном отсутствии седловидных пятен
и коричневых полос по бокам живота. Подвес на нижней стороне шеи
у самок не развит, и нижняя сторона последней имеет более светлую
окраску по сравнению с верхом.
Следует отметить, что у баранов западной части ареала преобладают
более яркие рыжевато-желтые тона. У восточных рас, наоборот, яснее
выступают коричневато-бурые оттенки. У баранов Кызылкумов, напри-
мер, преобладание буроватых тонов и буровато-серую окраску туловища
вообще следует считать более типичной.
Мозговая часть черепа (рис. 106) в среднем несколько укорочена
по сравнению с таковой у европейского муфлона, но у большинства рас
хорошо развита; длина мозговой коробки от заднего выступа затылоч-
ного бугра (acrocranion) до заднего края глазницы составляет от 43 до
47.4% основной длины черепа и от 62.4 до 73% длины морды (от prosthion
до переднего края глазницы). Зароговая часть черепа значительно вы-
ступает назад позади рогов и сравнительно мало отогнута книзу; наруж-
ные слуховые проходы (meatus auditorius externus) расположены на уровне
задних краев оснований роговых отростков или несколько позади по-
следних. Длина морды составляет от 63.1 до /0.5% основной длины че-
репа. Межглазничная ширина черепа, за крайне редкими исключениями,
OVIS
503
составляет не больше двух третей длины морды от prosthion до точки
слезно-скулового шва на краю глазницы.1 Длина роговых отростков
значительно превосходит обхват их в основаниях. Носовые кости сред-
Рис. 106. Череп Ovis orientalis Gmel.
ней ширины или узкие; ширина их составляет от 29 до 44% собственной
длины. Межчелюстные кости обычно (хотя не всегда) соприкасаются
с^носовыми. Костное нёбо сравнительно короткое; при горизонтальном
1 У самок она всегда значительно меньше.
504
BOVIDAE
его положении передний край хоан всегда располагается заметно впе-
реди уровня передних краев глазниц. В области крыловидного гребня
(crista pterygoidea), на границе височной впадины и глазницы, теменная
кость у взрослых самцов, за редкими исключениями, не соприкасается
с алисфеноидом; орбитосфеноид, наоборот, обычно входит в контакт
с височной костью (рис. 103, Б). У самок одинаково часто встречаются
оба типа сочетания костей в области crista pterygoidea. В области попе-
речного затылочного гребня у взрослых животных мастоидная кость,
как правило, соприкасается с теменною, оттесняя чешую височной от
затылочной кости (рис. 104, А).
История и географическое распространение (рис. 107). Азиатские
муфлоны, вероятно, произошли от общего предка с европейским. На
о. Кипре до сих пор обитает форма морфологически весьма близкая к ги-
потетическому предку обоих видов. Ископаемые остатки муфлонооб-
разных баранов из областей, близких к современному их ареалу, очень,
редки.
В настоящее время ареал О. orientalis в том объеме, как этот вид при-
нимается в данной работе, охватывает Малую Азию, Иран, Афганистан,
Кашмир, Пенджаб, Белуджистан, а также западные и южные части
советской Средней Азии: Устюрт, Копет-Даг, горы южного Узбекистана
и Таджикистана, включая юго-западный угол Памира.
В СССР крайним западным районом распространения азиатского
муфлона является южное Закавказье в пределах Армении и Нахиче-
ванской АССР. Основными местами его обитания здесь являются Сарай-
Булагский и Западно-Даралагезский хребты, а также Приараксинские
горы к востоку от Ордубада. А. Саркисов (1944) северным пределом рас-
пространения барана в Закавказье считает возвышенность Айдохмая
Вединского района Армении. С. К. Даль (1954) приводит этот вид также
для южного отдела Зангезурского хребта и отрогов последнего (хребтов
Баргушатского и Мегринского). Далее, по данным этого автора, в Арме-
нии дикие бараны встречаются во многих местах Даралагезского (Айоц-
дзорского) хребта в ряде пунктов Азизбековского района: в окрестно-
стях Хыдзорута, Амагу, Гнишика и Хачика.1 В небольшом числе ба-
раны держатся также в Урусском хребте, по горным массивам до р. Азата.
А. А. Саркисов западной границей в Армении считал р. Веди, но, по
С. К. Далю, муфлоны зимой появляются еще дальше на склонах ске-
летных гор в Арташатском районе. К югу ареал арменийского муфлона
уходит в пределы смежных районов Турции и Ирана. Внутри очерчен-
ного ареала в Закавказье численность муфлонов возрастает в юго-во-
сточном направлении. Больше всего их в районах Каракуша, Чалхана.
и на Сары-Даге (Саркисов, 1944). В коллекциях ЗИН АН СССР имеется
череп барана, доставленный в 1911 г. из с. Мастора (№ 11504), к юго-
западу от горы Алагез. Из этого же района имеется пара рогов, датиро-
ванных 1849 г. (№ 946). Однако А. Саркисов сомневается в точности ука-
зания на местонахождение этих экземпляров: «По крайней мере, — пи-
шет он (1944), — жители этого района не имеют понятия о диком баране
и никаких конкретный данных, подтверждающих его наличие там, не
имеется».
В Закаспии районами местообитания муфлонообразных баранов яв-
ляются Устюрт, Копет-Даг, Большие и Малые Балханы.
1 А. А. Саркисов (1944) северную границу здесь проводит на широте сел Микоян
и Кюки.
OVIS
505
Устюрт населяет сравнительно крупная раса степного барана (О. orien-
talis areal Eversm.). В конце прошлого столетий Г. С. Карелин (цит. по
Н. В. Насонову, 1923) находил диких баранов на северных уступах Устю-
рта близ побережья залива Мертвый Култук. В настоящее время они
здесь распространены главным образом по Западному чинку Устюрта
и в горах полуострова Мангышлак (Бажанов, 1945). Северные пределы
распространения точно не известны, но в качестве мест обитания и этот
автор указывает чинки Устюрта вблизи Мертвого Култука.
Главные места концентрации баранов на Устюрте — окрестности
сора Тузбаир. Встречаются и к югу от этих мест, в частности у Ак-Булака
Рис. 107. Распространение Ovis orientalis Gmel.
1 — XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше.
(около 43° с. ш.), в районе залива Кендирли. Многочисленны они по
побережью Кара-Богаз-Гола, уже в пределах Туркменской ССР.
На полуострове Мангышлаке еще недавно степной баран водился
на Каратау, но теперь^ по В. С. Бажанову (1945), его в этих горах нет.
Сохранился в незначительном числе в горах северного Актау, недалеко
от побережья залива Сарыташ и в южном Актау в окрестностях Таучик.
Восточной границей распространения О. orientalis areal Eversm. Н. В. На-
сонов (1923) считал Аральское море, но конкретных указаний об оби-
тании его к востоку от указанных ранее мест в литературе нет.
В пределах Туркмении, как уже говорилось, бараны до сих пор дер-
жатся на Западном чинке Устюрта вдоль восточного побережья Кара-
Богаз-Гола. По данным Н. М. Михеля (Антипин, 1941), в 30-х годах
этого столетия они встречались изредка по северному берегу этого залива
между Чагала и Сартасом, по обрывам Южно-Мангышлакского плато.
В эти же годы был добыт баран на Южном чинке Устюрта в районе ко-
лодца Сыкиз-Хан.
Местами обитания дикого степного барана до сих пор являются возвы-
шенности северо-западных Каракумов. Встречаются бараны, по А. К: Ру-
стамову (1950), в районе колодца Чагыл, в Горах Ирсарыбаба и на Туаркыре.
5p6
BOVIDAE
В конце прошлого столетия дикие бараны водились на побережье
Красиоводского залива, в окрестностях Красноводска, в горах Куба-
Даг, но, в связи с проведением железной дороги и колонизацией края,
в этих районах они вскоре были истреблены.1
В Больших и Малых Балханах распространена уже другая раса
азиатского муфлона — копетдагский, или афганский баран (О. orien-
lalis cycloceros Hutt.). На Больших Балханах, где бараны были много-
численны еще около 30 лет назад, они теперь стали редки, а на Малых
Балханах за последние два десятилетия как будто истреблены вовсе.
Редки сейчас бараны в западном Копет-Даге. По Г. П. Дементьеву
(1945), их нет уже около Кизыл-Арвата и Казанджика, но встречаются
еще к югу от последнего в Кюрендаге, а также в горах между рр. Сум-
баром и Чандыром. Г. П. Дементьев (1951) видел вс. Шарлоук ручного
барана, пойманного на хребте Сонгудаг. В небольшом числе есть они
у Куйляра, к юго-западу от Кизыл-Арвата. На востоке от последнего
нет баранов у Нохура, но имеются еще в горном массиве Супча к западу
от Бахардена, а также в горах у аулов Кодж и Заулу.
В центральном Копет-Даге, по личным исследованиям В. И. Цалкина,
«бараны распространены практически повсеместно, встречаясь везде,
где есть для них подходящие стации». Но сокращение ареала здесь обо-
значается еще более отчетливо, чем в западной части Копет-Дага. К во-
стоку от Гяурса бараны сравнительно малочисленны, встречаются юж-
нее станций Ахсу и Бабадурмаз в хребте Зиранев.
На крайнем юге Туркмении областью распространения барана является
Бадхыз, особенно часть его, прилегающая к р. Теджену с востока.
Здесь они распространены в узкой полосе от Акар-Чешме до Чамчаклы-
Чанга. Много баранов М. П. Розанов (1937) встречал по хребту Гёз-
гельды (у Акар-Чешме), особенно на вершине последнего и у родников
в ущелье Кизыл-Гер, где попадались табуны их по 40—50 голов. Б. П. Сквор-
цов (1948) трех баранов добыл на южных склонах Эльбирик-Кыра. В срав-
нительно большом числе бараны наблюдались в ущелье Кызыл-Джар
и на северном обрыве впадины Ер-Ойлан-Дуза (в центре Бадхыза). Дальше
к востоку они встречаются в Фисташковом хребте к востоку от Кушки
(Громов, 1937), а также в других местах пограничной полосы между
Кушкой и Мургабом, в частности у застав Рабат-Кашан, Караул-Хана
и в районе р. Кашана (Флеров, 1932). К востоку от Мургаба обыкновенны
бараны в горах Кермили, в урочище Беш-Тешек (около 30 км южнее
Тахта-Базара), у источников Ходжа-Гаурдек и Екимбай в 100—120 км
к востоку за Мургабом (Цалкин, 1948).
Несколько изолированных очагов обитания диких баранов (подвид
О. orientalis severtzovi Nas.) расположены в возвышенностях централь-
ных Кызылкумов. Здесь бараны были установлены еще в конце прошлого
века в горах Букантау, Актау, на возвышенностях между последними
и Тамдытау (в районе колодца Ак-Кудук), в Нуратау, а также, вероятно,
в Джетымтау. В далеком прошлом они обитали, вероятно, также в горах
Султантау к северу от Турткуля (Цалкин, 1951). Во всех ли перечислен-
ных местах бараны сохранились до сих пор и в каком числе, удовлетво-
рительных сведений нет. Известно, что они есть в Тамдынском районе
Бухарской области, в частности в хребте Букантау. Единичные экземп-
ляры встречаются на горе Актау юго-западнее колодца Тамды (Бажанов,
1 Эти и приводимые ниже сведения о распространении баранов в Туркмении даны,
в основном, по В. И. Цалкипу (1948).
OVIS
507,
1945). В Нуратау, по данным Р. Н. Мекленбурцева (1937), бараны, со-
хранились лишь в западной части начиная от кишлака .Синтаб. Срав-
нительно многочисленны они здесь только у кишлаков Кискен и Янгалы.
В не столь далеком прошлом возвышенности Кызылкумов, по-видимому,
составляли часть сплошного ареала азиатского муфлона, сливавшуюся
раньше с ареалом его в Таджикистане, в частности в Туркестанском и Зе-
равшанском хребтах.
Крайний юго-восток Туркмении, южная часть Узбекской и почти вся
Таджикская ССР (кроме Памира), населены следующей расой азиат-
ского муфлона — бухарским бараном, или уриалом (О. orientalis bocha-
riensis Nas.). По собранным А. А. Гуреевым (1937) сведениям, бараны
встречаются в горах Карлюка и Кугитангтау (крайние юго-западные
отроги системы Гиссарского хребта). В южном Узбекистане дикие бараны,
до сих пор водятся, кроме пограничного с Туркменией хребта Кугитанг-
тау, также в расположенных к северу от последнего горах Танги-Дувал
(хребты Зиралик и Куюк), в горах Сан-Муллы (Ширабадский район
Сурхан-Дарьинской области) и в пограничном хребте Кызырык-Дари
(Султанов, 1939). Судя по коллекционным экземплярам ЗИН АН СССР,
обитают они также в Байсунтау и Бабатаге.
В Таджикистане северной границей распространения барана до по-
следнего времени считалась р. Зеравшан (Федченко, 1873). Однако ока-
зывается, что до сих пор бараны встречаются даже в северных предгорьях
Туркестанского хребта. Так, по совсем недавним данным В. Б. Дубинина
(1954), они являются основным промысловым животным в предгорьях
Хавастского района Ташкентской области на границе с Таджикистаном.
Два экземпляра этого зверя в 1942 и 1943 гг. были добыты южнее кишлака
Верхний Сармыч.
С. Ю. Мадаминов (1953) в списке добытых им в Ленинабадской области
зверей называет О. orientalis, но не указывает, к сожалению, точного
местонахождения. К югу от Зеравшана дикий баран водится повсеместно
в горах вплоть до р. Пянджа. Автору лично удалось в 1951 г. убедиться
в наличии его в горах, окружающих Дангаринскую равнину (Каратау
на Вахше, Джилантау, Сарсариак). К. К. Флеров и И. М. Громов (1935)
в огромном числе встречали их летом 1932 г. между Кировабадом (Са-
раем) и Пархаром. Из других мест нахождения этого вида в Таджикистане,
судя по коллекциям ЗИН АН СССР, можно назвать еще Бальджуан,
Тахта-Куватские и Кокульские горы, «горы Каратау на Пяндже»1 (горы
Тавакчи или Тарвакчи) и левый берег р. Кафирнигана у кишлака Ка-
бадиан Шаартузского района.
Восточной границей распространенияуриала вТаджикистанеК. К. Фле-
ров и И. М. Громов (1935) считают долготу Куляба. По заслуживающим
доверия данным В. П. Лаздина (Насонов, 1923), отсутствуют бараны
в Дарва.зском хребте.
Подвиды. До настоящего времени выделены и описаны 14 рас азиат-
ского муфлона, некоторые из них иногда рассматривались как самостоя-
тельные виды.
Н 1) О. orientalis anatolica Valenciennes — Анатолийский баран. Раз-
меры средние; основная длина черепа около 225 мм. Рога самцов гете-
ронимные, лобовая грань сильно скошена наружу. Самки комолые.
Седловидные пятна у самцов обычно имеются. Подвес на нижней стороне
1 Речь, по-видимому, идет о хребте Каратау, на правой стороне р. Вахша.
508
BOVIDAE
шеи не доходит до головы. Распространение: южная часть
Малой Азии (горы Тавра и Антитавра).
2) О. orientalis armeniana Nasonov — Закавказский, или Армений-
ский муфлон. Размеры средние; основная длина черепа у самцов 215—
235 мм. Рога самцов гетеронимные. Лобовая грань сильно скошена на-
ружу, иногда совсем слита с боковой. Самки встречаются как комолые,,
так п с рогами. Седловидные пятна у самцов имеются, но выражены раз-
лично. Отличием от предыдущей расы являются якобы слабое развитие
передне-наружного ребра и более значительная разница в ширине между
внутренней и наружной поверхностями рогов. Распростране-
ние: Армения, восточная Турция, северо-западный Иран. Целесооб-
разность отделения от предыдущей расы вызывает большие сомнения^
Цалкин (1951) относит к этой же расе баранов, описанных Блайсом (Blyth,
1840) из окрестностей Эрзурума, которых Насонов считал принадлежа-
щими к следующему подвиду.
3) О. orientalis urmiana Guenter — Урмийский баран. Размеры несколько
меньше, чем у предыдущих рас; основная длина черепа у самцов состав-
ляет 204—221 мм. Рога самцов слабо первертированные или гетероним-
ные. Передне-наружное ребро слабо выражено и лобовая грань скошена
наружу. Самки имеют рога не всегда. Седловидные пятна у самцов вы-
ражены слабо. Подвес не доходит до головы. Распространение:
остров Коюн-Дагы на оз. Урмия (Иран). Есть предположение, что это
не исконные обитатели острова, а потомки баранов, завезенных из разных
областей Армении и Ирана (Насонов, 1923; Саркисов, 1944).
4) О. orientalis isphaganica Nasonov — Исфаганский баран. Размеры
небольшие; основная длина черепа самца, по-видимому, не превышает
220 мм. Передне-наружное ребро на рогах самцов выражено более от-
четливо". Кроме того, от предыдущей расы отличается якобы более крутым
загибом рогов. Имеют ли рога самки — неизвестно. Распростра-
нение: западный и юго-западный Иран к югу от Курдистана и Ирак-
Аджеми.
5) О. orientalis orientalis Gmelin — Восточноэльбурсский баран. Раз-
меры средние; основная длина черепа самцов около 225 мм. Рога самцов
первертированные; наружное ребро более или менее отчетливое. Лобо-
вая грань немного скошена наружу, но, в отличие от предыдущих рас,
на всем протяжении плоская. Седловидные пятна отсутствуют. Самки
имеют рога не всегда. Отличием от предыдущих рас является также бо-
лее мощный подвес на нижней стороне шеи у. самцов, достигающий го-
ловы. Распространение: восточная часть Эльбурсского хребта.
6) О. orientalis erskinei Lydekker — Западноэльбурсский баран. От
предыдущего подвида отличается, по Насонову (1923), более узкими ро-
гами, выпуклой лобовой гранью и отсутствием пучков удлиненных во-
лос по бокам головы. Распространение: Эльбурсский хребет.
В. И. Цалкин (1951) объединяет эту расу с предыдущим подвидом.
7) О. orientalis cycloceros Hutton — Копетдагский баран (рис. 105).
Размеры, сравнительно, крупные; основная длина черепа самцов от
231 до 265 мм. Рога самцов сильно первертированные или гомонимные.
Передне-наружное ребро их резко выражено. Лобовая грань плоская
или даже вогнутая и не имеет скоса наружу. Самки, как правило, имеют
рога. Седловидные пятна отсутствуют. Подвес у самцов хорошо развит
и достигает головы. Распространение: Туркмено-Хоросан-
ские горы, юго-восточная Туркмения, Большие и Малые Балханы, Аф-
ганистан (границы распространения в последнем не известны).
OVIS 509
8) 0. orientalis dolgopolovi Nasonov — Баран Долгополова. От пре-
дыдущего подвида отличается лишь меньшей степенью перверзии рогов
у самцов, и поэтому В. И. Цалкиным (1951), по-видимому, справедливо
объединяется с последним. Распространение: горы между
Эльбурсом и Копет-Дагом.
9) О. orientalis areal Eversmann — Устюртский баран. От предыду-
щего подвида отличается еще более мощным развитием подвеса, более
узкой мордой, а также тем, что самки всегда рогаты. Распростра-
нение: Устюрт, полуостров Мангышлак, побережье залива Кара-
Богаз-Гола.
10) О. orientalis punjabensis Lydekker — Пенджабский баран. Размеры
сравнительно небольшие; основная длина черепа самцов около 210—
215 мм. Рога самцов слабо первертированные. Передне-наружное ребро
не всегда резкое. Лобовая грань сильно скошена наружу. Самки всегда
дмеют рога. Седловидные пятна отсутствуют. Подвес у самцов хорошо
развит и доходит до головы. Распространение; Соляной хре-
бет и некоторые другие районы Пенджаба.
11) О. orientalis, blanf or di Нише — Белуджистанский баран, или баран
Бланфорда. Очень плохо изученная раса. По-видимому, очень близка,
если не идентична, с предыдущей. ,Р а с п р о ст ранение: восточный
Белуджистан.
12) О. orientalis vignei, Blyth — Баран Малого Тибета, или баран
Винье. Размеры средние; основная длина черепа самцов. 226—228 мм.
JPora у самцов слабо первертированные. Передне-наружное ребро ясно
выражено, но лобовая грань скошена наружу. Самки имеют рога,. Седло-
видное пятно отсутствует. Подвес у самцов хорошо развит и доходит
до головы. Распространение: Ладак, Малый Тибет, Вахан
и Ишкашим. < .
13) О. orientalis bochariensis Nasonov — Таджикский баран. Размеры
сравнительно небольшие; основная длина черепа у самцов от 202 до
225 мм. Рога у последних первертированные, изредка даже гомонимные.
Поперечное ребро, обычно, выражено слабо. Лобовая грань скошена
наружу. Самки всегда имеют’ рога. Седловидные пятна отсутствуют.
Подвес у самцов хорошо развит и доходит до головы. Распростра-
нение: Таджикистан на восток до Дарваэа и на север до р. Зеравшан.
14) О. orientalis severtzovi Nasonov — Баран Кызылкумов, или баран
Северцова. Размеры сравнительно крупные; основная длина черепа сам-
црв около 240 мм. Рога у самцов всегда гомонимные. Передне-наружное
ребро на рогах самцов всегда резко выражено. Лобовая грань плоская
или вогнутая и наружу не скошена. Самки имеют рога. Седловидные пятна
отсутствуют. Подвес развит слабо и до основания головы не доходит.
Отличием от других подвидов является также более темная, серовато-
буроватая окраска. Распространение:. возвышенности Кы-
зылкумов.
Биология. Азиатский муфлон, как и другие виды рода Ovis, — зверь
горных ландшафтов. Даже зимой, когда условия в высокогорье стано-
вятся для них мало благоприятны, бараны только в немногих местностях
спускаются в предгорные равнины. Пределы вертикального распростра-
нения баранов колеблются от 100 до 150 м над ур. м., как это наблюдается
В северо-западных Каракумах (Цалкин, 1948), и до 3130 м—предела,
указываемого С. К. Далем (1954) для арменийского муфлона.
Однако утверждение С. И. Билькевича (Насонов, 1913), что бараны
в Коиетдаге занимают «совершенно одинаковые области» с безоаровыми
510
BOVIDAE
козлами и что «чем выше горы и больше ущелий, скалистых обрывов,
тем вероятней найти баранов», совершенно не верно. В противополож-
ность козлам, бараны в нормальных условиях не являются обитателями
скалистых гор. Только настойчивое преследование и занятие подходящих
для них стаций заставляет муфлонов уходить в зону высокогорий и в мало-
доступные скалы. В норме бараны таких мест явно избегают.
Более характерны открытые горные стации со спокойным рельефом:
плато, пологие склоны, закругленные вершины. Правда, бараны не из-
бегают и даже имеют привычку держаться в таких местах, где участки
спокойного рельефа сочетаются с ущельями, глубокими оврагами или
выступами скал. Но ущелья и обрывы служат баранам лишь местами
отдыха и укрытия от зноя и зимней непогоды. В качестве мест постоян-
ного обитания и выпаса даже небольшие лужайки между скал их, как
правило, не удовлетворяют. При преследовании бараны редко ищут
спасения в неприступных скалах и больше рассчитывают на быстроту
своих ног, приспособленных к бегу, чем на значительно слабее развитую,
по сравнению с козлами, способность карабкаться по крутым камени-
стым склонам. «Часто, — пишут С. И. Огнев и В. Г. Гептнер (1929), —
бараны проводят жаркую пору дня, не скрываясь в ущельях, но отды-
хая в тени густых арчей. Из этого уже видно, что барану ущелье само
по себе, как чисто механическое условие безопасности, нужно в меньшей
степени, чем козлу, важна, главным образом, тень для дневного отдыха.
Главное условие — пастбище достаточно широкое, чтобы заметить из-
далека опасность. В этом коренная разница его биологии от биологии
козла». Необходимым условием местообитания муфлонов, кроме нали-
чия хорошего пастбища и широкого кругозора, является также близость
источника водопоя. В таких условиях бараны живут в Закавказье. Ле-
том они держатся в полосе горных степей с разнотравно-злаковой и ксе-
рофитной растительностью, зимой спускаются к подножью хребтов,
вплоть до равнин в долине среднего течения Аракса. Избегают не только
глубоких ущелий и скал, но и альпийских лужаек (Саркисов, 1944; Даль,.
1954).
Сходные стации занимают бараны в Копетдаге. Это — горные плато,
изрезанные долинами и ущельями, пологие склоны гор с растительным
покровом, состоящим из типчака, ковылей и пыреев с примесью разно-
травья. Излюбленные места — открытые, ровные плато, пересеченные
рядом сравнительно пологих ущелий и покрытые богатой растительностью'
степного типа с разбросанными местами зарослями и отдельными деревьями
арчи (Огнев и Гептнер, 1929; Цалкин, 1948).
В Таджикистане местообитаниями баранов являются склоны гор,,
порой довольно крутые, но обычно имеющие сглаженный рельеф, по-
крытые растительностью полупустынного и степного типа. Горы Кара-
тау на Пяндже, где К. К. Флеров и И. М. Громов (1935) встречали много
баранов, представляют ряд сильно изрезанных и пересеченных оврагами
горных цепей с крутыми склонами округлых очертаний, поросшими
редкой фисташкой, образующей местами рощи и небольшие лесочки.
Такой же рельеф, по моим наблюдениям, имеют горы Сарсариак на Вахше,
также являющиеся местами обитания уриалов. Летом бараны здесь дер-
жатся в верхней полосе гор на более богатых разнотравной зеленой рас-
тительностью участках вблизи родников. Осенью спускаются ниже, до-
ходя до высоких предгорий, растительность которых носит полупустын-
ный характер. Значительная часть зверей, впрочем, и на лето остается
в среднем поясе гор, если там нет стад домашних овец. В узких долинах?
OVIS
511
и в фисташковых рощах они находят укрытие от зноя, а питаются высох-
шей травой и зеленой листвой фисташек.
На пологих горных склонах и мягких вершинах гор держатся бараны
в Нуратау. Только в жаркую дневную пору они укрываются в скалах
(Меклеибурцев, 1937). На Устюрте предпочитают, по В. С. Бажанову
(1945), места ближе к верхнему краю обрывов, выходят даже на самый
край плато. Однако, как и в Нуратау, к подножью гор, на равнины, ба-
раны даже зимою спускаются очень редко.
В большинстве районов распространения азиатского муфлона сезон-
ные миграции выражены слабо или отсутствуют вовсе. Обычно происхо-
дят лишь незначительные вертикальные перемещения популяций. Как
уже отмечалось, на лето бараны поднимаются более высоко в горы, при-
влекаемые, очевидно, более прохладным климатом и лучшей обеспечен-
ностью сочными зелеными кормами. На зиму они спускаются в нижнюю
полосу гор и даже в предгорья. В Закавказье, например, по А. А. Сар-
кисову (1944), муфлоны зимой подходят иногда к населенным пунктам
и используют в качестве выпасов озимые поля.
Очень слабо выражены сезонные миграции в Центральном Копет-Даге;
в одних и тех же местах баранов можно встретить в течение всего года
(Насонов, 1913). Однако и здесь, по В. И. Цадкину (1948), такие мигра-
ции имеют место. При этом выше всех летом поднимаются в горы самцы,
тогда как самки с Молодыми держатся заметно ниже. На Устюрте бараны
ведут практически совсем оседлый образ жизни, придерживаются, обычно,
небольших урочищ, имеющих родник, и далеко от него не уходят.
Ясно выраженные регулярные сезонные миграции муфлонов не только
в вертикальном, но и широтном направлении, наблюдаются, по-видимому,
только в Закавказье. Здесь, по А. А. Саркисову (1944), значительная
часть их откочевывает на зиму в пределы Турции и Ирана, не останав-
ливаясь перед таким препятствием, как р. Араке. Миграции проходят
определенными путями. Большие стада баранов, особенно в суровые
зимы, переселяются с запада на восток; часть их направляется через
Карчеванский проход недалеко от с. Джейранлы и, переправившись
через Араке, уходит в пределы Ирана на Дарашамские горы. Другой
путь миграции проходит через Волчьи Ворота у ст. Велидаг и через Араке
ведет, с одной стороны, на Кареинское плато в Иране, а с другой —
в Турцию, на южные склоны Арарата и в пределы Баязетского санджака.
Из возвышенностей советского Бадхыза большая часть баранов от-
кочевывает в соседние более высокие районы Афганистана (Громов, 1937).
Кроме регулярных сезонных перемещений, имеют место нерегуляр-
ные перекочевки баранов, главным образом летом, связанные с выго-
ранием растительности в засушливые годы и недостатком по этой причине
корма и питьевой влаги.
В зависимости от численности в той или иной местности, муфлоны
держатся летом группами от-5 до 15—20 голов. Летние табунки состоят,
в основном, из самок с приплодом текущего года, но иногда вместе с ними
держатся и один или даже два самца (Флеров, 1932). Самцы чаще дер-
жатся отдельными небольшими группами; вместе с ними можно видеть
и холостых самок. Старые самцы часто держатся в одиночку.
В конце осени группки баранов начинают объединяться и образо-
вывать табуны, размеры которых зимою доходят до 100—150 и даже
до 200 голов. Особенно велики стада бывают во время зимних перекоче-
вок (Саркисов, 1944). Табуны распадаются весною перед окотом; от них
отделяются самки и держатся в одиночку, пока не окрепнет молодняк.
512
BOVIDAE
Вожаком в смешанном стаде является взрослый самец. Последний,
когда стадо находится на отдыхе или при пастьбе, стоит на возвышенности
или камне и зорко смотрит по сторонам. Увидев опасность, он топает
ногой и первый пускается в бегство. При беге впереди находится вожак,
за ним следуют самки с молодыми, а позади всех старые бараны. Сторожа,
по-видимому, непостоянны и время от времени сменяют один другого.
А. А. Саркисов (1944) наблюдал табун арменийских муфлонов в течение
четырех часов, за время которых сторожевые бараны сменялись шесть раз.
В южной Туркмении, цо сообщению Л. Морица (1931), бараны исполь-
зуют случайное соседство кекликов {Caccabis chucar schestoperovi Sush.);
последние предупреждают зверей о появлении опасности своим криком.
Бегают муфлоны быстро. По словам С. И. Огнева и В. Г. Гентнера
(1929), бег их настолько быстр и ловок, что «не видно как животное каса-
ется земли». В случае необходимости они делают высокие, до 1.5 м, и длин-
ные прыжки, легко перемахивают через кусты и камни. Нередко спры-
гивают вниз с высот до 10 м; при прыжке голову и рога закидывают назад,
передние и задние ноги смыкают вместе, но приземляясь падают не на
рога,. как нередко описывается в охотничьей литературе, а на копыта
широко расставленных ног (Мекленбурцев, 1937).
В пределах избранного местообитания муфлоны ведут сравнительно
оседлый образ жизни, придерживаются определенных мест отдыха, кор-
межки и водопоев. При переходах пользуются одними и теми же путями,
в результате чего в местностях, где баранов много, ими вытаптываются
хорошо заметные тропы. Этими тропами иногда пользуются и другие
звери.
В Больших Балханах бараны ежедневно, по утрам, поднимаются
на самый верх, а к вечеру спускаются в пониженные места, выбирая
пологие, легкие спуски (Лаптев, 1934).
Днем, в жаркие солнечные часы, бараны укрываются в ущельях, под
навесами скал или в тени крупных арчевых и фисташковых деревьев.
На жировку (пастьбу) летом выходят, когда спадает жара. Кормятся
до наступления сумерек. Пьют на закате или в начале ночи. Ночью,
по крайней мере некоторое время, отдыхают. На рассвете снова пьют
и направляются в горы, где пасутся вблизи мест дневного отдыха до
наступления жары. .
Лежки баранов, по-видимому, постоянны; в Таджикистане они, по
К. К. Флерову и И. М. Громову (1935), имеют вид вытоптанных довольно
глубоких, до 1.5 м, ям, иногда даже нор, уходящих под скалы, корни
кустарников и деревьев или просто под нависающие склоны. Рытье глу-
боких лежек имеет целью, видимо, не столько маскировку, сколько за-
щиту от вредного действия высокой температуры.
В зимнее время бараны пасутся все светлые часы суток. В сильные
холода и ненастье укрываются в глубоких, защищенных от ветра ущельях
или в скалах. .
Основу питания азиатского муфлона в летний период составляют
разнообразные травы. Видовой состав пищи зависит от района обитания,
но в основном — это злаки: ковыли, типчак и пырей. Ковыли, типчак?,
пыреи и астрагал считаются В. Й. Цалкиным (1948) основными кормами
копетдагских баранов, хотя составленный им список поедаемых расте-
ний включает 17 видов. Охотно поедаются и такие виды разнотравья,
как крестовница, бурачок, эспарцет, зверобой и некоторые другие, но
они имеют меньшее значение, хотя бы потому, что составляют незначи-
тельную долю в общем травостое.
OVIS
613
В Закавказье бараны охотно едят люцерну, в Нуратау — тюльпаны
и особенно крупные листья Megacarpala (Мекленбурцев, 1937). Полыни,
как и другими копытными, муфлонами поедаются лишь зимою. Травя-
нистый корм поедается как в зеленом, так и в сухом виде. В Бадхызе
и Таджикистане при питании летом сухой травой, в качестве зеленого
корма употребляется листва и побеги фисташки (Громов, 1937; Флеров
и Громов, 1935). В. С. Бажанов (1945), исследуя желудки баранов на
Устюрте, обнаружил в них четыре вида солянок, из которых основу со-
держимого составляла Anabasis turgaica Iljin etKrasch., затем карагану,
колючую курчавку, морскую полынь (Artemisia maritima L.). В неболь-
шом количестве содержался житняк и тростник.
Зимой бараны питаются, в основном, также остатками сухой травы,
торчащими из-под снега, и пасутся на бесснежных участках. Откапывать
траву из-под снега муфлоны, по-видимому, мало способны (Цалкин,
1948). При недостатке другой пищи зимой они объедают тонкие ветки
кустарников и даже грызут кору. Последнюю муфлоны иногда предва-
рительно. сбивают ударами рогов о стволы деревьев (Насонов, 1913).
И. М. Громов (1937) находил в желудках баранов остатки членистоно-
гих: жуков, фаланг и даже скорпионов.
Хотя в литературе и встречаются указания, что муфлоны могут по-
долгу обходиться без питья и что их часто находят вдали от источников
водопоя, ' все же наличие воды, по-видимому, является необходимым
условием возможности существования их в том или ином районе.
В засушливые годы они бывают вынуждены откочевывать из безвод-
ных районов (Бажанов, 1945; Насонов, 1913). При недостатке или от-
сутствии пресной воды муфлоны пьют соленую, и не только из ручьев,
но и из сильно соленых луж (Флеров, 1932).
Голос азиатского муфлона сходен с голосом домашней овцы, но более
грубый и короткий. Пользуются им бараны редко, — он слышится
обычно, когда звери скликают друг друга. Самцы-сторожа, заметив опас-
ность, издают звук, сходный с «кии . . .к» (Флеров и Громов, 1935), а во
время бега, по М. К. Лаптеву (1934), звуки, похожие на «тох-тох».
У муфлонов хорошо развиты как слух, обоняние, так и зрение. Наи-
более остро обоняние. Муфлоны — очень чуткие и осторожные звери.
Считается, что с подветренной стороны подойти к ним ближе, чем на 300
шагов, невозможно. Нередко даже, не видя человека, они могут учуять
его по ветру за 300—400 шагов и дальше (Насонов, 1913; Саркисов, 1944).
Особенно осторожны самки с ягнятами. С другой стороны, муфлоны
проявляют нередко признаки любопытства. Завидя человека, если он
движется спокойно, они иногда смотрят на него, не двигаясь, и подпу-
скают шагов на двести. Были случаи, когда на автомашине удавалось
приблизиться к неподвижно стоящим баранам на 50 шагов (Саркисов,
1944). При беге иногда останавливаются и оглядываются назад.
Линька в Закавказье и южной Туркмении у диких баранов начинается
в конце марта и тянется около двух месяцев. К концу мая у взрослых
самцов она заканчивается. У самок, в связи с деторождением, она затя-
гивается, и еще в июне можно видеть не вылинявших животных. При
возможности, бараны в это время трутся о деревья и кусты,‘-J оставляя
на них клочья старой шерсти. Осенью смены волосяного покрова не про-
исходит. Со средины или конца сентября начинает лишь отрастать пух
и более быстро, чем летом, расти остевой волос. Отрастание волоса за-
канчивается и полный зимний наряд звери получают перед.. началом
гона, к ноябрю (Саркисов, 1944; Цалкин, 1948).
33 Фауна СССР, И. И. соколов
314
BOVIDAE
Половой зрелости азиатские муфлоны, по В. И. Цалкину (1948), до-
стигают и начинают участвовать в размножении на третьем году жизни.
У отдельных животных течка наступает в конце октября. Массовый же
гон у баранов в большинстве районов протекает в период с середины
ноября по первую половину декабря.
В это время животные держатся табунками до 10—15 голов, в кото-
рых находятся по одному-два, а то и больше взрослых самцов. Изгнания
самцами друг друга из стада, по-видимому, не происходит, но между
ними происходят поединки. Разойдясь метров на двадцать, они стреми-
тельно сближаются и с силой ударяются основаниями рогов, так что
звук удара в горах бывает слышен за 2—3 км. Иногда самцы сцепляются
рогами, сцепившись водят друг друга, возятся, падают, издают стону-
щие звуки. Однако, в противоположность, например, оленям, утомлен-
ные самцы прекращают борьбу и мирно остаются оба в стаде, так что
участвовать в покрытии самок могут все находящиеся при стаде бараны.
Через некоторое время борьба может возобновиться. Случаев тяжелых
увечий или убийства во время драк не известно. Но самцы в это время
теряют обычную осторожность и чаще, чем обычно, становятся жертвой
охотника или хищного зверя.
Самки во время течки и драк самцов ведут себя спокойно. Ухажи-
вание диких баранов за самками сходно с тем, что наблюдается у домаш-
них овец: самец с тихим блеянием ходит за самкой, трется шеей о ее бока,
пытается покрыть (Саркисов, 1944). По окончании полового сезона самцы
не отделяются от табунов и остаются с самками до весны.
Беременность у диких муфлонов, как и у домашних овец, длится
около пяти месяцев. Первые случаи окотов могут иметь место уже в конце
марта, но в основном рождение молодняка происходит во второй поло-
вине апреля и в первой половине мая.
Перед окотом самки отделяются от стад, в одиночку уходят в глубо-
кие ущелья или каменистые россыпи, где в укромных местах родят яг-
нят. Приносят большею частью по два ягненка, реже по одному или по
три. Известны, правда, очень редкие случаи, когда было даже четыре
ягненка (Флеров и Громов, 1935).
Ягнята азиатских муфлонов, в противоположность европейскому,
первое время довольно беспомощны, следовать за матерью не могут и
при опасности не бегут, а затаиваются (Мориц, 1931). Когда они доста-
точно окрепнут, что происходит через педелю-полторы после рождения,
самки выходят с ними в более открытые места и вновь объединяются
в небольшие группы.
Кормятся молоком матерей ягнята до сентября или октября, но зе-
леный корм начинают понемногу употреблять раньше, с месячного воз-
раста. Голос муфлонят мало отличается от голоса домашнего ягненка.
К годовалому возрасту молодые муфлоны достигают несколько больше
двух третей роста взрослых и около одной трети их веса. Полного роста
в высоту достигают к 4—5 годам, но увеличение длины тела и живого
веса продолжается до 7 лет и, быть может, даже дольше (Цалкин, 1948).
Продолжительность жизни в естественной обстановке зависит от того,
сколь интенсивно их преследуют; по некоторым данным не превышает
12 лет (Огнев и Гептнер, 1929).
В Тбилисском зоопарке самка арменийского муфлона, по сообщению
А. А. Саркисова (1944), жила в течение 10 лет и в течение всего этого вре-
мени давала здоровый приплод. В Лондонском зоологическом саду самка
уриала прожила 11 лет 7 месяцев и 20 дней (Flower, 1931).
OVIS
515
Врагами азиатского муфлона из мира млекопитающих являются,
в первую очередь, волки и леопарды. Однако вред и от тех, и других
не так велик. Волк с трудом догоняет взрослого и не слишком старого
барана. Кроме того, на Больших Балханах, например, волков в горах,
по М. К. Лаптеву (1934), нет, и вообще они в горы заходят менее охотно.
В питании леопарда (Pardus pardus L.) дикие бараны занимают важное
место, но в силу малой численности этого хищника, вред от него также
незначителен. Волки охотятся на баранов стаями нагоном. Леопарды
же обычно подкарауливают их в одиночку на путях переходов, подпол-
зая к ним из-за прикрытий или в высокой траве“йа животе, как это на-
блюдал С. И. Билькевич (Насонов, 1913). На молодых могут нападать
крупные хищные птицы, например бородач (Gypaetus barbatus L.), неко-
торые из более мелких диких кошек, даже гепард (Acinonyx jubatus
Schreb.), а на новорожденных лисицы (Мориц, 1931). Однако мас-
сового характера гибель от хищников не носит- Основной причиной
снижения численности диких баранов является нерациональный промы-
сел, в еще большей степени — освоение излюбленных ими стаций под
выпас домашних животных.
Массовых заболеваний среди азиатских муфлонов до сих пор не на-
блюдалось. Паразитофауна их изучена еще недостаточно. Из эктопара-
зитов Л. Мориц (1931) находил на копетдагских муфлонах муху-крово-
соску (Melophagus ovinus L.). В. И. Цалкин (1948) упоминает о подкож-
ном оводе на копетдагских муфлонах без указания их видовой принадлеж-
ности. К. Я. Грунин (1951) описал новый вид носоглоточного овода с
уриала, добытого в Кулябской области Таджикской ССР, — Rhinoest-
rus tshernyshevi Grun.
На муфлонах в Закавказье установлено два вида клещей: Наета-
phisalis sulcata Can. et Fane, и Ripicephalus turanicus B. Pom. (Саркисов,
1944).1
Г. А. Григорян (1951) при вскрытии 6 муфлонов, добытых в Армении,
обнаружил 21 вид паразитических червей, 15 из которых оказались об-
щими с безоаровыми козлами и 15 общими с местными домашними ов-
цами. В наибольшем числе у всех исследованных зверей находились
и, по-видимому, наибольшее патогенное значение имеют два вида легоч-
ных нематод — Protostrongylus davtiani Ssav. и Р. muraschkinzewi Dawt.,
вызывающих поражение легочной ткани.
Хозяйственное значение. Азиатский муфлон имеет промысловое
значение главным образом как мясной зверь. До сих пор в некоторых
районах существует организованный промысел муфлонов. Только в цен-
тральном Копетдаге, по расчетам В. И. Цалкина (1948), ежегодно до-
бывается около 1000 баранов.
Мясо муфлонов обладает хорошими вкусовыми качествами, доста-
точно нежно, жирно и ценится, по словам С. И. Билькевича, выше мяса
джейрана (Насонов, 1913). Наилучшей упитанностью бараны обладают
с сентября по ноябрь, перед началом гона, во время которого самцы
вновь худеют.
Кожа, хотя особенной ценности и це представляет, безусловно мо-
жет служить сырьем для выработки шевро и хрома.
Одна или несколько рас азиатского муфлона несомненно явились
радоначальниками домашних овец. Скрещивание с последними легко
1 В литературе еще встречаются требующие проверки сведения о находках Во-
ophilus calcaratus Bir. (сообщение Г. В. Сердюковой).
33*
516
BOVIDAE
происходит и и настоящее время. Пойманные в молодом возрасте,
муфлоны легко приручаются, привязываются к ухаживающему за ними
человеку. Случаи поимки и выращивания их в неволе в районах рас-
пространения и сейчас не так редки. С. И. Огнев и В. Г. Гептнер (1929)
в южной Туркмении видели жившую уже несколько лет прирученной
самку, которая не делала никаких попыток к бегству, паслась вместе
со стадом домашних овец и вместе с ними сама возвращалась в селение.
В г. Тавризе трехлетний ручной самец свободно разгуливал по улице,
но его пришлось позднее посадить на привязь, так как он стал нападать
на встречных людей (Саркисов, 1944).
В зоологических садах муфлоны хорошо размножаются. При скре-
щивании с домашними овцами дают во всех случаях вполне плодовитое
потомство. Получены также плодовитые гибриды от скрещивания копет-
дагского барана и уриала с муфлоном. Гибридные самки в обоих слу-
чаях имели рога (Gray, 1954). В Ереванском зоопарке получены гибриды
от скрещивания самки европейского муфлона с самцом арменийского
муфлона (Саркисов, 1953).
Имеются требующие проверки сведения о получении гибридов от
скрещивания барана Винье (О. о. vignei Blyth) с аргали (Ewart, 1914;
Саркисов, 1953). О плодовитости гибридов аргали с уриалом ничего не
известно.
Охота. Из способов добычи муфлонов наиболее распространенным
в настоящее время является скрадывание. Это трудный, но увлекатель-
ный спорт. Он требует от охотника много выносливости и выдержки.
Баран, особенно там, где сильно подвергается преследованию, очень
осторожен. Необходимо подкрадываться против ветра, тщательно пря-
чась за камни, кусты и другие укрытия. Более успешным бывает скра-
дывание баранов-самцов во время гона, когда они, во-первых, легко
обнаруживают себя по далеко слышному стуку рогов, а во-вторых, ув-
леченные дракой, позволяют охотнику подойти иногда очень близко.
Можно подкарауливать баранов на тропах у переходов и на местах во-
допоев. Стрелять лучше пулей, а при хорошем заряде пороха — и круп-
ной картечью.
В некоторых местностях ловят баранов в капканы, расставляя по-
следние на постоянных тропах и местах отдыха животных (Мекленбур-
цев, 1937).
Ovis ammon Linnaeus — Архар, или аргали (рис. 108—112)
Linnaeus С., 1758, Syst. Natur., ed. X, t. I : 70 (первооп.); П а л л а с П. С.,
1788, Путешеств. по разным провинциям Российского госуд., ч. 1, полов. 1 : 312, 315,
319, 320(распр.); М а а к Р., 1859, Путешеств. на Амур, СПб. : 112 (ист.); R ad de G.,
1861, Beriehte u. Reisen im Siiden von Ost-Sibirien, St.-Petersb. (Beitr. z. Kenntn.
d. Russ. Reich., 23) : 433—434 (ист.); Северцов H. A., 1873a, Изв. Общ. любит,
естествозн., антропол. и этногр., VIII, 2 : 85 (сист., распр.); Северцов Н. А.,
18736, Природа, 1 : 144—245 (общ., изобр.); Brooke V., 1875, Proc. Zool. Soc.
London : 509—526 (морф.); Иванов Д., 1885, Природа и охота, 1 : 1—48 (распр.,
биол.); Lydekker R., 1898, Wild oxen, sheep a. goats of all lands, London : 174—
203 (общ., изобр ); De m ido f f E., 1900, After wild sheep in the Altai and Mongolia,
London : I—XII-j-1—324 (общ., изобр.); ГоловнинД. H., 1901, Природа и охота,
III: 1—11, IV : 1—9, V : 137—149, VI : 95—104 (общ., изобр.); Туркин Н. В. и К. А. Са-
тунин, 1904, Звери России, т. IV, Bovidae, М. : 560—582-J-672—753 (общ., изобр.);
Яблонский Н. И., 1907, Природа и охота, XI : 7 (распр.); Lydekker R.,
1913, Catal. Ungulate Mammals Brit. Mus., I, London : 92—109 (сист., синон., изобр.);
Насонов Н. В., 1914, Изв. Акад, наук : 695—726 (морф., сист., распр., изобр.);
Rowland Ward, 1914, Records of Big. Game, London : 396—400 (porLj,
Ewart J. C., 1914, Trans. Highl. Agricul. Soc., Scotl., 26 : 74—101 (цит. no Gray,
OVIS
617
1954) (гибр.); Чернавин В. В., 1916, Ежегодн. Зоол. муз., XXI, 2/3 : 200—213
(биол., распр.); Дорогостайский В. Ч., 1918, Ежегодн. Зоол. муз., XXIII,
1 : 32—37 (распр., биол.); Н а с о н о в Н. В., 1923, Географии, распростр. диких ба-
ранов Старого Света, Птг., изд. Акад, наук : 69—123 (сист., распр., синон., изобр.);
Селевин В. А., 1925, Охотник, 6—7:28—29 (распр.); Morden W., 1928,
Natur. history, XXVIII : 486—500 (биол., ох., изобр.); Северцов С. А., 1921,
Землеведение, XXXI, 2/3 : 180—181 (распр., биол.); Teilhard de Cha г din Р.
et J. Piveteau, 1930, Ann. Paleont., 19 : 3—132 (эвол.); Булавин A., 1934,
Охотник и рыбак Сибири, 3 : 24 (распр.); Б у т а р и и Н. С., 1935, Тр. Киргизск.
компл. экспед., IV, 3 : 59—76 (волос); Громова В. И., 1935, Докл. Акад, наук
СССР, IV, 1/2 : 97—99 (ист.); Денисов В. Ф., 1935, Тр. Киргизск. компл. экспед.,
IV, 3 : 81—113 (хоз. знач.); Розанов М. П., 1935, в кн. : Матер, по млекопит. и
птицам Памира, Л., изд. Акад, наук СССР : 34—37 (распр.); Румянцев Б. Ф.,
Н. С. Б у т а р и н и В. Ф. Д е н и с о в, 1935, Тр. Киргизск. компл. экспед., IV,
3 : 15—58 (морф., сист., гибр., изобр.); Ф л е р о в К. К., 1935, в кн.: Виноградов Б. С.,
Е. Н. Павловский и К. К. Флеров, Звери Таджикистана, Л., изд. Акад, наук СССР :
127—130 (общ., изобр.); Gromova V., 1936, Neue Forsch. in Tierzucht und Abstam-
mungslehre (Festschr. z. 60. Geburtstag v. prof. J. U. Duerst), Bern : 81—91 (сист.);
Шнитников В. H., 1936, Млекопит. Семиречья, М.—Л., изд. Акад, наук СССР :
134—137 (распр., биол.); Громова В. И. и В. И. Г р о м о в, 1937, Тр. Советск,
секц. Международн. ассоц. по изуч. четверт. периода, 1 : 55—57 (ист.); В о h 1 i п В.,
1938, Nov. Acta Soc. Sci. Upsala, ser. IV, v. XI, № 2 : 1—54 (эвол., ист.); К о л о-
с о в А. М., 1939, Учен. зап. Моск. гос. унив., 20 : 176—181 (распр., биол.); Pen Н.,
1939, Bull. Fan Memorial Inst. Biol., Zool. ser., IX, 2 : 71—219 (морф.); Соколов С. С.,
1939, Зоол. журн., XVIII, 3 : 344—450 (биол.); Б у т а р и н Н. С., 1940, Тр. Инет,
генет. Акад, наук СССР, 13 : 175—247 (морф., гибр., изобр.); Машковцев А. А.,
1940, Боец-охотник, № 7 : 24—28 (распр.); Огнев С. И., 1940, Матер, к позн.
фауны и флоры СССР, н. с., Отд. зоол., 3 : 79 (распр.); Антипин В. М., 1941,
Млекопит. Казахстана, т. III, Копытные, Алма-Ата, Каз. ОГИЗ : 77—93 (общ.,
изобр.); Schaub S., 1943, Ecl. Geol. Helvet., XXXVI, 2 : 282 (эвол., ист.);
Афанасьев А. В., 1945, Изв. Акад, наук Казахск. ССР, 5 : 29 (распр.);
Б а ж а н о в В. С., 1945, Изв. Акад, наук Казахск. ССР, сер. зоол., 5 : 75—78 (распр.);
Микулин М. А. и А. Я. Исаева, 1945, Изв. Акад, наук Казахск. ССР, 5 : 22
(распр.); Антипин В. М., 1946, Вести. Акад, наук Казахск. ССР, 7/8 : 49—50
(распр.); Антипин В. М., 1947, Изв. Акад., наук Казахск. ССР, сер. зоол., 6 :
3—22 (биол., эвол.); Афанасьев А. В. и А. А. Слудский, 1947, Изв. Акад,
наук Казахск. ССР, сер. зоол., 6 : 57 (распр.); Зверев М. Д.и А. А. Келей-
ников, 1947, Тр. Алма-Атинск. заповеди., IV : 57—58 (мигр.); Cobb Е., 1947,
Field, London : 189 (рога); Белослюдов Б. А., 1948, Изв. Акад, наук
Казахск. ССР, сер. зоол., 8 : 61 (распр.); Боев С. Н., И. Б. С о к о л о в а и
В. И. Бондарева, 1948, Изв. Акад, наук Казахск. ССР, сер. паразитол., 6:
85—98 (параз.); Громов В. И., 1948, Тр. Инет. геол, наук, 64 : 433—434 (ист.);
Громова В. И., 1948, Изв. Акад, наук СССР, сер. биол., 5 : 528—529 (ист.);
Зверев М. Д., 1948а, Тр. Алма-Атинск. зоопарка, I : 94—116 (биол.); 3 в е-
р е в М. Д., 19486, Тр. Алма-Атинск. гос. заповеди., VII : 146 (быстрота бега); Куз-
нецов Б. А., 1948, Звери Киргизии, М., изд. Моск. общ. испыт. прир. : 115—118
(общ.); Мекленбурцев Р. Н., 1948, Бюлл. Моск. общ. испыт. прир., отд.
биол. LIII, 5 : 65—84 (общ.); Шульпин Л. А., 1948, Изв. Акад, наук Казахск.
ССР, сер. зоол., 7 : 67 (распр., биол.); Абдилдабеков К. А., 1950, Изв. Кир-
гизск. фил. Акад, наук СССР, I—IX : 77—78 (распр., биол.); Б у т а р и н И. С.,
1950, Сов. зоотехния, I : 35—48 (гибр., хоз. знач.); Петров О. В., 1950, Тр. Ле-
нингр. общ. естествоисп., LXX, 4 : 155—193 (распр.); Пидопличко И. Г.,
1951, О ледниковом периоде, II, Киев, изд. Акад, наук УССР : 58—59 (ист.); Ц а л-
к и н В. И., 1951, Горные бараны Европы и Азии, М., изд. Моск. общ. испыт. прир. :
255—265 (сист., синон., распр., изобр.); Ellerman J.a. Т. Morriso n-S со 11,
1951, Checklist of palaerctic a. indian mammals, London : 413—416 (сист., синон.);
Янушевич А. И., 1952, Фауна позвоночн. Тувинской области, Новосибирск,
изд. Зап.-Сибирск. фил. Акад, наук СССР : 62 (распр.); Афанасьев А. В.,
В. С. Бажанов и др., 1953, Звери Казахстана, Алма-Ата, изд. Акад, наук Каз.
ССР : 493—497 (общ., изобр.); Верещагины. К., 1953, Тр. Естеств.-иет. муз.
им. Зардаби, Баку, 6 : 302—308 (ист.); Мадам ин ов С. Ю., 1953, Тезисы докл.
на выездной сессии Акад, наук Таджикской ССР в Ленинабаде, Сталинабад : 175
(распр.); Саркисов А. А., 1953, Природа, 2 : ИЗ (гибр.); Андрушко А. М.,
1954, Уч. зап. Лен. гос. унив., 181, сер. биол. наук, 38 : 17 (распр.); Банников А. Г.,
1954, Млекопит. Монгольск. Народи. Республ., М., изд. Акад, наук СССР : 266—289
(общ.); Боев С. Н., 1954, Зоол. журн., XXXIII, 4 : 779—787 (параз.); Gray А.,
518
BOVIDAE
1954, Mammalian hybrids, London : 76—77 (гибр.); Дементьев Д. П. и П. С. Т ю-
р и н, 1954, Тр. Инет. зоол. и паразитол. Киргизок, фил. Акад, наук СССР, II : 154
(распр.); Насимович А. А., 1955, Роль режима снежн. покрова в жизни копыт-
ных, М., изд. Акад, наук СССР : 144—157 (распр., биол.); Шапошников Ф. Д.,
1956, Природа, 1 : 109—110 (мигр.).
Описание. Баран очень крупных размеров; длина тела взрослых
самцов от 177 до 200 см и даже выше, высота в холке от 106 до 125 см,
косая длина туловища (от плечелопаточного сочленения до заднего вы-
ступа седалищного бугра) от 102 до 114 см. Живой вес вполне взрослых
самцов составляет от НО до 170 кг, а в исключительных случаях, по-
видимому, достигает 200 кг.1 Основная длина черепа у самцов 258—349 мм.
Самки значительно меньше самцов; высота в холке у них колеблется
от 95 до 112 см, косая длина туловища от 85 до 95 см, живой вес от 59
до 90—100 кг. Основная длина черепа у взрослых самок 232—302 мм.
Телосложение крепкое. Сравнительно короткое, массивное туловище
покоится на высоких, не очень тонких ногах. Обхват груди позади ло-
паток у самцов составляет в среднем 120 см, максимально до 135 см. Вы-
сота в крестце на 2—7 см превышает высоту в холке. Линия спины пря-
мая, несколько наклонная вперед. Длина хвоста около 11—18 см. Го-
лова массивная, у самцов увенчанная мощными рогами. Профиль носа,
как правило, прямой, но иногда слегка горбоносый. Уши длиной около
9—13 см, очень подвижные. Шея у самцов сравнительно короткая и тол-
стая. Копыта относительно общей величины животного небольшие;
длина их на передних конечностях составляет 70—78 мм, на задних
59—67 мм. Высота передних копыт 41—45 мм, задних на 2—4 мм меньше.
Самки более легкого сложения. Голова у них легкая, с тонкой прямой
мордой. Ноги и шея производят впечатление длинных и тонких, особенно
в летней шерсти. .
Рога у самцов аргаловидных баранов отличаются мощным развитием,
особенно толщиной. Длина их по изгибу передней поверхности у вполне
взрослых самцов колеблется в пределах 74—155 см, а по некоторым дан-
ным (Rowland Ward, 1914), достигает даже 190 см. Длина четырех сег-
ментов (прирост второго, третьего, четвертого и пятого года жизни) со-
ставляет от 67 до 115 см (Цалкин, 1951). Обхват рогов в основаниях ко-
леблется от 28.5 до 50 см (25.2 — 54.9% длины всего рога), но очень редко
бывает менее 30 см. Продольный диаметр сечения рога в основании (от
заднего ребра до наиболее выдающейся точки передне-внутреннего)
составляет 103—107 мм; наибольший поперечный диаметр рога — от
65 до 138 мм. Средние величины объема четырех сегментов у разных рас,
по В. И. Цадкину (1951), составляют, в среднем, от 1300 до 2500 см3. Рога
поставлены полого по отношению к продольной оси черепа, мало высту-
пают над плоскостью лба. С профилем носовых костей они образуют
угол, близкий к 180°.
Изгиб рогов у самцов всегда гомонимный. При этом рога, описывая
в пространстве более или менее растянутую спираль в полный оборот,
концами направлены вверх и в стороны или в стороны и слегка вниз.
Передне-наружное ребро на рогах хорошо выражено; передняя и наружная
грани рога образуют угол не более тупой, чем передняя с внутренней
гранью. Передняя (лобовая) грань плоская, расположена перпендику-
лярно продольной оси поперечного сечения рога, а нередко даже скошена
1 Размеры тела и живой вес приводятся по данным Б; Ф. Румянцева с соавторами
(1935), В. Ф. Денисова (1935), Н. С. Бутарина (1939) и Р. Н. Мекленбурцева (1948).
OVIS
51ff
заметно внутрь. Наружная грань рога у вершины слабо вогнутая, к ос-
нованию становится плоской или даже слегка выпуклой. Внутренняя
грань в дистальной половине сильно вогнута, но к основанию рога ста-
новится плоской или даже выпуклой. Поперечное сечение рога в осно-
вании имеет форму равнобедренного треугольника с более острым углом,
соответствующим заднему ребру рога. Описанная форма рогов типична
для баранов Каратау, Тянь-Шаня, Казахского нагорья. У самых круп-
Рис. 108. Архар (Ovis аттоп L.).
ных рас с Памира, Алтая и Монголии наблюдается вторичное притуп-
ление передне-наружного ребра рогов, особенно ближе к их основаниям.
Часто сильно закруглено также передне-внутреннее и в меньшей сте-
пени заднее ребро рога. Передняя грань рога становится при этом более
или менее выпуклой, но и в этих случаях она, как правило, остается
скошенной внутрь, а не наружу. Наружная грань у верхушек плоская,
к основаниям становится сильно выпуклой. Выпуклой становится и вну-
тренняя грань рога, только ближе к вершинам вогнутость ее остается
ясно заметной. Поперечное сечение основания рога, в силу отмеченных
особенностей, у крупных рас имеет округло-треугольную форму. Во
всех случаях вершины рогов сильно уплощены, лобная грань постепенно
суживается и сходит на нет, так что на конце рога фактически имеется
только два ребра — переднее и заднее, и две грани — наружная и внут-
ренняя.
Поперечная морщинистость на рогах аргаловидных баранов хорошо
выражена, особенно на лобовой грани. Отдельные складки (морщины)
обычно широкие и высокие, хотя, в противоположность козлам, вплот-
520
BOVIDAE
ную прилегают друг к другу. У некоторых рас, например у алтайского
аргали, глубокие поперечные борозды разделяют высокие и узкие мор-
щины.
Самки аргаловидных баранов всегда рогатые. Рога у них более корот-
кие (от 23 до 53 см), лишь изредка (у алтайской и центральноазиатских
рас) достигают полуторной длины черепа; часто имеют ясно выражен-
ный гомонимный изгиб; сильно сжаты с боков. Более или менее отчет-
ливы на рогах лишь передне-внутреннее и заднее ребро, причем оба
обычно закруглены. Поперечное сечение рогов у самок продольно-оваль-
ное, с плоской или вогнутой внутренней и выпуклой наружной гранью.
Морщинистость более мелкая, чем у самцов, но всегда имеется. Цвет
рогов светлый, желтовато-серый, реже темно-бурый.
Вполне развитый шерстный покров аргали состоит из грубых ломких
остевых волос и тонкого извитого подшерстка (пуха) (Бутарин, 1935).1
Остевые волосы слабо волнистые, на боках, шее и спине несколько бо-
лее мягкие. Очень толстые и грубые волосы на крупе около хвоста, на
животе и на шее. Длина остевых волос на туловище в зимнем меху —
около 5—6 см. Подвеса на нижней стороне шеи и бакенбард в том виде,
в каком они имеются у муфлонов, у аргаловидных баранов нет. Однако
в большинстве случаев удлиненные до 9—10 см и более грубые волосы
снизу и с боков шеи образуют подобие ошейника или манишки, рас-
пространяющейся на переднюю сторону груди и плечи. Удлинены до
8—9 см волосы в виде пучка на затылке и прилегающей части верха шеи.
Морда впереди глаз, а также ноги ниже запястных и скакательных су-
ставов покрыты коротким блестящим волосом, плотно прилегающим
к коже. Более редкий и мягкий волос, особенно в летнее время, покры-
вает подмышки, пахи, область молочной железы и наружных половых
органов.
Окраска аргаловидных баранов, так же как и у муфлонов, подвер-
жена географической и индивидуальной изменчивости. В одних случаях
она более насыщенная и темная, в других более бледная и светлая. Не
одинаково выражены и контрасты между темно и светло окрашенными
участками тела. Вместо рыжевато-желтых оттенков, характерных для
муфлонов, особенно в западной части ареала, в окраске баранов восточ-
ной части Средней Азии, южной Сибири, Монголии и Китая преобладают
коричнево-бурые тона.
Остевые волосы с боков туловища и шеи от основания светло-бурые,
кверху постепенно темнеют. Перед вершиной обычно, хотя и не всегда,
имеется светлое коричневое или желтоватое кольцо. Заостренные кон-
чики волос, пока не обломаны, черно-бурого цвета. Тот или иной отте-
нок окраске придают степень и характер посветления предвершинной
части ости. Окраска пуховых волос (подшерстка) однотонная, светло-
бурая или грязно-белая.
Общая окраска туловища буро-коричневая, иногда с желтоватым
или рыжеватым оттенком. Темной продольной полосы вдоль средней
линии спины в зимнем меху нет, но вообще верхняя сторона туловища
темнее, чем бока, постепенно светлеющие книзу. Седловидных пятен
по бокам, свойственных многим расам муфлонов, у аргали нет. Лишь
иногда на их месте и вообще на боках туловища и спине имеется незна-
чительная примесь белых волос, придающих окраске седоватый отте-
1 Утверждение Н. А. Северцова (18736 : 197), что у О. a. polii Blyth и О. а. ка-
relini Sev.. нет подшерстка и зимой, ошибочно.
OVIS
521
нок. По бокам туловища от локтевых к коленным суставам тянутся темно-
коричневые полосы, отграничивающие бока от светлой окраски живота.
Иногда эти полосы, впрочем, слабо выражены, почти незаметны, или
представлены пятнами позади локтей и впереди коленных суставов.
Шея, особенно с боков и снизу, имеет более светлую буровато-жел-
тую, иногда почти белую окраску. Низ груди и живота чаще желто- или
грязно-белый, но иногда бывает буро-коричневый, но более светлый,
чем бока и верх тела. Пахи, внутренняя сторона бедер, область молоч-
ной железы и наружных половых органов светлые, грязно- или желто-
белые. Светлая, часто с охристо-рыжим оттенком окраска захватывает
ягодицы и распространяется на круп по сторонам и выше хвоста. Иногда
она довольно четко отделяется от соседних темно окрашенных участков
бедер и крупа, но чаще незаметно переходит и сливается с окраской по-
следних. Хвост грязно-белый, иногда на конце темный, или по верху
его проходит темно-бурая полоса, как продолжение темной окраски
средней части крупа.
Голова, как правило, светлее туловища, буровато-коричневая или
желтовато-бурая. Лоб, а иногда и щеки, несколько темнее. Вокруг глаз
часто имеется темное коричневое кольцо. Темя и затылок темно-бурые, даже
темнее туловища. На носу позади ноздрей обычна темная коричневая
поперечная полоса или пятна по обеим сторонам носа. Конец морды,
верхняя и нижняя губа, а также низ головы светлые, грязно- или желто-
белые. Уши сверху буро-коричневые или буровато-серые, внутри грязно-
белые.
Ноги выше запястий и скакательных суставов более или менее одно-
тонные с боками туловища, часто имеют значительную примесь белых
волос, придающих ногам серовато-коричневую окраску. Внутренняя
сторона предплечий и голеней беловатая, сливающаяся со светлой окрас-
кой груди и живота. Ниже запястий и скакательных суставов, а иногда
и с половины предплечий ноги светлые, желто- или грязно-белые, но
часто передняя и наружная стороны этих частей светлые, буровато-ко-
ричневые, благодаря смеси белых и коричневых волос. Резкая гра-
ница между темно и светло окрашенными частями ног у аргаловидных
баранов отсутствует.
Во второй половине зимы, в результате обламывания темно окрашен-
ных верхушек волос и общего выцветания, окраска баранов светлеет,
становится желто-бурой или даже белесой. Уменьшаются контрасты
между светло и темно окрашенными участками тела.
Летняя окраска не слишком сильно отличается от зимней. Только
в некоторых случаях в ней проявляются ясно рыжие тона, вдоль средней
линии шеи и спины часто появляется отчетливая темно-бурая полоса,
да темные полосы по бокам туловища иногда вовсе не заметны.
Половой диморфизм выражен очень слабо. Самки окрашены немного
светлее самцов, менее отчетливы у них, обычно, полосы по бокам туло-
вища и нет ясно выраженного удлинения волос на шее; удлиненные же
волосы на затылке и по верху шеи часто имеются и у самок. Молодые
первые месяцы жизни имеют светлую серовато-желтую окраску, с более
темной коричнево-бурой головой. Потемнение окраски у них наступает
к осени, на 5—6-м месяце жизни.
В черепе (рис. 109) мозговая часть еще более укорочена, чем у муф-
лонов. Длина мозговой коробки от заднего выступа затылочного бугра
(acrocranion) до заднего края глазницы составляет редко более 46%
(от 40.7 до 46.7%) основной длины черепа и от 58.0 до 69 % длины морды
522
BOVIDAE
(от ptosthion до переднего края глазницы). Зароговая часть черепа слабо,
особенно у самцов, выступает назад позади роговых отростков и сильно
отогнута книзу. Дорзальная ее поверхность круто спадает вниз позади
рогов и у самцов нередко становится почти в одну плоскость с затылком.
Наружные слуховые проходы (meatus auditorius externus), даже у самок,
редко расположены позади уровня задних краев роговых отростков;
у самцов же они всегда находятся целиком под основаниями последних.
Лицевая часть черепа, наоборот, относительно длинная; длина морды
составляет ие менее 64% (и до 75%) основной длины черепа. Межглаз-
ничная ширина черепа, даже у самцов, редко превышает две трети рас-
стояния от prosthion до точки слезно-скулового шва на краю глазницы.
Роговые отростки и абсолютно и относительно более толстые, чем у муф-
лонов, но более тонкие и длинные, чем у снежного барана. Обхват их
в основаниях не превышает собственной длины. Носовые кости средней
ширины или узкие; ширина их составляет от 33 до 43% собственной длины.
Межчелюстные кости всегда достигают носовых и соприкасаются с ними,1
а иногда глубоко вклиниваются между носовыми и верхнечелюстными
костями. Костное нёбо сравнительно короткое; при горизонтальном его
положении передний край хоан расположен впереди уровня передних
краев глазниц. В области крыловидного гребня (crista pterygoidea) на
границе височной впадины и глазницы у самцов, а также, за редкими
исключениями, и у самок, теменная кость не соприкасается с алисфе-
ноидом; зато орбитосфеноид входит в контакт с височной костью
(рис. 103, Б). В области поперечного затылочного гребня у взрослых самцов
мастоидиая кость всегда соприкасается с теменной, разъединяя височ-
ную и верхнюю затылочную кости (рис. 104, А). У самок мастоидная"
кость иногда не достигает теменной и височная кость входит в контакт
с чешуей затылочной кости (рис. 104, Б).
История и географическое распространение (рис. 110). В настоящее
время область распространения аргаловидных баранов включает горные
системы Средней и Центральной Азии (к востоку от Каратау и Памира
и на юг до Гималаев включительно), а также горы южной Сибири — Ал-
тай и Саяны. Первоначальный ареал их был гораздо шире. Остатки круп-
ных баранов обнаружены в верхнеплиоценовых отложениях западной
Европы (Франция; Schaub, 1943) и Китая (Bohlin, 1938; Teilhard de
Chardin et Piveteau, 1930). В Европе аргаловидные бараны прослежи-
ваются до верхов плейстоцена. Остатки их найдены, например, в средне-
и верхнепалеолитических стоянках Крыма, частично уже вместе с остат-
ками баранов типа муфлона (Громова, 1935; Громова и Громов, 1937).
На Кавказе находки аргаловидных баранов известны из битуминозных
песков Бинагадинского местонахождения четвертичной фауны на Ап-
шеронском полуострове (средний или верхний плейстоцен, хозарский
ярус; Верещагин, 1953) и в пещерных отложениях верхнеплейстоцено-
вого возраста в бассейне р. Квирилы (северо-восточная Колхида, пе-
щера Мгвимеви, и близ Чиатур; Громова, 4948), Задний коренной зуб
крупного барана найден также в комплексе верхнеплейстоценовой фауны
в окрестностях города Мараги в северо-западном Иране (Громова, 1948)? 1 2
1 Кроме случаев, когда между носовыми и верхнечелюстными костями сохра-
няется открытая щель.
2 Ссылка И. Г. Пидопличко (1951) на В. И. Громова (1948) о том, что в Мезинской
палеолитической стоянке на территории Черниговской области найден аргаловидный
баран, является явным недоразумением. Речь идет об О. argaloides Nehring, относя-
щемся, как указывает и В. И. Громов, к типу муфлонов.
Рис. 109. Череп Ovis аттоп L.
524
BOVIDAE
Следует, однако, отметить, что отношения европейских ископаемых
аргаловидных баранов с современными аргали Средней и Центральной
Азии совершенно не ясны. Защищаемая В. И. Громовой точка зрения,
что муфлонообразные бараны являются измельчавшими потомками круп-
ных аргалоподобных, нуждается в более солидном подтверждении, так
как находится в противоречии с представлениями о направлении эволю-
ции морфологических особенностей рода.
В Азии, кроме упомянутых находок, аргали в ископаемом состоянии
известен из верхнепалеолитической стоянки под Красноярском и из
Забайкалья (Громов, 1948). В ряде мест Сибири и Средней Азии, как
будет показано ниже, бараны исчезли уже в историческое время.
Очерк современного и былого распространения аргали в пределах
СССР удобнее начать с Казахстана. Здесь бараны, хотя и спорадически,
встречаются во многих местах Казахского нагорья. Наиболее западным
пунктом их обитания оказываются горы Улутау (Антипин, 1941; Афа-
насьев и Слудский, 1947). Восточнее Улутау бараны в небольшом числе
живут в настоящее время в Жана-Аркинском районе по правому берегу
р. Сарысу (Цалкин, 1951). На северо-западе Казахского нагорья бараны
указываются для гор Ортау (Насонов, 1923). В 1948 г. на территории
б. заповедника Боровое Кокчетавской области было обнаружено три
случайно зашедших архара (сборн. «Заповедники СССР», т. II : 309).
К юго-востоку от Акмолинска в Казахском нагорье бараны встре-
чаются в большинстве возвышенностей до широты Каркаралинска, в
частности в Каркаралинских и Кентских горах. Б. А. Белослюдов (1948)
встречал их в гранитном мелкосопочнике высот Сары-Арка: на горках
Мунглы, Кзылтау и Актау. Обычны архары в районе горы Кызылрай
(Цалкин, 1951). Раньше на север они доходили до Баян-Аульских гор,
где были истреблены лишь в 1914—1918 гг. (Селевин, 1925). По данным
последнего автора, в 20-х годах настоящего столетия архары еще довольно
многочисленны были в Дельбегетейских горах, в 80—85 км к юго-востоку
от Семипалатинска, а также в других группах гор: Аркате, Кокони,
Доголан и Урдатау. Далее к югу и юго-востоку область распространения
архара охватывает хребты Калбинский, Чингиз-Tay, Тарбагатай, Семис-
тау и Саур.1 Имеются они, судя по коллекционным экземплярам, в районе
Кокпекты, в горах Монрак, а в 60-х годах прошлого столетия водились
на возвышенности Сар-Тологой (на северной окраине Тарбагатая) и к за-
паду от нее, в горах Аркарлы (Насонов, 1923). В последнем пункте, судя
по недавним данным Б. А. Белослюдова (1948), бараны не редки и до
сих пор.
Как показали наблюдения автора во время экспедиции 1956 г. в Бет-
Пак-Далу, до сих пор архары являются обычными обитателями невысо-
ких горушек Байгара и Джамбул. Здесь экспедицией было собрано не-
сколько черепов крупных самцов и добыты две самки архара. Наблю-
дались нами архары даже на мелких сопках к северо-востоку от Байгары
в окрестностях метеорологической станции в урочище Тюкен. Отдель-
ные экземпляры, говорят, проникают еще дальше вглубь пустыни вплоть
до западного побережья оз. Балхаш. По данным Б. А. Белослюдова (1948),
дикие бараны появлялись в урочищах Каракамыш и Когашик (в окрест-
ностях гидрометстанции Бет-Пак-Дала). Для С. Н. Боева (1948) архары
отстреливались в районе Булаттау (46.5° с. ш. и 72° в. д.), где, по его сло-
вам, эти звери держатся круглый год.
1 О распространении баранов к востоку от Иртыша и оз. Зайсан см. ниже, стр. 528.
OVIS
526
Областью почти сплошного распространения архаров является хре-
бет Каратау на Сыр-Дарье. Здесь они многочисленны по всему хребту
вплоть до северной оконечности его Кара-Мурун (Бажанов, 1945). Не
исключена возможность более северных заходов; самка архара в 1934 г.
была встречена в урочище Кендерлык, в западной части пустыни Бет-
Пак-Дала (Антипин, 1941).
Из Каратау ареал архара переходит в систему гор западного Тянь-
Шаня. На Таласском Алатау в пределах Казахстана бараны держатся
на территории заповедника Аксу-Джебаглы, где, впрочем, не так много-
Рис. 110. Распространение Ovis аттоп L.
1 — XX в. (области постоянного обитания); 2 — XIX в.; 3 — XVIII в. и раньше;
4 — отдельные заходы в настоящее время.
численны (Шульпин, 1948). Встречаются они также по Угамскому хребту
(Антипин, 1941). Далее к востоку бараны обитают на северных склонах
Киргизского хребта, в частности в районе Мерке (Северцев, 1873), а также
в Чу-Илийских горах. В последних они до сих пор довольно многочисленны
и даже заходят отсюда в прилегающие возвышенности Бет-Пак-Далы,
В Заилийском Алатау бараны водятся по всему хребту, но в средней
части очень редки и держатся только по южному склону. Более обычны
в западной половине хребта, в Кастекских горах (Антипин, 1941). В во-
сточной части Заилийского Алатау в настоящее время они, по С. С. Со-
колову (1939), постоянно встречаются по Сарытау с прилегающими вер-
ховьями рр. Тургени, Асы и Дженешке, а также в урочищах Боскуль,
Ортатау и Пейрама. Далее к востоку они временами бывают, но постоянно
не живут на отрогах Заилийского Алатау — Турайгыр, Тюре-Джайляу,
на Бакайтюбе. Кроме того, они держатся в урочище Бакалы-Карабастау
и в Богутах. По собранным С. И. Огневым (1940) сведениям, живут в го-
рах Сары-Булак (в Джаланашской лесной даче) и на хребте Турайгыр.
В б. Алма-Атинском заповеднике зимой нередко спускаются в Сюгатин-
скую долину, пересекая ее в самом узком месте между хребтами Турайгыр
и Богуты (Зверев, 1948).
626
BOVIDAE
Распространены бараны также на Кунгей-Алатау, где они обычны
в верховьях рр. Большой Кебени и Тау-Чилика (Антипин, 1941).
Еще далее к востоку архары населяют и весьма многочисленны в хребте
Кетмень, оттуда спускаются зимой в долину Чекулдысу, а также в неболь-
ших соседних хребтах Каратау, Ельчин-Буйрюк и Жабыр (Микулин и
Исаева, 1945; Антипин, 1946).
В Джунгарском Алатау архары до сих пор многочисленны не только
в самом хребте, но и в его предгорьях. Появляются, например, в Капчагое
на р. Или (Афанасьев и др., 1953). Н. В. Насонов (1923) западной гра-
ницей джунгарской расы (О. аттоп littledalei Lyd.) считал р. Чилик,
левый приток Или.
В коллекциях ЗИН АН СССР имеется экземпляр барана, добытый
на сопке Калкантау по правому берегу р. Или между рр. Чилик иЧарын.
В западной части Джунгарского Алатау архары весьма обычны в отдель-
ных небольших хребтах Катутау, Актау, Ортатау, Дулантау и
Калкантау (Антипин, 1941). В верховьях Биже (левый приток Ка-
ратала) бараны очень редки и встречаются лишь в небольшом числе на
хребте Чаган (Афанасьев, 1945). В 60-х годах прошлого столетия баранов
встречали еще в районе Копала, но, по Н. А. Северцову (1873а), они
заходили туда только поздно осенью во время перекочевок. В. Н. Шнит-
ников (1936) сообщал о стаде архаров, наблюдавшемся в 1910 г. в очень
низких горках Кок-Кия, к северу от тракта между Копалом и Арасаном.
В его распоряжении были также заслуживающие доверия сведения о том,
что архары держались в изолированных хребтиках Коракой и Буракой
вблизи Талды-Кургана, расположенных среди низменной полынной
степи в нескольких десятках километров от подножий главного хребта
Джунгарского Алатау. Во время путешествия Н. А. Северцова (18736), да
вероятно и сейчас, архары обычны в верховьях рр. Коксу, Каратала,
Саркана и Лепсы.
Широко, хотя и не повсеместно, бараны распространены в Киргиз-
ской ССР. На Киргизском (б. Александровском) хребте они водятся на
всем его протяжении, но в более низкой западной части, имеющей высоту
около 1600 м, бывают, главным образом, осенью и зимой, в восточной же
держатся в течение всего года, преимущественно на южном склоне (Чер-
навин, 1916). По сообщению А. А. Машковцева (1940), многочисленны
бараны на Киргизском хребте в районе Карабалты. Севернее Киргизского
хребта Н. А. Северцовым (1873) они отмечались в районе Токмака. На
Таласском Алатау, по В. В. Чернавину (1916), бараны держатся, главным
образом, по южному склону, тогда как на северном, по крайней мере
от г. Аулиэ-Ата и до отрога, которым Таласский Алатау соединяется
с Киргизским хребтом, их, по-видимому, нет. О спорадическом распро-
странении архаров в западной части Таласского Алатау не так давно
сообщал К. А. Абдилдабеков (1950).
Местами обитания являются также северные склоны Джумгольского
и Сусамырского хребтов. В последнем больше всего баранов в вер-
ховьях р. Арамсы (Чернавин, 1916).
По собранным Б. А. Кузнецовым (1948) данным, архары в сравнительно
небольшом числе встречаются вокруг оз. Сонкёль, на северных склонах
Кавакского и на Джумгольском хребте, а также в горах, окружающих
Сусамырское плато. На Чаткальском хребте, по данным того же автора,
бараны, по-видимому, не встречаются. Нет их также в горах Ташкумыр-
ского района и в возвышенностях к югу от Кетмень-Тюбинской котло-
вины. Не приводятся они и в составленном Д. Н. Кашкаровым списке
OVIS
521
млекопитающих окрестностей курорта Арслан-Боба (Кузнецов, 1948).
Не известно, есть ли архары в Кураминском хребте, но в тезисах доклада
С. Ю. Мадаминова (1953) о млекопитающих Ленинабадской области,
в числе добытых лично автором видов упоминается, впрочем без указания
точного местонахождения, наравне с азиатским муфлоном и архар. К югу
от оз. Сонкёль Б. А. Кузнецов указывает баранов для урочища Арпа
Ат-Башинского района Тянь-Шаньской области.
Судя по коллекционным экземплярам ЗИН АН СССР, бараны дер-
жатся в районе пос. Гульчи и в горах Ошского района Киргизской ССР.
Распространены они также на Алайском и Заалайском хребтах. Особенно
многочисленны на последнем, в частности в районе Бардабы и Иркеш-
тама. Бригадой пограничников в течение трех зимних месяцев здесь было
заготовлено 7 т архарьего мяса, а рога добытых баранов с трудом были
вывезены на полуторатонной машине (Кузнецов, 1948). Зимой со склонов
окружающих гор архары нередко спускаются в равнинную часть Алай-
ской долины (Андрушко, 1954).
На северной стороне Иссык-Куля бараны распространены на Кунгей-
Алатау. На последнем они, впрочем, держатся только в западной поло-
вине к востоку до бассейна р. Чон-Аксу или немного дальше (Петров,
1950; Дементьев и Тюрин, 1954). К югу от Иссык-Куля они встречаются,
хотя и немногочисленны, в Терскей-Алатау. Еще Н. А. Северцов (18736)
встречал много баранов в бассейне Нарына в верховьях рр. Атбаши и
Улана. Экспедиция М. В. Певцова, проходившая из Пишпека в Каш-
гар между Иссык-Кулем, Сонкёлем и Чатыркёлем, на своем пути встре-
чала много баранов (Цалкин, 1951). В большом числе водились в то время
архары в пограничном хребте Кокшаал-Тау. Множество баранов Н. А. Се-
верцов (1873) находил по Аксаю, в частности в горах Бос-Адыр, между
рр. Кыны и Межерюм, а также между Курткой и оз. Чатыркёль. Долину
Аксая, Большой Кебени, верховья Нарына и сырты по Сарыджазу
В. Н. Шнитников (1936) относит к числу тех мест Семиречья, где эти звери
были наиболее многочисленны и в те годы. До последних лет обычны
архары в районе Хан-Тенгри, в частности в районе водораздела рр. Те-
кеса, Баянкола, Капки и Сарыджаза (Антипин, 1947).
К югу от Заалайского хребта архары населяют среднюю и восточную
части Памира. Нет их на Дарвазе, а также в западных частях Рушанского
и Шугнанского хребтов. На р. Шахдаре крайним западным пунктом,
где встречаются архары, считался кишлак Джаухангоз «в пяти километ-
рах от Хорогского поста» (Наносов, 1923). По К. К. Флерову (1935),
западная граница распространения этого вида на Памире, «пересекая
р. Кызылсу (верхнее течение Вахша), идет на юго-восток; пересекая
р. Мухсу и через ледники Федченко и Шереметьева — к р. Мургабу,
в 10 верстах ниже Ташкургана. Далее на юг, пересекая р. Гупт, его при-
ток Тагузбулак и через перевалы Кок-Бо и Мас, к слиянию рр. Памира
и Вахандарьи, затем вверх по Памиру до восточной оконечности оз. Зор-
куль». Однако Р. Н. Мекленбурцев (1948), на основании собственных
данных, считает, что в южной части очерченная К. К. Флеровым граница
должна быть довольно существенно сдвинута к западу. Так, группа ар-
харов наблюдалась на горах по правому берегу Риджеса (приток Шах-
дары). Наблюдались они также на берегах оз. Турумтайкуль и на впа-
дающей в Шахдару у развалин Кала-и-Джаушангоз р. Саукташ, где
бараны живут круглый год.
В долине р. Шахдары архары регулярно спускаются не ниже Кок-
бая, по Токузбулаку не ниже Койтекеза, но по хребту между ними ареал
528
BOVIDAE
их выступает длинным языком на запад. Несколько черепов старых сам-
цов этого вида Р. Н. Мекленбурцеву пришлось видеть в куче рогов Capra
sibirica у кишлака Шош-Булат. М. П. Розанов (1935) западной границей,
где еще в массе встречаются бараны, называет Хоргуш, но единичные
экземпляры встречаются до р. Мац.
К востоку от очерченной границы архары встречаются и местами весьма
многочисленны во всех подходящих для них районах восточного Памира:
в урочище Маркансу в окрестностях оз. Кара-Куль у Памирского поста,
у перевала Кызыл-Рабат, у оз. Зоркуль (Розанов, 1935). Д. Л. Иванов
(1885) встречал архаров в верховьях Оксу, на Аличуре, Рянгкуле, Кокуй-
Бели и Беляндкиике. Большое число их он встречал в верховьях р. Истыка
у водораздела с р. Памир.
В пределах Алтая в прошлом столетии аргали были обычным зверем
в верховьях рр. Катуни и Бухтармы, в Нарымском и Курчумском хреб-
тах. Сохранились они здесь, по-видимому, и сейчас, но стали очень редки
(Антипин, 1941; Афанасьев и др., 1953).1 В средине XVIII в. бараны встре-
чались, по И. Гмелину (цит. по Цалкину, 1951), в окрестностях Усть-
Каменогорска и в горах по р. Бухтарме. Во время путешествия П. С. Пал-
ласа, в конце того же века они были распространены во всех безлесных
частях Алтая, в частности на Тигирецких белках, но уже в 20-х годах
XIX в. они были отсюда оттеснены и сохранялись лишь в горах по Катуни
и Чуе (Северцов, 18736). К этому же времени бараны исчезли на Аргуте.
Остатки их были доставлены Геблером (Колосов, 1939) из пещер по р. Ча-
рышу. К средине XIX в. бараны перестали водиться в горах по Катуни,
но все еще многочисленны были на р. Чуе.
К началу нынешнего столетия область распространения аргали на
Алтае сократилась еще больше; экспедиция Холлистера (цит. по Колосову,
1939) не нашла их в Чуйской степи севернее Кош-Агача. В 30-х годах,
по А. Булавину (1934), они были немногочисленны, но встречались в уро-
чищах Бора-Бургазы и Узун, в верховьях Уландрыка, Большие Ши-
беты и Аксае. В незначительном числе держатся в лесной части Коша-
гачского аймака на территории Джассатерского сельсовета. Судя по кол-
лекционным экземплярам, сохранились бараны в Чуйских Альпах,
в верховьях рр. Чуи, Черной речки, Башкауса и Чулышмана. По-види-
мому, немало их в междуречье Чаган-Бургазы и Тархатты. А. М. Колосов
(1939) при поездке на р. Чаган-Бургазы отмечал аргали в урочищах
Байн-Чаган, Саржематы, Тозунду. Имеются, по данным этого автора,
бараны по р. Уландрыку, Малой Шибетты, в урочище Себестей и восточнее
на границе высокогорного плато Укок, в урочище Джюмалы. Широко
распространены они также в Сайлюгемском хребте. Черепа баранов на-
ходили в устье р. Калбакая (приток Башкауса), на перевале из верховий
Башкауса в Кош-Агач. Неоднократно наблюдались в окрестностях
оз. Джулу-Куля, а А. М. Колосов (1939) даже пользовался тропами их в исто-
ках Чулышмана. Здесь они распространены к северу до р. Шавлы (Цал-
кин, 1951).
В Тувинской области, по данным А. И. Янушевича (1952), аргали
не редки в западной части Танну-Ола; возможно, держатся и в восточной
части Тувинского нагорья. В западной половине Тувы они распространены
до бассейна р. Кемчика включительно (Цалкин, 1951). Бараньи тропы
и помет архаров В. Ч. Дорогостайский (1918) видел на южном склоне
Танну-Ола в истоках р. Дзайгал-Гол.
1 В верховье р. Курчума баранов находил Н. К. Яблонский (1907), но в окрестно-
стях оз. Маркаколь их, по данным этого автора, нет.
OVIS
529
В пределах Западных и Восточных Саян, по крайней мере в настоящее
время диких баранов нет. Только В. Ч. Дорогостайский (1918) слышал,
будто бы встречаются они на Нуху-Дабане в верховьях р. Иркута, но
проверить самому эти сведения ему не пришлось.
В Монгольской Народной Республике, по суммированным А. Г. Бан-
никовым (1954) данным, бараны распространены по всему Монгольскому
и Гобийскому Алтаю, по хребту Хурхэ-Ула и по ряду мелких поднятий
до 110° в. д. Далее, они населяют большую часть заалтайской Гоби,
кроме обширных котловин и больших межгорных сухих долин, магистраль-
ный хребет Хангай от истоков Онгин-Гола и Орхона на востоке до Бол-г
ная на западе и Хан-Хухей. Кроме того, до сих пор бараны встречаются
в горах к западу от оз. Убсу-Нур, в предгорьях Танну-Ола, на правом
берегу р. Тэс, у Шабарин-Гола и в западном Прикосоголье, на поднятиях,
отделяющих Дархатскую котловину от оэ. Косогол. По свидетельству
Радде (цит. по Банникову, 1954), сотню лет назад они водились в неболь-
шом числе в восточном Прикосоголье и Хэнтэе, но в настоящее время
ни в этих районах, ни на Большом Хингане в пределах Монгольской
Народной Республики диких баранов нет.
Сравнительно недавно бараны были широко распространены в южном
Забайкалье. Черепа их найдены в бассейне р. Джиды, а также близ с. Усть-
Кяхты по левому берегу р. Савы и на Селенге (Цалкин, 1951). Мессер-<
шмидт (цит. по Насонову, 1923) в 1724 г. получил туши диких баранов
от охотников с. Галкина на р. Ингоде. Паллас (1788) нашел много баранов
на невысоких горах Одон-Чолон между р. Ингодой и Снопом, а также
к юго-востоку от р. Борзи вдоль границы до Абагайтуя. Однако уже в 50-х
годах прошлого столетия местное население Даурии знало их только,
по названию. Последние бараны, до этого времени жившие незначитель-
ными группами на голых скалистых горах к югу от пограничного поста
Соктуй (Тутхалтус, Шире и Соктуе), были уничтожены в суровую снежную
зиму 1831/32 года (Radde, 1861). Эвенки, кочевавшие по рр. Шилке и
Онону, сообщали Р. Мааку (1859), что они охотились в то время за бара-
нами, державшимися небольшими группами в горах по этим рекам. Однако
нет уверенности, что в данном случае речь шла не о толстороге {О- ni-
vicola Eschsch.), а об аргали. Известен случай добычи в 1918 г. аргали
в пограничном хребте по р. Онону в 20 км от Мапгута (Цалкин,:
1951).
Подвиды. В пределах вида О. аттоп L. в том объеме, как он прини-
мается в настоящей работе, Н. В. Насонов (1923) признавал наличие
следующих подвидов.
1) О. аттоп аттоп L. — Алтайский баран, аргали. Самая крупная
раса баранов. Высота в холке достигает у крупных самцов 125 см, основ-
ная длина черепа 350 мм, живой вес до 200 кг. Рога у самцов не очень
длинные, но толстые, с притупленным передне-наружным ребром и вы-
пуклой наружной гранью. Удлинение волос на шее и груди слабо выра-
жено. Распространение: советский (северный) Алтай.
2) О. аттоп hodgsonii Blyth — Гималайский баран. Размеры крупные:
Заметно выражено удлинение волос на шее. Крутой изгиб рогов, но сами
рога менее массивные, чем у типичной расы. Р а с.п ространение:
Гималаи от Кашмира па западе и до Сиккима на востоке.
3) О. аттоп dalai-lamae Przevalskij — Баран западного Тибета. От пре-
дыдущих двух рас отличается более короткими рогами, слабо развитым
удлинением волос на шее и темно-бурыми, иногда даже черными пятнами
на боках возле нахов. Распространение: западный Тибет от
34 Фауна СССР, И. И. Соколов
530
BOVIDAE
Алтынтага, Токуздавана и Русского хребта на севере до хребта Прже-
вальского на юге. На восток доходит до оз. Кумкуль.
4) О. аттоп jubata Peters — Баран восточного Тибета. По размерам
приближается к типичной расе. От последней отличается сильно развитым
удлинением волос на шее, образующих у ее основания и на плечах по-
добие свисающей вниз манишки. Распространение: северо-во-
сточный Китай, горы Алашань, Наныпань, восточная часть Куньлуня
(Цалкин, 1951).
5) О. аттоп kozlovi Nasonov — Ябарайский баран. Размеры сравни-
тельно небольшие. Высота в холке, даже у самцов, немного больше 100 см.
В черепе, по Цалкину (1951), характерна короткость носовых костей.
Распространение: горы Ябарай в южной Гоби; возможно также
обитание этой расы в возвышенностях заалтайской Гоби.
6) О. аттоп mongolica Severtzov — Баран северной Монголии. Сходен
с типичной расой; отличается от последней трудно определимыми на глаз
особенностями изгиба рогов. Распространение: горы северной
Монголии, хребет Танну-Ола, южное Забайкалье до р. Селенги на во-
стоке. В. И. Цалкин (1951) объединяет в один подвид с алтайской
расой.
7) О. аттоп przevalskii Nasonov — Сайлюгемский баран. Сходен с ти-
пичной расой; отличается от последней «особым характером кривизны
рогов». Распространение: Сайлюгемский хребет. В. И. Цадки-
ным (1951) включается в один подвид с алтайской расой.
8) О. аттоп nigrimontana Severtzov — Каратауский баран. Размеры
по сравнению с другими расами аргаловидных баранов небольшие; основ-
ная длина черепа у самцов не превышает 285 мм. На рогах хорошо раз-
вито передне-наружное ребро. Передняя и наружная грани рога плоские.
По Цалкину (1951), особенностью данной расы является также более
темная окраска. Распространение: хребет Каратау (правобе-
режье р. Сыр-Дарьи).
9) О. аттоп karelini Severtzov — Тянь-шаньский баран. Размеры более
крупные, чем у предыдущего подвида; основная длина черепа у взрослых
самцов доходит до 310 мм. Окраска более светлая. Переднее ребро рога
у основания часто притуплено. Распространение: Киргизский
и Угамский хребты, Таласский Алатау, Чу-Илийские горы, Заилийский
и Кунгей-Алатау, область к югу от Иссык-Куля до Нарына.
10) О. аттоп littledalei Lydekker —Баран Литльдаля. Отличается от
предыдущих рас более слабым изгибом и сильно расходящимися в стороны
нисходящими ветвями рогов. Распространение: Джунгарский
Алатау. Цалкин (1951) объединяет эту расу в один подвид с баранами
Памира.
11) О. аттоп coIlium Severtzov — Североказахстанский баран. Сходен
с тянь-шаньским подвидом (О. a. karelini Sev.). Отличается от последнего
некоторыми деталями изгиба рогов (Насонов, 1914), а по Цалкину (1951)
также длинными межчелюстными костями, почти достигающими слезных.
Распространение: Саур, Джаир, Тарбагатай, Семитау, Чингиз-
Тау, возвышенности Казахского нагорья на северо-восток до Иртыша,
на запад до 35° в. д., на юг до Балхаша, Сасыккуля и Алаколя (Цал-
кин, 1951).
12) О. аттоп polii Blyth — Памирский баран, или архар. Крупная
раса, немного уступающая по величине алтайской, а по длине рогов даже
превосходит последнюю. От тянь-шаньского подвида отличается, в сред-
нем, более темной окраской, а также формой рогов с сильно отогнутыми
OVIS 531
в стороны концами. Распространение: Памир, Кокшаал, горы
северной Индии приблизительно до Хунзы.
13) О. аттоп heinsii Severtzov. — Отличием тянь-шаньской расы
является, якобы, более сильное расхождение в стороны нисходящих частей
рогов. Описан из б. Токмакского уезда (ныне Фрунзенская область Кир-
гизской ССР). В. И. Цалкин (1951), по-видимому, справедливо считает
особенности, приписываемые этой «расе», проявлением индивидуаль-
ной изменчивости и включает ее название в синонимы О. a. karelini
Sev.
14) О. аттоп sairensis Lydekker. — По мнению описавшего подвид
автора, отличиями от других рас является массивность рогов, делающих
изгиб более полного оборота. В качестве области распространения ука-
зываются горы Саур и Джаир (местности к юго-востоку от Зайсана и
западная Джунгария). Насонов (1923) и Цалкин (1951) сомневаются
в реальности существования данной расы, и последний из этих авторов
включает ее в синонимы О. a. coIlium.
Кроме того, В. И. Громова (Gromova, 1936) установила подвид:
15) О. a. intermedia Gromova. — Близок к алтайской расе. Характе-
ризуется короткими, но массивными рогами, относительно узким черепом
с короткими и широкими носовыми костями. Распространение:
центральная Гоби, горы Наин-Богдо.
Биология. На обширной территории своего ареала с разнообразными
ландшафтно-климатическими условиями архары живут в неодинаковой
обстановке. Они обитают на высотах от 300 и до 5500 м над ур. м., от
незначительных возвышенностей Бет-Пак-Далы до высочайших сыртов
Хан-Тенгри и- плоскогорий Памира. Общей и наиболее типичной особен-
ностью архаров является привязанность к гористому, но сглаженному,
мягкому рельефу. Они избегают как совершенно ровных, открытых рав-
нин, так и гор с сильно- выраженным скалистым рельефом и вообще об-
ширных скальных массивов. Сравнивая местообитания семиреченского
архара с таковыми сибирских козерогов, Н. А. Северцов (1873а) пишет:
«Он [архар ] не довольствуется, подобно им, скудными клочками паст-
бищ на выступах скал, а требует пастбищ более обширных, степи или
широких травянистых склонов». Говоря о памирской расе, он также ука-
зывает, что «это животное, собственно, есть житель не горных хребтов,
а высоких холмистых площадей, где растут альпийские представители
степной растительности: кипец (Festuca), мелкая полынь (Artemisia), даже
солянки (Salsoleae), составляющие его корм; в горах он только укрывается,
и то избегая слишком скалистых хребтов, например Кок-кия у Аксая,
где я нашел только Capra skym~>. По этой причине, несмотря на то, что
ареалы сибирского козерога и архара во многом совпадают и оба вида
являются безусловно жителями гор, встречать в одних и тех же местах
козлов и баранов приходится сравнительно редко.
Типичными стациями О. аттоп являются обширные открытые про-
странства по пологим безлесным склонам гор и широким межгорным
долинам, покрытые растительностью степного типа, а также обширные
горные плато, имеющие поблизости выходы скал, каменистые россыпи
или ущелья. Лесных склонов и даже густых кустарниковых зарослей
архары избегают.
Если для козлов скалы являются главным и необходимым условием
существования, то для баранов они имеют второстепенное значение, —
лишь как места укрытий; даже в случае опасности архары спасение ищут
часто не в скалах, а в быстром беге.
34*
532
BOVIDAE
В центральном Казахстане архары живут в наиболее возвышенных
частях Казахского мелкосопочника, покрытых растительностью степного
типа. «Только на этих безлесных пастбищах, — пишет Н. А. Северцов
(18736), — и водятся архары; везде, избегая лесов, держатся они в самых
скалистых частях гор, но не постоянно, а часто уходя на травянистые
склоны и даже на предгорные степные пастбища, где встречаются с сай-
гаками».
В высокогорной части Джунгарского Алатау бараны придерживаются
альпийских лугов и пологих склонов, лишенных леса. В предгорьях его,
например в Катутау, они живут на мелкосопочном горном плато, из-
резанном саями и неглубокими обрывистыми ущельями. Здесь на обед-
ненных участках бараны находят корм, а в ущельях и саях — укрытия
в жаркие дневные часы суток. В течение круглого года бараны живут
в невысоком (800—1000 м над ур. м.) хребтике Актау в юго-западных
предгорьях Джунгарского Алатау. Здесь нет хороших пастбищ, а вер-
шины вообще почти лишены растительности, но зато имеются убежища
в виде многочисленных пещерок и крутых тенистых саев; кормиться же
звери выходят на межгорную тырсово-полынную степь (Антипин, 1941).
В Сары-Арка и Бет-Пак-Дале баранов неоднократно встречали в пу-
стынной обстановке, вообще мало напоминающей горы; в этих случаях
необходимые элементы горной среды архары находили, по-видимому,
в обрывах чинков, в ущельях и саях (Белослюдов, 1948).
На Улутау бараны держатся среди почти голых, с редкими пятнами
арчи гранитах по уступам верхней части хребта (Афанасьев и Слудский,
1947).
В западном Тянь-Шане, по В. В. Чернавину (1916), архары предпо-
читают горы с мягкими контурами и высокие плато; однако оттесняемые
стадами домашних животных летом «бараны уходят на самые вершины
гор и лазают по скалам не хуже горных козлов. Но и тут все-таки они
держатся таких гор, где скалы отвесны не на всем протяжении, где долины
не так узки, где всегда можно выйти из скал на более или менее ровное
место». Вероятно, вытеснение и преследование явилось причиной того,
что в центральной и южной части Каратау типичными местообитаниями
архаров в настоящее время являются крутые каменистые склоны и ущелья
(Антипин, 1947).
В Заилийском Алатау архары распространены в большом числе в во-
сточной, несколько пониженной части хребта, где на сглаженных его
складках преобладают высокогорные луговые и степные ландшафты
(Соколов, 1939).
В центральном Тянь-Шане, в частности в районе между главным
хребтом и хребтом Кетмень, типичными местообитаниями являются поло-
гие склоны последнего и мелкосопочные горы, подымающиеся на 400—
800 м над высокогорным плато. Мягкий рельеф и хорошие пастбища соз-
дают благоприятные условия для существования архара. Стацию архаров
образуют и сырты горного узла Хан-Тенгри, расположенные на высоте
4000 м над ур. м. Здесь, как и на северных склонах центрального Тянь-
Шаня, архары бывают, главным образом, летом, на зиму спускаясь
в более низкие малоснежные места. На крутых северных склонах Тянь-
Шаня архаров летом можно видеть в зарослях арчи и других кустарников
по соседству с еловым лесом (Антипин, 1947).
В горах Киргизии, по Б. А. Кузнецову (1948), «излюбленными местами
обитания архара являются слабо всхолмленные пространства сыртов,
площади альпийских лугов, участки высокогорной степи на высоких
OVIS
533
плоскогорьях, холмы предгорий, ограничивающих высокогорные долины
и котловины. Реже держатся на травянистых склонах горных хребтов».
На Памире, по данным Р. Н. Мекленбурцева (1948), наиболее под-
ходящими стациями для архаров являются места, где ксерофитные расти-
тельные группировки сочетаются, с одной стороны, с альпийскими лугами,
а с другой — с совершенно лишенными растительности участками на гор-
ных склонах. Последние бывают покрыты щебнем или щебневатым мелко-
земом и служат местами отдыха в дневное время. Хотя места обитания
Рис. 111. Горки Джамбул, восточная Бет-Пак-Дала, стация архаров. (Фото автора).
памирского архара представляют горную сторону со сглаженными кон-
турами, однако, по мнению этого автора, для него необходимы и скалы.
«Правда, — пишет Р. Н. Мекленбурцев (1948), — эти животные при спо-
койной кормежке, на отдыхе, при переходах, могут встретиться в скалах
только в исключительных случаях. Зато они постоянно находят здесь
надежное убежище в случае настойчивой погони каких-нибудь хищни-
ков».
Наконец, на Алтае, по наблюдениям А. М. Колосова (1939), летом
бараны держатся в нагорной степи полупустынного типа, но иногда под-
нимаются выше, до линии вечного снега.
Сезонная смена мест обитания у архаров выражена в разной степени.
Там, где их мало беспокоят (например, в районе Хан-Тенгри, в некоторых
районах Памира), а на пастбищах, благодаря малому количеству осад-
ков и сдуванию ветром, не образуется глубокого снега, бараны ведут
почти оседлый образ жизни. «Везде, где только был наблюдаем О. polii, —
пишет Н. А. Северцов (1873а), — он на своих высокогорьях держится
и летом, и зимой, которая там холодна, но малоснежна». Впрочем, ио
последним данным Р. Н. Мекленбурцева (1948), и на Памире наблюдается
534
BOVIDAE
сезонная смена мест обитания и перемещения архаров. Летом они пред-
почитают альпийские лужайки по склонам гор на высотах от 4500—5500 м.
Зимой спускаются на расположенные на 500—600 м ниже пастбища и
даже в широкие долины, где зимою меньше снега, более доступны злаки,
а весною раньше, чем на склонах гор и альпийских лугах, появляется пер-
вая зелень.
Практически оседлый образ жизни ведут и бараны, населяющие Ка-
захское нагорье,, возвышенности Сары-Арка и Бет-Пак-Далы. Летом
они придерживаются наиболее возвышенных участков, а к зиме спус-
каются к подножью гор в степи, покидаемые к этому времени домашними
стадами.
В районах, где архарам приходится жить в условиях более пересе-
ченного рельефа, у них наблюдается резко выраженная сезонная смена
мест обитания, приобретающая характер, хотя и не слишком обширных,
но регулярных миграций. Основными причинами, побуждающими зве-
рей совершать сезонные миграции, служат зимой различная глубина
снежного покрова, затрудняющего добычу корма, летом—-гнус и вы-
сыхание травянистой растительности, составляющей основу питания
архаров, а также непосредственное преследование и вытеснение диких
баранов с их исконных пастбищ стадами домашних животных.
Чаще всего сезонные перемещения совершаются в вертикальном
направлении. Лётом; бараны поднимаются в верхний пояс гор вплоть
до альпийских лугов и линии снега. Здесь они находят сочный зеленый
корм, защиту от зноя и насекомых; кроме того, стада домашних живот-
ных не всегда добираются до высокогорий. В начале зимы звери вновь
спускаются в субальпийскую зону, на южные солнцепечные склоны горы
и даже в предгорные и межгорные долины, где снега мало или он отсут-
ствует вовсе.
Вертикальное направление сезонные миграции баранов носят в За-
илийском и Джунгарском Алатау, Тарбагатае, но особенно резко они
выражены в высокогорном центральном Тянь-Шане. Здесь, как уже
отмечалось, бараны лето проводят на сыртах, альпийских лугах и степных
участках северных склонов, а в сентябре, когда в высокогорье начинаются
обильные снегопады, они спускаются ниже на открытые и почти лишен-
ные снега небольшие хребты Ельчин-Буйрюк, Каратау и мелкосопоч-
ник Жабьер. Весною, начиная с февраля, по мере таяния снега и по-
явления свежей зелени в горах, происходят обратные перемещения,
заканчивающиеся к маю (Антипин, 1947). Первыми и особенно высоко
в горы уходят взрослые самцы, тогда как самки с только что родившимся
молодняком дольше задерживаются на более низких высотах.
Иногда, впрочем, в зависимости от местных условий, вертикальные
миграции имеют противоположный только что описанному характер.
На Алтае, например, зиму бараны проводят не в долинах, где выпадает
много снега, а высоко в горах, где на обнаженных постоянными ветрами
участках они находят необходимый им минимум корма (Северцов, 18736).
Впрочем, в особо снежные зимы аргали и здесь спускаются с гор в Чуй-
скую степь, в которой в нормальные годы пасутся только осенью и вес-
ною (Колосов, 1939).
В северо-западной Монголии, по данным В. Ч. Дорогостайского (1918),
летом бараны держатся в степных долинах вдали от гор, на чистых местах,
и только осенью, с наступлением заморозков, откочевывают на склоны
гор, где дольше сохраняется зелень; зимой же ведут бродячую жизнь
в,поисках мест, где засохшая трава не занесена глубоким снегом.
OVIS 535
В некоторых районах имеют место миграции баранов в широтном
направлении. Например на Алтае, кроме вертикальных миграций, час-
тично наблюдается перекочевка их на зиму из СССР в малоснежные
районы Монголии, а весной обратно, связанная, вероятно, с тем, что
в Чуйских Альпах и на Сайлюгемском хребте снег, по-видимому, слишком
глубок, чтобы здесь могли прокормиться эти звери (Колосов, 1939).
В восточной части Заилийского Алатау из района Тау-Чилика бараны
ежегодно — с ноября по январь — мигрируют в горы Большие и Ма-
лые Богуты, а в феврале или марте, в зависимости от времени стаивания
снега, переселяются в обратном направлении, пересекая степную часть
Сюгатииской долины (Зверев и Келейников, 1947).
В западной части Таласского Алатау часть популяции баранов на лето
поднимается высоко в горы, а другая, наоборот, спускается в мелкосо-
почник, где звери находят оптимальные кормовые условия, и благодаря
более открытым ветру местам, защиту от насекомых (Северцов, 1929).
Известны также регулярные миграции архаров из западной части Талас-
ского Алатау в хребет Каратау и обратно (Шапошников, 1956).
Архар — стадное животное; размеры образуемых стад зависят как
от общей численности зверя в том или ином районе, так и от времени
года. Взрослые самцы летом держатся отдельными группами от 4 до 12
толов, в число которых могут входить и один-два молодые. Реже, но все же
встречаются и одиночно держащиеся самцы. Самки с ягнятами, за ис-
ключением периода окота, а также молодняк обоего пола до 3 лет, обра-
зуют летом смешанные стада, доходящие до нескольких десятков голов.
В Монголии и на Памире стада самок с молодняком и летом могут
достигать 150—200 голов. Первое время после окота самки с только что
родившимся молодняком держатся в одиночку или образуют неболь-
шие группы по нескольку особей вместе.
Яловые самки и молодняк прошлых лет образуют иногда самостоя-
тельные стада. Изредка и летом в стаде самок можно видеть одного-двух
взрослых самцов, обратная же картина наблюдается крайне редко.
Самцы в массе присоединяются к самкам только осенью на период
гона; по окончании последнего они, как правило, снова держатся отдельно.
Крупные смешанные стада, насчитывающие по нескольку сотен эк-
земпляров, образуются осенью и в начале зимы. Зимой массовые скопле-
ния носят часто временный характер, вызываются недостаточностью
зимних пастбищ со слабым снеговым покровом и при наступлении бла-
гоприятных кормовых условий легко распадаются на более мелкие группы.
Вожаком в стаде является обычно старый и крупный самец или
самка. Вожак следует впереди стада, первым поднимается на гребень
или вершину горы и осматривает местность. Специальных сторожей в ста-
дах баранов, по-видимому, нет. Естественно, что чаще первыми замечают
опасность более взрослые и осторожные бараны.
При пастьбе архары не расходятся широко; стадное чувство развито
у них сильнее, чем у козлов. В противоположность последним, бараны
при испуге не бросаются врассыпную, а сбиваются в кучу и бегут плотной
группой.1
1 Автору, однако, пришлось быть свидетелем следующего случая в горах Джам-
бул в юго-восточной Бет-Пак-Дале 28 мая 1956 г. Вспугнутое близкими выстрелами
А. А. Слудского стадо архаров, состоявшее из двух десятков голов самок с молодня-
ком, понеслось по гребню горы. Вскоре от него в разных местах отделились по-
одиночке молодой самец и холостая самка; на моих глазах, по-видимому не заметив
меня, они спустились по склону горы и залегли между камней, тогда как остальной
табун исчез н горах.
536
BOVIDAE
Пасутся бараны в течение большей части суток, но наиболее активны
в утренние и вечерние часы. Поднявшись с ночлега, они начинают пас-
тись одновременно и, как правило, поднимаются вверх по склонам. В жар-
кие полуденные часы архары отдыхают и пережевывают жвачку где-
нибудь у гребня горы или на открытом альпийском склоне. Часов с 4—5
дпя, когда спадает жара, звери вновь выходят на кормежку, на этот раз
медленно спускаются вниз по склонам, доходя иногда до верхней границы
леса. Таков летний суточный ход жизни баранов в высоких горах; в низ-
ких предгорьях и мелкосопочнике звери на день уходят не вверх, а, на-
оборот, вниз к подножью гор и в ущелья, где находят защиту от зноя
в тени пещер, выступов скал и отдельных камней. В верхние участки
звери в этих условиях уходят пастись на ночь. По В. М. Антипину (1947).
бараны летом деятельны и ночью, но очень осторожны. Р. Н. Меклен-
бурцев (1948), впрочем, считает, что архары пасутся только вечером и ут-
ром; ночь же проводят лежа, и утром их можно найти там, где они кор-
мились вечером накануне. В пасмурную погоду нет столь резкого де-
ления светлой части суток на часы отдыха и кормежки, но все же боль-
шая часть зверей в середине дня отдыхает.
Для отдыха бараны располагаются на открытых местах с широким
кругозором. В восточном Памире, по Р. Н. Мекленбурцеву (1948), и в Бет-
Пак-Дале, по моим наблюдениям, такими местами служат пятна щебнистых
россыпей по склонам гор, где окраска зверей отлично сливается с окру-
жающим фоном.
Перед тем, как лечь, баран ’ выбивает копытами неглубокую ямку,
в которую и ложится. Места лежек часто бывают постоянны, около них
скапливается значительное количество помета.
В высокогорье бараны не ищут тени, а, наоборот, любят, особенно во
время линьки, греться на солнцепеке, принимать «солнечные ванны»,
подставляя солнцу не только бока, но и живот (Соколов, 1939). По наблю-
дениям В. Ч. Дорогостайского (1918), в северо-западной Монголии в ясную
и теплую погоду бараны имеют обыкновение днем посещать русла пересы-
хающих в это время речек и часами валяться в песке. Здесь они, по-ви-
димому, не только греются, но и освобождают себя от насекомых.
В отношении посещения архарами водопоев сведения противоречивы.
В. Ч. Дорогостайский (1918) считает, что специально на водопой они не
ходят, и пьющими их он вообще ни разу не видел. Другие авторы, наблю-
давшие архаров, говорят о регулярном и даже двукратном в день посеще-
нии водопоев, а наличие последних считают одним из необходимых усло-
вий существования в той или иной местности этого вида (Кузнецов, 1948;
Мекленбурцев, 1948).
В зимний период архары активны и пасутся только в светлые часы
суток, для чего выходят на солнцепечпые склоны, в открытую межгор-
ную степь и даже в речные долины (Антипин, 1947). Днем отдыхают
мало. На ночь зимой уходят в укрытые от ветра места, для чего иногда
вынуждены подниматься в горы. Ложась отдыхать, выкапывают в снегу
глубокие лежки.
В бураны, равно как и в сильно дождливую погоду, бараны пасутся
мало и неохотно, предпочитая отлеживаться в укрытиях, под защитою
скал или в пещерах. Однако холода, как такового, не боятся. Во всяком
случае, даже в снегу, ложатся не тесно, а на некотором расстоянии друг
от друга.
В пределах избранного на данный сезон района бараны, если их систе-
матически и настойчиво не тревожат, стойко придерживаются определен:-
OVTS
них местообитаний. В определенных местах они жируют, польиуютсп
одними и теми же водопоями, местами отдыха и даже одними и темп жп
путями идут с отдыха на пастбище и обратно. На путях переходов ими
часто вытаптываются ясно обозначенные тропы (рис. 112), которые иногда
используются человеком, так как проходят обычно по наиболее удобным
местам гор и перевалов.
В спокойной обстановке архары передвигаются шагом. Будучи нотро
вожены, но не испуганы, уходят ускоренным шагом или рысью. Ноожщ
Рис. 112. Тропа архаров, горы Байгара, восточная Бет-Пак-Дала. (Фото автора).
данно и сильно испуганные, несутся вскачь, сбиваясь в кучу и не разби-
рая пути. Отбежав некоторое расстояние, звери несколько раз останав-
ливаются, смотрят в сторону, откуда угрожает опасность, после чего или
успокаиваются, или уходят еще более быстрым аллюром.1 На скаку сильно
вскидывают кверху зад, как это делают в подобных случаях домашние
овцы. В. В. Чернавин (1916) пишет, что ему на ровном месте удавалось
на верховой киргизской лошади сравнительно легко догонять архаров.
Р. Н. Мекленбурцев (1948) сомневается в этом и утверждает, что на
Памире даже лучшие из киргизских собак не могут догнать архара ни
в горах, ни на равнине; при этом последний выдерживает преследование,
не сбавляя хода в течение часа. Преследованием с автомобиля М. Д. Зве-
рев (1948) установил, что крупные большерогие самцы О. а. ka.reli.ni
в Алма-Атинском заповеднике не могут бежать быстрее, чем 50 км в час,
тогда как более легкие, молодняк и самки, могут развивать скорость до
1 В результате наблюдений в Бет-Пак-Да ле, у меня сложилось убеждение, что
архары гораздо пугливее джейранов. В противоположность последним, они почти
никогда не остановятся после выстрела по ним, пока не скроются из вида.
538
BOVIDAE
60 км. С большой легкостью и быстротой бараны передвигаются по крутым
скалистым горам, «легко лазают по таким скалам, куда не добраться ни
собаке, ни волку». Одинаково легко бегут как вверх, так и вниз по та-
ким крутым склонам, где лошадь не пройдет и шагом (Чернавин, 1916;
Мекленбурцев, 1948).
Основную пищу архаров составляют разнообразные, в том числе и
жесткие травы, произрастающие в местах их обитания. Почти повсе-
местно постоянными кормами являются типчак (Festuca sulcata Hack.),
ковыли (Stipa) и некоторые другие злаки.
В Джунгарском Алатау, по исследованиям В. М. Антипина (1947),
летом питание состоит, кроме злаков, из листьев таволги (Spiraea hype-
ricifolia L.), курчавки (Atraphaxis frutescens Eversm.) и сочных солянок
(Anabasis sp.). Осенью первое место в рационе занимает лук (Allium sp.),
а весной предпочтение отдается ковылю и курчавке. Ковыль главную роль
в питании архаров играет не только летом, но и зимой. В Заилийском
Алатау бараны летом охотно поедали разнотравье: виды горцов (Poly-
gonum), герани (Geranium) и др. (Соколов, 1939).
На Памире, по Р. Н. Мекленбурцеву (1948), излюбленными летними
пастбищами архаров являются альпийские лужайки, основу раститель-
ного, покрова которых составляют осоки (Carex) и кобрезии (Cobresia).
Виды осок и злаков (Роа, Stipa, Festuca), пока снег не глубок, служат
кормом и зимой. По мере углубления снегового покрова звери переходят
на питание более высокими растениями, торчащими из-под снега (Ног-
deum, Roegneria). Основным же зимним кормом архаров здесь являются
терескен (Eurotia ceratoides С. А. М.) и различные виды полыней (Arte-
misia).
В конце весны бараны с большой охотой поедают цветущие примулы
(Primula nivalis Pall.) и веточки шпорника (Delphinium dasyanthum Kar.
et Kir.). В юго-западной части Джунгарского Алатау известно поедание
зимой веток белого саксаула, а на Алтае — лишайников и мхов. В. М. Ан-
типин (1947) наблюдал баранов, пасущихся на озимых посевах.
Аргали — чуткий зверь, но не пугливый и мало сообразительный. На
открытом месте замечает человека на расстоянии 2—3 км, но редко обра-
щается немедленно в бегство.
Стадо, даже испуганное выстрелами, не всегда далеко убегает и иногда
на виду у охотника вновь начинает кормиться.
Все чувства у баранов развиты хорошо, но полагаются они, по-види-
мому, главным образом на обоняние. Заметив издали приближающегося
человека, архары сбиваются в кучу, но даже идущего прямо на них при
безветрии или против ветра подпускают на 500—800 м, а двигающегося
стороной — еще ближе (Мекленбурцев, 1948). Однако, почуяв запах че-
ловека, даже не видя его, бараны немедленно обращаются в бегство. По-
видимому, они способны зрением воспринимать издали ощущение движу-
щегося предмета, но разобраться в том, представляет ли он опасность или
нет, не в состоянии. То же и со слухом. Их можно испугать за 300 м лег-
ким шорохом шагов или щелканьем взводимого курка, но, с другой сто-
роны, известны случаи (Дорогостайский, 1918), когда бараны не могут ра-
зобрать—откуда идут звуки и бросаются на выстрел. Обнаружив опасность,
баран издает свистящий звук, похожий на «чшу-ую-ю», независимо от
того, находится ли он один или в стаде.
Смена волосяного покрова у архаров происходит один раз в году —
весной. Сроки начала и окончания линьки не одинаковы в разных частях
ареала, а кроме того зависят от состояния организма животных. В разных
OVIS
539
районах она тянется со средины марта до средины июля. Архары Ка-
ратау вылинивают, по-видимому, уже в мае (Антипин, 1947). На Памире,
по Р. Н. Мекленбурцеву (1948), линька начинается в апреле, а в середине
мая все звери ходят уже в летней шерсти. В северо-западной Монголии
архары вылинивают только к началу июля (Дорогостайский, 1918). У са-
мок, имеющих ягнят, линька заканчивается дней на 20—25 позднее, чем
у самцов и холостых самок. В конце мая 1956 г. в горках юго-восточной
части Бет-Пак-Далы основная масса архаров вылиняла, но нам встретился
молодой самец, у которого в бинокль было ясно видно много еще не вы-
павшей старой шерсти. У добытой 30 мая там же взрослой самки,
имевшей ягненка, следы старой шерсти были только на шее.
В первую очередь старая шерсть выпадает на лопатках, морде, боках
туловища. Дольше всего она удерживается на затылке и в виде отдельных
разрозненных пучков на верхней стороне шеи, спины, а также на нижней
стороне груди и передней части живота (Дорогостайский, 1918; Антипин,
1947).
Половой зрелости самки архаров, по мнению В. М. Антипина (1947),
могут достигать уже на втором году жизни, но Р. Н. Мекленбурцев (1948)
не мог установить беременности ни у одной из добытых им полуторалет-
них самок; поэтому он считает их способными к размножению с возраста
не моложе двух лет. Самцы достигают половой зрелости также в возрасте
2—2У2 лет, так как у убиваемых полуторалетних зверей в ноябре семен-
ники еще лишены спермы (Румянцев и др., 1935). Самцы начинают активно
участвовать в размножении позднее — с 4 или даже 5 лет, так как до
этого отгоняются более взрослыми крупными зверями.
Перед началом гона взрослые самцы, до этого державшиеся отдельно,
присоединяются к стадам самок. Сроки протекания гона не одинаковы
в разных частях ареала О. аттоп. В Киргизии, по данным Б. Ф. Румян-
цева с соавторами (1935), он происходит в течение ноября и декабря.
Первых самцов в стадах самок они начали встречать во второй половине
октября. В западном Тянь-Шане течка начинается раньше, около сере-
дины октября (старого стиля), и тянется с месяц (Чернавин, 1916; Абдилда-
беков, 1950). Судя по времени окота, еще раньше, в среднем, протекает гон
у баранов Каратау (на Сыр-Дарье). На Памире, наоборот, время гона
смещается на более поздние сроки. В ноябре большинство самцов держится
еще отдельно от самок, а спаривание происходит в декабре и первой поло-
вине января (Мекленбурцев, 1948). Такие же сроки гона приводит А. М. Ко-
лосов (1939) для Алтая.
Взрослые самцы-бараны создают себе гаремы от 8 до 25 самок, вместе
с которыми находится известное число неполовозрелых животных, вклю-
чая и самцов. Взрослых самцов в стаде самок, по собранным В. В. Черна-
виным (1916) опросным данным, может быть несколько, например по три.
По данным же Р. Н. Мекленбурцева (1948), в гареме находится только
один половозрелый самец. Половозрелые, но слабые и молодые, не подпу-
скаемые сильными соперниками самцы, в это время бродят отдельными
группами неподалеку от гаремов (Румянцев и др., 1935).
Самцы в период спаривания сильно возбуждены, гоняются за самками,
становятся менее осторожны. Подойти на выстрел к ним в это время го-
раздо легче. Между самцами происходят турнирные поединки. Расходясь
и снова сближаясь, они с огромной силой ударяются с разбега основаниями
рогов и лбами. Звуки ударов рогов, говорят, слышны в горах за несколько
километров. Достоверных случаев гибели одного из дерущихся соперни-
ников не известно и, по-видимому, вообще таковые бывают крайне редко.
540
BOVIDAE
По окончании гона самцы вновь отделяются от самок и, собираясь в не-
большие группы, уходят в горы.
Продолжительность беременности у архаров точно не известна.
Б. Ф. Румянцев с соавторами (1935) считают, что она такова же, как и
у домашних овец, т. е. около 5 месяцев. Однако, судя по срокам гона и
окота, Р. Н. Мекленбурцев (1948) справедливо определяет ее в полгода.* 1
Рождение молодняка в Казахском нагорье, южном Алтае, Сауре и
Тарбагатае происходит в первых числах мая, в западном Тянь-Шане и
Заилийском Алатау — в марте и апреле, а в Каратау, по В. М. Антипину
(1947), первые ягнята появляются в феврале и заканчивается окот к концу
марта. На Памире первые ягнята появляются не раньше конца мая или.
начала июня.
Перед окотом самки удаляются от стада в более глухие каменистые или
кустарниковые участки, где и приносят одного, реже двух ягнят. Тройни
наблюдаются очень редко. Первые дни ягнята слабы и беспомощны, мо-
гут быть пойманы руками и обычно лежат, затаившись между камней
или в кустах. Однако уже на 3—4-й день они следуют за матерью и не
могут быть пойманы даже собакой (Мекленбурцев, 1948). Самки, держа-
щиеся первые дни после окота одиночно и в одном месте, дней через 10—
15, когда ягнята окрепнут, начинают бродить более широко и объединяются
в небольшие группы, к которым в дальнейшем присоединяется молодняк
прошлого года. Матери кормят ягнят до сентября—октября, но у отдель-
ных, даже беременных самок Р. Н. Мекленбурцев (1948) обнаруживал
молоко в вымени до первых чисел февраля. Молоко архариц по вкусу и
химическому составу существенно не отличается от молока домашних
овец. Зеленый корм ягнята начинают щипать уже в первые недели жизни,
к осени большая часть их переходит на самостоятельное питание.
Родятся архарята (Б. Ф. Румянцев с соавторами, 1935) мелкими,
около 4.5 кг, мало отличаясь по величине от местных домашних ягнят.
К осени они достигают половины роста взрослых. Половой диморфизм
в размерах при рождении выражен слабо. Самочки-ягнята лишь немного
меньше самцов; только впоследствии они начинают сильно отставать от
самцов в росте. Растут архары очень долго, особенно самцы, по крайней
мере до 10 лет (Бутарин, 1940). Последнее, вместе с более высокой интен-
сивностью роста, объясняет факт, почему при одинаковом с домашними
овцами весе при рождении дикие архары достигают огромных размеров
во взрослом состоянии. Б. Ф. Румянцев с соавторами (1935) и Н. С. Бу-
тарин (1940) показали, что прирост в весе у архара от рождения до взрос-
лого состояния превышает таковой у курдючных овец и новокавказских
мериносов в несколько раз.
Максимальная продолжительность жизни архаров в естественной
обстановке, по-видимому, не превышает 12 лет, а у самок даже меньше
(6—7 лет). Большинство находимых в горах черепов баранов принадле-
жит животным в возрасте 4—5 лет, реже 5—10-летним (Мекленбурцев,
1948).
Из врагов архара на первом месте по значению стоит волк. Волки ба-
ранам наносят больший ущерб, чем козлам, так как бараны держатся
на более ровных и открытых местах, и спасение часто ищут пе в скалах,
а в быстроте своих ног, которая не всегда спасает от не слишком быстрого,
1 Сам Б. Ф. Румянцев (1935) пишет, что оплодотворение самок происходит около
1 ноября, а молодые архарята в возрасте 1—2 дней были пойманы только 30 апреля,
т. е. через 6 месяцев.
OVIS
541
но более выносливого волка. Охотятся волки на архаров стаями, часами
гоняют их до утомления, стараясь не дать уйти в скалы или отбить одиноч-
ного зверя, с которым управиться гораздо легче. Иногда часть хищников
прячется в засаде, а другая старается гнать на них преследуемых баранов
(Мекленбурцев, 1948). В первую очередь при этом гибнут старые больше-
рогие самцы, особенно зимой, не способные быстро бежать по хотя бы
сколько-нибудь глубокому снегу. В западном Тянь-Шане таким же обра-
зом охотятся на архаров красные волки (Суоп alpinus Pall.) (Черна-
вин, 1916).
Барс (Felis uncia Schr.) еще легче справляется с бараном, но в силу
своей малочисленности и приверженности к скальному ландшафту, ко-
торого архары избегают, существенного вреда их поголовью не наносит.
Другие хищные млекопитающие (рысь, медведь) и птицы практического
значения, как враги архара, не имеют.
Сведения об эпизоотиях среди архаров имеются только в отношении
Памира (Мекленбурцев, 1948). Здесь в конце прошлого столетия наблю-
далась установленная ветеринарным персоналом сибирская язва. Кроме
того, наблюдалась какая-то неизвестная эпизоотия, носящая у местного
населения название «кара-юпка» (черное легкое) по патологическим изме-
нениям, наблюдающимся в легких. Внешне это заболевание выражалось
в тяжелом учащенном дыхании, удушливом кашле с кровью, угнетении
и слабости, доходящей до того, что больные животные почти не реагиро-
вали на приближение человека. Часть животных выздоравливала, но мно-
гие погибали. В 1886—1887 гг. «кара-юпка» охватила обширные районы
Памира и унесла около половины всего населявшего их поголовья
архаров. Болели ею также сибирские козероги и домашний скот: козы,
овцы, верблюды, но ни люди, ни хищные звери не заражались.
В многоснежные зимы значительное число архаров гибнет от бескор-
мицы. П. П. Сушкин (цит. по Насонову, 1923) видел кладбище, около 70
бараньих черепов, на перевале через Сайлюгемский хребет, которое он
считал остатками жертв бурана. А. М. Колосов (1939) упоминал о скопле-
ниях скелетов аргали в Чуйских Альпах, например в районе Чагах-Бур-
гизы и в Сайлюгеме, которые могли иметь только такое же происхождение.
Много ослабленных голодом зверей в такие годы гибнет от волков, в пер-
вую очередь опять-таки крупные старые самцы, которые из-за больших
рогов не могут опускать глубоко в снег морду и передвигаться по глубо-
кому снегу.
Из эктопаразитов на архарах известны подкожный и носоглоточный
оводы. Видовая принадлежность первого до сих пор не установлена.
Личинки подкожного овода на архарах отмечены в Джунгарском Алатау,
Каратау, центральном Тянь-Шане (Антипин, 1947) и на восточном Памире
(Мекленбурцев, 1948). Р. Н. Мекленбурцев пишет, что, спасаясь от ово-
дов, архары летом поднимаются к линии вечных снегов и, тем не менее,
«редкий архар не имеет под кожей личинок овода, которые находятся
только на одном, строго определенном месте — на пояснице». Выпадение
личинок подкожного овода на Памире происходит в марте и апреле. В кол-
лекциях Зоологического института АН СССР имеется шкура самца О. ат-
топ nigrimontana Sev., добытого в горах Каратау 1 февраля 1910 г.
(№ 14239), со множеством свищей на спинной части от личинок под-
кожного овода. Попутно следует отметить, что у двух архаров, добы-
тых в последних числах мая 1956 г. в горах Джамбул (юго-восточная
Бет-Пак-Дала), личинок подкожного овода и их следов нам обнаружить
не удалось.
542
BOVIDAE
Носоглоточных оводов на архарах зарегистрировано два вида: Oestrus
ovis L. в Терскей- и Кунгей-Алатау (Гребенюк и Сартабаев, 1955) и Rhi-
noestrus tshernyshevi Grun. в горках Джамбул в Бет-Пак-Дале.1 Кроме того,
паразитизм личинок носоглоточного овода без установления их видовой
принадлежности отмечен на восточном Памире (Мекленбурцев, 1948),
Джунгарском Алатау, Каратау и центральном Тянь-Шане (Антипин, 1947).
Нападают на архаров также клещи, из которых с достоверностью уста-
новлены Ilyalomma plumbeum Panz., Ixodes persulcatus P. Sch. и Dermacen-
tor pavlovskyi 01. (данные Г. В. Сердюковой). Р. Н. Мекленбурцев (1948)
находил на памирских архарах в начале зимы присосавшихся к коже
в области грудины паразитов, по внешности напоминавших личинку
овода, но более плоских и широких в средней части. К какому не только
виду и роду, но и классу, относятся эти паразиты — не известно.
До настоящего времени у архаров зарегистрировано 32 вида паразити-
ческих червей (Боев, Соколова и Бондарева, 1948). Одних легочных нема-
тод, паразитирующих у этих баранов, известно 7 видов (Боев, 1954).
Гельминтами поражены поголовно все, по крайней мере взрослые звери.
Однако специфичных для О. аттоп гельминтов не известно. Известный
состав фауны паразитических червей архаров близок к таковому совме-
стно с ним обитающих диких и домашних видов.
Хозяйственное значение. Самым ценным продуктом, получаемым от
убитого архара, является мясо. От взрослого самца архара получается
туша (без нижних частей ног и головы) от 55 до 96 кг, от самки 34—36 кг.
Выход чистого мяса (убойный вес) у взрослых самцов составляет около
56% (максимально 63.6%) живого веса, у самок несколько меньше (49.2—
54.8%), а у молодняка, наоборот, несколько больше — до 63.5% (Дени-
сов, 1935; Бутарин, 1940).1 2
Мясо диких баранов по вкусу напоминает мясо домашних, но имеет
специфический привкус дичи и более жестко. На вид оно темно-красного
цвета, без мраморности (без прослоек жира между мышцами), свойствен-
ной мясу домашних баранов. Разваривается мясо даже молодых архаров
труднее и дольше, чем мясо домашних.
Химический состав мяса самца 8 лет, по исследованию В. Ф. Денисова
(1935), был следующий: сырого протеина 84.0%, сырого жира 6.26%,3
минеральных веществ 4.52%, безазотистых экстрактивных веществ 5.22%.
Подкожный жир, по данным того же автора, у архаров совершенно
не развит. Основная масса жировой ткани откладывается в брюшной и
грудной полости. От взрослых, хорошо упитанных зверей, внутреннего
сала получается свыше 3 кг. По питательным и вкусовым достоинствам
жир архара близок к жиру домашней овцы, такой же тугоплавкий (темпе-
ратура плавления 48—53° С).
В ряде районов промысел архара имеет важное значение для населе-
ния. Так, по Р. Н. Мекленбурцеву (1948), на Памире каждый средней
руки охотник ежегодно добывает 20—25 архаров, и в общем архарятина
составляет не менее половины всего потребляемого киргизами мяса.
Шерсть архаров содержит около 85% мертвого волоса и поэтому шкуры
как овчины употребляются редко. Безусловно, они годны для выделки
хрома, а от молодых зверей — даже на шевро. Шерсть, вероятно, могла бы
1 В последнем случае определение произведено К. Я. Груниным.
2 Приведенные цифры относятся к зверям, добытым в период с сентября по ноябрь,
когда они упитаны более, чем в другое время.
3 Все в процентах к абсолютно сухому веществу.
OVIS
543
служить для набивки мебели и матрацев. Однако товарный выход шкур
диких архаров до сих пор незначителен. Частично местным населением
они применяются в быту в качестве ковриков на пол, для чего лучше под-
ходят шкуры крупных самцов с густым мехом. Иногда из шкур делают
бурдюки для хранения молочных продуктов. Для этой цели употребляется
снимаемая целиком шкура, начиная с ноги или шеи. Немалая доля шкур
от добываемых на Памире архаров бросается на месте (Мекленбурцев,
1948).
В большинстве случаев бесцельно бросаются и рога вместе с отрезае-
мой на месте добычи тяжелой головой архара, хотя при плановой заго-
товке они могли бы служить для разных поделок (Чернавин, 1916).
Неволю архары переносят плохо, в зоопарках долго не живут. В Ле-
нинградском зоопарке две пойманные в сентябре 1953 г. полувзрослыми
самки памирского архара пали в феврале 1955 г. от туберкулеза, не про-
жив в неволе и полтора года.
В литературе имеются ссылки на получение гибридов при скрещивании
архара с бараном Винье (О. orientalis vignei Blyth) и с арменийским муф-
лоном (О. orientalis armeniana Nas.). Однако о внешнем виде и плодови-
тости полученных гибридов ничего не известно (Саркисов, 1953; Ewart,
1914). Известны многочисленные случаи получения вполне плодовитых
гибридов при скрещивании архара с различными породами домашних
овец (Gray, 1954).
В Советском Союзе работы по гибридизации аргаловидных баранов
с домашними овцами ведутся в плановом порядке с начала 30-х годов на-
стоящего столетия и имеют своей целью выведение новых, более совершен-
ных пород. Была разработана методика искусственного осеменения до-
машних овцематок спермой из придатков семенников от убитых самцов-
рахаров, давшая весьма положительные результаты.
Спермой архара осеменялись киргизские курдючные овцы (Румянцев
и др., 1935). Были получены гибридные ягнята, которые по внешнему виду
(экстерьеру) и темпераменту напоминали больше архарят, чем ягнят кур-
дючных овец. По общим размерам они не отличались от ягнят исходных
форм; по пропорциям телосложения, в частности по характеру головы,
занимали промежуточное положение. Масть у гибридных ягнят была более
близка к домашней форме, зато хвост и уши приближались к архарьему
типу. Характер (структура) шерстного покрова также был средний между
исходными формами, но более близкий к дикому виду. Гибридные ягнята,
родившиеся от домашних маток, были очень дики и по сравнению с чисто-
кровными домашними весьма подвижны. Дальнейшая судьба гибри-
дов не известна. Опыты по гибридизации архаров с курдючными овцами,
вероятно вследствие их хозяйственной нецелесообразности, были прекра-
щены.
Гораздо более плодотворной оказалась плановая гибридизация архара
с тонкорунными овцами, проводимая с 1934 г. последовательно Институ-
том генетики Академии наук СССР и Академией наук Казахской ССР.
Спермой, полученной от убитых самцов-архаров, осеменялись овцематки
мериносов новокавказского типа. Гибриды первого поколения по внешнему
виду напоминали архаров, а по шерсти — мериносов. От последних они
отличались живым темпераментом и большой подвижностью, способ-
ностью передвигаться по крутым горным склонам. От дикого вида гиб-
риды первого поколения унаследовали тип роста, в результате чего были
значительно крупнее одновозрастных мериносов. В отношении пропор-
ций телосложения, головы и ушей наблюдалась мозаичная наследствен-
544
BOV ID АН
ность, а в отношении широтных промеров — даже гетерозис. Шерстные
качества, в основном, унаследовались от мериносов.
Гибриды первого поколения скрещивались повторно с мериносовыми
матками, а гибриды второго и третьего поколения, в дальнейшем, разво-
дились в себе. В результате систематического отбора и подбора, к 1950 г.
была создана новая культурная порода овец — казахский архаро-мери-
нос, совмещающая в себе высокую шерстную и мясную продуктивность
с прекрасной приспособленностью к круглогодичному пастбищному со-
держанию в горах (Бутарин, 1940, 1950).
Охота. Способы охоты на архаров описали Д. Иванов (1885), Д. Н. Го-
ловин (1901), Н. В. Туркин и К. А. Сатунин (1904), а в последние годы
Р. Н. Мекленбурцев. Наиболее распространенным является скрадывание,
охота с подхода, в принципе ничем не отличающаяся от таковой на других
копытных животных. Главное условие — уметь скрытно подойти к зверю
против ветра, так как запах человека архар чует за километр. Так как
бараны живут на обширных открытых пространствах, то для разыски-
вания стад в местах, где водятся бараны, охотники отправляются верхом
на лошадях.
Подкарауливают баранов за прикрытиями на местах кормежек, во-
допоев и на постоянных путях суточных переходов. Ловят и капканами,
расставляя последние на тропах.
На Памире широко практикуется охота с собаками. Последние, зави-
дев баранов, начинают преследовать их. Догнать их собаки не могут,
но, преследуя часами, заставляют утомленных зверей становиться на от-
стой где-нибудь на выступе скалы, куда преследователи забраться не
могут. Окружив скалу, собаки громким лаем дают знать об этом охотнику
и до прихода его не позволяют зверям скрыться. Однако и в этом случае
к баранам надо подходить скрытно, чтобы раньше времени не испугать
зверя, и стрелять из-за камня.
Второй способ охоты с собаками, применяемый на Памире, тоже до--
бычлив, но требует хорошо дрессированных собак. Сущность его заклю-
чается в том, что собаки не дают уйти обнаруженному стаду архаров,
забегают с противоположной от охотников стороны и, умело маневрируя,
лаем и бросками заставляют зверей набежать почти вплотную на распо-
ложившихся в засаде стрелков (Мекленбурцев, 1948).
В западной части Таласского Алатау для охоты за архарами приме-
няют борзых, а изредка устраивают облавы (Абдилдабеков, 1950).
Ovis (Pachyceros) nivicola Eschscholtz — Снежный баран,
или толсторог, чубук (рис. ИЗ—115)
Eschscholtz F., 1829, Zool. Atlas, I : 1 (первооп., и->обр.); Е г m a n n А.,
1838, Reise um die Erde, I, 2 : 350 (распр.); Middendorf A., 1853, Reise in die
auRer. Norden u. Ost°n Sibiriens, II, 2 : 116—118 (распр.); Нейман К. К.,
1871, Изв. Русек, географ, общ., 1:19 (распр.); Schmidt F., 1872, Mem. Acad. Sci.
St.-Petersb., ser. VII, t. XVIII, No. 1 : 40 (распр.); Поляков И. С., 1873, Зап.
Рус.ск. географ, общ. по общей географ., III : 72—73 (распр.); Северцов Н. А.;
1873а, Изв. Общ. любит, естествозн., аптропол. и этнограф. VIII. 2 : 86, 153 (распр.,
морфол.); Северцов Н. А., 18736, Природа, 1 : 208—215 (распр., биол.); Си-
до р о в М. К., 1881, Изв. Общ. любит, естествозн., антропол. и этнограф., XXXVII,
1 : 45 (распр.); Майдель Г., 1894, Путешеств. по сев.-вост, части Якутской
области 1868—1878 гг., СПб., Изд. Акад, наук : 200 (распр.); Иохельсон В. И.,
1898, Очерк зверопромышл. и торговли мехами в Колымском округе, СПб. : 102
(распр.); Lydekker R., 1898, Wild oxen, sheep and stoats of all lands, London : 221 —
226 (общ., синон., изобр.); Lydekker R., 1902, Proc. Zool. Soc. London, II : 84
(сист., морф.); Allen J., 1904, Bull. Amer. Mus. Natur. Hist., XX : 293—298 (сист.,
морф.); Demidoff E., 1904, A shooting trip to Kamchatka, London:!—302
ovis
545
'(распр., биол., ох.); Туркин Н. В. и К. А. Сатунин, 1904, Звери России,
т. IV, Bovidae, М. : 560—563, 716—723 (общ., изобр.); Matschie Р., 1907, в кн.:
NiedieckP. Kreuzfahrten im Beringmeer, Berlin: 235—236 (сист., изобр.); Neumann L,
1908—1909, Notes Leyden Mus., XXX. : 86—88 (параз.); Pocock R., 1910, Proc.
.Zool. Soc. London : 861 (морф.); Толмачев И. IL, 1911, По Чукотскому побе-
режью Ледовитого океана, СПб. : 84—85 (распр.); Бутурлине. А., 1913, Дневн.
Зоол. отд. Общ. любит, естествозн., антропол. и этнограф., нов. сер, 1,5: 253 (распр.);
Kowarzik R., 1913, Zool. Anzeig., XLI, 10 : 439—445 (сист.); Lydekker R.,
1913, Catal. Ungulate mammals Brit. Mus., I, London : 121—122 (сист., синон.); H a-
сонов H. В. и В. Ч. Дорогостайский, 1915, Изв. Акад, наук : 1599—
1616 (морфол., сист., биол., изобр.); Толстов С. М., 1916, Наша охота, 18 : 16
(распр.); Насонов Н. В., 1923, Географич. распр. диких баранов Старого Света,
Птг., изд. Акад, наук : 124—138 (сист., синон., распр., изобр.); Suschkin Р.,
1925, J. mammals, vol. 6, No. 3 : 150—154 (сист., распр.); Караев А. И., 1926,
Эконом, жизнь Дальн. Востока, 4 : 143 (распр.); Огнев С. И., 1926, Млекопит.
сев.-вост. Сибири, Владивосток : 139—141 (распр.); Тассовский Г. Н., 1927,
Произв. силы Дальн. Востока, IV : 530+534—535 (распр.); Дулькейт Г., 1927,
Охотник, 1 : 13 (распр.); Сокольников Н. П., 1927, Бюлл. Моск. общ. испыт.
прир., отд. биол., XXXVI, 1/2:131—133 (распр., биол.); Яковлев Е., 1930,
Тр. Зоол. секц. Среднесибирск. отд. Русск. географ, общ., 1:59; Белополь-
ский Л. О., 1933, Тр. Зоол. инет. Акад, наук СССР, 1 : 184—185 (распр.); Афа-
насьев А. В., 1934, Тр. Совета по изуч. произв., сил, сер. дальневост., 2 : 262
(распр.); Золотарев Н. Т., 1934, Тр. Совета по изуч. произв. сил, сер. дальне-
вост., 3:161 (распр.); Наумов Н. П., 1934, Тр. Полярн. комисс. 17:66—67
(распр.); Флеров К. К., 1935, в кн.: Адлерберг Г. П., Б. С. Виноградов,
Н. А. Смирнов и К. К. Флеров, Звери Арктики, Л., изд. Главсевморпути : 224—236
(общ., синон., изобр.); Gromova V., 1936, Neue Fortschr. in Tierzucht u. Abstam-
mungelehre (Festschr. z. 60. Geburtstag v. Prof. J. U. Duerst), Bern : 84 (морф., сист.);
Подаревский В. Б., 1936, Пробл. охотхозяйств, акклим. в вост. Сибири, Ир-
кутск, ОГИЗ : 47 (распр.); Фаворский В. П., 1936, в сборн. «Охотничье про-
мысл. хозяйство Вост.-сибирск. края», Иркутск, ОГИЗ : 39 (распр.); Туров G. С.’,
1937, Снежный баран, М.—Л., КОИЗ : 1—109 (биол.); Frick Ch., 1937, Bull. Amer.
Mus. Natur. Hist., 69 : 554—555 (ист.); П ле ч e в E. И., 1939, Изв. Русск. географ,
общ., 71 : 1380—1381 (распр., биол.); Самородов А. В., 1939, сборн. тр. гос.
зоол. музея при Моск. гос. унив., V : 18 (распр.).; Шнакенбург П. Б., 1939,
Сов. север, 3 : 90—92 (распр.) (цит. по Портенко, 1941); Портенко Л. А., 1941,
Фауна Анадырского края, Л., изд. Главсевморпути : 89—91 (распр.); С к а л о н В. Н.,
1941, Природа и соц. хозяйство, VIII : 278—279 (распр.); Harper F., 1945, Extinct
and vanishing mammals of Old World, N. Y. : 595—599 (синон., распр.); Бажа-
нов В. С., 1946, Бюлл. Моск. общ. испыт. прир., Отд. биол., LI, 4/5 : 100 (распр.);
Г а в р и л о в В. Т., 1947, Зоол.журн.,XXVI, 6 : 561—562 (биол.); Громова В. И.,
1947, Докл-. Акад, наук СССР, LVII, 5 : 493—495 (ист.); Аверин Ю. В., 1948,
Тр. Кроноцкого заповеди., 1:175—189 (морф., биол., изобр.); Павлов Е. И.,
1949, Промысл, звери Читинской обл., Чита, Облизд. : 112—116 (общ., изобр.);
Скалой В. Н., 1949, Охрана природы, 8 : 118—121 (распр.); Аверин Ю. В.,
1951, Бюлл. Моск. общ. испыт. прир., отд. биол., нов. сер., LVI, 1 : 15—26 (распр.,
биол.); Цалкин В. И., 1951, Горные бараны Европы и Азии, М., изд. Моск. общ.
испыт. прир. : 265—-275 (сист., синон., распр., изобр.); Е 1 1 е г m a n J. а. Т. Мог
rison-Scott, 1951, Checklist of palaearctic a. indian mammals, London : 413
(сист., синон.); Насимович A. A., 1955, Роль снежн. покрова в жизни копытных,
М., изд. Акад, наук СССР : 141—144 (распр., биол.).
Описание. Баран сравнительно крупной величины; длина тела взрос-
лых самцов от 162 до 178 см; высота в холке от 97 до 105 см. Основная длина
черепа 228—255 мм. Живой вес взрослых самцов составляет, в зависи-
мости от упитанности, 79—129 кг. Самки значительно мельче; длина тела
у них колеблется от 127 до 145 см, высота в холке от 84 до 94 см, основная
длина черепа от 205 до 236 мм, живой вес от 45 до 55 кг.1 Встречаются
изредка и более крупные экземпляры. Так, Ю. В. Аверин (1951) полагает,
что вес хорошо упитанных крупных самцов может достигать 140 кг, а са-
мок — до 60—65 кг.
1 Цифры размеров тела и живого веса приводятся по Б. Т. Гаврилову (1947) и
Ю. В. Аверину (1951). . . ,
35 Фауна СССР, И. И. Соколов
546
BOVIDAE
Телосложение более массивное и несколько более неуклюжее, чем
у других баранов. Конечности сравнительно толстые и производят впечат-
ление более коротких. Высота в крестце на несколько сантиметров пре-
восходит высоту в холке. Линия спины прямая, наклоненная вперед или
выпуклая. Хвост равен, или немного длиннее уха, около 9—11 см. Голова
не длинная, но широкая, особенно в глазницах. Морда относительно ко-
роткая. Профиль носа прямой. Уши сравнительно короткие; длина их
не превышает 9—10 см. Шея короткая и толстая. Копыта крупные, черно-
бурого цвета. Длина их у самцов на передних конечностях составляет
66—80 мм, на задних 59—69 мм. Высота передних копыт 37—46 мм, зад-
них на 1—3 мм меньше. Телосложение самок более легкое; но голова и
у них более широкая, а ноги более толстые, чем у самок других видов ба-
ранов.
Рога снежного барана сравнительно короткие, но относительно своей
длины толстые, откуда и происходит название толсторога. Длина рогов
по изгибу передней поверхности у взрослых самцов колеблется в пределах
43—111 см, но длиннее 1 м бывают редко. Длина четырех сегментов (при-
рост второго, третьего, четвертого и пятого годов жизни) составляет от
36 до 75 см (Цалкин, 1951). Обхват рогов в основаниях колеблется в пре-
делах от 25 до 36 см (29—59.8% длины всего рога). Продольный диаметр
сечения рога в основании (от заднего ребра до наиболее выдающейся точки
передне-внутреннего) составляет 91—111 мм; наибольший поперечный
диаметр рога от 67 до 97 мм. Рога поставлены круто и сильно выступают
над поверхностью лба. С профилем носовых костей они образуют угол
около 140—150°. Особенностью рогов толсторога является слабое расхо-
ждение их в стороны, под углом не более 70° (Gromova, 1936).
Изгиб рогов всегда гомонимный. У взрослых самцов рога описывают,
хотя и не всегда, крутую спираль в полный оборот и концами обращены
несколько в стороны и одновременно вверх, реже вперед или вниз. Передне-
наружное ребро на рогах хорошо развито. Передне-внутреннее также
ясно выражено, но несколько притуплено. Передняя (лобовая) грань рога
образует с боковой гранью угол значительно более острый, чем с внутрен-
ней. Отличительной от других видов особенностью рогов снежного барана
является то, что лобовая грань выступает в сторону и нависает над боко-
вой гранью рога, в результате чего на последней вдоль передне-наружного
ребра образуется продольный желобок, особенно резко выраженный
у основания рога. Лобовая грань плоская или слегка выпуклая, обычно
в той или иной мере скошена внутрь. Внутренняя грань ближе к вершине
вогнутая, к основанию становится сильно выпуклой. Наружная грань от
основания до вершины рога выпуклая, если не считать желобка вдоль
передне-наружного ребра рога. Заднее ребро рогов толсторога, в противо-
положность другим видам баранов, сильно закруглено. Вследствие этого,
поперечное сечение рога в основании имеет форму четырехугольника
с сильно выпуклой стороной, соответствующей заднему ребру рога. Вер-
шины рогов, как и у других баранов, уплощены; лобовая грань постепенно
суживается и сходит на нет, так что конец рога имеет только два притуп-
ленные ребра — переднее и заднее, и соответственно две грани — наруж-
ную и внутреннюю.
Характерной особенностью рогов снежного барана является слабо
развитая и мелкая морщинистость. Поверхность роговых чехлов несет
мелкие, тесно прилегающие друг к другу, низкие и слабо дифференци-
рованные морщины. Иногда морщинистость столь сглажена, что стано-
вится почти незаметной, поверхность же рогов несет только поперечные
OVIS
547
круговые борозды (кольца) на границах участков годичного прироста
рога.
Самки снежного барана всегда имеют рога. Рога у них, по сравнению
с самцами, очень короткие; длина колеблется в пределах от 13 до 32 см,
обычно не превышает общую (наибольшую) длину черепа, а если превос-
ходит, то меньше, чем в полтора раза. Изгиб их дугообразный, с концами,
загнутыми назад и в стороны. Рога сжаты с боков, имеют два закруглен-
ные ребра. Поперечное сечение их продольно-овальное, с выпуклой на-
Рис. ИЗ. Снежный баран (Ovis nivicola Eschsch.).
ружной и несколько вогнутой внутренней поверхностью. Морщинистость
рогов у самок тоже мелкая но ясно выражена. Цвет рогов у толсторогов
светлый, желтовато-бурый, реже темно-бурый.
Волосяной покров снежного барана во вполне развитом состоянии
очень густой, по сравнению с другими видами баранов, более длинный и
сравнительно мягкий, состоит из прямой, длинной ости и тонкого извитого
подшерстка. Остевые волосы слабо волнистые, легко обламываются, осо-
бенно на концах. Длина их на боках туловища около 7—9 см. Удлинены
волосы на шее, особенно на верхней ее стороне, а также на боках выше
подмышек и иногда над пахами, где они могут достигать 12—13 см. Более
короткий упругий волос покрывает голову и нижние части конечностей
ниже запястных и скакательных суставов. Подвес на нижней стороне
шеи и бакенбарды у основания головы в том виде, как они имеются у муф-
лонов, у толсторогов отсутствуют. Нет у них и подобия манишки на пле-
чах и на передней стороне груди, как у некоторых рас аргали. Особо
толстый и грубый волос покрывает нижнюю сторону груди и пространство'
на крупе вокруг хвоста. Более редкие и мягкие волосы растут в пахах и
на задней части живота.
35'*
548
BOVIDAE
Окраска снежного барана подвержена возрастной, сезонной и геогра-
фической изменчивости. Характерным является отсутствие красноватых
и рыжих тонов. Преобладают коричневый и бурый оттенки. Представи-
телям типичной расы (с Камчатки) свойственна более темная однотонная
окраска. Верх и бока туловища зимой — от коричнево-бурого до буровато-
желтого цвета. Иногда круп немного темнее или, наоборот, светлее, но
ясных посветлений на боках туловища, подобно седловидным пятнам муф-
лонов, нет. Местами, особенно на плечах и шее, заметен легкий желто-
ватый налет. К концу зимы окраска сильно выцветает, иногда становится
совсем светлой (Цалкин, 1951). Отдельные волосы ости туловища от осно-
вания светло-бурые, часто почти белые, кверху постепенно приобретают
коричнево-бурую расцветку. Верхушечные части имеют желтоватое кольцо,
а самые приостренные кончики волос, пока не обломаны, черно-бурые.
Пуховые волосы (подшерсток) желтовато-белые. От степени потемнения
срединной части остевых волос и от размеров светлой предвершинной зоны
зависит общий тон окраски животного. Нижняя сторона груди и перед
живота коричнево-бурый. Остальной живот, пахи, промежность, область
наружных половых органов желтовато- или грязновато-белые. Белая
окраска распространяется также на ягодицы и прилегающие к хвосту
части крупа, образуя довольно четко отграниченное околохвостовое. зер-
кало, разделенное срединной полосой у корня хвоста. На границе темных
боков и светлой окраски живота от локтевых к коленным суставам тянутся
более или менее ясные коричневые полосы; часто они, впрочем, почти не
заметны. У баранов Станового хребта позади лопаток на уровне плече-
лопаточных суставов имеется с каждой стороны по белому пятну. Для
баранов Якутии (подвид О. п. lydekkeri Kow.) характерно общее посвет-
ление окраски туловища. Последняя зимою может становиться желтовато-
белой или очень светлой буро-коричневой, а в' отдельных случаях приоб-
ретает дымчатый оттенок. От основной буро-коричневой окраски остается
лишь темная полоса из смеси буро-коричнезых и белых волос вдоль сред-
ней линии спины от холки до корня хвоста, да иногда неясные коричне-
вые пятна на боках или крупе. Светлое околохвостовое зеркало в этих
случаях сливается незаметно с окраской крупа, равно как нет резкой гра-
ницы между боками и животом. Пограничная с животом полоса по бокам
туловища или вовсе отсутствует, или от нее сохраняются лишь небольшие
коричневые пятна на подмышках и впереди коленных суставов. Окраска
низа груди образована смесью буро-коричневых и желтоватых волос.
Шея у темно окрашенных рас одноцветна с туловищем; у светло окра-
шенных она снизу, с боков, равно как и передняя сторона плеч коричнево-
бурые. На верхней стороне шеи в основной коричневой окраске имеется
значительная примесь белых волос.
Голова сверху одного цвета с туловищем, коричнево-бурая или желто-
вато-белая. Белая окраска в последнем случае распространяется также
на темя и затылок. Конец морды и передняя сторона нижней губы грязно-
илй желто-белые. Позади ноздрей на переносье у светло окрашенных осо-
бей имеется поперечная коричневая полоса, переходящая на бока морды.
Стороны и низ головы буровато-коричневые или серовато-бурые с при-
месью желтых волос. Уши буро-коричневые, к верхушкам беловатые,
внутри грязно-белые. Волосы по бокам-головы, ниже ушей, коричневые,,
равно1 как иногда и полосы между глазами й углом рта. Хвост сверху и.
с боков буро-коричневый, темнее туловища, снизу белый.
Наружная и внутренняя стороны предплечий и голеней одного цвета
с туловищем. Передняя сторона их, особенно ниже запястных и скака-
OVIS
549
тельных суставов, более темная, интенсивного темно-коричневого цвета,
иногда с золотистым оттенком. Задняя сторона ног желтовато-белая или
чисто белая, причем, в противоположность другим баранам, темная окраска
передней стороны резко отграничена от светло окрашенной задней стороны
ног. Узкие белые полосы вдоль задней стороны ляжек доходят до около-
хвостового зеркала и сливаются с ним. Светлая окраска около копыт
переходит на наружную сторону бабок, а с внутренней стороны, наоборот,
темная коричневая окраска переходит и охватывает заднюю сторону ба-
бок между главными и добавочными копытами.
Летняя окраска отличается от зимней более тусклым серовато-бурым
оттенком верха и боков туловища и наличием, хотя и не всегда, темной
буро-коричневой полосы вдоль средней линии шеи и спины. У светло окра-
шенных рас летом окраска светло-буроватая. Только полоса вдоль сред-
ней линии туловища, шея и низ груди сохраняют более темную буро-ко-
ричневую расцветку, да конечности, как и зимой, интенсивного темно-
коричневого цвета. Половой диморфизм в окраске выражен слабо. Самки
мало отличаются от самцов. Молодые вскоре после рождения желтовато-
бурые, позднее приобретают буровато-серую окраску. Серый оттенок
особенно отчетливо выражен на голове и ушах. Вдоль средней линии спины
имеется темно-коричневая полоса. Хвост темно-коричневый. Низ живота,
околохвостовое зеркало и задние стороны ног желто- или грязно-белые.
В черепе (рис. 114) характерна большая ширина, особенно мозговой
части. Ширина в глазницах всегда превышает расстояние от prosthion
до заднего края верхнего зубного ряда.1 Мозговая часть черепа хорошо
развита. Длина мозговой коробки от заднего выступа затылочного бугра
(acrocranion) до заднего края глазницы составляет не менее 49% (от 49.3
до 53.7%) основной длины черепа и от 71.8 до 83.8% длины морды (от
prosthion до переднего края глазницы). Зароговая часть, особенно у сам-
цов, сравнительно слабо выступает позади рогов и с затылочной поверх-
ностью образует очень тупой угол. Наружные слуховые проходы у сам-
цов располагаются целиком под основаниями роговых отростков, у са-
мок — на уровне задних краев или несколько позади последних. Лице-
вая часть черепа умеренной длины; длина морды составляет от 62.9 до
69.7% основной длины черепа. Межглазничная ширина черепа даже
,у самок всегда больше двух третей (от 67.6 до 85% у самцов) расстояния
от prosthion до точки слезно-скулового шва на краю глазницы.
Отличительной особенностью черепа толсторогов является также ко-
роткость и необычайная толщина роговых стержней. Обхват в основаниях
всегда больше их длины, нередко превосходя последнюю больше, чем
в полтора раза. В отличие от других видов баранов, предглазничные ямки
на черепе развиты очень слабо, иногда едва заметны.1 2 * Носовые кости
короткие и широкие; их ширина составляет от 40 до 60% собственной
длины и редко бывает меньше 43% последней. Межчелюстные кости не
всегда достигают носовых костей и соприкасаются с ними.
Костное нёбо сравнительно короткое; при горизонтальном положении
черепа передний край хоан расположен впереди уровня передних краев
глазниц. В области крыловидного гребня (crista pterygoidea), на границе
височной впадины и глазницы, как правило, лобная кость входит в сопри-
1 Из других видов баранов только у большерогих самцов О. аттоп L. ширина
черепа иногда немного (не более чем на половину длины М8) превосходит указанное
расстояние. У самок же ширина черепа всегда меньше.
2 В соответствии с этим, и предглазничные железы у толсторогов, по-видимому,
рудиментарны (Рососк, 1910).
550
BOVIDAE
косновение с алисфеноидом, что особенно хорошо заметно на черепах
молодых животных. Орбитосфеноид не соприкасается ни с теменной,
ни с височной костью (рис. 103, В). В области поперечного затылочного
гребня мастоидная кость не доходит до теменной, а височная входит в кон-
такт с верхней затылочной костью (рис. 104, Б). Особенностью зубной
системы толсторогов является отсутствие у 40—45% зверей переднего
премоляра в нижней челюсти (Р2), а у самцов иногда и в верхней, или ру-
диментарное его состояние.
История и географическое распространение (рис. 115). Снежные
бараны северо-восточной Сибири генетически тесно связаны с североаме-
риканскими толсторогами (О. canadensis Shaw) и, несомненно, происхо-
дят с последними от общего корня. Время обособления их от общего ствола
рода Ovis L. не известно. Со среднего плейстоцена толстороги известны
на Аляске. В Азии остатки их известны лишь в верхнепалеолитической
стоянке Мальта под Иркутском (конец плейстоцена) и в отложениях того же
возраста на р. Барзас (левый приток Томи, в Кузнецком Алатау, 86° в. д.
и 56° с. ш.; Громова, 1947). Современный ареал О. nivicola включает горы
восточной Сибири от Енисея до побережья Охотского моря и от побе-
режья Ледовитого океана до южных склонов Станового хребта и южной
оконечности Камчатки. В деталях распространение его представляется
в следующем виде.
Изолированный очаг обитания толсторога находится на севере Красно-
ярского края в горах Путорана к востоку от Норильска. А. Ф. Мидден-
дорф (Middendorf, 1853) нашел баранов в горах Сыверма на 67° с. ш. и
«особенно у истоков р. Хеты». Ф. Шмидт (Schmidt, 1872) указал его для
Медвежьих гор к востоку от Норильских озер. М. К. Сидоров (1881) доста-
вил Московскому обществу любителей естествознания, антропологии
и этнографии рог чубука с «низовьев Енисея». По-видимому, существенно
не изменилась эта часть ареала О. nivicola и до настоящего момента. В 30-х
годах, по В. Б. Подаревскому (1936), район обитания снежного барана
здесь шел, «по-видимому, сплошной полоской от верховьев р. Романихи
в бассейне Хеты на верховья р. Маймечи и затем к западу на истоки
р. Аякли. Здесь он охватывает верховья Котуя, Кочечумы, Хеты, водораз-
дел Курейка — Н. Тунгуска и через водораздел Хантайка — Пясина
перебрасывается в скалистый район озер Кита, Глубокое, Ляма». По
Н. П. Наумову (1934), к востоку от этого района бараны встречаются
еще в верховьях р. Тукалана, но, по-видимому, повсеместно редки и до-
бываются случайно. Сообщение С. М. Толстова (1916) о том, что область
распространения отсюда доходит до р. Оленека, по-видимому, ошибочно.
Ошибочным является и указание К. К. Флерова (1935 : 234) о распро-
странении баранов на Таймыре. В. М. Сдобников, проведший на Таймыре
3 года, сообщил мне, что этого зверя здесь нет, а по условиям природной
обстановки и быть не может. К югу от Полярного круга бараны на Туру-
ханском севере не заходят (Яковлев, 1930). В пределах Якутии снеж-
ные бараны держатся к востоку от Лены. Известен единственный случай
добычи толсторога на левобережье в 40 км от Лены, между населенными
пунктами Сиктях и Говорова (Скалой и др., 1941). Распространены ба-
раны по большей части Верхоянского хребта. В конце прошлого столетия
они встречались до северной его оконечности и устьев Лены, но в настоя-
щее время их как будто в этой части хребта уже нет. Есть бараны в Хара-
улахских горах. Лайдекер (Lydekker, 1902) описал экземпляр, добытый
в 40 км от устьев Яны. В коллекциях ЗИН АН СССР имеются экземпляры,
добытые в горах за 450 км к северу от Верхоянска. Водятся толстороги
OVIS
551
в большинстве гор северной Якутии к востоку от р. Яны: на Тас-Хая-
тахе, хребте Черского, Томус-Хае, Андрей-Тасе, Момском и ряде дру-
гих .^Однако распространены они здесь не повсеместно и неравномерно.
Рис. 114. Череп Ovis nivicola Eschsch.
По сравнительно свежим данным В. Н. Скалона с соавторами (1941),
в пределы б. Верхоянского округа заходят только по правым притокам
р. Нельчехе и служат лишь случайной добычей. Попадаются бараны также
изредка в правобережье р. Адычи (приток Яны), ниже впадения в нее
р. Чарки. В Абыйском районе их нет вовсе. В Оймяконском районе ветре-
552
BOVIDAE
чаются только в частях, пограничных с Момским районом. Последний
в настоящее время является центром распространения этого зверя в Якут-
ской АССР. Здесь баран является обычным зверем по всему Момскому
хребту; наибольшая плотность наблюдается по хребту Илинь-Тас, по
правобережью р. Ожигина и в бассейне р. Зырянки. В этих местах толсто-
рог является основным промысловым зверем. Севернее р. Ожигина в лево-
бережье Колымы баранов нет. Южнее указанных районов бараны дер-
жатся в Томно неком районе, в Брюнгядинских горах, а также по верхнему
и среднему течению р. Томпо.
По данным тех же авторов, имеются отдельные очаги обитания чубука
в южной Якутии. Так, в промысловых количествах бараны держатся в рай-
оне оз. Токо, по р. Алгаме от горы Горугун до вершины р. Ундетина. До-
бываются они здесь также по р. Утука, Талла и в окрестностях оз. Хуто.
В Учурском районе бараны встречаются по Алдано-Учурскому хребту,
на водоразделе рр. Алдана и Гыныма, по среднему течению р. Гонама
и по р. Негускан (левому притоку Учура) водятся до 58° с. ш. Еще во вто-
рой половине прошлого столетия довольно широко были распространены
в Олекмо-Чарском нагорье. Несомненно о чубуке слышал в 1866 г. от
эвенков И. С. Поляков (1873), хотя и называл его козлом (Aegoceros sp.):
«Тунгусы, кочующие теперь при устье Муи, — писал он, — несколько
десятков лет назад убивали его в большом количестве в системе рр. Чары
и Олекмы. . . Прежде он был в тех местностях весьма обыкновенным
животным; всякий промышленник убивал его, а наиболее ловкие и про-
ворные клали в один день более чем по полудесятку. . . Полагают, что этот
козел еще и теперь там встречается. . . Из местностей, лежащих недалеко-
от Муи и в которых этот козел также в настоящее время встречается,
тунгусы указывали на окрестности оз. Орона и именно на долины рр. Си-
гикты, Чульбана и Ахукана; здесь козел убивался в недавнее время».
Небольшой очаг обитания баранов сохранился до сих пор в Каларском
районе Читинской области. По свидетельству В. Н. Скалона (1949), ба-
раны обитают в окрестностях оз. Нечатка, по впадающим в него рр. Абсат,
Суломат Ельгер, а также на притоках р. Чары—Большому иМалому Тора.
По слухам, они имеются также в верховьях Чары, на р. Саллыкыт. Вниз
по Чаре бараны держатся до устья р. Сен, где в 1929 г. былиубиты2 барана.
По данным В. П. Фаворского (1936) и В. Б. Подаревского (1936), сохра-
нились толстороги в хребте Кодар в верховьях рр. Меркулак (впадает
в оз. Лепринда), и Сегикта до р. Апсада. На хребте Кодар экземпляр
барана был добыт в 1946 г. (Павлов, 1949). По словам В. Н. Скалона (1949}
в верховьях р. Токо бараны были истреблены лет около 30 тому назад.
В. Н. Скалой категорически утверждает, что в б. Витимо-Каренгском и
Тунгиро-Олекминском районах этого зверя нет.
Раньше ареал О. nivicola простирался еще значительно южнее. Об этом
свидетельствует уже упоминавшаяся находка ископаемого толсторога
в Кузнецком Алатау, а также указания В. Н. Скалона (цит. по Цалкину,
1951) о том, что в XVII в. дикие бараны, возможно, водились по Апгаре.
Нет полной уверенности также в том, что не были толсторогами те бараны
по рр. Шилке и Онону, об охоте на которых сообщал Р. Маак (стр. 529)
в середине прошлого века. Во всяком случае, где-то в Прибайкалье в свое-
время смыкался ареал О. nivicola и О. аттоп или произошла дивергенция
между ними.
Нет удовлетворительных сведений о распространении толсторога в Ста-
новом хребте. По собранным Г. Тассовским (1927) данным, водятся
бараны в б. Гилюй-Ольдойском районе (Амурская область), главным
OVIS
553
образом в гольцах Типтур (верховья Гонама и Сутама). Известно также
(Насонов и Дорогостайский, 1915; Туров, 1937), что встречаются они в не-
большом числе на южных склонах восточной его части в верховьях р. Зеи
Большого и Малого Оконона. В большом количестве В. Ч. Дорогостай-
ский встретил их в этом районе на северных склонах хребта, в частности
в горах по рр. Малой и Большой Тускани, Много баранов, по полученным
им данным, держится в горах, расположенных километрах в 100 к северо-
западу от Окононской озерной впадины и повсюду на восток от оз. Оконон.
На запад и юго-запад от озера бараны заходят лишь случайно. Нет их
Рис. 115. Распространение Ovis nivicola Eschsch.
1 — XX в. (области постоянного обитания); 2— XIX в.; 3 — XVIII в. и раньше.
в горном кряже Дихтанрэ, расположенном километрах в 60—65 к югу
от Главного Станового хребта, равно как и в хребте Тукурингра.
Областью распространения толсторога являются горные хребты край-
него северо-востока Сибири к востоку от р. Колымы, Охотское побережье
и полуостров Камчатка.
В 60-х годах прошлого столетия дикие бараны, по словам К. К. Ней-
мана (1871), были распространены «по всей Чукотской земле» и доходили
.до побережья Ледовитого океана. Сам он встретил их в большом числе
в горах около р. Каменной, притока р. Орловки (бассейн Б. Анюя).
Г. Майдель (1894) отмечал значительное количество толсторогов на Боль-
шом и Малом Барановых камнях, на побережье к востоку от Колымы
и в горах к востоку от р. Белой. По словам И. П. Толмачева (1911),раньше
баран местами встречался в изобилии по Чукотскому побережью Ледо-
витого океана.
Некоторыми деталями современного распространения толсторога в пре-
делах так называемого Анадырско-Чукотского края мы обязаны Н. П. Со-
кольникову (1927), Л. О. Белопольскому (1933), Л. А. Портенко (1941)
и некоторым другим авторам. По словам первых двух, бараны обыкновенны
554
BOVIDAE
в горах Анадырского хребта и распространены по нему от Берингова
пролива до Ледовитого океана. В наибольшем числе они держатся в Пе-
кульнейских горах между рр. Танюрер и Белой, т. е. там, где находил
их Майдель, а также в горах левого истока р. Белой. На эти же места ука-
зывал Л. О. Белопольскому И. В.. Друри. Водятся бараны в горах к се-
веру от зал. Большого Креста, близ вершины Матачингай, в верховьях
р. Амгуемы. Известно, что есть они в верховьях рр. Чауна,
Еропола и их притоков (Маракуам, Умкувэем), на водоразделах
рр. Окапайте, Маракуама и Керальгуама (Яблона). Встречались
бараны также на р. Малый Пеледон, на перевале между Б. Пеледоном и
Саламихой, в горах на водоразделе рр. Ленли и Тымкыли (69°10' с. ш.
и 169°00' в. д.). В 30-х годах добывались они на сопках у р. Ворожеи.
Водятся в Щучьем хребте близ р. Бараньей (левый приток Анадыря).
Изредка добываются они в окрестностях с. Еропола, а также в отрогах
Русского хребта, хребта Пал-Пал, в верховьях р. Маи. Л. А. Портенко
(1941), со слов ламутов, указывает их для хребта Рарыткин. Далее, место-
нахождения баранов известны по Корякскому хребту, в частности по р. Ха-
тырке, а также в районе мыса Наварина и к югу от последнего (Караев,
1926). Водятся эти звери в горах по побережью от мыса Наварина до Олю-
торского, в частности в районе бухт Шлюпочной, Дежнева, Анастасии,
Наталии, Глубокой, в районе озер Мимельгытьгын и Колан (Шнакен-
бург, 1939) и горах Олюторского полуострова. В б. Олюторском округе
Камчатской области, по словам А. В. Самородова (1939), О. nivicola явля-
ется обычным промысловым зверем. Как сообщил В. И. Цалкину (1951)
М. Г. Волков, «в Олюторском районе бараны многочисленны в прибреж-
ной зоне, в особенности на мысе Говена. Внутри страны их меньше. Были
неоднократно встречены Волковым бараны как на западных, так и на
восточных склонах Корякского хребта. Обыкновенны они также на Ма-
метчинском, Пенжинском, Палгинском, Ушнылемском, Ушканьем, Тай-
гоносском, Колымском и других хребтах Пенжинского района. В северо-
западной части Пенжинского района один из отрогов Колымского хребта
носит название хребта Кытепнэй (Бараньи горы). В этих хребтах баранов
успешно промышляют охотники ламуты и чукчи Аянского сельсовета».
В 1930 г. три барана были добыты на западном берегу Пенжинской губы
в 10—12 км от устья р. Парени (Портенко, 1941). В 1935 г., по В. С. Ба-
жанову (1946), в небольшом числе баранов встречали в верховьях Пен-
жины по правобережью Нинчеваяма, в окрестностях Красного мыса,
а также близ истоков р. Сухой Аянки, Б. Мургаля, обоих Куйбовеемов
и в некоторых местах по правобережью Пенжины. Довольно много в то
время было их в горах Пал-Пал; встречались и на Понтонейском хребте
(северное продолжение Камчатского хребта) и на хребтиках, соединяю-
щихся с ним в левобережье среднего течения Пенжины. Встречались они
также по горам полуострова Тайгоноса и на Каменском хребте по ниж-
нему течению Пенжины. В последние годы, добавляет В. С. Бажанов,
баранов, как говорят, уже не стало в правобережье Пенжины и вообще
их реже видят во многих пунктах района.
Л. О. Белопольский (1933), на основании материалов Приполярной
переписи 1926—1927 гг., приводит, кроме того, следующие места добычи
толсторогов: в Анадырском районе — р. Чиринай и ее приток р. Научи-
ринай, а в Чукотском районе пункты — Каленьмаум, Коконай, Мульх-
тикан и Уськамынаыргын.
К сожалению, нет сведений о распространении снежного барана в пра-
вобережье среднего течения Колымы, на западных склонах Колымского
OVIS
555
хребта, по Северному и Южному Анюйским хребтам, а также в бассейне
р. Омолона. Надо, однако, по аналогии с районами, лежащими к востоку,
полагать, что бараны должны водиться и здесь.
Обстоятельную сводку о прошлом и современном распространении
толсторогов на Камчатке дал недавно Ю. В. Аверин (1951). Здесь бараны
до сих пор распространены по большей части хребтов и сопок полуострова.
Обитают, по-видимому, по всему Срединному хребту, хотя нет достовер-
ных сведений о северной его части, от Парапольского дола до р. Паланы.
К югу от последней местонахождения баранов указывались в истоках
р. Верной (сопка Айнелькан, гора Шишель), в верховьях р. Тигиль (между
горой Шишель и сопкой Анаун), в истоках р. Козыревки, на сопке Ичин-
ской, в районе к югу от горы Камень, а также в бассейнах рр. Белой Анд-
риановки и Колпаковой, для Озеровской и Яковской падей. В южной
части Срединного хребта широко распространены в Малкинских горах
(верховья рр. Быстрой и Камчатки). В средине прошлого и начале нынеш-
него столетия толстороги встречались местами также по западному по-
бережью полуострова, изолированно от основного ареала, например,
на высоких мысах Ухтолокском и Омгоне, а также в окрестностях с. Ти-
гиль. Ю. В. Аверин полагает, что в настоящее время здесь баранов уже
нет.
К востоку и югу от р. Камчатки местами обитания являются Ганаль-
ский, Валагинский хребты и хребет Тумрок. Указывают их также для
бассейнов Левой Авачи, Кехкуй и Тополевки. В Валагинском хребте
они наблюдались в районе г. Попутной, у Кроноцкого перевала и в исто-
ках р. Валагинской. Нет сведений о баранах на хребте Кумроч, но по-
скольку они живут на соседних с хребтом Ключевских вулканах и на Ши-
велуче, автор полагает, что должны быть толстороги и здесь.
Далее бараны распространены на Камчатке по большей части вулка-
нического высокогорья, от мыса Лопатки до вулкана Шивелуч включи-
тельно. К северу от м. Лопатки1 бараны отмечались на Камбальной и
Кошелевой сопках, в горах вокруг Курильского озера, на сопке Опале,
по рр. Плотниковой и Карымчина. Наблюдались также на вулканах
Ксудач, Мутновской, Горелом, Корякской, Жупановском, Карымском,
Березовом, Малом и Большом Семячиках, в бессейне р. Вахиль.
В пределах Кроноцкого заповедника бараны весьма обыкновенны
на всех сопках около Кроноцкого озера, а именно: на вулканах Тауншиц,
Унана, Узон, Кихпиныч, Крашенинникова, Кроноцком, Конради, По-
путном, Гамчен и группе Гамченских вулканов, выходящих к вершине
р. Сторож. В центре заповедника они в небольшом числе обитают в исто-
ках р. Богачевки. Многочисленны также в средних и верхних частях бас-
сейнов всех рек Кроноцкого полуострова.
К северу от Кроноцкого заповедника бараны живут на вулкане Кизы-
мен, в бассейне рр. Щапиной и Сторож, на хребте Тумрок, в истоках
р. Тобалчик, на вулкане Малый Тобалчик, а также в горах между рр. Сто-
рож и Малой Чажмой. Далее местами обитания является группа Ключев-
ских вулканов (сопки Удина, Зимина, Камень, Плоская, Ключевская,
Средняя). Живут снежные бараны и на береговых скалах восточного по-
бережья Камчатки. В числе мест обитания на побережье Ю. В. Аверин
(1951) называет: основание мыса Лопатки, побережье между мысами Ин-
канющ и Хадутка, мысы Пиратков и Асача, бухты Лиственичная и Ахом-
1 Таким образом, мыс Лопатка (около 51° с. ш.) является крайним южным пунк-
том ареала О. nivicola Eschsch., как этот вид принимается в настоящей работе.
556
BOVIDAE
тен. На мысе Асача бараны теперь истреблены (Плечев, 1939), равно как
вытеснены они на всем побережье между бухтой Ахомтен и Авачинским
заливом, заходя сюда лишь случайно с вулкана Вилючик. К северу от
Авачинского залива бараны встречаются в районе бухт Халыгер, Биче-
винский, Моржовой и на мысах Налачевом и Шипунском. Далее к северу’
наблюдаются на мысах Аргали, Среднем, Кронье и по побережью до
устья р. Жупановой. Охотились за ними по побережью Шипунского по-
луострова, но теперь здесь эти звери стали редки. Встречаются чубуки по-
всему обрывистому побережью от р. Тюшевки до мыса Кроноцкого и от-
сюда на север до мыса Чажма. Изолированная от остального камчатского
ареала, небольшая группа снежных баранов живет на береговых скалах
и хребтах Начикинского полуострова вблизи Укинской бухты.
В заключение Ю. В. Аверин подчеркивает, что «за пределами пере-
численных выше горных районов, побережья и его изолированных йысов,.
снежные бараны на Камчатке, по-видимому, не встречаются».
Следует отметить, что в средине XVIII в. бараны водились на Куриль-
ских островах, а по некоторым данным (Цалкин, 1951) даже на Алеутских.
Несомненно распространены бараны в южной половине Колымского
хребта, в верховьях Колымы, Нерско-Колымском плоскогорье (Скалой
и др., 1941) и на большей части Охотского побережья. По словам В. Йохель-
сона (1898), в бывшем Колымском округе чубук распространен по всем
хребтам к востоку от Колымы и по размерам добычи занимал тогда второе-
место среди других крупных животных. О распространении толсторога
в верховьях Колымы в начале настоящего столетия писал С. А. Бутур-
лин (1913). Однако детали распространения его в этой части ареала из-
вестны очень мало.
По А. Эрманну (Ermann, 1838), бараны живут на р. Аллах-Юнь (при-
ток Алдана) между Гарнастахом и ст. Аллах-Юнь. По сведениям, сообщен-
ным К. К. Флерову (1935) «Л. В. Бианки и другими последними исследо-
вателями Якутии, снежные бараны населяют почти все горные районы
этой страны». В большом числе толстороги водятся в окрестностях Ейри-
нейской губы (Насонов, 1923). Известны бараны из окрестностей Охотска.
Бэкстон (цит. по С. И. Огневу, 1926) в числе других мест обитания толсто-
рогов называет Аян, окрестности Охотска, Олу, Ямск и Нояхинск. Насе-
ляют они также хребет Джугджур, в частности отмечены их местонахож-
дения вблизи Ляна, на перевале между Аяном и Нельканом (Насонов,.
1923). Наличие баранов на аянской стороне хребта отмечал Г. Дулькейт-
(1927). В средине прошлого столетия они водились в горах бассейна
Учура; особо много их было в горах Шапча (или Чапча) к востоку от вер-
ховий р. Половинной (Маи), притока р. Уды (Миддендорф, 1853). Послед-
няя, по-видимому, является южным пределом распространения О. nivi-
cola на Охотском побережье. По сведениям, полученным на месте Н. Т. Зо-
лотаревым (1934), толстороги живут в гольцах Янканского нагорья (во-
дораздел Маи и Уды), главным образом в верховьях р. Чогор, где иногда,
собираются в значительные стада. Как показали исследования Л. Шренка,
(цит. по Цалкину, 1951) и А. В. Афанасьева (1934), в горах к югу от Уды,,
в частности в хребте Дуссе-Алинь, снежных баранов нет.
Подвиды. Общепринятыми являются следующие подвиды О. nivicola..
1) О. nivicola nivicola Eschscholtz — Камчатский снежный баран..
Характеризуется темной, однотонной окраской и особо массивными ро-
гами. Распространение : Камчатка.
Описанный И. Алленом (Allen, 1904) как самостоятельный вид, О. stor-
cki из западной Камчатки представляет одно из индивидуальных уклоне-
ovis
557
лий и последующими исследователями не принимается даже в качестве
подвида.
2) О. nivicola alleni Matschie — Тайгоносский снежный баран. Отли-
чается от типичной расы некоторыми особенностями изгиба рогов, в ча-
стности направлением концов вперед. Распространение: Тай-
гоносский полуостров, Колымский хребет, Охотское побережье, хребет
Джугджур.
3) О. nivicola potanini Nasonov —- Снежный баран Станового хребта.
Особенностью этого подвида является наличие белого пятна на боках по-
зади плеч. В. И. Цалкин (1951) объединяет эту расу с предыдущим под-
видом и наличие упомянутого пятна считает характерной чертой для обоих
подвидов.
4) О. nivicola lydekkeri Kowarzik — Якутский снежный баран. От
предыдущих подвидов отличается, во-первых, сильным развитием в ок-
раске туловища светлого тона, часто подавляющего основную буро-ко-
ричневую расцветку. Особенностью рассматриваемой расы являются
также более короткие и тонкие, по сравнению с типичным подвидом,
рога. Распространение: горы северной Якутии, возможно
также бассейн р. Анадыря.
5) О. nivicola borealis Severtzov — Норильский снежный баран. Изучен
слабо. По Н. В. Насонову (1923), особенностью данного подвида является
удлинение волос на боках выше пахов. Распространение: горы
Путорана в верховьях рр. Пясины, Хеты, Котуя (междуречье Енисея и
Лены).
Биология. Местообитания и образ жизни снежнего барана изучены
совершенно не достаточно. По-настоящему не известны даже пределы
вертикального распространения. В отношении предпочитаемых местооби-
таний толсторог заметно отличается от других баранов, проявляет черты
сходства с козлами и занимает сходную с последними экологическую нишу.
Повсеместно снежные бараны населяют наиболее скалистые, трудно до-
ступные участки гор, даже если мало преследуются человеком. В При-
енисейском севере, по словам Н. П. Наумова (1934), они являются обита-
телями «местностей с сильно расчлененным гористым рельефом, богатыми
скалами и голыми безлесными горами». Также характеризует С. А. Бу-
турлин (1913) местообитания толсторогов в Колымском крае.
Описывая специфические трудности охоты на снежных баранов в Ябло-
новом (Становом) хребте, В. Ч. Дорогостайский (Насонов и Дорогостай-
ский, 1915) подчеркивает, что добыча их значительно более трудна, чем
других диких баранов. «Прежде всего, — пишет он, — невозможность
пользоваться какими бы то ни было вьючными животными сильно сокра-
щает район охотничьих экскурсий; пробраться в места, обитаемые бара-
нами, на лошадях нечего и думать, даже на оленях не везде можно про-
ехать, поэтому приходится охотиться исключительно пешком, а при пе-
шем способе охоты на ночлег необходимо возвращаться к лагерю, так как
при страшной крутизне гор нет возможности носить с собой достаточно
провианта и теплой одежды, чтобы ночевать в горах... Мне приходилось
охотиться за горными баранами на степных плоскогорьях Монголии.
Там я всегда пользовался лошадьми, и благодаря этому в день мы объез-
жали в общей сложности верст 30—40, тогда как в Яблоновом хребте,
если сделаешь в день верст 9—10, то это нужно считать очень хорошим».
Лучше изучены стации толсторогов на Камчатке (Гаврилов, 1947;
Аверин, 1948, 1951). Здесь бараны встречены от морского побережья до
^высоты 2570 м, где никакой растительности уже нет; основная же масса;
558
BOVIDAE
их постоянно живет на высотах от 1000 до 1600—1800 м. Для всех место-
обитаний баранов обязательно наличие открытых низкотравных выпасов
по соседству с неприступными скалами, служащими местом спасения.
Ю. В. Аверин (1951) все местообитания толсторогов сводит к двум
основным типам.
Первый тип — высокогорные стации, расположенные выше верхней
границы леса и кустарникового сланца (800—900 м). Они представляют
сочетание открытых пастбищ на склонах, хребтах и в долинах с прилегаю-
щими к ним скалистыми обрывами и отдельными скалами. Пастбищами
здесь служат летом альпийские лужайкии богатые растительностью склоны,,
а зимой — бедные растительностью, но малоснежные, обдуваемые ветрами
хребты и скалы. На вулкане Шмидта (Кроноцкий заповедник), обитае-
мом баранами, основными стациями баранов являются скалистые зубцы
верхних разрушенных частей вулкана и радиально расходящиеся от
вершины гребни барранкосов.1 Первые лишены растительности и служат
убежищами на случай опасности и непогоды, тогда как кормовыми уго-
дьями являются гребни барранкосов, покрытые ниже 1000 м над ур. м.
сомкнутым травянистым покровом. Чаще всего бараны держатся на па-
стбищах, имеющих скалистые выступы, выходящие к обрывам или глубо-
ким оврагам (Гаврилов, 1947).
Второй тип местообитаний толсторога на Камчатке — это береговые-
уступы с обрывистыми скалами. Они представляют крутые склоны под-
ходящей к морю высокой суши, высотой 100—300 м, идущие параллельно-
морскому берегу и иногда отделенные от него узкой лентой побережья,,
но часто подходящие вплотную к морю. Склоны покрыты кустарниковыми:
сланцами, спускающимися почти до моря. Пастбищами на береговых усту-
пах служат небольшого размера травянистые площадки, расположенные-
на вершинах мысов, склонах берегового уступа или лужайках среди
скал. Небольшие группы толсторогов местами живут в полосе леса между
побережьем и высокогорьем на скалистых участках невысоких хребтов.
(500-800 м).
. Как на Камчатке, так и на севере у Ледовитого океана (Бутурлин,
1913), баранов летом нередко можно видеть на побережье у самого моря.
Снежные бараны ведут сравнительно оседлый образ жизни и сколько-
нибудь значительных сезонных миграций не совершают. Можно говорить-
лишь о смене мест обитания по сезонам в пределах незначительной терри-
тории, занимаемой популяцией, о смене, вызываемой, в основном, усло-
виями залегания снежного покрова и связанной с ним возможностью
добычи корма. Зимовками служат места с неравномерным или неглубоким
снегом, в общем расположенные ниже, чем места летнего обитания. На
Становом хребте зимой бараны пасутся на склонах гор, где ветер сдувает
снег и дает возможность животным кормиться сухим травянистым кормом;
иногда в эту пору бараны спускаются в полосу кустарников, где едят по-
беги и кору молодых деревьев. К весне толстороги концентрируются на
южных склонах хребта, где почва раньше освобождается от снега и по-
является первая молодая зелень. Весной и летом они живут в самых вы-
соких частях гор и почти совсем не спускаются в долины. Особо недо-
ступных . скалистых мест придерживаются летом самцы, тогда как са-
мок с молодыми можно встретить на более открытых склонах, террасах
и седловинах между отдельными горами. Осенью звери иногда спускаются
1 Барранкосы — узкие овраги, прорезывающие склоны конуса вулкана от вер-
шины до его основания.
OVIS
559
в полосу леса, чтобы полакомиться грибами, но долго здесь не остаются и,
насытившись, вновь возвращаются в скалы (Насонов и Дорогостайский,
1915; Туров, 1937).
Примерно такой же характер носят сезонные перемещения толсторо-
гов, населяющих хребты Камчатки. Места зимнего обитания здесь также
располагаются, как правило, ниже летовочных территорий, однако выше
полосы равномерно глубоких снегов; обычно они представляют участки,
где группы скал («стойбища»), служащие местами укрытий, сочетаются
с выдувами по хребтам и склонам, служащими местами зимних пастбищ.
С вулкана Кихниныч (на территории Кроноцкого заповедника) бараны
уходят зимовать в долину р. Гейзерной, до самого побережья, где на ак-
тивных гейзерных площадках растительность зимой свободна от снега,
а весной начинает вегетировать намного раньше, чем в других местах
(Аверин, 1948). Летовки толстороги и здесь проводят выше, заходя иногда
к безжизненным вершинам вулканов.
Несколько своеобразный характер носят сезонные передвижки тол-
сторогов, обитающих вблизи побережий. С высоких прибрежных мысов,
где звери проводили зимовки, они в мае, а иногда и раньше, спускаются
вниз к морю, где в это время уже появляется зелень. По мере появления
растительности выше по склонам, бараны совершают обратные перемеще-
ния в горы (Гаврилов, 1947; Аверин, 1948). В сентябре они вновь начинают
постепенно приближаться к морю (Плечев, 1939).
Держатся снежные бараны сравнительно небольшими группами,
больших стад не образуя. В Олюторском районе на Камчатке средняя
величина стада 5—7 голов (Самородов, 1939). Стада в 2—3 десятка голов
в опромышляемых районах составляют уже редкость. Максимальное по
численности стадо, наблюдавшееся в Кроноцком заповеднике в конце
сентября, состояло из 60 голов (Аверин, 1951). Смешанные стада, состоя-
щие из животных разного возраста и пола, наблюдаются поздней осенью,
зимой и ранней весной. В мае от смешанных стад отделяются беременные
самки. В июне обособляются и взрослые самцы, которые держатся груп-
пами до десятка голов или единично. Отдельные смешанные табунки
толсторогов можно встретить уже в сентябре, но массовое табунение
происходит в октябре.1 В летний период чаще всего можно встретить
табунки, состоящие из самок с ягнятами-сеголетками, яловых самок и
молодняка прошлых лет. Впрочем, как отмечает В. Ч. Дорогостайский
(Насонов и Дорогостайский, 1915), одиночно держащимися зверями
бывают и самки, и молодые.
Вожаком в смешанном стаде обычно бывает старая самка. Есть ли
у толсторогов специальные сторожа во время отдыха стада — не известно.
В обычных условиях снежные бараны ведут дневной образ жизни.
Летом выходят на кормежку с восходом солнца и пасутся до 7—8 часов
утра. После этого направляются к солонцу или на отдых. Вторично
выходят на кормежку часа в четыре после полудня и пасутся до наступле-
ния темноты. Иногда и вечером посещают солонцы. В жаркие дни, в це-
лях защиты от кровососущих насекомых, для отдыха выбираются хорошо
обдуваемые ветром места: выступы скал, площадки около горных вершин
или седловины между двумя цепями гор. В прохладные дни бараны на
пастбищах появляются позднее, для отдыха выбирают укрытые от ветра
места, держатся вообще несколько ниже, а во время дождя укрываются
1 В. Ч. Дорогостайский (Насонов и Дорогостайский, 1915) отмечает, что на Ста-
новом хребте самцы и зимой ходят отдельно от самок.
560
BOVIDAE
под навесами скал. На ночлег бараны уходят наверх в скалы. В зимние корот-
кие дни они не отдыхают и все время проводят в пастьбе, чтобы собрать
необходимый минимум поддерживающего корма.
Наблюдается ясно выраженная привязанность к избранным местам
обитания. На места отдыха, ночлега и на солонцы бараны ходят опре-
деленными путями, обозначенными иногда хорошо заметными тропами.
Для дневных лежек, укрытий от непогоды и врагов, в спокойной обста-
новке пользуются в течение длительного времени одними и теми же ме-
стами. На этих стойбищах и лежках зачастую скапливается значительное
количество помета, по которому узнают о наличии в данном районе снеж-
ных баранов.
Снежные бараны легко карабкаются и передвигаются по крутым
склонам и скалам, но очень быстро бегать не могут. Об этом можно судить
по тому, что иногда их, особенно старых отяжелевших самцов, легко
догоняют собаки (Насонов и Дорогостайский, 1915). Обычный аллюр
в спокойной обстановке — шаг; при этом животные идут гуськом и ста-
вят ноги след в след, так что получается впечатление, будто прошел
один баран, оставив нечеткий расплывчатый след (Аверин, 1948). Впереди
всегда идут более старые звери — самцы или самки.
Основу питания толсторогов в течение всего года составляют различ-
ные травы. Так как вегетационный период в районах обитания О. nivi-
cola очень короток, всего 4—4% месяца, то большую часть года бараны
вынуждены питаться «сеном на корню». Не отказываются бараны при
возможности и от древесно-веточного корма: от листвы кустарников и
невысоких деревьев, а зимой — от коры и тонких веток.
Ю. В. Аверин (1951) приводит список поедаемых снежными баранами
растений, состоящий из 38 видов. Наиболее разнообразен ассортимент
кормовых растений, естественно, в бесснежный период. В это время
первостепенными по кормовому значению видами являются 17 видов,
такие, как ситник (Juncus beringianus Buch.), осоки (Carex atrofusca
Schkuhr и C. pyrenaica Wahlenb.), два вида ив (Salix pseudopolaris Wahlb.
и S. berberifolia Pall.), из бобовых несколько видов остролодок (Oxytro-
pis), из злаков мятлик Комарова (Роа komarovi Boshev) и вейник (Cala-
magrostis monticola Petr, et Korn.). Из разнотравья охотно и в больших
количествах поедаются полынь Комарова (Artemisia komarovi Р. Pol.)
и камнеломка Нельсона (Saxifraga nelsoniana D. Don.). На Становом
хребте толстороги охотнее посещают места, покрытые альпийской то-
локнянкой (Arctostaphylos alpina L.); остатки последней В. Ч. Дорого-
стайский (Насонов и Дорогостайский, 1915) неизменно находил в желуд-
ках убитых им баранов. Кроме того, со слов орочен, охотно едят они
листву и побеги ползучей ивы (Salix baicalensis Turcz.) и полярной бе-
резы (Betula fruticosa Pall.). По Е. И. Павлову (1949), прекрасным кормом
является рано зацветающее на солнечных склонах гор растение, носящее
название «ургуя» (вероятно, Pulsatilla sp.). Как уже отмечалось, при
случае бараны не прочь полакомиться и грибами. На Анадыре и Чукотке
главной пищей толсторогов является кочедыжник, или горная рута,
Asplenium ruta muraria L. (Сокольников, 1927).
Зимой кормовая база снежного барана сильно суживается не только
количественно, но и качественно. Во-первых, становится весьма ограни-
ченной площадь пригодных для пастьбы мест, благодаря образованию
глубокого или слишком плотного снежного покрова. Во-вторых, звери
вынуждены довольствоваться остатками тех растений, обычно более
грубых, которые не разрушаются от дождя, снега, мороза и ветра. По-
ovis
561
прежнему одним из основных кормов является ситник, некоторые злаки
и побеги ив. Снижается удельный вес в питании осок, но зато сильно
возрастает роль некоторых видов вересковых, как подбела (Cassiope
lycopodioides D. Don), азалеи (Loiseleuria procumbens Desv.), гаррима-
неллы (Harrimanella stelleriana Cov.). Важное знамение в зимнем питании
толсторогов имеют лишайники, в частности «олений мох» (Cladonia rangi-
ferina Web.), затем плаун (Lycopodium sp.) и даже зеленые мхи. В ка-
честве второстепенного корма зимой поедаются листья и побеги голубики
(Vaccinium uliginosum L. и V. vulcanorum Кот.), брусники (Vaccinium
vitis idaea L.) и шикши (Empetrum nigrum L.).
Зимой баранам приходится нередко добывать корм из-под плотного
снега, разбивая и разгребая его копытами. В раскопанных лунках трава
выедается вплотную к земле, нередко с корнями и приставшими к по-
следним частицами почвы, обнаруживаемой в кале. Вес содержимого
желудка у взрослых зверей летом может доходить до 10—11 кг, зимой
обычно не более 4—5 кг (Аверин, 1951).
Потребность в минеральном питании толстороги Станового хребта
удовлетворяют посещением естественных солонцов и поеданием на них
почвы. В качестве солонцов служат выходы вулканического туфа красно-
вато-бурого цвета со включенными в него кристаллами повареной соли.
Жадно лижут и грызут в этих местах бараны землю, выгрызая в ней
углубления и даже пещеры. Посещение солонцов начинается весной
с появлением молодой зелени; первую половину лета бараны ходят на
них часто, с половины июля реже, а в августе солонцевание прекращается
вовсе. Больше пользуются солонцами самки и молодые, тогда как самцы
посещают их сравнительно редко (Насонов и Дорогостайский, 1915).
О солонцевании толсторогов на Камчатке ничего не известно..
Потребность в воде удовлетворяется в бесснежное время года влагой,
содержащейся в зеленом корме, или из попадающихся на пути водоемов.
Во всяком случае, даже в Становом хребте, где места обитания баранов
характеризуются почти полным отсутствием воды, по словам В. Ч. Доро-
гостайского, они не нуждаются в особых местах для водопоя и вообще
пьют редко (Насонов и Дорогостайский, 1915).
Там, где толстороги не подвергаются преследованию, они не пугливы.
В Кроноцком заповеднике, например, к ним, по словам Ю. В. Аверина
(1951), легко можно подойти на несколько десятков метров, а нередко
и вплотную. Особенно доверчивы самки и молодые. Однако, утверждение
В. Н. Скалона (1949) о том, что «классический прием [охоты] — увидев
барана, повесить одежду и обходить зверя — удается с ним отлично»,
следует считать явным преувеличением. В местах, где на них охотятся,
бараны очень чутки и осторожны (Бутурлин, 1913; Портенко, 1941;
Туров, 1937). По словам орочен, они очень боятся лошадей, палаток,
а особенно дыма костров и человеческих фигур; обнаружив их, немедленно
уходят, даже не проявляя любопытства (Насонов и Дорогостайский,
1915).
Из чувств у толсторогов лучше развито зрение. «Подкрадываясь
к стаду, — пишет В. Ч. Дорогостайский, — достаточно чуть высунуть
голову из-за камня, как животные уже замечают опасность и обращаются
в бегство». Гораздо слабее у них обоняние. По словам того же автора,
они не могут отличить старые следы охотника от свежих, если даже тот
прошел несколько минут назад. Слух у толсторогов, возможно, острый,
но они мало полагаются и на него, благодаря множеству звуков в горах.
Во всяком случае, по В. Ч. Дорогостайскому, от шума падающих из-под
36 Фауна СССР, И. И. Соколов
562
BOVIDAE
ног неосторожного охотника камней, бараны убегают редко, так как при-
выкли к звукам произвольно падающих в горах камней. В том же духе
об остроте чувств анадырского толсторога высказывался Н. П. Соколь-
ников (1927).
Смена волосяного покрова происходит один раз в году — весной и
в начале лета. Сроки начала ее не установлены. Известно, что на Кам-
чатке разгар падает на май и июнь. Заканчивается она к началу—сере-
дине августа (Аверин, 1951). На Становом хребте, по данным В. Ч. Дорого-
стайского (Насонов и Дорогостайский, 1915), линька заканчивается
раньше; 20 июня (старого стиля) старые самки были наполовину еще
в зимней шерсти; молодые 2—3-летние животные старую шерсть имели
только на ногах, затылке, нижней стороне грудной клетки, и отдельные
старые волосы, разбросанные по всему туловищу; в начале июля все
добытые им животные были уже полностью в летней шерсти. Начинают
линьку раньше и быстрее проделывают ее взрослые здоровые самцы,
несколько позднее — холостые самки и молодняк предшествующих лет.
Позже других групп линяют самки, имевшие в данном году приплод.
Сильно растягивается линька у больных и истощенных животных. В про-
тивоположность сведениям В. Ч. Дорогостайского, Ю. В. Аверин (1951)
пишет, что ноги у толсторогов вылинивают первыми, так же как и живот;
дольше же всего старая шерсть удерживается на голове между рогами.
Вскоре после смены старой шерсти (на Камчатке уже в августе), на-
чинается отрастание и зимнего меха. В конце ноября можно встретить
самцов, имеющих вполне развитый зимний волосяной покров. До конца
года мех сохряняет свою пышность и целость, но уже в феврале осте-
вой волос становится ломким и сильно сечется (Аверин, 1948).
Половой зрелости снежные бараны-самки достигают на третьем году
жизни, в возрасте около 2% лет; самцы, возможно, несколько раньше
(Аверин, 1951).1 Сведения о сроках размножения несколько противоре-
чивы. Так, на Туруханском севере, по Н. П. Наумову (1934), гон на-
чинается во второй половине ноября и тянется до средины декабря. Окот
беременных самок происходит около половины июня. Сходные сроки
гона (ноябрь—декабрь) для Камчатки приводит Ю. В. Аверин (1951),
но добавляет, что случай спаривания наблюдался также в конце октября.
Окот на Камчатке, по его данным, имеет место в конце мая—июне. На
Становом хребте, по собранным В. Ч. Дорогостайским данным (Насонов
и Дорогостайский, 1915), спаривание у баранов начинается во второй
половине сентября или в начале октября (старого стиля); рождение
молодняка у большинства самок происходит в мае (Туров, 1937). Со-
здается впечатление, что у О. nivicola период беременности длится более
5 месяцев.
В период гона, как и у других баранов, происходят поединки самцов,
причем соперники иногда, по-видимому, сбрасывают один другого в про-
пасть (Насонов и Дорогостайский, 1915). Мясо самцов в период течки
приобретает специфический неприятный запах. Перед окотом беременные
самки для родов на некоторое время удаляются из стада. Приносят, как
правило, по одному ягненку. Достоверных случаев окота толсторогов
двойнями не известно. Яловость наблюдается у 7—10% взрослых самок.
К концу июля или началу августа ягнята достигают веса 13—20 кг. Про-
1 Е. И. Плечев (1939), впрочем, указывает на более ранний срок полового созре-
вания — вторую осень.
OVIS
563'
должительность питания молоком не установлена. Ю. В. Аверин (1951)
наблюдал сосущего ягненка 18 сентября.
Продолжительность жизни снежного барана в естественных условиях
не известна. В коллекции Зоологического Института АН СССР не имеется
экземпляров старше 13—14 лет. Вероятно, этот возраст, судя по степени
стертости зубов, недалек от предельного для этого зверя.
Из естественных врагов толсторога на первом месте по значению стоят
волк и росомаха. Росомаха, подкарауливая, поймает его скорее, чем волк,
и находки задранных этим хищником баранов на Камчатке представляют
обычное явление. Волк менее приспособлен к охоте в скалах, но благодаря
своей многочисленности вообще наносит больший ущерб популяции бара-
нов. Сильнее всего страдают толстороги от обоих хищников зимой. По сло-
вам Ю. В. Аверина (1948, 1951), на зимовках экскременты росомахи
и волка состоят почти исключительно из остатков баранов. Летом баранам
спасаться от хищников в скалах легче; кроме того, и росомаха, и волк
в это время спускаются для размножения в полосу леса, куда бараны не
заходят. Как редкое исключение толсторог может стать добычей медведя.
Молодые, недавно родившиеся ягнята, по-видимому, не так редко стано-
вятся жертвой орлана-белохвоста (Heliaeetus albicilla L.), а по А. В. Само-
родову (1939), и белоплечего орлана (Heliaeetus pelagicus Pall.). Но вред от
пернатых хищников не велик.
Весьма существенную роль в снижении численности толсторогов играют
неблагоприятные кормовые и климатические условия зимовок. Как уже
отмечалось, звери зимой недоедают, вследствие ограниченности и бедности
кормами пастбищ. Нередки, особенно вблизи побережий, гололедицы и
образование под действием ветра слишком плотного снега, который звери
пробить копытами не в состоянии. Положение усугубляется тем, что сви-
репые снежные бури, сопровождаемые сильными холодами, иногда не
позволяют животным по нескольку дней выходить из укрытий и заставляют
их голодать. Вероятно, немалое число баранов гибнет зимой от бескормицы.
Кроме того, ослабленные звери легче становятся добычей хищников.
Е. И. Плечев (1939) сообщает, что крайне осторожные в другое время,
весной же сильно истощенные за зиму толстороги слабо реагируют даже на
приближение человека. В первую очередь зимой гибнет неокрепший молод-
няк от последнего приплода. Ю. В. Аверин (1951) определил, что почти
половина (около 45.7%) молодняка погибает, не достигнув годовалого»
возраста. Поэтому прирост популяции лимитируется, прежде всего, не-
благоприятными условиями зимовок.
Паразиты и болезни толсторогов не изучены совершенно. Известно
только (Туров, 1937), что на Становом хребте они страдают от носового,
овода. О паразитировании носового овода на камчатском баране сообщал
автору также В. Т. Гаврилов. По-видимому, с дикого барана Камчатки
Л. Нейманом (Neumann, 1908—1909) описан подвид иксодового клеща.
Dermacentor variegatus kamschadalus Neum.
Хозяйственное значение. Снежный баран имеет важное значение как
мясной промысловый зверь. В ряде районов северо-восточной Сибири
(в Якутии, Чукотском национальном округе, на Камчатке) промысел его
издавна играет не последнюю роль в питании местного населения. Ю. В. Аве-
рин (1951) считает, что только на Камчатке добывается ежегодно 800—1000
баранов. Баранина считается здесь лучшим мясом, особенно мясо самок
и молодых прошлогодних зверей. По Н. П. Сокольникову (1927), мясо
толсторога вкусом напоминает таковое домашней овцы, но по качеству ему
уступает. Лучшую упитанность бараны имеют поздней осенью и в начале
36*
564
BOVIDAE
зимы перед гоном. Особенно жирны бывают в это время самцы. Взрослый
самец дает в среднем около 65, максимально до 72 кг мяса. Кроме того,
получается около 10—12 кг жира, а от одного крупного самца весь собран-
ный жир весил 17.2 кг (Гаврилов, 1947). К весне (апрелю-маю) бараны
становятся сильно истощенными («сухими») и совершенно теряют запасы
жира.
Шкуры идут на изготовление одежды, спальных мешков, в качестве
ковриков, на подстилку. На Камчатке из шкур баранов шьются незамени-
мые на зимнем охотничьем промысле спальные мешки-кукули, по качеству
(теплоте) превосходящие оленьи и собачьи. Лучшим временем для убоя
баранов на кукули считается конец сентября—начало октября, пока
шерсть еще не слишком отросла и крепко держится в коже (Аверин, 1951).
В январе—феврале волос сильно сечется, ломкий, и такие шкуры для
пошивки одежды и спальных мешков уже не годятся. На Анадыре шкуры
толсторогов употребляются для выделки замши, более тонкой и мягкой,
чем оленья (Сокольников, 1927).
Рога снежных баранов повсеместно используются населением для
изготовления ложек, гребешков, пуговиц, рукояток к ножам и других
мелких поделок.
Опытов по скрещиванию снежного барана с другими представителями
рода Ovis и домашними овцами до сих пор, по-видимому, не предпринима-
лось. Если гибридизация с домашней овцой возможна, то была бы, ве-
роятно, перспективной в отношении возможности выведения домашней
породы овец, приспособленной к суровым условиям Заполярья.
Охота. Способы охоты на толсторогов разнообразны. Наиболее обыч-
ной является охота скрадом. Подкарауливают баранов на солонцах, у троп
на переходах. Эти способы в принципе ничем не отличаются от этих приемов
добычи других копытных (см. раньше). В некоторых районах охотятся
с собаками, ставящими неспособных много и быстро бегать зверей «на от-
стой». Вот как описывает эту охоту у орочен В. Ч. Дорогостайский (Насо-
нов и Дорогостайский, 1915): «Охотники отправляются вдвоем, причем
один идет впереди и высматривает баранов, а другой ведет сзади собак.
Стараются подойти к баранам на выстрел, стреляют и спускают собак.
Собаки с лаем бросаются преследовать баранов, и если попались старые
зажиревшие самцы, то быстро их настигают и останавливают. Самок же
и молодых догнать собакам далеко труднее, так как они убегают значи-
тельно быстрее. Охотники по доносящемуся до них лаю определяют место,
где собаки задержали барана, спешат к ним и, незаметно подкравшись,
удачным выстрелом завершают охоту. Хорошо держащие баранов на
месте собаки очень редки, почему и ценятся очень дорого — до 100 и более
рублей».
По словам того же автора, осенью орочены подкарауливают баранов
у верхней границы леса, куда звери приходят лакомиться грибами.
Подсемейство BOVINAE — БЫКОВЫЕ
Характеристика подсемейства. Рога у большей части родов и видов
имеют как самцы, так и самки, причем у последних они обычно немного
меньше, чем у самцов. Основаниями рога в большинстве случаев сдвинуты
назад за глазницы, иногда располагаются на вершине черепа над затылком.
Внутри роговых стержней, за немногими исключениями, имеются внутрен-
ние полости, нередко доходящие до вершины. Форма рогов различная:
BOVIANE
565
в виде простых спичек, гетеронимно свернутые или скрученные, гомонимно
изогнутые, серповидные. Поверхность рога обычно несет от одного до трех
продольных ребер (килей).
Носовое зеркало большое, занимает все пространство между ноздрями
и переднюю сторону верхней губы. Ноздри широко расставлены; расстоя-
ние между ними больше расстояния между краем верхней губы и ноздрей.
Межпальцевые железы отсутствуют, но у многих форм имеются отсут-
ствующие в других подсемействах особые железы около дополнительных
копыт.. У подавляющего большинства видов отсутствуют также иредглаз-
ничные и паховые железы. Хвост средней длины или длинный, достигаю-
щий концом скакательного сустава. Молочная железа всегда имеет четыре
соска.
Зароговая часть черепа у большей части родов и видов короче лба.
Лобные кости средней длины или длинные, редко составляют меньше 36%
основной длины черепа. Передние края глазниц не сдвигаются назад
дальше уровня последнего верхнего моляра. Этмоидальные щели иногда
имеются, но предглазничные ямки, за исключением рода Tetracerus Leach,
отсутствуют. Основная затылочная кость расширена в задней и сужена
в передней части (рис. 123). Передний край хоан расположен впереди
или позади уровня заднего края верхних моляров. Слуховые пузыривздуты,
вытянуты и сжаты с боков. Резцы широкие, но равномерно убывающие от
первой пары к клыкам или первая их пара расширена в форме асимметрич-
ных лопаток, а остальные резко убывают в ширине от первой пары к клы-
кам. Заднекоренные зубы (по крайней мере нижние) имеют добавочные
столбики или бугорки. Последний премоляр нижней челюсти (Р4) имеет
открытыми и переднюю, и заднюю долинки или замкнутую переднюю
долинку.
Географическое распространение. Вся Африка, средняя и южная
Европа, южная и юго-восточная Азия, Северная Америка.
Классификация. Подсемейство Bovinae включает средних и крупных,
в пределах своего семейства, животных (исключение — мелкий Tetracerus
quadricornis Blainv.). Большинство их характеризуется массивностью
телосложения, короткостью конечностей, в скелете которых пястная кость
иногда короче не только лучевой, но и плечевой кости. Все без исключения
представители данной группы в той или иной мере в образе жизни связаны
с лесом или парковым ландшафтом.
Роды данного подсемейства группируются в две трибы.
1) Bovini — Буйволы и быки. Рога прямые, серповидные или гомонимно
изогнутые. Паховых желез нет. В черепе передний край хоан расположен
позади уровня боковых выемок заднего края костного нёба и почти всегда
позади уровня заднего края последнего верхнего моляра (М3). Верхние
моляры имеют добавочные столбики. Первая пара резцов в виде широких,
почти симметричных лопаток; остальные резцы постепенно убывают в ши-
рине от первой пары к клыкам (рис. 61, В). Распространен, и е —
то же, что и всего подсемейства. Триба включает современные роды: Bos L.
(быки), Poephagus Gray (яки), Bison Н. Smith (бизоны), Bibos Hodgson
(лобастые быки), Bubalus Н. Smith, (азиатские буйволы), Апоа Н. Smith
(целебесский карликовый буйвол — аноа), Syncerus Hodgson (африкан-
ские буйволы), Boselaphus Blainville (нильгау) и Tetracerus Leach, (четырех-
рогая антилопа).
2) Tragelaphini — Винторогие антилопы. Рога гетеронимно свернутые
или скрученные. Паховые железы, как правило, имеются (исключение —
род Taurotragus Wagn. и подрод Boocercus Thom.).
566
BOVIDAE
В черепе передний край хоан расположен впереди уровня заднего
края последнего верхнего моляра (М3) и почти всегда впереди боковых
выемок заднего края костного нёба. Верхние моляры не имеют ни добавоч-
ных столбиков, ни.бугорков. Первая пара резцов расширена в виде асим-
метричных лопаток. Остальные резцы резко убывают в ширине от первой
пары к клыкам. Распространение: Африка, к югу от Сахары.
Триба включает три современные рода: Tragelaphus Blainville (лесные
антилопы), Strepsiceros Frisch (куду) и Тaurotragus Wagn. (канны).
В современной дикой фауне СССР подсемейство Bovinae представлено
родом бизонов (Bison); кроме того, уже в историческое время вымер пред-
ставитель рода Bos, первобытный бык, или тур (Bos primigenius Boj.).
ТАБЛИЦА ДЛЯ ОПРЕДЕЛЕНИЯ РОДОВ И ВИДОВ ПОДСЕМЕЙСТВА
BOVINAE ФАУНЫ СССР
1 (2). Холка приподнята в виде горба. Высота в холке значительно превос-^
ходит высоту в крестце. Линия спины заметно спадает кзади. На ниж-
ней стороне головы, шеи и груди имеется подвес из длинных грубых
волос (рис. 116). На черепе ширина лба между рогами и глазницами
превосходит ширину его в скуловых дугах. Затылочная часть черепа
выступает позади рогов. Последние отходят от черепа приблизительно
на средине расстояния между глазницами и поперечным затылочным
гребнем (рис. 117). В скелете 14 пар ребер^. /. Род. Bison=<(cTp. 566).
В фауне СССР один вид: Bison bonasus L. — Зубр (стр. 568).
2 (1). Холка не выступает вверх в виде горба. Высота в крестце мало усту-
пает, а иногда даже равна высоте в холке. Линия спины почти прямая.
Волосы на нижней стороне головы и шеи не удлинены и не образуют
подвеса (рис. 122). На черепе ширина лба между рогами и глазницами
меньше ширины черепа в скуловых дугах. Рога расположены на
макушке черепа и затылочная часть последнего не видна позади
рогов (рис. 123). В скелете 13 пар ребер . . . Род Bos (стр. 599).
В фауне СССР в домашнем состоянии один вид: Bos taurus L. — крупный рогатый
скот. В исторические времена вымер один из диких предков последнего: Bos primi-
genius Boj. — Первобытный бык, или тур (стр. 601).
Род BISON Н. SMITH—БИЗОНЫ
Falconer Н., 1868, Fauna antique sivalensis, London : 555 (эвол.); A 1 1 e n J.
1876, Mem. Mus. Ccmparat. Zool. Harvard Coll., Cambridge, IV, № 10 : I—IX-|-1—246
(морф., сист., изобр.); Teilhard de Chardin P. etJ. Piveteau, 1930,
Ann. Paleontol., XIX : 3—132 (эвол.); Громова В. И., 1935, Тр. Зоол. инет.,
II : 77—204 (морф., эвол); Garretson М., 1938, American bison, N. Y : 1—254
(морф., сист.); Skinner М. and О. Kaisen, 1947, Bull. Amer. Mus. Natur.
Hist., vol. 89, № 3 : 123—256 (морф., эвол., сист.); Бурча к-А брамовичН. О.,
1949, Докл. Акад, наук Азербайдж. ССР, V, 11 : 474—479 (ист.); Roe F., 1951,
The north american buffalo, Toronto :I—VIII-J-1—957; Соколов И. И., 1953,
Тр. Зоол. ивст. Акад, наук СССР, XIV : 78—80 (морф., эвол.); Бурчак-Абрамо-
в и ч Н. И., 1957, Иск. быки Ст. Света, I, Баку, изд. АН Азерб. ССР: 17—65 (морф.).
Характеристика рода. Животные крупной величины. Холка высокая,
приподнятая в виде горба, вследствие удлинения остистых отростков
первых 3—5 грудных позвонков. Высота в крестце поэтому заметно меньше
высоты в холке. На нижней стороне головы и шеи подвес из длинных волос.
Череп широкий (рис. 117), с резко выступающими в стороны трубко
BISON
567
образными глазницами. Наибольшая ширина его составляет не менее 60%
основной длины. Лоб позади глазниц суживается в направлении рогов.
Заглазничная ширина лба (в самом узком месте между рогами и глазницами)
превышает ширину черепа в скуловых дугах. Лобная поверхность черепа
без резкой границы переходит в теменную. Межроговой валик, свойствен-
ный многим другим видам трибы Bovini, отсутствует.|3ароговая часть
черепа сравнительно хорошо развита и выступает позади рогов. Дорзаль-
ная ее поверхность расположена под очень тупым углом к плоскости лба
и почти прямым к плоскости затылка, принимая заметное участие в обра-
зовании крыши мозговой коробки. При взгляде на череп сверху задние
выходы височных впадин хорошо видны позади роговых отростков. Рас-
стояние между ними больше расстояния между наружными краями заты-
лочных мыщелков (отличие от рода Bibos Hodgson). Рога расположены
приблизительно на середине расстояния между глазницами и попереч-
ным затылочным гребнем (у живого, спокойно стоящего зверя, следова-
тельно, ниже макушки головы). На черепе расстояние между глазницей
и основанием рогового стержня лишь немного больше передне-заднего
диаметра глазницы. Поперечное сечение рогов почти идеально круглое,
иногда лишь (у некоторых ископаемых форм) немного сплюснутое в дорзо-
вентральном направлении. Поверхность роговых стержней без продольных
ребер (килей; отличие от Bubalus Н. Smith, Апоа Н. Smith, Syncerus
Hodgson, Bibos Hodgson, Boselaphus Blainville Tetracerus Leach.).
В скелете характерно наличие 14 грудных позвонков и 14 пар ребер
(отличие от всех других родов Bovini, кроме Poephagus Cray).
История и географическое распространение. В историческое время
ареал рода Bison в Старом Свете ограничивался некоторыми областями
Европы. С 20-х годов нашего столетия в естественной обстановке истреблен.
В настоящее время сохранился лишь в полудиком состоянии в ряде
заповедников и зоопарков. Другой очаг обитания — Северная Америка,
где бизон хотя сохранился (Roe, 1951), но за последнее столетие также
почти целиком истреблен. В доисторические времена ареал был гораздо шире.
Корни рода Bison не известны. Судя по направлению специализации
в черепе и ряду косвенных палеонтологических доказательств (Соколов,
1953), филогенетическая ветвь бизонов отделилась от общего ствола под-
семейства, вероятно, еще в миоцене. Остатки, достоверно относящиеся
к этому роду, известны, начиная с верхнего плиоцена Индии, в лице при-
митивного Bison sivalensis Falc. (Falconer, 1868). Последнего можно рас-
сматривать как одного из прямых предков современных европейского
зубра (В. bonasus L.) и американского бизона (В. bison L.). Находки зубров
верхнеплиоценового возраста известны также из северного Китая
(Teilhard de Chardain et Piveteau, 1930), Англии и Азербайджана (Бурчак-
Абрамович, 1949).
С нижнего плейстоцена многочисленные формы рода Bison широко
распространены по большей части Европы и Азии вплоть до побережья
и островов Северного Ледовитого океана (Громова, 1935). В среднем плей-
стоцене или даже раньше представители рода проникли из северо-восточ-
ной Сибири через «Берингов мост» в Северную Америку, где дали вторичную
вспышку формообразования (Skinner and Keisen, 1947). Начиная с конца
ледниковой эпохи, в Евразии, в силу не достаточно еще установленных
причин, началось вымирание зубра. В течение почти всего послеледнико-
вого периода в Азии и большей части Европы зубр уже не водился. Прибли-
зительно в это же время произошло разобщение ареалов литовского и кав-
казского зубров.
-568
BOVIDAE
Классификация. Современные формы рода Bison группируются в два
Рида.
' 1) Bison bison Linnaeus — Бизон. От европейского зубра отличается;
а) сравнительно слабо развитым и низким задом, б)красновато-коричневым,
вместо серовато-коричневого, оттенком окраски, в) черно-коричневыми
ушами, сильным удлинением волос на лбу и на предплечьях, ^сравнитель-
но коротким хвостом и д) более короткими рогами. Распростране-
ние: Северная Америка.
2) Bison bonasus Linnaeus —Зубр. Характеристику см. ниже. Рас-
пространение: Европа, западная часть Главного Кавказского
хребта.
Bison bonasus Linnaeus —Зубр (рис. 116—121)
Linnaeus С., 1758, Syst. Natur. ed. X, 1 : 71 (первооп.); Volborth A.,
1825, De bobus uro, amo et caffro, Berlin : I—IV-J-1— 26 (общ.); Baer C., 1836,
Bull. Sci. Acad. Sci. St.-Petersb., 1.1, ser. I, 20 : 153—155 (морф.); P u a c h G., 1837,
Polens Palaeontologie, Stuttgart : 195—209 (ист.); Owen R., 1848, Proc. Zool. Soc.
London : 126—133 (анат.); Усов С. A., 1865, Зубр, M., изд. Моск. зоол. сада :
1—114 (общ.); Brandt J., 1867, Verh. Russisch. keiserl. Mineralog. Gesellsch.,
St.-Petersburg, Ser. 2, Bd. 2 : 133—184 (распр.); Allen J., 1876, Mem. Mus. Comparat.
Zool. Harvard Coll., Cambridge, IV, 10:41—46 (морф.); Lydekker R., 1898,
Wild oxen, sheep and goats of all lands, London : 64—79 (общ., изобр.); Рузский M.,
1898, Уч. зап. Казанск. ветеринарн. инет., XV :1—49 (общ.); Websterg G.,
1899, Arb. Naturforsch.-Vereins, Riga, N. F., 9 : 1—80 (распр.; цит. по Башкирову,
1939); К а р ц о в Г. П., 1903, Беловежская пуща, СПб. : 131—238 (общ., изобр.);
Туркин Н. В. и К. А. Сатунин, 1904, Звери России, т. IV, Bovidae, М : 754—
799 (общ., изобр.); Mertens А., 1906, Abhandl. u. Berichte d. Museum f. Natur-
п. Heimatkunde zu Magdeburg, Bd. I, H. II : 64 (ист.); Hilzheimer M., 1909,
Jahresheft. Vereins f. Vaterl. Naturk., Wurtemberg, LXV : 241—269 (морф., сист.);
Динник H. Я., 1910, Звери Кавказа, I, Тифлис : 138—158 (общ.); Фила-
тов Д. П., 1910, Ежегодн. Зоол. муз. Акад, наук, XV, 4 : 0171—0217, табл. IX—X
(биол., изобр.); Врублевский К., 1912, Арх. ветеринарн. наук, 8 : 746—778
(пит.); Lydekker R., 1912, The ox and its kindred, London : 215 (общ., изобр.);
Филатов Д. П., 1912, Зап. Акад, наук, сер. VII, т. XXX, 8 : 1—39 (биол.);
Эккерт Н. И. иВ. В. Феддерс, 1912, Сборн. работ в память проф. И. М. Са-
довского, СПб. : 260—346 (бол.); Lydekker R., 1913, Catal. Ungulate mammals
Brit. Mus., I, London : 35—37 (общ., синон.); Сатунин К. А., 1914, Кавказский
зубр, Тифлис : 1—16 (общ.); Иванов И. И. и Ю. А. Филиппенко, 1915,
Описание гибридов между зубром, бизоном и рогатым скотом, Птг., : 1—33; К о ж е в,-
ников Г. А., 1917, Охотничий вестник, 9—10 : 162 (охр.); Соловьев Д. К.,
1918, Типы организаций, способствующих охране природы, Птг., : 24, 45 (акклим.);
Кулагин Н. М., 1919, Зубры Беловежской пущи, М., изд. Моск. науч. инет. :
I—VI-j-1—166 (общ.); П о р те н ко Л. А., 1919, Висти природничо! секцп Украшьск.
паукового товариства, I, 3—4:140—146 (акклим.); Hilzheimer М., 1920,
Arch. f. Naturgesch., Abt. A, LXXXIV, 6 : 41—87 (морф., эвол., сист.); D e m e 1 K.,
1922, Ochrana Przyrody, Krakow, 3 : 5 (акклим.); Фортунатов Б. К., 1922,
Изв. Гос. запов. Аскания-Нова, I : 1—66 (морф., акклим.); Фортунатов Б. К.,
1924, сборн. «Аскания-Нова», М. : 319—341 (гибр.); Васильковский П. Е.,
1927, Охрана природы и краеведы, М.—Л. : 7 (охр.); Догель В. А., 1927, Природа,
11 : 905 (охр.); J i с k е 1 i С., 1927, Вег. Internat. Gesellsch. f. Erhaltung d. Wisents,
II, 2 : 163—170 (ист.); Koch W., 1927, Ber. Internal. Gesellsch. Erhalt. d. Wisents.,
II, 2 : 175—183 (морф., сист.); Беднарский M., 1928, Охрана природы, I : 15
(охр.); Фортунатов Б. К., 1929, Изв. Гос. степи, запов. Чапли (б. Аскания-Нова),
VII : 69—87 (гибр., разв.); Calinescu R., 1931, Zeitschr. f. Saugetierk., VI :
78—84 (ист.); Розанов М. П., 1931, Сборн. работ по изуч. фауны Крымск, запов.,
М.—Л. : 60—62 (акклим.); Groeben G. (red.), 1932, Das Zuchtbuch, Berlin : 7—
50, Taf. I—X (разв., изобр.); Koch W., 1932, Abh. Mathem—Naturwiss. Abt. Bayer.
Akad. d. Wissensch. Bd. 15, Supplem. : 553—678 (морф.); Флеров К. К., 1932,
Изв. Акад, наук СССР, Отдел матем. и естеств. наук, 10 : 1579—1590 (сист.); М о h г Е.
(red.), 1933, Bericht d. Internat. Gesellsch. z. Erhaltung d. Wisents., Ba. 5, H. 2 : 53—60
(плем. книга); Андреева E. Г., 1935, сборн. «Гибридиз. и акклиматиз. животн.»,
II : 112—135 (гибр.); Боголюбский С. Н., 1935, сборн. «Гибридиз. и аккли-
матиз. животн.», II : 89—110 (гибр.); Громова В. И., 1935, Тр. Зоол. инет., II :
BISON
569
77—204 (морф., эвол.); Кос h W.,1935, Anat. Record,61 : 371—376 (морф.); M о h rE.,
1937, Bericht d. Internat. Gesellsch. z. Erhaltung d. Wisents, Bd. 5, H. 3 : 6'1—74 (илем,
книга); Заблоцкий M. А., 1938, Зоол. журн., XVII, 5 : 233—238 (разв.); Б а-
ш к и р о в И., 1939, сборн. «Кавказский зубр», М., изд. Главн. управл. по заповеди,
зоопаркам и зоосадам : 3—72 (общ., изобр.); Заблоцкий М. А., 1939а, сборн.
«Кавказский зубр», М., изд. Главн. управл. по заповеди, зоопаркам и зоосадам : 73—
144 (акклим., гибр.); Заблоцкий М. А., 19396, Научно-метод. зап. Главн. управл.
по заповеди., III : 141—159 (разв.); Рухлядев Д. П., 1939, Научно-метод. зап.
Главн. управл. по заповеди., IV : 79—83 (бол.); Schertz Е., 1939, Senckenbergiana,
Bd. 18, № 1/2 : 37—71 (морф.); Зубарева В. И., 1940, Природа, 8 : 82—86 (ист.);
Кулагин Н. М., 1940, Фауна БССР, т. I, вып. IV, Копытные, Минск, изд. Акад,
наук БССР : 5—И (общ.); Северцов С. А., 1941, Дин. насел, и приспособ, эвол.
жив. : 111 (биол.); KalelaO. иМ. Salmi, 1944, Luonnon Ystava, Helsinki, vol. 48,
№ 1 : 49—52 (цит. no Zool. Rec. 83) (ист.); Harper F., 1945, Extinct and vanishing
mammals of Old World, N. Y. : 531—538 (ист., изобр.); К r e t z о i M., 1946, Ann.
Mus. Natur. Hungarici, vol. 39, №5/6 : 105—107 (ист.); Glower R./1947, J.Mammal.,
vol. 28, № 4 : 333—342 (общ.); Z a h i n s k i J. (red.), 1947, Ksiegi rodowodowe zuhrow,
Warszawa : 1—32, tabl. I—VIII (плем. книга); Крайнова Л. В., 1947, сборн.
«XX лет Кавказск. гос. заповеди.», М. : 61—73 (акклим.); Заблоцкий М. А.,
1948а, Охр. прир., 3 : 22—48 (ист.); Заблоцкий М. А., 19486, Природа, 2 : 40—45
(ист. разв.); Заблоцкий М. А. 1948в, Наукаижизнь, 5 : 32—34 (ист.); Mohr Е.,
1948, Natur u. Volk, 78 : 146—151 (рога); Zabinski J., 1949а, J. Soc. Preserv.
Fauna Empire, London, LIX : 11—28 (разв.); Zabinski J. (red.), 19496, Ksiegi
rodowodowe zubrow, Warszawa : 33—52, tab. IX—XX (плем. книга); Заблоцкий M. А.,
1949, Научно-метод. зап. Главн.управл. по заповеди.,XIII : 128—146 (биол.,восстановл.);
Б и б и к о в а В. И., 1950, Билл. Моск. общ. испыт. прир., Отд. биол., LV, 5:35—43 (ист.);
Zabinski J., 1950, Ochrona przyrody, Krakow, XIX : 102—115 (восстанови.); Lut-
tschwager H., 1950, Anat. Anzeig., Bd. 97, H. 18/20 : 385—387 (морф.); Bur-
ton M., 1951, Ill. London News, 219 : 1078 (морф., изобр.); Zabinski J., 1951,
Ksiegi rodowodowe zubrow, Warszawa : 53—73, tab. XXI—XXVI (плем. книга);
Крайнова Л. В., 1951, Билл. Моск. общ. испыт. прир., Отд. биол., LVI, 6 : 3—17
(пит.); Пидопличко И. Г., 1951, О ледниковом периоде, II, Киев, изд. Акад, наук
УССР : 50—53 (ист.); Цалкин В. И., 1951, Докл. Акад, наук СССР, LXXV1I,
2 : 349—352 (ист.); Заблоцкий М. А., 1952, Болып. советск. энциклопед., 2-е изд.,
17 : 230—231 (общ., изобр.); Кириков С. В., 1952, Изв. Акад, наук СССР, сер.
географ., 6 : 31—48 (ист.); Mohr Е., 1952, DerWisent., Leipzig : 1—75(общ., изобр.);
Жабинский Я., 1953, Природа, 3 : 85—87 (восстановл.); Бурчак-Абра-
м о в и ч Н. О. и В. И. Н а н и е в, 1954, Тр. Естеств.-историч. музея им. Зардаби,
Баку, VIII : 78—186 (ист.); Gray А., 1954, Mammalian hybrids, London : 62 (гибр.);
Zabinski J., 1954, Wszechswiat, Krakow, 7 : 169—172 (восстанови.); C e n-
k i e г S., 1954, Chronmy przyrode ojczista, X, 1 : 34—42 (разв.); Fuller W., 1954,
J. mammal., vol. 85, № 3 : 454—456 (зубы); Ganasinski R., 1955, Med. We-
teryn., II, 9 : 526—530 (восстановл.); Соколов И. И., 1955, Природа, 3 : 48—55
(одомаш.); Заблоцкий М. А., 1956, Государств, плем. книга зубров и бизонов,
I, М., изд. Мин. сельск. хоз. СССР : 1—116 (морф., ист. восстанови., изобр.).
Описание. Зубр — наиболее крупный зверь современной фауны Евро-
пы. Лишь в росте он иногда уступает крупному лосю, но по весу значительно
превосходит последнего. Длина тела у взрослых самцов составляет
от 234 до 350 см; высота в холке от 158 до 195 см. Основная длина черепа
самцов колеблется в пределах 449—489 мм. Живой вес от 430 до 1000 кг.
Обычный вес зубра-самца среднего возраста и средней упитан-
ности Г. Карцов (1903) определяет в 32 пуда (512 кг). По-видимому, встре-
чались отдельные самцы еще более крупного, чем приведенные цифры,
веса. Так, Г. Карцов (1903) определял вес убитого при нем и выпотрошен-
ного старого крупного быка около 800 кг (50пудов); живой его вес (вместе
с внутренностями), следует поэтому полагать, был около тонны. Приводи-
мые иногда еще более высокие цифры, до 1900 кг (Башкиров, 1939; Websterg,
1899), по-видимому, не всегда достоверны. Достоверно известный случай
живого веса 12-летнего самца зубробизона «Ермыша» (3/4-кровный по зубру)
в 1200 кг, сообщенный автору М. А. Заблоцким, возможно, связанс явле-
нием гетерозиса, обычного при межвидовой гибридизации животных.
570
BOVIDAE
Живой вес отстрелянного в моем присутствии в октябре 1958 г. 58/64-
кровного зубра самца 4 лет в Кавказском заповеднике составил 589 кг.
Самки заметно мельче самцов. Г. Карцов (1903) живой вес их определяет
на 30% меньше одновозрастных самцов. Приводимая этим автором цифра
живого веса 10-летней зубрицы в 528 кг, по-видимому, относится к живот-
ному выше средних размеров. Всё же самки весом 370—400 кг, по-видимому,
довольно обычны. Длина тела у взрослых самок составляет около 240—
290 см, высота в холке до 160 см. Основная длина черепа у них колеблется
в пределах 420—449 мм.
Общее телосложение зубра тяжелое, массивное, вместе с особенностями
волосяного покрова придающее животному свирепую внешность. В туло-
вище обращает внимание непропорциональное развитие передней и задней
части. Передняя часть необычайно массивная, особенно у самцов. Холка
очень высокая, благодаря удлинению составляющих ее скелетную основу
остистых отростков позвонков, и образует подобие горба, еще более увели-
чиваемого вертикально стоящими на его гребне волосами. Мощно развитая
мускулатура плечевого пояса и лопатки усиливает массивность передней
части тела зубра. Грудная клетка очень глубокая, но плоская, вследствие
слабой изогнутости ребер. Задняя часть туловища зубра производит впе-
чатление недоразвитой, хотя и не в такой сильной степени, как у американ-
ского бизона. Таз представляется узким, по сравнению с плечевым поясом,
и посаженным низко. Высота в крестце у самцов на 20—25 см, у самок
на 12—15 см меньше высоты в холке. Впечатление слабости и недоразвито-
сти зада усиливается более коротким и более плотно прилегающим волосом
в этой части тела. Линия профиля спины круто падает кзаду, часто образуя
при этом заметный волнообразный изгиб. Живот поджарый, еще более
подчеркивающий глубину грудной клетки. Хвост в сечении круглый,
длиною 50—60 см с кистью длинных волос на конце—80—100 см, до-
стигающий или даже спускающийся несколько ниже скакательного су-
става, если только не вытерт и не оборван. При возбуждении и беге
хвост часто изгибается и поднимается кверху. Шея у зубра короткая,
толстая и широкая, вследствие высокой холки кажется низко посаженной
на туловище. Верхняя линия ее профиля круто спадает кпереди, еще
больше оттеняя горбообразную высокую холку. На нижней стороне
шеи имеется кожная складка с бахромой из длинных грубых волос.
Голова длиной 65—75 см, очень широкая, с выпуклым лбом, выступаю-
щими глазницами, относительно небольшими глазами, широко расставлен-
ными рогами и притупленной на конце мордой. Большая ширина лба
и густой длинный волос придают голове округлый вид, скрывают границу
между нею и шеей. «Челка сверху, а борода и нижняя шейная грива,
составляющая ее продолжение, снизу настолько округляют общий вид
головы в профиль, что выдающейся от черепа морды совершенно не видно
и даже на близком расстоянии не сразу возможно определить, где нахо-
дится рот зверя» (Карцов, 1903). Благодаря низкой посадке шеи голова
также опущена книзу и при спокойном состоянии животного ее макушка
(затылочный бугор) расположена ниже уровня корня хвоста. Носовое
зеркало занимает передний край верхней губы, значительную часть про-
странства между ноздрями и верхние края последних; цвет его темный,
свинцово-серый. Ноздри продольно-овальные, расположены по бокам
переднего края морды. Зрачок глаз вертикально-продолговатый (Карцов,
1903). Радужная оболочка темно-коричневая. Уши сравнительно короткие,
около 16—17 см, но густо обросшие косматой шерстью, торчащие позади
рогов в стороны и кверху.
BISOiX
571
Ноги, относительно общей величины и массивности зверя, у зубра не
толстые. Благодаря большой глубине грудной клетки, передние ноги
производят впечатление коротких. Копыта, по сравнению с американским
бизоном, широкие полукруглые и слабо изогнутые на внутренней стороне.
Длина копыт на передних ногах 139—173 мм, на задних 117—158 мм; вы-
сота передних копыт 90—106 мм, задних 78—107 мм.
Рис. 116. Зубр (Bison bonasus L.).
Самки отличаются от самцов зубров, помимо меньшей общей величины,
легкостью сложения. Туловище у них, по Г. Карцову (1903), относительно
длиннее; холка выделяется меньше, линия спины менее покатая, голова
сравнительно небольшая; общий облик несколько напоминает коровий.
Еще больше напоминает коровий облик склад молодых зубрят на первом-
втором году жизни, с их слабо выступающей холкой, легкой головой и
тонкими рогами.
Иллюстрацией пропорций телосложения зубров может служить при-
водимая нами таблица их промеров, составленная по литературным дан-
ным (зубробизон отстрелян нами в Кавказском заповеднике в 1958 г.).
Рога у зубров имеют и самцы, и самки. Изредка наблюдаемые случаи
отсутствия их (Карцов, 1903) объясняются, вероятно, потерей их в резуль-
тате травм в молодом возрасте. Рога у зубров, сравнительно с другими
572
BOVIDAE
Размеры и вес беловежских и кавказских зубров
Зубробизон i 58/64-крови 4 лет Беловежские Кавказские
<5 30 лет £ 10 лет 1 £ 8 мес. <5 3 лет 2 старая <5 взрослый £ старая $ 3 лет
Длина от верхней губы до конца хвоста (см) .... 381 375 350 162 298 309 306 (без 306 293
Обхват грудной клетки (см) 238 234 200 104 202 206 хвоста) 253 205
Длина головы (см) .... 73 64 60 31 71 61 — —. •—1
Обхват головы над глазами (см) 129 102 61 110 99 98 109
Расстояние между основа- ниями рогов (см) .... 28 24 13 24 20 20 24.5
Длина шеи (см) — 39 44 18 47 — — — —
Обхват шеи на уровне сре- дины длины (см) .... 130 92 53 117
Высота в холке (см) . . . 183 178 166 95 159 — 179.6 — 158
Высота в крестце (см) . . 183(?) 178 171 106 159 166 — 164 158
Обхват пясти (см) .... 23 24 20 12 —— 25 25.3 — .—
Длина хвоста без волос (см) 57 63 58 30 64 — — — —
Длина хвоста с волосами (см) 89 80 88 44 101 — — — —
Вес без внутренностей (кг) 437 576 416 224 320 — — — —
Живой вес (кг) 589 672 528 288 432 — — — —
быками, небольшие, но все же длиннее, чем у бизона; длина их по изгибу
у самцов от 33 до 51 см, меньше общей (наибольшей) длины черепа. Роговые
чехлы круто гомонимно изогнуты. Отходя от головы, рога направляются
в стороны и слегка вниз, затем загибаются вверх и назад, а вершинами обра-
щены назад и навстречу друг другу. Сравнительно толстые у оснований (от 28
до 36 см), рога быстро утончаются к вершинам. Поперечное сечение круг-
лое. Поверхность роговых чехлов гладкая; только у оснований она попе-
речно морщинистая. Степень выражения морщинистости хоть сколько-
нибудь точным критерием возраста животного служить не может.
У самок рога немного короче, чем у самцов, более тонкие и сильнее
загнуты внутрь концами. Цвет рогов и копыт у зубра черно-бурый.
У самцов, а нередко также и у самок, от трения и ударов о деревья,
а также вследствие привычки самцов рыть рогами землю, роговые чехлы
на вершинах начинают лупиться; иногда при этом образуется подобие
венца из волокон расщепленного рога, сквозь который выступает вновь
образованная острая верхушка. Таким образом происходит частичная
регенерация роговой ткани, как бы смена чехлов рога (Mohr, 1948, 1952).
К старости рога сильно притупляются и делаются короче.
Волосяной покров зубра очень густой. Состоит он, как и у большинства
других копытных, из более грубой толстой ости и тонкого извитого подшер-
стка. Остевые волосы, принимая во внимание величину животного, у зубра
сравнительно мягкие и неломкие, по длине весьма неравномерные. Более
длинный волос покрывает голову и переднюю часть туловища. Особенно
длинные, до 25—26 см, свешивающиеся на лоб и виски волосы растут на
темени и между рогами, образуя подобие шапки или чолки. Последняя,
у зубра, впрочем, не достигает той пышности, которая столь характерна
BISON
573
для бизона. Длинные, до 7—8 см, довольно грубые, направленные вперед
волосы покрывают лоб и верх морды почти до ноздрей, свешиваясь над
покрытым коротким волосом передним концом рыла. Уши и снаружи,
и внутри одеты мягкой, но длинной и лохматой шерстью. Длинные
грубые волосы на нижней стороне головы образуют свешивающуюся
вниз бороду длиной 35—40 см. Такие же волосы длиною до 20—
25 см образуют подвес на нижней стороне груди и шеи. В противополож-
ность бизону, борода у зубра не четко отделена от подвеса на шее и иногда
сливается с последним. У самцов пучок длинных волос окружает отверстие
препуция. Отличием зубра является также то, что волосы на предплечьях
мало удлинены (не более 10 см), и не образуют чулок, характерных для
бизона. На верхней стороне шеи и холки волосы также удлинены в стоя-
чую, а зимою, иногда, даже слегка свешивающуюся на сторону гриву.
На задней стороне туловища позади лопаток волосы короче. Длина их
на боку зимой составляет 3—4 см. У беловежского зубра волосы на туло-
вище волнистые или слабо извитые; у кавказского шерсть курчавая, зави-
тая иногда довольно мелкими завитками. В конце зимы, перед началом
линьки шерсть взлохмачивается и даже сбивается в войлок. Короткий,
плотно прилегающий волос покрывает конец морды вокруг ноздрей и носо-
вого зеркала, небольшое кольцо вокруг глаз и ноги ниже запястья и ска-
кательных суставов. Хвост от основания покрыт одинаковой с туловищем
короткой шерстью. Отступя 40—50 см от корня, волосы на нем постепенно
удлиняются и грубеют, образуя на конце кисть длиною 40—50 см. В зим-
нем меху волосы и на хвосте удлиняются; поэтому граница между кистью
и корневой частью становится менее заметной.
Окраска зубра, сравнительно с многими другими копытными нашей
фауны, довольно однотонная, буро-коричневая.1 Передняя часть тела,
по сравнению с задней, несколько темнее. Особенно темны бока и низ головы
а также борода и подвес на шее, имеющие черно-коричневый оттенок.
Темнее других частей туловища окрашенживот. Темно-коричневая окраска
свойственна и нижним частям ног, но волосы, непосредственно прилегающие
к венчикам основных и добавочных копыт, обычно, более светлого
желтовато-коричневого цвета. Темная буро-коричневая полоса часто
тянется вдоль средней линии шеи и спины. Проксимальная часть хвоста
однотонна с туловищем, кисть же состоит из черно-коричневых с неболь-
шой примесью белых волос. Более светлую желтовато-коричневую окраску,
с дымчатым налетом, имеют плечи и холка, а иногда также бока шеи и
верхняя сторона головы. «При ярком солнечном освещении — пишет
Г. Карцев (1903), — зубры кажутся как бы осыпанными по горбу и плечам
серебристым порошком». В противоположность бизону, уши одного тона
с туловищем. Светлый беловато-коричневый цвет имеют волосы на самом
конце морды и на нижней губе, тогда как глаза окружаются кольцом
чисто коричневого цвета. Волосы подшерстка более светлого чем ость,
желтовато-бурого цвета.
Летом, вскоре после линьки, шерсть на зубре более короткая, плотно
прилегающая к коже, особенно на задней части тела. Окраска летом свет-
лее, чем зимою, нередко приобретает серовато-желтый оттенок.
Половой диморфизм в волосяном покрове и окраске у зубров заметен
очень слабо.
1 Непонятно, почему в большинстве описаний зубра окраска его называется тем-
но-бурой или серовато-бурой. На основании просмотра значительного числа шкур
,в коллекциях ЗИН АН СССР и живых зубров в зоопарках, Центральном зубровом
питомнике и Кавказском заповеднике преобладает не бурый, а коричневый оттенок.
574
BOVIDAE
Молодые в первые месяцы после рождения желтовато-коричневые.
Только заметно удлиненные мягкие волосы на месте будущего подвеса
имеют темно-коричневую или темно-бурую окраску. К зиме первого года
жизни шерсть у молодняка становится красновато-коричневой с пепель-
ным налетом.
Молочная железа у зубриц, как и у всех быков, имеет четыре соска,
но она очень невелика и, даже в период лактации, снаружи не видна из
окружающей ее шерсти.
Характерную особенность зубра составляет свойственный и самцам,
и самкам мускусный запах. Этот запах ощущается в течение круглого года,
но особенно силен у самцов в период течки (яра), а также зимою в сырую
погоду и летом в сильную жару, когда стадо укрывается в тенистых влаж-
ных зарослях леса. Происхождение мускусного запаха зубра не выяснено.
Так как запах усиливается после бега, можно предполагать, что он связан
с деятельностью потовых желез. Впрочем, мускусным запахом обладает
будто бы и молоко зубриц. О силе запаха можно судить по тому, что по
ветру он ощущается нередко на расстоянии 150 шагов от пасущегося стада
и в продолжение часа после ухода зубров с мест жировки (Карцов, 1903).
Не менее сильно чувствуется мускусный запах и от убитого зубра.
Характеристика краниологических особенностей рода Bison была дана
раньше (стр. 566). Отличиями от американского бизона в черепе зубра
(рис. 117) являются следующие его особенности: а) несколько меньшие,
в среднем, общие размеры; б) более длинные и абсолютно, и относительно
роговые стержни, сильнее изогнутые у самцов назад и к тому же заметно
скрученные к вершинам. Кроме того, череп зубра и абсолютно, и относи-
тельно уже, имеет более укороченную лицевую часть, чем у бизона. Лоб-
ная поверхность черепа зубра имеет больше впадин и бугров, чем у бизона;
особенно ярко выражены впадины у переднего края межтеменных костей
и в средней части лба, между глазниц. Отличительным признаком является
форма слезной кости, более узкой и имеющей обособленный передне-внут-
ренний узкий зубец, вклинивающийся между верхнечелюстной и носовой
костями (Hilzheimer, 1909; Громова, 1935). Следует, однако, иметь в виду,
что большинство приведенных отличий (кроме, быть может, относительной
длины и массивности роговых стержней) имеет не абсолютный, а лишь
относительный характер, и только по совокупности признаков, и то не
всегда, можно отличить череп зубра от черепа бизона.
История и географическое распространение (рис. 118). Как установлено
исследованиями В. И. Громовой (1935), современный зубр является из-
мельчавшим потомком первобытного зубра {Bison prisons Boj.) — вида,
широко распространенного в Евразии с нижнего и до конца верхнего плей-
стоцена. Первобытный зубр явился одновременно предком и американского
бизона. Мельчание ствола зубра, вследствие ухудшихся к концу леднико-
вой эпохи климатических условий, началось, по мнению В. И. Громовой,
с конца последнего межледниковья (рисе—вюрма). К этому времени отно-
сится находка в Германии фрагмента черепа с сильно укороченными,'‘но
еще массивными роговыми стержнями, относимого по размерам уже
к виду Bison bonasus (Hilzheimer, 1920). Форма, промежуточная между
короткорогими расами первобытного зубра конца плейстоцена и современ-
ным В. bonasus, найдена в отложениях верхнепалеолитической стоянки
Ильской на Северном Кавказе (Громова, 1935).
Расцвет европейского зубра приходится на послеледниковое время.
Остатки, несомненно относящиеся к В. bonasus, известны из голоценовых
отложений ряда мест западной Европы. В противоположность первобыт-
BISON
575
ному зубру (В. priscus Boj.), в Азии современный зубр, по-видимому,
никогда не водился. В бостонной Европе в голоцене распространение его
было более ограниченным по сравнению с плейстоценовым предком.
Восточная граница, по-видимому, никогда не заходила дальше побережья
Ладожского озера, нынешней Новгородской и соседних с нею областей,
Рис. 117. Череп Bison bonasus L.
Белоруссии и западной части Украины.1 Остатки зубра найдены в неоли-
тических стоянках на южном берегу Ладожского озера и около Новгорода.
В. И. Громова (1935) считает, что уже к концу ледникового периода про-
изошло и разобщение некогда единого ареала рода Bison Н. Smith в Ев-
ропе и на Кавказе. Остатки мелкого зубра найдены на берегу оз. Севан
в слоях, относимых к эпохе бронзы или ассирийской, т. е. за 3000 лет до
1 Имеется, правда, указание на находку субфоссильного черепа зубра в Фин-
ляндии (Kalela и Salmi, 1944).
576
BOVIDAE
нашей эры. Раскопки древнего города Ольвии на территории нынешней
Николаевской области показывают, что здесь зубры водились еще в начале
нашей эры (Пидопличко, 1951).
Уже в историческую эпоху зубрбылраспространенивзападнойЕвропе,
и на территории юго-запада европейской части Советского Союза в местах,
где в настоящее время его нет уже в течение многих десятилетий. Во вре-
мена Юлия Цезаря зубры населяли леса Германии и Бельгии, откуда
доставлялись в римские амфитеатры. Сведения об обитании его во Франции
относятся к VI в. (Harper, 1945). В Семиградье (Трансильвании), зубры
являлись предметом охоты еще в VII в. Последние экземпляры их на воле
здесь были уничтожены в самом конце XVIII в. (Jickeli, 1927). В Венгрии,
будто бы, жили до 1790 г. (Kretzoi, 1946). В Восточной Пруссии последние
зубры были истреблены в 1755 г. (Lydekker, 1913), в Саксонии — в 1793 г.
(Harper, 1945). Дольше они сохранялись в лесах на территории Польши
(Pusch, 1837).
На территории СССР распространение зубра, в свое время, охватывало
современную Литовскую ССР, Белоруссию, юго-западную и правобереж-
ную Украину и даже Молдавию. Заходил ли в исторические времена зубр
в пределы Латвии и Эстонии — не известно. При раскопках древнего
Гродно в слоях, относящихся к XII—XVI вв. н. э., среди костей других
животных обнаружено значительное число их, принадлежащих зубру,
по размерам существенно превосходившему современных (Цалкин, 1951).
В XII в. зубры служили объектом охоты киевских князей (Зубарева,
1940; Пидопличко, 1951). По свидетельству современников, племянник
византийского императора Андроник Комнен, гостивший у галицкого
князя Ярослава Остросмысла (XII в.), охотился у него на зубров. Боль-
шое число письменных свидетельств о зубре относится к XVI в. (Кириков,
1952). Наиболее ценным, но еще не редким зверем в это время он являлся
в лесах Великого Княжества Литовского. По данным Мстибогского ста-
росты Воловича, в XVI в. зубры жили в Гродненской, Кнышинской,
Бельской, Беловежской, Перестунской и других крупных литовских пу-
щах. К тому же времени относятся указания Герберштейна о зубрах в По-
лесье и аналогичные данные Кромера об обитании их в Подолии. В XVI в.
в большом числе зубры обитали в правобережном лесо-степном Подне-
провье, а также в окрестностях Брацлавского замка, но в опубликованном
в 1650 г. «Описании Украины» Боплана, при перечислении водящихся
в ней зверей, упоминания о зубрах уже нет.
Имеется неопределенное указание о том, что в начале XVIII в. в Во-
ронежской губернии якобы встречались «зубры или дикие быки, подобные
ауроксам» (Кириков, 1952); сведения эти, впрочем, не были в дальнейшем
никем подтверждены; в лесостепном междуречье Днепра и Дона зубры,
по-видимому, никогда не водились. В лесах Молдавии, по словам Канте-
мира, зубры жили еще в 80-х годах XVIII в. В Карпатах достоверно они
существовали до 1762 г. (Galinescu, 1931). В Буковине небольшое число
зубров сохранялось, будто бы, до начала XIX в. (Harper, 1945); однако
достоверность этих сведений вызывает сомнения. В западной Европе
зубр исчез, по-видимому, в конце XVIII в. Последний зубр в Восточной
Пруссии был убит между Лавиау и Тиместом в 1755 г. (Mertens, 1906).
Основной причиной уменьшения численности зубра и исчезновения во
многих местах, где еще сравнительно недавно он был обычным зверем,
явилось прямое истребление человеком. Такое животное, как зубр, всегда
являлось заманчивой добычей. Сохранились неопровержимые доказатель-
ства того, что уже палеолитические люди с их примитивными орудиями
BISON
577
устраивали коллективные охоты на первобытного зубра (Бибикова, 1950)»
Не меньшее значение имела косвенная, вытесняющая деятельность чело-
века. Вырубание лесов, основной стации зубра, распашка и использование
под выпас домашнего скота лесных пастбищ — все это влекло за собою
сокращение угодий, пригодных для существования этого зверя.
Рис. 118. Распространение Bison bonasus L.
1 — XX в.; 2 — XIX в.; 3 — XVIII в. и раньше; 4— места
акклиматизации.
К началу XIX в. единственными местами, где сохранялись в диком
состоянии зубры, были Беловежская пуща на территории Гродненской
губернии 1 и западная часть Большого Кавказа.
Беловежская пуща издавна являлась местом охоты лиц царствовавшего
в России дома Романовых. Наряду с прочими охранительными мероприя-
тиями, с 1803 г. здесь категорически воспрещался отстрел и ловля зубров
1 В настоящее время частично входит в состав Польской республики, частично —
в состав Брестской области Белорусской ССР.
37 Фауна СССР, И. И. Соколов
578
BOVIDAE
без «высочайшего» на то разрешения. Для зверей проводилась заготовка
и раздача зимних кормов в виде сена и раскладываемого в кормушки
овса, картофеля и овощей (Карцев, 1903; Заблоцкий, 1948а). Изредка,
раз в несколько лет проводившиеся царские охоты не влияли отрицательно
на поголовье зубров в пуще, и последнее в первые 50 лет с момента уста-
новления запрета неуклонно численно возрастало. Если в 1810 г. количество
их исчислялось в 310 голов, то в 1857 г. оно достигало цифры 1898 голов
(Заблоцкий, 1948; Mohr, 1952). С начала 60-х годов поголовье стало резко
уменьшаться и долгое время колебалось в пределах 400—600 зубров.
Причину создавшегося положения стали усматривать в происходящем,
якобы, процессе естественного «вырождения» зубра, не приспособленного,
будто бы, к условиям современной эпохи. Признаки вырождения зубра
усматривали в пониженной интенсивности размножения, повышенной
смертности и измельчании. Не без основания одной из причин вырождения
беловежского зубра считали близко родственное размножение в пределах
одного стада. Однако, как выяснилось позднее, главной причиной приоста-
новки роста поголовья и снижения численности зубров в Беловеже яви-
лась перенаселенность ее копытным зверем. В погоне за увеличением чис-
ленности и многообразия дичи в Беловежскую пущу, начиная с 1865 г.,
многократно завозились благородные олени и лани, сильно размножив-
шиеся к началу XX в. Возникшая острая межвидовая конкуренция из-за
пищи повела к резкому ухудшению естественных пастбищ пущи, к истреб-
лению лесного подроста и местами даже к прекращению процесса естест-
венного возобновления леса (Врублевский, 1912). Хроническое голодание
животных влекло за собою снижение плодовитости, понижение сопротив-
ляемости к заболеваниям, повышенную смертность животных.
В результате некоторых мер по снижению перенаселенности и улуч-
шению подкармливания животных, в начале нынешнего столетия число
зубров стало вновь возрастать и к 1910 г. достигло 1200 голов. Губитель-
ной для беловежского зубра оказалась Первая мировая война. 6 августа
1915 г., перед приходом немцев в Беловежскую пущу, в ней насчитывалось
739 зубров. При первом же учете их, проведенном немцами после занятия
пущи, оказалось только триста. В 1916 г. зубров оставалось 150—200,
в 1917—1918 гг., по одним данным 200, а по другим — только 68 голов.
9 февраля 1921 г. бывшим лесным сторожем Беловежской пущи Варфоло-
меем Шпаковичем был застрелен последний вольный зубр Беловежа (За-
блоцкий, 1948а; Mohr, 1952).1
Другим, с дальних времен изолированным очагом обитания зубра яв-
лялся северо-западный Кавказ. О существовании здесь этого зверя цивили-
зованный мир узнал практически лишь в прошлом столетии, в то время,
когда зубр по тем же причинам, что и его беловежский собрат, находился
на пути к исчезновению. Первые документальные данные о кавказском
зубре в виде его шкуры, описанной академиком К. Бэром (Baer, 1836),
поступили в 1836 г. Но и после этого видовая принадлежность жившего
на Кавказе дикого быка до средины 60-х годов подвергалась сомнению
(Усов, 1865). Только в 1867 г., когда в Московский зоосад был доставлен
с Кавказа живой зубр, многолетний спор был окончательно решен.®
В доисторическое время область распространения зубра на Кавказе
была значительной и некогда, вероятно, сливалась с ареалом беловежского 1 2
1 По другим данным (Заблоцкий, 19486) последние беловежские зубры были уни-
чтожены еще в 1919 г.
2 Историю исследования кавказского зубра — см. Башкиров, 1939.
BISON
579
зубра. Уже упоминалась находка, свидетельствующая об обитании мелкого
зубра в бронзовую или ассирийскую эпоху в районе Севана. Необходимо
иметь также в виду неоднократно поступавшие, хотя до сих пор и непрове-
ренные сведения о том, что в провинциях Ирана Мезандеран и Гилян
(в районе г. Решта, около южного побережья Каспийского моря) до сих
пор водятся зубры (Башкиров, 1939).
Изменение ареала и ход последовательного истребления зубра на
Кавказе рисуется в следующем виде.1 Судя по многочисленным черепам,
снесенным в свое время охотниками в древние святилища-дзауры Север-
ной Осетии (в бассейне р. Уруха), зубры здесь водились до конца XVIII
или даже начала XIX в. (Бурчак-Абрамович и Наниев, 1954). По сведе-
ниям, относящимся к XVII в., в то время они обитали на границе Абха-
зии и Мингрелии. В XVIII в. экземпляр зубра был добыт у горы Бештау
севернее Пятигорска.
В начале прошлого столетия зубры водились на водоразделе Кубани
и Терека, от р. Агара до Чегема. Но уже к 50-м годам они, по-видимому,
не заходили на восток от Кубани, основная же масса была сосредоточена
к западу от р. Теберды. Северную границу распространения зубра в сре-
дине прошлого столетия, к моменту окончательного завоевания рус-
скими Кавказа, И. С. Башкиров (1939) проводит по р'р. Курджипсу и
Белой, на широте станиц Дагестанской и Абадзехской, далее к востоку
на р. Лабу километров сорок севернее Псебая, на р. Уруп и Кубань около
устья Малого Зеленчука. Зубры в это время населяли все пространство
между Малой и Большой Лабой, водились по рр. Карамыху, Большому
Тегеню, Ходзь, Карибижгону, Куве, Кунчь, Яскыке, Псекенч. На за-
паде зубр доходил в это время, вероятно, до р. Пшехи, а возможно, даже до
Пшиш. На Пшехе эти звери изредка встречались до конца 90-х годов XIX в.
К югу от Главного Кавказского хребта в прошлом столетии зубр оби-
тал в верхнем течении рр. Шахи, Сочи, Мзымты, Псоу, Бзыпи, возможно
также в верховьях Кодори и Ингура. В качестве пунктов местонахожде-
ния зубров в Абхазии в конце прошлого и начала нынешнего века назы-
ваются: район оз. Рица, водораздел рр. Лашипсе и Гега, бассейн р. Бзыпи.
Известен случай единичного захода зубра в 1915 г. в район г. Сухуми.
Предполагается, что ко времени окончательного покорения Кавказа
поголовье кавказского зубра исчислялось в 2000 голов.
В связи с военными действиями против горцев, колонизацией пред-
горных районов, в результате вырубания лесов, проникновения на на-
горные пастбища домашнего скота и усовершенствования огнестрельного
оружия, во второй половине XIX в. быстро стала сокращаться как об-
ласть распространения зубра, так и его численность. Некогда сплошной
ареал стал распадаться на изолированные участки, что, в свою очередь,
грозило вырождением вследствие близко родственного разведения.
Учреждение в 80-х годах великокняжеской «Кубанской охоты», с за-
претом добычи, более или менее налаженной охраной и даже частичной
подкормкой несколько замедлило процесс истребления, но не приоста-
новило вытеснения зубров с обжитых угодий и общего процесса сниже-
ния их численности. Если еще в 80-х годах зубры встречались на Зелен-
чуках и в верховьях Урупа, то к концу XIX в. основная область их оби-
тания ограничивалась пространством от верховьев рр. Белой и Пшехи
на западе до р. Большой Лабы на востоке, причем к северу дальше устьев
р. Киши и Уруштена она уже не распространялась. Вне границ означен-
1 В основном, по И. С. Башкирову (1939) и Н. Я. Диннику (1910).
37*
580
BOVIDAE
ной области и к югу, и к северу наблюдались только отдельные заходы
зубров. В 80-х годах число зубров на Кавказе определялось цифрой
800—1000 голов, а в 90-х годах разными лицами назывались цифры от
400 до 700 голов. К началу XX в., в результате мер охраны, число зубров,
возможно, несколько возросло и составляло к 1910 г. около 600 голов.
Однако область обитания, в силу непрекращающейся колонизации края,
к этому времени еще больше сократилась, и к востоку от Малой Лабы
зубры уже больше не встречались (Филатов, 1912).
Несколько расширилась область обитания в годы Первой мировой
войны, когда к востоку зубры стали встречаться вновь до Большой Лабы.
Еще в 1911 г. созданная при Академии наук специальная Комиссия
для выработки мер к охране кавказского зубра внесла в Государствен-
ную Думу проект организации в горной полосе Кавказа государствен-
ного заповедника, так как к тому времени стало ясно, что «Кубанская
охота», преследовавшая в основном удовлетворение охотничьих интересов
великих князей, не способна предохранить зубра от окончательного
истребления. Однако проект, предложенный Академией наук, даже не
был рассмотрен Государственной Думой.
Поставленный вторично через два года по инициативе Природоохра-
нительной комиссии Русского географического общества вопрос об орга-
низации Кавказского заповедника в 1913 г. был рассмотрен, но отклонен
Советом Министров. Новый проект предусматривал отчуждение под за-
поведник части земель кубанского казачества, а по мнению царского
правительства «охрана редких зоологических пород не отвечает понятию
общегосударственной полезной меры, ради осуществления которой можно
поступиться неприкосновенным вообще правом частной собственности»
(Заблоцкий, 1948).
Вопрос об организации зубрового заповедника положительно, хотя
и с запозданием, был решен лишь при Советской власти.
К 1917 г., когда великокняжеская «Кубанская охота» была национа-
лизирована, число зубров на Кавказе еще определялось цифрой около
500 голов. Губительными для зверя оказались последующие годы Гра-
жданской войны и разрухи. Зубров уничтожали укрывавшиеся в горах
банды. Еще больше звери страдали от пользовавшихся бесконтрольностью
того времени местных браконьеров. К 1920 г. оставалось не больше 100 зуб-
ров, уходивших под влиянием преследования с обжитых мест и попадав-
ших зачастую в тяжелую обстановку. Много животных погибло от эпизо-
отии, занесенной в горы домашним скотом. К 1921 г., когда окончательно
установилась Советская власть на Кубани и Черноморье, в живых на
Кавказе оставалось не более 50 зубров.
Ряд принятых местными органами постановлений по охране зубра не
дал надлежащих результатов. Число зверей катастрофически убывало.
12 мая 1924 г. декретом Совнаркома РСФСР был, наконец, учрежден
Кавказский государственный заповедник с основной задачей — сохра-
нить кавказского зубра. Однако недостаточность штатов на огромной
территории заповедника не позволила сразу поставить надлежащим
образом учет и охрану исчезающего зверя. Обследовавшая заповедник
в 1925 г. Комиссия не смогла даже точно установить, сохранились ли
к тому времени на Кавказе зубры, или уже исчезли (Заблоцкий, 1948).
Практически, к 1925 г. зубры, по-видимому, действительно были истреб-
лены. Позднее отмечались лишь отдельные встречи зубров и следов их,
главным образом на территории заповедника и вблизи его границ;, не-
многих зверей,- уходящих, вероятно, от преследования, находили в ме-
BISON
581
стах, где они исчезли уже несколько десятилетий назад, например в вер-
ховьях Зеленчуков (Башкиров, 1939). В восточной части Кавказского
заповедника, на г. Алоусе, в 1927 г. был убит последний кавказский
зубр.
Таким образом, к 1928 г. в мире не осталось ни одного вольно живу-
щего зубра. Только в некоторых частновладельческих парках и зооло-
гических садах разных стран Европы сохранилось пять-шесть десятков
экземпляров, с которыми предстояло начать работу по восстановлению
исчезнувшего в природе вида.
Подвиды. Подавляющим большинством исследователей беловежский
и кавказский зубры принимаются за подвиды одного вида Bison bonasus L.1
1) В. bonasus bonasus Linnaeus — Беловежский, или литовский зубр.
Размеры более крупные; высота в холке взрослого быка до 185 см; жи-
вой вес, в среднем, 512 кг. Сложение тяжелое. Окраска сравнительно
светлая, серовато-бурая с охристо-коричневым оттенком. Волосы почти
прямые или слабо завитые, на боках таза более длинные. Длина волос
в бороде у быка достигает 370—400 мм. Копыта удлиненные.1 2 Распро-
странение: в доисторические времена и до конца XVIII в. —запад-
ная Европа, Польша, Литва, Полесье, правобережная Украина, Молда-
вия, Румыния. С начала XIX в. и до 1921 г. — только Беловежская пуща.
В настоящее время — только в некоторых зоопарках и заповедниках
Европы в полуприрученном состоянии.
2) В. bonasus Caucasians Satunin — Кавказский зубр. Размеры более
мелкие; высота в холке взрослого быка около 160 см; живой вес, в сред-
нем, около 450 кг. Сложение более легкое; конечности, сравнительно,
тонкие. Окраска более темная коричнево-бурая, с шоколадным оттенком.
Шерсть на всем теле сильно курчавая. Длина волос на бороде не превы-
шает 250 мм. Копыта короткие, высокие и округлые. Распростра-
нение: северо-западный Кавказ. В настоящее время в чистом виде
ни в природе, ни в зоопарках не сохранился.
Следует отметить, что надлежащего сравнительно морфологического,
даже остеологического исследования зубров до сих пор не проведено.
В связи с тем, что обе расы, возможно, произошли независимо от общего
или разных предков и что ареалы их разъединены с конца ледникового
периода, с общебиологической точки зрения, быть может, правильнее
рассматривать диких беловежского и кавказского зубров как самостоя-
тельные виды.
Биология. Можно говорить о двух экологических типах зубра: рав-
нинном и горном. Первым является беловежский зубр, обитатель равнин-
ных смешанных лесов Европы, тогда как кавказский считается типичным
горно-лесным зверем. Обшим для обеих форм является привязанность
к лесу. Как и большинство других представителей подсемейства Bovinae,
виды рода Bison — первично лесные звери. Приспособление американ-
ского бизона (Bison bison bison L.), а также, вероятно, некоторых рас перво-
бытного зубра (Bison priscu2 Boj) к жизни в условиях степи — явление,
несомненно, вторичного порядка. Лес для вольного зубра являлся не
только местом отдыха и укрытия от врагов, но предоставлял животному
также весьма существенную и количественно и качественно долю пищи.
1 Отличия даются по К. К. Флерову (1932).
2 Приводимые К. К. Флеровым (1932) размеры копыт вызывают сомнения: у боль-
шинства парнопалых, в том числе и у зубра, передние копыта длиннее задних, а не
наоборот, как следует из цифр цитируемой работы.
>82
BOVIDAE
Однако сплошных лесных массивов, особенно хвойных и лишенных
молодого подроста, зубры также избегают. Излюбленными стациями, при
прочих равных условиях, являются смешанные леса с хорошо развитым
подлеском из лиственных пород или участки леса, чередующиеся с лес-
ными лужайками и открытыми полянами.
На Кавказе зубры держались на высотах от 900 до 2100 м. Летом,
в спокойных условиях, они предпочитали держаться вблизи верхней
границы леса, среди зарослей березняка, рододендронов и горного клена,
по соседству с субальпийскими лугами (Динник, 1910). Здесь звери на-
ходили не только в изобилии сочный зеленый корм, но, вследствие
большей прохлады, и защиту от кровососущих насекомых. В дождливую
и холодную погоду они скрывались под покровом густого хвойного леса.
На зиму зубры спускались вниз в лесные долины, где незначительная
глубина, а иногда и полное отсутствие снега облегчали животным добычу
пищи и передвижение, а лес предоставлял защиту от непогод в трудное
для животных время. Иногда звери доходили даже до крайних предгор-
ных селений. Известны случаи, когда в Абхазии зубры на зимовки спу-
скались к морскому побережью. Часть животных зимой оставалась в го-
рах, используя для этого долины таких рек, как Киши, Умныря, Малой и
Большой Лабы (Башкиров, 1939).
Передвижки кавказских зубров весной, по мере таяния снега, из
долин в горы, а с наступлением зимы в обратном направлении, еще 70—
90 лет назад имели характер регулярных сезонных миграций. По мере
вырубания лесов звери лишались обжитых зимовок, а с приходом на
высокогорные пастбища стад домашнего скота, вынуждены были уходить
и с альпийских лугов, довольствуясь пастьбой на опушках и лужайках
внутри леса.
Если их не преследуют, зубры отличаются большой консервативностью
и привязанностью к местам обитания. Бывший владелец соседней с Бело-
вежской Свислочской пущи не мог изгнать живших в ней зубров, несмотря
на то, что не проводил для них заготовки на зиму корма (Усов, 1865).
Известны лишь единичные случаи дальних заходов отдельных животных:
уже упоминавшийся случай добычи зубра в окрестностях Сухуми или
заходы из Беловежской пущи в леса б. Волынской и Минской губерний.
В пределах избранного на данный сезон местообитания они стойко при-
держиваются своих мест отдыха, кормежек, водопоя. В местах, где про-
водится подкормка, зубры привыкают держаться вблизи стогов сена,
кормушек и других мест раздачи корма и далеко от них не уходят.
Кормятся вольные зубры ранним утром и вечером. Ночью и в дневные
часы, обычно, отдыхают. В нежаркую погоду зубры выходят на кормежку
и днем. Иногда пасутся и ночью, особенно, лунной. Для отдыха выбирают
укрытые места, где животные спасаются не только от дождя и ветра, но
и от солнца. Места лежек, так называемые «точки», более или менее по-
стоянны и представляют собою большей или меньшей величины (в за-
висимости от числа пользующихся ими зверей) площадки с выбитой ро-
гами и копытами травой и превращенной в пыль почвой. Здесь зубры
не только отдыхают, но и принимают «песочные ванны», валяются на
боку, вытянув ноги, хотя перекатываться через хребет, как это делают
лошади, они не в состоянии. Зубры хорошо переносят сырость и холод,
зимой любят кататься в снегу. Гораздо хуже переносят жару. Особенно
чувствительны к перегреванию телята зубров, в противоположность
бизонам, как показал опыт разведения их в Аскании-Нова. Забираться
в воду от жары и насекомых, как делают некоторые другие копытные,
Рис. 119. Зубро-бизоны в Кавказском заповеднике.
584
BOVIDAE
зубры не имеют обыкновения. Летом, при сухой почве, заходят иногда
в болота, по-видимому, ради прохлады. Но, вообще, мест, сырых и богатых
водой, особенно зимой, зубры избегают. Осенью и ранней весной изредка
выходят на хлебные поля, очевидно, привлекаемые молодыми всходами
посевов (Mohr, 1952). В Беловежской пуще, в свое время, они любили
валяться на вспаханной рыхлой почве.
В сведениях о питании зубра много противоречивого. Правы, по-
видимому, те, кто утверждает, что пищей ему служит почти все то, что
он находит в богатом подростом лесу, на опушках и лесных полянах.
Принципиально важен вопрос о потребности зубра в древесно-веточ-
ном корме. К. Врублевский (1912), например, утверждает, что при до-
статочном количестве травянистого корма зубры могут обходиться во-
обще без поедания частей кустарников и деревьев. Однако теперь можно
считать установленным, что это не так. Типично лесной зверь, зубр,
в противоположность степному бизону, нуждается в обязательном уча-
стии древесно-веточного корма в рационе в течение всего годй. Многолет-
ний опыт разведения зубров в Аскании-Нова показал, что 50% всех
случаев падежа этих зверей, лишенных здесь древесного корма, проис-
ходили от желудочно-кишечных заболеваний (Заблоцкий, 1939).1 Ис-
следования Л. В. Крайновой (1951) по питанию вольно пасущихся зубро-
бизонов в Кавказском заповеднике показали, что «в условиях прекрасных
луговых пастбищ древесные корма зубры поедают постоянно. . . Древесный
корм необходим зубрам в течение круглого года. В летний период зубры
нуждаются в нем наряду с травянистым. Лес — не только жилище, но
и постоянный источник корма в виде листьев, побегов и коры деревьев».
Практически, в условиях Кавказского заповедника, в той или иной
мере поедаются все древесные породы, произрастающие в районе выпаса
зубров, причем почти половина является основным древесным кормом.
Излюбленными видами считаются ильм и рябина (Динник, 1910). Охотно
поедаются также разные виды ив. осина, ясень, граб и некоторые другие.
Поедаемыми частями этих пород являются листья, тонкие ветви, кора.
Кора поедается в течение всего года, но особенно охотно весной, когда
она сочна и легко отделяется от древесины. Зубры подрезают кору рез-
цами на высоте морды и движением головы или пятясь назад отдирают
снизу вверх длинные, до 3—4 м, ленты (у тех, конечно, пород, у которых
кора может таким образом отдираться). Лакомой осенней пищей являются
желуди, буковые орехи, а на Кавказе также плоды дикой яблони и груши,
ради которых зубры «дежурят» под деревьями и даже откапывают опав-
шие фрукты и желуди из-под снега.
Побеги хвойных деревьев (сосны и, особенно, ели) при наличии дру-
гих кормов, практически не поедаются вовсе. В Беловежской пуще,
по С. Усову (1865), зубры не употребляли коры тополя и липы.
Л. В. Крайнова (1951) в списке кормов зубра перечисляет свыше 100 ви-
дов травянистых растений, из которых около сорока являются основными.
На первом месте по значению для зубра из травянистых растений стоят
виды разнотравья, злаков и некоторые из бобовых, например восточный
козлятник (Galega orientalis Lam.). Одним из излюбленных и поедаемых
в большом количестве растений является борщевник (Heracleum pubes-
cens МВ.), а в Беловежской пуще, по старым данным, зубровая трава
(Hierochloe australis В. et Sch.).
1 У бизонов только 20%.
BISON
585
Летом основу питания зубра составляют травянистые растения,
в первую очередь крупное сочное разнотравье, у которого поедаются
не только листва и верхушки, но и сочные, еще не одеревяневшие, стебли.1
Зубр без вреда поедает ядовитую для других животных чемерицу (Veral-
rum lobelianum Bernh.). В работах старых авторов (Динник, 1910) встре-
чается также указание на поедание зубрами листьев папоротников, ш*
употребляемых в пищу домашним скотом. Наравне с травой зубры и ле-
том охотно едят листья, тонкие ветки, а также кору ив, лещины, граба,
яблони и ряда других деревьев. При вполне достаточном количестве дру-
гого, более питательного корма, употребление зубрами коры летом сле-
дует поставить в связь с потребностью в горьких и дубильных веществах,
возбуждающих пищеварительные процессы.
С наступлением осени, когда многие травы грубеют, меняется видовой
состав травянистого корма, еще составляющего основу питания. Поедаются
в первую очередь сочные зеленые части растений, отава на месте летних
покосов. По мере усыхания трав, все большую роль в питании начинают
играть древесно-веточные корма, на которые почти целиком звери пере-
ходят в октябре или ноябре.
Зимой главную пищу зубров составляют ветки и кора различных
кустарников и деревьев. Откусывают ветви толщиною до 1.5—2 см. Осо-
бенно охотно зубры кормятся около срубленных и поваленных ветром
деревьев, на вершинах которых звери в большом количестве находят
не обкусанные ранее ветки и тонкую кору. В сильные морозы (ниже 15е)
кору сгрызать трудно, и поэтому зубры в такие дни кормятся больше
ветвями. Один из излюбленных зимних кормов — древесные лишайники,
собираемые со стволов и низких ветвей деревьев. Известны случаи по-
едания наземных лишайников типа кладоний (Заблоцкий, по Л. Край-
новой, 1951). После зимних бурь зубры собирают сорванные ветром ветки
и лишайники прямо со снега. В качестве зимних зеленых кормов исполь-
зуются ежевика и овсяница, а по И. С. Башкирову (1939) также падуб
(Ilex aquifolium L.) с его вечнозелеными листьями, торчащими из-под
снега. Не отказываются зубры, пока снег не свыше 30 см, и от засохшей
травы, опавших листьев, желудей и фруктов. Доставая этот корм, зубры
разрывают снег копытами, а круговыми движениями морды образуют
в нем лунки.
В начале весны звери хотя, в основном, питаются еще древесно-
веточным кормом, но с жадностью набрасываются на первую зелень:
пролеску, первоцвет, лапчатку, осоки. Охотно обрывают в это время
концы веток с набухшими почками и распускающейся листвой.
По мере развития растительности, зелень принимает все большее уча-
стие в питании, и в конце мая—начале июня зубры окончательно пере-
ходят на летний рацион, состоящий, в основном, из трав и зеленых ли-
стьев.
Основу зимнего питания зубров Беловежской пущи в последние десяти-
летия их существования составляло специально заготавливавшееся для
них сено. В сене зубры выбирали, главным образом, бобовые и разно-
травье, втаптывая и разбрасывая значительную его часть по снегу.
Интересно, что дикие кавказские зубры на территории бывшей «Кубан-
ской охоты» упорно отказывались от заготовляемого для них сена (Дин-
ник, 1910). В Аскании-Нова привезенные туда зубры привыкли есть
1 Однако, как сообщил автору М. А. Заблоцкий, предпочтительное поедание зуб-
рами трав не подтверждается последними наблюдениями в Беловежской пуще.
586
BOVIDAE
степное сено, но это плохо отражалось на пищеварении и служило одной
из основных причин гибели животных.
В условиях паркового полувольного содержания зубры быстро при-
выкают и хорошо поедают концентрированные корма: зерно (особенно
раздавленное, так называемую дерть), отруби, корнеплоды, в первую оче-
редь свеклу. Последнюю они способны выгрызать из земли. В Кавказ-
ском заповеднике реакклиматизируемые зубры охотнее собирают и едят
картофель, выкапываемый ими из земли, чем раздаваемый в кормушках.
Известны случаи объедания зубром стеблей и початков кукурузы (Край-
нова, 1951).
На Кавказе и в прежнее время, и теперь зубры охотно посещают есте-
ственные и искусственные солонцы, а также минеральные и сероводород-
ные источники.
Зубров к источникам привлекает, по-видимому, не столько вода,
сколько грунт, так как, обычно, они пьют предварительно взмученную
ими самими воду, лижут камни и даже грызут землю по берегам солонце-
ватых водоемов (Динник, 1910). В Беловежской пуще естественных со-
лонцов, по-видимому, нет, но здесь так же, как и на Кавказе, зубры
охотно лижут и едят соль, разложенную в кормушках и высыпанную на
землю. Как естественные солонцы, так и задаваемую соль звери употреб-
ляют в течение круглого года, но наиболее интенсивное «солонцевание»
происходит весной, при переходе на зеленые корма, а также в августе
и сентябре, возможно, в какой-то связи с наступлением брачного сезона.
На Кавказе тяга к минеральному питанию у зубров настолько сильна, что
к солонцам тянулись на многие километры хорошо, из года в год про-
топтанные зверями тропы.
На воле зубры держатся небольшими группами, не превышающими,
обычно, 10 голов. Часто подобная группа является семейной и состоит из
одной или двух самок с их потомством за последние 3—4 года, в число
которого входят и молодые 2—3-летние самцы. Взрослые самцы вне пе-
риода полового сезона держатся отдельными группами по нескольку
особей. Группами держатся и временно прогнанные от стада старым
быком молодые самцы. «Одинцами» являются, и то, по-видимому, сравни-
тельно редко, старые самцы. Одиночками иногда держатся также старые
бесплодные самки. В условиях вольного обитания более крупные стада,
до 20—40 голов, по-видимому, получались в результате случайного
схождения нескольких групп на водопое, солонцах и на более удобных
зимовках. Роль вожака в стаде зубров, вне периода брачного сезона,
выполняет старая самка.
В условиях паркового содержания, вследствие недостаточности пло-
щади обитания, а возможно также вследствие привыкания к человеку,
зубры часто держатся более крупными группами, по нескольку десятков
голов. При этом в состав групп могут входить животные разного пола и
возраста, включая взрослых самцов. Только молодые самцы на период
яра отгоняются взрослыми быками и обособляются в отдельные группы,
да побежденный взрослый самец может стать временно одиноким бро-
дягой (Mohr, 1952).
Если не считать периода спаривания, зубры ведут себя по отношению
друг к другу мирно. Только самка отважно защищает новорожденного от
подходящих к нему зубров. Изредка, впрочем, и вне брачного сезона
могут происходить стычки; несколько зверей могут напасть на од-
ного, обычно более слабого или молодого и даже убить его (Mohr,
1952).
BISON
587
В спокойной обстановке зубры передвигаются шагом, рысью же поль-
зуются редко и неумело. Однако при внешней неуклюжести они отли-
чаются легкостью и даже порывистостью движений. На коротких дистан-
циях галопом они могут развивать большую скорость. Зубрята очень
игривы, напоминая в этом отношении домашних телят. Иногда признаки
игривости проявляют и взрослые звери. Испуганные или преследуемые
зубры легко перепрыгивают через поваленные деревья и другие препят-
ствия. Охотнее бегут под гору (Динник, 1910). В зоопарках, становясь
на задние ноги или с разбега, перебираются через высокие, 2—3-метровые
заборы (Усов, 1865; см. фотографию у Е. Mohr, 1952 : 20). Известны не
единичные случаи, когда зубр, чтобы достать высоко расположенную
ветку с листьями, становился на задние ноги, передними упирался
в ствол дерева и вытягивал шею. На Кавказе, по Диннику (1920),
зубры свободно передвигаются по очень крутым склонам, но скал избе-
гают.
Голос зубра — глухое, несколько растянутое хрюканье, которым
звери перекликаются друг с другом. Храп или сопение, обычно, служат
признаком раздражения. Зубрята мычат сходно с домашними телятами,
но более грубо. Изредка и взрослые зубры, раненные или пойманные
в клетку, издают громкий, протяжный рев.
У зубра, несомненно, хорошо развиты слух и, особенно, обоняние.
На зрение же он полагается гораздо меньше и, по-видимому, различает
не столько сам предмет, сколько его движение. Бывали случаи, когда,
заметив приближающегося человека, звери обращались в бегство, только
восприняв его обонянием. По ветру могут услышать звук шагов или
почуять запах человека за 500 шагов (Карцов, 1903).
О характере зубра и его отношении к человеку мнения противоречивы.
С одной стороны, подчеркивается его необыкновенная «чуткость», даже
при полувольном парковом содержании. Заметив издали приближение
человека, звери стараются бесшумно скрыться в лесной чаще. Внезапно
испуганные, наоборот, бросаются с шумом и треском сквозь ломаемые
кусты и деревья. В качестве примера чуткости приводится неудачная
охота известного английского охотника-спортсмена Литльделя в его
первую и вторую поездки за кавказским зубром в 90-е годы: «Следы
зверя удавалось видеть часто, в особенности по низменным лесистым
местам. . . Несколько ' раз Литльдель находил даже совершенно свежие
следы зверя, слыхал хряст шагов по сучьям, видел обглоданные деревья,
кал, но за 24 вылазки ему ни разу не удалось приблизиться настолько,
чтобы видеть зверя, — до такой степени он чуток и осторожен» (Туркин
и Сатунин, 1904).
С другой стороны, известны случаи, когда тот же дикий кавказский
зубр подпускал человека на 7—8 шагов и позволял сделать подряд не-
сколько фотоснимков (Динник, 1910). Во всяком случае, панического
страха перед человеком зубр не испытывает и, даже в случае явной опас-
ности, держит себя часто сравнительно спокойно.
На человека дикий зубр, даже раненный, нападает редко. Недавно
отелившаяся самка при приближении человека делает только вид, что
обороняет, а в конце концов убегает, покидая теленка (Динник, 1910).
В условиях паркового содержания и при организованной подкормке
зубры, чувствуя безопасность, ведут себя по отношению к человеку
значительно смелее, чем на воле. Нередко в этих случаях они проявляют
даже признаки агрессивного поведения. Не рекомендуется, например,
входить в зубровый загон без хорошей палки, которой можно отмахнуть
588
BOVIDAE
агрессивно настроенного зверя, или удаляться далеко от ограды, на ко-
торую, в случае опасности, можно быстро забраться. Особенно прихо-
дится оберегаться самок с молодым зубренком. В Кавказском запо-
веднике зубрицы не раз заставляли людей взбираться на деревья.
Агрессивно держатся иногда и самцы. С. Усов (1865), со слов Жиль-
берта, приводит случай, правда, мало вероятный, когда бык-одинец жил
около дороги, проходящейчерез Беловежскую пущу, и нападал на проез-
жавшие подводы, если с последних не сбрасывали ему сена.
Зубры обладают сообразительностью и достаточно развитым интел-
лектом. Они неплохо ориентируются в новой обстановке, научаются
распознавать грозящую опасность или неприятность (Mohr, 1952). Срав-
нительно быстро вырабатывают условные рефлексы. Автору в Централь-
ном зубровом питомнике около Серпухова удалось наблюдать, как со
всех концов зубрового загона звери стремглав мчались на рожок сторожа
к корытам с разложенным кормом.
Взятые для воспитания в раннем возрасте, телята хорошо привыкают,
даже привязываются к человеку и становятся ручными. Однако полное
приручение, а тем более одомашнивание зубра, по-видимому, никогда
не удавались. Причиной, очевидно, является особый склад нервной
системы этого зверя (Соколов, 1955). Даже выращенный с раннего воз-
раста около человека, зубр взрослым становится строптивым и опасным.
В зоопарках зубры также привыкают и отличают от других ухаживаю-
щих за ними людей, но М. А. Заблоцкий рассказывал, что в Московском
зоопарке зубр убил служителя, убиравшего дворик.
Смена волосяного покрова происходит один раз в году. Линька начи-
нается с средины февраля и затягивается до средины июля. Как и у других
копытных*, ранее и быстрее вылинивают хорошо упитанные быки и хо-
лостые самки. Запаздывают с линькой самки, имеющие телят, и больные
или истощенные звери. Сначала выпадает подшерсток, пластами висящий
и постепенно сползающий по дольше сохраняющейся ости. Вероятно,
испытывая зуд от раздражения мертвым волосом, зубры трутся боками
и головой о деревья, ускоряя процесс линьки. В первую очередь старая
шерсть выпадает на углу нижней челюсти и одновременно в промеж-
ности. Дольше всего она остается на брюхе и бедрах, где даже осенью
можно видеть редкие зимние волосы, хотя подшерсток и на этих местах
в это время уже выпал.1 Смена волоса происходит быстрее на задней
части туловища, чем на передней. В летней шерсти у взрослых самцов
бороды и подвеса нет, но у самок и молодых самцов он только становится
более редким. На туловище шерсть первое время после линьки очень
короткая, плотно прилегает к коже. Заметное удлинение ее происходит
в сентябре; тогда же отрастают вновь борода и подвес на нижней стороне шеи.
Растут зубры медленно, полного развития достигают на 7—8-м году
жизни, а в весе прибывают до 8—9 лет. Половое созревание наступает
значительно раньше. Самки обычно впервые оплодотворяются на 3-м
или даже на 4-м году жизни, и телят, следовательно, они приносят не
раньше 3—4 лет. Известны, однако, случаи оплодотворения самок на 2-м
году жизни, вызываемые или совместным содержанием самцов и самок
в парках и зоосадах, или стремлением владельцев максимально форсировать
воспроизводство зубров. Полноценное потомство в таких случаях, обычно,
не получалось, телята рождались недоразвитыми, а у самок-первотелок
часто не оказывалось молока (Заблоцкий, 1939). Самцы также способны
1 Данные М. А. Заблоцкого.
BISON
589
к оплодотворению часто на втором году жизни, но даже при полуволыюм
содержании отстраняются от размножения до 4—5-летнего возраста более
сильными взрослыми быками.
В условиях Беловежской пущи в последнее столетие существования
там зубров считалось нормой, когда самка приносила телят только через год
или даже раз в три года. Это явление рассматривалось даже как один
из показателей якобы естественного вымирания зубров. Как выяснилось
позднее, истинной причиной пониженной плодовитости беловежских
зубров являлись неудовлетворительные условия их существования,
в частности, недостаток полноценных кормов. В той же Беловежской
пуще, в Грибовском зверинце, где зверям обеспечивалось нормальное
питание, большинство взрослых самок приносили телят ежегодно. Нор-
мально самки могут плодоносить до 18-20 лет и даже позднее. Э. Моор
(Mohr, 1952) приводит случай выдающейся племенной продуктивности
зубрицы Каука: первого теленка она родила в возрасте 4 лет и затем,
с 1925 г. по 1945 г., принесла 20 телят (8 самцов и 12 самок), последнего
уже в возрасте 24 лет. Срок продуктивной половой деятельности самцов —
тоже около 20 лет, но половое влечение у некоторых из них сохраняется
чуть ли не до 30 лет. Возможность добиться почти ежегодной плодови-
тости у зубриц доказывает также практика некоторых заповедников
Советского Союза, и в частности, Центрального зубрового питомника
около Серпухова под Москвой.
Брачный сезон у зубров и в Беловежской пуще, и на Кавказе про-
исходит в период с августа по октябрь включительно. Продолжительность
его в нормальных условиях — от 2 до 3 недель. У отдельных самок течка,
по Н. М. Кулагину (1919), длится около 6—7 дней.
Продолжительность беременности у зубрицы — около 9 месяцев,
так что отелы происходят в мае и июне, растягиваясь в норме даже и у жив-
ших в вольных условиях зверей до полутора месяцев. При полувольном
же или загонном содержании эти сроки, вследствие измененных условий
существования, нарушаются еще больше. Хотя нормально максимальное
число (около 30—40%) отелов происходит в мае, все же процесс растя-
гивается с апреля по декабрь, а отдельные случаи могут быть в январе,
феврале и даже марте (Заблоцкий, 1949; Mohr, 1952). В Аскании-Нова
отел и гон, в соответствии с более теплым климатом, происходят, в сред-
нем, несколько раньше, а в Швеции, наоборот, пик кривой отелов сдви-
гается с мая на июнь (Заблоцкий, 1949). Нормальные сроки размножения
нарушаются при ухудшении кормовых условий и связанных с этим про-
пусках беременности, а также при старении самок. Иногда, если зубрица
потеряла теленка или последний отнят для искусственного выпаивания,
она вскоре может придти в охоту, оплодотвориться и успеть принести
в том же году второго теленка (Башкиров, 1939 ': 43).
Самцы, в том числе и одинцы, в период брачного сезона присоединя-
ются к стадам и создают около себя гаремы от двух до восьми самок.
Процесс яра (гона) у зубров проходит спокойнее, чем у оленей. Нет ни-
какого рева, изредка самцы только хрюкают или пыхтят. Более сильные
самцы иногда просто отгоняют от стада молодых и более слабых соперни-
ков, которые пасутся в этом случае группами отдельно. Но в Беловеже
и некоторых других местах, где самцы и самки не содержались отдельно,
между быками происходили поединки, иногда кончавшиеся для одного
из соперников увечьем или смертью. Самки нередко также бодаются
с самцами, но, вероятно, в данном случае речь идет не о борьбе, а о за-
игрывании или своеобразном ухаживании самца. У вольных кавказских
590
BOVIDAE
зубров драки между самцами были мало заметны, а некоторыми отрица-
лись вовсе (Динник, 1910; Туркин и Сатунин, 1904). Самцы в период
гона худеют. Кроме того, у них, как и у самок, в это время столь уси-
ливается мускусный запах, что мясо иногда становится непригодным
в пищу.
Дней за семь до отела самка удаляется от стада и в укромном
сухом месте приносит теленка. Теленок всегда один. Единственный до-
стоверно известный случай рождения двойни у чистокровных зубриц при-
водит для Плесского парка (верхняя Силезия) Э. Моор (Mohr, 1952),
но оба близнеца оказались мертворожденными.1 Вес новорожденного
зубренка около 22—26 кг, при росте около 70 см. Первые 2—3 дня теленок
лежит, вставая лишь для того, чтобы пососать молока. К 7—8-му дню
он настолько крепнет, что может следовать за матерью, и та вновь воз-
вращается к своей группе. Мать защищает теленка не только от человека
и врагов, но и от других зубров, которые иногда одним взмахом головы
могут убить шныряющего под ногами зубренка. По старым данным,
рождается будто бы больше самцов, чем самок.
Теленок сосет мать не меньше 8—10 месяцев, а если она не забере-
менела в этом году, то может пользоваться материнским молоком свыше
года. Количество выделяемого за сутки зубрицей молока не известно, но
есть данные, что выкармливаемый искусственно зубренок, будто бы, спо-
собен выпить молоко от двух средней удойности коров (Карцов, 1903).
И. С. Башкиров (1939), исходя, вероятно, из незначительной величины
вымени, считал, что молока у зубрицы немного, но оно более жирно, чем
у коровы. С двух-трех недель теленок начинает щипать нежные листочки,
но полностью на растительный корм переходит только в конце осени
или зимою.
Рога у зубрят начинают расти в возрасте около полутора месяцев.
Полного развития, как уже упоминалось, зубры достигают около 7—8
лет. Продолжительность жизни определяют в 30—40 лет и даже до
50 лет (Усов, 1865; Карцов, 1903). Однако максимальный, достоверно
установленный их возраст — 27 лет (Mohr, 1952). На самом деле естест-
венная продолжительность жизни, вероятно, несколько больше, так
как престарелых зубров обычно отстреливают раньше наступления естест-
венной смерти.
Единственным врагом зубра из млекопитающих, если не считать
человека, является волк. Теоретически возможно нападение и медведя,
но достоверные случаи подобного рода не известны. Волки могут пред-
ставлять угрозу практически лишь зимою, при глубоком снеге. В этом
случае группа в 3—4 волка может загнать на лед или в глубокий снег
даже взрослого самца и одолеть его. Чаще жертвой становятся молодые
и самки (Башкиров, 1939). При нападении волков зубры применяют
круговую защиту, укрывая внутри замкнутого круга слабых и молодых
животных (Карцов, 1903).
На Кавказе известны единичные случаи гибели зубров, срывавшихся
со скал, попадавших случайно в щели, откуда звери не могли выбраться,
и т. п. Случаи такого рода происходили, обычно, при вынужденной
зимовке в горах и в глубокоснежные зимы.
Из эпизоотических заболеваний среди зубров известны ящур и сибир-
ская язва, которыми звери заражаются через пастбища от домашних
животных. Вспышки ящура на Кавказе зарегистрированы в 90-х годах
1 О случае мертворожденных двоен сообщал М. А. Заблоцкий (1939).
BISON
591
прошлого столетия, затем в 1907 г. Большая эпизоотия, унесшая значи-
тельную часть и без того уже почти уничтоженного кавказского зубра,
была в 1919 г. (Башкиров, 1939). По имеющимся данным, в 1919 г. зубры
падали не только от ящура, но и от сибирской язвы. От ящура погибло
значительное число зубров за последние годы в Польше.
В 1910 г. в Беловежской пуще среди диких копытных имела место
эпизоотия геморрогической септицемии, от которой погибло около
50 зубров (Эккерт и Феддерс, 1912). Вспышка геморрагической септицемии
наблюдалась среди реакклиматизированных на Кавказе зубров в 1941 г.
Поголовье удалось спасти только благодаря своевременно проведенным
предохранительным прививкам (Заблоцкий, 1948). Из других заболеваний
отмечались случаи падежа от туберкулеза в Аскании-Нова вследствие
неблагоприятных климатических условий (Заблоцкий, 1939).
Из глистных инвазий у зубров зарегистрированы фасциолез, дикро-
целиоз, эхинококкоз и диктиокаулез (легочно-глистная болезнь) (Эккерт
и Феддерс, 1912; Кулагин, 1919; Заблоцкий, 1939).1 У зубробизонов
в Аскании-Нова, а раньше, по-видимому, также в Крымском заповеднике,
наблюдались случаи телязиоза — заболевания глаз, вызываемого нема-
тодой Thelazia rhodesi Desm. и ведущего к потере зрения на один или оба
глаза (Рухлядев, 1939). Интересно замечание Г. Карцова (1903) о том,
что ему ни разу не удалось наблюдать на коже зубров дыр от подкожных
оводов. По устному сообщению Г. В. Сердюковой, в Беловежской пуще
на зубров нападает клещ Ixodes ricinus L.
Хозяйственное значение. Уже несколько десятилетий перед окончатель-
ным истреблением на воле зубр имел только спортивно-охотничий инте-
рес, как крупный и поэтому особенно ценный зверь для высокопоставлен-
ных охотников-спортсменов. В настоящее время он имеет для человечества,
прежде всего, культурное значение, как редкий, почти исчезнувший, но
вновь восстанавливаемый вид крупного животного, как пример творческой,
преобразующей природу деятельности человека.
В перспективе зубр имеет, несомненно, и важное практическое зна-
чение. При успешном размножении, в будущем им могут быть населены
леса многих областей нашего Советского Союза, и народное хозяйство
получит дополнительный источник мясных ресурсов.
Мясо зубров, вообще, довольно грубо, волокнисто и по качеству
уступает таковому домашнего скота. Особенно жестко оно у взрослых
быков и старых зверей. Иногда оно по этой причине, а также вследствие
сильного мускусного запаха в период брачного сезона, совершенно не-
пригодно в пищу. При варке приобретает темный синеватый или багровый
цвет и имеет привкус дичи. Лучшими вкусовыми качествами обладает
мясо самок и молодых зубров. Мясо взрослых зверей рекомендуется,
перед приготовлением из него пищи, вымачивать в уксусе или молоке
(Карцов, 1903).
Кожа зубров подвергалась выделке и в Беловежской пуще, и на Кав-
казе. К сожалению, ее технические свойства в выделанном виде не известны.
Возможно, она пригодна в качестве подошвенного материала. В Беловеже
из зубровой кожи сыромятной выделки изготовлялась конская сбруя.
1 У вольных зубров Беловежской пущи А. К. Мордвилко обнаружил около 15 ви-
дов сосальщиков, круглых и ленточных червей. Патогенное значение, по-видимому,
имела лишь печеночная двуустка (Fasciola hepatica L.), вызывавшая поражение желчных
ходов печени и нарушение желчеотделения. Поголовно у всех зубров в печени обнару-
живалось от нескольких десятков до 300—600 штук двуусток (Кулагин, 1919).
592
BOVIDAE
Рога убитых зубров с давних пор употреблялись в качестве украшения
и для разных мелких поделок.
Из шерсти зубра могут изготовляться теплые перчатки. Однажды из
нее был изготовлен вязаный жилет. Но он, во-первых, сохранял мускус-
ный запах, свойственный зубру, а кроме того, хотя на ощупь и был мяг-
ким, настолько раздражал кожу даже через белье, что к носке оказался
непригодным (Mohr, 1952).
Гибридизация зубра. Зубр и бизон являются примером сравнительно
легкого получения межродовых гибридов, обладающих к тому же, хотя
и ограниченной, плодовитостью. И зубр, и бизон не только свободно
скрещиваются между собою, но образуют также ограниченно плодови-
тые помеси с домашним крупным рогатым скотом.
Первые гибриды были получены Балицким от скрещивания бело-
вежского зубра со скотом еще в 1848 г. (Карцов, 1903). Русским исследо-
вателям принадлежит и приоритет получения гибридов зубра с бизоном
(зубробизонов), имеющих такое большое значение для сохранения и
восстановления зубра, а также получение сложных гибридов (зубр X
бизонХ крупный рогатый скот). В наиболее широких масштабах гибриди-
зация проводилась в Аскании-Нова, начиная с 1902 г. (Иванов и Филип-
ченко, 1915; Фортунатов, 1922, 1924; Заблоцкий, 1939).
При гибридизации с крупным рогатым скотом обычно скрещивали
домашнюю корову с самцом зубра. Гибриды обладали более крупным
ростом. По пропорциям телосложения они носили промежуточный ха-
рактер. Зад туловища становился более высоким и в то же время более
широким. Гибрид зубра со швейцарским скотом в опытах Балицкого
имел голову, шею и перед туловища, сходные с зубром. У помесей зубра
с серым украинским скотом голова часто приобретала коровий облик;
но и в этом случае удлинение волос на лбу, а также развитие бороды
и подвеса на нижней стороне шеи и подгрудка, хотя и не в такой сильной
степени, как у чистокровного зубра, имели место. Рога приближались
к бычьему типу изгиба, более длинные и толстые в основаниях. Горб,
обычно, значительно менее выражен у гибридов, чем у чистокровных
зубров. Волосяной покров на туловище короткий, прямой и плотно при-
легающий к коже. Только иногда наблюдалась легкая курчавость. Окра-
ска при гибридизации с серым украинским скотом получалась двух типов:
а) темная, почти чисто черная, лишь с просвечивающим на лбу и верх-
ней стороне шеи рыжеватым оттенком, б) буро-коричневая, типичная
для зубра с проступающим через нее серым, свойственным украинской
породе цветом, который на спине иногда доминировал над основной
зубровой мастью. Хвост был среднего между зубром и домашним скотом
типа.1 Самец-гибрид в опытах Балицкого не мычал, а «хоркал» и обладал
большей подвижностью, силой и ловкостью, по сравнению с домашними
быками.
Самцы-гибриды между зубром и серым украинским скотом всегда
были бесплодны, а самки, наоборот, плодовиты. В опытах же Балицкого
по скрещиванию зубра со швейцарским скотом плодовитыми были не
только гибридные самки, но и самцы.
Гибриды зубра с серым украинским скотом обладают ценными рабо-
чими качествами. Воспитанные под наблюдением человека и кастриро-
ванные, они могут быть использованы в упряжке. Гибриды очень сильны
1 По краниологии гибридов см. работы С. Н. Боголюбского (1935) и Е. Г. Андрее-
вой (1935).
BISON
593
и выносливы. «При совместной работе полукровных бизонов с серыми
волами приходилось сменять в день три упряжки домашних волов, тогда
как полукровки не выказывали никакой усталости» (Фортунатов, 1922).
Однако практического значения гибридизация зубра с домашним быком
не получила. Помимо технических трудностей, связанных со скрещива-
нием и осеменением, очень часто наблюдались трудные отёлы или нерастёлы,
сопровождающиеся гибелью маток, а также большой процент мертво-
рождения. Мясо гибридов по качеству уступает таковому крупного ро-
гатого скота.
Совершенно легко происходит гибридизация зубра с бизоном. Во всех
отношениях гибриды носят промежуточный между исходными формами
характер, но, как правило, наблюдается ясно выраженный гетерозис
(увеличение размеров) и повышенная жизненность гибридного потомства.
Гибридизация с бизоном имеет большое значение в деле увеличения зубро-
вого поголовья путем поглотительного скрещивания, а также для осве-
жения крови зубра в целях повышения конституциональной крепости
животных.
Восстановление поголовья зубров в СССР и за границей.1
Начиная с 1821 г., зубров, отловленных в Беловежской пуще, неодно-
кратно вывозили за границу. Там их содержали в зверинцах (Констан-
тинопольский в Турции, Скерневицкий в Польше, Шеннбруннв Австрии)
или в охотничьих парках (Вобурн в Англии, Плесе в Верхней
Силезии). Позднее, когда в крупных европейских городах появились
зоологические сады, многие из них также получили зубров непосредст-
венно из Беловежской пущи.
В России беловежские зубры были завезены в Гатчину (Петербург-
ская губерния) и на Украину (Пилявино, Крым, Аскания-Нова).
За 40-летний период (1861—1901 гг.) в Гатчину было привезено 15 зуб-
ров (8 быков и 7 коров). Первый приплод в «Зверинце»получили в 1864 г.
С 1878 г. звери имели возможность использовать подножные корма не
только на территории «Зверинца», но и в окружавших его лесах, в связи
с чем их начали подкармливать только зимой одним сеном. До 1902 г.
рост стада происходил замедленно. В 1902 г. в Гатчине насчитывалось
19 зубров — 13 коров и 6 быков (Карцов, 1903). Такое соотношение
полов обеспечило более быстрое нарастание численности в течение после-
дующих 15 лет, и в 1917 г. в Гатчине было уже 36 зубров (Соловьев, 1918).
В годы гражданской войны стадо, насчитывавшее уже 40 голов, подверг-
лось истреблению браконьерами (Васильковский, 1927). В 1918 г. осталось
только 11 зубров, затем пало еще 2. Вскоре они были окончательно ис-
треблены (Фортунатов, 1929).
Аналогичная судьба постигла и зубров, завезенных в Крым и Пиля-
вино. В Крым зубры были доставлены в 1913 г. Г. А. Кожевников (1917)
указывает, что из Беловежской пущи в Крым было отправлено 5 зубров —
4 самки и 1 самец. М. П. Розанов (1931) замечает, что исходная группа
состояла из 1 быка и 3 коров. Помещенные первоначально в долине
р. Урдуклы, звери вскоре перекочевали на обширные лесные поляны
горы Синоб-Даг, с которых спускались к кормушкам в долину р, Урдуклы
только на зимнее время. Зубры размножались медленно, поэтому в 1917 г.
стадо состояло только из 9 голов. В этом же году началось истребление
зубров браконьерами, и в конце 1919 г. был убит последний зубр —
старый бык, привезенный из Беловежской пущи (Розанов, 1931).
1 Настоящий раздел по просьбе автора написан М. А. Заблоцким.
38 Фауна СССР, И. И. Соколов
594
BOVIDAE
О Пилявском охотничьем парке упоминает Лайдекер (Lydekker, 1912),
Л. А. Портенко (1919) и Демель (Demel, 1922). Зубры, содержавшиеся
в лесу на огороженной территории, давали приплод. В 1908 г. в стаде были
1 бык и 2 коровы, полученные из Беловежской пущи, и 1 теленок, родив-
шийся в Пилявино. Вместе с зубрами находились бизоны, что привело
к рождению 2 гибридов — зубробизонов, первый из которых появился
на свет в 1909 г. В 1917 г. в Пилявино имелось 22 зубра и 9 бизонов.
Л. А. Портенко, посетивший Пилявино в 1918 г., уже не застал ни зубров,
ни бизонов. Звери были выбиты браконьерами в 1917 г.
Наибольший размах работы по разведению зубров получили в Аскании-
Нова. Правда, акклиматизацию чистокровных зубров здесь следует
считать не удавшейся. В 1902 г. из Беловежской пущи доставили самца
и самку, а в 1910 г. еще 2 самки. Звери выпасались в открытой степи.
В течение 10 лет, с 1904 по 1913 г., родилось 12 зубрят (7 самок и 5 сам-
цов), но половина из них погибла в возрасте до 6 месяцев, чему способ-
ствовал, безусловно, и ранний отъем их от матерей. От зубров сумели
получить только 2 поколения (Заблоцкий, 1939). Неудача, прежде всего,
была обусловлена полным отсутствием в Аскании-Нова необходимого
зубру древесного корма. Как и в Пилявино, зубров содержали вместе
с бизонами, однако первый гибрид-зубробизон в Аскании-Нова был
получен на 2 года раньше, чем в Пилявино, — в 1907 г. От самок гибридов
бизона с крупным рогатым скотом при спаривании с самцами-зубрами
получили сложных гибридов (зубрXбизонXкрупный рогатый скот). На-
чиная с 1921 г. гибридные самки были использованы для скрещивания
сначала с зубробизонами, а затем с дополнительно завезенными в 1925—
1933 гг. 4 чистокровными зубрами. В результате проведения поглоти-
тельного (преобразовательного) скрещивания намечалось вывести чисто-
породных зубров. Асканийские гибридные зубры неоднократно выво-
зились в зоопарки СССР и за границу, где также оставили приплод от
скрещивания с чистокровными зубрами (Заблоцкий, 1938, 1939, 19486).
Эти же животные в количестве 10 голов (2 группы по одному самцу и
четыре самки) были переселены из Аскании-Нова в горные леса Крыма
и Кавказа, где хорошо акклиматизировались (Крайнова, 1947, 1951; Заб-
лоцкий, 19486).
Кавказское гибридное стадо (рис. 119, 120), пополненное в послевоен-
ные годы 8 чистокровными самцами, ныне (1958 г.) насчитывает 164 головы;
будучи разделено на 2 группы, оно обитает в бассейнах рр. Киши и
Малой Лабы — там, где 30 лет тому назад еще встречались дикие кав-
казские зубры. Как известно, только один из них, пойманный в 1907 г.,
был вывезен в Германию, где скрещивался с беловежскими зубрицами.
Имя этого зубра «Кавказ № 100 Р. Ц. Р.»1 встречается в родословных
многих современных чистокровных и гибридных зубров.
Асканийские гибридные зубры в количестве 56 голов, так же как и
крымские, размножившиеся до 14 голов, были уничтожены оккупантами
в 1941—1942 гг.
После окончания Великой Отечественной войны в Советском Союзе
было организовано еще 2 новых зубровых питомника — в государствен-
ном заповеднике Беловежская пуща (1946 г.) и в Приокско-террасном
государственном заповеднике (1948 г.). Исходное поголовье для этих
питомников предоставила дружественная Советскому Союзу Польская
Народная республика. За период с 1946 г. по 1956 г. в СССР было заве-
1 Р. Ц. Р. — Reinzuchtregister, племенная книга зубров (см. ниже).
BISON
59з
зено 20 чистокровных зубров (12 самцов и 8 самок) и 2 гибридных (чисто-
породных) самки.
Стадо Центрального зубрового питомника в Приокско-террасном
заповеднике было пополнено также 3 бизонами (1 самец и 2 самки) и 3
зубробизонками (сложными гибридами), доставленными из Рижского, Алма-
Атинского и Ростовского н/Д зоопарков. Особенно успешно размно-
жение идет в Центральном зубровом питомнике, где за 9 лет работы (1948—
1957 гг.) число животных увеличилось в восемь раз - с 4 до 34 особей
(рис. 121). За это время сюда было завезено 18 животных (7 самцов и 11 са-
Рис. 120. Самец зубро-бизон Кавказский заповедник.
мок). В питомнике выращено 56 телят (25 бычков и 31 телочка). Быстрый
рост стада позволил в течение 9 лет существования Центрального зубро-
вого питомника вывезти отсюда в зоопарки и заповедники 35 чистокровных
и гибридных зубров (21 самца и 14 самок) и организовать еще 2 новых
пункта вольного разведения зубров — в Мордовском и Хоперском го-
сударственных заповедниках. В Хоперском заповеднике работа по раз-
ведению зубров была начата в 1955 г., в Мордовском — в 1956 г. В пер-
вом из этих заповедников имеется 16 голов зубров (самцы чистокровные,
самки — сложные гибриды), во втором-—12.
В целом по СССР за послевоенный период (1946—1958 гг.) стадо вы-
росло с 29 до 280 особей, что уже на 150 голов превышает число животных,
имевшихся в 1941 г. до начала Великой Отечественной войны. На 1 ян-
варя 1958 г. в СССР имелось 79 зубров, 19 бизонов, 141 зубробизон и 41
сложный гибрид. Кроме пяти упомянутых заповедников, приплод
получают, правда, менее регулярно, зоопарки Алма-Атинский (сложные
гибриды), Рижский и Аскания-Нова (бизоны). В прочих 15 зоопарках
эти виды представлены преимущественно самцами, в числе которых
в Москве, Ленинграде, Риге, Алма-Ате, Гродно, Одессе, а также и в Аска-
нии-Нова живут чистокровные зубры.
38*
596
ВО VI DAB
Проблему восстановления зубра можно будет считать решенной лишь
в том случае, когда на территории СССР будет создано несколько само-
стоятельных популяций вольноживущих зубров, общей численностью
в несколько тысяч голов. При правильной организации работы эта за-
дача может быть решена в течение ближайших 15 лет. К концу VIII пяти-
летки, учитывая темпы роста зубрового стада за последние 10 лет, мы
можем иметь в СССР не менее 2500 зубров (из них чистокровных 385).
Поэтому основной задачей в деле восстановления зубра на ближайший пе-
риод времени по-прежнему остается всемерное наращивание численности
этого редкого, исчезающего вида и возможно широкое его расселение
как в местах прежнего обитания, так и в других лесных и лесогорных,
относительно малоснежных районах нашей страны, подходящих по своим
физико-географическим и кормовым условиям для существования зубра
(заповедники Окский, Воронежский, Крымский, Тебердинский, Судзу-
хинский). Уже сейчас необходимо приступить к отысканию и обследованию
подходящих мест на южном Алтае, в Карпатах, в южной части Саян,
на Тянь-Шане и в приморских лесах Дальнего Востока. Не менее 300 зуб-
ров должно сохраняться в питомниках, для чего необходимо органи-
зовать еще не менее трех питомников в дополнение к двум существующим.
При расселении зубров сначала необходимо заселять заповедники и
только затем уже охотничьи хозяйства и территории лесхозов. Много-
летний опыт разведения зубров показывает, что эти звери не хуже, чем
лось, приспосабливаются к жизни в культурном ландшафте, с правильным
ведением лесного хозяйства, что позволяет перейти к вольному разве-
дению зубров и в населенных областях европейской части СССР.
Вторая задача, о которой нельзя забывать, заключается в принятии
необходимых мер к сохранению зоологического подвида равнинного
беловежского зубра {Bison bonasus bonasus L.). Среди 95 чистокровных
зубров, имеющихся в СССР, беловежских только 28 (16 самцов и 12 самок).
Остальные 67 (35 самцов и 32 самки), как и все гибридные зубры Кавказ-
ского заповедника, будучи потомками упомянутого выше кавказского
зубра «Кавказа», имеют прилитие крови кавказского горного подвида
{Bison bonasus caucasicus Sat.), в связи с чем их следует рассматривать
как подвидовые (внутривидовые) помеси. Прежде всего обособлен-
ное разведение беловежского подвида нужно обеспечить в самой Бело-
вежской пуще — родине беловежского зубра, где должны остаться только
представители беловежского подвида. Для того, чтобы исключить воз-
можность стихийной гибридизации, всех зубров с прилитием крови кав-
казского зубра нужно в ближайшее время оттуда перевести в другой за-
поведник (например, в Окский или Воронежский). В зубровом питомнике
«Беловежа», организованном еще в 1929 г., а также в лесном массиве
Беловежской пущи в резервате того же названия Польской Народной
Республики с 1951 г. оставлены только представители беловежского
равнинного подвида. Работу этих двух пограничных заповедников (Бело-
русского и Польского) необходимо координировать. Важно отметить,
что в результате продолжающихся спариваний беловежских зубриц
с самцами, имеющими прилитые крови кавказского зубра, число бело-
вежских зубров во всех странах мира за 30 лет не увеличилось. В 1921 г.
их было 45 (20 самцов и 25 самок), в 1950 г. — 46 (20 самцов и 26 самок).
В то же время число особей с прилитием крови кавказского зубра уве-
личилось более чем в 10 раз; в 1921 г. — 8, в 1950 г. — 82.
В 1921 г., когда в Советском Союзе были начаты работы по восстанов-
лению поголовья зубра, во всех странах мира оставалось всего лишь
BISON
5У7
53 зубра.1 Большая часть зверей находилась в зоологических садах, где они
не размножались или размножались плохо. Выращиваемый приплод
не покрывал гибели взрослых животных. В связи с этим к 1927 г. числен-
ность зубров снизилась до 48 особей. Этот год ознаменовался выходом
в свет первого выпуска «Племенной книги зубров» (ред. Groeben, 1932;
Mohr, 1933, 1937), которую нашло необходимым издавать Международное
общество сохранения зубра, основанное передовой научной обществен-
ностью на Парижском международном конгрессе деятелей охраны при-
Рис. 121. Беловежские зубры в Центральном зубровом питомнике. Слева на-
право: Побуг, Пожога, Помета, Морошка (зубренок). Июль 1956 г.
(Фото А. Г. Сергеева).
роды в 1923 г. (Беднарский, 1928). В числе основателей Общества были
русские ученые — профессора Г. А. Кожевников и Н. Д. Российский,
известный польский зоолог Ян Штольцман и др. В 1927 г. доклад о дея-
тельности Международного общества сохранения зубра был заслушан
на Будапештском международном зоологическом конгрессе (Догель,
1927). В результате деятельности Международного общества, в дополнение
к уже имевшимся в то время зубровым питомникам (Шеннбрунн, Плесе,
Вобурн, Визеград, Шарбов, Бойтценбург, Сканзен, Лонгсйон) возник
целый ряд новых: Шпринге (1928 г.), Беловежа (1929 г.), Шорфхейде
(1933—1945 гг.),1 2 Смардзевице (1934 г.), Неандерталь (1936—1945 гг.),
Дарсс (1936—1945 гг.), Неполомице (1938 г.), Авеста (1938 г.), Хелла-
брунн (1938 г.), Златовесь (1938—1946 гг.), Роминтен (1941—1945 гг.),
Горце (1950—1954 гг.).
1 Если не считать некоторого количества вскоре уничтоженных вольных зубров
на Кавказе.
2 В скобках после тире указан год ликвидации питомника.
598
BOVIDAE
В зоологических садах условия содержания и кормления зубров
были улучшены, подобраны пары или племенные группы. Поголовье
зубров начало расти более быстро и к 1944 г. достигло цифры в 146 голов.
Последние годы Второй мировой войны, когда европейские страны стали
местом ожесточенных боев, в зону военных действий попали многие из
перечисленных зубровых питомников (Glower, 1947; Zabinski, 19496,
1950). В результате войны поголовье зубров упало почти до уровня 1937 г.,
когда во всех странах мира было 84 зубра. За пять послевоенных лет
(1946—1951 гг.) численность зубрового стада снова стала расти и увели-
чилась с 90 до 143 голов. Этому немало содействовало возобновление
деятельности Польского отдела Международного общества сохранения
зубра (г. Варшава), взявшего на себя инициативу продолжения деятель-
ности общества и по изданию «Племенной книги зубров» (ред. Ян Жабин-
ский, — Jan Zabinski, 1947, 19496, 1951). В последнем (третьем после-
военном) выпуске опубликованы списки всех чистокровных зубров, имев-
шихся на 1 января 1951 г. Анализ опубликованных сведений, проведен-
ный М. А. Заблоцким (1952 : 230—231), показывает, что различные
страны обладали в то время следующим количеством зубров: Польша —
65, Швеция — 23, СССР — 22, Германская Федеративная Республика —
18, Голландия — 8, Дания — 3, Англия — 2, Чехословакия — 1.
В 1953—1954 гг. ряд заграничных зоосадов и зубровых питомников
постигло большое несчастье. В результате неожиданно вспыхнувшей эпи-
зоотии ящура пало около 40 чистокровных зубров, в том числе в Поль-
ских резерватах погибло 32 головы (10 самцов и 22 самки). Причины и
характер заболевания, повлекшего за собою массовую гибель зубров
и приведшего к опустошению и ликвидации 2 зубровых резерватов (Горце
и Плесе) описаны Ганасинским (Ganasinski, 1955). Упомянутая эпизоотия
свела почти на нет результаты работы 2 последних лет. Если за период
с 1951 по 1953 г. поголовье зубров во всех странах мира увеличилось
более чем на 40 голов (с 143 до 184), то в течение следующих 2 лет (1953—
1955 гг.), в результате эпизоотии, абсолютный прирост поголовья зуб-
ров составил лишь 30 голов.
В 1955 г. мировое поголовье зубров определялось цифрой в 214 голов,
из которых в Польше было 71. На второе место вышел Советский Союз,
имевший тогда уже 48 зубров.1 Количество зубров в других странах
Европы, по сведениям Международного общества сохранения зубра
(Zabinski J., 1955), видно из следующего перечня: на 1 января 1955 г.
в Швеции было 29 зубров, в Германии — 38, в Голландии — 11, в Да-
нии и в Бельгии по 4, в Чехословакии — 3, в Австрии — 2 и в Венгрии,
Англии, Швейцарии и Италии — по 1 зубру.
Третья экологическая конференция, созванная в 1954 г., в связи с край-
ней малочисленностью зубров в настоящее время, отметила необходи-
мость установить обмен информацией о разведении зубров, их происхо-
ждении, размещении и т. п. между всеми странами, где имеются зубры
и прежде всего между Советским Союзом и странами народной демокра-
тии (Заблоцкий, 1954). Параллельно с изданием «Государственной пле-
менной книги зубров СССР» (Заблоцкий, 1956) важно продолжать также
и выпуск через каждые 2 года «Племенной книги зубров» Международ-
ного общества сохранения зубра, отражающих ход разведения зубров
во всех странах мира.
1 На 1 января 1959 г. в СССР имеется 95 чистокровных зубров, т. е. за 4 года
поголовье удвоилось.
BOS
599
Род BOS LINNAEUS—БЫКИ, ИЛИ ТУРЫ
Во janus L., 1827, Nov. Acta Physico-Medica Acad. Caes. Leop. Carol., XIII,
2 : 411—476 (морф., сист.); Meyer H., 1835, Nov. Acta Physico-Medica, XVII,
1 : 152—155 (сист.); Owen R., 1846, A history of. Brit, fossil mammals and birds,
London : 508 (сист.); Falconer H., 1859, Catal. fossil Vertebr. As. Soc., Calcutta :
232 (сист.); Lydekker R., 1788, Palaeontol. indica, ser. X, vol. I : 112 (ист., сист.);
Thomas Ph., 1881, Bull. Soc. Zool. de France, VI : 125—135 (сист.); P о m e 1 A.,
1894, Carte Geol. de 1’Algerie, Paleontol. Monogr., IV : 1—106 (ист., сист.); M a 1 s-
b u rg K.., 1911, Bull. Intern. Acad. Sci. Crackvie, ser. В, № 5B : 340—348 (сист.);
A d a m e t z L., 1925, Bull. Int. de Г Acad. Polon. Sci. et Lettr., cl. mathem. et natur.,
ser. B, 5/6 : 591—622 (ист., сист.); Leithner О., 1927, Ber. Internal. Gezellsch.
Erhalt. d. Wiesents, Bd. 2, H. 1/2 : 1—140 (морф., сист., ист.); Вассоевич Н.,
1929, Азербайдж. нефтян. хоз., 8/9 : 16—38 (цит. по Бурчак-Абрамовичу, 1952) (ист.,
распр.); Громова В. И., 1931, Ежегодн. Зоол. музея Акад, наук СССР, XXXII :
293—366 (морф., сист., ист.); Авакян Л. А., 1946, Изв. Акад, наук Армянск. ССР,
естеств. науки, 9 : 59—65 (ист.); La Baume W., 1947, Eclogae geologicae helvetiae,
vol. 40, № 2 : 308—316 (ист., сист.); Громов В. И., 1948, Тр. Инет. геол, наук,
64 : 432—433 (ист.); Вурчак-Абрамович Н. О., 1950, Докл. Акад, наук
СССР, LXX, 5 : 875—878 (эвол.); Даль С. К., 1950, Докл. Акад, наук Армянск.
ССР, XI, 4:133—137 (ист., распр.); Вурчак-Абрамович Н. И., 1952,
Тр. Ест.-ист. муз. им. Зардаби, Баку, V : 181—202 (сист.); Верещагин Н. К.,
1953, Бюлл. Комис, по изуч. четвертичн. периода, 18 : 39—50 (ист.); Соколов И. И.,
1953, Тр. Зоол. инет. Акад, наук СССР, XIV : 71—74 (морф. эвол.).
Характеристика рода. Животные крупной величины. Холка не вы-
сокая и не приподнята в виде горба. Остистые отростки первых грудных
позвонков удлинены, по сравнению с другими, незначительно. Высота
в крестце лишь немного меньше высоты в холке, а иногда равна послед-
лей. На нижней стороне шеи и головы подвес из длинных волос отсутствует.
Череп сравнительно узкий и вытянутый в длину. Глазницы умеренно
выступают в стороны. Наибольшая ширина черепа составляет меньше
60% основной длины черепа. Лобная поверхность, если не считать за-
глазничного сужения, имеет форму вытянутого прямоугольника, ширина
ее перед основаниями роговых стержней приблизительно равна ширине
лба в области глазниц (рис. 123). Заглазничная ширина лба (в самом
узком месте между рогами и глазницами) меньше наибольшей ширины
черепа в скуловых дугах. Задний край лба в виде хорошо развитого ва-
лика сильно выдается назад и резко отграничен от теменной поверхности
черепа. Последняя совершенно не принимает участия в образовании
крыши мозговой коробки. Дорзальная поверхность зарогового черепа
оттеснена назад, поставлена под острым углом к плоскости лба и под
тупым к плоскости затылка, нависая над последним. При взгляде на
череп сверху задние выходы височных впадин не видны. Расстояние
между ними больше расстояния между наружными краями затылочных
мыщелков (отличие от рода Bibos Hodgson). Рога отходят от задних углов
лба по краям межрогового валика (у живого, спокойно стоящего
зверя на самой макушке головы). Расстояние между глазницей и осно-
ванием рогового отростка значительно больше диаметра глазницы. По-
перечное сечение роговых отростков приближается к круглому, но всегда
заметно уплощено у оснований в дорзовентральном направлении. Про-
дольные ребра (кили) на поверхности роговых стержней, в отличие от
ряда других родов подсемейства, отсутствуют.
В скелете 13 грудных позвонков и 13 пар ребер.
История и географическое распространение. Корни рода Воз ведут
к роду Urmiabos Bartsch. (Вурчак-Абрамович, 1950), известному из нижне-
плиоценовой фауны Мараги в Иранском Азербайджане и обладавшему
600 BOVIDAE
совокупностью черт, которые позволяют считать его предковой формой не
только для быков, нои для яков (Poephagus Gray). Остатки, заведомо отно-
сящиеся к роду Bos, появляются только в верхнем плиоцене. В это время
в Индии жил длиннорогий В. acutifrons Lyd. (Lydekker, 1878), имевший
еще слабо развитый межроговый валик. Остатки того же геологического
возраста, не оставляющие сомнения в принадлежности их к роду быков,
известны из северной Африки (Thomas, 1881).
Время первоначального появления туров на территории Европы до-
стоверно не известно, но, по-видимому, они проникли сюда через Малую
и Среднюю Азию также не позднее верхнего плиоцена или нижнего плей-
стоцена. Н. Вассоевич (1929) среди других находок из нижнечетвертич-
ных отложений Таманского полуострова упоминает роговой отросток
Bos sp. Однако описания этого рогового отростка не было дано, место-
нахождение его в настоящее время не известно и нет уверенности, что
он принадлежал быку, а не найденному в таманской фауне первобытному
зубру (Bison schoetensacki Freudenb.).
Остатки настоящих быков известны из доледникового квартера ниж-
ней Волги и из пограничных с плиоценом или верхнеплиоценовых отло-
жений р. Псекупса на Северном Кавказе (Громов, 1948).
Ареал рода Bos был весьма обширным. В свое время туры населяли,
кроме северной Африки, большую часть Евразии, включая Британские
острова и южную часть Швеции. На север область распространения про-
стиралась до 57—60° с. ш. В противоположность первобытному зубру,
первобытные быки никогда не проникали на территорию Нового Света.
По-видимому, не было их также в Ирландии.
Классификация. Систематика рода Bos запутана. Описано значитель-
ное число форм, принимаемых то за подвиды, то за самостоятельные
виды. В. И. Громова (1931) произвела обстоятельную ревизию рода и
все многообразие четвертичных форм туров свела к двум видам: круп-
ному ледниковому Bos trochoceros Meyer (Meyer, 1835) и несколько измель-
чавшему его потомку, позднеплейстоценовому и голоценовому В. primi-
genius Boj. (Bojanus, 1827). Последний существовал на территории
средней и восточной Европы, а возможно также Средней и Малой
Азии уже в исторические времена и окончательно вымер в начале
XVII в. (см. ниже). Н. И. Бурчак-Абрамович (1952) описал из верхнеплей-
стоценовой фауны Бинагадов на Апшеронском полуострове (вюрм) новый
вид быка В. mastan-zadei Burtsch., по краниологическим признакам близ-
кий к плейстоценовому же индийскому виду В. namadicus Falc. (Falco-
ner, 1859). Однако видовая самостоятельность описанной Н. И. Бур-
чаком формы вызывает сомнение, поскольку В. И. Громова принимает
В. namadicus лишь за подвид В. trochoceros.
Находки остатков дилювиальных туров (Bos trochoceros) на террито-
рии СССР единичны (Громова, 1931:330; Авакян, 1946).
Спорным и неясным остается вопрос о карликовых формах туров:
В. longifrons Ow. (Owen, 1846), В. minutus Malsb. (Malsburg, 1911), В. bra-
chyceros europaeus Adam. (Adametz, 1925). Размеры черепа у этих форм
в ряде случаев не превосходят таковые черепа мелких рас домашнего
скота. Однако принадлежность к последнему исключается в одних слу-
чаях геологическим возрастом находок, в других — морфологическими
особенностями карликовых туров. Некоторыми исследователями (Leith-
пег, 1927; La Байте, 1947) мелкие черепа принимаются за черепа самок
В. primigenius Boj. Однако следует иметь в виду, что некоторые из нахо-
док карликовых туров имеют плейстоценовый возраст. Следовательно,
BOS
601
даже при принятии сильно выраженного у туров полового диморфизма
в размерах, трудно признать эти находки за черепа самок при гигант-
ских размерах черепа дилювиальных самцов.
В пределах Советского Союза остатки карликовых туров найдены
в Армении в донных отложениях оз. Севан (Даль, 1950) и в бассейне р. Урала
(Верещагин, 1953). Вопрос о систематическом положении карликовых
туров важен в связи с проблемой происхождения некоторых групп домаш-
него крупного рогатого скота.
В настоящее время род Bos представлен только домашней формой,
крупным рогатым скотом В. taurus L., но уже в исторические времена на
территории СССР водился в диком состоянии первобытный бык, или тур
В. primigenius Bojanus.
Bos primigenius Bojanus — Первобытный бык, или тур
(рис. 122—124)
Herberstain S., 1556, Rerum moscoviticarum. . . —см. Герберштейн.
1908; V i g е в е г В., 1573, Le chroniques et annales de Poloigne, Paris : XXIII (опис.,
биол.); Gesner C., 1606, Historiae animalium, I, Francoforti : 141—145 (опис.,
биол., изобр.); Pallas P., 1777, Act. Acad. Sci. Petropol., pars poster. : 223 (ист.);
Bojanus L., 1827, Nov. Act. Physoco-Medica Acad. Caes. Leop. Carol., t. 13, pars
2 : 411—476 (морф., сист., эвол., изобр.); Smith Н., 1827, Griffith’s Cuvier Animal
kingdom, V : 411—417 (общ., изобр.); Б о п л а и, 1832, Описание Украины, СПб. :
94 (ист.); Cuvier G., 1835, Recherclees sur les ossemets fossiles, 4 edit., VI, Paris :
228—238 (ист.); Беляев И., 1865, Зап. Русск. общ. акклим., I : 67—80 (ист.,
изобр.); Brandt J., 1867, Verhandl. Russ. Keiserl. Mineral. Gesellscb., St.-Petersb.,
2 ser. Bd. II : 185—227 (распр.); Wrzesniowski A., 1878, Ztschr. f. -wissensch.
Zool., Bd. XXX, Supplem., Leipzig : 492—555 (ист.); Иностранцев A., 1882,
Доисторический человек каменвого века побережья Ладожского озера, СПб. : 42'—43
(морф., ист.); Nehrins А., 1888, Sitzber. Gesellscb. Naturf. Freunde, Berlin : 54
(морф.); N e h r i n g A., 1889, Sitzber. Gezellsch. Naturf. Freunde, Berlin : 5—7 (сист.,
морф.); В иж e н e p Б., 1890, Мемуары, относ, к южн. России, I, Киев : 59—88
(опис., биол.); Сатунин К. А., 1895, Природа и охота, XI : 141—149 (ист.); Слов-
цов И., 1895, Тобольск, губ. ведом., 7 : 3 (ист.); N е h г i п g А., 1896, Landvirtsch.
Jahrb. : 915—933 (изобр.); N е h ri ng А., 1898, Globus, LXXIV, 3 : 44—47 (ист.);
D u e r s t J., 1899, Die Binder von Babylonien, Assyrien and Agypten, Berlin : 8—11
(ист-.); Долгих И., 1905, Мвимый единорог, риму и реэм Востока, Рига : 1—85
(общ., изобр.); N о а с k Th., 1905, Zool. Anzeig., XXVIII, 23 : 749—760 (изобр.);
Mertens А., 1906, Abhandl. u. Bericht. Mus. f. Naturk., Magdeburg, Bd. I, H. II :
45—119 (общ., изобр.); Герберштейн С., 1908, Записки о московских делах,
СПб. : 174-—175 (общ. изобр.); La Baume W., 1909, Ztschr. Naturk. Gesellscb.,
Danzig., N. F., 12 : 62—76 (морф., сист.); Hilzheimer M., 1910, Jahrbuch f.
wissensch. u. prakt. Tierzucht, V : 74 (морф.); Lydekker R., 1912, The ox and
its kindred, London : 37—67 (общ., изобр.); Leithner О., 1927, Ber. Internat.
Gesellscb. f. Erbalf. d. Wisents, Bd. 2, H. 1 /2 : 1—140 (морф., сист.); Г ромова В. И.,
1930, Природа, 7/8 : 755—770 (ист.); S z а 1 а у А., 1930, Zool. Garten., N. F., Ill,
9—10 : 255—263 (окраска); Громова В. И., 1931, Ежег. Зоол. музея Акад, ваук
СССР, XXXII : 293—366 (морф., сист., ист.); Heck L., 1934, Ber. Internat. Gezellsch.
f. Erhalt. d. Wisents, 3 : 287 (восстанови.); Me ховский M., 1936, Трактат о двух
Сарматиях, М.—Л., изд. Акад, наук СССР : 111 (ист.); J ennison G., 1937, Ani-
mals for show and pleasure in ancient Rome, Manchester : 167 (ист.); Зубарева В. И.,
1940, Природа, 8 : 82—86 (ист.); Harper F., 1945, Extinct a. vanishing mammals
of the Old World, N. Y. : 511—513 (ист.); La Baume W., 1947, Ber. scbvciz. Pa-
laontol. Gesellsch., 40 : 299 (сист.); Колесник H. H., 1949, Эволюция крупного
рогатого скота, Сталинабад, изд. Акад, наук СССР : 1—329 (одом.); Lehman U.,
1949, Neue Jahresberg. f. Mineral, e. t. c. Abhandl., Abt. B. 2 : 163—266 (морф., сист.,
ист.); Б у рч а к -А б p амо в и ч Н. О., 1951, Изв. Акад, наук Армянск. ССР,
IV, 2 : 135—145 (морф.); Пидопличко И. Г., 1951, О ледниковом периоде, II,
Киев, изд. Акад, наук УССР : 53—55 (ист.); Heck Н., 1951, Oiyx, I, 3 : 117—122
(восстанови.); Бу рчак-Абрамович Н. И., 1952, Тр. Естеств.-ист. музея им.
Зардаби, V : 181—202 (морф., сист.); Lukaszewicz К., 1952, Ochrona przyrody,
20 : 1—33 (общ., изобр.); Heck L., 1952, Jahrb. d. Nassauischen Ver. f. Naturk.,
90 : 107—124 (восстанови.); Цалкин В. И., 1952, Бюлл. Моск. общ. испыт. при-
роды, Отд. биол., LVII, 2 : 81—83 (ист.); Верещагин Н. К., 1953, Бюлл. Комисе.
602
BOVIDAE
по изуч. четвертичн. периода, 18 : 39—50 (ист.); Lengerken Н., 1953, Der Ur,
Leipzig : 1—80 (общ., изобр.); Н е г re W., 1953, K.osmos, Bd. 49, № 11 : 504—507
(ист., морф., восстановл., изобр.); Picard Р., 1954, Naturalia, 4:41—43 (ист.);
Le n g е г к е n Н., 1955, Ur, Hausrind und Mensch, Berlin:!—191 (ист., одомашн.,
изобр.).
Имеется ряд свидетельств современников о существовании с ранних
исторических времен до конца средних веков на территории Европы,
Передней и Средней Азии крупного дикого быка, весьма сходного по общему
облику с домашним крупным рогатым скотом и известного под названиями;
ур, тур, ауэрокс, риму или реэму. Имеются даже описания и изображения
этого зверя, сделанные лицами, видевшими его, например известным
средневековым историком и дипломатом С. Герберштейном (Herberstain,
1556).
Однако одновременное существование на той же территории другого
близкого вида, европейского зубра {Bison bonasus L.), и, в особенности,
большая редкость тура в последние столетия его существования послу-
жили причиной того, что представления и сами названия этих двух ви-
дов стали путаться даже современниками. Не случайно С. Герберштейн
был вынужден под изображением тура сделать подпись: «Я—ур, по-
польски тур, по-немецки аурокс, невежды дали мне имя бизона».
Позднее эта же причина послужила основанием для ряда исследова-
телей, начиная с Бюффона, поставить под сомнение вообще существова-
ние в историческую эпоху дикого быка-тура. Паллас (Pallas, 1777), на-
пример, считал, что зубр и тур — два названия одного и того же живот-
ного, что Герберштейн был просто введен в заблуждение и что изображен-
ный им безгорбый бык не что иное, как одичавший буйвол. Такой же точки
зрения придерживались польский палеонтолог Пуш (Mertens, 1906) и
даже Боянус (Bojanus, 1827), впервые описавший Bos primigenius в каче-
стве самостоятельного вида, якобы вымершего еще в доисторический пе-
риод.
В настоящее время накопилось достаточное количество неопровержи-
мых доказательств в пользу бесспорного существования дикого быка-
тура уже в историческую эпоху. Из числа этих доказательств, кроме
самого убедительного свидетельства С. Герберштейна, приведем лишь
немногие.1 Как показал И. Долгих (1905:50), древнеславянский язык
различал названия зубра и тура. Плиний ясно отмечал тура и зубра,
когда писал: «Скифия очень бедна животными, и в Германии их немного,
но замечательны два вида быков: именно, украшенный гривою бизон и
отличающийся силою и быстротою ур». Живший, примерно, в это же время
монах Эксгарм, перечисляя пищу, которой питались в его монастыре,
называет отдельно зубра (Wisent) и тура (Ur) (Сатунин, 1895). На мировой
карте Эбшторфа, изданной в 1284 г., среди других изображений живот-
ных, характерных для разных местностей, показаны как Bonacus (зубр),
так и Urns (тур).
Лаврентий Суров (цит. по Сатунину, 1895) в 1564 г. писал: «Заблужда-
ются те, которые уров (Uros) зовут бизонами (Bisontes), ибо бизоны мно-
гим отличаются от уров». Наконец следует помнить, что еще недавно
(а кое-где и сейчас) в замках, церквях и гостиницах хранились, то в ка-
честве сосудов для вина, то просто в качестве украшений, рога, которые,
судя по размерам, могли принадлежать только дикому быку-туру.
1 Из ряда сводок по истории Bos primigenius Boj. лучшими являются -работы
А. Вр.кэсниовского (Wrzesniowski, 1878), И. Долгих (1905), А. М. Мэртенса (Mertens,
1906), К. Лукашевича (Lukaszewicz, 1952) и Г. Ленгеркена (Lengerken, 1953).
BOS
603
Описание. Дать описание Bos primigenius Boj. в той же полноте и
последовательности, как для современных животных, невозможно. По
существу более или менее удовлетворительные и внушающие доверие
сведения о его наружности даны лишь лично видевшим, по крайней
мере труп, тура С. Герберштейном (Herberstain, 1556) и известным зо-
ологом конца XVI — начала XVII в. К. Геснером (Gesner, 1606). С. Гер-
берштейном же дано и первое, сколько-нибудь удовлетворительное изо-
бражение тура. Существуют споры — видел ли Герберштейн живых ту-
ров и сделаны ли рисунки тура и зубра для его «Московии» с натуры или
с чучел, которые стояли в его доме в Вене и впоследствии исчезли бес-
следно. Более вероятно последнее предположение. Бесспорно, однако, то,
что польским королем Сигизмундом-Августом был подарен Герберштейну,
-бывшему в то время послом при его дворе, выпотрошенный труп тура,
покрытый кожей, кроме лба, с которого она была срезана раньше. Ри-
сунки тура и зубра, сделанные, по-видимому, с неумело набитых чучел,
очень несовершенны. Животные выглядят неуклюжими и в весьма при-
нужденных позах. Вероятно, искажены некоторые пропорции телосло-
жения. Но, во-первых, изображение тура здесь гораздо лучше, чем на
упомянутой карте Эбшторфа. Во-вторых, ценность его заключается в том,
что оно изготовлено по заказу лица, которое видело зверя в натуре и,
следовательно, могло предупредить грубые искажения. Уже на этих
несовершенных изображениях переданы наиболее характерные отличи-
тельные черты тура и зубра; только рога у обоих видов показаны одина-
ковыми, причем в большей степени страдает против истины изображение
зубра. По-видимому, лучшим из всех известных является так называемое
«аугсбургское» изображение тура (рис. 122), впервые воспроизведенное
Г. Смитом (Smith, 1827). По словам этого автора, оно представляет копию
обнаруженной им у одного торговца в Аугсбурге картины посредственного
качества, сделанной маслом на дереве, и судя по технике выполнения,
относящейся к первой четверти XVI в., т. е. лет за сто до окончательного
вымирания тура. Изображенный на картине бык не имел подвеса на шее,
но был покрыт довольно грубыми волосами, обладал большой головой,
толстой шеей и небольшим подгрудком. Окраска его «черная, как сажа»
(sooty black), кроме белого подбородка. Рога имели характерный для
румынских и подольских домашних быков изгиб и светлую окраску,
кроме темных верхушек. В углу картины были видны остатки геральди-
ческих знаков и, кроме того, написанное золотыми готическими буквами
почти стертое слово «Thur». Дальнейшая судьба оригинала не известна —
он исчез бесследно, а сделанная с него и опубликованная Г. Смитом копия
в дальнейшем многократно воспроизводилась.
Все другие описания и изображения первобытного быка отрывочны,
несовершенны, а главное во многом противоречивы.
Критическое сопоставление имеющихся описаний современников и
найденных в земле почти полных скелетов (число последних в настоящее
время больше десяти) позволяет сделать заключение, что тур представлял
собой крупного быка, значительно превосходившего размеры зубра и
домашнего скота, даже крупных пород. Высота взрослых самцов в холке
составляла 175—200 см, а по некоторым данным даже больше (Негге,
1953).
Величина черепа туров сильно варьирует. Это зависит, помимо сравни-
тельно высокой личной изменчивости, от недостаточно выясненных поло-
вых отличий и геологического возраста отдельных экземпляров. Обще-
известен факт измельчания ряда крупных млекопитающих, в том числе
604
BOVIDAE
и быков, во вторую половину четвертичного периода. А. Неринг (Neh-
ring, 1889) приводит максимальную длину самого крупного из виденных
им черепов в Лондонском музее — 912 мм; вероятно, то был череп ледни-
кового тура, В. trochoceros Meyer.
По О. Лайтнеру (Leithner, 1927), передняя длина черепа (от prostion
до середины межрогового гребня) самцов аллювиальных туров, относя-
щихся к виду В. primigenius Boj., колеблется от 633 до 738 мм. Основиая
длина черепа, по данным того же автора и В. И. Громовой (1931), состав-
ляет у самцов 505—593 мм.
Все авторы, описывавшие тура, утверждают, что общим обликом и
телосложением он был сходен с домашним крупным рогатым скотом.
Если считать, что высота относилась к длине тела как 3:5, то последняя,
оудя по скелету, следовательно была в пределах 290—330 см. Длина
туловища от плечелопаточного сустава до заднего выступа седалищного
бугра (косая длина) составляла у самцов 210—220 см. Высота в крестце,
по крайней мере у самцов, была в большинстве случаев несколько ниже
высоты в холке, но в отдельных случаях, как например, на аугсбургском
изображении, почти равна ей и линия верхнего профиля спины и поясницы
была почти прямой, и горизонтальной. Столь характерный для зубра
«горб» (высокая холка) у тура совершенно не был развит.
Голова, в противоположность мнению Г. Смита (Н. Smith, 1827), не
производила впечатления тяжелой. Уши, по-видимому, были сравнительно
короткие. Глаза, наоборот, крупные, возможно слегка навыкате. Шея,
судя по дошедшим изображениям и сохранившимся остаткам скелета,
была короткой и широкой, а верхняя линия ее профиля, в противополож-
ность зубру, более или менее горизонтальной. Подгрудок в виде складки
кожи на нижней стороне шеи, судя по аугсбургской картине и изображе-
нию у Герберштейна, имелся, но был очень небольшим. Голова в спокой-
ном состоянии держалась сравнительно низко.1
Конечности были толстые, крепкие, но, по сравнению с зубром, более
высокие. Общее телосложение было, по-видимому, несколько более лег-
ким, чем у зубра. Хвост сравнительно длинный с кистью длинных во-
лос на конце, больше, чем у зубра, спускавшихся ниже скакательных
(голеностопных) суставов.
Рога были и у самцов, и у самок. И абсолютные, и относительные их
размеры значительно больше, чем у позднечетвертичных и современных
зубров. Длина их, по изгибу большой кривизны, в исключительных слу-
чаях доходила до 198 см (Lydekker, 1912). Роговой чехол молодого самца,
найденный в торфянике у Руммельсбурга, имел длину по большой кри-
визне 76 см, по прямой от основания до вершины 45 см и обхват в основа-
нии около 38 см (Mertens, 1906). У крупных экземпляров обхват, судя
по толщине роговых стержней, в отдельных случаях доходил до 60—65 см.
О величине рогов можно судить по тому, что некоторые из роговых чех-
лов, употреблявшихся в качестве кубков, вмещали жидкости до 4 л. Хотя,
как показали исследования В. И. Громовой (1931), форма рогов тура
весьма изменчива, изгиб их весьма характерен: отходя от верхушки
головы, рога направлялись в стороны и одновременно несколько назад,
затем круто загибались вверх и вперед, а верхушками несколько внутрь
и иногда снова вверх, как на аугсбургской картине. Поверхность роговых
чехлов у оснований шероховатая, перешнурованная кольцевидными взду-
1 В этом отвошении, по-видимому, более близко к истине изображение Гербер-
штейна, по сравнению с аугсбургским.
BOS
605
тиями и круговыми бороздками между ними; выше она, по аналогии с длин-
норогими домашними быками, гладкая, блестящая. Цвет рогов светлый
(желтоватый или светло-бурый), кроме темных, вероятно, черно-бурых
концов (см. аугсбургское изображение; Н. Smith, 1827).
У самок рога были и абсолютно и относительно не только короче,
но и тоньше. Кроме того, крутизна изгиба в дистальной половине внутрь
у самок меньше (Leithner, 1927).
В противоположность зубру, большая часть тела тура была покрыта,
хотя и довольно грубым, но сравнительно коротким и неизвитым воло-
сом. Кроме кисти на конце хвоста, только на лбу до уровня передних
Рис. 122. Первобытный бык — тур {Bos primigenius Boj.). (Аугсбургское изо-
бражение, — по Н. Smith, 1827).
краев глазниц волосы были длинные, курчавые, спутанные наподобие
войлока, и придавали зверю вместе с огромными рогами якобы свире-
пый вид. В летнее время после линьки короткий блестящий волос на
туловище плотно прилегал к телу. Зимой же, подобно породам домашнего
скота, пользующимся круглогодичным пастбищным содержанием, он был
более длинным и отчасти лохматым.
Сведения об окраске первобытного быка противоречивы. В описаниях
большинства современников (Herberstain, 1556; Vigener, 1573; Gesner,
1606), а также на аугсбургской картине она называется черной. Харак-
терным для польского тура считается наличие неширокой светлой по-
лосы вдоль хребта, по-видимому, состоявшей из смеси темных и белых
волос. С. Герберштейн (1556) писал о турах, что «это настоящие лесные
быки, ничем не отличающиеся от домашних быков, за исключением того,
что они совершенно черные и имеют вдоль спины белую полосу, наподо-
бие линии». Однако, как справедливо полагает А. Сцалай (Szalay, 1930),
в данном случае слово «черный» следует понимать не буквально, а в смысле
очень темный. Взрослые самцы, по-видиМому, на самом деле имели черно-
бурую окраску. Конрад Геснер (Gesner, 1606) подтверждает черную окраску
туров, но добавляет, однако, что теленок-самец имеет черно-коричневую
масть, к возрасту одного года становится совсем черным, за исключением
«черноватой» (вероятно, более светлой) полосы на хребте, шириною около
606
BOVIDAE
двух пальцев. Самки, по его данным, сохраняют эту черно-коричневую
окраску в течение всей жизни и черными бывают редко. Следует иметь
в виду, что К. Геснер, хотя и называет тура сходным с черными домаш-
ними быками, но в немецком издании своего «Thierbuch» (1583: 128) изо-
бражает его, по существу, бурым, с желтоватым низом шеи, груди и жи-
вота. Копыта у европейского тура были светлее, чем у домашнего быка.
Имеются в литературе указания и на иную расцветку первобытного
быка. Не следует из-за несовершенства техники живописи того времени
придавать большого значения тому факту, что на упомянутой выше карте
Эбшторфа конца XIII века «Urns» имеет окраску красно-желтого цвета.
Около Шонебекка в Германии был найден фрагмент черепа с остатками
волос у оснований роговых отростков; спереди волосы были белые, сзади
красно-бурые. Во-первых, цвет волос мог измениться за время пребыва-
ния черепа в земле. Во-вторых, сильно оспаривалась принадлежность
этого фрагмента дикому быку (Mertens, 1906).
Указание Мукантеса, что подаренный в 1596 г. польским королем
папскому нунцию кардиналу Гаэтано тур был серым, Лайдекер (Lydek-
ker, 1912), объясняет трудно доказуемым предположением, что этот
экземпляр не был исконно диким, а имел примесь домашнего скота, по-
тому что происходил не из Якторовского леса (последнего убежища тура),
а из королевского резервата вблизи Варшавы.
Весьма вероятно, имелась и достаточно ярко выраженная географиче-
ская изменчивость в окраске туров. Есть основания предполагать, что туры
южной и юго-восточной Европы были более светлыми. На одной римской
мозаике изображен дикий бык темной шиферно-серой окраски. Во дворце
легендарного короля Миноса на о. Крите обнаружено изображение,
по-видимому, дикого быка светло-бурого с оттенком сепии цвета и почти
белыми рогами (Lydekker, 1912). На гробницах египетских фараонов
имеются цветные картины с изображениями красно-бурых быков, частично
пятнистых, со светлым животом и внутренними сторонами ног (Долгих,
1905; Szalay, 1930). На некоторых из этих изображений рога и копыта
показаны черными, что А. Сцапай (Szalay, 1930) склонен рассматри-
вать, наравне с красно-бурой общей окраской, как расовые отличия афри-
канских от европейских туров.
Остеологические отличия тура и зубра в посткраниальном скелете
изучены слабо и, кроме наличия у первого только 13 пар ребер (к тому же
более широких и плоских), 6 поясничных позвонков вместо 5 и более
коротких остистых отростков грудных позвонков, вообще незначительны.
Зато достаточно беглого взгляда, чтобы отличить череп тура (рис. 123) от
черепа зубра. Основные краниологические отличия туров и зубров были
приведены раньше. Здесь уместно привести лишь некоторые дополнитель-
ные особенности.
Характерен сравнительно узкий, вытянутый лоб; длина заглазничной
его части составляет, в противоположность зубру, не меньше 90%, а иногда
превосходит его ширину. Лобная поверхность более или менее ровная
(плоская), но в той или иной мере волнистая, иногда с гребешком вдоль
сагиттального лобного шва. Глазницы, по сравнению с зубром, меньше
выступают в стороны. Профиль линии межрогового гребня прямой, иногда
в средней части несколько выпуклый или, наоборот, вогнутый. Роговые
отростки, отходя от задних углов лба, направляются в стороны и слегка
назад, затем загибаются вверх,1 вперед и вершинами внутрь. Поверхность
1 При горизонтальном положении черепа.
Рис. 123. Череп Bos primigenius Boj,
608
BOVIDAE
роговых стержней обычно несет продольные бороздки, по направлению
которых хорошо заметно легкое гомонимное их скручивание вокруг оси
рога. Межчелюстные кости часто, хотя и не всегда, соприкасаются с носо-
выми костями.
Мнения относительно полового диморфизма у тура расходятся. А. Мер-
тенс (Mertens, 1906), например, считает, что по крайней мере в отношении
величины рогов половой диморфизм у В. primigenius меньше выражен,
чем у большей части пород домашнего скота. Если же принимать черепа
карликовых туров за таковые самок В. primigenius, то резко выраженный
половой диморфизм должен, наоборот, считаться характернейшей особен-
ностью этого вида. В. Херре (Негге, 1953), например, полагает, что самцы
тура превосходили на одну треть или даже на половину размеры современ-
ных домашних быков, тогда как самки по величине приближались к совре-
менному домашнему скоту.
История и географическое распространение. Наиболее правдоподоб-
ным является предположение В. И. Громовой (1931) о том, что вид В. pri-
migenius Boj. произошел в результате постепенного мельчания с конца
ледникового периода плейстоценового В. trochoceros Meyer. И морфологи-
ческую грань, и границу во времени между этими двумя видами провести
очень трудно. При этом мельчание четвертичных туров происходило неза-
висимо одновременно и в Европе, и в Азии, где потомком верхнеплиоцено-
вых и нижнечетвертичных туров был В. namadicus Falc., остатки ко-
торого в Индии найдены вместе с орудиями палеолитического чело-
века.
Считается, что в западной Европе, в частности в Германии, туры появи-
лись лишь перед или в начале рисского времени, а как вид В. primigenius
жил с последней межледниковой эпохи или даже начиная с низов голоцена
(Громова, 1931; La Вашпе, 1947; Lehman, 1949).
Несомненно, палеолитический человек сталкивался с туром, охотился
на него. Наскальные изображения быков типа тура встречаются в ряде
пещер верхнего палеолита Франции, Испании, северной Африки, Сицилии
и некоторых других мест (Lengerken, 1953). Известны находки в Дании
и Англии черепов с кремневыми наконечниками от стрел и копий, вонзив-
шихся в лоб. В Англии найден даже полный скелет тура с пробитым лбом,
а в Дании скелет со следами повреждения каменным орудием ребер (Lydek-
ker, 1912).
Следует отметить, что тур, по крайней мере в Европе, по-видимому,
никогда не был особенно многочислен. Об этом свидетельствует не только
редкость его находок в ископаемом состоянии, но и малочисленность пе-
щерных его изображений по сравнению с зубром.
В Англии остатки диких быков (некоторые из них достигали огромных
размеров) обнаруживаются, начиная с эпохи мустье (средний палеолит).
Когда тур вымер на Британских островах — точно не известно, во вся-
ком случае — еще в доисторическое время. Вслед за отступанием ледника
он в .постплейстоцене заселил Данию и юг Швеции. В свайных построй-
ках неолита Швеции кости дикого тура найдены даже в большем числе,
чем кости зубра (Mertens, 1906). Однако и здесь тур рано вымер. Встре-
чающиеся иногда указания о существовании этого вида в Англии и Шве-
ции в историческую эпоху (Долгих, 1905 : 46), по-видимому, основаны
на недоразумении.
Имеются многочисленные свидетельства о существовании первобыт-
ного быка в предисторическую и раннеисторическую эпоху в северной
Африке, Передней Азии и юго-восточной Европе. Уже упоминались
BOS
609
изображения быков на гробницах египетских фараонов, относящиеся
к началу IIГ тысячелетия до н. э.
В библейские времена ареал тура включал Сирию. Изображения
сцен охоты на диких быков имеются на древних памятниках Ассирии
и Вавилона. В библейских писаниях тур фигурирует под названием
«риму» или «реэм» (Долгих, 1905).
Имеются неопровержимые доказательства существования тура на
о. Крите в среднеасинойский период (энеолит или бронза). При раскопках
дворца легендарного царя Миноса в Кносее, сооруженного за 2000 лет
до н. э., найдены обожженные черепа туров, а также уже упоминавшаяся
фреска с изображением быка светло-бурой масти. Келлер пытался пока-
зать, что легенда о минотавре основана на поимках и содержании во
дворце Миноса диких быков, обитавших в те времена в лесах о. Крита
(Lydekker, 1912). Упоминание о каких-то диких быках встречается
в древнекитайской и индийской литературе, но в данном случае нет уве-
ренности в том, что речь идет не о других представителях Bovini, напри-
мер о яке или гауре (Долгих, 1905).
Проследим основные черты истории первобытного быка, начиная
с эпохи бронзы в Европе. В поэмах Гомера нет упоминания о диком быке,
равно как нельзя узнать тура и у Аристотеля в «Истории животных».
Однако общеизвестны прекрасные изображения ловли и приручения
быков на вазе из гробницы Вафио при Амикле на Пелопонесе, относя-
щейся к микенскому времени (середина II тысячелетия до н. э.). Известны
две римские статуэтки, одна из которых изображает зубра, а другая
тура. Изображения сцен охоты на диких быков, по И. Долгих (1905),
встречаются на барельефах Помпеи, а также на одной римской мозаике,
доказывая этим, что тур в это время еще населял южную Европу.
Первые письменные указания современников о существовании в исто-
рическое время в Европе диких быков-туров принадлежат Юлию Цезарю
и относятся к I в. до и. э. Юлий Цезарь, описывая в «De Bello Gallico»
Герцинский лес, занимавший в то время всю современную Германию
и пространство к востоку от нее, указывал, что в этом лесу водятся быки,
«называемые Uri, величиною только немного меньше слонов, но по внеш-
нему виду, сложению и окраске — настоящие быки». В самой Италии,
похоже, в то время туров не было, но, привозимые издалека, они изредка
появлялись на арене римского цирка (Jennison, 1937). О быках в Германии,
как мы уже видели, в I в. н. э. писали Плиний и Сенека. О турах в Вогез-
ском лесу («Wasgenwald», возможно в Вогезскпх Альпах), а также в Майн-
ской провинции во Франции упоминается в литературных источниках
VI в. Однако уже в то время эти животные, по-видимому, были здесь
редки, так как право охоты на них сохранялось только за королем,
а виновные в незаконном убийстве тура наказывались смертной казнью
(Долгих, 1905 : 45).
В IX в. Карл Великий устраивал для знатных гостей охоты на туров
в лесу около Aix-la-Chapelle.
О мясе тура, как пище монахов Сентгаленского монастыря (Швейца-
рия) в XI в., писал Эксгарм (Сатунин, 1895). К этому же периоду отно-
сятся сведения, что во время Первого крестового похода, проходя Герма-
нию, крестоносцы встречали туров.
В «Песне о Нибелунгах», написанной около 1200 г., говорится, что
Зигфрид в числе другой дичи убил четырех туров и одного зубра около
Вормса. К XII в. относятся сообщения о том, что около 1170 г. в Рейн-
ской области время от времени на этих зверей охотились знать и короли.
39 Фауна СССР, И. И. Соколов
ею bovidae
Охота на туров уже в то время почиталась как нечто выдающееся, и трофеи
в виде рогов, а иногда и черепов тщательно сохранялись.
Несомненно, водились туры в Германии в XIII и по крайней мере
половине XIV в. Так, по сообщению Лукаса Давида, герцог Отто Браун-
швейгский в 1240 г. дарил братьям туров и зубров. В 1364 г. в Хинтер-
поммерне был убит зубр, который, «как думают, был крупнее тура»
(Mertens, 1906).
Записи в счетной книге Мариенбургского ордена с несомненностью
свидетельствуют, что в период с 1399 по 1409 г. дикие быки-туры были очень
редки и ценны, но еще встречались в Восточной Пруссии (около Кенигс-
берга) и в Литве, герцог которой Виттовит подарил в 1409 г. гроссмей-
стеру ордена четырех молодых туров (Nehring, 1898). Судя по всему,
в остальной Германии, как и вообще в западной Европе, к концу XIV в.
туры были уже истреблены; с этого времени даже название этого живот-
ного, постепенно стираясь в памяти людей, начало путаться с названием
зубра. По-видимому, не позднее середины ХАЛ в. тур исчез и в Пруссии
(Долгих, 1905).
Последним убежищем этого вида на земле осталась Польша, где он
влачил существование в полуприрученном состоянии до начала XVII в.
Основной областью обитания туров служила здесь западная часть ста-
рой Польши, Мазовия, расположенная по рр. Висле, Бугу и Нареву. Все
известные источники подтверждают, что и здесь они были редки. Уже
в 1298 г. герцог Мазовии Болеслав запретил охоту на туров. Позднее
запреты были подтверждены герцогом Зимовичем (1359) и королем Польши
Владиславом Ягелло. О ценности и редкости туров уже в то время го-
ворит тот факт, что Зимович разрешал охоту в своих владениях герцо-
гине Вышгорода на всех зверей, кроме тура.
В начале XVI в. туры оставались, и то, по-видимому, в небольшом
числе, в Якторовском (или Вискитском) лесу, расположенном в 55—60 км
к юго-западу от Варшавы, между деревнями Вискитки и Мищезова,
к северу до Сохачева.1 Здесь на пространстве около 200 кв. км звери
находились под строгой охраной и, по существу, уже в полувольном
состоянии.
Король Польши Сигизмунд III Ваза, видя угрозу полного исчезнове-
ния туров, число которых к тому времени исчислялось всего несколькими
десятками экземпляров, издал уже запоздалый указ об охране их
мест обитания (Harper, 1945). Крестьяне дер. Якторовки были освобож-
дены от налогов и других повинностей, но должны были заготовлять
для туров сено и все необходимое другое, а также подкармливать их в зим-
нее время. Специальные егеря должны были вести наблюдения за живот-
ными, не давать им широко разбредаться, а вышедших за пределы охран-
ной зоны возвращать обратно. В их же обязанности входил, будто бы,
отстрел экземпляров, относительно которых было известно, что они скрещи-
вались с домашним скотом (Lukaszewicz, 1952).
Однако принятые меры, по-видимому, не достигали цели и не смогли
приостановить прогрессировавшего процесса вымирания этого замеча-
тельного зверя. По свидетельству Яроцкого (цит. по Долгих, 1905),
в 1564 г. в Якторовском лесу еще было стадо из 30 туров (22 старых самки,
3 молодых самца и 5 телят), да, кроме того, было около 8 старых, держав-
шихся одиночно самцов. В 1599 г. оставалось уже 24 тура, в 1602 г.
только 5 голов (по другим данным 4: 3 самца и 1 самка); в 1620 г. в живых
1 Теперь от этого леса уже ничего не осталось.
BOS
611
оставалась всего одна самка, которая, как выяснилось, пала в 1627 г.
в возрасте 30 лет.
Лукашевич (Lukaszewi.cz, 1952) полагает, что хранящийся в Livru-
stkammaren в Стокгольме и датированный 1620 г. рог тура, заделанный
в серебро, принадлежал последнему из якторовских самцов-туров.
Следует отметить, что некоторое количество туров в конце XVI в.
содержалось еще в королевском резервате вблизи самой Варшавы. Этот
резерват в 1596 г. посетил, хотя живых туров сам и не видел, секретарь
папского нунция при польском дворе Муканте, и отсюда происходил
подаренный его шефу труп тура. По-видимому, в каком-то количестве
еще в 1610 г., а быть может и позднее, сохранялись туры в известном зве-
ринце графа Замойского в Замостье (Wrzesniowski, 1878).1
Считается, что последний экземпляр тура пал в 1627 г. Следует,
однако, иметь в виду, что кое-где в зоопарках и зверинцах туры могли
сохраниться несколько дольше. Не исключена возможность, что дольше,
чем в Якторовском лесу, они жили в зверинце Замойского. Имеется
требующее проверки указание, будто бы еще в 1669 г. туры жили в зооло-
гическом саду Кенигсберга (теперь Калининград), если только не имело
здесь места обычное смешение названий тура и зубра (Lengerken, 1953).
На территории СССР большинство ископаемых находок тура (рис. 124)
относятся к западным и северо-западным областям европейской части
Союза, т. е. близко к тем местам, где дольше всего удерживался этот вид
и в историческое время (Громова, 1931). Область распространения В. pri-
migenius и в наших пределах никогда не заходила к северу от 57—60° с. ш.
Неизвестно ни одной находки на территории СССР, достоверно от-
носящейся к палеолиту. Находки тура в неолитических стоянках также
не многочисленны. Самая северная из них происходит из стоянки на
южном берегу Ладожского озера (Иностранцев, 1882). Остатки дикого
быка, отнесенного Ю. Дюрстом (цит. по Громовой, 1931) к В. namadicus,
обнаружены в южных слоях древнего городища Анау (2000—3000 лет
до н. э.) под Ашхабадом.
Обитали туры на равнинах европейской части СССР, несомненно,
и в историческую эпоху. Об этом свидетельствуют многочисленные упоми-
нания этого животного в произведениях устного народного творчества —
в сказках, песнях, былинах. Ясное указание на тура имеем, например,,
в былине о Добрыне Никитиче, в сказании о Георгии Храбром и других
произведениях. О каком-то полумифическом быке говорится в «Калевале».
Свидетелями былого распространения тура являются отдельные сохра-
нившиеся слова, обороты, а также названия рек, урочищ, населенных
пунктов.1 2 По всем этим данным, тур водился не только в Литве, Волыни
и Приднепровье, но, если правильно происхождение этих географиче-
ских названий, то и дальше к северу и востоку вплоть до Сибири и степей
по Дону. Так, названия селений и урочищ «Турово» имеются в Новго-
родской, Псковской, Калининской, Московской, Костромской, Ярослав-
ской, Вологодской областях и даже в Красноярском крае.3
В западной Сибири на р. Туре был найден фрагмент черепа тура,
упоминаемый И. Словцовым (1895). Имеются, хотя и немногочислен-
ные, документальные данные о существовании диких быков на территории
СССР в историческую эпоху. Прежде всего, следует еще раз напомнить
указание Плиния об обитании туров в Скифии, т. е. на юге Русской рав-
1 См. историю тарпана, стр. 49.
2 Сводка этих данных имеется в работах И. Беляева (1865) и И. Долгих (1905).
3 См. указатель к Атласу мира, изд. 1954 : 457.
39*
612
BOVIDAE
нины в I веке н. э. Общеизвестно указание Владимира Мономаха в «По-
учении детям» о том, что в Черниговской земле «по Рови ездя. . . тура
два метала мя на рбзех и с конем». Уже упоминалось, что на грани XIV
и XV вв. туры были под Кенигсбергом ив Литве. Матвей Меховский (1936),
чей «Трактат о двух Сарматиях» появился впервые в 1517 г., писал о Литве
и Московии: «Так как леса там большие, то во множестве попадаются
и ловятся крупные звери: буйволы и лесные быки, которых они на своем
языке зовут турами или зубрами». Хотя здесь и имеет место смешение
названия тура и зубра, но факт наличия двух видов быков в это время
(«буйвола» и «лесного быка») остается несомненным.
И. Долгих (1905 : 50) цитирует место из «Жития святого Кассиана
Угемского», где говорится, что на грани XV и XVI вв. под Угличем будто
бы еще охотились на зубров и «буйволов», под которыми автор справед-
ливо подозревает туров. Ссылка же И. Г. Пидопличко (1951) на Боплана
(1832), что будто бы в начале XVII в. туры водились в северной части
Украины, основано на недоразумении; Боплан, во-первых, пишет о буй-
волах, а во-вторых, из дальнейшего совершенно ясно, что в данном абзаце
речь идет вообще о домашних животных («крупном скоте», длинношер-
стных и жирнохвостых валахских овцах, «белых зайцах»).
Имеются и палеонтологические доказательства обитания на Руси
в средние века тура. При исследовании костных остатков, найденных
при раскопке древнего Новгорода, в слоях, относящихся к XIII—XVвв.,
В. И. Цалкин (1952) обнаружил два стержня рога, принадлежавшие,
судя по величине и структуре, даже не очень мелкому туру. Этот же автор
сообщает, что, согласно летописям, в конце XV в. архиепископ Новгород-
ский Геннадий подарил псковичам оправленный в золото рог тура.
Остатки тура были обнаружены при раскопке городища у Вышгорода
в слоях, датируемых XI—XIII вв. н. э. (Зубарева, 1940). Таким образом,
возможно, что и на нашей территории последние туры вымерли в конце
XV—XVI в. ™
Биология. Сведения по биологии тура немногочисленны и отрывочны.
В описаниях современников мы находим их лишь у Свенцицкого, Муканте,
Виженера и Геснера. Наиболее полное описание образа жизни и привычек
тура дал К. Геснер (Gesner, 1606). Он использовал не только данные
С. Герберштейна (Herberstain, 1556), но и наблюдения двух из своих
друзей — Бонаруса и врача Шнеебергера, по-видимому имевших возмож-
ность непосредственно наблюдать этого зверя. В разных частях своего
ареала и в разные периоды своей истории биологические особенности туров
менялись.
Все источники периода последних столетий существования тура
в Европе сходятся в том, что он был типично лесным зверем и часто даже
назывался лесным быком. В свое время он населял обширнейший Герцин-
ский лес, а под конец — остаток последнего, Якторовскую лесную пущу.
Якторовский лес, по данным Лукашевича (Lukaszewicz, 1952), состоял
из сосны, дуба, ясеня, ольхи, клена, вяза, граба, березы. Окрестности
леса составляли марши (болота). При этом, по свидетельству современ-
ников, тур населял наиболее глухие участки этой пущи.
Анализируя указания русской народной поэзии о туре, И. Беляев
(1865) приходит к выводу, что он «был животным травоядным, любил дер-
жаться в местностях сырых, болотистых и лесистых, как привольных для
корма и уединенных».
По-видимому, правильны указания, что тур придерживался в основном
более влажных биотопов. Быть может с этим связана сравнительная часто-
Рис. 124. Карта ископаемых находок Bos primigenius Boj. (из В. И- Громовой, 1931). ® — места отдельных
находок.
614
BOVIDAE
та его ископаемых находок в западной половине восточной Европы и то
обстоятельство, что Прибалтика была последним убежищем этого вида.
Однако представление о туре, как об обитателе «самых густых мест леса»
(Gesner, 1606), мне кажется не совсем верным. Лесные чащи, особенно
в последний период существования тура, служили лишь местами укрытий.
Строение зубной системы и общее сложение говорят о том, что это был ти-
пично травоядный зверь, который при своей большой величине не мог
довольствоваться листвой и скудной травянистой растительностью под
пологом густого леса.
Правы, вероятно, Р. Лайдеккер (Lydekker, 1912) и другие авторы,
предполагавшие, что, по крайней мере в спокойной обстановке, первобыт-
ный бык должен был придерживаться более светлых участков леса, а кор-
мился на лесных лужайках или прилегающих к ним участках степи.
К этому же выводу пришла, анализируя былое распространение В. primi-
genius, В. И. Громова (1931), считавшая его жителем «смешанных и лист-
венных лесов и открытых степей, но не тайги». Впрочем, обитателем откры-
тых сухих степей тур, мне кажется, также не являлся, ибо строение корен-
ных зубов показывает, что последние не были приспособлены к пережевы-
ванию жестких степных растений.
Зимой туры жили, по свидетельству Шнеебергера, небольшими стадами,
летом же поодиночке. Взрослые самцы держались всегда одинцами и при-
соединялись к стадам только на период брачного сезона. Возможно, что,
как и буйволы, туры в жаркие дневные часы любили забираться в воду,
спасаясь от жары и насекомых.
Летом туры питались, вероятно, в основном травою, не брезгая листвой
и молодыми побегами некоторых кустарников и деревьев. Осенью, по
Шнеебергеру (Gesner, 1606), ели желуди, от которых становились очень
жирны. Зимой питались сухими листьями и ветками, возможно грызли
кору некоторых деревьев (резцы их к этому способны). В Якторовском
лесу кормились и специально заготовляемым для них сеном. Летом из
леса выходили на открытые поля, пожирали сжатые хлеба (а вероятно, и
на корню) и даже разбрасывали их рогами.
Все свидетельства современников и народного эпоса сходятся в том, что
тур был очень сильным, быстрым и временами свирепым животным. В ста-
ринных русских сказаниях и былинах он представлялся образцом силы,
храбрости и быстроты. Рассерженный, он легко поднимал на рогах и сбра-
сывал на землю всадника с конем. Невзирая на массивность, легко и быстро
бегал. Ни зверей, ни людей не опасался и даже не уступал дороги (пос-
леднее, вероятно, относится к периоду, когда звери под охраной на-
ходились уже в полувольном состоянии). Легко приходил в раздражение.
Особенно опасны были старые самцы-одинцы, иногда без всякого повода
набрасывавшиеся на человека. По свидетельству Остророга (цит. по Дол-
гих, 1905), туров нельзя было содержать в одном зверинце с зубрами, во
избежание постоянной вражды и столкновений.
Еще Юлий Цезарь сообщал, что туры, даже пойманные в молодом воз-
расте, не поддаются приручению. Шнеебергер также подтверждает, что
неоднократные попытки воспитать турят под домашней коровой всегда
оканчивались неудачей (гибелью дикого теленка).
Голос тура, вероятно, походил на голос домашнего быка. Иероним
Морщтин сообщал, что зубр «хрюкает», а тур «стонет», что, судя по извест-
ному нам голосу зубра, соответствует действительности (Долгих,
1905 : 56).
BOS
615
Спаривание у туров происходило в сентябре. Между самцами в это
время происходили ожесточенные поединки, кончавшиеся нередко смертью
обеих сторон.
Отел происходил в мае. Перед отелом самки удалялись в самую чащу,
где оставались с новорожденными около 20 дней. После этого выходили
на более открытые места. Матери заботливо оберегали телят от волков и
человека. Изредка наблюдались случаи позднего спаривания и отела
(в сентябре), но, как правило, родившиеся осенью телята зимой погибали
(Gesner, 1606).
Известны были неоднократные случаи спаривания диких туров с до-
машним скотом, но гибридное потомство оказывалось нежизнеспособным
и почему-то погибало, особенно в суровые зимы (Герберштейн, 1908;Luka-
szewicz, 1952).
По словам Шнеебергера (Gesner, 1606), туры были недолговечны и
будто бы редко проживали более 15 лет. Однако известно, что последняя
в мире турица пала в Якторовском лесу в возрасте 30 лет (Lukaszewicz,
1952).
Врагов, кроме человека, у тура практически не было. Волки могли пред-
ставлять опасность только для недавно родившегося молодняка. «Очень
часто наблюдалось, — писал Свицкий (цит. по Долгих, 1905 : 57), — как
один бык одерживал победу над несколькими волками, убив и растерзав их,
сам оставшись невредимым».
Хозяйственное значение. Как мясной зверь, тур, по-видимому, боль-
шого значения для человека не имел, вследствие своей малочисленности,
хотя, как мы уже отмечали, в одном из швейцарских монастырей мясо
туров фигурировало в качестве пищи монахов. Мясо тура, убитого в коро-
левском резервате под Варшавой, пробовал уже упоминавшийся раньше
секретарь папского нунция Муканте. По его словам, оно очень напоминает
мясо домашнего скота, только немного суше и тверже.
Несомненно, что при редких случаях убоя диких быков использовалась
их толстая кожа. Наиболее, говорят, ценилась кожа со лба, покрытая
длинными вьющимися волосами, особенно если она была вырезана еще
у живого зверя. По представлениям XVI в., даже ношение куска такой
кожи предохраняло от преждевременных родов, облегчало последние.
По свидетельству Свицкого (цит. по Долгих, 1905 : 57), такой же силой
обладали пояса из кожи, сдираемой с умирающих быков. Два таких пояса
мать польского короля Бона подарила в свое время за оказанную услугу
С. Герберштейну, а последний один из этих поясов преподнес жене авст-
рийского короля Фердинанда.
По словам Плиния, в его время рога туров шли на отделку острий копий,
употреблялись для изготовления сигнальных рожков в армии, а также,
разрезаемые на тонкие пластинки, вставлялись в фонари вместо стекол.
Солекопами Велички в Польше рога тура применялись в качестве светиль-
ников в рудниках (Lukaszewicz, 1952).
Позднее, когда тур стал редкостью и охота на него превратилась в при-
вилегию именитых особ, рога, помимо значения в качестве украшений,
тщательно сохранялись иногда вместе с черепами как очень ценные тро-
феи, отделывались золотом и серебром, преподносились в качестве ценных
подарков, употреблялись как кубки для вина и т. п.
В эпоху Юлия Цезаря, когда туры, по-видимому, были менее редки,
их ловили в ямы, вероятно выкапываемые и маскируемые на путях перехо-
дов. Позднее применялись только групповые охоты. Охота на диких быков
всегда представлялась как образец необычайного мужества, силы, ловкости
£16 BOVIDAE
и отваги. Если принять во внимание характер и силу этого зверя, а также
несовершенство оружия того времени, которое состояло, в основном, из
копий, луков и бомбард, то легко представить, что охота на него действи-
тельно была нелегким и опасным делом. Вот как описывает ее, в изложении
К. Геспера (Gesner, 1606), современник тура Шнеебергер:
«Охотники намечают одного наиболее сильного, постоянно вызываю-
щего других на борьбу (самца) и по разрешению короля охотятся на него. .
«. . . На охоту выходят много конных и пеших со стрелами, бомбардами и
большим числом собак. Вскоре того тура, на которого они решили охотиться,
криками и собачьим лаем заставляют отделиться от остальных, в то время
как всадники, бомбардисты и другие прячутся частью во рвах, частью за
большими деревьями, дабы при виде большой толпы, бой не сделался бы
слишком ожесточенным. По отделении тур преследуется бомбардами;
сам он, несмотря на все это, нападает даже на большую толпу и бегает
иногда в продолжение целого дня и больше, пока его не поразят в грудь,
после чего он, наконец, сваливается. Крестьяне тотчас рубят бревна и
валят на него с боков, чем и хотят удержать его, чтобы он не мог снова
подняться для сопротивления и настойчивого продолжения борьбы.
Удерживая его таким образом, они еще при жизни сдирают кожу с того
места, где на лбу, между рогами, находится закрученная кудрявая шерсть,
придающая животным страшный вид, и, удалив ее, они, наконец, убивают
его. Лишь только они убьют его, как вынимают сердце и, разрезав его
пополам, вынимают маленькую косточку формы креста; эту косточку,
вместе с кожей, содранной со лба, они посылают королю: то и другое це-
нится очень высоко. Некоторые говорят, что этот хрящик полезен бере-
менным и сильно страдающим природах. Мясо в коже посылается королю,
то в свежем виде, то соленое. Я слышал, что блаженной памяти король
Сигизмунд посылал несколько раз соленое мясо тура, в виде ценного по-
дарка, императору Карлу Пятому».
Главное же значение В. primigenius для человека заключалось в том,
что он явился родоначальником ряда пород домашнего крупного рогатого
скота. Целая их группа, на основе общности происхождения от европей-
ского тура, объединяется в тип или подвид В. taurus primigenius.1
Одомашнение тура происходило, по-видимому, в разных местах и в раз-
ное время, вероятно, от неолита до бронзовой эпохи (2000—3000 лет до н. э.).
Главными очагами одомашнения считаются восточное Средиземноморье,
Передняя Азия, Месопотамия, Иран, а также Придунайские и Приднепров-
ские степи.
Причиной вымирания тура явилось не прямое истребление, а косвен-
ное, вытесняющее влияние человеческой культуры. Житель леса и лесо-
степи, он не мирился с результатами хозяйственной деятельности человека.
По мере освоения, распашки и использования под выпас домашних жи-
вотных участков степи, вырубания лесов, сокращались пригодные для него
биотопы. Дикий зверь должен был уходить в леса, где он не находил
необходимых для него условий.
Особенно страдали туры в суровые и многоснежные зимы. Геснер
(Gesner, 1606) сообщает о том, что в 1557 г. в Якторовском лесу значитель-
ное число их погибло от сильного холода. Лукашевич (Lukaszewicz, 1952),
использовавший все архивные материалы, утверждает, что суровая зима
1 Предками остальных типов (В. taurus brachyceros Biitim., В. taurus turano-mongo-
licus Koi., B. taurus indicus L.) считаются другие подвиды или виды тура (Колесник,
1949).
BOS
617
1598/99 годов также была причиной резкого сокращения (до 24 голов)
оставшихся в живых туров. Причиной быстрого вымирания их в Якторов-
ском лесу этот автор считает и непрекращающиеся рубки леса, выпас до-
машнего скота, особенно свиней осенью. Последние становились конку-
рентами по использованию основного нажировочного корма — желудей.
Немало способствовала вымиранию и плохая, по его данным, организация
охраны, плохой уход за животными, эпизоотии и, наконец, беспокойство,
причиняемое турам от регулярных охот в пуще, заставлявших животных
удаляться в наиболее глухие, необжитые места леса.
Еще в начале прошлого века в Польше была высказана идея о жела-
тельности восстановления прообраза тура путем скрещивания примитив-
ных рас домашнего скота (Lukaszewicz, 1952). С ЗО-х годов нашего века
эта работа осуществляется в зоопарке Мюниха братьями Г. и Л. Гекк
(Heck, 1934, 1952). Сущность ее сводится к тому, что проводятся в различ-
ных комбинациях скрещивания разных примитивных пород домашнего
скота (корсиканского, испанского, паркового и т. д.), в результате которых
стремятся получить животных, которые по типу сложения и окраске при-
ближались бы к идеалу польского тура.
И хотя авторами этого начинания был выброшен хвастливый лозунг
«Тур вновь живет!», ни о каком восстановлении вымершего 300 лет назад
животного речи быть не может — эволюция не обратима. Получены живот-
ные, сходной с В. primigenius окраски, с формой рогов, сходной с таковой,
приписываемой туру, но не больше. Среди популяции «восстановленного
тура» очень нередки случаи появления пестрой масти.
И прав В. Херре (Нёгге, 1953), когда, оценивая результаты опытов
братьев Гекк, пишет: «Тур не восстановлен! Не имеющие научного значе-
ния скрещивания домашнего скота дают, благодаря сходной окраске и
сходной форме рогов, только бледное представление об этом импозантном
диком звере».
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ1
acteon, Hypoderma 230
acutifrons, Bos 600
Addax 300, 301*
Adenota 299, 305
adolescens, Sus 71
advena, Sus 71
aegagrus aegagrus, Capra 412
aegagrus blythi, Capra 412
aegagrus, Capra 7, 402, 404—416, 405*,
409*, 411*, 418, 420, 424, 444, 460, 474
aegagrus, Capra aegagrus 412
aegagrus cretica, Capra 412
aegagrus turcmenicus, Capra 412
Aegoceros 552
aenigmatica, Przevalskiana 340
Aepyceros 305, 306
affinis, Cervus elaphus 218
alaiana, Capra sibirica 430
alaschanicus, Cervus elaphus 218
albicilla, Heliaes tus 563
albirostris, Cervus (Przewalskium) 174
Alcelaphinae 300, 301*
Alcelaphini 301*
Alcelaphus 302
Alces 7, 140—142, 144, 173
alces, Alces 151*, 234—267, 237*, 239*,
241*, 243*, 255*, 263*
alces, Alces alces 247
alces americanus, Alces 248
alces bedfordi, Alces 248
alleni, Ovis nivicola 557
Allohippus 36
alpinus, Cyon 439, 541
altaica, Gazella gutturosa 336
Alticamelinae 100
Alticamelus 100
altus, Alticamelus 99
americana, Antilocapra 119
americanus, Alces alces 248
americanus, Oreamnos 388
Ammcdorcas 305
ammon am mon, Ovis 529
ammon coIlium, Ovis 530, 531
ammon dalailamae, Ovis 480, 529
ammon heinsii, Ovis 531
ammon hodgsoni, Ovis 480, 529
ammon intermedia, Ovis 531
ammon jubata, Ovis 530
ammon karelini, Ovis 480, 520, 530, 531
537
ammon kozlovi, Ovis 530
ammon littledalei, Ovis 526, 530
ammon mongolica, Ovis 530
ammon ngrimontana, Ovis 530, 541
ammon, Ovis 7, 481, 482, 489*, 516—544,
519*, 523*, 525*, 533*, 537*, 549, 552
ammon, Ovis ammon 529
ammon polii, Ovis 479, 480, 520, 530
ammon przewalskii, Ovis 530
ammon sairensis, Ovis 531
Ammotragus 476, 478
Amphimerycidae 118
Amphitragulus 141*
anatolica, Ovis orientalis 507
Anchitherium 31
angustirostris, Rangifer 280
Anoa 565, 567
Anthracotheriidae 68
Anthracotherium 21
Antidorcas 307
Antifer 141*
antiqua, Ovis 488
antiquus, Cervus elaphus 208
Antilocapridae 66, 119, 296
Antilope 300, 304, 305
antilopum, Pallasiomyia 371
antipini, Parafilaria 371
appendiculata, Damalinia 327
areal, Ovis orientalis 505, 509
arcar 480
Archaeohippus 31*
Archaeomeryx 118
arcticus, Moschus moschiferus 131*
argaliformes 478, 480
argaloides, Ovis 522
armeniana, Ovis orientalis 508, 543
Artiodactyla 6, 9, 10, 22, 23, 63, 65
asiatica, Rupicapra rupicapra 393
asiaticum, Hyalomma 327
Asinus 31*, 32, 34
attila, Sus scrofa 87, 88
aurelianense, Anchitherium 23
auribarbis, Cephenomyia 230
Axis 141*, 173, 174
1 Жирными цифрами обозначены страницы, где помещено описание или харак-
теристика данной формы. Звездочки указывают на страницы с рисунками названной
формы..
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ 619
Babirussa 71
bactrianus, Cervus elaphus 213, 217*, 218
bactrianus, Camelus 102, 103
bactrianus domesticus, Camelus 108
bactrianus ferus, Camelus 6, 102, 103—
115, 105*, 107*, 110*
balcanica, Rupicapra rupicapra 393
barbarus, Cervus elaphus 218
barbarus, Sus scrofa 88
barbatus, Gypaetus 515
barbatus, Sus 71, 96
bedfordi, Alces alces 248
bedfordi, Capreolus capreolus 161
berezovskii, Moschus 121, 122, 126
Bibos 565, 567, 599
Bison 7, 565, 566—568, 574, 575, 581
bison Bison 567, 568, 581
blanfordi, Ovis orientalis 509
Blastomeryx 141*, 272
blythi, Capra aegagrus 412
bochariensis, Ovis orientalis 453, 509
boeticus, Sus scrofa 88
bonasus, Bison 7, 25*, 301*, 566, 567, 568—
598, 571*, 575*, 577*, 583*, 595*, 602
bonasus, Bison bonasus 581, 596, 597*
bonasus bonasus, Bison 581, 596
bonasus caucasicus, Bison 581, 596
Boocercus 565
borealis, Ovis nivicola 557
Bos 7, 565, 566, 599, 600, 601
Boselaphina 301*
Boselaphus 302, 565, 567
Bovidae 3, 7, 10, 13, 21, 64, 66, 117—121,
137, 140, 295—299, 301*, 303
Bovina 301*
Bovinae 7, 301*—304, 564—566, 581
Bovini 301*, 565, 567, 609
bovis, Cysticercus 341
Bovoidea 119
brachyceros, Bos taurus 616
brachyceros europaeus, Bos 600
brauneri, Cervus 219
Brontotheriidae 26
Bubalina 301*
bubalis, Bubalus 299
Bubalus 565, 567
Budorcas 302, 344
caballus, Equus 32, 54
caballus gmelini, Equus 6, 32, 33, 45—
52, 47*, 49*
Calliforidae 134
Calobatippus 31*
Camelidae 6, 9, 66, 99, 100
Camelinae 6, 100, 101
Camelini 101
Camelops 98, 99, 101
Camelopsini 101
Camelus 6, 98, 100, 101, 102
canadensis, Cervus elaphus 217, 218
canadensis, Ovis (Pachyceros) 478, 550
Capra 7, 344, 345, 400, 401, 403, 405, 408,
431, 450, 456—458, 465, 477
capreoli, Hypoderma 170
capreoli, Theileria 170
Capreolus 6, 140, 141,143,144,151,160, 173
capreolus bedfordi, Capreolus 161
capreolus, Capreolus 6, 144, 145—172,
147*, 149*, 151*, 153*,
capreolus, Capreolus capreolus 161
capreolus melanotis, Capreolus 161
capreolus, Pelea 305
capreolus pygargus, Capreolus 161
Capricornis 343, 375
Caprina 301*
Caprinae 7, 301*—303, 342, 343, 352
Caprini 344
Carnivora 98
carpatica, Rupicapra rupicapra 394
cartusiana, Rupicapra rupicapra 393
cashmiriensis, Capra falconeri 450
castilianus, Sus scrofa 88
caucasica, Capra (Turns) 7, 301*, 394,
403,455—475,457*, 459*, 461*, 463*,
467*
caucasica, Capra caucasica 465
caucasica cylindricornis, Capra 296, 459*,
462, 465
caucasica dinniki, Capra 465
caucasica, Rupicapra rupicapra 393
caucasica severtzowi, Capra 403, 430,
457*, 464, 465
caucasicus, Bison bonasus 581, 596
caucasicus, Oestrus 440
caudatus caudatus, Nemorhaedus 380
caudatus griseus, Nemorhaedus 381
caudatus, Nemorhaedus 7, 375—384, 377*,
379*, 381*
caudatus, Nemorhaedus caudatus 380
caudatus raddaeanus, Nemorhaedus 381
celebensis, Sus 72
Cephalophinae 300, 301*
Cephalophus 297, 300
Cervalces 141*, 143
Cervavitus 141*, 144, 174, 240
cervi, Lipoptena 170, 182, 230, 258
Cervidae 6, 10, 21, 64, 66, 118, 120, 121,
137, 138—141*, 142, 174, 183, 191,
204, 264, 268, 272, 296
Cervinae 6, 140, 141*, 143
Cervulinae 140, 141*, 143, 151, 264
Cervus 6, 140, 141*, 143, 144, 148, 150,
173, 174, 175, 177
Chalicotheriidae 26
chapmani, Equus (Hippotigris) 62
chialtanensis, Capra falconeri 450
chirodontus, Sus scrofa 88
Choeropotamus 70
coIlium, Ovisa mmon 530, 531
comes, Sus 71
Condilarthra 10, 26, 64
Connochaetes 302
Connochaetini 301*
continentalis, Sus scrofa 88
coreanus, Sus scrofa 88
corinnae, Crivella 327
corsicanus, Cervus elaphus 218, 226
Creodonta 10
cretica, aegagrus Capra 412
cristatus, Sus 72
cycloceros, Ovis orientalis 480, 506,
508
620 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
cylindricornis, Capra caucasica 296, 403,
459*, 462, 465
dahli, Camelus dromedarius 102
dalailamae, Ovis аттоп 480, 529
Dama 141*, 144, 173, 175, 180
dama, Cervus (Dama) 6, 138, 175, 176—
183*, 177*, 179*, 186, 208
dama fossilis, Cervus 180
dama, Gazella 307
Damaliscus 302
davidianus, Elaphurus 144
davtiani Protostrongylus 415, 515
dementievi, Capra sibirica 431
Diacodexis 64
diana, Hypoderma 170, 230, 258
Dichobune 64
Dicoryphochoerus 71
Dicrocerus 141*, 143, 144
dinniki, Capra caucasica 465
dolgopolovi, Ovis orientalis 509
domesticus, Camelus bactrianus 108
Dremotherium 141*
dromedarius, Camelus 102
dromedarius dahli, Camelus 102
Dromomerycinae 139, 141*
dzerenae, Pharyngomyia 340
Elaphodus 140, 141*
Elaphurus 140, 141*, 144
elaphus affinis, Cervus 218
elaphus alashanicus, Cervus 218
elaphus antiquus, Cervus 208
elaphus bactrianus, Cervus 213, 217*, 218
elaphus barbarus, Cervus 218
elaphus canadensis, Cervus 217, 218
elaphus, Cervus 6, 138, 139*, 141*, 176,
204—234, 207*, 209*, 211*, 213*,
217*, 221*, 223*, 227*, 233*
elaphus, Cervus elaphus 207*, 216, 227*
elaphus corsicanus, Cervus 218, 226
elaphus elaphus, Cervus 207*, 216, 227*
elaphus hippelaphus, Cervus 218
elaphus maral, Cervus 216
elaphus sibiricus, Cervus 201, 213, 214,
217, 218, 221*, 223*, 227*
elaphus songaricus, Cervus 219
elaphus wallichi, Cervus 218
elaphus xanthopygus, Cervus 215, 218
elaphus yarkandensis, Cervus 218
elongatus, Metastrongylus 95
Entelodontidae 68
Eohippus 30, 31*
Eostylocerus 141*
Epieuryceros 141*
Epihippus 30, 31*
Equidae 6, 13, 26, 30, 31*
equiferus, Equus 56
Equus 6, 26, 27, 30, 31*, 32, 34, 46, 53
erskinei, Ovis orientalis 508
Eschatius 98
etuerianus, Cervus 208
Eucladocerus 141*, 180, 240
Eumeryx 139
europaeus, Bos brachiceros 600
falconeri, Capra falconeri 450
falconeri, Capra (Orthaegoceros) 7, 442—
455, 443*, 445*, 447*, 449*
falconeri cashmiriensis, Capra 450, 451
falconeri chialtanensis, Capra 450
falconeri falconeri, Capra 450, 451
falconeri megaceros, Capra 450
falconeri heptneri, Capra 450
falconeri jerdoni, Capra 450
falconeri ognevi, Capra 450
falzfeini, Sus 72
Ferae 10
Ferungulata 10
ferus, Camelus bactrianus 6, 102, 103—
115, 105*, 107*, 110*, 111*
fossilis, Cervus dama 180
Gazella 7, 299, 300, 304, 305*—307
Gazellinae 7, 300, 301, 303, 304
Gazdlini 301, 305
Gelocidae 118
Gelocus 118, 297
Gigantocamelus 101
Giraffidae 66, 119
gmelini, Equus caballus 6, 32, 33, 45—
52, 47*, 49*
gmelini silvaticus, Equus 49
goral, Nemorhaedus 375
Gorgon 302
granti, Gazella 307
granulosus, Echinococcus 340
griseus, Nemorhaedus caudatus 381
gutturosa altaica, Gazella 336
gutturosa, Gazella 7, 307, 308, 311*,
317*, 331—342, 333*, 335*, 337*
hagenbeckii, Capra sibirica 430
heinsii, Ovis ammon 531
Helicoportacina 301*
Helohyus 70
Hemionus 31*
hemionus, Equus (Asinus) 6, 33—44,
35*, 37*, 39*, 41*, 54
hemionus, Equus hemionus 40, 42
hemionus hemionus, Equus 40, 42
hemionus hemippus, Equus 42
hemionus kiang, Equus 40
hemionus onager, Equus 40, 42
hemippus, Equus hemionus 42
Hemitragus 344
hepatica, Fasciola 327
heptneri, Capra falconeri 450
hidruntinus, Equus 36
hillieriana, Gazella subgutturosa 319
Hipohippus 31*
Hipparion 26, 31*
Hippomorpha 27, 30
Hippopotamidae 66, 69
Hippotigris 31*, 32
Hippotraginae 300, 301
Hippotragus 300, 301
hircus, Capra 402, 415, 453, 474
hispidium, Gnathostoma 95
hodgsoni, Ovis ammon 480, 529
hodgsoni, Pantholops 343
Homacodon 64
АЛФАВИТНЫЙ УКАЗА ТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ 621
hominis, Gasterodiscoides 95
hortulorum, Cervus nippon 188
Hydropoles 137, 141
Hydropotiriae 140, 143
Hyemosclius 119
Hylochoerus 70, 71
Hyopsodus 64
Hyotherium 70
Hyperlragulidae 118
Hyracoidea 9, 22
Hyracotherium 30
Ibex 402
ibex Capra 402, 424, 430
ibex, Capra ibex 430
ibex ibex, Capra 430
indicus, Bos taurus 616
inermis, Hydropotes 139, 140
inexpectata, Cordilobia 134
intermedia, Ovis ammon 531
isphaganica, Ovis orientalis 508
jerdoni, Capra falconeri 450
jubata, Ovis ammon 530
jubatus, Acinonyx 515
kamschadalus, Dermacentor variegatus 563
karelini, Ovis ammon 480, 530, 531, 537
kiang, Equus hemionus 40
knoblochi, Camelus 102, 109
Kobus 299, 304, 305
kopschi, Cervus nippon 188
kozlovi, Ovis ammon 530
Lama 98—101
latffrons, Alces 240
lervia, Ovis (Ammotragus) 477, 478
leucomistax, Sus 72
leucomistax, Sus scrofa 88
Linognathoides 440
Lithocranius 305, 306
littledalei, Ovis ammon 526, 530
longicornis, Trichodectes 182
longifrons, Bos 600
Longirostromeryx 141*
Lophiomeryx 118, 138
lupus, Canis 439
lybicus, Sus scrofa 88
lydekkeri, Ovis nivicola 548, 557
Machaeomeryx 141*
magnus, Cervus 186
majori, Sus scrofa 88
maral, Cervus elaphus 216
marsupiaiis, Gazella (Antidorcas) 306, 307
mastan-zadei, Bos 600
matsehiei, Ovis musimon 490*
Mazama 141*, 264
megai-eros, Capra falconeri 450
Megaloceros 141*
megalot is, Madoqua 305
meiuerlzh'ageni, Hylochoerus 70
melampus, Aepyceros 305
mi lanotis, Capreolus capreolus 161
Melophagus 340, 440
meridionalis, Sus scrofa 88
Merycbippus 31*, 32
Merycopotamus 68
Mesohippus 30, 31*
mesopotamica, Cervus (Dama) 177
Metacervulus 141*
Micromeryx 141*
multispinosa, Cephenomyia 182
minutus, Bos 600
Miohippus 30, 31*
Miolabis 100
Madoqua 304, 305
mongolica, Ovis ammon 530
mongolica, Saiga tatarica 360
Morenelaphus 141*
moschatus, Ovibos 299
Moschidae 6, 66, 119 120, 121
moschiferus Moschus 6, 120, 121,122—136,
123*, 127*, 129*
moschiferus, Moschus moschiferus 130
Moschus 6, 121
moupinensis Sus scrofa 88
mubaschkinzewi, Protostrongilus 515
mufloniformes 478—480
Muntiacinae 140
Muntiacus 140
muraschincevi, Protostrongilus 415
musimon, Ovis 7, 480, 481, 482—497,
483*, 487*, 489*
musimon pallasii, Ovis 490
nahoor, Ovis (Pseudois) 117, 477, 478
namadicus, Bos 600, 608, 611
namadicus, Equus 36
namadicus, Sus 71
N anger 307
necrophorus, Proactinomyces 295
Nemorhaedini 301*, 343
Nemorhaedus 7, 343, 345, 374, 375
Neocervinae 7, 141*, 143, 263
Neotragini 299, 301*, 305
Neotragus 64, 296, 305
nigrimontana, Ovis ammon 480, 530, 541
nigripes, Sus scrofa 88
nippon, Cervus 6, 141*, 151*, 176, 183—
204, 185*, 187*, 189*
nippon, Cervus nippon 188
nippon hortulorum, Cervus 188
nippon kopschi, Cervus 188
nippon nippon, Cervus 188
nippon taioanus, Cervus 188
nivicola alleini, Ovis 557
nivicola borealis, Ovis 557
nivicola lydekkeri, Ovis 548, 557
nivicola nivicola, Ovis 556
nivicola, Ovis (Pachyceros) 7, 299, 478,
480—482 , 489*, 529 , 544—564, 547*,
551*, 553*
nivicola, Ovis nivicola 556
nivicola potanini, Ovis 557
Nonruminantia 6, 66, 67, 68
Notoungulata 10
nubiana, Capra 402, 455
Odocoileinae 141*
Odocoileus 141*, 264
ognevi, Capra falconeri 450
622 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
Oioceros 301
olympica, Rupicapra rupicapra 393
onager, Equus hemionus 40, 42
ophion, Ovis 477,481
Oreamnos 344
Oreotragus 305
orientalis anatolica, Ovis 507
orientalis areal, Ovis 505, 509
orientalis armeniana, Ovis 508, 543
orientalis blanfordi, Ovis 509
orientalis bochariensis, Ovis 453, 507, 509
orientalis cycloceros, Ovis 506, 508
orientalis, Dicrocoelium 135
orientalis dolgopolovi, Ovis 509
orientalis erskinei, Ovis 508
orientalis isphaganica, Ovis 508
orientalis ophion, Ovis 488
orientalis orientalis, Ovis 508
orientalis, Ovis 453, 482, 493, 497—516,
499*, 503*, 505*
orientalis, Ovis orientalis 508
orientalis punjabensis, Ovis 509
orientalis severtzovi, Ovis 506, 509
orientalis, Sus 72, 87
orientalis urmiana, Ovis 508
orientalis vignei, Ovis 479, 480, 509,
516, 543
ornata, Rupicapra rupicapra 393
Orohippus 30, 31*
Orthaegoceros 403
Oryx 300, 301
otophila, Haemaphysalis 170, 415
Ourebia 305
Ovibos 301*, 344
Ovibovini 301*, 344
ovinus, Melophagus 515
Ovis 7, 344, 345, 475, 477, 478, 479, 488,
500, 509, 550, 564
ovis, Oestrus 440, 542
Oxydactylus 98, 100
Pachyceros . 478
Palaeochoerus 70
Palaeodorita 65, 118
Palaeomerycinae 121, 139, 141*
Palaeomeryx 141*
Palaeotheriidae 26, 31
pallasii, Ovis musimon 490
Pantholopini 343
Pantholops 301*
papuensis, Sus 72
Paracamelus 99, 101
paragutturosa, Gazella 334
Parahippus 31*
Paranchitherium 31 *
Paratylopus 100
pardus, Pardus 515
parva, Rupicapra rupicapra 393
parvipes, Moschus moschiferus 131
pavlovskyi, Dermacentor 440, 542
pearsoni, Rangifer tarandus 279
Pecora 118, 119, 121
pecorum, Gastrophilus 44, 62
pelagicus, Heliaeetus 563
Pelea 301*
Peleini 305
Peregrinus, Sus 71
Perissodactyla 6, 9, 10, 22, 23—25
perrieri, Cervus 208
persulcatus, Ixodes 135, 258, 542
Phacochoerus 70, 71
Phenacodus 11
phylarchus, Rangifer tarandus 280
picta, Pharyngomyia 170, 182, 230, 258
picticaudata, Gazella 306, 307
pictus, Dermacentor 258
platyrhynchus, Rangifer 265
Piesippus 31*, 32, 36
Pliocervus 141, 151
Pliohippus 31*, 32
pliotarandoides, Eucladocerus 240
plumbeum, Hyalomma 327, 542
Poebrotheriinae 100
Poephagus 565, 567, 600
polii, Ovis ammon 479, 480, 520, 530, 533
Porcula 72
Potamochoerus 70
potanini, Ovis nivicola 557
primigenius, Bos 7, 299, 566, 601—617,
605*, 607*, 613*
primigenius, Bos taurus 616
prisca, Capra 402, 408
priscus, Bison 574, 575, 581
priscus, Sus 77
Proboscidea 9, 22
Procapra 307, 334
Procapreolus 141*, 143, 151
Prodorcas 307
Prodremotherium 118
Propalaeochoerus 70
Protoceratidae 118
Protohippus 31*
Protolahinae. 100
Protolabis 98
Protoungulata 10
Protylopus 98
przevalskii, Ovis ammon 530
przewalskii, , Equus 6, ,32, 33,, 46—48,
50, 52—63; 55*, 57*—59*
przewalskii, Gazella 307
Przewalskium 141* .
Pseudalces 141*
Pseudois 476, 478
Pseudolabidinae 100 , ,
Pseudolabfe 9Я,. .
Pudu 141*, 264
pumilio, Rhipicephalus 327
punjabensis, Ovis orientalis 509
pygargus, Capreolus capreolus 161
pyrenaica, Capra (Orthaegoceros) 403
pyrenaica,, Rupicapra rupicapra 393
quadricornis, Tetracerus 565
raddeanus, Nemorhaedus caudatus 381
raddeanus, ,,Sus scrofa 88
raddei, Capra 465
Racomylus 99
Rangifer 7, 137, 141*, 142, 173, 263—
265, 272
Raphicerus 305
Reaunca 305
Reduncini 301*,' 305
АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ 623
reginae, Gazella subgutturosa 319
reiseri, Sus scrofa 88
Rhinocerotidae 26
ricinus, Ixodes 170, 258, 399, 591
rossicus, Rhipicephalus 327
Ruminantia 6, 13, 66, 67, 98, 101, 115,
118
Rupicapra 7, 344, 345, 384, 390
rupicapra asiatica, Rupicapra 393
rupicapra balcanica, Rupicapra 393
rupicapra carpatica, Rupicapra 393
rupicapra cartusiana, Rupicapra 393
rupicapra caucasica, Rupicapra 393
rupicapra olympica, Rupicapra 393
rupicapra ornata, Rupicapra 393
rupicapra parva, Rupicapra 393
rupicapra pyrenaica, Rupicapra 393
rupicapra, Rupicapra 7, 301*, 384—400,
387*, 389*, 391*
rupicapra, Rupicapra rupicapra 393
rupicapra rupicapra, Rupicapra 393
Rupicaprina 301*
Rusa 141, 173, 174
sachalinensis, Moschus moschiferus 131
Saiga 7, 301, 343—345
saiga, Skrjabinodera 371
Saigini 343
sairensis, Gazella subgutturosa 319
sairensis, Ovis ammon 531
salvanus, Sus (Porcula) 71, 72
sanmeniensis, Equus 36, 56
schoetensacki, Bison 600
schantungensis, Ovis 477
scrofa attila, Sus 87, 88
scrofa barbarus, Sus 88
scrofa boeticus, Sus 88
scrofa castilianus, Sus 88
scrofa chirodontus, Sus 88
scrofa continentalis, Sus 88
scrofa coreanus, Sus 88
scrofa leucomystax, Sus 88
scrofa lybicus, Sus 88
scrofa majori, Sus 88
scrofa meridionalis, Sus 88
scrofa moupinensis, Sus 88
scrofa nigripes, Sus 88
scrofa raddeanus, Sus 88
scrofa reiseri, Sus 88
scrofa scrofa, Sus 87, 88
scrofa sennariensis, Sus 88
scrofa, Sus 6, 68, 69, 72—97, 75*, 79*,
81*, 91*
scrofa, Sus scrofa 87, 88
scrofa taivanus, Sus 88
seistanica, Gazella 319
sennariensis, Sus scrofa 88
severtzovi, Ovis orientalis 506, 509
severtzowi, Capra caucasica 403, 457*, 464
sibirica alaiana, Capra 430
sibirica, Capra (Ibex) 7, 402, 404, 416—
442, 419*, 421*, 423*, 425*, 448, 450,
474, 528
sibirica, Capra sibirica 430
sibirica dementievi, Capra 430
sibirica formosovi, Capra 430
sibirica hagenbeckii, Capra 430
sibirica sibirica, Capra 430
sibiricus, Cervus elaphus 201, 213, 214
217, 218, 227*
sibiricus, Moschus 122
sibiricus, Moschus moschiferus 131
sibiricus, Rangifer tarandus 280
sifanicus, Moschus 121, 122, 125, 126
silvaticus, Equus gmelini 49
sinaitica, Capra 402
sinensis, Gazella 334
Sirenia 9, 22
sivalensis, Bison 567
sivalensis, Camelus 102
skrjabini, Marschallagia 399
skyn, Capra 531
soemmeringi, Gazella 307
solium, Taenia 95
songaricus, Cervus elaphus 219
spekei, Tragelaphus 299
spiralis, Trichinella 95
Stenomylinae 100
Stenomylus 99
Stephanocemas 141*
stimulator, Cephenomyia 170
storcki, Ovis 556
Strepsiceros 301*, 566
strozzii, Sus 71
subgutturosa, Gazella 7, 301*, 307, 308—
331, 309*. 311*, 315*, 317*, 327*, 332
subgutturosa, Gazella subgutturosa 319
subgutturosa reginae, Gazella 319
subgutturosa sairensis, Gazella 319
subgutturosa subgutturosa, Gazella 319
subgutturosae, Pavlovskiata 326
Subungulata 9
Suidae 6, 66, 68, 69, 70
Suiformes 66, 67, 68
Suina 10, 12
sulcata, Haemaphysalis 515
Sus 6, 66, 70, 71
Syncerina 301*
Syncerus 565, 567
Tabanidae 370
taioanus, Cervus nippon 188
taivanus, Sus scrofa 88
Tapiridae 26
tarandi, Oedemagena 290
tarandus pearsoni, Rangifer 279
tarandus pbylarchus, Rangifer 280
tarandus, Rangifer 7, 138, 151*, 265,
266—295, 269*, 271*, 275*
tarandus, Randifer tarandus 279
tarandus sibiricus, Rangifer 280
tarandus tarandus Rangifer 279
tarandus valentinae, Rangifer 279
tatarica mongolica, Saiga 360
tatarica, Saiga 7, 343, 345—374, 348*,
349*, 351*, 353*, 355*
tatarica, Saiga tatarica 360
tatarica tatarica, Saiga 360
Taurotragus 301*, 565, 566
taurus, Bos 566, 601
taurus brachyceros, Bos 616
taurus indicus, Bos 616
624 АЛФАВИТНЫЙ УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИЙ ЖИВОТНЫХ
taurus primigenius, Bos 616
taurus turano-mongolicus, Bos 616
taxicolor, Budorcas 299
Tayassu 66
Tayassuidae 66, 69
tshernyslievi, Rhinoestrus 515, 542
tenuicollis, Cysticercus 341
Tetracerus 302, 565, 567
thomasi, Camelus 102
tibialis, Linognathus 327
timoriensis, Sus 72
Titanotheriidae 26
Tossunnoria 402, 477
Trachelocele 307
Tragelaphini 301*, 565
Tragelaphus 301*, 566
Tragocerina 301*
Tragulidae 64, 66, 116, 118, 138, 297
Tragulus 119
trochoceros, Bos 600, 604, 608
trompe, Ceplienomyia 290
Tubulidentata 9
turanicus, Rhipicephalus 327, 515
turano-mongolicus, Bos taurus 616
turcmenicus, Capra aegagrus 412
turgaicus, Lophiomeryx 138
Turocapra 403
turovi, Moschus moschiferus 131
Turus 403
Tylopoda 5, 6, 64—67, 98, 99, 101
ulrichii, Cephenomyia 258
uncia, Felis 439, 541
Ungulata 4, 6, 9, 10, 13, 21
Urmiabos 599
urmiana, Ovis orientalis 508
Uros 602
Urus 602
valentinae, Rangifer tarandus 279
variegatus kamschadalus, Dermacentor
563
Vermipsilla 440
verrucosus, Sus 71, 72, 75
vignei, Ovis orientalis 479, 509, 516, 543
vittatus, Sus 72
vsevolodovi, Cystocaulus 440
walburtoni, Haemapbysalis 440
wali, Capra 403
wallichi, Cervus elaphus 218
xanthopygus, Cervus elaphus 215, 218
Xenopsylla 440
Xiphodontidae 99
yarkandensis, Cervus elaphus 218
yarkandensis, Gazella 319
zdanskii, Ovis 477
АЛФАВИТНЫЙ УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Абиссинский козел 403
Азиатская антилопа 319
Азиатские ослы 34, 36, 40, 42
Азиатский буйвол 565
Азиатский муфлон 7, 482, 493, 497—
516, 499*, 501*, 503*, 505*, 527
Аксисы 174
Алашанский вапити 218
Аллювиальный тур 604
Алтайский аргали 520, 529
Алтайский баран 529
Альпийский козерог 402, 424, 430, 464
Американская снежная коза 344, 388
Американские мозоленогие 100
Американские олени 7, 141, 263, 272
Американский баран 478, 479, 493
Американский бизон 567, 568, 570, 571,
574, 581 '
Американский лось 248, 257
Американский толсторог — см. аме-
риканский баран
Амурский горал 7, 375—384, 377*, 379*,
381*, 386
Анадырский толсторог 562
Анатолийский баран 507
Аноа 565
Аноплотерии 65, 66
Антилопа-косуля 305
Антилопа-прыгун 307
Антилопы 66, 119, 297, 299
Антракотерии 65', 66, 68
Аравийский козерог 403
Аргали (Архар) 7, ИЗ, 479, 480, 482,
489*, 516—544, 519*, 523*, 525*,
533*, 537*, 547
Аргаловидные бараны 479—481, 488,
518, 519—522, 524, 530, 543
Аркар 478
Арменийский муфлон 504, 508, 509, 512,
514, 516, 543
Архар — см. Аргали
Архаро-меринос 544
Асканийский марал 212
Афганский баран (см. также копетдаг-
ский баран) 506, 508
Африканские серые ослы 34
Африканский буйвол 565
Африканский гривистый баран 477, 478
Африканский тур 606
Ауэрокс 602
Бабируссы 70, 71
Бактриан — см. двугорбый верблюд
40 Фауна СССР, И. И. Соколов
Бараны 7, 13, 43, 66, 119, 299, 344, 345,
376, 390, 401, 402, 435, 475—481,
483, 485, 489, 492
Баргузинский олень 280
Барс 94, 398, 439, 541
Бегемоты 13, 65, 66, 69
Безоаровый козел 7, 402, 404—416, 405*,
409*, 411*, 418, 420, 422, 436, 442,
455—457, 458, 460, 463, 464, 474,
509, 515
Беловежский зубр 572, 573, 578, 581,
592, 593, 596, 597*
Белоголовый сип 372
Белонлечий орлан 563
Белохвост, орлан 563
Белуджистанский баран 509
Березовского кабарга, 122
Беркут 170, 329
Бизон 565—568, 572—574, 582, 584, 592—
595, 602
Бизоны 7, 565, 566
Благородные олени 140, 144
Благородный олень 6, 138, 139*, 150,
172, 176, 181, 184, 204—234, 207*,
209*, 211*, 213*, 217*, 221*, 223*,
227*, 233*, 248, 251, 254, 255, 260,
268, 286, 474, 496, 497, 578
Блохи 440
Бландфорда баран 509
Болотный козел 305
Бородавочники 70
Бородатая свинья 71, 96
Бородач, орёл 439, 515
Бронтотерии 26
Бубалы 300
Буйвол 565, 602, 612, 614
Бурый гриф 372 ,
Бухарский баран 507, см. также тад-
жикский баран
Бухарский олень (хангул) 213, 216,
217*j 221, .226, 230
Быки 7, 16, 66, 119, 299, 565, 599, 600,
601, 603, 604, 608, 609, 615
Быковые 7, 302—304, 564
Бычий солитер 341 . ,
Валахская овца 612
Варварийский олень 218 i
Вапити 205, 216, 218
Верблюд Даля 102
Верблюдовые 6,. 9, 63, 66, 99, 100
Верблюды 6, 16, 65,. 66, 99, 101, 103, 104,
106, ИЗ, 116, 390
626 АЛФАВИТНЫЙ УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Вилорог 66, 119, 296, 298
Винторогие антилопы 565
Винторогий козел (мархур) 7, 401, 403,
442—456, 443*, 445*, 447* 449*, 458
Виргинские пампасовые олени 141, 264
Власоед 182
Водяной козел 299, 304, 305
Водяные олени 140
Волк 43, 94, 166, 170, 171, 182, 202, 203,
230 , 258 , 289 , 326 , 338 , 340 , 370,
382, 383, 398, 415, 454, 473, 493,
496, 515, 538, 540, 541, 563, 590, 615
Восточно-кавказский тур — см. даге-
станский тур
Восточно-сибирский лось 248
Восточно-эльбурский баран 508
Восточного Тибета баран 530
Вошь 327, 440
Высшие жвачные 118, 119, 121
Газелевые 7, 300, 303, 304
«Газелеобразные» верблюды 99, 100
Газели 7, 299, 305, 310, 318, 332
Гаур 609
Гепард 515
«Гигантские свиньи» 68
Гималайский баран 529
Гималайский горал 375
Гипертрагулиды 118
Гиппарионы 26, 31*, 32
Гиппопотамы 66, 68, 69
Гнус 92, 172, 370
Голубой баран — см. нахур
Горалы 7, 343, 344, 345, 374, 375, 378,
380
Горный меринос 494
Гривистый баран — см. африканс-ий гри-
вистый баран
Гуэмалы 141, 264
Давида олень 140, 144
Дагестанский тур — 459*, 462, 464, 465,
467*, 468, 470, 472, 494, 496
Даманы 9
Даниэль 176
Двугорбый верблюд 101—103, 105, 109,
112
Делювиальный тур 600
Джейран 7, 43, 301*, 307, 308—333, 309*,
311*—315*, 317*, 327*, 338, 339, 348,
360, 364, 365, 367, 371, 515, 537
Джигетай — см. кулан
Дзерен 7, 43, 308, 311*, 317*, 319, 331—
342, 333*, 335*, 337*, 360, 364
Дикая коза — см. косуля
Дикая лесная лошадь — см. лесные тар-
паны
Дикая лошадь — см. кулан, лошадь
Пржевальского, тарпан
Дикая свинья — см. кабан
Дикий буйвол 299
Дикий бык 576, 578, см. также перво-
бытный бык.
Дикий двугорбый верблюд 4, 6, 102
103—115, 105*, 107*, 111*
Динника тур 465
Дождевые черви 90—92, 95, 96
Долгополова баран 509
Домашние бактрианы — см. домашние
двугорбые верблюды
Домашние верблюды 44, 104, 106, 108,
110, 114, 541
Домашние двугорбые верблюды 102, 106,
108, ИЗ, 114
Домашние лошади 34, 44, 45, 46, 49—52.
54, 57, 60, 62
Домашний бактриан •— см. домашний
двугорбый верблюд
Домашний бык 115, 592, 593, 605, 606,
608, 614
Домашний верблюд 104, 109, 112
Домашний двугорбый верблюд 102—106.
109, 114
Домашний осел 34, 44
Домашний северный олень 273, 276, 281,
286____293 295
Домашняя коза 323, 401, 402, 415, 440,
466, 474, 541
Домашняя лошадь 32, 44, 46, 47, 51, 52,
54, 62, 538, 544, 557, 561
Домашняя монгольская лошадь 57, 62
Домашняя овца 13, 39, 323, 348, 364,
415, 466, 471, 474, 480, 490, 491, 493,
494, 497, 510, 513—516, 537, 540—
543, 564
Домашняя свинья 64, 72, 75, 93, 95, 96,
108
Дромедар 103, — см. также одногорбый
верблюд
Дрофа 339
Дукеры 300
Европейская дикая лошадь — см. тарпан
Европейская косуля 161, 163*, 171
Европейский благородный олень 205,
207*, 212, 216
Европейский лось 239*, 247, 257, 259
Европейский муфлон 7, 480, 481, 482—
497, 483*, 487*, 489*, 501—503, 514,
516
Европейский олень 145, 176, 204, 228
Жвачные 3, 6, 13, 15, 29, 66, 67, 101,
104, 115, 116—119, 121, 139
Желудочный овод 44, 62
Жирафовые 119 |
«Жирафообразные» верблюды 99, 100
Жирафы 65, 66, 137, 139
Жиряки 9, 13
Жуки 90
Забайкальский олень — см. баргузин-
ский олень
Закавказский муфлон 508
Замбары 174
Западно-кавказский тур 430 , 457*, 464,
470, 472
Западно-китайская кабарга 122.
Западно-эльбурский баран 508
Западного Тибета баран 529
Заяц 133
Зебра Чепмана 62
АЛФАВИТНЫЙ УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ 627
Зебры 32
Зобатая газель — см. дзерен
Зубр 3, 5, 7, 17, 25*, 222, 232, 301*, 566,
567, 568-601, 571*, 57'5*, 577*, 597*,
602—606, 608—612, 614
Зубробизон 583*, 584, 591, 592, 594, 595*
Изюбрь 191, 192, 205, 215, 216, 218,
221, 224, 231—234, 256, 258
Иксодовые клещи 170, 440, 563
Испанский козел 403, 455
Испанский тур — см. испанский козел
Исфаганский баран 508
Кабан 6, 72—97, 75*, 79*, 81*, 91*, 108,
162, 171, 258, 259
Кабарга 6, 116, 121, 122—136, 123*,
127* 129* 297
Кабарги 3, 4,’ 6, 21, 66, 119, 120, 121,
123, 132, 134, 136, 138—140, 146,
176, 184, 205
Кабарговые 120
Кавказские каменные козлы (туры) 403
Кавказский зубр 567, 572, 573, 578—
581,582, 585, 587, 589—591, 594, 596
Кавказский каменный козел (тур) 7, 301*,
403, 412, 441, 455—475, 457*, 459*,
461*, 463*, 467*
Кавказский олень 212, 225, см. также
каспийский благородный олень.
Кавказский тур — см. кавказский ка-
менный козел.
Камчатский снежный баран 556, 563
Канны 566
Каратаусский баран 530
Карликовые антилопы 299, 304, 305
Карликовый тур 600, 601., 608
Каспийский благородный олень 216,
см. также кавказский олень
Кашмирский хангул 218
Кианг 34, 40, 42
Кипрский муфлон 481, 488
Киргизская лошадь 537
Киргизская собака 537
Кистеухие свиньи 70
Китайская кабарга 121,124,126, см. также
западно-китайская кабарга
Китайский водяной олень 139, 140
Китайский горал 375
Клещи 135, 230, 258, 327, 370, 383, 399,
415, 515, 542, 591
Кожный овод 170, 230, 258, 290, 327,
340, 370, 383, 541
Козерог 402, 420, 425, 428
Козлообразные 7, 303, 342
Козлы 7, 66, 119*, 299, 344, 345, 383,
390, 400—403, 491, 510, 519, 531,
535, 540, 552, 557.
Комары 170, 258, 290
Кондилартры 10, 64
Коне-куланы 44
Копетдагский баран 506, 508, 512, 515, 516
Копытные 3, 6, 9—17, 19—24, 29, 43,
60, 63, 64, 66, 73, 74, 103, 117, 118,
123, 125, 128, 129, 132, 135, 146,
184, 202, 205, 326, 327, 341, 398
Коровьи антилопы 300
Корсак 340
Корсиканский муфлон — см. европейский
муфлон
Корсиканский олень — 218, 226
Косули 6, 140, 143, 144
Косуля 6, 145—172, 147*, 149*, 151*,
153*, 163*, 173, 176, 204, 226, 232,
474, 496, 497
Красный волк 439, 541
Креодонты 10
Кровососущие насекомые — см. гнус
Кролик 98, 299
Круглые черви 95, 170
Крымский благородный олень 222
Крымский муфлон 496, 497
Крымский олень 219, 228, 494
Ксифодонты 99
Куду 566
Куку-яман ИЗ, см. также нахур
Кулан 3, 6, 32, 33—44, 35*, 37* 39*, 40,
41*, 61
Куланоид 44
Куница 134, 170
Курдючная овца 540, 543
Кызылкумов баран 509
Левретка 304
Ламы 99
Лани 144, 175
Лань 3, 6, 138, 145, 175, 176—183, 177*,
179*, 183*, 204, 269, 578
Легочные нематоды 440, 515
Ледниковый тур 604
Ленточные черви 95, 170
Леопард 94, 230, 398, 415, 473, 515
Лесная антилопа 566
Лесной северный олень 279—281, 293
Лесные свиньи 70
Лесные тарпаны 49, 50, 52
Лисица 118, 134, 170, 182, 230, 340, 370,.
439, 496, 515
Литльдаля баран 530
Литовский зубр — см. беловежский зубр
Лобастые быки 565
Лоси 7, 140, 142, 144, 173, 234
Лось 205, 232, 234—263*, 237*, 239*,
241*, 243*, 255*, 268, 269, 272, 569
Лошади 6, 13, 26, 27—30, 31*, 32, 99, 115
Лошадиные 6, 26, 27, 31, 42, 45
Лошадиные антилопы 300
Лягушки 91
Мазамы 141, 264
Малого Тибета баран 509
Мамонт 109
Мансарорварский олень 218
Маньчжурская косуля 161
Марал 17, 201, 205, 212—216, 217—221*,
223*—225, 227*, 230—233*, 234, 260
Мархур — см. винторогий козел
Медведь 134, 166, 170, 258, 399, 439, 473,
541, 563, 590
Мендес 300
Меринос 493, 494, 543, 544
Месопотамская лань 177
Метастронгипюсы 95
40*
628 АЛФАВИТНЫЙ УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ
Мечезубые 99 '
Мозолёйогие 5, 6, 6'5—67, 98—100, 116
Монгольская сайга 360
Мошки 290
Мулы 44
Мунтжакй 140, 143, 144, 151, 174
Мускусный бык 13, 299
Муфлонообразные бараны 477—481, 488,
504, 520, 521, 524
Муха 134
Муха-кровососка 230, 258, 340, 341, 384,
440, 512
Мышевидные грызуны 91
Мышечная трихина 95
Насекомые 96, 103, 190, 290
Настоящие бараны 478
Настоящие копытные 9, 10
Настоящие олени 6, 140, 143, 144, 173,
175, 240
Настоящие ослы 34, 43
Нахур ИЗ, 477, 478
Нежвачные 6, 64, 66, 67, 68
Нематоды 371, 399
Непарнокопытные 9
Ненарноналые 6, 9, 12, 16, 23, 24, 26, 27,
30, 63, 64, 65
Нильгау 565
Новоземельский олень 279
Новокавказский меринос 540
Норильский снежный баран 557
Носовой овод — см. носоглоточный овод
Носоглоточный овод 170, 182, 230, 258,
290, 340, 383, 515, 541, 542, 563
Носороги 26, 29, 30
Нубийский козел 402, 455
Обыкновенная косуля 144
Обыкновенная лань 177
Обыкновенный кабан 72
Овцебыки 344
Одногорбый верблюд 102, см. также
дромедар
Олени 3, 4, 6, 10, 21, 65, 66,119*, 121, 123,
137 138, 139, 140, 141*—144, 146,
148, 164, 173, 174, 176, 178, 184, 205,
238 256 514
Оленьки бб’ 116, 118, 121, 125, 139, 140,
297
Оленьковые 116—118
Оленья муха — кровососка 170, 182
Онагр 33, 40— см. также кулан
Оронго 343
Осло-куланы 44
Ослы 32, 34, 42, 54, 62
Охотский северный олень 280
Палеодонты 68, 70, 118
Палеомерицины 139
Палеотерии 26, 27
Памирский архар — см. памирский баран
Памирский баран 530, 533, 542, 543
Парнопалые 3, 6, 9, 10, 12, 20, 23, 25, 63—
66, 68, 69, 118
Парнокопытные 9, ем. также парнопалые
Пекари 66, 69
Пенджабский баран 509
Первичнокопытные 10, 26, 64
Первобытная коза 402
Первобытный бык (тур) 4, 7, 108. 299.
566, 599, 600, 601—617, 605*,'607*.
613*
Первобытный зубр 13, 581, 600
Персидский онагр 42
Печеночная двуустка 327
Подкожный овод — см. Кожный овод
Полорогие 3, 7, 10, 14, 21, 65, 66, 117, 118,
119, 120, 125, 137, 138, 140, 295, 297,
298, 301*, 302, 303
Полукопытпые 9
Полуослы 32, 34, 36, 46, 47, 48, 53, 54, 56
Пржевальского лошадь 4, 6, 25*, 32, 33,
46, 50, 52—55*—58*—63*
Пржевальского олень 174
Приматы 16
Протостронгилиды 415
Протоцератиды 118
Пуду 141, 264
Пятнистые олени 190—192, 201—204
Пятнистый олень 3, 6, 176, 183—204,
185*, 189*, 192, 201, 208, 231, 233, 234
Речные свиньи 70
Риму 602, 609
Россомаха 134, 170, 230, 258, 563
Рысь 134, 170, 182, 230, 258, 383, 398,
415, 439, 473, 541
Рээму 602, 609
Сайга 7, 43, 109, 319, 324,' 328, 343, 344,
345—374, 348*, 349*, 350, 351*, 353*,
355*
Сайгак — см. сайга
Сайлюгемский баран 530
Саранча 90
Сахалинская кабарга 131
Свиные 7, 68, 69
Свиные олени 174
Свиньи 6, 9, 12, 16, 17, 65, 66, 69—71.
72, 76, 77, 82, 89, 90, 92—95, 116
Северной Монголии баран 530
Северные олени 7, 12, 137, 141*, 142,
173, 264, 265, 273, 277, 282, 283, 284,
289, 294, 296—298
Северный олень 7, 13, 138, 151*, 266, 268.
269*, 270, 271*, 273—275*, 276, 280,
281, 286, 291, 293, 295
Североамериканский толсторог 550
Североказахстанский баран 530
Северцова баран 509
Северцова тур 456, 457*, 464, 474
Серао 343, 344, 375
Серна 7, 89, 301*, 344, 345, 383, 384—400,
387*, 389*, 391*, 412, 413, 442, 463,
465, 468, 473—475, 491
Сернобыки 300
Серны 7, 384
Серые ослы 34
Серый вол 593
Серый -волк 438
Сибирская кабарга 121, 123, 124, 126, 131
Сибирская косуля 145, 146, 161, 163*. 171
АЛФАВИТНЫЙ УКАЗАТЕЛЬ РУССКИХ НАЗВАНИЙ ЖИВОТНЫХ 629
Сибирский благородный олень — см. ма-
рал
Сибирский козел — см. сибирский козе-
рог
Сибирский козерог 7, 402, 404, 408, 416—
442, 419*, 421*, 423*, 425*, 448, 450,
457, 464, 474, 531, 541.
Сиреновые 9
Сирены 9
Сирийский онагр 42, 43
Ситатунга 299
Слепни 170, 258, 290, 370
Слоны 9
Снежная коза — см. Американская снеж-
ная коза
Снежный баран 7, 299, 477, 478, 480—
489*, 522, 529, 544—564, 547*. 551*,
553*
Соболь 134
Собственно свиньи 70
Соколы 329
Сосальщики 95, 170
Сохатый — см. лось
Среднеевропейский олень 218, 219
Станового хребта снежный баран 548,
557, 561
Степной баран 505, см. также устюртский
баран.
Степной бизон 584
Степной тарпан 50
Сычуанская косуля 161
Таджикский баран 509, см. также уриал.
Тайгоносский снежный баран 557
Такин 299, 344
Тапиры 24—26, 29, 30
Тарпан 4, 6, 32, 33, 45—52, 47*, 49*
54—56, 112
Тары 344
Тигр 94, 134, 205, 230, 258
Титанотерии 26, 27
Толсторог — см. снежный баран.
Тонкорунная овца 543
Тонкошейная финна 341
Трагулиды 65, 121, 138
Трематоды 135
Трихостронгилиды 399
Трубкозубы 9
Тундровый домашний олень 293
Тундровый олень 279, 280, 282
Тур (бык) — см. первобытный бык
Тур (козел) — см. кавказский каменный
козел
Туро-коза 474
Тэк 402, см. также сибирский козерог
Тянь-шаньский баран 530
Тянь-шаньский марал 219
Ур 602
Уриал 507, 516, см. также таджикский
баран
Урмийский баран 508
Уссурийский лось 238, 248, 251, 257
Устюртский баран 509, см. также степной
баран
Филин 134, 170
Хабтагай — см. дикий двугорбый верблюд
Халикотерии 26, 27
Хангул 213, 216, 217*, 221—223, 225,
228, см. также бухарский олень
Харза 134, 383
Хищные птицы 134, 170
Хорек 170
Хохлатые олени 140 ’ .
Хрущи 90
Целебесский карликовый буйвол (аноа)
565
Центральнокавказский тур 462. 464, 465
Чайка 370
Черная лань 178
Чернотелки 90
Чиру 343
Четырехрогая антилопа 565
Чубук — см. снежный баран
Шерстистый носорог 109
Шу — 218
Энтелодоны 65, 68
Эхинококки 340
Южноамериканские копытные 10
Ябарайский баран 530 ’
Ягнятник — орел 454, 473
Як 565, 600, 609
Якутский снежный баран 548, 557
Яркендский олень 218
Ястреба 329
Ястреб-тетеревятник 134
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
Абдилдабеков К. А. 417, 426, 517, 526,
539, 544
Абрамов К. Г. 3, 20, 74, 86, 146, 160,
183, 184, 188, 190, 192, 235, 252, 254,
258, 259, 375, 376, 378,' 380, 382—384
Авакян Л. А. 599, 600
Авербург В. В. 100, 101
Аверин К). В. 146, 183, 202, 267, 277,
282, 285, 288, 291, 545, 555—564
Агеенко Ilj Д. 236
Адлерберг Г. П. 21, 71—73, 122, 266,
545
Адольф Т. А. 346, 347, 352—354, 356,
358, 359, 373
Акаевский А. И. 267, 268
Алекперов X. М. 314, 323, 410
Алекперова Н. А. 347, 354
Александров Л. 266, 274
Александрова В. Д. 267, 283, 284
Аллен Г. 42, 58, 104, 332
Аллен И. 556
Алфераки С. Н. 72, 83, 346, 354, 416
Андреева Е. Г. 568, 592
Андрушко А. М. 517, 527
Антипин В. М. 20, 33, 38, 73, 85, 123,
145, 157, 158, 169, 308, 316, 320, 326,
346, 357, 359, 417, 426, 427, 432, 436,
498, 505, 517, 524—528, 532, 534, 536,
538—542
Анучин А. В. 73
Анучин Д. Н. 45, 53, 58, 60, 346, 356
Аргентов А. 266
Арсеньев В. А. 183, 202
Асадов С. М. 146, 170, 308, 327, 385,
399, 455, 473
Афанасьев А. В. 73, 74, 84, 86, 122, 133,
134, 145, 146, 157, 158, 160, 168, 205,
214, 308, 316, 322, 323, 325, 347, 357,
361, 417, 427, 517, 524, 526, 528, 532,
545, 556
Бабенышев В. П. 346, 354
Бабушкина В. А. 101, 102
Баврин Б. 73, 205
Бажанов В. С. 73, 84, 89, 145, 158, 204,
205, 213, 235, 245, 308, 316, 346, 356,
417, 498, 505, 506, 511, 513, 517, 525,
545, 554
Базыкин М. 123, 145, 235, 267
Банников А. Г. 4, 20, 34, 40, 43, 53, 58,
60, 61, 74, 86, 87, 104, 112, 114, 130,
146, 160, 205, 216, 247, 267, 277,
278, 308, 318, 320, 322, 323, 326, 331,
336—341, 346, 347, 359, 360, 363, 366,
369, 370, 417, 429, 517, 529
Барабаш-Никифоров И. И. 145, 155
Баранчеев Л. М. 20, 74, 86, 146, 160,
268, 278
Барминцев Ю. Н. 101, 102
Башкиров И. 266, 276, 569, 578, 579,
581, 582, 585, 589—591
Беднарский М. 568, 597
Бедный С. Н. 347
Белке Г. 82
Белопольский Л. О. 122, 130, 235, 247,
266, 277, 545, 553, 554
Белослюдов Б. А. 517, 524, 532
Беляев И. Д. 601, 611, 612
Беляева Е. И. 346, 356
Берг Л. С. 346, 358, 364, 365, 367
Берггрин А. П. 101, 114, 346, 367
Бианки Л. В. 159, 556
Бибикова В. И. 569, 577; см. также
Зубарева В. И.
Билькевич С. И. 323, 404, 509, 515
Бихнер Е. А. 20, 53, 482
Благовещенский Д. И. 327
Благовещенский И. 146, 157, 205, 214,
236, 246
Бобринский Н. А. 20, 204, 213, 267, 276,
308
Богачев В. В. 308, 313
Богданов М. 145, 156, 266, 276
Богданов О. П. 443, 449, 453
Богданов П. К. 74, 82
Боголюбский С. Н. 101, 103, 104, 106,
568, 592
Боев С. Н. 204, 417, 440, 517, 524,
542
Боль Б. К. 266, 267, 283, 287
Бондарева В. И. 308, 327, 517, 542
Боплан Г. 45, 50, 346, 353, 601, 612
Борисяк А. А. 306
Бородин Л. П. 235
Боянус Л. 602
Брандт А. Ф. 45
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
631
Браунер А. А. 45, 73, 82, 104, 108, 112,
145, 154, 165, 204, 482, 493, 494
Броме-Скунке Ф. 289
Бромлей Г. Ф. 184, 188, 190
Брэм А. Э. 38, 69
Брюн Л. А. 267, 274
Буйновский Н. А. 101, 102, 104, 109
Букстон Е. 492
Булавин А. 331, 334, 427, 428, 517, 528
Буль М. 109
Бурчак-Абрамович Н. И. 566, 567, 569,
579, 599, 600, 601
Бутарин Н. С. 417, 441, 517, 518, 520,
540, 542, 544
Бутурлин С. А. 235, 236, 239, 240, 242,
247, 251—253, 255, 257, 259, 261, 545,
556—558, 561
Бэр К. 578
Вальтер А. 38, 413
Варенцов П. 33
Басенко Е. П. 308, 346, 349, 362, 363,
365—369, 371, 372
Васильев Я. 384, 392
Васильковский П. Е. 568, 593
Вассоевич Н. 599, 600
Вебер М. 102
Вендланд О. В. 183, 191
Верещагин Н. К. 73, 83, 95, 146, 155,
159, 205, 212, 236, 245, 267, 276, 308,
313—315, 320, 322, 323, 326, 385,
391, 392, 404, 408, 410, 455, 456,
462—474, 517, 522, 599, 601
Ветуляни Т. 45, 50
Виженер Б. 601, 612
Вильдермет Г. 266
Виноградов Б. С. 21, 73, 145, 266, 416,
417, 425, 442, 475, 498 517, 545
Витович В. В. 308, 314
Волков М. Г. 554
Волкова И. П. 273
Волкопялов Б. 93
Воробьев К. А. 346, 367
Воршев В. Н. 347, 373
Врангель Ф. 266, 291, 292
Вржесниовский А. 602
Врублевский К. 72, 95, 145, 164, 176,
180, 204, 210, 222, 223, 232, 568,
578, 584
Вялов О. С. 306
Гааке В. 72
Гавеман А. В. 236, 249
Гаврилов В. Т. 545, 557—559, 563, 564
Гагенбек 60
Ганасинский Р. 598
Тассовский Г. Н. 73, 86, 122, 129, 145,
204, 215, 266, 278, 545, 552
Геблер Ф. 427, 528
Гекк Г. 617
Гекк Л. 617
Гептнер В. Г. 34, 43, 45, 51, 52, 123,
205, 219, 267, 282, 346, 372, 385, 392,
404, 408, 410—414, 443, 455, 466, 469,
472, 474, 475, 498, 510, 512, 514, 516
Герберштейн С. 346, 352, 601—605, 612,
615
Герн Б. Ф. 416, 427
Геснер К. 603, 605, 606, 612, 616
Гладков Н. А. 346, 358
Глич Л. 347, 354, 363
Гмелин И. 359, 528
Гмелин С. 45, 50, 51
Годнев Е. Д. 236, 245, 260
Головнин Д. Н. 416, 516, 544
Горбачев С. Н. 234, 244, 256
Горбунов Е. И. 267, 288
Граве Г. Л. 73, 146
Граменицкий М. И. 183
Гребельский С. Г. 267
Гребень Л. К. 482, 494
Гребенюк Р. В. 542
Греве К. 182, 353
Григорьев Н. 266, 276
Григорьева Т. А. 183
Григорян Г. А. 404, 415, 498, 515
Гримм О. 266
Громов В. И. 73, 78, 235, 267, 272, 308,
313, 346, 352, 353, 417, 424, 482, 483,
488, 517, 522, 524, 599, 600
Громов И. М. 236, 240, 308, 317, 404, 411,
443, 448, 449, 498, 506, 507, 510—514
Громова В. И. 26, 27, 31, 33—35, 45, 47,
48, 52, 53, 55, 56, 104, 109, 346, 354,
400—402, 417,'424, 443, 448, 455, 475,
477, 479, 480, 482, 488, 517, 522,
524, 531, 545, 550, 566—568, 574, 575,
599—601, 604, 608, 611, 613, 614
Грум-Гржимайло Г. Е. 52—54, 58, 59,
61—63, 103, 110, ИЗ
Грум-Гржимайло М. Е. 52—54, 58, 59,
61—63, 110
Грунин К. Я. 44, 62, 123, 135, 170, 182,
230, 258, 308, 309, 327, 331, 340, 346,
358, 370, 384, 440, 455, 473, 498, 515,
542
Гуреев А. А. 152, 416, 424, 443, 448,
498, 507
Гусев В. М. 347
Гутникова 3. И. 184
Гюльмсирян Н. Г. 346, 369
Дагва Э. 104, 105, 114
Даль С. К. 34, 36, 145, 162, ю4, 166, 169,
347, 361, 371, 393, 404, 405, 408, 410,
413—415, 498, 504, 509, 510, 599, 601
Данилов Д. Н. 20, 73, 80, 235, 241, 242,
244—247
Данилов Н. Н. 235
Данилович А. П. 73, 82
Даршкевич Я. 73, 83, 146, 156, 183, 235,
245
Двойченко Г. 184, 202
Демель К. 594
Дементьев Г. П. 73, 145, 159, 167, 168,
171, 308, 317, 318, 320, 322, 323, 325,
329, 331, 346, 404, 417, 426, 498, 506
Дементьев Д. П. 518, 527
Демин В. С. 417, 422
Денисов В. Ф. 416, 440, 441, 517, 518, 542
Деревягин П. Я. 308, 321
Джанашвили А. Г. 455, 474
632
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
Динник Н. Я. 20, 72, 74, 76, 83, 87, 89—
91, 93, 94, 145, 162, 167, 170, 204,
212, 220, 221, 224, 228, 230, 231, 233,
308, 311, 313, 314, 346, 354, 374, 384—
386, 390—399, 404, 406, 408, 410, 412—
414, 416, 455, 462, 464—466, 468, 470—
475, 497, 568, 579, 582, 584—587, 590
Дмитриев В. В. 3, 20, 145, 157, 165, 204,
214, 219, 220, 222—224, 229, 230, 232,
235, 246, 248, 250, 251, 267, 282, 286,
416, 427, 428, 431—433, 435—439
Добротворский М. И. 266, 273, 276
Доброхотов В. И. 346, 354
Догель В. А. 568, 597
Долгих И. 601, 602, 606, 608—612, 614,
615
Долгушин И. А. 169
Долгушин Л. Д. 236, 242, 250
Донауров С. С. 3, 73, 74, 90, 92—96
Доппельмаир Г. Г. 122, 128, 132, 134,
204, 215, 266
Доржин Э. Д. 53, 60, 61
Дорогостайский В. Ч. 183, 201, 202, 416,
429, 516, 528, 529, 534, 536, 538, 539,
545, 553, 557, 559—562, 564
Дорофеев В. 159
Доценко В. 126
Друри И. В. 267, 273, 276, 277, 281, 282,
291, 294, 554
Друри С. И. 160
Дубинин В. Б. 74, 95, 308, 317, 417, 425,
498 „ 507
Дулькейт Г. 545, 556
Егоров Н. Н. 236, 246
Егоров' О. В. 154, 158, 417, 422, 426,
431—441
Елпатьевский В. С. 146, 156
Емельянов А. А. 122, 136, 204, 266, 278
Емельянов М. А. 73
Ендржеевский А. 455
Ергольский К. Р. 236
Ерещенко В. И. 349
Жабинский Я. 569, 598
Жарков И. В. 20, 205, 212, 236, 252
Железняков Д. Ф. 443, 448, 450—452
Жильберт 588
Житков Б. М. 176, 180, 204
Заблоцкий М. А. 3, 5, 568, 569, 578, 580,
584, 585, 588—594, 598
Завадовский М. М. 176, 182, 204, 228
Зайцев С. В. 205, 234
Заленский В. 53
Залесский П. М. 20, 73, 85, 235, 427, 428
Залетаев В. 74, 83
Зверев М. Д. 308, 324, 328, 346, 359,
366, 367, 417, 432, 433, 517, 526, 535,
537
Золотарев Н. Т. 73, 90, 122, 132, 135,
145,160, 266, 275, 278, 380, 545, 556
Зубарева В. И. 568, 576, 601, 612, см.
также Бибикова В. И.
Зубаровский М. 266, 273
Иваненко Б. И. 146, 172
Иванов А. И. 122, 129, 145
Иванов В. В. 347, 373
Иванов Д. Л. 416, 516, 528, 544
Иванов И. И. 568, 592
Иванов М. Ф. 482, 483, 494
Ильина Г. И. 184, 202, 203
Иностранцев А. 601, 611
Иохельсон В. И. 544, 556
Исаева А. Я. 417, 427, 517, 526
Исаков ГО. А. 73, 83, 95, 267, 273
Исмагилов М. Н. 309
Ишунин Г. И. 33, 308, 309, 326, 328
Каверзнев В. Н. 73, 96, 97, 308, 331,
346, 375, 385.
Каденаций А. Н. 376, 383, 482, 497
Казаченко Б. 236, 245
Казнаков А. Н. 53, 59
Казневский П. Ф. 205, 212
Кайзер Г. А. 346, 356
Калниньш А. И. 73, 78, 92, 146, 205,
235
Капланов Л. Г. 73, 80, 145, 205, 216, 221,
222, 224, 225, 229, 231, 235, 236,
247_____259
Караев А. И. 545, 554
Карачаровский В. В. 346, 356
Кардымович Э. И. 475
Карелин Г. С. 33, 36, 214, 505
Карцов Г. П. 568—571, 573, 574, 577,
587, 590, 592, 593
Кашкаров Д. Н. 426, 449, 526
Кащенко Н. Ф. 53, 60, 122, 126
Келейников А. А. 417, 432, 433, 517, 535
Кемберленд 106
Кеппен Ф. П. 45, 51
Кесслер К. Ф. 72, 82
Кириков С. В. 20, 45, 50, 74, 78, 83, 146,
156, 169, 171, 176, 180, 212, 236, 244,
245, 267, 274, 276, 347, 353, 569, 576
Клеменц Д. 53, 59, 61—63
Клюшкин Е. А. 34, 44, 205, 213, 222, 223
Кнорре Е. П. 235, 236, 250, 251, 253—
257, 259, 261—263
Ковалевский В. О. 11, 21, 23, 115, 295,
297
Коварцик Р. 490
Кожевников Г. А. 568, 593, 597
Козлов В. В. 183, 184, 202
Козлов П. К. 53, 62, 104, 106, 110, 112,
115, 156
Козлова-Пушкарева Е. В. 122, 132—134,
136
Колесник Н. Н. 601, 616
Колесников И. И. 74, 84, 308, 316, 329,
346, 356, 371
Колесников И. М. 346
Колосов А. М. 204, 214, 331, 334, 337,
338, 340, 341, 346, 359, 427, 428, 442,
517, 528, 533—535, 539, 541
Колюшев И. И. 267, 286, 287
Копылов И. П. 21, 73, 85, 123, 128, 132,
146, 159, 205, 215, 235, 254, 267, 278,
417, 429
Копырин С. 331, 342
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
Корнеев А. П. 21,74, 82, 146, 154, 236, 244
Коровин Е. П. 33, 308, 328
Костин В. П. 309, 316, 320, 321, 324—
326, 328
Котов В. А. 455, 473
Котовщикова М. А. 204, 212, 226, 230
Кравченко П. 53, 62
Крайнова Л. В. 569, 584—586, 594
Красовский В. П. 236, 245
Кромер 576
Круликовский Л. 266, 274
Круликовский П. 145, 156
Кузнецов А. 204
Кузнецов Б. А. 20, 73, 74, 82, 84, 146, 155,
158, 167, 176, 180, 205, 214, 231—
233, 266, 267, 278, 417, 426, 517, 526,
527, 532, 536
Кузнецов Н. 205
Кузякин А. П. 20
Куклин С. А. 145, 156, 235, 267, 274, 276
Кулагин Н. М. 21, 53, 73, 145, 235, 568,
569 589 591
Кульпин П. И. 309, 316, 321, 324, 347,
364, 365, 367, 371
Куражсковский Ю. Н. 74, 82, 205, 267,
273, 274, 283
Куроптев А. 266, 273
Кювье Ж. 99
Лавров А. 145, 154
Лавров Н. П. 145, 156—158, 162, 164, 170
Лавровский А. А. 346, 354, 361, 364, 369,
371 372
Ладыгин В. Ф. 104, 112, ИЗ, 115
Лаздин В. П. 507
Лайдекер Р. 188, 307, 319, 430, 492,
550, 594, 606, 614
Лайтнер О. 604
Лаптев И. П. 123, 128, 146, 157, 236, 245
Лаптев М. К. 204, 213, 404, 411, 414,
498, 512, 513, 515
Лебле Б. Б. 236, 242, 267, 273
Лев Д. Н. 417, 425
Леваневский М. 33
Левиев П. 417, 424
Ленский А. А. 184
Лентовский М. Н. 145, 158
Линг X. И. 74, 78
Литльдейл Г. 106
Лихачев А. И. 184, 201, 236, 257, 259
Лихачев Г. Н„ 235, 251
Лобачев С. В. 266, 274, 276
Логинов В. В. 73
Лодыженская В. И. 347
Лукашевич К. 602, 611, 612, 616
Лукашкин А. С. 331, 336, 337, 339—341
Лус Я. Я. 21, 475, 480
Лэкке В. 105, 106
Лэттимор О. 112
Любимов М. П. 184, 205, 234
Лялицкая С. 417, 429
Ляпунов И. 73, 80
Маак Р. 159, 183, 187, 516, 529, 552
Мадаминов С. Ю. 74, 96, 498, 507, 517,
527
Майдель Г. 266, 291, 544; 553, ,554
Макридин В. П. 236, 242
Максимов А. Н. 266, 292
Маленков И. К. 309, 317 ,
Марвин М. Я. 267, 273
Марков Е. Л. 73, 74, 96, 97, 146, 170,
498
Мартынов Н. М. 235,, 236, 245, 251, 252,.
256
Матюшкин 291
Мачульский С. Н. 235, 259, ,331, 341
Машковцев А. А. 267, 268, 517, 526,
430, 517, 526
Мейсель М. Н. 183
Мекленбурцев Р. Н. 417, 425, 431, 438,
439, 498, 507, 511—513, 516—518,
527, 528, 533, 536—544
Мельникова Т. Г. 497
Менард Г. А. 183
Мензбир М. А. 73, 145
Мертенс А. М. 602, 608
Мертц П. А. 205, 212, 222
Мессершмидт 529
Меховский М. 601, 612
Мигулин О. О. 73, 82
Миддендорф А. Ф. 204, 214, 266, 274,
276, 292, 550, 556
Микулин М. А. 417, 427, 517, 526
Мил Д. 145, 152
Мильков Ф. Н. 267, 276
Минервин В. Н. 308, 320, 322, 323
Миролюбов И. И. 183, 184, 186, 190, 192
Митюшев П. В. 183, 205, 226, 229, 230,
234
Михель Н. М. 505
Мишин И. П. 267, 278
Моор Э. 589, 590
Морщтин И. 614
Мордвилко А. К. 591
Мориц Л. 498, 512, 514, 515
Моррисон-Скотт Т. 307, 319, 430
Му канте 612
Мурзаев Э. М. 34, 104, 110,^112,5114
Мэтью В. 99, 100
Назарова И. В. 236, 260
Наниев В. И. 569, 579
Нарышкин Д. К. 234, 252—254
Насимович А. А. 3, 13, 21, 73, 74, 80,
85, 86, 89, 123, 126, 146, 154, 156, 184,
204, 205, 222, 236, 246, 268, 376, 385,
391, 392, 394—396, 398, 399, 405, 417,
427, 443, 455, 462, 466, 468—473, 498,
518, 545
Насонов Н. В. 475, 478, 479, 482, 488—
490, 497, 498, 505, 507—509, 511, 513,
515—517, 524, 526, 527,529—531,541,
545, 553, 556, 557, 559—562, 564
Наумов Н. П. 122, 128, 235, ' 246, 266,
277, 281—284, 287—291, 545, 550,
557, 562
Наумов С. П. 122, 128, 134, 266, 276,
281, 287
Нейман К. К. 544, 553, 563
Неринг А. 604
Никитин В. Н. 100, 235, 244
634
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
Никитин К. Н. 73, 80, 146
Николаевский А. Д. 266, 267, 283, 287,
295
Никольский А. М. 122, 204, 231, 266,
278, 285
Никольский Г. В. 346, 358
Новиков В. К. 184, 205
Новиков Г. А. 73, 82, 86, 122, 129, 136,
145, 159, 204, 222, 234, 267, 278
Новиков И. И. 101, 103
Носилов К. 234
Огнев С. И. 308, 316, 404, 414, 417, 427,
498, 510, 512, 514, 516, 517, 525, 545,
556
Орлов Е. И. 346, 356
Осадчий П. 73, 205
Осокорев С. 236, 244
Остророг 614
Паавер К. 74, 78
Павленко С. М. 183
Павлинин В. Н. 235
Павлов Е. И. 123, 133, 267, 278, 331,
336, 340, 545, 552, 560
Павловский Е. Н. 21, 73, 145, 416, 442,
475, 498, 517
Паллас П. С. 50, 56, 132, 145, 155, 276,
307, 334, 340, 345, 356—358, 366, 516,
528, 529, 602
Паровщиков В. Я. 482
Пастернак Ф. А. 146, 160, 164
Пачоский И. 346, 350
Певцов М. В. 53, 58, 60, 103, 105, 527
Петров О. В. 417, 426, 517, 527
Пидопличко И. Г. 73, 78, 236, 240, 244,
267, 272, 346, 352, 400, 402, 404, 475,
477, 517, 522, 569, 576, 601, 612
Плечев Е. И. 545, 556, 559, 562, 563
Плигинский В. Г. 346, 353
Подаревский В, В. 73, 86, 122, 128, 129,
132, 145, 159, 160, 204, 215, 267, 278,
331, 336, 416, 429, 545, 550, 552
Подсосов Л. А. 235, 256
Поляков И. С. 52, 54, 105, 106, 544, 552
Попов В. А. 146, 156, 236, 260, 357
Попов С. П. 235, 253, 261—263
Портенко Л. А. 235, 247, 250, 267, 277,
545, 553, 554, 561, 568, 594
Поршняков С. Н. 80, 152, 274
Потанин Г. 58
Потоцкий Я. 104
Похилевич Л. 234, 244
Пржевальский Н. М. 33, 52, 53, 58, 60,
86, 103, 105, 106, 110, 112—114, 183,
191, 331, 336, 338—341, 426
Птушенко Е. С. 73, 82, 145, 155, 235, 244
Пузанов И. И. 235
Пузарг Е. 104
Пуш 602
Пфиценмайер Е. В. 404, 415, 455, 464
Пхакадзе Г. М. 101, 102
Радде Г. 313, 337, 413, 529
Раевский В. 73, 80, 145
Разевиг В. А. 145, 455
Разоренова А. П. 417, 442
Райский А. П. 146, 156
Ралль Ю. М. 146, 155, 347, 354
Регель 449
Реутт Н. 455
Риттер К. 112
Роборовский В. И. 53, 104—106, 110,
113, 115
Розанов М. П. 33, 39, 425, 426, 498, 506,
517, 528, 568, 593
Российский Н. Д. 597
Роуланд Вард 444, 457
Рудинский К. 482, 491
Рузский М. 568
Румянцев Б. Ф. 45, 52, 53, 57, 517. 518,
539, 540, 543
Румянцев Н. В. 33, 39
Рустамов А. К. 308, 317, 498, 505
Рухлядев Д. П. 21, 74, 95, 145, 170, 204,
230, 482, 483, 497, 569, 591
Рыжиков К. М. 123, 135, 146, 170
Рычков П. И. 45, 50
Рябова Т. И. 183, 191
Рященко Л. П. 183, 184, 204
Сабанеев Л. П. 145, 152, 154, 156, 164,
165, 172, 204, 212, 266, 274
Саблина Т. В. 21, 74, 95, 146, 176, 180,
205, .236
Саверкин Н. П. 183
Салганский А. А. 184, 203
Салихбаев X. 308, 317
Самородов А. В. 74, 83, 96, 267, 277,
545, 554, 559, 563
Саркисов А. А. 183, 306, 385, 392, 393,
404, 405, 414, 416, 455, 483, 493, 498,
504, 508, 510—517, 543
Сартабаев С. К. 542
Сатунин К. А. 3, 21, 122, 124, 133, 135,
137, 145, 156, 168, 176, 180—183, 204,
212, 226, 232—234, 252, 255, 257, 259,
266, 274, 276, 295, 308, 313, 346, 375,
384, 404, 414, 416, 427, 437, 441, 455,
465, 474, 497, 516, 544, 545, 568, 587,
590, 601, 602, 609
Свенцицкий 612
Свицкий 615
Сдобников В. М. 267, 272, 281, 288, 289,
291 550
Северцов Н. А. 204, 214, 220, 234, 475,
478—480, 516, 517, 520, 525—528, 531—
535, 544
Северцов С. А. 45, 137, 145, 152, 235,
295, 416, 569
Седельников А. 158
Селевин В. А. 33, 36, 38, 73, 85, 416,
427, 517, 524
Семенов-Тян-Шанский О. И, 235, 242,
249, 251—253, 256—258, 263, 267,
273, 281—289
Сергеев А. М. 266, 273
Сердюкова Г. В. 152, 258, 327, 399, 415,
440 М5 ^49
Сержанин Й. Н.’ 74, 80, 176, 181, 235,
242 251
Сиверцов И. И. 183
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
635
Сидоров М. К. 544, 550
Симпсон Г. 10, 98—100
Симуков А. Д. 104, 105, 112, 114, 115
Скалой В. Н. 123, 129, 130, 132, 145,
146, 157, 159, 172, 204, 205, 215, 235,
246, 247, 267, 284, 331, 416,428, 438,
439, 545, 550—552, 556, 561
Скворцов В. П. 73, 83, 404, 411, 417,
424, 498, 506
Скотт В. 99, 100
Словцов И. 33, 38, 601, 611
Слудский А. А. 21, 33, 34, 38, 43, 74, 146,
171, 236, 241, 246, 259, 308, 309, 325,
326, 328, 331, 340, 346, 347, 357, 359,
361, 362, 364, 365, 368, 370—372,517,
524, 532, 535
Смирнов Н. А. 21, 73, 95, 122, 129, 145,
204, 231, 267, 545
Смит Г. 603, 604
Соколов И. И. 3, 13, 21, 146, 168, 170,
204, 224, 230, 233, 265—268, 270, 273,
280, 287, 289, 293, 295, 297, 299, 306,
308, 320, 343, 347, 358, 374—376,
385, 400, 475, 566, 567, 569, 588, 599
Соколов С. С. 346, 359, 361, 364, 366,
370, 517, 525, 532, 536, 538
Соколова И. В. 308, 327, 346, 371, 417,
440, 517, 542
Сокольников Н. П. 545, 553, 560, 562—564
Сокур И. Т. 74, 82, 205, 210, 221
Соловьев Д. К. 122, 204, 215, 232, 234,
255, 266, 268, 287, 416, 429, 431, 439,
568, 593
Станюкович К. В. 308, 322
(jTRTTHflTC 33
Страутман ф. И. 73, 96, 322, 325
Строганов С. У. 73, 80, 145, 152, 154
Строганова А. С. 74, 83, 235, 245, 347,
356, 363, 369
Судариков В. Е. 123, 135, 146, 170
Султанов Г. С. 308, 317, 443, 448, 450—
454, 498, 507
Суров Л. 602
Сушкин П. П. 541
Сцалай А. 605, 606
Сысоев В. П. 73, 74, 80, 123, 130, 133,
136, 146, 160, 165, 205, 216, 236, 247, 260
Таежник 375, 378, 384
Татаринов К. А. 73, 96
Тейлар де Шардэн П. 109
Теплов В. П. 3, 21, 73, 74, 90, 92—96,
235, 255, 256, 258
Тесдорпф О. 491—493
Тимофеев В. В. 146, 171, 417
Тихомиров А. А. 53, 54
Тихонов 53
Толмачев И. П. 545, 553
Толстов С. М. 234, 246, 545, 550
Толстой А. 346, 358
Толстой Н. Н. 73, 97, 204, 233
Томилин А. Г. 146, 156, 267, 274
Топачевский В. А. 236, 240, 244, 267, 272
Троицкий В. П. 266
Троицкий В. Н. 278, 416, 428
Трубецкой П. С. 442, 449—452
Тугаринов А. Я. 122, 129, 145
Туркевич В. Г. 346
Туркин Н. В. 3, 21, 122, 124, 133, 135,
137, 145, 156, 168, 176, 180—183, 204,
212, 226, 232—234, 252, 255, 257, 259,
266, 274, 276, 295, 346, 375, 384, 404,
414, 416, 427, 437, 441, 455, 474, 497,
516, 544, 545, 568, 587, 590
Туров И. С. 236
Туров С- С. 145, 154, 235, 244, 455, 471,
474, 545, 553, 559, 561—563
Тюрин П. С. 417, 426, 518, 527
Тэви А. С. 183, 204, 231
Усов С. А. 568, 578, 582, 584, 587, 588,
590
Успенский Г. А. 176, 181
Ушков С. Л. 183, 202
Фаворский В. П. 545, 552
Феддерс В. В. 568, 591
Федорович ф. ф. 145, 155, 234, 244
Федосов А. В. 73, 80, 146, 235, 244
Федченко А. П. 497, 507
Федюшин А. В. 73, 145
Фетисов А. С. 145, 146, 157, 164—166,
170—172, 267, 278, 331, 336
Филатов Д. П. 568, 580
Филипченко Ю. А. 72, 73, 87, 568, 592
Флауэр С. 398
Флеров К. К. 3, 4, 21, 27, 33, 39, 73,
83, 84, 87, 121—123, 125, 128, 131, 133,
135, 137, 138, 141, 143—146, 151, 158,
161, 168, 174, 176, 180, 184, 186, 188,
204, 205, 208, 214, 218, 219, 222, 225,
226, 230, 235, 236, 240, 247, 263, 265—
267, 272, 280, 306, 308, 317, 318, 326,
404, 411, 412, 414, 416, 424, 430, 442,
443, 449, 475, 480, 498, 506, 507, 510—
514, 517, 527, 545, 550, 556, 568, 581
Фоканов В. А. 74, 84
Формозов А. Н. 13, 21, 73, 78, 146, 152,
154, 156, 205, 209, 210, 213, 214, 235,
267, 276, 283, 284, 331, 346, 372, 373,
385, 392, 404, 408, 410, 413, 455, 466,
469, 472, 474, 475
Формозова Л. Н. 267, 283
Фортунатов В. К. 204, 482, 568, 592, 593
Фрик Ч. 100
Хавесон Я. И. 5, 98—104, 106, 108—
НО, 112
Харпер Ф. 59
Хахлов В. А. 308, 316
Хедин С. 105, 106, 110, 112, 113
Херре В. 608, 617
Хильцеймер М. 492
Хоменко И. П. 346, 352
Храневич В. 73, 235, 244
Хрусцелевский В. Н. 331, 336
Цагарев П. 205, 214
Цалкин В. И. 34, 74, 82, 122, 123, 125,
205, 210, 219, 225—227, 229, 346, 353,
404,-417, 421, 424, 426, 427, 430, 437,
443, 445, 448, 449, 455, 464, 475, 479,
636
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
480, 483, 484, 498, 499, 506, 508—
515, 517, 518, 524, 527—531, 545, 546,
548, 552, 554, 556, 557, 569, 576, 601,
612
Цедтвич Ф. 398
Черкасов А. А. 123, 132—134, 136, 205,
231, 235, 254, 331
Чернавин В. В. 517, 526, 532, 537—539,
541, 543
Черский И. Д. 45, 52, 204, 209, 346, 357
Читян С. А. 347
Чуди 384, 396, 399, 400
Шапошников Л. В. 235, 443, 453, 454
Шапошников Ф. Д. 21, 205, 221, 223,
267, 268, 277, 518, 535
Шарлемань Н. В. 34, 36, 205, 210, 236, 244
Шатерникова Т. М. 33, 39
Шатилов И. Н, 45, 51
Шварц С. С. 235, 246
Шерешевский Э. И. 455, 464, 482. 485,
491, 492, 494—496
Шиллингер Ф. Ф. 73, 76, 145, 176, 180—
183, 204, 206
Шильдер В. 455, 464, 474
Шкургин А. В. 34
Шмидт Ф. 550, 558
Шнакенбург П. В. 545, 554
Шнаревич И. Д. 417, 427, 435, 437, 438,
440
Шнеебергер 614—616
Шнитников В. Н. 33, 38, 73, 84, 145,
170, 308, 316, 324, 328, 346, 359, 374,
416, 427, 431, 436, 439, 517, 527
Шренк Л. 556
Штегман В. К. 73, 91, 94, 146, 162, 166,
169, 170
Штольцман Я. 597
Шульпин Л. А. 417, 436, 437, 517, 525'
Шульц Р. С. 376, 383
Шухов И. Н. 145, 158, 204, 266, 276
Щеголев В. Н. 266, 274
Щекин В. А. 34
Щербаков А. Н. 123
Щербаков И. 123, 145, 235, 266
Эванс Г. 381—383
Эверсманн Э. А. 33, 45, 46, 50, 266, 276,
308, 316, 346, 357, 361, 369
Эверт В. К. 308, 328
Эквтимишвили 3. С. 385, 392, 395, 405,
414, 455, 466—468, 472
Эккерт Н. И. 568, 591
Эллерманн И. 307, 319, 430
Эндрюс Р. 338, 340
Эрманн А. 556
Юдин К. А. 215
Юнатов А. А. 60, 61, 104, 112
Юргенсон П. В. 122, 123, 126, 132, 145,
163
Яблонский Н. И. 516, 528
Яковлев Е. 122, 128, 145, 158, 235, 245,
246, 545, 550
Янушевич А. И. 74, 85, 123, 127, 146,
157, 205, 214, 215, 236, 246, 267, 277,
331, 334, 417, 517, 528
Янушко П. А. 205, 219, 221, 222, 226,
228, 232, 483, 491, 492, 494—497
Яроцкий 610
Adametz L. 400, 402, 599, 600
Adlerberg G. 374, 375
Allen G. 33, 43, 53, 58, 104, 121, 123,
308, 319, 331—333, 336, 375, 381,
383
Allen J. 544, 556, 566, 568
Altmann M. 236
Amon R. 71, 73, 78, 87
Andrews R. 33, 331, 338, 340
Antonins H. 204
Antonins O. 21, 27, 45, 53, 56, 60, 443, 454
Baer G. 568, 578
Beaux O. 400
Belke G. 72, 82
Beninde I. 204
Bennett 218
Benson B. 295, 296
Berger K. 146
Blainville M. 305, 565, 566, 567
Blanford W. T. 103, 105, 218, 404
Blasius I. 204, 226
Bley F. 234, 260
Blyth E. 455, 475, 497, 508, 509, 529, 530
Bogdaschew N. 100, 101
Bohlin B. 306, 331, 334, 401, 475, 477,
517 522
Bojanus L. 599, 600, 601, 602
Bolkay St. 385
Bonaparte Ch. 385
Bouchud I. 267
Boule M. 101, 102, 104, 109, 308, 313,
384, 390
Bourdelle E. 27, 53
Brandt F. 122, 234, 568, 601
Brehm 21, 23, 33, 45, 53, 69, 385, 399,
400, 404, 442, 443, 455, 475, 482, 490,
491, 492, 498
Bromee-Skuncke F. 236, 258, 267, 289
Brook C. 234, 261
Brooke V. 137, 305, 308, 331, 516
Bruhin H. 21
Buchner E. 122, 455
Burton M. 569
Butler R. 404
Buxton A. 176
Buxton E. 482
Cabrera A. 267, 385
Calinescu R. 568, 576
Carette E. 266, 272
Carter T. 22
Cenkier S. 569
Chardin P. 313, 331, 334; см. также
Teilhard de Chardin P.
Charlemagne 219
Cobb E. 385, 391, 443, 517
Colbert E. H. 69, 71, 101, 115
Cope E, 63
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
637
Couturier М. 385, 386, 390, 400
Cumberland 105, 106
Cuvier G. 218, 601
Danek Z. 385, 391
Danford C. 404
Dechambre E. 267, 293
Degerbol M. 341
Demel K. 568, 594
Demidoff E. 516, 544
Dombrowski R. 137, 145
Diierst J. 295, 296, 490, 601
Dyk V. 483
Eichwald 72, 80
Eidmann H. 204, 235
Ellermann J. R. 21, 71, 73, 146, 184,
205, 235, 267, 306, 307, 308, 319,
374, 375, 385, 400, 404, 417, 430, 443,
455, 464, 475, 478, 498, 517, 545
Erdbrink D. 236, 240
Ermann A. 544, 556
Erxleben F. 218, 404
Eschholz F. 122
Eschscholtz F. 544, 556
Evans G. 375, 381, 382, 383
Eversmann E. 509
Ewart I. C. 45, 497, 516, 543
Falconer H. 101, 566, 567, 599, 600
Falz-Fein F. 45, 51
Findley J. 235
Fitzinger L. I. 71
Flerow C. 266
Flower S. 21, 73, 94, 235, 258, 266, 455,
474, 482, 493, 498, 514
Flower W. 21, 122, 295, 308, 326, 375,
383, 385, 398, 404, 415, 430
Frechkop S. 22, 27, 63
Friant M. 22
Frick Ch. 346, 352
Frisch 566
Fuller W. 569
Ganasinski R. 569, 598
Garretson M. 566
Garrod A. 121, 122
Gesner C. 601, 603, 605, 612, 614—616
Girtanner H. 384, 399
Glitsch L. 346, 347, 354, 363
Glower R 569, 598
Gmelin I. 331, 345
Gmelin S. 497, 508
Granger W. 115, 118, 137
Gray A. 22, 74, 385, 400, 417, 441, 455,
483, 493, 498, 516, 517, 543, 565,
567, 569
Gray I. E. 99
Gregory W. K. 22
Greve K. 72, 78, 176, 180, 182, 204, 346,
352, 353, 357
Groeben G. 568, 597
Gromowa V. 475, 479—482, 484, 488,
498, 517, 531, 545, 546.
Giienter 508
Giildenstaedt A. 308, 455
Had wen S. 266
Hagenbeck 53
Harper F. 22, 33, 45, 53, 59, 346, 352,
443, 448, 483, 485, 545, 569, 576,
601, 610.
Harrison R. 176
Haughton S. 26
Heck H. 45, 601, 617
Heck L. 204, 482, 490, 492, 493, 601, 617
Hedin S. 104, 105, 113
Herberstain S. 601, 602, 603, 605, 612
Herre W. 267, 292, 475, 483, 488, 602,
603, 608, 617
Heude P. 308, 374, 375
Hilzheimer M. 53, 56, 400, 404, 415, 416,
442, 455, 568, 574, 601
Hodgson B. 122, 218, 305, 400
Hodgson I. 565, 567
Hollister N. 122, 266, 331, 336
Hornaday W. 266, 290
Hume 450, 509
Hutton Th. 442, 450, 497, 508
Hyett A. 176
liliger J. K. 98, 99
Ivrea 404
Jacobi A. 145, 234, 240, 265, 266, 286
Jaczewski Z. 146, 168, 205, 228
Janicowski T. 45
Jennison G. 601, 609
Jickeli C. 568, 576
Kaisen O. 566, 567
Kalela O. 569,. 575
Karpinski I. 235
Keibel 145
Keilbach K. 22
Keller F. 384, 385, 400
Kiesseling W. 204
Kinloch A. 442
Koch A. 482, 488
Koch W. 568, 569
Koller O. 385, 393
Kolster R. 266
Kostron K. 235, 258
Kowalewskie W. 21
Kowarzik R. 475, 490, 545, 557
Kretzoi M. 400, 569, 576
La Baume W. 599—601, 608
Lantis M. 267, 295
Lataste ML 305
Lattimore O. 104, 105, 112
Leche W. 101, 102, 104—106
Lederer G. 22
Lehman U. 601, 608
Leister C. W. 346
Leithner O. 599—601, 604, 605
Leizewitz W. 416
Lengerken H. 602, 608, 611
Lesbre F. X. 101, 102
Linke W. 204, 205
Linnaeus C. 101, 122, 145, 176, 204,
234, 266, 345, 384, 400, 516, 568, 581
638
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
Littledale G. 103, 106
Lonnberg Е. 266, 295, 297
Loomis F. 26, 27
Lorenz-Liburnau L. 404, 416
Lubicz-Niezabitowski E. 235, 240
Lukaszewicz K. 601, 610—612, 615—617
Lutschwager H. 569
Lydekker R. 21, 22, 23, 27, 33, 53, 69,
71, 72, 137, 145, 183, 188, 217, 234,
264—266, 295, 299, 306—308, 319,
331, 333, 346, 374, 375, 384, 400, 404,
406, 416, 424, 430, 442, 450, 455,
475, 482, 492, 497, 508, 509, 516,
530, 531, 544, 545, 550, 568, 576,
594, 599, 600, 601, 604, 606, 608,
609, 614.
Macewen W. 137
Major F. 71, 295, 296
Malsburg K. 599, 600
Mann C. 384, 399
Matheson C. 123, 135
Matschie P. 53, 60, 72, 455, 465, 545, 557
Matthew W. D. 26, 27, 63, 98, 99, 115,
118, 137
Maxim J. 400, 402
Meerwarth H. 385
Menzbier M. 455
Merk-Buchberg M. 385
Mertens A. 568, 576, 601, 602, '604, 606,
608, 610
Meyer F. 416
Meyer H. 599, 600
Middendorf A. 544, 550
Millais J. 475
Milne-Edwards A. 122, 218, 375
Mochi U. 22
Modell W. 137
Mohr E. 346, 352, 568, 569, 572, 578,
584, 586—590, 592, 597
Momberg H. 74, 95
Morden W. 517
Morrison-Scott T. C. 21, 71, 73, 146, 184,
205, 235, 267, 306, 307, 308, 319, 374,
375, 385, 400, 417, 430, 443, 455, 464,
475, 478, 498, 517, 545
Morrison W. 234
Muller S. 71
Miiller-Liebenswalde L. 145
Murie J. 346
Murie O. 205, 265, 266, 282
Murray A. 266
Nasonov N. 490, 497, 508, 509, 530, 557
Nehring A. 101, 346, 482, 488, 601, 604,
610
Neumann O. 384
Neumann L. 545, 563
Newton E. 400, 402
Noack Th. 53, 416, 601
Noback Ch. 137
Nordmann 308, 314
Ogilby 216
Oloff H. 73
Omsted A. 235
Osborn H. 26
Osternburg W. 235
Owen R. 568, 599, 600
Pallas P. S. 33, 45, 122, 132, 331, 334,
340, 482, 601, 602
Palmer L. 266
Pearson H. 69
Pen H. 517
Peters 530
Peterson R. 235, 236
Picard P. 602
Pilgrim G. E. 69, 71, 115, 295, 297, 306
Piro A. 104, 108
Piveteau R. 331, 334, 475, 477, 517, 522,
566, 567
Pocock R. I. 69, 115, 122, 125, 137, 145,
150, 234, 253, 266, 295, 297, 306, 331,
333, 374, 375, 385, 388, 545, 549
Pomel A. 101, 599
Pommerol F. 482, 488
Potocki I. 103, 104
Pousargues E. 104
Przevalskij N. 529
Pusch G. 568, 576
Radde G. 33, 36, 38, 72, 308, 317, 331,
337, 375, 404, 413, 455, 516, 529
Reymond M. 53
Reynolds S. 346, 352
Riney Th. 176
Ritter C. 103, 112
Robl H. 74
Roc F. 566, 567
Romer A. S. 22, 63, 98, 99, 137
Rouiller C. 455
Rowland Ward 416, 418, 442, 444, 455,
457, 516, 518
Riitimeier L. 295
Sabanejew L. 475
Sakurai T. 145, 169
Salesski P. 122, 126
Salmi M. 569, 575
Satunin K. 455, 581
Schaeffer B. 63
Schaub S. 374, 375, 475, 477, 483, 488,
517, 522
Schertz E. 569
Sehin z H. 400
Schlegel H. 71
Schlosser M. 115, 118
Schmidt F. 544, 550
Schneider K. 482, 492
Schrenk L. 375
Sebroter H. 72
Schulz H. 235
Schwarz E. 33, 295, 306, 400
Sclater Ph. 22, 137, 295, 308, 331, 346. 400
Scott W. B. 98, 99, 100
Selve I. 26
Seton E. 265, 267, 282
Severtzov N. 217, 219, 530, 531, 557
Shitkow B. 475
Simpson G. 10, 22, 23, 63, 98, 99, 100
115, 118, 295
АЛФАВИТНЫЙ УКАЗАТЕЛЬ АВТОРОВ
в39
Skinner М. 566, 567
Skuncke F. 235
Smith Н. 272, 565, 567, 575, 601,
603—605
Soffel К. 346
Spottel W. 53
Stehlin H. G. 69
Stirton R. 26, 27
Stive H. 146, 169
Strahl H. 204
Suschkin P. 545
Swanson C. 205
Szalay A. 601, 605, 606
Szaflarski I. 74, 95
Tandler I. 266
Tegner H. 146
Teilhard de Chardin P. 101, 102, 104, 109,
308, 475, 477, 517, 522, 566, 567
Temminck 183
Tesdorpf O. 482, 490—493
Thomas O. 22, 72, 290, 306, 308, 331, 346
Thomas P. 599, 600
Thorington J. 385, 399
Tong E. 53
Toula F. 482, 488
Trassaert M. 475, 477
Vetulani T. 45, 50
Vigener B. 601, 605
Vittoz R. 22
Vogt T. 205
Volborth A. 568
Wagner I. 442, 450, 565, 566
Waldo C. 137
Walter A. 33, 38, 72, 308, 317, 404, 413
Weber M. 22, 23, 63, 69, 101, 102, 295
Websterg G. 568, 569
Wettstein O. 146
Whitehead G. 385
Wilscki B. 137
Wimans W. 176, 204
Wolf B. 404
Wollebaeck A. 265, 266
Wood H. 23
Wortman I. L. 99
Wrzesniowski A. 601, 602, 611
Yarham E. R. 385
Young A. 234
Zabinski J. 569, 598
Zakin V. 450
Zedtwitz F. 385, 398
Zittel K. A. 99, 100, 295
Zukowsky L. 234
СОДЕРЖАНИЕ
Предисловие ........................................................... .3
Систематический указатель видов копытных фауны СССР .................. 6
Введение ................................................................ 9
Характеристика надотряда копытных..................................... 9
Череп копытных .................................................. 13
Строение черепа .......................................... . . 13
Измерения черепа ................................................ 17
Зубные формулы .................................................. 17
Измерение тела животных ................................................ 19
Основная литература по копытным......................................... 20
Систематическая часть ........................................... , . . 23
Таблица для определения отрядов копытных млекопитающих.................. 23
Отряд Perissodactyla — Непарнопалые .................................... 23
Семейство Equidae — Лошадиные ................................. 26
Род Equus Linnaeus — Лошади............................... 27
Отряд Artiodactyla — Парнопалые......................................... 63
Подотряд Nonruminantia (= Suiformes) — Нежвачные.................. 67
Семейство Suidae — Свиные ..................................... 69
Род Sus Linnaeus — Свиньи ................................ 71
Подотряд Tylopoda — Мозоленогие .................................. 98
Семейство Camelidae — Верблюдовые ............................. 99
Подсемейство Camelinae . .................................. 100
Род Camelus Linnaeus — Верблюды.......................... 101
Подотряд Buminantia — Жвачные ................................... 115
Семейство Moschidae — Кабарговые.............................. 120
Род Moschus Linnaeus — Кабарги........................... 121
Семейство Cervidae — Олени ................................... 137
Подсемейство Cervinae — Настоящие олени ................... 143
Род Capreolus Gray — Косули ............................. 144
Род Cervus Linnaeus — Настоящие олени ................... 173
Род Alces Gray •— Лоси .................................. 234
Подсемейство Neocervinae — Американские олени.............. 263
Род Rangifer Н. Smith — Северные олени................... 265
Семейство Bovidae — Полорогие ................................ 295
Подсемейство Gazellinae — Газелевые ....................... 304
Род Gazella Blainville — Газели.......................... 305
Подсемейство Caprinae — Козлообразные ..................... 342
Род Saiga Gray — Сайга................................... 345
Род Nemorhaedus Flower et Garson — Горалы................ 374
Род Rupicapra Blainville — Серны ........................ 384
Род Capra. Linnaeus — Козлы ............................. 400
Род Ovis Linnaeus — Бараны............................... 475
Подсемейство Bovinae — Выковые ............................ 564
Род Bison Н. Smith — Бизоны.............................. 566
Род Bos Linnaeus — Быки, или туры ....................... 599
Алфавитный указатель латинских названий животных....................... 618
Алфавитный указатель русских названий животных......................... 625
Алфавитный указатель авторов .......................................... 630